/























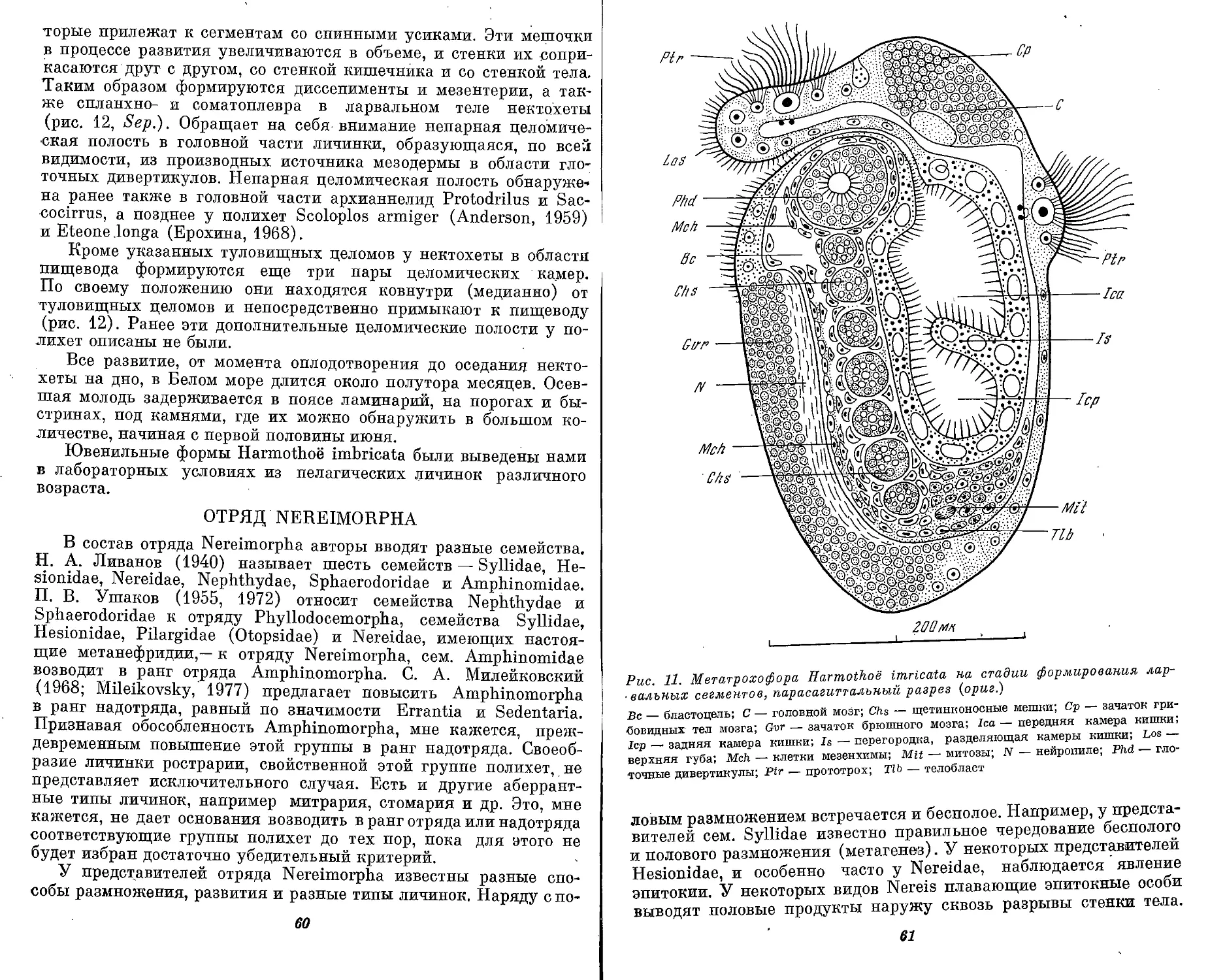
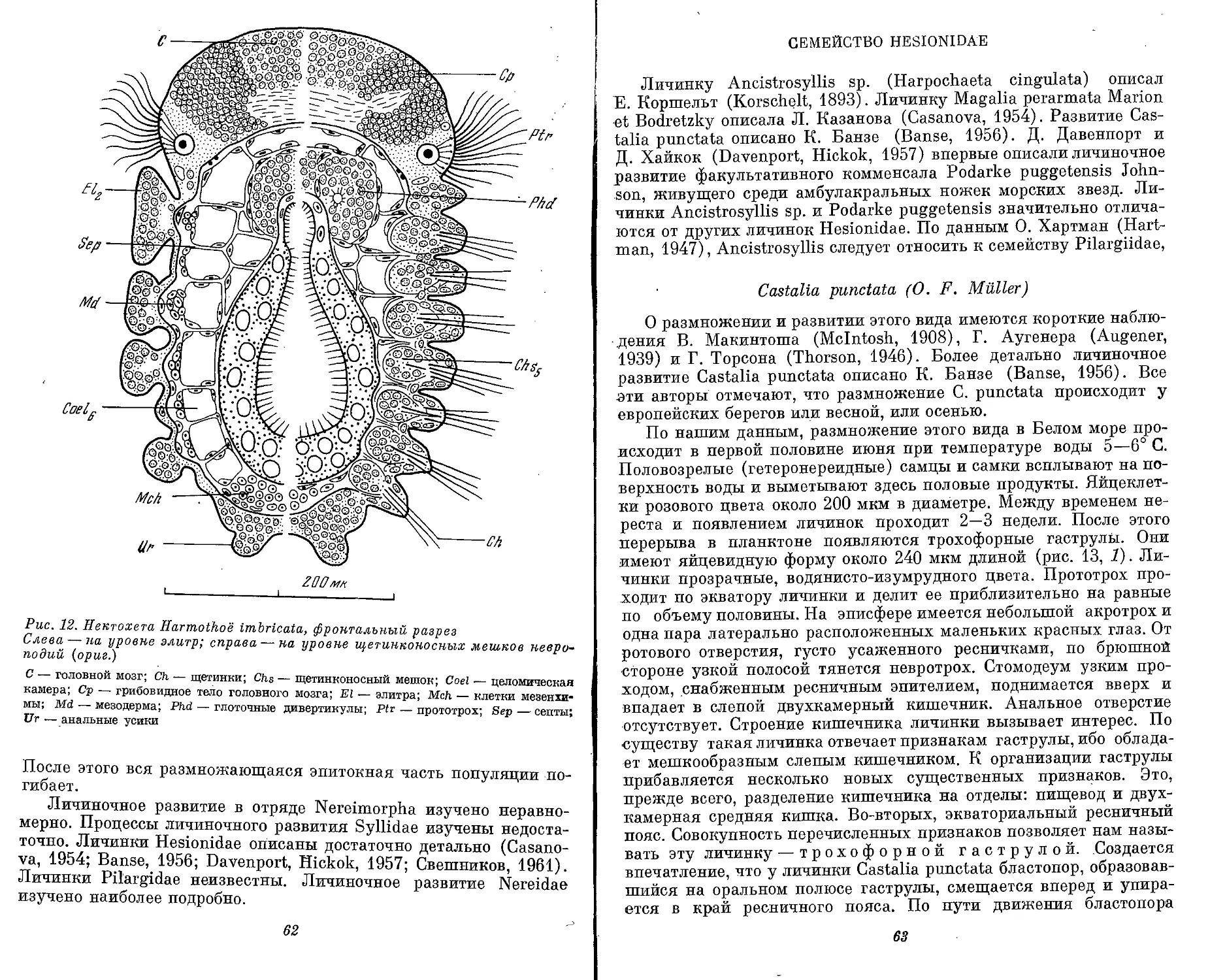
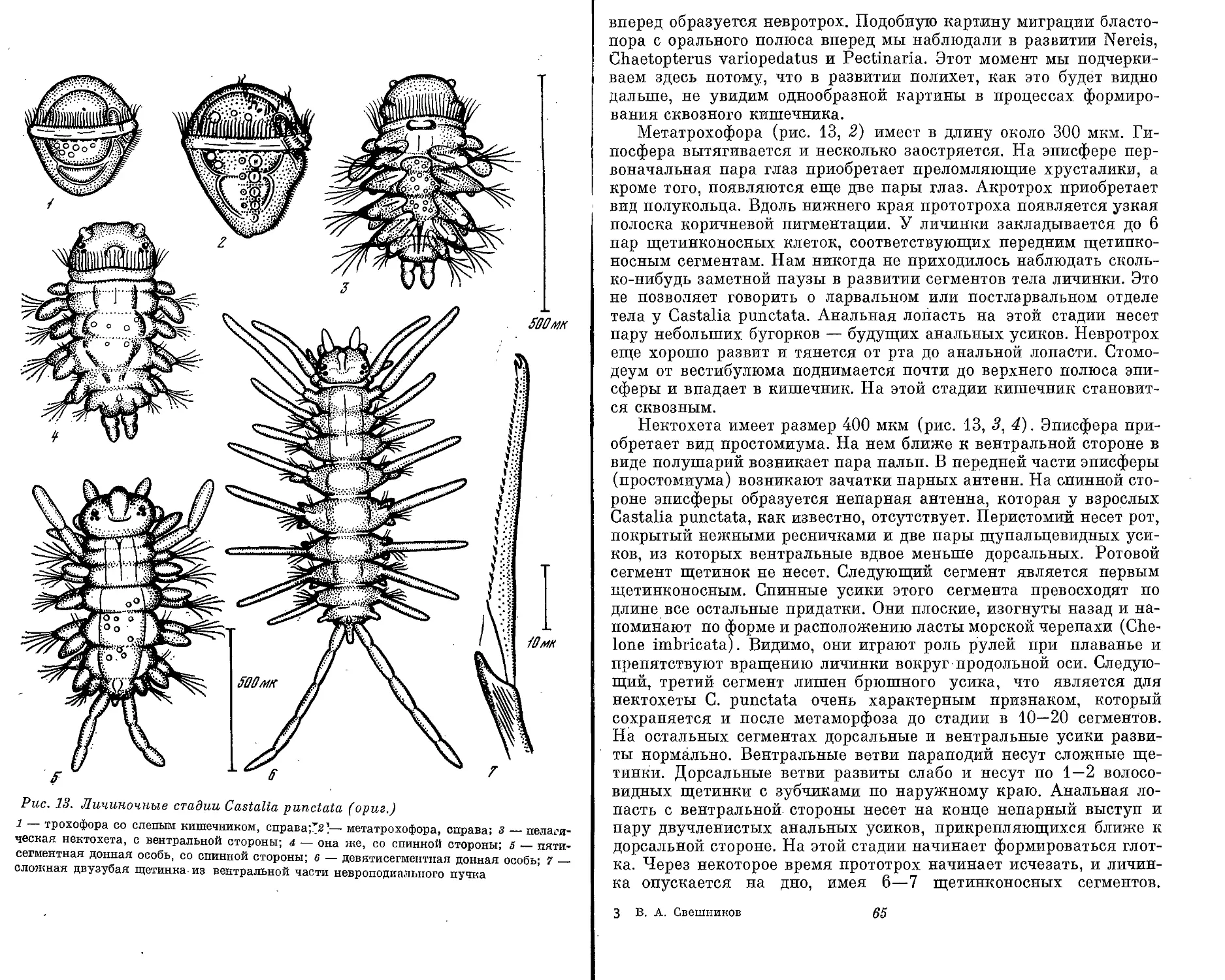


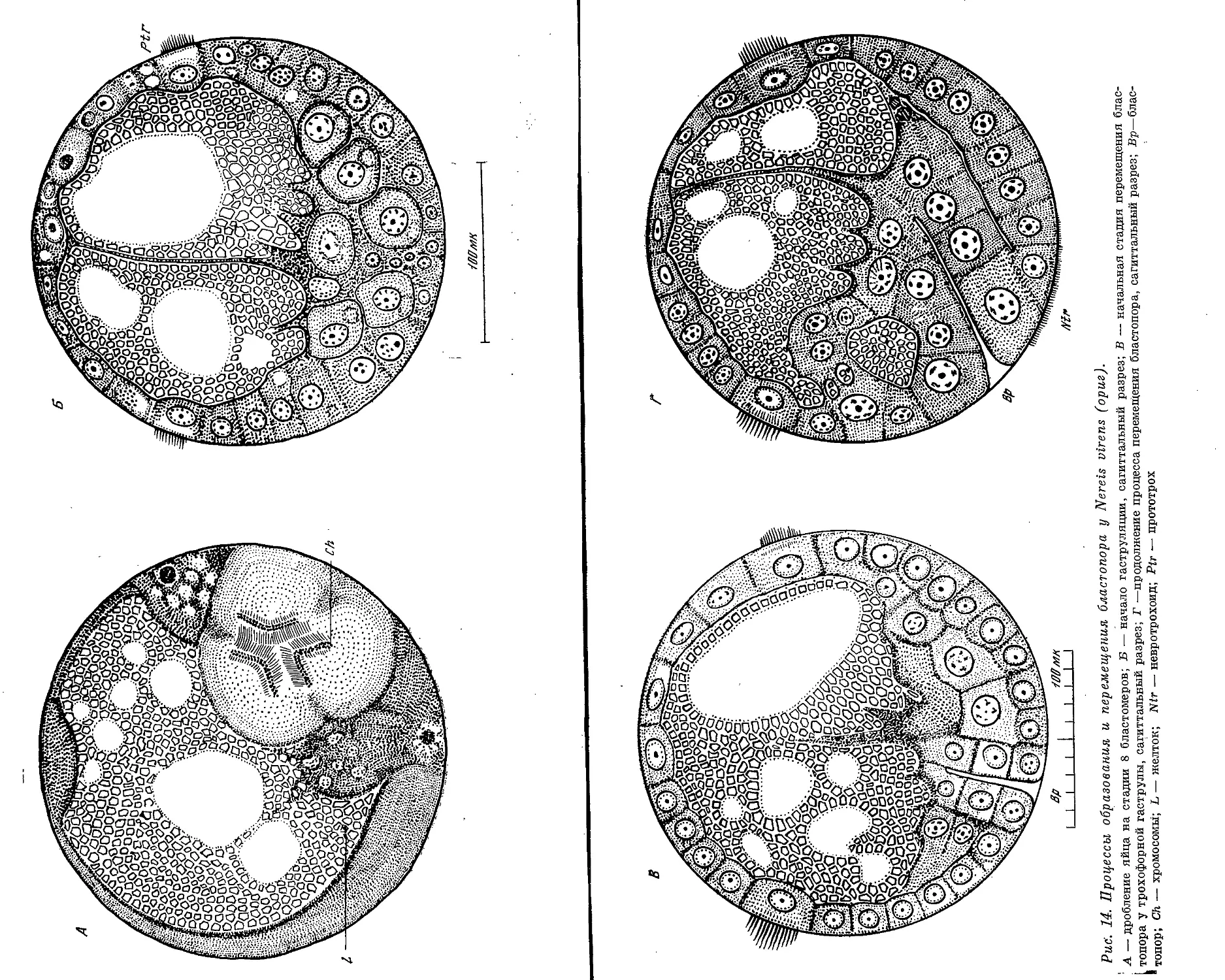
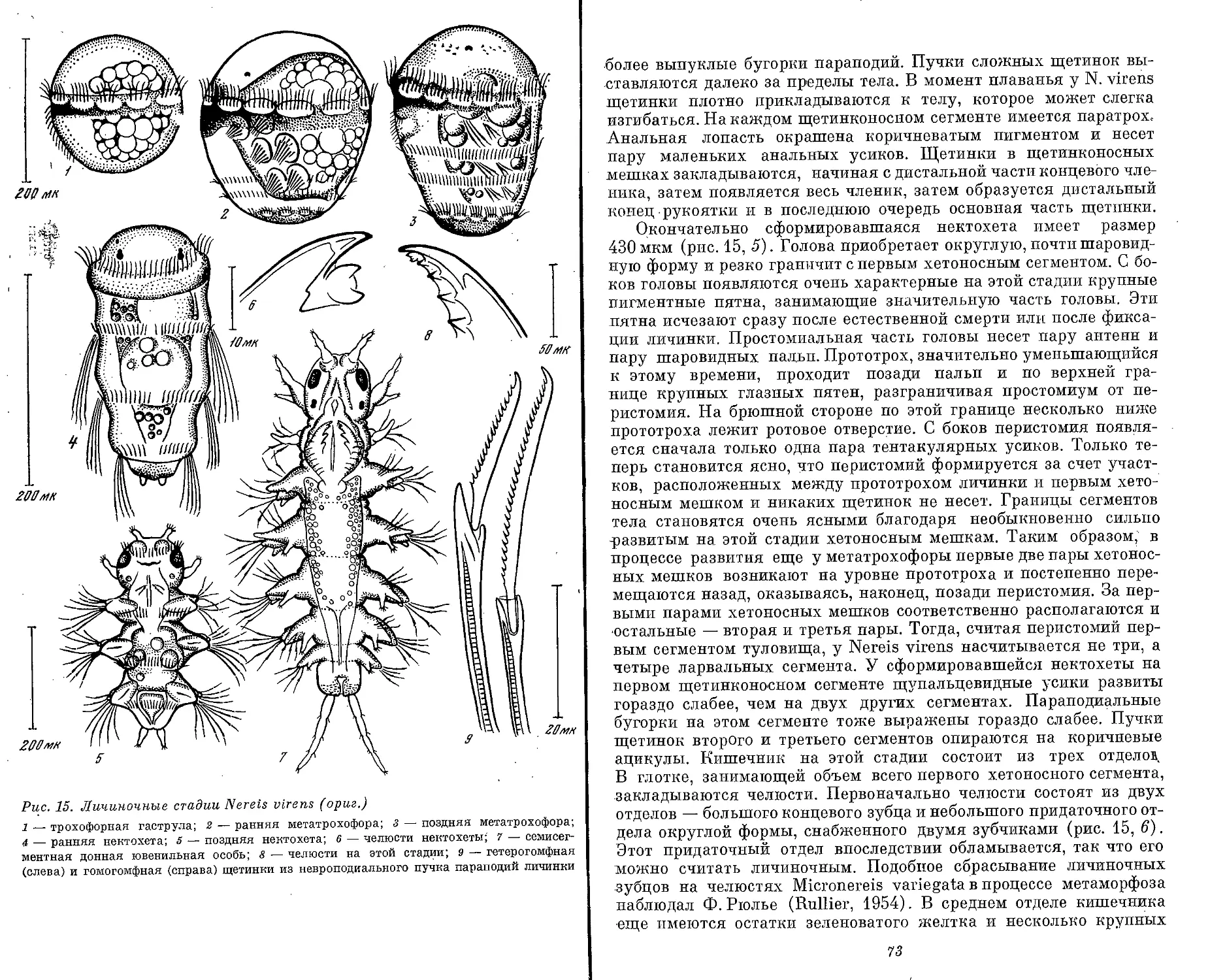

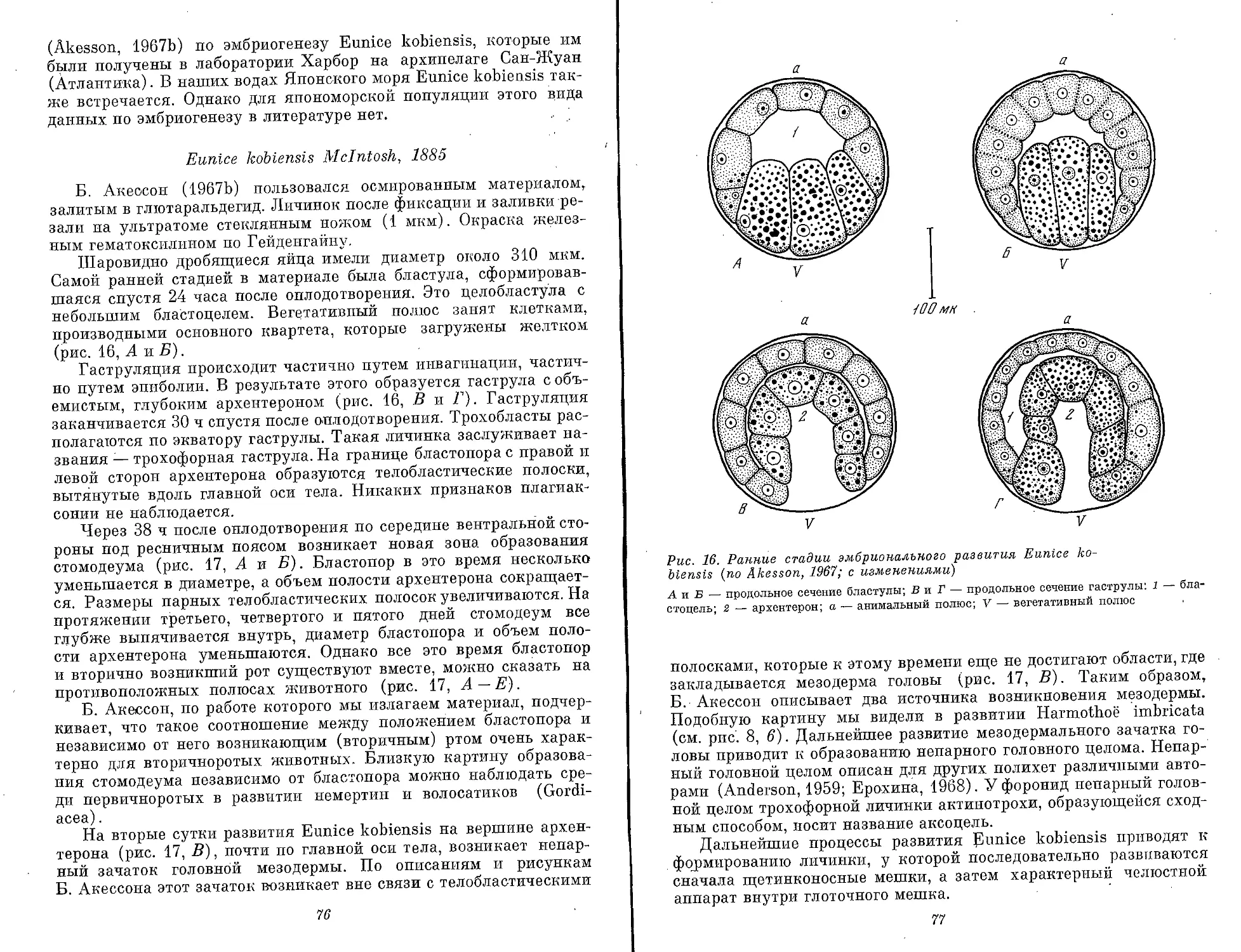
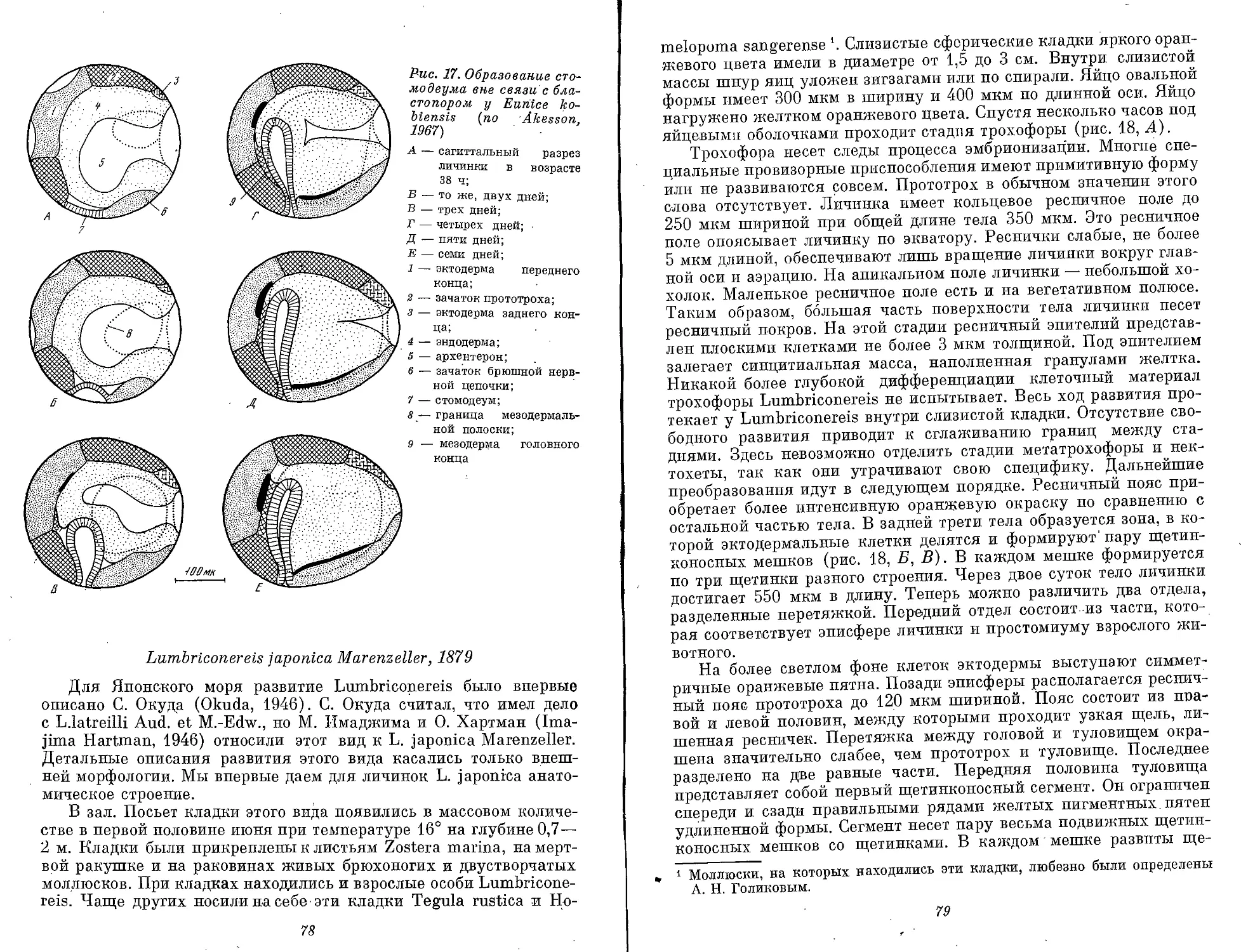
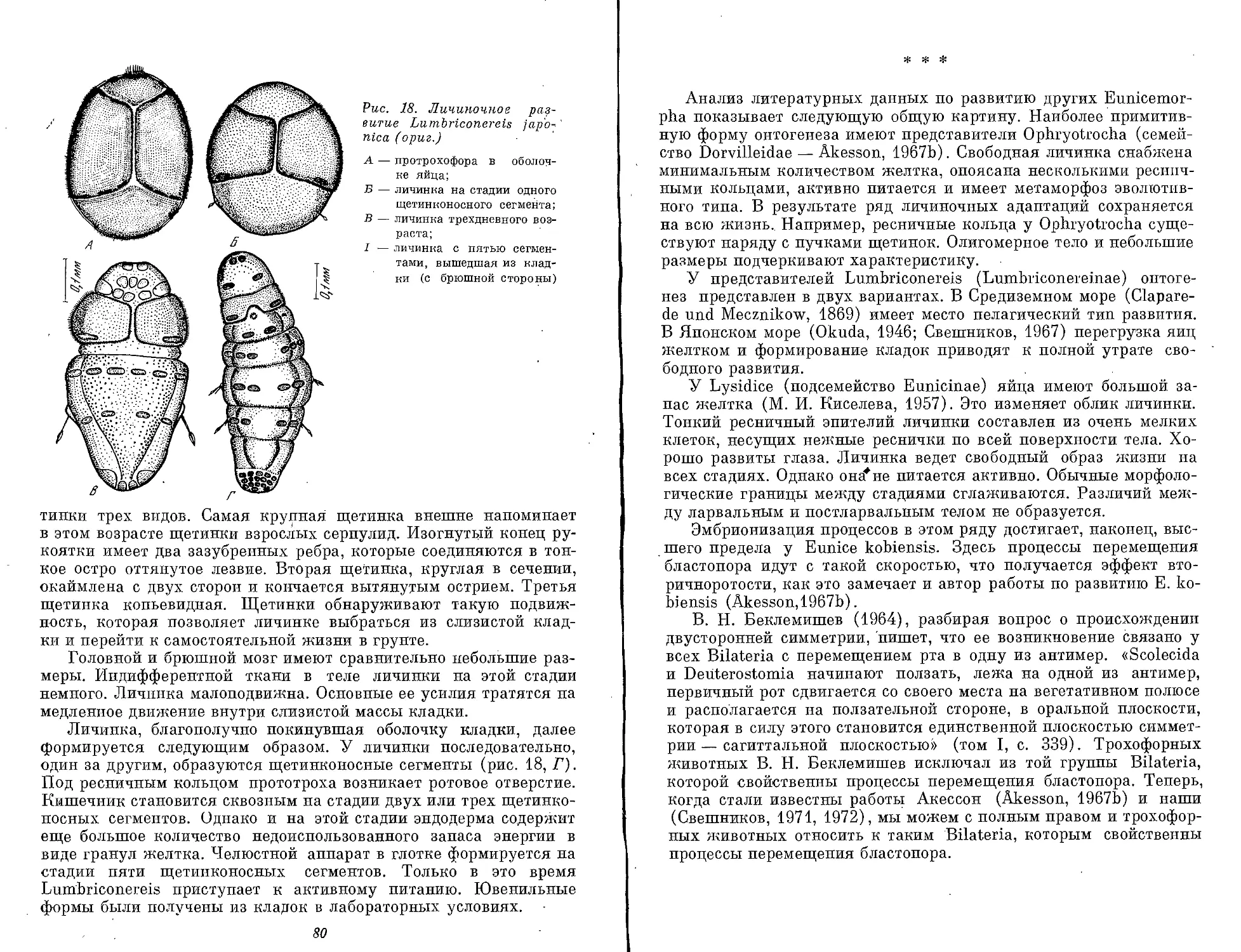





