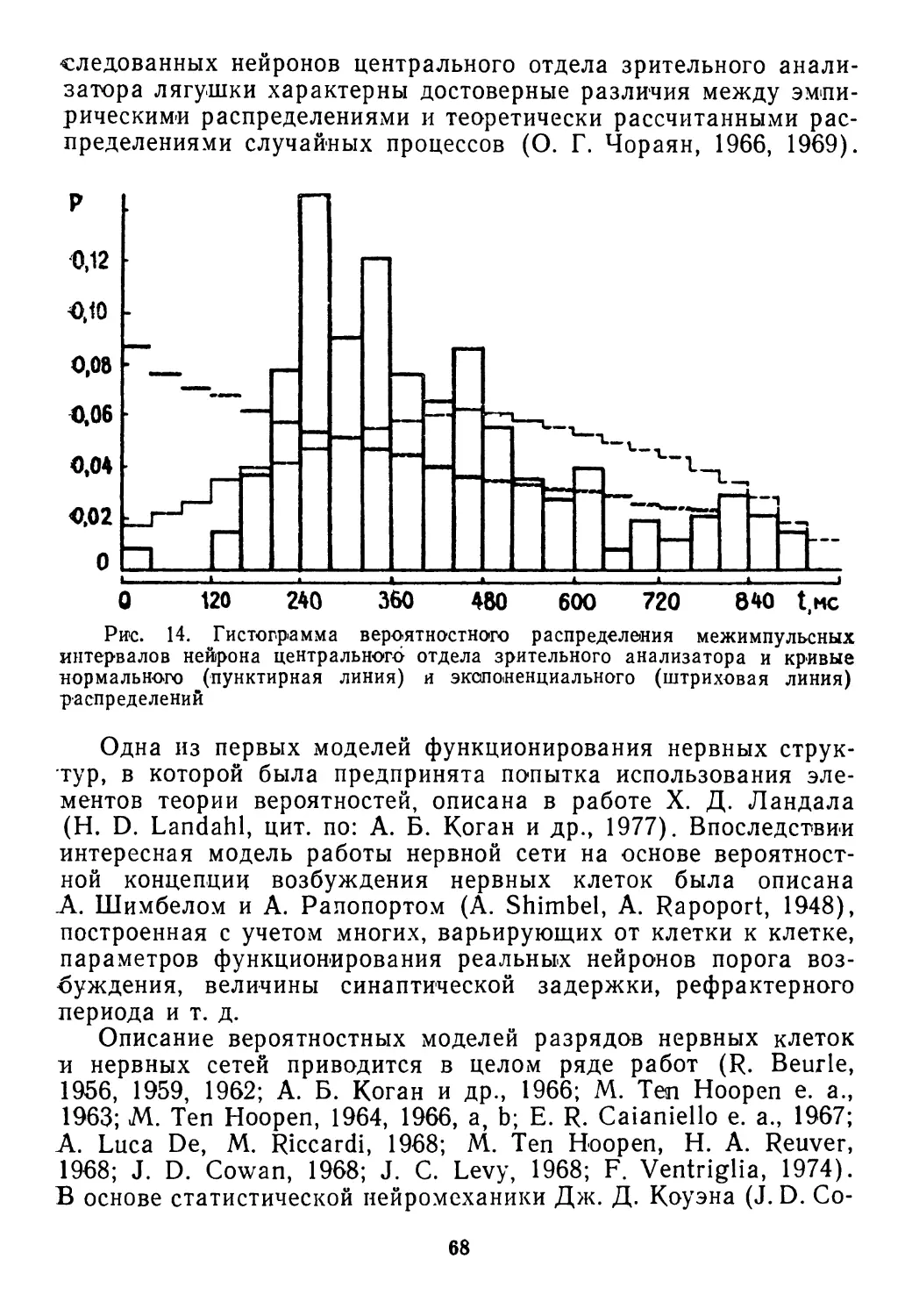
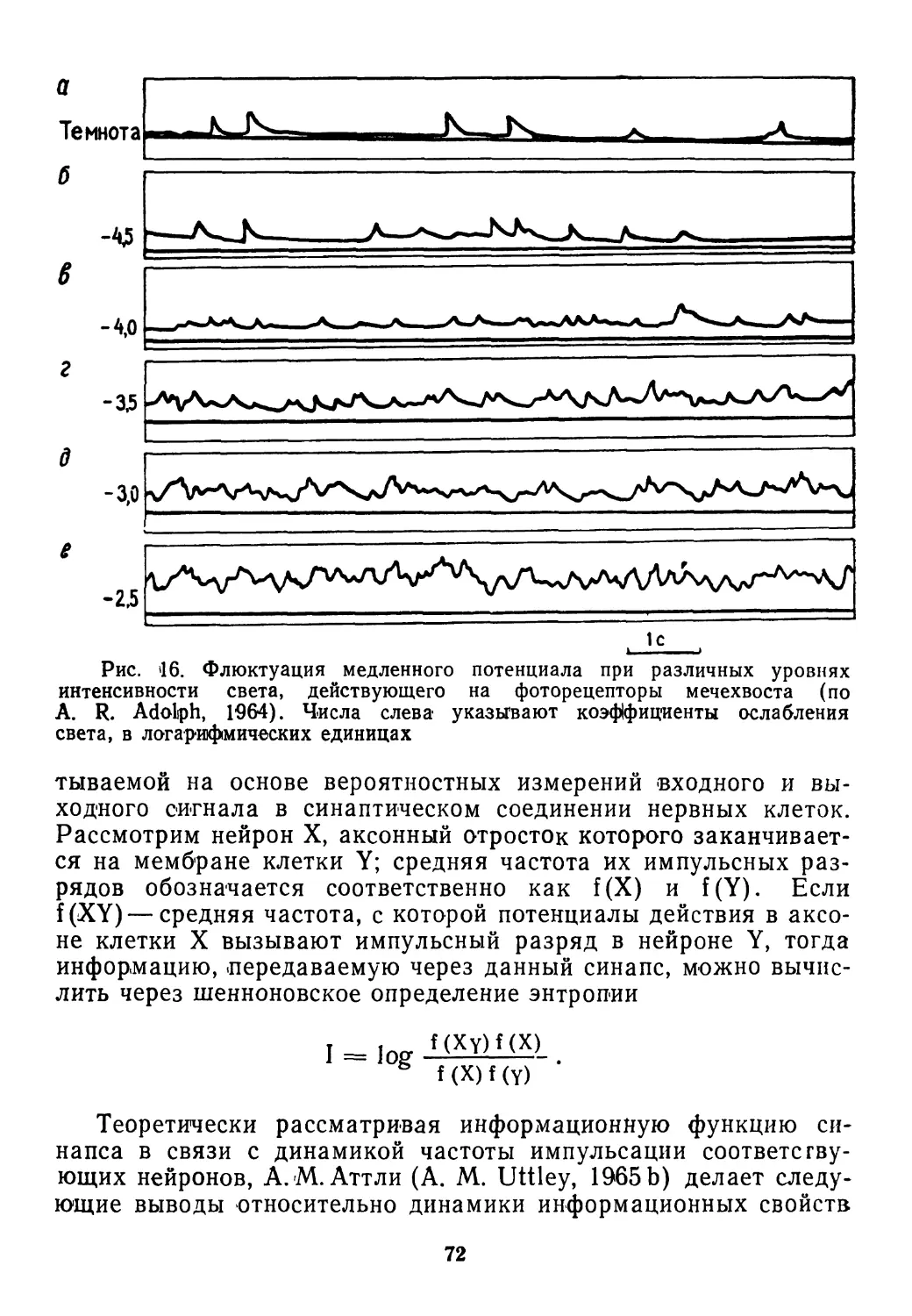
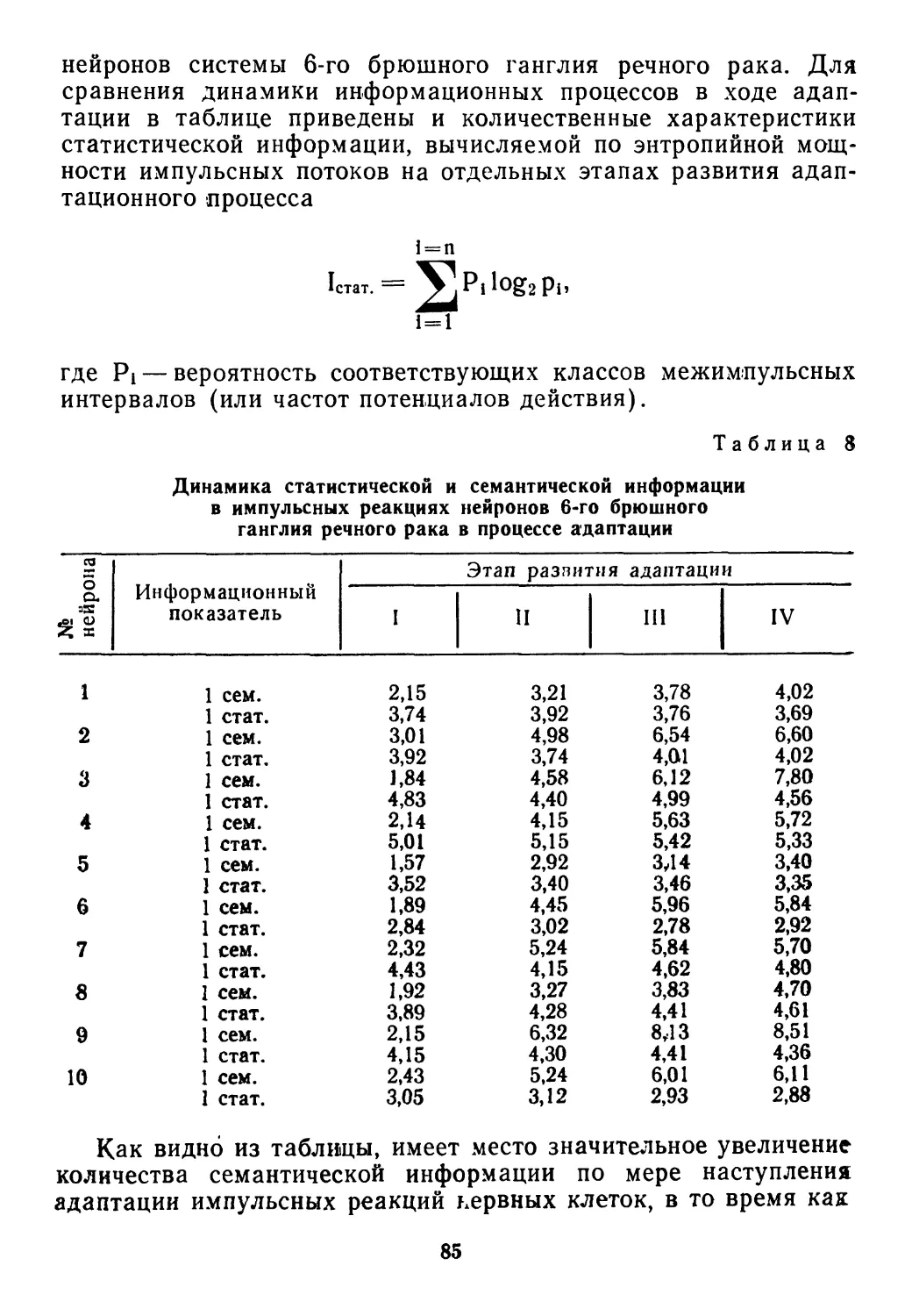
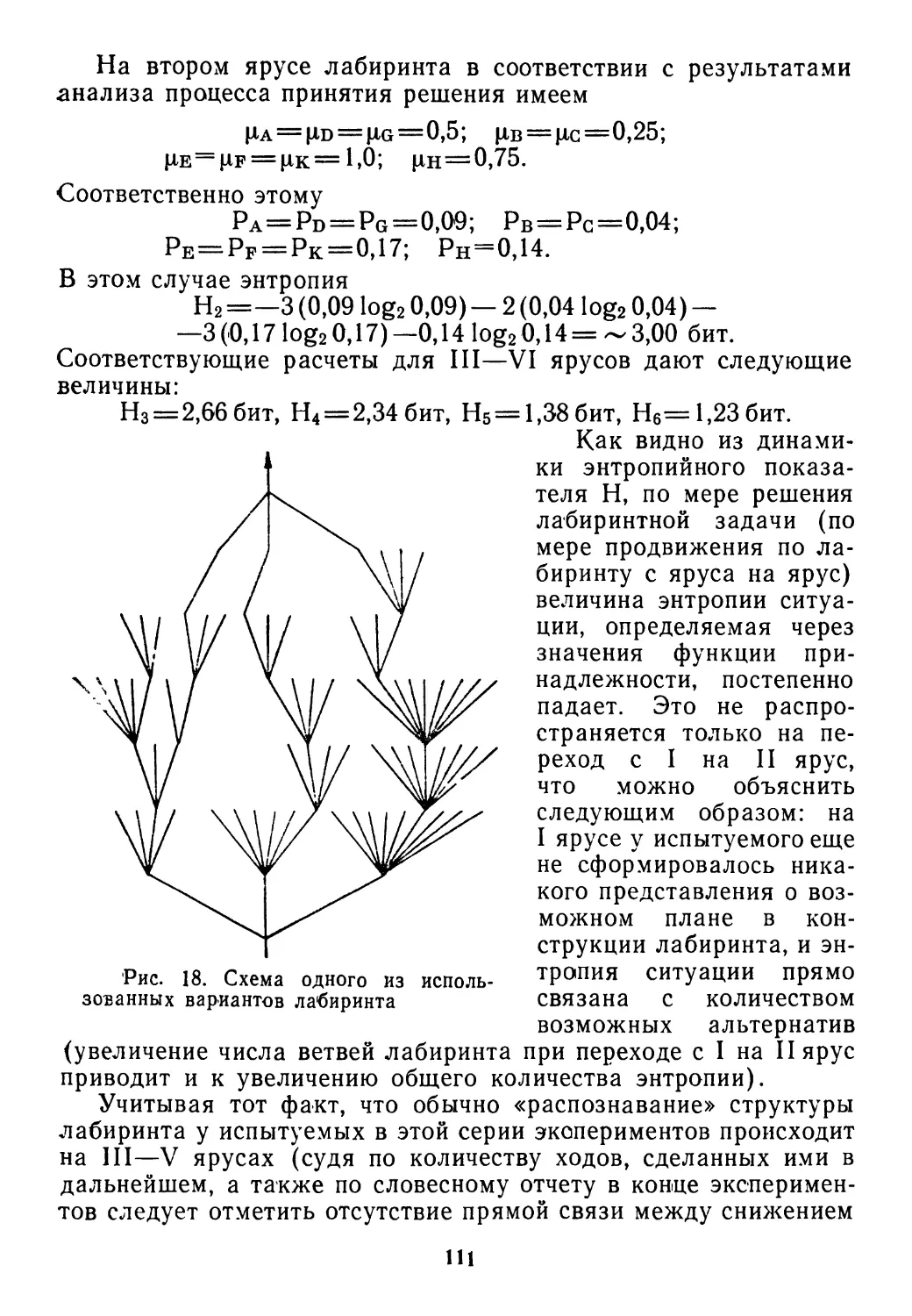
/































































































































































Author: Коган А.Б. Чораян О.Г.
Tags: медицина нервная система патогенез нервная деятельность
Year: 1980




Text
северо-кавказский научный центр высшей школы
ростовский-на-дону ОРДЕНА ТРУДОВОГО КРАСНОГО знамени
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
А. Б. К О Г А Н, О. Г. ч О Р А я н
ВЕРОЯТНОСТНЫЕ
МЕХАНИЗМЫ
НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Ответственный редактор
кандидат медицинских наук Г. А. Кураев
ИЗДАТЕЛЬСТВО РОСТОВСКОГО УНИВЕРСИТЕТА
1980
Печатается по решению Совета биологического учебно-
научного комплекса Ростовского-на-Дону государственного
университета
Рецензенты доктора биологических наук К. Е. Бугаев,
Н. В. Жуковская.
Коган А. Б., Ч о р а я н О. Г. Вероятностные механизмы
нервной деятельности. Издательство Ростовского универси-
тета, 1980.
176 с.
В монографии приведены результаты экспериментальных исследова-
ний вероятностных механизмов деятельности нервной системы: субкле-
точных структур, нервной клетки, нейронных ансамблей, функциональных
систем мозга. На каждом уровне конструкции нервной системы рас-
смотрено соотношение жесткого и вероятностного обеспечения функ-
ции как результат оптимальной организации информационно-управляю-
щей деятельности структур мозга. На базе теоретических обобщений
результатов собственных исследований и литературных данных выдви-
гается и развивается оригинальное представление о вероятностных
нейронных ансамблях как основе структурно-функциональной органи-
зации нервной системы, обеспечивающей свойства высокой ее надеж-
ности и пластичности. Описан ряд оригинальных приемов информацион-
ного анализа вероятностных механизмов и размытой логики адаптивно-
го поведения целостного организма и его функциональных систем, обу-
словленного регуляторной деятельностью нервных структур.
Монография представляет интерес для студентов, врачей, физиологов,
психологов, специалистов в области бионики, биофизики, биологической
и медицинской кибернетики.
20.7.2—020
К М 175'703)-80 без объявления
© Издательство Ростовского
университета, 1980
Причинность есть нечто, могущее
присутствовать в большей или
меньшей степени, а не только
просто быть или не быть.
Н. Винер
Гл ав а I. ДЕТЕРМИНИЗМ В ЖИВЫХ СИСТЕМАХ
1. КОНЦЕПЦИЯ ОДНОЗНАЧНОГО
И СТАТИСТИЧЕСКОГО ДЕТЕРМИНИЗМА
В любой научной дисциплине определение детерми-
нации изучаемых явлений составляет один из цен-
тральных (разделов, ибо выявление причинно-след-
ственных отношений изучаемых процессов, наблюдае-
мых феноменов и составляет основу научного познания:
закономерностей той или иной формы движения мате-
рии. Вместе с тем полнота решения проблемы детерми-
низма может служить и служит показателем зрелости?
данной научной дисциплины. Прогресс в научном реше-
нии проблемы и заключается в последовательном пере-
ходе от созерцательного, описательного подхода к
вскрытию внутренних закономерностей, механизмов,
протекания процессов, имеющих определенную про-
странственно-временную протяженность, в которой
состояния системы в разные временные промежутки и
разная их пространственная локализация всегда оказы-
ваются тесно взаимосвязанными, хотя эта связь может
носить чрезвычайно сложный опосредованный харак-
тер. Вполне справедливым является утверждение, что-
детерминизм как принцип всеобщей закономерной
связи явлений представляет собой одну из наиболее*
важных основ научного мировоззрения .
Как подчеркивает Б. М. Кедров (1968), детерми-
низм есть общее учение, признающее существование*
универсальной закономерной связи всех явлений мира
и отрицающее существование каких-либо явлений и ве-
щей в мире вне этой универсальной связи. Взаимосвязь
явлений может носить как прямой, непосредственный"
характер, так и быть более или менее опосредованной
через ряд промежуточных звеньев, число которых может-
з
возрастать практически до бесконечности. Поэтому в системе на-
ших естественнонаучных знаний об окружающем материальном
.мире концепция детерминизма занимает особое место как прин-
цип универсальной связи явлений, охватывающий все много-
образие форм движения материи.
Сущность принципа детерминизма определяется следующи-
ми положениями: 1) любое явление имеет свою причину;
2) развитие любого явления подчиняется некоей закономер-
ности; 3) возможно познание любого явления и тем самым
предсказание любого материального процесса. Несмотря на то,
что философским и естественнонаучным основам причинно-
следственных отношений в мире материальных явлений пос-
вящено значительное число публикаций, и сегодня нельзя счи-
тать, что формулировка диалектического принципа детерминиз-
ма во всем его многообразии завершена или близка к заверше-
нию. Новые открытия во всех областях науки выявляют но-
вые своеобразные черты детерминации, наглядно демонстрируя
«неисчерпаемость» материального мира.
В ходе развития концепции детерминизма о природе при-
чинности явлений материального мира, о возможности ее поз-
нания высказывались и аргументировались различные пред-
ставления, многие из которых порой носили диаметрально про-
тивоположный, взаимоисключающий характер. Так, в свое вре-
мя известный философ Б. Рассел (В. Russel, 1913) утверждал,
что все философы любой философской школы воображают, что
причинение является одной из наиболее фундаментальных ак-
сиом или постулатов науки. Однако, .как это ни странно, в наи-
более развитых науках, таких, как гравитационная астрономия,
слово «причина» никогда не встречается... Закон причинности
точно так же, как многое из того, что выдержало испытание в
рамках философии, является пережитком прошлого века, ко-
торый сохраняется, подобно монархии, только потому, что
ошибочная его поддержка не причиняет вреда. Почти пол-
ностью с ним солидаризуется один из видных специалистов в
области кибернетики Г. Цопф, который писал в 1966 году,
что он предубежден против предположения о высокой упоря-
доченности во Вселенной и против допущения, что какой-бы
то ни было порядок познаваем в рамках ныне доступных нам
методов.
Марксистско-ленинское диалектическое представление о де-
терминизме как отражении объективно существующей и позна-
ваемой причинно-следственной связи явлений и процессов ма-
териального мира дано в ряде работ советских философов
4
(И. Т. Фролов, 1961, 1973; Б. М. Кедров, 1968, 1973; В. С. Ви-
шаренко, 1971, 1974, 1975; В. С. Готт, А. Д. Урсул, 1971;
А. С. Кравец, 1971; Ю. В. Сачков, 1971; А. И. Филюков, 1972;
и др.).
Научное представление любого явления предполагает нали-
чие всеобщей (причинной связи явлений и предметов материаль-
ного мира. Однако в целом это верное представление в прошлом
науки было доведено до категорического и поэтому неверного
утверждения, что всегда за определенной причиной с необходи-
мостью должно наступить определенное следствие.
Форма детерминации, причинной обусловленности явления
в каждом конкретном случае бпределяется характером взаимо-
действия тех структур, в которых они происходят. В настоящее
время общепринятым является представление о двух формах
детерминизма явлений реального мира: жесткий однозначный
(или лапласовский) детерминизм и статистический, вероятност-
ный, детерминизм. Жесткий детерминизм характерен для про-
цессов, протекающих в сравнительно простых и высокостабиль-
ных системах, когда каждый последующий этап в развитии про-
цесса оказывается однозначно определенным предыдущим эта-
пом, состоянием. Об этом в свое время писал Лаплас (1908),
считающий, что любое настоящее состояние Вселенной одно-
значно определяется ее предшествующим состоянием. Наличие
случайности Лаплас считал связанным, обусловленным только
субъективным характером нашего знания, следствием нашего
незнания, полностью отрицая возможность объективного суще-
ствования категории случайного в окружающем нас материаль-
ном мире. В лапласовской форме детерминизма признается
только определенность параметров явлений природы и пол-
ностью отрицается их неопределенность. Лапласовский механи-
ческий детерминизм хотя и признает возможность проявления
случайности, но не придает ей существенного значения. Соглас-
но этой концепции, случайность отражает лишь ограниченность
наших знаний об изучаемых процессах и не является принци-
пиальной, характерной чертой самого рассматриваемого явле-
ния. Жесткий лапласовский детерминизм отражает процессы,
закономерности которых либо однозначны, либо очень близки
к однозначным. Таким образом, механический детерминизм, при-
знавая всеобщую причинную необходимость в материальном ми-
ре явлений, исключает диалектическую связь причины, необхо-
димости со случайностью, о которой говорил Ф. Энгельс: «...то,
что утверждается как необходимое, слагается из чистых слу-
чайностей, а то, что считается случайным, представляет собой
форму, за которой скрывается необходимость...»
5
Философские аспекты ограниченности сферы действия лап-
ласовского детерминизма рассмотрены в ряде работ (В. С. Ви-
шаренко, 1971; А. С. Кравец, 197-1; и др.). Так, например, оста-
навливаясь на недостатках жесткого детерминизма, В. С. Ви-
шаренко справедливо замечает, что лашласовский детерминизм,
признавая множественность причинных факторов, .предполагает
их число ограниченным (<в противном случае невозможно было
бы полное познание всех детерминирующих агентов). Однако та-
кое представление противоречит основным положениям совре-
менной науки о неисчерпаемости материи вглубь и бесконечно-
сти ее в пространстве и во времени. Поэтому в принципе в ма-
териальном мире ни одно событие, ни один процесс не могут
быть полностью изолированы от внешних и внутренних случай-
ных возмущений и описаны однозначно.
В отличие от механического однозначного статистический де-
терминизм присущ процессам, носящим вероятностный характер.
Результаты таких вероятностных процессов не могут быть про-
гнозированы однозначно. Каждое отдельное взаимодействие,
отклады-вающее определенный отпечаток на конечный результат,
является в статистически детерминированном процессе относи-
тельно случайным, однако при достаточно большом числе вза-
имодействующих явлений в результате может быть получен
вполне определенный эффект (В. С. Вишаренко, 1971). Процес-
сы, протекающие в статистически детерминированных системах
с большим числом взаимозаменяемых элементов, будучи веро-
ятностными по форме, тем не менее могут обеспечить получе-
ние достаточно определенного конечного результата. В. С. Ви-
шаренко (1971) предлагает различать среди всего разнообразия
видов статистической детерминации 2 различные формы. Одна
характеризует процессы вероятностные как по своей форме, так
и по результату, другая — процессы вероятностные по форме, нс
близкие к однозначным по результату. При первой форме стати-
стического детерминизма процессы принципиально неопределен-
ны по отношению к вызывающим их факторам. Вторая форма
статистической детерминации наблюдается обычно в иерархиче-
ских процессах, в которых случайные взаимодействия нижеле-
жащего уровня благодаря большому своему числу могут выра-
зиться в относительно однозначном результате. Таким образом,
сущность вероятностного детерминизма заключается в том, что
в статистически детерминированном процессе каждое отдельное
взаимодействие наступает случайно, но при большом числе та-
ких взаимодействий в итоге возникает определенный результат.
По словам Н. Винера (1976), нежесткий статистический харак-
тер связи между элементами системы позволяет уяснить ее из-
6
менчивость, понять, что мир вероятностных отношений — это мир
процесса. Вероятностные системы со статистической детермина-
цией явлений отличаются отсутствием строгой регулярности,
жесткой связанности элементов, жесткой упорядоченности про-
цессов, протекающих в системе. При этом случайной называет-
ся такая переменная системы, которая неопределенным образом
изменяет свое значение, но относительная частота проявления
каждого из возможных значений является достаточно посто-
янной.
Признание статистической детерминации явлений означает,
что вероятность -служит не субъективной характеристикой пред-
метов материального мира, а реальным объективным свойством
систем, в которых протекают эти явления, т. е. вероятность ста-
тистического результата системы — производное специфики са-
мой системы, ее структуры.
Вероятность принадлежит к числу важнейших общенаучных
и многогранных понятий, она может рассматриваться как не-
кая «субкатегория», отображающая существенные аспекты диа-
лектической связи причины и следствия, возможности и дей-
ствительности, случайности и необходимости. Вероятность как
математическая категория (в этом значении она чаще всего ис-
пользуется в различных научных дисциплинах) выступает в ви-
де численной характеристики степени возможности появления
определенного события в тех или иных условиях, т. е. служит
количественной характеристикой объективно существующей свя-
зи между различными событиями универсального множества
возможных событий.
Статистический детерминизм как одна из форм неопределен-
ности явлений окружающего мира -имеет свою определенную
специфику в каждой отрасли науки. Так, в квантовой механике
хорошо известна объективно существующая неопределенность в
виде соотношения неопределенностей В. Гейзенберга (1932).
Принцип Гейзенберга отражает объективную неопределенность
между пространственно-временным и импульсным энергетиче-
ским составляющими микрочастиц, проявляющуюся в такой
зависимости <их друг от друга, когда определенность одних со-
стояний порождает неопределенность других. Полная определен-
ность состояния движения частицы означает совершенную не-
определенность ее положения в пространстве. В работах Гей-
зенберга и Бора по анализу процесса физического измерения
убедительно было показано, что ни одно измерение не может
дать результатов, противоречащих соотношению неопределен-
ностей: состояние движения частицы при измерении оказыва-
лось тем более неопределенным, чем точнее было измерено ее
7
положение в пространстве. Поскольку, как подчеркивал Л. де
Бройль (1963), любую материальную систему можно, с точки
зрения современной физики, свести к набору частиц, то нужно
знать координаты и скорости различных частиц системы в один
и тот же момент времени. Сущность же соотношений неопреде-
ленностей заключается в том, что точное и одновременное зна-
ние этих величин (невозможно.
Опираясь на высказывание В. Гейзенберга (1932): «Закон
причинности в его классической форме может быть определен,
согласно его сути только для замкнутых систем», — некоторые
зарубежные философы пытались представить принцип неопре-
деленности Гейзенберга как отрицание детерминизма, как при-
знание абсолютной свободы пространственно-временной дина-
мики процессов и явлений материального мира. Вскрывая суть
этих заблуждений, Б. М. Кедров (1973) подчеркивал, что не
было бы недоразумения (что неопределенность Гейзенберга
опровергает детерминизм), если бы созданию квантовой меха-
ники (объясняющей процессы микромира) не предшествовал
взгляд, что детерминизм вообще — это лапласовский механиче-
ский, что причинность, закономерность вообще всегда должны
носить характер механический, однозначно определенный. На
необходимость более широкой интерпретации категории причин-
ности указывал и Н. Винер, который писал: «Причинность есть
нечто, могущее присутствовать в большей или меньшей степени,
а не только просто быть или не быть» (Н. Винер, 1967).
Диалектико-материалистическое представление принципа де-
терминизма, как пишет И. Т. Фролов (1973), позволяет понять
однозначные отношения механической однозначной причинной
обусловленности явлений и процессов лишь как один из видов
причинности, возможный в случае искусственной изоляции объ-
ектов от тех условий, в которых они функционируют.
2. ДЕТЕРМИНАЦИЯ ПРОЦЕССОВ
В БИОЛОГИЧЕСКИХ СИСТЕМАХ
Подчеркивая существенные различия в детерминизме физи-
ческих и биологических процессов, известный физик М. Дель-
брок (М. Delbruck, 1949) писал: «...зрелого физика, впервые
сталкивающегося с проблемами биологии, ставит «в тупик то об-
стоятельство, что в биологии нет «абсолютных явлений». Каж-
дое явление представляется иным в разных местах и в разное
время».
Как и в других областях естествознания, в биологии можно
выделить: 1) закономерности, отражающие строго однозначные
8
причинно-следственные отношения рассматриваемых явлений, и
2) (закономерности, проявляющиеся в статистической (взаимосвя-
зи явлений, которые благодаря большому числу причинных, об-
условливающих факторов в совокупности могут определить не-
которую общую тенденцию в развитии процесса (А. А. Мали-
новский, 1973). Хотя в принципе в биологических системах име-
ют место те же 2 типа детерминизма: жесткий, механический
и статистический, вероятностный, последний занимает несрав-
ненно более важное место в живых системах.
Развитие теоретических основ детерминизма я>влений в жи-
вых системах имеет большую историю. Многие примеры при-
чинной обусловленности ряда биологических и физиологических
процессов в свете лапласовского детерминизма мы встречаем в
работах крупнейшего физиолога XIX века К. Бернара. Автор
считал возможным представить все формы развития и реали-
зации функций животного организма на языке господствующего
тогда в физике и механике принципа жесткого лапласовского
детерминизма, случаи же, которые не укладывались в эту схе-
му, связывались с отсутствием соответствующих знаний (К. Бер-
нар, 1896). В последующих иоследованиях было установлено,
что жесткая детерминация действительно имеет место в таких
биологических процессах, где каждый последующий этап опре-
деляется однозначно предшествующим; такими процессами, на-
пример, являются процесс развертывания генотипической ин-
формации или процессы функционирования отдельных физиоло-
гических систем (в их основных проявлениях). Исследование
физиологических механизмов гомеостатичеакой регуляции функ-
ций животного организма, их развития в онтогенезе и передачи
по наследству основных и специфических механизмов функцио-
нирования свидетельствует о том, что без высокой стабильно-
сти, жесткой детерминированности специализированных систем
живого организма было бы невозможным не только его про-
грессивное развитие, но и вообще существование, нормальная
жизнедеятельность животного организма.
По мере развития вероятностных статистических идей, даль-
нейшего накопления экспериментальных данных о специфике в
организации и функционировании животного организма, накоп-
ления большого фактического материала в различных областях
биологии (в особенности в генетике, в эволюционной биологии
и др.) вероятностные понятия, представления об объективном
существовании неопределенности стали применяться и в науках
о живых системах.
Рассматривая различные факторы, обусловливающие нали-
чие неопределенности в сложных биологических системах,
9
Э. Майр (1970) считает необходимым указать на следующие.
1. Исключительная сложность организации живого. В этом
он -солидаризуется с В. Эльзассером (W. Elsasser, 1958), под-
черкивающим, что самой отличительной особенностью живых
организмов является их неограниченная структурная и динами-
ческая сложность. Поскольку специфические закономерности
живого не могут быть выведены из физико-химического взаимо-
действия, Эльзассер приходит к выводу о необходимости посту-
лирования для -биологических систем биотонической причинной
связи явлений. Эта особенность организации структуры и функ-
ции живого делает возможным определить причину возникнове-
ния некоторого явления в -биологической системе как условие
необходимое, но недостаточное, т. е. как одно из множества в
совокупности достаточных условий, без которого данное явле-
ние не могло бы произойти.
2. Неопределенность событий, не связанных с их значением.
В качестве примера можно привести спонтанную мутацию, обус-
ловленную «ошибкой» в репликации ДНК.
3. Уникальность живых систем на высоких уровнях биологи-
ческой организации, ибо в мире уникальных явлений не могут
успешно действовать общие законы, подобные законам, функ-
ционирующим в классической механике. Уникальность биоло-
гических систем рассматривается как основной фактор, опреде-
ляющий различия между биологической и физической науками.
4. Возникновение новых качеств, свойств на высших уровнях
интеграции биологической системы. Хорошо известно, что если
два существа объединяются на более высоком уровне интегра-
ции, то не все свойства нового объединения можно логически
вывести, прогнозировать на основании свойств, составляющих
его компонентов. Это свойство, в какой-то мере идентичное прин-
ципу неаддитивного сложения, в процессе (построения сложных
систем является производным понятия организации. А. Селт-
Дьердьи (4964) организацию называл одним из основных прин-
ципов жизни. Он писал: «... мы понимаем под этим, что при объ-
единении двух вещей рождается нечто новое, качества которого
неаддитивны и не могут быть выражены через качества состав-
ляющих его компонентов».
Э. Майр (1970) признавал существование в органическом
мире двух различных типов процессов, один из которых свой-
ствен системам, обладающим высокой целостностью (клетки,
организмы, колонии), развитие и функционирование которых
происходит по относительно жестким программам. Другой тип
представлен процессами, протекающими в дискретных живых
системах (популяции, виды, биоценозы), имеющими статистиче-
10
ский характер. Эти два различных типа процессов обусловли-
вают в -свою очередь наличие двух соответствующих, не своди-
мых друг к другу форм причинно-следственных отношений, одна
из которых свойственна объектам функциональной биологии,
другая характерна для эволюционных процессов.
Анализируя философские аспекты причинно-следственных
отношений в биологических системах, В. С. Вишаренко (1971,
1976) приходит к .заключению, что детерминизм здесь склады-
вается из одновременного взаимодействия физико-химических
процессов (инфрадетерминация), процессов, факторы взаимо-
действия которых локализованы внутри самих живых систем
(интердетерминизм) и воздействия супра-систем — биосферы
Земли и экосферы Солнца (супрадетерминизм).
Статистический и жесткий типы детерминизма в органиче-
ской природе тесно взаимосвязаны. Неразрывное единство и
взаимодействие статистических и жесткодетерминированных
процессов обеспечивают специфическое единство устойчивости
и изменчивости как основы существования и развития живого
мира. Без статистических процессов естественного отбора в по-
пуляциях животных организмов невозможно было бы создание
новой биологической информации, а без жесткодетерминирован-
ных систем и процессов в животном мире было бы невозможным
закрепление и сохранение полезных для вида и индивидуума
отдельных свойств и качеств живого организма (В. С. Вишарен-
ко, 1975). По сути дела об этом же говорит А. Д. Урсул (1971)\
когда он указывает на глубокую взаимосвязь и взаимообуслов-
ленность неопределенности и определенности в механизмах
функционирования биологических систем: определенность явле-
ний природы существует только на основе их неопределенно-
сти, возникает из нее> неопределенность — это определенность
в ее. становлении.
Как указывает В. С. Вишаренко (1971, 1973), для процес-
сов, протекающих в живых системах, характерен чрезвычайно
тонкий баланс между жесткими и статистическими процессами.
Само взаимодействие жестких и статистических процессов в
биологическом организме -не остается постоянным. В процессе
прогрессивной эволюции животных организмов степень стати-
стичности процессов в надорганизменных' системах может сни-
жаться. Так, по данным К. М. Завадского (1968), у видов, стоя-
щих на низкой ступени эволюции, в процессе естественного от-
бора для выработки какой-либо адаптации требуется гигантское
количество материала. У более высокоорганизованных видов
животных необходимая адаптация достигается в более короткий
11
срок при меньшем значении статистичности (И. И. Шмаль-
гаузен, 1961).
При эмпирическом подходе к познанию закономерностей
функционирования биологических систем, связанных с особен-
ностями детерминизма биологических явлений, важным пред-
ставляется возможность 'предсказания результата этого дина-
мического процесса, возможность прогнозирования его хода раз-
вития. Сравнивая особенности статистической детерминации в
физическом и биологическом эксперименте, Н. В. Тимофеев-Ре-
совский и Р. Р. Ромпе (1959) пишут: «С точки зрения статисти-
ческих закономерностей число идентичных единиц в биологиче-
ских опытах надо считать незначительным. Это ведет к тому,
что в биологии даже в тех случаях, когда математическое ожи-
дание числа прореагировавших элементарных частиц прибли-
жается к единице и когда, кроме того, мы имеем еще дело с
одновременным (протеканием нескольких конкурирующих реак-
ций, результат опыта является практически неопределенным и
не может быть (предсказан с достаточной вероятностью».
Статистический вероятностный характер прогнозирования
биологических процессов отмечает и Э. Майр (1970), считаю-
щий, что, как и во многих других областях науки (если не счи-
тать некоторые основные химические или физические процес-
сы), предсказание биологических явлений почти всегда носит
статистический характер. «Мы, — пишет он, — можем с большой
точностью сказать, что среди 1000 новорожденных будет немно-
гим более 500 мальчиков, но мы -не можем предсказать пол дан-
ного, еще не родившегося ребенка».
Таким образом, причинно-следственные отношения в биоло-
гических системах существенным образом отличаются от при-
чинности в физике и механике. Своеобразие детерминации био-
логических процессов заключается в том, что оно либо вообще
не дает возможности предсказывать, либо в лучшем случае по-
зволяет получить лишь вероятностный прогноз.
3. СТАТИСТИЧЕСКИЕ ЗАКОНОМЕРНОСТИ
В ЭВОЛЮЦИОННОМ ПРОЦЕССЕ
Основные закономерности эволюционного процесса, выявлен-
ные Ч. Дарвином, выражаются в непрерывном приспособлении
организмов к среде, в которой они обитают. Любое эволюцион-
ное изменение приобретается организмом в результате стати-
стических процессов мутагенеза и естественного отбора.
Процессы прогрессивного развития животного мира в значи-
тельной мере содержат элемент статистичности. Хотя вероят-
12
постный, статистический характер закономерностей изменения:
отдельных признаков и свойств живого организма является осно-
вой эволюционного процесса, в последнем важная роль принад-
лежит и жесткодетерминированным механизмам, обеспечиваю-
щим путем реализации жестких генетических программ закреп-
ление результатов такого эволюционного развития и видоизме-
нение функций и структуры организма. Эволюционный процесс
развития и совершенствования живых организмов основан, с од-
ной стороны, на статистических .процессах мутагенеза, с дру-
гой— на реализации жестких генетических программ.
Как справедливо указывал И. И. Шмальгаузен (1968), со-
временные представления об источниках мутаций далеки от
индетерминизма. Сложные проявления приспособительной из-
менчивости в мире живых существ часто нельзя объяснить не-
посредственной детерминацией их внешними условиями, они но-
сят опосредованный характер, учитывающий внутреннюю спе-
цифику организмов. Это означает, что факторы внешней среды
могут дойти до наследственного аппарата лишь через сложную
систему взаимозависимостей в живом организме. Такое опосре-
дование определяет вероятностный характер влияния среды на
организм и проявляется в случайностях мутационного процесса
(И. И. Шмальгаузен, 1968).
Закономерности передачи и преобразования наследственной
информации в свете современной концепции биогеоценоза доста-
точно полно рассмотрены И. И. Шмальгаузеном. В экосистеме
в качестве управляемого объекта рассматривается популяция»,
а в качестве регулятора — биогеоценоз. Поскольку биогеоценоз
играет роль регулятора эволюционного процесса отдельных по-
пуляций животных организмов (по линии прямой связи влия-
ния биогеоценоза на вид), в системе должны присутствовать и
присутствуют каналы обратной связи (для передачи информа-
ции о состоянии популяции). Как подчеркивает И. И. Шмаль-
гаузен (1968), нет прямой связи между наследственной инфор-
мацией по первому каналу (от биогеоценоза к виду) и обратной
информацией по второму каналу (от популяции к биогеоценозу).
Передача информации по этим каналам связи происходит на
разном уровне: наследственная информация передается на вну-
триклеточном (молекулярном) уровне организации, а обрат-
ная—только на уровне организации отдельной особи. В такой
пространственной разнесенности механизмов передачи разно-
родной информации — одна из причин вероятностных законо-
мерностей, характерных для эволюционного процесса.
Наследственная информация преобразуется в процессах ин-
дивидуального развития в средства передачи обратной инфор-
13
мадии в фенотипе особи. В биогеоценозе через (процессы есте-
ственного отбора и процессы размножения -происходит новое
преобразование этой информации © наследственную с переходом
от уровня организации особи (в фенотипе) на уровень органи-
зации клетки (половые клетки, зиготы). Этим замыкается пол-
ный круг преобразований в элементарном цикле эволюционного
процесса (см. схему).
Искажения передачи наследственной информации, происхо-
дящие под влиянием случайных факторов внешней среды (чаще
всего радиационных и химических воздействий), проявляются
в виде дискретных наследственных изменений, носящих случай-
ный характер. Эти изменения известны под названием мутаций.
Мутации иногда возникают без каких-либо видимых причин
только у отдельных особей данного вида. Это следует рассмат-
ривать как следствие индивидуально повышенной чувствитель-
ности данной особи к мутагенным воздействиям (к необычным
стимулам, к необычной интенсивности привычных стимулов —
температуры, влажности и т. д.).
Помехи
I
Передача наследственной информации
через зиготу и клеточное деление
Жизненные
средства
Преобразование ин-
формации в биогео-
ценозе (естественный
отбор и половой про-
цесс)
t
I
Преобразование ин-
формации в индиви-
дуальном развитии
(реализация феноти-
пов)
____________I
Жизненные
средства
Передача обратной информации через фенотипы
(активность особей)
t
Помехи
Общая схема регулирующего механизма эволюции (по И. И. Шмальга-у-
гзену, 1968)
Таким образом, мутации являются причиной изменения жи-
вых существ, зависящих от наследственных свойств организма
и от факторов внешней среды. В эволюционном процессе фак-
торы внешней среды играют роль агентов, освобождающих те-
чение формообразовательных процессов в животном организме.
Прогрессивная эволюция в мире живых существ достигается
благодаря освобождению развивающегося организма из-под
влияния случайных факторов внешней среды. Освобождение ор-
14
ганизма от прямой детерминирующей роли факторов среды вме-
сте с тем означает установление, усиление системы внутренних*,
факторов развития, определяющих специфическое течение фор-
мообразовательных процессов.
Разнообразные проявления мутаций, изменяющих исходные*
признаки и свойства организма в различных направлениях, со-
ставляют основу классической формы «неопределенной измен-
чивости» Ч. Дарвина (Н. В. Тимофеев-Ресовский и др., 1969).
Говоря об отрицательной и положительной роли мутаций в-
эволюционном процессе, И. И. Шмальгаузен (1968) подчерки-
вал, что мутации, в общем виде представляющие собой нару-
шения нормальной организации и нормального течения функ-
ций организма, при бесконтрольном их накоплении всегда при-
водят к распаду организации и дезинтеграции организма. Хотя
никакие помехи как случайные внешние воздействия на орга-
низм сами по себе не являются факторами прогрессивной эво-
люции, мутационные изменения путем естественного отбора при-
нимают определенное участие в эволюционном процессе, спо-
собствуя развитию частных адаптаций.
Конечный положительный результат мутационных измене-
ний структуры и функции организма, статистических по своей
природе, А. А. Малиновский (1973) объясняет следующим обра-
зом. Мутации как результат случайных изменений наследствен-
ного кода, как правило, являются вредными. Но, поскольку име-
ет место определенное (хотя и относительно малое) количество*
полезных мутаций, последние благодаря особенностям живых:
организмов, заключающихся в расширенной репродукции, ока-
зываются способными в случае удачных для естественного от-
бора вытеснить и нормальную форму, и многочисленные вред-
ные мутации. Неудачные мутации, хотя и многочисленны в пла-
не их разнообразия, но каждая из них в отдельности никогда*
не достигает в обычных условиях высокой численности. Полез-
ные же мутации (малочисленные в отношении их разнообразия)
размножившись, занимают преобладающее большинство числен-
ного состава данного вида животного организма. Поэтому есть,
все основания считать, что эволюционный процесс является про-
цессом закономерным, основанным на статистических предпо-
сылках, но протекающих в строго определенном направлении:
повышения видовой жизнеспособности.
Итак, эволюционный процесс является прекрасным приме-
ром, иллюстрирующим возможность получения вполне опреде-
ленного конечного результата в сложной системе, характери-
зуемой высоким удельным весом статистических вероятностных?
взаимоотношений между составляющими ее компонентами.
15
4. ДИНАМИЧЕСКИЙ ХАРАКТЕР АДАПТИВНЫХ
ИЗМЕНЕНИЙ ФУНКЦИЙ ОРГАНИЗМА
Адаптивное поведение организма, приспособление его к окру-
жающим условиям среды, обеспечиваемое соответствующими
реакциями физиологических систем, определяется характером
влияний внешних факторов среды. Против неблагоприятных
факторов среды организм вырабатывает средства массивной или
активной защиты. Благоприятные факторы среды способствуют,
облегчают реализацию биологических функций организма.
Для изучения механизмов взаимодействия организма и сре-
ды в экспериментальной физиологии используется классическая
методика «стимул — реакция». В детерминированном процессе
при постоянстве стимула будет иметь место и постоянство ре-
акций. Однако в многочисленных экспериментах установлено,
что реакции физиологических систем организма даже при иден-
тичных стимулах не остаются постоянными, а носят вероятност-
ный характер. В ответ на идентичные стимулы отдельные пара-
метры реакции (латентный период, интенсивность, форма реак-
ции) варьируют в широком диапазоне. Такая вариабельность
индивидуальных реакций отдельных компонентов физиологиче-
ских систем лишь на первый взгляд представляется контрасти-
рующей со значительной устойчивостью, стабильностью физио-
логически важных констант организма (артериального давле-
ния, содержания сахара в крови, содержания СО2 и О2 в крови
и т. д.). Одна из возможных причин вероятностных характери-
стик элементарных реакций организма заключается в особен-
ностях физиологического обеспечения регуляторных механизмов
в животном организме. Это прежде всего наличие множества
параллельно функционирующих контуров регуляции каждой от-
дельной физиологической функции или физиологически важной
константы. Например, функция теплопродукции в организме
обеспечивается процессами окисления в мышцах (при движе-
нии) и во внутренних органах. Функция теплоотдачи обеспечи-
вается изменением величины поверхности тела, учащением ды-
хания, потоотделением, изменением интенсивности кроветока в
коже. Наличие многих факторов, регулирующих эти два проти-
воположно направленных процесса регулирования температуры
тела, обусловливает возможность достижения одного и того же
конечного результата за счет различных их комбинаций, напри-
мер, при определенном фиксированном уровне процессов окис-
ления в мышцах один и тот же конечный итог в регуляции тем-
пературы тела организма может достигаться один раз преиму-
щественной активацией дыхания, в другой раз — препмуществен-
16
ным -изменением интенсивности кроветока в коже и т. д. И то-
гда становится понятным, что если за (параметр, по которому
оценивается влияние того или иного внешнего воздействия на
организм, выбрать частоту дыхательных движений, очевиден ва-
риабельный характер эффекта -идентичных стимулов.
К такому же выводу о вероятностном характере реакций от-
дельных компонентов физиологических 'систем организма можно
прийти и при анализе влияний факторов среды, действующих на
систему углеводного обмена в организме (по параметру содер-
жания сахара в крови). Повышение количества глюкозы в крови
достигается усилением процесса всасывания сахара в тонком
кишечнике, ферментативным распадом гликогена до глюкозы
(процессы гликогенолиза), образованием сахара из белков и
жиров. Противоположный эффект имеют распад глюкозы (гли-
колиз) в мышцах и нервной ткани в процессе их работы, фер-
ментативный синтез гликогена из глюкозы, образование жиров
из глюкозы. В условиях такой многофакторной регуляции двух
•противоположно направленных процессов повышения и пониже-
ния содержания сахара в крови физиологически константное
значение его в крови (на уровне 90—ПО мг %) может обеспе-
чиваться различными комбинациями и разной выраженностью
отдельных компонентов многозвенного регуляторного механизма
углеводного обмена в животном организме. Для физиологиче-
ского организма важен конечный результат, способы же его до-
стижения могут быть весьма различными -за счет значительной
вариабельности функциональных параметров отдельных физио-
логических систем организма. В этом находит свое выражение
широко распространенный в мире живых систем принцип экви-
финальности, принцип, который, по мнению Л. Берталанффи,
основоположника общей теории систем, присущ всем сложным
системам с иерархическим принципом организации.
Эквифинальность в различных проявлениях жизнедеятельно-
сти живых существ обусловлена термодинамическими особенно-
стями животного организма как открытой многокомпонентной
сложной системы». Открытой называется такая система, которая
обменивается с внешней средой и веществом и энергией (закры-
тая система в процессе взаимодействия со средой обменивается
только' энергией, а замкнутая, изолированная система не обме-
нивается ни веществом, ни энергией). Все открытые системы (а
они составляют большинство), как отмечает Л. Берталанффи,
являются эквифинальными, способными из различных началь-
ных положений переходить в одинаковое конечное состояние, в
противоположность замкнутым системам, у которых конечное
состояние однозначно определяется начальным (каждому конеч-
17
ному состоянию соответствует свое начальное состояние).
В соответствии со вторым законом термодинамики состоя-
ние системы характеризуется энтропией — мерой неопределен-
ности (S):
S=k InW,
где к — постоянная Больцмана;
W — термодинамическая вероятность системы.
Согласно этому закону, в замкнутой системе энтропия мо-
жет либо оставаться неизменной (при обратимых реакциях),,
либо возрастать (в случае необратимых реакций). Из приведен-
ного выше уравнения видно, что изолированная система само-
произвольно переходит .из менее вероятного в более вероятное
состояние, так как при этом энтропия системы возрастает.
В закрытой системе (обменивающейся со средой только энер-
гией) ход реакций ограничен количеством участвующих в ней
веществ, и по окончании реакции устанавливается постоянство
системы во времени (И. Пригожин, 1960). Напротив, в откры-
той системе — биологическом организме, в котором происходит
непрерывный обмен со средой не только энергией, но и веще-
ством, термодинамическое равновесие проявляется в некотором
стационарном состоянии системы (типа динамического равнове-
сия), характеризующемся тем, что скорости химических реак-
ций и диффузии метаболитов постоянны. В стационарном состоя-
нии энтропия открытой системы остается постоянной (но не
принимает максимально возможное значение). Постоянство
энтропии в этом случае поддерживается за счет того, что систе-
ма непрерывно извлекает свободную энергию из среды. И. При-
гожин (1960) характеризует динамику энтропии в открытой си-
стеме следующим образом:
dS dSj . dSe
"dt dt ~dT ’
dSe
где — характеристика обмена энтропией между биологиче-
ской системой и окружающей средой;
dSj
— —характеристика энтропии необратимых процессов*,
внутри -самой системы.
По мере приближения необратимых процессов к стационар-
dS
ному состоянию величина — стремится .к минимально возмож-
ному при данных условиях положительному значению.
В биохимической основе клеточных механизмов функциони-
рования физиологических систем животного организма важное
18
место занимают диссипативные структуры, характеризующиеся
различным отклонением от равновесного состояния (I. Prigo-
gine е. а., 1972). Они наблюдаются в процессах гликолиза, син-
теза белков в .клетке и т. д. Диссипативные структуры по-раз-
ному ведут себя в зависимости от того, насколько они близки
к состоянию (к точке) термодинамического равновесия. Вблизи
термодинамического равновесия диссипативные структуры* стре-
мятся к максимальной неупорядоченности, вдали от него — пре-
обладает стремление к организованности. При этом в окрест-
ностях точки термодинамического равновесия поведение систе-
мы жестко детерминировано (малые флюктуации затухают),
вдали от нее — является стохастическим (конечное состояние в
сильной мере зависит от вероятности образования флюктуаций
данного типа).
Образование диссипативных структур и их роль в протека-
нии физиологических процессов обмена веществ означает но-
вый принцип организации — «организацию через флюктуации».
Такие структуры могут служить хорошей иллюстрацией слож-
ной формы причинной обусловленности явлений — вероятност-
ного детерминизма на уровне обменнохимических реакций в жи-
вых .клетках. Вместе с тем организация через флюктуации во
многом напоминает широко распространенный метод регулиро-
вания физиологических функций животного организма, где в ка-
честве регулируемых величин выступают такие физиологиче-
ские константы, как артериальное давление, осмотическое дав-
ление, pH крови и т. д. Физиологический механизм такой регу-
ляции, поддерживания численных значений этих констант в до-
пустимых пределах функциональной нормы, известный под на-
званием гомеостаза—стабилизирующей1 саморегуляции функций,
.активируется благодаря наступающим флюктуациям жизненно
важных параметров физиологических систем организма, улав-
ливаемым, воспринимаемым специализированными рецепторны-
ми образованиями и вызывающим комплекс реакций, направ-
ленных на восстановление исходных значений этих физиологи-
ческих констант.
