/








































































































































































































































































































Text
АКАДЕМИЯ НАУК СССР
фауна
ссср
ЧЕШУЕКРЫЛЫЕ
том
хп
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА * ЛЕНИНГРАД
ЗООЛОГИЧЕСКИЙ И НСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ № 42
ФАУНА СССР
Главный редактор акад. Е. Н. ПАВЛОВСКИЙ
Редактор А. А. ШТАКЕЛЬБЕРГ
НАСЕКОМЫЕ ЧЕШУЕКРЫЛЫЕ
Том XII
И. В. КОЖАНЧИКОВ
ВОЛНЯНКИ (OR.GYIDAE)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА . 1950 . ЛЕНИНГРАД
Редактор издания А, А. Штакельберг
ПРЕДИСЛОВИЕ
Волнянки Orgyidae (=Liparidae, Lymantriidae) представляют группу
чешуекрылых, исключительно интересную в теоретическом отношении.
Вместе с тем особенности биологии этих видов определяют высокую зна-
чимость их для сельского хозяйства и в экономии природы в целом. Среди
волнянок, есть ряд вредителей лесов и лесных полезащитных посадок,
плодово-ягодных культур, причем некоторые из них являются массо-
выми видами, причиняющими периодически весьма существенный ущерб
народному хозяйству.
Для теоретических работ по систематике чешуекрылых волнянки
использованы слабо; в большей мере виды этого семейства затронуты
исследованиями по генетике, биологии и экологии. Слабая степень иссле-
дованности систематики семейства привела к тому, что сколько-нибудь
правильное понимание положения волнянок в системе отряда чешуекры-
лых установилось лишь в конце прошлого столетия, но и до сих пор оно
оценено лишь в специальной литературе. В сводках систематическое
положение сем. Orgyidae не обосновано. Состав семейства, по разным
авторам, очень различен, а классификация внутри семейства и ныне
еще совершенно не разработана.
В предлагаемом исследовании дается более обширное, но вполне со-
гласующееся с современными воззрениями представление о филогенетиче-
ских связях сем. Orgyidae в отряде чешуекрылых, а система родов и групп
внутри семейства приводится, в сущности говоря, впервые. Она, конечно,
не может расцениваться как не подлежащая изменениям, но, так как
ц основу ее, помимо внешней морфологии, положены также особенности
анатомического строения, биоэколотии и географического распростране-
ния Orgyidae, это позволяет надеяться на правильную оценку эволюции
этих насекомых.
В общей части дан ряд теоретических положений по биологии, эколо-
гии и географическому распространению волнянок, имеющих общее
значение. Положения эти разработаны на основе многих личных исследова-
ний, как наблюдательных, так и экспериментальных. Эксперименталь-
ными исследованиями, помимо многих вредных видов, затронуты также
некоторые относительно редкие на территории СССР Orgyidae, причем
ряд видов изучен подробно. Многие виды были присланы мне живыми
моим покойным отцом из южной Сибири (Минусинск). Обширная спе-
циальная литература по биологии волнянок, в частности касающаяся
вредных видов, использована с возможной полнотой; ряд положений
проверен экспериментально и новыми наблюдениями в природе. В связи
с этим внесено много дополнений в данные по биологии даже обычных
вредных видов, а также исправлен ряд ошибочных представлений, сло-
жившихся в литературе.
4
ПРЕДИСЛОВИЕ
Вопросы распространения волнянок разработаны с возможной полно-
той. Новые, добытые для волнянок факты послужили для дальнейшего
развития ряда теоретических положений по биогеографии и экологии,
высказанных мною в работе по фауне Agrotinae СССР (1937) и в ряде спе-
циальных работ (1935—1946).
Мною использованы, помимо коллекций палеарктических видов Orgyi-
dae, хранящихся в Зоологическом институте Академии Наук СССР и поло-
женных в основу этого исследования, также многие экзотические предста-
вители. Последние привлечены только для понимания филогенетических
отношений семейства и особенностей морфологии его представителей,
а также для построения системы внутри семейства. Филогенетические
отношения семейства и классификация видов в его пределах основаны
на специальном изучении морфологии, кроме имаго, также' личиночных
фаз. Некоторое число гусениц волнянок мне было передано А. М. Гера-
симовым. Все эти материалы как по имагинальной фазе, так и по личин-
кам были дополнены изучением коллекций Ленинградского университета,
Всесоюзного Института защиты растений, Высших курсов прикладной
зоологии и фитопаталогии, а также сборами частных лиц, в особен-
ности А. И. Куренцова, Н. Н. Филипьева и моего отца, В. Д. Кожан-
чикова. Всех упомянутых здесь лиц я' вспоминаю с глубокой благодар-
ностью.
Рисунки, карты, графики и схемы, исключая тотальные рисунки,
выполнены мною; последние как для имаго, так и для личиночных фаз
изготовлены с натуры художником Ф. И. Гуняевым. Заимствованные
рисунки оговорены.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
Сем Orgyidae
I. Подсем. Orgyinae
1. Род Olene Hb.
1. О. olga Obth. . ’...................................................... 225
2. О. fascelina L........................................................ 227
2. Gynaephora Hb.
1. G. selenitica Esp.................................................... 234
2. G. rossii Curt......................................................... 237
3. G. lugens Kozh........................................................ 241
4. G. pumila Stgr......................................................... 244
5. G. selenophora Stgr.................................................... 245
6. G. alpherakii Gr.-Gr............................................... 246
7. G. sincera, sp. nov. ............................................' . 248
3. Род Dasychira Steph.
1. D. lunulata Btl........................................................ 252
2. D. virginea Obth. . ................................................... 253
3. D. consanguis Wil...................................................... 254
4'. D. pseudabietis Btl................................................... .256
5. D. albodentata Brem.................................................... 257
6, D. abietis Schiff...................................................... 259
7. D. pudibunda L. . ................................................... 261
7a. D. pudibunda modesta Kirby............................................ 265
4. Род Laelia Steph.
1. L. coenosa Hb. .................................................... 267
5. Род Orgyia Ochs.
1. 0. thyellina Ochs..................................................... 276
2. O. antiqua L........................................................... 278
3. 0. gonostigma P....................................................... 281
4. 0. dubia Tausch....................................................... 285
5. 0. ericae Germ. ..................................................... 287
6. 0. trigotephras Bd..................................................... 289
7. 0. prisca Stgr......................................................... 290
8. O. aurolimbata Gn...................................................... 291
9. 0. ochrolimbata Stgr................................................... 292
10. 0. flavolimbata Stgr................................................... 293
6. Род Penthophera Germ.
1. P. morio L............................................................. 295
tn. P. morio caucasica Heyl. . ............................................ 298
6
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
7. Род Cifuna Wlk.
1. С. locuples Wlk......................................................... 300
2. С. eurydiee Btl.......................................................... 301
3. C. jankowskii Obth....................................................... 303
8. Род Nomens Wlk.
1. N. disparilis Stgr........................................................ 306
9. Род Artaxa Wlk.
1. A. subflava Brem......................................................... 310
2. A. piperita Obth........................................................ 312
3. A. niphonis Btl............<............................................. 313
10. Род Euproctis Hb.
1. E. similis Fuessly........................................................ 320
2. E. chrysorrhoea L........................................................ 323
3. E. karghalica Moore....................................................... 327
4. E. melania Stgr. :............~........................................... 330
5. E. susanna Stgr........................................................... 330
И. Род Arctornis Germ.
1. A. alba Brem.............................................................. 332
2. A. 1-nigrum Mull.......................................................... 333
12. Род Ivela Swinh.
1. 1. auripes Btl............................................................ 336
2. 1. ochropoda Ev........................................................... 338
' 13. .Род Leucoma Steph.
1. L. salicis L............................................................. 342
2. L. Candida Str.......................................................... 345
3. L. flavosulphurea Ersch................................................. 347
4. L. sartus Ersch......................................................... 350
5. L. surtur В. H. ........................................................ 350
14. Род Ocneria Hb.
1. O. monacha L............................................................. 355
2. O. grisescens Stgr....................................................... 362
3. O. matura Moore .......................................................... 362
4. O. dispar L.............................................................. 365
5. O. rubea F. .............................................................. 374
15. Род Ocneropsis, gen. nov.
1. 0. relictus, sp. nov............................................. . . . 377
16. Род Parocneria Dyar
1. P. detrita Esp............................................................ 381
2. P. lapidicola H. S....................................................... 381
3. P. komarovi Chr........................................................... 385
4. P. amabilis Chr........................................................... 386
5. P. griseipennis, sp. nov................................................. 387
6. P. signatoria Chr......................................................... 389
7. P. terebynthi Frr........................................................ 390
8. P. terebynthina Stgr...................................................... 392
9. P. raddei Chr..................................................... • . 393
II. Подсем. Acronictinae
1. Триба MOMINI
17. Род Panthauma Stgr.
1. P. egregia Stgr........................................................... 402
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ 7
18. Род Trisuloides Btl.
1. Т. eornelia Stgr........................................................... 404
19. Род Anacronicta Warr.
i. A. caliginea Btl........................................................... 407
2. A. nitida Btl.............................................................. 409
3. A. obscura Leech...................................................... . 409
20. Род Acronictoides, gen. nov.
1. A. lichenodes Graes....................................................... 410
21. " Род Diphthera Ochs.
1. D. coenobita Esp......................................................... 413
22. Род Calocasia Ochs.
1. C. coryli L. . . .......................................................... 418
2. C. mus Obth............................................................... 420
23. Род Raphia Hb.
1. R. hybris Hb. ................................................... _ . . . 423
2. R. approximate Alph....................................................... 424
3. R. peusteria Piingl...............................................’. . . . 425
4. R. obsolete, sp. nov....................................................... 426
24. Род Episema Ochs.
1. E. coeruleocephala L...................................................... 428
25. Род Belciana Wlk.
1. B. staudingeri Leech...................................................... 431
26. Род Мота Hb.
1. M. ludifica L............................................................ 433
2. M. champa Moore ........................................................ 437
27. Род Daseochaeta Warr.
1. D. alpium Osh............................................................. 441
28. Род Belciades, gen. nov.
1. B. niveola Motsch. . . ..................................'. ............ 444
29. Род Euromoia Stgr.
1. E. mixta Stgr. ............................................................ 446
30. Pog Subleilconyda, gen. nov.
1.. S. palshkovi Fil...................................................... • • 447
31. Род Саппа Wlk.
1. C. malachitis Obth....................................................... 449
2. C. pulchripicta Wlk........................................................ 450
32. Род Thyatirides, gen. nov.
1. T. trimaculata Brem.............................................. • • • 452
2. Триба ACRONIOTINI
33. Род Subacronida, gen. nov.
1. S. megacephala Schiff...................................................• 458
2. S. centralis Ersch....................................................... 460
3. S. grumi Alph........................................................... 461
8
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
34. Род Acronicta Ochs.
1. A. major Brem................................................................. 468
2. A. aceris L................................................................... 469
3. A. elaeagni Alph........... ,................................................ 472
4. A. consanguis Btl. ........................................................... 473
5. A. leporina L............................................................ • 475
6. A. senica Ev..............................................................\ • 478
7. A. alni L................................................................... 480
8. A. leucocuspis Btl.......................................................... 483
9. A. incretata Btl............................................................ 485
10. A. cuspis Hb; ...................................................... • 487
11. A. tridens Schiff.......................................................... 489
12. A. psi L. ......................................‘............................ 492
13. A. angustimacula, sp. nov.................................................- 495
14. A. bellula Alph............................................................. 497
15. A. hercules Feld........................ . .............................. 499
16. A. phaedriola Drd.........................................•................. 500
17. A. phaedra Hamps........................................................ . 501
18. A. terrigena Graes........................................................... 503
19. A. strigosa Schiff.......................................................... 504
10. A. catocaloida Graes......................................................... 508
11. A. carbonaria Graes.......................................................- 509
12. A. digna Btl................................................................. 511
23. A. raphael Obth............................................................. 513
24. A. menyanthidis View................................................... . .514
25. A. auricoma Schiff......................................................... 517
26. A. euphorbiae Schiff........................................................ 520
27. A. abscondita Tr............................................................ 523
28. A. orientalis Mann......................................................... 525
29. A. pasiphae Drd............................................................. 526
30. A. lutea Brem. et Grey . ................................................... 526
31. A. rumicis L............................................................... 529
32. A. pulverosa Hamps. . . . . ............................................. 533
33. A. cinerascens, sp. nov................' .................................... 533
34. A. cubitata Warr............................................................. 534
35. Род Hampsonia, gen. nov.
1. H. jankowskii Obth. . ....................................................... 535
2. H. albonigra Herz. ......................................................... 536
36. Род Craniophora Snell.
1. C. transversa, sp. nov..................................................... 540
2. C. niveosparsa Mts. ...................................................... 541
3. C. praeclara Graes........................................................ 542
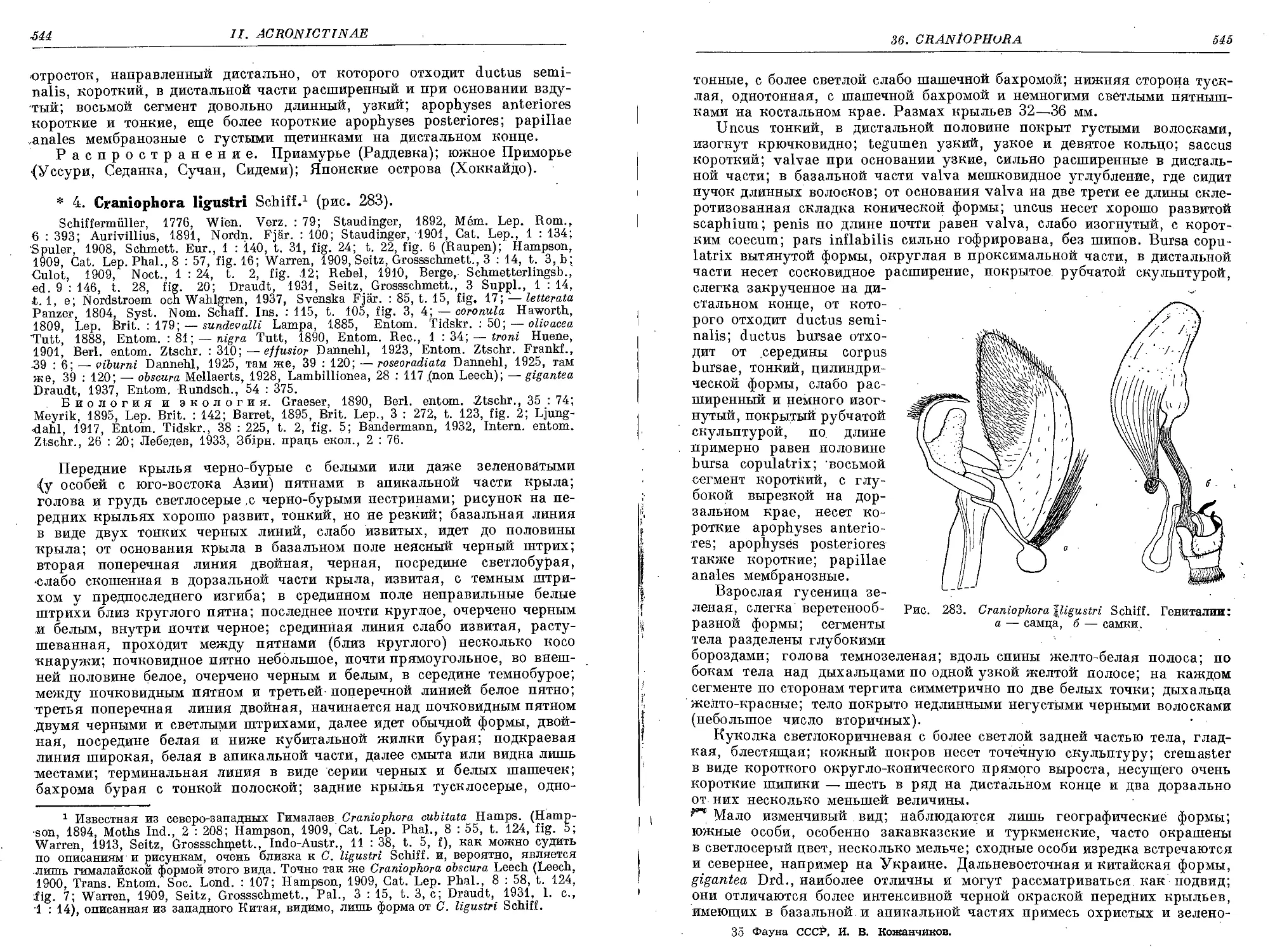
4. C. ligustri Schiff......................................................... 544
5. C. pontica Stgr............................................................ 546
6. C. hemileuca Piingl........................................................ 548
37. Род Arsilonche Ld.
1. A. albovenosa Goeze. ........................................................ 549
38. Род Ozycesta Bd.
1. O. geographica F. . . .................. . ............................. . 552
39. Род Eogena Gn.
1. E. contaminei Ev......................................,................... 555
40. Род Simyra Ochs.
1. S. dentinosa Frr. .......................................................... 558
2. S. nervosa Schiff............................................................ 561
41. Род Leiometopon Stgr.
1. L. simyrides Stgr....................................................... .’ 563
ОБЩАЯ ЧАСТЬ1
Глава I
ВНЕШНЕЕ СТРОЕНИЕ ТЕЛА ВЗРОСЛЫХ НАСЕКОМЫХ
Строение тела взрослых Orgyidae обнаруживает, наряду со многими при-
митивными чертами, черты специализации и нередко редукции и руди-
ментации. Это связано с биологией Orgyidae и цикликой их развития, кото-
рые являются специализованными и характеризуются у большинства
видов рудиментацией имагинальной фазы — укорочением длительности
жизни, афагией и обычно смещением процессов формирования половой
системы и созревания половых продуктов на преимагинальные фазы.
Рудиментация имагинальной фазы Orgyidae требует специальной оценки
особенностей строения этой группы чешуекрылых не только для разреше-
ния вопросов филогении, но и для понимания этих особенностей строения
с точки зрения сравнительной морфологии.
Голова Orgyidae представляет компактную, обычно короткую череп-
ную коробку со слабо дифференцированным лобным склеритом и налич-
ником. Глазки отсутствуют или развиты в числе двух. Глаза развиты
хорошо и могут быть разными по форме; обычно они округлы, но нередко
имеют эллипсоидную форму или даже заострены с дорзальной стороны
(Orgyia Ochs., Gynaephora НЬ.). Глаза слагаются из многих тысяч фасе-
ток; поверхность их, как правило, голая, но для некоторых родов харак-
терно наличие на поверхности их волосков (Мота Hb., Calocasia Ochs.).
Усики'построены из большого числа члеников,1 2 или простых (щетинковид-
ные), или несущих зубцы и выросты (пильчатые и гребенчатые). У самцов
чаще встречаются гребенчатые (в разной мере) усики, а у самок слабо
пильчатые или щетинковидные, несущие лишь реснички на члениках.
Редко оба пола имеют усики последнего типа, но в этом случае реснички
на члениках у самцов более длинны; у непарного шелкопряда, по
Емельяновой (1924), длина отростка членика (среднего) составляет у самки
3—-4% длины усика, у самца —20—28%. На гребенчатых и пильчатых
усиках самцов, на их* гребешках, располагаются многочисленные обоня-
тельные органы (сенсиллы), слабо развитые у самок.
Ротовые части или развиты и тогда вполне типичны для чешуекрылых,
или рудиментарны. Можно констатировать в пределах семейства все
1 Для более полного ознакомления с морфологией Orgyidae, помимо рисунков,
приведенных в общей части, необходимо использовать описания и рисунки специаль-
ной части по соответствующим родам, названия которых здесь приведены в скобках.
2 Число члеников в усиках Orgyidae в пределах вида не постоянно. Обычно оно
равняется 40—60. Число члеников различно у разных полов (несколько больше
у самцов); так, у самцов непарного шелкопряда оно равно 44.0 при колебаниях их
числа в пределах 39—49, у самок — 41.8. при колебаниях в пределах 37—45.
10
ОБЩАЯ ЧАСТЬ
этапы рудиментации их до полной утраты челюстей, верхней и нижней
губ и зарастания ротовой полости (Ocneria Hb., Orgy id Ochs.). Нижне-
губные щупальца при этом всегда развиты и имеют три членика (рис. 1);
нередко они даже большие (Ocneria Hb.), хотя в ряде случаев при руди-
ментации ротовых органов и они уменьшаются в размерах и становятся
слабыми (Gynaephora Hb., Orgyia Ochs.). В случае нормально развитых
ротовых органов есть верхняя губа, несущая epipharynx; по сторонам
ее расположены piliferi; длинные galeae образуют хоботок, нередко по
размерам превышающий в 5—6 раз голову; дорзально от них располо-
жены маленькие одночлениковые нижнечелюстные щупальца; нижняя
губа короткая и несет всегда трёхчлениковые щупальца, у которых вто-
рой членик наибольших размеров; третий членик губных щупалец нередко
короток и иногда рудиментарен (Orgyia Ochs., Мота Hb., Daseochaeta
Warr.). Губные щупальца могут быть направлены прямо вперед (у боль-
Рис. 1. Самка Gynaephora lugens Kozh. р. а — papillae anales;
р. g — papillae genitales.
шинства родов), свисать (Gynaephora Hb., Calocasia Ochs.) или быть под-
нятыми вверх (Belciana Stgr., Euromoia Stgr.).
Рудиментация ротовых органов касается прежде всего galeae и их
длины; хоботок прекращает функционировать еще задолго до их полной
утраты (Calocasia Ochs., Arsilonche Ld.). Начало рудиментации хоботка
знаменуется слабым его скручиванием — рудиментацией мускулатуры;
на этом этапе хоботок уже утрачивает значение органа питания. При этом
ротовые органы сохраняют еще челюстные щупальца, хотя и слабые,
а также нормальную верхнюю губу. Следующим ^тапом рудиментации
ротовых органов является утрата верхней губы и челюстных щупалец
при наличии еще довольно длинных galeae (Leucoma Steph., Artaxa Wlk.,
Eogena Bd.). Наконец у некоторых форм (Olene Hb., Oxycesta Hb.) руди-
менты хоботка ничтожны и прозрачны, но сохраняются еще рудименты
верхней губы; нижняя губа также еще есть. При крайней степени руди-
ментации ротовых органов (Orgyia Ochs., Ocneria Hb., Parocneria Dyar)
можно видеть лишь складочки на месте нижних челюстей и верхней губы;
нижняя губа при этом также почти сливается с черепной коробкой, а губ-
ные щупальца укрепляются на последней.
В пределах рода Orgyia Ochs, у самок наблюдается дальнейшая руди-
ментация ротовых органов, причем она идет в связи с рудиментацией
СТРОЕНИЕ ТЕЛА
11
черепной коробки и ее придатков в целом. Усики становятся короткими,
но не за счет уменьшения числа члеников, а их размеров. В преде-
лах рода есть все переходы от нормального строения головы до полной
ее утраты. У самок О. thyellina Btl. строение головы мало отличается
от такового самцов; самки О. antiqua L., О. gonostigma F., О. trigoteph-
ras Bd., О. ericae Germ, еще имеют хорошо развитую черепную коробку,
но заметно рудиментарные глаза и усики. Самки О. dubia Tausch., О. och-
rolimbata Stgr. характеризуются крайней рудиментацией головы (рис. 2).
Строение груди характеризуется слабым развитием переднегруди,
всегда несущей patagia, сильным развитием среднегруди, тергит которой
всегда имеет хорошо развитый щиток и несет tegulae, и слабым развитием
заднегруди. Расчленение сегментов груди мало типично для Orgyidae,
оставаясь однообразным во всяком случае в пределах всей серии Сага-
drinina. Для Orgyidae типична лишь в некоторых группах рудиментация
грудного отдела, например у видов Orgyia и у самок некоторых других
родов изучаемого семейства. Уместно отметить, что при рудиментации
груди, при почти полной утрате крыльев и при сильном ослаблении ног,
Рис. 2. Самка Orgyia dubia Tausch. с — рудименты головы; р —
рудименты ног; р. а — papillae anales; р. g— papillae genitalis.
склериты груди не утрачиваются, но становятся лишь мельче, оставаясь
дифференцированными. Слияние склеритов груди характеризует лишь
крайние степени рудиментации при полной утрате ног и крыльев (О. du-
bia Tausch., О. ochrolimbata Stgr.).
Строение ног в пределах семейства в отношении общего их типа одно-
образно и неоригинально; развиты все отделы (тазик, вертлуг, бедро, го-
лень, лапка), причем лапка всегда пятичлениковая. Различна пропорция
отдельных частей ног; так, голени могут быть более длинными (Ocneria
Hb., Leucoma Steph.) и более короткими (Orgyia Ochs.), но изменения про-
порций не велики. Разнообразно строение эпифиза, который может иметь
вид большой и широкой лопасти, по длине равной голени (Orgt/га Ochs.,
Arsilonche Ld.), или, будучи такой же длины, может быть узким, почти
шпоровидным (Olene Hb., Leucoma Steph.), или, наконец, может быть
очень коротким, но широким (Acronicta Ochs., Calocasia Ochs.). Шпоры
всегда есть на средних и задних голенях, но на последних одна пара
может исчезать и остается лишь последняя пара (Leucoma Steph., Ivela
Btl., Orgyia Ochs., Gynaephora Hb.). Размер шпор также варьирует —они
то большие (Daseochaeta Warr., Acronicta Ochs.), то короткие (у большей
части родов подсем. Orgyinae) или почти рудиментарны (Parocneria Dyar,
Leucoma Steph., Ivela Btl.). Малый размер шпор типичен для родов, ли-
шенных одной их пары на задних голенях, почему этот факт позволительно
12
ОБЩАЯ ЧАСТЬ
рассматривать как случай общей рудиментации шпор. Когти варьируют
по форме и являются или простыми (Diphthera Ochs., Cynaephora Hb.,
Ocneria Hb.), или снабженными отростками; отросток часто развивается
на вентральной стороне когтя в виде большой (Ciiuna Wlk., Numens
Wlk., Artaxa Wlk.) или малой лопасти (Gynaephora Hb., Orgyia Ochs.).
В ряде случаев отросток расположен на боковой (внутренней) стороне
когтя, короткий (Lichnoptera Н. S.) или длинный (Acronicta Ochs.). Prae-
tarsus характеризуется наличием paranychia под когтями (Ocneria Hb.,
Leucoma Steph.)или едва прикрывающих основание когтей (Orgyia Ochs.).,
Между когтями всегда хорошо развит эмподий, довольно значительно
склеротизованный и сильно волосистый. Наоборот, пульвиллы всегда
отсутствуют.1
Рудиментация ног у самок рода Orgyia характеризуется крайним
уменьшением всех их частей, особенно члеников лапки, которые в ряде
случаев теряют границы. Рудиментируется и претарзальный отдел,
утрачивается зубец на когтях, и они становятся очень слабыми; при край-
них степенях рудиментации когти исчезают вообще (О. dubia Tausch.).
Строение крыльев обнаружи-
вает прежде всего изменчивость
в жилковании (особенно ради-
альной системы) обеих их пар,
а также в размерах, пропорци-
ях и форме. Жилкование пе-
редних крыльев характеризу-
ется свободной, неветвящейся
субкостальной жилкой, пяти-
ветвистой радиальной жилкой,
отчасти рудиментарной, имею-
щей три ветви медиальной жил-
кой, первая ветвь которой
сближена основанием с пятой
кубитальной. Последняя всегда
полная и имеет две ветви. Анальная жилка имеет две самостоятельные
ветви, первая из которых обычно несколько рудиментарна, а вторая развита
вполне (третья ветвь всегда' рудиментарна). Вазальная половина меди-
альной жилки не всегда полностью утрачена, но у видов Eloria Wlk-,
Penthophera Germ, и особенно у самок рода Ocneria Hb. она довольно хо-
рошо развита и вполне видна в поле дискоидальной ячеи. Можно видеть
ее первое ветвление и отхождение третьего ствола (рис. 3), а затем второе
ветвление, которое сохраняется всегда в форме дискальных жилок, свя-
зывающих первый и второй медиальные стволы.
Жилкование задних крыльев близко таковому передних, но характе-
ризуется анастомозом или лишь сближением субкостальной и радиальной
жилок и образованием полузамкнутой (Leucoma Steph., Dasychira Steph.)
или вполне замкнутой базальной ячеи (Penthophera Germ., Calocasia
Ochs.). Радиальная жилка (единственный ствол) обычно соединена с пер-
вым медиальным стволом или очень короткой дискальной (Мота НЬ.,
Acronicta Ochs., Subacronicta Kozh.), или анастомозом (Euproctis Hb.);
в последнем случае первая ветвь медиальной жилки укорочена. Две дру-
гие ветви медиальной жилки обычно сильно сближены основаниями
Рис. 3. Жилкование переднего крыла Ocneria
dispar L.
ветвью радиальной жилки, а третья с
1 Терминология структур имаго по Кузнецову (1915) и Снодграссу (Snodgrass,
СТРОЕНИЕ ТЕЛА
13
и сдвинуты к кубитальной, с которой связаны или помощью дискальной,
или третья ветвь анастомозирует с первой кубитальной частично или пол-
ностью {Euproctis Hb., Artaxa Wlk.). Кубитальная жилка имеет две
ветви и развита вполне; две ветви имеет и анальная жилка (рис. 7).
Строение радиальной ячеи на передних крыльях различно. Обычно
она образуется частичным анастомозом и развитием короткой дискальной
жилки между пятой радиальной ветвью и стебельком третьего и четвер-
того радиальных стволов; второй радиальный ствол при этом ответвляется
от костального края ячеи на разных уровнях. Первый медиальный ствол,
анастомозируя с последним радиальным, отходит от дорзального края
ячей также на разном уровне. Иногда первый медиальный ствол связан
с радиальной системой лишь короткой дискальной жилкой. Ветвление
второго радиального ствола и в связи с этим положение радиальной ячеи
могут быть различны. Обычно второй радиальный ствол ветвится сразу
после отхождения первого радиального, и потому добавочная, радиальная
ячея обособлена от него полностью
{Dasychira Steph., Cynaephora Hb.,
Orgyia Ochs., Lae.Ua Steph., Cifuna
Wlk.), но в некоторых случаях,
например у некоторых особей Arct-
ornis Brem., второй радиальный
ствол отходит от первого в преде-
лах ячей (рис. 4).
Возникновение и исчезновение
радиальной (добавочной) ячеи в
общем связано с полнотой анасто-
моза радиальных стволов; в пре-
делах некоторых родов ячея не
только изменчива по размерам
{ArctornisGerm.,Dasychira Steph.),
но может и отсутствовать вовсе
Рис. 4. Различные типы ветвления ради-
альной системы переднего крыла у Arct-
ornis alba Brem.
в связи со степенью развития ана-
стомоза радиальных стволов; при полном анастомозе их она у некото-
рых особей одного и того же вида может исчезать {Penthophera Germ.).
Отсутствие радиальной ячеи у ряда видов Orgyidae {Ocneria Hb., Par-
ocneria Dyar, Leucoma Steph., Euproctis НЪ., ArZaxa Wlk.) есть, невидимому,
следствие полноты анастомоза радиальных стволов (рис. 5, 6). Ветвле-
ние радиальных жилок наблюдается всегда в области вершины крыла
с преобладанием окончания большинства их по костальному {Ocneria
Hb., Ivela Swinh., Dasychira Steph., Olene Hb., Gynaephora Hb., Orgyia
Ochs., Cifuna Wlk.) или по терминальному краю {Leucoma Steph., Arctor-
nis Germ.).
Форма крыльев дает обычно слабые изменения (чаще порядка видо-
вых особенностей) ширины их, более тупого или острого апекса. Лишь
в пределах трибы Acronictini у некоторых видов родов Pharetra Hb. {obli-
nita Sm.-Abb., carbonaria Graes., lanceolaria Grt.), Arsilonche Ld.
крылья очень узки, а у видов последнего рода они характеризуются
также заостренным терминальным краем. Форма крыльев меняется также
при рудиментации их у самок Orgyia Ochs. У нормально крылатых самок
О. thyellina Btl. крылья сильно выгнуты по костальному краю; сходны
по форме крылья самок рода Olene Hb. У большинства видов рода Orgyia
Ochs, крылья самок имеют вид коротких, нефункционирующих ланцето-
видных лопастей, несущих по краям длинные реснички и лишенных жил-
14
ОБЩАЯ ЧАСТЬ
кования. Задние крылья в этих случаях, будучи еще более короткими,
имеют вид округлых лопастей (рис. 8).
Брюшной отдел тела Orgyidae слагается из десяти сегментов, но по-
следний (десятый) сегмент всегда рудиментарен и в ряде случаев исчезает
вообще; рудиментарен обычно и девятый сегмент. Строение брюшных
сегментов просто и мало характерно для Orgyidae. Может быть лишь от-
мечено, что у самок тергальная и стернальная области слабы, а плей-
риты тонки, перепончаты и сильно растяжимы; такое строение сегментов
брюшка приводит к возможности сильного его увеличения в связи с разра-
станием гонады (Ocneria Hb., Orgyia Ochs.).
У самок Orgyidae десятый сегмент сохраняется всегда в виде рудимента,
сливаясь с девятым; он несет papillae anales. У самцов он почти всегда
утрачен, но и у видов Dasychira Steph, anus окружен полным или частич-
ным коротким кольцом (рис. 9), лежащим дистально от девятого сегмента.
Этот отдел брюшка самцов Dasychira Steph, утратил типичный облик
сегмента, так как обычно лишен ясно
выраженных плейритов, а в ряде слу-
чаев также и стернального отдела, но
он несет волосяной покров. Сегмент
этот отделен мембраной от девятого сег-
мента и является самостоятельным от-
делом. Сохранение столь обособленного
и полного десятого сегмента брюшка
у самцов рода/)asychira Steph, связано
с отсутствием у них uncus и tegumen..
Девятый сегмент в этом случае корот-
кий и нередко несколько рудиментар-
ный (в дорзальной половине). Функции
uncus выполняет вырост восьмого сег-
мента— superuncus, в связи с чем дор-
зальная сторона восьмого сегмента у
видов Dasychira Steph, приобретает
замещает tegumen. Этот тип строения
восьмого сегмента и обусловливает сохранение столь примитивных черт
архитектуры брюшка самцов в пределах этого рода. В пределах рода
Artaxa Wlk. uncus также может быть неразвит, но у видов этого рода
superuncus не констатирован, а, напротив, в примитивной форме разви-
вается настоящий, еще парный uncus.1
Строение tegumen и uncus у Orgyidae обнаруживает примитивные
черты. Часто uncus сохраняет парное строение — слагается из двух
латеральных половин, еще не слитых вместе (Gynaephora Hb., Artaxa
Wlk., некоторые Parocneria Dyar) или имеет ясные следы срастания из
двух половин в виде дорзального жолоба или гребня (Olene Hb., Orgyia
Ochs.). Кроме того, Orgyidae имеют недлинные латеральные выросты
Рис. 5. Строение добавочной ячеи
у Penthophera morio L. а — Венгрия;
б — Кавказ.
массивность и функционально
1 Uncus в копулятивном аппарате самцов чешуекрылых служит для прижимания
конца брюшка самки к valvae во время копуляции. Он может функционально заме-
щаться аналогичным придатком восьмого сегмента, как то описано для видов Dasy-
chira.. Тем не менее придаток восьмого сегмента (superuncus) может иметь и другую
функцию. Так, у Colias aurorina В. S. развиты одновременно как tegumen и uncus
на девятом сегменте, так и superuncus на восьмом сегменте. В этом случае superuncus
служит лишь покрышкой и защитой для uncus. Строение superuncus у Colias и Dasy-
chira близко, но у видов последнего рода он более острый и по форме напоминает
uncus, тогда как у Colias он имеет вид склеротизованного широкого и короткого
(округлого) выроста.
СТРОЕНИЕ ТЕЛА
по сторонам uncus, которые сидят на общем с ним основании, так назы-
ваемые subunci. Также очень характерно наличие шва между основанием
uncus (с subunci) и tegumen, т. е. еще отсутствие полного слияния девятого
и десятого тергитов брюшных сегментов. Лишь у наиболее специализо-
ванных представителей семейства (Penthophera Germ., Acronicta Ochs.,
Sirnyra Ochs, и некоторых других) наблюдается полное срастание этих
элементов; из высших Orgyidae (Acronictinae) они не слиты лишь у немно-
гих (Calocasia Ochs., Episema Ochs, и Diphthera Ochs.). Полное слияние
девятого и десятого сегментов обычно все же не ведет еще к утрате sub-
Рис. 6. Ветвление радиальной системы переднего крыла. У —
Panthauma Stgr., Diphthera Ochs.,' Trisuloides Wlk., Charadra
Wlk., Daseochaeta yVarr.; 2— Calocasia Ochs., Episema Ochs.,
Raphia Hb.; 3 — Мота Hb., Agriopodes Hamps., Euromoia Stgr.,
Belciades, gen. nov., Ganna Wlk.; 4— Craniophora Snell., Suba-
cronicta, gen. nov., Acronicta Ochs., Hampsonia, gen. nov., Eogena
Gn., Sirnyra Ochs., Arsilonche Ld., Oxycesta Hb.; 5 — Numens
Wlk.; 6 — Penthophera Germ., Olene Hb.'; 7 —Ivela Wlk.; 8 —
Ocneria Hb.
unci — они заметны почти у всех представителей в виде больших выростов
или небольших бугорков (Acronicta Ochs., Subacronicta Kozh., Craniophora
&ae\\.,\Eogena Gn.).
Тергальная область девятого и десятого сегментов брюшка обнаружи-
вает некоторые изменения в строении^ нетипичные для семейства в целом,
но характеризующие лишь отдельные группы родов. Так, для ряда видов
рода Orgyia Ochs, типично разрастание основания uncus, т. е. тергальной
области десятого сегмента (отделенной швом от девятого сегмента), столь
значительное, что оно вытесняет почти нацело тергит девятого сегмента.
У других родов, напротив, наблюдается рудиментация десятого сегмента
при срастании его с девятым (Craniophora Snell., Acronicta Ochs.). У мно-
гих Orgyinae и видов Momini uncus имеет сложное строение, часто являясь
широким, желобчатым, гребенчатым или снабженным сложными структу-
рами (Diphthera Ochs.). У специализованных представителей семейства
16
ОБЩАЯ ЧАСТЬ
uncus мало изменчив — он является тонким, заостренным, -несущим
щетинки с дорзальной стороны.
Строение девятого сегмента брюшка самцов Orgyidae в общем мало
характерно; он узок (если видны его границы на tegumen) и образует
на вентральной стороне saccus. Последний может быть очень мал (Gynae-
phora Hb.) или рудиментарен; напротив, у некоторых видов saccus может
быть очень большим (Craniophora Snell.), служа вместилищем для андро-
коний. Valvae, являясь латеральными придатками девятого сегмента,
развиты всегда. Строение их примитивно; дистальный конец valva никогда
не имеет хватательного аппарата (cucculus), который типичен для Noctui-
dae. Обычно дистальный конец valva перепончатый и покрыт волосками.
Лишь у видов Ocneria Hb. и Parocneria Dyar он вытянут шпагообразно
и несет на дистальном конце хватательный шип. У большинства родов
подсем. Orgyinae harpe отсутствует (Gynaephora Hb., Olene Hb., Orgyia
Ochs., Ocneria Hb.) или очень слабо развита (Dasychira Steph.). Среди
подсем. Acronictinae harpe отсутствует лишь у немногих примитивных
Рис. 7. Жилкование заднего крыла, а — Euproctis chrysor-
rhoea L.; б — E. similis Fuessly.
родов (Calocasia Ochs., Episema Ochs., Саппа Wlk., Belciana Stgr.),
а также у некоторых специализованных, где, вероятно, утрачена
(Craniophora Snell.);- у большинства родов этого подсемейства harpe
сложно развита и характерна. В роде Craniophora Snell., структурно
очень специализованном, утрата harpe, вероятно, вызвана превращением
valvae в органы ношения андроконий, так как в них развивается кармано-
образное углубление, где разрастаются очень длинные и многочисленные
железистые волоски, сидящие в виде больших пучков. Valvae Cranio-
phora Snell, покрыты тонкой мембраной и не имеют значения основного
хватательного аппарата. Примитивные Hampsonia Kozh., близкие к Cra-
niophora Snell., сохраняют еще harpe; она есть также у наиболее прими-
тивных видов Craniophora Snell, (transversa Kozh.). Субанально, в полости
девятого сегмента, разрастаются склеротизованные пластинки. Почти
всегда есть fultura inferior обычно большая, нередко слагающаяся из двух
латеральных половин (Acronicta Ochs., Ocneria Hb.), большею частью
отделенная от penis, реже срастающаяся с ним в проксимальной части
(Cifuna Wlk.). Fultura superior развивается у меньшего числа форм, но
типична, например, для видов Calocasia Ochs., у которых она к тому же
несет на поверхности сложную (в виде шипов) скульптуру и достроена
из двух латеральных половин. Чаще этот склерит развит слабее и даже
слабо дифференцирован. Стенки анального соска редко склеротизуются,
СТРОЕНИЕ ТЕЛА
17
напротив, они обычно мембранозны, не образуя scaphium или siibscaphium.
Лишь у некоторых видов, например у Artaxa Wlk., anus имеет плотные '
стенки и покрыт тонкими волосками. Недоразвитие у этих видов uncus
и tegumen говорит за примитивность строения их анальной области и за
возможное наличие у них рудимента десятого сегмента, слитого со стен-
ками anus. В связи с этим, может быть, развит волосяной покров на по-
верхности anus у видов Artaxa Wlk.
Строение penis мало характерно для сем. Orgyidae и в общем совпадает
с тем, что известно для большинства чешуекрылых (исключая Jugata).
Он образован более или менее прочной трубкой, несущей в проксималь-
ной части мешковидный отдел (соесшп penis) разной длины у разных
видов. Pars inflabilis penis несет большею частью инкрустаций в виде
разнообразных шипов, зубцов, пластинок, очень различных по форме
в пределах семейства. Реже инкрустации penis возникают за счет его на-
ружной стенки (Dasychira Steph., Euromoia Stgr.).
Рис. 8. Самка Orgyia gonostigma F. p. a — papillae anales;
p. g— papillae genitales.
Дистальный конец брюшка самок также разнообразно видоизменен в
пределах семейства, но у всех исследованных мною видов развитого десятого
сегмента не имеет: он слит с рудиментарным девятым, образуя с ним
единый комплекс. Значительно видоизменяется вагинальная область вось-
мого сегмента, на которой часто возникает вырезка {Euproctis Hb., Gynae-
phora Hb.) или пластинка ‘ (lamella postvaginalis). Реже развивается
предвлагалищная пластинка (lamella antevaginalis), как, например,
у Euproctis Hb., Artaxa Wlk. Восьмой сегмент брюшка самки всегда несет
передние апофизы, которые очень варьируют по длине (являясь харак-
терными лишь для видов, реже, для родов). Девятый сегмент неполный,
обычно сохраняется в виде небольших склеритов при основании papil-
lae anales; обычно эти склериты сливаются с основаниями задних апо-
физов;* последние сильно варьируют по длине и нередко бывают очень
длинны (Numens Wlk., Cifuna Wlk., Artaxa Wlk., Euproctis Hb., некото-
рые виды Olene Hb., Ocneria Hb., Orgyia Ochs.).
У некоторых представителей семейства (Ocneria Hb., Parocneria Dyar)
последние сегменты брюшка самок сильно видоизменены в связи с образо-
нанием яйцеклада. Для этих форм характерно сильное .разрастание ин-
терсегментальных мембран, которые образуют множество складок. Papil-
lae anales видоизменяются в этом случае в твердые длинные пластинки,
2 Фауна СССР, И. В. Кожанчиков.
18
ОБЩАЯ ЧАСТЬ
а передние и задние апофизы достигают весьма большой длины; восьмой
сегмент также становится узким и длинным (рис. 10). Яйцеклад, т. е. вытя-
нутый восьмой сегмент и длинные papillae anales, способен втягиваться
глубоко в предыдущие сегменты.
Рис. 9. Анальные сегменты мужского брю-
шка Dasychira pudibunda L. VI—X — сег-
менты брюшка; s — superuncus.
Такое строение конца брюшка ти-
пично для видов, откладывающих
яйца под чешуйки коры, и особен-
но характерно для монашенки
и розового (японского) непарного
шелкопряда. В противополож-
ность этому у большинства видов
семейства papillae anales явля-
ются короткими и мягкими и по-
крыты многими мелкими волос-
ками.
Эйдман (Eidmann, 1935) спе-
циально изучал мускулатуру кон-
цевых сегментов брюшка самки
монашенки, у которой яйцеклад
особенно велик. Он Описывает
шесть групп специальных мышц,
приспособленных к работе яйце-
клада и укрепленных на апофизах
и отчасти на тергитах седьмого
и восьмого сегментов. Часть их
является ретракторами (рис. 10, 1, 2), другие—протракторами (4, 5,
6). Специальная мышца управляет поворотами конца брюшка (3). Сле-
дует подчеркнуть, что в яйцекладе Orgyidae не наблюдается развития
специальных органов, но лишь
специфичное развитие обычных
структур — apophyses, papil-
lae — и усиление всегда при-
сутствующих мышц.
Совокупительная сумка (bur-
sa copulatrix) образует разной
формы мешковидный орган,
снабженный то коротким и мем-
бранозным, то длинным и неред-
ко склеротизованным протоком
(ductus bursae). Стенки сово-
купительной сумки обычно мем-
бранозные, без инкрустаций, но
у некоторых видов на ее поверх-
ности возникают laminae den-
tatae (Cifuna Wlk., Leucoma
Steph., Artaxa Wlk., Arctornis
Germ., чаще у низших Orgyidae).
Форма совокупительной сумки
в наибольшей мере меняется
в связи с величиной и степенью
Рис. 10. Яйцеклад монашенки (Ocneria mo-
nucha L.) (По Эйдману, 1935). 1, 2 —мышцы-
ретракторы; 3— мышца, управляющая яй-
цекладом; 4—6 —-мышцы-протракторы; р. а—
papillae anales; d. gl. s — ductus glandulae
sebaceae; d. r — ductus receptaculi; о. c —
oviductus communis; d. s — ductus seminalis;.
d. b — ductus bursae.
разрастания ее к ductus seminalis; это
разрастание часто очень велико и нередко сильно склеротизовано
(Hyboma Hb., Acronicta Ochs., Craniophora Snell., Мота Hb.). Редко»
совокупительная сумка очень мала (Orgyia Ochs., Penthophera Germ.).
СТРОЕНИЕ ТЕЛА
19
Адаптация концевых сегментов брюшка Orgyidae к специализованной
яйцекладке ведет к возникновению вторичных структур. Тем не менее
присутствующие всегда у самок Orgyidae papillae genitales — рудименты
яйцеклада низших групп насекомых — никогда не принимают участия
в формировании яйцеклада!. Существование papillae genitales у Orgyi-
dae впервые отмечено Гольдшмидтом (Goldschmidt, 1912), Гольдшмидтом
й Поппельбаумом (Goldschmidt u. Poppelbaum, 1914), а позже описано
Кузнецовым (1916; рис. 197, 199). Органы эти, как оказалось, имеют
большой сравнительно-морфологический интерес, так как являются го-
мологом valvae.
Экспериментальными работами Мейзенгеймера (Meisenheimer, 1907),
Гольдшмидта (1912, 1934), Голдшмидта и Поппельбаума (1914), Космин-
ского (1929, 1935а, 19356) и некоторых других авторов по гинандромор-
физму Orgyidae, преимущественно Ocneria dispar L., показана гомология
частей полового аппарата самцов и самок. Их данные вполне согласуются
с чисто морфологическими сопоставлениями и предположениями, выска-
занными ранее [Петерсен (Petersen, 1900—1904а); Цандер (Zander, 1903);
Кузнецов, 1916], но расширяют их. Здесь я даю таблицу гомологии поло-
вых придатков самцов и самок Orgyidae (табл. 1), которую я заимствую
из работы Гольдшмидта (1934); в ней я изменил лишь терминологию при-
менительно к общему изложению.1 * *
Таблица 1
Гомология частей полового аппарата самцов и самок Ocneria dispar L,
(цо Гольдшмидту, 1934)
Самки
1. Papillae anales
2. Apophyses posteriores
3. Ostium yaginale
4. Ductus bursae
5. Papillae genitales
6. Uterus
7. Bursa copulatrix
Самцы
1. Uncus
2. Девятый сегмент 4
3. Ostium ductus ejaculatori
4. Penis
5. Valvae
6. Ductus ejaculatorius
7. Нет гомолога
В отношении элементов, приведенных в табл. 1, требуется отметить
лишь, что uterus — область, относящаяся к oviductus communis (Снод-
грасс, 1935), едва ли является гомологом ductus ejaculatorius. Напротив,
этот последний наверное гомолог bursa copulatrix, если penis является
гомологом ductus bursae. Uterus скорее гомолог дистальных частей
брюшка самцов.
Гомологии полового аппарата самок и самцов, приведенные в табл. 1,.
принимает большая часть авторов, работавших над половым аппаратом
Orgyidae, в частности Ocneria dispar L., в связи с изучением интерсексов
|Косминский, 1929, 1935; Гольдшмидт и Сегучи (Goldschmidt u. Segiichi,
1922); Дюбуа (Du Bois, 1931)]. Гомология гаметоцитов не вызывает сомне-
ний, так как у непарного шелкопряда констаГированы переходы спер-
миев в яйца и наоборот. Несомненна также гомология фолликулов семен-
ников и яйцевых трубочек яичников, точно так же как vasa deferentia
и педицеллярных отделов яйцевых трубочек. Далее, не вызывает сомне-
1 Вопрос о гомологии частей полового аппарата чешуекрылых в целом выходит
им пределы изучения сем. Orgyidae, но он должен быть здесь затронут, так как виды
итого семейства послужили основным материалом для проведения указанных гомо-
логий.
2*
20
ОБЩАЯ ЧАСТЬ
ний гомология uncus полового аппарата самцов и papillae anales самок,
valvae и papillae genitales, apophyses и девятого сегмента брюшка самцов.
Для указанных органов во всех случаях доказано превращение гомоло-
гичного органа одного пола в орган другого пола у интерсексов. Напро-
тив, гомологии bursa copulatrix, ductus bursae, penis, ductus ejaculatorius
и oviductus communis далеко не столь определенны. По Дюбуа (1931,
с некоторыми поправками Косминского, 1935а, 19356) penis является
гомологом bursa copulatrix, a ductus ejaculatorius и кожный чехол вокруг
penis (Penistasche) — гомологом oviductus communis («vagina» и «uterus»).
Это толкование кажется вероятным и в известной части близко к выска-
занному выше. Но Косминский (1935а, 19356) склонен видеть гомологию
penis и vagina, т. е. для penis находит гомологии в дистальной части
oviductus communis; он не находит гомологии для receptaculum seminis,
bursa copulatrix и uterus самок в половом аппарате самцов. Точно так же,
по Косминскому (1935а, 19356), нет гомолога в половом аппарате самок
и для ductus ejaculatorius. Работы Косминского, Гольдшмидта, Дюбуа
и других авторов не осветили пока гомологии этих органов, хотя Космин-
ский склонен видеть подтверждение своей точки зрения об отсутствии
для них гомологий в отсутствии изменчивости penis и ductus ejaculatorius
у мужских интерсексов в тех случаях, когда другие части полового аппа-
рата их обнаруживают ясные переходы в структуры другого пола.
Развитие половых придатков до конечных этапов протекает в фазе
куколки, хотя закладываются они раньше. Для непарного шелкопряда
наличие зачатка penis в виде выроста гиподермы (герольдова органа),
образованного девятым сегментом, отмечено уже на первой личиночной
стадии. Этот зачаток сильно увеличивается и дифференцируется к моменту
окукления. Точно так же и у самок зачатки половых придатков намечены
уже у гусениц в первой стадии в виде имагинальных дисков восьмого
и девятого сегментов; но они разрастаются лишь при росте гусениц и окон-
чательно формируются в фазе куколки. Начальная дифференциация
половых структур, нужно полагать, наступает еще в эмбриональный
период развития Orgyidae.
Приведенные факты, подтверждающие гомологию половых придатков
Orgyidae, дают некоторые исходные точки для понятий примитивности
или специализации структур полового аппарата вйдов этого семейства,
что очень важно для выяснения вопросов систематики группы. Так,
парный uncus Artaxa Wlk., Parocneria Hb. и некоторых других видов се-
мейства указывает на примитивность этих структур.1 Точно так же отсут-
ствие uncus и наличие рудиментов десятого сегмента у видов Dasychira
Ochs, есть признак, характеризующий примитивные черты строения
этого рода.
Сроки дифференциации имагинальных структур в фазе куколки изу-
чены подробно у непарного шелкопряда Дюбуа (1929), Косминским
(1935а, 19356) и другими авторами. Сравнительное исследование полов
этого вида показало запаздывание формирования имагинальных струк-
тур самок. Следовательно, у самок диморфных Orgyidae имеет место
примитивность структур в связи с их недоразвитием. Половые продукты
самцов непарного шелкопряда формируются в личиночной фазе, у самок—
в фазе куколки. Развитие копулятивных органов у сампов (герольдов
орган) происходит в личиночной фазе, а дифференцировка и формиро-
1 За это же говорит и возникновение парного uncus в случае недоразвития его
у гинандроморфиых особей или при недоразвитии структур полового аппарата при
рентгенизации (Кузнецов, 1916; Бобров, 1930; Косминский, 1935а, 19356).
СТРОЕНИЕ ТЕЛА
21
вание их — в первые моменты развития куколок. Формирование половых
органов самок наблюдается только в фазе куколки, причем не в первые
моменты развития. В таб^и. 2 даются сроки дифференцировки органов
разных полов непарного шелкопряда, причем весь период развития куко-
лок разбит на восемь отрезков.
Таблица 2
Сроки дифференцировки имагинальных структур разных полов непарного
шелкопряда в куколочной фазе (по Косминскому, 1935)
Стадии развития куколок
Название органа
1. Образование выростов на
усиках:
самцов................
самок.................
2. Образование чешуи па
крыльях:
самцов................
самок.................
3. Образование рисунка и
окраски на крыльях:
самцов................
самок ................
4. Копулятивный- аппарат
самцов: I
penis . ’.............
valvae................
5. Копулятивный аппарат са-
мок:
papillae anales et geni-
tales ..............
apophyses et lamellae
Примечание. Знак —нет дифференцировки: знак + дифференцировка
имеется, О — окончание дифференцировки.
Большая часть органов имагинальной фазы у непарного шелкопряда,
как, например, крылья, penis и ряд других кожных структур, имеет
период предварительной дифференцировки перед переходом в куколку.
Ряд органов формируется, тем не менее, в фазе куколки. Это касается
чешуйчатого покрова, усиков. Пигментация покровов происходит в фазе
куколки лишь частично, а в основном протекает в момент выхода имаго
и в первые моменты его жизни. Аналогично формирование имагинальных
структур и у других видов.
Чешуйчатый и волосяной покров, исключая немногие формы (Реп-
Ihophera Germ.), развит у Orgyidae хорошо. Тело обычно покрыто воло-
сками, иногда очень длинными (Olene Hb., Gynaephora Hb.), и небольшим
числом чешуй. Развитие чешуй на голове, груди и брюшке у низших
Orgyidae незначительно; если чешуи и развиваются, то имеют вид расши-
ренных волосков (Artaxa Wlk.). Развитие чешуй у специализованных
22
ОБЩАЯ ЧАСТЬ
видов {Acronictinae) значительно даже в тех случаях, когда тело покрыто
длинными и густыми волосками {Calocasia Ochs.). На конце брюшка
самок многих видов семейства, как специальное биологическое приспо-
собление, развивается нередко толстый.слой волосков, покрывающих седь-
мой и восьмой сегменты в виде подушки {Euproctis Hb., Ocneria Hb.,
Artaxa Wlk., Penthophera Germ., OleneHb., ( ynaephoraJlb., EpisemaOchs.).
Крылья Orgyidae покрыты почти всегда только чешуей разной величины
и формы и имеют лишь небольшую примесь волосков при основании.
У низших форм семейства {Euproctis Hb., Artaxa Wlk.) чешуи на крыльях
разнообразны по форме, причем на поверхности крыльев имеется большое
количество волосков, расположенных между чешуйками в виде редкого
слоя {Leucoma Steph., Euproctis Hb.). Редко крылья покрыты только
волосками, без чешуй (самки Penthophera Germ., Orgyia Ochs), что связано
скорее с явлениями редукции.1
Рисунок на крыльях (и на теле) Orgyidae часто развит слабо и у ряда
родов их, в частности у некоторых богато представленных в тропиках,
отсутствует совершенно {Euproctis Hb., Laelia Steph., Eloria Wlk.). У неко-
торых из этих представителей нередко возникает скопление пигмента
(темного) вдоль жилок, например у видов Ivela Swinh., Arctornis Germ.,
Eloria Wlk., Penthophera Germ., в виде частичных или полных полос
или возникают отдельные пигментированные пятна на них. Формирование
поперечных пигментных полос и пятен на крыльях Orgyidae протекает
лишь отчасти в связи с жилками и идет, повидимому, через этап структур-
ных элементов рисунка, структурных полос. У видов Caviria Wlk. попе-
речные полосы на крыльях (как и крыло) белого цвета, но образованы
скоплением более широких и толстых чешуй, резко отличных от осталь-
ных чешуй крыла (тонких, волосковидных). Эти структурные различия
в чешуйчатом покрове на крыльях Caviria Wlk. и придают им полосатый
облик, кажущуюся гофрированность. Рисунок крыльев у Artaxa Wlk.
и Euproctis Hb. также обусловлен отчасти структурой чешуй; темные
чешуи являются более крупными по сравнению с окружающими их бе-
лыми.
Таким образом рисунок крыльев Euproctis Hb. и Artaxa Wlk. предста-
вляет следующую ступень развития структурного рисунка Eloria Wlk.
У специализованных видов- семейства пигментные элементы рисунка
безусловно преобладают над структурными и часто окрашенные чешуи
не отличаются по размерам и форме от бесцветных. У генерализованных
представителей семейства часто пигментный рисунок не возникает, но
окраска имеет характер разлитых общих тонов или представляет непра-
вильно разбросанные пятна, не образующие еще системы поперечных
полос {Aroa Wlk., Numens Wlk.).
Виды Orgyidae умеренных широт и тропические представители, насе-
ляющие открытый ландшафт, характеризуются сложным рисунком,
построенным из поперечных зубчатых полос, идущих параллельно одна
другой; таковы виды Orgyia Ochs, Ocneria Hb., Dasychira Ochg.; в преде-
лах рода Ocneria Hb. наблюдается формирование пятен, может быть из
обрывков поперечных линий. Формирование их, в частности почковид-
ного пятна, возможно и иным путем, как то ясно можно видеть на примере
1 Тем не менее нет уверенности, что в волосяном покрове самок Orgyia Ochs-
ицеет место проявление примитивных черт, как то видно, например, в строении чешуй-
чатого покрова у нормально крылатых самок. Так, самки непарного шелкопряда
характеризуются не только присутствием узких, волосковидных кроющих чешуй,
которых нет у самцов, но вообще узкой чешуей на крыльях.
СТРОЕНИЕ ТЕЛА
23
Ocneria Hb., у которых имеется скопление пигмента вдоль дискальной
;килки. Подобные скопления пигмента налицо также у видов, лишенных
поперечных полос и имеющих, следовательно, зачаток пятна, образо-
вавшегося, безусловно, без участия поперечных линий (Arctornis Germ.,
drtoa Wlk.). Круглое пятно возникает на месте ветвления первой ме-
диальной жилки (первого ее ветвления), возможно также в связи с пигмен-
тацией этой жилки. В дальнейшем она исчезает, а пятно сохраняемся.
Таким образом у Orgyidae формирование рисунка, как и первичное распре-
деление пигмента у Eloria Wlk. или Ivela Swinh., в известной мере свя-
зано с жилкованием.1
Рисунок, слагающийся из поперечных линий и двух пятен, круглого
и почковидного, типичен для большинства видов семейства и особенно
для всех высших групп — Acronictinae. Развитие третьего, клиновидного,
пятна в пределах семейства не наблюдается. В некоторых случаях имеется
лишь зачаток его {Trisuloides Wlk.). Среди высших Orgyidae есть предста-
вители с недоразвитым еще почковидным пятном, например виды Мота
lib., Belciana Stgr., Charadra Wlk. Среди этих групп существует ряд
видов, у которых круглое и почковидное пятна представлены не в виде
скопления темного пигмента, но в виде сложных, многократно очерченных
семными линиями пятен. Среди этих же групп тип рисунка с продоль-
ными полосками по жилкам менее распространен, но наблюдается напри-
мер, в роде Acronicta Ochs., ряд видов которого характеризуется присут-
ствием Штрихов вдоль кубитальной, анальной и медиальной жилок.
Распределение темного пигмента вдоль жилок типично также для видов
Arsilonche Ld. и Simyra Ochs.
В пределах сем. Orgyidae нередки случаи редукции рисунка. Кроме
многочисленных случаев частичной его утраты у отдельных видов {Epi-
sema Ochs., Acronicta Ochs.), имеются целые роды, характеризующиеся
полной утратой не только линий, но и пятен {Subleucoriycta Kozh., Eogena
Gn., Simyra Ochs., Arsilonche Ld.). Редукция рисунка наблюдается также
всегда при редукции крыльев, как то можно видеть на примере рода
Orgyia Ochs. Редукция рисунка у Orgyidae является лишь частным слу-
чаем рудиментации структур половой фазы. В частности, все упомяну-
тые случаи полной редукции рисунка {Simyra Ochs., Eogena Gn. и др.)
характеризуются также утратой и редукцией ротовых частей и сокраще-
нием длительности жизни половой фазы. Редукция рисунка идет тем
не менее не всегда параллельно редукции ротовых частей, обнаруживая
известную самостоятельность.
Общее развитие пигментации покровов в пределах сем,- Orgyidae раз-
лично. Среди архаичных представителей обращает внимание распростра-
нение белых окрасок {Leucoma Steph., Ivela Swinh., Eloria Wlk., Gavi-
ria Wlk., и многие другие), которые обусловлены отсутствием или бед-
ностью пигментов. Часто наблюдается замещение белых окрасок желтыми
и наоборот как в пределах близких видов {Artaxa Wlk., Euproctis Hb.),
так и в пределах одного вида (например у Acronicta lutea Brem. et Grey,
Le.ucoma flavo sulphur ea Ersch.). Изменение белой окраски в желтую и обрат-
но в связи с условиями развития специально изучено мною на примере
1 За важную роль жилкования в формировании первичных элементов рисунка
говорят и наблюдения при возникновении окрасок у интерсексов. Развитие черного
пигмента в крыльях интерсексов наблюдается, по Гольдшмидту (1922), так, как
если бы поток пигмента растекался от основания крыла в направлении жилок. Это же
Подтверждает и Косминский (1935а, 19356) своими экспериментами с непарным шел-
копрядом.
24
ОБЩАЯ ЧАСТЬ
Acronicta lutea Brem. et Grey. Рост гусениц этого вида при температуре
25—30° Ц сопровождается обильным развитием охристых и желтых пиг-
ментов у них и затем появлением бабочек с яркожелтыми задними крыль-
ями. При пониженных температурах (ниже 25°) желтые пигменты разви-
ваются слабо или полностью отсутствуют. Желтые пигменты этого вида
принадлежат к типу липохромов (легко растворимы в спирту, эфире
и хлороформе); они пропитывают покровы тела гусениц, а затем и имаго
и концентрируются также в желтке яиц. Наблюдается тесная связь
распространения типичной желтой формы, Acronicta lutea Brem.
et Grey, — с южными широтами (Китай, Маньчжурия) и белой ее формы,
Acronicta lutea leucoptera Btl., с северными частями ареала (Амур, Забай-
калье, южная Сибирь). Подобную же связь распространения с южными
и северными частями ареала и с разными стациями дают желтая и белая
формы Leucoma flavosulphurеа Ersch. Желтая (типичная) форма этого
вида распространена в предгориях Тянь-шаня-и Ала-тау, белая форма,
cretacea Stgr., известна из Семиречья (Нарын) и из высокогорий (запад-
ный Памир). Точно так же и в пределах рода Euproctis Hb. обращает вни-
мание почти чисто белая окраска типично палеарктических видов (Е. si-
mi li s Fuessly, chrysorrhoea L.) и охристая или желтая-—видов, распростра-
ненных в тропиках или на юге Палеарктики (Е. susanna Stgr.). Для близ-
кого к Euproctis Hb. рода Artaxa Wlk., распространенного в тропиках
и в южных широтах Палеарктики, типична желтая и охристая окраски
почти для всех видов. Желтые и охристые окраски распространены очень
широко и в других родах Orgyidae, распространенных в тропиках.
Желтые и охристые пигменты замещают белые окраски лишь при от-
сутствии меланинов; при развитии последних эти окраски вытесняются,
ограничиваясь областями тела, бедными темными пигментами (задние
крылья), или являясь окраской покровов личиночной фазы.
Развитие зеленых пигментов значительно менее интенсивно в пределах
семейства, но также типично для южных, преимущественно субтропи-
ческих, форм (Саппа Wlk., Belciana Stgr., Daseochaeta Warr., Мота Hb.).
Распределение зеленых пигментов, в отличие от охристых, характери-
зуется концентрацией их на передних крыльях и связью с меланинами;
черные и зеленые окраски передних крыльев нередко замещаются (Сга-
niophora Snell.). На примере Craniophora ligustri Schiff, можно видеть
различную степень развития зеленых пигментов у разных географических
форм. Китайская форма Craniophora ligustri gigantea Drd. характеризуется
яркой окраской и наличием большого количества зеленого пигмента на
передних крыльях, тогда как европейская, типичная форма окрашена
более бледно и лишена зеленого пигмента. Развитие зеленых окрасок
у гусениц Orgyidae незначительно, главным образом за счет прозрачности
кожи и просвечивания зеленой окраски крови (Craniophora Snell., Ну-
Ьота НЬ.).
Наибольшего развития, особенно у видов Orgyidae умеренных широт,
достигают меланистические окраски. С развитием меланинов связано
и развитие рисунка. Меланины лишь отчасти замещаются желтыми
и зелеными пигментами, чаще же они приобретают доминирующее значе-
ние. Развитие меланинов несомненно стимулируется пониженными тем-
пературами, сопровождающими рост. Это можно видеть как из географи-
ческого распределения окрасок, так и из экспериментов. Так, опыты,
поставленные мною над Acronicta leporina L., показали, что рост гусениц
этого вида при температуре 20—25° сопровождается развитием белой
или желтоватой окраски кожи и волосяного покрова, а при низкой темпе-
МОРФОЛОГИЯ ПРЕИМАГИНАЛЬНЫХ ФАЗ 25
ратуре — развитием черных волос. Бабочки, развивающиеся в таких усло-
виях, имеют в. первом случае белую, во втором — серую окраску. Имеется
связь распространения пигментных форм A. leporina L. с термическим
режимом мест обитания; более темные формы (leporella Stgr., bradypo-
rina Tr.) свойственны северным широтам и северо-восточной Азии.
Наличие изменений окраски и, в частности, развитие меланисти-
ческих черт при низких температурах отмечено для гусениц Orgyia
antiqua L. Бобровым (1930). Это же констатировано и мною, кроме
отмеченных выше случаев с Acronicta leporina L., также для О. antiqua L.,
О. gonostigma F., Calocasia coryli L., Daseochaeta alpium Osb. Наряду
с этим Бобровым описаны формы гусениц О. antiqua L. с белой и черной
окраской дорзальных щеток; эти признаки удерживаются в ряде поколе-
ний и дают свои изменения. Точно так же для непарного шелкопряда
описаны Гольдшмидтом (1934) разные географические цветовые наслед-
ственные формы; некоторые из них' (японские) имеют светлую окраску и
напоминают гусениц ивовой волнянки.
Цветовые формы бабочек Orgyidae, описанные большею частью как
аберрапии (формы -неизвестного генезиса), многократно указывались
в систематической литературе. Гейнрих (Heinrich, 1916) описал форму
златогузки (A. plumbociliata Hein.), характеризующуюся затемненным
терминальным краем крыльев. Урбан (Urbahn, 1935) описал ряд темных
форм от Laelia coenosa Steph.; у этого вида меланистические формы могут
иметь как затемненную периферию крыльев, так и диффузное (по жил-
кам) распределение темного пигмента (L. nigrociliata Urb.). Известна
меланистическая форма Leucoma salicis L. с черной бахромой и подобная же
форма Arsilonche albovenosa Goeze. Возникновение меланистических форм
Orgyidae может быть связано с дефектами циркуляции крови в крыловой-
пластинке в период развития ее. Они получены также под действием
лучей рентгена Бобровым (1930).
Глава II
ВНЕШНЯЯ МОРФОЛОГИЯ ПРЕИМАГИНАЛЬНЫХ ФАЗ
Строение преимагинальных фаз Orgyidae обнаруживает большое разно-
образие. Яйца обычно довольно крупны, диаметр их около 0.5—1.0 мм.
Они всегда имеют плотный хорион, скульцтированный, чаще в виде ра-
диальных ребрышек, или несущий очень мелкую, точечную скульптуру,
редко гладкий, блестящий. Хорион более плотен у видов подсем. Orgy-
inae, нередко настолько, что труден для разрезания скальпелем (.lene
Hb., Gynaephora Hb., Orgyia Ochs.). Форма яиц разнообразна, но можно
наметить два основных их типа. Гладкие яйца, нескульптированные,
шаровидные, нередко слегка вдавленные на дорзальной стороне или
вытянутые, типичны для видов подсем. Orgyinae и для многих Momini
из Acronictinae (Мота Hb., Diphthera Ochs.). Уплощенные или полушаро-
видные яйца, несущие обычно скульптуру в виде радиальных ребрышек,
типичны для большинства видов трибы Acronictini. Из. последних лишь
у Oxycesta Hb. яйца почти круглые, хотя несут обычную для Acronictini
рубчатую скульптуру. Полукруглые или почти круглые яйца, но также
несущие рубчатую скульптуру, свойственны большинству видов Momini
(Calocasia Ochs., Daseochaeta Warr.).
Окраска яиц Orgyidae или связана с пигментацией хориона, или,
При более прозрачном и тонком хорионе, зависит от пигментов желтка
26
ОБЩАЯ ЧАСТЬ
яиц, что свойственно, например, большинству видов Acronictinae. В пер-
вом случае окраска яиц матовобелая или светлосерая (Ocneria Hb., Olene
Hb., Dasychira Steph., Gynaephora Hb., Orgyia Ochs.), редко зеленоватая
(Diphthera Ochs); во втором случае окраска их зеленоватая (Calocasia
Ochs., Subacronicta Kozh.), беловато-желтая (Daseochaeta Warr., Acronicta
Ochs.), охристая или оранжевая (Acronicta lutea Brem. et Grey). Яйца
второго типа обычно меняют окраску при развитии в связи с возникно-
вением пигментов (чаще охристых или коричневых) в серозной оболочке
(виды Acronicta Ochs., Subacronicta Kozh.). При толстом хорионе измене-
ния окраски яиц при развитии почти или совсем незаметны; они обнару-
живаются обычно лишь в течение первых часов или даже минут после
откладки и незадолго до выхода гусениц. Так, у непарного шелкопряда
или у Orgyia antiqua L. свежеотложенные яйца — зеленовато-желтого
цвета, но спустя несколько часов они, в связи с твердением хориона,
приобретают типичную окраску: беловато-желтую у первого вида ц мато-
вобелую— у второго. У непарного шелкопряда
и у монашенки окраска яиц меняется перед
выходом гусениц, чего у видов Orgyia Ochs, не
наблюдается. По Христенсен (1937), у яиц
Orgyia antiqua L. хорион спустя несколько
часов после откладки сильно твердеет и ста-
новится более хрупким. У этого вида яйца,
уходящие в зимовку, имеют хорион, пропитан-
ный коричневым пигментом, что не характерно
для них в течение лета. Микропилярный аппа-
рат яиц Orgyidae специально изучен лишь для
Orgyia antiqua L. Христенсеном (1937). Микро-
пилярные отверстия в числе 3—5 у этого вида
располагаются на микропилярной пластинке,
ппагтиикя йи^Р°Рил?Рная имеющей вид розетки. Эта пластинка распо-
пластинка яйца Orgvia ап- г г
tiqua L. (По Христенсену, ложена на верхней стороне яйца, на месте
1937). ’ вдавления (рис. 11).
Структуры гусениц, в частности структуры
их кожных покровов очень разнообразны. У молодых, только что отродив-
шихся из яиц гусениц некоторых Acronictinae (Лcronicta Ochs., Triaena
Hb., Daseochaeta Warr.) наблюдается развитие пупилл, или склероти-
зованных щитков при основании щетинок, при слабом развитии вторичных
волосков и кожных бугров (бородавок). Редко, например у Episema Ochs.,
такое строение кожных покровов остается и у взрослых гусениц, или
вместо волосков на бородавках или пупиллах развиваются многочисленные
шипики (Triaena Hb.). У Acronicta alni L. с ростом наблюдается рудимента-
ция некоторого числа щетинок и более сложное строение остающихся
(рис. 12). Подавляющее большинство видов Orgyidae характеризуется обиль-
ным развитием вторичных волосков и развитием больших, нередко сложных
бородавок на коже уже с первой стадии гусениц (рис. 13). Часто обиль-
ное появление вторичных волосков наблюдается после линьки во вторую
стадию. Вторичные волоски у гусениц Orgyidae располагаются большею
частью только на бородавках, редко на коже, преобладая лишь на кож-
ных буграх (неполных бородавках), как то типично для взрослых гусе-
ниц Acronicta Ochs, и Triaena Hb. (leporina L., aceris L., psi L., tri-
dens Schiff., cuspis Hb.).
Строение головы гусениц довольно однообразно. Изменчивы размеры
головы; в некоторых случаях она заметно крупнее, чем обычно (Subacro-
МОРФОЛОГИЯ ПРЕИМАГИНАЛЬНЫХ ФАЗ
27
nicta Kozh.). Форма головы также несколько различна. У одних видов
она угловатая (Daseochaeta Warr.), у других — округлая (Acronicta Ochs.)
или со слабо выраженными углами полушарий. Эпикраниальный шов
то глубоко вдавлен (Daseochaeta Warr.), то едва заметно углублен
(рис. 14). Лобный склерит по форме и размерам однообразен; несколько
меньше он у видов Orgyinae и Momini и крупнее у видов Acronictini',
clypeus обычно пропорционален лбу. Верхняя губа всегда с вырезкой
на вентральной стороне, более глубокой у видов Orgyinae. По сторонам
головы по шесть глазков. Черепная коробка покрыта вторичными щетин-
ками, в большинстве случаев обильными (Gynaephora Hb., Acronicta Ochs,
и многих других родов семейства), лишь иногда их немного или совсем
нет, как у некоторых видов рода Acronicta Ochs, (alni L.).
Рис. 12. Типы волосков и бородавок гусеницы Orgyidae. 1 — Ocneria dispar L.;
2—Orgyia antiqua L.; 3— Acronicta alni L.; 4—Gynaephora lugens Kozh.
Строение ног, исключая хетотаксию, однообразно. Грудные ноги
не имеют специфичных черт. обычного для гусениц типа. Они несут один
коготь, снабженный зубцом. Последний может отсутствовать у некото-
рых видов. Форма когтей мало характерна. Брюшные ноги всегда в числе
пяти пар на брюшных сегментах, расположены по обычному для всех
Caradrinina типу (на третьем, четвертом, пятом, шестом и десятом сегмен-
тах). Подошвы их, имеющие вид плоской складки, несут однорядные
и одноярусные, длинные, загнутые на концах крючья, расположенные
в виде длинной, посредине несколько выгнутой полосы. У видов подсем.
Orgyinae крючья и подошва ног несколько длиннее (рис. 15).
Кожа гусениц Orgyidae несет железы или типа осматерий, или откры-
вающиеся на поверхности кожи каналами. Железы типа осматерий харак-
терны для видов подсем. Orgyinae. Они располагаются в подавляющем
большинстве случаев на тергитах шестого и седьмого брюшных сегмен-
тов и снаружи заметны в виде окрашенных (красных, желтых) вороно-
чек. Редко (у Leucoma Steph, и Ocneria Hb.) такие же железы есть также
на двух первых брюшных сегментах. У видов Ocneria Hb., кроме этих
непарных сегментарных железок, есть еще мелкие парные, расположенные
также на тергитах брюшных сегментов. Активное состояние желез, сопро-
28
ОБЩАЯ ЧАСТЬ
вождает состояние возбуждения гусеницы. Секрет желез не изучен, Желе-
зистый характер носят также у некоторых видов Orgyidae {Euproctis Hb.)
волоски, покрывающие кожу. Эти волоски внешне не отличимы от обыч-
ных. Основание их снабжено железистой клеткой. Секрет этих однокле-
точных железок выделяется по каналу волоска и является ядовитым
для человека, вызывая у него воспаление кожи [Павловский и Штейн
(Pawlowsky u. Stein, 1927)].
Расположение бородавок на коже гусениц Orgyidae вполне соответ-
ствует расположению первичных щетинок, как то было показано Дай-
ром (Dyar, 1894) для гусениц вообще. Возникновение бородавок из пер-
вичных волосков на гусеницах Orgyidae легко проследить цри изучении
стадийных изменений их структур. Они возникают всегда за счет усиле-
ния щетинки и образования пупиллы при ее основании. Далее наблюдается
разрастание вздутия- до небольшой бородавки, покрытой мелкими шипи-
ками или тонкими волосками, сидящими рядом с основной, первичной
Рис. 13. Гусеница Dasychira albodentata Brem. (Рис. Ф. Гуняева).
щетинкой. Заканчивается образование бородавки развитием длинных,
вторичных волосков, равных но размерам первичной щетинке или даже
более длинных.
Расположение бородавок и первичных щетинок на сегментах гусениц
Orgyidae следующее (рис. 16). На переднегруди щетинки а, р, у и S распо-
лагаются обычно на общем щитке; у взрослых гусениц они редко предста-
влены одиночными щетинками (Episema Ochs., Acronicta alni L.).1 Часто
1 Я пользуюсь терминологией Фракера (Fracker, 1915) для обозначения хето-
таксии Orgyidae.
Для хетотаксии гусениц чешуекрылых предложено несколько терминологических
схем [см. Герасимов (Gerasimov, 1935,1937)]; все они сводятся к трем. Одни авторы
[Дайр, 1901 ;Квэл (Quail, 1904); Джерман (Garman, 1921); Герасимов, 1935] используют
цифровые обозначения, принимая твердые понятия для первой, второй и т. д. щети-
нок на каждом сегменте. Система каждого автора оригинальна, и первая или пятая
щетинка одного имеет иной номер у другого. Пользование этими обозначениями обя-
зывает не только постоянно помнить автора, по которому ведется счет, но также не-
удобно в случае смещения щетинок. Другие авторы [Ширбэк (Schirbeek, 1917); Кузне-
цов, 1915; Герасимов, 1935] используют словесное обозначение щетинок, пытаясь
характеризовать и закрепить их положение на сегменте •— subdorsale, supraventrale
и т. д. Эти обозначения также неудобны при сильных смещениях щетинок и затруд-
нительны в связи с большим числом систем. Третьи авторы [Тсу (Tsou, 1914): Фракер,
1915] используют для обозначения щетинок алфавит (латинский или греческий),
не претендуя на территориальное закрепление щетинок или их групп цифровым
порядком или словесно. Эта терминология кажется мне наиболее практичной, к тому
же она наименее запутана, так как греческий алфавит использован только одним
автором.
МОРФОЛОГИЯ ПРЕИМАГИНАЛЕНЫХ фаз
29
эти группы сидят в виде пучков волосков, слабо отделенных друг от друга,
например, у видов Orgyia Ochs., Olene Hb., Diphthera Ochs., Мота Hb,
У большинства видов Orgyidae у взрослых гусениц, сливаясь, они обра-
зуют общее скопление волосков на щитке переднегруди, редко сохраняя
некоторые следы самостоятельности (Dasychira Ochs.). Лишь у немногих
видов Orgyidae (Ocneria rubea Hb., Parocneria Dyar, Daseochaeta Warr.)
на переднегруди наблюдается возникновение самостоятельных борода-
вок на месте первичных щетинок а, (3, у и сидящих на общем, но
не сильно склеротизованном щитке. В первой стадии гусениц Acronic-
tinae эти группы очень часто представлены одиночными щетинками.
В этом случае на переднегрудном щитке с каждой стороны располагается
всего по четыре щетинки (Daseochaeta Warr., Acronicta aceris L., A. psi L.,
A. lutea Brem. et Grey). Чаще переднегрудной щиток несет более четырех
одиночных щетинок, например пять у Calocasia coryli L., семь у Subac-
ronicta Kozh. (рис. 236), или пучки волосков, как то типично для всех
видов подсем. Orgyinae.
Рис. 14. Голова гусениц Orgyidae. а—Daseochaeta alpium Osb.;
б — Ocneria monacha L.
Группа p на переднегруди всегда самостоятельна. В старших стадиях
гусениц она почти всегда состоит из пучка щетинок (минимум из двух);
одной щетинкой она представлена лишь у взрослых гусениц Episema
coeruleocephala L. и Acronicta alni L., но у первого вида бородавка при
основании этой щетинки несет также короткие волоски. Эта бородавка
у Orgyidae является комплексной, возникающей за счет слияния с груп-
пой г, причем в подавляющем большинстве случаев е мигрирует к группе
р; последняя остается на своем месте. Лишь у Daseochaeta Warr, слияние
этих бородавок обусловлено миграцией щетинок р. В первой стадии
можно констатировать часто только две щетинки — р и е. Редко наблю-
дается одна, тогда это р так как г утрачивается. Присутствие пучка
волосков на месте этих двух щетинок есть вторичное явление, но не ред-
кое для первой стадии гусениц Orgyidae.
Перед стигмой на переднегруди располагается большая комплексная
бородавка, возникающая за счет бородавок х и -с. Очень редко она несет
только две щетинки (Episema Ochs.) или одну (Acronicta alni L.); в послед-
нем случае одна из них (?)) рудиментарна. Утрата этой щетинки взрос-
лыми гусеницами Acronicta alni L. несомненна, так как в первой стадии
гусениц у них на переднегруди перед стигмой всегда имеется две щетинки.
30
ОБЩАЯ ЧАСТЬ
У подавляющего большинства видов Orgyidae группа х с первой личиноч-
ной стадии несет большое количество щетинок, нередко очень длинных,
торчащих вперед перед головой в виде кистей (Orgyia Ochs., Laelia Steph.,
Dasychira Ochs.). У Acronictinae, наоборот, в первой личиночной стадии
очень часто группа х переднегруди несет только две щетинки (х и z).
Число щетинок возрастает лишь после первой линьки. Редко число их
уменьшается после линяния (Л. alni L.).1
Вентрально от стигм на переднегруди располагается бородавка
группы к, которая у взрослых гусениц очень редко несет одну (Episema
Ochs.) или две щетинки (Acronicta alni L.). Две щетинки в группе я
очень типичны для первой стадии многих Acronictinae. Обычно группа
тг несет пучок волосков и превращена в большую бородавку. При основа-
нии передней ноги расположены щетинки групп а и т, в первой стадии
гусениц в числе немногих щетинок, в старших стадиях в виде пучков
волосков, сидящих на склеротизованных щитках.
Рис. 15. Строение брюшных ног гусениц Orgyidae.
а—Acronicta (Pharetra) rumicis L.; б — Ocneria monacha L.
Хетотаксия средне- и заднегруди почти тождественна. Здесь с каждой
стороны сегмента располагается не более пяти групп щетинок, не считая
щитков, лежащих по сторонам основания ноги — групп т и а. Щетинки <т
и [В средне- и заднегруди обнаруживают среди Orgyidae тенденцию к сбли-
жению и объединению. Для родов подсем. Orgyinae типично положение
их в виде обособленных бородавок; лишь у некоторых родов (Olene Hb.,
Gynaephora Hb.) они сильно сближены и даже отчасти слиты ( у видов
последнего рода). У Penthophera Germ., Parocneria Dyar. в области распо-
ложения этих двух бородавок находится лишь одна; она, возможно,
-образовалась в результате слияния двух. У видов Leucoma Steph, и Ocne-
ria Hb. группа а представлена одной первичной щетинкой, в то время
как (3 развита в виде большой бородавки; у предыдущих родов есть осно-
вание считать вероятным слияние этой одной щетинки с бородавкой р.
1 На переднегруди группа & видимо рудиментарна, так как во всех случаях,
когда перед стигмой располагаются только первичные щетинки, они сидят в числе
двух. Развитие вторичных щетинок ведет к возникновению пучка волосков
у Orgyidae', третья щетинка в этой группе одна не появляется.
МОРФОЛОГИЯ ПРЕИМАГИНАЛЬНЫХ ФАЗ
31
В пределах подсем. Acronictinae слияние бородавок а и IS на средне-
и заднегруди является правилом.
У взрослых гусениц Orgyidae бородавка а и 8 ра средне- и заднегруди
редко бывают в виде простых щетинок; это наблюдается, например,
у Subacronicta megacephala Schiff. и Acronicta alni L. У большинства видов
они с первой личиночной стадии представлены пучками щетинок, сидя-
щими на бородавках. У гусениц Acronictinae в первой стадии они часто
Рис. 16. Схема расположения щетинок и бородавок на пе-
редне- и среднегруди и на брюшных сегментах (первом — вось-
мом) для первой (У) и для последней (2) стадии гусениц
Orgyidae.
представлены самостоятельными одиночными (первичными) щетинками,
сидящими на пупиллах (Subacronicta Kozh., Triaena Hb., Acronicta Ochs.);
часто они объединяются основаниями и после первой линьки образуют
одну общую бородавку. У гусениц Calocasia coryli L. они уже в первой
стадий полностью слиты, причем есть лишь одна большая щетинка на этой
бородавке, вероятно первичная, тогда как прочие — мелкие, вероятно
вторичные; одна из первичных щетинок этой комплексной бородавки
у Calocasia рудиментируется.1
1 Слияние групп а и 3 на средне- и заднегруди у Calocasia coryli L. несколько
отлично от того, что наблюдается у других Acronictinae, ибо у последних нет редук-
ций первичных щетинок при слиянии этих двух групп. Факт рудиментации одной из
первичных щетинок этих групп сближает процесс слияния а и в у Calocasia и видов
подсем. Orgyinae.
32
ОБЩАЯ ЧАСТЬ
Группа f средне- и заднегруди представлена всегда самостоятельной
бородавкой или пупиллой. В первой стадии у видов подсем. Acronicti-
пае, включая и Calocasia coryli L., она несет одну первичную щетинку
и лишь после первой линьки становится бородавкой. Средне- и задне-
грудь редко несет я обнаружил эту щетинку лишь у Subacronicta Kozh.,
у некоторых видов рода Acronicta Ochs, (leporina L., aceris L., rumicis L.,
menyanthidis View.), у Eogena Gn., Simyra Ochs., Arsilonche Ld., Oxy-
cesta Hb. У видов подсем. Orgyinae ее нет. Группа х на средне- и заднегруди
есть всегда. По Фракеру (1915), у Arctiidae и Orgyidae она является ком-
плексной, образованной слиянием групп х и 7). Изучение первой стадии
гусениц Orgyidae не дает основания для таких заключений. Это особенно
отчетливо на видах подсем. Acronictinae, у которых в первой стадии
гусениц эта группа нередко представлена всего одной щетинкой (Calo-
casia Ochs., Acronicta Ochs., Daseochaeta Warr.). Группа л всегда есть
в виде самостоятельной пупиллы или бородавки, чаще она несет пучок,
волосков, редтхО у взрослых гусениц одну щетинку (Acronicta alni L.).
У гусениц первой стадии обычно есть только одна первичная щетинка
в группе х.
Хетотаксия первых восьми брюшных сегментов однообразна; обнару-
живают отличия лишь два первых сегмента, а также седьмой и восьмой,
но только в строении вентральной их стороны. Дорзальная сторона
каждого сегмента характеризуется присутствием трех групп с каждой
стороны — а, р и о. Группы а и 13 очень часто располагаются рядом
и превращаются в плоские, большие, равные одна другой по размерам
бородавки. Они особенно велики на первых пяти или четырех брюшных
сегментах, когда несут дорзальные щетки волосков (Dasychira Ochs.,
Orgyia Ochs., Laelia Steph., Cynaephora Hb.). Часто также плоские боро-
давки а и ji развиваются на восьмом брюшном сегменте, когда последний
несет кисть волосков. Редко у Orgyidae на всех сегментах бородавки а
и р большого размера и плоские (Acronicta aceris L.). У ряда видов
Orgyidae (Euproctis Hb., Penthophera Germ., Parocneriq Dyar) бородавки
а и 13 всех брюшных сегментов сливаются, образуя общие волосистые
площадки; у видов Artaxa Wlk. они слиты на всех сегментах, кроме
первого брюшного; у видов Мота НЬ., наоборот, они слиты лишь на вось-
мом брюшном сегменте.
Группа х почти всегда есть на брюшных сегментах, но может утра-
чиваться и отсутствует, например, у Penthophera Germ. Обычно группа х
представлена первичной щетинкой не только в первой стадии, но
и у взрослых гусениц (Laelia Steph., Dasychira Ochs., Orgyia Ochs., Artaxa
Wlk., Calocasia Ochs., Episema Ochs., Eogena Gn. и у ряда видов Acro-
nicta Ochs.). Реже она представлена одной щетинкой лишь на некоторых
сегментах брюшка, тогда как на других несет пучок волосков, как, напри-
мер, у взрослых гусениц Acronicta aceris L., Simyra Ochs., Oxycesta Hb.
У многих видов Orgyidae (Subacronicta Kozh., Diphthera Ochs., Мота
Hb., Daseochaeta Warr., Gynaephora Hb., Leucoma Steph.) на всех сегмен-
тах брюшка группа х представлена пучком щетинок. Типично положе-
ние этой группы на брюшных сегментах прямо за стигмой, на ее уровне
(Gynaephora Hb., Artaxa Wlk., Мота Hb., Calocasia Ochs.). Нередко
она имеет такое положение на первых шести брюшных сегментах
и на восьмом, тогда как на седьмом сегменте она смещена вентрально
и сближена (Diphthera Ochs., Episema Ochs.) или слита (Daseochaeta
Warr., Acronicta О., Oxycesta Hb., Arsilonche Hb., Eogena Gn.) с группой tj.
Реже группа x сдвинута* дорзально и слита (Ocneria Hb., Parocneria Dyar,
МОРФОЛОГИЯ ПРЕИМАГИНАЛЬНЫХ ФАЗ
33
группа располагается
Я Л
Рис. 17. Хетотаксия девя-
того и десятого сегментов
брюшка гусеницы Episema
coeruleocephala L.
Leucoma Steph.) или сильно сближена {Subacronicta Kozh.) с группой р.
последнем случае смещение щетинок z наблюдается на всех сегментах
брюшка, кроме седьмого.
Под стигмой брюшных сегментов всегда располагаются две боро-
давки — -си у, тогда как положение групп v и <т на сегментах брюшка,
несущих ноги или лишенных их, различно. Группы z и у в первой личи-
ночной стадии часто несут всего одну щетинку {Acronicta Ochs.), а у неко-
торых видов на сегментах, несущих ноги, р. не развивается, например,
на третьем, четвертом, пятом и шестом сегментах у Acronicta lutea Brem.
et Grey, Acronicta rumicis L.; у этих видов у- появляется лишь после пер-
вой линьки; v присутствует всегда, но на сегментах, несущих ноги,
она располагается на щитке на внешней стороне ноги; на первом, вто-
ром, седьмом и восьмом сегментах брюшка э
свободно на коже. Это же относится и к группе
т, которая на сегментах брюшка, несущих
ноги, располагается на внутренней стороне
пог. Обе последние группы часто представ-
лены первичными или лишь, немногими еще,
вторичными щетинками на сегментах, лишен-
ных ног, но на брюшных ногах обычно они
представлены пучками волосков, даже в тех
случаях, когда наблюдается сильная общая
редукция вторичных и отчасти первичных во-
лосков {Episema Ochs.). В первой личиночной
стадии эти обе группы часто представлены
лишь первичными щетинками не только у
Acronictinae, но и у Orgyinae {Olene Hb.,
Gynaephora Hb.).
Девятый и десятый сегменты брюшка гу-
сениц Orgyidae как в отношении размеров,
так и хетотаксии обнаруживают некоторые
черты рудиментации; особенно это касается
последнего сегмента, который также очень силь-
но модифицирован. Девятый сегмент (рис. 17)
уменьшен, но на его дорзальной стороне с боков, дорзально, как обычно,
располагаются щетинки а, (Зир. Латерально расположены щетинки z и т;,
а с вентральной стороны <т и v; группа у. на этом сегменте не развита.
Группы эти, исключая группу а, большею частью несут по нескольку
щетинок каждая; одни первичные присутствуют лишь в первой стадии.
В некоторых случаях щетинки а и [В могут сливаться, например у Laelia
Steph., и тогда несут длинный пучок волосков. Десятый сегмент сильно
изменен и несет на тергите, превращенном в анальный щиток, с каждой
стороны по четыре группы щетинок: <z., j3, р и, вероятно, /., сдвинутую
к р; При основании ноги, в виде щитка с вторичными щетинками, раз-
растается группа л, а с внутренней стороны ноги небольшой щиток,
несущий пучок щетинок с. Бородавки девятого сегмента обычно мало
отличаются пр размерам от предыдущих, но на десятом сегменте они
часто рудиментарны и на анальном щитке нередко слиты одна с дру-
гой.
Волосяной покров гусениц различен в отношении густоты, длины
волосков и формы их. Эти особенности, хотя и в разной мере каждая,
сильно связаны с возрастными изменениями гусениц. Некоторые общие
моменты этих изменений при росте кожи могут быть отмечены. Длина
3 Фауна СССР, И. В. Кожанчиков.
34
ОБЩАЯ ЧАСТЬ
волосков гусениц Orgyidae. большею частью значительна; часто она более
ширины их тела; в ряде случаев (Acronicta rumicis L., A. auricoma Schiff.}
пропорции к размерам тела при росте остаются одинаковыми, но у мно-
гих видов они сильно меняются по возрастам (Acronicta lutea Brem. et
Grey, A. alni L.). Обычно волоски, покрывающие тело гусеницы, щети-
нистые, реже тонкие, шелковистые (Acronicta leporina L., A. aceris L.,
Daseochaeta alpium Osb.); наиболее типичная форма волосков пильчатая
(рис. 12). Волоски несут по сторонам небольшие щетинки. Такие волоски,
типичны для первой стадии гусениц всех видов Orgyidae. Простые волоски
редки; напротив, обычны сильно перистые волоски или со многими боко-
выми отросточками, как, например, на дорзальных щетках (ynaephora.
Hb., или с длинными вильными отростками, образующими булаву, как,,
например, на бородавках z переднегруди гусениц Orgyia Ochs. Для
взрослых гусениц Acronicta alni L. очень характерны длинные, на концах
расширенные в форме лопаточек волоски. Для первой стадии гусениц
Ocneria Hb. типичны волоски, вздутые .при основании или в базальной
половине,' которые Вахтль и Корнаут (Wachtl u. Kornauth, 1893) оши-
бочно описали как аэрофоры. Холодковским (Cholodkowsky, -1894) было1
показано, что полость этих волосков наполнена жидкостью; позже Инге-
ницкий предложил для них название «токсофор». Коллинз (Collins, 1915)
описал эти волоски для непарного шелкопряда как везикулярные, руко-
водствуясь тем, что он не нашел специальных железистых клеток при
основании этих волосков, хотя секрет в полости волосков он обнаружил;
им также была обнаружена большая трихогенная клетка в основании
волоска. Роль аэрофоров у мелких гусениц непарного шелкопряда играют,,
как выяснилось, длинные и перистые волоски, в большом количестве
покрывающие тело; они характерны, кроме монашенки и непарного
шелкопряда, и для других видов рода Ocneria Hb. и прочих Orgyidae
(Orgyia Ochs., Dasychira Steph.).
Изменения строения тела гусениц при росте затрагивают преиму-
щественно кутикулярные структуры и в первую очередь размеры боро-
давок, число и форму волосков на них. В меньшей мере затрагивается
положение отдельных щетинок (или бородавок) на сегментах тела. В пре-
делах подсем. Orgyinae изменения при росте гусениц меньшие, чем у гусе-
ниц Acronictinae. В пределах рода Euproctis Hb.. изменений в хетотаксии
при росте гусениц нет; у видов Ocneria Hb. наблюдается лишь смена
вздутых волосков и длинных пильчатых щетинок, типичных для первой
стадии, более короткими, но также пильчатыми волосками, свойствен-
ными всем старшим стадиям гусениц. У видов Orgyia Ochs, бородавки
st и р в первой стадии на первых четырех сегментах брюшка свободны
и не несут перистых волосков в форме щеток; после второй линьки эти
бородавки, разрастаясь, сближаются и почти срастаются в плоскую пло-
щадку (между ними остается еще шов, исчезающий у Euproctis Hb.).
У видов Orgyia Ochs, кисточки перистых волосков на щетинках z передне-
груди и на а и (3 восьмого сегмента появляются также в третьей стадии;
в первой личиночной стадии волоски на этих бородавках пильчатые
такие же, как на прочих бородавках тела. Слияние бородавок а и {3 при
росте гусениц, подобное тому, что описано для Orgyia Ochs., наблюдается
также у Dasychira Steph, и Olene Hb., тогда как у С ynaephora Hb. имеет
место лишь их разрастание и сближение в старших стадиях, но не сраста-
ние. Срастание бородавок а и [3 на брюшных сегментах наблюдается в стар-
ших стадиях гусениц также у некоторых видов рода Acronicta Ochs.
(eporina L.).
МОРФОЛОГИЯ ПРЕИМАГИНАЛЬНЫХ ФАЗ
35
В пределах подсем. Acronictinae изменения кутикулярных структур
при росте более разнообразны и сложны. Характерно резкое увеличение
числа вторичных волосков после первой линьки, хотя наблюдается уве-
личение числа их также и в старших стадиях. У некоторых видов Acro-
nictinae (Triaena) развитие вторичных волосков в старших стадиях слабое
и наблюдается даже их редукция (Acronicta alni L.). Меньшее развитие
иторичных волосков у гусениц первой стадии видов Acronictinae по срав-
нению с Orgyinae, вероятно, связано с выходом их из яиц на более ранней
стадии онтогенеза. Это видно хотя бы из того, что гусеницы Acronictinae
п именно видов, для которых характерно недоразвитие вторичных щети-
нок в первой стадии, линяют большее число раз, чем Orgyinae. Так, напри-
мер, у видов Acronicta Ochs., Triaena Hb. наблюдается не менее шести
линек, а в ряде случаев семь и даже восемь, тогда как у Orgyinae —
четыре, пять и лишь у самок некоторые видов шесть. Виды Acronictinae,
ныходящие из яиц с обильным развитием вторичных волосков (Diphthera
Ochs., Calocasia Ochs., Pharetra Hb.), линяют меньшее число раз, не более
шести. Недоразвитие вторичных волосков у видов Acronicta Ochs, и
Triaena Hb. в эмбриональном развитии может быть связано с малыми
размерами яиц и бедностью их желтком, по сравнении со всеми осталь-
ными видами этого подсемейства и тем более видами Orgyinae. У видов
Triaena Hb. и Acronicta Ochs, наблюдаются наибольшие изменения струк-
тур при росте. Факт выхода из яиц недоразвитых гусениц Acronictinae
рисуется мне как частность биологии этих видов, вызвавшая модифи-
кацию личиночных структур некоторых биологических специализован-
пых форм.1
Изменения структур кутикулы при росте Acronictinae могут быть
разбиты на две группы. Из числа изученных мною форм у Calocasia Ochs,
и Diphthera Ochs, изменения кутикулярных структур при росте очень
близки к тому, что типично для Orgyinae. Здесь молодые, только что
отродившиеся из яиц гусеницы имеют вполне сформированные бородавки
но только на переднегруди (на ее щитке), но и на всех сегментах тела,
при наличии, впрочем, известного числа щетинок, сидящих поодиночке
на пупиллах. У этих видов нет увеличения числа линек при росте или
оно не велико (Pharetra Hb.). Резкие изменения в облике гусениц насту-
кают после первой линьки, когда на месте каждой группы щетинок воз-
никают большие бородавки, несущие большие пучки волосков. У Calo-
easia Ochs, первичные щетинки сохраняют большую величину по сравне-
1 Из излагаемых фактов можно легко видеть, что Оедность желтком яиц Triaena
lib. и Acronicta Ochs, является следствием высокой специализации биологии има-
гииальной их фазы. Существенно отметить, что из яиц мелких и бедных желтком,
гппичных для ряда видов Acronictinae, выходят гусеницы, имеющие небольшое число
пгоричных щетинок. Развитие вторичных волосков компенсируется дополнительными
птьками. У Noctuidae, яйца которых также мелки и бедны желтком (хотя в ряде слу-
чнои, например у Hadeninae и не столь бедны, как у Acronictinae), выходящие из яиц
। уышицы всегда имеют лишь первичные волоски, и этот тип личиночных структур
остпотся неизменным до окукления. Совки линяют обычно шесть, редко семь (Agro-
Itn) раз. Таким образом для Noctuidae можно говорить о первичной простоте личи-
ночных структур; она является для них. окончательной, не требующей компенсации
при росте и сопровождает нормальное развитие, тогда как у Acronictinae появление гу-
ПЧ11Щ с малым числом вторичных щетинок — явление вторичное, требующее компен-
< пц|1И и компенсируемое в процессе роста добавочными линьками. У Noctuidae, в экспе-
риментальных условиях даже если и возникают добавочные линяния (например при
। vкости у гусениц озимой совки восемь линек), то они никогда не ведут к изменениям
1ИЧИПОЧНЫХ структур и появлению вторичных щетинок; они являются компенсатор-
ными лишь к. условиям роста в целом. Отсутствие значения добавочных линек в струк-
ivpni.rx изменениях отмечено также для A. ypsilon Rott. [Риплей (Ripley, 1923)].
3*
36
ОБЩАЯ ЧАСТЬ
нию с прочими. Облик взрослой гусеницы начинает проявляться обычно
с третьей стадии, например, пучки волосков, торчащие вперед, на боро-
давках [з среднегруди у Calocasia Ochs., увеличенные в размерах боро-
давки [В на первом брюшном сегменте у Acronicta rumicis L. Окончатель-
ный облик взрослых гусениц приобретается в четвертой стадии; это
касается главным образом окраски и размеров волосков. В некоторых
случаях, впрочем, окончательной является пятая стадия, например
у Acronicta lutea Brem. et Grey, приобретающей лишь в последней стадии
длинные шелковистые волоски, белую боковую линию и более плоские,
несколько меньшего размера, чем в предыдущих стадиях, бородавки.
У видов второй группы {Daseochaeta Warr., Subacronicta Kozh.,
Triaena Hb.) гусеницы первой стадии характеризуются отсутствием боро-
Рис. 18. Особенности хетотаксии первого, брюшного
сегмента гусениц Calocasia coryli L. по стадиям I—V.
второй группы обладают увеличенным числом личиночных стадий, неко-
торые до восьми {Acronicta psi L.). Первая линька у них характерна появ-
лением пучков волосков и небольших бородавок или на всех сегментах
тела, или лишь на некоторых (на переднегруди у Subacronicta megacephala
Schiff.). В третьей стадии наблюдается сильное увеличение бородавок,
а затем, до пятой и шестой стадий, происходит постепенное изменение
их размеров и числа волосков, покрывающих их. Облик взрослой гусе-
ницы приобретается или в пятой стадии {Acronicta alni L.), или в шестой
{Daseochaeta alpium Osb., Subacronicta Kozh.). При сильно усложненных
структурах некоторых видов {Acronicta aceris L., A. leporina L.) -облик
взрослой гусеницы проявляется лишь при крайне увеличенном числе
линек в седьмой и даже в восьмой стадии {Acronicta psi L.).
Более мелкие изменения при росте в структурах личиночной фазы
Orgyidae можно видеть из следующих кратких характеристик основных
типов.
У гусеницы первой стадии Calocasia coryli L. все бородавки, распо-
ложенные ниже стигм, а также бородавки х и (3 на брюшных сегментах
несут только первичные щетинки; прочие бородавки — комплексные.
Первичные щетинки имеют лишь т) и v на среднегруди. Длина всех щети-
нок в первой стадии однообразна. После первой линвки все группы,
МОРФОЛОГИЯ ПРРИМАГИНАЛЬНЫХ ФАЗ
37
кроме х сегментов брюшка, а также кроме v, с и у на седьмом брюшном
сегменте, на восьмом сегменте эти же плюс т, и а на первом и втором сег-
ментах брюшка, имеют вторичные щетинки. Щетинки х переднегруди
превращаются в большие выпуклые бородавки и несут длинные пучки
волосков. В третьей стадии все группы и на всех сегментах тела стано-
вится многощетинковыми, исключая группу х брюшных сегментов, кото-
рые и во всех остальных личиночных стадиях имеют первичные щетинки.
В третьей стадии на среднегруди развиваются мощные бородавки (3, несу-
щие длинные пучки волосков. Окраска тела в первых трех стадиях
ве меняется; волоски и кутикула серого цвета. В четвертой стадии разви-
ваются еще больших размеров пучки волосков на бородавках [В среднегруди
в появляются всюду на бородавках частично рыжие и черные волоски. Этот
тип гусеницы, но с большей примесью рыжих волосков остается и в пятой
стадии (рис. 18).
Рис. 19. Особенности хетотаксии первого брюшного сег-
мента гусениц Daseochaeta alpium Osb. по стадиям I—VI.
У Daseochaeta alpium Osb. в первой стадии все группы являются одно-
щетинковыми (рис. 19), а изменения хетотаксии при росте подобны
только что описанным.
У Subacronicta megacephala Schiff, смены личиночных структур при
росте иные. В первой стадии переднегрудной щиток несет, кроме обычных
шести щетинок с каждой стороны, еще по две вторичных. Группы х передне-
груди и второго брюшного сегмента несут по две щетинки, тогда как
все прочие щетинки сидят поодиночке на пупиллах на всех сегментах,
в большинстве являясь первичными. Во второй стадии группа х. передне-
груди несет пучок щетинок, тогда как прочие группы остаются такими же,
кик и в первой стадии. В третьей стадии на переднегруди, кроме много-
щетинковой группы х, возникает многощетинковая группа тг; на брюш-
вых сегментах также появляются многощетинковые группы х и т„ Группа
z па брюшных сегментах в этой стадии смещается дорзально, к основа-
нию группы р, исключая седьмой сегмент, где она смещается вентрально
к т). В последующих стадиях число вторичных щетинок возрастает на
группах, лежащих вентрально от стигм, тогда как дорзальные группы
остаются однощетинковыми. Облик взрослой гусеницы проявляется
। пятой стадии (рис. 20).
38
ОБЩАЯ ЧАСТЬ
У Acronicta aceris L; и других видов подрода Acronicta Ochs, изме-
нения структур при росте специфичны. Первая стадия характеризуется
большинством первичных щетинок, сидящих на пупиллах; небольшие
бородавки, несущие две или три щетинки, расположены лишь на дор-
зальной стороне тела (группа а); есть небольшие волосистые площадки
па дорзальной стороне брюшных сегментов. Со второй стадии группы а
превращаются в настоящие бородавки, они несут обычные пильчатые
волоски. В третьей стадии группы а, 3 и р брюшных сегментов приобре-
тают вид выпуклых бородавок; ниже стигм располагаются лишь первич-
ные волоски, сидящие на пупиллах. В четвертой стадии облик гусениц
резко меняется, так как бородавки дорзальной стороны тела становятся
плоскими и несут многочисленные тонкие волоски; тонкие же волоски
покрывают и вентральную сторону тела, скрывая первичные щетинки.
В пятой и последующих стадиях (до седьмой) бородавки остаются такими
Рис. 20. Особенности хетотаксии первого брюшного
сегмента гусениц Subacronicta megacephala Schiff,
по стадиям I—VI.
же плоскими, как и в четвертой, но, кроме того, они сдвигаются так,
что а. и р располагаются почти на одном уровне. В шестой стадии возни-
кает очень большое количество волосков на коже между бородавками
и приобретается вполне облик взрослой гусеницы (рис. 21). Окраска
первых пяти стадий одинакова: кутикула серая, волоски и пупиллы
или бородавки черные или черно-серые, голова тоже черная. Волоски
на первом и втором, а также на шестом и седьмом брюшных сегментах
более светлые. С пятой стадии появляются охристые и оранжевые пучки
волосков и на дорзальной стороне тела рисунок в виде колец.
У видов Triaena Hb. группы psi L. изменения структур гусениц при
росте иного типа. Гусеницы первой стадии имеют одиночные щетинки,
сидящие на пупиллах, исключая сближенные на средне- и заднегруди
щетинки гири парные щетинки х'и ~ переднегруди; развитие вторичных
щетинок очень слабо (рис. 22). После первой линьки возникают бородавки
с сидящими на них отдельными сильными щетинками (первичными)
и большим числом мелких (вторичных) щетинок. Бородавки а на первом
брюшном сегменте увеличиваются больше, чем другие. В третьей стадии
возникают плоские кожные щитки на месте бородавок, а бородавки а
переднего брюшного сегмента сильно увеличиваются в размерах и сли-
ваются в один непарный вырост, покрытый мелкими шипиками. В чет-
МОРФОЛОГИЯ ПРЕИМАГИНАЛЬНЫХ ФАЗ
39
вертой и пятой стадиях увеличиваются лишь размеры выроста на первом
абдоминальном сегменте, когда он достигает предельных размеров.
На этих стадиях увеличивается до предела число вторичных волосков.
Последующие личиночные стадии не вносят изменений в хетотаксию.
Рис. 21. Особенности хетотаксии первого брюшного сегмента
гусениц Acronicta aceris L. по стадиям I—VII.
Окраска при росте гусениц меняется следующим образом. В первой
.и второй стадиях окраска кожи беловатая с выделяющимися на ней чер-
ными щетинками и черными пятнами на первом грудном и первом, чет-
вертом, пятом и восьмом брюшных сегментах. В третьей стадии окраска
Рис. 22. Особенности хетотаксии первого брюшного сегмента
гусениц Acronicta psi L. по стадиям I—VIII.J
более однотонная и темная, а вдоль спины появляется желтая полоса.
В четвертой и пятой стадиях окраска остается примерно одинаковой
и лишь усиливается дорзальная полоса. В шестой стадии появляются
красные пятна по сторонам дорзальной полосы, последняя же становится
40
ОБЩАЯ ЧАСТЬ
более светлой. Этот тип окраски и кожных структур остается в после-
дующих стадиях до окукления.
У Acronicta alni L., вида очень бедного вторичными щетинками.,
в течение всего развития гусениц не появляется бородавок. Изме-
нения структур первых личиночных стадий заключаются в усилении
щетинок, пупилл и склеротизованных площадок, на которых они сидят.
Вторичные щетинки появляются лишь в предпоследней стадии. Четвер-
тая стадия гусениц характеризуется появлением толстых пластинчатых
волосков на бородавках а и [3 переднегруди. В пятой стадии появляются
пластинчатые волоски на всех бородавках |3 и развиваются сильные скле-
ротизованные пластинки на тергитах сегментов. Окраска первых трех
стадий однообразна — светлосерая с черным; гусеницы щетинистые,
щетинки сильно выделяются на кутикуле. Четвертая стадия типична
черной окраской кутикулы с чисто белыми пятнами на ней, а в пятой:
Рис. 23. Особенности хетотаксии первого брюшного
сегмента гусениц Acronicta lutea Brem. et Grey no
стадиям I—V.
стадии появляются характерные желтые или желто-белые поперечные-
широкие площадки на тергитах.1
У Pharetra lutea Brem. et Grey и других видов этого подрода Acro-
nicta Ochs, гусеницы первой стадии имеют только дорзальные бородавки'
(а), несущие вторичные щетинки; прочие щетинки сидят поодиночке
на пупиллах (рис. 23). После первой линьки все группы образуют боро-
давки, обильно покрытые вторичными волосками; особенно крупны!
бородавки а, р и т, а также и другие, только х и S остаются с одной пер-
вичной щетинкой. В третьей и четвертой стадиях все бородавки увели-
чиваются, но сохраняют пупиллы, хотя и приобретают много волосков..
Бородавки х и <т на брюшных сегментах становятся также многощетин-
ковыми, впрочем первая лишь на некоторых. В пятой стадии наблюдаются:
уменьшение размеров бородавок и более плоская их форма; меняется;
также тип волосков •— они становятся более длинными и тонкими..
1 По Холодковскому (1903), окраска этих площадок на тергитах обусловлена,
пигментом, расположенным в клетках гиподермы, тогда как кутикула, покрываю-
щая их, прозрачна. Волоски гусениц Acronicta alni, имеющие форму лопаточек,,
должны быть отнесены к типу железистых, ибо трихогенные клетки таких волосков-
имеют железистый характер и связаны каналом с полостью волоска.
МОРФОЛОГИЯ ПРЕИМАГИНАЛЬИЫХ ФАЗ
41
Группы z и а на брюшных сегментах вновь становятся бедны волосками.
В пятой стадии приобретается типичный облик взрослой гусеницы и появ-
ляются яркие белые боковые полосы.
В пределах подсем. Orgyinae интенсивное развитие первичных волос
наблюдается с первой стадии, причем рост гусениц вносит незначитель-
ные изменения в хетотаксию, как и у видов подрода Pharetra Hb. из Acro-
nictinae. Специфичные черты появляются обычно уже после первой
линьки, а типичный облик взрослой гусеницы данного вида часто прояв-
ляется уже после второй линьки (в третьей стадии). В качестве примера
изменений хетотаксии при росте Orgyinae может служить схема изме-
нения структур при росте гусениц Orgyia antiqua L. (рис. 24).
При переходе в пронимфу у гусениц Orgyidae часто наблюдаются
изменения в окраске кутикулы, исключая
немногие формы, например непарный шелко-
пряд, монашенка. Окраска кутикулы или
бледнеет (большинство Orgyinae), или возни-
Рис. 24. Особенности хетотаксии первого
брюшного сегмента гусениц Orgyia antiqua
L. по стадиям I—IV.
Рис. 25. Куколка Leucoma
salicis L. (Рис. Ф. Гуня-
ева).
кает более яркая окраска, например красная окраска пронимф Calocasia
coryli L., оранжевая Acronicta leporina L. и A. aceris L. Часто в этот
период меняется также окраска волосков, например у Acronicta leporina
L. волоски чернеют. Нередко происходит также уменьшение (втяги-
вание) бородавок.
Куколки Orgyidae характеризуются присутствием волосяного покрова
в виде пучков волосков по бокам брюшка за стигмами и пучков их на дор-
зальной стороне тела и на голове. Редко, например у с ynaephora Hb.,
волоски развиваются в виде густого слоя, войлока, покрывающего дор-
зальную сторону куколки. В ряде случаев, именно у Acronictinae, степень
развития волосков на кутикуле куколок не велика. Например, у видов
Triaena Hb. и Hyboma Hb. волоски сидят в виде коротких пучков только-
за стигмами. Пучки волосков у куколок имеют в основе то же положение,
что и бородавки на кутикуле гусениц (рис. 25).
Кутикула куколок несет разную скульптуру в различных частях
тела; Она всегда есть в области cremaster и очень часто развита на сег-
42
ОБЩАЯ ЧАСТЬ
ментах брюшка, особенно на их дистальных концах и в базальной поло-
вине. Реже скульптура развита на кожном покрове головы и в торакаль-
ной области. Скульптура кутикулы различна; обычно она в виде ямок
или бугорков (например в базальной части брюшных сегментов), имею-
щих разный диаметр, разную глубину или выпуклость; на грудном
отделе скульптура развита обычно в виде складочек. Голова и конеч-
ности тела или гладкие, или несут шипики и выросты, иногда значи-
тельные, например у Pharetra Hb. У ряда видов кутикула куколок совер-
шенно гладкая и блестящая {Daseochaeta Warr.).
Строение cremaster очень разнообразно, обычно характеризует роды
и подроды, относительно редко — виды. Cremaster Orgyidae, являясь,
как всегда, придатком десятого брюшного сегмента, имеет вид кониче-
ского выроста, несущего на конце крючья или шипы; поверхность его
обычно имеет глубокие борозды или ямки. Расположение крючьев или
щетинок на cremaster особенно типично для крупных групп. У видов
Рис. 26. Cremaster куколок Orgyidae.
1—• Leucoma salicis L.; 2—Diphthera coenobite. Esp.; 3— Acronicta cuspis Hb.;
4 —- A. tridens Schiff.
Pharetra Hb. он в виде короткого округлого выроста, несущего пучок
густых щетинок; у видов Triaena Hb. дистальный конец cremaster несет
небольшое число длинных крючьев, обычно шесть или восемь. У Diph-
thera coenobita Esp. имеется один длинный, направленный дистально шип,
несколько притупленный на конце; у Calocasia coryli L. cremaster в виде
нетолстого морщинистого выроста, несущего короткие крючья; у видов
Orgyia Ochs, cremaster тонкий, конический, с короткими шипами на конце
(рис. 26). По форме и общему типу строения cremaster могут быть выде-
лены группы родов, близкие и по прочим признакам. Так, например,
виды Acronicta Ochs., Craniophora Snell., Eogena Bd., Simyra Ochs., Arsi-
lonche Ld. имеют общий облик cremaster в виде короткого тупого выроста,
несущего пучок щетинок. Общий тип строения cremaster свойствен Dasy-
chira Steph., Olene Hb., Orgyia Ochs, и Gynaephora Hb. — родам, близким
также по прочим структурам как личинок, так и имаго.
Форма куколок также различна в пределах разных родов и нередко
может характеризовать родовые группировки (группы близких родов).
Так, куколки Orgyia Ochs, сильно сплющены дорзовентрально, сильно
расширены посередине и сужены к концам; такой же тип куколок у Dasy-
chira Steph., Gynaephora Hb., Olene Hb. Куколки Ocneria Hb. типичны
сильно суженным коническим брюшком и вальковатой формой. Это же
АНАТОМИЯ
43
наблюдается у куколок Leucoma Steph, и Calocasia Ochs. Куколки Acro-
nicta Ochs, почти цилиндрической формы; это же типично для Craniophora
Snell., Simyra Ochs., Arsilonche Ld.
Глава III
ОСОБЕННОСТИ АНАТОМИЧЕСКОГО СТРОЕНИЯ
Из черт анатомического строения Orgyidae требуется характеризовать
лишь некоторые и прежде всего изменения анатомического строения,
происходящие в метаморфозе, а также некоторые особенности анатоми-
ческого строения имагинальной фазы. Анатомия личиночной фазы мало
характерна и совпадает во всех существенных чертах с общими схемами
описания анатомии гусениц, приводимыми в сводках. Из анатомических
черт личиночной фазы следует отметить лишь сильное развитие скелет-
ной мускулатуры, а также соединительной ткани (жирового тела) и всегда
значительное развитие прядильных желез. У гусениц Orgyidae кишеч-
ный тракт по объему занимает наибольшую часть полости тела; причем
весьма значительные размеры имеет средняя кишка. Нервная система
слагается из подглоточного и надглоточного головных ганглиев, трех
грудных и восьми брюшных. Трахейная система во всех без исключения
случаях перипнейстического типа (одна пара мезоторакальных стигм,
сдвинутых на переднегрудь, и восемь пар брюшных) с шестью продоль-
ными стволами трахей и сильным ветвлением их отростков. Кровеносная
система представлена сердцем, построенным из десяти нерезко дифферен-
цированных камер и короткой аорты.
Наибольшие изменения в строении претерпевают при развитии Orgyi-
dae пищеварительные органы и половая система, причем наибольшее
разнообразие в строении наблюдается у имаго. Специализованная биоло-
гически имагинальная фаза Orgyidae, помимо изменений скелета, всегда
характеризуется большей или меньшей рудиментацией пищеварительных
органов и относительной гипертрофией половой системы. Для такого
развития половой системы материалом являются личиночная мускула-
тура и соединительная ткань. Требуется подчеркнуть, что у Orgyidae
в имагинальной фазе налицо не только рудиментация органов, но также
и недоразвитие их, что ведет в ряде случаев к сохранению примитивных
черт их строения.
В строении пищеварительных органов наблюдаются изменения, парал-
лельные тому, что описано для ротовых органов имаго. У видов Orgyinae
характеризующихся полной рудиментацией ротовых частей (Ocneria Hb.,
Orgyia О.), наблюдается также полная рудиментация средней кишки
и слюнных желез в имагинальной фазе. Больших размеров зоб является
у них продолжением почти нормального пищевода; 1 он срастается по-
средством узкого эктодермального канала, непосредственно с задней киш-
кой. Последняя у Orgyinae.развита вполне и имеет вид тонкой цилиндри-
ческой трубки, у выхода снабженной большим слепым резервуаром
(соесшп). Слепой мембранозный резервуар имеет .очень тонкую кутикулу
и открывается коротким ректальным отделом (rectum) задней кишки
наружу. В личиночной фазе Orgyidae сильно развита, средняя кишка
1 По своей форме и положению относительно пищевода зоб Orgyinae очень напо-
минает зоб Jugata, Zeuzeridae, некоторых низших Tineidae и Psychidae [Петерсен
(Petersen, 1900)].
44
ОБЩАЯ ЧАСТЬ
и недоразвиты прочие. Число мальпигиевых сосудов у Orgyidae в личи-
ночной и имагинальной фазах одинаково (рис. 27).
Подбирая разные виды Orgyidae по степени редукции кишечника*
имаго, можно видеть все переходы от сильно рудиментированных пище-
варительных органов к нормальным для чешуекрылых. Так, у Calocasia
coryli L. (рис. 28) в имагинальной фазе средняя кишка утрачена не вполне:
от нее остается рудимент в виде короткого, несколько вздутого отдела.
Положение зоба в этом случае также более типично; он отделен от пище-
вода и от основания средней кишки протоком, хотя коротким и широким,
но все же самостоятельным. Строение прочих частей пищеварительных
Рис. 27. Пищеварительные Рис. 28. Пищеварительные органы имаго Diphthera
органы имаго Orgyia anti- coenobita Esp. (1) и Calocasia coryli L. (2).
qua L.
органов сходно с тем, что описано для Orgyinae. Меньшая рудиментация
средней кишки налицо у Diphthera Ochs., у которой она может быть
даже функционирует, например для всасывания воды (рис. 28). Наконец
у видов Мота Hb., Acronicta Ochs, и Daseochaeta Warr, пищеваритель-
ный аппарат вполне типичен для Lepidoptera. Передняя кишка большая,
длинная и толстая, снабженная восемью пилорическими отростками
в проксимальной области (рис. 29). Хорошо развиты слюнные железы;
зоб вполне отделен от пищевода. Слюнные железы у видов Orgyinae,
Calocasia Ochs., Diphthera Ochs., а также, вероятно, у Simyra Ochs.,
Oxycesta Hb., Eogena Bd. и других видов Orgyidae с рудиментарными
ротовыми частями, утрачены. Для видов Ocneria. Hb., в частности мона-
шенки и непарного шелкопряда, установлено отсутствие пищеваритель-
ных ферментов в кишечнике, что связывается с рудиментацией секретор-
ных отделов пищеварительного аппарата средней кишки и слюнных
желез.
АНАТОМИЯ
45
Рис. 29. Пищеварительные
органы имаго Acronicta aurico-
та Schiff.
то они не дозревают,
гонада самок формируется
У Orgyidae обращает внимание разная степень развития и дифферен-
циации задней кишки, в частности ее слепого отдела. Параллельно с руди-
ментацией средней кишки размеры слепого отдела задней кишки возра-
стают и достигают максимума при полной утрате средней кишки. С уве-
личением слепой кишки стенки ее становятся все более тонкими; у видов,
имеющих короткую слепую кишку, стенка ее имеет ячеистую скульптуру.
Изменения размеров слепой кишки носят адаптивный характер. Являясь
резервуаром для экскрементов в период метаморфоза, слепая кишка
дает параллельное увеличение размеров с усилением процессов гистолиза
и гистогенеза. Изменения строения тела в период метаморфоза у Orgyidae
касаются не только перестройки органов вегетативных функций, но
(у видов с рудиментарными ротовыми частями) главным образом гистоло-
гической структуры органов в связи с использованием всех резервных
материалов и продуктов гистолиза тканей
для формирования половых продуктов и раз-
вития гонады. У всех непитающихся в
имагинальной фазе видов Orgyidae формиро-
вание половой системы и полное развитие
главной массы яиц заканчивается на фазе
куколки. Таким образом слепая кишка в
этом случае служит местом изоляции про-
дуктов обмена, при таком типе метаморфоза
очень обильных. У самцов Orgyinae слепая
кишка несколько меньше, чем у самок, осо-
бенно при наличии полового диморфизма
(Orgyia Ochs.), что, вероятно, связано с .бо-
лее слабым развитием гистолитических про-
цессов у них по сравнению с самками.
Формирование половой системы у боль-
шинства видов Orgyidae сдвинуто на ранние
фазы онтогенеза.. У видов Orgyinae оно проте-
кает в последних стадиях личиночной фазы
(рис. 30), тогда как развитие куколок
характеризуется созреванием яиц. Выходя-
щие из куколок самки Orgyia Ochs, имеют
вполне развитые и зрелые яйца, сконцентри-
рованные в педицеллярных отделах яйцевых
трубочек (рис. 31). Область вителлярия
яичников у видов этого рода в фазе имаго
вичтожна и не содержит созревающих яйце-
клеток, а если таковые там образуются,
У видов Ocneria Hb. и Leucoma Steph.
также в личиночной фазе, но с меньшей полнотой, чем у Orgyia Ochs,
(в частности не образуются яйцевые камеры), а созревание яиц охваты-
вает не только период развития куколок, но и первые часы жизни имаго.
< '.вежевышедшие самки ивовой волнянки имеют 30—50 незрелых яйце-
клеток в каждой яйцевой трубочке и такое же количество зрелых, гото-
вых к откладке. В период имагинальной жизни лишь небольшая часть
незрелых яиц (наиболее крупных) дозревает в течение первых суток,
тогда как 20—30 мелких ооцитов большею частью так и остаются несо-
зревшими после гибели самки (рис. 32). Степень дозревания ооцитов
в имагинальной фазе у видов Orgyidae с редуцированным питанией, воз-
можно, стоит в связи с упитанностью особи в личиночной фазе, но полное
46
ОБЩАЯ ЧАСТЬ
созревание всего запаса ооцитов, по всей вероятности, осуществляется
редко.1 2
По Шпренгелю (Sprengel, 1928) у самок непарного шелкопряда
сразу после выхода из куколок количество зрелых яиц в яйцевых
трубочках менее половины общего их числа. Автором приводятся сле-
дующие данные постметаморфного созревания яиц у непарного шелко-
пряда (табл. 3)?
Таблица 3
Постметаморфное развитие яиц непарного шелкопряда
(по Шпренгелю, 1928)
Возраст самок (в часах) Число яиц в яйцевых трубочках Общее число яиц % зрелых яиц
зрелых незрелых
3 148 252 400 37.0
5 . 200 250 450 44.4
7 178 233 411 42.5
8 181 361 542 33.5
12 308 238 546 56.4
15 501 135 630 77.1
18 486 107 593 80.7
24 564 63 627 90.0
У видов Acronicta Ochs, и других Acronictinae с хорошо развитым
имагинальным питанием формирование гонады и развитие яйцевых камер
происходит в фазе куколки, но созревание яиц падает на имагинальную
фазу. Яйца созревают в течение всей жизни особи, обычно в течение
20—30 суток, причем первые зрелые яйца появляются на 2—6-е сутки
после выхода самки из куколки. Период созревания яиц сильно зависит
от термического режима при жизни особи. Строение гонады у видов
этой группы несколько отлично от предыдущей (рис. 33). Яйцевые тру-
бочки имеют относительно небольшой педициллярный отдел и в це-
лом слагаются из громадного вителлярия, содержащего обычно в каж-
1 Изложенные факты, касающиеся характера созревания ооцитов Orgyidae,
имеют большой интерес для вопросов влияния условий среды на половую продук-
цию самок при росте и развитии Orgyidae. Из них можно видеть, что даже для Or-
gyinae не только условия личиночного роста могут влиять на плодовитость, хотя для
видов Orgyinae они очень важны. Могут влиять на плодовитость и условия жизни
имаго. Условия, определяющие плодовитость самок Acronictinae, еще более, сложны
в связи с большим сроком формирования и созревания ооцитов. Они близки к тем,
которые известны и описаны для большинства чешуекрылых [см.: К о ж а н ч и-
к о в. 1938. Плодовитость чешуекрылых в зависимости от экологических условий.
Зоол. журн., 16 (4): 644—663].
2 Несмотря на то, что значительная часть яиц у самок непарного шелкопряда
не развита в первые моменты после выхода из куколок, оплодотворение (спаривание)
самок наблюдается сразу после выхода их из куколок и в подавляющем большинстве
случаев в природе происходит в первые сутки. Это типично для всех Orgyinae и мно-
гих Acronictinae {Calocasia Ochs., Oxycesta Hb.). Фактическое оплодотворение яиц
происходит в момент прохождения яиц мимо ostium ductus receptaculi, т. е. в течение
Всего периода яйцекладки. Недостаточно ясен характер оплодотворения яиц у неко-
торых видов рода Orgyia Ochs, {dubia Tausch:, prisca Stgr., ochrolimbata Stgr. и неко-
торых других).
АНАТОМИЯ
47
дой трубочке около 50 крупных, ооцитов (близких к полному созре-
ванию) и большое число мелких; мелкие ооциты характеризуются малым
количеством желтка.1
Гонада самок Orgyidae построена, как и у большинства Frenata,
из восьми трубочек по четыре в каждой половине яичника. Яйцеводы
и общий яйцевод короткие. В общий яйцевод в средней или нижней его»
части впадает проток семеприемника, снабженного
железой. Рядом с ним впадает в общий яйцевод
проток совокупительной сумки. Близ выхода общего
яйцевода в него впадает проток парной клеевой же-
лезы (glandula sebacea). Резервуары клеевой железы
вливаются почти непосредственно в короткий проток. i
Они представляют расширение каждого из выводных
Рис. 32. Вершинная
область яйцевых тру-
бочек яидника Leuco-
ma salicis L. 1 — сра-
зу после выхода из
Рис. 30. Яичник (левая
половина) Orgyia gono-
stigma F. на стадии
взрослой гусеницы.
Рис. 31. Половая система сам-
ки Orgyia antiqua L. сразу
после выхода из куколки (пра-
вый яичник удален).
куколки; 2 — после
окончания яйцеклад-
ки, незадолго до ги-
бели (/ — вершинная
нить; II— гермарий ;
III — вителлярий).
Steph.). Сама придаточная
(Ocneria Hb., Leucoma
железа является трубчатой. В некоторых слу-
чаях, например у Leucoma Steph., дистальная часть
общего яйцевода заметно изменена. Она расширена,
мускулиста (рис. 34) и образует небольшой, слепой
вырост, направленный проксимально,' В эту расширенную часть
общего яйцевода впадает большая придаточная железа. Железа
1 Снодграсс (1935) считает, что у чешуекрылых яйцевые трубочки всегда харак-
н'ризуются большими педицеллярными отделами и малыми вителляриями. Это, как
видно, справедливо лишь для видов, не питающихся в имагинальной фазе, во всяком
I'.iyrae в пределах изучаемого семейства.
48
ОБЩАЯ ЧАСТЬ
Рис. 33. Яичники Acronictinae (правая половина). 1 — Acronicta ps-i
U. сразу после выхода из куколки при нормальной зимовке; 2 — А.
lutea Brem. et Grey, развившейся без диапаузы (при 27°).
Рис. 34. Дистальный отдел половой системы самки Leucoma
salicis L. gl. s — glandulae sebaceae; r. s— receptaculum
seminis; о. c — oviductus communis; ov — oviductus; d. s —
ductus seminalis.
БИОЛОГИЯ
49
своеобразна, имеет короткие прямые пальцевидные отростки, но сама,
кик правило, трубчатая.
Строение половой системы самцов Orgyidae мало характерно. Строе-
ние семенников обычное для Frenata —оба они одеты общей оболочкой
н образуют комплексный непарный орган. Каждый семенник слагается
из четырех камер. Семепроводы отходят или отдельно от каждой половины
семенника (обнаруживают более примитивное положение), как то типично
для Orgyinae, или при выходе также объединены общей обкладкой и идут
раздельно лишь ниже комплексного семенника (Acronicta Ochs., Subac-
ronicta Kozh.).
Г л а в a IV
БИОЛОГИЯ
Смена поколений Orgyidae у разных их видов в связи с различиями
циклов развития проходит неодинаково. Между общими условиями
обитания и цикликой развития Orgyidae имеется связь, однако на харак-
тер циклов развития непосредственно влияют также особенности погод-
ных условий данного года, наличие или отсутствие зимовки, пищевой
режим и, в частности, ритм роста кормовых растений и ряд других фак-
торов.
В тропиках виды Orgyidae населяют главным образом лесные терри-
тории, т. е. районы с устойчивым, как сезонным, так и суточным термо-
гисрорежимом. Будучи полифагами, Orgyidae дают в тропиках смену
поколений в течение года примерно с одинаковым ритмом. Саванны
п тропические степи заселены видами Orgyidae значительно беднее.
Таким образом поливольтинизм для видов подсем. Orgyinae является
безусловно основной и исходной формой цикла развития. Виды подсем.
Acronictinae в тропиках представлены бедно; они заселяют преимуще-
ственно горные территории. Биология их не изучена, циклы развития
lie известны.
В умеренных широтах Orgyidae населяют также лесные территории,
причем численность видов подсем. Orgyinae здесь ниже, чем Acronictinae.
Для лесных видов Orgyidae умеренных широт типичен моновольтинизм
г наличием диапаузы в той или иной фазе развития, в которой наблю-
дается зимовка данного вида. Бивольтинизм среди лесных Orgyidae
редок, а поливольтинные виды отсутствуют. Кувада (Kuwada, 1929)
указывает для Dasychira strigata Moore и D. lunulata Btl. на Хондо в год
дна поколения; лёт одного происходит в апреле, другого — в августе.
В материковой полосе лесной зоны Евразии встречаются лишь моно-
польтинные виды Orgyinae (Dasychira Steph., Ocneria Hb.). Моноволь-
niiiiibi также большинство видов Acronictinae, распространенных в зоне
хвойных лесов Евразии (Diphthera Ochs., Daseochaeta Warr., многие
пиды Acronicta Ochs.). Напротив, Orgyidae, распространенные в степной
полосе Палеарктики, в известной части являются би- и поливольтин-
пыми. Таковы, например, виды Orgyia Ochs, (gonostigma F., antiqua L.),
Purocneria Dyar, многие виды Pharetra Hb. Тем не менее и здесь большая
часть Orgyinae и многие виды Acronictinae моновольтинны, например
пиды Eogena Gn., Oxycesta Hb., отчасти Sirnyra Ochs. Поливольтинизм
палеарктических видов Orgyidae представляет явление вторичное, так как
пи связан у этих видов с диапаузой. В табл. 4 сопоставлены длительность
развития фаз и циклы некоторых видов Orgyidae, преимущественно
4 Фауна СССР, И. В. Кожанчиков.
50
ОБЩАЯ ЧАСТЬ
из числа изученных мною. Длительность развития приводится для сред-
них термических условий порядка 15—25°; для зимующих стадий дана
длительность развития включая также и период зимовки.
Табл ица 4
Сезонное распределение и длительность развития фаз некоторых видов Orgyidae
Название вида Длительность развития Фаза зимовки
яйца гусеницы куколки имаго
*Orgyia postica Wlk 7—11 16—20 7— 9 3—6 Нет
О. antiqua L 9—10 м 25—50 5—20 3—8 Яйцо
0. gonostigma F 8—30 . 9—10 м 6—20 3—8 Гусеница
**Parocneria, amanda Stgr. . . < 7—10 24—26 8—11 — »
Olene fascelina L 8—40 9—10 м 10—30 6—12 »
Dasychira pudibunda L 9—30 35—60 8— 9 м 6—12 Куколка
Euproctis chrysorrhoea L. . . . 10—20 9—10 м 10—35 6—10 Гусеница
Ocneria dispar L 9—10 м 30—70 10—50 5—12 Яйцо
0. monacha L. . 9—10 м 49—120 8—45 5—15 »
Leucoma salicis L 8—40 9—10 м 10—30 6—10 Гусеница
Diphthera coenobita Esp. . . . 6— 7 32—56 9—10 м 6—12 КукОлка
Calocasia coryli L. . 7—21 35—70 9—Юм 5—14 »
Daseochaeta alpium Osb. . . . 5—15 17—50 9—10 м 20—25 ' »
**Subacronicta megacephala Schiff. 4—15 20—60 9—10 м 15—20 »
Acronicta alni L 4—16 26—70 9—10 м 15—20 »
A. leporina L 4—15 27—70 9—10 м 15—20 »
A. aceris L 4—16 30—80 9'—10 м 15—20 »
**A. psi L 5—14 26—60 9—10 м 20-25 »
A. menyanthidis View 6—15 26—75 9—Юм 15—20 »
**A. auricoma Schiff 4—20 20—90 9—10 м 20—30 »
**A. lutea Brem. et Grey . . . 5—16 17—60 9—10 м 2 О'— 30 >>
**A. rumicis L . . . 5—15 15—40 9—10м 20—30 >)
Примечание. Длительность развития дана в сутках, исключая те случаи,
когда поставлена буква «м» — тогда она дана в месяцах, включая и месяцы зимовки.
Одной звездочкой отмечены первично-поливольтинные виды, двумя звездочками —
вторичн'о-поливольтинные.
Обращает внимание (табл. 4) более быстрое развитие всех фаз поли-
вольтинных видов по сравнению с моновольтинными. Это справедливо
не только для исходных поливольтинных типов, например тропической
Orgyia postica Wlk., но и для вторично-поливольтинных, например Acro-
nicta rumicis L. по сравнении с моновольтинным Acronicta menyanthidis
View.
Исходным циклом Orgyidae является • поливольтинизм с одинаковыми
сроками, затрачиваемыми на развитие каждого поколения, т. е. то, что
наблюдается сейчас у видов, населяющих влажные тропики. Переход
к жизни в широтах с переменным климатом приводит первоначально
к возникновению обязательной диапаузы в личиночных фазах. Для видов
подсем. Acronictinae характерен этот второй тип циклов, так как это
подсемейство, видимо, сформировалось в условиях переменного климата.
Дальнейшая адаптация циклов'развития к переменному и неустойчи-
вому климату ведет к возникновению лабильной диапаузы, которая про-
является лишь в определенных условиях роста и развития особи и допу-
БИОЛОГИЯ
51
<кает, в зависимости от условий, моно-, ди- и поливольтинизм. Этот вто-
ричный поливольтинизм типичен для некоторых групп Acronictinae.
Зимовка у Orgyidae и, в частности, у видов подсем. Orgyinae, распро-
страненных в умеренных широтах, наблюдается в разных фазах цикла
развития, но никогда не происходит в имагинальной фазе. Часто зимовка
и той или иной фазе онтогенеза типична для видов отдельных родов.
Так, у видов рода Ocneria Hb. (monacha L., dispar L., matura Btl.) зимовка
типична в фазе яйца, причем диапаузирует уже вполне сформирован-
ная, почти готовая к выходу, пигментированная гусеница; зимовка иных
фаз никогда не наблюдается. Это же известно для некоторых Orgyia
Ochs, (antiqua L., dubia Tausch.), но здесь развитие яйца идет весною.
Ниды Olene Hb. и Gynaephora Hb. (rossii Curt., lugens Kozh., groenlandica
Hom., selenitica Esp.) зимуют в стадии полувзрослой гусеницы (четвертая
стадия). Средние же возрасты гусениц, но более молодые, чем у предыду-
щих родов, зимуют у. видов Euproctis Hb. и Arctornis Germ, и, вероятно,
также у Artaxa Wlk.; обычно зимует третья стадия сразу после линяния.
Мелкие гусеницы (вторая стадия) зимуют у Leucoma Steph, и, вероятно,
у видов Parocneria Dyar. Гусеницы (неизвестно какой стадии) зимуют
также у видов Numens Wlk. и Cifuna Wlk. Из видов рода Orgyia Ochs,
лишь О. gonostigma F. зимует в фазе гусеницы (третья и четвертая ста-
дии), тогда как остальные виды—в фазе яйца (antiqua L., thyellina Btl.,
ericae Germ., prisca Stgr.); 0. gonostigma F. принадлежит к наиболее гене-
рализованным видам рода. Также и видыDasychira Ochs.: часть их зимует
и фазе гусенипы (abietis Schiff., albodentata Brem.), часть в фазе куколки
(pudibunda L., lunulata Btl., strigata Moore). Диапауза куколок Dasychira
< )chs. не прочна и может протекать без воздействия отрицательных тем-
ператур.
Основной фазой зимовки видов Orgyinae является гусеница. Можно
считать, что зимовка и диапауза в фазе гусеницы для видов, заселяющих
широты с переменным климатом, являются исходным типом цикла.
За это говорит зимовка в фазе гусеницы тропических видов Orgyidae,
находящих в пограничные районы Палеарктики. В этой фазе у Orgyidae
происходит и формирование диапаузы в связи с адаптацией к зимовке.
Диапауза яиц видов рода Ocneria Hb. также, по существу, затрагивает
личиночную фазу, так как диапаузирует уже закончившая свое эмбрио-
нальное развитие гусеница. Диапауза яиц видов Orgyia Ochs., биологи-
чески высокоспециализованных, наблюдаемая на ранних стадиях эмбрио-
генеза [формирование мезодермы, по Христенсену (1937)], может рас-
сматриваться как дальнейший сдвиг диапаузы с личиночной фазы. Эта
трактовка тем более вероятна, что в роде Orgyia Ochs, есть виды (О. gono-
stigma F.), зимующие в фазе гусеницы. Диапауза куколок немногих
гпециализованных видов Dasychira Ochs. (D. pudibunda L. и некоторых
других) точно так же является вторичной, возникшей за счет сдвига
диапаузы с поздних личиночных возрастов на фазу куколки. У Dasy-
ehira abietis Schiff., например, зимуют гусеницы последней стадии.
Виды подсем. Acronictinae характеризуются зимовкой и диапаузой
и фазе куколки; это точно установлено для видов Calocasia Ochs., Оху-
I'esta. Hb., Subacronicta Kozh., всех изученных видов рода Acronicta Ochs.,
Diphthera Ochs., Daseochaeta Warr., Craniophora Snell., Eogena Bd.,
Simyra Ochs., т. e. для всех известных в отношении биологии видов Асго-
nietinae. Не изучены со стороны биологии маньчжурские и гималайские
нпдм этого подсемейства; возможно, что среди них, например видов
I'risulbides Hamps, или Anacronicta Warr., есть случаи зимовки в фазе
4*
52
ОБЩАЯ ЧАСТЬ
гусеницы. Диапауза куколок Acronictinae очень прочна, исключая виды
Calocasia Ochs.,- дающие, как и Dasychira Steph., вылет нормальных
бабочек из куколок без охлаждения их ниже нуля. Влияние отрицатель-
ной или близкой к 0° температуры необходимо для нормального течения
диапаузы и для вылета нормальных и плодовитых имаго у громадной
массы видов Acronictinae. Редко у видов Acronictinae наблюдается двух-
летняя диапауза {Acronicta aceris L.).
Очень своеобразен и интересен цикл развития Episema coeruleocephala
Esp. — он связывает циклы развития Orgyinae и Acronictinae. У Episema
Ochs., как и у всех видов Acronictinae, диапаузируют куколки, но диапауза
их протекает в течение лета (как у Malacodea из С eometridae). Бабочки
вылетают осенью и всегда погибают в этот же сезон, но яйца диапау-
зируют и зимуют, как ?о наблюдается у Ocneria Hb. и Orgyia Ochs.
[Голлас (Hollas, 1935)1.
Зимовка Orgyidae протекает большею частью близ поверхности почвы,
на лесной подстилке или непосредственно в ней, реже в почве. В част-,
ности, большинство видов, зимующих в фазе гусеницы, уходит в под-
стилку на поверхность почвы, зимуя под снегом, например виды Olene
Hb., Gynaephora Hb., Dasychira Ochs. Гусеницы некоторых видов, напри-
мер гусеницы Leucoma Steph/и некоторых видов Euproctis Hb. (Е. similis
Fuessly), изготовляют шелковые чехлики в щелях коры деревьев, где
и происходит их зимовка. Редко зимовка протекает в гнездах (Е. chry-
sorrhoea L.), изготовленных гусеницами из паутины и построенных из мно-
гих ее слоев, которыми они оплетают еще зеленые листья кормового
растения. Гнезда остаются висеть в течение всей зимы на ветвях деревьев.
Зимовка яиц происходит часто открыто, как, например, яиц Orgyia Ochs.,
зимующих на поверхности коконов самок, которые находятся в щелях
коры; в течение зимы яйца остаются под непосредственным воздействием
ветра, осадков и колебаний температуры. У видов Ocneria Hb. яйца боль-
шею частью откладываются в щели коры кучками, где и зимуют, причем
они покрываются или пенистыми выделениями, или, как у непарного
шелкопряда, слоем волосков с конца брюшка самки. Зимовка куколок
происходит всегда в приземном слое, обычно в лесной подстилке (Calo-
casia Ochs., Pharetra Hb., Daseochaeta Warr.), реже в старой, загнившей
древесине (Acronicta Ochs., Triaena Hb.), обычно в пнях, упавших
деревьях или в прикорневой части стволов ослабленных растущих деревьев
(Subacronicta Kozh.). Куколки зимуют обычно под защитой снегового
покрова.
Период размножения в цикле развития Orgyidae большею частью
охватывает начало или середину лета, так что период личиночного роста
совпадает с моментом наиболее интенсивного роста растений, т. е. охва-
тывает весну и начало лета. У моновольтинных видов Orgyidae период
роста и размножения тесно связан с фенологией древесных лиственных
пород, а период твердения листвы и ее усыхания совпадает с воз-
никновением диапаузы. У поливольтинных видов и эврифагов этой связи
нет, но и у нйх наиболее энергично размножение протекает в начале
лета, тогда как последующие поколения частично или полностью уходят
в диапаузу и остаются на зимовку. Редко лёт имаго падает на осень;
это наблюдается у поливольтинных Parocneria Dyar; из Acronictinae это
известно для Episema coeruleocephala Esp.
Суточный ритм жизни Orgyidae у разных видов и разных их фаз раз-
личен. Имаго большинства видов активны в ночные часы; таковы, напри-
мер, Dasychira Ochs., Olene Hb., Calocasia Ochs., Acronicta Ochs., Diph-
БИОЛОГИЯ
53
I hero, Ochs., Daseochaeta Warr. В других случаях, напротив, активность
надает на дневные часы; таковы Orgyia Ochs., (ynaephora Hb., отчасти
Ocneria Hb. В некоторых случаях бабочки активны в сумерки и перед
заходом солнца, например Laelia Steph., Leucoma Steph., Ocneria Hb.
Гусеницы имеют, невидимому, менее выраженный суточный ритм пове-
дения, хотя и здесь могут быть отмечены ночные виды, например Раг-
oeneria Dyar (Ramachandra, 1922), Calocasia Ochs. Большая часть Orgyi-
dae в гусеничной фазе активна в течение всех суток; периоды питания
прерываются лишь краткими периодами покоя или моментами линяния.
Может быть отмечена некоторая связь дневной активности Orgyidae
< зональным и широтным их распределением. Виды, распространенные
в горной тундре или Арктике, активны днем (f ynaephora Hb.). Днев-
ная же активность типична для некоторых степных и лесостепных видов
{Orgyia Ochs., Ocneria Hb., Ocnerorgyia Stgr.). Лесные виды являются
преимущественно ночными.
Образ жизни Orgyidae характеризуется некоторыми деталями, заслу-
живающими быть отмеченными. Откладка яиц самками происходит
по двум основным типам. По первому типу яйца откладываются кучками
сразу все или почти все в течение короткого срока. Этот тип яйцекладки
отмечен для видов Orgyinae и для некоторых Acronictinae {Oxycesta Hb.,
Sirnyra Ochs., Diphthera Ochs., Episema Ochs.); он наблюдается при
отсутствии необходимости отыскания специфичного пищевого субстрата
для гусениц. Яйца кучками откладываются на кору деревьев {Ocneria
Hb., Dasychira Ochs.), на ветви кустарников {-ynaephora Hb., Olene
I lb.), на подстилку в лесу {Dasychira Ochs.) или на кокон самки {Orgyia
Ochs.). Этот тип яйцекладки распространен преимущественно у видов
с рудиментарным имагинальным питанием, при малой подвижности
самок. Крайний случай представляет яйцекладка видов Orgyia Ochs.,
когда яйца откладываются на том месте, где выходит самка, а у некоторых
видов даже в полость кокона {Orgyia dubia Tausch., О. ochrolimbata Stgr.).
По второму типу яйцекладка происходит поодиночке или неболь-
шими кучками (по 30—20 яиц); она типична для активных самок, боль-
шею частью способных к питанию {Subacronicta Kozh., Acronicta Ochs.,
Craniophora Sn.), но наблюдается также у некоторых видов, не питаю-
щихся в имагинальной фазе, например у Calocasia Ochs. При кладке
поодиночке яйца откладываются почти всегда на нижнюю сторону
листьев кормовых растений, причем в ряде случаев {Subacronicta Kozh.,
Triaena Hb., Acronicta Ochs.) на листья определенного возраста.
Существует связь этих двух типов яйцекладки не только с актив-
ностью самок, но и с разной активностью гусениц, выходящих из яиц.
Самки, яйцекладка которых проходит по первому типу, мало активны,
но выходящие из яиц гусеницы характеризуются высокой способностью
к расселению, большею частью олиго- или полифаги с первых же дней
жизни. Самки с яйцекладкой второго типа очень активны, но выходящие
из яиц гусеницы очень слабо приспособлены к самостоятельной жизни,
могут питаться только строго определенными растениями, нередко только
паренхимой нижней стороны листа определенного возраста {Subacronicta
megacephala Schiff., Acronicta alni L.). Они чрезвычайно чувствительны
к толщине стенок клеток, слагающих паренхиму, и к толщине кутикулы
листа.
Различия в типах яйцекладки коррелированы также с запасом желтка
в яйцах, с дальнейшим темпом роста и типом развития гусениц. При
яйцекладке первого типа яйца богаты желтком, крупны, эмбриональное
54
ОБЩАЯ ЧАСТЬ
развитие протекает длительный срок, а выходящие гусеницы характери-
зуются развитием многих вторичных структурных и биологических черт.
При яйцекладке второго типа яйца бёдны желтком (исключая Calocasia
Ochs.), эмбриональное развитие коротко, а выходящие гусеницы харак-
теризуются не только слабым развитием вторичных черт, но обнаружи-
вают в ряде случаев недоразвитие органов, в связи с чем у этих видов
стоят удлиненный период роста и увеличенное число личиночных стадий.
Формирование типичного облика гусениц Orgyidae отнесено в этом слу-
чае на более поздние стадии (четвертую или третью).
Наличие у самок Orgyidae яиц, богатых желтком, приводит к чрез-
вычайному увеличению веса и размеров гонады, что, естественно, ограни-
чивает активность самок. Созревание большого количества крупных
яиц требует также больших количеств питательного материала, который
не может быть получен в имагинальной фазе прежде всего в связи с пони-
женной активностью самок. Это направление биологической специали-
зации ведет у Orgyidae к смещению периода созревания половых продук-
тов на более ранние стадии онтогенеза, когда созревание их осуще-
ствляется полностью за счет личиночного питания, отчего период личи-
ночного роста самок часто удлиняется (за счет последней стадии
гусениц). В связи с подобным созреванием половых продуктов стоит
рудиментация имагинальной фазы не только в отношении структур',
но также в отношении общей активности и длительности жизни. Яйце-
кладку поодиночке (разбросанно), казалось бы, естественно считать
примитивной, но в пределах сем. Orgyidae она обнаруживает черты спе-
циализации, не меньшие, чем откладка всего запаса яиц в одном месте.
Самки Subacronicta Kozh. и некоторых видов Acronicta Ochs, отыскивают
листья определенного возраста и определенных видов растений, расту-,
щих в специфичных условиях- (например при отсутствии прямого сол-
нечного освещения). Самки этих видов в высшей мере активны и биоло-'
гически полно приспособлены к специализованной яйцекладке; для них
характерны растянутый период созревания половых продуктов и зна-
чительная длительность жизни (около месяца, против пяти—шести
суток, максимум полмесяца, у видов с редуцированным имагинальным
питанием). У некоторых видов Pharetra Hb. наблюдается промежуточный
тип яйцекладки, при котором самки откладывают яйца небольшими куч-
ками (по 30—50 яиц), причем без особого выбора частей растения, но
всегда на зеленые растения.
Таким образом, рассматривая разные типы яйцекладки Orgyidae,
можно считать характерным для всех групп семейства наличие высокой
биологической специализации. Исключением является подсем. Orgyinae,
где специализация несколько меньшая вследствие отсутствия выбора
кормового растения; свойственный видам этого подсемейства тип яйце-
кладки вызвал возникновение ряда частных моментов биологической
специализации: обогащение яиц желтком, значительную и, в конце
концов, полную рудиментацию активности и питания самок, эври-
бионтность и, в частности, эврифагию выходящих из яиц гусениц.
Структурно, по циклам развития и по поведению виды подсем. Acro-
nictinae (Acronicta Ochs., Subacronicta Kozh.) обнаруживают большую
степень сложности. Характерен чрезвычайно специализованный тип
яйцекладки со строгим выбором не только кормового растения и его
части, но даже степени зрелости его листьев. Для этих видов налицо
сохранение примитивных чёрт первых личиночных стадий в связи с недо-
развитием гусениц в эмбриогенезе.
БИОЛОГИЯ
55
Количество яиц, продуцируемых самками разных видов Orgyidae,
различно и в значительной мере коррелировано с размерами яиц, типом
яйцекладки и степенью активности самок. В табл. 5 приведены данные,
характеризующие плодовитость разных видов Orgyidae в связи с длитель-
ностью жизни самок.
Таблица 5
Плодовитость и длительность жизни самок некоторых
видов Orgyidae
Название вида
Длитель-
ность
жизни
(в днях)
самки
Плодови-
тость
одной
самки 1
Olene fascelina L.................
Orgyia antiqtua L.................
O. gonostigma F...................
Ocneria dispar L..................
O. monacha L.2....................
Leucoma salicis L.................
Calocasia coryli L................
Diphthera coenobita Esp, . .. . ,
Daseochaeta alpium Osb............
Subacronicta megacephala Schiff. .
Acronicta aceris L........... . .
auricoma Schiff................
Я. rumicis L......................
.4. lutea Brem. et Grey...........
5—7
4—6
4—6
9—12
9—12
8—12
8—14
8—12
20—25
15—20
15—25
20—30
20—30
20—30
300—350
200—450
200—450
400—600
150—300
200—500
100—250
400—800
200—500
300—500
200—400
100—500
200—600
800—1300
Из данных табл. 5 видно, что чамки видов Orgyidae, которые не пи-
таются в имагинальной фазе и яйца которых очень крупны, откладывают
псе же по 300—600 яиц каждая. Это наблюдается всегда при крайне сни-
женной активности самок. Из Acronictinae примерно такое же число яиц
откладывают самки Diphthera coenobita Esp., — биологически очень близ-
кие Orgyinae. Наоборот, самки Calocasia coryli L., вида, близкого к. Diph-
thera coenobita Esp., но очень активные, откладывающие яйца поодиночке
пн листья кормовых растений, характеризуются намного меньшей пло-
довитостью, несмотря на то, что яйца их мельче и имеют более тонкий
хорион. Значительно меньшую плодовитость обнаруживают самки мона-
шенки, по сравнению с непарным шелкопрядом также значительно более
активные. Монашенка откладывает яйца небольшими кучками и прячет
их в щелях коры. Это требует более высокой активности самок этого
вида по сравнению с непарным шелкопрядом, самки которого отклады-
вают все яйца в одну кладку.
Самки активных, питающихся в имагинальной фазе видов Acronictinae
обладают высокой плодовитостью, например виды Subacronicta Kozh.,
Pharetra НЬ., но они откладывают мелкие яйца, бедные желтком. Из Асго-
iiietinae выделяется несколько видов, именно Acronicta rumicis L.
и A. lutea Brem. et Grey, которые откладывают яйца кучками (по 10—
30 шт.) и характеризуются наибольшей плодовитостью. Самки их менее
1 Указаны пределы обычных, средних количеств зрелых яиц.
2 Данные Цвёльфера (Zwolfer, 1934).
56
ОБЩАЯ ЧАСТЬ
активны, чем самки видов, отыскивающих места для яйцекладки и откла- -
дывающих яйца поодиночке.
Выход гусениц Orgyidae из яиц, в смысле темпов их отрождения,
у разных видов различен. У видов Orgyinae наблюдается всегда более
растянутое отрождение гусениц по сравнению с большинством Acronic-
tinae. Отчасти это может быть связано с длительностью эмбрионального
развития, большей у Orgyinae. Об этом свидетельствует большая растя-
нутость сроков отрождения гусениц у видов, зимующих в фазе яйца,
эмбриональное развитие которых охватывает многие месяцы. При выходе
хорион яйца поедается гусеницами не всегда, но все же часто, например
у Acronictinae. В некоторых случаях и именно у видов Orgyia Ochs, (на-
пример, по литературным данным, у
О. dubia Tausch.) выходящие из яиц
гусеницы поедают остатки самки.
Личиночное развитие Orgyidae ха-
рактерно многими своеобразными чер-
тами. Помимо связи структур первой
стадии гусениц с общим типом яйце-
кладки и с обилием или бедностью яиц
желтком, с этим также связано число
линек при росте гусениц. У видов
Orgyinae число линяний в общем мень-
ше, чем у Acronictinae. Среди первых
виды Orgyia Ochs, характеризуются-
четырьмя линьками у самцов и пятью
-у самок.1 У Ocneria Hb. самцы линяют
четыре раза, самки пять, но большая
часть видов Orgyinae и некоторые Mo-
mini (Calocasia Ochs.) имеют пять линек
у обоих полов. У Acronictinae большая
часть видов характеризуется шестью
Рис. 35. Рост гусениц непарника
(Ocneria dispar L.) при питании листь-
ями черемухи (Prunus padus).
1 — самки; 2 — самцы.
линьками у обоих полов. Ряд видов
Acronictinae и именно те, у которых
гусеницы отрождаются особенно недо-
развитыми, а взрослые характерны
ооильным развитием вторичных волос
(Acronicta aceris L., A. leporina L.), линяют семь и восемь раз. У этих
видов наблюдается различное число линек. Среди Acronictinae есть
виды, имеющие пять (Acronicta lutea Brem. et Grey) и даже четыре
линьки (Episema coeruleocephala Esp.). Эти виды характеризуются или
недоразвитием вторичных волос во взрослом состоянии (Episema Ochs.),
или, как Acronicta lutea Brem. et Grey, обильным развитием их сразу
после выхода из яйца, подобным тому, что известно для видов Orgyinae.
Личиночный рост Orgyidae распадается на две фазы (рис. 35): подго-
товительный период, который можно было бы назвать фазой формиро-
вания, когда наблюдается не столько увеличение размеров особи,
сколько возникновение типичного или почти типичного ее облика, и пе-
риод собственно роста, когда вес и размеры особи обычно в течение корот-
1 Бобров (1930) считает, что лишняя (по сравнению с самцами) линька самок
не является простым удлинением периода их роста, как то принято считать [Дайр,
(Dyar, 1890)], но является добавочной стадией, удлиняющей определенные средин-
ные этапы развития гусеницы (III—IV стадии). Данные хетотаксии не говорят
в пользу точки зрения Боброва.
БИОЛОГИЯ
67
кого срока сильно возрастают. У Orgyinae первая фаза охватывает пер-
вые две-три личиночные стадии; это же наблюдается у ряда видов Асго-
nictinae (Calocasia Ochs., Pharetra Hb.). У видов Acronictinae, у которых
из яиц выходят недоразвитые гусеницы (Acronicta Ochs., Subacronicta
Kozh.), наблюдается более длительный подготовительный период, охва-
тывающий три или четыре первые личиночные стадии, за которыми уже
следует быстрый рост особи. Характерной чертой кривых роста Orgyidae
(рис. 35) является всегда сильное падение веса после максимума кривой.
Нго падение веса особи в конечной фазе роста в значительной мере обу-
словлено потерей воды и изменением химизма тканей. Оно намного
больше у видов, зимующих в фазе куколки, чем у тех, у которых куколки
развиваются в течение летнего времени (имеют короткий период разви-
тия). Исключением, повидимому, является Episema coeru eocephala Esp.-,
куколки которой хотя и развиваются летом, но диапаузируют.
Рост поверхности тела Orgyidae, так называемый линейный рост,
также сложен. Для его изучения наиболее удобны твердые отделы кожи,4
а частности кутикулярный чехол черепной коробки (Дайр, 1890). Темпы
роста черепной коробки различны у видов, обладающих разным числом
линек. В первых личиночных стадиях индекс прироста черепной коробки
(табл. 6) примерно одинаков, порядка 1.5, у разных видов семейства
(Olene Hb., Orgyia Ochs., Leucoma Steph., Ocneria Hb., Diphthera Ochs.,
Calocasia Ochs., Daseochaeta Warr., Acronicta Ochs.). У видов, линяющих
большее число раз, в частности у всех Acronictinae, индекс прироста
черепной коробки остается таким до последней стадии. Подобные же
величины дает коэффициент прироста черепной коробки у видов Orgyinae
Таблица 6
Рост черепной коробки (наибольший ее диаметр, в Миллиметрах) по стадиям
у некоторых видов Orgyidae
Название вида Стадии развития
I II Ш IV V VI VII VIII
Olene jascelina L 0.54 0.84 1.20 1.80 3.60 —
Orgyia gonostigma F. .... 0.42 0.66 1.08 2.00 2.501 — —
O. antiqua L 0.42 0.72 1.20 2.10 2.70 — —
!,высота salicis L 0.48 0.66 1.08 2.10 4.00 — — —
Ocneria dispar ‘L 0.60 1.10 2.80 3.00 4.20 6.201 — —
O. monacha L 0.62 1.08 1.53 2.34 3.30 4.32 — —
Diphthera coenobita Fsp. . . . 0.54 0.84 1.20 1.80 3.00 4.50 — —
Calocasia coryli L 0.36 0.66 1.44 2.97 4.00 — — —
Daseochaeta alpium Osb. . . . 0.36 0.60 0.84 1.32 2.20 3.60 — —
Subacronicta megacephala Schiff. 0.42 0.68 1.08 1.60 2.40 4.00 — —
Acronicta aceris L 0.42 0.54 1.03 1.62 2.97 3.63 4.50 —
A, leporina L. 0.42 0.60 1.02 1.92 3.00 4.50 — —
.1. alni L 0.42 0.60 1.02 1.80 3.25 4.50 — —
.1. psi L 0.36 0.54 0.96 1.38 1.80 2.70 3.30 4.80
.1. rumicis L 0.30 0.50 0.80 1.44 2.00 3.30 — —
A. auricoma Schiff 0.30 0.48 0.75 1.32 1.92 2.88 —
,1. lutea Brem. et Grey . . . 0.30 0.72 1.26 2.10 3.25 — —
1 Последняя линька приводится только для самок; для монашенки все цифры
даны по Цвёльферу (1934).
;>8
ОБЩАЯ ЧАСТЬ
и случло добавочной линьки у самок (Ocneria Hb.). При сокращении
числи стадий характерны коэффициенты порядка 2.00 для последней
или двух последних стадий. Это свидетельствует о том, что здесь изме-
нились последние стадии личиночного развития.
Вообще может быть отмечено, что соотношение первичных и вторич-
ных признаков гусениц и число их линяний связаны не столь простой
зависимостью, как то рисовалось, например, Дайру (1890). Точно так же
но просты соотношения и коэффициентов прироста особей от стадии
к стадии [Келер (Keler, 1934)].
Личиночный рост в случае диморфизма полов различается не только
по наличию добавочной линьки у самок, как отмечено выше, но и по интен-
сивности прироста живой массы. У самок видов Orgyinae интенсивность
•прироста всегда выше, в ряде случаев в несколько раз, чем у самцов
(рис. 35). Различия темпов роста полов прямо пропорциональны степени
полового диморфизма в имагинальной фазе и особенно размерам особей.
> По характеру поведения гусеницы Orgyidae распадаются на две основ-
ные группы. Главная масса видов обоих подсемейств характеризуется
высокой активностью гусениц с первых моментов их выхода из яиц.
Они многоядны, не связаны тесно с кормовым субстратом, легко мигри-
руют. В некоторых случаях, например у видов рода Ocneria Hb. (monacha
L., dispar L., matura Btl.), зимующих в фазе яйца в состоянии диапаузы,
выходящие из яиц гусеницы в течение нескольких первых суток не рас-
ползаются и не питаются, но сидят близ места выплода друг возле друга.
Этот период жизни «в зеркалах» характеризуется пигментацией и скле-
'ротизацией покровов и перевариванием желтка, остающегося еще в ки-
шечнике гусеницы. Спустя известный срок (обычно 5—10 суток, в зависи-
мости от погодных условий) они расползаются и живут далее пооди-
ночке, так же как и все остальные виды этого семейства, которые почти
сразу покидают место выплода, например, виды Orgyia Ochs. У видов
Leucoma Steph, период жизни в зеркалах длится несколько часов.
Мелкие гусеницы Orgyidae обычно подвижны и светолюбивы. Для
питания они всползают на кроны деревьев или на облиственные побеги.
Для многих их видов, в частности почти для всех видов с редуцированным
питанием имаго, первая личиночная стадия является периодом расселе-
ния вида. У некоторых видов, например, Ocneria Hb., гусеницы первой
стадии обладают даже специальными приспособлениями к пассивному
расселению, например разносу ветром. Гусеницы, висящие на паутинках,
подхватываются ветром и уносятся за десятки километров от места вы-
плода [Холодковский, 1894; Burges, 1913; Щербаков, 1914]. Большее
или меньшее пассивное расселение ветром мелких гусениц наблюдается
у всех видов подсем. Orgyinae и у некоторых Acronictinae, нетесно свя-
занных с субстратом (например Calocasia Ochs., Pharetra Hb.).
Среди видов Orgyinae и Acronictinae имеются формы со специализо-
ванной биологией, например златогузка или виды Simyra Ochs. У зла-
тогузки выходящие из яиц гусеницы одного выводка не расползаются,
хотя и весьма активны, но питаются вместе и после первой линьки начи-
нают изготовлять общее гнездо из паутины, стягивая для этой цели по
нескольку листьев. В этом гнезде гусеницы одного выводка зимуют все
вместе. После зимовки некоторое время они питаются также вместе,
затем расползаются и живут поодиночке. Момент расселения их падает
на конец весны. Уместно отметить, что у этого вида довольно активны
самки и расселение вида происходит как в личиночной, так и в имаги-
нальной фазе. У близкого к златогузке Euproctis similis Fuessly рассе-
БИОЛОГИЯ 59
шшие гусениц происходит, как и у большинства других видов Orgyinae,
и первой личиночной стадии; зимуют молодые гусеницы третьей стадии,
пи к и у златогузки, но поодиночке. У видов Simyra Ochs, и Oxycesta Hb.
гусеницы живут гнездами, подобно златогузке, но не только в первых
। тпдиях, а в течение всего периода роста, расползаясь лишь незадолго
до окукления.
Небольшое число видов Orgyidae, относящихся к подсем. Acronicti-
пче, характеризуется малой активностью гусениц, в особенности моло-
дых, только что отродившихся. Выходящие из-яиц гусеницы Subacro-
fiicta Kozh. и некоторых Acronicta Ochs. (Triaena. Hb., Acronicta Ochs.)
« разу после выхода укрепляются брюшными ногами при помощи
шелковинок на поверхности листа и, не передвигаясь, питаются вокруг
моста прикрепления в течение всей первой стадии, до момента линьки.
Особь покидает место питания только в крайнем случае, при сильном
беспокойстве или при полном высыхании листа. Это отрицательно влияет
nit ее жизненные функции. Нормально смена места питания особи в пер-'
пых стадиях наблюдается при каждой линьке, когда сразу после линя-
ния гусеница мигрирует на свежий лист, где вновь укрепляется и остается
питаться до новой линьки. Только после третьей или четвертой стадии
гусеницы этих видов Acronictinae переходят к более подвижному образу
жизни и свободно сидят на листьях, чаще на пластинках, реже на череш-
ки х. Особенно тесная связь с кормовым субстратом в первых личиночных
стадиях характерна для гусениц Subacronicta megacephala Schiff, и Acro-
nie/.a alni L. Такой и близкий к нему образ жизни типичен для узких
олигофагов из Acronictinae. Расселение этой группы видов почти нацело
связано с активностью самок и их миграциями и лишь в слабой мере —
с распространением гусениц последних стадий.
Кормовые растения Orgyidae относятся почти исключительно к видам
древесных лиственных растений. Большая часть видов Orgyidae много-
ндпа, причем нередко наблюдается известное расширение многоядности
г. ростом.
Более многоядны виды Orgyinae',. виды Acronictinae — нередко оли-
гофаги и иногда почти монофаги. Тропические виды Orgyidae связаны
главным образом с Могасеае (Ficus, Morus), Sapotaceae, Euphorbiaceae,
Leguminosae, Bombaceae, что наблюдается в тропических влажных лесах.
II саваннах наблюдаются связи Orgyidae с Leguminosae (Mimosaceae),
Euphorbiaceae, Rutaceae (Citrus). В умеренной зоне Голарктики особенно
обычны кормовые связи Orgyidae с Fagaceae (Fagus, Quercus, Castanea),
Hosaceae (Malus, Prunus), Rutaceae (Rhus, Phellodendron), Betulaceae
(Hotula, Carpinus, Alnus), Salicaceae (Salix, Populus) и отчасти c Juglan-
ilncoae, Myricaceae и Coniferae. В степях Голарктики и в полупустынях
обычны кормовые связи Orgyidae с Terebynthaceae, Euphorbiaceae и
отчасти с Gramineae. Ниже дан подробный разбор кормовых отношений
Orgyidae. Сейчас я хочу лишь подчеркнуть пищевую связь видов этого
гомойства с растениями, относящимися к исходным типам покрыто-
семянных, как в тропиках, так и в умеренных широтах. Многоядность
Orgyidae и легкость замещения одних кормовых растений другими харак-
теризуют семейство в целом.1
1 Гусеницы Orgyidae являются типичными фитофагами и хищничество и кани-
Гшлизм среди них редки, являясь следствием жизни в искусственных (лабораторных)
условиях, голодания и скученности. Мне никогда не приходилось наблюдать кани-
Гшлизма у Orgyidae, но Бобров (1930) указывает на случаи канибализма у Orgyia ап-
60
ОБЩАЯ ЧАСТЬ
Пронимфальная фаза Orgyidae коротка и типична лишь немногими'
чертами поведения. Задержка метаморфоза (диапауза) у Orgyidae в про-
нимфальной фазе (в противоположность многим другим чешуекрылым)
никогда не наблюдается. Гусеница после окончания питания подыскивает
Место для окукления, заплетая листья, делая кокон в щелях коры, вгры-
заясь в древесину или уходя в подстилку. Наблюдается два биологи-
ческих типа пронимф у Orgyidae. Один тип наблюдается у
видов Orgyinae, исключая немногих Dasychira Ochs. Пронимфы этого
типа окукляются чаще на кормовых растениях и не уходят далеко
от места питания. Длительность развития пронимф в этом случае
очень мала—до двух-трех суток, а нередко даже только несколько
часов.
Другой тип очень активных пронимф наблюдается у видов, зимующих
в фазе куколки, т. е. почти у всех Acronictinae и некоторых Dasychira
Ochs. Пронимфы этих видов уходят в припочвенный слой, где в подстилке
делают кокон, Сплетая его из растительных остатков и шелка (Diph-
theraOchs., Daseochaeta Warr., Calocasia Ochs., Simyra Ochs., Pharetra Hb.,
Eogena Bd., Dasychira Ochs.), или вгрызаются в слабую, полугнилую
древесину {Subacronicta Kozh., Acronicta Ochs.). У этих видов пронимфа.
живет до 5—10 суток, обычно сильно меняя окраску и форму по сравне-
нию с гусеницей. Например, пронимфы Acronicta leporina L. приобретают
черную или желто-бурую окраску волос и кутикулы.
Развитие куколок протекает в коконах или в колыбельках, если оку-
кление происходит в древесине. Коконы обычно плотны, но у ряда видов,
например у Calocasia Ochs., Leucoma Steph;,* 1 Ocneria Hb., они построены
из немногих нитей, которыми стянуты листья или оплетена щель в коре.
Виды Orgyinae характеризуются короткими сроками развития куколок
(как правило 6—12 суток при средней температуре 20—25°). У видов
Acronictinae и некоторых Dasychira Ochs., напротив, длительность раз-
вития куколок обычно очень велика и равняется нескольким месяцам,
что связано с наличием диапаузы и зимовки. Для куколок Orgyinae может
быть отмечен половой диморфизм в сроках развития. Куколки самок
развиваются всегда в течение значительно более короткого времени,
чем куколки самцов. Как видно, соотношения здесь обратны тому, что
описано для гусениц: в фазе гусеницы рост самок характеризуется
большей длительностью. Если более длительный срок роста самок обус-
ловлен увеличенной ассимиляцией, вызванной развитием половых про-
дуктов, то укорочение срока развития куколок у самок некоторых Orgy-
inae обусловлено, без сомнения, редукцией имагинальных структур
(табл. 7).
Из данных табл. 7 видно, что укороченные сроки развития куколок
самок наблюдаются у видов Orgyia Ochs., для которых характерна силь-
ная рудиментация имагинальных структур. У непарного шелкопряда,
tiqua L. В своих опытах я наблюдал и у этого вида предпочтение питания корой вето-
чек березы (при полном объедании листвы) канибализму, несмотря на чрезмерное
скопление особей разных возрастов.
1 Указания [Дегенер (Deegen,er, 1913); Уигглесуорс (A igglesworth, 1933, 1939)]
на вымазывание пронимфами (гусеницами) Leucoma salicis L. коконов порошковид-
ными выделениями мальпигиевых сосудов являются недоразумением, вошедшим
в сводки и на русском языке (Уигглесуорс). Коконы, изготовляемые пронимфами
Lerucoma Steph., очень слабы, состоят из немногих нитей и по типу близки к коконам
непарного шелкопряда. Эта особенность биологии характерна для видов сем. коконо-
прядов (Lasiocampidae), и ее можно наблюдать, например, у кольчатого шелкопряда.
БИОЛОГИЯ
61
\ которого рудиментация самок значительно меньше, различия сроков
развития полов у куколок меньше (при высоких температурах они почти
т'и'зают).
Таблица 7
Длительность развития разных полов некоторых Orgyidae в личиночной и
куколочной фазах
Длительность развития (в сутках)
' || Ocneria dispar L. Orgyia antiqua L. Orgyia gonostigma К.
гусеницы' куколки гусеницы куколки гусеницы куколки
у? И н й И IS Й й й н И
п ЕГ к й ЬГ к tr к й Д’
g S S 8 S я S я S S S
а Сб СВ СВ сб сб сб сб сб
1 -•« О Q о о о о О о о о о
Ь>,0 77.7 60.0 2,4.2 29.0 49.3 44.3 12.0 22.0 14.0 22.3
",0.0 51.2 42.0 16.7 21.1 39.0 32.4 8.0 13.0 35.0 29.0 7.0 12.5
2|>.о 38.0 32.6 10.3 11.2 27.0 23.2 G.5 10.0 26.0 22.5 5.2 9.1
'.17,5 36.7 31.5 8.9 9.0 28.0 24.0 5.5 8.0 23.0 19.5 5.7 7.5
У видов подсем. Acronictinae, у которых половой диморфизм выражен
слабо* различия в сроках развития куколок разных полов отсутствуют.
Имагинальная фаза Orgyidae биологически характеризуется резко
риаличной активностью полов. У семейства в целом очень отчетливо
выражены направление рудиментации активности самок и компенса-
торное возрастание активности самцов. В подсем. Acronictinae большая
часть видов обладает в общем одинаковой активностью полов. Некоторое
снижение ее у самок может быть отмечено для Diphthera Ochs. Степень
снижения активности их примерно такая же, как и у Euproctis Hb. из
Orgyinae, самки которых, впрочем, активны еще в такой мере, что оты-
скивают необходимые условия для питания гусениц. Яйца этих видов
Orgyidae откладываются на листья кормовых растений. Среди Orgyinae
у видов Leucoma Steph., Ocneria Hb. и Parocneria Dyar активность самок
уже заметно снижена. Самки Leucoma salicis L. летают по вечерним зо-
рим, особенно в тихую погоду. Откладка яиц происходит не всегда на
листья кормовых растений, но часто также на стволы их. У видов Ocneria
lib. яйцекладка нормально производится на стволы и ветви деревьев,
причем самки их летают редко, например у Ocneria matura Btl., или со-
всем не летают, как у непарного шелкопряда. Самки последнего вида
летают только в очень тихие вечера, делая небольшие перелеты обычно
но опушкам леса. Все или почти все яйца откладываются видами Ocne-
ria Hb. в одно место. Дальнейшая редукция активности, сопровождае-
мая некоторой редукцией крыльев, типична для самок Penthophera Germ.
< аники Penthophera morio L. никогда не летают, откладывая все яйца близ
места выплода. Та же степень активности самок наблюдается у Orgyia
thyellina Btl., у которых крылья или развиты нормально, или редуци-
рованы. Для большинства видов Orgyia Ochs, отмечается крайняя руди-
ментация активности самок вместе с сильнейшей редукцией органов
движения» У одних из них самки выползают из кокона и кладут яйца
пи ого поверхность (О. antiqua L., О. gonostigma L., О. ericae Germ.),
62
ОБЩАЯ ЧАСТЬ
у других они остаются в коконе, куда откладывают и яйца (О. dubia
Tausch., О. prisca Stgr., О. ochrolimbata Stgr.).
В связи с рудиментацией активности самок стоят повышение актив-
ности самцов и специализация их поведения. Уже при небольшой руди-
ментации активности самок резко возрастают активность самцов и при-
влекаемость их специфическим запахом самок. При сильной рудимен-
тации самок наблюдается почти круглосуточная активность самцов,
например у видов Orgyia Ochs, и Ocneria Hb. Самцов непарного шелко-
пряда привлекают не только живые самки, но также вытяжка из при-
даточных желез половой системы самок или просто отрезанный конец
брюшка самки [Коллинз' и Пэте (Collins a. Paths, 1932)]. Улавливание
запаха самки самцами непарного шелкопряда констатировано на рас-
стоянии 2.38 км.1 В то время как мономорфные виды Orgyidae являются
только ночными, диморфные часто активны в течение почти круглых суток,
но особенно в дневные часы. Преимущественно ночными являются слабо
диморфные Dasychira Ochs., Olene Hb., Leucoma Steph, и даже некоторые
виды Ocneria Hb. (0. monacha L.). Самцы сильно диморфных видов Orgyia
Ochs, летают в течение почти всех суток, прекращая лёт только в самое хо-
лодное время (обычно в ночные и предутренние часы), а днем при силь-
ных грозах, ветрах и т. п. Для этих видов типично некоторое смещение
максимума активности на более теплые и даже жаркие часы суток.
Спаривание Orgyidae связано с состоянием готовности самок к яйце-
кладке. Внешне это выражается характерной позой самки (раздвинуты
крылья и выставлен конец брюшка) и ритмическим движением papillae
anales. У видов Orgyidae с длительным периодом созревания, самки йото-
рых выходят с незрелыми половыми продуктами (многие Acronictinae),
спаривание наступает обычно через несколько суток после выхода из
куколок. Самки видов Orgyidae, выходящие из куколок зрелыми (Calo-
casia Ochs., Diphthera Ochs., Oxycesta Hb.), спариваются сразу после
выхода.2 Можно наблюдать, например, спаривание самок непарного шел-
копряда или видов Orgyia Ochs., едва лишь вышедших из куколок, у не-
парного шелкопряда с еще неразвитыми крыльями. В наибольшей сте-
пени самцы этих видов привлекаются самками через несколько часов
после выхода последних из куколок. Оплодотворенные самки Orgyidae
уже не привлекают самцов. Неоплодотворенные самки привлекают сам-
цов в течение всей жизни, но после трех, четырех и более суток эта спо-
собность самок слабеет.
1 Коллинз и Пэте (1931, 1932)-указывают, что вещества, привлекающие самцов
непарного шелкопряда, извлекаются из желез конца брюшка самки ксилолом,
бензином и спиртом. Алкогольная вытяжка сохраняет привлекающие свойства
в течение первых трех лет. Для вытяжки лучше брать конец брюшка самки на второй
день после выхода из куколки. По исследованиям Барта (Barth, 1937), проведенным
на огневках (Plodia), привлекающие самцов вещества выделяются, железистыми
клетками интерсегментальной мембраны восьмого и девятого сегментов брюшка
самки. Выделение секрета происходит выпотеванием его сквозь хитин. Его отдаче
способствуют движения papillae anales. Для неоплодотворенных самок Orgyidae, при-
влекающих самцов, особенно для видов с рудиментарными крыльями, характерно-
сильное движение (пульсация) этих papillae.
2 Поданным Румянцева( 1936), спаривание бабочек Leucoma salicis L. наступает
через два-три дня после выхода из куколок. По моим данным, спаривание этого вида,
как и у других видов подсем. Orgyinae, может наступать вслед за выходом из куколок
как самцов, так и самок. Нормально, т. е. при достаточном обилии бабочек этого вида
в природе, спаривание их наступает в течение первых суток (в первую ночь) после
выхода из куколок как самцов, так и самок.
БИОЛОГИЯ
63
Длительность созревания половых продуктов самок, выходящих из
куколок незрелыми, равна 3—5 суткам, как у Acronictinae (Daseochaeta
Warr., Acronicta Ochs). У этих видов самки питаются нектаром. При
отсутствии нектарного питания в некоторых случаях, например у Calo-
casia Ochs, и Diphthera Ochs., период созревания охватывает тот же срок.
У видов Orgyinae сроки созревания половых продуктов у самок короче;
например, у непарного шелкопряда весь запас яиц, по Шпренгелю (1928),
созревает в течение первых суток жизни, а значительная часть зрелых
яиц есть уже в момент выхода из куколки. То же типично для ивовой
волнянки, златогузки и многих других видов Orgyinae. У видов
Orgyia Ochs, выходящие из куколок самки имеют весь запас яиц
зрелыми.
У большинства видов Orgyidae яйцекладка начинается почти сразу
вслед за окончанием копуляции (Orgyia Ochs., Ocneria Hb., Leucoma
Steph.), когда еще не закончено развитие всех ойцитов. У видов Acro-
nictinae начало яйцекладки наступает на два-три дня позднее момента
выхода из куколок, а процесс созревания полного запаса яиц самки растя-
гивается на всю жизнь. Увеличение общего числа зрелых яиц за счет
постметаморфного созревания их у большинства видов подсем. Orgyinae
незначительно. Для некоторых же его видов (Orgyia Ochs, или Leucoma
Steph.) его нет вообще: у этих форм запас яиц определен уже при выходе
особи из куколки.
Длительность спаривания Orgyinae не велика.1 В обычных условиях
длительность копуляции непарного шелкопряда, видов Orgyia Ochs,
или -Leucoma Steph, охватывает период 20—40 минут, реже 1—1.5 часа
(при пониженной температуре). Несколько более длительно спаривание
у видов Acronictinae, но и у них оно обычно ограничено сроком в 1 —
2 часа. Нередки повторные спаривания, хотя нормально при копуляции
со свежевышедшим самцом одно спаривание достаточно для оплодотво-
рения всех яиц, которые Имеет самка Ocneria Hb. или Orgyia Ochs. Мак-
симальное число копуляций для одного и того же самца Orgyia Ochs,
или непарного шелкопряда — пять-восемь, но последние копуляции
часто дают количество половых продуктов, недостаточное для оплодо-
творения всех яиц самки. В некоторых случаях спаривание бывает свя-
зано с некоторыми особенностями поведения самцов, например про-
никновения их в коконы самок, что связано с полной редукцией органов
движения самок. Это описано для Orgyia dubia Tausch. (Сахаров, 1914).
Самец при этом проделывает передними ногами в коконе отверстие.
Самки Orgyia dubia Tausch. вне кокона не оплодотворяются вообще и не
привлекают самцов.
Высокая активность самцов Orgyinae является условием относительно
легкой гибридизации этих видов, во всяком случае в искусственных усло-
виях. В природе я никогда не наблюдал межвидовых спариваний Orgyi-
dae, хотя такие случаи описаны для видов Orgyia Ochs. В лабораторных
условиях межвидовые скрещивания и гибридное потомство получены для
видов, не соприкасающихся ареалами или редких в пределах общего
ареала. Гибридные особи, доведенные до взрослого насекомого, получены
1 Рейф (Reiff, 1909) указывает, что у златогузки копуляция длится в течение
целой ночи. Он даже наблюдал особей златогузки в состоянии копуляции днем на
гом месте, куда насекомые слетались вечером на свет. Мои наблюдения над копуляцией
итого вида (в лаборатории) показали действительно большую длительность ее по
сравнению с копуляцией других видов Orgyidae-. она длится 8—10 часов вместо обыч-
вых 1—2 часов у большинства Orgyinue и 20—40 минут у Orgyia Ochs, и Ocneria Hb.
64
ОБЩАЯ ЧАСТЬ
от скрещивания Orgyia antiqua L. с О. thyellina Btl. и О. antiqua L. с
О. vetusta Bd. и с О. leucostigma Sm. et Abb. Для этих видов получены
гибриды как при скрещивании самцов одного вида с самками другого,
так и наоборот. Мои попытки получения гибридов Orgyia antiqua L. и
О. gonostigma F. не дали положительных результатов, хотя спаривания
самцов О. antiqua L. с самками О. gonostigma F. наблюдаются довольно
часто. Яйца во всех случаях (было использовано более десяти самок
Orgyia gonostigma F.) были неоплодотворенными. Обратные спаривания,
т. е. самцов Orgyia gonos igma F. с самками О. antiqua L., вообще не уда-
ются, может быть в связи с относительно малой активностью самцов
О. gonostigma F. Неудача гибридизации этих видов в моих опытах, воз-
можно, связана с очень большими различиями их циклов развития.
Гибридизация Orgyia thyellina Btl. и О. antiqua L. — видов, структур-
но довольно далеких, но имеющих однообразные циклы, — удается
легко.
Гольдшмидт (1920, 1934) в работах по гибридизации географических
форм непарного шелкопряда показал условия получения так называ-
емых интерсексов. Полученные гибридные особи имеют разную степень
развития промежуточных вторично-половых признаков, приближаю-
щих их в большей мере то к самцам, то к самкам. Условия формирования
половой системы таких особей, часто построенных мозаично в отношении
признаков обоих полов, изучены очень неполно. В природе известны мно-
гочисленные находки так называемых гинандроморфов, например у видов
Ocneria Hb. и Leucoma Steph. Структурно эти особи нередко представляют
почти то же, что и получаемые экспериментально интерсексы (Кузнецов,
1916). Условия возникновения гинандроморфов в природе не известны.
Но, без сомнения, они не всегда йвляются продуктом скрещивания раз-
ных географических форм. Скорее возникновение гинандроморфов
в природе связано с действием факторов среды, что доказано и экспери-
ментально для видов Ocneria Hb. и Leucoma Steph. (Емельянова, 1924; Хир-
шлерова, 1934; Косминский, 1934).
Для Orgyidae есть ряд указаний, отчасти старых, отчасти современных
или недавних [Гентнер (Gentner, 1915); Гепп (Нерр, 1936)], на наличие
партеногенеза. Эти указания касаются непарного шелкопряда и видов
Orgyia Ochs, (antiqua L., gonostigma F., ericae Germ.., dubia Tausch.). Почти
во всех случаях они носят характер лишь попутных замечаний. Спе-
циальных исследований по возможным условиям партеногенеза Orgyidae
нет, в,некоторых случаях имеются указания явно сомнительного свойства.
Я сделал много попыток получения партеногенетического развития
у видов Orgyia Ochs, (antiqua L., gonostigma F., отчасти ericae Germ.,
dubia Tausch.) и Ocneria dispar L., но во всех случаях (на сотнях самок)
получил отрицательные результаты. Отсутствие партеногенеза у Orgyia
dubia Tausch. подчеркивает и Сахаров (1914). Бошковская (Boszkowska,
1933) указывает на отсутствие партеногенеза у Laelia coenosa Hb. Цвёль-
фер (Zwolfer, 1933) установил это же для монашенки на основе изучения
400 самок. Для непарного шелкопряда Вейенберг (Weijenbergh, 1870)
описывает случай развития двух партеногенетических поколений; это же
подтверждает Пирс (Pearce). Фернальд (Fernaid), как и я, получил отри-
цательные результаты в специально поставленных опытах. Для непар-
ного шелкопряда вопрос о партеногенезе, в связи с завозом этого вида
в Америку, мог иметь практический интерес, но и в полевых условиях
там он разрешился в отрицательном смысле.
КЛА ССИФИКАЦИЯ
65
' Г л а в а V
КЛАССИФИКАЦИЯ И ФИЛОГЕНИЯ
В этой главе разбираются вопросы о положении изучаемой группы
в системе Lepidoptera и об ее составе; оба вопроса тесно связаны с раз-
бором филогенетических связей семейства в целом и родов, в него вхо-
дящих.
Впервые Буадюваль (Boisduval, 1840) очертил признаки Orgyidae,
выделив трибу Liparides (подсем. Orgyinae), которую помещал по сосед-
ству с Arctiidae и со сборной группой Bombycini (Lemoniidae, Lasiocam-
pidae, Thaumetopoeidae). Буадюваль руководствовался признаками личи-
ночной фазы, в частности наличием у гусениц кожных желез на дорзаль-
ной стороне брюшных сегментов, наличием волосатых куколок и связью
видов Liparides с древесными растениями. Признаки имаго им не были
разработаны. Он отметил лишь сильно гребенчатые усики самцов и актив-
ность их днем. Буадюваль (1840) впервые дал диагноз этой трибы (сов-
падающей по своему таксономическому значению с семейством);1 * * * 5 он
сближал своих Liparides с Bombycidae, но не с Noctuidae.
I До Буадюваля не только Линней (Linne, 1759), но и Шифермюллер
{Schiffermuller, -1775) и другие авторы не видели общности между разными
группами и родами Orgyidae, выделяя некоторые из них в самостоятель-
ные комплексы и помещая их среди видов Arctiidae и Lasiocampidae.
У Шифермюллера виды Orgyia Ochs., Dasychira Ochs, и Calocasia Ochs,,
выделены в группу Bombyces Tendipedes, виды Leucoma Steph., Euprb-
ctis Hb. и Ocneria Ochs. — в группу Bombyces Albidae, род EpisemaOchs,
отнесен к Bombyces Capitones. Таксономическое значение этих групп
не ясно. В работе Гюбнера (Hubner, 1805) положение видов Orgyidae
также не ясно, но у этого автора они объединены в целостную группу;
хотя эта группа еще не названа. Это же Гюбнер сделал и позже (1816).
Существенным шагом вперед после работы Буадюваля (1840) явилось
исследование Геррих-Шефера (Herrich-Schaeffer, 1850), который ввел
признаки жилкования в диагноз Liparidina (Orgyidae), специально отме-
тив положение второго медиального ствола передних крыльев и строе-
ние базальной ячеи на задних крыльях, образованной субкостальной
и радиальной жилками. Геррих-Шефер указывает, что виды этого се-
мейства по жилкованию далеки от Nolodontidae и Bombycidae и особенно
от Saturniidae и несколько ближе к Endromididae и Geometridae. Наиболее
близки Liparidina, по мнению этого исследователя, к Arctiidae (Chelo-
niidae) и Noctuidae. От Noctuidae их отличает лишь слабый хоботок, раз-
личия в числе шпор и слабость их, а также отсутствие глазков. На осно-
1 вании этого он присоединяет Diph her a coenobita Esp. к Liparidina, тогда
как виды Calocasia Ochs, и Мота НЬ. оставляет в сем. Noctuidae. Род
Penthophera (Hypogymna) Germ., по мнению Геррих-Шефера, является
переходным к Psychidae.
Уокер (Walker, 1855) выделил сем. Liparidaeno признакам, указанным
Буадювалем, использовав почти без изменения его диагноз. Он суще-
1 Диагноз Буадюваля гласит: «Larvae pigrae, saepium arboricolae, pilis fasci-
culato-tuberculato vestitae; tuberculo primo laterali alteris magis producto, longius
piloso; dorso postico yesicularis duabus retractibus praedito. Pupae laxae folliculatae,
pilis fasciculatis saepius sparsae. Alae semi-deflexae. Antennae maris valde, feminae
obsoletius pectinatae vel dentatae. Abdomen maris mediocre, feminis obesum. Volatis
maris saepium diurnus». Я привожу его как первую по времени (1840) характеристику,
очерчивающую облик изучаемой группы.
5 Фауна СССР, И. В. Ко'жанчиков.
66
ОБЩАЯ ЧАСТЬ
ственно расширил объем семейства за счет экзотических его представите-
лей. Систематическое положение семейства в целом у Уокера менее-
правильно, чем у Геррих-Шефера, так как он отделяет Orgyidae (Lipari-
dae) от Noctuidae, выделяя трибу Bombycites, состав которой очень пестрый
(Psychidae, Hepialidae, Arctiidae). Сем. Liparidae помещено им близ Arcti-
idae, Notodontidae и Lasiocampidae.
Ауривиллиус (Aurivillius, 1891) объединил Orgyidae c Arctiidae-
в одно семейство, влив в него также Dilobidae, Acronictinae, Lithosiinde,
Cymbinae, Nyctaeolinae и Nolinae. Он базируется на признаках жилко-
вания и структурах личинок, но подробно не разбирает этих групп. Его
система не была оценена вполне, хотя она является весьма интересной
и в некоторых отношениях очень правильной.
Существенным шагом вперед, в смысле общего положения Orgyidae-
в системе Lepidoptera, является классификация Мейрика (Meyrick, 1895).
Использовав основные положения Геррих-Шефера, Мейрик впервые-
Рис. 36. Хетотаксия среднегруди гусениц первой стадии. 1 — Noctuidae (Agrotis):
2 — Lithosiidae (Lithosia); 3—Syntomidae (Syntomis); 4—-Orgyidae (Calocasia)-.
5 — Orgyidae (Olene): 6 — Orgyidae (Leucoma); 7 — Orgyidae (Acronicta aceris L.);
8—-Lasiocampidae (Macrothylacia); 9 — Lemoniidae (Lemonia); 10 — Notodontidae
(Phalera).
(йе считая попытки Уокера) серьезно обосновал крупные группировки
семейств (серии). Его серия Caradrinina, которая включает семейства
Arctiidae, Caradrinidae (Noctuidae), Plusiidae и Ocneriidae, объединяет
близко родственные группы и отражает в сильной степени также взгляды
Ауривиллиуса. Позже (1927) он расширил круг родственных групп этой
серии, включив сюда также Nolidae и Hylophilidae, выделив, впрочем,
оба эти семейства из входивших ранее в эту же серию семейств Arctiidae
и Caradrinidae. Виды серии Caradrinina Мейрик (1895) сближает с Lasio-
campina (Lasiocampidae, Endromididae) и Notodontina (Geometridae,
Notodontidae, Sphingidae и др.). Эти же в общем филогенетические сближе-
ния принимает и Гэмпсон (Hampson, 1898), который отбросил, однако,
понятие серий (онпринимает лишь семейства); Гэмпсон дополнил филоге-
нетические построения Мейрика присоединением к серии семейств Cara-
drinina еще двух —Syntomidae и Agaristidae, считая их специализован-
ными ветвями Lymantriidae (Orgyidae). Эта последняя схема была принята
Кузнецовым (1915) для серии Noctuodea (Caradrinina Meyrik).
Разработка признаков личиночных фаз Дайром (Dyar, 1892), а позже
Фракером (Fracker, 1915) в целом подтвердила правильность сближения
упомянутого выше ряда семейств Геррих-Шефером и Мейриком в серию
КЛАССИФИКАЦИЯ
67
Caradrinina. Наряду с этим выяснилась связь Orgyidae с Notodontidae
которые, по Дайру, вместе с некоторыми другими семействами относятся
и одну серию — Bombycoidea. К этой же серии семейств Фракер (1915)
относит и Geometridae.
В обширном круге позднейших работ по имагинальной фазе чешуе-
крылых, вышедших в течение текущего столетия, например работ Штау-
дингера (Staudinger, 1901), Голланда (Holland, 1903), Шпулера (Spuler,
1903), Ребеля (Bebel, 1908), Берге (Berge, 1910), Зейца (Seitz, 1913),
Горинга (Hering, 1932), вопросы филогении чешуекрылых вообще и,
и частности, изучаемого семейства выпали из поля зрения. Эти работы,
носят характер каталогов и фаунистических сводок. Они дали много
нового в отношении состава сем. Orgyidae, в особенности тропических его
представителей, именно работы Геринга (1926), Шауса (Schaus, 1927)
и Колленетт (Collenette, 1932, 1938). Большая часть видов семейства
объединена недавно в каталоге Брика (Bryk, 1934), содержащем также
некоторый синонимический материал, при составлении каталога спе-
циально, впрочем, не оцененный.
Во всех каталогах, включая и последний, географическое распростра-
нение видов освещено слабо; только отдельные виды (монашенка, непар-
ный шелкопряд) затронуты специальными исследованиями.
В связи с недавними работами Эггерса (Eggers, 1919, 1940) по тимпа-
нальным органам чешуекрылых вновь возник интерес к филогении чешуе-
крылых и, в частности, сем. Orgyidae.
Бёрнер (Borner, 1939) предложил новую систему чешуекрылых и дал
новую трактовку филогении Orgyidae, в целом, впрочем, близкую к тому,
что давалось Геррих-Шефером и Мейриком. Он сблизил Noctuidae
с Notodontidae, но не с Orgyidae и Arctiidae, хотя все эти семейства поме-
щаются им в одну серию Noctuodea вместе с Syntomidae и Dioptidae.
Им использованы признаки строения тимпанальных органов [Эггерс,
1927; Эггерс и Горбанд (Eggers u. Gohrband, 1938)].
Систематическое положение сем. Orgyidae может быть обосновано
двумя группами признаков. К первой группе наиболее заметных и всегда
используемых признаков относятся преимущественно отрицательные
черты строения имагинальной фазы — рудиментация ротовых частей,
глазков, нередко крыльев, чешуйчатого покрова. Этими признаками,
и также некоторыми чертами жилкования и наличием осматерий у гу-
сениц на шестом и седьмом брюшных сегментах очерчены в литературе
границы сем. Lymantriidae (Orgyinae). Другая группа признаков, из
которых практически использовано лишь жилкование, включает основ-
ной тип строения головы, груди и брюшного отдела (тимпанальные ор-
ганы, половые придатки), жилкование крыльев, строение личинок и их
хетотаксию и тип строения яиц. Этот комплекс признаков имеет боль-
шую ценность в характеристике семейства, так как не только очерчивает
ого границы, но проливает свет и на филогенетические связи его в боль-
шой мере, чем отрицательные черты строения имаго, имеющие скорее
частное значение.
Признаки хетотаксии гусениц (рис. 36, 37, 38, 39), и именно первой
стадии, строение грудного и брюшного отделов, а также особенности
жилкования объединяют семейства Orgyidae, Noctuidae (-\-Agaristidae),
Arctiidae и Syntomidae в целостный комплекс. Для них типичны сильное
сближение и М% основаниями, та или иная степень анастомоза суб-
костальной и радиальной жилок на задних крыльях, наличие тимпа-
нальных органов на заднегруди, положение групп щетинок a, [i и р
68
ОБЩАЯ ЧАСТЬ
и легкость объединения двух первых групп, положение группы т под
стигмой и близ нее и слабое развитие хватательных задних' брюшных
Рис. 37. Хетотаксии первого брюшного сегмента гусениц
Orgyidae. Взрослых: Ocneria rubea Hb. (7); Subacronicta mega-
cephala Schiff. (2); Dasychira pudibunda L. (3); Artaxa sp. (4)j
Parocneria detrita Hb. (6) Acronicta leporina L. (7); и первой
стадии 'Euproctis karghalica Moore (5).
строения личиночных покровов и структур имаго в отношении указан-
ных признаков. Notodontidae, сближаемые Мейриком с семействами
серии Sphingodea, обнаруживают в хетотаксии гусениц первой стадии
большое сходство с тем, что известно для Garadrinina. С семействами
Рис. 38. Хетотаксии шестого брюшного сегмента гусениц первой стадии. 1 — Noc-
tuidae (Agrotis); 2 — Lithosiidae (Lithosia): 3—Syntomidae (Syntomis); 4— Orgyi-
dae (Daseochaeta): 5 — Orgyidae (Olene); 6 — Orgyidae (Calocasia); 7—Orgyidae
(Acronicta aceris L.); 8 — Lasiocampidae (Macrothylacia); 9 — Lemoniidae (Lemonia);
10 —- Notodontidae (Phalera).
этой же серии сближает Notodontidae также и положение тимпанальных
органов на заднегруди, но не на первом брюшном сегменте, как у Geo-
metridae.1 Отличия жилкования Notodontidae, в частности медиальной
1 По данным Мартина и Котнера (Martin a. Cotner, 1934), между Noctuidae и
Sphingidae также намечается родство белков, улавливаемое серодиагностически,
тогда как, например, между Noctuidae и Nymphalidae уловить его не удается. Если
связь химизма белков Noctuidae с Sphingidae достоверна, то легче всего она рисуется
через Notodontidae.
КЛА ССИФИКАЦИЯ
69
системы, от того, что известно для семейств серии Garadrinina, хотя
и значительны, но не принципиальны и едва ли могут расцениваться как
первостепенно важный признак. Thaumetopoeidae, как то было показано
<’иком (Sick, 1940), не могут быть отделены от Notodontidae, так же как
Рис. 39. Хетотаксия седьмого брюшного сегмента взрослой гусеницы (7) и первой
стадии (2) Agrotis gladiaria Morr. (по Фракеру, 1935), гусеницы первой стадии Litho-
sia lurideola Zinck. (3) и взрослых гусениц Calocasia coryli L. (4), Dasychira pudibunda
I.. (5), Penthophera morio L. (6), Мота ludifica L. (7), Diphthera coenobita Esp. (8),. Ac-
ronicta leporina L. (9), Acronicta lutea Brem. et Grey (10).
и от Dioptidae, которые по личиночным структурам очень близки к Noctu-
idae (Фракер, 1915). Может быть подчеркнуто большое габитуальное
сходство как имаго (например Cnetodonta), так и личинок (например
Pygaera) с бабочками и гусеницами Orgyidae. Lasiocampidae, напротив,
Рис. 40. Схема филогенетических отношений Orgyidae в преде-
лах серии семейств Caradrinina (Noctuodea).
обнаруживают лишь отдаленные связи с Orgyidae и Notodontidae, давая
крупные отличия в хетотаксии (положение ц на брюшных сегментах
и хи р на среднегруди) и в жилковании. Намеченные филогенетические
связи Orgyidae иллюстрирует прилагаемая схема (рис. 40).
70
ОБЩАЯ ЧАСТЬ
Ближайшие филогенетические связи сем. Orgyidae несомненно имеются
с Noctuidae, Arctiidae и Syntomidae. Оценка их на основании признаков
какой-либо одной из фаз невозможна. Как я отмечал при описании био-
логии Orgyidae, примитивность структур одной из фаз цикла развития
может быть обусловлена высокой специализацией другой фазы и на-
оборот. В связи с этим сближения указываемых семейств возможны лишь
на основании признаков всех фаз или во всяком случае двух основных —
имагинальной и личиночной. Для этих сближений необходима оценка
типа адаптации к условиям размножения и расселения видов данной
группы, т. е. формы жизни данной группы.
По типу адаптации к расселению и размножению семейства, родствен-
ные Orgyidae, могут быть разбиты на три группы. Первая группа, куда
можно отнести Orgyidae, характеризуется малой активностью имагиналь-
ной фазы, часто кратковременной, и отсюда процессами редукции и руди-
ментации многих анатомических черт имаго, но высокой специализацией
полового процесса. Для нее характерны богатство яиц желтком и высокая
активность личиночной фазы, личинок первой стадии и как исключение
высокая активность личинок лишь со второй или третьей стадии. Харак-
терной чертой гусениц, которая может быть связана с приспособлением
к расселению младших стадий и к жизни гусениц открыто на растениях
(приспособление к уменьшению отдачи воды), является развитие густого
волосяного покрова (вторичных волосков).
Вторая группа видов, в которую могут быть отнесеныNoctuidae, харак-
теризуется обратной ролью фаз цикла развития в решении той же жиз-
ненной задачи. Здесь расселение обусловлено полностью или почти пол-
ностью активностью имагинальной фазы. С этим связаны бедность яиц
желтком и слабость развития личиночных структур, в частности наличие
только первичных волосков в течение всего периода роста гусениц. Адап-
тация личиночной фазы к расселению в этом случае не велика.
Возникновение этих двух основных биологических типов в серии
Garadrinina можно трактовать как результат адаптации к условиям раз-
ных, типичных для каждого типа ландшафтов. В первом случае — адап-
тации к закрытому ландшафту, во втором —; к открытому, степному. «
Степные условия, благоприятные для питания имаго и расселения вида
в имагинальной фазе, требуют одновременно приспособления к сухости
и в связи с этим к специфичным условиям обитания личинок 1 (в почве,
стеблях, корнях, плодах, свернутых листьях и т. д.). Эти условия оби-
тания гусениц защищают их также от неустойчивого термического ре-
жима, как суточного, так и сезонного.
Третья группа, к которой можно отнести Arctiidae и Synto-
midae, менее ясна в силу худшей изученности биологии этих
групп, но она характерна, как и первая, значительной активностью
личиночной фазы. Активность имаго в этом случае большая, чем
у Orgyidae, причем нередко (например у Syntomidae) сохраняется
имагинальное питание. Выведение какого-либо одного из этих типов
из другого затруднительно. Напротив, все они могут быть выведены лишь
из архаичной группы чешуекрылых, имевших примерно одинаковую
активность к расселению как в личиночной, так и в имагинальной фазе.
В наибольшей мере примитивные черты циклов развития сохранили
1 Непостоянство гидротермического режима в условиях открытого ландшафта
умеренных широт таково, что адаптация к этим условиям, видимо, не может осуще-
ствляться путем изменения кожных покровов (например развитием волосяного по-
крова на них), но требует глубоких перестроек в биологии и циклах развития видов.
КЛАССИФИКАЦИЯ
71
Syntomidae и отчасти Arctiidae, тогда как Noctuidae и Orgyidae представ-
i яются специализованными группами.
Трудность выведения одной какой-либо из этих групп из другой
видна из предложенных классификаций. По Гэмпсону (1903), Syntomidae
могут быть выведены из Arctiidae на том основании, что имеют более
'цециализованные отношения субкостальной и радиальной жилок на
одних крыльях. Orgyidae расцениваются на этом основании как наи-
более примитивная группа. Тем не менее по структурам tegumen Syn-
loinidae очень примитивны и почти тождественны с Orgyidae, но не с Arcti-
idae, точно так же как и по присутствию papillae genitales, утраченных
в других группах серии Caradrinina. Для Penthophera Germ, характерно
строение тимпанальных органов, очень примитивное и почти тожде-
ственное с Syntomis (Эггерс и Горбанд, 1938). Гусеницы первой стадии
Syntomidae структурно очень близки к Noctuidae, но не к Orgyidae. Габи-
гуально виды Orgyidae сходны то с Syntomidae {Penthophera), то с Noc-
tiiidae {Acronicta), то с Arctiidae {Numens). Признаки жилкования заднего
крыла дают как бы прямой ряд от примитивных Orgyidae к Noctuidae
и Arctiidae и далее к высокоспециализованным в этом признаке Synto-
midae. Этот один признак, принадлежащий лишь одной из фаз развития,
как видно из изложенного, не может быть серьезной основой для клас-
сификации и понимания филогенетических отношений семейств серии
(laradrinina.
Состав Orgyidae определяется двумя основными группами: Orgyinae
и Acronictinae, причем среди видов последнего подсемейства выделяются
\domini, по многим признакам переходные. Состав первого подсемейства
ко вызывает в настоящее время сомнений, но Acronictinae всеми авторами,
исключая Ауривиллиуса, относились к Noctuidae. Основанием для при-
соединения Acronictinae к Noctuidae служат признаки, принадлежащие
преимущественно роду Acronicta (и близкими нему): наличие хоботка,
тыкнутая базальная ячея на задних крыльях и наличие простых глаз-
ков. Тем не менее близкие роды, в частности относящиеся к трибе Momini,
вызывали у многих авторов сомнения в принадлежности к Noctuidae.
Горрих-Шефер относил Diphthera Ochs. {Panthea) к Liparides. Шпулер
(1.908) отмечает большее сходство Caloca ia Ochs, с Lymantriidae, чем
с Noctuidae. Гроте (Grote) выделил особое семейство Apatelidae {Acroni-
<’tinae), не решаясь объединять эти виды с Noctuidae. Только Дайр (1899)
и категорической форме утверждает полную идентичность Acronictinae
по личиночным признакам с N octuidae.
Прежде всего необходимо подчеркнуть очень большую близость се-
мейств Orgyidae и Noctuidae в целом как по признакам личинок, так и
имаго. Хетотаксия личинок резко различна лишь в связи с присутствием
вторичных- волос, тогда как основной тип расположения первичных
волос для обоих семейств очень близок. Дайр (1899), впрочем, склонен
видеть кардинальные отличия этих семейств в положении щетинок х
па брюшных сегментах, которые у Noctuidae имеет тенденцию смещаться
вентрально и на седьмом сегменте всегда сдвинуты вентрально, тогда
как у Orgyidaeони располагаются за стигмойили даже сдвинуты дорзально.
Кроме того, он указывает на сближение и даже слияние щетинок а и
па средне- и заднегруди у Noctuidae и на раздельное положение их у
Orgyidae (s. str.). По строению имаго эти два семейства разделяются на
основании признаков рудиментации ротовых органов и по наличию
незамкнутой базальной ячеи на задних крыльях у Orgyidae (s. str.) и про-
тивоположных признаков у Noctuidae. В литературе господствует мне-
72
ОБЩАЯ ЧАСТЬ
ние, что признаки Acronictinae сближают их с Noctuidae, но не с Orgyi-
dae.
Изучение хетотаксии гусениц и признаков имаго Noctuidae и Orgyi-
dae показывает, что провести резкую грань между этими семействами
невозможно. Напротив, намечаются более сложные отношения между
ними; по ряду признаков виды Acronictinae более близки к Orgyidae,
но не к Noctuidae. Бородавка х на брюшных сегментах у Orgyidae дей-
ствительно расположена за стигмой, включая и седьмой сегмент, .но
положение ее у Orgyidae не устойчиво: х нередко редуцирована и даже-
утрачена, например у некоторых Orgyia Ochs, и Euproctis Hb., а у Pen-
thophera Germ., вероятно, слита с тр Сдвиг х у видов Ocneria Hb., Paroc-
neria Dyar и Leucoma Steph, дорзально и слияние ее с р отнюдь не ти-
пичны для семейства в целом. Это наблюдается лишь у специализованных
видов семейства и является частным случаем неустойчивого положения х.
Существенно отметить, что у некоторых из них, именно у Parocneria
Dyar, на седьмом брюшном сегменте наблюдается более вент-
ральное положение х по сравнению с предыдущими сегментами и
отсутствие слияния ее ср. Для Noctuidae типично смещение х на седь-
мом брюшном сегменте вентрально, но в первой личиночной стадии сдвиг-
этот в ряде случаев не велик. Точно такая же картина характеризует-
Arctiidae и Syntomidae. Таким образом признак смещения х вентрально
на седьмом сегменте возник в пределах серии Caiadrinina. У генерали*-
зованных представителей этой серии положение ее примитивно и измен-
чиво, а у специализованных, наоборот, определенно и характеризуется
одним из положений, наблюдаемых у Orgyidae.
Положение групп а. и |3' на грудных сегментах не характерно для Noc-
tuidae та Orgyidae, но у последних опять-таки более изменчиво. У Dasy-
chira Steph, или Ocneria Hb. группы а и [3 на грудных сегментах обособлены
и значительно разъединены, но они у Gynaephora и Leucoma Steph,
сближены, а у Penthophera Germ, и Parocneria Dr. полностью
слиты.1 На брюшных сегментах положение этих бородавок очень измен-
чиво. То они лишь сближены, как, например, у Dasychira Steph., то час-
тично слиты, как у Artaxa Wlk.; их полное слияние типично для Eu-
proctis Hb., Penthophera Germ, и Parocneria Dyar. Таким обра-
зом для Orgyidae в отношении положения первичных щетинок и боро-
давок есть несколько частных типов, но в основе хетотаксия у них та же,,
что у Noctuidae. Для Noctuidae, как кажется, характерен один из част-
ных типов хетотаксии — сдвиг бородавки х вентрально на брюшных
сегментах и сближение или слияние сс и [3 на последних грудных. Этот
же частный тип хетотаксии характерен для Arctiidae и Syntomidae.
Хетотаксия гусениц Acronictinae обнаруживает значительную бли-
зость их к Orgyidae. Помимо факта развития вторичных волосков, начиная
с первой личиночной стадии, эти группы сближает неустойчивое поло-
жение х брюшных сегментов, легкость слияния а и р грудных и брюшных
сегментов и отсутствие е на брюшных сегментах. Как и у Orgyidae,
1 Бэкстон (Buxton, 1921), впервые отметивший для Orgyidae на примере Parocne-
ria Ocnerorgyia) наличие лишь двух бородавок надх на средне- и заднегруди, считает
их р ир, полагаях исчезнувшей. Мнение Бэкстона основано лишь на сравнении Оспег-
orgyia Stgr. и Ocneria Hb. Если же учесть очень сильное сближение а и р на сред-
негруди у Leucoma Steph., то более вероятным становится, напротив, слияние а и р.
Исчезновение этих первичных щетинок в пределах всего сем. Orgyidae не констати-
ровано, тогда как сближение характерно, кроме Leисота. Steph., также для Gynae—
phora Hb. и всех Acronictinae.
КЛА ССИФИКАЦЙЯ
7^
у Acronictinae х на брюшных сегментах.может оставаться прямо за стиг-
мой не только в первой личиночной стадии, как то наблюдается у пода-
вляющего большинства видов, но и в старших (Мота НЬ.). Большею
частью х на седьмом брюшном сегменте не сливается с rt, но лишь сдви-
гается вентрально. Лишь у наиболее специализованных групп (Phare-
Ira Hb., Simyra Ochs, Eogena Gn.) она сливается c rt. Смещение x вен-
трально на седьмом брюшном сегменте у Acronictinae является вторичным,,
возникшим недавно и в пределах семейства. Об этом свидетельствует
хетотаксия первой личиночной стадии, где бородавка х на седьмом сег-
менте расположена прямо за стигмой. В некоторых случаях у Acronic-
linae, как и у некоторых видов Orgyinae (Ocneria Hb., Leucoma Steph.),
z брюшных сегментов смещается дорзально, располагаясь над стигмой
л почти объединяясь с р. Это наблюдается у Subacronicta Kozh., гусе-
ницы которой напоминают виды подсем. Orgyinae также формой тела
и размерами переднегрудных х. Наоборот i на брюшных сегментах у гу-
сениц Acronictinae никогда не развита в противоположность видам совок
(Noctuidae'), у взрослых гусениц которых она всегда есть.
Положение групп а и |3 грудных и брюшных сегментов также сбли-
жает виды Orgyinae и Acronictinae. В типичном случае эти две группы
щетинок на брюшных сегментах гусениц Orgyinae обособлены, но у Dasy-
chira Steph., Gynaephora Hb. и Orgyia Ochs, они сближены, a у Euproctis
Hb., Penthophera Germ., Leucoma Steph, и Parocneria Dyar — слиты.
У Leucoma Steph, слияние этих бородавок можно видеть в первой личи-
ночной стадии. У Artaxa Wlk., рода, очень близкого к Euproctis Hb.,
у взрослых гусениц группы аир первого брюшного сегмента еще не слиты
полностью, хотя и соприкасаются, тогда как на всех остальных сегмен-
тах брюшка они слиты полностью. Точно так же и у Acronictinae а и р
на брюшных сегментах обособлены, но в ряде случаев, сближаясь, дают
большие плоские бородавки, наподобие таковых у Dasychira Steph,
п Laelia Steph. Это наблюдается у Acronicta aceris L. и A. leporina L.
У Acronicta leporina L. эти бородавки сливаются полностью на брюш-
ных сегментах, так же как у Euproctis Hb. или Artaxa Wlk., и могут даже
нести кисточки волосков. У Мота ludifica L. из Acronictinae бородавки
а и р на восьмом брюшном сегменте слиты, они образуют конический"
вырост, как у Penthophera Germ, или Leucoma Steph.
Наконец, наиболее заметный признак, выделяющий Orgyinae, —при-
сутствие железок на дорзальной стороне шестого и седьмого (реже всех)
сегментов брюшка, является у видов Orgyinae вторичным, возникшим
подавно, выраженным не у всех видов (Dasychira Steph.) и в онтоге-
незе возникающим лишь после первой личиночной стадии. Физиологи-
ческая роль этих железок не ясна, но несомненно их провизорное
значение, характеризующее вторичные черты обмена старших личиноч-
ных стадий. Первая личиночная стадия для видов обоих подсемейств чрез-
вычайно близка и у некоторых видов не отличима.
Приведенные здесь факты, а также ряд других данных, описанных:
в главе о морфологии личиночных фаз, не оставляют сомнения в цело-
стности всей массы видов Orgyinae и Acronictinae, имеющих.примитивные
черты строения по сравнению с прочими семействами серии Caradrinina.
Оба подсемейства сближают даже детали личиночных структур (например
присутствие пильчатых волосков у гусениц первой стадии, глубоко выре-
занная верхняя губа и др.).
Черты биологии и структуры куколок также очень близки у Orgyi-
nae и Acronictinae. Окукление этих видов всегда происходит на поверх-
-74
ОБЩАЯ ЧАСТЬ
ности почвы в коконах. Это отличает виды Orgyinae в целом от Noctuidae.
Гусеницы всех видов собственно совок (Trifinae), с которыми только
и могут быть сближены Acronictinae, уходят всегда для окукления
в почву; лишь Quadrifinae (Phytometrinae, Noctuinae)1 окукляются
наземно, но в коконах, очень отличных по строению от того, что типично
для Orgyidae. Напротив, детали строения коконов и общий их тип у Ог-
,gyinae и Acronictinae очень близки. Рыхлые коконы Calocasia Ochs.,
изготовленные из листьев, стянутых редкой паутиной, очень напоминают
по деталям постройки то, что типично для Ocneria Hb. или Leucoma
Steph. Точно так же плотные коконы Diphthera Ochs., Мота Hb. и Da-
seochaeta Warr., имеющие сигаровидную форму, совершенно сходны с ко-
конами Gynaephora Hb. или Dasychira Ochs. Лишь виды рода Acronicta
Ochs, имеют в некоторых случаях своеобразное строение коконов; так,
виды подродов Acronicta Ochs, и Triaena Hb. окукляются в древесине
и строят очень рыхлые коконы, расположенные в колыбельке из огрыз-
ков древесины.
Куколки Acronictinae даже таких видов, как Acronicta alni L., имеют
волоски за стигмами на брюшных сегментах. Обычно эти волоски распо-
лагаются пучками (Diphthera Ochs., Calocasia Ochs., Мота Hb., Daseo-
chaeta Warr., Simyra Ochs., большая часть видов Acronicta Ochs.), лишь
у некоторых видов рода Acronicta Ochs, они одиночные и короткие. При-
сутствие волосков на кожных покровах куколок Orgyidae не может рас-
сматриваться только как следствие обилия волосков у гусениц, но
является специфичным признаком, так как, например, у видов Arctiidae,
гусеницы которых обильно покрыты вторичными волосками, накожном
покрове куколок волосков нет. Из семейств, родственных Orgyidae, лишь
Lasiocampidae имеют короткие волоски на коже куколок, тогда как ку-
колки Noctuidae голы.
Черты строения имагинальной фазы также в сильной степени сбли-
жают Orgyinae и Acronictinae. Ротовые органы видов Orgyinae рудимен-
тарны, но не в равной степени у всех представителей. У видов Euproc-
tis Hb., Ocneria Hb., Parocneria Dyar, Orgyia Ochs., Penthophera Germ,
•ротовые органы утрачены полностью, но у видов Artaxa Wlk., Gynaephora
Hb., Leucoma Steph., Dasychira Steph, они еще присутствуют и нередко
даже хоботок этих видов довольно длинен и закручен, спирально (Artaxa
Wlk.). Из Acronictinae значительная часть родов также характеризуется
рудиментацией ротовых органов — Panthauma Stgr., Diphthera Ochs.,
•Calocasia Ochs., Charadra Wlk., Raphia Hb., Simyra Ochs., Eogena Gn.,
вплоть до полной их утраты у Oxycesta Hb., но многие имеют нормально
развитые ротовые части. Часть представителей Acronictinae, у которых
наблюдается рудиментация ротовых органов, характеризуются также
утратой глазков (Diphthera Ochs., Episema Ochs.), что типично для
Orgyinae.2 Жилкование передних крыльев Orgyinae и Acronictinae
идентично. Заднее крыло Orgyinae характеризуется большой, не вполне
1 Виды Quadrifinae представляют, без сомнения, наиболее генерализованных
совок, несмотря на развитие черт специализации гусениц хотя бы в виде редукции
частей брюшных ног. Признаки жилкования, строение головы и распространение
•обличают глубокую древность этих видов. Наземное окукление гусениц, конечно,
также является примитивным.
2 В другом, также примитивном семействе чешуекрылых, у Hesperiidae, как и у
видов Orgyidae, наблюдаются различия в развитии глазков. У одних видов (Lerema)
они есть, у других, довольно близких видов, они отсутствуют [Скёддер (Scudder, Proc.
Bost. Soc. Nat. Hist., XVI: 165—166)]. Этот признак и у Hesperiidae, как и у видов
^изучаемого семейства, не может служить основанием для его разделения.
КЛАССИФИКАЦИЯ
75
шмкнутой базальной ячеей между основанием субкостальной и радиаль-
ной жилок, но у ряда видов Orgyia Ochs, и у Penthophera Germ, эта ячея
нполне замкнута анастомозом субкостальной и радиальной жилок, даже
оолее полным, чем у видов Acronictinae, у которых она в ряде случаев
но замкнута {Diphthera Ochs., Panthauma Stgr.). Наконец у Acronictinae,
так же как и у Orgyinae, девятый сегмент по сторонам uncus несет sub-
uii.ci, которые, хотя быв рудиментарном состоянии, есть у всех видов.
S Calocasia они очень большие. Для Noctuidae subunci не типичны. Точно
гик же у ряда видов Acronictinae {Calocasia Ochs., Raphia Hb., Episema
Ochs., Diphthera Ochs.) uncus отделен мембранозным швом от тергита
i,святого сегмента. Строение tegumen примитивно, он двойной. Это очень
типично для Orgyinae, но совершенно не характерно для Noctuidae.
Cfuno _
Olene
Penthophera
Dasychira
Artaxa
Jvela
Lichnoptera
Panthauma
Diphthera
Episema
Mama
Eogena
Acronicta
Craniophora
Arsilonche
Oxycesta
Рис. 41. Схема филогенетических отношений подсемейств, триб
и некоторых родов сем. Orgyidae.
Наконец, рисунок видов Acronictinae характерен отсутствием клиновид-
ного пятна, что свойственно также Orgyinae, но не типично для Noctuidae,
у видов которых это пятно есть.
Приведенные факты и соображения по биологии и структурам Acro-
nictinae и Orgyinae позволяют, по моему мнению, объединить их в одно
семейство, оставляя самостоятельность за ними в ранге подсемейств.
Их. необходимо считать близко родственнымй группами, причем виды
Acronictinae являются дериватом архаичных типов Orgyinae. Признаки
Orgyidae (обоих подсемейств): их личиночные структуры, черты жилко-
вания и строение полового аппарата, отделяют их от прочих семейств
серии Caradrinina и, в частности, достаточно отчетливо от ближайшего
семейства Noctuidae. Сходство Acronictinae с Noctuidae только внешнее;
Acronictinae в действительности имеют с ними мало общего как струк-
турно, так и биологически. На рис. 41 я привожу схему, иллюстрирую-
щую филогенетические связи обоих подсемейств Orgyidae и их родов.
76
ОБЩАЯ ЧАСТЬ
Схема требует некоторых замечаний. Подсем. Orgyinae распадается
на две группы родов, тесно связанных, — это ветви Artaxa Wlk. и Dasy-
chira Steph. Оба указанных рода являются наиболее генерализованными
из изученных мною, связывая все остальные роды подсемейства, распро-
страненные в Голарктике. В тропиках, без сомнения, есть еще свои гене-
рализованные ветви семейства, но они не заходят в Голарктику. Обе
ветви обособлены не резко и не могут быть выделены в трибы. Их при-
знаки наиболее легко уловимы в личиночной фазе. Группа Artaxa Wlk.
характеризуется гусеницами с гладкой дорзальной поверхностью, без
кисточек, тогда как для видов группы Dasychira Ochs, характерно их
наличие. В имагинальной фазе есть мелкие признаки в строении valvae,
специфичные для этих групп. Род Penthophera Germ, занимает промежу-
точное положение. Он очень своеобразен и обособлен от обеих ветвей,
но в то. же время сочетает признаки их обеих — в гусеницах признаки
ветви Artaxa Wlk., в имагинальной фазе—Dasychira Steph. Деление
видов Orgyinae на Areolatae и Inareolatae, как то предлагает Штранд
(Strand, 1910), не может быть использовано, так как возникновение или
утрата добавочной ячеи на передних крыльях у Orgyinae происходит легка
и наблюдается в пределах родов (Arctornis Germ.) или даже видов (Arct-
ornis l-nigrum Miill., Penthophera morio L.).
Acronictinae распадаются на две трибы: генерализованные Momini,
целиком дендрофильные, удерживающие черты Orgyinae в сильной сте-
пени, и Acronictini, более специализованные, но в некоторых случаях
также очень полно сохраняющие черты строения Orgyinae, особенно
в личиночной фазе (Subacronicta Kozh.). Внутри трибы Momini затруд-
нительно дать более мелкие группировки. Роды этой трибы являются
очень обособленными, часто мономорфны или олигоморфны; они'пред-
ставлены обрывками, разрозненными видами (родами), реликтами тре-
тичного времени. В трибе Acronictini намечается две группы: роды, груп-
пирующиеся около полиморфного рода Acronicta Ochs., и мономорфные
роды группы Arsilonche Ld., стоящие несколько в стороне от прочих.
Представление об объеме родов сем. Orgyidae дает табл. 8.
Состав родов, особенно тропических, нельзя считать, конечно, точна
установленным, но палеарктические роды изучены значительно полнее.
Тем не менее общее сравнение объема родов может быть произведено без
боязни впасть в большую ошибку. В подсем. Orgyinae как раз наименее-
полно изучены тропические роды, которые являются полиморфными и,
несомненно, наиболее обильны видами.1 Дальнейшее изучение состава
этих родов может только увеличить и развить картину их полиморфизма
(Dasychira Steph., La'elia Steph., Orgyia Ochs., Artaxa Wlk.,. 'Euproctis
Hb., Leucoma Steph., Ocneria Hb.). Напротив, роды, распространенные
в широтах умеренного климата, изучены значительно лучше, они все
бедны видами (Ivela Swinh., Numens Wlk., Penthophera Germ., Gynae-
phora Hb., Cifuna Wlk., Arctornis Germ.) и являются специализованными
формами, дериватом тропических генерализованных Orgyidae. Сущест-
венно отметить, что эти роды представлены в Палеарктике как в области
лесов с опадающей листвой (Numens Wfk., Ivela Swinh., Arctornis Germ.),
1 Найти, например, несколько десятков новых видов рода Penthophera Germ.,
единственный представитель которого был описан еще Линнеем, невероятно, так как
двухсотлетнее изучение фауны Голарктики, конечно, дало бы указание на это. Это же
касается в той или иной мере и других голарктических олиго- и мономорфных родов
Orgyidae, представители которых были описаны еще Линнеем, Фабрициусом, Буа-
дювалем и другими авторами.