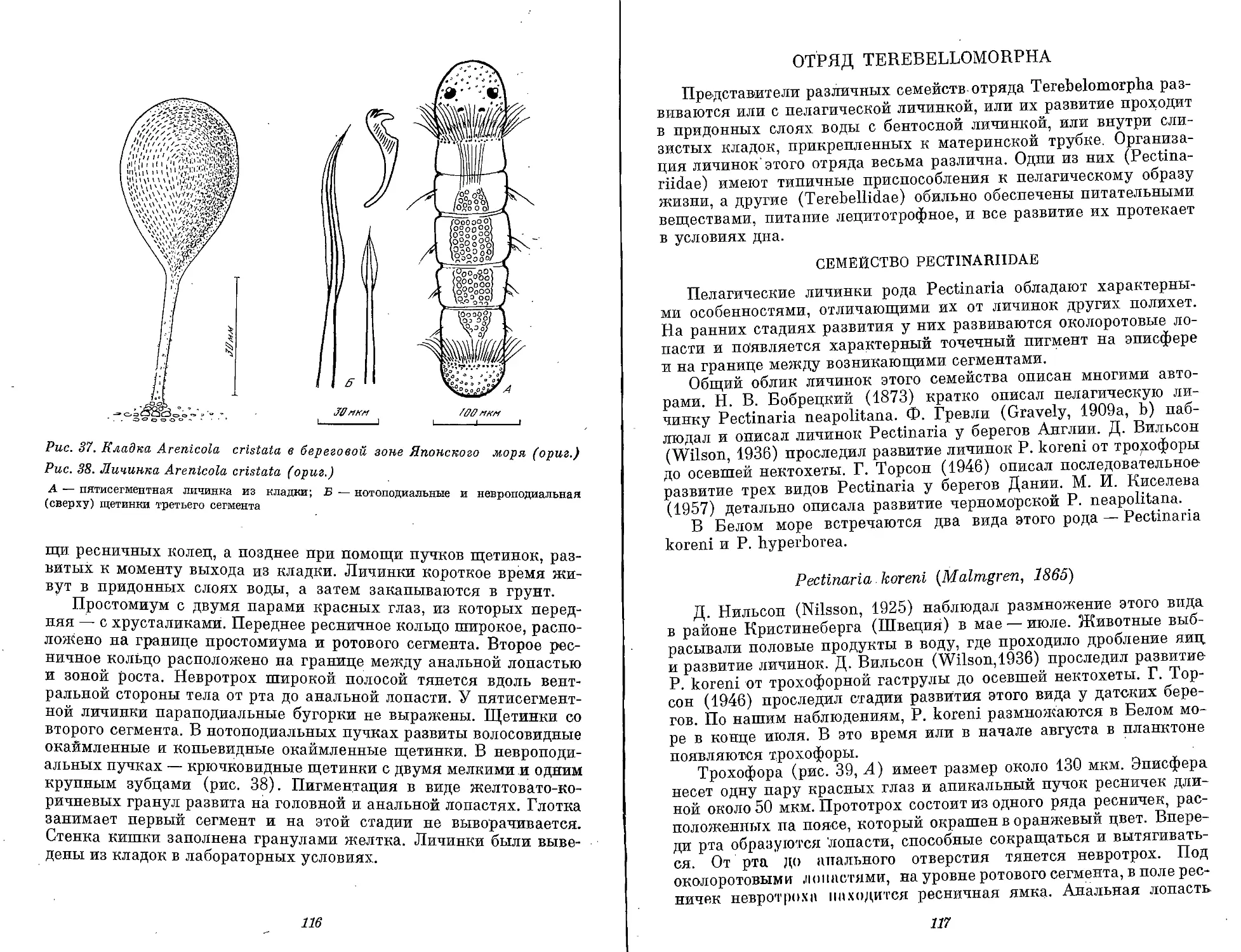
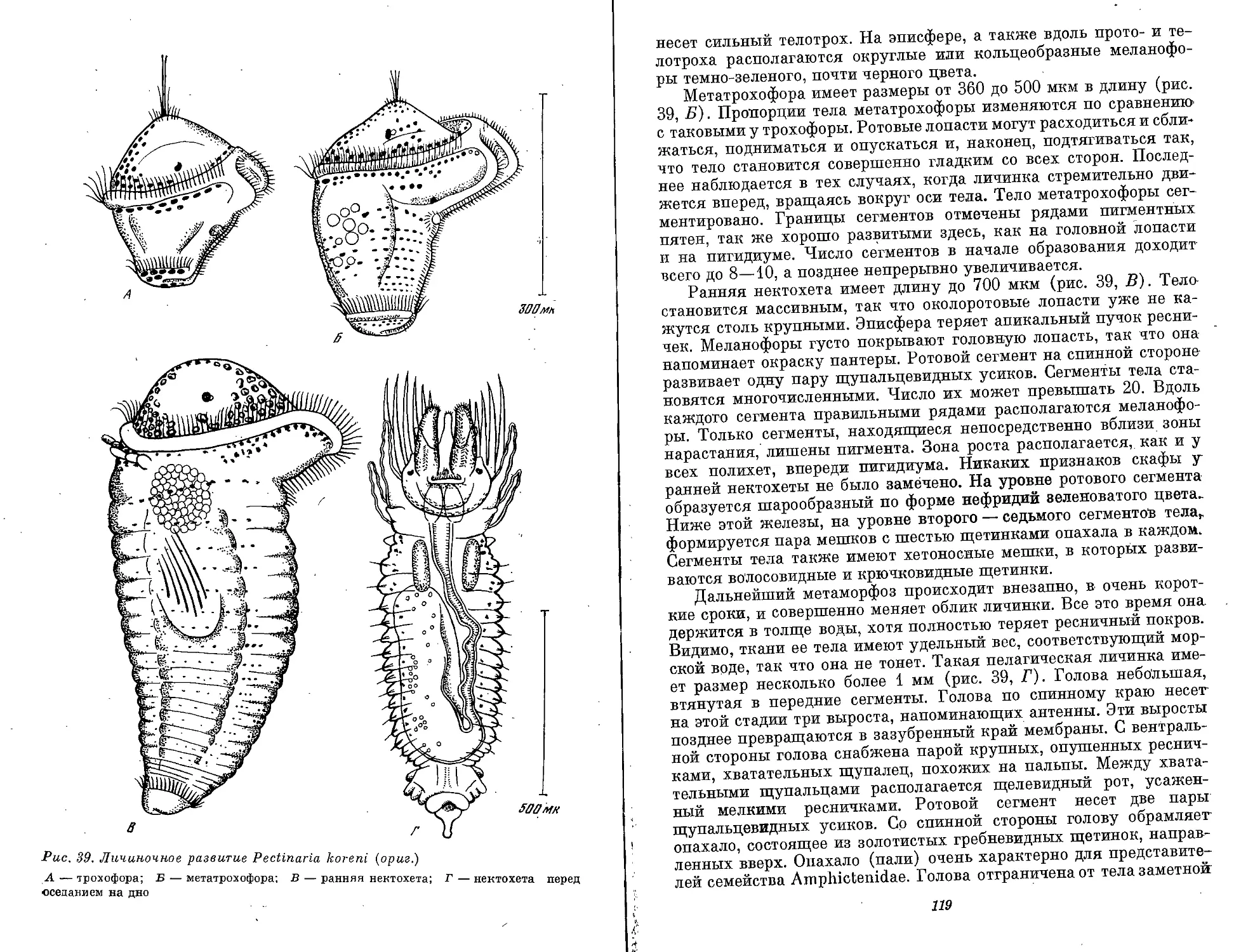




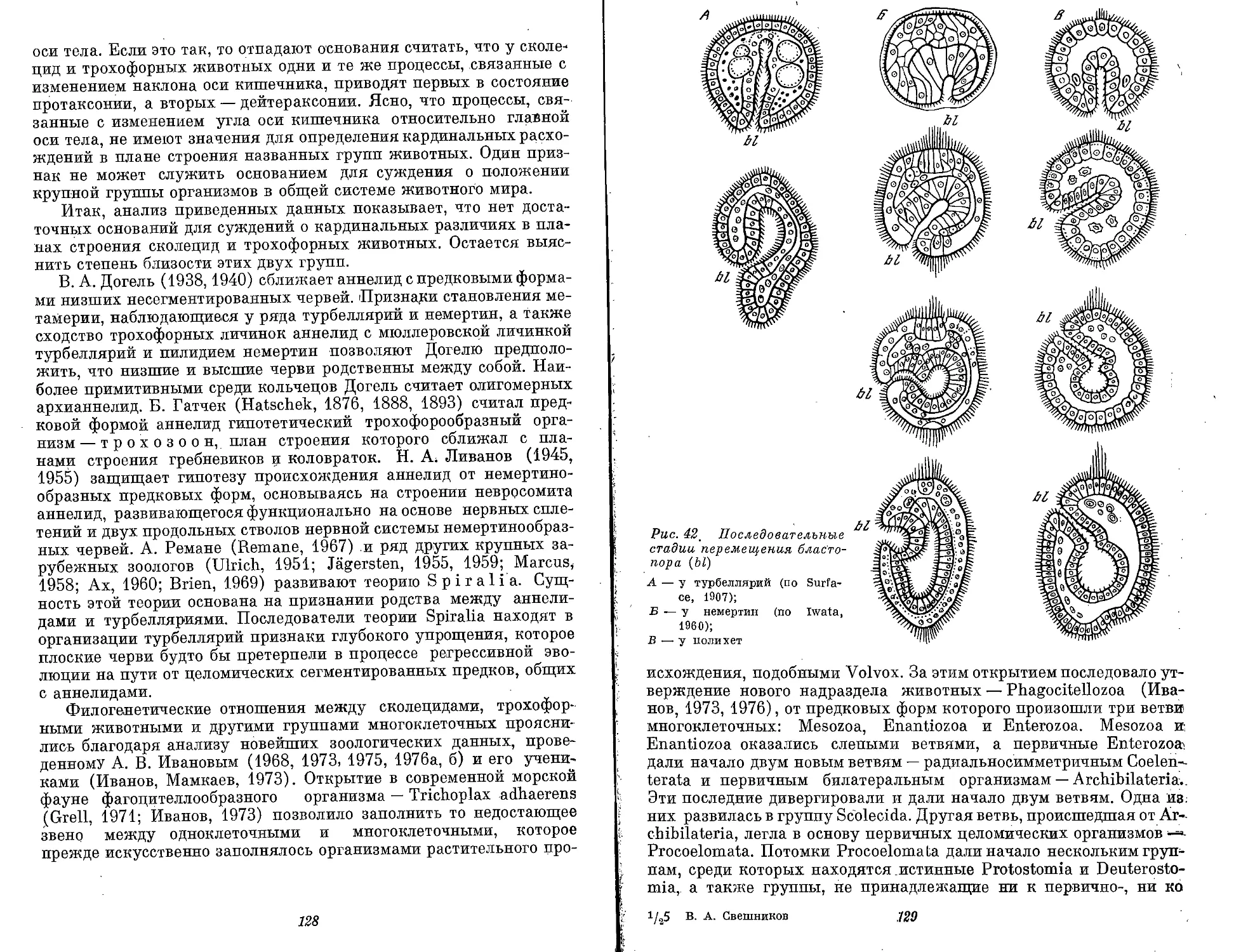
















Text
В. А.СВЕШНИКОВ
МОРФОЛОГИЯ
ЛИЧИНОК ПОЛИХЕТ
ИЗДАТЕЛЬСТВО «НАУКА
АКАДЕМИЯ НАУК СССР
Институт эволюционной морфологии
и экологии животных
им. А. Н. Северцова
МОРФОЛОГИЯ
ЛИЧИНОК ПОЛИХЕТ