Рассматривая биологический процесс эмбриогенеза,
К С. Тринчер (1964) приходит к выводу, что суть теоремы При-
гожина (открытая термодинамическая система стремится к
устойчивому стационарному состоянию) не может быть приме-
нена к эмбриональному периоду развития животного организма,
характеризуемому непрерывным нарастанием энтропии. Рост
удельного производства энтропии в ходе эмбриогенеза служит
энергетическим проявлением развития животного мира, форми-
рующего все более сложные органы для выполнения целесооб-
19
разных специфических функций. При этом возрастание энтро-
пии в непосредственном окружении организма связано с интен-
сивным возрастанием отрицательной энтропии (негэнтропии) в
самом организме, служащей мерой структурного усложнения
и дифференцировки живой системы.
Постоянный обмен веществ как обязательный элемент жиз-
ни приводит .к непрерывному росту энтропии вне организма. Во
взаимодействующей -системе «живой организм — внешняя ере-
да» такой процесс роста энтропии вне организма сопровождает-
ся возникновением негэнтропии внутри его (Э. Шредингер,
1947). Энтропийная стационарность открытых систем — живот-
ных организмов (и их подсистем) — обеспечивается равенством
скорости возникновения энтропии в результате процессов обме-
на веществ и скорости возникновения негэнтропии. Иначе го-
воря, стабильность животного организма достигается благодаря
негэнтропийным процессам в результате расхода макроэргических
высокоструктурированных пищевых веществ, потребляемых жи-
вотными. Процесс жизнедеятельности биологического организ-
ма есть негэнтропийный процесс структурообразования за счет
химической энергии и низкой энтропии высокоструктурирован-
ных макроэргических соединений пищевых веществ, поступаю-
щих в организм.
Таким образом, животный организм представляет собой от-
крытую термодинамическую систему, преобразующую потенци-
альную химическую энергию пищевых веществ в энергию био-
логических процессов, протекающих в организме, и выделяю-
щую из организма во внешнюю среду некоторое количество
энергии в виде тепла. Поэтому состояние биологического орга-
низма можно охарактеризовать количеством негэнтропии, име-
ющей противоположный смысл, чем энтропия, определяющей
степень упорядоченности биологической системы
1 = — klnW6 = kin—,
6 w6
где I — негэнтропия системы;
Wo — термодинамическая вероятность биологического орга-
низма.
С уменьшением во времени величины Wo растет величина
1/Wo, а это означает увеличение степени упорядоченности -струк-
туры животного организма в ходе биологической эволюции.
Важнейшей специфической особенностью организации и дея-
тельности живых организмов является то обстоятельство, что
внешняя энергия (энергия, поступающая в организм) прежде
чем трансформируется в работу, энергетическую деятельность
20
животных, предварительно превращается в энергию структуры
организма, определяя неравновесное его состояние. Деятельность
всех живых систем, а следовательно, и физиологических систем
животного организма в любых условиях среды 'направлена про-
тив равновесия, которое должно1 было бы наступить в данной
среде три данном первоначальном состоянии системы. Всеоб-
щий закон биологии обычно формулируется так (Э. Бауэрг
1935): «все и только живые системы никогда не бывают в рав-
новесии и исполняют за счет своей -свободной энергии постоян-
ную работу против равновесия, требуемого законами физики и
химии при существующих внешних условиях» (принцип устой-
чивого неравновесия Бауэра).
Хорошо извеоно, что почти все физиологические константы
организма по сути дела не являются, строго говоря, констант-
ными величинами, а подвержены определенным колебаниям
в некоторых физиологических пределах. Это привело к тому,
что известная концепция Кэннона о гомеостазе (W. В. Каппой,
1932) в последнее время была дополнена понятием гомеокине-
тической регуляции (У. Мак-Каллок и А. С. Айбералл, 1967).
Сущность представления о гомеокинетическом регулировании
физиологических функций заключается в утверждении, что ор-
ганизм в процессе своей жизнедеятельности стремится не к
абсолютной стабилизации физиологических констант, а к дина-
мической их регуляции, к удержанию их в определенных преде-
лах при наличии непрерывных небольших колебаний регулиру-
емого параметра. Эту концепцию успешно развивает у нас в
стране В. А. Шидловский с сотрудниками (В. А. Шидловский,
В. <Н. Новосельцев, 1973; В. А. Шидловский, 1978). Авторы со-
вершенно справедливо считают ошибочным предположение о
том, что устойчивость целого организма как сложной мульти-
параметрической системы должна иметь в своей основе высокую
устойчивость каждой составляющей организм физиологической
системы, стабилизацию каждой физиологической функции в от-
дельности. Сложная неравновесная система организма имеет
гораздо большие возможности обеспечения конечного устойчи-
вого* состояния, чем жесткая стабилизация составляющих его
элементов.
Гомеостаз (точнее — гомеокинез) целостного организма обес-
печивается путем мультипараметрической регуляции, не требу-
ющей устойчивости каждого параметра в отдельности. Для жи-
вотного организма обязательным является лишь требование ста-
бильности лишь конечного результата деятельности системы. В
качестве примера, иллюстрирующего это представление,
В. А. Шидловский рассматривает перестройки вегетативных
21
.функций организма, обеспечивающих приспособительные реак-
ции организма 1на «различные воздействия внешней среды. При-
способительные изменения вегетативных функций в формиро-
вании адаптивного поведения животного организма при этом со-
ставляют промежуточное звено между метаболическими процес-
сами организма, его «метаболическим котлом» и внешней сре-
дой. Отсюда и значение комплексного изменения метаболиче-
ских функций, связанных с обеспечением необходимого уровня
тканевого дыхания (изменение внешнего дыхания, работы сердца
и органов кровообращения). Соответствующие изменения много-
связной респираторно-гемодинамической системы обеспечивают
необходимый уровень тканевого дыхания за счет транспорта га-
зов из внешней среды к клеточным элементам. Такие приспосо-
бительные реакции вегетативных функций могут достигаться
разными формами взаимно-согласованных изменений функции
внешнего дыхания, обмена газов в альвеолах, дыхательной функ-
ции крови, центрального и периферического кровообращения и
микроциркуляции.
Поддерживание интенсивности тканевого дыхания на опре-
деленном уровне не требует обязательной стабилизации частоты
и глубины дыхательных движений организма, количества эри-
троцитов в крови, содержания гемоглобина в крови и т. д., хотя
каждый из этих факторов, взятый в отдельности, оказывает весь-
ма существенное влияние на уровень тканевого обмена в орга-
низме. Потребности организма в кислороде могут быть удов-
летворены при самых разных количественных взаимоотношениях
между параметрами внешнего дыхания, гемодинамики и дыха-
тельной функции крови. В этом и заключается сущность мульти-
параметрической регуляции уровня кислородного снабжения тка-
ней организма, обусловливающей значительные отклонения, ва-
риации в стабилизирующей регуляции каждого из этих пара-
метров в отдельности. Как подчеркивают В. А. Шидловскнй и
В. Н. Новосельцев (1973), животный организм не стремится
жестко соблюдать физиологические константы, а легко сдвигает
их в процессе мультипараметрического регулирования. Это про-
исходит в интересах организма, функционирование которого
определяется не отдельными параметрами, а скоординированной
деятельностью функциональных систем, обеспечивающей необ-
ходимое перераспределение физиологических констант соответ-
ственно условиям взаимодействия организма с внешней средой.
Многосвязное (мультипараметрическое) регулирование означа-
ет, что в соответствующей функциональной системе конечный
адаптивный результат представлен множеством составляющих
его параметров.
22
Интересны в этом отношении попытки объяснения стабильно-
сти конечного результата в сложной иерархической структуре
животного организма с позиции теории функциональной системы
П. К. Анохина (1970, 1971) как следствие механизмов саморе-
гуляции целостного организма. Так, в работе В. А. Полянцева
(1973) утверждается, что при экспериментальном анализе, про-
веденном на электронной модели системы, удовлетворяющей
требованиям к функциональной системе, можно показать, что
стабильный по своим параметрам результат соответствует пуас-
соновскому потоку. Это рассматривается как достижение детер-
минированного результата случайным потоком действия.
Функциональная система 'представляет собой динамически
формирующуюся организацию, избирательно объединяющую
разнородные центральные и периферические аппараты для по-
лучения полезного для организма в данный момент результата.
В соответствии с системным представлением одна и та же нерв-
ная структура может принимать участие в реализации разных
функций. В этом можно усмотреть определенную преемствен-
ность концепции функциональной системы П. К. Анохина при-
менительно к структурам мозга и (представления И. Н. Фили-
монова (1949) о мультифункциональности структур старой и
новой коры мозга.
В исследованиях О. С. Андрианова (1977) о роли пусковых и
исполнительных механизмов условнорефлекторного поведения
животных шолучены новые экспериментальные данные, свиде-
тельствующие о значительном динамизме процессов мобилиза-
ции определенных структур мозга в реализации одного и того
же эффекта. Из этих результатов делается заключение о том,
что организация каждого из узловых этапов функциональной
системы, лежащей в основе сложного приспособительного пове-
дения, не носит характера жесткой приуроченности этих этапов
к какой-либо одной структуре мозга. Различные нейронные ан-
самбли неравной системы могут участвовать в реализации не од-
ного, а нескольких механизмов системной деятельности на по-
веденческом уровне, и степень участия той или иной ансамбле-
вой структуры в каждом из этапов системных /процессов явля-
ется различной, изменчивой по мере образования и закрепления
адаптивных поведенческих навыков.
5. СТАТИСТИЧЕСКИЙ ХАРАКТЕР НОРМЫ И ПАТОЛОГИИ
Вероятностные проявления физиологических систем организ-
ма в значительной степени определяются тем обстоятельством,
что функционирование отдельных органов, тканей протекает, как
23
правило, в некотором достаточно широком диапазоне колебаний
основных (параметров, изменения в пределах которых не ска-
зываются отрицательно в конечном результате адаптивного по-
ведения организма. Этот рабочий диапазон колебаний, откло-
нений отдельных функциональных параметров от их среднеста-
тистической .величины известен *в медицине и ib ее теоретической
основе — биологии — в виде понятия нормы.
В истории развития медицины вопросы о норме и патологии,
о границах между «нормой» и «ненормой», о содержании поня-
тия нормы всегда были предметом оживленных дискуссий, в
ходе которых выдвигались и обосновывались крайне противо-
положные представления (как полностью отрицающие объек-
тивную основу этого понятия, так и рассматривающие норму
как механическую среднестатистическую величину анализиру-
емых параметров без учета функциональных особенностей ор-
ганизма, изменений внешних условий, адаптивного приспособи-
тельного изменения целостного организма и отдельных его фи-
зиологических систем). Эти опоры не утихли и в настоящее вре-
мя. В соответствующей медицинской и биологической литерату-
ре в последнее время появился ряд работ монографического пла-
на, в которых рассматриваются различные аспекты проблемы
нормы и патологии, включая и гносеологические корни идеали-
стических, субъективных представлений о норме (Р. Уильямс,
I960; Г. И. Царегородцев, 1963; И. В. Давыдовский, 1962;
А. Д. Адо, Г. И. Царегородцев, 1970; А. Д. Степанов, 1975;
А. А. Корольков, В. П. Петленко, 1977; и др.).
Идеалистические субъективные .представления о норме сво-
дятся к утверждению, что это понятие является лишь резуль-
татом единого общепринятого мнения ученых о допустимых пре-
делах отклонений физиологических параметров, наблюдаемых
при функционировании животного организма. С этой точки зре-
ния объективной оценки понятия нормы просто не существует и
•не может существовать (Д. Мелмент, 1948; L. King, 1954; и др.).
Так, например, Д. Мелмент утверждает, что никакая статистика
не в состоянии отделить границы нормальных и патологических
процессов в организме.
Материалистический подход к анализу проблемы основан на
представлении, что норма характеризуется определенными де-
маркационными границами, в пределах которых имеющие мес-
то различные колебания изучаемых параметров не приводят к
качественным изменениям ни в морфологическом, ни в физиоло-
гическом состоянии организма или отдельных его частей. «Нор-
ма,—как пишут А. Д. Адо, Г. И. Царегородцев (1970), —гар-
моническая совокупность структурно-функциональных данных
24
организма, адекватных окружающей его среде и обеспечива-
ющих организму оптимальную жизнедеятельность». При таком
подходе к проблеме становится возможным понятие нормы на-
делить объективным содержанием — определенным уровнем ре-
ально наблюдаемого функционального состояния организма,
применительно <к которому и оценивать количественные харак-
теристики изучаемых параметров.
Критикуя подход к проблеме научного определения термина
нормы, широко распространенного в различных науках, вклю-
чая и социологические, Э. В. Ильенков (1968) весьма образно
писал: «О нормальных человеческих условиях развития гово-
рить можно и нужно точно так же, как можно и нужно говорить
о «норме» в медицине и нейрофизиологии. И хоть «норму»
эту нелегко установить и там, и тут, хоть границы между «нор-
мой» и ее нарушением трудно прочертить с такой же строгостью,
с какой ее можно прочертить между «гением» и «идиотом», нор-
ма эта все же есть. Иначе ни к чему была бы медицина и со-
циальные науки тоже. И не надо... увиливать от этого обстоя-
тельства разговорами об «относительности» всяких норм. От-
носительность относительностью, а здоровый человек все же от-
личается от больного».
Справедливо стараясь связать понятие нормы с функцио-
нальным состоянием, точнее с функциональным назначением
нормальных величин функциональных показателей, некоторые
авторы (Г. И. Царегородцев, 1963; А. А. Корольков, В. П. Пет-
ленко, 1977) считают наиболее рациональным определение нор-
мы живых организмов с точки зрения функционального опти-
мума. В целом положительно оценивая такой подход, тем не ме-
нее следует заметить, что и он не лишен некоторых недостатков.
Так, например, повышение артериального давления до 159—170
мм рт. ст. при атеросклеротических поражениях сосудов, явля-
ющееся для данного функционального состояния организма оп-
тимальным, носит компенсаторный характер, обеспечивает преж-
ний нормальный уровень метаболизма в тканях (для проталки-
вания крови через суженный просвет сосудов требуется более
высокое давление). Однако такие величины артериального дав-
ления, естественно, не могут считаться нормальными.
Аналогичное положение имеет место и с многими другими
физиологическими константами, значительные отклонения ко-
торых в ту или другую сторону могут носить компенсаторный
характер и поэтому должны рассматриваться как оптимальные,
хотя количественные значения их при этом могут выходить да-
леко за пределы физиологической нормы.
Поскольку в плане обмена энергией животные организмы ха-
25
рактеризуются способностью сохранять неравновесное состояние
с окружающей средой, важнейшей функцией, лежащей в основе
жизнедеятельности животного организма, следует считать обе-
спечение этого неравновесного состояния. В неравновесном со-
стоянии живых систем флюктуирующим показателям нормы при-
надлежит важная роль как индикатора динамического характе-
ра нормопоказателей, так и неотъемлемого элемента в механиз-
мах автоматической саморегуляции — начального фактора, за-
пускающего систему регуляторных гомеостатических механиз-
мов.
Физиологические колебания среднестатистической величины
нормы для многих показателей имеют достаточно широкие пре-
делы. В табл. 1 приведен диапазон колебаний некоторых неор-
ганических компонентов в кропи человека.
Таблица 1
Колебания содержания некоторых неорганических ионов
в крови человека (по Е. С. Albritton, 1952)
Элемент Источник Границы колебаний, мг-экв/1000 мл
Натрий Цельная кровь Сыворотка Форменные элементы 72—91 132—144 8,7—28,6
Калий Цельная кровь Сыворотка Форменные элементы 39—62 3,6—4,8 89—101
Кальций Плазма 4,3—5,2
Хлориды Цельная кровь 71—87
Диапазон колебаний количественных характеристик многих
органических и неорганических компонентов крови у человека
приводится в работе Р. Уильямса (1960), посвященной изложе-
нию проблемы биохимической индивидуальности. Приведенные
автором данные свидетельствуют о весьма значительном разма-
хе колебаний этих величин (в несколько раз). Анализ материала
привел автора к выводу, что практически каждый человек пред-
ставляет собой в том или ином отношении отклонение от нор-
мы. Выдвинутое Р. Уильямсом положение о значительных инди-
видуальных различиях организма — сложной системы со мно-
жеством составляющих ее подсистем и компонентов — обосно-
вывается наряду с большим количеством фактического матери-
ала также и следующими расчетами. Если, например, 0,95 попу-
ляции «нормально» в отношении какого-либо одного измеряе-
26
мого признака, то только 0,902^ (0,95)2 популяции будет «нор*
мальным в отношении обоих измеряемых признаков, а в отно-
шении 10 и 100 независимых признаков—только 0,60^ (0,95)10
и 0,0059^ (0,95) 10°. Поэтому уловить и однозначно определить
границу между нормой и патологией часто очень трудно и на ос*
нове оценки лишь одной характеристики, одного параметра в
отрыве от комплекса других не менее важных 'параметров прак-
тически невозможно. Научно обоснованное определение границы
между нормой и патологией возможно лишь при системном комп-
лексном подходе к оценке множества функциональных пара-
метров живого организма на основе вычисления некоторой сред-
нестатистической величины «нормы», учитывающей индивиду-
альные колебания отдельных физиологических констант.
Весьма часто значительные вариации наблюдаются не только
в самих функциональных параметрах, но и морфологических
характеристиках обеспечивающих их структур. Так, например,
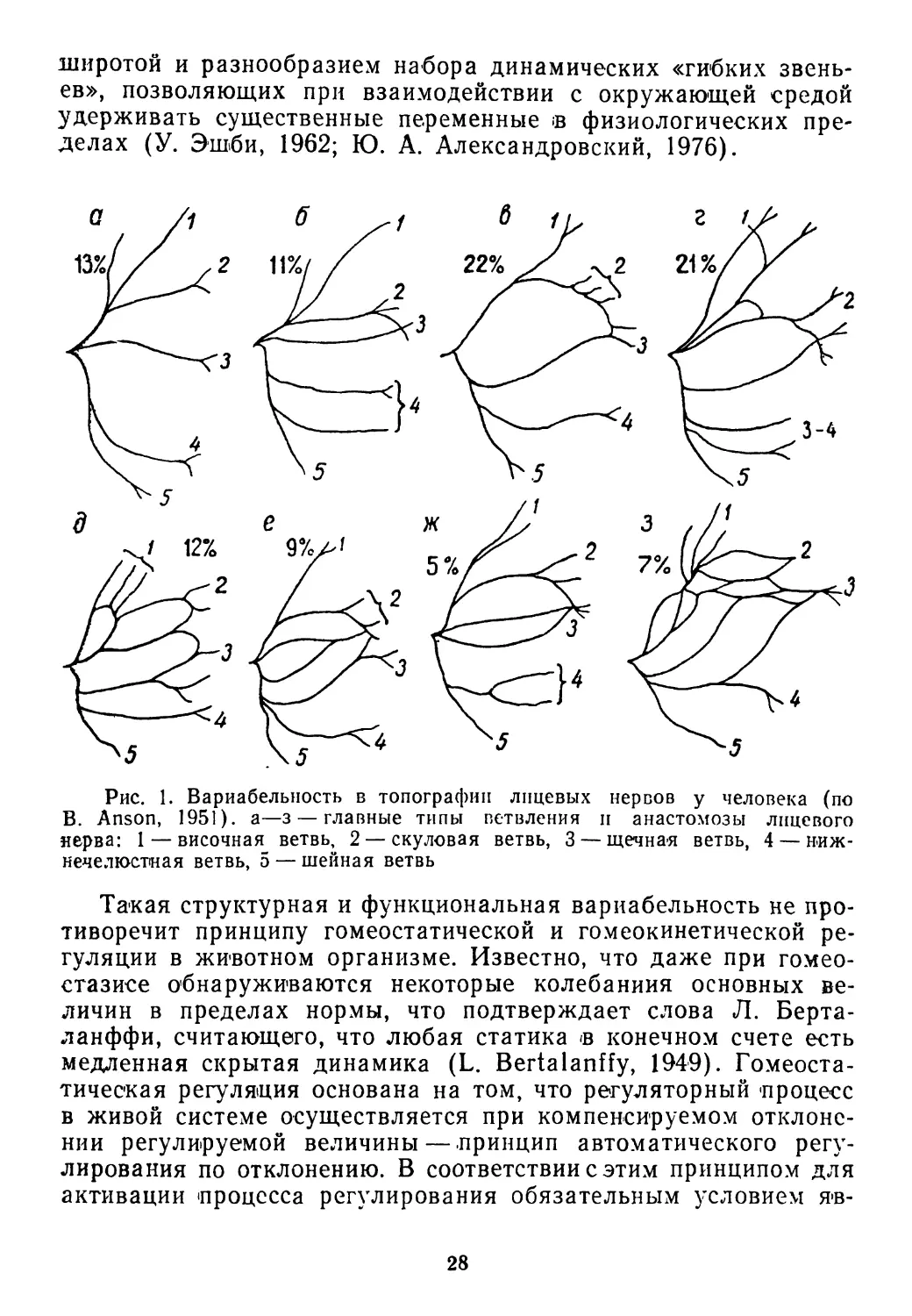
в атласе Б. Энсона (В. Anson. 1951) приводится изображение 8
различных типов расположения лицевых нервов, встречающихся
у человека в норме (рис. 1). Как и в отношении других анато-
мических структур (костной, мышечной, сосудистой систем), ав-
тор не находит какого-либо единого типа строения. Значитель-
ной морфологической изменчивости подвержены и другие нерв-
ные структуры: не существует стандартной нижней границы
спинного мозга, широко варьирует расположение места вхожде-
ния различных нервов в спинной мозг. Примерно у 15% людей
вообще отсутствует прямой пирамидный тракт в спинном моз-
гу (Р. Уильямс, I960).
В работе У. X. Растеда (W. Н. Rustad, 1954) по исследованию
топографии и структуры возвратных нервов гортани на 100 тру-
пах было показано, что в 57% случаев нерв входит в гортань
без всяких разветвлений, а в 43% ствол нерва разветвляется до
вхождения в гортань на несколько ветвей (от 2 до 6). У 35%
трупов оба возвратных нерва были одноствольными, у 24% оба
нерва имели разделенный ствол.
Анализ нейрофизиологических механизмов психических явле-
ний позволил Н. П. Бехтеревой (1971) высказать предположе-
ние, что в основе реализации психической деятельности челове-
ка лежит сложная взаимно координированная работа корково-
подкорковой структурно-функциональной системы со звеньями
различной степени жесткости. По мнению автора, увеличение
количества «гибких звеньев» в системе реализации психической
деятельности является основным принципом функционального
усложнения рабочих конструкций мозга. Возможности адапта-
ционного поведения человека в значительной мере определяются
27
широтой и разнообразием набора динамических «гибких звень-
ев», позволяющих при взаимодействии с окружающей средой
удерживать существенные переменные ® физиологических пре-
делах (У. Эшби, 1962; Ю. А. Александровский, 1976).
Рис. 1. Вариабельность в топографии лицевых нервов у человека (по
В. Anson, 1951). а—з — главные типы ветвления и анастомозы лицевого
нерва: 1 — височная ветвь, 2 — скуловая ветвь, 3 — щечная ветвь, 4 — ниж-
нечелюстная ветвь, 5 — шейная ветвь
Такая структурная и функциональная вариабельность не про-
тиворечит принципу гомеостатической и гомеокинетической ре-
гуляции в животном организме. Известно, что даже при гомео-
стазисе обнаруживаются некоторые колебаниия основных ве-
личин в пределах нормы, что подтверждает слова Л. Берта-
ланффи, считающего, что любая статика <в конечном счете есть
медленная скрытая динамика (L. Bertalanffy, 1949). Гомеоста-
тическая регуляция основана на том, что регуляторный -процесс
в живой системе осуществляется при компенсируемом отклоне-
нии регулируемой величины — -принцип автоматического регу-
лирования по отклонению. В соответствии с этим принципом для
активации процесса регулирования обязательным условием яв-
28
ляется начальное отклонение регулируемой величины под влия-
нием сигналов среды. По этому поводу П. К. Анохин (1962) пи-
сал, что само отклонение от конечного приспособительного эф-
фекта служит стимулом возвращения, достижения системой это-
го эффекта.
В плане приспособления организма к окружающей среде
амплитуда колебаний физиологических параметров организма
и его подсистем является важным элементом обеспечения
устойчивости нормальной деятельности организма. На это обра-
щал внимание еще Н. Винер (1958), который писал, что, с точ-
ки зрения кибернетики, любая система, в том числе и биологи-
ческая, может устойчиво функционировать только благодаря от-
сутствию определяющих ее жестких рамок.
Нарушение регуляторного процесса в ряде случаев приводит к
патологическому состоянию организма, проявляющемуся в раз-
личных типах фиксированных стереотипных реакций (Г. В. За-
левский, М. С. Роговин, 1970).
Стереотипия — это крайняя форма ригидности, неспособ-
ности изменить поведение, патологическое возвращающееся,
повторяющееся движение, поза, .произнесенное слово или фраза,
имеющие мало общего со стимулом или реальной ситуацией
(Н. В. English, А. С. English, 1959; W. J. Coville, цит. по:
Г. В. Залевский, М. С. Роговин, 1970).
Характеристика нормы реакции организма на внешние сти-
мулы в определенной мере связана и с генетическими особен-
ностями развития, формирования свойства реактивности биоло-
гической системы. Как подчеркивает М. Е. Лобанов (1956), фе-
нотипичесткая изменчивость животного организма в процессе
индивидуального развития в значительной мере коррегируется
рамками, границами нормы реакции, определяемой генотипом
индивидуума. В данном случае под нормой реакции подразуме-
вается способность генотипа обеспечивать ограниченную измен-
чивость онтогенетических процессов под влиянием различных
стимулов внешней среды. Поскольку норма реакции изменяется
в генетически детерминированном диапазоне, по словам
И. И. Шмальгаузена (1964), генотип — это унаследованная нор-
ма реакции, определяющая число возможных фенотипов при
различных условиях среды. Таким образом,, норма — это рабо-
чий диапазон функционирования живых систем, детерминиро-
ванный как генотипом (по сравнительно жестким однозначным
законам), так и фенотипом (по статистическим, вероятностным
закономерностям). Чем шире раздвинуты эти границы, тем, оче-
видно, и больше возможностей для адаптивного поведения жи-
вотного организма.
29
В науке должно искать идеи. Нет идеи — нет
науки. Знание фактов только потому и
драгоценно, что в фактах скрываются идеи:
факты без идей — сор для головы и памяти.
В. Г. Белинский
Глава II. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ
ФУНКЦИОНИРОВАНИЯ НЕРВНОЙ СИСТЕМЫ
Среди физиологических систем нервная система за-
нимает особое положение, так как она объединяет всех
их в организм как единое целое и устанавливает его
взаимоотношения с окружающим миром. Она осуще-
ствляет непрерывную обработку информации о текущих
событиях, оценку их значения для своей жизни, приня-
тие решений о необходимых в данной ситуации дейст-
виях и гибко управляет этими действиями. Ее роль ин-
формационно-управляющей системы организма наложи-
ла особый отпечаток на строение и функции нервных
механизмов, определила замечательные свойства их ис-
ключительной надежности, чрезвычайной .пластичности,
высокой эффективности и сделала нервную систему уни-
кальной по своей функциональной организации. Эта уни-
кальность в значительной степени связана с требования-
ми приспособления к стохастичности окружения, в ко-
тором надо было поддерживать свое существование.
Эволюция мира животных происходила в условиях
бесконечно разнообразных, непрерывно изменяющихся
факторов среды. В сложном изменчивом суровом мире
жизненно важные события нельзя предсказывать одно-
значно, а можно лишь ожидать их наступления с боль-
шей или меньшей вероятностью. Поэтому в процессе
естественного отбора преимущества получили организмы,
способные строить свое поведение с учетом этих вероят-
ностей.
1. ВЫЖИВАНИЕ В СТОХАСТИЧЕСКОЙ СРЕДЕ
Чтобы выжить в условиях чрезвычайного разнообра-
зия, часто гибельных непредсказуемых воздействий
окружающего мира, организм должен обладать способ-
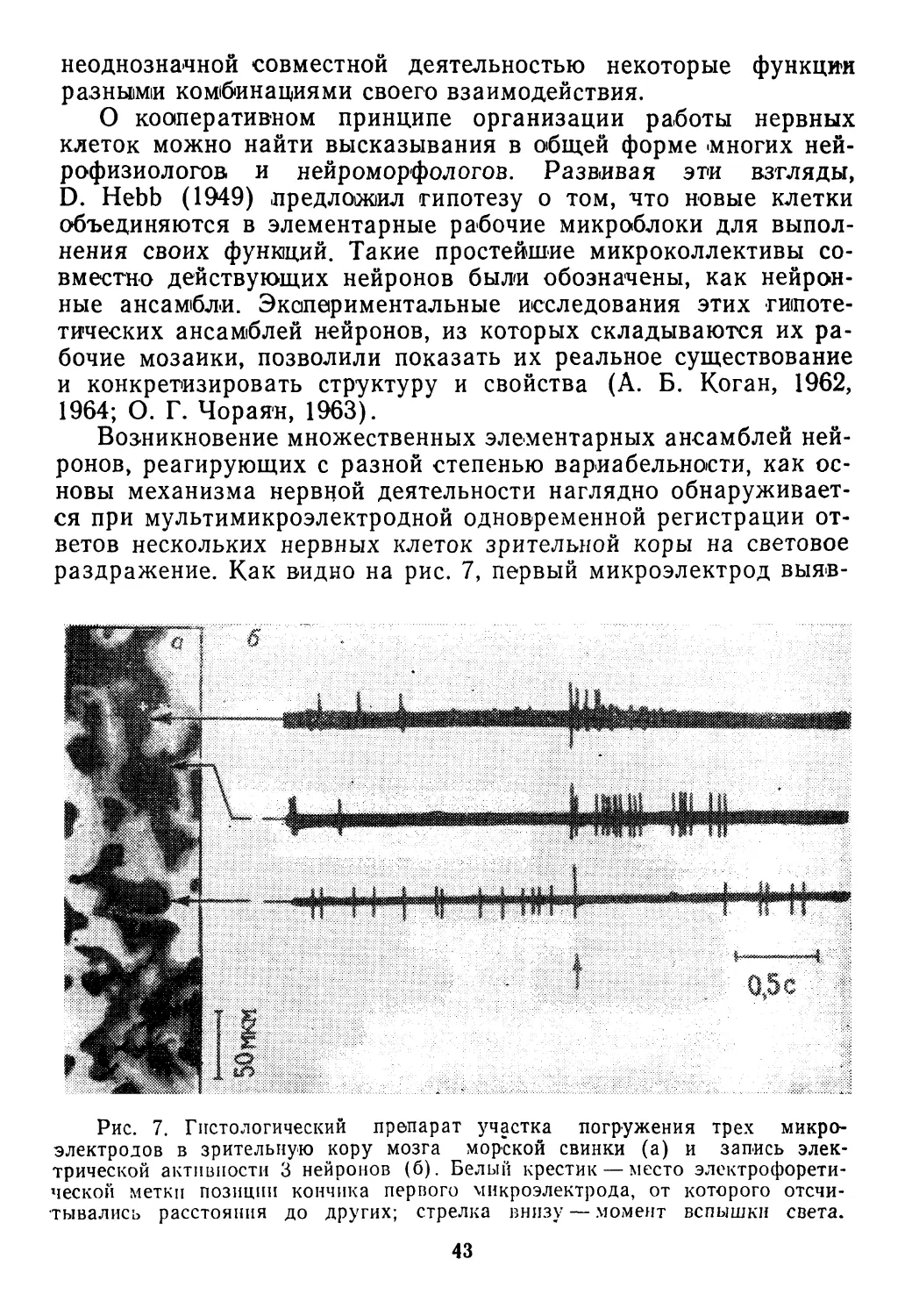
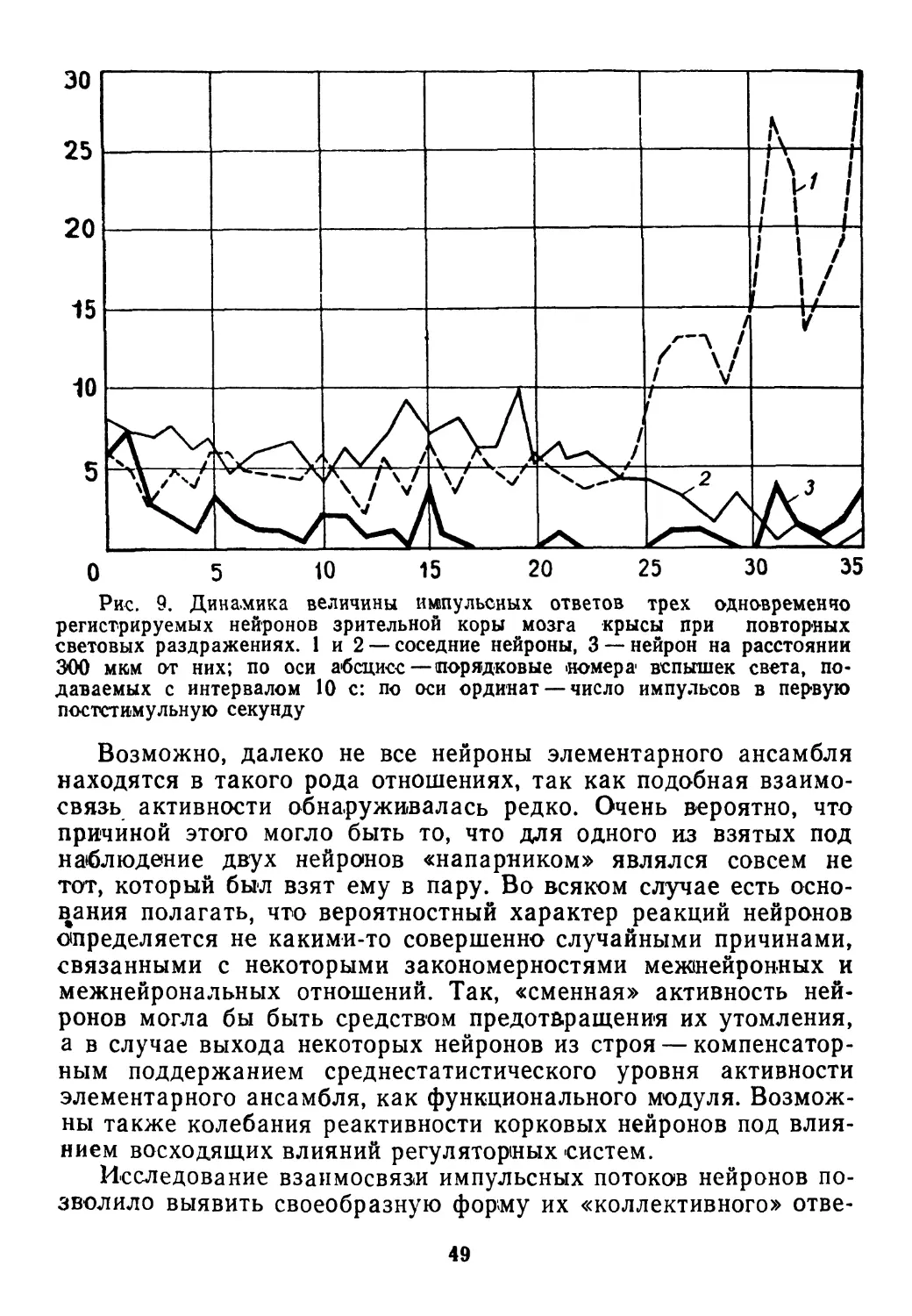
30
ностыо адекватно реагировать на все эти воздействия. Такая
способность достигла высокого совершенства в свойствах нерв-
ной системы. Наиболее общая характеристика этих свойств
связана с требованиями, которым должно удовлетворять управ-
ление целесообразным поведением объекта в обстановке слу-
чайных событий. Эти требования в абстрактном виде исследует
теория статистических решений с учетом особенностей регули-
рования в биологических системах, обеспечивающих выживание.
Необходимость принимать последовательные решения о дей-
ствиях в ответ на непрерывные изменения обстановки позволяет
рассматривать приспособительное поведение в некотором отно-
шении как процесс игры, где одним из игроков является орга-
низм, управляемый нервной системой, а в качестве другого вы-
ступает природа. Так как вредные и 'полезные воздействия по-
следней с этой точки зрения выглядят как «случайные ходы», то
процессы обеспечения выживания могут быть описаны в поня-
тиях теории статистических игр (Б. Блекуэлл, М. Гиршик, 1958).
При этом выбор альтернатив ответных реакций приобретает
вероятностный характер.
Определяя 'выживание как сохранение существенных пере-
менных функций организма в определенных границах при нали-
чии непрерывных возмущений, У. Эшби (1959) рассматривает
деятельность нервной системы как адаптивного регулятора, ко-
торый разнообразию возмущений противопоставляет разнооб-
разие регулируемых им состояний. Если обозначить изменчи-
вый мир событий через Т, а раз-
нообразные. происходящие в нем
возмущения — через D, то, что-
бы обеспечить выживание, нерв-
ная система в роли регулятора R
должна воздействовать на Т так,
чтобы осуществленные перемен-
ные организма Е не выходили
за границы, совместимые с его
жизнью (рис. 2). По такому «за-
кону необходимого разнообразия»
для выживания в мире событий,
наступление которых можно ожи-
дать лишь с некоторой вероят-
ностью, необходим регулятор, спо-
собный перекрыть разнообразие
событий вероятностным характе-
ром своего функционирования.
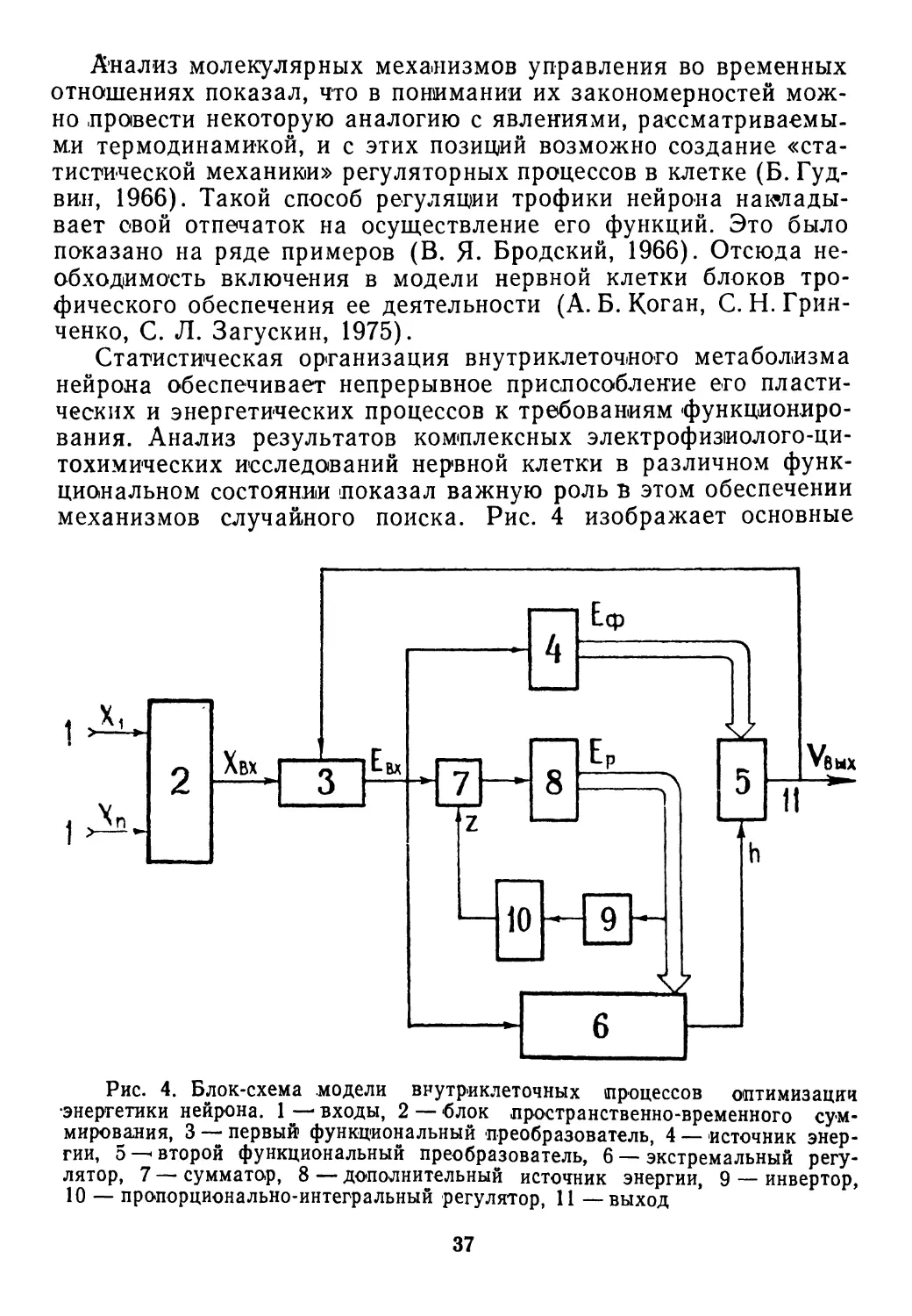
•Рис. 2. Место нервной системы
как регулятора в отношениях ор-
ганизма и среды (по У. Эшби,
1959)
31
Многие характерные черты и закономерности .процессов це-
лесообразного поведения, обеспечивающего выживание в стоха-
стической среде, раскрываются при их исследовании с исполь-
зованием понятий теории автоматов, особенность которых со-
стоит в том, что они подвергаются случайным воздействиям и
должны выбирать оптимальную стратегию реакций. Здесь также
речь идет о некоторой своеобразной игре с природой. Однако
в отличие от классической теории игр, предполагающей знание
игроками системы платежных функций, в этом случае априорная
информация о вероятности наступления тех или иных событий,
отсутствует (М. Л. Цетлин, 1969). Если при известных априор-
ных вероятностях выбора из множества А апостериорная вероят-
ность определяется по формуле Байеса
Р = (А/Х) =-------Р(А)Р(Х/А)------
v ’ Ss(A) Р (А) Р (Х/А) d2 (А)
и означает такой выбор, при котором апостериорная плотность
вероятности максимальна, то при отсутствии априорной инфор-
мации используется метод максимума правдоподобия. Тогда
выбор между двумя альтернативами ai и а2 определяется их
апостериорными вероятностями
р (а,/Х) =--------PiZWli)------..
Рх Р (Х/ах) Н-р2 Р (Х/а,)
р (а,/Х)=-------р2 Р (Х/а?)---.
PiР (X/ai) + р2 Р (Х/аО
Их отношение даст коэффициент правдоподобия Н(Х) =
_ P(X/aQ
Р(Х/а2) ’
значение которого (больше единицы) приводит к решению в
пользу альтернативы аь а меньше единицы — в пользу а2
(А. Р. Шахнович, Д. И. Шапиро, 1973).
Несмотря на свою стохастичность среда существования изо-
билует ситуациями, повторяющимися с большей или меньшей
частотой в большей или меньшей степени. Это обстоятельство
определило исключительное значение такого свойства нервной
системы, как память в выборе альтернатив поведения, обеспе-
чивающего выживание. Более подробно о вероятностных меха-
низмах обучения речь будет идти в следующей главе. Здесь
можно лишь упомянуть о некоторых наиболее общих аспектах
32
стратегии «приспособительного поведения с использованием про-
шлого опыта. Способность выбрать 1из многих вероятных реше-
ний наиболее целесообразное характеризует «интеллигентные-
системы» (Н. Oestreicher, 1974). В таком выборе учет прошлого-
опыта резко (повышает шансы на выживание. Исчисление веро-
ятностей полезных и вредных эффектов действий, предпринимае-
мых чисто случайно или учитывая результаты предыдущих, по-
зволяет количественно оценить выгоду последней стратегии:
(С. В. Фомин, М. Б. Беркинблит, 1973).
Эволюция мира животных сменяется историческим разви-
тием человеческого общества, где вместо биологического вы-
живания ставятся задачи общественно полезной деятельности.
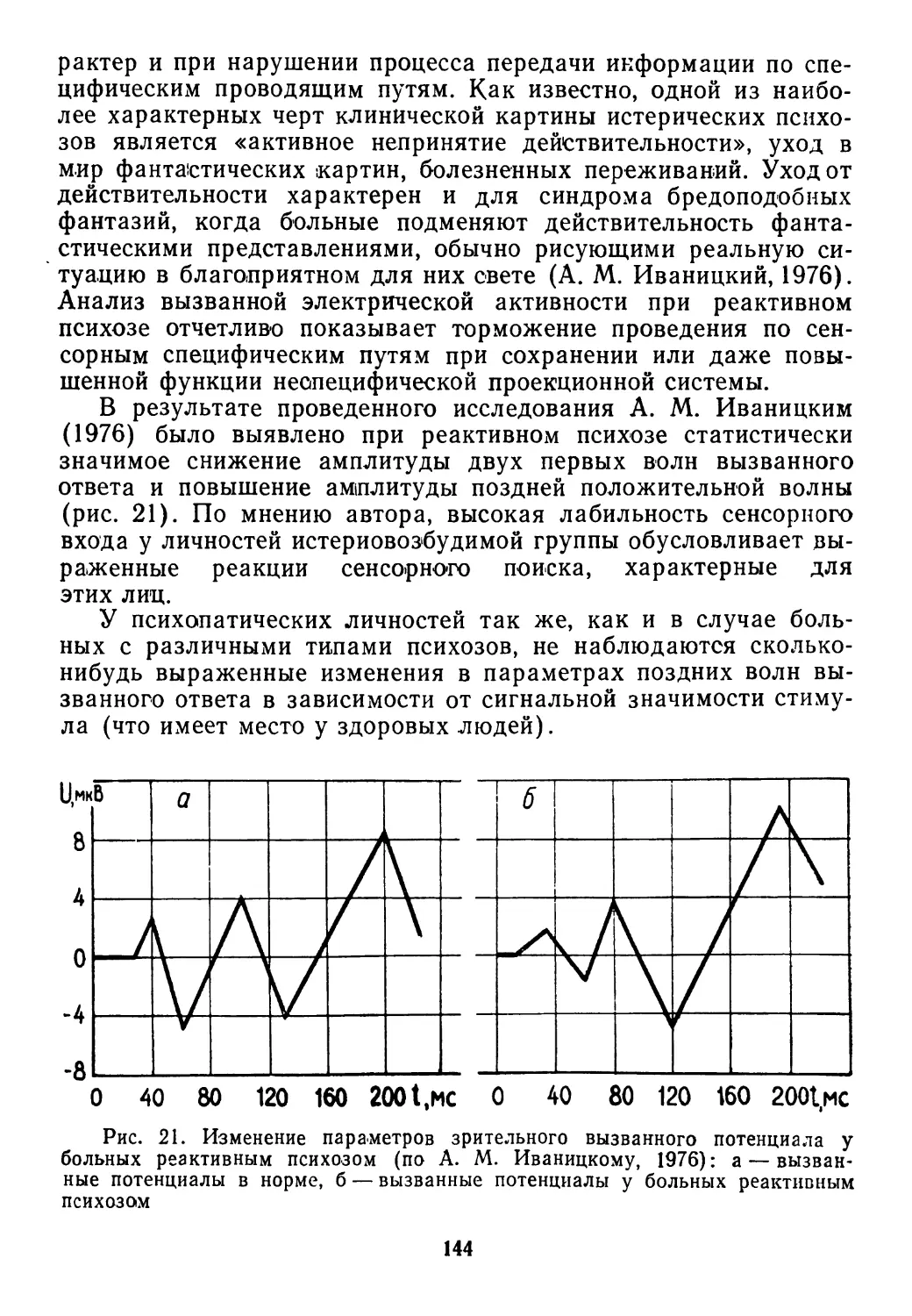
Здесь действует язык «размытой» логики, которая оперирует
лингвистическими переменными, имеющими не количественное^
а качественное выражение, и распространяется на оценки веро-
ятности (Л. Заде, 1976). При этом такие основные акты мышле-
ния человека, как процессы принятия решений, определяющих
успех его профессиональной и иной деятельности, включают в-
себя вероятностный выбор в нечетких множествах альтернатив,
умозаключений и действий (О. Г. Чораян, 1979).
2. ЧЕРТЫ СТРУКТУРНОЙ ОРГАНИЗАЦИИ МОЗГА
Мозг как орган «выживания» в стохастической среде разви-
вался в направлении совершенствования своей способности
управлять целесообразнЫхМ поведением организма в самых раз-
нообразных, неожиданных ситуациях. Непредвиденное множе-
ство таких ситуаций, в которых всегда много варьирующих об-
стоятельств, затрудняющих однозначное определение их значе-
ния в процессе выживания, делает невозможным обеспечение
поведения в каждой из них фиксированными программами дей-
ствия. Поэтому нервная система приспосабливалась своим строе-
нием и свойствами к задаче адекватного реагирования на такие
ситуации, учитывая неоднозначность каждой из них и оцени-
вая их биологическое значение в категориях вероятности.
По-видимому, наиболее общая черта морфологической орга-
низации нервной системы, дающая максимальные возможности
текущего варьирования ее формирующихся и расформировыва-
ющихся функциональных структур, заключается в наличии пу-
•тей, по которым любая комбинация рецепторов, воспринима-
ющих событие во внешнем мире (или в самом организме) мо-
жет в ответ на это событие привести в действие эффективные
органы в любой, необходимой в данном случае комбинации.
Эту черту строения нервной системы Г. И. Поляков (1965) рас-
33
сматривает как основной «биологически обусловленный прин-
цип универсальной взаимосвязи всех воспринимающих раздра-
жения элементов — рецепторов — со всеми элементами, реали-
зующими ответные реакции на раздражения — эффекторами, ко-
торыми обладает организм». Анатомическая реализация такого
принципа в общей связи осуществляется путем множественного
перекреста нервных путей, где в точках их пересечения нахо-
дятся нервные клетки, направляющие потоки импульсов в ту
или иную сторону (рис. 3).
Рис. 3. Структурная организация всеобщего перекреста нервных путей
(по Г. И. Полякову, 1965): а — схема перекрещивающихся путей возбужде-
ния от воспринятых рецепторами воздействий (верхние стрелки) к эффек-
торам, осуществляющим ответные действия (нижние стрелки); б—сеть ней-
ронов, возникающих на пересечениях нервных путей; г — рецептор, э — эф-
фектор, п — нейрон
Множественные пересечения путей проведения импульсных
потоков, в которых узлы их конвергенции и дивергенции обра-
зованы нервными клетками, обусловливают своеобразную кон-
струкцию рабочих механизмов мозга в виде нейронных сетей.
В такой сети, как показывает само название, множество петель
представляет широкие возможности выбора разнообразных пу-
тей прохождения импульсов. Если исходить дальше из предель-
но упрощенной схематической нейронной сети, представленной
на рис. 3, то каждый нейрон в узлах пересечения уже ставит
задачу выбора из двух возможностей. В реальной нейронной се-
ти все оказывается несравненно более сложным, так как каж-
34
дый нейрон имеет много входов и выходов на другие нейроны.
Учитывая результаты специальных исследований нейроно-
архитектоники коры мозга (D. Sholl, 1956; J. Szentagothai, 1969;.
Г. И. Поляков, 1973), приходится признать, что колоссальное
изобилие входов и выходов каждого коркового нейрона делает
нереальной задачу однозначного описания его реакции, исходя
из сведений о состоянии всех его входов и 'выходов. Об этом
убедительно говорит расчет, приведенный Б. Бернсом (1969).
Исходя из того, что на каждый квадратный миллиметр зритель-
ной коры мозга кошки приходится 25000 входящих волокон при
75000 выходящих, а также учитывая, что каждое афферентное
волокно от наружного коленчатого тела снабжает своими терми-
налями около 5000 корковых клеток и что плотность нейронов,
в коре составляет 72000 на квадратный миллиметр при ее тол-
щине 1,75 мм, он определил число путей (N), по которым им-
пульсные потоки могут .поступать к каждому нейрону
жт 25000-5000
N— 72000 • .1,75 ~ 000‘
Так как из коры выходят аксоны лишь половины корковых:
нейронов, то, по-видимому, полученную цифру следует еще-
удвоить. Конечно, состояние тысяч независимых входов, актив-
ность каждого из которых зависит в свою очередь от множе-
ства влияний, не может быть однозначно учтено, и конечный
результат в виде реакций нейрона определяется лишь с неко-
торой вероятностью.
Такая избыточность нейронных элементов и межнейронных,
связей, являясь характерной особенностью структурной органи-
зации, особенно высших отделов мозга, дает возможность про-
являться мультифуйкциональности (И. Н. Филимонов, 1949),
которая обеспечивает пластичность нервных механизмов и ком-
пенсацию их нарушений (О. С. Адрианов, 1976). При этом пере-
стройки используемых путей и центров происходят под влия-
нием столь большого числа в значительной мере неучитываемых
факторов, что характеристику новой формирующейся структу-
ры можно .предсказать лишь с известной долей вероятности.
Вероятностный характер таких перестроек усугубляется тем, что
болыпая избыточность нейронных элементов дает широкие воз-
можности их осуществления разными способами с использова-
нием различных наборов клеток и организацией разнообразных
коммуникаций между ними.
Особое значение среди множественных межнейронных ком-
муникаций, придающих вероятностный характер взаимодейст-
виям нейронов в их популяциях, приобретают обратные связи.
35
Они создают морфологическую основу для возвратных влияний
и их распространения на другие отделы мозга, а также для
возникновения замкнутых цепей реверберации импульсных по-
токов, что еще более усложняет и обусловливает неоднознач-
ность структур, реализующих определенные функции.
3. ОРГАНИЗАЦИЯ ВНУТРИКЛЕТОЧНЫХ ПРОЦЕССОВ
Вероятностная природа субклеточных процессов наглядно
иллюстрируется феноменами броуновского движения протоплаз-
мы. Бесчисленное множество случайных соударений не подда-
ется индивидуальному контролю, но обусловливает закономер-
ный статистический результат. Приводя броуновское движение в
качестве одного из примеров, известный физик Э. Шредингер
(1947) рассматривает статистическую природу физических и
химических процессов в живой клетке как важный фактор ее
упорядоченной организации. Он считает «многоатомность» орга-
низма необходимым условием достаточной определенности и
точности вероятностной детерминации происходящих в нем про-
цессов. Подчеркивая статистический характер всех закономер-
ностей в явлениях жизни, Шредингер утверждает, что «всякий
другой род закономерностей и упорядочностей, который можно
себе представить, непрерывно нарушается и делается недействи-
тельным вследствие безостановочного теплового движения ато-
мов».