КЛАССИФИКАЦИЯ
77
Таблица 8
Число видов в пределах разных родов сем. Orgyidae
Название рода Число видов Название рода Число видов
I. Orgyinae t. Dasychira Steph 70 8. Episema Ochs 1
.2. Laelia Steph 104 9. Саппа Wlk 5
3. Orgyia Ochs 54 10. Acronictoides Kozh. . . . . 1
4. Olene Hb 44 (?) 11. Belciana Wlk . 3(?)
5. Cifuna Wlk. ...... 3 12. Belciades Kozh 1
6. Gynaephora Hb 7 • 13. Bathyra Wlk 2
7. Penthophera Germ 1 14. Gaujonia Dogn 4
8. Artaxa Wlk 225? 15. Lichnoptera H. S 12
9. Euproctis Hb 161 16. Euromoia Stgr. 1
10. Arctornis Germ 2 (?) 17. Raphia Hb. 5
11. Iveta Wlk 2 18. Daseochaeta Warr 12
12. Leucoma Steph 57 19. Subleuconicta Kozh .... 1
13. Ocneria Hb 116 20. Agripodes Hamps 4
14. Parocneria Dyar 20 21. Thyatirides Kozh. . . . . 1
15. Numens Wlk 8 22. Cosmodes Gn. ...... 1
II. А с г о ri i c t i n a e 1) Momini 1. Panthauma Stgr 1 2) Acronictini 23. Subacronicta Kozh. . . . 24. Acronicta Ochs 25. Hampsonia Kozh, .... 5 90 2
2. Trisuloides Butl. 15 26. Craniophora Snell. . . . 8
3. Anacronicta Warr 5 27. Eogena Gn 1
4. Diphthera Ophs 8 28. Sirnyra Ochs 3
5. Calocasia Ochs 7 29. Arsilonche Ld 1
6. Charadra Wlk 7 30. Oxycesta Hb 3
’.7. Мота Hb 12? 31. Leiometopon Stgr 1“
так и в степях (Ocneria Hb., Penthophera Germ.) и даже в арктоальпийской
•области (Gynaephora Hb.) без большого преобладания на лесных терри-
ториях. Этот факт говорит за то, что дифференциация этих специализо-
панных форм из тропических представителей шла более или менее одно-
временно в разных ландшафтах. Тем не менее должно констатировать
несколько более легкое проникновение Orgyidae в зону широколиствен-
ных лесов Голарктики, куда только и заходят почти все известные в уме-
ренных широтах представители тропических родов.
Виды подсем. Acronictinae почти целиком связаны с умеренными широ-
тами, и именно с лесами с опадающей листвой Голарктики, особенно с ле-
сами третичного типа (амурские и китайские широколиственные леса). Эти
виды тесно связаны с условиями мягкого, переменного климата. Как
видно, подавляющее большинство родов этого подсемейства олигоморфны
или мономорфны, включая также и немногих представителей, перешед-
ших к жизни в степях и полупустынях (Oxycesta Hb., Eogena Gn.). Только
один род Acronictinae — Acronicta Ochs. — является полиморфным,
далеко уступающим, впрочем, по числу видов полиморфным тропическим
родам семейства, несмотря на худшую изученность тропических родов
Orgyidae и относительно полную исследованность этого голарктического
рода. Все подсем. Acronictinae должно рассматриваться как дериват
тропических Orgyidae, как возникшее в условиях третичных широколист-
78
ОБЩАЯ ЧАСТЬ
венных лесов; без сомнения, оно было более широко распространено»в тре-
тичное время, в дальнейшем как число его видов, так и территории оби-
тания в связи с послетретичными переменами климатического режима
Голарктики сильно сократились. В последующем изложении я даю
оценку филогенетических и экологических связей в распространении
Orgyidae, что имеет значение для понимания изложенных общих филоге-
нетических связей семейства.
Глава VI
ЭКОЛОГИЯ
Orgyidae в целом могут быть характеризованы как гигрофильные
и стенотермичные виды; для них типичны черты адаптации к жизни
в условиях устойчивого термического режима (табл. 9). Основным типом
стаций обитания семейства являются широколиственные леса в умеренных
широтах и вечнозеленые широколиственные леса (гилеи) влажных тро-
пиков. Стадиальные условия для большинства тропических видов Orgyi-
dae сколько-нибудь подробно не изучены. Однако, рассматривая условия
среды на территориях, обильно заселенных Orgyidae, все же можно вы-
нести некоторые суждения по экологии тропических видов. Для Каме-
руна, например, известно несколько сотен видов изучаемого семейства,
среди которых очень большое число — виды эндемичные, а также общие-
с видами Конго и других территорий влажных тропиков Западной
Африки. Леса Камеруна и вообще западной Африки характеризуются
высокой устойчивостью климата как в течение года, так и суток [Ричардс
(Richards, 1939); Эванс (Evans, 1939)]. Возникновение полициклии и
полифагии, столь характерных для сем. Orgyidae в целом, нужно связывать
с возможностью круглогодового развития при обязательной смене кор-
мового режима в разных поколениях. С условиями же гилей, именно-
с бедностью там нектарного питания, может быть связана рудиментация
ротовых органов имагинальной фазы.1
Широколиственные леса умеренных широт отличают сезонная смена
климатических условий и значительная неустойчивость термического
режима в течение суток (минимальная в период вегетации лесов). Сезон-
ные термические смены здесь достигают нескольких десятков градусов,
причем типично ежегодное падение температуры зимой ниже нуля. Го-
дичная амплитуда, например, в Цзинсене (Корея) —28°, в Чундзине
(Голубая река) — 22.3°, во. Владивостоке — 34.3°, в Симле (западные
Гималаи) — 15.6°, в Линце — 21.7°. Суточная смена термического режима,
в широколиственных лесах наименьшая (порядка 5—8°) в течение летних
месяцев и наибольшая весной, до наступления полного облиствления
деревьев. Более устойчива температура в смешанных хвойно-широколи-
ственных лесах, где облиствение лишь увеличивает ее стабильность, но
экоклимат кардинально не меняет. В хвойных, и именно темнохвойных
лесах, устойчивость термического режима в течение суток максимальная,
но здесь средние суточные температуры понижены,2 часто в июле 12—14°,
что в свою очередь неблагоприятно.
1 Рудиментация ротовых органов у дендрбфильных чешуекрылых представляет
явление, характеризующее разные и совсем не родственные группы (Sphingidae,.
Orgyidae, Saturniidae, Lasiocampidae, Notodontidae, Drepanidae, Aegeriidae).
2 Я не упоминаю здесь об изменчивости режима влажности воздуха, так как
влажность воздуха тесно связана с термическим режимом среды и им обуслов-
лена. Устойчивый режим влажности только и может характеризоваться стойким
термическим режимом (например в гилеях).
экология
79'
Экологические условия обитания Acronictinae, населяющих леса,
широт умеренного климата и субтропиков, характеризуются сезонной
цикличностью не только термического режима, но и развития растений,
причем значительным синхронизмом развития разных видов древесной
растительности. С этим может быть связан и основной тип циклов раз-
нития видов Orgyidae, распространенных в Голарктике и заселяющих
леса с опадающей диствой. Для их циклов характерен моновольтинизм
с диапаузой, наступающей в период окончания вегетации древесных по-
род (август—сентябрь в северных широтах, октябрь—ноябрь на юге),,
и моно- или олигофагия. Диапауза обеспечивает значительную или высо-
1’ис. 42. Климограммы некоторых территорий Евразии и Африки. А: 1 — западная-:
Африка (Дебунджа); 2 — Гималаи (Симла); В : 1 — южное Приморье (Пост Ольга);
2— средняя Европа (Мюнхен); 3 — южная Сибирь (Минусинск); 4— Аральские
степи (Казалинск).
кую холодостойкость зимующих особей, а олиго- или монофагия допу-
скает максимальную приспособленность цикла насекомого к циклу раз-
вития кормового растения. Для развития зимующих фаз и облиствения их
кормовых растений типична близость критических низких и высоких
|(!мператур и сумм тепла (Кожанчиков, 1940). Это известно даже для поли-
фагов, например непарного шелкопряда. Для характеристики сезонной
циклики гидротермического режима основных условий обитания Orgyi-
dae. (гилеи, широколиственные леса, хвойные леса, степи) я привожу серию
климограмм (рис. 42).
Orgyidae, распространенные в умеренном климате, но относящиеся
к родам, широко распространенным в тропиках, и в Голарктике также
часто связаны с условиями обитания, имеющими максимально устой-
чивый гидротермический режим. Так, для монашенки в средней Европе
характерны стации хвойно-широколиственных лесов (буково-еловых)
30
ОБЩАЯ ЧАСТЬ
склонов Альп зоны 150—600 метров. Для Olene olga Obth., видов Cifuna
Wlk., Artaxa Wlk. и Ocneria matura Btl. типична связь с кедрово-широко-
лиственными лесами. Эти виды в наибольшей мере несут черты тропи-
ческих Orgyidae.1
Главная масса видов Orgyinae умеренных широт, а также почти все
Acronictinae, свойственные Голарктике, распространены как в хвойно-
широколиственных, так и в широколиственных и даже мелколиственных
лесах, но заметно преобладают в первых. Немногие виды Acronictinae,
например Diphthera Ochs., связаны с хвойными лесами (виды этого
рода связаны в Евразии преимущественно с островными борами). Не-
большое число видов Acronictinae и Orgyinae характерно для кустарни-
ковых стаций, в частности для опушек леса и окраин болот (виды Calo-
casia Ochs., Acronicta menyanthidis View., A. rumicis L., A. lutea Brem.
Рис. 43. Термический режим
приземного слоя воздуха в
течение дня в сосновом лесу
с подлеском (1) и на соседнем
лугу (2).
et Grey, A. alscondita Tr., Olene fascelinaL.,
отчасти виды Orgyia Ochs., Dasychira pudi-
bunda L.). Некоторые представители Orgy-
idae- (Leucoma Steph., Subacronicta Kozh.)
связаны с уремами речных долин.
Переход видов Orgyidae в открытые ста-
ции и,- в частности, даже на опушки лесов
и болот связан с адаптацией их к жизни в
условиях неустойчивого гидротермического
режима. Это особенно необходимо подчерк-
нуть для видов, распространенных в степях
на засушливых и жарких стациях. Нарис.43
приведены синхронные кривые температуры
для смежных участков леса, опушки и луга,
где я наблюдал развитие в природе многих
видов Orgyidae—в лесу Diphthera coenobita
Esp., Daseochaeta alpium Osb., Dasychira
pudibunda L., на опушке по кустарникам
березы— Calocasia coryli L., видов Acronicta
Ochs, (auricoma Scjiift., rumicis L.) и Orgyia
Ochs, (antiqua L., ericae Germ., gonostigma
F.) Переход в открытый ландшафт связан
с адаптацией к новым пищевым условиям
(в частности к травянистой растительности) и к неустойчивому терми-
ческому режиму. В связи с последним моментом переход этот у Orgyidae
осуществляется через промежуточные стации — кустарники опушек,
окраин болот и болотную растительность, уремы речных долин (Кожан-
чиков, 1941).
Адаптация Orgyidae к условиям обитания в открытом ландшафте до-
стигается двумя путями. Известная часть видов (их немного), относя-
щихся к роду Acronicta Ochs. (Pharetra rumicis L., euphorbias Schiff.)
и отчасти к Orgyia Ochs, (ericae Germ., dubia Tausch.), характеризуется
поливольтинизмом, высокой степенью эвритермии и эврихумидности
и полифагией. Значительная степень приспособленности этих видов
1 Может быть отмечено, что эндемики из Orgyidae, филогенетически тяготеющие
к видам, заселяющим тропики Старого Света или горные леса южной Америки, столь
обильные в лесах южного Приморья, присутствуют здесь, без сомнения, в связи
с сохранением специфичных условий, в частности устойчивого гидротермического
режима этих лесов.
экология
81
к обитанию в открытом ландшафте иллюстрируется данными табл. 9
и рис. 44—48.
В противоположность этой группе видов другие виды Orgyidae, оби-
тающие в открытом ландшафте, достигают адаптации к неустойчивому
Таблица 9
Термические пределы и оптимумы развития некоторых видов сем. Orgyidae
Оптимумы Пределы
Название вида развития . развития Условия
яйца обитания
гус. кук. яйца гус. кук.
I группа
Ocneria dispar L. . . . 10—25 20—26 18—28 6—33 5—32 10—32
О. monacha L 10—23 17—25 15—25 6—30 6—31 8—32 Широколиствен-
Acronicta psi L 20—27 15—25 18—26 8—34 5—30 8—31 ные и хвойно-
4. aceris L 15—25 20—25 18—22 9—34 9—30 — широколиствен-
Daseochaeta alpium ные леса
Osb 15—25 15—25 15—25 9—32 9—31 6—31
II группа
Olene fascelina L. . . . 18—25 15—25 — 10—32 10—32
Leucoma salicis L. . . 15—25 — 15—25 10—32 — 10—32
Diphthera coenobita Esp. Subacronicta megacepha- 20—25 21—25 — 15—28 12—26 — Зона березы и боры
la Schiff . 18—25 20—27 15—25 9—30 7—31 7—32
Acronicta leporina L. 18—25 15—22 15—20 9—30 7—30 8—30
.1. alni L 15—20 16—25 15—25 12—30 6—31 9—32 1
Ill группа
Orgyia ericae Germ. . . 20—28 — — — 1
O. antiqua L 15—25 15—22 15—25 10—29 6—31 5—32
<). gonostigma F. ... 15—25 15—25 15—22 10—29 5—31 5—32
Calocasia coryli L. . . 15—20 13—22 15—20 8—28 9—31 6—31 Зона березы,
Acronicta Lutea Brem. et кустарники,
Grey 18—28 15—27 20—27 8—32 8—34 10—32 опушки болот
.4. menyanthidis View. . 15—25 17—24 18—24 8—30 7—32 8—30
.1. auricoma Schiff. . . 15—27 16—27 18—25 9—34 8—34 9—32
.1. rumicis L 15—28 14—30 18—26 9—34 7—35 7—33
гидротермическому режиму путем специализации цикла и сезонного
распределения фаз развития, строго соответствующих сезонной смене
климатических условий и вегетации их, кормовых растений. Эти виды
характеризуются моновольтинизмом и редко дают два поколения в се-
нов (как и лесные); для них характерна моно- или олигофагия личиноч-
ных фаз при рудиментации половой фазы {Simyra Ochs., Oxycesta Hb.,
I'logena Gn.). Первый тип адаптации к жизни в открытом ландшафте ведет
к возрастанию эврибионтности вида и его значительному экологическому
и географическому распространению. Он встречается у Orgyidae реже,
чем второй. Второй тип адаптации обусловливает специализацию стаци-
luii.noro распределения видов и ограничивает их ареалы распространения.
Данные, приведенные в табл. 9, требуют некоторых пояснений.
В каждой из трех групп, несмотря на общую связь с тем или иным био-
топом, наблюдается пестрота в смысле приуроченности видов к стациям.
(j Фауна СССР, И. В. Кожанчиков.
82
ОБЩАЯ ЧАСТЬ
Она наименьшая в первой группе; во второй группе виды, связанные
с зоной березы, отчасти связаны с березняками {Acronicta alni L., Olene
Daseochaeta alpium Osb.; 2— Di
coenobita Esp.
fascelina L.), отчасти с. осинниками
и уремами {Leucoma salicis L., Subac-
ronicta megacephala Schiff.), тогда
как некоторые, как, например, Diph-
thera coenobita Esp., приурочены к
старым борам и связаны со стациями,
обладающими очень стойким гидро-
термическим режимом. Третья груп-
па видов не резко отделена от вто-
рой, но эти виды, кроме леса, жи-
вут также на опушках и даже на
совершенно открытых территориях,
как Acronicta rumicis L., Orgyia ericae
Germ. Для большинства из них ха-
рактерны расширение области тер-
мического оптимума и повышение
этой зоны на термической шкале.
У некоторых видов этой группы,
кроме того, наблюдается расширение
области пригодных для жизни тер-
мических условий за счет повыше-
ния верхнего термического предела,
как то известно для Acronicta auricoma F., A. lutea Brem. et Grey. У ви-
дов, живущих на разных,
хотя и смежных стациях, различие терми-
ческих оптимумов и пределов
может быть очень большим
(рис. 44, 45). Так, для Acronicta
menyanthidis View, типичны уз-
кий термический оптимум и
сильно ограниченные пределы
шкалы температуры, пригодной
для развития преимагинальных
фаз. Этот вид распространен
на севере и заселяет влажные
и холодные стации (березняки
на мшистых болотистых почвах
по понижениям рельефа). Си-
стематически и экологически
ближайшие виды Pharetra {ru-
micis L., auricoma Schiff., lutea
Brem. et Grey), живущие на
Рис. 45. Смертность гусениц при развитии
в разных термических условиях. 1— Acro-
nicta rumicis L.; 2—Daseochaeta alpium
Osb.; 3—Diphthera coenobita Esp.
и пределов и более широкими их гра-
б Orgyidae с разными биотопами видна
сухих опушках и окраинах
торфяников или даже на лугах
(по суходолам), поросших ку-
старником (ивовым и березо-
вым), характеризуются более
высокой температурой оптимума
ницами. Связь подсемейств и трг
из табл. 10.
Виды подсем. Orgyinae концентрируются преимущественно в гилеях,
главным образом на равнинах, но встречаются еще в довольно большом
экология
83
Таблица 10
Распределение видов Orgyidae 1 по биотопам
Число видов
Биотоп
Гилеи ....................
Саванны ..................
Тропические степи . . . , .
Субтропические широколи-
ственные леса ............
Широколиственные леса уме-
ренных широт Л............
Зона березы и хвойных лесов
умеренных широт...........
Степи умеренных широт . . .
Пустыни умеренных широт .
Тундры, (горные и равнин-
ные) ............. . . . .
Orgyinae Momini Acroni- ctini
1207 .29 2 3
316 ИЗ 0
66 1 0 0
313 44 17
187 43 121
15 8 25
30 0 12
7 0 1
6 0 0
числе и в саваннах, а также в субтропических широколиственных лесах.
Меньшее число их известно из бореальных широколиственных лесов
(несмотря на лучшую изученность их по сравнению с субтропиками).
Хвойные леса бедны видами этого подсемейства, несколько богаче ими
степи (сухие), особенно тропические. Пустыни умеренных широт и тундры
имеют лишь единичных представителей этого подсемейства. В подсем.
Acronictinae виды Momini представлены еще довольно богато в гилеях,
по лишь в горных, с более низкой и изменчивой температурой. Из саванн
их почти не известно, но широколиственные леса как субтропиков, так
и умеренных широт населены ими обильно. Хвойные леса бедны видами
Momini, а из степей, пустынь и тундр виды этой трибы не известны со-
всем. Виды трибы Acronictini дают еще больший сдвиг максимума обилия
в умеренные широты, причем также почти нацело в широколиственные
леса, хотя более обильно по сравнению с Momini они представлены
в зоне березы и хвойных лесов, отсутствуя, как и последние, .в тундрах.
1 Числа видов Orgyidae, распространенных в саваннах и сухих тропических сте-
пях, без сомнения, несколько выше, чем истинные, так как ссылки в литературе на
моста обитания этих видов очень общи и говорят лишь о нахождении данного вида
па той или иной большой территории, которая хотя в основном и занята степью или
си ванной, но имеет также участки влажного тропического леса или широколиствен-
ных лесов. _ Для ряда видов (формально распространенных в степях) на'основании
специальной литературы мною было установлена несомненная связь с лесами, куда
оци и отнесены в таблице. Большая часть местонахождений не поддается более точ-
ной расшифровке в экологическом смысле. В отношении всей таблицы необходимо
отметить, что число видов тропических Orgyidae еще далеко не исчерпано. Фауна
многих территорий тропиков еще не затронута специальными исследованиями.
Напротив, для умеренных широт инвентаризация видов подсем. Orgyinae почти пол-
ностью закончена. Степень изученности видов подсем. Acronictinae значительно пол-
ное, так как они почти нацело ограничены умеренными широтами.
2 Более половины (17) видов распространены в горных гилеях Старого Света
(Зондские острова). К ним относятся Мота Hb., Belciana Stgr., Trisuloides Hamps.,
rimlatha Wlk. Большая часть остальных населяет горные гилеи Южной Америки
(Колумбия), в частности виды Lichnoptera Н. S. и Gaujonia Dogn.
3 Виды Momini, распространенные в саваннах, заселяют главным образом тер-
ритории Южной Америки (Венецуэлу, Колумбию).
6*.
84
ОБЩАЯ ЧАСТЬ
Как в тропиках, так и в умеренных широтах биотопы открытого
ландшафта заселены видами Orgyidae слабо. Необходимо подчеркнуть,
что саванны и степи заселяются видами Orgyidae. обычно при сильном
развитии травянистой и кустарниковой растительности. Пустынные
степи в умеренных широтах, где распространение видов Orgyidae изучено
более точно, населены немногими, преимущественно специфичными пред-
ставителями (Parocneria Dyaf, Albarracina Stgr., Sirnyra Ochs., Eogena
Gn.). Обычно в пустынях (особенно тропических) виды Orgyidae связаны
с оазисами, исключая отмеченные пустынные виды.
Суточная ритмика жизни видов Orgyidae, населяющих крайние усло-
вия — биотопы пустынь и тундры, приспособлена к их своеобразному
гидротермическому и световому режиму. Так, в пустынях активность
всех фаз развития имеет место
в ночные часы, а переживание
жаркого и сухого времени суток
осуществляется в тени растений
под сухими листьями и в поверх-
ностном слое почвы. Типичны
суточные миграции гусениц ночью
на деревья для питания, днем в
поверхностный слой почвы для пе-
реживания жаркого времени.1 Точ-
но так же и в аркто альпийском
ландшафте немногие обитающие
виды Orgyidae характеризуются
определенными чертами поведения.
Распространенные там виды рода
Gynaephora Hb. ведут гелиофиль-
ный образ жизни, используя в вы-
соких широтах Арктики тепло
прямой круглосуточной радиации
солнца. Из видов, ведущих ночной
образ жизни и нормально избега-
ющих прямого солнечного осве-
щения, в Арктику проникает лишь
Рис. 46. Смертность яиц при развитии
в разных условиях влажности при темпе-
ратуре 20°. 1—Daseochaeta alpium Osb.;
2—Orgyia gonostigma F.; 3— O. antiqua
L.; 4—Leucoma salicis L.' 5—Orgyia eri-
cae Germ.; 6—Ocneria dispar L.
один — utene jascettna ь. иораз
жизни этого вида в Арктике, впрочем, не известен; для личиноч-
ной фазы не исключен гелиофильный образ жизни, для имаго он сомни-
телен. Настоящие ночные виды и виды, избегающие прямого солнечного
света, каковы все Acronictinae, в Арктику не проникают, хотя из горной
лесотундры еще известны.
Распределение видов Orgyidae по биотопам показывает в целом их
высокую гигрофильность, термофильность и мегатермичность. Хвойные
леса и тем более тундры, несмотря на высокую влажность и, в первом
случае, на очень устойчивый термический режим, населены ничтожным
числом видов этого семейства. Степи, несмотря на сухость и изменчивый
режим влажности, имеют не меньшее число видов Orgyidae, чем хвойные
леса, исключая стенохумидные виды трибы Momini, отсутствующие
в степях совершенно. Для степей и полупустынь характерно на-
1 Требуется отметить, что и для видов Orgyidae умеренных широт, например для
монашенки, отмечены (Кулагин) суточные миграции. На жаркий период суток в юж-
ных частях ареала гусеницы этого вида спускаются с крон по стволам деревьев
в более прохладный ярус, вновь поднимаясь для питания вечером на кроны деревьев
экология
85
личие не только широко распространенных видов или видов, относя-
щихся к широко распространенным родам, следовательно эвритопных,
каковы Orgyia Ochs., но, главное, наличие большого числа специфичных
представителей, распространенных нередко локально (Leiometopon Stgr.,
Hogena Gn., Parocneria Dyar, Albarracina Stgr., Oxycesta Hb.). Это об-
стоятельство подчеркивает высокую потребность в тепле Orgyidae, труд-
нее приспособляющихся к недостатку тепла в зоне хвойных лесов, чем
к изменчивому гидротермическому режиму степей, где количество тепла
достаточно и даже избыточно.
Зональное распространение Orgyidae, ограниченное обычно одной
зоной или двумя родственными (например широколиственные леса суб-
1’ис. 47. Термогигрограмма-оптимум
(отсутствие смертности) для яиц мо-
нашенки (Ocneria monacha L.) — пунк-
тир (по Цвёльферу, 1934) и непарника
(О. dispar L.) — сплошная линия,
и осенне-летний период развития
(а) и весной (Ь).
Влажность Воздуха
Рис. 48. Термогигрограмма для разви
тия яиц Acronicta rumicis L.
1 — оптимум (отсутствие смертности);
2 — граница полного вымирания яиц
при развитии.
тропиков и широколиственные леса умеренных широт, гилеи, субтропи-
ческие леса и т. д.), характеризует малую их физиологическую пластич-
ность. Лишь отдельные виды семейства, например Olene fascelina L.,
Ocneria dispar L. и немногие другие, заселяют несколько зон; первый
вид, например, распространен и обычен как в хвойных лесах, так
и в зоне широколиственных лесов и даже в тундре и пустынях (Монголии).
Представление о потребности видов Orgyidae в тепле дает рассмотре-
ние сумм тепла, необходимых для полного развития разных их видов
и разных фаз их (табл. 11; рис. 49—51). Приводимый материал был изу-
чен путем постановки многочисленных экспериментов с палеарктическими
видами семейства.1 Тропические виды семейства не затронуты экспери-
1 Для экспериментов и наблюдений над каждым видом использовано в среднем
1000—2500 особей, а для некоторых видов даже больше. Эксперименты охватили
икт.тилетний период, а наблюдения— около 10 лет; для всех видов было исследовано
86
ОБЩАЯ ЧАСТЬ
ментами за отсутствием материала. Литературные сведения эксперимен-
тального характера для них также отсутствуют. И для палеарктических
представителей семейства литературные данные очень скудны и касаются
всего трех вредных представителей — монашенки, непарного шелкопряда
и златогузки, причем для последних видов эти сведения очень неполны
[Али (Ali, 1933); Сэндерсон и Пэре (Sanderson a. Pairs, 1913)].
Таблица 11
Суммы тепла и нижние термические пределы для развития некоторых видов
Orgyidae
Название вида t Нижние пороги развития в °Ц Сумма тепла ДЛЯ развития Сумма тепла для всего цикла Условия обитания
яйца гус. кук. яйца гус. кук.
I группа
Ocneria monacha*L. * 1 . . . 6.0 6.0
О. dispar L 6.0 5.0
Acronicta psi L 8.5 5.0
Daseochaeta alpium Osb. 8.5 9.0
Acronicta aceris L 9.5 8.8
II группа Diphthera coenobita Esp. . 15.0 12.5
Leucoma salicis L 9.0 ——
Olene fascelina L 10.5 —
Subacronicta rnegacephalo. Schiff 9.0 7.0
Acronicta leporina L. . . . 9.0 7.5
A. alni L 12.0 6.0
Ill группа Orgyia ericae Germ. . . . 13.5
0. antiqua L . 10.0 4.0
0. gonostigma F 11.0 8.0
Calocasia coryli L 8.5 6.0
Acronicta lutea Brem. et Grey 8.0 8.0
A. menyanthidis View. . . 8.0 7.5
A. auricoma Schiff. . . . 9.0 8.0
A. rumicis L 9.0 7.5
8.5 305 804 130 1239 Широколи-
10.0 416 714 153 1283 ственные или
.8.5 87 600 555 1242 хвойно-широ-
6.0 97 385 239 721 колиственяые
10.0 82 551 200 833 леса
— 50 175 364 — — Зона березы;
155 отчасти хвой-
ные леса
7.0 90 396 183 669 (боры) и ши-
8.2 80 512 211 803 роколиствен-
. 9.5 65 471 120 656 ные
109 —
5.0 9.5 6.0 253 135 98 440 417 600 166 ‘120 152 859 672 850 Кустарники и болота зоны березы;
9.0 100 532 177 809 отчасти широ-
8.5 . 100 457 233 790 ; колиственяые
9.0 90 463 225 778 леса
7.5 92 390 243 725
Адаптация к неустойчивому термическому режиму и к недостатку
тепла при расселении Orgyidae к северу достигается одним и тем же
путем — уменьшением потребной для развития суммы тепла. Об этом,
свидетельствует рассмотрение сумм тепла более чем 20 видов семейства.
около 50 000 особей. Кривые, характеризующие зависимость развития разных фаз
исследованных видов от термического режима, я привожу выборочно (в силу их боль-
шого числа), для наиболее характерных случаев.
1 Для монашенки я привожу данные по Цвёльферу (1933, 1934). По развитию
куколок и гусениц видов Orgyia Ochs, и Ocneria Hb. приведены средние данные обоих
полов.
экология
87
Характерно также снижение суммы тепла у видов эвритермичных, засе-
ляющих биотопы с наименее устойчивым термическим режимом. Эври-
Рис. 49. Влияние термических условий на длительность
развития яиц. 1 — Orgyia antiqua L.; 2 — 0. ericae Germ.;
3 — Leucoma salicis L.; 4 — Acronicta psi L.
термичные же виды, в нашем случае виды Orgyia Ochs, и некоторые Асго-
rdcta Ochs., распространены к северу в наибольшей мере. Нижние терми-
Рис. 50. Влияние термических условий на длитель-
ность развития гусениц. 1 — Ocneria dispar L.;
2— Orgyia antiqua L.; 3—-Acronicta rumicis L.
чсские пороги развития отличаются у разных видов слабо и менее опре-
деленно, оставаясь примерно одной величины как у видов, связанных
г широколиственными лесами, так и у заселяющих зону хвойных лесов.
Приспособление к использованию для развития крайних низких темпе-
88
ОБЩАЯ ЧАСТЬ
ратур у видов Orgyidae выражено слабо,1 в большей мере, невидимому,
у бореальных видов подсем. Orgyinae, а также у видов трибы Acronictini.
Может быть отмечена связь уменьшения сумм тепла с уменьшением раз-
меров особей этих видов и побледнением их окраски. Правило это дей-
ствительно при сравнении видов Orgyidae, относящихся к одному роду,,
или форм одного вида, распространенных в пределах разных зон.
Разнообразие сумм тепла связано не только с особенностями зональ-
ного и стадиального распределения Orgyidae, но отражает также и осо-
бенности онтогенетического развития и, следовательно, циклику разных
видов. Так, мегатермность видов Orgyinae стоит в связи с большой потреб-
ностью к теплу их яиц. Этот факт вяжется с наличием большого коли-
Рис. 51. Влияние термических условий на длительность развития
куколок. 1 — Ocneria dispar L.; 2—Daseochaeta alpium Osb.; 3 —
Acronicta alni L.‘, 4 —A. psi L.
чества желтка в яйцах этих видов и с возникновением многих вторичных
личиночных черт в эмбриональном их; развитии. Напротив, потребность
в тепле куколок Orgyinae меньшая, чем у Acronictinae, и, без сомнения,
стоит в связи с рудиментацией половой фазы этих видов и потому с более
быстрым ее развитием. Мегатермность ряда видов подсем. Acronictinae
(Subacronicta Kozh., Calocasia Ochs., Acronicta leporina L.), напротив,
связана с большими суммами тепла, необходимыми для развития гусениц
и особенно куколок, которые обладают диапаузой и дают в высокой сте-
пени развитых, специализованных имаго. Яйца видов Acronictinae бедны
желтком и дают недоразвитых гусениц; они характеризуются быстрым
развитием и малыми суммами тепла. Обращают внимание малые суммы
1 В пределах других групп насекомых, подвергавшихся специальному изуче-
нию в направлении адаптации их к распространению на север, наблюдается иное.
Например, у листоедов Phaedon распространение к северу достигается значитель-
ным снижением порогов развития, использованием крайне низких (еще положитель-
ных) температур для развития преимагинальных фаз (Кожанчиков. 1939. Роль
тепла в развитии и распространении овощных листоедов Phaedon cochleariae F. и
Gastroidea viridula Deg. Изв. Высш. курс, прикл. зоол. и фитопатол., IX: 5—39).
экология
89
тепла, необходимые для развития гусениц Acronicta alni L. и Sabacronicta
megacephala Schiff., видов, у которых в личиночном развитии вторичные
черты выражены слабо.
Связь величин суммы тепла со спецификой онтогенеза бросается
в глаза и при рассмотрении их для разных фаз диморфных видов
(табл. 12). Так, суммы тепла, необходимого для развития самок и самцов
в личиночной и куколочной фазах у видов Ocneria Hb. и Orgyia Ochs.,
очень различны. Суммы тепла в личиночной фазе для самок значительно
больше, чем для самцов, что связано с большей интенсивностью процес-
сов ассимиляции самок, накопляющих громадные количества жировой
ткани и развивающих более богатую мускулатуру, что далее исполь-
зуется организмом для образования желтка яиц. Гусеницы самок, как
указывалось, имеют добавочную линьку. Напротив, большая рудимен-
тация имагинальной фазы самок по сравнению с самцами ведет к значи-
тельному сокращению сроков развития и сумм тепла для куколок самок.
Тем не менее следует специально подчеркнуть, что особенности онтоге-
неза, если их учитывать, не затемняют отмеченной выше географической
закономерности в величинах сумм тепла. Уменьшение сумм тепла у видов,
эвритермичных и распространенных далеко к северу, проходит красной
нитью. Так, виды Acronicta rumicis L. и A. auricoma Schiff., вполне разви-
вающие вторичные черты в личиночной фазе, одновременно характери-
зуются широкой адаптацией к термическому режиму и малыми суммами
тепла, необходимыми для развития личиночных фаз и цикла в целом.
Таблица 12
Суммы тепла для личиночной и куколочной фаз некоторых диморфных видов Orgyidae
Название Суммы тепла для: 1
гусениц куколок
вида самцы | самки самцы самки
Ocneria dispar L. . Orgyia antiqua L. . Orgyia gonostigma F. . 615.4 440.5 394.0 781.7 539.0 440.0 159.2 201.0 157.3 148.0 132.5 87.7
В табл. 13 приведены общие суммы тепла и средние термические
(нижние) пороги для развития зимующих фаз Orgyidae. Величины эти
1 Приводимые здесь факты еще раз говорят о том, что сумма тепла, представляя
специфичный экологический показатель вида, должна тем самым являться и спе-
цифичной физиологической величиной (К ozhantshikov. 1936. Zool. Anz.,
113:7). В связи с этим может быть отмечено еще одно положение. Адаптация к жизни
и умеренных широтах с изменчивым . суточным и сезонным термическим режимом,
где налицо недостаток тепла как в единицу времени (сутки), так и в целом, в тече-
ние всего цикла развития, ведет к возникновению физиологических типов, более
экономно расходующих тепло при развитии, в частности более полно использующих
единицу тепла (калорию). Частным случаем этого, без сомнения, является отмечен-
ное выше уменьшение размеров и обеднение пигментации бореальных и северных
видов изучаемого семейства по сравнению с субтропическими, что я отмечал выше.
Факт более экономного использования тепла как физиологическая особенность се-
верных видов животных (во всяком случае беспозвоночных) в широком обсуждении
выходит за рамки настоящей работы [И. Кожанчиков. 1946. О лабиль-
ности процессов развития насекомых в отношении термических влияний. Зоол. жури.,
25 (1) : 27—35).]
SO
ОБЩАЯ ЧАСТЬ
для периода развития фаз до диапаузы и после нее могут быть очень
отличны, в зависимости от момента, на который падает диапауза в раз-
витии данного вида. У видов Orgyidae, у которых диапауза яиц наступает
в самых ранних фазах эмбрионального развития, сумма тепла опреде-
ляется почти полностью весенним периодом их развития. Напротив,
у видов рода OcneriaHb., у которых диапаузирует в яйце почти завершив-
шая свое эмбриональное развитие гусеница (покрытая кутикулой и имею-
щая вполне развитыми все органы), сумма тепла для весеннего периода
развития составляет одну четвертую или одну пятую часть от общего
ее объема. Для личиночной фазы эти отношения также разнообразны.
Таблица 13
Величины сумм тепла и нижних порогов развития для
некоторых видов Orgyidae до и после диапаузы
Название вида До диапаузы После диапаузы
нижний предел сумма тепла нижний предел сумма тепла
Ocneria dispar L. . . 6.8 310.3 5.7 106.3
Ocneria monacha L. . 6.8 240.0 4.9 65.0
В табл. 13 нет данных для куколок Acronictinae, также обладающих
диапаузой. Диапауза их проходит на очень ранних стадиях развития,
во всяком случае еще до гистогенеза, хотя гистолиз (в значительной
мере за счет развития пронимфы) в момент Диапаузы уже почти закон-
чен. Каковы суммы тепла для развития куколок до и после диапаузы,
определить очень трудно. Косвенно о продолжительности обоих периодов
развития можно судить по сравнению сроков развития куколок летнего
и зимующего поколений данного вида, как то возможно у поливольтин-
ных видов рода Acronicta Ochs.; у Acronicta rumicis L. и A. auricoma
Schiff., например, длительность развития куколок зимующего и летнего
поколений почти равна или даже для летнего поколения превышает
величины для зимующего. Из этого можно заключить, что почти все
развитие куколок у этих видов протекает весной, после диапаузы
(табл. 14).
Приведенные выше величины (табл. 11—14) сумм тепла и нижних
порогов развития разных видов Orgyidae характеризуют отчасти также
фенологию разных их фаз и групп. Яйца видов подсем. Orgyinae разви-
ваются большею частью в середине лета {Laelia Steph., Arctornis Germ.,
Dasychira Steph, и др.). У видов Orgyia Ochs., несмотря на то, что зимуют
яйца, выход гусениц наблюдается в начале лета, так как яйца требуют
больших сумм тепла в весенний период, когда проходит почти все их раз-
витие. Лишь у видов Ocneria Hb. требуемые ими для развития большие
суммы тепла обеспечиваются осенним периодом развития, а выход гусе-
ниц весною наблюдается с первыми теплыми днями, так как весенний
отрезок эмбрионального развития очень короток. Развитие яиц видов
подсем. Acronictinae, исключая Episema Ochs., протекает в течение ран-
него лета, охватывая,очень короткие сроки, и слабо влияет на феноло-
гию видов этого подсемейства.
экология
91
Таблица 14
Длительность развития куколок Acronicta Ochs,
зимующего и летнего поколений
Темпе- ратура (°Ц) Длительность развития куколок (в сутках)
Acronicta rumicis L. Acronicta auricoma Schiff.
зимующее поколение летнее поколение зимующее поколение летнее поколение
32.0 8.0 8.0
30.0 9.2 7.9 10.0 8.4
27.5 10.5 8.0 10.5 • —
25.0 12.0 11.2 12.0 —
20.0 17.4 24.0 24.0 —
Форма кривых зависимости развития яиц разных видов семейства
от термического режима специфична (рис. 49). Специфичны и термиче-
ские кривые для развития личиночной фазы. На рис. 50 можно видеть
разный объем термической шкалы, в пределах которой возможен рост
гусениц некоторых, наиболее типичных представителей Orgyidae. При
разделении личиночного периода на два — до диапаузы и после нее —
появление гусениц весной и дальнейшее их развитие и питание начи-
наются сразу вслед за наступлением стойкой теплой погоды. Это наблю-
дается еще до облиствения кормовых растений {Euproctis Hb., Olene Hb.,
('ynaephora Hb., Arctornis Germ., Leucoma Steph.), когда гусеницы пи-
таются почками. Лишь после известного срока они переходят на питание
молодыми листьями. Кривые зависимости развития куколок от терми-
ческого режима (рис. 51) также отражают специфичные Черты разных
видов. По характеру этих кривых можно выделить две большие группы
видов. В одной группе куколочное развитие проходит быстро полностью
в течение летнего периода. Это наблюдается почти у всех, видов подсем.
Orgyinae. Оно подобно также у большинства Acronictinae (.Diphthera Ochs.,
Calocasia Ochs., Мота Hb., Daseochaeta Warr., Acronicta Ochs, группы
rumicis L.) и некоторых Orgyinae (Dasychira Steph.), хотя и зи-
мующих в фазе куколки, но имеющих, короткий период развития,
Протекающего почти исключительно весной. Вторая группа сла-
гается из видов Acronicta Ochs, группы Triaena (psi L., cuspis
lib., tridens Schiff.) и некоторых других видов этого подсемейства (Epi-
sema Ochs.), развитие куколок которых длится большой срок, а появле-
ние имаго наблюдается в середине лета или даже под осень. Эти виды
требуют больших сумм тепла для развития куколок.
Влияние термических условий, выходящих за пределы температур,
Пригодных для развития, возможно в природе в связи с сильными терми-
ческими колебаниями осенью и весной. Оно даже в пределах небольших
сроков (порядка часов) может вести к глубоким и часто необратимым
изменениям обмена веществ и структур. Необходимо подчеркнуть, что
подобные изменения наступают лишь при отсутствии диапаузы. При ее
наличии как кратковременные температурные влияния (шоки), уак
и длительные проходят, не отражаясь на дальнейшем развитии. В связи
с этим при повышении температуры поздним летом осенние заморозки
слабо или вообще не отражаются на развитии зимующих фаз. Весен-
92
ОБЩАЯ ЧАСТЬ
ние же заморозки, падающие на развитие тех же фаз, скорее могут пред-
ставлять опасность для развития особей после диапаузы.1
При воздействиях, даже кратковременных, крайней низкой или край-
ней высокой температуры при развитии куколок видов подсем. Orgyinae
наблюдается массовая гибель особей. Выходящие немногие имаго почти
всегда бесплодны и нередко уродливы. В опытах Емельяновой (Emelja-
noff, 1924) с непарным шелкопрядом было установлено, что при одно-
трехкратном воздействии на куколок в течение 24 часов температуры
37—40° или 6—8° у выходящих бабочек наблюдается дегенерация
не только спермиев или яиц, но даже утрата у самок целых яйцевых
трубочек и, конечно, бесплодие особей. Наряду с этим наблюдалось
недоразвитие у самцов вторичных половых признаков — гребешков
усиков и сенсилл на них, а также гонапофизов (valvae, saccus), часто
присутствие недоразвитого парного uncus. У самок, напротив, наблюда-
лись некоторые удлинения выростов на члениках усиков и большая,
по сравнению с обычной, величина papillae genitales. У обоих полов
отмечалось изменение окраски, у самцов в сторону посветления, у самок
в сторону потемнения. Таким образом у обоих полов изменения структур
под влиянием термических шоков представляют инверсию (по существу,
просто недоразвитие) вторичных половых признаков. Косминским (Kos-
minsky, 1924а) параллельно с Емельяновой (1924) описано появление
подобных же дефектных индивидов (интерсексов) в потомстве особей
непарного шелкопряда, подвергавшихся действию термических шо-
ков при развитии куколок. Им же (1924b) описывается недоразвитие
усиков у самцов ивовой волнянки при влиянии низкой температуры,
порядка 3°, на куколок. Структуры усиков совпадали с теми, что
наблюдались у самок.
Orgyidae в целом гигрофильны, но обнаруживают специфичное отно-
шение к влажности; некоторые виды их очень устойчивы к сухости,
как, например, некоторые Orgyia Ochs., Parocneria Dyar, Albarracina Stgr.
Представление об отношении разных видов к условиям гигрорежима
дает рассмотрение кривых зависимости смертности преимагинальных
фаз от условий влажности (рис. 46, 47, 48, 52). Оптимум для развития
яиц большинства видов лежит при 60—90% относительной влажности
воздуха, но степень выносливости их к пониженой влажности у видов,
населяющих открытый ландшафт, значительна. Так, у лесных Daseo-
chaeta alpium Osb. и Orgyia gonostigma F. вымирание яиц при развитии
на 50% наступает при падении влажности воздуха до 50%, тогда как
у видов, заселяющих кустарниковые стации, каковы Leucoma salicis L.,
Orgyia antiqua L. и особенно Orgyia ericae Germ., заселяющего верещат-
ники, развитие яиц даже при влажности воздуха в 6% ведет к отмира-
нию лишь 25% особей. Высокая устойчивость яиц к сухости наблюдается
и у эвритермного и вообще эврибионтного непарного шелкопряда, засе-
ляющего преимущественно лиственные леса, но живущего также
и в кустарниковых степных стациях. Напротив, у близкого, но чисто
лесного вида — монашенки — развитие яиц ограничено значительно
более узкими пределами влажности (рис. 47). Влияние условий влаж-
- 1 Явление эстивации, т. е. перенесения высоких температур и засухи в состоя-
нии диапаузы, у Orgyidae наблюдается редко. Оно вероятно для куколок Episema
coeruleocephala L. и для отдельных особей Pharetra Hb. (rumicis L., auricoma Schiff.).
Также вероятно наличие эстивации при развитии яиц Orgyia dubia Tausch. и гусениц
некоторых видов рода Parocneria Dyar.
экология
93
пости на темны развития яицы Orgyidae не велико. Длительность развития
их в условиях разной влажности дает различия порядка часов, редко
суток, возрастая в условиях сухости и избыточной влажности.
Помимо специфичных отличий реакции разных видов Orgyidae в пе-
риод эмбрионального развития на влияние влажности, реакция их
у одного и того же вида меняется в зависимости от стадии эмбриональ-
ного развития, на которую падает данное влияние. Яйца непарного
шелкопряда в летний период развития (до диапаузы) относительно легко
переносят как предельную высокую влажность, так и крайнюю сухость
воздуха, но стенотермны. После диапаузы, весной, эмбриональное разви-
тие непарного шелкопряда в большей мере ограничено условиями влаж-
ности и в меньшей термическим режимом (рис. 47). У монашенки также
устойчивость яиц к условиям влажности большая в период развития
их до диапаузы (в осенне-летний период) по сравнению с весенним перио-
дом развития (после диапаузы). Стойкость яиц к условиям влажности
слабо зависит от толщины хориона, будучи, повидимому, в большей
мере обусловлена его физико-химическими свойствами (может быть
составом). Так, толщина хориона яиц Acronicta rumicis L. очень не
велика (сквозь него можно наблюдать развитие эмбриона), тогда
как стойкость яиц этого вида к сухости значительна и большая, чем
многих других видов Orgyidae, имеющих толстый хорион яйца; разви-
тие яиц Acronicta rumicis L. возможно еще при влажности воздуха
н 5—6% (рис. 48).
Стойкость личиночной фазы (гусениц) Orgyidae к условиям влажности
разнообразна и специфична, но в общем велика. Гусеницы монашенки,
по Цвёльферу (1934), имеют оптимум развития при 90% относительной
влажности воздуха, но нормальное развитие их возможно в пределах
влажности воздуха 100—50%. Для непарного шелкопряда границы эти
шире (100—40%), но оптимум условий влажности для личиночного раз-
вития этого вида также лежит близ 80—90%. Гусеницы Orgyidae, несу-
щие густой волосяной покров, более устойчивы к сухости, чем те, кожа
которых несет редкие и короткие волоски. Менее устойчивы к сухости
также младшие возрасты гусениц. Слабая сухостойкость личиночной
фазы типична для Diphthera Ochs., Daseochaeta Warr., и большинства
видов Acronicta Ochs, (кроме видов Pharetra Hb.), развитие которых тор-
мозится уже при влажности воздуха в 75—60%. В природе в пределах
Голарктики в южных ее частях и в континентальных областях режим
влажности выступает в качестве ограничивающего фактора вместе с коле-
баниями температуры.
Наибольшую устойчивость к влиянию влажности обнаруживают
куколки Orgyinae (рис. 52), что может быть связано с развитием у них
плотного кутикулярного покрова и с присутствием у многих видов также
плотных шелковых коконов. Последний момент, видимо, имеет второсте-
пенное значение, так как куколки ивовой волнянки, непарного шелко-
пряда и ряда других видов, почти лишенные кокона, тем не менее очень
устойчивы к колебаниям влажности. В пределах подсем. Acronictinae
устойчивость к изменениям влажности обусловливается также наличием
у них диапаузы. Для куколок непарного шелкопряда установлена обшир-
ная область влажности в пределах 10—100%, пригодная для развития
куколок при температуре 12—30°. Волее чувствительны к сухости
куколки Leucoma salicis L., Olene fascelina L., Orgyia gonostigma F., Dasy-
ehira pudibunda L., а из Acronictinae — виды Acronicta Ochs, (кроме
Pharetra Hb. и Subacronicta Kozh.).
94
ОБЩАЯ ЧАСТЬ
Адаптация к жизни в широтах умеренного климата с холодной зимой
выражается у Orgyidae возникновением диапаузы и холодостойкости
зимующих их фаз. Холодостойкость бореальных Orgyidae специфична
и нередко очень значительна. В ряде случаев она обнаруживает большую
индивидуальную изменчивость. Она связана, как то было установлено
специальными исследованиями (Кожанчиков, 1938, 1939), с наличием
диапаузы и присутствием у холодостойких особей термостабильного
дыхания, не подавляемого низкими температурами (рис. 53). Общее
представление о холодостойкости бореальных видов семейства, и именно
зимующих их фаз, дает табл. 15.
Холодостойкость видов Orgyidae, будучи специфичной, в известной
•мере зависит также от условий среды. Одно из них —это длительность
действия отрицательной температуры. Это видно хотя бы из опытов Эше-
риха (Цвёльфер, 1934) с яйцами монашенки, который показал, что при
Рис. 52. Смертность куколок Acronicta
rumicis L. в разных условиях влаж-
ности.
коротких экспозициях они переносят
охлаждение до —70°. Точно так же
Рис. 53. Дыхание яиц непарника
(Ocneria dispar L.) в разных тер-
мических условиях. 1 — поглоще-
ние кислорода; 2 — выделение
углекислоты; 3 — дыхательный
коэффициент.
яйца непарного шелкопряда выносят
получасовое охлаждение ниже.— 55°. В природе зимовка яиц мона-
шенки при падении температуры до —40° еще возможна [Кнохе (Knoche,
1929)], но какова степень вымирания яиц при этом, не учтено. По моим
наблюдениям над непарным шелкопрядом несомненно, что значитель-
ные, но кратковременные охлаждения до —40, —45° (в пределах
суток) переносятся яйцами этого вида легко, а длительные действуют
губительно.
Другим существенным условием являются момент развития и состоя-
ние особи, когда действуют отрицательные температуры. При действии
их в период диапаузы стойкость зимующих фаз Orgyidae велика, при
запоздалом действии мороза в начале весны (конец февраля, март или
апрель) холодостойкость этих же зимующих особей значительно сни-
жается. Последнее обстоятельство может быть поставлено в связь с физио-
логическими изменениями при развитии (рис. 54).
экология
95
Таблица 15-
Холодостойкость зимующих фаз некоторых видов
Orgyidae
Название вида Фаза зимовки Зона смертель- ных температур (В °Ц) при кратко- временном влиянии 1
Ocneria dispar L Яйца 40—55
О. monacha L. » 40—45
Dasychira pudibunda L Куколки 25—30
Olene jascelina L Гусеницы Ниже 40
Euproctis chrysorrhoea L » 20—25
Orgyia antiqua L Яйца Ниже 40
Daseochaeta alpium Osb Куколки 30—35
Calocasia coryli L . » 35—40
Acronicta rumicis L » 40—55
A. lutea Brem. et Grey .... » 40—45
A. auricoma Schiff ' . » 35—40
A. psi L » Ниже 40
A. aceris L » 25—30
время взятий яиц из природы после 1 шевцаля/всутках)
Рис. 54. Длительность развития яиц непарни-
ка {Ocneria dispar L.), взятых из природы в
период 23 февраля — 15 апреля; после 15 ап-
реля—начало развития яиц в природе.
Независимо от сказанного и после окончания диапаузы холодостой-
кость особей Orgyidae, развивающихся весной, может быть относительно
большой. Так, по Фернальду (Fernaid, 1898), яйца непарного шелко-
пряда после окончания диапа-
узы еще способны переносить
охлаждение до —28°. Вышед-
шие из яиц гусеницы мона-
шенки в «зеркалах» также до-
вольно холодостойки. По Ар-
ниму (Arnim, 1936), они спо-
собны переносить сильные за-
морозки, до —7°. Нольте (Nolte,
1939) указывает, что вышедшие
из яиц гусеницы монашенки
могут переносить, не погибая
в дальнейшем, температуру —1,
—2° в течение 6 суток, вымирая
нацело при этих условиях лишь
через 21 день. Замечательно,
что длительное охлаждение
задерживает
развитие гусениц (например
не происходит пигментации покровов) лишь на срок действия холода,
давая слабое последействие, во всяком случае незаметное в последующем
развитии их (в природных условиях) по сравнению с особями, не под-
1 Кратковременные охлаждения, которые были использованы для этих изме-
рений, определялись сроками в 12—24 часа. При длительных охлаждениях порядка
мпогих суток эта зона несколько смещается вверх. В природе, при наличии суточ-
ной ритмики температуры, действие крайних температур обычно порядка полусуток.
96
. ОБЩАЯ ЧАСТЬ '
’I
вергавшимися длительному охлаждению. По моим данным (1940), яйца
непарного шелкопряда в момент выхода из них гусениц, перенесенные
в условия температуры от —6° до —9°, способны переносить задержку
развития до двух месяцев. Спустя этот срок наблюдается нормальное
отрождение гусениц при наступлении необходимых условий температуры.
Очень важным условием холодостойкости Orgyidae является режим
влажности при зимовке. Де-Флитер (De Fluiter, 1934) указывает, что
за период 1917—1932 гг. в Голландии (Бермонд) дождливые и влажные
зимы неизменно вызывали снижение численности златогузки, тогда как
значительные отрицательные температуры (порядка —20° как предел),
типичные для сухих зим, не влияли отрицательно на зимовку гусениц.
По Нольте (1939), отродившиеся весной гусеницы монашенки при повы-
шенной влажности вымирают при температуре —1, —2° скорее, чем
в сухом воздухе. По моим наблюдениям, гусеницы Olene fascelina L.
под Ленинградом сильно вымирают за период зимовки во влажные, хотя
и теплые зимы (с многочисленными и длительными оттепелями). Напро-
тив, сухую, хотя и очень суровую зиму 1939/1940 г., когда наблюдались
падения температуры до —50°, гусеницы этого вида пережили вполне
благополучно и появились весной в массе'. Сухость в период зимовки
особенно необходима для видов, зимующих в личиночной фазе. Это нахо-
дит отражение и в поведении зимующих гусениц Orgyidae, которые, как то
известно, например, для (ynaephora selenitica Esp., Arctornis l-nigrwn
Mull., Leucoma Steph., отыскивают для зимовки сухие места. В частности,
первые два вида, уходящие на зимовку в поверхностный слой почвы,
отыскивают сухие ее участки, обычно песчаные. Другие виды, как ивовая
волнянка или златогузка, зимуют в шелковых коконниках поодиночке
(первый) или в общем гнезде. Шелковая ткань й расположение коконов
йли гнезд высоко над почвой обеспечивают сухость при зимовке.
В связи с наличием шелковых коконов у куколок, часто очень плот-
ных, как то характерно для подавляющего большинства видов Acronic-
tinae, исключая Calocasia coryli L., и для видов Orgyidae (Dasychira Ochs.),
зимующих куколкой, роль влажности в зимовке куколок менее велика.
Так, куколки Dasychira pudibunda L., вообще относительно мало холо-
достойкие, хорошо переносят влажные й даже дождливые, но теплые
зимы, вымирая почти нацело в сухие, но холодные (например в
1939/1940 г. в Прибалтике). Напротив, защитная роль коконов от действия
низких температур (со стороны теплопроводности их) очень не велика.
Отсутствует подобная же защитная роль и волос, покрывающих кладки
непарного шелкопряда, златогузки, синеголовки и других видов волня-
нок. Защитное значение их несомненно в отношении действия влажности.
По Кулагину (Kulagin, 1897), гибель очищенных от волос яиц непарного
шелкопряда наступает уже при —15° Р (—18.5° Ц), тогда как яйца,
не очищенные от волос, при этих же условиях выносят температуру
до —50° Ц. Рейф (Reiff, 1910) специально проверял эти данные и пока-
зал, что разница в холодостойкости яиц непарного шелкопряда, покры-
тых волосками или оголенных при охлаждении их до —21.5°, отсут-
ствует. Яйца как в том, так и в другом случае переносят в сухом воздухе
такое охлаждение, не погибая. Это же было подтверждено и мною, с тою
лишь разницей, что были испытаны значительно более низкие темпера-
туры. В природных условиях в холодостойкости Orgyidae большую роль
играет снеговой покров, так как зимовка гусениц и подавляющего боль-
шинства куколок протекает в лесной подстилке. Точно так же и яйца,
..например видов Orgyia Ochs., Ocneria Hb., откладываются на прикор-
экология
iff
новые части древесных стволов или даже на корни, камни и т. д., где
они оказываются защищенными от действия мороза слоем снега.
Холодостойкость видов Orgyidae в ряде случаев, видимо, также раз-
лична у локальных географических форм. Специальных экологических
исследований в этом направлении нет, но есть некоторые попутные наблю-
дения Гольдшмидта, который указывает на связь холодостойкости яиц
географических форм непарного шелкопряда с условиями их обитания.
При влиянии пониженной температуры (до —12°) в период с октября
но январь лучший выход дали яйца форм, распространенных на терри-
ториях, имеющих холодную зиму. Необходимо отметить, что во всех опы-
тах этого автора смертность яиц была очень высокой (минимум 75%).
Это указывает на слишком раннее перенесение яиц в тепло (20° в январе).
Таблица 16
Выход гусениц разных географических форм непарного
шелкопряда после охлаждения до—12° (по Гольдшмидту,
1934; сокращено)
« Географическое происхождение Темпера- тура январ- ской изо- термы (в°Ц) Число яиц Общий процент выхода
Хоккайдо — 5 61 25
Гензен (Корея) — 4 32 13
Берлин — 3 33 18
Нигата (северный Хоккайдо) . — 3 51 13
Бухара — 1 50 2
Фукуа } (сРеднее х°нд°) . + 3 + з 26 37 0 6
Портачи (Италия) ..... + .9 35 8
Диапауза большинства видов Orgyidae, распространенных в умерен-
ных широтах, является обязательным звеном цикла развития. Она воз-
никает в каждом поколении в определенной фазе развития данного вида.
Лишь у немногих видов она проявляется (также на определенной стадии
цикла) в зависимости от условий, сопровождающих развитие особи
(рис. 55). Этот второй тип диапаузы представляет биологическую специ-
ализацию и определяет возможность существования вторичного поли-
польтинизма у Orgyidae, населяющих умеренные широты. Она возникает
как видоизменение диапаузы первого типа. Диапауза Orgyidae характери-
зуется полной остановкой процессов морфогенеза, глубокими биохими-
ческими изменениями организма и рядом особенностей обмена веществ.
Для диапаузирующих яиц непарного шелкопряда, куколок Acronictinae
пли гусениц Olene Hb., Gynaephora Hb., Euproctis Hb. и ряда других
ппдов типична высокая устойчивость к наркозу, асфиксии и действию
низких температур. Это отличает их от развивающихся и растущих фаз
этих же видов, для которых указанные влияния губительны. Так, по Фран-
клину (Franklin, 1917), яйца непарного шелкопряда в диапаузе устойчивы
к длительному затоплению. По Миннот (Minnott, 1922), гибель яиц непар-
ного шелкопряда от затопления наступает лишь при длительности его,
например с декабря по май. По моим данным (1938, 1939), хлороформный
7 Фауна СССР, И. В. Кожанчиков.
{)Я
013.1ЦАЯ ЧАСТЬ
наркоз но убивает диапаузирующих куколок Acronictinae (Л. rumicis L.,
Л. auricoma Schiff.), даже при увеличении срока действия и дозы по срав-
55. Возникновение диапаузы
Рис.
некоторых видов Orgyidae при развитии
гусениц в разных термических усло-
виях. 1 — Leucoma salicis L.;2 — Orgyia
gonostigma F.; 3 — Acronicta rumicis L.;
4 — Acronicta rumicis L. при развитий
куколок в разных термических
условиях.
нению с теми, что достаточны для
гибели развивающихся особей, в 20—
30 раз. По Рейфу (1910), диапаузи-
рующие яйца непарного шелкопряда
устойчивы к пищеварительным сокам
птиц, например Turdidae. Они про-
ходят через их кишечник неповре-
жденными .
В пределах сем. Orgyidae прояв-
ление диапаузы у разных видов спе-
цифично. Специфична прежде всего
глубина изменений обмена веществ
в состоянии диапаузы. В связи с
этим у одних видов нормальное про-
текание диапаузы требует охлажде-
ния и именно действия отрицатель-
ных или близких к нулю положи-
тельных температур, тогда как' у
других видов это не является обя-
зательным. Разнообразие и слож-
ность отношений здесь очень велики,
но изучены еще неполно. Во всяком
случае в пределах семейства могут
быть намечены два типа диапаузы
(вероятно крайние),. Одна — длительная (прочная) диапауза, нормально
прекращающаяся лишь после воздействия отрицательных температур
(рис. 56, 57), причем тре-
бующая много месяцев (4—5)
их влияния. Она не про-
ходит нормально при высо-
кой температуре (порядка
25° и выше), при которой
погибают все особи, но при
умеренных и низких положи-
тельных температурах у неко-
торых особей прекращается
тем скорее, чем ниже тем-
пература. Диапауза этого
типа наблюдается, например,
у гусениц Leucoma salicis L.,
Olene fascelina L., Orgyia
gonostigma F. Второй тип —
короткая и нестойкая диа-
пауза , дающая сокращение
срока остановки развития 1
с повышением температуры
и проходящая также при по-
вышенных и высоких тем-
У
Рис. 56. Длительность диапаузы (в сутках)
в разных термических условиях у куколок.
1 — Acronicta auricoma Schiff.; 2 — A. lutea Brem.
et Grey; 3 — Calocasia coryli L.; 4— Daseochaeta
alpium Osb. Сплошная линия — длительность
диапаузы, пунктир (только для 1 и 4) — процент
особей, диапауза которых вообще не прекра-
тилась.
1 Остановка морфогенетических процессов в состоянии диапаузы для самых
разных видов и фаз развития Orgyidae несомненна, В связи с этим позволительно
сослаться на специальные цитологические исследования процессов морфогенеза яиц
экология
99
ш'ратурах, хотя развитие особей в этих условиях не нормально.
Действие отрицательных температур и в этом- Случае безусловно
благоприятно, как можно судить по более полному выходу нормально
плодовитых особей после этих влияний, но не обязательно. В табл. 17
дана характеристика реакции нескольких видов Acronictinae в состо-
нпии диапаузы на влияние разных положительных температур. Из нее
легко усмотреть специфичную реакцию их.
Таблица 17
Состояние диапаузы куколок Acronictinae при постоянных положительных
температурах с августа по май 1
Темпе- ратура в °Ц Название вида
Calocasia coryli L. Daseochaeta alpium Osb. Acronicta auri- coma Schf. Acronicta lutea Brem. et Grey
диапауза гибель диапауза гибель диапауза гибель диапауза гибель
в процентах
27.5 25.0 20.0 17.5 15.0 11.0 0 0 0 0 0 0 80 29 23 0 0 0 0 0 92 26 15 0 100 32 8 20 58 44 0 0 0 40 75 . 87 53 53 50 47 25 17 0 . 57 9 7 17 25 / 80 22 75 90 83 75
Цвёльфер (Zwolfer, 1934) исследовал длительность состояния диапаузы
ниц монашенки в разных условиях тепла и влажности воздуха (без пред-
варительного действия отрицательных температур). Лишь в пределах
Н -15° и 35—100% относительной влажности воздуха наблюдался выход
гусениц, причем через 7—8 месяцев после откладки яиц (начало экспери-
мента) и при смертности не ниже 50%, а в ряде серий достигавшей 95%;
а остальных сериях наблюдалась полная смертность эмбрионов. У яиц
монашенки состояние диапаузы, следовательно, скорее первого типа.
Требуется подчеркнуть, что прекращение диапаузы при тех или иных
условиях и переход особи в следующую фазу развития еще сами по себе
но характеризуют эти условия как вполне благоприятные для развития.
Тик, состояние диапаузы куколок Acronicta lutea Brem. et Grey хотя
и может проходить при положительных температурах, но вышедшие
пн куколок бабочки бесплодны и обладают недоразвитыми яичниками
(рис. 33, 2), которые, кроме того, и сильно деформированы. При всяких
условиях дальнейшей жизни особь так и погибает бесплодной. Это же
отмечается для Acronicta menyanthidis View, и многих видов этого рода
(leporina L., rumicis L., euphorbias F., alni L.), а также для других родов
(Subacronicta Kozh.). Тем не менее у Daseochaeta alpium Osb., вида сдли-
гпрпичевых, проведенные Слайфер (S 1 i f е г. 1931. Mitotic activity in the grasshop-
per embryo. Journ. Morph, and Physiol., 51: 613—618); Слайфер с несомненностью
понизала отсутствие размножения клеток (митозов) у диапаузирующих яиц саранчи
(Melanoplus differentialis Tbos.).
1 Предварительное охлаждение куколок отсутствовало. Вычитание из 100 суммы
процентов для диапаузирующих и погибших куколок дает возможность знать и
грогыо величину — процент развившихся и вышедших в этих условиях имаго,
7*
100
ОБЩАЯ ЧАСТЬ
тельной диапаузой, прекращение ее без охлаждения длительным влия-
нием положительной температуры (25°) не нарушает плодовитости бабо-
чек. Яйца, отложенные ими, нормально развиваются и дают жизнеспо-
собных, нормально растущих гусениц. Напротив, для непарного шелко-
пряда значение положительной температуры в период диапаузы яиц небла-
гоприятно. Диапаузирующие гусеницы Orgyia gonostigma^., Olene fasce-
lina L. и Leucoma salicis L. без предварительного охлаждения погибают на-
цело при положительных температурах. Последействие условий, в которых
протекала диапауза, очень сложно и специфично и, помимо вымирания
особей после диапаузы и бесплодия тех, которые развились, может выра-
Рис. 57. Сроки эмбрионального развития у форм непарника (Ocneria
dispar L.) из' Массачузеттса (М) и Портичи (П) у родительских осо-
бей и гибридов первого поколения (А) и у гибридов второго
поколения (В). (По Гольдшмидту, 1934).
жаться еще многими другими ненормальностями, например нарушением
секреторной деятельности акцессорных желез самцов,1 неправильно-
стями их половой деятельности и рядом других еще мало исследованных
моментов. Таким образом диапауза, являясь результатом адаптации
цикла развития вида к ежегодным сменам пищевого и гидротермического
режима, в свою очередь определяет возможность процветания данного
вида только в условиях соответствующего климатического типа.
Приспособление к сезонным сменам гидротермического режима у неко-
торых видов сем. Orgyidae усложнено благодаря наличию локальных
географических форм, максимально приспособленных к гидротермиче-
1 Дефекты плодовитости, как показала впервые Норрис, у чешуекрылых в ряде
случаев обязаны ненормальностям половой системы самцов, но не самок. Последнее
связано с нарушениями секреции акцессорных желез полового аппарата самцов и
может быть вызвано неблагоприятным режимом температуры при развитии куколок.
Позже этого же порядка дефекты плодовитости были найдены мною и описаны Н. По-
спеловым для мельничной огневки и совок озимой и хлопковой. (См.: М. Norris.
1934. Adult nutrition, fecondity and longevity in the genus Ephestia. Proc. Zool. Soc.
Lond: 903—934; И. К о ж а н чи к о в. 1938. Плодовитость чешуекрылых в зави-
симости от экологических условий. Зоол, жури,, 16 (4): 643—663).
экология
101
(иим сменам данного местообитания. Так, значение охлаждения в период
диапаузы может быть в разной мере обязательно для разных географи-
ческих форм, как то вероятно для непарного шелкопряда и ряда видов
рода Acronicta Ochs, (leporina L., rumicis X^.,psi L.). Некоторые сомнения
ость лишь в том, связаны ли эти различия с существованием прочно сло-
жившихся и имеющих свой ритм развития географических форм, как то
утверждает Гольдшмидт,1 или эти различия должны быть отнесены
за счет особенностей онтогенеза особей, развившихся в разных условиях,
т. е. за счет адаптаций, даже ограниченных одним поколением. Из Orgyidae
лишь для непарного шелкопряда (Goldschmidt, 1934) имеются специаль-
ные экспериментальные данные, показывающие существование разной
глубины диапаузы яиц у особей различного географического происхо-
ждения (табл. 18).
Таблица 18
Длительность развития''(включая диапаузу) яиц непарного шелкопряда
разного географического происхождения при температуре 11°, без охлаждения
(по Гольдшмидту, 1934)
Географическое происхождение Температура ; январской изотермы (В °Ц) % выхода гусениц по дням Общий про- | цент выхода
71 81 91 101 111 121
Хоккайдо — 5 0 0 0 0 0 0 0
Гоул (Корея) — 4 0 0 0 0 0 0 0
Огандай (Маньчжурия) . . . — 0 0 0 5 0 0 5
Ниигата } (севеРное Хо=Д°) . —3 — 3 0 0 0 0 0 0 0 0 0 0 . 0 0 0 0
Токио 0 5 0 5 0 45 0 55
Нива ) +3 0 0 0 0 0 0 0
Напалма 1 <сРеднее Хондо) + з • + 3 0 , 0 15 0 25 0 5 0 25 15 15 0 85 15
Гифу J + 3 . 0 0 0 0 10 0 10
Нухара — 1 0 0 0 0 0 0 0
Самарканд . . . . — 1 0 0 0 ° 0 0 0
Вопрос о географической изменчивости диапаузы в пределах вида
сложен прежде всего потому, что не известно, в какой мере получаемые
различия в реакции на охлаждение (или на другие воздействия) в период
диапаузы следует относить за счет особенностей, часто значительных,
данных особей и что принадлежит географической форме как таковой.
По Гольдшмидту, различия длительности диапаузы яиц разных геогра-
фических форм непарного шелкопряда сохраняются в потомстве при
скрещивании. Географические формы являются, следовательно, доста-
точно обособленными по признаку диапаузы (рис. 57). В то же время
также вполне достоверно установлено, что в части ареала распростране-
нии непарного шелкопряда наблюдается регулярный выход части гусе-
1 Наличие локальных географических форм внутри вида, кроме непарного шел-
Коирида, известно для многих видов Acronictinae (см. ниже главу о географическом
распространении).
102 ОБЩАЯ ЧАСТЬ
лиц из яиц в тот же сезон, как то известно для особей, завезенных в Новую
Англию, или для особей из Марокко [де-Лепиней (De Lepiney, 1930).
Диапауза у одних особей проходит здесь без влияния низких температур,
тогда как у других особей, живущих здесь же, влияние холода обяза-
тельно. Возможно, что это различие в большей мере обусловлено особен-
ностями климата, чем физиологией особей, относящихся к данным геогра-
фическим формам. Фернальд (1898) специально указывает, что полное
развитие яиц непарного шелкопряда в тот же сезон наблюдается в "слу-
чае неустойчивого термического режима осенью и наличия волн тепла
после заморозков. В Марокко также наблюдается выход гусениц непар-
ного шелкопряда в тот же сезон в случае падения температуры и затем
нового длительного потепления. Необходимо специально подчеркнуть,
что, несмотря на развитие яиц непарного шелкопряда в ряде мест в тот же
сезон, нигде в пределах ареала распространения непарного шелкопряда
не наблюдается второго поколения. Следовательно, отмеченные случаи
развития гусениц этого вида в тот же сезон не могут оцениваться как
нормальное явление.
В связи с описанным следует указать, что выход в тот же сезон осо-
бей, нормально диапаузирующих, отмечен не только для непарного шел-
копряда, но также и для златогузки [Tuleschkov (Тулешков, 1935)].
Мною констатирован выход отдельных особей Calocasia coryli L. в тот же
сезон без диапаузы или со слабо выраженной диапаузой как от особей,
собранных в природе, так и от воспитанных в лаборатории. Точно так же
без диапаузы выход бабочек наблюдался мною в отдельных случаях
и у Acronicta alni L.- и Daseochaeta alpium Osb., хотя основная масса куко-
лок оставалась в состоянии диапаузы в этих же условиях и уходила
нормально в зимовку.
Кормовые отношения видов сем. Orgyidae сложны. Кормовые растения
тропических видов и видов Orgyidae, распространенных в субтропиках,
известны, к сожалению, неполно. Из них основными являются семейства
растений, группирующиеся близ Amentaceae-Rosaceae. Важнейшими
кормовыми растениями являются Moraceae, Rutaceae, Magnoliaceae,
Myricaceae, Myrtaceae, Lauraceae, Zingiberaceae, Rubiaceae, Meliaceae,
Theaceae, Euphorbiaceae, Verbenaceae, Lythraceae, Leguminosae, Erica-
ceae, Sterculiaceae, отчасти Rhamnaceae и Casuarinaceae. За счет этих,
в общем очень разнообразных, растительных комплексов живет главная
масса тропических представителей семейства, видов Dasychira Steph.,
Artaxa Wlk., Euproctis Hb., Olene Hb., Ocneria Hb., Orgyia Ochs, и ряда
других. Часть видов, и именно или специализованные, или, напротив,
многоядные, является потребителями Palmae (Orgyia Ochs.), Gramineae
и Cyperaceae (Laelia Steph.) и Malvaceae (Orgyia Ochs., Olene Hb.). Важ-
нейшими кормовыми растениями тропических видов Orgyidae, насколько
можно судить по числу указаний, являются Moraceae, Euphorbiaceae,
Leguminosae, Rutaceae, Theaceae и Magnoliaceae. Тропические виды
сем. Orgyidae, будучи представлены архаичными типами, по питанию
также связаны с примитивными растительными группами. Кормовые
растения их включают преимущественно древние типы покрытосемянных.
В умеренных широтах главная масса видов Orgyidae живет за счет
Fagales, Rosales и Salicales. Потребителями Fagales являются Dasychira
Steph., Gynaephora Hb., Olene Hb., Orgyia Ochs., Numens Wlk., Artaxa
Wlk., Euproctis Hb., Arctornis Steph., Ocneria Hb., Parocneria Dyar, Calo-
casia Ochs., Charadra Wlk., Daseochaeta Warr. Из Fagales главнейшими
кормовыми группами являются виды родов Quercus, Castanea, Fagus,
экология
103
Carpinus, Ostryia, Corylus, Betula. Видами Alnus, несмотря на широкое
распространение их в Голарктике, питается небольшое число видов
семейства, отчасти специализованных (Acronicta alni L.). За счет Rosales
питаются частью те же представители семейства, которые указаны по-
требителями Fagales, частью специфичные — Мота Hb., Episema Hb.
Большая часть родов Orgyidae потребляет как виды Fagales, так и Rosa-
les; таковы Dasychira Steph., Gynaephora Hb., Olene Hb., Orgyia Ochs.,
Euproctis Hb., Arctornis Germ., Ocneria Hb., Parocneria Dyar., Charadra
Wlk., Acronicta Ochs. Важнейшие кормовые растения из Rosales — это
Malus, Pirus, Prunus, Crataegus, Spiraea, Rubus — древесные породы
и кустарники. Травянистые представители Rosales, даже встречающиеся
в лесах, слабо поедаются видами Orgyidae. Видами Salicales питаются
все те представители Orgyidae, которые указаны потребителями Rosales
и Fagales. Специфичными потребителями Salicales являются виды Leu-
сота Steph, и Subacronicta Kozh. Специализованные на питании иво-
цветными представители Orgyidae связаны в основном с видами Populus,
хотя могут питаться также и ивами (Salix). Виды ив служат, наоборот,
пищей главной массе многоядных Orgyidae. Кроме трех указанных основ-
ных растительных групп, голарктические виды Orgyidae питаются видами
Juglandaceae, Hamamelidaceae, Rutaceae, Lauraceae, Myricaceae, Mora-
ceae, Terebynthaceae. Этими и некоторыми другими растениями отмечено
питание лишь отдельных видов семейства, что, возможно, обусловлено
слабым распространением этих растений .в Голарктике. Немногие виды
Orgyidae специализованы на питании некоторыми из них, например
Parocneria terebynthi Frr. на Pistaceae. Наконец растения из семейств
Tiliaceae, Aceraceae, Elaeagnaceae, Rhamnaceae, Oleaceae, Caprifoliaceae,
Ulmaceae, хотя и широко распространенные в Голарктике, служат пищей
немногим, преимущественно многоядным видам семейства, наиболее
многоядным из тех, которые указаны в качестве потребителей Fagales,
Rosales и Salicales (Ocneria Hb., Orgyia Ochs.). Они являются второсте-
пенными кормовыми растениями, служащими преимущественно заменой
основных групп при известных экологических условиях. Лишь за счет
Oleaceae живут своеобразные и специфичные потребители, этого семей-
ства— Craniophora Snell., способные также питаться видами Caprifo-
liaceae. Специализованными потребителями Aceraceae, Ulmaceae и Elae-
agnaceae являются лишь отдельные виды, способные, впрочем, питаться
и другими растениями, но несомненно предпочитающие эти специфичные
растения. Таковы виды рода Acronicta Ochs. — A. aceris L., A. elaeagni
Alph. и A. americana Grt. Первостепенное значение голосемянных и в пер-
вую очередь хвойных (Pinaceae) в качестве кормовых растений видов
Orgyidae несомненно лишь для видов умеренных широт. В Голарктике
виды рода Diphthera Ochs, целиком специализованы на питание Pinaceae,
главным образом на видах Pinus и отчасти Abies, Picea и Larix. Из Orgyi-
nae специфичными потребителями хвойных являются лишь немногие
виды Dasychira Steph., связанные с Abies и Larix, это D. abietis Schiff,
и D. albodentata Brem. Хвойными могут питаться также и некоторые
i из наиболее многоядных видов семейства — монашенка, непарный шел-
копряд, Gynaephora selenitica Esp., виды Orgyia Ochs. Некоторые много-
ядные Orgyidae могут питаться также видами Тахасеае и Cupressaceae,
например непарный шелкопряд, но питание этими растениями оказывает
неблагоприятное влияние на развитие.1 Связь видов Acronictinae с хвой-
1 Монашенка, как известно, является вредителем хвойных пород, в частности
оли и сосны. Вредность этого вида для лиственных деревьев значительно ниже. Спе-
104
ОБЩАЯ ЧАСТЬ
ними, без сомнения, более первична по сравнению с видами Orgyinae,
если среди первых возникли специализованные на Pinaceae особые роды,
имеющие архаичные черты строения. Среди видов Orgyinae, напротив,
специализировались на питании хвойными немногие отдельные виды,
относящиеся к родам, широко распространенным в тропиках и в целом
являющимся потребителям^ лиственных пород. В ряде случаев обнару-
живают питание хвойными полифаги из Orgyidae, например представи-
тели родов Orgyia Ochs, и Ocneria Hb.
Сопоставление кормовых связей голарктических видов сем. Orgyidae
дано в табл. 19.
Таблица 19
Распределение видов Orgyidae (распространенных в Голарктике) по группам
кормовых растений
Семейство или порядок растений ') Общее число ви- ' дов потребителей Число видов Семейство или порядок растений ! Общее число ви- дов потребителей Число видов
j полифагов I олигофагов монофагов полифагов ; олигофагов i i ! монофагов
Pinaceae .... 17 3 4 10 Ulmaceae .... 10 4 5 1
Тахасеае .... 1 1 — Tiliaceae .... 9 6 з
Cupressaceae . . . 1 1 — — Ericaceae .... и '11 — —
Magnoliaceae . . . 1 1 — —— Leguminosae . . . 5 5 — —
Lauraceae' .... 1 1 —— Euphorbiaceae . . 7 4 2 1
Hamamelidaceae 1 1 Oleaceae 7 2 4 1
Saxifragaceae . . 3 3 — — Caprifoliaceae . . 2 1 1 . —
Rutaceae .... 3 2 1 — Ampelidaceae . . 2 2 — —
Anacardiaceae. . . 2 1 — 1 Gramineae .... 6 4 1 2
Platanaceae . . . 1 1 — — . Cyperaceae .... 2 1 1 —
Aceraceae 8 5 2 1 1 Typhaceae . . . 1 1 — —
Moraceae .... 8 8 — —. Liliaceae .... 1 1 — —
Rosaceae .... 52 25 23 4 P lantaginaceae . . 2 2 —— —
Fagaceae .... 58 27 31 — Polygonaceae . . . 6 6 — —
Betulaceae .... 57 24 33 —— Gentianaceae . . . ’2 2 — —
Salicaceae .... 58 36 16 6 Scrophulariaceae 2 2 — —
Juglandaceae . . 3 3 — — Pinaceae 1 1 — —
Myricaceae . . . 7 7 — — Plumbaginaceae . . 4 1 2 1
Elaeagnaceae . . . 2 1 — 1(’) Compositae .... 5 — —
Из данных табл. 19
Salicaceae и Coniferae
можно видеть, что Rosaceae, Betulaceae, Fagaceae,
характеризуются не только наибольшим общим
циальными исследованиями [Маркс (Maercks, 1935)] установлено, что хвойные расте-
ния и, в частности, ель (тем более сосна) являются значительно менее благоприят-
ным кормом для этого вида по сравнению с лиственными, в частности буком. Появле-
ние монашенки как вредителя хвойных нужно рассматривать на фоне уничтожения
старых буковых и вообще лиственных насаждений Евразии (особенно Западной
Европы). Климатический режим разреженных лиственных лесов неблагоприятен
для жизни монашенки, и она здесь, несмотря на оптимум питания, не может успешно
размножаться. Напротив, устойчивый гидротермический режим спелых еловых лесов
благоприятен для жизни этого вида: в этих условиях, несмотря на худший пище-
вой режим, монашенка может не только сохраняться как вид, но и размножаться
в массах. Общее распространение этого вида указывает также на связь с распростра-
нением лиственных, в частности широколиственных пород, слабо перекрывая ареалы
последних, что говорит [Вилке (Wilke, 1931)] также за первичную связь монашенки
с лиственными породами,
экология
105
числом потребителей из видов Orgyidae, но, главное, наличием большого
числа олигофагов, специализованных на питании немногими родствен-
ными группами растений. Для этих же растительных групп характерно
наличие монофагов (родов и видов). Монофаги особенно типичны для
хвойных. Наоборот, первичные кормовые растения видов подсем. Orgyi-
nae, каковы Magnoliaceae, Lauraceae, Hamamelidaceae, Myricaceae и ряд
других, в условиях Голарктики имеют потребителями лишь полифагов.
Видами Juglandaceae, Elaeagnaceae, Tiliaceae, Ulmaceae, Aceraceae,
Ericaceae и Ampelidaceae питается вообще немного видов семейства,
преимущественно полифагов. Родов же, специализованных на питании
ими, нет совсем. Oleaceaeи Caprifoliaceae, как правило, избегаются видами
Orgyidae, за исключением специализованных на питании этими растениями
некоторых видов Craniophora Snell. Saxifragaceae также имеют второ-
степенное значение как кормовые растения Orgyidae и потребляются
также по преимуществу полифагами. Лишь для арктической Gynaephora
rossii Curt, камнеломки отмечены в числе основных кормовых растений,
хотя и в этом случае, видимо, большее значение имеют виды Salicaceae,
Betulaceae и Ericaceae [Дайр (Dayr, 1896)].
Особый интерес как кормовые растения Orgyidae имеют однодольные
и, в частности, Gramineae и Сурегасеае, за счет которых живет хотя и не-
большое число голарктических видов семейства, но для них типична
высокая степень кормовой специализации их потребителей. Интересно
также, что виды Penthophera Germ, и Laelia Steph., специализованные
на питании видами Glumiflorae, наряду с многими чертами специализации
строения, обнаруживают многие анцестриальные черты. Питание видами
Glumiflorae отмечено также и для своеобразных Arsilonche Ld. из Acro-
nictinae, видов, обнаруживающих многие черты специализации в строе-
нии тела и биологии, для которых эти кормовые связи могут считаться
вторичными.
В условиях открытого ландшафта (степи, пустыни) голарктические
виды Orgyidae являются потребителями Euphorbiaceae, Plumbaginaceae,
Polygonaceae, Gentianaceae, Scrophulariaceae Leguminosae, Compositae,
Rosaceae и Linaceae, причем первые два семейства растений характерны
для видов Acronictinae, тогда как остальные — для видов Orgyinae. Для
видов же Orgyinae в условиях открытого ландшафта типична тесная связь
с Glumiflorae, тогда как виды Acronictinae хотя и питаются злаками,
как и перечисленными выше другими видами растений, но олигофаги
из Acronictinae, заселяющие открытый ландшафт, известны лишь для
Euphorbiaceae и Plumbaginaceae. Кормовые связи в условиях открытого
ландшафта Голарктики для видов обоих подсемейств различны. Для
видов Orgyinae питание как однодольными, так и другими видами расте-
ний, свойственными открытому ландшафту, в ряде случаев носит первич-
ный характер {Laelia Steph., Penthophera Germ., Orgyia Ochs, и др.),
являясь в Голарктике отзвуком кормовых связей этих родов в тропиках,
где многочисленные представители этих родов (исключая Penthophera
Germ.) имеют те же кормовые отношения. Напротив, для видов Acro-
nictinae питание травянистыми двудольными, включая виды Plumbagi-
naceae и Euphorbiaceae, на которых наблюдается наибольшая их кормо-
вая специализация, не говоря о Glumiflorae, следует рассматривать как
вторичное. Эти кормовые связи возникли, в результате перехода с дре-
весных растений типа Rosales и Amentaceae (с ними нередко еще не по-
рвана связь, как у Arsilonche Ld.), который совершился на территории
Голарктики,
106
ОБЩАЯ ЧАСТЬ
Рассмотрение классификации Orgyidae в связи с системой цветковых
растений приводит к картине, очень далекой от параллелизма в развитии
этих групп организмов. Напротив, здесь налицо очень сложные взаимо-
отношения. Кормовые связи Orgyidae формировались не только парал-
лельно филогенетическому развитию видов этого семейства чешуекрылых
и потребляемых ими растений, но и на фоне распространения как Orgyi-
dae, так и их кормовых растений. Уловлена может быть лишь общая
связь питания сохраняющих анцестральные черты видов Orgyidae (Olene
Hb., Dasychira Ochs.) с древними растительными типами (Moraceae,
Magnoliaceae, Rutaceae) как для тропиков, так и для Голарктики. Наибо-
лее архаичные Diphthera Ochs, обнаруживают кормовые связи даже
с голосемянными (Coniferae). В целом специализованные представители
Orgyidae обнаруживают кормовые связи с высшими порядками, цветковых
растений (Oleaceae, Compositae, Plumbaginaceae). Эти связи могут выра-
жаться двояко. В одном случае специализация на них приводит к потере
связей с исходными типами кормовых растений, как то известно для
Craniophora Snell., Eogena Gn. В другом, напротив, расширение круга
кормовых растений осуществляется за счет полифагии, как то типично
для большинства высших групп семейства, например для видов Ocneria
Hb., Acronicta Ochs., Orgyia Ochs., Euproctis Hb. Специально нужно
отметить переход на питание Rosales, Fagales, Salicales и Coniferae в пре-
делах Голарктики видов Orgyidae, относящихся к родам и сейчас живу-
щим в тропиках за счет Moraceae, Magnoliaceae, Lauraceae, Theaceae,
каковы Dasychira Steph., Olene Hb., Euproctis Hb.1 Это является показа-
телем того, что расселение по территории является более мощным факто-
ром в смене кормовых растений, чем параллельная дифференциация
видов кормовых растений и их потребителей Orgyidae на одной и той же
территории.
Наконец необходимо отметить почти полное отсутствие потребления
видами Orgyidae травянистых Rosales, тогда как древесные Rosaceae
(Pomaceae, Prunoideae) являются важнейшими их кормовыми растениями.
Особое положение занимают также виды Coniferae и Euphorbiaceae как
кормовые растения видов разных подсемейств Orgyidae. Для Acronictinae
голосемянные являются первичными кормовыми группами (с ними свя-
заны древнейшие типы подсемейства), тогда как для Orgyinae — вторич-
ными (с ними связаны, напротив, наиболее специализованные. формы).
В отношении Euphorbiaceae положение обратное. Потребители их из Acro-
nictinae (Simyra Ochs., Eogena Bd.. Oxycesta Hb.) — всё виды в высо-
кой мере специализованные. Основные черты смены кормовых ра-
стений в эволюции сем. Orgyidae иллюстрирует прилагаемая схема
(рис. 58).
Приведенные факты показывают, что смена кормовых растений в эво-
люции Orgyidae идет при сильном ограничении выбора их групп эколо-
гическими условиями, в частности климатическим режимом. Адаптация
к климатическим условиям является для Orgyidae, следовательно, более
стойкой эколого-физиологической чертой по сравнению с пищевыми отно-
шениями. Последние более лабильны. Об этом свидетельствует, напри-
мер, питание монашенки елью — вида, связанного первично с листвен-
ными древесными породами, в частности с Fagales. Будучи видом стено-
1 Существенно, что у этих голарктических представителей, несмотря на иные
кормовые связи, сохранилась еще возможность питаться и этими исходными расти-
тельными типами, как, например, шелковицей у Euproctis similis Fuessly,
экология
107
термным, в современных условиях средней Европы монашенка
необходимый экоклимат лишь в спелых еловых насаждениях.
находит
Compositae
Рис. 58. Схема распределения ботанических групп растений по кормовым
отношениям Orgyidae. Группы. растений, с которыми обнаруживаются пер-
вичные связи Orgyidae, обведены; стрелки указывают направление смены
кормовых растений.
Смены кормовых растений в эволюции Orgyidae, протекающие в усло-
виях закрытого (лесного) ландшафта, наиболее закономерны и понятны
108
ОБЩАЯ ЧАСТЬ
с точки зрения биохимических отношений. Переход Orgyidae в открытый
ландшафт связан с очень сильными изменениями кормовых связей, при-
чем травянистые формы основных кормовых растений (например Rosales)
могут быть в этих условиях и не использованы. Питание в открытом
ландшафте осуществляется за счет других растений, в частности кустар-
ников из Saliacaceae, Ericaceae или сочных травянистых растений из
Euphorbiaceae, Polygonaceae, Plumbaginaceae, Compositae. Ведущее зна-
чение в формировании новых кормовых отношений получают локальные
условия в связи с экоклиматом. Заселяются стации окраин болот, поймы
рек, влажные луга.
Влияние пищевых условий на развитие Orgyidae не менее разнооб-
разно и сложно, чем физических факторов. В природе влияние пищи
сказывается прежде всего при зимовке в фазе яйца, когда выходящие ран-
ней весной гусеницы сразу сталкиваются с быстрыми изменениями химизма
кормовых условий в период ранневесеннего, быстрого роста растений.
Велико влияние кормового режима весною также и для видов зимую-
щих гусеницей.
Одним из важных моментов, определяющих успешное развитие
видов, зимующих яйцом и гусеницей, каковы Ocneria Hb., Orgyia
Ochs., Gynaephora Hb., Olene Hb., является выход с мест зимовки,
синхронный с появлением свежих почек или листочков их кормовых
растений. Позднее или слишком раннее их появление весной может быть
губительным для видов этой группы в связи с недостаточной питатель-
ностью кормовых растений. Для непарного шелкопряда случаи несвое-
временного выхода гусениц весной отмечены для .северной Африки
(Марокко), где этот вид является потребителем вечнозеленых дубов
(Quercus suber), вегетация которых начинается довольно поздно (в мае).
На зимних листьях этих дубов развитие молодых гусениц непарного
шелкопряда невозможно. По Лепинею, в некоторые годы, например
в 1928 г., в связи с преждевременным выходом гусениц весною до начала
роста первых листочков дуба, они погибают в количестве до 50%. В зоне
лесов с опадающей листвой, наоборот, всегда наблюдается выход гусениц
непарного шелкопряда, синхронный с развитием листвы кормовых расте-
ний. Аналогично гусеницы монашенки в еловых насаждениях появляются
весной большею частью одновременно с ростом молодой хвои, которая
необходима для питания их в первой стадии. Наоборот, в сосновых наса-
ждениях, прогревание которых весною наступает раньше, выход из яиц
гусениц монашенки нередко наступает еще до начала развития молодой
хвои на соснах. Многочисленными эксперимептами Майера (Mayer, 1940)
по кормлению гусениц этого вида листьями разного возраста устано-
влено, что нормальное развитие их возможно лишь при синхронном
появлении весною гусениц и молодых листьев бука, еловой хвои и т. д.
и дальнейшем параллельном развитии листьев и хвои и потребляющих
их гусениц монашенки. Расхождение в сроках выхода гусениц и появле-
ния листьев в два-три дня может иметь уже значение для жизнеспособ-
ности особей и дальнейшего их роста. При запоздалом выходе гусеницы
встречают затвердевшие покровы листьев и иной химизм. Это затруд-
няет их питание, вызывает дальнейшее отставание в росте и,, наконец,
гибель.
Фактором, вызывающим несвоевременный выход гусениц Orgyidae
с мест зимовки, является термический режим. Таким образом косвенное
влияние климатических условий, через пищу, очень велико. Особенно
опасен для развития Orgyidae запоздалый выход. Питание подросшими
экология
109
и загрубевшими листочками губительно не только для гусениц мона-
шенки, но также, по моим наблюдениям, неблагоприятно для многих
видов Acronictinae, например для Subacronicta megacephala Schiff. Прежде-
временный выход гусениц Olene Hb., Arctornis Germ., Euproctis Hb.,
Orgyia gonostigma P. и других видов, зимующих полувзрослой гусеницей,
менее губителен, так как крупные гусеницы более стойки к изменениям
химизма их кормовых растений, но и здесь роль неадэкватных условий
питания несомненна.
Orgyidae, зимующие в фазе яйца или гусеницы, в весенний период
очень устойчивы к голоданию. Для монашенки или непарного шелко-
пряда отмечена возможность голодания только что отродившихся гусениц
без вреда для дальнейшего развития их в течение 8—10 суток. Высокая
устойчивость к голоданию известна и для перезимовавших гусениц Eupro-
ctis Hb. (в частности златогузки), Olene Hb., Orgyia Ochs. У видов, зиму-
ющих в фазе полувзрослой гусеницы, такая устойчивость менее выражена.
Наоборот, в период роста недостаток и тем более отсутствие пищи влияют
резко отрицательно. Эксперименты показывают слабую приспособлен-
ность растущих взрослых и полувзрослых гусениц Orgyidae к голоданию.
Гусеницы непарного шелкопряда II—IV стадии вымирают нацело через
4—5 суток голодания. Сильное (порядка суток-двух) голодание в период
роста сказывается очень вредно на дальнейшем развитии гусениц. Наблю-
дения над развитием Orgyia antiqua L. (Парфентьев, 1925), непарного
шелкопряда [Гофман (Hoffman, 1933); Трейман, 1938] и разных видов
Acronictinae (Acronicta rumicis L., A. auricoma Schiff., A. lutea Brem.
et Grey, Daseochaeta alpium Osb., Calocasia coryli L. и др.), проведенные
мною, показывают у голодавших особей уменьшение темпов роста, раз-
меров и веса куколок, задержку окукления,1 разную степень снижения
плодовитости голодавших особей и появление бесплодных особей.
По Трейману (1938), следствием голодания гусениц непарного шелко-
пряда являются не только указанные явления, но и повышенная смерт-
ность эмбрионов в яйцах, отложенных самками, недостаточно питавши-
мися в личиночной стадии. По Копечу (Кореё, 1924), голодание гусениц
непарника ведет к глубоким нарушениям плодовитости, в частности сам-
цов, с чем связано неполное оплодотворение самок и появление в потом-
стве дефектов развития и повышенной смертности. В естественных усло-
виях голодание растущих гусениц Orgyidae является редким и даже
исключительным случаем, например при массовом размножении того
или иного вида, как то отмечалось для монашенки и непарного шелко-
пряда (Кеппен, Мокржецкий), но и здесь чаще наблюдается питание
необычными растениями, т. е. смена кормового режима (на худший),
чем прямое голодание.
Более общим в природной обстановке является влияние особенностей
химизма пищи на рост и развитие Orgyidae. Оно связано прежде всего
с широко распространенной среди них многоядностью и потому с отно-
сительной легкостью замены одних кормовых растений другими. Вместе
с тем оно обусловлено пестротой химизма кормовых растений, даже
одних и тех же видов, выросших в разных условиях среды. В отношении
многоядности нужно оговориться, что даже наиболее широкие полифаги,
как непарный шелкопряд или Acronicta rumicis L., в природе в общем
питаются специализованно, предпочитая и нормально используя ограни-
1 Голодание в последние дни личиночного развития ускоряет окукление.
по
ОБЩАЯ ЧАСТЬ
ченный круг кормовых растений. Для.полифагов при недостатке основ-
ных кормовых растений доступны для питания представители многих
других цветковых растений, с видами которых они в природе нормально
не связаны.
Представление о значении качества пищи для роста поли- и олиго-
фагов из Orgyidae дает табл. 20 и рис. 59 и 60.
Из приведенных в табл. 20 данных видно, что такие многоядные виды,
как представители родов Ocneria Nb. и Acronicta Ochs. (Pharetra Hb.),
не говоря об олифагах Calocasia Ochs., Daseochaeta Warr., Subacronicta
Kozh. и некоторых Acronicta Ochs. (A. alni L.), дают полную выжи-
ваемость особей при росте лишь на немногих (оптимальных) растениях.
Для полифагов это определенно выражено лишь при влиянии пищевого
режима в течение всего периода роста, тогда как для олигофагов оно
уже вполне ясно с первой личиночной стадии. Минимальное вымирание
монашенки, для которой, к сожалению, нет достаточно разнообразных
экспериментов, констатировано при питании буком. По Мэрксу (1935),
питание этого вида яблойей также благоприятно, хотя смертность при
росте все же достигает 10—15%. Замечательно, что при питании яблоней
особи монашенки достигают наибольшего веса, какой известен для этого
вида при питании разными растениями. Питание монашенки хвойными
деревьями (Pinaceae) в целом менее благоприятно, чем лиственными.
Из хвойных наилучшие пищевые качества обнаруживает ель, затем
лиственница, наихудшие — сосна. При питании монашенки хвойными
деревьями в младших возрастах гусениц всегда наблюдается высокая
смертность, порядка 50%. Пережившие особи в дальнейшем почти не вы-
мирают.
Несомненно, что питание видами Fagales и Rosales наиболее благо-
приятно для развития монашенки. Для питания Daseochaeta alpium
Osb. благоприятны только виды Fagales. Виды сем. Betulaceae опти-
мальны для питания видов Calocasia Ochs, и некоторых Acronicta Ochs.
(Л. leporina L.). Наоборот, для питания видов Triaena Hb. и Acronicta
Ochs, оптимальны как растения этого семейства, так и Rosales, а для
Pharetra Hb. {rumicis L., lutea Brem. et Grey) также и Salicales. Наконец
для Subacronicta megacephala Schiff, оптимальны только Salicales. Для
непарного шелкопряда, как и для монашенки, оптимальные кормовые
качества обнаруживают Fagales и именно Fagaceae, но также благопри-
ятно питание Rosaceae (Prunus, Mains, Pirns) и некоторыми Salicaceae
(Salix caprea). Кормовые отношения, сходные с тем, что известно для
непарного шелкопряда, обнаруживает и златогузка, однако с большей
специализацией на Rosales.
Пищевые качества и в пределах оптимальных для данного вида расти-
тельных групп меняются как в зависимости от условий их произрастания,
так и от фенологического момента их использования. Различия особенно
велики для древесных растений с опадающей листвой, имеющих строго
сменяющуюся по сезонам стадийность роста и цикл превращения пище-
вых материалов. Они менее выражены у травянистых, особенно однолет-
них растений, вегетирующих короткий срок. В этом, без сомнения,
и нужно искать условия для возникновения в пределах сем. Orgyidae
вторичной полициклии при переходе на питание травянистыми расте-
ниями {Acronicta Ochs.). Один факт полифагии сам по себе еще недостато-
чен, как то видно на примере дендрофильных монашенки и непарного
шелкопряда. Изменчивость кормовых качеств растений затрудняет оценку
их питательности, а сравнение близких видов растений требует большого
а
РГ
Н
ко
сб
1 Данные для монашенки приведены на основании работ Пите (Pictet 1920), Маркса. (1935), Затлера (Sattler, 1939) и Майера (1940). Следует
специально отметить сведения, сообщаемые Трейманом (1938) по кормлению непарного шелкопряда дубом. В опытах этого автора смертность гусе-
ниц в норме (?) достигала 99.3-98.8J/o, что может быть отнесено только за счет неопытности его в работе с этим видом и за счет негигиеничных
условий развития гусениц. В нормальных условиях смертность непарного шелкопряда при питании дубом практически отсутствует.
112
бВЩАЯ ЧАСТЬ
материала. Например, до сих пор, несмотря на большое число работ
по непарному шелкопряду, неясно, обнаруживают ли более благоприят-
ные пищевые свойства Fagales (Castanea, Quercus, Fagus) или виды Rosa-
ceae (Pirns, Mains). К тому же нередко влияние среды так велико, что
растение, будучи по природе оптимальным, может в данных условиях
Длительность роста (В сутках.)
Рис. 59.
на темпы роста и вес гусениц Daseo-
chaeta alpium Osb. 1 — бук (Fagus
sylvatica); 2 — дуб (Quercus robur);
3 — береза (Betula pubescens); 4 —
береза, но в конце лета и осенью.
Влияние кормовых условий
оказаться совершенно негодным для питания.
В питательности кормовых растений, особенно древесных, наиболее
общее значение имеет фенологический момент их использования. Макси-
мально питательными для Orgyidae, зимующих яйцом или гусеницей,
особенно для моновольтинных видов и олигофагов, являются листья
деревьев в период формирования пластинки. Окончившие рост и твер-
деющие листья питательны для гусениц тех видов, которые зимуют в фазе
куколки. Гусеницы старших стадий
этих видов нередко обнаруживают
даже специальную приспособлен-
ность к питанию листьями разного
возраста, не исключая и желтеющие
осенние листья, что отмечено, напри-
мер, для Dasychira pudibunda L. и
Acronicta psi L. Эти виды способны
завершать рост, начиная с четвер-
той стадии, при питании желтыми
листьями, причем питание ими мо-
жет быть даже благоприятно. У по-
ливольтинных видов и полифагов
приспособление к питанию листь-
ями разной степени зрелости выра-
жено еще более резко. Например,
гусеницы Acronicta rumicis L. нор-
мально растут, начиная с первой
личиночной стадии при питании жел-
теющими, осенними листьями ивы
(Salix fragilis).
В течение вегетационного сезона
условия освещения, влажности воз-
духа и почвы, термический и солевой
режим почвы обусловливают очень большую пестроту пищевых условий
у дендрофильных Orgyidae. Они оказывают сильнейшее влияние на
alpium Osb. 1 — бук (Fagus
развитие древесины и других углеводов в листьях, на количество в них
белка и других веществ, что в свою очередь сказывается на росте их по-
требителей.
Для многих видов Orgyidae характерно возрастание многоядности
гусениц с ростом. Это отмечено преимущественно для полифагов, как
непарный шелкопряд, Acronicta rumicis L., но наблюдается также
и у олигофагов, например у Calocasia coryli L. В связи с этим обстоятель-
ством нередко полувзрослые и взрослые гусеницы питаются необычными
для них растениями. Так, для непарного шелкопряда не раз отмечалось
питание взрослых гусениц злаками, отчасти культурными (Брянцев,
1928; Талицкий, 1928). Специальные эксперименты показали (Келус,
1939), что для непарного шелкопряда питание злаками, в частности
пыреем, хотя и ведет к сильному падению веса и плодовитости бабочек,
но не обусловливает значительной смертности. Подобные резкие укло-
нения в питании взрослых гусениц не часты и обычно вызваны массц-
экология
из
природе
выми размножениями. Питание многих Orgyidae в старших стадиях
не характерными для них растениями наблюдается и при наличии основ-
ных кормовых растений. Для Calocasia coryli L., например, не раз отме-
чалось в природе питание взрослых гусениц осиной, ивой, ольхой, хотя
рост этого вида с первых стадий при питании этими растениями проте-
кает ненормально и сопровождается очень высокой' смертностью, по срав-
нению с питанием на березе (табл. 20). Питание гусениц Calocasia coryli L.
первой стадии какими-либо иными растениями, кроме березы, в
наблюдается редко.
Для большинства олигофагов и особенно для монофагов из Orgyidae
изменения кормовых отношений
для полифагов. С ростом на-
блюдается повышение кормовой
специализации. Так, напри-
мер, у Daseochaeta alpium
Osb. — олигофага — старшие
стадии гусениц не могут питать-
ся орешником или восковником
и погибают, в то время как мо-
лодые способны расти на такой
диэте. Точно так же монофаг
Diphthera coenobita Esp. обна-
руживает меньшую требова-
тельность к возрасту хвои в
первых личиночных стадиях,
будучи в состоянии питаться
как молодой, так и старой хвоей
сосны, тогда как полувзрослые
и взрослые гусеницы растут
при питании лишь старой сос-
новой хвоей.
с ростом обратны тому, что указано
Рис. 60. Влияние кормовых условий на тем-
пы роста и вес гусениц Acronicta rumicis L.
1 — манжетка (Alchemilla vulgaris); 2 —
ломкая ива (Salix fragilis); 3 — черный то-
поль [(Populus nigra); 4 — береза (Betula
pubescens).
Все факторы среды, влияю-
щие на процессы роста и мета-
морфоз Orgyidae, в значитель-
ной степени влияют и на жизнь
имаго, затрагивая в первую
очередь длительность жизни и
плодовитость особей. Помимо монашенки и непарного шелкопряда, раз-
носторонне изученных в этом отношении многими авторами, мною спе-
циально исследован ряд видов подсем. Acronictinae (Daseochaeta Warr.,
Calocasia Ochs., Acronicta Ochs., Subacronicta Kozh.) и Orgyinae (Leucoma
.Steph., Gynaephora Hb., Ocneria Hb.).
Все виды семейства в отношении влияния условий среды на жизнь
имаго распадаются на две группы. В одну входят виды с утраченным
имагинальным питанием, у которых формирование и отчасти созревание
половых продуктов сдвинуты на куколочную и личиночную фазы. Для
птой группы отношения более просты и обусловлены главным образом
влиянием среды на личиночное развитие. Вторая группа включает виды
с наличием имагинального питания. В этом случае формирование и во вся-
ком случае созревание половых продуктов падает на имагинальную фазу
(лишь отчасти на фазу куколки). Здесь отношения более сложны и свя-
заны как с воздействием условий среды в течение жизни и развития
преимагинальных фаз, так и имаго.
8 Фауна СССР, И. В. Кожанчиков.
114
ОБЩАЯ ЧАСТЬ
Для первой группы видов Orgyidae и, в частности, для непарного
шелкопряда и монашенки работами Цвёльфера (1933), Гофмана (1933),
Левитта (1935), Затлера (1939) и некоторых других авторов установлена
тесная связь между изменениями веса особей, их плодовитостью и дли-
тельностью жизни. Особи, у которых развитие преимагинальных фаз
прошло в условиях оптимума, имеют наибольший вес (он может быть
принят за нормальный), характеризуются высокой плодовитостью и дли-
тельностью жизни, близкими к максимальным. Цвельфер (1933) показал,
что максимальная плодовитость самок монашенки наблюдается при
развитии гусениц в условиях температуры 22—25°. В этих же условиях
наблюдается наибольший вес особей и развивается нормальное (50%)>
число самок. Развитие при пониженной температуре ведет к сильному
вымиранию самок в личиночной фазе, 'тогда как при повышенных темпе-
ратурах вымирают в большей
мере самцы. И в тех и в дру-
гих условиях, кроме нару-
шения соотношения полов,
наблюдается падение веса
особей, по сравнению с оп-
тимумом (рис. 61). При оп-
тимуме наблюдается отсут-
ствие смертности монашенки:
при росте и примерно равное-
соотношение полов.
Цвёльфер (1933) и Затлер
(1939) показали на большом
материале для монашенки,
что депрессивные условия
развития, чем бы они ни были
вызваны, ведут к падению
веса особей, особенно самок,
и к снижению плодовитости..
Есть тесная связь между
этими величинами, которую
указанные авторы считают-
возможным выразить прямой
Рис. 61. Влияние термических условий при ро-
сте монашенки {Ocneria monacha L.) на вес ку-
колок самок (1), плодовитость самок (2), соотно-
шение полов (3) и вес куколок самцов {4). (По
Цвёльферу, 1934).
(рис. 62). Для самок непарного шелкопряда получена подобная же кор-
реляция между величинами веса и плодовитостью на материале, собран-
ном в природе (Левитт, 1935; Пятницкий, 1935). Она может быть выражена
также прямой, но имеющей иной тангенс угла к оси ординат (рис. 62)..
Зависимость плодовитости от веса самок для непарного шелкопряда была
обнаружена Пятницким (1935) на материале, собранном в лесах Крыма
на разных расстояниях от центра очага -размножения этого вида. По
мере удаления от очага вес и плодовитость особей возрастают. Ранее-
этих авторов Парфентьев (1925) при исследовании влияния голодания
на развитие Orgyia antiquaL. указывал на тесную связь между степенью
истощения гусениц при голодании, величиной развивающихся самок
и их плодовитостью. Им была указана здесь обратная связь.1
1 Вопрос о связи между весом самок и их плодовитостью был разработан на сос-
новой совке {Panolis flammea L.) Цвёльфером, его данные были подтверждены Ильин-
ским. После этих работ для еще более обширных исследований были использованы.
Orgyidae. Эта связь в той простой форме, как она рисуется Цвёльфером (1931), Ильин-
экология
115
По существу эти простые отношения между весом самок и их плодо-
витостью могут быть получены лишь в эксперименте при воспитании
в одинаковых условиях всех фаз развития. Но плодовитость самок зави-
сит от воздействия среды как в период роста, так и в период развития
куколок и жизни самок. Таким образом фактически плодовитость особей
разного веса может быть нивелирована условиями среды. Наоборот,
у особей одного веса, попавших в имагинальной фазе или куколкой в раз-
ные условия, она может быть разной (рис. 63). У видов с редуцирован-
ным питанием имаго длительность жизни этой фазы очень коротка, почему
и воздействия на плодовитость могут быть минимальны. К тому же они
ограничены лишь конечными моментами формирования половых про-
дуктов.
У видов Orgyidae, для которых типично имагинальное питание и боль-
шая длительность жизни имаго, условия развития преимагинальных фаз
Рис. 62. Плодовитость самок и вес куколок монашенки
(Ocneria monacha L.). 1 — при росте в разных термиче-
ских условиях. (По Цвёльферу, 1934); 2 — при влия-
нии кормовых условий. (По Саттлеру, 1939); 3 — то
же для особей непарника, взятых из природы. (По
Левитту, 1934).
в значительно меньшей степени определяют фактическую плодовитость
самок. Влияние условий среды в период личиночного развития принци-
пиально то же, что описано для предыдущей группы, но слабее, а усло-
вия имагинальной жизни могут кардинально Менять плодовитость особей,
совершенно нивелируя и изменяя величины плодовитости особей, ожи-
даемые на основании условий личиночного развития, . На рис. 63 даны
изменения плодовитости в зависимости от термического режима при
жизни имаго для трех видов Orgyidae, имеющих разную степень рудимен-
тации питания имаго. Можно видеть, что у Daseochaeta alpium Osb., вида
с энергичным имагинальным питанием, термический режим в период
жизни имаго в широчайших пределах меняет плодовитость, независимо
от потенциальных возможностей особей. Сходная картина наблюдается
у видов Acronicta Ochs, и Subacronicta Kozh. У Calocasia coryli L. нет
питания имаго, но вышедшие из куколок самки должны дозревать извест-
ный срок (двое суток или сутки). Для этого вида значение термиче-
ским, Пятницким (1935) и другими, имеется лишь для видов с редуцированным има-
гинальным питанием, но и здесь она все же не так определенна (Левитт, 1935). Она
может быть выражена лишь средними величинами.
8*
116
ОБЩАЯ ЧАСТЬ
Рис. 63. Плодовитость самок Daseochaeta alpium
Osb. (I), Calocasia coryli L. (2) и Orgyia gonostigma
F. (3) при жизни их в разных термических усло-
виях.
ского режима при жизни имаго значительно меньше, а на изменениях
плодовитости в большей мере сказываются условия развития личиночных
фаз. Наконец у самок Orgyia gonostigma F., у которых яйцекладка начи-
нается в день выхода имаго, зависимость плодовитости от термических
условий в период жизни имаго почти отсутствует, а изменения плодови-
тости почти полностью связаны с условиями личиночного развития и,
в частности, питания гусениц. У Orgyia gonostigma F. уловить влияние
термического режима при жизни имаго на плодовитость почти невоз-
можно .
Плодовитость и длительность жизни видов, питающихся в имаги-
нальной фазе, в большой мере обусловлены самими условиями имагиналь-
ного питания, в частности
составом нектарного пита-
ния и количеством пот-
ребляемого пищевого ма-
териала. Лучшим пище-
вым материалом является
глюкоза, но вполне пита-
тельны также фруктоза,
сахароза и раффиноза.
Примесь органических ки-
слот (яблочной, молочной,
щавелевой) вредна. Крах-
мал как примесь допу-
стим, но влияние его не-
благоприятно . В чистом
виде крахмал перевари-
вается слабо. Зависимость
от условий имагинального
плодовитости и длительности жизни имаго
питания при использовании разных пищевых веществ иллюстрируется
табл. 21.
Таблица 21
Влияние пищевых условий на длительность жизни и плодовитость
самок Acronicta rumicis L.
Диэта Число особей Длительность жизни (в сутках)1 Плодовитость (число отложен- ных яиц)
Голод 18 10.0 ( 4—13) Нет
Вода 15 11.2 ( 6—16) 65 (0—230)
Щавелевая кислота 10 9.0 ( 7—10) Нет
Глицерин j 20 12.1 ( 4—18) 27 (9—273)
Маннит 18 16.3 ( 6—22) 95 (0—409)
Глюкоза 20 20.1 ( 8—32) 260 (0—632)
Фруктоза 20 %-й раствор 21 17.0 (11—20) 94 (0—220)
Сахароза 19 16.8 ( 5—24) 94 (0—360)
Лактоза 20 9.2 ( 5—14) 9 (0—89)
Раффиноза 14 17.0 ( 6—32) 237 (0—638)
Крахмал 3 %-й раствор 15 12.0 ( 6—18) 28 (0—215)
1 В скобках приведены минимальные и максимальные величины.
экология
117
Факторы среды в широчайшей мере влияют на поведение, а в связи
с этим и на расселение Orgyidae. Условия обитания видов этого семейства
в закрытом ландшафте (в лесу) очень сложны и пестры. Изменения пове-
дения, естественно, имеют характер не только циклических, суточных
изменений активности, но комплекс сложных реакций обусловливает
выбор на данный момент наиболее благоприятных мест обитания.
Влияние световых условий на поведение Orgyidae очень велико, особенно
в весеннее время..Только что отродившиеся из яиц или вышедшие с зи-
мовки гусеницы, как то установлено для златогузки [Гесс (Hess, 1919)],
монашенки [Брандт (Brandt, 1936, 1937а, 1937в); Гундертмарк (Ннп-
dertmark, 1936, 1937а)] и непарного шелкопряда, очень активны и харак-
теризуются резко выраженным предпочтением яркого света. Это же изве-
стно и для некоторых (далеко не всех) Acronictinae, например видов
Pharetra Hb. По Гессу (1919), гусеницы златогузки не только особенно
сильно реагируют на ультрафиолетовые лучи, но и привлекаются ими
в наибольшей мере, чем всеми остальными лучами спектра. По Гундерт-
марку (1936в, 1937в), только что отродившиеся гусеницы монашенки
обнаруживают хорошо развитое цветное зрение и зрение формы. Они
предпочитают голубой и фиолетовый цвета (вероятно и ультрафиолето-
вый, который не был испытан). Предпочитаемой формой являются узкие
прямоугольники и треугольники, по сравнению с круглыми (шарообраз-
ными) формами. Для гусениц Orgyidae, и именно только что отродив-
шихся, типичен отрицательный геотаксис- (Гесс, 1919; Гундертмарк,
1937в; Брандт, 1937в). Реакции на свет молодых гусениц Orgyidae, соче-
таясь с отрицательным геотаксисом весною, ведут к быстрому переме-
щению их из прикорневой части стволов, где обычно они выводятся,
на кроны деревьев и к отысканию там необходимых пищевых условий.,
Отыскание пищи весною облегчается значительной быстротой движения/
молодых гусениц и выносливостью их к голоданию. По Брандту (1936),
только что отродившиеся и еще не питавшиеся гусеницы монашенки
способны покрыть расстояние в 100 метров. По мере роста гусеницы
Orgyidae теряют столь резко выраженную реакцию на разность освещения
и миграции их теперь связаны скорее с термическим режимом.
Обонятельные реакции, во всяком случае на большом расстоянии,
в отыскании пищи гусеницами Orgyidae играют подчиненную роль или
даже вовсе не имеют значения [Гётц (Gotz, 1936, 1938)]. Тем не менее
при даче гусеницам видов Acronicta Ochs., златогузке, непарному шел-
копряду и другим на выбор растений, нормально не служащих им пищей
и не поедаемых ими охотно, они долгое время ползают по ним, не пробуя
их. Напротив, при наличии среди любого количества несъедобных или
мало съедобных растений хотя бы одного пригодного для питания, они
в течение короткого срока (часто нескольких минут) скопляются именно
на нем, пожирая его. Таким образом, на близком расстоянии роль хеморе-
цепции гусениц и химизма питающих растений в выборе кормового суб-
страта несомненна. Роль зрительных и тактильных реакций в этом слу-
чае, без сомнения, на втором плане (Гундертмарк, 1936а, 1937а). По Гётцу
(1936), вкусовые реакции гусениц монашенки играют очень ограниченную
роль. Автор склонен видеть основу в выборе пищи гусеницами этого
вида в использовании физических свойств субстрата. Роль хеморецепто-
ров у гусениц Orgyidae в выборе пищи несомненна во всяком случае при
непосредственном контакте с пищей. Несомненна также их высокая сте-
пень специализации. Неясно лишь, с какого момента в выборе пищи
наступает переход от реакции на физические особенности субстрата
118
ОБЩАЯ ЧАСТЬ
(Брандт, 1936; Гётц, 1936, 1938) к реакциям на его химизм, хотя также
нет сомнения, что это происходит еще до разгрызания тканей.
Миграции молодых гусениц Orgyidae на кроны деревьев в поисках
пищи играют большую роль и в распространении их, в частности с по-
мощью ветра. Факт разноса молодых гусениц непарного шелкопряда
ветром был известен еще в прошлом столетии (Холодковский, 1894).
Специальными исследованиями американских авторов [Бёрджес (Bur-
gess, 1913); Коллинз (Kollins, 1917)] внесена некоторая ясность в этот
вопрос. Для разноса ветром гусениц непарного шелкопряда благопри-
ятна погода с температурой воздуха порядка 18° и с теплым ветром.
Разнос гусениц ветром возможен начиная с температуры воздуха 10°.
Кроме того, для разноса гусениц благоприятны средние скорости ветра,
порядка 4—8 м/сек. Сильные и резкие ветры не благоприятны. Разнос
гусениц может происходить и при слабом ветре, порядка 10 миль в час.
Висящие на паутинках гусеницы непарного шелкопряда подхватываются
ветром с еще голых в это время (весною) ветвей деревьев и уносятся вос-
ходящими токами воздуха до высот 100—300 метров и как максимум (?),
повидимому, до 600 метров [Коллинз и Бэкер (Collins a. Baker, 1934)].
Расстояние, на которое констатирован разнос их ветром, достигает 10 ки-
лометров. У собранных методом клеевых щитов и решеток гусениц непар-
ного шелкопряда обнаружен пустой кишечник, причем экспериментально
доказано, что после переноса ветром они способны питаться и нормально
расти. В лесах Северной Америки установлена чрезвычайно большая
роль ветра в распространении непарного шелкопряда, и именно за счет
разноса мелких гусениц. Значительная роль ветра в расселении непар-
ного шелкопряда установлена в лесах Крыма Пятницким (1935) и Пар-
хоменко (1936).
Для монашенки роль ветра в расселении молодых гусениц установ-
лена в принципе’та же, что и для непарного шелкопряда, но, видимо,
меньшая (Гундертмарк, 1938). Несомненна роль ветра в распространении
молодых гусениц и многих других видов Orgyidae, и именно тех, у кото-
рых первая стадия гусениц имеет большое число вторичных волос, как,
например, у видов Euproctis Hb., Orgyia Ochs., Pharetra Hb., Calocasia
Ochs. Для златогузки, тем не менее, роль ветра в расселении гусениц
первой стадии второстепенна или просто отсутствует, так как гусеницы
живут выводком в шелковом гнезде до весны и расползаются лишь после
зимовки. Первые стадии гусениц этого вида недоступны действию ветра
благодаря ряду особенностей их биологии — сильному плетению пау-
тины и изготовлению гнезда, появлению их в середине лета, когда деревья
вполне облиствены и движение воздуха в кронах ослаблено, и ряду
других.
Роль ветра в расселении Orgyidae несомненна также при разносе им
бабочек, как то указано, например, для монашенки [Лоос (Loos, 1927)].
Бабочки монашенки при посредстве воздушных токов способны пре-
одолевать высоты до 100Q метров и многие десятки километров расстоя-
ния. Биологическое значение ветра в разносе бабочек Orgyidae во многих
случаях второстепенно, потому что оплодотворенные самки их часто почти
не летают (или вообще лишены возможности полета) и потому подхва-
тывание их ветром или вообще исключено, или имеет вероятность значи-
тельно меньшую, чем для самцов (златогузка, монашенка). Лишь для
некоторых видов семейства возможно ожидать значительную роль ветра
в распространении имаго (у видов Acronictinae), но она ближе не изу-
чена.
ЭКОЛОГИЯ 119
Факторы среды и особенно термический режим в сильной степени
влияют на выбор мест окукления, а отсюда нередко и на выбор мест для
яйцекладки самками. Так, для непарного шелкопряда типична яйцекладка
в щели коры на прикорневые части деревьев, но при повышенной темпе-
ратуре яйцекладка часто наблюдается в корни деревьев, по трещинам
почвы, иногда даже под уровнем почвы (Бёрджес, 1913; Пархоменко,
1936), на камни, лежащие в тени на почве, и в трещины камней и скал
в глубине (Бёрджес, 1913; Бей-Биенко, 1924; Пятницкий, 1935). Дей-
ствию же высокой температуры следует приписать также окукление
и яйцекладку непарного шелкопряда на кронах деревьев, где движение
воздуха сильнее, отчего температура ниже и более изменчива, чем в при-
корневой части деревьев (Бёрджес, 1913; Щелкановцев,' 1926; Пархо-
менко, 1936). На кронах яйцекладка часто наблюдается в районе гнезд
птиц, например на вороньих гнездах (Бёрджес, 1913). Различия мест
яйцекладки непарного шелкопряда отчасти связаны с миграциями ба-
бочек (Щелкановцев, 1926), отчасти с миграциями гусениц (Бёрджес,
1913; Бей-Биенко, 1924). Гусеницы всегда отыскивают места для окук-
ления, куда не проникают прямые солнечные лучи. Часто яйцекладка
непарного шелкопряда и всегда у видов Orgyia Ochs, происходит в ме-
стах выплода самок. У видов, самки которых подвижны, каковы боль-
шинство Acronictinae, выбор мест яйцекладки более сложен. В одних
случаях он определяется просто присутствием кормового растения, на
которое, часто даже без выбора определенной его части, откладываются
яйца, как то известно для видов Pharetra Hb., кладущих яйца кучками.
В других случаях место яйцекладки определяется, помимо термического
режима и присутствия кормового растения, еще очень многими другими
условиями (освещенностью места, возрастом листа и пригодностью его
для питания молодых гусениц). Это наблюдается, когда яйца отклады-
ваются поодиночке, как, например, у видов Acronicta Ochs., Subacronicta
Kozh., Calocasia Ochs, и ряда других. В этих случаях молодые гусеницы
строго ограничены в использовании кормового субстрата и начинают
питаться нормально лишь на месте выплода, не отыскивая пищи.
Биоценотические связи Orgyidae чрезвычайно многообразны и сложны.
Виды Orgyidae во всех фазах развития являются пищевым субстратом для
разнообразных паразитов и хищников из насекомых и для насекомояд-
ных птиц. Паразиты в свою очередь служат пищей и уничтожаются пара-
зитами второго порядка, а последние нередко служат пищей для парази-
тов третьего порядка. Это явление сверхпаразитизма, когда один и тот
же вид может быть и паразитом и сверхпаразитом, чрезвычайно услож-
няет биоценотические отношения Orgyidae. Хищные жуки и хоботные
(клопы) также часто питаются за счет Orgyidae. Это прежде всего связано
с обилием особей многих видов Orgyidae и заметной ролью их в ланд-
шафте. Птицы в меньшей мере связаны с видами Orgyidae, так как воло-
систые и нередко ядовитые гусеницы этого семейства пожираются отно-
сительно немногими видами птиц.
Часто наблюдаемые массовые размножения ряда видов Orgyidae почти
всегда влекут за вобою изменения характера растительности в связи
с угнетением роста деревьев. Ослабленные деревья служат в дальнейшем
субстратом для короедов, усачей и златок, а также других древесинных
насекомых и часто, усыхая, погибают. Известна гибель больших площа-
дей леса и смена их новыми формациями как следствие массовых раз-
множений монашенки, непарного шелкопряда, златогузки и других
Drgyidae. На тесную связь очагов короедов и массовых размножений мо-
120
ОБЩАЯ ЧАСТЬ
нашенци, а также ряда других Orgyidae есть много указаний в литературе-
[Ратцебург (Ratzeburg, 1838—1840); Эшерих (Escherich, 1914); Комарек
(Komarek, 1926); Прель (Prell, 1925), и многие другие]. Отмечена зави-
симость массовых появлений долгоножек {Tipula nubeculosa Mg.) [Дин-
глер (Dingier, 1924)] на следующий год после массовых размножений
монашенки. Это, возможно, связано с изменениями химизма почвы и
экоклимата, но, повидимому, главным образом с появлением большого
количества пищи для Tipula в виде отмирающих корней древесных расте-
ний и обилия в почве мицелия грибов. Orgyidae являются субстратом для:
симбиотических и патогенных бактерий, вирусов (полиэдренная болезнь),,
грибков и простейших. Наконец сами виды Orgyidae могут способствовать
возникновению эпизоотий среди растений и животных или даже обуслов-
ливать развитие таковых среди них. Отмечен разнос грибка — паразита,
видов Ribes (Cronartium ribicola) гусеницами непарного шелкопряда.
|Траватт и Позей (Gravatt a. Posey, 1918)]. Несомненен разнос патогенных
грибков и бактерий гусеницами Orgyidae как с экскрементами, так и на
поверхности тела. Равным образом и выпадающие волоски гусениц
Orgyidae, прилипая к листьям растений и попадая с пищей гусеницам
или личинкам других насекомых, могут вызывать среди них повальные-
заболевания. Это отмечено, например, для гусениц тутового шелкопряда
при массовых размножениях желтогузки {Euproctis similis Fuessly)*
[Иамамасу (lamamasu, 1926)].
Из паразитов Orgyidae главное число падает на многоядные виды,
паразитирующие в личиночных фазах. Это паразиты преимущественно-
из перепончатокрылых, виды Ichneumonidae {Ichneumon, Cryptus, Pimpla,
Ophiori) и Braconidae {Apanteles, Microgaster, Meteorus), а также из дву-
крылых— Larvivoridae.1 Некоторые виды паразитов, как, например,
Apanteles melanoscelus Ratz., довольно полно специализованы на питании:
определенными видами Orgyidae, в частности указанный вид связан
с гусеницами непарного шелкопряда. Он дает два поколения в период,
их роста, диапаузирует и зимует в фазе пронимфы в кокончиках. Боль-
шею частью паразиты из перепончатокрылых поливольтинны, в связи
с чем наблюдается смена хозяев у полифагов. Так, Apanteles liparidis
Rche. и A. porthetriae Meusbk., развиваясь в двух поколениях на непар-
ном шелкопряде, затем переходят на златогузку и даже гусениц сос-
нового шелкопряда, в которых и зимуют. У паразитических двукрылых,
например у Compsilura concinnata Mg., Sturmia scutellata R. D., 5. incon-
spicua Mg., Phorocera agilis R. D., Larvivora larvarum L., Lydella nigripes
Fall., Carcellia, Zenillia и ряда других, точно так же циклы развития
лишь соприкасаются с Orgyidae, гусеницы которых служат им в те или
иные моменты года пищевым субстратом. Некоторые из паразитических
мух, как, например, первые три вида, являются массовыми паразитами
непарного шелкопряда и златогузки. Неспециализованные циклы раз-
вития имеются и у паразитов куколок Orgyidae, часть которых парази-
тирует также и в гусеницах, например Monodontomerus. Тем не менее
есть виды, специализованные целиком на паразитировании в куколках,
как Brachymeria, но они не вполне ограничены рамками сем. Orgyidae.
Паразиты яиц {Chalcididae) обнаруживают наибольшую связь с определен-
ной фазой развития Orgyidae, будучи ограничены лишь яйцом, но, как
и другие паразиты, обнаруживают высокую степень полифагии, парази-
1 Подробнее о паразитах см. в Специальной части, где списки их даны для каж-
дого вида сем. Orgyidae.
экология
121
тируя не только на Orgyidae, как, например, Schedins, но и на яйцах
многих других чешуекрылых. Напротив, некоторые виды яйцеедов
(Anastatus disparis Ruschka) обнаруживают высокую степень пищевой
специализации, будучи ограничены, как упомянутый вид, только одним
хозяином (непарным шелкопрядом) и имея циклы развития, вполне
приспособленные к циклу хозяина. В целом циклы развития паразитов
Orgyidae изучены далеко не достаточно и степень связи их с циклами
хозяев пока не ясна.
Взаимоотношения паразитов Orgyidae очень сложны. Помимо связи
их с определенными хозяевами из Orgyidae и некоторой или значительной
(редко полной) кормовой специализации на них, ряд паразитов обна-
руживает также связь с другими хозяевами. Последние частично из
чешуекрылых, но из других семейств, в особенности Notodontidae, Lasio-
campidae, частично связь эта имеет место даже с другими группами насе-
комых из перепончатокрылых и двукрылых. Некоторые из паразитов,
в частности многие виды двукрылых, являются паразитами паразити-
рующих на Orgyidae же наездников, т. е. сверхпаразитами.
Существенное значение имеет сильная конкуренция между парази-
тами Orgyidae, особенно в случае слабо летающих форм, например неко-
торых Chalcididae, как то отмечено для яйцеедов непарного шелкопряда.
Так, Anastatus disparis Ruschka и Schedins kuwanae Haw. оба паразити-
руют в яйцах непарного шелкопряда, причем нередко одно и то же яйцо
оказывается зараженным личинками обоих видов. Количество питатель-
ного материала в яйце непарного шелкопряда недостаточно для питания
личинок обоих видов паразитов, и часто оба паразита погибают от голода.
Сильно распространен гиперпаразитизм. Например, на Apanteles melano-
sceles, паразите гусениц непарного шелкопряда, паразитирует более
десятка видов Ichneumonidae (Hemiteles, Gelis, Mesochorus, Acrolyta,
Thysiotorus,Ephialtes), несколько ресятков видов Chalcididae (Eupelmus,
Haltichella, Spilochaicis, Eusagyia, Monodontomerus, Eupteromalus, Elas-
mus, Ditrachys, Coelophistia, Habrocytus, Hypopteromalus, Dimmockia,
Cirrospilns, Plenrotropis, Symbiosis'). Часть из них является в то же время
паразитами непарного шелкопряда и паразитами сверхпаразитов, на-
пример живущих на паразитах Apanteles melanosceles, т. е. обнаруживают
паразитизм третьего порядка. Наконец куколки тахин пожираются хищ-
ными жуками из Elateridae (Corymbites) [Вихман (Wichmann, 1925)].
Существенным фактором в жизни Orgyidae являются хищники из насе-
комых. Из них на первом месте стоят виды жужелиц-красотелов Calosoma
(sycophanta L., inquisitor L.). Личинки и жуки этих видов пожирают,
ползая по сучьям и стволам деревьев, куколок и гусениц Orgyidae (мона-
шенку, непарного шелкопряда, златогузку) и для некоторых их видов
являются очень существенным фактором динамики популяции. По Эска-
лера (Escalera, 1924), в Испании в уничтожении гусениц непарного шел-
копряда существенную роль играют (кроме жужелиц) также мертвоеды
(Xylodrepa quadripunctata Schrb.), хорошо лазящие по деревьям. Истре-
бление гусениц непарного шелкопряда этими жуками было отмечено
также и в других частях ареала распространения этого вида (Венгрия,
Чехословакия). В Марокко яйца непарного шелкопряда уничтожаются
жуками Trogoderma versicolor Crtz. из Dermestidae (Лепиней, 1930).
Но они уничтожаются также, хотя может быть менее регулярно, чем
предыдущим видом, другими видами жуков, например Tenebroides
marrocanus Reitt., Anthrenns verbasci L., Dermestes lardarius L., Dermes-
tes erichsoni Gyllh., Oeypus olens Mull., Cantharis rustica Ball., Steropus
122
ОБЩАЯ ЧАСТЬ
globosus F., Cardbus latus Deg. var. gougeleti Reiche. Отмечено питание
яйцами непарного шелкопряда также некоторых Staphylinidae, но здесь
степень связи не ясна. Указано также, что гусеницы Tinea fuscipunctella
Haw. пожирают яйца непарного шелкопряда. Яйца разных видов Orgyi-
dae, а также и их куколки уничтожаются муравьями (Прель, 1925). Нако-
нец клещики (Trombidiidae) высасывают и уничтожают яйца непарного
шелкопряда.
Гусеницы Orgyidae (Dasychira Steph., Calocasia Ochs, и ряд других)
•часто служат пищей хищным клопам, например Picromerus bidens L.
Гусениц непарного шелкопряда уничтожают клопы Podisus serieventris
Bryce. В Австралии (Виктория) клопы Dindymus versicolor Н. S. часто
высасывают гусениц Orgyia anartoides Wlk. Для тропической южной Аф-
рики есть указания на питание хищных гусениц огневок (Phycitinae) 1
гусеницами (преимущественно окукляющимися) и куколками Orgyidae
(Dasychira Steph.).
И здесь биоценотические взаимоотношения очень сложны. Хищные
насекомые являются врагами не только одних Orgyidae, но и многих
других видов насекомых, в том числе хищников и паразитов. Кроме
того, насекомоядные позвоночные часто предпочитают питаться хищ-
никами и паразитами, живущими за счет Orgyidae, но не последними,
как то указывалось в отношении птиц. Несомненна лишь значительная
роль кукушек и сорокопутов в уничтожении личиночных фаз Orgyidae,
но куколки уничтожаются более обширным кругом насекомоядных птиц,
в особенности в тех случаях, когда покровы их мягки и слабо волосисты
(Leucoma Steph., ряд видов Acronictinae). Ритзема-Бос (Ritzema-Bos,
1914) указывает, что яйца Dasychira 'pudibunda L. уничтожаются, по-
мимо кукушек, также сойками (Garrulus) и синицами (Parus). Истребле-
ние яиц Orgyidae птицами, повидимому, незначительно, так как они или
имеют очень твердую оболочку, как, например, яйца Olene Hb., Dasy-
chira Ochs., Orgyia Ochs, и многих других видов, или покрыты густым
слоем волосков (Ocneria Hb., Euproctis Hb.), затрудняющих пожирание
кладок. Наконец яйца Acronictinae, мягкие и незащищенные, мелки,
откладываются часто поодиночке и мало заметны, к тому же развиваются
короткий срок. В отношении яиц Orgyidae, имеющих даже не слишком
твердую скорлупу, каковы яйца монашенки и непарного шелкопряда
(освобожденные от войлока волосков), исследованиями Рейфа (Reiff,
1910а) показана трудность переваривания их в желудках птиц. Так,
яйца, съеденные дроздами, вьюрками, голубями и совами (Turdidae,
Fringillidae, Columbidae, Bubonidae), совершенно не перевариваются
и дают в значительном проценте нормальный выход гусениц после про-
хождения через кишечник птицы. Таким образом, при пожирании яиц
Orgyidae, имеющих твердый хорион, возможно, придется учитывать также
роль птиц в расселении этих видов и разносе их яиц по территории, а
не только в уничтожении их.
Роль земноводных в уничтожении Orgyidae не ясна, но, повидимому,
она не велика, хотя бы в силу их дендрофильного образа жизни. Отме-
чены случаи пожирания личиночных фаз видами Hyla, Rana и Bufo,
но не часто. Из млекопитающих связь с Orgyidae обнаружена для скунса
(Mephotis mephitica) в Америке, где указано пожирание этими живот-
ными куколок непарного шелкопряда. Для северной Африки (Алжир)
отмечена косвенная связь лисиц с Orgyidae через уничтожение лисами
1 Без более точного определения.
экология
123
жужелиц, хищничающих за счет Orgyidae. Пожирание жужелиц лиси-
цами в этих условиях столь сильно; что лисы являются фактором, за-
трудняющим применение биологического метода борьбы с непарным
шелкопрядом (в частности использование видов Calosoma).
Биоценотические связи Orgyidae осложняются также наличием пато-
генных микроорганизмов, частью специфичных только для этих видов
Lepidoptera, частью патогенных также для окружающих животных
и растений. Orgyidae могут получить микроорганизмы или с пищей, или.
через паразитов, прокалывающих покровы при откладке яиц в гусениц,
или, наконец, при контакте разных особей одного вида при еде, при спа-
ривании. Следует отметить также большое число не паразитических,
случайных, но частью облигаторных микроорганизмов — обитателей
•кишечника Orgyidae, например, различных Bacillus, описанных Пайо
(Palliot). Наряду с индифферентными микроорганизмами несомненно
наличие также болезнетворных для Orgyidae, иногда вызывающих поваль-
ные заболевания ряда их видов (непарного шелкопряда, краснохвоста,
монашенки и др.). Таков вирус,1 вызывающий полиэдренную болезнь
очень многих видов Orgyidae', он распространен чрезвычайно .широко.
Затем, по-Янишу (Janisch, 1938), грибок Aspergillus versicolor Vuill.
в высокой мере патогенен для гусениц монашенки. Далее, микроб Strep-
tococcus disparis Gias. [Глезер (Glaser, 1918)] патогенен для гусениц
непарного шелкопряда. Наряду с этим из споровиков (Microsporidia)
Plistophora schubergi Zwolf. может вызывать массовую гибель гусениц
непарного шелкопряда и златогузки.
Наибольшее распространение и несомненно громадное экологическое
значение, в силу развития грандиозных эпизоотий, имеет полиэдренная
болезнь. Она давно уже отмечалась для монашенки и непарного шелко-
пряда, для которых эффективен один и тот же вирус. Кроме того, она ши-
роко распространена среди других видов Orgyia Ochs., из которых особенно
подвержена заболеванию Orgyia antiqua L. и в меньшей мере, повидимому,
Orgyia gonostigma F. Полиэдренная болезнь констатирована также для
гусениц Dasychira pudibunda L. и Olene fascelina. L., реже для видов
Euproctis Hb. и Calocasia Ochs. Специальным бактериологическим и
достаточно точным исследованием эта болезнь изучена при эпизоотиях
монашенки и непарного шелкопряда [Чэпман и Глезер (Chapman a. Gla-
ser, 1916; Комарек и Брейндль, 1924; Прель, 1926]. Для других видов
она установлена только по внешним, впрочем очень типичным призна-
кам.
В развитии заболеваний большую роль играют условия среды.
В случае полиэдренной болезни условия среды играют даже ведущую
роль, так как инфекция вирусом возможна всегда. При оптимальных
гидротермическом режиме и условиях питания развитие эпиздотий по-
лиэдренной болезни не наблюдается. Факторами, способствующими раз-
витию ее, являются температура, сухость и неполноценное питание.
Развитие эпизоотий полиэдренной болезни наблюдается в очагах раз-
множения монашенки или непарного шелкопряда после оголения гусе-
ницами деревьев, когда качество листьев, развивающихся на них, сильно
ухудшается, и возрастают сухость воздуха и его температура.
1 По данным Комарека и Брейндля (1924), полиэдренная болезнь вызывается
возбудителями типа кокков. Эта точка зрения не разделяется большинством микро-
биологов, работавших над полиэдренной болезнью; она является устаревшей.
124
ОБЩАЯ ЧАСТЬ
Глава VII
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Рассмотрение географического распространения Orgyidae в известной
мере резюмирует предыдущие главы, так как понимание географических
фактов требует привлечения всех данных, касающихся жизни Orgyidae.
Должны быть использованы материалы как по био-экологии и физиоло-
гии Orgyidae, так и в неменьшей мере по их морфологии и филогении,
причем все они должны быть рассмотрены в разрезе эволюции Orgyidae.
Естественно, что этот круг вопросов требует привлечения обширной
литературы.
Географическое распространение видов, родов- и даже подсемейств
Orgyidae обнаруживает тесную связь с условиями обитания. Так, границы
ареалов распространения обнаруживают легко уловимые связи с кли-
матом. Они фактически обусловлены или отдельными ведущими его
факторами, преимущественно условиями термо- и гигрорежима, или ком-
плексом их. Связь границ ареалов Orgyidae с пищевыми условиями менее
резка, но достаточно ясна у узких олигофагов, например у Leucoma
salicis L., Eogena contaminei Ev. У многоядных видов лишь общая кон-
фигурация ареалов совпадает с распространением их основных кормовых
растений. Напротив, всегда ясную связь дает обилие особей данного вида
с условиями наиболее полного роста кормовых растений (рис. 64). Рас-
пространение Orgyidae, будучи ныне, в голоцене, нацело обусловлено
экологическими связями и условиями среды, специфичными для разных
видов, сколько-нибудь полно может быть понято лишь при рассмотрении
его в историческом разрезе.
Распространение сем. Orgyidae обширно. Orgyidae встречаются почти
на всей суше земного шара, но количественно резко преобладают в тро-
пиках и субтропиках Старого Света. Из них виды подсем. Orgyinae доми-
нируют в тропиках, а виды Acronictinae — в субтропиках и умеренных
широтах. Из всех континентов Северная Америка наиболее бедна видами
Orgyidae. Не многочисленны виды этого семейства и в Южной Америке.
Общее представление р распределении числа видов подсемейств и триб
Orgyidae по разным территориям земного шара дает рассмотрение цифр
в табл. 22 и карт (рис. 64, 65, 67).1
Наибольшую самостоятельность фауны Orgyidae при наибольшем
обилии видов обнаруживают эфиопская и индомалайская области, имею-
щие по характеру фауны этого семейства много общего между собою
(табл. 22). Для эфиопской области типичны полиморфные роды {Eu-
proctis Hb., Laelia Steph.) и ряд небольших, целиком эндемичных родов
(Cimola Wlk., Pirga Auriv., Marbla Swinh., Marbloides Her., Bracharoa
Hamps., Lomadonta Holl., Lacipa Wlk., Nyctimera Hb и др.). Для индо-
малайской области характерны из полиморфных родов Artaxa Wlk.,
Leucoma Steph., Ocneria Hb. {Lymantria Hb.) и большое число мелких
эндемичных родов {Etobema Wlk., Ponderia Swinh., Cispia Wlk., Dura
1 В качестве подразделений суши, для которых приводится число видов семей-
ства, в табл. 22 использованы обычные зоогеографические подразделения суши, при-
чем для Палеарктики — согласно моей схеме (Кожанчиков, 1937). Для других терри-
торий даются общепринятые подразделения, но более схематизованные, чем для
Палеарктики. Исходя из специфичности фауны Orgyidae, степени изученности ее
на данной территории и важности этой территории для понимания формирования
фауны Orgyidae СССР, в одних случаях используется более дробное деление суши,
в-других-—менее дробное и более схематичное.
Рис. 64. Территории, обильно заселенные видами Acronictinae (точки), и области распространения их основных
кормовых растений: 1 — Betulaceae, 2—-Fagaceae.
126
ОБЩАЯ ЧАСТЬ
Moore). Обе области связаны наличием многих общих полиморфных
родов. Из них прежде всего могут быть названы обширные роды Dasy-
chira Ochs, и Olene Hb., обильно представленные как на Зондских остро-
вах, так и в западной Африке. Кроме того, виды Laelia Steph, и Euproc-
tis Hb. хотя и резко преобладают в Африке, но представлены также'
и в индомалайской фауне значительным числом видов. Наоборот, виды
Ocneria Hb., будучи характерными для индомалайской фауны, богато
представлены также на Мадагаскаре и есть в Африке. Это же, но в мень-
шей мере касается видов Artaxa Wlk., а также некоторых мелких родов,
например Inans Moore. Может быть отмечена большая близость мадага-
скарской фауны Orgyidae к индомалайской, чем к африканской. Так,
род Numens Wlk., будучи типичен для Гималаев и Индии, имеет предста-
вителей и на Мадагаскаре, но не в Африке. Это же может быть отмечено
и для Thalatha (malagassica Hamps.) —рода, распространенного на Зонд-
ских островах, отчасти в Гималаях, в западном Китае и в северной Ав-
стралии. Кроме этого рода, из видов подсем. Acronictinae для индомалай-
ской фауны характерны виды Мота Hb., Belciana Stgr. и Trisuloides
Btl., неизвестные из эфиопской области. Необходимо подчеркнуть нали-
чие в тропиках и, в частности, в индомалайской фауне генерализованных
представителей подсем. Acronictinae, столь характерного для умерен-
ных широт (Голарктики). Эфиопская фауна почти не имеет представи-
телей этого подсемейства.
Австралийская фауна Orgyidae, будучи близкой к индомалайской,
обнаруживает тем не менее несколько большую обособленность от нее,
чем африканская. Для Австралии и Тасмании характерны большой
полиморфный род Anthela Wlk. и некоторое число мелких эндемичных
родов (Munichryia Wlk., Pterolocera Wlk., Chelepteryx Gr. R. Grey, Notaxa
Wlk., Aposita Turn., Gephyroneura Turn., Chennala Swinh.). Все эти роды
близки к йндомалайским, хотя и выделяются в особое подсемейство.1
Но в Австралии нет представителей полиморфного рода Olene Hb. и почти
отсутствуют Dasychira Steph, и Ocneria Hb., столь типичные для индо-
малайской и африканской фауны Orgyidae. Виды Artaxa Wlk., Eu-
proctis Hb. и Laelia Steph, также очень немногочисленны в Австралии
и наблюдаются только в северной И северо-восточной ее частях. Лишь
род Orgyia Ochs, представлен несколько большим числом видов (anarto-
ides Wlk., australis Wlk.), близких к африканским типа О. tricolor
Н. S. и евразийской О. dubia Tausch., т. е. степным видам этого рода,
но не лесным его представителям типа О. antiqua L. Acronictinae почти
отсутствуют в австралийской фауне; там есть (Квинслэнд) лишь один
эндемичный вид из рода Thalatha Wlk., затем своеобразный представи-
тель монотипического рода Cosmodes Gn. (С. elegans Gn.), близкий к во-
сточноазиатским Саппа Wlk. Особо необходимо отметить, что еще до-
вольно богатая фауна Orgyidae папуасской подобласти (почти исключи-
тельно Новой Гвинеи) является по составу почти нацело индомалай-
ской. Ее слагают не только общие с индомалайской фауной роды Orgyi-
.1 Подсем. Antheliinae Strd., в которое объединяются эндемичные или характер-
ные для Австралии виды Orgyidae, морфологически почти не характеризовано. Оно
выделено скорее на основании габитуальной обособленности этих видов и априорных
суждений об автономности австралийской фауны вообще. Требуется отметить, что
признаки, положенные в основу классификации Orgyidae Strand, как, например,
деление всего семейства на два отдела — виды, имеющие добавочную ячейку (Агео-
latae), и не имеющие ee(lnareolatae),— очень ненадежны. Этот признак может при-
сутствовать или отсутствовать не только в пределах родов (Arctornis Germ.), но и видов
(Penthophera morio L.).
РАСПРОСТРАНЕНИЕ
12Т
Таблица 22:
Географическое распространение видов Orgyidae на территории
земного шара
Название территории Число видов из под- семейства :
Orgyinae Acronictinae
Momini Acronictini
I. Эфиопская область 1. Восточноафриканская 197 0 0
2. Западноафриканская 477 2 0
3. Южноафриканская 95 2 1 (?)•
4. Мадагаскарская 107 4 0
П. Индомалайская область 1. Индийская 107 3 4
2. Малайская 317 17 2
3. Филиппинские острова 54 0 0
III. Неотропическая область 1. Центральноамериканская 37 12 1
2. Бразильская 121 15 0
3. Чилийская .... 5 0 0
ДУ. Австралийская область 1. Папуасская ; 145 5 1
2. Австралийская 153 3 1 (?>
3. Тасмания 18 0 0
4. Новая Зеландия 0 1 0
V. Голарктическая область 1. Арктическая 4 0 0
2. Бореевразийская: а) Европейская (западная и восточная) . . 13 5 20
б) Западносибирская 8 3 12.
в) Восточносибирская 12 4 11
3. Маньчжурская: а) Гималайская . . 154 30 16
б) Китайская (средняя и южная) .... 174 17 37
в) Японо-корейская 52 11 32-
г) Амурская 23 16 30
д) Монголо-тибетская 4 1 4
4. Ту рано-африканская: а) Североафриканская и ирано-туран- ская . . ' 24 0 9-
б) Нагорно-туркестанская . 11 1 12.
5, Средиземноморская . 11 1 1
6. Канадская 5 2 31
7. Подобласть Скалистых гор 5 10 53
8. Аллеганская 20 9 62
Рис. 65. Сферы влияния индомалайской (I), африканской (2) и центральноамериканской (3) фаун на фауну Orgyidae
Голарктики. Черным закрашены территории, обильно заселенные видами Orgyidae, с обилием анцестральных форм.
РАСПРОСТРАНЕНИЕ
129
dae, но очень часто и виды. Эндемичные представители папуасской фауны
Orgyidae, как, например, виды Dasychirides Beth.-Bek., обнаруживают
большую близость к индомалайским Dasychira Steph. Напротив, харак-
терный для Австралии род Anthela Swinh. известен с Новой Гвинеи всего
в числе нескольких видов. И в отношении фауны Orgyidae Австралии
несомненна тесная генетическая связь ее с индомалайской фауной, но
она сильно обеднена влиянием ксерофильных условий. Там, где еще
сохраняются условия обитания, пригодные для гигрофильных лесных
видов этого семейства, фауна эта имеет почти неизменный индомалайский
тип и довольно обильна видами. Заселяющие эту территорию виды
сильно изменены габитуально, исключая эврибионтных представителей
рода Orgyia Ochs.1
•Фауна Orgyidae Южной Америки, будучи еще более бедной видами,
чем австралийская, по сравнению со всеми рассмотренными областями
представляется в систематическом отношении оригинальной и сильно
обособленной. Для Южной Америки эндемичны очень своеобразные
роды Eloria Wlk., Caviria Wlk.,2 * * * * * * 9 Thagona Moeschl., Sarsina Wlk., Staethe-
rinia Btl., Desmoloma Feld., Phiditia Moeschl., Rolepa Wlk., Bazia *
Schr., Doa N. et Dyar, Tepilia Wlk., Leucalodes Dyar из Orgyinae uBathyra
Wlk., Gaujonia Dogn. и Lichnoptera H. S. из Acronictinae. Большая часть
родов олигоморфны, лишь некоторые полиморфны, например Eloria
Wlk., Thagona Moeschl. Возможно говорить о двух, примерно одинаково
обособленных группах Orgyidae. Одна — малайско-африканская, вклю-
чающая фауну Австралии, а также, как показано далее, фауну большей
части Евразии и Северной Америки. Вторая группа, по самостоятель-
ности вполне равнопенная первой, — неотропическая, представленная
типичными или близкими формами лишь в небольшой части Северной
Америки, на ее юго-западе, и на востоке Азии. В неотропической области
не известно ни одного рода Orgyidae, распространенного также в ма-
лайско-африканской фауне. Нужно, впрочем, отметить, что систематиче-
ская изученность африканских Orgyidae еще не настолько полна, чтобы
можно было настаивать на полном отсутствии связей в виде общих родов
между неотропической и эфиопской фаунами этого семейства. В запад-
ной Африке есть ряд представителей Orgyidae, имеющих во всяком слу-
чае большое габитуальное сходство с неотропическими Eloria Wlk. и
Caviria Wlk. Наоборот, филогенетическая связь неотропических энде-
мичных Orgyidae с эндемичными для Хондо и юго-восточного При-
морья Ivela Btl. несомненна. Более того, виды Ivela Btl. (ochropoda Ev.,
auripes Btl.) по многим морфологическим особенностям очень близки
к неотропическим Eloria Wlk. Они более близки к этим неотропическим
представителям, чем к какому-либо другому роду изучаемого семейства.
Точно так же эндемичный уссурийский Panthauma Stgr. из Acronictinae
обнаруживает сходство в структурах с неотропическими Bathyra Dogn.,
1 Есть сомнения даже в самостоятельности австралийской О. australis Wlk.
и африканской О. tricolor Н. S. Эта последняя в свою очередь почти не отличима
от евразийской О. dubia Tausch.
2 Колленетт (Collenette, 1934, Stylops, 3 : 113 описывает для юго-восточ-
ной Азии виды Caviria. Wlk., например С. chrysoscela Coll., но принадлежность их
к этому неотропическому роду сомнительна. Дайр (Dyar, 1897, Canad. Entom.,
29 : 16) давно уже отмечал, что азиатские виды, относимые к этому роду, по существу
далеки от типичных неотропических его представителей. Бывшие в моих руках нео-
тропические Caviria Wlk. по структурам гениталий обнаруживают очень значитель-
ную обособленность. Азиатские виды, относимые к этому роду, я не имел воз у<и<-
пости исследовать.
9 Фауна СССР, И. В. Кожанчиков.
Рис, 66, Области-царства «арктогейское» (Z-рЗ) и «нотогейское» (2) для фауны гигрофильных Orgyidae и обособление
(пунктир) области «цеогейской» для ксерофильных Orgyidae (3),
РА СПРОС ГРАНЕНИЕ
131
но Panthauma Stgr. также близок и к индомалайским Trisula Moore.
Эндемичные неотропические Orgyidae сосредоточены на территории гви-
ано-бразильского щита, в Колумбии и по длине Амазонки. Более широко
распространенные их виды проникают в центральную Америку. Связь
общими родами неотропической фауны с Северной Америкой ничтожна.
Лишь род Charadra Wlk. из Acronictinae распространен как в центральной
так и, более широко, в Северной Америке. Он несет черты строения нео-
тропических Bathyra Wlk. и Lichnoptera Н. S., являясь, без сомнения,
филогенетически более молодым производным этих родов.
Влияние неотропической фауны Orgyidae на фауну Голарктики несом-
ненно и в пределах подсем. Acronictinae довольно велико. Оно более
сложно, чем влияние индомалайской фауны. В частности несомненно,
что генерализованные представители подсем. Acronictinae сосредоточены
в неотропической фауне, где единственно и представлены большим чис-
лом видов. Виды Momini, обнаруживающие наиболее примитивные
черты строения среди всех известных голарктических представителей
подсем. Acronictinae, филогенетически тяготеют к неотропическим родам
этой трибы, причем как восточноазиатские (Мота Hb., Belciana Stgr..,
Panthauma Stgr., Belciades Kozh., Саппа Wlk,), так и североамериканские
их представители (Charadra Wlk., Diphthera Wlk.).
Фауна Orgyidae Голарктики и особенно неарктического ее отдела
бедна видами подсем. Orgyinae. Тем не менее она обнаруживает значи-
тельную специфичность, хотя в то же время и несомненное, иногда тес-
ное, родство с фауной южных областей. Для Голарктики эндемично боль-
шинство родов подсем. Acronictinae, тогда как роды Orgyinae в подавляю-
щем большинстве являются общими с тропическими и имеют здесь лишь
эндемичные виды или группы (Dasychira Steph., Olene Hb., Laelia Steph.).
Обычны случаи распространения видов этого подсемейства как в уме-
ренных широтах Голарктики, так и в пограничных южным областям
субтропиках, например Ocneria matura Moore, Laelia coenosa Hb. В тро-
пиках голарктические виды семейства не представлены совершенно.
Из подсем. Acronictinae распространены только в Голарктике Acronicta
Ochs., Subacronicta Kozh., Craniophora Snell., Diphthera Ochs., Calocasia
Ochs., Panthauma Stgr., Sirnyra Ochs., Arsilonche Ld., Hampsonia Kozh.,
Eogena Bd., Oxycesta Hb., Leiometopon Stgr., Agriopodes Hamps., Sub-
leuconycta Kozh., Daseochaeta Warr., Thyatirides Kozh., Euromoia Stgr.,
Anacronicta Warr., Belciades Kozh. Преимущественно голарктическими-
родами являются Trisuloides Btl., Саппа Wlk., Charadra Wlk., Belciana
Stgr. и Мота Hb. Из подсем. Orguinae эндемичны для Голарктики лишь
Gynaephora Hb., Penthophera Germ., Arctornis Mull. (?), Animomyia Dyar,
Parocneria Dyar, Ivela Btl., Cifuna Wlk. и Albarracina Stgr. Многие
голарктические роды последнего подсемейства обнаруживают близость
к тропическим, филогенетически являясь их дериватом. Таковы Paroc-
neria Dyar и Ocneria Hb., Gynaephora Hb. и Olene Hb., Penthophera
Germ, и Orgyia Ochs, и некоторые другие.
Могут быть намечены три территории в пределах Голарктики, где
особенно легко можно обнаружить связи голарктических Orgyidae с тро-
пическими (рис. 65). Фауна Orgyidae области Средиземного моря и Перед-
ней Азии (имея в виду ксерофильные территории и в первую очередь
турано-африканскую подобласть) обнаруживает связи с эфиопской фау-
ной не только в пределах близких и общих родов, но также и видов.
Так, Laelia impura Her. распространен в Абиссинии и Марокко. Этот вид
является также наиболее близким к евразийскому Laelia coenosa L.
9*
132
ОБЩАЯ ЧАСТЬ
Фауна Orgyidae юго-востока Палеарктики, и именно маньчжурской ее
подобласти, обнаруживает тесную связь с индомалайской фауной мно-
гими общими родами и видами из родов Olene Hb. (например О. mendosa
Hb.), Dasychira Steph., Ocneria Hb., Мота Hb. Третья область генети-
ческих связей фауны Голарктики — в южной Неарктике и в юго-восточ-
иой Палеарктике (Хондо, южное Приморье, отчасти Хинганы и
Забайкалье) с неотропической фауной. Здесь связи более отдаленные,
лишь близкими, но не общими родами, как, например, Ivela Btl., Diph-
thera Ochs., Daseochaeta Warr., Мота Hb. в Азии, Eloria Wlk., Charadra
Wlk., Agriopodes Hamps, в Америке. Общие виды между этими терри-
ториями (юго-восточная,Азия, юг Северной Америки и Южная Америка)
отсутствуют. Дериватом неотропической фауны Orgyidae являются широко
распространенные в Голарктике Diphthera Ochs, и Calocasia Ochs, более
богато в видовом отношении представленные в Неарктике и сложив-
шиеся, без сомнения, на территории Голарктики, вероятно в ее неаркти-
ческом отделе.
Сравнение изложенной общей картины распространения Orgyidae
с тем, что известно для других групп животных и, в частности, для наи-
более хорошо изученных разных групп позвоночных обнаруживает ряд
сходных черт. Это требуется подчеркнуть в связи с огромными разли-
чиями в систематическом положении, геологическом возрасте и даже
в современных условиях обитания тех и других. Фауна подсем. Orgyinae
(не всего семейства) Голарктики тяготеет к индо-африканской фауне.
Здесь мы имеем общую конфигурацию «северного царства» в понимании
Гадова (Gadow, 1898, 1901) для амфибий, «северной зоны» Блэкфорда
(Blanford, 1890) и Берга (1910) для рыб, «арктогеи» Ньютона (Newton,
1893) и Бианки (1905) для птиц, то же Склетера (Sclater, 1893), Осборна
(Osborn, 1900), Лидеккера (Lydekker, 1896) и Циттеля (Zittel, 1893) для
млекопитающих.1 Требуется подчеркнуть это сходство лишь для Orgyi-
nae, но и здесь несомненно влияние на состав фауны Голарктики нёотро-
пической фауны, особенно в ее неарктическом отделе, а также на востоке
Азии.
Далее, в согласии с тем, что описано упомянутыми авторами для
позвоночных, для Orgyidae может быть выделено царство «нотогеи»
(рис. 66), обладающее хотя и не богатой, но весьма своеобразной и обо-
собленной фауной. Голарктика в сильной мере заимствовала свою фауну
хвойных лесов и лесов с опадающей листвой, в частности видов подсем.
Acronictinae, из «нотогеи». В то же время налицо несомненная связь
архаичных групп индомалайской и неотропической фаун Orgyidae.2
Таким образом, выделение «нотогеи» для Orgyidae может быть проведено,
но должны быть подчеркнуты связь этих архаичных форм и с индомалай-
1 Н. Gadow. 1898. Bronn’s Klassen und Ordnungen des Tier-Reichs. VI (4) :
296. Vogel, II, Leipzig; A. Newton. 1893. Geographical distribution. Dictionary
of Birds, London : 314; В. Л. Бианки. 1905. Основные понятия о зоогеографи-
ческих единицах вообще и о внепалеарктических Старого Света в частности. Есте-
ствозн. и география, 7; Н. G a d o w. 1901. Amphibia and reptiles. Cambridge. Nat.
Hist., 8 : 74; w. Blanford. 1890. The anniversary adress of the president. Quart.
Journ. Geol. Soc. Lond., 46 (Proc.) : 76; Л. Верг. 1910. Рыбы бассейна Амура.
Зап. Акад. Наук, 8 сер., 24 (9) : 269; Р. Sclater. 1893. The nearctic region and
its mammals. Nat. Sci., 3 : 288; H. Osborn. 1900. Correlation between tertiary
mammals horizons of Europe and America. Ann. New York Acad. Sci., 13 : 46; R. L y-
. de kker. 1896. A geographical history of mammals. Cambridge; R. Zittel. 1893.
Die geologische Entwicklung, Herkunft und Verbreitung der Saugetiere. Sitzungsber.
mat.-phys. Classe Akad. Wiss., Miinchen 23(2) : 195.
2 Аналогичная тому, что известно, например, для тапиров из млекопитающих.
РАСПРОСТРАНЕНИЕ
133
ской фауной и влияние «нотогеи» на формирование фауны лесов с опа-
дающей листвой Голарктики.
Распространение видов сем. Orgyidae в Голарктике не равномерно.
Оно характеризуется преобладанием их числа и обилием эндемизма на
территориях, пограничных южным областям, связывающим эфиопскую,
индомалайскую и неотропическую фауны с Голарктикой. Средиземно-
морье, область Передней Азии и прилежащих засушливых территорий
при общей бедности видами Orgyidae имеют специфичную фауну с боль-
шим числом эндемиков. Из эндемичных родов могут быть названы Albar-
racina Stgr., Penthophera Germ., Eogena Bd., Oxycesta Hb., Leiometopon
Stgr. и Parocneria Dayr. Есть ряд эндемичных видов из широко распро-
страненных родов, например виды Ocneria Hb. (rubea Hb., atlantica Rbr.)T
Parocneria Dyar (detrita Esp.), Craniophora Snell, (pontica Stgr.) и ряд
других. Эндемичные роды семейства дают или ясные филогенетические
связи с малайско-африканскими представителями семейства (например
Parocneria Dyar, Penthophera Germ.), или менее ясные, как у некото-
рых Acronictinae.1 Область юго-востока Азии охарактеризуется как оби-
лием видов сем. Orgyidae вообще, так и числом эндемиков. Необходимо •
подчеркнуть, что это относится к обоим подсемействам. Виды Orgyidae
и в особенности эндемики концентрируются на юго-востоке Палеарктики
в пределах маньчжурской подобласти, на территориях ее гималайской,
китайских, амурской и японо-корейской провинций, тогда как монголо-
тибетская провинция бедна видами Orgyidae, хотя и типична наличием
эндемизма. Фауна гигрофильных Orgyidae Голарктики (во всяком случае
зоны широколиственных лесов) обнаруживает связи к этой территории,
если иметь в виду филогенетические отношения слагающих ее видов. ,
Для маньчжурской подобласти эндемичны роды Iveta Btl., Panthau-
та Stgr., Hampsonia Kozh., Anacronicta Warr., Euromoia Stgr., Bel-
ciades Kozh., Subleucony eta Kozh., Acronictoides Kozh., Thyatirides Kozh.
Кроме того, ряд родов, распространенных в тропиках, в индомалайской
и эфиопской областях, в Палеарктике представлен только в маньчжурской
подобласти. К ним относятся Саппа Wlk., Belciana Stgr. и др., а также
виды Numens Wlk., распространенные там же, но известные и с Мада-
гаскара. Число эндемичных видов в фауне маньчжурской подобласти
Палеарктики очень велико..
Своеобразным очагом является фауна Хондо и южного Приморья
в значительной мере обособленная от прилежащих территорий мань-
чжурской подобласти и связанная преимущественно с западным Китаем
и отчасти с Гималаями. Об этом свидетельствует распространение видов.
Саппа Wlk., Numens Wlk., Ocneria matura Btl. и ряда других. В то же
время территория эта частично связана, хотя и более отдаленно, с нео-
тропической фауной. На это указывают ареалы Ivela Btl., Panthauma
Stgr. Ранее (1937) мною отмечались подобные же связи для Agrotinae.
Семенов-Тян-Шанский (1899) отметил наличие среди Cerambycidae, т. е.
1 Филогенетические связи эндемичных для этой территории Acronictinae с индо-
малайской фауной в ряде случаев вполне определенны, особенно когда это касается
эндемичных видов, относящихся к широко распространенным родам. Так, эндемич-
ная для Крыма и Передней Азии Craniophora pontica Stgr. наиболее близка не
к широко распространенной, как в Европе, так и на востоке Азии Craniophora
ligustri L., но к эндемичным восточноазиатским видам этого рода — к тибетской
С. hemileuca Piingl. и японской С. niveosparsa Mts. Это же относится и к некоторым
другим видам. Для многих эндемичных Acronictinae, известных из Средиземноморья
и Передней Азии, особенно для олиго- и мономорфных родов этого семейства, фило-:
генетические отношения очень сложны и обнаруживают обширные связи.
134
ОБЩАЯ ЧАСТЬ
опять дендрофильных насекомых, очень заметного представителя нео-
тропической фауны — Callipogon (Eoxenus) relictus Sem. — в фауне юж-
ного Приморья. Караджа (Caradja, 1935, 1938) для западного Китая
приводит ряд примеров, показывающих связь лесной западнокитайской
фауны Lepidoptera с южноамериканской. Так, по его мнению, южно-
китайский Polygrammodes priscalis Gar. наиболее близок к’ неотропиче-
скому Polygrammodes ostrealis Gn., причем он более архаичен, по сравне-
нию с последним видом. Вторая часть утверждения Караджа кажется
мне недостаточно обоснованной. Он не изучал достаточно углубленно
роды Pyralidae, поэтому филогенетические отношения их едва ли доста-
точно ясны. Отмеченные выше виды Orgyidae и роды их, дающие связи
с неотропической фауной, эндемичны для южного Приморья, как
и известный давно Callipogon (Eoxenus) relictus Sem.1
Из приведенных выше фактов видно, что фауна Orgyidae хвойных
лесов Голарктики гетерогенна и обнаруживает связи с двумя достаточно
удаленными и самостоятельными центрами — восточноазиатским и нео-
тропическим. Указанное .наиболее резко выражено в подсем. Acronic-
tinae, где часть родов (Саппа Wlk., Euromoia Stgr., Belciana Stgr., Belci-
ades Kozh., Anacronicta Warr., Мота Hb., Acronictoides Kozh.) тяготеет
к индомалайской фауне, часть (Acronicta Ochs., Diphthera Ochs., Calo-
casia Ochs., Raphia Hb., Craniophora Snell.) —к неотропической. В подсем.
Orgyinae большая часть родов является дериватом индомалайской фауны
или даже общими ей и только виды Ivela Btl. представляют дериват нео-
тропической фауны.2 Этот же характер имеет и гигрофильная (дендрофиль-
ная) фауна горных районов турано-африканской подобласти, куда лес-
ные элементы Голарктики или проникают неизменными, как, например,
Leucoma Candida Stgr. и Ocneria dispar L. в Джунгарский Ала-тау, Olene
fascelina L. туда же и в северную Монголию, или представлены эндемич-
ными видами. Последнее может быть отмечено для некоторых видов
Leucoma «Steph, (flavo sulphur еа Ersch., sartus Ersch., surtur В. H), Eupro-
ctis Hb. (karghalica Moore) и ряда других родов. Из гигрофильных же
представителей Orgyidae дериватом индомалайской фауны являются
альпийская и арктическая фауны. Фауны эти вообще очень бедны и по-
тому, может быть, столь целостны по происхождению. Напротив, высо-
кая степень гетерогенности проявляется в рамках ксерофильных фаун
Голарктики, виды Orgyidae которых большею частью эфиопского про-
исхождения,. но смешаны также с дериватами индомалайской (Simyra
Ochs.) и неотропической (Eogena Gn., Oxycesta Hb., Raphia Hb., Leio-
metopon Stgr.) фаун.
Дериватом индомалайской фауны и, в частности; фауны юго-восточ-
ной Азии является также фауна Orgyidae Северной Америки, исключая
1 А. Семенов-Тян - Ша некий. 1899. Callipogon (Eoxenus) relictus,
sp. nov., представитель неотропического рода дровосеков в русской фауне. Труды
Русек. Энтом. общ., 32 (1) : 562—580; A. Caradja. 1935. Uber die zoogeographi-
schen Verhaltnisse in den westchinesischen Provinzen. Intern, entom. Ztschr., Guben,
29:105—109; A. Caradja. 1938. Materialien zu einer Microlepidopteren-Fauna Nord-
Fukiens. Stett. Ent. Ztg., 99 : 254—257; A. Caradja. 1938. Materialien zu einer
Lepidopteren-Fauna des Taipeischan-Massivs (Tsinlingshan), Provinz Shensi. Iris, 58 :
103—111; A. Caradja und R. M e у г i c k. 1937, 1938. Materialien zu einer
Microlepiopterenfauna des Julingshanmassivs. Iris, 51 : 137—180, 52 : 1—30.
2 В связи с изложенным уместно отметить работы Эйдмана (Е i d m a n n. 1938.
Beitrage zur Kentnis der Fauna von Slid. Labrador, insbesondere des Flussgebietes des
Matamek River. Zoogeographica, 2 (2) : 204—245. Эйдман очень ярко рисует гетеро-
генность фауны южного Лабрадора и специально отмечает довольно большое участие
в ней неотропических элементов.
РАСПРОСТРАНЕНИЕ
135
вида Charadra Wlk. и некоторых других представителей Acronictinae.
Из Orgyinae в Северной Америке есть лишь четыре рода: Orgyia Ochs.,
Olene Hb., Gynaephora Hb. и неясный’ малоизученный, эндемичный
калифорнийский род Animomyia Dyar, видимо близкий к Orgyia Ochs.
Первые три рода лишь заходят в Неарктику, будучи богато представлен-
ными в индомалайской фауне (первые два) и в умеренной и северной
Азии (третий). В Неарктике они представлены эндемичными видами,
большею частью близкими к видам, распространенным в маньчжурской
подобласти, например видами Orgyia Ochs, типа thyellina Btl., видами
Olene Hb. типа mendosa Hb., olga Obth. Напротив, виды подсем. Acro-
nictinae более богато представлены в Северной Америке, где много
видов рода Acronicta Ochs., в подавляющем большинстве эндемичных.
Виды poponDiphthera Ochs., Calocasia Ochs., Subacronicta Kozh. представ-
лены в Америке также более богато, -чем в Евразии, причем здесь, кроме
генерализованных эндемичных Charadra Wlk., есть еще близкий к евра-
зийским Daseochaeta Warr. Agriopodes Hamps.1 Из других представителей
семейства в фауне Северной Америки распространены голарктический
монотипичный Arsilonche Ld. и (эндемичные) представители Raphia Hb. —
рода, более богато представленного в фауне Евразии.
Характеристика фауны Orgyidae Средиземноморья2 более сложна,
так как истинно средиземноморская фауна (дендрофильные виды) очень
трудно отделима от пустынно-степной тур ано-африканской фауны этого
семейства, фауны бедной, но также очень своеобразной. Тем не менее
ряд элементов фауны Средиземноморья имеется и в сем. Orgyidae. Почти
эндемичен для Средиземноморья род Parocneria Dyar, лишь один вид
которого распространен от берегов Средиземного моря до Тихого океана,
встречаясь пятнами в Средиземноморье, на Кавказе и в Средней Азии.
Это связанный с Anacardiaceae и особенно с Pistacea Parocneria terebyn-
thi Frr. Другие виды этого рода сосредоточены в Средиземноморье, при-
чем они характерны пятнистым и прерывчатым распространением. Так,
Parocneria raddei Chr. известен с Эльдара и из Ирака, terebynthina Stgr. —
с Кавказа и из Сирии. Ряд видов рода Ocneria Hb., вообще типичного
для индомалайской фауны, ограничен в распространении Средиземно-
морьем. Таковы Ocneria atlantica Rbr., известный из северной Африки
(Атлас), rubea НЬ., ограниченный в распространении Пиренейским полу-
островом и южной Францией. Для Канарских островов эндемичен Olene
fortunata Rgfr., близкий к индомалайской О. mendosa Hb. и к ряду видов
этого рода, распространенных в юго-восточной части Северной Америки.
Для южного Крыма и Малой Азии эндемична Craniophora pontica Stgr.,
близкая к тибетской hemileuca Piingl. и восточноазиатской niveosparsa
Mts. (—pacifica Fil.). Часть восточноазиатских видов Orgyidae известна
как с юго-востока Азии, так и из Средиземноморья (а также западной
Европы). Таковы Craniophora ligustri L., Daseochaeta alpium Osb.3 Эти
1 Кроме Agriopodes Hamps., несомненно относящихся к Acronictinae и близких
к Daseochaeta Warr., что я лично мог проверить на фактическом материале, полу-
ченном мною от Мэк Доннофа (Оттава), Гэмпсон (1901) относит к Acronictinae
и к этой же группе родов виды Leuconycta Hamps. Род этот, изученный мною на доста-
точном материале, должен быть отнесен к Noctuidae (Metachrostinae).
2 Средиземноморская подобласть в моем понимании (см. И. Кожанчиков,
1937. Фауна СССР, Agrotinae), т. е. литторальная зона Средиземного моря с ее релик-
товой древесно-кустарниковой растительностью.
3- Из сказанного выше видно, что эндемы средиземноморской фауны сильно напо-
минают своими связями общеизвестный пример голубой сороки {Cyanopica), пиреней-
ском {С: cooki Вр.) и китайской {С. суапа Pall.). Ряд примеров сходных связей, но уже
136
ОБЩАЯ ЧАСТЬ
виды тяготеют к фауне Orgyidae юго-востока Азии, особенно к китайско-
гималайской фауне.
Для средиземноморской фауны Orgyidae следует считать характер-
ными и других представителей, дающих более сложные географические
отношения и обнаруживающих связи с центральной и Южной Америкой.* 1
Таковы виды рода Raphia Hb. —R. hybris Hb., ограниченная в распро-
странении Пиренейским полуостровом, но близкая к среднеазиатской
R. approximata Alph., к описываемой мною ниже R. obsoleta, sp. nov. *
из- Ирана, к локально распространенной на востоке Азии R. peusteria
Piingl. и к R. frater Grt., известной из Северной и Южной Америки (Кор-
дильеры). Аналогичны географические связи своеобразной Episema еое-
ruleocephala L., хотя и распространенной за пределами Средиземноморья,
но характерной также для этой подобласти. Episema coeraleocephala L.
филогенетически тяготеет к южноамериканским Charadra Wlk. и Lich-
noptera Н. S. Это же справедливо в отношении Anilonche albovenosa
Goeze, прерывчато, но несколько более широко распространенного,
подобно видам Raphia Hb., и сохранившего видовое единство в пределах
всего ареала как в Европе, так в Америке и Азии. Этот представитель
также тяготеет к центрально американской фауне, в частности к своеоб-
разным видам рода Acronicta Ochs, группы oblonga Sm. К этим же видам
и особенно к Arsilonche Ld. близки виды рода Sirnyra Ochs., хотя и распро-
страненные далеко за пределы Средиземноморья (S. nervosa Schiff, известна
из всей Средней Азии, Монголии, Тибета и из многих точек восточной
Азии), но представленные эндемичным средиземноморским видом S. den-
tinosa Schiff., характерным для северной Африки, Малой Азии и Кав-
каза. Наконец к этим же своеобразным представителям средиземно мор-
ской фауны Orgyidae должны быть отнесены и виды рода Oxycesta Hb. —
О. geographica F. и О. chamesices Gn., известные с Иберийского полуост-
рова, из южной Франции, Далмации, Украины и Кавказа, а также моно-
типический Eogena contaminei Ev., распространенный в южном Крыму,
на Кавказе, в южной Украине, на северном Кавказе, и в южном По-
волжье. Отпрыском этой же фауны, ныне уже совершенно не связанным
кавказских и отчасти альпийских видов с восточноазиатскими из чешуекрылых дают
Agrolinde, например виды подрода Chersotis—albufurca Ersch., multifida ~Ld.,sanctmorizi
В. H. (I. Kozhantschikov). 1935. Eurasiatische Unterbrechung der Verbreitungsbereiche
einiger Agrotinen-Arten in Beziehung auf ihre Geschichte und Bildung der Arten. Zool
Jahrb., Syst., LXVI. H. И. Кузнецов в своей работе «Нагорный Дагестан и его зна-
чение в истории развития флоры Кавказа» (Изв. Русек. Географ, общ., XIIIVI (6—7) :
213—260) подчеркивает подобные же связи для ряда растений, например берез (Betula
raddeana Trautw. и Betula medvedevi Rgl.), соответствующих маньчжурским, китай-
ским, дальневосточным и отчасти североамериканским видам этого рода.
1 В отношении приведенных здесь фактов очень важны указания Невского (Мате-
риалы к флоре Кугитанга и его предгорий. Труды Бот. инет. Акад. Наук СССР,
сер. 1, вып. 4,1937 : 199—346), который обращает внимание на самобытность среди-
земноморской кустарниковой флоры (Pistacia, Nitraria, Glycyrrhiza, Ephedra). Он
называет ее «флорой эфедры», отличая от самобытного средиземноморского элемента,
но обнаруживающего связь с южными континентами — «гондванско-средиземномор-
ской флорой» или «флорой Вельвичии» Попова (1937). Отмеченные мною элементы
фауны Orgyidae Средиземноморья стоят в прямой (пищевой) связи с «флорой эфедры»
и ни в коем случае не могут быть поставлены в аналогию с «флорой Вельвичии»,
хотя они генетически и тяготеют к Южной Америке. В связи с этим важно отметить
также работу Сетчелла (S е t с h е 1 1. 1935. Geographic elements of the marine
flora of the Pacific Ocean. Amer. Nat., 59 (725): 560—577), который указывает,
что эндемичные средиземноморские рдестовые (Potamogetonaceae) обнаруживают связь
с флорой Вест-Индии. Сетчелл (1935) склонен рассматривать область древнего Тетйса
колыбелью этих видов, а присутствие их в Вест-Индии как результат расселения.
РАСПРОСТРАНЕНИЕ
137
с Средиземноморьем, является и своеобразный Leiometopon simyrides
Stgr., пятнисто распространенный в Средней Азии.1 Существенно отме-
тить, что какие-либо специфичные виды Orgyidae, связанные с влажными
лесами субтропического типа (Черноморское побережье Кавказа, Кол-
хида, Талыш), в Средиземноморье не известны.
Фауна турано-африканской подобласти может быть охарактеризована
немногими эндемичными Orgyidae. К таковым относятся виды своеобраз-
ного рода Albarracina Stgr., ограниченные в распространении северной
Африкой и Сирией. Очень характерна также для фауны турано-афри-
кпнской подобласти Orgyia dubia Tausch., распространенная вообще
несколько шире. Дериватом этой же фауны является своеобразный Реп-
thophera morio L., сохранившийся ныне лишь за пределами этой подоб-
ласти в условиях сухих полупустынных степей. Ими и ограничиваются
типичные ксерофильные представители турано-африканской подобласти.
II число их можно было бы по формальным признакам добавить еще
/.eiometopon Stgr., но, как упоминалось, этот род представляет дериват
средиземноморской фауны, хотя и ограничен в распространении Средней
Азией. Столь слабое развитие типичных для турано-африканской подоб-
ласти ксерофильных элементов среди Orgyidae, вероятно, связано с труд-
ностью адаптации гигрофильных и дендрофильных видов этого семейства
к условиям пустыни. Косвенно это подтверждает факт значительно боль-
шего обилия эндемичных горных Orgyidae турано-африканской подоб-
ласти, обитающих мезофильные стации (кустарники, леса) гор, предгорий
и речных долин (тугаи). Эти виды являются, впрочем, мало" характер-
ными для турано-африканской подобласти. Помимо присутствующих
гноеобразных эндемичных видов Raphia Hb. [approximata Alph., obso-
lete Kozh.), здесь известны и другие. Из лесной зоны Тянь-шаня, Копет-
дага и Ала-тау известны Orgyia prisca Stgr., О. ochrolimbata Stgr., Leu-
eotna flavosulphurea Ersch., L,. sartus Ersch., L. surtur В. H., Euproctis
karghalica Moore, ряд эндемичных видов Acronicta Ochs, [elaeagni Alph.)
и Subacronicta Kozh. [centralis Ersch., grumi Alph.). Из Гиссарского
хребта описаны характерные архаичные Acronicta angustimacula,
нр. п., и Ocneropsis relictus, gen. et sp. n. Следует подчеркнуть про-
никновение в горные районы турано-африканской подобласти мно-
гих широко распространенных в бореевразийской подобласти видов,2
которые при этом не дают даже подвидов. Так, проникают в Джунгарский
Ала-тау и Тянь-шань виды Leucoma Steph. [Candida Stgr., salicis L.),
1 Как видно, распространение приведенных здесь видов Orgyidae имеет много
оГйцого с тем, что известно для моллюсков Geomalacus (Sharff. 1907. European ani-
iiiiiIh). Распространение видов Sirnyra Ochs., Eogena Bd. и Oxycesta Hb. сходно с тем, что
обнаруживают выхухоли (Myogale), но степень сохранения ареалов первых полней.
Наконец аналогичные связи обнаруживают и ареалы угрей (Anguilla) — европейского
(Anguilla anguilla L.), чрезвычайно близкого к вестиндскому (Anguilla rostrata
I л Sueur).
и Это характерно для гигрофильных групп, заселяющих лесные территории.
Факт проникновения широко распространенных лесных гигрофильных «таежных»
иломоптов в горные условия Средней Азии в форме хотя и обедненного, но все же лес-
ного ценоза отмечен уже давно. Он оценен еще Вилькинсом в его работах по Средней
Лики и, в частности, при обсуждении выделенной им «туранской» зоогеографической
провинции (A. Wilkins. 1890. Les Cicindeles Touraniennes. Horae Soc. Entom.
Ilona,, 24 : 86—119). Примеров этого типа достаточно среди всех животных и расти-
гпльпых групп. В известном числе они собраны Вилькинсом и особенно• подробно
Моппбиром (1914) в работе «Зоологические участки Туркестанского края и вероятное
происхождение фауны последнего». Уместно отметить, что виды таежных птиц в горах
Сродной Азии дают лишь эндемичные формы, не обособленные до видов.
138
ОБЩАЯ ЧАСТЬ
широко распространенный непарный шелкопряд (Ocneria dispar L.),
Orgyia antiqua L., Olene fascelina L., ряд видов Acronicta Ochs, (rumicis
L., euphorbiae Schiff., psi L., strigosa Schiff.). Существенно отметить, что
в горной зоне турано-африканской подобласти из Orgyidae есть лишь
эндемичные виды, но нет родов. Ряд видов представлен очень близкими
к распространенным в бореевразийской подобласти викарирующими
последним. Так, Acronicta elaeqgni Alph. чрезвычайно близка к Acro-
nicta aceris L., которая не известна из Средней Азии, Subacronicta mega-
cephala Schiff, заменена в горах Средней Азии S. centralis Erschi и
S. grumi Alph. Некоторые из этих видов, например виды Leucoma Steph.,
Orgyia Ochs, и Acronicta Ochs., населяют даже довольно сухие террито-
рии, приникая в условия полупустыни 1 по долинам рек. Таким образом,
фауна Orgyidae турано-африканской подобласти сформирована двумя
основными группами видов: пустынно-степными, очень характерными,
пришедшими с юга и дифференцировавшимися до ранга родов, древними
элементами турано-африканской фауны, отчасти сложного генезиса
(если причислить к ним также роды Leiometopon Stgr., Raphia Hb.), и
более молодыми, мезофильными, горно-лесными элементами, проник-
шими с севера и востока, преимущественно распространенными в горах
в лесной зоне. Эти виды уже довольно полно приспособлены к ксерофиль-
ным условиям турано-африканской подобласти.2
Фауна Orgyidae северных подобластей Палеарктики также своеобразна
и сложна. Для бореевразийской подобласти, точнее для зоны хвойных
лесов и березы Голарктики, могут быть названы несколько эндемичных
родов и ряд видов Orgyidae. Таковы Calocasia Ochs, и Diphthera Ochs.,
связанные первые с лиственными породами (Fagales и отчасти Salic ales
и Rosales), вторые —с хвойными (Pinaceae). Число эндемичных видов очень
1 В других группах насекомых среди лесных элементов горной фауны Средней
Азии выделяются эндемы, которые при этом распространены обычно лишь в северных
горных системах (Тянь-шань, Ала-тау, Тарбагатай) и представлены также на. юге
Сибири (южный Алтай, редко южный Саян). На юго-западе эти эндемы достигают
Памира и даже Кавказа. Примером таких видов могут служить некоторые Agrotinae,
например Еихоа deserticola Kozh., распространенный в Памире, Семиречье и на южном
Алтае (И. К о ж а н ч и к о в. 1937. Фауна СССР, Agrotinae). Аналогичные черты
распространения обнаруживает ряд других видов Agrotinae — Ochropleura pudica
Stgr., О. plumbea Stgr. Среди Pyralidae Ancylosis /lamella Rag. обнаруживает подоб-
ное же распространение (Н. Филипьев, 1930, Ежегодн. Зоол. Муз. Акад.
Наук СССР: 223—226). Дифференциация этих представителей горной фауны Средней
Азии наблюдается лишь за счет ксерофильных элементов турано-африканской фауны,
каковы Agrotinae, Pyralidae. Поэтому подобные эндемы, вероятно, неслучайно отсут-
ствуют в сем. Orgyidae.
2 Участие лесных элементов в формировании эндемичной ксерофильной фауны
Средней Азии известно не только для одних Orgyidae, но также и для жуков. Харак-
терно при этом, что генезис родов тот же, что и для Orgyidae. Наблюдается внедрение
в фауну Средней Азии по хребтам китайско-гималайской фауны (роды Leucoma Steph.,
Orgyia Ochs., Ocneria Hb. все наиболее богато представлены в Индо-Малезии и в ки-
тайско-гималайской фауне). Из Coleoptera, по Семенову-Тян-Шанскому и Боровской
(1935), виды дровосеков Apatophyses Cherv., обитающие солонцеватые, довольно влаж-
ные еще стации полупустынь (с наличием тополей), в фауне Средней Азии являются
дериватом горных гигрофильных Protapatophyses Sem. et Ваг., обитающих субтропи-
ческие леса западных Гималаев, Кашмира и юго-западного Китая (см. Энтом. обозр.,
24 (1—4) : 59—90). Аналогично присутствие в Средней Азии ряда видов уховерток,
ограниченных также почти исключительно горной зоной (А. Семенов-Тян-
Шанский. 1935. Изв. Акад. Наук СССР: 825—830; Бей-Биенко. 1936.
Фауна СССР, Кожистокрылые). Для элементов китайско-гималайской и индомалай-
ской фауны в Средней Азии можно обнаружить все переходы от гигрофильных усло-
вий — лесов Средней Азии, где они наиболее обильны, — к пустыням, где налицо
лишь немногие, наиболее специализованные виды.
РАСПРОСТРАНЕНИЕ
139
велико, это прежде всего многие Acronicta Ochs.—auricoma Schiff., meny-
anthidis View., aceris L., cuspis Hb., psi L., tridens Schiff., leporina L.,
senica Ev. и ряд других видов этого рода, затем Subacronicta megacephala
Schiff., многие виды Orgyinae (Ocneria monacha L., Euproctis similis Fues-
sly, Dasychira abietis Schiff., Olene fascelina L., Orgyia gonostigma E., Leu-
coma salicis L., L. Candida Stgr.). Ряд видов, в том числе некоторые
Orgyidae бореевразийского типа распространены циркумполярно по зоне
хвойных лесов. Таково распространение Acronicta leporina L., встреча-
ющейся от берегов Атлантического океана до Тихого в Евразии и всюду
по лесной зоне Северной Америки. Зона хвойных лесов и березы Северной
Америки характеризуется тем не менее большим числом эндемичных ви-
дов Orgyidae и несколькими эндемичными родами (Agriopodes Hamps.),
почему естественно должна быть выделена в особую (канадскую) под-
область.
Фауна Orgyidae арктической подобласти Палеарктики, будучи крайне
бедной, представляется, пожалуй, даже более обособленной от прочих,
чем фауна бореевразийская. Она характеризуется наличием почти энде-
мичного рода Gynaephora Hb., по существу аркто-альпийского, так как
ряд видов (G. pumila Stgr., G. grumi Alph., G. selenophora Stgr.) заселяет
горные тундры Центральной Азии. Один вид, Gynaephora selenitica
Schiff., распространен в лесной зоне, в бореевразийской подобласти
(преимущественно в Прибалтике). Тем не менее для фауны Арктики
виды Gynaephora Hb. очень характерны. Эндемичны для этой фауны
Gynaephora rossii Curt., G. lugens Kozh., G. groenlandica Hom. Все эти
виды очень обычны в Арктике, являясь существенным элементом ланд-
шафта. Из других видов Orgyidae, свойственных бореевразийской под-
области, в арктическую проникают очень немногие. Наиболее далеко,
вплоть до крайних широт Арктики (о. Врангеля), распространена Olene
fascelina L. — вид, эврибионтный и заселяющий, кроме лесной области
Евразии, также лесостепные, пограничные с полупустынями северной
Монголии (Танну-ола, Хангай, Кентей) и Средней Азии (Тянь-шань).
Этот вид связан с ивоцветными (Salicales). Из Acronictinae же лишь Acro-
nicta menyanthidis View, и A. euphorbiae Schiff, отмечены для европейской
тундры (Лапландия), хотя оба вида связаны с низкорослыми кустарни-
ками и травянистыми растениями (Rosales, Ericaceae). В азиатскую тундру
кроме Acronicta menyanthidis View, возможно проникновение Acronicta
auricoma Schiff.; вид этот собран был мною в зоне криволесья Саян. Воз-
можно проникновение в Арктику также эврибионтного Acronicta rumi-
cis L. В Сибири в горно-арктической тундре Станового хребта распро-
странена своеобразная Acronicta senica Ev. (=pfilzenmayeri Herz), изве-
стная также из Забайкалья, западной Сибири и с южного Урала. Факт
слабого проникновения видов Orgyidae в Арктику является замечатель-
ным, так как кормовые растения их в виде разнообразных ив, вересковых,
розоцветных и берез проникают далеко в тундру и идут до ее крайних
пределов.
Распространение Orgyidae при учете филогенетических связей разных
групп позволяет наметить пути и время формирования фауны этого се-
мейства для разных областей суши, в частности для СССР. Могут быть
намечены следующие важные связи и разъединения в пределах ареала
семейства:
1) филогенетическая близость западноафриканской и центрально-
африканской фаун к индомалайской в пределах общих родов; 2) бблыпая
(чем первых двух) обособленность неотропической фауны, причем бли-
140
ОБЩАЯ ЧАСТЬ
зость к индо малайской; 3) несамостоятельность австралийской фауны
и подчиненность ее индомалайской; 4) бедность, но высокое своеобразие
фауны центральной Америки и северной части Южной Америки (Боли-
вия); 5) значительная обособленность фауны Голарктики (до ранга под-
семейств) от южных областей, но несомненное ее развитие за счет фауны
последних; 6) наличие трех источников для формирования фауны Голар-
ктически— африканского, индомалайского и неотропического; 7) наи-
большее богатство в Голарктике фауны маньчжурской подобласти и тес-
ная связь ее с индомалайской фауной; 8) наличие связи маньчжурской
фауны (в пределах близких родов) с неотропической; 9) наиболее замет-
ное влияние маньчжурской фауны на формирование фауны хвойных
лесов Голарктики; связь ее с аллеганской и калифорнийской подобла-
стями; 10) резкое различие генезиса ксерофильной и гигрофильной фаун
турано-африканской подобласти Палеарктики; связь первой с африкан-
ской, второй— с индомалайской; 11) отсутствие близости между фауной
широколиственных лесов Европы с богатой их фауной Северной Америки,
но общая связь той и другой с индомалайской; 12) большая близость
полярных видов восточноамериканской Арктики к видам европейской
Арктики, чем к видам азиатской и большая близость последних к аркти-
ческим видам западной Америки; 13) тесная связь фауны Orgyidae хвой-
ных лесов и березы Го’ларктики с неотропической фауной; 14) связь
средиземноморской фауны с неотропической (в пределах близких родов)
и связь ее с индомалайской фауной общими родами.
Приведенные факты прежде всего свидетельствуют о значительной
древности основных типов сем. Orgyidae. Время формирования подсе-
мейств и родов возможно относить не ранее, чем к мезозою (юра, мел).
К этим же выводам о возрасте отдельных представителей изучаемого
семейства пришли Караджа (Caradja, 1931) и Варнеке (Warnecke, 1938).
Первый подверг специальному обсуждению распространение Laelia
coenosa Hb. и Orgyia ericae Germ, в Румынии и некоторых других видов
этого рода в южной и средней Европе. Он считает необходимым относить
существование этих видов в Европе еще к дотретичному времени. Вар-
неке (1938), занимавшийся специально ареалом и стациальным распре-
делением Laelia coenosa Hb. в Европе, пришел совершенно независимо
от Караджа к тому же мнению.
Изучение обилия видов современной фауны Orgyidae и областей сохра-
нения анцестральных форм позволяет наметить возможные территории
формирования обоих подсемейств, не касаясь пока истории формирования
отдельных фаун. Было бы трудно сказать определенно, которое из под-
семейств Orgyidae является более древним. Нет сомнения, что время
формирования подсем. Acronictinae, как и Orgyinae, нужно относить
к мезозою, вероятно к юре. Территории и условия формирования обоих
подсемейств существенно различны. Территория формирования подсем.
Orgyinae в основе охватывает малайско-африканскую сушу с некоторыми
прилежащими областями центральной и Южной Америки (Боливия,
Венецуэла, Гвиана, Вест-Индия). На этих территориях (рис. 67) доныне
сохранились архаичные типы этого подсемейства, причем более обильно
в малайско-африканском секторе. Здесь и ныне Orgyinae достигают наи-
большего разнообразия видов. Отсутствие архаичных типов подсем.
Orgyinae на большей части территории Южной Америки, в Австралии,
Северной Америке и в умеренной зоне Евразии говорит против возмож-
ности его формирования здесь. Жизненные циклы видов Orgyidae, столь
однообразные у громадного числа тропических видов, говорят за воз-
142
ОБЩАЯ ЧАСТЬ
мощность формирования этого подсемейства в условиях теплого, влаж-
ного и устойчивого в течение круглого года климата, подобного современ-
ному климату гилей. Какие-то условия, может быть суровость климата
Южной Америки в конце мезозоя, помешали обильному развитию видов
Orgyinae на этом материке, где по существу сохранились лишь реликты,
тогда как малайско-африканская суша характеризовалась наличием
пригодных для развития этой группы условий с конца мезозоя в течение
всего третичного времени и доныне.
Область формирования подсем. Acronictinae ограничивается почти
нацело юго-восточной Азией и севером Южной Америки (рис. 68). В Аф-
рике нет не только анцестральных типов этого подсемейства, но почти нет
его представителей вообще. Мадагаскар занимает промежуточное поло-
жение, так как на этом острове есть представитель архаичного рода
Thalatha Wlk. (malagassica Hamps.), типичного для индомалайской фауны.
С Мадагаскара же описан один вид рода Craniophora Snejl. Есть все
основания считать, что дифференциация подсем. Acronictinae происхо-
дила от тропических типов, может быть по облику имагинальной фазы
близких к видам Orgyinae. Дифференциация видов Acronictinae рисуется
в связи с адаптацией к умеренному климату, характеризующемуся се-
зонным распределением погодного режима. Архаичные типы этого под-
семейства сохранились ныне также в тропиках, например виды Trisu-
loides Btl., Мота Hb., Belciana Stgr., Thalatha Wlk., Cannes Wlk. виндо-
малайской фауне и виды Lichnoptera Н. S. и Gaujonia Dogn. — в нео-
тропической, причем в обоих случаях в горных условиях, на высотах
порядка 1500—2500 метров над уровнем моря. Можно представить себе
формирование Acronictinae на территориях с умеренным климатом, близ-
ким к климату гиЬей.1 Может быть, наиболее правильно рассматривать
возникновение Acronictinae в связи с орогенетическими процессами.
Широкое распространение видов этого подсемейства на территории се-
верных и умеренных широт Голарктики, вероятно, есть уже вторичное
явление — заселение благоприятных обширных территорий раз возник-
шими более эврибионтными типами семейства. За это говорит отсутствие
в Голарктике примитивных эндемичных типов Acronictinae, которые
обильны в горных условиях тропиков. Тем не менее и в Голарктике,
но опять в горных условиях,' обнаружен, ряд архаичных типов этого
подсемейства на юго-востоке Азии (Гималаи, Китай, южное Приморье
таковы виды Panthauma Stgr., Enromoia Stgr., Thyatirides Kozh. Таким,
образом территория формирования этого подсемейства оказывается сдви-
нутой несколько севернее, по сравнению с тем, что обнаруживают виды
подсем. Orgyinae (рис. 68). Кроме гор тропиков, территория формирова-
ния видов (родов) подсем. Acronictinae охватывает площадь запад-
ного Китая, южного Приморья, Японских островов, Кореи, Тайвана,
южных отрогов североамериканских Кордильер, Аллеганских гор и
отчасти площадь Бразильского нагорья (Боливия).
Время и территории формирования обоих подсемейств находят обос-
нование и в их кормовых связах. Виды Orgyinae являются потребителями
1 Филогенетические отношения равнинных северных видов Acronictinae к высо-
когорным (архаичным) видам этого подсемейства имеют аналогию среди других групп
животных. Так, высокогорный кавказский тетерев (Lirarus mlocosievriczi Тнгс.), рас-
пространенный локально на Кавказе, по многим признакам должен быть признан
первичным (линька, строение рулевых, по сравнению с широко распространенным
косачом (Lirurus tetrix L.). Подобные же отношения обнаруживает высокогорная
Prometheomys Sat. к равнинным видам полевок (К. С а т у н и н. 1910. Некоторые
соображения о происхождении фауны Кавказского края. Тифлис.).
Рис 68. Распространение видов подсем. Acronictinae. Черным показаны территории обилия анцестральных форм',
сплошная линия ограничивает территорию современного обилия видов Acronictinae точечный пунктир — общее
распространение Acronictinae.
/// ОБЩАЯ ЧАСТЬ
многочисленных и разнообразных лиственных древесных пород, типич-
ных для тропиков, преимущественно семейств Могасеае, Lauraceae,
Euphorbiaceae, Leguminosae, отчасти Fagaceae. Растения эти типичны
для ровного и мягкого климата. Acronictinae специализованы на питании
отчасти теми же группами растений (Могасеае, Fagaceae), именно ви-
дами их, заселяющими' территории с переменным климатом, но также,
а в некоторых случаях и главным образом на питании типичными для
Голарктики, близкими к Fagaceae, видами Betulaceae, Salicaceae и Ro-
saceae. Пищевые связи частного порядка для Acronictinae (с Myricaceae,
Juglandaceae, Ericaceae, Coniferae) дополняют общую картину. Дифферен-
циация семейств древесных покрытосемянных растений датируется палео-
ботаниками второй половиной мезозоя (юра, мел), а формирование
современных их широко распространенных представителей (берез, видов
ольхи) — началом третичного времени (эоцен, олигоцен), как о том сви-
детельствуют данные Палибина, Краснова, Сьюорда, Кеппена и Веге-
нера. Не случайна, вероятно, связь видов Arsilonche Ld., Raphia Hb.,
Мота Hb., Beleiana Stgr. и других архаичных Acronictinae, имевших
прерывистое распространение, с этими группами покрытосемянных
(Salicaceae, Rosaceae).1
Деление сем. Orgyidae на два подсемейства и дифференциация их на
основные родовые группировки может быть связана с моментом дифферен-
циации низших типов покрытосемянных (древесных) растений. Таким
образом для древних групп, сохраняющих архаичные черты и связан-
ных с Salicaceae, Fagaceae, Rosaceae, есть все основания относитьпериод
дифференциации к мезозою.
Изучение сложения и распространения флор Голарктики в прошлом
показало наличие двух их основных типов. Данные Криштофовича, Эндо,
Вульфа 2 показывают существование в Голарктике двух типов
флоры: одного — тропического, сложившегося на территории южной Азии,
Африки и Средиземноморья, по Криштофовичу — «полтавского», пред-
ставленного в настоящее время реликтами средиземноморской (вероятно
горной) флоры и богатой тропической растительностью южной и юго-
восточной Азии; другого — «тургайского» (по Криштофовичу), сформи-
рованного растительностью с сезонно опадающей листвой, характеризую-
щегося, в частности, широколиственными древесными породами (Fagales,
Salicales, Rosales). Этот последний тип сложился на территории
западного Китая и Маньчжурии (по Криштофовичу — «ангарского»
материка) и характеризует в основе флору Голарктики. Ныне он доми-
нирует во флоре гор западного Китая, а его дериватом является гигро-
фильная флора Евразии. Время дифференциации и формирования этих
двух флор датируется меловым периодом и даже концом юрского. В тре-
тичном периоде признается существование двух флористических обла-
стей: одной «полтавской», охватывавшей часть Африки, юг Европы
(берега Тетиса) и южную Азию, и другой, более обширной, «тургайской»,
1 Связь всего сем. Orgyidae с древесными растениями имеет глубокие истори-
ческие корни. Есть основания считать, что в конце мезозоя и в начале третичного
времени (эоцен, олигоцен) гигрофильные ценозы были бедны травянистыми расте-
ниями. За это говорит факт существования в начале третичного времени большого
числа листоядных млекопитающих, например Dinoceros, Diceraterium и многих других,
подобно современным слонам, связанных с древесной, но не травянистой раститель-
ностью. (М. Мензбир. 1934. Очерки истории фауны Европейской части СССР.
ГИЗ : 1—223).
2 Имеется в виду обобщающая работа Вульфа (1939. Очерк истории флоры Восточ-
ной Азии. Изв. Гос. Геогр. общ., 10 : 1447).
РАСПРОСТРАНЕНИЕ
145
простиравшейся от восточной Азии через всю Сибирь до полярной (ныне)
Европы и Азии. Из приведенного не трудно видеть, что в распространении
Orgyidae в виде их подсемейств мы имеем соответствие тому, что описано
палеоботаниками.
Утверждение о былом более обширном распространении в Голарк-
тике видов Acronictinae, связанных с широколиственными лесами, так же
как и геологическое время их происхождения (формирования), имеет
доказательства в условиях распространения и генезисе других групп
насекомых, связанных с этими лесами в умеренных широтах. Особенно
полную аналогию тому, что описано выше, обнаруживают тли (Aphi-
dodea). Будучи специализованы на питании древесными растениями
(Rutaceae, Могасеае, Fagales, Rosales, Salicales), тли точно так же обна-
руживают обилие видов в областях наиболее полного сохранения ныне
(в голоцене) широколиственных древесных пород. Но для тлей возмож-
ность анолоциклии дает условия для сохранения их на территориях,
где основные кормовые растения уже исчезли. Территории, занятые
этими анолоцикличными формами, могут быть, конечно, вторично засе-
лены ими, но в ряде случаев присутствие таких форм указывает на нали-
чие здесь в прошлом их основных кормовых растений. В этом случае,
следовательно, неполноциклые формы являются реликтами когда-то
существовавших здесь более полных дендрофильных ценозов. Таково
распространение Geoica Hart., связанных с фисташкой (Pistacea), а в виде
неполноциклых форм на корнях злаков, распространенных до Британских
островов, Финляндии и средней Сибири. Это же известно для Proci-
philus Koch., связанных с видами Fraxinus, Eriosoma Leech., связанных
с Ulmaceae, и многих других. Время формирования указанных родов
тлей должно быть отнесено к концу мезозоя (юра, мел), а видов — к на-
чалу третичного времени.1
Следует специально отметить в отношении Orgyidae, а также и других
дендрофильных насекомых (например многих жуков), что тип фауны,
который мог бы быть назван «тургайским», т. е. тип форм, приспособлен-
ных к сезонным, циклическим сменам климата и пищевого режима, оби-
тающих в лесах с опадающей листвой, сформировался не только на тер-
ритории юго-восточной Азии, но в начальные периоды формирования
(вероятно в юре и меловом) складывался также на территории севера
Южной Америки (Боливия) и центральной Америки. Особенно в этом
отношении большую роль сыграли горные территории центральной
Америки, Боливии и Индо-Малезии.2 На этих территориях и ныне оби-
1 А. Мо р двилко. 1901. Естественные условия существования тлей и стоя-
щие в связи с ними некоторые особенности их жизненного цикла и строения. Тр.
Русск. Энтомолог, общ., 33 : 162—302; А. Мордвилко. 1925. Экология циклов
и происхождения гетереции (миграции) у тлей. Защ. раст., 2 (7) : 1—9; A. Mord-
v i 1 к о. 1934. On the evolution of Aphids. Arch. Naturgesch. (N. F.), 3 (1) : 1—60;
A. Mordvilko. 1935. Die Blattlause mit unvollstandiegm Generationszyklus
mid ihre Entstehung. Ergebn. Zool., VIII : 35—328; F. R о 11 e s and Th. F r i-
н о n. 103. The plant lice, or Aphidae, of Illinois. Div. Nat. Hist. Surv., 19 : 121—447.
2 Давнюю связь территории юго-восточной Азии и центральной Америки и важ-
ную роль этой области в формировании фауны умеренных широт Голарктики можно
подтвердить, помимо приведенных фактов по Acronictinae и Orgyinae, также замеча-
тельным ареалом рода Juglans L. Растения эти, как отмечалось, принадлежат к группе
основных кормовых растений видов Orgyidae. Ареал рода Juglans L. (см. Н. И. Куз-
нецов, 1925. Ботанико-географический атлас. Ореховые— Juglandaceae. Вып. 4;
И. Н е к р а с о в а. 1928. Род Juglans L. в Туркестане. Труды по прикл. бот.
и сел., 18 (2) : 303—330) поразительно соответствует территориально тому, что отме-
•чопо для архаичных типов Acronictinae. Аналогичные связи в отдаленном прошлом.
Ю Фауна СССР, И. В. Кожанчиков
146
ОБЩАЯ ЧАСТЬ
тают наиболее архаичные типы Acronictinae (МотаТАЪ., Belciana Swinh.,..
Thalatha Wlk., Trisuloides Wlk., Lichnoptera H. S., Gaujonia Dogn.).
В связи с указанным территорию юго-восточной Азии (горы Китая,..
Гималаи) и гор Северной Америки следует рассматривать как вторич-
ную, но весьма важную область, .сыгравшую весьма существенную роль-
в развитии фауны Orgyidae, большую, чем горы тропиков, давших лишь-
первичные типы Acronictinae. В то же время территория юго-восточной!
Азии явилась также убежищем для этой фауны в период плейстоценовых
оледенений.
Доминирующее положение Acronictinae в Голарктике, по сравнению
с Orgyinae, повидимому, связано лишь отчасти с былым, более широким
распространением этих элементов «тургайского» ландшафта (как об этом
свидетельствуют реликтовые нахождения в ряде мест Голарктики видов-
Acronictinae), но, вероятно, главным образом с расселением этого подсе-
мейства по территории Голарктики в конце третичного времени (миоцен,,
плиоцен) как видов, более приспособленных к условиям переменного-
климата, по сравнению с видами Orgyinae. Из Orgyinae также возник ряд.
типов, приспособленных к жизни в Голарктике, причем из родов наиболее-
обширно распространенных в тропиках. Приспособленными к жизни
в широтах с переменным климатом ныне являются, например, виды
Dasychira Ochs., Olene Hb., Laelia Steph., Orgyia Ochs, и др., составляю-
щие все же ничтожный процент от общего числа видов этих родов, известных
из тропиков. Требуется подчеркнуть, что приспособление видов Orgyinae
к жизни в условиях климата умеренных широт происходило на тех же-
пограничных с тропиками территориях, как и видов Acronictinae, т. е..
прежде всего в горах западного Китая и горных областях Северной
Америки (Кордильеры, Аллеганы), но в меньшем масштабе, чем первых..
Генезис фауны Orgyidae арктической подобласти Голарктики имеет
известную связь с тем, что дает анализ фауны лесов. Фауна эта пред-
ставлена немногими эварктами из рода Gynaephora Hb.1 (рис. 69) и одним
видом Olene Hb. {fascelina L.). Эваркты из рода Gynaephora Hb. тесно
связаны, с горно-альпийскими видами этого рода, заселяющими Сред-
нюю Азию {G. pumila Stgr., G. selenophora Stgr.), и имеют родственные
формы в высокогорье Гималаев (Lachana ladakensis Moore). Род Gynae-
phora Hb. очень близок к роду Olene Hb., почти нацело тропическому.
Из Olene Hb. дифференцировалось в общем лишь небольшое число энде-
мичных представителей в Голарктике в областях распространения широ-
колиственных лесов юго-восточной Азии (западного Китая, Маньчжу-
рии, южного Приморья) и Северной Америки (Калифорния, Аллеганы).
Немногие виды этого рода эврибионтны, заселяют тропики и широко-
лиственные леса, например Olene mendosaWh., широко распространенный:
в тропиках, на Тайване, в континентальном Китае и Японии. Таким образом,
арктическая фауна Orgyidae генетически непосредственно связана с тропи-
ческой фауной этого семейства, причем фауной гигрофильной (гилей).
Представители сухих тропиков из 'Orgyidae не дали производных, засе-
ливших Арктику, хотя в фауне Голарктики в целом эти формы представ-
_________________ •
между юго-восточной Азией и центральной Америкой доказывает также и распро-
странение замечательных Atyidae (Decapoda) из группы Xiphocaris, в частности видов-
Xiphocardinella.Один из видов этой группы известен из пещер Закавказья (Кутаиси) —
Xiphocardinella kutaissiana Sadowsk., тогда как другие локализованы в индомалай-
ской фауне, в Австралии и Вест-Индии (А. Садовский. Xiphocardinella kutais-
siana, nov. gen. etsp. (Atyidae) из подземной пещеры под Кутаисом. Отд. оттиск: 1—12).
1 Полнее по распространению видов рода Gynaephora Hb. см. КоЖанчикоа
(1.948). . .
148
ОБЩАЯ ЧАСТЬ
лены. Можно было бы сказать, что фауна Orgyidae Арктики является
осколком «полтавского», по Криштофовичу, ландшафта третичного вре-
мени. Отсутствие в арктической фауне представителей Acronictinae
(видов «тургайского» ландшафта) должно рассматриваться не более как
частный случай. Отсутствие их в Арктике может быть связано с дендро-
фил ьным образом жизни их, с высокой пищевой специализацией (часто
олигофагией) и в связи с этим с очень высокой специализацией циклов
развития. Среди Agrotinae виды Triphaenini по типу распространения
совершенно соответствуют Acronictinae, но они как раз являются основ-
ным контингентом арктической фауны Agrotinae. Различия биоэкологии
видов Triphaenini из Agrotinae'TX. Acronictinae из Orgyidae заключаются
в отношении к субстрату й термогигрорежиму, а также к кормовым ра-
стениям. В то время как Acronictinae в основном являются видами ден-
дрофильными, лазящими по растениям, не способными к жизни на по-
верхности почвы, часто специфично приспособленными к циклам жизни
их кормовых растений, нередко моно- или олигофагами, виды Triphae-
nini из Agrotinae — полифаги, живут на поверхности почвы и в поверх-
ностном ее слое и не связаны тесно с растениями. Эти особенности биоло-
гии Agrotinae создали условия для проникновения их в Арктику. Не слу-
чайно, повидимому, что из Acronictinae в горную тундру проникают виды
Pharetra Hb. {menyanthidis View., auricoma Schiff., euphorbiae Schiff.),
гусеницы которых связаны не только с деревьями и кустарниками, но
также с травянистыми растениями и могут жить на поверхности почвы.
Точно так же отсутствие в Арктике пустынно-степных Orgyidae не
является ни в коей мере общим фактом, но имеет специфику в биоэко-
логии семейства. Из Agrotinae виды рода Еихоа Hb., в целом заселя-
ющие зону полупустынь Голарктики, распространены также в аркти-
ческой подобласти {Е. islandica Stgr., Е. drewseni Stgr., Е. dissona
Moschl.), где представлены даже эндемичными видами.
Что касается территории формирования арктических Orgyidae, то
не случайно, вероятно, присутствие в роде Gynaephora Hb., на ряду
с1 чисто арктическими представителями {G. rossii Curt., G. groenlandica
Hom.) видов, распространенных в Арктике и в высокогорьях хребтов
восточной Азии (G. lugens Kozh.), эндемов горной фауны южной Сибири
{G. relicta В. Н.) и эндемов высокогорий южного Алтая, Ала-тау, Алая,
Памира и восточного Тибета (G. selenophora Stgr., G. pumila Stgr.,
G. alpherakii Gr. Gr.). Горные эндемы обнаруживают наибольшее сход-
ство с видами Olene Hb., характеризуясь при этом архаичными чертами
строения, по сравнению с арктическими представителями рода, более
специализованными. Есть основания считать, что территорией форми-
рования первичных типов рода Gynaephora Hb. явились высокогорные
области Тибета, пограничные с субтропическими территориями запад-
ного Китая (Юньнань). Формирование (рис. 69, 70, 71) этих высокогор-
ных тибетских и центральноазиатских архаичных типов Gynaephora Hb.
{alpherakii Gr. Gr., selenophora Stgr., pumila Stgr.) могло осуществиться
за счет форм типа Olene Hb., распространенных в горах западного Ки-
тая. Время формирования их вероятнее всего связывать с моментом под-
нятия Гималаев и Сибирского плато, т. е. с третичным временем (олиго-
цен, миоцен, плиоцен), синхронным периоду формирования альпийских
ценозов этих горных систем. Время формирования эндемов, распро-
страненных в Арктике, может быть тем же, учитывая, что эндемы эти
сосредоточены в- ее азиатско-американском секторе {G. rossii Curt.,
G. groenlandica Hom.), где в миоцене также было значительное охлажде-
Рис. 70. Азиатско-американская и европейско-американская (заштриховано) области формирования арктических
Orgyidae территории распространения анцестральных форм (закрашено черным).
150
ОБЩАЯ ЧАСТЬ
ние. Для некоторых аркто-альпийских видов Gynaephora Hb. (lugens
Kozh.), как раз характеризующихся многими вторичными структурами,
вероятно более позднее происхождение, совпадающее, может быть,'с пли-
оценом или постплиоценом, т. е. временем расселения альпийских цено-
зов к северу и арктических к югу. Для Orgyidae, следовательно, налицо
теснейшая генетическая связь альпийской восточноазиатской фауны
с арктической циркумполярной. Для другой, ближе изученной мною
группы, видов Agrotinae, эта связь также несомненна, с тем лишь добав-
лением, что генетические связи с арктической фауной обнаруживают
также и эндемы европейских Альп и альпийской зоны североамерикан-
ских горных систем. В отношении последних, в частности Белых Гор,
следует отметить присутствие на них арктических (Dyar, 1894), но' не
эндемичных видов Gynaephora Hb. (rossii Curt., groenlandica Hom.)
Фаунистические группировки Orgyidae не исчерпываются наличием
тропических (индомалайского и неотропического), китайско-гималайского
й аркто-альпийского типов (рис. 67, 68, 70, 71). Есть необходимость выде-
лить еще ряд других и прежде всего калифорнийско-японский тип фауны
Orgyidae, представленный обоими подсемействами. Могут быть названы
как Orgyinae (Ivela Swinh.), так и Acronictinae (Calocasia Ochs., Daseo-
chaeta Warr., Diphthera Ochs.), представленные тождественными родами
или викарирующими типами в восточноазиатской и западноамериканской
фаунах. Западноамериканская фауна более богата этими группами, фило-
генетически связанными с неотропическими. Этот тип фауны Orgyidae
является американским аналогом только что разобранной азиатской
китайско-гималайской лесной фауны. Он дал ряд широко распространен-,
ных групп Orgyidae лесной зоны Голарктики. Формирование этих свя-
зей следует относить к концу мезозоя, а развитие широкораспространен-
ных типов — к началу третичного времени. Несомненна связь этого типа
фауны с горно-лесным типом ее, формировавшимся на территории Бра-
зильского нагорья.
С неотропической фауной Orgyidae, но уже исключительно видов
Acronictinae, обнаруживает связь и европейская, в частности средизем-
номорская, их фауна. Этот тип фауны мог бы быть назван лигурийским
(рис. 72), но я предпочитаю для него более полный термин — «бразиль-
ско-лигурийский».
Я упоминал уже, что роды Episema Ochs., Oxycesta Hb., Eogena Bd.
и Leiometopon Stgr. эндемичны для фауны Средиземноморья, южной Ев-
ропы и Средней Азии. Наряду с этим они филогенетически близки к неко-
торым неотропическим родам, в частности к Lichnoptera Н. S. и Gaujonia
Dogn. Близки к упомянутым евразийским родам и представленные лишь
различными видами в Евразии и в центральной и Южной Америке Raphia
Hb. Отдельные элементы этой фауны в виде Arsilonche Ld. и Simyra Ochs,
обнаруживают остаточное, пятнистое распространение в южной Сибири,
северной Монголии, в Забайкалье, на Дальнем Востоке, в восточном
Тибете и северном Китае. Замечательно полное отсутствие этих пред-
ставителей или близких к ним видов в юго-западном Китае, а также
в индомалайской и африканской фаунах.
Более молодым типом фауны Orgyidae, характерным для Средней Азии
и Средиземноморья, формирование которого все же необходимо относить
к началу третичного времени (эоцен, олиго цен), следует считать западно-
азиатский тип. Он представлен видами обоих подсемейств, причем более
богато видами Orgyinae. Распространение этого типа фауны видно на
примере ареалов видов Ocneria Hb., Parocneria Dyar и некоторых Ей-
Рис. 71. Китайско-гималайский тип фауны Orgyidae. Черным закрашена территория обилия анцестральных форм;
ареал непарника; 2 — видов Olene Hb. (в северном полушарии).
152
ОБЩАЯ ЧАСТЬ
proctis Hb. (рис. 73). Главная масса видов рода Ocneria Hb. заселяет индо-
африканскую сушу, причем они особенно обильны на территории Индо-
Малезии. Значительное число их известно с территорий юго-востока
Азии, занятых лесами с опадающей листвой. В Голарктике виды Ocne-
ria Hb. представлены немногими эндемами, известными из гор Азии (Гима-
лаи), например Ocneria grisescens Stgr., О. maturaEiA. Широко распростра-
ненными видами в Палеарктике являются лишь два — Ocneria dispar L. и О.
monacha L. В Средиземноморье из видов Ocneria Hb. известен лишь один,
эндемичный О. гидеаНЬ. Может быть не случайно, что именно этот вид обна-
руживает наибольшее число признаков, сближающих его с родом Parocneria
Dyar (близким к Ocneria Hb.). Виды Parocneria Dyar локализованы почти
все на территории восточного Средиземноморья, именно в горах Малой:
Азии, Сирии, Кавказа и Ирана (Р. lapidicola Н. S., komarovi Chr., grisei-
pennis, 'sp. n., therebynthina Stgr., raddei Chr., signatoria Chr., amabilis?
Chr.). Лишь немногие виды известны из западного Средиземноморья
(Р. atlantica Stgr.) и из южной Европы (Р. detrita Hb.). Только один
вид — Parocneria terebynthi Err. — хотя и спорадично, но распространен:
на значительной территории, встречаясь как в южной Европе, так и
в ряде мест Передней и Средней Азии, а также известен с некоторых
территорий восточной Азии (западный Китай, южное Приморье). Сходное-
распространение обнаруживают также виды Orgyia Ochs, группы О. аи-
rolimbata Н. S., которые богато представлены в китайской фауне ((?. im-
maculata Gaede, parallela Gaede), один вид распространен в горах Средней
Азии (О. о chr о limb ata Stgr.), другой в альпийской зоне Кавказа (O.fla-
volimbata Stgr.), а третий ограничен Пиренеями (О. aurolimbataEi. S.).
Наконец этого же- типа распространение обнаруживают виды Euproctis-
Hb- (bipunctapex Hamps., chibiana Mis., karghalica Moore, susannaSbgv'.).
Leucoma Steph, (jlavosulphurea Ersch., sartus Ersch.) и Acronicta Ochs,
группы aceris L. В этих родах на территории восточного Средиземноморья
есть эндемичные виды, близкие к восточноазиатским.
Все последние роды {Euproctis Hb., Leucoma Steph.) являются гигро-
фильными и распространены так же или сходно с тем, что дает род.
Ocneria Hb., но они не дифференцировали в восточном Средиземноморье-
родов или подродов. Напротив, некоторые представители востбчноазиат-
ских родов проникают в Средиземноморье и южную Европу неизмен-
ными, как, например, Craniophora ligustri L., или дают здесь лишь под-
виды, как то известно для Daseochaeta alpium Osb., Dasychira pudibunda L.
Это характеризует недавнее проникновение южноазиатских форм в Сре-
диземноморье, а наличие в этих же родах (Craniophora Snell.) эндемичных
ксерофильных сильно дифференцированных видов в Средиземноморье-
говорит за длительный период формирования западноазиатского типа
фауны Orgyidae. Лишь этим можно объяснить наличие в нем таких древ-
них типов, как Parocneria Dyar, вместе с слабо дифференцированными
видами и подвидами. Независимо от времени дифференциации все пред-
ставители Orgyidae западноазиатского типа фауны являются дендро-
фйльными видами, связанными с теми же основными кормовыми расте-
ниями, как и восточноазиатские (Fagales, Rosales, Salicales). Все эти
виды обнаруживают очень определенные черты адаптации к жизни в усло-
виях пустынно-степного ландшафта.
Связь ксерофильных элементов средиземноморской фауны, в частности,
восточного Средиземноморья, с фауной восточного Тибета и западного-
Китая отмечал еще Караджа. Он полагает, что формирование элементен
этого фаунистического типа осуществлялось в раннетретичное время, а тер-
Рис. 72, Бразильско-лигурийский тип фауны Orgyidae (Acronictinae). Черным показано распространение
арцестральных форм, штрихом — отдельные их нахождения,
.154
ОБЩАЯ ЧАСТЬ
риториально связывает формирование его с областью Тетиса. Эта точка зре-
ния может быть всецело поддержана и дополнена еще многими приме-,
рами не только из распространения чешуекрылых, но и из других групп
насекомых, а также из распространения позвоночных. Ранее (1935,
1937) мною было описано распространение видов Chersotis Bd. из Agrotinae
(Noctuidae), например видов: С. hahni Chr., C.multifida Ld., C. calorica
Crt., С. bonza Piingl., C. capnistis Stgr., C. senna Piingl., C. agalma Piingl.
и многих других.
Филогенетические связи ксерофильных элементов среднеазиатской
и североафриканской полупустынной и пустынной фауны Orgyidae иные.
На прилагаемой карте (рис. 74) я привожу распространение видов Ог-
gyia Ochs, группы dubia Tausch. Распространение этого типа показывает
отчетливые связи и у ксерофильных Orgyidae с пустынной фауной южной
Африки и Австралии. Ранее (1937, 1941) для видов Agrotinae мною были
показаны филогенетические связи пустынных турано-африканскйх ви-
дов этой группы чешуекрылых с Южной Африкой, восточной Африкой,
-Австралией и восточным Индостаном. Для Orgyidae филогенетические
связи пустынно-степных элементов турано-африканской фауны установ-
лены (Orgyia Ochs., Albarracina Stgr., Penthophera Germ.) лишь в пределах
Старого Света и Австралии. Это говорит о более позднем формирова-
нии пустынно-степной фауны Orgyidae, по сравнению с Agrotinae (Noctui-
dae), видами Pyralidae (Lepidoptera), Acridiidae (Orthoptera), Myrrneleon-
tidae (Neuroptera), обильно представленными не только в Старом Свете,
но и в Южной Америке. Замечательно, что в то время как пустынно-
степные ^виды Agrotinae или Pyralidae связаны питанием с Ghenopodia-
сеае, пустынно-степные виды Orgyidae специализованы на Leguminosae
и Gramineae. Анцестральные типы пустынно-степных видов Orgyidae,
в частности видов Orgyia Ochs., представлены в фауне Индо-Малезии. Фор-
мирование ксерофильной фауны видов Orgyidae можно представить лишь
на осколках «гондваны» в эоценовое или олигоценовое время, на террито-
риях юго-восточной Африки, южной Азии и Австралии. В связь с этим
относительно молодым возрастом ксерофильных элементов изучаемого
семейства может быть поставлено отсутствие в нем обособленных родов,
что типично для Agrotinae, Acridiidae и ряда других пустынных групп
насекомых.
Виды Orgyidae турано-африканской фауны, распространенные в пу-
стынях и полупустынях, представляют большой зоогеографический инте-
рес. Они подчеркивают и для этого типа фауны значительную протяжен-
ность его формирования во времени, так как нет сомнения, что они
являются относительно молодыми его элементами. В других группах
насекомых (и многих других членистоногих), представленных в турано-
африканской фауне обособленными родами, время дифференцировки
пустынных элементов должно быть отнесено к мезозою (юра, мел). Об
этом свидетельствуют обширные филогенетические связи пустынно-степ-
ных групп фауны Средней Азии с южными континентами. Кроме видов,
Ochropleura НЬ. и Agrotis Ochs, из Agrotinae (Кожанчико.в, 1937), может
быть назван ряд примеров, показывающих принципиальное значение этих
связей для фауны Средней Азии. Туркестанские термиты—• Hoioter-
mes (Isoptera) являются дериватом ксерофильных, тропических видов
этой архаичной группы насекомых. Скорпионы и сольпуги из пауко-
образных обнаруживают подобные же филогенетические связи и ареалы.
Этого же типа ареалы и филогенетические связи имеются среди пчелиных
у видов Diphoglossinae, для которых, как и для видов Agrotinae, несом-
Рис. 73. Западноазиатский тип фауны Orgyidae (точки). Черным показаны территории распространения ближайших
родственных форм.
156
ОБЩАЯ ЧАСТЬ
ненно присутствие архаичных типов на южных материках и их дери-
ватов в бореальных пустынях Голарктики (Средняя Азия, Колорадо,.
Калифорния).
Сказанное выше позволяет дать следующую характеристику форми-
рования фауны Orgyidae разных подобластей Палеарктики. Маньчжурская
подобласть сложена в основе древним, аутохтонным типом фауны
(китайско-гималайским). Известную роль в северных и восточных про-
винциях* подобласти играют элементы калифорнийско-японского типа,
а в южных и восточных — индомалайского. Турано-африканская под-
область сформирована пустынной, аутохтонной, хотя и не богатой фауной
Orgyidae. Более многочисленны, но не типичны для этой подобласти
ксерофильные элементы бразильско-лигурийского типа, особенно обиль-
ные в западных провинциях подобласти, тогда как восточные и горные
провинции заселены видами западноазиатского типа фауны Orgyidae.
В горах же характерно присутствие широко распространенных элемен-
тов китайско-гималайского типа фауны или дериватов последнего. Из
них дифференцировалось известное число также мезо- или даже ксеро-
фильных эндемов среднеазиатской фауны. Средиземноморская подобласть,
в основе сложена ксерофильными элементами бразильско-лигурийского
типа фауны; элементы эти особенно многочисленны в западных провин-
циях подобласти. Восточные провинции Средиземноморья обильно засе-
лены элементами западноазиатского типа фауны. Всюду, но в особенности
в южных и восточных провинциях, значительна примесь элементов ту-
рано-африканского типа фауны. В горных условиях налицо распростра-
нение дериватов или широко распространенных видов китайско-гималай-
ского типа фауны. Эндемичные виды представлены главным
образом за счет первых двух типов и немногими представителями
третьего. Бореевразийская подобласть в основном сформирована гигро-
фильными лесными элементами китайско-гималайского типа фауны Or-
gyidae. В восточных провинциях налицо примесь калифорнийско-япон-
ского типа фауны, распространенных вообще в известном числе по всей
подобласти. На юге борее'вразийской подобласти в горных условиях
заметна примесь индомалайского типа фауны, а в высокогорных областях
и в Прибалтике — присутствие элементов (эндемов) альпийско-арктиче-
ского типа фауны. На юге западных провинций подобласти значительное
распространение имеют элементы турано-африканского, бразильско-лигу-
рийского и западноазиатского типов фауны Orgyidae. Арктическая под-
область в основе сложена аутохтонными, альпийско-арктическими ви-
дами Orgyidae. Местами в эту подобласть проникают некоторые более-
эврибионтные элементы китайско-гималайского типа фауны.
Приведенная сжатая характеристика подобластей Палеарктики пока-
зывает высокую степень гетерогенности их фауны Orgyidae, из обзора кото-
рой можно вынести изЛстное суждение о генезисе этой фауны в целом.
Резюмируя изложенные материалы о территориях формирования
и филогенетических связях Orgyidae, можно наметить для этого семей-
ства следующий ряд фаунистических типов.