Статистический характер результата межмолекулярных вза-
имодействий усложняется тем, что рабочие механизмы активных
клеток, в том числе и нервных, построены из макромолекул, кон-
формационные свойства которых обусловливают их широкую
изменчивость в формировании на их основе более сложных
структур (М. В. Волькенштейн, 1965). При чрезвычайном раз-
нообразии комбинаций взаимодействия молекул, индивидуаль-
ное поведение которых непредсказуемо, «эквифинальность» та-
кого статистического процесса обеспечивает определенное на-
правление превращений метаболического цикла. В свою очередь
различные метаболические циклы участвуют в формировании
внутриклеточных процессов более высокого уровня. Гибкость та-
кой многоуровневой статистической организации определяет ее
способность к далеко идущим приспособительным перестройкам
при изменении функционального состояния и условий деятель-
ности клетки. Этому соответствует и чрезвычайная вариабель-
ность параметров ее существенных переменных, с которой свя-
заны особенности управления внутриклеточными процессами
{Ф. Гродинз, 1966).
36
Анализ молекулярных механизмов управления во временных
отношениях показал, что в понимании их закономерностей мож-
но провести некоторую аналогию с явлениями, рассматриваемы-
ми термодинамикой, и с этих позиций возможно создание «ста-
тистической механики» регуляторных процессов в клетке (Б. Гуд-
вин, 1966). Такой способ регуляции трофики нейрона наклады-
вает свой отпечаток на осуществление его функций. Это было
показано на ряде примеров (В. Я. Бродский, 1966). Отсюда не-
обходимость включения в модели нервной клетки блоков тро-
фического обеспечения ее деятельности (А. Б. Коган, С. Н. Грин-
ченко, С. Л. Загускин, 1975).
Статистическая организация внутриклеточного метаболизма
нейрона обеспечивает непрерывное приспособление его пласти-
ческих и энергетических процессов к требованиям функциониро-
вания. Анализ результатов комплексных электрофизиолого-ци-
тохимических исследований нервной клетки в различном функ-
циональном состоянии показал важную роль в этом обеспечении
механизмов случайного поиска. Рис. 4 изображает основные
Рис. 4. Блок-схема модели внутриклеточных процессов оптимизации
энергетики нейрона. 1-— входы, 2 — блок пространственно-временного сум-
мирования, 3 — первый функциональный преобразователь, 4 — источник энер-
гии, 5 —• второй функциональный преобразователь, 6 — экстремальный регу-
лятор, 7 — сумматор, 8 — дополнительный источник энергии, 9 — инвертор,
10 — пропорционально-интегральный регулятор, 11 — выход
37
звенья организованной таким образом системы регулирования
трат энергии (А. Б. Коган, С. Н. Гринченко, С. Л. Загускин,
1975). Входные воздействия Xi ... суммируются в Хвх, и под
влиянием выходного сигнала нейрона УВых. преобразится в сиг-
нал «входной энергии» Евх. В то время как функциональная со-
ставляющая энергии Еф расходуется на выработку выходных
сигналов нейрона, регуляторная составляющая Ер затрачивает-
ся на работу поискового экстремального регулятора, который
изменяет динамический порог возбуждения нейрона h, влияет
на его импульсный разряд, а по обратной связи минимизирует
траты энергии Еф. Когда минимизация Еф достигнута, выходной
сигнал Z, суммируясь с сигналом Евх, .приводит к нулю Ер и тем
самым прекращает поисковые операции.
Именно статистическая организация, обусловливающая гиб-
кость метаболической основы выполнения функций нервной
клетки в разных условиях и в разном состоянии, обеспечивает
оптимизацию ее деятельности. Различают проявляющиеся при
этом биохимические механизмы: в первом случае «эксплуататив-
ные», обеспечивающие приспособление к изменениям среды, а
во втором случае — «компенсаторные», восстанавливающие на-
рушенные функции клетки (П. Хочачка, Дж. Сомеро, 1977). По-
следнее представляет собой интерес с точки зрения оценки на-
дежности осуществления нейроном своей функции. Для количе-
ственной оценки надежности молекулярного механизма обеспе-
чения нормы метаболических процессов предлагается вычисле-
ние вероятности безотказного действия при производстве опре-
деленных микромолекул с учетом дублируемости этого произ-
водства (А. В. Савич, М. И. Шальнов, 1977).
Наличие обратных связей в многоуровневой организации
внутриклеточных процессов при возрастании ее сложности и
участия достаточно большого числа компонентов делает все бо-
лее вероятным переход этих процессов в циклические (Н. Ви-
нер, 1958). При этом в каждом данном локусе метаболической
системы они приобретают колебательный характер. Такие ос-
цилляторы обнаруживают свойства нелинейности и могут всту-
пать между собой во взаимоотношения, результат которых стро-
ится на статистической основе. По-видимому, некоторые свойства
внутриклеточных осцилляторов проявляются в ритмоводящих,
или пейсмекерных потенциалах нейронов (Е. Н. Соколов, 1975).
В этом случае ритмическая генерация принципов обусловливает-
ся гипер- и деполяризацией возбудимой мембраны й результате
колебательных процессов, главным образом изменений метабо-
лического обеспечения активного транспорта ионов и проницае-
мостью клеточной мембраны.
38
Другим проявлением свойств внутриклеточных осцилляторов
при их активности может служить явление так называемого за-
хватывания ритма, когда частота колебаний одного из них при-
ближается к частоте колебаний другого (Б. Гудвин, 1966). Та-
кое захватывание ритма, например, наблюдалось пр.и регистра-
ции импульсного ответа нейрона растяжения рака на электри-
ческую стимуляцию нарастающей частотой (А. Б. Коган, Б.Г. Ре-
жабек, О. С. Васильева, 1973). На рис. 5 показана зависимость
частоты импульсного разряда от частоты раздражения, наноси-
мого через микроэлектрод, установленный у аксонного холмика
нейрона. В определенных диапазонах частот импульсные раз-
ряды точно следуют частоте раздражения. Если исходить из
концепции захватывания ритма сближающимися по частоте ге-
нераторами, то можно предположить, что здесь проявились свой-
ства по крайне мере трех внутренних осцилляторов. Первые два
полностью захвачены ритмом раздражения, третий проявляет
некоторое отставание. Статистическая природа формирования
внутренних осцилляторов позволяет нервной клетке гибко менять
свои резонансные ха-
рактеристики, приспо-
сабливаясь к текущим
условиям деятельности.
Статистические за-
кономерности лежат в
основе деятельности
ионных механизмов воз-
буждения и торможе-
ния нейрона. Обосно-
вывая ионную теорию
возбуждения, П. П. Ла-
зарев (1916) исходил
из представления о мас-
совых потоках ионов.
Электрическая актив-
ность нервной клетки
возникает на мембра-
Рис. 5. Захватывание частотой разд-
ражения частоты следования импульсов
нейрорецепторой клетки речного рака. По
оси абсцисс — частота раздражения, по
оси ординат — частота импульсации ней-
ронов
нах в результате соз-
дания разностей ионных концентраций, лишь глобальные отноше-
ния которых выражаются соответствующими формами (А. Ходж-
кин, 1965). А так как эти разности концентраций ионов не-
прерывно поддерживаются работой «ионных насосов» метабо-
лической природы, то совершенно естественно, что они имеют
статистическое происхождение. В механизме деятельности воз-
будимой мембраны нейрона важную роль играют изменения ее
39
проницаемости. Здесь также мы сталкиваемся с невозможностью
учитывать индивидуальное поведение элементов этого механиз-
ма—предполагаемых «пор» мембраны, и все заключения осно-
вываются на статистических оценках. Вместе с тем делаются
попытки связать такие глобальные статистические оценки с про-
цессами на молекулярном уровне (А. А. Лев, 1976).
Основная функция отдельной клетки в нервной системе за-
ключается в получении, переработке и передаче информации.
Поэтому особое значение приобретают процессы, разыгрываю-
щиеся в межклеточных соединениях — синапсах. Электронный
микроскоп позволил увидеть в структуре синапса на участках
пресинаптической мембраны сети пузырьков — везикул, напол-
ненных медиатором. Когда их содержимое изливается в синапти-
ческую щель, происходит раздражение постсинаптической мем-
браны. Эти выбросы медиатора представляют -собой дискретные,
независимые друг от друга события. Поэтому они возникают в
случайной последовательности, имеют вероятностный характер,
и лишь их статистическая совокупность определяет закономер-
ности поддержания некоторого суммарного уровня поляризации
постсинаптической мембраны и однозначного эффекта ее депо-
ляризации.
Миниатюрные потенциалы имеют амплитуду, составляющую
лишь сотые доли вызванного потенциала, и они возникают в
случайной последовательности, указывая на вероятностный ха-
рактер опорожнения синаптических везикул. В свою очередь
вариабельность амплитуды указывает на то, что разные везику-
лы содержат разное количество молекул медиатора. Следова-
тельно, совместное действие таких элементарных единиц действия
медиатора имеет статистическую природу и неоднозначно приво-
дит к достижению порогового значения генераторного потенциала.
Статистический характер того, как возбудительный постои-
наптический потенциал, вызываемый нервным импульсом, обра-
зуется из множества миниатюрных единиц, убедительно про-
явился в экспериментах с подавлением активности медиатора,
уменьшающим число невыброса из везикул. Так, снижение кон-
центрации калия и повышение содержания магния уменьшало
величину вызванных потенциалов ступенеобразно, каждый раз
на величину, лежащую в пределах варьирующих значений ми-
ниатюрных потенциалов. Когда количество участвующих в ре-
акции миниатюрных единиц становилось достаточно малым, то
гистограммный анализ выявлял, что распределение амплитуд
вызванных потенциалов имело мультиноминальный вид с подъ-
емами на участках, кратных среднему значению амплитуд ми-
ниатюрных потенциалов (рис. 6).
40
Закономерные отношения стимулов и ответов рецепторов,
обеспечивающие правильность отражательной деятельности сен-
сорных систем, проявляются лишь на уровне статистического
усреднения их элементарных реакций, которые оказываются
весьма вариабельными. Так, даже при постоянной нагрузке на
мышцу в потоке проприоцептивных импульсов, сохранявших
0 2 4 6 8
Число выделенных порций ацетилхолина (число МПКП)
Рис. 6. Миниатюрные потенциалы и их отношение к вызванным по-
тенциалам нервно-мышечного синапса: а — запись миниатюрных потенциа-
лов (по Р. Fatt, В. Katz, 1962); б — гисто-граммы распределения ампли-
туды вызванных потенциалов и их отношение к распределению амплитуд
миниатюрных потенциалов 1М1ГВКП (по Y. Boyd, A. Martin, 1966). По оои
ординат — число наблюдаемых событий: 1 — теоретическое число «отказов>;
2 — экспериментальные данные; 3 — теоретическое распределение; 4 — Гаус-
сову плотность вероятности амплитуд МГВДП
определенную среднюю частоту, наблюдался разброс межим-
пульсных интервалов, происхождение которого связывали с «теп-
ловым движением ионов» и другими предсказуемыми причинами
(A. Buller, Y. Nicholls, J, Strom, 1953). Еще более выражена
статистическая природа процессов рецепции в зрительной систе-
ме. Исключительно высокая чувствительность палочек к свету
имеет в своей основе случайную 'последовательность независи-
мых дискретных воздействий световых квантов на родопсин, ко-
торые подчиняются распределению Пуассона и носят стохасти-
ческий характер (Н. Barlow, 1974). Эти воздействия запускают
молекулярные механизмы, подобные недавно описанным для
возбудимых мембран в виде «одиночных каналов» (G. Ehren-
stein, Н. Legar, 1977). Было высказано предположение, что
функции таких молекулярных триггеров, включающих ионные
потоки, под действием единичных квантов света могут выпол-
нять макромолекулы, изменяющие свое конформационное со-
стояние (С. Э. Шноль, 1979).
4. ОРГАНИЗАЦИЯ ЭЛЕМЕНТАРНЫХ
НЕЙРОННЫХ ОБЪЕДИНЕНИИ
«Мозг — сложная система, слишком сложная, чтобы описать
ее в деталях, и слишком детальная, чтобы описать ее статисти-
чески» (М. Conrad, 1974). В этом .парадоксе выражено главное
затруднение, возникающее при изучении принципов организации
и конструкций рабочих механизмов мозга. С одной стороны, ис-
ключительная множественность элементов вариабельных реак-
ций, из которых складываются функциональные системы мозга,
делает нереальной возможность понять способ их работы и свой-
ства деятельности. Отсюда попытки описать эту деятельность,
исходя из статистических закономерностей организации взаимо-
действия нейронов, обозначаемых однородными элементами в
случайных нервных сетях (Р. Берл, 1966; Б. Бернс, 1969). С дру-
гой стороны, функциональная и морфологическая гетероген-
ность нейронов, получающая все более далеко идущие подтверж-
дения и уточнения (И. С. Беритов, 1969; Г. И. Поляков, 1973),
лишает оснований такой простой стохастический подход, требу-
ющий однородности взаимодействующих элементов.
Попытка выйти из этих затруднений была предпринята в
представлении о вероятностной ансамблевой организации мо-
заик нейронной активности как основе рабочих механизмов
мозга (А. Б. Коган, 1962—1979). Здесь предполагается своего
рода «относительная статистичность» функционирования, при
которой микрогруппы нейронов могут однозначно реализовать
42
неоднозначной совместной деятельностью некоторые функции
разными комбинациями своего взаимодействия.
О кооперативном принципе организации работы нервных
клеток можно найти высказывания в общей форме «многих ней-
рофизиологов и нейроморфологов. Развивая эти взгляды,
D. Hebb (1949) .предложил гипотезу о том, что новые клетки
объединяются в элементарные рабочие микроблоки для выпол-
нения своих функций. Такие простейшие микроколлективы со-
вместно действующих нейронов были обозначены, как нейрон-
ные ансамбли. Экспериментальные исследования этих гипоте-
тических ансамблей нейронов, из которых складываются их ра-
бочие мозаики, позволили показать их реальное существование
и конкретизировать структуру и свойства (А. Б. Коган, 1962,
1964; О. Г. Чораян, 1963).
Возникновение множественных элементарных ансамблей ней-
ронов, реагирующих с разной степенью вариабельности, как ос-
новы механизма нервной деятельности наглядно обнаруживает-
ся при мультимикроэлектродной одновременной регистрации от-
ветов нескольких нервных клеток зрительной коры на световое
раздражение. Как видно на рис. 7, первый микроэлектрод выяв-
Рис. 7. Гистологический препарат участка погружения трех микро-
электродов в зрительную кору мозга морской свинки (а) и запись элек-
трической активности 3 нейронов (б). Белый крестик — место электрофорети-
ческой метки позиции кончика первого микроэлектрода, от которого отсчи-
тывались расстояния до других; стрелка внизу — .момент вспышки света.
43
ляет совместную реакцию двух нейронов, по-видимому, располо-
женных по соседству, о чем свидетельствует перекрытие элект-
рических полей их импульсации. В этой реакции участвует и
нейрон, активность которого регистрировал второй микроэлек-
трод, кончик которого находился на расстоянии 50 мкм от пер-
вого. Вместе с тем нейрон, расположенный еще дальше, на рас-
стоянии более 100 мкм от первого, не только не участвует в
совместной реакции, но даже тормозится. Таким образом, вы-
рисовываются контуры элементарного объединения нейронов в
проекционном поле зрительной коры в виде компактной груп-
пы синергично возбуждающихся клеток, окруженных тормозя-
щимися. Характерно, что при повторных световых раздраже-
ниях характер и последовательность импульсных ответов иссле-
дуемых нейронов каждый раз оказывается несколько иной.
Группирование корковых нейронов в элементарные ансамб-
ли для совместного выполнения своих функций можно рассмат-
ривать как одно из проявлений кооперативного способа их дея-
тельности. Они образуют первый уровень иерархической орга-
низации рабочих механизмов мозга. При этом вариабельность
ответов отдельных клеток усредняется и формирует коллектив-
ную реакцию ансамбля, как рабочей единицы коркового функ-
ционального модуля.
Когда индивидуальное участие нейрона становится не одно-
значно обязательным, а вероятностным и возможна относитель-
ная взаимозаменяемость элементов, повышается надежность
нервных механизмов. В результате функционирование системы
становится малозависящим от состояния той или иной отдель-
ной нервной клетки. С другой стороны, подвижная структура
таких рабочих единиц, формируемых вероятностным участием в
них нервных клеток, обусловливает большую гибкость межней-
ронных связей и легкость их перестроек, которые определяют
свойства пластичности, характерные для нервных механизмов
высших отделов мозга.
Объединение нейронов в элементарные вероятностные ан-
самбли и возникновение их мозаик в ответ на афферентное
раздражение были обнаружены в различных экранных структу-
рах мозга разных животных: крыше среднего мозга (О. Г. Чо-
раян, 1963, 1969; М. Эль Гохари, 1974) и коре мозжечка (Фам
Хыу Хоан, 1967) лягушки, в больших полушариях мозга птиц
(В. Г. Савина, 1971), соматосенсорной (А. Г. Сухов, 1969) и
зрительной (А. Э. Тамбиев, 1971; П. Н. Ермаков, 1978) коре
мозга крысы, зрительной коре мозга морской свинки (Л. Н. Под-
44
ладчикова, 1978; С. А. Чебкасов, 1972), зрительной (Н. И. Пав-
ловская, 1970) и сенсомоторной (А. А. Буриков, 1969) коре моз-
га кролика, теменной коре мозга кошки (Г. Л. Фельдман, 1972).
Статистическая природа ансамблевой организации делает очень
условными все ее .параметры, в частности пространственные ха-
рактеристики. Однако при достаточном числе наблюдений усред-
ненные значения могут дать некоторый материал для их сравни-
тельной оценки. Такие оценки выводились на основании экспе-
риментального определения степени отставления одного микро-
электрода от другого, при котором сохранялось преобладание
синергизма реакций отводимых ими нейронов. Учитывая вероят-
ность захвата микроэлектродами разных частей ансамбля и его
окружения, можно было понять, что резкое падение процента
синергично реагирующих пар означало расхождение микроэлек-
тродов на величину, превышающую его размеры.
Несмотря на вероятностный характер участия нейронов в
формировании элементарных ансамблей, усредненные показа-
тели пространственных характеристик последних с достаточной
определенностью отражают различия поступающих афферент-
ных сигналов и свойства исследуемых проекционных структур.
Например, в соматосенсорной коре мозга кролика резкое паде-
ние синергизма реакций нейронов наступало в ответ на раздра-
жение специфического вентролатерального ядра таламуса при
отставлении микроэлектродов на расстояние более 100 мкм, а
при раздражении неспецифического центрально-медиального
ядра для этого потребовалось увеличить расстояние между ми-
кроэлектродами более 200 мкм. Из вариабельных реакций ней-
ронов статистически складываются элементарные нейронные ан-
самбли определенных пространственных характеристик. Их вре-
менные характеристики также имеют статистическую природу,
проявляющуюся, например, неодновременным вовлечением в ак-
тивность нейронов, синергичные реакции которых образуют
микроочаги возбуждения ансамблей.
Статистический способ формирования рабочих механизмов
мозга из нейронов с реакциями разной степени вариабельности
занимает тем более видное место, чем выше организована нерв-
ная система. Это проявляется по возрастанию в эволюционном
/ряду нестабильности главным образом таких показателей реак-
ций, как латентный период, количество импульсов в ответе, по-
рядок их следования. Например, большинство нейронов коры
мозга крысы отвечали на афферентное раздражение с отклоне-
ниями числа импульсов в ответе, составляющими 0,1—0,3 его
среднего значения, а вариации ответов нейронов коры мозга
кошки превышали этот уровень. О закономерном возрастании
45
’Вариабельности латентных периодов нейронных реакций в эво-
люции нервной системы дает представление табл. 2.
Таблица 2
Вариабельность латентных периодов ответов нейронов
на афферентное раздражение
Нервное образование Число исследован- ных нейроноз | Отклонение от средн, значения, %
до 0,1 0,1 —0,3 J свыше 0,3
Подглоточный ганглий улитки 31 58 36 6
.Ретикулярная формация среднего мозга лягушки 49 33 55 12
Зрительные доли лягушки 74 28 18 54
Зрительная кора морской свинки 34 13 30 57
Закономерное увеличение доли вероятностных процессов в
механизмах нервной деятельности по мере их эволюционного со-
вершенствования подтверждает значение этого способа обеспе-
чения их свойств пластичности и надежности. Вместе с тем на-
растание нестабильности реакций может достичь степени, при
которой потеряется определенность узора ансамблевой мозаики,
в котором кодируется обрабатываемая информация. Такая край-
няя степень нестабильности достигается при полном выпадении
того или иного из очередных ответов нейрона, т. е. их нерегу-
лярности. Оценка свойств вероятностных реакций нейронов по
этим показателям выявила, что возрастание нестабильности от-
сюда не означает повышения нерегулярности ответов, которые
при этом даже могут становиться более регулярными. Из 100
случайно выбранных нейронов зрительной коры крысы в 76%'
случаев имели место нестабильные реакции (более 0,1 среднего
значения ответа), в том числе 43% —нерегулярные ответы. А в
выборке 80 нейронов зрительной коры кошки нестабильность
реагирующих нейронов несколько возросла (80%). Следователь-
но, можно думать, что вероятностная организация ансамблевой
мозаикн, делая ее динамичной, вместе с тем обеспечивает инва-
риантность основных черт кодового узора.
Необходимо подчеркнуть, что вероятностный способ органи-
зации рабочих механизмов мозга не означает рандомизации ак-
тивности всех нейронов, участвующих в деятельности этих меха-
низмов. Не говоря о том, что степень нестабильности реакций,
46
Рис. 8. Одновременно зарегистрирован-
ные реакции (а, б, в,) двух соседних ней-
ронов зрительной доли мозга лягушки на
свет и вызванные потенциалы в отведении
от поверхности. Горизонтальная линия—
отметка светового раздражения
как уже было показа-
но, могла быть различ-
ной, но часть нейронов,
отвечала настолько ста-
бильно, что по всем ха-
рактеристикам повтор-
ных реакций нельзя бы-
ло обнаружить каких-
либо отклонений. На
рис. 8 показаны такие
ответы, стабильные по>
латентным периодам
числу импульсов, их
распределению и дру-
гим характеристикам
одного нейрона, в то-
время как соседний ней-
рон обнаруживает чрез-
вычайную нестабиль-
ность своих реакций.
Обращает на себя внимание то обстоятельство, что одновре-
менно регистрируемый суммарный вызванный потенциал ока-
зался совершенно стабильным по своей форме в ответе на каж-
дое раздражение, несмотря на вариабельность индивидуальных
реакций большинства нейронов, участвующих в его формирова-
нии. В этом наглядно проявляется способность вероятностной
организации формировать однозначные функциональные струк-
туры из статистически вероятностных элементов.
Вскрытие интимных процессов межнейронного взаимодей-
ствия, в результате которых из множества вариабельных реак-
ций нервных клеток образуется определенный акт системной дея-
тельности, является сложнейшей задачей, близкой в своем фи-
лософском аспекте к проблеме перехода количества в новое ка-
чество. Однако ряд фактов, полученных при изучении деталей
внутренней структуры ансамблевой мозаики, позволяет выска-
зать по этому поводу некоторые предположения. Оказалось, что
однозначно или вероятностно реагирующие нервные клетки рас-
полагаются в образованном ими элементарном нейронном ан-
самбле в определенном порядке, а именно: первые сосредото-
чены в центральных отделах его микроочага возбуждения, а
вторые — преимущественно на периферии. Так, например, иссле-
дование 64 нейронов сенсомоторной коры мозга показало, что
у тех из них, которые были зарегистрированы микроэлектрода-
ми, находящимися в фокусе возбуждения, т. е. в центре зоны
47
синергичных возбудительных реакций, коэффициент вариации
числа импульсов в ответе на тактильное раздражение составлял
в среднем 4,6, а у тех, которые регистрировались микроэлектро-
дом, отставленном на 100 мкм, он возрос до 5,8.
Так как расположенные в центральной эоне элементарного
ансамбля более стабильные по своим реакциям нейроны оказа-
лись и коротколатентными (22 мс в среднем) по сравнению с
расположенными более периферически (25 мс в среднем), то
можно предположить, что они первыми активируются термина-
лями афферентных волокон. Именно эти первично активируемые
нервные клетки образуют определенную основу, своего рода ске-
лет каждого элементарного ансамбля, и создают в своей сово-
купности канву рабочей мозаики, От них получают импульсы
и «вторично активируются, естественно, с гораздо меньшей ста-
бильностью нейроны, образующие периферию микроочагов воз-
буждения. Эти вторичноактивирующиеся нервные клетки и со-
ставляют основную массу вероятностных участников текущего
формирования и переформирования динамической мозаики ней-
ронной активности. По-видимому, они являются тем подвижным
резервом, который позволяет осуществлять непрерывно идущие
перестройки рабочих механизмов мозга и обеспечивает пластич-
ность его приспособительной деятельности.
Оказалось также, что между нейронами вероятностного уча-
стия в формировании ансамблевой мозаики могут возникнуть
довольно сложные отношения, проявившиеся в некоторых фор-
мах взаимосвязи их активности. Одна из наиболее интересных
форм такой взаимосвязи заключалась в том, что некоторые па-
ры нейронов, .принадлежащие к одному элементарному ансамб-
лю, временами начинали работать как бы «по очереди», усили-
вая и ослабляя овои ответы на афферентные раздражения. На
рисунке 9 представлена динамика реакций трех одновременно
регистрируемых нейронов зрительной коры мозга крысы на .мно-
гократно предъявляемое световое раздражение. Начиная с 10-го
предъявления и почти по 25-е, первый из пары соседних нейро-
нов увеличивал свои ответы, когда второй их уменьшал, и
уменьшал их, когда второй усиливал свои реакции. Однако по-
сле 25 применений светового стимула второй нейрон по какой-то
причине начал выходить из строя вплоть до полного прекраще-
ния ответов, и тогда первый нейрон резко увеличил реакции, как
бы компенсируя дефицит активности, созданный отказом от ра-
боты своего «напарника». Активность третьего нейрона, принад-
лежащего ввиду далекого расположения к какому-то другому
микроочагу возбуждения, оставалась независимой от всех этих
событий.
48
30
Рис. 9. Динамика величины импульсных ответов трех одновременно
регистрируемых нейронов зрительной коры мозга крысы при повторных
световых раздражениях. 1 и 2 — соседние нейроны, 3 — нейрон на расстоянии
300 мкм от них; по оси абсцисс — порядковые «номера вспышек света, по-
даваемых с интервалом 10 с: по оси ординат — число импульсов в первую
постстимульную секунду
Возможно, далеко не все нейроны элементарного ансамбля
находятся в такого рода отношениях, так как подобная взаимо-
связь активности обнаруживалась редко. Очень вероятно, что
причиной этого могло быть то, что для одного из взятых под
наблюдение двух нейронов «напарником» являлся совсем не
тот, который был взят ему в пару. Во всяком случае есть осно-
вания полагать, что вероятностный характер реакций нейронов
определяется не какими-то совершенно случайными причинами,
связанными с некоторыми закономерностями межнейронных и
межнейрональных отношений. Так, «сменная» активность ней-
ронов могла бы быть средством предотвращения их утомления,
а в случае выхода некоторых нейронов из строя — компенсатор-
ным поддержанием среднестатистического уровня активности
элементарного ансамбля, как функционального модуля. Возмож-
ны также колебания реактивности корковых нейронов под влия-
нием восходящих влияний регуляторных систем.
Исследование взаимосвязи импульсных потоков нейронов по-
зволило выявить своеобразную форму их «коллективного» отве-
49
та jia афферентное раздражение, характерную для кооператив-
ной организации. При этом нейроны не реагировали на аффе-
рентное раздражение. В это время резко повышалась теснота
связи импульсами. Незаметная в активности отдельных нейро-
нов реакция нейронного ансамбля заключалась в экстренной
перестройке межнейронных отношений, одним из проявлений
которой была синхронизация импульсных потоков. Пример та-
кой реакции в табл. 3.
Таблица 3
Импульсная активность двух одновременно регистрируемых нейронов
(X и Y) зрительной коры морской свинки
Характеристика импульсных потоков Тип нейрона Фон Свет Достоверность различия (при 95 % уровне)
Средняя частота (гц) X
Средняя длительность меж- X
импульсного интервала (мс) Y
Функция распределения X
длительности межимпуль-
сных интервалов Y
Коэффициент корреляции X
импульсных потоков Y
4,5 5,4 Недостоверно
2,3 2,0 »
227 182 »
464 492 »
По критерию однородностей
к-выборок
Достоверных различий нет
0,63 0,88 Достоверно
«Коллективный» ответ нейронной популяции проявлялся в
изменившемся соотношении потоков аритмичных импульсов,
представляющих собой для каждого нейрона фоновую актив-
ность. Ее аритмичность обусловлена главным образом перемен-
ной афферентацией от рецепторов, циркуляцией импульсов по
цепочкам вставочных нейронов и колебаниями поляризации кле-
точных мембран (П. Г. Костюк, А. И. Шаповалов, 1964). Влия-
ние этих факторов по мере усложнения нервной системы стано-
вится все более значительным, и поэтому аритмизация фоновой
импульсации возрастает в эволюционном ряду. Однако несмотря
на случайный порядок следования импульсов в потоке, их вза-
имоотношения выявляют, как в случае «коллективного» ответа,
определенные статистические закономерности, которые могут
свидетельствовать о некоторой определенной организации их
взаимоотношений. Наличие такой организации, определяющей
характер взаимосвязи реакций нейронов, следует также из со-
поставления коэффициентов корреляции импульсных потоков
пар нейронов при их фоновой и вызванной активности (табл. 4).
50
Таблица 4
Коэффициенты корреляции импульсных потоков фоновой
и вызванной активности нейронов зрительной коры
мозга морской свинки
Исследованный показатель Положительная корреляция I Отрицательная корреляция
Средняя величина коэффициента кор- реляции при фоновой активности 0,13 + 0,02 0,07 ±0,03
при реакции на свет 0,42 + 0,16 0,34 ± 0,09
Число нейронов, исследованных при фоновой активности 286 114
Из них сохранили тот же знак корреляции при реакции на свет 248 112
По-видимому, важным фактором такой начальной организа-
ции, дающей определенное направление взаимодействию веро-
ятностно реагирующих нервных клеток, может быть распределе-
ние терминалей афферентов, по которым они активируются, и
структура межнейронных связей. В этой связи могут иметь
определенное значение пространственные характеристики эле-
ментарных нейронных ансамблей, размеры которых соответству-
ют зонам ветвления вторичных специфических афферентов
(D. Sholl, 1956). Одним из способов осуществления взаимосвя-
зи активности нейронов в пределах элементарного ансамбля мо-
жет быть показанное на рис. 10 их компактное расположение
и переплетение апикальных дендритов, в своеобразный жгут
(С. А. Шибкова, Г. С. Клепач, 1969).
В понимании тончайших и сложнейших механизмов вероят-
ностной организации кооперативной деятельности нервных кле-
ток мозга делаются лишь первые шаги. Однако эта проблема
привлекает, особенно за последнее время, все больше внимание
исследователей. Накапливаются факты, подтверждающие взгляд
на пластичность и надежность функционирования мозга как про-
явление свойств его «|бессхемной» конструкции. Подчеркивается
значение «нежестких» компонентов нейронных механизмов дея-
тельности мозга человека (Н. П. Бехтерева, 1971). Кооператив-
ный принцип объединения нейронов »в элементарные микропопу-
ляции, из которых образуются вероятностные мозаики, полу-
чает как морфологические, так и функциональные обоснования.
Морфологи уже давно обратили внимание на множественные
группировки нервных клеток, которые В. А. Бец назвал «гнез-
51
Рис. 10. Гистологический препарат участка поля 17 зрительной коры
мозга морской свинки, в .котором микроэлектродным исследованием опреде-
лялись элементарные нейронные ансамбли. (Серебрение по Глиссу-Камп-
сону. Об. 40X6,3)
дами» крупных пирамид в моторной коре, А. С. Догель — «ко-
лониями» ганглиозных клеток в сетчатке, Д. Шолл — «агрега-
тами», а Г. И. Поляков — «гроздьями» корковых клеток. За по-
следнее время физиологическое исследование подобных элемен-
тарных нейронных объединений раскрыло многие свойства та-
ких функциональных «модулей» (А. С. Батуев, 1978), «микро-
систем» (М. М. Хананашвили, 1972), «нейроценозов» (С. А. Куз-
нецов, 1979). Дальнейшее изучение нейрональных механизмов
деятельности мозга >и статистических закономерностей форми-
рования его функциональных структур может способствовать
пониманию природы замечательных свойств гибкого приспособ-
ления к непрерывно меняющимся условиям работы и своему
состоянию, которое характерно для мозга как высоко совершен-
ной живой информационно-управляющей системы.
5. МЕХАНИЗМЫ УСЛОВНОРЕФЛЕКТОРНОЙ
ДЕЯТЕЛЬНОСТИ ОРГАНИЗМА
Характерной особенностью многих (Приобретенных навыков
является то, что они формируются в условиях стохастической
внешней среды, когда вероятность одновременного наступления
52
во времени (и в пространстве) двух разных стимулов почти все-
гда бывает меньше 1, и тем не менее через некоторое время в
цёнтральных нервных структурах, отвечающих за определенные
поведенческие реакции, образуются соответствующие ассоциа-
ции. Это в полной мере относится и к механизму образования
условных рефлексов-, наиболее распространенной ситуацией об-
разования которых (не в эксперименте, а в реальных условиях
жизни животного организма) является положение, когда веро-
ятность подкрепления условного стимула безусловным почти ни-
когда не достигает единицы, когда сама последовательность
подкрепляемых условных стимулов носит случайный характер.
Вероятностный компонент реакции занимает значительное ме-
сто на всех этапах условнорефлекторного акта, состоящего из
ряда последовательных процессов в периферических, афферент-
но-эфферентных и центральных ассоциативных системах.
При этом выраженность вероятностных соотношений в услов-
норефлекторной деятельности во многом определяется эволю-
ционной динамикой соотношения вероятностно-детерминирован-
ных и жестко-детерминированных механизмов в приспособитель-
ных реакциях организма. С нарастанием в ходе эволюции веро-
ятностного компонента взаимодействия элементов нервной си-
стемы (А. Б. Коган и др., 1969, 1975) происходит увеличение
приспособительных возможностей организмов. Чем ниже на
эволюционной лестнице развития находится животный организм,.,
тем более выражены у него жестко детерминированные фиксиро-
ванные связи, а следовательно, и фиксированные стереотипные
реакции и тем беднее динамические, пластичные формы услов-
норефлекторной деятельности.
Особое значение вероятностные механизмы нервной деятель-
ности приобретают в функционировании высших отделов мозга
человека, обусловливая ведущее значение его «нежестких си-
стем» (Н. П. Бехтерева, 1971).
Широкие возможности вероятностного выбора путей реализа-
защии нервного акта возникают в ходе образования условного
рефлекса. Весьма наглядно это демонстрируется в опытах
А. Б. Когана, Н. М. Николаевой (1958) с выработкой условных
рефлексов на прямое электрическое раздражение коры голов-
ного мозга. Поскольку в этих опытах было точно известно место-
положение коры, от которого распространяется условное возбуж-
дение, то методом хирургического выключения различных об-
ластей мозга можно было оценить степень необходимости тех
или иных путей для проведения сигнала к пополнительным нерв-
ным центрам. Впоследствии эти опыты с некоторой модифика-
цией были повторены Г. М. Глумовым (1964), который у разных.
53
животных попытался ис- п
ключить все возможные пу-
ти движения сигнала от оча-
га условного возбуждения
(естественно, что у разных
животных выключаются раз-
ные пути). В результате бы-
ло установлено, что услов-
ный рефлекс мог реализо-
ваться после любой из этих
операций. Эти результаты,
очевидно, свидетельствуют о
том, что условное возбужде-
ние в структурах мозга рас-
пространяется не по какому-
то одному строго определен-
ному пути, а может пользо-
ваться многими нервными
путями.
Вероятностная природа
закономерностей формирова-
ния условнорефлекторной
деятельности четко показана
в опытах с нерегулярным
подкреплением условного
стимула безусловным (Д. Н.
Меницкий, В. В. Трубачев,
1974). Анализ результатов
образования условных реф-
лексов, выработанных на
раздражения, подкреплен-
ных стереотипно или стоха-
стически, выявил отсутствие
сколько-нибудь существен-
Рис. 11. Динамики образования
условного оборонительного рефлекса у
рыб при разной вероятности подкреп-
ления безусловным раздражителем (по
Д. Н. Меницкому, 1966). По оси орди-
нат — число предъявленных условных
стимулов; по оси абсцисс — вероятность
подкрепления; в скобках — информаци-
онная неопределенность, в битах
ных различий в скорости
формирования этих приспособительных ответов, причем услов-
норефлекторные связи, возникающие при случайном подкрепле-
нии, оказываются часто более устойчивыми (их труднее уга-
шать) , чем при стереотипном подкреплении.
В экспериментах по выработке двигательных условных реф-
лексов с частичным подкреплением (только часть предъявлен-
ных условных стимулов подкреплялась безусловным стимулом,
причем порядок -следования подкрепляемых стимулов носил слу-
чайный характер ib разных сериях экспериментов) Д. Н. Мениц-
54
ким (1966) было установлено, что рефлекс вырабатывался тем
лучше, чем выше была вероятность подкрепления условного раз-
дражителя безусловным (рис. 11). Анализ полученных кривых,
характеризующих степень эффективности образования условного
рефлекса от величины вероятности подкрепления начального ин-
дифферентного стимула безусловным, показал, что успешность об-
разования условнорефлекторной реакции прямо пропорциональ-
на логарифму вероятности (подкрепления очередного сигнала и,
следовательно, связана и с динамикой энтропийного показателя.
В работе В. В. Трубачева (1968) также было обнаружено,
что неподкрепление части условных раздражителей безуслов-
ным не препятствует образованию условного рефлекса, а в ча-
сти случаев это даже (приводит к повышению скорости формиро-
вания двигательного условного рефлекса. В работе этих авторов
(Д. Н. Меницкий, В. В. Трубачев, 1974) установлено, что при
вероятностном подкреплении в случае экологической адекватно-
сти применяемых сигнальных раздражителей конечный резуль-
тат образования условного рефлекса был не менее успешным,
чем при регулярном подкреплении, а часто условнорефлекторные
реакции, выработанные в таких условиях, оказывались более
устойчивыми к процедуре угашения.
Неопределенность ситуации не позволяет принимать одно-
значные решения, и они становятся в той или иной степени ве-
роятностными. При этом для выбора наиболее оптимальных в
данной обстановке форм поведения информационно-управляю-
щая система мозга работает по закономерностям вероятностной
логики. Чем больше неопределенность, тем больше необходимо
выполнять логических операций выбора. Поэтому время осу-
ществления реакции на сигналы с их выбором существенно удли-
няется. Оно оказывается в прямой линейной зависимости от ко-
личества информации, которое несет каждый сигнал (W. Hick,
1952). Время реакции на сигнал, который нужно выбрать из
нескольких сигналов, можно вычислить по формуле
А=К log2(n+ 1),
где А — время реакции с выбором, К — время реакции без вы-
бора,, п— число сигналов, из которых производится выбор.
Вероятностная природа организации сложных произвольных
движений человека обеспечивает пластичность их непрерывно
совершающихся перестроек и надежность осуществления в са-
мых разнообразных условиях. В этом отношении чрезвычайно
характерно, что тренировка к выполнению заданного упражне-
ния лишь усиливает возможность его выполнения каждый раз
55
несколько иной комбинацией мышечных сокращений в зависи-
мости от множества случайных обстоятельств (С. В. Гурфинкель,
Я. М. Кац, М. Л. Шик, 1965). Поэтому возникает противоречие
между срочностью и точностью выполнения двигательного акта,
так как выбор оптимальной комбинации взаимодействующих
мышц из многих вероятных требует времени, о чем свидетель-
ствует удлинение латентного периода более точных двигатель-
ных реакций (М. П. Иванова, 1978). Еще более наглядно веро-
ятностный характер нервных процессов деятельности высших
отделов мозга проявляется в организации механизмов речевых
функций (Т. А. Аминев, 197'2). Так, эксперименты с воспроиз-
ведением рядов случайных чисел показали, что объем кратко-
временной памяти представляет собой случайную функцию вре-
мени, и вероятность запоминания может быть вычислена по фор-
муле интегральной функции распределения объема памяти. Ве-
роятностный характер ассоциативных реакций проявляется как
в варьировании ответов на один и тот же словесный стимул,
так и в колебаниях времени ассоциативного ответа. Здесь дает
о себе знать множественность путей ассоциативных связей и
многоэтапность их нервных механизмов. С этим связана вариа-
бельность речевого выражения понятий и их восприятия.
Разработка микроэлектродной техники регистрации импульс-
ных потенциалов отдельных нервных клеток позволила просле-
дить на клеточном уровне особенности вероятностной динамики
условнорефлекторных реакций. Проблема нейронных механиз-
мов условнорефлекторного поведения рассмотрена во многих
работах (F. Morrell, 1961; Г. Джаспер и др., 1962; J. Bures,
О. Buresova, 1965; Н. Н. Василевский, 1968; Б. И. Котляр, 1970;
А. Б. Коган и др., 1976, 1977; Г. И. Шульгина, 1978; и др.).
Условнорефлекторные изменения импульсной активности пре-
имущественно проявляются в развитии реакций, имитирующих
ответы клетки на безусловный стимул (К. Kamikawa а. о., 1964;
И. Н. Книпст, Г. И. Шульгина, 1966; J. Bures, О. Buresova, 1965;
Н. Н. Василевский, 1968; Г. И. Шульгина, 1978; и др.). Почти во
всех работах отмечается, что одной из характерных черт клеточ-
ных реакций при выработке условного рефлекса является раз-
нообразие параметров индивидуальных ответов нервных клеток.
Риччи и др. (G. Ricchi а. о., 1957) выделили 9 типов нейронных
реакций на действие условных стимулов. Аналогичные резуль-
таты получены и в исследованиях Ф. Моррелла (F. Morrell,
1961), Камикавы и др. (К. Kamikawa а. о., 1964), Я. Буреша
и О. Бурешовой (J. Bures, О. Buresova, 1965). В работе Г. Джас-
пера и др. (1962) показано, что в двигательной области коры
мозга у обезьян примерно 15% клеток снижали, а 18% повы-
56
шали частоту своей импульсной активности при действии 40—50
последовательно предъявляемых стимулов.
По-видимому, значительная вариабельность клеточного про-
явления условнорефлекторных реакций в значительной мере
связана с полифункциональностью нервных элементов в смысле
возможного их включения в разные системы, обеспечивая раз-
личные ответы на один и тот же стимул (в зависимости от про-
шлого опыта). Причем эта полифункциональность и вытекающая
из нее индивидуальность условнорефлекторных клеточных реак-
ций связана с многообразием .морфологических связей, с систем-
ными перестройками в их активности (П. К. Анохин, 1968;
Н. Н. Василевский, 1968; А. Б. Коган и др., 1977; Г. И. Шуль-
гина, 1978; и др.).
Очевидно, одной из первых математических моделей, описы-
вающих вероятностные отношения между различными факто-
рами, определяющими образование условного рефлекса, являет-
ся модель, предложенная сотрудником И. П. Павлова Н. А. Ро-
мановым (il935). Автором было предложено описывать интен-
сивность вырабатываемого условного рефлекса R S (на услов-
ный стимул С, подкрепляемый безусловным раздражителем U)
следующим образом:
где М — число предъявлений условного стимула, подкреплен-
ного безусловным;
N—число неподкрепленных предъявлений условного сти-
мула;
К, L и е — константы, имеющие определенные положительные
значения (>0) для различных раздражителей (при
этом необходимо соблюдать одно условие, чтобы ве-
личина L была больше, чем е).
Кривые угашения и восстановления условного рефлекса, рассчи-
танные по этой формуле, достаточно хорошо удовлетворяли ре-
зультатам экспериментальных исследований.
Вероятностные математические модели условнорефлектор-
ных реакций впоследствии были описаны в ряде работ (Н. Lan-
dehl, цит. по: А. Б. Коган и др., 1977; R. Solomon, L. Wynne,
1953; A. Uttley, 1956; Р. Буш, Ф. Мостеллер, 1962; А. Рапопорт,
цит. по: А. Б. Коган и др., 1977; Б. Д. Бернс, 1969; А. А. Ляпу-
нов и др., цит. по: А. Б. Коган и др., 1977).
Вероятностная модель А. Аттли (A. Uttley, 1956)—машина
условных вероятностей — представляет собой систему переклю-
57
Двигательные реакции
5
6
00000000111110111001100001000001000000100000001
00000000000000000000000111110111001100001000001
Рис. 12. Элементы условно-вероятно-
стной машины, необходимые для обучения
(по Б. Берн-су, 1969). 1 и 3—сенсорное
раздражение звонком, условный раздражи-
тель; 2 и 4 — вид мяса, безусловный раз-
дражитель; 5— поведение xi; 6 —пове-
дение Х2
чений, при помощи которых любая комбинация входных сигна-
лов может быть поставлена в однозначное соответствие с состоя-
нием машины; в модели обеспечивается возможность выявления
общих элементов в двух сложных событиях и оценки вероятно-
сти одного из этих событий в зависимости от статистических ха-
рактеристик появления у него элементов, общих с другим собы-
тием. В качестве таких событий выступают безусловная и услов-
ная реакции, а в виде классифицируемых элементов—сигнальные
раздражители. А. Аттли рассматривает гипотетический экспери-
мент ио образованию условного рефлекса и предлагает вычис-
лять условную вероятность подкрепления как отношение числа
подкрепленных предъявлений условного раздражителя к обще-
му числу подкрепленных и неподкрепленных предъявлений
условного стимула. В этом отношении предлагаемая формула
А. Аттли во многом напоминает формулу, приведенную в рабо-
те Н. А. Романова (1936).
В условно-вероятност-
ной модели условного реф-
лекса Б. Бернса (1969)
находит отражение про-
цесс образования условно-
рефлекторной связи в
динамике взаимодействия
потоков нервных импуль-
сов, поступающих от ре-
цепторов условного и
безусловного раздражите-
лей (рис. 12).
В естественных усло-
виях жизни, полной слу-
чайностей, обучение про-
исходит на основе веро-
ятностной оценки значе-
ния событий и принятия
решений. При этом пер-
вые реакции на сигналы
осуществляются в услови-
ях неопределенности ожи-
их подкрепления или не-
даемых результатов, и лишь по мере
подкрепления эта неопределенность начинает уменьшаться, а
вероятность правильного выбора возрастать. Эти закономерно-
сти обучения получают наибольшее выражение в стратегии
«проб и ошибок».
58
Регуляция всех звеньев материальной
динамики живой системы и управление
ими опираются на информационные про-
цессы.
В. А. Энгельгардт
Глава III. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ
КОДИРОВАНИЯ И ДЕКОДИРОВАНИЯ
ИНФОРМАЦИИ В НЕРВНОЙ СИСТЕМЕ
1. ПОНЯТИЕ О КОДИРОВАНИИ ИНФОРМАЦИИ
В информационно-управляющей функции нервной си-
стемы значительное место занимают процессы кодирова-
ния. Кодом считается система знаков, -символов, которая
служит для передачи, переработки и хранения информа-
ции, т. е. код предполагает наличие в коммуникационной
системе особого способа отображения информации, при
котором соблюдаются некоторые правила соответствия
между получаемым и передаваемым сообщением и сово-
купностью символов, его обозначающих. Процесс коди-
рования заключается в операции отождествления симво-
лов, знаков одного кода с символами, знаками другого
кода -в ходе восприятия, переработки и передачи сооб-
щения в информационных каналах связи, образующими-
ся отдельными нервными клетками, нейронными сетями,
цепочками последовательно соединенных нейронов. На-
личие различных способов кодирования позволяет пере-
давать сообщения, выраженные в разной форме, но име-
ющие одно и то же информационное содержание.
Необходимость .кодирования информации определяет-
ся потребностями приспособления формы сообщения к
системе, к каналу связи. Так, форма подавляющего боль-
шинства сообщений, которые воспринимает животный ор-
ганизм> такова, что их невозможно непосредственно пе-
редавать по нервным каналам связи. Для создания опти-
мальных условий информационной деятельности нервной
системы необходимо эти сообщения преобразовать опре-
деленным образом, т. е. закодировать их.
Рассматривая различные методы изучения процессов
кодирования информации в нервной системе, А. И. Фес-
cap (А. Е. Fessard, 1971) выделяет 2 основных подхода
59
к проблемам кодирования. Первый, получивший название функ-
ционального подхода, основан на экспериментальном анализе
характеристик входного сообщения и 1выявлении структурно-
функционального их отображения в реальных воспринимающих
структурах. Только при таком в определенном смысле взаимно-
однозначном отношении между входным сигналов — стимулиру-
ющей энергией — и выходным сигналом системы связи, являю-
щимся отражением воспринимаемого сообщения, оказывается
возможным «прямое непосредственное «изучение процесса транс-
формации одних кодовых сигналов в другие при инвариант-
ности информационного содержания сообщения (S. Wiersma,
R. Т. Adams, 1950; S. Hagiwara, К. Morita, 1963, 1965; A. Herz
е. а., 1964; Т. Н. Bullock, S. Chichuku, 1965; Н. Ratliff а. о., 1968;
L. Stark, 1968; и др.).
Второй подход, известный как эвристический, основан на тео-
ретическом или модельном рассмотрении множества возможных
кодов в поисках оптимального (в отношении какого-либо вы-
бранного критерия), который и считается функционирующим в
данной системе коммуникации (R. Stein, 1967; G. Farber, 1968;
Е. Harth а. о., 1970; G. С. Cheng а. о., 1970; Y. Kulikowski, 1971;
D. Slepian , Y. К. Wolf, 1973).
Оба эти подхода имеют свои недостатки, но, взаимно допол-
няя друг друга, позволили в настоящее «время выявить основные
принципы информационной деятельности нервных структур. В
тех случаях, когда имеют место достаточно полные знания как
о структуре, так и функции отдельных элементов, образующих
рассматриваемый нервный контур связи, возможно использова-
ние функционального, экспериментального подхода, который мо-
жет быть дополнен для выведения окончательного суждения о
реальности предполагаемого способа кодирования информации
результатами теоретической модельной проверки выдвигаемой
гипотезы. Однако там, где эти знания отсутствуют, тем не менее
существенные результаты могут быть получены и при исполь-
зовании эвристического подхода, не говоря уже о том, что ре-
зультаты эвристических приемов могут помочь исследователю в
поиске новых нетривиальных методов более рационального экс-
периментирования.
Считая процесс кодирования центральным механизмом в ин-
формационной деятельности нервных структур, Д. X. Перкел и
Т. X. Баллок (D. Н. Perkel, Т. Н. Bullock, 1968) предложили весь-
ма широкое рабочее определение кодирования как процесса
представления и трансформации информации в нервной систе-
ме. При таком определении кодирования последнее включает в
себя все основные элементы информационной функции нервной
60
системы. Действительно, выделяемые авторами формальные
аспекты кода: референт, трансформация, передача трансформи-
рованной информации, интерпретация,— в сущности почти исчер-
пывают все содержание информационного процесса, протекаю-
щего в нервной системе. Каждый из компонентов этого сложного
информационного процесса отличается определенными коли-
чественными характеристиками информационных показателей,
таких как энтропийная мощность сообщения, информационная
емкость, пропускная способность канала связи, избыточность
информационная (сообщения) и канальная (недозагруженность
нервного канала связи), помехоустойчивость, надежность и т. д.
2. ЭЛЕМЕНТЫ КОДИРОВАНИЯ ИНФОРМАЦИИ
В НЕРВНЫХ КЛЕТКАХ
В качестве кодовых элементов в информационной функции
нервных клеток могут выступать многие лабильные характери-
стики их деятельности, находящиеся в определенных силовых и
временных отношениях со входными воздействиями. Это могут
быть как импульсные распространяющиеся потенциалы дейст-
вия, генерируемые в аксонном холмике (или в начальном сег-
менте аксона) — наиболее распространенная форма генерации
сигналов в нейронных системах связи, так и неимпульсные фор-
мы проявления деятельности нейронов: градуальные изменения
мембранного потенциала, синаптические потенциалы, пейсмекер-
ные потенциалы и др. В структурах нервной системы выявлено
большое количество короткоаксонных нейронов, амакриновых
клеток, нейронов с трудно дифференцируемыми аксонными от-
ростками, у которых, по-видимому, распространение возбужде-
ния (передача информации) происходит без импульсных потен-
циалов (С. Н. Bullock, 1969; Y. D. Vincent, В. Duffy, 1972).
Роль того или иного вариабельного параметра функциональ-
ной активности нейронов в информационной деятельности опре-
деляется особенностями структурной их организации, топогра-
фическими характеристиками, функциональным назначением
и т. д. Так, у рецепторных клеток, выполняющих функцию сен-
сорных датчиков в организме, различные виды внешней стиму-
лирующей энергии вызывают образование градуальных местных
генераторных электрических потенциалов, которые, в свою оче-
редь, становятся основой формирования распространяющихся
потенциалов действия, поступающих в дальнейшем в централь-
ные нервные структуры соответствующей анализаторной систе-
мы. Хотя у разных рецепторов на входе информация представ-
лена различными сигналами, на выходе их, как правило, наблю-
61
дается одна и та же форма сигналов — импульсные потенциа-
лы. Рецепторные образования у высших млекопитающих, фор-
мирующиеся в результате длительного эволюционного развития
животных организмов в процессе совершенствования их приспо-
собительного поведения, отличаются чрезвычайной чувствитель-
ностью к адекватным раздражителям. Достаточно' отметить, что
возбуждение фоторецепторной клетки может быть достигнуто
одним фотоном, обонятельной рецепторной клетки — 1—4 моле-
кулами пахучего вещества, слуховых рецепторов — колебаниями
с амплитудой, равной 0,1 диаметра атома водорода.
Многим рецепторным нейронам свойственно частотное коди-
рование, при котором интенсивность стимула отображается,
трансформируется в разную частоту генерации импульсных по-
тенциалов в афферентном нервном волокне. В экспериментах
это впервые было показано в работе И. Эдриана (Е. Adrian,
1932). Частота потенциалов действия в афферентных волокнах
обычно находится в логарифмической зависимости от силы раз-
дражения. Этой логарифмической зависимости придается боль-
шое значение в эффективном функционировании рецепторных
образований нервной системы. Так, в работе У. Раштона (1964}
было показано, что постоянство дискриминационного отноше-
ния сигнал/шум обеспечивается в периферической рецепторной
системе зрительного анализатора у рыбы мечехвоста именно ло-
гарифмической зависимостью между интенсивностью стимула и
реакцией рецепторной клетки.
В исследованиях С. Хагивары и X. Мориты (S. Hagiwara,
Н. Morita, 1963), Т. Сцабо (Т. Szabo, 1967), Т. Сцабо и С. Ха-
гивары (Т. Szabo, S. Hagiwara, 1967), Т. X. Баллока и С. Чичуку
(Т. Н. Bullock, S. Chichuku, 1965) установлено, что в электриче-
ском органе у рыб имеют место 2 типа рецепторных клеток, ко-
дирующих интенсивность внешнего сигнала либо латентным пе-
риодом (фазовым скачком), либо изменением вероятности цик-
лов разряда клеток электрического органа. Как известно, сущ-
ность работы электрического органа, которым обладают некото-
рые виды рыб, заключается в том, что разряды клеток электри-
ческого органа образуют некоторое электрическое поле, интен-
сивность которого изменяется в связи с нахождением в воде
различных объектов. Эти изменения приводят к перераспределе-
нию плотности тока в различных участках поверхности тела ры-
бы, что и становится сигналом, определяющим особенности пере-
распределения импульсных потенциалов в нервном волокне
«электрорецептора» рыбы. Исследованиями С. Хагивары и X. Мо-
риты (S. Hagiwara, Н. Morita, 1963) было установлено, что в
нервных волокнах боковой линии рыб кодирование информации
62
происходит либо градуальным изменением частоты импульсных
потенциалов, либо изменением вероятности наступления импуль-
са в нервном волокне после каждого раздражающего стимула
(«вероятностное» кодирование). Интересные результаты об осо-
бенностях функционирования фазовых и вероятностных кодеров
электрочувствительных волокон нерва латеральной линии у не-
которых электрических угрей (Eigenmannia virescens) описаны
X. Шайгом и др. (Н. Scheigh е. а., 1973) в опытах с блокирова-
нием разрядов электрического органа при помощи кураре
(рис. 13). Показано, что при вероятностном кодировании актив-
дб -40 -30 -20 -10 0 Дб -40 -30 -20 -10 0
Рис. 13. Схема работы «фазового» и «вероятностного» кодеров (по
Н. ScheUh а. о, 1973). По оси абсцисс — интенсивность стимула, по оси
ординат—фазовый скачок (а) и вероятность пропуска импульса (б). Сплош-
ная линия характеризует диапазон функционирования «фазового», пре-
рывистая — «вероятностного» кодера
ность электрорецепторов находится в вероятностной зависимости
от импульсных разрядов электрического органа. Вероятностное
кодирование заключается в данном случае в том, что в состоя-
нии покоя электрорецепторы в ответ на разряды электрического
органа генерируют импульсы нерегулярно; .по мере увеличения
интенсивности стимула вероятность появления импульсных по-
тенциалов в электрорецепторах плавно повышается, в резуль-
тате чего животное получает возможность оценивать интенсив-
ность стимула.
Для большинства центральных и эфферентных нейронов ос-
новными носителями информации являются импульсные потоки,
а в качестве кодового элемента могут выступать любые стати-
63
стические «параметры импульсной активности, градуально изме-
няющиеся под влиянием афферентных стимулов разной интен-
сивности. Многочисленные экспериментальные исследования в
этой области показали, что различные статистические характе-
ристики импульсных потоков нервных клеток имеют важное ин-
формационное значение в процессе функционирования нейронов
и удовлетворяют условиям, требованиям, предъявляемым к кан-
дидатам в кодовые элементы, согласно определению Д. X. Пер-
кела и Т. X. Баллока (D. Н. Perkel, Т. Н. Bullock, 1968). По
мнению авторов, для того чтобы какой-либо статистический па-
раметр импульсного потока считать возможным кандидатом ко-
да, необходимо раскрыть его формальные свойства и, во-вторых,
выявить условия, при которых такое кодирование практически
используется в системе связи, образованной нервными структу-
рами. В качестве кодового элемента, информационного показа-
теля, могут выступать средняя частота импульсации, средний
межимпульсный интервал, симметрия моды, картина узора меж-
импульсных интервалов и др. (S. Hagiwara, 1954; S. Wiersma,
R. Т. Adams, 1950; O.-J. Grusser e. a., 1962; G. Werner, V. B. Mo-
untcastle, 1963, 1965; R. W. Rodieck e. a., 1962; J. P. Segundo e. a.,
1963, 1966; G. P. Poggio, L. J. Viernstein, 1964; A. Herz e. a.,
1964; В. Я. Пятигорский, 1966; J. C. Levy, 1968; M. А. Куликов,
1968; F. Ratliff, 1968; A. B. Kogan, O. G. Ghorayan, 1973;
G. P. Moore e. a., 1966; G. Gestri e. a., 1966; О. Г. Чораян,
1975—1976, В. Д. Глезер и др., 1968; и др.). Постоянство ампли-
туды и длительности отдельных импульсных потенциалов сви-
детельствует о том, что многие нейроны попользуют частотное
кодирование информации (A. Herz е. а., 1964; Е. Adrian, 1932;
Н. В. Barlow, 1959, 1961; Н. Т. Herman, Z. Stark, 1962; Н. Н. Ва-
силевский, 1966, 1968; С. A. Terzuolo, Е. J. Bayly, 1968; Р.М. Ме-
щерский, 1972; R. Stein, 1967; и др.), при котором интенсивность
стимула отражается в количестве распространяющихся потен-
циалов действия, генерируемых нервной клеткой в фиксирован-
ном отрезке времени.
Результаты нейрофизиологических исследований Ф. Рэтлифа
и др. (F. Ratliff, 1968) указывают на то, что при одной и той же
средней частоте импульсации дисперсия межимпульсных интер-
валов в разряде темноадаптированной эксцентрической клетки
рыбы мечехвоста (Limulus) больше, чем у адаптированной к
свету клетки. Из этого факта делается вывод о том, что инфор-
мация об адаптации (о степени адаптации) в зрительной систе-
ме животного может передаваться, кодироваться величиной ко-
эффициента вариации импульсного потока, отражающего сте-
пень дисперсии межимпульсных интервалов.
64
Возможность использования определенного узора межим-
пульсных интервалов в качестве кодовых элементов при восприя-
тии и обработке информации в нервной системе впервые была
установлена С. Вирсма и Р.Т. Адамсом (S.Viersma, R. Т. Adams,
1950). В опытах этих авторов было показано, что нервно-мышеч-
ное соединение ракообразных обладает значительной чувстви-
тельностью к распределению во времени импульсных потен-
циалов: более сильные сокращения мышцы чаще наступали при
чередующихся коротких и длинных сериях импульсных потен-
циалов, чем при одинаковых межимпульсных интервалах. Ана-
логичные результаты были получены в работе Дж. П. Сегундо-
с сотрудниками (J. Р. Segundo е. а., 1966), обнаружившем вы-
сокую чувствительность к временному узору импульсных потен-
циалов у ганглионарных клеток в центральной нервной системе-
аплизии: три импульсных потенциала в фиксированном времен-
ном отрезке с варьирующим положением среднего импульсного
потенциала вызывали значительные различия в постсинаптиче-
ских эффектах. Авторы обнаружили также существенные разли-
чия в эффекте стохастически распределенных во времени пачек
импульсных потенциалов с постоянньш количеством импульсных
сигналов в пачке и фиксированным общим временем действия
этих импульсных потенциалов.
Даже в экспериментально контролируемых условиях постоян-
ства информационного входа нервной клетки (или отсутствия
каких-либо стимулов) регистрируемый от нейрона импульсный
поток является нерегулярным, что послужило основанием для
разработки стохастических моделей генерации импульсных раз-
рядов (S. Hagiwara, 1954; A. R. Adolph, 1959, 1964; G. В. Arden,.
U. Soderberg, 1961; W. I.-Mc Gill, 1963; Н. Н. Преображенский,
Н. В. Яро-вицкий, 1963; D. Н. Perkel е. а., 1964; М. Теп Ноореп
е. а., 1963; G. L. Gerstein, В. Mandelbrot, 1964; А. Б. Коган
и др., 1966; М. Теп Ноореп, Н. A. Reuver, 1968; А. Р. Адольф,
1970).
Анализируя статистику импульсных потоков, генерируемых
проприорецепторами портняжной мышцы лягушки, С. Хагивара
(S. Hagiwara, 1954) пришел к выводу, что генерация распро-
страняющихся потенциалов действия обусловливается 2 незави-
симыми процессами. Один из них, доступный прогнозированию,—
динамика восстановления величины порога клетки (Тр), кото-
рый можно определить по формуле
ТР=А exp(C/t),
где А — величина реобазы;
65
t — отрезок времени после предыдущего импульсного потен-
циала.
С — константа для данной клетки.
Второй непредсказуемый фактор заключается в небольших
флюктуациях величины порога Тп, определяемых как некоторая
функция от времени: Tn=f(t), где f(t) — случайный процесс.
Флюктуации латентных периодов импульсных разрядов на
идентичные стимулы связывались А. А. Вервееном и X. И.Дерк-
соном (A. A. Verveen, Н. Е. Derksen, 1965) с соответствующими
колебаниями мембранного потенциала. Нерегулярные узоры
импульсных разрядов, генерируемые в ответ на постоянные
•стимулы клетками слухового анализатора кошки, описаны
Л. Дж. Вирнштейном и Р. Г. Гроссманом (L. J. Viernstein,
R. G. Grossman, 1961), Н. Дж. Кянгом (N. Y-S. Kiang, 1966). На
математических моделях флюктуации возбудимости нервных кле-
ток, описанных в работах М. Тен Хупена и др. (М. Теп Ноореп
<е. а., 1963) и Э. В. Нарушевичус и В. А. Магинскас (1966), по-
казано, что величина порога возбуждения клетки является объ-
ектом шумоподобных вариаций.
Физиологической основой таких колебаний порога возбужде-
ния нервной клетки являются небольшие (0,5 мв) деполяриза-
ционные изменения мембраны нейронов. Деполяризационные из-
менения мембраны клеток соматосенсорной коры кошки описаны
Ч. Ли (С.—L. Li, 1959, 1961). Р. Б. Штейн (R. В. Stein, 1967)
считает, что вариабельность разряда нервной клетки связана с
флюктуациями мембранного потенциала, которые в свою очередь
обусловлены изменением проницаемости нейрональной мем-
браны.
В работе А. Р. Мартина и К. Л. Бренча (A. R. Martin,
С. L. Branch, 1958) показано, что частота встречаемости любых
межимпульсных интервалов в активности бецовских пирамидных
клеток кошки описывается экспоненциальным распределением
F = N-At/Texp
где N —общее число наблюдаемых интервалов;
Т —। длина среднего межимпульсного интервала.
Согласно Р. Г. Гроссману и Л. Дж. Вирнштейну (R.G. Gross-
man, L. J. Viernstein, 1961), распределение межимпульсных ин-
тервалов большинства нейронов слухового ядра кошки хорошо
аппроксимируется кривой экспоненциального распределения
P(t)=re~r/t,
66
где P(t)—вероятностная плотность межимпульсных интерва-
лов длительностью t;
г — средняя частота разряда нейрона.
По данным Г. В. Ардена и У. Зедерберга (G. В. Arden,
U. Soderberg, 1961), А. Герца и др. (A. Herz а. о., 1964), им-
пульсные потоки фоновой активности волокон зрительного нер-
ва представляют собой стационарный пуассоновский процесс с
распределением вида
Pk(t)= nFe-xt’
где X — интенсивность потока импульсов.
Экспоненциальная форма большинства интервальных распре-
делений импульсных потоков, регистрируемых из волокон зри-
тельного тракта, а также низкие значения сериальной корреля-
ции последовательных межимпульсных интервалов, по мнению
Дж. О. Фастера и др. (J. О. Faster а. о., 1965), указывают на
случайный характер последовательности импульсных потенциа-
лов, генерируемых ганглиозными клетками сетчатки. В цент-
ральных нервных структурах импульсные потоки клеток стано-
вятся более упорядоченными и значительно отличаются от слу-
чайного распределения (A. Herz е. а., 1964; О. Г. Чораян, 1966г
1975; и др.).
Для оценки степени стохастичности в структуре импульсных
потоков клеток центральных нервных структур можно исполь-
зовать метод сравнения кривых вероятностного распределения
межимпульсных интервалов активности центральных нейронов
и сравнение их с известными кривыми наиболее часто встреча-
ющихся распределений (нормального и экспоненциального). Для
определения вероятности распределения гауссовского потока ис-
пользуется уравнение функции Лапласа (О. Г. Чораян, 1966)
позволяющее определить вероятность нахождения любой jnop-
мально распределенной величины в интервале Xj<X<X2 (X —
средний интервал, о — среднее квадратическое отклонение, Ф —
значение интегральной функции Лапласа). Для построения кри-
вой экспоненциального распределения было использовано урав-
нение, примененное А. Р. Мартином и К. Л. Бренчом (A. R. Mar-
tin, С. L. Branch, 1958). Такой анализ, пример которого приве-
ден на рис. 14, показал, что для подавляющего большинства ис-
67
следованных нейронов центрального отдела зрительного анали-
затора лягушки характерны достоверные различия между эмпи-
рическими распределениями и теоретически рассчитанными рас-
пределениями случайных процессов (О. Г. Чораян, 1966, 1969).
Рис. 14. Гистограмма вероятностного распределения межимпульсных
интервалов нейрона центрального отдела зрительного анализатора и кривые
нормального (пунктирная линия) и экспоненциального (штриховая линия)
распределений
Одна из первых моделей функционирования нервных струк-
тур, в которой была предпринята попытка использования эле-
ментов теории вероятностей, описана в работе X. Д. Ландала
(Н. D. Landahl, цит. по: А. Б. Коган и др., 1977). Впоследствии
интересная модель работы нервной сети на основе вероятност-
ной концепции возбуждения нервных клеток была описана
А. Шимбелом и А. Рапопортом (A. Shimbel, A. Rapoport, 1948),
построенная с учетом многих, варьирующих от клетки к клетке,
параметров функционирования реальных нейронов порога воз-
буждения, величины синаптической задержки, рефрактерного
периода и т. д.
Описание вероятностных моделей разрядов нервных клеток
и нервных сетей приводится в целом ряде работ (R. Beurle,
1956, 1959, 1962; А. Б. Коган и др., 1966; М. Теп Ноореп е. а.,
1963; М. Теп Ноореп, 1964, 1966, a, b; Е. R. Caianiello е. а., 1967;
A. Luca De, М. Riccardi, 1968; М. Теп Ноореп, Н. A. Reuver,
1968; J. D. Cowan, 1968; J. С. Levy, 1968; F. Ventriglia, 1974).
В основе статистической нейромеханики Дж. Д. Коуэна (J. D. Со-
68
wan, 1968) лежит выведенное автором «уравнение движения»,
являющееся нелинейным дифференциальным уравнением, отра-
жающим, воспроизводящим многие изменения в активности
нервных сетей как результат стохастического взаимодействия
внутри и между ансамблями нервных клеток. В некоторых слу-
чаях в этих уравнениях может быть представлена осцилляторная
активность нейронов, характеризуемая средними значениями
функциональных характеристик ансамбля нервных клеток.
Математическая модель работы нервных сетей, основанная
на использовании статистической нейромехапики, описана в ра-
боте Ф. Вентриглия (F. Ventriglia, 1974), в которой нейронные
сети рассматриваются как газ, в состав которого входят как ча-
стицы с фиксированным положением (нейроны), так и свобод-
ные частицы (нейронные импульсы). В основе модели лежит до-
пущение, что путь прохождения нервных импульсов не обяза-
тельно связан с физической структурой нервной сети. Утвержда-
ется, что для макроскопического модельного описания достаточ-
но рассмотреть распределение скорости (и положения) импуль-
сов, а также положения нейронов в каждый момент времени.
Приведенные автором дифференциальные уравнения удовлетво-
рительно описывают динамику изменения распределения этих
параметров нейронной сети.
Вероятностные аспекты частотного кода рассмотрены в ра-
боте Р. Б. Штейна (R. В. Stein, 1967). На рис. 15 представлен
идеализированный разряд из серии импульсных потенциалов со
средней частотой vmin. В течение периода времени t на клетку
Р(х) Р(у/Х) Р(у)
Рис. 15. Схематизированный импульсный разряд со средней частотой,
изменяющейся под влиянием раздражения, приложенного к клетке в тече-
ние времени t (по R. В. Stein, 1967)
69
действует стимул (генераторный потенциал, пресинаптический
импульсный разряд и т. д.), изменяющий частоту спонтанных
потенциалов действия v. Среднее число импульсных потенциалов,
являющееся результатом действия стимула, есть X. Действи-
тельное (реально наблюдаемое в ответ на каждый стимул) ко-
личество импульсных потенциалов — Y, вызванное отдельным
стимулом, является случайной вариабельной серией. Три вида
вероятностных величин ассоциируются с X и Y:
Р(Х)—вероятность того, что ответ на данное приложение
стимула есть X;
iP(Y)—вероятность ответа Y;
P(Y/X)—вероятность того, что данный стимул X генерирует
Y импульсов.
Эта последняя вероятность может быть использована для ха-
рактеристики информационного канала в нейронных системах
связи.
•Стохастический компонент вызванного импульсного потока
нервных клеток имеет множественную природу: случайные изме-
нения последовательных межимпульсных интервалов могут быть
результатом неконтролируемых изменений локального метабо-
лизма нервной клетки, результатом спонтанного освобождения
квантов медиатора в синаптической щели из окончания пресинап-
тического волокна и т. д.
П. Фет и Б. Катц (Р. Fatt, В. Katz, 1950) в опытах с внутри-
клеточной регистрацией микроэлектродом местных деполяриза-
ционных изменений в области концевой пластинки в волокнах
скелетной мышцы лягушки в отсутствие возбуждения двигатель-
ного нерва впервые наблюдали случайное распределение локаль-
ных электрических потенциалов. Мышечная концевая пластин-
ка — специфическое нервно-мышечное синаптическое образование
с химической формой передачи возбуждения. «Биологический
шум» в нервно-мышечном препарате П. Фет и Б. Катц (Р. Fatt,
В. Katz, 1952) характеризовали как случайную последователь-
ность миниатюрных потенциалов концевой пластинки с ампли-
тудой порядка 1/100 нормальной величины потенциала действия
в ответ на импульсные потенциалы в двигательном нерве. Ми-
ниатюрные потенциалы постоянной амплитуды возникают в не-
предсказуемые моменты времени со средней частотой от I до
100 гц. В последующих работах (J. Del Castillo, В. Katz, 1954;
R. Birks, F. Me Intosh, 1957; P. Fatt, 1974; B. Katz, J962) было
установлено, что миниатюрные потенциалы концевой пластинки
являются результатом спонтанного квантованного освобождения
ацетилхолина в пресинаптических волокнах, происходящего от-
дельными порциями по несколько тысяч молекул и в случайные
70
моменты времени независимо от времени появления предыдущих
потенциалов. По данным П. Фета и Б. Катца (Р. Fatt, В. Katz,
1952), распределение интервалов между спонтанными миниатюр-
ными потенциалами в нервно-мышечном синапсе соответствует
приведенной ранее формуле экспоненциального распределения
F=N At/T exp (—t/T).
Спонтанные миниатюрные потенциалы концевой пластинки
наблюдали также А. В. Лили (A. W. Liley, 1956), Дж. Дудел и
С. Кафлер (J. Dudel, S. Kuffler, 1961).
Случайно распределенные во времени электрические потен-
циалы были зарегистрированы в фоторецепторах фасеточного
глаза рыбы мечехвоста (S. Yeandle, 1958; М. G. Fuortes, 1959;
A. R. Adolph, 1964), саранчи (J. Н. Scholes, 1964), пиявки
(J. В. Walther, 1966). Разновидностью случайной нейроэлектри-
ческой активности в центральной нервной системе как позвоноч-
ных (L. G. Brock е. а., 1952; S. Nishi, К. Koketsu, 1960), так и
беспозвоночных (G. A. Kerkut, R. Thomas, 1963; К. Kusano,
S. Hagiwara, 1961) являются спонтанные синаптические потен-
циалы. Важно отметить, что такая случайная нейроэлектриче-
ская активность в функциональном отношении не тождественна
информационному шуму, она часто выполняет важную интегра-
тивную функцию в работе нервной клетки. Непосредственное
участие в информационных процессах случайных синаптических
потенциалов проявляется в том, что вызываемые ими флюктуа-
ции суммарного мембранного потенциала клетки являются по-
казателем интегральной функции нейрона. В работе А. Р. Адоль-
фа (A. R. Adolph, 1964) показано, что после нескольких минут
темновой адаптации омматидия микроэлектродом, расположен-
ным в эксцентрической клетке, регистрируются чаще всего под-
пороговые флюктуации мембранного потенциала, которые имеют
случайное распределение как во времени, так и по амплитуде
(рис. 16), но тем не менее, суммируясь с афферентными воздей-
ствиями, могут оказывать существенное влияние на распреде-
ление во времени распространяющихся потенциалов действия.
Информационное значение вероятностной организации пере-
дачи возбуждения через синаптическое образование рассмотрено
в работе А. М. Аттли (А. М. Uttley, 1959, 1965 а, Ь). Предпола-
гается, что синаптическое соединение характеризуется значи-
тельной вариабельностью в отношении передачи через него им-
пульсного сигнала, которую можно оценить по величине пост-
сина.птического потенциала. Величина последнего в свою оче-
редь может быть определена шенноновской энтропией, рассчи-
71
Рис. 16. Флюктуация медленного потенциала при различных уровнях
интенсивности света, действующего на фоторецепторы мечехвоста (по
A. R. Adolph, 1964). Числа слева указывают коэффициенты ослабления
света, в логарифмических единицах
тываемой на основе вероятностных измерений входного и вы-
ходного сигнала в синаптическом соединении нервных клеток.
Рассмотрим нейрон X, аксонный отросток которого заканчивает-
ся на мембране клетки Y; средняя частота их импульсных раз-
рядов обозначается соответственно как f(X) и f(Y). Если
f(XY) — средняя частота, с которой потенциалы действия в аксо-
не клетки X вызывают импульсный разряд в нейроне Y, тогда
информацию, передаваемую через данный синапс, можно вычис-
лить через шенноновское определение энтропии
I = log -МО.
Теоретически рассматривая информационную функцию си-
напса в связи с динамикой частоты импульсации соответству-
ющих нейронов, А.М. Аттли (А. М. Uttley, 1965 b) делает следу-
ющие выводы относительно динамики информационных свойств
72
синапса, ассоциируемых с известными физиологическими фено-
менами: 1) если повторное идентичное возбуждение в клетке X
приводит к усилению возбуждения, то улучшается процесс пе-
редачи информации в данном синапсе (физиологический фено-
мен облегчения); 2) если возбуждение нейрона X не сопровож-
дается возбуждением клетки Y (или приводит к более ослаблен-
ному возбуждению), трансинформационная функция синапса па-
дает (феномен физиологической адаптации после начальной ак-
тивации).
Большое число синаптических контактов каждой нервной
клетки с другими (число таких контактов, по данным Д. А. Шол-
ла (D. A. Sholl, 1956), Дж. Бока (J. Т. Вок, 1959), доходит
до 10000) создает широкий выбор пути движения информации
по нейронным каналам связи. При этом реализация каждого
конкретного пути зависит от различных факторов, многие из ко-
торых носят случайный непредсказуемый характер. Вместе с
тем анализ физиологических процессов, лежащих в основе си-
наптической передачи сигналов, позволяет рассматривать синап-
тическое образование как вероятностную систему. Поэтому сто-
хастичность фунционирования каждой отдельной нервной клетки
может в значительной степени обусловливаться неопределен-
ностью синаптической передачи через множество синаптических
соединений в полисинаптических каналах связи. При большом
числе независимых возбуждающих входов, что действительно
имеет место в центральных нервных структурах, становится по-
нятным случайный характер наступления момента генерации
импульсного ^потенциала в нервной клетке, какова бы ни была
временная и пространственная организация этих независимых
входных импульсных потоков (число таких независимых источ-
ников возбуждения для коркового нейрона составляет, по дан-
ным Б. Бернса (1969), примерно 2000). Нервные клетки — источ-
ники дискретных сообщений и вместе с тем каналы передачи этих
сигналов—генерируют последовательности распространяющих-
ся потенциалов действия, порядок следования которых часто
случаен, непредсказуем. Оценка генерации и передачи этих сиг-
налов в нервной системе, ino-видимому, возможна лишь на язы-
ке теории вероятностей, вероятностных соотношений взаимосвя-
занных нервных клеток. В частности, такой взгляд на природу
информационных процессов в нервной системе нашел отражение
в представлениях о непропорциональности и неаддитивности
в работе мозга (Д. О. Вальтер и В. Р. Адей, 1968).
Таким образом, вероятностный характер информационной
деятельности нейрона определяется множеством экзо- и эндоген-
ных факторов: случайной пространственно-временной мозаикой
73
поступающих к клетке возбуждающих и тормозящих импульсов,
спонтанным выделением квантов медиатора из окончаний пре-
синаптического волокна, флюктуацией порога возбуждения ней-
рона под влиянием вариабельных процессов локального метабо-
лизма нервной клетки и т. д.
3. МЕХАНИЗМЫ ОБЕСПЕЧЕНИЯ НАДЕЖНОЙ
ПЕРЕДАЧИ ИНФОРМАЦИИ
Проблема организации надежного функционирования инфор-
мационно-управляющих систем мозга привлекает большое вни-
мание не только биологов и медиков, но и широкий круг спе-
циалистов в области техники, для которых удивительные приме-
ры эффективной и надежной работы центральных нервных струк-
тур служат идеальным образцом, эталоном при конструирова-
нии и создании искусственных систем автоматического управле-
ния и связи.
Хорошо известны клинические и экспериментальные данные,
свидетельствующие о чрезвычайной гибкости и надежности си-
стем мозга, огромные компенсаторные возможности нервных
структур. Достаточно 'вспомнить, что всемирно известный фран-
цузский ученый Л. Пастер сделал все свои открытия, владея
лишь «половиной мозга» (одно полушарие головного мозга бы-
ло полностью поражено после болезни). Или другой пример с
Финеасом Гейджем, мастером дорожников-строителей, получив-
шим серьезную травму: железная палка диаметром около 3 см
насквозь пронзила головной мозг, войдя через его левую щеку
и выйдя около темени. После такой серьезной травмы он про-
жил еще 12 лет, причем все это время он был дееспособной лич-
ностью с хорошей памятью (Д. Вулдридж, 1965).
В соответствующей литературе описаны многие примеры бла-
гополучного исхода клинических операций удаления обширных
областей мозговой ткани при новообразованиях и различных
других поражениях структур головного мозга. Еще более мно-
гочисленные примеры известны из практики нейрофизиологиче-
ских исследований, в которых экстирпация или функциональное
выключение значительных объемов мозговой ткани не приводи-
ли к сколько-нибудь заметным функциональным изменениям,
нарушениям.
Традиционным, получившим распространение в различных
областях техники, способом обеспечения надежности является
структурное или функциональное дублирование, создание опре-
деленного вида избыточности сообщения, канальной избыточно-
сти и т. д. Однако примеры дублирования в целях обеспечения
74
надежности известны не только (а может быть, и не столько) в
технике, но и в биологии, медицине. Отмечая известный факт,
что работа многих жизненно важных физиологических систем
организма обычно дублируется рядом других систем или органов,
и придавая этому большое функциональное значение в деятель-
ности животного организма, Баркрофт (1937) писал: «Дубли-
рование встречается так часто, что едва ли является случайным.
Очевидно, оно составляет определенное свойство архитектуры
функции и производит тем большее впечатление, что оно осу-
ществляется столь различными путями. Но я часто удивляюсь,
почему не дублировано само сердце. Меня всегда удивляет, что
в организме только одно сердце».
Многие свойства клеточной организации центральной нерв-
ной системы указывают на наличие частичного дублирования и
резервирования в процессе функционирования рабочих механиз-
мов мозга. Биологическое значение такого дублирования заклю-
чается в повышении надежности деятельности мозга за счет опе-
ративного введения, по мере надобности, в систему дополнитель-
ных элементов, т. е. путем использования принципа избыточно-
сти. Объем информационной памяти нейронных элементов мозга
оценивается примерно в 1013—1020 битов (Дж. Нейман, 1956).
ЭВМ, использующая в качестве памяти 1010 ячеек (что пример-
но соответствует количеству нервных клеток мозга), не может
иметь память, превышающую 1010 битов (Б. И. Крюков, П. О. Ма-
каров, 1963). Следовательно, надо допустить, что в основе орга-
низации рабочих механизмов мозга лежит принцип, который
может обеспечивать высокую точность и надежность нейронных
конструкций мозга при гораздо меньшей избыточности его эле-
ментов. В литературе описан ряд модельных исследований раз-
личных способов более экономичного использования рабочих
элементов системы, обеспечивающих высокую надежность ее
функционирования. Так, в работе Дж. Неймана (1956) описан
способ построения органа высокой надежности, заключающийся
в- том, что информация передается не одиночной линией — ана-
логом нервного волокна,— а пучком из большого числа линий
связи. Такой механизм мультиканальной передачи идентичной
информации, по мнению автора, должен лежать в основе кон-
струкции высоконадежного автомата. Однако при таком спосо-
бе ’повышения надежности системы необходимо очень большое
число параллельно организованных каналов связи. Так, при на-
чальной ненадежности элемента порядка 0,005 для получения
вероятности ошибки 10-10 при передаче информации в системе
необходимы 10000 линий. В модельных исследованиях Дж. Коу-
эна (J. Cowan, 1968) модульная избыточность искусственной
75
нейронной сети обеспечивает относительно высокую надежность
при гораздо меньшем числе избыточных элементов, чем это тре-
буется в модели Неймана.
Многочисленные нейрофизиологические исследования, ре-
зультаты которых приведены в главе 2, дают основание пола-
гать, что подобный принцип уже давно используется в живой
природе в виде одного из проявлений кооперативного принципа
организации и функционирования в нервной системе — формиро-
вания вероятностных нейронных ансамблей (А. Б. Коган, 1964—
1976; О. Г. Чораян, 1963—1976; Н. П. Бехтерева, 1971; М. М. Ха-
нанашвили и др., 1975; О. С. Адрианов, 1976; и др.). Когда эво-
люционное совершенствование мозга достигает такой степени,
при которой гибкость, пластичность, надежность его функцио-
нирования уже не может быть обеспечена простым дублирова-
нием и резервированием, возникает, формируется новый принцип
функциональной организации нейронов в структурно-функцио-
нальные объединения — вероятностные нейронные ансамбли,
обеспечивая тем самым в каждой конкретной ситуации опера-
тивную избыточность нервных клеток, каждая из которых в дру-
гое время при другой ситуации может быть включена в другой
клеточный ансамбль.
Четко наблюдаемое нарастание аритмии импульсной актив-
ности в филогенезе (А. Б. Коган, О. Г. Чораян, 1965; О. Г. Чо-
раян, 1976) направлено на расширение рабочего диапазона сиг-
нальных значений статистических параметров импульсной актив-
ности, а следовательно, и к увеличению информационной емко-
сти таких нейронных каналов связи. Вместе с тем в рандомиза-
ции временной структуры импульсной активности, по-видимому,
проявляется механизм обеспечения надежности и адаптивных
свойств нервной системы, совершенствующихся в ходе эволю-
ционного развития животного мира. Как подчеркивал П. К. Ано-
хин (1971), природа развивалась в сторону увеличения степеней
свободы элементов функциональной системы, в сторону повы-
шения числа возможных взаимодействий между ними. Физиоло-
гической основой этих последних на примере отдельных нервных
клеток, очевидно, и является нарастание стохастичности времен-
ной организации генерируемых ими импульсных потоков.
Хорошо известно, что, используя избыточную информацию,
всегда можно добиться надежности передачи даже в присутствии
шума. Поэтому одним из основных способов борьбы с помехами
в каналах с шумами является повторение сообщения, которое
наряду с организацией коррелятивных взаимозависимостей..м£Ж-
ду кодовыми элементами сообщения способствует повышению
надежности передачи информации в сложных многоканальных
76
системах связи. Необходимое соотношение между уровнем шу-
мов и избыточной информацией приведено в теореме К. Шенно-
на о канале с шумами (К. Шеннон, 1963): если некоторое коли-
чество информации нужно передать с идеальной надежностью-
в присутствии шума, то необходимо по крайней мере такое ко-
личество избыточной информации, которое будет равно количе-
ству неоднозначности, обусловленной шумом (Г. Кастлер, 1960;
К. Шеннон, 1963). Количество эффективно передаваемой инфор-
мации равно общему количеству передаваемой информации
минус количество информации, теряемой вследствие шума.
Поэтому источник сообщения может надежно передать при-
емнику некоторое количество информации в присутствии шума
лишь при условии, что передается намного большее количество^
информации.
В информационных системах принцип избыточности опреде-
ленным образом связан с экономичностью работы, в результате
чего эффективность функционирования системы в целом в раз-
ные моменты времени может ассоциироваться в большей мере
то с одним, то с другим критерием (избыточностью или эконо-
мичностью). Применительно к деятельности нервной системы
это означает, что в разных ситуациях (в разном функциональ-
ном состоянии, при действии повторных идентичных стимулов,,
на разных стадиях адаптации, при формировании определенных
навыков целесообразного поведения и т. д.) эффективное функ-
ционирование рабочих конструкций мозга (как и отдельных нерв-
ных клеток) определяется разной динамикой, разным соотно-
шением показателей избыточности и экономичности. Так, напри-
мер, начально избыточная реакция любого возбудимого элемен-
та (как на уровне клеточной реакции, так и системной, напри-
мер, ориентировочной реакции животного организма) сменяется
впоследствии более экономичной формой ответа, характеризу-
ющейся уменьшением показателя избыточности. Это хорошо»
можно видеть на . примере анализа динамики избыточности фо-
новых и вызванных импульсных потоков, рассчитываемой по»
энтропийным показателям зависимости последовательных меж-
импульсных интервалов. Известно, что максимальную энтропию'
имеют независимые сигналы, в случае же статистической зави-
симости между сигналами, генерируемыми источником, энтропия
сигналов уменьшается параллельно с повышением тесноты связи
между ними. Представим импульсный поток нервной клетки в..
виде последовательности чисел 0 и 1 (0 означает отсутствие,.
1 — наличие импульсного потенциала) в квантованных во време-
ни равных отрезках (величина этого кванта должна быть мень-
ше минимального межимпульсного интервала анализируемой им-
77
пульсной серии. Тогда Pt и Pj означают соответственно полные
безусловные вероятности этих 2 различимых состояний нервной
клетки, a Pi/i; Pi/j; Pj/j и Pj/i — 4 условные вероятности, соответ-
ствующие разным комбинациям зависимости последовательных
состояний (0/0, 0/1; 1,0, 1/1). По значениям полных и условных
вероятностей можно рассчитать величину энтропии
Н = [Pi (P|/l 10g2 Pl/I + Pi/j 10g2 Pl/j) +
+ Pj (F*j/j l°g2 Pj/j + Pj/1 log2 Pj/1 )].
Избыточность (D) равна разности 1 и величины относитель-
ной энтропии (тт-):
по
D = 1 - — ,
Но
где Но — максимальная энтропия при равновероятном распре-
делении.
Вычисленные таким способом величины показателя избы-
точности для фоновых и вызванных импульсных потоков нерв-
ных клеток приведены в табл. 5.
Таблица 5
Динамика избыточности импульсных потоков нервных клеток
центрального отдела зрительного анализатора лягушки при их возбуждении
№ нейрона Величина избыточности в фоновой импульсной активности Величина избыточности в вызванной импульсной активности
реакция на 1 — 15 световое раздражен. реакция на 16—20 световое раздражен.
1 0,89 0,68 0,49
2 0,85 0,71 0,60
3 0,72 0,63 0,48
4 0,70 0,53 0,39
5 0,68 0,60 0,54
6 0,67 0,52 0,47
7 0,63 0,55 0,49
8 0,61 0,50 0,38
9 0,56 0,47 0,40
10 0,51 0,44 0,38
Данные, приведенные в таблице, указывают на существенно
более высокие значения избыточности в фоновых импульсных
потоках (по сравнению с вызванной активностью). По мере на-
несения повторных идентичных стимулов величина избыточности
падает, стабилизируясь к 10—20 стимулу.
78
X. Б. Барлоу (Н. В. Barlow, 1968), рассматривая избыточ-
ность как полезное свойство информационного потока, повыша-
ющее его помехоустойчивость, предлагает различать 2 вида из-
быточности: внутреннюю, связанную с особенностями структур-
ной организации системы связи, и внешнюю избыточность вход-
ного импульсного потока. Среди множества способов определе-
ния помехоустойчивости системы связи наиболее простым и ши-
роко распространенным является вычисление логарифма вели-
чины, обратно пропорциональной вероятности ошибки при пере-
даче сигнала (регистрируя выходной сигнал по сравнению со
входным)
S-log!A,
где Ро — вероятность ошибки.
Некоторые результаты такого анализа на примере нейронных:
систем связи в узловой нервной системе беспозвоночных живот-
ных и зрительном анализаторе позвоночных ^приведены в табл. 6.
Из таблицы видно, что мера надежности передачи информации
прямо пропорциональна величине избыточности входного им-
пульсного потока, хотя в некоторых случаях такая связь обна-
Таблица 6-
Динамика показателя надежности передачи информации
и избыточности в различных нервных структурах (по О. Г. Чораяну, 1976)
Избыточность информа- ционной емкости нейронов зрительного анализатора лягушки Мера надеж- ности । Избыточность информа- ционной емкости нейронов речного рака Мера надеж- ности
ганглиозные клетки сетчатки клетки кры- ши сред- него мозга рецептор- ные нейроны 1 промежу- точные ней- роны 6-го брюшного ганглия
0.80 0,53 3,4 0,84 0,63 2,7
0,77 0,62 3,2 0,71 0,51 2,4
0,86 0,59 3,3 0,70 0,61 2,1
0,54 0,41 1,6 0,56 0,29 2,0
0,50 0,29 1,9 0,49 0,34 1,8
0,35 0,42 1,6 0,48 0,37 1,8
0,27 0,39 1,6 0,44 0,33 1,6
0,29 0,61 2,2 0,54 0,38 1,9
0,37 0,77 2,7 0,54 0,41 1,8
0,41 0,74 3,4 0,38 0,36 1,5
79
руживается и между мерой помехоустойчивости и избыточностью
выходного импульсного потока. Последнее может свидетельство-
вать о существовании обратной связи механизма обратной
афферентации, обеспечивающего динамическое перераспреде-
ление афферентной импульсации в реализации оптимальной,
для организма, целесообразной реакции системы (П. К. Ано-
хин, 1968).
Необходимость избыточного числа центральных компонентов
реакций нервной системы для обеспечения надежности ее функ-
ционирования обосновывается в работе -М. Веллера и Р. Берда
(М. Weller, R. Bird, 1973). О значительной структурной избыточ-
ности в реализации функций центральных нервных структур
свидетельствуют и результаты исследований С. М. Блинкова и
М. В. Пуцилло (1968), основанные на хирургических вмешатель-
ствах в структуры мозга — удалении некоторых участков цент-
ральных ядерных структур.
Основным критерием оценки надежности функционирования
нервной клетки является вероятность проведения возбуждения
от синаптических контактов на входе нейрона до концевых раз-
ветвлений его аксонного отростка (Б. С. Сотсков и др., 1972).
Ненадежность нервной клетки проявляется в виде генерации
ложного импульса или в «пропадании» отдельных импульсов в
ходе передачи информации. По данным Б. С. Сотскова и др.
(1972), надежность нейрона в среднем составляет 0,9—0,99.
Сравнение этой величины с показателями надежности элементов
современных радиоэлектронных систем (0,9999) показывает, на-
сколько низка надежность реальных нервных клеток. Тем не ме-
нее рабочие конструкции мозга, включающие тысячи и десятки
тысяч подобных ненадежных элементов, функционируют доста-
точно надежно и эффективно. Это может быть связано с неко-
торыми особенностями кодирования и передачи информации.
Во многих исследованиях (A. Rapoport, W. Horvath, 1960;
У. Розенблит, 1961; У. Раштон, 1964; В. Д. Глезер и др., 1968;
Т. Osawa, 1969; и др.) показано, что эффективным способом ко-
дирования информации при ее передаче является группирование
се в пачки.
Математическое обоснование природы надежной передачи
пачки импульсных потенциалов приводится в работе Л. С. Гам-
баряна и Ю. М. Гаспаряна (1968). Надежность функционирова-
ния системы связи, представленной соединением рецептора с
афферентным нервным волокном, авторы предлагают оценивать
вероятностью того, что из m импульсных потенциалов, последо-
вательно поступающих на вход, хотя бы п импульсов достигают
выхода системы. Тогда вероятность передачи информации по
80
волокну Pn, m, определяющую надежность системы рецептор —
волокно, можно вычислить следующим образом:
т—1
Pn,m = 1 - V РПК.
к=0
Это означает, что надежность системы прямо пропорциональ-
на п и обратно пропорциональна ш. Например, если вероятность
передачи одиночного импульса P(N)=0,5, п = 10, то РПт=
= 0,96875.
Следовательно, несмотря на достаточно низкую вероятность
передачи одиночного импульса P(N) в результате специфиче-
ского кодирования сообщения в пачку последовательных им-
пульсных потенциалов вероятность передачи информации в та-
кой системе значительно повышается. Эти результаты подтверж-
даются данными, полученными в модельных экспериментах на
ЭВМ И. Хартом и Г. -Пертиле (Е. Harbh, G. Pert’ile, 1972). Авто-
ры установили высокую надежность передачи пачки импульсов
в цепочку элементов, в то время как отдельные потенциалы пло-
хо воспринимаются, воспроизводятся на выходе такой цепочки.
Таким образом, надежность работы нервной системы являет-
ся одним из следствий ее организации, она вытекает из кон-
структивных особенностей составляющих нервную систему кле-
ток и ансамблей клеток, обусловлена многими факторами: нали-
чием большого числа входных каналов распространения возбуж-
дения, значительной структурной, функциональной и информа-
ционной избыточностью, резервированием нервных элементов
и т. д.
4. СЕМАНТИЧЕСКИЕ АСПЕКТЫ ИНФОРМАЦИОННОЙ
ДЕЯТЕЛЬНОСТИ НЕРВНОЙ СИСТЕМЫ
Вероятностный характер функциональных проявлений рабо-
чих конструкций мозга и образующих их отдельных нервных кле-
ток ставит вопрос о том, в какой мере наблюдаемая вариабель-
ность касается существенно важных, информационно значащих
параметров нервного кода. Вполне понятно, что допустимый диа-
пазон вариаций в различных формах реакций отдельных нерв-
ных клеток будет существенно различным для основных функ-
ционально важных и второстепенных, менее важных для реали-
зации данной функции статистических характеристик информа-
ции, закодированной в рассматриваемом импульсном потоке.
Понятно, что одни и те же вероятностные характеристики эле-
81
ментов сообщения в разном функциональном состоянии системы
связи или при решении разных задач могут иметь существенно
различное значение.
Решение этого вопроса оказывается в свою очередь тесно
связанным с проблемой выделения полезной информации в сум-
марном информационном потоке, циркулируемом в нервных ка-
налах связи. Многие авторы весьма скептически относятся к
статистической теории связи при попытке практического исполь-
зования ее в анализе информационной деятельности нервной си-
стемы, поскольку она не дает возможности оценить содержа-
тельную сторону сообщения, семантику информации. Статисти-
ческая теория информации не может выявить различия между
заурядной информацией и ценной информацией, если статисти-
ческая структура сообщения (содержащего эту информацию) и
свойства канала связи в обоих случаях идентичны. Информа-
ционная емкость сообщения в статистической теории связи за-
висит лишь от числа символов (от алфавита кода) и вероятности
каждого из символов. Это обстоятельство является одним из
основных факторов, объясняющих весьма скромные успехи ста-
тистической теории связи в биологии, в частности в нейрофизио-
логии, центральный объект которой (информационная деятель-
ность структур мозга) казалось бы, должен разрабатываться
именно на основе статистической теории связи К. Шеннона
(1963).
Отмечая слабые стороны статистической теории информации
применительно к анализу сложных закономерностей функциони-
рования биологических организмов, А. Сент-Дьердьи (1964) пи-
сал следующее: «Если какой-либо процесс 999 раз происходит
одним путем и только один раз — другим, то физик, не колеб-
лясь, скажет, что первый путь и есть истинный. Биология — это
наука о невероятном, и думаю, что в принципе для организма
существенны только статистически невероятные реакции». Эту
же особенность информационной деятельности живых систем
имел в виду И. И. Шмальгаузен (1968), когда он писал, что в
биологии основное значение приобретает качественная оценка
информации, в то время как количество информации, хотя и
имеет определенное значение, оказывается на втором месте.
Одним из подходов к количественному определению полезной
информации может служить вычисление информативности кле-
точных реакций, информационного содержания вызванных им-
пульсных разрядов нейрона. Импульсную реакцию нервной клет-
ки можно охарактеризовать через изменение вероятности на-
ступления определенных классов событий, в качестве которых
могут выступать частоты потенциалов действия или классы меж-
82
импульсных интервалов. Определив вероятности этих парамет-
ров фоновой и вызванной импульсной активности, можно вычис-
лить информативность импульсных реакций нервных клеток сле-
дующим образом (О. Г. Чораян, 1976):
i1 = siog2!k,
где Рн — вероятность класса событий в фоновой активности;
Рк — соответствующая вероятность в вызванной импульсной
активности.
Значения этих вероятностных величин определяются из постсти-
мульных гистограмм. В табл. 7 приведены некоторые результаты
такого анализа импульсных реакций клеток центральных нерв-
ных образований у разных животных.
Та б лица 7
Величина информативности клеточных реакций
Вид нервных клеток Величина показателя информативности
Нейроны висцерального ганглия виноградной улитки Нейроны 6-го брюшного ганглия речного рака Нейроны центрального отдела зрительного ана- лизатора лягушки Нейроны центрального отдела зрительного анали- затора кролика 0,54 + 0,11 0,50 ± 0,07 0,62 + 0,15 0,51+0,16
Данные, приведенные в таблице, свидетельствуют о значи-
тельных вариациях в величине информативности реакций нейро-
нов даже в пределах одной и той же нервной структуры у одно-
го и того же животного.
Как видно из формулы, метод выявления информативности
реакции определяет в сущности общий прирост информации по
сравнению с фоном по значениям соответствующих вероятностей,
однако он, естественно, не может характеризовать свойство по-
лезности, т. е. семантику передаваемой информации. Более или
менее разработанным приемом количественной оценки семанти-
ческой информации является тезаурусный подход. Согласно это-
му методу, количество информации определяется не как мера
неопределенности выбора, а как характеристика приобретенного
знания под действием данного сообщения (Ю. А. Шрейдер,
83
1974). Поскольку в основе семантического подхода к оценке ин-
формации лежит представление о решающей роли информаци-
онного тезауруса и учет ситуационного фактора (R. Carnap,
J. Bar-Hilel, 1952; Р. Уэлз, 1965; Р. Г. Пиотровский, 1975;
Ю. А. Шрейдер, 1975; Д. А. Поспелов, 1976; и др.), то методы
количественного определения количества полезной информации
оказываются связанными в той или иной мере с вычислением
условных вероятностей. Вместе с тем для определения конкрет-
ного информационного содержания того или иного сообщения
важно знать, для выполнения какой функции, для решения какой
цели используется данное сообщение. Иначе говоря, количествен-
ное определение семантической информации возможно лишь в
целенаправленных процессах с определенной динамикой инфор-
мационного состояния системы (в -плане приближения к цели
в результате получения положительной информации или удале-
ния от нее — в случае дезинформации).
Один из таких методов, описанный ранее (О. Г. Чораян,
1977) и используемый в данной работе, заключается в следу-
ющем. На разных стадиях анализируемого целенаправленного
процесса по гистограмме распределения межимпульсных интер-
валов (или частот потенциалов действия) вычисляются вероят-
ности отдельных классов событий (символов, которыми коди-
руется сообщение), что дает возможность определить суммарную
величину семантической информации следующим образом:
Ic.- = Siog2-^,
Ро
где ро — вероятность данного класса символов, рб<1) — апостери-
орная байесовская вероятность каждого класса, определяемая
по формуле
п
Ур.Р.
1=1
где Pi — вероятность символа до и pi — после осуществления со-
бытия, выражение в знаменателе — сумма произведений веро-
ятностей символов сообщения до и после наступления события —
какого-либо целенаправленного действия.
В табл. 8 приведены некоторые результаты такого анализа
семантической информации в импульсных потоках афферентных
84
нейронов системы 6-го брюшного ганглия речного рака. Для
сравнения динамики информационных процессов в ходе адап-
тации в таблице приведены и количественные характеристики
статистической информации, вычисляемой по энтропийной мощ-
ности импульсных потоков на отдельных этапах развития адап-
тационного процесса
1=п
1стат. = Рi Pi»
где Pi — вероятность соответствующих классов межимпульсных
интервалов (или частот потенциалов действия).
Таблица 8
Динамика статистической и семантической информации
в импульсных реакциях нейронов 6-го брюшного
ганглия речного рака в процессе адаптации
№ нейрона Информационный показатель Этап развития адаптации
I II Ш IV
1 1 сем. 2,15 3,21 3,78 4,02
1 стат. 3,74 3,92 3,76 3,69
2 1 сем. 3,01 4,98 6,54 6,60
1 стат. 3,92 3,74 4,01 4,02
3 1 сем. 1,84 4,58 6,12 7,80
1 стат. 4,83 4,40 4,99 4,56
4 1 сем. 2,14 4,15 5,63 5,72
1 стат. 5,01 5,15 5,42 5,33
5 1 сем. 1,57 2,92 ЗД4 3,40
1 стат. 3,52 3,40 3,46 3,35
6 1 сем. 1,89 4,45 5,96 5,84
1 стат. 2,84 3,02 2,78 2,92
7 1 сем. 2,32 5,24 5,84 5,70
1 стат. 4,43 4,15 4,62 4,80
8 1 сем. 1,92 3,27 3,83 4,70
1 стат. 3,89 4,28 4,41 4,61
9 1 сем. 2,15 6,32 8,13 8,51
1 стат. 4,15 4,30 4,41 4,36
10 1 сем. 2,43 5,24 6,01 6,11
1 стат. 3,05 3,12 2,93 2,88
Как видно из таблицы, имеет место значительное увеличение
количества семантической информации по мере наступления
адаптации импульсных реакций нервных клеток, в то время как
85
показатели статистической информации практически не изменя-
ются.
Одно из возможных объяснений наблюдаемым фактам за-
ключается в следующем. Уменьшение, ослабление импульсной
реакции афферентных нейронов при адаптационных явлениях
могло бы значительно снизить способность адекватной оценки
организмом силовых характеристик стимулов (которые при
идентичных раздражениях, естественно, остаются постоянными).
Однако компенсаторное повышение семантического содержания
получаемых сообщений, повышение ценности каждого из симво-
лов, которыми закодирована информация, позволяет организму
нейтрализовать отрицательный эффект адаптационных измене-
ний импульсных реакций афферентных нейронов.
Другой интересный факт, проливающий некоторый свет на
природу семантической информации, формируемой как под влия-
нием информационного содержания входного импульсного пото-
ка, так и информационного тезауруса приемника информации,
состоит в особенностях динамики соотношения величин статисти-
ческой и семантической информации в импульсных реакциях на
повторные адекватные раздражения. Если в начальной стадии
действия стимулов величина статистической информации почти
всегда превышает количество семантической информации, то по
мере предъявления повторных стимулов величина семантической
информации растет и начинает превышать величину статистиче-
ской информации, содержащейся в данном сообщении. Этот
факт, очевидно, объясняется тем, что содержательная оценка
любого сообщения, осуществляемая его адресатом, обеспечива-
ется не только восприятием определенного объема информации
из общего объема статистической информации входного сообще-
ния, но и путем извлечения, преобразования части информации,
хранящейся в памяти системы и определенным образом связан-
ной, когерентной с поступающим извне сообщением» т. е. привле-
чением информационного тезауруса нервной клетки.
Интересные особенности перераспределения вероятностной
структуры импульсного потока выявлены при вычислении коли-
чества семантической информации, содержащейся в вызванных
импульсных сериях в ответ на действие условного и безусловно-
го стимулов при образовании условного рефлекса (табл. 9). Об-
наружено, что количество семантической информации, содержа-
щейся в вызванных на безусловный стимул импульсных пото-
ках (в особенности до выработки и упрочения условного реф-
лекса), как правило, .превышает информационный объем реакции
на условный стимул. Однако в динамике роста статистической
и семантической информации на разных стадиях формирования
86
Таблица 9
Динамика семантической информации в импульсных реакциях
корковых нейронов кошки на безусловные и условные раздражители
№ нейрона Количество семантической информации в вызванных импульсных потоках (в битах)
на 1 — 5 сочетания на 6 — 10 сочетания на 11 —15 сочетания на 16 — 20 сочетания
1 1,12 _1,76_ 2,64 _2,9,1_
2,42 2,50 2,38 2,60
2 1,82 2,31 3,01 3,43
2,24 2,43 2,54 2,71
3 1,99 2,63 3,23
3,02 ' 2,84 2,70 2*51
4 2,09 _з,оо_ 3,96 4,83_
2,56 2,60 2,71 2,58
5 1,84 1,99_ 2,74 3,44
2,91 2,64 2,50 *2,71
6 2,33 3,82 4,76 5,11
2,74 2,89 2,95 3,04
7 2,08 2,74 3,48 4,72_
2,73 2,69 2,71 3,15
8 2,43 3,94 4,73 5,60
2fi2 3,11 3,40 3,54
9 1,73 3,02 4,76 5,44_
здГ 2,99 3,16 3,42
10 2,94_ 3i98_ 4,56 5,08__
3,90 4,12 4,30 4,21
Примечание: в числителе — значение для условного раздражителя,
в знаменателе — для безусловного.
условного рефлекса наблюдаются обратные отношения между
ответами на условный и безусловный раздражители: уже в на-
чальной фазе образования условнорефлекторной связи в корко-
вых анализаторных структурах прирост полезной (семантиче-
ской) информации в реакциях на условные стимулы обычно пре-
вышает величину прироста семантической информации на без-
условные стимулы. Прирост величины семантической информа-
ции в вызванных импульсных потоках в ответ на повторные
предъявления безусловного стимула (если он имеет место) вы-
ражен намного слабее, чем в случае импульсных реакций на
условные стимулы. Следует также заметить, что в случаях, когда
последовательные предъявления условных стимулов не приводи-
ли к существенным изменениям в величине семантической ин-
87
формации (а следовательно, и не вызывали значительных изме-
нений в вероятностной структуре соответствующих импульсных
потоков), абсолютная величина последней обычно выше, чем в
импульсной реакции, вызванной безусловным стимулом.
Полученные результаты позволяют рассматривать вероят-
ностный механизм превращения ранее индифферентного стимула
в сигнальный значащий — основу условнорефлекторного поведе-
ния— как процесс извлечения корковыми анализаторными ней-
ронами полезной сигнальной информации из поступающего сум-
марного информационного потока, превышение которой по абсо-
лютным значениям над соответствующей величиной импульсных
реакций на 'безусловный раздражитель свидетельствует о боль-
шей пластичности, о большем диапазоне возможных вариаций
реакции нейрона на условные стимулы, а следовательно, и о
большем удельном весе вероятностного компонента в ответах
нервных клеток на условные стимулы по сравнению с реакциями
на безусловные раздражители.