1. Индомалайский (и африканский) тропический тип, представленный
почти исключительно видами Orgyinae, экологически связанный с. гиле-
ями и биологически представленный поливольтинными и полифагич-
ными видами, полиморфный, широко распространенный в тропиках
Старого Света. Вероятное время его формирования в мезозое (юра-^мел).
2. Неотропический (горный), представленный обоими подсемействами,
но в большем объеме анцестральными видами Acronictinae, биоэкологи-
«1 C e cn О
«г i . »- . g
(£> { bCTr NJ \ a о
>0 9i |z V о
о Q
о tSA > ed О
о (Ф g
i о 1 &Ч, !> «с Г ~т~^ i -4<i cfi® ' ! « ? \ ( 9
А- CT Ы о
? — n O
D 61 / »
1 31 го f < ^3) 1 fir J_J h Й1 c> Q £
—F < f 1 e c c> G « .e
о Э 1
Рис. 74. Распространение видов группы Orgyia dubia Tausch. В Австралии — О. anartoides Wlk., О. athlophora Turn,
О. retinopepla Turn; в южной Африке — О. tricolor Н. S., в Палеарктике — О. dubia Tausch.
158
ОБЩАЯ ЧАСТЬ
чески характеризующийся моно- и олиговольтинизмом, распространенный
в горной зоне Кордильер Южной Америки и в Вест-Индии. Вероятное время
его дифференцировки — в конце мезозоя (мел.).
3. Китайско-гималайский, в основном производный от, индомалай-
ского, имеющий анцестральные формы в горах тропиков Старого Света,
но наиболее обильно представленный в фауне юго-восточной Азии и дав-
ший основной (гигрофильный) тип фауны Голарктики, слагающийся из
видов, экологически связанных с лиственными лесами, в развитии-моно-
вольтинных. Вероятное время дифференцировки — в 'конце мезозоя
и в начале третичного времени (эоцен, олигоцен).
4. Калифорнийско-японский, производный от неотропического, пред-
ставленный обоими подсемействами, распространенный на юго-западе
Северной Америки и в восточной, умеренной Азии, слагающийся из видов,
экологически связанных с хвойными и лиственными лесами, в развитии
сезонно-циклических, моно- и олиго вол ьтинных. Вероятное время диффе-
ренцировки этого типа — в мезозое и в начале третичного времени (оли-
гоцен).
5. Бразильско-лигурийский, производный от неотропического, пред-
ставленный видами Acronictinae, распространенными в ксерофильных
условиях, в особенности в литоральной Зоне Средиземного моря и в Сред-
ней Азии, экологически связанный с травянистой и кустарниковой расти-
тельностью, моно- и олиговольтинный, строго цикличный. Дифференци-
ровка его вероятна в конце мезозоя.
6. Западноазиатский, производный от китайско-гималайского, пред-
ставленный в горной зоне южных систем от Гималаев до берегов Атланти-
ческого океана, почти нацело слагающийся из видов Orgyinae, экологи-
чески характерных связью с древесными растениями, сезонно-цикличных,
моно-и олиговольтинных. Дифференцировка его —в середине третичного-
времени (олигоцен, миоцен).
7. Турано-африканский, для Orgyinae производный африканского-
сектора индомалайской фауны (типа), слагающийся из пустынных видов,
связанных с пустынной, кустарниковой растительностью, сезонно-циклич-
ных, моно- и олиго вол ьтинных. Дифференцировка его вероятна в мезо-
зое (мел) и в начале третичного времени (эоцен, олигоцен).
8. Альпийско-арктический, для Orgyinae производный от первого-
типа, его индомалайского сектора, дифференцировавшийся в горной зоне
юго-восточной Азии и северо-восточной Америки, представленный только-
видами Orgyinae, экологически ограниченный горной или арктической
тундрой, слагающейся из моновольтинных видов, заселяющих высоко-
горья восточноазиатских и североамериканских горных систем и два
сектора Арктики — берингийский и гренландский. Вероятное время
дифференцировки этого типа фауны Orgyidae — в середине третичного
времени (миоцен).1
1 Штегман (1936, 1938) сделал попытку характеристики авиафауны Палеарктики
отдельными фаунистическими типами. Он принимает следующие типы: арктический,
сибирский, европейский, средиземноморский, монгольский, тибетский и китайский.
Эти типы целиком установлены на основе формы ареалов (без учета филогенетических
отношений), почему представляются сложными, ценотическими типами и, конечно,
гетерогенны. Автор не вышел за пределы Палеарктики и потому не уловил генети-
ческих типов, ей свойственных. Палеарктические типы авиафауны Штегмана совер-
шенно совпадают с обычными биогеографическими делениями Палеарктики, что есте-
ственно и ожидать, так как и те и другие отражают только современные условия
обитания в Палеарктике и основаны на сравнении распределения организмов без
учета условий возникновения ареалов во времени и филогенетических отношений видов.
РАСПРОСТРАНЕНИЕ
159'
Границы ареалов распространения Orgyidae обусловлены климатическим
режимом, причем распределение видов, приспособленных к условиям среды;
данной территории, и создает ее фаунистический облик. Голарктика
в целом характеризуется наличием сезонно переменного климата. Южной
ее границей является территория выпадения снега (рис. 75). Она совпа-
дает с прохождением изотермы средних абсолютных минимумов в О0,
и изотермы средней годовой термической амплитуды порядка 20°. Следо-
вательно, южная граница Голарктики является территорией, где хотя
и слабо, но ежегодно, выражена зима. Фауна Orgyidae Голарктики сла-
гается из видов (или родов), приспособленных к сезонным, циклическим
сменам климата. Этот биологический тип в фауне Голарктики является,
абсолютно доминирующим.
Формирование фауны Orgyidae Голарктики и отдельных ее подобластей
шло параллельно с изменениями климата в прошлом. Оно шло в первую'
очередь за счет адаптации физиологических особенностей видов и их.
циклов развития к более суровым и неустойчивым условиям среды. Эти
экологические особенности видов Orgyidae, так же как й форма их ареалов,
распространения, дают все переходы от очень неполно приспособленных
к жизни в континентальных условиях, как, например, Dasychira pudi-
bunda L. (рис. 76), к другим, хорошо приспособленным к жизни в этих
условиях сурового, переменного климата. Примером последних может
служить Olene fascelina L., распространенная по всей Палеарктике до са-
мых крайних северных широт.
Подобласти Палеарктики распадаются на два основных климатиче-
ских типа. Один тип представлен исходным климатическим циклом маньч-
журской подобласти с обильным осадками летом и сухой зимой. Этот же
климатический тип имеется в бореевразийской подобласти, с той разни-
цей, что лето здесь более бедно осадками, а годичная термическая ампли-
туда увеличена за счет зимних минимумов (рис. 77). Близок климатиче-
ский цикл и в арктической подобласти, но здесь он модифицирован распре-
делением освещения, круглосуточного летом. Для Арктики также специ-
фичны бедность осадками и термическая динамика в течение суток. Есть
все основания считать, что в этих подобластях мы имеем развитие одного
климатического цикла. Не случайно, вероятно, и сходство фауны этих
трех подобластей.
Динамика условий влажности обратна в условиях климата Средиземно-
морья и пустынь. Более устойчивы и потому несомненно исходны для
Orgyidae условия гидротермического режима в средиземноморской под-
области. Развитие определенных и сходных со Средиземноморьем черт
гидротермического режима обнаруживает турано-африканская под-
область. В связи с этим не случайно сходство фаун этих двух подобластей.
Адаптация Orgyidae к физико-географическим условиям определяет-
дифференциацию фаун и возникновение типов ареалов, но в‘ряде случаев
также приводит и к внутривидовой дифференциации. У непарного шелко-
пряда, у Orgyia dubia Tausch. и у многих видов Acronictinae наблюдается.
Методика картографирования, предложенная Штегманом (1936), лишь наглядно,
но чрезвычайно схематично и потому, пожалуй, даже произвольно рисует сложный,
по экологическим признакам мозаичный, состав авиафауны разных частей Палеарк-
тики. Она лишь подчеркивает общеизвестную стадиальную (экологическую) мозаику
каждой биогеографической единицы. (Б. Ш те гм ан. 1936. О принципах зоогео-
графического деления Палеарктики на основе изучения типов орнитофауны. Изв.
Акад. Наук СССР, мат. и ест.: 523—563; Б. Ш те гм ан. 1938. Фауна СССР. Птицы,.
(н. с.), 1 (2) : 1—76.
ц Фауна СССР, И. В. Кожанчиков.
Рис. 75. Климатологическая характеристика территории Палеарктики. Сплошные линии — изотермы средней годовой
амплитуды температуры; пунктир—изотермы абсолютных минимумов; пунктир с точкой—-июльские изотермы; пунктир
с двумя точками — северная граница средиземноморского климата; точечный пунктир — граница пустынного климата.
Рис. 76. Распространение Dasychira pudibunda L. (1) и Orgyia antiqua L. (2).
162
OB 7/АН ЧАСТЬ
«мозаичное» распределение форм в пределах ареала. Разнообразие струк-
тур Orgyidae в зависимости от распространения сравнительно. не велико.
Бросается в глаза значительно меньшая изменчивость имагинальноп
фазы, чем это наблюдается среди многих других групп чешуекрылых.
Этот факт может быть в известной мере связан с кратковременностью
ее существования у многих видов. Возможно, что в целом малая изменчи-
вость Orgyidae определяется малой экологической разностью условий,
заселяемых большинством их видов, и, кроме того, во многих случаях
небольшими их ареалами. Ничтожны по площади ареалы, например, у мно-
гих видов Parocneria Dyar, Cifuna Wlk., у многих видов OrgyiaOchs., Dasy-
Рис. 77. Два типа годичного цикла осадков в Палеарктике.
Сплошные линии — закрытый (лесной) ландшафт, пунктир —откры-
тый (степной и пустынный). 1—Шанхай; 2—Пенза; 3—Верхо-
янск; 4—Селегард; й—Малага; 6—Александрия; 7—Новая
Бухара; 8— Джедда.
chira Ocli., OZeneHb., Euproctis Hb., Ocneria Hb., Leucoma Steph., Acronicta
Ochs., Craniophora Snell. Очень велико число олиго- и монотипических
родов, ареалы видов которых ничтожно малы, как то известно для Panthau-
та Stgr., Euromoia Stgr., Thyatirides Kozh., Subleuconycta Kozh., Pentho-
phera Germ., Eogena Bd., Ozycesta Hb., Leiometopon Stgr. и некоторых
других. Обычное представление о широком распространении видов Orgyi-
dae связано с обширными и иногда громадными ареалами немногих из них,
очень заметных и часто массовых видов, как, например, Ocneria monacha
L., О. disparL., Orgyia antiquaL., О. gonostigmaPDasychira pudibunda L.,
Olene fascelina L., Leucoma salicis L., L. Candida Stgr., Calocasia coryli
L., Daseochaeta alpium Osb., видов рода Acronicta Ochs, (leporina L.,ace-
ris L., tridens Schiff., psi L., euphorbias Schiff.), Arsilonche albovenosa
Goeze, Simyra nervosa Schiff, и других. Тем не менее и эти виды чаще
всего ограничены в обитании определенными биотопами, а широкое
распространение их связано с фактом широкого распространения
РАСПРОСТРАНЕНИЕ
163
последних, как, например, зарослей Salicaceae по долинам рек для ивовой
волнянки, .зарослей березового кустарника (поросли Betula pubescens)
для Orgyia antiqua L., О. gonostigma F., Calocasia coryli L., Acronicta rumi-
cis L., A. auricoma Schiff. Возможно, в связи с этим находится слабая
изменчивость этих широко распространенных видов. Эврибионтные виды
среди Orgyidae представлены относительно небольшим числом.
Из числа широко распространенных видов некоторые обнаруживают
заметные различия в пределах своих ареалов. Степень их не одинаковау
разных видов,а географические формы наблюдаются двух типов.Могут быть
названы следующие виды Orgyidae, у которых констатировано наличие гео-
графических форм: Ocneria monacha L., О. disparF., Orgyia dubia Tausch.,
0. trigotephras Bd., Olene fascelina. L., Dasychira pudibunda L., Euproctis
chrysorrhoea L., E. similis Fuessly, Parocneria lapidicola H. S., Acronicta
leporina L., A. menyanthidis Niew., A. auricoma Schiff., A. euphorbias Schiff.,
A. tridens Schiff., A. rumicus L., Arsilonche albovenosaGoeze, Simyra nervosa
Schiff. Orgyidae в подавляющем большинстве случаев характеризуются не
столько цветовыми и структурными различиями (имаго или личиночных
фаз) в пределах их ареалов, сколько различиями циклов развития
и биоэкологии в целом. Это установлено для географических форм
непарного шелкопряда Гольдшмидтом (1911, 1934), отмечено Беляевым
для Orgyia antiqua L. и мною констатировано для ряда видов
рода Acronicta Ochs. Морфологически эти формы могут быть очень
слабо дифференцированы. Нет сомнения, что этот тип географической
изменчивости у Orgyidae преобладает. Другие случаи изменчивости,
когда можно говорить лишь о цветовых или структурных различиях
(в личиночной или имагинальной фазе), также известны, например, у Acro-
nicta menyanthidis View., A. auricoma Schiff., A. leporina L., A. lutea
Brem. et Grey, однако большею частью нет уверенности, что одни только
морфологические различия географических форм обязаны просто недоста-
точному знанию их биоэкологии.
Следует подчеркнуть, что хотя между размерами ареалов и сте-
пенью изменчивости видов Orgyidae есть известная связь, но гео-
графическая изменчивость их не может быть поставлена в прямую зави-
симость от площади ареала. Нет также простой связи между изменчиво-
стью и разнообразием условий среды обитания. Так, на юго-востоке
Азии непарный шелкопряд, обнаруживает на смежных, довольно одно-
типных биотопах обилие форм, а в горных условиях Средней Азии,
где условия обитания не менее, но, может быть, даже более разнообразны,
непарный шелкопряд не обнаруживает сколько-нибудь резких форм. Нет
сомнения, что наибольшее разнообразие географических форм у многих изу-
чаемых видов наблюдается на территориях, наиболее длительно обитаемых
ими (Дарвин, 1859).1 Это, например, несомненно для непарного шелкопряда.
У него распределение форм в целом несет общие черты распределения видов
в роде Ocneria Hb. Так, наибольшее разнообразие видов рода Ocneria
Hb. в Голарктике наблюдается в юго-восточной Азии; здесь же обнару-
жено и наибольшее обилие форм непарного шелкопряда. В этом типе гео-
графической изменчивости мы, наверное, имеем дело с отзвуком
глубокой истории вида. Формы непарного шелкопряда, известные
с востока Азии, ныне уже настолько дифференцированы, что даль-
нейшее развитие их, вероятно, в недалеком геологическом будущем
должно дать вполне обособленные виды. Не случайно гибридиза-
1 Ч. Дарвин. 1859. Происхождение видов. Изд. АН СССР, 1939.
11*
164
ОБЩАЯ ЧАСТЬ
ция подобных форм, как то показали многочисленные эксперименты
с непарным шелкопрядом, обнаружила громадный процент неправильно
развитых особей и бесплодных гибридов.
Указанный тип географической изменчивости с обилием внутривидо-
вых географических форм на территориях ареала, наиболее длительно
обитаемых видом, далеко не может быть обобщен на все случаи внутриви-
довой географической изменчивости Orgyidae. Это может быть обосновано
экспериментальным материалом и многими полевыми наблюдениями
(Acronicta lutea Brem. et Grey, Acronicta rumicis L., Leucoma salicis L.,
Ocneria dispar Hb. и другие). Обитание более яркой, крупной
и имеющей яркожелтые задние крылья, типичной китайской формы
Acronicta lutea Brem. et Grey на юге ареала вполне вяжется с необходи-
мостью формирования желтого и охристых пигментов этого вида при вы-
сокой температуре. Это доказано экспериментально. Наоборот, распро-
странение на севере ареала этого вида более мелкой, бледной, белокрылой
формы — Acronicta lutea leucoptera Butl. также вполне соответствует
реакции этого вида на температуру. Таким образом дифференциация
форм в других случаях может быть совершенно не связана с территорией
формирования данного ’ вида, но обилие их может быть обусловлено
разнообразием условий обитания. При этом как частный случай
наибольшая разность этих условий может наблюдаться как раз в области
формирования вида, и распределение форм в этом случае может быть
совершенно аналогичным тому, что известно, например, для непарного
шелкопряда. Для случаев географической изменчивости Orgyidae нет со-
мнения в легкой физиологической обратимости ряда молодых географи-
ческих форм, например цветовых, которые, кстати, совершенно не
обнаруживают биологических различий в циклах развития и дают при
скрещивании вполне плодовитых и нормальных по реакции на среду
особей. Кроме Acronicta lutea Brem. et Grey это обнаружено у Acronicta
rumicis L., A. leporina L.
Наиболее разносторонне изучена географическая изменчивость непар-
ного шелкопряда (Ocneria dispar L.). Географическое распределение
его форм на юго-востоке Азии видно из карты (рис. 78). Общее распреде-
ление географических форм в Палеарктике иллюстрируется рис. 79. Цве-
товые и структурные признаки географических форм непарного шелко-
пряда как в имагинальной, так и в личиночной фазах не велики и очень
изменчивы.
Можно достоверно различать лишь две группы географических форм:
1) более крупных, имеющих темно окрашенных (почти шоколадного цвета)
самцов восточноазиатских Ocneria dispar L., распространенных до Забай-
калья, но особенно характерных для Дальнего Востока, и 2) более мелких
и светлых, имеющих желтоватых самцов, форм, распространенных . от
Забайкалья до берегов Атлантики. Рейнигом справедливо отмечено
уменьшение размеров особей непарного шелкопряда с востока на запад.
Уменьшение размеров особей не является тем не менее постепен-
ным, как полагает Рейниг, считающий, что наибольшими размерами
характеризуются О. d. japonica Motsch., меньшими амурские О. d. fumida
Btl. (—praeterea Kardak.), еще меньшими типичный евразийский О. dis-
par L. и наименьшими — иберийский О. d. bordigalensis Mab. (=andalu-
siaca Rein.). Различия размеров достоверны лишь для первых трех форм,
причем первые две практически неразличаются, но обе отличны от третьей.
Четвертая, андалузская карликовая форма, описанная Рейнигом, характе-
ризуется иными признаками, но карликовые особи непарного шелкопряда
РАСПРОСТРАНЕНИЕ
165
наблюдаются решительно всюду в связи с условиями развития и пи-
тания.
Восточные формы непарного шелкопряда (О. dispar japonica Motsch.,
O.d. fumidaBtl.) характеризуются в личиночной фазе значительно большим
развитием светлого пигмента в тергальной области брюшных сегментов,
по сравнению с западными. В некоторых случаях, например у особей,
взятых из Кумамато (Япония), расположение и форма брюшных тергаль-
Рис. 78. Географическое распространение восточноазиатски± форм непарника
(Ocneria dispar L.). (По Гольдшмидту, 1934). 1 — маньчжурско-китайская; 2 — ко-
рейская; 3 — форма Киу-Шиу; 4 — южнояпонская; 5 — форма Гифу; 6 — примор-
ская; 7 — горная (Кофу); 8 — северояпонская; 9 — форма Хоккайдо.
ных пятен у гусениц чрезвычайно напоминает окраску гусениц видов
Leucoma Steph, группы salicis L. Уместно отметить, что род Leucoma
Steph, (см. специальную часть) в большей мере сохраняет анцестральные
черты во всей организации, равно как и в распространении. Восточные
формы непарного шелкопряда характеризуются также более медлен-
ными темпами роста. Ускоренный рост их есть признак специали-
зации. Это приспособление к более короткому периоду вегетации
кормовых растений (мелколиственных пород). Таким образом, можно
присоединиться к мнению Рейнига, что восточноазиатские формы
120
Рис. 79. Географическое распределение основных комплексов форм непарника (Ocneria dispar L.). 1 — восточные формы
типа japonica Motsch.; 2 — амурские формы типа praeterea Kard.; 3 — комплекс форм типа dispar L.; 4 — пиренейские формы
типа bordigalensis Mab, (=andalusiaca Rein.).
РАСПРОСТРАНЕНИЕ
167
непарного шелкопряда более древни и имеют более архаичные черты,
как в структурах, так и в биологии в целом. Из западных форм лишь
средиземноморские обнаруживают в личиночной фазе светлую окраску
тергальных отделов брюшных сегментов, но окраска эта совершенно
иного типа и во всяком случае более специализована, чем у восточных
форм. Нельзя отрицать, однако, возможности наличия и в Средиземно-
морье более древних форм непарного шелкопряда, чем заселяющие зону
хвойных лесов и березы Палеарктики.
При ближайшем знакомстве с этими географическими формами, которые
структурно слабо дифференцированы, как, например, L. dispar japonica
Motsch. и L. d. fumida Btl. (==praeterea Kard.) или типичная евразийская,
все же они.оказываются далеко не однородными. По мнению Гольдшмидта,
едва ли не каждой географически обособленной территории соответ-
ствует своя, локальная форма с комплексом наследственных структур-
ных и биологических признаков. Это едва ли справедливо для всего
ареала непарного шелкопряда, но вероятно для его восточной части.
Только на востоке Азии (не включая Амур и Приморье) Гольдшмид-
том обнаружены,, по крайней мере, девять локальных форм, также едва ли
достаточно однородных. Гольдшмидт (1934) различает «китайско-маньч-
журскую форму», «корейскую», «форму Киу-шиу», «южнояпонскую»,
«форму Гифу», «приморскую», «горную» (Кофу), «северояпонскую» и
«формуХоккайдо». В пределах этих форм горно-японские особи из Кофу,
Токио, Аомори и ряда других мест Хондо обнаруживают наибольшую
степени домиантности признаков, по сравнению как с более северными —
Хоккайдо, так с западными — из Кореи и Маньчжурии -г- и с южными —
с Киу-Шиу. Это, может быть, говорит за то, что в горных японских
формах непарного Шелкопряда налицо более древние типы этого вида
с признаками, сложившимися давно и прочно. За это же говорит и харак-
тер цветовых и структурных признаков гусениц и имаго восточных форм
непарного шелкопряда.
Все эти формы непарного шелкопряда обнаруживают высокую степень
биологической адаптации к местным условиям обитания. Они характе-
ризуются специфичностью общего цикла развития и прежде всего
длительностью и глубиною диапаузы яиц. Диапауза более глубока и дли-
тельна у северных и западных форм. У них развитие яиц требует
охлаждения, что не обязательно для южных. Признаки' форм выра-
жаются также . темпами роста и числом личиночных стадий, следо-
вательно длительностью личиночного периода. Для всех форм специфичны
темпы развития гонад. Различия темпов роста,.а следовательно и интен-
сивности ассимиляторного процесса в целом, находят выражение в разме-
рах особей и в размерах клеток тела, в частности половых, и в количестве
в них хроматина. Западные формы более богаты хроматином ядра, по
сравнению с восточными. Все эти признаки в их конкретных величинах
сочетаются в разнообразных и сложных комбинациях. Характер призна-
ков не оставляет сомнения в постепенном возникновении их во времени
в связи с адаптацией непарного шелкопряда к новым условиям в процессе
завоевания более суровых, по условиям обитания, территорий. Они
говорят весьма убедительно за то, что формы эти ныне имеют свои пути
эволюции.
Аналогичную изменчивость на территории ’ареала распространения
обнаруживает Orgyia dubia Tausch-. У этого вида она совершенно
по затронута экспериментальным анализом. Наибольшее обилие форм
Orgyia dubia Tausch. наблюдается в Средиземноморье (рис. 80). Из Пире-
Рис. 80. Географическое распространение форм Orgyia dubia Tausch. 1 — O.d. splendida Rbr; 2 — 0. d. arcerii Rag.; 3 — O.
d. orana Pow.; 4 — O. dubia var.; 5 — O.d. / osephina Aust, и О. d. /udaeaStgr.'. 6 — O.d. turcica Ld.; 7 — O. d. dubia Tausch.;
8 — O.d, interrupta G .-G:’.; 9 — O.d. heplapolamica, f. nov.; 10 — 0. d. baicalensis, f. nov.
РАСПРОСТРАНЕНИЕ
169
леев и из северо-западной Африки известна Orgyia dubia splendida Rbr. —
крупная и яркая форма. Близкая к ней, но несколько более мелкая Orgyia
dubia arcerii Rag. заселяет южную Францию, Корсику, Сардинию, Сици-
лию и Апеннинский полуостров. Южнее, в Тунисе, обитает светлая
форма — Orgyia dubia orana Pow. На Балканском полуострове распро-
странена близкая к типичной, неназванная форма. Египет, Сирию и Па-
лестину заселяет Orgyia dubia josephina Aust.; здесь же встречается
несколько более светлая и яркая форма, напоминающая западную
(О. splendida Rbr.) Orgyia dubia judaea Stgr. Восточнее и севернее
в Малой Азии распространена Orgyia dubia turcica Ld., а еще севернее,
в Закавказье, на Северном Кавказе и в южной Украине —Orgyia dubia
typica Tausch., ыкоторой весьма близка восточная, туркестанская (Турк-
мения, северный Иран) Orgyia dubia interrupta Gr. Gr. (=transcaspica¥Av&.).
Восточнее распространены локальные более темные формы, Orgyia dubia
heptapotamica Kozh. в Семиречье и Orgyia dubia baicalensis Kozh. в При-
байкалье.
Orgyia dubia Tausch. не обнаруживает уменьшения размеров в направле-
нии удаления от области обилия форм. Восточные формы имеют лишь
несколько более темную окраску. Обилие форм наблюдается опять-таки
в областях, наиболее длительный период обитаемых этим видом. Общий
тип ареала Orgyia dubia Tausch. указывает на его деградацию в современ-
ную эпоху. Неясно, насколько формы Orgyia dubia Tausch. обнаружи-
вают адаптацию к местным условиям обитания,, но вероятность такой
адаптации велика, если учесть, что территории, к которым привязаны
отмеченные формы, даже в условиях Средиземноморья, отличаются весьма
резко выраженными местными особенностями климата и, в ча-
стности, его годичной циклики. Географические формы Orgyia dubia Tausch.
обнаруживают большую лабильность признаков, по сравнению с непар-
ным шелкопрядом. Для них типичны значительные различия в резкости
рисунка и в окраске, причем в пределах одной и той же области имеются
разные цветовые формы, например в Передней Азии —Orgyia dubia
josephina Aust, и Orgyia dubia judaea Stgr., может быть возникшие в связи
с экологическими особенностями местообитаний.
Того, же типа географическая изменчивость, что описана для непар-
ного шелкопряда и Orgyia dubia Tausch., обнаружена и для ряда других
видов Orgyidae, например для О. trigotephras Bd., которая дает ряд локаль-
ных форм в Средиземноморье (О. anceps Obth., transiens Stgr., sicula
Stgr., Corsica Bd., orientalis Stgr.), для О. ericae Germ., у которой в центре
ареала много слабо дифференцированных форм, а на востоке описана
локальная О. ericae zimmermani Graes. (Амур). Две формы — восточную
(амурскую) и западную (европейскую) обнаруживает Dasychira pudi-
bunda L.; Laelia coenosa Hb. дает восточную, более ограниченную в распро-
странении (Китай) форму L. Candida Leech, более крупную и яркую, по
сравнению с широко распространенной типичной формой. Аналогичным
образом Ocneria matura Moore, распространенный в Гималаях, представ-
лен в Китае, Маньчжурии, ю. Приморье и на Японских островах, более
яркой формой — О. aurora Btl. (= jusca Leech). Parocneria lapidicola
H. S. дает ряд форм в Средиземноморье — темную в Малой Азии (Р. urbi-
cola Stgr.), однотонную сирийскую, известную также с Кипра (Р. phoe-
nissa Rghfr.) и распространенную в. Ираке (Р. mardina Stgr.). Parocneria
signatoria Chr. представлена в Палестине локальной формой Р. phoeni-
tens Stgr. Сходные отношения географической изменчивости обнаруживает
Acronicta euphorbiae Schiff., пятнисто распространенная по болотистым
170
ОБЩАЯ ЧАСТЬ
и песчаным территориям Прибалтики, Украины, Средиземноморья и за-
падной Сибири (A. montivaga Gn., myricae Gn., eaphrasiae Brahm,
esulae Hb.).
Характерной чертой всех подобных географических форм Orgyidae
является та или иная степень изоляции их друг от друга и прерывчатое
распространение, а для видов, имеющих подобные географические формы,
та или иная степень деградации и распада ареала. Возникновение подоб-
ных форм есть основания ставить в связь с постепенным ухудшением
условий обитания этих видов на всей территории ранее сплошного
ареала.
В' ряде случаев географическая изменчивость Orgyidae является более
сложной, как то несомненно для Acronicta ramicis L., Olene fascelina L.
и некоторых других видов. Здесь при обширных сплошных ареалах
и обилии особей данного вида на большей части территории его распро-
странения несомненно наличие , двух типов географических форм. Одни
из них территориально не разобщены, но постепенно переходят одна
в другую, другие более или менее географически изолированы. Возникно-
вение первых связано с влиянием среды в текущий момент. Подобные
формы обособляются на территориях с сильно отличными условиями
обитания (в частности климата). Эти формы могли бы быть названы моло-
дыми, продуктом ныне действующих влияний. -Морфологические р1злщ-
чия этих форм не стойки и обратимы, как то констатировано с несомнен-
ностью для ряда форм Acronicta ramicis L., lutea Brem. et Grey и некото-
рых других видов этого рода. Географические формы второго типа
у этих же видов являются в значительной мере остаточными, результа-
том изменения данного вида в процессе длительной в прошлом адаптации к
меняющимся условиям обитания. Следует добавить, что первые формы воз-
никают в результате освоения видом новых территорий. Не случайно они
наблюдаются в областях сплошного распространения вида и сопрово-
ждают его насыщенные популяции (рис. 79). Напротив, вторые формы
этих же видов отражают утрату необходимых для них общих условий
обитания.
Распространение Acronictaramicis L. охватывает обширную территорию
от Японских островов и Гималаев до берегов Шотландии и Гибралтара
(рис. 81). На востоке ареала Acronicta ramicis L. редка. Из Китая (Юнь-
нань) описана крупная темная форма—Acronicta ramicis regifica Drd.
Эта же или близкая к ней форма населяет и другие территории западногоКи-
тая. Из Гималаев описана столь же редкая и обособленная, еще более круп-
ная, сиккимская форма—Acronicta ramicis indica Moore. Обе формы яв-
ляются аналогом японских и китайских форм непарного шелкбпряда. Они
сильно дифференцированы структурно и являются-остаточными формами
Acronicta ramicis L. Этого же типа форма A. ramicis L. известна с Даль-
него Востока; она заселяет также Японские острова, Корею и северный
Китай, включая Маньчжурию. Территория от Забайкалья до Пиренеев
занята наиболее обычной, типичной формой A. ramicis L. Из Средней Азии
известна светлая форма —A. ramicis turanica Stgr., очень обычная, мас-
совая по оазисам. Она распространена также в Закавказье и в, южном
Поволжье, где отмечена как вредитель садов." Аналогичные светлые
формы с менее ясными ареалами обитания описаны из южной Европы —
Acronicta ramicis distincta Zub. et Vuk. из Румынии и Бессарабии,
А. г. meridionalis DanHehl из южной Европы и А. г. pallida Roths, из север-
ной Африки. В Шотландии обитает однотонная, серая форма этого вида —
А. г. salicis Curt.
Рис. 81. Распределение географических форм Acronicta rumicis L. 1 — А. г. rumicis L.; 2 — А. г. turanica Stgr.; 3 A. ramicis
var.; 4 —A. r. regifica Drd.; 5 —A. r. indica Moore; 6 — A. r. salicis Curt.; 7 — A. r. pallida Roth.; 8 A. r. distincta
Zub. et Vuk.; 9 — A. r. meridlonalis Dannehl.
172
ОБЩАЯ ЧАСТЬ
Все последние формы — иного типа. Они сопровождают насыщенные
популяции, нередко являются массовыми, но заселяют территории, сильно
отличные от тех, где распространена типичная форма. Они распространены
в пустынях, сухих степях, по литторали Средиземного моря или в области
северного безлесья (верещатники, лесо-тундры). Эти формы в основе,
вероятно, обязаны влиянию климата. Вместе с тем Acronicta rumicis L.
является многоядным видом, связанным по питанию как с многолетними,
так и с однолетними растениями.
Сходную картину внутривидовой изменчивости обнаруживает Olene
fascelina L., для которой несомненьо наличие ряда остаточных форм на
юго-востоке ареала (О. /. nivalis Stgr., О. /. angelis Tschetw.). В то же
время широко распространенная субарктическая форма Olene fascelina
obscura Zett., вероятно, обязана влиянию климата (рис. 82). Для этого
вида, олигофага, специализованного на питании древесными растениями
преимущественно из Fagales, Salicales и Rosales, описаны пищевые формы,
специализованные (?) на питании мотыльковыми (О. fascelina medicagi-
nis Hb.) и хвойными (О. f. laricis Schille).
Глава VIII
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
Хозяйственное значение Orgyidae как отрицательного фактора в жизни
человека велико. В пределах семейства насчитывается несколько десят-
ков видов, которые отмечены в литературе как вредители разнообразных
культурных растений или как потребители полезных растений дикой
флоры. Среди Orgyidae есть ряд так называемых массовых вредителей,
например монашенка, непарный шелкопряд/ златогузка. Они известны
для умеренных широт северного полушария, но и для тропиков и субтро-
пиков отмечены массовые формы—Olene mendosa Hb., Orgyia postica
Wlk., вредящие там разнообразнейшим культурным растениям.
Экономическое значение вредных видов Orgyidae определяется не только
наличием постоянного ущерба от Них хозяйственным растениям, но, глав-
ное, угрозой возникновения массовых размножений вредных видов, когда
наносимый ими вред принимает катастрофические размеры и далековыходит
за рамки одногодового ущерба или грозит уничтожением целых лесных
или садовых массивов, как то имело место, например, в Северной Аме-
рике. Массовые размножения вредных Orgyidae влекут за собой уничтоже-
ние не только обычных кормовых растений, для охраны которых ежегодно
применяются различные мероприятия, но приводят к порче и утрате
многих других. Например в 1927 г. в период массового размножения не-
парного шелкопряда в Европейской части СССР отмечалось не только
сплошное уничтожение листьев древесной растительности и кустарни-
ков, но также уничтожение гусеницами этого вида хлебов —пшеницы
(Брянцев, 1928) и озимой ржи (Талицкий, 1928), причем они поедали
не только листья злаков, но и их стебли (солома), т. е. гибли и колосья
растений. Это отмечалось многократно для непарника (Кеппен, 1883).
Точно так же массовые размножения непарного шелкопряда в Север-
ной Америке ознаменовались уничтожением не только обширных про-
странств леса и территорий, занятых плодовыми садами, но также
и огородов.
Наряду с этим при массовых размножениях обнаруживается ряд
растений, совершенно не поедаемых вредными Orgyidae. Так, Прелль
Рис. 82. Распределение географических форм Olene fascelina L. 1 — О. f. fascelina L.; 2 — О. f. obscura Zett.; 3 — 0. f. nivalis
Stgr.; 4 — O. f. angelis Tschetw.; 5—0. fascelina var.; 6 — O. fascelina var.
174
ОБЩАЯ ЧАСТЬ
(Prell, 1924) указывает, что среди еловых насаждений часть елей была
нетронута монашенкой при ее массовом размножении. И в лабора-
торных условиях гусеницы не поедали хвою с этих растений, но
эта. же хвоя, вымоченная в воде в течение двух суток, делалась
пригодной для питания гусениц монашенки. По мнению Прелля,
причиной, тормозившей питание гусениц монашенки этой хвоей,
было повышенное содержание терпентина у этих особей ели. Для
непарного шелкопряда зафиксировано и при массовых размножениях
избегание питания Pinus rigida, хотя произраставшие в этих же условиях
Pinus strobus и Pinus silvestris поедались и часто были оголены [Гейль
(Heil, 1937)]. Для этого же вида характерно, по Гейлю, в случае массовых
размножений избегание видов дрока (Genista) и почти полное отвергание
белой акации (Robinia pseudacacia), как то справедливо приводится
Вукасовичем (Vukasovic, 1936) для Югославии и давно уже отмечалось
для нашего юга.
Массовые размножения вредных Orgyidae наблюдаются в самых различ-
ных областях ареала распространения семейства и характеризуют раз-
ные, хотя и немногие виды его. Так, размножения монашенки (Ocneria
monacha L.) и златогузки (Euproctis chrysorrhoea L.) часты в Западной
Европе, там же, в восточной Европе и Сибири — непарного шелкопряда
(Ocneria dispar L.), в юго-восточной Азии этого же вида и розового непар-
ного шелкопряда (Ocneria matura Btl.), а также Olene mendosa Hb. В Ги-
малаях и субтропиках южной Азии, кроме двух последних видов, наблю-
даются размножения также Orgyia postica Wlk. и многих видов Dasychira
Ochs. В Северной Америке необычайно сильны размножения завезенных
туда непарного шелкопряда, златогузки и ивовой волнянки (Leucoma
salicis L.), в Африке на севере известны размножения непарного
шелкопряда, а в тропиках — многих видов Dasychira Ochs, и Eup-
roctis Hb.
Массовые размножения Orgyidae не только приносят прямой ущерб,
но . являются отрицательным фактором, обусловливающим размножение
так называемых вторичных вредителей. Они влекут появление короедов
на ослабленных деревьях, что наблюдалось уже давно в лесной практике
[Рацебург (Ratzeburg, 1838)]. Комарек (Komarek, 1931) отмечает, что
в Чехословакии за размножением монашенки следует размножение корое-
дов (Hylastes palliatus Gyll., Xyloterus lineatus 01., Ips cembrae Heer,
I. amitinus Fich.), которое ведет к гибели елей, а также размножение
и других менее опасных короедов, каковы: Hylastes cuniculariusEr., Cry-
phalus abietis Rtz., Ips laricis F., Pityogenes chalcographus L., Pityophthorus
micrographusL., P. pityographus Rtz. Распространение Ips typographus L.
на очагах массовых размножений монашенки меньшее, так как на елях,
поврежденных монашенкой, кора высыхает слишком быстро, что ведет
к гибели личинок типографа. Связь Ips typographus L. с распространением
монашенки, по Комареку (1933), заключается в том, что он является,
так же как и монашенка, горным видом, связанным с еловыми насажде-
ниями. Напротив, размножения Ips cembrae Heer, по Преллю (1925а),
стоят в тесной связи с размножениями монашенки. Нехлеба (Nechleba,
1927) наблюдал миграции Ips cembrae Heer с лиственницы на ель вслед
за размножениями монашенки. Размножение короедов вслед за размно-
жениями монашенки на Дальнем Востоке отмечал Куренцов. Динглер
(Dingier) отмечает массовое появление личинок комара-долгоножки (Ti-
pula nubeculosa Mg.) в лесах на следующий год после появления
монашенки; он ставит это в связь с повышением содержания азота в почве
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
175
в связи с большим количеством падающих на почву экскрементов. Мас-
совые размножения Euproctis similis Fuessly оказывают большое влияние
на питание других листоядных насекомых, например в Японии гусениц
тутового шелкопряда, которые отравляются ядовитыми волосками гусе-
ниц Euproctis Hb., прилипающими к листьям.
Таким образом массовые размножения Orgyidae глубоко влияют
на состояние леса или сада. Отрицательное значение их совершенно
не ограничивается тем периодом времени, пока наблюдается размножение,
но оно ведет, помимо прямых потерь, еще к потерям от последующего
ослабления прироста растений. Так, по Парамонову (1934), прирост дубов
в Крыму в связи с недавним (1930—1932) размножением там непарного
шелкопряда был снижен на 30—50%. Развитие вторичных вредителей
часто завершает отрицательную роль листоядных Orgyidae . Кроме того,
отрицательное значение массовые размножения Orgyidae имеют в связи
с разносом грибных и вирусных болезней растений, как то установлено
для непарного шелкопряда в отношении ржавчины (Coronarium
ribicola), которой гусеницы заражают виды Ribes (Граватт и Позей,
1918).
В пределах семейства распределения вредных видов по родам очень
характерно (табл. 23). Наиболее важны в экономическом отношении
виды подсем. Orgyinae, особенно в Голарктике. В тропиках вредные
виды из Orgyinae, пожалуй, более многочисленны, но экономическое зна-
чение их, видимо, меньше, чем видов этого подсемейства, распространен-
ных в умеренных широтах. Точно так же меньшее экономическое значение
видов подсем. Acronictinae связано отнюдь не с слишком малым числом
вредных видов. Напротив, почти все Acronictinae по характеру питания
должны быть отнесены к числу вредных, так как кормовыми растениями
их являются или ценные древесные лесные породы или плодовые деревья.
Тем не менее малочисленность большей части видов этого подсемейства
исключает отрицательную роль большинства их в жизни человека.
Это же касается и тропических видов подсем. Orgyinae, разнообразие ви-
дов которых очень велико, а характер питания мог бы определить роль,
их как хозяйственно важных вредителей. Тем не менее на деле роль их
в местном ландшафте относительно второстепенна. Таким образом факто-
ром, определяющим значение видов Orgyidae как вредителей сельскохо-
зяйственных растений, является потенциальная возможность давать
очажные или сплошные, на больших территориях, массовые размножения.
По своему систематическому положению вредные виды Orgyidae могут быть
названы специализованными, а географически могут быть определены
как расширяющие ареалы распространения, заселяющие новые террито-
рии. Высокая или значительная плотность популяций (конечно не
на всей территории ареала и не ежегодно) типична для вредных видов
Orgyidae.
Вредные Orgyidae обнаруживают и другие особенности биологического
и географического характера. Прежде всего все они относятся к поли-
морфным группам. Они сосредоточены в полиморфном подсемействе
и в большем числе относятся к полиморфным родам (Orgyia Ochs., Dasy-
chira Steph., Euproctis Hb., Artaxa Wlk., Ocneria Hb., Acronicta Ochs.).
Массовые вредители из Orgyidae все относятся к* морфологически измен-
чивым или полиморфным видам. Этот факт указывает, что возможность
массовых размножений связана с изменчивостью видов Orgyidae, может
быть, со способностью их приспособляться к условиям обитания, а также
с их физиологической пластичностью.
Таблица 23
Систематическое и географическое распределение вредных видов Orgyidae
Систематическое положение
и название вида
Ареал вреда
Подсем. Orgyinae
1. Olene Hb.
О. fascelina L..............................
О. mendosa Hb...............................
О. dudg'eoni Swinh..........................
2. Gynaephora Hb.
G. selenitica Esp....................... .
3. Orgyia Ochs.
0. thyellina Btl. ..........................
O. antiqua L.................................
O. gonostigma F.............................
0. dubia Tausch.............................
O. vetusta Hamps.............................
0. postica Wlk..............................
O. viridescens Wlk...........................
O. turbata Btl. . . .........................
0. anartoides Wlk...........................
O. prisca Stgr..............................
4. Dasychira Steph.
D. pudibunda L..............................
D. abietis Schiff...........................
D. albodentata Brem.........................
D. pseudabietis Btl.........................
D. seouris Hb...............................
D. twaitesi Moore...........................
D. horsfieldi Saund.........................
D. endophaea Hamps. ........................
D. niobe Weym.'.............................
D. georgiana Faw............................
5. Hemerocampa Dyar
H. leuco stigma Wlk.........................
6. Penthophera Germ.
P. morio L. . . •...........................
7. Pseudodura Strd.
P. dasychiroides Strd.......................
8. Redoa Moore
R. cygna Moore..............................
9. Arctornis Hb.
A. alba Brem.................................
A. (?) rubricosta Faw........................
10. Euproctis Hb.
E. chrysorrhoea L...........................
E. similis Fuessly .........................
E. producta Wlk.............................
E. karghalica Moore.........................
E. mediosquamosa Beth.-Bek..................
Европа, западная Азия
Индия, Формоза, Суматра, западный
Китай
Тайван
Скандинавия (Финляндия)
Япония
Западная Европа
» »
Поволжье, юго-восточная Европа
Восточная Африка, Занзибар
Цейлон, Тайван, южная и северная
Австралия
Тайван
Малайский архипелаг
Австралия
Тянь-шань
Западная Европа
» » Западная Сибирь
Забайкалье
Япония
Западные Гималаи (Пенджаб), Бирма
Цейлон
»
Центральная Африка (Конго)
» » »
Уганда
Северная Америка (восточные штаты)
Средняя Европа
Тайван
»
»
Уганда
| Западная и средняя Европа
Европа и юго-восточная Азия
' Восточная и центральная Африка
Средняя Азия (Тянь-шань).
i Конго, Уганда
Продолжение
Систематическое положение
и название вида
Ареал вреда
Е. taiivana Shir........................ , .
Е. flexuosa Veen.............................
Е. fraterna Moore.............................
Е. varians Wlk................................
E. lunata Wlk.................................
E. puloerea Leech............................
E. pseudoconspersa Strd.1 ........
E. sericea Wilem............... .' ...........
E. xanthorrhoea Kollar
E. atomaria Wlk. ..........................
.E. terminalis Wlk. . ........................
E. subfasciata Wlk..........................
11. Artaxa Wlk.
.A. scintillans Wlk...........................
A. latifascia Wlk.............................
.A. flava Brem................................
.A. bipunctapex IJamps..........:.............
12. Leucoma Steph.
E. salicis L. . ..............................
E. Candida Stgr...............................
E. flaoosulphurea Ersch......................
13. Cijunu Wlk.
C. locuples Wlk..............................
14. Ocneria Hb.
•O. dispar L..................................
Тайван
Восточная Индия, Малайский архи-
пелаг
Гималаи, Индокитай
Тайван
Западные Гималаи (Пенджаб)
Япония
Тайван
»
Западные Гималаи (Пенджаб)
Цейлон
Южная Африка (Трансвааль)
Индия •
O. monacha L...............................
•O. matura Btl..............................
‘O. ampla Wlk...............................
O. seroa. F................................:
O. xylina Swinh..........................>
O. nebulosa Wilem. ......................' |
• O. reducta Wlk.............
15. Parocneria Dyar }
E. lapidicola H. S.........................|
P. detrita- Esp.
E. amanda Stgr..............................
Подсем. Acronictinae
1. Eiphthera Ochs.
D. coenobita Esp............................
2. Calocasia Ochs.
C. coryli L. ..............................
3. Daseochaeta Warr.
D. alpium Osb..........................
4. Episema Ochs.
E. coeruleocephala L...................
Цейлон
Малайский архипелаг
Цейлон, Япония
Индокитай, южный Китай
Европа, западная Сибирь
Сибирь
Семиречье
Южное Приморье
Европа, северная Африка, западная
Сибирь, восточная Азия, Северная
Америка
Европа, западная Сибирь, ю. Приморье
Гималаи, Япония, ю. Приморье
Индия, Бирма, Цейлон
Тайван
»
»
Австралия
Сирия, Палестина
Поволжье
Ирак
Средняя Европа
Западная и южная Европа
Западная Европа, Украина
» » , Кавказ
_ 1 Указываемая для Тайваня Euproctis conspersa Btl. является синонимом вредя-
щей там же Е. pseudoconspersa Strd. (F. Bryk, 1934 : 221); точно так же Ocneria olso-
leta Wlk.—синоним О. seroa F. (F. Bryk, 1934 : 168).
12 Фауна СССР, И. В. Коисанчиков.
178
ОБЩАЯ ЧАСТЬ
Продолжение?
Систематическое положение
и название вида
Ареал вреда
5. Subacronicta Kozh.
S. megacephala Schiff......................
6. Acronicta Ochs.
A. rumicis L. . . . '.......................
A. tridens Schiff...........................
A- Psi L....................................
A. aceris L.................................
A. major Brem...............................
A. consanguis Btl...........................
A. dactylina Sm.............................
A. longa Gn.................................
A. incretata Btl............................
7. Arsilonche Ld.
A. albovenosa Goeze ........................
8. Simyra Ochs.
S. nervosa Schiff...........................
Украина
Европа, Средняя Азия, Япония
Поволжье, Закавказье
Европа, Поволжье, Кавказ, Средняя
. Азия
Западная Европа, Украина
Китай
Корея, Япония
Северная Америка
» »
Западный Китай, Корея, Япония
Поволжье
Поволжье, сев. Кавказ
Связь изменчивости Orgyidae с плотностью популяций находит под-
тверждение и в территориальном распределении вредных видов. Высокая
плотность популяций отмечена у видов, заселяющих области малого
разнообразия видов Orgyidae. Так, область гилей, где видовое разнообра-
зие Orgyidae велико, характеризуется наличием малого числа и при этом
локально вредящих видов. Виды Orgyidae, относящиеся к тропическим
группам, которые могут быть названы массовыми вредителями, например
широко распространенные Orgyia postica Wlk., Olene mendosa Hb., виды
Artaxa Wlk., обнаруживают высокую численность в субтропиках —
в западном Китае, на Т айване, на Японских островах, но не в тропиках, ?
т. е. в местах видового разнообразия групп, к которым они принадлежат.
Точно так же субтропические группы Orgyidae, например многие группы
видов Ocneria Hb., Dasychira Ochs., Euproclis Hb., характеризуются вы-
сокой плотностью популяций не в субтропиках, но в умеренных широтах
Европы и Азии. Acronictinae, в видовом отношении богато представлен-
ные в широколиственных лесах Северной Америки и юго-восточной Азии,
имеют там мало вредных видов и, наоборот, число их больше в Европе
и Средней Азии. Наконец в пределах вида высокую численность обнару-
живают не те популяции, для которых характерно большое количество
локальных форм, но популяции, заселяющие территории, где полимор-
физм выражен слабо. Так, высокая численность. непарного шелкопряда
или монашенки наблюдается в Европе и Западной Сибири, где нет разно-
образия форм этих видов, но не на юго-востоке Азии, где имеется поли-
морфизм этих видов. Точно так же Acronicta rumicis L. обнаруживает
высокую численность не в местах обилия близких видов и локальных
форм, т. е. на юго-востоке Азии или в Европе, но в Закавказье и Средней
Азии. Характеризуя эти факты в целом, можно высказать предположе-
ние о том, что вредные виды Orgyidae относятся к полиморфным груп-
пам, являясь структурно изменчивыми видами, высокая плотность попу-
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
179
ляций которых наблюдается на заселяемых ими (осваиваемых) терри-
ториях, где не возникла еще полная адаптация к новым условиям, но'
где особи данного вида находят достаточно благоприятные условия для
существования.1
В этом отношении интересны факты акклиматизации Orgyidae. Заве-
зенные в новые условия в Северную Америку непарный шелкопряд,
златогузка и ивовая волнянка в течение короткого времени размно-
жились и стали видами первостепенного хозяйственного значения. Роды
эти—Ocneria Hb., Euproctis Hb. и Leucoma Steph. —не представлены
в американской фауне вообще. Завоз многих видов из родов, типичных
для Америки, например Acronicta Ochs, {rumicis L., auricoma Schiff.) и
Orgyia Ochs, {antiqua L., gonostigma F.), несмотря на многократные слу-
чаи, не сопровождался размножением их там.
Круг повреждаемых видами Orgyidae хозяйственных растений обши-
рен, но он включает главным образом древесные растения и кустарники,
тогда как травянистые растения представлены меньшинством. В связи
с этим Orgyidae выступают прежде всего как вредители лесного хозяйства
и садовых культур, тогда как луговым растениям, огородным и полевым
культурам они вредят' реже.
В лесном хозяйстве Orgyidae наносят очень значительный вред как
хвойным (Pinaceae), так и лиственным (Fagales, Salicales, Juglandales,
Aceraceae, Platanaceae, Tiliaceae) древесным породам. Хвойным массо-
вый вред наносит монашенка {Ocneria monacha L.), реже вредят обыкновен-
ный непарник {Ocneria dispar L.) и розовый {О. matura Btl.), Ocneria
reducta Wlk., виды Dasychira Steph, {abietis Schiff., albodentata Brem.),
Euproctis terminalis Wlk., Orgyia antiqua L. и Diphthera coenobita Esp.
Лиственные деревья повреждаются часто на громадных площадях
обыкновенным {Ocneria dispar L.) и розовым {О. matura Btl.) непарником,
Ocneria serva F., O. xylina Swinh., видами Euproctis Hb. {similis Fuessly,
chrysorrhoea L., scintillans Wlk.), Dasychira Steph, {pudibunda L., pseud-
abietis^A.YLeucorna'SVi^- {salicis L., Candida Stgr., flavosulphurea Ersch.),
Olene Hb. {mendosa Hb., fascelina L.), Acronicta Ochs. {acerisL., rumicis L.,
tridens Schiff.), Subacronicta megacephala Schiff., Calocasia coryli L., Daseo-
chaeta alpium Osb.
Плодовые деревья и кустарники (Rosales, Saxifiagaceae, Rutaceae,
Anacardiaceae) очень сильно повреждаются обыкновенным {Ocneria dis-
1 Это положение, мне кажется, не ограничивается одними Orgyidae, но касается
и многих других вредных насекомых, позвоночных животных и даже растений. Напри-
мер, азиатская саранча совсем не столь многочисленна в тропиках Старого Света,
где она представлена рядом форм, но обильна в умеренных широтах, где условия
для ее существования в какой-то мере новы. Капустная белянка относительно мало-
численна в Средиземноморье, где обнаруживает разнообразие форм, но является мас-
совым видом в Прибалтике, близ окраины ареала. Точно так же озимая совка (Agrotis-
segetum Schiff.) является массовым видом в Европе, где она осваивает новые терри-
тории и не обнаруживает разнообразия форм, но не на юго-востоке Азии, где формы
этого вида разнообразны. Обыкновенный кролик не является массовым видом в местах
природного ареала (Европа), но он сделался массовым вредителем сельского хозяй-
ства на территориях, заново им освоенных (Австралия, Британские острова). Это
же касается филлоксеры {Phylloxera vastatrix Planch.), кровяной тли (Eriosoma
lanigerum Hausm.) и многих других животных, завезенных из Америки в Европу.
Также и элодея (Elodea canadensis) размножилась в Европе в массовом количестве,
будучи завезенной из Америки. Напротив, завезенные в Америку непарный шелко-
пряд, златогузка, японский жук и ряд других видов животных стали там многочис-
ленными и более обильными, чем на территориях основного ареала.
12*
180
ОБЩАЯ ЧАСТЬ
par L.) и розовым (О. matura Btl.) непарником, многочисленными видами
Euproctis Hb. (chrysorrhoea L., similis Fuessly, pulverea Leech, montis Leech,
varians Wlk., conspersa Btl., karghalica Moore, fraterna Moore, xanthor-
rhoea Koll.), Orgyia Ochs, (antiqua L., postica Wlk. thyellina Btl., gono-
stigma F.,prisca Stgr., oetusta Hamps., anartoides Wlk.), Artaxa Wlk. (flava
Brem., flavinata Wlk.), Olene Hb. (mendosa Hb., fascelina L.), Parocneria
lapidicola H. S., P. amanda Stgr., Redoa cygna Moore, Arctornis alba
Brem., Pseudodura dasychiroides Strd., Episema coeruleocephala L.,
Calocasia coryli L., видами Acronicta Ochs, (rumicis L., aceris L., tridens
Schiff., incretata Btl.).
Субтропические древесные и кустарниковые технические ' растения,
как, например, шелковица, чайный куст, Cinchona, Erythrina, некоторые >
виды Lyihraceae, повреждаются видами Euproctis Hb. (similis Fuessly,
atomaria Wlk., fraterna Moore), Artaxa bipunctapex Hamps, видами Orgyia
Ochs, (antiqua L., postica Wlk., turbata Btl.), Olene Hb. (mendosa Hb.,
fascelina L-), Dasychira Steph. (h>rsfieldi Strd., pudibunda L.) и Acronicta
major Brem. В тропиках отмечен постоянный вред культуре технических
пальм (Palmae), которые повреждаются видами Dasychira Steph, (niobe
Weym., endophaea Hamps.), Euproctis Hb. (mediosquamosa Beth.-Bek.,
fraterna Moore).
Огородным и бахчевым культурам (Cruciferae, Polygonaceae, Legumi-
nosae, Euphorbiaceae, Cannabinae) виды Orgyidae вредят главным образом
в тропиках. Из них здесь отмечены виды Orgyia Ochs, (postica Wlk., ve-
tusta Hamps., turbata Btl.), Olene mendosa Hb., Euproctis Hb. (producta
Wlk., conspersa Btl., 1ипа1аУЕ\к.), Arctornis rubricostaFaw., Artaxa bipunct-
apex Hamps. В умеренных широтах вред этим растениям, большей частью
случайный, отмечен лишь от Acronicta rumicis L. и Gynaephora selenitica
Esp.
Хлопок в тропиках постоянно повреждается Olene mendosa Hb. и Or-
gyia postica Wlk., а в умеренных широтах отмечены повреждения видами
Acronicta Ochs, (rumicis L., longa Gn.).
Культурные злаки (Graminae) повреждаются наиболее редко видами
Orgyidae. Здесь отмечены Dasychira securis Hb., Arsilonche albovenosa
Goeze \ Penthophera morio L. и —как случайный объект — непарник
(Ocneria dispar L.).
Вредят указанным хозяйственным растениям как многоядные, так
и специализованные виды Orgyidae. Из числа хвойных растений
повреждаются сосны (Pinus), ели (Picea), пихты (Abies), лиственницы
(Larix), кедры (Gedrus), Tsuga, Thuya, можжевельники (Juniperus), кипа-
рисы (Cupressus), особенно сильно — четыре первых рода.
Из многоядных видов Orgyidae хвойными древесными породами
питаются монашенка, непарный шелкопряд и редко Orgyia antiqua L.
На хвойных же и, в частности, на ели и пихте живет специализованная
'Dasychira abietis Schiff., а на даурской лиственнице—Dasychira albo-
dentata Brem. Из лиственных древесных пород в особенности сильно '
повреждаются дубы (Quercus), буки (Fagus), грабы (Carpinus), березы
(Betula), каштаны (Castanea), клены (Acer), липы (Tilia), вязы (Ulmus),
орехи (Juglans), орешники (Gorylus), конские каштаны (Aesculus), чере-
муха (Prunus), рябина (Sorbus), причем преимущественно многоядными
1 Повреждения лугов и хлебных злаков в умеренных широтах тремя последними
видами являются случайными, местными. Виды Arsilonche Ld. и Penthophera Germ,
вообще в норме редки. Напротив, тропическая Dasychira securis Hb., видимо,
постоянно вредит сахарному тростнику.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
181
видами: Ocneria Hb.r. Euproctis Hb., Acronicta Ochs., Orgyia Ochs., Da-
sychira Steph., уничтожающими также и различные ивоцветные (Salix,
Populus). С последними связаны виды Subacronicta Kozh., Leucoma Steph. '
Другие вредные Orgyidae преимущественно вредят лишь отдельным груп-
пам растений. Так, непарный шелкопряд повреждает преимущественно
дубы и грабы, златогузка — дубы и плодовые деревья, Acronicta aceris L.—
клены, Calocasia coryli L. — березы.
Из плодовых деревьев и ягодных кустарников в особенности повре-
ждаются яблони (Malus), груши (Pirns) и косточковые — абрикосы (Ргц-
nus armenica), вишни (Prunus cerasus), сливы (Prunus domestica), черешни
(Prunus avium, P. mahaleb). В меньшей мере Orgyidae вредят другим
плодовым — персику (Amygdalus persica), миндалю (Amygdalus com-
munis), мушмуле (Mespilus germanica), орехам (Juglans), фигам (Ficus
carica), цитрусовым (Citrus nobilis, C. aurantium), фисташкам (Pistacia
terebinthus, P. vera). Вредят преимущественно многоядные виды .
из тех же родов, что упомянуты для лиственных лесных пород, часто
те же виды (непарный шелкопряд, златогузка), причем как на террито-
рии Европы и Азии, так и на других континентах. Так, даже для Австра-
лии отмечен вред от местных видов Orgyidae (Orgyia anartoides Wlk.,
О. postica Wlk.) ныне разводимым там яблоням и косточковым (сливы),
где специализованные виды Orgyidae — вредители садовых деревьев —
почти отсутствуют. Из более специализованных видов могут быть наз-
ваны Episema coeruleocephala L. и златогузка, связанные преимущественно
с розоцветными, а в Сирии — Parocneria lapidicola Н. S. Другие виды
Parocneria Dyar вредят фисташке. В Ираке фигам вредит, может быть
специализованная на этой группе (Moraceae), Parocneria amanda Stgr.
Вместе с тем и субтропические плодовые растения, как цитрусы, повреж--
даются всеми теми же многоядными видами Orgyidae — Ocneria Hb.,
Orgyia Ochs., Dasychira Ochs., Euproctis Hb.
Ягодные кустарники — виды Saxifragaceae, Rosaceae, Ericaceae и ряд
других, в частности разные виды смородины (Ribes nigrum, R. rubrum,
R. petraeum), крыжовник (Ribes grossularia), малину (Rubus idaeus),
землянику и клубнику (Fragaria vesca), шиповник (Rosa canina), чернику,
голубику и бруснику (Vaccinium vitis-idaea, V. uliginosum, V. myrtil-
lus), облепиху (Hippophae rhamnoides), барбарис (Berberis vulgaris),
лох (Elaeagnus), калину (Viburnum), разные виды винограда (Vitis vini-
fera, V. amurensis) — повреждают все .те же многоядные, обычные у нас
непарный шелкопряд, златогузка, краснохвост, виды Orgyia,Ochs. Для
Средней Азии на этих же растениях, в частности на лохе, отмечены Eu-
proctis karghalicaMooren Acronicta elaeagni Alph., повреждающие, впрочем,
также плодовые деревья и специфичные скорее географически, чем в отно-
шении кормовых растений. Для южного Приморья отмечен вред ягодным
кустарникам из Saxifragaceae от видов Artaxa flava Brem. и Euproctis
Hb., а также Cifuna locuples Wlk. (Куренцов; 1939), но специфичность
этих видов как потребителей Saxifragaceae сомнительна.
К многоядным Orgyidae принадлежат и вредители декоративных расте-
ний и городских парков. Розам, сирени, жимолостям (Lonicera), кизиль-
нику (Соtoneaster), бирючине (Ligustrum), азалиям (Azalea) вредят
те же многоядные виды Orgyia Ochs., Dasychira Steph., Acronicta Ochs..
Ocneria Hb. Парковым и иным городским насаждениям тополей и ив
в ряде случаев очень серьезно вредили ивовые волнянки, в Европе — Leu-
coma salicis L., в Сибири и на Дальнем Востоке — Leucoma Candida Stgr.,
а в горной Средней Азии — Leucoma flavosulphurea Ersch.
182
ОБЩАЯ ЧАСТЬ
Технические-, растения в умеренных широтах: повреждаются слабо,
большей частью случайно и не специфичными видами Orgyidae, обычно
теми же многоядными видами, которые вредят плодовым и лесным поро-
дам. Так, в Испании плантациям пробкового дуба вредят непарный шелко-
пряд и златогузка. На Украине хондрилла (Parthenium argentatum)
повреждается Acronicta rumicis L. Как вредитель чайного куста в Закав-
казье отмечена Episema coeraleocephala L. В Англии хмелю и конопле
вредят обычные у нас виды Orgyia Ochs., Dasychira pudibundaL., Acronicta
rumicis L.
Конопля на Японских островах повреждается Acronicta consanguis
Btl. В Югославии шелковице (Morus) вредит Acronicta aceris L., а на
Японских островах эти же растения повреждают Euproctis similis Fuessly
и Acronicta rumicis L. В тропиках значение Orgyidae как вредителей тех-
нических растений, напротив, очень велико. Чайный куст, какао, дадап
. (Theobroma), масляничная пальма (Elaeis), хинное дерево (Cinchona),
сахарный тростник, клещевина, коф'ейное дерево, Zingibar officinalis,
Hibiscus, Cassia fistula, Uncaria gambar, Zizyphus jujuba, Hevea brasi-
liensis сильно повреждаются на Тайване, в Индокитае, на Малайском
архипелаге, в Индии, на Цейлоне, в Африке рядом видов Orgyidae (Or-
gyia turbata Btl., О. postica Wlk., О. vetusta Bd., Olene mendosa Hb.,Artaxa
scintillans Wlk., A. flava Brem., Euproctis mediosquamosa Beth.-Bek.,
Dasychira niobe Weym., D. endophaea Hamps., Ocneria ampla Wlk.) и рядом
других видов. Некоторые из них, как, например, Redoa cygna Moore,
повреждающая на Тайване чайный куст, могут быть специфичны. Точно
так же хлопчатник в тропиках повреждается очень многими видами
Orgyidae и прежде всего многоядными Orgyia postica Wlk. О. producta
Wlk., О. vetusta Bd., Euproctis Hb., ArZaxa Wlk., Olene mendosa Hb.B уме-
ренных широтах в Средней Азии и в Закавказье отмечены как вредители
хлопка лишь Acronicta rumicis L. (Васильев, 1914), а в Северной Аме-
рике — Acronicta longa Gn.
Полевые культуры и огородные растения повреждаются видами
Orgyidae преимущественно случайно. Помимо отмеченных выше случаев
для непарного шелкопряда известны случаи повреждения капусты
в Индии Euproctis subfasciata Strd., бобов в Китае Artaxa bipunctapex
Hamps., лука и люцерны в Бирме Olene mendosa Hb. и Dasychira securis
Hb., капусты и бобов на Малайском Архипелаге Orgyia turbata Btl.,
Artaxa scintillans Wlk. В наших широтах даже случайный вред огород-
ным и полевым культурам от Orgyidae редок. Специфичным вреди-
телем щавеля, ревеня, шпината можно считать Acronicta rumicis L. (Кеп-
пен, 1883), а бобовых — клевера, эспарцета, гороха — Gynaephora sele-
nitica Esp. (Linnaniemi u. Hukkinen, 1921). В тропиках специфичным вре-
дителем полевых культур — кукурузы, риса, сорго, сахарного тростника
(в Индии) — является Dasychira securis Hb. Наконец случайный вред
от Orgyidae отмечен в тропиках Casuarina, мимозам, акациям. Последние
растения, возможно, специфично повреждаются некоторыми видами,
например Orgyia anartoides Wlk. в Австралии.
Главная масса вредных видов Orgyidae сосредоточена на территории
Евразии и Африки в умеренных широтах, субтропиках и тропиках этих
континентов и на близлежащих островах (Малайский архипелаг, Мада-
гаскар). В Австралии вредные Orgyidae очень немногочисленны, преиму-
щественно те же, что распространены в Индо-Малезии. Очень бедна
вредными видами Orgyidae Северная Америка, где наиболее серьезными
вредителями являются завезенные из Евразии. Заслуживает быть отме-
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
183
ненным, что ввезенный в Северную Америку олигофаг Leucoma salicis L.
'повреждает там в первую очередь ввезенные туда европейские виды то-
полей — Populus alba, Р. nigra, Р. deltoides,. но не туземные — Populus
trichocarpa, Р. tremuloides, и точно так же — ввезенные виды ив и их
гибриды, но не туземные американские ивы. Условия массовых размно-
жений ивовой волнянки в Северной Америке тесно связаны с наличием
ввезенных видов Salicaceae. Напротив, для завезенных в Америку много-
ядных видов Orgyidae, таких, как непарный шелкопряд и златогузка,
пищевой базой явились туземные широколиственные леса, которым они
и наносят в связи с массовыми размножениями грандиозный ущерб. В Юж-
ной Америке среди туземных Orgyidae вредные виды не отмечены вообще.
Для понимания причин массовых размножений Orgyidae, условий
их вредоносности и условий, ограничивающих размножения, а отсюда
и возможности активного влияния на размножения, необходимо рас-
смотреть некоторые основные черты биоценотики этих видов. Для семей-
ства в целом характерны прежде всего высокая степень устойчивости
(стабильности) биоценетических отношений и прочная территориальная
приуроченность видов. Для дендрофильных видов при этом учитывается
постоянство кормовых источников. Это создают многолетние кормовые
растения, при питании отдельными индивидуумами которых могут раз-
виться многие сотни поколений их потребителей видов Orgyidae, как,
например, потребителей буков, дубов, каштанов и других деревьев. Это
'-обусловливает чрезвычайно высокую степень постоянства условий пита-
ния и микроклиматического режима. Последнее ведет к высокой степени
биологической специализации видов, к обеднению ценотических связей
при отборе наиболее полно приспособленных к жизни в данных локальных
условиях популяций. В процессе существования и развития дендро-
•фильных видов Orgyidae естественно возникновение очень мелких попу-
ляций, так как это следует зй адаптацией к жизни на небольших терри-
ториях, адаптацией к деталям основной среды обитания. В связи с этим
в естественных и давно сложившихся ценозах, например в девственных
.лесах тропиков (гилеях) или в лесах субтропиков (например в юго-вОс-
точной Азии), наблюдается пятнистое распределение не только видов,
но и популяций. Такое стабильное во времени и на территории существо-
вание дендрофильных Orgyidae ведет к возникновению стабильных популя-
ций и живущих за их счет паразитических животных, в первую очередь
паразитических и хищных насекомых, которые из года в год находят
в одних и тех же местах, на одних и тех же деревьях их хозяев —особей
Orgyidae. Эти стабильные ценотические отношения Orgyidae находят
выражение во многих частностях биологии, обличающих черты адаптации
к условиям обитания как у хозяев — видов Orgyidae, так и у паразитов —.
тахин, наездников и других. Они выражаются в типе яйцекладки, в ме-
-стах и характере питания, в условиях дневки и ночевки особей, в типе
уничтожения паразитом или хищником жертвы и в ряде других особен-
ностей.
Стабильные условия жизни дендрофильных Orgyidae и их прочные
ценотические отношения характеризуются медленной, отчасти цикличе-
ской, сменой условий обитания. Смена эта стоит в связи с естественным
старением пищевого субстрата — древесных растений, ведущим к воз-
никновению неподходящих для данной популяции условий существования
вообще и питания в частности. Последнее обстоятельство, вызывая необ-
ходимость отыскания новых, благоприятных условий жизни, и ведет
ж постепенной (во времени) смене мест обитания отдельных популяций
184
ОБЩАЯ ЧАСТЬ
данного вида. Кроме того, смена мест обитания обусловливается объеди-
нением и вымиранием популяций на территориях, ставших непригодными
для жизни. Таким образом, в старых лесах наблюдается постоянное, хотя
и медленное, цикличное движение популяций Orgyidae. Вместе с тем
наблюдается постоянное мелкоочажное размножение их на отдельных
благоприятных для жизни территориях на растениях, находящихся в опти-
мальном экологическом состоянии. Параллельно с этим идет процесс даль-
нейшей адаптации особей к изменяющимся условиям — одних, остав-
шихся на старых территориях, на стареющих кормовых растениях,,
других, напротив, переселившихся на новые, более молодые или в
видовом отношении иные кормовые растения. Лишь часть особей попадает
в условия питания, сходные с теми, что' она имела вначале.
Указанные особенности биоценотики дендрофильных Orgyidae благо-
приятствуют возникновению локальных популяций, тем более дробных,
чем старше.по возрасту лесные насаждения, заселенные ими. В девствен-
ных лесах, например в гилеях тропиков, в горных лесах Гималаев, запад-
ного Китая, Маньчжурии или южного Приморья, фактически и наблю-
дается большое число локальных популяций Orgyidae. Здесь наблюдается
большое число локальных форм у широко распространенных видов
и обилие эндемов. При столь большой дробности видовых популяций
в условиях старых лесов, в связи с пестротой кормового, светового и гидро-
термического режима их, естественно, нет места для высокой общей плот-,
ности населения данного вида, а следовательно, нет условий и для мас-
совых размножений. Однообразные условия эти на обширных террито-
риях могут возникнуть лишь при наличии однотипных лесонасаждений,
например при возобновлении леса, при наличии лесных посадок. Они мо-
гут возникать также цосле пожарищ, вырубок.
В связи с изложенным необходимо подчеркнуть отмеченное выше чисто
эмпирическое положение, что высокая плотность популяций у видов:
Orgyidae наблюдается в полиморфных группах и у полиморфных видов,
т. е. у пластичных форм, при соприкосновении их с обширными,
площадями однообразных и благоприятных (в общем) условий оби-
тания. Такое соприкосновение может быть вызвано разными причи-
нами. Оно может быть следствием естественного расселения физиологи-
чески пластичных видов, например видов Gynaephora Hb. в Арктику.
Точно так же контакт этот может возникнуть под влиянием человека —
или в связи с созданием обширных лесных площадей — лесокультурными
работами, закладкой садовых массивов, или, напротив, в связи с заносом
эврибионтных видов на новые незаселенные ими территории. Первый;
случай имеет место с монашенкой в Западной Европе, где она размно-
жается в искусственных еловых насаждениях, и на ibre СССР со злато-
гузкой, которая часто размножается в массах в плодовых садах. Второй
случай дают примеры завоза и размножения в Северной Америке непар-
ного шелкопряда и златогузки. Указанное определяет, тем не менее,
лишь территорию, возможную для массовых размножений Orgyidae.
Темпы размножений, ритм их и периодика различны не только у разных
видов, но и у популяций одного и того же вида и определяются многими,
другими условиями, также требующими анализа.
Прежде чем характеризовать ритм и объем массовых размножений,
необходимо еще дополнить изложенную схему ценотических отноше-
ний Orgyidae. У разных видов имеется различная степень устойчи-
вости связей их с условиями среды, что, конечно, непосредственно влияет
на частоту и объем массовых размножений. В частности резко различна.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
185
прочность ценотических отношений у видов Orgyidae, специализованных
на питании древесными и травянистыми растениями. Малая длительность
вегетации и жизни последних не может обеспечить той степени устойчи-
вости кормовых отношений их потребителей, что дает древесная расти-
тельность. В том случае, если кормовые отношения все же остаются проч-
ными, вид становится высокоспециализованным в отношении цикла разви-
тия, будучи целиком приспособлен к циклу развития кормового растения..
Естественно, что распределение этих видов еще более локально, чем
распределение олигофагов из дендрофильных Orgyidae. Примером видов
Orgyidae, специализованных на питании травянистыми растениями, явля-
ются Penthophera Germ., Oxycesta Hb., Eogena Bd., Simyra Ochs.,
Leiometopon Stgr. Для них возможность массовых размножений требует
сочетания очень многих обстоятельств.
Но питание травянистыми (часто однолетними) растениями может
приводить и к обратному — к высокой степени лабильности ценотических
отношений. Возникает полифагия, как, например, у видов Pharetra Hb.
(rumicis L., auricoma Schiff., euphorbiae Schiff.) или Orgyia Ochs., и чрез-
вычайно расширяются, становясь непрочными, ценотические отношения.
Эти виды имеют условия для массовых размножений, подобные описанным
выше.
Может быть легко прослежена разная степень устойчивости и обшир-
ности ценотических отношений Orgyidae разных климатов. Виды Orgyi-
dae, заселяющие устойчивые условия климата умеренных широт часто
моновольтинны и приурочены к питанию одним или немногими родствен-
ными видами растений. Распространение видов Orgyidae в неустойчивый
климат, в пустынный и степной ландшафты или в горную и арктическую
тундру ведет к расширению кормовых связей и к увеличению объема цено-
тических отношений. При этом возникают приспособления к пережива-
нию неустойчивого термического режима.
Изложенное показывает, что для Orgyidae можно ожидать разные
и специфичные территории для массовых размножений у разных видов
и разную степень их массовости. Для одних видов территории массовых
размножений обширны и постоянны в пределах их ареалов обитания,,
у других они неустойчивы и могут иметьочажный характер, как, например,
у Acronicta rumicis L. Точно так же ритмы размножений, периодика
и характер течения массового размножения, без сомнения, обнаруживают
специфичность. Среди Orgyidae фактический материал позволяет пока
наметить лишь некоторые экологические типы их по характеру града-
ционных кривых и массовых размножений (рис. 83).
Два момента требуют специального обсуждения при рассмотрении
периодики массовых размножений Orgyidae — тип градационной кривой,
которая определяется потенциалом размножения вида, и возникновение
необходимой для размножения экологической ситуации. Потенциал -раз-
множения вредных Orgyidae определяется интенсивностью размножения
(числом яиц от пары особей), устойчивостью особей при развитии в раз-
ных фазах к условиям среды (степенью их эврибионтности), устойчивостью-
их к болезням и паразитам и темпами развития, т. ,е. числом поколений
в год. Для Orgyidae можно говорить о некоторой специфичной максималь-
ной устойчивости особей при развитии к условиям среды и о максимальной
плодовитости как об основных условиях, определяющих величину потен-
циала размножения. Темпы развития в пределах почти всего семейства
однообразны (моновольтинизм), исключая немногие виды (Pharetra Hb.).
Конкретные величины плодовитости и устойчивости особей к условиям
186
ОБ'ЦАЯ ЧАСТЬ
«среды меняются в зависимости от многих условий (см. главу по эколо-
гии), и полная величина потенциала размножения достигается, как част-
ный случай, в условиях оптимума.
Благоприятная экологическая ситуация для размножений Orgyidae
складывается также во всяком случае из двух основных групп факторов —
условий питания и климатического режима. Условия произрастания
основных кормовых растений не
Вое.мя (годы) -----
Рис. 83. Типы градационных кривых. 1 —
монашенка (Ocneria monacha L.); 2 — напар-
ник (Ocneria dispart,.): 3 — щавелевая стрель-
чатка (Acronicta rumicis L.j. А и Б — нор-
мальные годы до и после размножения; I и
Та — подготовительные годы первой и второй;
II — продромальная стадия; III — максимум
(эруптивная стадия) на третьем или четвер-
том году размножения; IV —то- же на чет-
вертом или пятом году размножения; V —
кризис на втором (стрельчатка), пятом
"(непарник) или шестом-седьмом (монашенка)
/году размножения, а — при арабских цифрах
обозначает случаи более длительного или
более кратковременного размножения.
(1838—1840) для Западной Европы и Эверсмана (1844) для СССР. Для
•СССР есть уже более точные данные, за пол столетие, начиная с работ
Кеппена (1883).
Массовые размножения монашенки наблюдаются преимущественно
в северо-западной и центральной Европе — на территории Германской
низменности, в Прибалтике, на северных склонах Альп и в Карпатах
до высоты 657 метров (Комарек, 1931). Это же в южной Швеции. В СССР
случаи массовых размножений монашенки значительно реже. Они наблю-
дались .на территории Белоруссии, северной Украины, в Московской
области, в Горьковском крае, на среднем и южном Урале. Несколько
всегда и не во всех случаях
характеризуют оптимальный
климатический режим. Связь
эта кажется первоначально сама
собой разумеющейся, но на при-
мере монашенки можно видеть
иное. Для этого вида оптималь-
ным кормовым растением яв-
ляется бук, тогда как ныне
климатические условия в бу-
ковых лесах Западной Европы
неблагоприятны. Напротив, в
еловых лесах Европы с их
устойчивым гидротермическим
режимом монашенка находит
необходимые климатические ус-
ловия для развития, несмотря
на менее благоприятные усло-
вия питания. Различие клима-
тических условий по годам и
по сезонам года в разных ча-
стях ареала распространения
дает бесконечное разнообразие
сочетанйй, благоприятных и не-
благоприятных для размноже-
ния. Отсюда понятна чрезвычай-
ная запутанность вопросов мас-
совых размножений Orgyidae,
в частности, например, непар-
ного шелкопряда, златогузки,
монашенки. Тем не менее фак-
тический материал по массовым
размножениям довольно велик,
ибо он накоплен более чем за
столетие, протекшее с момента
опубликования работ Рацбурга
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
187
случаев размножения этого вида было отмечено на Алтае, а также в лесах
южного Приморья (Куренцов, 1939). Известны случаи массовых размноже-
ний монашенки в Японии, где они были отмечены в еловых лесах из Picea
jocoensis и Picea sachalinensis. В средней Европе и Прибалтике, а также
в Белоруссии и на Украине монашенка вредит преимущественно еловым
насаждениям, на востоке Европы северо-восточнее линии Витебск—
Орел, по Шевыреву (1894а, 18946), монашенка чаще повреждает сосновые
насаждения. На Дальнем Востоке вред от нее обнаружен опять-таки
в еловых лесах. Повреждения лиственных пород монашенкой возможны
всюду, но они носят спорадический характер. Случаи размножения на
лиственных породах отмечены в брянских лесах и в грабовых лесах
близ Винницы. В Чехословакии, по Комареку (1931), в начале прошлого
«столетия и до 1892 г. размножения монашенки наблюдались преимущест-
венно в сосновых, а в течение всего остального времени — преимущественно
в еловых лесах. За период с 1892 по 1930 г. в Чехословакии наблюдались
размножения монашенки в еловых насаждениях 418‘раз, в смешанных
хвойных лесах 125 раз, в хвойно-лиственных лесах 70 раз, в со'сно-
вых лесах 24 раза, в пихтовых лесах 2 раза и в березовых 2 раза. Размно-
жения монашенки отмечаются преимущественно в спелых (средневозраст-
ных) еловых насаждениях, полных, порядка полноты 1.0—0.6, редко
в разреженных до 0.4. В средней Европе в старых сосновых лесах до конца
прошлого столетия монашенка имела необходимые условия для размно-
жения. С вырубкой сосновые леса утратили необходимые условия для
жизни этого вида. Они более благоприятны в условиях культуры ели
в средней Европе. Приуроченность монашенки к разным насаждениям
«определяется не столько пищевыми условиями, сколько гидротерми-
ческим режимом. В северных областях ареала этого вида, характеризую-
щихся большей'влажностью, и ныне сосновые леса более благоприятны
для размножения монашенки.
Размножения монашенки следуют через длительные промежутки
времени, порядка 30—40 лет.1 2 Они продолжаготся обычно 5—7 лет
и охватывают очень обширные территории. Например, в 1890—1894 гг.
размножением монашенки в России была охвачена площадь свыше мил-
лиона га. Размножение монашенки в средней Европе в период с 1852
по 1919 г. охватило площадь свыше двух с половиной миллионов га.
Градационная кривая для монашенки характеризуется несколькими
(двумя)годами подготовки размножения, после чего в течение.трех или
четырех лет развивается массовое количество особей, а затем в течение
одного или двух леу наступает снижение численности особей до обыч-
ного числа (рис. 83).
Массовые размножения непарного шелкопряда (рис. 84)а наблюдаются
на территории почти всей Европы, кроме широт выше 60° с. ш., на севере
Африки, в Крыму, на Кавказе, в Западной Сибири (до Енисея) и в гор-
ной Средней Азии. Наиболее часты они в районе Альп и Карпат, где
отмечены начиная с 1720 года. Для России массовые размножения не-
парного шелкопряда указаны Кеппеном (1856—1879), Авериным (1913),
1 В Чехословакии массовые размножения монашенки наблюдаются последнее
время часто, через 7—10 лет. Лейтольд (Leuthold, 1931) полагает увеличение частоты
размножения монашенки за счет экстенсивной культуры ели.
2 Подробно литература по размножениям непарного шелкопряда как . в Старом,
так и в Новом Свете собрана Шедлем [S с h е d 1, 1936. Der Schwammspinner (Por-
thetria dispar L.) in Euroasien, Afrika und Neuengland. Monogr. angew. Entom., 12 :
1—242)],
188
ОБЩАЯ ЧАСТЬ
Камышным (1924), Римским-Корсаковым (1913). Они более редки
в Средиземноморье, но отмечены для Испании; очень редко массовые
размножения наблюдаются в северной Африке. Более часты в северном
Средиземноморье, где отмечены для Италии, для Крыма. На северном
Кавказе, на южном Урале, в северном Казахстане, на Алтае и в Сая-
нах размножения непарного шелкопряда значительно реже. В южном
Приморье и в Японии непарный шелкопряд обычен, но размножения
его редки и не достигают той степени, что наблюдается в Европе.
Всюду для непарного шелкопряда кормовыми растениями являются
лиственные древесные породы, прежде всего дубы. В Крыму важную
роль в питании непарного шелко-
пряда имеет грабинник (Carpinus
orientalis Mull.), тогда как обыч-
ный граб повреждается слабо
(Пархоменко, 1936; Пятницкий,
1935). Разные виды дубов повре-
ждаются также в разной степени
(см. таблицу 25), причем некото-
рые виды, например Quercus lan-
uginosa Lam., повреждаются почти
столь же значительно, как и во-
сточный граб (грабинник).
В Сибири, на Алтае, Бей-Би-
енко (1924) наблюдал размноже-
ние непарного шелкопряда в со-
сновых (кедровых) насаждениях
(Pinus Silvestris, Р. sibirica).
Рис. 84. Динамика численности непарни-
ка {Ocneria dispar L.) в Крыму в течение
периода массового размножения 1929—
1934 гг. (1) и число яиц в одной кладке
по годам (2). (По Пархоменко, 1936).
В Средней Азии размножения
непарного
Мною в западных Саянах (1923)
наблюдалось массовое размноже-
ние непарного шелкопряда в пой-
ме Енисея (р. Тукеек-Кем) в че-
ремухово-березовых насаждениях,
шелкопряда наблюдались в
плодовых садах и лесах предгорий, состоящих из диких плодовых
(яблоня, груша, терн, грецкий орех, лох и др.). Размножения в
хвойных лесах наблюдаются редко, причем только в сосновых. Но и в
лиственных лесах поражаются средневозрастные или даже молодые,
часто разреженные насаждения полнотою порядка 0.2—0.5 (рис. 85).
Таблица 25
Степень повреждения непарным шелкопрядом в Крыму
некоторых древесных пород (по Пархоменко, 1936).
Название породы Число проб % повреж- дений
Quercus sessiliflora Ehrh 57 18
Quercus lanuginosa Lam 52 53
Carpinus orientalis Mull. ..... 50 62
Carpinus betulus L 25 10
Fagus orientalis L 13 0.4
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
189
Массовые размножения непарного шелкопряда в местах наибольшего
обилия вида следуют, например в Крыму, через 10—20 лет1 и продол-
жаются обычно 2—4 года (рис. 83). Массовым размножением обычно ох-
ватываются обширные территории в несколько сотен тысяч гектаров,
хотя обычно не столь значительные, как при размножениях монашенки.2
Часты местные, локальные размножения. Градационная кривая (рис. 84)
характеризуется одним подготовительным годом и следующими за ним
одним, двумя, редко тремя годами массового размножения. Падение
плотности популяции до нормального уровня наступает обычно в те-
чение одного года кризиса.
Массовые размножения златогузки имеют много общего с тем, что
описано для непарного шелкопряда, но площадь, где они наблюдаются
в пределах ареала распространения, значительно меньшая, чем для
последнего. Они отмечены
для средней Европы, При-
балтики, Украины, южного
Приуралья, Кавказа и По-
волжья. Обычно, и в ряде
мест очень часто, наблюда-
ются локальные размноже-
ния этого вида в садах и лес-
ных полезащитных полосах.
В лесных массивах, и при
этом только лиственных ле-
сах, размножения златогузки
наблюдаются реже и на ме-
нее обширных территориях,
чем то констатировано для
непарного шелкопряда. Та-
кие случаи отмечены, на-
пример, для Поволжья, где в
1921-—1922 гг. златогузка
размножилась на площади
в несколько десятков квад-
ратных километров дубового
леса. Массовые размножения
Рис. 85. Плотность населения непарника в
Крыму в зависимости от полноты ' насаждения.
(По Пархоменко, 1936). 1 — степень поврежде-
ния леса; 2 — число яйцекладок в среднем на
одно дерево при разной полноте насаждения.
в дубовых лесах отмечены
и для окраин ареала этого вида, например для северной Африки
и Иберийского полуострова. Периодика массовых размножений злато-
гузки изучена недостаточно полно. Они наблюдаются, по всей вероят-
ности, несколько чаще, чем у непарного шелкопряда, но охватывают
меньшие площади. Часто они имеют очажный характер. Невидимому,
аналогична картина массовых размножений и ритма их для краснохво-
ста {Dasychira pudibunda L.), ивовой волнянки (Leucoma salicis L.) и видов
1 Для Крыма, например, массовые размножения отмечены в следующие годы
(Кеппен, 1883; Кулагин, 1923; Мокржецкий и Щеголев, 1914; Пятницкий, 1935;
Пархоменко, 1936): 1842, 1861—1863, 1869—1871, 1884—1885, • 1911—1913, 1920—
1921, 1929—1932. В условиях Крыма по Пятницкому (1935) дикая груша избегается
непарным шелкопрядом. Это давно еще отмечал Кеппен (1883) для б. Харьковской
губернии. В то же время в Средней Азии дикие плодовые потребляются непарником
•охотно.
2 Исключая, конечно, случай массового размножения непарного шелкопряда
при колонизации им Северной Америки, но и здесь периоды более интенсивного раз-
множения сменялись депрессиями.
190
ОБЩАЯ ЧАСТЬ
Orgyia Ochs, {antiqua L., gonostigma F.). Для всех этих видов типичны
значительно меньшие территории массовых размножений, чем для
непарного шелкопряда, большая очажность их и более короткий период
градационной кривой. Массовые размножения краснохвоста {Dasychira
pudibunlaD.) отмечены для средней Европы и для зоны широколиственных
лесов СССР. В течение последних лет они наблюдались в дубовых лесах в
Харьковской области. В 1926—1928 гг. и особенно в 1932 г. массовое раз-
множение этого же вида, также в дубовых лесах, наблюдалось в Воронеж-
ской области на территории свыше 30 тысяч га. В 1933 г. здесь же и в.
Харьковской области наблюдалось размножение златогузки на площади
свыше 10 тысяч га. Массовые размножения ивовой волнянки наблюдались
в разных местах ареала —в Ленинграде в 1939—1940 гг., в 1943 г.
в парковых насаждениях Ленинграда. В парковых же насаждениях они
отмечены для Москвы, Северного Кавказа, Западной Сибири, Среднего
Поволжья. Они носят локальный характер, так как в сильной мере
связаны с питомниками, парковыми насаждениями, а в нетронутой при-
роде с уремами. Размнсжения видов Orgyia Ochs, нередко очень значи-
тельны, например размножение Orgyia antiqua L. в березовых лесах
окрестностей Ленинграда в 1938—1939 гг. Несмотря на это, они никем
сколько-нибудь специально не изучались, хотя, по видимому, явление
это наблюдается не редко в лиственных лесах средней Европы, Европей-
ской части СССР и, вероятно, в Западной Сибири.
Из вышеизложенного видно, что между частотой размножений,
объемом их (площадью, охватываемой размножением), длительностью
размножений и типом градационной кривой данного вида имеется связь.
Так, у видов Orgyidae ритм массовых размножений определяется не
только сочетанием благоприятных для вида условий среды, но и биоло-
гическими особенностями, в частности специфичной лабильностью про-
цессов размножения, длительностью цикла развития и величинами потен-
циала размножения в целом. Виды с длительными циклами (годичными
у Orgyidae), с малым потенциалом размножения, со стойкими ценоти-
ческими отношениями характеризуются более редкими периодами массо-
вых размножений. Для них типична многолетняя градационная кривая.
Виды поливольтинные, биологически лабильные, с короткими циклами
развития, эврибионтные, с обширными, но неустойчивыми ценотическими
отношениями характеризуются частыми массовыми размножениями иодно-
летней или во всяком случае немноголетней градационной кривой. Приме-
ром первых может служить монашенка, примером вторых—Acronicta rumi-
cis L. Сравнение типов градационных кривых (рис. 83) и биологических
особенностей видов Orgyidae (табл. 24) показывает, что у моновольтинных
видов с малым потенциалом размножения массовые размножения опре-
деляются возникновением длительной благоприятной экологической
ситуации (оптимум). Чем ниже потенциал размножения и чем устойчивее
ценотические отношения, тем длительнее должен быть подготовительный
период к размножению и тем длительнее будет период размножения,
а следовательно, тем обширнее будут территории, охваченные массовым
размножением (монашенка, непарный шелкопряд). При высоком потен-
циале размножения и высоких темпах развития отношения эти обратны,
условия для массового размножения могут возникать часто, но размно-
жения, естественно, носят очажный характер и они кратковременны.
Благоприятная экологическая ситуация для размножения Orgyidae
оценивается с двух сторон: со стороны физических условий среды и со
стороны биотических факторов, в особенности паразитов и хищников,
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
1ST
живущих за счет данного вида. В начальный период изучения массовых
размножений вредных Orgyidae (в связи с завозом непарного шелкопряда
и златогузки в Северную Америку) большая часть авторов была склонна
оценивать положительную экологическую ситуацию почти исключительно-
в пользу биотического фактора — численности паразитов. Этому способ-
ствовало успешное применение биологического метода борьбы с непарным?
шелкопрядом и златогузкой в лесах Северной Америки, где виды эти.
в течение многих лет давали, как казалось, стойкое размножение. При-
менение специфичных паразитов (Apanteles, некоторых тахин — Сот-
psilura) и в особенности яйцеедов [Schedins, Anastatus), сцециализован-
ных на Orgyidae, устранило опасность массовых размножений этих видов?
в Северной Америке. Существенную роль сыграли также хищники из
Carabidae (Calosoma). Тем не менее, более внимательное изучение плот-
ности населения непарного шелкопряда в Северной Америке указывает
на наличие и там периодичности размножений этого вида на фоне его-
все увеличивающегося ареала (рис. 86).
Таблица 24?
Связь периодики массовых размножений и типов градационных кривых . Orgyidae с характером и величиною потенциала . размножения
Название вида Длительность периода между размноже- ниями (лет) Длитель- ность градацион- ной кривой (лет) Вольтинизм Плодови- тость 1 самки (кол. яиц)?
Монашенка (Ocneria то- nacha L.) 30—40 5—7 Моновольтин- 15’)—300-
Непарный шелкопряд (Oc- neria dispar L.) 10—20 2—4 ный Моновольтин- 400— G001'
Щавелевая стрельчатка (Acronicta rumicis L.) . . Часты, почти 1 ный Поливольтин- 200—600
ежегодны ный
Попытки применения местных паразитов в борьбе с монашенкой?
и непарным шелкопрядом в Европе не дали положительных результатов..
Интродукция новых паразитов и в Евразии давала лишь некоторое,,
иногда заметное снижение численности.2 Вместе с тем параллельное
изучение динамики численности вредных Orgyidae (в особенности мона-
шенки) в естественных ценозах Евразии в течение всего периода массо-
вого размножения показало отставание увеличения плотности населения
паразитов в годы подъема численности Orgyidae. Таким образом увели-
чение плотности населения вредных видов Crgyidae не может быть,
поставлено в связь только с паразитизмом, оно должно иметь и иные при-
чины. Лишь в случае падения численности в завершающий (или завер-
шающие) год отмечается относительное увеличение плотности населен
1 У отдельных особей непарника в особо благоприятных условиях плодовитость
может достигать 1800 яиц, но средние ее величины обычно колеблются в указанных
пределах.
2 Положительные результаты в борьбе с непарным шелкопрядом биологический,
метод дал в пределах естественного обитания на окраинах ареала.
192
ОБЩАЯ ЧАСТЬ
ния паразитов.1 Увеличение процента зараженности паразитами может
быть обусловлено в этом случае также и уменьшением числа жертв
(хозяина).2 Наблюдение физиологического состояния особей вредных
Orgyidae в течение периода размножения показывает, особенно в пе-
риоды падения и подъема численности, большие отличия. Происходит
увеличение веса, большая плодовитость (большее число потомков), более
равномерное соотношение полов, полное оплодотворение всех яиц,
меньшая смертность преимагинальных фаз в течение развития. По Фёль-
Рис. 86. Динамика численности непарника в широколиственных лесах Север-
ной Америки за период 1905—1933 гг. по числу яйцекладок (оплошна линия)
и гусениц (пунктир). (По Шедлто, 1936).
келю (Voelkel, 1930), это особенно типично в первые два года массо-
вого размножения (подготовительные). В завершающий период размноже-
ния, наряду с повышением смертности при развитии, наблюдается также
1 Комарек в начальный период своих работ по массовым размножениям мона-
шенки в Чехословакии признавал паразитов, в частности тахин, единственным серьез-
ным регулятором размножений этого вида. Тем не менее, после ряда лет специальных
изысканий этот автор (1931—1937) переменил мнение и считает роль паразитов как
регуляторов размножений второстепенной. По его мнению, роль их в нарастании
численности отсутствует (или очень невелика). Еще ранее Ружичка (Ruzicka,
1923—1931) защищал это положение, основываясь на своих работах по монашенке,
также в Чехословакии. (См. также сноску на стр. 199).
2 Интересно наблюдение Смите ван-Бёрджеса (Smiths van Burgess, I, 1924), кото-
рый указывает для Голландии факт заражения тахинами почти каждой гусеницы
монашенки в конце массового размножения (в период кризиса). Но, по его наблюде-
ниям, многие особи, несмотря на это, дают нормальных имаго. Автор приходит к за-
ключению, что в период массовых размножений возникает повышение иммунитета
к паразитам.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ 193
снижение плодовитости особей, неправильноё соотношение полов,
наличие неоплодотворенных яиц. Все это показывает, что факторы,
регулирующие размножение Orgyidae, в частности специально изученных
монашенки, непарного шелкопряда, златогузки и ивовой волнянки,
лежат вне сферы влияния паразитов. Действие последних является лишь
частным моментом, накладывающимся на общий комплекс факторов,
влияющих на изменение численности в период размножения. Условия
для нарастания численности создает весь комплекс среды смещением
к оптимуму для данного вида (Ружичка, 1923—1931; Яниш, 1936 —
1939).
Роль климата в массовых размножениях Orgyidae ныне оценена всеми.
Не случайно, конечно, что зона массовых размножений монашенки не
выходит как в широтном, так и в высотном направлениях (в Альпах,
Карпатах) за пределы июльской изотермы 16°Ц. Влияние климата
только на физиологическое состояние особей и биологические моменты
в их жизни чрезвычайно велико и сложно. Оно возможно в отношении
снижения или увеличения плодовитости как путем непосредственного
воздействия условий тепла и влажности, а также радиации, так и через
питательность кормовых растений. Значение погодных условий перво-
степенно в смертности особей при росте и развитии. Не менее велика
их роль в поведении особей, в активности полов, в спаривании, а отсюда
и в полноте оплодотворения. Не менее важно их влияние на отыскание
пищи. Чрезвычайно велика роль термогигрорежима в период зимовки
и диапаузы Orgyidae, а также влияние этих условий на необходимый
•синхронизм в развитии кормовых растений и появлении питающихся
фаз Orgyidae. Наконец чрезвычайно многообразно влияние климата на
жизнь комплекса паразитов, сверхпаразитов и хищников-потребителей
Orgyidae. Здесь, кроме упомянутых влияний, в той же мере действитель-
ных, как и для Orgyidae, возникает еще новое условие—нарушение
синхронизма в появлении фаз развития паразита и жертвы. В целом
признается большая чувствительность паразитов к климатическим
влияниям и меньшая их эврибионтность по сравнению с хозяевами —
видами Orgyidae, например большая чувствительность Tachinidae к све-
товым и термическим условиям, по сравнению с Orgyidae.
Для наиболее заметных и экономически важных видов Orgyidae —
монашенки, златогузки, непарного шелкопряда — ныне выявились уже
направления погодных изменений, благоприятные для возникновения
размножений. Ружичка (1931) считает, что в обычные годы в условиях
средней Европы местами резерваций монашенки являются смешанные
буково-еловые или широколиственные леса, реже боры, но они всегда
характеризуются недостаточно устойчивым режимом влажности. Еловые
леса, благоприятные в отношении влажности в обычные годы, имеют
в условиях средней Европы слишком низкую температуру. В теплые
годы в еловых лесах, при их высокой и стойкой влажности, наблюдается
также благоприятный термический режим и, таким образом, создаются
наилучшие условия для размножения монашенки. В эти (тепцые) годы
смешанные или широколиственные леса, а также боры оказываются
слишком сухими и жаркими, и монашенка в них исчезает. Непосредствен-
ными наблюдениями в природе показано, что нормальная активность
имаго (в частности спаривание бабочек) возможна лишь при темпера-
туре выше 15°, т. е. в теплые ночи [Седлачек (Sedlaczek, 1909—1911)].
Седлачек (1918), а за ним Лейтольд (1931), Кнохе (Knoche, 1929): и дру-
гие авторы признают, что теплая (даже жаркая)'погода в июле и августе,
13 Фауна СССР, И. В. Коисанчиков.
194
ОБЩАЯ ЧАСТЬ
при условии теплого мая1 в.течение двух, трех лет подряд, является
основным фактором для размножения монашенки в еловых лесах Европы.
Теплая весна — необходимое условие для нормального выхода из яиц
гусениц, теплое лето благоприятно для роста и размножения. Непосред-
ственное доказательство значения климатических условий в размноже-
ниях монашенки (в связи с влиянием их на активность имаго) дает Кнохе
(1929). Он показал, что в 1922 г. (в Альпах) на высоте до 350 метров яйца;
монашенки встречались почти во всех кладках нормально оплодотворен-
ные (неоплодотворенных яиц было только 8%), тогда как на высоте-
630 метров в тех же местах неоплодотворенными были все яйца. По Сед-
лачеку (1909—1918), для нормальной активности имаго монашенки
благоприятны температура 18° и влажность воздуха порядка 70%.
Полное оплодотворение всех яиц происходит при условии наличия более-
12 теплых вечеров (ночей с температурой близ 18°) в период лёта,
имаго —в августе.
Независимо от приведенных наблюдений в природе, потребовавших
много внимания и опыта со стороны исследователей, эксперимен-
тальные исследования Цвёльфера (1934) показали существование в цикле-
развития монашенки двух биологических периодов, особенно чувстви-
тельных в отношении влияния климата. Первый из них совпадает с раз-
витием яиц и с первыми днями жизни гусениц в весеннее время. В при-
роде он протекает в конце апреля и в мае. Нормальное развитие в этот-
период возможно при 10—25° для выхода гусениц2 и 18—25° для их раз-
вития в первой стадии. Наиболее благоприятна температура порядка 18°..
Второй критический период в развитии преимагинальных фаз мона-
шенки совпадает с периодом максимального роста гусениц и с разви-
тием куколок. Оптимальными условиями являются термический режим
порядка 20—27°. Температура ниже 20° заметно ухудшает рост, а ниже-
16° резко неблагоприятна. Условия увлажнения благоприятны в преде-
лах высокой, но стойкой влажности, около 70—90%. Такие же-
условия благоприятны и для жизни имаго.3 В целом, как видно, данные,,
полученные экспериментальным методом, вполне подтверждают и разъ-
ясняют наблюдения, сделанные в природе. Мнение Седлачека (1909,
1911) и Цедербауера (1911), обоснованное чисто эмпирическими, поле-
выми материалами, имеет глубокую физиологическую основу.
Значение пищевых условий в массовых размножениях Orgyidae.
менее определенно, что, может быть, , стоит в связи с многоядностью-
1 Сопоставления Цедербауера (Zederbauer, 1911) показали, что массовые размно-
жения монашенки в средней Европе в зоне с 400—1000 мм осадков следуют за теплыми
и засушливыми периодами. Им были использованы данные Брюкнера по периодиче-
ским изменениям климата в Европе.
2 Наличие в природе большого числа погибших яиц отмечалось многими авто-
рами [Тубёф (Tubeuf, 1892); Эшерих (Escherich, 1911, 1912); Кнохе, 1912, 1932]. При-
чины гибели их различны, и она обусловлена не только отсутствием оплодотворения.
Она может быть вызвана также дефектами его. Так, развитие куколок при повышен-
ной температуре ведет к неправильностям оплодотворения и к появлению бесплодных
яиц. Цвёльфер (1934) различает три типа погибших яиц, которые можно обнаружить,
весною. Первый тип характеризуется гибелью в самые начальные периоды развития,,
что связано с влиянием среды поздним летом и осенью. Второй тип характеризуется
гибелью при вполне развившемся эмбрионе. В этом случае гибель происходит в тече-
ние зимы. Третий тип характеризуется гибелью при наличии вполне развившейся
гусеницы. В этом случае гибель наступает весной, в связи с неблагоприятными усло-
виям? для выхода гусениц.
з По Ребелю (Rebel, 1921), сильные, особенно ночные, холодные дожди губи-
тельны для бабочек монашенки .
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
195
Рис. 87. Значение экспозиции скло-
нов в плотности населения непар-
ника в Крыму. (По Пархоменко,
1936).
гусениц, а в младших стадиях также в связи с выносливостью их к го-
лоду. Вахтль (Wachtl, 1910) считал важным фактором размножений
монашенки неприспособленность к питанию молодых (первой стадии)
гусениц этого вида прошлогодней хвоею ели. Предполагая возможность
гибели гусениц монашенки от голода весною, при позднем развитий
еловой хвои, Вахтль (1910) предложил культуру поздней, зеленой
разновидности ели (Picea excelsa v. chlorocarpa) вместо обычной (Picea
excelsa v. erythrocarpa), так как первая дает молодую хвою на месяц
позже. Седлачек (1910) еще тогда же указывал на крайнюю растяну-
тость выхода .гусениц монашенки, что, при стойкости их к голоду
и в связи с возможностью питания почками, делает сомнительным
большое значение позднего развития зеленой разновидности ели.
Исследования последнего времени по значению растений (в разной ста-
дии роста) в росте гусениц монашенки заставляют осторожно относиться
к утверждению Седлачека, хотя эти данные говорят пока лишь о воз-
можности запаздывания выхода гусениц
монашенки, но не о возможности преж-
девременного их выхода, на что ука-
зывал Седлачек. В сосновых лесах
роль асинхронизма в развитии хвои
в питании гусениц монашенки имеет
меньшее значение, так как ими по-
едается и старая хвоя. Тем не менее
имеются указания, что чисто еловые
насаждения в пищевом отношении ме-
нее благоприятны для жизни мона-
шенки, чем смешанные елово-буковые.
Положительное значение примеси бука
в еловых лесах можно ожидать и на
основании экспериментов Мэркса (1935).
Ряд авторов и особенно Мокри
(Mokry, 1922, 1924) склонны вообще
отрицать роль пищевых условий в мае- •
совых размножениях Orgyidae, в частности монашенки, но они трак-
туют влияние пищи слишком абстрактно.
Степень изученности условий массовых размножений непарного
шелкопряда значительно меньшая, чем. для монашенки. Для Крыма
отмечено (Мокржецкий, 1914; Пятницкий, 1935), что массовые размно-
жения следуют за теплыми сухими годами. В целом в пределах ареала
этого вида замечено, что более высокая плотность популяции наблюда-
ется в сухих лиственных лесах. В гористых местностях, например на
Крымской Яйле, размножения непарного шелкопряда наблюдаются
на южных, юго-восточных и юго-западных склонах (рис. 87), как на это
специально указывал Пархоменко (1936). Анлогичные наблюдения
сделаны Бей-Биенко (1924) на Алтае, а в Саянах мною (1923). Размноже-
ния непарного шелкопряда в хвойных лесах редки; они наблюдаются
лишь в сосновых или кедровых (Pious sibirica).
На основе всего комплекса материалов по непарному шелкопряду
могут быть даны лишь общие соображения о благоприятном погодном
режиме для размножений. Весной благоприятна теплая и сухая погода
с дружным развитием листвы, при средних суточных температурах выше
15° до 18—20°. Для роста гусениц важна умеренная влажность, но не
слитком высокая температура, порядка средней суточной 20—22°.
13*
t96 ОБЩАЯ ЧАСТЬ
Падение температуры (ниже 18° средней суточной), особенно длительное,
при сочетании с высокой влажностью влияет на рост резко отрицательно.
Для жизни имаго и развития куколок также благоприятна не дожд-
ливая теплая погода, при средней суточной температуре порядка 22—
25°. В этих условиях оплодотворение яиц протекает быстро и полно,
а начальные моменты их развития — успешно. Эти же условия при влаж-
ности порядка 75% благоприятны и для развития куколок.
Условия массовых размножений златогузки, ивовой волнянки,
краснохвоста, видов Orgyia Ochs, {antiqua L., gonostigma F.) ждут еще
специальных исследований. Сейчас могут быть сделаны лишь некоторые
замечания. Для размножения златогузки (в условиях широколиствен-
ных лесов) благоприятны сухое и теплое лето и сухая зима. Влажная,
хотя бы и теплая, зима с оттепелями и дождями крайне неблагоприятна
для зимовки. Наоборот, для размножения краснохвоста {Dasychira
pudibunda L.) благоприятна теплая, хотя бы и влажная зима, при теплом
и сухом (не дождливом) лете. Влажное, хотя бы и теплое, лето харак-
теризуется сильным развитием заболеваний гусениц (Ритзема-Бос, 1914).
Для размножения Orgyia antiqua L. благоприятны эти же условия,
особенно засушливое и теплое лето с ясной погодой в период лёта имаго.
Для размножения Orgyia gonostigma F. благоприятны такие же летние
условия, но необходима сухая зима. По отношению к условиям зимовки
виды Orgyidae можно разделить на две группы. Для одной группы видов,
зимующих в личиночной фазе (даже в гнездах), благоприятна сухая
погода в течение зимы; это справедливо для Dasychira abietis Schiff.,
Olene fascelina L., Orgyia gonostigma F., Euproctis chrysorrhoea L., Gynae-
phora selenitica Esp. и для видов, зимующих в фазе яйца с вполне сфор-
мированной гусеницей, как Ocneria Hb. {dispar L., monacha L.). Для дру-
гой группы видов, наоборот, благоприятна мягкая, хотя бы и влажная
или даже отчасти дождливая (с оттепелями), зима. Сюда относятся
Dasychira pudibunda L., Calocasia coryli L., Subacronicta Kozh., Acronic-
ta Ochs., некоторые виды Orgyia Ochs., т. e. виды, зимующие в фазе
куколки или яйца на ранних стадиях развития последнего.
Угасание массовых размножений у всех видов Orgyidae имеет общие
черты. Возрастание плотности населения ведет к плохому росту, раз-
реживанию и частичной (или даже полной) гибели кормовых растений.
За этим уменьшается общее количество корма и падает его питательность.
Слабо растущие листья растений становятся более бедны белком и более
богаты древесиной, сильно меняется их химизм в целом. Сильно меня-
ется и общий физический режим среды в сторону большей изменчивости
его. Воздух в таких разреженных лесах сильно нагревается днем и осты-
вает ночью, более сух и более подвижен. Голодание гусениц и каче-
ственно недостаточное питание их ведут к заболеваниям. В частности,
среди Orgyidae широко распространяются эпидемии полиэдренной болезни,
желтухи. Развитие ее типично для кризиса (завершающих лет) при
размножениях монашенки, непарного шелкопряда, Dasychira pudibunda
L., Orgyia antiqua L. Возрастает процент особей, пораженных парази-
тами. Недоедание и заболеваемость гусениц ведут к снижению плодови-
тости особей, переживших период массового размножения (рис. 88).
Голод ведет к большей смертности самок, требующих у Orgyidae всегда
большего количества пищи и большего срока для развития. В целом
завершение йассового размножения характеризуются пониженной плодо-
витостью особей, высокой смертностью, развитием эпизоотий, миграцией
и расселением (выселением из очагов размножения) оставшихся в живых
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
197
особей.1 Миграция обусловливается возникновением непригодных усло-
вий для жизни данной популяции на территории, где произошло мас-
совое размножение, в течение ближайшего отрезка времени. Развитие
эпизоотий (полиэдренной болезни, флашерии), возможно, обусловлено
еще и ролью паразитов, в частности наездников, переносящих инфекцию
от одной особи к другой.
Для борьбы с вредными Orgyidae могут быть применены профилак-
тические и истребительные меры, причем для проведения первых важен
прогноз возникновения размножений. Лишь при этом условии возможно
сколько-нибудь полное и рациональное использование профилактиче-
ских мероприятий. В целом оно возможно в садах, в лесных и садовых
питомниках, лесных полезащитных посадках, в парковом и городском
хозяйстве и лишь в исключительном случае в лесных массивах. Истре-
бительные меры применяются всегда в экстренных случаях обычно после
того, как размножение уже произошло. Эти меры проводятся всюду,
где они оправдываются экономичес-
ки. Они могут быть механическими,
биологическими и химическими. Для
проведения истребительных меро-
приятий, так же как и для прове-
дения профилактических, необходи-
мы сведения о плотности населения,
т. е. нужны обследования заражен-
ности территории данным, вредным
видом Orgyidae. Такие сведения
необходимы и для прогноза измене-
ний численности, а также для учета Ю0% 7У% периферия новый
эффективности проведенных меропри- °43г 1/433 04333 0433
ятий.
В целом методы учета плотности
населения Orgyidae те же, что и для
других насекомых, —учет числа осо-
Рис. 88. Зависимость размеров куколок
самок непарника от плотности на-
селения гусениц. (По Пятницкому,
1935).
бей на единицу площади или на
растение (дерево). Не имея возможности изложить их подробно,
я здесь затрагиваю те из них, которые специфичны для Orgyidae.
Прежде всего необходим учет зимующих фаз (зимующего запаса)
данного вида—яиц непарного шелкопряда, розового непарника, мона-
шенки, античной волнянки, синеголовки (Episema coeruleocephala L.).
Учет производится по местам яйцекладки — на коре деревьев, на лесной
подстилке, на корнях в их припочвенном слое, на камнях и т. д. Точно
так же производится учет зимующих гусениц златогузки, желтогузки
и туркестанской златогузки (Euproctis karghalica Moore), ивовых волня-
нок, лунницы (Cynaephora selenitica Esp.), Dasychira abietis Schiff., Orgyia
gonostigma F., тоже зимующих в щелях коры, на ветвях, в гнездах, в лес-
ной подстилке или в верхних слоях почвы. Наконец зимующий запас
куколок должен быть учтен для краснохвоста (Dasychira pudibunda L.),
Acronictinae (Acronicta Ochs., Calocasia Ochs., Subacronicta Kozh., Daseo-
chaeta Warr.), зимующих в лесной подстилке или в старой древесине.
1 Например, по наблюдениям Мокржецкого и Брагиной (1917) в Крыму в 1913 г.,
при массовом размножении непарного шелкопряда в 1914 г. наступило резкое ослабле-
ние особей. Самки были мелки и откладывали вместо обычных 500—600 яиц лишь
60—200 яиц, при этом вообще наблюдалось резкое преобладание самцов.
198 ОБЩАЯ ЧАСТЬ
Отыскание гусениц и куколок наиболее трудно и требует больших
навыков.
Кроме того, в течение вегетационного периода учет всех фаз развития
возможен при помощи сбора гусениц, учета числа падающих экскремен-
тов на светлые щиты, подставленные под деревья, сбора яиц или ловли
бабочек. Для Orgyinae есть специфический метод учета сампов по вылову
их на самок [Бёрджес, 1924; Лемари (Lemarie, 1933)]. Может быть также
применен метод самоловки на свет или активный сбор самцов, прилетев-
ших к самке. Этот метод может дать оценку площади, зараженной непар-
ным шелкопрядом или Orgyia antiqua L., наиболее резко реагирующих
на присутствие самок.1 Не менее важен метод учета числа молодых гусе-
ниц при помощи клеевых щитов, на которые приклеиваются разносимые
ветром гусеницы первой стадии. Он позволяет не только судить о плот-
ности населения данного вида, но и оценить опасность заселения данной
территории вредными Orgyidae.
Из профилактических мер борьбы с вредными Orgyidae могут быть,
упомянуты прежде всего мероприятия по уничтожению зимующего
запаса, например сбор и уничтожение яиц непарного шелкопряда, мона-
шенки и античной волнянки соскабливанием их ножом с коры деревьев
в ведра и сжиганием; сбор кладок монашенки Производить труднее в силу
того, что они менее заметны. Яйца непарного шелкопряда в ряде случаев
проще уничтожать смазыванием кладок керосином с добавкой дегтя,
чтобы обработанные кладки были легко заметны по их черному цвету.
.Эффективно уничтожение зимующих гнезд златогузки сбором и сжиганием
их осенью, после опадения листвы (в садах мера эта особенно эффективна,
так как проходит параллельно , со сбором гнезд боярышницы). Сбор
и уничтожение гусениц желтогузки и ивовой волнянки, зимующих пооди-
ночке, возможен лишь в исключительных случаях, на особо ценных расте-
ниях и при особенно обильном размножении. Точно так же уничтожение
куколок Acronictinae, Dasychira pudibunda L., гусениц Dasychira abietis
Schiff., Gynaephora selehitica Esp., зимующих в лесной подстилке может
быть достигнуто использованием домашних животных — выпасом их
(куры, свиньи). Изоляция древесных крон от зимующих в подстилке
и на прикорневых частях стволов личиночных фаз Orgyidae достигается
со значительной полнотой применением клеевых колец из гусеничного
клея. Неимеющие доступа к кронам гусеницы вредных Orgyidae погибают
с голоду и уничтожаются паразитами и хищниками-. Клеевые кольца
накладываются на деревья возможно выше, перед началом первых крупных
ветвлений, обычно на высоте груди. Для накладки клеевых колец сначала
сглаживается скобелем шероховатая кора дерева, а затем лопаточкой
накладывается полное кольцо клея шириною 3—5 см и толщиною 3—5 мм.
При изоляции отдельных сильно зараженных площадей могут быть
использованы для взрослых гусениц (и для средневозрастных) загради-
тельные канавки, которыми окапывается зараженная площадь. В некото-
рых случаях, например при заносе массовых видов (монашенка, непарный
шелкопряд, златогузка) на новые благоприятные для размножения терри-
тории, как то случилось в Северной Америке, уместно применение барьер-
1 То обстоятельство, что этим методом можно собрать только одних самцов, ума-
ляет его практическое значение, так как пропорция полов может быть весьма различ-
ной (Цвёльфер, 1934, 1935; Пятницкий, 1935; Пархоменко, 1936). Тем не менее рас-
пределение самцов дает достаточное понятие о плотности популяции при больших
масштабах сбора и на обширных территориях, когда влияние отдельных, локальных
условий обитания выпадает.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕИЕ
19ST
•ной зоны, где полностью уничтожаются все особи данного вида всеми
доступными способами. Наконец, общей мерой,, имеющей профилакти-
ческое значение, является охрана й привлечение, особенно в сады,
парки, питомники, но также и в леса, насекомоядных птиц. В частности,
для Orgyidae следует использовать не мелких птиц (пеночек, славок и т. п.),
но соек, кукушек, поползней. Точно так же уместна охрана паразитических
и хищных насекомых —Ichneumonidae, Braconidae, Chalcididae, Proctotru-
pidae из Hymenoptera, Laroiooridae из Diptera, Carabidae из Coleoptera,
некоторых хищных клопов (Picromerus} из Rhynchota, клещиков (Trombi-
dium) и некоторых других беспозвоночных.
Существеннейшим профилактическим мероприятием является форма
.лесоиспользования и лесоводства, а также форма садоводства. Могут
быть указаны следующие принципиальные моменты, важные со стороны
борьбы с вредными Orgyidae. Должны быть организованы лесорубки так,
чтобы не создавать одновозрастных и однообразных по составу насажде-
ний. Наличие последних, как показал опыт лесоводства в средней Европе
в отношении монашенки (Седлачек, 1916, 1917) и опыт лесопользования
в Крыму (Пятницкий, 1935), является существенным условием для массо-
вых размножений Orgyidae.
Проведение профилактических мер, вместе с достаточной осведомлен-
ностью о численности вредных Orgyidae и о возможности размножения
их в ближайшее время на данной территории, представляется наиболее
эффективным в условиях леса, крупных массивов садов и т. д. Истреби-
тельные мероприятия в таких условиях, помимо крайней дороговизны,
часто просто невозможны по условиям времени. Нейтрализация вредо-
носной деятельности таких видов, как монашенка, непарный шелкопряд,
златогузка, в случае массовых размножений часто сводится лишь к макси-
мальной нейтрализации ущерба от них, как то было, например, при
массовых размножениях монашенки у нас и в средней Европе. В усло-
виях сада и небольших лесных массивов (парки, питомники и др.) может
быть использован сбор всех фаз развития вредных Orgyidae, причем
•особенно яиц, куколок и бабочек, когда уничтожение может быть про-
изведено до нанесения вреда. Сбор гусениц вредных Orgyidae, например
гусениц краснохвоста, в летнее время может быть осуществлен также
•стряхиванием их на матерчатые щиты. Лов бабочек возможен на свето-
ловушки (златогузка) и на самок (непарник).
Метод применения болезней и паразитов Orgyidae с истребительными
целями требует специальной оценки условий обитания и размножения
вредного вида в каждом случае особо. Размножение в лабораторных
условиях яйцеедов —Anastatus disparis Ruschka, Schedius kuwanae Haw.,
Apanteles julvipes Hal. и др., а также ряда видов тахин —Compsilura
concinnata Mg., Sturmia scutellata R. D., S. nidicola Towns., Carcellia
gnava Mg., хищных жуков, например жужелиц Calosoma 1 {sycophanta L.,
inquisitor L.) и затем выпуск их могут при известных условиях дать поло-
жительный результат. Положительными сторонами этого метода являются
•относительная дешевизна и длительное последействие в связи с размно-
1 По Нехлеба (1927, 1928), роль хищников и, в частности, видов жужелиц в сни-
жении численности вредных Orgyidae (например непарного шёлкопряда) ничтожна.
Он считает, что угасание массовых размножений стоит в связи не с деятельностью
паразитов и хищников, но зависит от снижения плодовитости после размножений
{см. выше). Роль же хищников и паразитов этот автор видит в условиях нормального
•существования популяции, т. е. в начальные периоды, перед размножением (может
быть в подготовительные годы).
20Q ОБЩАЯ ЧАСТЬ
жением паразитов в природе. Использование бактериальных (флашерия)
и вирусных (полиэдренная болезнь) заболеваний личиночных фаз Orgyi-
dae требует больших знаний экологической ситуации размножения,,
а в то же время развитие эпизоотий этих возбудителей еще не ясно со сто-
роны физиологических условий развития болезни. Метод этот, являясь,,
может быть, очень перспективным, до сих пор, несмотря на многочислен-
ные попытки, не дал положительных результатов в полевой обстановке..
Он требует еще специального изучения.
Использование химического метода в борьбе с вредными Orgyidae г
буручк в принципе просто, практически громоздко и дорого из-за боль-
ших площадей в случае обработки лесных массивов. Более рентабельно
использование его в садах, парках, особенно в питомниках. Наиболее
употребительными при этом методе являются кишечные яды — париж-
ская зелень, мышьяковистокислый свинец (джипсин), мышьяковокислый
кальций, мышьяковистокислый натрий, кремнефтористый натрий и
органические и синтетические препараты — дихлордифенил трихлорметил-
метан (ДДТ), гексахлорцгкло гексан (ГХЦГ). Гусеницы вредных Orgyidae-
отличаются высокой выносливостью к кишечным ядам. Минимальная
летальная доза на 1 г живого веса гусеницы непарного шелкопряда равна
0.9 мг фтористого натрия, то же для арсената натрия — 0.10 мг. Для
златогузки эти же отношения 0.8 мг кремнефтористого натрия, 0.10 мг
арсената натрия и 0.07 мг парижской зелени (Додонов, 1936). В связи
только с этим обстоятельством расход яда на обработку лесных (и садо-
вых) площадей при борьбе с вредными Orgyidae очень велик. При боль-
ших площадях работа приобретает грандиозный объем. Расход арсената
кальция, например, на 1 га зараженного гусеницами монашенки или
непарного шелкопряда леса выражается в 8-—12 кг.
Химический метод эффективен не только своим непосредствен-
ным действием, но и последействием,- которое было установлено именно
для Orgyidae. Оказалось, что сублетальные дозы мышьяковистых препа-
ратов вызывают дефекты в развитии половых продуктов бабочек, пере-
несших отравление в стадии гусениц, приводящие к бесплодию
нормально оплодотворенных самок. Этот факт повышает значение
химического метода в истреблении массовых вредителей из Orgyidae.
Нанесение ядов на большие площади осуществляется с самолетов
опыливанием, причем против вредных Orgyidae с успехом используются
и порошковидные контактные яды. Обычные технические приемы обра-
ботки растений помощью ручных и даже конных опрыскивателей
или опыливателей мыслимы лишь в садовом хозяйстве и в питом-
никах. Уже в парковом хозяйстве необходимо применение моторных
машин, а в лесном хозяйстве химическая обработка больших массивов;
реальна лишь при участии авиации (Коротких, 1932; Новосельский^
1931).
ЛИТЕРАТУРА
К I г л а в е
Бобров А. 1930. Экспериментальные исследования над шелкопрядом Orgyia
antiqua L. Русск. зоол. журн., 10 : 71—116.
Дербенева Н. 1926. Изменение мускулатуры у Stilpnotia salicis L. под влия-
нием холода на куколок. Русск. зоол. журн., 6 : 20—32.
Ем ельянова Н. 1924. Влияние температуры на вторичные половые признаки
у Limantria dispar L. Русск. зоол. журн., 4 (1—2).
ЛИТЕРАТУРА
201
К ос минский П. 1924. Некоторые данные по развитию усиков у непарного шелко-
пряда в связи с явлением интерсексуальности. Русск. зоол. журн., 4 (3—4) :
134—140.
Косминский П. 1924. Получение интерсексуальных форм у Stilpnotia salicis L.
под влиянием холода на куколку. Там же, 4 : 186—197.
Косминский П. 1930. Исследование над интерсексуальностью непарного шел-
копряда. Limantria' dispar, ч. I. Там же, 10, 2 : 1—50.
Косминский П. 1935. Морфологическое исследование интерсексуальных сам-
цов. Там же, 14 : 271—310.
Косминский П. 1935. Исследование над интерсексуальностью у непарного
шелкопряда, ч. II. Там же, 14 (4) : 621—636.
Косминский П. и Н. Варшаве р. 1935. Получение интерсексуальных
изменений у непарного шелкопряда под влиянием температуры. Там же, 14 (3) :
439—464.
Кожанчиков И. 1937. Совки Agrotinae. Фауна СССР, Чешуекрылые, XIII,
2 : XVI + 679.
Кузнецов Н. 1915. Чешуекрылые. Введение. 1. Фауна России, 1 : 1—336.
Кузнецов Н. 1916. Морфология полового аппарата у чешуекрылых. Русск.
энтомол. обозр., 16 : 1—42.
Brandt Н. 1936. Untersuchungen uber die Morphologie der Kopulationsorgane der
Nonne, Lymantria monacha L. Zool. Anz., 114 : 161—168.
Dubois A. 1929. Morphologische Untersuchungen uber die Variation Und Abnor-
mitaten des Kopulationsapparates intersexueller Weibchen von Lymantria dis-
par L; Ztschr. Morph. Oekol. Tiere, 15 : 447—459.
Dubois A. 1931. Morphologische Untersuchungen uber die Entwicklung des Kopu-
lationsapparates der intersexuellen Weibchen von Lymantria dispar L. Arch
Entwicklungsmech., 124 : 93—137.
Eidmann H. 1935. Uber die Morphologie der Legerohre der Nonne, Lymantria
monacha L. (Lepidoptera, Lymantriidae). Zool. Anz., Ill (3—4) : 104—108.
Goldschmidt B. 1912. Erblichkeitsstudien an Schmetterlingen. Ztschr. Abst.
Vererb.-I.ehre, 7 : 1—62.
Goldschmidt B. 1934. Lymantria. Bibliographia Genetica, 11 : 1—186.
Goldschmidt B. und H. Poppelbaum. 1914. Erblichkeitsstudien an
Schmetterlingen. Ztschr. Abst. Vererb. Lehre, 11 : 280—316.
Goldschmidt B. und S. S a g u c h i. 1922. Die Umwandlung des Eierstocks
in einen Hoden beim intersexuellen Schwammspinner. Ergebn. anat. Entwick-
lungsgesch., 65 : 226—253.
Kosminsky P. 1930. Erhaltung mannlicher und weiblicher Intersexe in ein und
derselben Zucht. Biol. Zentralbl., 50 : 351—357.
Kosminsky P. und X. G о lo w ins ka ja. 1929. Zur Morphologie des Gesch-
lechtsapparates der Lepidopteren. Ztschr. Morph. Oekol. Tiere, 15 : 459—473.
Meisenheimer J. 1907. Ergebnisse einiger Versuchsreihen uber Extirpation
und Transplantation der Geschlechtsdriisen bei Schmetterlingen. Zool. Anz.,
XXXII : 393—400.
Petersen W. 1900. Beitrage zur Morphologie der Lepidopteren. Зап. Акад. Наук,
СПб., 9 : 1—144.
Petersen W. 1904a. Die Morphologie der Generationsorgane der Schmetterlinge
und ihre Bedeutung fur die Artbildung. Там же, 14 : 1—84.
Petersen W. 1904b. fiber Dendrolimus pini L. und D. segregatus В. H. Русск.
Энтом. обозр., 4 : 163—166.
Beichelt M. 1925. Schuppenentwicklung und Pigmentbildung auf den Flugeln
'von Lymantria dispar L. unter besonderer Berucksichtigung des sexuellen Dimor-
phismus. Ztschr. Morph. Oekol. Tiere, 3 : 477—525.
Sato H. 1932. Die postembryonale Differenzierung der Gonaden von Lymantria dis-
par. Ztschr. Zellforsch., 16 (1) : 63—88.
Snodgrass W. 1935. Principles of insect morphology: IX + 667. London.
Urbahn E. 1935. Geschwarzte Lymantriiden. Intern, entom. Ztschr., 29 : 399—400.
Zander E. 1903. Beitrage zur Morphologie der mannlichen Geschlechtsanhange
an Lepidopteren. Ztschr. wiss. Zool., 74 : 4.
Ко II главе
Кулагин H. 1897. Строение кожных желез личинок непарного шелкопряда.
.Изв. Моск. с.-х. инет., 3, 1 : 12—13.
Buxton Р. 1921. Liparid-moth (Ocnerorgyia amanda Stgr.) destructive to figs in
Mesopotamia. Bull. Entom. Bes., 11 : 181—186.
.202
ОБЩАЯ ЧАСТЬ
Chapman Г. 1893. Some neglected points in the pupae of the heterocerous Lepi-
doptera. Trans. Entom. Soc.. bond. : 97—119.
Cholodkowsky N. 1894. Uber die sogenannten Aerophoren der Nonnenraupe.
Forstl. naturw. Ztschr., 3 : 240—242.
Cholodkow s'k у N. 1903. Ueber die gelben Flecke und die kolbenformigen Haare
der Raupe von Acronicta alni L. Zool. Jahrb., Syst., 19 (5) : 554—556.
Collins C. 1915, Dispersion of gipsy-moth larvae by the wind. U. S. Dep. Agr.,
Washington, Bull. 273 : 1—23.
Doting E. 1936. Beitrage zur Kenntnis der Noctuideneier. Entom. Ztschr. Frankf.,
49 : 445—448.
Dyar H. 1894. A classification of Lepidopterous larvae. Ann. Amer. Acad. Sci. New
York, 8 : 194—232.
(Dyar H. 1895. Additional notes on the classification of Lepidopterous larvae. Trans.
Amer. Acad. Sci., 14 : 49—62.
Dyar H. 1901. A century of larval descriptions. Entom. Rec., 13 : 40.
’F о r b e s W. 1910. A structural study of some caterpillars. Ann. Amer. Entom. Soc.,
3 : 94—144. .
<F о r b e s W. 1911. A structural study of the caterpillars. Ann. Amer. Entom. Soc.,
4 : 261—279.
’Forbes W. 1916’. On certain caterpillar homologies. Journ. N. Y. Entom. Soc.,
24 : 137—142.
Tracker S. 1915. The classification of lepidopterous larvae. Ulin. Biol. Monogr.,
2 (1) : 1—161.
Garman H. 1921. Some neglected setae of lepidopterous larvae. Ann. Entom. Soc.
Amer., 14 : 145—149.
Gerasimov A. 1935. Zur Frage der-Homodynamie der Borsten von Schmetterlings-
raupen. Zool. Anz., 112 : 177—194.
Gerasimov A. 1937. Beitrag zur Systematik der Psychiden auf Grund der Erfor-
schung der Raupen (Lepidoptera). Zool. Anz., 120 : 7—17.
Gerasimov A. 1937. Bestimmungstabelle der Familien von Schmetterlingsraupen.
Stett. entom. Ztg., 98 : 281—300.
Hinton H. 1946. On the homology and nomenclature of the setae of lepidopterous
larvae, with some notes on the phylogeny of the Lepidoptera. Trans. Roy. Entom.
Soc. Lond., 97 (1) : 1—37.
K. u w a d a A. 1929. The eggs, larvae and pupae of Dasychira strigata Moore and D.
lunulata Btl. Kontyu, 3 (2) : 85—88. . .
Kuwada A. 1931. The adult, eggs, larva and pupa of Nygmia piperita Obth. Idem,
5 : 125—128 (японск.).
Mosher E. 1916. A classification of the Lepidoptera based on the characters of the
pupa. Bull. Ulin. State Labor., 12 : 1—160.
Pawlowsky E. und A. Stein. 1927. Experimentelle Untersuchungen uber die
Wirkung der Gifthaare der iiberwinternden Goldafterraupen (Euproctis chrysor-
rhoea L.) auf die Menschhaut. Ztschr. Morph. Oekol. Tiere, 9 : 615—-637.
Poulton E. 1891. The external morphology of the lepidopterous pupa. Trans. Linn.
Soc. London, Zool. Ser., 2 (5) : 245—263.
Quail A. 1904. On the tubercles of thorax and abdomen in first larvae stage of Lepi-
doptera. Entomologist, 37 : 269.
R i p 1 e у L. 1923. The external morphology and postembryology of Noctuid larvae.
Ulin. Biol. Monogr., 8 : 7—87.
S c h i e r b e e k A. 1917. Over het setale patroon der rupsen. Proc. Sci. Acad. Wet.
Amsterdam, XIX : 1156—1161. .
Tong A. 1932. The ova of British Lepidoptera; Noctuidae. Proc. Lond. Entom. and
Nat. Hist. Soc., 4 : 1—15.
T s о u Y. 1914. The body setae of lepidopterous Larvae. Trans. Amer. Mier. Soc., 33 :
233—260, 110—113',
К III главе
i
Поспелов В. 1910. Постэмбриональное развитие и имагинальная диапауза
у чешуекрылых. Зап. Киевск. общ. естественен., 21 : 163—418.
Тихомиров А. 1882. История развития тутового шелкопряда в яйце. Изв. Общ.
люб. естествозн., антроп. и этногр., 32 (4) : 1—80.
X о лодковский Н. 1886. Мужской половой аппарат чешуекрылых (сравни-
тельно-анатомическое исследование). Прил. к 42 тому Зап. Акад. Наук, СПб.,
4 : 4+1—130.
ЛИТЕРАТУРА
203
Berlese А. 1909. Gli insetti. Milano, 1 : 1—1004.
E i d m a n n H. 1931. Morphologische und physiologische Untersuchungen am weibli-
chen Genitalapparat der Lepidopteren. 2. Physiologischer Tail. Ztschr. ang.
Entom., 18 (1) : 57—112.
S prengel L. 1928. Untersuchungen liber Zustand und Entwicklung der Eier in den
Ovarien geschlupfter Lepidopteren. Anz. Schadlingsk., 4 : 25—30.
Weber H. 1933. Lehrbuch der Entomologie. Jena : 7—726.
К IV г л а в e
Бей-Биенко Г. 1924. Материалы к биологии непарного шелкопряда. Тр. Омск,
с.-х. акад., 3 : 155—160.
Добровольский Б. 1949. Златогузка на Дону и Северном Кавказе. Уч. зап.
Гос. Унив. им. Молотова, Ростов н/Д, 15 (6) : 35—45.
Кожанчиков И. 1947. Зимовка и диапауза у чешуекрылых семейства Orgyidae.
Изв. Акад. Наук СССР, сер. биол., 6 : 653—673.
Лучник В. 1914. Ивовый шелкопряд. Защ. раст., 4 (22) : 14.
Мориц П. 1921. К биологии златогузки. Тр. Ставроп. с.-х. инет., 1 ."23—30.
Румянцев П. 1925. К биологии непарного шелкопряда. Изв. Ставроп. станции
защ. раст., 1 : 23—26.
Р у м я ац ев П. 1936. Биология ивовой волнянки (Stilpnotia salicis L.) в город-
ских условиях в Москве. Бюлл. Моск. общ. исп. прир., биол. (н. с.),
45 : 271—278.
•Сахаров Н. 1914. Несколько данных к биологии Orgyia dubia Tausch. Русск.
энтом. обозр., 14 : 391—-397.
Шевырев И. 1893. Описание вредных насекомых степных лесничеств. СПб. :
1—31.
Шевырев И. 1901. Непарный шелкопряд. Лесной журнал, 31 (6) : 1132—1137*
Щелкановцев Я. 1926. Из летних и зимних наблюдений над насекомыми в лес-
ных дачах Воронежского с.-х. института. Зап. Ворон, с.-х. инет., 6 : 219—224.
Щербаков Т. 1914. Заметка о непарном шелкопряде. Тр. Естественноист. музея
Таврич. губ. земства, 3 : 1—34;
Шрейнер Я. 1908. О некоторых бабочках, вредящих плодовым садам. Тр. Бюро
антомол. II, 16 : 16—37.
Шугуров А. 1907. Этюды по экологии непарного шелкопряда в русских лесах.
Лесной журнал, 37 : 1218—1234.
Айе А. 1926. Der Schwammspinner. Entom. Ztschr. Frankf., 39 : 145, 149; 153, 158.
Boczkowska M. 1933. On the life history of Laelia coenosa Hb. (Lepidoptera,
Lymantriidae). Ann. Mus. Zool. Polon, Wars., 10 : 41—50.
Christensen P. 1937. Zur Histologie und Embryologi'e der uberwinterten Eier
von Orgyia antiqua L. Zool. Jahrb., Anat., 62 : 567—582.
Collins C. and S. Paths. 1931. Attractants for the male gipsy-moth. Journ.
Econ. Entom., 24 : 561—562.
Collins C. and S. Paths. 1932. Attractants for the flying gipsy-moth as an aid
in locating new infestations. Techn. Bull. U. S. Dep. Agr. Wash., 336 : 1—43.
Dyar. H. 1890. The number of moults in lepidopterous larvae. Psyche, 5 : 175, 420.
Eckstein K. 1933. Der Buchanrotschwanz (Dasychira pudibunda L.). Anz. Schad-
lingsk., 9 : 52—54.
Escherich K. 1931. Die Forstinsekten Mitteleuropas. Bd. 3: XI+ 825.
Gleichauf R. 1937. Arctornis 1-nigrum Mull. Entom. Ztschr. Frankf., 51 : 29—32.
G о e t z e H. 1936. Hemerocampa leucostigma Sm. and Abb. eihe Aufzucht aus dem
Ei. Entom. Ztschr. Frankf., 49 : 515—518.
Grevillius A. 1905. Zur Kenntnis der Biologie des Goldafters (Euproctis chry-
sorrhoea L.) und dutch denselben verursachten Beschadigungen. Beihefte Bot.
Centralbl., 18 (2) : 222—322.
H a n n о R. 1929. Anlockversuche bei Lymantria monacha L. Ztschr. angew. Entom.,
25 : 628—641.
Hepp A. 1936. Parthenogenesis bei Orgyia antiqua L. Entom. Rundsch., 53 :
294________295
HJo 11 a s F. 1935. Eine Eizucht von Diloba coeruleocephala L. Entom. Anz. Wien,
15 : 180—183,-
К e 1 e r S. 1934. Uber die Kopfindices der Larven und die Dayrsche Hypothese. Polsk.
• Pismo. Ent., 12 : 173—180.
L a m p a S. 1900. Lofskogsnunnan (Ocneria dispar L.). Entom. Tidskr., 21 : 39—48.
Lederer G. 1923. Handb. prakt., Entom., Bd. I—II—III. Frankfurt.
204 ОЯЩАЯ ЧАСТЬ
I, innaniemi W. und J. Hukkinen. 1921. Zur Biologie und Verbreitung
der Dasychira selenitica Esp. mit besonderer Beriicksichtigung ihres Massen-
auftretens in Finnland. Acta Soc. Fauna et Flora Fennica, 48 (7) : 1—27.
Pabst M. 1888. Die Entwicklungsgeschichte von Panthea coenobita Esp. nebst Mit-
teilungen uber das Aufsuchen des Schmetterlings sowie liber Erziehung der Raupe..
Iris, 1 : 115—118.
P r iif f e r J. 1937. Weitere Untersuchungen liber die Mannchenanlockung bei Lyman-
tria dispar L. Zool. Polon., Lwow, 2 : 43—66.
Ramachandra R. 1922. Notes on the life histories of two Mesopotamian moths.
Bull. Entom. Res., 12 : 477—479.
S chne i der-0 rell i 0. 1913. Uber Schwammsp inner und Goldafter. Schweiz.
Ztschr. Obst- und-Weinb., Frauenfeld, 22 : 18—22* 38—41.
Standfuss M. 1896. Handbuch der palaarktischen Grossschmetterlinge. 2 AufL,
Jena : 1—392.
S ylven E. 1942. Die Biologie des Buchenspinners, Dasychira pudibunda L. Ztschr.
angew. Entom., 30 : 119—142.
Weijenbergh H. 1870. Quelques observations de parthenogenese chez les Lepi-
dopteres. Arch. Neerl. : 258—264.
W о h n i g A. 1927. Orgyia thyellina Btl., zwei Aberrationen dieser Art und eine Kreu-
zung mit Orgyia antiqua L. Intern, entom. Ztschr., 21 : 36—40.
К V г л а в e <
Aurivillius Ch. 1891. Nordens Fjarilar. Stockholm.
Bang-Haas. 0. 1926—1930. Novitates Macrolepidopterologicae. I—V.
Bang-Haas 0. 1927. Horae Macrolepidopterologicae, I : XXXVIII+128.
Barnes W. and J. McDunnough. 1917. Check list of the Lepidoptera of
Boreal America. Herald Press.
Boisduval J. 1840. Genera et index methodicus europaeorum Lepidopterorum.
VII-J-238.
Borner G. 1939. Die Grundlagen meines Lepidopterensystems. Verh. VII Intern.
Kongress Entomol. Berlin, II : 1372—1424.
Bryk F. 1934. Lymantriidae. Junk’s Lep. Cat., 62 : 3—441.
Butler A. 1881, 1889. Illustrations of typical specimen of Lepidoptera Heterocera
in the collection of the British Museum. V : XII+74; VII : IV+124.
Collenette C. 1935. Genera and types of the Lymantriidae. Stylops, IV : 241—246.
C u 1 о t J. 1909. Noctuelites et geometres d’Europe. Acronyctinae. Geneve : 17—27.
Dyar H. 1897. A generic revision of the Hypogymnidae. Canad. Entom., XXIX :
12—16.
Dyar H. 1899. On the fluctuation of the postspiracular tubercle in Noctuid larvae..
Proc. Entom. Soc. Washington : 370—373.
Eggers F. 1919. Das thoracale tympanale Organ einer Gruppe der Lepidoptera Hete-
rocera. Zool. Jahrb., Anat., 41 : 273—376.
Eggers F. und J. . Gohr b and. 1938. Hypogymna morio L. — ein Sonderfall
in der Gesetzmassigkeit phyletischer Korrelation? Zool. Jahrb., Syst., 71 :
265—276.
Esper E. 1777—1815. Die Schmetterlinge in Abbildungen nach der Natur. Erlangen,.
I-V. ’ .
Fabricius J. 1787. Mantissa Insectorum. II. Hafniae.
FabriciusJ. 1793—1794. Entomologia systematica emendata et aucta. Ill (1—2)..
Hafniae.
F г e у e r C. 1831—1858. Neuere Beitrage zur Schmetterlingskunde. Augsburg. I—VIE
, Guenee A. 1852. Species general des Lepidopteres. Noctuelites. I—III.
Hampson G. 1898. Catalogue of the Syntomidae. London, I : 1—20.
Hampson G. 1909. Catalogue of the Noctuidae in the collection of the British Mu-
seum. VIII : 1—178 (Acronictinae).
Hampson G. 1913. To же, XIII : 337—400 (Mominae).
Haworth A. 1803—1829. Lepidoptera Britannica. London : 1—609.
Hemming F. 1937. A bibliographical and systematical account of the entomolo-
gical works of J. Hiibner. London, 1:39 + 605; II : 1—274.
Herrich-Schaeffer G. 1845. Systeinatische Bearbeitung der Schmetterlinge
von Europa zugleich als Text, Revision und Supplement zu Hiibner. II. Spinner
und Eulen : 1—450.
Hoffmann E. 1894. Die Gross-S chmetterlinge Europas. Stuttgart : XXXXI -|- 240.
Hormusaki C. 1904. Analytische Ubersicht der palaarktischen Lepidopteren-
familien. Berlin : 1—68.
ЛИТЕРАТУРА 205
Hiibner J. 1805. Sammlung europaeischer Schmetterlinge, II, III (Bombyces).
H ii b n e r J. 1816. Verzeichniss bekannter Schmetterlinge. Augsburg : 158—-162.
Kennel C. und F. Eggers. 1933. Die abdominalen Tympanalorgane der Lepi-
dopteren. Zool. Jahrb., Anat., 57 : 1—103.
Kirby W. 1892. A synonymic catalogue of Lepidoptera Heterocera (Moths). London :
XII + 951.
Lederer J. 1857. Die Noctuinen Europas. Wien : XV + 251.
Lempke B. 1936. Lymantria dispar L. et ses forms. Lambillionea, 36 : 227—236,
257—263.
Linn e C. 1758. Systema Naturae. Editio decima. Holmiae.
Linn ё C. 1761. Fauna Suecica. Editio altera. Stockholmiae.
Lins tow V. 1913. Die neue lepidopterologische Nomenclatur und die Hiibner’schen
Gattungsnamen, besonders der Noctuinen. Berl. entom. Ztschr., 58 : 21—29.
Martin S. and F. Cotner. 1934. Serological studies of moth proteins with spe-
cial reference to phylogenetic significans (Noctuidae). Ann. Entom. Soc. Amer.,
27 : 372—383.
M cDunnough J. 1916. On the types of' certain Noctuid-genera occuring in North
America. Entom. News, Philad., 27 : 393—400.
M e у r i с к E. 1895, 1927. A Handbook of British Lepidoptera. London — New York,
. ed. I : 1—843; ed. II : 1—914.
Nordstroem F. and E. Wahlgren. 1937. Svenska Fjarilar. Stockholm.
Ochsenheimer F. 1810. Die Schmetterlinge von Europa. Ill—V : 1—360.
Pierce F. 1909. The genitalia of the group Noctuidae of the Lepidoptera of the Bri-
tish Islands. XII + 88.
Prout M. 1924. Revision of the genus Trisuloides Btl. Bull. Hill Mus., I (3) : 400—418.
Schiffermiiller J. 1776. Systematisches Verzeichnis der Schmetterlinge der
Wiener Gegend. Wien.
Sick H. 1935. Die Bedeutung der Tympanalorgane der Lepidoptera fur die Systema-
tic Verh. Deutsch. Zool. Ges. : 131—135.
Smith J. 1893. Catalogue of the lepidopterous subfamily Noctuidae found in Boreal
North America. Bull. U. S. Nat. Mus., 44 : 1—424.
Staudinger O. 1901. Catalog der Lepidopteren des palearktischen Faunengebietes.
I : XXX + 334.
Stephens J. 1829. Illustrations of British Entomology. Haustellata. II : 1—202.
Treitschke F. 1825—1835. >Die Schmetterlinge von Europa (Fortsatzung des
Ochsenheimerschen Werkes). Leipzig, V—X.
К VI главе
E м ч у к Е. 1938. Динамика каталазы у непарного и соснового шелкопрядов за время .
их развития. Зб1рн. праць екол. : 161—180.
К е л у с О. 1939. О роли кормовых растений в развитии непарного шелкопряда.
Зоол. журн., 18 (6) : 1010—1019.
Кожа нчик ов И. 1939. Термостабильное дыхание как условие холодостойкости
насекомых. Зоол. журн., 18 : 86—98.
Кожанчиков И. 1940. Значение физических условий среды в развитии яиц
непарного шелкопряда. Вести, защ. раст., 3 : 3—16.
Кожанчиков И. 1941. Об условиях смены кормовых растений у дендрофиль-
ных насекомых. Зоол. журн., 20 (3) : 382—397; Природа, 1941, 1 : 78—79.
Ко ж ан чиков И. 1949. Значение сезонных изменений листьев кормовых
растений в развитии непарного шелкопряда (Ocneria dispar L.). Докл. АН
СССР, 66,6 : 1203—1206.
Левитт М. 1935. Изменчивость куколок и плодовитость бабочек непарного шелко-
пряда. Тр. инет. зоол. и биол., Киев, 2 : 135—170.
Пархоменко В. 1936. Непарный шелкопряд в лесах Крыма. Киев. ВУАН :•
1—117.
Парфентьев И. 1925. Опыт с неполным голоданием гусениц Orgyia antiqua L. ,
Защ. раст., 1 (6) : 201—206.
СавицкаяЗ. 1939. Динамика количества воды и жира в теле насекомых в связи
с холодостойкостью. Зб1рн. праць екол. : 145—160.
Сахаров Н. 1928. К изучению холодостойкости насекомых. Журн. опытн. агрон.
юго-вост., Саратов : 1—20.
Тарануха М. 1937. Методы определения интенсивности питания Lymantria
dispar L. Тр. инет. зоол. и биол., Киев, 14 : 100—109.
Т рейман Ф. 1938. Количество гусениц, выходящих из яиц непарного шелко-
206
ОБЩАЯ ЧАСТЬ
пряда й их жизнеспособность в первые дни в связи с условиями питания мате-
ринского поколения. 36ipH. праць екол., 5 : 117—144.
А 1 i М. 1933. Experimentelle Untersuchungen uber den Einfluss von Temperatur und,
Luftfeuchtigkeit auf Entwicklung des Schwammspinners (Lymantria dispar L.).
Ztschr. angew. Entom., 20 (3) : 355—381,
Arnim F. 1936. Zur Kenntnis der Temperaturabhangigkeit der Nonne (Lymantria
monacha L.). Ztschr.' angew. Entom., 22 : 533—557.
Brandt H. 1936. Die Abhangigkeit der Bewegungsgeschwindigkeit der Nonnenraupe-
(Lymantria monacha L.) von der Temperatur. Ztschr. vergl. Physiol., 23 :
715—720.
Brandt H. 1937a. Untersuchungen fiber die Aenderung des Geschlechtverhaltnisses
bei der Nonne (Lymantria monacha L.) und ihre Uhrsachen. Ztschr. angew. Entom.,
24 : 87—94.
Brandt H. 1937b. Untersuchungen liber die Aenderung der photo- und geotakti-
schen Reaktionen der Nonnenraupe, Lymantria monacha L. im Verlaufe des
Raupenlebens. Ztschr. vergl. Physiol., 24 : 188—197.
Burgess A. 1913. The dispersion of the gipsy-moth. U. S. Dept. Agr. Bur. Entom.r
Bull., 119 : 1—62,
Chapman J. and R. Glaser. 1916. Further studies on wilt of gipsy-moth cater-
pillars. Journ. Econ. Entom., 9 : 149—169.
Collins C. 1917. Methods used in determining wind dispersion of the gipsy-inoth
and some other insects. Journ. Econ. Entom., X : 170—177.
Collins C. and W. Baker. 1934. Exploring the upper air for windbona gipsy-
moth larvae. Journ. Econ. Entom., 27 : 320—327.
De Lepiney J. 1927, 1928. Les insectes nuisibles du chene liege dans la foret de
la Mamora (Maroc.). Ann. Epiphyt., XIII : 145—174; XIV : 313—321.
De Lepiney J. 1930. Contribution a 1’etude du complex de Lymantria dispar.
Mem. Soc. Nat. Maroc., 23 : 1—400.
D у a r H. 1896. The arctic Lymantriid larvae from Mt. Washington, Psyche, 7 :
328—329.
E i d m a n n H. 1938. Neue Untersuchungen fiber die Nonne. Anz. Schadlingsk., 14 :
13—18.
Emel j an off N. 1924. Intersexualitat bei Lymantria dispar L. unter Einwirkung
de- Temperatur. Biol. Zentralbl., 44 : 106—109. ,
Escalera M. 1924. Note sobre dos Coleopteros que atacan a la Lymantria dispar
у al Tortrix viri'dana en el Escurial. Boll. Soc. Esp. Hist. Nat., XXIV : 273—274.
Escherich K. 1914. Die Forstinsekten Mi.tteleuropas. Bd. I : 186—390.
Evans G. 1939. Ecological studies on the rein forest of south Nigeria. II. The atmo-
spheric environmental conditions. Journ. Ecol., 27 (2) : 436—482.
F lu i ter H. 1934. On the brown-tail moth and the factors reducing its numbers
during hibernation. Tijdschr. Plantenziekten, XL : 1—35.
Franklin H. 1917. Report of the cranberry substation for 1916. Mass. Agric. Exper.
Sta., Amherst, Bull. 180—183—234.
Glaser R. 1913. The polyedral virus of insects with a theoretical consideration of
filtrable viruses generally. Science, Lancaster, XLVIII : 301—302.
G о t z B. 1936. Beitrage zur Analyse des Verhaltens von Schmetterlinsraupen beim
Aufsuchen des Futters und Verpuppungsplatzes. Ztschr. vergl. Physiol.,
Gotz B. 1938. Sinnesphysiologische Untersuchungen an Schmetterlingsraupen und
ihre praktische Bedeutung. Anz. Schadlingsk., 14 : 92—93.
Hess C. 1919. Neue Untersuchungen fiber den Lichtsinn bei wirbellosen Tieren. Arch,
ges. Physiol., Pfliiger, 136.: 282.
H irschlerowa Z. 1934. Uber sexuelle Umstimmung bei Lymantria dispar L.
unter dem Einflusse chemischef Faktoren. Kosmos, 58 : 327—339.
Hoffman C. 1933. Der Einfluss von Hunger und engem Lebensraum auf das Wachs-
tunigUnd die Fortpflanzung der Lepidopteren. Ztschr. angew. Entom., 20 :
Hundertmark A. 1936a. Die Orientierung der Eiraupen der Nonne, Lymantria
monacha L. Arb. physiol, angew. Entom., 3 : 221—226.
Hundertmark A. 1936b. Helligkeits- und Farbenunterscheidungsvermogen der
' Eiraupen der Nonne (Lymantria monacha L.). Ztschr. vergl. Physiol., 24 :
42—57.
Hundertmark A. 1937a. Das Formenunterscheidungsvermogen der Eiraupen
der Nonne (Lymantria monacha L.). Ztschr. vergl. Physiol., 24 : 563—582.
Hundertmark A. 1037b. Verbreitungsmoglichkeiten- der Nonne, Lymantria
monacha L. durch die Eiraupen. Ztschr. angew. Entom., 24 : 118—128.
литература
20~
Janisch Е. 1933. Untersuchungen iiber die Oekologie und Epiderniologie der Nonne.
Arb. biol. Reichsanst., 20 : 269—290.
Janisch E. 1938a. Das klimatische Optimum der Nonnenraupe. Mitt. Forstwirtsch..
und Forstwissensch., 3 : 372—386.
Janisch E. 1938b. Eine neue Pilzkrankheit bei Nonnenraupen. Arb. physiol, angew..
Entom:, V : 1—21.
Kopec S. 1924. Studies on the influence of inanition on the development and the dura-
tion of life in insects. Biol. Bull., 46 : 1—21, 22—34.
К n о che E. 1929. Schadling, Klima und Bekampfung. Arb. Biol. Reichsanst. Land-
und Forstwirtsch., XVI : 705—775.
К n о che E. 1932. Klima und Nonne. I. Die Entwicklung des Embryo. Arb. Biol.
Reichsanst. Land- und Forstwirtsch., XX (2) : 193—235.
Komarek J. 1933. Wichtige Neubeobachtungen aus der Biologie der Nonne. Anz..
Schadlingsk., 9 : 77—82.
Komarek J. 1937. Kritisches iiber die Bedeutung der Insektenparasiten der Nonne..
Ztschr. angew. Entom., 24 : 95—117..
Komarek J. und V. В r e i n d 1. 1924. Die Wipfelkrankheit der Nonne und der-
Erreger derselben. Ztschr. angew. Entomol., X : 99—162.
Kozhant. shikov I. 1938. Physiological conditions of coldhardiness in insects...
Bull, entom. Res., 29 (3) : 253—262.
Kulagin N. 1897. Zur Biologie von Ocneria dispar L. in Russland. Illustr. Wochen-
schrift Entomol, 2 :418—420.
Loos K. 1920—1921. Zur eines machtigen Nonnenschwarmes iiber das Glatzer-Gebierge-
in die Tschechoslowakei. Vereinschr. Forst-, Jagd- und Naturk., Prag : 60—62.
Loos K. 1927. Die Wanderung der Nonnefalter. Sudetendeutsche Forst- und Jagd-
ztschr., XXVIII : 33—34.
Ludwig W. 1933. Seitenstatigkeit niederer Tiere in Ein- und Zweilichtversuch...
I. Lymantria dispar-Raupen. Ztschr. wiss. Zool., 144 : 469—495.
M a e г к s H. 1935. Der Einfluss der Nahrung auf die Entwicklung der Nonnenraupe..
Arb. physiol, u. angew. Entom., 2 (3) ; 175—195.
Mayer A. 1940. Ernahrungsphysiologische Untersuchungen an Nonnenraupen (Ly-
mantria monacha L.). Ztschr. angew. Entom., 27 (2) : 157—208; (3) : 408—449..
M innott C. 1922. The gipsy-moth on cranberry logs. U. S. Dept. Agr., Bull., 1093 :
1—19.
M о s h e r F. 1915. Food plants of the gipsy-moth in America. U. S. Dept. Agr., Bull.,.
250 : 1—39.
Nolte H. 1939. Die Wiederstandsfahigkeit der Spiegelraupen der Nonne (Lymantria
monacha L.) gegen Kalte. Anz. Schadlingsk., 15 : 11—12.
Nishikawa J. 1918. The causes of «Patch» disease of silkworm and the terrible-
urticating hairs of the gold-tail moth caterpillars, Porthesia similis Fuessly.
Journ. Silk Industr., Tokyo : 998—1003.
Petkoff P. 1939. Die Einfluss des Futters auf die Entwicklung des Goldafters, Nyg--
mia phaeorrhoea L. und des Ringelspinners, Malacosoma neustria L. Verb..
VII Intern, entom. Kongr., Berlin, IV : 2388—2396.
Pictet A. 1905. Des diapauses embryonnaires, larvaires et nymphales chez les insec-
tes lepidopteres. Bull. Soc. Lepidopter., Geneve, I : 98—153.
Pictet A. 1919. Recherches experimentales sur 1'adaptation de Lymantria dispar L.
aux Coniferes et de 1’autres essences. Mitt. Schweiz, entom. Ges., XIII : 20-’
—54.
Pictet A. 1920. Observations biologiques sur Psilura monacha L. Mitt. Schweiz,
entom. Ges.,. XIII : 63—64.
P r e 11 H. 1924. Uber die Immunitat von Fichten gegen Nonnenfrass und ihre Uhrsa-
chen. Tharandter Forstl. Jahrb., LXXV : 58—71.
Reiff W. 1909. The relation of sex to heliotropism in the browntail moth. Psyche,.
XVI, 6 : 115—119.
Reiff W. 1910a. Some experiments on the resistance of gipsy-moth eggs to the dige-
stive fluids of birds. Psyche, XVII, 4 : 161—164.
Reiff W. 1910b. On the resistance of gipsy-moth eggs (Liparis dispar L.) to cold and',
other conditions. Psyche, XVII, 2 : 69—73.
San d e rson E. and L. P e a i r s. 1913. The relation of temperature to insect .
life. New Hempshire Coll. Agr. Exper. Stat. Techn. Bull., 7 : 1—125..
Sattler H. 1939. Die Entwicklung der Nonne, Lymantria monacha L. in ihrer Abhan-
gigkeit von der Nahrungsqualitat. Ztschr. angew. Entom., 25 : 543—583.
S c h e d Г К. 1936. Der Schwammspinner (Porthetria dispar L.) in Euroasien, Africa
und Neuengland. Monogr. angew. Entom., 12 : 1—242.
Sedlaczek W. 1915. Einflusse der Witterung auf die ortsweisen Lebenserscheinun--
208
ОБЩАЯ ЧАСТЬ
gen der Nonne (Lymantria monacha L.). Ztschr. ges. Forstwesen, 41 :
321—342.
Sedlaczek W. 1916. Einwirkung des Klimas auf die Entwicklung der Nonne (Ly-
mantria monacha L.). Verh. zool.-hot. Ges. Wien, 66 : 28—33.
Sumners J. 1922. Effect of low temperature on the hatching of gipsy-motb eggs.
U. S. Dept. Agr. Bull.„1080 : 1—14.
Tuleschkov R. 1935. Uber die Uhrsachen der Uberwinterung der Lymantria
dispar L., Lymantria monacha L. und anderen Lymantriiden im Eistadium.
Ztschr. angew. Entom., 22 : 97—117.
Tuleschkov R. 1936. Sur les differences physiologiques pendant le developpe-
ment des oeufs des Lepidopteres, specielement chez la famille Lymantriidae.
Mem. Soc. Sci. Boheme, Prague, 10 : 1—24.
Van der M e r w e C. 1921. A note on Dasychira extorta and its lepidopterous para-
site. South Afric. Journ Sci., XVII (2) : 192—193.
Zwolfer W. 1927. Die Pebrine des Schwammspinners und Goldafters—neue wirt-
schaftlich bedeutungsvolle Infektionskrankheit. Ztschr. angew. Entom., 12 :
498—500.
Zwolfer W. 1933. Studien zur Oekologie und Bevolkerungslehre der Insekten. 2.
Die Nonne, Lymantria monacha L. Ztschr. angew. Entom., 20, 1 : 1—50:
Zwolfer W. 1934. Die Temper aturabhangigkeit der Entwicklung der Nonne (Lyman-
tria monacha L.) und ihre bevolkerungswissenschaftliche Auswertung. Ztschr.
angew. Entom., 21 : 333—334.
К VII главе
Алфера к и С. 1876. Чешуекрылые Северного Кавказа. Тр. Русск. Энтом. общ.
10 : 3—34.
А л ф е р а к и С. 1908. Чешуекрылые окрестностей Таганрога. Тр. Русек. Энтом.
общ., 38 : 558—618.
Б р е м е р О. 1864. Lepidopteren Ostsibiriens. Зап. Акад. Наук, СПб., 8 (7) : 1—104.
Гильтебранд.т В. 1915. К фауне Macrolepidoptera Тверской губернии. Русск.
энтом. обозр., 15 : 596—603.
Гюнтер А. 1896. Список чешуекрылых, найденных в Олонецкой губернии. Изв.
СПб. биол. лаборат., 1 : 21—33.
.Ершов Н. 1874. Чешуекрылые. Путешествие в Туркестан Федченко А., 2 (5) :
1—128.
Ершов Н. 1876. Список чешуекрылых из окрестностей Шахруда в Персии. Тр.
Русск. Энтом. общ., 8 : 321.
Ершов Н. и А. Ф и л ь д. 1870. Каталог чешуекрылых Российской Империи.
Тр. Русск. Энтом. общ., 4 : 130—204.
Зайцев Ф. 1906. К фауне Macrolepidoptera Новгородской губернии. Тр. Пресн.
биол. ст. СПб. общ. естествоисп., 2 : 44—62.
Кавригин В. 1894. Verzeichniss der im St. Petersburger Gouvernement gefun-
denen Schmetterlinge. Изд. Русск. Энтом. общ. : 1—57.
Кожанчиков В. 1923. Материалы к фауне чешуекрылых Минусинского края.
Ежегодн. Гос. музея им. Мартьянова, 1 : 1—50.
Кожанчиков И. 1937. Совки Agrotinae. Фауна СССР, Чешуекрылые,
XII, 3:1-678.
Кожанчиков И. 1941. Об условиях смены кормовых растений у дендрофиль-
ных насекомых. Зоол. журн., 20 (3) : 382—397.
-Кож. анчиков И. 1946а. Гипотеза ангарского происхождения фауны Голарктики.
Природа (5) : 72—75.
Кожанчиков И. 19466, Экологические предпосылки для зоогеографических
делений Евразии. Журн. общ. биол., 7 (1) : 35—48.
Кожанчиков Й. 1948. Род Gynaephora Hb., нго распространение и филогене-
тические связи. Тр. Зоол. инет. Акад. Наук СССР, 7 : 149—-161,
К р у л и к о в с к и й Л. 1899. Опыт каталога чешуекрылых Казанской губернии.
Тр. Московок. Общ. исп. прир., 13 : 157—219.
К руликовский Л. 1909. Чешуекрылые Вятской губернии. Мат. позн. фауны
и флоры Росс, имп., зоол., 9 : 48—250.
Кузнецов Н. 1900.. К фауне Macrolepidoptera Псковской губернии. Тр. Русск.
Энтом. общ., 33 : 85—131.
Куренцов А. 1936. Чешуекрылые хребта Сихотэ-алиня и вопрос о. происхо-
ждении его фауны. Вестн. Дальневост. фил. Акад. Наук СССР, 20 : 137—172.,
Куренцов А. 1939. Бабочки — Macrolepidoptera — вредители деревьев и кустар-
ников Уссурийского края. Тр. Горнотаежн. ст., 3 : 107—205.
ЛИТЕРАТУРА
209
Петерсен В. 1924. Lepidopteren-Fauna von Estland. Изд. 2, Таллин, 1 : 1—317;
2 : .318—590,
П о р т н я г и и Д., 1919. Чешуекрылые, собранные на протяжении Обь-Енисей-
ского канала на границе Томской и Енисейской губерний. Изв. Томск, унив.,
68 : 1—21.
Шапошников X. 1904. Заметки о Macrolepidoptera центральной части северо-
западного Кавказа. Ежегодн. Зоол. муз. Акад. Наук, СПб., 9 : 189—259.
Ш у г у р о в А. 1906. К лепидоптерофауне Херсонской губернии. Зап. Новоросс.
общ. ест., 29 : 35—82.
•Э в е р с м а н Е. 1844. Fauna lepidopterologica Volgo-Uralensis. Казань : 1—633.
Зверем ан Е. 1854. Beitrage zur. Lepidopterologie Russlands und Beschreibung
einiger anderen Insecten aus den siidlichen Kirgisensteppen, den ndrdlichen Ufern
des Aral Sees und des Sir-Darjas. Билл. Моск. общ. исп. прир., 27 (2) : 174—
205.
Bang-Haas О. 1930. Register der faunistischen Literalur der palaarktischen
Grossschmetterlinge. Novit. Macrolep., V : 1—128.
Barret Ch. 1900. The Lepidoptera of the British Islands. Vol. 1—7.
Bremer O. 1864. Lepidoptera Ost-Sibiriens, insbesondere des Amur-Landes. Mem.
Acad. Sci. Petersb., VIII (1) : 1—103.
Bremer O. und м . Grey. 1853. Beitrage zur Schmetterlingsfauna des nordli-
chen Chinas. 28 pp.
C a r a d j a A. 1934. Herkunft und Evolution der palaarktischen Lepidopteren. Intern,
entom. Ztschr., 28 : 133, 161, 187, 267, 361, 381.
Cara d j a A. 1937. Materialien zu einer Mikrolepidopterenfauna Julingshanmassivs
. (Provinz Junnan). Iris, 51 : 138—182.
Carmin J. and D. S c h e n k i n. 1933. The fauna of Palestinian plants. 3. Alhagi
maurorum D. C. Bull. Soc. Ent. Egypte, Cairo, 16 : 204—217.
Collenette C. 1932a. The Lymantriidae of the Malay Peninsula. Novit. Zool.,
Tring, 38 : 49—102.
•Collenette C. 1932b. Fauna buruana, Lepidoptera, Fam. Lymantriidae. Treubia, .
Buitenzorg, 7 : 385—387.
Collenette C. 1932c. New African and Indo-Australian Lymantriidae. Novit.
Zool., Tring, 37 : 159—180.
Collenette C. 1932d. Dr. K. Jordan’s expedition to South West Africa and Angola.
New Lymantriidae. Mem. Mus. Hist. Nat. Belg., 4 : 47—51.
Collenette C. 1933a. Notes on the genus Lymantria Hb. with des riptions of new
species. Novit. Zool., Tring, 39 : 21—33.
Collenette C. 1933b. New Lymantriidae (Lepidoptera, Heterocera) in the British
Museum collection. Ann. Mag. Nat. Hist., (10) 11 : 33—48.
Collenette C. 1933c. New Indo-Australian Lymantriidae (Lepidoptera). Stylops,
2 : 43—45. •
Collenette C. 1936a. On a collcetion of Lymantriidae (Heterocera) from North
Junnan. Ann. Mag. Nat. Hist., (10), 17 : 329—346.
Collenette C. 1936b. New African Lymantriidae. Entomologist, 69 : 178—181.
Collenette C. 1936c. Lymantriidae from North Junnan. Entom. Monthly Mag.,
72 : 90—91. ' '
Collenette C. 1936d. New Lymantriidae from Madagascar. Novit. Zool., Tring,
40 : 153—169.
Collenette C. 1936e. New Lymantriidae from the Belgian Congo. Rev. Zool. Bot.
Afr., Bruxelles, 28 : 249—252.
Collenette C. 1936f. New Lymantriidae from the Belgian Congo. Rev. Zool. Bot.
Afric., Bruxelles, 28 : 404—405.
Collenette C. 1937a. New African Lymantriidae (Lepidoptera, Heterocera). Ann.
Mag. Nat. Hist., (10), 29 ; 604—617.
Collenette C. 1937b. Lymantriidae from the Belgian Congo. Ann. Mag. Nat.
Hist., (10), 29 : 421—425.
Collenette C. 1937c. Some new Lymantriidae from the Belgian Congo. Там же,
(10) 30 : 280—286.
Collenette C. 1938a. On the collection of Lymantriidae (Heterocera) from China.
Proc. Entom. Soc. Lond. (B), 7 : 211—221.
Collenette C. 1938b. Some new and little known Malayan Lymantriidae. Journ.
F. M. S. Mus., Kuala Lumpur, 18 : 263—266.
Collenette C. 1938c. Some Lymantriidae from Abyssinia; Entom. Monthly Mag.,
74 : 152—153.
Collenette C. 1938d. New Palearctic and Indo-Australian Lymantriidae in the
British Museum Collection. Ann.' Mag. Nat. Hist., (11) 2 : 318—387.
14 Фауна СССР, И. В. Кожанчиков.
210
ОБЩАЯ ЧАСТЬ
Collenette С. 1938е. New. species of Lymantriidae (Lepidoptera, Heterocera) from,
the Kataya and other localities. Там же, (11) 2 : 209—223.
С о 1 1 e n e t t e C. and G. Carpenter. 1933. New species and descriptions of
larvae of ‘ Lymantriidae from Uganda. Entom. Monthly Mag., 69 : 258—270.
Cotes E. and C. S w i n h о e. 1887—1889. Catalogue of the mothes of India. Cal-
cutta : 1—812.
Draudt M. 1931. Noctuidae, Acronyctinae. Seitz, Grossschmett., III. Suppl.
1 : 5—14.
Gaede M. 1932. Lymantriidae. Seitz, Grossschmett., II, Suppl. 1 : 95—106.
Graeser L. 1892. Beitrage zur Kenntnis der Lepidopteren-Fauna des Amur-Landes,
B'erl. entom. Ztschr., 37 : 209—234.
Hampson G. 1892. The fauna of British India including Ceylon and Burma. Moths,.
I : 432—494.
Hampson G. 1901. Catalogue of the Lepidoptera Phalenae in the British Museum,
3 : 19—690.
Hampson G. 1912. The Moths of India. Suppl. paper to the volume in the fauna
of British India. Series IV, parts III—V. Bomb. Journ. Nat. Hist. Soc., 21 :
411—466, 868—911, 1232—1272.
Hering M. 1926. Lymantriidae. Seitz, Grossschmett., Fauna Africana, 14:
129—204.
Hering M. 1932. Die Schmetterlinge (Lepidoptera). Tierw. Mitteleur., VI (3) : 1—94.
Hormusaki C. 1933. Neue Erliiuterungen zu meinen Ansichten liber die geogra-
phische Evolution der europaischen Lepidopteren. Entom. Rundsch., 50 :
285—287.
Hudson G. 1928. New Zealand Moths and Butterflies. London, ed. I : 1—140; ed.
II : 1—386.
Kozhantschikov I. 1935. Eurasiatische Unterbrechung der Verbreitungs-
bereiche einiger Agrotinen-Arten in Beziehung auf ihre Geschichte und Bildung'
der Arten. Zool. Jahrb., System., 66 (6) : 549—563.
LindrothC. 1931. Die Insektenfauna Islands und ihre Probleme. Zool. Bidrag,.Upp-
sala, XIII : 105—589.
Matsumura S. 1933. Lymantriidae of Japan-Empire. Ins. Matsum., 7 : 111—152.
Moore F. 1879—1888. Descriptions of the Indian Lepidopterous Insects from the
collection of the late Mr. W. S. Atkinson. Heterocera, I—III, Calcutta.
Moore F. 1882—1883. The Lepidoptera of Ceylon. London, 2 : 1—162.
N a m u r a K. 1937. Studies on the Moth fauna of Mandchuria. Festschr. Strand, III :
467—505.
Osthelder L. 1925. Die Schmetterlinge Siidbayerns. Mitt. Munch, entom. Ges.,.
XV : 1—376.
Rothschild N. 1917. Supplemental notes to the Mr. Charles Oberthiires <Faune-
de Lepidopteres de la Barbarie», with lists of the specimens contained in the-
Tring Museum. Novit. Zool., Tring, XXIV : 325—373.
S chaus W. 1927. Lymantriidae. Seitz, Grossschmett., Fauna Americana (N. und S.
America), VI:535—564.
Schultze A. 1934. Neue und bekannte Lymantriiden aus Aequatorialafrika. Iris.
48 : 104—121.
Strand E. 1911. Lymantriidae. Seitz, Grossschmett., Fauna Palearctica., II: »
109—141.
Strand E. 1925. Lymantriidae. Fauna Indo-Australia. Там же, 10 : 291—388.
Turner A. 1921. Revision of Australian Lepidoptera. Liparidae. Proc. Linn. Soc_
N. S. W., Sydney, 45 : 474—499.
Warnecke G. 1938. Laelia coenosa Hb. Stett. entom. Ztg., 99 : 231—235.
W a 1 1 e n g r e n H. 1863—1881. Lepidoptera Scandinaviae. Heterocera. I—III.
К VIII главе
Бр янцев Б. 1928. Непарный шелкопряд, Lymantria dispar L., как вредитель-
посевов. Защ. раст., 5 (3—4) : 370—371.
Васильев И. 1914. Насекомые и другие вредители хлопка в Ферганской области/
наблюдавшиеся в 1914 г. Тр. Бюро энтом., 11(6) : 1—23.
Додонов Н. 1936. О сравнительной устойчивости некоторых видов насекомых
к мышьяковистым и фтористым препаратам. Тр. защ. раст., 3 сер., 7 : 55—78..
Голубева М., С. О б о л е н с к и и, Н. Мейер и В. Поспелов.. 1933.
Биологический метод борьбы с вредителями в сельском хозяйстве. Сельхозгиз.
Москва—Ленинград.
ЛИТЕРАТУРА
211
Кеппен Ф. 1878—1883. Вредные насекомые. Томы 1, 2, 3, СПб.
Коротких П. 1932. Применение самолетов для борьбы с вредителями сельского
и лесного хозяйства. Сельхозгиз.
Кулагин Н. 1894. Вредное для лесов насекомое, монашенка (Ocneria monacha L.)
и главнейшие меры борьбы с ним. Москва.
Кулагин Н. 1898. Непарный шелкопряд. Изд. 2-е.
Кулагин Н. 1923. Вредные насекомые, т. 2. Изд. 3-е, Москва—Ленинград.
Мокржецкий С. 1898. Вредные животные и растения в Таврической губ.
по наблюдениям в 1898 г. и меры борьбы с ними. Симферополь.
Мокржецкий С. 1913. Энтомологический календарь для садоводов. Симферо-
поль : 1—53.
Мокржецкий С. и И. Щеголев. 1914. Вредные насекомые и болезни расте-
ний, наблюдавшиеся в Таврической губернии в течение 1913 года;
Мокржецкий С. и А. Брагина. 1917. Вырождение непарного шелкопряда
в Крыму. Журн. прикл. энтомол., 1 : 21—22.
Новосельский А. 1931. Применение авиации и аэрофотосъемки в борьбе
с вредными для леса насекомыми. Тр. и исслед. по лесн. хоз. и леей, пром., 15.
Парамонов А. 1934. К вопросу о влиянии повреждений непарного шелкопряда
в отношении прироста дубов в Крыму. Зб1рн. праць Зоолог. Муз., Кшв,
13 : 97—110.
Парамонов А. 1935. К вопросу о лесохозяйственном значении Краснохвостого-
шелкопряда, Dasychira pudibunda L. Там же, 14 : 81—89.
Прозоров С. 1939. Вредители и болезни леса, их учет и борьба с ними. Красно-
ярск : 1—132.
Пятницкий Г. 1935. Факторы, способствующие и ограничивающие массовые
вспышки непарного шелкопряда в Крыму. Вопр. экол. и биоценол 2 :
100—120.
Римский-Корсаков М. 1935 (ред.). Лесная энтомология. Гостехиздат.
Ленинград.
Руднев Д. 1936. Вплив якост! харчу на пл!дшсть Lymantria dispar L. Зб1рн.
праць сект. екол. наземн. тварпн, Ки1'в, 3.
Сахаров И. 1893. Вредные для лесных и садовых насаждений бабочки. Изв.
Сарат, Гос. инет. с.-х. и мелиор., 4.
Силантьев А. 1895. Монашенка (Ocneria monacha L.) в лесах Царства Поль-
ского в 1893 г., ее образ жизни, распространение и борьба с нею. Москва.
Соболев А. 1897. История массового размножения непарного шелкопряда в Туль-
ской губернии в 1892—1896 гг. СПб.
Старк В.'1931. Вредные лесные насекомые. Сельхозгиз.
Старк В., И. Старк и И. Б е л а н о в с к и й. 1932. Методы и техника
борьбы с вредными насекомыми. I. Сельхозгиз.
Талицкий В. 1928. Непарный шелкопряд, Lymantria dispar L. как вредитель
посевов. Защ. раст., 5 (5—6) : 677—678.
Чу гунин Я. 1949. Очаговая цикличность массовых размножений непарного
шелкопряда. Зоол. журн., 28 (5) : 431—438.
III е в ы р е в И. 1894а. Шелкопряд-монашенка или шелкопряд-монах и способы
борьбы с ним. Изд. Лесн. департ. : IV + 72..
III е в ы р е в И. 18946. Монашенка и спутники ея в лесах Средней и Восточной
Европы летом 1894 г. СПб., : 1—32.
Шевырев И. 1908. О вреде, причиненном в Беловежской Пуще Монашенкой,
Ocneria monacha L. Русск. энтом. обозр., 8 : 26.
Щелкановцев Я. 1932. Очерки по биологии лесных вредных насекомых и мерам
борьбы с ними. Изд. 2-е.
Яцентковский А. 1931. Обследование лесов, зараженных вредными бабоч-
ками. Ленинград.
В а е г W. 1923. Beitrage zur Lebensweise der Nonne und Versuche mit deren Bekiimp-
fung. Tharandter Forstl. Jahrb., LXXIV : 240—247.
Brown R. 1931. Observations on the satin-moth and its natural enemies in Central
Europe. Bull. Circ. U. S. Dept. Agric., 176 : 1—19.
Burgess A. 1924. Methods of estimating abundance and demage caused by the gipsy-
moth. Journ. Econ. Entom., XVII : 180—183.
Burgess A. 1931. The satin-moth, an introduced enemy of Poplars and Willows.
U. S. Dept. Agric., Wash., Bull., 167 : 1—16.
D e 1 a s s u s. 1923. Le Liparis dispar en Algerie. Rev. Agric. Afric. Nord, XXI :
520—524.
Delassus 1925. La lutte contre le Liparis dispar dans le massif de 1'Edough. Rev.
Agric. Afric. Nord, XXIII : 334—336, 348—352.
14*
212
ОБЩАЯ ЧАСТЬ
Eckstein К. 1894. Zur genaueren Kenntnis der Nonneneier. Forstl. Naturw.
Ztschr., 3 : 191—192.
Escalera M. 1925. Noticia biologica sobre Lymantria dispar. Boll. R. Soc. Esp.
Hist. Natur., XXV : 337—340.
Escherich K. 1911. Tote Nonneeier. Naturw. Ztschr. Forst-und Landwirtsch.,
9 : 237—246.
Escherich K. 1912. Nonneprobleme. Там же, 10 : 65—85. *
G r a w a 11 G. and G. Posey. 1918. Gipsy-moth larvae as agents in the dissemi-
nation of the white-pine blister-rust. Journ. Agric. Res., Wash., D. С., XII :
429-439.
Howard L. 1924. Insect Parasites of Insects. Proc. Entom. Soc. Wash., XXVI :
27—46.
Howard L. 1930. A history of applied Entomology. Smithsonian Inst. Misc. Coll. :
1 -I- 564.
Janisch E. 1936. Physiologische Grundlagen der Nonneprognose. Anz. Schadlingsk.,
12 : 77.
Janisch E. 1939. Di; Bedeutung des Optimums fur den Massenwechsel forstschad-
licher Insekten. Verh. VII Intern. Entom. Kongr., Ill : 1974—1990.
Knoche E. 1922. Die Wipfelk’ankheit der Nonnenraupen. Nachrichtenbl. deutsch.
Pflanzenschutzdienst., II : 98—99.
К о m a г e к J. 1931. Mniskova kalamita v letech 1917—1927. Rec. Trav. Inst. Rech,
agron. Tschecosl., 78 : 1—-256.
Komarek J. 1933. Wichtige Neubeobachtungen aus der Biologie der Nonne. Anz.
Schadlingsk., 9 : 93—96.
Komarek J. 1937. Kritisches Wort iiber die Bedeutung der Insektenparasiten der
Nonne. Ztschr. angew. Entom., 24 : 95—117.
L e m a r i e J. 1933. Neue Kontrollmethode des Nonnenvorkommens. Anz. Schad-
lingsk., 9 (3) : 43—44.
Lepiney J. de. 1930. Contribution a 1'etude du complex de Lymantria dispar. Mem.
Soc. Sei. Natur. Maroc., 23 : 1 —100.
Leuthold. 1931. Fichtenkultur und Massenvermehrung der Nonne. Tharandter
forstl. Jahrb., LXXXII (1) : 53—81.
Loos K.. 1915—1916. Einige В eob a ch tungen, Versuche und Untersuchungen iiber die
Lebensweise der Tachine, Parasetigena seg egata Rd. auf dem Libacher Herr-
schaftgebiet. Vereinschr. Forst- und Naturkunde, Prag, 12 : 537—563.
Maleno t t i E. 1931. Le polveri arsenicali contro il bombice dispari. Primo experi-
ment in Italia con distributore di polvere a motoro contro gl’insetti dei boschi.
Riv. mens. Touring. Cl. ital., «d’Alpe».
M о к г у T. 1922. The true reasons for the increase of the nun-moth in Central Europe
in recent years. Ztschr. Pflanzenkr. und Gallenkunde, Stuttg., XXXII : 72.
M о к г у T. 1924. From my experiences of the nun-moth. Centralbl. Bakter. Parasit.
Infektionskr., LXII : 7—12. •
Nechleba A. 1927. Verkiimmern und Verderben von Bruten forstschadlicher Insek-
ten. Anz. Scnadlingsk. , III : 115—117.
Nechleba A. 1928. Beschrankte Niitzlichkeit des Calosoma sycophanta L. Anz.
Schadlingsk., IV : 89.
Niisslin O. and L. Rhumbler. 1927. Forstinsektenkunde. 4 Aufl., Berlin.
P r e 11 H. 1924. Uber die Immunitat von Fichten gegen Nonnenfrass und ihre Uhrsa-
chen. Tharandter forstl. Jahrb., 75 : 58—71.
Ratzeburg J. 1838—1840. Die Forstinsekten. I—II, Berlin.
Rebel H. 1921. Nonnenvermehrung im Hofoldinger Forst 1899—1902. Ztschr. angew.
Entom., VII : 311—333.
R e h L. 1925. Tierische Schadlinge an Nutzpflanzen. Sorauer, Handb. Pflanzenkr.,
IV (1) : 433—445. Berlin.
Ritzema-Bos J. 1914. Eine Belangrijke Vretetij van de Banken-Borstel rups
of den Foodstaart (Dasychira pudibunda L.) in het Elspeter Bosch. Tijdschr.
Plantenzickten, Wageningen, XX : 115—140.
R u z i ck a J. 1923. Uber den Einfluss der Niederschlage auf die Vermehrung und
den Frass der Nonne. Ztschr. Forst- und Jagdtwirtsch., XLI : 98—99.
R u z i с к a J. 1924. Die neusten Erfanrungen iiber die Nonne in Bohmen. Zentralbl.
ges. Forstwesen, 1 : 33—67, 159—185, 315—336.
R u z i e к a J. 1925. Das- Verhalten der Vogel zur Nonne in Bohmen und Mahren im
Zeitraume 1884—1924. Ztschr. angew. Entom., 11 : 147—148. >
R u z i с к a J. 1931. Einige Gedanken zur Nonnenforschung. Anz. Schadlingsk., VII :
81—82.
Sedlaczek W. 1909. Die Nonne, Lymantria monacha L. (Studien uber ihre Lebens-
ЛИТЕРАТУРА
213
weise und Bekampfung). Centralbl. ges. Forstwes., 35 : 145—164 193—213
242—261. ..
Sedlaczek W. 1910. Uber das Verhalten der Nonnenraupen auf friih- und spat-
treibenden Fichten. Там же, 36.
Sedlaczek W. 1916. Einwirkung des Klimas auf die Entwicklung der Nonne.
Verhandl. zool.-bot. Ges., Wien 66 : 28—33.
Sedlaczek W. 1917. Relation between climate and life-cycle of the tussock moth
(Lymantria monacha L.). Monthly Bull. Agric. Intell, and Pl. Dis., Rome,
VIII : 1200.
Sedlaczek W. 1918. Das Auftreten der Nonne in Bohmen im Jahre 1918. Centralbl.
Forstwesen., : 219—227.
Smits van Burgest C. 1924. A note on the immunity in insects from parasiti-
sation by other insects. Entom. Ber. Ned. Entom. Vereen., II : 240.
Trag&rdh J. 1920. Undersonninger over mumians upptradande i Gualov 1915—
1917. Medd. Statens Skogsoksanstalt, Stockholm, 17 : 301—328.
T u b e u f C. 1892. Weitere Beobachtungen iiber die Krankheiten der Nonne. Forstl.
Naturw. Ztschr., 1 : 277—279.
Voelkel H. 1930. Vorlaufge Mitteilung iiber den gesetzmassigen Ablauf der Mas-
senvermehrung von Insekten. Nachrichtenbl. Deutschen Pflanzenschutz, 10 :
1—44.
Vukasovic P. 1936. Contribution a I’etude de Lymantria dispar L. en 1935. Arh.
Min. Pol. Poprivr., 3 : 1—42.
W a c h 11 F. 1910. Neue Gesichtspunkte iiber die Entstehung der Nonnenkalamitaten
und die Mittel zu ihren Abwehr. Centralbl. Ges. 'Forstwesen.
Wilke S. 1931. Uber die Bedeutung tier- und pflanzengeographischer Betrachtungen
fur den Forstschutz.. Arb. Biol. Reichsanst. Land- und Forstwirtsch 18 :
583—675.
Wolff M. und A. К r a u s s e. 1922. Die forstlichen Lepidopteren. Jena.
Zederbauer E. 1911. Klima und Massenvermehrung der Nonne, Lymantria mo-
nacha L. Mitt. Forstl. Versuchsw. Oesterr., 36 : 1—19.
Z w б 1 f e r W. 1925. Eine Polyederseuche als Uhrsache des Erloschens einer lokalen
Goldafterkalamitat. Die kranke Pflanze, II (12) : 239—240.
Z wolf er W. 1935. Klima und Nonne. Forstwissensch. Zentralbl. : 23—24. 753 -
767, 795—800.
СПЕЦИАЛЬНАЯ ЧАСТЬ
Сем. ORGYIDAE.1 — ВОЛНЯНКИ
Boisduval, 1840, Gen. Index Meth. Eur. Lep. : 66; Walker, 1855, Cat. Lep. Het.
Brit. Mus., 4 : 7-77; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 429; Staudinger, 1901, Cat.
Lep., 1 : 113; Spuler, 1908, Schmett. Eur., 1 : 125; Strand, 1910, Seitz, Gross-
schmett., 3 : 109; Strand, 1915, там же, 10 : 291; Stauder, 1924, Entom. Anz., 4 ; 73;
Jordan, 1924, Nov. Zool., 31 : 217; Hering, 1926, Seitz, Grossschmett., 14 : 129; Schaus,
1927, там же, 6 : 536; Hering, 1932, Tierw. Mitteleur. : 482; Gaede, 1932, Seitz, Gross-
schmett., Suppl., 2 : 95; Collenette. 1933, Nov. Zool.,. 29 : 31; Bryk, 1934, Lep. Cat.,
62 : 1—327; Collenette, 1935, Stylops, 4 ; 241.
Крупные или средних размеров бабочйи; крылья вполне покрыты
чешуей (редко прозрачные); рисунок на крыльях у низших представи-
телей семейства часто не развит совсем (Orgyinae); крылья одноцветные,
чаще белые или желтоватые, совсем без черных полос или с немногими;
ряд родов, особенно высших Orgyidae (Acronictinae), имеет вполне разви-
тый рисунок в виде системы поперечных полос, отчасти или вполне обра-
зующих пятна (круглое и почковидное); клиновидное пятно не развито
(некоторый намек на возникновение его можно видеть у представителей
Trisuloides Btl.). Окраска у низших представителей однотонная — белая,
желтая, реже зеленая, у высших — серая; жилкование передних крыльев
характерно соединением основания второго и третьего медиальных ство-
лов; радиальная жилка или образует добавочную ячею, или ветвится
просто; образование добавочной ячеи связано с частичным анастомозом
1 Название семейства в литературе запутано. По приоритету должно быть поста-
влено название Liparidae, данное впервые этому семейству Гюбнером (Hiibner, 1816)
по одному из старейших родов — Liparis Ochs. (1810). Название рода Liparis Ochs.
(1810) является преоккупированным, так как Артеди (Artedi, 1738) давно уже описал
рыб под этим названием, прочно вошедшим в литературу. По Свайно (Swinhoe, 1922,
Ann. Mag. Nat. Hist. : 449) и Колленетт (Collenette, 1935, Stylops : 241—246), назва-
ние Артеди (1738) не может быть принято как данное не по бинарной номенклатуре
до десятого издания «Systema Naturae» (1758) Линнея. Но, как оказалось, Скополи
(Scopoli, 1777, Instr. Hist. Nat. : 453) переописал рыб Liparis Artedi под двойным
названием, вследствие чего название Liparidae (Artedi) Scopoli все же остается
за рыбами.
Широко используемое в литературе название Lymantriidae также не может быть
использовано, потому что род lymantria Hb. (1816) описан позже, чем Ocneria Hb.
(1816), к которому нужно, как оказывается, относить теперь все виды бывшего рода
Lymantria Hb. Дайр (1897), за ним Кузнецов (1915) принимают для этого семейства
название Hypogymnidae. Тем не менее' название Hypogymna Hiibner не было опубли-
ковано, но лишь приведено в рукописи и потому не принято в литературе вообще.
Из всех остальных родов этого семейства Orgyia Ochs. (1810) и Arctornis Germ. (1810)
являются старейшими. Ауривиллиус (1891) использовал первое название для подсем.
Orgyinae, которое он относил к Arctiidae. Это подсемейство по смыслу соответствует
первому подсемейству системы, принятой мной в настоящем издании, почему я
принимаю это название для описываемого семейства в целом.
СПЕЦИАЛЬНАЯ ЧАСТЬ
215
второго и третьего радиальных стволов; наличие ее констатировано
„для самых низших Orgyidae (Dasychira Ochs.), утрата связана с полным
анастомозом основания (более половины длины) второго и третьего ради-
альных стволов, которые в этом случае сидят на длинном стебельке (Eu-
proctis Hb., Ocneria Hb., Parocneria Dyar.). Наличие стебелька характери-
зует специализованные роды; на задних крыльях типична большая ба-
зальная ячея, образованная субкостальной и радиальной жилками;
ячея или открытая, когда нет анастомоза субкостальной и радиальной
жилок — у большинства низших Orgyidae, или замкнутая полным анасто-
мозом этих жилок, как это наблюдается у высших Orgyidae', у некоторых
специализованных родов Orgyinae (Penthophera) и у большинства родов
подсем. Acronictinae вторая медиальная жилка на задних крыльях (так же
как и на передних) сдвинута основанием к третьей медиальной и нередко
с ней анастомозирует основанием или даже сливается полностью (ряд
видов рода Euproctis Hb.). Типична редукция ротовых частей, которые
или рудиментарны настолько, что остатки их полностью скрыты в воло-
сяном покрове головы (большинство Orgyinae и Oxycesta Hb. из Acronic-
tinae), или. хотя и рудиментарны, но заметны в виде еще хорошо разви-
того хоботка (Leucoma Steph., Artaxa Wlk., Diphthera Ochs., Panthauma
Stgr., Simyra Ochs., Eogena Bd., Leiometopon Stgr.); реясе ротовые органы
развиты вполне и функционируют (Мота Hb., Daseochaeta Warr., Acro-
nicta Ochs.). Ноги без шипов на голенях (исключая Daseochaeta Warr.)
и бедрах; когти простые или с зубцом. Количество сегментов брюшка
у самцов равно девяти, но у низших представителей (Dasychira Steph.)
есть остаток десятого сегмента в виде почти полного, но слабого кольца
(покрытого волосками) вокруг anus; у этих представителей uncus не раз-
вит и функционально замещен superuncus восьмого сегмента; у боль-
шинства видов uncus развит вполне, причем от девятого сегмента часто
отделен швом и лишь у высших представителей (родов трибы Acronictini)
он с ним вполне спаян; при основании uncus по сторонам его имеются
короткие, реже длинные отростки (subunci), сохраняющиеся в виде
небольших выступов и у высших Orgyidae (Acronicta Ochs., Craniophora
Snell.).
Преимагинальные фазы развития Orgyidae характеризуются рядом
признаков. Яйца почти всегда несколько уплощенные в дорзовентраль-
ном направлении, иногда очень плоские. Гусеницы типичны развитием
бородавок на коже, на месте первичных щетинок и возникновением вто-
ричных волосков, локализованных на этих бородавках и в меньшей мере
разрастающихся просто на коже (некоторые виды Acronicta Ochs.); есть
•случаи недоразвития бородавок и образования только пупилл (Acronicta
alni L.) или, напротив, редукция их (Episema coeruleocephala Esp.); перед-
негрудь типична слитыми бородавками х и ч\ и разрастанием этой
комплексной большой бородавки перед стигмой; обычны, во всяком случае
у многих родов, слияние [i и а. брюшных сегментов и редукция а или
слияние ее с р на средне- и заднегруди; бородавка х на брюшных сегмен-
тах утрачена или сдвинута и даже слита с и, или, реже, сдвинута и слита
(не полностью) с р; на брюшных ногах развиты склеротизованные пластин-
ки (щитки), несущие большое число щетинок. Возникновение бородавок
наблюдается с первой стадии, в последующих стадиях возможно их уси-
ление; возникновение их со второй стадии — реже; развитие вторичных
щетинок начинается всегда с первой стадии, хотя в последующих число
.их,обычно сильно возрастает. Куколки типичны наличием скульптиро-
: ванных дистальных частей брюшных сегментов и пучков волосков (иногда
216
ORGYIDAE
негустых и коротких) за стигмами и на вентральной и дорзальной сторо-
нах брюшка, а также (не всегда) на спинке и на голове.
Биологически Orgyidae характеризуются кормовой связью с древес-
ными растениями, в основном с лиственными породами — Salicales,.
Fagales, Rosales; питание хвойными возможно и в ряде случаев специ-
фично (например, для некоторых неарктических видов Olene Hb.), но
не характерно для видов Orgyidae. Питание травянистыми растениями
также не характерно, хотя ряд видов живет за счет травянистых Legu-
minosae и Rosaceae и есть виды, специализованные на питании Euphor-
biaceae; питание травянистыми растениями наблюдается также у некото-
рых полифагов (Ocneria Hb., Acronicta Ochs.). Яйцекладка производится
кучками по несколько десятков или сотен яиц, и расселение гусениц
в молодом возрасте имеет место у большинства видов Orgyidae с руди
ментарной имагинальной фазой. Среди Acronictini у ряда видов, питаю-
щихся в половой фазе, наблюдаются случаи откладки яиц поодиночке
(виды подродов Acronicta Ochs, и Triaena Hb.) на листья кормовых расте-
ний и высокая специализация молодых гусениц, почти неспособных
к отысканию пищи. Окукление, как правило, происходит на листьях
растений, в щелях коры, в развилках ветвей и реже в сухой древесине
или в лесной подстилке на поверхности почвы. Период перед окукле-
нием характеризуется сбрасыванием волосков (часто также изменением
их окраски) и использованием их для кокона. Коконы плетутся из редких
шелковых нитей (у тех видов, у которых фаза куколки падает на летний’
период) или они более плотны и готовятся из многих шелковых нитей,
куда вплетаются волоски или сухие листья (у зимующих в фазе куколки)..
Orgyidae распространены почти по всему свету (рис. 89), но численное
распределение видов дает очень ясное указание - на концентрацию их
в тропиках Старого Света, именно на территории тропической Африки,
Индостана, Индокитая и Малайского архипелага. Видами этого семейства,
богаты влажные тропики (Конго, Нигерия, Зондские острова) и в мень-
шей мере саванны (восточной Африки, Индостана, Зондских островов);
на территории тропической (центральной и отчасти Южной) Америки’
виды Orgyidae малочисленны, но своеобразны. Обильно населены субтро-
пики Азии (Гималаи, Китай, Япония), где есть ряд эндемичных родов.
Неарктика бедна видами Orgyidae, в частности известны немногие энде-
мичные роды. В Австралии и на океанских островах виды Orgyidae редки
и эндемичные роды не многочисленны. Виды подсем. Orgyinae концен-
трируются на территориях влажных тропиков и субтропиков, тогда как-
виды подсем. Acronictinae наиболее обильны в субтропиках и в зоне широ-
колиственных лесов умеренных широт Голарктики. Хвойные леса бедны-
видами Orgyidae, а в степях и полупустынях виды этого семейства пред-
ставлены ничтожным числом часто высоко специализованных форм..
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
1 (2). Хоботок большею частью очень сильно редуцирован (не виден
при рассмотрении объекта в лупу без специальной препаровки^
или, редко, заметен в лупу, но рудиментарен; передние крылья
с добавочной ячеей или без нее; базальная ячейка на задних крыльях
большая и большей частью незамкнутая; крылья большею частью-
с слабо развитым, рисунком, нередко одноцветные; у гусениц
на шестом и седьмом брюшных сегментах, редко также на первомг
и втором сегментах есть выворачивающиеся железы (красные,.
Рис. 89. Общее распространение видов сем. Orgyidae.
Черным обозначены территории с числом видов 500—300; сеткой — 200—100; вертикальной штриховкой — 50—30;
горизонтальной штриховкойgo—10 и пунктиром — 5—1,
218
I. ORGYINAE
желтые и т. д.); бородавки и т. на средне- и заднегруди свободны,
но часто слиты на первых брюшных сегментах. I. Подсем. Orgyinae
2 (1). Хоботок почти всегда виден в лупу на объекте; он развит нормально
или рудиментарен и слабый (у Oxycesta Hb. не виден); добавочная
ячейка на передних крыльях всегда развита; базальная ячейка
разных размеров, нередко небольшая и замкнутая; рисунок на пе-
редних крыльях большею частью развит хорошо, окраска нередко
(если рисунок.редуцирован) яркая (зеленая, желтая); выворачи-
вающиеся железы на брюшных сегментах у гусениц не развиты;
бородавки fl и а на средне- и заднегруди слиты, на первом и втором
брюшных сегментах они всегда свободны...........................
.....................................II. Подсем. Acronictinae
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ ПО ЛИЧИНОЧНОЙ
ФАЗЕ (ПО ВЗРОСЛЫМ ГУСЕНИЦАМ)
1 (2). Шестой и седьмой брюшные сегменты с дорзальной стороны несут
непарные, выворачивающиеся железы, обычно видные в виде воро-
ночек или мясистых выступов. Редко такие железки есть на первых
двух брюшных сегментах. Бородавки а и S на средне- и заднегруди
почти всегда не слиты; лишь у . Penthophera Germ, они слиты;
с каждой стороны груди по пять бородавок. Бородавки 7. и 13 на всех
или только на первых и последних брюшных сегментах часто слиты.
Бородавка х на переднегруди (перед стигмой) несет пучки волосков,
направленные вперед .................................I. Orgyinae
2 (1). Железки на дорзальной стороне брюшных сегментов отсутствуют;
бородавки р и а на средне- и заднегруди всегда слиты; с каждой
стороны этих грудных сегментов по четыре бородавки (редко есть
небольшая бородавка рядом с ъ—&) на брюшных сегментах боро-
давки а и р на первом — седьмом сегментах всегда свободны (три
над стигмой); на переднегруди нет больших пучков волосков перед
стигмой; редко есть также латеральные пучки на среднегруди . .
......................................... II. Acronictinae
I. Подсем. ORGYINAE
Виды крупные или средних размеров. Ротовые части редуцированы
большею частью очень сильно; рудименты их совершенно скрыты в воло-
сяном покрове головы; реже ротовые органы довольно длинные, и руди-
.менты видны в форме закрученного короткого хоботка; губище щупальца
большею частью слабые; глаза всегда голые; задние ноги с одной или двумя
парами шпор на голенях; радиальная ячея или развита, или утрачена; ба-
зальная ячея на задних крыльях большая, большею частью не замкнутая
анастомозом субкостальной и радиальной жилок; рисунок или не развит
совсем (крылья однотонные), или развит в виде поперечных полос; почко-
видное пятно часто недоразвито; клиновидное пятно всегда отсутствует;
uncus почти всегда отчленен швом от девятого сегмента, нередко двой-
ной, иногда не развит; в последнем случае есть остатки десятого сегмента;
щевярый сегмент часто несет subunci в виде длинных выростов по сторо-
нам основания uncus или в виде бугорков; редко они вполне утрачены.
Гусеницы в общем характерны отсутствием слияния бородавок fl и а
на средне- и заднегруди (исключая роды Penthophera Germ., Leucoma
.Steph., Parocneria Dyar); бородавка z большею частью прямо за стигмой
ТАБЛИЦА РОДОВ
219
несет небольшой пучок волосков (редко одну щетинку); иногда она слита
с р и сдвинута дорзально, в нескольких случаях утрачена; волоски гусе-
ниц всегда пильчатые и нередко перистые. Куколки несут густые волоски
в виде пучков на дорзальной стороне тела, за стигмами и в меньшем
количестве на вентральной стороне тела.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ ПО ИМАГИНАЛЬНОЙ ФАЗЕ
1 (18). Радиальная ячейка на передних крыльях развита; если она
недоразвита, то к этой тезе все же отходят виды с черными полу-
прозрачными крыльями, имеющие самок с укороченными крыльями,
или особи с одноцветно белыми крыльями и черной дискальной
жилкой, или, наконец, белые особи с длинными губными щупаль-
цами.1
2 (13). Задние голени с двумя парами шпор; полы не сильно диморфны,
самки всегда крылаты.
3 (6). Задние голени (а также голени других пар ног) покрыты густыми
длинными пушистыми волосками; губные щупальца всегда корот-
кие, свешиваются вниз, покрыты торчащими волосками.
4 (5). Дорзальная сторона заднегруди и первых брюшных сегментов
без пучков волосков; uncus не развит, он замещен (функционально)
superuncus восьмого сегмента; есть остатки десятого сегмента в виде
латеральных, покрытых волосками пластинок по сторонам аналь-
ного сосочка .................................3. Dasychira Steph.
5 (4), Заднегрудь и второй брюшной сегмент несут с дорзальной стороны
по непарному короткому густому пучку черных коротких волос;
uncus развит вполне, простой или со слабой дорзальной бороздой;
десятый сегмент утрачен........................... . 1. Olene Hb.
6 (3). Задние голени (а также голени других ног) покрыты прижатыми
волосками; губные щупальца покрыты чешуйками или гладкими
прижатыми волосками.
7 (10). Передние и задние крылья без рисунка чисто белые или желто-
белые; иногда они с черным штрихом на месте дискоидальной жилки.
8 (9). Губные щупальца длинные, косо направленные дистально и вниз,
второй членик их большой, третий членик хорошо виден, занимает
около 1/3 второго; коготки на всех ногах простые; harpe не развита
................................................4. Laelia Steph.
9 (8). Губные щупальца короткие, третий членик их почти полностью
, утрачен; коготки на лапках с сильным вентральным зубцом; harpe
с сильным стержневидным выростом. . . . . .11. Arctornis Germ.
10 (7). Крылья с хорошо развитым рисунком из полос и нередко из хо-
рошо развитых пятен; окраска яркая — желтая, черная или корич-
невая.
11 (12). Передние крылья с хорошо развитыми пятнами (особенно почко-
видными); есть две поперечных линии на передних крыльях, не ши-
рокие и темные; valva с острым сильно склеротизованным дисталь-
ным концом ................ ....................7. Cifuna Wlk.
12 (11). Передние крылья без обычных пятен; линии на них широкие
в виде косых белых или желтых полос на черном фоне; окраска
. 1 Некоторые особи Penthophera Germ., Arctornis Germ., Laelia Steph, иногда
не имеют радиальной ячейки на передних крыльях; в этих случаях должно быть обра-
щено внимание на дополнительные признаки.
220
I. ORGYINAE
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
черная или светложелтая; valva короткая с многими мелкими зуб-
цами по дистальному скошенному краю............8. Numens Wlk.1
(2). Задние голени с одной парой шпор; полы часто сильно диморфны,
самки короткокрылы или бескрылы.
(15). Крылья бедно покрыты чешуей, полупрозрачные, с черным
хорошо заметным жилкованием; голени всех ног покрыты прижа-
тыми волосками; базальная ячея на задних крыльях замкнута
вполне; uncus плотный, без шва............6. Penthophera Germ.
(14). Крылья густо покрыты чешуей, голени всех ног или только
передних покрыты длинными пушистыми волосками; базальная
ячея на задних крыльях не замкнута, uncus с бороздой на дор-
зальной стороне.
(17). Тело более или менее массивное при относительно пропорцио-
нальных крыльях; самки крылаты или с рудиментарными крыльями;
глаза круглые или слабо суженные с боков; голени всех ног покрыты
длинными пушистыми волосками; valva короткая и широкая,
с продольной складкой от основания...........2. Gynaephora Hb.
(16). Тело тонкое, стройное; крылья большие и широкие; самки почти
или совсем бескрылы и даже безноги (лишь у одного вида кры-
латы); глаза заметно сужены с боков; голени, во всяком случае
задних ног, покрыты прижатыми волосками; valvae заостренные,
без продольной складки, часто короткие..........5. Orgyia Ochs.
(1). Радиальная ячейка на передних крыльях всегда отсутствует.
(24). Задние голени с двумя парами шпор.
(21). Когти на всех ногах простые; крылья с хорошо развитым рисун-
ком в виде черных или коричневых полных (или частичных) полос
на светлом фоне; полы сильно диморфны, но самки крылаты . . .
................................................14. Ocneria Hb.
(20). Когти на всех ногах с большим вентральным зубцом; передние
крылья или совсем без рисунка, одного цвета с задним, большей
частью белые или светложелтые, или несут рисунок в виде охри-
стых полос и точек.
(23). Uncus двойной, разделен на две симметричных латеральных
половины; anus несет многочисленные мелкие волоски; тело тонкое,
крылья пропорционально большие, широкие и округлые; окраска
большей частью желтая или охристая . . . . . .9. Artaxa Wlk.
(22). Uncus целостный; отделен швом от десятого сегмента; anus
мембранозный, без волосков; тело довольно толстое; крылья про-
порционально короче, чем у предыдущего рода, и острее, а также
уже; окраска большей частью белая............10. Euproctis Hb.
(19). Задние голени с одной парой шпор.
(26). Когти всех ног с сильным вентральным зубцом; valva без выроста
с дорзальной стороны или на дистальном конце, на внутренней
стороне несет склеротизованную бородавку, усаженную многочис-
ленными, разно направленными шипами; передние ноги охристые
................................................12. Ivela Swinh.
(25). Когти всех ног простые; valva с более или менее значительным
выростом на дорзальном крае или на дистальном конце, без боро-
давки на внутренней стороне; все ноги одного цвета.
(28). Передние крылья без рисунка, .одного цвета с задними: белые,
желтоватые или черно-серые, реже черные; хоботок у некоторых
] К этой тезе относится также Qcneropsis Kozh. (см. стр. 376).
ТАБЛИЦА РОДОВ
221
видов заметен в виде небольшого рудимента; valva в дистальной
частй округлена, дорзальный край ее несет сильный, клерсотизо-
ванный вырост ...........................13. Leucoma Steph.
28 (27). Передние крылья с хорошо заметным рисунком или его руди-
ментами, редко однотонные, обычно отличного и более темного
цвета, по сравнению с задними; хоботок утрачен и не виден
на объекте; valva с острым, сильно склеротизованным дистальным
концом........................................16. Parocneria Dyar.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА БРОДОВ ПО ЛИЧИНОЧНОЙ ФАЗЕ
. (ПО ВЗРОСЛЫМ ГУСЕНИЦАМ)
1 (16). Над стигмами брюшных сегментов (1—8) простые (непарные)
бородавки; на дорзальной стороне первых четырех или пяти брюш-
ных сегментов и на восьмом брюшном сегменте или только на пер-
вом и втором брюшных сегментах непарные кисточки или пучки
щетинистых или курчавых волосков; если пучков волосков нет,
то на средне- и заднегруди с каждой стороны по четыре бородавки
(у. и р слиты).1
2 (15). Средне- и заднегрудь имеет с каждой стороны по пять борода-
вок; на первых четырех, пяти или двух брюшных сегментах непар-
ные дорзальные пучки (щетки) волосков; восьмой сегмент большею
частью также несет пучок волосков.
3 (12). Над стигмами брюшных сегментов (1—8) по три бородавки; две
верхних аир если и сближены и соприкасаются, то полностью не
слиты, и даже при крайнем их сближении виден шов (эти бородавки
сильно сближены на первых четырех-пяти брюшных сегментах).
4 (7). Бородавки <х и р на шестом и седьмом брюшных сегментах при-
мерно равных размеров; р не сильно увеличена; х на брюшных
сегментах всегда очень бедна волосками, несет только один длинный
волосок или рудиментарна; волоски, слагающие щетки, слабо зуб-
чатые или перистые.
5 (6). Бородавки а и р на первых четырех брюшных сегментах и на
восьмом брюшном сегменте почти равны по размерам; на.конце
брюшка только один пучок волосков на восьмом его сегменте;
по сторонам переднегруди нет сильных пучков волосков (кисточек
перистых волосков) ...».......................3. Dasychira Steph.
6 (5). Бородавка а на первом брюшном сегменте значительно больше р,’
которая лежит прямо под ней (не в стороне, как у Dasychira Steph.);
на восьмом брюшном сегменте а значительно меньше р, на конце
брюшка две кисти волосков, одна на восьмом, а другая на девятом
сегменте; по сторонам первого грудного сегмента длинные черные
кисточки перистых волосков....................... . 4. Laelia Steph.
7 (4). Бородавка р на шестом и седьмом брюшных сегментах значительно
больше, чем а (в пять-шесть раз); бородавка х на брюшных сегмен-
• тах большею частью несет пучок волосков; если на этой бородавке
одна щетинка или она рудиментарна, то на переднегруди длинные
кисточки сильно перистых волосков.
8 (11). Первые пять брюшных сегментов несут непарные кисточки
(щетки) волосков (иногда не сильно развиты); бородавка х на брюш-
ных сегментах всегда несет пучок волосков; волоски, сидящие
1 Обозначение хетотаксии (бородавок) для гусениц я принимаю по Фракеру
(S. Fracker, 1930 (1915), Illinois Biol. Monogr., 2 : 1—161).
222
Г. ORGYINAE
на z переднегруди, слабо перистые или пильчатые, но не перистые
и не булавовидные; тело густо покрыто длинными щетинистыми
волосками.
9 (10). Бородавка а на среднегруди сильно сближена с (3, соприкасается
с ней; на бородавке р по сторонам тела сильные и длинные щети-
нистые волоски (пучки) далеко торчат в стороны; дорзальные щетки
на первых пяти брюшных сегментах короткие и нередко несут
сильно перистые волоски; перистые же волоски нередко имеются
и на дорзальной стороне груди и последних брюшных сегментах;
тело несколько уплощено........................2. Gynaephora НЬ.
10 (9). Бородавки а и на среднегруди не сильно сближены; тело более
или менее равномерно покрыто длинными волосками; дорзальные
щетки на брюшных сегментах длинные и слагаются из слабо пиль-
чатых волосков; тело вальковатое......................1. Olene НЬ.
И (8). Дорзальные щетки располагаются лишь на.первых четырех брюш-
ных сегментах, всегда вполне развиты; переднегрудь несет по сто-
ронам длинные торчащие вперед пучки перистых головчатых воло-
сков, сидящих на бородавке х; волоски, сидящие на бородавках,
не густые; z на брюшных сегментах несет одну или две щетинки.
...............'...................................5. Orgyia Ochs.
12 (3). Над стигмами брюшных сегментов по две бородавки на всех
восьми брюшных сегментах или лишь начиная со второго; лате-
ральные кисточки на переднегруди и дорзальные щетки не развиты.
13 (14). На первом брюшном сегменте « и f не слиты, но сильно сбли-
жены (соприкасаются); а несколько крупнее, чем (3. На остальных
сегментах брюшка бородавки эти слиты вполне . .9. Artaxa Wlk.
14 (15). На всех брюшных сегментах над стигмами по две бородавки с каж-
дой стороны (а и р) слиты; на дорзальной стороне первых двух брюш-
ных сегментов короткие курчавые волоски. . . 10. Euproctis НЬ.
15 (2). На средне- и заднегруди по четыре бородавки с каждой стороны,
аир слиты; эти бородавки слиты и на всех брюшных сегментах
(над стигмами брюшных сегментов по две бородавки); тело покрыто
короткими жесткими всюду примерно равными по длине волосками
, .......................................... 6. Penthophera Germ.
16 (1). Над стигмами первого—восьмого брюшных сегментов двойные
бородавки, х частично слита с р, но видны хорошо обе; на дор-
зальной стороне тела нет густых1 кисточек волосков; гусеницы
нередко несколько уплощенной формы.
17 (18). На средне- и заднегруди с каждой стороны по четыре бородавки;
аир слиты; эти бородавки на первых восьми сегментах брюшка
также слиты; над стигмами на брюшных сегментах по три боро-
давки (считая две над стигмами). Пучки щетинок на дорзальных
бородавках (а) первого брюшного сегмента длиннее других ....
.........................................16. Parocneria Dyar1
18 (17). На средне- и заднегруди по сторонам по четыре больших боро-
давки, а кроме того обособлена а в виде одной сильной щетинки,
сидящей на пупилле; бородавка а есть также и на брюшных сег-
ментах.
19 (20). Непарные выворачивающиеся железки, кроме шестого и седь-
мого брюшных сегментов есть также на первом и втором брюшных
1 К этой тезе подходит также род Ocnerorgyia Stgi’,, являющийся, невидимому,
синонимом Parocneria Dyar.
I. OLENE
223'
сегментах; щетинка z на среднегруди сильно сближена ср...
.......................................... 13. Leucoma Steph.
20 (19). Непарные выворачивающиеся железки есть только на шестом
и седьмом брюшных сегментах, как у всех видов Orgyidae щетинки
х и р на среднегруди расположены как обычно; нередко а несет
пучок волосков (не только одну сильную щетинку). 14. Ocneria Hb.
1. Род OLENE НВ.
Hiibner, 1827, Zutr. Exot. Schmett., 2 : 19 (typus mendosa Hb.); Kirby, 1892, Syn..
Cat. Lep Het., 1 : 488; Barnes and Me Dunnough, 1913 Contrib. Nat. Hist. N.
Amer., 2 : 52; Schaus, 1927, Seitz, Grossschmett., 6 : 537; Bryk, 1934, Lep. Cat., 62 : 8..
Тип рода О. mendosa Hb.1
Хоботок рудиментарен; губные щупальца слабые, тонкие и короткие,
торчащие вперед; последний членик их очень короток; голова и ноги
покрыты длинными пушистыми волосками; передние ноги очень длинные-
(нормально, в покойном состоянии особей, они торчат далеко вперед
головы насекомого); коготки на всех ногах с широким основанием, обра-
зующим маленький выступ (зубчик) с вентральной стороны; голова,,
грудь и брюшко покрыты тонкими пушистыми волосками без примеси
чешуи; глаза большие, почти круглые, окруженные длинными волосками,
нависающими над ними; крылья вытянутые, сравнительно узкие (осо-
бенно у самок); радиальная ячейка большая, вытянутая, широкая; тре-
тий, четвертый и пятый радиальные стволы отходят на общем стебельке,
или пятый радиальный ствол отходит свободно от конца ячеи рядом
(из одной точки) с первыми двумя стволами; второй радиальный ствол
отходит от костального края ячеи; первый медиальный ствол от ее дор-
зальной стенки к базальной половине; базальная ячейка на задних
крыльях узкая, незамкнутая; субкостальная и радиальная жилки на зад-
них крыльях не анастомозируют; третий медиальный ствол отходит
на общем стебельке с первым кубитальным; дискоидальная ячея на зад-
них крыльях не вполне замкнутая.
Uncus хорошо развит, сложной формы (разделенный вдоль или взду-
тый); saccus также развит; valvae без harpe, с небольшой складкой в ба-
зальной половине; ductus bursae длинный; apophyses длинные. Полы
диморфны; самки более крупны с округлыми, более нежными, часто
полупрозрачными крыльями.
Гусеницы густо волосистые, после первой линки приобретающие черты
взрослых. Бородавках переднегруди несет короткие пучки волосков, слабо
выдающиеся вперед; на брюшных сегментах (взрослых гусениц) она несет
немногие волоски в виде пучков. Бородавки аир первых пяти брюшных сег-
ментов расположены рядом, почти слиты, большие, несут пучки недлинных
пильчатых волосков. Бородавки а шестого и седьмого брюшных сегментов
очень малы, несут небольшие'пучки волосков. Восьмой сегмент несет непар-
ный пучок слабо перистых волос, сидящих на бородавках а и р. Эти же
бородавки на среднегруди небольшие, заметно сближены (рис. 93). У куко-
лок хорошо развиты лабиальные щупальца, они равны примерно ширине
1 Описание родов во всех случаях,' если это специально не оговорено, дано на
основании родовых типов с учетом Признаков других видов, входящих в данный род.
В тех случаях, когда тип рода относится к тропической (или иной экзотической фауне)
и не мог быть мною исследован (это имело место в одном случае), диагноз рода состав-
лен на основании близких видов, что в соответствующем месте оговорено.
T. ORGYINAE
максиллы; дорзальная сторона тела густо покрыта волосками; усики
при основании очень широкие; дорзальная сторона пятого и шестого
брюшных сегментов не сужена; верхняя губа и clypeus выше уровня
.прочих частей ротового аппарата; cremaster направлен дорзально.
Обширный полиморфный род, сосредоточенный главным образом
в тропиках и богато представленный в юго-восточной Азии, а также
в восточной части Северной Америки (восточные штаты). Наиболее
•обильны виды Olene Hb. в экваториальной западной Африке (Золотой
Берег, Сиерра-Леоне, Нигерия) на территории влажных тропических
лесов; отсюда известны О. seitzi Her., pheosia Hamps., basalts Wlk., quin-
quepunctata Moeschl., pulcherrima Her.,, gloveroides Her., omissa Her.,
cedestis Her., mediofascia Her., pais Her., proletaria Holl., albosignata Hb.
Значительное число видов описано с Малайского архипелага (О. misana
Moore, albiplaga Swinh., costalis Wlk., basinigra Hegl., mendosa Hb.)
также с территорий тропического леса, причем последний вид широко
заселяет прилегающие тропические острова (Филиппины, Тайван),
Индию, Гималаи, Китай и Японские острова, являясь в ряде мест вре-
дителем сельскохозяйственных растений -г- хлопчатника, Zizyphus, Тег-
minalia. Из Индии и южных Гималаев известны также эндемичные
виды (О. dudgeoni Swinh., orirnba Swinh., magnalia Swinh., cymotaSwinh.,
nigra Hamps.); два эндемичных вида описано из Японии (О. inclusa Wlk.,
tarawanensis Mts.), тогда как в Китае распространена только О. men-
dosa Hb. Один эндемичный вид (О. olga Obth.) описан из южного При-
морья; один (О. fascelina L.) широко распространен в лесной зоне Ев-
разии от Тихого до Атлантического океана и заходит в тундру, и один
эндемичный вид описано Канарских островов (О. fortunataRghir.), нередко
здесь массовый и вредящий лесу.
В Северной Америке виды Olene Hb. довольно многочисленны на юго-
востоке (Каролина, Арканзас, Массачузеттс, Ныо-Хэмпшайр, Нью-Джер-
сей, южная Канада), —О. basiflava Pack., kervillei Barn, et MacD., vagans
Barn, et McD., dorsipennata Barn, et McD., atomaria Wlk., achatina Sm.
et Abb., cinnamomea Grt. et Rob., manto'Streck., montana Beut., /nreiDyar,
lagiata Wlk., griseofacta Dyar и bonivelli Barn. etBenj.; один вид {О. stix
Barn, et McD.) описан из юго-западной Канады (Британская Колумбия).
Виды Olene Hb. очень гигрофильны, а большинство их, кроме того,
и термофильны; они связаны с древесной растительностью; основными
кормовыми растениями являются Fagales (Betulaceae,. Fagaceae), Rosales,
включая и низкорослые виды, и Salicales (Populus, Salix); ряд видов
•связан с Pinaceae (Abies, Pinus), например, О. montana Beut., lagiata
Wlk., pint Dyar. Для большинства видов специфична многоядность,
а также питание взрослых гусениц, кроме указанных древесных расте-
ний, многими другими видами растений, например Malvaceae, Могасеае,
Theaceae, Cactaceae, Rutaceae, Euphorbiaceae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Самцы
1 (2). Вторая и третья поперечные линии тонкие; в срединном поле
у костального края большое белое пятно треугольной формы; основ-
ной цвет черно-коричневый (шоколадный); uncus сильно вздут
при основании; valva небольшая, треугольная. .* 1. О.olga Obth.
2 (1). Вторая и третья поперечные линии очень широкие, интенсивно
черные; срединное пятно одного цвета с общим фоном крыла, кото-
1. OLENE
225
рый большею частью черный, но может быть бело-желтым; uncus
плоский; valva широкая, с небольшим выступом на дорзальном
крае.......................................... *2. О. fascelina L.
Самки
11 (2). Передние крылья светлые, желто-серые, с двумя нерезкими зиг-
загообразными поперечными линиями; в костальной половине их
широкое белое с неправильными контурами пятно в срединном
и базальном полях; совокупительная сумка мешковидная . . . .
. ...........................................* 1. О. olga Obth.
2 (1). Передние крылья чаще темные, пепельно-серые или, реже, свет-
лые, почти белые, однотонные, с двумя почти прямыми, широкими
черными или темными (иногда охристыми) полосами; совокупитель-
ная сумка сложной формы.....................,.. ..* 2. О. fascelina L.
* 1. Olene olga Obth. (рис.. 90, 91).
Oberthiir, 1881, Et. Entom., 5 : 34, t. 2, fig. 1, 2; Graeser, 1888, Berl. entom. Ztschr.,
32 : 124; Staudinger, 1901, Cat .Lep., 1 : 115 Oyn.); Strand, 1910, Seitz, Grossschmett.,
2 : 112 (syn.), t. 19, g; Gaede, 1932, там же, Suppl. : 95 (syn.), t. 8, c; Bryk, 1934, Lep.
Cat., 62 : 9 (syn.); Куренцов, 1939, Tp. Горно-таежн. ст., 3 : 145.
Резко диморфный вид; самец коричнево-серый, короткокрылый, рисунок
нерезкий; голова и грудь шоколадно-серые, покрыты слегка утолщен-
ными волосками; брюшко светлосерое, на конце несколько затемненное;
на дорзальной стороне первого и третьего сегментов брюшка есть пучки
больших блестящих черных чешуй, резко выделяющихся на светлом
^юне; базальная линия на передних крыльях утрачена, базальное поле
однотонное; вторая поперечная линия в виде широкой полосы, расту-
шеванная, слабо заметная в дорзальной половине крыла; в костальной
части срединного поля широкое почти треугольное белое пятно, захва-
тывающее также часть костального поля за третьей поперечной линией,
на фоне которого отдельные, неправильно разбросанные черные чешуйки;
почковидное пятно слито со светлым костальным краем, растушеванное,
в середине тусклокоричневое; третья поперечная линия у костального
края широкая, черная на белом фоне, под костальным краем выгнута
кнаружи, образует острый зубец, далее скошена базально, тонкая, про-
стая, слабо волнистая; срединное поле сильно сужено в дорзальной
половине; подкраевая линия серая с немногими белыми штрихами, вол-
нистая, не резко выраженная на однотонном внешнем поле; терминаль-
ная линия в виде тонкой коричневой полоски; бахрома шоколадно-корич-
невая с немногими неправильными белыми пестринами; задние крылья
однотонные шоколадно-серые, несколько более светлые в середине, с тон-
кой терминальной полоской; дискоидальные пятна на них слабо заметны
с верхней стороны; бахрома одного цвета с общим фоном; нижняя сто-
рона светлая, серо-шоколадная, с тонкой белой терминальной полоской
и неясными дискоидальными пятнами.
Самка крупнее, с вытянутыми, слегка округленными крыльями;
:голова и грудь бело-серые, брюшко почти чисто белое; на дорзальной
стороне заднегруди и второго сегмента брюшка (как и у самца, но боль-
ших размеров) пучки блестящих черных чешуй; дистальный конец брюшка
покрыт шелковистыми черными волосками (со слабой примесью белых воло-
сков); передние крылья бело-коричневые с черным рисунком; костальная
половина крыла от основания до третьей поперечной линии белая с немно-
15 Фауна СССР, И. В. Кожевников.
226
1. ORGYINAE
гими неправильно разбросанными темными чешуйками; в средней части
крыла от его основания черный штрих, почти достигающий второй попе-
речной линии; дорзальная половина крыла и поле между третьей попе-
речной и подкраевой линиями желто-коричневое с неправильно разбро-
санными черными чешуйками; вторая поперечная линия черная, дает
широкую петлю под костальным краем, в дорзальной половине смыта;
третья поперечная линия тонкая, дает два широких изгиба, далее пря-
мая, скошенная базально; подкраевая линия белая и коричневая, дает
Рис. 90. Olene olga Obth. Самец. (Рис. Ф. Гуняева).
три широких изгиба; внешнее поле белое с нерезкой белой терминаль-
ной линией; бахрома белая; задние крылья почти чисто белые с широкой
нерезкой, тусклой полосой; нижняя сторона одноцветно белая с непра-
вильно разбросанными одиночными черными чешуйками; усики самцов
сильно гребенчатые, у самок с очень короткими гребешками. Размах
крыльев самцов 30—37 мм, самок 45—55 мм.
Uncus вздут в базальной части в виде пузыря, дистальный конец его
острый, направлен косо вниз; латерально от его основания расположены
лопасти; девятое кольцо широкое, тергальная его область отделена
от основания uncus швом; вентральная половина этого сегмента узкая;
saccus вытянут, треугольной формы; есть scaphium; valva почти тре-
угольной формы, выгнута дорзально, при основании широкая, с продоль-
ной складкой от основания; fultura inferior полукруглая, с широкой
вырезкой в дорзальной части; penis тонкий и длинный с несколько рас-
ширенным coecum. Bursa copulatrix мешковидная, мембранозная, с руб-
чатой скульптурой; ductus bursae длинный, изогнутый, склеротизован
в дистальной половине; восьмой сегмент длинный, несет толстые расши-
ренные в дистальной части и довольно длинные apophyses anteriores;
apophyses posteriores еще более длинные; papillae anales заостренные..
Гусеница-, по Палшкову, желто-зеленая с тремя длинными и густыми
кисточками волосков на первых трех брюшных сегментах и черным пят-
ном на последующих; кисточки первого грудного сегмента длинные,
на концах черные; кисточка волосков на восьмом брюшном сегменте-
длинная, 7келто-зеленая; голова и грудные ноги коричневато-жел-
тые.
1. OLENE
227
Близка, как то правильно указывал Обертюр (Oberthiir, 1881), к Olene
fascelinah., особенно самки; неправильное сведение в синонимы к этому
виду Dasychira albodentata Brem. является следствием плохой сохран-
ности типа последнего вида (см. ниже Dasychira albodentata Brem.).
Распространение. Южное Приморье (Владивосток, Аскольд.
Сучан, Сица).
Биология. Зимующая стадия не известна; лёт бабочек, по Ку-
ренцову (1939), в сентябре. Гусеницы питаются (по Палшкову) черемухой
и (по Грезеру) лещиной и
кленом; окукление в листьях ----
в рыхлом коконе, куда впле- / 'х
таются и опавшие волоски. ' \ Ч
Стацией являются кедрово- С . р. I
ширкоолиственные леса. \ ’> )
1х (
* 2. Olene fascelina L. I \
(рис. 92, 93). /у 'ч \ А / • •xJ '
Linne, 1758, Syst. Nat., ed. A'.'l V |
10 : 503 (Bombyx)\ Aurivillius, I 1.1
1891, Nordn. Fjar. : 93, t. 25, \E;;7 1J I
fig. 4; Staudinger, 1901, Gat. Lep., /'I
1 : 115; Spuler, 1908. Schmett. zA::l гЧа£йц|'\<дд
Eur., 1:128, 351, t. 25, fig. 22; fi'iw-i W/i гй
Wahlgren, 1908, Entom. Tidskr.', /•'./ VW a
29 : 133; Strand., 1910, Seitz,Gross- \ '// б
schmett., 2 : 112, 120, t. 19, fig.; v
Rebel, 1910, Berge, Schmetter-
lingsb. : 115, t. 21, fig. 9; Vorbrodt
und Muller-Rutz, 1912, Schmett.