В целенаправленных реакциях организма, когда конечное
целевое состояние достигается в ходе эксперимента, возможно
определение количества прагматической информации на разных
стадиях формирования конечного результата. В этих случаях по
картине вероятностной структуры различных электрографиче-
ских проявлений адаптивного приспособительного поведения жи-
вотного организма можно судить о степени оптимальности выра-
ботанной и реализуемой стратегии информационно-управляю-
щей системы в ходе сбора, избирательной фильтрации и перера-
ботки сенсорной информации. Хотя прагматическая информация,
как и семантическая, характеризует содержательную ценность
циркулируемой и перерабатываемой в нейронных системах свя-
зи, между ними есть и определенные различия, сводящиеся к
мере универсальности, полифункционального назначения этой
информации. Если семантическую информацию в каком-то при-
ближении можно условно определить как информацию, имею-
щую отношение практически ко всем реализуемым при помощи
данной системы функциям (информация «на все случаи жизни»),
то прагматическая информация характеризует лишь ту часть се-
мантической информации, которая имеет отношение к каждой
отдельной, рассматриваемой в данный момент, реализуемой в
данной конкретной ситуации функции.
В основе ранее описанного (О. Г. Чораян, 1975) и исполь-
зуемого здесь метода вычисления количества прагматической ин-
формации лежит способ, предложенный В. А. Стуловым (1973)
для определения относительной и суммарной ценности отдель-
ных стимулов в сообщении. При этих расчетах за символы, ко-
88
торыми кодируется сообщение, принимаются разные частоты по-
тенциалов действия или классы межимпульсных интервалов в
постстимульной гистограмме
Qc = P i’Q,
где Qc — суммарная ценность (по всем классам гистограммы);
Pi — вероятность определенных символов (классов частот
потенциалов действия или межимпульсных интерва-
лов) ;
Q —относительная ценность, определяемая по постстимуль-
ной гистограмме в стадии достижения конечного ре-
зультата, в состоянии достижения цели.
1=п
1=1
— коэффициент, характеризующий изменение количества инфор-
мации при учете ее ценности, п — число классов в гистограмме.
Некоторые результаты таких расчетов на примере анализа
динамики количества суммарной ценности сообщений и прагма-
тической информации, отражающей особенности вероятностных
узоров межимпульсных интервалов в вызванных импульсных по-
токах нервных клеток в процессе образования условного рефлек-
са или адаптации, приведены в табл. 10.
Таблица 10
Динамика прагматической информации в вызванных
импульсных потоках клеток 6-го брюшного ганглия
речного рака в процессе адаптации
№ нейрона Период начальной реакции Период выраженных признаков адаптации
1* 0,084 0,096
2* 0,091 0,132
3* 0,047 0,092
4* 0,074 0,102
5* 0,065 0,095
6* 0,062 0,108
7* 0,088 0,110
8* 0,056 0,083
9* 0,026 0,043
89
№ нейрона Период начальной реакции Период выраженных признаков адаптации
10* 0,031 0,059
। ** 3,24 4,37
2** 3,82 5,13
3** 2,85 4,39
4** 4,01 4,83
5** 3,11 4,32
6** 3,82 5,11
у** 2,99 4,01
g** 3,88 4,73
9** 3,74 4,57
10** 2,81 3,82
Примечание: * — Qc; ** — 1Прагм
Как видно из таблицы, в ходе развития адаптационного про-
цесса наблюдается существенное повышение величины суммар-
ной ценности сообщения, приводящее к соответствующему уве-
личению количества прагматической информации в вызванных
импульсных потоках. Это означает, что введение фактора значи-
мости приводит к существенному увеличению скорости перера-
ботки информации, т. е. к интенсификации процесса восприятия
и переработки информации в системе связи, образованной нерв-
ными клетками.
Данные, приведенные в табл. 11—12, свидетельствуют о том,
что у «обучаемых» нейронов (т. е. у клеток, у которых насту-
Таблица 11
Динамика величины суммарной ценности сообщений,
выраженных узорами импульсных потоков корковых
нейронов кошки при образовании условного рефлекса
№ нейрона № сочетания
1 — 5 6—11 12—17 18 — 23 24 — 29
1 0,18 0,19 0,24 0,25 0,27
2 0,09 0,10 0,47 0,23 0,25
3 0,27 0,36 0,49 0,57 0,56
4 0,23 0,39 0,40 0,41 0,43
5 0,08 0,1,1 0,23 0,25 0,28
6 0,17 0,22 0,24 0,24 0,27
7 0,23 0,29 0,37 0,41 0,40
8 0,11 0,13 0,26 0,27 0,29
9 0,14 0,23 0,31 0,41 0,43
10 0,15 0,18 0,23 0,36 0,38
90
Таблица 12
Динамика прагматической информации в импульсных реакциях
корковых нейронов кошки в ответ на условные стимулы
К-во прагматической информации в реакциях (в битах)
Xs нейрона на 1 — 5 сочетания на 6— 11 сочетания на 12— 17 сочетания на 18—23 сочетания на 24 — 29 сочетания
1 3,49 3,58 3,73 4,01 4,13
2 3,01 3,11 3,54 3,82 3,99
3 2,74 3,72 4,12 4,52 4,58
4 3,01 4,08 4,16 4,22 4,30
5 3,42 3,57 3,89 3,98 4,12
6 1,96 2,28 2,36 2,36 2,49
7 2,37 2,58 2,91 3,08 3,05
8 2,84 3,08 4,11 4,22 4,30
9 3,09 4,15 4,48 4,96 5,08
10 2,73 2,92 3,19 3,59 3,68
пают условнорефлекторного типа перестройки вероятностного
узора межимпульсных интервалов), имеет место четкое повыше-
ние суммарной ценности сообщения, сопровождаемое уменьше-
нием энтропийной мощности импульсного потока и увеличением
количества прагматической информации, по мере образования
и упрочения условного рефлекса.
Лишь мысли свет, не скованный числом,
осветит тайны и достигнет правды.
Д. Байрон
Г л а в а IV. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ
ИНФОРМАЦИОННОЙ ДЕЯТЕЛЬНОСТИ
1. ХАРАКТЕР ПРОЦЕССОВ ПРИНЯТИЯ РЕШЕНИЯ
В НЕОПРЕДЕЛЕННОЙ СИТУАЦИИ
Принятие решения как органическая часть любого
поведенческого акта тесно связано с мерой неопределен-
ности ситуации, в которой принимается то или иное ре-
шение. Процесс принятия решения является универсаль-
ным процессом в различных формах деятельности жи-
вотного и человеческого организма. Об этом хорошо
сказал Дж. Коуэн (J. Cohen, 1964), который писал, что
«когда бы мы ни посмотрели на человека, в какую бы
то ни было эпоху и при любой социальной форме орга-
низации общества, мы видим, что от рождения и до
смерти он постоянно чувствует себя в неопределенности;
поэтому жизнь может быть описана как непрерывная
последовательность операций выбора». Управление про-
изводством, руководство большим коллективом людей,
регулирование уличного движения, пилотирование воз-
душных кораблей и многие другие операции, которые вы-
полняет современный человек, являются наглядным при-
мером принятия решения в многосвязных сложных проб-
лемных ситуациях. Принятие решения играет важную
роль и в жизни животных. Животные также должны при-
нимать решения, когда они ищут пищу, партнера, при
смене сна и бодрствования и т. д., причем эти решения
они должны принимать таким образом, чтобы максими-
зировать свои шансы на выживание и продолжение ви-
да (D. J. Me Farland, 1977).
В случае полной определенности ситуации нет никакой
проблемы принятия решения—практически решение при-
нимается однозначно и автоматически. Проблема в при-
нятии решения возникает лишь в том случае, когда в си-
стеме, в ситуации выбора имеет место неопределенность.
92
Принятие решения означает поиск всех возможных способов
достижения цели, сравнение их предпочтительности, полезности
(с целью выбора наиболее предпочтительного), на основе чего
и выполняется операция принятия решения — выбора определен-
ной альтернативы, альтернативного действия. Выбор альтерна-
тивы как центральный механизм принятия решения является ве-
роятностным механизмом, основанным на процессе устранения
избыточных степеней свободы действия субъекта, процессом, в
котором вероятность выбранной альтернативы есть функция по-
лезности альтернативы.
В процессе принятия решения различаются 2 принципиально
различные фазы: 1) генерация разнообразия (в которой из уни-
версального многообразия действий выбирается класс возмож-
ных допустимых путей решения, удовлетворяющих условиям ре-
шаемой задачи) и 2) ограничение этого разнообразия с целью
определения отдельного варианта действия (с точки зрения эф-
фективности этого способа достижения цели). Сущность процес-
са принятия решения можно представить следующим образом:
лицо, принимающее решение, вначале выдвигает взаимно исклю-
чающие альтернативы, затем производит их сравнительную оцен-
ку, на основе которой собственно и осуществляет операцию
выбора. Операция выбора, определение сравнительной ценности
альтернатив практически выполняется в условиях сбора разно-
образной информации, характеризующей различные стороны рас-
сматриваемого альтернативного действия, явления или предмета,
и обычно выражается либо в количественных мерах субъектив-
ной вероятности, либо в численных значениях функции принад-
лежности, принимающей промежуточные значения в интервале
0-4.
Структуру и последовательность действий, характеризующих
механизм принятия решения, обычно представляют схематически
в виде некоторого древовидного процесса, в котором по мере
решения проблемы — принятия решения в широком смысле это-
го слова — отсекаются бесперспективные ветви. Такими беспер-
спективными ветвями являются действия, приводящие к повто-
ряемости промежуточного результата, нарушению условий зада-
чи и т. д., т. е. явления, которые определяют приемы эвристи-
ческого поиска решения задачи. Принять правильное решение
означает выбрать из числа рассматриваемых альтернатив такую,
которая с учетом многих факторов (успешность суждения, целе-
сообразность, экономичность и т. д.) будет оптимизировать об-
щую ценность стратегии и тактики решения задачи (D. V. Lind-
ley, 1971; М. Де Гроот, 1974).
Решение, принятое на некотором уровне п, можно рассматри-
93
вать в качестве стратегического по отношению к решению на
уровне n + 1 и тактического в отношении решения, принимаемо-
му на уровне п — 1 (Б. И. Аронович, 1974). Эффективность ко-
нечного решения проблемы в сложной многоступенчатой задаче
в разной степени зависит от эффективности применяемой такти-
ки или стратегии: при плохой стратегии даже самая лучшая, оп-
тимальная тактика не сможет помочь в общем решении пробле-
мы, в то время как хорошая стратегия даже при плохих такти-
ческих приемах обеспечивает достижение конечного результата
(хотя естественно далеко не оптимальным способом).
Следует заметить, что элемент неопределенности присутствует
на каждом из описанных выше этапов решения проблемной си-
туации. На этапе сбора информации составления структурно-
информационной модели среды у субъекта неопределенность во
многом связана с практической невозможностью выделения и
учета полного множества альтернативных действий, путей реше-
ния рассматриваемой задачи. Никогда нельзя быть уверенным,
что в рассматриваемом наборе не упущен ряд альтернативных
действий (или хотя бы одна альтернатива), имеющих отношение
к решаемой задаче. На следующем этапе принятия решения —
сравнительной оценке альтернатив — наличие существенной не-
определенности вытекает уже из самого механизма такой оценки
по принципу «более предпочтительно», «менее предпочтительно»,
из отсутствия общепринятых объективных показателей меры по-
лезности. Следует также заметить, что любая классификацион-
ная задача, решаемая обычно в отсутствии точных количествен-
ных операций, осуществляется, как правило, на языке неопреде-
ленных, нечетко сформулированных понятий.
Сама неопределенность, размытость, присущая в той или иной
мере всем процессам принятия решения, как указывает Г. С. По-
спелов (1976), может быть двух видов: а) обусловленная вероят-
ностной организацией внешней среды и вероятностным характе-
ром взаимодействия организма со средой в процессе развития
его приспособительного поведения, б) связанная с неточностью,
приблизительностью представлений лица, принимающего реше-
ние, о конечных целях, задачах, а также о способах и путях ре-
шения проблемных задач.
В логической основе решения задач различают обычно 3 раз-
личных типа выведения заключительных результатов из некото-
рых исходных начальных условий: дедуктивные, абдуктивные и
индуктивные решения. Каждому из этих типов свойственно опре-
деленное соотношение строгих, точных и приблизительных эври-
стических компонентов.
Дедуктивные решения (выведение заключительного утверж-
94
дения из одного или нескольких исходных) относятся к числу
строгих решений и характеризуются полной определенностью.
Абдуктивные решения — выявление наиболее вероятных исход-
ных утверждений из некоторых заключительных — отличаются
значительной неопределенностью, хотя в ряде случаев и могут
быть отнесены к числу строгих решений. Большей неопределен-
ностью отличаются индуктивные решения, направленные на вы-
явление наиболее вероятных закономерностей на основе анализа,
сопоставления ряда исходных утверждений.
С этими особенностями различных типов умозаключений свя-
заны особенности человеческого мышления — субъективная пред-
почтительность одних логических приемов рассуждения по
сравнению с другими, детально исследованные в работах многих
психологов (A. Tversky, D. Kahneman, 1977; D. Kahneman,
A. Tversky, 1971; P. Slovic e. a., 1977; и др.). Большинство лю-
дей считают более легким, отдают предпочтение задаче выведе-
ния результата, следствия по известным исходным причинам пе-
ред обратной задачей — выявлению причины исхода из известно-
го результата, следствия. Результаты психологических экспери-
ментов (A. Tversky, D. Kahneman, 1977) показывают, что боль-
шинство испытуемых более уверены в предсказании результата,
зная причину, нежели в определении причины по известному ре-
зультату, хотя объективно в эксперименте эти две вероятности
были равны друг другу.
Неопределенность ситуации выбора в процессе принятия ре-
шения можно оценить, используя энтропийную меру оценки мно-
жества альтернатив:
H = log2n,
где п — число альтернатив.
Это означает, что энтропия ситуации прямо пропорциональ-
на числу альтернатив. В случае, когда возможно нахождение
числовых значений вероятностей этих альтернатив, неопределен-
ность ситуации можно вычислить через классическую меру шен-
ноновской энтропии
i=n
1=4
где Pi — вероятность каждой из п альтернатив.
Ясно, что максимальная энтропия ситуации будет наблюдать-
ся в том случае, когда рассматриваемые альтернативы имеют
равную вероятность. Напротив, энтропия ситуации равна 0, т. е.
95
имеет место полная определенность, когда вероятность одной из
альтернатив равна 1. Таким образом, вероятностный характер
процесса решения обусловлен мерой неопределенности ситуации:
чем больше степень энтропии, т. е. чем меньше информации,
тем меньше оснований для однозначно определенного решения,
тем более вероятностным становится решение. В этих случаях
проблемных ситуаций информационно-управляющая система моз-
га возмещает, компенсирует дефицит информации использова-
нием более тонкого и сложного аппарата оценки вероятности
того или иного события (А. Б. Коган и др., 1977), требующего
более продолжительного времени для осуществления процесса
принятия решения. Так, в опытах М. А. Алексеева и др. (1962)
было установлено, что латентный период двигательной реакции
на световой стимул, предваряемый предупредительным звуковым
стимулом, находится в прямой линейной зависимости от количе-
ства требуемой информации, вычисляемой как логарифм средне-
квадратического отклонения интервалов времени между этими
двумя сигналами. Неопределенность ситуации может изменяться
и при изменении числа предъявляемых сигналов. В опытах
В. Хика (W. Hick, 1952) было продемонстрировано, что с увели-
чением числа таких сигналов, т. е. с увеличением неопределен-
ности ситуации, растет и величина скрытого периода реакции,
что можно было описать следующей формулой:
A = Klog2(n+l)>
где А — латентный период реакции в ситуации выбора:
К — латентный период реакции без выбора;
п — число дифференцируемых сигналов.
Следует, однако, заметить, что в целом ряде случаев, когда
увеличение числа альтернатив приводит к такой ситуации, в ко-
торой лицо, принимающее решение, перестает надеяться на об-
наружение, выявление оптимальной альтернативы, такой зависи-
мости между числом альтернативных действий и временем, за-
траченным на операцию выбора, не наблюдается. В этих слу-
чаях лицо, принимающее решение, делает ряд действий (выбо-
ров) наугад, не пытаясь сразу отыскать лучший вариант, чтобы
впоследствии на основе анализа ошибочных ходов построить,
выработать определенную стратегию целенаправленного поиска.
Этому соответствует определенная кривая динамики временных
интервалов, идущих на операцию выбора. Так, например, в опы-
тах нашего сотрудника В. Н. Кироя (1979) было показано
(рис. 17), что увеличение времени, затраченного испытуемым на
производство первого хода в решении лабиринтной задачи, на-
блюдавшееся при увеличении числа альтернатив до 6—7, сме-
96
17. Динамика времени, затрачен-
ного на производство первого хода на
разных ярусах лабиринта. По оси ор-
динат — время, в с, по оси абсцисс —
номер яруса
нялось в последующем (по
мере увеличения альтерна-
тив— возможных ветвей ла-
биринта до 10—14) умень-
шением его. При увеличении
числа альтернатив более чем
до 7—8 испытуемый как бы
теряет надежду уже на пер-
вых шагах решения лаби-
ринтной задачи (поскольку
анализируется динамика ла-
тентного периода только пер-
вого хода) найти, отыскать
вариант, максимизирующий
шанс обнаружения нетупи-
кового хода в лабиринте; поэтому испытуемый в таких случаях:
выбирает очередной ход наугад без предварительного анализа
ситуации, следствием чего является сокращение времени на осу-
ществление операции выбора. Тот факт, что уменьшение латент-
ного периода реакции выбора происходит при увеличении числа
альтернатив до 7—8, напоминает известное правило «магическо-
го числа 7», характеризующего объем оперативной памяти чело-
века (Дж. Миллер, 1964). Очевидно, такое совпадение нельзя
считать случайным, оно в какой-то мере указывает на тесную
связь операции выработки плана действия человека в проблем-
ной ситуации с объемом краткосрочной оперативной памяти его.
В нейрофизиологической литературе описан ряд эксперимен-
тальных работ, посвященных проблеме эффективности вероят-
ностных раздражений в производстве операции выбора. Это
прежде всего относится к проблеме вероятностного подкрепления
в условнорефлекторной методике исследования закономерностей
высшей нервной деятельности в несколько ином аспекте приме-
нительно к условнорефлекторному поведению животных, рас-
смотренной уже в предыдущей главе. Актуальность таких иссле-
дований обусловливается тем, что в реальной, естественной жиз-
ни как животных, так и человека вероятность подкрепления
условного раздражителя безусловным всегда меньше 1, к тому
же случайным является и номер условного стимула, подкреп-
ляемого безусловным. К числу первых работ такого плана сле-
дует отнести исследования П. К. Анохина (1968) с использова-
нием методики выработки двигательной реакции («побежки»)
животного в условиях опыта с двумя кормушками.
Эта же цель — анализ эффективности целенаправленных дей-
ствий организма в условиях ситуационной неопределенности —
97
преследовалась и в экспериментальном исследовании М. А. Алек-
сеева с сотрудниками (1962). Авторы показали, что человек ре-
шает задачу выбора вполне удовлетворительно при вероятност-
ном подкреплении его действий. Сами количественные характе-
ристики решения такой задачи были достаточно близки к теоре-
тически рассчитанным ожидаемым результатам: чем выше была
вероятность подкрепления, тем больше было нажатий на «мак-
симальную кнопку», т. е. на ту кнопку, с которой вероятность
подкрепления была больше. Суть экспериментов заключалась в
том, что перед испытуемыми ставилась задача нажать на одну
из 2 кнопок для получения звукового сигнала, случайно подклю-
чаемого в разной последовательности то к одной, то к другой
кнопке. Тот факт, что испытуемые в этом эксперименте преиму-
щественно нажимали на «максимальную кнопку», указывает на
то, что несмотря на случайность подкрепления у испытуемых все
же создается некоторое общее впечатление о возможности под-
крепления с той или другой кнопки. Авторы приходят к выводу,
что рыхлость, нечеткость вырабатываемого при этом в структу-
рах мозга стереотипа определяется тем, что реализация каждого
движения зависит от очень многих, не поддающихся учету пере-
менных, меняющихся по закону случайного распределения.
Одним из основных положений теории принятия решения
является утверждение, что не существует решения, наилучшего
в абсолютнОлМ смысле. Решение может считаться наилучшим для
данного лица в отношении данной конкретной цели. Поэтому
обычно принятием решения считается отыскание некоторого ком-
промиссного варианта, не являющегося оптимальным ни для
одной отдельной цели, но оказывается наиболее приемлемым для
всей совокупности, множества целей (В. Л. Волкович, 1971).
В задачах принятия решения используются различные реша-
ющие правила, критерии, определяющие тот или иной выбор.
Эти решающие правила определяют порядок предпочтения воз-
можных альтернативных действий, помогают выделить оптималь-
ное подмножество альтернатив из числа возможных. В качестве
таких критериев хорошо известны минимаксный критерий, кри-
терий минимаксного риска, критерий показателя пессимума —
оптимума и т. д. В теории антагонистических игр основным прин-
ципом оптимальности является принцип максимина (miny шах
f(x, у) функции двух переменных f(x, у): человек, принимающий
решение и использующий при этом правило максимина, стре-
мится максимизировать свой выигрыш при наиболее неблагопри-
ятном действии противника. При интуитивном подходе к множе-
ству альтернатив обычно используются аксиомы сравнения веро-
ятностного отношения по принципу «не более вероятно, чем»
98
(L. J. Savage, 1954; D. Davidson, P. A. Suppes, 1956; D. Kahne-
man, A. Tversky, 1972).
Степень уверенности лица, принимающего решение при вы-
боре определенной альтернативы, описывается величиной субъ-
ективной вероятности этого альтернативного действия. Субъек-
тивной вероятностью называется оценка объективной вероятно-
сти человека, его субъективное представление о шансах на-
ступления того или иного события. Связь между субъективными
оценками вероятности и реализацией, воспроизводимостью пред-
сказанного результата в различных жизненных ситуациях осно-
вана на следующих 3 эмпирически выведенных постулатах
(П. Линдсей, Д. Норман, 1974):
1) люди обычно переоценивают встречаемость событий, име-
ющих низкую вероятность, и недооценивают встречаемость со-
бытий, характеризующихся высокими значениями вероятности;
2) люди считают, что событие, не наступившее в течение не-
которого времени, имеет большую вероятность наступления в
ближайшем будущем;
3) люди переоценивают вероятность благоприятных для них
событий и недооценивают вероятность неблагоприятных.
В большинстве проблемных ситуаций человек старается дей-
ствовать таким образом, чтобы, исходя из субъективной вероят-
ности и полезности, можно было бы максимизировать ожи-
даемый выигрыш. Комбинация субъективной вероятности с по-
лезностью называется субъективной ожидаемой полезностью
(СОП) события. Формула для вычисления субъективной ожи-
даемой полезности похожа на формулу математического ожида-
ния; отличие заключается в том, что вместо объективной веро-
ятности Р здесь фигурирует субъективная вероятность Ps, а вме-
сто ценности V — полезность U (П. Линдсей, Д. Норман, 1974).
Если событие Е может привести к результату А с субъективной
вероятностью Ps(A) или к результату В с субъективной вероят-
ностью Ps(B), то
COn(E) = [U(A)Ps(A)] + [U(B)+Ps(B)].
Важным решающим правилом статистической теории реше-
ния является сравнение между собой апостериорных вероятно-
стей различных гипотез с целью выбора той, апостериорная ве-
роятность которой выше. Иначе говоря, основной смысл реша-
ющего правила заключается в том, что с учетом результатов на-
блюдения принимается более вероятная гипотеза. Так, в случае
двухальтернативной простой гипотезы решающее правило запи-
сывается следующим образом. Из двух гипотез hi и h2 выбирает-
99
ся hi, если Р(hi/et) >Р(h2/ei), где ei — полученная информация.
Получение субъектом некоторой информации ei изменяет его ин-
формационный тезаурус, при этом априорные вероятности Р> пе-
реходят в апостериорные (Ю. П. Леонов, 1977).
Смысл групповых экспертных оценок, часто привлекаемых к
решению проблемных ситуаций, заключается в том, что оценки,
получаемые от группы экспертов, будут более надежными, бо-
лее «объективными», чем оценки, даваемые отдельным лицом,
отдельным экспертом (Н. Я. Виленкин, Ю. А. Шрейдер, 1977).
Поскольку жизнь и развитие человеческого вида на Земле
протекает в вероятностном мире, то нервная система как основ-
ной аппарат обеспечения приспособительного поведения уже с
момента своего возникновения, по мнению Г. И. Полякова (1964),
формировалась как орган, способный учитывать статистические
закономерности изменений, происходящих в окружающей дей-
ствительности. «Можно сказать, что в деятельности нервной си-
стемы всем ходом предшествующей эволюции предусмотрен
большой запас неопределенности в отношении влияний внешней
среды. Имеются основания полагать, что переработка информа-
ции в биологических системах животного типа основана на функ-
циональном единстве детерминированного и вероятностного
принципов действия», — писал Г. И. Поляков (1964).
Жесткодетерминированный и вероятностный компоненты по-
веденческих актов, связанных с принятием решения, характери-
зуются определенной динамикой в ходе формирования, станов-
ления приспособительных реакций организма. В теории функ-
циональной системы П. К. Анохина (1970) принятие решения
рассматривается как результат афферентного синтеза, произ-
водимого организмом на основе ведущей мотивации. Принятие
решения освобождает организм от большого количества степе-
ней свободы и обеспечивает формирование интеграла эфферент-
ных возбуждений, необходимых для приспособительного поведе-
ния организма в данный момент и в каждой конкретной си-
туации.
2. ПРОГНОЗИРОВАНИЕ
Эффективность адаптивного поведения человека в значитель-
ной степени связана с его уникальной способностью предвидеть,
прогнозировать наступление определенных событий и, следова-
тельно, соответствующим образом подготовиться к ним. Это за-
мечательное свойство человеческого ума уже давно было из-
вестно психологам, в работах которых анализ психофизиологи-
ческих механизмов прогнозирования занимает значительное ме-
100
сто. С. Л. Рубинштейн (1958) писал, что, обладая аппаратом
мышления, человек способен устанавливать такие связи и отно-
шения между предметами и явлениями материального мира, на-
ходить способы решения таких задач, с которыми он встречает-
ся впервые. Во многих случаях решения проблемных ситуаций
со значительным элементом неопределенности человек как бы
выхватывает из общего разнообразного потока информации не-
которые отдельные сообщения, группы сигналов, играющие роль
матричных элементов в распознавании образов, отношений меж-
ду реальными предметами и явлениями. Хорошим примером то-
му является механизм чтения (особенно того вида, который из-
вестен под названием «партитурного», чтения «по диагонали»,
когда глаза скользят по тексту не горизонтально по каждой
строчке, а по диагонали, пропуская значительную часть слов в
каждой строчке), при котором нет необходимости последователь-
но и раздельно воспринимать и анализировать структуру мно-
жества знаков, образующего соответствующий текст.
Аналогичным образом осуществляется практически любой
случай распознавания отдельных предметов или обстановки в
целом. Для идентификации опознаваемого предмета нет необхо-
димости в создании в мозгу изоморфной модели объекта — созда-
нии точной копии, модели, достаточно восприятия лишь немно-
гих наиболее существенных характеристик его структуры. Прак-
тика показывает, что степень адекватности распознаваемого об-
раза — построения в мозгу его модели — определяется не с точ-
ностью до изоморфизма, а с точностью до узнавания. Отсюда
становится понятным, что конечный приспособительный резуль-
тат любого более или менее сложного акта человеческой дея-
тельности может быть осуществлен лишь с определенной огра-
ниченной степенью вероятности. Достижение биологической це-
ли-организма требует учета вероятностных отношений различ-
ных сигналов, определение меры вероятности появления того
или иного сигнала (Г. И. Поляков, 1964). На языке психологи-
ческих понятий это означает формирование субъективных моде-
лей, адекватных статистической структуре опознаваемых пред-
метов. Прогнозирование событий, экстраполирование момента
наступления сигнала, по словам А. Н. Леонтьева и Е. П. Крин-
чик (1964), способствует оптимизации процесса восприятия раз-
новероятных сигналов, поскольку оно является выражением
установки испытуемого на активное ожидание.
По-видимому, одной из первых работ по изучению психофи-
зиологических характеристик вероятностного прогнозирования
следует считать исследование И. Брунсвика (Е. Brunswick,
1939), который собственно и ввел в психофизиологическую ли-
101
тературу термин «вероятностное ожидание». В процессе обучения,
по мнению И. Брунсвика, происходит формирование отношений
вероятностного ожидания между возможной формой стимуляции
и формой ответной активности. Он пишет, что вероятностная
структура внешней среды воспринимается и фиксируется в па-
мяти системы, обеспечивая тем самым положительный опыт ре-
шения аналогичных задач (Е. Brunswick, 1943). Это свойство
человеческого ума определяет эффективность экстраполяцион-
ной деятельности его.
Различные аспекты проблемы вероятностного прогноза рас-
смотрены в различных работах П. К. Анохина (1968) под на-
званием опережающего возбуждения, Е. Н. Соколова (1960)
под названием нервной модели стимула, Н. А. Бернштейна
(1966) — программы двигательной активности, Л. В. Крушинско-
го (1960) — экстраполяционного рефлекса. П. К. Анохин (1977)
писал, что опережающее отражение действительности является
основой всей нашей психической деятельности и составляет сущ-
ность предсказания, что предсказание, прогнозируемая оценка
будущего составляет основу всей интеллектуальной деятельно-
сти человека.
Есть все основания утверждать, что в деятельности человека
практически нет ситуаций, в которых вероятностное прогнозиро-
вание (если понимать его в широком смысле со всеми упомяну-
тыми выше аспектами) не имело бы важного значения. Почти
все реакции организма основаны на сопоставлении в мозгу мо-
дели вероятного будущего, которое определяется протекающими
независимо от организма внешними событиями, и модели того,
к чему он стремится (Г. И. Поляков, 1964). В этом плане автор
считает, что ориентировочные реакции организма должны быть
отнесены к классу реакций на расхождение или рассогласование
прогноза (реакция на низкую вероятность наступления ожидае-
мого события).
Образование условного рефлекса — один из ведущих приемов
формирования приспособительного адаптивного поведения жи-
вотного и человеческого организма представляет собой физио-
логический феномен преобразования неопределенной информа-
ции в определенную, т. е. реакцию на уменьшение неопределен-
ности в среде. Как указывает Г. И. Поляков (1964), узловым мо-
ментом в образовании условного рефлекса является оценка ве-
роятности подкрепления условного стимула безусловным. Ско-
рость образования условного рефлекса прямо пропорциональна
вероятности подкрепления, хотя чувствительность к вероятност-
ной структуре событий и явлений внешней среды широко варьи-
рует у разных животных. Чем выше стоит животный организм
102
на эволюционной лестнице, тем выше его чувствительность к
«вероятностной структуре среды», тем точнее он оценивает бу-
дущее. Так, например, по данным И. Брунсвика (Е. Brunswick,
1939), крысы не могли различать вероятностное отношение 75:25
от равновероятного (50:50). Следует, однако, заметить, что по
данным Д. А. Бирюкова (1963, цитир. по Г. И. Полякову, 1964)
в несколько другом варианте экспериментов (в опытах на Т-об-
разном лабиринте) крысы могут определять, воспринимать раз-
личную степень вероятности подкрепления уже при разнице в
5—10% между количеством подкреплений в правом и левом раз-
ветвлениях лабиринта.
Достаточно высокая способность детей к дифференцированию
числовых характеристик вероятностной структуры стимулов по-
казана в работе С. Дж. Мессика и С. М. Сэли (S. J. Messick,
С. М. Saley, 1957). Авторы изучали процесс развития вероят-
ностного ожидания у детей в возрасте от 3 до 8 лет, предъявляя
им картинки с изображением больших и маленьких животных и
предлагая угадать очередность их появления. Вероятность по-
явления больших изображений варьировала от 0,6 до 0,9. В экс-
периментах было установлено, что уже через несколько минут
(через 6—8 минут) у детей проявлялись признаки усвоения, диф-
ференцирования разных вероятностей появления больших и ма-
леньких картинок. Способность человека усваивать вероятност-
ную структуру стимулов была подтверждена в ряде работ
(М. А. Цискаридзе, 1969; W. К. Estes, 1976; В. А. Иванников,
1977; и др.).
В опытах нашего сотрудника В. Н. Ковалевского со случай-
ным предъявлением на световом табло знаков «плюс» и «минус»
было установлено, что уже после 20—30 таких предъявлений
число правильных предсказаний, угадываний оказывалось суще-
ственно более высоким, чем при начальных предъявлениях сиг-
налов. При введении в схему определенной закономерности «рас-
познавание», правильное предсказание очередного предъявления
знака наступало еще раньше. При этом параллельно наступало
сокращение времени двигательной реакции нажатия на соответ-
ствующую кнопку (испытуемому предлагалось определить на-
ступление определенного знака соответствующим нажатием на
одну из кнопок: если ожидается знак «плюс», нажать левую
кнопку, если «минус»—правую кнопку).
М. А. Цискаридзе (1969) на основании своих эксперимен-
тальных исследований приходит к выводу, что человек, опираясь
на прошлый опыт, может прогнозировать наступление определен-
ных событий, строить вероятностную модель будущего, в соот-
ветствии с которой и осуществляет преднастройку своих функ-
103
циональных систем для реализации наиболее оптимального, це-
лесообразного поведенческого акта. Этот процесс проявляется в
уменьшении времени реакции в тех ситуациях, которые прогно-
зируются с высокой вероятностью. При изменении статистиче-
ской структуры среды, приводящей к рассогласованию прогно-
за с реальной картиной среды происходит приспособление к но-
вой среде посредством перестройки вероятностного прогноза.
В последние годы проблемы вероятностного прогнозирования
особенно интенсивно разрабатываются И. М. Фейгенбергом и
его сотрудниками (И. М. Фейгенберг, 19бЗ; И. М. Фейгенберг,
В. А. Иванников, 1969; И. И. Гуревич, И. М. Фейгенберг, 1977).
Авторы справедливо утверждают, что прогнозирование на основе
прошлого опыта не может быть абсолютным, прогнозирование
всегда носит вероятностный характер. Экстраполировать, прогно-
зировать дальнейшие изменения среды на определенный отрезок
времени помогает человеку прошлый опыт. Реагирование орга-
низма в соответствии с таким вероятностным прогнозом позво-
ляет ему резко уменьшить число ошибочных реакций, следова-
тельно, оно является наиболее оптимальным для активного при-
способления к окружающей среде. Оптимальность такого пове-
дения, основанного на результатах вероятностного прогноза, свя-
зана с тем, что последний дает возможность реализовать адап-
тивную реакцию раньше, чем появляется пусковой стимул. В
условиях неопределенного прогноза организм выполняет работу
по подготовке к нескольким возможным действиям. Это соответ-
ствует ориентировочной реакции организма па неопределенность
ситуации. Чем больше неопределенность прогноза, тем больше
физиологических систем вынужден подготовить к действию орга-
низм, тем более сильную ориентировочную реакцию мы наблю-
даем (И. М. Фейгенберг, В. А. Иванников, 1969). Напротив,
условнорефлекторную реакцию авторы рассматривают как от-
вет па стимул, организуемый на базе индивидуального опыта
человека и позволяющий прогнозировать появление в будущем
некоторой определенной ситуации. Угашение ориентировочной
реакции, сопровождаемое образованием условного рефлекса сво-
дится к ряду реакций организма на прогнозируемую ситуацию,
когда прогноз становится все более и более определенным. .От-
сюда авторы делают следующий вывод: условнорефлекторная
реакция организма проявляется всегда в ситуации определен-
ного прогноза, ориентировочная реакция — в условиях неопреде-
ленного прогноза (И. М. Фейгенберг, В. А. Иванников, 1969).
Следует, однако, заметить, что способность к вероятностному
прогнозированию, составление такого вероятностного прогноза
еще не означает, что человек всегда действует сообразно этому
104
прогнозу. Так, например, М. И. Ярвик (М. Jarvick, 1951) на ос-
нове анализа результатов экспериментов приходит к заключению,
что человек в реальной обстановке часто действует не по вероят-
ностным законам, а поступает вопреки им, хотя оценка ситуации
на основе учета вероятностной структуры среды и, следователь-
но, вероятностное научение бесспорно имеет место. Иначе го-
воря, человек, усваивая вероятностную структуру среды, борет-
ся с прогнозом, основанным только на вероятности появления
событий.
3. ЭЛЕМЕНТЫ РАЗМЫТОЙ ЛОГИКИ В РАБОТЕ .МОЗГА
Большинство классов явлений и предметов реального мира
в противоположность понятию класса или множества в матема-
тике не имеют четких границ, которые отделяли бы их от пред-
метов, не входящих в эти классы (Р. Беллман, Л. Заде, 1976;
К. Asai а. о„ 1977).
Сложным системам, функционирующим в реальном мире при-
сущи 2 различных вида неопределенностей: случайность и раз-
мытость; первая соответствует случайности, неопределенности
появления объекта в пространстве и во времени, вторая указы-
вает на смысловую неопределенность объекта. Основное разли-
чие между случайностью и расплывчатостью заключается в том,
что случайность характеризуется неопределенностью, касающей-
ся принадлежности или непринадлежности некоторого объекта
к нерасплывчатому четкому множеству. Расплывчатость, нечет-
кость— свойство таких классов предметов и явлений, в которых
имеются различные градации, степени принадлежности, проме-
жуточные между полной принадлежностью и непринадлежностью
объектов к данному классу. С этой особенностью и связаны
принципиальные различия между центральными понятиями —
функцией принадлежности в теории размытых множеств и функ-
цией распределения вероятностей в теории вероятностей. Как
подчеркивает Г. С. Поспелов (1976), если функция распределе-
ния вероятностей имеет объективную основу в виде накопления
статистических данных, то функция принадлежности (численные
значения ее) является субъективной по самой своей сущности,
что сближает их с понятием субъективных вероятностей, кото-
рые столь широко используются в психологических исследова-
ниях.
Указывая на принципиально важную роль элементов нечет-
кости, размытости в сложных системах, Л. Заде (L. Zadeh, 1969)
пишет, что сложность организации и функционирования систем
приводит к признанию существенной степени размытости как
105
отличительной особенности живых систем. Решения многих задач
повседневной жизни человека, таких как распознавание обра-
зов, разработка оптимальной стратегии поведения, различные
формы мыслительной деятельности человека и т. д., происходят,
как правило, при нечетких условиях, в ситуациях, недоступных
точному количественному описанию. Теория размытых множеств
и размытых алгоритмов, успешно развиваемая в последние 15 лет
в работах Л. Заде и многочисленных его последователей (L. Za-
deh, 1965—1976; М. Shimura, 1975; М. Kokawa е. а., 1975;
С. V. Negoita, Р. A. Raleseu, 1975, 1977; S.-K. Chang, 1972;
С. L. Chang, 1975; М. Kochen, 1975; R. R. Yager, 1977; К. Asai
e. a., 1975, 1977; N. Naplatanoff, D. Lakow, 1977; V. D. Dimitrov,
1977; Орловский C. A., 1971; А. Б. Коган, О. Г. Чораян, 1977;
О. Г. Чораян, 1977, 1978; В. С. Радзиевский и др., 1976; Д. И. Ша-
пиро, 1975—1978), в сущности есть попытка создания концеп-
туальной основы для оперирования размытыми понятиями, раз-
мытыми представлениями в количественном или квазиколичест-
венном плане.
Н. Винер (1966) в свое время называл способность человека
оперировать нечеткими понятиями —главным преимуществом
его перед машиной. Это замечательное свойство человеческого
мозга — оперирование, манипулирование расплывчатыми кате-
гориями— во многом обусловлено ролью, которую играет в жиз-
ни человека язык — ведущая форма кодирования информации.
Характерной чертой человеческого языка как основного сред-
ства коммуникации является полифункциональный характер со-
ставных его элементов, словесных конструктов, свидетельству-
ющий о неоднозначных вероятностных отношениях между зна-
ком и обозначаемым им предметом. По-видимому, постоянная
практика использования натурального языка с его вероятност-
ным отношением между знаком и денотатом (обозначаемым)
послужила прекрасной тренировкой для человеческого ума в
манипулировании нечеткими понятиями, что дает возможность
человеку оперировать размытыми множествами и размытой ло-
гикой. Именно размытая логика мыслительной деятельности че-
ловека обеспечивает ему возможность эвристического решения
многих сложных проблем, которые невозможно решить обычны-
ми алгоритмическими методами. Очевидно, следует согласиться
с Л. Заде (1976), когда он пишет, что мы не можем развдть
большую точность наших количественных измерений, чем это
позволяют нам наши качественные концепции. А качественные
оценки в силу своей природы не могут быть очень точными.
Центральным понятием в теории размытых множеств являет-
ся понятие о функции членства, принадлежности, с помощыр ко-
106
торой задается расплывчатое множество. Размытое множество
(или подмножество) — это класс с нерезкими, нечеткими грани-
цами, в котором переход отдельных элементов от принадлежно-
сти к непринадлежности — градуальный процесс. Размытое под-
множество А универсального множества U характеризуется
функцией принадлежности ц: и-^[0,1], которая описывает эле-
мент ueU некоторым числовым значением функции рА(и) в ин-
тервале [0,1].
Эта величина функции принадлежности характеризует сте-
пень принадлежности данного элемента к размытому подмно-
жеству А. Когда пространство (подмножество) А содержит толь-
ко 2 вида точек со значением функции принадлежности 0 и 1,
множество А является неразмытым, четким, и функция принад-
лежности сводится к соответствующей характеристической функ-
ции неразмытого множества.
Основные определения и операции над размытыми множест-
вами сводятся к следующему.
1. Размытое множество является пустым тогда и только тог-
да, когда его функция принадлежности равна 0.
2. Два размытых множества А и В равны (А = В), если и
только если fA(x)=fB(x) для всех хеХ.
3. Комплемент размытого множества А, обозначаемый как А',
определяется следующим образом:
fA'(x) == 1 — fA(x) для всех хеХ.
4. Размытое подмножество А содержится в размытом под-
множестве В (или что одно и то же: А есть подмножество В
или А^В), если и только если
fA(x)^fB(x) для всех хеХ.
5. Объединение двух размытых множеств А и В с соответ-
ствующими функциями принадлежностей fA(x) и fB(x) есть раз-
мытое множество С, значение функции принадлежности в кото-
ром определяется как
fG(x) =max[fA(x), 1в(х)] для всех хеХ.
Это выражение характеризует наименьшее размытое множе-
жество, содержащее и А, и В.
6. Интерсекция двух размытых множеств А и В с соответ-
ствующими функциями принадлежности fA(x) и fB(x) есть раз-
мытое множество С, функция принадлежности которого опреде-
ляется следующим образом:
fG(x) =min[fA(x), fB(x)] для всех xdX.
Это выражение характеризует самое большое размытое мно-
жество, содержащееся и в А, и в В.
7. Концентрация размытого множества Con АДА2 (равенство
по условию) результируется либо в уменьшении вероятности
107
лринадлежности (т. е. уменьшение соответствующей, функции
лринадлежности) в случае а>1, либо в увеличении численных
значений функции принадлежности при а<1.
Хотя в ранних работах по теории размытых множеств неод-
нократно подчеркивалось принципиальное отличие ее от теории
вероятностей, как вполне справедливо отмечают Б. Р. Гейнес
и др. (В. R. Gaines, 1976; В. R. Gaines, L. J. Kohout, 1977), тео-
рия вероятностей имеет много разных аспектов и утверждение
об отсутствии связи между любым из них и теорией размытых
множеств следует считать сильным преувеличением. Это верно
хотя бы потому, что форма задания численных значений функ-
ции принадлежности в интервале 0—1- уже есть некоторая фор-
ма теории вероятностей. В действительности несмотря на суще-
ственные различия между ними есть и формальные, и логические
связи между теорией размытых систем и вероятностной теорией.
Иначе говоря, хотя концепция размытого множества не обяза-
тельно является вероятностной, каждое размытое множество, за-
даваемое определенными значениями функции принадлежности,
уже имеет тем самым вероятностную меру, а сами правила опе-
рации над размытыми множествами являются такими же, как
и над вероятностными явлениями, предметами (Е. Н. Ruspini,
1970). На возможность перевода численных значений функции
принадлежности в вероятностные величины указывает и сам
Л. Заде (L. Zadeh, 1968)
m(A) = 1Р(А)’ f XBA(x)dP,
Rn
где ца — функция принадлежности элемента х к множеству А
в пространстве Rn, а Р(А) — фактор нормирования.
Понятием «размытый алгоритм» обозначается упорядоченное
множество размытых инструкций, обеспечивающих приблизи-
тельное решение проблемы. Неточная инструкция как элемент
размытого алгоритма может приобрести точное значение (в рам-
ках необходимого масштаба точности для решения проблемы)
благодаря использованию численных значений функции принад-
лежности размытого множества.
Между размытыми алгоритмами и эвристическим поиском,
широко используемым при решении сложных проблемных си-
туаций в условиях дефицита информации о среде, есть тесная
связь. Стратегию эвристического поиска можно свести к вычис-
лению значений функции принадлежности элементов в размы-
тых множествах (S.-K. Chang, 1972). Теорию размытых мно-
жеств можно рассматривать в качестве математической модели
108
неточных концепций, в которой неточность является не помехой,
а единственно возможным способом описания, формализации
сложной проблемной ситуации.
Перспективным и многообещающим направлением теории
размытых множеств является разработка возможных ее прило-
жений в принятии решений, основанных на неполной, неточной
информации об изучаемом явлении или процессе. Одна из пер-
вых работ в этом направлении опубликована Р. Веллманом и
Л. Заде (R. Bellman, L. Zadeh, 1970), в которой задача приня-
тия решения в расплывчатых условиях формулируется в виде
задачи достижения нечеткой цели, определяемой как размытое
подмножество множества всех исходов. Расплывчатой целью на-
зывается такая цель, которую можно описать как расплывчатое
множество в соответствующем пространстве. При расплывчатых
целях и ограничениях конечное решение определяется как пере-
сечение множества расплывчатых целей и ограничений. Выбор
результирующего решения заключается в этом случае в нахож-
дении так называемого максимизирующего решения, при кото-
ром функция принадлежности пересечения множества целей и
множества ограничений принимает максимальное значение. В
этом плане результирующее решение может рассматриваться
как оптимальное (Г. С. Поспелов, 1976). Использование основ-
ных положений теории размытых множеств при теоретико-мно-
жественных операциях интерсекции и концентрации позволяет
получить объективную меру оценки оптимальности выбора той
или иной альтернативы в проблемной ситуации (Л. А. Заде, 1976;
R. R. Yager, 1977).
Поскольку многие проблемы, решаемые человеком в усло-
виях неопределенности, достаточно хорошо можно аппроксими-
ровать одной из модификаций широко используемой в экспери-
ментальной психологии лабиринтной задачи, представляет зна-
чительный интерес рассмотрение процесса решения лабиринта
человеком, используя основные положения теории размытых
множеств. Для этой цели была поставлена специальная серия
экспериментов на испытуемых — студентах университета. Испы-
туемому предъявлялся лабиринт поэтапно—по отдельным ярусам
(число ярусов лабиринта в разных экспериментах варьировало
в пределах 7—10). Испытуемым предлагалось найти выход из
лабиринта, сделав минимальное число ходов. На каждом ярусе
лабиринта есть только один выход на следующий ярус, все
остальные выходы лабиринта тупиковые. В структуре каждого
из типов лабиринта имеется определенная закономерность в рас-
положении нетупикового выхода.
Кроме словесного отчета и фиксации сделанных ходов (вы-
109
боров) в ходе эксперимента у испытуемого регистрировали ЭЭГ
от симметричных точек лобной, теменной и затылочной областей
головного мозга с целью определения электрографических кор-
релятов активности различных структур мозга на разных этапах
принятия решения (фон, стадия решения лабиринтной задачи,
момент принятия решения (выбора), последействие). Оценка эф-
фективности процесса принятия решения на разных его этапах
по мере обучения испытуемого, развития способностей к вероят-
ностному прогнозированию структуры решаемого лабиринта
проводилась по показателям информационного анализа ЭЭГ по
описанной ранее методике (О. Г. Чораян, 1978; О. Г. Чораян,
В. Н. Кирой, 1978), по динамике времени, затраченного на про-
изводство первого хода на каждом ярусе, а также по количеству
сделанных ходов.
Как показал анализ результатов этих экспериментов, много-
ярусная структура использованных лабиринтов обеспечивала у
большинства испытуемых «распознавание» общей закономерно-
сти в расположении выходов, вероятностное прогнозирование то-
пографии последующих элементов лабиринта уже к 3—5 ярусу.
На этом этапе решения лабиринтной задачи уже наблюдались
статистически достоверные изменения анализируемых парамет-
ров: наблюдалось сокращение времени, затраченного на произ-
водство первого хода на каждом ярусе, более выраженные из-
менения наступали в статистической структуре параметров ЭЭГ,
приводящие к увеличению количества семантической инфор-
мации.
Численные значения функции принадлежности определяли по
количеству сделанных испытуемым ходов по отдельным ветвям
разных ярусов. Максимальное число ходов по отдельным вет-
вям принималось за верхний предел функции принадлежности
(р = 1,0), по отношению к нему определялись другие значения р
(для остальных ходов лабиринта) в интервале 0,1. Полученные
таким образом значения функции принадлежности на первом
ярусе лабиринта (рис. 18) имеют вид: рд = 0,9; рв=1,0; рс=0,9;
p,D=0,9; рЕ=0,9. Это означает, что ветвь А выбирается с вероят-
ностью 0,9/4,6 (Рд=0,19), ветвь В —с вероятностью 1/4,6 (Рв =
= 0,24), ветви С, D и Е с вероятностью Pc=Pd = Pe=0,19. Эн-
тропия такой ситуации Hi, согласно статистической теории связи
К. Шеннона (1963), равна
i-^n
Н, = - ^ Pi log2 Pi=- 4 (0,19 log2 0,19) -
i = l
—0,24 log2 0,24 = ~ 2,33 бит.
110
Рис. 18. Схема одного из исполь-
зованных вариантов лабиринта
На втором ярусе лабиринта в соответствии с результатами
анализа процесса принятия решения имеем
|Xa = |1d = |1g=0,5; цв = цс=0,25;
це=pf = цк= 1,0; цн=0,75.
Соответственно этому
Ра=pD = pG=0,09; Рв = Pc=0,04;
Pe=Pf = Pk = 0,17; Рн=0,14.
В этом случае энтропия
Н2=—3 (0,09 log2 0,09) — 2 (0,041og2 0,04) —
—3 (0,17 log2 0,17) —0,14 log2 0,14 = ~ 3,00 бит.
Соответствующие расчеты для III—VI ярусов дают следующие
величины:
Н3 = 2,66 бит, Н4 = 2,34бит, Н5 = 1,38 бит, Н6= 1,23 бит.
Как видно из динами-
ки энтропийного показа-
теля Н, по мере решения
лабиринтной задачи (по
мере продвижения по ла-
биринту с яруса на ярус)
величина энтропии ситуа-
ции, определяемая через
значения функции при-
надлежности, постепенно
падает. Это не распро-
страняется только на пе-
реход с I на II ярус,
что можно объяснить
следующим образом: на
I ярусе у испытуемого еще
не сформировалось ника-
кого представления о воз-
можном плане в кон-
струкции лабиринта, и эн-
тропия ситуации прямо
связана с количеством
возможных альтернатив
(увеличение числа ветвей лабиринта при переходе с I на II ярус
приводит и к увеличению общего количества энтропии).
Учитывая тот факт, что обычно «распознавание» структуры
лабиринта у испытуемых в этой серии экспериментов происходит
на III—V ярусах (судя по количеству ходов, сделанных ими в
дальнейшем, а также по словесному отчету в конце эксперимен-
тов следует отметить отсутствие прямой связи между снижением
111
величины энтропии и моментом решения задачи. Резкое сниже-
ние величины энтропийного показателя наступает при переходе
с IV на V ярус (мера информации, вычисляемая по количеству
устраненной неопределенности Н4 — Н5, в этом случае макси-
мальна: 2,34— 1,38=0,96 бит).
В психофизиологических исследованиях показано, что в слож-
ных проблемных ситуациях при операции выбора лицо, прини-
мающее решение, обычно склонно переоценивать значения ча-
сто встречающихся вариантов (т. е. имеющих большую вероят-
ность) и, напротив, уменьшать ценность редко встречающихся
вариантов, тем самым суживая область поиска в диапазоне воз-
можных альтернатив (Р. Restle, 1961; A. Tversky, 1972). Иначе
говоря, альтернативы, вероятности которых (точнее значения
функции принадлежности их к размытому множеству желаемых
альтернатив) для испытуемого ниже некоторого порогового зна-
чения практически им не принимаются во внимание и исключа-
ются из общего множества возможных альтернатив. Это обстоя-
тельство и объясняет относительно быстрое принятие решения
человеком в проблемной ситуации с неполной неточной инфор-
мацией о среде. Видимо, в условиях значительной неопределен-
ности (при примерно равных объективных характеристиках
энтропии ситуации) единственно возможным рациональным при-
емом выбора альтернативного действия является стремление
субъекта, принимающего решение, максимизировать величину
функции принадлежности, доводя значение его до величин, близ-
ких к 1, а затем выбрать альтернативу с численным значением
функции принадлежности, близким к 1, напротив отбрасывая
альтернативы, величины функции принадлежности которых близ-
ки к 0. Поэтому в рассмотренной выше серии экспериментов
имела определенный смысл имитация пороговой логики решения
человеком лабиринтной задачи путем направленной фильтрации
отдельных альтернатив.
Рассмотрим динамику энтропийного показателя при порого-
вом значении функции принадлежности ц^0,5. Тогда для I яру-
са имеем прежние значения:
Н1=2,33 (Pa = Pc = Pd = Pe = 0,19; Рв=0,24).
Для II яруса:
PA = PD = PG = 0,l; Рн = 0,43;
рк = р = рЕ=0,19 (Н2=—3(0,1 log20,1) — 3(0,19logo0,19) —
—0,13 log2 0,13 = 2,77 бит.
Соответственными расчетами для II—VI ярусов получаем
следующие величины энтропийного показателя:
Н3=2,44 бит, Н4= 1,5 бит, Н5=0,93 бит, Н6=0.
Эти результаты показывают, что при пороговой логике операции
112
над размытыми множествами и соответственно над вероятност-
ными мерами функции принадлежности имеет место тесная кор-
реляция между динамикой энтропийного показателя и момен-
том распознавания (решения) лабиринтной задачи. Максималь-
ное падение энтропии наблюдается при переходе от III к IV яру-
су, т. е. там, где испытуемый (судя по его действиям и после-
дующему словесному отчету) распознал схему лабиринта Н3—
—iH4=0,94 бит. Подобные результаты получены и на других ва-
риантах лабиринтных задач. Некоторые результаты такого ана-
лиза на одном из вариантов лабиринтов, по данным 10 испы-
туемых, приведены в табл. 13.
Таблица 1$
Динамика энтропийного показателя на разных ярусах
лабиринтной задачи до и после введения пороговой
логики выбора альтернатив
№ Величина энтропии до введе-
яруса ния порогового значения р.
Величина энтропии при пороговом
значении р.>0,5
I 3,05 ± 0,35
II 3,20 ± 0,28
III 2,75 ± 0,31
IV 2,36 ± 0,28
V 1,68 ±0,30
VI 1,18 ±0,26
2,75 ±0,31
2,41 ±0,32
2,09 ±0,21
1,25 ±0,18
1,01 ±0,28
0,88 ±0,18
Эти данные позволяют считать, что человек оперирует чис-
ленными значениями функции принадлежности на базе порого-
вой логики отбора альтернатив с переоценкой их величин при
изменении проблемной ситуации (по мере продвижения по ла-
биринту).
Многочисленные психофизиологические и психологические ис-
следования указывают на то, что в человеческом сознании ха-
рактеристика взаимосвязи между предметами реального внеш-
него мира проводится в терминах, трудно поддающихся точному
количественному измерению. В то же время эти отраженные, мо-
делируемые в структурах мозга образы предметов и их связей
служат основанием для формирования альтернатив, рассматри-
ваемых при операции принятия решения. Сама оценка образо-
ванных таким способом альтернатив также производится на язы-
ке неточных размытых множеств с использованием размытой:
логики индуктивных и дедуктивных умозаключений (по типу
«более предпочтительно», «менее предпочтительно» и т. д.).
пз
Эти особенности человеческой логики операции размытыми
понятиями хорошо проявляются во многих случаях решения
проблемных ситуаций, требующих мультипараметрической оцен-
ки альтернативных действий. В ходе трудовой деятельности че-
ловеку часто приходится принимать решения в отношении пла-
нирования и реализации некоторых действий в условиях нечет-
кого определения, нечетких характеристик ситуации, при нали-
чии нескольких одновременно действующих оценочных показа-
телей, возможных результатов. Например, в условиях производ-
ственной деятельности частыми являются задачи выбора тех или
иных действий из множества возможных при одновременном уче-
те двух (или более) основных показателей: эффективности (по-
лезности) и рентабельности (экономичности); при этом каждое
из альтернативных действий может иметь и имеет практически
разное субъективное оценочное значение. Такие задачи можно
решить, используя операции нечеткой оптимизации, т. е. оптими-
зации в размытой среде с нечеткими ограничениями.
Рассмотрим одну из таких типовых задач, решаемых чело-
веком— задачу выбора места работы — на одном из 5 возмож-
ных предприятий Пь П2, П3, П4 и П5. Оценивая привлекатель-
ность каждого из этих предприятий, человек принимает во вни-
мание лишь следующие 4 фактора: возможность творческой ра-
боты (Qi), наличие друзей на этом предприятии (Q2), размер
заработной платы (Q3), близость расположения предприятия к
дому, где он живет (Q4). В пределах шкалы 0—1 условно оце-
нивается каждое из этих предприятий в отношении выбранных
критериев. В результате мы получаем условную меру, являю-
щуюся числовым значением функции принадлежности каждого
предприятия к размытому множеству предприятий с заданными
свойствами, обеспечивающими творческую работу — размытому
множеству Qi, наличие друзей — Q2, высокую заработную пла-
ту— Q3, к совокупности предприятий близкорасположенных к
дому — Q4.
При таком способе задания численных значений функции
принадлежности jiq:u->[0,1] пусть мы имеем следующие раз-
мытые множества:
JOJ 0$, 0,7 0^
Qi=l п/ п2' п3 Ч Ч г
гад, ад, ад,ад, адд
Q‘2 = t п/ п/ п3Ч’ п5);
114
г оз, II, 11, 111
Qs=t Пх’ n3’ n3’ n4’ n5J
(• 0J5, (X6, 0J, 0,6 ОД!
Q4 = l п/ na ’ Пз ’ Щ ’n5 J
Тогда предприятие с максимальным значением функции при-
надлежности (ino совокупности учитываемых характеристик)
можно определить по операции интерсекции размытых множеств.
(L. Zadeh, 1965, 1975; R. Yager, 1977), сущность которой заклю-
чается в нахождении максимального размытого множества, со-
держащегося в любом из рассматриваемых размытых множеств:
Г 0,5 0,6 0,5 0,4 0,5)
Q=QiAQ2AQ3AQ4=[ П1’ па ’ П3 ’ П4 ’ П5 J ’
Это означает, что таким предприятием, отличающимся по сово-
купности значений, максимальной привлекательностью, обладает
предприятие П2, характеризующееся максимальным значением
функции принадлежности.
Описанную выше процедуру определения оптимальности вы-
бора при мультипараметрической оценке альтернатив можно ис-
пользовать лишь в случае равнозначной, одинаковой ценности
принимаемых во внимание характеристик. Однако в реальной
жизни более частыми являются случаи, когда разные параметры
альтернативных действий имеют разную ценность. Эту относи-
тельную ценность разных параметров можно учесть при теоре-
тико-множественной операции концентрации размытых мно-
жеств СопА=Аа.
Как известно, операция концентрации размытых множеств
приводит к возрастанию величины функции принадлежности при
значении а<3 и к уменьшению при а>1. Причем эти изменения,
незначительные при величинах, близких к 1, носят весьма суще-
ственный характер по мере уменьшения величины функции при-
надлежности.
Для вычисления величины показателя степени в операции
'концентрации размытого множества необходимо построение со-
ответствующей матрицы на основе попарного сравнения выбран-
ных, рассматриваемых параметров альтернатив. Пусть для лица,
принимающего решение, рассмотренные выше характеристики
предприятия имеют следующие коэффициенты ценности: Q1==5;
Q2=Q3=3; Q4= 1. Попарно сравнивая эти параметры и выра-
зив конечные результаты в вероятностных величинах, получаем
следующую n-размерную матрицу (в данном случае п=4):
115
Qi Q2 Q3 Q4
Qi 0,22 0,04 0,07 0,09
Q2 0,13 0,03 0,04 0,05
Q3 0,13 0,03 0,04 0,05
Q< 0,04 0,01 0,01 0,02.
Построчно суммируя значения вероятностей 0 и перемножив
на размерность матрицы (п=4), можно определить численные
значения коэфициента а:
Q, =0,42-4 = 1,68;
Q2=0,25-4 = 1,00;
Q3=0,25-4 = l,00;
Q4=0,08-4=0,32.
Тогда, выполнив теоретико-множественную операцию концентра-
ции размытых множеств, получаем новые множества:
Q?=
Q?=
q:=
(ОД)1*68 (0,9)1368 (0,5) *-88 (0,7) *-»8 (°Д)1>68
п, п2 Пз Пз П5
( 0,54 0,83 0,32 0,54 0,43 )
> — > — —} — — > — — >
1 П1 П2 Пз 1П4 П5 J
( (0,6)*-» (0,6)*.» ., (°-9) 1з° ? (ОД)1»0 (0,5)1>0 ]
( П1 Пг Пз 1П4 Пз J
_ ( 0,6 0,6 0,9 , °’7 , °»5 '
= 1 п, п2 Пз П4 П5 , 1 >
' (0,8) ° (-0,8)2^ , (0Д)1>0 (0,4)1,0 (0,6)м_) _
m Пг Пз Пз П5 J ~
f 0,8 °,8 0,7 0,4 0,6
= t П1 Иг Пз п4 ’ п5
( (0,5) °.»2 т (0,6) °>32 , (0,7) 0,32 (0,6)0,32 (Ю,8)°>32 ]
1 П1 •Пг Пз Пз Пз Г
( 0,79 0,85 0,89 0,85 0,93 I
J — “ 1 > ! — — »
— 1 п, Пг Пз П4 П5 J
Предприятие с максимальным значением функции принад-
лежности (по совокупности учитываемых характеристик) можно
определить по операции интерсекции размытых множеств
116
Q* = Q?nQ?nQ3enQ: =
( 0,54 0,6 0,32 0,4 0,43 1
I П1 Пг Пз П4 П5 J *
Таким предприятием, как и раньше, остается предприятие П2,
хотя последовательность расположения рассматриваемых пред-
приятий (по мере убывания значения функции принадлежности)
существенно изменяется: П2—Fli—П5—П4—П3 вместо П2—(П3 =
= П] =Пб) — Пь
4. СЕМАНТИЧЕСКИЕ АСПЕКТЫ СИТУАЦИОННОЙ
ИНФОРМАЦИИ
Значительная степень неопределенности в информационных
процессах, образующих основу сложных актов мыслительной
деятельности человека, связана с индивидуальными вариациями
в оценке содержательной ценности, воспринимаемой и перера-
батываемой в нервной системе информации. Информационное
содержание раздражающего агента, информативность сущест-
венных параметров стимула вместе с тем зависит и от его био-
логической значимости, «сигналыюсти» (А. Б. Коган, О. Г. Чо-
ран, 1970).
Одним из самых замечательных свойств адаптивного приспо-
собительного поведения животных является их способность рас-
познавать и выделять из общего входного информационного по-
тока полезную актуальную информацию. Как верно подметил
Г. И. Поляков (1964), животный организм, и в частности чело-
век, в отличие от искусственных технических систем умеет реаги-
ровать не только на количество информации, содержащейся в
сообщениях, но и на ее качество (измерять и оценивать содер-
жательность информации). Физиологическим механизмом, обес-
печивающим эту замечательную способность организма, являет-
ся образование условного рефлекса, фиксирующего, закрепля-
ющего отношения организма к сигналам, несущим недостаточно
определенную информацию о том, что где-то что-то меняется,
т. е. информацию лишь о возможности наступления чего-то, что
может иметь существенное значение для организма (Г. И. По-
ляков, 1964). Такого же мнения придерживается В. А. Звягин-
цев (1965), подчеркивающий, что значение всегда производно от
коммуникативного намерения. Общение человека с машиной, пи-
шет он, оказывается возможным лишь в тех случаях, когда че-
ловек спускается до «интеллектуального» уровня машины и нала-
гает на язык формальные ограничения, которые лишают его мно-
117
гих существенных качеств и превращают его в искусственный
препарат. Мышление машины, по словам В. А. Звягинцева, име-
ет особый характер, оно оперирует не понятиями, как человек,
а значимостями. Значимости же в машине далеко не равнознач-
ны человеческим понятиям, они представляют только их мертвый
слепок, лишенный творческой энергии и обобщающей силы.
Экспериментальные данные об изменении информативности
стимула в связи с различиями в его вероятностных характери-
стиках были получены в исследованиях Саттона и др. (S. Sutton,
1969). Авторы считают, что информация, содержащаяся в факте
предъявления стимула, тем больше, чем меньше вероятность по-
явления его среди других раздражений, иначе говоря, чем не-
ожиданнее он для испытуемого. Авторы установили также стро-
гую математическую зависимость между информативностью сти-
мула и амплитудой поздней положительной волны электриче-
ского ответа (волны РЗОо в ЭЭГ). В простых экспериментальных
исследованиях наблюдалась прямая связь между этими двумя
показателями стимула и реакции, в сложных эксперименталь-
ных условиях эта корреляция носила более сложный характер:
при низкой вероятности стимула амплитуда волны РЗОо увеличи-
валась, при высокой вероятности — уменьшалась. В эксперимен-
тах, где испытуемому предлагалось предсказать появление по-
следующего сигнала, было установлено, что при большой вероят-
ности стимула амплитуда волны Р303 возрастает, если предска-
зание было неправильным, и уменьшалось — если оно верное.
Эксперименты с вероятностным подкреплением выявляют суще-
ственные изменения в параметрах условной отрицательной вол-
ны в записи ЭЭГ, описанной Г. Уолтером (1966). Многие авторы
считают волну РЗОо положительным колебанием электрического
потенциала, развивающимся после условного отрицательного ко-
лебания.
Значительная вариабельность различных волн вызванных
электрических потенциалов мозга показана во многих работах
(R. Jones е. а., 1965; Е. Callaway е. а., 1970). Особенно вариа-
бельными, судя по этим данным, являются амплитудные харак-
теристики различных электрографических феноменов, латентные
периоды вызванных электрических потенциалов характеризова-
лись обычно большей устойчивостью, стабильностью, Л.Лайфер
с сотр. (L. Leifer е. а., 1973), используя матрицу из 4 различ-
ных стимулов, чередующихся с разной степенью вероятности, и
предлагая испытуемым угадывать следующий) раздражитель, по-
лучили данные, подтверждающие предположение, что амплитуда
условной отрицательной волны прямо коррелирует с вероят-
ностью ожидаемого события, в то время как амплитуда волны
118
Рзоо находится в отрицательной корреляционной связи с вероят-
ностью прогнозируемого события.
Биологическая значимость сообщения является характеристи-
кой семантического и прагматического аспектов информации. В
статистической теории информация рассматривается как харак-
теристика случайного исхода опыта, как реализация некоторого
случайного выбора из широкого множества возможностей. Од-
ной из первых попыток использования теоретико-информацион-
ных моделей при описании поведения человека являются работы
В. Хика (Hick W., 1952) и Р. Хаймана (R. Hyman, 1953), в ко-
торых рассматривалась связь между временем, затраченным на
выполнение дизъюнктивной реакции и количеством информа-
ции, вычисляемой по энтропийной оценке стимула. Информаци-
онное содержание стимула изменялось путем варьирования об-
щего числа символов, алфавита источника информации, вероят-
ности предъявления отдельных символов и статистики межсим-
вольных связей. Количество информации в статистической теории
связи определяется через меру неопределенности или энтропии
выбора, которая в свою очередь характеризует сообщение как
совокупность знаков (символов) и частоту появления данного
знака или комбинации знаков. Напротив, количество семанти-
ческой информации менее связано со статистикой символов, об-
разующих сообщение, оно представляет собой содержательную
оценку сообщения (Е. К. Войшвилло, 1966).
Прагматический аспект информационного содержания вход-
ного сообщения связан с двойственным характером информаци-
онного тезауруса, который, с одной стороны, можно оценить в
плане обеспечения картины полного описания внешней среды, с
другой — сточки зрения действий субъекта, изменения поведения
его при получении данного сообщения. В этом последнем случае
полезная информация будет характеризовать прагматическую
ценность информации (R. Wells, 1961; Ю. А. Шрейдер, 1963;
В. И. Николаев, 1968; В. И. Николаев, М. В. Кузнецова, 1977).
В общем виде величина информации, содержащейся в неко-
тором высказывании А, выражается следующей формулой:
1(A) =—logP(A),
где Р(А) — вероятность (степень правдоподобия) А. Поэтому
два текста Tj и Т2, состоящие из множества высказываний по-
добных А, могут считаться равнозначными в семантическом ас-
пекте (т. е. содержащими одинаковое количество семантической
информации), если они изменяют на одинаковую величину ин-
формационный тезаурус приемника этих сообщений, текстов.
Критикуя предлагаемые многими авторами приемы измерения
семантической и прагматической информации текста, В. Н. Пес-
119
тунова и Р. Г. Пиотровский (1970) считают невозможным их
практическое использование, поскольку невозможно определить
весь ряд альтернативных логических вариантов и выразить их
через вероятностные меры. Поэтому, по мнению авторов, един-
ственно реальным способом остается оценка семантико-прагма-
тической информации через синтаксическую информацию, изме-
ряемую с помощью классической шенноновской методики вычис-
ления энтропии.
На существенно важную роль объема прагматической инфор-
мации в определении эмоционального поведения организма ука-
зывают исследования П. В. Симонова (1975). Автор связывает
это с самой природой прагматической информации, определяе-
мой как изменение вероятности достижения цели организмом
благодаря полученному сообщению. При дефиците прагматиче-
ской информации, по мнению автора, возникает отрицательная
эмоция, активно минимизируемая субъектом. Превышение по-
лученной информации над имевшимся ранее прогнозом ведет к
возникновению положительных эмоциональных состояний, мак-
симизируемых субъектом.
Как было уже отмечено ранее (см. гл. III), семантическая
информация, как и прагматическая, будучи субъективной по
своей природе, в большей мере обусловливается свойствами ин-
формационного тезауруса приемника, индивидуума, чем стати-
стикой символов, знаков, которыми образовано сообщение. По-
скольку информационный тезаурус человека отличается значи-
тельными индивидуальными различиями, становятся понятными
существенные вариации количественных характеристик полезной
(семантической и прагматической) информации. Трудности в
построении единой общей теории информации, охватывающей
все три ее вида (статистическую, семантическую и прагматиче-
скую), связаны, во-первых, с тем, что понятие ценности является
весьма многогранным, неоднозначным, во-вторых, оно несет на
себе черты субъективности, является производным полноты и
широты информационного тезауруса приемника, адресата дан-
ного сообщения. Ситуационный фактор, модифицирующий про-
цесс восприятия и переработки информации, оказывает влияние
на информационные механизмы через соответствующие измене-
ния оперативного уровня информационного тезауруса приемни-
ка сообщения.
Существуют различные способы количественной оценки се-
мантической информации информационного тезауруса системы и
его изменений под влиянием афферентного сообщения. Одним из
них является информационный анализ следовых процессов на
примере статистической взаимосвязи количественных парамет-
120
ров реакций организма. Рассмотрим динамику количества семан-
тической информации в последовательно взятых отрезках ЭЭГ
реакции, информационного выражения следовых процессов как
одного из основных компонентов информационного тезауруса,
предопределяющего способность организма извлекать из данно-
го сообщения максимальное количество семантической инфор-
мации. Предметом анализа были ЭЭГ испытуемых (в разных
отведениях — лобных, теменных, затылочных и париетальных)
при решении ряда тестовых заданий, требующих принятия ре-
шения в многоальтернативной ситуации. В процессе решения
таких задач дифференцировались следующие стадии: фон (‘пред-
решение), стадия решения, момент решения (ответ испытуемо-
го) и последействие. Прежде всего вычисляется количество ста-
тистической информации в ЭЭГ по энтропийной мощности ста-
тистических ее параметров
Нэмп:= 2 Р1 1о§2 Рь
где Pi — вероятность соответствующего параметра.
Показатель статистической упорядоченности, организованности
вычисляется как отношение между эмпирической и максималь-
ной энтропией (последнее характерно для равновероятного рас-
пределения) :
р = 1 — ^эмп
Нмакс
где Нэмп —эмпирическая энтропия;
НМакс — максимальная энтропия.
Данные табл. 14, в которой приведены результаты такого
анализа, показывают, что энтропийная мощность и количество
относительной энтропии (Нэмп/НМакс) в стадии решения несколь-
ко повышается (следовательно, мера организованности соответ-
ственно падает). Эти изменения особенно проявляются в момент
решения, в последействии наблюдаются обратные изменения.
Уменьшение степени упорядоченности в момент наиболее ак-
тивной мыслительной деятельности (при операции принятия ре-
шения), видимо, соответствует реакции десинхронизации, кото-
рая рассматривается обычно в качестве наиболее характерного
проявления активности структур мозга.
В описанном выше способе анализа меры организованности
ЭЭГ в качестве событий (символов, которыми, возможно, зако-
дирована информация в ЭЭГ) принимается факт пересечения
кривой ЭЭГ «изоэлектрической линии». В этом случае запись
ЭЭГ- можно разделить на равные промежутки времени, в кото-
рых текущее значение статистических параметров ЭЭГ оценива-
лось двоичным кодом: 1 — наличие пересечения изоэлектриче-
121
Таблица 14
Динамика относительной энтропии вычисляемой в ЭЭГ картине
левой фронтальной зоны испытуемых на разных стадиях решения задач
Испытуе- мые Величина относительной энтропии
фон стадия решения момент решения последействие
к. с. 0,124 0,141 0,199 0,164
В. А. 0,382 0,510 0,632 0,524
С. Р. 0,483 0,532 0,610 0,600
в. к. 0,492 0,563 0,578 0,532
А. И. 0,513 0,572 0,639 0,621
А. Р. 0,610 0,724 0,819 0,742
С. Е. 0,643 0,704 0,892 0,901
Г. И. 0,792 0,843 0,862 0,837
Б. 0. 0,832 0,884 0,903 0,812
С. А. 0,848 0,883 0,886 0,844
ской линии в выбранном временном кванте 0 — непересечение.
При таком представлении ЭЭГ можно вычислить 2 полные ве-
роятности Pi и Pj (i — пересечение, j — непересечение) и 4 услов-
ные вероятности, характеризующие следовые процессы: Pi/i, Pi/j>
Pj/j, Pj/i- Тогда энтропию (Н'), характеризующую степень вза-
имосвязи параметров ЭЭГ в последовательно взятых квантах
времени, вычисляют следующим образом:
Н' = — [Pi (Pi/i 10g2 Pi/i 4" Pi/j l°g2 Pi/j) +
+P1 (Pj/j l°g2 Pj/j + Pj/i 10g2 Pj/i) ] •
Некоторые результаты анализа динамики вычисляемой та-
ким образом условной энтропии приведены в табл. 15.
Наблюдаемое падение величины условной энтропии по мере
решения задачи может служить косвенным доказательством, с
одной стороны, усиления тесноты связи между электрическими
потенциалами мозга в последовательно взятых интервалах вре-
мени, с другой — повышения роли информационного тезауруса
системы в процессе формирования и реализации некоторой так-
тики решения задачи.
Результаты этих исследований ставят вопрос о том, в какой
мере наблюдаемое изменение энтропийной мощности ЭЭГ — ре-
акций, вычисляемое на основе условных вероятностей следова-
ния разных компонентов суммарной ЭЭГ, связано с динамикой
семантической информации, содержащейся в этих же электриче-
ских потенциалах (в предположении, что информационно зна-
122
Таблица 15
Динамика условной энтропии кривой ЭЭГ испытуемых
на разных стадиях решения задач
Испытуе- мые Величина условной энтропии
фон стадия решения момент решения последействие
В. А. 0,87 0,80 0,69 0,74
В. к. 0,85 0,69 0,58 0,61
С. А. 0,83 0,73 0,65 0,71
Б. О. 0,76 0,69 0,62 0,63
к. с. 0,75 0,70 0,66 0,66
С. Р. 0,71 0,66 0,63 0,60
А. И. 0,69 0,68 0,64 0,65
С. Е. 0,66 0,61 0,53 0,57
Г. И. 0,64 0,53 0,41 0,48
А. Р. 0,63 0,56 0,40 0,42
чащими параметрами электрической активности мозга является
частота фазовых переходов волн ЭЭГ — пересечение изоэлектри-
ческой линии).
Определение количества семантической информации в ЭЭГ
проводилось по соответствующему алгоритму, описанному в гл. 3,
с той лишь разницей, что определение доопытной и апостериор-
ной вероятностей событий производилось из гистограмм частот
фазовых переходов волн ЭЭГ.
Динамика семантической информации в ЭЭГ, зарегистриро-
ванной от левой лобной области мозга, приведена в табл. 16.
Таблица 16
Динамика семантической информации на разных стадиях решения криптографических задач
Испытуе- мые Кол-во семантической информации
фон стадия решения момент решения последействие
к. с. Я ,74 2,13 2,41 2,08
С. С. 1,35 2,14 2,28 1,71
С. Р. 1,04 1,42 1,73 1,52
А. И. (1,08 1,31 1,38 1,36
Г. А. 1,15 1,43 1,40 1,30
123
В табл. 17 иллюстрируются особенности изменений количе-
ства семантической информации в ЭЭГ, регистрируемой от раз-
ных зон головного мозга на разных стадиях решения испытуемы-
ми логических задач. Как видно из результатов, приведенных в
обеих таблицах, у испытуемых имеет место существенное увели-
чение количества семантической информации в стадии решения
и в момент решения задачи. В последействии наблюдается, на-
против, уменьшение количества семантической информации.
Таблица 17
Динамика семантической информации в картине ЭЭГ
на разных стадиях решения испытуемым логических задач
(по О. Г. Чораяну, 1978)
№ задачи Отведе- ние Кол-во семантической информации
фон решение момент решения последейст- вие
1 Fr. s. 1,2 1,7 1,8 —
Fr. d. 1,27 1,36 .1,38 —
Par. s. 0,54 1,47 0,48 —
Осс. s. 2,6 3,13 2,42
2 Fr. s. — 0,68 1,34 1,16
Fr. d. — <1,39 2,06 0,57
Par. s. — 0,58 0,48 0,74
Occ. s. — 1,61 1,82 2,46
3 Fr. s. 0,32 1,09 1,49 1,58
Fr. d. 1,42 1,53 4,7 1,67
Par. s. 0,31 1,1 1,45 1,00
Occ. s. 1,79 2,00 2,13 0,33
4 Fr. s. 1,38 1,39 2,58 1,11
Fr. d. 0,8 0,58 .1,13 0,33
Par. s. 0,47 1,18 1,7 0,26
Occ. s. 1,57 1,47 2Д 2,46
5 Fr. s. — 0,5 1,27 0,76
Fr. d. — 0,66 '1,17 1,27
Par. s. — 0,62 0,58 0,66
Occ. s — 1,89 1,39 1,89
6 Fr. s. 1,2 — 2,1 2,15
Fr. d. 1,27 — 1,47 1,18
Par. s. 0,54 — 0,86 0,69
Occ. s. 2,6 — 2,62 3,09
Примечание: прочерком отмечены случаи, в которых анализ не про-
водился.
В экспериментах с использованием стандартных интеллек-
туальных тестовых заданий (Г. Айзенк, 1972) было показано,
что наиболее заметные изменения при решении таких задач име-
ли место в показателях семантической информации ЭЭГ, реги-
124
Рис. 19. Динамика семантической информации в ЭЭГ разных отведений:
а — Fr. s., б — Fr. d., в — Par. s., г — Par. d. По оси абсцисс: Ф — фон,.
Р — стадия решения, М — момент решения, П — последействие; по оси ор-
динат— количество семантической информации (среднее из Г5 опытов)г
в битах
стрируемой от лобных областей головного мозга (рис. 19). Эти
изменения в большинстве случаев сводятся к следующему: вна-
чале при переходе от фона к решению количество семантической
информации растет, этот рост принимает еще более крутой ха-
рактер при переходе от начала решения до момента «принятие
решения», после чего (в последействии) наблюдается падение
величины семантической информации, хотя она редко достигает
фонового уровня на протяжении всего наблюдаемого периода
последействия (до 20—40 сек).
Аналогичные результаты были получены при анализе дина-
мики семантической информации в кривых ЭЭГ, испытуемых при
решении ими лабиринтных задач; значительное возрастание ко-
личества семантической информации ЭЭГ, отводимой от лобных
долей мозга (особенно, от левой фронтальной зоны), наблюда-
лось на стадии формирования стратегии, соответствующей «про-
движению» испытуемого на 1—3 ярусах многоярусной структуры
лабиринта. На последующих ярусах, т. е. когда у испытуемого
сформировался некоторый вероятностный прогноз структуры ла-
биринта, рост семантической информации значительно замедлял-
ся (рис. 20). В ЭЭГ, отводимой от теменных областей (чаще
из левой теменной области), изменения количества семантиче-
125
€кой информации носили менее выраженный характер на ста-
дии формирования вероятностного прогноза, хотя на последую-
щих ярусах (4—7) лабиринта на этапе реализации выработан*
бит
ной стратегии эти изменения
были более существенными.
Постоянно растущая роль
машинного мира с увеличени-
ем числа и усложнением авто-
матических технических уст-
ройств, ставит перед современ-
ным человеком задачу оптими-
зации работы сложных био-
технических систем. В целях
оптимизации путей решения за-
дач управления широким клас-
сом больших систем, у кото-
рых нельзя предусмотреть все
возможные ситуации в ходе
их функционирования, для ко-
торых характерна большая раз-
мерность алгоритмов управле-
ния, в последние годы предла-
3
2
1
О
Ф Р м п
Рис. 20. Динамика семантической
информации, вычисляемой по ЭЭГ, ле-
вой лобной области в ходе решения
лабиринтной задачи. По оси орди-
нат — семантическая информация, в
битах; по оси абсцисс — этапы (Ф —
фон, Р — решение, М — момент при-
нятия решения, П — последействие)
гается метод ситуационного
управления (Д. А. Поспелов,
1969, 1976; Л. Б. Нисенбойм, Л. С. Загадская, 1975; Ю. И. Клы-
ков, Ю. И. Лобанов, 1976; Г. С. Поспелов, 1977; А. Б. Карунин
и др., 1977; и др.). Метод ситуационного управления в значи-
тельной мере учитывает, имитирует многие элементы как раз-
мытой логики, так и семантической основы мыслительной дея-
тельности человека.
Д. А. Поспелов (1969) считает, что единственной возмож-
ностью расширения способностей машины в решении творческих
задач, решаемых человеком, является создание внутри нее се-
миотической системы внешнего мира. В такой машине, названной
автором гироматом, отображается и реализуется структурно-си-
стемная модель внешней среды. Принцип ситуационного управ-
ления использован при построении ряда семиотических моде-
лей: управления объектов в системах диспетчерской службы мор-
ского порта, при проектировании и возведении промышленных
зданий и т. д. (А. В. Колесников, В. Ф. Пономарев, 1977;
А. Б. Карунин и др., 1976). Семиотической системой называется
языковая модель объекта, отражающая основные черты, зако-
номерности реально протекающих процессов.
В ситуационном управлении различают 3 основных момента:
126
1) описание модели объекта производится на естественном язы-
ке и носит семантический характер; 2) основу модели управле-
ния составляет процесс обучения; 3) построение модели объекта
осуществляется на основе изучения его поведения в различных,
условиях.
Построение модели проблемной ситуации является централь-
ным в ситуационном управлении. В случае ситуационного управ-
ления человек строит структурную модель проблемной ситуации,,
выделяя в исходной ситуации наиболее важные элементы и от-
ношения. Поэтому сама ситуационная модель и ее элементы:
оказываются тесно связанными с прошлым опытом, с информа-
ционным тезаурусом человека, принимающего решение. Как ука-
зывают Д. А. Поспелов, В. Н. Пушкин (1972), творческая дея-
тельность, продуктивное мышление заключается в формировании:
семантической модели проблемной ситуации и целенаправлен-
ном ее преобразовании.
Процесс распознавания ситуации, начинающийся с восприя-
тия отдельных элементов, составляющих сложный образ, есть
процесс отражения в ассоциативных структурах мозга отдель-
ных свойств рассматриваемого образа и сравнение их с соответ-
ствующими «эталонными» элементами хранящегося в памяти:
человека образа этого предмета. Но как сам образ предмета в.
информационном тезаурусе человека, так и восприятие свойств,
объекта носит вид абстрактных в количественном отношении, не-
детализированных элементарных информационных процессов.
Эталонная модель, без которой невозможно распознавание ка-
кого бы то ни было предмета или явления, в психологии извест-
на под названием схемы, прототипа (Р. Клацки, 1978). Под схе-
мой психологи подразумевают набор правил для создания или
описания прототипа, которым в свою очередь обозначают аб-
страктную фигуру, отображающую основные элементы некото-
рого множества стимулов.
В распознавании образа большое внимание психологами и
психофизиологами уделяется концепту «семантического контек-
ста». Хорошо известно, что в своей практической деятельности
человек решает многие задачи распознавания образов с доста-
точно высокой скоростью, что трудно себе представить при на-
личии большого числа потенциальных возможностей выбора и
сравнительно низкой скорости элементарных информационных:
процессов в нейронных системах связи у человека. Экономия
времени, и таким образом повышение скорости операций распо-
знавания образа, может достигаться процессами параллельной
обработки информации, использованием семантического смысло-
вого аспекта информации, которая поступает в организм в про-
127
цессе перцепции. Например, в одной из главных операций в рас-
познавании образа — классификации — знание перечня стимулов,
которые могут встретиться в данной ситуации (т. е. ограничение
универсального множества стимулов до подмножества действи-
тельно встречаемых в этих условиях стимулов), позволяет ис-
ключить из рассмотрения большое множество других образов.
.Хорошим примером этому являются результаты исследования
Г. Райчера (G. Reicher, 1969) и Д. Уиллера (D. Wheeler, 1970),
в которых было показано, что при зрительном восприятии буква
легче идентифицируется, если она предъявляется не изолирован-
но, а входит в состав некоторого слова. Распознавание буквы,
находящейся в слове, резко сокращает число рассматриваемых
значений остальных букв в слове, позволяет ограничить область
поиска и анализа небольшим числом принимаемых во внимание
букв. Это дает возможность ограничиться проверкой лишь не-
которых признаков, отбросив многие другие.
Учет именно фактора семантического контекста лежит в ос-
нове составления различных анаграмм, 'криптографических за-
дач, ребусов, головоломок, когда авторы стараются выбрать
наименее вероятные комбинации букв, событий явлений, тем са-
мым затрудняя решение этих задач «стандартно» мыслящим че-
ловеком, привыкшим к широкому использованию своего инфор-
мационного тезауруса.
В качестве другой четкой иллюстрации роли семантического
контекста в распознавании является описанная многими автора-
ми способность шахматных мастеров воспроизводить позицию
«фигур на шахматной доске при достаточно краткой их экспози-
ции. Так, Де Гроот (De Groot, 1965, 1966) отмечает, что шах-
матные мастера могут безошибочно воспроизвести позицию при
-ее экспозиции в течение не более чем 5 с. Однако если вместо
реальной позиции те же шахматные фигуры расположить на дос-
ке случайным образом, мастера и опытные шахматисты запо-
минают и воспроизводят картину гораздо хуже и ничуть не луч-
ше начинающих шахматистов.
На использовании принципа семантического контекста осно-
ваны известные приемы улучшения мнемонической деятельности
человека: так для запоминания набора бессмысленных слов или
длинных числовых рядов человек часто использует приемы свя-
зывания, отождествления их с другими известными, значащими
для него понятиями.
Вместе с тем тот факт, что многие предметы, образы могут
распознаваться и вне специального контекста, свидетельствует
о том, что в нашей эталонной системе допускаются некоторые
вариации, некоторая размытость входящих в нее элементов.
128
Человек, должен верить, что непонятное
можно понять: иначе он не стал бы раз-
мышлять о нем.
И. Гете
Г л а в а V. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ
ФОРМИРОВАНИЯ ПАТОЛОГИЧЕСКИХ
СОСТОЯНИЙ НЕРВНОЙ СИСТЕМЫ
1. СООТНОШЕНИЕ ЖЕСТКОДЕТЕРМИНИРОВАННОГО
И ВЕРОЯТНОСТНОГО КОМПОНЕНТОВ РЕАКЦИЙ
В ПАТОЛОГИЧЕСКОМ СОСТОЯНИИ
Многим патологическим состояниям организма и его
отдельных физиологических систем свойствен статисти-
ческий характер процессов, протекающих в органах и
тканях. Так, например, процесс канцерогенеза представ-
ляет собой процесс формирования малигнизированных
клеток, протекающий независимо от целостного организ-
ма (Д. Гексли. Цит. по: В. П. Петленко, Г. И. Царего-
родцев, 1976). Первично малигнизация развивается в со-
матических клетках как результат нарушения генетиче-
ской информации. Последнее объясняет значительную
независимость малигнизированных клеток от системных
интегральных реакций организма, ускользание их из-под
контроля интегрирующих организменных реакций и без-
удержный рост и размножение их.
К фиксированным формам поведения, составляющим
значительную часть известных форм расстройств нор-
мального протекания физиологических процессов в орга-
низме, относится широко распространенная в психиатри-
ческой клинике форма патологии—навязчивые состояния.
Термином навязчивость обозначается широкий круг не-
произвольно наступающих и многократно повторяемых
действий или состояний, неадекватных внешней реальной
ситуации и поэтому являющихся нежелательными и не-
целесообразными. Навязчивые состояния разной степени
выраженности характерны для различных неврозов.
Одним из проявлений навязчивых состояний может
служить стереотипия — патологические стереотипные ре-
акции. Г. В. Залевский и М. С. Роговин (1970) считают,
129
что патологические стереотипии развиваются в ряде случаев из
начально целесообразных поведенческих актов, имевших ранее
в жизни человеческого организма (или вида) определенное зна-
чение (например, крестное знамение у религиозных людей), од-
нако впоследствии потерявших этот смысл, ставших механиче-
скими, автоматическими и вышедшими из-под контроля созна-
ния и воли. Авторы полагают, что такого типа нарушения объяс-
няются выпадением активной регулирующей роли механизма
обратной связи, призванной оценивать и направлять приспосо-
бительные реакции организма в плане формирования его адап-
тивного поведения (путем усиления, поддержания адекватных,
полезных и исключения, подавления неадекватных реакций). На-
личие таких коррегирующих влияний механизма обратной связи
на многие поведенческие реакции организма определяет высо-
кий динамический характер последних, поскольку реальное мно-
гообразие ситуаций внешней среды должно найти отражение и
в соответствующем многообразии ответных реакций организма,
чтобы последний оказался приспособленным к этой среде. Сте-
реотипия, фиксированность реакций организма в этих условиях
становится препятствием для адаптивного поведения организма
и является уже сама по себе проявлением патологии. Поэтому
следует присоединиться к мнению Г. В. Залевского и М. С. Ро-
говина (1970), считающих всякую действительно фиксированную
форму поведения патологией, поскольку она подрывает основы
физиологических механизмов обеспечения необходимого органи-
ческого взаимодействия организма и среды, в результате чего
любой поведенческий акт организма реализуется независимо от
внешней среды и вне плана целенаправленного приспособитель-
ного его поведения.
В современной медицине общепринятым является представ-
ление о том, что каждый человек представляет собой четко
обособленную индивидуальность, что индивидуальные поведен-
ческие реакции человека широко варьируют как в норме, так
и в патологии. Однако это резко контрастирует с удивительным
сходством поведения психических больных (П.П. Волков, 1977).
Как подчеркивает автор, в большой психиатрической больнице
всегда можно найти группы больных, страдающих различными
заболеваниями, но почти тождественных по своему поведению.
Не отрицая клинической ценности учета особенностей симптома-
тики отдельных психических заболеваний, следует тем не менее
подчеркнуть, что в принципе любой симптом и синдром может
наблюдаться при определенных условиях при любом заболева-
нии. Это хорошо согласуется и с другим известным в психиатрии
фактом, что при психических заболеваниях общий «набор» раз-
130
личных вариантов патологического поведения невелик, хотя, учи-
тывая многообразие патологических нарушений высших нервных
функций, этот набор мог быть чрезвычайно большим. Статисти-
ческий, вероятностный характер патологических проявлений
высших функций мозга определяется 2 факторами: с одной сто-
роны, любой этиологический фактор может вызвать практиче-
ски любой ответ из неограниченного разнообразия возможных
индивидуальных реакций организма на него, с другой стороны —
один и тот же симптом, комплекс симптомов (реакция организ-
ма на болезнетворное начало) могут иметь место при различ-
ных заболеваниях.
В современной медицине все более и более склоняются к
мнению, что в этиологии различных заболеваний нельзя прида-
вать абсолютное значение какому-либо одному болезнетворному
фактору. Особенности поражения организма данным болезне-
творным агентом обусловлены разнообразными факторами внеш-
ней и внутренней среды организма, связаны с индивидуальными
особенностями организма. При действии одной и той же причи-
ны реакция организма может быть далеко неодинаковой. Хоро-
шо известны случаи, когда родственники или обслуживающий
инфекционного больного персонал несмотря на тесный контакт
не заболевают. Различная индивидуальная чувствительность на-
блюдается и в случае профессиональных заболеваний: несмотря
на одинаковые условия работы лишь у небольшой части работ-
ников отмечаются признаки профессионального заболевания,
причем клинические признаки болезни оказываются выражен-
ными в разной степени у разных больных. Наряду с ведущей
причиной (специфическим фактором), определяющей наступле-
ние и протекание данного заболевания (на последнее могут ока-
зывать и реально оказывают влияние многие факторы), внешние
проявления любого отдельного заболевания характеризуются
значительной случайностью. По этому поводу весьма убедитель-
но высказался Н. К. Боголепов (1967), который писал, что пато-
логические процессы всегда детерминированы и в патогенезе бо-
лезней всегда можно проследить определенную закономерность,
прокладывающую себе путь через ряд случайностей.
Проблема соотношения жесткодетерминированного и случай-
ного в этиологии различных заболеваний в свое время была
предметом оживленной дискуссии, отчасти в связи с выходом
в свет во многом спорной, проблематичной, хотя и весьма ори-
гинальной в медико-философском плане, книги одного из .круп-
нейших отечественных патологоанатомов академика АМН СССР
И. В. Давыдовского ('«Проблема причинности в медицине». М.,
1962). Знаменательным событием в этом научном споре стала
131
открытая дискуссия во II МОЛМИ в 1965 г., в которой приняли
участие многие известные медики (инфекционисты, физиологи,
патофизиологи, патологоанатомы, терапевты, невропатологи, пси-
хиатры и др.). Материалы дискуссии были опубликованы в спе-
циальном издании («О проблеме причинности в медицине». М.,
1965).
Основные выводы книги И. В. Давыдовского и его выступле-
ния на этой дискуссии сводились к следующему: 1) болезнь —
это форма приспособления организма к вероятностно организо-
ванной среде; болезнь — это такой же необходимый атрибут жиз-
ни, как питание, дыхание и т. д.; 2) вероятность заболевания
определяется индивидуальностью, которая и детерминирует кон-
кретное проявление заболевания; индивидуальность организма
порождает загадочную для врача неопределенность, поскольку,
казалось бы, совершенно одинаковые условия (видовые, возраст-
ные, половые, социальные и т. д.) порождают неодинаковый ре-
зультат. Подчеркивая важную роль в поражении организма бо-
лезнетворным началом, И. В. Давыдовский приводит известные
слова великого русского хирурга Н. И. Пирогова: «Случай — вот
творческое начало». В практической медицине есть много пре-
красных иллюстраций случайности заболевания, того, что веду-
щий болезнетворный фактор может привести к заболеванию ор-
ганизма лишь при наличии многих дополнительных условий.
Так, например, из 1000 носителей вируса полиомиелита 990 им
не болеет вовсе, остальные 10 болеют по-разному, и из них толь-
ко 1—<2 заболевают паралитической формой.
Характеризуя соотношение обязательного и дополнительного,
модифицирующего факторов в определении болезни, выступив-
ший в дискуссии известный специалист в области психиатрии
О. В. Кербиков (1965) образно высказался так: «Причинность —
такая королева, выход которой происходит всегда в сопровож-
дении свиты: свита — это условия... Причина всегда необходима,
но она не всегда достаточна для действия. Достаточной причина
становится лишь в надлежащих условиях».
Четкие изменения в мыслительных процессах, наступающие
при различного рода органических поражениях центральной
нервной системы, также связаны с изменением соотношения меж-
ду жесткодетерминированными и вероятностными компонентами
сложных актов интеллектуальной деятельности человека. Так,
например, у больных, страдающих выраженным общим атеро-
склерозом, поражающим и мозговые сосуды, обычно преобла-
дают проявления более простого способа действия, элементар-
ного конкретно-ситуационного, нежели абстрактно-логического
(тем более многозначно-логического). При этом больные обыч-
132
но используют привычные, более часто встречающиеся в обыч*
ной жизни алгоритмы поведения, хотя они и могут быть далеко
не оптимальными, адекватными для данной реальной ситуации.
Иначе говоря, у этих больных затруднена оценка смыслового,
прагматического содержания воспринимаемой и перерабатывав*
мой информации, в значительной степени обусловленная нару-
шением достаточно успешно функционирующего у здорового
человека механизма усвоения, учета и практического использо-
вания вероятностной картины пространственно-временной струк-
туры -ситуаций, внешней среды на основе привлечения прошлого
опыта. Для таких больных практически более сильными и зна-
чимыми становятся тривиальные стандартные композиции, и по-
этому они без особого труда решают типичные тестовые задания,
требующие лишь тривиального стандартного мышления с резким
сужением диапазона поисковых действий. Поэтому в некоторых
случаях внешние проявления поведения таких больных как бы
производят впечатление об их большой способности к абстрак-
ции (в рамках резко ограниченной зоны эвристического поиска)
по сравнению со здоровыми людьми. Иначе говоря, у них исклю-
чается или резко затрудняется возможность решения сложных,
редко встречающихся проблемных ситуаций, но зато как бы об*
легчается решение небольшого числа сравнительно простых сте-
реотипных реакций.
2. .ИЗМЕНЕНИЕ ПРОГНОЗА ПРИ ФОРМАХ ПСИХОПАТОЛОГИИ
Сложная совокупность физиологических и психических про-
цессов, составляющая нормальный психический статус здорово-
го человека, есть результат сбалансированной деятельности не-
скольких уровней сознания, обеспечивающей адекватность пси-
хической реакции на окружающую обстановку в соответствии
с сенсорным настоящим и прошлым человека. При таком сба*
лансированном скоординированном состоянии регуляторных
уровней у здорового человека психический мир выступает как
гармоническое целое. «Раздробленность» психики, фрагментар-
ность проявлений низших и высших уровней психики — одна из
наиболее часто встречающихся форм психопатологии. В целом
ряде психических болезней в результате изменения нормально-
го соотношения уровней мышления нарушается нормальное
взаимодействие организма со средой, человек как бы замыкает-
ся в узком кругу своего внутреннего мира, который порой мало
в чем оказывается скоррегированным с внешним миром.
Психопатологические синдромы, наблюдаемые у маниакаль-
ного или депрессивного больного, хотя и отличаются значитель-
на
ной сглаженностью, полиморфизмом и динамичностью своих
проявлений, в целом характеризуются «невозможностью уравно-
вешиваться с настоящим и держаться в нем» (Т. Ф. Пападопу-
лос, 1975). Мышление такого больного все больше и больше
теряет свои связи с объективным миром, с данными органов
чувств, оно перестает контролироваться и коррегироваться про-
шлым опытом. Для больных являются типичными неадекватные
гиперболизированные реакции, характерные для промежуточных
фаз развития парабиотического, гипнотического состояния, па-
радоксальные ответы на стимулы, не коррегирующие с интен-
сивностью этих стимулов. Так, например, стук в дверь таким
больным воспринимается «как канонада начавшейся войны»,
при прикосновении к плечу — «появляются крылья и он превра-
щается в летающего ангела», любой резкий запах ассоциирует-
ся с ядами, с отравлением живых существ, в том числе и самого
больного. При развитии маниакально-депрессивного психоза на-
ступают сложные многозвеньевые расстройства как в восприя-
тии внешнего мира, так и в самоощущении и осознании себя
как личности.
Менее выраженные изменения характерны для психопатий —
врожденных или сформировавшихся в течение жизни аномалий
характера, занимающих ^промежуточное положение между пси-
хической нормой и болезнью. Психопатии характеризуются пре-
имущественно сдвигами в эмоционально-волевой сфере. Психо-
патическим личностям свойствен недоучет прошлого опыта, в ре-
зультате их поведение представляется неадекватным реальной
ситуации; психопаты склонны к неожиданным импульсивным по-
ступкам с резко выраженной эмоциональной окраской. Хотя пси-
хопаты в какой-то мере с годами меняются в своем характере
(психопатии — патологические характеры), внутренняя стерж-
невая структура их душевного склада сохраняется. Психопати-
ческие личности произвольно оперируют с объективными мера-
ми вероятности тех или иных альтернативных действий, руко-
водствуясь в своих практических действиях показателями субъ-
ективной вероятности, мало обращая внимание на величину
объективно существующей вероятности того или иного альтер-
нативного действия. Так, истерическая личность в определенных
условиях может использовать свой аффект для достижения це-
ли, хотя априори достижение этой цели характеризуется ма-
лой вероятностью. Как указывает А. М. Иваницкий (1976), исте-
рик может подметить тонкие детали обстановки и некоторые
нюансы настроения окружающих (если они идут в плане его
доминирующей мотивации) и, умело ими пользуясь, достигает
поставленной цели.
134
Важным синдромом, часто встречающимся в картине различ-
ных психозов и занимающим ведущее место в определении пси-
хического состояния человека является синдром психического
автоматизма. Синдром психического автоматизма Кандинского-
Клерамбо характеризуется большим многообразием расстройств,
хотя с нарастанием органического дефекта, определяющего этот
синдром, психические проявления становятся все более элемен-
тарными, низшие и негативные психические автоматизмы начи-
нают преобладать над высшими и позитивными, пока не исчез-
нут полностью при глубокой деменции (Л. К. Хохлов, 1969).
Основные проявления психического автоматизма в виде соответ-
ствующих псевдогаллюцинаций, явлений насильственного мыш-
ления в наиболее четкой форме выявляются в состоянии, ха-
рактеризующемся преобладанием тормозного процесса.
Более дифференцированные изменения в сенсорно-перцептив-
ной и логико-мнестической деятельности наблюдаются при ши-
зофрении. При шизофрении, если нет продуктивных галлюци-
наторных синдромов, отсутствие заметных изменений в перцеп-
тивной деятельности таких больных дает основание говорить об
интактности, сохранности процессов восприятия при шизофре-
нии (Ю. Ф. Поляков, 1974). Известно, что в норме у здорового
человека характер перцептивной деятельности определяется
сложностью системы альтернативных действий и вероятностны-
ми характеристиками этих альтернатив: чем выше сложность
системы альтернатив, тем труднее процесс распознавания си-
туации, «образа» и т. д. Вместе с тем при разных вероятностях
альтернатив легче и лучше распознаются те сигналы, которые
имеют высокую субъективную вероятность.
Существенно иначе ведет себя больной шизофренией. По
данным Ю. Ф. Полякова (1974), больные шизофренией в 4 раза
чаще, чем здоровые, воспринимают маловероятные сигналы (в
опытах с предъявлением через слуховой анализатор отдельных
слов и предложений, в которых отсутствуют окончания). В этих
опытах было установлено, что больные шизофренией более ус-
пешно по сравнению со здоровыми испытуемыми воспринима-
ют маловероятные по прошлому опыту речевые сигналы и «ме-
нее успешно — более вероятные. Автором показано, что если здо-
ровые испытуемые преимущественно руководствуются вероят-
ностью той или иной гипотезы по контексту фразы, учитывая
статистические связи слов внутри предложения или букв в
слове, то для больных шизофренией большое значение приобре-
тают внешние акустические характеристики сигналов (число
слогов, ударная гласная и т. д.). Аналогичные результаты были
получены и для зрительного восприятия. Больные 'шизофренией
135
в процессе распознавания образов в большей мере, чем здоро-
вые люди, используют формальные признаки сигналов, они бо-
лее точны в текущей перцептивной деятельности (без коррек-
ции ее прошлым опытом).
Это свойство коррекции настоящего (текущей информации
в результате непосредственного процесса восприятия) прошлым
опытом, которое более всего оказывается нарушенным у боль-
ных шизофренией, Ю. Ф. Поляков называет свойством «актуали-
зации» событий, признаков, предметов. Многочисленные резуль-
таты экспериментальных исследований, приведенные в книге
автора, подтверждают его вывод о том, что у больных шизофре-
нией вследствие нарушения актуализации, привлечения близких,
родственных сведений из прошлого опыта имеет место сущест-
венная перестройка и процесса восприятия, возникает необходи-
мость учета и анализа большого объема характеристик, свойств
стимула для обеспечения верного восприятия объекта. Естествен-
но, что такая перестройка процессов перцепции приводит к ухуд-
шению, к нарушению оптимального характера протекания про-
цессов распознавания образов. В результате мы видим при не-
котором выигрыше в редких случаях (когда распознаются ма-
ловероятные, редко встречающиеся образы) существенный про-
игрыш в большинстве обычных жизненных ситуаций.
Это находит подтверждение и в том факте, что при решении
задач, связанных с выдвижением и последующим перебором
многих альтернатив, больные шизофренией рассматривают гораз-
до больше вариантов обобщения и различения сравниваемых
предметов, чем здоровые. Так, в работе Ю. Ф. Полякова (1974),
где приводится изложение результатов сравнительного исследо-
вания на здоровых и больных шизофренией, описан случай, ко-
гда здоровые испытуемые находили 203 способа для обобщения
и различения предложенных 12 пар сравниваемых предметов,
в то время как больные шизофренией рассматривали 556 раз-
личных вариантов (опыты ставились на 50 здоровых и 50 боль-
ных шизофренией). Больные шизофренией с гораздо большей
легкостью находили возможности для сравнения, хотя предла-
гаемые ими обобщения, сравнения могут произвести впечатление
«странных»: например, при нахождении общих свойств у тарел-
ки и лодки больные приводят следующее: «не пропускают жид-
кости», «являются несъедобными»; при другой предъявляемой
паре слов «ботинок — карандаш» больные говорят «они издают
звуки» и т. д. Следует при этом заметить, что с точки зрения
формально-логической умозаключения больных не являются
ошибочными, они вполне удовлетворяют требованиям задачи.
По-видимому, у больных шизофренией имеет место четко вы-
136
раженная тенденция вычленять в процессе сравнения рассмат-
риваемых предметов наряду со стандартными, часто встречаю-
щимися признаками, нестандартные, редко повторяющиеся при-
знаки, низкая вероятность встречаемости которых у здоровых
людей является основанием отбрасывать, не принимать их во
внимание. Употребление нестандартных признаков, оперирова-
ние маловероятными характеристиками, очевидно, лежит в осно-
ве нетривиальной логики мыслительной деятельности больных
шизофренией, что позволяет им в некоторых случаях находить
нетривиальные решения проблемных задач легче и лучше, чем
это могут делать здоровые люди.
По мнению Ю. Ф. Полякова (1974), основной патологией в
мыслительных процессах у больных шизофренией, проявляю-
щейся при решении классификационных задач (а эти задачи со-
ставляют обязательное звено любой более или менее сложной
творческой задачи), служит расширение круга используемых
свойств, признаков за счет актуализации латентных характери-
стик, малозначимых с точки зрения здоровых людей, общепри-
нятой практики применения предметов и явлений. Очень инте-
ресными и наглядными в этом плане представляются результа-
ты экспериментов с «творческой задачей Секея». Суть экспери-
мента заключается в следующем. На столе лежат различные
предметы: весы, ключи, электрический фонарик, баночка с солью,
счетные палочки, грузы, различные гири и т. д. Среди этих пред-
метов находится обычная восковая свеча. Испытуемым пред-
лагается, используя какие-то из этих предметов, уравновесить
весы так, чтобы через некоторое время они сами вышли из рав-
новесия без дополнительных вмешательств. Опыты ставились
на здоровых испытуемых и больных шизофренией. Оказалось,
что больные шизофренией в среднем легче, чем здоровые люди,
находили правильное решение задачи, основная трудность кото-
рой была связана с необходимостью актуализации свойства, яв-
ляющегося благодаря влиянию прошлого опыта «латентным»
(правильный ответ, заключенный в помещении на одну чашку
весов свечи, которую зажигают, основан на достаточно второ-
степенном признаке — свойстве свечи гореть и уменьшаться в
весе; обычно здоровые испытуемые в процессе решения предла-
гают более известные, часто встречающиеся в повседневной
практике приемы — помещение на одну из чашек весов легко
испаряющихся растворов и т. д.).