Schweiz, 1 : 230; Brundin, 1917,
Entom. Tidskr., 37 : 216; Nean- Рис. 91. Olene olga Obth. Гениталии: a — самца;
der, 1918, там же, 42 : 43; Brun- б — самки.
din, 1918, там же, 42 : 66;
Marschner, 1932, Entom. Rundsch., 49 : 246; Gaede, 1932, Seitz, Grossschmett., Suppl.,
2 : 95; Matsumura, 1931, 6000 Insects Japan : 698, fig. 413; Matsumura, 1933,
Ins. Matsum., 7 : 116; Bryk, 1934, Lep. Cat., 62 : 11; Nordstroem und Wahlgren, 1937,
Nordn. Fj r.: 65, t. 13, fig. 9; — medicaginis Hiibner, 1800, Schmett. Eur., 2 : fig. 81
(Bombyx); —• obscura Zetterstedt, 1840, Ins. Lapp. : 42; — nivalis Staudinger, 1888,
Stett. Entom. Ztg., 48 : 97; — laricis /Schille, 1899, Soc. Entom., 14 : 73; — obscurata
Staudinger, 1900, Iris, 12 : 337; — angelus Tschetwerikov, 1904, Русск. энтом. обозр.,
4 : 77; —helgelandiae Strand, 1906, Zool. Zentralbl. ; 798; —callunae Peets, 1908,
Jahresber. Naturf. Ges. Hannover, 55 : 214; — proletaria, Strand, 1910, Seitz, Gross-
schmett., 2 : 112; — unicolor Schulz, 1910, Entom. Ztschr. Stuttgart, 24 : 35; — cauca-
sica Scheljuzhko, 1919, Neue Beitr. Ins.-Biol., 1 : 131; — alpina Kitt, Verh, zool.-bot.
Ges. Wien, 74/75 : 69; — nigrotecta Dannehl, 1928, Mitt. Munch, entom. Ges., 19 : 103;—
karafutonis Matsumura, 1933, Ins. Matsum., 7 : 116.
Биология и экология. Васильев, 1913, Хозяйство: 16; Reh, 1925,
Sorauer, Handb. Pflanzenkrankh., 4 : 443; Белозор, 1931, Зб1рн. праць Зоол. муз.,
10 : 152; Лебедев, 1933, Зб1рн. праць екол., 1 .: 72.
Голова и грудь пепельно-серые, заднегрудь в проксимальной части
несет охристые чешуйки; второй и третий сегменты брюшка несут большие
пучки черных чешуй, из них больший пучок на втором сегменте; брюшко
пепельно-серое, дистальный конец его черный; усики самцов сильно
гребенчатые, у самок они несут короткие гребешки на члениках; димор-
физм полов в размерах и рисунке не резок. У самок примесь охристых
и черных чешуй больше; передние крылья у самцов относительно корот-
кие и широкие, у самок вытянутые и округлые, пепельно-серые, с двумя
резкими слабо развитыми черными и охристыми поперечными линиями
15*
228
1. ORGY IN Ah
и неясным остальным рисунком; у самцов рисунок полнее; базальное
поле с ясной короткой черной базальной линией у самого корня крыла;
вдоль срединной жилки от основания белый штрих; по всему полю непра-
вильно разбросанные крупные охристые и черные чешуйки; вторая попе-
речная линия в виде густо расположенных крупных чешуй, оттенена
охристым, особенно обильно у самок, изогнута под костальным краем,
далее прямая; третья поперечная линия почти прямая, идет несколько
косо параллельно терминальному краю крыла; она особенно интенсивная
у|костального края крыла и имеет примесь охристых чешуй, особенно
обильную в дорзальной
половине крыла; внешнее
поле уже срединного или
одной с ним ширины;
подкраевая линия белая,
грубо волнистая, с внут-
ренней стороны оттенен-
ная черным, в виде не-
правильно разбросанных
чешуй и штрихов; терми-
нальная линия в виде
черной полосы, прерван-
ная; бахрома серая, одно-
тонная; задние крылья
однотонно серые, несколь-
ко более темные в базаль-
ной половине, светлые по
краю; у некоторых особей
(особенно самцов) заметна
растушеванная краевая
линия; бахрома светлая,
почти белая; нижняя сто-
рона светлосерая, по
костальному краю непра-
вильно разбросанные бе-
лые чешуйки; есть широ-
кая неполная поперечная линия, затушевывающая дискоидальные пятна;
самки бледнее и имеют большую примесь охристых чешуй. Размах
крыльев самцов 34—42 мм, самок 40—55 мм-
Uncus широкий и плоский, с глубокой двойной борбздой с дорзаль-
ной стороны; дистальный конец его тонкий, уплощен и сильно склеро-
тизован; основание uncus большое, образует небольшие латеральные
вздутия; девятое кольцо короткое, отделено швом от основания uncus,
в вентральной части несет небольшой saccus; вокруг anus разрастается
склеротизованная пластинка, разделенная с дорзальной стороны; fultura
inferior в. виде плоского склерита, с вентральной стороны овального;
valva при основании широкая, довольно длинная, с дорзальной стороны
несет при основании короткий вырост в виде ступеньки; penis почти
прямой с тонким и коротким coecum; pars inflabilis без инкрустаций.
Bursa copulatrix короткая, сложной формы, несет рубчатую скульптуру;
ductus seminalis отходит от ее дистального конца при основании ductus
bursae; последний тонкий, складчатый, в вагинальной области сильно
склеротизован; восьмой сегмент длинный, в дистальной половине слабо
склеротизован; apophyses anteriores тонкие и длинные; apophyses poste-
1. OLENE
229
riores длиннее первых; papillae anales небольшие, короткие, в виде сла-
бых мембранозных лопастей, несущих многочисленные щетинки.
Яйца матовые, серо-беловатые, овальной формы; наибольший диаметр
около 1 мм, наименьший, примерно, 0.7—0.8 мм.
Гусеницы в младших стадиях зеленовато-желтые с черным, в старших
стадиях черно-белые или черно-желтые; тело покрыто бородавками,
несущими пучки черно-зелено-
ватых и черно-белых волосков;
на дорзальной стороне первых
пяти брюшных сегментов гус-
тые и толстые кисти черно-
белых волосков; седьмой и
восьмой брюшные сегменты
с дорзальной стороны почти
голые и несут лишь вывора-
чивающиеся железки; на дор-
зальной стороне восьмого брюш-
ного сегмента длинная черно-
белая или черно-зеленая кисть
волосков; по сторонам первого
грудного сегмента небольшие
(не длиннее прочих) и негу-
стые пучки волосков; по бокам
под стигмами неширокая охри-
стая полоса, особенно ясная
на брюшных сегментах; вен-
тральная сторона тела охри-
стая, посередине (между ног)
черно-серая; грудные и брюш-
ные ноги черные; голова черная
с немногими охристыми пят-
нами .
Куколка блестящая, корич-
нево-черная, обильно покрыта
волосками лишь по дорзальной
стороне тела; вентральная сто-
рона даже брюшных сегментов
почти голая; кожа брюшных
сегментов почти не скульптиро-
вана; cremaster в виде гладкого
Рис. 93. Olene fascelina L. Хетотаксии перед-
не- и среднегруди и брюшных сегментов
I, VI, VII и VIII гусеницы I стадии (а)
и взрослой (б).
почти цилиндрического стерж-
ня, на дистальном конце несущего недлинные-крючья.
.Относительно однообразный вид, во всяком случае в отношении
рисунка; окраска меняется от почти черной до белой. Однотонная черно-
серая форма с несколько редуцированным рисунком, известная с севера
ареала из разных мест арктической Европы и Сибири, названа О.
obscura Zett. С юга — из гор Средней Азии (Тянь-шань) — известна
светлая, почти белая форма с несколько более гребенчатыми усиками
самцов — nivalis Stgr.; подобная же форма, но с несколько более интен-
сивными черными поперечными полосами на крыльях описана из северной
Монголии как angelus Tschetw. Последняя форма (светлая) прини-
мается Штаудингером, без достаточных оснований, как самостоятель-
ный вид.
230
1. ORGYINAE
Распространение. Средняя Европа к северу от Пиренеев
{Бильбао), всюду в Альпах (до северной Италии), по Германской низ-
менности, по Висле, в Карпатах; Скандинавия (Швеция, Норвегия,
Лапландия); Финляндия (всюду по Лапландии до горной и приморской
тундр); в Европейской части СССР, начиная от тундры, всюду по лесной
зоне, лесостепи и отчасти по степи (Хибины, Усть-Сысольск, Петрозаводск,
Котлас, Вологда, Каунас, Рига, Таллин, Тарту, Ленинград, Калинин,
Москва, Ярославль, Псков, Могилев, Брянск, Харьков, Киев, Чернигов,
Винница); Поволжье (Сарепта, Саратов); Приуралье (Чкалов, Губерли,
Уральск); Северный Кавказ (Ставрополь, Эльбрус, Казбек, Ларс, Кис-
ловодск); в Сибири также всюду по лесной зоне и отчасти тундре и степи
(Акмолинск, Кокчетав, Минусинск, Красноярск, Новосибирск, Омск,
Томск, Тобольск), Алтай (Чемал, Онгудай, Иртыш, Семипалатинск),
Тарбагатай, Тянь-шань, Семиречье, Алай, Заалайский хребет, Гиссар-
ский хребет (2080 м); Саяны; Прибайкалье (Иркутск, Вилюй, Яна);
Яблоновый хребет; северо-восточная Азия (Яна от Адыча до Иктырхая,
Якутск); о. Врангеля, бухта Роджерса 1 (сборы Портенко); Сахалин
{Нуиво); Индигирка; Приамурье (Хабаровск); южное Приморье (Уссури,
Черниговка); северная Монголия (Улан-Батор, Уляссутай, Орок-нор,
Халха).
Биология и экология. Зимует полувзраслая гусеница
(III—IV стадии) в лесной подстилке и мхе; весной появляется с повыше-
нием температуры воздуха до 5—б°; гусеницы многоядны, питаются
Pinaceae (Larix europaea), Fagaceae (Quercus, Fagus), Rosaceae (Pirns,
Sorbus, Rubus), Salicaceae (Salix, Populus), Betulaceae (Betula, Corylus),
Leguminosae (Vicia, Medicago, Trifolium), Compositae (Lactuca, Taraxa-
cum), Ericaceae (Cassandra, Andromeda, Calluna), Euphorbiaceae (Euphor-
bia). Из них предпочитаются Betulaceae, Fagaceae и Rosaceae; с осени
до зимовки на кустарниках и травянистых растениях (Leguminosae,
Ericaceae и др.), весной на древесных, особенно на Rosaceae (Sorbus,
aucuparia) и Betulaceae (Betula pubescens), летом — преимущественно
на кустах березы (Betula pubescens). Окукление в июне, в рыхлом
коконе из рыхло сплетенных нитей и волосков, в подстилке или в корнях.
Лёт бабочек в конце июня, в июле, происходит только ночью; днем бабочки
прячутся в травянистом покрове и очень редко остаются на стволахдеревьев.
Яйцекладка кучками по несколько десятков яиц на ветви растений или
на кору стволов; яйца густо покрываются волосками черного цвета
с конца брюшка самки. Развитие яиц в июле в течение 8—15 суток; гусе-
ницы диапаузируют. Дает всегда одно поколение. Наилучшими условиями
зимовки является устойчивая, сухая, морозная погода. Отмечен как вре-
дитель лесных пород, например орешника, а на юге даже как вредитель
люцерны. Известны паразиты — Microgaster sp. (Braconidae), Ichneumon
bellipes Wesm., Pimpla ‘instigator F., Anilasta ebenina Gr., Sagaritis
ebenina Tscliek. (Ichneumonidae) и Larvivora larvarum L. (Larvivo-
ridae)
1 Особи этого вида с о. Врангеля (бухта Роджерса) темнее обычных, более одно-
тонные и с редуцированным рисунком; самцы имеют лишь остатки темной третьей
поперечной линии и полулунный штрих вместо почковидного пятна; самки однотон-
ные с слабым развитием охристых чешуй. Повидимому, это крайняя северная форма
О. fascelina L., наиболее близкая к О. obscura Zett. По структурным признакам (кроме
гениталий, которые не могли быть мною изучены) особи эти тождественны с типичной
формой.
1. OLENE, 2. GYNAEPHORA
231
4 2. Род GYNAEPHORA HB.
Hiibner, 1816, Verz. bekannt. Schmett.: 16 1; Staudinger, 1901, Cat. Lep., 1 : 114;
115 {Dasychira, Dasorgyia); Strand, 1910, Seitz, Grossschmett., 2 : 120 {Dasychira, Dasor-
gyia); Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 : 293 {Dasorgyia)', Fracke/. 1930,
Illinois Biol. Mon., 2 (1), ed. 2 : 98; Gaede, 1932, Seitz, Grossschmett., 2 Suppl. :
99; Bryk, 1934, Cat. Lep., 62:83 {Dasychira, Dasorgyia);—Dasorgyia Staudinger,
1881, Stett. entom. Ztg., 42 : 406 (typus pumila Stgr.); — Byrdia Schaus, 1927, Seitz,
Grossschmett., 6 : 537 {typus rossii Crt.).
Тип рода G. selenitica Esp.
Хоботок рудиментарный; лоб гладкий, покрытый густыми короткими
волосами; глаза голые, небольшие, эллиптической формы, окружены
короткими волосками; губные щупальца короткие, тонкие и слабые,
свисающие вниз, последний членик их рудиментарный, едва заметный,
покрытый длинными пушистыми волосками; усики самцов сильно гребен-
чатые, у самок гребешки очень короткие; передние голени значительно
длиннее первого членика лапки; голени задних ног с одной парой шпор;
коготки всех лапок с небольшим острым выростом с вентральной стороны
при широком основании когтя; радиальная ячейка маленькая, узкая
и короткая; второй радиальный.ствол отходит от наружной стенки ячеи,
третий и четвертый на стебеАьке, пятый свободный, отходит от вершины
ячеи из одной точки с двумя предыдущими; первый медиальный ствол
отходит от середины ячеи с помощью дискальной жилки; базальная ячея
на задних крыльях ланцетовидной формы, замкнутая (субкостальная
и радиальная жилки анастомозируют); третий медиальный ствол слит
основанием с первым кубитальным и анастомозирует на известном рас-
стоянии (на одной шестой длины ствола).
Uncus хорошо развит, несет с дорзальной стороны или небольшой
шов, или глубокую борозду так, что разделен на две части вдоль; saccus
хорошо развит; valvae без harpe, мембранозные, с узкой склеротизован-
ной складкой в вентральной трети, идущей от основания до половины
длины valva; ductus bursae длинный, в дистальной половине склероти-
зован; apophyses длинные. Голова, грудь и брюшко покрыты длинными
пушистыми (или короткими) волосками без примеси чешуй. Крылья
короткие и широкие. Полы диморфны — самки крупнее и у бореальных
и арктических видов типичны большими крыльями, тогда как для высо-
когорных видов указывается (Штаудингер) наличие несколько укорочен-
ных крыльев у самок.
Гусеницы густо волосистые, с длинными боковыми волосками и отно-
сительно короткими дорзальными; брюшная сторона почти голая; харак-
терны отдельные длинные щетинки, торчащие по бокам, из густого воло-
сяного покрова; щетинки х (рис. 94) переднегруди в виде большой круглой
бородавки, несущей пучок выдающихся вперед перистых волосков.
Бородавки а и р средне- и заднегруди сильно сближены (соприкасаются);
эти же бородавки на первых пяти брюшных сегментах большие, распо-
ложены рядом, несут густые щетки сильно перистых волос. Бородавки а ше-
стого и седьмого сегментов брюшка маленькие, но [3 на этих же сегментах
очень большие. На бородавках р всех сегментов тела длинные щетинковид-
ные (щетинистые) торчащие в стороны волоски; а и р на восьмом брюшном
сегменте крупные (но неравных размеров), несущие пучок волосков.
1 Тип рода G. selenitica Esp. указан Кирби (1892); первый вид в роде Gynaephora
Hb. G. antiqua L., но в этот же род входит G. selenitica Esp.; Гюбнером тип рода не от-
мечен.
232
1. ORGYINAE
ментов I VI, VII и VIII взрослой гусе-
ницы.
Щетинки x на всех сегментах брюшка в виде маленькой бородавочки,
несущей небольшой пучок волосков. Куколки короткие, уплощенные-
дорзовентрально, густо волосистые, на дорзальной стороне несущие
буквально войлок волосков; вентральная сторона тела почти голая.
Яйца почти круглые, с толстым гладким хорионом, крупные, до 1 мм
диаметром; откладываются кучками, покрываются короткими волосками:
в виде войлока; волоски эти обычно черного цвета.
Олиготипический род, пятнисто распространенный в бореальной,. '
альпийской и арктической зонах Голарктики; очень близок к предыду-
щему (Olene Hb.), характер-
ный, тем не менее, рядом при-
знаков, отделяющих его (строе-
ние глаз, одна пара шпор на
задних голенях, короткие val-
vae, редукция крыльев у са-
мок), но сближающих с видами
Orgyia Ochs.
Gynaephora Hb. известны из
альпийской зоны гор Централь-
нс1й Азии (Тибет, Памир, Тянь-
шань) — G. selenophora Stgr,,.
pumila Stgr., alpherakii Gr. Gr.,.
sincera Kozh.; один вид распро^
странен в сибирской Арктике-
—G. lugens Kozh. — , а близкий
к нему G. rossii Curt, заселяет
арктическую Америку (Грен-
ландия, Лабрадор, Аляска). В арктической Америке распростра-
нен второй эндемичный вид — G. groenlandica Нош., известный
из Гренландии и с о. Мелльвиль. Наконец в Европе распространена
G. selenitica Esp., втречающаяся спорадически на сухих стациях по воз-
вышенностям средней Европы, Скандинавии и Прибалтики.1
Кормовые растения видов этого рода изучены недостаточно. Отме-
чены кустарники (ивы, березы, вересковые) и многолетники из травя-
нистых. Биология видов Gynaephora НЬ. почти не изучена; для боль-
шинства видов специфична дневная активность бабочек.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (6). Бахрома на передних крыльях черная и желтая или белая, ша-
шечная; на задних крыльях она также шашечная, но менее резкая;
на месте почковидного пятна тонкий полулунный штрих; крупные-
виды — 20—50 мм. -
2 (5). Передние крылья пепельно-серые, бело-серые или почти чисто
белые; дискоидальные пятна хорошо видны с вентральной стороны;
задние крылья тусклосерые или чисто белые с широкой черной,
каймой; uncus плоский с глубокой бороздой.
1 По Дайру (Dyar, 1897), к роду Gynaephora Hb. принадлежит также и Lachana-
ladakensis Мооте (Hampson, 1892, Moths Ind., I : 435), известная из Кашмира (Ладак).
Я не имел материала по этому виду, но по описанию и изображению он действительно
близок к среднеазиатским горным видам этого рода. Этот вид является, типом рода
Lachana Moore (1888, Proc. Zool. Soc. bond.: 397) и, если указание Дайра справедливо,
этот род уходит к Gynaephora Hb. в синонимы.
2. GYNAEPHORA
233
3 (4). Передние крылья белые или бело-серые; срединное поле узкое
(за счет сдвинутой дистально второй поперечной линии); попереч-
ные линии интенсивно черные, широкие; подкраевая линия четкая,
белая, сильно зубчатая; задние крылья чисто белые или слабо серо-
ватые с слабым желтоватым оттенком при основании или без него
..................................„•.........* 3. G. lugens Kozh.
4 (3). Передние крылья пепельно-серые с беловатыми штрихами; сре-
динное поле широкое; поперечные линии, особенно третья, не-
сколько редуцированы, тонкие; подкраевая линия не сильно зуб-
чатая; задние крылья желто-серые..................2. G. rossii Curt.
5 (2). Передние крылья коричнево-серые с коричневыми штрихами;
задние крылья однотонно серые или черные с немногими коричне-
выми мазками; дискоидальные пятна с нижней стороны не развиты;
uncus высокий, с узкой и неглубокой продольной бороздой на дор-
зальной стороне..............................* l.G. selenitica Esp.
6 (1). Бахрома на передних крыльях однотонная или очень слабо ша-
шечная; почковидное пятно широкое, в виде растушеванного штриха;
мелкие виды — 18—25 мм.
7 (10). Задние крылья в середине светлые, желтые или серые, с широ-
кой черной, четко отграниченной краевой каймой.
8 (9). Передние крылья со стушеванными, оттененными светлым попе-
речными линиями; срединное поле посредине с белым пятном.
...................................* 5. G. selenophora Stgr.
9 (8). Вторая и третья поперечные линии на передних крыльях резко
выражены, четкие, слабо извитые; срединное поле однотонное
...............................................* 4. G. pumila Stgr.
10 (7). Задние крылья однотонно черные или несколько осветленные
у основания.
И (12). Передние крылья лимонно-желтые с интенсивно черным рисун-
ком или серые с широкими желтыми мазками; задние крылья жел-
тые с черной каймой........................6. G. alpherakii Gr. Gr.
12 (11). Передние крылья серые или желтовато-серые, с не резким чер-
ным рисунком; задние крылья серые с черной каймой.................
......................................* 7. G. sincera, sp. nov.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПО ВЗРОСЛЫМ ГУСЕНИЦАМ
1 (2). На переднегруди по сторонам длинные торчащие вперед кисточки;
на дорзальной стороне первых пяти брюшных сегментов широкие
двухцветные кисточки (черные и белые по сторонам); на восьмом
брюшном сегменте длинная хорошо видная кисточка...................
..............................................* 1. G. selenitica Esp.
- 2 (1). Кисточки на первом грудном сегменте и на восьмом брюшном
слабо дифференцированы; дорзальные щетки на первых пяти брюш-
ных сегментах распушенные; подобные же короткие пучки воло-
сков на дорзальной стороне грудных и последних брюшных сег-
ментов.
3 (6). Щетки на дорзальной стороне первых пяти брюшных сегментов
слагаются из сильно перистых волосков, имеющих сероватый (седой}
оттенок; все волоски этих щеток примерно одной длины; длинные
волоски очень редки.
4 (5). По сторонам бородавок р (с нижней стороны их) на брюшных сег-
ментах и при основании всех пучков грудных сегментов яркожел-
,234
1. ORGYINAE
тые пучки волосков; боковые пучки волосков черные, щетинистые;
желтые и черные волоски слабо перистые, серые сильно перистые
.................................................2. G. rossii Curt.
-5 (4). Дорзальная сторона тела несет серые или серо-желтые и серовато-
коричневые сильно перистые волоски; бока тела покрыты однотон-
ными коричнево-черными или коричнево-желтыми волосками (чер-
ных и яркожелтых волосков нет) .... * 3. G. Ingens Kozh.
6 (3). Щетки на дорзальной стороне первых пяти брюшных сегментов
слагаются из слабо перистых волос; эти волоски не имеют седого
тона; на дорзальной стороне тела есть примесь большого коли-
чества длинных волос.
7 (8). Щетки на первых двух брюшных сегментах интенсивно черные;
на третьем—пятом брюшных сегментах — темнооранжевые; черные
щетки несколько более длинные, чем оранжевые; на восьмом сег-
менте брюшка пучок интенсивно черных волосков........................ . .
...........................................G. groenlandica Hom.
•8 (7). Все щетки на дорзальной стороне брюшных сегментов черно-
коричневые; бока несут щетинистые желтые волоски........................
...................................* 5. G. selenophora Stgr. (?) 1
* 1. Gynaephora selenitica Esp.2 (рис. 94, 95).
Esper, 1783, Schmett. Eur., 3(2) : 18, t. 82, fig, 5; p. 49, t. 88, fig. 2 (Bombyx'y, Auri.-
villius, 1891, Nordn. Fjar. : 93, t. 25, fig. 3; Staudinger, 1901, Cat. Lep., 1 : 115; Spu-
ler, 1908, Schmett. Eur., 1 : 128, t. 25, fig. 21; Strand, 1910, Seitz, Grossschmett., 2-: 111,
t. 19, e; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 115, t. 21, fig. 8; Vorbrodt und
Muller-Rutz, 1912, Schmett. Schweiz : 230; Petersen, 1924, Lep. Fauna Estl.,ed. 2,
(1) : 140; Hering, 1932, Tierw. Mitteleur. : 483; Bryk, 1934, Lep. Cat., 62 : 8; —pa-
radoxa Fabricius, 1787, Mantissa Insectorum, 2 : 130 (Bombyx)', — lathyri Hiibner.
1800, Eur. Schmett., Bomb., t. 20, fig. 79, 80; — lathyra Swinhoe, 1923, Ann. Mag. Nat.
Hist., 11 (9) : 297 (Bombyx).
Биология и экология. Linnaniemiund Hukkinen, 1921, Acta Soc. Fauna
et Flora Fenn., 48 : 1—27; Lederer, 1923, Handb. prakt. Entom., 3 : 142; Reh, 1925,
Sorauer, Handb. Pflanzenkrank., 4 : 443; Seitz, 1929, Entom. Anz., 9 : 19; Romanizyn,
1930, Prazn. Monogr., 6 : 166; Frey, 1932, Not. Entom., 12 : 64; Gronbloem, 1936, Acta
Soc. Fauna et Flora Fenn., 58 : 12; Wimmer, 1938, Entom. Ztschr. Frankf,, 52 : 33.
1 Принадлежность этой формы, собранной в числе трех взрослых гусениц в Тянь-
шане, к роду Gynaephora Hb. несомненна; принадлежность именно к виду G. seleno-
phora Stgr. очень вероятна, но доказана пока быть не может, так как эти особи не вы-
ведены из яиц и из них не выведены бабочки.
2 Gynaephora groenlandica Hom. (Homeyer, 1874, Zweite Deutsch. Nordpol. Fahrt,
2 : 409; Schaus, 1927, Seitz, Grosschmett., 6 : 537; Gaede, 1932, там же, 2 Suppl. : 96,
t. 8 с) является самостоятельным видом, повидимому наиболее близким к G. seleni-
tica Esp., но не к G. rossii Curt., с которой ее сравнивали и даже- иногда пу-
тали. Имаго описано и изображено неузнаваемо (описан только самец плохой сохран-
ности). Тем не менее Дайр (Dyar, 1897, Psyche, 8 : 153) дает точное описание гусеницы
этого вида по особям, собранным в Гренландии (Уилкокс-Хид), одна из которых дове-
дена до имагинальной фазы — получен самец. Описание это не оставляет сомнения
в близости G. groenlandica Hom. к G. selenitica Esp. Я привожу следующие выдержки
из этого описания.
«Голова 3.4 мм шириною, округлая, тусклого черного цвета, густо покрытая
длинными вторичными волосками. Тело совершенно черное, покрытое большими зер-
нистыми бородавками, расположение которых вполне нормально: первая и четвертая
маленькие. Я не в состоянии точно описать их на свернувшихся особях. Железки
на 10 и И сегментах (члениках) беловатые. Дорзальные пучки есть на члениках 5—9
и 12, как у selenitica Esp., но пучки на 5—6 сегментах и на 12 черные, а на 7, 8 и 9
темнооранжевые. Черные пучки волос несколько длиннее и торчат несколько выше
оранжевых и несколько выше среднего уровня дорзальных волосков. Все волоски
тонко шиповатые, но не перистые. Меньшая особь имеет голову 2.5 мм ширины.
«Куколка напоминает rossii Curt., но длинные дорзальные волоски более сильно тор-
чат и черные, а не беловатые; кутикула тоньше, черная и несет выступы в числе
2. GYNAEPHORA
235
Передние крылья, голова и усики коричнево-серые, прочие части
тела и задние крылья у самцов черно-серые, у самок серые; передние
крылья широкие, несколько выгнутые по костальному краю, задние
крылья сильно расширены; базальная линия почти неясная, растуше-
ванная; базальное поле черно-охристое; вторая поперечная линия в виде
широкой черно-коричневой полосы, слабо извитая; срединное поле узкое,
вильно суженное в дорзальной половине, почковидное пятно широкое,
белое, со слабыми темными штрихами; третья поперечная линия начи-
нается растушеванной полосой за почковидным пятном, сдвинута далеко
к внешнему краю, дает резкие и грубые зигзаги, ниже почковидного’
пятна сильно скошена базально; между почковидным пятном и третьей
поперечной линией охристое пятно; за третьей поперечной линией
на этом же уровне желтый штрих; подкраевая линия белая и серая,
частично прерванная, неправильно извилистая, в дорзальной ее части
белое пятно; терминальная линия тонкая, коричневая; бахрома шашеч-
ная— черная и охристая; задние крылья черные со слабым охристым
оттенком, дискоидальные пятна желто-охристые или вообще не развиты;
бдхрома жёлто-белая; нижняя сторона передних крыльев черная лишь
в базальной части, во внешней половине коричнево-желтая; нижняя
сторона задних крыльев черная с охристой бахромой; самки серого цвета,
рисунок на крыльях тот же, но резче выражена белая подкраевая линия
и белое почковидное пятно; охристые тона у самок не развиты; задние
крылья чисто серые; чешуйчатый покров не густой; усики самцов сильно
гребенчатые, у самок гребешки почти отсутствуют, усики тонкие. Размах
крыльев самцов 20—^25 мм, самок 30—35 мм.
Uncus короткий и широкий, с глубокой продольной бороздой, деля-
щей его на две половины; дистальный конец его простой; вокруг anus
склеротизованный валик, образованный разрастанием его стенки; девя-
тый сегмент Отделен швом от основания uncus, с вентральной стороны
он узкий, не образует saccus; valvae короткие и широкие, почти квадрат-
ные, с небольшой складкой, идущей от основания в вентральной трети
valva; fultura inferior в виде простой пластинки, расширенной в дорзаль-
ной части; penis неправильно изогнут и в дистальной части расширен;
coecum тонкий и короткий; pars inflabilis без инкрустаций. Bursa copu-
трех пар на месте брюшных личиночных ног на 8, 9 и 10 члениках; этих придатков
я не нашел у куколок rossii Curt.».
По Уокеру (1927), в Оксфордском музее существует еще один экземпляр G. groen-
landica Нош., найденный на крайнем севере Америки, на о. Меллевиль в Полярном
море; при сборе отмечено (найдена Фишером в 1819—1820 гг.), что бабочка летала
днем у куста ивы (Salix arctica и S. dpposjtifolia).
В последнее время Кнабен (Knaben, 1927, Skriften от Svalbard og Ischavet,
71: 19) сообщил ряд новых сведений по Gynaephora groenlandica Нот. Он указы-
вает, что вид этот живет в личиночной фазе на ивах и очень обычен в восточной
и западной Гренландии. По наблюдениям в 1930 г. Gynaephora groenlandica Hom.
появлялась в первой половине августа: 5 августа в Moskusoksefjorden, 9 августа в Ап-
tarctichamna, 9 августа в Керр Petersens, 13 августа в Dicksonfjorden, но имаго он
не нашел, почему и полагает, что лёт имаго происходит в конце июля. Он наблюдал
очень много гусениц, пестрых по размерам и, вероятно, по возрасту, по не сумел вы-
вести ни одной бабочки.
Упоминание о Gynaephora groenlandica Hom. можно найти в следующих работах:
Packard, 1877, Amer. Nat. : 52; McLachlan, 1879, Journ. Linn. Soc., Zool., 14 : 112;
Aurivillius, 1890, Groenl. InsektenL, 15 : 12, 1137; Skinner and Mengel, 1892, Proc.
Nat. Sci. Phil. : 158; Bang-Haas, 1895, Medd. Groenl., 19 : 111; Bang-Haas, Lep. Groenl.
Vid. Meddel. Dansk. Foreinig., Kobenhavn : 180; Vanhoffen, 1897, Groenl. Exped. :
145; Strand, 1905, Rep. Nord Exped. «Fram», 3 : 12; Nielsen, 1907, Medd. om Groenl.,
29 : 402; Nielsen, 1910, 1. c., 43 : 34; Henriksen om Lundbeck, 1917, Conspectus Fau-
nae Groenlandiae, Pars 2, Medd. om Groenl., 22 : 675.
236
1. ORGYINAE
Рис. 95. Gynaephora selenitica Esp. Гениталии:
а — самца, б — самки.
latrix небольшая, мешковидная, слабо склеротизованная, с мембраноз-
ным и довольно длинным ductus bursae; восьмой сегмент короткий,
несколько более длинный в вентральной половине; apophyses anteriores
и posteriores почти равной длины, тонкие и длинные; papillae anales
поч'ги прямоугольные, небольшие, слабо суженные в дистальном напра-
влении .
Яйца 1.1—0.9 мм диаметром, почти круглые, сплюснутые дорзовен-
трально; поверхность хориона гладкая и блестящая; сразу после откладки '
окраска зеленовато-желтая
или коричневатая, вскоре-
затем тускнеющая; на дор-
зальной стороне яйца' тем-
ное пятнышко.
Взрослая гусеница черно-
коричневая, покрыта черно-
бурыми длинными щетини-
стыми волосками, более длин-
ными по сторонам тела на
бородавках р и т;, торчащими
в стороны и несколько вверх;
на переднегруди эти пучки
(на бородавках х) длиннее,
имеют вид тонких, торчащих
вперед кисточек; на дорзаль-
ной стороне второго и
третьего грудных сегментов
на бородавках аир также-
щетинистые волоски, торча-
щие вверх и* назад; эти
щетинистые волоски заметно-
пильчатые, отростки их
раза в два-три превышают диаметр волосков; на дорзальной стороне
первых пяти брюшных сегментов на бородавках аир широкие щетки
черных и беловато-серых волосков; белые расположены по сторонам
щеток на краях бородавок р; эти белые волоски заметно более пери-
стые, чем боковые щетинистые, их отростки превышают диаметр во-
лоска в 3—6 раз; на дорзальной стороне последних брюшных сегментов
после пятого черно-бурые щетинистые волоски, торчащие косо назад;,
на восьмом сегменте они образуют неясно дифференцированную кисть;
вентральная сторона тела почти голая, темнокоричневая, почти черная,
несет небольшие и короткие пучки волосков; грудные и брюш-
ные ноги одного цвета с кожей; голова черная, густо волоси-
стая.
Куколка длиною 13—16 мм, блестящая, желто-коричневая; глаза
черные; крылья и усики черно-коричневые; конец брюшка красно-корич-
невый; дорзальная сторона тела покрыта густыми и длинными нежными
желто-коричневыми волосками.
Как то было давно уже установлено, этот вид близок к лабрадорской
G. rossii Curt, и G. groenlandica Hom. По приведенным материалам видно
также, что он близок к среднеазиатским горным мелким видам, относи-
мым по недоразумению в отдельный род Dasorgyia Stgr. Мало измен-
чивый вид; форма lathyri Hb. является синонимом и не может быть харак-
теризована никакими отличительными признаками.
2. GYNAEPHORA
237
Распространение. Средняя Европа—-Альпы, восточная Фран-
ция, Бавария; средняя Германия; Саксония; Судеты; Карпаты; долина
Вислы (Радом); Прибалтика (Вильнюс, Каунас, Рига, Таллин, Нарва,
Луга, Псков, Ленинград); Карельский перешеек (Выборг, Приозерск,
Токсово); Карелия (северный берег Ладожского озера, Петрозаводск);
Финляндия (Тавастия, Савотия); в Финляндии и Карелии идет на север
до 62—63° с. ш.; в Европейской .части СССР, помимо северо-запада,
локально в лесной зоне и лесостепье. (Валдай, Москва, Можайск, Калуга,
Мичуринск, Казань, Полтава).
Биология и экология. Зимует полувзрослая гусеница;
весной выход с мест зимовок сразу после таяния снега; гусеницы сразу
после зимовки линяют (до начала питания); питаться начинают зелеными
частями перезимовавших растений; окукление в апреле—мае, в черно-
коричневом коконе, изготовленном из шелковых нитей и волосков; раз-
витие куколок около месяца. Вылет бабочек в мае, на севере в июне
(первая половина), сначала преимущественно самцов, затем самок. Самки
мало активны, сидят на травинках и побегах; самцы очень подвижны
и летают днем; спаривание наступает сразу после выхода из куколок,
а яйцекладка — следом за окончанием спаривания. Яйца откладываются
кучками на листья и побеги кормовых растений по 50—25 шт. и покры-
ваются черными волосками с конца брюшка самки; плодовитость самок
не велика — 150—200 яиц. Развитие яиц около 25 суток. Гусеницы
очень активны и многоядны. Указаны следующие кормовые растения:
Pinaceae (Pinus, Larix, Picea), Salicaceae (Salix), Betulaceae (Carpinus,
Betula), Fagaceae (Quercus), Rosaceae (Rosa, Rubus, Fragaria), Aceraceae
(Acer), Tiliaceae (Tilia), Ericaceae (Calluna, Erica, Vaccinium), Polygo-
naceae, Leguminosae (Sarothamnus, Hedysarum, Onobrychis, Lathyrus),
Compositae; преимущественно питание травянистыми растениями —
видами Leguminosae и Polygonaceae; из древесных — Fagales. До зимовки
линяет несколько раз (повидимому четыре раза); зимовка в поверхност-
ном слое почвы; для зимовки благоприятны легкие песчаные почвы.
Всегда дает одно поколение. Распространена локально, но местами
обычна. Отмечена как вредитель бобовых кормовых трав в Финляндии;
отмечены повреждения молодых сеянцев лиственницы.
Известны паразиты: из Braconidae — Microgaster spurius Wesm.; из
I chneumonidae — Ichneumon fabricator Gr., I. insidiosus Wesm.; из Larvioo-
ridae—Pales pavida Mg., Larvivora larvarum L.
2. Gynaephora rossii Curt.1 (рис. 96, 97).
Curtis, 1835, Descr. App. Narr. Ross. II Voy. N. W. Pass. : 70, t. fig. 10; Moesch-
ler,1870, Stett. entom. Ztg.: 252; Packard, 1876—1877, Entom. Monthly Mag., 13 : 228
{Lariap, Aurivillius, 1885, Lep. Vega Exped. : 78, t. 1, fig. 2; Staudinger, 1901, Cat.
Lep., 1 : 115; Strand, 1910, Seitz, Grossschmett., 2 : lll;Schaus, 1927, там же, 6:537,
t. 72, a ; Nordstroem, 1927, Entom. Tidskr., 48 : 56; Scheljuzhko, 1927, Ann. Soc. En-
tom. France, 91 : 285, t. 5, fig. 1; Walker, 1927, Entom. Monthly Mag., 63 : 126; Gaede,
1932, Seitz, Grossschmett., 2 Suppl., : 96; Bryk, 1934, Lep. Cat., 62 : 10; Ermolajev,
1935, Folia Zool., 8 : 153.
Биология. Dyar, 1896, Psyche, 7 : 328; Fracker, 1930, Classif. Lep. Larvae,
ed. 2 : 99; Clark, 1935, Rep. Smithsonian Inst. : 267—296.
1 Гедэ (1932) ставит синонимом к Gynaephora rossii Curt. Konokareha daisetsu-
zana Mts. (1927, Ins. Matsum., 2 : 120, Lasiocampidae; Matsumura, 1932, 6000 Insects
Japan : 120, fig. 3, Liparidae). Брик (1934) считает самостоятельным этот вид,
а также род Konokareha, установленный Матсумура (1927). Мне К. daisetsuzana Mts.
известен только по описанию и изображению (не очень хорошему); по литературным
.данным и описанным признакам, его следует считать самостоятельным видом во вся-
ком случае.
238
1. ORGYINAE
Голова, грудь, брюшко и передние крылья пепельно-серые с черным
и белым рисунком; усики самцов с длинными гребешками на члениках;,
гребешки серо-черные, членики белые; усики самок тонкие, с. короткими,
но также двусторонними гребешками; половой диморфизм не резок;
самки несколько крупнее, с брюшком большего размера и всегда серыми,
задними крыльями; задние крылья самцов желтоватые; базальная линия
в виде широкой черной полосы, с базальной стороны оттененная белым;
базальное поле однотонное, бело-серое; вторая поперечная линия в виде
широкой черной, почти прямой полосы (иногда, если более тонкая, мелко
извитая); срединное поле однотонно серое, равное по ширине в косталь-
ной и дорзальной частях; почковидное пятно в виде черного полулуния
на пепельно-белом фоне, иногда почти не заметно и видно лишь белое
Рис. 96. Gynaephora rossii Curt. Самец. (Рис. Ф. Гуняева).
пятно; третья поперечная линия начинается далеко за почковидным пятном,
слабо выгнутая к внешнему краю и очень слабо зубчатая в средней части,
оттенена белым по внутреннему краю, особенно сильно у костального
края и с наружной стороны у дорзального края, где нередко видно белое
пятно; внешнее поле однотонное, серое с тонкими темными полосками
на месте жилок; подкраевая линия в виде белой густо зубчатой частично
стушеванной полосы с базальной стороны у костального и дорзального
краев крыла, оттененной черным; терминальная линия тонкая, черная;
бахрома серая и белая; задние крылья самок тусклосерые, несколько
затемненные по.внешнему краю; у самцов они желто-серые с широким
черным краем и бело-желтой бахромой; нижняя сторона тусклосерая
у самцов с примесью желтых тонов и резкой широкой просвечивающей
черной каймой на задних крыльях; на передних крыльях рисунок просве-
чивает д виде неясных полос и пятен. Размах крыльев самцов 32—35 мм,
самок 40—45 мм.
Uncus широкий, глубоко разделенный продольной бороздой на две
половины, дистальный конец его раздвоен; anus включен в склеротизо-
ванную капсулу, образованную расширенными scaphium и subscaphium;
девятый сегмент узкий, слабо разросшийся в вентральной части, saccus
недоразвиты; fultura inferior в виде большой разросшейся вокруг penis
пластинки; valvae широкие и короткие, в дистальной части косо срезан-
ные, слабо выемчатые по дорзальному и вентральному краям; в базальной
трети valva склеротизованная складка; penis изогнут и в дистальной
2. GYNAEPHORA
239‘
части расширен; coecum короткий; pars inflabilis не несет инкруста-
ций. Bursa copulatrix короткая и широкая; ductus bursae широкий, отхо-
дит от ее правой стороны, мембранозный в проксимальной половине,
посредине несколько сужен, в дистальной части склеротизован; ductus
seminalis отходит от широкой, мембранозной половины ductus bursae
(проксимальной его части); apophyses posteriores значительно длиннее,
чем apophyses anteriores; papillae anales короткие и мембраноз-
ные.
Взрослая гусеница, по Дайру (1896), «имеет округленную голову,
бархатисто-черного цвета, густо волосистую; наличник черный, гладкий;.
Рис. 97. Gynaephora rossii Curt. Гениталии: а — самца,,
б — самки.
задняя часть головы с примесью коричневого; ширина головы 3.2 мм..
Бородавки типичны для Orgyidae — три над стигмой на члениках 3
и 4; бородавка I на брюшке умеренно крупная; бородавка IV очень ма-
ленькая, позади дыхальца; пластинки на ногах (брюшных) ясные. Вы-
ворачивающиеся бородавки (я.елезки) на члениках 10 и 11 обычные,
беловатые, более или менее скрытые в волосяном покрове, часто совер-
шенно. Тело черное с седоватым серым оттенком; волоски толстые, все
перепутанные, некоторые сильно перистые, но не превращены в перышки.
Пучки волос на бородавках I на члениках 5—9 и на 12 несколько более
густые, но не образуют настоящих кисточек или щеток. Волоски серые
с примесью черных и с наличием яркожелтых волос на нижней стороне
бородавки II на члениках от 5 до 13 и при основании всех пучков волос
на грудных сегментах. Брюшные (вентральные) пучки волос маленькие.
Серые волоски густо перистые (сильно перистые) на бородавках от I
до V, черные и желтые волоски только шиповатые. Волоски не очень
длинны большею частью, но не все».
«Куколка с дорзальной стороны покрыта густыми беловатыми (белыми)
волосками; брюшные сегменты не несут придатков, остатков брюшных,
ног, на 8—10 сегментах».
240
1. ORGYINAE
Слабо изученный вид, распространенный в трудно доступных местах;
американские особи довольно многочисленны (с Лабрадора) и имеют
однообразный облик (описаны выше).
Банг-Гааз (Bang-Haas, 1927)1 описывает по одной самке из Саян
(Туран) форму от G. rossii Curt, под названием relictus В.-Н. Описание
очень кратко; оно гласит: «Отличия от типичной самки с Лабрадора из
колл. Штаудингера не значительны. Рисунок хорошо совпадает. Основ-
ной рвет всех крыльев темнее; перед внутренней поперечной линией на
передних крыльях более светлая перевязь. Четкое светлое пятно у зад-
него угла переднего крыла. Задние крылья почти одноцветно черные;
краевая перевязь едва заметна. Нижняя сторона черно-серая с отчетливо
отмеченными срединными точками на всех крыльях». Фото описанной сам-
ки характеризует какую-то однотонную форму, несомненно относящуюся
к роду Gynaephora, но более стройную, чем rossii Curt, или тем более
lugens Kozh. Описывать эту форму как самостоятельный вид невоз-
можно за полным отсутствием материала. Известен только тип. На-
личие черных задних крыльев сближает relictus В.-Н. с балтийско-
скандинавской G. selenitica Esp. Банг-Гааз (цит. раб., стр. 117) упоминает
о сообщении Л. Шелюжко, который отмечает еще два нахождения этой
•формы на северо-востоке Азии. Одно в Коньям (Чукотский п-ов), где
была взята самка экспедицией «Вега», и другое по сборам В. Кожан-
чикова на северном склоне Яблонового хребта, где в горах Да<угджур
(р. Джелинда) были взяты в июне 1915 г. самки. Первая находка была
опубликована Ауривиллиусом и дано изображение этой самки (см.
G. lugens Kozh.), вторая находка мне известна в натуре. Эти особи
принадлежат к G. lugens Kozh. и имеют массивное тело и светлые зад-
ние крылья с темной каймой. Таким образом сближение этих особей
с формой relicta В.-H., которое сделано Шелюжко по фотографиям, не
обосновано. Если саянская форма — relicta В.-Н. действительно имеет
черные задние крылья, то она может быть сближаема с selenitica Esp.
и тогда этот случай пополнит еще список викарирующих видов чешуе-
крылых Скандинавии и Саяна. Все остальные указания на наличие
G. rossii Curt, в Азии относятся к G. lugens. Из двух особей, соб-
ранных В. Д. Кожанчиковым на Яблоновом хребте (р. Джелинда, приток
Алдана) и определенных Шелюжко (1927) как G. rossii Curt., одна была
мною исследована. Эта особь несколько более коричневая, чем собранные
в Арктике, но, без сомнения, принадлежит к G. lugens Kozh. Указания
Ермолаева на наличие G. rossii Curt, на р. Пясине также относятся к этому
виду. Ермолаев имел лишь куколки и из одной извлек совершенно сформи-
рованную особь. Детальное исследование гениталий этой особи им произве-
дено не было, но я имел материал Бесчетнова, служивший также и Ермо-
лаеву; все особи, бывшие в моих руках с Пясины, относятся к G. lugens.
Точно так же изображенная Ауривиллиусом (1887) самка, добытая
на восточной оконечности Чукотского полуострова (зал. Коньям), при-
надлежит к описываемой ниже G. lugens Kozh., как то явствует
из даваемого им изображения. Ауривиллиус не имел материала
с Лабрадора по G. rossii Curt., чем и объясняется неточность определе-
ния. Указание Нордштрёма (1927) о находке G. rossii Curt, на о. Врангеля
основано на его личном мнении и на мнении Н. Я. Кузнецова, прини-
мавшего участие в определении этой особи; мне эта особь не известна,
1 Bang-Haas О. 1927. Horae Macrolep. 1:77, t. 10, fig. 6.
2. GYNAEPHORA
241
ко принадлежность ее к G. rossii Curt, очень сомнительна; вероятно
и здесь имело место неправильное определение.
Распространение. Гренландия (Полярис Бай); Лабрадор
(Гудсонов пролив 62°35' с. ш. и 77°22' з. д., пролив Ферри 72°40' с. пь
и 9Г53' в. д.); Уошингтон Моунт (гольцовый пояс); Аляска (п-ов Барро).
Биология. Слабо изучен в природе, но есть ряд интересных
попутных замечаний Кёртиса; бабочки этого вида на Лабрадоре очень
•обычны. Например, в проливе Ферри за один день 16 июля 1832 г. было
собрано около сотни особей. Гусеницы холодостойки и выносливы к про-
мораживанию. Кёртис указывает, что они твердеют, но выносят пониже-
ние температуры до —40°. Кормовые растения в полярной Америке,
по Кёртису, камнеломки.Saxifraga tricuspidata и S. oppositifolia. В горах
на Уошингтон Моунт, по Дайру (1897), гусеницы были собраны на Vacci-
nium и Betula. Из гусениц, собранных в арктической Америке, выведен
паразит Ichneumon lariae Curt.
* 3. Gynaephora lugens Kozh. (рис. 98, 99).
Кожанчиков, 1948, Тр. Зоол. инет. АН СССР, 7: 151.
Значительно массивнее G. rossii Curt., ярче, рисунок резче, окраска
<белее; крылья по форме близки к G. rossii Curt., но несколько шире;
грудной отдел широкий. Размах крыльев самцов примерно 40 мм, самок
45 мм.1
Передние крылья бело-серые, без примеси диффузного черно-серого
тона, как у G. rossii Curt.; базальная линия интенсивно черная, широкая,
идет до половины крыла; вторая поперечная линия также широкая,
несколько скошенная базально, слабо выгнутая посередине, черная,
оттененная белым; срединное поле узкое, значительно уже, чем у G. rossii
Curt., за счет того, что вторая поперечная линия сильно отодвинута
от основания крыла; на месте почковидного пятна черный штрих, широко
обведенный белым; подкраевая линия зубчатая, черная и белая, дает
два зигзага к термену, белая, оттенена темным; у дорзального края крыла
между этой и третьей поперечной линией большое белое пятно; бахрома
шашечная —черная и белая; задние крылья почти чисто белые, очень
слабо желтоватые у основания, имеют интенсивно черную резко отграни-
ченную кайму; бахрома на них белая; нижняя сторона белая с просве-
чивающими черными элементами рисунка; дискоидальные пятна сверху
слабо видны на задних крыльях, с нижней стороны четкие, в виде точек;
на передних крыльях они не развиты; тело покрыто бело-серыми воло-
сками; членики усов белые, гребешки усиков тонкие, усажены короткими
ресничками. Самки более тусклые, с полупрозрачными крыльями, с не-
сколько редуцированным рисунком.; нередко рисунок совершенно тот Hie
(не утрачен совсем), что и у самцов; задние крылья обычно не имеют
резкого черного ободка; на конце брюшка самки на восьмом и девятом
сегментах слой густых черных волос.
1 Описание дается преимущественно на основании спиртового материала; кроме
того, я имел возможность просмотреть расправленные особи этого вида, которые
в числе примерно десятка определены И. Я. Кузнецовым как G. rossii Curt. Одна
особь этого вида (самка изображена Ауривиллиусом (Aurivillius, 1887, Nordenskjold
Vega Exped., IV : 78, t. I, fig. 2). Изображение (рисунок) не является удач-
ным, но вполне узнаваемо. К этому же виду, вероятно, относится и особь с
о. Врангеля, определенная Нордстрёмом и Кузнецовым как G. rossii Curt.
16 Фауна СССР, И. В. Кожанчиков.
212
1. ORGYINAE
Uncus широкий, плоский, конический, с глубоким вырезом (жолобом).
на дорзальной стороне и несколько расширенным концом, покрыт корот-
кими волосками; девятый сегмент вполне отчленен от uncus, узкий, saccus-
ие образует; стенки anus образуют широкое склеротизованное кольцо,
не вполне сомкнутое на дорзальной стороне; valvae широкие и короткие,
с косо срезанным дистальным краем; базальная половина valva несколько
утолщена и склеротизована, дистальная половина мембранозная и воло-
систая; fultura inferior в виде изогнутой неширокой пластинки с неболь-
шими складочками по сторонам; penis короткий, слабо выгнут дорзалЬно
и не сильно расширен в дистальном направлении; pars inflabilis без инкру-
стаций. Bursa copulatrix мембранозная, небольшая, почти яйцевидной
формы; ductus bursae изогнут
под прямым углом, на месте
изгиба сужен, в базальной
части (более половины его
длины) широкий и мембраноз-
ный; дистальная часть ductus
bursae . расширена к ostium
и сильно склеротизована;
вагинальная область выреза-
на широко; волоски в не-
большом количестве есть сра-
зу под vagina и на мембране
под восьмым сегментом;
восьмой сегмент широкий,
сильно распиренный в дор-
зальной половине, где несет
небольшое число пластинок;
apophyses anteriores доволь-
но длинные, длиннее скле-
рртизованной части ductus
bursae; apophyses posteriores
длиннее anteriores, почти рав-
ны по длине ductus bursae;
Рис. 98. • Gynaephora lugens Kozh. Гениталии:
а — самца, б — самки
papillae anales широкие, несколько косо срезанные по дорзальному краю,,
черные, густо покрыты короткими и редкими длинными щетинками. Весь.'
копулятивный аппарат самки сильнее склеротизован, чем у G. rossii Curt.
Яйца очень плоские, диаметром около 1 мм и высотою 0.4—0.5 мм;
хорион тусклобелого цвета, шероховатый.
Взрослая гусеница темной или светлой коричнево-желтой окраски;
дорзальные пучки волосков, сидящие на бородавках а и (3, более корот-
кие, все равной длины (точно обрубленные), светлее прочих, с серым,
седоватым оттенком; степень развития этого оттенка несколько варьи-
рует (я имел для обозрения около двух десятков гусениц); серая окраска
их стоит в связи с сильной перистостью волосков, отростки волосков
превышают их диаметр в 10—20 раз и торчат в стороны; эти перистые
волоски особенно обильны на указанных бородавках первых пяти брюш-
ных сегментов, где они образуют сильно распушенные щетки; на дор-
зальной стороне грудных и последних брюшных сегментов волоски также
перистые, но несколько длиннее и менее равных размеров; на восьмом
брюшном сегменте они особенно неравномерны и образуют подобие непра-
вильного пучка (отдаленно напоминающего пучок восьмого сегмента
G. selenitica Esp.); боковые пучки волосков, расположенные на бородав-
2. GYNAEPHORA
243
ках ниже стигм, несколько более темно окрашены, более каштановые
или коричневатые и менее перистые, хотя выросты на них вполне заметны
(простым глазом); волоски, слагающие эти пучки, разной длины, щети-
нистые, торчат в стороны; боковые пучки переднегруди обычно совер-
шенно не выдаются относительно других, но у некоторых особей они
слегка длиннее; кожа, как и волосяной покров, варьирует в окраске
в отношении яркости тона; у
особей, имеющих яркую окраску
волосков, она черная, даже бар-
хатистая, у светлоокрашенных
особей она коричневая; брюш-
ная сторона тела черно-корич-
невая или желто-коричневая.,
почти голая, пучки волосков
мелкие; брюшные и грудные
ноги несколько светлее или од-
ного тона с кожей; голова тем-
нокоричневая, с более светлыми
прилобными участками; гусени-
цы самок несколько крупнее,
имеют более темную окраску и
менее перистые волоски на дор-
зальных щетках.
Куколка черно-коричневая
с вентральной стороны и светло-
коричневая с дорзальной; дор-
зальная сторона тела густо по-
крыта короткими, примерно
равной длины, волосками жел-
то-коричневого цвета; пучки
волосков по бокам редкие; вент-
ральная сторона почти голая с
четырьмя рядами небольших
пучков волосков, из которых
особенно короткие средние;
скульптура на брюшных сегмен-
тах в виде косых или попереч-
ных тонких складочек и очень
мелких точек; cremaster в виде
почти цилиндрического сильно
а
Рис. 99. Gynaephora lugens Kozh. Хето-
таксия передне- и среднегруди и брюшных
сегментов I, VI, VII и VIII гусеницы
I стадии (а) и взрослой (б).
склеротизованного выроста, покрытого тонкими бороздками и складоч-
ками, несет на дистальном конце небольшой пучок крючковидных волосков.
Систематическое положение вида ясно; он является близким к G. ros-
sii Curt., но сохраняет также ряд черт, сближающих его с G. selenitica'^]).
Распространение. Полярный Урал (Зайцев); низовья Ени-
сея, Дудинка (Трлстой), р. Волчанка (Рычков), р. Пясина (Бесчетнов);
западный Таймыр, зал. Гафнера, зал. Дика (Старокадомский), р. Лидия
(Мизин); восточный Таймыр, р. Яну-Тарида — устье р. Балахын, Куту-
Конг (Толмачев); Хатангский залив, мыс Нордвик (Старокадомский);
устье р. Лены, Быков полуостров (Бруснев); Селяхская губа, р. Селях;
Новосибирские острова, Большой Ляховский остров (Смесов), Малый
Ляховский остров (Оленкин); низовья Индигирки, с. Хянгастай, Колым-
ская протока; Медвежьи острова; о. Врангеля; Чукотский- полуостров,
16*
24:4
1. ORGYINAE
Уэллен (Портенко); побережье Берингова моря, бухта Эмма (Старокадом-
ский); Камчатка; Яблоновый хребет, р. Джелинда (Кожанчиков).1
Биология и экология. Некоторые сведения по феноло-
гии и биологии этого вида, а также по условиям обитания дают резуль-
таты сборов личиночных стадий; известные мне по спиртовому материалу.
На западном берегу Таймырского полуострова, на берегу зал. Дика
(76°48' с. ш. и 100°50' в. д.), обнаружены взрослые гусеницы 28 щоня
1915 г.; несколько ранее, 19 июня, здесь же собраны из желудков поморника
(Stercorarius parasiticus) куколки и бабочки (Старокадомский). Несколько
южнее, на побережье зал. Гафнера (76°30' с. ш. и 100°50' в. д.), найдены
пронимфы в коконах, в сухой траве (злаках) 4 июня 1915 г. на низком
берегу, где только что стаял снег, и тундра еще не просохла (Старокадом-
ский). Таким образом, окукление происходит вскоре после таяния снега,
может быть еще на проталинах. Зимует гусеница, вероятно, несколько
р.аз; это видно из сборов взрослых и полувзрослых гусениц в конце лета
на Медвежьих островах и на Чукотском полуострове. Окукление в плот-
ных коконах, которые сотканы из шелковых нитей и волосков.. Основной
лёт бабочек падает на июль (главная масса находок с 15 июля) и август
(до двадцатых чисел), но есть находка (на Таймыре) в последней декаде
июня. Это дает возможность полагать период окукления растянутым,
а период развития куколок коротким (около двух недель).. Яйца откла-
дываются на растения кучками и покрываются черными волосками с конца
брюшка самки. Кормовые растения, вероятно, главным образом —. ивы;
Зайцевым найдена взрослая гусеница на иве на полярном Урале; Старо-
кадомский указывает на нахождение гусениц на Таймыре в «траве».
- *4. Gynaephora pumila Stgr.
Staudinger, 1881, Stett. entom. Ztg., 42 : 405 (Dasychira)', Kirby, 1892, Syn. Cat.
Het. Lep., 1 : 485; Staudinger, 1901, Cat. Lep., 1 : 114; Strand, 1910, Seitz, Gross-
schmett., 2 : 120; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 293; Bryk, 1934,
Lep. Cat., 62 ; 83.
Этот вид мне в натуре не известен, поэтому я привожу описание
Штаудингера (1881): «Две пары из Зайсана, которые, вероятно, были
выведены из куколок. Обе самки (особенно одна) имеют своеобразные,
может быть несколько уродливые крылья; являются ли они от природы
укороченными у самок этого вида, что я склонен принять, утверждать
этого не могу. Самцы в размахе 22 и 25 мм. Передние крылья серо-корич-
невые, с грубыми черными чешуйками и растушеванным черным диско-
идальным пятном и с двумя рудиментарными поперечными’ линиями, наме-
ченными на одной трети их длины и у.наружного края. Задние крылья
черноватые с желто-серой бахромой. Этот мелкий вид имеет сходство
в рисунке и общем габитусе с fascelina Esp., только мельче этого ..вида
1 Распространение этого вида выяснено путем изучения как коллекционного
сухого материала, так и спиртовых сборов, в которых мною обнаружены все фазы
развития этого вида. Принадлежность бабочек к описанным куколкам установлена
извлечением сформированных особей (покрытых имагинальной кутикулой) из описан-
ных куколок и идентичных по гениталиям с описанными бабочками. В таких же коко-
нах и из тех же материалов мною обнаружены пронимфы, имевшие строение волосков
и кожи то же, что и гусеницы, собранные в разных местах Арктики. Яйца могли быть
изучены на основании зрелых и одетых хорионом, но еще неотложенных, найденных
в половых путях самок. Этот материал мною обнаружен среди неопределенных
сборов Зоологического института Академии Наук СССР.
2. GYNAEPHORA
245
в два или даже три раза. Усики самцов также сильно гребенчаты, но щу-
пальца значительно короче, чем у fascelina Esp. Волосяной покров груди
и строение ног и брюшка кажутся у обоих видов одинаковыми. Окраска
этих частей, как и передних крыльев, коричневая или желто-серая. По-
следние (крылья) с грубыми черными штрихами, как у fascelina Esp.,
но, исключая упомянутые выше элементы, почти без рисунка. Небольшое
черное пятно на конце ячейки наиболее заметно. От темных базальной
и наружной поперечных линий остаются только следы. С нижней стороны
крылья светлее, с темными чешуйками по костальному краю. Задние
крылья черные, при основании светлее (серые), с серой бахромой. Одна
самка имеет довольно сильно развитые крылья (размах 23 мм), такой же
длины, как тело. У другой самки они едва достигают одной трети длины
тела. У этой последней можно заметить ясный рисунок, напоминающий
таковой у длиннокрылой самки, который находится на желто-сером
фоне с примесью разбросанных черных чешуек. Задние крылья черно-
коричневые со светлым. наружным краем. Усики коротко пильчатые.
Брюшко более сильно покрыто волосками, чем у видов Dasychira Steph.,
почти так же как у самок Orgyia Ochs.; только у Olene fascelina L. послед-
ний сегмент несет такие же волоски. Эта pumila кажется объединяет
рба рода (ранее Boisduval объединял их) и заслуживает, повидимому,
выделения в особый род, который может быть назван Dasorgyia»'.
Не касаясь видовых признаков в описании Штаудингера, требуется
отметить, что строение цог этого вида было просмотрено невнимательно
и не отмечена одна пара шпор, свойственная всем видам Gynaephora Hb.
Распространение. Алтай — Зайсан.
5. Gynaephora selenophora Stgr.1 (рис. 100).
Staudinger, 1887, Stett. entom. Ztg., 48 : 96; Grum-Grzhimailo, 1890, Mem. Lep.
Rom., 4 : 556; Kirby, 1892, Syn. Cat. Het. Lep., 1 : 485; Staudinger, 1901, Cat. Lep.,
1 : 114; Strand, 1910, Seitz, Grossschmett., 2 : 120; Swinhoe, 1923, Ann. Mag. Nat.
Hist., 10 (9) : 293; Bryk, 1934, Lep. Cat., 62 : 83; — pumila. Grum-Grzhimailo, 1890,
Mem. LepARom., 4 : 556, t. 19, fig. 4 (non Staudinger); — pamiricola Strand, 1910,
Seitz,' Grossschmett., 2 : 120, t. 22, b (=purnila Gr. Gr., nom. nov.).
Голова, грудь и брюшко серо-оливковые с просвечивающими серыми
тонами, покрыты длинными шелковистыми волосками; усики и ноги
того же цвета; передние крылья черно-серые с бело-желтоватым рисунком;
базальная линия не развита, в базальном поле серо-оливковые штрихи
между жилками (участки, покрытые длинными волосками); внешняя
часть базального поля светложелтая; вторая поперечная линия в виде
широкой полосы, растушеванная, в костальной половине крыла скошена
1 Из Киргизского хребта (р. Тегарек) из сборов И. Чернавина (24 июня 1913 г.)
в спиртовых материалах Зоологического института Академии Наук СССР есть три
гусеницы Gynaephora Hb. По хетотаксии они тождественны с теми, что описаны и изоб-
ражены для G. selenitica Esp. и G. rossii Curt., так что нет сомнений в принадлежности
их к этому роду (ранее они были определены как Dasorgyia Stgr. Герасимовым). Они
несколько мельче, обликом похожи скорее на арктических Gynaephora Hb., чем на
G. selenitica Esp. Очень вероятна принадлежность их к G. selenophora Stgr.
Дорзальная сторона несет пучки черно-коричневых сильно перистых волосков;
на первых пяти сегментах брюшка дорзальные щетки слабо дифференцированные,
распушенные; бока желто-коричневые, несут торчащие в стороны пучки щетинистых
волосков разной длины (сидящих на р и '<); на переднегруди эти пучки не выдаются
больше, чем на других сегментах; на восьмом брюшном сегменте есть несколько неяс-
ных дорзальных пучков; брюшная сторона почти голая, коричневая; боковые щетини-
стые волоски слабо перистые.
246
1. ORGYINAE
кнаружи, в дорзальной половине крыла идет перпендикулярно его про-
дольной оси; в дорзальной половине крыла эта линия растушевана и слита
•с нижней частью третьей поперечной линии; круглое пятно не развито;
ячейка между второй поперечной линией и почковидным пятном бело-
желтая; костальный край в срединном поле желтовато-серый; срединное
поле в костальной половине почти в три раза шире, чем в дорзальной;
почковидное пятно в виде черного полулуния, резко заметно на фоне
крыла, с внешней стороны оттенено желто-белым; третья поперечная
линия в виде тонкой черной полоски, тонкб зубчатой по жилкам, несколько
более широкой на костальном крае, широко окаймленной снаружи бело-
желтой полосой; она начинается над почковидным пятном, близко оги-
бает его снаружи и сильно сдвинута базально в дорзальной половине
крыла; внешнее поле черно-
серое с обрывками белой
подкраевой линии; терми-
нальная линия в виде тонкой
черной полоски; бахрома с
внешней стороны темная,
оливково-серая; задние
крылья черно-серые с серо-
вато-оливковым волосистым
внутренним краем и неболь-
шим белым срединным полем;
где выделяется черное диско-
идальное пятно; бахрома
selenophora Stgr. Генита- желто-белая; нижняя сторо-
самца. на обеих пар крыльев бело-
желтоватая, края их затем-
крыле ясное дискоидальное пятно. Известны только
Рис. 100. Gynaephora
лии
иены; на каждом
самцы. Размах крыльев 20—22 мм.
Uncus короткий и толстый, с глубокой продольной бороздкой, деля-
щей его вдоль, отделен швом от девятого сегмента; последний —узкий;
saccus короткий; anus склеротизован вокруг; valvae короткие, почти квад-
ратной формы, со склеротизованной складкой от основания; penis слабо
изогнут, расширяющийся в дистальном направлении, с коротким соесшп
и без инкрустаций на pars inflabilis.
По Грум-Гржимайло, встречается на Памире среди снеговых пятен
на талых полянках, поросших Ranunculaceae и Gentiana. Самцы летают
днем. Очень близка к G. pumila Stgr.
Распространение. Средняя Азия Ферганская долина
(Маргелан), Алай (Арча-бахи), Заалайский хребет (Кизил-арт), Памир
(Кара-кузун, р. Беик, 4200 м), Гиндукуш.
6. Gynaephora alpherakii Gr. Gr. (рис. 101, 102).
Grum-Grzhimailo, 1891, Тр. Русск. энт. общ., 25 :464: Kirby, 1892, Syn. Cat.
Het. Lep., 1 : 923; Staudinger, 1901, Cat. Lep., 1 : 115; Strand, 1910, Seitz, Gross-
schmett., 2 : 120, t. 22 b; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 293; Bryk,
1934, Lep. Cat., 62 : 83; — seme novi Grum-Grzhimailo, 1891, Тр. Русск. Энт. общ., 25 :
464; —grumi Staudinger, 1901, Cat. Lep., 1 : 115; — haulberti Oberthur, 1911, Et. Lep.
Comp., 5: 337, t. 83, fig, 782, 783 (Trichosona); — staudingeri Bang-Haas, 1938, Entom.
.Ztschr. Frankf., 52. : 179.
Голова, грудь и брюшко оливково-серые с примесью желтых тонов
от окраски вершин волосков в желтый цвет; волоски покрова тела густые
2. GYNAEPHORA
247
и длинные; усики самцов очень сильно гребенчатые, гребешки усиков
вдвое большей длины, чем у среднеазиатских видов; ноги оливково-
серые, на концах желтые, покрыты длинными волосками; передние крылья
желтые, с черным рисунком; базальное поле желто-серое с примесью
охристых тонов; первая поперечная линия не развита; вторая попереч-
ная линия в виде прямой широкой по краям, особенно по жилкам, размы-
той полосы; у костального края эта линия дает длинный конический
штрих черного цвета вдоль радиальной системы жилок, достигающий
почковидного пятна; костальный край‘(в срединном поле) того же цвета;
круглое пятно не развито; почковидное пятно в виде широкой растуше-
ванной черной лунки на дискальных жилках; третья поперечная линия
Рис. 101. Gynaephora alpherakii С г. Gr. Самец.
(Рис. Ф. Гуняева).
черная, неширокая, растушеванная, зубчатая по жилкам, далеко огибает
почковидное пятно, оставляя между ним и линией желтую ячейку; эта ли-
ния оттенена снаружи широкой желтой полосой; внешнее поле желто-
серое с темными жилками, с намеченной желтым слабой подкраевой
полосой или однотонное; терминальная линия в виде растушеванной
серой полоски; бахрома при основании серая, с внешней стороны жел-
тая; задние крылья желтые при основании, по внешнему краю затемнен-
ные; бахрома желтая; нижняя сторона однотонная, желтая, несколько
затемненная при основании, дискоидальные пятна не развиты. Размах
крыльев 25 мм. Известны только самцы.
Uncus широкий и короткий, с глубоким желобом на дорзальной сто-
роне; девятый сегмент отделен широким швом от основания uncus; saccus
не развит; valvae широкие и короткие, со складкой на уровне одной трети
от вентрального края, идущей от основания valva; penis короткий и широ-
кий, изогнутый, в дистальной части расширенный; соесшп короткий,
изогнут вентрально.
Форма semenovi Gr. Gr. имеет тот же рисунок иа крыльях, но более
темно окрашена, с немногими желтыми полосами и пятнами; задние
крылья одноцветно черные с желтой бахромой; нижняя сторона желтая
лишь на передних крыльях, черная на задних; дискоидальные пятна раз-
виты. Описана из одних мест с типичной.
248
1. ORGYINAE
Распространение. Тибет, Сининские горы, Куку-нор, Амдо,
Китай (Сычуань, Ганьсу).
* 7. Gynaephora sincere Kozhantshikov, sp. nov.
Голова и грудь оливково-серые, покрытые нежными тонкими волос-
ками; брюшко с дорзальной стороны и с боков черное, покрыто корот-
кими волосками; с вентральной стороны несет негустые, светлосерые-
волоски; ноги рыжевато-коричневые; усики самцов сильно гребенчатые^
в той же мере, как у G. selenophora Stgr., менее, чем у G. alpherakii Gr..
Gr.; передние крылья желто-серые с растушеванным, неясным рисунком:;
базальное поле черно-серое, более темное при основании крыла; вторая;
. поперечная линия широкая, растушеванная, бело-серая, несколько-
извитая в дорзальной части крыла, идет перпендикулярно его продольной
оси; круглое пятно не раз-
вито; на его месте неясное-
скопление темного пигмента,
слитое со второй поперечной
линией; почковидное пятно-
в виде размытого черного-
полулунного пятна, слитого-
с третьей поперечной линией;
последняя начинается косым;
штрихом,черная, ниже поч-
ковидного пятна, сильно выг-
нута базально, но не слита,
со второй линией (как у G.
Рис. 102. Gynaephora alpherakii Gr. Gr. Гени- selenophora Stgr.), зубчатая
талии самца. п0 жилкаМ1 оттенена белым
с внешней стороны; под-
краевая линия очень резкая, контрастная, бело-желтая и темная;
терминальная линия в виде тонкой серой полосы; бахрома шашеч-
ная — бело-серая и черная; задние крылья по внешнему краю чер-
ные, в середине серые, с едва намеченными дискоидальными пятнами;
бахрома бело-желтая; нижняя сторона бело-серая; основание крыльев-
черное, черный широкий ободок на задних крыльях; дискоидальные-
пятна развиты в виде слабых тонких изогнутых штрихов на обеих парах
крыльев; терминальная линия в виде тонкой, желтоватой полоски; бах-
рома бело-серая. Размах крыльев 24 мм.
Распространение. Один самец взят на р. Мац (Памир)
29 VI 1909 (3600 м).
Новый вид отличается от алайских и памирских G. selenophora Stgr.
прямой второй поперечной линией, хорошо развитой подкраевой линией
и темными задними крыльями; от тибетских G. alpherakii Gr. Gr. отли-
чается строением усов и третьей поперечной линии. Этот вид наиболее-
обособлен из всех горных видов Gynaephora Hb.
3. Род DASYCHIRA STEPH.1
Stephens, 1829, Ill. Brit.Entom., Haustell., 2 : 58; Aurivillius, 1891, Nordn,. Fjar. :
92; Hampson, 1892, Fauna Brit. Ind., 1 : 433; Staudinger, 1901, Cat. Lep., 1 : 115;
Spuler, 1908, Schmett. Eur., 1 :128; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 114;
1 Стефенс (1829) принимает название Dasychira по Гюбнеру (Hiibner, 1810, Ten-
tamen : 1), который дал это родовое название только для D. pudibunda L.; этот, вид:
3. DASYCHIRA
24 9
Strand, 1910, Seitz, Grossschmett., 2 : 109; Hering, 1932, Tierw. Mitteleur. : 483;.
Nordstroem und Wahlgren, 1937, Svenska tjar. : 66, t. 13, fig. 8; — Elkneria Bor-
ner, 1932, Fauna Deutschl., ed. 4 : 393 (typus pudibunda L.).
Тип рода D. pudibunda L.
Хоботок рудиментарный; губные щупальца торчат вперед, довольно
длинные, второй членик их с вентральной стороны густо опушен, послед-
ний членик маленький; глаза большие, голые, окружены короткими волос-
ками; усики самцов сильно гребенчатые, у самок гребешки короткие;
лоб гладкий; голени передних ног не менее чем вдвое превышают по длине
первые членики передних лапок, дистальный конец голеней простой;
коготки всех лапок простые; радиальная (добавочная) ячея большая,,
вытянутая, сильно сдвинута к костальному краю крыла; Р2 свободный,
отходит от ее наружной стороны; третий, четвертый и пятый радиальные
стволы отходят на стебельке; отходит от верхней половины ячеи; ба-
зальная ячея на задних крыльях большая, полузакрытая; Мй на задних
крыльях выходит из одной точки с кубитальным стволом не анастомозируя.
Uncus не развит; есть остаток десятого сегмента в виде неполного или<
полного кольца вокруг anus; saccus не развит; восьмой сегмент широкий,,
или простой, или (почти у всех видов) несет superuncus; valvae большие,
имеют склеротизованную лопасть, снабженную зубцами; ductus bursae
длинный. Полы диморфны, но самки всегда крылатые, крупнее самцов,
бледнее окрашены (очень похожи у разных видов), имеют более широкие-
крылья и слабо гребенчатые усики.
Признаки личиночных фаз следующие. Гусеницы покрыты длинными
густыми волосками, слабо пильчатыми, не щетинистыми; кожа несет
крупные бородавки; гусеницы после первой линьки приобретают облик
взрослых. Бородавки х на переднегруди большие, но несут короткий,
пучок волос. Бородавки а и |3 на первых четырех брюшных сегментах
почти равны по размерам, тесно сближены (почти слиты) и несут пучки'
коротких, щетинистых волосков (щетки), торчащих дорзально. Эти же
бородавки на восьмом брюшном сегменте несут непарный пучок волосков
(кисть), направленных косо дистально. На шестом и седьмом брюшных
сегментах бородавки а и [3 почти равных размеров, но значительно меньше,
чем на прочих сегментах брюшка (рис. 103). Бородавка х на брюшных
сегментах маленькая, с небольшим числом волосков. Бородавки а и (3
на средне- и заднегруди разделены. Выворачивающиеся желёзки есть
только две, на шестом и седьмом брюшных сегментах. Куколки валько-
ватой формы, волосистые на дорзальной стороне более, чем на вентраль-
ной, но не столь своеобразно, как у видов предыдущего рода. Губные
щупальца по размерам примерно равны ширине максилл, хорошо развиты.
Усики с широким основанием и заметно выпуклые. Пятый и шестой
сегменты брюшка заметно сужены на дорзальной стороне. Яйца круп-
ные, примерно в 1 мм диаметром, имеют плотный хорион, матовые, про-
долговатые округлые, не покрываются волосками при откладке.
является, следовательно, типом рода, хотя род Dasychira описан Стефенсом (1829)^.
но не Гюбнером (1810). В роде Dasychira Стефенс (1829) приводит два вида: D. fasce-
lina L. и D. pudibunda L.
Брик (1932) приводит к роду Dasychira Steph., большое число синонимов исклю-
чительно из тропической фауны; состав этих синонимов очень пестрый, и они никем не
обоснованы. Поскольку изученность этих видов ничтожна и многие из них, как мне;
кажется, представляют самостоятельные группы, а также в связи с тем, что обзор этих’
родов не может быть дан в пределах фауны СССР, синонимы эти мною не приводятся..
250
1. ORGYTNAE
Полиморфный род, широко распространенный в тропиках; из тропи-
ческой Африки и с Малайского архипелага описано более 200 видов,'
многие из которых были выделены при описании в особые роды и лишь
после (без достаточных оснований) слиты с Dasychira Steph. Хотя все
авторы относят очень большое число тропических видов к роду Dasy-
chira Steph. (Штранд, 1919; Геринг, 1926; Брик, 1934), тем не менее мно-'
гие из них очень своеобразны, некоторые из них принадлежат: к роду
Olene Hb., другие требуют выделения в особые роды.
Распространение видов Dasychira Steph, наибольшее в юго-восточной
Азии и на Малайском архипелаге, тогда как из Северной и Южной Аме-
рики виды этого рода не описаны. В Африке ътарът. Dasychira Steph, концен-
трируются в области Золотого Берега, Камеруна и Нигерии, хотя известны
также из восточной и южной Африки, но во всех случаях населяют влаж-
Рис. 103. Dasychira pudibunda L. Хетотаксия
передне- и среднегруди и брюшных сегментов
I, VI, VII и VIII взрослой гусеницы.
ные, преимущественно лесис-
тые территории. Из Африки
несомненными Dasychira Steph,
являются D. extorta Dist.,
georgiana Fawc., tessmanni Her.,
polia Her., thanatos Her., nebu-
lifera pluto Her., rhopalum
Her., melochlora Her., cangia
Dre., macrodonta Her., greeni
Janse. Несколько очень типич-
ных видов описано с Мадагас-.
кара (D. butleri Swinh., lemuria
Her., zena Her., ania Her.,
aurantiaca Her.). Значительное
число видов Dasychira Steph,
описано с Новой Гвинеи (D.
minor Bet.-Bak., kenricka Bet.-
Bak., subnigra Bet.-Bak., subnigropunctala Bet.-Bak., brunnea Bet.-
Bak., grossa Pag., horsfieldi Saund.), часть из которых распростра-
нена по Малайскому архипелагу и один вид заходит в Австралию —
D. horsfieldi Saund.; эндемичный D. vitensis Bet.-Bak. известен с
Фиджи.
Индия и Гималаи также богаты видами Dasychira Steph. (D. dalber-
giae Moore, grotei Moore, horsfieldi Saund., feminula Hamps., thwaitesi
Moore, cinctata Moore, albescens Moore, bohna Moore). Некоторые из этих
видов проникают в западный Китай, Корею и Японию. (D. dalber-
giae Moore, grotei Moore), где (в Японии и Корее) многочисленны эндемич-
ные представители (D. aurifera Scriba, maculosa Mts., hirafamae Mts., su-
:zukii Mts., conjuncta Wil., semicirculosa Gaede, albibasalis Mts., angulosa
Hamps., arakawae Mts., atomariana Mts., baibarana Mts., citrineffa Mts.,
cyanea Mts., flaoa Mts., horishanella Mts., kagina Mts., kibarae Mts., kika-
chii Mts., nachiensis Mts., postfusca Swinh., sachalinensis Mts., sationella
Mts., sationis Mts., taiwanensis Mts., takamukuana Mts.). Южное При-
морье, Приамурье и Маньчжурия также довольно богаты видами Dasy-
chira Steph., отчасти эндемичными (D. conjuncta Wil., pseudabietis Btl.,
lunulata Btl., oirginea Obth., abletisSchiff., pudibunda svdosp. modesta Kirby).
Один вид (D. albodentata Brem.) распространен в Забайкалье, Якутии
и северной Монголии. В средней Сибири зарегистрирован (как редкость)
только D. abietis Schiff., а в Европе, кроме этого вида, распространен
еще типичный D. pudibunda L.
3. DASYCKIRA
251
Видь! Dasychira Steph, связаны с древесной растительностью и преиму-
щественно с широколиственными лесами; для евразийских видов типична
связь с Fagales (Betulaceae, Fagaceae), Rosales и отчасти с Pinaceae (Picea,
Larix). Тропические виды Dasychira Steph, слабо изучены в отношении
кормовых растений, но для них есть указания на питание лиственными
породами, специфичными для тропиков. Биологически Dasychira Steph,
характеризуются в умеренных широтах моновольтинизмом и зимовкой в
личиночной (гусеницы средних возрастов) фазе и редко в фазе куколки.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (10). Бахрома шашечная; передние крылья с несколькими часто
сильно зубчатыми и интенсивными поперечными линиями, окра-
шены в серый или черно-серый цвет. Мельче 30—60 мм.
2 (5). Penis с вентральной стороны имеет стержневидный, на конце
несколько вздутый отросток; поперечные линии на передних крыльях
в виде четких сильно зубчатых полос.
3 (4). Рядом со второй поперечной линией, двойной и широко
расставленной, проходит интенсивно черная поперечная средин-
ная полоса; базальное поле короткое, почти чисто белое; основной
цвет темнобурый; вентральный отросток penis короткий .....
.............................................*4. D. pseudabietis Btl.
4 (3). Срединная линия нерезкая или редуцирована; вторая попереч-
ная линия простая, интенсивно черная, разделенная вдоль лишь
у дорзального края крыла; базальное поле большое, светлосерое;
вентральный отросток penis длинный, вздутый на дистальном
конце..........................................*6. D. abietis Schiff.
5 (2). Penis с вентральной стороны без отростка; поперечные линии
четкие, но не сильно зубчатые.
6 (7). Вторая и третья поперечные линии бархатисто-черные, тонко
очерченные белым, почти без зубцов; в срединном поле у косталь-
ного края неправильное белое пятно . . . *3. D. conjuncta.Wil.
7 (6). Вторая и третья поперечные линии зубчатые или волнистые;
срединное поле однотонное.
8 (9). Срединное поле несколько затемнено; подкраевая линия свет-
лая, но не резкая; superuncus большой; penis на дистальном конце
• с вентральной стороны несет плоский вырост; зубцы harpe круп-
ные .............................................*7. D. pudibunda L.
9 (8). Срединное поле не затемнено; подкраевая линия белая, четкая,
сильно зубчатая; superuncus очень мал; penis на дистальном конце
простой; зубцы на harpe очень мелкие . . *5. D. albodentata Brem.
10 (1). Бахрома однотонная; передние крылья с сильно редуцирован-
ным рисунком, светлосерые или белые; крупные виды 45—80 мм.
11 (12). Передние крылья светлосерые с короткой, в виде четкого пятна
у костального края, двойной, второй поперечной линией; superun-
cus двойной; penis без выроста; harpe с двумя сложными бугорча-
тыми выростами..................................*1. D. lunulata Btl.
12 (И). Передние крылья чисто белые с двумя тонкими поперечными
коричневыми полосками; superuncus простой, короткий; penis
с коротким косым выростом с вентральной стороны близ дисталь-
ного конца; harpe с правильным рядом зубцов.........................
..........................................*2. D. virginea Obth.
252
1. ORGYINAE
*1. Dasychira limiilata Btl. (рис. 104, 105).
Butler, 1877, Ann. Mag. Nat. Hist., 20 (4) : 403; Butler, 1878, 111. Lep. Het. Brit.
Mus., 2 : 11, t. 24, fig. 8; Oberthiir, 1881, Et. Entom., 5 : 35, t. 5, fig. 7; Kirby, 1892,
Syn. Cat. Lep. Het., 1 : 483; Staudinger, 1892, Mem. Lep. Rom., 6 : 307; Leech, 1899,
Trans. Entom. Soc. Lond. : 125; Staudinger, 1901, Cat. Lep., 1 : 116; Strand, 1910,
Seitz, Grossschmett., 2 : 114; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 : 96, t. 8, a;
Matsumura, 1925, Joum. Coll. Agric. Sapporo, 15 : 111; Matsumura, 1931, 6000
Insects Japan : 700, fig. 420; Matsumura, 1933, Ins. Matsum., 7 :118; Bryk, 1934,
Lep. Cat., 62 : 18; — acronycta Oberthiir, 1881, Et. Entom., 5 : 35, t. 5, fig. 7,
8; — solitaria Staudinger, 1887, Mem. Lep. Rom., 3:202, t. 12, fig. 1; — lunulata
Kirby, Syn. Cat. Lep. Het., 1 : 483; — solitalana Matsumura, 1933, Ins. Mat-
sum., 7 : 118.
Передние крылья, грудь, голова и брюшко пепельно-серые или буро-
серые; задние крылья также буро-серые; половой диморфизм выражен
Рис. 104. Dasychira. lunulata Btl. Самец. (Рис. Ф. Гуняева).
лишь в строении усиков и в размерах, тогда как рисунок одинаков у обоих
полов; у самок лишь основной цвет без примеси пепельного; базальная
линия не выражена, базальное поле однотонное, в нижней половине
слитое со срединным; вторая поперечная линия намечена двумя широ-
кими штрихами на костальном крае, в нижней части соединенными и обра-
зующими петлю у costa; у самок и некоторых самцов промежуток между
этими полосами затемнен, и тогда на месте начала поперечной линии
развито черное пятно; третья поперечная линия ясная, в виде одной
полосы, слабо выгнутая посредине, где нерезкая и зубчатая по жилкам,
идет почти перпендикулярно продольной оси крыла; за третьей попе-
речной линией у костального края черная извитая полоса; внешнее поле
однотонное, без подкраевой и терминальной полос, или они бывают слабо
развиты (особенно у самок); в последнем случае подкраевая линия в виде
простой извилистой сильно сдвинутой к терминальному краю темной
полоски; терминальная линия в виде тонкой серой полосы; бахрома се-
рая; задние крылья буровато-серые, слегка желтоватые при основании,
с растушеванным широким дискальным пятном, широкой серой полосой
поперек крыла и нерезкой темной терминальной полоской; бахрома
светлосерая, почти белая; нижняя сторона буровато-серая у самок и бе-
ловато-серая у самцов; дискальные пятна в,виде размытых полулунных
пятен; дискальная область передних крыльев густо волосистая; усики
3. DASYCHIRA
253
самцов сильно гребенчатые, гребешки желтые, членики усиков белые;
у самок гребешки короткие,.но окраска усов та же. Размах крыльев
самцов 55—45 мм, самок —80—65 мм.
Uncus не развит; девятое кольцо очень узкое, не образует saccus;
superuncus на восьмом сегменте в виде двух больших крючкообразно изо-
гнутых выростов, массивных при основании, заостренных; десятый сег-
мент хорошо развит, расположен дорзально от anus, несет мелкие волоски
с боков, вентральная сторона его не развита; valva вытянутая, мембра-
нозная, с хорошо выраженной
базальной лопастью в виде
склеротизованной пластинки, .
несущей сложно расположен-
ные выросты, особенно силь-
ные по сторонам; fultura inferior
почти полулунной формы с вы-
емкой с дорзальной стороны;
penis простой, слабо изогнутый
в дистальной части, несет мел-
кие шипики на pars inflabilis.
Bursa copulatrix очень неболь-
шая, мембранозная; снабжена
коротким и широким сильно
склеротизованным ductus bur-
sae; ductus seminalis отходит
от ее нижней части; восьмой
сегмент короткий, несколько
большей величины при основа-
нии apophyses; последние длин-
ные, *в проксимальной части
расширенные; papillae anales
широкие, почти прямоугольные.
Наиболее близок k D. virgi-
пеа Obth. и D. grotei Moore, рас-
пространенным в Северной Рис. 105. Dasychira lunulata Btl. Гениталии:
Индии) западном Китае и на « — самки, б — самца.
Тайване.
Распространение. Приамурье (Усть-Майская); южный Са-
халин (Южно-Сахалинск); южное Приморье (Сучан, Сидеми, Океанская,
Владивосток, Ворощилов-Уссурийский); Японские острова (Хоккайдо);
западный Китай (Шенси).
*2. Dasychira virginea Obth. (рис. 106).
. Oberthiir, 1879, Diagn. Lep. Askold : 7; Oberthiir, 1881, Et. Entom., 5 : 23, t. 1.
fig. 9.; t. 5, fig. 5; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 483; Staudinger, 1901, Cat, Lep.,
1 : 116; Strand, 1910, Seitz, Grossschmett., 2 : 114, t. 19, f; Wileman, 1911, Trans,
Entom. Soc. Lond. : 271; Swinhoe, 1923, Ann. Mag. Nat. Hist., 10 (3) : 297; Gaede,
1934, Seitz, Grossschmett., 2 Suppl. : 96; Bryk, 1934, Lep. Cat., 62 ; 19.
Голова, грудь и передние крылья у самцов белые, брюшко и задние
крылья серые, последние по внешнему краю светлые; у самок тело и обе
пары крыльев белые или бело-сероватые; рисунок на передних крыльях
слабо развит; базальное поле широкое, одноцветное, базальная линия
не развита; вторая поперечная линия двойная, черная, с примесью охри-
стых чешуй, местами слитая в одну линию, слабо извитая, почти перпен-
254
1. ORGYINAE
тые, греоешки яркожелтые, членики
Рис. 106. Dasychira. virginea Obth. Генита-
лии: а —^гамца, б — самки.
дикулярная продольной оси крыла; третья поперечная линия в виде
одной неширокой черной полосы, слабо извитой, выгнутой кнаружи
у дорзального края крыла; срединное поле узкое, уже базального и внеш-
него; последнее одноцветное, без линий; бахрома белая, задние крылья
без рисунка, у самцов затемнены в середине, у самок чисто белые с очень
тонкой темной терминальной полосой; нижняя сторона крыльев чисто
белая с нечетким рисунком и полулунным дискоидальным пятном на зад-
них крыльях или одноцветная (у самок); усики самцов сильно гребенча-
белые; у самок гребешки очень
короткие. Размах крыльев самцов
43—49 мм, самок 60—Об мм.
Uncus не развит; восьмой сег-
мент образует на дорзальной сто-
роне короткий superuncus с двумя
латеральными лопастями; девятый
сегмент узкий, слабый, не обра-
зует saccus; есть остатки десятого
сегмента, образующие - небольшие
пластинки, сидящие по сторонам
anus; fultura" inferior в виде двух,
при основании сросшихся, острых
выростов, направленных дорзаль-
но; базальная часть valva сильно
склеротизована, несет на вен-
тральной стороне не длинные, но
сильные зубцы, основание вздуто,
дистальный конец ее мембраноз-
ный; penis почти цилиндричцркий,
несколько изогнутый, на вентраль-
ной стороне несет длинный цилин-
дрический вырост; дистальный
конец penis’несет многочисленные,
мелкие шипики. Bursa copulatrix
мешковидная, мембранозная, несет
тонкую рубчатую скульптуру;
ductus bursae в проксимальной
части мембранозный, в дистальной части сильно склеротизован; восьмое
кольцо узкое несколько более сильно склеротизованное в дистальной
половине, несет длинные и тонкие apophyses anteriores; apophyses ро-
sterires также тонкие и длинные; и те и другие короче ductus bursae;
papillae anales почти квадратной формы.
Изменчивость окраски незначительна; наблюдается изменчивость сте-
пени развития рисунка; есть особи совсем белые; у некоторых особей,
напротив, линии рисунка, расширяясь, затемняют базальное и средин-
ное поле. Наиболее близка к китайской/), grotei Moore.
Распространение. Амур; Южное Приморье (Уссури,
Аскольд, Владивосток, Сучан, Сидеми); западный Китай (Шенси).
* 3. Dasychira conjuncta Wil. (рис. 107, 108).
Wileman, 1911, Trans. Entom. Soc. Lond. : 270; Gaede, 1932, Seitz, Grossschmett.,
2 Suppl. : 95, t. 8, c.
Голова и грудь черно-серые, покрыты не только волосками, но и уз-
кими чешуйками; брюшко черно-серое с толстыми и короткими пучками
3. DASYCHIRA
2&5
волосков на дорзальной стороне первых брюшных сегментов; нижняя
сторона брюшка светлосерая, ноги и нижняя сторона груди того же
цвета; членики лапок черные, кольчатые; усики самцов сильно гребенча-
тые, гребешки серые, членики черные, у самок гребешки короткие; перед-
ние крылья узкие, несколько вытянутые и округлые в апикальной части;
базальная линия в виде двух косых штрихов, наружный из которых ин-
Рйс. 107. Dasychira conjuncta Wil. Самец. (Рис.
Ф. Г.уняева).
тенсивно черный; базальное поле однотонное; вторая поперечная линия
черная и светлосерая, не извитая, посредине слабо выгнутая кнаружи;
в срединном поле темный штрих вдоль анальной жилки, соединяющий
эту линию с третьей поперечной линией; между второй поперечной линией
и почковидным пятном белое пятно, несколько затемненное по краям,
окаймляющее почковидное
пятно в виде полулуния;
почковидное пятно наме-
чено несколькими черными
штрихами в середине свет-
лое ; срединное поле в сред-
ней части сужено; внеш-
нее поле однотонное с бе-
лыми и темными клиновид-
ными штрихами и четкой,
черной подкраевой линией,
посредине сильно извитой,
Рис. 108. Dasychira conjuncta Wil. Гениталии самца.
сильно сдвинутой к краю
крыла; терминальная линия в виде тонкой черной полоски; бахрома
черно-серая; задние крылья однотонно серо-желтоватые с неясной рас-
тушеванной поперечной полосой и размытыми дискальными пятнами.
Полы слабо диморфны, самки лишь несколько крупнее. Размах крыльев'
самцов 37—42 мм, самок 50 мм.
Uncus не развит;. девятый сегмент узкий, saccus очень маленький;
восьмой сегмент и особенно его тергит расширены, но superuncus на нем
не развит; есть рудименты десятого сегмента, располагающиеся по сторо-
нам anus в виде пластинок, несут короткие волоски; fultura inferior кони-
ческой формы, по вентральному краю несет три небольших выроста
256
1. ORGYINAE
и один вырост на дистальной стороне, дорзальный ее край несет мягкий
волосистый вырост; penis конический, расширенный в проксимальной
части, несет большое количество мелких шипиков на pars inflabilis.
Описана по серии особей, из которых только одна была самка; измен
чивость очень мала; очень характерный, стоящий особняком в роде вид,
габитуально наиболее похож на D. pseudabietis Btl., но структурно далек.
Распространение. Южное Приморье (Аскольд, Владивосток,
Виноградовка, Яковлевка, Сучан, Сидеми); Японские острова (Хондо).
. * 4. Dasychira pseudabietis Btl. (рис. 109, 110).
Butler, 1885, Cist. Entom., 3 : 118 (Calliteara)', Leech, 1899, Trans. Entom. Soc.
Lond., 37 124; Swinhoe, 1903, Trans. Entom. Soc. Lond., 41 : 472; Strand, 1910, Seitz,
Grossschmett., 2 : 113, t. 19, h; Strand, 1914, Entom. Mitt., 3 : 114; Strand, 1914,
Suppl. Ent., 3 : 36; Swinhoe, 1923, Ann. Mag. Nat. Hist. 10 (3) : 296; Matsumura, 1932,
6000 Insects Japan : 700, fig. 422; Matsumura, 1933, Ins. Matsum., 7 : 120; Gaede, 1934,
Seitz, Grossschmett., 2 Suppl. : 282; Bryk, 1934, Lep. Cat., 62 : 15;— pryeri Butler,
1885, Cist. Entom., 3 : 119; — coreana Matsumura, 1927, Journ. Coll, agric. Sap-
poro, 19 : 36, t. 2, f. 10.
Голова й грудь светлосерые, покрытые толстыми волосками, имею-
щими облик почти чешуй; вентральная часть тела и брюшка светлосерая;
Рис. 109. Dasychira pseudabietis Btl. Самец. (Рис. Ф. Гуняева).
усики самцов сильно гребенчатые, гребешки желто-коричневые, членики
белые; лапки серые с черными колечками; передние крылья черно-серые
с белым и черным рисунком; базальное поле белое с неправильно раз-
бросанными черными чешуйками и ясной первой поперечной линией,
слабо извитой, несколько выгнутой кнаружи в дорзальной части крыла;
вторая поперечная линия почти прямая, черная, ординарная, довольно
.широкая, проходит косо базально, давая штрихи по жилкам к основанию
крыла; срединная линия сильно сдвинута базально и идет почти рядом
со второй поперечной линией в дорзальной части крыла, выгибаясь кна-
ружи; срединное поле серое, однотонное с несколько затемненными жил-
ками, широкое, шире внешнего и базального попей, почковидное пятно
маленькое, в виде полулунного штриха, очерчено черным, в середине
светлосерое; третья поперечная линия далеко отставлена от почковидного
пятна, так что оно находится почти в середине срединного поля, слабо из-
витая, идет косо к дорзальному краю; внешнее поле светлосерое с черной
и белой подкраевой линией, сильно зубчатой; терминальная линия тон-
кая, черная, слабо извитая, проходит на близком расстоянии от терми-
3. DASYCHIRA
257
Дальнего края; бахрома черно-серая, шашечная; задние крылья тускло-
серые, волосистые по внутреннему краю, со слабой растушеванной внеш-
ней полосой; дискоидальное пятно сверху выражено слабо; терминаль-
ная полоска тонкая, черная; бахрома серая; нижняя сторона черно-серая
с ясным темным просвечивающим дискал ьным пятном; задние крылья
светлосерые, дискоидальное пятно ясное; через оба крыла проходит
косая серая линия. Самки крупнее, светлосерого цвета, с черным тонким
рисунком; подкраевая линия более ясная; задние крылья светлосерые;
передние крылья более округлой формы. Размах крыльев самцов 42—
48 мм, самок 45—55 мм.
Uncus не развит; девятый сегмент узкий; saccus не развит; восьмой
сегмент несет superuncus и латерально от него две коротких лопасти;
остатки десятого сегмента в виде
очень маленьких лопастей лате-
рально от anus; fultura inferior
в виде двойного, при основании
сросшегося заостренного склерита;
valva в базальной части расши-
ренная, со склеротизованной боль-
шой лопастью по вентральному
краю; эта лопасть несет сильные,
Рис. НО. Dasychira pseudabietis Btl. Ге
ниталии самца.
но короткие шипы в дистальной
половине; дистальная половина
valva мембранозная; penis рас-
ширен и изогнут в проксимальной
части; coecum направлен почти прямо вентрально; в дистальной части
penis на pars inflabilis большое число мелких шипиков; вентральный
край penis несет недлинный округленный в дистальной части напра-
вленный косо дистально отросток.
Наиболее близка по облику и структуре полового аппарата к D. albo-
denlaia Brem., но все же резко отлична. Известна в небольшом числе
особей, в общем однообразных; варьируют лишь интенсивность (резкость)
рисунка и степень развития белого цвета.
Распространение. Приамурье; южное Приморье; (Виноградов.-
ка); Маньчжурия (Мукден); Корея; Японские острова (Хондо). \
*5. Dasychira albodentata Brem. (рис. Ill, 112).
Bremer, 1864, Lep. Ost.-Sib. : 102, t. 8, fig. 13; Kirby, 1892, Syn. Cat. Lep. Het.,
1 : 483; Staudinger, 1901, Cat. Lep., 1 : 115; Strand, 1909, Seitz, Grossschmett., 2 :
112; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 : 404; Gaede, 1932, Seitz, Grossschmett.,
2 Suppl. : 95; Bryk, 1934, Lep. Cat., 62 ; 9.
Голова, грудь и передние крылья черно-серые; брюшко и задние
крылья светлосерые, тусклые; лоб, нижняя часть головы и передние
ноги черные; остальные две пары ног светлосерые; усики самцов сильно
гребенчатые, гребешки коричнево-серые, членики усиков белые; базаль-
ная линия интенсивно черная, в середине крыла дает резкий изгиб кна-
ружи; базальное поле почти по всей ширине бело-серое с неправильно
разбросанными черными чешуйками и темным пятном при основании у на-
ружного края; дорзальная часть базального поля темиосерая; у некоторых
особей и в частности у типа базальное поле однотонное; срединное поле
узкое, темное, одинаковое по ширине у костального и дорзального краев,
скошенное; почковидное пятно узкое, серповидной формы, очерчено черным,
17 Фауна СССР, И. В. Кожанчиков.
258
1. ORGYINAE
особенно интенсивное с внешней стороны, в середине светлосерое; третья
поперечная линия черная, тонкая, грубо зубчатая по жилкам; внешнее
поле тусклое, черно-серое, однотонное; подкраевая линия тонкая, белая;
терминальная линия в виде тонкой черной полоски; бахрома серая, ша-
шечная; задние крылья черно-серые, однотонные, тусклые; бахрома
на них бело-черная; нижняя сторона серо-черная, дискоидальное поле
передних крыльев более темное; самки более однотонные, светлая косталь-
ная часть базального поля у них стушевана, стушеваны также вторая
и третья поперечные линии; одна самка почти белая, но рисунок у нее
тот же; самки более ширококрылы и круглокрылы. Размах крыльев сам-
цов 32—-42 мм, самок 40—50 мм.
Рис. 111. Dasychira albodentata Brem.
Самец. (Рис. Ф. Гуняева).
Uncus не развит; девятый сегмент узкий; saccus рудиментарный;
superuncus в виде очень маленького выроста восьмого тергита; по сторо-
нам его две небольшие боковые лопасти; valva мембранозная в дисталь-
ной половине; базальная половина ее в вентральной части утолщена,
сильно' склеротизована и несет многочисленные короткие, но сильные
зубцы; fultura inferior с двумя острыми длинными выростами, направлен-
ными дорзально; penis расширен в проксимальной части,, соесшп корот-
кий; pars inflabilis несет многочисленные, тонкие и довод но длинные ши-
пики. Bursa copulatrix мешковидная. Несет тонкую рубчатую скульптуру;
ductus bursae длинный, в дистальной половине сильно склеротизован;
ductus seminalis отходит от верхней, мембранозной половины ductus bursae;
восьмой сегмент короткий, с короткими (менее половины длины ductus
bursae) apophyses anteriores; apophyses posteriores более длинные и тон-
кие; papillae anales широкие и короткие, несколько конические, покрыты
густыми короткими волосками.
В литературе этот вид смешивался с Olene olga Obth. Синонимика
впервые была установлена Штаудингером, который не видел типа и свел
синонимику по указаниям Герца и Грум-Гржимайло, которые не разобра-
лись в этих видах в должной мере. Штранд (1910) изображает самца
О. olga Obth. под названием D. albodentata Brem.;Тедэ (1932), хотя и пи-
шет, что изображает самку 7). albodentata Brem., тем не менее опять рисует
О. olga Obth. Мною внимательно изучен тип Бремера из Кяхты (он
3. DASYCHIRA
259
(Кяхта, p. Аргунь,
Монголия (оз. Дзергу-
не имеет брюшка и вообще сильно облетан, брюшко отсутствовало еще
при описании) и подобрана серия особей из разных мест; D. albodentata
/ Brem. представляет хороший вид, наиболее близкий к широко распро-
страненным D. pudibunda L. и D. abietis Schiff.; на Японских островах
и в южном Приморье известны виды, более близкие к D. albodentata
Brem.-—например, D. sachalinensis Mts.
Распространение. Забайкалье
р. Урюмкан, Троицкосавск, Ботый); Якутск;
ческое).
Биология и э к о л о -
гия. Заселяет холодные тер-
ритории, на которых еще в июне
регулярно наблюдаются замо-
розки. Встречается по сухим,
южным склонам, поросшим
даурской лиственницей (Larix
dahurica) с подлеском из даур-
ского рододендрона (Rhododen-
dron dahuricum). Гусеницы
зимуют в средних стадиях.
Питание изучено недостаточно,
но известно, что гусеницы
питаются хвоей даурской лист-
венницы. Окукление на ство-
лах деревьев и на скалах.
Отмечена как вредитель да-
урской лиственницы в Забай-
калье (Теплоухов, 1937), где
наблюдалось массовое размно-
жение этого вида на протяже-
Рис. 112. Dasychira albodentata Brem. Гени-
талии: а — самца, б — самки.
нии нескольких десятков кило-
метров по Аргуни.
* 6. Dasychira abietis Schiff.
(рис. ИЗ).
Schiffermuller, 1776, Syst. Verz. Schmett. Wien, 2 : 55; Aurivillius, 1891, Nordn.
Fjar. : 93, t. 25, fig. 6, a, b; Spuler, 1908, Schmett. Bur., 1 : 123, t. 25, fig. 23;
Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 115, t. 21, fig. 10, a,b; Strand,
1910, Seitz, Grossschmett., 2 : 113, t. 19, g; Mewes, 1914, Entom. Tidskr., 35 : 11; Strand,
1914, Entom. Mitt., 3 : 331; Carpelan, 1921, Not. Entom., 1 : 17; Swinhoe, 1923, Ann.
Mag. Nat. Hist., 11 : 296; Petersen, 1924, Lep. Fauna Estl., ed. 2, 1 : 140; Hering, 1932,
Tierw. Mitteleur. : 483; Bryk, 1934, Lep. Cat., 62 : 14; —medioobscura Schult. 1910,
Entom. Zeitschr. Stuttgart, 24:35;—caligata Lumma, 1932, Intern. Entom.
Zeitschr., 26 : 249.
Биология и экология. Lederer, 1923, Handb. prakt. Entom., 3 : 143;
Osthelder, 1926, Schmett. Siidbayerns, 1 (2) : 197; Lumma, 1932, Intern. Entom. Ztschr.,
26 : 75; Куренцов, 1936, Тр. Дальневост. фил. Акад. Наук СССР, 1 : 146.
Голова, грудь, брюшко и передние крылья черно-серые и белые; самки
значительно светлее, почти белые; губные щупальца торчат вперед,
последний членик их маленький, тонкий; усики самцов сильно гребенча-
тые, членики их белые, гребешки желто-коричневые; ноги белые с чер-
ными колечками; первые сегменты брюшка с дорзальной стороны у сам-
цов несут короткие пучки волосков; передние крылья с ясным и резким
рисунком; базальное поле белое, особенно в костальной половине; базаль-
17*
260
1. orgyinae
ная линия интенсивно черная, широкая, с сильным зубцом под косталь-
ным краем и у дорзального края, связана штрихом с основанием крыла;
вторая поперечная линия в виде широкой сильно скошенной полосы,
дает зубец к основанию крыла; срединное поле неширокое, уже внешнего
и базального, в базальной половине затемнено; почковидное пятно в виде
узкого треугольного пятна; третья поперечная линия начинается над
почковидным пятном, тонкая, черная, зубчатая по жилкам, дает глубокий
зубец к почковидному пятну, где влита в темное пятно, у дорзального
края крыла дает резкий выгиб кнаружи; подкраевая линия белая, широ-
кая, дает мягкие изгибы, резко заметная на темном фоне; терминальная
линия в виде коротких, черных штрихов; бахрома бело-серая; задние
Рис. 1'13. Dasychira. abietis
а — самца, б —•
Schiff. Гениталии:
самки.
крылья тусклосерые, не-
сколько более темные в сере-
дине, бахрома белая у самок
задние крылья белые; ниж-
няя сторона коричнево-серая
у самцов и почти белая у са-
мок; оба крыла пересекает
нерезкая поперечная пере-
вязь; на заднем крыле рас-
тушеванное дискоидальное
пятно. Размах крыльев сам-
цов 40—45 мм, самок 45—
55 мм.
Uncus не развит; девятый
сегмент узкий, не образует
saccus, восьмой сегмент об-
разует короткий superuncus,
имеющий по сторонам не-
большие лопасти; valvae
мембранозные с утолщенной
и склеротизованной вентраль-
ной частью, несущей мелкие
многочисленные крепкие зуб-
цы; fulturainferior в виде
двух острых и длинных выростов, сросшихся при основании; penis
с очень длинным изогнутым и утолщенным на дистальном конце вы-
ростом, направленным косо дистально; pars inflabilis несет многочис-
ленные мелкие зубцы. Bursa copulatrix почти яйцевидной формы,
покрыта тонкой рубчатой скульптурой; ductus bursae длинный,
в нижней половине склеротизован; ductus seminalis тонкий, отходит
от мембранозной части ductus bursae; восьмой сегмент узкий, несколько
расширенный в проксимальной части; apophyses anteriores длинные и
тонкие; apophyses posteriores также тонкие и длинные; papillae anales
слабо конические.
Взрослая гусеница (по Шпулеру) светлозеленая с интенсивно черными
межсегментальными промежутками, покрыта коричневатыми волосками;
дорзальная сторона несет белые и черные пятна; по сторонам тела —по
белой с чёрными пятнами полосе; четыре спинных кисточки коричнево-
желтые, пара черных кисточек первого грудного сегмента и одна
несколько изогнутая кисточка, на восьмом сегменте брюшка, длинная,
коричнево-желтая; дыхальца белые, окружены черным; голова зеленая
с двумя темными штрихами.
3. DASYCHIRA
261
Куколка светлокоричневая, с вентральной стороны покрыта ред-
кими волосками, на дорзальной стороне несет пучки недлинных волос-
ков; вентральная сторона брюшных сегментов покрыта скульптурой
в виде мелких морщинок, дорзальная сторона их гладкая; cremaster
в виде толстого конического выроста, несколько желобчатого; дистальный
его конец несет недлинные крючья.
Мало изменчивый вид; особи с Амура несколько мельче и более одно-
тонны; довольно редок. Более обычен на востоке ареала; на Хондо
отмечен вред Cryptomeria.
Распространение. Средняя Европа — Альпы, Бавария, Чехо-
словакия, южная Германия, Прибалтика; Карпаты, Висла; южная
Скандинавия, южная Финляндия; в Европейской части СССР только
в зоне хвойных лесов (Ленинград, Витебск, Москва, Калуга, Казань,
Чебоксары); Сибирь (Красноярск, Иркутск); Хингано-Архаринский р-н.;
южное Приморье (бухта Самарга, р. Сама, Сучан), обычна, по Куренцову,
в елово-пихтовой зоне; южный Сахалин (Ишиносава); Хондо (Токио).
Биология. Зимует гусеница в лесной подстилке; кормовые ра-
стения только Abietinae (Picea excelsa, Р. obovata, Abies sibirica); окукле-
ние в мае—июне в лесной подстилке в рыхлом коконе из желтых нитей
и волосков; лёт бабочек в июле; период развития гусениц охватывает
июль—май; зимовка гусениц обычно сопровождается высокой смерт-
ностью.
*7. Dasychira pudibunda L,—Краснохвост (рис. 103, 114, 115).
Linne, 1758, Syst. Nat., ed 10 (1) : 503 (Bombyx); Linne, 1761, Fauna Suecica, 2 :
296; Aurivillius, 1891, Nordn. Fjar. : 93, t. 25, fig. 5; Staudinger, 1901, Cat. Lep., 1 :
116; Tullgren, 1907, Entom. Tidskr., 28 : 209; Spuler, 1908, Schmett. Eur.,1 : 129,
t. 25, fig. 24, a, b; Nielsen, 1908, Sexuell. Dim. Lep.: 11—15, fig. 5—10; Rebel, 1910,
Berge, Schmetterlingsb., ed. 9 : 115, t. 21, fig. 11; Strand, 1910, Seitz, Grossschmett.,
2 : 114; Wermelin, 1912, Entom. Tidskr., 32 : 138; Meves, 1914, там же, 34 : 11;
Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 : 295; Petersen, 1924, Lep. Fauna EstL, ed. 2,
1 : 140; Zerny, 1927, Eos, 3 : 350; Vorbrodt, 1930, Tessin-Misox, Schmett. : 267; Mar-
schner, 1932, Entom. Rundsch., 49 : 246; Hering, 1932, Tierw. Mitteleur., Schmett. :
483; Bryk, 1934, Lep. Cat., 62 : 15; — scopularia Linne, 1758, Syst. Nat., ed. 10 : 521
(Geometra);— justice. Muller, 1764, Fauna Friedrichsd. : 47; — porrecta-alba Retzius,
1783, Gen. Spec. Ins. : 38 (JPhalena); — juglandis Hiibner, 1800, Eur. Schmett., Bomb.:
fig. 84, 85 (Bombyx); — concolor Staudinger, 1871, Cat. Lep., ed. 1, 1 : 29; —• jutica
Kirby, 1892, Syn. Cat. Lep. Het., 1 : 482 (Notolophus); — albescens Lempke, 1937,
Tijdschr. Entom., 80 : 278; —jusca Lempke, 1937, там же, 80 : 278; — unilineata Lempke,
1937, там же, 80 : 278; — bicolor Cockayne, 1940, Entom. Rec., 52 : 86; — nesiotes
Bryk, 1942, Iris, 52 : 25. 1 '
Биологияи экология. Escherich, 1914, Forstins. Mitteleur., 1 : 133,
fig. 123; Roman, 1917, Entom. Tidskr., 38 : 262; Schille, 1921, Ztschr. Oesterr. entom.
Ver., 6 : 45; Wolf und Krausse, 1922, Wichtigste Forstins. : 132, fig. 160—162; Hollas,
1922, Lepidopt. Rundsch., 2 : 13; Led rer, 1923, Handb. prakt. Entom, 3 : 142, 145;
Reh, 1925, Sorauer, Handb. Pflanzenkrankh., 4 : 442; Самойлович, 1925, Защ. раст.,
1 : 214; Старк, 1926, там же, 3 : 17; Nusslin-Rhumbler, 1927, Forstins.: 463; Гросс-
гейм, 1928, Тр. Млеев. ст., 5 : 25; Eidmann, 1929, Ztschr. angew. Entom., 15 : 30,
fig. 26—28; Bertram, 1929, Entom. Ber., 7 : 409; Белозор, 1931, 36ipn. праць Зоол.
муз. Украшськ. Акад, наук, 10 : 152; Лебедев, 1933, 36ipii. праць екол. 1 : 62;
Eckstein, 1933, Anz. Schadlingskunde, 9 : 52; Auel, 1935, Entom. Anz., 15 : 15, 2;
Heese, 1935, Intern, entom. Ztschr., 29 : 306; Rometctsch, 1935, Entom. Ztschr.
Frank., 49 : 87.
* Описанная Лоркевичем (Lorkewitz, 1930, Verb, zool.-bot. Ges. Wien, 80 : 7, 10)
Dasychira grundi из средней Европы чрезвычайно близка к D. pudibunda L.; автор
дает только цветовые отличия этого вида и по существу совершенно не обосновывает
его; отличия в копулятивном аппарате D. pudibunda L. и D. grundi Lork. ничтожны
и укладываются в пределы изменчивости первого вида.
262
1. ORGYINAE
Голова и грудь оливково-серые, покрыты волосками, без примеси
чешуй; брюшко и нижняя сторона тела желто-серые; усики самцов сильно
гребенчатые, гребешки желто-коричневые, членики усиков белые; у са-
мок усики с короткими, но также желто-коричневыми гребешками; губ-
ные щупальца коричнево-серые; передние крылья самцов светлосерые,
самок желто-серые; рисунок черно-коричневый, довольно ясный; базаль-
ное поле бело-серое с неправильно разбросанными черными чешуйками;
базальная линия в виде тонкой черной полоски; вторая поперечная ли-
ния идет перпендикулярно продольной оси крыла, прямая двойная,
темносерая, посередине бело-серая; круглое пятно не развито; средин-
ная линия сдвинута базально, растушеванная и почти слитая со второй
Рис. 114. Dasychira pudibunda L. Самец. (Рис. Ф. Гуняева).
линией; почковидное пятно вытянутое, несколько суженное с дорзаль-
ного края; третья поперечная линия начинается над почковидным пят-
ном, несколько выгнута кнаружи под костальным краем, далее идет прямо
к дорзальному краю, неправильно извитая, простая темносерая; подкрае-
вая линия в виде неправильных белых и серых штрихов; терминальная
линия в виде тонкой серой, в нижней части извитой полосы; бахрома ша-
шечная— темная и серая — или одноцветно серая; задние крылья
светлосерые, покрытые нежными волосками; на них есть темное растуше-
ванное дискоидальное пятно и тусклая растушеванная полоса по краю;
бахрома светлосерая; нижняя сторона белая, слегка затемненная на пе-
редних крыльях в области дискоидальной ячеи, имеет неполную почти
прямую поперечную линию; дискоидальные пятна ясные в виде широких
растушеванных мазков. Самки крупнее, более светлой окраски, базаль-
ное поле слабо выделяется на общем фоне крыла, линии рисунка несколько
редуцированные, хорошо заметны вторая и третья поперечные линии.
Размах крыльев самцов 35—45 мм, самок 45—60 мм.
Uncus не развит; девятое кольцо очень узкое; saccus отсутствует;
восьмое кольцо образует superuncus в виде короткого крючкообразного
выроста, снабженного по сторонам лопастями; десятый-сегмент сохранен
в виде узких лопастей по сторонам anus; valva мембранозная, базальная
ее часть с вентральной стороны имеет утолщенную лопасть, несущую
по краю короткие, но сильные зубцы, расположенные в ряд; fultura infe-
rior в виде двух конических выростов, сросшихся при основании; penis
3. DASYHIRA
263
•сильно выгнут дорзально, несет с вентральной стороны на дистальном
конце лопасть; pars inflabilis penis покрыта многочисленными мелкими
щетинками. Bursa copulatrix мешковидной формы, мембранозная, покрыта
слабой рубчатой скульптурой; ductus bursae длинный, две трети его мем-
бранозные, дистальная часть склеротизована; от его середины отходит
ductus seminalis; восьмой сегмент короткий, несет длинные apophyses
anteriores, по длине почти равные ductus bursae; apophyses posteriores
значительно короче первых; papillae anales короткие, суженные в дисталь-
ной части.
Рис. 115. Dasychira pudibunda L. Гениталии: а — самца,
б — самки.
Яйца почти шаровидной формы (слабо овальные) с темной точкой
с дорзальной стороны, гладкие, бело-серые, матовые; диаметр 1.0—1.2 мм.
Взрослая гусеница желтовато-зеленая с дорзальной стороны, с вентраль-
ной черно-серая; голова и грудные ноги желтые, брюшные ноги черно-
бурые при основании и желто-зеленые на концах; тело покрыто желто-
зелеными волосками, сидящими на небольших бородавках; встречаются
гусеницы с темными, почти черными волосками на теле; на дорзаль-
ной стороне первых четырех брюшных сегментов (первого—четвертого)
по большой короткой непарной щетке волосков желто-охристого цвета;
на дорзальной стороне восьмого сегмента непарный направленный косо
дорзально и назад розово-красный или розово-черный пучок волосков;
интерсегментальные мембраны между сегментами первым—пятым барха-
тисто-черные; между другими сегментами брюшка лишь слабо черные
полоски; на тергитах пятого—восьмого сегментов двойные черные по-
лоски, частью прерванные; железка развита хорошо лишь на седьмом
сегменте брюшка, на шестом она слабая или не развита совсем.
Куколка светлокоричневая, несет большое число пучков волосков
на дорзальной стороне тела (всех сегментов); вентральная сторона почти
264
1. ORGYINAE
голая; скульптура на брюшных сегментах в виде мельчайших точек;
cremaster в виде толстого конического гладкого (блестящего) выроста,
несущего на дистальном конце многочисленные мелкие крючья. Изменчи-
вость этого вида незначительна; светлая форма с примесью белого на груди,
голове и на передних крыльях, особенно в базальном поле, получила, -
название fuglandis Hb.; темная форма одноцветная, почти без рисунка,
описана как concolor Stgr.; обе формы описаны из средней Европы.
Распространение. Средняя и западная Европа к северу
от Пиренеев (Бильбао)—Альпы до северной Италии, Германская низ-
менность, долина Дуная, Карпаты; в Средиземноморье указана лишь
для Сардинии (Чеккони, 1924); Швеция. (Упланд, Эланд, Даларнэ),
Норвегия, Финляндия (Ниландия); в СССР всюду распространен в При-
балтике (Рига, Таллин, Каунас), далее к востоку идет от Карелии (Петро-
заводск) и линии Вологда—Молотов (вероятно, несколько севернее) всюду
по лесной зоне на юг, проникая местами в степную полосу (Ленинград,
Псков, Новгород, Москва, Калуга, Ярославль, Тула, Киев, Чернигов,
Курск, Харьков, Полтава, Винница, Таганрог, Херсон, Каменец-Подольск,
Бахмут, Кйшенев); Крым (указан для южного берега); Северный Кавказ
(Ставрополь, Туапсе); Закавказье (Боржоми, Тбилиси, Лагодехи, Киров-
абад, долина Аракса); Поволжье (Казань, Вольск); Приуралье (Чкалов,
Свердловск); западная Сибирь (Омск); горная Средняя Азия.
Биология и экология. Зимует куколка в корнях, в ще-
лях коры и лесной подстилке, в желто-сером плотном двойном коконе;
диапауза наблюдается непрочная и нарушается в условиях повышенной
температуры в течение немногих месяцев; лёт бабочек в июне, в конце
мая; только ночью; яйца откладываются кучками по 10—100 шт. в один
ряд, на стволы деревьев на высоте трех-четырех метров и не покрываются
волосками; при массовых размножениях яйца кладутся в разных местах,
часто даже на почву. Гусеницы появляются во второй половине июня —
начале июля и растут в течение всего лета; окукление продолжается со
второй половины августа и до середины и даже конца сентября (в лесостепи |
- кончается в октябре). Дает одно поколение, как исключение наблю- i
дается частичный вылет второго. Гусеницы многоядны, но живут преиму- i
щественно на лесных породах, особенно на Betulaceae (Betula, Carpinus,
Corylus, Ostryia), Fagaceae (Quercus, Fagus, Castanea), Rosaceae (Pirus,
Prunus, Crataegus, Rosa, Rubus), Salicaceae (Salix, Populus) и Ulmaceae
(Ulmus); очень редко взрослые гусеницы на PinaceaA (Pinus) и Euphor-
biaceae (Euphorbia).
Вредит садам и чаще лиственным лесам (в средней Европе — буку,
на востоке — дубу). Вред, помимо западной и средней Европы, отме-
чался в Харьковской, Киевской и Смоленской областях (Гусев, 1935). Мас-
совые размножения и вред отмечались в Голландии, Франции, Германии,
Бельгии и Болгарии. В садах вредит груше, яблоне и косточковым.
Отмечены многочисленные паразиты из перепончатокрылых и двукры-
лых: из Proctotrupidae — Ceraphron albipes Rtzb., Teleas zetterstedti Rtzb.,
Telenomus dalmani Rtzb., T. truncatus Ns.; из Braconidae— Eulophus
ramicornis Geoffr., E. bombycicornis Ns., Microgaster triangulator
Wesm., M. gracilis Curt., M. affinis Ns., M. spur ius Wesm., Rhogas genicu-
lator Ns.; из Ichneumonidae— Ichneumon alboguttatus Gr., Campoplex
mixtus Brischke, Glypta flaaolineata Gr., G. brischkei Hgr., G. teres Gr.,
Hemiteles areator Gr., Trogus alboguttatus Gr., Pimpla instigator F.,
P. pudibundae Rtzb., Ichneumon balticus Rtzb., I. fabricator F., I. flavi-
femur Thbn., I. sexlineatus Rtzb., I. bilunatus Gr., I. computatorius Mill—
3. DASYCHIRA, 4. LAELIA 265
ler, I. deliratorius L., Stenichneumon militarius Thnb., P. instigator F
Cratichneumon sicarius Gr., - M elanichneumon bimaculatus Schrk., Bari-
chneumon bilunulatus Gr., Paniscus cephalotus Holmgr., Campoplex
falcator F., Pimpla spuria Gr.; из двукрылых — Carcelia gnava Mg.,
C. excisa Fall., C. lucorum Mg., Compsilura concinnata Mg., Exorista glauca
Mg., E. affinis Fall., Leskia aurea Fall., Phorocera assimilis Fall., Phryxe
vulgaris Fall., Winthemia quadripustulata F.
* 7a. Dasychira pudibunda modesta Kirby.
Kirby, 1892, Syn. Cat. Lep. Het., 1 : 483 (nom. nov.); Strand, 1910, Seitz, Gross-
schmett., 2 : 114, t. 22, а; Куренцов 1922, Зап. Южн.-уссур. отд. Геогр. общ., 1 : 23;
Gaede, 1932, Seitz, Grossschmett., 2 Suppl. : 96; Bryk, 1934, Lep. Cat., 62 : 17; —
pudica Staudinger, 1887, Mem. Lep. Rom., 3 : 204 (pudibunda var. pudica, nomen prae-
occ.); — immunda Kardakoff, 1928, Entom. Mitt., 17 : 416, t. 8, fig. 13; — pfjeminata
Kardakoff, 1928, там же, 17 : 416, t. 8, fig. 14.
Хорошо дифференцированы от западного подвида лишь самцы; этот
подвид типичен значительным развитием белого цвета на передних
крыльях и широкой черной перевязью в срединном поле; базальное поле
бело-серое с хорошо заметной первой поперечной (базальной) линией;
срединное поле в костальной части лишь частично занято широкой, раз-
мытой, расширяющейся к дорзальному краю черно-серой с примесью
бурых тонов полосой, которая у дорзального края крыла - захватывает
все срединное поле и часть базального; третья поперечная линия оттенена
с внешней стороны также широкой бурой растушеванной каймой; внеш-
нее поле белое (сероватое), с немногими черными штрихами, слабой под-
краевой полосой и тонкой терминальной линией; бахрома шашечная;
задние крылья с широкой черно-серой краевой полосой, темным основа-
нием и размытым дискоидальным пятном; бахрома белая и черная,
белые же концы жилок. Нижняя сторона бело-серая, более темная на пе-
редних крыльях, с черными дискоидальными пятнами на обеих парах
крыльев. Строение усиков и структуры гениталий не различаются у за-
падного и восточного подвидов; нет отличий и в личиночных стадиях.
Этот подвид ограничен в распространении восточной Азией.
Распространение. Приамурье (Хабаровск, Раддевка); юж-
ное Приморье (Сучан, Яковлевна, Седанка, Владивосток, Аскольд, Тигро-
вая, Адими, Черная речка, Суйфун, Ворошилов-Уссурийский); южный
Сахалин; Курильские острова; Корея; Японские острова (Хоккайдо,
Хондо).1
4. Род LAELIA STEPH.
Stephens, 1827, Cat. Brit. Ins., 2 : 52; Stephens, 1828, Brit. Entom., Haustell.,
2 : 62; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 459; Hampson, 1892, Fauna Brit. Ind.,
Moths, 2 : 44; Staudinger, 1901, Cat. Lep., 1 : 117; Swinhoe, 1903, Trans. Entom. Soc.
Lond. : 439; Aurivillius, 1904, Ark. Zool., 2 (4) : 68; Strand, 1910, Seitz, Grossschmett.,
2 : 121; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 116; Strand, 1915, Seitz, Gross-
schmett., 10 : 306; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 89; Hering, 1926, Seitz,
Grossschmett., 14 : 158; Collenette, 1934, Ann. Mag. Nat. Hist., 13 : 210; Bryk, 1934,
Lep. Cat., 62 : 95; — Cycnia Walker, 1855, Cat. Lep. Het. Brit. Mus., 3 : 683 (typus
testacea Wlk.); — Ricine .Vulker, 1855, там же, 4:824 (typus sufjusa Wlk.); — Char-
dinesSNsYkeir, 1855, там же, 4 : 797 (typus htura Wlk.);— Pro codec a Walker, 1855, тамже,
1 С Курильских островов Брик (Bryk, 1942, Iris, 56 : 25) описал форму от D. pu-
dibunda L. под названием D. hesiotes. Особи D. pudibunda L. из сборов Н. Н. Кона-
кова с Курильских островов вполне сходны с дальневосточными D. pudibunda mode-
sta Kirby (pudica Stgr).
266
1. ORGYINAE
4 : 812 (typus obsolete. F.=quadrata Wlk.); —Odagra Walker, 1855, там же, 32 : 401
(typus devistita Wlk.); — Нагара Moore, 1878, Descr. Ind. Lep. Atkins. : 47 (typus
atestacea Hamps. = testacea Moore). — Laelioides Moore, 1883, Lep. CeyL, 2 : 83
(typus fasciate Moore).
Тип рода L. coenosa Hb.
Хоботок редуцирован, слабый; губные щупальца длинные, торчащие
вперед или слабо свешивающиеся вниз, второй членик их покрыт длин-
ными волосками, длинный; третий их членик короткий и слабый; усики
самцов очень сильно гребенчатые, гребешки на концах несут по три длин-
ных щетинки; передние крылья с тупыми углами; первый членик перед-
них лапок более половины длины передних голеней; задние голени с двумя
парами шпор; коготки всех ног простые; радиальная ячейка на передних
крыльях небольшая; второй радиальный ствол отходит от ее наружной
стороны; третий и четвертый радиальные стволы на стебельке; пятый
радиальный ствол свободный от-
ходит почти из одной точки с
третьим и четвертым; первый ме-
диальный ствол отходит от сере-
дины ячеи; дискальные жилки на
передних крыльях редуцированы;
задние крылья с очень узкой и
длинной базальной ячеей, дис-
кальные жилки на задних кры-
льях также недоразвиты.
Uncus отделен швом и мем-
браной от девятого сегмента; sac-
Рис. 116. Laelia coenosa Hb. Хетотаксия
иередне-и среднегруди и брюшных сегмен-
тов I, VI, VII и VIII взрослой
гусеницы.
cus недоразвит; valva без harpe;
penis с инкрустациями на pars
inflabilis; bursa copula trix без
инкрустаций на поверхности; apo-
physes длинные.
Гусеницы густо волосистые, покрыты длинными волосками; кожа не-
сет небольшие плоские бородавки. Бородавки х на переднегруди крупные,
несут пучки перистых волосков, торчащие вперед. На прочих сегментах
бородавки х несут лишь одну щетинку. Бородавки а на первых четы-
рех брюшных сегментах крупные, плоские, несут короткие щетки тор-
чащих дорзально густых волосков; бородавки [3 на этих же сегментах
небольшие (рис. 116), лежащие вентрально от а, сближены с ними. На
восьмом и девятом брюшных сегментах бородавки [3 значительно крупнее
а и несут пучки сильно перистых, кистеобразных волосков, направленных
косо дистально (сидят один за другим). Выворачивающиеся железки
как обычно на шестом и седьмом брюшных сегментах.
Политипический род, сосредоточенный во влажных тропиках Старого
Света; главная масса — свыше семидесяти видов Laelia Steph. — известна
из тропической Африки (Сиерра-Леоне, Либерия, Слоновый берег, Золо-
той берег, Нигерия, Огове, Конго, северная Родезия и восточная эквато-
риальная Африка г), немногие виды известны из южной Африки и есть
1 Из Марокко описана форма от абиссинского Laelia impura Her. (Audegud,
1935, Mitt. Schweiz, entom. Ges., 16 : 419—420) — brauni Aud.
Судя по изображению это очень своеобразный представитель описываемого рода,
далекий от евразийского L. coenosa Hb. Наличие этого вида в Палеарктике весьма
интересно в отношении связей фауны. О близости описанной особи Laelia Steph, из
Марокко к абиссинской L. impura Her. судил не только. Одегю, но также Лесерф и
Коллепетт.
L LAELfA
267
два эндемичных для Мадагаскара (L. diaphanella Mab., croperoides Her.);
один вид (Л. melaxantha Wlk.) распространен в Африке и известен из Ин-
дии. Большинство африканских видов Laelia Steph, сильно отличается
от азиатских и индомалайских и во всяком случае образует особую группу
в пределах рода. Тем не менее некоторые из африканских Laelia Steph,
достаточно типичны и несомненно относятся к индомалайской группе
этого рода (A. incerta Her., leucoptera Her., sericaria Her. и ряд других).
В индомалайской фауне виды Laelia Steph, распространены преиму-
щественно в Индии и на Цейлоне (A. adalia Swinh., ju.lv ala Hamps., ит-
brina Moore, atestacea Hamps., sufjusa Wlk., lilacina Moore, prolata Swinh.,
calamaria Hamps., venosa Moore, coenosa Hb., exclamationis KolL, litura
Wlk., testacea Wlk., heterogyna Hamps.); часть этих видов эндемична для
Индии, но некоторые из них проникают также на Малайский архипелаг
(A. sufjusa Wlk., venosa Moore, coenosa Hb.), где есть несколько эндемиков
с Явы (A. buana Moore, adora Moore) и с Суматры (L. moszkowskii Strd.).
Только два вида Laelia Steph, известны из Австралии: с севера—L. ob-
soleta F. и из Квинслэнда — L.. ostracina Turn. Напротив, к северу от
Малайского архипелага и к востоку от Индии распространение Laelia
более значительно — ряд видов известен с Тайвана (A. striata Wilem.
devestita Wlk., costalis Mts., ochripalpis Strd.), один вид указан для Гон-
конга (L. coenosa Hb.) и один описан из Японии (A. japonibia Strd.); нако-
нец один вид (A. coenosa Hb.) широко распространен в Китае и Японии
и далее на запад во многих местах умеренной Евразии от Тихого до Атлан-
тического океана. Из Северной и Южной Америки виды Laelia Stph.
или сколько-нибудь близкие к ним формы не известны.
Как видно из распространения, виды Laelia Steph, гигрофильны;
единственный несколько ближе известный вид A. coenosa Hb. связан
с влажными лугами и живет за счет однодольных растений, преимуще-
ственно Gramineae. Кормовые связи и биология тропических видов не
изучены.
* 1. Laelia coenosa Hb. (рис. 116, 117, 118).
Hiibner, 1804, Eur. Schmett., Bombyces : 120, t. 51, fig. 218; t. 77, fig. 323, 324,
325 (Bombyx)', Kirby, 1892, Syn. Cat. Lep. Het., 1 : 459; Leech, 1899, Trans. Entom.
Soc. Lond. : 120; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 116, t. 21, fig. 12; Strand,
1910, Seitz, Grossschmett., 2 : 122, t. 19, i; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) :
93; Joannis, 1929, Ann. Soc. Entom. France, 98 : 415; Matsumura, 1931, 6000 Ins.
Japan : 710, fig. 468, a; Matsumura, 1931, Ins. Matsum., 4 : 136, t. 22, fig. 21; Gaode,
1932, Seitz, Grossschmett., 2 Suppl : 105; Collenette, Nov. Zool'., 37 : 179; Matsu-
mura, 1933, Ins. Matsum., 7 : 132; Amsel, 1933, Zoogeogr., 2 : 99; Bryk, 1934, Lep.
Cat., 62 : 97; Urbahn, 1935, Intern, entom. Ztschr., 29 : 199; Warnecke, 1936, Entom.
Rundsch., 53 : 123; Warne.cke, 1938, Rev. Fr. Lep., 9 (10) : 173;— sinensis Walker,
1855, Cat. Lep. Het. Brit. Mus., 4 : 829; — brevicornis Walker, 1858, там же, 7 : 1729; —
sangaica Moore, 1877, ,Ann. Mag. Nat. Hist., 4 (9) : 92;—gigantea Butler, 1885, Cist.
Entom., 3 : 117; — Candida Leech, 1899, Trans. Entom. Soc. Lond. : 121;—paucipuncta
Strand, 1910, Seitz, Grossschmett., 2 : 122, t. 19, i; —formbsana Strand, 1914, Suppl.
Entom., 3 : 37; —fuscolimbata Cerva, 1918, Rovart. Lap., 25 : 101; — nigrociliata
Urbahn, 1935, Intern, entom. Ztschr., 29 : 199, fig. 1—3.
Крылья, грудь и голова белые или беловато-желтые; рисунок на
передних крыльях редуцирован; вдоль жилок на передних крыльях
светлые и темные штрихи, по жилкам светлые и между ними темные;
в терминальной области на передних крыльях неправильно разбросан-
ные черные точки (чешуйки), задние крылья однотонные, желто-белые;
нижняя сторона желто-белая, костальный и терминальный края перед-
них крыльев у самцов дымчато-желтые, у самок чисто белые; усики
268
1. ORGYINAE
самцов сильно гребенчатые, членики белые, отростки их серо-черные;
губные щупальца покрыты длинными волосками с вентральной стороны;
ноги желто-белые; брюшко того же цвета; самки более светлой окраски,
почти или чисто белые. Размеры очень изменчивы; размах крыльев 25—
50 мм, главная масса особей 25—40 мм; самки несколько крупнее самцов.
Uncus плоский с боков, узкий, при основании расширенный, отде-
лен швом и мембраной от девятого сегмента; тергит девятого сегмента
расширен, стернит узкий, не образует saccus; fultura inferior большая,
почти круглая, в ' середине с большим почти круглым вырезом, края
которого несут многочисленные мелкие зубчики; valvae почти треуголь-
ной формы, сильно выгнутые вентрально, вентральный угол острый;
Рис. 117. Laelia coenosa Hb. Самец. (Рис. Ф. Гуняева).
дистальная часть valva покрыта многочисленными волосками; penis
короткий, толстый, к дистальному концу конический, несет в дисталь-
ной части мелкие шипики. Bursa copulatrix большая, мешковидная,
мембранозная, несколько расширенная в проксимальной части, почти
цилиндрическая; ductus bursae мембранозный и узкий, длиннее apophy-
ses, стенка его несет многочисленные продольные складки; восьмой
сегмент короткий, несет длинные apophyses anteriores; apophyses poste-
riores также длинные; papillae anales короткие и широкие, несут мел-
кие щетинки; есть сильно склеротизованная lamella postvaginalis в виде
складки кожи.
Яйца сферические, уплощенные дорзовентрально, белые с серым
колечком и серой точкой на месте вдавления.
Взрослая гусеница беловатая или серовато-желтая, покрыта волос-
ками того же цвета и отчасти черными; вдоль спины темная прерванная
полоса (иногда не развита); по бокам на уровне стигм по одной тонкой
темной полоске; бородавки одного цвета с прочими частями кожи; красно-
желтые бородавки (Ламперт, Шпулер, Зейтц), по данным Божовской
(BoczowsKa, 1933), не характерны для гусениц этого вида (живых);
мною на спиртовых материалах красно-желтые бородавки также не
констатированы; на переднегруди на х. с каждой стороны по кисточке
черных перистых волосков, торчащих вперед; на первых четырех брюш-
ных сегментах с дорзальной стороны по одной густой короткой щетке
волосков; на шестом и седьмом брюшных сегментах железки в виде
вороночек; на восьмом брюшном сегменте непарный дорзальный пучок
4. LAELIA
269
перистых волосков, направленных косо, дистально; на девятом сегменте
несколько более короткий пучок такого же типа; грудные и брюшные
ноги желто-серые; голова желтая с темными ротовыми частями.
Куколка 16—20 мм, коричневая, покрытая шелковистыми белыми
или желтыми волосками.
Изменчивость, как отмечалось, велика в размерах и меньше в окраске;
очень крупные особи с юго-востока Азии описаны как Laelia gigantea
Btl., они ничем не отличаются от обычных, кроме размеров. Совершенно
белые без примеси желтых тонов и черных чешуй особи из Китая опи-
саны как Candida Leech; форма sangaica Moore, описанная из Китая
и Японии, с примесью охристых тонов; особи с примесью темных серых
тонов, также известные из Ки-
тая, описаны как sinensis Wlk.
Последняя, а также может быть
предыдущая не достаточно изу-
чены и могут оказаться особыми
видами. Изменчивость европей-
ских особей касается также
лишь цветовых признаков, при-
чем развития темных тонов;
описаны меланистические фор-
мы— nigrociliata Urb. и fusco-
limbata Cerva.
P а с п p о с т р а нение.1
Пиренейский полуостров (Ка-
талония, Барселона); Франция;
Германская низменность (Мек-
ленбург, Штеттин, Берлин,
Эмден, Гарц); Голландия; Ют-
ландия; Британские острова
(где ныне исчез, по Варнеке);
Карпаты (Буковина); долина
Вислы (Сарны); долина Дуная
(ряд точек); Македония; До-
бруджа; Малая Азия (Троя);
Палестина; Сирия; северная
Африка (Рифф); Молдавия (Ки-
шинев); Украина (Винница,
Полтава, Чернигов, Волынь); Крым (Керчь); Закавказье (Поти, устье
Куры); восточный Казахстан (Алма-ата); южный Алтай (Колываиь);
Приамурье (Зея, Уркан, Хабаровск, Благовещенск); Шантарские остро-
ва, Сахалин; южное Приморье (Уссури, Владивосток, Сучан, Суйфун,
Тигровая, Океанская, Сидеми, Яковлевка, Виноградовка, Ворошидов-
Уссурийский); Японские острова (Хоккайдо, Хондо); Корея; Китай
(Гон-конг, Ханг-янг, Шанхай, Нинг-по); Вьетнам (Тонкин).
Биология. Зимуют гусеницы в средних возрастах, как то было
установлено для средней Европы [в лабораторных условиях (Boczkowzka,
1933)]есть указания на зимовку яиц, но они не подтверждены исследо-
ваниями. Появление весной не изучено; питающие растения —Сурега-
сеае и Gramineae, виды Carex (limosa, chordorrhizza, acuta, riparia, pseudo-
Рис. 118. Laelia coenosa Hb: Гениталии:
a—• самца, 6 — самки.
1 Дополнения к распространению этого вида в средней Европе могут быть най-
дены в работе Варнеке на прилагаемой в ней схематичной карте. '
270
1. ORGYINAE
cyperus, lasiocarpa), Eriophorum, Agrostis, Phragmites, Calamagrostis,
Baekmannia, Luzula; питание Роа и Phleum неблагоприятно. Окукление
в июне — июле на стеблях растений; коконы удлиненной формы.укреп-
ляются вдоль стеблей, часто на Equisetum. Длительность развития
пронимф (в коконах) около двух-трех суток. Развитие куколок в тече-
ние двух-трех недель. Выход бабочек в конце июня. Бабочки летают
перед заходом солнца. Спаривание наблюдается один раз; самки откла-
дывают яйца в течение трех-четырех суток; общее количество яиц 200—
250 штук.Яйца откладываются цепочками по 10—20 вдоль стеблей
кормовых растений. Развитие яиц продолжается 10—15 суток. Стацией
являются влажные луга (нередко заливные); в уссурийской тайге отме-
чены тростниковые сосняки; не частый вид и всегда распространен-
ный спорадически. Отмечены случаи (в средней Европе) значительных
размножений. В СССР массовое размножение этого вида отмечено на
Кавказе (в Аланском районе) В. Смольянниковым в 1937 г. Гусеницы
в массе встречались на тростнике в плавнях рек Кубани и Аланки и после
уничтожения тростника мигрировали на поля, где уничтожали по пре-
имуществу злаки.
5. Род ORGYIA OCHS.1
Ochsenheimer, 1810, Schmett. Eur., 3 : 208; Aurivillius, 1891, Nordn. Fiar. : 91;
Kirby, 1892, Syn. Cat. Lep. Het., 1 : 492 (Notolophus)' Staudinger, 1901, Cat. Lep.,
1 : 113; Swinhoe, 1903, Trans. Entom. Soc. Lond. : 458; Aurivillius, 1904, Ark.
Zool., 2 (4) : 67; Spuler, 1908, Schmett. Eur., 1 : 126; Rebel, 1910, Berge, Schmetter-
lingsb., ed. 9 : 114; Strand, 1910, Seitz, Grossschmett., 2 : 109, 116; Chapman, 1919,
Entom. Rec., 3 : 41; Turner, 1921, Proc. Linn. Soc. N. S. Wales, 45 : 493; Gaede, 1932,
Seitz, Grossschmett., 2 Suppl. : 97; Hering, 1932, Tierw. Mitteleur. : 482; Bryk, 1934,
Lep. Cat., 62 : 61; — Notolophus Germar, 1812, Prodromus, 3 : 35 (typus antiqua L.);—
Tnchiosoma Lucas, 1849, Exped. Alger.: 376 (typus algyricum Lucas); — Acyphas Wal-
ker, 1855, Cat. Lep. Het. Brit. Mus., 4 : 797 (typus fusca Wlk.); — Lacida Walker,
там же, 4 : 803 (typus anartoides Wlk.); — Teia Walker, 1855, там же, 4 : 803 (typus
anartoides Wlk.); — Microptero'gyna Rambur, 1866, Cat. Syst. Lep. Andal., 2 : 281
(typus antiqua L.); — Thylacyna Rambur, 1866, там же, 2 : 283 (typus ericae Germ. sec.
Kirby, 1892); — Apterogynis Guenee, 1875, Stat. Sci. d’Eure-Loire : 78 (typus
antiqua L.);—Hemerocampa Dyar, 1897, Canad. Entom., 29:29 (typus leucostigma
Sm. et Abb.).
Тип рода О. an'iqua L.
Хоботок редуцирован, есть лишь невидимые под лупой очень малень-
кие остатки челюстей, заметные лишь у самцов, у самок полная редукция;
глаза маленькие, голые, овальной формы, у некоторых видов очень
узкие (хорошо отличны по этому признаку и по голеням задних ног
от Cifuna Wlk.); губные щупальца слабые, у многих видов очень укоро-
чены и почти не видны в волосяном покрове, последний членик их всегда
утрачен; усики самцов сильно гребенчатые; у самок, если есть усики,
1 Оксенгеймер (1810) принимает состав Orgyia Ochs, следующим: О. pudibunda L.,
abietis Schiff., fascelina L., selenitica Esp., gonostigma F., antiqua L., указывая, что
два последних вида отличаются от остальных тем, что самки их бескрылы и самцы
держат в покое крылья распластанными. Гюбнер (Hiibner, 1816, Verz. : 161) прини-
мает тот же состав рода Orgyia Ochs., исключая два последние вида, которые он вводит
в род Gynaephora Hb. ; ни один из авторов не назвал родового типа. Кёртис (Curtis
1816, Brit. Entom., 8 : 378) впервые указал тип рода — Orgyia antiqua L. Все после-
дующие авторы принимали род Orgyia Ochs, по Кёртису, исключая Кирби (1892), кото-
рый для О. antiqua L. принимает название Notolophus, a Dasychira Steph, сводит в
синонимы-к Orgyia Ochs. В связи с тем, что тип рода Orgyia Ochs. — О. antiqua L. —
был указан 'Кёртисом (1816), я принимаю состав рода по этому автору.
5. ORGYIA
271
то они с ничтожными гребешками или ресничками; передние голени
длиннее первого членика этих же лапок; голени задних ног с одной
задней парой шпор; коготки всех ног у самцов с зубцом на внутренней
стороне; у самок коготки простые, исключая самок О. thyellina Btl.,
antiqua L., gonostigma F.; радиальная ячейка на передних крыльях
большая, широкая, ромбическая; третий и четвертый радиальные стволы
на стебельке, отходят от вершины ячеи; пятый радиальный ствол сво-
бодный, отходит также от конца ячеи; первый медиальный ствол отходит
от середины ячеи; дискальная жилка на передних крыльях утрачена;
базальная ячея на задних крыльях замкнутая неполным анастомозом;
второй медиальный ствол рудиментарный; третий ствол на общем
стебельке с первым кубитальным.
Uncus или широкий и простой, или разделенный вдоль глубокой
бороздой и с двойным концом; tegumen разделен швом; penis расширен
в дистальной части. Bursa copulatrix маленькая, нежная, мешковидная.
Тело покрыто волосками, без примеси чешуи. Полы диморфны; самки
или с нормальными крыльями (несколько больших размеров, чем у сам-
цов), короткокрылы, или совершенно бескрылы; у некоторых видов они
лишены также ног и даже головы. Кожные покровы самок нежны, тело
покрыто шелковистыми волосками.
Гусеницы покрыты неравномерно длинными волосками; волоски, сла-
гающие пучки, перистые; бородавки х переднегруди большие, несут
пучки длинных сильно перистых волосков, направленные вперед
(рис. 122, 124); на брюшных сегментах группа х в виде небольшой
бородавочки или рудиментарна, несет одну щетинку или лишена ее;
бородавки аир на средне- и заднегруди сильно разобщены. На первых
четырех сегментах брюшка эти же бородавки очень большие, почти
полностью слиты (есть лишь шов между ними), несут короткие пиль-
чатые белые или иначе окрашенные волоски в виде щеточек. Бородавки
р первых двух сегментов могут нести кисти сильно перистых волосков.
Бородавки а. шестого и седьмого брюшных сегментов маленькие, во много
раз мельче, чем бородавки [3 на этих же сегментах. На восьмом брюшном
сегменте бородавки а большие, но вдвое меньше, чем [3 на этих же
сегментах. Куколки блестящие, черные или коричневые, с редкими
небольшими пучками волосков, сильно суженные к дистальному концу;
куколки самок уплощенные дорзовентрально. Яйца крупные, до 0.7—
0.6 мм диаметром, с плотным матовым хорионом, не покрываются воло-
сками с конца брюшка самки.
Широко распространенный политипический род, связанный в распро-
странении преимущественно с индомалайской подобластью и богато пред-
ставленный в Голарктике (рис. 119); в Африке, Австралии и Южной
Америке виды Orgyia Ochs, представлены бедно. В Африке они известны
из Нигерии (О. vaporata Her., albocostata Bet.-Bek., vetusta Bd.), из юж-
ной Африки (0. tricolor Н. S.), из восточной Африки (О. mixta Snell.,
nigricostata Joie, et Taib.); два вида описаны с Мадагаскара (О. aurantia
Mab., malagassica Kenr.); все эти виды эндемичныдля Африки, но, исклю-
чая О. tricolor Н. S., не совсем ясны в отношении систематического
положения.
С Малайского архипелага известно большое число видов Orgyia Ochs.
(О. varia Wlk., dimidiata Wlk., nebulosa YN\k,,oss&ata\N\k., jlavittata Pag.,
nigriplaga Swinh., postica Wlk., nigrocrocea Wlk., tisdala Swinh., turbata
Btl.); многие из них очень напоминают евразийские виды; один энде-
мичный вид Orgyia Ochs, описан с Новой Гвинеи (Ch dawara Swinh.)
Рис. 119. Распространение видов рода Orgyia Ochs. Черным указаны территории обилия видов.
5. ORGYIA
273
и туда же проникает О. postica Wlk.;1 несколько очень характерных
видов описано из Австралии (Н. Ю. Уэллс, Виктория, юго-восточная
Австралия — О. australis Wlk., retinopepla Lower, hemicalla Lower, anar-
toides\N\k.), очень напоминающих евразийскую О. dubia Tausch. и афри-
канскую О. tricolor Н. S. Два малайских вида (О. turbata Btl. и О. postica
Wlk.) проникают на Малакку и в Индию, где есть два эндемичных вида
(О. nucula Swinh. и О. senica Hamps.); один эндемичный вид описан
с Цейлона (О. viridescens Wlk.).
Умеренная Азия также типична эндемичными видами или распростра-
ненными локально, как, например, виды, распространенные на юго-востоке
Азии (О. immaculata Gaede, parallela Gaede, thyellina Btl.) или связан-
ные с горными системами (О. aurolimbata Gn., jlavolimbata Stgr., ochro-
limbata Stgr., prisca Stgr., panlacroixii Obth.), или распространенные
широко ((?. dubia Tausch., trigotephras Bd., ericae Germ., gonostigma F.),2
Один вид (О. antiqua L.) распространен по всей лесной и'лесостепной
зонам Евразии (как и О. gonostigma F.) и проникает в Северную Америку,
где представлен очень обособленным подвидом О. badia Edw. В Северной
Америке есть большое число эндемичных видов Orgyia Ochs.,3 концен-
трирующихся в южных штатах и Мексике. Отсюда известны (включая
также и более северные виды из Канады и атлантических штатов) О. 1еи-
costigma Sm. et Abb, vetusta Bd., inornate. Beut., oslari Barn., pseudotsu-
gata Barn, et MacD., definite Pack., falcataSchaus, coresia Dre., /aZiscaSchs.,
bipuncta Dre.; многие из этих видов очень напоминают японскую О. thyel-
lina Btl. (например pseudotsugata Barn, et MacD.). Самки неарктических
видов изучены слабо, но у тех видов, у которых они известны, они корот-
кокрылы или бескрылы. Несколько видов Orgyia Ochs, описано из
центральной Америки (Гватемала, Сальвадор, Коста-Рика — О. guate-
malica Schs., costaricensis Schs., povera Schs., salvadora Schs.), один вид
1 Принадлежность вида О. postica Wlk. к роду Orgyia Ochs, в последнее время
подвергнута сомнению. Гарднер (Gardner, Indian Forest Rec., 6 : 299), первоначально
относивший, как и другие авторы, этот вид к Orgyia Ochs., недавно (Gardner, 1946,
Trans. Roy. Entom. Soc. Lond., 96 (4) : 61—72) высказался за принадлежность этого
вида к подсем. Acronictinae. Этот отказ мотивируется ссылкой на работу Колленетт
(Collenette, 1935, Stylops : 242), которая считает О. postica Wlk. типом рода Thiaci-
das. Кроме того, гусеницы О. postica Wlk., по Гарднеру (1946), лишены выворачиваю-
щихся железок на шестом и седьмом сегментах брюшка, что столь типично (хотя
не абсолютно) для Orgyinae. Колленетт (1935) не приводит никаких данных для под-
тверждения своего мнения о принадлежности О. postica Wlk. к Acronictinae, но
ссылается лишь на работу Дайра (Dyar, 1897, Canad. Entom., 29 : 12). Между том
Этот автор приводит О. postica Wlk. (цит. раб., стр. 15) в числе видов Notolophus Germ.,
т. е. Orgyia Ochs.
2 Кроме упомянутых видрв, описаны еще два вида Orgyia Ochs, из Средиземно-
морья, являющиеся, по моему мнению, лишь формами от О. trigotephras Bd. Это пирега
Trti. и rupestris Rbr.
3 Дайр (189.7) указывает, что американские виды Orgyia Ochs, по признаку числа
шпор на задних голенях распадаются на две группы. О. vetusta Bd. имеет одну пару
шпор на задних голенях и' потому совпадает по признакам с видами Orgyia Ochs.,
тогда как О. leucostigma Sm. et Abb. и О. definita Pack, имеют две пары шпор. Дайр
(1897) выделяет их в особый род — Hemerocampa Dyar. Шаус (1927) ошибочно указы-
вает, что все виды Hemerocampa Dyar, куда он включает и О. vetusta Bd., характери-
зуются одной парой шпор, чем отличны от Orgyia Ochs. Просмотр видов О. vetusta
Bd. и О. leucostigma S. А., произведенный мною, подтверждает указание на различие
в числе шпор у них. Требуется отметить, что американские видь! Hemerocampa Dyar,
в частности именно О. leucostigma Sm. et Abb. — тип рода, обнаруживают очень боль-
шую близость к евразийским Orgyia Ochs., например к О. antiqua L., с которой дает
гибриды. Вероятно правильнее пока принимать эти виды с двумя шпорами за под-
род от Orgyia Ochs.
18 Фауна СССР, И. В. Кожанчиков
274
I. ORGYINAE
известен из Колумбии — О. columbiensis Dogn.; далее на юг, из других
частей Южной Америки, виды Orgyia Ochs, не известны.
Биологически видыOrgyia Ochs, характеризуются высокой активностью
самцов и малой активностью или даже почти полной неподвижностью'
самок; самцы летают днем, отыскивая самок по запаху. Яйцекладка на
поверхность кокона или в кокон; яйца кладутся или в один ряд и остав-
ляются голыми (О. antiqua L., ericae Germ.), или откладываются кучкой
и густо покрываются волосками (О. gonostigma F.). Степень редукции
крыльев и органов движения самок в известной мере связана с место-
обитаниями—виды, населяющие влажные леса (например О. thyellina
Btl.), имеют крылатых, еще довольно активных самок; степные виды
и Особенно полупустынные (О. dubia Tausch.) имеют самок не только
бескрылых, но безногих и безголовых. Зимовка в умеренных широтах
протекает в фазе яйца или в личиночной; в тропиках развитие проте-
кает весь год, и для О. postica Wlk., например, на о. Тайван отмечено
до шести поколений. Виды Orgyia Ochs, многоядны, но связаны преиму-
щественно с кустарниками из Salicales, Fagales и Rosales, а также с травя-
нистыми и кустарниковыми Leguminosae и Mimosaceae; возможно также
питание многими другими растениями (Theaceae, Palmae, Geraniaceae,
Euphorbiaceae и др.). Известны массовые виды и вредители лесных и
сельскохозяйственных культур; особенно значительный вред отмечался
от некоторых субтропических и тропических видов (О. turbata Btl.,pos-
tica Wlk.) чайц'бму кусту, какао (Theobroma), Erythrina lythosperma
и клещевине.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Самцы
1 (14). Передние крылья темнее задних;. рисунок хорошо развит или,
если линии рисунка редуцированы, с неправильными пятнами
и полосами: бахрома на обеих парах крыльев более или менее-
одного цвета с общим фоном крыла.
2 (3). Передние и задние крылья желтые; рисунок на передних крыльях
в виде широких черных полос; задние крылья желтые с широкой
черной каймой; uncus вздут, со слабой бороздой на дорзальной
стороне...................................*4.0, dubia , .Tausch.
3 (2). Окраска крыльев коричневая или бурая, рисунок в виде тонких
полос.
4 (5). У окончания третьей поперечной линии, у дорзального края
крыла, нет белого пятна; передние крылья с примесью фиолёто-
вых тонов; почковидное пятно в середине белое, четко очерчено;
оба пола крылаты.............................1.0. thyellina Btl.
5 (4). На передних крыльях у дорзального их края при третьей попе-
речной линии белое пятно; почковидное пятно более или менее
стушевано.
6 (9). Задние крылья однотонно черные или коричнево-черные, перед-
ние крылья темнокоричневые с светлыми пятнами у костального
края. -
7 (8). Внешнее поле на передних крыльях с неправильными желтыми
пятнами у костального края; первая и вторая поперечные линии
четкие, тонкие; uncus с глубокой бороздой на дорзальной стороне,
на дистальном конце не расщеплен; valva коническая, с простым
дорзальным краем . . •.....................* 3. О. gonostigma L-
5. ORGYIA
27 5
8 (7). Внешнее поле у костального края фиолетово-серое; линии рисунка
несколько стушеваны; в срединном поле у костального края свет-
лый фиолетово-серый треугольник; uncus с гребнем на дорзальной
стороне; valva с короткой ступенькой на дорзальном крае . . .
............................................6. О. trigotephras Bd.
9 (6). Задние крылья охристые и желтоватые или желто-коричневые;
передние крылья желто-коричневые.
10 (И). Передние крылья с черными растушеванными поперечными
линиями; срединное поле однотонное; uncus плоский; valva с длин-
ным направленным дистально выростом на дорзальном крае . . . .
.................................................* 2. О. antiqua L.
И (10). Передние крылья с нерезкими коричнево-желтыми линиями;
в срединном поле у костального края светлое неправильное или
треугольное пятно; valva без длинного дорзального отростка; uncus
высокий, на дистальном конце расщеплен.
12 (13). Почковидное пятно тонко очерчено светлым; valva короткая,
простая; penis на дистальном конце слабо расширен............. . .
.........................................* 7. О. prisca Stgr.
13 (12). Почковидное пятно растушевано; penis на дистальном конце
расширен; valva с небольшим отростком на дорзальном крае ....
...............................................* 5. 0. ericae Germ.
14 (1). Передние и задние крылья однотонные с светлой бахромой или
светлым внешним краем, реже с неясной срединной полосой; ли-
нии рисунка и пятна редуцированы совершенно.
15 (16). Крылья светлые охристо-коричневые, в середине несколько
более светлые; широкие при тонком стройном теле, бахрома на них
желтая; девятый сегмент с небольшим subunci.................... . .
....................................*9.0. ochrolimbata Stgr.
16 (15). Крылья шоколадно-коричневые или темные, без охристых тонов,
одноцветные, с бледножелтой или охристой яркой бахромой и не-
редко также с охристым терминальным краем.
17 (18). Передние крылья оливково-коричневые с растушеванной поло-
сой в середине; бахрома на них светложелтая или яркая рыже-
охристая, причем охристый цвет захватывает и терминальный край.
...........................................* 10. О. flavolimbata Stgr.
18 (17). Передние крылья шоколадно-коричневые с бело-желтой бахро-
мой, однотонные.............................*8. О. aurolimbata Gn.
Самки ’
1 (2), Самки крылаты или имеют лишь наполовину укороченные крылья
........................................... 1. О. thyellina В11.
2 (1). Самки с рудиментарными крыльями или совсем без крыльев;,
иногда редуцированы ноги и голова.
3 (6). Крылья хорошо заметны, в виде небольших лопастей, по раз-
мерам почти равны ногам; ноги вполне развиты, функционируют;
коготки несут хорошо видные вентральные зубцы.
4 (5). Крупный, 15—20 мм; брюшко широкое, почти прямоугольное;
тело покрыто черными, желто-серыми или чисто белыми шелкови-
стыми волосками.............................*3. О. gonostigma L.
5 (4). Мельче, 10—18'мм; брюшко овальное или вытянуто-овальное,
узкое; волоски, покрывающие тело, желто-белые................'. .
..............................................* 2. О. antiqua L.
18*
276
I. ORGYINAE
6 (3). Крылья утрачены вполне или очень малы, по размерам меньше
длины ног и нередко спрятаны в волосках тела; ноги есть или их
нет совсем, так же как и головы.
7 (10). Ноги развиты вполне; крылья заметны в виде очень маленьких
рудиментов; коготки на ногах развиты; длина тела 10—-15 мм.
8 (9). Окраска желтоватая или серо-желтая; тело покрыто желто-
серыми волосками................................*5.0. ericae Germ.
9 (8). Тело покрыто чисто белыми или бело-серыми волосками ....
........................................6.0. trigotephras Bd.
10 (7). Крылья утрачены вполне; ноги если есть, то не функционируют,
слабые; голова нередко утрачена; самки не выходят из коконов.
11 (12). Ног нет, голова рудиментарна; волоски, покрывающие тело,
желто-серые; длина 18—20 мм.....................*4.0. dubia Tausch.
12 (И). Ноги есть, хотя и слабые; голова недоразвита или развита
вполне.
13 (14). Голова рудиментарна; ноги слабые; тело широкое и толстое,
длиною примерно 15 мм; кожа покрыта темносерыми волосками.
..........................................* 9. 0. ochroiimbata Stgr.
14 (13). Голова развита вполне, маленькая; тело узкое, овальное; длина
тела .примерно 10—15 мм; кожа покрыта серебристыми белыми
волосками.1........................................*7. 0. prisca Stgr.
1. Orgyia thyellina Btl. (рис, 120).
Butler, 1881, Trans. Entom. Soc. Lond. : 10; Kirby, 1892, Syn. Cat. Lep. Het.,
1 : 495; Leech, 1899, Trans. Entom. Soc. Lond. : 118; Strand, 1910, Seitz, Grossschmett.,
2 : 120, t. 19, c; t. 22, b; Seitz, 1914, Entom. Rundsch., 31 : 91; Klotz, 1915, Intern,
entom. Ztschr., 8 : 177; Albrecht, 1917, Entom. Ztschr., 28 : 60, fig. 1; Wileman, 1918,
Philipp. Journ. Sci., 13 : 152, t. 1, fig. 3; Swinhoe, 1923, Ann. Mag. at. Hist., 11 (9):
418; Gaede, 1932, Seitz, Grossschmett., 2 .Suppl. : 98; Bryk, 1934, Lep. Cat., 62 : 73; —
flavescens Wohnig, 1927, Intern, entom. Ztschr., 21 : 38;— grisea. Wohnig, 1927, там
же, 21 : 39.
Биология. Wohnig, 1927, Intern, entom. Ztschr., 21 : 36; Clausen, 1931,
Bull. U. S. Dep. Agric., Giro., 168 : 7, 20.
Передние крылья у самцов коричнево-серые с черным и фиолетовым
рисунком; голова и грудь светлосерые; губные щуцальца снизу черные,
усики сильно гребенчатые; брюшко черное с кисточкой на третьем сег-
менте; ноги покрыты длинными желтыми волосками; базальная линия
темнокоричневая и белая, короткая, почти прямая; базальное поле
однотонно коричневое, несколько более темное в костальной, половине;
вторая поперечная линия в вйде широкой коричневой полосы, оттененной
по сторонам фиолетово-белым, слабо изогнутая; срединное поле шире
внешнего и базального, в костальной половине черное, черный цвет
распределен неравномерно; черное пятно в срединном поле окаймлено
неправильно разбросанными бело-фиолетовыми чешуйками; дорзальная
половина срединного поля коричневая; почковидное пятно неясное,
в виде темного вытянутого диффузного штриха, частично покрытого
фиолетово-белыми чешуйками; третья поперечная линия темнофиолето-
вая и коричневая, посредине почти черная, мягко зубчатая по жилкам;
внешнее поле с черными пятнами, подкраевая линия слабо волнистая,
оттенена черным в костальной половине, у дорзального края крыла она
расплывается в белое пятно; по терминальному краю нерезкие фиолетово-
1 К этому же виду близка и самка О. jlavolimbata Stgr., видимо несколько более
рудиментарная; мне известны особи лишь очень плохой сохранности.
5. ORGYIA
277
черные пятна; терминальная линия в виде тонкой полоски, бахрома
желто-коричневая; задние крылья однотонно черные с беловатой бахро-
мой; нижняя сторона черно-желтоватая, более светлая по внешнему
краю и жилкам. Самки желто-белого цвета с немногими черными пят-
нами на крыльях; встречаются две формы — вполне крылатая и с сильно
укороченными (до одной трети) крыльями; у первой типичны крылья
вытянутые, несколько серповидно загнутые (типа самок Olene. Hb.); рису-
нок не резок, но по типу тот же, что и у самцов; под костальным краем
во внешнем поле черное пятно; подкраевая линия черная, растушеван-
ная; задние крылья однотонные, желто-белые; на заднегруди и на третьем
сегменте брюшка черная кисточка волос; нижняя сторона желто-белая;
на передних крыльях просвечивают черные пятна. Размах крыльев сам-
цов 25—30 мм, самок 40—45 мм.
Uncus плоский, почти прямой, неширокий, слабо изогнутый лишь
в дистальной части; основа-
ние uncus большое, с парой
латерально расположенных
отростков, отделено швом от
девятого сегмента; послед-
ний узкий, стернит его не *•
образует saccus; valvae вы-
тянутые, в базальной части /
широкие; у вентрального /
угла valva небольшой вы-
Рис. 120. Orgyia. thycllina'Bll. Гениталии самца.
рост, покрытый волосками,
и другой большой гладкий
вырост на дорзальном крае,
правленный дистально, слегка загнутый внутрь; дистальная половина valva
узкая, мембранозная, несущая многочисленные мелкие волоски; fultura
inferior в виде мембранозной пластинки, имеющей остроугольный вырост
на дорзальной стороне; penis сильно выгнут дорзально, длинный, кони-
ческий, имеющий складочки; дистальная половина valva сильно расши-
рена, срезана косо, конец закруглен.
В систематическом отношении представляющий большой интерес вид,
наиболее сохранивший среди других голарктических видов Orgyia Ochs,
примитивные черты. По ряду признаков — форма крыльев и рисунок
у самок, кисточки на брюшке гусеницы — он сближается с видами Olene
Hb., являясь тем не менее несомненной Orgyia Ochs. Другие виды Or-
gyia Ochs, утратили эти признаки вполне.
Распространение. Корея; Японские острова (Хондо);
Тайван.
Биология. Зимует яйцо; гусеницы живут на листьях растений,
многоядны, питаются видами Salicaceae (Salix, Populus),. Betulaceae
(Betula, Alnus, Carpinus, Corylus), Fagaceae, Могасеае, Rosaceae (Pirus,
Prunus, Crataegus); предпочитаемыми растениями являются Salicaceae
и Rosaceae (Pirus). Дает несколько поколений в течение года. Самцы
весьма активны. Дает гибриды с О. antiqua L., к которому в системати-
ческом отношении наиболее близок; известны, как гибриды О. antiqua L.
(самка) и О. thyellina Btl. (самец) [Клееман (Kleemann, 1933, Intern,
entom. Ztschr., 27:23, t. 3)], называемые thyelantiqua Kleem., так и гиб-
риды 0. thyellina Btl. (самка) и О. aitiqua L. (самец), называемые ап-
thyellina John [Ион (John, 1922, Entom. Ztschr., 30 :34)]. Гибриды бес-
плодны.
278
Г. ORGYINAE
Вредит в Японии и Корее сельскохозяйственным растениям, пре-
имущественно садовым — яблоне, груше, шелковице, вишне.
*2. Orgyia antiqua L.— Античная волнянка (рис. 121, 122).
Linne, 1758, Syst. Nat., ed. 10 : 508; Linne, 1761, Fauna Suecica, ed. 2 : 297;
Aurivillius, 1891, Nordn. Fjar. : 92, t. 25, fig. 2, b; Staudinger, 1901, Cat. Lep., 1 : 114;
Spuler, 1908, Schmett. Eur., 1 : 126, fig. 81, t. 25, fig. 15; 1910, Berge, Rebel, Schmet-
terlingsb., ed. 9 : 114; Strand, 1910, Seitz, Grossschmett., 2 : 117, t. 19, a; Lagerberg,
1911, Entom. Tidskr., 32 : 28; Wahlgren, 1912, там же 33 : 86; Swinhoe, 1923, Arm.
Mag. Nat. Hist., 11 (9) : 415; Petersen, 1924, Lep. Fauna EstL, ed. 2, 1 : 140; Gaede,
1932, Seitz, Grossschmett., 2 Suppl. : 97; Hering, 1932, Tierw. Mitteleur. : 483; Bryk,
1934, Lep. Cat., 62 : 62; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 64, t. 13,
fig. 5; — paradoxa Retzius, 1783, Gen. Spec. Ins. : 36 (Bombyx); — nova Fitsch, 1865,
Rep. Ins. New Jork, 8 : 193; — badia Edwards, 1874, Proc. Cal. Acad., 5 : 186; — con-
finis Grum-Grzhimailo, 1891, Тр. Русск. Энтом. общ., 25 : 463; — modesta Heyne,
1899, Soc. Entom., 14 : 98; — infernalis Rebel, 1910, Berge, Schmetterlingsb., ed. 9 :
114; — bukovina Strand, 1910, Seitz, Grossschmett., 2 : 117;—dilutior Schultz, 1910,
Entom. Ztschr., Stuttgart, 24 : 35; —grisea Denso, 1912, Iris, 26 : 132; — bukourna.
Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 415;—mandzhurica Matsumura, 1933,
Ins. Matsum., 7 : 112, t. 3, fig. 13; — septentrionalis Rangnow, 1935, Entom. Rundsch.,
52 : 189, t. 1, fig. 10; — mongolica Alpheraky, in litt.
Биология. Tullgren, 1907, Entom. Tidskr., 28 : 209; Spuler, 1908, Raupen,
t. 15, fig. 25; Ridsen, 1908, Berl. Entom. Ztschr.,'52 : 32; Peyron, 1909, Kgl. Sv. Vet-
Akad. Handl., 44 : 92, t. 4, fig. 5; Albrecht, 1913, Entom. Ztschr. Frankf., 26 : 207;
Escherich, 1914, Forstins. Mitteleur., 1 : 250; Wolff und Krausse, 1922, Wichtigste
Forstins: 131; Lederer, 1923, Handb. prakt. Entom., 3: 148—150; Парфентьев, 1925,
Защ. раст., 1 : 201—206; Reh, 1925, Sorauer, Handb. Pflanzenkarnkh., 4 : 444;
Sprengel, 1928, Anz. Schadlingsk., 4 : 25; Энгельгардт, 1928, Защ. раст., 5 : 51; Луч-
ник, 1929, Изв. Ставр. Ст, защ. раст., И : 23; Козякина, 1930, Защ. раст,, 7 : 88;
Kuskov, 1933, Beitr. Kunde EstL, 18 : 124; Лебедев, 1935, 36ipH. праць. екол., 1 : 62
Голова, грудь и брюшко у самцов серо-коричневые; ноги покрыты
длинными пушистыми волосками; передние крылья желто-коричневые
с серо-коричневым рисунком; базальная линия почти не развита; вторая
поперечная линия в виде двух темных полос, слитых в костальной поло-
вине крыла; срединное поле равно по ширине базальному и внешнему,
слабо расширенное в середине; почковидное пятно несколько светлее
общего фона крыла, тонко очерчено коричневым; третья поперечная
линия в виде растушеванной, слабо зубчатой по жилкам полосы; у дор-
зального ее края с внешней стороны ярко белое пятно, окруженное
темно коричневым; внешнее поле несколько светлее общего фона крыла
с разбросанными охристо-желтыми пятнами; подкраевая линия в виде
тонкой темной и светлой полоски, мелко волнистая, в дорзальной части
нередко имеющая белый треугольный штрих; бахрома шашечная —
темно коричневая и охристая; задние крылья однотонные коричнево-
рыжие с слабыми штрихами вдоль жилок; бахрома на них охристая
с немногими желтыми штрихами; нижняя сторона светложелтая с чер-
ной и Темной Поперечными полосами; дискальное поле и костальный
край черные; бахрома желтая; брюшко того же цвета. Самки с сильно
укороченными крыльями серого цвета с примесью желтоватых тонов;
тело покрыто нежными волосками; усики короткие с короткими же-
сткими гребешками; крылья в виде ничтожных придатков, задние короче
передних; ноги вполне развиты, покрыты желтыми волосками, коготки
с коротким зубцом на вентральной стороне.' Размах крыльев самцов
25—30 мм, длина тела самки 10—20 мм.
Uncus тонкий и длинный, слабо изогнутый, сидит на широком осно-
вании, разрастающемся латерально, отделен широким швом от девятого
5. ORGYIA
279
Рис. 121. Orgyia antiqua L. Гениталии:
a :— самца б — самки.
сегмента; тергит последнего довольно длинен; fultura inferior почти
округлая, с дорзальной стороны имеет вырез почти треугольной формы;
valva довольно длинная, базальная половина ее более сильно склероти-
зована, широкая, с дорзальной стороны несет длинный отросток, направ-
ленный дистально, искривленный дорзально; дистальная половина valva
мембранозная; penis расширенный в дистальной части. Bursa copulatrix
мембранозная, почти шаровидная, маленькая, без скульптуры на поверх-
ности; ductus seminalis отходит от ее дистальной части; ductus bursae
короткий и тонкий, со складчатыми стенками; ostium ductus bursae
окружен склеротизованным валиком; восьмой сегмент очень короткий,
несет длинные и тонкие apophyses • anteriores, превышающие длину
ductus bursae; apophyses pos-
teriores также тонкие и длин-
ные, короче первых; papillae
anales короткие и узкие.
Яйца беловато-желтые, при-
плюснутые в дорзовентральном
направлении; поверхность их
матовая, несет скульптуру в
виде мелких точек; дорзальная
поверхность ворончато вдавле-
на, с точкой посредине; диа-
метр 0.3—0.4 мм.
Гусеницы старших стадий
•серого цвета с красными бо-
родавками по спине и бокам;
по сторонам первого сегмента
по пучку длинных и перистых
волосков, сидящих на красных
бородавках; первые четыре
брюшных сегмента несут с дор-
зальной стороны недлинные
пучки желтых волосков, слабо
перистых, сидящих на склеро-
тизованных темных пластинках;
по сторонам первого брюшного
сегмента по одному пучку белых
перистых волосков, сидящих на красных бородавках; на втором брюш-
ном сегменте по сторонам большие пучки сильно перистых темных волос-
ков, торчащие в стороны; с дорзальной стороны восьмого брюшного
сегмента большой непарный пучок перистых: волосков разной длины,
более длинных в дистальной части пучка; вентральная сторона тела свет-
ложелтая, ноги желто-белые, голова черно-коричневая.
Куколка темнокоричневая, покрытая негустыми желтыми волосками;
поверхность кожи блестящая, со скульптурой на брюшных сегментах
в виде небольших рубчиков; cremaster в виде конического выступа, несу-
щего на дистальном конце недлинные, но многочисленные крючья и
имеющего на поверхности продольные бороздки.
Изменчивы размеры особей и окраска; последняя варьирует от желто-
до темнокоричневой; наиболее светлые особи, имеющие почти белое
срединное поле, известны из северной Монголии — форма mongolica
Alph. (in litt.). Темные формы описаны — одна из Средней Азии (confinis
Gr. Gr.) и еще более темная, черно-коричневая dilutior Schultz и infer-
280
I. ORGYINAE
nalis Rbl., также черно-коричневая, но без охристых штрихов, известны
из средней Европы.
Распространение. Средняя и Западная Европа (Пиреней-
ский полуостров, Альпы, Германская низменность, Карпаты); Сканди-
навия (Швеция, Лапландия, Норвегия, Финляндия); Британские острова;
Исландия; в Европейской части СССР по всей лесной зоне и степи
(Выг-озеро, Петрозаводск, Рига, Таллин, Вильнюс, Нарва,' Ленин-
град, Псков, Ярославль, Москва, Тула, Орел, Курск, Полтава, Харьков,
Херсон, Одесса, Таганрог, Ки-
шинев); Крым (Севастополь,
Бельбек, Альма, р. Аксу); Се-
верный Кавказ (Ставрополь,
Ростов); Поволжье (Саратов,
Рис. 122. Orgyia antiqua L. Хетотаксия перед-
не- и среднегруди и брюшных сегментов I,
VI, VII и VIIL гусеницы I стадии (а)
и взрослой (б).
Ростов); Поволжье (Саратов,
Сарепта, Куйбышев, Казань);
Закавказье (Боржоми, Атехури,
Лагодехи, Батуми, Ленкорань);
Приуралье (Чкалов, Уфа, Гу-
берли). В Сибири на севере
распространение неясно, но,
судя по некоторым нахожде-
ниям (Обь-Енисейский канал.
Подкаменная Тунгуска, Улус,
Якутск), — вероятно обширно.
По лесостепи и югу лесной зо-
ны Сибири всюду (Омск, Томск,
Тайшет, Чулым, Канск, Бар-
наул, Минусинск); Алтай; се-
верный Казахстан (Семипала-
тинск Акмолинск, Кокчетав);
Кузнецкий Ала-тау (Тельбес);
Саяны; Прибайкалье (Витим,
Иркутск); Забайкалье; При-
амурье (Хабаровск, Нико-
лаевск, Благовещенск); южное
Приморье (Ворошилов-Уссу-
рийский, Владивосток, Сучан);
северная Монголия (Хангай,
Улан-батор); Японские острова
(Мацуяма); Маньчжурия (Кван-
тунг); Корея; Тибет (Куку-
нор, Амдо).
Биология и экология. Зимуют яйца на поверхности ко-
кона, открыто, на стволах деревьев; отрождение гусениц проходит растя-
нуто, в мае—июне. Вначале гусеницы питаются почками, позже листь-
ями; гусеницы многоядны, питаются различными древесными растениями
и кустарниками — Salicaceae (Salix, Populus), Betulaceae (Betula, Alnus,
Corylus, Carpinus), Fagaceae (Fagus, Quercus), Rosaceae (Pirus, Prunus,
Malus, Crataegus etc.), Tiliaceae, Aceraceae, Ericaceae, реже травянистыми;
отмечено питание хвойными растениями — Pinaceae (Picea, Pinus).
Оптимум роста гусениц — при температурах ниже 20°; при высоких
температурах наблюдается сильное развитие эпизоотий — заболевание
полиэдренной болезнью. Предпочитаемыми растениями, рост на кото-
рых сопровождается наименьшей смертностью от болезней, являются
5. ORGYIA
281
виды берез. Развитие гусениц до июля; окукление в щелях на стволах
и ветвях деревьев и в листьях в рыхлом коконе; лёт бабочек в июле —
начале августа; самцы отыскивают самок днем. Обычно дает второе поко-
ление, а в южных частях ареала и третье, всегда частичные. Лёт бабочек
второго поколения в начале сентября. Яйца диапаузируют и зимуют.
Дает гибриды с американскими видами Orgyia Ochs, и с азиатской
О. thyellina Btl. (см. предыдущий вид); с последним видом, кроме упомя-
нутых выше случаев, известен гибрид wintersteini Wohnig (1927, Intern,
entom. Ztschr., 21 : 38) —О. thyellina Btl. (самец) — О. antiqua L. (самка).
Из американских видов гибридизирует О. vetusta Bd.; получен гибрид
от самца этого вида и самки О. antiqua L., называемый vetantiqua Kleem.
(Kleemann, 1937, Entom. Ztschr., 23 : 348). Известен гибрид c leucostigma
Sm. et Abb.: самец этого вида и самка antiqua L., называемый leucanti-
qua Kleem. (Kleemann, 1929, Intern, entom. Ztschr., 23 : 348; Kleemann,
1933, там же, 27 : 22). Гибриды бесплодны, хотя получены в имагиналь-
ной фазе. Все гибриды получены только в лабораторных условиях.
Результаты гибридизации этого вида интересны для систематики, так
как гибридизация удается только в пределах определенной группы ви-
дов — дальневосточного О. thyellina Btl. и американских, к которым,
включая также и китайские виды parallela Gaede и immaculata Gaede,
О. antiqua L. наиболее близок морфологически. Многочисленные попытки
гибридизации О. antiqua L. с О. gonostigma L., не говоря уже про другие
виды, не дали положительных результатов; не были получены даже
оплодотворенные яйца, несмотря на то, что спаривание (в многочислен-
ных моих опытах) постоянно наблюдалось, но только самцов О. gono-
stigma L. с самками О. antiqua L., а не наоборот.1
Вредит лесным породам, плодовым садам и Декоративным посадкам;
отмечался вред не только лиственным, но и хвойным породам — ели
и сосне [Нюслин и Румблер (Niisslin und Rhumbler, 1927)], в Америке
в наибольшей мере вредит декоративным растениям [Трегёрне (Treherne,
1916)]. В садах сильный вред приносит абрикосам, яблоне, сливе на юге
СССР, где специально отмечен на Украине (Аверин, 1913), в Крыму
(Мокржецкий, 1898), в Закавказье (Уваров, 1919) и на Дальнем Востоке
(Энгельгардт, 1928); отмечен как вредитель ивы в средней полосе Евро-
пейской части СССР (Плигинский, 1915).
Известны многочисленные паразиты, особенно для гусениц: из Рго-
ctotrupidae — Telenomus dalmani Rtzb.; из Braconidae — Apanteles for-
mosusWesm., Eulophus ramnicornis Geoffr., Microgaster solitarius Rtzb.,
M. affinis Ns., BJiogas geniculator Ns.; из Ichneumonidae — Campoplex
carbonarius Rtzb., C. nigripes Gr., C. cultrator Gr., Ichneumon semirufus
Wesm., Pezomachus cursitans F., P. agilis Fst., Sagaritis raptor Zett.,
Pimpla instigator Pz., P. stercorator Gr., P. examinator F., Epiurus inqui-
sitor Scop., Casinaria nigripes Gr., Mesochorus stigmaticus Br., Ani-
lasta ebenina Gr.; из Diptera — Carcelia gnava Mg., C. excisa Fall.,
C. lucorum Mg., Exorista sp., Phryxe vulgaris Fall., Larvivora impotens
Rd., L. vidua Mg.
* 3. Orgyia gonostigma L. (рис. 123, 124).
Linne, 1767, Syst. Nat., add. (2), Г (2) : 826 (Phalena); Aurivillius, 1891, Nordn.
Fjar., 1 : 92, t. 25, fig. 2; Staudinger, 1901, Cat. Lep., 1 : 114; Spuler, 1908, Schmett.
1 По исследованию Беляева (1930), эти виды Orgyia Ochs, имеют значительные
отличия в числе хромозом. Беляев дает следующие числа для гаплоидного числа хро-
мозом: thyellina Btl. — 11, antiqua L. — 14, leucostigma Sm. et Abb. — 28, gonostigma
F. — 30 и ericae Germ. — 30.
282
I. ORGYINAE
Eur., 1 : 126, t. 25, fig. 14; Matsumura, 1909, Thous. Ins. Jap. Emp., Suppl., 1 : 66,
t. 10, fig. 20; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 114, t. 21, fig. 6; Strand,
1910, Seitz, Grossschmett., 2 : 117, t. 19; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) :
416; Petersen, 1924, Lep. Fauna Estl., ed. 2, 1 : 139; Osthelder, 1926, Schmett. Siid-
bayerns, 1 (2) : 196; Hering, 1932, Tierw. Mitteur. : 483; Bryk, 1934, Lep. Cat., 62 :
64; Warnecke, 1936, Entom. Rundsch., 53 : 133; Nordstroem och Wahlgren, 1937, Svenska
Fjar. : 65, t. 13, fig. 6; — antiqua Scopoli, 1763, Entom. Carn. : 198 (non Linne); — re-
cens Hiibner, 1816, Verz. bekannt. Schmett. : 161; — approximans Butler, 1881, Trans.
Entom. Soc. Lond. : 10; —etrusca Verity, 1904, Boll. Soc. Entom. ItaL, 36 : 187 (Orgyia
prisca Bd., forma); — approximans Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 416;—
jlaoescens Maslowsky, 1923, Polsk. Pismo ent., 2 : 133.
Биология. Spuler, 1908, Raupen, t. 15, fig. 24; Turati, 1914, Atti Soc. Sci.
Nat. ItaL, 53 : 541; Lederer, 1923, Handb.prakt. Entom., 3 : 148; Reh, 1925, Sorauer,
Handb. Pflanzenkrankh., 4 : 444; Энгельгардт, 1928, Защ. раст., 5 : 51; Бассель, 1929,
Изв. Сиб. ст. защ. раст., 3—6 ; 150; Щеголев, 1930, Насек, вред. масл. культ. : 116;
Щеголев, 1931, Насек, поврежд. масл. культ.: 124, 209.
Голова, грудь и брюшко серо-коричневые; ноги покрыты длинными
волосками; передние крылья темнокоричневые с черными и белыми
элементами рисунка и охристыми неправильными пятнами; рисунок
четкий; базальная линия в виде темной Прямой полосы, оттененной
белым; базальное поле темное, коричневое с неправильно разбросанными
белыми чешуйками в костальной половине, дорзальная его половина
с нерезкими желтыми штрихами; вторая поперечная линия в виде тонкой
белой и черной полосы, плавно выгнутая к внешнему краю, слабо вол-
нистая; срединная линия в виде почти прямой сильно растушеванной
по середине полосы, сливающаяся в темной части срединного поля; ко-
стальная часть этого последнего с многочисленными белыми чешуйками;
почковидное пятно ясное, очерченное белым, посредине черно-коричневое
с тонким желтым штрихом; третья поперечная линия в виде черной диф-
фузной полосы, слабо и неясно зубчатой по жилкам; у дорзального края
в срединном поле белое пятно в форме запятой; костальная часть средин-
ного поля охристо-желтая; подкраевая линия у костального края (белая
далее коричневая, слабо волнистая; терминальная линия в виде тонкой
черной полоски; бахрома коричневая; снаружи черно-рыжая с немно-
гими белыми штрихами; задние крылья однотонные черно- или шоколадно-
коричневые с светлой желто-серой бахромой; нижняя сторона черно-
коричневая; внешнее поле передних крыльев светложелтое, почти белое
в апикальной части; усики самцов сильно гребенчатые, на концах гре-
бешков коротенькие щетинки в числе двух. Самки очень короткокрылы;
тело покрыто густыми короткими шелковистыми волосками, темносерое,
желтоватое или почти белое; усики очень тонкие с короткими выростами
на члениках; ноги покрыты белыми волосками, развиты хорошо; коготки
с зубцом на вентральной стороне. Размах крыльев самцов 25—35 мм,
длина тела самки 17—22 мм.
Uncus плоский с глубокой бороздой на дорзальной стороне, имеет
вид двух гребней, покрытых недлинными волосками, короткий, загнутый,
сросшийся с двумя лопастями, расположенными под ним и образованными
разрастанием латеральных выростов девятого сегмента и subscaphium,
отделен швом от девятого сегмента; tegumen короткий; saccus треуголь-
ный; valva почти треугольной формы, простая, усаженная в дистальной
части грубыми волосками, в базальной части образует складку, несущую
зубчики; fultura inferior небольшая, в виде сильно склеротизованной
небольшой пластинки; penis слабо расширен в дистальной части : соесшп
уплощен; pars inflabilis penis без инкрустаций. Bursa copulatrix мешко-
видная, расширенная в проксимальной части и суженная в дистальной,
5. ORGYIA
283
мембранозная, маленькая; ductus bursae короткий и мембранозный, уз-
кий, постепенно переходит в corpus bursae; ductus seminalis отходит
от дистальной части совокупительной сумки посредством небольшого
расширения цилиндрической формы; lamella antevaginalis короткая
и мягкая, густо усаженная волосковидными длинными шипиками чер-
ного цвета; lamella postvaginalis маленькая, голая; восьмой сегмент
узкий, неправильной формы, наиболее широк при основании; apophyses
anteriores длинные, изогнутые; длинные же и apophyses posteriores;
papillae anales короткие, в дистальной части округленные, покрытые
густыми щетинками.
Рис. 123. Orgyia gonostigma L. Гениталии:
а — самца, б — самки.
Яйца матово-белые, слабо блестящие, почти круглые, сплющенные
несколько дорзовентрально, вдавленные на дорзальной стороне, с точ-
кой; диаметр 0.3—0.4 мм.
Взрослая гусеница желто-серая, желтый цвет кожи располагается
в виде полос; вдоль спины широкая темная полоса; нижняя сторона тела
однотонно желто-серая; бородавки, покрывающие тело, черные, несут
пучки белых слабо перистых волосков; дорзальные бородавки, исклю-
чая первые четыре сегмента брюшка, несут короткие пучки чисто белых
сильно перистых волосков; первые четыре брюшные сегмента несут
четыре щетки коротких, желто-серых волос; первый сегмент груди несет
по сторонам длинные и тонкие пучки перистых на концах черных волос-
ков; восьмой сегмент брюшка с дорзальной стороны несет непарный та-
кой же пучок; брюшные и грудные ноги черные и желтые.
Куколка светлокоричневая с более светлыми межсегментальными
участками; склеротизованные пластинки покрыты недлинными желтыми
волосками; cremaster. в виде недлинного остроконечного выроста с ко-
роткими крючковидными выростами; кожа гладкая, блестящая, без
заметной (в лупу) скульптуры.
284
I. ORGYINAE
a
Изменчивость не велика; заметно меняется лишь размер особей в связи
с условиями личиночного питания. На востоке Азии известна темная
несколько более крупная и несколько более острокрылая форма, аррго-
ximans Btl.
Распространение. Средняя Европа к северу от Пиренеев (Биль-
бао)—Альпы, Германская низменность, Карпаты, долина Дуная, Висла;
локально в Средиземноморье (средняя Италия, Корсика); Британские
острова; южная Швеция;
Финляндия; запад Европей-
ской части СССР (Вильнюс,
Рига, Тарту, Нарва); далее
на восток к югу от линии
Петрозаводск — Молотов
всюду по лесной зоне, лесо-
степи и отчасти по степи
(Ленинград,Псков, Калинин,
Москва, Калуга, Брянск,
Винница, Каменеп-Подольск,
Бендеры); Крым (точно не
известно и сомнительно);
Северный Кавказ; Закав-
казье (Ленкорань); Повол-
жье (Казань, Куйбышев);
Приуралье (Чкалов, Губер
ли); в Сибири северная гра-
ница не ясна, видимо всюду
по лесостепи и в южной по-
ловине лесной зоны (Омск,
Новосибирск, Томск, Мину-
синск, Тайшет,'Канск, Бар-
наул, Шадринск, Иркутск);
Саяны (Казыр-сук, Тукеек-
кем); Алтай (Немал, Томь);
Забайкалье (Сретенск); При-
амурье и Приморье (Раддев-
ка, Рейново-Джалинда, Ха-
Рис. 124. Orgyia gonostigma L. Хетотаксия пе-
редне- и среднегруди и брюшных сегментов
I, VI, VII и VIII гусеницы I стадии (а) и
взрослой (б).
баровск, Сидеми, Аскольд,
Сучан); южный Сахалин;
Японские острова (Хоккайдо,
Хондо); Корея.
Биология. Зимует гу-
сеница обычно в третьей стадии; зимовка поодиночке в шелковых
чех ликах, в сухих листьях; весной питание с наступлением тепла
(температура выше 5—10°); гусеницы многоядны, питаются—Sali-
caceae (Salix— разные виды, Populus tremula), Betulaceae (Betula, Car-
pinus, Alnus, Corylus), Fagaceae (Fagus, Quercus), Rosaceae (Rosa, Pirus,
Malus, Prunus, Sorbus, Rubus, Amelanchier, Crataegus); заметно пред-
почитаются ивы и розоцветные (плодовые деревья); окукление в конце
июля; самцы летают днем, отыскивая самок по запаху; менее активны,
чем у предыдущего вида; яйцекладка кучкой на поверхность кокона,
яйца лежат в несколько слоев и покрываются волосками с конца брюшка
самки; развитие яиц в июле в течение 14—-20 суток; выходящие из яиц
гусеницы расползаются и первое время интенсивно питаются и растут, за-
5. ORGYIA
285
тем после первых двух линек питаются слабо и впадают в диапаузу, оста-
ваясь на зимовку; при росте в условиях повышенной температуры (выше
25°) часть особей (на юге ареала всегда) дает второе поколение ине пере-
стает интенсивно питаться в третьей стадии; диапаузирующие гусеницы,
помимо поведения (завивки коконника), отличаются окраской; у них серая
окраска вместо желтой у растущих, не диапаузирующих особей. Вредит
плодовым деревьям — груше (Витковский, 1914) на Украине и яблоне
и сливе (Энгельгардт, 1928) на Дальнем Востоке.
Известны паразиты преимущественно личиночной фазы: из Ргос-
iotrupidae — Telenomus dalmani Btzb.; из Braconidae —Microgaster sp.;
из Ichneumonidae —Campoplex cultratdr Gr., Casinaria seniculaGr., Ichneu-
mon sp., Sagaritis declinator Gr.; из Diptera — Carcellia excisa Fall.,
Exorista affinis Fall., Larvivora larvarum L.
* 4. Orgyia dubia Tausch. (рис. 125).
Tauscher, 1806, Mem. Soc. Nat. Mose., 1 : 176, t. 13, fig. 3 (Bombyx); Kirby, 1892,
Syn. Cat. Lep. Het., 1 : 492; Staudinger, 1901, Cat. Lep., 1 : 114; Spuler, 1908,
Schmett. Eur., 1 : 128, t. 25, fig. 20; Strand, 1910, Seitz, Grossschmett., 2 : 119, t. 19 c.;
Powell, 1916, Oberthiir, Et. Lep. Comp., 12 : 272; Oberthiir, 1917, Et. Lep. Comp.,
13 : 37, t. 419, fig. 3553, 3555, 3556; Rothschild, 1920, Nov. Zool., 24 : 356, t. 1, fig. 1,
2, 3, 7, 9, 10, 42, 43; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 422; Schwingeschuss,
1931, Verh. zool.-bot. Ges. Wien, 81 : 18; Gaede, 1932, Seitz, Grossschmett., 2 Suppl.:
99; Hering, 1932, Tierw. Mitteleur. : 436; Amsel, 1933, Zoogeographica, 2 ; 99;
Rothschild, 1933, Nov. Zool., 38 : 324; Zerny, 1933, Iris, 47 : 63; Bryk, 1934, Lep. Cat.,
62 : 70; Hordhammer, 1934, Intern, entom. Ztschr., 24 : 87; — seleniaca Fischer-Wald-
heim, 1823, Entom. Imper. Ross., 2 : 249, t. 10, fig. 3; — alhagi Fischer-Waldheim,
1839, Bull. Soc. Nat. Mose., 1 : 117) t. 4, fig. 2; — splendida Rambur, 1842, Fauna
Andal., 1.15, fig. 3—6; — algiricum Lucas, 1849, Exped. Alg., 3 : 376, t. 3, f; — turcica
Lederer, 1853, Verh. zool.-bot. Ges. Wien, 2 : 117; — josephina Austaut, 1880, Natu-
raliste : 202, 212; — interrupta Grum-Grzhimailo, 1890, Mem. Lep. Rom., 4 : 555; —
judaea Staudinger, 1901, Cat. Lep., 1 : 114; — transcaspica Krulikowsky, 1906, Soc.
Entom., 21 : 50; — medionigra Ribbe, 1909, Iris, 23 : 219; — pricta Ribbe, 1909, там же,
23 : 219; — mancha Ribbe, 1909, там же, 23 : 219; — obliterella Strand, 1910, Seitz,
Grossschmett., 2 : 119; — isolatella Strand, 1910, там же, 2 : 119, t. 19, c; — umbripen-
nis Strand, 1910, тамже, 2 : 119, t. 19, c; — algirica Strand, 1910, также, 2 : 119; —
deserticola Powell, 1916, Oberthiir, Et. Lep. Comp., 12 : 165; — intermedia Powell,
1916, тамже, 12 : 267; — orana Powell, 1916, тамже, 12 : 267; — aurea Oberthiir, 1922,
тамже, 12 : 201, t. 534, fig. 4443, 4444; — arcerii Ragusa, 1923, Boll. Soc. Entom. Ital.,
55 : 19; — lunulata Hordhammer, 1934, Intern, entom. Ztschr., 24 : 87. fig. 6.
Биология. Rangnow, 1912, Intern, entom. Ztschr., 5 : 334; Сахаров, 1914,
Русск. энтом. обозр., 14 : 391, рис. 1—2; Chretien, 1916, Ann. Soc. Entom. France,
85 : 374; Lederer, 1923, Handb. prakt. Entom., 3 : 149; Reh,1925, Sorauer, Handb.
Pflanzenkrankh., 4 : 444.
Голова и грудь покрыты длинными оливково-серыми волосками,
сквозь которые просвечивает темный цвет кожи; этого же цвета волоски
покрывают и тергиты базальной половины брюшка; ноги покрыты длин-
ными волосками того же цвета; дистальная часть брюшка покрыта свет-
ложелтыми волосками; губные щупальца сильно укорочены и спрятаны
в густом волосяном покрове головы; усики сильно гребенчатые, серые,
членики их темносерые; передние крылья желто-черные, полосатые,
рисунок сильно видоизменен; базальное поле оливково-черное с жел-
тыми неправильно разбросанными чешуями, более темное посредине;
вторая поперечная линия в виде очень широкой черной полосы, почти
прямая, посредине слабо выгнутая и дающая у дорзального края крыла
сильный изгиб кнаружи, достигающий третьей поперечной линии; сре-
динное поле желтое, неправильной формы, посредине узкое; почковидное
пятно черное, почти прямоугольное, резко выделяется на желтом фоне;
•третья поперечная линия в виде широкой полосы, черная, плавно изо-
286
I. ORGYJNAE
гнутая, особенно широкая в дорзальной половине крыла, окаймлена сна-
ружи желтым; подкраевая линия в виде тонкой мелкозубчатой по жил-
кам полоски; терминальная линия не развита; бахрома желтая; задние
крылья желтые с черным костальным краем и черной каймой по внеш-
нему краю, не доходящей до заднего угла крыла; базальная часть зад-
него крыла несколько затемнена; бахрома желтая; нижняя сторона
крыльев желтая с черными костальным и внешним краями задних
крыльев, черным внешним краем на передних и черными дискоидаль-
ными пятнами на обеих парах крыльев. Самки бескрылые, лишены даже
рудиментов крыльев, без ног и нормально развитой головы; тело по-
крыто короткими, серовато-желтыми волосками. Размах крыльев сам--
цов 20—30 мм, длина тела самки 10—20 мм.
Uncus гладкий, очень вздутый в средней части, сильно выгнут, ди-
стальный конец его заострен; девятый сегмент короткий, особенно,
в области pleurae; saccus хорошо развит; fultura inferior в форме плоского
склерита дугообразной формы; valva почти треугольной формы, сильно
выгнута вентрально; дистальный
r-щ ----X к°нец ее приподнят; базальная
\ А —-у / J половина valva широкая, на дор-
---------------—==«£^-.-.. у J зальном крае имеет небольшое
утолщение; вся поверхность valva
С -------------------------------— Xхнесет мелкие волоски; penis имеет
ш Д™нный (около ’/;( части penis)
J[r Ж coecum, изогнутый вентрально;
AyJA г '-Jr. дистальная половина penis расши-
рена, инкрустации на pars inflabi-
lis в виде мельчайших шипи-
Рис. 12а. Orgyia dubia Tausch. Гениталии ков
самца. ' ,
Яйца почти круглые, слабо-
сплюснутые дорзовентрально, чисто белые, матовые; диаметр 0.3—Я.4 мм.
Взрослые гусеницы имеют желто-серую кожу, нередко переходящую
в черно-серую, покрыты красными бородавками; железы седьмого и вось-
мого брюшных сегментов карминно-красные; на первых четырех брюш-
ных сегментах по большой кисточке густых волосков черных или жел-
тых, 'бархатистых; часть волосков в этих кистях белые, перистые; на
дорзальной стороне восьмого брюшного сегмента непарный длинный
пучок белых и черных перистых волосков; первый грудной сегмент
несет на борадавке v. с каждой стороны по латеральному длинному
пучку белых перистых волосков; грудные ноги черно-серые, брюшные
черно-красные.
Наблюдается значительное разнообразие в окраске самцов и, в част-
ности, в размерах (в ширине) окраски черных и желтых полос на крыльях;
желтые особи с узкими черными полосами описаны как orana Powell,
judaea Stgr., turcica Ld., algirica Lucas, темные формы — как josephina
Aust.; темные же особи известны из Иркутска —baicalensis forma nov.
и из Семиречья —heptapotamica, forma nov. Вариации окраски велики
в пределах одних и тех же мест, что отмечено, например, для Средизем-
номорья — формы obliterella Strd., isolatella Strd., почему выделение под-
видов на основании этих признаков затруднительно; часто Основой вариа-
ций являются, видимо, местные экологические влияния. Тем не менее не-
сомненно наличие ряда географических, локальных, хотя и мелких форм.
Распространение. Средиземноморье — Испания (Андалу-
зия), южная Франция, Апеннинский.'полуостров, Далмация, Балканский.
5. ORGYIA
287
полуостров (Греция, Болгария), острова Средиземного моря (Сицилия,
Корсика), северная Африка, Алжир (Батна, Оран), Тунис, Марокко,
Египет (Александрия, Аскван), Сирия (Хайфа, Лизан), Палестина;
Малая Азия (Аксехир); северный Иран; Закавказье (Армения: Ордубад;
Азербайджан, Дербент); южное Поволжье (Астрахань, Садовое, Сара-
тов, Сарепта, о. Бирючий); южная Украина (Ворошиловград, Полтава);
Ростов, Таганрог; Северный Кавказ (Ставрополь), Туркмения (Ашха-
бад, Атрек); Мангышлак; Кара-кумы; Семиречье (Или); южный Алтай
(Зайсан); Прибайкалье (Иркутск); по сведениям Таушера (1806), этот
вид встречается под Москвой, но никаких более поздних указаний на его
наличие здесь нет; фактический материал из-под Москвы также мне
не известен.
Биология. Зимуют яйца, отложенные внутрь кокона; гусеницы
выходят в течение апреля—мая; питание гусениц разными растениями,
но преимущественно мотыльковыми Leguminosae (Alhagi camelorum,
Glycyrrhysa, Hedysarum alhagi), возможно также Salicaceae (Salix, Popu-
lus), Fagaceae (Quercus ilex), Rosaceae (Cornus), Chenopodiaceae (Kochia
sedoides, K. prostrata, Salsola, Atriplex, Helimocnemis), Compositae
(Artemisia fragrans). Окукление на растениях в большом желтоватом
коконе; развитие куколок продолжается около двух недель. Самцы ле-
тают днем, отыскивая самок по запаху. Развитие яиц лётом в течение
9—16 суток; дает несколько поколений в течение года. По указаниям
Рангнова (1912), у этого вида наблюдается партеногенез; проверка мною
этого факта на нескольких самках дала отрицательный результат; не-
сомненно, что естественный партеногенез у этого вида отсутствует.
Отмечены паразиты из I chneumonidae— Gambrus adustus Gr. и
Pimpla instigator F.
* 5. Orgyia ericae Germ. (рис. 126).
Germar, 1818, Ins. Eur., 8 : 17; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 494; Auri-
villius, 1891, Nordn. Fjar. : 92; Spuler, 1908, Schmett. Eur., 1 : 127, t. 25, fig. 15;
Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 114; Strand, 1910, Seitz, Grossschmett.,
2 : 118, t. 19, b; Eckstein, 1915, Schmett. Deutsch., 32 : 41; Swinhoe, 1923, Ann. Mag.
Nat. Hist., 11 (9) : 417; Petersen, 1924, Lep. Fauna Estl., ed. 2, 1 : 140; Cleyhills, 1928,
Not. Eptom., 8 : 62; Hering, 1932, Tierw. Mitteleur.: 483; Bryk, 1934, Lep. Cat., 62 :
69; Gronblom, 1936, Acta Soc. Fauna et Flora Fennica, 58 : 12; — antiquoides Hiibner,
1818, Eur. Schmett., 2, fig. 279, 280; — intermedia Frivaldsky, 1866, Magy Tud. Acad.
Eskon., 11 (4) : 149, t. 5, fig. 1; —zimmermani Graeser, 1888, Berl. entom. Ztschr.,
22:122;—prisca Leech, 1890, Entomologist, 23:111 (non Staudinger); — leechi
Kirby, 1892, Syn. Cat. Lep. Het., 1 : 495 (nom. nov.).
Биология. Bartel, 1914, Mitt. Munch, entom. Ges., 5:17; Lederer, 1923,
Handb. prakt. Entom., 3 : 149; Kuskov, 1927, Beitr. Kunde Estl., 11 : 80; Kuskov,
1931, там же, 17 : 67; Kuskov, 1933, там же, 18 : 131; Nordmann, 1937, Acta Soc.
Fauna et Flora Fennica, 60 : 648, fig. 17, 18.
Голова, грудь и брюшко коричнево-охристые и серые; усики желто-
коричневые, членики почти чисто желтые; тело покрыто длинными гус-
тыми желто-коричневыми волосками; передние и задние крылья одно-
тонные охристо-коричневые с очень слабым рисунком; при хорошо раз-
витом рисунке базальная линия намечена в виде темного штриха; базаль-
ное поле однотонное; вторая поперечная линия в виде широкой расту-
шеванной полосы, слабо выгнутая и извитая в дорзальной части крыла,
дает штрих к третьей поперечной линии; срединное поле узкое,' уже
базального и внешнего, в костальной части его фиолетово-серое тре-
угольное пятно; почковидное пятно намечено темным полулунным штри-
хом, окружено серо-седой тонкой полоской; третья поперечная линия
288
I. ORGYINAE
в виде неширокой темной полосы, дает двузубчатый выгиб в области
медиальной системы; подкраевая линия в виде слабой растушеванной
полоски, несколько затемненной в области кубитальной системы, ватой
части крыла снаружи линии четкое белое пятнышко; терминальная полоса
очень слабая; бахрома желто-коричневая; задние крылья темнее перед-
них, однотонные коричнево-желтые с светлой желтой бахромой; нижняя
сторона желто-коричневая с широкой растушеванной поперечной линией.
Самки имеют ничтожные рудименты крыльев; усики в виде нитевидных
придатков со слабо дифференцированными члениками; ноги короткие
со слабыми рудиментарными лапками, несущими простые коготки; тело
покрыто короткими пуши-
- л стыми белыми волосками.
Ч . ;} Размах самцов 24—30 мм,
Л длина самок 10—15 мм.
к — —___,—Uncus большой, сильно
А. /Ж / ' 1 изогнутый, на дистальном
-----(/ \л I х—конце расщеплен, с вент-
\у. v ральной стороны несет
короткие щетинки; scap-
V hium сильно развит, изо-
\/[ А,' гнутый, треугольный, на
дистальном конце зао-
стренный; tegmnen вздут,
Рис. 126. Orgyia ericae Germ. Гениталии самца. большой, со-слаоым швом
посередине; saccus малень-
кий; valva очень маленькая, в виде короткой лопасти, с небольшим
отросточком на дорзальном крае; penis длинный, сильно расширенный
в дистальной части, в середине более тонкий, в проксимальной части
округлен, вздут; соесиш не длинный.
Яйца очень похожи на яйца Orgyia antiqua L., но мельче их.
Взрослая гусеница шафранно-желтого цвета с черными продоль-
ными полосами; голова черная; первый грудной сегмент несет по бо-
кам два пучка перистых волосков черного цвета; первые четыре
брюшных сегмента несут по одному непарному пучку; кисти волос
беловато-желтого цвета, железы на шестом и седьмом сегментах брюшка
желтого цвета; восьмой сегмент брюшка несет длинный непарный
дорзальный пучок волосков черного цвета, перистых; ноги черные; боро-
давки желто-серые, покрыты волосками того же цвета.
Рисунок на крыльях самцов часто утрачивается, особенно бело-фио-
летовое пятно (под костальным краем в срединном поле, а также белое
пятнышко); такие однотонные особи с обедненным рисунком являются
наиболее обычными; из средней Европы описана форма intermedia Friv.,
имеющая ясный рисунок, но несколько более темная; описанная Гре-
зером (1888) О. zimmermani Graes. из Благовещенска представляет яркую
форму ericae Germ. — вида, распространенного в Китае и Забайкалье,
хотя и не частого на территории восточной Азии, — но не синоним
antiqua L., как полагал Брик (1934).
Распространение., Северо-западная Европа (Германская
низменность, Штеттин, Померания, преимущественно по верещатникам);
южная Скандинавия (Швеция); Финляндия; в Европейской части СССР
на севере локально (Каунас, Таллин, Рига, Нарва, Ленинград, Псков,
Луга); на юге Европы чаще — долина Дуная, Молдавия, Укра-
ина (Чернигов, Каменец-Подольск, Полтава, Харьков); Крым (Алупка);
5. ORGYIA
289
Северный Кавказ (Кисловодск, Ларс); Закавказье (Дербент, Баку);
Поволжье (Казань); Приуралье (Уральск, Чкалов); Алтай (Онгудай);
Сибирь (Тобольск, Обь-Енисейский канал); Забайкалье; Приамурье
(Благовещенск); Китай (Дадзянлу, Квансиен).
Биология. Зимуют яйца, отложенные, на поверхность кокона;
выход гусениц весной, растянут; гусеницы растут в течение июня и июля,
питаясь разными растениями, преимущественно кустарниками и травя-
нистыми — Ericaceae (Erica tetralix, Calluna vulgaris, Vaccinium uli-
’inosum, Andromeda polyf lia), Salicaceae (Salix, Populus), Myricaceae
(Myrica gale), Betulaceae (Betula, Alnus), Fagaceae (Quercus), Rhamnaceae
(Rhamnus pallasi), Leguminosae (Alhagi camelorum, Onobrychis), Tama-
ricaceae (Tamarix). Окукление в рыхлом желто-белом коконе тут же на
растениях. Дает на севере одно, на юге ареала два поколения в сезон.
Заселяет открытые территории, на севере преимущественно верещатники
и торфяные болота. Распространение локальное. Известны паразиты
Apantelus liparidis Bche и Microgaster sp. из Bracoaidae, Anilasta
ebenina Gr. и Campoplex sp. из Ichneumonidae и Pales pavida и Lar-
vivora larvarum L. из Larvivoridae.
6. Orgyia trigotephras Bd. (рис. 127).
Boisduval, 1929, Index Meth. Eur. Lep. : 46; Boisduval, 1834, leones Lep., 2 : 140,
t. 61, fig. 3—5; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 494; Staudinger, 1901, Cat. Lep.,.
1 : 114; Spuler, 1908, Schmett. Eur., 1 : 127, t. 25, fig. 17; Ribbe, 1909, Iris, 23 : 218;
Strand, 1910, Seitz, Grossschmett., 2 : 117, t. 19, e; Rothschild, 1917, Nov. Zoo’l.,
24:354; Wagner, 1921, Intern, entom. Ztschr., 15 : 18, fig. 1; Swinhoe, 1923, Ann. Mag.
Nat. Hist., 11 (9) : 416; Rothschild, 1933, Nov. Zool., 38 : 325; Reisser, 1933, Eos,
9 : 216; Schneider,» 1936, Entom. Ztschr., 50 : 191; — Corsica Boisduval, 1834, leones
Lep., 2 : 142, t. 61, fig. 6—8; — ramburi Mabille, 1866, Ann. Soc. Entom. France,
6 (4) :557, t. 6, fig. 6; — anceps Oberthiir, 1884, Et. Entom. Comp., 9 : 37, t. 3, fig. 5; — si-
cula Staudinger, 1901, Cat. Lep., 1 : 114; orientalis Staudinger, 1901, там же, 1:114 —
transiens Stauginger, 1901,' там же, 1 : 114; — calabra Stauder, 1916, Ztschr. wiss.
Insektenbiol., 12 : 110, t. 5, fig. 8; — sicula Turati, 1916, там же, 12 : 110; — sebdo-
riensis Oberthiir, 1916, Et. Lep. Comp., 12 : 276; — nobli Oberthiir, 1916, там же,
12 : 276; —sebdonensis Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 419; — dilu-
tior Babacek, 1926, Ztschr. Oesterr. entom. Ver. Wien, 11 : 35.
Голова и вентральная сторона груди фиолетово-серые, дорзальная
сторона груди желто-серая; передние и задние крылья коричнево-серые
и коричнево-черные, рисунок редуцирован; базальная линия в виде сла-
бого штриха; вторая поперечная линия в виде немногих тонких, разроз-
ненных серо-фиолетовых штрихов; в костальной части срединного поля
почти треугольное неправильно растушеванное серо-фиолетовое пятно,
лежащее в костальной части крыла и почти сливающееся со светлым
остатком почковидного пятна; ниже этого пятна в срединном поле тем-
ный растушеванный штрих; третья поперечная линия в виде тонкой ко-
ричнево-черной полосы, выгнутой* кнаружи под костальным краем, далее
почти прямой; подкраевая линия начинается фиолетово-белым пятном
под костальным краем, далее идет, прерываясь в виде тонкой бело-фиоле-
товой полоски; терминальная линия тонкая темнокоричневая; бахрома
однотонная тоже темнокоричневая; задние крылья однотонные шоко-
ладного цвета с бахромой того же цвета; нижняя сторона однотонная,
желто-коричневая. Самки с ничтожными рудиментами крыльев; усики
очень тонкие с волосками на члениках вместо гребешков; ноги рудимен-
тарны, но членики еще есть, с небольшими рудиментарными коготками
без зубцов на них; тело покрыто белыми шелковистыми волосками. Раз-
мах самцов 20—25 мм, длина самок 10—15 мм.
19 Фауна СССР, И. В. Кожанчнков.
290
1. ORGYINAE
Uncus толстый, с гребнем на дорзальной стороне, сильно загнут на-
дистальном конце, с раздвоенным концом и с бороздой на дорзальной
стороне; tegumen очень большой, разделен швом лишь при основании
unciis; девятое кольцо очень короткое, в вентральной части образует
очень маленький треугольный saccus; scaphium большой, сильно склеро-
тизован и срастается с основанием uncus; fultura inferior в виде до-
вольно широкой пластинки, изогнутой дугообразно; valvae очень корот-
кие и маленькие, конические, с небольшим выростом на дорзальной
стороне, покрыты мягкими и короткими волосками, особенно обильно
в дистальной части; penis почти прямой, длинный, расширенный на
дистальном конце и косо срезан; coecum около одной трети длины penis.
Наибольшее внешнее сходство и сходство структур гениталий
с О. ericae Germ.
.----Распространение.
,..<1 -Ju Пиренейский полуостров (Испа-
ния, Португалия); южная Фран-
- ция (Монпелье); Апеннинский
полуостров (Тоскана, Флорен-
<ч——----ция, Калабрия); острова Среди-
АЖУ земного моря (Сицилия, Кор-
сика); северная Африка (Ма-
рокко, Тунис); Сирия, Пале-
Рис. 127. Orgyiatrigotephras Bd. Гениталии стина; Малая Азия; возможно
самца. нахождение в Закавказье.
Биология. Зимуют
яйца; гусеницы многоядны, преимущественно в июне—июле на травя-
нистых растениях и кустарниках — Leguminosae (Genista Corsica, G.
myrtifolia, Spartium lobelii), Rosaceae (Rosa), Fagaceae (Quercus ilex,
Q. coccifera); окукление на растениях.
* 7. Orgyia prisca Stgr. (рис. 128).
Staudinger, 1887, Stett. entom. Ztg., 48 : 95; Grum-Grzhimailo, 1890, Mem. Lep.
Rom., 4 : 556; Staudinger, 1901, Gat. Lep., 1 : 114; Strand, 1910, Seitz, Grossschmett.,.
2 : 118, t. 22, c; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 417; Bryk, 1934, Lep.
Cat., 62 : ,68 (syn.). .
Более ширококрылая, чем О. trigotephras Bd. и О. ericae Germ, и имеет
более стройное тело; голова, грудь и брюшко серо-охристые; усики сам-
цов сильно гребёнчатые, гребешки по длине более ширины головы; ноги
покрыты длинными густыми волосками; передние крылья серо-коричне-
вые, рисунок нерезкий, нередко почти редуцирован; базальная линия
не развита; базальное поле с примесью большого количества белых че-
шуй; вторая поперечная линия в виде очень широкой выгнутой полосы
посредине темной, по сторонам бело-серой; срединное поле узкое в ко-
стальной половине и у дорзального края; светлое пятно сливается с неяс-
ным почковидным пятном, имеющим темные штрихи; третья поперечная
линия выгнута к внешнему краю, светлосерая, нерезкая, слабо зубчатая
по жилкам; подкраевая линия очень слабая, сильно растушеванная,
постепенно переходящая в общий тон внешнего поля; у дорзального края
крыла с внешней стороны этой линии белое нерезкое пятно; терминаль-
ная линия в виде очень тонкой коричневой полоски; бахрома темно-
коричневая, однотонная, темнее общего фона крыла; задние крылья
значительно светлее передних, рыже-коричневые; бахрома желтая; ниж-
няя сторона желто-рыжая, одного цвета на обеих парах крыльев, свет-
5. ORGYIA
291
лое пятно просвечивает под костальным краем в области срединного
поля. Самки имеют рудиментарную голову, усики почти совершенно
утрачены, слабо развиты также глаза; ноги есть, но очень маленькие
и слабые, лапки имеют очень маленькие членики, коготки есть, простые;
крылья почти совершенно утрачены, имеют вид ничтожных лопастей,
несущих щетинки; тело покрыто тонкими беловато-серыми волосками.
Размах самцов 25—28 мм, длина самок 10—15 мм.
Uncus толстый, высокий, с глубокой бороздой с дорзальной стороны,
дистальный конец его острый и двойной; основание uncus широкое, раз-
растается рядом с девятым сегментом; девятый сегмент отделен от uncus
глубоким швом; saccus мембранозный, небольшой; anus несет scaphium
в виде двух располо-
женных латерально пла- ---X
стинок; valvae очень \ \ А i
короткие, в виде про- Vjx.
стых, слегка заострен- ----------
ных лопастей, не имею- \. _______ \\А
щих инкрустаций; ful-
tura inferior в видедуго-
образно изогнутой пла- Л/ г Д
стирки, лежащей с вен-
трал' ной стороны и с
боков enis; penis впро- п .оо си.
„ r г Рис. 128. Orgyia prisca Stgr. Гениталии самца.
КСИМалЬНОЙ части ТОН- г а ... .
кий, в дистальной ча-
сти расширен; соесшп длинный, округлый; на pars inflabilis тонкие
склеротизованные пластинки, ' несущие очень мелкие шипики,:
Брик (1934) ошибочно синонимизирует этот вид с О. trigotephras Bd.
Это же без всяких оснований было сделано Штрандом (1910); О. prisca
Stgr. сильно дифференцированный вид, характерный значительно боль-
шей рудиментацией самок, чем О. trigotephras Bd.
Распростра не н и е. Средняя Азия — Ферганская долина
(Ош, Маргелан, Ургенч). .
Биология. Зимуют яйца внутри старого кокона самки; выход
гусениц в конце апреля; гусеницы питаются листьями .яблони (Malus
domestica) и, вероятно, многими другими растениями; включая и дикие
розоцветные. Окукление в июне, выход бабочек в конце июня. Самцы
летают днем, отыскивая самок; самки не выходят из коконов и там же
откладывают яйца. Отмечен вред яблоне в Средней Азии, —в Фергане
(Плотников, 1926; Архангельский, 1925). Наиболее обычна в горах.
* 8. Orgyia auroiimbata Gn.
Guenee, 1885, Ann. Soc. Entom. France, 4 : 635, t. 8, fig. 4; Kirby, 1892, Syn.
Cat. Lep. Het., 1 : 493; Staudinger, 1901, Gat. Lep., 1 : 113; Chapman, 1903, Entom.
Rec., 15 : 113; t. 4, 5; Spuler, 1908, Schmett. Eur., 1 : 126, t. 25, fig. 13; Strand, 1910,
Seitz, Grossschmett., 2 : 117, t. 19, b; Oberthiir, 1923, Et. Lep. Comp., 20 : 77; Swin-
hoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 416; Bryk, 1934, Lep. Cat., 62 : 66; —gua-
darramensis Staudinger, 1871, Cat. Lep., ed. 1 : 65.
Голова, грудь и брюшко однотонные, шоколадно-коричневые; крылья
того же цвета с желтоватым отблеском; нижняя сторона крыльев и
брюшка несколько светлее (желтее); крылья без рисунка; бахрома перед-
них и задних крыльев светложелтая, на внутренней стороне задних
крыльев желтая; усики самцов сильно гребенчатые; гребешки тонкие
19*
292
Л. ORGYINAE
и длинные, покрыты тонкими ресничками; голова густо волосистая;
глаза черные; ноги серо-коричневые, желтые к концам; лапки охристо-
коричневые. Самки имеют ноги и голову почти голые; ноги слабые, но
членики лапок еще сохранены, развиты и коготки; крылья в виде ничтож-
ных (около 0.5 мм) рудиментов; тело покрыто серебристыми желтоватыми
волосками. Размах крыльев самцов 21—27 мм, длина самок 8—12 мм.
Распространение. Пиренейский полуостров (Испания,
Кастилия, Каталония, Сиерра Гвадаррама); Штаудингером (1901) ука-
зан для Средней Азии, Семиречья (Или, Иссык-куль); фактический
материал, подтверждающий это, в коллекции Зоологического института
Академии Наук СССР отсутствует; указание на наличие этого вида
в Средней Азии внушает сомнение.
* 9. Orgyia ochroiimbata Stgr. (рис. 129, 130).
Staudinger, 1881, Stett. entom. Ztg., 42 : 405; Romanoff, 1889, Mem. Lep. Rom.,
2 : 8; Radde, 1899, Колл. Кавк, муз., 1 : 435; Staudinger, 1901, Cat. Lep., 1 : 113;
Strand, 1910, Seitz, Grossschmett., 2 : 117, t. 19, b; Swinhoe, 1923, Ann. Mag. Nat.
Hist., 11 (9) : 416; — aurolimbata var. Christoph, 1885, Mem. Lep. Rom., 2:8; — chri-
stophi Alpheraky, 1899, Mem. Lep. Rom., 9 : 202, t. 13, fig. 1.
Голова, грудь и брюшко покрыты рыже-коричневымй волосками;
глаза черные; усики сильно гребенчатые, гребешки по длине равны ши-
Рис. 129. Orgyia ochroiimbata Stgr. Самец. (Рис. Ф. Гуняева). '
рине головы, членики желтые; ноги покрыты короткими желтыми волос-
ками; передние и задние крылья без .рисунка, однотонные, коричнево-
охристые, несколько более светлые в средней части; светлая окраска
на передних крыльях располагается в виде неясной широкой полосы
в середине крыла; бахрома яркая охристая на обеих парах крыльев;
нижняя сторона охристо-коричневая с темными жилками. Самки имеют
рудиментарную голову, усики очень слабые, рудиментарные глаза; ноги
очень короткие и слабые; тело покрыто короткими густыми шелкови-
стыми волосками. Размах крыльев самцов 28—32 мм, длина тела, самок
10—15 мм.
5. ORGYIA
293
Uncus короткий, толстый, сильно изогнутый, на дистальном конце
коротко раздвоенный, не покрытый волосками; valva в базальной части
широкая с небольшим склеротизованным заворотом по дорзальному краю;
дистальная половина valvae коническая, сильно вздернутая дорзально,
мембранозная, слабо покрыта волосками; есть широкий subscaphium;
penis сильно вытянутый, посередине утолщен и расширен в дистальной
части; соесшп длин-
ный..
Распростра-
нение. Северный Кав-
каз, р. Зея (субальпий-
ская зона); Закавказье
(Боржоми, Бакуриани);
Армения (?). Последнее
указание по Штаудин-
геру (1901); я не имел
в руках материала из
этих мест.
Рис. 130. Orgyia ochroiimbata Stgr.. Гениталии самца.
* 10. Orgyia flavolimbata Stgr. (рис. 131).
Staudinger, 1881, Stett. entom. Ztg., 42 : 404; Alpheraky, 1882, Тр. Русск. энтом.
общ., 17 : 35, t. 1, fig. 35; Staudinger, 1901, Cat. Lep., 1 : 113; Strand, 1910, Seitz,
Grossschmett., 2 : 117, t. 19, b; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 416;
Draeseke, 1926, Iris, 40 : 94; Bryk, 1934, Lep. Cat., 62 : 66; — tristis Grum-Grzhimailo,
1890, Mem. Lep. Rom., 4 : 554, t. 20, fig. 6.
Голова, грудь и передние крылья оливково-охристые; брюшко и
задние крылья оливково-серые; голова покрыта длинными шелковистыми
волосками, скрывающими глаза; усики сильно гребенчатые, членики их
желтые; ноги покрыты длинными оливково-серыми волосками; передние
и задние крылья однотонные; на передних
Рис. 131. Orgyia flavolimbata Stgr. Гениталии
самца.
крыльях темная диффузная
полоса в области дискаль-
ных жилок; бахрома на
передних крыльях бледно-
желтая; на задних кры-
льях бахрома серо-жел-
. тая; нижняя сторона одно-
тонная, оливково-серая;
ноги и усики у самок
очень короткие, недораз-
витые; голова рудимен-
тарная; тело покрыло ко-
роткими густыми воло-
сками. Размах крыльев самцов 25—30 мм, длина самок 8—15 мм.
Uncus крючкообразно изогнут, на дистальном конце раздвоенный,
имеет широкое основание; девятый сегмент короткий, особенно в вен-
тральной половине, образует небольшой saccus; anus несет по сторонам
две склеротизованные пластинки; fultura inferior в виде узкой пластинки,
изогнутой дугообразно; valvae маленькие, в базальной половине широ-
кие, сидят на широком основании, дорзальный край их сильно склеро-
тизован, дистальная половина узкая, коническая; penis очень длинный,
утолщенный в дистальной части; его соесшп занимает около одной трети
длины; pars inflabilis не имеет инкрустаций.
294
J. ORGYINAE
Гусеница, по Грум-Гржимайло (1890), светлооранжевой окраски,'
снизу желтоватая, бока серые; имеет три полосы —дорзальную черную
и две боковых, желтоватые, снизу оттененные черным; голова черная;
ноги оранжевые; первые четыре сегмента брюшка несут густые кисти
волосков охристого цвета; передний грудной сегмент несет по бокам
по кисти черных перистых волосков, такой же непарный пучок находится
на дорзальной стороне брюшного восьмого сегмента; тело покрыто боро-:
давками, несущими желто-серые пучки волосков.
Окраска крыльев самцов варьирует в сторону черной; изменчив
также цвет краев крыльев — то бледножелтый, то охристо-оранжевый,
причем варьирует также и ширина этой желто-охристой каймы; темные,
почти черные особи из Тянь-шаня описаны как tristis Gr. Gr.
Распространение. Средняя Азия —• Ферганская долина
(Гульча, Ош); Тянь-шань; Памир (?); Семиречье (Джунгарский Ала-тау,
Алма-ата, Кульджа); Тарбагатай; южный Алтай (Зайсан); северный
Китай —Сычуань (Сунпантин); последнее указание Дрезеке (1926) вну-
шает сомнения; мне материал из Китая не известен.
6. Род PENTHOPHERA GERM.* 1
Germar, 1812, Syst. Gloss. Prodr. : 33; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 490;
Staudinger, 1901, Cat. Lep., 1 : 113; Spuler, 1908, Schmett. Eur., 1 : 125; Strand, 1910,
Seitz, Grossschmett., 2 : 109, 116; Rebel, 1910, Rerge, Schmetterlingsb., ed. 9 : 113;
Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 422; Hering, 1932, Tierw. Mitteleur. :
482; Rryk, 1934, Lep. Cat., 62 : 81; — Liparis Hiibner, 1816, Verz. bekannt. Schmett.,
158, non Ochsenheimer (typus morio L.);—Penthophera Speyer, 1856, Deutsch.