Эти представления хорошо согласуются с результатами ис-
следований И. М. Фейгенберга (1963—1977), особенностей на-
рушений механизмов вероятностного прогнозирования у боль-
ных шизофренией. Анализ характера протекания ориентировоч-
137
ного рефлекса у больных шизофренией, отличающихся резко вы-
раженной картиной дефекта с преобладанием в клинической
картине негативной симптоматики, но без отчетливых бредовых
или галлюцинаторных расстройств, привел автора к гипотезе
о преимущественном нарушении у этих больных механизма ве-
роятностного прогнозирования.
Поведение больного шизофренией таково, что создается впе-
чатление, будто бы при сохранности в его памяти прошлого опы-
та тем не менее информация о вероятности этих событий дезор-
ганизована, в результате чего возрастает неопределенность про-
гноза настоящих событий. У больных почти не угасают ори-
ентировочные реакции, имеет место повышенная чувствитель-
ность к обычным стимулам, резко затруднена адаптация, при-
способление к изменяющимся условиям среды и т. д. В целом
поведение больного определяется неадекватностью, неэкономич-
ностью реакций нервной системы на различные стимулы, след-
ствием чего становится быстрое ее «истощение». Видимо, в этом
плане неосознаваемой защитной реакцией больного является
стремление резко сузить круг возможных воздействий, как бы
стремление уйти в искусственно созданный бедный мир с огра-
ниченным числом активных элементов, с жестко установленным
порядком в нем.
Как уже указывалось выше, прошлый опыт, память больно-
го в целом существенно не изменена, нарушена лишь способ-
ность практически руководствоваться этим опытом, знание веро-
ятностной организации встречавшихся в прошлом аналогичных
событий, использование прошлого опыта для решения реально
стоящих перед больным задач. Складывается впечатление, что
все прошлые события для больного приобретают одинаковую
значимость, он теряет способность улавливать различия в их
вероятностях. Наступающее для больного сглаживание различий
между вероятностями разных ситуаций, событий, сигналов, при-
водящее к вероятностной дезорганизации среды, приводит к ро-
сту энтропии в мозговых системах переработки информации
(И. М. Фейгенберг, 1977). В результате мы видим большую
«рыхлость» ассоциаций больного, ему одинаково легко (или оди-
наково трудно) извлекать из памяти высоковероятную или мало-
вероятную ассоциацию. Этим обстоятельством определяется
«оригинальность» мышления шизофренического больного, не отя-
гощенного какими бы то ни было предубеждениями. Нарушения
вероятностного прогноза у больных шизофренией хорошо прояв-
ляются при решении многих тестовых заданий (предъявление
необычных стимулов, тест на иллюзию Шарпантье и 'Т. д.). Если
испытуемым дать в руки 2 одинаковых цилиндра, но разных
138
объемов, то здоровые обычно полагают, что цилиндр с меньшим
объемом тяжелее (в действительности оба цилиндра имеют оди-
наковый вес). Больные шизофренией в этом опыте лучше здоро-
вых испытуемых оценивают вес цилиндров, поскольку у них пер-
цептивный аппарат мозга «не отягощен» прошлым опытом, для
них мало значения имеют такие моменты, как преднастройка,
установка, т. е. психофизиологические феномены, опять-таки яв-
ляющиеся следствием прошлого опыта.
При предъявлении шаблонных типичных изображений, явля-
ющихся высоковероятными, для правильного распознавания
больными шизофренией их приходилось представлять в гораздо
более четкой, сфокусированной форме, чем здоровым испытуе-
мым. Напротив, предъявление необычных стимулов оказывается
для больных при распознавании более легкой задачей, чем для
здоровых испытуемых. Такие же результаты были получены при
тахистоскопическом предъявлении различных изображений: для
верного распознавания высоковероятных изображений больным
требовалось больше времени, чем здоровым, и, напротив, при
предъявлении маловероятных изображений им требовалась мень-
шая экспозиция, чем здоровым.
Это объясняется тем, что у здорового человека для распозна-
вания образов нет необходимости в подробном рассмотрении
всех деталей предмета: он воспринимает лишь некоторые основ-
ные детали, а остальное достраивает благодаря прошлому опы-
ту, иначе говоря, дефицит текущей информации у него компен-
сируется вероятностным прогнозом. В результате для опознания
обычных шаблонных предметов здоровому человеку достаточна
лишь короткая экспозиция, позволяющая воспринять лишь не-
большое число его признаков. Механизм распознавания образов
у больного шизофренией совсем иной: он не может пользоваться
вероятностным прогнозом, не может достраивать текущую ин-
формацию ассоциативной информацией, извлекаемой из памяти,
и поэтому он вынужден собирать достаточно полные сведения,
усваивать все детали предъявляемого образа. Естественно, на
это уходит гораздо больше времени, чем у здорового человека,
оперирующего вероятностным прогнозом.
Иное мы видим при предъявлении незнакомых, необычных,
нестандартных стимулов и образов. Для больного шизофренией
механизм распознавания остается тем же: он детально рас-
сматривает весь предмет, объект опознания, пока из такого по-
дробного анализа не получает цельное представление об образе,
предмете. Необходимое время экспозиции существенно не изме-
няется (по сравнению с экспозицией стандартных изображений).
У здорового человека возникает необходимость увеличения вре-
139
мени экспозиции для сбора дополнительной, более детальной ин-
формации об изображаемом предмете. Однако при этом вероят-
ностный прогноз вместо положительной роли начинает играть
роль отрицательного фактора: родственная, ассоциируемая по
прошлому опыту, информация не только не помогает процессу
верного распознавания уникального предмета, но ухудшает его,
увеличивая общую продолжительность необходимого времени
экспозиции опознаваемого предмета. Иначе говоря, вероятност-
ное прогнозирование, нарушенное у больных шизофренией, недо-
статочно помогает ему при предъявлении обычных шаблонных
изображений, но оно и мало мешает ему при необходимости рас-
познавания необычных, нестандартных предметов.
Такое представление о природе расстройств в механизме ве-
роятностного прогнозирования у больных шизофренией находит
подтверждение и в результатах исследования И. М. Фейгенбер-
га (1975), в котором был отмечен факт выравнивания у больных
времени реакции на разновероятные стимулы при анализе вре-
мени, затраченного на выполнение дизъюнктивных реакций.
Таким образом, наиболее важным моментом в патологии выс-
ших функций мозга при шизофрении является увеличение ин-
формационной неопределенности, приводящей к ухудшению
адаптационного приспособительного поведения таких больных,
как результат уменьшения негэнтропийного эффекта моделиру-
ющей деятельности мозга, в частности проявляющейся в глубо-
ких нарушениях нейро- и психофизиологических механизмов ве-
роятностного прогнозирования (И. М. Фейгенберг, 1963, 19*77;
И. М. Фейгенберг, В. Л. Леви, 1965; В. М. Михлин, 1973; и др.).
Существенные нарушения в механизме вероятностного про-
гнозирования отмечены также у бол*ьных с поражением лобных
долей головного мозга и v больных-олигофренов (Е. Ф. Бажин,
1973).
3. ХАРАКТЕРИСТИКИ НЕЙРОФИЗИОЛОГИЧЕСКИХ
ПРОЯВЛЕНИИ РАБОТЫ СТРУКТУР МОЗГА
ПРИ ПСИХОПАТОЛОГИЧЕСКИХ ПРОЦЕССАХ
Принцип причинности в патологии означает, что всякое изме-
нение в состоянии живой системы детерминировано материаль-
ными взаимодействиями этой системы с факторами среды. Это
означает, что психопатологические процессы должны найти от-
ражение в особенностях нейрофизиологических характеристик
функционирования структур мозга.
Данные патофизиологии свидетельствуют о том, что шизо-
140
френия характеризуется отчетливой функциональной слабостью
коры головного мозга, повышенной склонностью к тормозным
реакциям. При этом торможение часто неполное, с элементами
парадоксальных реакций на стимулы разной интенсивности име-
ет преимущественную локализацию в высших корковых центрах
мыслительной деятельности человека. Так, А. Абаскулиев (1967)
пишет, что в результате нарушения интермедиарного обмена у
больных, страдающих шизофренией, имеет место токсическое по-
ражение высокомолекулярными белковыми ядами, носящими
специфический характер и эффективно повреждающими «специ-
ально человеческие области головного мозга».
Нарушения познавательных процессов у больных шизофре-
нией прежде всего связаны с ухудшением, расстройством уже
описанного ранее механизма избирательного извлечения из па-
мяти и использования прошлого опыта индивидуума. Результаты,
исследований Ю.Ф. Полякова (1974) свидетельствуют о том, что*
в процессе познавательной деятельности больных нарушается
лишь процесс актуализации, выбора сведений из памяти при со-
хранности самого объема этой памяти. В случае шизофрении
патологический процесс в сущности почти не затрагивает те ви-
ды умственной деятельности, которые имеют четкую, однознач-
но детерминированную схему операций, жесткую полностью фор-
мализуемую программу реализации.
Для понимания физиологической природы нарушения меха-
низмов вероятностного восприятия сенсорной информации, из-
влечения из памяти ранее фиксированной информации, вероят-
ностного прогнозирования динамического процесса важным
представляется анализ различных компонентов вызванного по-
тенциала при образовании условного рефлекса с привлечением
и отвлечением внимания (А. М. Иваницкий, 1976). В то время
как у здоровых испытуемых использование этой процедуры, как
правило, приводит к соответствующим направленным измене-
ниям компонентов вызванного потенциала, у больных шизофре-
нией наблюдаемые изменения отдельных волн вызванного по-
тенциала носили хаотический характер и не были связаны с
сигнальной значимостью применявшихся стимулов. Отмечено
также, что изменения ответа в теменной и затылочной областях
у больных носили разнонаправленный характер и также не име-
ли четкой корреляции между собой.
Интересные результаты о вариабельной природе различных
компонентов вызванного потенциала у больных шизофренией
были получены И. Каллавеем с сотр. (Е. Callaway е. а., 1965),
Р. Т. Джонсом с сотр. (R. Jones е. а., 1966). В этих эксперимен-
тах было установлено, что в вызванных потенциалах на 2 при-
141
менявшихся в случайном порядке звуковых сигналах (тоны вы-
сотой 600 и 1000 Гц) у больных, страдающих шизофренией, име-
ет место гораздо меньшая корреляция, чем у здоровых испытуе-
мых. Поскольку испытуемым предлагалось не обращать внима-
ния на звуковые раздражения, авторы этой работы на основе
полученных результатов делают вывод о том, что больные шизо-
френией склонны акцентировать свое внимание на несуществен-
ных сигналах. В последующих исследованиях авторов (Е. Calla-
way, 1969; R. Jones, Е. Callaway, 1970) было показано, что при
применении двух одинаковых тонов различия между отдельны-
ми ответами у больных шизофренией были выше, чем у здоровых
испытуемых. При этом авторами отмечен факт непостоянства
амплитудных и временных параметров вызванного потенциала
на фоне общего падения амплитуды ответов на звуковые стиму-
лы (тоны разной высоты).
Значительная вариабельность параметров вызванного потен-
циала наблюдалась и в случае применения зрительных сигналов-
(К. Lifshitz, 1969). Коэффициент корреляции между двумя по-
следовательно зарегистрированными вызванными ответами у
больных составлял 0,428 (у здоровых испытуемых он состав-
лял 0,785). Соответствующие значения стандартного отклонения
составляли 0,362 и 0,167. Эти результаты автором рассматрива-
ются как свидетельство более высокой неопределенности сенсор-
ной информации у больных шизофренией. Аналогичные результа-
ты в отношении вызванных потенциалов на зрительные сигналы
у больных шизофренией описаны и в работе А. А. Кирпиченко
(1970).
Уменьшение корреляционной связи между вызванными потен-
циалами, зарегистрированными из разных областей коры голов-
ного мозга, при шизофрении сопровождается и существенным
затруднением в выработке условного рефлекса.
Обсуждая результаты экспериментов И. Каллавея и К. Лиф-
шиц о повышении вариабельности вызванных потенциалов у
больных шизофренией, И. М. Орниц (Е. Ornitz, 1969) приходит
к заключению, что рост вариабельности параметров вызванных
ответов отражает неустойчивость регуляции сенсорного входа,
результатом чего и является нарушение константности восприя-
тия, чувство неуверенности и изменение реактивности, опреде-
ляющие особенности поведения больного, страдающего шизо-
френией.
Значительная вариабельность вызванных потенциалов, отсут-
ствие их закономерных изменений, определяемых условиями экс-
перимента, очевидно, могут быть связаны с неадекватностью
поступков больного.
142
С таким представлением о природе и возможной нейрофи-
зиологической основе поведения больного шизофренией согласу-
ются и результаты исследований Т. И. Вековица и П. Б. Блевета
(Т. Е. Weckowicz, Р. Blewett, 1959), П. Шакова (Р. Shakow,
1963). По мнению Т. И. Вековица и П. Б. Блевет (Т. Е. Wecko-
wicz, Р. В. Blewett, 1959), для больного шизофренией характер-
на неспособность к селективному отбору информации. Как счи-
тают авторы, механизм, оценивающий значимость раздражений,
работает у этих больных на основе ложной «теории вероятно-
сти». По данным П. Шакова (Р. Shakow, 1963), фрагментарность
восприятия, наблюдаемая у больного, страдающего шизофре-
нией, проявляется во внимании к незначительным деталям при
затрудненной оценке внутреннего смысла, значимости события
и ситуации в целом.
К таким же выводам о механизме нарушения свойства адек-
ватной оценки больным шизофренией реальной ситуации при-
ходит и В. В. Плотников (1968), описавший случаи, когда боль-
ные при знакомстве с репродукциями художественных картин не
могли правильно объяснить смысл изображенного на них собы-
тия. Так, при рассмотрении репродукции картины И. Е. Репина
«Иван Грозный убивает своего сына» для больного наиболее
значимым сигналом оказался узор рисунка на ковре (или тако-
го же плана второстепенные детали картины). Даже когда его
внимание привлекли к крови на голове царевича, он сказал, что
«здесь, очевидно, пролилась краска».
Сравнительный анализ изменений ранних и поздних компо-
нентов вызванного потенциала показал, что при шизофрении в
основном нарушено проведение по неспецифическим проекцион-
ным путям, что в свою очередь приводит к изменению соотно-
шения между 2 видами информации (прямая, непосредственная
информация от источника и опосредованная через неспецифиче-
скую систему мозга). Таким образом, симптомокомплекс пато-
логических изменений в организме, наступающий при шизофре-
нии, характеризуется снижением уровня афферентного потока
по неспецифическим путям (оно проявляется в уменьшении ампли-
туды поздних волн вызванного потенциала, в повышении ва-
риабельности ответа, замедлении циклов восстановления и в
парадоксальном эффекте на раздражение высокой интенсивно-
сти). Все это вместе взятое определяет те особенности перцепции
и использования текущей и зафиксированной в памяти инфор-
мации, которые делают поступки больного шизофренией не со-
ответствующими реальной ситуации, поведение его перестает
быть адаптивным, приспособительным.
Поведение больного теряет адаптивный целесообразный ха-
143
рактер и при нарушении процесса передачи информации по спе-
цифическим проводящим путям. Как известно, одной из наибо-
лее характерных черт клинической картины истерических психо-
зов является «активное непринятие действительности», уход в
мир фантастических картин, болезненных переживаний. Уход от
действительности характерен и для синдрома бредоподобных
фантазий, когда больные подменяют действительность фанта-
стическими представлениями, обычно рисующими реальную си-
туацию в благоприятном для них свете (А. М. Иваницкий, 1976).
Анализ вызванной электрической активности при реактивном
психозе отчетливо показывает торможение проведения по сен-
сорным специфическим путям при сохранении или даже повы-
шенной функции неспецифической проекционной системы.
В результате проведенного исследования А. М. Иваницким
(1976) было выявлено при реактивнохм психозе статистически
значимое снижение амплитуды двух первых волн вызванного
ответа и повышение амплитуды поздней положительной волны
(рис. 21). По мнению автора, высокая лабильность сенсорного
входа у личностей истериовозбудимой группы обусловливает вы-
раженные реакции сенсорного поиска, характерные для
этих лиц.
У психопатических личностей так же, как и в случае боль-
ных с различными типами психозов, не наблюдаются сколько-
нибудь выраженные изменения в параметрах поздних волн вы-
званного ответа в зависимости от сигнальной значимости стиму-
ла (что имеет место у здоровых людей).
Рис. 21. Изменение параметров зрительного вызванного потенциала у
больных реактивным психозом (по А. М. Иваницкому, 1976): а — вызван-
ные потенциалы в норме, б — вызванные потенциалы у больных реактивным
психозом
144
У психопатов обычно наблюдаются изменения амплитуды
ранних волн ответа (увеличение — у психопатов истериовозбуди-
мой группы, уменьшение — у психопатов тормозной группы).
Таким образом, при различных формах психозов, как и при
шизофрении, очевидно, имеет место блокада одной из двух вос-
ходящих проекционных систем, что приводит к постоянному ин-
формационному дефициту, в свою очередь препятствующему
адаптивному поведению такого больного.
4. НЕКОТОРЫЕ ХАРАКТЕРИСТИКИ ДЕТЕРМИНАЦИИ
НЕЙРОФИЗИОЛОГИЧЕСКИХ ПРОЦЕССОВ ПРИ ЭПИЛЕПСИИ
Симштомокомплекс патологических проявлений деятельности
мозга, известный под названием эпилептического синдрома, на-
блюдается при разных неврологических и психических заболе-
ваниях и отличается значительной неоднородностью. Это объяс-
няет тот факт, что многие авторы и сейчас еще придерживают-
ся той же точки зрения, которую более 40 лет назад высказал
М. Б. Кроль (цит. по: А. И. Болдырев, 1976): «На эпилепсии*
нельзя смотреть как на самостоятельную болезнь, а лишь как
на синдром, могущий проявляться при разных заболеваниях:
центральной и периферической нервной системы». Однако по-
давляющее большинство специалистов в области психиатрии и:
невропатологии в настоящее время склонны выделять эпилепти-
ческую болезнь как отдельную нозологическую единицу со спе-
цифической (хотя и в значительной степени варьирующей) симп-
томатикой. Основными чертами эпилепсии являются взрывча-
тый характер приступов, чрезвычайная сила возбуждения и пе-
риодичность этих пароксизмов.
Для эпилепсии характерен полиморфизм пароксизмов, обус-
ловленный как разной интенсивностью возбуждения, исходящего
из эпилептического очага, так и одновременным сосуществова-
нием нескольких очагов (А. И. Болдырев, 1971, 1976). Соотно-
шения моно- и полиморфизма у больных не остаются постоян-
ными на разных этапах развития болезни. По мере развития эпи-
лептического приступа наблюдается общая тенденция: умень-
шение мономорфных и нарастание полиморфных приступов,
(А. И. Болдырев, 1971).
В основе этиологии болезни у взрослых лежат различные, ра-
нее перенесенные воспалительные поражения мозга, в особенно-
сти воспалительные процессы арахноидальной оболочки мозга.
В сфере психических проявлений болезни на первый план:
выступают такие признаки как замедленность и инертность мыс-
145
лительных процессов, чрезмерная патологическая обстоятель-
ность, вязкость мышления. Одним из наиболее важных нейро-
физиологических признаков эпилептического поражения являет-
ся патологическая концентрация возбудительного процесса в
структурах мозга. В поведении больного эпилепсией это прояв-
ляется высокой инертностью его мышления — больного эпилеп-
сией чрезвычайно трудно свернуть с избранной им темы.
Об отсутствии генерализованного общего возбуждения всех
структур мозга в случае эпилепсии, хотя клиническая картина
самого эпилептического пароксизма наводит на такую мысль,
говорил еще И. П. Павлов: «Ясное дело, что какого-нибудь воз-
буждения сплошного в этой болезни (эпилепсии. — А. К., О. Ч.)
отнюдь нет».
Анализируя ЭЭГ, зарегистрированную во время судорожных
приступов, Пенфилд и Джаспер (1958) обратили внимание на
сложную природу судорожных пиков в ЭЭГ. По мнению авто-
ров, развитие судорожного разряда обусловливается как увели-
чением общего числа активных нейронов, степенью их синхро-
низации, так и увеличениехМ уровня деполяризации мембраны
отдельной клетки и определенной направленностью электриче-
ских потенциалов клетки относительно регистрирующих элек-
тродов.
Одно из кардинальных свойств эпилептического разряда кор-
ковых нейронов — высокая частота импульсации (до 500 и более
импульсов в секунду)—было впервые отмечено Баумгартнером
(Baumgartner G., 1954). По данным Фессара (Fessard А., 1958)
и Ф. Моррела (F. Morrell, 1961), в зоне эпилептического очага
в корковой ткани создаются условия, благоприятствующие изо-
ритмичности; электрические поля, возникающие во время актив-
ности нервных клеток, создают условия для синхронизации ней-
ронов благодаря эфаптическим влияниям, что благоприятствует
распространению эпилептического возбуждения на соседние
участки мозга.
Даже в состоянии эпилептического приступа нервные клетки
в очаге пароксизмального возбуждения не представляют гомо-
генную структуру. В работе Ч. Л. Ли (С. L. Li, 1955) было по-
казано, что после аппликации стрихнина на кору мозга в эпи-
лептическом очаге были обнаружены два типа клеток: одни ней-
роны генерировали пачки импульсов только во время «стрихнин-
ных» волн поверхностной электрограммы, другие — в интерва-
лах между ними, круто обрывая свой импульсный разряд с по-
явлением на электрокортикограмме «стрихнинных» волн. По мне-
нию автора, нейроны первой группы выполняют специфические
функции в коре, связанные с проекционными корковыми зона-
146
ми, в то время как нейроны второй группы выполняют функции
регулятора уровня возбудимости корковых нейронов.
Процесс «эпилептизации» нервной клетки под влиянием эпи-
лептогенного фактора X. Мацумото и С. Марсан (Н. Matsu-
moto, S. Marsan, 1964, а, в) делят на 2 стадии: в первой стадии
действия такого фактора, например пенициллина, в корковых ней-
ронах возникают пароксизмальные деполяризационные сдвиги,,
следующие ритмически и прерывающиеся гиперполяризацион-
ными волнами амплитудой 5—10 мВ и длительностью 400—
600 мс; потенциал покоя при этом не менялся. По мере усиления
действия пенициллина при тонической фазе судорожного при-
падка происходит переход ко второй стадии, выражающейся в
значительной и устойчивой деполяризации сомы с ритмическими
осцилляциями мембранного потенциала.
В наших «экспериментах» с эпилептизацией центральных ней*
ронов под влиянием поверхностной аппликации стрихнина
(О. Г. Чораян, 1969) мы не обнаружили заметного увеличения
числа нейронов с фоновой импульсной активностью. Но пример*
но в 50% случаев наблюдались определенные изменения в ха*
рактере фоновой и вызванной импульсной активности нервных
клеток. Они заключались в значительном увеличении количе*
ства нейронов с групповой пачечной импульсной активностью
(табл. 18). Далее увеличивалось число синхронизированных воз*
буждений нервных клеток, увеличивался диаметр зоны синергии*
но работающих нейронов (с 200—300 мкм в интактном состоя-
нии до 600—1000 мкм в «эпилептическом» состоянии). На рас-
стоянии между кончиками регистрирующих микроэлектродов
100—1000 мкм в центральном отделе зрительного анализатора
лягушки в подавляющем большинстве случаев (до 85%) наблю-
дались синергичные отношения как в условиях вызванной им-
пульсной активности, так и при «спонтанно» возникающих эпи-
лептических пароксизмах. Некоторые результаты этих исследо-
ваний приведены в табл. 19.
Полученные результаты свидетельствуют о том, что специфи-
ческие изменения в импульсной активности, имеющие место при
развитии судорожных приступов, обусловливаются как измене-
ниями электрических потенциалов отдельных нервных клеток,,
так и особенностями клеточной организации возбуждения в эпи-
лептическом очаге.
Наблюдавшееся нами уменьшение амплитуды потенциалов,
действия в пачке при внеклеточной регистрации, очевидно, яв-
ляется проявлением различного уровня поляризации мембраны
клетки, феномена катодической депрессии, наступающего в ре-
зультате характерной для эпилептического приступа цикличе-
147
Таблица 18
Изменение частоты встречаемости нейронов с различным типом
фоновой импульсации при действии стрихнина
Тип активности Число нейронов (в %)
до действия стрихнина после действия стрихнина
Непрерывный аритмический 74 65,5
Пачечный, групповой 19 31
Отсутствие фоновой активности 7 3,5
Таблица 19
Взаимосвязь нейронов при световом раздражении
в состоянии эпилептиформных пароксизмов
(по данным корреляционного анализа)
Статистические
параметры
Положительная корреляция Отрицательная корреляция
фоновая вызванная фоновая вызванная
импульсная импульсная импульсная импульсная
активность активность активность активность
Среднее арифметическое
Среднее квадратическое
отклонение
Достоверность различия
0,23 0,42
0,09 0,47
(Р < 6,01)
ОД 9 0,38
ОДЗ 0,21
4,3
(р < 0,01)
ской пароксизмальной деполяризации клеточной мембраны.
Результаты, приведенные в таблице 20, показывают, что при
развитии судорожного состояния у животного наступает увели-
чение числа реагирующих на свет нейронов (преимущественно
за счет возбудительных реакций). Такая повышенная активность
нервных клеток на адекватные стимулы является результатом
некоторого ослабления тормозных постсинаптических потенциа-
лов, сопровождаемого возрастанием удельного веса возбудитель-
ных постсинаптических потенциалов.
По-видимому, преимущественная роль электротонических эк-
страсинаптических влияний и определяет увеличение удельного
веса жесткодетерминированных фиксированных форм взаимо-
148
Таблица 20
Изменение частоты встречаемости разных типов реакций нейронов
на световое раздражение до и после введения стрихнина
Число реакций (в %)
Тип реакции
нейронов
до введения
стрихнина
после введения
стрихнина
Возбуждение на включение
света Торможение на включение 13 24
света Возбуждение на выключе- 9 4,6
ние света Торможение на выключение 15,8 23
света Возбуждение в течение 8,4 7,6
всего действия света Торможение в течение всего 14 16,9
действия света 10 2
Отсутствие реакции 29,2 22,5
связи как импульсных потенциалов исследуемых нейронов, так
и потенциалов действия отдельных нейронов и эпилептических
волн суммарной поверхностной электрограммы (О. Г. Чораян,
1969). Такое увеличение жесткодетерминированных форм вза-
имосвязи различных типов электрической активности отдельных
нейронов и их совокупности было отмечено и в работе С. А. Мар-
сапа (S. Marsan, 1965).
Если сопоставить эти результаты нейрофизиологических ис-
следований и клинических проявлений эпилепсии с патологиче-
скими проявлениями шизофрении, следует согласиться с мне-
нием Б. Г. Будашевского и Д. Н. Меницкого (1973), полагаю-
щих, что у больных эпилепсией нарушается прежде всего про-
цесс сличения внутренних моделей с внешней средой, тогда как
для шизофрении более характерно нарушение гипотезообразова-
ния, процесса перестройки внутренних моделей из-за снижения
уровня мотивации.
5. ВОСПРИЯТИЕ ВЕРОЯТНОСТНОЙ СТРУКТУРЫ
СИГНАЛОВ ВНЕШНЕЙ СРЕДЫ
ПРИ НЕВРОТИЧЕСКИХ состояниях
Неврозы — определенный класс психогенных заболеваний —
формируются как следствие неблагоприятных, патогенных усло-
вий жизни индивидуума. Неврозы чаще всего возникают в ре-
149
зультате конфликта человека с окружающей действительностью.
Невроз, как указывает В. Н. Мясищев (1960), представляет со-
бой болезнь личности, и в первую очередь болезнь развития лич-
ности, невроз — это сложный продукт социального развития че-
ловека, складывающийся на основе как развития социально-
исторической среды, так и индивидуального развития в усло-
виях этой среды. К такому мнению приходит и П. В. Симонов
(1968), который считает невроз чисто человеческим заболева-
нием, обусловленным индивидуальными особенностями данного
человека, условий его воспитания, формирующих влияний об-
щественной среды.
Невротическое состояние, формируемое у человека, как пра-
вило, является отражением развиваемого конфликта между же-
лаемым и реально достигаемым. Неразрешимость конфликта ос-
нована на соотношении объективной необходимости и субъек-
тивных качеств индивидуума. Патогенная ситуация, приводящая
к психоневрозу, характеризуется индивидуальной и относитель-
ной неразрешимостью — неразрешимостью для данного индиви-
дуума и в данной обстановке.
Невроз возникает в результате столкновения потребностей,
желания индивидуума с реальной действительностью, которая
его не удовлетворяет. Такие ситуации часто возникают у чело-
века в различных условиях производственной деятельности, ко-
гда он длительное время вынужден выполнять определенные обя-
занности, с которыми не справляется (так называемый невроз*
относительной недостаточности). Такую ситуационную недоста-
точность, приводящую к развитию невротического состояния, ин-
дивидуум пытается компенсировать мобилизацией усилий с мак-
симальным использованием внешних возможностей. Но даже
при этих условиях мобилизации всех своих возможностей чело-
век, не сумев справиться с решением стоящей перед ним зада-
чи, впадает в болезненное невротическое состояние (В. Н. Мя-
сищев, 1960).
По образному выражению П. В. Симонова (1968), невроз —
это болезнь ненайденных выходов, подавленных стремлений, не-
разрешенных проблем, последствие драматической беспомощно-
сти живого существа. Роль эмоционального переживания чело-
века в развитии невротического состояния хорошо видна из из-
вестной формулы, предложенной автором (П. В. Симонов, 1968):
Э=П(Н — С), где Э —эмоция; П — потребности индивидуума;
Н—информация, необходимая для удовлетворения потребностей;
С—реально существующая информация, которая может быть ис-
пользована для организации целенаправленных действий.
В учении И. П. Павлова неврозы делятся на чисто человече-
150
ские патологические состояния — истерии и психастении — и об-
щие у человека и у животных — неврастении. Симптомокомплекс
неврастении отражает картину перевозбуждения и истощения
нервной системы. Истерический синдром ассоциируется с уско-
ренным, упрощенным импульсивным аффективным разрядом,
проявляющимся в своей крайней форме в виде истерического
припадка. Исследования физиологических механизмов, лежащих
в основе развития истерического невроза, привели И. П. Павло-
ва к утверждению, что этот тип неврозов характерен для слабой
нервной системы, склонной к разлитому пассивному торможе-
нию коры головного мозга. Истерическая реакция является ре-
зультатом того, что возникшее в некоторой зоне коры мозга воз-
буждение в силу слабости данной области мозга и торможения
в других ее зонах сопровождается сильным возбуждением под-
корковых структур мозга, что поддерживает возбуждение одного
участка коры и торможение других. При истерии становятся
более выраженными, выступают на первый план реакции пер-
вой сигнальной системы.
Обязательным условием развития неврастении является не-
умение рационально организовывать психические и физические
средства для достижения цели. Нерациональное использование
сил объясняет неудачу индивидуума в достижении поставленной
задачи, что приводит к развитию более сильного возбуждения,
к чрезмерному беспокойству в обычной ситуации. Обостренная
озабоченность, лихорадочное усилие, песоразмеримое объектив-
ной реальностью, влекут за собой, по словам В. Н. Мясищева
(I960), функциональную дезорганизацию и нервное истощение.
Психастеническая форма невроза проявляется в поведенче-
ских реакциях в виде признаков неуверенности, нерешительно-
сти, слабой воли. В нейрофизиологическом плане неврастения
выступает как проявление слабости корковой деятельности, сла-
бости реакций первой сигнальной системы.
Как видно из этих характеристик и истерической, и невра-
стенической формы невроза, оба эти патологические состояния
в конечном итоге представляют собой четко выраженное нару-
шение нормальных адекватных реакций организма и его отдель-
ных систем на различные внешние стимулы разной интенсивно-
сти. Организм в целом, прежде всего его информационно-управ-
ляющая система мозга, утрачивает способность градуальной и
адекватной вероятностной характеристики ситуации, реакции на
стимулы. При этом часто наблюдаются четко выраженные при-
знаки парабиотических реакций. Внешнее проявление невроти-
ческих состояний во многом напоминает феномен истериозиса
Н. Е. Введенского. Как известно, истериозис Н. Е. Введенского,
151
формируемый при длительном тетаническом раздражении аффе-
рентного нерва, обусловливается реакцией неспецифической сен-
сибилизации. Сам Н. Е. Введенский подчеркивал связь между
явлением истериозиса и некоторыми формами невротических ре-
акций человека, в особенности сенсибилизации нервных структур.
В лаборатории И. П. Павлова его сотрудниками (М. П. Пет-
рова и др.) было достаточно четко показано, что эксперимен-
тальные неврозы возникают, как правило, вследствие перена-
пряжения процесса раздражения под действием сильных, слож-
ных или необычных стимулов, вследствие перенапряжения преж-
де всего тормозного процесса, столкновения в структурах мозга
двух противоположно направленных процессов. Эти принципи-
ально важные установки в плане выяснения нейрофизиологиче-
ских основ нарушения нормальной деятельности мозговых струк-
тур нашли дальнейшее подтверждение и развитие в работе
М. М. Хананашвили (1978), вскрывшего некоторые информаци-
онные механизмы невротических реакций как на основе соб-
ственных экспериментальных исследований, так и анализа со-
временных исследований патофизиологической природы экспери-
ментальных неврозов.
Для понимания нейрофизиологических механизмов развития
и становления неврозов важным представляются результаты ис-
следований по выработке условного рефлекса на разные без-
условные раздражители. В опытах И. А. Ветюкова (1962) при
чередовании условных рефлексов, образованных на пищевой и
электрокожный стимулы, наблюдались сложные межцентральные
взаимоотношения, сопровождающиеся повышением возбудимо-
сти, возникновение очага с повышенной возбудимостью, пара-
биотическими стадиями и т. д. Дальнейшее длительное приме-
нение этих стимулов приводило к уменьшению работоспособно-
сти клеток коры головного мозга и к повышению возбудимости
элементов нижележащих структур центральной нервной систе-
мы, к наступлению срыва в деятельности центральной нервной
системы, развитию невротических состояний.
В экспериментах К. Гехта и др. (1977) с обучением, точнее
с переобучением при вероятностном подкреплении, и обучением
с вероятностным стрессом показано, что при смене постоянного
подкрепления подкреплением с разными вероятностями у боль-
шинства исследованных животных (собак) имело место развитие
выраженного эмоционального напряжения с последующим невро-
тическим срывом.
В работе А. Я. Мехедовой (1975) было показано, что при
введении вероятностного пищевого подкрепления (формирова-
ние вероятностного стереотипа реакции) вслед за постоянным
152
подкреплением условных сигналов, т. е. в ситуации информаци-
онного дефицита, у собак, фиксированных в экспериментальном
станке, нередко возникали эмоциональные расстройства с после-
дующим развитиехМ невротического состояния разной длительно-
сти. На этом основании автором (А. Я. Мехедова, 1975) делает-
ся заключение, что, по-видимому, замена .постоянного подкреп-
ления вероятностным у животных приводит к срыву в деятель-
ности центральной нервной системы. Учитывая широкую рас-
пространенность аналогичной ситуации в производственной и по-
вседневной деятельности человека, не следует особенно удивлять-
ся весьма частым случаям невротических заболеваний человека
при длительных неблагоприятных условиях его жизни.
Заключение
Принцип детерминизма как характеристика закономерной
связи явлений в многообразии форм движения материи зани-
мает центральное место в системе наших представлений об окру-
жающем материальном мире. Лапласовский жесткий детерми-
низм— наиболее известная и исторически ранняя форма при-
чинно-следственных отношений в системе естественно научных
знаний — отражал состояние естествознания своего времени, ко-
гда среди научных дисциплин на первое место выступила меха-
ника и механическое понимание материальной структуры, харак-
теризуемое однозначными причинно-следственными отношения-
ми, стало распространяться на многие области знаний об окру-
жающей природе.
Однако последующее развитие естествознания, в частности
становление, формирование квантовой механики, показало прин-
ципиальную неоднозначность в микромире. Н. Бор установил ве-
роятностный характер перехода электронов с одного энергетиче-
ского уровня на другой. В. Гейзенберг обосновал знаменитый
принцип неопределенности, утверждающий невозможность одно-
значного определения положения в пространстве и в то же вре-
мя энергетического состояния микрочастицы.
Принцип неопределенности Гейзенберга отражает объектив-
ную неопределенность, складывающуюся между пространствен-
ным и импульсно-энергетическим состоянием микрочастиц и вы-
ражающуюся в их зависимости друг от друга. Эта связь состоит
в том, что определенность одних состояний с необходимостью по-
рождает неопределенность других состояний. Дж. Лэмб и
Дж. Изаакс (J. Lamb, J. Р. Isaacs, 1969) полагают, что реаль-
153
ная гейзенберговская неопределенность проявляется и в деятель-
ности биологической клетки, включая молекулярный уровень ее
функционирования и уровень межнейронных соединений — си-
напс.
Анализ процессов, развивающихся в одиночном синапсе, по-
зволяет рассматривать эту основную структуру соединений нерв-
ных клеток как вероятностную систему. Стохастическое поведе-
ние клеток в нервной системе создается неопределенностью си-
наптической передачи через множество межнейронных соедине-
ний. Схема жесткой детерминации оказалась несостоятельной
в науках, имеющих дело со сложными и развитыми объектами,
чем объекты исследования классической физики, прежде всего
при анализе биологических и социальных явлений.
Вероятностные характеристики отвергают не детерминизм во-
обще, а лапласовское представление о детерминизме как одно-
значной определяемости событий начальными условиями. В каж-
дой конкретной ситуации, в данном функциональном состоянии
биологической системы формируются совершенно определенные
адекватные этим условиям действия. Однако эти определенные
детерминированные процессы целостной системной деятельности
образуются не на основе раз и навсегда определенных жестких
отношений между элементами, компонентами системы, а путем
вероятностного их взаимодействия.
Результаты морфофизиологического изучения взаимоотноше-
ний одновременно регистрируемых реакций нейронов привели к
представлению о вероятностно-статистической организации нерв-
ных клеток в рабочие конструкции мозга (А. Б. Коган, 1962—
1979; О. Г. Чораян, 1963—1979; А. Б. Коган, О. Г. Чораян, 1965—
1979). Этими исследованиями показано, что единицами функ-
циональной системы анализаторной, управляющей и информа-
ционной деятельности мозга являются, как правило, не отдель-
ные нервные клетки, а их объединения—вероятностные нейронные
ансамбли со статистической реализацией их функций. Были по-
лучены многочисленные факты, свидетельствующие о том, что
участие отдельных нейронов в каждОхМ ансамбле не фиксирован-
ное, а вероятностное, в результате чего в каждый конкретный
момент времени в мозгу образуются и распадаются динамиче-
ские мозаики возбужденных и заторможенных клеток, паттерны
которых служат элементами позиционного пространственного ко-
дирования информации в структурах мозга.
Вариабельность нейронных реакций означает существенную
изменчивость мозаики ансамблевой активности. В филогенети-
ческом ряду имеет место рост доли нейронов с вариабельными
реакциями. Очевидно, вариабельность нейронных реакций явля-
154
ется не вредным, «шумом», а эволюционно-прогрессирующим
приспособлением.
Установленная способность центральных нервных механизмов
к значительным компенсаторным перестройкам указывает на
отсутствие в большинстве случаев жесткого закрепления функ-
циональных свойств отдельных нейронов и межнейронных связей
и возрастание в ходе эволюции статистических и вероятностных
принципов функциональной организации центральных нервных
механизмов.
Эволюционное преимущество статистической организации по
сравнению с жесткой структурой заключается в том, что конеч-
ная реализация определенного акта нервной деятельности на
основе динамических мозаик возбужденных и заторможенных
нейронных ансамблей становится мало зависящей от состояния
отдельных функциональных единиц и допускает гибкие и опера-
тивные перестройки центральных нервных механизмов. Стати-
стический характер реализации функции во многом обеспечивает
их высокую надежность, а вероятностный характер включения
нервных клеток в ту или иную функциональную структуру в
значительной мере определяет их приспособительную пластич-
ность— свойства, которые получили особенное развитие именно
в аналитико-синтетических процессах высшей нервной деятель-
ности (А. Б. Коган, 1979). Чрезвычайная пластичность и надеж-
ность биологических систем основана на сочетании вероятност-
ных и однозначно действующих механизмов, составляющих спе-
цифику живого. Эволюционное развитие и совершенствование
механизмов адаптивного поведения животных в стохастических
условиях внешней среды связано, как указывает И. И. Шмаль-
гаузен (1961), с тем, что в них ведущую роль начинают играть
процессы статистической природы. Иначе говоря, переход от пре-
имущественно «схемных», жесткоопределенных, к преимущест-
венно «бессхемным», вероятностным, конструкциям является за-
кономерным результатом эффективности эволюционных пере-
строек в центральных нервных механизмах животных. Вероят-
ностно-статистическая природа механизмов нервной деятельно-
сти и пластичности высших функций мозга .получает подтверж-
дение и в результатах исследования различных форм психиче-
ской деятельности и функционирования второй сигнальной си-
стемы (Г. А. Аминев, 1972; Н. П. Бехтерева, 1971; Н. П. Бехте-
рева, П. В. Бундзен, 1976; Н. П. Бехтерева и др., 1977). Вероят-
ностные свойства проявляются и в приспособительной деятель-
ности физиологических систем при компенсации нарушенных
функций (В. А. Полянцев, 1974).
При осуществлении целостных актов нервной деятельности в
155
качестве вероятностных компонентов выступают уже не отдель-
ные нервные клетки, а функциональные блоки совместно рабо-
тающих нервных центров. Это хорошо проявляется в локомотор-
ных координациях, когда вся кинематика двигательного акта
определяется статистическим результатом действия приложен-
ных сил и выполнение одного и того же движения может про-
исходить за счет сокращения различных групп мышц. Так, даже
у тренированных спортсменов хорошо выработанным автоматиз-
мом движений при повторных выполнениях упражнения каждый
раз оно осуществляется несколько иной комбинацией мышечных
сокращений (В. С. Гурфинкель и др., 1965).
Представление о том, что закономерности высшей нервной
деятельности обусловлены вероятностными механизмами, выте-
кает уже из классических экспериментов К. Лешли (К. Lashley,
1933), в которых была установлена высокая степень взаимоза-
меняемости корковых структур. Возможность использования раз-
ных путей при частичном их выключении для движения нервных
процессов в структурах мозга была показана на примерах рас-
пространения реакции десинхронизации фоновой ритмики коры
мозга (А. Б. Коган, 1962), эпилептиформной активности (Г. А.Ха-
савов, 1978) и иррадиации зрительных вызванных потенциалов
на соматосенсорную кору мозга (Н. Н. Ткаченко, 1963). Идея
о вероятностном принципе организации отражательной функции
мозга разделяется многими крупными нейрофизиологами А. Фес-
сар (1962), Б. Бернс (1969), Г. Уолтер (1966) и др.
Итак, лишь на первый взгляд вероятностный детерминизм
представляется противоречащим причинно-следственной опреде-
ленности в организации и функционировании мозга. При более
глубоком рассмотрении проблемы вероятностная организация
выступает как проявление неоднозначного статистически гибко-
го детерминизма—более высокой формы детерминации явлений
в материальном мире (Ashby W., 1962; Коган А. Б., 1964, 1979;
Кравец А. С., 1971; Берг А. И. и др., 1971; Вишаренко В. С.,
1971; Чораян О. Г., 1974, 1979). Определенность системных ре-
акций мозга обеспечивается статистическими результатами мно-
жественного вероятностного взаимодействия нервных клеток,
образующих иерархию уровней структурно-функциональной ор-
ганизации мозга. Мозг представляет собой сложную вероят-
ностную систему, в которой определенность функции достигает-
ся статистическим путем на основе неоднозначного вероятност-
ного взаимодействия миллиардов разнородных нервных клеток.
ЛИТЕРАТУРА
'Маркс К. Дебаты о свободе печати. — Маркс К., Энгельс Ф. Соч.„
т. 1, с. 64.
Энгельс Ф. Людвиг Фейербах и конец классической немецкой фило-
софии.— Маркс К., Энгельс Ф. Соч., т. 21, с. 303.
Абакумов А. О некоторых патогенетических механизмах шизофре-
нии. М., 1907.
Абаскулиев А. О некоторых патогенетических маханизмах шизо-
френии. М., 1967.
Адольф А. Р. Стохастические подпороговые процессы в фоторе-
цепторах и нервно-мышечных соединениях и модель дробового шума.—
В кн.: Модели нейронных структур. М., 1970, с. 64.
Адрианов О. С. О принципах организации интегративной деятель-
тельности мозга. М., 1976.
Адрианов О. С. О роли различных систем мозга в организации:
церебральных функций. — В кн.: Проекционные и ассоциативные системы
мозга. М., 1977, с. 4.
Адо А. Д., Царегородцев Г. И. Борьба материализма и идеализ-
ма в учении о здоровье и болезни человека. М., 1970.
Айбералл А. С., Мак-Калок У. Гомеокинез — организационный
принцип сложных живых систем. — В кн.: Философские вопросы биологии иг
биокибернетики. М., 1970, с. 141.
Айзенк Г. Проверьте свои способности. М., 1972.
Александровский 10. А. Состояние психической дезадаптации и
ее компенсация. М., 1976.
Алексеев М. А., 3 а л к и н д М. С., Кушнарев В. М. Решение-
человеком задачи выбора при вероятностном подкреплении двигательных,
реакций. — В кн.: Биологические аспекты кибернетики. М., 1962, с. 198.
Аминев Г. А. Вероятностная организация центральных механиз-
мов речи. Казань, 1972.
Анохин П. К., Ст ре ж Е. Изучение динамики высшей нервной
деятельности. — «Физиол. журн. СООР.» 1933, № 46, с. 280 — 293.
Анохин П. К. Теория функциональной системы как предпосылка к
построению физиологической кибернетики. — В кн.: Биологические аспекты
кибернетики. М., 1962, с. 198.
Анохин П. К. Общие принципы формирования защитных приспо-
соблений организма. — «Вестник АМН СССР», 1962, № 4, с. 16.
Анохин П. К- Биология и нейрофизиология условного рефлекса..
М., 1968.
А н о х и н П. К. Теория функциональной системы. — «Усп. физиол. наук»,.
1970, т. 1, с. 19.
Анохин П. К. Физиологические аспекты функциональной системы.—
«В®пр. философии», 1971, № 3, с. 55.
Анохин П. К. Теория функциональной системы и ее место в по-
строении теоретической биологии. — В кн.: Эволюция темпов индивиду-
ального развития животных. М., 1977, с. 9 — 18.
Аронович Б. И. Системы отображения информации и инженерная
психология. Л., 1974.
Бажин Е. Ф. Об особенностях вероятностного прогнозирования при
некоторых психопатологических и нейропсихологических синдромах. — В
кп.: Шизофрения и вероятностное прогнозирование. М., 1973, с. 68.
157
Баркрофт Дж. Основные черты архитектуры физиологических
функций. М., 1937, с. 289.
Батуев А. С. Кортикальные механизмы интегральной деятельности
мозга. Л., 1978.
Бауэр Э. С. Теоретическая биология. М. — Л., 1935.
Беллман Р., Заде Л. Принятие решений в расплывчатых условиях.—
В кн.: Вопр. анализа и процедуры принятия решений. М., 1976, с. 172.
Берг А. И., Бирюков Б. В., Новиков И. Б. Методологические
аспекты кибернетики. — «Коммунист», 1971, № 18, с. 86.
Беритов И. С. Структура и функции коры большого мозга. М., 1969.
Б ер л Б. Функциональная организация в случайных сетях. — В кн.:
Принципы самоорганизации. М., 1966, с. 358—386.
Бернар К. Введение к изучению опытной медицины. Спб., 1896.
Бернс Б. Неопределенность в нервной системе. М., 1969.
Бернштейн Н. А. Очерки по физиологии движения и физиоло-
гии активности. М., 1966.
Бехтерева Н. П. Нейрофизиологические аспекты психической
деятельности человека. Л., 1971.
Бехтерева Н. П., Бундзен П. В. Информационная детермина-
ция пространственно-временной структуры нервного кода вербальной па-
мяти. — В кн.: Вопр. кибернетики. От нейрофизиологии к нейрокибернети-
ке. М., 1976, с. 30.
Бехтерева Н. П., Бундзен П. В., Го Голицын Ю. Л. Моз-
говые коды (психической деятельности. Л., 1977.
Блекуэлл Б., Гиршик М. Теория игр и статистических реше-
ний. М., 1953.
Блинков С. М., Пуцило М. В. О зависимости компенсации функ-
ций от количества нейронов. — «Вопр. нейрохирургии», 1968, № 3, с. 15.
Боголепов Н. К. Философский аспект проблемы невропатологии,
1967. М., 1967.
Болдырев А. И. Эпилепсия у взрослых. М., 1971.
Болдырев А. И. Эпилептические синдромы. М., 1976.
Бродский В. Я. Трофика клетки. М., 1966.
Бройль Л. де. Революция в физике. М., 1963.
Будашевский Б. Г., Меницкий Д. Н. Особенности отображе-
ния вероятностной структуры внешней среды при некоторых патологи-
ческих состояниях высшей нервной деятельности. — В кн.: Шизофрения и
вероятностное прогнозирование. М., 1973, с. 5.
Буриков А. А. Нейронная активность двигательной коры при вы-
сокочастотном раздражении таламуса. — Материалы XVII конф, физиол.
юга РСФСР, т. 1. Ставрополь, 1969, с. 18— 19.
Буш Р., >Мо с т е л л е р Ф. Стохастические модели обучаемости.
М., 1962.
Вальтер Д, О., Ад ей В. Р. Линейные процессы в мозгу. — Тез.
доклада. Международный енмпоз. ИФАК по техн, и биолог, пробл. упр.
Ереван, 1968, с. 7.
Василевский Н. Н. Фоновая и вызванная импульсная активность
нейронов. — В кн.: Механизмы деятельности центрального нейрона. М.—
Л., 1966, с. 149.
Василевский Н. Н. Нейрональные механизмы коры больших по-
лушарий. Л.., 1968.
В е т ю к о в. И. А. Истериозы и неврозы. — В кн.: Проблемы ла-
бильности, парабиоза и торможения. М., 1962, с. 57.
158
Виленкин Н. Я., Шрейдер 10. А. Мажоритарные пространства и
квантор «большинства». — В кп.: Семиотика и информатика, вып. 8. М.,
1977, с. 45.
Винер Н. Кибернетика. М., 1958.
Ви н ер Н. Творец и робот. М., 196'6.
Винер Н. Я — математик. М., 1967.
В и ш а р е н к о В. С,. Об особенностях детерминизма в живых систе-
мах— В кн.: Методологические проблемы биологии. Л., 197'1, с. 4.
В иш арен.ко В. С. Типы и формы детерминации. — В кн.: Мето-
дологические проблемы биологии. Л., 1971, с. 65.
В и ш а р е н к о В. С. Детерминизм и особенности биологической орга-
низации. — В кн.: Логика и методология науки. М., 1973, с. 210.
В и ш а р е н ко В. С,. Детерминация в биологических процессах.
Л, 1975.
Войшвилло Е. К. Попытка семантической интерпретации статисти-
ческих понятий информации и энтропии.—В кн.: Кибернетику на службу
коммунизма. М. — Л., 1966, с. 275.
Волков П. П. О природе психопатологических явлений. — В кн.:
Проблемы системного подхода в психиатрии. Рига, 1977, с. 15.
В о л к о в и ч В. Л. Экспертные оценки в задачах принятия сложного
решения. Киев, 1971.
В о л ь к е н ш т е й н М. В. Молекулы и жизнь. М., 1965.
Вулдридж Д. Механизмы -мозга. М., 1965.
Гамбарян Л. С., Г а сп а р я н Ю. М. Некоторые вопросы передачи
информации по нервному волокну. — ДАН, 1968, т. 178, с. 745.
Гейзенберг В. Физические принципы квантовой теории. М. —
Л, 1932.
Гехт К , М е х е д о в а А. Я., Т р е и т о в К. Значение неопределенности
в процессе обучения для уровня эмоционального напряжения и развития
экспериментального невроза. — «Журн. высш, нервн. деят.», 1977, № 2,
с. 328.
Глезер В. Д. и др. О кодировании сигналов в импульсных последо-
вательностях.— В кн.: Статистическая электрофизиология. Вильнюс, 1968,
с. 141.
Глумов Г. М. О роли межкорковых и корково-подкорковых путей
в осуществлении условного рефлекса. Автореф. канд. дис. Ростов н/Д, 1964.
Готт В. С., Урсул А. Д. О категориях определенности и неопре-
деленности.— «Вопр. философии», 1971, № 6, с. 55.
Гродинз Ф. Теория регулирования и биологические систе!мы. М., 1966.
Г у д в и и Б. Временная организация клетки. М., 1966.
Г уревич И. И., Фейгенберг И. М. Какие вероятности «рабо-
тают» в психологии. — В кн.: Вероятностное прогнозирование в деятель-
ности человека. хМ., 1977, с. 9.
Г у р ф и н к е л ь В. С., К о ц Я. М., Шик М. Л. Регуляция позы че-
ловека. М., 1965.
Давыдовский И. В. Проблема причинности в медицине. М., 1962.
Де Гроот М. Оптимальные статистические решения. М., 1974.
Джаспер Г., Риччи Г., Доун Б. Микроэлектродный анализ
разрядов корковых нейронов клеток при выработке условных оборонитель-
ных рефлексов у обезьян. — В кн.: Электроэнцефалографические исследования
высшей нервной деятельности. М., 1962, с. Г29.
Ермаков П. Н. О функциональной организации нейронов зритель-
ной коры мозга крысы при локальной и диффузной световой стимуляции.
Автореф. канд. дис. Ростов н/Д, 1978.
159
3 а в а д с к и й К. М. Вид и видообразование. Л., 1968.
Заде Л. Понятие лингвистической переменной и его применение к
принятию приближенных решений. М., 1976.
Зале в ск ий Г. В., Роговин М. С. Фиксированные формы поведе-
ния и их значение для неврологической и психиатрической клиники.
I. Персеверация. — «Журн. невропатологии и психиатрии», 1970, т. 70,
с. Г255.
За леве кий Г. В., Р о говн и М. С. Фиксированные формы поведения
и их значение для неврологической и психиатрической клиники. II. Навяз-
чивости, ритуалы, стереотипии. — «Журн. невропатологии и психиатрии»,
1970, т. 70, с. 1725.
Звягинцев В. А. Значение и понимание с точки зрения машины.—
В кн.: Теоретические проблемы прикладной лингвистики. М. 1965, с. 38.
Иванова М. П. Электрофизиологическое исследование произвольных
движений человека. М., 1978.
Иваницкий А. И. Мозговые механизмы оценок сигналов. М., 1976.
Иванников В. А. Поведение человека в ситуации выбора. — В кн :
Вероятностное прогнозирование в деятельности человека. М., 11977, с. 112.
Ильенков Э. В. Проблема абстрактного ц конкретного в свете
«Капитала» Маркса. — В кн.: «Капитал» Маркса, философия и современ-
ность. М., 1968, с. 186.
К а р у н и н А. Б., Клыков Ю. И., Куликов Ю. А. Модели ситуа-
ционного управления в проектировании организации строительства.— «Изв.
АН ClOOP. Техн, кибернетика», 1976, вып. 2.
К а р у н и н А. Б., Крюков А. А., Клыков Ю. И. Задача обучения
з системах ситуационного управления. — В кн.: Вопр. кибернетики (теория
и практика ситуационного управления), вып. 18. М., 1977, с. 28.
Кастлер Г. Азбука теории информации. — В кн.: Теория информа-
ции в биологии. М., 1960, с. 8.
Кедров Б. М. О детерминизме. — «Философские науки», 1968, № 1,
•с. 41.
Кедров Б. М. Научная концепция детерминизма. — В кн.: Современ-
ный детерминизм. Законы природы. М., 1973, с. 6.
Кербиков О. В. Выступление в дискуссии. — В кн.: О проблеме при-
чинности в медицине. М., 1965.
К и р п и ч е п к о А. А. Исследование нейрофизиологических нарушений у
больных шизофренией. Докт. дне. М., 1970.
Кирой В. Н. О некоторых нейрофизиологических проявлениях процес-
са решения человеком мыслительных задач. Автореф. канд. дис. Ростов н/Д,
J979.
К л а ц к и Р. Память человека. Структуры и процессы. М., 1978.
Клыков Ю. И., Лобанов IO. И. Принципы построения семиотиче-
ских систем решения задач. — В кн.: Общая теория систем и интеграция
знаний. М., 1976, с. 90 — 93.
К н и п с т И. Н., Шульгина Г. И. Электроэнцефалоскопические ис-
следования образования двигательного оборонительного условного рефлек-
са.— В кн.: Нервные механизмы двигательной деятельности. М, 1966, с. 317.
Коган А. Б., Николаева Н. И. О соотношении электрических
потенциалов и возбудимости нейронов коры мозга. — Тез. докладов конф,
по вопр. электрофизиологии центр, нервн. системы. М., 1958, с. 65.
Коган А. Б. Выражение процессов высшей нервной деятельности в
электрических потенциалах коры мозга при свободном поведении живот-
ного.— В кн.: Электроэнцефалографическое исследование высшей нервной
деятельности. М.» 1962, с 42.
160
Коган А. Б. О некоторых принципиальных особенностях организации
и свойств информационного аппарата мозга. — В кн. Проблемы нейрокибер-
нетики. Ростов н/Д, 1962, с. 16—‘17.
Коган А. Б. Вероятностно-статистический принцип нейронной органи-
зации функциональных систем мозга. — ДАН, 1964, т. 154, с. 123-1.
Коган А. Б., Чораян О. Е Центральная организация нервных про-
цессов и основная активность нейронов. — «Журн. высш, нервн. деят.», 1965,
т. Г5. с. 225.
Коган А. Б., Петунии Ю. И., Чораян О. Г. Исследование им-
пульсной активности нейронов методами теории случайных процессов. —
«Биофизика», 1966, выл. 5, с. 887.
Коган А. Б., Чораян О. Г. Физиологические механизмы анализа
сигналов. — В кн.: Пространственная ориентация животных. М., 1970,
с. 13—16.
Коган А. Б., Карпенко Л. Д., Чораян О. Г. Эволюционные ас-
пекты формирования гомеостатической системы нейронных ансамблей. —
В кн.: Проблемы бионики. М., 4973, с. 280.
Коган А. Б., Режабе к Б. Г., Васильева О. С. Нелинейное
захватывание ритма при раздражении изолированной нейрорецептор ной клет-
ки. — ДАН, 1973, т. 216, с. 493—495.
Коган А. Б., Гринченко С. Н., 3 а г у с к и н С. Л. Устройство для
моделирования нервной клетки. Авт. свид. № 494752 — «Бюл. изобрет.»,
1975, № 6.
Коган А. Б., Кураев Г. А., Соболева И. В. О нейрональной
организации очага условного возбуждения при выработке условного рефлек-
са на электрическую стимуляцию сенсорной коры кошки. — «Журн. высш,
не.рвн. деят.», 1976, т. 26, с. 59.
Коган А. Б., Кураев Г. А., Чораян О. Г. Информационные ха-
рактеристики импульсной активности нейронов сенсорной коры кошки при
выработке оборонительного условного рефлекса. — «Журн. высш, нервн.
деят.», 1976, т. 26, с. 1151.
К о г а н А. Б. и др. Биологическая кибернетика. М., 1977.
Коган А. Б., Чораян О. Г. Элементарные нейронные ансамбли моз-
га. — «Журн. высш, нервн. деят.», 1977, т. 27, с. 922.
Коган А. Б., Чораян О. Г. Вероятностные нейронные ансамбли с
позиции теории размытых множеств. — «Физ ио л. журн. QCIOP», 1977, т. 63,
с. 125)1.
Коган А. Б. Функциональная организация нейронных механизмов моз-
га. Л., 1979.
Коган А. Б., Чораян О. Г. Элементы размытых алгоритмов интел-
лектуальной деятельности. — «Физнол. человека», 1979, т. 5, № 3, с. 552.
Колесников А. В., Пономарев В. Ф. Построение языка описа-
ния объекта диспетчерского управления. — В кн.: Автоматизированные си-
стемы управления в рыбной промышленности. Калининград, 1977, с. 83.
Корольков А. А., Петленко В. П. Философские проблемы тео-
рии нормы в биологии и медицине. М., 1977.
Ко ст юк П. Г., Шаповалов А. И. Электрофизиология нейрона.—
В кн.: Современные проблемы электрофизиологических исследований нерв-
ной системы. М., 1964, с. 3'1.
Котляр Б. И. Ми кроэлект родные исследования условно-рефлекторных
реакций.— В кн.: Нейронные механизмы обучения. М., 1970, с. НО.
Коуэн Дж. Вероятностная логика и синтез надежных организмов из
ненадежных компонентов. — В кн.: Автоматы. М., ,1956, с. 68.
Кр а в е ц А. С. Вероятность и системы. Воронеж, 1970.
161
Кравец А. С. О философском аспекте понятия вероятности. — «Вонр.
философии», 1971, т. 1, с. 57.
Крушинский Л. В. Формирование поведения животных в корме
и патологии. М., I960.
Крюков Б. И., Макаров П. О. О проблеме двойной сигнализации
в вопросе моделирования нервной деятельности. — В кн.: Применение ма-
тематических методов в биологии. М., 1963.
Куликов М. А. Алгоритмы комплексного исследования фоновой им-
пульсации нейронов. — В кн.: Статистическая электрофизиология, т. 2. Виль-
нюс, 1968, с. 254.
Кузнецов С. А. Функциональная организация нейронов и нейронных
популяций моторной коры. Кишинев, 1979.
Лазарев П. П. Исследования по ионной теории возбуждения. М., 1916.
Лаплас П. Опыт философии теории вероятностей. М., 1908.
Лев А. А. Моделирование ионной избирательности клеточных мембран.
М., 1976.
Леонов Ю. П. Теория статистических решений и психофизика.
М., 1977.
Леонтьев А. Н., Кринчик Е. П. Переработка информации чело-
веком в ситуациях выбора. — В кн.: Инженерная психология. М., 1964,
с. 295.
Линдсей П., Норман Д. Переработка информации у человека.
М., 1974.
Лобанов М. Е. Генетика и условный рефлекс. — «Изв. АН СССР.
Биология», 1956, № 2, с. 29.
Майр Э. Причина и следствие в биологии. М., 1970, с. 47.
Малиновский А. А. О причинности в биологии. — В кн.: Совре-
менный детерминизм. Закон природы. М., 1973, с. 468.
Мак Кал л ок У., Айбералл А. Гомеокинез — принцип организа-
ции сложных живых систем. — Тез. докл. Международного симпоз. ИФАК
по техн, и биол. пробл. управл. Ереван, 1967, с. 9.
хЧе л мент Д. Курс количественной медицины. М., 1948.
Меницкий Д. Н. Моделирование структуры и функции центральной
нервной системы. — В кн.: Механизмы деятельности центрального нейрона.
М. — Л., 1966, с. 203.
Меницкий Д. Н., Трубачев В. В. Информация и проблемы выс-
шей нервной деятельности. Л., 1974.
Мехедова А. Я. Вероятностное подкрепление и неврологические сры-
вы в условиях свободного поведения. — «Журн. высш, нервн. деят.», 1975,
вып. 6, с. 1188.
Мещерский Р. М. Анализ нейронной активности. М., 1972.
Миллер Дж. Информация и память. — В кн.: Кибернетика и живой
организм. Киев, 1964, с. 7—46.
Михлин В. М. Некоторые личностные особенности при шизофрении в
свете нарушений вероятностного прогнозирования. — В кн.: Шизофрения и
вероятностное прогнозирование. М., 1973, с. 78.
Моррелл Ф. Данные микроэлектродных исследований и изучение ста-
бильного потенциала, указывающие на локализацию процесса замыкания на
дендритах. — В кн.: Электроэнцефалографическое исследование высш, нервн.
деят. М. — Л., 1958, с. 54.
Мясищев В. Н. Личность и неврозы. Л., 1960.
Нарушевичус Э. В., Магинскас В. А. Флуктуация межспай-
ковых интервалов у автонейронов. — В кн.: Динамика нейронных сетей.
Вильнюс, 1966, с. 67.
162
Нейман Дж. Вероятностная логика и синтез надежных организмов
«з ненадежных компонентов. — В кн.: Автоматы. М., 19*5в, с. 68.
Николаев В. И. Об одном методе определения ценности инфор-
мации. — В кн.: Проблемы инженерной психологии, вып. ,1. М., 1968,
с. 124—129.
Николаев В. И., Кузнецова М. В,. О прагматических характе-
ристиках информации. — В кн.: Автоматизированные системы управления.
Л., 1977, с. 10.
Нисенбойм Л. Б., Загадская Л. С. О понятийном аппарате и
некоторых вопросах организации ситуационных моделей1 управления. —
«Вопр. кибернетики», вып. 14. М., 1975, с. 30.
Нисенбойм Л. Б., Загадская Л. С. Семиотические модели: успе-
хи и .перспективы. — «Кибернетика», 1976, № 6, с. 114.
Орловский С. А. Принятие решений. — «Автоматика и телемехани-
ка», 1971, № 11, с. 106.
Павловская Н. Н. Нейронная структура реакции десинхронизации
электрокортикограммы кролика. Автореф. канд. дис. Ростов н/Д, 1970.
Павловские клинические среды, т. 2. М.—Л., 1955, с. 374.
Пападопулус Т. Ф. Острые эндогенные психозы. М., 1975.
Пенфилд В., Джаспер Г. Эпилепсия и функциональная анатомия
головного мозга человека.. М., 1958.
Пестун ова В. Н., Пиотровский Р. Г. Измерение смысловой и
прагматической информации в тексте. — В кн.: III симшоз. по психолингви-
стике. М., 1*97’0, с. 30.
Петленко В. П., Царегородцев Г. И. О двух формах детер-
минации биологических процессов как причине их дискретности и непрерыв-
ности.— В кн.: Патологическая физиология и экспериментальная терапия.
М, 1976, № 6, с. 80.
Пиотровский Р. Г. Текст, машина, человек. Л., 1975.
Плотников В. В. Об одном виде нарушения зрительного восприя-
тия ври шизофрении. — «Вопр. психологии», 1968, вып. 4, с. 123.
Подладчикова Л. Н. Динамика функциональной организации ней-
ронов зрительной коры при изменении уровня ее возбудимости. Автореф.
канд. дис. Ростов н/Д, 1978.
Поляков Г. И. Проблема происхождения рефлекторных механизмов
мозга. М., 1964.
Поляков Г. И. О принципах нейронной организации мозга. М., 1965.
Поляков Г. И. Основы систематики нейронов новой коры большого
мозга человека. М., 1973.
Поляков Ю. Ф. Патология познавательной деятельности при шизо-
френии. М., 1974.
П о л я н ц е в В. А. Особенности методологии аналитического и систем-
ного подходов. — В кн.: Принципы системной организации функций. М.,
1973, с. 91.
Полянцев В. А. Изучение приспособительного поведения с позиций
функциональной системы. Автореф. докт. дис. М., 1974.
Поспелов Г. С. Предисловие. — В кн.: Вопр. анализа и процедуры
принятий решений. М., 1976.
Поспелов Г. С. Искусственный интеллект и АСУ. — В кн.: Вопр. ки-
бернетики. М., 1977, с. 71.
Поспелов Д. А. «[Сознание», «самосознание» и вычислительные ма-
шины.— В кн.: Системные исследования. М., 1969, с. 17-8.
Поспелов Д. А., Пушкин В. Н. Мышление и автоматы. М., 1972.
Поспелов Д. А. Большие системы (ситуационное управление).
М, 1975.
163
Поспелов Д. А. Семиотические модели: успехи и перспективы. —
«Кибернетика», 1976, т. 6, с. 1'14.
Преображенский Н. Н., Яровицкий Н. В. Применение ма-
тематических методов исследования импульсной активности центральных ней-
ронов мозга. — «Биофизика», 4963, т. 8, с. 387,.
Пригожин И. Введение в термодинамику необратимых процессов.
М.. 1960.
Пятигорский В. я. О стационарности спонтанной активности ней-
ронов дорзального спиномоэжечкового тракта. — «Биофизика», 1966, т. 11,
с. 116.
Радз невский В. С. Использование концептуального подхода при
описании размытых множеств. — VIII Всесоюзный симпоз. по кибернетике.
Тбилиси, 1976, с. 137—140.
(Раштон У. Периферическое кодирование в нервной системе. — В кн.:
Теория связи в сенсорных системах. М., 1964, с. 53.
Розенблит У. О некоторых количественных аспектах электрической
активности центральной нервной системы. — В кн.: Пробл. современной био-
физики, вып. 2. М., Г961, с. 282.
Романов Н. А. О возможности контакта между теорией вероятно-
стей и учением акад. И. П. Павлова об условных рефлексах. — ДАН, 1935,
т. 1, с. 193.
Рубинштейн С. Л. О мышлении и путях его исследования. М., 1958.
Савина В. Г. О динамике нейронных реакций крыши среднего мозга
на световые раздражения в пре- и постнатальном онтогенезе кур. — Докл.
XVIII конф, физиол. Юга РСФСР. Воронеж, 1971, с. 247—248.
Савич А. В., Ш а л ь н о в М. И. Молекулярные механизмы обеспече-
ния надежности клеток. — В кн.: Системы надежности клетки. Киев, 1977,
с. 46—56.
Сачков Ю. В. Информация и вероятность.—«Вопр. физиол.», 1971,
т. 6, с. 45.
Сачков Ю. В. Введение в вероятностный мир. М., 1971.
Сент-Дьердьи А. Введение в субмолекулярную биологию. М., 1964,
с. 22.
С и м о н о в П. В. Болезнь неведения. М., 1968.
Симонов П. В. Высшая нервная деятельность человека. Мотивацион-
но-эмоциональные аспекты. М., 1975.
Соколов Е. Н. Нервная модель стимула и ориентировочный реф-
лекс. — «Вопр. психологии», I960, № 4, с. 128.
Соколов Е. Н. Пейсмекерный потенциал в организации ритмической
активности нейрона. — В кн.: Пейсмекерный потенциал нейрона. Тбилиси,
1'975, с. 13.
Соте ко в Б. С., Гамбарян Л. С., Мкртчян С. О. К вопросу о
надежности мозга. — В кн.: Функциональные модели биологических систем.
Ереван, 1972, с. 7.
Степанов А. Д. Норма, болезнь и вопросы здравоохранения. Горь-
кий, 1975.
Стулов В. А. Количество информации.—.В кп.: Разработка и кон-
струирование средств автоматизации и систем управления. Киев, 1973, с. 24.
Сухов А. Г. О функциональной организации корковых нейронов крысы
при нормальной и эпилептической активности. Автореф. канд. дис. Ростов.
н/Д, 1969.
Тамбиев А. Э. О межнейронных отношениях в центральной коре
мозга крысы при нормальной и эпилептической активности. Автореф. канд.
дис. Ростов н/Д, 1971.
164
Тимофеев-Ресовский Н. В., Ромле Р. Р. О статистичности
и принципе усилителя в биологии, вып. 2. М., 1959, с. 213.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Ябло-
ков А. В. Краткий очерк теории эволюции. М., 1969.
Ткаченко Н. Н. О путях распространения первичных зрительных
ответов по коре мозга кошки. — Материалы IV конф, по электрофизиол.
нервн. системы. Ростов н/Д, 1963', с. 382.
Т р и н ч ер К. С. Биология и информация. М., Г964.
Уильямс Р. Биохимическая индивидуальность. М., 19*60.
Уолтер Г. Живой мозг. М., 1966.
У рсул А. Д. Информация. М., 197'1.
Уэлз Р. Мера субъективной информации. — В ки.: Новое в лингвисти-
ке. М., 1965, с. Г67.
Фам X ы у Хоан. Функциональная организация нейронных систем
коры мозжечка. — «Физиол. журн. OQGP», 1967, т. *53.
Фейгенберг И. М. Вероятностное прогнозирование в деятельности
мозга. —«Вопр. психологии», 1963, № 2, с. 59.
Фейгенберг И. М. Нарушение вероятностного прогнозирования при
шизофрении. — В кн.: Шизофрения и вероятностное прогнозирование. М.,
1973, с. 5.
Фейгенберг И. М., Иванников В. А. Взаимоотношение услов-
ной и ориентировочной реакций. — «Вопр психологии», 1969, № 5, с. 39.
Фейгенберг И. М., Леви В. Л. Вероятностное прогнозирование и
экспериментальное исследование его при патологических состояниях (иллю-
зия Шарпантье у больных шизофренией). — «Вопр. психологии», 1965, № 1,
с. 42.
Фельдман Г. Л. К модели нейронной перестройки при развитии
сна. — В кн.: Пробл. нейрокибернетики. Ростов н/Д, 1972, с. 250—255.
Фесс ар А. Анализ замыкания временных связей на уровне нейро-
нов.— В кн.: Электроэнцефалографическое исследование высш, нервн. деят.
М., 1962, с. 147.
Филимонов И. Н. Сравнительная анатомия коры большого мозга
млекопитающих. М., 1949.
ФилюковА. И. Эволюция и вероятность. Минск, 197*2.
Фицнер Л. Н. Биологические поисковые системы. М., 1977.
Фомин С. В., Берки нблит М. Б. /Математические проблемы в
биологии. М., 1973.
Фролов И. Т. О причинности и целесообразности в живой природе.
М, 1964.
Философия и современная биология. Под ред. Фролова И. Т.
М, 1973.
X а н а н а ш в и л и М. М. Механизмы нормальной п патологической услов-
норефлекторной деятельности. Л., 1972.
X а н а н а ш в и л и М. М., 3 а р к е ш е в Э. Г., Усова И. П. Микроси-
стема как структурно-функциональная основа интеграции нейронов при за-
мыкании временной связи. — В кн.: XII съезд Всесоюзного физиол. об-ва.
Л., 1975, т. 1, с. 59.
Ха на наш вил и М. М. Экспериментальная патология высшей нервной
деятельности. М., 1978.
Хасабов Г. А. Нейрофизиология связей коры больших полушарий
приматов. М., 1978.
ХоджкинА. Нервный импульс. М., 1965.
Хохлов Л. К. К психопатологии, клинике и нозологии синдрома Кан-
динского -Клерамбо. — В кн.: Синдром психического автоматизма. М., 1969,
с. 19.
165
Хочачка П., Семеро Дж. Стратегия биохимической адаптации.
М., 1977.
Царего.родцев Г. И. Диалектический материализм и медицина. М.,
1963, с. 432.
Ц е т л и н М. Л. Исследования по теории автоматов и моделированию
биологических систем. М., 1969.
Цискаридзе М. А. Зависимость реакций человека на стимул от
некоторых вероятностных характеристик ситуаций. Автореф. канд. дпс.
М., 1909.
Ц о п ф Г. Отношение и контекст. — В кн.: Принципы самоорганизации.
М., 1966, с. 309.
Чебкасов С. А. Пространственное распределение вызванных потен-
циалов и организация импульсных ответов нейронов зрительной коры мозга
морской свинки. Автореф. канд. дис. Ростов н/Д, 1972.
Чораян О. Г. К функциональной нейроноархитектонике. — «Физиол.
журн. СООР», 1963, т. 49, с. 1026.
Чораян О. Г. Взаимосвязь фоновой и вызванной импульсной актив-
ности нейронов КОМ лягушки. — «Физиол. журн. ССОР», 1965, т. 54,
с. 1024.
Чораян О. Г. Гистограммный метод анализа импульсной активности.—
В кн.: Пробл. нейрокибернетики. Ростов н/Д, 1966, с. 90.
Чораян О. Г. Нейронная организация центрального отдела зрительно-
го анализатора лягушки. Ростов н/Д, 1969.
Чораян О. Г. Вероятностный детерминизм в деятельности нервной
системы. — В кн.: Вероятностно-статистическая организация нейронных ме-
ханизмов мозга. Ростов н/Д, 1974, с. 20.
Чораян О. Г. Кибернетика нервных клеток. Ростов н/Д, 1975.
Чораян О. Г. Информативность импульсных реакций нервных кле-
ток. — «|Изв. СКНЦ BiUI. Бстеств. науки», 1976, № 2, с. 101.
Чораян О. Г. К методике определения количества полезной инфор-
мации в импульсных потоках нервных клеток. «Биофизика», 1977, т. 22,
с. 738.
Чораян О. Г. Концепция размытых алгоритмов и некоторые вопросы
высшей нервной деятельности человека. — В кн.: XXV совещ. по пробл.
высш, нервн. деят., посвященного памяти И. П. Павлова. Горький, 1977,
т. 2, с. 170—471.
Чораян О. Г. Полезная информация. Ростов н/Д, 1978.
Чораян О. Г. Информационные функции нейронов в свете теории
размытых множеств. — II Международная конф, стран-членов СЭВ по основ-
ным пробл. бионики. — «Бионика-78». Реф. докл. М. — Л., 1978, т. 1,
с. 68—70.
Чораян О. Г., Кирой В. Н. Методический подход к анализу по-
лезной информации в ЭЭГ человека. — «Физиол. человека», 1978, № 3,
с. 564—567.
Чораян О. Г. Размытые алгоритмы мыслительных процессов. Ро-
стов н/Д, 1979.
Шапиро Д. И. О методах принятия решений в сложных системах
управления. — В кн.: Вопр. кибернетики, вып. 8. М., П975, с. 123—134.
Шапиро Д. И. О решении одного класса многокритериальных за-
дач. — В кн.: Многокритериальные задачи принятия решений. М., 1978,
с. 169—183.
Ш а х н о в и ч А. Р., Шапиро Д. И Математические методы в иссле-
довании биотехнических систем. М., 1973.
Шеннон К. Работы по теории информации и кибернетики. М., 1963.
Ш и б к о в а С. А., Клепач Г. С. К архитектоническим особенностям
166
поля 17 большого мозга морской свинки.. — «Арх. анат. гистол. эмбриол.»,
1969, т. 56, с. 56—60.
Шидловский В. А. Мультивариантная адаптивная регуляция веге-
тативных функций.—В кн.: Вопр. кибернетики, вып. 37. М., 1978, с. 3.
Ш и д ло в с ки й <В. А., Новосельцев В. Н. Мультипараметрическое
обеспечение гомеостаза и гомеокинеза. — В кн.: Принципы системной орга-
низации функций. М., 1973, с. 81.
Шмальгаузен И. И. Интеграция биологических систем и их само-
регуляция.— «Бюл. МОИП», 1961, вып. 2, с. 104.
Шмальгаузен И.. И. Регуляция и формообразование в индивиду-
альном развитии. М., 1964, с. 106.
Шмальгаузен И. И. Кибернетические вопросы биологии. Новоси-
бирск, 1968.
Ш н о л ь С. Э. Физико-технические факторы биологической эволюции.
М., 1979.
Шредингер Э. Что такое жизнь с точки зрения физики. М., 1947.
Шрейдер Ю. А. О количественных характеристиках соматосенсорной
информации. — «Бюл. НТИ», 1963*, № 10, с. 3*3—38.
Шрейдер Ю. А. Семантические основы информатики. М., 1974.
Шрейде.р Ю. А. Семантические аспекты информатики (лекции).
М., 1975.
Шульгина Г. И. Биоэлектрическая активность головного мозга и
условный рефлекс. М., 1978.
Эль Гохари М. О вариабельности фоновой и вызванной импульсной
активности нейронов крыши среднего мозга лягушки. Автореф. канд. дис.
Ростов н/Д, 1974.
Эшби У. Введение в кибернетику. М„ 1959.
Эшби У. Конструкция мозга. М., 1962.
Adolph A. R. Feedback in physiological sistems: An application of
feedback analysis and stochastic models to neurophysiology. — Bull. Math.
Biophys., 1969, v. 21, p. 133.
Adolph A.R. Spontaneous slow potential fluctuations in the Limulus
photoreceptor. — J. Gen. Physiol., 1964, v 48.
Adrian E. The mechanism of nervous action. Electrical studies on the
neuron. L., Oxford Univ. Press, 1932.
Albritton E. C. Standard values in blood. W. B. Saunders Co.
Philadelphia a. L., 1952.
Anson B. Atlas of human anatomy. W. B. Saunders Co. Philadelphia a.
L., 1951.
Arden G. B., Soderberg U. The transfer of optic information
through the lateral geniculate body of the rabbit— In: Sensory Communication,
1961, p. 521. 4
Asai K., Tanaka H., Oku da T. Decision-making and its goal in
a fuzzy environment. — In: Fuzzy sets and their applications to cognitive and
decision processes. N. Y, San Francisco, L., 1975, p. 257—277.
Asai K., Tanaka H., О k u d a T. On discrimination of fuzzy states
in probability space. Kybernetes, 1977, v. 6, p. 185—192.
Barlow H. B. Sensory mechanisms, the reduction of redundancy and
intelligence, — In: Mechanization of thought processes. L., 1959, p. 537.
Barlow H. B. Three points about lateral inhibition. In: Sensory
communication. N. Y, L. Mass. Technol. Press, 1961, p. 782.
Barlow H. B. Sensory mechanisms; the reduction of redundancy and
intelligence. — In: Cybernetics. Baltimore a. o. Univ. Park Press, 1968, p. 183.
Barlow H. B. Visual sensitivity. —In: Physic's and Mathematics of the-
167
Nervous System. Springer — Verlag — Heidelberg—Berlin—N. Y. 1974, p. 198—
210.
Baumgartner G. Microelectrode recordings from single cortical neurons
in the normal state and during epileptic discharges. — EEG a. Clin. Neurophysiol
1954, v. 6, p. 520.
Bellman R., Zadeh L. Decision-making in fuzzy environment. —
Monog. Sci., 1970, v. 17, p. 141.
Bertalanffy L. Das Biologische. Weltbild. Bern, 1949.
В e u r 1 e R. L. Properties of a mass of the cells capable of regenereting
pulses. — Phil. Trans. Roy. Sos., 1956, B. 240, v. 55.
В e u r 1 e R. L. Storage and manipulation of information in the brain. —
IEEE, 1959, v. 5, d. 75.
В e u r 1 e R. Storage and manipulation of information in random networks. —
In: Aspects Theory Artificial Intelligence. N. Y., 1962, p. 19.
Birks R. I., Me I n t о sh F. C. Acetylcholine metabolism of nerve
endings. — Brit. med. Bull., 1957, v 13, p. 157.
В о к J. T. Histonomy on the cerebral cortex. Amsterdam, L., N. Y., 1959.
Boyd J., Martin A. The end-plate potential in mammalian muscle. — J.
Physiol., 1956, v. 132, p. 74—91.
Brock L. G., С о о m b s J. S., E с с 1 e s J. C. The recording of potentials
from motoneurones with an intracellulare electrode. — J. Physiol, 1952, v. 117,
p. 431—460.
Brunswick E. Probability as a determiner of rat behavior.—J. Exp
Psychol., 1939, p. 25.
Brunswick E. Organismic achievement and environmental probability.
Psychol. Rev., 1943, p. 50.
Buller A., Nicholls Y., Storm Y. Spontaneous fluctuations of
ecxitability in the muscle spindle of the frog. — J. Physiol., 1953, vol. 122, p. 409.
Bullock T. H. Species differences in effect electro receptor input on
electric organ pacemakers and other aspects of behaviour in electric fish. —
In: Brain, Behavior and Evolution, 1969, v. 2, p. 85.
Bullock T. H., C h i c h i к u S. Further analysis of sensory coding
in electroreceptors of electric fish. — Proc. Nat. Acad. Sci., 1965, v. 54, p. 422.
Bures J., Buresova O. Relationship between spontaneous and evoked
unit activity in the inferior colliculus of rat. — J. Neurophisiol., 1965, v. 28,
,p. 641.
Burns B. D. The mammalian cerebral cortex. Edward Arnold. L., 1958.
C a i a n i e 11 о E. R., L u c a J., Riccardi L. M. Reverberations and
•control of neural networks. Kybernetik, 1967, v. 4, p. 216.
Callaway E. Diagnostic uses of the average evoked potential. — In:
Average evoked potentials. Methods, results and evaluations. Washington, 1969,
p. 299—332.
Callaway E., Jones R., Layne R. S. Evoked response and seg-
mental set of schizophrenia. — Arch. Gen. Psychiat., 1965, v. 12, p. 83.
Callaway E., Jones R. T., Layne R. S. Evoked responses and
segmental set of schizophrenia.—EEG a. Clin. Neurophysiol., 1970, v. 29,
p. 421—428.
Cannon W. B. The wisdom of body. Oxford. 1932.
Carnap R., В a r - H i 1I e 1 Y. An outline of theory of semantic infor-
mation. — Tech. Rep. 247, MYT, 1952.
Chang S. - K. On the execution of fuzzy programs using finite machi-
nes. — IEEE Trans, on Computers. 1972, v. C—21, N 3, p. 241.
Chang C. L. Interpretation and execution of fuzzy program. In: Fuzzy
sets and their application to cognitive and decision processes. N. Y., San
Francisco, L., Acad. Press, 1975, p. 191.
168
Cheng G. C., Ledley R. S., Onyang B. Pattern recognition
with time interval modulation information coding. IEEE Trans. Aerospace and
Electron Syst., 1970, v. 6, p. 226.
Cohen J. Behaviour in uncertainty and its social implications. L., 1964.
Conrad M. Molecular information processing in the central nervous
system. Part П. — In: Physics and Mathematics of the Nervous System.
Berlin — Heidelberg — N. Y. — Springer — Verlag. 1974, p. 108—127.
Cowan J. D. Statistical mechanics of nervous nets. — In: Neural networks.
Springer—Verlag—Berlin—Heidelberg—N. Y., 1968, p. 181.
Davidson D., Suppes P. A. Finitistic axiomatization of subjective
probability and utility. — Econometrics, 1956, v. 24.
De Groot A. D. Thought and choice in chess. The Hague, Mouton,
1965.
De Groot A. D. Perception and memory versus thinking. — In: Problem
solving. N. Y., Wiley, 1966.
Del Castillo J., Katz B. Quantal components of the endplate
potential. —J. Physiol, 19154, v. 124, p. 569.
Del b ru ck M. Trans. Conn, Acad. Arts Sci., 1949,v. 33, p. 173.
Dimitrov V. D. Social choice and self-organization under fuzzy
management.—Kybernetes, 1977, v. 6, p. 153.
D u d e I Y., Ku f f 1 er S. W. Presynaptic inhibition at the crayfish ne-
uromuscular junction. — J. Physiol., 1961, v. 155, p. 543.
Ehrenstein G., Legar H. Electrically gated ionic channels in lipid
bilayers. — Quart. Rev. Biophys., 1977, v. 10, p. 1
Elsasser W. The physical foundation of biology. L. — N. Y., 1958,
p. 19:
English H. B., English A. C. A comprehensive dictionary of
psychological and psychoanalitical terms. N. Y., 1959.
Estes W. K. The cognitive side of probability learning. — Psychol. Rev.r
1976, v. 83, p. 37.
Farber G. Berechung und Messung des Informationflusses der Nerven-
faser. — Kybernetik, 1968, v. 7, p. 17.
Fatt P., Katz B. Some observations on biological noise.—Nature,
1950, v. 166, p. 597.
Fatt P., Katz B. Spontaneous subthreshold activity of motor nerve
endings. — J. Physiol., 1952, v. 117, p. 109.
Fatt P. Presynaptic cell characteristics determining membrane potential
changes. — In: Lecture Notes in Biomathematics. Berlin, 1974, v. 4, p. 150.
Fessard A. Les mechanismes de synchronization interneuronique et
leur intervention deus la crise epileptique. — In: Bases Psychol, et Aspects Clim
de I'Epilepsie. Paris, 1958, p. 37.
Fessard A. E. Les problemes du code nerveux. — In: Physique Theorie
a. Biologie, Paris, 1971, p. 170.
F u о r t e s M. G. F. Discontinuous potentials evoked by sustained il-
lumination in the eye of Limulus. — Arch. Ital. Biol., 1959, v. 97, p. 49.
Fuster J. O., Creutzfeldt O, Straschill O. Intracellular
recording of neuronal activity in the visual system. — Z. vergl. Physiol., 1965*
v. 49, p. 605.
Gaines B. R. Foundations of fuzzy reasoning.— Int. J. Man-Machine
Studies, 1976, v. 8, p. 623.
Gaines B. R., Kohout L. J. The fuzzy decade: a bibliography of
fuzzy system and closely related topics. — Int. J. Man-Machine Studies, 1977*
№ 9, p. 23.
Gerstein G. L., Mandelbrot B. Random walk models for the
spike activity of a single neuron. — Biophys. J., 1964, v. 4, p. 41.
169
Gestri G., Maffei L., Petrachi D. Statistical independence of
events in neighbouring retinal units. — Brain Res., 1966, p. 397.
G г о s s m a n B. G., Viernstein L. J. Discharge patterns of neurons
in cochlear nucleus. — Science, 1961, v. 134, p. 99.
Gruss er O.-J., Hellner K. A., Grusser—Cornehls U.
Die Information subertragung im afferenten visuellen System. — Kybernetik,
1962, v. 1, p. 175.
Hagiwara S. Analysis of interval fluctuation of the sesory nerve fibres.
Jap. — J. Physiol, 1954, v. 4, p. 234.
Hagiwara S., Morita R. Coding mechanisms of elcctroreceptor
fibers in some electric fish. — J. Neurophysiol., 1963, v. 26, p. 551.
Hagiwara S. e. a. Electroreceptor mechanisms in a frequency weakly
electric fish. — J. Neurophysiol., 1965, v. 28, p. 784.
H a r t h E., Beck В., P e r t i 1 e G., Young F. Signal stabilization
and noise suppression in neural system. — Kybernetik, 1970, v. 7, p. 112.
Harth E., Pertile G. The role of inhibition and adaptation in sensory
information processing. — Kybernetik, 1972, v. 9, p. 12.
H e b b P. O. The organization of behavior. N. Y., 1949.
Herman H. T., Stark Z. Information role and capacity in a cray-
fish photoreceptor structure in the crayfish tail ganglion.—Anat. Rec., 1962,
v. 147, p. 209.
Herz A., Creutzfeldt O., Fuster J. Statistische Eigenschaften dor
Neuronaktivitat im ascendierendcn visuellen System. — Kybernetik, 1964, v. 2,
p. 61.
Hick W. H. On the role of gain of information. Quart.—J. Exp. Psychol,
1952, v. 4, p. 11.
Hyman R. Stimulus information as a determinant of reaction time. —
J. Exp. Psychol., 1953, v. 45, p. 3.
J a r v i с к M. Probability learning and negative recency effect in the
serial anticipation of alternative symbols. — J. Exp. Psychol., 1951, v. 41.
Jones R. T., Blacker К- H., Callaway E. Perceptual disfunction
in schizophrenia: clinical and auditory evoked responses findings. — Am. J. Psy-
chiatr., 1966, v. 123, p. 639.
Jones R. T., Callaway E. Auditory evoked responses in schizo-
phrenia. Biol. Psychiatr., 1970, v. 2, p. 632.
Kahneman D., Tversky A. The belief in the «law of small num-
bers».— Psychol. Bull., 1971, v. 76, p. 105.
Kahneman D., Tversky A. Subjective probability: a judgement of
representativeness. — Cognitive Psychology, 1972, v. 3, p. 430.
Kamikawa K-, Me II wain J. T., A de у W. R. Response of tha-
lamic neurons during classical condition. — EEG a. Clin. Neurophysiol., 1964,
v. 17, p. 485.
К a n del E. R., T au c L. Mechanism of heterosynaptic facilitation in
the giant cell of the abdominal ganglion of Aplysia depilans. — J. Physiol.,
1965, v. 181, p. 28.
Katz B. The transmission of impulses from nerve to muscle and the
subcellular unit of synaptic action. — Proc. Roy. Soc., B, 1962, v. 155, p. 455.
Katz B., Miledi R. Membrane noise produced by acetylcholine.—
Nature, 1970, v. 226, p. 962.
Kerkut G. A., Thomas R. Acetylcholine and the spontaneous IPSP
in snail ganglion neuron. — Compar. Biochem. a. Physiol., 1963, v. 8, p. 84.
Kiang N. Y.-S. Discharge pattern of single fibers in the cat auditory
nerve. — MIT, Cambridge, 1966.
King L. What is disease? — Philosophy of Science, 1954, N 3, p. 192.
Kochen M. Applications of fuzzy sets in psychology. — In: Fuzzy sets
170
and their applications to cognitive and decision processes. N. Y., San Francisco,
1975, p. 395.
Kogan А. В., C h о г а у a п O. G. Some information theory application
to the physiology of nervous cells. — Kybemetes, 1973, v. 2, p. 77.
Kok a w a M., Nakamura K-, Ola M. Experimental approach to
fuzzy simulation of memorizing, forgetting and inference processes. — In: Fuzzy
sets and their applications to cognitive and decision processes. N. Y., San Fran-
cisco, 1975, p. 409.
Kulikowski J. Information channels of the senses. — In: Bionics.
Warszawa, 1971, p. 97.
Kusano K., Hagiwara S. On the integrative synaptic potentials of
Onchidium nerve cell. — Jap. J. Physiol., 1961, v. 11, p. 23.
Lamb J. C., Isaacs J. P. Indeterminacy, the synapse, the mnemonic
microstate and the psyche. — In: Condition Reflex, 1969, p. 1.
Lashley K. S. Brain mechanisms and intelligence. N. Y., L., 1963.
L e i f e г L. J., Otto D. A., Huff E. M. Evoked potential correlates
of a probability learning task-topographic analysis. — EEG a. Clin. Neurophysiol.»
1973, v. 349, p. 741.
Levy J. C. Statistical approach to the study of neural networks. — In:
Neural Networks. Berlin—Heidelberg—N. Y., 1968, p. 145.
L i C.-L. Functional properties of cortical neurons with particular reference
to strychninization. — EEG a. Clin. Neurophysiol. 1955, p. 475.
Li C.-L. Cortical intracellular potentials and their responses to the
strychnine. — J. Neurophysiol., 1959, v. 22, p. 4.
Li C.-L. Cortical intracellular synaptic potentials. — J. Cell. Comp.
Physiol., 1961, v. 68, p. 153.
Lifshitz K. An examination of evoked potentials as indicators of
information processing normal and schizophrenia subjects. — In: Average Evoked
Potentate. Methods and Evaluations. Washington, 1969, p. 357.
L i 1 e у A. W. An investigation of spontaneous activity at the neuromuscu:
lar junction of the rat. — J. Physiol., v. 132, p. 68.
Lindley D. V. Making decisions. L., N. Y., Sydney, Toronto, 1971.
Luca de A., Riccardi M. Probabilistic description of neurons. —
In: Neural Networks. Berlin a. o., 1968, p. 100.
Mars an S. A. Electrical activity of the brain: slow waves and neuronal
activity. — Israel J. Med. Sci., 1965, v. 1, p. 104.
Martin A. R., Branch C. L. Spontaneous activity of Betz cells
in cat's with midbrain lesions. — J. Neurophysiol., 1958, v. 21, p. 368.
Matsumoto H., M a r s a n A. C. Cortical cellular phenomena in
experimental epilepsy: interictal manifestations. — Exptl. Neurol., 1964, v. 9,
p. 286.
Matsumoto H., Marsan A. C. Cortical cellular phenomena in
experimental epilepsy: ictal manifestations. — Exptl. Neurol. 1964 b, v. 9, p. 305.
Me Farland D. J. Decision making in animate. — Nature, 1977, v. 269,
p. 15—21.
Me Gill W. J. Stochastic latency mechanisms. — In: Handbook of
Mathematical Psychology. N. Y., Wiley, 1963, p. 309.
Messick S. J., Salley С. M. Probability learning in children some
exploratory studies. — J. Genet. Psychol., 1957, p. 90.
Morrell F. Microelectrode studies in chronic epileptic foci. Epilepsia,
1961, v. 2, p. 81.
Moore G. P., P e г к e 1 D. H., Segundo J. P. Statistical analysis
and functional interpretation of neuronal spike data. — Ann. Rev. Physiol., 1966,
v. 28, p. 499.
171
Naplatanoff N., Lakov D. Decision-making in vaque conditions. —
Kybernetes, 1977, v. 6, p. 91—93.
N e g о i t а С. V., R a 1 e s c u D. A. Applications of fuzzy sets to systems
analysis. Basel a. Stuttgart, 1975.
N e g о i t а С. V., R a 1 e s c u P. A. On fuzzy optimization. — Kyberne-
tes, 1977, v. 6, p. 193.
Nishi S., Koketsu K. Electrical properties and activities of single
sympathetic neurons in frogs. — J. Cell, a Comp. Physiol., 1960, v. 55.
Oestreicher H. On the structure of intelligent system. — In: Kyber-
netik and Bionik. Munchen Wien. R. Oldenburger. 1974, v. 5. p. 232—249:
О r n i t z E. M. A note on the AEP of antistic children during sleep. —
In: Average evoked potentials. Methods, results and evaluutions. Washington,
1969, p. 355.
О s a w a T. Information properties of a neuron. Mem. Fac. Engng. Kyoto
Univ., 1969, v. 31, p. 95.
Perk el D. H. e. a. Pacemaker neurons: effects of regularly spaced
synaptic input. Science, 1964, v. 145, p. 61—63.
Perkel D. H., Gers tain G. L, Moore G. P. Neuronal spike
trains and stochastic point processes. 1. The single spike train. Biophys. J.,
1967, v. 7, p. 419.
Perkel D. H., В u 11 о с к T. H Neural coding. Neurosci. Res. Progr.,
1968, v. 6, p. 2.
P о g g i о G. P., V i e r n s t e i n L. J. Time series analysis of impulse
influences of thalamic somatic sensory neurons. — J. Neurophysiol., 1964, v. 27,
p. 517.
Prigogine I., Nicolis G., Babloyantz A. Thermodynamics of
evalution. — Phis. Today, 1972, v. 25, p. 23.
Rapoport A., Horvath W. Y. The theoretical channel capacity of
single neuron as determined by various coding systems.— Inform, a. Control,
1960, v. 3, p. 335.
Ratliff F. On fields of inhibitory influence in a neural networks.—
In: Neural networks. Springer—Verlag— Berlin, 1968, p. 6.
Ratliff F., Hartline N. K., Lange D. Variability of interspike
intervals in optic nerve fibers of Limulus: effect of light and dark adaptation.—
Proc. Nat. Acad. Sci., USA, 1968, v. 60, p. 464.
R e i c h e г G. M. Perceptual recognition as a function of meaningfulness
of stimulus material. — J. of Exper. Psychol., 1969, v. 81, p. 275—280.
R e s 11 e P. Psychologv of judgement and choice a theoretical essay.
N. Y., L., 1961.
Ricchi G., Doane B., Jasper H. Congr. intern, sci. neurol. I-st.
Congr. Brussels, 1957, p. 401.
Rodieck R. W., Kiang N. J. - S., Gerstein G. L. Some quanti-
tative methods for the study of spontaneous activity of single neurons. —
Biophys. J., 1962, v. 2, p. 351.
Ruspini E. H. Numerical methods for fuzzy clustering.—Inform. Sci.,
1970, v. 2, p. 319.
Russel B. On the notion of conscionsness. — Proc, of the Aristotelian
Society, 1913, v. 13, p. 1.
R u s t a d W. H. J. Clin. Endocrinol. Metabolism, 1954, v. 14, p. 87:
Savage L. J. The foundations of statistics. N. Y., 1954.
S c h e i g h H., Bullock T. H., H a m s t г a R. H. Coding properties
of two classes of afferent nerve fibers: high — frequency electroreceptors in the
electric fish Eigenmania. — J. Neirophysiol., 1973, v. 36, p. 39.
Scholes J. H. Discrete subthreshold potentials from the dimly let
insect eye. — Nature, 1964, v. 202, p. 1173.
172
Segundo J. P., Moore G. P., Stensaas L., Bullock T. H.
Sensitivity of neurons in Aplysia to temporal patterns of arriving impulses. —
J. Exp. Biol., 1963, v. 40, p. 643.
Segundo J. P., P e г к e 1 D. H., Moore G. P. Spike probability
in neurons influence of temporal structure in the train of synaptic event*.—
Kybernetik, 1966, v. 3, p. 67.
S h а к о w P. Psychological deficit in scizophrenia. — Behav. Sci., 1963,
v. 8, p. 275.
Shimbcl A., Rapoport A. A statistical approach to the theory of
the central nervous system. — Bull. Math. Biophys., 1948, v. 10, p. 41.
S h i m u r a M. An approach to pattern recognition and associative memo-
ries using fuzzy logic. — In: Fuzzy sets and their applications to cognitive and
decision processes. N. Y. — San Francisco, 1975, p. 395.
S h о 1 1 D. A. The organization of the cerebral cortex. L. — N. Y., 1956.
S 1 e p i a n D., Wolf J. K. Noiseless coding of correlated information
sources. — IEE Trans. Inform. Theory, 1973, v. 19, p. 471.
Slovic P., Fischoff B., Lichtenstein S. Behavioral decision
theory. — Ann. Rev. Psychol., 1977, v. 23, p. 1—39.
Solomon R., Wynne L. Psychol. Monogr., 1953, v. 67, p. 4.
Stark L. Neurological control systems. N. Y., 1968.
Stein R. The information capacity of nerve cells using a frequency
code. — Biophys. J., 1967, v. 7, p. 797.
Sutton S. The specification of psychological variables in an average
evoked potential experiment. — In: Average evoked potentials. Methods, results
and evaluations. NASA, Washington, 1969, p. 237—262.
Szentagothai J. Architecture of the cerebral cortex. — In: Basic
mechanisms of the epilepsie. Boston, 1969, p. 13 — 28.
Szabo T. Activity of peripheral and central neurons involved in electro-
reception. — In: Lateral Line Detectors. Bloomingtons, 1967, p. 295.
Szabo T., Hagiwara S. A latency change mechanism involved in
sensory coding of electric fish. — Physiol, a. Behav., 1967, v. 2, p. 331.
Ten Ноореп M. On an inpulse interval generating mechanism. — Jap. J.
Physiol., 1964, v. 14, p. 607.
Ten Ноореп M. Probabilistic firing of neurons considered as a first
pass age problem. — Biophys. J., 1966 a, v. 6, p. 435.
Ten Ноореп M. Impulse of the thalamic neurons. An attempted theo-
retical interpretation. — Brain Res., 1966 b, v. 3 (2), p. 123.
Ten Ноореп M., Hertog A., Reuver H. A. Fluctuation in
excitability of nerve fibers — a model study.— Kybernetik, 1963, 2, p. 1.
Ten Ноореп M., Reuver H. A. Recurrent point processes with
reference to neuronal spike trains.— Math. Biosci., 1968, v. 8, d. 1.
T e r z u о 1 о С. А., В а у 1 у Е. J. Data transmission between neurons. —
Kybernetik, 1968, v. 5, p. 83.
Tversky A. Elimination by aspects: a theory of choice. — Psychol. Rev.,
1972, v. 79, p. 281.
Tversky A., Kahneman D. Causal schemate in judgements under
uncertainty. — In: Progress in Social Psychology, 1977, p. 143.
U111 e у A. M. A theory of the mechanism of learning based on ttie
computation of conditional probabilities. Proc. I st Int. Congr. on Cybernetics,
Namur, Paris, 1956.
U 11 1 e у A. M. Conditional probability computing in a nervous system. —
In: Mechanisms of thought processes., 1959, v. 1, p. 121.
U 111 e у A. M. Does a cortical synapse transmit Shannon information. —
In: The anatomy of memory. Palo Alto, Calif., 1965, a p. 318.
173
Uttley A. M. The probability of neural connexions. — Proc.Roy. Soc.,
1965, b, 144, p. 229.
Ventriglia F. Kinetic approach to neural systems. — Bull. Math. Biol.,
1974, v. 36, p. 535.
Verveen A. A., Derksen H. E. Fluctuations in membrane potential
of axons and the problem of coding. — Kybernetik, 1965, v. 11, p. 152.
Viernstein L. J., Grossman F. Neural discharge pattern in the
transmission of sensory information. Information theory. IV London Sympos.,
1961, p. 252.
Vincent J. D. Duffy B. Les codages Nerveux. Bordeaux Med., 1972,
v. 5, p. 93.
Walther J. B. Single cell responses from the primitive eyes of an
annelid. — In: The functional organization of the compound eye. Oxford, Pergam.
Press., 1966.
Weckowicz T. E., Blewett P. B. Size constancy and abstract
thinking in schizophrenic patients. — J. Ment. Sci., 1959, v. 105, p. 441.
Weller M., Bird R. Economy in the brain. Lancet, 1973, p. 7817.
Wells R. A measure of subjective information. Proc, of Symp. AppL
Math., 1961, N. 12, p. 14.
Werner G., Mountcastle V. B. The variability in central neuron
activity in a sensory system and its implications for the central reflection of
sensory events. — J. Neurophysiol., 1963, v. 26, p. 958.
Werner G., Mountcastle V. B. Neural activity in mechanoreceptive
cutaneous afferents: stimulus — responses relations, Weber functions and in-
formation transmission. — J. Neurophysiol., 1965, v. 28, p. 359.
Wheeler D. D. Processes in word recognition. Cognitive Psychology,
1970, v. 1, p. 59—85.
Wiersma S., Adams R. T. The influence of nerve impulses sequence
on the contractions of different crustacean muscles.P h у s i о 1. Comp., 1950, v.
2, p. 20.
Yager R. R. Multiple objective decision—taking using fuzzy sets.
Int.—J. Man—Machine Studies, 1977, v. 9, p. 375—382.
Y e a n d 1 e S. Electrophysiology of the visual system. Discussion. Am.
J. Ophthalmol., 1958, v. 46.
Young Y. Z. A model of the brain. Oxford, Clarendon, 1964.
Zadeh L. Fuzzy sets and systems. Procc. Symp. System Theory, N. Y.,
1965, p. 29.
Zadeh L. Probability measures of fuzzy events. — J. Math. Analysis
and Applications. 1968, v. 23, p. 421.
Zadeh L. Biological application of the theory of fuzzy sets andsystems. —
In: Biokybernetics of the central nervous system. Boston, 1969, p. 199.
Оглавление
Глава I. ДЕТЕРМИНИЗМ В ЖИВЫХ СИСТЕМАХ 3
1. Концепция однозначного и статистического детерминизма 3
2. Детерминация процессов в биологических системах 8
3. Статистические закономерности в эволюционном процессе 12
4. Динамический характер адаптивных изменений функций организма 16
5. Статистический характер нормы и патологии 23
Глава II. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ ФУНКЦИОНИРО-
ВАНИЯ НЕРВНОЙ СИСТЕМЫ 30
1. Выживание в стохастической среде 30
2. Черты структурной организации мозга 33
3. Организация внутриклеточных процессов 36
4. Организация элементарных нейронных объединений 42
5. Механизмы условнорефлекторной деятельности организма 52
Глава III. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ КОДИРОВАНИЯ
И ДЕКОДИРОВАНИЯ ИНФОРМАЦИИ В НЕРВНОЙ
СИСТЕМЕ 59
1. Понятие о кодировании информации 59
2. Элементы кодирования информации в нервных клетках 61
3. Механизмы обеспечения надежной передачи информации 74
4. Семантические аспекты информационной деятельности нервной си-
стемы 81
Глава IV. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ ИНФОРМАЦИОН-
НОЙ ДЕЯТЕЛЬНОСТИ 92
1. Характер процессов принятия решения в неопределенной ситуации 92
2. Прогнозирование 100
3. Элементы размытой логики в работе мозга 105
4. Семантические аспекты ситуационной информации 117
Глава V. ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ ФОРМИРОВАНИЯ
ПАТОЛОГИЧЕСКИХ СОСТОЯНИЙ НЕРВНОЙ
СИСТЕМЫ 129
1. Соотношение жесткодетерминированного и вероятностного компо-
нентов реакций в патологическом состоянии 129
2. Изменения прогноза при формах психопатологии 133
3. Характеристики нейрофизиологических проявлений работы струк-
тур мозга при психопатологических процессах 140
4. Некоторые характеристики детерминации нейрофизиологических
процессов при эпилепсии 145
5. Восприятие вероятностной структуры сигналов внешней среды при
невротических состояниях 149
Заключение 153
Литература 157
КОГАН Александр Борисович
ЧОРАЯН Ованес Григорьевич
ВЕРОЯТНОСТНЫЕ МЕХАНИЗМЫ
НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Редактор К л и м ч ук Т. М.
Технический редактор Кислова Т. М.
Корректоры К о и ч а н и н а 3. Р.,
Хубиева 3. М.
Обложка Морозова Д. А.
ИБ № 461.
Изд № 20/1269. Сдано в набор 18 04 80 Подписа-
но в печать 17 10 80 ПК 18128 Формат 60x84'/i6.
Бумага тип. № 1. Гарнитура литературная. Вы-
сокая печать Физ. пл И Усл п л 10,23 Уч -
изд п. л. II. Тираж 700 экз Заказ № 77. Це-
на 1 р. 10 К.
Издательство Ростовского университета, г. Ро-
стов-на-Дону, 6, ул Пушкинская, 160.
Типография им. М И. Калинина Ростовского
областного управления по делам издательств, по-
лиграфии и книжной торговли, г. Ростов-на-До-
ну, ул. 1-я Советская, 57.