Schmett. : 119 (typus morio L.).
Тип рода P. morio L.
Усики самцов сильно гребенчатые, ротовые части редуцированы;
губные щупальва короткие, последний членик их очень короток, почти
не виден; второй членик их очень тонкий; щупальца свисают вниз; глаза
небольшие, мелко фасеточные, Голые, круглые; ноги покрыты короткими
волосками; первый членик лапки передних ног короче передних голеней;
голени задних ног с одной (последней) парой шпор; коготки на всех
лапках простые; радиальной ячейки на передних крыльях или нет
(у европейских особей), или она ромбическая, широкая (у кавказских);
третий, четвертый и пятый радиальные стволы в последнем случае от-
ходят на стебельке и ветвятся в апикальном углу крыла; жилкование
в области добавочной ячеи очень изменчиво, то ячейка образована непол-
ным анастомозом третьего и четвертого радиальных стволов, то они обра-
зуют стебелек, то пятый радиальный ствол выходит самостоятельно от
стенки ячеи, то, наконец, этот ствол находится на общем стебельке
с двумя предыдущими; первый медиальный ствол отходит от середины
ячеи; дискоидальные жилки утрачены; базальная ячея на задних крыльях
ланцетовидной формы, она образована полным слиянием радиальной
и субкостальной жилок и вполне замкнута, слияние жилок на большем
расстоянии; первый медиальный ствол анастомозирует с радиальным;
1 Используемое часто родовое название Hypogymna Hiibner (1810, Tentamen:
1, typus morio L.) не может быть принято, так как «Tentamen» Гюбнера не был напе-
чатан; позже (1816) Гюбнер ставит название Hypogymna как «штамм» (Stamm), куда
входят два рода — Liparis Ochs, и Ocneria Hb. (detrita L.). Liparis имеет первым видом
morio L., но содержит также много других видов (dispar L., monacha L., similis Fues-
sly, riibea F.). Впервые тип morio L. был фиксирован за родом Penthophera Germ. Рам-
буром (Rambur, 1866, Cat. Syst. Lep. Andalus., 2 : 273).
6. PENTHOPHERA
295
Рис. 132. Penthophera morio L. Хетотаксия
передне- и среднегруди и брюшных сег-
ментов.. I, VI, VII и VIII взрослой
гусеницы.
крылья самцов покрыты редкими и длинными, ланцетовидной формы,
чешуйками и волосками, крылья самок — только волосками. Полы
сильно диморфны; самки короткокрылы и узкокрылы, брюшко по форме
напоминает таковое самок Orgyia Ochs.
Гусеницы несут на коже негустые короткие волоски (см. рис. 132);
бородавки, покрывающие тело, плоские и круглые; больше других
выдается бородавка х. на переднегруди, она также несет короткие щетинки.
Бородавки а и [з на средне- и заднегруди слиты и образуют большую,
вытянутую, овальной формы бородавку. На брюшных сегментах боро-
давки а и р также слиты; х на брюшных сегментах утрачена, z брюшных
сегментов вытянутая, большая.
Есть только две выворачиваю-
щиеся железки на шестом и седь-
мом брюшных сегментах.^
Монотипический род, ограни-
ченный в распространении только
средней Европой и восточным
Средиземноморьем; систематиче-
ски обособлен, но наиболее бли-
зок к Orgyia Ochs. Налицо специ-
фичные черты высокой степени
адаптации и специализации—утра-
та чешуйчатого покрова на кры-
льях, полная изоляция базальной
ячеи на задних крыльях, коротко-
крылость самок. Замечательным
фактом является изменчивость ра-
диальной ячейки в пределах ви-
да — факт, показывающий высокую степень лабильности признаков
жилкования в пределах Orgyidae.
* 1. Penthophera morio L. (рис. 132, 133, 134, 135).
Linne, 1767, Syst. Nat., ed. 12, 1 (2) : 828 (Bombyx)', Kirby, 1892, Syn. Cat. Lep.
Het., 1 : 490; Staudinger, 1901, Cat. Lep., 1 : 113; Spuler, 1908 Schmett. Eur., 1 :
126, t. 25, fig. 12; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 113, t. 21, fig. 5, 6;
Strand, 1910, Seitz; Grossschmett., 2 : 116, t. 19, a; Swinhoe, 1923, Ann. Mag. Nat.
Hist., 11 (9) : 422; Osthelder, 1926, Schmett. S. Bayerns, 1 (2) : 196; Ankinsini, 1931,
Mem. Acad. Rom., 7 : 199; Hering, 1932, Tierw. Mitteleur. : 482; Bryk, 1934, Lep.
Cat., 62 : 82; — caucasica Heylaerts, 1887, Compt. Rend. Soc. Entom. Belg., 31 : 88;
Heyi e t«, 1889, Mem. Lep. Rom., 5 : 201, t. 9, fig. 4; — natlyi Aigner-Abafi, 1906,
Rov. Lap., 13 : 69; — illustreciliata Stauder, 1921, Iris, 35 : 182; — omninolimbata
Stauder, 1921, там же, 35 : 182; — eburnina Stauder, 1921, там же, 35 : 182; —• jalcata
Stauder, 1921, там же, 35 : 182; — feminicolorata Stauder, 1921, там же, 35 : 182; —°-
aureomicans Stauder, 1921, там же, 35 : 182;—dejuncta Stauder, 1921, там же, 35 : 182.
Биология. Sajo, 1894, Ztschr. Pflanfcenkrankh., 4 : 100; Aigner-Abafi, 1900,
Illustr. Ztschr. Entom., 5 : 201; Hoffman, 1916, Entom. Jahrb., 25 : 145; Rozsypal,
1922, Ochr. Rosl., 2 : 17; Lederer, 1923, Handb. prakt. Entom., 3 : 146; Stauder, 1924,
Entom. Anz., 4 : 73; Romanizyn, 1930, Prazn. Monogr,, 3 : 167.
Среднеевропейская форма, крупнее и менее волосистая; передние
крылья типичны отсутствием добавочной ячеи; голова и грудь покрыты
густыми черными волосками, несколько рыжеватыми по сторонам груди;
ноги покрыты плоскими черно-серыми волосками; задние ноги имеют
большую примесь желтых волосков; усики самцов сильно гребенчатые,
гребешки черные, членики усиков серые; глаза и губные щупальца чер-
ные; брюшко серое с темными'основаниями сегментов, кольчатое; перед-
296
I. ORGY/NAP
ние и задние крылья прозрачные, однотонные, без рисунка, дымчато-
серые, бахрома черно-серая, жилки резко выделяются черным; нижняя
сторона того же цвета, также без рисунка. Самки с очень длинным и тол-
стым брюшком и укороченными крыльями; окраска крыльев и брюшка
Рис. 133. Penthophera morio L. Самец. (Рис. Ф. Гуняева).
серая; крылья прозрачные, без рисунка, покрыты только длинными
волосками, особенно по краю; усики с очень маленькими гребешками
на члениках; ноги серые. Размах крыльев самцов 22—25 мм, самок 19 —
21 мм, длина тела самок 14—16 мм.
Uncus конический, гладкий, вальковатой формы, слабо изогнутый
вентрально, тупой; tegumen без швов; вентральная половина девятого
Рис. 134. Penthophera morio L. Самка. (Рис. Ф. Гуняева).
сегмента узкая, образует узкий треугольный saccus; valvae короткие,
лопастевидные, расширенные в дистальной части; с дорзальной стороны
у основания valva неглубокая и узкая выемка; вдоль дорзального края
valva на уровне одной трети ее высоты склеротизованная складочка;
в. PENTHOPHERA
297
есть складочка и при основании valva; penis короткий, почти прямой,
сильно расширенный в дистальной части, с коротким и заостренным
соесшп. Bursa copulatrix маленькая, мембранозная, без скульптуры на
поверхности, в- средней части слабо расширенная;, ductus bursae корот-
кий, мембранозный; ductus seminalis отходит от средней части совоку-
пительной сумки, толстый и короткий; apophyses anteriores короткие,
расширенные в проксимальной части; восьмой сегмент недлинный, уко-
рочен в вагинальной области, где образует валик; apophyses posteriores
короче первых; papillae anales широкие и короткие, несколько более
длинные в середине, в проксимальной части несут венчик длинных ще-
тинок.
Рис. 135. Penthophera morio L. Гениталии: а — самца,
б — самки.
Взрослые гусеницы желто-серые с черным рисунком; рисунок в виде
продольных черных полос как на дорзальной, так и на вентральной
стороне; на дорзальной стороне тела полосы распадаются на черные
прямоугольные отрезки; узкая дорзальная полоса оторочена двумя
широкими латеральными полосами; тело покрыто желто-серыми боро-
давками, несущими пучки недлинных волос бело-желтого цвета; брюш-
ные ноги желто-серые, грудные черно-серые; голова черная; бородавки
на всех сегментах равных размеров, не высокие; железки на шестом
и седьмом сегментах брюшка заметны в виде бугорков.
По наблюдениям Штаудера (1921), типичная форма в общем мало
изменчива; этот автор выводил сотни особей из куколок и только на гро-
мадном материале, может быть не безупречном в отношении условий
воспитания, отметил наличие цветовых форм; им подобраны цветовые
формы от типичной черной разных оттенков до коричнево-рыжей; часть
их названа (см. синонимы). В природе окраска этого вида очень устой-
чива.
Распространение. Средняя Европа (южная Германия, вос-
точные Альпы, южные Альпы и северная Италия, долина Дуная, Висла,
Карпаты, Босйия); северные Балканы, Румыния; Молдавия; Украина
(Смела, Умань, Черкассы, Звенигородка, Каменец-Подольск, Винница,
298
1. ORGYINAE
Могилев-Подольский, Киев); указана для северной части лесной зоны—'
Кама, Елабуга.
Биология. Зимуют гусеницы в средних стадиях развития; появ-
ление весной в апреле—мае; питание гусениц различными злаками
(по Штаудеру, очень многоядны, но предпочитают злаки), особенно ви-
дами Lolium (Lolium perenne, Lolium tomulentum); окукление в середине
лета среди растений в очень рыхлом коконе; куколка развивается около
трех недель; самки не летают, самцы очень активны; яйцекладка куч-
ками на ветки и сухие стержни трав. Дает одно полное поколение в сезон,
а при наличии теплого (или жаркого) лета частичное второе. Отмечен
вред озимым хлебам в средней Европе (Венгрия).
* la. Penthophera morio caucasica Heyl.
Heylaerts, 1887, Compt. Rend. Soc. Entom. Belg., 31 : 88; Heylaerts, 1889, Mem.
Lep. Rom., 5 : 201, t. 9, fig. 4; Staudinger, 1901, Oat. Lep., 1 : 113; Bryk, 1934, Lep.
Cat., 62 : 82.
Кавказские особи дымчато-черные с некоторой примесью коричневых
волосков, более волосистые и меньших размеров; крылья покрыты гу-
стыми волосками по жилкам; тело густо волосистое; волоски, покрываю-
щие тело, интенсивно черные; усики самцов сильно гребенчатые, гребешки
черные, членики бело-серые. Устойчивым различием подвидов является
жилкование; кавказские особи все без исключения имеют хорошо раз-
витую добавочную ячейку. -Строение гениталий обеих форм не . дает
отличий.
Распространение. Дагестан (Куруш, Шах-даг); Закавказье
(Баку); Армения; последнее указание по Штаудингеру, без большей
точности обозначения места, возможно ошибочно; фактический материал
из Армении мне не известен.
Биология этой формы не известна.
7. Род CIFUNA WLK.
Walker, 1855, Cat. Lep. Het. Brit. ?Mus., 5 : 1172; Hampson, 1892, Fauna Brit.
Ind., Moths, 1 : 446; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 458; Staudinger, 1901, Cat.
Lep., 1 : 115; Swinhoe, 1903, Trans. Entom. Soc. Lond. : 479; Strand, 1910, Seitz,
Grossschmett., 2 : 121; Strand, 1914, Entom. Mitt., 3 : 329; Strand, 1915,Seitz, Gross-
schmett., 10 : 305;-Swinhoe, 1923, Ann. Mag. Nat. Hist., 10 (9) : 413; Hering, 1926,
Seitz, Grossschmett., 14 : 188; Joannis, 1929, Ann. Soc. Entom. France, 98 : 416; Bryk,
1934, Lep. Cat., 62 ; 92; —Barynza Moore, 1879, Descr. Lep. Atk. : 45 (typus cervina
Moore).
Тип рода C. Locuples Wlk.
Хоботок рудиментарный; глаза голые, большие, круглые или чуть
суженные; губные щупальца торчат вперед, второй членик их густо
опушен длинными волосками, третий членик очень маленький; усики
самцов сильно гребенчатые, на конце гребешков по три щетинки; лоб
гладкий; первый членик передних лапок короче передних голеней;
голени задних ног с двумя парами шпор; коготки на всех лапках с ши-
роким вентральным зубцом; голова и грудь покрыты длинными волос-
ками, почти без примеси чешуй; примесь волосков на крыльях ничтожна;
радиальная ячейка большая, ромбической формы; третий и четвертый
радиальные стволы на очень длинном стебельке (ветвление у самого
apex); первый медиальный• ствол отходит от середины ячеи; дискальные
жилки рудиментарны, базальная ячея, на задних крыльях замкнута
7. Cl FUN A
299
вполне, длинная, ланцетовидная; второй медиальный ствол на задних
крыльях отходит посредством дискальной жилки.
Uncus сложной формы, часто с бороздой на дорзальной стороне; val-
vae со склеротизованной лопастью в базальной части, на которой нахо-
дятся сильные шипы и нередко сложный вырост; дорзальный край valva
с выростом. Bursa copulatrix несет laminae dentatae; apophyses тонкие
и длинные. Полы слабо диморфны,, самки всегда крылаты.
Олиготипический род, ограниченный в распространении юго-восточ-
ной Азией, причем только один вид (С. locuples Wlk.) распространен более
или менее широко, заселяя Индию, Гималаи, западный Китай, Япон-
ские острова и южное Приморье, тогда как три остальных вида энде-
мичны, один (С. eurydicae Btl.) ограничен только Китаем, Японскими
островами и южным Приморьем, один (С. jankowskii Obth.) известен
только из южного Приморья и один (С. cervina Moore) — только из
Гималаев (может быть форма от locuples Wlk.).
С других территорий виды Cifuna Wlk. не известны; описанные как
Cifuna Wlk. из Африки С. nigriplagata В. Век. и С. marginenotata Her.,
без сомнения, к Cifuna Wlk. не принадлежат. Очень сомнительна также
принадлежность к Cifuna Wlk. индийского biundulalis Hamps.
Виды Cifuna Wlk. наиболее близки к видам Orgyia Ochs.; наибольшее
сходство с видами группы О. antiqua L. дает С. locuples Wlk. как по ри-
сунку, так и по структурам гениталий; два другие вида Cifuna Wlk.
обнаруживают, наряду с локализацией географического распростране-
ния, значительно выраженные черты специализации морфологических
структур.
Биологически виды Cifuna Wlk. не изучены; отмечена лишь связь
их с широколиственными лесами, основывающаяся, помимо питания
лиственными древесными породами и кустарниками—Bosaceae, Vita-
ceae, Saxifragaceae (Bibes),—также на высокой гигрофильности видов
этого рода. Род Cifuna Wlk. следует рассматривать как осколок какой-то
древней группы китайско-гималайского комплекса форм типа Dasychira
Steph. — Orgyia Ochs.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). 1/ костального края передних крыльев во внешнем поле широкий
черный треугольник; окраска коричневая или серая; дорзальный
край valva несет короткий вырост; penis тонкий, конический; duc-
tus bursae короткий; uncus плоский.
2 (3). Передние крылья серые, задние одноцветные черные или серые;
дорзальный край valva несет короткие конические выросты; ди-
стальный конец ее загнут крючковидно; lamina dentata с дорзаль-
ной стороны bursa copulatrix.................* 2. С. eurydice Btl.
3 (2). Передние крылья коричневые или черные, задние коричнево-
серые; valva несет крючковидный вырост у дорзального края;
дистальный конец ее прямой; lamina dentata расположена на вен-‘
тральной стороне совокупительной сумки ..........................
.....................................* 3. С. jankovskii Obth.
4 (1). Передние крылья желто-коричневые, без черного треугольника
во внешнем поле у костального края; задние крылья светлые; valva
вытянутой формы, с длинным изогнутым отростком на дорзальном
крае; penis примерно одной толщины по всей длине;ипсиз компакт-
ный, высокий, с глубокой бороздой на дорзальной стороне; duc-
tus bursae длинный . . ................... * 1. С. locuples Wlk.
300
ORGYINAE
* 1. Cifuna Iocupies Wlk. (рис. 136, 137).
Walker, 1855, Cat. Lep. Het. Brit. Mus., 5 : 1173; Hampson, 1892, Fauna Brit.,
Ind., Moths, 1 : 446; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 458; Leech, 1899, Trans.
Entom. Soc. Lond., 37 : 122; Staudinger,.1901, Cat. Lep., 1 : 115; Swinhoe, 1903, Trans.
Entom. Soc. Lond., 41 ; 479; Nagano, 1909, Konch. Sek., 13 : 411; Strand, 1909, Seitz,
Grossschmett., 2 : 121; Wileman, 1918, Philipp. Journ. Sci., 13(4) : 154; Swinhoe,
1922, Ann. Mag. Nat. Hist., 10(9) : 413; Draeseke, 1926, Iris, 40 : 54; Gaede, 1932,
Seitz, Grossschmett., 2 Suppl. : 99, t. 8, b; Bryk, 1934, Lep. Cat., 62 : 93; Куренцов,
1939, Вести. Дальневост. фил. Акад. Наук СССР, 10 : 130; —confusa Bremer, 1861,
Bull. Acad. Sci. St.-Petersb., 3 : 479 (Artaxa)’, Bremer, 1864, Lep. Ost-Sib. : 42, t. 4,
fig. 5. . ‘
Передние крылья рыже-коричневые, у самок бурые с небольшой
примесью рыжих тонов в базальной части крыла и в области почковид-
ного пятна; базальное поле в дорзальной половине охристо-рыжее, ко-
Рис. 136. Cifuna Iocupies Wlk. Самец. (Рис. Ф. Гуняева).
стальная его половина буро-коричневая; вторая поперечная линия широ-
кая, двойная, дающая зигзаги к внешнему краю у costa, посередине этой
линии зигзаг, направленный базально, а в нижней части —направлен-
ный дистально; срединное поле равной ширины на всем протяжении,
шире, чем базальное или внешнее; круглое пятно не развито, почковидное
пятно узкое, вытянутое, сильно выгнутое в средине, желтое или желто-
коричневое, четко очерченное коричневым; костальная половина сре-
динного поля желтая, иногда все срединное поле серовато- или фиолетово-
коричневое; третья поперечная линия в виде неизвитой и лишь слабо
выгнутой кнаружи простой коричневой полосы, в костальной половине
внешнего поля охристо-желтое пятно; подкраевая линия началом сдви-
нута почти к apex, волнистая, скошенная базально, извитая, коричневая
и желтая, отмеченная белыми чешуйками у костального края; часть
внешнего поля между подкраевой линией и третьей поперечной линией
затемнена, черно-коричневая; терминальная линия коричневая, с вну-
тренней стороны осветленная, несколько отступающая от терминального
края у заднего угла крыла, бахрома рыже-коричневая; задние крылья
одноцветно желтые с слабыми сероватыми дискоидальными пятнами, тон-
кой коричневой треминальной полоской и коричнево-желтой бахромой;
нижняя сторона желтая с многими черными чешуйками, особенно по
7. CIFUNA
301
костальному краю крыла; дискоидальные пятна развиты; заметна по-
перечная темнокоричневая полоса; самки темнее, но рисунок у них тот
же; усики самцов сильно гребенчатые, гребешки серые, членики желтые;
у самок гребешки усиков короткие. Размах крыльев самцов 35—40 мм,
самок 45—50 мм.
Uncus толстый и короткий, загнутый вентрально, с глуоокой бороз-
дой на дорзальной стороне; основание его широкое, образует по сторонам
два вздутия, отделено неполным швом от девятого сегмента; последний —
короткий, образует с вентральной стороны хорошо развитой saccus; по
сторонам две латеральных пластинки, сильно склеротизованные; ful-
tura inferior в виде склероти-
зованной пластинки, имеет тре-
угольное основание, свернута
в полутрубку; valvae вытяну-
тые, в базальной части имеют
склеротизованную лопасть, не-
сущую по сторонам зубцы (не
образующую еще выроста, как
у других видов этого рода);
дорзальная сторона valva имеет
длинный плоский и узкий (не-
сколько расширенный в ди-
стальной части) вырост; penis
длинный, расширенный в ди-
стальной части, с длинным со-
есшп, изогнутым вентрально и
со сложной зубчатой пластин-
кой на pars inflabilis. Bursa
copulatrix мешковидная, несет
рубчатую скульптуру, а на
дорзальной стороне поперечно
расположенную, имеющую ха-
рактер склеротизованной склад-
ки lamina dentata; ductus bur-
sae окружен склеротизованной
складкой, несущей многочислен-
Рис. 137. Cijuna Iocupies Wlk. Гениталии:
а — самца, б — самки.
ные хитиновые ворсинки и шипики; восьмой сегмент короткий, склероти-
зован в проксимальной части, несет длинные и тонкие apophyses ante-
riores; apophyses posteriores также тонкие и длинные, короче первых;
papillae anales в виде плоских лопастей, мембранозные.
Распространение. Приамурье (Бурея, Хабаровск); Шан-
тарские острова; южное Приморье (Океанская, Янчихе, р. Адими,
Виноградовка, Сучан, Сидеми, Владивосток, Аскольд, р. Мангугай,
Уссури); Сахалин; Корея; Японские острова (Хоккайдо, Хондо); Тай-
ван; Китай; восточные Гималаи (Кхазиа-Хиллс); северо-восточная
Индия (северная Бенгалия).
* 2. Cifuna enrydice Btl. (рис. 138).
Butler, 1885, Cist. Entom., 3 : 118 (P orthetriay, Leech, 1888, Proc. Zool. Soc. Lond. :
632; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 459; Leech, 1899, Trans. Entom. Soc. Lond. :
123; Staudinger, 1901, Cat. Lep., 1 : 115; Swinhoe, 1903, Trans. Entom. Soc. Lond. :
483; Strand, 1910, Seitz, Grossschmett., 2:121, t. 19,d; Swinhoe, 1923, Ann. Mag.
Nat. Hist., 11 (9) : 413, 424; Scriba, 1928, Entom. Rundsch., 35 : 14; Bryk,
1934, Lep. Cat., 62 : 93; Куренцов, 1936, Tp. Дальневост. фил. Акад. Наук СССР,
302
Л ORGYINAE
Btl. Гениталии: a —
самки.
Рис. 138. Cifuna eurydice
самца, б —
1:152; Куренцов, 1939, Тр. Горн.-таежн. ст., 3:146; — anata Staudinger, 1887,-
Мёш. Lep. Rom., 2 : 206, t. 12, fig. 2 {Dasychira)', — ornata Swinhoe, 1923, Ann. Mag.
Nat. Hist., 10 (9) : 413.
Голова, грудь и передние крылья зеленовато-серые (оливковые),
задние крылья и брюшко самцов черные, у самок темносерые; базальная
линия хорошо развита,, двойная, черная, слабо волнистая, посредине
оливковая; базальное поле между этой линией и второй поперечной ли-
нией черно-оливковое, черный тон распространен за пределы второй
линии в срединное поле; вторая поперечная линия черная и белая, за-
метна в виде извитой белой полоски, черный тон крыла сливается с за-
темненным базальным полем; срединное поле темнооливковое, однотон-
ное, без пятен и рисунка;
третья поперечная линия
в виде светлой растуше-
ванной полосы у косталь-
ного края, черная, более
резкая; под костальным
краем широкий черный
штрих; у начала подкрае-
вой линии черный полу-
лунный штрих, подкрае-
вая линия нерезкая, замет-
на в виде отдельных чер-
ных штрихов неправиль-
ной формы, соединенных
слабой светлой полоской;
терминальная линия в ви-
де узкой оливковой по-
лоски; бахрома светло-
оливковая; задние крылья
самцов одноцветно черные
с оливково-охристой бах-
ромой; у самок внешний
край задних крыльев ши-
роко окаймлен оливково-
черная с оливковосерым внешним краем перед-
серым; нижняя сторона
них крыльев и просвечивающими черными пятнами; у самок нижняя
сторона желто-черная с широкой черной лимбальной линией; усики
самцов очень сильно гребенчатые, длина гребешков более ширины голо-
вы; у самок гребешки усиков очень короткие. Размах крыльев сам-
цов 35—40 мм, самок 40 —45 мм.
Uncus короткий, плоский, треугольной формы с небольшими высту-
пами у основания, которое широкое и образует два латеральных выроста,
идущих параллельно девятому сегменту; последний отделен швом, корот-
кий, с вентральной стороны образует saccus; valvae очень широкие при
основании, дистальный конец их заострен; в узкой дистальной трети их
две складки, образующих на дистальном конце загнутый внутрь вырост;
весь дистальный конец valva покрыт волосками, сидящими на бородав-
ках; в базальной части valvd два выроста — один у вентрального ее
край, несколько извитой, шиповатый, волосистый, другой — на дор-
зальном крае, короткий, гладкий, изогнутый внутрь; fultura inferior
в виде широкого склеротизованного тяжа, укрепленного основанием на
saccus и дистальной частью на основании penis; penis длинный, изогну-
7. СI FUGA
303
тый, конический, более тонкий к дистальному концу, где также уплощен,
с коротким соесиш. Bursa copulatrix изогнутая, почти веретеновидной
формы, несет рубчатую скульптуру и с дорзальной стороны в форме
склеротизованной складки, расположенной поперек, lamina, dentata;
ductus bursae короткий, мембранозный, склеротизован лишь у ostium;
ductus seminalis отходит на границе corpus и ductus bursae; восьмой
сегмент короткий, широкий, склеротизован в проксимальной части,
несет длинные и тонкие -apophyses anteriores; apophyses posteriores не-
сколько более длинные; papillae anales широкие, короткие и мембраноз-
ные.
Взрослая гусеница (по Куренцову, 1939) почти черная, щетки на спине
красные; по бокам пучки длинных светлых волос; на первом и втором
сегментах черные кисточки; голова большая, черная. Куколка с жел-
тыми брюшными кольцами, лежит в просторном буром коконе.
Распространение. Южное Приморье (Уссури, Владивосток,
Аскольд, Супутинка, Седанка, Сучан, Сидеми); Японские острова
(Хондо); Вьетнам (Тонкин).
Биология. Биология изучена слабо; зимуют, невидимому, яйца
(может быть молодые гусеницы); взрослые гусеницы обычно наблю-
даются в июне; окукление в июле; выход бабочек в конце июля; окукле-
ние на растениях в просторном буром коконе; гусеницы многоядны,
питаются Vitaceae (Vitis amurensis), по Грэзеру (1888), и Crataegus, по
Куренцову (1939). Очень обычна в кедрово-широколиственных, лесах;
вредит лесным породам.
* 3. Cifuna jankowsxii Obth. (рис. 139, 140).
Oberthiir, 1884, Et. Entom. Comp., 10 : 13, t. 1, fig. 9 (Orgyia); Graeser, 1888, Berl.
Entom. Ztschr., 32 : 123; Staudinger, 1892, Mem. Lep. Rom., 6 : 305; Kirby, 1892,
Syn. Cat. Lep. Het., 1 : 459; Staudinger, 1901, Cat. Lep., 1 : 115; Strand, 1910, Seitz,
Grossschmett., 2 : 121, t. 19, d; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 413;
Matsumura, 1933, Ins. Matsum., 7 : 112; Bryk, 1934, Lep. Cat., 62 : 94; Куренцов,
1936, Тр. Дальневост. фил. Акад. Наук СССР, 1 : 152; Куренцов, 1939, Тр. Горно-'
таежн. ст., 3 : 146.
Голова и грудь серо-коричневые; передние крылья коричнево-охри-
стые, темные; самки имеют общий тон крыльев и груди, желто-серый,
более светлый, чем самцы; базальная линия в виде прямой пепельно-
серой полосы; вторая поперечная линия очень широкая, сильно ско-
шенная кнаружи, черная, с многочисленными неправильно разбросан-
ными чешуйками и белой внешней подкраевой полосой; срединное поле
почти без рисунка; слабо намечено темнокоричневым узкое почковидное
пятно; ширина срединного поля примерно равна ширине внешнего и
базального; третья поперечная линия белая с тонкой черной полоской
посредине, заметно расширенной у костального края, S-образно изви-
тая, мелко и слабо зубчатая по жилкам; между этой и подкраевой лини-
ями под костальным краем черное пятно; подкраевая линия мелко из-
витая, темная, местами смытая, идет параллельно внешнему краю крыла;
терминальная линия в виде тонкой зубчатой полоски, несколько отстав-
ленной от терминального края; бахрома серо-коричневая; задние крылья
черно-коричневые с неясными черными дискоидальпыми пятнами и ши-
рокой неясной черной лимбальной линией; терминальная линия узкая,
черная; бахрома серо-коричневая; нижняя сторона охристо-желтая с чер-
ным пятном в дискоидальном поле передних крыльев, с хорошо замет-
ными почти полулунной формы дискальными пятнами на передних и
304
1. ORGYINAE
задних крыльях и с прерванной лимбальной линией; усики самцов -
сильно гребенчатые, у самок гребешки короткие. Размах 30—-35 мм самцы,
35—-40 мм самки.
Рис. 139. Cifuna jankowskii ОЫЬ. Самец. (Рис. Ф. Гуняева).
Uncus ^треугольной формы, заостренный, с бороздой на дорзальной
стороне; основание его широкое, с латеральными выростами, отделено
швом от девятого сегмента; последний короткий, не образует большого
saccus; Valvae длинные, в базаль-
Рис. 140. Cijuna jankowskii Obth. Гени-
талии: а —: самца, б — самки.
ной половине широкие, в дисталь-
ной сильно суженые; в базальной
части valva склеротизованная ло-
пасть с большим шиповатым от-
ростком у дорзального края; на
дорзальном крае этой лопасти ко-
роткий сильно загнутый отросток;
дистальный конец valva со скле-
ротизованной складкой, образую-
щей на конце короткий зубец,
покрытый волосками: fultura in-
ferior срастается с вентральной
стороной девятого сегмента и осно-
ванием penis; последний плотно
укреплен на ней, длинный, изо-
гнутый дорзально, сильно склеро-
тизован, конический, заострен-
ный. Bursa copulatrix не длинная,
почти цилиндрическая, мембра-
нозная, несет в вентральной части
тонкую, в виде складки, распола-
гающуюся поперек corpus bursae,
lamina dentata; ductus bursae
короткий, мембранозный в ваги-
нальной области, несет склеротизованную пластинку часто с отростками;
ductus seminalis отходит от основания ductus bursae; восьмой сегмент
склеротизован сильно по проксимальному краю, широкий и короткий;
7. CIFUNA, 8. NUMENS
305
-apophyses anteriores тонкие и длинные, такие же и apophyses posteri-
•ores; papillae anales в виде плоских мембранозных довольно больших
лопастей.
Взрослая гусеница (по Куренцову, 1939) золотисто-желтая; в отличие
от предыдущего вида имеет щетку светлых волосков на спине, а кисточки
па первом и втором сегментах черные и значительно более длинные, чем
у Cifuna eurydice Btl. Куколка черная.
Распространение. Южное Приморье (Уссури, Владиво-
сток, Яковлевна, Евгениевка, Сучан, р. Сица, р. Супутинка, р. Бикин,
>Сидеми).
Биология. Зимуют, повидимому, яйца; взрослые гусеницы
^попадаются в июне, питаются разными лиственными породами, отме-
чены Vitis amurensis, по Грэзеру (1888), Ribes mandzhurica, Malus mand-
zhurica, по Куренцову (1939); окукление в середине июля; выход
бабочек в конце июля, гусеницы предпочитают яблоню. Отмечен как
вредитель на Дальнем Востоке.
8. Род NUMENS WLK.
Walker, 1855, Cat. Lep. Het. Brit. Mus., 3 : 662; Kirby, 1892, Syn. Cat. Lep. Het.,
1 : 455; Semper, 1898, Schmett. Philipp., 2 : 460; Staudinger, 1901, Cat. Lep., 1 : 116;
Swinhoe, 1903, Trans. Entom. Soc. Lond. : 432; Strand, 1910, Seitz, Grossschmett.,
2 : 109, 126; Strand, 1915, там же, 10 : 316;'Swinhoe, 1922, Ann. Mag. Nat. Hist., 11 (9) :
•82; Hering, 1926, Seitz, Grossschmett., 14 : 155; van Eecke, 1928, Zool. Meded.Mus.
Leid., 11 : 97; van Eecke, 1930, Heteroc. Sumatr. : 247; Matsumura, 1933, Ins. Mat-
sum., 7 : 141; Gaede, 1934, Seitz, Grossschmett., 2 Suppl. : 283; Bryk, 1934, Lep.
Cat., 62 : 138; — Pseudomesa Walker, 1855, Cat. Lep. Het. Brit. Mus., 4 : 923 (typus
quadriplagiata Wlk.).
Тип рода N. siletti Wlk.
Хоботок рудиментарный; губные щупальца маленькие, слабые, тор-
чащие вперед, с коротким последним члеником; они покрыты чешуйками
и короткими волосками; усики самцов гребенчатые, гребешки на ди-
стальных концах несут очень короткие тонкие щетинки; усики самок
с укороченными гребешками; первый членик передней .лапки более поло-
вины длины передней голени; голени задних ног с двумя парами шпор;
.коготки всех ног с широким зубцом на вентральной стороне; глаза оваль-
ные, небольшие, голые, окруженные волосками; дистальный конец
брюшка самки без волосяной подушки; передние крылья с заостренным
верхним углом; радиальная ячейка на передних крыльях есть; ниже нее
отходит лишь первый радиальный ствол, второй ствол отходит от ее
костальной стенки или на стебельке с третьим и четвертым, которые всегда
стебельчатые и отходят от вершины ячеи; пятый ствол свободный, отходит
от внутренней стенки ячеи или из одной точки с предыдущими; первый
медиальный ствол отходит от нижней четверти ячеи; второй медиальный
ствол на задних крыльях вполне сближен основанием с третьим; ба-
зальная ячейка узкая, почти замкнутая.
Uncus короткий, отделен швом от девятого сегмента; последний обра-
зует латеральные выросты по сторонам основания uncus; saccus недоразвит;
penis несет многочисленные шипы на pars inflabilis; apophyses длинные.
О лиготипический род, представленный немногими видами в индо-
малайской фауне и на Мадагаскаре. Наибольшее число видов известно
для Индии и Гималаев (ДГ. patrana Moore, siletti Wlk., fragrans Thoms.,
contrahens Wlk.); из этих видов N. siletti Wlk. заселяет восточную Индию
(Бирму), о-ва Малайского архипелага (Борнео, Ява) и известен с Филип-
20 Фауна СССР, И. В. Кожанчиков.
306
I. ORGYINAE
пинских островов, откуда описан один эндемичный вид — A'. insolita
Schultze, тогда как с Малайского архипелага эндемиков из рода Numens-
Wlk. не известно. Севернее, в Китае, Корее и Японии, а также в южном
Приморье распространен N. disparilis Stgr., очень близкий к гима-
лайским Numens Wlk.; один эндемичный вид известен с Тайвана—N. 1а-
kamukui Mts. Наконец два вида Numens Wlk. описаны с Мадагаскара
(A. staudingeri Saalm. и N. praestana Saalm.), очень похожие на гималай-
ские виды и на маньчжурского N. disparilis Stgr. (особенно первый вид).
Из Африки виды Numens Wlk. не известны, хотя несколько видов;
Геринг (1926) отнес в этот род, несмотря на то, что они давно уже были
выделены Голландом в специальный род Heteronygmia, так как действи-
тельно скорее напоминают виды из рода Artaxa Wlk., чем виды Numens
Wlk. Не известны виды Numens Wlk. и из Австралии, а также из Аме-
рики, как Южной, так и Северной, где нет к ним даже близких форм.
Биология и кормовые связи тропических видов Numens Wlk.
не известны, хотя, без сомнения, это все виды, дендрофильные; N. dispa-
rilis Stgr. связан с Betulaceae (Carpinus, Ostryia). Виды Numens Wlk.
представляют осколок какой-то древней группы, тяготеющей к Artaxa.
Wlk. и Euproctis Hb.
*1. Numens disparilis. Stgr. (рис. 141, 142). '
Staudinger, 1887, Mem. Lep. Rom., 3 : 200, t. 1, fig. 2 a, b; Kirby, 1892, Syn. Cat.
Lep. Het., 1 : 456; Leech, 1899, Trans. Entom. Soc. Lond., 37 : 629; Staudinger, 1901,
Cat. Lep., 1 : 116; Swinhoe, 1903, Trans. Entom. Soc. Lond., 41 : 432; Strand, 1910,
Seitz, Grossschmett., 2 : 126, t. 20, c; Swinhoe, 1923, Ann. Mag. Nat. Hist.. 11 (9) : 82;
Bang-Haas, 1927, Hor. Macrolepidopt., 1 : 76; Gaede, 1932, Seitz, Grossschmett., 2,
Suppl: 283; Bryk, 1934, Lep. Cat., 62 : 139; —albofascia Leech, 1888, Proc. Zool. Soc.
Lond. : 629, t. 31, fig. 8; — separata Leech, 1890, Entomologist, 23 : 112; — biseparata
Strand, 1910, Seitz, Grossschmett., 2 : 126, t. 22, g; — hayashii Matsumura, 1911,
Thous. Insects Jap., Suppl. 3 : 2, t. 30, fig. 2.
Полы диморфны. Голова, грудь и передние крылья самцов бархатисто-
черные, с примесью бурых тонов, особенно на голове и груди; жилки на
передних крыльях белые, тонко очерченные, хорошо видные на черном1
фоне; от костального края поперек крыла косо к tornus широкая и пря-
мая белая или желтоватая полоса, суживающаяся к дорзальному краю;
задние крылья буро-черные с буро-белыми жилками, с размытым белым1
пятном в средней части и светлобурой внутренней стороной; нижняя сто-
рона красно-бурая с тонкими беловатыми жилками; белые пятна просве-
чивают на нижней стороне; лоб, нижняя сторона груди и ноги рыже-бурые;
брюшко с дорзальной стороны черное, с вентральной белое, с боков бурое;
усики сильно гребенчатые. Самки имеют более округлую форму крыльев;
голова, грудь и передние крылья черные; жилки на передних крыльях
желтые; у основания крыла проходит широкая желто-белая полоса, почти
прямая; от базальной трети крыла косая широкая желто-белая полоса,
идущая кобо к tornus, вливается в широкую желтую полосу, идущую
вдоль терминального края крыла; задние крылья оранжево-желтые с не-
правильными большими черными пятнами по терминальному краю, иногда
сливающимися в неправильной формы перевязь; нижняя сторона желто-
оранжевая с круглым черным дискоидальным пятном на задних крыльях
и тремя черными круглыми пятнами — двумя на передних и одним
на задних крыльях; брюшко желтое; губные щупальца и ноги желто-
бурые. Размах крыльев самцов 42—46 мм, самок 50—55 мм.
Uncus короткий, слабо изогнутый вентрально, сидит на щироком ос-
новании, сросшемся с anus; девятый сегмент узкий, отделен швом от
S. NUMENS
307
uncus, saccus не образует; valvae короткие и очень широкие; по дисталь-
ному скошенному краю расположены зубцы; на дорзальном крае valva
выемка и два больших зубца; fultura inferior в виде слабой мембраны;
Рис. 141. Numens disparilis Stgr. Самец. (Рис. Ф. Гуняева).
142. Numens disparilis Stgr. Гениталии?1
а — самца, б — самки.
penis сильно расширен в проксимальной части с большим мешковидным
coecum и тонким дистальным отделом; pars inflabilis несет многочислен-
ные расположенные пучком мелкие шипики и сидящие отдельно длинные
шипы. Bursa copulatrix меш-
ковидная, по краям слабо
выемчатая, несет глубокую
рубчатую скульптуру и <,с
вентральной стороны узкую
lamina dentata, усаженную
большим числом мелких, но
сильных выростов; ductus
bursae при основании узкий,
далее выгнутый, посредине
сильно расширен, в дисталь-
ной части вновь несколько
сужен, равномерно склеро-
тизован; apophyses anteriores
расширенные в дистальной
части, длинные, но короче
ductus bursae; apophyses pos-
teriores несколько короче;
восьмой сегмент короткий,
несколько конический: papil-
lae anales мягкие, покрыты рис
короткими волосками.
Распространение.
Амур; южное Приморье (р. Викин, Уссури, Владивосток, Аскольд);
Корея; Японские острова (Охайяма); центральный и западный Китай.
Биология не изучена; кормовые растения — Betulaceae (Garpi-
nus, Ostryia); гусеницы в июне—июле.
20*
308
I. ORGYINAE
9. Род ARTAXA WLK.i
\yalker, 1855, Cat. Lep. Het. Brit. Mus., 4 : 794; Moore, 1882, Lep. CeyL,
2 : 85; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 451; Bryk, 1934, Lep. Cat., 62:215
(syn.); — Cispidia Walker, 1855, Cat. Lep. Het. Brit. Mus., 4 : 857 (typus plagiata
Wlk.); — Ama Walker, 1855, там же, 4 : 1176 (typus apicalis Wlk.);—Adlullta
Walker, 1865, там же, 32 : 394 (typus oreosauria Swinh.); — Themaca Walker,
1865, там же, 32 : 394 (comp ar ata Wlk.); —Orvasca Walker, 1865, тамже, 32 : 502
(typus subnotata Wlk.).
Тип рода A. flava F. (non Bremer).2
Хоботок есть в виде рудимента или даже закручен спирально, недлин-
ный; губные щупальца, торчащие вперед, длинные; второй членик их
особенно длинный, третий членик около одной четверти длины второго;
глаза большие, круглые, голые; усики самцов очень сильно гребенчаты,
гребешки на концах несут две-три длинных щетинки; первый членик
лапки передних ног более половины длины голени этих ног; ноги боль-
шею частью покрыты густыми волосками; голени задних ног с двумя
парами шпор; коготки всех ног с широким вентральным зубцом; голова
и грудь покрыты плоскими, узкими и длинными чешуйками и неболь-
шим количеством волосков,; крылья покрыты широкими чешуйками
с очень малой примесью волосков; первый радиальный ствол свободный;
второй, третий, четвертый и пятый ветвятся позже отхождения первого
медиального ствола, два первых в костальный край и два вторых в тер-
минальный; второй и третий медиальные стволы отходят посредством
дискальных от первого кубитального; базальная ячея на задних крыльях
большая, ланцетовидная, незамкнутая (очень редко почти полный ана-
стомоз и замкнутая ячея — у Artaxa piperita Obth.); третья медиальная
жилка на задних крыльях обнаруживает разную степень анастомоза
с первым кубитальным стволом — или свободна, как и вторая медиаль-
ная, и они обе отходят с помощью дискальных {Artaxa niphonis Btl.),
или на стебельке, или, наконец, полностью слита с первым кубитальным
стволом {A. piperita Obth.); брюшко покрыто мелкими волосками, конец
его у самок несет пучок темноокрашенных (коричневых или рыжих)
волосков.
Из южного Приморья из сборов Нассовского и Волкова (1940) мною
получены гусеницы Orgyidae (от А. М. Герасимова), среди которых ока-
залось два вида Artaxa Wlk. Они, без сомнения,1 2 относятся к этому роду,
хотя при них и не было выведенных бабочек; одни, более крупные,
принадлежат, повидимому, к виду A. niphonis Btl., другие —кА. sub-
jlava Brem. (ниже я привожу их описания при соответствующих видах
рода). Сравнение их с рисунками гусениц Artaxa Wlk. с Цейлона в ра-
боте Мура (1882)- дает очень большое сходство; ожидать в южном При-
морье гусениц этой группы можно только для двух указанных видов
(A. piperita Obth. мельче остальных и встречается значительно реже).
1 Описание этого рода дано на основании изученйя палеарктических видов
и, в частности, A. subflava Brem., близкого к flava F. — типу рода Artaxa Wlk.
2 Типом рода Artaxa Wlk. Кирби (1892) указывает ВопгЪух flava F.; по Штранду
(1910), ВопгЪух flava F. является nomen nudum, и название это должно быть отброшено.
К. виду A. flava F., отмеченному как тип рода Artaxa Wlk., Кирби (1892) в этом же
каталоге ставит синонимом digramma Boisduval (Icon. Beg. Anim. Ins., t. 86, t. 4,
1829), распространенную на Цейлоне и описанную Муром (1882). Таким образом,
если считать название A. flava F. названием без описания, то типом рода Artaxa
Wlk., по Кирби (1892), должен остаться A. digramma Bd., близкий к A. subflava
Brem.
9. ARTAXA
309
Хетотаксия обоих видов одинакова (рис. 143) и очень близка к тако-
вой видов Euproctis Hb.; передне-, средне- и заднегрудь по хетотаксии
не отличаются от этих же сегментов Euproctis Hb., но первый абдоми-
нальный сегмент обнаруживает большую примитивность, чем у Eupro-
ctis Hb.; бородавка (3 на этом сегменте лишь сильно сближена с а, но
не слита с ней. Таким образом, меньшая специализация видов Artaxa
Wlk., по структурным признакам в имагинальной стадии (двойной
uncus), совпадает й с меньшей структурной специализацией личиночной
фазы. Тело покрыто двумя типами волосков — длинными, зубчатыми,
негустыми и курчавыми, сидящими на больших (двойных) бородавках
на спине гусеницы.
Большой полиморфный род, состав которого ясен лишь в общих чер-
тах; многие тропические виды требуют еще переописания; трудность
Рис. 143. Artaxa subjlava Brem. Хетотаксия
передне- и среднегруди и I, VI, VII и
VIII брюшных сегментов взрослой гусеницы.
характеристики состава рода
связана с тем, что после Мура
(1882) и Кирби (1892) род Arta-
xa Wlk. перестали отличать
от Euproctis Hb. и Porthesia
Steph., хотя два последних
рода различали. Изучения жил-
кования видов, вводимых в эти
два рода, тем не менее не про-
изводилось; так, например,
О. piperita Obth. по жилко-
ванию задних крыльев при-
надлежит к роду Porthesia
Steph., как его понимают Шпу-
лер (1908), Штранд (1910),
Геринг (1932), между тем этот
вид относят (Штранд, 1910;
Брик, 1934) в род Euproctis Hb. По существу родовые признаки ни
для Artaxa Wlk., ни для Euproctis Hb. или Porthesia Steph, не были даны,
и распределение видов по родам производилось по внешнему облику
на глаз.
На основании просмотра описаний и изображений разных видов
Euproctis Hb. и Porthesia Steph., а также некоторых видов в натуре можно
считать, что род Artaxa Wlk. является почти нацело индомалайским.
Наибольшее число видов этого рода (свыше полутораста) распространено
в Индии, Гималаях, в Индокитае и на островах Малайского архипелага.
Значительно распространение видов Artaxa Wlk. на юго-востоке Азии —
в Китае, Корее и Японии, где есть ряд эндемов (A. straminea Leech, con-
spersa Btl., staudingeri Leech, flavinata Wlk., mantis Leech, bipunctapex
Hamps., inconspicua Leech, recurvata Leech, seitzi Strd., tarasan Holl.,
riukiuana Mts., aurata Wlk., curvata Wil., sakaguchii Mts., magniplaga
Gaede, parviplagiosa Strd.); три приведенных ниже вида из этой группы
заходят в пределы СССР в южное Приморье, будучи распространенными
в Китае, Корее и Японии.
На Новой Гвинее виды Artaxa Wlk. еще многочисленны, но из Австра-
лии (почти исключительно из ее восточной части) известно не более двух
десятков видов этого рода; для Тасмании отмечен один вид, а для Новой
Зеландии нет указаний на наличие не только видов Artaxa Wlk., но во-
обще близких видов типа Euproctis Hb. Виды Artaxa Wlk. отсутствуют
также в Африке, на Мадагаскаре и в Южной и Северной Америке.
310
I. ORGYINAE
Биологически виды этого рода совершенно не изучены, хотя некото-
рые отмечены как вредители культурных растений, например чая, в тро-
пиках.1
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Передние крылья с большим количеством черного цвета, заливаю-
щим дорзальный край их или с размытым черным пятном у дор-
зального края; на месте дискоидальной жилки черная точка; зад-
ние крылья самцов черные; uncus очень короток.....................
. . . ....................................* 3. A. niphonis Btl.
2 (1). Передние и задние крылья однотонно желтые или с желтыми, ко-
ричневыми или охристыми элементами рисунка; черный рисунок
не развит; на месте дискоидальных жилок нет черных точек; uncus
длинный.
3 (4). Передние крылья с коричнево-желтой перевязью сложной формы
и четырьмя черными точками во внешнем поле; реже передние
крылья однотонные; valva с отростком с вентральной стороны;
bursa copulatrix узкая и длинная..............* 2. A. piperita Obth.
4 (3). Передние крылья с узкой почти прямой слабоохристой или жел-
товатой полосой, реже они одноцветно желтые; valva без выроста;
совокупительная сумка короткая и широкая..........................
.......................................* 1 • A. subfl a va Brem.
* 1. Artaxa subflava Brem. (рис. 143, 144).
Bremer, 1861, Bull. Acad. Sci. Pet., 3 : 479 (jlava Brem., nomen praeoccupatum); Bre-
mer, 1864, Lep. Ost.-Sib. : 41, t. 3,fig. 19; Staudinger, 1901, Cat. Lep., 1 : 116: Swinhoe,
1903, Trans. Entom. Soc. Lond. : 415; Matsumura, 1909, Thous. Ill. Ins. Jap., Suppl.
1 : 58, t. 10, fig. 4; Strand, 1910, Seitz, Grossschmett., 2 : 134, t. 21, a; Swinhoe, 1922.
Ann. Mag. Nat. Hist., 11 (9) : 52; Куренцов, 1922, Зап. Южно-уссур. отд. Географ.
общ., 1 : 23; Matsumura, 1931, 6000 Ins. Japan : 705, fig. 444; Matsumura, 1933, Ins.
Matsum., 7 : 125; Bryk, 1934, Cat. Lep., 62 : 218; Collenette, 1934, Stylops, 3 : 116; —
flava Bremer (non Fabricius), 1861, Bull. Acad. Sci. Pet., 3 : 479 (Area); — intense.
Butler, 1877, Ann. Mag. Nat. Hist., 20(4) : 402 (Artaxa); — bipunctigera Strand,
1910, Seitz, Grossschmett., 2 : 136.
Голова, грудь' и крылья светложелтые, слегка охристые; передние
крылья покрыты чешуйками без примеси волосков; базальная часть передо
них крыльев охристо-желтая; срединное поле с широкой изогнутой тем-
ной полосой, которая в костальной половине широкая, в дорзальной
сужена, она образована крупными коричнево-черными чешуйками; по
терминальному краю несколько (два—четыре) больших бурых пятен —
скоплений крупных бурых чешуек; бахрома желтая; дорзальный край
передних крыльев несет длинные, ланцетовидные чешуи и волоски в виде
широкой бахромы; задние крылья одноцветные, желтые; нижняя сторона
одноцветно желтая, у самок однотонная, у самцов костальный и терми-
нальный края передних крыльев бурые; усики, губные щупальца и ноги
одноцветно желтые; усики сильно гребенчатые, гребешки члеников несут
длинные острые щетинки; брюшко одноцветно желтое, дистальный конец
1 Гусеницы тропических видов рода Artaxa Wlk. не описаны сколько-нибудь
подробно, но изображены, например у Мура (1882); по габитусу они очень напо-
минают евразийских Euproctis similis Fuessly и chrysorrhoea L., но по рисункам бро-
сается в глаза отличие выростов (бородавок) на первом и втором брюшных сегмен-
тах; они бблыпих размеров, чем'у Euproctis Hb. Сходство их с дальневосточными
видами Artaxa Wlk. очень велико.
9. ARTAXA
311
брюшка у самцов слабо затемнен, у самок несет подушку буро-желтых
волосков. Размах крыльев самцов 25—30 мм, самок 30 —40 мм.
Uncus двойной, в виде пары тонких слабо изогнутых в дистальной
части округлых отростков; tegumen широкий; anus несет scaphium,
покрытый мелкими волосками; вентральная половина девятого сегмента
узкая, не образует saccus; valvae при основании широкие, изогнутые,
в дистальной части суженные, покрыты мелкими густыми волосками;
penis почти цилиндрической формы с двумя выростами, в дистальной
"части несколько утолщен. Bursa copulatrix мембранозная, мешковидная,
покрыта очень тонкой рубчатой скульптурой; восьмой сегмент сложной
•формы, длинный, в вагинальной области образует складку, с дорзальной
стороны сужен, несет длинные
•apophyses anteriores; они длин-
нее ductus bursae; apophyses
posteriores длинные; papillae
anales короткие, несколько
заостренные.
Гусеницы этого вида не
•описаны.' Мною были получены
четыре особи (через А. М. Ге-
расимова) из сборов Кассов-
ского в Сучане. Они были най-
дены на орешнике в паутинном
гнезде; гусеницы полувзрослые.
Они, без сомнения, относятся
к роду Artaxa- Wlk. и, вероят-
но, к виду A. subflava Brem.
Я даю следующее их описание.
Голова черная, блестящая, по-
крыта негустыми темными вто-
ричными волосками; основание
усиков, вентральная часть на-
личника и отдельные части
нижней губы матово-белые;
тело уплощенной формы
Рис. 144. Artaxa sabjlava Brem.. Гениталии:
а — самца, б — самки.
кое, почти веретеновидное (при
рассмотрении сверху), так как первые три брюшных сегмента рас-
ширены; дорзальная сторона грудных сегментов черная, с тонкой ох-
ристой срединной полоской на передне- и среднегруди, расширяющейся
в неправильное дорзальное пятно на заднегруди; первые два брюшных
сегмента с дорзальной стороны черные, несколько вздуты и несут по две
больших бородавки черного цвета (и, кроме них, еще по две маленьких (3);
на дорзальной стороне третьего и четвертого брюшных сегментов непра-
вильные охристые пятна; маленькое желтое пятнышко на тергите пятого
сегмента; прочие сегменты брюшка черные; железки на шестом и седь-
мом брюшных сегментах матово-белые; восьмой сегмент несколько вздут
и несет бородавки большей величины; по бокам тела на уровне стигм
и ниже их неправильная охристо-желтая полоса; стигмы черные, хорошо
заметны; бородавки по бокам тела желтые; вентральная сторона тела
черная с очень мелкими бородавками, несущими мелкие и немногие
волоски; ноги, как брюшные, так и грудные, черные со светлыми колеч-
ками; кожа покрыта двумя типами волосков — длинными, слабо пери-
стыми и короткими, более сильно перистыми, но не курчавыми.
312
I. ORGYINAE
Распространение. Амур; южное Приморье (Ворошилов-
Уссурийский, Сучан, Аскольд, Сидеми, Супутинка, р. Сица, Седанка,
Лянчихе); Корея; Японские острова (Хондо, Хоккайдо); северный не
западный Китай.
Биология и экология. Гусеницы в первой половине лета;:
отмечено питание Rubus melanolasius (Куренцов, 1939); гусеницы редки..
В южном Приморье в уремных лесах. Лёт бабочек в июле—августе..
* 2. Artaxa piperita Obth. (рис. 145, 146).
Oberthiir, 1880, Et. Entom., 5 : 35 (Leucoma); Kirby, .1892, Syn., Cat. Lep. Het.,.
1 : 451 (Artaxa)', Leech, 1899, Trans. Entom. Soc. Lond. : 135; Staudinger, 1901, Cat.
Lep., 1 : 116; Strand, 1910, Seitz, Grossschmett., 2 : 136, t. 21, c; Swinhoe, 1922, Ann.
Mag. Nat. Hist., 10 (9) : 481 (Nygmia); Kawada, 1931, Kontyu, 5 : 125; Matsumura,
1931, 6000 Ins. Japan : 708, fig. 456; Gaede, 1932, Seitz, Grossschmett-., 2. SuppL:
Рис. 145. Artaxa piperita Obtn. Самка. (Рис. Ф.
Гуняева).
104, t. 10, a; Matsumura, 1933, Ins. Matsum., 7 : 147 (Porthesia); Bryk, 1934, Lep. Cat.,
62:220; Collenette, 1934, Stylops, 3:11; — snelleni Staudinger, 1887, Mem. Lep..
Rom., 3 : 207, t. 12, fig. 3; — staudingeri Matsumura, 1909, Thous. Ill. Ins. Jap., SuppL,
1 : 60, t. 10, fig. 8.
Грудь, крылья и голова лимонно-желтого цвета; рисунок на передних
крыльях редуцирован; на светложелтом фоне крыла расплывчатое-
желто-бурое пятно, идущее от основания крыла и расширяющееся в сред-
ней части крыла в неправильную размытую широкую перевязь; на тер-
минальном поле передних крыльев четыре небольших пятна (того же-
цвета, как и перевязь); перевязь и пятна несут крупные неправильно-
разбросанные чешуйки, выделяющиеся в виде мелких точек черного-
цвета; бахрома передних крыльев желтая; дорзальный край крыла несет
длинные, ланцетовидные чешуйки, свисающие на заднее крыло; задние
крылья чисто желтые; нижняя сторона желтая с просвечивающей темной
неясной перевязью на передних крыльях; губные щупальца, ноги и ос-
нование брюшка одноцветно желтые; конец брюшка у самцов светложел-
тый, у самок последние сегменты несут подушку рыже-желтых волос,
усики самцов сильно гребенчатые, с тонкими гребешками и на их конце
9. ARTAXA
31,
щетинками; у самок гребешки усиков очень укорочены. Размах крыльев
самок 30—35 мм, самцов 25—30 мм.
Uncus плоский, в виде пластинки, в дистальной части выемчатый,
с двумя выростами (двойной); девятое кольцо в дорзальной половине
вздутое, широкое, в вентральной части образует конический и длинный
saccus; fultura inferior короткая и широкая, выемчатая по дорзальному
краю; valvae вытянутые, но широкие, с коротким коническим вентраль-
ным отростком в дистальной части; penis в дистальной части расширен
и склеротизован. Bursa copulatrix мембранозная, длинная, расширенная
• в проксимальной части; ductus bursae короткий и широкий, склероти-
зован по всей длине, соединен
перетяжкой с bursa copulatrix;
восьмо.! сегмент сильно расши-
рен в вентральной части, в
дистальной половине несколько
сильнее склеротизован и пунк-
тирован; apophyses anteriores
и posteriores узкие и длинные;
papillae anales с небольшими
острыми выростами в дисталь-
ной части.
Распространение.
Приамурье (Хабаровск); южное
Приморье (Уссури, Владивос-
ток, Аскольд, Сйдеми, Сучан,
Виноградовка, Седанка); Ко-
рея; Япония (Хондо); север-
ный и средний Китай.
* 3. Artaxa niphonis Btl.
(рис. 147, 148).
Butler, 1881, Trans. Entom. Soc.
Lond. : 9 (Chaerotricha)', Kirby, 1892,
Syn. Cat. Lep. Het., 1 : 447 (Nygmia)',
Leech, 1899, Trans. Entom. Soc.
Lond. : 133; Staudinger, 1901, Cat.
Lep., 1 : 116; Swinhoe, 1903, Trans.
Entom. Soc. Lond. : 418; Strand,
1910, Seitz, Grossschmett., 2 : 136;
Swinhoe, 1922, Ann. Mag. Nat.
Hist., 10 (9): 481; Куренцов, 1922,
Рис. 146. Artaxa piperita Obth. Гениталии:
а — самца, б — самки.
Зап. Южно-уссур. отд. Географ, общ., 1 : 24;
Matsumura, 1931,6000111. Ins. Japan : 707, fig. 454; Matsumura, 1933, Ins. Matsum.,
7 : 128; — squamosa Butler, 1881 Trans. Entom. Soc. Lond. : 9 (Chaerotricha); —
raddei Staudinger, 1887, Mem. Lep. Rom., 3-: 207, t. 17, fig. 3 (Porthesia)', — coreana
Staudinger, 1892, там же, 6 : 311 (Artaxa)',—melanostigma Moltrecht, 1929, Зап.
ВлаДив. отд. Географ, общ., 3 (2) : 50; — karonis Matsumura, 1933, Ins. Matsum.,.
7 : 128, t. 3, fig. 2. .
Основной цвет лимонно-желтый; грудь у обоих полов желтая, у сам-
цов с значительной примесью серого тона; голова желтая; глаза чер-
ные с немногими желтыми пятнами; усики самцов сильно гребенчатые,
у самок гребешки более короткие, усики покрыты белыми чешуйками;
щупальца покрыты желтыми и черными чешуйками, последний членик
их короткий; ноги желтые, на лапках продольные ряды черных чешуй;
брюшко у самца с дорзальной стороны темносерое, с боков и вентральной
стороны желтое; у самок брюшко чисто желтое; передние крылья у сам-
All
I. ORGYINAE
цов желтые лишь по костальному краю и вдоль терминального края, -
прочие части шоколадно-черного цвета с немногими неправильно разбро-
санными желтыми чешуйками; у самок они почти чисто желтые с боль-
Рис. 147. Artaxa niphonis Btl. Самец. (Рис. Ф. Гуняева).
длим размытым и стушеванным темным пятном у дорзального края;
в области дискоидальной жилки круглое черное пятно, более резкое
У самца; задние крылья самцов кофейно-черные с желтой бахромой,
у самок' одноцветно жел-
тые; в области дискоидаль-
ной жилки на задних
крыльях темное круглое
пятно, более яркое у сам-
ца; нижняя сторона пе-
редних крыльев у самца
черная с желтым косталь-
ным краем, желтыми жил-
ками и бахромой; нижняя
сторона задних крыльев
желтая с немногими жел-
тыми полями между жил-
ками; на задних крыльях
более ясные дискоидаль-
ные пятна; у самок ниж-
няя сторона обеих пар
крыльев светложелтая с
коричневыми круглыми
дискои дальними пятнами
на каждом крыле. Размах
крыльев самцов 32—36 мм,
самок 36—42 мм.
Uncus короткий, двойной; девятое кольцо короткое, saccus недораз-
вит; anus разрастается в виде конуса, сильнее обычного склеротизован,
покрыт короткими волосками; valvae короткие и широкие, при основании
выгнутые, с зубцами по вентральному краю и с одним большим зубцом
л средней части; дистальный конец valva несет длинный склеротизован-
9. ARTAXA, 10. EUPROCTIS
315
ный крючкообразный отросток; penis расширен в проксимальной части,
с недлинным coecum, дистальная часть penis несет многочисленные мел-
кие шипики; на pars inflabilis небольшие склеротизованные пластинки.
Bursa copulatrix большая, мешковидная, мембранозная, с небольшой
почти треугольной lamina dentata на внешнем крае; ductus bursae скле-
ротизован, расширен в дистальной части, от его основания отходит duc-
tus seminalis; восьмой сегмент неширокий, суженный в дистальной части,
несет короткие apophyses anteriores; apophyses posteriores короткие;
papillae anales почтй прямоугольные, мембранозные.
Гусеницы этого вида не известны. Повидимому, к нему относятся
четыре особи, полученные мною из сборов Волкова; они были собраны
на розах, орешнике, смородине и алтайском крыжовнике на ст. Океан-
ской близ Владивостока 9 июля 1940 г. Принадлежность их к роду
Artaxa Wlk. несомненна и очень вероятна именно к этому виду. Голова
черная и блестящая, бедна вторичными щетинками; ротовые части чер-
ные; вдоль спины на грудных сегментах широкая оранжево-охристая
полоса, окаймленная с боков черным; дорзальная сторона первых трех
брюшных сегментов черная, несет более крупные бородавки, чем на про-
чих; тергиты четвертого, пятого, шестого, седьмого, а также отчасти
третьего сегментов брюшка желто-охристые; окраска располагается
в виде неправильной полосы, окаймленной черным; бока тела желто-
охристые; брюшная сторона черная; ноги, как брюшные, так и грудные,
черно-бурые; тело покрыто желто-охристыми волосками; бородавки тела
небольшие, черные, с охристыми пятнышками; на восьмом сегменте они
охристо-желтые, охристые они и на дорзальной стороне грудных сегмен-
тов и по бокам тела; волоски, покрывающие тело, сильно перистые,
двух типов — одни более длинные, редкие, с редкими недлинными рес-
ничками и другие, сидящие густо, короткие, курчавые и сильно пери-
стые, особенно обильные на дорзальной стороне тела, на больших круг-
лых и плоских бородавках.
Изменчивость бабочек не велика; в наибольшей мере варьирует чер-
ная окраска крыльев, которая то почти совсем исчезает, то, напротив,
крылья почти чисто черные; наиболее темные особи описаны как согеапа
Stgr., но такие темные особи известны и из южного Приморья и с Амура;
редко бывают самки с примесью черной окраски, как и самцы, или со-
вершенно черные.
Распространение. Приамурье (Раддевка, Хабаровск);
южное Приморье (Виноградовка, Седанка, Владивосток, Сидеми, р. Сица,
р. Суйфун, Сучан, Ворошилов-Уссурийский); Корея; Японские острова
(Хондо); северный Китай.
Биология. Гусеницы питаются листьями Ostryia и Carpinus
cordata (по Грэзеру).
10. Род EUPROCTIS НВ.
Hiibner, 1816, Verz. bekannt. Schmett. : 159; Aurivillius, 1891, Nordn. Fj’.r. : 94;
Kirby, 1892, Syn. Cat. Lep. Het., 1 : 442; Hampson, 1892, Fauna Brit. Ind., Moths,
1 : 470; Staudinger, 1901, Cat. Lep., 1 : 116; Swinhoe, 1903, Trans. Entom. Soc. Lond. :
395; Spuler, 1908, Schmett. Eur., 1 : 132; Rebel, 1910, Berge, Schmetterlingsb., ed.
9 : 116; Strand, 1910, Seitz, Grossschmett., 2 : 135; Swinhoe, 1922, Ann. Mag. Nat.
Hist., 10 (9) : 435; Hering, 1926, Seitz, Grossschmett., 14 : 143; Bryk, 1934, Lep.
Cat., 62 : 215; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 67, t. 14, fig. 13; —
.Nygmia. Hiibner, 1816, Verz. bekannt. Schmett. : 193 (typus phaeorrhoea Don., sec.
Kirby, 1892);—Porthesia Stephens, 1829, Ill. Brit. Entom. Haustell., 2 : 65 (typus
similis Fuessly); — Antipha Walker, 1855, Cat. Lep. Het. Brit. Mus., 4 : 806; (typus
316
1. ORGYINAE
costalis Wlk.); — Urocoma Herrich-Schaeffer, 1855, Aussereurop. Schmett., 1 : 82 (typug
limbalis H. S.); :— Chianophasma Butler, 1886, Trans. Entom. Soc. Lond. : 384 (typus
paradoxa Btl.).
Тип рода E. chrysorrhoea L.
Хоботок утрачен вполне; губные щупальца слабые, свисающие вниз
или косо дистально; третий членик их довольно длинный, второй корот-
кий, около одной трети длины второго; лоб гладкий, покрытый воло-
сками; глаза большие, голые; усики самцов очень сильно гребенчатые;
гребешки несут на концах две длинные щетинки и одну короткую; пер-
вый членик передней лапки около половины длины передней голени;,
голени задних ног с двумя парами шпор; коготки сильно изогнуты,,
с большим острым щироким шипом на вентральной стороне; голова и
грудь покрыты тонкими пушистыми волосками; крылья, кроме чешуи,
покрыты волосками; радиальная ячейка на передних крыльях не раз-
вита: второй, третий, четвертый и пятый радиальные стволы ветвятся
позже отхождения от этого ствола первой медиальной жилки, которая
отходит с помощью дискальной лишь немного выше ветвления первого
радиального ствола; базальная ячея на задних крыльях большая, вытя-
нутая, заостренная, незамкнутая или вполне замкнутая, когда (как,
например, у Е. chrysorrhoea L.) радиальная и субкостальная жилки ана-
стомозируют на известном участке вполне; третий медиальный ствол
и первый кубитальный на задних крыльях на стебельке или слиты вполне,,
второй медиальный ствол отходит посредством дискальной жилки; на
передних и задних крыльях в дискоидальной ячее остаток медиального
ствола в виде складочки; брюшко покрыто тонкими волосками; послед-
ние сегменты брюшка часто окрашены более ярко и у самок несут подушку
густых коротких волос (золотистых, коричневых или иных).
Степень анастомоза третьей медиальной и первой кубитальной жилок
на задних крыльях различна даже у одного и того же вида; так, напри-
мер, среди серии Е. similis Fuessly я обнаружил особей, у которых эти
жилки были разделены в дистальной части на самостоятельные стволы
примерно на одной шестой их длины, в то же время у Е. chrysorrhoea L.
и Е. karghalica Moore эти жилки нередко слиты на протяжении более
трети их длины. Наконец у этих же видов и ряда других слияния этих
жилок может совсем не быть. Обращает внимание связь распространения
на север со степенью редукции покровных образований (чешуйчатого
покро'ва), ротовых частей и слияния жилок; повидимому, наиболее
специализованные виды рода могут иметь это слияние большим, чем
генерализованные. По всем другим признакам виды, относимые к Рог-
thesia Steph, (rnelania Stgr., similis Fuessly), представляют один комплекс
с видами Euproctis Hb. и филогенетически от них неотделимы.
Гусеницы покрыты неравномерной длины волосками, часть из которых
сложной, курчавой и перистой формы; окраска тела яркая с примесью охри-
стых и красных тонов. Первыйи второй сегменты брюшка заметно шире про-
чих. Бородавка х переднегруди большая, несет пучки коротких волосков, не
выдающихся вперед; на сегментах брюшка х или несет одну щетинку, или
полностью утрачена. Бородавки а и (3 на средне-и заднегруди далеко расста-
влены. Эти же бородавки на всех сегментах брюшка слиты и несут короткие-
нередко курчавые волоски, особенно часто на первых сегментах. Бородавки
о несут щетинистые волоски. Часто есть ядовитые волоски. Куколки чер-
ные, блестящие, сильно конической формы; губные щупальца равны
примерно половине ширины максилл; усики самцов широкие по всей
длине. Волоски, покрывающие тело куколки., негустые, отдельными
10. EU PROCT is
317
небольшими пучками. Яйца крупные — 0.7—0.8 мм диаметром, покры-
ваются обычно золотистыми волосками с конца брюшка самки, отклады-
ваются кучками.
Большой, широко распространенный род, наиболее богато представ-
ленный в африканской фауне, где виды Euproctis Hb. обитают не только
в области влажных тропических лесов (Золотой Берег, Камерун, Огове),
но главным образом в саваннах восточной Африки и степях южной
(Кения, Родезия, Капская Земля). В тропической Африке распростра-
нено около ста видов этого рода; некоторые из них напоминают по обли-
ку европейскую chrysorrhoea L. или сирийскую melania Stgr., ио боль-
шая часть сходны с малоазийской susanna Stgr. Ряд видов известен с Ма-
дагаскара.1
В Индии, на Цейлоне и в Гималаях виды Euproctis Hb. еще довольно
многочисленны (Е. semisignata Wlk., postincisa Moore, semivitta Moore,
flavipuncta Hamps., endoplagia Hamps., divisa Wlk., albescens Swinh.,
oirgo Swinh., costalis Wlk., xanthorrhoea Ko 11., subfasciata Wlk., signata
Blanch., venosa Moore, cervina Moore); значительное число видов есть на
Новой Гвинее (Е. virginea Beth.-Bek., aroa Beth.-Bek., alba Beth.-Bek.,
meeki Beth.-Bek., ekeikei Beth.-Bek., awgarra Beth.-Bek., gracilior Pag.)
и в восточной Австралии (Е. aliena Btl., panabra Turn., galactopsis Turn.,
enthusana Turn., hololeuca Meyr.); три вида указаны для Тасмании (Н. 1еи-
comelas Wlk., mixta Btl., anacausta Meyr.).
На островах Малайского архипелага виды Euproctis Hb. менее много-
численны (F. tenuis Swinh., civitta Swinh., virguncula Wlk., isabellina
Heyl., biplagatana Strd., signiplaga Wlk.); немногие виды проникают
в Китай (Е. albescens Swinh., divisa Wlk.), на Филиппинские острова
{Е. albescens Swinh., nigrofasciata Semp.); один вид описан из Японии
(Н. karapind Strd.), где распространен также евразийский Е. similis
Fuessly.
В умеренной полосе (лесостепи) Евразии распространен только один
вид — Е. similis Fuessly, но в Средней Азии известно два эндемичных
вида (Е. laciaea Moore и Е. karghalica Moore). Один эндемичный вид опи-
сан из Аравии и Сирии (Е. melania Stgr.), один из Малой Азии (Е. susanna
Stgr.) и два из северной Африки (Тунис, Алжир — Е. boulifa Dum. и
Е. charmentanti Vuill.); один вид (Е. chrysorrhoea L.) распространен почти
всюду в Европе и на Кавказе (рис. 149).
В Северной и Южной Америке нет видов Euproctis Hb.; в восточные
штаты в конце прошлого столетия был занесен европейский Е. chrysor-
rhoea L., акклиматизировавшийся там и ставший бичом культурных
садов и лесов (широколиственных), значение которого, однако, сейчас
значительно уменьшилось благодаря успешному применению пара-
литов.
Биологически хорошо изучены только два евразийские вида, которые
связаны с Fagales и Bosales; они характеризуются одним поколением
в сезон и зимовкой в фазе гусеницы. Циклика и биология тропических
видов не изучены; связь их с древесной и'кустарниковой растительностью
несомненна,.
1 Состав рода в африканской фауне см. у Геринга (Hering, 1926, Seitz, Gross-
schmett., 14 : 143); отсутствие там видов Artaxa Wlk. избавляет от необходимости
специального их перечисления, тем более, что число видов этого рода там очень
велико (более ста).
Рис. 149. Распространение палеарктических видов рода Euproctis Hb.
златогузка (Е. chrysorrhoea L.), черным отмечен ареал вредной деятельности, 2 — Е. karghalica Moore, 3 — Е; similis Fuessly.
10. EUPROCTIS
319
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ.
1 (4). Передние крылья чисто белые или с небольшими темными стуше-
ванными пятнами или штрихами у дорзального края, в середине
и во внешней половине крыла.
2 (3). Третий медиальный ствол на задних крыльях свободный или
на общем стебельке с первым кубитальным, но не слит с ним пол-
ностью; uncus на дистальном конце с бороздой; valva с гладкими,
краями; penis в дистальной части с зубчиками с вентральной сто-
роны; девятый сегмент несет subunci; bursa copulatrix без lamina
dentata; ductus bursae снабжен lamella antevaginalis.............
......................................* 2. E. chrysorrhoea L.
3 (2). Третий медиальный ствол на задних крыльях слит с первым куби-
тальным, редко на очень длинном стебельке; uncus острый па ди-
стальном конце; valva с гофрированными краями; subunci не
развиты; penis на дистальном конце с выростом, направленным вен-
трально; bursa copulatrix с вентральной стороны несет lamina den-
tata; ductus bursae в середине склеротизован, без lamella anteva-
ginalis ....................................* 1. Е. similis Fuessly
4 (1). Передние крылья белые с черным и охристым рисунком или жел-
тые с таким же рисунком.
5 (8). Крылья белые с охристым и черным рисунком.
6 (7). Крупный, 30—40 мм; во внешнем поле черные и охристые пятна
не слиты в сплошную полосу; на месте дискальной жилки охристое
или черно-охристое пятно.................* 3. Е. karghalica Moore
7 (6). Мельче 25—30 мм; дискальная жилка белая; во внешнем поле
черные и охристые штрихи слиты или почти слиты в полосу . . .
...............................................4. Е. melania Stgr.
8 (5). Крылья желтые с охристыми тонами; на передних крыльях две-
поперечных охристых полных или неполных полосы...................
..........................................5. Е. susanna Stgr.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПО ГУСЕНИЦАМ (ВЗРОСЛЫМ
И ПОЛУВЗРОСЛЫМ)
1 (4). На первом и втором брюшных сегментах небольшие утолщения,
несущие короткие густые и курчавые волоски; курчавые волоски
находятся также нй брюшных бородавках, расположенных над
стигмами на каждом сегменте (р).
2 (3). Вдоль спины яркая красная полоса, разделенная тонкой черной
полоской; курчавые волоски, покрывающие бородавки, чисто белые;
нижняя сторона тела с яркими красными полосками.................
....................................* 1. Е. similis Fuessly
3 (2). Вдоль спины узкая темная полоска; курчавые волоски, покры-
вающие бородавки, охристые; брюшная сторона тела с охристо-
серыми полосками; красный цвет на коже отсутствует..............
.................................... * 2. Е. chrysorrhoea L.
4 (1). На дорзальных бородавках первого и второго брюшных сегментов
щетинистые (зубчатые) волоски; курчавых волосков нет на боковых
брюшных бородавках над стигмами (р); вдоль спины широкая
яркая охристая полоса; грудные сегменты с дорзальной стороньг
желтые .............................. . * 3. Е. karghalica Moore
320
I. ORGYINAE
*1. Euproctis similis Fssl. —Желтбгузка (рис. 150, 151).
Fuessly, 1775, Verz. Schweiz. Ins. : 35 (Phalaenay, Fuessly, 1778, Mag. Iris.,
2 : 287; Aurivillius, 1891, Nordn. Fjar. : 94, t. 25, fig. 5 {Euproctis)', Kirby, 1892, Syn.
Cat. Lep. Het., 1 : 445; Standfuss, 1896, Handb. Pal. Schmett., 1 : 168; Leech, 1899,
Trans. Entom. Soc. Lond. : 40; Staudinger, 1901, Cat. Lep., 1 : 116; Swinhoe, 1903,
Trans. Entom. Soc. Lond. : 392; Spuler, 1908, Schmett. Eur., 1 133, t. 25, fig. 29;
Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 116, f. 37, t. 24, f. 4; Strand, 1910,
.Seitz, Grossschmett., 2 : 134, t. 24, i; Petersen, 1924, Lep. Fauna EstL, ed. 2 : 150;
Matsumura, 1931, 6000 Ins. Japan : 723; Matsumura, 1933, Ins. Matsum., 7 : 148;
Bryk, 1934, Lep. Cat., 62 : 203 (Porthesia)', Nordstroem und Wahlgren, 1937, Svenska
Fjar. : 68, t. 14, fig. 14; — auriflua Fabricius, 1787, Mantissa Ins., 2 : 125 (Bombyx)-, —
nyctaeq Grum-Grzhimailo, 1891, Tp; Русск. Энтом. общ., 25 : 464; — rebeli Haberhauer,
1902, Soc. Entom., 17 : 82; — trimaculata Strand, 1910, Seitz, Grossschmett., 2 : 134; —
quadrimaculata Strand, 1910, там же, 2 : 134; — coreacola Matsumura, 1933, Ins. Mat-
sum., 7 : 148; —wilczinskia Wize, 1934, Polsk. Pismo Entom., 13 : 108; Wize, 1936, Lam-
billionea, 36 : 183;—punctillata Lempke, 1937, Tijdschr. Entom., 80 : 284.; — sjoquisti
Bryk, 1942, Iris, 56 :26.
Биология и экология. Шевырев, 1890, Сельск. и лесн. хоз., 165 :
13; Eltringham, 1913, Trans. Entom. Soc. Lond.: 423, t. 12; Pictet, 1921, Bull. Soc.
Lep. gen., 4 : 186; Wolff und Krausse, 1922, Wichtigste Forstins. : 128; Lederer, 1923,
Handb. prakt. Entom., 3 : 167; Reh, 1925, Sorauer, Handb. Pflanzenkrankh., 4 : 440;
Somamasu, 1926, Ins. World, 39 (2) : 53; Niisslin und Rhumbler, 1927, Forstinsekten-
kunde : 461, fig. 382; Белозор, 1931, 36ipn. праць Зоол. муз., 10 : 144; Лебедев, 1933,
Зб1рн. праць 1нст. биол., 1 : 62; Филипьев, 1932, Тр. защ. раст., 5 : 368, Куренцов,
1939, Тр. Горнотаежн. ст., 3 : 150.
Голова, грудь, передние крылья и задние крылья белые; рисунок
редуцирован; у самцов на передних крыльях есть остатки второй
и третьей поперечных линий в виде широких растушеванных полос,
образующих бурые неправильной формы пятна у дорзального края кр^ла;
передние крылья покрыты широкими чешуйками и волосками лишь
в базальной части; вдоль дорзального края передних крыльев редкая
бахрома из длинных и очень широких чешуй и волосков; нижняя сторона
передних крыльев у самцов вдоль костального края затемнена, он черно-
бурый, далее к дорзальному краю крыла слабая примесь бурых чешуй;
у самок нижняя сторона крыльев чисто белая, рисунок верхней стороны
также большею частью совершенно утрачен; губные щупальца снизу
и с внутренней стороны белые, снаружи черные; ноги белые, несколько
затемнены по внутренней стороне голеней; базальная часть брюшка
белая, не густо покрытая волосками, дистальные сегменты, начиная
с седьмого, золотисто-желтые; у самки эти волоски образуют густую
подушку на конце брюшка; усики самцов сильно гребенчатые, белые,
гребешки желтоватые; у самок гребешки усиков короткие. Размах крыльев
самцов 30—40 мм, самок 35—45 мм.
Uncus тонкий, но довольно высокий, с плоским дистальным концом,
сидит на широком основании, отделен швом от девятого сегмента; тергит
девятого сегмента довольно длинный, стернит его образует длинный
треугольной формы плоский saccus; fultura inferior образует склероти-
зованное кольцо, сквозь которое проходит penis; valva при основании
широкая, дистальный конец ее сужен и вздернут дорзально; вентраль-
ный край valva несет небольшие выросты; penis в проксимальной части
расширен, соесшп короткий; на дистальном конце penis короткий отхо-
дящий в сторону вырост; pars inflabilis не несет инкрустаций. Bursa
copulatrix расширена в проксимальной части, длинная, мембранозная,
с тонкой рубчатой скульптурой, несет с вентральной стороны в прокси-
мальной части lamina dentata, почти круглую, с двумя короткими выро-
'гтами; ductus bursae короткий и широкий в дистальной части, в средней
10. RU PROCT IS
321
части склеротизован; ductus seminalis отходит от его проксимального
отдела; восьмой сегмент равномерной длины всюду, не широкий и корот-
кий, несет длинные и тонкие apophyses anteriores; apophyses posteriores
также тонкие и длинные; papil-
lae anales короткие и широкие.
Яйца круглые, серой окрас-
ки, хорион тусклый, диаметр
0.5—0.4 мм. Взрослая гусени-
ца черная с коричневым и
красным рисунком; вдоль спины
неширокая карминно-красная
полоса, прерванная па первом
и втором брюшных сегментах
и разделенная вдоль тонкой
черной полоской, несколько
расширенной между сегмента-
ми; на переднегруди дорзаль-
ная полоса почти редуцирова-
на; на тергитах первого и вто-
рого сегментов мясистые невы-
сокие бородавки, покрытые
короткими черными волосками
и более длинными курчавыми
седыми волосками, несущие
также небольшое . число длин-
ных черных щетинок, такие же, Рис. 150. Euproctis similis Fuessly. Гепита-
но меньших размеров бородавки лии: а —'самца, б — самки.
сидят с дорзальной стороны
двумя рядами латерально от дорзальной линии; под стигмами боковая
красная полоса и ряд больших красных бородавок, несущих серые
волоски, а также маленьких красных бугорков; ниже этих рядов боро-
давок располагается над уровнем
брюшных ног ряд серых борода-
вок, несущих светлосерые волоски;
вентральная сторона черно-серая
с красными неправильно разбро-
санными пятнами; голова черпая
с желтыми адфронтальными поля-
ми; ноги черные, слегка сероватые
на концах; брюшные ноги серые;
стигмы черные, почти круглые,
слабо выделяются на коже.
Куколка покрыта негустыми
короткими оелыми и серыми воло-
сками, неравномерно распределен-
ными на коже; кутикула черно-
Рис. 151. Euproctis similis Fuessly. Хето-
таксия’ передне- и среднегруди и брюш-
ных сегментов I, VI, VII и VIII
взрослой гусеницы.
ких;
щего
ков.
коричневая, на брюшных сегмен-
тах несет скульптуру в виде мелких
бугорков, очень острых и корот-
cremaster в виде гладкого конического недлинного выроста, несу-
на дистальном конце большое количество коротких крючоч-
Изменчивость как взрослой, так и личиночных фаз ничтожна.
21 Фауна СССР, И. В. Кожанчиков.
322
I. ORGYINAE
Распространение. Центральная и южная Европа, от Пире-
неев (Бильбао) всюду по Германской низменности, по долине Дуная,
в Альпах, Карпатах (Буковина) и Балканах (в горы идет далее злато-
гузки); на юге граница идет по средней Италии и северной Греции;
на островах Средиземного моря (Корсика, Сардиния) редка (Чеккони);
Британские острова; южная Швеция (Эланд, Готланд); Финляндия; в Евро-
пейской части СССР от Прибалтики (Рига, Таллин, Тарту) и линии Ленин-
град—Вологда—Молотов всюду по лесной зоне, лесостепи и отчасти
по степи (Псков, Москва, Тула, Серпухов, Мичуринск, Калуга, Брянск,
Харьков, Полтава, Одесса, Киев, Чернигов, Таганрог, Кишинев, Нико-
лаев); Поволжье (Казань, Куйбышев, Саратов, Сарепта); Приуралье (Чка-
лов, Уральск); Крым (всюду);Северный Кавказ (Ахты, Ставрополь); Закав-
казье (Боржоми, Тбилиси, Лагодехи, Талыш); северная Турция (Кази-
копорань); в Сибири от границы Тобольск—Томск частью в лесостепи
на равнине (Кунгур, Омск, Тюмень), но большею частью в горах— Алтай
(Усть-Бухтарма, Усть-Каменогорск), Саяны (Сухая Гора, Казыр-сук);
северный Казахстан (Семипалатинск); Прибайкалье (Иркутск); При-
амурье (Тукурингра, Зея, Благовещенск, Хабаровск); южный Сахалин
(Ишиносава); Курильские острова; южное Приморье (Владивосток,
Сидеми, Сучан, Иман, Ворошилов-Уссурийский, Виноградовка, Тигро-
вая); Маньчжурия (Хинганы); Японские острова (Хоккайдо, Хондо);,
северный и западный Китай (Пекин); Тибет (Амдо).
Биология и экология. Зимуют гусеницы в третьей стадии
в специально сплетенных шелковых коконниках, поодиночке в щелях
коры или под корой; весной появляются с наступлением тепла и растут
до июля—августа; многоядны, питаются видами Salicaceae (Salix, Popu-
lus), Betulaceae (Betula," Corylus, Alnus), Fagaceae (Fagus, Quercus), Rosa-
ceae (Rosa, Prunus, Pirus, Crataegus, Sorbus), Saxiphragaceae (Ribes),
Ericaceae (Calluna), Rhutaceae (Phellodendron), Caprifoliaceae (Lonicera),
Rhamnaceae (Paliurus); предпочитаемыми растениями' являются ивы,
березы и розоцветные, а на юго-востоке Азии Rhutaceae и Могасеае;.
окукление на западе ареала в июне—июле, в южном Приморье в июле —
августе; окукление в беловатом коконе, куда вплетаются волоски; коконы
помещаются на коре, в щелях ее, на сучьях и реже в поверхностном:
слое подстилки; лёт бабочек в июле—августе; откладка яиц кучками
по 100—150 шт. на листья растений; яйца покрываются подушечкой
золотистых волосков с кон(?а брюшка самки; развитие яиц продолжается
12—14 суток; гусеницы по выходе расползаются и живут поодиночке
с первой стадии. Дает почти всегда одно поколение; на юге ареала
(по Шпулеру, 1908) в жаркие годы дает частичное второе, всегда мало-
численное.
Вредит преимущественно садам — яблоне, груше, косточковым,
серьезный вредитель плодовых деревьев на Украине и Северном Кав-
казе; севернее вред отмечен в Киевщине, в Горках; вредит в ряде мест
средней Европы, а также в Англии; на востоке ареала вредит в Китае-
и Японии садам и шелковице; кроме косвенного вреда шелководству,
отмечен прямой вред в связи с заболеванием и смертностью гусениц туто-
вого шелкопряда, вызываемыми отравлением их ядовитыми волосками
желтогузки, попадающими с пищей в желудок гусениц (при размноже-
нии желтогузки на культурах шелковицы). Лесохозяйственное значение
не велико, но многократно отмечался вред лиственным породам.
Указан ряд паразитов для личиночной стадии преимущественног
для куколок и в меньшем количестве паразитов яиц: из Chalcididae —•
10. EUPROCTIS
323
Pteromalus puparum L., P. processioneae Rtzb., Torymus obsoletus F.;
из Braconidae —Microgaster lactipennis Rtzb., M. consularis Br., M. posti-
cus Ns., M. dilutus Rtzb., M. fulvipes Hal., M-. inclusus Rtzb., M. ultdr
Rhd., M. similis Ns., Rhogas testaceus Hal., Rh. tenuis Rtzb., Rh. pulchri-
pes Wesm.; из Ichneumonidae — Automalus alboguttatus Gr., Banchus
compressus F., Ichneumon consimilis Wesm., Mesochorus stigmator Thunb.,
Stenichneumon multicinctus Gr., S. alboguttatus Gr., Pimpla instigator
Pz., P. examinator F. Из двукрылых указаны следующие виды: Carcelia
sp., Ceromasia ferruginea Mg., Compsilura concinnata Mg., Erycia fatuaMg.,
Exorista westermanni Zett., Lydella nigripes Fall., Masicera sp., Pales'paoida
Mg., Plagia elataMg., I.aroioora larvarum L., Tricholyga sp., Zenillia liba-
trix Pz.1
* 2. Euproctis chrysorrhoea L. — Златогузка (рис. 152, 153).
Linne, 1758, Syst. Nat., ed. 10 (1) : 502 (Bombyx)', Aurivillius, 1891, Nordn. Fjiir. :
94, t. 26, fig. 4; Kirby, 1892, Syn. Cat. Lep. Het., 442; Staudinger, 1901, Cat. Lep.,
1 : 116; Spuler, 1908, Schmett. Eur., 1 : 132, t. 25, fig. 28; Rebel, 1910, Berge, Schmot-
terlingsb., ed. 9 : 118, t. 24, fig. 3; Strand, 1910, Seitz, Grossschmett., 2 : 110, 135,
t. 21, e; Vorbrodt und Muller-Rutz, 1912, Schmett. Schweiz, 1 : 234; Oberthiir, 1916,
Et. Lep. Comp., 12 : 281; Rothschild, 1917, Nov. Zool., 24 : 355; Oberthiir, 1922, там же,
19 : 205; Oberthiir, 1923, там же, 20 : 76; Petersen, 1924, Lep. Fauna Estl., ed. 2 (1) : 141;
Gaede, 1932, Seitz, Grossschmett., 2 Suppl. : 104; Hering, 1932, Tierw. Mitteleur.. : 484;
Bryk, 1934, Lep. Cat., 62 : 217; — auriflua Esper, 1785, Schmett. Eur., 3 : 207, t. 39,
fig. 6; — phaeorrhoea Donovan, 1801, Brit. Ins., 10 : 555 (ВотЪух'у. — nigrita Kruli-
kovsky, 1895, Тр. Общ. естественен. Сарат. унив., 1 : 38; Iris, 21 : 218; — puncti-
gera Teich, 1899, Corr. Nat. Ver. Riga, 41 : 87; — nigrosignata Bandermann, 1906,
Entom; Ztschr., 20 : 97, e, f; — punctella Strand, 1910, Seitz, Grossschmett., 2 : 135,
t. 21, e; —flavescens Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 116; — abdominala
Strand, 1910, там же, 2 : 135; —plumbociliata Heinrich, 1916, Deutsch, entom. Ztschr. :
510; — punctifera Oberthiir, 1916, Et. Entom. Comp., 12:281;—xanthorrhoea Ober-
thiir, 1916, там же, 12 : 282.
Биология и экология. Мокржецкий, 1899, Зап. Симф. общ, сад.,
7 : 29; 9 : 42; 14 : 31; Pictet, 1906, Bull. Soc. Lep. Gen., 1 : 114; Шрейпер; 1908,
Тр. Бюро прикл. энтом., 2(16) : 31, рис. 6; Schultze, 1914, Ztschr. wiss. Insekleiihiol.,
9 : 13; Deegener, 1919, Deutsch, entom. Ztschr. : 81; Мориц, 1921, Тр. Ставр, c.-x.
инет., 1 : 23; Wolff und Krausse, 1922, Wichtigste Forstins., 1 : 122, fig. 148; Lede-
rer, 1923, Handb. prakt. Entom., 3 : 169; Zwolfer, 1926, Verb. Deutsch, entom. Ges.
angew. Entom. : 98; Reh, 1925, Sorauer, Handb. Pflanzenkrankh., 4 : 441; Сахаров,
1926, Защ. раст., 3 : 180; Engel, 1927, Ztschr. Morph. Oekol. Tiere, 9 : 615; Niisslin
und Rhumbler, 1927, Forstins., : 460, fig. 382, 383; Pawlovsky und Stein, 1927, / oils hr.
A orj h. Oekol Tiere. 9 : 615; Павловский и Штейн, 1928, Защ. раст., 4 : 556; Ban
dermann, 1930, Entom. Anz., 10 : 224; Weis, 1931,'Ztschr. angew. Entom., 20 : 502;
Лебедев, 1933, 36ipn. праць Муз. зоол. УАН, 1 : 62; Gosswald, 1934, Ztschr.
angew. Entom., 20 : 502, Martin, 1934, Tijdsc.hr. Entom., 77 : 239,
Грудь и крылья белые; рисунок на передних крыльях редуцирован,
они одноцветно белые или с немногими черными пятнами; крупные'
чешуйки черного цвета образуют скопления в виде двух небольших поло-
сок около дискальной жилки; такие же чешуйки могут давать скопления в
виде небольших пяти-шести пятен по внешнему краю крыла; задние крылья
чисто белые; бахрома белая на обеих' парах крыльев; передние крылья
густо покрыты недлинными волосками, закрывающими чешуйки; но дор-
зальному краю передних крыльев бахрома из длинных волосков; нижняя
сторона крыльев у самок чисто белая, у самцов передние крылья затем-
нены по костальному краю коричнево-черными неправильно разбросан-
ными чешуйками; голова у самок почти чисто белая; у самцов по сто-
1 Список паразитов приведен по Ледереру (Lederer, 192 3, Handb. prakt.
Entom., Ill : 168—169).
21*
324
I. ORGYINAE
ронам лба и глаз расположены коричнево-рыжие волоски; губные
щупальца у самцов коричнево-рыжие, более светлые в дистальной части,
у самок почти чисто белые; ноги белые, у самцов голени передних ног
рыже-коричневые; лапки всех ног желтоватые; брюшко у самца белое
лишь при основании и с вентральной стороны, дорзальная сторона всех
сегментов, начиная с третьего, рыже-коричневая; у самки все сегменты
брюшка бело-серые и лишь на последних, начиная с седьмого, сидит
большая подушка ярких коричнево-рыжих коротких волос; усики обоих
полов гребенчатые; у самцов гребешки очень длинные, членики усов
белые, гребешки желтые. Размах крыльев самцов 26—32 мм, самок
32—40 мм.
Uncus толстый и длинный, несколько утолщенный в дистальной части;
основание uncus разрастается в виде
Рис. 152. Euproctis chrysorrhoea L. Гени-
талии: а — самца, б—самки.
латеральных больших склеритов,
несущих в вентральной части ко-
роткие дистально направленные
выросты; девятый сегмент отде-
лен швом от основания uncus;
тергит длинный, несколько взду-
тый, стернит образует длинный
несколько конический и плоский
saccus; valvae почти треугольной
формы, сидят на узком основании;
дистальный конец valva сильно
конический, округлен; penis поч-
ти цилиндрический с коротким
coecum и мелкими шипиками на
дистальном конце; pars inflabilis
без инкрустаций. Bursa copulatrix
небольшая, вытянутая, расширен-
ная в проксимальной части, мем-
бранозная, покрытая тонкой руб-
чатой скульптурой, постепенно
переходит в ductus bursae, кото-
рый мембранозный и длинный;
duclus seminalis отходит от ди-
стальной части ductus bursae; есть
lamella antevaginalis в виде выре-
занной пластинки; восьмой сег-
мент короткий и широкий, равной длины в разных частях; apophyse-
an'eriores и posteriores длинные (длиннее due us bursae); papillae anales
узкие, довольно длинные и заостренные. Отличается от Е. similis Fues-
sly по отсутствию у самок lamina dentata, наличием lamella anteva-
ginalis, местом отхождения ductus seminalis острыми и удлиненными
papillae anales.
Яйца круглые, слегка приплюснутые в дорзовентральном направле-
нии, серые, темнеющие с развитием; хорион гладкий; диаметр 0.5—.0.4 мм.
Взрослые гусеницы черно-серые с коричнево-рыжими бородавками,
точками и такими же волосками; вдоль спины две тонкие охристые по-
лоски, особенно заметные на тергитах и смытые между сегментами, раз-
деленные узкой черной полоской; на первом и втором брюшных сегмен-
тах сидят сильно сближенные парные дорзальные бородавки, несущие
редкие длинные щетинки и многочисленные короткие курчавые коричнево-
рыжие волоски; на месте этих бородавок дорзальные охристые полоски
10. EUPROCTIS
325
прерваны; прочие брюшные сегменты несут подобные же бородавки,
но меньших размеров, расставленные шире; грудные сегменты несут
небольшие едва выдающиеся бородавки, покрыты неправильной формы
красными пятнами; железы на шестом и седьмом брюшных сегментах
красного цвета; боковые линии (по одной с каждой стороны) над стиг-
мами белые, прерванные между сегментами; ниже этих линий сидит ряд
бородавок, несущих с дорзальной стороны ряд курчавых коричнево-
рыжих волосков и небольшое число рыжих щетинок; стигмы черные;
ниже их сидят два ряда бородавок, несущих пучки желто-белых щети-
нок; голова красно-коричневая; ноги коричнево-черные; волоски на всем
теле зазубренные; общая величина волосков относительно небольшая.
Куколка черно-коричневая, обильно покрытая короткими рыже-
коричневыми волосками; скульптура брюшных сегментов в виде тонких
мельчайших выростов кутикулы;
дистальный конец брюшка вытя-
нутой формы, причем вытянут
частично девятый и весь десятый
сегменты; эта вытянутая часть
дистального конца брюшка кукол-
ки несет многочисленные короткие
крючковидные волоски. Степень
изменчивости окраски не велика;
варьирует степень развития чер-
ного пигмента на передних крыль-
ях преимущественно самцов; осо-
би с черными пятнышками на
крыльях названы punctella Strd.,
punctigera, Teich.; особи с темными
пятнышками, слившимися в непол-
ные полосы, — nigrosignata Ban-
derm.; особи с желтоватым оттен-
cremaster слабо дифференцирован,
Рис. 153. Euproctis chrysorrhoea L. Хето-
таксия передне- и средпегруди и брюш-
ных сегментов I, VI, VII и VIII по-
лувзрослой гусеницы. .
ком и с примесью серого тона на
передних крыльях описаны как jlavescens Rbl.; самцы с затемненным
более обычного брюшком названы abdominata Strd.
Распространение. Средняя и южная Европа, от северной
и средней Италии всюду в Альпах, Карпатах и на низменности до побе-
режья Атлантического океана; восточные Пиренеи; Балканы; острова
Средиземного моря (Сардиния, Корсика, Сицилия); северная Африка
(Мавритания, Марокко); Британские острова; Швеция (Эланд, Готланд);
в Европейской части СССР от Прибалтики (Вильнюс, Рига, Каунас, Тар-
ту) и линии Ленинград (чаще в Луге) — Молотов всюду по лесной зоне, лесо-
степи и по степи (Москва, Серпухов, Калуга, Пенза, Курск, Орел, Воро-
неж, Полтава, Харьков, Чернигов, Киев, Николаев, Одесса, Таганрог,
Каменец-Подольск, Кишинев); Крым (Феодосия, Севастополь, Бельбек,
Керчь, Симферополь); Северный Кавказ (Железноводск, Новороссийск,
Ставрополь, Красная Поляна, Кубань, Ростов, Владикавказ, Кисло-
водск, Эльбрус); Поволжье (Казань, Куйбышев, Ульяновск, Саратов,
Сарепта, Астрахань, Ахтуба); Приуралье (Уральск, Чкалов); Закавказье
(Тбилиси, Артвин, Батуми, Лаходехи, Кировабад, долина Аракса, Орду-
бад, Нахичевань); Малая Азия (Карс, Казикопорань, Анатолия, Анкара,
Марат, Аксехир).
Биология и экология. Зимуют' молодые гусеницы третьей
стадии в гнездах, обычно особи одной кладки в общем гнезде; гнезда
32b
J. orgyinae
сплетаются из шелковых нитей, причем размер гнезда сильно зависит
от породы дерева (размеров листа); гнездо имеет верхний рыхлый слой
шелка и внутренний плотный слой (или слои), где и сидят гусеницы
между слоями; гнезда остаются висеть в течение всей зимы на ветвях;
весной питание гусениц начинается с появлением вздутых почек и при
наличии температуры (средней суточной) порядка 8—10°; для весеннего
выхода гусениц из гнезд благоприятны не длительные, но обильные
дожди; питание гусеницы продолжается и с распусканием листвы, кото-
рая при сильном размножении златогузки уничтожается нацело, равно
как цветы и завязи. Гусеницы многоядны, они питаются видами Sali-
caceae (Salix, Populus), Fagaceae ( agus, Quercus, Castanea), Betulaceae
(Carpinus, Corylus, Betula), Rosaceae (Pirus, Prunus, Rosa, Sorbus, Cydo-
nia, Crataegus), Ulmaceae (Ulmus), Aceraceae (Aesculus, Acer), Tiliaceae
(Tilia); предпочитаемыми являются Fagales и Rosales, в лесах — дуб,
бук, в садах —• яблоня, груша. Окукление в конце июля — в августе
в рыхлом яйцевидной формы коконе, в щелях коры, в развилинах ветвей.
Лёт бабочек в июле—августе; бабочки живут недолго. Яйцекладка
ночью на листья и побеги кормовых растений кучками по 100—300 шт.;
яйца покрываются подушечкой золотистых волосков с конца брюшка
самки.
Выходящие гусеницы не расползаются, а живут гнездом, сначала
питаются энергично и первый . раз быстро линяют, а затем после
второй линьки плетут гнездо, впадают в диапаузу и зимуют.
Дают всегда одно поколение. Диапаузирующие гусеницы не очень
холодостойки; по Сахарову, переносят температуру до —20°; судя
по ареалу, нижний предел холодостойкости около —40°.
Серьезный вредитель плодовых и лесных деревьев; вредная деятель-
ность зарегистрирована во всей средней Европе; в СССР преимущественно
в средней полосе и на юге Европейской части, в Крыму, на Северном
Кавказе, в Закавказье, на Украине, в Поволжье, на южном Урале.
Йз плодовых деревьев особенно сильно повреждаются яблоня, груша,
слива, абрикос, черешня, айва, вишня; из лесных пород наиболее, повре-
ждаемыми являются дуб, бук, осина, ива, клен и вяз, в меньшей мере
повреждается береза, тогда как ясень не повреждается совсем. Указания
на вред от златогузки на Дальнем Востоке (Куренцов, 1926) относятся
к желтогузке {Euproctis similis Fssl.).
Вред особенно велик при массовых’ размножениях, когда деревья
совершенно оголяются в связи с объеданием листвы гусеницами.
В СССР массовые размножения златогузки отмечались в Молдавии,
в Крыму,, во многих местах Украины, в Ставрополье, в Туапсе, в ряде
мест средней полосы — Орел, Воронеж, Сызрань, по Средней и Нижней
Волге и в Башкирии. Особое значение вредная деятельность златогузки
имеет в лесокультурных полосах южного Урала, Среднего и Нижнего
Поволжья (Гусев, 1935; Парфентьев, 1938). Массовые размножения злато-
гузки отмечались также в Испании (Каталония); в окрестностях Мадрида,
близ Сарагоссы (Aullo, 1918); в Италии, Франции, Югославии, Болгарии,
Германии, Швейцарии, Голландии, в Сирии, в окрестностях Дамаска.
Златогузка завезена в Северную Америку (восточные штаты), где раз-
множилась и стала одним из серьезнейших вредителей садов и лесов.
Отмечены случаи прямого вреда человеку при сильных размножениях;
волоски гусениц, а по Ледереру (1923), и бабочек (с грудного отдела),
ядовиты и, разносясь по воздуху, при попадании в органы дыхания
человека могут причинять воспаление их.
10. EUPROCTIS
327
Известны многочисленные паразиты златогузки;1 из Ichneumonidae —
Campoplex conicus Rtzb., Cryptus moschator F., S tenichneumon scutellator
Gr., Protichneumon disparis Poda, Hemiteles sociaZts Rtzb., Pimpla insti-
gator F.’, P. examinator F., P. (Apechtes) brassicariae Poda, Exetastes cla-
valor Gr., E. inquisitor Gr., Itoplectis alternans Gr., Ichneumon disparis
Poda, I. consimilis Wesm., I. sarcitorius L., I. scutellator Gr., I. termina-
iorius Gr., Idiolispa atripes Gr., Theronia atalantae Poda, Th. flavicans F.,
Limneria difformis Thoms., Mesochorus dilutus Rtzb., M. pallidus Brisch.,
M ..pallipes Brisch., M. pectoralis Rtzb., Omorus difformis Gmel.; из Bra-
•conidae — Bracon geniculator Ns., Habrobraconbreoicornis Wesm., Meteorus
versicolor Wesm., M. ictericus Ns., Microgaster difficilisNs., M. disparis
Bche., M. connexus Hal., M. calceatus Hal.’, M. consularis Hal., M. dilutus
Rtzb., M. inelusus Rtzb., M. fuloipes Hal., M. lactipennis Rtzb., M. lac-
teicolor'Vier., M. posticus Ns., M. solitarius Rtzb.,7l/. ultor Rhd., M. vitri-
pennis Hal., Rhogas geniculator Ns., Rh. testaceus Spin., Rh. pulchripes
Wesm., Rh. tenuis Rtzb.; из Chalcididae—Chalets scirripoda Forst., Mono-
dontomerus aereus Wlk., M. dentipes Boh., Psychophagus omnivora Wlk.,
Oophthora semblidis Aur., Pteromalus processioneae Rtzb., P. puparum L.,
P. boucheanus Rtzb., P. nidul.ans Thoms., Torymus anpphelus Rtzb., T. obso-
letus Ns., T. regius Ns., Trichogramma evanescens Westw. Из двукрылых
число паразитических форм меньше, но также довольно значительно;
могут быть указаны: Carcelia cheloniae Rd., Ceromasia ferruginea Mg.,
Compsilura concinnata Mg., Cyclotophrys anser Towns., Digonochaeta seti-
pennis Fall., Echinomyia praeceps Mg., Exorista westermanni Zett., Eudo-
romyia magnicornis Zett., Erycia fatua Mg., Larvivora larvarum L., Lydella
nigripes Fall., Masicera silvatica Fall., Pales pavida Mg., Phryxe vulgaris
Fall., Tachina latifrons Tnd., Thyella floralis Mg., Tricholyga sorbillans
Wied., Zenillia libatrix Pz., Z. fauna Rd., Zygobothria nidicola Towns.
Паразиты обнаруживают сложные отношения, и ряд из них является
не только паразитами златогузки, но также и паразитами ее паразитов
(сверхпаразиты). Есть большое число специфичных сверхпаразитов.
Из перепончатокрылых часть видов, например виды Mesochorus, Eutodon,
JDibrachys, могут являться сверхпаразитами различных других перепон-
чатокрылых.
* 3. Euproctis karghalica Moore (рис. 154, 155, 156).
Moore, 1878, Ann. Mag. Nat. Hist., 5 : 231; Moore, 1879, Second Jark.Miss., Lepid.:
7, t. 1, fig. 18; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 443; Staudinger, 1901, Cat. Lep.,
1 : 1.16; Strand, 1910, Seitz, Grossschmett., 2 : 138, t. 21, i; Swinhoe, 1923, Ann. Mag.
Nat. Hist., 11 (9) : 48; —'kargalica Grum-Grzhimailo, 1886, Mem. Lep. Rom., 4 : 559
{Porthesiay, — karghalika. Strand, 1910, Seitz, Grossschmett., 2:134;—flavosulphu-
rea Grum-Grzhimailo, 1886, Mem. Lep. Rom., 4 : 559.
Биология. Суворов, 1894, Зап. Зап.-сиб. отд. Русск. Географ, общ., 17 :
5; Herz, 1900, Ежегодн. Зоол. муз. Акад. Наук, 5 : 448; Reh,' 1925, Sorauer, Handb.
Pflanzenkrankh., 4 : 442; Архангельский, 1926, Защ. раст., 2 : И; Плотников, 1926,
Вреди, насек. Средн. Азии : 109.
Грудь и крылья чисто белые, серебристые; рисунок на передних
крыльях утрачен, представлен в виде немногих пятен; в области дискои-
дальных жилок круглое охристое с черным пятно, слагающееся из круп-
ных охристых и черных чешуек; в терминальной части крыла восемь
круглых черно-охристых пятен, слагающихся из таких же чешуй; задние
1 Я привожу список их по Ледереру (Lederer, 1923, Handb. prakt. Entom.,
Ill : 170—171) с дополнениями; для Ichneumonidae — по Мейеру (1937).
328
1. ORGYINAE
крылья чисто белые; передние крылья, кроме чешуй, покрыты слоем
недлинных белых волосков; нижняя сторона крыльев чисто белая,
костальный край передних крыльев затемнен, у самки слабо, у самца
сильно; голова белая с примесью коричнево-охристых волосков по краям
лба и вокруг глаз; глаза большие, черные; губные щупальца коричнево-
охристые в основании (первые два членика) и белые на конце; голени
передних ног у самцов охристые, лапки и бедра белые; основные членики
ног покрыты длинными редкими волосками; две другие пары ног белые;
у самок все ноги белые; брюшко белое у обоих полов лишь при основании,
первые три сегмента; дистальные сегменты брюшка рыже-бурые, более
светлые, почти белые, с вентральной стороны у самок; три последних
сегмента брюшка окрашены особенно интенсивно и у самки несут тол-
стую подушку охристых волосков; усики самцов сильно гребенчатые,
Рис. 154. Euproctis karghalica Moore. Самка. (Рис. Ф. Гу-
няева).
членики усиков белые, гребешки желтые; отростки члеников на дисталь-
ных концах несут длинные острые щетинки; у самок отростки члеников:
короче. Размах крыльев самцов 30—35 мм, самок 35—40 мм.
Uncus толстый, длинный, слабо изогнутый, почти цилиндрический;
его основание разрастается вентрально, образуя небольшие subunci;
девятый сегмент отделен от основания uncus швом; вентральная часть
несет saccus в виде плоского узкого мешочка; fultura inferior в виде
округлой пластинки с большими вырезами с дорзальной стороны и двумя
недлинными, острыми латеральными отростками; valvae вытянутые
в дистальной части, расширенные, с косо срезанным дистальным концом,
покрытым короткими волосками; penis расширен в базальной части
с коротким соесшп; дистальный конец его вздут и сильно склеротизован.
Bursa copulatrix мембранозная, расширена в проксимальной половине,
дистальная часть ее почти цилиндрическая с слабо рубчатой скульптурой;
ductus seminalis отходит от дистальной части совокупительной сумки;
ductus bursae очень короткий, мембранозный; восьмой сегмент сильно
склеротизован, с большой выемкой на дорзальной стороне, удлиненный
в основании apophyses anteriores; последние длинные, расширенные
в проксимальной части; apophyses posteriores длинные, тонкие; papillae
10. EUPROCTIS!
329'
anales вздутые, конической формы, сильно склеротизованные, довольно-
длинные.
Яйца около 0.3—0.4 мм диаметром, почти круглые, сплюснутые дор-
зовентрально, блестящие, коричневато-желтые; хорион их гладкий;
помещаются в толстом слое
золотистых волосков.
Взрослые гусеницы белова-
то-серые с черными полосами
и коричнево-рыжим рисунком;
волоски, сидящие на бородав-
ках, беловато-желтые и светло-
серые; дорзальная сторона
грудных сегментов желто-ох-
ристая с тонкими поперечными
полосками; дорзальная сторо-
на первых двух брюшных сег-
ментов черная с рыжеватыми
бородавками; последующие
пять сегментов брюшка с дор-
зальной стороны охристые с
черными полосками и тонкой
прерванной срединной черной
полоской; выворачивающиеся
железки на шестом и седьмом
сегментах брюшка охристые;
тергиты восьмого и анального
сегментов черные со слабыми
охристыми полосками; по сто-
ронам окрашенных тергитов
Рис. 155. Euproctis karghalica Moore. Генита-
лии: а — самца, б — самки.
вдоль тела по широкой светлосерой полосе; под ней над стиг-
мами ряд темных бородавок, заметных на светлом фоне; бока желто-
серые, желто-серые же и бородавки, лежащие под стигмами; стигмы1
черные; нижняя сторона тела желто-серая с немногими черными попе-
речными линиями; брюшные
ноги желтосерые, грудпыо —
желтоватые; голова светло-
желтая, более сильно (темно)'
пигментированная в области
глазков и ротовых частей.
Распространение.
Ферганская долина (Гульча,
Наманган, Маргелап); Талас-
ский Ала-тау (Ташкент, Чим-
ган); Зеравшанский хребет (Са-
марканд); Гиссарский хребет
(Кондара, Кзыл-там, Сталин-
абад, Вахш); Памир (Хорог,
Канд-там, Шугпап); Семиречье
(Нарыи, Кульджа, Чилик, Ка-
ра-турук, Пишпек, Алма-ата, Иссык-кулъ, Или, р. Кара); Тарбагатай
(Зайсан); южный Алтай (Иртыш, Усть-Каменогорск, Малокрасноярск,
Семипалатинск).
Биология. Зимуют гусеницы, невидимому, поодиночке, так как-
.330
J.. ORGYINAE
зимующие гнезда не отмечены; рост гусениц в первой половине лета
и весной; гусеницы многоядны, но, невидимому, главным образом на Rosa-
ceae и Elaeagnaceae (Elaeagnus), на которые падает главная масса указа-
ний. Яйцекладка кучками по несколько сотен яиц, которые покрываются
•слоем золотистых волосков с конца брюшка самки; яйца кладутся
на листья и побеги кормовых растений.
4. Euproctis melania Stgr.
Staudinger, 1891, Iris, 4 : 255; Staudinger, 1901, Cat. Lep., 1 : 116; Strand, 1910,
Seitz, Grossschmett., 2 : 134, t. 22, h; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10 (9) :
476; Daniel, 1932, Mitt. Munch, entom. Ges., 22 : 159; Bryk, 1934, Lep. Cat., 62 : 205;—
melaniodes Strand, 1910, Seitz, Grossschmett., 2 : 134; t. 22, h.
Голова, грудь и передние крылья чисто белые; на передних крыльях
во внешнем поле широкая растушеванная и частично разорванная полоса,
•более интенсивная у дорзального края; эта прерывистая полоса (или
ряд пятен) образована скоплением крупных черных и коричневых чешуй;
иногда эта полоса совершенно не развита; у некоторых особей, кроме
внешней полосы, есть еще черное пятно в средней части крыла у дорзаль-
ного края; у самок развитие пигмента на передних крыльях значительно
слабее; задние крылья одноцветно белые; бахрома белая на обеих парах
крыльев; кроме чешуй, передние крылья обильно покрыты белыми воло-
сками, задние крылья имеют только чешуйки; нижняя сторона крыльев
белая; передние крылья по костальному краю и у основания коричневые,
задние крылья при основании также коричневые; у самок нижняя сто-
рона чисто белая; усики самцов сильно гребенчатые, желтоватые; глаза
большие, черные; второй членик губных щупалец черный с белым колеч-
ком на дистальном конце; третий членик маленький, покрытый желто-
вато-белыми волосками; ноги покрыты длинными белыми волосками
с примесью небольшого количества черных волосков; брюшко у самцов
одноцветное, беловато-желтое или слегка затемненное на дистальном
конце; у самок дистальные сегменты несут толстую подушку коричневых
волосков. Размах крыльев 26—30 мм.
Распространение. Малая Азия, Мараш; Эль-Джезире; Кур-
дистан. Возможно наличие этого вида в Закавказье.
5. Euproctis susanna Stgr.
Staudinger, 1894, Iris, 7 : 258, t. 9, fig. 9; Staudinger, 1901, Gat. Lep., 1 : 116;
Strand, 1910, Seitz, Grossschmett., 2 : 138, t. 21, i'; Hering, 1926, там же, 14 : 143;
Gaede, 1932, там же, SuppL, 2 : 104, t. 8, h; Bryk, 1934, Lep. Cat., 62 : 227; —fasciata
Swinhoe, 1922, Ann. Mag. Nat. Hist.. 10Д9) : 484.
(^Голова, грудь и передние крылья желтые с примесью красно-рыжих
тонов, задние крылья желто-охристые; крылья покрыты крупными широ-
кими чешуйками; базальное поле передних крыльев желтое с небольшой
примесью чешуек ржавого цвета, несущее крупные взъерошенные
чешуйки; по середине крыла две широких поперечных ржаво-желтых
извитых полосы, по середине крыла выгнутых одна к другой и -частично
смытых; поле между этими полосами покрыто крупными черными чешуй-
ками, имеющими характер точек; вдоль терминального края крыла
черные чешуйки, образующие скопления между жилками в виде непол-
ных и неправильных восьми пятен; бахрома охристо-желтая; задние
крылья одноцветные охристо-желтые, более яркие у самца; нижняя сто-
рона обеих пар крыльев желто-охристая с охристо-рыжим костальным
10. EUPROCTIS, 11. ARCTORNIS
331
краем; лоб и основание усиков охристо-красные; усики самцов сильно
гребенчатые, желтые; гребешки на концах несут острые щетинки; перед-
ние ноги почти целиком охристо-ржавые; две другие пары ног желтые
с примесью небольшого числа охристых волосков; на конце брюшка
у самки подушка ржаво-желтых волосков; у самца конец брюшка
такого же цвета. Размах крыльев самцов 20—25 мм, самок 30—35 мм.
Распространение. Северная Африка — Египет; Палестина,
долина Иордана; Аравия.
11. Род ARCTORNIS GERM.
Germar, 1810, Syst. Gloss. Prodromus : 18; Kirby, 1892, Syn. Cat. Lep. Het., 1 :
432; Staudinger, 1901, Cat. Lep., 1 : 117; Rebel-, 1910, Berge, Schmetterlingsb., ed.
9 : 117; Strand, 1910, Seitz, Grossschmett., 2 : 109, 123; Bryk, 1934, Lep. Cat., 62 : 111.
Тип рода A. l-nigrum Mull.
Хоботок редуцирован, едва заметен; губные щупальца короткие,
покрытые чешуей, направлены вперед и вверх, последний членик их
очень короток, почти не виден; лоб гладкий; глаза большие, голые, окру-
женные волосками; первый членик передних лапок не более четверти
или одной пятой длины передней голени; голени задних ног с двумя
парами шпор; лапки передних ног очень укорочены и в целом не превы-
шают длину передних голеней; коготки всех ног сильно выгнуты и имеют
широкий зубец на вентральной поверхности; усики обоих полов сильно
гребенчатые; у самцов гребешки более длинные. Передние крылья по-
крыты волосками лишь при основании; волоски и здесь не скрывают
чешуйчатого покрова; радиальная ячея на передних крыльях развита,
но может и отсутствовать, узкая и длинная; третий и четвертый ради-
альные стволы на стебельке, пятый радиальный ствол отходит от внутрен-
ней стенки этой ячеи; первый медиальный ствол отходит почти от осно-
вания ячеи посредством дискальной; ячея может отсутствовать; субко-
стальная и радиальная жилки на задних крыльях образуют болыпухо
базальную ячею, но не сливаются, а лишь подходят близко; второй
медиальный ствол на задних крыльях развит вполне и соединен с прочими
хорошо развитой дискальной жилкой.
Uncus плоский со следами продольного шва; valvae со сложной harpe,
несущей зубчатые отростки; penis короткий, без инкрустаций на pars
inflabilis. Bursa copulatrix с большой lamina dentata на вентральной
стороне; apophyses короткие.
Олиготипический род, представленный только двумя видами, огра-
ниченными в распространении Палеарктикой и преимущественно юго-
восточной Азией; один из них распространен более широко и заселяет
почти все территории, занятые в Евразии широколиственными лесами,
встречаясь и в Европе. Есть сомнения в отношении слабой изученности
признаков многочисленных видов Orgyidae, известных из тропикохз, где
возможно ожидать наличие видов Arctornis Germ., особенно в ипдома-
лайской фауне.
Систематически наиболее близок к Leucoma Steph, и Artaxa Wlk.,
особенно к первому роду, от которого, однако, отличается рядом хоро-
ших признаков.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Дискальная жилка на передних крыльях окрашена в черный
цвет; окраска образует фигуру на подобие буквы «Ь», выделяю-
щуюся на белом фоне, или дает простой черный штрих; губные
332
I. ORGYINAE
щупальца в дистальной части черные; на ногах черные колечки.
...........................................*2. A.i-nigrum Mull.
2 (1,). Передние крылья чисто белые, одноцветные; губные щупальца
в дистальной части охристые; голени всех ног с небольшими охри-
стыми пятнами.....................................* 1, A. alba В rem.
* 1. Arctornis alba В rem. (рис. 157).
Bremer, 1861, Bull. Acad. Sci. Pet., 3 : 478 (Aroa); Bremer, 1864, Lep. Ost-Sib. :
41, t. 3, fig. 18; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 432; Staudinger, 1892, Mem. Lep.
Rom., 6 : 308; Leech, 1899, Trans. Entom. Soc. Lond. : 143 (Leucoma)', Staudinger,
1901, Cat. Lep., 1 : 117; Swinhoe, 1903 Trans. Entom. Soc. Lond. : 380; Strand, 1910,
Seitz, Grossschmett., 2 : 123; Swinhoe,
1922, Ann. Mag. Nat. Hist., 10
(9) : 476;Bryk, 1934, Lep. Cat., 62 :
112; — sinensis Moore, 1877, Ann.
Mag. Nat. Hist., 4 : 92 (Redoa);—
l depuncta Strand, 1910, Seitz, Gross-
\ schmett., 2 : 123; — nigripunctata.
A Warnecke, 1924, Iris, 38: 154.
' Грудь, брюшко и крылья
белого цвета; передние крылья
молочно белые, без рисунка, с
маленькой охристо-черной точ-
кой на месте дискальной жил-
ки; эта темная точка нередко
не развита; крылья покрыты
блестящими чешуйками; задние
6 крылья одноцветно белые;
нижняя сторона обеих пар
крыльев .белая со слабым жел-
товатым оттенком при основа-
нии и по костальному их краю;
голова охристо-белая, темя и
основание усиков охристые;
охристые последние членики
Губных шупалец; передние го-
лени и лапки покрыты длин-
ными белыми волосками, дс не-
большим количеством охристых
со слабым желтоватым оттенком;
Рис. 157. Arctornis alba Brem. Гениталии:
a — самца, 6 — самки.
волосков; средние и задние ноги белые
усики самцов с очень длинными гребешками, желтоватые, членики,
с вентральной стороны светложелтые. Размах крыльев самцов 32—
37 мм, самок 40—45 мм.
Uncus треугольной формы, плоский, сочленен посредством шва с девя-
тым сегментом; девятый сегмент расширен в дорзальной половине, в вен-
тральной половине образует небольшой плоский saccus; valvae слабо
склеротизованы, длинные, выгнутые дорзально; в базальной части valva
склеротизованная лопасть, несущая в основании двойной зубчатый отро-
сток; penis очень короткий, несет на pars inflabilis мелкие зубчики. Bursa
copulatrix почти цилиндрическая, мешковидная, с большой lamina den-
tata, усаженной зубчиками и расположенной на вентральной поверх-
ности совокупительной сумки; ductus bursae короткий; восьмой сегмент
длинный, образует вокруг ostium ductus bursae валик, покрытый мно-
гими мелкими складочками; apophyses anteriores очень короткие; apophy-
11. ARCTORNIS
333
ses posteriores вдвое длиннее; papillae anales короткие, несут густые
тонкие волоски.
Изменчивость признаков не велика; окраска остается всегда белой,
но охристые и черные пигменты могут быть развиты в большей или мень-
шей мере; особи с хорошо развитыми черными точками на передних
крыльях названы A. nigripunctata Warnecke.
Распространение. Приамурье (Хабаровск); Шантарские
острова (Б. Абрек); южное Приморье (Уссури, Яковлевка, Виногра-
довка, р. Супутинка, р. Суйфун, Сучан, р. Сица, Владивосток, Аскольд);
Корея; центральный и северный Китай (Маньчжурия, Квансиен).
Биология почти не изучена; по указаниям Грэзера (1888), гусе,
ницы живут на монгольском дубе (Quercus mongolica Fisch.); по Курен-
цову (1939), бабочки встречаются в смешанных лесах в июле.
*2. Arctornis 1-nigrum Mull. (рис. 158 ,159).
Muller, 1764, Fauna Friedrichsdal. : 40 (Rombi/ж); Aurivillius, 1891, Nordn. Fjar.:
93, t. 26, fig. 6; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 432; Staudinger, 1901, Cat. Lep.,
1 : 117; Spuler 1908, Schmett. Eur., 1 : 130, t. 25, fig. 26; Rebel, 1910, Berge, Schmet-
terlingsb., ed. 9 : 117, t. 24, fig. 1; Strand, 1910, Seitz, Grossschmett., 2 : 123, t. 20, a;
Vorbrodt und Miiller-Rutz, 1912, Schmett. Schweiz, 1 : 231; Swinhoe, 1922, Ann. Mag.
Nat. Hist., 10 (9) : 478; Petersen, 1924, Lep. Fauna Estl., ed. 2 (1) : 141; Gaede, 1932,
Seitz, Grossschmett., 2 Suppl. : 99; Hering, 1932, Tierw. Mitteleur. : 483; Bryk, 1934,
Lep. Cat., 62 : 111; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 66, t. 14, fig. 11; —
v-nigrum Fabricius, 1775, Syst. Entom. : 577, — nervosa Schiffermiiller, 1776, Syst.
Verz. Wien. Lep. : 52 (Bombyx)-, — vau-nigrum Hampson, 1892, Fauna Brit. Ind.,
Moths, 1 : 487; (Leucomc); — costa-nigra Lambillion, 1908, Rev. Namur., 8 : 43; —
evanescens Schultz, 1910, Entom. Ztschr. Frank!., 24 : 35; — pusillata Dannehl, 1925,
Entom. Ztschr. Frank!., 39 : 107.
Биология. Graeser, 1888, Berl. entom. Ztschr., 33:124; Eckstein, 1915,
Schrift. Lehrerversamml. Naturk., 32 : 46, t. 24, fig. 4, a, b; Lederer, 1923, Handb.
prakt. Entom., 3 : 152; Stauder, 1924, Entom. Anz., 4 : 74; Куренцов, 1936, Tp. Даль-
невост. фил. Акад. Наук СССР, 1 : 146; Куренцов, 1939, Тр. Горнотаежн. ст., 3 : 146.
Голова, грудь и крылья чисто белые; передние крылья имеют утра-
ченный рисунок, одноцветные, дискальная жилка полностью или ча-
стично окрашена в черный цвет; в связи с изломом дискальных жилок
черный пигмент распределяется в виде углообразной (L-образной) фигуры;
чешуйчатый покров передних крыльев густой, чешуйки двух типов —
прижатые к плоскости крыла и узкие, торчащие вверх; жилки окрашены
в желтоватый цвет, слабо выделяются на белом фоне; задние крылья
того же типа, как и передние; бахрома на обеих парах крыльев чисто
белая; нижняя сторона крыльев чисто белая; усики самцов сильно гре-
бенчатые, гребешки желтые, членики усиков белые; у самок гребешки
усиков короче, но также желтые; губные щупальца при основании
и с вентральной стороны белые, дорзальная их сторона и дистальный
конец черные; голени передних и средних ног с черным пятном у вершины;
лапки всех пар ног имеют черные первый и последний членики; брюшко
покрыто белыми волосками не густо, просвечивают желтоватые кожные
покровы; дистальный конец брюшка самки простой. Размах крыльев
самцов 35—40 мм, самок 40—50 мм.
Uncus в виде тупого, разделенного бороздой на две половины выроста,
отделен швом от девятого сегмента; девятое кольцо короткое, не образует
saccus; valvae мембранозные, при основании расширены, со склеротизо-
ванной лопастью, образующей кармапообразное углубление; при осно-
вании лопасти harpe со стержневидным зубчатым отростком; шипики
расположены на дистальном конце отростка; дистальный конец valva
334
I. ORGYINAE
мембранозный, покрытый мелкими волосками; penis короткий, слабо
изогнутый, без инкрустаций. Bursa copulatrix перепончатая, изогнутая,
в проксимальной части расширенная, на вентральной стороне несет
Рис. 1а8. Arctornis l-nignun Mull. Самец.
(Рис. Ф. Гуняева).
Рис. 159.
Arctornis l-nigrum Mull. Гениталии:
а — самца, б — самки.
большую lamina dentata с зубцами, направленными в полость совокупи-
тельной сумки; ductus bursae очень короткий; восьмой сегмент короткий,
образует складку на вентральной стороне вокруг ostium ductus bursae;
apophyses anteriores и posterio-
res короткие; papillae anales
узкие, слабо выгнутые по вент-
ральному краю.
Яйца почти круглые, не-
сколько уплощенные в дорзо-
вентральномнаправлении, свет-
лозеленые, блестящие.
Взрослая гусеница черная
с желтыми или ржаво-желтыми
боками; на дорзальной стороне
первых пяти брюшных сегмен-
тов кисточки красно-рыжего
цвета на третьем и четвертом
брюшных сегментах и белые
на остальных; первый и второй
грудные сегменты несут белые
кисточки; восьмой- брюшной сег-
мент несет пучок белых волос-
ков; волоски, покрывающие те-
ло, сидят на небольших бо-
родавках; они желтоватые или
красно-желтые; голова черная (описана по Шпулеру, 1908; Ламперту,
1912; Куренцову, 1939; все описания слишком кратки и недостаточны).
Распространение. Средняя Европа (Альпы, средняя Ита-
лия, долина Дуная, Германская низменность); Швеция; Прибалтика
11. ARCTORNIS, 12. 1VRLA
33N
(Висла, Рига, Каунас); восточнее преимущественно на юге — Румы-
ния, Молдавия, Украина (Каменец-Подольск, Киев, Харьков, Пол-
тава, Винница); на севере пятнами; в зоне широколиственных лесов
(Могилев—Горки, Калуга, Курск, Серпухов, Казань); местами в По-
волжье (Саратов, Вольск), на южном Урале (Чкалов), на Северном
Кавказе (Ставрополь) и в Закавказье (Ареш, Боржоми); указан для
Финляндии (Шпулер, 1908), но ошибочно [Грёнблом (Gronblom, 1924)];.
Приамурье (от Шилки до Николаевска, Раддевка, Хабаровск); южное-
Приморье (Суйфун, Ворошилов-Уссурийский, Сучан, Седанка, Уссури,
Сидеми, р. Мангугай); Корея; Японские острова.
Биология. Зимуют гусеницы третьей стадии свободно, в свер-
нутых листьях; весной благоприятно увлажнение талой водой или
дождями для активизации развития гусениц; выход из мест зимовки-
через 12—15 суток после наступления теплой погоды с дождем; рост-
в весеннее время протекает до половины июня; гусеницы многоядны,
но преимущественно на широколиственных древесных породах — Faga-
ceae (Fagus, Quercus, Castanea), Betulaceae (Carpinus, Ostryia, Corylus,
Betula), Rosaceae (Malus, Crataegus), Ulmaceae (Ulmus), Salicaceae (Popu-
lus, Salix); часто отмечаются белые тополя как основные кормовые расте-
ния; на безлесных местах, по Шпуйеру, гусеницы встречаются на кустар-
никах— Ericaceae (?); по Куренцову (1939), на Дальнем Востоке
гусеницы встречаются на травянистых Rosaceae (Filipendula); окукление
в конце‘июня; лёт бабочек в июле; яйца откладываются на растения;
развитие яиц 8—10 суток. Всегда дает одно поколение.
12, Род IVELA SWINH.
Swinhoe, 1903, Trans. Entom. Soc. Lond. : 388; Strand, 1910, Seitz, Grossschmett,,..
2 : 124; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10 (9) : 469; Bryk, 1934, Cat. Lep.,
62 : 121.
Тип рода I. auripes Btl.
Хоботок очень короток; губные щупальца очень слабые, с очень,
коротким, почти утраченным последним члеником, направлены вперед;,
глаза круглые, голые; усики самцов двусторонне-гребенчатые, гребешки
на концах несут небольшие одиночные щетинки; первый членик передних
ног вдвое короче голени этих же ног; коготки всех ног с отростком (шипом)
на вентральной стороне; поверхность крыльев покрыта чешуей с неболь-
шой примесью волосков лишь у дорзального края крыла; тело стройное;
крылья широкие; добавочная ячейка не развита; третий, четвертый и пятый
радиальные стволы на длинном стебельке, ветвление .радиальных жилок в
костальный край и в область apex; первый медиальный ствол отходит с по-
мощью ди скальной жилки от радиального ствола; базальная ячейка на зад-
них крыльях вытянутая, почти изолированная неполным анастомозом суб-
костальной и радиальной жилок; второй медиальный ствол на задних
крыльях отделен дискальной от третьего ствола; uncus при основании
двойной, в дистальной половине сплошной, заостренный; valvae округ-
лые, со сложной складчатой harpe, несущей бородавку с сильными
шипами и длинными щетинками; ductus bursae склеротизован, короткий;
apophyses длинные; есть небольшая lamina dentata на вентральной сто-
роне совокупительной сумки.
Род близкий к Leucoma Steph. (Stilpnotia Westw.), отличающийся
тем не менее целым комплексом признаков — еще не вполне сросшимся
при основании uncus, строением valvae, ветвлением радиальных жилок:
336
I. ORGYINAE
•по костальному краю, но не по терминальному, строением ногтевого
аппарата. Род Ivela Swinh. следует рассматривать как исходный по отно-
шению к Leucoma Steph.
К роду Ivela Swinh. в литературе относят (Брик, 1934) всего два
вида — I. auripes Btl. и I. eleuteroides Semper, известный также с юго-
востока Азии (Лузон); кроме того, к этому рЬду относится I. ochropoda
Ev., ранее считавшаяся видом Leucoma Steph. Тем не менее было бы
.преждевременно считать род Ivela Swinh. ограниченным этими немно-
гими видами. Обитающий в центральной (южная Мексика, Панама,
Коста-Рика) и в Южной Америке (Венецуэла, Гвиана, Амазонка, Колум-
бия, Перу) очень характерный, но слабо изученный род Eloria Wlk.
s(Schaus,' 1927, Seitz, Grossschmett., 6 : 539),.число видов которого рав-
няется нескольким десяткам, по комплексу признаков (ширококрылость,
наличие лишь одной пары шпор, отсутствие добавочной ячеи и тип жил-
кования) и по габитусу видов очень близок Ivela Swinh. (рис. 160).
Недостаточная изученность этого рода (в частности, отсутствие данных
по строению копулятивных органов) мешает пока объединению обоих
родов. В центральной и Южной Америке обитает другой, без сомнения
самостоятельный род Caviria Wlk., также близкий к Ivela Swinh.
В тропиках Старого Света и в других частях земного шара не известно
столь близких к Ivela Swinh. родов, как Eloria Wlk., но в индомалайской
фауне роды Sitvia Wlk. и Cispia Wlk. напоминают Ivela Swinh., хотя
дают отличия в числе шпор на задних голенях. Африканские' роды —
Cropera Wlk., Crorema Wlk. и Ruanda Strd. —хотя и дают значитель-
ное габитуальное сходство видов с Ivela Swinh., но отличаются струк-
турой ног и признаками жилкования.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Передние крылья тусклого цвета, полупрозрачные, с темными
жилками; усики самцов с примесью большого количества черных
.чешуй; valva с небольшой бородавкой в средине, усаженной корот-
кими шипами; более крупный, 45—50 мм ... 1. I. auripes Btl.
.2 (1). Передние крылья блестяще белые, с большими и густо сидящими
чешуйками; жилки белые; valva несет посредине большую боро-
давку сложной формы, усаженную длинными и сильными шипами;
мельче, 25—40 мм................................*2. I. ochropoda Ev.
1. Ivela auripes Btl. (рис. 161, 162).
Butler, 1877, Ann. Mag. Nat. Hist., 4 : 402 (Leucoma); Kirby, 1892, Syn. Cat.
Lep. Het., 1 : 432 (Arctornis); Strand, 1910, Seitz, Grossschmett., 2 : 124, t. 20,b;Swin-
Ihoe, 1922, Ann. Mag. Nat. Hist., 10 (9).: 469; Matsumura, 1931, 6000 Ins. Japan : 709,
..fig. 466; Matsumura, 1933, Ins. Matsum., 7 : 131; Bryk, 1934, Lep. Cat., 62 : 121; —
.denudata Swinhoe, 1892 (non Walker), Cat. Het. Lep., 1 : 202 (Sitvia).
Передние крылья белые с дымчатым налетом вдоль жилок, особенно
-сильным, почти черным, у самцов; костальный край затемнен; темная,
дымчато-серого цвета и бахрома; передние крылья широкие, по облику
несколько напоминают дневных бабочек; задние крылья меньше затем-
нены, бахрома на них дымчатая, жилки почти белые; нижняя сторона
.крыльев такая же, как и верхняя; усики самцов сильно гребенчатые,
черные; глаза круглые, черные; губные щупальца сверху черные, с боков
..и снизу белые; передние ноги (их голени и лапки) желтые (охристые);
150
Рис. 160. Распространение видов рода Ivela Wlk. (в Азии) и Eloria Wlk. (в Америке).
22 Фауна СССР, И. в. Кожанчиков.
338
1. orgyinae
грудь и брюшко покрыты белыми волосками и узкими чешуйками. Раз-
мах крыльев самцов 45—55 мм, самок 50—60 мм.
Uncus тонкий, уплощенный с боков, почти прямой, сидит на широ-
ком двойном основании, с небольшой бороздой на дорзальной стороне;
Рис. 161. Ivela auripes Btl. Самец. (Рис.
Ф. Гуняева).
девятый сегмент отделен широким швом от основания uncus; девятый
стернит расширен,'имеет небольшие выросты, не образует saccus; fultura
inferior в виде пластинки, имеющей округлый дорзальный и плоский,
и угловатый вентральный края; valva короткая и толстая, сильно скле-
ротизованная, густо волосистая; harpe в виде складки с небольшой воло-
Рис. 162. Ivela auripes Btl. Гениталии самца.
систой и покрытой шипиками бородавкой; penis длиннее valva, его сое-
cum недлинный, изогнут вентрально; в месте срастания penis с vallum,
большая склеротизованная складка; дистальный конец penis заострен.
Распространение. Японские острова (Хондо); Корея; север-
ный Китай (Сычуань); западный Китай (Омисиен).
* 2. Ivela ochropoda Ev. (рис. 163).
Eversmann, 1847, Бюлл. Моск. общ. исп. прир., 20 (2) : 76, t. 5. fig. 1—3; Graeser,.
1888, Berl. entom. Ztschr.,32'.125; Staudinger, 1892, Mem. Lep. Rom., 6: 309; Kirby, 1892,
Syn. Cat. Lep. Het., 1 : 443; Staudinger, 1901, Cat. Lep., 1 : 117; Strand, 1910, Seitz,
Grossschmett., 2 : 124, t. 22, d; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10 (9) : 462;
12. IVELA, 13. LEUCOMA
339
Matsumura, 1931, 6000 Ins. Japan : 712, fig. 479; Matsumura, 1933, Ins. Matsum.,
7 : 151; Bryk, 1934, Lep. Cat., 62 : 117 (Stilpnotia). .
Передние крылья чисто белые; жилки их особенно обильно покрыты
чешуей; рисунок не развит; задние крылья также одноцветно белые;
голова и грудь белые, покрыты длинными пушистыми волосками, в глу-
бине которых, непосредственно покрывая кожу, сидят чешуйки; на- го-
лове очень небольшая примесь дымчатых волосков; глаза большие, чер-
ные; губные щупальца покрыты охристыми волосками, усики самцов
с большими двусторонними гребешками; членики их покрыты белыми
волосками, отростки черные; голени передних ног и их лапки охристые;
охристые же лапки и частично голени средних и отчасти задних ног;
усики самок с довольно
большими гребешками.
Размах крыльев самцов
25—35 мм. самок 30—
40 мм.
Uncus острый, сильно
конический, в основании
двойной, плотно срастает-
ся с tegumen; девятый
сегмент недлинный, в вен-
тральной части образует
плоский небольшой saccus;
valva почти округлая,
несет сложную harpe в
виде изогнутой складки;
в ее средней и дистал ьной
частях шаровидные выро-
сты, усаженные многочис-
ленными шипами и волос-
ками, направленными в
разные стороны; penis ко-
роткий, почти цилиндри-
ческий, В дистальной час- Рис. 163. Ivela ochropoda Ev. Гениталии: а — сам-
ти заостренный,- с доволь- ца, б — самки,
но широким и вздутым
coecum, Bursa copulatrix мембранозная, тонкая, с очень маленькой lamina
dentata на вентральной стороне; ductus bursae короткий, вздутый, сильно
склеротизован, срастается широко с восьмым сегментом; последний
очень короткий, расширенный лишь при основании apophyses, которые
длинные; apophyses posteriores короткие; papillae anales большие, покры-
тые мягкими длинными волосками.
Распространение. Южное Приморье (Сучан, Суйфун,
Аскольд, Владивосток, Виноградовка); Забайкалье (Кяхта, Даурья);
Прибайкалье (Иркутск); есть указание для Томска (Внуковский, 1935),
которое я не имел возможности проверить по фактическому материалу.
Японские острова (Хондо, Хоккайдо); северный Китай; Маньчжурия.
13. Род LEUCOMA STEPH.
Stephens, 1828, Ill. Brit. Entom. Haustell.,'2 : 64; Aurivillius, 1891, Nordn. Fjar. :
94; Kirby, 1892, Syn. Cat. Lep. Het., 1 :433; Staudinger, 1901, Cat. Lep., 1 : 117; Spu-
ler, 1908, Schmett. Eur., 1 : 130; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 :Д17;
Strand, 1910, Seitz, Grossschmett., 2 : 123; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10
340
1. ORGYINAE
(9) : 461; Hering, 1932, Tierw. Mitteleur. : 483; Bryk, 1934, Lep. Cat., 62 : 113; Nord-
stroem och Wahlgren, 1937, Svenska Fjar. : 66; — Laria Schrank, 1802, Fauna Boica',
2 (2) : 152, nomen praeoccupatum (typus salicis L.); — Stilpnotia Westwood and Hamphe-
reys, 1841, Brit. Moths, 1 : 90 (typus salicis L.); — Leucosia Rambur, 1866, Cat. Syst.
Lep. d’Andalus., 2 : 266 (typus salicis L.); — Redoa Moore, 1878, Proc. Zool. Soc.
Lond. : 401 (typus cygna Moore); —Nymphyxis Grote, 1895, Mitt. Mus. Hildesheim.,
1 : 4 {typus salicis L.).
Тип рода L. salicis L.1
Хоботок слабый, но довольно длинный; глаза большие, голые, окру-
жены волосками, с малой примесью чешуй; усики обоих полов сильно
гребенчатые, у самок отростки члеников короче: крылья, кроме чешуй,
покрыты волосками, у некоторых видов волосяной покров на крыльях
очень густ; лоб гладкий; губные щупальца слабые, направленные прямо
вперед или несколько свисающие вниз; третий членик их хорошо виден,
второй довольно большой; голени передних ног вдвое длиннее первого
членика лапки этих же ног; голени задних ног с одной парой шпор;
коготки всех ног простые; радиальная ячея на передних крыльях не раз-
вита; третий, четвертый и пятый радиальные стволы на длинном стебельке
открываются в область apex или в терминальный край; первый медиаль-
ный ствол отходит с помощью дискальной жилки от этого стебля; у неко-
торых особей есть остатки медиального ствола в дискоидальной ячее;
базальная ячея варьирует от малых размеров до очень больших, но
всегда незамкнута; второй и третий медиальные стволы на задних
крыльях сильно сближены основаниями; uncus целостный, сплошной,
при основании его латеральные сильные выросты tegumen; valva сильно
выгнута, ковшевидной формы, несет сильный склеротизованный вырост
на дорзальном крае; ductus bursae склеротизован; есть lamina dentata;
apophyses длинные. Полы слабо диморфны.
Строение личиночных фаз следующее. Гусеницы уплощенной формы,
с длинными пучками волосков на бородавках р по сторонам тела; имеют
сложный белый рисунок на тергальной стороне сегментов; выворачиваю-
щиеся железки, кроме обычных на шестом и седьмом брюшных сегмен-
тах, есть также на первом и втором сегментах брюшка. Бородавка •/.
переднегруди большая, несет пучок волосков, направленный вперед
(рис. 164). Бородавка а на средне- и заднегруди несет только две щетинки;
на брюшных сегментах <z несет только одну щетинку. Бородавка р на
брюшных сегментах слита с /, и лежит за стигмой. Куколки черного
цвета с блестящей кутикулой (черной) и светлыми волосками; брюшко
сильно суженное в дистальном направлении; усики самцов широкие.
Яйца крупные — .0.8—1.0 мм диаметром, почти круглые, имеют матовый
желто-зеленый хорион; покрываются пенистыми выделениями придаточ-
ных желез полового аппарата самки.
Объем и распространение в тропиках рода Leucoma Steph, сколько-
нибудь точно установлены быть не могут; тропические виды, близкие
к евразийским Leucoma Steph., разбиваются разными авторами по двум
родам — Leucoma Steph, и Stilpnotia Westw., несмотря на то, что для
обоих родов типом является один и тот же вид — L. salicis L.; различия
родов не даются, а описания как видов, так и родов очень кратки, поверх-
ностны и не пригодны для определения их; должно добавить, что с ними
смешиваются также виды Arctornis Germ. Приводимая ниже характери-
1 Тип рода Leucoma salicis L. фиксирован за родом Leucoma Steph, в 1840 г.
Вествудом (Westwood, Intern. Glass. Insect., 2 : 92).
13. LEUCOMA
341
стика распространения дается на основании изучения оригинальной
литературы.
Кирби (1892) относит к роду Leucoma Steph., кроме двух евразийских
видов (L. salicis L. и L. flavosuphurea Ersch.), один вид с Мадагаскара
{L. cretosa Saalm.), два вида из Индии (L. sericea Moore и L. ochripes Moore)
и четыре сомнительных вида из центральной Америки.
Род Leucoma Steph. {Stilpnotia Westw.), насколько сейчас можно
судить по литературе (по описаниям), является более богатым видами и рас-
пространен преимущественно в Азии, причем представлен наибольшим чис-
лом видов в тропиках на юго-востоке Азии и на Малайском архипелаге.
Виды, относящиеся к этому роду с Малайского архипелага (преимуще-
ственно с Явы, Борнео и Целебеса), следующие: L. semihyalina Swinh.,
riguata Snell., lepta Coll., submarginala Wlk., niveata Wlk., egens Feld.,
Coll., rujimarginata Swinh., ecnomoda
wmutissima Swinh., monobalia
Swinh., micacea Wlk., gigantea
Auriv., perfecta Wlk., divisa Wlk.
Один вид с Новой Гвинеи (А.
intacta Wlk.) и один сомнитель-
ный вид описан из Австралии
{L. semiochrea Н. S.). Некото-
рые виды, известные на Малай-
ском архипелаге, заселяют так-
же Малакку, где есть и свои
виды {L. phrika Coll., poecilo-
nipha Coll., nivosa Wlk., fla-
vescens Moore, egerina Swinh.,
camurisquama Coll., singaporen-
sis Strd., discirufa Swinh., phas-
matodes 'Coll., hipp'aria Swinh.,
niphobola Coll., primula Swinh.).
Некоторые из этих видов, а так-
же свои известны с Филип-
Рис. 164. Leucoma salicis L. Хстотаксия пе-
редне- и средпегруди и брюшных сегментов
I, VI, VII и VIII гусеницы I стадии.
пинских островов {L. divisa Wlk., submarginata Wlk., marginalis Wlk.)
и с Тайвана, где есть часть своих видов {L. niveata Wlk., jormosensis
Strd., flavicostata Mts.). Из Индии известно меньшее число видов, чем
с Малайского архипелага {L. comma Hutt., silhetica Wlk., divisa Wlk.,
diaphana Moore, phasmatodes Coll., flavescens Moore, submarginata Wlk.);
еще меньшее число видов распространено в западном Китае, Корее и на
Японских островах {L. submarginata Wlk., diaphana Moore, moorei Leech,
suigensis Mts., doii Mts., chichibensis Mts.).
Два вида {L. salicis L. и L. Candida Stgr.) широко распространены
в умеренной зоне Евразии, а три вида эндемичны для гор Средней Азии
{L. flavosulphurea Ersch., sartus Ersch. и surtur В.-H.).
Из Африки виды Leucoma Steph. {Stilpnotia Westw.) достоверно
не известны, но один вид (не считая ранее указанного L, crelosa Saalm.) —
L. brontonepha Coll. —описан недавно с Мадагаскара. Из Южной Аме-
рики виды этого рода также не известны, равно как и из Северной Аме-
рики, но в южную Канаду и восточные штаты занесен европейский
L. salicis L., размножившийся и расселившийся там. Из Южной Америки
известны близкие роды —£aviria Wlk. и Eloria Wlk.
Евразийские виды Leucoma Steph, зимуют в личиночной фазе и свя-
заны преимущественно с Salicaceae, в особенности с видами тополей;
кормовые связи и биология тропических видов не изве&тны.
B42
1. ORGYINAE
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (6). Крылья и тело белые или желтые, но не черные.
2 (5). Крупные виды — 35—60 мм; передние крылья чисто-белые без
всякой примеси желтых тонов, нередко полупрозрачные; valva
с зубчатым завернутым вентральным краем, без сильного одного
зубца.
3 (4). Передние крылья тусклобелые, покрыты узкими чешуйками
с большим количеством волосков; valva с немногими зубцами по за-
вернутому вентральному краю; lamina dentata расположена ближе
к polus bursae................................ • . * 1. L. salicis L.
4 (3). Передние крылья мелового цвета, не прозрачны (даже у самок),
покрыты широкими чешуйками; valva с многочисленными зубцами
по вентральному краю; lamina dentata сдвинута в дистальную
часть совокупительной сумки.................' . . * 2. L. Candida Stgr.
5 (2). Мельче, 32—37 мм; передние крылья желтые или, реже, белые
со слабым желтоватым оттенком; bursa copulatrix без lamina den-
tata; вентральный край valva несет один сильный вырост . . . .
................................* 3. L. flavosulphurea Ersch.
6 (1). Крылья и тело черные, коричневые или черно-бурые.
7 (8). Шоколадно-коричневый или коричнево-черный; размах крыльев
30—35 мм..................................* 4. L. sartus Ersch.
8 (7). Чисто черный, несколько крупнее; размах крыльев 39 мм . . .
..........................................* 5. L. surtur В. Н.
* 1. Leucoma salicis L.—Ивовая волнянка (рис. 165).
Linne, 1758, Syst. Nat., ed. 10 (1) : 502 (Bombyx); Aurivillius, 1891, Nordn. Fjar. :
04, t. 26. fig. 1; Staudinger, 1901, Cat. Lep., 1 : 117; Spuler, 1908, Schmett.- Eur.,
1 : 130, f. 83, t. 25, f. 27; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 117, t. 24, fig. 2;
Strand, 1910, Seitz, Grossschmett., 2 : 123, t. 20, a; Vorbrodt und Muller-Rutz, 1912,
Schmett. Schweiz, 1 : 232; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10 (9) : 461; Petersen,
1924, Lep. 'Fauna EstL, ed. 2 (1) : 142; Zerny, 1927, Eos, 3 ; 350; Engelhardt, 1928,
Bull. Brookl. Entom. Soc., 23 : 250; Hering, 1932, Tierw. Mitteleur..: 484; Bryk, 1934,
Lep. Cat., 62 : 113; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 66, t. 14, fig. 10; —
schesti Capronnier, 1879, C. R. Soc. Entom. Belg., 21 : 129; — ni.gripehnata Staudinger,
1899, Iris, 12 : 338; — nigrociliata Fuchs, 1900, Jahrb. Nass. Ver. Naturk., 56 : 71; —
rubicunda Strand, 1901, Schrift. Nat. Ges.'Danzig, N. F., 10 : 285; — neumanni Ban-
dermann, 1929, Entom. Anz., 9 : 340.
Биология. Standfuss, 1896, Handb. pal. Grossschmett., ed: 2 : 168; Король-
ков, 1913, Насек, поврежд. сады; Лучник, 1914, Люб. природы : 14; Reh, 1926,
Sorauer, Handb. Pflanzenkrankh., 4 : 440; Hepp, 1928, Lep. Rundsch., 2 : 53.
Чисто белый, с густо волосистыми головой и грудью; усики сампов
сильно гребенчатые, гребешки покрыты тонкими волосками и на концах
несут тонкие щетинки; лоб белый; глаза большие, черные, окруженные
с боков и,снизу черными волосками; губные щупальца покрыты черными
волосками; хоботок заметен; ноги белые с черными колечками, покрыты
длинными волосками, лапки черные с немногими белыми штрихами;
брюшко покрыто редкими белыми или желтоватыми волосками, кожа
просвечивает черным; передние крылья без рисунка, покрыты редкими,
узкими чешуйками, полупрозрачны; базальная половина крыльев и ко-
стальный край их покрыты, кроме чешуи,, волосками; жилки слегка
желтоватые, выделяются на фоне крыла; самки несколько крупнее;
гребешки члеников их усиков значительно короче; крылья имеют более
округлую форму, особенно их верхний угол, чешуйчатый покров на них
13. LEU COM A
343
Рис. 165. Leucoma salicis L. Гениталии:
а — самца, б — самки.
беднее, они полупрозрачны. Размах крыльев самцов 35—45 мм, самок
45 —55 мм.
Uncus тонкий, сидит на широком двойном основании; девятый сегмент
неширокий (равномерной ширины), образует небольшой saccus; valvae
ковшеобразной формы с завернутым и сильно склеротизованным внеш-
ним краем; в дистальной части valva имеет два зубца — вентральный
короткий, сильно конический и дорзальный длинный, сидящий на широ-
ком основании, на дистальном конце с коротким шипиком; fultura infe-
rior в виде сильно склеротизованной пластинки с округлым дорзальным
вырезом; penis короткий, почти прямой, с коротким coecum, заостренный
в дистальной части; на pars inflabilis сильно склеротизованная пластинка
с коротким шипом. Bursa copulatrix мешковидная, несколько расширен-
ная проксимально, мембра-
нозная, несет в верхней по-
ловине lamina dentata в виде
небольшой почти круглой
пластинки; ductus bursae
короткий, сильно склероти-
зован, есть lamella anteva-
ginalis в виде короткой почти
•сердцевидной пластинки;
восьмой сегмент широкий,
но очень короткий, большей
длины при основании apo-
physes anteriores; последние
короткие, но длиннее ductus
bursae, расширенные в прок-
симальной части; apophyses
posteriores длиннее восьмого
сегмента; papillae anales
очень короткие и широкие.
Яйца круглые, крупные,
почти до 1 мм диаметром,
зеленоватой окраски; хорион
гладкий, матовый.
Взрослая гусеница серо-коричневая; по спинной поверхности белая
или желтоватая полоса, разделенная рыже-серыми перемычками на тер-
гитах, особенно брюшных сегментов; кожные железки находятся, кроме
шестого и седьмого брюшных сегментов, также на тергитах первого и вто-
рого брюшных сегментов; по сторонам широкой дорзальной полосы две
прерванных узких белых или желтоватых полоски, оттененные черным;
на тергитах по паре красных бородавок, сидящих на темных перемычках,
разделяющих дорзальную линию; по бокам желто-серые бородавки;
волоски, покрывающие тело, бело-желтые; голова черная; грудные ноги
черные, брюшные желтые.
Куколка блестящая, черная, покрытая длинными желто-белыми мяг-
кими курчавыми волосками, сидящими пучками на небольших вздутиях
кожи; кожа гладкая без скульптуры; cremaster длинный, почти стержне-
видный, покрытый глубокими продольными бороздами, на дистальном
конце его сидят многочисленные короткие закрученные спирально
крючья.
Изменчивость как во взрослой, так и в личиночной фазах не велика;
гусеницы при развитии в повышенных температурах характеризуются
I. ORGYINAE
более сильным развитием железок на тергитах брюшных сегментов.
Имаго варьирует в окраске; особи с рыжеватым оттенком, очень редкие-
впрочем, описаны как rubicunda Strd., особи серовато-желтого цвета —
как soheSti Casp., особи с темной, почти черной бахромой и краем крыльев,
очень редкие,—как nigrociliata Fuchs.; в наибольшей мере изменяется
размер особей в связи с условиями личиночного питания.
Распространение. Вся средняя и западная Европа от Пире-
неев до побережья Балтийского и Средиземного морей; северная и сред-
няя Италия, Альпы, Карпаты и северные Балканы; северная Африка
(Б. Атлас); острова Средиземного моря (Корсика, Сардиния); Швеция.
(Эланд, Готланд, Дарланд, Азеле, Ликзеле, Торне); Финляндия (Алан-
дия, Ниляндия); в Европейской части СССР от Мурманского побе-
режья (Александровск) и Архангельска всюду по лесной зоне, лесо-
степи и отчасти по степной полосе (Каунас, Рига, Таллин, Петро-
заводск, Вологда, Молотов, Ленинград, Москва, Калинин, Тамбов, Орел,
Брянск, Воронеж, Киев, Полтава, Херсон, Одесса, Таганрог); Крым
(Севастополь, Бельбек, Керчь); Поволжье (Казань, Куйбышев, Улья-
новск, Саратов, Сарепта, Астрахань); Приуралье (Чкалов, Красноуфимск,
Курган); Северный Кавказ (Ставрополь, Новороссийск, Дзауджикау,
Майкоп, Гуанчхир, -Дагестан); Закавказье (Лагодехи, Армения); Малая;
Азия; в Сибири от линии Березов—Обь-Енисейский канал—Вилюй
всюду по лесной зоне и степи (Омск, Барабинская степь, Томск, Мину-
синск, Бунбуй, Канск, Иркутск, Байкал, Якутск); Алтай (Телецкое
озеро, Томь, Онгудай); Саяны (Можарские озера, Казыр-сук); Казахстан
(Семипалатинск, Акмолинск); Средняя Азия. (Джунгарский Ала-тау,.
Тарбагатай, Тянь-шань); северная Монголия (Хангай, Кентей).
Биология и экология. Зимуют гусеницы второй стадии
в щелях коры, в сухих листьях или в подстилке, поодиночке, в малень-
ких белых кокончиках; Нюсслин (1927) указывает на зимовку яиц, как
исключение—гусениц; Гусев (1935) также полагает зимовку этого вида,
в фазе яйца; многочисленные наблюдения в окрестностях Ленинграда,
проводившиеся мною в течение нескольких лет, подтвердили указания;
на зимовку только гусениц второй стадии; всегда наблюдалась одна,
линька и только после этого диапауза; зимовка яиц в условиях лесной
полосы и вообще для данного вида мало вероятна. Весной гусе-
ницы начинают питаться с появлением листвы (может быть почек); они.
связаны в природе только с видами Salicaceae (Populus, Salix), пред-
почитают белый тополь, но обычны также на осине и черном тополе, а так-
же на некоторых видах ив (Salix саргеа); для взрослых гусениц отмечено--
питание лещиной и ольхой; как редкость отмечено питание плодовыми
деревьями (Корольков, 1913); окукление в конце июня, на севере в июле,,
в рыхлом коконе из белых шелковых нитей, в щелях коры и на ветвях;
лёт бабочек в конце июля по вечерам; они живут не долго — 5—10
суток — и откладывают яйца на кору и листья кормовых растений;
яйца откладываются кучками по 20—30 шт., покрываются белой пени-
стой массой; общее количество откладываемых самкой яиц 200—300 шт.;
гусеницы вскоре после выхода из яиц расползаются и живут поодиночке,,
сначала энергично питаясь, а после линьки впадают в диапаузу и зимуют..
Встречается в долинах рек, в уремах, в степной полосе, в балках,
часто в городах — в парках — и в лесных питомниках. В СССР вред от-
мечен в ряде мест: в Ленинграде, Москве, Курске, Астрахани, Харькове,
западной Сибири. Вредит посадкам ив и тополей; вред отмечен в Ис-
пании (Сарагосса) тополям и ивам, в Голландии, Швейцарии, Бель-
13. LEUCOMA
34o
гии, Люксембурге, Германии, Франции, Прибалтике и южной Швеции.
После завоза в Америку (около 1920 г. в восточную Канаду) там сильно-
размножилась и стала серьезным вредителем Salicaceae, особенно ввезен-
ных видов — Populus nigra, Р. alba; туземные виды тополей — Populus-
tremuloides, Р. trichocarpa или совсем не повреждаются или повре-
ждаются слабо; из ив вредит Salix basia'ndra и. S babylonica. Несмотря
на недавний в общем завоз, в Америке наблюдались много раз массовые
размножения этого вида в Вашингтоне, Нью-Фаундлэнде, в штате Мэн,
в восточной Канаде и других местах, сопровождавшиеся опустошениями
больших территорий; в пределах СССР массовое размножение этого вида
отмечено в Поволжье (Гусев, 1935).
Отмечены различные паразиты как яиц, так гусениц и куколок;
из Chalcididae — Eutedon oinidae Rtzb., Pteromalus boucheanus Rtzb.,
P.eticeros Rtzb., P.halidayanus Rtzb., P.pini Htg., P. puparumL.; из Bra-
conidae — Bracon circumscriptus, Meteorus formosus Rhd., M. bimaculatus
Rtzb., M. scutellator Ns., M. obscuratus Ns., M. ruficrus Ns., M. dijjicilis-
Ns., M. delectus Hal., M. melanoceles Rtzb., M. salebrosus Mrsch., M. so-
litarius Rtzb., Bhogas unicolor Wesm., Rh. praerogator Ns.; из Proctotrupi-
dae — Telenomus punctiventris Thms., T. nitidulus Thins.; из Ichneumo-
nidae — Cryptus analis Gr., Limneria assimilis Gr., Protichneumon disparts
Poda, Cratichneumon fabricator F., C. nigritarsus Gr., Mesochorus
semirufus Hgr., M. ater Rtzb., M. politus Gr., M. splendidus Gr., Pezo-
machus terebrator Rtzb., P. fasciatus Gr., Pimpla instigator F., P. ster-
corator F., P. arctica Zett, P. examinatbr F., Theronia atalantae Poda,
Ichneumon haglundi Holmgr., Apechta brassicariae Poda, Epiurus inqui-
sitor Scop., P.eltopius anxius Wesm.; из двукрылых известно меньшее'
число паразитов — Carcelia cheloniae Rd., С. excisa Fall., C. gnava
Mg., Compsilura concinnata Mg., Lydella stalulans Mg., Micropalpus haemor-
rhoidalis Fall., Nemoria mirabilis B., Pales pavida Mg., Plagia elata Mg.,
Agria affinis Fall. (Мейер, 1937; Ледерер, 1923).
* 2. Leucoma Candida Stgr. (рис. 166, 167).
Staudinger, 1892, Mem. Lep. Rom., 6 : 308; Staudinger, 1901, Cat. Lep., 1 : 117;
Spuler, 1908, Schmett. Eur., 1 : 130; Strand, 1910, Seitz, Grossschmett., 2 : 123; Nagano,.
1916, Bull. Nava ent. Lab., 1 : 12, t. 4, fig. 20—24; t. 9, fig. 24; Swinhoe, 1922, Ann.
Mag. Nat. Hist., 10 (9) : 462 (bona species); Matsumura, 1933, Ins. Matsum., 7 : 151;
Bryk, 1934, Lep. Cat., 62 : 115; Куренцов, 1935, Вести. Дальневост. фил. Акад. Наук-
СССР, 2 : 68; Куренцов, 1939, Тр. Горнотаежн. ст., 3 : 147.
Близок к предыдущему виду, но отличается, помимо структурных
признаков в гениталиях, более густым чешуйчатым покровом и, главное,
более крупными чешуйками, покрывающими крылья, почему крылья
не прозрачны (как бы более толстые); они имеют чисто белый цвет, бле-
стящие; усики с черными колечками; ноги черные с небольшими белыми
колечками; брюшко густо волосистое, кожные покровы не просвечивают;
самки этого вида отличаются от предыдущих еще более самцов; в проти-
воположность предыдущему виду крылья их совсем не прозрачны; ука-
зание Штаудингера на меньшие размеры L. Candida»Stgr. по сравнению
с L. salicis L. ошибочно. Размах крыльев самцов 32—38 мм, самок
45—60 мм.
Uncus в основании широкий, в дистальной части несколько сужен
и на конце изогнут и несколько утолщен, сочленен швом с tegumen;
девятый сегмент широкий, образует небольшой конической формы saccus;
fultura inferior в виде двух округлых лопастей, сросшихся посредине;.
:Uf>
1. ORGYINAE
valvae сильно склеротизованы; в дистальной половине valva заострена,
на дорзальной ее стороне коротких!, но сильный и изогнутый внутрь
вырост, вентральный край valva завернут, склеротизован и зубчат:
penis сложно изогнут с длинным coecum, несет на pars inflabilis мелкие
щетинки. Bursa copulatrix почти цилиндрической формы, мембранозная,
имеет короткий, широкий и прямой мембранозный ductus bursae; ваги-
нальная область снабжена lamella antevaginalis в виде сильно двузуб
чатой пластинки; восьмой сегмент широкий, удлиненный лишь в дор-
зальной части; apophyses anteriores короткие, posteriores тонкие, длиннее
первых; papillae anales очень большие, несколько суженные в дистальной
части, мембранозные.
Рнс. 166. Leucoma Candida Stgr. Гениталии:
а — самца, б —• самки.
Личиночные фазы этото вида изучены еще неполно. Мне известны
одна гусеница и несколько куколок, собранные Черновским 8 августа
1949 г. близ Семипалатинска. Они очень характерны, хотя и напо-
минают обычную Leucoma salicis L. Взрослая гусеница беловато жел-
тая с широкой, желто-белой спинной полосой, прерванной на сегмен-
тах и по сторонам с двумя узкими, прерванными на сегментах, корич-
невыми линиями. Эти линии представлены на сегментах короткими
штрихами, отделенными друг от друга бородавками и р. Послед-
ние желтоватого цвета и серые, большие, резко выделяются на коже.
Все бородавки несут мягкие желто-серые волоски. Голова черная;
грудные ноги черные со светлыми колечками; брюшные ноги одного
цвета с кожей. Стигмы в середине темносерые, по краям черные, мел-
.кие, круглые на первом сегменте брюшка, на прочих овальные. Же-
лезки на первом и втором брюшных сегментах двойные, черные, выво-
рачиваются в виде двух расходящихся трубочек; железки на шестом
и седьмом брюшных сегментах брюшка непарные, белые в виде вороно-
чек. Куколки беловато желтые с ярким, черным рисунком; голова,
13. LEU COM A
347
ротовые части и усики черные или желто-коричневые; темные концы
лапок; тергиты грудных сегментов разделены вдоль черной полосой
с неправильными, черными пятнами по ее сторонам; жилки на крыльях
черные; черные стигмы на брюшных сегментах в виде узких щелей,
черные полоски на межсегментальных мембранах и черные пятна снизу
на сегментах брюшка; cremaster черный, длинный, почти стержневид-
ный, с продольными бороздками и с многими спирально закрученными
крючьями на дорзальной стороне, сходен с тем, что описано для L. sali-
cis L. Кожа куколок несет пучки беловато-желтых, густых и довольно
длинных волосков.
Распространение. Южное Приморье (Иман, Сучан, Супу-
тинка, Бикин, Суйфун, Сица, Сидеми, Хуалаза); Йриамурье (Зея, Благо-
вещенск, Хабаровск, Николаевск, Усть-Майская, хуторРейново); Якутск;
Яблоновый хребет; Витим; Забайкалье (Даурья, Сретенск, Троицкосавск);
Улан-удэ; южный Енисей (Минусинск); Алтай (Онгудай, Ойротия, Киреев-
ское);. Иртыш (Семипалатинск); возможно нахождение в Тарбагатае и
Ала-тау, Японские острова (Хондо, Хоккайдо); Корея, северный Китай;
Маньчжурия; северная Монголия (рис. 167).
Биология. Зимуют Мелкие гусеницы; распределение фаз разви-
тия в течение вегетационного времени сходно с таковым у L. salicis L.
Гусеницы питаются тополями (Populus nigra, Р. alba, Р. tremula), реже
ивами; в южном Приморье отмечены виды, питающиеся, кроме ив и топо-
лей, также Chasemia macrolepis Кош. Заселяет речные уремы иногда
в массовом количестве (по Уссури; Куренцов, 1939). Вредит,, как и пер-
вый вид, ивам и тополям; вред указан для южного Приморья (Курен-
цов, 1935, 1939) и Японских островов (Хоккайдо).
* 3. Leucoma flavosulpburea Ersch. (рис. 168, 169).
Erschov, 1872, Тр. Русск. энтом. общ., 8 : 316; Erschov, 1874; Fedtschenko, Reise
Turkest. : 35, t. 3, fig. 34 (Leucoma); Grum-Grzhimailo, 1890, Mem. Lep. Rom., 4 : 558,
t. 20, fig. 7; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 434; Staudinger, 1901, Cat. Lep.,
1 ; 117; Strand, 1910,. Seitz, Grossschmett., 2 : 124; Swinhoe, 1922, Ann. Mag. Nat.
Hist., 10 (9) : 462; Gaede, 1932, Seitz, Grossschmett., 2 Suppl. : 99, t. 8, k; — cretacea
.StEiudinger, 1901, Cat. Lep., 1 : 117; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10 (9) : 462
(bona species).
Передние крылья, голова, грудь, брюшко и задние крылья темного
шафранно-желтого цвета; передние крылья без рисунка, однотонные,
с жилками, несколько более светлыми; крылья покрыты узкими чешуй-
ками и волосками; нижняя сторона крыльев бело-желтая; голова и грудь
покрыты нежными пушистыми волосками без примеси чешуй; усики
самцов сильно, двусторонне гребенчатые, членики уСиков светложелтые,
отростки их черно-серые; глаза большие, черные; губные щупальца
покрыты длинными черными волосками снизу и желто-белыми сверху;
ноги покрыты бело-желтыми длинными волосками с небольшой примесью
черных волосков; передние годени с внутренней стороны черные; самки
окрашены бледнее, белые, со слабым желтоватым оттенком; членики
усиков с короткими отростками. Размах крыльев самцов 35—45 мм,
самок 35—50 мм.
Uncus заостренный, при основании плоский и широкий, с двумя
бороздками по дорзальной стороне; отделен швом от девятого сегмента;
последний с дорзальной стороны длинный, с вентральной не образует
saccus; fultura inferior широкая, в виде пластинки, с внутренней стороны
шесколько выемчатая; valvae округлой формы, вздутые, несут по два
Рис. 167. Распространение видов рода Leucoma Steph, в Палеарктике.
1 — L, salicis L.; 2— L, Candida Stgr.; 3—L. flavosulphurea Ersch,
13. LEU COM A
349
отростка, направленных косо и вверх; penis сложно изогнут с длинным
coecum, почти равным его длине. Bursa copulatrix мешковидная, без
скульптуры; ductus bursae короткий, снабженный па поверхности склад-
Рис. 168. Leucoma jlavosulphurea Ersch. Самец. (Рис.
Ф. Гуняева).
ками; восьмой сегмент короткий, несколько длиннее в месте укрепления,
apophyses anteriores; последние заостренные и довольно длинные; apo-
physes posteriores короче первых; papillae anales широкие и короткие,
в дистальной части округ-
лые.
Изменчивость признаков
не велика; варьирует окрас-
ка особей; известна чисто бе-
лая форма — cretacea Stgr.,
более мелкая — 32—37 мм,
описанная из Семиречья и
из северной Ферганы; воз-
можна связь этой формы с
севером ареала вида.
Распро странение.
Горная Средняя Азия: Фер-
ганская долина (Ош, Воа-
дил, Маргелан, Юрга, Лян-
гар, Вагаман); Семиречье
(Нарын, Фрунзе, Чилик,
Алма-ата, Талгар, Джасил-
куль); Алайский хребет
(Гульча); Памир (Вахан,
Калай-вамар), Зеравшан- рис. ^gg. £еисота jlavosulphurea Ersch. Геиита-
скии хребет (Самарканд, На- лии: а — самца, б — самки,
ра-тюбе).
Биология. Вид обычный в оазисах Средней Азии на тополях
(Грум-Гржимайло, Вельтищев); видимо дает (по Грум-Гржимайло) два
поколения в сезон, так как бабочки были найдены в апреле и в августе;
около Оша в мае, близ Самарканда — в июне.
350 I
I. ORGYINAE
* 4. Leucoma sarhis Ersch.
Erschov, 1874, Fedtschenko, Reise Turkest. : 36, t. 2, fig. 32; Kirby, 1892, Syn.
Cat. Lep. Het., 1 : 466; Staudinger, 1901, Cat. Lep., 1 : 117; Strand, 1910, Seitz, Gross-
schmett., 2 : 124, t. 21, d; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10 (9) : 462; Gaede,
1932, Seitz, Grossschmett. 2 Suppl. : 99; Bryk, 1934, Lep. Cat., 62 : 117.
Передние крылья кофейно-серые, почти шоколадного цвета; этого же
цвета голова и грудь; голова имеет примесь рыжего цвета на лбу и уси-
ках; передние крылья покрыты длинными густыми волосками, прикры-
вающими сверху чешуйчатый покров; рисунок на передних крыльях
отсутствует; задние крылья менее волосистые, покрыты узкими чешуй-
ками и редкими толстыми волосками, окрашены светлее, особенно при
основании; бахрома как передних, так и задних крыльев ‘коричнево-
серая; брюшко волосистое; усики самцов с длинными гребешками, по-
крыты волосками, концы гребешков несут по одной длинной толстой
щетинке; глаза большие, черные; губные щупальца недлинные, торча-
щие вперед, покрытые черными и коричневыми волосками; последний
членик их очень мал; передние голени покрыты длинными пушистыми
волосками, с внутренней стороны черные; черный цвет несут на них
чешуйки и плоские волоски; лапки всех ног черные и коричневые; самки
окрашены бледнее, светлошоколадного цвета, менее волосисты (брюшко,
крылья и ноги); губные щупальца и ноги также имеют примесь черных
чешуй и волосков; усики с более короткими гребешками, концы гребеш-
ков несут также по одной сильной щетинке, более короткой, чем у сам-
цов. Размах крыльев самцов 30—32 мм, самок 34 мм. Близок к преды-
дущему виду; известен в небольшом количестве особей.
Распространение. Горная Средняя Азия: Таласский Ала-
тау (Ташкент); Зеравшанский хребет (Самарканд); Семиречье (Кульджа,
Или, Иссык-куль); по Штаудингеру (1901), также Фергана.
* 5. Leucoma surtur В. Н.
Bang-Haas, 1912, Iris, 26 : 107; Gaede, 1932, Seitz, Grossschmett., 2 Suppl. : 99.
Известен только тип этого вида, очень свежий самец; изображения
его не дано, описание очень кратко. Мне вид в натуре не известен, при-
вожу оригинальное описание.
«Хотя есть только один самец (очень свежий), я вынужден описать
новый вид на основании этого замечательного экземпляра. Этот новый
вид наиболее близок к L. sartus Ersch., но отличается равномерным чер-
ным цветом как верхней, так и нижней сторон крыльев и тела (примерно
как самка Ocneria detrita Esp.); все части тела также черные. Передние
крылья несколько шире, чем у L. sartus Ersch., и усики имеют несколько
более сильные гребешки. Размах крыльев 39 мм.» (Банг-Гааз).
Распространение. Средняя Азия (Юлдус).
14. Род OCNERIA НВ.
Hiibner, 1816, Verz. bekannt. Schmett. : 153 (typus rubea F.); Aurivillius, 1891,
Nordn. Fjar. : 94; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 477; Hampson, 1892,
Fauna Brit. Ind., Moths, 1 : 459; Pagenstecher, 1899, Lep. Bism. Arch, in Zool., 29 : 42;
Staudinger, 1901, Cat. Lep., 1 : 117; Swinhoe, 1903, Trans. Entom. Soc. Lond. : 403;
Spuler, 1908, Schmett. Eur., 1 : 130; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 117;
Strand, 1910, Seitz, Grossschmett., 2 : 126; Strand, 1915, Seitz, Grossschmett., 10 :
320; Strand, 1916, Arch. Naturgesch., A, 3 : 80; Hering, 1927, тамже, 14 : 193; Hering,
1932, Tierw. Mitteleur. : 484; Collenette, 1933, Nov. Zool., 39:21; Bryk, 1934, Lep.
Cat., 32 : 144; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 67; — Liparis Ochsenl
14. OCNERIA
351
heimer,1 1810, Schmett. Eur., 3 : 186; — Lymantria Hiibner, 1816, Verz. bekannt.
Schmett. : 160 (typus monacha L.); —Porthetria Hiibner, 1816, тамже : 160 (typus dis-
par L.); —Psilura Stephens, 1822, Ill. Brit. Entom. Haustell., 2 : 57 (typus monacha
L.); — Erasta Gistl, 1848, Nat. Thierr. : 9 (typus monacha L.); — Enome Walker, 1855,
.Cat. Lep. Het. Brit. Mus., 4 : 883 (typus ampla Wlk.); —Pagella Walker, 1866, тамже,
35 : 1922. (typus cure ifer a Wlk.); —Nagunda Walker, 1879, Lep. Atk. : 53 (typus semi-
cincta Wlk.); —Pyramocera Butler, 1880, Journ. Linn. Soc. Lond., Zool., 15 : 86 (typus-
juliginea Btl.). '
Тип рода О. rubea F.
Хоботок редуцирован вполне; усики самцов сильно гребенчатые;
глаза большие, голые, круглые, окружены волосками и чешуей; губные-
щупальца развиты вполне, второй членик их длинный, третий членик
очень маленький; ноги покрыты длинными волосками; первый членик
передних лапок короче передних голеней; голени задних ног с двумя
парами шпор; коготки на лапках всех ног простые; радиальная ячея
на передних крыльях не развита, жилки радиальной системы располо-
жены по форме ячеи; первый и второй радиальные стволы подходят
близко один к другому; третий и четвертый радиальные стволы на длин-
ном стебельке; дискоидальные жилки развиты; на задних крыльях базаль-
ная ячея большая, в дистальной части замкнутая коротким анастомозом
радиальной и субкостальной жилок; вторая и третья медиальные жилки,
на задних крыльях сильно сближены основаниями; дискальные жилки
развиты вполне.
Uncus простой, плотно спаян с девятым сегментом; лишь редко есть
следы шва при основании uncus; tegumen часто несет subunci; valvae-
конические, с острым дистальным концом. Bursa copulatrix маленькая,
мембранозная; последние сегменты брюшка' у самок вытянутые. Полы
обычно сильно диморфны; самки больших размеров и светлее окрашены.
Гусеницы уплощенной формы; в первой стадии несут длинные волоски
«аэрофоры» и короткие, колбовидно вздутые при основании; характерны
длинные волоски по сторонам тела. Первые четыре брюшных сегмента не-
сут небольшие парные выворачивающиеся железки; на дорзальной стороне-
шестого и седьмого брюшных сегментов по одной срединной железке.
Бородавка /. на переднегруди большая, торчащая вперед, с большим
пучком волос, направленным вперед. Бородавка а на втором и третьем'
грудных сегментах, а также на сегментах брюшка несет по две щетинки
или небольшой пучок мелких; бородавка р во много раз меньше [i на
этих же сегментах. Бородавка z. на брюшных сегментах сдвинута дор-
зально, располагается над стигмами и почти слита с р; редко, например
у Ocneria rubea F., бородавка z. только сдвинута дорзально, но не слита
с р на восьмом брюшном сегменте.
Куколки черно-коричневого цвета, сильно суженные дистально; кожа
покрыта отдельными негустыми пучками волосков. Дыхальца располо-
жены в головной половине на подвижных брюшных сегментах. Головной
отдел несет такие же большие пучки волос, как и прочие отделы тела...
Губные щупальца очень малы, нередко почти незаметны. Яйца крупные,
1.0—0.9 мм диаметром, с плотным матовым хорионом. Яйцекладка куч-
ками; яйца покрываются или волосками с конца брюшка самки, или
выделениями придаточных желез полового аппарата..
Обширный политипический, но в общем целостный род, обильно пред-
ставленный на юге и юго-востоке Азии (рис. 170); наибольшее число
1 У Оксенгеймера (1810) первым видом рода Liparis стоит!.. morio L. (nonScopoli,
1777), но род этот включает также ряд других видов и среди них L. monacha L.; тип рода
не отмечен автором. Впервые как тип рода указан Кирби (1892) — L. monacha L...
„К 2 ° О.
20 90 60 31 <67 ^5^ V ** / Хл-, йЕ ту 6т \ 0 » \ z**\ 1 0 } —. <0 0 o\ 0
- . о 1 ч i 0
о ts» / 0 00
о ш ? 0
- 4 . £ О -'°5 : ? ! 4 x К L 4
гЖ т FZ 0
<5> ' вС2^Г z 0
СО \ ’ Ч "Тст^ ^<[5 ~~' * -"**1 /*ур г 5*l I 0
IC с ц- ,,j (_Т • J 0 s ? 0 0 Oj Q
t=/4z О e a 0 О
* “* "“d § ‘ eo
Рис. 170. Распространение видов рода Ocneria Hb.
Черным обозначены территории с числом видов 10—15; сеткой с числом видов 4—8; штриховкой с числом видов
14. OCNERIA 353
видов концентрируется в Гималаях, Индии и на Малайском архипелаге.
Часть индо-гималайских видов эндемичны (О. marginata Wlk., semicincta
Wlk., obfuscata Wlk., todara Moore, concolor Wlk., incerta Wlk., lepcha
Moore, phmbalis Hamps., grisea Moore, ampla Wlk., rhodina Wlk., bivit-
tata Moore), ограничены только Гималаями и Индией. Известное число
эндемиков описано также для Цейлона (О. viola Swinh., fuliginosa Moore,
grandis Wlk., postfusca Swinh.). Многие индо-гималайские эндемы, а также'
некоторые цейлонские очень напоминают по габитусу ’ евразийскую
О. monacha L.; меньшее число сходно с О. dispar L. Часть индо-гималай-
ских видов заселяет также Цейлон (О. .subrosea Swinh., ampla Wlk.),
а некоторые распространены шире и заселяют также южный Индокитай,
Малакку и острова Малайского архипелага (О. obsoleta Wlk., similis
Moore, ganara Moore, golinaria Swinh.). На островах Малайского архи-
пелага, особенно на Борнео, Яве и Целебесе, есть ряд эндемичных видов
Ocneria Hb. (strigata Auriv., dubiosa Auriv., ganaha Swinh., beatrix Stoll.,
asoetria Hb., brunneiplaga Swinh., loacana Semp., narindra Moore, margi-
nala Wlk., maculata Semp., simplex Pag., rosina Pag., brotea Cr., inordi-
nata Wlk.); некоторые из видов, населяющих Малайский архипелаг,
распространены шире, например О. lunata Stoll., и заселяют также Филип-
пинские острова и Новую Гвинею. G последнего острова известен ряд энде-
мичных видов (О. silca Swinh., ekeikei Bet.-Bek., ninayi Bet.-Bek., nova-
guinensis Bet.-Bek., ganaroides Strd.); немногие эндемики известны с Фи-
липпинских островов (О. tagalica Auriv., curvijera Wlk.) и с Малакки
(О. singapura Swinh.). Небольшое число видов Ocneria Hb. известно
из Австралии (О. anlennata Wlk., turneri Swinh.); они ограничены только
севером этого материка (Норк, Мортон).
Индомалайские виды распространены также в Западном Китае {О. si-
nica Moore, concolor Wlk., obsoleta Wlk.); там же есть эндемичные виды
(О. di’ssoZwtaSwinh., nigriplagiata Gaede, apicibrunnea Gaede); большее число
эндемичных видов известно для Тайвана (О. kosempornis Strd., melanopygon
Strd., umbrifera Wil., nebulosa Wil., xylina Swinh.); сюда же проникает
широко распространенный О. dispar L. Севернее, на Японских островах
и в южном Приморье распространено небольшое число эндемичных видов —
О. sakaguchii Mts., lutescens Btl. в. Японии, О. grisescens Stgr. в южном
Приморье. Один вид (О. malura Moore) известен для Индии, но не рас-
пространен в Китае, а широко распространен в южном Приморье и на
Японских остравах. Два вида (О. dispar L. и О. monacha L.) широко
распространены в лесной зоне Евразии. В Северной Америке нет тузем-
ных видов Ocneria Hb., но евразийский О. dispar L. был занесен в восточ-
ную ее часть и акклиматизировался там вполне; в Южной Америке совсем
нет видов Ocneria Hb.
Большое число эндемичных видов Ocneria Hb. известно с Мадагаскара
(atdla Swinh., rebuti Pouj., velutina Mab., nigricostata Kenr., dulcinea
Btl., rosea Btl., paroula Kenr., rufofusca Mab., joanisi Le Cerf, melanocera
Mab., vitrina Mab., heptasticta Mab., rhodophora Mab., fuliginea Btl., barica
Mab., fumosa Saalm., uxor Saalm.); значительное число также типичных
Ocneria Hb. описано из восточной тропической Африки (bananae Btl.,
eddela Swinh., pruinosa Her., carriola Swinh., gondona Swinh., melete Fawc.,
taurina Her., menecles awe., vrete Fawc., conspersa Her., melissa Fawc.).
Меньшее число и в ряде случаев не вполне типичных по габитусу видов
Ocneria Hb. описано из западной тропической Африки — Конго, Каме-
руна, Того (rusticana Her., vacillans Wlk., bananoides Her., bananae Btl.,
multiscripta Holl.).
23 Фауна СССР, И. В. Кожанчиков.
334
I. ORGYINAE
Палеарктические виды увязаны с древесными растениями и преиму-
щественно с широколиственными породами — Fagales, Rosales и отчасти
с Coniferae. Кормовые растения тропических видов мало известны, но
в ряде случаев отмечают или эти же группы растений или древесные
Leguminosae. Изученные палеарктические виды гигрофильны и стенотер-
мичны; судя по ареалу рода это же типично и для большинства видов
Ocneria Hb.
Большая часть видов биологически характеризуется малой активностью
самок; зимовка палеарктических видов протекает в фазе яйца; в тро-
пиках размножение видов Ocneria Hb. и их развитие протекают круглый
год; диапауза для тропических видов не описана. Широко распростра-
ненные в Палеарктике виды характеризуются массовым количеством
особей и большим экономическим значением как вредители сельского
и лесного хозяйства. Массовые тропические виды и вредители культур-
ных растений среди тропических видов Ocneria Hb. не описаны.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). Передние и -задние крылья однотонные — серые или охристо-
розоватые, без рисунка совсем или с немногими его рудиментами;
дорзальный край valva несет один дистально направленный вырост.
2 (3). Серый; передние крылья тусклого цвета с неясным, стушеванным
рисунком; задние крылья однотонно серые с узкой растушеванной
темносерой каймой; бахрома слабо шашечная или однотонная;
дорзальный отросток valva широкий и большой ...................
................................... * 2. О. grisescens Stgr.
3 (2).- Охристо-розоватый, бедно покрытый чешуей; на месте диско-
идальной жилки светлый штрих; бахрома на крыльях простая;
задние крылья однотонные, без каймы; однотонные же и передние
крылья, без рисунка; дорзальный отросток valva не длинный .
.............................................5. О. rubea F.
4 (1). Передние крылья белые, серые или иной окраски с хорошо разви-
тым рисунком; нередко есть примесь чисто розовых тонов на брюшке
и задних крыльях самок; бахрома шашечная: valva или не имеет
боковых выростов (несет только острый конец), или имеет два
выроста по дорзальному краю и один по вентральному.
5 £6). Передние крылья белые или светлые с четким черным рисунком
в виде сильно зубчатых линий; иногда рисунок сильно растушеван
и линии сливаются между собою, тогда крылья почти черные; зад-
ние крылья тусклосерые; брюшко самок с длинным яйцекладом
(который образован седьмым—девятым сегментами брюшка и может
втягиваться в предыдущие сегменты); valva без выростов на боковых
сторонах, острая; девятый сегмент с subunci ... .......... .
.............................. . . .* 1. О. monacha L.
6 (5). Рисунок бурый или желтый, нерезкий, слабо зубчатый; крылья
коричнево-желтые или желто-серые, но не белые; если есть subunci,
то valva несет отростки по дорзальному и вентральному краям,
если нет subunci, то нет и отростков на valvae, которая острая.
7 (8). Передние крылья с белыми продольными штрихами вдоль жилок;
светлые — белые и серые — штрихи есть также на груди (спинке)
. и на голове; брюшко самок сильно розоватое или розовое, розовые
и крылья: valva с тремя отростками (не считая острого дистального
конца); девятый сегмент несет subunci...........................
........................................* 3. О. matura Moore
14. OCNERIA
355
8 (7). Передние крылья желто-коричневые без продольных светлых
штрихов; линии рисунка сильно растушеваны; крылья самок желто-
белые; valva без боковых выростов, только с острым дистальным
концом; subunci не развиты; конец брюшка самок несет подушку
коричневых или желтых коротких густых волосков .....................
............................................. * 4. О. dispar L.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПО ГУСЕНИЦАМ (ВЗРОСЛЫМ
И ПОЛУВЗРОСЛЫМ)
I (2). Бородавки, покрывающие тело, рыже-желтые; вдоль спины две
латеральных полосы темного цвета; голова светложелтая, одно-
тонная; на переднегруди бородавка х небольшая, несет короткие
пучки волосков; щетинки, покрывающие тело, рыжие....................
................................................5. О. rubea F-
2 (1). Бородавки, покрывающие кожу, серо-желтые или красные и си-
ние; бородавка х (перед стигмой) на переднегруди сильно развита
и несет длинные пучки. волосков, направленных вперед; голова
желто-серая с черными полосами и черным рисунком; щетинки,
покрывающие тело, черные.
3 (4). На тергите среднегруди трапециевидное черное бархатистое
пятно (несколько стушеванное по заднему краю); бородавки на
всех сегментах желто-серые, не резко выделяются на фоне кожи
.....................’.............................*10. monacha L.
4 (3). Тергит среднегруди окрашен одинаково с прочими частями тела;
грудные сегменты, а также два первых брюшных с дорзальной сто-
роны несут по две синих или синеватых бородавки; тергиты прочих
брюшных сегментов несут красные бородавки; все бородавки резко
выделяются на фоне кожи.............................* 4. О. dispar L.
*1. Ocneria monacha L. —Монашенка (рис. 171, 172, 173).
Linne, 1758, Syst. Nat., ed. 10 : 501 (Bombyx); Aurivillius, 1891, Nordn. Fjar.,
94, t. 26, fig. 3; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 477; Staudinger, .1901, Cat. Lep.,
1 : 117; Spuler, 1908, Schmett. Eur., 1 : 131, t, 25, fig. 30; Rebel, 1910, Berge, Schmet-
terlingsb., ed. 9 c 118, t. 24, fig. 5; Strand, 1910, Seitz, Grossschmett., 2 : 110, 128,
t. 20, g; Brundin, 1916, Entom. Tidskr., 37 : 102; Petersen, 1924, Lep. Fauna EstL,
ed. 2, 1 : 142; Osthelder, 1926, Schmett. Siidbayerns, 1 (2) : 200; Marschner, 1932,
Entom. Rundsch., 49 : 246; Gaede, 1932, Seitz, Grossschmett., SuppL, 2 : 100; Hering,
1932, Tierw. Mitteleur. : 482; Bryk, 1934, Lep. Cat., 62 : 151; Brandt, 1926, Zoo],
Anz., 114 : 161; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 67, t. 14, fig. 5; —
heterociliata Muller, 1764, Fauna Ins. Friedrichsd. : 47; —eremita Hiibner, 1804, Eur.
Schmett., 2 : f. 246; — nigra Freyer, 1833, Neuere Beitr. Schmett., 2 : 5, t. 98, fig. 2,
3; — transiens Mieg, 1886, Le Naturalists, 8 : 238; — atra Linstow, 1907, Entom. Ztschr.
Stuttg., 21 : 96; — lutea Auel, 1908, Ztschr. wiss. Insektenbiol., 4 : 39; —flavivei
ter Krulikovsky ,1909, Русск. Энтом. обозр., 9, : 303; — dbsoleta Schultz, 1910, Entom.
Ztschr. Stuttgart, 24 : 36; — subjusca Schultz, 1910, там же, 24 : 36; — flaviabdomino-
lis Schultz, 1910, там же, 24: 36;—gracilis Krulikovsky, 1911, Русск. Энтом.
обозр., И : 445;—fasciata Hanneman, 1916, Intern, entom. Ztschr., 10 : 37; —
mediojusca Lambillion, 1919, Rev. Mus. Soc. Namur : 39;—brunnea Stipan, 1923,
Entom. Ztschr., 37 : 40; — kusnezovi Kolossov, 1928, Entom. Ztschr., 41 : 481; —
yunnanensis Collenette, 1933, Nov. Zool., 39 : 23, t. 3, fig. 3.
Биология и экология. Judeich und Nitzsche, 1895, Lehrb. Forstins.,
2 : 803; Standfuss, 1896, Handbuch Grossschmett. : 57, 162, 204, 227, 261, 307, 309,
316; Lampa, 1899, Entom. Tidskr., 20 : 5, 81; Aurivillius, 1899, тамже, 20 : 279; Auel,
1900, Ztschr. wiss. Insektenbiol., 5 : 158, 183; Brake, 1909, Entom. Ztschr., 23 : 156;
Eckstein, 1910, Deutsch. Forstzeitschr., 25 : 428; Scheidter, 1910, Naturw. Ztschr. Land-
wirtschaft, Stuttgart : 373; Коппер, 1916, Леси.” жури., 45 : 1421; Escherich, 1914,
Forstins. Mitteleur., 1 : 183, 200, 243, 250, 302, 312, 314, 325, 351; Wolff und Krausse.
23*
ЗЬ6
1. ORGYINAE
1922, Wichtigste Forstins : 124, fig. 150—156; Fischer, 1925, Entom. Ztschr. Frankf.’,
38 : 39, 73, 78, 83; Escherich, 1925, Anz. Schadlingsk., 1 : 68; Старк, 1926, Защ. раст.,
3 : 17; Knoche, 1929, Arb. Biol. Reichsanst., 16 : 700; Knoche, 1932, Arb. Biol.Reichs-
anst., 20 : 269; Zwolfer, 1933, Ztschr. angew. Entom., 20 : 1; Zwolfer, 1934, тамже,
21 : 33; Eidmann, 1935, Anz. Schadlingsk., 11 : 80; Eidemann, 1935, Zool. Anz., Ill :
104; Куренцов, 1936, Тр. Дальневост. фил. Акад. Наук СССР, 3 : 50; Janisch, 1936,
Anz. Schadlingsk., 12 : 77; Куренцов, 1939, Тр. Горнотаежн. ст., 3 : 150.
Передние крылья, голова и грудь белые с черным резким рисунком;
ноги белые с черными колечками и пестринами, покрыты. длинными
пушистыми волосками; брюшко серое и черное с розоватыми или розо-
Рис. 171. Монашенка (Ocneria monacha L.). Самец.
(Рис. Ф. Гуняева).
выми волосками; базальная линия на передних крыльях начинается
черным штрихом на костальном крае, далее идет в виде интенсивной
черной полосы, обрывающейся в дорзальной трети крыла; базальное поле
широкое с неправильно разбросанными крупными черными пестринами;
вторая поперечная линия также простая, черная, сильно извитая, начи-
нается широким черным почти четыреугольным пятном на костальном
крае, идет перпендикулярно продольной оси крыла; срединное поле
очень неправильной формы, сильно расширенное в середине; круглое
пятно в виде черной точки; почковидное пятно подковообразной формы,
черное; третья поперечная линия сильно зубчатая, начинается двумя
черными пятнами на костальном крае, далее идет двойная, сильно ско-
шенная базально; подкраевая линия черная, сильно зубчатая, зубцы
достигают почти терминального края; терминальная линия стушевана;
бахрома шашечная — белая и черная; задние крылья тусклосерые, одно-
тонные, несколько затемненные по внешнему краю; бахрома шашечная —
белая и серая; нижняя сторона тусклосерая с нерезкими пятнами на ко-
стальном крае, чередующимися с белыми штрихами; на плоскости крыла
неясные, растушеванные поперечные, линии; дискоидальные пятна не
развиты; бахрома шашечная; самка более светлой окраски с сильно рас-
тушеванным рисунком’; брюшко самки розовое с черными полосами,
несколько крупнее. Размах 40—45 мм самцы, 50—55 мм самки.
Uncus сплющен с боков и выгнут, дистальный конец его острый, осно-
вание uncus широкое, отделено хорошо заметным швом от девятого сег-
мента; последний короткий, образует по сторонам основания uncus не-
14. OCNERIA
357
большие конические выросты; есть хорошо развитойвассиэ, треугольный,
несколько заостренный; valvae сильно изогнуты; базальная половина
valva широкая, несет небольшой плоский острый отросток у дорзального
края; дистальная половина ее узкая, сплющенная дорзовентрально,
с округленным дистальным концом; fultura inferior почти треугольной
формы, широкой стороной расположена дорзально, с двумя небольшими
выростами по сторонам, почти плоская; penis заостренный в дистальном
направлении, в проксималь-
ной части широкий, имеет
небольшой заостренный сое-
cum. Bursa copulatrix неболь-
шая, почти цилиндрическая,
мембранозная, без инкруста-
ций; ductus seminalis отходит
от ее дистальной части почти
от основания ductus bursae;
последний почти цилиидри--
ческйй, склеротизован по
всей длине; ostium его снаб-
жен небольшими двумя la-
mellae; между седьмым и
восьмым сегментами длинная
складчатая membrana itner-
segmentalis; восьмой сегмент
узкий и длинный, вентраль-
ная- его сторона мемраноз-
ная, складчатая; apophyses
anteriores тонкие и длинные;
длинная складчатая мемб-
рана между восьмым сег-
ментом и papillae anales;
последние длинные, склеро-
тизованные, усаженные тон-
Рис. 172. Монашенка (Ocneria monacha L.).
Гениталии: а — самца, б — самки.
кими длинными . волосками;
apophyses posteriores тонкие, более длинные, чем первые.
Яйца полушаровидные, несколько сплющенные дорзовентрально;
хорион гладкий, покрытый мельчайшими точками и шипиками; диа-
метр 0.9—1.2 мм.
Взрослая гусеница желто-серой окраски с тонким штриховым, поло-
сатым рисунком; вдоль спины располагаются тонкие чёрно-серые полоски;
на дорзальной стороне второго грудного сегмента черное трапециевид-
ное пятно, брюшная сторона несет черные неправильной формы точки;
сегменты тела покрыты круглыми небольшими бородавками; бородавки
по сторонам первого грудного сегмента значительно больше других,
несут длинные пучки серых волосков; прочие бородавки синие и крас-
ные, несут также пучки серых волосков; брюшные и грудные ноги жел-
тые; голова желто-коричневая.
Куколка темнокоричневая, сильно суженная в дистальной части;
брюшные сегменты, тергиты груди и головы покрыты длинными жел-
тыми волосками; cremaster в виде конического выступа последнего сег-
мента, покрытого короткими сильно загнутыми крючьями; сегменты
брюшка имеют точечную скульптуру.
Разнообразие окраски бабочек довольно велико; у гусениц она ме-
358
I. ORGYINAE
няется лишь в отношении интенсивности; для бабочек обычны мелани-
стические формы — совершенно черная почти без рисунка —atra Linst.,
коричнево-черная с резким черным рисунком — eremita Hb., серая форма
с резкими расплывающимися у костального и дорзального краев крыла
линиями — nigra Frey; светлые формы реже; описана светлая форма
с редуцированным рисунком — lutea Au., имеющая желтое брюшко
вместо обычного красного. Все формы встречаются в одних и тех же
географических областях и связаны с изменениями экологических усло-
вий во времени, но не с определенными территориями.
Распространение. Западная и средняя Европа (вся северная
часть Пиренейского полуострова, Германская низменность, Альпы до
северной Италии, Карпаты, Балканы до северной Греции); Британские
острова (до широты Дублин — Кэмбридж —Ньюкэстль); Ютландия; Шве-
ция (Эланд, Упланд); Норвегия (до широты Христиании); Финляндия;
северо-запад Европейской части СССР (Литва, Латвия,Эстония); далее
к востоку от линии Ленинград—Вологда, Киров, Молотов всюду
по лесной зоне (Псков, Ярославль, Москва, Калуга, Витебск, Тамбов,
Рязань, Владимир, Горький, Орел, Курск, Харьков, Киев, Полтава);
есть указания для юга (Каменец-Подольск, Херсон, Винница), где редка;
Поволжье (Казань, Ульяновск, Куйбышев, Саратов); Северный Кавказ
(Краснодар, Кубань, Ставрополь, Дзауджикау); Закавказье (Боржоми,
Бакуриани, Лагодехи); Приуралье (Челябинск, Чкалов, Свердловск,
Уфа); Крым (Алушта, Козьмодемьянский монастырь); в Сибири по лес-
ной зоне и лесостепью не часто до Байкала (Омск, Томск, Каинск, Бара-
бинск, Курган, Барнаул, Карачи, Колывань); Алтай (Чемал, Бийск,
Семипалатинск); Прибайкалье (Черемхово, Иркутск); Приамурье (Бу-
рея, Хабаровск, Николаевск, Яма-Линь); южный Сахалин (Ново-Саха-
линская); южное Приморье (Иман, Уссури, Сучан, Хуалаза, Пан-Дынза,
Колумбэ, Сица, Лаболаза, Сидеми); Япония (Хоккайдо); есть одна особь
из Гималаев — Сикким на высоте 2100 метров (рис. 173). _
Биология и экология. Зимуют яйца с вполне сформиро-
ванной и уже пигментированной, но диапаузирующей гусеницей. Выход
гусениц происходит весной после того, как термический режим (10—15°)
держится в течение нескольких суток; нижний предел развития яиц
при 5°; свежёвышедшие гусеницы первое время (от 1 до 15 суток, смотря
по погоде) сидят так называемым «зеркалом», не питаясь; продолжи-
тельность неактивного состояния молодых гусениц зависит от терми-
ческого режима весны. После периода покоя молодые гусеницы присту-
пают к питанию, всползая на кроны деревьев и расползаясь по подлеску;
в этот период они очень стойки к голоданию. Молодые гусеницы питаются
распускающимися листочками или хвоей (молодой); в сосняках они
питаются старой хвоей, так как к моменту выхода гусениц молодая хвоя
еще не успевает развиться; последний факт неблагоприятен для жизни
монашенки в сосновых насаждениях. Часто уничтожаются молодые
почки, начавшие надуваться, еще под чешуйками; наконец пищей
молодым гусеницам могут служить пыльца и ткани сережек сережкоцвет-
ных растений (ива, лещина, береза и др.).
Гусеницы многоядны, но выбор их кормовых растений обнаруживает
большую сложность, чем обычно у Orgyidae и даже близких видов Ocne-
ria Hb. Предпочитаемыми являются лиственные породы из Be ulaceae,
Fagaceae, Salicaceae и некоторые хвойные. Оптимальными кормовыми
растениями являются Fagaceae (Fagus sylvatica, Quercus robur, Q. ses-
sili flora), Betulaceae (Carpinus bet ulus, Corylus avellana), Rosaceae (Cydo-
Рлс.-ПЗ. Распространение монашенки (Ocneria -monacha L.). Черным обозначена территория, где она вредит.
360
I. ORGYINAE
nia vulgaris, Crataegus monogyna, C. oxyacantha, Pirus malus), Salicaceae
(Salix viminalis, S. alba), Ericaceae (Vaccinium uliginosum, V. myrtil-
lus), Pinaceae (Larix decidua, Picea excelsa, Pinus Silvestris, P. strobus).
Развитие возможно также при питании некоторыми другими расте-
ниями — Pirus torminalis, Rosa canina, Prunus serotina, Prunus spinosa,
P. padus, Sorbus aucuparia, Alnus incana, Betula verrucosa, Pinus cembra,
P. banksiana, Abies alba, Picea pungens, Pseudotsuga douglasi.
. Питание грушей, тополями, терном, некоторыми ивами (Salix aurita),
пушистой березой, вязами, кленами и рядом других растений, близко
родственных оптимальным, ведет к полному вымиранию особей или до
начала линяния во вторую, стадию, или вскоре после него. Монашенка,
видимо, очень чувствительна к каким-то соединениям, выключающим
эти расстения из числа кормовых; так, Salix viminalis является хорошим
кормовым растением, но гибриды этой ивы с несъедобными видами ив
уже непригодны для питания гусениц. Растения из Oleaceae, Caprifolia-
ceae, Juglandaceae, Rhamnaceae, Leguminosae избегаются. Известны
случаи поедания взрослыми гусеницами травянистых растений. Гусеницы
едят преимущественно ночью, не экономно, теряя значительные коли-
чества несведенных верхних частей листьев или хвои, т. е. начинают
грызть их с середины; днем гусеницы часто сидят, спрятавшись в щелях
коры.
Основными стациями являются спелые еловые насаждения с ровной
температурой, около 15—20°, в течение периода развития и высокой
стойкой влажностью, почему в природе этот вид размножается в хвойных
лесах реже, являясь вредным в лиственных. В южном Приморье мона-
шенка связана также с зоной хвойных лесов, заселяя елово-пихтовые
и елово-кедровые стации (Куренцов, 1939). Нижний термический порог
развития гусениц близ 7—8°, оптимум близ 22—24°. Период роста про-
должается обычно 50—70 суток. Окукление происходит в конце июня,
в июле; обычно гусеницы окукляются в щелях коры или в сухих лисТьях
и’хвое, стянутых паутиной, в рыхлом коконе. Куколка развивается в те-
чение .15—20 суток. Лёт бабочек происходит ночью, в июле—августе;
копуляция и яйцекладка происходят сразу после выхода, обычно близ
мест выплода на стволах деревьев. Яйца откладываются в щели коры и
под ее чешуи кучками по 10—50 шт., располагаясь там, где кора^еще
шероховата и образует много щелей. Общее число откладываемых самкой
яиц —100—400 шт. Эмбриональное развитие почти нацело протекает
в течение летнего, времени; пигментированная и почти готовая к выходу
гусеница диапаузирует и зимует в яйце. По окончании эмбриональцого
развития, в состоянии диапаузы, яйца холодостойки и переносят охла-
ждение до —40°, —50°. В редких случаях (при неизученных пока усло-
виях развития предыдущих фаз) диапауза яиц не наступает и выход гу-
сениц происходит в тот же год, но второе поколение этого вида никогда
не развивается.
Является одним из серьезнейших вредителей леса, наносящим ущерб
преимущественно хвойным породам, особенно ели, реже размножается
в елово-сосновых, чисто сосновых и сосново-лиственных лесах; в чисто
лиственных насаждениях размножения очень редки; в 1923—1924 гг.
отмечено размножение монашенки в Брянских лиственных лесах;
в 1927 г. Белозор наблюдал размножение этого вида в грабовых лесах
Украины (Винница). Территория вредной деятельности вида охватывает
зону хвойных лесов преимущественно средней Европы (Германия, Гол-
ландия, Силезия, Прибалтика), охватывая также и горные системы
14. OCNERIA
361
(Альпы, Карпаты), где массовые размножения наблюдаются до высоты
450—600 метров. В северной Европе размножения монашенки наблюда-
лись в Швеции, также в еловых лесах; для южной Европы указаны слу-
чаи размножения монашенки в Испании, в Пиренеях, где вред, однако,
не был велик, во Франции (Савоя), где были сильно повреждены еловые
насаждения; были отмечены размножения монашенки также в Альпах,
в Швейцарии. На востоке Европы размножения монашенки наблю-
даются реже, но указаны для СССР в Московской и Смоленской обла-
стях, на северо-западной Украине и в южной части Ивановской области,
а также в южной части Горьковской области, в Приуралье (Свердловск,
Челябинск) и в Среднем Поволжье. Размножение монашенки отмечено
также на западном Алтае. В пределах СССР вредит как еловым, так
и сосновым лесам; территории' юго-западнее линии Витебск—Орел ха-
рактерны повреждением ели, в прочих местах (включая и Алтай) вред
преимущественно сосновым лесам. В южном Приморье монашенка хотя
и многочисленна, но массовым вредителем пока не отмечена, умеренно
повреждает елово-кедровые и елово-пихтовые насаждения; вред От нее
на южном Сахалине и на Японских островах тем не менее значителен.
Известны многочисленные хищники и паразиты, уничтожающие мо-
нашенку в разных стадиях развития. Из первых яйцами питаются ли-
чинки верблюдки (Raphidia)', гусениц поедают хищные жужелицы
Calosoma и высасывают клопы Picromerus bidens L.
Особенно многочисленны паразитические перепончатокрылые и дву-
крылые, уничтожающие в разных фазах монашенку. Из перепончато-
крылых известны 1 из Ichneumonidae — Automalus alboguttatus Gr.,
Barichneumon nigritarius Gr., Co' lichneumon sugillatorius L., Eurylabus
dirus Wesm., Ichneumon insidiosus Wesm., I. melanocerus Rtzb., I. rap-
torius Gr., Protichneumon disparis Poda, Pimpla alternates Gr., P. bras-
sicariae Poda, P. capulifera Krchb., P. examinator F., P.'inquisitor Scop.,
P. instigator F., P. quadridentata Thoms., P. rufata Gmel., P. turionellaeL.,
P. arctica Zett., P. varirornis F., Ephialtes tuberculatus Forcr., Theronia
atalantae Poda, Xylonomus irrigator F., Anilastes rapax Gr., Aphanistes
ruficornis Gr., Casinaria claviventris Hl mgr., Omorgus difformis Gmel.,
Ophion luteus L., Trophocampa scutdlaris Tschk., Mesochorus silvarum
Curt., M. s'igmafor Thubg., M. vitticollis Holmgr.; из Braconidae —
Apanteles difficilis Ns., A. glomeratus L., A. inclusus Rtzb., A. melanos-
celis Rtzb., A. nigriventris Ns., A. solitarius L., Macrocentrus colldris
Spin., Meteorus bimaculatus Wesm., M. scutellator Ns. M. versicolor
Wesm.; из Chalcididae — Cratotechus larvarum L., Trichogramma evanescens
Westw., T. fasciatum Perk., T.preciosa Kolub.; из Proctotrupidae — Teleag
laeviusculus Rtzb., Теlenomus bombycis Mayr. Из двукрылых (Larvivoidea)
паразитируют следующие виды: Agria affinis Fall., A. monachae Kram.,
A. ruralis Fall., Carcelia excisa Fall., C. lucorum В. В., C. sussurans
R. D., Ceromasia florum Macq. (Rond.), C. ferruginea Mg., Compsilura
concinnata Mg., Ernestia rudis Fall., Erycia festinans Mg., I arvivora lar-
varum L., L. rustiea Film, Masicera silvatica Fall., Pales pavida Mg.,
P. pumicata Mg., Parasetigena segregate R. D. (=silvestris R. D.), Pelle-
tieria nigricornis Mg., P. prompta Mg., Phorocera assimilis Rond., Ph. caesi-
frons Macq., Phryno vetula Mg., Prosopaea fugax Rd., Redtenbacheria
insignis Egg., Sarcophaga aratrixPand., S. atropos Mg., S. falculate Rond.,
1 Я привожу список паразитов монашенки по Колубаеву (Kolubajev, 1938,
Ztschr. angew. Entom., 24 : 102—117).
362
1. ORGYTNAE
S. priyigna Rond., S. schutzei Kram., S. tuberose, Pand., S. uliginosa Kram.,
Sturmia bimaculala Htg., S. scutellala R. D., Tachina fera L., Thelaira
nigripes F., Tricholyga sorbillans Wied., Gymnochaeta viridis Fall., Micro-
lachina nympharum Rond., Thelymorpha vertiginosa Fall.
Вторичные паразиты, т. e. паразиты, живущие за счет паразитов мо-
нашенки, также известны в большом числе, но в меньшем, чем то описано
для непарного шелкопряда. Они указаны из перепончатокрылых, в ча-
стности из семейств Ichneumonidae, Braconidae, Aphidiidae, Chalcididae,
и один вид известен из мух-жужжал — Hemipenthes (Anthrax) morio L.
*2. Ocneria grisescens Stgr. (рис. 174).
Staudinger, 1887, Mem. Lep. Rom., 3 : 289; Kirby, 1892, Syn. Cat. Lep. Het.,
1 : 476; Staudinger, 1901, Cat. Lep., 1 : 118; Strand, 1910, Seitz, Grossschmett., 2: 129,
t. 22, f; Swinhoe, 1923, Ann. Mag.
Nat. Hist., 11 (9) : 425; Bryk, 1934,
Lep. Cat., 62 : 156; —albescens Stau-
K-X dinger, 1887, Mem. Lep. Rom., 3 :
Ц ' '289, t. 12, fig. 4.
"'1AГолова, грудь и брюшко
с—• I_' X?- ' желто-серые; грудь и голова
///л покрыты волосками и чешуей,
чешуйки узкие; брюшко по-
крыто только волосками; глаза
круглые, серо-зеленые; усики
Рис. 174. Ocneria 8^^ Stgr. Гениталии сильно гребенчатые, гребешки
желто-серые, членики покрыты
бело-серыми чешуйками; передние крылья желто-серые, однотон-
ные, почти без рисунка; дискальные жилки несут черные че-
шуйки и образуют ломаную черную полоску; ниже этой полоски
неправильные черные штрихи в виде отдельно разбросанных пятен; по
терминальному краю крыла остатки подкраевой линии в виде неправиль-
ных пятен; бахрома желто-коричневая с редкими темными штрихами;
задние крылья без рисунка, желто-серые с бахромой того же цвета; ноги
серо-желтоватые, передние ноги розоватые или розовые; усики самок
с короткими гребешками. Размах крыльев 40 мм самцы, 53 мм самки.
Uncus большой, конический, постепенно загнут крючковидно, ди-
стальный конец его острый; основание uncus отделено швом от девятого
тергита, который сильно склеротизован и образует два дистально направ-
ленных выроста по сторонам uncus; saccus очень небольшой; valvae
длинные, плотно соприкасающиеся основаниями; базальная половина
valva широкая, несет два широких слегка изогнутых острых выроста —
более короткий и широкий дорзальный и более длинный сильнее изо-
гнутый несколько более узкий вентральный; penis длинный, почти прямой,
с слегка отогнутым дорзально дистальным концом; соесшп длинный,
несколько тоньше дистальной части penis, по длине равный ей; penis
укреплен сильно разросшимся vallum.
Очень своеобразный вид, близкий к monacha L., но дающий ряд отли-
чий не только в рисунке и окраске, но и в структуре гениталий.
Распространение. Южное Приморье (Уссури, Владивос-
ток, Сидеми, Аскольд).
*3. Ocneria matura Moore. —Розовый непарный шелкопряд (рис. 175,
176).
Moore, 1865, Proc.. Zool. Soc. Lond. : 85; Hampson, 1892, Fauna Brit. Ind., Moths,
.1 : 464; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 479; Staudinger, 1901, Cat. Lep., 1 : 117;
14. OCNERIA
36'3
Swinhoe, 1903, Trans. Entom. Soc. Lond. : 128; Strand, 1910, Seitz, Grossschmett.,
2 : 128; t. 20, e; Swinhoe, 1923, Ann. Mag. Nat. Hist,, 11 (9) : 429; Joannis, 1929,
Ann. Soc. Entom. France, 98 : 420; Matsumura, 1931, 6000 Ins. Japan : 714; Collenette,
1932, Nov. Zool., 38 : 95; Matsumura, 1933, Ins. Matsum., 7 : 137; Bryk, 1934, Lep.
Cat., 62 : 151; — aurora. Butler, 1877, Ann. Mag. Nat. Hist., 4 (20) : 403;—fusca
Leech, 1888, Proc. Zool. Soc. Lond. : 629, t. 31, fig. 9.
Биология. Graeser, 1888, Berl. entom. Ztschr., 32:125; Nissigay, 1918,
Ins. World, 22 : 13; Clausen, 1931, Bull. U. S. Dep. Agric., 168 : 20; Филиппов, 1932,
Tp. загц. раст., 5 : 318. (
Полы сильно диморфны; усики самцов сильно гребенчатые с желто-
серыми гребешками, членики черные; голова и грудь желто-серые с се-
рыми пестринами; брюшко желтое с пучками серых волосков на тергитах;
нижняя сторона брюшка и груди желтая; передние крылья желто-
серые с серым и белым рисунком в виде многочисленных поперечных
Рис. 175. Розовый пепарник (Ocneria matura Moore). Самец.
(Рис. Ф. Гупяева).
полос; жилки светложелтые; базальная линия в виде двух черных полос,
отороченных белым; между полосами пучок желтых волосков; вторая
поперечная линия бело-желтая и серая, слабо изогнутая в дорзальной,
части крыла; круглое пятно в виде растушеванного серого пятна; почко-
видное полулунной формы, серое, сильно сближенное с круглым; средин-
ная тень в виде широкой растушеванной полоски; третья поперечная
линия двойная, очень широко расставленная, белая и серая; подкраевая
линия в виде неправильных белых зубцов на сером фоне; терминальная
линия в форме темных пятен по краю крыла; бахрома желто-серая, розо-
ватая;; задние крылья тусклые, серо-желтые, с неясным темным диско-
идальным пятном и растушеванной краевой линией, волнистой, местами
прерывистой, бахрома светложелтая; нижняя сторона желтая, слегка
розоватая, однотонная, с круглыми дискоидальными пятнами на задних
крыльях. Самки бело-розовые; задние крылья, брюшко, основание уси-
ков, губные щупальца, ноги и концы жилок па передних крыльях ярко-
розовые, прочие части тела розоватые; рисунок несколько редуцирован
и напоминает самок непарного шелкопряда; вторая и третья попереч-
ные линии редуцированы или даже вполне утрачены; последние сегменты
брюшка вытянуты в форме яйцеклада, как у монашенки. Размах крыльев
самцов 40—50 мм, самок 70—90 мм. .
364
I. ORGYINAE
Рис. 176. Розовый непарник (Ocneria matura
Moore). Гениталии: а — самца, б —'самки.
Uncus тонкий, округлый, изогнутый крючковидно, острый; базаль-
ная его половина плоская, отчленена швом от девятого сегмента; послед-
, ний образует по сторонам основания uncus острые дистально направ-
ленные выросты; saccus отсутствует; valvae короткие, на дистальном
конце несут три гладких заостренных выроста, два из которых (дорзаль-
ные) соединены основанием, причем один меньших размеров; дорзальный
край valva несет короткий конический вырост; penis длиннее valva с длин-
ным гладким соесшп, ди-
стальная часть его расши-
рена; pars inflabilis несет
слабые склеротизованные
тяжи. Bursa copulatrix меш-
ковидная, покрыта скульпту-
рой в виде многочисленных
склеротизованных складо-
чек, постепенно переходит
в ductus bursae, проксималь-
ная часть которого мембра-
нозная, складчатая; дисталь-
ная его часть расширенная,
сильно склеротизована; duc-
tus seminalis отходит от ди-
стальной части совокупи-
тельной сумки; девятый и
отчасти восьмой сегменты
вытянуты, с вентральной
стороны мембранозные, силь-
но [.складчатые; [ apophyses
anteriores и posteriores очень
длинные; мембрана при осно-
вании papillae anales.склад-
чатая, сильно растягиваю-
щаяся; papillae anales длин-
ные и плоские; последние
сегменты образуют яйцеклад.
Изменчивость не велика. Синонимические названия относятся к не-
сколько более розовым и мелким особям aurora Btl. и к более темным
fusca Leech.
Распространение. Южное Приморье (Владивосток, Аскольд,
Сучан, Тигровая, Бикин, Сица, Супутинка, Хуалаза, Лунза). Гималаи,
Кашмир (Кангра), Сикким; Индия, Бенгалия; западный Китай, Омисиен;
Японские острова (Хондо); о. Лоохао.
Биология. Зимуют яйца с почти закончившей развитие гусени-
цей; выход гусениц в мае; первые 4—5 суток- гусеницы не питаются и
не расползаются, остаются в «зеркальцах»; период роста охватывает май,
июнь и отчасти июль; гусеницы многоядны, но ограничены в природе
лиственными породами, особенно Quercus dentata в Японии и Quercus
mongolica в южном Приморье, а также древесными розоцветными, осо-
бенно яблоней; в отличие от японской формы непарного шелкопряда
(О. dispar fumida Btl.), этот вид размножается и вредит в Японии
садам; круг кормовых растений розового непарного шелкопряда вклю-
чает Salicaceae, Rosaceae, Betulaceae, Fagaceae и, вероятно, ряд других
древесных растений. С весны гусеницы питаются почками, позже листь-
14. OCNERIA
365
ями, предпочитают держаться на листьях, но не на ветвях. Окукление
в листьях в рыхлом коконе и на ветвях-; лёт бабочек в конце июля, в ав-
густе. Самки этого вида более активны, чем самки непарного шелко-
пряда, тогда как самцы менее активны. Откладка яиц на стволы деревьев
под чешуйки коры кучками; яйца обильно покрываются выделениями
(самка имеет очень большие придаточные железы); по Нишигата (1918),
самки охотно кладут яйца на кору елей. Яйца концентрируются на при-
корневой части стволов.
*4. Ocneria dispar L. —Непарник, непарный шелкопряд (рис. 177,
178, 179, 180).
Linne, 1758, Syst. Nat.,ed. 10 : 501 {ВотЪух); Aurivillius, 1891, Nordn. Fjar. : 94,
t. 26, fig. 2; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 475; Staudinger, 1901, Cat. Lep.,
1 : 117; Spuler, 1908, Schmett. Eur., 1 : 131, t. 26, fig. 5; Rebel, 1910, Berge,
Schmetterlingsb., ed. 9 : 117, t. 24, fig. 6; Strand, 1910, Seitz, Grossschmett., 2 : 127;
Oberthiir, 1916, Et. Lep. Comp. : 289; Swinhoe, 1922, Ann. Mag. Nat. Hist., 10 (9) :
426; Petersen, 1924, Lep. Fauna Estl., ed. 2 (1) : 142; Goldschmidt, 1924, Arch. Mikrosk.
Anat. Entw., 101 : 92; Osthelder, 1926, Schmett. Siid-Bayerns, 1 (2) : 199; Schaus, 1927,
Seitz, Grossschmett., 6 : 553; Hering, 1932, Tierw. Mitteleur. : 484; Gaede, 1932,
Seitz, Grossschmett., 2 Suppl. : 101; Amsel, 1933, Zoogeographica, 2 : 99; Bryk, 1934,
Lep. Cat., 62 : 145; Nordstroem und Wahlgren, 1937, Nordn. Fjar. : 67, t. 14, fig. 4; —
disparina Muller, 1802, Fauna Schles;, 3, fig. 1; — nigra Selys, 1857, Ann. Soc. Entom.
Belg., 1 : 52; — japonica Motschulsky, 1860, Et. Entom. : 31; — bordigalensis Mabille,
1876, Bull. Soc. Entom. France, 6 (5) : 9; — disparoides Mabille, 1876, Ann. Soc. Entom.
France, 6 (5) : 521; —fumida Butler, 1877, Ann. Mag. Nat. Hist., 4 (20) : 402; — hadina
Butler, 1881, Trans. Entom. Soc. Lond. : 11; — semiobscura Mieg., 1881, Naturaliste,
.8 : 237; —erebus Mieg, 1881, там же, 8 : 237; — major Fuchs, 1899, Jahrb. Nassau Ver.,
52 : 132; —variegata Lambillion, 1907, Rev. Mansuel Ent. Namur, 7 : 51; —fasciata
Lambillion, 1907, там же, 7 : 51; — brunnea Schultze, 1910 Internal. Entom.
2efts hr., Guben 4 : 26; —submarginalis Schultz, 1910, Entom. Ztschr. S u gart, 24 :
36; — insignata Schultz, 1910, там же, 24 : 35; — angulijera Schultz, 1910, там же,
.24 : 35; — unifasciata Schultz, 1910, там же, 24 : 36; — fragarius Ribbe, 1910, Iris,
;23 : 221; — wladiwostokensis Strand, 1910, Seitz, Grossschmett., 2 : 127, t. 20, e; —
.destrigata Klemensiewiez, 1911, Spraw. fiz. Kom., 46 : 6; — suffusa Schultz, 1912,
Intern, entom. Ztschr. Guben, 6 : 263, 319; — unicolor Lambillion, 1913, Cat. Lep.
Belg., Add. : 441; — alba Stauder, 1914, Iris, 28 : 16; — atra Schultze, 1917, Suppl.
Entom.,-6 : 24 — eremita Bandermann, 1924, Intern, entom. Ztschr., 18 : 30 {dispar X
japonica); — lactea Manon, 1926, Rev. Zool. Agric. Appl., 25 : 55; —dealbata Manon,
1926, там же, 25 : 55; — radiata Manon, 1926, там же, 25 55; — asiatica Wnukowsky,
1926, Русск. Энтом. обозр., 20 : 79; — obsoleta Wnukowsky, 1926, там же, 26 : 79; —
.unicolor Wnukowsky, 1926, там же, 20 : 80; ochracea Wnukowsky, 1926, там же,
20 : 80; — ochracea-obsoleta Wnukowsky, 1926, там же, 20 : 80; ochracea-unicolor
Wnukowksy. 1926, там же. 20 : 8>>; — albescens Matsumura, 1927, Journ. Coll. Agric.
.'Sapporo, 19 : 25; —.praeterea Kardakoff, 1928, Entom. Mitt., 17 : 416, t. 8, fig. 15,
16; — examinata Kardakoff, 1928, там же, 17 : 417; — fasciatella Strand, 1934, Bryk, Lop.
Cat., 62 : 148; — bremenskyi Knop, 1937, Entom. Ztschr., 51 : 262; — mediterranea Gold-
schmidt, 1940, Mat. Basis Evol. : 59; —bocharae Goldschmidt; 1940, там же : 59; —
hdkkaidensis Goldschmidt, 1940, там же : 59; — japonica Goldschmidt, 1940, там же:
.59; — obscura Goldschmidt, 1940, там же : 59; — cho'senensis Goldschmidt, 1940,
там же : 60; — nesiobia Bryk, 1942, Iris, 56 : 25.
Би ология и экология. Шевырев, 1890, Сельск. леей, хоз., 125 : 1—8;
Judeich und Nitzsche, 1895, Lehrb. Forstins., 2 : 794, t. 5, fig. 1; Standfuss, 1896,
Handb. Schmett., ed. 2 : 49, 59, 157, 168, 227, 308, 318; Lampa, 1900, Entom. Tidskr.,
21 : 39, 1.1, fig. 1—6; Кузнецов, 1904( Русск. Энтом. обозр., 4 : 203; Pictet, 1906, Bull,
:Soc. Lep. Gen., 1 : 139; Шугуров, 1907, Лесн. журн., 37 : 1205; Шрейнер, 1908, Тр.
Бюро энтом., 2 : 14; Rebel, 1913, Ztschr. wiss. Insektenbiol., 9 : 148; Escherich, 1914,
Forstins. Mitteleur., 1 : 122, 231, 240, 338, fig. 131, 234, 93, И; Кузнецов, 1916, Русск.
Энтом. обозр., 15 : 109; Ljungdahl, 1917, Entom. Tidskr., 38 : 227; Klatt, 1920, Biol.
Zentralbl., 11 : 539; Wolff und Krause, 1922, Wichtigste Forstins. : 130, fig. 157—
159; Lederer, 1923, Handb. prakt. Entom., 3 : 157; Kosminsky, 1924, Biol. Zentralbl.,
44 66; Emeljanov, 1924, там же, 44 : 106; Reichert, 1925, Ztschr. Morph. Oekol. Tiere,
3 : 477; Косминский, 1925, Русск. зоол. журн., 5 : 1; Escalera, 1925, Bol. Soc. Esp.
Hist. Nat., 25 : 337; Румянцев, 1925, Изв. Ставр.-энтом. общ., 1 : 23; Niisslin und
366
1. ORGYINAE
Rhumbler, 1927, Forstinsektenk. : 459, fig. 381; Zwolfer, 1927, Ztschr. angew. Entom.'
1.2 : 498; Kopec, 1927, Biol. Gener., 3 : 375; Косминский, 1928, Русск. зоол. жури.,
9 : 3—61; Kosminsky, 1930, Biol. Zentralbl., 50 : 351; Goldschmidt, 1933, Bibliogra-
phia Genet., 11 : 5—174; Левитт, 1935, Тр. Инет. биол. зоол. ВУАН, 4 : 135; Ali,
1933, Ztschr. angew. Entom., 20 : 354; Hoffman, 1933, там же, 20 : 70; Лебедев, 1935.
Зб1рн. працз бюл. 1 екол., 2 : 118; Пятницкий, 1935, Вопр. биоц. экол., 2 : 100;
Пархоменко, 1936, Непарный шелкопряд, 1—117; Shedl, 1936, Der Schwamms,pin-
ner. Monogr. angew, Entom., 12 : 1—242; Schwingenschuss, 1938, Entom. Rundsch. ,
55 : 161; Кожанчиков, 1940, Вести, защ. раст., 3 : 3.
Полы сильно диморфны; передние крылья у самцов коричнево-серые,
этого же цвета голова и грудь; задние крылья желтовато-коричневые
или желто-серые; брюшко этого яте цвета с черными штрихами на терги
Рис. 177. Непарник {Ocneria dispar L.). Самец.
(Рис. Ф. Гуняева).
тах; вентральная сторона груди и брюшка светлая, желто-серая; ноги
того же цвета; усики самцов сильно гребенчатые, гребешки серые, чле-
ники желто-серые; рисунок на передних крыльях обычно вполне ясный;
базальная линия в виде короткого черного штриха; базальное поле темно-
серое с черными штрихами; вторая поперечная линия простая, черная,
мелко извитая; круглое пятно в виде черной точки близ второй попереч-
ной линии; почковидное пятно в форме черного полулуния; срединная
линия широкая, растушеванная; третья поперечная линия начинается
интенсивным черным штрихом, далее она зубчатая, тонкая, простаяг
темнокоричневая; подкраевая линия буро-коричневая, слабо волнистая,
растушеванная; поле между подкраевой линией и внешним краем темное;,
бахрома шашечная — коричневая и белая; задние крылья при основании
желтоватые, покрыты длинными волосками, внешнее поле их темное,
коричневатое, в виде широкой краевой полосы; бахрома светложелтая;,
среди самцов нередки особи темнокаштанового цвета или коричневые;
рисунок остается тем же, но несколько темнее; задние крылья в этом-
случае почти однотонные лишь с несколько более темным внешним краем;
нижняя сторона однотонная, темнобурая или бледножелтая; дискоидаль-
ные пятна ясные на передних и задних парах крыльев; у светлых особей
просвечивают линии рисунка и затемненный край передних крыльев..
Самки значительно крупнее, белые или желтовато-белые, или, редко,
коричневато-белые; рисунок тот же, что у самцов, почти черный, тонкий.
14. OCNERIA
367
несколько редуцирован; основание крыльев покрыто желтоватыми волос-
ками; брюшко самки желтое, на конце несет подушку темных коричнево -
черных или коричнево-желтых волосков; усики черные, слабо гребен-
чатые. Размах крыльев самцов 40—50 мм, самок 60—90 мм; размеры
обоих полов сильно изменчивы.
Uncus длинный и тонкий, полого загнут вентрально; основание его
отделено швом от девятого сегмента, слабым, почти исчезнувшим; saccus
довольно большой, в виде узкого почти треугольного мешка; fultura
Рис. 178. Непарник (Ocneria dispar L.). Гениталии:
а — самца, б — самки.
inferior в форме продолговатой пластинки, выгнута дугообразно; valvae
большие, сильно изогнутые, базальные части их сильно сдвинуты, почти
соприкасаются; базальная половина valva широкая, образует небольшой
выступ в дистальной части на вентральном углу; дистальная половина
valva штангообразная, узкая, коническая, слабо заостренная; penis
длинный, в средней части слабо выгнутый, в дистальной части срезан
косо; coecum довольно длинный, тонкий; инкрустдций на pars inflabilis
нет. Bursa copulatrix мембранозная, в проксимальной части расширена,
в середине имеет небольшую перетяжку, дистальная ее половина ци-
линдрическая; поверхность совокупительной сумки покрыта тонкой
рубчатой скульптурой; ductus bursae короткий, широкий, мембранозный;
вагинальная область имеет вокруг широкую склеротизоваиную рамку;
ductus seminalis отходит от нижней четверти совокупительной сумки;
восьмой сегмент короткий, равномерной длины во всех частях; apo-
physes anteriores длинные и толстые, расширенные в проксимальной части;
apophyses posteriores также расширенные на концах, более короткие;
papillae anales длинные, конические, несущие в основании венчик длин-
ных щетинок.
Рис. 179. Непарник (Ocneria dispar L.). Географическое распространение. Черным отмечен ареал, где он вредит.
14. OCNERIA
369
Яйца сначала желтые, затем желто-серые, гладкие, почти круглой
формы, слегка приплюснутые в дорзовентральном направлении; диа-
метр яиц 1.0—1.2 мм.
Взрослые гусеницы желто-коричневые со сложным мелким рисунком
из тонких светло желтых и черно-коричневых, бархатистых полосок;
вдоль спины узкая белая полоса, на которой йа шестом и седьмом сегмен-
тах брюшка расположены же-
лезки коричнево-желтого света;
по сторонам этой полосы вдоль
спины по ряду больших боро-
давок — синих на грудных и
двух первых брюшных сегмен-
тах и красных на остальных
шести брюшных сегментах; они
несут пучки черных волосков:
латерально от бородавок рас-
полагаются узкие белые поло-
сы, проходящие по всем сегмен-
там тела насекомого; по бокам
большие серые бородавки, не-
Рис: 180. Непарник (Ocneria dispar L.). Хе-
тотаксия передне- и среднегруди и брюшных
сегментов I, VI, VII и VIII гусеницы
первой стадии (а) и взрослой (б).
сущие пучки длинных желтых
волосков; по сторонам первого
грудного сегмента по одной
очень крупной борода ке, несу-
щей пучок волосков, направ-
ленных косо вперед; вентраль-
ная сторона тела желто-серая;
брюшные и грудные ноги жел-
тые.
Куколка темноко ричневая;
брюшные тер гиты, тер гиты
грудных сегментов и голова
несут пучки негустых рыжева-
тых волос; брюшные сегменты
покрыты мельчайшей точечной
скульптурой и более крупными
точками; cremaster в виде не-
длинного конического выроста,
пбйрытого бороздами, несет многочисленные утолщенные на копнах
крючья.
Изменчивость цвета и отчасти рисунка (большая или меньшая
его редукция) как во взрослой, так и в личиночной стадиях велика,
о чем свидетельствуют названия, характеризующие разные формы
непарного шелкопряда. Большая часть названий относится к незна-
чительным отклонениям, но некоторые заслуживают быть отмеченны-
ми. Черные и темные северо-западные формы описаны (из Германии
и Англии) как erebus Mieg., atra Schultz, semiobscra Mieg., восточно-
азиатские темные формы непарного шелкопряда несколько крупнее —
japonica Motsch., /uznidaBtl.; самки этих форм могут достигать, по Личу
(1899), 114 мм в размахе. В средней Сибири распространена желтоватая
светлая форма — asiatica Wnuk. Известны также как формы с редуци-
рованным рисунком — insignata Schultz, obsoleta Wnuk., так и формы
с очень резким рисунком — fasciata Rol., bordigalensis Mab. Гусеницы
24 Фауна СССР, И. В. Кожанчиков.
370
I. ORGYINAE
также дают изменчивость окраски: то встречаются особи без спинных
полос и имеющие лишь пятнышки, то, напротив, особи с сильным разви-
тием полос. Структурные признаки изменчивы слабо.
Изменчивость непарного шелкопряда, ’ как показали экспери-
менты Нитэ (1906) и многочисленные мои наблюдения, помимо форм,,
описанных Гольдшмидтом (1922—1934), стоит в связи с условиями
роста и развития, в частности сильно зависит от пищи гусениц; так,
например, особи, питавшиеся Alchemilla vulgaris, имеют облик, очень
напоминающий asiatica Wnuk., такие же особи получены при питании ’
орешником; окраска сильно меняется и от термического режима. В связи
с этим попытки систематизации цветовых форм едва ли имеют науч-
ный смысл, если проводятся только формально по коллекционному
материалу, всегда при этом недостаточному. По данным Гольдшмидта,
восточно-азиатские формы в наибольшей мере обособлены и отли-
чаются некоторыми биологическими особенностями, например менее дли-
тельной диапаузой; морфологически они также наиболее отличны от про-
чих, тем не менее выделение их в самостоятельные виды (Брик, 1934).
лишено основания. К тому же степень изученности их биологических
признаков очень слаба (Гольдшмидт, 1933), а морфологически (струк-
турно) они неотличимы от европейских. Выделение их в подвиды также
затруднительно, так как в пределах одного ареала (Японские острова,
Китай, южное Приморье) обитает несколько в разной мере дифферен-
цированных биологически и слабо отличимых морфологически форм.
Таким образом, внутривидовая изменчивость непарного шелкопряда ждет
еще внимательного и всестороннего исследования.
Распространение. Европа от Пиренеев до Прибалтики
и Кавказа (Пиренеи, Альпы, вся Италия и острова Средиземного моря,
Карпаты, Балканы, Германская низменность, Прибалтика).; Британские
острова (до широты Глэсго), южная Скандинавия; южная Финляндия;
северная Африка, Марокко, Алжир (в Египте и на Канарских островах
нет); северный Иран (Астрабад); Малая Азия (Анкара); Сирия (Ливан);
Палестина (Иерусалим); в Европейской части СССР от линии Ленинград,
(в Луге более обычен) — Вологда —Молотов всюду по лесной и частично
по степной полосе; наиболее обилен в лесостепье (Москва, Ярославль, Пен-
за, Рязань, Воронеж, Полтава, Киев, Чернигов, Таганрог, Одесса, Нико-
лаев, Бендеры, Кишинев, Измаил); Крым (Бельбек, Севастополь, Керчь,
Феодосия, Судак, Алушта); Северный Кавказ (Туапсе,, Эльбрус, Дзау-
джикау, Железноводск, Куруш, Кума, Ставрополь); Дагестан (Дер-
бент); Закавказье (Гагры, Геленджик, Поти, Боржоми, Тбилиси, Лен-
корань, Казах, Лагодехи); Армения (Араке, Ордубад, Ареш); Поволжье
(Куйбышев, Саратов, Камышин, Астрахань); Приуралье (Чкалов,
Уральск, Челябинск, Уфа, Свердловск); западная Сибирь (Кустанай,
Тюмень, Шадринск); Алтай (Бухтарма, Усть-Каменогорск, Чемал,
Онгудай, Барнаул, Ульбинское, Бийск, Семипалатинск, Баян-аул,
Павлодар); Тарбагатай; Семиречье (Алма-ата, Джасилькуль, Копал);
северный Тянь-шань (Самарканд, Ташкент, Джизак, Сары-булак, Кир-
гизский хребет); Ферганская долина (Наманган), Копет-даг (Сумбар);
Саяны (Ус, Тукеек-Кем, Тунка), Минусинск; Прибайкалье (Иркутск,
Байкал); Забайкалье (Шилка, Троицкосавск, Даурский хребет); При-
амурье (Раддевка, Рейнова, Благовещенск, Хабаровск, Буреинские горы,
Николаевск); южное Приморье (Иман, Улахэ, Яковлевка, Сучан, Вино-
градовка, Ворошилов-Уссурийский, Седанка, Океанская, Владивосток,
Сица, Супутинка, Суйфун, Судзухэ); северная Монголия (Кобдо, Улас-
14. OCNhRlA
371
сутай, Танну-ола, Судзуктэ, Улан-Батор); Маньчжурия (Хинган); Япон-
ские острова (Хоккайдо, Хондо, Сикок, Киу-Шиу); Тайван; Корея;
северный Китай; южный и западный Китай.
Биология и экология.' Зимует в фазе яйца, где лежит
вполне развитая покрытая кутикулой и пигментированная гусеница;
зимовка проходит в состоянии диапаузы разной глубины у разных
географических форм — южные формы характеризуются более корот-
ким периодом диапаузы, не требующим охлаждения ниже нуля,
для северных форм охлаждение в период диапаузы обязательно для
нормального дальнейшего развития; весенний период развития гусениц,
в яйце характеризуется перевариванием желтка; выход из яиц весной
с наступлением стойкой теплой погоды, когда полуденные температуры
превышают 10—15°, а средние суточные 5—6°; нижний порог развития
яиц при 6°. Сразу после выхода из яиц гусеницы несколько суток сидят
вместе, не расползаясь, в «зеркальцах», пигментируются и затем распол-
заются. Мелкие гусеницы ползут к вершинам деревьев и в этот период,
легко подхватываются ветром (повисая на паутинах, на ветвях) и раз-
носятся; период расселения непарного шелкопряда падает именно на
это время. Сразу же после выхода из яиц гусеницы в высокой мере много-
ядны; кормовыми растениями (включая питание и взрослых гусениц)
могут являться Taxaceae (Taxus baccata), Cupressaceae (Juniperus com-
munis, J., sabina, J. virginiana, Thuja occidentalis, Cupressus sempervi-
rens, C. limetta), Pinaceae (Pinus silvestris, P. cembra, P. thunbergj,
P. densifolia, P. rigida, P. resinosa, P. banksiana, P. monticola, P. strobus,
P. maritima, Picea excelsa, P. rubens, P. canadensis, Larix europaea,
L. laricina, L. decidua, L. dahurica), Salicaceae (Salix viminalis, S. alba.
S. fragalis, S. caprea, S. discolor, S. interior, S. pentandra, Populus nigra,
P. tremula, P. alba, P. tremuloides, P. grandidentata, P. balsamifera,
P. deltoides), Myricaceae (Myrica gale, M. carolinensis), Juglandaceae
(Juglans nigra, J. cinerea, J. regia), Hamamelidaceae (Hamamelis virgi-
niana, Liquidambar stiraciflua), Betulaceae (Betula verrucosa, B. pubes-
cens, B. lenta, B. lutea, B. populifolia, B. nigra, B. patyrifolia, Alnus
incana, A. glutinosa, Carpinus betulus, C. orientalis, C. carolineana, Ostrya
virginiana, 0. carpinifolia, Corylus avellana), Fagaceae (Fagus sylvatica,
F. orientalis, F. grandifolia, Castanea vesca, C. dentata, Quercus robur,
Q. serrata, Q. cerris, Q. ilex, Q. pubescens, Q. suber, Q. coccifera, Q. glan-
dulifera, Q. prinus, Q. prinoides, Q. macrocarpa, Q. palustris, Q. stellata,
Q. rubrum, Q. umbricaria, Q. bicolor, Q. alba, Q. velutina), Anacardiaceae
(Bhus capalluna, Bh. glabra, Rh. hirta, Phellodendron amurense), Lau-
raceae (Liriodendron tupulifera), Platanaceae (Platanus occidentalis),
Aceraceae (Acer platanoides, A. tataricum, A. pseudoplatanus, A. negundo,
A. platanus, A. saccharum, A. rubrum, A. saccharinum, A. spicarum, A. pseu-
dosylvanicum, Aesculus hippocastanum), Могасеае (Morus alba, M. nigra,
M. rubra), Ulmaceae (U. campestris, U. effusa, U. pedunculata, U. subrosa,
U. americana, U. fulva, Celtis occidentalis, C. glabrata), Berberidaceae
(Berberis vulgaris), Tiliaceae (Tilia americana, T. cordata), Rosaceae (Pru-
nus domestica, P. occidentalis, P. armeniaca, P. spinosa, P. avium, P. lau-
rocerasus, P. padus, Amygdalis persica, Padus virginiana, P. nana, Crataegus
pyrocantha, C. oxyacantha, Sorbus aucuparia, Pirus malus, P. communis,
P. americana, Rosa canina, R. centifolia, R. virginiana, R. rubiginosa,
Cornus sp., C. stranifera, Amelanchier canadense, Spiraea tomentosa, S. sa-
licifolia, Geum urbanum, Alchemilla vulgaris, Fragaria vesca), Ericaceae
(Erica meladenca, Calluna vulgaris, Vaccinium angustifolium, V. vacillanss
24*
373
I. ORGYINAE
V. cotymbascqm, Oxycoccus macrocarpus, Azalia), Oleaceae (Olea europaea,
Fraxinus excelsior, F. nigra, F. quadrangula, F. pennsylvanica, F. ameri-
cana, Ligustrum vulgare, Syringa vulgaris), Saxifragaceae (Ribes uva-crispi,
R. alpinum, R. rubrum, R. vulgare,. R. nigrum) и Polygonaceae (Rheum
rhaponticum, Rumex confertus, Polygonum sp.). Излюбленными из этих
растений, на которых рост с первой стадии протекает быстро и не со-
провождается вымиранием, являются: Rhutaceae, Hamamelidaceae, Sali-
caceae, Fagaceae, Rosaceae, Tiliaceae; виды Coniferae, Lauraceae, Ole-
aceae поедаются неохотно. Травянистые растения (исключая розоцветные)
также поедаются неохотно, но при массовых размножениях отме-
чалось питание Pisum sativum, Lupinus albus, Onobrychis sativa из
Leguminosae; Brassica oleracea из Cruciferae; Gonium, аси'аиз Umbel-
liferae; Cucumis sativus из Cucurbitaceae; Taraxacum officihale, Lactuca
sativa из Compositae; Secale cereale из Gramineae, а также Vitis vinifera
из Ampelidaceae и Viburnum, Sambucusиз Caprifoliaceae. По указаниям
Маршаля (1912), некоторые растения — лютиковые, из злаков Actaea
spicata, Impatiens, Barbarea из крестоцветных, а из сложноцветных —
полыни не поедаются гусеницами непарного шелкопряда даже при
голодании.
Гусеницы в природе растут в течение 50—80 суток, термофильны, '
имёют нижний термический порог при 10°, а оптимум примерно при
22—25°; самцы линяют четыре раза, самки пять раз; окукление в июне—
июле в щелях коры, в развилинах ветвей или в листьях; на юге окукле-
ние может наблюдаться в расщелинах скал, в лесной подстилке у осно-
вания стволов и в корнях (Крым, Алтай); развитие куколок 15—25 су-
ток. Выходящие бабочки активны днем, причем копулировать начинают
вскоре же по выходе (через несколько часов), нередко самки копулируют
с еще не вполне расправившимися крыльями; самки мало активны, часто
не летают совсем или летают немного в тихие вечера. Яйцекладка следует
почти сразу за окончанием спаривания, обычно близ места выплода, наи-
более часто на кору деревьев на высоте нескольких метров от основания;
на юге отмечена яйцекладка на ветви деревьев в кронах и в расщелины
скал, на камни, на корнях деревьев под подстилкой и т. д.; на Дальнем
Востоке отмечена кладка на.ульи, на стены домов, на заборы. Развитие
яиц происходит сразу после откладки и заканчивается на 20—25-е сутки,
когда наступает диапауза. Иногда (а в Алжире даже часто) наблюдается
выход гусениц в тот же год, но второго поколения нигде в пределах ареала
вида не бывает. Откладка яиц производится большими кучками; самка,
если ее не тревожить, откладывает весь запас яиц в одно место, покры-
вая их густым слоем волосков охристо-желтого цвета с конца брюшка;
число яиц в кучке колеблется от 100 до 650; плодовитость сильно меняется
в зависимости от питания в личиночной фазе. Диапаузирующие яйца
очень холодостойки и способны переносить температуры до —50°;
холодостойкость яиц разных географических форм различна.
Непарный шелкопряд заселяет смешанные и лиственные леса и сады,
являясь серьезнейшим вредителем сельского и лесного хозяйства. Объе-
дение листвы ведет , к потере годичного прироста или к усыханию де-
ревьев. Область вредной деятельности обширна и охватывает всю сред-
нюю Европу, Кавказ, Крым, восточную Европу, западную Сибирь,
Алтай, южное Приморье, Японию и Китай. В садах вредит яблоне,
груше, айве, косточковым (вишня, черешня, абрикос, персик), в лесу
лиственным породам, особенно вязу, дубу, липе, грабу, буку и ряду дру-
гих деревьев; при сильном размножении могут повреждаться хвойные
14. OCNERIA
373
(соСна, кедр, лиственница) и редко, при полном объедании раститель-
ности — кустарниковой и древесной, уничтожаются травянистые расте-
ния, например культурные злаки (Брянцев, 1928). Массовые размножения
наблюдаются на обширной территории — в северной Африке в Алжи-
ре и в ряде мест Испании, где вредит вечнозеленым и пробковым дубам,
во Франции, в Швейцарии, Италии, особенно в северной и средней,
в Сицилии, в ряде мест Югославии, в Болгарии, всюду в Германии.,
в Голландии. В СССР массовые размножения наблюдались всюду в лесо-
степье, в широколиственных лесах Европейской части и в садовых райо-
нах степной полосы, а также в лесных полезащитных посадках; они
отмечены в Приуралье на Кавказе, в Закавказье, в Крыму, в западном
Казахстане, на Алтае. Массовые размножения отмечены также в Японии.
После заноса в 70-х годах прошлого столетия в Америку достиг там к
началу настоящего грандиозной численности и сделался первостепенным
вредителем лиственных лесов и садов; вредная деятельность и размноже-
ние вида приостановлены ввозом паразитов с территории природного
ареала (из Евразии):
Известны многочисленные хищники и паразиты, имеющие выдающе-
еся значение в регуляции численности непарного шелкопряда. Из жестко-
крылых важнейшими видами, хищничающими за счет разных фаз разви-
тия и преимущественно за счет гусениц, являются Calosoma sycophanta
L., С. inquisitor L., Habrocarabus latus Dej., Xylodrepa quadripunctata
Schreib., Dermestes erichsoni Gyll., D. lardarius F., Attagenus sp., Tene-
broides maroccanus Reitt., Steropus globosus F.; яйца уничтожаются
также клещиками Trombidium, высасывающими их.
Специальными многолетними исследованиями очень многих авторов
установлены следующие виды паразитических насекомых, связанных
с непарным шелкопрядом.1 Из перепончатокрылых Ichneumonidae: Тго-
gus lutorius F., Ichneumon disparis Poda, 1. pictus Gmel., I. leucocherrus
Wesm., I. sarcitorius L., I. rubens'Wesm., Cryptus cyanator Gr., C. liparidis
Rnd., C. amoenus Gr., Pimpla inquisitor F., P. porthetriae S. E., P. insti-
gator F., P. disparis S. E., P. examinator F., P. flavicans F., P. .brassicariae
Poda, P. holmgreni Schmidekn., P. pluto Ashm., P. pedalis Cress., P. coh-
quisitor Say, P. tenuicornis Cress., Ophion merdarius Gr., Campoplex
conicus Rtzb., C. rapax Gr., Omorgus difformis Gmel., Theronia atalantae
Poda, Th. flavescens Cress., Casinaria tenuiventris Gr., Limneria disparis
Vier., L. difformis Gr., L. validum S. E., Opheltes glaucopterus L., Pri-
stomerus vulnerator Pz., Pezomachus agilis S. E., P. hortensis Br., Paniscus
testaceus Gr., P. cephalotus Holmgr., Hyposter disparis Vier., Barylipa
perspicillator Gr., Ichnus assertorius Gr., I. flavus Rd., Eurylabus dirus
Wesm., Hemiteles nanus Gr., H. schaffneri Schmiedkn., H. bicolorinus
Gr., H. areator. Parez., H. bipunctator Thunb., H. pictipes Gr., H. aestivalis
Gr., H. an'hracinusForst., H. hortensis Gr., Trichomma enecator Rossi., Enic-
ospilus merdarius Gr., Barylypa delica^or Thunbg., Mesochorus semirufus
Holmgr., M. gracilis Br., M. viticollis Holmgr., M. pecloralis Rtzb.;
Braconidae: Meteorus japonicus Ashm., Apanteles fulvipes Hal., A. lipara-
dis Bche., A. porthetriae Muesb., A. melanoscelus Rtzb., A. tenebrosus
Wesm., A. ocneriae Stanov, A. glomeratus L., A. vitripennis Hal., A. lac-
teicolor L., Microgaster tibialis Brischke, M. punctiger Rd., M. pubescens
1 Список паразитических насекомых дан ио Шедлю (1936) для Евразии, т. е.
естественного ареала непарного шелкопряда.
374
1. orgyinae
Rtzb., M. calcaetus Hal., M. nigerrimus Rd., M. brevicornis S. E., M.
fulvipes Brischke, Meteorus scutellator Wesm., M. versicolor Wesm., M.
pulchricornis Wesm., Habrobracon brevicornis Wesm.; Chalcididae: Chaicis
flavipes Rz., Ch. callipus Kby., C. obtasatas Wlk., M onodontomerus aereus
Wlk., Anastatus disparis Ruschka, Schedins (Oencyrtus) kuwanae How.,
Schedius masii Mercet, Oophthora semblidis Aur.., Pteromalus pini Hart.,
P. halidayanus Rtzb., Atoposomoidea ogimae How., JBrachymeria intermedia
Ns, Encyrtus tardus Rtzb., Melittobia acasta Wlk., Perisopterus javensis
How.; Proctotrupidae: Hadronotus howardi Mokrz., Telenomus phalaenarum
Ns. Паразитические двукрылые также многочисленны: Agria affinis
Fall., A. monachae Kram., Carcelia affinis Fall., C. cheloniae Rnd.,
C. excisa Fall., C. lucorum Mg., C. sussurans Rd., C. recussata Pand.,
C. separata Rd., Chaetomyia crassiseta Rnd., Compsilura concinnata Mg.,
Eudoromyia magnicornis Zett., Ernestia consobrina Mg., Exorista blanca S.
E., E. westermanni Zett., E. erythrostoma Hrt., Gaurax anchora S. E.,
Histochaeta marmorata F., Lydella nigripes Fall., L. crassiseta Taschenb.,
Macheiria serriventris Rd., Masicera fatua S. E., Meigenia bisignata Mg.,
Pales pavida Mg., Phorocera agilis R. D., Rhacodineura antiqua
Mg., Sarcophaga affinis Fall., S. albiceps Mg., S. uliginosa Kr., Sturmia
atropivora R. D., A. bimaculata Rtt., S. inconspicua Mg., S. scutellata R.
D., Larvivora conjugata Rd., L. festiva S. E., L. larvarum L., L. larvicola
Rtzb., L. monachae Rtzb., L. noctuarum Rd., L. rusticaMg., L. bella Mg., L.
pinivora Rtzb., L. quinquevtata Rtzb., L. moretii R. D., Tachina fera L.,
T. japonica Tows., Telymorpha vertiginosa Fall., Tricholyga grandis Zett.,
T. segregata Rd., T. sorbillans Wied., Winthemia speciosa Egg., Zenillia
libatrix Panz., Crossocosmia sericariae Coronalia.
В Северной Америке непарный шелкопряд в связи с колонизацией
на новой территории приобрел ряд новых паразитов, перешедших из мест-
ной фауны, например, виды Achaetoneura (Diptera)-H др. Комплекс пара-
зитов, связанных с непарным шелкопрядом, обнаруживает очень боль-
шую сложность отношений. Одни из них являются монофагами и связаны
только с непарным шелкопрядом, другие многоядны в пределах самых
различных семейств чешуекрылых. Наконец некоторые многоядные виды
обнаруживают способность паразитировать как на непарном шелко-
пряде, так и на его паразитах, например Schedius kuwanae How. Но
наряду с этим обнаружен и ряд так называемых сверхпаразитов (вто-
ричных паразитов), связанных исключительно с паразитами непарного
шелкопряда, но не с самим непарным шелкопрядом. Таких видов, живу-
щих на паразитах непарного шелкопряда, насчитывается большое число,
как из перепончатокрылых (Ichneumonidae, Chalcididae), так (в меньшем
числе) и из жуков (Hydnocera).
5. Ocneria rubea F. (рис. 181, 182).
Fabricius, 1878, Mani. Ins., 2 : 117 (Bombyx'y, Staudinger, 1901, Cat. Lep., 1 : 118;
Spuler, 1908, Schmett. Eur., 1 : 132, t. 26, fig. 18, 14; Strand, 1910, Seitz, Grossschmett.,
2 : 132, t. 21, c; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 219, t. 27, fig. 8; Ober-
thiir, 1916, Et. Lep. Comp., 12 : 274; Rothschild, 1917, Nov. Zool., 24 : 358; Oberthiir,
1922, Et. Lep. Comp., 19 (2) : 204; Oberthiir, 1923, там же, 20 : 76; Aue, 1926, Entom.
Ztschr. Frankf., 80 : 25; Lederer, 1923, Handb. prakt. Entom., 3 : 16; Gaede, 1932,
Seitz, Grossschmett., 2 Suppl. : 102; Hering, 1932, Tierw. Mitteleur. : 484; Bsyk,
1934, Lep. Cat., 62 : 178; — rubrior Fuchs, 1900, Jahrb. Nassau Ver., 53 : 45; — brun-
nea Rocci, 1923, Mem. Soc. Entom. Ital., 2:9; — hillmani Dannehl, 1925, Entom.
Ztschr., 39 : 108; — obscura Babacek, 1927, Ztschr. Oesterr. ent. Ver., 12 : 65.
Голова,'грудь, брюшко и крылья коричнево-серые с сильным розовым
оттенком, особенно на задних крыльях; тело покрыто длинными шелко-
14. OCNERIA
375
Рис. 181. Ocneria rubea F. Гениталии:
а — самца, б — самки.
вистыми волосками; рисунок редуцирован; базальная линия в виде
слабого штриха при основании крыла; вторая поперечная линия широ-
кая, растушеванная (слагается из крупных черных чешуй), сильно и
грубо извитая посредине и по внешнему краю, сильно выгнутая кнаружи;
третья поперечная линия сильно сдвинута к терминальному краю, сла-
гается из узких черных штрихов, идет почти параллельно внешнему
краю, незубчатая; внешнее поле крыла очень узкое; терминальная линия
на передних крыльях отсут-
ствует ; почковидное пятно
внутри белое, маленькое,
тесно приближено к дискаль-
ной жилке, овальной фор-
мы, оттенено коричнево-розо-
вым; костальный край перед-
них крыльев затемнен; зад-
ние крылья , с очень ясной
параллельной наружному
краю темной полосой, слабо
извитой; терминальная ли-
ния в виде очень слабо
заметной полоски; бахрома
светлее крыла; нижняя сто-
рона без рисунка; внешний
край крыльев охристо-розо-
вый; усики самцов сильно
гребенчатые, членики их
покрыты светлыми, почти
белыми чешуйками; усики
самок с короткими гребеш-
ками; рисунок на крыльях
самок сильно редуцирован,
крылья несколько уже, мень-
ше и более прозрачны, чему самцов; чешуйчатый покров беднее. Раз-
мах крыльев самцов 30—32 мм, самок 28—32 мм.
Uncus плоский, несколько выемчатый по сторонам с боков, на дисталь-
ном конце заострен, слабо загнут, плотно срастается с девятым сегмен-
том, шва нет; tegumen короткий, saccus большой, широкий; fultura in-
ferior большая, желобчатая, слабо вырезана с дорзальной стороны, почти
сердцевидной формы; valva в базальной половине широкая, в дисталь-
ной узкая, прямая, штангообразная; на дистальном ее конце небольшой
лубец, загнутый внутрь; базальная (широкая) половина несет два вы-
роста — дорзальный, сидящий при основании тонкого конца valva,
и вентральный, сидящий на расширенной части, несколько сдвинутый
к основанию valva; penis тонкий, конический, сужающийся к дисталь-
ному концу, более сильно склеротизован по вентральной стороне, coecum
примерно равен одной трети .его длины. Bursa copulatrix маленькая,
яйцевидной формы, мембранозная, без скульптуры; ductus seminalis
отходит от ее левой стороны со средней части; ductus bursae короткий,
широкий, цилиндрический, сильно склеротизован; восьмой сегмент
узкий и длинный; apophyses anteriores тонкие и очень длинные, более
чем в три раза превышают длину ductus bursae; apophyses posteriores
тонкие, но несколько короче первых; papillae anales небольшие, мембра-
нозные.
376
I. ORGYINAE
Гусеница рыже- или желто-серая; спинная сторона ее светлая с се-
рыми латеральными полосками, прерванными между сегментами; на
брюшных сегментах при начале каждого из них по паре латерально рас-
положенных черных точек; передне- и среднегрудь серые; того же цвета
и два последних брюшных сегмента; бока и нижняя сторона тела светло-
желтые; голова желтая с черными глазками и темно коричневыми рото-
выми частями; грудные и брюшные ноги желто-серые; тело покрыто
волосками, сидящими на плоских желтых бородавках.
Распространен и ё.
Южная Европа — Пиренеи (юж-
ная Андалузия), Альпы (Ти-
роль, Тербан), Апеннины (се-
верная Италия); северная Аф-
рика— Марокко, Б. Атлас.
Биология. Зимует гу-
сеница; кормовые растения
Fagaceae (Quercus) мелкие дуб-
ки; яйцекладка на листья и
ветки дуба.
15. Род OCNEROPSIS KOZHAN-
TSIHKOV, gen. nov.
Тип рода О. relictus, sp.»
nov.
Хоботок рудиментирован вполне. Голова маленькая с большими
голыми глазами. Щупальца короткие, торчат вперед, второй членик их
длинный, последний очень короткий. Усики менее половины длины перед-
него крыла, двусторонне гребенчатые. Передние голени длиннее первого
членика лапки с хорошо развитым большим epiphysis. Толени средней
пары ног с одной парой пшор, голени задних — с двумя парами, сильно
сближенными. Тело покрыто тонкими волосками и узкими чешуйками.
Радиальная ячейка на передних крыльях узкая и короткая, образована
коротким анастомозом J?3+i и пятого радиального стволов. Третий и чет-
вертый радиальные стволы на длинном стебельке и ветвятся лишь
в области apex крыла. Второй медиальный ствол расположен по середине
между первым и третьим. Первый медиальный ствол отходит непосред-
ственно от радиального ствола. Субкостальный и радиальный стволы
на задних крыльях свободные. Uncus толстый и расширенный на дисталь-
ном конце с бороздой на дорзальной стороне. Subunci длинные, искрив-
ленные дорзально. Saccus не развит. Valvae в основании лопастевидно
расширены, в средней части со склеротизованной складкой. Дистальный
конец valva несет лопасть, покрытую копоткими волосками.
Личиночные признаки описываемого рода не известны. Признаки жил-
кования сближают новый род Ocneropsis с Ocneria Hb., тогда как струк-
туры гениталий очень своеобразны и не обнаруживают сходства ни
с одним из известных родо^ сем. Orgyidae. Габитуально Ocneropsis скорее
напоминает из нашей фауны виды Parocneria Dyar типа terebynthi Frr.,
но не представителей рода Ocneria Hb.
Ряд признаков рода Ocneropsis, gen. nov., можно оценивать как при-
митивные, особенно положение второго медиального ствола на передних
крыльях (между Му и М2 по середине расстояния). Такое положение этой
жилки не типично для видов Orgyidae, распространенных в умеренных
15. OCNEROPSIS, 16. PAROCNERIA
377
широтах, НО’ не редко у более примитивных представителей семейства,
распространенных в тропиках. Вероятно, описываемый ниже О. relictus
представляет остаточный вид фауны палеогеновых или даже мезозойских
лесов Средней Азии.
* 1. Ocneropsis relictus Kozhantshikov, sp. nov. (рис. 183).
Габитуально несколько напоминает виды Parocneria Dyar типа tere-
bynthi Frr., но обнаруживает ряд отличий в форме крыльев и в рисунке
на них.
Тело и крылья покрыты черными, серыми и белыми
ками. На груди, особенно на tegulae, а также
на передних крыльях в области срединного поля
немногочисленные неправильно разбросанные жел-
тые чешуйки, не образующие рисунка. Рисунок на
передних крыльях развит слабо. Первая попереч-
ная линия едва намечена на костальном крае у
самого основания крыла. Вторая поперечная линия
начинается лишь немного отступя от основания
крыла, растушеванная, черная, идет косо кнаружи,
но несколько выше анальной жилки резко повора-
чивает почти прямо к дорзальному краю крыла.
Срединная линия в виде сильно растушеванной ши-
рокой, черной полосы, образующей в средней части
неясное полулунное пятно. Она слабо выгнута
под костальным краем кнаружи, а у дорзального
края, напротив, сдвину-та базально, причем здесь
сливается со второй поперечной линией. Третья
поперечная линия начинается на костальном крае
почти в дистальной трети крыла, черная, оторочена
снаружи белым, идет сначала слабо заметная,
изгибаясь, кнаружи, а от М3 изогнута базально.
В срединном поле в области дискальных жилок,
между R и Си, темный полулунный штрих, остаток
почковидного пятна, с ним почти сливается третья
поперечная линия. Подкраевая линия начинается
волосками и чешуй-
Рис. 183. Ocneropsis
relictus Kozh. Гени-
талии самца.
двумя черными расположенными косо базально штрихами в обла-
сти apex, затем идет тонкая, отороченная белым, слабо изви-
тая. Терминальная линия слабо заметная, тонкая, серая. Зад-
ние крылья тусклого серого цвета с более темными жилками. Нижняя
сторона крыльев одноцветная, серая, с неясной просвечивающей попе-
речной полосой. Грудь более темного цвета, чем брюшко, последнее одно-
цветное, серое. Усики желтовато-серые, недлинные, лишь немного более
одной трети длины переднего крыла. Размах крыльев 29 мм.
Очень характерный вид, известный пока по одному облетанному
самцу, взятому В. Гуссаковским в ущелье Кондара, 110Q м, в долине
р. Варзоб (Гиссарский хребет), 5 июля 1946 г.
Распространение. Средняя Азия — Гиссарский хребет
(ущелье Кондара).
16. Род PAROCNERIA DVAR
Dyar, 1897, Can. Entom., 29 : 13 (typus detrita Esp.); Staudinger, 1901, Cat. Lep.,
1 : 118 (Ocneria part.); Spuler, 1908, Schmett. Eur., 1 : 131 (Ocneria part.); Rebel, 1910,
378
1. ORGYINAE
Рис. 184. Ocnerorgyia amanda Stgr. Хетотаксия
передне- и среднегруди и брюшных сегментов
I, VI, VII и VIII взрослой гусеницы.
(Схематизовано по Buxton, 1924).
Berge, Schmetterlingsb., ed. 9 : 118 (Ocneria part.); Strand, 1910, Seitz, Grossschmett.,
2 ; 131 (Ocneria part.); Bryk, 1934, Lep. Cat., 62 : 176.
Тип рода P. detrita F.
Хоботок редуцирован вполне; глаза голые, часто большие, круглые;
усики самцов сильно гребенчатые; на конце гребешков по одной длинной
щетинке; губные щупальца длинные, несколько свисающие вниз, послед-
ний членик их длинный, кони-
ческий, по длине около одной
трети второго; голени передних
ног длиннее первого членика
лапки; коготки всех ног прос-
тые; голени задних ног с одной
(второй) парой шпор; тело по-
крыто только длинными воло-
сками; радиальная ячея на
передних крыльях не развита;
третий и четвертый радиальные
стволы на длинном стебельке;
первый медиальный ствол отхо-
дит с помощью дискальной
жилки от общего радиального
ствола; базальная ячея на зад-
них крыльях вытянутая, почти
ланцетовидной формы, в ди-
стальной части замкнутая не-
полным анастомозом радиальной и субкостальной жилок; второй и третий
медиальные стволы и первый кубитальный на задних крыльях сильно
. сближены, но разделены дискальными жилками.
Uncus плоский, иногда на дис-
тальном конце расщепленный
вдоль, плотно спаян с девятым
сегментом, без шва; tegumen без
subunci; valvae с длинным ди-
стальным выростом. Bursa сори- -
latrix очень маленькая, мембра-
нозная. Полы диморфны: но в
меньшей мере, чем у Ocneria Hb.,
самки несколько крупнее и значи-
тельно бледнее окрашены.1
Личиночные признаки видов
этого рода следующие. Гусеницы
покрыты короткими волосками.
Бородавка х переднегруди боль-
шая, несет пучок волосков, тор-
чащих вперед (рис. 185); а и т;
на переднегруди в виде само-
стоятельных (лишь отчасти) слитых бородавок; а и средне- и заднегруди,
1 Ocnerorgyia amanda Stgr. (Iris, 6 : 242, 1891) не был у меня в руках, почему
я не могу вынести окончательное решение относительно синонимики этого рода
с Parocneria Dyar (1897) и восстановить приоритет названий. По очень хорошему
описанию Бэкстона (Buxton, 1921, Bull. Entom. Res., 11 : 181—186) гусеницы
О. amanda Stgr. можно видеть почти полное совпадение того, что мне известно для
Parocneria detrita Esp. (рис. 184). Я почти не сомневаюсь, что Р. amanda Stgr. при-
надлежит к группе видов, объединяемых сейчас мною в род Parocneria Dyar.
Рис. 185. Parocneria detrita Esp. Хето-
таксия передне- и среднегруди и брюш-
ных сегментов I, VI, VII и VIII взро-
слой гусеницы.
120
Рис. 186. Распространение видов рода Parocneria Dyar. Черным обозначена территория обилия видов этого рода.
380 I. ORGYINAE
а также всех брюшных сегментов слиты и образуют большие бородавки;
бородавки р брюшных сегментов сдвинуты вентрально и слиты или сбли-
жены (на седьмом и восьмом сегментах) на брюшных сегментах с боро-
давкой х. Бородавка ц на шестом брюшном сегменте большая, узкая
и длинная. Непарные выворачивающиеся железки есть лишь на шестом
и седьмом брюшных сегментах.
Целостный олиготипический род, близкий к Ocneria Ochs.; заселяет
засушливые степи, полупустыни и пустыни Средиземноморья. Главная
масса видов концентрируется в Средиземноморье на его засушливых
территориях — в северной Африке (Алжир, Марокко, северная Сахара,
Ливийская пустыня) —Р. mus Obth., uniformis Roth., flavipalpata Stgr.,
в северной Африке и на Пиренейском полуострове (Андалузия) — Р. atlan-
tica Rbr. и по северному побережью Средиземного моря (Р. detrita Esp.),
Наибольшее число видов концентрируется на территории Сирии, Пале-
стины, Ирака, Малой Азии и Закавказья (Р. samarita Stgr., nora Stgr.,
signatoria Chr., flavipalpata Stgr., raddei Chr., terebynthina Stgr., komarovi
Chr., lapidicola H.S., abbleta Stgr.); один вид известен с Сардинии (Р. кгие-
geri Trti.), и два вида заселяют южную Европу (Р. detrita Esp. и Р. tere-
bynthi Frr.), причем последний вид распространен также пятнами в юж-
ной Сибири (на Алтае), в Китае и южном Приморье. Два вида известны
из Средней Азии — Р. amabilis Chr. и Р. signatoria Chr. (рис. 186).
Биологически виды Parocneria Dyar почти не изучены; у известных
биологически видов зимует гусеница (в средних стадиях); у некоторых
видов возможно ожидать зимовку в фазе яйца (например у Р. grisei-
pennis, sp. п.). Заселяя сухие территории, ча'сто пустыни, виды Parocneria
Dyar все же связаны с группами древесных растений — видами Fagaceae
и Anacardiaceae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Передние крылья окрашены одинаково с задними, черно-серые,
однотонные, почти совершенно без рисунка (есть слабый остаток
третьей поперечной линии); чешуйчатый покров на крыльях негу-
стой; valva имеет два выроста на вентральном крае й лишена их на
дорзальном крае...............................* 1. Р. detrita Esp.
2 (1). Передние и задние крылья разной окраски; если окраска их
одинакова, то они белые и имеют рисунок или светлосерые и очень
широкие, но не черные или черно-серые.
3 (4). Тело стройное; крылья большие, тупые и широкие, серой окраски;
почковидное пятно почти в форме круга; valva с одним длинным
дорзальным выростом . . ................... .* 6. Р. signatoria Chr.
4 (3). Тело более или менее толстое; крылья неширокие с острым или
заостренным apex; valva с двумя или более выростами.
5 (10). Задние крылья белые или серые; брюшко того же цвета, но не
желтое или розовое; передние крылья светлосерые или белые; мел-
кие виды — 20—35 мм.
6 (7).’Чисто белой окраски с очень слабым рисунком (несколько штри-
хов) на передних крыльях или без него . . . .* 9. Р. raddei Chr.
7 (6). Передние крылья желто-серые или светлосерые с хорошо разви-
тым рисунком.
8 (9). Задние крылья серые, передние бело-серые; по вентральному
краю vaRa два зубца, кроме длинного дистального; более круп-
ный — 23—36 мм................................* 7.Р. terebynthi Frr.
16. PAROCNERIA
381
9 (8). Задние крылья белые, передние желто-серые; по вентральному
краю valva нет зубцов, но в средней части ее с внутренней стороны
есть один короткий, кроме длинного дистального; мельче —• 23—
25 мм....................................... * 8. Р. terebynthina Stgr.
10 (5). Задние крылья желтые, охристые, желтоватые, серо-желтоватые
или розоватые; брюшко серое, желтоватое или розоватое; передние
крылья черно-серые; крупные виды — 35—60 мм, если мельче,
то задние крылья яркожелтые.
11 (12). Брюшко, голова и грудь с значительной примесью розового;
задние крылья бело-серые с розовым внутренним краем; рисунок
на передних крыльях несколько стушеван . .* 2. Р. lapilicola Н. S.
12 (И). Розовый цвет на теле и крыльях не развит; рисунок на передних
крыльях заметен хорошо, если стушеван, то задние крылья желтые.
13 (16). Задние крылья желтые (нередко яркие); uncus целостный, острый.
14 (15). Более крупный — 35—60 мм; передние крылья несколько одно-
тонные; вентральный (лежащий в плоскости valva) отросток направ-
лен дорзально, слабо конический....................* 3. Р. komarovi Chr.
45 (14). Мельче — 35—45 мм; передние крылья с резким рисунком;
вентральный отросток valva направлен дистально, толстый ....
..................................................* 4. Р. amabilis Chr.
16 (13). Задние крылья светлосерые со слабым желтоватым оттенком
или без него; uncus на дистальном конце расщеплен.....................
......... ...........................* э. Р. griseipennis, sp. nov.
* 1. Parocneria detrita Esp. (рис. 185, 187).
Esper, 1785, Schmett. Eur., 3 : 229; t. 44, fig. 6; Kirby, 1892, Syn. Cat. Lep. Het.,
1 : 466; Staudinger, 1901, Cat. Lep., 1 : 118; Spuler, 1908, Schmett. Eur., 1 : 132, t. 26,
fig. 2; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 118; Strand, 1910, Seitz, Gross-
schmett., 2 : 131, t. 21, e; Eckstein, 1915, Schrift Deutsch Lehrervers., 32 : 58; Swin-
hoe, 1923, Ann. Mag. Nat. Hist., И (9) : 435; Lederer, 1923, Handb. prakt. Entom.,
3 : 166; Bryk, 1934, Lep. Cat., 62 : 176; Лебедев, 1935, 36ipn. праць екол., 2 : 32; —
orientalis Daniel, 1939, Entom. Rundsch., 56 : 360.
Голова, грудь, брюшко и крылья. черно-серые с примесью коричне-
вых тонов; ноги того же цвета; нижняя сторона тела более серая; окраска
крыльев более светлая, чем тела; рисунок слабо развит; слабо заметны
остатки второй поперечной линии и хорошо видна растушеванная мелко
зубчатая по жилкам третья поперечная линия; костальная половина
передних крыльев темнее прочих их частей; бахрома передних и задних
крыльев черно-серая, более темная при основании; терминальная линия
более заметна на задних крыльях; нижняя сторона крыльев однотонная,
тусклая, темносерая, более темная по костальному краю; усики самцов
с длинными гребешками (длина средних гребешков равна одной пятой
или одной шестой длины всего уса); членики усика покрыты светлыми,
почти белыми чешуйками; усики самок с короткими гребешками, глаза
черно-серые; ноги покрыты длинными шелковистыми волосками; брюшко
самок сильно увеличено по сравнению с брюшком самцов. Размах крыльев
27—32 мм, у обоих полов без особых различий.
Uncus короткий, слабо изогнутый, заостренный, сидит на широком
основании, плотно слитом с девятым сегментом, без шва; saccus большой,
очень широкий, заостренный; fultura inferior в виде прямоугольника,
заостренная и рассеченная в дорзальной части; valvae широкие и корот-
кие, с завернутым внутрь острым внешним краем, несущим три выроста —
два крайних коротких (дистальный округлый) и средний длинный; дор-
382
I. ORGYINAE
зальный край valva несет длинный, крючковидный, загнутый вентрально,
гладкий отросток; отростки valva сильно склеротизованы; penis длинный,
почти прямой, в дистальной части косо срезанный, короткий; pars infla-
bilis не несет инкрустаций. Bursa copulatrix мембранозная, короткая,
без скульптуры; ductus bursae почти прямой, сильно склеротизован,.
в дистальной части расширен; ductus seminalis отходит от дистальной
части совокупительной сумки; восьмой сегмент короткий, с вентральной
стороны мембранозный; papillae
anales лопастевидные, почти
прямоугольные; apophyses an-
teriores недлинные, несколько
длиннее восьмого сегмента;
apophyses posteriores короткие.
Общий тип гениталий обоих
полов очень близок к terebyn-
thi Frr.
Гусеница сизовато-серая,
покрытая короткими серыми
волосками, сидящими пучками
на бородавках; вдоль спины
широкая белая полоса, прерван-
ная на первом брюшном сег-
менте (который целиком темно-
серый) и стушеванная на пос-
ледних; бока тела светлее
спины, почти того же цвета,
как спинная полоса; брюшная
сторона светлосерая; ноги, как
брюшные, так и грудные, желто-
серые; бородавки, покрываю-
Рис. 187. Parocneria detrita Esp. Гениталии: Щие тело, небольшие, плоские,
а — самца, б — самки. голова черная с желтоватыми
ротовыми частями и лобным
треугольником (clypeus).
Распространение. Центральная и южная Европа — южная
Франция, Альпы, Германская низменность (Берлин, Гамбург); Польша
(Ченстохов, Радом, Варшава), Судеты, Карпаты (Буковина); долина
Дуная (Венгрия); Балканы; Украина (Харьков, Киев, Чернигов); По-
волжье (Казань, Тетюши, Куйбышев, Камышин, Сарепта); Малая Азия
(Аксехир); Крым; данные по Крыму есть лишь в литературе (Брик, 1934),
.и нахождение вида там не может быть мною подтверждено фактическим
материалом, хотя вероятно.
Биология. Зимует полу взрослая гусеница в лесной подстилке;
выход с мест зимовки поздней весной (в мае); кормовые растения гусе-
ниц Fagaceae (Quercus); окукление в рыхлом коконе в листьях и на ство-
лах; лёт в июле-августе; яйца откладываются кучками на листьях
кормовых растений и не покрываются волосками с брюшка самки. В по-
следнее время отмечены размножение и вред дубам в лесных полеза-
щитных посадках в Поволжье. Констатирован паразит Exochilum
circumflexum L. (Ichneumonidae).
*2. Parocneria lap!dicola H. S. (рис. 188).
Herrick-Schaeffer, 1851, Aussereuropaeische Schmett., 6 : 52, fig. 158, 159; Kirby,
1892, Syn. Cat. Lep. Het., 1 : 477; Staudinger, 1901, Cat. Lep., 1 : 118; Strand, 1910,
16. PAROCNERI A
' 383
Seitz, Grossschmett., 2 : 129, t. 23, e; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 425;
Gaede, 1932,Seitz, Grossschmett., 2, Suppl. ; 181; Zerny, 1933, Iris, 47 : 63; Gaede,
1934, Seitz, Grossschmett., 3, Suppl. 2 : 283; Bryk, 1934. Lep. Cat., 62 : 156; Schwin-
genschuss, 1938, Entom. Rundsch., 55 : 162; — atlantica Herrich-Schaeffer, 1850,
Aussereuropaeische Schmett., 6:51, fig. 144—146 (non Rambur); — urbicola Stau-
dinger, 1871, Cat. Lep. Pal., 1 : 28 (ed. 1); Staudinger, 1878, Tp. Русск. энтом.
общ., 14 r 353; — phoenissa Rogenhofer, 1891, Sitzungsber. zool.-hot. Ges. Wien,
41 : 85; — destituta Staudinger, 1891, Iris, 4 : 256; —mardina Staudinger, 1891, Iris,
4 : 337; — libanicola Staudinger, 1899, Iris, 12 : 161; —maraschi Daniel, 1932, Mitt.
Munch, entom. Ges., 22 : 15.
Биология. Bodenhei-
mer, 1930, Schadlingsfauna
Palaest., Monogr. angew. En-
tom., 10 : 226, 227.
. Грудь, голова и брюш-
ко, особенно с боков,
желто-розовые с примесью
серых и черных волосков,
многочисленных на голове
и груди и в меньшем числе
на дорзальной стороне
брюшка; розовые волоски
на теле особенно много-
численны у самок; усики
самцов сильно гребенча-
тые, у самок гребешки
короткие; передние кры-
лья желто-серые или ро-
зово-серые, почти без ри-
сунка, особенно у самок;
базальная линия в виде
широкого черного штриха,
по костальному краю рас-
тушеванного ; вторая попе-
речная линия хорошо за- _ .оо „ . , „ с г
r г „ Рис. 188. Parocneria lapidicola Н. S. Гениталии,
метна' в костальной поло- а - самца, б - самки. .
вине крыла, далее размы-
тая и неясная; срединное поле без рисунка, в области дискальных жилок
неясное светлое пятно; третья поперечная линия начинается интенсив-
ным черным штрихом, далее темносерая, слабо и мягко зубчатая; внеш-
нее поле однотонное, без полос; бахрома желто-серая; задние крылья
бледножелтые с широкой черной растушеванной полосой по краю и чер-
ным костальным краем, у самок розовато-желтые с желто-серой каймой
и светлосерой бахромой; нижняя сторона крыльев желто-белая с затем-
ненными терминальными краями, у самок она однотонно серая; косталь-
ный край передних крыльев несет широкие большие чешуйки, что отли-
чает этот вид от Р. komarovi Chr. Размах крыльев самцов 35—40 мм,
самок 55—60 мм.
Uncus на длинном широком слегка коническом основании, тонкий,
крючковидный, направлен почти дорзально относительно расположения
девятого сегмента; saccus небольшой, широкий; fultura inferior большая,
слегка овальной формы, расширенная в вентральной половине, в дор-
зальной половине заостренная, с треугольным вырезом на одну треть ее
высоты; valvae при основании широкие, в дистальной половине тонкие,
384
I. ORGYINAE
штангообразные, с крючком на дистальном конце; базальная широкая
половина valva несет очень маленький острый вырост на дорзальном
крае и больший вырост близ ее вентрального угла; вдоль valva небольшая
складочка; penis длиннее valva, почти прямой, сильно расширенный
в дистальной части, с коротким округлым coecum. Bursa copulatrix не-
большая, почти шаровидная, мембранозная, без скульптуры; ductus
bursae короткий, сильно извитой, тонкий, складчатый; ostium ductus
bursae открывается на склеротизованной пластинке, расположенной на
сильно разросшейся складчатой мембране между седьмым и восьмым
сегментами; седьмой сегмент узкий, еще более узкий восьмой сегмент,
они образуют яйцеклад; восьмой сегмент несколько длиннее в дорзаль-
ной половине, несколько укорочен и мембранозный в вентральной; длин-
ная и сильно гофрированная мембрана между этим сегментом и papillae
anales; последние плоские, короткие, мембранозные; apophyses ante-
riores и posteriores тонкие и длинные, слабо расширенные.
Ocneria destitata S;gr., описанная из Ирака (Мардин), очень близка
к Р. lapidicola Н. S., известной из этих же мест; какие-либо отличия,
которые можно считать за видовые, при описании и позже никем не даны, I
тогда как известна большая изменчивость Р. lapidicola Н. S.; в связи
с этим, пока не будет доказано, что О. destitata Stgr. является самосто-
ятельным видом, правильнее считать эти названия синонимами.
Изменчивость этого вида касается главным образом основного цвета,
того или иного-его тона, развития розового тона и степени развития ри-
сунка; сравнение закавказских destitata Stgr. с типичной малоазийской
формой Р. lapidicola Н. S. показало невозможность их разграничения не
только как видов, что до сих пор принималось, но и как подвидов;
никаких различий между этими формами я обнаружить не мог как
в ‘структурах гениталий, так и в признаках цвета и рисунка; форма
libanicola Stgr., описанная из Палестины, несколько более однотонная,
с более резким рисунком; задние крылья одноцветные, желтобелые,
без затемненного внешнего края.
Распространение. Сирия и Палестина (Яффа, Ливия);
Ирак (Мардин); Малая Азия (Анатолия, Анкара, Аксехир, Мараш,
Смирна); Кипр. Литературные указания на наличие этого вида в Закав-
казье (Романов, 1885) касаются описываемого ниже нового вида — grisei-
pennis, sp. n.; тем не менее, по устному сообщению М. Рябова, ему изве-
стна одна особь этого вида с Малого Кавказа (Зангезур); мне закавказ-
ские особи этого вида не известны.
Б и о л о гия. Зимуют гусеницы в щелях коры и под камнями
(Боденгеймер, 1930); период роста в Палестине охватывает февраль й
март; в апреле окукление, в середине апреля вылет бабочек и в конце
апреля, в мае появление гусениц нового поколения; в течение года
в Палестине появляются три поколения, второе заканчивает развитие
в конце июля, а третье — в сентябре—октябре; гусеницы третьего поко-
ления уходят в зимовку. Яйцекладка кучками, причем яйца покрыва-
ются волосками с конца брюшка самки; гусеницы живут первое время
совместно по сто и более особей, с ростом постепенно расползаюся. Кор-
мовые растения — Rosaceae (Prunus, Лт'gdalus). Плодовитость самок до
250 яиц; яйца розоватые. Указанная Боденгеймером биологическая осо-
бенность— покрытие яиц слоем волосков—вызывает некоторые сомнения,
как в определении им вида, так и наблюдения; самки Р. lapidicola
И. S., как и других близких видов Ocneria, имеют яйцеклад, но не
подушку волос на конце брюшка.
16. PAROCNERIA
385
Серьезный вредитель миндаля в Палестине, где отмечен для ряда
мест.
* 3. Parocneria komarovi Chr. (рис. 189).
Christoph, 1882, Тр. Русск. энтом. общ., 17 : 109; Romanoff, 1885, Mem. Lep.
Rom., 2 : 10, t. 1, fig. 2; Kirby, 1892, Syn. Cat. Lep. Het., 1 : 477; Strand, 1910, Seitz,
Grossschmett, 2 : 129, t. 21, a; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 425; Bryk,
1934, Lep. Cat., 62 : 157.
Голова и грудь серо-желтого цвета, вентральная сторона охристо-
желтая; ноги покрыты желтыми
глаза черные: усики самцов
сильно гребенчатые, гребеш-
ки желто-серые, более интен-
сивно окрашенные по бокам;
передние крылья желто-се-
рые со слабо развитыми эле-
ментами рисунка; косталь-
ный край их затемнен скоп-
лением мелких темных че-
шуек; базальная линия в
виде растушеванной простой
темносерой полоски; базаль-
ное поле светлосерое с при-
месью большого количества
узких чешуек желто-сер.ого
цвета; вторая поперечная ли-
ния в виде слабо извитой,
идущей перпендикулярно оси
крыла, полосы, несколько ;
более узкая в дорзальной
половине крыла; срединное
поле широкое, шире, чем
базальное и внешнее; сре-
r Р
динная линия широкая, из-
витая, сильно растушеван-
волосками с примесью серых и черных;
189. Parocneria komarovi Chr. Гениталии:
а — самца, б — самки.
ная, теряющаяся в дорзаль-
ной части крыла; третья поперечная линия в виде тонкой слабо зубчатой
полоски, слабо выгнутая посредине, начинается черным пятном у кос-
тального края; внешнее поле однотонное, желто-серое, со следами светлой
подкраевой линии; терминальная линия не развита; бахрома шашеч-
ная — черная и желто-серая; задние крылья оранжево-желтые с черно-
серым ободком, идущим только по костальному и внешнему краям; бах-
рома желто-охристая; нижняя сторона охристо-желтая, передние крылья
затемнены по костальному краю и в терминальной области; самки имеют
более стушеванный рисунок; задние крылья и брюшко светложелтые;
примеси розовых тонов как у самца, так и у самки нет. Размах крыльев
самнов 35—40 мм, самок 55.-60 мм.
Строение гениталий обоих полов очень близко к таковому у Р. lapidi-
cola Н. S.; valvae и penis почти идентичны, но есть небольшие отличия
в верхнем отростке valva, который у Р. komarovi Chr. почти не развит;
отличен также у Р. komarovi Chr. более длинный и широкий uncus. Раз-
личия самок несколько более значительны; совокупительная сумка
у Р. komarovi Chr. крупнее и имеет прямой короткий ductus bursae, более
2а Фауна СССР, И. В. Кожанчиков.
386
1. ORGYINAE
широкий, чем у Р. lapidicola Н. S.; отлична также вагинальная пла-
стинка, меньших размеров, чем у Р. lapidicola Н. S. Прочие части поло-
вого аппарата очень близки у обоих видов.
Распространение. Закавказье — долина Куры (Кировабад),
долина Аракса (Мигры, Нювады, Ордубад).
* 4. Parocneria amabilis Chr. (рис. 190, 191).
' Christoph, 1887, Mem. Le.p. Rom., 3 : 60, t. 3, fig. 8 (Orgyia); Staudinger, 1901,
Cat. Lep., 1 : 118; Strand, 1910, Seitz, Grossschmett., 2 : 133, t. 21, d; Swinhoe, 1923,
Ann. Mag. Nat. Hist., 11 (9) : 436; Gaede, 1932, Seitz, Grossschmett., 2 Suppl. : 103;
Bryk, 1934, Lep. Cat., 62 : 179; — autamnalis Wiltshire, 1943, Journ. Bomb. Nat.
Hist. Soc., 43 : 622.
Голова светло-желтая, особенно лоб; грудь сверху желто-серая
с вентральной стороны светложелтая; брюшко желто-серое; усики серые,
Рис. 190. Parocneria amabilis Chr. Самец. (Рис. Ф. Гуняева).
особенно их гребешки, членики бело-серые: передние крылья серо-желтые
с сильно редуцированным рисунком; базальная линия не развита, базаль-
ное поле желто-серое, несколько более темное в костальной части; вторая
поперечная линия черно-серая, тонкая, нерезкая, более широкая в ко-
стальной половине; срединное поле в середине затемнено широкой расту-
шеванной полосой; круглое пятно неясное, стушеванное; почковидное
пятно в виде светложелтого неправильной формы участка, лежащего
на более светлом фоне наружной половины срединного поля; третья
поперечная линия двойная, посредине желтая, слабо зубчатая; внешнее
поле однотонное, желто-серое, терминальная линия в виде тонкой полоски;
бахрома желто-серая; задние крылья оранжево-желтые с четко отграни-
ченной черной каймой по костальному и внешнему краям; внутренний
край задних крыльев желтый; бахрома желтая; нижняя сторона светло-
желтая, более интенсивно желтая на задних крыльях; на них заметны
слабые дискоидальные пятна; внешний и костальный края крыльев затем-
нены; самки желто-серые с еще более сильно стушеванным рисунком,
с тусклыми желто-серыми задними крыльями и того же цвета брюшком;
усики самцов сильно гребенчатые, усики самок с очень короткими отро-
стками на члениках. Размах крыльев самцов 30—35 мм, самок 43 мм.
Очень близок по строению гениталий к двум предыдущим видам —
Р. lapidicola Н. S. и Р. komarovi Chr. Uncus конический, длинный, не
16. PAROCNERIA
387
отделен швом от основания, почти прямой, с загнутым острым дистальным
концом; saccus треугольной формы, широкий; девятый сегмент короткий;
fultura inferior в виде большой почти прямоугольной пластинки с широ-
ким треугольным дорзальным вырезом; valva имеет широкую базальную
половину и гладкую узкую, в виде искривленного конического выроста,
дистальную; базальная половина на вентральном внешнем углу несет
широкий большой конический вырост, направленный дистально; penis
почти прямой со вздутым дистальным концом; coecum широкий и округ-
лый, длинный, по длине почти равен половине penis; pars inflabilis без
инктрустаций.
Этот очень характерный вид входит в состав группы Parocneria Dyar
типа lapidicola Н. S., komarovi Chr., представляя, невидимому, самый
северный вид этой группы, заселяющей полупустынную зону северной
Африки, Малой Азии
и Кавказа.
Распростране-
ние. Туркмения (Аш-
хабад, Ахал-текке, Сум-
бар, Айдере); Иран.
* 5. Parocneria gri-
seipennis Kozhantshi-
kov, sp. nov. (рис. 192).
Голова, грудь и пе-
редние крылья тускло- Рис. 191. Parocneria amabilis (Ьг, Гениталии самца„
го черно-серого цвета
с желтоватым оттенком
или желто-серые; голова и грудь покрыты тонкими шелковистыми
волосками; на крыльях желтые чешуйки располагаются вдоль жилок
и по линиям рисунка; базальная линия намечена коротким расту-
шеванным штрихом на костальном крае крыла; вторая поперечная линия:
идет почти прямо перпендикулярно продольной оси, широкая, слабо
волнистая, черная; срединная линия в виде широкой размытой черной
полосы, затемняющей или- костальную половину срединного поля, или
почти все поле; внешняя часть поля почти всегда не затемнена; третья
поперечная линия черная, сильно растушеванная, оттененная желтым
с внешней стороны (или однотонная), слабо выгнутая в середине крыла
кнаружи; почковидное пятно, а также круглое не развиты; жилки во внеш-
нем поле несколько осветлены желтым, или внешнее поле вполне одно-
тонное, желто-серое; подкраевая линия не развита; вдоль терминального
края крыла тонкая желтая полоска; бахрома шашечная — черпая
и желто-серая; задние крылья желтовато-белые с примесью черных,
чешуй по жилкам и с черной каймой; бахрома желтая с немногими чер-
ными штрихами; нижняя сторона светложелтая с серым или желто-белая
с серым; костальный и терминальный края крыльев темносерые; третья
поперечная линия и срединная линия в костальной части крыла, просве-
чивают темносерым; брюшко желто-серое. Самки очень похожи па самок
Р. komarovi Chr., несколько крупнее самцов, имеют более растушеван-
ные линии рисунка и большую примесь бледножелтых тонов; от самок
Р. komarovi Chr. отличаются более бледной окраской, в частности окраской
задних крыльев без примеси оранжевых тонов. Размах крыльев сампов=
35—40 мм, самок 40—45 мм.
25*
388
J. orgyinae
дифференцированным
Рис. 192. Parocneria griseipennis, sp. nov. Гени-
талии: a — самца, 6 — самки.
Uncus плоский, на дистальном конце расщеплен, степень расщепле-
ния варьирует; девятый сегмент несет короткие выступы — редуциро-
ванные subunci; saccus короткий; valva узкая с острой изогнутой и кони-
ческой дистальной частью; в средней части valva, ближе к ее вентральному
краю, узкий конический отросток; penis длинный, сложно изогнут,
несет склеротизованную складку на дистальном конце; pars inflabilis
без инкрустаций. Строение гениталий самок ближе к Р. komarovi Chr.,
чем к Р. lapidicola Н. S. Bursa copulatrix более мешковидная, со слабо
ductus bursae, широким и коротким; apophyses
и papillae anales длинные.
Своеобразный вид, при-
нимавшийся частью за' Р.
lapidicola Н. S. (самцы),
частью за Р. komarovi Chr.
(самки); более близок ко
второму виду, но как в
окраске и рисунке, так и
по структурам гениталий
дает ряд отличий от обоих
видов. Характерно полное
отсутствие розовых тонов
на брюшке и крыльях,
чем этот вид резко отли-
чается от Р. lapidicola Н.
S.; отсутствие оранжевых
тонов на крыльях отличает
от Р. komarovi Chr. Самки
этих трех видов менее
дифференцированы, чем
самцы, но габитуально
характеризуются этими же
различиями тонов окрас-
ки, имея, как и самцы,
отличия в структуре по-
лового аппарата.
Особи Р. komarovi Chr. известны в больших количествах из разных
мест долины Аракса, откуда этот вид был описан впервые (Ордубад);
западнее.—.в долине Куры —- Р. komarovi Chr. встречаются, видимо,
реже, так как известно лишь небольшое число их из Кировабада; Р. gri-
seipennis, sp. nov., собран мною в конце сентября в Грюнфельде (долине
Куры) и известен также из Кировабада.1
Распространение. Закавказье — долина Куры (Грюнфельд,
Кировабад).
1 Точных данных по фенологии Р. komarovi Chr. и Р. griseipen: it:. sp. nov., нет.
Если судить по времени сбора обоих видов, то, видимо, в Закавказье каждый из них
дает два поколения; лёт одного в конце июня, лёт второго в сентябре. Для Р. koma-
rovi Chr. есть особи, пойманные и летом, и осенью, причем они не дают отличий и,
в частности, оранжевый цвет сохраняется у всех; Р. griseipennis, sp. nov. был ловлей
мною лишь осенью, тогда как летом при тех же методах лова не встречен; по устному
сообщению М. Рябова, этот вид также летает и летом; летние Р. komarovi Chr. известны
и меньшем числе, чем осенние, может быть, летнее поколение этих видов малочис-
леннее осеннего.
16. PAROCNERIA
389
!i: 3. Parocneria signatoria Chr. (риС. 193).
Christoph, 1883, Iris, 6 : 88; Alpheraky, 1897, Mem. Lep. Rom., 9: 203, t. 8,
fig. 5a, b; Staudinger, 1901, Cat. Lep., 1 : 118; Strand, 1910, Seitz, Grossschmett.,
2 : 132, t. 21, b; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 436; Dumont, 1927,
Bull. Soc. Entom. France : 306; Gaede, 1932, Seitz, Grossschmett., 2 Suppl. : 102;
Amsel, 1933, Zoogeograph., 2 : 99; В yk. 1934, Cat. Lep., 62 : 177; — poenitens Stau-
dinger, 1899, Iris, 12 : ,162; — nisseni Rothschild, 1912, Nov. Zool., 19 : 125; Gaede,
Seitz, Grossschmett., Suppl., 2 : 102, t. 8, k; — militaris Oberthiir, 1914, Bull. Soc.
Entom. France : 386; — algeriea Oberthiir, 1916. Et. Lep. Comp., 11 : 22 t. 330,
fig. 4727, 4728.
Ширококрылый, тело cl ройное, мало пропорционально крыльям; голова
и передние крылья серые с примесью очень слабых бурых тонов; базаль-
ная линия в виде тонкого
черного штриха, растянутого
также вдоль кубитального
ствола; базальное поле несет
многочисленные черные че-
шуи; вторая поперечная ли-
ния в виде двойной черной
широкой полосы, частично
прерванной; срединное поле
очень широкое, несколько
более расширенное в ко-
стальной половине; почко-
видное пятно круглое или
овальное, внутри серо-корич-
невое, очерчено черным, ле-
жит за дискальной жилкой;
третья поперечная линия
черная, широкая, двойная,
широко расставленная по-
средине, светлосерая, слабо
зубчатая, прерванная на
жилках, идет почти парал-
лельно внешнему краю, остав-
ляя очень узкое наружное
поле; последнее серое, одного
цвета со срединным, не более
Рис. 193. Parocneria signatoria Chr. Гениталии:
а — самца, б — самки.
одной трети ширины последнего; терминальная линия в виде тонкой полоски
при основании бахромы; бахрома серая, разделенная вдоль тонкой полос-
кой; задние крылья при основании более светлые, по внешнему краю имеют
неправильную, размытую полосу; бахрома на них серая; нижняя сторона
желто-серая со слабо просвечивающей двойной третьей линией; глаза
большие, очень заметно выдаются; усики самЦов имеют очень тонкие
членики и длинные гребешки с одним волоском на конце каждого; брюшко
серое, у самок толстое, заметно утолщенное на дистальном конце; самки
крупнее самцов. Размах крыльев самцов 24—30 мм, самок 37—41 мм.
Uncus широкий, плоский, почти прямой, загнут лишь на дистальном
конце, плотно слит с девятым сегментом, без шва; tegumen слабо склеро-
тизован, короткий; fultura inferior щитовидная, с расщепленным дор-
зальным краем, в виде склеротизованного выступа; valva почти ромби-
ческой формы, по внешнему краю несет мелкие шипики, усаженные
щетинками; в базальной части ее с дорзальной стороны длинный искрив-
ленный отросток с небольшими шипиками на дистальном конце; penis
390
I. ORGYINAE
длинный и тонкий, сложно искривлен, coecum не более четверти его
длины. Bursa copulatrix очень маленькая, мембранозная, без скульптуры;
ductus bursae короткий,- широкий, прямой, складчатый, открывается
на большой овальной вагинальной пластинке; apophyses anteriores
и posteriores длинные и тонкие; восьмой сегмент недлинный, довольно
широкий; papillae anales мембранозные, большие.
По рисунку и габитусу этот вид стоит особняком среди рода; в строе-
нии гениталий также есть признаки (отсутствие вентрального отростка
на valva). отличающие этот вид от прочих в пределах рода, но структуры
генитального аппарата обнаруживают определенную связь с видами
группы Р. terebynthi Frr.
Распространение. Закавказье (Эльдар); Туркмения (Сумбар,
Балханы, Узун-су), Таджикистан (Куляб). Северная Африка, Алжир,
Марокко; Сирия, Эремийский хребет; Палестина (долина Иордана);
Ирак (Тигр, Багдад);
*7. Parocneria terebynthi Frr. (рис. 194, 195).
Freyer, 1839, Neuere Beitr. Schmett., 3 : 110, t. 272, fig. 1 (Bombyx)\ Freyer, 1845,
там же, 5 : 59, t. 419, fig. 1; Kirby, 1892, Syri. Cat. Lep. Het., 1 : 466; Staudinger, 1901,
Cat. Lep., 1 : 118; Spuler, 1908, Schmett. Eur., 1 : 132, t. 26, fig. 1; Strand, 1910,
Seitz, Grossschmett., 2 : 131, t. 21, b; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 119;
Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 435; Gaede, 1932, Seitz, Grosschmett.,
2 Suppl. : 102; Schwingenschuss, 1938, Entom. Rundsch., 55 : 162; — unicolor Stau-
dinger, 1894, Iris, 7 : 260; — terebinth! Spuler, 1908, Schmett. Eur., 1 : 132; — stotz-
neri Draeseke, 1926, Iris, 40 : 5 3.
Голова, грудь и передние крылья светлосерые, у самок почти чисто
белые; брюшко и задние крылья одноцветно серые; вокруг глаз желтые
волоски; на передних крыльях большая примесь светлосерых, пепель-
ного цвета чешуй; базальная линия в виде двух черных тонких штрихов;,
вторая поперечная линия черная, в виде сильно разрозненных штрихов,
выгнутая посредине кнаружи; базальное поле в дорзальной половине
белое, в костальной темное; третья поперечная линия идет почти парал-
лельно внешнему краю, слабо выгнутая, тонкая, зубчатая по жилкам;
срединная линия широкая, растушеванная, идет в виде косой полосы
через срединное поле, почти параллельная третьей поперечной линии;
подкраевая линия сильно волнистая, особенно хорошо развита у косталь-
ного края; жилки в срединном поле затемнены; почковидное пятно белое,
узкое по внешней стороне дискальной жилки, сильно выгнутое в середине,
у самцов часто редуцированное; терминальная линия в виде тонкой по-
лоски, серая; бахрома шашечная — серая и белая; задние крылья одно-
цветные, тусклосерые с тонкой темносерой терминальной полоской,
бахрома их чисто белая; нижняя сторона одноцветно серая с белыми
неправильно разбросанными чешуями по костальному краю и со слабо
просвечивающим белым почковидным пятном; усики самцов сильно гре-
бенчатые; длина гребешков посредине усика равна одной четверти его
длины; самки крупнее самцов, с выгнутыми по костальному краю перед-
ними крыльями; усики самок с заметными, но значительно более корот-
кими гребешками, чем у самцов. Размах крыльев самцов 23—28 мм,
самок 33—36 мм.
Uncus короткий, конический, загнутый крючкообразно, сидит на ши-
роком основании, полностью слитом с девятым сегментом; saccus боль-
шой, треугольной формы, широкий; fultura inferior длинная, в средней
части несколько суженная, склеротизованная в дорзальной части, заост-
16. PAROCNERIA
391
ренная, с узким вырезом; valvae широкие и выгнутые; вентральный край
valva острый, несет три зубца, из которых средний наиболее длинный,
Рис. 194. Parocneria terebynthi Frr. Самец. (Рис. Ф. Гуняева).
тупой; на дорзальном крае сильно склеротизованный гладкий длинный
вырост, изогнутый вентрально; penis тонкий, очень длинный, почти пря-
мой, с недлинным соесшп,
отогнутым вентрально. Bursa
copulatrix небольшая, мем-
бранозная, несет рубчатую
скульптуру, имеет большой
сильно склеротизованный
due'us bursae, образующий
кармановидное расширение в
дистальной половине; ostium
ductus bursae без инкруста-
ций, открывается в складча-
тую большую мембранозную
пластинку; восьмой сег-
мент короткий, склеротизо-
ван лишь на дорзальной сто-
роне; стерпит его мембраноз-
ный; apophyses anteriores и
posteriores тонкие и длинные,
последние несколько более
короткие; последние сегмен-
ты брюшка вытянутые и об-
разуют яйпеклад; есть боль-
шие придаточные железы;
papillae anales в виде до-
вольно толстых мембраноз-
ных волосистых лопастей. Рис. 195. Parocneria terebynthi Frr. Гениталии:
Форма uni col or Stgr. из « — самца,, б — самки.
Ирака почти белая, без
рисунка, особенно самки; stotzneri Draes. из Китая также со слабым
рисунком, тусклосерая; может быть, самостоятельный, вид; изучена
недостаточно.
392
T. ORGYINAE '
Распространение. Закавказье (Тбилиси, Закатали, Муганди);
южный Алтай, Семипалатинск (пос. Ульбинский); южное Приморье
(Сидеми); Малая Азия (Аксехир, Анкара); Ирак (Тигр); Сирия; Север-
ный Китай (Пекин).
Биология не изучена; указаны лишь кормовые растения, из
которых отмечена фисташка Pistacea terebynthus (Anacardiaceae).
*8. Parocneria terebynthina Stgr. (рис. 196).
Staudinger, 1894, Iris, 7 : 2r9; Staudinger, 1901. Cat. Lep., 1 : 118; Strand, 1919,
Seitz, Grossschmett., 2 : 132, t. 22, h; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9)
435; Bryk, 1934, Lep. Cat., 62 ; 177.
Голова, грудь и передние крылья желто-серые; последние сильно вытя-
нутые, выгнутые по костальному краю, с резко скошенным терминальным
краем; глаза большие, круглые, очень резко выдающиеся; базальная
линия на передних крыльях
в виде короткого черного
штриха; вторая поперечная
линия черная, двойная, ши-
роко расставленная, скошен-
ная базально; базальное поле
светложелтое со слабо • за-
темненным костальным кра-
ем; срединное поле широкое,
темное, с немногими желты-
Рис. 196. Parocneria terebynthina Stgr. Гениталии ми чешуйками; срединная
самца. линия в виде косой расту-
шеванной полосы, идущей
параллельно третьей поперечной линии; почковидное пятно очень узкое,
желтое, расположено на месте дискальной жилки; третья поперечная
линия идет параллельно внешнему краю, резко извитая, тонкая в ко-
стальной половине, растушеванная у дорзального края; срединное поле
темное, резко заметное на фоне крыла; внешнее поле узкое, почти равно-
мерное по ширине, с многочисленными чешуйками, неправильно разбро-
санными по крылу; подкраевая линия в виде растушеванной, неясной
полосы, заметной лишь в середине крыла; бахрома шашечная — темная
и желтая; задние крылья чисто белые с тонкой терминальной полоской
по краю и чисто белой бахромой; нижняя сторона передних крыльев
тусклосерая с просвечивающими светлыми элементами рисунка; задние
крылья чисто белые; терминальная линия на передних и задних крыльях
четкая; усики самцов сильно гребенчатые, членики их светлосерые.
Размах крыльев 23—25 мм.
Uncus короткий, плоский и почти прямой, загнут лишь на дистальном
конце; anus большой, мембранозный; девятый сегмент короткий, вполне
слит с основанием uncus, без шва; saccus длинный, конический; valva
короткая, со слабо зубчатой дистальной частью, несколько вогнутая
с дорзальной стороны, несет длинный, сильно склеротизованный отро-
сток и короткий почти прямой направленный косо дорзально другой,
вентральный отросток; fultura inferior высокая, плоская с вентральной
стороны и рассеченная с дорзальной стороны, с боков выемчатая; penis
тонкий и длинный, на конце косо срезан; соесшп короткий.
Для описания служили особи из Эльдара и Алазани; особи из
vИрака в натуре мне не известны; по рисунку и по описанию они вполне
сходны с эльдарскими.
16. PAROCNERIA
393
Распространение. Закавказье (Эльдар, Алазань); Ирак
(Мар дин).
* 9. Parocneria raddei Chr. (рис. 197).
Christoph, 1885, Mem. Lep. Rom,, 2 : 20, t. 1, fig. 3, 4; Kirby, 1892, Syn. Cat.
Lep. Het., 1 : 466; Staudinger, 1901, Cat. Lep., 1 : 118; Strand, 1910, Seitz, Gross-
schmett., 2 : 132, t. 21, b; Swinhoe, 1923, Ann. Mag. Nat. Hist., 11 (9) : 435; Bryk,
1934, Lep. Cat., 62 : 177; Zerny, 1935, Mem. Soc. Nat. Maroc., 42 : 47; — vestilina
Staudinger, 1894, Iris, 7 : 260.
Голова, грудь и передние крылья белые с примесью небольшого коли-
чества желтых волосков; передние крылья вытянутые, с сильно скошен-
ным внешним краем; базальная линия в виде слабого черного штриха
или совсем утрачена; вторая поперечная линия частично прерванная,
черно-коричневая, простая, почти по всей длине сильно скошенная ба-
Рис. 197. Parocneria raddei Chr. Самец.
(Рис. Ф. Гуняева).
зально, в костальной части более широкая; срединное поле широкое,
чисто белое, с хорошо развитой или частично редуцированной срединной
линией, наиболее ясно видной у костального и у дорзального краев;
кубитальный ствол в срединном поле несколько затемнен; третья попереч-
ная линия слабо извитая по жилкам, более широкая в промежутках между
ними; внешнее поле неширокое, вдвое более узкое, чем срединное; тер-
минальная линия в виде тонкой коричневатой полоски; бахрома желтая
и желто-белая; задние крылья чисто белые с тонкой коричневатой терми-
нальной полоской и белой бахромой. Нижняя сторона передних крыльев
светлосерая с неясно просвечивающими белыми и темными элементами
рисунка; нижняя сторона задних крыльев чисто белая. Брюшко у самки
все чисто белое, у самца слабожелтоватое на конце (указание Штранда
на желтое брюшко самки не соответствует тому, что имеется на типах Хри-
стофа). Размах крыльев самцов 29 мм, самок 35 мм.
Типичная форма этого вида известна мне только по типам (две особи
из Армении); форма vestilina Stgr., описанная из Ирака, отличается почти
одноцветными белыми передними крыльями; две особи, определенные
Штаудингером как vestilina Stgr. из Эльдара, мне известны по коллекции
Зоологического института Академии Наук СССР. Габитуально эти особи
заметно отличаются от типичных; они были собраны в августе (1 VIII 1886,
две самки), тогда как армянские особи — в мае. Недостаток материала
не позволяет решить вопрос о систематическом положении этих двух
394 II. ACRONICTINAE
форм; я пока оставляю их, как то было принято авторами, их описав-
шими, как формы одного вида.
Распространение. Закавказье — Армения (Шарофан, Бар-
тис), долина Куры (Кировабад), Эльдар; Ирак (Мардин).
II. Подсем. ACRONICTINAE
Виды средних размеров, редко крупные; ротовые органы или развиты
вполне и видны в виде скрученного хоботка, или рудиментарны; в одном
случае они почти полностью утрачены и остатки их скрыты в волосяном
покрове головы (Orycesta Hb.); бедра, голени и лапки всегда без шипов;
задние голени с двумя парами шпор; радиальная ячея развита; базальная
ячея на задних крыльях большая или маленькая, в последнем случае
замкнутая анастомозом субкостальной и радиальной жилок; рисунок
на передних крыльях обычно развит вполне, в виде поперечных полос;
круглое и почковидное пятна или недоразвиты (Diphthera Ochs., Мота
Hb., Belciana Stgr.), или развиты вполне (Acronicta Ld., Subacronicta
Kozh. и Craniophora Snell.); реже рисунок не развит (Arsilonche Ld.,
Eogena Gn., Simyra Ochs.), рудиментарен; в последнем случае рудимен-
тарны также и ротовые органы. Десятый сегмент всегда отсутствует;
uncus почти всегда слит полностью с девятым сегментом (исключая Calo-
casia Ochs., у которого он ясно отчленен), редко широкий, но никогда
не двойной, обычно тонкий и острый; по сторонам основания uncus
выросты девятого сегмента (subunci) иногда очень короткие, даже слабо
заметные, в виде бугорков.
Гусеницы типичны слиянием бородавок а. и [3 на средне- и заднегруди;
редко наблюдается слияние этих бородавок на брюшных сегментах (Acro-
nicta Ochs.). Куколки покрыты небольшими пучками коротких (иногда
редких, одиночных) волосков за стигмами и на дорзальной стороне послед-
них брюшных сегментов.
Виды Acronictinae распространены преимущественно в умеренных
широтах Голарктики (рис. 198); главная масса видов концентрируется
в зоне широколиственных лесов Азии (Гималаи, Китай, Япония); боль-
шое число видов и отчасти эндемичных родов, но меньше, чем в юго-восточ-
ной Азии, связано с широколиственными лесами Северной Америки.
Широколиственные леса Европы не имеют эндемичных родов и бедны
видами; для зоны хвойных лесов Голарктики эндемичен только один
род (Diphthera Ochs.), она бедна и видами Acronictinae. Степи и полу-
пустыни Средиземноморья и Средней Азии имеют несколько эндемичных
родов Acronictinae, в степной и полупустынной зонах Северной Америки
эндемичных родов нет. Арктика и зона тундры не имеют эндемичных ви-
дов Acronictinae’, виды лесной зоны лишь заходят в Арктику в самом огра-
ниченном количестве. В тропиках Acronictinae представлены бедно — нет
эндемичных родов; китайско-гималайские роды Trisuloides Wlk., Саппа
Wlk., Belciana Stgr., Мота Hb. распространены в тропиках почти исклю-
чительно в горах в определенной зоне их (Индостан, Цейлон, Зондские
острова, Новая Гвинея), где дают ряд эндемичных видов.
Экологически Acronictinae —• типичные лесные насекомые, связан-
ные с древесными растениями групп Salicales, Fagales, Rosales и близ-
кими к ним; в открытом ландшафте они представлены вторичными, произ-
водными и высокоспециализованными формами.
Подсемейство распадается на две целостных трибы — Momini и
Acronictini.
ТАБЛИЦА ТРИБ, ТАБЛИЦА РОДОВ ПО ГУСЕНИЦАМ 395
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ТРИБ
1 (2). Вторая медиальная жилка на задних крыльях развита вполне;
рисунок на передних крыльях часто только в виде поперечных
полос, которые или не образуют еще пятен, или образуют лишь
неполное почковидное и круглое пятна; окраска крыльев и особенно
передних часто, помимо серой, также охристая, зеленая или жел-
тая; базальная ячейка на задних крыльях большая, часто не замк-
нутая полным анастомозом субкостальной и радиальной жилок;
uncus часто сильно утолщен или весь, или лишь при основании,
в средней части или на дистальном конце; иногда tegumen сильно
расширен, и основание uncus отчленено от него швом.............
...........................................1. Триба Momini
2 (1). Вторая медиальная жилка на задних крыльях сильно редуциро-
вана и часто заметна лишь в виде слабой складочки; базальная
ячея на задних крыльях небольшая, большею частью замкнутая
анастомозом субкостальной и радиальной жилок; рисунок на перед-
них крыльях представлен, кроме поперечных линий, также пятнами—
круглым и почковидным; окраска крыльев, особенно передних,
почти всегда серая или черно-серая, редко лишь с примесью корич-
невых, охристых или зеленых тонов; uncus всегда тонкий; tegumen
короткий, прочно спаян с основанием uncus ... .................
.......................................2. Триба Acronictmi
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ ПОДСЕМ. ACRONICTINAE
ПО ЛИЧИНОЧНОЙ ФАЗЕ (ВЗРОСЛЫМ ГУСЕНИЦАМ)
i (2). Грудные сегменты и первый брюшной (первые четыре сегмента
тела) несут с дорзальной стороны длинные и густые пучки волос-
ков; дорзальная сторона средне- и заднегруди бархатистая, черно-
синяя; бородавка х на седьмом брюшном сегменте не слита с -и,
несет пучок волосков.........................21. Diphthera Ochs.
2 (1). Первые сегменты тела (грудь и первый брюшной) с дорзальной
стороны несут пучки волосков той же длины, что и прочие сегменты;
окраска их одинакова с прочими; если первый и второй грудные
сегменты затемнены, то они несут большие бородавки, покрытые
курчавыми волосками; бородавка х на седьмом брюшном сегменте
имеет разное положение.
3 (4). На восьмом брюшном сегменте над стигмой две бородавки —
большая (слитые а и р) и маленькая (р); большие бородавки вось-
мого сегмента сближены и образуют конический вырост; тело по-
крыто небольшими черными бородавками, несущими черные волоски;
бородавки х на седьмом сегменте почти на том же уровне, что и па
прочих..........................................26. Моша НЬ.
4 (3). На восьмом брюшном сегменте над стигмой с каждой стороны
по три бородавки; если на этом сегменте есть конический вырост,
то есть также вырост и на первом брюшном сегменте; бородавка х
на седьмом сегменте может быть свободной или слитой с т;.
5 (6). На среднегруди два длинных латеральных пучка волосков, сидя-
щих на бородавках р (вторых сверху); на первом, втором и вось-
мом брюшных сегментах непарные кисточки волосков на дорзальной
стороне (на бородавках (3); тело плоское; бородавка х на седь-
мом брюшном сегменте не слита с и, несет только одну щетинку;
396
II. ACRONICTINAE
бородавка z на всех остальных сегментах брюшка также с одной
сильной щетинкой...........................22. Calscasia Ochs.
6 (5)^ Кисточек волосков по сторонам Среднегруди нет; нет и трех кисто-
чек на дорзальной стороне брюшных сегментов; редко есть неболь-
шие дорзальные кисточки, но тогда они образованы плоскими воло-
сками, или если есть одна кисть на первом брюшном сегменте, то
она сидит на большой бородавке.
7 (8). Бородавки небольшие, черные, резко выделяются на светлом
голубовато-зеценом фоне кожи; они несут по одной или по две
длинных и сильных черных щетинки и усажены множеством мелких
шипиков; бородавка т) на брюшных сегментах с двумя сильными
щетинками; щитки на брюшных ногах несут большое количество
щетинок; бородавка х на седьмом сегменте брюшка не слита с z . .
.................................................24. Episema Ochs.
8 (7). Бородавки выделяются на фоне кожи не резко, часто они одного
цвета с ней; обычно они крупные и несут (во всяком случае три
низших) более двух щетинок; если на бородавках по одному волоску,
то они в виде лопаточек и тергиты воскового, желто-белого цвета.
9 (Ю). Черепная коробка глубоко вдавлена теменным швом и сильно
угловатая (образует углы с боков по сторонам ротовых частей);
на тергитах первого, третьего и шестого брюшных сегментов желто-
белые пластинки, бородавки на тергитах на этих пятнах желто-
белые; волоски, покрывающие тело, пушистые, коричневые; боро-
давка х на седьмом брюшном сегменте слита с ц . , . . . . . . .
............................................27. Daseochaeta Warr.
10 (9). Черепная коробка округлая; тергиты окрашены в один цвет
с прочими частями сегментов, или тергиты всех сегментов желто-
белые; в последнем случае на них сидят волоски в форме лопаточек.
И (12). Дорзальная половина тела покрыта лишь пупиллами, на которых
сидят по одному длинному простому волоску; на боках бородавки
несут много волосков; бородавка р на брюшных сегментах почти
слита с х, которая сильно сдвинута дорзально и расположена над
стигмой; на седьмом сегменте бородавка х вентрально от стигмы,
но не слита ст;..........................33. Subacronicta, gen. nov.
12 (11). Тело покрыто пучками волосков, сидящими или на бородавках,
или прямо на коже; если на коже мелкие бородавки несут по од-
ному волоску, то волоски в форме лопаточек.
13 (18). Бородавка х на седьмом брюшном сегменте большею частью
(исключая виды Pharetra Hb.) не слита с -и; на прочих сегментах
брюшка (первом—шестом и восьмом) х сидит прямо за стигмой
или слабо сдвинута вентрально; бородавка т; на среднегруди
обычно не развита (исключая виды Pharetra Hb.); верхняя губа
несет острый или округлый глубокий вырез ..................... .
..........................................34. Acronicta Ochs.
14 (17). Верхняя губа имеет глубокий острый вырез; тело покрыто
небольшими бородавками несет. пупиллы или просто пучки
волосков на коже; бородавка х на седьмом сегменте брюшка боль-
шею частью не слита с ц.
15 (16). Вся кожа покрыта тонкими шелковистыми волосками, образую-
щими скопления (пучки), где они более длинны; дорзальная сторона
тела не несет выростов на первом и восьмом сегментах; пластин-
чатые волоски, если они есть, многочисленные, простые..............
...................................... Подрод Acronicta Ochs.
1. M0MIN1
397
16 (15). Пучки длинных волосков сидят на плоских бородавках, резко
дифференцированы от мелких волосков, покрывающих тело; если
пучков волосков нет, то на пупиллах сидят толстые и длинные
волоски в форме лопаточек; на первом и восьмом сегментах брюшка
есть мясистые выросты, или тергиты всех сегментов несут желто-
белые участки.............................'. . Подрод Triaena Hb.
17 (14). Верхняя губа несет глубокий, но округлый вырез; тело покрыто
крупными выпуклыми бородавками, развитыми уже со второй ста-
дии; бородавка х. на седьмом брюшном сегменте слита с •/;; иногда
есть бородавка & на среднегруди................Подрод Pharetra Hb.
18 (13). Бородавка х на седьмом брюшном сегменте всегда слита с ц;
b на среднегруди всегда есть; бородавка х на прочих сегментах
брюшка (первом—шестом и восьмом) сидит на уровне до дорзального
края стигмы; верхняя губа имеет всегда широкий вырез.
19 (20). Тело покрыто короткими, примерно равной длины на всех сег-
ментах, жесткими волосками; верхняя губа хотя и с округлым,
но довольно глубоким вырезом (около половины длины) ....
.............•................................37. Arsilonche Ld.
20 (19). Волоски, покрывающие тело, разной длины; если они примерно
равной длины, то длинные.
21 (22). Все тело покрыто пушистыми длинными светложелтыми волос-
ками; кожа светложелтая; вдоль спины широкая оранжево-желтая
с черными штрихами (редко почти черная) полоса; голова желтая
................................................. 39. Eogena Gn.
22 (21). Тело покрыто короткими и длинными волосками; длинные волоски
часто расположены преимущественно в передней части тела; вдоль
спины черная полоса.
23 (24). Стигмы черные, хорошо видны на светлом фоне (или скрыты
на черном фоне); голова желтая или желто-бурая с светлыми при-
лобными участками; крючья на брюшных ногах желтые.................
.......................................... 40. Simyra Ochs.
24 (23). Стигмы белые с черным ободком, лежат на черном фоне; голова
черная с резко заметными желтыми прилобными участками; крючья
на брюшных ногах черные...................... . 38. Oxycesta Hb.
1. Триба Momini
Aurivillius, 1891, Nordn. Fjar. : 96 (Dilobidae); Smith, 1899, Proc. Un. St.
Nat. Mus.’ : 21 : 3 (Pantheini)’, Hampson, 1913, Cat. Lep. Phal., 13 : 335 (Mominae).
Крупные или средних размеров виды, обычно ширококрылые; окраска
крыльев часто яркая — желтая, зеленая, охристая, иногда бурая, серая
или черная; рисунок развит, слагается из многих самостоятельных полос
и недоразвитых пятен или образует сложный орнамент {Саппа Wlk.,
Thyatiri'dp.s Kozh.), или редуцирован {Sublauconycla Kozh.). Глаза волоса-
тые или голые; губные щупальца большею частью длинные, изогнутые,
торчащие прямо или вверх перед лбом; последний членик их хорошо раз-
вит, нередко длинный; редко щупальца короткие и свешиваются вниз
(тогда глаза волосатые); хоботок очень часто рудиментарный, не заметен
в волосяном покрове головы или заметен в виде рудимента; нередко
он развит вполне и закручен спирально; Мх на передних крыльях от-
ходит в»болыпинстве случаев от низа или от нижней части радиальной
ячеи, которая всегда развита; на задних крыльях отходит в большин-
стве случаев от места слияния с Си, развита хорошо, реже редуцирована;
39S
II. ACRONICTINAE
коготки на ногах простые или с отростком на внутренней стороне; голени
всех ног без шипов или (у Daseochaeta Warr.), редко, несут шипы; усики
самцов редко гребенчатые {Panthauma Stgr.), чаще простые и несут лишь
более длинные реснички, чем у самок. Uncus отделен от tegumen швом
или отчленен, но нередко полностью слит с ним основанием; есть .subunci
или их небольшие рудименты в виде бугорков по сторонам uncus; valvae
без cucculus, обычно с простой harpe, редко сложно инкрустированы.
Личиночные стадии для большинства видов не известны, но в тех
случаях, когда описаны, они всегда близки к Acronictini, хотя нередко
габитуально напоминают в сильной степени представителей Orgyinae.
Представители Momini распространены почти исключительно в северном
полушарии (рис. 198), причем они особенно многочисленны и разнообразны
на юго-востоке Азии в зоне широколиственных лесов Гималаев и Китая.1
В Центральной и Южной Америке есть эндемичные роды этой трибы,
представленные небольшим числом видов. В Северной Америке известны
лишь немногие эндемичные роды Momini.
В Евразии эндемичными родами являютсяEpisema Ochs., ограниченный
Европой, Panthauma Stgr., Euromoia Stgr., Belciades Kozh. vt Subleu cony eta
Kozh., локально распространенные в южном Приморье, Acronictoides Kozh.
и Thyatirides Kozh., распространенные в южном Приморье и в западном Ки-
тае, Elynodes Hamps., ограниченный западным Китаем, Thyracona Moore
и Geony ~ta Hamps., распространенные в Гималаях. Кроме того, в Гималаях
и юго-восточной Азии распространены роды, выходящие за пределы
Азии — на территорию Зондских островов, Новой Гвинеи и, редко, Австра-
лии, где известны обычно немногие представители их в горах (на высотах
1500—2100 метров), тогда как главная масса их видов распространена
в субтропиках Азии. Так, Trisuloides Moore богато представлены в Гима-
лаях, ряд видов заходит в Китай и один достигает южного Приморья;
из тропиков известны только два вида этого рода (с Явы и Новой Гвинеи).
Такое же распространение типично для видов Саппа Wlk., Trisula Moore,
Daseochaeta Warr., Anacronicta Warr., BelcianaWlk., ThalatlaWXk. Разли-
чия в распространении этих родов сводятся к степени расселения к северу
и в тропиках, но все они дают наибольшее число видов в Гималаях и за-
падном Китае. Так, виды Thalatha Wlk. не идут на север далее западного
Китая; в тропиках виды этого рода спорадически распространены довольно
широко (Мадагаскар, Цейлон, Зондские острова, Новая Гвинея, Северная
Австралия). Виды других родов, например Belciana Wlk., Саппа Wlk.
и ряд других, на севере достигают южного Приморья и Хондо, но в тропи-
ках распространены слабо. Один род этой группы —Мота НЬ., давая
тот же ареал, что и предыдущие, распространен на север шире и имеет
одного представителя в северо-западной Европе, а на севере Азии дохо-
дит до Енисейска. Наконец к этому же комплексу родов принадлежит
и близкий к Саппа Wlk. Cosmodcs Gn., эндемичный для тропиков и извест-
ный с Новой Гвинеи и из восточной Австралии.
Три рода Momini имеют голарктические ареалы; виды Raphia Hb.
распространены спорадически по полупустынной зоне Евразии и Северной
1 Вид Cetola dentata Wlk., типичный для северной Индии, по Гарднеру (Gardner,
1946, Trans. Roy. Entom. Soc. Lond., 96 (4) : 61—72), по структурам гусениц отно-
сится не к Amphipyrinae, как то считал Гэмпсон, но также является представителем
Acronictinae. Мне этот вид в натуре не известен, ни по имаго, ни по гусенипам, почему
вопрос о его систематическом положении я выяснить не мог. Гарднер указывает, что
гусеницы этого вида довольно сильно отличаются от типичных Acronictinae развитием
вторичных щетинок.
I
Рис. 198. Распространение видов трибы Momini.
Черным обозначены территории с числом видов 20—30; сеткой с числом видов 10—20; вертикальной штриховкой
с числом видов 5—10; горизонтальной штриховкой с числом видов 1—4.
400 77. ACRONICTINAE (1. MOMINI)
Америки (Пиренеи, Тянь-шань, Тибет, Техас); виды Calocasia Ochs,
и Diphthera Ochs, распространены шире по лесной зоне Евразии и Север-
ной Америки; оба рода дают большее обилие видов в Северной Америке.
В Северной Америке есть только два эндемичных рода — Agriopodes
Hamps., близкий к евразийским Daseochaeta Warr., распространенный
на западе (Британская Колумбия, Калифорния, Аризона), и другой
род — Charadra Wlk., родственный Calocasia Ochs, и Diphthera Ochs.,
очень своеобразный и сильно обособленный, может быть близкий к цен-
тральноамериканским родам этой трибы, распространенный в восточ-
ных штатах и Мексике, заселяющий широколиственные леса Северной
Америки и проникающий в Центральную Америку (Ch. higracreta Edw.,
nitens Schaus.).1
Центральная Америка имеет три рода трибы Momini — Lichnoptera
Н. S., распространенный на территории Колумбии, Коста-Рики, Гвате-
малы, южной Мексики, откуда известна главная масса видов; некоторые
виды этого рода доходят до Калифорнии и Флориды.на север и один вид
распространен в Перу, на юге; Gdujonia Dogn., известный также, с этой
территории, и Bathyra Wlk., два вида которого распространены в Колум-
бии и Мексике. Эти три рода близко родственны между собою, но очень
обособлены от азиатских; из родов, распространенных в Азии, лишь
Panthauma Stgr. и Diphthera Ochs, близки к ним. Четвертый, центрально-
американский, монотипический род Polyonicta Hamps, стоит особняком
от прочих и имеет, повидимому, некоторое сходство с уссурийским Еиго-
moia Stgr.
Кроме этих родов, к Momini, по Гэмпсону (1913), из африканской
фауны принадлежат виды Elaeodes Hamps., очень своеобразные и совсем
не изученные формы, и монотипический мадагаскарский род Epicausis
Btl., также очень своеобразный. Эти роды если и принадлежат к этой
трибе, то представляют сильно обособленные ветви. Мне они в натуре
не известны.
Все известные в отношении биологии виды Momini являются дендро-
фильными формами, не исключая и видов Raphia Hb., которые хотя
и распространены в полупустынной зоне, но связаны в питании с Sali-
•cales. Из всей трибы лишь виды Diphthera Ochs, питаются хвойными
(Pinus, Larix, Picea), причем и евразийский, и американские виды.
Прочие Momini связаны с лиственными породами и особенно с широко-
лиственными Fagales, Rosales, Salicales.
В пределах трибы широко распространены роды, имеющие во взрос-
лой фазе рудиментарные ротовые органы; это особенно типично для древ-
них, генерализованных представителей Panthauma Stgr., Diphthera
Ochs.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ ПО ИМАГИНАЛЬНОЙ ФАЗЕ
1 (12). Глаза покрыты густыми или, реже, редкими, но всегда хорошо
видными волосками.
2 (7). Хоботок рудиментарный, слабо закрученный или висящий.
1 Кроме этих двух родов, для американской фауны Фракер (1915) указывает
еще для Acronictinae и, конечно, для трибы Momini род Polyogramma Hb. (typus heb-
raicti Hb.). Монотипический род, известный только из лесной зоны Северной Америки
(Мас.сачузеттс, Нью-Йорк, Техас, Флорида, Британская Колумбия), пожалуй наибо-
лее близкий по рисунку к дальневосточной Subleuconycta Kozh. Мне этот род известен
только по описанию; Фракер имел для исследования гусениц Polyogramma hebraica Hb.
ТАБЛИЦА РОДОВ
401
3 (4). Valvae без harpe; tegumen несет по сторонам отростки (subunci)
...............................................32. Calocasia Ochs.
4 (3). Harpe на valva в виде складки, несет отростки разной длины
и формы.
5 (6). Передние крылья белые с черным рисунком, состоящим из линий
(без пятен); tegumen со сложными выростами; penis с тремя боль-
шими зубцами на pars inflabilis................21. Diphthera Ochs.
6 (5). Передние крылья тусклосерые с яркими, светлыми пятнами
на темном фоне; линии рисунка недоразвиты; penis без зубцов . . .
........................’...................... 24. Episema Ochs.
7 (2). Хоботок развит вполне.
8 (9). Усики самцов гребенчатые; отростки harpe лежат вдоль вентраль-
ного канта по середине valva; задние крылья яркожелтые . . .
.....................,...................... . 18. Trisuloides Btl.
9 (8). Усики самцов простые; отросток harpe лежит по вентральному
канту valva; задние крылья серые или белыё, если есть примесь
желтого цвета, то слабая.
10 (И). Передние крылья бледнозеленые или белые с пестрым черным
рисунком; линии широкие, посредине светлые; девятое кольцо
не имеет выростов типа subunci.......................26. Мота НЬ.
11 (10). Передние крылья серые, с обычными узко сдвинутыми линиями;
девятое кольцо имеет выросты по сторонам (subunci) .................
........................................ 19. Anacronicta Warr.
12 (1). Глаза голые, окружены ресничками или чешуйками, или только
чешуйками.
13 (16). Хоботок рудиментарный, маленький.
14 (15). Отросток harpe направлен дистально, косо, вдоль вентрального
края valva; усики самцов в базальной половине с длинными гре-
бешками, в дистальной с короткими; крупные бабочки — 55—
65 мм..........................................17. Panthauma Stgr.
15 (14). Отросток harpe направлен дорзально; усики самцов равномерно
гребенчатые по всей длине; бабочки меньших размеров — 30—
40 мм......................’.........................23. Raphia Hb.
16 (13). Хоботок развит вполне, закручен спирально.
17 (22). Губные щупальца длинные, последний членик их длинный, за-
остренный, около половины длины второго членика.
18 (19). Линии рисунка многочисленные, не слитые в парные линии;
круглое и почковидное пятна не развиты; valva без отростка на
harpe, широкая, с утолщенным вентральным краем......................
.............................. 25. Belciana Wlk.
19 (18). Линии рисунка в виде обычных полос; пятна, особенно круглое,
развиты; valva несет harpe с отростком.
20 (21). Valva с острым отростком на вентральном крае; отросток направ-
лен вниз; отросток harpe направлен косо и дорзально; окраска
передних крыльев зеленая, линии рисунка черные, прерван-
ные ..........................................27. Daseochaeta Warr.
21 (20). Valva без отростка, с гладким вентральным краем; harpe с двумя
отростками — коротким, торчащим дорзально, и длинным, направ-
ленным дистально под valva . .................28. Belciades, gen. nov.
22 (17). Губные щупальца менее длинные, также торчащие вверх перед
лбом, но последний членик их короткий, не более трети или чет-
верти длины второго.
23 (24). Коготки на всех лапках простые; передние крылья изумрудно-
26 Фауна СССР, И. В. Кожанчиков.
402
II. ACRONICTINAE
зеленого цвета с белым или охристым рисунком; valva без harpe
или каких-либо отростков........................31. Саппа Wlk.
24 (23). Коготки всех лапок с зубцом; передние крылья не изумрудно-
зеленые, обычно серые, черные, коричневые или белые; строение
гениталий иное.
25 (26). Отросток harpe направлен назад, широкий, усаженный зубцами;
задние крылья желтые, передние с продольными желтыми маз-
ками .............................................29. Euromoia Stgr.
26 (25). Отросток harpe направлен дорзально или наискось дистально:
окраска крыльев белая, серая или коричневая.
27 (28). Девятое кольцо имеет небольшие отростки в дорзальной половине
(subunci); valva снабжена отростком на дорзальном крае; отросток
harpe направлен дорзально; передние крылья темносерые с широ-
ким белым пятном посредине...............20. Acronictoides, gen. nov.
28 (27). Девятое кольцо простое; valva без отростка на дорзальном крае;
отросток harpe направлен дорзально; прочие признаки иные.
29 (30). Valva широкая, расширенная в дистальной части; отросток harpe-
лежит в плоскости valva по середине valva; тергит девятого кольца
очень широк; передние крылья коричневые, с тремя яркими жел-
тыми пятнами...............................32. Thyatiridest gen. nov.
30 (29). Valva узкая, несколько заостренная; отросток harpe лежит у вен-
трального канта valva; тергит девятого кольца узкий; передние
крылья бело-серые почти без рисунка, с черным четыреугольником
под костальным краем.....................30. Subleuconycta, gen. nov.
17. Род PANTHAUMA STGR.
Staudinger, 1892, Mem. Lep. Rom., 4 : 404 (typus egregia Stgr.); Staudinger,
1901, Gat. Lep., 1 : 130; Warren, 1909, Seitz, Grossschmett., 3 : 18.
Тип рода P. egregia Stgr.
Хоботок сильно редуцирован, но заметен в виде рудимента; лоб сильно-
выпуклый; глаза большие, голые, округлой формы; усики самцов сильно
и двусторонне гребенчатые, от половины длины их гребешки сильно уко-
рочены; губные щупальца слабо выдаются вперед с коротким последним
члеником; первый членик передних лапок не более половины длины
голени этой же ноги; голени задних ног в дистальной половине резко
и сильно утолщены; коготки на всех ногах с коротким зубцом па внутрен-
ней стороне; радиальная ячейка На передних крыльях узкая и длинная;
от ее основания отходит и от конца Р5; М2 на задних крыльях развита
вполне, вполне развиты и дискальные; по бокам брюшка пучки желези-
стых волосков.
Монотипичный, видимо реликтовый род, отчасти близкий к; южно-
американским Bathyra Wlk., Lichnoptera Н. S., Gaujonia Dogn., отчасти
к индийским Trisula Moore и голарктическим Diphthera Ochs. Неясно-
также отношение к видам рода Anepholcia Prout. Наибольшее сходство
Panthauma Stgr. имеют с видами Trisula Moore (typus variegata Moore, non
Trisuloides variegata Moore).
Известен только один вид, локально распространенный в южном
Приморье.
* 1. Panthauma egregia Stgr. (рис. 199).
Staudinger, 1892, Mem. Lep. Rom., 6 : 404 (Panthauma Stgr.); Staudinger, 1901,
Gat. Lep., 1 : 130; Warren, 1909, Seitz, Grossschmett., 3 : 18, t. 4, а; Филипьев, 1927,
Ежегодн. Зоол. муз. Акад. Наук СССР, 27 : 230; Куренцов, 1929, Зап. Общ. изуч..
Амурск, края, 2 : 47.
17. PANTHAUMA, 18. TR1SUL0IDES
403
Передние крылья бело-серые с темным бурым рисунком и неясными
размытыми желтоватыми пятнами; голова и грудь того же цвета; чешуй-
чатый покров грубый, блестящий, негустой; базальное поле однотонное,
темнобурое, несколько осветленное в дорзальной половине; первая попе-
речная линия редуцирована; вторая поперечная линия двойная, широко
расставленная, посредине бело-серая, нерезко, но заметно извитая;ъкруг-
лое пятно отсутствует, не развито и клиновидное; .почковидное пятно
в виде неясной темной лунки при дискоидальной жилке; срединное поле
однотонное, слегка желтоватое; третья поперечная линия двойная, начи-
нается двумя широко расставленными штрихами черногоЩвета на косталь-
ном крае, далее внутренняя
половина ее редуцирована,
но от кубитальных жилок до
дорзального края крыла она
рновь идет двойная, слабо
зубчатая по жилкам; внеш-
нее поле узкое, у костально-
го края близ третьей попе-
речной линии затемнено, да-
лее бело-серое с неясными
желтыми штрихами; подкрае-
вая линия в виде тонкой
полосы, слагающейся из тре-
угольных штрихов; бахрома
бело-серая; задние крылья
тусклые, бело-серые, с тем-
ными жилками и двумя рас-
тушеванными темными поло-
сами, хорошо заметными
лишь у внутреннего края
крыла; внешнее поле их за-
темнено; терминальная линия
в виде слитых черных тре-
угольников четкая; бахрома Рис. 199. Panthauma egregia Stgr. Гениталии:
i r a — самца, б — самки.
бело-серая; нижняя сторона
бело-серая с темными жилка-
ми, слегка затемнена по. внешнему краю, с ясными лунными пятнами,
на переднем и заднем крыльях. Размах крыльев 55—80 мм.
х Valva широкая, лопастевидная, утолщенная в базальной части; harpe
несет недлинный, слабо изогнутый и направленный дистально отросток;
uncus простой; penis несет мелкие шипики и два больших шипа на pars
inflabilis; bursa copulatrix узкая, вытянутая, без скульптуры; ductus
bursae неширокий, сильно изогнутый; ductus seminalis отходит от осно-
вания ductus bursae; apophyses короткие.
Распространение. Южное Приморье (Владивосток, Сучан).
Биология вида не известна; стацией являются смешанные леса
(маньчжурская зона); питающими растениями, видимо, лиственные по-
роды, так как бабочки сидят только на коре лиственных деревьев (Курен-
цов, 1929).
18. Род TRISULOIDES BTL.
Butler, 1881, Ann. Mag. Nat. Hist., 7 : 36 (typus sericea Btl.); Staudinger, 1901,
Gat. Lep., 1 : 131; Warren, 1909, Seitz, Grossschmett., 3 : 18 (Xanthomantis Warr.);
Hampson, 1913, Cat. Lep. Phal., 13 : 339; Warren, 1913, Seitz, Grossschmett., Indo-
26*
404 II. ACRONICTINAE
Austr., 11 : 40; Strand, 1914, Arch. Naturgesch., A, 80 : 142; Prout, 1924, Bull. Hill
Mus., 1 (3) : 400—412, t. 13, 17; — Tambana Moore, 1882, Lep. Ark. Coll. : 155 (typus
variegata Moore); — Xanthomathis Warren, 1909, Seitz, Grossschmett., 2:18 (typus
Cornelia Stgr.); —Disepholcia Prout, 1924, Bull. Hill Mus. : 404 (typus coerulea Btl.); —
Smilepholcia Prout, 1924, там же ; 406 (typus luteifascia Hamps.).
Тип рода T. sericea Btl.1
Глаза большие, круглые, покрыты волосками; хоботок хорошо развит;
лоб гладкий; усики самцов двусторонне гребенчатые и несут на члениках
пучки ресничек; ноги покрыты густыми волосками; передние голени вдвое
длиннее первого членика лапки этой ноги; коготки на всех ногах про-
стые; губные щупальца короткие, торчащие вперед, с коротким передним
члеником; голова и грудь покрыты недлинными волосками почти без
примеси чешуй; радиальная ячея узкая и длинная; отходит от осно-
вания ячеи; Р5 отходит от ее дистального конца; Р3 и Pi на стебельке;
ТИ2 на задних крыльях развита вполне, связана хорошо развитой дискаль-
ной жилкой с прочими.
Valva коническая; harpe несет один отросток, направленный дистально
и наискось; penis с многочисленными мелкими шипами на pars inflabilis.
Bursa copulatrix мешковидная, без инкрустаций.
Полиморфный род, близкий к Anacronicta Warr.; часть видов неясна
в систематическом отношении; неясна также самостоятельность родов
Anepholcia Prout и Trisula Moore, особенно последнего. Распространение
ограничено преимущественно Гималаями, лишь немногие виды известны
из Китая, а из СССР только один Т. Cornelia Stgr. из южного Приморья;
из тропиков также известны немногочисленные виды, именно с Явы —
Т. calocalina Moore, известный также из Индии, и Т. papuensis Warr,
с Новой Гвинеи (рис. 200). В Гималаях виды Trisuloides Btl. особенно
многочисленны в Сиккиме и Кашмире (luteifascia Hamps., sericea Btl.,
coerulea Btl., albiplasa Warr., catocalina Moore, albitessellata Hamps.,
entoxantha Hamps.); в западном Китае встречаются отчасти эти же виды,
отчасти эндемики (sericea Btl., subflava Wilem., c-albwn Leech, conta-
minata Drd., chekiana Dr.). Из Африки, а также с материков Нового Света
виды Trisuloides Btl. не известны.
* 1. Trisuloides cornelia Stgr. (рис. 201).
Staudinger, 1888, Stett. entom. Ztg. : 246 (Acronicta); Staudinger, 1892, Mem. Lep.
Rom., 6 : 393, t. 6, fig. 2 (Acronicta)', Staudinger, 1901, Cat. Lep., 1 : 133 (Acronicta)',
Warren, 1909, Seitz, Grossschmett., 3:4, t. 4, a; Hampsom, 1913, Cat. Lep. PhaL,
13 : 345 (Trisuloides)', филипьев, 1927, Ежегодн. Зоол. муз. Акад. Наук СССР, 28 : 231;
Куренцов, Д929, Зап. Общ. изуч. Амурск, края, 9 : 52;— honrathi Graeser, 1888, Berl.
entom. Ztschr. : 315 (Diphthera).
Голова, грудь и передние крылья шоколадно-черные; рисунок черный,
нерезкий; базальное поле в средней части с многими бархатистыми чер-
ными чешуйками; вторая поперечная линия черная, простая, очень сильно
изогнутая в средней части к внешнему краю и соединена посредством
белого клиновидного пятна с третьей поперечной линией; круглое пятно
правильной формы, в центре с черной точкой, выполнено коричнево-
серым; почковидное пятно почти прямоугольной формы, очерчено черным,
в середине с черными колечками; срединная тень заметна лишь на косталь-
1 Структуры типичного вида, sericea Btl., включая и гениталии, кратко описаны
Проут (1924); гениталии этого вида изображены Тальботом (1924). Строение Tri-
suloides sericea Btl. очень близко к нашему Т. cornelia Stgr., принадлежность которого,
следовательно, к роду Trisuloides Btl. несомненна.
е> <4 о Г to / "Я® s о U i fjX; 7 ! 4 1 □ CT kvZS 7-^b; H) □ CT о — ' ICO . —6 g Q 0> — .— .... § C5
о \ ' ^ ' t 2
о = у о PJ ——— Y1 * / 1—" ’’ 1 v V rl 7; ——Ж •fc J M t c. ——d >0 120 150 I
О) . Ч ЗЪ \ ЬО Л e( ° V 1? ? "\ Ст к £ b „<*<?- о — . '.i e X x. * о
X4- / tp1 / : ' о C3 M
к 0 H? i~K.v О a о о Л| о
яйSI g o ”
Рис. 200. Распространение видов рода Trisuloides Btl.
i
I
406
JI. ACRONICTINAE
Valva
Рис. 201. Trisuloides Cornelia Stgr. Гениталии: a —
самца, 6 — самки.
ном крае, сливается с почковидным пятном и далее теряется; срединное
поле очень узкое, в середине перехваченное, более узкое, чем внешнее
поле, которое занимает почти половину крыла; подкраевая линия черная,
неширокая, с тремя сильными зигзагами; задние крылья желтые со сла-
бым оранжевым оттенком, костальная и апикальная части черные; темное
лунное пятно слито с темным костальным краем, слегка затемнены концы
жилок; терминальная линия и бахрома черные; нижняя сторона желто-
серая, передние крылья по костальному и терминальному краям черные;
задние крылья с тонкой черной терминальной полоской; лунные пятна
на передних и задних крыльях заметны. Размах крыльев 35—50 мм.
Valva коническая с заостренным дистальным концом; костальный
и вентральный края ее
утолщены; harpe узкая,
несет наискось дистально
направленный отросток,
несколько зубчатый на
конце; uncus несколько
вздут; девятое кольцо про-
стое; penis несет много
тонких и длинных зубцов
на pars inflabilis. Bursa
copulatrix мешковидно
вздутая, большая, дисталь-
ный угол ее склеротизо-
ван, склеротизован (и
желто окрашен) также, и
короткий ductus bursae;
ductus seminalis отходит
от сосковидного расшире-
ния совокупительной сум-
ки с правой стороны;
apophyses anteriores и
posteriores небольшие.
Мало изменчивый вид,
очень близкий к китай-
ским и гималайским
представителям рода; описанный из Китая Т. contaminata Draudt (Entom.
Rundsch., 54 : 40, 1937) чрезвычайно напоминает Т. Cornelia Stgr. и, воз-
можно, представляет лишь географическую форму последнего.
Распространение. Южное Приморье (Тигровая, Сучан,
Уссури, Яковлевка, Виноградовка, Суйфун, Владивосток, о. Аскольд).
Виология не известна, не известны и личиночные стадии; стацией
являются смешанные леса; биологически связана, вероятно, с широко-
лиственными породами.
19. Род ANACRONICTA WARR.
Warren, 1909, Seitz, Grossschmett., 3 : 18 (typus caliginea Btl.); Prout, 1924, Bull.
Hill Mus., 1 (3) : 407.
Тип рода A. caliginea Btl.
Хоботок вполне развит; глаза большие, круглые, покрыты редкими
недлинными волосками; губные щупальца большие, слабо приподнятые
вверх, направленй почти прямо вперед, последний членик их малень-
19. AN ACRON ICT A .
407
кий, второй длинный, покрыт волосками и чешуей; первый членик перед-
ней лапки около половины длины голени той же ноги; коготки на всех
лапках с небольшим зубцом на внутренней стороне; губные щупальца
большие, слабо приподнятые вверх, направлены почти прямо вперед,
последний членик их маленький, второй покрыт длинными волосками
и чешуей; усики обоих полов простые, покрытые недлинными реснич-
ками; радиальная ячея узкая, недлинная, отходит от нижней четверти
ячеи с помощью дискальной или непосредственно от нее; Т1/2 на задних
крыльях развита вполне; дискальная жилка или редуцирована как
на передних, или развита нормально.
Valva лопастевидная с небольшой harpe, от которой отходит отросток
вдоль дорзального края, направленный дистально; девятое кольцо с не-
большими отростками типа subunci, покрытыми короткими волосками;
uncus и penis обычного типа. Bursa copulatrix мешковидная, без инкру-
стаций.
Олиготиппческий род, ограниченный в распространении юго-восточ-
ной Азией; главная масса видов концентрируется в северо-западных
Гималаях (Кашмир, Сикким—A. obscura Leech, infausta Wlk., glauca
Hamps., flavala Moore); один из этих видов (A. infausta Wlk.) проникает
в тропики (Бирма), кроме того, с Явы известен эндемичный вид с гор
(до 1500 метров) —A. pallidipennis Warr. Большое число видов населяет
западный Китай, где также частью налицо гималайские виды (A. obs-
cura Leech, caliginea ВЫ. и nitida Btl.), распространенные в Корее и дости-
гающие южного Приморья; последний вид проникает в Японию, где есть
также эндемик A.. plumbea ВЫ.
Все представители рода Anacronicta Warr, однообразны, но север-
ные виды отличаются темной серой окраской, тогда как южные имеют
примесь охристых и желтых тонов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (1). Крупный, 50—60 мм; передние крылья светлосерые; пятна
частично очерчены интенсивно черным; круглое пятно большое;
светлое пятно между почковидным пятном и третьей поперечной
линией большое, вдвое больше, чем почковидное пятно.................
.............................................* 2. A. nitida Btl.
2 (1). Меньших размеров, 45—55 мм; передние крылья темнокоричневые;
прочие признаки иные.
3 (4). Между почковидным пятном и третьей поперечной линией светлое
пятно такой же величины, как и почковидное; внешнее поле между
подкраевой линией и краем крыла белое . . . * 1. A* caliginea Btl.
4 (3). Срединное поле однотонное, с хорошо развитой срединной линией
и ясно видными пятнами; внешнее поле однотонное, темное ....
................................................3. A. obscura Leech.
* 1. Anacronicta caliginea ВЫ. (рис. 202).
Butler, 1881, Trans. Entom. Soc. Lond., : 185 (Acronicta)', Staudinger, 1901, Cat.
Lep., 1 : 131; Herz, 1904, Ежегодн. Зоол. Муз. Акад. Наук, 9 : 269; Warren, 1909,
Seitz, Grossschmett., 3 : 18, t. 3, k; Hampson, 1913, Cat. Lep. PhaL, 13 : 352 (Trisuloi-
des); Prout, 1924, Bull. Hill Mus., 1 (3) : 408; Филипьев, 1927, Ежегодн. Зоол. муз..
Акад. Наук, 23 : 230; Draeseke, 1928, Iris, 42 : 297; — nitida Graeser, 1888, Berl.'
entom. Ztschr. : 316 (Мота Hb., non Btl.).
Голова и грудь серо-коричневые с примесью черных чешуй на patagia; пе"
редкие крылья буро-серые с немногими охристыми мазками, рисунок не-
408
11. ACRONICTINAE
резкий, базальная линия в виде едва заметной двойной, черной и серой
полоски на костальном крае; базальное поле более темное в костальной
половине; вторая поперечная линия двойная, посредине темносерая,
базальная половина ее черная, дистальная охристо-коричневая; клино-
видное пятно не развито, круглое пятно небольшое, овальной формы,
в середине серое, очерчено черным; срединная линия в виде почти прямой
растушеванной полосы, расплывчатой в средней части крыла, между
пятнами; почковидное пятно с базальной стороны четко и широко ото-
рочено в виде черного полулунья, в середине коричнево-белое, открытое
в терминальный и костальный края и переходящее в белое, неправильной
формы пятно, занимающее
Рис. 202. Anacronicta caliginea Btl. Гениталии:
а— самца, б — самки.
рая, двойная; нижняя сторона буросерая
остальную часть поля до
третьей поперечной линии;
последняя двойная, чер-
ная, посредине светлосе-
рая, зубчатая по жилкам,
слабо выгнутая к терми-
нальному краю; срединное
поле широкое, шире внеш-
него, более темное в ко-
стальной половине; под-
краевая линия состоит из
двух сильно волнистых
серебристо-белых полосок,
идущих- близ терминаль-
ного края, с базальной
стороны отороченных чер-
ными штрихами; терми-
нальная линия в виде
прерывистой черной поло-
ски; бахрома серая; зад-
ние крылья тусклосерые,
слегка охристые по внут-
реннему краю, с широкой
темной полосой, терми-
нальная линия в виде тон-
кой полоски; бахрома се-
с двумя растушеванными
полосами и тонкой терминальной полоской; на месте почковидного
пятна светлое пятно; лунное пятно на задних крыльях развито. Размах
крыльев 40—45 мм.
Valva широкая, лопастевидная, несущая на внутренней поверхности
немногочисленные волоски; harpe небольшая, занимающая лишь часть
базальной половины, прерванная посредине и несущая в дистальной
части отросток, на конце заостренный и загнутый; тергит девятого кольца
с небольшими отростками, расположенными латерально и покрытыми
короткими волосками; uncus тонкий, покрытый негустыми волосками;
penis без зубцов на pars inflabilis. Bursa copulatrix мембранозная, мешко-
видной формы, с небольшим расширением, от которого отходит ductus
seminalis, и небольшим слепым расширением с вентральной стороны;
ductus bursae короткий; apophyses anteriores и posteriores также корот-
кие.
Наиболее близок к японскому A. nitida Btl.
19. ANACRONICTA
409
Распространение. Приамурье (Хабаровск); южное Приморье
(Суйфун, Владивосток, Ворошилов-Уссурийский, Сидеми); Япония (Токир,
Иокагама); Корея (Гензан); Китай.
* 2. Anacronista nitida Btl.
Butler, 1878, Ann. Mag. Nat. Hist., 1 : 197; (Acronicta)-, Butler, 1879.Ill. Types
Brit. Mus., Het., 3 : 16, t. 45, fig. 1; Warren, 1909, Seitz., Grossschmett., 3 : 18, t. 3,
k; Hampson, 1913, Cat. Lep. Phal., 13 : 351, fig. 90; Matsumura, 1925, Journ. Coll.
Agric. Univ. Sapporo, 20 : 49; Matsumura, 1927, Ins. Matsum., 1 : 110; Draeseke,
1928, Iris, 42 : 297; —moupiensis Leech, 1900, Trans. Entom. Soc. Lond. : 527.
Голова, переднегрудь и tegulae пепельно-серые, заднегрудь и брюшко
темносерые; передние крылья светлосерые с ясным рисунком; базальная
линия намечена у основания крыла двумя штрихами; под костальным
краем вдоль радиальной жилки черный штрих; базальное поле светло-
серое, в костальной половине несколько затемнено; вторая поперечная
лйния широкая, двойная, посредине светлосерая, слабо извитая, дает
два изгиба к внешнему краю крыла, почти перпендикулярная оси крыла;
серединная тень, размытая в костальной половине серединного поля, сли-
вается с краями почковидного и круглого пятен; круглое пятно вытянутое,
продольной осью лежит вдоль крыла, очерчено черным, с внешней стороны
интенсивно черное, в средней части коричнево-серое, а в базальной части
светлосерое; почковидное пятно вытянутое, почти овальное, четко очер-
чено черным лишь с базальной стороны, снаружи грань его стушевана,
и оно открыто в большое бело-серое пятно, лежащее близ третьей попереч-
ной линии; от нижнего угла почковидного пятна длинный черный штрих
вдоль Сир пересекающий третью поперечную линию; светлый участок
за почковидным пятном резко выделяется на фоне темного серединного
поля; внешнее поле однотонное, серое с зигзагообразной белой подкрае-
вой линией, отороченной с базальной стороны черным; терминальная
линия в виде серии черных и белых штрихов; бахрома серая; задние
крылья тусклосерые, однотонные, с тонкой светлой полоской по краю
и с белосерой бахромой; нижняя сторона тусклосерая, однотонная. Размах
крыльев 50—60 мм. '
Распространение. Южный Сахалин; Япония (Токио, Иока-
гама); западный Китай.
3. Anacronicta obscura Leech.
Leech, 1900, Trans. Ent. Soc. Lond. : 527 (Acronicta)-, Warren, 1900, Seitz,
Grossschmett, 3 : 18, t. 3, c; Hampson, 1913, Gat. Lep. Phal., 13 : 354, t. 233, fig. 13;
Prout, 1924, Bull. Hill Mus., 1 : 408.
Голова, грудь и передние крылья темнокоричневые; рисунок хорошо
развит; базальная линия в виде короткого черного штриха; базальное
полуоднотонное; вторая поперечная линия черная, двойная, посредине
светлая, слабо извитая; срединная тень широкая, черная, растушеванная
особенно интенсивно в костальной половине крыла, проходит между
пятнами; круглое пятно правильной формы, небольшое, очерчено черным,
в середине светлое, хорошо видное на фоне крыла; почковидное пятно
небольшое, овальное, несколько стушеванное в дистальной половине,
с темными колечками в середине, несколько светлее общего фона; третья
поперечная линия двойная, черная, зубчатая по жилкам, посредине
Светлая; подкраевая линия черная, грубо зубчатая, с внешней стороны
белая; терминальная линия в виде серии черных штрихов в основании
410
II. ACRONICTINAE
бахромы; бахрома темнокоричневая; задние крылья темносерые,
несколько затемненные по внешнему краю, у самки однотонные, темные;
терминальная линия тонкая, черная; бахрома серая; нижняя сторона
с неясной просвечивающей третьей поперечной линией.
Распространение. Западный Китай; Индия (Пенджаб).
20. Род ACRONICTOIDES KOZHANTSHIKOV, gen. nov.
Тип рода A. lichenodes Graes.
Хоботок развит вполне; лоб гладкий, сильно выпуклый; глаза боль-
шие, круглые, голые, окруженные чешуйками; губные щупальца длин-
ные, торчащие вверх перед лбом, до уровня темени, последний членик
довольно длинный—-около трети длины второго; усики обоих полов
нитевидные, покрытые короткими ресничками; первый членик лапки
более одной трети длины голени; коготки на всех ногах с широким отрост-
ком на внутренней стороне; голова и грудь покрыты чешуей и волосками;
радиальная ячея вытянутая, ромбической формы, от нижней ее трети
или от половины с помощью дискальной жилки отходит Мл, Ps и Р4 на
длинном стебельке; М2 на задних крыльях несколько редуцирована;
дискальные жилки на передних и задних крыльях недоразвиты.
Valva с выростом на дорзальном крае; harpe с широким отростком,
направленным дорзально; девятое кольцо с небольшими subunci; penis
несет многочисленные шипы на pars inflabilis. Bursa copulatrix снабжена
длинным изогнутым ductus bursae.
Систематическое положение A. lichenodes Graes. в литературе неопре-
деленно; Штаудингер (1901) помещает этот вид в Acronictinae, Грэзер
(1892) в Agrotinae, Лич (1900) в Sarrothripinae, Гэмпсон (1903) и Уоррен
(1912) в Hadeninae. Уоррен (1909) объединяет lichenodes Graes. вместе
с angusta Wlk. и ypsilon Btl. в особый род Gerbathodes Warr., принимая
типом вид angusta Wlk. Этот последний, однако, близок к видам Oligia
Hb. (Hadeninae).1
* 1. Acronictoides lichenodes Graes. (рис. 203).
Graeser, 1892, Berl. entom. Ztschr., 37 : 216 (Graphiphora Hb.); Staudinger, 1901,
Cat. Lep., 1 : 131 (Craniophora)', Hampson, 1908, Cat. Lep. PhaL, 7 : 368, t. 117,
fig. 17 (Oligia.)', Warren, 1909, Seitz, Grossschmett., 3 : 175, t. 41, b (Gerbathodes)', Hamp-
son, 1913, Gat. Lep. PhaL, 13 : 400 (Trisuloides Btl.); — paupera Staudinger, 1892,
Mem. Lep. Rom., 6 : 394 (Craniophora Snell.); — connexa Leech, .1900, Trans. Entom.
Soc. Lond. : 516 (Gerbatha).
Голова и грудь покрыты серыми и черными чешуйками; крылья светло-
серые с белыми и черными пятнами и черным рисунком; базальная линия
в виде короткой черной полосы при основании крыла; базальное поле
серое с неправильно разбросанными белыми чешуйками; вторая попереч-
ная линия в виде интенсивной черной полосы, нерезко отороченной белым,
слабо извитой, выгнутой в средней части крыла в виде треугольника
к термену, срединное поле в нижней половине интенсивно черное (местами
буроватое), в костальной половине белое; круглое пятно на белом фоне
в виде неясного силуэта овальной формы; серединная линия намечена
на костальном крае растушеванным треугольным черным пятном, пре-
1 Бутлер (Butler, 1879, Ill. Тур. Het. Brit. Mus., 3 : 24, 47, fig. 1, 2) дал изобра-
жение типов этих видов (angusta Wlk. и ypsilon Btl.); оба вида очень далеки от опи-
сываемого и принадлежат действительно к Hadeninae.
20. ACRONICTOIDES, 21. DIPHTHERA
411
рванная и заметная лишь ниже, в дорзальной половине крыла на темном
фоне срединного поля в виде темной неясной полоски; почковидное пятно
в виде неясной светлосерой, очерченной черным лунки, растушеванной
к костальному краю; третья поперечная линия в виде прерванной черной
полосы, сильно скошенной базально и идущей параллельно терминаль-
ному краю; внешнее поле неширокое, уже серединного; подкраевая линия
в виде слабой, местами прерванной полоски; терминальная линия в виде
черной прерывистой узкой полосы; бахрома темносерая; задние крылья
темносерые, несколько более светлые при основании; терминальная
полоска тонкая, серая с
немногими темными точ-
ками; бахрома серая; ниж-
няя сторона желто-серая,
несколько более темная
на передних крыльях, с
неправильно разбросанны-
ми черными чешуйками, с
неясной поперечной поло-
сой и штриховидными дис-
кал ьными пятнами на пе-
редних и задних крыльях.
Размах крыльев 34—
38 мм.
Очень характерный и
своеобразный вид, напо-
минающий обликом неко-
торых Acronicta Ochs., на-
пример A. alni L.
Распр остран е-
н и е. Южное Приморье
(Сучан, Виноградовка,
Яковлевка); Приамурье
(Хабаровск); западный
Китай (котип соппеха
Leech).
Рис. 203. Acronictoides lichenodes Graes. Гениталии
а — самца, б — самки.
21. Род DIPHTHERA OCHS.1
Ochsenheimer, 1816, Schmett. Eur., 4 : 62 (typus coenobite. Esp.); Aurivillius, 1891,
Nordn. Fjar. : 95 (Panthea)’, Staudinger, 1901, Gat. Lep., 1 : 130; Spuler, 1908, Schmett.
Eur., 1 : 134 (Panthea)’, Warren, 1909, Seitz, Grossschmett., 3 : 10 (Panthea)’, Culot,
1 В связи с тем, что родовое название Diphthera Ochs, долгое время неправильно
относилось к роду Мота НЬ., для первого рода было принято в литературе название
Panthea НЬ. (1827), которое является синонимом Diphthera Ochs., так как типом обоих
родов является один и тот же вид — coenobita Esp. У Оксенгеймера (1816) род Diph-
thera включает три вида: coenobita Esp., ludifica L. и alpium Osb. (= orion Esp.). Bee
три вида относятся к разным родам. Вид ludifica L. был выделен Гюбнером (1818)
в род Мота, а вид alpium Osb. принадлежит к роду Daseochaeta Warr., хотя долгое
время он принимался ошибочно за вид рода Мота НЬ. Неправильное название родов
особенно укрепилось после работы Ледерера (1857), который отнес к роду Мота НЬ.
вид alpium Osb., а к роду Diphthera Ochs вид ludifica L. Гэмпсон (1913) впервые ука-
зал типы этих родов — для Diphthera Ochs, тип coenobita Esp., а для Мота НЬ. —
тип ludifica L. Проут (1922) считает неправильным указание Гэмпсона, ссылаясь
на Ледерера (1857), который будто бы фиксировал ранее типы родов. Фактическая
справка показывает, что Ледерер (1857) типов не указывал, но по примеру прежних
412
II. ACRONICTINAE
1909, Noct., 1 : 17; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 141 (Panthea)',
Hampson, 1913, Cat. Lep. Phal., 13 : 365; Nordstroem und Wahlgren, 1937, Svenska
Fjarl. : 78; —Diphthera Treitschke, 1825, Schmett. Eur., 5 : 47 (typus coenobita Esp.);—
Panthe'a Hiibner., 1827,- Verz. bekannt. Schmett. : 202 (typus coenobita Esp.); — Andela
Walker, 1861, Can. Nat. and Geol., 6 : 37 (typus acronyctoides Wlk.); —Platycerura
Packard, 1864, Proc. Entom. Soc. Philad., 3 : 374 ( typus furcilia Pack.).
Тип рода D. coenobita Esp.
Хоботок рудиментарный; лоб гладкий; губные щупальца очень корот-
кие, с очень маленьким, почти незаметным последним члеником; усики
самцов гребенчатые; глаза покрыты короткими густыми волосками, боль-
шие, круглые, окружены ресничками; голова и грудь покрыты длинными
волосками с примесью чешуй; на дорзальной стороне первых сегментов
брюшка пучки волосков; первый членик передних лапок более чем вдвое
короче передних голеней; задние голени утолщены в дистальной половине
' (напоминают строение у
Рис. 204. Diphthera coenobita Esp. Хетотаксия пе-
редне- и среднегруди и брюшных сегментов I,
VI, VII и VIII взрослой гусеницы.
Panthauma Stgr.); коготки
на всех лапках простые;
радиальная ячейка узкая
и длинная; Mt отходит от
ее нижней четверти; Р31Ч
и Р5 на стебельке; М2 на
задних крыльях развита
вполне;дискальная жилка
нормальная лишь в дор-
зальной половине, кос-
тальная часть ее редуци-
рована.
Valva с небольшим от-
ростком на harpe; tegumen
со сложными выростами.
Bursa copulatrix с лопа-
стевидным отделом в ди-
стальной части и с шаровидным расширением, от которого отходит
ductus seminalis; apophyses короткие.
Личиночные фазы этого рода характеризуются следующими призна-
ками. Гусеницы (рис. 204) уплощенной формы с большими пучками боко-
вых щетинок; торакальные сегменты тела имеют бархатистую кожу,
синеватые. Бородавка х на переднегруди большая, несет длинные ще-
тинки, торчащие вперед. Щетинки а, [3, у и 8 на переднегруди сидят
на общем щитке; бородавка р обособлена; а и [3 на среднегруди слиты
в целостную бородавку; на среднегруди есть Бородавка х на брюшных
сегментах прямо за стигмой, имеет несколько щетинок; на седьмом брюш-
ном сегменте х сдвинута вентрально; бородавки а брюшных сегментов
несут кисточки волосков.
авторов давал после рода список видов. Оба эти рода, названия которых были заим-
ствованы Ледерером, им описаны —Diphthera Ochs, для ludifica L. и Мота Hb. для
alpium Osb. Точно так же Геррих-Шэффер (1843) еще до Ледерера (1857) описывал
род Diphthera Ochs, только для вида ludifica L., но тоже не указывал типа. По пра-
вилам номенклатуры типом рода принимается названный тип. Для обоих родов тины
впервые были указаны Гэмпсоном (1913), причем как раз это виды, стоящие первыми
в списках впервые выделенных видов: Оксенгеймером (1816) для рода Diphthera —
coenobita Esp. и Гюбнером (1818) для Мота — ludifica L., где этот вид является единст-
венным. Таким образом принимаемая мною синонимика, по Гэмпсону (1913), яв-
ляется обоснованной.
21. DIPHTHERA
4/3
Куколки вальковатой формы, блестящие, коричневой окраски, с ред-
кими короткими волосками за стигмами; cremaster направлен дистально.
Яйца крупные, почти правильной округлой формы с „матовым хорионом,
откладываются открыто кучками на ветки деревьев.
Представители этого рода родственны, с одной стороны, Panthauma
Stgr., с другой — видам Charadra Wlk., Calocasia Ochs., Raphia Hb.
Олиготипический род, дающий наибольшее разнообразие видов на
востоке (Великие Озера, восточные штаты) и на западе (Калифорния)
Северной Америки; здесь распространены D. acronyctoides Wlk., furcilia
Pack., gigantea French., virginaria Grt., portlandia Grt., palata Grt.
В Евразии широко распространен только один вид (D. coenobita Esp.).
Другой, D. grisea Wilem., описанный с Тайвана, эндемичен и сильно
отличается от всех остальных представителей этого рода; возможно
должен быть выделен в самрстоятельный род. Как американские виды,
так и евразийский вид являются бореальными формами, не заходящими
даже в субтропики; связаны с хвойными лесами. Они заселяют зону
хвойных лесов Голарктики и связаны преимущественно с видами рода
Pinus как в Евразии (с Pinus silvestris), так и в Северной Америке, где
виды Diphthera Ochs, распространены частью в Скалистых Горах (Кали-
форния), частью в Аллеганах.
* 1. Diphthera coenobita Esp. (рис. 204, 205, 206).
Esper, 1775, Schmett. Eur., 3 : 196, t. 37, fig. 7; t. 82, fig. 2; Aurivillius, 1891,
Nordn. Fjar. : 95, t. 26, fig. 7; Staudingbr, 1901, Cat. Lep., 1 : 130; Spuler, 1908,
Schmett. Eur., 1 : 134, t. -32, fig. 1; Culot, 1909, Noct., 1 : 17, t. 1, fig. 1;
Warren, 1909, Seitz, Grossschmett., 3 : 19, t. 2, a; Rebel, 1910, Berge, Schmctter-
• lingsb., ed. 9 : 141; Hampson, 1913, Cat. Lep. Phal., 13 : 366, fig. 95; Petersen, 1924,
Lep. Fauna Estl., ed. 2 : 143; Draudt, 1931, Seitz, Grossschmett, Suppl., 3 : 5; John,
1931, Folia Zool. Hydrobiol., 3 : 272; Gronblom, 1936, Acta Soc. Fauna, et Flora Pen-
nies, 58 : 12; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 78, t. 15, fig. 1; —
latefascia Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 141; — ussuriensis Warnecke,
1916, Neue Beitr. Syst. Insekt., 1 : 32; — immaculata, Scheljuzhko, 1920, Ztschr. wiss.
Insektenbiol., 15 : 188.
Биология и экология: Kirby, 1895, Butt. Moths. Eur. : 115; Pabst,
1888, Iris, 1 ; 115; Slevogt, 1903, Arch. Kurl. Livl. : 66, 123; Spuler, 1908, Raupen,
t. 22, fig. 13; Kuskov, 1931, Beitr. Kunde Estl., 17 : 68.
Голова и грудь белые с черными пестринами, покрыты плоскими воло-
сками; усики самцов двусторонне-гребенчатые, передние крылья заострен-
ные, вытянутые, черные и белые, с ясными растушеванными линиями;
базальная линия в виде недлинной черной и белой полосы; базальное поле
темное, во внешней половине белое; вторая поперечная линия черная,
грубо извитая, в средней части крыла дает изгиб, достигающий срединной
линии; круглое пятно в виде неправильной черной точки на белом фоне;
срединная линия в виде широкой черной линии, дающей три изгиба
н проходящей через все крыло; почковидное пятно узкое, полулунной
формы, очерчено черным, в середине белое; третья поперечная линия на-
чинается широким черным штрихом на костальном крае, слабо выгнутая
к,,внешнему краю, далее скошенная базально, сильно волнистая по жил-
кам, черная, с внешней стороны отороченная широкой белой полосой;
внешнее поле широкое, значительно более широкое, чем срединное,
с сильно зигзагообразной белой подкраевой линией, отороченной с вну-
тренней стороны черным; терминальная линия в виде серии длинных
черных штрихов; бахрома шашечная — белая и черная; задние крылья
тусклосерые с темными жилками и двумя неясными растушеванными поло-
сами; терминальная линия в виде тонкой черной полосы, бахрома белая
414
11. ACRONICTINAE
С тонкими черными штрихами; нижняя сторона белая с просвечивающими
линиями на крыльях; терминальная линия ясная; ноги черные с белыми
колечками на концах члеников. Размах крыльев 40—55 мм.
Рис. 205. Diphthera coenobita Esp. Самка. (Рис. Ф. Гуляева).
Рис. 206. Diphthera coenobita Esp. Гениталии:
а — самца, б — самки.
Valva короткая, широкая, несет узкую harpe с недлинным торчащим
вверх отростком; saccus большой, вытянутой формы; tegumen с мно-
гими сложными выростами; uncus в виде прямого стержня; penis с тремя
большими зубцами на pars
inflabilis. Bursa copulatrix
мешковидная, без скульпту-
ры, в нижней части склероти-
зована до светлокоричневой
окраски, образует лопасте-
видный вырост и шарообраз-
ное расширение, от которого
отходит ductus seminalis;
apophyses короткие.
Яйца крупные, около1
0.5—0.8 мм диаметром, ша-
рообразной формы, голубо-
вато-зеленые, покрытые ра-
диальными бороздками ;после
двух-трех суток ,развития
несколько темнеют.
Взрослые гусеницы серо-
коричневой окраски с при-
месью желтых тонов; пучки
коричневых волосков сидят
на круглых бородавках; осо-
бенно длинные пучки волос-
ков сидят на первом и восьмом
брюшных сегментах ; грудные сегменты черные с синеватыми бархатистыми
полосами между ними; гусеницы сидят, согнув переднюю часть тела, так что
синие полосы выставлены и хорошо видны; вдоль спины широкая желто-
белая полоса, частично прерванная и видная на каждом сегменте в виде
двух штрихов; по бокам две охристо-желтых полоски, образующие, ско-
21. DIPHTHERA 22. CALOCASIA
41o
пления желтого пигмента над дыхальцами; последние белые с черным
ободком; по бокам ниже дыхалец по широкой белой полосе.
Куколка толстая и относительно короткая; брюшные сегменты по-
крыты короткими волосками; верхняя губа в виде треугольной пластинки,
сильно выемчатая с вентрального края; кожа брюшных сегментов гладкая,
блестящая, имеет скульптуру в виде редких мелких точек; cremaster
в виде короткого и толстого сильно морщинистого выроста, несущего
четыре длинных тонких стержня на конце и много мелких стержней, за-
гнутых крючкообразно, сидящих по сторонам и вентрально от больших.
Уссурийские особи (D. ussuriensis Warn.) несколько крупнее (45—
60 мм) и более белые; особенно светлы передние крылья; есть некоторые
отличия в гениталиях самцов; у уссурийских особей отросток harpe
несколько длиннее и дистальный конец valva более косо срезан; строение
penis то же.
Распространение. Западная Европа от северной Италии
до северного побережья; на Пиренейском полуострове и на Британских
островах отсутствует; Альпы, Карпаты; Германская низменность; южная
Швеция и южная Финляндия; северо-запад Европейской части СССР,
далее к востоку от линии Петрозаводск—Молотов спорадически распро-
странен в лесной зоне (Ленинград, Москва, Калуга, Чернигов, Киев,
Волынь); Сибирь (Барнаул, Минусинск, Иркутск); Саяны (Казыр); Саха-
лин; южное Приморье (Сучан, Тигровая, Виноградовка, Ворошилов-
Уссурийский); Курильские острова; Риу-Киу (Цусима). Всюду встре-
чается локально. .
Биология и экология. Яйца откладываются на хвою
кучками по 30—50 шт.; вышедшие гусеницы держатся в первой стадии
вместе, затем расползаются; развитие яйца длится 5—6 суток; гусеницы
питаются старой хвоей сосны (Pinus silvestris) и, по некоторым сведениям,
также Abies alba, Picea excelsa, Larix ецгораеа; развитие гусениц проте-
кает в июле—августе; окукление в старой хвое, в плотном коконе из ко-
ричневых нитей; куколка диапаузирует и зимует; наблюдается всегда
одно поколение. Бабочки появляются весной в мае—июне и держатся
у вершин крупных сосен, где копулируют, кладут яйца и днюют; плодо-
витость самки 500—800 яиц; длительность жизни 5—10 суток.
22. Род CALOCASIA OCHS.
Ochsenheimer, 1816, Schmett. Eur., 4 : 63 (typus coryli L.); Hiibner, 1827, Verz.
bekannt. Schmett. : 201; Aurivillius, 1891, Nordn. Fjar. : 95; Staudinger, 1901, Cat'.
Lep., 1 : 131; Spuler, 1908, Schmett. Eur., 1 : 135; Culot, 1909, Noct., 1 : 18; Rebel,
1916, Berge, Schmetterlingsb., ed. 9 : 142; Warren, 1909, Seitz, Grossschmett., 3 : 11,
t. 2, c; Hampson, 1913, Cat. Lep. Phal., 13 : 360; Draudt, 1931, Seitz, Grossschmett.,
Suppl., 3 : 5; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 78; —Demos Stephens,
1829, П1. Brit. Entom. Haustell., 2 : 59 (typus coryli L.).
Тип рода C. coryli L.
Хоботок рудиментарен, едва заметен; лоб гладкий; губные щупальца
свисают вниз, слабые, тонкие, с коротким последним члеником, покрыты
длинными редкими волосками; глаза круглые, небольшие, покрыты
волосками и окружены длинными волосками — ресничками; усики сам-
цов гребенчатые; первый членик лапки около половины длины голени;
коготки всех лапок с коротким зубцом на внутренней стороне; радиальная
ячейка узкая, маленькая; Мг отходит с помощью дискальной от радиуса
416
II. ACRONICTINAE
ниже радиальной ячейки; М2 на задних крыльях развита вполне и свя-
зана дискальной с М3.
Uncus отчленен от tegumen; fultura superior большая, с зубцами;
девятое кольцо имеет subunci; valva без отростков; penis без инкрустаций.
Bursa copulatrix имеет laminae dentatae.
Строение личиночных фаз следующее. Гусеницы уплощенной формы,
покрыты короткими волосками; в первой стадии бородавка х передне-
груди несет пучок волосков, как у видов Orgyinae, но после линьки во
второй стадии х этого сегмента несет небольшие пучки волосков; напро-
тив, комплексная бородавка х—п на среднегруди имеет пучок волосков,
торчащих вперед. Бородавка х
всех брюшных сегментов на
уровне стигм за стигмами, на
седьмом сегменте сдвинута вент-
рально. Расположение борода-
вок типично для Acronictinae
(рис. 207 а, б). Гусеницы сразу
после выхода из яйца имеют
облик почти типичный; после
первой линьки приобретаются
признаки взрослых. Куколки
имеют блестящую гладкую ко-
жу, сильно расширены в тора-
кальной области, с тонким зао-
стренным брюшным отделом;
Рис. 207. Calocasia coryli L. Хетотаксия перед-
не- и среднегруди и брюшных сегментов I,
VI, VII и VIII взрослой гусеницы (б) и гу-
сеницы первой стадии (а).
волоски короткие и тонкие, за
стигмами. Яйца почти круглые
с приплюснутой вентральной
поверхностью; хорион их несет
тонкую скульптуру в виде реб-
рышек; откладка поодиночке, на
нижнюю поверхность листьев.
Близок к североамерикан-
ским Charadra V\flk., но также
несомненна его связь с Episema
Ochs. {—Diloba Bd.) и Raphia
Hb., как на то указывал еще
Смит (1899); Charadra Wlk.
отличаются от всех этих родов
наличием хоботка; они менее
близки к Calocasia Ochs., чем
. два последних рода.
Олиготипичный род; виды Calocasia Ochs, распространены по зоне
лиственных лесов Голарктики (рис. 208), причем особенно богато пред-
ставлены в северо-восточной Америке (Великие Озера, восточные штаты)
и на востоке’Азии. В Северной Америке распространены С. electa Sm.,
jlavicollis Sm., infanta Sm., propinquilinea Grt.; по зоне лиственных лесов
Евразии распространен широко лишь С. coryli L., на Амуре и по Уссури
его замещает С. mus Obth., а на Японских островах и на Тайване С. suzukii
Mats, и С. umbrosa Wilem. Виды Calocasia Ochs. — бореальные формы,
в Азии не идущие даже в Гималаи, а в Америке не выходящие за пре-
делы Аллеганских гор на юге. Ареал рода связан с распространением
видов Betulaceae.
Рис. 208. Распространение видов рода Calocasia Ochs.
27 Фаjна СССР, И В Кожанчиков
418
II. ACRONICTINAE
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Subunci короткие и широкие; fultura superior с небольшим числом
зубцов на вентральной стороне; окраска передних крыльев серо-
стального цвета.................................... * 1. С. coryli L.
2 (1). Subunci довольно длинные и узкие, заостренные; fultura superior
усажена шипами на всей дистальной поверхности; окраска перед-
них крыльев фиолетово-серая.........................* 2. С. mus Obth.
1. Calocasia coryli L. (рис. 207, 209).
Linne, 1758, Syst. Nat., ed. 10 : 10; Aurivillius, 1891, Nordn. Fjar. : 95; Staudin-
ger, 1901, Cat. Lep., 1 : 131; Spuler, 1908, Schmett. Eur., 1 : 136, t. 31, fig. 8; Warren,
1909, Seitz, Grossschmett., 3 : 11, t. 2, c; Culot, 1909, Noct., 1 : 18, t. 1, fig. 4; Rebel,
1910, Berge, Schmetterlingsh., ed. 9: 142, t. 28, fig. 11; Hampson, 1913, Cat. Lep.
PhaL, 13 : 362, fig. 94; Draudt, 1931, Seitz, Grossschmett., 3 Suppl. : 5, t. 1, a; Groen-
blom, 1936, Acta Soc. Fauna et Flora Fennica, 58 : 12; Nordstroem och Wahlgren,
1937, Svenska Fjar. : 78, t. 15, fig. 3; — avellanae Heune, 1901, Berl. entom. Ztschr. :
309;—melanotica Haverkampf, 1906, Ann. Soc. Entom. Belg., 1 : 158; —- weymeri
Holdhaus, 1910, Intern, entom. Ztschr., 3 : 240; —medionigra Vorbrodt, 1912, Schmett.
Siidbayerns : 237; — betulae Lenz, 1917, JMitt. Munch, entom. Ges., 8 : 45; — uni-
formis Turati, 1919, Natur. SiciL, 23 : 265; — umbrosissima Turati, там же, 23 : 26; —
wautersi Dufrane, 1925, Lambillionea, 25 ; 31; — alba Derenne, 1926, там же, 26 : 42.
Б и о л огия и экология. Slevogt, 1903, Arch. Kurl. Livl. : 66, 124;
Spuler, 1909, Raupen, t. 50, fig. 6; t. 21, fig. 20; Ljungdahl, 1918, Entom. Tidskr., 39 :
217, fig. 1; Romaniszyn, 1930, Prazn. Monogr., 6 : 235; Lebedev, 1933, 36ipn. прац.ь
екол., 1 : 66; Лебедев, 1935, тамже, 2 : 35; Aue, 1937, Entom. Rundsch., 55 : 80.
Передние крылья светлосерые с слабой примесью белого; голова
и грудь того же цвета, покрыты длинными пушистыми волосками;
брюшко светлосерое; базальная линия на передних крыльях редуциро-
вана; базальное поле у основания крыльев серое, во внешней половине
буро-черное; вторая поперечная -линия перпендикулярна передней оси
крыльев, дает в средней части крыла треугольный изгиб кцаружи, широ-
кая, черная, ординарная; клиновидное пятно не развито; круглое пятно
большое, сильно сдвинуто к почковидному, очерчено черным, в середине
темнобуроё, срединная линия растушеванная, идет через круглое пятно
поперек крыла, заметна по всей длине, слабо волнистая; почковидное
пятно неполное, в виде лунки на месте дискальной жилки, слагается из
двух темных штрихов, разделенных светлым, связано темным штрихом
со срединной линией; срединное поле неширокое, равное по ширине
в костальной и дорзальной частях, уже, чем внешнее поле; третья попереч-
ная линия в виде черной полосы, снаружи несколько осветленной, очень
слабо выгнута кнаружи у костального края, далее почти прямая, очень
слабо зубчатая; внешнее поле светлее срединного, с белой, подкраевой
линией, отороченной изнутри серым, слабо волнистой; терминальная
линия в виде тонкой непрерывакицейся полоски; бахрома серая; задние
крылья тусклосерые с темной терминальной полосой; нижняя сторона
серая со слегка просвечивающими пятнами и полосами. Размах .крыльев
30—35 мм.
Valvh узкая, в средней части выгнутая вентрально, имеет более сильно
склеротизованные дорзальный и вентральный края; harpe не развита;
fultura superior состоит из двух пластин, большая, кровлеобразной формы,
в дистальной части с зубцами; fultura inferior треугольная, также выгнута
кровлеобразно; uncus отчленен, сильно заостренный, голый; девятое
кольцо несет короткие и широкие отростки; penis с многочисленными
мелкими шипиками на pars inflabilis. Bursa copulatrix мешковидная,
J
22. CALOCASIA 419
Рис. 209. Calocasia coryli L. Гениталии:
а — самца, б — самки. .
ру в виде густых мелких точек в начале
мембранозная, имеет две слабо склеротизованных laminae dentatae;
ductus bursae короткий и широкий; apophyses почти не развиты.
Яйца круглые, покрытые радиальными бороздками, беловато-желтой
окраски, диаметром около 0.3—0.5 мм; через два-три дня после откладки
под хорионом появляется темнокоричневый пигмент, располагающийся
в виде колечка на дорзальной стороне яйца.
Гусеницы первых стадий плоские, покрыты короткими серыми воло-
сками равной длины на всей поверхности тела; старшие стадии, начиная
с третьей, приобретают бело-
ватую или желтоватую окраску
и покрыты более редкими воло-
сками; волоски сидят на не-
больших бородавках; боковые
пучки волосков второго груд-
ного сегмента длинные и гус-'
тые, торчат в виде кисточек;
на дорзальной стороне первого
и второго грудных сегментов
сидят непарные пучки рыжих
волосков; такой же пучок
сидит на восьмом брюшном
сегменте; голова желто-белая;
особи, развивающиеся в тени-
стых и сырых, местах, темнее;
перед окуклением гусеницы
краснеют.
Куколка с толстыми груд-
ными сегментами и сильно кони-
ческим тонким брюшком; брюш-
ные сегменты покрыты корот-
кими, многочисленными воло-
сками; они гладкие и блестящие
в средней части и несут скулыг
и в конце сегмента; cremaster в виде неширокого конического покрытого
глубокими бороздами выроста, несущего на- дистальном конце много-
численные крючко видно изогнутые недлинные щетинки.
Мало изменчив как в отношении величины, так и структур; .от С. mus
Obth. отличается более широким срединным полем; вторая и третья попе-
речные линии у этого вида проходят в стороне от пятен; у С. mus Obth.
они проходят через пятна; в срединном поле нет штриха, соединяющего
вторую и третью поперечные линии; меланестические особи описаны как
weymeri Hold. „
Распространение. Лесная и лесостепная зоны Евразии
от Альп до Байкала (рис. 208), крайние местонахождения на севере —
Британские острова, южная Швеция; Финляндия, Лапландия; Карелия
(Петрозаводск); Молотов, Енисейск, Иркутск; на юг идет далеко за пре-
делы лесостепья, по горам и мелким лесным участкам; крайние местона-
хождения на юге — северная Италия; южные Карпаты; Крым (Бельбек);
Таганрог; Северный Кавказ (Ставрополь); Дон; Закавказье (Боржоми);
Армения (?); южный Урал; северный Казахстан (Петропавловск); южная
Сибирь (Барнаул, Сталинск, Минусинск).
Биология, и экология. Яйца откладываются поодиночке
на нижнюю сторону листьев, в лесной зоне преимущественно на листья
27*
430
ACRONICTINAE
берез (Betula pubescens, Betula humilis). Гусеницы живут поодиночке,
с первых стадий склеивая листья паутиной, старшие стадии делают из
листьев плотный домик, при линьке совсем заплетаемый. Рост всех стадий
возможен только на видах Fagales (Betula, Quercus, Carpinus, Fagus и Co-
rylus; виды Alnus не пригодны для питания младших стадий) и на некото-
рых Salicaceae (Salix fragilis). Лучшим кормовым растением является
береза, именно Betula pubescens; максимальное число особеи”"как в ста-
дии яйца, так и молодых гусениц встречается на низкорослых березках
по окраинам болот. Старшие стадии более многоядны и встречаются
Рис. 210. Calocasia mus Obth. Гениталии:
а — самца, б — самки.
на осине, тополе (черном),
рябине, яблоне, ольхе. Окук-
ление между несколькими
стянутыми паутиной листья-
ми (как у непарного шелко-
пряда). Куколки диапаузи-
руют и зимуют. В зоне хвой-
ных лесов дает одно поко-
ление, хотя всегда наблю-
дается вылет бесплодных
особей осенью. Для южных
частей ареала канстатиро-
вано частичное второе поко-
ление. Бабочки живут 3—
7 суток, при пониженной
температуре до 15 суток и
откладывают до 300—
500 яиц; оптимумом темпе-
ратуры для яйцекладки
является 15—16°.
Днем бабочки прячутся в
кустарнике, на стволах де-
ревьев сидят редко. Отмечены
паразиты — Anomalon rufi-
corne Gr., Ophion luteus L.,
Compsilura concinnata Mg.
* 2. Calocasia mus Obth. (рис. 210).
Oberthiir, 1884, Et. Entom., 10 : 17, t. 12, fig. 4 (Dilobap, Staudinger, 1901, Cat.
Lep., 1 : 131; Hampson, 1913, Cat. Lep. Phal., 13 : 361; — ussuriensis Kardakov,
1928, Entom. MitteiL, 17 : 418, t. 2, fig. 20;— grisescens Kardakov, там же, 17 :
418, t. 2.
Передние крылья одноцветно серые с седый, фиолетово-белым нале-
том; базальная линия в виде слабой полоски, редуцирована; вторая попе-
речная линия сильно сдвинута дистально, проходит, касаясь круглого
пятна, идет почти прямая до дорзального края, где выгнута кнаружи;
от середины этой линии темный штрих, соединяющий ее с третьей попереч-
ной линией; вторая поперечная линия здесь несколько выгнута; круглое
пятно большое, очерчено черным, светлосерое в середине; срединная
линия нерезкая, заметная лишь у костального края, далее редуциро-
вана; третья поперечная линия темная, слабо выгнутая у костального
края, идет у самого почковидного пятна, ниже его слабо изгибаясь ‘ба-
зально, черная, слабо зубчатая по жилкам, оттененная светлым с внешней
стороны; подкраевая линия светлая, в базальной части оттененная чер-
22. CALOCASIA, 23. RAPHIA
421
ним, слабо извитая; терминальная линия в виде тонкой полоски, тем-
ная; бахрома темносерая; задние крылья одноцветные, тусклосерые,
терминальная линия слабая; бахрома серая; нижняя сторона серая с не-
правильно разбросанными светлосерыми чешуйками и слабо просвечиваю-
щим рисунком. Размах крыльев 35—40 мм.
Valva при основании широкая, конической формы, заостренная;
базальная часть ее утолщена, отделена от утолщенного вентрального конца
дистальной части мембранозным участком; fultura superior двустворча-
тая, обильно покрытая шипами, как на вентральной, так и на дорзальной
сторонах; fultura inferior широкая, сердцевидная; девятое, кольцо узкое,
с сосковидными направленными вентрально отростками; uncus отчленен
от девятого кольца, толстый, с выступом на вентральной стороне; penis
с многочисленными шипами (в виде коротких игл) на pars inflabilis. Bursa
copulatrix мешковидная, узкая, мембранозная, с небольшими штрихо-
образными laminae dentatae; ductus bursae широкий, покрытый рубчатой
скульптурой; apophyses короткие.
Распространение. Приамурье (Тукурингра, Хабаровск,
Николаевск); южное Приморье (Яковлевка, Владивосток, Тигровая,
Седанка, Ворошилов-Уссурийский, Суйфун); Сахалин; Япония (Хок-
кайдо); северный Китай (Пекин).
Питающие растения — Alnus, -Betula (Graeser, 1888).
23. Род RAPHIA НВ.
Hiibner, 1822, Verz. bekannt. Schmett. : 212 (typus hybris Hb.); Staudinger, 1901,
Gat. Lap., 1 : 179; Spuler, 1908, Schmett. Eur., 1 : 230; Gulot, 1909, Noct., 1 : 180;
Prout, 1928, Bull. Hill. Mus., 2 : 11.
Тип рода hybris Hb.
Хоботок редуцирован, короткий, примерно в половину обычной
длины; глаза голые, круглые; усики самцов двусторонне гребенчатые;
щупальца небольшие, торчащие вперед, последний членик их маленький;
лоб гладкий, с пучком волосков, торчащих вперед; первый членик лапки
короче голени этой же ноги в дв.а—два с половиной раза, коготки всех
лапок простые; голова и грудь покрыты почти исключительно чешуй-
ками; брюшко без пучков волосков на тергитах, или с маленькими; Мх
на передних крыльях отходит от нижней четверти радиальной ячейки,
с помощью дискальной или непосредственно; Р3_4 на длинном стебельке;
на задних крыльях развита вполне, сохранены и дискальные жилки.
Valva коническая, несет отросток harpe, направленный прямо дор-
зально; penis без слепого участка. Bursa copulatrix мешковидная, мембра-
нозная.
Олиготипический род, ограниченный в распространении полупусты-
нями северного полушария (рис. 211). Из южной Франции и с Пиреней-.
ского полуострова известен R. hybris Hb., из северной Африки •—R. aethi- •
ops В. Н. (видимо лишь форма от первого вида). Из Средней Азии из-
вестны R. approximate, Alph. и R. obsoleta Kozh., а из Тибета и с Дальнего
Востока — R. peusteria Piingl.1 В Северной Америке виды Raphia Hb.
также связаны с сухими полупустынными районами (Техас, Мексика);
1 Описанная Бутлером (Butler, 1878, Ann. Mag. Nat. Hist., Ser. 5, 1 : 1931
там же, 1878, Ill. Types Brit. Mus., Lep. Het., 2 : 3, t. 31, fig. 5) из Японии Raphia fas-
ciata Btl. очень своеобразна и, судя по изображению (в красках), скорее принадлежит к
какому-либо из родов подсем. Hadeninae (например к Parastichtis), но не кэтомуроп?’.
1A О С К ta о с О
> 0 30 60 90 120 ISO (во ISO 120 90 60 S ° . 0 . до 60 90 120 150 (ВО 150 120 90 60 t
ЙЙТ kyZ\ \ f
л• -*~
) '(( с
/ 1 у ч t?s> ' » «
Д .. /
>-< 1 \ 1 5 Г\*Л te. 4>^D
у' —J *
et V Ь и\ / : а
° '’-> \ S **
*S-\! ч!. / S&* / s | sz/b^y ! ' "\ о 1 ? о о W
3^С’ о о Q О
ОС 0 ОС 09
Рис. 211. Распространение видов рода Raphia Hb.
23. RAPHIA
423
известны R. frater Grt. и R. piazzi Hill., стоящие ближе к восточноазиат-
ским видам, чем к европейским, особенно первый вид.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). В срединном поле передних крыльев большое белое пятно; почко-
видное пятно не развито; передние крылья черно-серые; valva ко-
роткая и широкая почти ромбической формы; отросток harpe далеко
не достает дорзального канта valva . . * 3. R. peusteria Piingl.
2 (1). Срединное поле передних крыльев однотонное, серое или с примесью
желтых тонов; почковидное пятно заметно; прочие признаки иные.
3 (4). Valva широкая, лишь несколько суженная в дистальной части;
отросток harpe короткий, не выдается за пределы дорзального края
valva, но почти достигает его............* 2. R. approximata Alph.
Рие. 212. Raphia hybris Hb. Гениталии самца.
4 (3). Valva конической формы, сильно суженная в дистальной части,
отросток harpe длинный, далеко выдающийся за пределы valva . . .
................................................ 1. R. hybris Hb.
1. Raphia hybris Hb. (рис. 212).
Hiibner, 1827, Noct. Eur., fig. 518; Staudinger, 1901, Cat. Lep., 1 : 179; Spuler’
1908, Schmett. Eur., 1 : 200, t. 31, fig. 7; Spuler, 1908, Raupen, t. 21, fig. 19; Culot,
1909, Noct., 1 : 180, t. 33, fig. 7; Zerny, 1927, Eos, 3 : 377.
Передние крылья светлосерые с нерезким, но четким рисунком, желто-
ватые или с примесью желтых тонов лишь при основании; вторая попе-
речная линия простая, черная, сначала прямая, затем изогнутая базально,
окаймленная с внутренней стороны белым; срединное поле беловатое
с маленьким, в виде точки, очерченным черным круглым пятном и расту-
шеванным почковидным пятном; последнее очерчено не полно, в- середине
темносерое, с внешней стороны нередко размытое; срединная линия
в виде широкой черной полосы, прерванной в средней части, волнистая;
третья поперечная линия начинается над почковидным пятном, далее
выгнута к внешнему краю крыла, ниже почковидного пятна сдвинута
базально, черная, оттенена с внешней стороны светлой полосой, зубчатая
по жилкам; срединное поле той же ширины, что и внешнее; последнее
обычно более светлое, с тонкой извилистой подкраевой линией; терми-
нальная линия в виде тонкой черной прерывистой полоски; задние крылья
тусклобелые с темным неясным пятном (обрывком срединной линии)
в базальном нижнем углу; терминальная линия тонкая, черная; нижняя
сторона светлосерая с крупными неправильно разбросанными белыми-
и. ACRONICTINAE
и черными чешуйками, особенно обильными в базальной части крыла;
на задних крыльях неясные лунные пятна. Размах крыльев 33—37^мм.
~ Valva коническая, в дистальной части сильно суженная; harpe в виде
толстой складки по вентральному канту valva, несет тонкий прямой от-
росток, направленный прямо дорзально и далеко выдающийся за пределы
valva; penis короткий, непосредственно переходящий в ductus ejacula-
torius, без coecum.
Распространение. Пиренейский полуостров (Андалузия,
Арагония); южная Франция; северная Африка (Мавритания).
Описание гусеницы
(Spuler, 1908) неудовле-
творительно; мне гусеница
в натуре не известна; пи-
тающее растение — то-
поль.
Рис. 213. Raphia approximata Alph.. Гениталии:
а — самца, б — самки.
* 2. Raphia approxi-
mata Alph. (рис. 213).
Alpheraky, 1887, Stett.
entom. Ztg. : 167; он' же,
1889, Mem. Lep. Rom., 5 : 124,
t. 6, fig. 1; Staudinger, 1901,
Cat. Lep., 1 : 179.
Передние крылья свет-
лосерые co слабожелтова-
тым налетом в базаль-
ной части, по линиям и
вокруг пятен; очень близ-
ка к Лг/Zn-zsHb.; базаль-
ное поле желто-серое с
неясной короткой темной
первой поперечной ли-
нией; вторая поперечная
линия двойная, в виде
растушеванных черных
выгнутая кнаружи, затем
не развито; круглое пятно
полос, посредине серая, широкая, сначала
направленная базально; клиновидное пятно
в виде точки, тонко очерчено черным, в середине желтое; срединная ли-
ния в виде широкой растушеванной полосы, прерванной посредине; почко-
видное пятно нерезкое, частично смытое, оторочено черным, в середине'
имеет темное пятно; третья поперечная линия начинается над почко-
видным пятном, далее резко выгнутая кнаружи, затем сдвинутая ба-
зально, зубчатая по жилкам, снаружи оттененная широкой желтой поло-
сой; внешнее поле однотонное, серое, с тонкой беловатой подкраевой
линией; терминальная линия в форме черных штрихов, прерванная;
бахрома серая; задние крылья тусклобелые с затемненными жилками
и темным нижним базальным углом; терминальная линия в виде тонкой
полосы; бахрома серая; нижняя сторона серая, с крупными неправильно
разбросанными белыми и черными чешуйками, особенно обильными
по костальному полю; просвечивает третья поперечная линия, и заметна
терминальная линия; дискоидальные пятна слабые, маленькие. Размах
крыльев 32—35 мм.
S3. ПАРНА A
Valva широкая, лишь в дистальной части сразу суженная; harpe несет
отросток, направленный дорзально, прямой, довольно толстый, не до-
стигающий дорзального края valva; penis с очень ма'леньким coecum
несет на pars inflabilis мелкие зубчики; uncus толстый и плоский. Bursa
copulatrix мембранозная, сильно вытянутая, изогнутая; расширение
ее к ductus seminalis сосковидное, направленное дистально; ductus bur-
sae короткий, с рубчатой скульптурой; apophyses короткие; papillae
anales широкие, усаженные многочисленными правильно расположен-
ными щетинками.
Распространение. Средняя Азия (Ташкент); Семиречье (На-
рын).
*3. Raphia ре asteria Piingl. (рис. 214, 215).
Piingeler, 1906, Iris, 19 : 216, t. 8, fig. 9; — illarioni Filipjev, 1937, Lambillionea,-
3 : 16, fig. 3.
Рис. 214. Raphia peusteria Piingl. f. illarioni Fil. Самец.
(Рис. Ф. Гуняева).
Передние крылья черно-серые с примесью белых чешуй и с белым
пятном в срединном поле; базальное поле широкое, большей ширины,
чем срединное,'темносерое с белым растушеванным пятном вдоль базаль-
ной жилки; базальная линия в виде короткого штриха, стушеванная;
вторая поперечная линия черная, простая, сначала идет почти перпенди-
кулярно продольной оси крыла, затем, ниже' кубитальной жилки, сдви-
нута базально; срединное поле сужено в середине, в костальной половине
белое с темным штрихом у костального края (остаток срединной линии);
дорзальная половина срединного поля темная с нерезкой срединной
линией, идущей параллельно третьей поперечной линии; круглое пятно
редуцировано, почковидное пятно заметно очень слабо в виде темного
штриха; третья поперечная линия начинается над серединой белого пятна,
резко изогнута кнаружи под костальным краем, далее сдвинута базально,
одинарная, черная, зубчатая по жилкам, снаружи окаймленная белым;
наружное поле уже срединного, черно-серое с тонкой растушеванной
белой подкраевой линией; терминальная линия в виде тонкой сплошной
темной полоски; бахрома серая; задние крылья белые с слегка затемнен-
ными жилками, тонкой темной терминальной полоской и тонким нижним
базальным углом; бахрома светлосерая: нижняя сторона светлосерая
426
I]. ACRO'NICTINAE
с просвечивающейся полосой и лунными пятнами на передних и задних
крыльях. Размах крыльев 32—36 мм.
Valva широкая,' почти ромбической формы; harpe небольшая с корот-
ким расширенным отростком; tegumen широкий, массивный; uncus отчле-
нен, сильно выгнутый и утолщенный в основании; penis в дистальной
части несет многочисленные мелкие шипы, сидящие на его стенке, но не на
pars inflabilis; coecum не развит.
Распространение. Южное Приморье (Виноградовка). Ти-
бет (Куку-нор).
Описанная Филипьевым (1937) R. illarioni Fil. из южного При-
морья по существу ничем не отличается от R, peusteria Piingl. с Куку-
нора; отличие, заключающееся в отсутствии желтого оттенка передних
крыльев у R. illarioni Fil., не может быть признаком даже для подвида,
так как у уссурийских особей легкий желтый оттенок все же есть.
Рис. 215. Raphia peusteria Piingl. Гениталии самца.
4. Raphia obsoleta Kozhantshikov, sp. nov.
Вид из группы hybris НЬ. —approximata Alph., мельче-этих видов,
светлее и имеет тонкий рисунок; характерны сильно вытянутые косо
вдоль крыла поперечные линии и суженное срединное поле.
Голова, грудь и передние крылья серо-стального цвета с примесью
желтоватых тонов, рисунок черный; задние крылья белые, брюшко,
ноги и усы жёлто-серые; первая поперечная линия в виде тонкой черной
в середине прерванной полосы, дает под костальным краем крыла глубо-
кий выгиб к основанию; вторая поперечная линия тонкая, интенсивно
черная, снаружи оттенена широкой светлой полосой и далее тонкой
темной Полоской; она начинается на костальном крае почти по середине
крыла, но в нижней части его дает резкий изгиб к его основанию
и в области анальной жилки почти подходит к первой поперечной линии;
круглое пятно не развито; почковидное пятно в виде темного почти круг-
лого пятна, оттенено светлым; третья поперечная линия начинается на
костальном крае, терминально от почковидного пятна, на его уровне
в плоскости крыла, сначала сильно выгнута кнаружи, затем от уровня
медиальной жилки сильно скошена базально, в дорзальной половине
крыла сильно зубчата по жилкам, снаружи вся окаймлена широкой свет-
лой полосой и тонкой темной полоской; срединная линия сильно расту-
шевана; срединное поле темнее других частей крыла; внешнее поле одно-
тонное с очень небольшими остатками светлой подкраевой линии; терми-
нальная линия в виде восьми черных штрихов; бахрома серая; задние
крылья чисто белые с очень слабым растушеванным пятном на нижнем
23. RAPHIA, 24. EPISEMA
427
базальном углу (признак рода Raphia Hb.); нижняя сторона обеих пар
крыльев светлая, почти белая, слегка затемнен костальный край '
крыльев, и слегка просвечивают тёмным третья поперечная и подкраевая
линии. Размах крыльев 34 мм.
Известен один самец этого характерного вида по сборам Зарудного
в 1901 г.
Распространение. Белуджистан (Калапошт-Сарбаз).
24. Род EPISEMA OCHS.1
Ochsenheimer, 1816, Schmett. Eur., 4 : 65 (typus coeruleocephala L.); Treitscke,
1825, Schmett. Eur.., 5 (1) : 111 (typus coeruleocephala L.); Aurivillius, 1891, Nordn.
Fjar. : 95 (Dilobidae)', Staudinger, 1901, Cat. Lep., 1 : 168; Spuler, 1908, Schmett. Eur.,
1 : 184' Warren, 1912, Seitz, Grossschmett., 2 : 332 (Cymatophoridae'y, Hampson,
1913, Cat. Lep. Phal., 13 : 593 (NoctuidaeY Closs-Hannemann, 1917, Suppl. Entom.,
4 : 10' Zerny, 1918, Verh. zool.-bot. Ges. Wien, 58 : 4Г — Diloba Boisduval, 1940,
Gen. Index Meth. Eur. Lep. : 88 (typus coeruleocephala L.).
Тип рода E. coeruleocephala L.2
Xo боток маленький, редуцирован, обычно висит, не закручен спирал ьно;
.губные щупальца торчат вперед, последний членик их тонкий и довольно
длинный; второй их членик покрыт длинными и густыми волосками; гла-
за голые, окруженные длинными ресничками; усики самцов двусторон-
не-гребенчатые; первый членик передней лапки менее чем в два раза ко-
роче передней голени; коготки всех лапок простые; лоб гладкий; голова
и грудь покрыты волосками и узкими чешуйками; на конце брюшка
самки густые коричневые волоски, образующие подушку; радиальная
ячея небольшая, узкая; Мл отходит от основания этой ячеи с помощью
маленькой*дискальной; М., на задних крыльях развита вполне, развиты
и дискальные жилки.
Valva узкая коническая, с узкой лежащей по вентральному канту
harpe; отросток harpe (как у Raphia Hb.) короткий, направленный прямо
дорзально; uncus отчленен от tegumen, толстый; penis цилиндрический,
со слабыми инкрустациями (мелкими шипиками) на pars inflabilis.
Гусеницы характерны слабым развитием вторичных щетинок, начи-
ная с первой стадии; они есть лишь при основании ног; крупные большею
частью одиночные щетинки сидят на крупных пупиллах черного цвета,
несущих мелкие зубчики (шипики); некоторые пупиллы, например р
переднегруди, несут короткие щетинки (рис. 216). Бородавки аир средне-
и заднегруди самостоятельны; бородавка х брюшных сегментов сидит
прямо за стигмой; на седьмом брюшном сегменте она сдвинута вентрально.
Монотипический род, ограниченный в распространении только Среди-
земноморьем; пищевыми растениями являются в основном розоцветные;
наиболее близок к Raphia Hb. и Calocasia Ochs., являясь в отношении
1 Из всех авторов лишь Ауривиллиус (1891) правильно помещал род Episema
Ochs. (Diloba Bd.), выделяя специальное подсем. Dilobinae, куда объединял Diphthera
Ochs., .Мота Hb,, Calocasia Ochs., Episema Ochs, и ошибочно Simyra Ochs. Это под-
семейство он включал в сем. Arctiidae, которое, кроме того, распадалось на Orgyinae
(Liparinae), Arctiinae, Nolinae и ряд других. Гэмпсон (1913) помещает род Episema
Ochs, в Noctuidae, в подсем. Phytometrinae, Уоррен (1909) помещает ого в подсем.
Hadeninae, Шпулер (1908) — также в Noctuidae в подсем. Trijinae.
2 Род Episema у Оксенгеймера включает, кроме первого вида coeruleocephala L.,
еще cincta Esp., trimacula Hb., tersa Hb.; первый вид указан типом рода Episema Ochs.
Hampson (1913). Ошибочное отнесение этого родового названия к виду trimacula Hb.
(Dertisa Wlk.) связано с привившимся к coeruleocephala L. синонимическим назва-
нием Diloba Bd.
428
II. ACRONICTINAE
строения гениталий как бы переходным между 4ними; сохраняет, однако,
ряд очень своеобразных древних черт, ^например в хетотаксии и в стро-
ении дистальных сегментов брюшка самок, сближающих его с подсем.
Orgyinae. Систематическое положение )рода до последнего времени не
было с достоверностью установлено.
*1. Episema coeruleocephala L. (рис. 216, 217).
Рис. 216. Episema coeruleocephala L. Хетотаксия
передне- и среднегруди и брюшных сегментов
I, VI, VII и VIII взрослой гусеницы.
Linne, 1758, Syst. Nat., ed 10 : 504 (Bombyx)’, Aurivillius, 1891, Nordn. Fjar. : 95;
Staudinger, 1901, Cat. Lep., 1 : 168; Spuler, 1908, Schmett. Eur., 1 : 184, t. 31, fig. 6’;
Culot, 1909, Noct., 1 : 137, t. 25, fig. 4; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 :
192—193, t. 28, fig. 9, b; Hampson, 1913, Cat. Lsp. Phal., 13 : 593, fig. 130; Warren,
1913, Seitz, Grossschmett., 3 : 332; Amsel, 1933, Zoogeogr., 2 : 101; Gronblom, 1936,
Acta Soc. Fauna et Flora Fennica, 58 : 17; — armena Staudinger, 1871, Cat. Lep.,
1 : 76; — bipartita Strand, 1903, Arch. Naturwissensch. Christian., 25 (9): 12;.—
orbimaculata Strand, 1903, там же, 25 (9) : 12; — separata Schultz, 1907, Soc. Entom.,
22 : 52; — coalita Meves, 1914, Entom. Tidskr., 35 : 14, 41; — infumata Schwingen-
schuss, 1918, Verh. zool.-bot. Ges. Wien, 68 : 150; — conferens Dammer, 1922, Entom.
Ztschr. Frankf., 35 : 100; —• cap-
nodes Dannehl, 1926, там же, 39 :
152; — nigrofasciata Hackrag, 1933,
Lambillionea, 33 : 54.
Биология и эколо-
гия. Spuler, 1908, Raupen, t. 21,
fig. 15; Ljungdal, 1918, Entom.
Tidskr., 39 : 232 fig. 19, 38; Са-
мойлович, 1925, Защ. раст.,1 : 215;
Bodenheimer, 1930, Schadlingsfa-
una Palaest., Mon. angew. Entom.,
10 : 236—239; Филипьев, 1932, Tp.
защ. раст., 5 : 374; Козякина,
1932, Изв. Инет, борьбы вред,
сельск. хоз., 2 : 133; Лебедев;
1933, Зб1рн. праць екол., 1 : 36,
Hollas, 1935, Entom. Anz., 15 :180;
Nickolson, 1935, Entom. Rec.,
97 : 52; Doring, 1937, Entom.
Ztschr., 51 : 115, t. 11, fig. 165.
Передние крылья вытянутые, с вытянутым верхним углом, серо-
фиолетовые, несколько более светлые в средней части; базальная линия
тонкая, темнокоричневая, несколько выгнутая кнаружи; базальное поле
в основной части рыжеватое; вторая поперечная линия тонкая, одинар-
ная, ступенчато изогнутая кнаружи; почковидное пятно сильно сбли-
жено с круглым, иногда сливается с ним, овальной или почковидной
формы, редко в виде восьмерки, широко очерчено белым, в середине желто-
бурое с немногими белыми чешуйками; круглое пятно в форме восьмерки
или двух почти слитых пятен, широко очерчено белым, в середине желто-
серое с немногими белыми чешуйками; третья поперечная линия начи-
нается за почковидным пятном, сначала она двойная, коричневая, посре-
дине беловатая, далее одинарная (редуцируется внешняя половина),
сильно скошенная базально; срединное поле в нижней части более чем
в четыре раза уже, чем по костальному краю; внешнее поле однотонное,
довольно широкое, равной ширины на костальном крае и на дорзальном;
подкраевая линия очень тонкая, едва намечена, белая, заметно волни-
стая; бахрома серая; задние крылья одноцветно серые с темными жил-
ками, дискальные пятна заметны, темные; бахрома серая; нижняя сто-
рона светлосерая с просвечивающими пятнами на передних крыльях,
с дискоидальными пятнами и поперечной растушеванной полосой на зад-
24. EPISEMA
429
них; терминальная линия на передних и задних крыльях в виде тонкой
полоски. Размах крыльев 35—40 мм.
Valva коническая, неширокая, дистальный конец ее несет мягкие
волоски; harpe с коротким, направленным дорзально выростом; uncus
толстый, отчленен от tegumen; penis цилиндрический, слабо изогнут.
Bursa copulatrix неширокая, мембранозная, несет сильную рубчатую
скульптуру; расширение ее к ductus seminalis имеет характер угловатого
выроста; ductus bursae широкий.
Яйца полушаровидные, серо-белые с зелеными ребрышками; покры-
ваются волосками с брюшка самки; откладываются кучками.
Гусеницы старших стадий голубовато- или зеленовато-белые с размы-
той желтоватой дорзальной линией и голубоватыми боковыми линиями;
Рис. 217. Episema coeruleocephala L. Гениталии:
a — самца, б — самки.
на грудных сегментах с
дорзальной стороны боль-
шие черные бородавки;
на других сегментах тела
имеются лишь пупиллы,
также черного цвета; как
на пупиллах, так и на бо-
родавках на всех сегмен-
тах всегда по одному боль-
. шому черному волоску,
вокруг основания которо-
го располагается много
мелких шипиков; ноги
(брюшные) волосистые и
несут по 12 щетинок;
брюшная сторона серо-
зеленоватая с небольшими
черными пупиллами; го-
лова голубая, лишь вер-
хушки полушарий черные.
Куколка покрыта ред-
кими коричневыми воло-
сками; кожа темнокорич-
невая, кремастер в виде
двух латеральных лопастей, каждая лопасть имеет три острых корот-
ких выроста.
Распространение. Южная и средняя Европа, включая
Пиренейский полуостров и Британские острова; в СССР — на север до
Латвии, Литвы, Ленинградской области (Ленинград, Луга), Калуги,
Казани, Горького, Уральска, на юг всюду до побережья Черного моря
(Одесса, Николаев, Анапа) и Каспия (Астрахань); всюду в Поволжье,
но на восток далее У рал ьска не известна; Крым — всюду; Кавказ '(Ставро-
поль); Закавказье (Кировабад, Тбилиси, Мцхет, Ереван); северный
Иран (Шахруд); Малая Азия (Анкара, Мараш); Сирия, Палестина (Ие-
русалим).
Биология. Зимуют яйца; есть указания на зимовку имаго
(Assmus, 1868); возможно, что зимуют и бабочки и яйца. Выход гусениц
ранней весной (в средней Европе в начале апреля). Гусеницы питаются
сначала почками, затем Листьями; многоядны, но связаны преимуще-
ственно с Rosaceae (Pirus, Prunus, Sorbus, Malus, Crataegus), Fagaceae
(Quercus, Corvlus), Salicaceae (Populus, Salix). У гусениц наблюдается
430
II. acronictinae
четыре линьки. Окукление на растениях, чаще в щелях коры, в плотных
коконах из шелковых нитей и кусочков листьев и коры, наступает в се-
рединеиюня. Пронимфа развивается около 10суток.' Куколка развивается
долго, по Голласу (Hollas, 1935) диапаузирует; диапауза прекращается
при падении температуры, к осени. Выход бабочек в сентябре. Бабочки
активны ночью; спаривание не длительно—час-полтора. Яйца откла-
дываются кучками на растения, покрываются черно-коричневым секре-
том, посредством которого.очень прочно держатся на субстрате, и войло-
ком коричневых волос; дает всегда одно поколение.
Вредит плодовым деревьям — яблоне, груше, миндалю, сливе, вишне,
персику, абрикосу, орешнику; наибольшая вредоносность в южных райо-
нах — Украина, Поволжье (Кеппен, 1883; Брамсон, 1883; Кулагин,
1928, Сахаров, 1915); есть указания на вред и для севера (Самойлович,
1925; Козякина, 1932). Указаны паразиты из Ichneumonidae—Ichneumon
sarcilorius L., Cryptus cyahator Gr., Hemiteles schaffneri Schmied.,
H. submar^inatus Bridgm., H. conformis- Gr., Anomalcn cerinops
Gr., A. fibulat r Gr., Mesochorus brevipetiolatus Rtzb., M. ftrmosus
Bridgm.
25. Род BELCIANA WLK.
Walker, 1862, Journ. Linn. Soc. N. H., 6 : 182 (typus biformis Wlk.); Hampson,
1894, Moths Ind.., 2 : 465, 471 {Polydesma); Herz, 1909, Ежегодн. Зоол. муз., 9 : 268;
Warren, 1909, Seitz, Grossschmett., 3 : 367; Draudt, 1937, Entom. Rundsch., 54 : 374
{Daseochaeta). #
Тип рода В. biformis Wlk.
Хоботок хорошо развит; лоб гладкий, покрыт чешуей и волосками;
глаза голые, большие, круглые; губные щупальца приподняты вверх,
почти серповидной формы, последний членик тонкий, довольно длинный,
около, половины длины второго; усики самцов простые, щетинковидные;
первый членик лапки передних ног более половины длины передней
голени этих же ног; коготки на всех лапках с зубцом на внутренней
стороне; Мг отходит от радиального ствола, далее ветвящегося на Р,,
Р3, Pi и Р5; радиальная ячейка отсутствует; М3 на задних крыльях раз-
вита вполне; рисунок на передних крыльях слагается из непарных линий,
и неполных пятен; линии самостоятельны, не образуют пар, обычных
для остальных родов подсемейства.
Valva мембранозная, широкая и большая;'вентральный кант образует
harpe в виде небольшой разделенной пополам складки без выростов;
девятое кольцо простое; penis несет инкрустации в виде многочисленных
мелких шипов на pars inflabilis.
Олиготипический род, ограниченный в распространении, юго-восточ-
ной Азией, повидимому наиболее примитивный из известных родов
в группе Мота НЬ. Главная масса видов концентрируется в северо-запад-
ных Гималаях (Сикким); отсюда известны В. thoracica Moore, В. tetraspila
Wlk., В. stratiovirens Moore, В. eurychlora Wlk., В. biformis Wlk. Из
тропиков известны виды Belciana Wlk.: с гор (высоты 600—2300 метров)
Суматры — В. kola Prout и В. sophronia Prout и Новой Гвинеи —
В. patricolor Prout, В. habroscia Prout, 'Х subserrata Prout. Один вид изве-
стен из западного Китая (5. trinubila Drd.), один вид найден в Корее
и в южном Приморье, но пока не найден в Японии {В. staudingeri Leech).
Этот род несомненно близок к М та НЬ., но более примитивен, что
видно из строения рисунка крыльев, в котором пятна развиты более
слабо, чем у Мота НЬ., а поперечные линии очень полны и лишь отчасти
25. BELCIANA
431
объединены в пары. Близки и структуры гениталий, которые также
обнаруживают примитивные черты по сравнению с-Мота НЬ. По литера-
турным данным положение рода неясно. Уоррен (1909) помещает его
близ Catocala, может быть из-за строения щупалец. Проут (1924) описы-
вает новые виды этого рода как представителей Ophiderinae (Noctuidae).
Несомненны изолированность этого рода и его древность, но также несом-
ненна его принадлежность к Momini и наибольшая близость к родам
Мота НЬ. и Belciades Kozh.
* 1. Belciana staudingeri Leech (рис. 218, 219).
Leech, 1900, Trans, Entom. Soc. Lond.. : 551; Warren, 1909, Seitz, Grossschmett.,
3 : 368.
Передние крылья голубовато-зеленые с тремя темными пятнами и чер-
ным рисунком; голова и грудь того же цвета, но голова более темной
окраски и затемнена штрихами; линии рисунка многочисленны, лишь
Рис. 218. Belciana staudingeri Leech. Самец,
(Рис. Ф. Гуняева).
отчасти объединены в обычные пары; базальная линия в виде^коротного
черного штриха у основания крыла; вторая поперечная линия в виде
двух широко расставленных полос, посредине темнокоричневая, особенно
в костальной половине, слабо выгнутая базально; сильно извитая средин-
ная линия также двойная, широко расставленная, охватывающая с обеих
сторон пятна; круглое пятно почти овальной формы, в середине голубо-
вато-зеленое, очерчено черным; почковидное пятно вытянутое, с внешней
стороны выемчатое, очерчено черным, особенно интенсивно с внешней
стороны; третья поперечная линия двойная, широко расставленная,
черная, посредине голубовато-зеленая, дает изгиб к внешнему краю;
внешнее поле затемнено в апикальной части и у торнуса; подкраевая
линия нерезкая, белая, с внутренней стороны темная; терминальная
линия двойная, черная, широко расставленная; бахрома черная; задние
крылья черные, у основания светлее, чем по краю, со слабым диско-
идальным пятном; нижний базальный угол их с двумя короткими белыми
и темными полосками; бахрома серая с тонкой черной полосой; нижняя
сторона тусклосерая с размытым дискоидальным пятном на передних
и задних крыльях; костальный и терминальный края передних крыльев
черные; терминальная полоска в виде тонкой линии. Размах крыльев
32—36 мм.
432
II. ACRONICTINAE
Valva широкая, толстая, загнутая вентрально, с заостренным и усе-
ченным концом; в дорзальной ее части при основании утолщение, уса-
женное волосками; обильно покрыт длинными волосками и ее вытянутый
дистальный конец; harpe в виде складки по вентральному канту valva,
разделенная посредине, без отростков; penis сильно изогнутый, несет
многочисленные шипики треугольной формы на pars inflabilis; uncus
тонкий и голый.
Распространение. Южное Приморье (Сидеми;; Корея (Ген-
зен).
Описанная из западных провинций Китая Драудом (Draudt, 1937)
В. trinubila Drd., известная мне только по изображению и описанию,
представляет, невидимому, синоним В. staudinger! Leech, с которой
сравнения произведено не было; вид этот неправильно отнесен Драудом
в род Мота НЬ.
26. Род МОМА НВ.
Hiibner, 1818, Verz. bekannt. Schmett. : 208 (typus ludifica L.); Aurivillius', 1891,
Nordn. Fjar. : 95 (Diphthera)’, Staudinger, 1901, Cat. Lep., 1 : 130; Spuler, 1908,
Schmett. Eur., 1 : 135 (Trichosea); Culot, 1909, Noct., 1 : 17; Warren, 1909, Seitz,
Grossschmett., 3 : 10; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 141; Warren, 1913,
Seitz, Grossschmett., Indo-Austr., 11 : 40; Hampson, 1913, Cat. Lep. Phal., 13 : 376;
Prout, 1924, Bull. Hill Mus., 1 : 439; Nordstroem och Wahlgren, 1937, Svenska Fjar. :
78; — Trichosea Grote, 1874, Bull. Buff. Soc. Nat. Hist. Sci., 2 : 213 (typus ludifica L.).
Тип рода M. ludifica L.
Хоботок хорошо развит, губные щупальца направлены косо вперед
и вверх, покрыты короткими волосками и чешуйками, последний членик
их короткий, круглый; усики самцов щетинковидные, слабо опушенные;
глаза .большие, круглые, густо покрыты волосками; передние голени
с наружной стороны с рядом щетинок; первый членик лапки менее чем
вдвое короче передней голени; коготки на всех ногах с очень маленьким
шипом на внутренней стороне; радиальная ячея большая, ромбическая;
Мг на передних крыльях отходит от середины радиальной ячеи с помощью
дискальной жилки; Ря w Pi на стебельке; М2 на задних крыльях развита
вполне; дискальные жилки на передних и задних крыльях редуциро-
ваны; рисунок близок к Belciana Wlk., линии рисунка слабо объединены
в пары; почковидное пятно неполное.
Valva вытянутая, усаженная грубыми щетинками; harpe несет длин-
ный крючковидно изогнутый отросток; penis с инкрустациями на pars
inflabilis. Bursa copulatrix без laminae dentatae.
26. MOM A
433
Гусеницы co струйчатым, сложным рисунком; кожа несет короткие .
мелкие волоски и пучки крупных щетинок, сидящих на. бородавках;
бородавки крупные; бородавка х переднегруди большая, несет пучок
щетинок, направленных вперед; а, (5 и р среднегруди слиты (но не теряют
самостоятельности) в узкую длинную поперечную бородавку в которой
р сохраняет наибольшую обособленность. На заднегруди слияние этих
бородавок менее полно, но также имеет место. Бородавка х брюшных
сегментов на дорзальном уровне стигм, причем на седьмом сегменте
на том же уровне, что и на прочих; st и восьмого брюшного сегмента
слиты и образуют большую бородавку, несущую кисточку волосков.
Олиготипический род, близкий к Belciana Wlk. и, может быть, отчасти
к южноамериканским Lichnoptera Н. S. Главная масса видов сосредото-
чена в Гималаях (71/. gypsochroa Hamps., obsolescent Warr., nigrescent
Hamps., champa Moore); часть этих видов проникает в Китай и один
из них (71/. champa Moore) — в южное Приморье, Корею и, повидимому,
в Сибирь (Енисейск). Из тропиков виды (рис. 220) Мота НЬ. известны
.лишь с гор, с высот 1500—2100 метров. Описаны два очень близких между
собою вида с Суматры (71/. androdes Prout и М. tamsi Prout), с о. Буру
описана 71/. leucotaenia Prout, может быть форма от обычной 71/. champa
Moore (Prout, 1924); из гор о. Серам описана 71/. nigricatena Prout —
также, вероятно, форма от обычной 71/. champa Moore; очень вероятно,
что суматранские виды, описанные Проутом, также не самостоятельны.
В Европе распространен обособленный географически и структурно
наиболее специализованный вид этого рода, 71/. ludifica L., заселяющий
широколиственные леса.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Основной цвет передних крыльев зеленовато-желтый, рисунок
менее резок, чем у следующего вида; почковидное пятно с дорзаль-
ной стороны большею частью замкнутое; valva узкая, длинная,
на конце заостренная; penis с немногими шипами на pars inflabilis.
. .......................................... * 1. М. ludifica L.
2 (1). Основной цвет розовато-белый, рисунок в виде интенсивно черных
линий; почковидное пятно1 открыто в нижней части и в костальный
край; valva более короткая и широкая, с расширенным дистальным
концом, penis с многими мелкими шипами на pars inflabilis . . .
. ......................................* 2. М. champa Moore.
’* 1. Мота ludifica L. (рис. 221, 222).
Linne, 1758, Syst. Nat., ed. 10 : 514; Aurivillius, 1891, Nordn. Fjar. ; 95, t. 33,
fig. 1; Staudinger, 1901, Cat. Lep., 1 : 130,; Spuler, 1908, Schmett. Eur., 1 : 135, t. 22,
fig. 12, a; t. 31, fig. 32; Warren, 1909, Seitz, Grossschmett., 3 : 10, t. 2, a; Culot, 1909,
.Noct., 1 : 17, t. 1, fig. 2; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 141; Hampson,
1913, Cat. Lep. Phal., 13 : 380, fig. 97; Prout, 1924, Bull. Hill Mus., 1 : 439—441,
t. 13, fig. 11—13; Petersen, 1924, Lep. Fauna Estl., ed. 2 (1) : 148; Draudt', 1931, Seitz,
Grossschmett., Suppl., 3 : 5, t. 1, a; Nordstroem und Wahlgren, 1937, Svenska Fjar. : 78,
t. 15, fig. 2; — lugens Culot, 1909, Noct., 1 : 17, t. 2, fig. 1; —difjusipuncta Strand,
1916, Arch. Naturgesch.,. 82, A (2) : 46.
Биология и экология. Spuler, 1908, Schmot. Eur., Raupen, t. 22,
fig. 12, a; Romanizyn, 1930, Prazn. Monogr., 4 : 234.
Голова, грудь и передние крылья беловато-зеленые (иногда слабо-
желтоватые) с четким и сложным черным рисунком; брюшко охристо-
желтое с черной дорзальной поверхностью: базальная линия в виде
23 Фауна СССР, И. В. Кожанчиков.
Рис. 220. Распространении видов -рода Мота НЬ, Черным обозначены области обилия видов.
26. MOM A
435
короткого черного штриха у костального края, дает несколько расту-
шеванный черный штрих ниже срединной жилки; вторая поперечная
линия в виде широкой двойной полосы, посредине такого же цвета, как
и все крыло, слабо зубчатая, выгнутая в средней части к внешнему краю
крыла; круглое пятно овальной формы, лежащее большой осью вдоль
крыла, очерчено черным, в середине бело-зеленое; срединная линия
четкая, намечена широким черным штрихом под круглым пятном, далее
выгнута кнаружи и редуцирована, но ниже круглого пятна в виде хорошо
заметной интенсивно черной полосы, местами прерывающаяся; вдоль
анальной жилки в срединном поле черный штрих, соединяющий средин-
ную линию со второй поперечной линией; почковидное пятно неполное,
очерчено черным лишь с базальной и терминальной сторон и открытое
в костальный и дорзальный края, средняя часть пятна одного цвета
с общим тоном крыла; третья поперечная линия черная, двойная, широ-
кая, слабо изогнутая, посредине бело-зеленая, у костального края затем-
ненная с внешней стороны, четко зубчатая по жилкам; ее внешняя поло-
вина обычно частично редуцирована и нередко остается в виде черных
штрихов по жилкам крыла; внешнее поле узкое, равно по ширине или
уже срединного, подкраев^я линия в виде волнистой тонкой черной
полосы, местами прерванной; внешняя часть наружного поля с черными
штрихами по жилкам; бахрома шашечная—черная и бело-зеленая;
задние крылья тусклобелые с темными, черноватыми жилками и бело-
желтоватым внутренним краем; бахрома белая с черными шашечками;
нижняя сторона белая с охристым внутренним краем задних крыльев
и желтым дискоидальным полем на передних; жилки черные, черные
штрихи у костального края передних и задних крыльев и черный верх-
ний угол передних. Размах 40—53 мм.
Uncus почти прямой, слабо заостряющийся, при основании несколько
утолщен; penis с длинной слепой частью, почти прямой, дистальная
часть его слабо изогнута; pars inflabilis несет небольшое число тонких
шипов (значительно меньше, чем у восточной — champa Moore); valva
сильно выгнута в средней части вентрально, длинная, несколько кони-
ческая, но не заостренная; дорзальный край ее несет негустые волоски;
не сильные, но многочисленные щетинки расположены на дистальном
конце; harpe в виде двух склеротизованных складок, из которых более
широкая лежит на вентральном крае valva; отросток harpe острый,
сильно искривленный, недлинный. Bursa copulatrix мешковидная, не-
сколько усложненной формы в дистальной части, с плоским склероти-
зованным расширением к ductus seminalis; последний тонкий; восьмой
сегмент короткий и широкий, в дистальной части несколько рассеченный
вдоль; apophyses anteriores и posteriores короткие; papillae anales мем-
бранозные, несколько заостренные.
Взрослая гусеница желто-коричневая с светло желтыми полосками;
вся кожа покрыта мелкими желтыми волосками; первый и второй груд-
ные сегменты с дорзальной стороны затемнены черно-коричневым и несут
большие желтые бородавки, покрытые короткими курчавыми желтыми
и белыми волосками; среднегрудь с дорзальной стороны также несет
симметричные площадки с охристыми штрихами; вдоль спины по всей
длине тела три узких частично прерванных желтых полоски; по сторо-
нам тела желтая волнистая боковая полоска на уровне стигм, и над ней
на каждом сегменте по четыре косых светлых штриха; брюшная сторона
. однотонно серая с примесью коричневых или желтых штрихов; бородавки
дорзальной стороны тела мелкие, черные, несут черные волоски; на боках
28s ,
436
II. acronictinae
бородавки желтые, с вентральной стороны тела их нет совсем;- грудные
ноги черные, брюшные желто-серые; восьмой брюшной сегмент вздут
Рис. 221. Мота ludifica L. Гениталии: а —
самца, б — самки.
и несет на дорзальной сто-
роне небольшой черный ко-
нический вырост—слившие-
ся дорзальные бородавки;
голова черно-бурая с жел-
товатой фронтальной частью;
окраска и тип грудных сег-
ментов несколько напомина-
ют таковые Diphthera coeno-
bita Esp.
Куколка широкая и пло-
ская (по форме напоминает
куколки Dasychira Steph.),
светлокоричневая; кожа
гладкая, блестящая; за стиг-
мами и вокруг них скульп-
тура в виде мелких коничес-
ких кутикулярных бугорков;
стигмы выдаются; грудной
отдел несет скульптуру в
виде полосок, шагренировки;
cremaster в виде плоского и
короткого выроста, черно-
коричневый, покрыт про-
дольными бороздками; ди-
стальный конец его усажен
мелкими короткими крючко-
видными волосками.
Рис. 222. Мота ludifica L. Хетотаксия
передне- и среднегруди и брюшных сегментов
I, VI, VII и VIII взрослой гусеницы.
Распространение. Средняя Европа — Германская низмен-
.ность, Альпы (Бавария, Швейцария, Французские Альпы), Карпаты,
Висла, северная Италия; до-
лина Дуная (Венгрия); Шве-
ция (Готланд, Даларне), Фин-
ляндия (Таушо); северо-запад
Европейской части СССР (Кау-
нас, Рига, Таллин, Нарва);
восточнее распространен в лес-
ной полосе пятнами (Ленин-
град, Новгород, Петрозаводск,
Москва, Молотов); есть указа-
ние на нахождение этого вида
в центральной Сибири (Ени-
сейск); последнее указание
может касаться и следующего
вида (что вероятнее); проверка
по материалам невозможна, так
как была найдена одна особь,
нахождение которой в настоящее время не известно.
Биология и экология. Зимует куколка; лёт бабочек
в июне—начале июля; гусеницы растут в течение всего лета и осени,
связаны преимущественно с Rosaceae (Sorbus aucuparia, Crataegus охуа-
26. MOM A
437
cantha, Pirus malus, Rosa canina); есть указание на питание Salicaceae
(Populus nigra, Salix caprea); чаще на рябине и яблоне. Окукление
в сухих листьях (в подстилке) в плотном коконе из шелковых желто-
коричневых нитей. Редкий и локально распространенный вид, более
обычный в средней Европе.
*2. Мота champa Moore (рис. 223, 224).
Moore, 1879, Proc. Zool. Soc. Lond. : 403, t. 33, fig. 2; Hampson, 1894, Moths
Ind., 2 : 435; Alpheraky, 1897, Mem. Lep. Rom., 9 : 158, t. 13, fig. 4; Staudinger, 1901,
Gat. Lep., 1 : 130; Warren, 1909, Grossschmet., 3 : 10, t. 2, a: Warren, 1913, Seitz,
Indo-Austral., 11 : 40, t. 5, h, i; Hampson, 1913, Gat. Lep. PhaL, 13 : 378, t. 234,
fig. 18; — ainu Wileman, 1911, Trans. Entom. Soc. Lond., 10 : 242, t. 30, fig. 18; —
diffusa Warren, 1912, Seitz,Grossschmett., Indo-AustraL, 11 : 40, t. 5.
Голова и грудь розово-белые с черным рисунком; основной членик
усика белый, жгутик черный; щупальца с дорзальной стороны черные
с вентральной белые; грудь белая с поперечными черными штрихами;
Рис. 223. Мота champa Moore. Самец. (Рис. Ф. Гуняева).
брюшко желто-белое с дорзальным рядом черных пучков волосков; пе-
редние крылья бело-розовые с резким и грубым черным рисунком; базаль-
ная линия в виде прерванных четыреугольных и ромбических штрихов,
более широких в дорзальной части крыла; вторая поперечная линия
в виде двух черных широко расставленных полос, дает два больших
изгиба и два больших черных штриха в срединном поле вдоль анальной
жилки; круглое пятно большое, овальной формы, расположено продоль-
ной осью вдоль крыла; срединная линия в виде прерванных широких
черных штрихов; почковидное пятно большое, неполно ограничено чер-
ным, вытянутое и изогнутое, открытое в костальный край; третья попе-
речная линия в виде двойной широко расставленной черной полосы,
посредине белой, дающей по жилкам черные штрихи; внешнее поло узкое,
уже срединного, с сильно зубчатой, состоящей из треугольников, черной
подкраевой линии; терминальная линия в виде недлинных конических
черных штрихов, упирающихся вершинами в терминальный край крыла;
бахрома шашечная — белая и черная; задние крылья при основании
желто-белые или охристые, к середине светлые, почти белые, по внеш-
нему краю с широкой черной полосой; бахрома белая с темными штри-
хами; нижняя сторона белая, основание крыльев и их дорзальный край
желтые; по костальному краю широкие черные пятна; передние крылья
затемнены сильнее, и пятна на них почти сливаются. Размах крыльев
50—60 мм.
438
IT. ACRONICTINAE
Valva более короткая и широкая, чем у предыдущего вида, изогнута
дорзально; по дорзальному краю и на дистальном конце большое число
сильных и длинных щетинок; базальная часть valva занята harpe, кото-
рая слагается из двух хитиновых полос — толстой вентральной и узкой
изогнутой дорзальной; при слиянии их короткий вырост, сильно изогну-
тый и направленный дистально; penis несет большое число расположен-
ных в два ряда мелких шипиков на pars inflabilis; дистальная часть
penis утолщена, соеспш более короткий, чем у М. ludifica L. Bursa copu-
latrix мешковидная, мембранозная, без инкрустаций, очень расширенная
Рис. 224. Мота champa Moore. Гениталии: а — сам-
ца, б — самки.
в проксимальной части, имеет сложной формы расширение к ductus semi-
nalis; ductus bursae короткий и конический, восьмой сегмент массивный.
Распространение. Южное Приморье (Сидеми, Владивосток,
Сучан, Уссури); Амур; южный Сахалин; северо-западные Гималаи
(Пенджаб); Северная Индия (Сикким, Ассам); западный Китай; Япон-
ские острова (Хондо).
Исследование уссурийских и гималайских особей показало, что все
они относятся к виду М. champa Moore и что настоящая М. ludifica L.,
видимо, не известна из юго-восточной Азии. Особь, добытая в Енисейске
(выведенная из гусеницы, найденной на рябине), мной не могла быть
проверена, и мне не совсем ясно, относится ли она к М. ludifica L.; более
вероятно, что она принадлежит к М. champa Moore.
27. Род DASEOCHAETA WARR.
Warren, 1909, Seitz, Grossschmett., 3 : 11 (typus viridis Leech); Aurivillius, 1891,
Nordn. Fjar. : 97 (Diphthera}', Staudinger, 1901, Cat. Lep., 1 : 130 (Diphthera)-,
Spuler, 1908, Schmett. Eur., 1 : 135; Hampson, 1909, Cat. Lep. Phal., 8 : 22; Rebel,
1910, Berge, Schmetterlingsb., ed. 9 : 141; Hampson, 1912, Journ. Bomb. Nat. Hist.
21. DASEOCHAETA
439
Soc., 31 : 420; Warren, 1913, Seitz, Grossschmett., Indo-Austr., 11 : 33; Draudt, 1931,
Seitz, Grossschmett., Suppl., 3 : 5; Nordstroem qoo Wahlgren, 1937, Svenska Fjar. :
80; — Diphtherocome Warren, 1909, Seitz, Grossschmett., 3 : 11 (typus pallida Moore).
Тип рода D. viridis Leech.
Хоботок хорошо развит, лоб гладкий, слабо выгнутый; губные
щупальца подняты вертикально вверх, торчат выше уровня темени;
последний их членик заостренный и длинный, более половины длины
второго (почти равен ему); глаза небольшие; первый членик передних
лапок более половины длины голени этих же ног, коготки на всех ногах
простые; грудь и голова покрыты чешуйками и волосками; радиальная
ячея на передних крыльях маленькая, узкая, от ее вершины, отходят
на общем стебельке Р2, Ps, Pi и Ps', Мг отходит от середины ячеи;
на задних крыльях развиты вполне; дискальные жилки на передних
и задних крыльях редуцированы.
Valva с острым выростом, направленным вентрально; дистальный
конец ее сужен; harpe широкая с коротким ложковидным отростком;
penis с инкрустациями,на pars inflabilis. Bursa copulatrix большая, мем-
бранозная, без laminae dentatae.
Гусеницы плоские, уплощенные дорзовентрально, несут длинные
щетинки по сторонам тела, сидящие на бородавках р брюшных сегментов
{рис. 227); прочие группы большею частью однощетинковые, сидящие
на пупиллах; х и р на среднегруди в первой личиночной стадии обосо-
блены, в старших стадиях слиты, как у большинства других родов под-
сем. Acronictinae. Среднегрудь несет бородавку т. Бородавка х брюшных
сегментов прямо за стигмой, на седьмом брюшном сегменте, в первой
•стадии слабо сдвинута вентрально, у гусениц старших стадий слита с щ
Гусеницы первой стадии несут все группы однощетинковые, сидящие
на пупиллах; характерные признаки взрослых приобретаются после
.двух линек, но бывают и после первой линьки. Куколки вальковатые,
с очень плотной и толстой кутикулой; брюшные сегменты у них плотно
соединены друг с другом; кожа гладкая, блестящая, несет небольшое
число очень тонких и коротких волосков, лишь на брюшных сегментах.
Яйца почти шаровидные, зеленовато-желтые, хорион их несет рубчатую
скульптуру, откладываются кучками по несколько десятков.
Олиготипический род; главная масса видов сосредоточена в северо-
западных Гималаях и северной Индии (D. pallida Moore, vigens Wlk.,
brevipennis Hamps., fasciata Moore, chrysochlora Hamps., marmorea Leech,
metaphae Hamps.), из которых некоторые проникают в Корею и Японию;
в Японии есть эндемичный D. viridis Leech (рис. 225). В Палеарктике
широко распространен только один вид (D. alpium Osb.), засоляющий
всю зону лиственных лесов Евразии и частично идущий в зону тайги.
Из тропической Индии эндемичные виды Daseochaeta Warr, не известны,
но туда проникает один гималайский вид (D. muscosa Hamps.). Из юж-
ного полушария известны два вида Daseochaeta Warr.; один (D. verbenata
Dist.) из южной Африки (Трансвааль) и второй (7). beryllodes Turn.)
из северной Австралии (Квинслэнд); оба вида не возбу?кдают сомнения
в принадлежности к роду Daseochaeta Warr.
В Северной и Южной Америке нет представителей рода Daseochaeta
Warr., но эндемичные североамериканские Agriopodes Hamps, близки
к палеарктическим Daseochaeta Warr. Близок к Daseochaeta Warr, также
и эндемичный индомалайский род Thalatha Wlk.1
1 Agriopodes Hampson, 1909, Cat. Lep. Phal., 8 ; 35 (typus fallax H. S.); Thalatha
Walker, 1862, Journ. Linn. Soc., Zool., 6 : 187 (typus sinens Wlk.).
27. DASEOCHAETA
44Г
* 1. Daseochaeta alpium Osb. (рис. 226, 227).
Osbeck, 1778, Gotheb. Samml. Handl. Vetensk. Afd., 1 : 52, t. 1, fig. 2; Aurivil-
lius, 1891, Nordn. Fjar. : 97, t. 33, fig. 2; Staudinger, 1901, Cat. Lep., 1 : 130; Spuler,
1908, Schmett. Eur., 1 : 135, t. 31, fig. 32; Hampson, 1909, Cat. Lep. Phal., 8 : 30,
fig. 7; Warren, 1909, Seitz, Grossschmett., 3 : 11, t. 2, c; Culot, 1909, Noct., 1 : 18,
t. 1, fig. 3; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 141, t. 29, fig. 5a, 6.; Turati,
1923, Atti Soc. Ital. Sci. Nat., 62, t. 13, fig. 7; Petersen, 1924, Lep. Fauna EstL, ed.
2 (1) : 149; Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 5, t. 1, a; Nordstroem.
und Wahlgren, 1937, Svenska Fjar. : 80, t. 15, fig. 2; — ludifica Sepp, 1762, Ins. Ned.,
1 (4) : 41, t. 9, fig. 1—8 (non Linne, 1758); — aprilina Schiffermiiller, 1776, Wien.
Verz. Schmett. : 70 (non Linne, 1758); —orion Esper, 1787, Europ. Schmett., 4 : 118,
fig. 4—-7; — runica Gmelin, 1788, Syst. Nat., 1 (5) : 2561; — murrhina Graeser, 1888,
Berl. entom. Ztschr. : 314; — rosea Tutt, 1891, Brit. Noct., 1 : 12;—glauca Turati,
1911, Ann. Mus. Zool. Univ. Napoli, 3 : 25; — designata Turati, 1919, Atti Soc. Ital.
Sci. Nat., 48 : 45; —fasciata Lenz, 1927, Osthelder, Schmett. Siidbayerris, 1 : 226.
Биология и экология. Meyrik, 1895, Brit. Lep. : 139; Barret, 1895,
Lep. Brit., 3 : 216, t. .119, fig. 3 : Sorhagen, 1917, Stett. entom. Ztg., 78 : 248; Kuskov,
1937, Beitr. Kunde EstL, 13 : 83; Romanizyn, 1930, Prazn. Monogr., 15 : 235; Белозор,
1931, Зап. Никои, on. ст., 2 : 96; Лебедев, 1933, Зб1рн. праць екол., 1 : 76; Cockayne,
1934, Trans. Entom. Soc. Lond., 67 : 170, t. 3, fig. 4; Doring, 1937, Entom. Ztschr.,
Frankfurt, 50 : 547, t. 2, fig. 11.
Голова ri грудь беловато-зеленые с черными пестринами; передние
крылья зеленые с черным рисунком и белыми штрихами; базальная
линия намечена у корня крыла слабым черным штрихом; вдоль дорзаль-
ного края в базальном поле черный штрих; вторая поперечная линия
в виде широкой черной полосы, слабо выгнутой к внешнему краю крыла
посредине; от основания крыла вдоль кубитальной системы растушеван-
ный белый штрих через все базальное и срединное поля; круглое пятно
не развито; в срединном поле вдоль радиальной системы белый штрих;
срединная линия в виде широкой волнистой, растушеванной полосы,
идущей через все крыло, почти прямой; в основании почковидного пятна
скопление черного пигмента, отграниченное от пятна белым штрихом;
почковидное пятно намечено полулунным черным штрихом, окаймлен-
ным белым; срединное поле очень широкое, вдвое шире внешнего; третья
поперечная линия редуцирована, заметна в виде черных и белых штри-
хов по жилкам; подкраевая линия в виде широкой черной полосы через
все крыло, с внутренней стороны оторочена белым, с наружной бело-
серая; терминальная линия в виде серии черных и белых штрихов; бах-
рома зеленая с немногими черными и белыми волосками; задние крылья
однотонные, серые, несколько более светлые у основания, нижний базаль-
ный их угол бело-серый с двумя темными поперечными полосками; бах-
рома серая с полоской, нижняя сторона бело-серая с неправильно раз-
бросанными черными и белыми чешуйками и двумя растушеванными
белыми полосами; бахрома шашечная — черная и белая. Размах крыльев
35—40 мм.
Valva широкая, резко суженная на дистальном конце, который покрыт
грубыми щетинками; harpe широкая, посредине разделенная, несет
короткий направленный наискось дистально вырост и сросшаяся с острым
выростом вентрального канта valva; penis суженный в средней части,
на дистальном конце несет большое число коротких треугольных шипи-
ков на pars inflabilis. Bursa copulatrix большая, мембранозная, без ин-
крустаций, в проксимальной части очень сильно суженная; ductus bursae-
короткий, широкий, сильно изогнутый; papillae anales очень маленькие.
Яйца почти шаровидные, несколько уплощенные снизу; бело-желто-
ватые, прозрачные, не меняющие окраски в течение всего развития;
хорион имеет тонкую рубчатую скульптуру в виде радиальных рубчиков-
442
IT. ACRONICTINAE
Гусеницы первой стадии желто-зеленые, вальцеватой формы, каждый
сегмент резко отделен от другого; волоски, покрывающие тело, желтые,
выделяются темными точками; гусеницы старших стадий коричнево-
серые с мелкими струйчатыми желтыми полосками, покрыты длинными
негустыми шелковистыми коричневыми волосками, сидящими на очень
слабых бородавках; первый, третий и шестой брюшные сегменты на тер-
гитах несут узкие поперечные желто-белые полосы (пятна); у некоторых
особей такие пятна бывают в' большем числе и располагаются почти
на всех брюшных сегментах; голова черная с сильно вдавленным темен-
ным швом и выдающими-
Рис. 226. Daseochaeta alpium Osb. Гениталии:
а — самца, о — самки.
ся полушариями.
Куколка красно-корич-
невая вальковатой формы,
по бокам брюшных сегмен-
тов покрытая очень коро-
тенькими рыжими волос-
ками; кожа брюшных сег-
ментов несет на вентраль-
ной стороне скульптуру в
виде ямок, на дорзальной
стороне она гладкая, но,
по краям сегментов в про-
ксимальной части каждого
сегмента расположён вы-
ступ в виде валика и рядом
желобок; cremaster слабо
развит; дистальный конец
брюшка округлен и уса-
жен шестью короткими
широкими зубцами, на-
правленными в разные
стороны.
^Изменчивость незначи-
тельна; почти в преде-
лах всего ареала от Бри-
танских островов до Амура распространена типичная форма, от которой
наблюдаются лишь более яркие или темные формы. К таким вариантам
относится большинство синонимических названий. На Амуре, по Уссури
и в южном Приморье распространена форма murrhina Graes., характер-
ная сильным развитием охристых тонов и металлическим блеском этих
охристых участков, которые располагаются во внешнем поле передних
крыльев вдоль подкраевой полосы; степень развития охристых тонов
различна у разных особей; задние крылья этих особей несколько темнее;
строение гениталий не отличается от строения типичной формы.
Распространение. Западная и средняя Европа, исключая
сухие степи южного побережья; Пиренеи, Альпы, Карпаты; на севере
Британские острова, южная Швеция, Финляндия; северо-запад Евро-
пейской части СССР, далее к востоку северная граница идет по линии
Петрозаводск—Молотов; на юг распространен всюду до границы лесо-
степи (Киев, Полтава, Куйбышев, Воронеж); известны находки с край-
него юга (Херсон); указан для Кавказа (Боржоми); в Сибири из-
вестны лишь отдельные точки (Минусинск, Омск, Томск, Енисейск);
обычен в южном Приморье (Владивосток, Сучан, Ворошилов-Уссурий-
27. DASEOCHAETA, 28. BELCIADES
443
ский); Приамурье (Хабаровск); Сахалин; Маньчжурия; северный Китай
(Пекин); западный Китай; Корея (Гензен, Пунг-Тунг); Японские острова
(Токио, Иокогама).
Биология и экология. Лёт бабочек в мае—июне, бабочки
питаются и живут 20—30 суток; яйца откладываются на листья кормовых
растений кучками по 30—40 яиц. Выходящие гусеницы держатся гнездом
и грызут лист (скелетируют в
первых стадиях) строем; по-
тревоженные, спускаются на
паутине; старшие стадии (IV—
V) держатся поодиночке; кор-
мовыми растениями являются
виды Fagaceae (Betula, Quercus,
Bagus); возможно также Carpi-
nus и Ostryia, из берез пред-
почитаются Betula pubescens
и В. humilis. В Европейской
части СССР и средней Европе
указан как вредитель дуба (Во-
ронихин, 1928). Окукление в
щелях коры в кокончике из
стадии (а) и взрослой (б).
волосками; губные щупальца изогну-
коричневых нитей, куда впле-
таются частички погрызенного
субстрата; куколки диапаузи-
руют и зимуют; почти всегда
одно поколение; второе бывает
очень слабое, и большая часть
вылетающих особей бесплодна.
Отмечен, паразит Aphanistes
armatus Wesm. (Ichneu monidae).
.28. Род BELCIADES KOZHANTSHI-
KOV, gen. nov.
Тип рода В. niveola Motsch.
Хоботок развит вполне,
лоб гладкий, но выпуклый;
глаза голые, большие, округ-
лые, окружены чешуйками и
ты* и направлены вверх, длинные, с очень длинным и острым послед-
ним члеником, который по длине.более половины второго членика; усики
обоих полов щетинковидные, слабо опушенные (покрыты короткими
ресничками); первый членик лапки передних ног около половины длины
передних голеней; голова и грудь покрыты чешуйками и волосками;
радиальная ячея на передних крыльях узкая; Мг отходит па передних
крыльях от середины ячеи; Р3 на стебельке, Pi и РБ свободно отходят
от вершины радиальной ячеи, почти из одной точки; М„ на задних
крыльях развита вполне и соединена короткой дискальной с Мх\ средняя
часть дискальной жилки утрачена; рисунок представлен парными поло-
сами и обычными пятнами.
Valva узкая, башмаковидной формы; harpe сильная, занимает вен-
тральную часть базальной части valva, несет два отростка; penis несет
инкрустации на pars inflabilis. Bursa copulatrix без laminae dentatae.
444
II. ACRONICTINAE
Монотипический род, известный только из южного Приморья-; един-
ственный представитель рода—В. niveola Mptsch. Этот вид был про-
пущен Гэмпсоном (1904—1913); Уоррен (1909) помещает этот вид в род
Belciana Wlk., в подсем. Noctuinae, и ошибочно в синонимы к staudingeri
Leech.
* 1. Belciades niveola Motsch. (рис. 228, 229).
Motschulsky, 1866, Бюлл. Моск. общ. исп. прир., 39 : 195; Staudinger, 1901,
Cat. Lep., 1 : 130; Warren, 1909, Seitz, Grossschmett., 3 : 368, t. 67, h; — virens-
Butler, 1881, Trans. Entom. Soc. Lond. : 192;— iapyx Staudinger, 1892, Mem. Lep.
Rom., 6 : 402, t. 7, fig. 1.
Грудь с дорзальной стороны серо-зеленая; переднегрудь и голова
более темные; лоб коричнево-серый; передние крылья серо-зеленые с чер-
ным пятном в базальном поле и с коричнево-черным с примесью зеленого
Рис. 228. Belciades niveola Motsch. Самец. (Рис. Ф. Гуняева).
внешним полем, базальная линия светлая, почти белая; между базальной
линией и второй поперечной линией черное пятно от костальной части
до середины базального поля; вторая поперечная линия белая и черная,
слабо волнистая, идущая почти перпендикулярно продольной оси крыла;
срединное поле однотонное, серо-зеленое с белым растушеванным пятном
вместо круглого пятна; на месте почковидного пятна размытое беловатое
пятно, почти четыреугольное; срединная линия ясная, в виде черной,
волнистой полосы, извитая, более широкая у костального края; третья
поперечная линия в виде тонкой черной полоски, идущей на фоне широ-
кой белой полосы; внешнее поле уже срединного, с черно-рыжими штри-
хами в апикальном углу и с немногими белыми пятнышками на косталь-
ном крае; подкраевая линия в виде растушеванной, волнистой зеленой
полосы на темном фоне; терминальная линия в виде коротких черных
штрихов, окаймленных зеленым; бахрома серая с белыми пестринами;
задние крылья тусклосерые с поперечными волнистыми линиями; терми-
нальная линия в виде тонкой полоски; бахрома светлосерая; нижняя,
сторона желто-серая с поперечной волнистой линией и лунными пятнами
на передних и задних крыльях; бахрома серая с белыми штрихами.
Размах крыльев 40—44 мм.
Valva узкая, вытянутая, с нескодько расширенным башмаковидным
дистальным концом, несущим большое количество нежных волосков;
harpe занимает вентральный кант базальной половины valva, она песет
28. BELCIADES, 29. EUROMOIA
445
два отростка — один короткий, направленный дорзально, и другой
длинный, направленный дистально, острый, выходящий за пределы valva;
uncus тонкий; девятое кольцо довольно длинное в дорзальной половине;
penis несет острые, но короткие шипы на pars inflabilis. Bursa copulatrix
вытянутая, ?узкая, мембранозная; ductus seminalis отходит от ее верхнего
полюса; ductus bursae короткий, изогнутый; apophyses длинные.
Распространение. Известен в большом количестве особей
из южного Приморья (Яковлевка, Виноградовка, Сучан, Владивосток,
Аскольд, Русский остров); есть указание для Амура (более не уточ-
ненное).
Рис. 229. Belciades niveola Motsch. Гениталии:
а — самца, б — самки. ' 1
29. Род EUROMOIA STGR.
Staudinger, 1892, Mem. Lep. Rom., 6 : 632 (typus mixta Stgr.); Staudinger, 1901,
Cat. Lep., 1 : 131; Warren, 1909, Seitz, Grossschmett., 3 : 12; —Euromoea Hampson,
1908, Cat. Lep. Phal., 7 : 425.
Тип рода Eu. mixta Stgr.
Хоботок развит вполне, лоб гладкий, слабо выпуклый; глаза боль-
шие, круглые, голые; щупальца длинные, изогнутые, торчат вверх перед
лбом, серповидной формы, третий членик их короткий, второй, напротив,
длинный и изогнутый; усики у обоих полов простые, покрытые короткими
ресничками, образующими бархатистую поверхность усика; первый
членик передних лапок более половины длины голени этих же ног;
коготки на всех ногах с большим зубцом на внутренней стороне; радиаль-
ная ячея на передних крыльях большая, ромбической формы (у одной
особи обнаружено отсутствие этой ячеи); Мг отходит от средней части
446
it. acronictinae
Рис. 230. Euromoia mixta Stgr. Гениталии:
а — самца, б_ — самки.
1
ком; базальное поле серое с охристым
ячеи, Ps и Pt от вершины ячей на длинном стебельке; М2 на задних
крыльях в виде очень слабой жилки; дискальные жилки как на перед-
них, так и на задних крыльях редуцированы; голова и грудь покрыты:
чешуйками и волосками; рисунок на крыльях в виде парных линий,
пятна недоразвиты.
Valva со сложным зубчатым отростком harpe; penis с многочисленными
сложными шипами как на pars inflabilis, так и на corpus penis; ductus
bursae длинный, изогнутый.
Монотипический род, наиболее близкий к группе Belciana — Мота,
но очень своеобразный; систематическое положение запутано; его относят
то к Hadeninae (Гэмпсон,
1908), то к Acronictinae
(Штаудингер, 1901); един-
ственный представитель —
Ей. mixta Stgr. ограничен в
распространении южным
Приморьем.
Окончательное решение
вопроса о систематическом
положении в пределах трибы,
как и для предыдущего рода,
может быть дано при изуче-
нии личиночной фазы, кото-
рая пока совершенно не из-
вестна и не описана.
* 1. Euromoia mixta Stgr..
(рис. 230).
Staudinger, 1892, Mem. Lep.
Rom., 6 : 632, t. 12, fig. 10;.
Hampson, 1908, Cat. Lep. Phal.,.
7 : 425, fig. 71; Warren, 1909,. •
Seitz, Grossschmett., 3 : 12, t. 2, d..
Голова и грудь серые с
примесью охристых тонов;,
передние крылья сёрые с не-
правильными охристыми маз-
ками и черным и белым рисун-
ютальным краем и охристым
мазком вдоль срединной жилки; базальная линия в виде тонкой белой:
полоски; вторая поперечная линия в виде черной полосы, очерченной
охристым/срединное поле очень широкое, особенно растянутое в сред-
ней части, с охристыми штрихами вдоль кубитального и радиального
стволов; круглое пятно не развито; почковидное пятно в виде растуше-
ванного белого пятна почти прямоугольной формы; срединная линия
растушеванная, в виде темной полосы, прерванной у почковидного пятна;
третья поперечная линия начинается над почковидным пятном, под
костальным краем резко выгнута кнаружи и далее идет вертикально
вниз на уровне последней четверти крыла, широко расставленная, двой-
ная, черная, посредине серая и охристая, зубчатая по жилкам, в дор-
зальной трети крыла скошена базально; внешнее поле уже срединного,
с слабо извитой золотисто-охристой подкраевой линией, окаймленной,
по внутренней стороне черными клиновидными штрихами; внешняя
половина наружного поля золотисто-охристая; терминальная линия.
29. EUROMOIA, 30. SUBLEUCONYCTA
.447
в виде охристой полоски, окаймленной темной полосой; бахрома серая;,
задние крылья охристо-желтые с черной каймой, темным дорзальны"
краем и черным основанием; бахрома желтая; терминальная линия в виде
желтой или охристой полоски; нижняя сторона охристо-серая, более
темная на передних крыльях; на передних крыльях просвечивает в ох-
ристо-желтый внешний край желтым почковидное пятно; на задних,
крыльях видна чёрная кайма. Размах крыльев 42—48 мм.
Valva вытянутая, несколько вздутая в дистальной части; harpe выем-
чатая по вентральному краю, несет широкий отросток, усаженный зуб-
цами; uncus тонкий; penis несет многочисленные треугольные зубцы
на pars inflabilis; поверхность penis также усажена зубцами. Bursa copu-
latrix с перехватом, покрыта рубчатой скульптурой; ductus seminalis
отходит от нижнего ее угла; ductus bursae склеротизован.
Распространение. Южное Приморье (Владивосток, Сучаи).
30. Род SUBLEUCONYCTA* KOZHANTSHIKOV, gen. nov.
' Тип рода S. palschkovi Fil.
Хоботок хорошо развит; глаза большие, голые, окруженные чешуй-
ками и короткими волосками; щупальца вздернуты вверх до уровня
темени, торчат перед лбом; последний членик их довольно длинный,
около трети длины второго, заостренный; второй членик также длинный,
изогнутый; лоб гладкий, плоский, первый членик передней лапки около
половины длины голени этой же ноги; коготки с отростками; отросток
коготка широкий, торчит почти прямо под коготком; голова и грудь
покрыты чешуйками и короткими волосками; добавочная ячейка большая,
ромбическая; Р3, Pi и Р- отходят на общем стебельке; отходит от се-
редины ячеи помощью дискальной; М2 на задних крыльях развита
вполне; развиты частично и дискальные. Valva вытянутая, узкая, с гру-
быми щетинками на дистальном конце; с одним дистально направленным
отростком.
С видами американского рода Leuconycta Hamps, имеет лишь габи-
туальное сходство, но даже в рисунке и окраске существуют большие
отличия. У описываемого вида рисунок сильно редуцирован, тогда как
у Leuconycta, Hamps, есть зачаток клиновидного пятна, которого нет
у волнянок, у Acronictinae и в том числе у описываемых представителей;
крылья у Leuconycta Hamps, широкие, покрытые грубой и крупной
чешуей, тогда как у Subleuconycta, gen. nov. чешуйки мелкие и крылья
относительно узкие, почти равные по форме видам рода Acronicta Ochs.
Отлично строение гениталий — у Leuconycta Hamps, нет harpe; отличны
также строение щупалец и форма их члеников.
Как отмечено выше, виды Leuconycta Hamps, очень обособлены и далеки
не только от видов Subleuconycta, gen. nov., но вообще от всех видов
изучаемого семейства.
* 1. Subleuconycta palschkovi Fil. (рис. 231).
Filipjev, 1937, Lambillionea, 37 : 64—66, fig.' 1, 2 (Leuconycta Hamps.).
Голова и грудь бело-серые с примесью немногих черных чешуй; перед-
ние крылья бело-серые с нерезким рисунком; базальная линия в виде
короткого двойного штриха; вторая поперечная линия намечена черным
и белым штрихами на костальном крае, доходит до кубитального ствола;,
базальное поле слито со срединным; круглое, почковидное и клиновидное
448
11. ACRONICTINAE
пятна не развиты; на костальном крае в области срединного поля расту-
шеванная срединная линия, вливающаяся в интенсивно черный четыре-
угольник между пятнами; третья поперечная линия намечена на косталь-
ном крае, далее редуцирована; подкраевая линия в виде тонкой и слабой
белой полоски; внешнее поле несколько затемнено; терминальная линия
в виде слабой полоски; бахрома серо-коричневая; задние крылья тускло-
белые с затемненными жилками; терминальная линия в виде темной,
ровной полоски; бахрома беловато-серая; нижняя сторона светлосерая
с примесью темносерых и черных чешуй, образующих скопление на перед-
них крыльях; на задних крыльях растушеванная поперечная линия
Рис. 231. Subleuconycta palschkovi Fil. (По Филипьеву, 1936).
а — самец (тип), б — гениталии самца.
и темные небольшие лунные пятна; четкая терминальная полоска на ниж-
ней стороне обеих пар крыльев. Размах крыльев 32 мм.
Valva узкая, длинной конической формы, несколько выгнутая дор-'
зально, дистальный конец округлен и несет грубые щетинки; вентраль-
ный край valva утолщен и несет harpe с коротким слабо изогнутым от-
ростком; направленным дистально; uncus утолщен в средней части; penis
вздут на дистальном конце.
Распространение. Южное Приморье (Сучан). Известен
только один самец.
31. Род CANNA WLK.
Саппа Walker, .1865, List. Lep. Brit. Mus., 23 : 790 (typus pulchripicta Wlk.);
Hampson, 1909, Cat. Lep. Phal., 8 : 19; Warren, 1909, Seitz, Grossschmett., 3 : 10;
Hampson, 1912, Journ. Bomb. Nat. Hist. Soc., 21 : 420. '
31. CANNA
449
Тип рода С. pulchripicta Wlk.
Хоботок вполне развит; глаза большие, голые, окруженные чешуей
и короткими волосками; лоб гладкий, невыпуклый'; губные щупальца
недлинные, приподняты вверх и прижаты ко лбу; последний членик их
короткий, примерно в четыре или пять раз короче второго; усики обоих
полов простые, покрытые очень короткими ресничками; первый членик
передней лапки более половины длины передней голени; коготки всех
лапок простые; голова и грудь покрыты чешуей с небольшой примесью
волосков. Добавочная ячея на передних крыльях большая, широкая,
ромбическая; Р2 отходит от ее костального края, Р3 и Р± на коротком
стебельке, Р5 свободна; Мг отходит примерно от половины высоты
ячеи; М., на задних крыльях несколько недоразвита; дистальные жилки
задних крыльев редуцированы, на передних крыльях они частично раз-
виты; рисунок специализован, линии слиты и затушеваны; все известные
виды имеют зеленую окраску.
Олиготипический род, главная масса видов которого концентрируется
в северо-западных Гималаях (С. malachitis Obth., pulchripicta Wlk.,
prasinaria Wlk); один из этих видов проникает в западный Китай, именно
С. malachitis Obth. Из Китая описан один эндемичный, но может быть
не самостоятельный вид — С. smaragdina Drd. Саппа malachitis Obth.
распространен и. далее на север до южного Приморья, откуда известен
также гималайский С. pulchripicta Wlk., пока не найденный в Китае.
Из тропиков (с Явы, повидимому с гор) известен только один вид —
С. ]'avensis Warr., напоминающий С. malachitis Obth., т.е. наиболее широко
распространенный вид рода Саппа Wlk.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Широкая коричневая поперечная линия на передних крыльях
слабо изогнута; затемненный дорзальный край с зеленым пят-
ном у основания крыла; на pars inflabilis длинные хитиновые
зубцы........................................* 1. С. malachitis Obth.
2 (1). Широкая поперечная линия на передних крыльях, посредине
сильно изогнутая кнаружи; дорзальный край переднего крыла
затемнен сплошной коричневой полосой; на pars inflabilis корот-
кие и широкие зубцы..........................* 2. С. pulchripicta Wlk.
*1. Саппа malachitis Obth. (рис. 232).
Oberthiir, 1881, Et. Entom., 5 : 80, t. 3, fig. 9; Hampson, 1894, Moths Ind., 2 : 294;
Staudinger, 1901, Cat. Lep., 1 : 130; Hampson, 1909, Cat. Lep. Phal., 8 : 20; Warren,
1909, Seitz, Grossschmett., 3 : 10, t. 2, b; —splendens Moore, 1888, Proc. Zool. Soc.
Lond. : 412 (non Hampson).
Голова и грудь желто-зеленого цвета, tegulae и заднегрудь коричнево-
желтые; передние крылья изумрудно-зеленые с золотисто-коричневым
и желтым рисунком; обычные линии рисунка редуцированы или слиты
(зеленая окраска головы, груди и крыльев со временем меняется в жел-
тую); у основания крыла широкая золотисто-коричневая полоса, снаружи
отороченная черным и белым, она занимает весь базальный отдел перед-
него крыла; дистально от нее светложелтое или зеленое пятно, слитое
с зеленым полем крыла; дорзальный край крыла за зеленым пятном
до половины затемнен, в виде широкой коричневой полосы, отороченной
(с костального края) черным и белым и в дистальной части слитой с широ-
29 Фауна СССР, И. В. Кожанчиков.
450
IT. ACRONICTINAE
черной терминальной полоской; бахрома
Рис. 232. Саппа malachitis Obth. Гениталии: а —
самца, б — самки.
кой коричнево-охристой поперечной полосой, идущей через середину
крыла; поперечная полоса слабо выгнута кнаружи посредине; она отте-
нена по сторонам белой и черной линиями; за этой полосой на зеленом
фоне остаток почковидного пятна; подкраевая линия в виде двух вытя-
нутых белых пятен в верхней и нижней частях крыла вдоль терминаль-
ного края, разделенных зеленым; на фоне белого внешнего поля два
черных полулуния у терминальной линии, с белыми точками внутри;
терминальная линия в виде тонкой штриховой черной полоски; бахрома
белая и зеленая с двумя черными пятнами; задние крылья белые с тем-
ным внешним краем, несколько суженные в базальной части, с тонкой
белая, слабо затемненная
в костальной половине
крыла; нижняя сторона
белая с темно бурыми,
почти черными пятнами во
внешней части крыльев;
заметна слабая терминаль-
ная полоска; ноги белые,
на дистальных концах
охристо-коричневые. Раз-
мах крыльев 32—35 мм.
Valva узкая и длинная,
почти с параллельными
краями, на дистальном ее
конце небольшая щеточка
из грубых щетинок; harpe-
не развита; penis вздут в
базальной части, изогнут
и сужен в дистальной,.
несет на pars inflabilis
немногочисленные длин-
ные шипы. Bursa copulat-
rix тонкая, мембранозная,
мешковидной формы, без
инкрустаций; ductus bur-
sae короткий, посредине
вздутый.
Близок к С. pulchripicta Wlk., но особенно к китайской С. smaragdina.
Drd. и зондской С. javensis Warr.
Распространение. Южное Приморье (Сучан, Владивосток,
Аскольд, Ворошилов-Уссурийский, Сидеми); северо-западные Гималаи:
(Пенджаб);, северная Индия (Сикким); восточный Китай.
* 2. Саппа pulchripicta Wlk. (рис. 233).
Walker, 1865, List. Lep. Brit. Mus., 23 : 790; Moore, 1861, Proc. Zool. Soc.
Lond. : 61, t. 6, fig. 10; Hampson, 1894, Moths Ind.,- 2 : 293; он же, 1909, Cat. Lep..
PhaL, 8 : 21; Warren, 1912, Seitz, Grossschmett., Indo-Austr., 11 : 33, t. 5, a.
Голова и переднегрудь изумрудно-зеленые; tegulae и заднегрудь,
охристо-коричневые; передние крылья изумрудно-зеленые с неправиль-
ным коричнево-охристым рисунком; как и у предыдущего вида, зеленый
цвет не стоек, окраска желтеет; основание переднего крыла золотисто-
коричневое; это пятно переходит далее в широкую полосу вдоль дор-
зального края крыла, тонко отороченную белым; от базальной трети
31. GANNA, 32. THYATIRIDES
451
крыла через его плоскость широкая поперечная золотисто-коричневая
косая полоса, окаймленная по обеим сторонам тонкой белой полоской; она
сильно изогнута (под прямым углом) в середине крыла и у дорзального
края крыла сливается с коричневой полосой, идущей вдоль дорзального
его края; от угла поперечной линии к апексу крыла косой интенсивно
зеленый растушеванный штрих; подкраевая линия в виде тонкой полоски
у самого терминального края крыла, оторочена белым и коричневым;
терминальная линия в виде тонкой полоски у самого терминального
края крыла, оторочена белым и коричневым; терминальная линия в виде
немногих черных штрихов по краю крыла; бахрома охристо-коричневая;
задние крылья чисто белые, слабо затемненные по внешнему краю; бах-
рома на них охристо-коричневая, нижняя сторона белая, слабо-охристо-
бурая по костальному краю; бах-
рома охристо-бурая; ноги также
охристо-бурые. Размах крыльев
32—36 мм.
Valva неширокая, длинная, на
конце заостренная, с параллель-
ными краями, на дистальном крае
с небольшой щеточкой щетинок;
harpe не развита; uncus большой,
крючковидный, почти голый; pe-
nis вздут в базальной части, ко-
нический, слабо изогнутый, с тон-
кими изогнутыми зубцами на pars
inflabilis.
Вид, хорошо отличимый от С.
malachitis Obth. как по генита-
Рис. 233. Саппа pulchripicta Wlk. Гени-
талии самца.
лиям, так и по рисунку, степени изгиба поперечной полосы и по отсут-
ствию зеленого пятна на дорзальном крае переднего крыла.
Распространение. Амур’ (без более точного указания —
коллекция Зоологического института Академии Наук СССР). Северная
Индия (Сикким).
32. Род THYATIRIDES KOZHANTSHIKOV, gen. nov.
Тип рода Th. trimaculata Brem.1
Хоботок развит, хотя менее длинен, чем у Саппа Wlk.; глаза боль-
шие, окруженные короткими волосками и чешуйками; губные щупальца
приподняты вверх, торчат перед лбом, второй членик их длинный, тре-
тий, напротив, очень короткий, в 6—7 раз короче второго; лоб гладкий
и плоский, первый членик лапки передней ноги около половины длины
передней голени; коготки с широким отростком на внутренней стороне;
усики у обоих полов простые, покрытые короткими ресничками; доба-
вочная ячея узкая, ромбическая; первый медиальный ,ствол отходит
от верхней ее трети помощью дискальной; второй медиальный ствол
на задних крыльях развит вполне; частично развиты также и дискаль-
ные. Рисунок сложный, в виде пятен, линии редуцированы и изменены;
1 Thyatirides trimaculata Brem. габитуально, по рисунку, обнаруживает значи-
тельное сходство с описанной из Африки Dasychira plbtzi Her., особенно с ее формой
cymatophorina Her. (Hering, 1926, Seitz, Fauna Africana, 14 : 186, 27, с). Последняя,
впрочем, едва ли принадлежит к роду Dasychira Steph., являясь как и ряд других
африканских представителей этого рода, очень своеобразной формой.
29*
452
II. ACRONICTINAE
valvae простые, мембранозные, несколько напоминают по форме valvae
видов Саппа Wlk., но несут небольшую harpe с одним отростком; гени-
талии самок также обнаруживают некоторое сходство с видами Саппа Wlk.
Своеобразный представитель Momini, ближайший к роду Саппа Wlk.
Слабо изучен, известна только имагинальная фаза, довольно редок.
Среди Orgyinae есть представители с таким типом рисунка в африканской
фауне. В связи с габитуальным сходством долго смешивался с видами
Thyatira Hb. (Cymatophoridae). Различия в жилковании этого вида
от Cymatophoridae впервые отмечены Н. Я. Кузнецовым.
* 1. Thyatirides trimaculata Brem. (рис. 234).
Bremer, 1861, Изв. Акад. Наук СПб.: 573, t. 5, fig. 5; Bremer, 1864, Зап. Акад.
Наук СПб., 8 : 47 (Thyatira); Staudinger, 1901, Cat. Lep., 1 : 259 (Thyatira)' War-
ren, 1912, Seitz, Grossschmett.,
2 : 324, t. 49, e (Thyatira)', Dalla-
Torre, 1921, Cat. Lep., Cymatopho-
ridae : 10 (Thyatira); — albomaculata
Leech, 1890, Entom.,
chinensis Leech, 1890,
23 ; 113; — tripunctata
1927, Horae Macrolep., 1 : 82, t. 10,
fig. 12.
Рис. 234. Thyatirides trimaculata Brem. Гени-
талии: а — самца, б — самки.
23 : 114; —
там же,
Bang-Haas,
Волова темнокоричневая с
черными волосками на темени;
грудь желтокоричневая с жел-
товатыми и розоватыми тонами;
брюшко бело-желтое с примесью
коричневых чешуй; передние
крылья коричневые с немно-
гими неправильными разбро-
санными желтыми чешуйками
и с тремя большими желто-
коричневыми с белым ободком
пятнами; рисунок редуцирован;
в базальной части крыла боль-
шое желто-коричневое с мелки-
ми коричневыми полосками
пятно, окруженное широким
белым ободком; в терминаль-
ной части крыла два подобных
пятна, из которых одно боль-
ших размеров у апекса, того же цвета, но оторочено белым, в форме полу-
луния; терминальный край крыла с тонкой белой полоской; остаток
терминальной линии в виде черных точек по краю крыла; бахрома в сред-
ней части крыла коричневая, в нижней и верхней частях белая; задние
крылья желто-серые с четкой темносерой полосой и широким темным
ободком по внешнему краю; бахрома белая; нижняя сторона крыльев
бледносерая с примесью бурых чешуй, более обильных на передних
крыльях, и растушеванной темной поперечной полосой. Размах крыльев
28—32 мм.
Valva в базальной половине узкая, далее расширенная в виде лопасти,
посредине имеет складку, сливающуюся с небольшой harpe, несущей
недлинный изогнутый и расширяющийся отросток; тергит девятого
сегмента сильно вздут, особенно в вентральной части; uncus большой;
32. THYATIRIDES
453
penis слабо изогнут, конический. Bursa copulatrix мембранозная, вздутая
в дистальной части, с перетяжкой посредине; ductus bursae короткий,
постепенно переходящий в тело сумки.
Слабо изменчивый вид; приведенные в синонимах названия касаются
мелких аберраций; темные особи из Китая описаны как tripunctata В. Н.
Распространение. Южное Приморье (Сучан, Владивосток,
Виноградовка, Яковлевка, Седанка); северный Китай (Пекин, Ганьсу);
Корея. .
2. Триба acronictini
Виды средних размеров, более узкокрылые, чем виды предыдущей
трибы; некоторые представители имеют острые ланцетовидные крылья 1
(Sirnyra Ochs.). Окраска обычно тусклая; яркие — зеленые, золотистые
и др. — окраски, характерные для многих родов предыдущей трибы
не типичны; рисунок слагается не только из поперечных линий, но также
из хорошо развитых пятен — круглого и почковидного, у некоторых
. видов, впрочем, недоразвитых; клиновидное пятно в рисунке и у этих
| t видов не развито; усики у обоих полов почти всегда простые (несколько
! более длинные реснички у самцов), лишь у видов рода Sirnyra Ochs.
; у самцов усики гребенчатые; М2 на задних крыльях обычно слабая,
[ иногда утраченная; голени всех ног всегда без шипов; глаза голые. Tegu-
men всегда вполне целостный, без поперечного шва; uncus заостренный
тонкий, при основании его на tegumen всегда или хорошо развитые,
или рудиментарные латеральные выросты; valvae большие, лишь в ред-
ких случаях простые, но тогда с андрокониями, чаще несут хорошо
развитую harpe, обычно с несколькими, часто разно направленными,
отростками; дистальный конец valva простой, без cucculus.
В пределах трибы у степных и полупустынных представителей наблю-
дается тенденция к рудиментации ротового аппарата, часто рисунка
и отчасти крыльев; этим характерны биологически крайне специализо-
ванные представители, имеющие имагинальную фазу кратковременной.
Виды трибы Acronictini распространены почти исключительно в Гол-
арктике. Наибольшее разнообразие наблюдается в Палеарктике, где
есть ряд эндемичных родов; два эндемичные рода есть в Неарктике.
За пределы Голарктики выходит один вид Craniophora Snell, (jasciata
Moore), известный из Индии (Зап. Гаты), северного Цейлона и с Новой
j Гвинеи, и немногие виды рода Acronicta Ochs., найденные в небольшом
числе в горах центральной Америки, на островах Малайского архипе-
' лага и в горах северной Австралии.
В Палеарктике распространены следующие эндемичные роды: Sub-
। acronicta Kozh., известный почти со всей территории Палеарктики и от-
I части Неарктики, но дающий наибольшее разнообразие видов в горах
Средней Азии; Harhpsonia Kozh., ограниченный в. распространении
только востоком Азии — южным Приморьем, Кореей, Маньчжурией
и Японскими островами; Craniophora Snell., распространенный и особенно
богатый видами на этой же территории, виды которого распространены
также в Китае, Гималаях, а два вида известны из Западной Европы;
Eogena Gn., монотипичный, распространенный в восточной Европе;
Sirnyra Ochs., особенно богато представленный на юго-востоке Европы,
но распространенный также в Средней Азии, Монголии, Сибири и Цен-
тральной Азии, и Leiometopon Stgr., эндемичный для полупустынь Сред-
в
Рис. 235. Распространение видов трибы Acronictini.
Черным обозначены территории с числом видов 40—50; сеткой — с числом видов 20—30; вертикальной штрихов,
кой — с числом видов 10—15; горизонтальной штриховкой — с числам видов 1—5.
ТАБЛИЦА РОДОВ
466
лей Азии. Роды эти дифференцированы очень полно, особенно степные
и полустепные (три последних).
Лишь два рода этой трибы распространены голарктически: моноти-
пичный Arsilonche Ld., выпадающий, однако, в центральной Сибири,
и обширный полиморфный род Acronicta Ochs., дающий наибольшее раз-
нообразие видов на юго-востоке Азии и на востоке Северной Америки. По
кормовым растениям и по стациальнрму распределению оба рода дают
однообразную картину как в Евразии, так и в Америке. В Северной Аме-
рике есть лишь два эндемичных рода этой трибы: Merolonche Gn. и Наг-
risemna Grt. — своеобразные олигоморфные роды, которые более всего
могут быть сближаемы с Acronicta Ochs, и являются, пожалуй, аналогом
Simyra Ochs, в Америке; они, однако, менее изменены и отдалены
от Acronicta Ochs. (рис. 235).
Виды трибы Acronictini экологически очень ясно распадаются на две
группы — дендрофильных Acronicta Ochs., Subacronicta Kozh., Hamp-
sonia Kozh., Craniophora Snell., живущих главным образом за счет Sali-
cales, Fagales и Rosales, и особенно обильных в местах сохранения тре-
тичной флоры, и степных родов, связанных с травянистыми растениями —
Arsilonche Ld., Oxycesta Hb., Eogena Gn., Simyra Ochs, и, вероятно,
Leiometopon Stgr., живущих преимущественно за счет Euphorbiaceae,
Plumbaginaceae, Scrophulariaceae, Glumiflorae, Cyperaceae и отчасти
Gompositae. Эти роды концентрируются в юго-восточной Европе и Сред-
ней Азии.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (8). Хоботок развит вполне, способен закручиваться, сильный; рису-
нок на передних крыльях развит хорошо, в виде линий и пятен;
от основания крыла часто черный штрих.
2 (3). Valva мембранозная, лишена harpe; при основании несет густую
кисть длинных волосков, сидящих в специальном кармановидном
углублении valva; bursa copulatrix закручена спирально...........
......................................36. Craniophora Snell.
3 (2). Valva несет harpe, которая имеет один или несколько отростков;
bursa copulatrix иной формы.
4 (5). Valva очень узкая, в дистальной части несет сильные щетинки;
harpe имеет один тонкий простой формы отросток, направленный
дистально; коготки на всех лапках с очень коротким зубцом на вну-
тренней стороне..........................35. Hampsonia, gen. nov.
5 (4). Valva большею частью широкая, нередко со сложной несущей
несколько отростков harpe; коготки на лапках с сильным зубцом
на внутренней стороне.
6 (7). Дистальный конец valva сильно склеротизован, несет крючко-
образный вырост, загнутый вентрально, заострен; отросток harpe
короткий..............................33. Subacronicta, gen. nov.
7 (6). Дистальный конец valva широкий, если острый, то без отростка
и не склеротизован, обычно несет волоски; harpe с многими отрост-
ками ........................................34. Acronicta Ochs.
8 (1). Хоботок редуцирован, виден только его рудимент; рисунок или
отсутствует совершенно, и крылья имеют лишь темные штрихи
по жилкам, или заметен в виде рудиментов и только у видов Oxy-
cesta Hb. и Leiometopon Stgr. развит.
456
II. acronictinae
9 (14). Рисунок редуцирован; на передних крыльях продольные штрихи
вдоль жилок, или крылья однотонные.
10 (11). Одноцветный, коричнево-красный; тело покрыто пушистыми
волосками; усики самцов простые; harpe с двумя отростками . . .
..............................................39. Eogena Gn.
11 (10). Крылья бельге или желтоватые, реже черно-серые, но не кир-
пичного цвета; усики самцов гребенчатые, или harpe с одним
отростком.
12 (13). Усики самцов гребенчатые; harpe с двумя отростками; penis
не несет зубцов на pars inflabilis.................40. Simyra Ochs.
13 (12). Усики самцов простые; harpe несет один длинный отросток,
выходящий далеко за пределы valva; penis несет сложную систему
зубцов на pars inflabilis; bursa copulatrix имеет на поверхности
laminae dentatae.................................37. Arsilonche Ld.
14 (9). Рисунок на передних крыльях развит в виде поперечных полос
и пятен или только полос.
15 (16). Рисунок мелкий, в виде многих поперечных полос; penis без зуб-
цов на pars inflabilis, с одним отростком; короткокрылые виды.
................................................ . 38. Oxycesta Hb.
16 (15). Рисунок в виде немногих поперечных полос; penis с зубцами
на pars inflabilis; harpe с тремя отростками; bursa copulatrix мем-
бранозная, мешковидная, маленькая .... 41. Leiometopon Stgr.
33. Род SUBACRONICTA KOZHANTSHIKOV, gen. nov.
Тип рода A. megacephala Schiff.
Хоботок хорошо развит, закручен спирально; лоб сильно вздут,
имеет округлый широкий выступ; щупальца прямые, тонкие, в базальной
части покрыты длинными волосками, второй и третий членики покрыты
гладкими чешуйками и волосками; последний членик щупиков очень
мал; голова и грудь покрыты узкими чешуйками и волосками; глаза
голые, большие; добавочная ячея на передних крыльях узкая, М± отхо-
дит от ее середины; Рз+ь на стебельке, Р5 отходит от анастомоза Р3+4;
и Ms связаны очень маленькой дискальной жилкой или лишены ее
и выходят из одной точки; Mz на задних крыльях редуцирована. Valva
несет хорошо развитую harpe в дистальной части; дистальный конец
valva сильно склеротизован и загнут крючковидно, острый; базальная
часть valva мембранозная и соединена узкой сильно склеротизованной
складкой с harpe. На передних крыльях между почковидным пятном
и третьей поперечной линией светлое пятно, как у Craniophora Snell.
Хетотаксия гусениц рода Subacronicta весьма интересна (рис. 236).
Дорзальная половина тела, исключая первый грудной и восьмой брюш-
ной сегменты, несет первичные щетинки; на восьмом брюшном сегменте
группа р несет пучок волосков и есть ряд вторичных щетинок. На передне-
груди вторичные щетинки есть уже со второй личиночной стадии.
Ниже стигм у взрослых гусениц бородавки х, tq и у несут пучки волосков.
Важно положение бородавки х; на первых шести брюшных сегментах
и на восьмом брюшном сегменте она сильно сдвинута дорзально и сбли-
жена, почтд сливается с р; будучи превращенной в бородавку х почти
стушевывает р, несущую всего одну длинную щетинку. На переднегруди
бородавка х сильно увеличена и выпукла, несет пучок сильных щетинок.
Положение ее на брюшных сегментах, исключая седьмой, где она сдви-
нута вентрально, очень напоминает некоторых Orgyinae (Leucoma Steph.,
33. SUBACRONICTA
457
Ocneria Hb.); напоминают их также уплощенная форма тела гусеницы
и тип бородавки /. переднегруди. Эти личиночные признаки очень инте-
ресны с филогенетической стороны.
Олигоморфный род, близкий к Acronicta Ochs. Обнаруживает прими-
тивные черты как«в структурах имаго, так и личиночной фазы (наличие
дорзальных бородавок только с первичными щетинками). Широко рас-
пространенным видом в Евразии является только один (S. megacephala
Schiff.); в горах Средней Азии
(A. grumi Alph. и S. centralis
Ersch.). Последний виц идет и
в предгорья (Таласский Ала-
тау); он наиболее близок к
5. megacephala Schiff. Из Китая
(юг провинции Шань-си) опи-
сан A. concerpta Drd., также
очень близкий к S. megacephala
Schiff, (может быть лишь форма
•этого вида). В Северной Амери-
ке известен вполне точно лишь
один представитель этого рода
(5. superans Gn.), распростра-
ненный в восточной Канаде
(Оттава), в восточных штатах
(Нью-Йорк) и в Скалистых Го-
рах (Вашингтон, Британская
Колумбия); структурно этот
вид наиболее близок к S.
grumi Alph.
По Смису (Smith, 1899), S.
superans Gn. относится к Acro-
nicta Ochs, группы lobelia, т. е.
к подроду Triaena по моей
классификации. Тем не менее,
просмотр структур гениталий
(Тянь-шань) распространены два вида
Рис. 236. Subacronicta megacephala Schiff. Хе-
тотаксия передне- и среднегруди и брюшных
сегментов I, VI, VII и VIII взрослой
гусеницы.
этого вида дает полное сход-
ство с видами Subacronicta, gen.
nov.; Дайр (1899), хотя не опи-
сывает подробно хетотаксию
гусениц S. superans Gn., все
же отмечает, что дорзальная сторона тела имеет мало щетинок, т. е.
вероятно только первичные, как то типично для Subacronicta, gen. nov.
Биологически изучен только один вид (A. megacephala Schiff.). Окук-
ление происходит в древесине (как видов Triaena НЬ.); цикл развития
и распределение фаз, как у Acronicta Ochs. Кормовые растения из евр-
азийских видов известны для A. megacephala Schiff., которая свя-
зана только с Salicales; американская S. superans Gn. связана с Betu-
laceae, Rosaceae и Aceraceae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). Дистальный конец valva с сильно склсротизоваиным, острым
отростком, загнутым вентрально; участок между почковидным
пятном и третьей поперечной линией выделяется светлым пятном
на фоне крыла.
458 II. ACRONICTINAE
2 (3). Окраска груди и передних крыльев бледносерая; задние крылья
чисто белые; отросток harpe длинный . . . * 2. S. centralis Ersch.
3 (2). Окраска груди и передних крыльев черно-белая; задние крылья
тусклосерые и беловатые, нередко с темной краевой полосой;
отросток harpe короткий....................* 1. S. megacephala Schiff.
4 (1). Дистальный- конец valva широкий, лопатовидный, без выроста,
сильно склеротизован; участок срединного поля между почковид-
ным пятном и третьей поперечной линией окрашен в один тон
с общим фоном крыла.................................* 3. S. grumi Alph.
* 1. Subacronicta megacephala Schiff, (рис. 236, 237).
Schiffermiiller, 1776, Wien. Verz. : 67; Aurivillius, 1891, Nordn. Fjar. : 99;
Staudinger, 1901, Cat. Lep., 1 : 131; Spuler, 1908, Schmett. Eur., 1 : 136, t. 31, fig. 11;
t. 21, fig. 23; Hampson, 1909, Cat. Lep. PhaL, 8 : 149; Warren, 1909, Seitz, Gross-
schmett., 3 : 15, t. 3, b; Culot, 1909, Noct., 1 : 19, t. 1, fig. 8, 9; Rebel, 1910, Berge,
Schmetterlingsb., ed. 9 : 143, t. 28, fig. 146; Bapg-Haas, 1927, Nov. Macrolep., 2 : 180;
Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 12, t. 1, c; Nordstroem und Wahlgren,
1937, Svenska Fjar. : 82, t. 15, fig. 8; — rosea Tutt, 1891, Brit. Moths, 1 : 16; — ochrea
Tutt, 1891, тамже, 1 : 16; — igdyrensis Teich, 1901, Korresp. Blatt. Nat. Ver. Riga,
44 : 17; — nigra Schawerda, 1902, Entom. Rec., 14 : 103; — pulla Strand, 1903, Arch.
Naturgesch., 25 : 9; — aethiops Krulikovsky, 1906, Soc. Entom., 21 : 50; — schlumbergi
Schultz, 1906, Entom. Ztschr., Guben, 20 : 73, fig. 1—2; — warpachowskii Krulikovsky,
1908, Soc. Entom., 23 : 123; — albidior Wagner, 1923, Ztschr. Oesterr. entom. Ver.,
8 : 21; — ankarensis Hering, 1923, Intern. Entom. Ztschr., 26 : 412; — dungerni Rang-
now, 1935, Entom. Rundsch., 52 : 233, t. 3, fig. 30.
Голова, грудь и передние крылья черно-серые, рисунок черный и
белый, четкий; передние крылья несколько округленной формы; базаль-
ная линия в виде двух черных штрихов, посредине серая, продолжается
ниже костального края в виде серого гптриха; вторая поперечная линия
двойная, широко расставленная, посредине светлосерая, сильно скошен-
ная, слабо волнистая; срединное поле в нижней части затемнено; между
почковидным пятном и третьей поперечной линией светлое пятно; средин-
ная линия растушеванная, нерезкая, заметная на костальном крае,
захватывает почковидное пятно; круглое пятно светлосерое, светлее
общего тона крыла, четко очерчено черным; почковидное пятно большое,
сильно растушеванное с неясными контурами, черное в базальной поло-
вине и светлое с внешней стороны; третья поперечная линия начинается
двумя штрихами на костальном крае, далее идет в виде одной черной
полосы, глубоко и четко зубчатой по жилкам; она очерчена с базальной
стороны белым, в дорзальной части крыла двойная, посредине белая;
внешнее поле однотонно серое со слабой прерывистой белой подкраевой
линией; терминальная линия в виде черных штрихов, заходящих на бах-
рому, последняя серая; задние крылья тусклосерые, коричневатые при
основании, жилки темные, по краю темная полоса; терминальная полоса
в виде тонкой линии; бахрома белая; нижняя сторона белая, в дискои-
дальном поле передних крыльев темносерая; терминальная полоса пре-
рывистая; по середине крыльев неясная разорванная перевязь, и стуше-
ванные дискоидальные пятна на задних крыльях. Размах крыльев
35—42 мм.
Uncus очень тонкий и недлинный, почти голый; tegumen более длин-
ный в дорзальной части; saccus сильно склеротизован, треугольной формы;
fultura inferior в виде щитовидной слабо склеротизованной пластинки;
valvae большие, сильно склеротизованные; базальная половина имеет
широкую складку, дистальный конец которой срастается с harpe; по-
следняя в виде косой складки, несет небольшой конический вырост, на-
33. SUBACRONICTA
459
Рис. 237. Subacronicta megacephalaSchiff.
Гениталии: а —• самца, б — самки.
правленный косо дорзально, не выходящий за плоскость valva; дисталь-
ный конец valva широкий, несет длинный сильно склеротизованный
острый вырост, направленный вентрально; penis почти равен по длине
valva, слабо изогнут; coecum равен одной четверти его длины; pars infla-
bilis несет многочисленные хитиновые шипы. Bursa copulatrix вытянутая
вздутая в проксимальной части, затем сужена и вновь расширена в ди-
стальной, где сильно склеротизована; поверхность ее несет рубчатую
скульптуру; ductus bursae тонкий, несет склеротизованные тяжи, корот-
кий, несколько более % части corpus bursae; восьмой сегмент короткий
и неширокий, несет apophyses anteriores, почти равные по длине ductus
bursae; apophyses posteriores короткие и острые; papillae anales слабо
дифференцированы от прочих частей
девятого сегмента, на дистальном
конце несут густые щетинки.
Яйца плоские, слабо выпуклые
посредине, диаметром около 1 мм,
желто-зеленого цвета, спустя два-три
дня меняют окраску, так как сероз-
ная оболочка приобретает красно-ко-
ричневый цвет, окрашена пятнисто.
Гусеницы габитуально меняются
мало с первой стадии до взрослых;
приобретаются лишь бородавки на
боках тела; окраска кожи коричнево-
бурая; середина сегментов темноко-
рйчневая, межсегментальные участки
светложелтые; дорзальная поверх-
ность покрыта первичными щетинка-
ми, сидящими на пупиллах, окрашен-
ных в желто-охристый цвет; бока и
вентральная сторона тела несут пло-
ские бородавки с пучками волосков
(у взрослых) светложелтой окраски.
Голова светлокоричневая с черным
рисунком; ноги желтые. Перед окуклением кожа темнеет и становится
охристого тона; волоски темнеют и сбрасываются.
Куколка темнокоричневая, покрытая короткими волосками по бокам;
поверхность брюшных сегментов несет скульптуру в виде неглубоких
почти круглых ямок; cremaster в виде толстого короткого выроста, не-
сколько конического, покрытого глубокими ямками и бороздами и несу-
щего большое число толстых щетинок, сидящих поодиночке па дисталь-
ном конце загнутых крючкообразно.
Распространение. Западная и средняя Европа от атланти-
ческого побережья (Пиренеи) до северной Италии; Альпы; Карпаты;
Британские острова; Скандинавия i(Швеция, Норвегия, Лапландия,
Финляндия); северо-запад Европейской части СССР, далее к востоку
распространена на юг от линии Петрозаводск— Великий Устюг всюду
по территории лесной зоны, лесостепи и отчасти степи (Ленинград,
Киров, Молотов, Кострома, Вологда, Москва, Рязань, Псков, Ярославль,
Казань, Тамбов, Ульяновск, Полтава, Могилев, Сарепта, Камышин,
Саратов, Астрахань, Таганрог, Одесса, Херсон, Каменец-Подольск);
Крым (южный берег); Северный Кавказ (Ставрополь); Приуралье
(Чкалов, Уральск); Закавказье (Борккоми, Куссари, Кировабад, Тби-
460
II. ACRONICTINAE
лиси, Лагодехи, Арарат, Идыр); северный Иран; Малая Азия (Анкара);
в Сибири от линии Обь-Енисейский канал — Иркутск (вероятно север-
нее) всюду (Омск, Томск, Минусинск, Барнаул, Канск); Саяны (Сухая
Гора); Алтай; Тянь-шапь; Приамурье (Хабаровск, Николаевск); южное
Приморье (Ворошилов-Уссурийский, Владивосток, Яковлевка).
Биология и экология. Вылет бабочек на севере ареала
в июне, на юге в мае; плодовитость самок 300—500 яиц; яйца отклады-
ваются на нижнюю сторону листьев кормовых растений (осина, тополь)
поодиночке; вначале они почти не видны, так как прозрачны. Кормовые
растения только Salicales, преимущественно виды Populus (nigra, alba,
balsamifera), реже ивы с гладкими листьями (Salix fragilis, S. daphnoi-
des), питание которыми возможно с первой стадии; гусеницы живут
поодиночке, днем обычно на поверхности листа, свернувшись полукру-
гом; известен каннибализм гусениц, особенно старших стадий. Окукле-
ние под корой или в начавшей загнивать древесине; гусеница делает
рыхлый кокон, предварительно выгрызая колыбельку в коре или дре-
весине. Куколка диапаузирует и зимует. В зоне хвойных лесов известно
одно поколение, в лесостепи два.
Отмечена как вредитель тополя на Дону (Зверезомб-Зубовский, 1917),
в Астрахани (Сахаров, 1916) и в Болгарии (Буреш, 1914). Из парази-
тов указаны следующие виды: С oel ichneumon centummaculatus Christ.,
Paniscus cephalot.us Holmgr., Paniscus melanurus Thoms. (Ichneumonidap).
*2. Subacronicta centralis Ersch. (рис. 238).
Erschov, 1874, Fedtschenko Reise : 37, t. 3, fig. 35; Herz, 1900; Ежегодн. Зоол.
муз. Акад. Наук, 5 : 449; Staudinger, 1901, Cat. Lep., 1 : 131; Spuler, 1908, Schmett.
Eur., 1 : 137, t. 31, fig. 11; t. 21, fig. 23 (Raupen); Hampson, 1909, Cat. Lep. Phal.,
8 : 148, t. 126, fig. 28; Warren, 1909, Seitz, Grossschmett., 3 : 15, t. 3, b; Draudt, 1931,
Seitz, Grossschmett., 3 Suppl., 1 : 12, t. 1, h; — persica Strand, 1915, Arch. Naturgesch.,
81, A (11) : 158.
Голова, грудь и передние крылья серо-белые, покрыты грубой чешуей,
блестящие; передние крылья остроугольные с сильно скошенным внеш-
ним краем и узким apex; базальная линия в виде двух неясных растуше-
ванных поперечных линий; базальное поле однотонное; вторая попереч-
ная линия двойная, очень широко расставленная, слабо волнистая,
растушеванная, сильно скошенная кнаружи; круглое пятно большое,
слабо заметное на общем фоне, очерченное серым, сильно сдвинуто к поч-
ковидному пятну, нередко соединено с ним штрихом; почковидное пятно
большое, однотонно серое, очерченное темносерым, узкое, очень глубоко
зубчатое с внешней стороны; срединная линия заметна слабым растуше-
ванным штрихом на костальном крае; третья поперечная линия сильно
зубчатая по жилкам, в виде темной полосы, начинается над почковидным
пятном, затем резко выгнута кнаружи и далее идет вниз, образуя ниже
кубитальной жилки изгиб к основанию крыла; срединное поле за почко-
видным пятном осветлено; внешнее поле однотонно серое со слабо замет-
ной подкраевой линией; терминальная линия в виде узких и не слитых
черных штрихов; задние крылья чисто белые с тонкой буроватой терми-
нальной полоской и белой бахромой; нижняя сторона белая с растуше-
ванной поперечной линией, отдельными неправильно разбросанными
черными чешуйками и тонкой терминальной полоской. Размах крыльев
38—42 мм.
По строению гениталий близок к S. megacephala Schiff.; uncus и saccus
одинаковы; базальная часть ча1ча|образует меньшей величины складку,
33. SUBACRONICTA
461
чем у S. megacephala Schiff.; harpe также более короткая; складка в дор-
зальной части valva выражена только у ее основания; отросток harpe
тонкий, более длинный, чем у S. megacephala Schiff, загнутый внутрь;
дистальный конен valva более широкий и несет менее длинный отросток,
чем у S. megacephala Schiff.; bursa copulatrix очень длинная, мембра-
нозная, лишь слабо расширенная в проксимальной части, сильно сужен-
ная в средней части и значительно расширенная в дистальной части,
где несет небольшой сосковидный отросток, от которого отходит ductus
seminalis; ductus bursae короткий, не более % длины bursa, имеет скле-
ротизованные тяжи; восьмой сегмент короткий, несет короткие apophyses
anteriores, менее половины длины ductus bursae; apophyses posteriores
очень короткие; papillae ana-
les как у предыдущего вида.-
Наиболее близок к 5.
megacephala Schiff., но отли-
чается как структурами ге-
ниталий, так и габитуально;
рисунок очень близок; для
обоих видов типично очень
узкое сильно выгнутое поч-
ковидное пятно. Особи габи-
туально сходные с S. centra-
lis Ersch. есть из Закавказья
(Ордубад, Кировабад, Бор-
жоми), с Северного Кавказа
(Кисловодск) и одна из Та-
ганрога. Исследование гени-
талий показало, что они
являются просто светлыми
S. megacephala Schiff.
Распространение.
Средняя Азия — Фергана
(Пенджикент, Самарканд),
Зеравшанский хребет; Гис-
сарский хребет (Кашка-
дарья, Китаб); Семиречье;
Тянь-шань (Кульджа); северо-западный Иран (Урумнах).
* 3. Subacronicta grumi Alph.
Alpheraky, 1897, Mem. Lep. Rom., 9:3 (Acronicta}; Staudinger, 1901, Cat.
Lep., 1 : 131; Warren, 1909, Seitz, Grossschmett., 3 : 15; Hampson, 1909, CuL. Lop.
Phal., 8 : 149..
Голова и грудь светлосерые, однотонные; передние крылья того же
цвета с черным рисунком; базальная линия в виде двух широко расста-
вленных темных штрихов, разделенных светлосерым; базальное поле
однотонное, серое; вторая поперечная линия широко расставленная,
двойная, посредине светлосерая, идет почти прямо через все крыло,
слабо волнистая; срединное поле широкое, вдвое шире, чем внешнее,
в базальной половине светлосерое, в терминальной темное; срединная
линия в виде широкой темной полосы, идет от костального края через
почковидное пятно и далее выгнута базально; круглое пятно правильной
формы, светлосерое, очерчено черным; почковидное пятно нерезкое,
в середине темное, очерчено темносерым; третья поперечная линия начи-
462
II. ACRONICTINAE
нается над почковидным пятном двумя штрихами, Далее резко выгнута
кнаружи, грубо зубчатая, ниже кубитального ствола выгнута базально;
внешнее поле темное, однотонное, с едва намеченной штрихами подкрае-
вой линией; терминальная линия в виде серии черных точек; бахрома
серая с темными штрихами; задние крылья чисто белые, с очень слабой
терминальной полоской, бахрома чисто белая; нижняя сторона однотон-
ная, светлосерая с несколько затемненной дискоидальной областью
на передних крыльях. Размах крыльев 44 мм.
Известен только один самец этого вида, по которому он был описан
как форма от S. megacephala Schiff. С. Четвериков впервые in lift, указал
на самостоятельность S. grumi Alph.; ближайшее исследование показало,
что это очень своеобразный вид. Гениталии могли быть изучены только
рассмотрением в лупу на объекте; дистальный конец valva сильно скле-
ротизован и построен так же, как и у S. megacephala Schiff., хотя
вырост имеет несколько иную форму; своеобразно также и строение
harpe.
Распространение. Средняя Азия — северный Тянь-шань.
34. Род ACRONICTA OCHS.
Ochsenheimer, 1816, Schmett. Eur., 4 : 62 (typus leporina L.); Aurivillius, 1891,
Nordn. Fjar. : 98; Staudinger, 1901, Cat. Lep., I : 131; Culot, 1909, Noct., 1 : 18; Hamp-
son, 1909, Cat. Lep. PhaL, 8 : 59; Warren, 1909, Setz, Grossschmett., 3 : 13; Rebel,
1910, Berge, Schmetterlingsb., ed. 9 : 142; Draudt, 1931, Seitz, Grossschmett.,‘SuppL,
3 : 7; Nordstroem und Wahlgren, 1937, Svenska Fjar. : £2; — Apratele Hiibner,
1806, Manuscr. Tentamen Lepid. : 1 (typus tritona Hb.); —Hyboma Hiibner, 1820,
Verz. bekannt. Schmett. : 200 (typus strigosa Schiff.); — Triaena Hiibner, 1820, там
же : 200 (typus psi L.); — losheaera Hiibner, 1820, там же : 201 (typus alni L.); —
Pharetra Hiibner, 1820, там же : 202 (typus auricoma Schiff.); — Arctomyscis Hiibner,
1820, там же : 202 (typus aceris L.); —Apatela Stephens, 1829, Ill. Brit. Entom.,
HaustelL, 3 : 34 (typus leporina L.); —Microcoelia Guenee, 1852, Noct., I : 33 (typus
fragilis Gn.); — Megacronycta Grote, 1873, Bull. Buff. Soc. Nat. Sci., 1 : 79 (typus
hastulifera Sm. et Abb.); — Lepitoreuma Grote, 1873, там же : 80 (typus ovata
Grt.);—Eulonche Grote, 1873, Bull. Buff. Soc. Nat. Sci., 1 : 81 (typus oblinita Sm.
et Abb.);’— Plataplecta Butler, 1878, Ann. Mag. Nat. Hist., (5), I : 195 (typus
pruinosa Gn.);—Mastiphanes Grote, 1882, New Check List : 23 (typus edolata
Grt.); — Viminia Chapman, 1890, Entom. Rec., I : 26 (typus rumicis L.); — Cuspidia
Chapman, 1890, там же, I : 26 (typus psi Ъ.у, — Pseudopunda Butler, 1890, Trans.
Entom. Soc. Lond. : 672 (typus bicolor Moore); — Aeronycta. Hampson, 1894, Fauna
Brit. Ind., Moths, 2 : 240 (typus leporina L.); — Tricholonche Grote, 1896,'Mitt.
Hildersheim Mus., 3 : 16 (typus afflicta Grt.); — Philorgyia Grote, 1896, там же, 3 : 17
(typus luteicoma Grt. et Rob.);.— Chamepora Warren, 1909, Seitz, Grossschmett.,
3 : 16 (typus auricoma Schiff.).
Биология. Rangnow, 1921, Ztschr. wiss. InsektenbioL, 16:135; Schulz,
1930, Intern, entom. Ztschr., 24 : 179—185; Кожанчиков, 1941, Зоол. журн., 20
(3) : 382—397; он же, 1941, Природа, (1) : 78—79.
Тип рода A. leporina L.
Хоботок хорошо развит, закручен спирально; щупальца приподняты
косо вверх, гладко покрыты • чешуйками и волосками; последний чле-
ник их маленький, заостренный, глаза голые, большие; лоб гладкий,
плоский или слабо выпуклый; усики обоих полов простые; брюшко
покрыто короткими волосками, и небольшим числом более длинных,
образующих пучки на дорзальной стороне первых брюшных сегментов;
передние крылья широкие, с хорошо развитым рисунком; Мг отходит
от середины добавочной ячейки; Mz и Mz связаны небольшой дискаль-
ной жилкой; Ps и Pi на стебельке, Р5 отходит от одной точки с ними;
М2 на задних крыльях редуцирована. Valva с хорошо развитой harpe,
34. ACRONICTA
463
связанной c basis valvae; задние крылья тусклобелые или серые, редко
желтоваты с или желтые.
Строение личиночных фаз следующее. Гусеницы в первой стадии
очень бедны вторичными щетинками, но всегда их имеют; особенно мало
вторичных щетинок у видов подродов Acronicta Ochs, и Triaena Hb.
в первой стадии. Развитие вторичных щетинок в первой стадии типично
для группы сс. Бородавка /. переднегруди не увеличена и не несет пучка
щетиной. Типичны сближение а и (3 на среднегруди уже в первой личи-
ночной стадии и слияние их в комплекс в следующих стадиях. Виды
некоторых подродов, в частности Acronicta Ochs, и Triaena Hb.,
характеризуются обособлением этих групп на среднегруди и в стар-
ших стадиях и даже у взрослых гусениц, например у Acronicta
al гг L.1 Есть случаи слияния а и fj брюшных сегментов, как у неко-
торых видов Orgyidae] это наблюдается, например, у Acronicta leporina
L. Группа д на среднегруди большею частью есть. Группа z брюшных
сегментов прямо за стигмой; на седьмом брюшном сегменте слабо сдви-
нута вентрально, но в ряде случаев даже слита с х- Куколки валько-
ватой формы, кутикула несет скульптуру в виде точек, шипиков или
ямок; волоски на коже сегментов очень короткие и тонкие. Яйца плоские
сплющенные дорзовентрально, имеют тонкий хорион, несущий рубчатую
скульптуру.
Полиморфный род, распространенный почти целиком в Голарктике
(рис. 239); Неарктика более богата видами, чем Палеарктика; в Пале-
арктике главная масса видов сосредоточена в северо-западном Китае
и отчасти в Японии, южном Приморье и северо-западных Гималаях;
лишь немногие виды распространены в бореальной зоне Евразии; в тро-
пики Старого Света тоже проникает очень немного видов (Л. albiorbis
Hamps., gastridia Swinh., nigralbata Warr., pruinosa Gn. и rubiginosa
Wlk.); они известны из Индии, с Цейлона и с Малайского архипелага
(Борнео); твердо установленных эндемичных видов Acronicta Ochs, для
тропиков не известно; все упомянутые виды распространены также в северо-
западных Гималаях. Эндемичные виды Acronicta Ochs, описаны — один
с Новой Гвинеи (Л. crenulata Beth.-Bek.), один из Австралии (A. phaeo-
cosma Turn.), два с Мадагаскара (A. mediovittata Roth, и A. pseudo-
bamra Roth.). Принадлежность этих видов к роду Acronicta Ochs, по
достоверна.
Главная масса видов Acronicta Ochs. Северной Америки концентри-
руется в восточной части материка (восточные штаты, Великие озера),
меньшее число видов распространено в бореальной зоне Северной Аме-
рики и в Скалистых Горах; в тропическую Америку проникает всего
два вида (A. velia Schaus и A. theodora Schaus—г Мексике, Гватемала).
Из Южной Америки виды Acronicta Ochs, не известны.
Биологически виды Acronicta Ochs, являются в общем целостной груп-
пой. Сезонное распределение фаз развития во всех известных случаях оди-
наково; зимует всегда куколка, причем обычно диапаузируот. Большин-
ство видов моно вол ьтинны, причем у некоторых (напримор у A. aceris L.)
частично наблюдается даже двухлетний цикл с двойной зимовкой ку-
колки; есть незначительное число видов. поливольтинвых, обычно
1 Самостоятельность групп а и 3 на среднегруди для Acronictinae является первич-
ным и исходным положением; этот исходный тип хототаксии является идентичным
с тем, что обнаруживают и Orgyinae. У Acronictinae признак этот сохраняется лишь-
у групп, несущих архаичные черты, или в первой стадии развития (всегда).
Рис. 239. Распространение видов рода Acronicta Ochs. Черным обозначены территории обилия - видов.
34. ACRONICTA
465
полифагов, распространенных не только в лесах (A. rumicis L., euphor-
bias Schiff.). Основными кормовыми растениями являются Salicaceae,
Betulaceae, Fagaceae, Rosaceae, Sapindaceae; отдельные виды связаны
с Euphorbiaceae, Ericaceae. По типу окукления наблюдается две группы
видов Acronicta Ochs.; виды подродов Acronicta Ochs, и Triaena Hb. для
окукления вгрызаются в древесину (обычно начинающую загнивать или
отмирающую), где делают колыбельку, слабо- используя шелкоотделе-
ние для постройки кокона; виды Pharetra НЬ. окукляются в плетеных
из шелка коконах; окукление их происходит в подстилке.1 2
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДЮДОВ
1 (6). На передних крыльях четкие черные стреловидные штрихи,
в числе трех (один у основания крыла, один у апекса и один у тор-
нуса); если штрихи слабо развиты, то рисунок редуцирован; окра-
ска белая или светлосерая, или передние крылья по дорзальному
краю интенсивно черные; задние крылья белые или серые, ио не
желтые и не желтоватые; harpe несет один, два или три отростка.
2 (3). Harpe с одним отростком, обычно тонким; на pars inflabilis два
ряда длинных шипов; рисунок на передних крыльях часто редуци-
рован.......................................... 1. AcronictaOchs.
3 (2). Harpe с двумя или тремя отростками; если с одним, то valva
заострена; рисунок обычно развит хорошо.
4 (5). Harpe большею частью с тремя отростками; если harpe имоот два
отростка (один редуцирован), то они направлены дорзалыю;
крупные и средних размеров виды, 35—50 мм.......................
........................................... 2. Triaena 11Ь.
5 (4). Harpe с двумя отростками, направленными косо дистально, иногда
загнутыми крючкообразно; мелкие или очень крупные виды,
30—35 мм и 50—65 мм...............................3. Hyboma Hb.
6 (.1). Черные штрихи на передних крыльях развиты слабо (обычно
резок один штрих при основании крыла); задние крылья белые
или желтовато-серые; harpe несет два отростка, расходящиеся
вилкообразно и направленные дистально; если отростки harpe
в числе трех (нижний раздвоенный), то задние крылья желтые . .
........................................... 4. Pharetra НЬ.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ПО ВЗРОСЛЫМ ГУСЕНИЦАМ^
1 (4). Волоски сидят густыми пучками непосредственно па коже; вся
кожа покрыта мелкими нежными волосками; бородавки или пу-
пиллы не развиты совершенно.
2 (3). Тергиты всех сегментов тела несут небольшие бело-розовые
почти круглые пятна, очерченные черным; тело покрыто большими
пучками белых и розовых волосков.................* 2. A. aceris L.
1 Не все известные в литературе виды Acron ida Ochs, могут быть определены
до подрода; в ряде случаев описания и изображения их недостаточны. I? числу видов,
которые я не мог отнести с уверенностью ни к одному из принимаемых мною подро-
дов, относятся lilacina Hamps., luteola Hamps., exempta Dyar, [ameola Dyar, polyporia
Dyar, jobasis Dyar, vanda Buch.
2 Определение подродов см. выше в таблице для родов Orgyidae. Но этой таблице
возможно определение гусениц последних трех стадий, т. е. тех, которые наиболее
заметны в природе.
30 Фауна СССР, И. В. Кожанчиков.
4 66
11. acronictinae
3
4
5
6
7
8
9
10
11
12
13
14
15
16
(2). Тергиты всех сегментов тела без рисунка, одного цвета с прочими
частями сегментов, без пятен; кожа покрыта шелковистыми нежно-
зелеными или желтоватыми волосками; редко на тергитах брюшка
непарные темносерые, почти черные кисточки (типичны для полу-
взрослых гусениц).............................* 5. A. leporina L.
(1). Волоски сидят на более или менее развитых бородавках; если
бородавки слабо развиты, то на первом брюшном сегменте есть
мясистый вырост, или тергиты несут мясистые желто-белые пло-
щадки. , •
(6). Тергиты всех сегментов тела утолщены, бело-желтого цвета,
резко выделяются на фоне черной кожи тела; по сторонам тергита
сидит по одному длинному расширенному лопаточкой волоску.
Молодые (до четвертой стадии) гусеницы имеют белую и черную
окраску, и тергиты их лишены мясистых площадок; волоски и
в предпоследних стадиях (III и V) расширены . . .*7. A. alni L.
(5). Тергиты всех сегментов тела одного цвета с прочими частями
сегментов, во всяком случае без мясистых площадок; прочие приз-
наки иные.
(12). Первый брюшной сегмент несет мясистый вырост; если этот
вырост не велик, то он несет длинный пучок волосков; бородавки,
покрывающие кожу, небольшие, плоские, правильные ряды боро-
давок отсутствуют.
(9). Вырост первого брюшного сегмента длинный, мясистый, покрыт
множеством коротких шипиков; длина этого выроста равна ширине
тергита -этого сегмента тела; вдоль спины широкая желтая полоса,
по сторонам которой (под нею) на каждом сегменте по два вытяну-
тых красных пятна................................* 12, A- psi L.
(8). Вырост первого брюшного сегмента короче, менее ширины этого
сегмента; если этот вырост очень короток, то он несет длинный
пучок волосков.
(11) . Вырост первого брюшного сегмента довольно большой,, без
длинного пучка волосков; вдоль спины яркая оранжево-желтая
полоса, по сторонам которой белые и оранжевые пятна.............
...................................... * 11. A. tridens Schiff.
(10). Вырост первого брюшного сегмента очень короткий, несет
длинный несколько изогнутый назад пучок щетинистых черных
волосков; вдоль спины желтая или зеленоватая широкая полоса
без пятен по сторонам.........................* 10. A. cuspis Hb.
(7). Первый сегмент тела не несет непарного дорзального выроста;
кожа покрыта правильными рядами бородавок; дорзальные боро-
давки (а также боковые) часто шиповатые.
(14). Кожа черная, покрытая круглыми черными бородавками,
которые несут пучки черных, длинных волосков; стигмы белые,
резко выделяются на черном фоне кожи; под стигмами проходит
яркая красная полоса.................* 24. A. menyanthidis View.
(13). Иной окраски; если черная, то со светлыми бородавками и без
красной боковой линии.
(16) . Бородавки светлее общего цвета кожи — светложелтые или
рыжие; бородавки круглые и крупные; кожа рыжая или светло-
коричневая; волоски, покрывающие бородавки, рыжие или черные
................................... * 25. A. auricoma Schiff.
(15). Бородавки одного цвета с прочими частями кожи, обычно они
несколько сплющены; вдоль спины заметная продольная полоса.
34. ACRONICTA
467
17 (20). Тергиты брюшных сегментов несут косые короткие белые
полосы; спинная полоса узкая, желтая.
18 (19). Кожа покрыта длинными и тонкими охристо-каштановыми
волосками; ниже дыхалец широкая ярко-белая и оранжевая
полоса; голова сильно округлая, темнокоричневая ...................
..................................* 30. A. lutea Brem. et Grey.
19 (18). Волоски, покрывающие кожу, недлинные, жесткие, рыжие или
охристо-коричневые; ниже дыхалец рыжая и белая неяркая
полоса..........................................* 31. A. rumicis L.
20 (17). Вдоль спины широкая черная полоса и по краям ее две белых
полосы; под стигмами прерванная ровная оранжево-красная
полоса......................-.............*26. A. euphorbiae Schiff.
1. Подрод Acronicta Ochs.
Рисунок развит вполне, есть черные продольные штрихи; он может
быть ослаблен или редуцирован; valva широкая, мембранозная в ди-
стальной половине; harpe узкая, в виде полосы, несет один длинный
отросток, направленный косо и вверх. Совокупительная сумка сильно
вытянута в длину; ductus seminalis отходит от. небольшого расширения;
ductus bursae короткий.
Виды этого подрода распространены в бореальной зоне Голарктики;
лишь один вид (A. leporina L.) свойствен Палеарктике и Неарктике,
остальные эндемичны для этих территорий. Число видов в Неарктико-
болыпе.
Из Палеарктики известны — A. novella Drd., aceris L., leporina L.,
eiaeagni Alph., gastridia Swinh., major Brem. {maxima Moore), senica Ev.
и omorii Mats. Для Неарктики описаны — A. rubricoma Grt., americana
Harris, hastulifera Sm. et Abb., hesperida Sm., dactylina Grt., jelina Grt.,
jrigida Sm., pacijica Sm., insita Wlk., cretata Sm., leporina L., populi Ril.,
lepusculina Gn., Cinderella Sm., transversa Sm., tota Grt., canadensis Sm.,
et Dyar, cyanescens Hamps., chinochroa Hamps., eldora Sm., similana Sm..
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). Рисунок на передних крыльях черный, редуцирован, есть лишь
обрывки линий и пятен; окраска белая или тусклосерая.
2 (3). Передние крылья чисто белые или бело-серые; линии рисунка
в виде обрывков; на месте почковидного пятна черная точка . . ..
........................................ * 5. A. leporina L.
3 (2). Передние крылья грязно-серые; остатки второй и третьей попе-
речных линий слиты в виде буквы X, расположенной в лежачем
положении........................................*6. A. senica Ev.
4 (1). Линии рисунка на передних крыльях развиты вполне или лишь,
несколько ослаблены, образуют резкие стрельчатые фигуры; три
черных штриха на передних крыльях развиты вполне.
5 (6). Крупный, около 50—65 мм; окраска тусклосерая; линии рисунка
частично редуцированы; задние крылья серые........................
......................................* 1. A. major Brem..
6 (5).' Мельче, 40—45 мм; окраска бело-серая или желто-сорая.
7 (8). Окраска бело-серая, без примеси желтых тонов; valva расширен-
ная в дистальной половине....................... . * 2. A. aceris L.
30*
468
II. ACRONICTINAE
8 (7). Окраска бело-желтоватая или серая с примесью охристых тонов;
valva слабо расширенная в дистальной половине.
9 (10). Harpe, кроме длинного косо и вверх направленного отростка,
несет небольшой второй отросток при основании первого; valva
в дистальной половине коническая; желтая окраска имеет диф-
фузное распределение..........................* 3. A. elaeagni Alph.
10 (9). Harpe несет только один длинный отросток; valva в дистальной
половине не сужена ; на передних крыльях мелкие охристые штрихи
.......................................* 4. A. consanguis Btl.
* 1. Acronicta (s. str.) major Brem. (рис. 240)
Bremer, 1868, Lep. Ost. Sib. : 48, t. 5, fig. 7; Oberthiir, 1880, Et. Entom., 5 : 69,
t. 7, fig. 5; Butler, 1881, Trans. Entom. Soc. Lond. : 119; Staudinger, 1892, Mem. Lep.
Rom., 6 : 384; Staudinger, 1901, Cat. Lep,, 1 : 131; Herz, 1904, Ежегодн. Зоол. муз.
Акад. Наук, 9 : 269; Hampson, 1909, Cat. Lep. PhaL, 8 : 117; Warren, 1909, Seitz,
Grossschmett., 3 : 15, t. 3, b; Куренцов, 1922, Зап. Южно-Уссур. отд. Географ, общ.,
1 : 27; Matsumura, 1927, Ins. Matsum., 1 : НО; Филипьев, 1927, Ежегодн. Зоол. муз.
Акад. Наук, 28:230; Draeseke, 1928, Iris, 42:297; — maxima Moore, 1881, Proc.
Zool. Soc. Lond. : 333; — anaedina Butler, 1881, Trans. Entom. Soc. Lond. : 19; —
defigurata Warren, 1909, Seitz, Grossschmett., 3 : 15; — anaedinella Strand, 1915, Arch.
Naturgesch., 85, A, 11 : 152.
Передние крылья острые, сильно скошенные по внешнему краю,
особенно у самцов; окраска их пепельно-серая с примесью небольшого
количества охристых тонов; от основания крыла вдоль срединной жилки
длинный черный штрих, дающий четыре ветвления; вторая поперечная
линия в виде двух широко расставленных черных штрихов на косталь-
ном крае и в средней части крыла у конца черного базального штриха,
нерезкая; клиновидное пятно не развито; круглое пятно небольшое,
правильной формы или слегка овальное, не четко очерчено черным,
в середине одного цвета с общим фоном крыла; срединная линия наме-
чена лишь на костальном крае темносерым штрихом; почковидное пятно
темное, почти треугольной формы, слабо выемчатое по внешнему краю,
очерчено черным, нередко растушеванное; третья поперечная линия
намечена на костальном крае двумя темными штрихами, далее резко
и далеко выгнута кнаружи, простая, мелкозубчатая по жилкам; внешнее
поле однотонное, несколько более темное, чем срединное; подкраевая
линия не выражена; в области торнуса тонкий черный штрих вдоль
анальной жилки, в базальной части отмеченный белым штрихом; терми-
нальная линия в виде серии черных точек; бахрома серая; задние крылья
тусклочерные, затемненные по внешнему краю, жилки темные; бахрома
светлосерая; нижняя сторона крыльев светлосерая, особенно по внеш-
нему краю, задние крылья с нижней стороны светлее; на нижней стороне
передних крыльев тусклое пятно; грудь сверху однотонная пепельно-
серая; брюшко светлосерое. Размах крыльев 50—65 мм.
Uncus большой, несет короткие и густые волоски, сильно изогнут
и вздут в средней части, острый; tegumen короткий, несколько более
длинный в боковых частях, образует при основании uncus, по сторонам,
небольшие выросты; anus несет длинный scaphium; saccus маленький,
округлый; valvae большие, почти прямые, несколько расширенные
посредине; в основании valva короткая, склеротизованная лопасть; harpe
в виде дву^х косых складок, несущих в месте срастания длинный отросток
и при основании его очень небольшой бугорок; дистальная половина
valva мембранозная, густо волосистая; penis короче valva, толстый,
с коротким coecum; pars inflabilis несет многочисленные длинные шипы
34. ACRONICTA
469
anteriores; apophyses postenores рудимен-
Рис. 240. Acronicta major Brom. Гои Италии:
a — самца, б — самки.
в средней части penis и группу более мелких в его дистальной части;
fultura inferior в виде щитовидной пластинки, заострена в вентральной
половине. Bursa copulatrix очень большая, вытянутой формы, в середине
суженная, более вздутая в дистальной части, чем в проксимальной,
мембранозная; ductus seminalis отходит от короткого сосковидного
расширения рядом с основанием ductus bursae с дорзальной стороны;
ductus bursae почти прямой, широкий при основании, по длине почти
равен трети corpus bursae; восьмой сегмент короткий и узкий, несет
короткие и тонкие apophyses
тарны; papillae anales мем-
бранозные с венчиком густых
щетинок на дистальных кон-
цах.
Распро стр анение.
Амур; южное Приморье (Во-
рошило в-Уссурийский, Сиде-
ми, Аскольд); Японские
острова (Хоккайдо, Хондо);
Корея; западный Китай;
восточные Гималаи (Ассам);
северо-западные Гималаи
(Пенджаб); Индия (Бирма).
Личиночные стадии не
изучены; отмечен как важ-
ный вредитель шелковицы
(Morus alba, М. nigra) в
Японии.
*2. Acronicta (s. str.)
aceris L. (рис. 241, 242).
Linne, 1758, Syst. Nat.,
ed. 10 : 514; Aurivillius, 1891,
Nordn. Fjar. : 99, t. 32, fig. 9;
Staudinger, 1901, Cat. Lep., 1:131;
Hampson, 1909, Cat. Lep. PhaL, 8 :
116 ;Warren, 1909, Seitz, Grossschmett., 3 :13, t. 2, g; Culot, 1909,Noct., 1 : 19, 1.1, fig. I 7 ;
Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 143, t. 28, fig. 13; Bang-Haas, 1926, Nov.
Macrolep.,, 1 : 159; Bang-Haas, 1927, там же, 2 : 179; Turner, 1927, Brit. Noct. : 51.; Dra-
udt, 1931, Seitz, i rossschmett., 3 SuppL, 1 : 11, t. 1, g; Gronblom, 1936, Acta Soc.
Fauna et Flora Fennica, 58 : 13; Nordstroem und Wahlgren, 1937, Svepska I' jiir. : 82, t.
15, fig. 7; — candelisequa Esper, 1796, Schmett. Eur., 4.: 191, fig. 1;.— iii/uscaln.
Haworth, 1809, Lep. Brit. : 177; — paradoxa Boisduval, 1829, Ind. Moth. : 60; - inter
media Tutt, 1891, Brit. Noct., 1 : 14; —judaea Staudinger, 1901, Cat. Lop., 1 : 131;
asignata Hirschke, 1910, Verh. zool.-bot. Ges. Wien, 60 : 413; — elineata Dufrene,
1925, Rev. Mus. Namur, 25 : 32; — calceata Dannehl, 1939, Mitt. Munch, eutom.
Ges., 19 : 104 — anthracina Burrau, 1950, Opusc. Entom., 15 (1) : 82.
Биология и экология. Meyrik, 1895, Brit. Lop. : 142; Barret, 1895,.
Lep. Brit., 3 : 231, t. 120, fig. 2; Bergner, 1913, Korrespondenzbl., Naturforsch. Ver.
H.iga, 56:23; Bandermann, 1917, Soc. Entom., 22:40; Israel, 1920, Mitt. Dout.
Dendrl. Ges. : 201; Reh, 1925, Sorauer, Handb. Pflanzonkrankh., 4 : 418; Engol, 1927,
Ztschr. Morph. Oekol. Tiere, 9 : 233; Eidmann, 1929, Ztschr. angew. Entom., 15 : 44;
Suite, 1929, Rev. Zool. Agric., 28 : 97; Oudemans, 1930, Entom. Boric,ht., 84 : 119;
Филипьев, 1932, Тр. защ. раст., 5 : 372; Warnecke, 1936, Entom. Jahrb., 45 : 106;
Doting, 1937, Entom. Ztschr., 50 : 547, t. 2, fig. 94.
Светлосерый с примесью черных и, роже, желтоватых чешуй; рису-
нок темный; от основания переднего крыла черный штрих с тремя вет-
влениями; первая поперечная линия в виде двух широко расставленных
170
II. ACRONICTINAE
Рис. 241. Acronicta. aceris L. Генита-
лии: а — самца, б — самки.
штрихов на костальном крае и ниже черного штриха в виде скобки;
вторая поперечная линия также двойная, идет косо; круглое пятно
овальной формы, вытянутое вдоль крыла, серое, оторочено черным;
срединная линия намечена темносерым, штрихом, идущим от костальною
края к почковидному пятну, слита с ним и далее идет слабая, волнистая
до дорзального края; почковидное пятно большое, широкое, оторочено
черным, внутри темнобурое, с темными штрихами у базального края;
третья поперечная линия двойная, широкая, посредине светлая, базаль-
ная половина этой линии стушевана, хорошо развита лишь в костальной
половине крыла, внешняя половина четкая, неправильно зубчатая по
жилкам; во внешнем поле вдоль кубитальной жилки черный штрих; вол-
нистая линия стушевана; терминаль-
ная линия в виде узких черных штри-
хов, захватывающих бахрому; под-
краевая линия не развита; бахрома
серая с полоской; задние крылья
тусклосерые с белой бахромой и темны-
ми жилками; нижняя сторона светлая
с неправильным темным рисунком и
. немногими неправильно разбросанными
темными чешуями; грудь светлосерая
с немногими черными чешуями; голова
того же цвета; губные щупальца не-
длинные, второй членик их черный,
третий короткий и белый. Размах
крыльев 40—45 мм.
Uncus толстый, несколько утолщен
в дистальной части и заострен, в сред-
ней части выгнут серповидно; tegumen
недлинный, образует небольшие лопасти
по сторонам у основания uncus; saccus
очень маленький, острый в прок-
симальной части, с гребнем на
дорзальной стороне; fultura inferior
щитовидной формы; valvae большие,
широкие, почти прямые, расширен-
ные дистально; базальная часть valva несет небольшую склероти-
зованную лопасть, в дистальной части срастающуюся с harpe;
последняя в виде двух косых в дистальной части срастающихся складок,
на месте срастания несущих длинный извитой косо направленный отро-
сток; дистальная часть valva мембранозная, несет мелкие волоски; penis
по длине равен valva, слабо расширен в дистальной части; coecum корот-
кий; pars inflabilis несет многочисленные длинные шипы и склеротизо-
ванные складки. Bursa copulatrix длинная, суженная в середине, с не-
сколько заостренным polus bursae, покрыта рубчатой скульптурой,
дистальная часть bursa склеротизована; ductus seminalis отходит от ди-
стальной части bursa (in toto) от небольшого сосковидного отростка;
ductus bursae довольно длинный, широкий, особенно в основании, склад-
чатый; восьмой сегмент короткий, несет короткие (не более половины
длины ductus bursae) apophyses anteriores; apophyses posteriores несколько
более длинные, тонкие, papillae anales мембранозные, короткие и широкие.
Яйца плоские с вентральной стороны и слабо выпуклые с дорзальной;
хорион несет скульптуру в виде ребрышек; сразу после откладки
34. ACRONICTA
471
окраска желто-белая; по мере развития сквозь хорион просвечивают
желтоватые линии, которые, сливаясь, оставляют лишь неправильные
окраска
они ста-
легко те- .
покровы.
Рис. 242. Acronicta aceris L. Хототаксия пе-
редне- и среднегруди и брюшных сегментов
I, VI, VII и VIII взрослой гусеницы.
светлые пятна.
Гусеницы старших стадий светложелтые, покрытые длинными жел-
тыми волосками, сидящими прямо на коже, пучками; на задних сегмен-
тах тела есть небольшие бородавочки, но и здесь волоски сидят преиму-
щественно на коже; первый, третий, четвертый и пятый сегменты
с дорзальной стороны несут пучки красных волосков, смешанных с жел-
тыми; примесь красных волосков варьирует, и иногда они отсутствуют;
грудные ноги черные; голова
темнокоричневая с желтыми ад-
фронтальными полями и черным
наличником; дыхальца черные,
очень заметные; на дорзальной
стороне тела на каждом сегмен-,
те по одному желтому, очер-
ченному черным, пятну; на
первом и втором сегментах гру-
ди эти пятна узкие, связанные
черной полосой и пронизанные
желтым штрихом, на последую-
щих сегментах пятна большие,
замкнутые, яркожелтые; с пре-
кращением питания
волосков меняется,
новятся черными и
ряются; темнеют и
Куколка светлокоричневая;
брюшные сегменты несут очень
редкие и короткие волоски; ба-
зальная половина брюшных
сегментов несет скульптуру в
виде плоских ямок; интерсег-
ментальная мембрана несет
скульптуру в виде тонких по-
лосок, образующих род ша тре-
нировки; cremaster в виде ко-
роткого плоского выроста,
поверхность его сильно морщи-
нистая, дистальный конец несет
два крючковидных выроста с дорзальной стороны и два латеральных
пучка крючьев (по 6 в каждом) с вентральной.
Распространение. Западная Европа — Пиренеи; средняя
Европа — Альпы, Германская низменность, Карпаты, Швеция, (Даларие,
Эланд, Готланд); южная Норвегия; Финляндия (Нилапдия); южная Ев-
ропа — Италия (Абруццы), Греция; северная Африка (Марокко); Пале-
стина (Ливан); Малая Азия (Аксехир, Витиния); Турция (Карс, Казико-
поран); в СССР от побережья Черного моря (Севастополь, Одесса, Херсон;
Новороссийск) всюду до северо-запада (Литва, Латвия, Эстония) и ли-
нии Ленинград (Петродворец, Дудергоф, Гатчина), Москва, Казань,
Чкалов, Уральск (указан для Новгорода, Пскова, Могилева, Горок,
.Днепропетровска, Полтавы, Таганрога, Чернигова, Винницы, Харькова,
Куйбышева, Воронежа); северовосточными форпостами ареала являются
4 72
II. ACRONICTINAE
Молотов и Кунгур; Закавказье (Боржоми, Тбилиси, Лагодехи, Куссари,
Ордубад); Талыш (Ленкорань); Туркмения (Айдере); Семиречье (Или,
Иссык-куль); северный Иран; Кашгар. Указание для Амура (Моль-
трехт, 1929) основано на неправильном определении.
Мало изменчив; форма candelisequa Esp. темнее, чем типичная,
с несколько усиленными черными штрихами, значительно реже, чем
типичная; известна из всех частей ареала вида.
Биология и экология. Зимует куколка; вылет бабочек
в мае — июне. Яйца откладываются поодиночке на нижнюю сторону
листьев кормовых растений; выходящие молодые гусеницы тут же скеле-
тируют листья, прикрепляясь паутиной к листу. Кормовые растения
включают только древесные породы — виды Acer (campestre и др.), Aes-
culus, Juglans, виды Quercus и некоторые Rosaceae; типичен для широко-
лиственных лесов; размножаясь, вредит клену и конскому каштану;
окукление в листьях,, на поверхности почвы, в плотном, сверху рыхло
сплетенном, коконе из желтых нитей, куда вплетаются также отпав-
шие с тела гусеницы волоски. Может наблюдаться частичное второе поко-
ление или частичная двухгодичная генерация (двойная зимовка куко-
лок). Специально отмечен как.вредитель клена на Дону и в Югославии:
(Сербии) как вредитель (?) шелковицы; есть указания на повреждения
плодовых деревьев. Из паразитов отмечены Ophion, costatus Rtzb. и
О. luteus L. (Ichneumonidae).
* 3. Acronicta (s. str.) elaeagni Alph. (рис. 243)
Alpheraky, 1889, Mem. Lep. Rom., 5 : 126, t. 6, fig. 2; онже, 1887, Stett. entom.
Ztg. : 167; Staudinger, 1901, Cat. Lep., 1 : 131; Warren, 1909, Seitz, Grossschmett.r
3 : 14, t. 2, i; Hampson, 1910, Cat. Lep. PhaL, 9 : 530.
Передние крылья, грудь и голова светлосерые с примесью бледного
желто-охристого тона, особенно сильного при основании крыльев; чешуй-
чатый покров грубый, многочисленны крупные черные чешуи; базаль-
ная линия лишь намечена двумя короткими штрихами на костальном
крае; от основания крыла интенсивно черный штрих, ветвящийся на три
отростка и кончающийся на уровне круглого пятна; вторая попереч-
ная линия намечена двумя широко расставленными черными штрихами
на костальном крае, сильно скошена (направлена к круглому пятну),
далее идет прерывистая; круглое пятно небольшое, правильной формы,
или овальное, четко очерчено черным, в середине светлосерое, желто-
ватое; почковидное пятно сильно сдвинуто базально, большое, в косталь-
ной половине широкое, несколько стушеванное, по краям и с дорзаль-
ной стороны тонко очерчено черным, в середине серое с немногими
черными штрихами; срединная линия лишь намечена интенсивно черным
штрихом на костальном крае; третья поперечная линия начинается на
костальном крае крыла сразу за почковидным пятном двумя интенсивно
черными штрихами, далее резко и далеко выгнута кнаружи, идет вначале
почти параллельно костальному краю, далее зубчатая по жилкам, местами
четко двойная у второй кубитальной жилки пересечена черным штри-
хом; расстояние от почковидного пятна до середины этой линии почти
вдвое более расстояния от нее до терминального края (у A. aceris эти
отрезки равны); подкраевая линия редуцирована; терминальная линия
в виде немногих точек, заходящих на бахрому; последняя серая с полос-
кой; задние крылья светлосерые с белой бахромой и темными жилками;
нижняя сторона крыльев светлосерая с темными штрихами. • Размах
крыльев 45—55 мм.
34. ACRONICTA
473
Uncus большой, раздут в средней части, с дорзальной стороны по-
крыт густыми короткими волосками, острый; tegumen короткий, несколько
более длинный по сторонам uncus; saccus маленький, треугольный; anus
имеет subscaphium; valvae почти прямые, широкие; базальная часть
valva несет небольшую склеротизованную лопасть по вентральному краю;
harpe в виде широкой складки, несет два отростка, один очень корот-
кий, направленный прямо дорзально, другой длинный, направленный
косо дистально, изогнутый; у базального верхнего угла valva склороти-
зованная складка; дистальная половина valva мембранозная, песет мел-
кие нежные волоски; penis посредине слабо изогнут дорзально, несколько
сужен; соесшп почти не развит; pars inflabilis несет многочисленные
длинные шипы. Bursa copulatrix длинная и узкая, в середине с пере-
тяжкой, несет грубую рубчатую
скульптуру; более широкая ее
дистальная часть склеротизована
и несет небольшой также скле-
ротизованный сосок, от которого
отходит ductus seminalis; ductus
bursae длинный и тонкий, склад-
чатый, по длине равен половине
corpus bursae; восьмой сегмент
довольно длинный, конический,
несет толстые apophyses anteri-
ores; apophyses posteriory более
короткие и острые; papillae anales
плоские и широкие, слабо склеро-
тизованы.
Ра сп ро ст ранение.
Средняя Азия — Таджикистан
(Ширабад); западный Памир (Ка-
лай-вамар, Хорог); Таласский
Ала-тау (Ташкент); Кара-Калпа-
кия (Турт-куль); северный Иран
(Мешхед). Рис. 243. Acronicta elaeagni Alph, Гонита-
Мало известный в литературе лии: а — самца, б~ самки,
вид; мною подобрана серия из раз-
ных мест, причем особенно большое число особей (несколько десятков) из
Калай-вамара (по сборам Лупповой). Сомнений в самостоятельности
elaeagni Alph. как вида нет. Гэмпсон (1909) с вопросом относил ого в сино-
нимы к tridens Schiff. Штаудингер (1901), а за ним Уоррен (1909) объеди-
няли asiatica Pouj. с этим видом, что мало вероятно; этот китайский вид,
по Гэмпсону, является синонимом consanguis Btl., с чем можно согла-
ситься. Описанная Уорреном (1909, Seitz, 3:17, t. 3, i), видимо по
какому-то уродливому экземпляру из Куляба (по одной самке), radiata
Warr., может быть, принадлежит сюда; к сожалению, тип остался мне
не известен и окончательное сведение синонимики сделано быть но может.
Биология. Установлено питание гусениц Elaeagnus hortensis;
кроме того, очень вероятно питание розоцвотными, особошю плодовыми;
сколько-нибудь полного описания личиночных стадий пет.
* 4. Acronicta (s. str.) consanguis Btl. (рис. 244).
Butler, 1879, Ann. Mag. Nat. Hist., 4(5) : 358; Staudinger, 1892, Mdm. Lep. Rom.,
4 : 390; он же, 1901, Cat. Lep., 1 : 133; Hampson, 1909, Cat. Lep. PhaL, 8 : 92, t. 125,
fig. 11; Warren, 1909, Seitz, Grossschmett., 3 ; 14, t. 3, h; Draudt, 1931, Seitz, Gross-
474
II. ACRONICTINAE
schmett., Suppl., 3 : 9;Филипьев, 1932, Труды защ. раст., 5 : 213; — asiatica Poujade,
1888, Bull. Soc. Entom. France, 8 (6) : 20 (sec. Hampson, 1909).1
Передние крылья, голова и грудь желто-серые с разбросанными
черными чешуйками и частично черным рисунком; базальная линия
в виде двух коротких черных штрихов на костальном крае, далее про-
должается в виде искривленного черного штриха, очерченного снаружи
белым; базальное поле желто-серое с неправильно разбросанными чер-
ными чешуйками; вторая поперечная линия в виде двух растушеванных
темных штрихов, широко разделенных желто-серым; круглое пятно
большое, эллиптическое, желто-серое, очерчено черным; почковидное
пятно также большое, светлее общего тона, желто-серое, неполно очерчено
черным, с внешней стороны слабо выемчатое; между пятнами диффузное
черное пятно, косое, слитое с почковидным; третья поперечная линия
начинается на костальном крае над почковидным пятном неясным штри-
хом, далее идет в виде мелко- и частозубчатой линии, несущей в зубцах
-белые штрихи; подкраевая линия в виде нерезкой извитой полосы, жел-
Рис. 244. Acronicta consanguis Btl. Гениталии самца.
той и белой, местами затемнена; терминальная линия состоит из черных
штрихов, заходящих на бахрому; рисунок неправильный, стрельчатый,
неясный; бахрома серо-желтая; задние крылья тусклосерые с белой
бахромой; нижняя сторона однотонно серая с нерезкими темными лун-
ными пятнами. Размах крыльев 42—48 мм.
Uncus большой, в базальной части толстый, несколько вздутый
в середине, в дистальной половине сильно конический; tegumen короткий
и узкий; saccus маленький, заостренный;, valvae большие, в проксималь-
ной части несколько более узкие, чем в дистальной; базальная часть
valva имеет склеротизованную лопасть, слитую с основанием harpe;
последняя в виде широкой складки, лежащей почти поперек valva, несет
изогнутый, несколько утолщенный в дистальной части отросток; дисталь-
ная часть valva (две трети ее длины) мембранозная, несет густые мелкие
волоски; penis несколько короче valva, сильно выгнут в средней части
дорзально, в дистальной части сужен; pars inflabilis несет многочислеи-
1 Известная из Гималаев (Кашмир, Сикким, Пенджаб), из северной Индии
(Ассам) и с Явы A. pruinosa Gn. (Noct., 1 : 53, 1852) очень близка к A. consanguis
Btl. и, очень вероятно, представляет лишь форму этого вида [см.: Hampson, 1909, Cat.
Lep. Phal., 8 : 91, t. 125,. fig. 10 и Warren, 1913, Seitz, >. rosss hmett., 11 (2) : 38,
t. 5, f]. Недавно описанная из Китая A. cavillatrix Draudt (Entom. Rundsch., 54 : 383,
fig. 2d) также, повидимому, представляет форму от A. consanguis Btl.; автор не дает
отличий , от этого вида и сравнивает свой вид лишь с alhistigma Ham ns.
34. ACRONICTA
475
ные мелкие шипики; fultura inferior в виде двух узких склеритов, срос-
шихся при основании.
Распространение. Южное Приморье (Уссури, Сучан,
Сидеми); Хондо (Токио, Иокогама); северо-западный Китай.1
Личиночные стадии и биология не изучены; отмечен как вредитель
конопли в Японии.
5. Acronicta (s. str.) leporina L.2 (рис. 245 , 246).
Linne, 1758, Syst. Nat., ed. 10 : 510; Aurivillius, 1891, Nordn. Fjar. : 99; Smith
and Dyar, 1899, Proc. U. S. Nat. Mus., ,21 : 59, t. 11, fig. 1, 2; Staudinger, 1901, Cat.
Lep., 1 : 131; Spuler, 1908, Schmett. Eur., 1 ; 136, t. 31, fig. 9; Culot, 1909, Noct.,
1 : 18, t. 1, fig. 7; Hampson, 1909, Cat. Lep. PhaL, 8 : 161, fig. 19; Warren, 1909, Seitz,
Grossschmett., 3 : 14, t. 3, a; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 142, t. 28,
fig. 12; Petersen, 1924, Lep. Fauna EstL, ed. 2 : 149; Draudt, 1931, Seitz, Grossschmett.,
3 SuppL, 1 : 13, t. 1, k; Nordstroem och Wahlgren, 1937, Svenska Fjar. : 82, t. 15,
fig. 6; — larose Engramelle, 1788, Ins. Eur. : 27, f. 297; — bradyporina Hiibner, 1818,
Schmett. Eur., Noct., fig. 570,'571; — bimaculosa Maasen, 1871, Stett. entom. Ztg. :
27; —vulpina Grote, 1883, Can. Entom., 15 : 8; — semivirga Tutt, 1888, Entomologist,
21 : 52; — leporella Standinger, 1888, Stett. entom. Ztg. : 245; — cineracea Graeser,
1888, Berl. entom. Ztschr., 32 : 310; — sancta Edwards, 1888, Entom. Amer., 3 : 185; —
rosea Tutt, 1891, Brit. Noct., 1 : 15; — maesta Dyar, 1904, Can. Entom., 34 : 29; —
melanocephala Mansbridge, 1905, Entomologist, 38:289;—• grisea Cochrane, 1906,
Entom. Rec., 18 : 102; — nigra Tutt, 1906 Entom. Rec., 18 : 147; — alba Gillmer,
1906, Insektenborse, 23 : 118; — melaleuca Culot, 1909, Noct., 1 : 19, t. 2, fig. 2; —
leucogaea Stichel, 1917, Ztschr. wiss. InsektenbioL, 13 : 290, t. 1, fig. 7; — f lave scans
Lempke, 1928, Entom. Ber., 9:128;—mesella Rangnow, 1935, Entom. Rundsch.,
52 : 223, t. 3, fig. 29; —grisescens Rangnow, там же, 52 : 223, t. 3, fig. 29, b; —
minor Rangnow, 1935, там же, 52 : 223, t. 3, fig. 29, c.
Светлосерого или чисто белого цвета с черным рисунком на передних
крыльях и груди или без него, однотонный; на передних крыльях неболь-
шой черный штрих на костальном крае, на месте начала второй попе-
речной линии; пятна редуцированы; срединная линия заметна в виде
слабого косого штриха на костальном крае и темного Полулуния у осно-
вания почковидного пятна; третья поперечная линия сильно выгнута
к внешнему краю крыла, заметна в виде слабых тонких черных штрихов
по жилкам, иногда редуцирована; терминальная линия в виде черных
шашечек (восьми), захватывающих часть бахромы; последняя чисто
белая; задние крылья чисто белые со слабо затемненными, тусклыми
жилками; нижняя сторона чисто белая с. темными лунными пятнами
на нижней стороне передних и задних крыльев и с тонкой темной поло-
ской по краю крыльев; в некоторых случаях рисунок развит полнее
и тогда есть полные вторая и третья поперечные линии и видны остатки
1 Описанная из Японии jozana Matsumura (1926, Ins. Mats., 1 : 1, I. 1, fig. 5),
по мнению автора, близкая к A. consanguis Btl., по даваемым фотографиям скорое напо-
минает какую-то форму, близкую к A.- rumicis L. (Draudt, 1931, Soitz, Grossschmett.,-
3 SuppL, 1 : 9, t. 1, d). Дрезеке (1928) указывает этот вид для сонорного Китая
Пекин).
2 В неарктической фауне среди семейства Notodontidae упоминается род
Apatelodes Pack., к которому относят, едва ли правильно объединяя, два вида —
A. torrefacta Abb. et Sm. и angelica (,.rt. Первый вид — Apatelodes torrefacta
Abb. et Sm. имеет гусениц, обнаруживающих большое впешпсо сходство с видами
Acronictinae и в особенности с гусеницами Acronicta leporina L. Габитус бабочек
обнаруживает отдаленное сходство в рисунке задних крыльев с видами Momini.
Жилкование имеет сходство с Notodontidae, куда и отнесен род Apatelodes (Pa-
ckard А., 1895, Mem. Acad. Sci. Wash., 7 : 95, t. 7, fig. 10, t. 9, fig. 1—8). Может
быть, в A. torrefacta Abb. et Sm. мы имеем одно из связывающих звеньев между
еемействами Orgyidae и Notodontidae.
476
it: acronictinae
пятен; рисунок полнее развит у особей с более темной окраской. Раз-
мах крыльев 35—45 мм.
Uncus длинный и толстый, в средней части вздутый, густо усажен-
ный мелкими волосками; tegumen короткий, образующий небольшие
выросты при основании uncus; saccus маленький, заостренный в прокси-
мальной части и изогнутый в дорзальной; valvae большие, толстое
и широкие, прямые; базальная часть valva несет небольшую склероти-
зованную лопасть, в дистальной части срастающуюся с harpe; последняя
несет длинный изогнутый отросток, направленный косо дорзально; при
основании отростка на harpe два небольших конических выроста; две
дистальных трети valva мембранозные, несущие мелкие короткие воло-
ски; penis равен по длине valva, в средней части слабо выгнут дорзально;
coecum короткий; pars inflabilis-
Рис. 245. Acronicta leporina L. Гениталии:
a — самца, б — самки.
несет многочисленные склероти-
зованные складки и в прокси-
мальной части многочисленные-
зубцы. Bursa copulatrix узкая и
длинная, мембранозная, несет
тонкую рубчатую скульптуру; в
дистальной трети corpus bursae
перетяжка; дистальная часть bursa
более, толстая, чем проксималь-
ная ; ductus seminalis отходит от
небольшого сосочка с правой
стороны от дистальной части
bursa; ductus bursae очень корот-
кий, складчатый; восьмой сегмент
короткий, широкий, несет корот-
кие apophyses anteriores; apophy-
ses posteriores также короткие,
равные по длине ductus bursae^
papillae anales мембранозные,
короткие и широкие.
Яйца с вентральной стороны
плоские, с дорзальной слабо вы-
пуклые, желтые; через три-четыре
дня после откладки темнеют в свя-
зи с пигментацией серозной оболочки, которая становится фиолетово-серой..
Гусеницы первых трех стадий светложелтые с темными поясками и немно-
гими черными кисточками на спине (стадии II и III), покрыты торчащими
недлинными волосками. Четвертая и пятая стадии имеют длинные шелко-
вистые волоски светложелтого или желто-зеленого цвета, изогнутые
и направленные в одну сторону; темные кисточки не всегда» остаются
у взрослых гусениц и всегда в малом числе (2—3); особи, растущие при
низкой температуре (порядка 12—15°), имеют зеленовато-черные волоски.
Перед окуклением волоски буреют и выпадают, а кожа принимает охри-
стую окраску. Грлова желто-зеленая, однотонная; ноги того же цвета;
дыхальца маленькие, овальные, желто-зеленые, в середине темнокоричне-
вые, слабо заметные.
Куколка темнокоричневая; кожа брюшных сегментов имеет скульп-
ТУРУ в виде колечек; cremaster булавовидной формы, усажен многочис-
ленными направленными в разные стороны изогнутыми крючьями.
Распространение. В Европе примерно от 66° с. ш., т. е.
34. acronicta
477
от северной Швеции и северной Финляндии на зап,аде и Петрозаводска,
Усть-Сысольска на востоке,— всюду по лесной зоне, лесостепи и отча-
сти степи. Британские острова, вся средняя Европа на запад до Пире-
неев; Альпы, Карпаты; Апеннинский полуостров; Европейская часть
СССР (Прибалтика, Ленинград, Псков, Ярославль, Казань, Москва,
Киров, Молотов, Куйбышев, Могилев, Киев, Полтава, Каменец-Подольск,
Таганрог); Северный Кавказ. Указан для южного берега Крыма (Мели-
•оранский, Тр, Русск. энтом. общ., 31:229), но фактический материал
мне не известен. Указан для Закавказья (Батуми, Боржоми, Кировабад)
и для Талыша (Христоф), где очень редок. В Сибири известны место- .
нахождения от широты Обь-Енисейского канала и Байкала всюду по
лесной и лесостепной зонам (Тюмень, Омск, Енисейск, Тайшет, Бар-
наул, Иркутск, Тобольск, Минусинск, Томск, Хабаровск, Николаевск
н/А.) и южнее—в Семиречье (Иссыккуль, Или), северной Монголии (Улан-
батор) и Тибете (Куку-нор,
Амдо); южное Приморье (Влади-
восток, Сучан, Аскольд, Сидеми);
Сахалин; Хоккайдо; Северная
Америка — Канада (Онтарио); во-
сточные штаты (Мэн, Нью-Хэмп-
шайр, Массачузеттс, северный
Иллинойс).
Слабо изменчивый вид, измен-
чива лишь окраска; по окраске и
рисунку можно выделить две
формы — leporina L. и leporella
Stgr., ограниченные также отча-
сти ареалами; первая характер- Рис. 246. Acronicta leporina L. Хототаксия
на для всего ареала вида, вторая передне- и среднегруди и брюшных сег-
14 А < м ’ 1 ментов I, VI, VII и VIII взрослой
ТОЛЬКО ДЛЯ Сибири; она 0ТЛИ- гусотгиттт.т.
чается беловато-серой окраской
и наличием хорошо заметных полос; структурных отличий от типич-
ной формы не дает. Есть основания рассматривать leporella Stgr.
как холодовую форму. Затем может быть отмечена bradyporina Hb., отлич-
ная от типичной присутствием черных полос рисунка (при сохранении
белой окраски), распространенная в пределах всего ареала вида. В Евр-
азии leporina L. является обособленным видом, не имеющим близких,
тогда как в Северной Америке есть ряд очень схожих не только по имаго,
но по личиночным стадиям {lepusculina Gn., americana Harr., populi Ril.,
rabricoma Gn., dactylina Grt. и jrigida Sm. et Dyar). Напротив, виды
группы aceris L. {major Brem., elaeagni Alph.) в Америке по предста-
влены.
Биология и экология. Зимует куколка; лёт бабочек в мае —
нюне. Яйца откладываются поодиночке на нижнюю поверхность листьев
кормовых растений, почти исключительно березы; яйца обливаются
секретом придаточных желез и очень слабо заметны. Гусеницы живут
поодиночке; первые стадии прикрепляются паутиной к листу лишь в период
линяния, старшие стадии менее подвижны. Кормовыо растопил ограничены
нормально лишь видами Betula (pubescens, ermani, humilis, реже verru-
cosa); из других Betulaceae возможно питание грабом (Carpinus) и ольхой
(Alnus); возможно развитие при питании видами ив (Salix) и даже топо-
лей (Populus); отмечается питание в природе некоторыми Rosaceae (Pirus,
Rosa). Типичным местообитанием являются средневозрастные березняки
4 78
JI. ACRONICTINAE
или березняки (молодые) окраин болот. Обычна также в смешанных ле-
сах. Окукление в загнившей древесине (пни, старые упавшие деревья)
или под корой; гусеница делает рыхлый кокон из зеленовато-белых нитей,
куда вплетает опавшие волоски и частички древесины или коры. На
севере (в зоне хвойных лесов) одно поколение, в лесостепье частичное
второе. Бабочки живут 20—25 суток; самка откладывает 200—400 яиц.
Из паразитов известны Ichneumonidae—Ophiot luteus L., Paniscus mela-
nurus- Thoms.
* 6. Acronicta (s. str.) senica Ev. (рис. 247).
Eversmann, 1856, Билл. Mock.- общ. исп. прир., 2, t. 3, fig. 7; он же, 1857, там
же, 1 : 85; Staudinger, 1901, Cat. Lep., 1 : 131, 133; Warren, 1909, Seitz, Grossschmett.t
3 : 15, t. 3, e, (literata Brem.,) p. 16 (x-signata Stgr.); Hampson, 1910, Cat. Lep. PhaL,'
9 : 530, 531; Draudt, 1931, Seitz, Grossschmett., SuppL, 3, 1 : 13; Kozhantshikov W.,
1936, Folia Zool. HydrobioL, 9 : 29; — literata Bremer, 1864, Lep. Ost-Sib. : 102, t.
8, fig. 14; — x-signata Staudinger, 1897, Iris, 10 : 329, t. 9, fig. 24.1
Основной цвет тела и крыльев пепельно-серый с многими мелкими
белыми и черными чешуйками; рисунок на передних крыльях редуци-
рован, характерны две черные линии, идущие наискось и отчасти вдоль
крыла и вместе дающие иксобразную фигуру (расположенную в лежа-
чем положении); базальная линия не развита; вторая поперечная линия
намечена черными штрихами на костальном крае, далее резко выгнута
кнаружи посредине и затем от середины также сильно выгнута базально
и оканчивается на дорзальном крае на одном уровне с началом ее на
costa; от середины этой линии резкий черный штрих к третьей попереч-
ной линии; круглое и почковидное пятна едва намечены неправильными,
кольчатыми фигурами; третья поперечная линия начинается над почко-
видным пятном черным штрихом на костальном крае, далее смыта и
затем резко выгнута базально, сливаясь с темным штрихом от второй
поперечной линии и дополняя цксообразную фигуру; во внешнем поле
отдельные черные штрихи вдоль жилок, не образующие определенной
линии; терминальная линия в виде длинных тонких радиальных штри-
хов, бахрома серая; задние крылья тусклосерые, более светлые у осно-
вания и затемненные по внешнему краю; бахрома белая; нижняя сторона
светлосерая с разбросанными темными чешуйками; передние крылья
снизу более темные. Размах крыльев 32—35 мм.
Uncus тонкий, длинный, почти голый; tegumen короткий, особенно
короткий в дорзальной части; saccus почти не развит; valvae большие,
широкие, в средней части сильно выгнутые вентрально и несколько
суженные; базальная часть valva у вентрального края несет небольшую-
складку, которая имеет короткий вырост; harpe слита с этой складкой,
небольшая, несет недлинный сильно заостренный отросток, направленный
косо дорзально; дистальная часть valva (две трети) мембранозная, по
канту несет ряд шипиков, направленных орально (cucculus), дистальный
конец valva густо покрыт мелкими шелковистыми волосками; penis по
длине равен valva, тонкий, в средней части выгнут дорзально, с корот-
ким coecum; pars inflabilis несет один длинный и острый шип и большое-
число тонких и длинных меньшей величины. Bursa copulatrix сильно-
1 Описанная Матсумура с японских островов (Хоккайдо, Хоншу) A. omorii
(Matsumura,' 1926, Ins. Mats., 1 : 3, t. 1, fig. 2), по мнению автора, близка к senica Ev.,
но имеет «совсем другой рисунок». По краткому описанию и прилагаемой плохой
фотографии нельзя составить впечатление об этом японском виде.
34. ACRONICTA
479
вытянута и несколько расширена в проксимальной части, в середине несет
немногочисленные ск'леротизованные тяжи; дистальная часть ее образует
склеротизованный сосковидный вырост, от которого отходит ductus
seminalis; ductus bursae короткий, тонкий в проксимальной половине
и широкий в дистальной, изогнутый; восьмой сегмент очень узкий, но
довольно длинный, тергит его мембранозный; apophyses anteriores тонкие
и длинные, по размерам около половины длины ductus bursae; apophyses
posteriores также тонкие, но более длинные; papillae anales мембранозные,
в дистальной части заостренных, по-
Южный Урал (Спасское); северный
в виде латеральных пластинок,
крыты редкими волосками.
Распространение.
Казахстан (Боровое); Алтай;
среднее течение Енисея (Ени-
сейск); Забайкалье (Нерчинск,
Олекминск, Кяхта); Яблоновый
хребет; Якутск.
Синонимика senica Ev., lite-
rata Brem. и x-signata Stgr.,
сведенная Уорреном и Драуд-
том, правильна. Я имел воз-
можность сравнить типы literata
Brem. (Кяхта), senica Ev. (Спас-
ское) и котип x-signata Stgr.
(Нерчинск); все эти особи, без
сомнения, принадлежат к одно-
му виду. Мало изученный вид
с неизвестной биологией и не-
известными фазами развития..
Систематическое положение яс-
но лишь в отношении принад-
лежности к роду (и подроду);
близкие виды как в Америке,
так и в Евразии не известны.
2. Подрод Triaena Hb.
Характерен стрельчатым ри-
сунком, наличием трех черных
штрихов по одному у apex,
Рис. 247. Acronicta senica Ev. Гениталии:
а — самца, б — самки.
basis и tornus крыла. Harpe сильно склеротизована, находится
на вентральном крае valva, несет два или три отростка. Bursa
copulatrix в дистальной части с хорошо развитым отростком, часто за-
крученным спирально. Обширный подрод, виды которого широко распро-
странены в лесной зоне Голарктики; наибольшее число видов связано
с широколиственными лесами юго-восточной части Северной Америки.
Из Палеарктики известно относительно небольшое число видов
[А. (Г.) alni L., leucocuspis Btl., cuspis Hb., subpurpurea Mts. incretata
Btl., tridens Schiff., psi L., jozensis Mts., sapporensis Mts. и bellula Alph.].
Неарктика богата видами Triaena Hb., которые американские энтомо-
логи (Smith and Dyar, 1899) объединяют в две группы — lobeliae Gn.
и persuasa Harv. Известны А. (п.) innotata Gn., betulae Ril., morula
Grt. et Rob., interrupta Gn., lobeliae Gn., fureifera Gn., hasta Gn., laetifica
Sm., manitoba Sm., thoracica Grt., strigulata Sva.,oin ula Grt., fragilisGm.,
paupercula Gtr., lithospila Grt., mansueta Sm., funeralis Grt. et Rob.,
ISO
11. ACRONICTINAE
tritona Hb., quadrata Grt., radcliffei Harv., falcula Grt., paralella Grt.,
rivellata Sm., grisea Wlk., connecta Grt., subochrea Grt., spinigera Gn.,
pruni Harr., afflicta Grt., liturata Sm., persuasa Harv., marmorata Sm.,
elisabeta Sm., theodora Schaus, velia Schaus. Оба последние вида
известны из центральной Америки (Мексика, Гватемала).
Палеарктические виды также частично проникают в тропики, хотя
большая часть их ограничена южными склонами Гималаев. Из видов,
заходящих в тропики, может быть отмечен А. (/\) rubiginosa Wlk., изве-
стный, кроме Гималаев, также с Борнео, и А- (Г.) albiorbis Hamps.
(= pruinosa Moore, non Guenee), известный с Цейлона.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Передние крылья с интенсивно черной широкой поперечной
полосой, заливающей.черным дорзальный край крыла; дорзальный
отросток harpe очень длинный и тонкий, далеко выдающийся за
дорзальный край valva................................* 7. A. alni L.
.2 (1). Окраска передних крыльев однотонно серая.
3 (4). Тело стройное при относительно широких крыльях; почковид-
ное пятно в верхней и нижней части редуцировано; ductus bursae
склеротизован по всей длине . . . * 13. A. angustimacnla, sp. nov.
4 (3). Тело массивное, пропорции крыльев и тела обычны; почковид-
ное пятно развито вполне; ductus bursae склеротизован лишь ча-
стично или полностью мембранозный.
5 (6). Передние крылья серо-фиолетовые,, рисунок тонкий, нерезкий;
harpe с двумя отростками; дорзальный отросток в виде широкой
лопасти, дистальный острый....................* 14. A. bellula Alph.
6 (5). Передние крылья бело-серые с резким рисунком; отростки harpe
разной длины.
7 (10). Вентральный отросток harpe очень короткий и тупой или со-
всем не развит; два других длинные, направлены дорзально.
В (9). Линии рисунка на передних крыльях простые; передние крылья
однотонные, нередко почти белые; расширение bursa, от которого
отходит ductus seminalis, слабо закручено............* 12. A. psi L.
9 (8). Линии рисунка четкие, двойные, в середине белые; расширение
bursa copulatrix, от которого отходит ductus seminalis, закручено
спирально....................................* 8. A. leucocuspis Btl.
10 (7). Вентральный отросток harpe длинный и .острый.
11 (12). Рисунок на передних крыльях резкий и грубый, черные штрихи
резкие; окраска пепельно-серая; harpe широкая, вентральный ее
отросток широкий и сильно конический ... * 10. A. cuspis Hb.
12 (11). Рисунок обычный; окраска передних крыльев тусклосерая;
harpe или узкая, или имеет полость в основании отростков.
13 (14). Harpe узкая; вентральный ее отросток отходит под острым
углом от дорзального; окраска передних крыльев серая с неболь-
шой примесью охристых тонов вдоль линий ............................
...................................... * 11. A. tridens Schiff.
14 (13). Harpe широкая с полостью при основании отростков; нижний
отросток отходит примерно под прямым углом от дорзального;
передние крылья серо-фиолетового цвета . . .........................
........................................* 9. A. incretata Btl.
* 7. Acronicta (Triaena) alni L. (рис. 248, 249).
Linn6, 1767, Syst. Nat., ed. 12 : 845; Graeser, 1888, Berl. entom. Ztschr. : 311;
Aurivillius, 1891, Nordn. Fjar. : 96, t. 32, fig. 6; Staudinger, 1892, Mem. Lep. Rom.,
34. ACRONICTA
481
•6 : 385; он же, 1901, Cat. Lep., 1 :132; Spuler, 1908, Schmett. Eur., 1 : 137, t. 31, fig. 12:
Hampson, 1909, Cat. Lep., 8 ; 123; Warren, 1909, Seitz, Grossschmett., 3 : 13, t. 2 h;
•Culot, 1909, Noct., 1 : 20, t. 1, fig. 10; Rebel, 19'10, Berge, Schmetterlingsb., ed. 9 : 143,
t. 28, fig. 15 a, b; Bang-Haas, 1927, Nov. Macrolep., 2 : 179; он же, 1929, там же,
4 : 40; Draudt, 1931, Seitz, Grossschmett., SuppL, 3, 1 : 11; Nordstroem och Wahlgren,
1937, Svenska Fjar. : 83, t. 15, fig. 19; —degener Schiffermiiller, 1776, Wien, Verz. :
70; — su.ffu.sa Tutt, 1890, Brit. Noct., 1 : 17; — obsoleta Tutt, 1890, там же, 1 : 17; —
-carola Phillips, 1898, Soc. Entom. : 50; — steinerti Caspar!, 1898, там же, 13 ;.3;
•Gaspari, 1898, Iris, 11 : 399; Caspari, 1899, тамже, 12, t. 2, fig. 5; — obscurior Caspari,
1910, Berge, Schmetterlingsb., ed-. 9 : 143; — nigromaculata Gelin, 1917, Cat. Lep.
-d’Ouest Fr. : 89; — eothina Dannehl, 1929, Entom. Ztschr. Frankf., 39:6; — inten-
jiva Draudt, 1937, Entom. Rundsch., 54 : 397; — nigromarginata Draudt, 1937, Nach-
richtenbl. Ber. zu Seitz’s SuppL, Bd. 3 : 238.
Тело и передние крылья светлосерые; рисунок на передних крыльях
резкий; характерна широкая черная перевязь на передних крыльях,
расплывающаяся в широкую
продольную полосу по дорзаль-
ному краю; базальная линия в
виде двух коротких темных
штрихов на костальном крае;
-от основания крыла интенсивно
черный широкий штрих, сли-
вающийся с затемненной ниж-
ней частью базального поля;
вторая поперечная линия двой-
ная, темцосерая, широкая, сла-
бо выемчатая кнаружи у ко-
стального края, сильно скошен-
ная, хорошо видная в темной,
.дорзальной части крыла; круг
.лое пятно большое, светлосерое,
очерченное черным и беловато-
серым, в середине серое; сре-
динная линия проходит между
пятнами и слита с упомянутой
выше поперечной перевязью;
.’почковидное пятно очерчено
белым, в середине черное; третья
поперечная линия намечена
Рис. 248. Acronicta alni L. Гопиталий:
а— самца, б — самки
двумя черными штрихами на костальном крае, далее идет в виде
тонкой полосы, слабо зубчатой, в дорзальной половине крыла
•сильно сдвинутой базально; срединное поле в нижней части очень сужено,
значительно уже, чем внешнее поле; на внешнем поле крыла два чорпых
штриха, один между Му и М2 и другой,'очень резкий, пересекает третью
поперечную линию у tornus; подкраевая линия в виде слабой белой вол-
нистой полосы; терминальная линия в виде серии черных штрихов,
заходящих на бахрому; бахрома серая, шашечная с полоской; нижняя
•сторона бело-серая, желтоватая в костальной половине передних крыльев
с растушеванной, волнистой темной полосой и темными штрихами по
костальному краю; дискоидальные пятна на всех четырех крыльях
в виде растушеванных штрихов; задние крылья чисто.белые или светло-
серые, по внешнему краю с темной терминальной полоской и шашечной
бахромой. Размах крыльев 35—40 мм.
Uncus тонкий, при основании выгнут вентрально, покрыт короткими
волосками; tegumen короткий и широкий, несколько более длинный
31 Фауна СССР, И. В. Кожанчиков.
482
II. ACRONICTINAE
в плевральной области, образует короткие округлые выросты при осно-
вании uncus; saccus маленький, плоский, округлый на проксимальном
конце; fultura inferior длинная, суженная в вентральной половине-
и вырезанная в дорзальной; valvae узкие, с параллельными краями,
в средней части выгнутые вентрально; в базальной части valva склеро-
тизованная лопасть; harpe большая, расположена на вентральном крае-
valva, несет три отростка, два направленных косо дистально, более
коротких, и один искривленный длинный отросток, направленный косо
дорзально; дистальная треть valva мембранозная, покрыта густыми
короткими волосками; penis короче valva, в средней части слабо изогнут
дорзально; co.ecum penis тонкий и короткий; pars inflabilis несет много-
численные острые недлинные шипы. Bursa copulatrix большая, расширен-
ная в проксимальной части, мембранозная, с очень слабой рубчатой
скульптурой; дистальная ее часть узкая, несет на дорзальной стороне
небольшой заостренный, сильно гофрированный вырост, от. которого
отходит ductus seminalis; ductus bursae короткий и широкий, скдероти-
зован в виде двух широких темных тяжей; восьмой сегмент узкий и длин-
ный; apophyses posteriores короткие и острые; apophyses anteriores
тонкие, по длине равны ductus bursae; papillae anales мембранозные,
короткие и широкие.
Яйца плоские с вентральной поверхности и слабо выпуклые с дор-
зальной; хорион несет радиальные ребрышки с тонкой поперечной насеч-
кой; сразу после откладки бело-желтые; с развитием (через 12—24 часа}
под хорионом, возникают охристо-коричневые полосы, образующие
крупно ячеистый рисунок; ячеи в базальной части яйца более крупные,
вытянутые.
Гусеницы младших стадий (II—IV) имеют черно-белую окраску;
голова и первый грудной сегмент черные с немногими беловатыми и буро-
ватыми пестринами; второй и третий грудные сегменты белые с непра-
вильными и немногочисленными черными точками; первые шесть брюшных
сегментов черные, седьмой сегмент белый, восьмой черный, девятый и
десятым белые; дорзальная часть восьмого брюшного сегмента вздута
и имеет форму конического выроста; волоски сидят на небольших кониче-
ских выступах, длинные и негустые; поверхность тела блестящая. Гусе-
ницы последней стадии черные с желтыми тергитами всех сегментов тела;.
межсегментальная мембрана и на спинной стороне черная; голова черная;
два волоска каждого сегмента, сидящие на желтых тергитах, длинные,,
на концах расширенные в виде лопаточек.
Куколка светлокоричневая, относительно тонкая; по сторонам стигм
сидят короткие и тонкие волоски на небольших вздутиях кожи; поверх-
ность брюшных сегментов покрыта редкими некрупными точками —
ячейками; дистальная половина каждого сегмента и интерсегментальная
мембрана покрыты большим числом точек; cremaster в виде сильно1
морщинистого темнокоричневого длинного выроста, на дистальном конце1
несет два сильных крючковидных и торчащих в стороны шипа; базально-
й вентрально от них с каждой стороны расположено по три сильно изо-
гнутых недлинных шипа (всего на cremaster восемь шипов).
Распространение. Средняя Европа,— Германская низмен-
ность, Альпы, Карпаты, Британские острова; Швеция (Упланд, Верм-
ланд, Даларне, Оланд); южная Финляндия; в Европейской части
СССР от Прибалтики (Литва, Латвия, Эстония) и линии Олонец—Мо-
лотов всюду по лесной зоне и лесостепи (Ленинград, Новгород, Псков,
Калуга, Москва, Серпухов, Казань, Саратов, Средняя Волга); в Сибири
34. ACRONICTA
483
известно немного нахождений, хотя несомненно распространение по
всей лесной и лесостепной зонам (Томск, Барнаул, Минусинск); Саяны
(Тукеек-кем); Приамурье (Хабаровск, Николаевск); южное Приморье
(Владивосток, Ворошилов-Уссурийский, Сучан, Тигровая, Сидеми, Се-
данка); южный Сахалин", Хоккайдо; западный Китай (Шанси). Наи-
более обычен в южном Приморье и в средней Европе. Указание
Штаудингера о нахождении этого вида в Армении сомнительно; факти-
ческого материала из Закавказья я не имел.
Мало изменчивый вид; замет-
но меняется лишь степень за-
темнения дорзального края пе-
редних крыльев и развитие
черной перевязи на них. Приво
димые синонимы относятся к
разным цветовым вариантам.
Крайние, наиболее темные,
формы описаны как casparii Snt.
для Европы и intensiva Drd. для
востока Азии.
Биология и эколо-
гия. Зимует куколка в состо-
янии диапаузы; вылет бабочек
в конце мая —в июне; бабочки
живут 15—20 суток; самка от-
кладывает 300—400 яиц; яйца
откладываются поодиночке на
нижнюю сторону листьев, пре-
имущественно ольхи и березы;
гусеницы младших стадий мало
подвижны, сидят на нижней
стороне листьев, прикрепив-
шись паутиной; кормовые рас-
тения в природе — почти исклю-
чительно виды берез (Betula
pubescens, В. ermani) и ольх
(Alnus incana, A. glutinosa).
Питание другими растениями
констатировано только для стар-
ших стадий гусениц, указывают-
ся Salixcaprea, Corylus avellana,
Pirus, Prunus. В зоне хвойных
сентября, гусеницы вгрызаются
колыбельку, которую оплетают редкими шелковыми нитями И отпавшими
волосками. В лесостепье дает частичное второе поколение. Типичными
местонахождениями являются старые березняки и олыпанники. Гигро-
фильна и требовательна к стойкому термогигрорежиму. Из паразитов
отмечен Campop lex unicinctus Gr. (Ichneumonidae).
Рис. 249. Acronicta alni L. ХетотиКсин перед-
не- и среднегруди и брюшных сегментов I,
VI, VII и VIII гусеницы первой стадии (а)
и взрослой (б).
лесов окукление в августе — начале
в гниющую древесину, изготовляют
* 8. Acronicta (Triaena) leucocuspis Btl. (рис. 250).
Butler, 1878, Aim. Mag. Nat. Hist., (5), 1 : 78; Butler, 1879, Ill. Het. Brit. Mus.,
3 :12, t. 44; Hampson, 1909, Cat. Lep. Phal., 8 : 111; Warren, 1909, Seitz, Grossschmett.,
3 : 14 (var. cuspis Hb.); Bang-Haas, 1926, Nov. Macrolep., 1 : 160; Draudt, 1931,
31*
484
II. ACRONICTINAE
Seitz, Grossschmett., 3 Suppl. (1) : 10, t. 1, f; — brunnior Strand, 1913, Arch. Natur-
geschichte, 81 (A) : 158, fig.obsuta Draudt, 1933, Entom. Rundsch., 50 : 157.* 1
Передние крылья светлосерые с неправильно разбросанными охристо-
серыми штрихами; рисунок резкий; от основания крыла черный штрих,
резкий или слабо заметный, достигающий второй поперечной линии
и дающий три ветвления; первая поперечная линия в виде двух черных
штрихов на костальном крае; вторая более ясная, двойная, черная,
широко расставленная, посредине светлая, начинается резким черным
штрихом на костальном крае, далее местами стушевана, скошена кнаружи;
круглое пятно сильно сближено с почковидным,- связано с ним узким
черным штрихом, овальное, тонкое очерченное черным; почковидное
выемчатое снаружи, также вполне очерчено черным;
пятно сил ьно
Рис. 250. Acronicta leucocuspis Btl. Генита-
лии: а — самца, б — самки.
срединная линия в виде черной
растушеванной полосы, более
заметной под почковидным пят-
ном, далее идет менее резкая,
с охристым оттенком, зубчатая,
параллельная третьей попереч-
ной линии; последняя тонкая,
черная у дорзального края,
двойная, сильно сдвинута к
внешнему краю, слабо зубча-
тая, пересеченная двумя попе-
речными черными штрихами —
одним против почковидного
пятна и другим близ кубиталь-
ной жилки; последний более
резкий, интенсивно черный;
подкраевая линия не развита;
терминальная линия в виде не-
скольких (7—8 шт.) темных
штрихов; бахрома шашечная —
белая и черная; задние крылья
тусклосерые с затемненным
внешним краем и темной серо-
охристой перевязью; терми-
нальная линия тонкая, темная; нижняя сторона передних крыльев
темносерая с охристо-серым костальным краем и белым дорзальным;
просвечивают некоторые элементы рисунка (темный штрих между пят-
нами, срединная и терминальная линии); задние крылья с нижней стороны
чистобелые с черным дискоидальным пятном, штриховой терминальной
линией и темной перевязью. Размах крыльев 34—42 мм.
Uncus тонкий, длинный, почти голый; tegumen довольно длинный,
образует небольшие лопасти по сторонам основания uncus; saccus боль-
шой, округленный в проксимальной части; fultura inferior подковообраз-
ная (в вентральной части узкая), разрастается по сторонам penis в виде
1 Описанные из Японии jezoensis Matsumura [1925, Journ. Coll. Agric. Univ.
Sapporo, 15 (3) : 114, t. 8, fig. 20] и sapporensis Matsumura (Insecta Matsumurana,
1 : 3, t. 1926, 1)близкие, по мнению автора, к A. cuspis Hb., едва ли являются само-
стоятельными видами; если судить по описанию и прилагаемым фото, это лишь формы
от leucocuspis Btl. Отсутствие описания структурных признаков и материала по этим
видам, относящимся к наиболее трудной группе Подсем. Acronictinae, лишает возмож-
ности суждения об их самостоятельности.
34. ACRONICTA
485.
двух узких тяжей, несущих мелкие шипики; есть scaphium в виде узкой
пластинки; valvae длинные, прямые, в дистальной части округлые;
базальная часть valva имеет склеротизованную лопасть; harpe в виде
широкой полосы несет два тонких более длинных отростка, слабо изо-
гнутых, направленных дорзально, и один вентральный, тупой, около
одной трети длины верхних, направленный косо 'вниз; penis толстый
с тонким coecum, выгнут в средней части дорзально, короче valva; pars
inflabilis несет многочисленные короткие, но сильные шипы. Bursa copula-
trix в проксимальной половине мембранозная, покрытая рубчатой
скульптурой, в дистальной половине сильно склеротизована, образует
широкий отросток, обрастающий основание ductus bursae и направлен-
ный в сторону (влево in toto); конец его, от которого отходит ductus
seminalis, повернут под прямым углом проксимально; ductus bursae
по длине более половины corpus bursae, широкий, складчатый; восьмой
сегмент узкий и короткий; apophyses anteriores острые и короткие, менее
половины длины ductus bursae, но вдвое длиннее apophyses posteriores;
papillae anales мембранозные, короткие и широкие.
Распространение. Амур (Николаевск); южное Приморье
(Владивосток, Сучан, Аскольд, Сидеми, Самарга); Сахалин (Нуиво,
Ишиносава); Хоккайдо (Хокодате, хребет Дайзетсу); Хондо (Иокогама);
Корея (Гензен); северный. Китай. Все известные мне указания на нахо-
ждение A. cuspis Hb. на Дальнем Востоке относятся к этому виду.
* 9. Acronicta (Triaena) incretata Btl. (рис. 251).
Butler, 1878, Ann. Mag. Nat. Hist. (5) 1 : 78; Butler, 1879, Ill. Het. Brit. Mus.',
3 : 12, t. 44, fig, 3; Staudinger, 1901, Cat. Lep., 1 : 132; Herz, 1904, Ежегодн. Зоол.
муз. Акад. Наук, 9 : 270; Hampson, 1909, Cat. Lep. PhaL, 8 : 109; Matsumura, 1925,
Journ. Coll. Agric., Sapporo, 15 : 119; Moltrecht, 1929, Зап. Владив. Отд. Географ,
общ., 3 : 52; Draudt, 1931, Seitz, Grossschmett., Suppl., 1 : 10, t. 1, f. — increta
Butler, 1878, Ann. Mag. Nat. Hist., 1 (5) : 78;.— intermedia Warren, 1909, Seitz, Gross-
schmett., 3 : 14, t. 2, k.
Биология и экология. Филипьев, 1932', Тр. защ. раст., 5:372;
Clausen, 1931, U. S. Dept. Agric. Circ., 168 : 7, 19.
Передние крылья тускло-серо-фиолетовые с примесью неправильна
разбросанных охристых тонов; от основания крыла вдоль срединной
жилки интенсивно черный штрих, дающий четыре ветви; первая попе-
речная линия неясная в виде двух косых штрихов; вторая зубчатая,
идет косо кнаружи, простая, двойная лишь у костального края; круглое
пятно овальное, большое или слито с почковидным или близко подходит
к нему; между пятнами черный штрих, иногда частично пронизывающий
и почковидное пятно; последнее тонко очерчено черным, в середине серо-
фиолетовое с примесью охристых тонов, большое, почти треугольной
формы; срединная линия заметна лишь у костального края охристо-
серая, далее смыта; третья поперечная линия начинается двумя штри-
хами над почковидным пятном, далее сильно выгнута кнаружи, простая,
дает слабый черный штрих против почковидного пятна и другой в области
кубитальной жилки, сильный, достигающий края крцла; внешнее поле
узкое, особенно в костальной половине, без линий с неправильными ох-
ристыми штрихами; терминальная линия в виде восьми черных треуголь-
ных штрихов; бахрома серая с черными штрихами. Задние крылья у самца
чисто белые с белой бахромой, у самки тусклосерые с неширокой серой
перевязью и слабой терминальной. линией, бахрома белая; нижняя сто-
рона серо-белая с ясной поперечной линией и дискоидальными пятнами
на передних и задних крыльях; терминальная линия просвечивает
486
11. ACRONICTINAE
в виде черных штрихов. Чешуйчатый и волосяной покров головы и груди
серо-фиолетовый; по бокам груди и головы черные полосы. Размах крыльев
40—45 мм.
Uncus тонкий, длинный и голый; tegumen короткий и широкий; sac-
cus острый и узкий; valvae по форме, как у A. leucocuspis Btl.; harpe
большая, более массивная, чем у A. tridens Schiff, в середине полая, несет
три отростка; дистальный верхний отросток особенно сильный, длиннее
других, нередко несколько зубчат; вентральный отросток сидит на широ-
ком основании, сильно конический, острый, направлен дистально, около
половины длины предыдущего; на месте срастания базальной лопасти
valva и harpe небольшая бородавка;
fultura inferior в виде подковы,
в вентральной части узкая, ее
боковые выросты сильно расши-
рены в дорзальной части и зуб -
чаты; penis толстый с коротким
соесшп, в средней части сильно
изогнут дорзально, несет много-
численные шипы на pars infla-
bilis. Bursa copulatrix очень
большая, сильно вытянутая,
изогнутая вправо (in toto),
расширенная в проксимальной
части, мембранозная; в дисталь
ной половине с левой стороны,
(in toto) склеротизованный ши-
рокий вырост, направленный
вентрально и в сторону (напра-
во); конец выроста направлен
орально; от этого выроста от-
ходит ductus seminalis; ductus
bursae не более одной трети
длины bursa, широкий, почти
прямой; восьмой сегмент узкий
и короткий; apophyses anteriores
тонкие/ не менее половины дли-
Рис. 251. Acronicta incretata Btl. Гениталии:
а — самца, б — самки.
ны ductus bursae; apophyses posteriores такой же длины, как и первые;
papillae anales узкие и короткие, мембранозные.
Распространение. Южное Приморье (Яковлевка, Влади-
восток, р. Сама, Ворошилов-Уссурийский, Аскольд, Сучан, Покровка);
Приамурье (Николаевск); южный Сахалин; Хоккайдо, Хондо (Иокогама);
Корея (Гензен); Китай (Пекин). Указан для Енисейска, но фактический
материал для этой территории мне не известен.
Наиболее близок к A. tridens Schiff., но хорошо отличается более
тусклым тоном передних крыльев; японские особи более крупны и имеют
более интенсивный фиолетовый тон.
Биология изучена слабо; гусеницы (не описаны) многоядны,
отмечено питание Betulaceae (Alnus, Betula) и Rosaceae (Pirus, Prunus,
Malus). В Китае, Корее и Японии вредит плодовым садам (яблоне, груше,
сливе, вишне).* 1
1 Описанный из Японии A. subpurpurea Mats. (Matsumura, 1926, Ins. Matsumur.,
1 : 2, t. 1, fig. 3) очень близок, по мнению автора, к A. incretata Btl.; автор указывает
только цветовые отличия между этими видами: строение гениталий им не исследовано;
мне этот вид известен только по литературе.
34. ACRONICTA
487
* 10. Acronicta (Triaena) cuspis Hb. Стрельчатка ольховая (рис.
.'252).
Hiibner, 1818, Noct. Eur., f. 504; Graeser, 1888, Berl. entom. Ztschr. : 311; Auri-
villius, 1891, Nordn. Fjar. : 98; Staudinger, 1892, Mem. Lep. Hom., 6 : 387; Staudin-
-ger, 1901, Gat. Lep., 1 : 132; Spuler, 1908, Europ. Schmett., 1 : 138, t. 31, fig. 16; Cu-
lot, 1909, Noct., 1 : 21, t. 1, fig. 14; Hampson, 1909, Gat. Lep. Phal., 8 : 110; Warren,
1909, Seitz, Grossschmett., 3 : 14, t. 2, h; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 :
147; Petersen, 1924, Lep. Fauna Estl., ed. 2 : 151; Кожанчиков, 1927, Русск. эптом.
обозр., 21: 137, рис. 1, 2, 7; Draudt., 1931, Seitz, Grossschmett., 3 Suppl., 1 : 10,
t. 1, f; Grosse, 1936, Entom. Ztschr., 50 : 219, 229; Nordstroem och Wahlgren, 1937,
.Svenska Fjar. : 84, t. 15, fig. 12; — caliginosa Schultz, 1908, Soc. Entom., 22 : 185;—
suffusa Spuler, 1908, Schmett. Eur., 1 : 138; — rosea Turati, 1911, Atti Soc. Nat.
Sci. Ital., 51 : 290 (Sardinia)', — taeniata Schultz, 1910, Intern, entom. Ztschr., 24 :
185; —obscurior Strand, 1915, Arch. Naturgesch., 81, A (11) : 158; —detyanea Strand,
1915, там же, 81 A (11) : 157; — belgica Draudt, 1931, Seitz, Grossschmett., 3 Suppl.:
10, t. 1,- f.
Биология и экология. Kirby, 1895, Butterfl. Moths Eur, : 158;
Kuskov, 1927, Beitr. Kunde Estl., 13 : 42.
Светлосерый co слабыми желтоватыми тонами на крыльях, груди
и голове; чешуйчатый покров блестящий; рисунок на крыльях резкий,
грубый, интенсивно черный; базальная линия в виде двух коротких штри-
хов на костальном крае; от основания крыла интенсивно черный широкий
штрих,- достигающий второй поперечной линии; последняя двойная лишь
в костальной половине крыла, далее внешняя ее половина редуцирована,
костальная часть ее имеет петлю кнаружи, косая; круглое пятно большое,
•светлосерое, неполно очерчено черным, сдвинуто к почковидному пятну
и соединено с ним черным штрихом; срединная линия растушеванная,
более интенсивна на костальном крае, идет косо к почковидному пятну,
.далее редуцирована; почковидное пятно большое, широкое, неполно
очерчено черным, выемчатое по внешнему краю, где нередко почти, или
совершенно стушевано; третья поперечная линия лишь на костальном
крае намечена двумя штрихами, далее внутренняя ее половина редуци-
рована, и она представлена тонкой черной полосой слабо зубчатой по
жилкам, снаружи затемненной, пересеченной двумя черными штрихами,
менее интенсивным в области медиальных жилок и более интенсивным
в области кубитальных; внешнее поле узкое с тонкой беловатой подкрае-
вой линией, почти смытой; терминальная линия в виде топких белых
и черных штрихов, заходящих на бахрому; последняя — серая с тонкой
полоской; задние крылья тусклосерые со слабо заметными жилками,
со слабой, размытой лимбальной линией и иногда заметной терминальной
полоской; бахрома светлая; нижняя сторона блестящая, светлосерая,
почти белая, затемненная в области дискоидальной ячеи на передних
крыльях, с неясной поперечной линией и растушеванными лунными пят-
нами на передних и задних крыльях. Размах крыльев 40—45 мм.
Uncus, tegumen, saccus и общая форма valvae близки к другим видам
этой группы (incretata Btl., tridens Schiff., psi L.); valva более широкая
чем у tridens Schiff., особенно в базальной части; базальная лопасть
valva длинная, сильно склеротизованная; harpe очень широкая; оба дор-
зальных отростка harpe одинаковых размеров, недлинные; вентральный
отросток имеет очень широкое основание, длинный, почти равен по длине
двум остальным, прямой, сильно конический, направлен косо вентрально;
fultura inferior такая же, как у A. tridens’, penis толстый, слабо изогнутый
со многими шинами на pars inflabilis. Bursa copulatrix по форме напо-
минает таковую A. tridens Schiff., но отросток ее в основании очень широк
.и вполне срастается с основанием ductus bursae; конец его завернут влево
488
II. ACRONICTINAE
Рис. 252. Acronicta cuspis Hb. Генита-
лии: а — самца, б — самки.
и направлен почти дистально; ductus, bursae короткий, менее половины
длины corpus bursae, изогнут в основании и расширен в дистальной части;
apophyses anteriores острые и короткие, менее половины длины ductus
bursae; apophyses posteriores более короткие.
Яйца плоские, слабо выпуклые посредине, диаметром около 1 мм;
поверхность хориона несет радиально расходящиеся рубчики; окраска
бело-желтая.
Гусеницы старших стадий (начиная с третьей) желто-серые или корич-
невые со сложным рисунком; вдоль спины широкая желто-голубая по-
лоса; бока коричневые с двумя оранжево-желтыми и черными пятнами
над каждым дыхальцем; брюшная сторона светложелтая; тело покрыто
небольшими бородавками, несущими пучки волосков, более длинных
и толстых на первых трех брюшных
сегментах; первый брюшной сегмент
несет на дорзальной стороне одну
небольшую бородавку (значительно-
более короткую, чем у A. psi L.),
густо покрытую волосками и несу-
щую длинный и густой пучок чер-
ных волосков; восьмой брюшной сег-
мент несколько вздут и несет на.
дорзальной стороне кисть волосков;
голова густо покрыта волосками.
Куколка вытянутой формы, с уз-
ким брюшком, светлокоричневая;
дистальная часть брюшка округлена,
не переходит постепенно в crema-
ster; последний сидит в виде при-
датка на гладком блестящем послед-
нем сегменте; брюшные сегменты в
дистальной части гладкие, в базаль-
ной части они несут скульптуру в.
виде неправильных точек; стигмы
выдаются; волоски за стигмами очень
короткие; cremaster в виде почти
цилиндрического вырюста, темноко-
ричневый, покрытый неправильными бороздами; на дистальном его-
конце восемь слабо изогнутых щетинок, по четыре на дорзальной и
вентральной сторонах.
Распространение. Средняя Европа от Альп (Ломбардия)-
и Карпат до Атлантического побережья и Прибалтики; Польша (Вар-
шава); южная Швеция (Даларне, Эланд, Готланд); южная Финлян-
дия (Савония, Ниландия); северо-запад Европейской части СССР
(Рига, Тарту, Вильнюс, Каунас); далее на восток от линии Петроза-
водск—Молотов всюду по лесной зоне и лесостепи (Ленинград, Нов-
город, Москва, Калинин, Калуга, Чернигов, Казань, Ярославль, Моги-
лев); Урал и далее на восток в лесной и лесостепной зонах в Сибири
(Омск, Томск, Минусинск). Указания Штаудингера (1901) для Семи-
речья (Или, Иссык-куль), так же как указания Гэмпсона (1909)
для Закавказья и Егорова для Северного Кавказа, не находят подтвержде-
ния; фактический материал из этих мест мне не известен; также сомни-
тельны указания на наличие A. cuspis Hb. в Николаеве и Каменец-По-
дольске. Указания ряда авторов (Грезер, 1888; Штаудингер, 1892; Моль-
34. ACRONICTA
489
трехт, 1929) на наличие A. cuspis Hb. в- южном Приморье и на Амуре
ошибочны и относятся к A. leucocuspis Btl.
Очень характерный по рисунку и строению гусениц вид; мало измен-
чив; мелкие и бледные особи могут быть смешаны тем не менее с psi L.
и tridens Schiff.
Биология и экология. Вылет бабочек в июне—-июле;
яйцекладка на листья кормовых растений; яйца откладываются пооди-
ночке. Развитие яиц 8—12 суток; главный период роста гусениц август—
сентябрь; кормовые растения только Betulaceae (Alnus incana, A. glu-
tinosa; Be'ula pubescens, B. verrucosa, B.'humilis); есть указания на пита-
ние гусениц лещиной (Gorylus avellana); по Тейху (Teich, 1888) пред-
почитаемым растением является ольха. Окукление в загнившей древесине
или во мхе; куколка диапаузирует и зимует. Почти на всей территории
ареала дает одно поколение; для Прибалтики указано частичное второе.
* 11. Acronicta (Triaena) tridens Schiff. Стрельчатка яблонная
(рис. 253).
Schiffermiiller, 1776, Wien. Verz. : 67; Aurivillius, 1891, Nordn. Fjar. : 98; Stau-
dinger, 1901, Cat. Lep., 1 : 132; Spuler, 1908, Schmett. Eur., 1 : 137, t. 31, fig. 4; Hamp-
son, 1909, Cat. Lep. PhaL, 8 : 108; Warren, 1909, Seitz, Grossschmett., 3 : 16, t. 3,f;
Culot, 1909, Noct., 1 : 20, t. 1, fig. 12; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 144,
t. 28, fig. 16; Petersen, 1924, Lep. Fauna Estl.,ed. 2 : 150; Кожанчиков, 1927, Энтом.
обозр., 21 : 139, fig. 3, 4; Draudt, 1931, Seitz, Grossschmett., S.uppl., 3, 1 : 10, t. 1,,
f; Nordtsroem och. Wahlgren, 1937, Svenska Fjar. : 83; — psi Hiibner, 1822, Schmett.,
Noct., fig. 5 (non Linne); — karghalica Moore, 1878, Ann. Mag. Nat. Hist. (5) 1 : 232;
Moore, 1878, Second Jarkend Mission, Lep. : 8, t. 1, fig. 9; — virga Tutt, 1888, EntomoL,
1 : 50; — rosea Tutt, 1891, Brit. Noct., 1:20; — bidens Tutt, 1891, там же, 1:20.
quinquedentata Tutt,' 1891, там же, 1:20;—juncta Tutt, 1891, там же, 1:20;—
asignata Spuler, 1908, Schmett. Eur., 1 : 137; —variegata, Strand, 1915, Arch. Natur-
gesch., 81 (A) : 157; — radoti Le Cerf, 1924, Bull. Soc. Entom. France : 25; — solto-
wensis Schultz, 1930, Intern, entom. Ztschr., Guben, 24 : 185; — obscurior Lattin, 1938,
Ztschr. Oesterr. entom. Ver., 23 : 96J
Биология: Meyrik, 1895, Brit. Lep. : 141; Barret, 1895, Lep. Brit., 3 : 247; t.
122, fig. 1; Spuler, 1908, Raupen, t. 21, fig. 26; Сахаров, 1915, Вредн. насек.: 14; Reh,.
1925, Sorauer, Handb. Pflanzenkrankh., 4 : 418; Сахаров, 1927, Журн, агрон. IO. В.:
385; Дехтярев, 1927/28, Зах. росл., 3/4 : 9; Филипьев, 1932, Труды защ. раст., 5 :
373; Romanizyn, 1935, Prazn. Monogr., 6 : 239; Лебедев, 1935, Зб1рн. праць екол.,.
2 : 34; Парфентьев, 1936, Итоги научно-исслед. раб. ВИЗР, 1935 : 212.
Голова и грудь бело-серые с боков с черной полосой; передние крылья
бело-серые с черным рисунком и неясными охристыми мазками; чешуй-
чатый покров блестящий; первая поперечная линия намечена черным
и белым штрихами на костальном крае, далее смыта; от основания крыла
интенсивный черный штрих ниже кубитального ствола; вторая попереч-
ная линия сильно скошена кнаружи; двойная, посредине бело-серая;
круглое пятно овальное, нередко большое, сдвинуто в терминальном
направлении, нередко открыто в костальный край и частично слито
с почковидным, в середине серое, очерчено черным; срединная линия
широкая, растушеванная, заметна лишь на костальном крае, далее сли-
вается с почковидным пятном и теряется; почковидное пятно большое,
неясное, в терминальной половине прерывисто очерчено черным, в сере-
дине серо-белое или темносерое; третья поперечная линия намечена
в виде двух темных штрихов лишь на костальном крае, далее идет в виде
1 Описанная Штаудингером (1901) как форма от cuspis Hb. A. taurica Stgr. изу-
чена слабо; по мнению Остгельдера и Гэмпсона это bona species, по ни тот ни другой
автор не дает обоснования этого вида; по доступным мне материалам (не типам) из
Закавказья, а также учитывая распространение A. cuspis Hb., A. taurica Stgr. следует
считать формой от tndens Schiff.
490
11. ACRONICTINAE
Рис. 253. Acronicta tridens Schiff.
Гениталии: а — самца, б — самки.
тонкой черной полосы, далеко выгнутой к внешнему краю крыла, слабо
и неправильно зубчатой по жилкам и пересеченной двумя черными штри-
хами — одним в области медиальных жилок и другим, более сильным,
в области кубитальных; вокруг второго темного штриха темное пятно;
подкраевая линия в виде тонкой волнистой полоски; терминальная линия
черная, состоит из серии черных штрихов, заходящих на бахрому; бах-
рома белая; задние крылья или чисто белые или слегка сероватые (у са-
мок) с тонкой терминальной полоской и белой бахромой; нижняя сторона
белая с четырьмя дискоидальными пятнами, просвечивающей черным
третьей поперечной линией; бахрома с четкой черной полоской. Раз-
мах крыльев 30—40 мм.
Uncus, tegumen и saccus такие же, как у A. psi L. или A. incretata
Btl.; valvae по форме также сходны, узкие, округлые в дистальной части;
на месте срастания базальной лопасти
и harpe нет склеротизованной бородав-
ки; harpe более узкая, чем у A. increta-
ta Btl., несет три отростка, более корот-
ких и тонких, чем у этого вида; вент-
ральный отросток сидит на узком осно-
вании, конический и острый, почти та-
кой же длины, как два первых (дорзаль-
ных); fultura inferior подковообразная,
дорзальные ее расширенные выросты
несут мелкие многочисленные шипики;
penis в средней части слабо изогнут,
толстый, с коротким coecum; pars infla-
bilis несёт многочисленные короткие и
сильные шипы. Bursa copulatrix рас-
ширенная в проксимальной части и за-
остренная в дистальной; проксимальная,
часть ее мембранозная, несет, рубчатую
скульптуру; дистальная часть склеро-
тизована и образует отросток, охваты-
вающий базальную часть ductus bursae,
загнутый улиткообразно; от конца его
отходит ductus seminalis; ductus bursae
широкий, сильно изогнутый в средней части, складчатый, несколько рас-
ширенный дистально; apophyses anteriores менее половины длины duc-
tus bursae, но вдвое длиннее apophyses posteriores; papillae anales мем-
бранозные.
Яйца плоские, слабо выпуклые с дорзальной стороны, беловато-жел-
тые, хорион несет радиальную неправильную рубчатую скульптуру; диа-
метр около 1 мм.
Взрослые гусеницы (начиная с третьей стадии) желто-серые со слож-
ным черным, желтым и оранжевым рисунком; вдоль спины широкая оран-
жевая полоса, разделенная вдоль тонкой черной полоской; по сторонам
-оранжевой полосы (иногда желтой) две черных прерванных полосы;
на сегментарных отрезках этих полос с каждой стороны симметрично
располагаются по одной (или две) белой и оранжевой точке; боковые
полосы желто-серые, однотонные; дыхальца черные, слабо выделяются
на общем фоне; на первом брюшном сегменте высокая (почти как у A. psi
,L.) черная бородавка, покрытая короткими и густыми черными волосками
и усаженная редкими и длинными черными щетинками; на восьмом брюш-
34. ACRONICTA
491
ном сегменте небольшой вздутый вырост; голова округлая, густо волоси-
стая.
Куколка стройная, светлокоричневая; брюшные сегменты несут то-
чечную скульптуру; точки более густые и грубые в основании сегментов,
у дистального канта они исчезают; вентральная часть последнего сегмента
округлена и не переходит в cremaster; последний короткий (почти вдвое
меньше, чем у A. cuspis НЬ.), покрыт глубокими и редкими продольными
косыми складками, несет восемь искривленных щетинок, две по дорзаль-
ному и шесть по вентральному краю.
Распространение. Средняя Европа — Альпы (Бавария),
Германия, долина Дуная (Вена); северная Италия; Далмация; Карпаты;
Финляндия (Ниландия, Аландия); южная Швеция; северо-запад Евро-
пейской части СССР (Рига, Таллин, Тарту), далее к востоку — от
линии Петрозаводск — Вологда—Молотов (вероятно несколько севернее)
всюду по лесной зоне, лесостепи и отчасти степной полосе (Ленинград,
Псков, Новгород, Москва, Калуга, Мичуринск, Воронеж, Ярославль,
Харьков, Полтава, Николаев, Херсон, Таганрог); Поволжье (Казань,
Ульяновск, Саратов); Приуралье (Чкалов, Спасское, Уральск); Северный
Кавказ (нижний Дон, Ставрополь, Кубань); Закавказье (Дербент); Си-
бирь от линии Тобольск—Томск—Иркутск (вероятно и севернее но
нет фактического .материала) всюду по лесной зоне и лесостепи (Омск,
Барнаул, Минусинск, Кузнецк, Тайшет, Канск, Бунбуй, Якутск);
Алтай (Усть-Бухтарма, Усть-Каменогорск, Тельбес, Онгудай); северный
Казахстан (Семипалатинск); Амур (Поярково); южное Приморье (Тигро-
вая, Сучан, Яковлевка, Виноградовка).
Характеристика ареала A. tridens Schiff, могла быть дана преи-
мущественно на основании фактического материала; фаунистические
списки не могли быть использованы за малыми исключениями, так
как отличие этого вида от A. psi L. без привлечения структур-
ных признаков не надежны. В общем ареалы обоих видов, невидимому,
близки, но A. tridens Schiff, более распространена на востоке Азии, тогда
как A. psi L. более обычна на юге Европы и редка в Сибири. Признаком,
отличающим по окраске A. tridens Schiff, от A. psi L., являются охристые
штрихи, почти всегда присутствующие у этого вида. Многочисленные
синонимы относятся к аберрантным формам, описанным или на основании
отличия общего тона окраски (Татт, 1888—1891) или на основании раз-
вития или редукции элементов рисунка.
Биология и экология. Зимует куколка в древесине
или в подстилке. Вылет бабочек в южной части ареала в мае—июне,
в северной — в июле. Яйца откладываются поодиночке на листья кормо-
вых растений. Гусеницы многоядны; питание преимущественно на Rosales
(Prunus padus, Р. domestica, Р. armeniaca, Р. cerasus, Pyrus doinestica,
Malus domestica, Crataegus oxyacantha, Sorbus aucuparia); в лесной зоне
гусеницы обычно на Fagaceae (Fagus sylvatica, Quercus robur), Betulaceae
(Betula pubescens, B. verrucosa, Alnus incana), Salicaceae (виды Salix
и Populus), Rhamnaceae (Rhamnus frangula). На севере дает одно поколе-
ние (лесная зона); южнее (в лесостепье и степи) дает частичное второе
и третье поколения (гусеницы до середины октября). На юге в ряде мест
отмечен как вредитель плодовых садов; указан для Астрахани Сахаровым
(1915), для Ставрополя Уваровым (1914, 1916), как вредитель яблони
на Дону Зверезомб-Зубовским (1917); на Украине (Харьков) как вре-
дитель садов указан Пачосским (1916), в Центральной черноземной по-
лосе (Воронеж) отмечен как вредитель лесных полезащитных посадок
492 IT. ACRONICTINAE
и плодовых — вишни и яблони Парфентьевым (1937); указания на
вредную деятельность касаются тех частей ареала, где развиваются два
и более поколений.
Из числа паразитов указаны перепончатокрылые: из Braconidae —
Apanteles fulvipes Hol., Chelonus irrorator F., Eutedon larvarum Rztb.
Etroxus stenogaster Wlk., Eulophus ramicornis Schff., Microgaster conge-
stus Wesm., M. connexusNs., M. consularis Hol.,7lf. difficilis Ns., M. ful-
vipes Wesm.-, M. fumipennis Msh., M. glomeratus L., M. irritator Msh.,
M. pallipes Wesm., M.ratzeburgi Rtzb., M.spinolae Ns.,M. spurius Wesm.,
M. subcompletus Ns., M. ultor Вт., M. rugulosus Ns., Rhitigaster irrorator
Ns., Taleas breviusculus Rtzb.; из Icbneumonidae —Amblyteles bellicosus
Wesm., Exetastes guttatorius Gr., E. tarsator F., Hemiteles bicolorinus Gr.,
H. conformis Gr., Mesochorus pictilis Rtzb., M. semirufus Hgr., Metopius
migratorius F., Ophion costatus Rtzb., O. luteus L., Paniscus cephalotus
Gr., P. testace.us Gr.; из Diptera — Compsitura concinnata Mg., Exorista
acronyctae R. D., E. affinis Fall., E. glauca Mg., E. glirina Rd., E. lota
Mg., E. noctuicida Rd., Pales pavida Mg., Sturmia atropivora R. D.,
Wint1 emia xanthogastra Rd.
* 12. Acronicta (Triaena) psi L. (рис. 254, 255).
Linne, 1758, Syst. Nat., ed. 10 : 514; Aurivillius,. 1891, Nordn. Fjar. : 98, t. 32,
fig. 7; Staudinger, 1892, Mem. Lep. Rom., 6 : 387; Staudinger, 1901, Cat. Lep., 1 : 132;
Spuler, 1908, Schmett. Eur., 1 : 138, t. 31, fig. 15; Hampson, 1909, Cat. Lep. Phal.,
8 : 106, fig. 18; Warren, 1909, Seitz, Grossschmett., 3 : 15, t. 3, d; Culot, 19’09, Noct.,
1 : 21, t. 1, fig. 13; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 144, t. 24, fig. 17; Peter-
sen, 1924, Lep.. Fauna Estl., ed. 2 : 151; Кожанчиков, 1927, Энтом. обозр., 21 : 140,
рис. 5,6; Grosse, 1936, Entom. Ztschr., 50 : 219, 224; Hoffmeyer, 1937, Entom. Ztschr.,
51 : 68, fig. 1—3; Heydemann, 1937, там же, 51 : 78, fig. 1—4; Lattin, 1937, там же,
50 : 517, fig. 1—6; Boursin, 1937, тамже, 51 : 70, fig. 1—4; Nordstroem och Wahlgren,
1937, Svenska Fjar. : 84, t. 15, fig. 11 (?); — cuspis Stephens, 1829, Entom. Haustell.,
3 : 40 (non Hiibner); — virga Tutt, 1888, Entomol. : 50; — bivirgae Tutt, 1888, там же:
50; — suffuse. Tutt, .1888, Entomol. : 50; — bidens Tutt, 1891, Brit. Noct., 1 : 22; —
rosea Tutt, 1891, Brit. Noct., 1 : 22; —juncta Tutt, 1891, там же, 1 : 21;— altaica
Staudinger, 1901, Cat. Lep., 1 : 132; — batnana Draudt, 1931, Seitz, Grossschmett.,
Suppl, 1 : 10, t. 1, e; — iliensis Draudt, 1931, тамже, 1 : 10, t. le; — psideleta Turner,
1935, Brit. Noct., 1 : 362; — crassistigma Lattin, 1938, Ztschr. Oesterr. entom. Ver.,
23 : 96; — solimana Draudt, 1938, Mitt. Munch, entom. Ges., 28 : 29.
Биология и экология. Meyrik, 1895, Brit. Lep. : 141; Barret, 1895,
Lep. Brit., 3 : 251, t. 122, fig. 2; Reh, 1925, Sorauer, Handb., 4 : 418; Лебедев, 1935,
36ipn. праць екол., 1 : 418; Aue, 1935, Entom. Anz., 15 : 125.
Голова и грудь пепельно-серые со слабым желтоватым оттенком,
по бокам черные; передние крылья тусклого серого цвета, также со сла-
бым желтоватым оттенком (не всегда); чешуйчатый покров неблестящий;
рисунок на крыльях тонкий, несколько редуцирован; базальная -линия
в виде двух черных штрихов, разделенных белым, заметная лишь на ко-
стальном крае; от основания крыла интенсивно черный штрих, достигаю-
щий второй поперечной линии, ветвящийся; вторая поперечная линия
черная, двойная, посредине светлая, сильно скошена, пронизана черным
штрихом; круглое пятно небольшое, почти правильное, неполно очерчено
черным, сильно сдвинуто к почковидному пятну, часто соединено с ним
черным штрихом или иногда сливается с ним; срединная линия лишь
намечена косым растушеванным штрихом на костальном крае, далее
теряется и не доходит до пятен; почковидное пятно во внешней половине
редуцировано, развито в базальной половине, четко очерчено черным,
внутри несколько темнее общего тона крыла, с немногими черными чешуй-
ками; третья поперечная линия начинается двумя черными штрихами
34. ACRONICT.A
493
на уровне дистального края почковидного пятна, далее идет в виде одной
четкой полосы, неправильно зубчатой, снаружи затемненной, особенно
в дорзальной половине крыла; она пересечена двумя черными штрихами —
одним в области медиальных, другим в области кубитальных жилок,
последний штрих достигает tornus; внешнее поле однотонное, почти ли-
шенное подкраевой линии; терминальная линия в виде восьми узких
черных штрихов, заходящих на бахрому; бахрома серая с тонкой поло-
ской; задние крылья тусклобелые, несколько темнеющие к внешнему
краю с тонкой терминальной полоской; жилки затемнены. Нижняя сто-
рона бело-серая с неправильно разбросанными темными чешуйками
и темными пятнами на костальном
дельных черных штрихов. Размах
крыльев 36—40 мм.
Uncus, tegumen, saccus и строе-
ние valva очень близки к tridens
Schiff.; базальная лопасть valva
в месте срастания с harpe обра-
зует складку; harpe узкая и корот-
кая, несет два длинных дорзаль-
ных отростка, третий вентральный
отросток не развит, или (у неко
торых особей) на его месте неболь-
шой бугорок; fultura inferior боль-
шая, сильно расширенная в дор-
зальной части и покрытая много-
численными шипами (более широ-
кая, чем у A. tridens Schiff.); penis
-слабо изогнут в средней части, не
-сет многочисленные шипы на pars
inflabilis. Bursa copulatrix, как у
A. tridens Schiff., расширена в
проксимальной части, но дисталь-
ная ее часть отделена небольшой
перетяжкой, менее сужена и сла-
бее склеротизована; отросток ди-
стального конца bursa не тесно ох-
ватывает основание ductus bursae,
крае; терминальная линия в виде от-
Рис. 254. Acr'onicta psi L. Гениталии:
а — самца, б — самки.
но, отходя с левой стороны (in toto) сначала на половину своей длины,
идет дистально, параллельно основанию ductus bursae, и лишь вторая
половина его повернута вентрально, и конец загнут проксимально; ductus
bursae сильно расширен при основании, длинный, в середине изогнут,
в дистальной части расширен; прочие части полового аппарата, как
у A. tridens Schiff. Различия строения гениталий самок A. tridens и A. psi,
выражающиеся, главным образом, в степени изгиба выроста bursa copu-
latrix, часто стушевываются и трудно улавливаются в связи с развитием
сперматофора и, может быть, с числом оплодотворений; различия в строе-
нии гениталий самцов очень постоянны.
Яйца плоские, слабо выпуклые с дорзальной стороны, бело-желтые;
хорион несет тонкую рубчатую скульптуру; рубчики располагаются
радиально и сходятся в середине; диаметр 0.8—1.0 мм.
Гусеницы младших стадий (I и II) светлосерые с крупными легко за-
метными негустыми черными щетинками и белыми волосками; старшие
•стадии (начиная с III) приобретают очень характерную внешность. Окра-
494
11. ACRONICTINAE.
ска серо-коричневая, вдоль спины широкая желтая полоса (исключая
переднегрудь); на каждом сегменте по сторонам этой полосы два больших
карминно-красных или (реже) желтых поперечных штриха; бока и брюш-
ная сторона желто-серые; голова и ноги черно-коричневые, голова густо
покрыта волосками; все сегменты тела покрыты мелкими пупиллами,
на которых сидят волоски; на первом брюшном сегменте очень длинный
непарный дорзальный вырост черного цвета, густо покрытым короткими
Рис. 255. Acronicta psi L. Хетотаксия передне-
и среднегруди и брюшных сегментов I, VI,
VII и VIII гусеницы первой стадии (а) и взрос-
лой (б).
черными волосками и усажен-
ный отдельными черными ще-
тинками; восьмой сегмент
брюшка также вздут в виде
небольшого треугольного вы-
роста; голова округлая.
Куколка очень близка.к
A. tridens Schiff, по форме те-
ла и по cremaster, отличается
несколько более тонкой
пунктировкой сегментов
брюшка; cremaster еще более
короткий, также покрыт гру-
быми бороздами; несет восемь
острых тонких слабо искрив-
ленных шипиков — шесть по
вентральному и два по дор-
зальному краю. ?
Сходство A. psi L. и А.
tridens Schiff, по имагиналь-
ной фазе находит подтверж-
дение и в структурах куко-
лок, также близких. Личи-
ночная фаза обоих видов силь-
но дифференцирована.
Распространение.
Вся Западная и средняя
Европа — Пиренеи, Альпы,
Карпаты, Средиземноморье
(северная Франция, Ита-
лия, Далмация, Греция),
острова Средиземного мо-
ря (Корсика, Сардиния);
северная Африка, — Алжир (Батна), Марокко (Пехер); Сирия (северный
Ливан); Малая Азия; Закавказье (Боржоми, Тбилиси, Лагодехи, Куссари,
Кировабад); северная Европа — Швеция, Финляндия (Ниландия, Або);
северо-запад Европейской части СССР (Прибалтика), далее на восток от ли-
нии Петрозаводск—Вологда—Молотов (вероятно несколько севернее)
всюду по лесной и лесостепной зонам и в степи (Ленинград, Москва, Псков,
Ярославль, Тула, Мичуринск, Могилев, Калуга, Волынь, Полтава, Киев,
Чернигов, Таганрог, Николаев); Поволжье (Казань, Ульяновск, Саратов,
Астрахань); Приуралье (Узян, Чкалов, Уральск); Северный Кавказ (Став-
рополь); Крым (Севастополь, Евпатория, Керчь); Сибирь, вероятно, всюду
по лесной зоне, но нахождения очень разрозненные (Тюмень, Томск,
Омск, Минусинск, Иркутск, Якутск, Верхоянск); Казахстан; Средняя Азия
(Ашхабад, Байрам-али). Из Приамурья и южного Приморья не известен.
34. ACRONICTA
495
Изменчивость выражается главным образом в изменении основного
тона окраски и отчасти резкости рисунка; с Алтая описана почти белая
форма этого вида — altaica Stgr.; сходные особи известны из других мест
Сибири—из Томска и Иркутска; особи с разным оттенком окраски
послужили Татту (1888—1891) для описания цветовых аберраций. Ха-
рактеристика ареала затруднена не внутривидовой изменчивостью, но
в большинстве случаев полным отсутствием внешних отличий от пре-
дыдущего вида; даваемое мною распространение основано на просмотре
коллекционного материала; указания Штаудингера на наличие темной
формы psi L. (suffusa Tutt.) на Дальнем Востоке и Амуре специальным,
исследованием больших материалов по видам этой группы с указанных
мест не подтвердились.
Биология и экология. Зимует куколка; вылет имаго
в южной части ареала (степная зона и лесостёпье) в мае—июне, в лесной
зоне в июле—августе; на юге наблюдается частичное второе поколение,
вылет которого происходит в июле—августе. Яйца откладываются по-
одиночке на нижнюю сторону листьев кормового растения. Гусеницы
многоядны, но наиболее обычны на березах и некоторых розоцветных
(древесных); круг кормовых растений включает Betulaceae (Betula pu-
bescens, В. verrucosa, Alnus incana, A. glutinosa, Gorylus avellana, Ost-
ryia, Garpinus), Fagaceae (Quercus, Fagus), Rosaceae (Sorbus aucuparla,
Rosa canina, Crataegus oxyacantha, Malus domestica, Prunus cerasus, P.
armeniaca, Pyrus domestica), указываются также Salicaceae (Salix, Popu-
lus). На севере преимущественно на березах, на юге на розоцветных и иво-
цветных. Окукление в древесине; куколка диапаузирует. Вредит плодо-
вым культурам; отмечена как вредитель в Астрахани, на Дону, в Рязани,
в Болгарии и Англии; повреждает абрикосы, черешни, вишни, груши
и яблоки; оказывается предпочтение косточковым; отмечалось поврежде-
ние роз в питомниках. Указаны паразиты: из Braconidae — Microplitis
fumipennis Rtzb., M. sordipes Ns., Rhogas pulchripes Wesm.; из Ichneu-
monidae — Coelichnewnon centummaculatus Christ., Ichneumon cessator
Gr., Hemiteles bipunctator Thunb., H. conformis Gmel., Eriobius brac-
catus Gmel., Paniscus cephalotus Holmgr., P. melanurus Thoms., Melopius
migratorius F.
* 13. Acronicta (Triaena) ang-nstimacula Kozhantshikov, sp. nov.
(рис. 256).
Габитуально напоминает виды подрода Hyboma и особенно обыкновен-
ную A. strigosa Schiff., но структурно сочетает признаки подродов Tri-
aena и Hyboma, причем более отчетливо выражены признаки первого под-
рода.
Окраска крыльев и тела светлосерая, рисунок черный и белый; крылья
относительно тела длинные и широкие, передние с заостренным apex;
тело тонкое* стройное; губные щупальца торчат прямо вперед, последний
членик их тонкий и длинный; рисунок на передних х^рыльях следую-
щий: первая поперечная линия короткая, намечена двойным штрихом
лишь на костальном крае крыла; вторая поперечная лилия в виде двух
широко расставленных штрихов и идет от костального края косо кнаружи
до середины крыла, слабо изгибаясь и сливаясь с широким черным штри-
хом, идущим от основания крыла; этот штрих проходит через все базаль-
ное поле, пересекает вторую линию, располагаясь отчасти в срединном
496
ll. ACRONICTINAE
поле; сначала он узкий, но на конце ложковидно расширен; срединное
поле в вентральной половине затемнено, широкое, особенно в середине,
так как третья поперечная линия сильно сдвинута кнаружи, к терминаль-
ному краю крыла; срединная линия намечена на костальном крае корот-
ким растушеванным штрихом между пятнами; круглое пятно овальной
формы, тонко и четко очерчено черным и оторочено изнутри белой поло-
сой, в середине серое; почковидное пятно по размерам лишь немного
более круглого, с неясными границами на костальной и дорзальной
сторонах, очерчено черным, в середине с темным полулунным штрихом;
оба пятна сильно сближены, так что почти не оставляют промежутка;
третья поперечная линия тонкая, слабо зубчатая, с внутренней стороны
оторочена тонкой белой полосой и пересечена маленьким черным и тонким
штрихом на уровне почковидного пятна; она начи-
нается на костальном крае над почковидным пятном,
но затем резко выгнута кнаружи, образуя широкую
петлю, тогда как в дорзальной половине крыла она
сильно сдвинута базально и слита с основанием про-
дольного черного штриха; внешнее поле очень узкое,
значительно менее половины ширины срединного поля,
более темное; подкраевая линия не выражена, от нее
есть лишь остатки в виде немногих белых пятнышек; в
нижней половине срединного поля широкий черный
штрих, заостренный в базальном направлении и ото-
роченный снизу белым; терминальная линия в форме
восьми черных штрихов; бахрома серая; задние крылья
однотонно серые с темными жилками и неясными
растушеванными поперечными полосами; терминальная
линия в виде прерванных черно-серых тонких штрихов;
бахрома белая; нижняя сторона крыльев белая, просве-
чивающая темным на передних крыльях и имеет немно-
гие черные чешуйки; заднее крыло с дискоидальным
пятном в форме точки и с неясной сильно зубчатой
полосой, идущей почти в средней части крыла; ноги
и нижняя часть тела светлосерые; глаза черные.
Рис. 256. Acronicta Размах крыльев 30 мм.
angustimacula, Строение гениталий самки напоминает виды Triaena.
sp. nov. Гениталии Bursa copulatrix мешковидная, вытянутой формы,
самки. с небольшим улиткообразно завернутым расши-
рением к ductus seminalis; расширение это имеет
короткий сосковидный отросточек, направленный проксимально;
ductus bursae широкий, как у видов НуЬота, частично склеротизован
и имеет много сложных складок; восьмой сегмент короткий с довольно
длинными apophyses anteriores; papillae anales узкие, мембранозные,
несущие густые короткие волоски; apophyses posteriores очень короткие
и слабо склеротизованные.
Описана по одной самке хорошей сохранности, взятой В. Гуссаков-
«ским в ущелье Кйндара в Гиссарском хребте 23 августа 1945 г. на высоте
1100 м. Описываемый вид представляется очень своеобразным. Вместе
с чертами специализации он обнаруживает много примитивных черт,
связывая два подрода в роде Acronicta Ochs. Вероятно это реликт третич-
ного времени лесной фауны Средней Азии.
Распространение. Средняя Азия —• Гиссарский хребет
(Кондара).
34. AC RON J СТ A
497
* 14. Acronicta (Triaena) bellula Alph. (рис. 257).
Alpheraky, 1895, Iris, 8 : 189; Staudinger, 1901, Cat. Lep., 1 : 132; Warren, 1909,
Seitz, Grossschmett., 3 : 13; Hampson, 1910, Cat. Lep. Phal., 9 : 530; Мольтрехт, 1929,
Зап. Владивост. отд. Географ, общ., 3 : 52.
Передние крылья, голова и грудь тусклой серо-фиолетовой окраски;
рисунок не резок; первая поперечная линия намечена на костальном
крае штрихом, далее почти редуцирована; от основания крыла вдоль куби-
тального ствола черный штрих, вначале очень резкий, далее тонкий,
достигающий второй поперечной линии; нижняя часть базального поля
черная; вторая поперечная линия двойная, черная, мало интенсивная,
посредине того же цвета, что и общий тон крыла, широкая; круглое
пятно большое, светлосерое, почти правильной формы, очерченное бурым,
сдвинутое дистально к почковидному пятну; срединная линия растуше-
ванная, более заметная у костального края, далее скошенная к почковид-
ному пятну и ниже растушеванная или совсем смытая; почковидное
пятно светлее общего тона
крыла с неясными контура-
ми, более четко и тонко очер-
ченное в базальной половине;
третья поперечная линия на-
мечена на костальном крае .
крыла над почковидным
штрихом двумя растушеван-
ными штрихами, далее внут-
ренняя линия редуцирована,
наружная полоска идет мел-
ко и неправильно зубчатая,
сильно выгнутая посредине,
пересеченная интенсивным
черным штрихом, достигаю- Рис 257 Acronicta ЪМи1а Alph. гениталии
щим tornus, идущим вдоль самца,
анальной жилки; внешнее по-
ле однотонное со слабо заметными светлыми штрихами, остатками подкрае-
вой линии; терминальная линия очень тонкая, буроватая, едва замет-
ная; бахрома светлосерая; задние крылья чисто белые самцов)
с тонкой терминальной полоской и белой бахромой; нижняя сторона
светлосерая, блестящая, с растушеванной срединной полосой и неясными
дискоидальными пятнами. Размах крыльев 30—32 мм.
Uncus тонкий и длинный, склеротизован на дистальном конце и за-
острен; tegumen вытянутый, образует небольшие выросты по сторонам
основания uncus; saccus большой, массивный, треугольный, закругленный
в проксимальной части; fultura inferior глубоко вырезанная па дорзаль-
ном крае, почти вся покрыта мелкими шипиками, ио скульптирована
лишь ее вентральная часть — узкая полоска; valvae большие и широкие,
слабо конические; базальная часть valva несет большую склоротизован-
ную лопасть, в дистальной части срастающуюся с harpe; последняя имеет
вид широкой поперечной склеротизованной складки; harpe сдвинута
к вентральному краю, несет короткий отросток, направленный дистально
и внутрь; дистальная треть valva мембранозная, несет многочисленные
волоски; penis в средней части выгнут дорзально, толстый; coocum корот-
кий и толстый; pars inflabilis несет многочисленные сильные шипы;
сильные шипы есть также на дорзальной стенке penis.
Распространение. Южное Приморье (Уссури, Сидеми, Сучан).
32 Фауна СССР, И. В. Кожанчиков.
498
II. ACRONICTINAE
Очень характерный вид Triaena, известный только по трем самцам.
Стоит обособленно среди видов подрода. Личиночные стадии не известны.
Среди евразийских видов Acronicta Ochs, нет близких видов к bellula
Alph., но среди американских radcliffei Harv. и особенно grisea Wlk.
близки к этому виду. Оба вида распространены в южной Канаде (Мани-
тоба, Албани), в области Великих Озер и в восточных штатах (Массачу-
зеттс, Нью-Йорк, Ныо-Гэмпшайр).1
4. Подрод Hyboma Hb.
Большая часть видов этого подрода более мелка, чем виды других
подродов (исключая hercules Feld.); передние крылья имеют выгнутый
костальный край, внешний край передних крыльев слабо скошен; черные
штрихи на передних крыльях выражены менее резко, чем у двух преды-
дущих подродов; valva неширокая, дистальный конец ее мембранозный
или заостренный; harpe в виде неширокой косой полосы, посередине
несет один или два коротких крючковидно изогнутых отростка.
Число видов, слагающих этот подрод, не велико; в Евразии широко
распространен только один А. (Н.) strigosa Schiff.; прочие палеарктиче-
ские виды этого подрода [А. (Н.) hercules Feld., terrigena Graes., chingana
Drd., phaedra Piingl., phaedriola Drd.] сосредоточены в зоне широколи-
ственных лесов южного Приморья, северного Китая, Маньчжурии, Кореи
и Японии. В Неарктике виды Hyboma Hb. известны в большем числе,
чем из Палеарктики, но также не многочисленны; они сосредоточены
в зоне широколиственных лесов склонов Скалистых Гор и в Аллеганах;
для Америки описаны А. (Н.) albarufa Grt., cwataGrt., modica Wlk., cla~
rescens Gn.. inclara Sm., increta Morr., retardata Wlk., exilis Grt., minella.
Dyar.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Крупный, 60—66 мм; передние крылья светлосерые с неправильными
охристо-серыми мазками, вытянутые, заостренные; valva сильно,
суждеа в дистальной половине, конец ее заострен и несет небольшой
конический вырост.......................* 15 А. (Н.) hercules Feld-
2 (1). Меньших размеров, 30-—45 мм; прочие признаки комбинируются
иначе.
3 (4). Передние крылья, грудь и голова темной тусклосерой окраски;
передние крылья покрыты неблестящей чешуей.....................
................................ * 18. А. (Н.) terrigena Graes.
4 (3). Передние крылья, голова и грудь светлосерые с многочисленными
черными и иногда слабоохристыми штрихами; чешуйчатый покров
блестящий.
5 (8). Harpe с двумя хорошо развитыми отростками; на pars inflabilis
penis нет сильных шипов.
1 Из сборов моего отца под Минусинском мною была получена одна куколка
Acronicta, из которой вылетела самка; она сильно обилась, почему не может быть,
описана достаточно полно. Она несомненно относится к Triaena', более бледной окраски,
чем A. psi L., по размерам такая же; цвет тусклосерый, слегка фиолетовый. По генита-
лиям наиболее близка к A. psi L., но отличается прямым отростком bursa copulatrix,
направленным прямо дистально. Куколка имеет десять шипов на cremaster, чем резко,
отлична от остальных Triaena НЬ.
34. ACRONICTA
499
6 (7). Размах крыльев 30—38 мм; круглое и почковидное пятна желто-
ватые; окраска передних крыльев, головы и груди серая; дорзаль-
ный отросток harpe покрыт волосками......................................
.....................................* 19. А. (Н.) strigosa Schiff.
7 (6). Размах крыльев 36—42 мм; пятна очерчены черным и в середине
окрашены в белый цвет; окраска передних крыльев, головы и груди
бело-серая с черными резкими штрихами и грубым рисунком; дор-
зальный отросток harpe длинный острый и не несет волосков . .
•...........................* 17. А. (Н.) phaedra (Piingl.) Hamps.
8 (5). Harpe с одним длинным дорзальным отростком; penis несет много-
численные шипы на pars inflabilis; рисунок тонкий........................
•....................................* 16. А. (Н.) phaedriola Drd.
* 15. Acronicta (Hyboma) hercules Feld. (рис. 258).
Felder, 1874, Reise Freg. Novara, t. 109, fig. 2; Staudinger, 1892, Mem. Lep. Rom.,
6 : 384; Staudinger, 1901, Cat. Lep., 1 : 131; Hampson, 1909, Cat. Lep. Phal., 8 : 114;
Warren, 1909, Seitz, Grossschmett., 3 : 14, t. 2, k; Draudt, 1931, Seitz, Grossschmett.,
3 SuppL, 1 : 11, t. 1, g; Куренцов, 1936, Тр. Дальневост. фил. Акад. Наук СССР, 1 :
147; —elongate, Oberthiir, 1884, Et. Entom., 10 : 20, t. 2, fig. 3 (var. luteicoma Grt.); —
aceroides Staudinger, 1892, Mem. Lep. Rom., 6 : 385; — (?) iria Swinhoe, 1899, Ann.
Mag. Nat. Hist., 3 (7) : 113.
Передние крылья, грудь и голова светлосерые с примесью охристый
и белых тонов; рисунок на передних крыльях нерезкий; от основания
крыла неясный, растушеванный черный штрих; первая поперечная линия
в виде двух косых темносерых штрихов на костальном крае; вторая попе-
речная линия в виде двух широко расставленных зигзагообразных полос,
сильно сдвинутых в дорзальной части крыла кнаружи; круглое пятно
большое, овальное, тонко и неполно очерчено черным, в середине охри-
стое ,и серое, во внешней части растушеванное, неясное; оно соединено
растушеванным черным штрихом с почковидным пятном; последнее боль-
шое, почти круглое, тонко очерченное черным, в середине серо-охристое,
также смытое во внешней части; срединная линия в виде косого черного
штриха на костальном крае, заметна лишь до почковидного пятна; третья
поперечная линия намечена двумя серыми, нерезкими штрихами на ко-
стальном крае, несколько позади почковидного' пятна, далее резко выгнута
к терминальному краю и затем идет почти параллельно ему двойная,
местами прерванная, сильно зубчатая; против почковидного пятна на этой
линии охристо-серое пятно, а против tornus ее пересекает нерезкий темно-
серый штрих, доходящий лишь до подкраевой линии; подкраевая линия
в виде серии треугольных серых штрихов, местами неясная; терминаль-
ная линия в виде черных неясных штрихов; бахрома бело-серая; задние
крылья белые, несколько тусклые по внешнему краю, жилки серо-охри-
стые; бахрома серая; нижняя сторона белая с неясной серой поперечной
полосой и небольшими дискоидальными пятнами на передних и задних
крыльях. Размах крыльев 50—66 мм.
Uncus недлинный, в средней части несколько вздут, слабо изогнут
и заострен, густо волосистый; tegumen короткий и широкий; есть sca-
phium; fultura inferior почти ромбической формы, с глубокой вырезкой
(двумя отростками) на дорзальной стороне; valvae большие, в средней
части сильно расширенные; в базальной части valva большая склероти-
зованная лопасть; в середине valva продолговатая бородавка, несущая
длинные волоски; harpe в виде складки, соединенной с базальной лопастью
valva, несет два коротких отростка — один направленный дистально,
32*
500
11. ACRONICTINAE
прямой и другой направленный дорзально и изогнутый назад; дистальный
конец valva сильно сужен, уплощен, склеротизован и несет щетку щети-
нок, а на вентральной стороне небольшой прямой вырост; penis длинный,
в средней части выгнут дорзально; pars inflabilis несет многочисленные
мелкие шипики. Bursa copulatrix большая, мембранозная, состоит из
двух половин, из которых правая (in toto) больше; ductus seminalis отхо-
дит с вентральной стороны правой половины от ее середины; основание
ductus bursae сильно склеротизовано; ductus bursae в проксимальной
половине узкий, в дистальной расширен, плоский и сильно склеротизо-
ван; восьмой сегмент узкий и довольно длинный, несет широкие, изогну-
тые apophyses anteriores, которые по длине больше восьмого сегмента;
apophyses posteriores руди-
ментарны; papillae anales
мембранозные, нерезко диф-
ференцированные, несут вен-
чик волосков на дистальном
конце.
Распространение.
Приамурье (Раддевка, Хаба-
ровск, Буреинские горы);
южное Приморье (Виногра-
довка, Седанка, Владивосток,
Аскольд, Ворошилов-Уссу-
рийский, Суйфун); Японские
острова (Хондо); северный
Китай (Пекин). •
Биология и эко-
логия. По Куренцову
(Тр. Горно-таежн. ст.,
3 :107—210, 1939), лёт бабо-
чек во второй половине лета
в ильмовошироколиственной
уреме рек южного Приморья;
гусеницы в августе — сентяб-
ре на листьях Ulmus japo-
Рис.
258. Acronicta hercules Feld. Гениталии:
а — самца, б — самки.
nica; гусеницы каштаново-коричневые, с жесткими черными к концу
заметно утолщенными щетинками, сидят на верхней стороне листьев вяза;
зимовка (по Грэзеру, 1888) в стадии куколки.
* 16. Acronicta (Hyboma) phaedriola Drd. (рис. 259).
Draudt, 1931, Seitz, Grossschmett., 3 Suppl., 1 : 8, t. 1, c.
Передние крылья пепельно-серые с неясными охристыми мазками
и с мелким черным рисунком; от основания крыла узкий черный штрих
до второй поперечной линии; первая поперечная линия в виде двух слабых
штрихов — черного и (снаружи) светлосерого; вторая поперечная линия
сильно скошена кнаружи, особенно в дорзальной половине крыла, широ-
кая, двойная, посредине пепельно-серая, менее ясная в костальной части;
круглое Пятно правильной формы в середине пепельно-серое, очерчено
черным, соединено тонким черным штрихом с почковидным пятном;
последнее частично смыто, особенно во внешней части, в середине охристо-
серое, сильно выгнутое, очерчено черным и белым; третья поперечная
линия сильно выгнута кнаружи, грубо зубчатая с внутренней стороны
пепельно-серая; в срединном поле неясные серые и охристые' штрихи;
34. ACRONICTA
501
внешнее поле узкое; подкраевая линия не развита; терминальная линия
в виде черных точек, заходящих также и на бахрому; бахрома серая.
Задние крылья однотонные тусклосерые с неясными поперечными поло-
сами; нижняя сторона тусклосерая с одной широкой поперечной полосой
и дискоидальными пятнами на передних и задних крыльях. Размах
крыльев 30—32 мм.
Uncus короткий, сосковидный, слабо загнутый, почти голый; tegumen
короткий, особенно в тергальной области; saccus длинный, треугольный,
широкий при основании; fultura inferior длинная, заостренная дорзально;
valvae большие, заостренные на дистальном конце; базальная половина
valva образует широкую склеротизованную лопасть; harpe в виде косой
складки, несущей у дорзаль-
ного края короткий’ посре-
дине изогнутый и направлен-
ный дорзально отросток; ди-
стальная половина valva не-
сет большое количество во-
лосков; дистальный конец ее
склеротизован и заострен;
penis по длине равен valva,
прямой в дистальной полови-
не и выгнут вентрально в
проксимальной; coecum pe-
nis длинный; pars inflabilis
в средней части несет много-
численные большие шипы и
в дистальной части склеро-
тизованную пластинку, уса-
женную мелкими шипиками.
Bursa copulatrix двойная,
левая часть ее мембраноз-
ная, мешковидной формы,
узкая, покрытая рубчатой
скульптурой, несколько су-
женная посредине, в дисталь-
ной части срастается с
большой правой частью,
Рис. 259. Acronicta phaedriola Drd. Гениталии:
.а — самца, б — самки.
сильно склеротизованной, сложно и сильно складчатой, покрытой точеч-
ной скульптурой, почти прямоугольной формы; от этой части bursa copu-
latrix, от ее проксимального конца, отходит ductus seminalis; ductus bur-
sae тонкий, суживающийся в дистальной части; восьмой сегмент длинный,
узкий; apophyses posteriores острые, короче, чем apophyses anteriores,
которые по длине несколько бол'ее половины ductus bursae; papillae
anales мембранозные, нечетко отчлененные, в дистальной части несут
мелкие волоски.
Распространение. Южное Приморье (Сучап, Випоградовка).
Биология не изучена; связан, повидимому, с розоцветными,
так как известна находка одной гусеницы (из которой выведена самка)
на яблоне в Сучане; гусеница не описана.
* 17. Acronicta (Hyboma) phaedra Hamps, (рис. 260).
Hampson, 1910, Cat. Lep. Phal., 9 : 520, t. 17, fig. 32 (Piingeler inlitt.); Филипьев,
1927, Ежегодн. Зоол. муз. Акад. Наук СССР, 27 : 230; Draudt, 1931, Seitz, Gross-
schmett., 3 Suppl., 1 : 8, t. 1, b.
502
II. ACRONICTINAE
Голова, грудь и передние крылья бело-серые (с разбросанными круп-
ными черными чешуями); рисунок передних крыльев резкий, белый
с черным; базальная линия в виде двух широко расставленных коротких
темных штрихов; от основания крыла тонкий черный штрих, слитый
с темнокоричневой нижней половиной базального поля; вторая попереч-
ная линия широкая, растушеванная, грубо зубчатая, посредине белая,
в нижней части дает темный штрих через срединное поле; круглое пятно
большое, белое, в центре слегка серое, очерчено черным; срединная ли-
ния в виде косой растушеванной полосы, идущей к почковидному пятну;
последнее четкое, большое, сильно выгнутое, очерчено черным, в середине
белое и серое; срединное поле в нижней части очень сильно сужено (менее
половины его в средней части), третья поперечная линия начинается
двумя размытыми штри-
хами на костальном крае,
далее очень резко выгну-
та к внешнему краю, гру-
бо зубчатая, отороченная с
базальной стороны белым,
затем резко скошена ба-
зально и делает крупные
зигзаги; наружное поле в
дорзальной части темно -
бурое с неправильно раз-
бросанными белыми чешу-
ями; срединная и верхняя
части серые; подкраевая
линия в виде черных штри-
хов; бахрома серая с по-
лоской; задние крылья бе-
лые, слегка затемненные
по внешнему краю, с тон-
кой коричневой терминаль-
ной полоской; бахрома
задних крыльев белая;
Рис. 260. Acronicta phaedra Hamps. Гениталии: нижняя сторона светлосе-
а — самца, б — самки. рая, блестящая, с тонкой
терминальной полоской.
Размах крыльев 36—42 мм.
Uncus тонкий, сильно загнут вентрально, покрыт короткими воло-
сками; tegumen в дорзальной части короткий, с боков более длинный;
saccus плоский, округленный, небольшой; valvae длинные, слабо выгну-
тые вентрально в середине, округленные в дистальной части; базальная
часть valva образует небольшую лопасть, срастающуюся у вентрального
края valva с основанием harpe; последняя имеет вид косой складки,
которая несет два отростка; оба отростка примерно равной величины,
острые, направленные косо дорзально; вентральный отросток зубчатый;
fultura inferior в виде почти шестигранной пластинки, слабо склероти-
зована посредине и у дорзального края; penis длиннее valva, .толстый,
слабо изогнутый, с коротким отогнутым дорзально coecum и с сильными
многочисленными шипами на pars inflabilis. Bursa copulatrix большая
и двойная; левая половина ее мембранозная, мешковидная, расширенная
в проксимальной части, покрытая рубчатой скульптурой, склеротиза-
ванная в дистальной части; правая половина также склеротизованная,
«
34. ACRONICTA
503
черно-коричневого цвета, несет точечную скульптуру, большая, образует
сосковидное расширение в проксимальной части, от которого отходит
ductus seminalis; ductus bursae широкий при основании, короткий,
несколько суженный в дистальной части; восьмой сегмент короткий с па-
раллельными краями, несет недлинные, острые apophyses anteriores;
apophyses posteriores похожи на первые, но несколько короче; papillae
anales мембранозные, в виде латеральных коротких лопастей с венчиком
щетинок на дистальных концах.
Распространение. Южное Приморье (Уссури, Яковлевка,
Виноградовка, Ворошилов-Уссурийский, Сучан, Тигровая, Владивосток,
Сидеми).
* 18. Acronicta (Hyboma) terrigena Graes.
Graeser, 1892, Berl. entom. Ztschr., 37 : 214; Staudinger, 1901, Cat. Lep., 1 : 132;
Hampson, 1910, Cat. Lep. Phal., 9 : 531 (=strigosa Schiff.); Warren, 1909, Seitz, Gross-
schmett., 3 : 16; Мольтрехт, 1929, Зап. Владивост. отд. Географ, общ., 3 : 53.
Известен только тип; габитуально очень напоминает strigosa Schiff,
и особенно форму bryophiloides Horm. (=casparii Stein.), но имеет ряд
особенностей, вследствие чего до накопления серии особей не может быть
безоговорочно сведён в синонимы к strigosa Schiff. Я даю описание
по типу Грэзера.
Передние крылья, грудь и голова тусклрсерыё; чешуйчатый покров
неблестящий [в отличие от А. (Н.) strigosa Schiff.], рисунок на передних
крыльях тонкий, нерезкий, черный и белый; от основания крыла широкий
черный штрих, несколько растушеванный; первая поперечная линия
намечена в виде слабого тонкого штриха, двойная, черная и светлосерая;
базальный черный штрих пересекает эту линию и достигает второй попе-
речной линии, которая на месте слияния со штрихом становится интен-
сивно черной; она двойная, широкая, суженная лишь у дорзального
края, сильно скошенная кнаружи; внешняя половина более зубчатая,
каждая из полос тонкая, темносерая, почти черная, между полосками
промежуток серый; от дорзальной половины этой линии вдоль анальной
жилки косой черный и светлосерый штрих, почти достигающий третьей
поперечной линии; круглое пятно очень большое, овальное, вытянутое,
не полностью очерчено черным, в середине светлосерое; срединная линия
намечена почти черным изогнутым штрихом на костальном крае, далее
не развита; почковидное пятно размытое, желто-белое, широкое, почти
полулунное, очерчено черным лишь с базальной стороны; расстояние
между круглым и почковидным пятнами малое; третья поперечная линия
тонкая, черная и светлая, начинается над почковидным пятном, затем
резко выгнута кнаружи, мелко, но четко зубчатая; срединное поло в мосте
выгиба третьей поперечной линии вдвое шире, чем у дорзального края;
от дорзального отдела третьей поперечной линии в наружном поло широ-
кий черный штрих вдоль второй кубитальной жилки, достигающий терми-
нальной линии; внешнее поле узкое, почти однотонное, со светлыми
штрихами, неправильно расположенными вдоль жилок; терминальная
линия в виде тонкой полосы, темносерая; бахрома двойная, с тонкой
серой линией посредине. Задние крылья тусклосерыо с неясными,более
темными радиальными штрихами; терминальная линия тонкая, серая;
в середине крыла неясная поперечная линия; бйхрома светлее общего
фона крыла. Нижняя сторона желто-серая с неправильно разбросанными
отдельными крупными чешуями; дискоидальные пятна только на задних
504
JI. ACRONICTINAE
крыльях, в виде темных точек; растушеванная поперечная линия через
оба крыла посередине. Глаза черные; щупальца темносерые. Размах
крыльев самки 30 мм.
Распространение. Приамурье (Благовещенск).
* 19. Acronicta (Hyboma) strigosa Schiff, (рис. 261).
Schiffermiiller, 1776, Wien. Verz. : 88; Aurivillius, 1891, Nordn. Fjar. : 98; Stau-
dinger, 1892, Mem. Lep. Rom., 6 : 385; Staudinger, 1901, Cat. Lep., 1 : 132; Spuler,
1908, Schmett. Eur., 1 : 137, t. 31, fig. 13; Hampson, 1909, Cat. Lep. Phal., 8 : 65;.
Warren, 1909, Seitz, Grossschmett., 3 : 16; Culot, 1909, Noct., 1 : 20, t. 1, fig. 11; Re-
bel, 1910, Berge, Schmetterlingsb., ed. 9 : 143; Chapman, 1914, Entomologist, 47 : 208;
Nordstroem och Wahlgren, 1937, Svenska Fjar : 83, t. 15, fig. 10; —favillacea Esper,,
1788, Schmett. Eur., 4, t. 127, fig. 4; — bryophiloides Hormusaki, 1891, Entom. Nach-
richtenbl., 17 : 145; — casparii, Steinert, 1897, Iris, 10 : 398; — nigrescens Barret,.
1896, Lep. Brit. Isl., 3 : 24; — adaucta Warren, 1909, Seitz, Grossschmett., 3 : 16,
t. 3, e.
Биология и экология. Graeser, 1888, Berl. entom. Ztschr. : 311;
Meyrik, 1895, Brit. Lep. : 141; Barret, 1895, Lep. Brit., 3 : 243, t. 121, fig. 2; Spuler,
1908, Raupen, t. 21, fig. 25; Kuskov, 1927, Beitr. Kunde Estl., 13 : 82.
Голова, грудь и передние крылья светлосерые; рисунок четкий, чер-
ный и белый; базальная линия в виде одной черной полосы, окаймленной
белым, заметна до половины ширины крыла; от основания крыла черный
штрих ниже кубитального ствола до второй поперечной линии; базальное
поле под штрихом темного цвета; вторая поперечная линия широкая,
по краям черная, в середине почти белая, сильно скошена, волнистая,
дрстигает дорзального края крыла на уровне круглого пятна; срединное
поле в нижней части затемнено; круглое пятно желто-белое в середине,
очерчено черным, почти круглое, большое; срединная линия черная,
растушеванная, заметная лишь у костального края; почковидное пятно
довольно большое, в середине однотонное, желто-белое, очерчено серо-
черной полоской, слабо выемчатое по внешнему краю; третья поперечная
линия начинается над почковидным пятном двумя темными косыми
штрихами; внутренняя половина ее редуцирована и за почковидным:
пятном идет в виде тонкой черной полоски, изнутри окаймленной белым,
слабо зубчатой, пересеченной одним черным штрихом под кубитальной
жилкой, достигающим tornus; внешнее поле однотонное, со следами слабо
волнистой белой подкраевой линии; терминальная линия в виде серии
треугольных неслитых между собою штрихов; бахрома серая с полоской;
задние крылья блестящие, белого цвета, с тонкой терминальной полоской;
бахрома белая; нижняя сторона светлосерая, более темная на передних
крыльях в области дискоидальной ячеи; дискоидальные пятна на задних
крыльях едва заметны, на передних не развиты; есть волнистая тонкая
линия, идущая поперек обоих крыльев. Размах крыльев 26—32 мм.
Uncus тонкий и длинный, почти голый; tegumen короткий в дорзаль-
ной части, расширен с боков; saccus маленький, слабо дифференцирован;
fultura inferior мембранозная; valvae большие, округло-эллиптические
в средней части выгнутые вентрально; базальная часть valva склероти-
зована и снабжена короткой лопастью; дорзальный край valva утолщен;
harpe в виде узкой поперечной складки с двумя выростами-— коротким
и округлым дорзальным и более длинным, острым вентральным, направ-
ленным дистально и загнутым внутрь; дистальная половина valva мем-
бранозная; penis короче valva, в средней части выгнут дорзально, толстый,
с коротким coecum; pars inflabilis несет многочисленные острые шипы.
Bursa copulatrix склеротизована, в проксимальной части сужена и по-
крыта рубчатой скульптурой; дистальная часть ее сильно расширена.
34. ACRONICTA
505
склеротизованаи дает мешковидный вырост, направленный проксимально,
от которого отходит ductus seminalis; ductus bursae при основании широ-
кий, в дистальной половине сужен, короткий — около одной трети
длины corpus bursae; apophyses anteriores тонкие, слабо изогнутые, по
длине равны восьмому сегменту; apophyses posteriores рудиментарны;
papillae anales мембранозные, несут мелкие щетинки в виде венчика
на концах.
Взрослая гусеница зеленая, не густо волосистая, с широкой непра-
вильной, коричнево-серой полосой, разделенной на две латеральные
половины; бородавки, покрывающие кожу, мелкие, несколько бблыпих
размеров лишь на втором грудном и первом брюшном сегментах; все они
несут по небольшому чис-
лу (5—9) волосков, сидя-
щих на светлых пупиллах,
окруженных черным коль-
цом; восьмой брюшной
сегмент вздут в виде ко-
нического выроста; дор-
зальная полоса на нем
сильно сужается и на по-
следних двух сегментах
крайне сужена; бородавки
восьмого сегмента также
увеличены в сравнении с
бородавками других брюш-
ных сегментов; нижняя
сторона тела светлозеле-
ная ; ноги зеленые; крючья
на ложных ногах розова-
тые; голова коричнево-
черная; размер взрослой
гусеницы около 2 см.
Рис. 261. Acronicta strigosa Schiff. Гениталии:
а — самца, б — самки.
Куколка светлоко-
ричневая , стройная, не-
сколько более короткая,
чем у видов Triaena Hb.; кожа гладкая; стигмы выдаются слабо; пучки
волосков по сторонам тела короткие, волоски тонкие; брюшные сегменты
несут в базальной части точечную скульптуру; в дистальной части сег-
мента точки мелкие и редкие; последний сегмент брюшка тупой, округ-
лый; cremaster в виде короткого склеротизованного сильно морщинистого
выроста, сидящего с дорзальной стороны последнего сегмента; он песет
восемь острых щетинок — две дорзальных, изогнутых крючковидпо,
и шесть прямых по вентральному краю.
Распространение. Средняя Европа — Альпы, Карпаты;
Британские острова; Ютландия; южная Скандинавия (Швеция); южная
Финляндия (Ниландия); северо-запад Европейской части СССР (Литва,
Латвия, Эстония), далее на восток от линии Ленинград — Молотов (не-
сколько севернее) по всей лесной зоне и лесостепью не часто (Ленин-
град, Псков, Киров, Москва, Калуга, Калинин) и редко в степи (Винница,
Таганрог, Херсон); Средняя Волга (Казань, Ульяновск); Северный
Кавказ; Крым (южный берег); Закавказье (Куссари); Приуралье (Спас-
ское); в Сибири (особенно южной) более обычен всюду по лесной зоне
(Минусинск, Канск, Иркутск); Алтай; Саяны (Тиберкуль, Сухая Гора);
506
JI. ACRONICTINAE
Яблоновый хребет; Забайкалье; Приамурье (Благовещенск); южное При-
морье (Уссури, Сучан, Покровка, Тигровая, Владивосток, Суйфун);
Корея (Гензен); Японские острова (Хоккайдо); Китай (Пекин); одна
особь известна из Средней Азии с этикеткой «Русский Туркестан».
Очень характерный и мало изменчивый вид; хорошо дифференциро-
ванная темная и мелкая форма bryophiloides Нотт, (известная из Буко-
вины, Румынии и южной Германии) может рассматриваться как подвид.1
Биология и экология. Зимует куколка; вылет бабочек
в лесной зоне в июле—начале августа;, рост гусениц в июле—сентябре;
питание гусениц преимущественно розоцветными, как дикими (Sorbus,
Prunus padus, Crataegus), так и культурными (Prunus domestica, Р. cera-
sus, Pirus, Malus); окукление в гнилой древесине или во мху. Куколка
диапаузирует. Отмечена как вредитель плодовых деревьев.
5. Подрод Pharetra Hb.
Виды Pharetra Hb. характерны относительно широкими и короткими
передними крыльями с тупым внешним и прямым костальным краями;
рисунок передних крыльев мелкий. Valva имеет хорошо развитую harpe,
Занимающую базальную ее половину; harpe несет два отростка, расхо-
дящихся и направленных дистально, имеющих различную форму у раз-
ных видов.
Виды этого подрода наиболее многочисленны во всем роде; они засе-
ляют не только лесную зону Голарктики, но также распространены в сте-
пях и даже полупустынях. Наиболее богато виды Pharetra Hb. представ-
лены в зоне широколиственных лесов. Из- Евразии известны A. (Ph.)
catocaloida Graes., metaxanthd Hamps., albistigma Hamps., subornata Leech,
carbonaria Graes., digna Btl., raphael Obth., menyanthidis View., auricoma
Schiff., euphorbiae Schiff., abscondita Tr., cinerascens sp. n. orientalis
Mann., lutea Brem., rumicis L., puloerosa Hamps., nigricans Leech, fas-
ciata Nag., expergita Drd., insitioa Drd., succidens Drd., bicolor Moore, albi-
stigma Hamps., punctinotata Hamps. В Северной Америке виды этого под-
рода представлены также богато, но менее обильны, чем в Евразии
[A. (Ph.) illitaSm., impleta Wlk. (=luteicoma Gtr. et Rob.), sperata Grt.,
noctioaga Grt., emaculata Sm., impressa Wlk., distans Grt., barnasi Sm.,
perdita Grt., edolata Grt., extricata Grt., xyliniformis Gn., oblinita Sm. et
Abb.,2 lanceolaria Grt., insolita Grt., dentata Grt., pyralis Sm., caesarea
Sm., lepetita Sm., hamamelis Gn., othello Sm., brumosa Gn.]. Из тропиков
виды этого подрода совершенно не известны, и лишь из северной Индии
описан A. (Ph.) pruinosa Gn., близкий к обычной A. rumicis L.
1 Описанная из Хингана Драудтом Acronicta chingana Drd. (in Seitz, Gross-
schmett., 3 Suppl., : 8, t. 1, c, 1931), по мнению автора, очень близка к Л. strigosa
Schiff. Отличия указаны лишь в окраске; A. chingana Drd. имеет примесь охристых
тонов у третьей поперечной линии и у темного штриха в дорзальной части средин-
ного поля; самка более крупная, темносерой окраски, с темными задними крыльями;
возможно, что chingana Drd. является синонимом terrigena Graes.; фактический мате-
риал по этому виду отсутствует, описание дано недостаточное.
2 Этот вид был выделен Гротэ (Grote, 1873, Bull. Buff. Soc. Nat. Hist. Sci.,
1 : 81) в особый род — Eulonche Grt.; этому же последовал и Гэмпсон (1909), самне изу-
чавший A. (Ph.) oblinita Sm. et Abb. По Смиту и Дайру (1899), это типичная Acro-
nicta Ochs, группы auricoma Schiff., как и два других вида, относимые Гротэ (1873)
к этому роду. Я имел для исследования A. (Ph.) oblinita Sm. et Abb. (из Виргинии)
и должен вполне присоединиться к мнению Смита. Необходимо указать, что ротовой
аппарат у этих видов[ A. (Ph.) oblinita Sm. et Abb., lanceolaria Grt., insolita Grt,] развит
вполне в отличие от видов Simyra Ochs., с которыми дает сходство форма крыльев
их и рядом с которыми Гэмпсон (1909) помещает род Eulonche Grt.
34. ACRONICTA
507
1
2
3
4
.5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
(4). Задние крылья охристо-желтые или тускложелтые; передние
крылья черные с белым.
(3). Дискоидальные пятна на задних крыльях выражены хорошо,
видны с верхней стороны; через заднее крыло темный штрих, охва-
тывающий и дискоидальное пятно; передние крылья черные с мел-
кими белыми штрихами.................* 20. A. catocaloida Graes.
(2). Дискоидальные пятна на задних крыльях или не развиты, или
заметны в виде слабых штрихов; нет темных продольных штрихов
. на задних крыльях.................* 30. A. lutea Brem. et Grey
(1). Задние крылья чисто белые, серые или со слабым палевым тоном,
но не охристо-желтые.
(8). Третья поперечная линия незубчатая, очень правильной формы,
образует лишь М-образную фигуру в средней части крыла, черная
и белая; передние крылья с сильными коричневыми тонами; задние
крылья палево-серые..
(7). Крупный, 38—52 мм; вентральный отросток harpe недлинный,
острый; penis изогнут слабо, дистальный конец его вздут ....
...........................................* 22. A. digna Btl.
(6). Меньших размеров, 30—35 мм; вентральный отросток harpe
широкий и тупой, короткий; penis сильно изогнут................
.......................................* 23. А. raphael Obth.
(5). Третья поперечная линия зубчатая по жилкам; передние крылья
серые или черные.
(10). Передние крылья узкие, интенсивно черные; задние крылья
чисто белые; отростки harpe длинные, расходятся под прямым
углом; harpe большая, занимает большую базальную часть valva;
penis с многими мелкими шипами на pars inflabilis..............
...................................* 21. A. carbonaria Graes.
(9). Передние крылья обычной ширины, серой или коричнево-серой
окраски; отростки harpe расходятся под острым углом.
(20). Дорзальный и вентральный отростки harpe равны по длине,
или вентральный отросток лишь несколько короче дорзального
(не более чем вдвое).
(13). Отростки harpe загнуты крючкообразно; сильно вздут в дисталь-
ной части............................* 26. A. euphorbiae Schiff.
(12). Отростки harpe прямые или слабо изогнутые, но не крючко-
образные.
(17). Отростки harpe достигают примерно половины длины valva;
лежат во всяком случае не у дистального ее конца.
(16). Дистальный конец valva узкий и длинный, тупой; harpe неболь-
шая, без выемки.............................* 31. A. rumicis L.
(15). Дистальный конец valva острый, дистальная половина ее
коническая; harpe большая, разделена выемкой...................
..................................* A. lutea leucoptera Btl.
(14). Отростки harpe лежат в дистальной трети valva; передние крылья
серого цвета без примеси (или с , небольшой примесью) черных
чешуй.
(19). Отростки harpe равной длины; вентральный отросток harpe
заострен, той же ширины, что и дорзальный; волосяной покров
на груди и брюшке длинный .... * 24. A. menyanthidis View.
(18). Отростки harpe разной длины, широкие; вентральный отросток
508 II. ACRONICTINAE
около половины длины дорзального, широкий, косо срезан на ди-
стальном конце; окраска крыльев однотонно серая, рисунок реду-
цирован; волосяной покров тела обычный...................' . . .
...............................................27. A. abscondita Тг.
20 (11). Вентральный отросток harpe не более четверти длины дор-
зального.
21 (22). Передние крылья бело-серые, задние крылья чисто белые; от-
ростки harpe тонкие, заостренные, несколько загнутые на кон-
цах .......................................* 28. A. orientalis Mann.
22 (21). Передние крылья темносерые, задние тусклосерые; отростки
harpe прямые, вентральный отросток широкий ...........................
........................................* 25. A. auricoma Schiff.
* 20. Acronicta (Pharetra) catocaloila Graes. (рис. 262).
Graeser, 1888, Berl. entom. Ztschr., 32 : 313; Staudinger, 1892, Mem. Lep. Rom.,
6 : 393; Staudinger, 1901, Gat. Lep., 1 : 133; Hampson, 1909, Gat. Lep. Phal.,
8 : 95, t. 125, fig. 13; Warren, 1909, Seitz, Grossschmett'., 3 : 13; Куренцов, 1922, Зап.
Южно-Уссур. отд. Географ, общ., 1 : 28; Draudt, 1921, Seitz, Grossschmett., 3 SuppL,
1 : 9; Куренцов, 1939, Тр. Горнотаежн. ст., 3 : 176.
Передние крылья черно-серые с примесью белых штрихов и отдельных
белых чешуек; базальная линия широкая, двойная, посредине серая,
скошенная; при основании крыла черный штрих не развит; вторая попе-
речная линия широкая, двойная, в середине почти черная, слабо волни-
стая, идущая косо кнаружи в направлении дорзального края; круглое
пятно почти правильной формы, тонко очерчено черным, в середине
белое и черное, белый цвет располагается в виде полумесяца; срединная
линия начинается двумя "растушеванными полосами на костальном крае,
которые далее сходятся, сливаясь, а затем расходятся вновь, сильно
выгнута кнаружи в середине крыла; почковидное пятно большое, во внеш-
ней половине сильно выгнуто (выемчатое), очерчено черным, в середине
черное и серое с черной точкой у внешнего края; подкраевая линия начи-
нается над почковидным пятном, далее резко выгнута к терминальному
краю, двойная, черная, зубчатая по жилкам, посредине светлосерая;
подкраевая линия зубчатая, белая и серая, большими зубцами достигаю-
щая терминальной линии; последняя в виде треугольных штрихов, рас-
положенных по внешнему краю; бахрома бело-серая. Задние крылья
тусклосерые и желтые, затемненные при основании и по краю, с большим
черным дискоидальным пятном; терминальная линия на них черная
и желтая; бахрома желтая; нижняя сторона желто-серая с неясными
черными полосами на передних крыльях и с черным дискоидальным
пятном; задние крылья с нижней стороны с широкой черной каймой.
Размах крыльев 40—52 мм.
Uncus слабо изогнут, цилиндрической формы, слабо покрыт корот-
кими волосками; tegumen короткий и широкий; saccus длинный, кони-
ческий, заостренный, слегка выгнутый дорзально; valvae широкие и боль-
шие; базальная часть valva имеет большую склеротизованную лопасть;
от базального верхнего угла valva косая складка, срастающаяся с пло-
ской harpe, которая несет три отростка — вентральный прямой, направ-
ленный дистально, тупой и широкий; средний широкий, конический, напра-
вленный косо дистально, и дорзальный отросток, тонкий, длинный, сильно
изогнутый; дистальная часть.valva мембранозная, несет густые волоски;
fultura inferior почти шестигранной формы; penis длиннее valva; coecum
penis длинный, в проксимальной части сильно выгнут вентрально; pars
34. ACRONICTA
509
Рис. 262. Acronicta catocaloida Graes. Гениталии:
а — самца, б — самки.
inflabilis penis имеет характер гофрированной склеротизованной трубки,
несущей сильные шипы в дистальной части. Bursa copulatrix мешковид-
ная, искривленная, несколько суженная в средней части, покрыта грубой
рубчатой скульптурой; дистальная часть ее сильно сужена и несет неболь-
шой конический искривленный отросток, от которого отходит ductus
seminalis; ductus bursae широкий и плоский, сильно склеротизованный,
расширенный в дистальной части, короче, чем половина длины corpus
bursae; восьмой сегмент очень короткий и узкий, несет длинные и тонкие
apophyses anteriores; apophyses posteriores рудиментарны; papillae ana-
les мембранозные, слабо дифференцированные от межсегментальной пере-
понки, с венииком щети-
нок в дистальной части.
Распростране-
ние. Приамурье (Хаба-
ровск); южное Приморье
(Уссури, Яковлевка, Вла-
дивосток, Русский остров,
оз. Ханка, Камень-Рыбо-
лов, Ворошилов-Уссурий-
ский); Японские острова —
Хоккайдо (Хакодате).
Очень характерный
вид, стоящий несколько
особняком в подроде и
имеющий признаки как
Pharetra НЬ. так и, от-
части, Triaena Hb.;из Ки-
тая и Японии описано не-
сколько видов, очень близ-
ких к A. (Ph.) catocaloida
Graes., может быть пред-
ставляющих синонимы
этого вида; имеющиеся
описания совершенно не-
достаточны, а фактический
материал по этим видам
мною ' не мог быть получен. Так, из западного Китая описан meta-
xantha Hamps. (Hampson, 1909, Cat. Lep. Phal., 8 : 139, t. 126, fig. 17),
по изображению крайне похожий на A. (Ph.) catocaloida Graes., с Япон-
ских островов описаны два вида A. (Ph.) subornata Leech (1888, Proc.
Zool. Soc. : 477, t. 1, fig. 6) и A. (Ph.) albistigma Hamps. (1909, Cat. Lep.
Phal., 8 :93, t. 125, fig. 12), также похожие на A. (Ph.) catocaloida Graes.
Распространение этих видов, по имеющимся данным, не идет за пре-
делы Китая и северной Японии.
Биология. Гусеницы питаются дубом (Quercus mongo lica); сидят
на верхней стороне листа, наполовину свернувшись. Гусеница темно-
зеленая с короткими коричневыми к концу утолщающимися щетинками.
Куколка зимует.
* 21. Acronicta (Pharetra) carbonaria Graes. (рис. 263).
Graeser, 1888, Berl. entom. Ztschr., 32 : 313; Graeser, 1889, тамже, 33 : 252; Stau-
dinger, 1892, Mem. Lep. Rom., 6 : 380; Staudinger, 1901, Cat. Lep., 1 : 133; Herz, 1904,
Ежегодн. Зоол. муз. Акад. Наук, 9 : 272; Hampson, -1909, Cat. Lep. Phal.,
510
I Г. ACRONICTINAE
8 : 85, t. 125, fig. 2; Warren, 1909, Seitz, Grossschmett., Pal., 3 : 13, t. 2, h; Draudt,
1931, тамже, Suppl., 1 : 8; — brumosa Leech, 1889, Proc. Zool. Soc. Lond. : 476 (non
Grote).
Передние крылья узкие, черно-серые с ясным, но не резким рисунком,
однотонные; первая поперечная линия двойная, несколько выгнутая,
зубчатая; вторая поперечная линия также двойная, широко расставлен-
ная, слабо волнистая, посредине черно-серая, слабо скошенная кнаружи;
круглое пятно неправильной формы, почти овальное, очерченное интен-
сивно черным; почковидное пятно большое, темносерое в середине, слабо
выемчатое снаружи, очерчено тонкой черной линией и пересечено средин-
ной растушеванной полосой; последняя хорошо развита ,;в»с редней части
Рис. 263. Acronicta carbonaria Graes. Гениталии:
а — самца, б — самки.
крыла выгнута кнаружи; третья поперечная линия начинается над почко-
видным пятном, далее резко выгнута кнаружи и затем идет параллельно
внешнему краю, зубчатая по жилкам, двойная, посредине светлосерая;
подкраевая линия с внутренней стороны белая, мелко зубчатая, слабо
волнистая; терминальная линия в виде мелких точек, широко расставлен-
ных; бахрома черно-серая; внешнее поле передних крыльев узкое, более
узкое, чем базальное; задние крылья чисто белые с тонкой коричнево-
серой (или черной) терминальной линией; бахрома белая. Нижняя сто-
рона белая с черно-серыми радиальными штрихами на передних крыльях
й слабыми дискоидальными пятнами, а также немногими темными штри-
хами на задних. Размах крыльев 36—40 мм.
Uncus длинный, изогнутый лишь при основании, заострен лишь
его дистальный конец; tegumen короткий; saccus массивный, в прокси-
мальной части тупой; fultura inferior в виде широкой пластинки, плоско
срезанной в дорзальной части и заостренной в вентральной; valvae уз-
кие и длинные, в середине выгнутые дорзально; базальная часть valva
образует большую склеротизованную лопасть, срастающуюся с harpe;
34. ACRONICTA
511
последняя находится в средней части valva, несет два отростка — вент-
ральный, направленный дистально, тупой, прямой и толстый, и дорзаль-
ный, направленный косо вверх, тонкий, сильно изогнутый; дистальная
половина valva тупая, мембранозная, несет густые волоски, более грубые
на дисталЬном конце; есть scaphium; penis длиннее valva, тонкий, слабо
расширяющийся дистально, с тонким недлинным coecum; pars inflabi-
lis penis несет многочисленные густо расположенные сильные шипы,
образующие (при покойном положении pars inflabilis) подобие чешуи.
Bursa copulatrix мембранозная, почти прямоугольной формы, несколько
суженная в дистальной части, где несет сосковидный отросток, направ-
ленный в сторону, от которого отходит ductus seminalis; ductus bursae
широкий и плоский, по длине не более половины corpus bursae, несет
рубчатую скульптуру; восьмой сегмент короткий, несет тонкие, длинные
и прямые apophyses anteriores; apophyses posteriores короткие и острые;
papillae anales мембранозные, в проксимальной части слабо дифференци-
рованные от сегментарной перепонки, несут густые волоски.
Распространение. Приамурье (Хабаровск); Японские ост-
рова — Хондо (Иокогама); Корея.
В литературе описано два близких к A. (Ph.) carbonaria Graes. вида —
из западного Китая (Дадзянлу, Вахан) nigricans Leech (1900, Trans. En-
tom. Soc. Lond., 37 : 104) и из Гималаев (Кашмир) bicolor Moore (1881,
Proc. Zool. Soc. Lond. : 332). Оба вида изучены очень слабо и известны
только в типах; по изображениям и описаниям есть все основания счи-
тать их синонимами carbonaria Graes.; структурные признаки их не опи-
саны и никем не изучались, вследствие чего окончательные выводы сде-
ланы быть не могут.
* 22. Acronicta (Pharetra) digna Btl. (рис. 264).
Butler, 1881, Trans. Entom. Soc. Lond., 19:176; Staudinger, 1892, Mem. Lep.
Rom., 6 : 389; Staudinger, 1901, Cat. Lep., 1 : 133; Herz, 1904, Ежегодн. Зоол. муз.
Акад. Наук, 9 : 271; Hampson, 1909, Cat, Lep. Phal., 8 : 116; Warren, 1909, Seitz,
Grossschmett., Pal., 3 : 14, t. 2, i; Draudt, 1931, тамже, SuppL, 1 : 12; — michael Ober-
thiir, 1884, Et. Entom., 10 : 18, t. 2, fig. 13.
Передние крылья пепельно-серые с черно-бурым, четким рисунком;
от основания крыла вдоль срединной жилки интенсивно черный штрих
до второй поперечной линии; первая поперечная линия намечена на ко-
стальном крае двумя черными штрихами; вторая поперечная линия двой-
ная, слабо извитая, почти перпендикулярная продольной оси крыла;
нижняя часть срединного поля затемнена; круглое пятно небольшое, пра-
вильной формы, очерчено черным, в середине серое с черной точкой
в центре) срединная линия ясная, неширокая, проходит через почковид-
ное пятно', извита параллельно третьей поперечной линии; почковидное
пятно большое, нередко стушеванное в верхней и нижней частях, неполно
очерчено черным, нередко размытое; третья поперечная линия очень
четкая, простая, бархатисто-черная, с внутренней стороны оторочена
белым, начинается темным над почковидным пятном, далее резко выгнута,
параллельно костальному краю и затем скошена вниз и базально, в сред-
ней части дает резкую М-образную фигуру; внешнее поле довольно широ-
кое, в базальной части затемнено, на костальном крае мелкие белые
пятна; подкраевая линия светлая, растушеванная, волнистая; терми-
нальная линия в виде мелких черных точек; бахрома коричнево-серая
с черными штрихами; задние крылья у самца белые, у самки серые; осно-
вание и внутренний край задних крыльев с тусклым палевым оттенком;
512 II. ACRONICTINAE
бахрома бело-серая. Нижняя сторона передних крыльев буро-серая с бе-
ловатым налетом по внешнему краю; задние крылья у самца белые,
у самки с темными перевязями — широкой по внешнему краю и узкой
посредине; дискоидальные пятна на задних крыльях не резкие, но ясные.
Размах крыльев 38—44 мм.
Uncus тонкий и длинный, почти голый, сильно загнут вентрально;
tegumen на дорзальной стороне сильно склёротизован, по сторонам uncus
образует маленькие сосковидные выросты; латеральные части tegumen
широкие; saccus недоразвит; fultura inferior щитовидной формы, с не-
большим гребневидным выростом в средней части; valvae длинные, слабо
Рис. 264. Acronicta digna Btl. Гениталии:
а — самца, б — самки.
конические; базальная половина valva несет широкую склеротизованную
лопасть; harpe в виде косой складки, идет от верхнего базального угла
valva до середины ее вентрального края, срастается с базальной лопастью;
она несет один длинный изогнутый внутрь отросток, направленный косо
дистально; при основании отростка harpe есть пучок волосков; дисталь-
ная половина valva сужена и склеротизована, несет многочисленные
волоски; penis короче valva, вздут в дистальной трети, почти прямой;
coecum короткий; pars inflabilis penis склеротизована и несет мелкие хити-
новые зубчики. Bursa copulatrix большая, мешковидная, мембранозная,
раздутая в средней части, в дистальной части несет сосковидный спи-
рально закрученный.вокруг основания ductus bursae отросток, покрытый
рубчатой скульптурой, от которого отходит ductus seminalis; ductus
bursae почти прямой, короткий, склеротизован в виде продольных тяжей;
восьмой сегмент короткий и узкий; apophyses anteriores тонкие и прямые,
около половины длины ductus bursae; apophyses posteriores тонкие и за-
остренные, короче первых; papillae anales мембранозные, короткие,
покрытые короткими волосками.
34. acronicta
613
Распространение. Южное Приморье (Яковлевна, Черни-
товка, Сидеми, Владивосток); Приамурье (Хабаровск); Японские ост-
рова — Хондо (Иокогама); Корея (Гензен).
* 23. Acronicta (Pharetra) raphael Obth. (рис. 265).
Oberthiir, 1884, Et. Entom., 10 : 19; t. 2, fig. 9; Staudinger, 1892, Mem. Lep. Rom.,
<6 : 388; Staudinger, 1901, Gat. Lep., 1 : 133; Herz, 1904, Ежегодн. Зоол. муз. Акад.
Наук, 9 : 271; Hampson, 1909, Gat. Lep. Phal., 8 : 147; Warren, 1909, Seitz, Gross-
schmett., 3 : 15, t. 3, e; —fixseni Graeser, 1888, Berl. entom. Ztschr.. 32 : 311; —
raphaelis Hampson, 1909, Gat. Lep. Phal., 8 : 133; — cubitata Warren, 1914, Nov.
Zool., 21 : 402.
Основной цвет передних крыльев, груди и головы коричнево-серый;
рисунок передних крыльев интенсивно черный, четкий; базальная линия
в виде двух широко расстав-
. ленных коротких, черных
штрихов; вдоль срединной
.жилки от основания крыла
'бархатисто-черный штрих,
достигающий второй попереч
ной линии; нижняя часть ба-
зального поля темная, почти
черная; вторая поперечная
линия прямая, слабо волни-
стая, двойная, широко рас-
ставленная, посредине серая;
круглое пятно большое,
•овальной формы, неполно
очерчено бархатисто-черным
ж белым, в середине темносе-
рое; срединная линия чер-
ная, растушеванная, выгну-
та кнаружи и слита' с почко-
видным пятном, далее идет
косо базально; почковидное
пятно в виде темного непра-
Рис. 265. Acronicta raphael Obth. Гениталии:
а — самца, б — самки.
вильной формы пятна, слабо
выемчатого с внешней стороны, неполно очерчено черным, в середине темно-
бурое, с немногими черными чешуйками; третья поперечная линия слабо
волнистая, но крупно-угловатая, начинается над почковидным пятном
тонкими белой и черной полосками, выгнута кнаружи, далее интенсивно
-бархатисто-черная, очерченная с внутренней стороны белым, посредине
дает неясную М-образную фигуру; внешнее поле в базальной половине
темное, во внешней светлосерое с тонкой неправильной белой подкрае-
вой полоской; терминальная линия в виде тонкой черной полосы, почти
непрерывная; бахрома серая; задние крылья тусклые, темносерые, более
темные по внешнему краю, с тонкой терминальной полоской; бахрома
желто-серая; нижняя сторона тусклосерая с неправильными темными
и светлыми полями, одной поперечной волнистой белой полосой и неяс-
ными лунными пятнами на каждом крыле. Самки более темные, нередко
со смытым, неясным рисунком. Размах крыльев 35—39 мм.
Uncus короткий, почти прямой, слабо расширенный в средней части,,
на конце заостренный; tegumen длинный, особенно в латеральных обла-
стях; saccus маленький, округленный в проксимальной части, прямой;
33 Фауна СССР, И. В. Кожанчиков.
514
II. ACRONICTINAE
valvae большие, выгнутые в середине вентрально; базальная половина
valva несет склеротизованную складку, срастающуюся с harpe; последняя
в виде длинной складки, идущей от дорзального края valva, покрыта
большим числом мелких волосков; она несет два отростка — вентраль-
ный, короткий и тупой, срастающийся с вентральным краем valva, направ-
ленный косо вентрально, и дорзальный отросток, направленный косо
дорзально, тонкий, выгнутый, конический; fultura inferior в виде пла-
стинки, несущей в средней части киль; penis короче valva, в середине
сильно выгнут дорзально, утолщающийся к дистальному концу; coecum
penis короткий; pars inflabilis слабо склеротизован, без зубцов. Bursa
copulatrix в виде большого мембранозного пузыря, расширенного в про-
ксимальной части, без скульптуры на поверхности; дистальная часть ее
несет сосковидный дистально направленный отросток, покрытый склеро-
тизованными складочками, от дистального конца которого отходит ductus-
seminalis; ductus bursae широкий,, расширенный в области ostium, по-
крыт склеротизованными складочками; восьмой сегмент короткий, несет
тонкие почти прямые apophyses anteriores превышающие длину кольца;
apophyses posteriores острые, сидят на длинном и широком основании;
papillae anales слабо дифференцированные, мембранозные, короткие
и широкие, с венчиком щетинок на дистальном конце.
Распространение. Яблоновый хребет (Усть-Майская); При-
амурье (Раддевка, Хабаровск, Вятское); южное Приморье (Уссури, Су-
чан, Яковлевка, Сидеми, Суйфун, Ворошилов-Уссурийский, Владивосток^
Аскольд); Малый Хинган; Корея.
Габитуально очень заметный вид, на первый взгляд стоящий особ®
в роде; по структурным признакам очень близок к A. (Ph.) menyanthidis
View. Территориально он замещает A. (Ph.) тепу ant'. IdisNiQ^. на востоке;
Азии. 1
* 24. Acronicta (Pharetra) menyanthidis View. (рис. 266, 267).
Vieweg, 1790, Tab. Verz. Brand. Schmett., 2, t. 2, fig. 1, 2; Aurivillius, 1891, Nordic
Fjar. : 99; Staudinger; 1901, Cat. Lep., 1 : 132; Spuler, 1908, Schmett. Eur., 1 : 138,
t. 31, fig. 17; Culot, 1909, Noct. Eur., 1 ; 21, t. 1, fig. 15; Hampson, 1909, Cat. Lep.
Phal., 8 : 134; Warren, 1909, Seitz, Grossschmett., 3 ; 17, t. 3, h; Rebel, 1910, Berge,
Schmetterlingsb., ed. 9 : 145; Petersen, 1924, Lep. Fauna Estl., ed. 2 (1) : 151; Draudt,,
1931, Seitz, Grossschmett., 3 Suppl., 1 : 12; Nordstroem och Wahlgren, 1937,.
Svenska Fjar. : 84; — obsoleta Tutt, 1891, Brit. Noct., 1 : 24; — scotica Tutt, 1891,.
тамже, 1 : 24; — sufjusa Tutt, 1891, тамже, 1 : 24; — sartorii Hockmeyer, 1904, Entom.,
Ztschr., Guben, 18 : 29, fig. 3, 4; — oirga Gillmer, 1905, тамже, 18 (32) : 128; — ardu-
enna Gillmer, 1905, там же, 18 (33) : 129; — jaeschkei Kujan, 1917, Intern, entom.
Ztschr., 10 : 141; — nigra Schaefer, 1925, Intern, entom. Ztschr., 18 ; 276..
Основной цвет пепельно-серый с примесью черных тонов; рисунок
на передних крыльях черный; базальная линия двойная, очень слабая,
посредине светлая; черный штрих при основании крыла не развит или
очень слабый; вторая поперечная линия широкая, двойная, посредине-
светлая, слабо скошенная кнаружи, слабо волнистая, в дорзальной части
крыла нередко смытая; круглое пятно очень маленькое, тонко обведено
черным, внутри светлое; срединная линия сильно растушеванная, широ-
кая, идет через почковидное пятно к третьей поперечной линии и далее
параллельно ей к дорзальному краю крыла; почковидное пятно неболь-
шое, неправильной формы, несколько расширенное в костальной поло-
вине, очерчено черным, внутри сначала светлосерое, в центре темносерое;
третья поперечная линия начинается на костальном крае на уровне дш
стального края почковидного пятна, сначала двойная, внутри светлая,.
34. ACRONICTA
515
далее простая, интенсивно черная, четко зубчатая по жилкам,с внутрен-
ней стороны оттенена широкой светлой полосой; в области кубитальных
жилок черный штрих, пересекающий третью поперечную линию; внеш-
нее поле однотонное или имеет светлую подкраевую полоску; терминаль-
ная линия очень тонкая й слабая; бахрома шашечная — черная и очень
тонкая и слабая; задние крылья однотонные, грязно-серые со светлой
бахромой; нижняя сторона однотонная, черно-серая с примесью немно-
гих беловатых чешуй при основании крыльев. Волосяной покров густой
Рис. 266. Acronicta menyanthidis View. Гениталии:
а — самца, б — самки.
и длинный на всем теле; волоски тонкие и нежные. Размах крыльев
35—40 мм.
Uncus тонкий, в средней части несколько утолщен, почти голый; tegu-
men длинный и узкий, при основании uncus по сторонам его дает по од-
ному небольшому выросту; saccus короткий и толстый, закругленный
в проксимальной части; valvae прямые, узкие, довольно длинные,
несколько заостренные на дистальном конце; основание valva несет ши-
рокую склеротизованную лопасть; harpe в виде двух косых складок —
дорзальной, идущей от верхнего базального угла valva, и вентральной,
идущей от базальной лопасти; на месте срастания этих складок находится
два отростка — один длинный, слабо изогнутый, направленный косо
33*
616
II. ACRONICTINAE
дорзально, второй почти прямой, более короткий, недостигающий конца
valva вентральный отросток; fultura inferior в виде округлой пластинки,
несущей в средней части киль;, penis короче valva, несколько утолщен
в дистальной части и выгнут дорзально в средней; coecum очень корот-
кий и тонкий; pars inflabilis penis несет многочисленные острые и силь-
ные шипы. Bursa copulatrix большая, расширенная в проксимальной
части, в дистальной половине образует вырост, дистальный конец кото-
рого подогнут и охватывает снизу (in toto) основание ductus bursae;
ductus seminalis отходит от конца этого отростка совокупительной
сумки; ductus bursae широкий, несколько изогнутый, складчатый; вось-
мой сегмент короткий и широкий, несет почти прямые короткие apophy-
ses anteriores, цо длине равные сегменту; apophyses posteriores, очень
тонкие и острые, слабо склеротизованные; papillae anales мембранозные,
слабо дифференцированные, не-
сут сильные щетинки в дисталь-
ной части.
Яйца полушаровидные, диа-
метром 0.5—0.8 мм, с радиаль-
ными бороздками на хорионе;
беловато-желтые сразу после
откладки, спустя три-четыре дня
покрываются неравномерными
пятнами темнокоричневого цве-
та.
Гусеницы первых трех ста-
дий серой окраски с немноги-
Рис. 267. Acronicta menyanthidis View. Хето- ми черными волосками; стар-
таксия передне- и среднегруди и брюшных шие стадии гусениц черной
сегментов I, VI, VII и VIII взрослой гусе- п j ^ г
ницы окраски, покрыты длинными
волосками, сидящими на боль-
ших бородавках; стигмы белые, резко выделяются на черном фоне
кожи; под дыхальцами идет широкая яркокрасная полоса; ноги и
голова черные^
Куколка почти черная, слабокоричневая между сегментами; голова
несет густую рубчатую скульптуру в виде шагренировки и на темени два
морщинистых бугра; среднегрудь несет скульптуру в виде тонких скла-
дочек; брюшные сегменты несут скульптуру в виде большого количества
острых шипиков и бугорков; дистальный край брюшных сегментов (одна
четверть длины сегмента) гладкий; cremaster в виде короткого выроста,
несущего два дорзальных и два вентральных конических бугорка; на се-
редине cremaster расположен пучок коротких прямых и острых щетинок,
направленных дистально.
Распространение. Средняя Европа (Альпы, Германская
низменность, Ютландия); Британские острова (Валлис, Шотландия);
Скандинавия (Швеция, Норвегия, Финляндия, Лапландия Шведская
и Финская); северо-запад Европейской части СССР (Эстония, Литва,
Латвия); далее к востоку только по зоне хвойных лесов (Архангельск,
Петрозаводск, Ленинград, Псков, Новгород, Тула, Чернигов); Урал; Ал-
тай (Джайдак); в лесной зоне Сибири немногие точки — Саяны (Буйба,
Казыр-сук), Иркутск, Витим; Амур (оз. Окупай); Шантарские острова
(Б. Шантар). Есть указание Дрезеке для западного Китая (Квансиен);
в моих руках материала из Китаяцо этому виду не было; указание может
быть*не точно в смысле определения вида.
34. ACRONICTA
517
Биология и экология. Лёт бабочек в мае—июне; яйца
откладываются кучками по 3—15 шт. на нижнюю сторону листьев;
наиболее часто яйцекладка на листья ивы. Гусеницы расползаются с пер-
вой же стадии и живут поодиночке. Кормовые растения для младших
стадий виды ив (Salix nigricans, S. triandra, S. repens, S. daphnoides),
Myrica gale, Betulaceae (Betula pubescens, B. verrucosa, B. humilis); для
старших стадий указывается также питание Rosaceae (Crataegus), Erica-
ceae (Vaccinium, Calluna), Compositae (Tussilago), Gentianaceae (Menyan-
thes). Окукление в плотном коконе в листьях; куколка диапаузирует
и зимует; дает всегда одно поколение. Типичной стацией обитания яв-
ляются окраины торфяников, поросшие ивовым и березовым кустарни-
ком; гигрофилен; подсыхающие болота заселяет слабо.
* 25. Acronicta (Pharetra) auricoma Schiff.1 (рис. 268, 269).
Schiffermulier, 1776, Verz. Schmett. Wien, : 67; Aurivillius, 1891, Nordn. Fjar.:
99, t. 32, fig. 9; Staudinger, 1892, Mem. Lep. Rom., 6 : 390; Staudinger, 1901, Gat.
Lep., 1 : 133; Spuler, 1908, Schmett. Eur., 1 : 138, t. 31, fig. 18; Culot, 1909, Noct.,
1 : 21, t. 1, fig. 16; Hampson, 1909, Cat. Lep. Phal., 8 : 132; Warren, 1909, Seitz, Gross-
schmett., 3 : 16, t. 3, f; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 145; Draudt,
1931, Seitz, Grossschmett., 3 SuppL, 1 : 11, t. 1, h; Nordstroem och Wahl-
gren, 1937, Svenska Fjar. : 84, t. .15, fig. 14; — lapathi Schrank, 1802, Fauna Boica :
308; — similis Haworth, 1809, Lep. Brit. ; 180;— menyanthidis Haworth, 1809, Lep.
Brit. : 180 (non Vieweg);—pepli Hiibner, 1818, Eur. Schmett., Noct., fig. 614; —
alpina Freyer, 1858, Neue Beitr. Schmett., t. 623; — pyhaevaarae Hoffman, 1893,
Stett. entom, Ztg., 54 : 126; — vernalis Frings, 1905, Soc. Entom., 20 : 73; — basi-
striata Kujan, 1931, Warnecke, Verh. Nat. Heimat Hamb. : 134.
Основной цвет головы, груди и передних крыльев черно-серый; ба-
зальная линия на передних крыльях двойная, черная, широко расстав-
ленная, посредине светлосерая; от основания крыла черный штрих,
несколько расплывчатый во внешней части; вторая поперечная линия
почти прямая, двойная, черная, в середине светлая, слабо волнистая,
слабо скошенная кнаружи; срединная линия проходит между пятнами,
черная, хорошо заметная на костальном поле, ниже слабо извитая, расту-
шёванная, иногда совсем редуцированная; круглое пятно светлосерое,
в середине с темной точкой; почковидное пятно темносерое, в середине
кольчатое; оба пятна очерчены черным; третья поперечная линия начи-
нается над серединой почковидного пятна двумя черными штрихами и
белым между ними, далее идет простая, черная, слабо зубчатая, дающая
М-образную фигуру в области М3 и С,, в дорзальной части крыла сильно
. скошена базально; срединное поле узкое по краям и расширенное в сере-
дине; подкраевая линия в виде неясной и неправильной светлой полоски,
1 По Смиту (Smith, 1893, Cat. Lep. Noct. Boreal America : 43) синонимом к пале-
арктической A. auricoma Schiff, является неарктическая brumosa, описанная Гена
(Guenee, 1852, Noct., 1 : 52). Это принимает и Штаудингер (Staudinger, 1901, Cat.
Lep. Pal., 1 : 133). Между тем brumosa Gn. представляет, по Гэмпсону (Hampson,
1909, Cat. Lep. Phal., 8 : 78, t. 124, fig. 27), синоним persuasa Harvey (1875, Bull.
Buffalo Soc. Natur. Sci., 2 : 271), считавшейся Смитом (1893) за самостоятельный
вид. Я не имел возможности проверить по фактическому материалу отношения пале-
арктической auricoma Schiff, к неарктическим brumosa Gn. и persuasa Harv., равно-
как и синонимику последних. В связи с этим brumosa Gn. мною исключена из форм
вида auricoma Schiff., тем более, что распространение этого вида в Палеарктике со-
всем не говорит за обязательное его присутствие в Северной Америке. В то же время
изображение brumosa Gn. (= persuasa Harv.) у Гэмпсона (1909) характеризует вполне
специфичную форму.
518
11. ACRONICTINAE
делящей пополам внешнее поле; терминальная линия в виде серии чер-
ных шашечек, заходящих на бахрому; последняя серая с тонкой полос-
кой; задние крылья тусклосерые, несколько более, светлые в базальной
части, бахрома белая или светлая; нижняя сторона, блестяще-серая,
костальный край передних крыльев несколько осветлен, с немногими
крупными черными штрихами; на задних крыльях темные дискоидальные
пятна; бахрома снизу чашечная — белая и черная. Размах крыльев
32—36 мм.
Uncus почти цилиндрический, несет короткие редкие волоски; tegu-
men узкий и недлинный, особенно короткий в тергальной области, обра-
зует два небольшие выроста по сторонам основания uncus; saccus плос-
кий, маленький, округлен в проксимальной части; fultura inferior в виде
овальной пластинки, выре-
занной на дорзальном
крае, в дорзальной поло-
вине несет на поверхности
мелкие шипики; valvae
большие, округлые в ди-
стальной половине, слабо
выгнутые вентрально в
средней части; базальная
часть valva несет склеро-
гизованную лонасть, ко-
торая в дистальной части
срастается с harpe; послед-
няя в дистальной части
valva образует два расхо-
дящихся отростка, направ-
ленных косо дистально;
вентральный отросток ко-
роткий, прямой и тупой,
дорзальный — длинный,
тонкий и изогнутый; ди-
стальный конец valva мем-
бранозный; penis короче
valva, с коротким округ-
лым coecum, в дисталь-
ной трети изогнутый дор-
зально, вздутый и склеротизованный на дистальном конце; pars infla-
bilis несет многочисленные зубцы. Bursa copulatrix короткая, широ-
кая, мешковидная, покрытая слабой рубчатой скульптурой, сужена
в дистальной части, где образует направленный дистально сосковидный
отросток, от которого отходит ductus seminalis; ductus bursae короткий
и широкий, складчатый; восьмой сегмент неширокий, короткий, его
тергит слабо склеротизован; apophyses anteriores тонкие, по длине рав -
ные ductus bursae; apophyses posteriores слабые, рудиментарные, короче
первых; papillae anales слабо дифференцированные, мембранозные.
Яйца полушаровидные, прикрепляются плоской стороной к субстрату;
выпуклая сторона несет рубчатую скульптуру в виде радиальных скла-
док; сразу после откладки яйцо серо-беловатое, спустя несколько суток
появляются охристые пятна.
Гусеницы первых трех стадий серые с немногими белыми и черными
пучками волосков; взрослые гусеницы изменчивы в окраске, но двух
34. ACRONICTA
519
•основных типов — рыжие, покрытые светложелтыми бородавками, и
черные, имеющие окраску волосков черную и черно-бурые бородавки;
окраска кожи и черепной коробки у первых (рыжих) светлая, желтова-
тая, ноги светложелтые, у черных гусениц кожа и черепная коробка,
а также ноги черные и бурые; есть особи, имеющие окраску волосков
желтую, но окраску кожи и бородавок темную или черную; иногда жел-
того цвета волоски и бородавки, а кожа темная. Признак окраски гусениц
•является наследственным и не связан с возрастными изменениями при-
знаков гусениц; особи, имевшие темную окраску гусениц, в моих экспе-
риментах в трех поколе-
ниях давали только чер -
ных гусениц (число особей
много сотен).
Куколка . черно-корич-
невая; брюшные сегменты
покрыты скульптурой в
виде неправильных и раз-
ных по величине ямок;
грудные сегменты несут
морщинистую скульптуру;
•cremaster в виде коротко-
го плоского выроста, не-
сущего по сторонам по две
маленьких пластинки; ди-
стальная часть cremaster
покрыта многочисленны-
ми короткими и остры-
ми незагнутыми щетин-
ками.
Распростране-
ние. Западная и средняя
Европа от Пиренеев (где
всюду до Португалии),
всюду в Альпах, Карпатах,
по Германской низменно-
сти, Ютландии и Сканди-
навии до Норвегии и Лап- Рис. 269, Acronicta auricoma Schiff. Хетотаксия пе-
•ландии; побережье Среди- редне- и среднегруди и брюшных сегментов I, VI,
земного моря (в Европе) VI1 и VIH гусеницы первой стадии (а) и взрос-
до средней Италии; Бри- лои ’’
танские острова; Фин-
ляндия; в СССР всюду в Прибалтике (Литва, Латвия, Эстония) и
далее на восток от южного берега Крыма и Северного Кав-
каза всюду по степной и особенно по лесной зонам до Архангельска,
Усть-Сысольска и Молотова (Одесса, Херсон, Чернигов, Москва, Яро-
славль, Казань, Петрозаводск, Псков, Ленинград, Новгород, Киров);
Северный Кавказ (Кубань, Кисловодск); Закавказье (Боржоми, Баку-
риани, Армения); Приуралье (Чкалов, .Тюмень, Свердловск); в Сибири
указана преимущественно для горных районов — Алтая (Чемал, Онгу-
дай, Бухтарма, Усть-Каменогорск), Саян (Буйба, Казыр-сук, Тиберкуль,
Сухая Гора), Яблонового хребта (Джелинда); в лесной зоне и лесо-
степье Сибири указана для Минусинска, Томска, Иркутска, Охотска;
Камчатка; Амур-(Николаевск); указание Штаудингера (1901) для Вороши-
520
JI. ACRONICTINAE
лова-Уссурийского не подтверждено сборами Куренцова, Филипьёва и.
Дьяконова и, может быть, ошибочно.1
Биология и экология. Зимует куколка (в состоянии
диапаузы); вылет бабочек в мае—июне; яйцекладка на нижнюю сторону
листьев кормовых растений (чаще ивы и березы) кучками по 3—10 яиц;,
гусеницы поодиночке сидят непрочно (легко стряхиваются) на листьях
кормовых растений; кормовые растения разнообразны, преимущественно’
виды берез (Betula pubescens, В. verrucosa, В. humilis) и ив (Salix nigri-
cans, S. fragilis, S. daphnoides); кроме того, отмечено питание видами
Populus, Carpinus, Quercus, Sorbus, Prunus, Ulmus, Myrica, Juglans,
Crataegus, Pirus и совсем редко Rosa, Rubus, Calluna, Vaccinium. Окук-
ление в сухих листьях в подстилке, гусеница плетет плотный кокон,
вплетая в него кусочки коры, сухие листья (которые обычно свертываются}
и отпадающие волоски. В зоне хвойных лесов дает одно, а на юге два.
поколения; в лесостепье и южнее всегда два и более. Типичной стацией
являются лесные опушки, окраины болот (старых торфяников), покрытые-
верещатниками и кустарником из березы и в меньшей мере ивы. Указаны,
паразиты из Ichneumonidae (Platylabus borealis).
* 26. Acronicta (Pharetra) euphorbiae Schiff.2 (рис. 270).
Schiffermiiller, 1776, Verz. Schmett. Wien : 67; Aurivillius, 1891, Nordn. Fjar.,.
99; Staudinger, 1901, Cat. Lep., 1 : 133; Spuler,• 1908, Schmett. Eur., 1 : 139.
t. 31, fig. 22; t. 22, fig. 3; Warren, 1909, Seitz, Grossschmett., 3 : 17, t. 3, g; Hamp-
son, 1909, Cat. Lep. Phal., 8 : 158; Pierce, 1909, Genit. Noct., t. 5; Culot, 1909, Noct.
Eur., 1 : 22, t. 1, fig. 18; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 145; Petersen,.
1924, Lep. Fauna Estl., ed. 2 (1) : 152; Wagner, 1921, Intern, entom. Ztschr., 24 : 471;.
Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 12, t. 1, i; Nordstroem och Wahl-.
gren, 1937, Svenska Fjar. : 84, t. 15, fig. 15; — obscura Strom, 1783, Dahsk. Vid. Selsk.
Skr. : 79; — euphrasiae Brahm, 1791, Ins. Kalend., 2 : 143; —esulae Hiibner, 1818,.
Europ. Schmett., Noct., fig. 613; — cyparissiae Hiibner, 1818, там же, fig. 615; — pa-
radoxa Boisduval, 1840, Gen. Ind. Meth. : 60; — abscondita Herrich-Schaeffer, 1845;-
Europ. Schmett., 2: fig. 87—89 (non Treitschke); — montivaga Guenee, 1852, Noct., 1 :
57; —glaucoptera Petersen, 1902, Fauna Estl., ed. 1 : 59 (sec. Petersen f. abscondita
Tr..); — parisiensis Culot, 1909, Noct., 1 : 23, t. 2, fig. 9; —fasciata Hanneman, 1916,.
Intern, entom. Ztschr., 10 : 63; — debilis Domaison, 1925, Bull. Soc. Entom. France :
308; — acerbata Schawerda, 1928, Ztschr. Oesterr. entom. Ver., 16 ; 31; — virgata Dan-
nehl, 1929, Mitt. Miinch. entom. Ges,, 19 : 104; — attomana Draudt, 1931, Seitz, Gross-
schmett., 3 SuppL, 1 : 13, t. 1, i — xanthomista Draudt, 1931, там же, 1 : 13; —
apennina Draudt, 1931, там же, 1 : 13; — korlana Draudt, 1931, там же, 1 : 13; —
farinosa Bytinsky-Salz, 1934, Intern, entom. Ztschr., 28 : 100; — andalusiaca Scha-
werda, 1934, Intern, entom. Ztschr., 28 : 416, fig. 7—10; — negrita Marten, 1934,.
Festschr. Intern, entom. Ver. : 46, fig. 8; — amasina Marten, 1934, там же : 46.
1 Описанная из Пиренеев (Арагония, Альбаррацин, Андалузия) Acronicta schivin-
genschussi Zerny (1927, Eos, 3 : 358, t. 9, fig. 5) очень близка к широко распространен-
ной A. auricoma Schiff.; она представляет, невидимому, просто пиренейскую форму
этого вида.
2 Описанная Пюнглером (Pungeler, 1906, Iris, 19 : 216, t. В, fig. 10) с Или и из-.
Александровского хребта tiena Piingl. — вид мало известной в литературе (Hampson,
1909, Cat. Lep. Phal., 8 : 150, t. 126, fig. 29; Draudt, 1931, Seitz, Grossschmett., Pal.,.
3, SuppL, 1 :1, t. И), близка к A. (Ph.) euphorbiae и возможно является просто формой,
что, по недостатку данных по этому виду, выяснить сейчас нельзя; она известна только-
в типах. Описание дано очень недостаточное, я привожу его полностью: «Размах
38 мм, длина переднего крыла 18 мм; близка к euphorbiae F:, крупнее, передние крылья
шире, темнее и более коричневые, с нижней стороны затемнены, только внешний-
край крыла и передний его край светлые, коричневато-серые; рисунок передних
крыльев неясный, похож на таковой euphorbiae, также и все остальное, как у этого-
вида». Описаны по двум самцам. По Драудту, может быть поставлена рядом с mega-
cephala Schiff., но по фотографии и по рисункам это, как полагает и Пюцглер (1906),
форма, близкая к euphorbiae Schiff.
34. ACRONICTA
521
Основной цвет головы, груди и передних крыльев тусклосерый; рису-
нок нерезкий, растушеванный; лишь у некоторых форм (особей) резкий;
базальная линия в виде двух широко расставленных штрихов на косталь-
ном крае, посредине светлая, короткая; вторая поперечная линия ско-
шена кнаружи, слабо волнистая, двойная, посредине светлая; круглое
пятно маленькое, обычно правильной формы или овальное, в середине
светлочерное, очерчено черным; почковидное пятно широкое, слаба
выемчатое с внешней стороны, в середине серое с немногими темными
штрихами; срединная линия редуцирована; третья поперечная линия,
начинается двумя широко расставленными штрихами на костальном
крае сразу за почковидным пятном, сильно выгнута к внешнему краю,
сильно зубчатая по жилкам, тонкая, простая; расстояние от края почко-
видного пятна до третьей поперечной линии в средней части крыла,
больше, чем ширина внешнего поля; внешнее поле темное с тонкой белой
линией, едва заметной, местами волнистой; тер’минальная линия тонкая,
состоит из серии сливающихся черных штрихов треугольной формы;
задние крылья чисто белые или тусклосерые с белой бахромой и темной
терминальной линией, жилки слабо затемнены; нижняя сторона тускло-
белая с неправильно разбросанными (больше в костальной части крыльев)
черными чешуйками, без рисунка. Размах крыльев 32—35 мм.
Uncus сильно загнут вентрально, почти цилиндрической формы,
покрыт небольшим количеством длинных волосков; tegumen узкий и
довольно длинный, образует небольшие конические выросты по сторонам
основания uncus; saccus маленький, конический, направлен косо вен-
трально; fultura inferior в виде большой в вентральной части округлой
пластинки, несет многочисленные мелкие шипики; valvae длинные и
широкие, в дистальной части округлые и сильно выгнутые дорзально;
вдоль вентрального края valva широкая склеротизованная harpe, сра-
стающаяся с большой склеротизованной лопастью при основании valva;
harpe несет два отростка, острых, сильно крючкообразно загнутых внутрь;
penis короче valva, сильно расширен в дистальной части, вздут перед,
дистальным концом, coecum короткий и узкий; pars inflabilis penis несет
многочисленные сильные зубцы. Bursa copulatrix большая, конической
формы, суженная в проксимальной части, с заостренными polus, тонкая,
мембранозная; дистальная часть ее несет вырост, загнутый под прямым
углом и направленный дорзально, от конца которого отходит ductus
seminalis; ductus bursae при основании вздут, короткий, расширенный
в. дистальной части; восьмой сегмент узкий и довольно длинный; apo-
physes anteriores тонкие и недлинные, острые, несколько короче ductus
bursae; apophyses posteriores тонкие, также острые, короче первых;
papillae anales короткие и широкие, с густыми короткими волосками
в дистальной части.
Яйца плоские, около 1 мм диаметром, посредине несколько кони-
ческие; сразу после откладки нежнооранжевой окраски, затем темнеют;
хорион несет радиальные рубчики на верхней стороне.
Гусеницы последних стадий желто-серые с черным рисунком; дор-
зальная сторона имеет черную срединную полосу, разделенную на пятна;
по сторонам ее по светлой желто-красной полосе; по бокам тела под ды-
хальцами желто-красная полоса, на которой сидит ряд желтых боро-
давок; брюшная сторона желто-серая; бородавки частью желтые, частью
черные; голова черная с желтыми адфронтальными полями; ноги черные,
черные же подошвы ложных ног; дыхальца белые, окруженные черным
валиком кожи.
522
11. ACRONICTINAE
Куколка черно-коричневая; голова и грудь густо морщинистые;, на
темени два небольших бугра; брюшные сегменты несут густую скульп-
туру в виде глубоких ямок и бугорков; особенно сильно развита скульп-
тура на дорзальной стороне сегментов; дистальная часть каждого сегмента
(около одной трети длины) несет очень мелкие точки, почти гладкая;
дыхальца жёлтые, выдаются; под ними расположены пучки коротких
рыжих волосков; cremaster в виде тупого короткого выроста ' резко от-
граничен от последнего сегмента лишь с вентральной стороны, морщи-
нистый; с дорзальной стороны точечный; по сторонам на дистальном конце
несет по одному коническому
выросту;
Рис. 270. Acronicta euphorbiae Schiff. Генита-
лии: а — самца, б — самки.
по. дорзальному краю имеет че-
тыре маленьких бугорка.
Распространение.
Западная и средняя Европа
(Пиренеи, Альпы, Герман-
ская низменность, долина
Дуная); европейское побе-
режье Средиземного мрря
(Испания, Италия, Далма-
ция, Греция); Корсика, Си-
цилия; Британские острова;
Ютландия, южная Сканди-
навия, Швеция (Готланд,
Эланд, Тюбру), Финляндия
(до Лапландии); в СССР
всюду в Прибалтике (Каунас,
Таллин, Рига) и далее на
восток от линии Ленинград
(Луга) — Казань всюду на
юг по лесостепной и степной
зонам, но встречается ло-
кально (Москва, Калуга, Пол-
тава, Киев, Воронеж, Винни-
ца, Николаев, Херсон); При-
уралье (Чкалов, Губер ли);
Поволжье; Северный Кавказ (Железноводск, горные степи в б. Терской
юбл.; склоны Эльбруса); Закавказье (Тбилиси; Боржоми, оз. Севан,
долина Куры); Закаспий (Мангышлак); Крым (Карадаг, Севастополь,
Бельбек); Малая Азия (Аксехир); распространение на восток неясно,
есть лишь отдельные точки — западная Сибирь: Омск (Давров), Бар-
наул (Внуковский); Алтай, Ульба (Внуковский); Тарбагатай (Гэмпсон).
Биология и экология. Зимует куколка; вылет бабочек
в апреле—мае на юге и в июне—июле на севере ареала; яйцекладка на
листья кормовых растений по одному яйцу. Гусеницы многоядны, но
преимущественно питаются травянистыми растениями и кустарниками;
наиболее часто на Euphorbiaceae (Euphorbia rigida, Е. cyparissias), но
также и многими другими растениями — Ericaceae (Erica carnea), Myri-
caceae (Myrica gale), Rosaceae (Alchemilla vulgaris), Betulaceae (Betula
pubescens, B. humilis), Salicaceae (Salix glabra, S. nigricans), Polygonaceae
(Rumex), Tamaricaceae (Myricaria), Scrophulariaceae (Euphrasia), Labi-
atae (Thymus serpylum), Gentianaceae (Gentiana asclepiades), Crassulaceae
(Sedum), Compositae (Achillea). На юге ареала два поколения; лёт вто-
рого в июле—августе. Окукление в сухих листьях в плотном коконе;
куколка диапаузирует. Стацией обитания на юге являются сухие песча-
34. ACRONICTA
523
ные участки степи, поросшие молочаем; на севере (в лесной зоне) вереща-
тники и окраины усыхающих торфяников.
* 27. Acronicta (Pharetra) abscondita Тг. (рис. 271).
Treitschke, 1835, Schmett. Eur., 10 (2) : 5; Staudinger, 1901, Cat. Lep., 1 : 133;
•Spuler, 1908, Schmett. Eur., 1 : 139, t. 31, fig. 19, 21; Hampson, 1909, Cat. Lep. Phal.,
8 : 159; Warren, 1909, Seitz, Grossschmett., 3 : 17 (syn. euphorbiae Schiff.); Culot,
1909, Noct. Eur., 1 : 23, t. 2, fig. 10; Rebel, 1910, Berge, Schmetterlingsb.,ed. 9 : 145;
Petersen, 1924, Lep. Fauna Estl., ed. 2 (1): 152; — Draudt, 1931, Seitz,Grossschmett.,
Pal. 3; SuppL, 1 : 12, t. 1, k;—myricae Guenee, 1852, Noct., 1 : 59; — eurasiae Tschet-
werikov, in litt.
Передние крылья узкие, почти ланцетовидной формы, тусклосерые,
•однотонные; рисунок не резок, линии стушеваны; первая поперечная
линия намечена нерезким черным штрихом на дорзальном крае; черного
штриха при основании крыла нет; вторая поперечная линия неясная,
в виде прерванного штриха на костальном крае, при основании круглого
пятна и на дорзальном крае, где она дает короткий штрих в дистальном
направлении; круглое пятно заметно в виде тонкого неясного колечка,
•середина которого одного цвета с общим тоном крыла; срединная линия
ясная, черная, растушеванная, широкая, выгнутая кнаружи до почко-
видного пятна и далее сильно сдивнутая базально (почти до второй по-
перечной линии) в дорзальной его половине; почковидное пятно непра-
вильной формы, затемнено в базальной части, в наружной размыто;
третья поперечная линия двойная, посредине белая, резко заметная
в костальной половине крыла, в дорзальной части его смыта; от начала
третьей поперечной линии по костальному краю белые точки; подкрае-
вая линия неясная, тонкая; терминальная линия в виде черных точек;
бахрома белая с черными штрихами; задние крылья тусклосерые или
беловатые со слабо заметными жилками, у самок темнее; бахрома белая;
дискоидальные пятна едва намечены на нижней стороне задних крыльев;
терминальная линия в виде тонкой полоски, коричнево-серая; нижняя
сторона как передних, так и задних крыльев тусклосерая с немногими
черными чешуями. Размах крыльев 30—32 мм.
Uncus при основании тонкий и заостренный на дистальном конце,
в средней части вздут; tegumen узкий на дорзальной стороне и расширен
с боков; saccus маленький, округленный; jultura inferior в виде неболь-
шой округлой с вентральной стороны пластинки; vallum penis несет
многочисленные мелкие шипики; valvae короткие, широкие, выгнутые
вентрально посредине; базальная часть valva несет небольшую склеро-
тизованную лопасть, усаженную волосками; по вентральному и дорзаль-
ному краям valva косые складки; harpe в виде складки с двумя слабо
изогнутыми отростками — вентральным, коротким и широким, и дорзаль-
ным, более длинным и тонким, направленным косо дорзально; penis
короткий и толстый, слабо изогнутый, с толстым coecum; pars inflabilis
penis несет небольшое число (10 шт) коротких но сильных шипов. Bursa
copulatrix почти яйцевидной формы с саключенным спирально неболь-
шим отростком на дистальном конце; dustus seminalis отходит от этого
отростка, с его дистального конца; поверхность bursa copulatrix покрыта
рубчатой скульптурой; dustus bursae, длинный, более половины длины
corpus bursae, широкий при основании, суженный к дистальному концу,
морщинистый; восьмой сегмент узкий; недлинный; apophyses anterio-
res конические, тонкие, недлинные, короче, чем восьмой сегмент; apo-
physes posteriores тонкие и рудиментарные; papillae anales мембранозные,
недифференцированные в проксимальной части.
524
11. ACRONICTINAE
Приуралье (Губерли, Уральск); одна
Рис. 271. Acronicta abscondita Тг. Гениталии:
а — самца, б — самки.
Гусеница (по Шпулеру, 1908) черно-коричневая с серией черных
пятен на дорзальной стороне, с рядом желтых бородавок и белым пятном
на каждом сегменте; по сторонам тела красные, а над ногами коричнева-
тые бородавки; ноги черные, покрытые белыми волосками; нижняя сто-
рона тела черно-коричневая; голова черная.
Распространение. Средняя Европа — Альпы (Шварц-
вальд), Польша (Варшава, Кельцы, Радом); Британские острова (Шотлан-
дия); Швеция (Лапландия), Финляндия (Ниландия); в СССР разрознен-
ные местонахождения — Прибалтика (Тарту, Таллин, Кингисепп), Ка-
релия (Петрозаводск), Ленинград, Псков, Могилев, Москва, Казань;
особь найдена мною в коллекции
Зоологического института Ака-
демии Наук СССР из Крыма
(Севастополь), совершенно сход-
ная по гениталиям с abscondita
Тг.; одна особь из Иркутска
(р. Качук) в коллекции Зоол.
института очень близка по об
лику к abscondita Тг., может быть
аберрация от auricoma Schiff.
Диагностически лишь не-
давно хорошо отграничен от
A. euphorbiae Schiff., вследст-
вие чего данные по распростра-
нению неудовлетворительны.
Принадлежность части форм к
euphorbiae Schiff, и до сих пор-
еще не окончательно ясна.
Мною по материалам Зооло-
гического института Академии
Наук СССР просмотрены все
основные формы euphorbiae
Schiff., и из всех только ту-
ricae Gn. может быть отнесена
к этому виду, тогда как glau--
coptera Peters., описанная как
форма от abscondita Тг., все же должна быть отнесена (по строению
гениталий) к euphorbiae Schiff., представляя мелкую и тускло окра-
шенную форму этого вида. Описанные в последнее время Драудтом,
Геннеманом, Мартеном и Шаверда формы от euphorbiae Schiff, из разных
мест средней Европы мною за отсутствием материала не могли быть
исследованы; очень вероятна, тем не менее, принадлежность их именно
к euphorbiae Schiff., как то считали и сами авторы.
Биология и экология. Зимует куколка; вылет бабочек
в июне—июле; яйцекладка на листья кормовых растений. Гусеницы
многоядны; как и для предыдущего вида, указано питание Salicacefte
(Salix caprea, S. fragilis, Populus tremula, P. nigra), Betulaceae (Betula
pubescens, B. humilis), Polygonaceae (Polygonum, Rumex), Ericaceae
(Calluna, Vaccinium), Euphorbiaceae (Euphorbia), Scrophulariaceae (Eu-
phrasia), Compositae (Hieracium, Achillea). Рост гусениц в июле—августе;
окукление в листьях в плотном коконе, изготовляемом в свернутых
листьях. Стацией обитания являются сухие луга или окраины торфя-
ников — верещатники. Отмечен паразит Rhogas rugosus Ns.
34. ACRONICTA
525
* 28. Acronicta (Pharetra) orientalis Mann. (рис. 272).
Mann, 1862, Wien, entom. Monatschr. : 370, t. 3, fig. 4; Lederer, 1865, Ann. Soc.
entom. Belg., 9 : 63, t. 3, fig. 7; Staudinger, 1901, Cat. Lep., 1 : 132; Hampson, 1909,
Cat. Lep. Phal., 8 : 111; Warren, 1909, Seitz, Grossschmett., 3 : 15, t. 3, c; Schawerda,
1914, Verh. zool.-hot. Ges. Wien, 64 : 359; Draudt, 1931, Seitz, Grossschmett.,
3 Suppl., 1 : 11, t. 1, g;—galvagnii Schawerda, 1916, Verh. zool.-hot. Ges. Wien,
66 : 483.1
Передние крылья светлосерые с примесью черных чешуй; от основания
крыла интенсивно черный штрих до второй поперечной линии; первая
поперечная линия почти не видна, едва замечена на костальном крае;
вторая поперечная линия в виде двух широко расставленных черных
полос, слабо скошена кнаружи, посредине серая, в нижней части не-
сколько размытая; клиновидное пятно не развито; круглое пятно неболь-
шое, овальной формы, расположено горизонтально, в середине серое;
срединная линия в виде косой черной полосы, в костальной части крыла
направлена косо к почко-
видному пятну, в нижней
части срединного поля
растушеванная и смытая;
почковидное пятно неболь-
шое, слабо выгнутое с
внешней стороны, в сере-
дине светлосерое с черной
полоской в центре; третья
поперечная линия двойная,
начинается над внешним
углом почковидного пятна, рис 272. Aeronicta orientalis Mann. Гениталии самца,
далее полого выгнута, силь-
но, но равномерно зубча-
тая по жилкам, четкая, посредине светлая, почти белая; против почко-
видного пятна на третьей поперечной линии нерезкий темный штрих,
такой же второй штрих в дорзальной ее части; подкраевая линия не раз-
вита; терминальная линия в виде серии черных треугольных штрихов;
бахрома серая с полоской. Задние крылья чисто белые с тонкой терми-
нальной полосой, бахрома белая; нижняя сторона белая с небольшим
количеством черных чешуй по костальному' краю передних и задних
крыльев и по внешнему их краю; дискоидальные пятна на задних крыльях
едва намечены, на передних не развиты. Размах крыльев 34—38 мм.
Uncus короткий, слабо изогнутый, конический в дистальной части,
•склеротизован, острый, tegumen узкий и довольно длинный; saccus
большой и широкий; fultura inferior в виде широкой щитовидной пла-
стинки; valvae конической формы, в средней части слабо выгнуты вен-
трально; базальная половина valva склеротизована, имеет небольшую
лопасть; harpe развита в дистальной половине valva, имеет вид короткой
•складки с двумя изогнутыми внутрь отростками — коротким вентраль-
ным и длинным коническим, направленным косо дорзально верхним;
дистальный конец valva мембранозный, несет негустые волоски; penis
по длине равен valva, сильно изогнут; соесшп короткий и толстый; pars
1 Описанная недавно Acronicta saadi Brand (1938, Entom. Rundsch., 55 : 498)
таз Малой Азии чрезвычайно близка к A. orientalis Mann. Автор не исследовал струк-
турных признаков и дает только описание рисунка; сведение синонимики за отсут-
ствием материала пока преждевременно.
526
II. ACRONICTINAE
inflabilis penis несет многочисленные недлинные шипы и дистально от
них сильно склеротизованную зубчатую пластинку.
Распространение. Герцеговина; Малая Азия (Лидия, Бит-
ния, Аксехир, Анкара).
29. Acronicta (Pharetra) pasiphae .Drd.
Draudt, 1936, Entom. Rundsch., 53 : 457, t. a, 2.
Голова, грудь и передние крылья бело-серые; первая поперечная:
линия намечена двумя черными штрихами на костальном крае крыла;
от основания переднего крыла черный штрих,, расширяющийся тре-
угольно близ второй поперечной линии; вторая поперечная линия косая,
двойная, посредине белая, на костальном крае крыла дает резкую петлю,
достигающую круглого пятна, и в дорзальной части крыла широкую
петлю наружу, как у A. orientalis Mann; круглое пятно овальное, вытя-
нутое, очерченное черным, в середине бело-серое; срединная линия расту-
шеванная, заметная лишь близ костального края; почковидное пятно
очерчено черным, слабо выемчатое с внешней стороны, в середине серое,,
к краям белое; третья поперечная линия зубчатая, двойная лишь у ко-
стального края, далее оторочена с базальной стороны белым, сильно изо-
гнута к внешнему краю, пересечена двумя черными штрихами — одним
слабым, между и М2 и другим более интенсивным, под С2; дорзальная
часть внешнего поля темная; подкраевая линия в виде серии черных
треугольных неслитых штрихов; бахрома бело-серая с тонкой поперечной
линией; задние крылья белые, полупрозрачные, темные по внешнему
краю, жилки более темные; бахрома белая; нижняя сторона на передних
крыльях серая, на задних белая; дискоидальные пятна едва заметны.
Размах крыльев 32 мм.
Этот вид известен мне только по рисунку и описанию немногих осо-
бей; очень напоминает orientalis Mann.
Распространение. Малая Азия (Дюльдюнь-даг, Аманус).
* 30. Acronicta (Pharetra) lutea Brem. et Grey (рис. 273, 274).
Bremer und Grey, 1852, Entom. Monatschr., 1 : 65; Bremer, 1864, Lep. Ost-Sib.,
t. 4, fig. 7; Oberthiir, 1884, Et. Entom., 10 : 18, t. 2, fig. 4; Staudinger, 1901, Cat. Lep.,
1 : 133; Hampson, 1909, Cat. Lep. Phal., 8 : 95; Warren, 1909, Seitz, Grossschmett.,
3 : 17, t. 3, h; Draeseke, 1928, Iris, 42 : 297; В. Кожанчиков, 1928, Ежегодн. Мину-
синск. муз., 6 (1) : 73; Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 9; Ermola-
yev, 1935, Entom. Anz., 15 : 283; — leucoptera Butler, 1881, Trans. Entom. Soc.
Lond., 19 : 395; — aurantior Strand, 1915, Arch. Naturgesch., 81 (11) : 157; — suigensis-
Matsumura, 1926, Ins. Matsum., 1 : 4, t. 1, fig. 4 a.
Голова, грудь и передние крылья черно-серые; рисунок черный,
резкий, особенно у типичной формы (с желтыми задними крыльями};
базальная линия двойная, идет до середины крыла, посредине серая;
ниже кубитальной жилки от базальной линии темный штрих, соединяю-
щий ее со второй поперечной линией; вторая поперечная линия двойная
и очень широкая, сильно скошенная кнаружи, слабо волнистая, посре-
дине светлосерая; срединное поле очень широкое, вдвое шире внешнего,,
более светлой окраски в области пятен; круглое пятно почти правильной
формы, большое, в середине светлосерое с черной точкой в центре; сре-.
данная линия широкая, растушеванная, в средней части крыла выгну-
тая и захватывающая почковидное пятно; последнее большое, почти круг-
лое, слабо выемчатое с внешней стороны, очерчено черным, в середине
темносерое; третья поперечная линия двойная, широкая, мелко зубча-
34. ACRONICTA
52 7
тая, посредине светлосерая; в ее нижней части с наружной стороны белое
пятно; внешнее поле однотонное, черное, с белой .подкраевой линией;
терминальная линия в виде черной полосы; бахрома темносерая с тонкой
полоской; задние крылья желтые с широким черным краем, на заднем
углу желтое пятно; бахрома желтая, снаружи белая; нижняя сторона
передних крыльев темносерая с немногими черными и желтыми пятнами:
на костальном крае; задние крылья желтые с темным внешним краем
и желто-белой бахромой. Размах крыльев 44—52 мм.
Uncus длинный, тонкий, в средней части несколько расширенный
и уплощенный, голый; tegumen узкий, но довольно длинный, на ди-
стальном крае несет при основании uncus два острых выроста; fultura
inferior килевидной формы,
сросшаяся из двух больших
пластинок, по дорзальному краю
несет точечную скульптуру;
valvae длинные, конические,
заостренные, в середине слабо
выгнутые вентрально; базаль-
ная половина valva несет боль-
шую склеротизованную лопасть,
срастающуюся с harpe; послед-
няя в виде двух складок, вместе
срастания которых находятся
два выроста — вентральный,
короткий и острый, сросшийся
с valva, и дорзальный, направ-
ленныйкосо дистально, извитой,
свободный; дистальная полови-
на valva склеротизована, глад-
кая; penis короче valva, силь-
но выгнут в средней части дор-
зально, расширен в дистальной
части; coecum penis короткий;
pars inflabilis сильно склероти-
зована и несет многочисленные
длинные и острые зубцы. Bursa
copulatrix очень сильно расширена в проксимальной части, резко сужена
в дистальной, склеротизована, несет точечную скульптуру; дистальный
конец ее образует на дорзальной стороне узкое сосковидное расширение,
завернутое на основание ductus bursae, от конца которого отходит ductus^
seminalis; ductu§ bursae короткий и складчатый; восьмой сегмент короткий
и узкий; apophyses anteriores тонкие, длиннее ductus bursae; apophyses
posteriores более короткие, но также тонкие; papillae anales короткие,
мембранозные.
Яйца плоские, выпуклые посредине, диаметром около 0.8—1.0 мм;
поверхность хориона несет слабую рубчатую скульптуру; сразу после
откладки светложелтого цвета, однотонные; с развитием принимают
темножелтую с охристым окраску, а затем коричневую, оставаясь одно-
тонными. •
Гусеницы младших стадий светлосерой окраски, очень близки к ма-
леньким гусеницам A. rumicis L. и Л. auricoma Schiff., начиная с третьей
стадии приобретают характерные черты. Тело покрыто длинными и тон-
кими желтыми волосками, которые на дорзальной стороне щетинистые’
Рис. 273. Acronicta lutea Brem. et Grey. Гени-
талии; a — самца, б — самки.
528
IT. ACRONICTINAE
и короткие, а по бокам шелковистые и длинные; кожа бархатисто-черная
со слабым коричневым оттенком; бородавки небольшие, почти круглые,
не сильно выдаются на поверхности кожи, несколько крупнее на вось-
мом сегменте; на вентральной стороне тела короткие волоски и мелкие
бородавки; ноги буро-коричневые; голова блестящая, черная, округлая,
^небольшая; сигмы одного цвета с кожей; ниже стигм и перед каждой
из них расположено яркобелое неправильной формы пятно;,пятна обра-
зуют прерванную боковую линию
на втором—восьмом сегментах
брюшка; на груди их нет.
Куколка черно-коричневая,
короткая и довольно широкая;
голова спереди гладкая и бле-
стящая; на темени округлый
вырост, покрытый густыми глу-
бокими ямками; переднегрудь
с дорзальной стороны глубоко
и редко морщинистая; средне-
грудь, крылья и ноги блестя-
щие; брюшные сегменты несут
скульптуру в виде бугорков и
ямок, особенно сильную с дор-
зальной стороны и только в
базальной части каждого сег-
мента (на 3/4 его длины); ди-
стальные сегменты брюшка мел-
ко скульптированы; cremaster
в виде слабо дифференцирован-
ного выступа, несет пучок ко-
ротких прямых и острых ще-
тинок, направленных дисталь-
но.
Этот вид дает две хорошо
разграниченные цветовые гео-
графические формы — типич-
ную, распространенную в Ко-
рее, по Уссури и доходящую
на север до Тукурингра, и 1еи-
coptera Btl., более мелкую, со
светлыми задними крыльями,
Форма leucoptera Btl. по внешним
L., настолько, что мелкие особи
.Рис. 274. Acronicta lutea Brem. et Grey. Хето-
таксия передне- и среднегруди и брюшных
сегментов I—V, VI, VII и VIII гусеницы пер-
вой стадии (а) и взрослой (б).
распространенную на запад до Енисея,
признакам очень напоминает rumicis
габитуально почти не отличимы; желтый цвет исчезает совершенно как
на задних, так и на передних крыльях, и отличием от rumicis L. является
лишь интенсивно черная окраска рисунка. Живые особи Л. lutea Brem.
et Grey любой из форм отличаются более энергичным поведением,
более пугливы и более стремительны в движениях (имаго) по сравне-
нию с A. rumicis L. Оба вида не скрещиваются ни при каких условиях.
Различия обеих» форм связаны с условиями роста; при росте при
повышенных температурах возникают более крупные и интенсивно окра-
шенные желтые особи, при пониженных (ниже 25°) наблюдаются мелкие
особи без желтого пигмента. В экспериментальных условиях из енисей-
ских белокрылых особей мною были получены крупные с желтыми зад-
ними крыльями особи в первом же поколении, типа китайских.
34. ACRONICTA 529
Распространение. Центральная Сибирь (Минусинск, Тай-
шет); Байкал (Иркутск); Забайкалье (Яблоновый хребет); Приамурье
(Буреинские горы, Тукурингра, Зея, Благовещенск, Хабаровск), южное
Приморье (Яковлевка, Владивосток, Ворошилов-Уссурийский, Уссури,
'Сидеми); северная Монголия (Кентей, Улан-Батор); западный Китай;
Корея; Япония.
Биология и экология. Зимует куколка; вылет имаго .
в июне—июле; яйцекладка кучками по 3—10 яиц на листья кормовых
растений; самки очень плодовиты и способны откладывать около
1500 яиц (максимум). Гусеницы многоядны; основные кормовые расте-
ния — Salicales (Salix fragilis, S. caprea, Populus nigra, P. tremula),
Betulaceae (Carpinus orientalis, Betula pubescens, B. humilis), Polygona-
ceae (Rumex, Polygonum aviculare), Myricaceae (Myrica gale), Rosaceae
(Crataegus, Alchemilla). Этот круг кормовых растений определен в экспе-
риментальных условиях; наблюдения в природе касаются лишь питания
гусениц восточным грабом (на Амуре) и тополем (на Енисее); в окрест-
ностях Минусинска гусеницы не редки на молодых тополях (2—3-лет-
иих), старые тополя не служат пищей. Типичной стацией на Енисее
являются окраины болот с зарослями мелких тополей и ив. Рост гусениц
в июле—августе; окукление в сухих листьях в плотном шелковом коконе
черного цвета; куколка диапаузирует. В природе дает одно поколение;
в экспериментальных условиях возможно получение частичного вылета
второго поколения лишь при 30°. Куколки очень холодостойки, пере-
носят температуры до —40, —50°.
* 31. Acronicta (Pharetra) rumicis L.1 (рис. 275, 276).
Linne, 1758, Syst. Nat., ed. 10 : 516; Aurivillius, 1891, Nordn. Fjar. : 99; Stau-
dinger, 1892, Mem. Lep. Rom., 6 : 390; Hampson, 1894, Moths Ind., 2 : 103; Hamp-
.son, 1896, тамже, 4 : 509; Staudinger, 1901, Cat. Lep., 1 : 133; Herz, 1904, Ежегодн.
Зоол. муз. Акад. Наук, 9 : 272; Spuler, 1908, Schmett. Eur., 1 : 139, t. 31, fig. 28;
.Hampson, 1909, Cat. Lep. Phal., 8 : 93; Warren, 1909, Seitz, Grossschmett., 3 : 17,
it. 3, i; Culot, 1909, Noct., 1 : 24, t. 2, fig. 11; Rebel, 1910, Berge, Schmetterlingsb., ed.
9 : 145, t. 22, fig. 156; Petersen, 1924, Lep. Fauna Estl., ed. 2 (1) : 152; Bang-Haas,
1926, Nov. Macrolep., 1 : 160; Bang-Haas, 1927, тамже, 2 : 180; Bang-Haas, 1928, там же,
3 : 40; Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 9, t. 1, d; Nordstroem och
Wahlgren, 1937, Svenska Fjar. : 85, t. 15, fig. 16; Куренцов, 1935, Вести. Дальневост.
фил. Акад. Наук СССР, 15 : 89; — salicis Curtis, 1829, Brit. Entom., 5, t. 136; —dif-
jusa Walker, 1857, Spec. Lep. Brit. Mus., 11 : 708; — indica Moore, 1867, Proc. Zool.
.Soc. Lond. : 47; —-bercei Saunders, 1870, Pap. France, 3 : 19; — turanica Staudinger,
1888, Stett. entom. Ztg., 48:65; Staudinger, 1889, Mem. Lep. Rom., 5, t. 8, fig.
-5; —lugubris Schultz, 1902, Soc. Ent., 16 (22) : 170; — alnoides Geest, 1903, Ztschr.
allg. Entom., 8 : 310;— suhrianna Gillmer, 1907, Entom. Rec., 19 : 91: — marginata Lam-
billion, 1909, Rev. Namur, 9 : 22; —grisea Warren, 1913, Seitz, Grossschmett., Indo-Aust-
.ral., 11 :.39, t. 5, g; — striata Meves, 1914, Entom. Tidskr., 35 : 14, 41; —priens Strand,
1915, Arch. Naturg., 81 (11) : 157; — polonica Priiffer, 1918, Bull. Acad. Polon. : 201; —
pallida Rothschild, 1920, Nov. Zool., 27 : 7; —prafferi Maslovsky, 1923, Polsk. Pismo
Ent.,2.’130, t. 4, fig. 10;— meridionalis Dannehl, 1925, Entom. Ztschr. Frankf.,
39 : 119; — distincta Zubowsky et Vuscinski, 1938, Bull. Mus. reg. Bessar., 8 : 26.
Биология и экология. Васильев, 1914, Тр. Бюро энтом., X, 10 : 19;
Уваров, 1914, Отч. Ставр. ст. защ. раст. : 26; Dubois, 1918, Vie agr. rur., 8 : 118;
Reh, 1925, Sorauer, Handb. Pflanzenkrankh., 4 : 418; Вогданов-Катьков, 1926, Защ.
раст., 4 : 398; Старк, 1926, там же, 3 : 18; Kuskov, 1927, Beitr. Kunde Estl., 13 : 82;
-Щеголев и Мамонов, 1928, Изв. on. дел. Сев. Кавк.’, 15; Щеголев, 1929, Защ. раст.,
6 : 403; Сиязов, 1930, Вред, хлопч. : 192; Щеголев и Струкова, 1930, Вред. масл.
культ. : 127, рис. 95; Айе, 1930, Entom. Anz., 10 : 231-; Лебедев, 1933, Зб1рн. праць
екол., 1 : 65; Нерреп, 1935, Entom. Rec., 47 : 38.
1 Описанная из Юньнани A. regifica Draudt (1937, Entom. Rundsch., 54 : 343,
Л. 4, fig. b) очень близка к A. rumicis L., являясь, вероятно, лишь { формой этого
.вида, более крупной и темной; рисунок очень плох.
34 Фауна СССР, И. В. Кожанчиков.
530
IT. ACRONICTINAE
Рис. 275. Acronicta rumicis L.
Гениталии: а — самца, б—
самки.
ненный в боковых частях
valvae конические, длинные,
Голова, грудь и передние крылья черно-серые; рисунок черный it
белый; базальная линия двойная, черная, посредине бело-серая, идег
через все крыло; базальное поле в костальной половине серое; от осно-
вания крыла черный штрих; вторая поперечная линия двойная, черная,
широкая, посредине черно-серая; срединное поле в области пятен свет-
лое, дорзальная часть его затемнена; круглое пятно правильной формы,
• маленькое, очерчено черным, в середине светлое с темным центром; сре-
динная линия широкая и ясная, начинается между пятнами, идет выг-
нутая кнаружи через почковидное пятно, далее выгнута базально,
теряется в темном дорзальном крае; почковидное пятно неправильной,
почти круглой формы, очерчено черным и далее белым, в середине серое;
третья поперечная линия начинается двумя черными и белым штрихами
на костальном крае над почковидным
пятном, далее резко выгнута кнаружи
(почти параллельна костальному краю),
далее идет параллельно терминальному
краю, черная, двойная, глубко зубчатан
по жилкам, посредине белая; между ку-
битальной и анальной жилками образует
белое, лунное пятно; внешнее поле тем-
ное с тонкой прерывистой подкраевой
линией; терминальная линия в виде серии
разделенных черных штрихов, захватыва-
ющих часть бахромы; задние крылья
однотонные, тусклосерые со светлой, почти
белой бахромой; нижняя сторона одно-
тонная с осветленным костальным краем,
имеющим ряд косых штрихов и с просве-
чивающей светлым тоном третьей попе-
речной линией; диско и дальнее пятно сла-
бое. Размах крыльев 35—45 мм.
Uncus длинный, в средней части
несколько расширен и уплощен, несет-
короткие волоски; tegumen узкий, ко-
роткий в тергальной области и удли-
saccus маленький, выгнут дорзально;
прямые; базальная половина valva несет
большую склеротизованную лопасть, слитую с harpe; последняя сла-
гается из двух срастающихся складок, несущих в месте срастания длин-
ный искривленный отросток, направленный косо дорзально;. дистальная,
половина valva в виде длинной лопаточки правильной формы, несущей
длинные густые волоски; fultura inferior килевидная, образованная двумя
сросшимися под углом пластинками; базальная ее часть зубчатая; penis
значительно короче valva, с коротким округлым coecum, расширенный
к дистальному концу, в дистальной половине сильно выгнут; pars infla-
bilis несет склеротйзованные складки и многочисленные острые склеро-
тизованные зубцы. Bursa copulatrix расширенная в проксимальной части,
искривленная, несет тонкую рубчатую скульптуру, местами склероти-
зована; ее дистальная, более узкая часть образует сосковидный отро-
сток, повернутый перпендикулярно продольной оси bursa; от его ди-
стального конца отходит ductus seminalis; ductus bursae равен примерно
половине длины corpus bursae, складчатый, неширокий; восьмой сегмент
длинный и узкий, тергальная его область мембранозная; apophyses ап-
34. ACRONICTA
531
teriores острые, около половины длины ductus bursae; apophyses pos-
teriores острые, несколько короче первых; papillae anales очень Короткие,
мембранозные, широкие, покрытые короткими густыми волосками.
Яйца плоские, полушаровидные, диаметром около 0.5 мм; хорион
покрыт радиальными бороздками; сразу после откладки яйца светлой,
бело-желтой окраски, спустя сутки или двое под хорионом появляются
коричнево-красные пятна неправильной формы.
Гусеницы младших стадий (до третьей) серые с немногими мелкими
черными и белыми пестринами; старшие стадии гусениц покрыты ры-
жими и черными волосками; вдоль спины проходит прерванная черная
дорзальная полоса, на каждом сегменте имеющая вид черного прямо-
угольника, у переднего края которого расположена желтая точка; по
сторонам дорзальной полосы проходят две других белого цвета, распа-
дающиеся на сегментах на самостоятельные
отрезки; по бокам на уровне стигм проходит
белая полоса в виде угловатых отрезков на
каждом сегменте; стигмы белые, овальной
формы, ясно заметные на фоне черных пятен,
где они располагаются; под стигмами и бе-
лой полосой дю бокам на каждом сегменте
охристые бородавки; первый брюшной сег-
мент несет пучки волосков, более длинные,
чем прочие сегменты. Голова черно-коричне-
вая с желто-красным эпикраниальным швом
и адфронтальными полями; нижняя сторона
тела оранжево-серая, ноги того же цвета.
Куколка темнокоричневая, покрытая не-
густыми волосками; брюшные сегменты
покрыты скульптурой в виде длинных ко-
нических выростов, особенно сильных с дор-
зальной стороны; грудные сегменты покрыты
морщинками; cremaster в виде плоского не-
длинного выроста с четырьмя короткими и
широкими шипами с каждой стороны, рас-
Рис. 276. Acronicta rumicis L.
Хетотаксия передне- и средпе-
груди и брюшных сегментов
I, VI, VII и VIII гусени-
цы первой стадии (а) и взрос-
лой (б).
положенными по два латерально; дистальный конец cremaster песет
многочисленные короткие и острые щетинки.
Распространение. Вся Европа от Пиренеев до Урала;
всюду в средней Европе (Альпы, Германская низменность), в Прибал-
тике и Скандинавии (Швеция, Норвегия, Финляндия, Лапландия); все
Средиземноморье (Пиренейский и Апеннинский пблуостровы, Далмация,
острова Средиземного моря — Корсика, Сицилия, Кипр); сонорная
Африка (Марокко); Сирия (Ливан), Палестина; Малая Азия (Бруссл,
Аманус, Мараш, Карс); Ирак (Багдад); в СССР от линии Архангельск—
Сыктывкар—Молотов (на востоке возможно севернее) всюду па юг до
побережья Черного моря и Закавказья (Петрозаводск, Псков, Ленин-
град, Вологда, Киров, Москва, Калинин, Калуга, Казань, Ярославль,
Воронеж, Киев, Белая Церковь, Полтава, Харьков, Чернигов, Николаев,
Херсон, Кишинев, Одесса, Таганрог, Ростов, Сарепта, Астрахань);
Северный Кавказ (Туапсе, Дагестан до 3000 м, Ставрополь); Крым
(Севастополь, Бельбек); Закавказье (Тбилиси, Лагодехи, Куссари, Бор-
жоми, Дербент, Ленкорань, Ордубад, Батуми, Кировабад); Иран (Астра-
бад); Средняя Азия — Фергана (Ош, Маргелап), Зеравшанский хребет
(Самарканд), Алма-ата, Кара-тау (Бала-мурун), Сыр-дарья (Байгакум.
34*
532
11. ACRONICTINAE
Ай-дарле), Тянь-шань, Семиречье '(Чилик, Зайсан); Урал (Свердловск,
Уфа, Челябинск); в Сибири от линии Тобольск—Обь-Енисейский ка-
нал— Иркутск всюду по лесной и лесостепной зонам (Омск, Томск,
Каинск, Тайшет, Бунбуй, Минусинск, Тибельти); Алтай; северный
Казахстан (Семипалатинск, Кокчетав, Акмолинск); Приамурье (Благо-
вещенск, Хабаровск, Николаевск); южное Приморье (Сучан, Суйфун,
Аскольд, Владивосток); Сахалин; Японские острова (Хакодате, Иоко-
гама, Токио, Киу-Сиу); северная Монголия (Кобдо, Суск, Ленсы, Агал-
каты); Корея (Гензан); западный Китай; северная Индия (Сик-
ким).
В основной части ареала слабо изменчивый вид; изменчивость обна-
руживает значительную сложность. Наиболее отличны по окраске сред-
неазиатские особи, описанные как A. turanica Stgr. [по Шпулеру (Spu-
ler, 1908), относящиеся к A. тепуanthidis View., что, конечно, совершенно
ошибочно]. Рисунок этих особей не отличим от типичных; идентично
и строение половых органов. Форма эта отличается светлосерой окра-
ской передних крыльев и чисто белой окраской задних. В экспериментах
такие особи я получал при развитии всего цикла в термическом режиме
30—32°. Эта форма является связанной с сухими и жаркими террито-
риями ареала. Вероятно, аналогичными же термическими формами
являются и некоторые другие, например A. alnoides Geeste, A. margi-
nata Lamb., A. pallida Roth., A. meridionalis Dann.
Гималайские и китайские особи выделяются в иные формы, вероятно
подвидового значения, обособленные значительно сильнее; таковы более
крупные, A. indica Moore и A. regifica Drd., известные пока в небольшом
чиспе экземпляров.
Биология и экология. Зимует куколка; вылет имаго на
юге ареала в апреле—мае, на севере в июне; яйцекладка на листья кор-
мовых растений кучками по 40—50 яиц, расположенных в один ряд,
несколько налегающих одно на другое, преимущественно на иву. Плодо-
витость самки до 700 яиц. Гусеницы многоядны, питаются видами Sali-
caceae (Salix), преимущественно с гладкими листьями, Betulaceae (Betula,
Carpinus, Ostryia), Fagaceae (Fagus, Quercus), Myricaceae (Myrica), Rosa-
ceae (Potentilla, Alchemilla, Sorbus, Crataegus, Malus, Pirus, Prunus,
Amygdalus) и Polygonaceae (Polygonum, Rumex), но могут питаться также
многими другими растениями — Oleaceae, Caprifoliaceae, Saxifragaceae,
Ericaceae, Graminae, Tiliaceae, Compositae, Linaceae, Cruciferae, но не едят
Labiatae, Urticaceae, Chenopodiaceae. Окукление в сухих листьях
в плотно сплетенном коконе, обернутом листом; куколка диапаузирует
и зимует. Диапауза 'куколок наступает всегда при росте и развитии
в условиях температуры 22° и ниже; развитие без диапаузы наблюдается
при росте в термических условиях выше этой зоны, особенно полное
развитие без диапаузы наблюдается при температурах выше 27°, но даже
при самых высоких температурах всегда остается небольшой процент
диапаузирующих особей. Дает одно поколение на севере ареала и два,
три и более на юге. Встречается в самых разнообразных стациях, но чаще
в зарослях кустарников —ивы и березы на севере и розоцветных на юге.
Вредит разным культурным растениям (Брамсон, 1881; Reh-Sorauer,
, 1925); указан как вредитель плодовых деревьев, ягодных кустарников,
декоративных растений, огородных и бахчевых культур и хлопчатника;
отмечен как вредитель хлопка в Фергане и в Закавказье, как вредитель
плодовых деревьев в Ставрополе и на Дону; отмечен как вредитель
хондриллы (Parthenium argentstum) на Украине и ревеня там же; ука-
34. ACRONICTA
533
зан как вредитель хмеля и плодовых деревьев в Англии и как вреди-
тель шелковицы в Японии; отмечено повреждение роз и сирени.
Из числа паразитов указаны перепончатокрылые: из Chalcididae —
Pterofnalus puparum L.; из Braconidae — Micrdplitis fumipennis Rtzb.,
Aleiodes nigriceps Rtzb., Microgaster congestusWesm., M. fulvipes Wesm.,
M. fuscicornis~Wesrii.', M. opavus Rbg., M.perspicuus Wesm., M. ruficornis
Wesm., M. rubens Ns., M. scutellator Ns., Apanteles congestus Rhd., A. ru-
micis Rhd., Bracon kollari Mrsh., Rhogas circumscriptus Ns., Rh. dimidi-
atus Ns.; из Ichneumonidae — Amblyteles fasciatorius Gr., A. vadatorius
Gr., Anomalon cerinops Gr., Colpotrochia elegantula Schr., Exetastes tar-
sator F., Limneria majalis Gr., L. unicincta Gr., Mesochorus pjctilis Hgr.,
M. semirufus Hgr., Paniscus testaceus Gr., Pimpla examinator F., P. insti-
gator E.,P. ornata Gr.,P. ooioora Rtzb., P. rufata Gmel., P. scandica Vill.,
Tryphon elegatulus Schr.; из Diptera — Compsilura concinnata Mg., Ly-
della nigripes Fall., Larvivora larvarum L., Zenillia fauna Rd.
* 32. Acronicta (Pharetra) pulverosa Hamps.
Hampson, 1909, Cat. Lep. Phal., 8 : 133, t. 126, fig. 9; Warren, 1909, Seitz, Gross-
schmett.,. 3 : 15, t. 3, d (как pruinosa Gn.); — pruinosa Leech, 1900, Trans. Entom.
Soc. Lond., 38 : 104 (nom. praeocc., non Guenee);—fuscosuffusa Strand, 1915, Arch.
Naturgesch., 81 (11) : 158; — sachalinensis Matsumura, 1925, Journ. Coll. Agric. Univ.
Sapporo, 15 (3) : 120, t. 11, fig. 13.
Основной цвет серо-беловатый с примесью бурых и желтых тонов;
базальная линия в виде двух черных штрихов, идущих от костального
края; от основания крыла ниже срединной жилки черный штрих, рас-
тушеванный в дистальной части; вторая поперечная линия двойная,
волнистая, скошенная кнаружи, посредине выгнутая; круглое пятно
большое, очерчено черным, в середине светлосерое со светлой полоской
по внутреннему краю, почти квадратное, несколько косое; срединная
линия в виде косого темного штриха, растушеванная, более заметная
в костальной части крыла; почковидное пятно большое, очерчено чер-
ным, внутри оторочено белым, тусклосерое в середине; в нижней части
срединного поля светлый штрих; третья поперечная линия двойная,
внутренняя часть ее видна лишь у костального края, посредине с белыми
полулунными штрихами, тонко зубчатая; в нижней ее части черный
штрих к termen и белое пятно (как у A. rumicis L.); подкраевая линия
в виде немногих белых несвязанных штрихов; терминальная линия в виде
серии черных штрихов; задние крылья тусклокоричневые, более светлые
при основании; бахрома на них белая; нижняя сторона коричневатая
с немногими темными чешуйками и просвечивающей поперечной линией.
Размах крыльев 34—38 мм.
Распространение. Сахалин; Хондо (Иокогама, Шима-
насаки); Киу-Сиу; западный Китай.
Очень близок к A. rumicis L., но напоминает также по габитусу и
А. тёпу an thidis View.; изменчивость этого вида незначительна, у некото-
рых особей наблюдается сильное затемнение передних крыльев (ab.
I, по Гэмпсону).
33. Acronicta (Pharetra) cinerascens Kozhantshikov, sp. nov.
Голова и грудь однотонные, желто-серые; передние крылья желто-
серые с черным нечетким рисунком, остроугольные, с прямым косталь-
ным краем и несколько вздернутыми верхними углами; базальная линия
в виде короткой черной полосы, отороченной светлосерым (желтоватым);
534 II. ACRONICTINAE
вторая поперечная линия стушеванная; круглое пятно в виде правиль-
ного или слегка овального пятна, небольшое, очерченное черным,
в середине желто-серое, с черной точкой в центре; почковидное пятно
узкое, растушеванное, в виде двух параллельных темных полосок, раз-
деленных желто-серым; третья поперечная линия начинается за почко-
видным пятном и идет в виде желто-серой полосы, шашечная — разделена
темными жилками; срединное поле вдвое шире внешнего; внешнее узкое,
по терминальному краю желтоватое; терминальная линия в виде тонкой
темной полоски; бахрома желто-серая; задние крылья желто-серые
с темными жилками и узкой желто-серой терминальной полоской; ниж-
няя сторона однотонно серая; бахрома светлее крыльев. Размах крыльев
29 ММ. г
Распространение. Северный Китай— Бейцегоу (Путята).
Известен в одном экземпляре; очень.характерный вид, напоминающий
в наибольшей мере A. euphorbiae Schiff.; есть известное сходство с A. au-
ricoma Schiff, и A. rumicis L.
* 34. Acronicta (Pharetra) cubitata Warr.
Warren, 1914, Nov. Zool., 21 : 403; Draudt, 1931, Seitz, Grossschmet., 3
Suppl., | : 8.
Малоизвестный вид; в литературе его изображения нет; известно
только оригинальное описание, сделанное очень шаблонно; привожу
перевод последнего.
«Переднее крыло: бледносерое, затемненное темносерым; темная тень
ниже наружной линии; срединная линия толстая и темная, затемняю-
щая почковидное пятно; суббазальная линия намечена двойным штри-
хом на костальном крае; внутренняя линия двойная, слегка косая кна-
ружи, выгнутая кнаружи у первой жилки; срединная тень выгнута
в месте слияния с почковидным пятном и далее идет косо базально, ка-
саясь выступа внутренней линии; круглое пятно правильной формы,
бледносерое, очерченное черным и с темным центром; почковидное пятно
^черноватое, очерченное черным; наружная линия выгнута резко кна-
ружи у костального края, затем идет вертикально слабо зубчатая
в области пятой жилки, затем искривлена й полого изогнута под почко-
видным пятном, снаружи белая и широко оторочена черным; подкраевая
линия неясная, заметная слабыми клинообразными темными штрихами;
бахрома черная и серая в базальной части и черная и белая.во внешней.
Заднее крыло: грязно-белое с желтовато-серым дискоидальным пятном,
наружной линией и терминальной полосой; терминальная линия чер-
ная; бахрома беловато-серая. Нижняя сторона: передние крылья темно-
серые; костальный край, термен и наружная линия беловатые; задние
крылья белые с серыми пятнами по костальному краю; дискоидальное
пятно, наружная линия и подкраевая тень темные. Голова, грурь и
брюшко бледного и темносерого цвета смешанной окраски; второй чле-
ник губных щупалец снаружи черный; ноги и pectus серые; лапки чер-
ные с белыми члениками. Размах крыльев самца 32 мм, самки 34 мм.
«Распространение. Приамурье, Хабаровск (июль, 1910, 1911, Бар-
сов); М. Хинган, Помпеевка (июнь, 1910, Манн).
«Усики желтые, гребенчатые и совершенно гладкие у самок; гребешки
тесные, короткие, булавчатые. Самка несколько светлее».
Полной уверенности в принадлежности этого' вида к Acronicta (Pha-
retra) Ochs, нет; мне этот вид совершенно не известен.
34. ACRONICTA, 35. HAMPSONIA
535
35. Род HAMPSONIA KOZHANTSHIKOV, gen. nov.
Хоботок -развит вполне; глаза большие, круглые, голые, окружены
волосками и чешуйками; губные щупальца длинные, торчащие косо
вверх; последний членик их длинный, конический, по длине почти равен
половине второго членика (у Craniophora Snell, этот членик короткий,
тупой, примерно равный четверти второго); первый членик передней
лапки по длине более половины длины передней голени этой же ноги;
коготки на всех лапках с очень маленьким зубчиком на внутренней сто-
роне; голова и грудь покрыты почти исключительно чешуей; добавочная
ячейка ромбический формы, Мг отхода от ее верхней трети; радиальные
стволы — третий, четвертый и пятый — на стебельке; М2 на задних
крыльях редуцирована; дискальные жилки слабые. Valva очень узкая,
в дистальной части несет грубые щетинки; harpe несет почти прямой
отросток, направленный косо дистально.
Пока известно только два вида этого рода, оба распространенные
в южном Приморья, северном Китае и на Японских островах — Hamp-
sonia jankowskii Obth. и Н. albonigra Herz.
Тип рода Н. jankowskii Obth.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Передние крылья широкие с косой темной поперечной перевязью,
с базальной стороны отороченной белым; harpe слабая, имеет
небольшой отросток, почти прямой. . . . * 2. Н. albonigra Herz
2 (1). Передние крылья узкие, округленные, без поперечной полосы,
- светлосерые; harpe сильная, занимает базальную треть valva, несет
большой сильно изогнутый отросток, торчащий косо дистально
..........................................* 1. Н. jankowskii Obth.
* 1. Hampsonia jankowskii Obth. (рис. 277).
Oberthiir, 1880, Et. Entom., 5 : 69, t. 7, fig. 1 (Acronicta Ochs.); Staudinger, 1892,
Mem, Lep. Rom., 6 : 395; Staudinger, 1901, Cat. Lep., 1 : 132; Herz, 1904, Ежсгодн.
Зоол. муз. Акад. Наук, 9 : 270; Hampson, 1909, Cat. Lep. Phal., 8 : 75; Warren, 1909,
Seitz, Grossschmet., 3 : 14, t. 2, k; Matsumura, 1925, Journ. Coll. Agric. Sapporo,
15 : 120; Draudt, 1931, Seitz, Grossschmett., 3 Suppl., 1 : 8, t. 1, c.
Голова, грудь и передние крылья серые со слабым фиолетовым от-
тенком; рисунок не резок; крылья вытянутые, узкие, непропорционально
большие по отношению к размерам брюшка; базальная линия в виде
двух штрихов, короткая, заметна лишь на костальном крае; базальное
поле большое, светлосерое; от основания крыла интенсивно черный
штрих, расширяющийся в дистальной части, четко очерчен; у дистальной
части этого штриха светлое пятно; вторая поперечная линия двойная,
сильно скошена терминально, довольно широкая, в середине светлосерая;
срединное поле затемнено сильно растушеванной срединной линией,
темный цвет заполняет нижнюю часть крыла; круглое пятно маленькое,
белое, очерчено черным; почковидное пятно овальное, небольшое очер-
нено черным, буро-охристое, в центре черное; поле между почковидным
пятном и третьей поперечной линией светлое с охристым оттенком;
третья поперечная линия начинается светлым штрихом на костальном
крае, далее выгнута, черная, тонкая, двойная, в середине бело-серая,
теряется в дорзальной части крыла; внешнее поле белое и серое с вол-
нистой белой подкраевой линией, дающей в нижней части белые пят-
536
II. ACRONICTINAE
нышки на темном фоне; терминальная линия в виде тонкой полоски,,
темная; бахрома серая с тонкой полоской; задние крылья темнобурые-
у обоих полов, бахрома на них светлая; нижняя сторона серая с неяс-
ной полосой и дискоидальными пятнами на задних крыльях. Размах,
крыльев 28—30 мм.
Uncus не длинный, тонкий, почти прямой, изогнут лишь при основа-
нии; tegumen короткий, особенно в дорзальной части, но довольно широ-
кий; saccus плоский и острый; fultura inferior в виде простой пластинки,
свернутой почти в трубку; anus несет в виде тяжа scaphium; valvae уз-
кие и длинные, выгнутые вентрально; базальная треть valva утолщена
и слита с harpe, которая несет сильно изогнутый большой отросток,
направленный дистально;
дистальная часть valva мем-
бранозная, несет на конце-
щеточку сильных щетинок:
penis’почти прямой, несколь-
ко короче valva; дистальный
конец его несет склеротизо-
ванную пластинку с двумя
большими и двумя маленьки-
ми зубцами; на pars inflabi-
lis сильный шип, сидящий
Рис. 277. Hampsonia jankowskii Obth. Генита- на сильном основании,
лии самца. • Распространение.
Южное Приморье (Суйфун,
Владивосток, Седанка, Русский остров, Сидеми); Сахалин; северный
Китай (Пекин); Хоккайдо; указан с Амура (Гемпсон, 1909), но мате-
риалов из этих мест нет.
* 2. Hampsonia albonigra Herz. (рис. 278). •
Herz, 1904, Ежегодн. Зоол. муз. Акад. Наук, 9 : 269, т. 1, рис. 3; Hampson,
1909, Cat. Lep. Phal., 8 : 53, t. 124, fig. 3; Warren, 1909, Seitz, Grossschmett., 3 : 13;
Draudt, 1931, тамже, SuppL, 1 : 14; Draudt, 1937, Entom. Rundsch., 54, t. 4, fig. 3, e.
Передние крылья черно-серые с белыми и охристыми мазками; база ль- (
ное поле черно-серое с интенсивно черной срединной жилкой; первая
поперечная линия выражена слабо; вторая в виде двух бархатисто-чер-
ных полос, посредине бело-серая, сильно скошена в нижней части крыла
кнаружи; поле между этой линией и очень резкой двойной черно-серой
срединной линией бело-охристое, несколько затемненное в нижней части
крыла; срединная линия широкая идет косо поперек крыла, почти пря-
мая; почковидное пятно почти круглое, нерезкое,.иногда почти смытое-
или, напротив, неполно очерчено черным, внутри охристо-серое; внеш- »
няя половина срединного поля за почковидным пятном почти белая,
в нижней части- с черными штрихами; третья поперечная линия наме-
чена двумя черными штрихами на костальном крае над почковидным
пятном, двойная, резко выгнутая к внешнему краю, посредине белая,
в нижней части крыла одинарная или совсем редуцирована; вдоль куби- i
тальной жилки тонкий верный штрих; внешнее поле тусклосерое с пре-
рывистой белой подкраевой линией; терминальная линия в виде круп-
ных бархатисто-черных штрихов; бахрома шашечная — белая и серая;
задние крылья тусклосерые, однотонные, с неясной перевязью; Нижняя
сторона серо-белая с неясным светлым рисунком; дискоидальное пятно
едва заметно на нижних крыльях. Размах крыльев 28—30 мм.
35. HAMPSONIA, 36. CRANIOPHORA
531'
Uncus длинный, покрытый короткими волосками, загнут почти под;
прямым углом вентрально; tegumen короткий и широкий; saccus неболь-
шой, мешковидный; valvae узкие и длинные, расширенные в дистальной
части, несут большие и густо расположенные волоски на дистальном
конце; в базальной части valva harpe в виде небольшой складки, несу-
щая небольшой, дистально направленный отросток; penis по длине почти
равен valva, прямой, на pars inflabilis несет сильные зубцы. Bursa copu-
latrix цилиндрической формы, покрыта рубчатой скульптурой, в ди-
стальной части имеет направ-
ленный дистально отросток,
закрученный спирально, от
конца которого отходит duc-
tus seminalis; ductus bursae
примерно равен по длине поло-
вине corpus bursae, отходит
от одного уровня с этим рас-
ширением, постепенно сужаю-
щийся к ostium; поверх-
ность . ductus bursae и ниж-
ней части bursa copulatrix по-
крыта рубчатой скульптурой;
восьмой сегмент короткий, не-
сет острые маленькие apophy-
ses anteriores; apophyses pos-
teriores очень короткие; pa-
pillae anales мембранозные,
плоские, покрытые короткими
волосками.
„ Распространение.
Южное Приморье (Влади-
восток, Аскольд); Корея, се-
верный Китай.
Рис. 278. Hampsonia albonigra Herz. Генита-
лии: а — самца, - б — самки.
36. Род CRANIOPHORA SNELL.
Snellen, 1872, Vlind. van Ned. : 262; Aurivillius, 1891, Nordn. Fjar. : 100; Stau-
dinger, 1901, Cat. Lep., 1 : 134; Hampson, 1909, Cat. Lep. Phal., 8 : 51; Warren, 1909,
Seitz, Grossschmett., 3 : 14 (Acronicta Ochs.); Culot, 1909, Noct., 1 : 24; Robel,
1910, Berge, Schmetterlingsb., ed. 9 : 140; Hampson, 1912, Journ. Bomb. Nat. Hist.
Soc., 21 : 424; Bang-Haas, 1926, Nov. Macrolep., 1 : 160; Bang-Haas, тамже, 3 : 61;
Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 13; Nordstroem och Wahlgren,
1937, Svenska Fjar. : 85; — Bisulcia Chapman, 1890, Entom. Rec., 1 : 28 (typus li-
gustri Schiff.).
Тип рода C. ligustri Schiff.
Хоботок развит вполне; глаза большие, круглые, голые, окруженные
волосками и чешуйками; щупальца недлинные, торчащие несколько'
косо вверх, покрытые прижатыми чешуйками и волосками, с коротким
последним члеником; усики самцов простые; голени передних ног вдвое
длиннее первого членика лапки этик же ног; коготки на всех йогах с очень
маленьким зубцом; лоб гладкий, покрытый чешуйками; голова и грудь
покрыты преимущественно чешуей; брюшко несет (не у всех видов)
пучки волосков на дорзальной стороне сегментов; жилкование очень
близко к предыдущим "родам трибы; Мг отходит от дистальной поло-
вины добавочной ячеи; Рэ+4 отходят на стебельке, Р& отходит от одной
ass
II. ACRONICTINAE
-точки с ними; М2 на задних крыльях редуцирована; на передних крыльях
между почковидным пятном и третьей поперечной линией светлое пятно;
окраска задних крыльев тусклобелая или серая.
Олигоморфный род; главная масса видов известна с юга Азии (Гима-
лаи, Китай, южное Приморье, Япония); в тропики заходят немногие
виды (рис. 279). Из Гималаев известны С. harmandi Рouj. (nigromaculata
Warr.), nigrivitta Hamps., nubilata Hamps, и fasciata Moore, последний
вид идет также в тропики (Бирма, Цейлон, Новая Гвинея). G Тайваня
известен С. picata Wilem.; из Китая известны частью те же виды, что
и из Гималаев, частью эндемики (С. obscura Leech, jactans Drd., ni-
aeosy?arsa Mts., /zemiZeaca Piingl., tigniumbra Drd. и fasciata Мооге);часть
этих видов, а также один гималайский известны с Японских островов,
откуда эндемикине описаны; отсюда известны С. fasciata Moore, niveosparsa
Mts. и pracclara Graes. В южном Приморье распространены два послед-
ние вида, эндемичная С. transversa, sp. п. и европейская С. ligustri
Schiff. В бореальной зоне Евразии нет видов Craniophora Snell., но
в южной Европе распространены С. ligustri Schiff, и С. pontica
.Stgr.; первый вид не идет далее зоны широколиственных лесов, а второй
ограничен восточным Средиземноморьем. Кроме этих видов, для, Кап-
ской земли Гэмпсон (1909) указывает Craniophora paragrapha Feld.;
^принадлежность этого вида к Craniophora Snell., впрочем, очень сом-
нительна; возможно даже, что этот вид не принадлежит вообще
к Acronictinae.
Кормовые растения известны только для С. ligustri L., который свя-
зан с видами Oleaceae (Ligustrum, Syringa, Fraxinus) и Caprifoliaceae
"(Viburnum).
Из стран Нового Света, а также из Австралии ‘виды Craniophora
Snell, не описаны; не известны они также и для Африки.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (10). Valva широкая, в базальной части имеет пазуху, в которой нахо-
дится пучок длинных направленных дистально волосков; corpus
bursae редуцирован и заменен сложно устроенным его рукавом, от
которого отходит с вершины ductus seminalis.
J2 (3). Передние крылья с примесью зеленого цвета, особенно при осно-
вании; uncus толстый, густо усаженный короткими волосками;
на дистальном конце valva у верхнего угла группа жестких корот-
ких щетинок, направленных проксимально; рукав bursae широ-
кий, неизвитой.............................* 3. С. praeclara Graes.
3 (2). Передние крылья серые или черно-серые, без примеси зеленого;
прочие признаки иные.
4 (5). Поперек переднего крыла широкая темная полоса, с базальной
стороны резко оттененная светлым; у дорзального края крыла два
резких белых пятна; valva с большой вырезкой в базальной части;
рукав совокупительной сумки перекручен спирально..................
......................................* 2. С. niveosparsa Mts.
5 (4). Передние крылья с более или менее равномерным распределе-
нием темного пигмента, осветлена лишь область близ почковидного
пятна; белые пятна у дорзального края крыла не выражены;
прочие признаки иные.
6 (7). Задние крылья тусклосерые, затемненные по краю; передние
крылья темносерые или черно-серые . . . * 4. С. ligustri Schiff.
Рис. 279. Распространение видов рода Craniophora Snel.
540
II. acronictinae
7 (6). Задние крылья чисто-белые или со слабым затемнением по крак>
и с темными жилками; передние крылья светлосерые.
8 (9). Круглое и почковидное пятна в середине окрашены в желто-ох-
ристый цвет; в базальной части крыла примесь охристых тонов . .
. .............................................* 5. С. pontica Stgr.
9 (8). Пятна выполнены черным или серым; базальная часть крыла,
темная, почти черная, без примеси охристых тонов ...................
.......................................6. С. hemileuca Piingl.
10 (1). Valva узкая, в базальной части без пучка волосков; дистальный
конец ее усажен в беспорядке грубыми щетинками; передние крылья
с темной, черно-коричневой полосой на пепельно-сером фоне . .
...................................* 1. С. transver sa, sp. nov.
Рис. 280. Craniophora transversa, sp. nov. Генита'
лии: a — самца, б — самки.
* 1. Craniophora trans-
versa Kozhantshikov, sp.
nov. (рис. 280).
Передние крылья свет-
лосерые с охристыми
штрихами и черным ри-
сунком; базальное поле
черное с небольшим чер-
ным штрихом вдоль сре-
динной жилки; первая по-
перечная линия неясная,,
но нередко с внутренней
стороны осветлена; вторая
поперечная линия неясная,
светлосерая, простая,
сильно скошенная к внеш-
нему краю, с наружной
стороны оттенена белым,
у дорзального края дает
черный штрих; клиновид-
ное пятно не развито; ба-
зальная половина средин-
ного поля белая с немно-
гими черными и охристы-
ми чешуйками; круглое'
пятно большое, оваль-
ной формы, неполно очерчено черным, иногда почти сливается со свет-
лой частью срединного поля, в середине бело-охристое; срединная линия
черно-серая, очень широкая, занимает всю среднюю часть срединного
поля, идет косо к tornus, затемняя также нижнюю часть внешнего поля;
почковидное пятно большое, овальное, в середине охристо-серое с по-
перечной серой полоской, очень слабо выемчатое с внешней стороны,
тонко очерчено черным; третья поперечная линия начинается черным1
штрихом над почковидным пятном, полого выгнута к внешнему краю,
двойная, тонкая, белая и черная, неглубоко зубчатая по жилкам; внеш-
нее поле бело-серое, затемненное в нижней и верхней частях; в нижней
его части интенсивно черный штрих; подкраевая линия белая, неясная;
терминальная линия в виде белых и черных штрихов; бахрома серая и,
охристая. Задние крылья одноцветно серые, несколько более темные во
внешней доловине, бахрома светлоохристая и серая. Нижняя сторона перед-
36. CRANIOPHORA
541
них крыльев темная, серо-охристая; задние крылья светлосерые с тем-
ной перевязью и дискоидальными пятнами. Размах крыльев 30—34 мм.
Uncus длинный, тонкий, конической формы, сильно загнут вентрально,
.дистальный конец его почти прямой; tegumen короткий с латеральными
короткими выростами; saccus небольшой, сильно конический; anus
несет хорошо развитый scaphium; valvae узкие и длинные, выгнутые
вентрально; дистальный конец и отчасти вентральный край valva несут
большое количество густо расположенных длинных щетинок; penis
несколько короче valva, почти прямой, с коротким соесшп, без инкруста-
ций на pars inflabilis. Bursa copulatrix вытянутая, несколько изогнутая,
расширенная в проксимальной части, несет в дистальной части вырост,
закрученный спирально и направленный проксимально, от конца кото-
рого отходит ductus seminalis; ductus bursae короткий и широкий; ди-
стальная половина совокупительной сумки, ее вырост и ductus bursae
покрыты рубчатой скульптурой; восьмой сегмент короткий, мембраноз-
ный в вентральной половине, несет короткие и острые apophyses ante-
riores; apophyses posteriores очень короткие, острые; papillae anales
узкие и длинные, мембранозные, в дистальной части несут густые длин-
ные щетинки.
Своеобразный, особо стоящий в роде вид; по структурам гениталий
он имеет известное сходство с видами Hampsonia Kozh., хотя valvae
и не имеют harpe. Габитуально напоминает также Н. albonigra Herz,
но имеет значительно большее сходство с Craniophora niveosparsa Mts.
Принадлежность к роду Craniophora Snell, не вызывает все же сомнений,
но этот вид является, повидимому, одним из наиболее генерализованных
в роде Craniophora Snell.
Распространение. Южное Приморье (Уссури, Сучан,
Седанка, истоки р. Сицы); Корея.
* 2. Craniophora niveosparsa Mts. (рис. 281).
Matsumura, 1926, Ins. Matsum., 1 : 4, t. 1, fig. 8; Draudt, 1931, Seitz, Gross-
schmett., 3 Suppl., 1:9, t. 1, d (niveosparsa Mts.); p. 14, t. 1, e (pacifica Fil.); —
pacifica Filipjev, 1927, Ежегодн. Зоол. муз. Акад.. Наук СССР, 28:37; — kalgana
Draudt, 1931, Seitz, Grossschmett., 3 Suppl. 1 : 14, t. 11.
Голова и грудь пепельно-серые с отдельными черными чешуйками;
передние крылья темносерые с немногими белыми пестринами, охри-
стыми пятнами и черным рисунком; базальная линия в виде двух недлин-
ных штрихов на костальном крае; базальное поле темное с тонким чер-
ным штрихом; вторая поперечная линия двойная, слабо извитая, широко
разделенная светлосерой срединной полосой; базальная часть средин-
ного поля светлая, внизу белая; круглое пятно большое, правильной
формы, очерчено черным и белым, в середине охристо-серое; под круглым
пятном ниже кубитальной жилки тонкий, черный штрих; срединная
линия очень ясная, широкая, слагающаяся из четырех полос, отчасти
захватывающих почковидное пятно, слабо волнистая, черная, контраст-
ная, с белой базальной частью срединного поля; почковидное пятно боль-
шое, почти квадратное, охристо-серое, очерчено, черно-бурым; область
между этим пятном и третьей поперечной линией выделяется светлым на
фоне крыла; третья поперечная линия в виде тонкой полоски, сильно
выгнутая, почти не зубчатая близ костального края крыла и сильно
зубчатая в дорзальной его части; внешнее поле узкое, по ширине равно
расстоянию от почковидного пятна до третьей поперечной линии; под-
краевая линия белая, тонкая, слабо волнистая, в нижней части внешнего
542
II. ACRONICTINAE
Рис. 281. Craniophora niveosparsa Mts
Гениталии: а — самца, 6—самки.
поля дает яркое белое пятнышко; терминальная линия в виде серии:
черных штрихов; бахрома серая; задние крылья тусклосерые, однотон-
ные, с несколько более светлой бахромой; нижняя сторона передних
крыльев темная, желто-серая, задних — блестяще-белая, с круглым,
дискоидальным пятнышком и неправильной, размытой поперечной ли-
нией. Размах крыльев 28—32 мм.
Uncus в базальной половине вальковатый, тонкий, в дистальной части
утолщенный, покрыт волосками, с заостренным и загнутым концом;
tegumen узкий, девятый сегмент очень короткий; saccus короткий и широ-
кий; valvae большие, лопастевидные, узкие в базальной половине и
сильно расширенные в дистальной, в базальной части с вентральной
стороны полость, в которой сидит большой пучок волосков; по середине
valva, почти от ее основания отходят складка и сильно ск лер отиз о ванный
тяж; на дистальной половине valva
короткие волоски; penis несколько
длиннее valva, слабо заостренный
в дистальной части: pars inflabilis
penis сильно гофрированная, частич-
но склеротизованная; fultura infe-
rior не отделена от vallum penis.
Bursa copulatrix несет большой вы-
рост, спирально закрученный, от
проксимального конца которого от-
ходит ductus seminalis; ductus bur-
sae сильно изогнут (дугообразно),
с многочисленными продольными
склеротизованными тяжами, осно-
вание его сильно гофрировано; вось-
мой сегмент недлинный, сильно выем-
чатый, более длинный при основа-
нии apophyses anteriores; послед-
ние — конические, довольно длин-
ные (длиннее восьмого сегмента);
apophyses posteriores короткие; pa-
pillae anales мембранозные.
Описанная Филипьевым (1927) Craniophora pacifica Fil. не была сопо-
ставлена с описанием и рисунком (фото) С. niveosparsa Mts. (1926); по-
следняя описана по рисунку и цвету; гениталии не исследованы. Срав-
нение С. pacifica Fil. с описанием С. niveosparsa Mts. говорит за полное
сходство обеих описанных форм и не вызывает сомнения в принадлеж-
ности обоих названий к одному виду.
Распространение. Южное Приморье (Уссури, Сучан,
Виноградовка, Владивосток, бухта Новик); северный Китай; западный
Китай; Японские острова — Хоншу (Киото).
* 3. Craniophora praeclara Graes. (рис. 282).
Graeser, 1890, Berl. entom. Ztschr., 35 : 74; Staudinger, 1901, Cat. Lep., 1 : 134;
Hampson, 1909, Cat. Lep. Phal., 8 : 54, t. 124, fig. 4; Warren, 1909, Seitz, Gross-
schmett., 3 : 15, t. 3, d; Matsumura, 1925, Journ. Coll. Agric. Sapporo, 15 : 23; Matsu-
mura, 1927, Ins. Matsum., 1 : 110; Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 14.
Голова и грудь пепельно-серые с немногими черными чешуйками;
передние крылья пепельно-серые с неправильными черными пятнами,
черным рисунком и серо-зелеными участками (отливающими зеленым;)
36. CRANIOPHORA
543
зеленоватый отлив есть у основания крыла, за почковидным пятном
и за третьей поперечной линией; базальная линия в виде двух разде-
ленных серым цветом штрихов; базальное поле в нижней части затем-
нено; вторая поперечная линия широкая, слабо извитая, посредине серая,,
по краям черная (двойная), ниже кубитального ствола дает размытый
желтый штрих; круглое пятно тонко очерчено темнобурым и широко
белым, в середине серое с зеленоватым блеском; срединная линия широ-
кая, двойная, растушеванная, идет в виде черной перевязи, в дорзаль-
ной части крыла исчезая и почти сливаясь с темной третьей поперечной
линией; почковидное пятно большое, но не широкое, слабо выемчатое,
очерчено черным, в середине однотонное черное; поле между этим пятном
и третьей поперечной линией светлое, однотонное, без рисунка; третья
282. Craniophora praeclara Graes. Генита-
лии: а —• самца, б — самки.
поперечная линия начинает-
ся двумя широко расставлен-
ными штрихами на косталь-
ном крае; далее остается
лишь одна полоса (наруж-
ная), она идет черная, освет-
ленная с базальной стороны,
слабо зубчатая по жилкам;
внешнее поле узкое, значи-
тельно уже расстояния от
почковидного пятна до
третьей поперечной линии,
серо-зеленоватое, с клино-
видным черным штрихом
близ второй и третьей меди-
альных жилок и ниже вто-
рой кубитальной, расплыв-
чатым, пересеченным тонкой
белой полоской; подкраевая
линия в виде сильно извитой
белой полоски; терминаль- Рис-
ная линия в виде серии чер-
ных штрихов; бахрома черно-
серая с ясной полоской и белыми штрихами (шашечная); задние крылья
тусклосерые, однотонные, с частично белой бахромой; нижняя сторона
серая с растушеванной серой полосой, слабыми дискоидальными пят-
нами и неправильными черными штрихами. Размах крыльев 40—44 мм.
Uncus сильно изогнут, в дистальной части расширяющийся, на конце
острый, отделен швом от tegumen; последний короткий, но широкий;
saccus, разросшийся в дистальной части вокруг основания valvae, слу-
жит вместилищем для пучка волосков; anus несет небольшой scaphium;
valvae узкие при основании, сильно расширены в дисталыюй половине;
от основания valva на две трети ее длины широкая, склеротизованная
складка; дистальная половина valva покрыта густыми тонкими волос-
ками, а на дистальном конце несет несколько сильных коротких щети-
нок; fultura inferior ромбической формы с двумя большими вырезками —
одной с дорзальной стороны, другой с вентральной; penis по длине
почти равен valva, в дистальной части слабо изогнут вентрально; соесшп
короткий; pars inflabilis penis гофрированная, в дистальной части склеро-
тизована и несет скульптуру в виде ямок. Bursa copulatrix мешковидная,
расширенная в проксимальной части, несет небольшой заостренный
544
II. acronictinae
«тросток, направленный дистально, от которого отходит ductus semi-
nalis, короткий, в дистальной части расширенный и при основании взду-
тый; восьмой сегмент довольно длинный, узкий; apophyses anteriores
короткие и тонкие, еще более короткие apophyses posteriores; papillae
anales мембранозные с густыми щетинками на дистальном конце.
Распространение. Приамурье (Раддевка); южное Приморье
(Уссури, Седанка, Сучан, Сидеми); Японские острова (Хоккайдо).
* 4. Craniophora ligustri Schiff.1 (рис. 283).
Schiffermiiller, 1776, Wien. Verz. : 79; Staudinger, 1892, Mem. Lep. Rom.,
6 : 393; Aurivillius, 1891, Nordn. Fjar. : 100; Staudinger, 1901, Cat. Lep., 1 : 134;
Spuler, 1908, Schmett. Eur., 1 : 140, t. 31, fig. 24; t. 22, fig. 6 (Raupen); Hampson,
1909, Cat. Lep. Phal., 8 : 57, fig. 16; Warren, 1909, Seitz, Grossschmett., 3 : 14, t. 3, b;
Culot, 1909, Noct., 1 : 24, t. 2, fig. 12; Rebel, 1910, Berge, Schmetterlingsb.,
ed. 9 : 146, t. 28, fig. 20; Draudt, 1931, Seitz, Grossschmett., 3 SuppL, 1 : 14,
>t. 1, e; Nordstroem och Wahlgren, 1937, Svenska Fjar. : 85, t. 15, fig. 17; — letterata
Panzer, 1804, Syst. Nom. Schaff. Ins. : 115, t. 105, fig. 3, 4; — coronula Haworth,
1809, Lep. Brit. : 179; — sundevalli Lampa, 1885, Entom. Tidskr. : 50; — olivacea
Tutt, 1888, Entom. : 81; — nigra Tutt, 1890, Entom. Rec., 1 : 34; — troni Huene,
1901, Berl. entom. Ztschr. : 310; — effusior Dannehl, 1923, Entom. Ztschr. Frankf.,
139 : 6; — viburni Dannehl, 1925, там же, 39 : 120; — roseoradiata Dannehl, 1925, там
же, 39 : 120; — obscura Mellaerts, 1928, Lambillionea, 28 : 117.(non Leech); — gigantea
Draudt, 1937, Entom. Rundsch., 54 : 375.
Биология и экология. Graeser, 1890, Berl. entom. Ztschr., 35 : 74;
Meyrik, 1895, Lep. Brit. : 142; Barret, 1895, Brit. Lep., 3 : 272, t. 123, fig. 2; Ljung-
dahl, 1917, Entom. Tidskr., 38 : 225, t. 2, fig. 5; Bandermann, 1932, Intern, entom.
Ztschr., 26 : 20; Лебедев, 1933, Зб1рн. праць екол., 2 : 76.
Передние крылья черно-бурые с белыми или даже зеленоватыми
<(у особей с юго-востока Азии) пятнами в апикальной части крыла;
голова и грудь светлосерые ,с черно-бурыми пестринами; рисунок на пе-
редних крыльях хорошо развит, тонкий, но не резкий; базальная линия
в виде двух тонких черных линий, слабо извитых, идет до половины
крыла; от основания крыла в базальном поле неясный черный штрих;
вторая поперечная линия двойная, черная, посредине светлобурая,
•слабо скошенная в дорзальной части крыла, извитая, с темным штри-
хом у предпоследнего изгиба; в срединном поле неправильные белые
штрихи близ круглого пятна; последнее почти круглое, очерчено черным
и белым, внутри почти черное; срединная линия слабо извитая, расту-
шеванная, проходит между пятнами (близ круглого) несколько косо
кнаружи; почковидное пятно небольшое, почти прямоугольное, во внеш-
ней половине белое, очерчено черным и белым, в середине темнобурое;
между почковидным пятном и третьей- поперечной линией белое пятно;
третья поперечная линия двойная, начинается над почковидным пятном
двумя черными и светлыми штрихами, далее идет обычдой формы, двой-
ная, посредине белая и ниже кубитальной жилки бурая; подкраевая
линия широкая, белая в апикальной части, далее смыта или видна лишь
местами; терминальная линия в виде серии черных и белых шашечек;
бахрома бурая с тонкой полоской; задние крылья тусклосерые, одно-
1 Известная из северо-западных Гималаев Craniophora cubitata Hamps. (Hamp-
son, 1894, Moths Ind., 2 : 208; Hampson, 1909, Cat. Lep. Phal., 8 : 55, t. 124, fig. 5;
Warren, 1913, Seitz, Grossschrpett., Indo-Austr., 11 : 38, t. 5, f), как можно судить
по описаниям и рисункам, очень близка к С. ligustri Schiff, и, вероятно, является
лишь гималайской формой этого вида. Точно так же Craniophora obscura Leech (Leech,
1900, Trans. Entom. Soc. Lond. : 107; Hampson, 1909, Cat. Lep. Phal., 8 : 58, t. 124,
fig. 7; Warren, 1909, Seitz, Grossschmett., Pal., 3 : 15, t. 3, c; Draudt, 1931, 1. c.,
1 : 14), описанная из западного Китая, видимо, лишь форма от С. ligustri Schiff.
36. CRANIOPHORA
545
тонные, с более светлой слабо шашечной бахромой; нижняя сторона туск-
лая, однотонная, с шашечной бахромой и немногими светлыми пятныш-
ками на костальном крае. Размах крыльев 32—-36 мм.
Uncus тонкий, в дистальной половине покрыт густыми волосками,
изогнут крючковидно; tegumen узкий, узкое и девятое кольцо; saccus
короткий; valvae при основании узкие, сильно расширенные в дисталь-
ной части; в базальной части valva мешковидное углубление, где сидит
пучок длинных волосков; от основания valva на две трети ее длины скле-
ротизованная складка конической формы; uncus несет хорошо развитой
scaphium; penis по длине почти равен valva, слабо изогнутый, с корот-
ким соесшп; pars inflabilis сильно гофрирована, без шипов. Bursa copu-
latrix вытянутой формы, округлая в проксимальной части, в дистальной
части несет сосковидное расширение, покрытое рубчатой скульптурой,
слегка закрученное на ди-
стальном конце, от кото-
рого отходит ductus semi-
nalis; ductus bursae отхо-
дит от середины corpus
bursae, тонкий, цилиндри-
ческой формы, слабо рас-
ширенный и немного изог-
нутый, покрытый рубчатой
скульптурой, по длине
примерно равен половине
bursa copulatrix; 'восьмой
сегмент короткий, с глу-
бокой вырезкой на дор-
зальном крае, несет ко-
роткие apophyses anterio-
res; apophyses posteriores
также короткие; papillae
anales мембранозные.
Взрослая гусеница зе-
леная, слегка веретенооб- Рис. 283. Craniophora Jligustri Schiff. Гениталии:
разной формы; сегменты а — самца, б — самки,
тела разделены глубокими
бороздами; голова темнозеленая; вдоль спины желто-белая полоса; по
бокам тела над дыхальцами по одной узкой желтой полосе; на каждом
сегменте по сторонам тергита симметрично по две белых точки; дыхальца
желто-красные; тело покрыто недлинными негустыми черными волосками
(небольшое число вторичных).
Куколка светлокоричневая с более светлой задней частью тела, глад-
кая, блестящая; кожный покров несет точечную скульптуру; cremaster
в виде короткого округло-конического прямого выроста, несущего очень
короткие шипики — шесть в ряд на дистальном конце и два дорзально
от них несколько меньшей величины.
У4 Мало изменчивый вид; наблюдаются лишь географические формы;
южные особи, особенно закавказские и туркменские, часто окрашены
в светлосерый цвет, несколько мельче; сходные особи изредка встречаются
и севернее, например на Украине. Дальневосточная и китайская формы,
gigantea Drd., наиболее отличны и могут рассматриваться как подвид;
они отличаются более интенсивной черной окраской передних крыльев,
имеющих в базальной и апикальной частях примесь охристых и зелено-
35 Фауна СССР, И. В. Кожанчиков.
546
II. ACRONICTINAE
ватых тонов, и большей яркостью рисунка; строение гениталий не отли-
чается от типичной формы. Задние крылья этой формы почти чисто белые
с тонкими черными штрихами по жилкам и тонким черным ободком по
краю; она крупнее, 40—42 мм. Очень напоминает индийскую С. nigro-
maculata Warr., возможно* являющуюся также лишь формой этого вида.1
Распространение. Средняя Европа (Германская низмен-
ность, Альпы, северная Италия); Британские острова (Шотландия);
южная Скандинавия (Упланд, Этланд, о. Готланд); Финляндия (Аландия);
юго-восточная Европа (долина Дуная, Карпаты, Герцеговина, Греция);
Румыния; Европейская часть СССР, кроме севера (Прибалтика, Ленин-
град, Москва, Казань, Калуга, Могилев, Херсон, Винница, Таганрог,
Одесса, Бендеры); Крым — южный берег; Северный Кавказ (Железноводск,
Кубань); Закавказье (Лагодехи, Истидера, Куссари, Ленкорань); Турк-
мения (Ашхабад); южное Приморье (Виноградовка, Яковлевна, Сучан);
северная Корея (Сидеми). Указание Суворова (1896) для Семипалатинска
мало, вероятно; материал из этих мест и даже с южного Урала отсут-
ствует; последним пунктом на восток, где этот вид достоверно встре-
чается, является Казань, но и здесь он-очень редок.
Биология и экология. Зимует куколка; вылет бабочек
в мае—июне; гусеницы живут поодиночке, питаются видами Oleaceae,
главным образом бирючины (Ligustrum), сирени (Syringa) и ясеня (Fra-
xinus); в природе встречаются также на калине (Viburnum). Окукление
в подстилке в плотном коконе; куколка диапаузирует; дает почти всегда
одно поколение и лишь в теплые годы в средней Европе два.
* 5. Craniophora pontica Stgr. (рис. 284, 285).
Staudinger, 1879, Тр. Русск. энтом. общ., 14 : 364; Christoph, 1885, Mem, Lep.
Rom., 2 : 27; Melioransky, 1898, Тр. Русск. энтом. общ., 31 : 239, t. 7, fig. 6;
Staundiger, 1901, Cat. Lep., 1 : 132; Hampson, 1909, Cat. Lep. Phal., 8 : 55, t. 124,
fig. 6; Warren, 1909, Seitz, Grossschmett., 3 : 15, t. 3, c, d; Culot, 1909, Noct., 1 :'2O,
t. 2, fig. 3; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 144; Филипьев, 1927, Еже-
годн. Зоол. муз. Акад. Наук СССР, 28 : 37, фиг. 6, 7; Draudt, 1931, Seitz, Gross-
schmett., 3 Suppl., 1 : 13, t. 1, b; Boursin, 1935, Intern, entom. Ztschr., 29 : 241,
fig. 8—12; — navasi Boursin, 1935, там же, 29 : 241, fig. 11.
Передние крылья светлосерые с неправильно разбросанными белыми
и желтыми штрихами; рисунок нерезкий; базальная линия в виде двух
косых штрихов, теряющихся в темном базальном поле; от основания
крыла темный тонкий штрих; нижняя часть базального поля с желтыми
чешуйками; вторая поперечная линия сильно скошена кнаружи, двойная,
черная, посредине серая, дает короткий треугольный изгиб к основа-
нию крыла; базальная часть срединного поля в виде светлой полосы;
круглое пятно правильной формы, тонко очерчено черным и широко
белым, в середине желто-серое; срединная линия в виде черно-серой
перевязи, косая, широкая, слагающаяся из трех параллельных полос,
в дорзальной части крыла сливается с третьей поперечной полосой;
почковидное пятно растушеванное, серо-желтое в центре и беловатое-
к краям, лишь частично очерчено темным, сливается со светлой частью
срединного поля у третьей поперечной линии; последняя — двойная,
1 Описанная Драудтом из северной Юньнани (западный Китай) С. jactans Dra-
udt (1937, Entom. Rundsch., 54 : 376, t. 4, fig. 1, d) очень близка к C. ligustri Schiff.;
автор дает дурной рисунок и краткое описание; изучение структурных признаков
сделано не было. Самостоятельность вида не ясна; мне он известен только по литера-
туре.
36. CRANIOPHORA
547
темная, посредине светлосерая, начинается у дистального края почко-
видного пятна, выгнута кнаружи, слабо зубчатая, в нижней части стуше-
Рис. 284. Craniophora pontica Stgr. Самка. (Рис. Ф. Гуняева).
ванная; внешнее поле сильно суженное, не шире расстояния между
третьей поперечной линией и почковидным пятном, тусклосерое, в ниж-
ней части с узким бе-
лым штрихом; подкрае-
вая линия неясная, в
виде разорванной белой
полоски; терминальная
линия в виде серии чер-
ных и белых штрихов,
заходящих на бахро-
му; последняя —серая;
задние крылья тускло-
белые с затемненными
слегка охристыми жил-
ками и немногими не-
правильными темными
растушеванными штри-
хами по внешнему краю;
бахрома белая; нижняя
сторона белая с темны-
ми пятнышками на ко-
стальном крае и в ди-
стальной части крыль-
ев. Размах крыльев
28—32 мм.
Uncus длинный, изо-
Рис. 285. Craniophora pontica Stgr. Гениталии: а —
самца, б — самки.
гнут, в дистальной по-
ловине утолщен, несет
большое количество во-
лосков; tegumen узкий; saccus небольшой, треугольный; fultura
inferior в виде пластинки неправильной формы, слабо склероти-
зованной; valvae в виде широкой лопасти, мембранозные, округлые-
35*
548
II. ACRONICTINAE
по дистальному краю; от базального верхнего угла косая склеротизо-
ванная складка, достигающая середины valva; дистальная половина
valva густо волосистая; от базального края valva с внешней его стороны
отходит пучок длинных тонких волосков; penis длиннее valva, слабо
изогнут дорзально; соесшп короткий; pars inflabilis penis без инкруста-
ций. Bursa copulatrix рудиментарна, но несет большой сильно расширен-
ный в проксимальной части отросток, закрученный спирально, от конца
которого отходит ductus seminalis; dustus biirsae тонкий и длинный,
сложно извитой, равномерной толщины, несет продольные склеротизо-
ванные складки на внутренней стороне; восьмое кольцо длинное, но
узкое, несет короткие apophyses anteriores; apophyses posteriores также
короткие; papillae anales плоские, мембранозные, слабо дифференциро-
ванные, дистальная часть их несет густые, но крепкие щетинки.
Распространение. Побережье Средиземного моря, Пире-
нейский полуостров (Испания); юго-восточная Венгрия (Геркулесбад);
северная Африка (Тунис); северная Сирия (Мараш); Малая Азия; Крым
(Алупка, Севастополь, Керчь); Молдавия; Закавказье (Кировабад, Тби-
лиси, Ордубад); северо-западный Иран (Шахруд); Курдистан.
6. Craniophora hemileuca Piingl.
Piingeler, 1899, Iris, 12 : 291, t. 8, fig. 6 (Acronicta Ochs.); Staudinger, 1901, Cat.
Lep., 1 : 132; Hampson, 1909, Cat. Lep. Phal., 8 : 64, t. 124, fig. 11; Warren, 1909,
Seitz, Grossschmett., 3 : 14, t. 2, h; Draudt, 1931, Seitz, Grossschmett., 3 SuppL,
1 : 8, t. 1, b; Draudt, 1937, Entom. Rundsch., -54 : 401, t. 4, e.
Малоизвестный вид; оригинальное описание очень кратко; я привожу
его полностью: «Размах 29 мм, длина переднего крыла 15 мм. Наиболее
близка к strigosa, но крупнее, передние крылья темнее, равномерного
красновато-черного цвета, у одной особи в середине более светлые. Рису-
нок как у strigosa, как у этого вида, светлое пятнышко на внутреннем
углу. Задние крылья чисто белые, только на конце несколько затемнены,
имеют прерванную темную краевую линию. Нижняя сторона как
у strigosa, нижние крылья белее; щупальца сильнее, покрыты волосками;
тело стройное».
Драудт (1937) впервые отнес этот вид К Craniophora Snell.; сравнение
со С. strigosa Schiff., проводимое Пюнгелером, основано на чисто габи-
туальном сходстве. Судя по фотографии и рисункам, этот вид наиболее
близок к С. pontica Stgr. Базальная часть крыла несколько затемнена
с очень слабым штрихом от основания крыла; линии рисунка двойные,
пятна хорошо развиты, в середине темные; срединное поле с примесью
охристых тонов, последние не заметны по дорзальному краю передних
крыльев. Принадлежность к Craniophora Snell, не вызывает сомнений.
Распространение. Тибет — Куку-нор.
37. Род ARSILONCHE LD.
Lederer, 1857, Noct. Eur. : 70; Staudinger, 1901, Cat. Lep., 1 : 134; Spuler, 1908,
Schmett. Eur., 1 : 141; Culot, 1909, Noct., 1 : 26; Warren, 1909, Seitz, Grossschmett.,
3 : 12; Hampson, 1909, Cat. Lep. Phal., 8 : 173 (Sirnyra Ochs.); Rebel, 1910, Berge,
Schmetterlingsb.’, ed. 9 : 147; Bang-Haas, 1926, Nov. Macrolep., 1 : 159; Bang-Haas,
1927, там же, 3 : 61; Bang-Haas, 1928, там же, 4 : 40.
Тип рода A. albovenosa Goeze.
Хоботок есть, но небольшой, слабый; губные щупальца короткие,
торчащие прямо вперед; последний членик их короткий, в 5—6 раз
37. ARSILONCHE
549
короче второго; второй членик покрыт длинными нежными волосками;
коготки всех ног простые, с небольшим утолщением при основании,
глаза большие, круглые, голые; усики самцов простые, такие же как
у самок; голова и грудь покрыты волосками, почти без чешуй; добавоч-
ная ячея узкая, почти ромбическая, отходит от основания верхней
ее четверти; 7?3 и Pi на длинном стебельке, Р$ соединен при основании
со стебельком; М2 на задних крыльях рудиментарна. Передние крылья
матовые.
Габитуально Arsilonche Ld. близок к Sirnyra Ochs., но, помимо струк-
турных отличий в имагинальной стадии, дают отличия также и гусе-
ницы — они имеют плоские бородавки и короткие щетинистые волоски.
По строению гениталий Arsilonche Ld. также близок к Sirnyra Ochs.,
но отличен строением головы, редукцией ротовых частей, наличием тон-г
кого волосяного покрова груди (без чешуй) и редукцией рисунка крыльев.
Хетотаксия гусениц близка тому, что описано для Acronicta Ochs.
Группы а и р на среднегруди взрослых гусениц слиты; есть ?) на этом же
сегменте. Группа ос на брюшных сегментах, исключая седьмой, прямо
за стигмой, на седьмом слита с п. Группы ос и на восьмом брюшном
сегменте на одном уровне (рис. 281).
Единственный вид этого рода A. albovenosa Goeze 1 распространен
очень широко, но пятнисто. Заселяет всю Европу и юго-восточную Азию
(отсутствуя в северо-восточной Европе и Сибири) и умеренную зону Север-
ной Америки от Калифорнии до Канады. Особи из Северной Америки
идентичны с евразийскими как по габитусу, так и структурно. Описанный
из Колорадо (Смит, 1900) Arsilonche colorada Sm. очень близок к A. albo-
venosa Goeze; отличия даны лишь в окраске, но не в структурах; очень
вероятно, что он представляет лишь форму голарктического A. albove-
nosa Goeze.
* 1. Arsilonche albovenosa Goeze (рис. 286, 287).
Goeze, 1781, Ent. Beitr., 3 (3) : 251 (Noctua); Aurivillius, 1891, Nordn. Fjar. : 98;
Staudinger, 1901, Cat. Lep., 1 : 134; Spuler, 1908, Schmett. Eur., 1 : 141, t. 31, fig. 3;
он же, 1908, Raupen, t. 21, fig. 17; add. : 351; Hampson, 1909, Cat. Lep. Phal.,
8 : 177, fig. 28; Warren, 1909, Seitz, Grossschmett., 3 : 12, t. 2, e, f; Culot, 1909,
Noct., 1 : 26, t. 3, fig. 2; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 147, t. 28, fig.
10; Kuhn, 1928, Intern, entom. Ztschr., 22 : 421, fig. 1; Draudt, 1931, Seitz, Gross-
schmett., 3 SuppL, 1 : 7, t. 1, b; Urbahn, 1932, Stett. entom. Ztg., 94 :141, fig. 6; Dra-
udt, 1937, Seitz, Grossschmett., 3, Nachtr.-Bericht., SuppL, 1 : 237; — pattens Sepp, 1777,
Ins. NederL, 2 (3) : 11, t. 3, fig. 1—9; — degener Sepp, 1777, Ins. NederL, 4, Corri-
genda; Hiibner, 1808, Europ. Schmett., Noct., fig. 380 (non Schiff.); — venosa Borkhau-
sen, 1792, Eur. Schmett., 4 : 716; — atomima Haworth, 1809, Lep. Brit. : 175; — cen-
tripuncta Herrich-Schaeffer, 1856, Neue Beitr. Schmett. : 4, fig. 24, 25; — henrici Grote,
1873, Bull. Buff. Soc., Nat. Sci., 1 : 10 (Leucania)',— eva'nida Grote, 1873, тамже, 1 : 10
(Leucania)', —fumosum Morrison, 1873, там же, 1 : 275 (Ablepharon)', —flavida Auri-
villius, 1880, Entom. Tidskr. : 38; — murina Aurivillius, 1880, там же : 37; — albida
Aurivillius, 1880, там же : 38; — ochracea Tutt, 1888, Entom. : 99; — argentea Tutt,
1888 : 100; — saepestriata Alpheraky, 1895, Iris, 8 : 188; — tanaica Alpheraky, 1904,
Тр. Русск. Энтом. общ., 38 : 589; — tristis Bang-Haas, 1907, Iris, 20 : 70; — neome-
laina Traub, 1928, Intern, entom. Ztschr., 22 : 188, 203; — cretacea Wagner, 1929, Mitt.
Munch, entom. Ges., 19 : 68, t. 2, fig. 8; — renimaculata Osthelder, 1932, там же,
22 : 83; — tjurana Draudt, 1936, Entom. Rundsch., 53 : 456, t. a, 1.
Биология и экология. Thaxter, 1877, Psyche, 1 : 188; Coquillett,
1880, Can. Entom., 12 : 45; Edwards, 1888, Entom. Amer., 3 : 171; Snyder, 1894, En-
1 Ледерер (1857) разделил род Sirnyra Ochs, на два и отнес виды A. dentinosa Frr.
и S. nervosa L. к роду Sirnyra Ochs., a A. albovenosa Goeze выделил в особый род. Из
последующих авторов лишь Гэмпсон (1909) не последовал этому и вновь объединил
эти разные формы в Sirnyra Ochs.
550
II. ACRONICTINAE
tom. News, 5 : 277; Meyrik, 1895, Brit. Lep. : 144; Barret, 1895, Lep. Brit., 3 : 275,
t. 123, fig. 3; Шембель, 1924, Отч. Астр. ст. защ. раст. : 29; Ильинский, 1927, Зап.
Астр. ст. защ. раст., 1 : 24; Айе, 1930, Entom. Anz., 10 : 232; Aue, 1932, там же,
12:245; Kuhn, 1928, Intern, entom. Ztschr., 28:421, fig. 1; Hoffman, ' 1916,
Entom. Ztschr. Frankf., 29 : 53.
Рис. 286. Arsilonchd alboeenosa Goeze. Гени-
талии: а — самца, б — самки.
Голова и грудь желтовато-белые с примесью немногих черных чешуй;
усики самцов слабо утолщенные; передние крылья округлые, слабо вытя-
нутые, неравномерно желто-охристой и белой окраски, с неправильно
разбросанными многочисленными черными чешуйками; основание перед-
них крыльев светлое; от дискоидальной ячеи и вдоль второй медиальной
жилки темный серо-желтый штрих; жилки светлее общего фона; терми-
нальная линия в виде очень тонких темных штрихов; бахрома желто-
белая; задние крылья чисто белые с белой бахромой; нижняя сторона
белая с охристо-серым косталь-
ным краем передних и задних
крыльев. Размах крыльев 30—
42 мм.
Uncus недлинный, сильно
изогнутый при основании, да-
лее почти прямой, почти ци-
линдрической формы, лишь у
дистального конца резко сужен;
tegumen очень короткий в тер-
гальной области, длинный в
плейральной; saccus очень не-
большой с округлым прокси-
мальным концом; valvae боль-
шие, слабо конические; базаль-
ная половина valva с вен-
тральной стороны утолщена,
срастается с harpe, имеющей
вид складки и несущей один
длинный конический отросток,
слабо изогнутый и направлен-
ный косо дорзально; дистальная половина valva мембранозная, несет
короткие густые волоски; fultura inferior в виде двух сросшихся пласти-
нок, образующих в месте срастания киль; penis короткий, расширяю-
щийся в дистальном направлении, изогнутый в дистальной трети; coecum
короткий; pars inflabilis несет несколько крупных сильно склеротизован-
ных пластин, слабо зубчатых. Bursa copulatrix большая, почти яйце-
видной формы; нижняя часть ее несет короткий отросток, закрученный
у основания ductus bursae; от конца этого отростка отходит ductus semi-
nalis; на вентральной стороне совокупительной сумки lamina dentata
в виде почти круглой, слабо склеротизованной пластинки, в центре
имеющей складку с зубцом; ductus bursae почти прямой, по длине при-
мерно равен половине длины corpus bursae, складчатый; восьмой сегмент
длинный и узкий, мембранозный с вентральной стороны; apophyses anterio-
res тонкие и длинные; apophyses posteriores короткие и слабые; papillae
anales широкие, слабо дифференцированные, несут венчик волосков
на дистальных концах.
Взрослая гусеница желто-коричневая с черным рисунком; голова чер-
ная или коричневая с желтыми, адфроптальнымй полями и желтыми
пятнами на прочих частях; тело несет желтые бородавки, покрытые
37. ARSILONCHE
551
короткими торчащими желтыми волосками; вдоль спины широкая черная
полоса, частью прерванная; бока желтые с многими мелкими темными
точками и черточками, иногда сливающимися и придающими кожному
покрову темный тон; под стигмами нерезкая желтая полоса; брюшная
сторона и ноги желто-серые; стигмы белые, окруженные черным.
Куколка черно-коричневая; брюшные сегменты имеют приподнятый
дистальный край, несущий скульптуру в виде круглых ямок; cremaster
в виде конического выроста, несет пучок прямых тонких щетинок.
Окраска передних крыльев и тела варьирует от желто-белой у типич-
ной формы до чисто белой (albida Auriv., argentea Tutt, creta-ea Traub.),
от темносерой и почти черной (tristis В. Н., neoivelaina Traub.) до сме-
шанной охристо-серой (irurina Auriv., j lav id a Auriv.); изменчивость
окраски не связана с ареалом распространения форм. Описанная Ост-
гельдером A. renimaculata Osth. из Малой Азии (Мараш) является, без
сомнения, лишь формой от обычной albovenosa Goeze, отличаясь полным
Рис. 287. Arsilonche albovenosa Goeze. Хёто-
таксия передне- и среднегруди и брюшных
сегментов I, VI, VII и VIII взрослой
гусеницы.
отсутствием желтых тонов и
развитием черных. Это тем бо-
лее несомненно, что позже из
Малой Азии была описана Дра-
удтом tjurana (1936) как форма
от европейской albovenosa Go-
eze. Учитывая большую измен-
чивость окраски этого вида,
нельзя считаться при описании
близких видов только с окра-
ской, но необходимо давать
описание структурных отличий,
что не было сделано Остгелде-
ром. Гэмпсон (1909) выделяет
американскую henrici Grt. как
самостоятельный вид без вся-
ких аргументов, в то время как Смит (1899) изучал структурные особен-
ности европейской albovenosa Goeze и американской henrici Grt. и объеди-
нил их; его рисунки гениталий дают право присоединиться к его
мнению.
Описанная Алфераки (1895) по одной самке из северной Монголии
saepestriata также выделена на основании лишь цветовых признаков,
в частности на основании наличия светлых штрихов вдоль крыльев
и более темных задних крыльев. Сколько позволяют судить признаки
строения головы, ног и чешуйчатого покрова, это обычная A. albovenosa
Goeze; может быть это северный, более мелкий подвид, о чем судить
сейчас невозможно. '
Распространение. Средняя Европа — Франция, Германия,
Польша (Варшава), Карпаты, Альпы; долина Дуная (Венгрия, Вена),
Далмация; Британские острова; южнаяЩвеция (Оланд, Готланд, Уп7
ланд, Блекинге); Финляндия (Ниландия, Або, Остроботтния); средняя
и южная полоса Европейской части СССР (Москва, Казань, Молотов,
Подольск, Чернигов, Таганрог, Херсон); Поволжье (Саратов, Сарепта);
южный Урал (Чкалов); Средняя Азия — Сыр-дарья (Ай-дарле), Зерав-
шанский хребет (Самарканд); Семиречье (Или, Кульджа); южный
Сахалин; Малая Азия (Мараш, Тогуз-Тюран); северная Монголия
(Улан-Батор); Иезо (Саппоро); Северная Америка — южная Канада,
Соединенные Штаты (Массачузеттс, Нью-Йорк, Калифорния).
552
11. ACRONICTINAE
Биология и экология. Зимует куколка; вылет бабочек
на севере ареала в мае—июне, на юге — в марте—апреле (в Калифорнии
в январе). Гусеницы живут поодиночке, многоядны, питаются видами
Salicaceae (Salix), Rosaceae (Rubus), Polygonaceae (Polygonum), Gramineae
(Phragmites, Elymus, Glyceria, Arundo), Iridaceae (Iris), Cyperaceae
(Carex), Typhaceae (Typha). Окукление в коконе из тесно сплетенных
листьев и шелковых нитей (собственно кокон —• из шелка). Стацией
обитания являются влажные луга и уремы, но на теплых территориях.
Б Нижнем Поволжье отмечена как вредитель сенокосов (Шембел, 1924).
38. Род OXYCESTA НВ.
Hiibner, 1827, Verz. bekannt. Schmett. : 144; Staudinger, 1901, Cat. Lep., 1 : 134;
Spuler, 1908, Schmett. Eur., 1 : 140; Hampson, 1909, Cat. Lep. Phal., 6 : 112 (Hadeni-
nae); Warren, 1909, Seitz, Grossschmett., 3 : 12; Rebel, 1910, Berge, Schmetter-
lingsb., ed. 9 : 146; Dalia-Torre, 1917, Intern, entom. Ztschr., 11 : 243; Bang-Haas,
1928, Nov. Macrolep., 3 : 61; — Clidia Boisduval, 1840, Index Method. : 68 (typus
geographica F'.).
Тип рода О. geographica F.
Хоботок редуцирован; щупальца направлены вперед и покрыты длин-
ными пушистыми волосками; последний членик щупалец довольно длин-
ный, заостренный; глаза большие, голые; усики самцов гребенчатые,
гребешки короткие, толстые, дистально расширенные; голова и грудь
покрыты длинными пушистыми волосками; брюшко покрыто короткими
волосками; крылья короткие, относительно широкие, костальный край
несколько выемчатый; apex вздернут; Р2 отходит близ вершины добавоч-
ной ячеи, Мг отходит от ее середины или несколько базальнее; Р3, Pi
й Р5 на коротком стебельке; Р5 отходит иногда почти без стебелька; Л/2
задних крыльев вполне развита, идет от дискоидальной ячеи.
Хетотаксия гусениц почти вполне соответствует тому, что описано
для Arsilonche Ld. и Acronicta Ochs., исключая восьмой брюшной сегмент.
На последнем бородавки ос и р слиты в одну (как у Мота НЬ.), что отли-
чает этот род весьма резко.
Олигоморфный род; известны всего два вида, очень близкие, распро-
страненные по полупустынной и степной зонам северного побережья
Средиземного моря и юго-восточной Европы от Пиренейского полуострова
(Арагония, Каталония) до южного Урала; распространение пятнистое.
Виды Oxycesta Hb. связаны с Euphorbiaceae (Euphorbia) и Scrophularia-
ceae (Linaria).
* 1. Oxycesta geographica F. (рис. 288, 289).
Fabricius, 1787, Mant. Ins., 2 : 167; Staudinger, 1901, Cat. Lep., 1 : 134; Hampson,
1906, Cat. Lep. Phal., 6 : 113, fig. 28; Spuler, 1908, Schmett. Eur., 1 : 140, t. 31, fig. 4;
Culot, 1909, Noct., 1 : 24, t. 2, fig. 13; Warren, 1909, Seitz, Grossschmett., 3 : 12,
t. 2, d; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 146; — austera Scriba, 1791, Beitr.
Ins. : 176, t. 7, fig. 5 (Bombyx); — sericina Hiibner, 1827, Beitr. Eur. Schmett., 2 (1),
t. 4, fig. 7.
Биология. Hoffman, 1893, Raupen : 74, t. 21, fig. 18; Kirby, 1895, Butt.
Moths Eur. : 155.
Голова и грудь покрыты длинными волосками, местами (на tegulae)
окрашенными более темно; у основания крыльев длинные волоски; перед-
ние крылья желто-коричневые с коричневым и серебристо-белым рисун-
ком; базальная линия в виде короткого коричневого штриха, с базальной
38. OXYCESTA
553
стороны осветленного; от основания крыла короткий треугольный сере-
бристый штрих, почти доходящий до второй поперечной линии; базальное
поле в костальной половине светложелтое, в дорзальной желто-серое;
вторая поперечная линия очень четкая, двойная, коричневая, посредине
серебристо-белая, особенно близ дорзального края крыла, в костальной
половине крыла она резко выгнута базально, почти к основанию крыла
и затем дистально; круглое пятно намечено неправильным диффузным
скоплением темного пигмента; срединная линия заметна в виде корот-
кого штриха на костальном крае; почковидное пятно почти намечено
в виде слабой луночки, растушеванное; третья поперечная линия очень
четкая, сначала идет прямо к дорзальному краю до М2, затем дает резкий
изгиб кнаружи, после чего опять почти по той же линии к дорзальному
краю крыла, у костального края двойная, далее в виде тонкой простой
черно-коричневой полоски, окайм-
ленной с внутренней стороны ши-
рокой серебристой полосой; внеш-
нее поле однотонное, тускло-
желтое, слегка светлеющее к терми-
нальному краю; подкраевая ли-
ния редуцирована; терминальная
линия в виде слабых светлых
штрихов; бахрома шашечная —
серая и желтая; задние крылья
светлые, желто-серые, несколько
светлее к основанию; у самок тем-
нее; бахрома на них желто-серая;
нижняя сторона серо-желтая с
неправильными светлыми пятнами,
осветленными, особенно в терми-
нальной части, жилками, со свет-
лой широкой размытой полосой
и шашечной бахромой. Размах
крыльев 20—26 мм.
Uncus короткий, слабо загнут
вентрально, слегка утолщен в ди-
стальной половине и заострен на конце; tegumen довольно длинный, но уз-
кий; saccus маленький; valvae в средней части изогнуты вентрально, ди-
стальный конец направлен дорзально; в базальной части valva у вентраль-
ного края склеротизованная складка, связанная швом с основанием harpe;
отросток harpe широкий при основании, конический, лежит в плоскости
valva; fultura inferior в форме диска, слабо склеротизована; penis короче
valva, в середине сильно выгнут дорзально; coecum узкий, около одной
четверти длины valva; pars inflabilis без инкрустаций. Bursa copulatrix
в виде суженного в дистальной части мешка, с коротким отростком в ди-
стальной части, от которого отходит ductus seminalis; поверхность bursa
copulatrix имеет рубчатую скульптуру; ductus bursae короткий, широкий,
суженный в дистальной части, имеет склеротизованные складки; восьмой
сегмент длинный, мембранозный с вентральной стороны; apophyses ante-
riores короче, чем восьмой сегмент; apophyses posteriores очень короткие,
заостренные; papillae anales мембранозные, почти прямоугольные, несут
короткие волоски в дистальной части.
Яйца почти правильной формы, круглые, несколько сплюснутые
в дорзовентральном направлении; хорион покрыт глубокими радиаль-
Рис. 288. Oxycesta geographica F. Генита-
лии: а — самца, б —• самки.
654
11. ACRONICTINAE
ними бороздками и ребрышками; сразу после откладки яйца бело-серова-
тые, диаметр их 0.5—0.7 мм.
Взрослая гусеница черно-коричневая с желтыми, оранжеватыми
кольцами (межсегментальными мембранами) и участками тергитов; вдоль
спины широкая черная полоса, слагающаяся из черных прямоугольни-
ков, расположенных на тергитах каждого сегмента; по краям этой полосы
по узкой желтоватой полоске; бока выше стигм серо-желтые со многими
мелкими неправильной формы черными пятнышками; ниже стигм
по бокам широкая желтая или желто-белая полоса; вентральная сторона
тела светложелтая, брюшные ноги на концах черные, передние ноги
черные целиком; голова черная, с желтыми адфронтальными полями;
на каждом брюшном сегменте по ряду бородавок, несущих пучки черных
волосков; стигмы черные или темнокоричневые, очерченные черными
колечками.
Куколка темнокоричневая, короткая и толстая, брюшные сегменты
несут скульптуру из острых коротких бугорков; все сегменты брюшка
покрыты волосками, более
густыми на боках, спинной
и брюшной сторонах; осо-
бенно волосисты последние
сегменты; cremaster корот-
кий, плоский в дистальной
части, на которой сидят гу-
стые щетинкообразные волос-
ки ; на краях cremaster шесть
больших коротких зубцов и
несколько мелких.
Мало изменчивый вид, цо
Рис. 289, Oxycesta geographica F. Хетотаксия описано две близких К нему
передне- и среднегруди и брюшных сегментов географических формы, одна
I, VI, VII и VIII взрослой гусеницы. из них может быть принята
за викарный вид — Oxycesta
chamaesices Н. S. (Herrich-Schaeffer, 1845, Europ. Schmett., fig. 568),
известна из южной Франции и Испании. Эта форма меньше размерами,
более узкокрылая и дает небольшие отличия в гениталиях; uncus не-
сколько толще, чем у О. geographica F.; valvae очень похожи, но более
прямые и острые; penis менее выгнут, имеет более короткий coecum и
на дистальном конце вырост; bursa copiulatrix более длинная, чем у
О. geographica F., менее склеротизована; восьмой сегмент несколько более
вырезан с дорзальной стороны. Описанная в последнее время из Испании
О. serratae Zerny (1927, Eos, 3 : 359, t. 9, fig. 17, 18) является лишь цве-
товой формой от О. chamaesices Н. S.
Распространение. Средняя Европа — Альпы (южный
Тироль), долина Дуная (Вена), Карпаты, Югославия, Далмация, южная
Румыния; Молдавия (Кишинев); Украина (Полтава, Таганрог, Черни-
гов, Воронеж, Кировоград, Николаев, Каменец-Подольск, Винница,
Харьков, Одесса); Крым (Керчь); Поволжье (Казань, Саратов, Вольск);
Северный Кавказ (Ставрополь, Кисловодск); Приуралье (Чкалов, Спас-
ское, Уральск); Закавказье; Малая Азия (Анатолия, Булгар-даг).
Биология и экология. Зимует куколка; вылет бабочек
в мае; яйцекладка сразу после вылета; яйца кладутся кучками на листья
кормовых растений. Гусеницы живут в первых стадиях в общем гнезде,
старшие стадии расползаются й живут поодиночке, но также далеко
38. ОХ УСЕЕТ A, 39. EOGENA
555
от гнезда не уходят. Основными кормовыми растениями являются виды
Euphorbiaceae (Euphorbia cyparissias), но отмечено питание также и Scro-
phulariaceae (Linaria). Дает два (может быть неполных) поколения;
вылет бабочек летнего поколения в июле—августе. Предпочитает теплые
и сухие, обычно песчаные стации.
39. Род EOGENA GN'.
, Guenee, 1852, Noct., 1 : 340; Staudinger, 1901, Cat. Lep., 1 : 134; Spuler, 1908,
Schmett. Eur., 1 : 140; Warren, 1909, Seitz, Grossschmett., 3 : 12; Hampson, 1909,
Cat. Lep. Phal., 8 ; 179; Culot, 1909, Noct., 1 : 25; Bang-Haas, 1926, Nov. Macro-
lep., 1 : 159.
Тип рода E. contaminei Ev. •
Хоботок редуцирован; щупальца короткие, покрыты короткими тор-
чащими волосками, направленными вверх и вперед; последний членик
щупалец короткий; глаза небольшие, голые, круглые; лоб гладкий,
покрытый волосками; усики самцов простые, тонкие; голова и грудь
покрыты только волосками, равномерно, без пучков; брюшко без пучков
волосков с дорзальной стороны; М1 отходит от середины добавочной
ячеи; Pi и Р3 на стебельке длиною около половины длины жилок; Р5
отходит из одной точки с двумя предыдущими, от конца ячеи; на зад-
них крыльях редуцирована, есть на ее месте лишь складка; крылья
широкие и короткие, почти без рисунка; valva очень похожа на valva
Sirnyra Ochs., несет два отростка harpe.
Хетотаксия гусениц совершенно того же типа, который описан для
Arsilonche Ld., но бородавки мельче (рис. 289). Типична большая (ком-
плексная) вытянутая бородавка ос—р на среднегруди, на которой обе
группы видны отчетливо.
Монотипический род, близкий к Sirnyra Ochs, и видам Acronicta Ochs,
подрода Pharetra Hb.; ограничен в распространении только юго-восточ-
ной Европой. Связан с видами Plumbaginaceae.
* 1. Eogena contaminei Ev. (рис. 290, 291).
Eversmahn, 1847, Вюлл. Моск. общ. исп. прир., 3 : 77, t. 5, fig. 6; Staudinger,
1901, Cat. Lep., 1 : 134; Spuler, 1908, Schmett. Eur., 1 : 140, t. 140, t. 28, fig. 16;
Warren, 1909, Seitz, Grossschmett., 3 : 12, t. 2, d; Hampson, 1909, Cat. Lep. Phal.,
8 : 179, fig. 29; Culot, 1909, Noct., 1 : 25, t. 2, fig. 15; Draudt, 1931, Seitz, Gross-
schmett., 3 SuppL, 1 : 36; — bombycina Menetries, 1948, Mem. Acad. Sci. Pet.,
6 : 285, t. 6, fig. 7 (Cosmia)', — eogene Freyer, 1852, Neue Beitr. Schmett., 6 : 53, t.
514; — contaminella Strand, 1915, Arch. Naturgesch., 81 (11) : 158.
Крылья, голова, грудь и брюшко рыже-кирпичного цвета нередко
с розоватым оттенком или с примесью серых тонов; рисунок редуциро-
ван; передние крылья более интенсивно окрашены по костальному краю,
в базальной части сероватые; базальная линия не развита; вторая попе-
речная линия у некоторых особей интенсивно коричневая, тонкая, в виде
одной полоски, неизвитая; круглое пятно утрачено; почковидное пятно
в виде луночки интенсивного темнокирпичного цвета, нередко растуше-
ванное; третья поперечная линия начинается над дистальной частью
почковидного пятна, идет в виде одной темной кирпичного цвета полоски,
слабо зубчатой по жилкам, нередко бывает редуцирована совершенно;
под почковидным пятном на этой линии темный штрих; прочие линии
отсутствуют; бахрома рыже-кирпичного цвета с тонкой темной полоской;
задние крылья кирпичного, розово-рыжего или серо-кирпичного цвета,
556
II. ACRONICTINAE
более светлые к основанию; бахрома на них рыжая; нижняя сторона
однотонная, рыже-серая. Размах крыльев 28—34 мм.
। Uncus длинный, постепенно утончающийся в дистальном направле-
нии, изогнут полого, почти голый; tegumen длинный, но узкий; saccus
небольшой, в проксимальной части загнут дорзально; fultura inferior
в виде слабо дифференцированной овальной пластинки; valvae узкие
и длинные, в базальной половине утолщены с вентральной стороны;
утолщенная часть срастается со складкой harpe, которая тянется от ба-
зального верхнего угла до внешнего края valva и несет один отросток,
длинный, изогнутый внутрь; дорзальная часть и дистальный конец valva
мембранозные, слабо волосистые; penis дугообразно выгнут дорзально,
имеет короткий coecum и несет на pars inflabilis большое число (более
10 шт.) длинных шипов, си-
Рис. 290. Eogena contaminei Ev. Гениталии:
а — самца, б — самки.
дящих в виде пучка, и оваль-
ную склеротизованную пла-
стинку. Bursa copulatrix
почти цилиндрической фор-
мы, густо покрыта рубчатой
скульптурой, в виде продоль-
ных тяжей; ductus bursae от-
ходит от ее дистального кон-
ца почти прямой, тонкий,
складчатый, по длине более
половины совокупительной
сумки; восьмой сегмент длин-
ный и широкий, несет длин-
ные и толстые (более длин-
6 ные, чем восьмой сегмент)
apophyses anteriores; apophy-
ses posteriores короткие, изо-
гнутые, тонкие; papillae ana-
les слабо дифференцирован-
ные, мембранозные, несут
густые волоски на дисталь-
ных концах.
Взрослая гусеница светло-
желтая или охристая с мелкими охристо
черными точками на кожном
покрове; вдоль спины широкая охристо-черная, шашечная полоса; на
каждом сегменте проксимальные участки полосы черные, равно как
и межсегментальные ее участки; вентральная сторона тела светложел-
тая; ноги желтые; голова желтая со скоплением темного пигмента
в области глазков и у лба; тело покрыто недлинными желтыми волосками,
сидящими на невысоких круглых, желтых бородавках и торчащих в
разные стороны; стигмы белые, ограниченные тонким черным валиком.
Слабо изменчивый вид; варьирует лишь интенсивность окраски.
Распространение. Южная Украина (Очаков, Аскания-
Нова); Крым (Симферополь, Камышлы, Керчь); Северный Кавказ (Сла-
вянская); Нижнее Поволжье (Саратов, Сарепта, Бирючий на Волге,
Баскунчак); южный Урал; Закавказье (Дербент, Эльдар).
Биология. Зимует куколка; вылет бабочек происходит растя-
нуто, в апреле—мае; гусеницы живут на видах Plumbaginaceae (Statice
gmelini, St. tomentella); окукление в сухих листьях, в августе (в Крыму).
Дает одно поколение. Связан с солончаками.
39. EOGENA, 40. SI MYRA
557
40. Род SIMYRA OCHS.
Ochsenheimer, 1816, Schmett. Eur., 4 : 81; Treitschke, 1825, Schmett. Eur., 2 (5) :
280; Aurivillius, 1891, Nordn. Fjar. : 96; Staudinger, 1901, Cat. Lep., 1 : 134; Spuler,
1908, Schmett. Eur., 1 : 141; Warren, 1909, Seitz, Grossschmett., 3 : 12; Hampson,
1909, Cat. Lep. Phal., 8 : 173; Culot, 1909, Noct., 1 :25; Rebel, 1910, Berge, Schmet-
terlingsb., ed. 9 : 146; Bang-Haas, 1926, Nov. macrolep., 1 : 159; Bang-Haas, 1927,
там же, 2 : 179; Bang-Haas, 1928, там же, 61; Draudt, 1931, Seitz, Grossschmett., 3,
SuppL, 1:6.
Тип рода S. dentinosa Frr., указан Ледерером (1857).
Хоботок есть, слабый, рудиментарный, но способный слегка закру-
чиваться; губные щупальца недлинные, почти прямо торчащие вперед,
слабо вздернутые, покрытые густыми короткими волосками и отдельно
сидящими длинными, последний членик их короткий и довольно тол-
стый; первый членик перед-
Рис. 291. Eogena contaminei Ev. Хетотаксия
передне- и среднегруди и брюшных сегментов
I, VI, VII и VIII взрослой гусеницы.
них лапок около половины
длины передних голеней;
коготки всех лапок с корот-
ким зубцом на внутренней
стороне; глаза голые, окру-
женные короткими волоска-
ми; голова и грудь покрыты
только волосками, без при-
меси чешуй, без пучков;
передние крылья вытянутой
формы, нередко остроуголь-
ные или ланцетовидные;
отходит от дистальной поло-
вины добавочной ячеи; Ps
и Pi на стебельке, кото-
рый менее половины их дли-
ны; Р5 отходит от одной точки со стебельком двух предыдущих ветвей
или на коротком стебельке; чешуйчатый и волосяной покров крыльев
и тела блестящий; harpe вытянутой формы, несет два отростка.
Хетотаксия гусениц идентична тому, что описано для Arsilonche Ld.
и Eogena Gn.; слияние бородавок аир среднегруди полнее, чем у Eogena
Gn.; аир восьмого сегмента на одном уровне, р сдвинута дорзально.
Бородавка х на седьмом сегменте слита с т) (рис. 295).
Олигоморфный род, распространенный (рис. 292) по степям и полу-
пустыням Евразии; кормовые растения преимущественно Euphorbiaceae.
Точно установлены лишь два вида 1 —; Simyra dentinosa Frr. и 6”. nervosa
Schiff.; из них от побережья Тихого океана по степям Тибета и Монго-
лии, южной Сибири, Средней Азии, юго-восточной Европы, Закавказья
и Малой Азии до Венгерских степей распространена только S. nervosa
Schiff., тогда- как S. dentinosa Schiff, ограничена только Средиземно-
морьем. Указываемые Гэмпсоном (1909) три вида Simyra Ochs, для Индии
(A. albicosta Hamps., conjusa Wlk. и conspersa Moore)2 внушают сомнения
1 Simyra oberthiiri Deckert, описанная Кюло (Culot, 1909, Noct. Eur., 1 : 26, t. 3,
fig. 1), по Ротшильду (Rothschild, 1920, Nov. Zool., 27 : 7), относится к роду Copi-
cucullia. Дюмон выделяет эту африканскую форму в особый род — Metalamia Dum.
(Dumont, 1928, Encycl. Entom., 3 : 21). Мне этот вид в натуре не известен.
2 Гарднер [Gardner, 1946, On larvae of the Noctuidae, Trans. Roy. Entom. Soc.
London, 96 (4) : 61—72] описывает гусеницу Simyra conspersa Moore и по всем призна-
кам сближает ее с прочими Acronictinae. Таким образом в отношении этого вида теперь
сомнения, высказанные выше, отпадают. Он относится, конечно, к подсем. Acronic-
tinae и роду Simyra Ochs.
558
1J. ACRONICTINAE
в принадлежности их вообще к Acronictinae и, возможно (насколько
можно судить по описаниям и изображениям), как то выяснилось недавно
для biittneri Нет., относятся к Hadeninae. Изученность их совершенно
недостаточная.
Из Неарктики виды Simyra Ochs, не известны; приводимые Гэмпсо-
ном S. henrici Grt., S. evanidum Grt. и S. fumosumMorr., по исследова-
ниям Смита (1899), являются синонимом евразийской A. albovenosa
Goeze. Виды Simyra Ochs, не описаны также и с других континентов
(исключая наличие их в средиземноморской Африке).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Передние крылья с хорошо развитым сложным темносерым рисун-
ком на белом и сероватом фоне; усики самцов с короткими гребеш-
ками; брюшко массивное даже у самцов; более крупных размеров;
передние крылья с закругленным верхним углом; размах 36—42 мм
.......................................... * 1. S. dentinosa Frr.
2 (1). Передние крылья без рисунка, чисто белые, серебристые или серые,
иногда с темными точками; усики самцов с длинными гребешками
на члениках; брюшко обоих полов тонкое; передние крылья за-
остренные, ланцетовидной формы, с острым верхним углом; размах
32—38 мм.................................* 2. S. nervosa Schiff.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
ПО ВЗРОСЛЫМ ГУСЕНИЦАМ
1 (2). Дорзальная поверхность тела окрашена в черный цвет; черная
окраска располагается в виде неправильных поперечных штрихов;
волоски, покрывающие кожу, сильные, щетинистые; боковые линии
нерезкие..................................* 1. S. dentinosa Frr.
2 (1). Вдоль спины неширокая прерванная черная полоса; боковые
части сегментов оранжевые и черные; щетинки, покрывающие тело,
более слабые.............................* 2. S. nervosa Schiff.
* 1. Simyra dentinosa Frr. (рис. 293).
Freyer, 1839, Neue Beitr. Schmett., 3 : 97, t. 203, fig. 2, 3; Staudinger, 1901,
Cat. Lep., 1 : 134; Spuler, 1908, Schmett. Eur., 1 : 141, t. 31, fig. 1; Warren, 1909, Seitz,
Grossschmett., 3 : 12, t. 2, e; Hampson, 1909, Cat. Lep. Phal., 8 : 175; Culot, 1909,
Noct., 1 : 25, t. 2, fig. 16; Rothschild, 1920, Nov. Zool., 28 : 8; Draudt, 1931, Seitz,
Grossschmett., 3 Suppl., 1:6; — tendinosa Herrich-Schaeffer, 1845, Europ. Schmett.,
Noct., 2 : 179, fig. 393—394; — leucaspis Fischer-Waldheim, 1840, Бюш. Моск,
общ. исп. прир. : 84, t. 3, fig. 2.1
Биология. Kirby, 1895, Butt. Moths Eur. : 161; Hoffman, 1893, Raupen :
73; Korb, 1915, Mitt. Munch, entom. Ges., 6:7; Buresch, 1918, Ztschr. wiss. Insecten-
biol., 23 : 275, t. 4.
Голова и грудь серо-беловатые с примесью рыжих тонов; усики и пе-
редние лапки охристо-желтые; усики самцов с короткими гребешками
на члениках; передние крылья узкие, с сильно вытянутым и округленным
apex; костальный край их почти прямой, слабо выемчатый; окраска
передних крыльев серая с неправильно разбросанными темными и охри-
1 Описанная из северной Африки (Тунис, Гафса) S. autumna Chretien (1910,
Ann. Soc. Entom. France, 79 : 497) очень близка к S. dentinosa Frr.; несмотря на дока-
зательства самостоятельности этой формы Ротшильдом (1920), ее следует рассматри-
вать лишь каК форму S. dentinosa Frr.
Рис. 292. Распространение видов рода Simyra Ochs.
560
11. ACRONICTINAE
Рис. 293. Sirnyra dentinosa Frr. Гениталии: а —
самца, б — самки.
стыми пятнами, жилки серо-желтые и окаймлены светлым (желто-серым);
терминальная линия в виде тонкой полоски, большею частью редуци-
рованная; бахрома белая; задние крылья белые, у самца при основании
затемненные, у самки однотонно тусклые с тонкой темной терминальной
полосой и белой бахромой; нижняя сторона светлосерая с затемненным
базальным отделом и срединной частью передних, а у самок задних
крыльев; на задних крыльях есть небольшое дискоидальное пятно. Раз-
мах крыльев 36—42 мм.
Uncus длинный, слабо конический, полого загнут, почти голый, слабо
заостренный; tegumen довольно длинный, особенно в латеральной обла-
сти; saccus маленький, выгнутый дорзально; valvae длинные, конические;
базальная часть valva более склеротизована, соединена складкой с осно-
ванием harpe; последняя в
виде складки имеет один
длинный прямой отросток,
направленный косо дорзаль-
но и другой направленцый
вентрально; fultura inferior
в виде плоской пластинки,
почти округлая; penis короче
valva, расширенный в ди-
стальной части, дистальный
конец его вздут и резко за-
гнут вентрально; coecum ко-
роткий и тонкий; pars infla-
bilis без инкрустаций. Bursa
copulatrix почти правильной
формы, овальная, густо по-
крыта рубчатой скульптурой;
ductus seminalis отходит от
ее дистальной части; ductus
bursae тонкий, почти прямой,
складчатый, длиною равен
примерно половине совоку-
пительной сумки; восьмой
сегмент короткий и широкий,
несет короткие заостренные . apophyses, anteriores; apophyses posteriores
слабые и тонкие; papillae anales слабые, мембранозные, в дистальной
части несущие длинные волоски.
Взрослая гусеница покрыта черными волосками; интерсегментальная
мембрана желтая; особенно интенсивно окрашены тергиты; по бокам
ниже стигм широкие светложелтые полосы; они проходят через ряд
бородавок, которые выделяются на светлом фоне; нижняя сторона желто-
коричневая; брюшные ноги желто-коричневые с темными склеротизован-
ными пластинками, несущими густые волоски на наружной стороне их;
грудные ноги черные; голова коричневая с неправильными черными
пятнами в области глазков, теменного шва и лба; тело покрыто большими
круглыми и плоскими бородавками, несущими пучки желтых волосков,
торчащих в разные стороны; стигмы черные.
Распространение. Юго-восточная Европа — Балканы и юг
Балканского полуострова, южная Болгария, восточная Румыния (Фили-
пополь), Фракия (Деде-Агач, Бодома), Македония. (Свети-Юро); южная
Украина (Харьков), Таганрог, Ростов; Поволжье (Бузулук, Саратов,
40. SI MYRA
561
Сарепта, Богдо, Хвалынск); Приуралье (Чкалов, Уральск, Губерли,
Кок-Джида, Эмба); Северный Кавказ (Дагестан); Туркмения; Алтай
(Усть-Каменогорск, Бухтарма); Закавказье (долина Куры, Акстафа,
Кировабад, Тбилиси, долина Аракса, Ордубад); Иран (Астрабад); Малая
Азия (Анкара, Аксехир, Мараш); Сирия (северный Ливан); северная
Африка (Тунис, Гафса).
Биолог и я и экология. Зимует куколка; вылет бабочек
в мае—апреле; яйца откладываются кучками на листья кормовых расте-
ний; гусеницы гнездами на видах Euphorbiaceae (Euphorbia gerardiana,
Е. esuloides); расползаются только для окукления; окукление в сухих
листьях в плотном коконе, сплетенном из шелковых нитей; куколки
диапаузируют или дают вылет второго поколения в то же лето. Распро-
странение локально по сухим песчаным степным и полупустынным
стациям.
* 2. Sirnyra nervosa Schiff, (рис. 294, 295).
Schiffermiiller, 1776, Wien. Verz. : 85; Staudinger, 1901, Cat. Lep., 1 : 134; Spu-
ler, 1908, Schmett. Eur., 1 : 141, t. 31, fig. 2; Hampson, 1909, Cat. Lep. Phal., 8 : 174,
fig. 27; Culot, 1909, Noct., 1 : 25, t. 2, fig. 17; Warren, 1909, Seitz, Grossschmett.,
3 : 12, t. 2, e; Rebel, 1910, Berge, Schmetterlingsb., ed. 9 : 146; — oxyptera Es-
per, 1788, Schmett. Eur., 4, t. 130, fig. 6 (Noctua)', — argentacea Herrich-Schaeffer,
1849, Noct. Eur., fig. 387, 388; — torosa Guen'ee, 1852, Noct., 1 : 62; — splendida Stau-
dinger, 1888, Stett. entom. Ztg., 49 : 245; — niveonitens Graeser, 1888, Berl. entom.
.Ztschr., 32 : 309; — argentea Spuler, 1908, Schmett. Eur., 1 : 141; — expressa Bang-Haas,
1912, Iris, 26 : 139; — rubribrunnea Strand, 1915, Arch. Naturgesch., 81 (A), 11 : 158; —
albicilia Strand, 1915, тамже, 81 (A), 11 : 158; — atrata Belling, 1922, Deutsch, entom.
Ztschr., 39 : 199.
Биология и. экология. Kirby, 1895, Butt. Moths Eur. : 161; Spuler,
1908, Raupen : 141, t. 21, fig. 16; Kuhn, 1934, Intern, entom. Ztschr., 28 : 420, fig.
3 Щеголев, 1929, Защ. раст., 6 : 403; Щеголев и Струкова, 1930, Насек, вред. масл.
раст. : 126, 127, 133, 210; Филипьев, 1932, Тр. защ. раст., 5 : 213.
Голова и грудь серовато-белые с примесью желтых тонов; усики сам-
цов гребенчатые, желтоватого цвета; передние крылья ланцетовидной
формы, без рисунка, серовато-белые с белыми продольными мазками;
три широких белых мазка идут от основания крыла: один, широкий
в области дискоидальной ячеи, сужающийся к терминальному краю
крыла, проходит между первом и вторым медиальными стволами, другой
в области кубитального ствола и третий по дорзальному краю крыла;
во внешнем поле между жилками короткие и узкие белые штрихи; жилки
белые, но окаймлены светлосерым; бахрома белая; задние крылья одно-
цветные, серовато-белые с белой бахромой; нижняя сторона передних
крыльев белая, жилки окаймлены темносерым, в виде продольных штри-
хов; задние крылья чисто белые. Размах крыльев 28—32 мм.
Uncus тонкий, длинный, почти голый; tegumen неширокий, довольно
длинный; saccus небольшой, правильной формы; fultura inferior в виде
цельнокрайней, почти прямоугольной пластинки, с дистально направлен-
ным выростом на дорзальном крае; valva продолговатая, несколько рас-
ширенная в дистальной трети; вдоль ее вентрального края от основания
склеротизованная складка, доходящая до harpe; последняя в виде складки
несет два отростка — один направленный вниз и дистально, другой на-
правленный косо дорзально, длинный, загнутый внутрь; penis по длине
примерно равен valva, в дистальной части сильно загнут вентрально;
еоесшп недлинный, узкий, прямой; pars inflabilis penis склеротизована,
без инкрустаций. Bursa copulatrix почти цилиндрической формы, слабо
«уженная, в середине без инкрустаций, с тонкой рубчатой скульптурой,
36 Фауна СССР, И. В. Кожанчиков.
562
II. ACRONICTINAE
образует небольшое расширение в дистальной части, отходящее в виде
соска дистально, от конца которого идет ductus seminalis; ductus bursae
короткий- и тонкий; восьмой сегмент
узкий, но довольно длинный; apophy-
ses anteriores тонкие, по длине равны
восьмому сегменту; apophyses posterio-
res очень короткие, заостренные; pa-
pillae anales мембранозные, слабо диф-
ференцированные, несущие в дисталь-
ной части мелкие густые щетинки.
Взрослая гусеница черно-желтая с
небольшими оранжевыми и черными
штрихами; вдоль спины широкая чер-
ная полоса, прерванная желтыми штри-
хами между сегментами; по сторонам
этой полосы по одной широкой желто-
охристой или желто-белой полосе; бо-
ка серо-охристые с перемежающимися
черными и охристыми штрихами; на
боках желтые бородавки; под стигмами
проходит широкая светлая желто-белая
полоса; вентральная сторона тела тем-
носерая или желто-серая; брюшные
ноги того же цвета; грудные ноги чер-
Рис. 294. Simyra nervosa Schiff. Ге- Hbie! стигмы черные; тело покрыто не-
ниталии: а — самца, б — самки. большими бородавками; голова желтая
с темно-коричневыми, почти черными
неправильными пятнами на черепной коробке в области глазков и по
сторонам головы.
Куколка светлокоричневая; брюшные сегменты несут, в дистальной
части скульптуру в виде шипиков и бугорков; по бокам брюшка за стиг-
мами, а на трех последних
сегментах также и с вент-
ральной стороны пучки ко-
ротких волосков; cremaster
в виде небольшого выступа
несет восемь сильных зуб-
цов, несколько мелких зуб-
чиков и пучок прямых густо
сидящих щетинок.
Изменчивость касается
окраски; основной белый
цвет то с примесью желтых
тонов(5. rubrobrunnea Strand),
то темных (V. air ata Belling)
или, напротив, без них (S.
argentea Spul., argentaeea Н.
S.); серебристая, чисто бе-
Рис. 295. Simyra nervosa Schiff. Хетотаксия пе-
редне- и среднегруди и брюшных сегментов I,
VI, VII и VIII взрослой гусеницы.
лая с темными задними крыльями сибирская форма splendida Stgr.,
принимавшаяся за самостоятельный вид, была изучена мною по материа-
лам из Минусинска, Сретенска и с Дальнего Востока; никаких отличий
в копулятивных органах и других структурах от типичных S. nervosa,
Schiff, мною не обнаружено. Существенно все же отметить, что обе формы —
40. SIMURA, 41. LEIOMETOPON
563
nervosa Schiff, и splendida Stgr. — встречаются в Минусинске вместе;
биологические различия между ними пока не отмечены.
Распространение. Средняя Европа — южная Германия
(Саксония), Альпы (Валлис), долина Дуная (Венгрия), Силезия, Кар-
паты, Польша (Варшава), северная Италия; юг Европейской части СССР
(Чернигов, Полтава, Таганрог, Херсон, Николаев, Одесса); Поволжье
(Казань, Сарепта); Приуралье (Чкалов, Уральск, Губерли, Спасское);
Северный Кавказ (Ставрополь, Ростов); Крым (Симферополь); Закавказье
(Дербент); Средняя Азия — Туркмения (Байрам-али, Ашхабад), Зерав-
шанский хребет (Самарканд), Кара-тау, Сыр-дарья (Байгакум), Тянь-
шань, Юлдус, Семиречье (Кульджа); Тарбагатай; южный Алтай (Онгу-
дай, Чемал, Усть-Бухтарма, Абакан); западная Сибирь (Минусинск);
Забайкалье (Сретенск); южное Приморье (Уссури, Владивосток); север-
ная Монголия (Улан-Батор); северный Китай; Тибет (Амдо); Малая
Азия.
Биология и экология. Зимует куколка; вылет бабочек
в мае; яйцекладка кучками на листья кормовых растений; гусеницы
питаются преимущественно видами Euphorbia, но едят также и ряд дру-
гих растений — Polygonaceae (Rumex), Plumbaginaceae (Statice Yati-
folia, S. sareptana), Baeckmannia.erycaeformis, а также отмечено питание
Juncus sp. и Agropyrum repens. Дает всегда одно поколение. Окукление
в сухих листьях в плотном коконе из шелка; куколка диапаузирует.
Стацией обитания являются сухие, часто песчаные, территорий. Отмечена
как вредитель подсолнечника и льна на Северном Кавказе (Щеголев и
Струкова, 1930).
Из паразитов отмечены из Ichneumonidae — Gambus ornatus Gr.,
Alexeter nebulator Thunb.
41. Род LEIOMETOPON STGR.
Staudinger, 1888, Stett. entom. Ztg. : 37; Staudinger, 1901, Cat. Lep., 1 : 134;
Warren, 1909, Seitz, Grossschmett., 3 : 13; Hampson, 1909, Cat. Lep. Phal., 8 :
172; Bang-Haas, 1926, Nov. Macrolep., 1 : 159.
Тип рода L. simyrides Stgr.
Хоботок редуцирован; лоб с большим округлым выростом, слабо
покрытым желтоватыми чешуйками; глаза большие, голые; коготки всех
ног тонкие, простые; губные щупальца недлинные, торчащие прямо
вперед; усики простые у обоих полов; передние крылья узкие, с выем-
чатым костальным краем и вздернутым верхним углом; и Р2 отходят
от добавочной ячеи почти на одном уровне с противоположных ее сторон,
от ее середины; Р3 и Pt на стебельке изменчивой длины, не более поло-
вины длины этих стволов, иногда коротком; Р6 отходит самостоятельно
от добавочной ячеи, рядом с двумя последними стволами его; М2 на зад-
них крыльях редуцирована; М3 сближена с первым кубитальным стволом,
но сидит на дискальной жилке; valva короткая; harpe с двумя отрост-
ками; bursa copulatrix простая, мешковидная, маленькая.
Мономорфный род, ограниченный в распространении полупустынями
Средней Азии.
* 1. Leiometopon simyrides Stgr. (рис. 296).
Staudinger, 1888, Stett. entom. Ztg. : 37; Staudinger, 1901, Cat-. Lep., 1 : 134;
Hampson, 1909, Cat. Lep. Phal., 8 : 172, fig. 26; Warren, 1909, Seitz, Grossschmett.,
3 : 13, t. 2, f; Draudt, 1931, там же, 3, Suppl., 1 : 7; — sincere. Warren, 1914, Nov-
Zool., 21 : 403.
36*
5164
II. ACRONICTINAE
Рис. 296. Leometopon simyrides Strg. Генита-
талии: а — самца, б— самки.
Голова, грудь и передние крылья светлосерой окраски с нерезкими
охристыми и белыми мазками; рисунок на передних крыльях редуци-
рован; вдоль радиального ствола светлый штрих, такой же штрих вдоль
кубитального ствола; от основания крыльев между кубитальной и аналь-
ной жилками тонкий черный штрих, окаймленный с костальной стороны
белым и дающий к tornus белый расширяющийся мазок; базальная линия
в виде едва заметного темного штриха на костальном крае, который
в базальном поле белый; базальное поле широкое, нижняя часть его
охристо-серая; вторая поперечная линия очень слабо заметна, в виде
отдельных растушеванных штрихов, тонкая, желто-серая, идет слабо
скошенная кнаружи; срединное поле в нижней половине светлосерое
без охристых тонов; круглое пятно утрачено; почковидное пятно наме-
чено в виде неправильного серого пятна при основании медиальных
жилок с белой точкой в середи-
не; от почковидного пятна к
apex широкий белый мазок,
затушевывающий третью попе-
речную линию; последняя на-
мечена в виде очень слабого
штриха на костальном крае и
под ним, далее смыта и хорошо
заметна лишь в дорзальной по-
ловине крыла в виде тонкой
темносерой полоски, окаймлен-
ной снаружи белым, слабо зуб-
чатой; внешнее поле узкое с
остатками белой подкраевой ли-
нии, растушеванной и расплыв-
шейся в виде мазков; терми-
нальная линия четкая, в виде
восьми темных штрихов; бахро-
ма светлосерая с полоской; зад-
ние крылья однотонно серые с
тонкой терминальной полоской
и белой бахромой; нижняя сто-
рона серая с неправильными белыми пятнами и тонкой четкой терми-
нальной полосой. Размах крыльев 30—32 мм.
Uncus почти прямой, короткий, резко суженый на дистальном конце;
tegumen длинный, особенно в тергальной области, обрастает основание
uncus, более короткий в латеральных отделах; saecus длинный,заострен-
ный, узкий; valvae короткие, почти эллиптической формы; базальная
часть valva утолщена и склеротизована по дорзальному краю, а также
с вентральной стороны; утолщенные отделы ее соединены тяжами со
складчатой harpe; последняя несет два отростка — дорзальный длинный,
направленный косо дистально, лежащий в плоскости valva, и вентраль-
ный короткий, тонкий, острый, торчащий в сторону отvalva; дистальная
половина valva мембранозная; penis почти прямой, подлине почти равен
valva; соесшп короткий; pars inflabilis несет сильные длинные шипы
в числе пяти. Bursa copulatrix длинная, расширенная в проксимальной
части и сильно суженная в дистальной, где несколько более сильно скле-
ротизована; проксимальная ее часть мембранозная, без скульптуры;
ductus seminalis отходит от ее дистального конца без специального рас-
ширения к нему; ductus bursae очень короткий, складчатый; восьмой
41. LEIOMUTOPON 565
сегмент короткий и широкий, заметно конический; apophyses anteriores
тонкие, по длине несколько превышают восьмой сегмент; apophyses poste-
riores более длинные, заостренные; papillae anales мембранозные, слабые,
несут короткие волоски, особенно густые в дистальной их части.
Плохо изученный в отношении изменчивости вид— известно всего
не более десятка особей; наблюдается различие между ними лишь в интен-
сивности белых штрихов на передних крыльях; самки светлее. Описан-
ная Уорреном (1914) L. sincera, по сборам В. Кожанчикова с Сыр-дарьи,
является только синонимом, но не цветовой формой. Очень своеобразный
вид, наиболее близкий к видам Simyra Ochs.
Распространение. Среднйя Азия — Сыр-дарья (Байгакум);
Семиречье (Сары-тогой, Чилик, Или, Ку'льджа).
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ СЕМ. ORGYIDAE
abdominata, Euproctis 323
abietis, Dasychira 250 , 251, 259
aboleta, Parocneria 380
abscondita, Acronicta 506, 508, 523
abscondita, Acronicta euphorbiae 520
acerbata, Acronicta 520
aceris, Acronicta 462, 465, 467, 459
aceroides, Acronicta 499
achatina, Acronicta 224
Acronicta 396, 455, 462
^Acronictinae 218, 394
Acronictini 395, 453
Acronictoides 402, 410
Acronycta 462
acronycta, Dasychira 251
acronyctoides, Diphthera 413
Acyphas 270
adalia, Laelia 267
adaucta, Acronicta 504
Adlullia 308
adora, Laelia 267
aethiopa, Subacronicta 458
aethiops, Raphia 421
afflicta, Acronicta 462, 480
Agriopodes 439
ainu, Moma 437 ;
alba, Acronicta 475
alba, Arctornis 332
alba, Calocasia 418
alba, Euproctis 317
alba, Ocneria 439
albacostata, Orgyia 271
albarufa, Acronicta 498
albescens, Dasychira 250
albescens, Dasychira pudibunda 261
albescens, Euproctis 317
albescens, Ocneria dispar 365
albescens, Ocneria grisescens 362
albibasalis, Dasychira 250
albicilia, Sirnyra 561
albicosta, Sirnyra 557
albida, Arsilonche 549
albidior, Subacronicta 458
albiorbis, Acronicta 463, 480
albiplaga, Olene 224
albiplaga, Trisuloides 404
albistigma, Acronicta 474, 506
albitesselata, Trisuloides 404
albodentata, Dasychira 227, 250, 251,
albofascia, Numens 306
albomaculata, Thyatirides 452
albonigra, Hampsonia 535, 536
albosignata, Olene 224
albovenosa, Arsilonche 548, 549
algerica, Parocneria 389
algirica, Orgyia 285
algiricum, Orgyia 270, 285
alhagi, Orgyia 285
aliena, Euproctis 307
alni, Acronicta 462, 466, 480
alnoides, Acronicta 529
alpherakii, Gynaephora 232, 246
alpina, Acronicta 517
alpina, Olene 227
alpium, Daseochaeta 439, 441
altaica, Acronicta 492
amabilis, Parocneria 381, 386
amanda, Ocnerorgyia 378
amasina, Acronicta 520
americana, Acronicta 467
ampla, Ocneria 351, 353
anacausta, Euproctis 317
Anacronicta 401, 406
anaedina, Acroni'cta 468
anaedinella, Acronicta 468
anartoides, Orgyia 270, 273
anata, Cifuna 302
ancarensis, Subacronicta 458
anceps, Orgyia 289
andalusiaca, Acronicta 520
And'ela 412
androdes, Moma 433
Anepholcia 402, 403
angelus, Olene 227, 229
angulifera, Ocneria 365
angulosa, Dasychira 250
angusta, Gerbathodes 410
angustimacula, Acronicta 480, 495
ania, Dasychira 250
ankarensis, Subacronicta 458
antennata, Ocneria 353
anthracina, Acronicta 469
anthyellina, Orgyia f. hybrida 277
antiqua, Orgyia 270, 275, 278
antiqua, Orgyia gonostigma 282
antiqaoides, Orgyia 287
Apatela 462
Apatele 462
1 Жирным шрифтом напечатаны названия таксономических единиц выше родов
и страницы, на которых помещены описания; курсивом напечатаны синонимы.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ* 567
apennina, Acronicta 520
apicalis, Artaxa 303
apicibrunnea, Ocneria 353
approximans: Orgyia 282
approximata, Raphia 421, 423, 424
aprilina, Daseochaeta 441
Apterogynis 270
arakawae, Dasychira 250
arcerii, Orgyia 285
Arctomyscis 462
Arctornis 219, 331
arduenna, Acronicta 514
argentacea, Sirnyra 561
argentea, Arsilonche 549
argentea, Sirnyra 561
armena, Episema 428
aroa, Euproctis 317
Arna 308
Arsilonche 396, 456, 548
Artaxa 220, 308
asiatica, Acronicta 474
asiatica, Ocneria 365
asignata, Acronicta 469
asignata, Acronicta tridens 489
asoetria, Ocnbria 353
atala, Ocneria 353
atestacea, Laelia 266, 267
Atipha 315
atlantica, Parocneria 380
atlantica, Parocheria lapidicola 383
atomaria, Olene 224
atomariana, Dasychira 250
atomima, Arsilonche 549
attomana, Acronicta 520
atra, Ocneria dispar 365
atra, Ocneria monacha 355
atrata, Simura 561
aurantia, Orgyia 271
aurantiaca, Dasychira 250
aurata, Artaxa 309
auratior, Acronicta 526
aurea, Orgyia 285
aureomicans, Penthophera 295
auricoma, Acronicta 462, 466, 508, 517
aurifera, Dasychira 250
auriflua, Euproctis chrysorrhoea 323
auriflua, Euproctis similis 320
auripes, Ivela 335, 336
aurolimbata, Orgyia 273, 275, 291
aurolimbata var. Orgyia 292
austera, Oxycesta 552
australis, Orgyia 273
autumna, Sirnyra 558
autumnalis, Parocneria 386
avellanae, Calocasia 418
awgarra, Euproctis 317
badia, Orgyia 273, 278
baibarana, Dasychira 250
baicalensis, Orgyia 286
bananae, Ocneria 353
bananoid.es, Ocneria 353
barica, Ocneria 353
barnasi, Acronicta 506
Baryaza -298
basalis, Olene 224
basiflava, Olene 224
basinigra, Olene 224
basistriata, Acronicta 517
batnana, Acronicta 492
Batyra, 400, 402
beatrix, Ocneria 353
Belciades 401, 443
Belciana 401, 430
belgica, Acronicta 487
bellula, Acronicta 479, 480, 496
bercei, Acronicta 529
beryllodes, Daseochaeta 439
betulae, Acronicta 479
betulae, Calocasia 418
bicolor, Acronicta 462, 506
bicolor, Dasychira 261
bidens, Acronicta psi 492
bidens, Acronicta tridens 489
biformis, Belciana 430
bimaculosa, Acronicta 475'
bipartita, Episema 428
biplagatana, Euproctis 317
bipuncta, Orgyia 273
bipunctapex, Artaxa 310
bipunctigera, Artaxa 310
biseparata, Numens 306
Bisulcia 537
biundulalis, Cifuna 299
bivirgae, Acronicta 492
bivittata, Ocneria 353
bocharae, Ocneria 365
bohna, Dasychira 250
bombycina, Eogena 555
bonivelli, Olene 224
bordigalensis, Ocneria 365
boulifa, Euproctis 317
bradyporina, Acronicta 475
brauni, Laelia 266
bremenski, Ocneria 365
brevivicornis, Laelia 267
brevipennis, Daseochaeta 439
brontonepha, Leucoma 341
brotea, Ocneria 353
brumosa, Acronicta 506, 517
brumosa, Acronicta carbonaria 510
brunnea, Dasychira 250
brunnea, Ocneria 365
brunnea, Ocneria monacha 355
brunnea, Ocneria rubea 374 '
brunneiplaga, Ocneria 353
brunnior, Acronicta 484
bryophiloides, Acronicta 504
buana, Laelia 267
bucovina, Orgyia 278
bukouma, Orgyia 278
butleri, Dasychira 250
buttneri, Leiicania 558
Byrdia 231
caesarea, Acronicta 506
calabra, Orgyia 289
calamaria, Laelia 267
c-album, Trisuloides 404
calceata, Acronicta 469
caligata, Dasychira 259
caliginea, Anacronicta 406, 407
568 АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
caliginosa, Acronicta 487
callunae, Olene 227
Calocasia 396 , 401, 415
camurisquama, Leucoma 341, 342, 345
canadensis, Acronicta 467
candelisequa, Acronicta 469
Candida, Laelia 267
Candida, Leucoma 341, 342, 345
cangia, Dasychira 250
Canna, 402, 448
capnodes, Episema 428
carbonaria, Acronicta 506, 507, 509
carola, Acronicta 481
carriola, Ocneria 353
casparii, Acronicta 504
catocalina, Trisuloides 404
catocaloida, Acronicta 506, 507, 508
caucasica, Olene 227
caucasica, Penthophera 295, 298
cavillatrix, Acronicta 474
Caviria 336
cedestis, Olene 224
centralis, Subacronicta 457, 458, 460
centripuncta, Arsilonche 549
cervina, Cifuna 298, 299
cervina, Euproctis 317
Chamepora, 462
chamesices, Oxycesta 554
Charadra 400, 413
Chardines 265
charmentanti, Euproctis 317
chekiana, Trisuloides 404
Chianophasma 316
chichibensis, Leucoma 341
chinensis, Thyatirides 452
chingana, Acronicta 506
chinochera, Acronicta 467
chosenensis, Ocneria 365
christophi, Orgyia 292
chrysochlora, Daseochaeta 439
chrysorrhoea, Euproctis 316, 319, 323
Cifuna 219, 298
cinctata, Dasycira 250
Cinderella, Acronicta 467
cineracea, Acronicta 475
cinerascens, Acronicta 533
cinnamomea, Olene 224
Cispia 336
Cispidia 308
citrinella, Dasychira 250
civitta, Euproctis 317
clarescens, Artaxa 498
Clidia 552
coalita, Episema 428
coenobita, Diphthera 412, 413
coenosa, Laelia 266, 267
coerulea, Trisuloides 404
coeruleocephala, Episema 427, 428
colorada, Arsilonche 549
columbiensis, Orgyia 274
comma, Leucoma 341
eomparata, Artaxa 308
concerpta, Subacronicta 457
concolor, Dasychira 261
concolor, Ocneria 353
conferens, Episema 428
corcfinis, Orgyia 278
confusa, Cifuna 300
confusa, Simyra 557
conjuncta, Dasychira 250, 251, 254
connecta, Acronicta 480
connexa, Acronictoides 410
consanguis, Acronicta 468, 473
conspersa, Artaxa 309
conspersa, Ocneria 353
conspersa, Simyra 557
contaminata, Trisuloides 404, 406
contaminei, Eogena .555
contaminella, Eogena 555
contrahens, Numens 305
coreacola, Euproctis 320
coreana, Artaxa 313
coreana, Dasychira 256
coresia, Orgyia 273
Cornelia, Trisuloides 404
coronula, Craniophora 544
Corsica, Orgyia 289
coryli, Calocasia 415, 418
Cosmodes 398
costalis, Euproctis 316, 317
costalis, Laelia 267
costalis, Olene 224 (
costa-nigra, Arctornis 333
costaricensis, Orgyia 273
Craniophora 455, 537
crenulata, Acronicta 463
cretacea, Arsilonche 549 s
cretacea, Leucoma 347
cretata, Acronicta 467
cretosa. Leucoma 341
Cropera 336
croperoides. Laelia 267
Crorema 336
cubitata, Acronicta 513, 514
cubitata, Craniophora 544
curvata, Artaxa 309
curvifera, Ocneria 351, 353
Cuspidia 462
cuspis, Acronicta 479, 480, 487
cuspis, Acronicta 492
cyanea, Dasychira 250
cyanescens, Acronicta 467
Cycnia 265
cygna, Leu coma 340
cymota, Olene 224
cyparissiae, Acronicta 520
dactylina, Acronicta 467
daisetsu2aha, Konokareha 237
dalbergiae, Dasychira 250
Daseochaeta 396, 401, 438
Dasorgyia 231
Dasychira 219, 248
dawara, Orgyia 271
dealbata, Ocneria 365
debilis, Acronicta 520
decyanea, Acronicta 487
defigurata, Acronicta 468
definite, Orgyia 273
defuncta, Penthophera 295
degener, Acronicta 481
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
5б9>
degener, Arsilonche 549
Demas 415
dentata, Acronicta 5Q6
dentata, Cetola 398
dentinosa, Simyra 557. 558
denudata, Ivela 336
depuncta,. Arctornis 332
deserticola, Orgyia 285
designata, Daseochaeta 441
destitute, Parocneria 383
destrigata, Ocneria 365
detrita, Parocneria 377, 380, 381
devestita, Laelia 266, 267
diaphana, Leucoma 341
diaphanella, Laelia 267
diffusa, Moma 437
diffusipuncta, Moma 433
digna, Acronicta 500, 507, 511
digramma, Artaxa 308
Diloba 427
dilutior, Orgyia antiqua 278
dilutior, Orgyia trigotephras 289
dimidiata, Orgyia 271
Diphthera 395, 401, 411
Diphthera 412
Diphtherocome 439
discirufa, Leucoma 341
Tiisepholcia 404
dispar, Ocneria 353, 355, 365
disparilis, Numens 305 , 306
disparina, Ocneria 365
disparoides, Ocneria 365
dissoluta, Ocneria 353
distans Acronicta 506
distincta, Acronicta 529
divisa, Euproctis 317
divisa, Leucoma 341
doii, Leu coma 341
dorsipennata, Olene 224
dubia, Orgyia 273, 274. 285
dubiosa, Ocneria 353
dudgeoni, Olene 224
dulcinea, Ocneria 353
dungemi, Subacronicta 458
eburnina, Penthophera 295
ecnomada, Leucoma 347
eddela, Ocneria 353
edolata, Acronicta 462, 506
effusior, Craniophora 544
efjeminata, Ocneria 265
egens, Leucoma 341
egerina, Leucoma 347
egregia, Panthauma 402
ekeikei, Euproctis 317
ekeitei, Ocneria 353
elaeagni, Acronicta 467, 472
Elaeodes 400
eldora, Acronicta 467
electa, Calocasia 416
eleuteroides, Ivela 336
elineata, Acronicta 469
elisabeta, Acronicta 480
Elkneria 249
elongata, Acronicta 499
Eloria 336
Elynodes 398
emaculata, Acronicta 506
endoplagia, Euproctis 317
Enome 351'
enthoxantha, 'trisuloides 404
entusana, Euproctis 317
Eogena 397, 456, 555
eogene, Eogena 555
eothina, Acronicta 481
Epicausis 400
Episema 396, 401, 427
Erasta 351 ,
erebus, Ocneria 365
eremita, Ocneria 355
eremita, Ocneria dispar f. hybrids 365
ereosaura, Artaxa 308
ericae, Orgyia 270, 273, 275, 287
esulae, Acronicta 520
etrusca, Orgyia 282
Eulonche 462, 506
euphorbiae, Acronicta 506, 507, 520
euphrasiae, Acronicta 520
Euproctis 220, 315
eurasiae, Acronicta 523
Euromoia 406, 445
eurychlora, Belciana 430
eurydicae, Cifuna 299, 301
evanescens, Arctornis 333
evanida, Arsilonche 549
examinata, Ocneria 365
exclamationis, Laelia 267
exempts, Acronicta 465
dxilis, Acronicta 498
expergita, Acronicta 506
expressa, Simyra 561
extorta, Dasychira 250
extricate, Acronicta 506
falcata, Orgyia 273
falcata, Penthophera 295
falcula, Acronicta 480
fallax, Agriopodes 439
fameola, Acronicta 465
farinosa, Acronicta 520
fascelina, Olene 224, 227
fasciata, Acronicta 506
fasciata, Acronicta euphorbiae 520
fasciata, Craniophora 538
fasciata, Daseochaeta 439
fasciata, Daseochaeta 441
fasciata, Euproctis 330
fasciata, Laelia 266
fasciata, Ocneria dispar 365
fasciata, Ocneria monacha 355
fasciata, Raphia 421
fasciatella, Ocneria 365
favillacea, Acronicta 504
felina, Acronicta 467
feminicolorata, Penthophera 295
feminula, Dasychira 250
fixeni, Acronicta 513
flava, Artaxa 308
Hava, Artaxa 308, 310
flava, Dasychira 250
flavala, Anacronicta 407
flavescens, Acronicta 475
-570
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
flavescens, Euproctis 323
flavescens, Leucoma 341
flavescens, Orgyia gonostigma 282
flavescens, Orgyia thyellina 276
flavicollis, Calocasia 416
flavicostata, Leucoma 341
flavida, Arsilonche 549
flavinata, Artaxa 309
flavipalpata, Ocneria 380
flavipuncta, Euproctis 317
flaviventer, Ocneria 355
flavittata, Orgyia 271,
flavoabdominalis, Ocneria 355
flavolimbata, Orgyia 273, 275, 293
flavosuphurea, Euproctis 327
flavosulphurea, Leucoma 341, 342, 347
formosana, Laelia 267
formosensis, Leucoma 341
fortunata, Olene 224
fragarius, Ocneria, 365
fragilis, Acronicta 462, 479
fragrans, Numens 305
frater, Raphia 423
frigida, Acronicta 467
fuliginea, Ocneria 351, 353
fuliginosa, Ocneria 353 .
fulvata, Laelia 267
fumida, Ocneria 365
fumdsa, Ocneria 353
fumosum, Arsilonche 549
funeralis, Acronicta 479
furcifera, Acronicta 412, 413
furcilla, Diphthera 412, 413
fusca, Dasychira 261
fusca, Ocneria 363
fusca, Orgyia 270
fuscolimbata, Laelia 267
fuscosuffusa, Acronicta 533
galactopsis, Euproctis 317
galvagnii, Acronicta 525
ganaha, Ocneria 353
ganara, Ocneria 353
ganaroides, Ocneria 353 ,
gastridia, Acronicta 463, 467
Gaujonia 400, 402
geographica, Oxycesta 552
Geonycta 398
georgiana, Dasychira 250
gigantea, Craniophora 544
gigantea, Diphthera 413
gigantea, Laelia 267
gigantea, Leucoma 341
glauca, Anacronicta 407
glauca, Daseochaeta 441
glaucoptera, Acronicta 520
gloveroides, Olene 224
golinaria. Ocneria 353
gonodona, Ocneria 353
gonostigma, Orgyia 273, 274, 281
gracilior, Euproctis 317
gracilis, Ocneria 355
grandis, Ocneria ,353
greeni, Dasychira 250
grisea, Acronicta 480
grisea, Acronicta leporina 475
grisea, Acronicta rumicis 529
grisea, Diphthera 413
grisea, Ocneria 353.
grisea, Orgyia thyellina 276
grisea, Orgyia antiqua 278
griseofacta, Olene 224-
griseipennis, Parocneria 380, 381, 387
grisescens, Acronicta 475
grisescens, Calocasia 420
groenlandica, Gynaephora 232, 234
grossa, Dasychira 250
grotei, Dasychira 250
grumi, Gynaephora 246
grumi, Subacronicta 457, 458, 461
grundi Dasychira 261
guadarramensis, Orgyia 291
guatemalica, Orgyia 273
Gynaephora 220, 231
gypsochroa, Moma 433
hahroscia, Belciana 430
hadina. Ocneria 365
hamamelis, Acronicta 506
Hampsonia 455, 535
Нагара 266
harmandi, Craniophora 538
Harrisemna 455
hastulifera, Acronicta 462. 467
haulberti, Gynaephora 245
hayashii, Numens 306
hebraica, Pplyogramma 400
helgelandiae, Olene 227
Hemerocampa 270
hemicalla, Orgyia 273
hemileuca, Craniophora 538 , 540 , 548
henrici, Arsilonche 549
heptapotamica, Orgyia 286
heptesticta Ocneria 353
hercules, Acronicta 498, 499
hesperida, Acronicta 467
heterociliata, Ocneria 355
heterogyna, Laelia 267
Heteronygmia 306
hillmani, Ocneria 374.
hipparia, Leucoma 341
hirajame, Dasychira 250
hokkaidensis, Ocneria 365
hololeuca, Euproctis 317
honrathi, Trisuloides 404
horishanella, Dasychira 250
horsfieldi, Dasychira 250
Hyboma 462, 465, 498
hybris, Raphia 421, 423
Hypogymna 294
Hypogymnidae 414
tgdyrensis Subacronicta 458
iliensis. Acronicta 492
illarioni, Raphia 425
illita. Acronicta 506
illustreciliata, Penthophera 295
immaculata, Orgyia 273
immaculata, Diphthera 413
immunda, Dasychira 265
impleta, Acronicta 506
impressa, Acronicta 506
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
571
impura, Laelia 266
incerta, Laelia 267
incerta, Ocneria 353
inclara, Acronicta 498
inclusa, Olene 224
inconspicua, Artaxa 309
increta, Acronicta 498
increta, Acronicta incretata 485 :
incretata, Acronicta 479, 480, 485
indica, Acronicta 529
infanta, Calocasia 416
infausta, Anacronicta 407
injernalis, Orgyia 278
infumata, Fpisema 428
injuscata, Acronicta 469
innotata, Orgyia 273
inordinata, Ocneria 353
inornata, Orgyia 273
insignata, Ocneria 365
insita, Acronicta 167
insitiva, Acronicta 506
insolita, Acronicta 506
insolita, Numens 306
intacta, Leucoma 347
intense, Artaxa 310
intension, Acronicta 481
intermedia, Acronicta incretata 485
intermedia, Acronicta major 469
intermedia, Orgyia dubia 285
intermedia, Orgyia ericae 287
interrupta, Acronicta 479
interrupta, Orgyia 285
loshaera 462
iria, Acronicta 499
isabellina. Euproctis 317
isolatella.' Orgyia 285
I vela 220, 335
jactans, Craniophora 538, 546
jaeschkei. Acronicta 514
jalisca, Orgyia 273
jankowskii, Cifuna 299, 303
/ankowskii, Hampsonia 535
japonibia, Laelia 267
japonica, Ocneria 365
japyx. Belciades 444
javensis, Canna 449
joanisi, Ocneria 353
jobasis, Acronicta 465
josephina, Orgyia 285
jozana, Acronicta 475
jozensis, Acronicta 479
judaea, Acronicta 469
judaea, Orgyia 285
juglandis, Dasychira 261
juncta, Acronicta psi, 492
juncta, Acronicta tridens 489
justica, Dasychira 261
jutica, Dasychira 261
kagina, Dasychira 250
kala, Belciana 430
kalgana Craniophora 541
karafutonis, Olene 227
karapina, Euproctis 317
kargalica, Euproctis 327
karghalica, Acronicta 489
karghalica, Euproctis 317, 319, 327
karghalika, Euproctis 327
karonis, Artaxa 313
kenriki, Dasychira 250
kervillei. Olene 224
kibarae, Dasychira 250
kikuchii, Dasychira 250
komarovi, Parocneria 380, 381, 385
Konokareha 237
koreana, Acronicta 520
kosempornis, Ocneria 353
kusnezovi, Ocneria 355
Lacida 270
lactaea, Euproctis 217
lactea, Ocneria 365
ladakensis, Lachana 232
Laelia 219, 265
Laelioides 266
laetifica, Acronicta 506
lagiata, Olene 224
lanceolaria, Acronicta 506
lapathi, Acronicta 517
lapidicola, Parocneria 380, 381, 382
Laria 340
laricis, Olene 227
larose, Acronicta 475
latefascia, Diphthera 413
lathyri, Gynaephora 234
lathyra, Gynaephora 234
leechi, Orgyia 287
Leiometopon 456, 463
lemuria, Dasychira 250
lepcha, Ocneria 353
lepetita, Acronicta 506
Lepitoreuma 462
leporella, Acronicta 475
leporina,, Acronicta 462, 466, 467, 475
lepta, Leu coma 347
lepusculina, Acronicta 467
letterata, Craniophora 544
leucantiqua, Orgyia f. hybrid a 281
leucaspis, Simyra 558
leucocuspis, Acronicta 479, 480, 483
leucogaea, Acronicta 475
Leucoma 221, 339
leucomelas, Euproctis 317
leucoptera-, Acronicta 507, 526
leucoptera, Laelia 267
Leucosia 340
leucostigma, Orgyia 270
leucotaenia, Moma 433
libanicola, ' Parocneria 383
lichenodes, Acronictoides 412
Lichnoptera 400, 402
ligustri, Craniophora 537, 538, 544
lilacina, Acronicta 465
lilacina, Laelia 267, 273
limbalis, Euproctis 316
Liparidae 214
Liparis Hb. 234
Liparis Ochs. 214, 350
literata, Acronicta 478
lithospila, Acronicta 479
litura, Laelia 265, 267
572
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
liturata, Acronicta 478, 480
1-nigrum, Arctornis 331, 332, 333
loacina, Ocneria 353
lobeliae, Acronicta 479
locuples, Cifuna 298, 299, 300
ludifica, Daseochaeta 441
ludifica, Mom'a 432, 433
lugens, Gynaephora 232, 241
lugens, Moma 433
lugubris, Acronicta 529
lunata, Ocneria 353
lunulata, Dasychira 250, 251, 252
lunulata, Orgyia 285
lutea, Acronicta 506, 507, 536
lutea, Ocneria 355
luteicoma, Acronicta 462, 506
luteifascia, Trisuloides 404
luteola, Acronicta 465
lutescens, Ocneria 353
Lymantria 350
Lymantriidae 214
macrodonta, Dasychira 250
maculata, Ocneria 353
maculosa, Dasychira 250
maesta, Acronicta 475
magnalia, Olene 224
magniplaga, Artaxa 309
major, Acronicta 467, 468
major. Ocneria 365
malachitis, Canna 449
inalagassica, Orgyia .271
mancha, Orgyia 285
mandzhurica, Orgyia 278
manitoba, Acronicta 479
mahsueta, Acronicta 479
manto, Olene 224
marashi, Parocneria 383
mardina, Parocneria 383
marginalis, Leucoma 341
marginata, . Acronicta 529
marginata, Ocneria 353
marginenotata, Cifuna 299
marmorata, Acronicta 480
marmorea. Daseochaeta 439
Mastiphanes 462
matura, Ocneria 353, 354, 362
maxima, Acronicta 467 468
medicaginis, Olene 227
mediofascia, Olene 224
mediofusca, Ocneria 355
medionigra, Calocasia 418
medionigra, Orgyia 285
medioobscura, Dasychira 259
mediovittata, Acronicta 463
mediterranea, Ocneria 365
meeki, Euproctis 317
megacephala, Subacronicta 456, 458
Megaeronicta 462
melaleuca, Acronicta 475
melania, Euproctis 317, 319, 338
melaniides, Euproctis 320
melanocephala, Acronicta 475
melanocera, Ocneria 353
melanopygon, Ocneria 353
melanostigma, Artaxa 313
melanotica, Calocasia 418
melaxantha, Laelia 267
melete, Ocneria 353
melissa, Ocneria 353
melochlora, Dasychira 250
mendosa, Olene 223, 224
menecles, Ocneria 353
menyanthidis, Acronicta 506, 507,514
menyanthidis, Acronicta auricoma 517
meridionalis, Acronicta 529
Merolonche 455
mesella, Acronicta 475
metaphae, Daseochaeta 439
micacea, Leucoma 341
michael, Acronicta 511
Microcoelia 462
Micropterogyna 270
militaris, Parocneria 389
minella, Acronicta 498
minor, Acronicta 475
minutissima, Leucoma 341
misana, Olene 224
mixta, Euproctis 317
mixta, Euromoia 445 , 446
mixta, Orgyia 271
modesta, Dasychira 250 , 255
modesta, Orgyia 278
modica, Acronicta 498
Moma 395 , 401, 432
Mominae 397
Momini 395, 397
monacha, Ocneria 353, 354, 355
mongolica, Orgyia 278
monobalia, Leucoma 341
moijtana, Olene 224
inontis, Artaxa 309
montivaga, Acronicta 520
moorei, Leucoma 341
morio, Penthophera 214, 215
morula, Acronicta 479
moszkowskii, Laelia 267
moupiensis, Anacronicta 409
multiscripta, Ocneria 353
murina, Arsilonche 549
murrhina, Daseochaeta 441
mus, Calocasia 418, 420
mus, Parocneria 380
muscosa, Daseochaeta 439
myricae, Acronicta 523
nachiensis, Dasychira 250
Nagunda 351
narindra, Ocneria 353
natlyi, Penthophera 295
naumanni, Leucoma 342
navasi, Craniophora 546
nebulifera, Dasychira 250
nebulosa, Ocneria 353
nebulosa, Orgyia 271
negrita, Acronicta 520
neomelaina, Arsilonche 549
nervosa, Arctornis 333
nervosa, Sirnyra 557, 558, 561
nesiobia, Ocneria 365
nesiotes, Dasychira 251
nigra, Acronicta leporina 475
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
573
nigra, Acronicta menyanthidis 514
nigra, Craniophora 544
nigra, Ocneria dispar 365
nigra, Ocneria monacha 355 ।
nigra, Olene 224
nigra, Subacronicta 458
nigralbata, Acronicta 463
nigrescens, Moma 433
nigrescens, Acronicta 504
nigricans, Acronicta 506
nigricatena, Moma 433
nigricostata, Ocneria 353
nigricostata, Orgyia 271
nigripennata, Leucoma 342
nigriplaga, Orgyia 271
nigriplagata, Cifuna 299
nigriplagiata, Ocneria 353
nigripunctata, Arctornis 332
nigrita, Euproctis 323
nigrivitta, Craniophora 538
nigrociliata, Laelia 267
nigrociliata, Leucoma 342
nigrocrocea, Orgyia 271
nigrofasciata, Episema 428
nigrofasciata, Euproctis 317
nigromaculata, Acronicta 481
nigromaculata, Craniophora 538
nigromarginata, Acronicta 481
nigrosignata, Euproctis 323
nigrotecta, Olene 227
ninayi, Ocneria 353
niphobola, Leucoma 341
niphonis, Artaxa 308, 310, 313
nisseni, Parocneria 389
nitida, Anacronicta 407, 409
nivalis, Olene 227, 229
niveata, Leucoma 341
niveola, Belciades 443, 444
niveonitens, Sirnyra 561
niveosparsa, Craniophora 538, 541
nivosa, Leucoma 347
nobli, Orgyia 289
noctivaga, Acronicta 506
nora, Parocneria 380
Notolophus 270
nova, Orgyia 278
novaguinensis, Ocneria 353
novella, Acronicta 467
nubilata, Craniophora 538
nucula, Orgyia 273
Numens 220, 305
nupera, Orgyia 273
nyctea, Euproctis 320
Nygmia 315
Nymphyxis 340
oberthiiri, Sirnyra 557
obfuscata, Ocneria 353
oblinita, Acronicta 462, 506
obliterella, Orgyia 285
obscusa, Acronicta 520
obscura, Anacronicta 407, 409
obscura, Craniophora 538
obscura, Craniophora ligustri 544
obscura, Ocneria dispar 365
obscura, Ocneria rubea 374
obscura, Olene 227, 229
obscurata, Olene 227
obscurior, Acronicta alni 481
obscurior, Acronicta cuspis 487
obscurior, Acronicta tridens 489
obsolescens, Moma 433
obsoleta, Acronicta alni 481
. * obsoleta, Acronicta menyanthidis 514
obsoleta, Laelia 266, 267
obsoleta, Ocneria 353
obsoleta, Ocneria dispar 365
obsoleta, Ocneria monacha 355
obsoleta, Raphia 421, 426
obsuta, Acronicta 484
ochracea, Arsilonche 549
ochracea, Ocneria 365
ochracea-obsoleta, Ocneria 365
ochracea-unicolor, Ocneria 365
ochrea, Subacronicta 458
ochripalpis, Laelia 267
ochripes, Leucoma 341
ochrolimbata, Orgyia 276, 292
ochropoda, Ivela 336, 338
Ocneria 220 , 350
Ocneropsis 376
Ocnerorgyia 378
Odagra 266
Olene 219, 223
olga, Olene 224, 225
olivacea, Craniophora 544
omissa, Olene 224
omninolimbata, Penthophera 295
omori i, Acronicta 467, 478
orana, Orgyia 285
orbimaculata, Episema 428
Orgyia 270, 220
Orgyidae 214
Orgyinae 218
oriens, Acronicta 529
orientalis, Acronicta 506, 508, 525
orientalis, Orgyia 289
orientalis, Parocneria 381
orimba, Olene 224
orion, Daseochaeta 441
ornata, Cifuna 302
Orvasca 308
oslari, Orgyia 273
ostracina, Laelia 267
othello, Acronicta 506
ovata, Acronicta 462, 498
Oxycesta 397, 456, 552
oxyptera, Sirnyra 561
pacifica, Acronicta 467
pacifica, Craniophora 541
Pagella 351
pais, Olene 224
palata, Diphthera 413
pollens, Arsilonche 549
pallida, Acronicta 529
pallida, Daseochaeta 439
pallidipennis, Anacronicta 407
palshkovi, Subleuconycta 447
pamiricola, Gynaephora 245
panabra, Euproctis 317
panlacroixii, Orgyia 273
674 АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Panthauma 401, 402
Panthea 412
Pantheini 397
papuensis, Trisuloides 404
paradoxa, Acronicta 469
paradoxa, Euproctis 316
paradoxa, Gynaephora 234
paradoxa, Orgyia 278
paragrapha, Craniophora 538
parallela, Acronicta 480
parallela, Orgyia 273
parisiensis, Acronicta 520
Parocneria 221, 377
parviplagiosa, Artaxa 309
parvula, Ocneria 353
pasiphae, Acronicta 526
patrana, Numens 305
patricolor, Belciana 430
paucipuncta, Laelia 267
paupera, Acronictoides 410
paupercula, Acronicta 479
Penthophera 220, 294
Penthophera 294
pepli, Acronicta 517
perdita, Acronicta 506
perfecta, Leucoma 341
persica, Subacronicta 460
persuasa, Acronicta 480, 517
peusteria, Raphia 421, 423, 425
phaedra, Acronicta 499, 501
phaedriola, Acronicta 499, 500
phaeorrhoea, Euproctis 315, 323
Pharetra 462, 465, 506
phasmatodes, Leucoma 341
pheocosma, Acronicta 463
pheonissa, Parocneria 383
pheosia, Olene 224
Philorgyia 462
phrika, Leucoma 341
piazzi, Raphia 423
picata, Craniophora 538
pini, Olene 224
piperita, Artaxa 308, 310, 312
plagiata, Artaxa 303
Plataplecta 462
Platy centra 412
plumbalis, Ocneria 353
plumbociliata, Euproctis 323
pluto, Dasychira 250
poecilonympha, Leucoma 341
poenitens, Parocneria 389
polia, Dasychira 250
polonica, Acronicta 529
Polyogramma 400
Polyonicta 400
polyporia, Acronicta 465
pontica, Craniophora 538, 540, 547
populi, Acronicta 467
porrecta-alba, Dasychira 261
Porthesia 315
Porthetria 350
portlandia, Diphthera 413
postfusca, Dasychira 250
postfusca, Ocneria 353
postica, Orgyia 271, 273
postincisa, Euproctis 317-
povera, Orgyia 273 '
praeclara, Craniophora 538 , 542
praestana, Numens 306
praeterea, Ocneria 365
prasinaria, Canna 449
pricta, Orgyia 285
primula, Leucoma 341
prisca, Orgyia 273, 275, 290
prisca, Orgyia ericae 287
Procodeca 265
prolata, Laelia 267
proletaria, Olene 224
proletaria, Olene fascelina 227
propinquilinea, Calocasia 416
pruinosa, Acronicta 462, 506
pruinosa, Acronicta albiorbis 480
pruinosa, Acronicta pulverosa 533
pruinosa, Ocneria 353
prufferi, Acronicta 529
pruni, Acronicta 480
pryeri, Dasychira 256
pseudabietis, Dasychira, 250, 251, 256
pseudobamra, Acronicta 463
P seudomesa 305
Pseudopunda 462
pseudotsugata, Orgyia 273
psi, Acronicta 462, 479, 480, 492
psi, Acronicta tridens 489
prideleta, Acronicta 492
Psilura 350
pudibunda, Dasychira 249, 251, 261
pudica, Dasychira 265
pulcherrima, Olene 224
pulchripicta, Canna 449, 450
pulla, Subacronicta 458
pulverosa, Acronicta 506, 533
pumila, Gynaephora, 232, 244
pumila, Gynaephora alpherakii 245
punctella, Euproctis . 323
punctifera, Euproctis 323
punctigera, Euproctis 323
punctillata, Euproctis 320
punctinotata, Acronic.ta 506
pusillata, Arctornis 333
pyhaevaarae, Acronicta 517
pyralis, Acronicta 506
Pyramocera 351
quadrata, Acronicta 480
quadrata, Laelia 266
quadrimaculata, Euproctis 320
quadriplagiata Numens 305
quinquedentata, Acronicta 489
quinquepunctata, Olene 224
radcliffei, Acronicta 480
raddei, Artaxa 313
raddei, Parocneria 380, 393
radiata, Ocneria 365
radoti, Acronicta 489
ramburi, Orgyia 289
raphael, Acronicta 506, 507, 513
raphaelis, Acronicta 513
Raphia 401, 421
rebeli, Euproctis 320
rebuti, Ocneria 353
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ 575
recens, Orgyia 282
recurvata, Artaxa 309
Redoa 340
relicta, Gynaephora 240
relictus, Ocneropsis 376, 377
renimaculata, Arsilonche 549
retardata, Acronicta 498
retinopepla, Orgyia 273
rhodina, Ocneria 353
rhodophora, Ocneria 353
rhopalum, Dasychira 250
Ricine 265
riguata, Leucoma 341
riukiuana, Artaxa 309
rivellata Acronicta 480
rosea, Acronicta cuspis 487
rosea, Acronicta leporina 475
rosea, Acronicta tridens 489
rosea, Daseochaeta 441
rosea, Ocneria 353
rosea, Subacronicta 458
roseoradiata, Craniophora 544
rosina, Ocneria 353
rossii, Gynaeohora 232, 237
Ruanda 336
rubea, Ocneria 351, 354, 355, 374
rubicunda, Leucoma 342
rubiginosa, Acronicta 463, 480
rubribrunnea, Simyra 561
rubricoma, Acronicta 467
rubrior, Ocneria 374
rufimarginata, Leucoma 341
rujofusca, Ocneria 353
rumicis, Acronicta 462, 467, 507, 529
runica, Daseochaeta 441
rupestris, Orgyia 273
rusticana, Ocneria 353
saadi, Acronicta 525
sachalinensis, Acronicta 533
sachalinensis, Dasychira 250
saepestriata, Arsilonche 549
sakaguchii, Artaxa 309
sakaguchii, Ocneria 353
salicis, Acronicta 529
salicis, Leucoma 340 , 342
salvadora, Orgyia 273
samarita, Parocneria 380
sangaica, Laelia 267
sapporensis, Acronicta 479
sartorii, Acronicta 514.
sartus, Leucoma 341, 342, 349
sationella, Dasychira 250
sationis, Dasychira 250
schlumbergi, Subacronicta 458
schwingenschussi, Acronicta 520 1
scopularia, Dasychira 261
scotica, Acronicta 514
sebdonensis, Orgyia 289
sebdoriensis, Orgyia 289
seitzi, Artaxa 309
seitzi, Olene 224
seleniaca, Orgyia 285
selenitica, Gynaephora 231, 232, 234
selenophora, Gynaephora 232, 245
semenovi, Gynaephora 245
semicincta, Ocneria, 351, 353
semicirculosa, Dasychira 250
semihyalina, Leucoma 341
semiobscura, Ocneria 365
semiochrea, Leucoma 341
semisignata, Euproctis 317
semivirga, Acronicta 475
semivitta, Euproctis 317
senica, Acronicta 467, 478
senica, Orgyia 273
separata, Episema 428
separata, Numens 306
septentrionalis, Orgyia 278'
sericaria, Laelia 267
sericea, Euproctis 341
sericea, Leucoma 341
sericea, Trisuloides 403, 404
sericina, Oxycesta 552
serratae, Oxycesta 554
sicula, Orgyia 289
signata, Euproctis 317
signatoria, Parocneria 380, 389
signiplaga, Euproctis 317
silca, Ocneria 353
siletti, Numens 305
silhetica, Leucoma 341
srmilana, Acronicta 467
similis, Acronicta 517
similis, Euproctis 315, 319, 329
similis, Ocneria 353
simplex, Ocneria 353
Simyra 397, 456, 557
simyrides, Leiometopon 563
sincera, Gynaephora 232, 248
sincera, Leiometopon 563
sinensis, Arctornis 332
sinensis, Laelia 267
sinensis, Thalatha 439
singaporensis, Leucoma 341
singapura, Ocneria 353
sinica, Ocneria 353
Sitvia 336
sjoquisti, Euproctis 320
smaragdina, Canna 312
Smilepholcia 404
snelleni, Artaxa 312
sohesti, Leucoma 342
solimana, Acronicta 492
solitalana, Dasychira 251
soltowensis, Acronicta 489
sophronia, Belciana 430 ,
squamosa, Artaxa 313
sperata, Acronicta 506
spinigera, Acronicta 480
splendens, Canna 449
splendida, Orgyia 285
splendida, Simyra 561
staudingeri, Artaxa 309
staudingeri, Artaxa piperita 312
staudingeri, Belciana 431
staudingeri, Gynaephora 245
staudingeri, Numens 306
steinerti, Acronicta 481
Stilpnotia 340
stix, Olene 224
stbtzneri, Parocneria 390
576
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
straminea, Artaxa 309
stratiovireris, Belciana 430
striata, Acronicta 529
striata, Laelia 267
strigata, Ocneria 353
strigosa, Acronicta 462, 499, 504
strigulata, Acronicta 479
Subacronicta 396, 455
subfasciata, Euproctis 317
subflava, Artaxa 308, 310
subflava, Trisuloides 404
subfusca, Ocneria 355
Subleuconycta 402, 447
submarginalis, Ocneria 365
submarginata, Leucoma 347
subnigra, Dasychira 250
subnigropunctata, Dasychira 250
subnotata, Artaxa 308
subochrea, Acronicta 480
subornata, Acronicta 509
subpurpurea, Acronicta 479, 486
subrosea, Ocneria 353
subserrata, Belciana 430
succidens, Acronicta 506
suffusa, Acronicta alni 481
suffusa, Acronicta cuspis 487
suffusa, Acronicta psi 492
suffusa, Acronicta menyanthidis 514
suffusa, Laelia 265, 267
suhrianna, Acronicta 529
suigensis, Acronicta 536
suigensis, Leucoma 341
sundevalli, Craniophora 544
superans, Subacronicta 457
surtur, Leucoma 341, 342, 350
susanna, Euproctis 3J.7, 319, 330
suzukii, Calocasia 416
suzukii, Dasychira 250
taeniata, Acronicta 487
tagalica, Ocneria 353
taiwanensis, Dasychira 250
takamukuana, Dasychira 250
takamuktii, Numens 306
Tambana 404
tamsi, Moma 433
tanaica, Arsilonche 549
tarawanensis, Olene 224
taurica, Acronicta 489
taurina, Ocneria 353
Teia 270
tendinosa, Simyra 558
tenuis, Euproctis 317
terebinthi, Parocneria 390
terebynthi, Parocneria 380, 390
terebynthina, Darocneria 380, 381, 392
terrigena, Acronicta 498, 503
tessmanni, Dasychira 250
testacea, Laelia 265, 267
testacea, Laelia atestacea 266
tetraspila, Belciana 430
Thalatha 398, 439
thanatos, Dasychira 250
Themaca 308
thoodora, Acronicta 463, 480
thoracica, Acronicta 479
thoracica, Belciana 430
thwaitesi, Dasychira 250
Thyatirides 402, 451
thyelantiqua, Orgyia f. hybrida 277
thyellina, Orgyia 273, 274, 276
Thylacyna 270
Thyracona 398
tiena, Acronicta 520
tigniumbra, Craniophora 538
tisdala, Orgyia 271
tjurana, Arsilonche 549
todara, Ocneria 353
torasan, Artaxa 309
torosa, Simyra 561
tota, Acronicta 467
transcaspica, Orgyia 285
transiens, Orgyia 289
transiens, Ocneria 355
transverse, Craniophora 538, 540
transverse, Acronicta 467
Triaena 462, 465, 479
Trichiosoma 270
Tricholonche 46 2
Trichosea 432
tricolor, Orgyia 271
tridens, Acronicta 479, 480, 489
trigotephras, Orgyia 273, 275, 289
trimaculata, Euproctis 320
trimaculata, Thyatirides 451, 452
trinubila, Belciana 430, 432
tripunctata, Thyatirides 452
tristis, Arsilonche 549
tristis, Orgyia 293
Trisula 398, 404
Trisuloides 401, 403
tritona, Acronicta 462, 480
troni, Craniophora 544
turanica, Acronicta 529
turbata, Orgyia 271, 273
turcica, Orgyia 285
turneri, Ocneria 353
umbrifera, Ocneria 353
umbrina, Laelia 267
umbripennis, Orgyia 285
umbrosa, Calocasia 416
umbrosissima, Calocasia 418
unicolor, Ocneria 365
unicolor, Olene 227
unicolor, Parocneria 390
unifasciata, Ocneria 365
uniformis, Calocasia 418
uniformis, Parocneria 380
unilineata Dasychira 261
urbicola, Parocneria 383
TJrocoma 316
ussuriensis, Calocasia 420
ussuriensis, Diphthera 413
uxor, Ocneria 353
vacillans, Ocneria 353
vagans, Olene 224
vanda, Acronicta 465
vaporata, Orgyia 271
varia, Orgyia 271
variegata, Acronicta 489
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
variegata, Ocneria 365
variegata, Trisula 402
variegata, Trisuloides 402
vau-nigrum, Arctornis 333
velia, Acronicta 463, 480
velutina, Ocneria 353
venosa, Arsilonche 549
venosa, Euproctis 317
venosa, Laelia 267
verbenata, Daseochaeta 4
vernalis, Acronicta
vestilina, Parocneria 393
vetantiqua, Orgyia f. hybrids 281
vetusta, Orgyia 271
viburni, Craniophora 544
vigens, Daseochaeta 439
Viminia 462
vinnula, Acronicta 479
viola, Ocneria 353
virens, Belciades 444
virga, Acronicta menyanthidis 514
virga, Acronicta psi 492
virga, Acronicta tridens 489
virgata, Acronicta 520
virginaria, Diphthera 413
virginea, Dasychira 250 , 252 , 253
virginea, Euproctis , 317
virgo, Euproctis 317
virguncula, Euproctis 317
viridescens, Orgyia 273
viridis, Daseochaeta 439
vitensis, Dasychira 250
vitrina, Ocneria 353
v-nigrum, Acrtornis 333
vulpina, Acronicta 475
> rete, Ocneria 353
warpachowskyi, Subacronicta 458
ivautersi, Calocasia 418
iveymeri, Calocasia 418
wilczinskia, Euproctis 320
wintersteini, Orgyia f, hybrids 221
wladiwostokensis, Ocneria 365
Xanthomantis 403, 404
xanthomista, Acronicta 520
xanthorrhoea, Euproctis 317
xanthorrhoea, Euproctis chrysorrhoea 323
x-signata, Acronicta 478
xylina, Ocneria 353
xyliniformis, Acronicta 506
ypsilon, Gerbathodes 410
yunnanensis, Ocneria 355
zena, Dasychira 250
zimmermani, Orgyia 287
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ПАРАЗИТИЧЕСКИХ
И ХИЩНЫХ НАСЕКОМЫХ
acasta, Melittobia 374
acronyctae, Exorista 492
adustus, Gambrus 287
aereus Monodontomerus .327, 374
aestivalis, Hemiteles 373
affinis, Agria 345, 361, 374
affinis, Carcelia 374
affinis, Exorista 265, 285, 492
affinis, Microgaster 264
affinis, Sarcophaga 374
agilis, Pezomachus 281, 373
agilis, Phorobera 374 .
albipes, Ceraphron 264
albipes, Sarcophaga 374
alboguttatus, Automalus 323, 361
alboguttatus, Ichneumon 264
alboguttatus, Stenichneumon 323
alboguttatus, Trogus 264
alternans, Itoplectes 327
alternatus, Pimpla 361
amoenus, Cryptus 373
analis, Cryptus 345
anchora, Gaurax 374
anopheles, Torymus 327
anser, Cyclotophrys 327
anthracinus, Hemiteles 373
antiqua, Rhacodineura 374
anxi, Peltopius 345
aratrix, Sarcophaga 361
arctica, Pimpla 345, 366
areator, Hemiteles 264, 373
armatus, Apanteles 443
assertorius, Ichnus 373
assimilis, Limneria 345
assimilis, Phorocera 265, 361
atalantae, Theronia 327, 345, 361, 373
ater, Mesochorus 345
atripes, Idiolispa 327
atropivora, Sturmia 374, 492
atropos, Sarcophaga 361
Attagenus sp. 373
aurea, Leskia 265
balticus, Ichneumon 264
bella, Sturmia 347
bellicosus, Amblyteles 492
bellipes, Ichneumon 230
bicolorinus, Hemiteles, 373, 492
bidens, Picromerus 361
bilunatus, Ichneumon 264
bilunulatus, Barichneumon 265
bimaculata, Sturmia 361, 374
bimaculatus, Meteorus 345, 361
bimaculatus, Melanichneumon 265
bipunctator, Hemiteles 373, 495
bisignata, Meigenia 374
blanca, Exorista 374
bombycis, Telenomus 361
bombycicornis, Eulophus 264
boucheanus, Pteromalus 327 , 345
braccatus, Eriobius 495
brassicariae, Apechta 345
brassicariae, Pimpla 327, 361, 373
brevicornis, Habrobracon, 327, 374
brevicornis, Microgaster 374
brevipetiolatus, Mesochorus 430
breviusculus, Taleas 492
brischkei, Glypta 264
caesifrons, Phorocera 361
calceatus, Microgaster 327, 374
callipus, Chaicis 374
capulifera, Pimpla 361
carbonarius, Campoplex 281
centummaculatus, Coelichneumon 495
cephalotus, Paniscus 265, 373, 492
cerinops, Anomalon 430, 533
cessator, Ichneumon 495
cheloniae, Carcelia 327, 345, 374
circumflexum, Exochilum 382
circumscriptus, Bracon 345
circumscriptus, Rhogas 532
clavator, Exetastes 327
claviventris, Casinaria 361
collaris, Macrocentrus 361
compressus, Banchus 323
computatorius, Ichneumon 264
concinnata, Compsilura 265, 323, 327,
345, 361, 374, 420, 492, 495, 533
concinnus, Campoplex, 327, 373
conformis, Hemiteles 430, 492, 495
congestus, Apanteles 533
congestus, Microgaster 492, 533
conjugata, Larvivora 374
connexus, Microgaster 327
conquisitor, Pimpla 373
consimilis, Ichneumon 323, 327
consobrina, Ernestia 374
consularis, Microgaster 327, 323
costatus, Ophion 492
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
579
crassiseta, Chaetomyia 374
cultrator, Campoplex j 281, 285
cursitans, Pczomachus 281
cyanalor, Cryplus 373, 430'
dalmani, Telenomus 264, 281, 285
declinator, Sagaritis 285
delectus, Meteorus 345
delicator, Barylypa 373
deliratorius, Ichneumon 265
dentipes, Monodontomerus 327 ' . 7
difficilis, Meteorus 345
difficilis, Microgaster 327
difformis, Limneria 327, 373
difformis, Omorgus 327, 361, 373
dilutus, Mesochorus 327
dilutus, Microgaster 323, 327
dimidiatus, Rhogas 533
dirus, Eurylabus 361, 373
disparis, Anastatus 374
disparis, Hyposter 373
disparis, Ichneumon 327, 373
disparis, Limneria 373
disparis, Microgaster 327
disparis, Pimpla 373
disparis, Protichneumon 327, 345/ 361
ebenina, Anilasta 230, 281, 289
ebenina, Sagaritis 230
elata, Plagia 323, 345
elegantula^ Calpotrochia 533
elegantulus, Tryphon 533
enecator, Trichomma 373
erichsoni, Dermestes 373
erythrosloma, Exorista 374
euceros, Pleromalus 345
evanescens, Trichogramma 327, 361
examinator, Pimpla 281, 323, 327, 345,
361, 373, 533
excisa. Carcelia 265, 281, 285, 345, 361,
374
fabricator,’ Cralichneumon 345
fabricator, Ichneumon 237, 264
falcator, Campoplex 265
falculata, Sarcophaga 361
fasciatorius, Amblyteles 533
fasciatum, Trichogramma 361
fasciatus, Meteorus 345
fatua,- Erycia 323, 327
fauna, Zenillia 327, 533
fera, Tachina 362, 374
ferruginea, Ceromasia 323, 327, 361
festinans, Erycia 361
festiva, Larvivora 374
fibulalor, Anomalon 430
flavescens, Theronia 373
flavicans, Pimpla 373
flavicans, Theronia 327
flavifemur, Ichneumon 264
flavipes, Apanteles 373, 492
flavipes Microgaster 374, 533, 323, 374
flavipes, Chaicis 374
flavolineata, Glypta 264
flavus, Ichnus 373
floralis, Thyella 327
florum, Ceromasia 361
formosus, Apanteles 281
formosus, Mesochorus 430
formosus, Meteorus 345
fugax, Prosopaea 361
fumipennis, Microgaster 492
fumipennis, Microplitis 533
geniculator, Bracon 327
geniculator, Rhogas 264, 281, .327
glauca, Exorista 265, 492
glaucopterus, Opheltes 373
glirina, Exorista 492
globosus, Steropus 373
glomeratus, Apanteles 361, 373
glomeratus, Microgaster 492
gnava, Carcelia 265, 281, 345
gracilis, Mesochorus 373
gracilis, Microgaster 264
grandis, Tricholyga 374
guttatorius, Exelasles 492
haemorrhoidalis, Micropalpus 345
haglundi, Ichneumon 345
halilayanus,' Pteromalus 345, 374
holmgreni, Pimpla 373
hortensis, Pezomachus 373 '
hortensis, Hemiteles 373
howardi, Hadronotus 374
ictericus, Meteorus 327
impotens, Larvivora 281
inconspicua, Sturmia 374
inclusus, Apanteles 361
inclusus, Microgaster 323, 327
inquisitor, Calosoma 373
inquisitor, Epiurus 281, 345, 361, 373
inquisitor, Exetasles 327
insidiosus, Ichneumon 237, 361
insignis, Redtenbacheria 361
instigator, Pimpla 264, 281, 287, 323,
327, 345, 361, 373, 533
intermedia, Brachymeria 374
irrigator, Xylonomus 361
irritator, Microgaster 492
irrorator, Chelonus 492
irrorator, Rhitegaster 492
japonica, Tachina 374
japonicus, Meteorus 373
javensis, Pcrisopterus 374
kollari, Bracon 533
ktrwanae, Schedins 374
lacteicolor, Microgaster 327, 373
lactipennis, Microgaster 323, 327
laeviusculus, Tesale 361
lardarius, Dermestes 373
lariae, Ichneumon 241
larvarum, Larvivora 230, 237, 285, 287,
323, 327, 361, 374, 533
larvarum, Cratotechus 361
larvarum, Eutedon 492
larvicola, Larvivora 374
latifrons, Tachina 327
580
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
latus, Habrobracon 373
leucocherrus, Ichneumon 373
libatrix, Zenillia 323, 327, 374
liparidis, Apanteles 289, 373
liparidis, Cryptus 373
lota, Exorista 402
lucorum, Carcelia 265, 281, 361, 374
lutorius, Trogus 373
magnicornis, Eudromyia 327, 374
majalis, Limneria 533
marmorata, Histochaeta 374
marrocanus, Tenebrioides 373
masii, Schedius 374
melanoceles, Meteorus 345
melanocerus, Ichneumon 361
melanoscelus, Apanteles 361, 373
melanurus, Paniscus 495
merdarius, Enicospilus 373
merdarius, Ophion 373
migratorius, Metopius 492, 495
militarius, Stenichneumon 265
mirabilis, Nemoria 345
mixtus, Campoplex 264
monachae, Agria 361, 374
monachae, Larvivora 374
moretti, Larvivora 374 .
morio, Hemipenthes 362
moschator, Cryptus 327
multicinctus, Stenichneumon 323
nanus, Hemiteles 373
nebulator, Alexeter 503
nidicola, Zygobothria 327
nidulans, Pteromalus 327
nigerrimus, Microgaster 373
nigriceps, Microgaster 533
higricornis, Pelletieria 361
nigripes, Campoplex 281
nigripes, Casinaria 281
nigripes, Lydella 323, 327, 374, 533
nigripes, Thelaira 362
nigritarius, Barichneumon 361
nigritarius, Cratichneumon 345
nigriventris, Apanteles 361
nitidulus, Telenomus 345
noctuarum, Larvivora 374
noctuicida, Exorista 492
nympharum, Microtachina 362
obscuratus, Chaicis 374
obsoletus, Torymus 323, 327
obtusatus, Chaicis 374
ocneriae, Apanteles 373
ogimae, Atoposomoidea 374
omnivora, Psychophagus 327
opavus, Microgaster 533
ornata, Pimpla 533
ornatus, Gambrus 563
ovivora, Pimpla 533
pallidus, Mesochorus 327
pallipes, Mesochorus 327
pallipes, Microgaster 492
pavida, Pales 237, 289, 323, 327, 345, 361
poctoralis, Mesochorus 327, 373
pedalis, Pimpla 373
perspicillator, Barylipa 373
perspicuus, Microgaster 533
phalaenarum, Telenomus 374
pictilis, Mesochorus 492, 533
pictipes, Hemiteles 373
pictus, Ichneumon 373
pini, Pteromalus 345, 374
pinivora, Larvivora 374
pluto, Pimpla 373
politus, Mesochorus 345
porthetriae, Apanteles 373
porthetriae, Pimpla 373
posticus, Microgaster 323
praeceps,' Echinomyia 327
preciosa, Trichigramma 361
privigna, Sarcophaga 362
processioneae, Pteromalus 323, 327
prompta, Pelletieria 361
prorogator, Rhogas 345
pubescens, Microgaster 373
pudibundae, Pimpla 264
pulchricornis, Meteorus 374 .
pulchripes, Rhogas 323, 327, 495
pumicata, Pales 361
punctiger, Microgaster 373
punciventris, Telenomus 345
puparum, Pteromalus 323, 327, 345, 533
quadridentata, Pimpla 361
quadripunctata, Xylodrepa 373
quadripustulata, Winthemia 2*65
quinquevittata, Larvivora 374
ramicornis, Eulophus 264, 281, 492
rapax, Anilastus 361
rapax, Campoplex 373
raptor, Sagaritis 281
raptorius, Ichneumon 361
ratzeburgi, Ichneumon 361
ratzeburgi, Microgaster 492
recussata, Carcellia 374
regius, Torymus . 327
rubens, Ichneumon 373
rubens, Microgaster 533
rudis, Ernestia 361
rufata, Pimpla 361, 533
ruficorne, Anomalon 420
ruficornis, Aphanistes 361
ruficornis, Microgaster 533
ruficrus, Meteorus 345
rugosus, Rhogas 524
rugulosus, Microgaster 492
rumicis, Apanteles 533
ruralis, Agria 361
rustica, Larvivora 361, 374
salebrosus, Meteorus 345
sarcitorius, Ichneumon 327, 373, 430
scandica, Pimpla 533
schaffneri, Hemiteles 373, 430
schutzei, Sarcophaga 362
scirripoda, Chaicis 327
scutellaris, Trophocampa 361
scutellata, Sturmia 362, 374
scutellator, Meteorus 345, 361, 373
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
scutellator, Microgaster 533
scutellator, Stenichneumon 327
segregate, Parasetigena 361
segregate, Tricholyga 374
semblidis, Oophthora 327, 374
semirufus, Ichneumon 281
semirufus, Mesochorus 345, 373, 492, 533
senicula, Casinaria 285
separata, Carcelia 374
sericariae, Crossocosmia 374
setipennis, Digonochaeta 327
sexlineatus,. Ichneumon 264
sicarius, Cratichneumon 265
silvarum, Mesochorus 361
silvatica, Masicera 327, 361
similis, Microgaster 323
socialis, Hemiteles 327
solitarius, Apanteles 361
solitarius, Meteorus 345
solitarius, Microgaster 286, 327
sorbillans, Tricholyga 327, 362, 374
sordipes, Microplitis 495
speciosa, Winthemia 374
spinolae, Microgaster 492
splendidus, Mesochorus 345
spuria, Pimpla . 265
spurius, Microgaster 237, 264, 492
stabulans, Lydella 345
stenogaster, Etroxus 492
stercorator, Pimpla 281, 345
stigmaticus, Mesochorus 281
stigmator, Mesochorus 323, 361
subcompletus, Microgaster 492
submarginatus, Hemiteles 430
sugillatorius, Coelichneumon 361
sussurans, Carcelia 361, 373
sycophanta, Calosoma 373
tardus, Encyrtus 374
tarsator Exetastes 492, 533
tenebrosus, Apanteles 373
tenuicornis, Pimpla 373
tenuis, Rhogas 323, 327
tenuiventris, Casinaria 373
teras, Glypta 264
terebrator, Pezomachus 345
terminatorius, Ichneumon 327
testaceus, Paniscus 373, 492, 533
testaceus, Rhogas 323, 327
tibialis, Microgaster 373
triangulator, Microgaster 264
Trombidium 373
truncatus, Telenomus 264
tuberculatus, Ephialtes 361
tuberosa, Sarcophaga 362
turionellae, Pimpla 361
uliginosa, Sarcophaga 361, 374
ultor, Microgaster 323, . 327, 492
unicincta, Limneria 533
unicinctus, Campoplex 483
unicolor, Rhogas 345
vadatorius, Amblyteles 533
validum, Limneria 373
varicornis, Pimpla 361
versicolor, Meteorus 327, 361, 373
vertiginosa, Telymorpha 374
vetula, Phryno 361
vidua, Larvivora 281
vinidae, Eutedon 345
viridis, Gymnochaeta 362'
vitripennis, Microgaster 327, 373
vitticollis, Mesochorus 361, 373
vulgaris, Phryxe 265, 281, 327
vulnerator, Pristomerus 373
westermanni, Exorista 323, 327, 374
xanthogastra, Winthemia 492
zetterstedti, Teleas 264
СОДЕРЖАЛИ Е
Стр.
Предисловие........................................................... 3
Систематический указатель............................................ 5
Общая часть............................................................ 9
Глава I. Внешнее строение тела взрослых насекомых............ 9
Глава II. Внешняя морфология преимагинальных фаз............. 25
Глава III. Особенности анатомического строения...............• 43
Глава IV. Биология .......................................... 49
Глава V. Классификация и филогения.......................... 56
Глава VI. Экология .......................................... 78
Глава VII. Географическое распространение................... 124
Глава VIII. Хозяйственное значение............................ 172
Литература . . . .............................................. 200
Специальная часть ................................................... 214
Сем. Orgyidae ............................................... 214
I. Подсем. Orgyinae ........................................ 218
II. Подсем. Acronictinae....................................... 394
Алфавитный указатель латинских названий сем. Orgyidae................ 566
Алфавитный указатель латинских названий паразитических и хищных насе-
комых ...................................................... 578
Печатается по постановленшд
Редакционно-издательского совета
Академии Наук СССР
ф
Технический редактор А. В. Смирнова
Корректоры А. П. Конради и А. Д. Копысова.
*
РИСО АН СССР № 3845. Подписано к пе-
чати 12 IX 1950 г. М. 40013. Бумага
70ХЮ8716. Бум. л. 187<. Печ. л. 3672.
Уч.-изд. л. 51.3. Тираж 2000. Зак. № 1678.
Цена в переплете 44 руб.
1-я тип. И'з-ва АН СССР. Ленинград, В. О.,
9 линия, дом 12.