Издательство «Наука»
Москва 1978
УДК 591.341.2; 595.14
Свешников В. А. Морфология личинок полихет. М.,
«Наука», 152 с.
Впервые проведен морфологический анализ п
для некоторых видов дана анатомия личинок поли-
хет морей Советского Союза. Методом искусственного
выведения идентифицированы многие виды личинок.
Составлены определительные таблицы. Показано зна-
чение морфологии личиночного развития для реше-
ния проблем морфологии, экологии и филогении
беспозвоночных. Монография рассчитана на специа-
листов зоологов широкого профиля, эмбриологов,
преподавателей и студентов биологических ВУЗов
и морских пр.актикумов.
Ил. 43, библ. 320 назв.
Ответственный редактор
доктор биол. наук
Н. Н. СМИРНОВ
„ 21008-098 „
С 055(02)—78 579~78> кн- 2
© Издательство «Наука», 1978 г.
ВВЕДЕНИЕ
Закономерности индивидуального развития животных приоб-
ретают большое значение в зоологии в связи с актуальными совре-
менными проблемами охраны и воспроизведения биологических
ресурсов. Планомерное восстановление биологических ресурсов
неразрывно связано с интенсивностью развития организмов в оп-
ределенных экологических условиях. Изучение индивидуального
развития в морской биологии в последние десятилетия открыло
ряд новых направлений.
Экология размножения и развития морских животных получила
многочисленные данные о значении пелагических личинок беспоз-
воночных в процессах трансформации энергии в океане. Ранее из-
вестные зависимости географической /принадлежности организмов
и сроков их размножения при строго определенных условиях и,
особенно, температурном режиме нашли новые подтверждения и
практические выходы. Биоценотические отношения между план-
ктоном и бентосом, характер внутривидовых и межвидовых отноше-
ний на протяжении жизненного цикла организмов — это весьма
актуальные и перспективные проблемы, без решения которых не
удается в полной мере обосновать практические положения о зако-
номерностях формирования донных и пелагических биоценозов,
прогнозирования кормовой базы рыб, организации промысла мор-
ских объектов и ведения марикультур (Зенкевич, 1947, 1951, 1963;
Strathmann, 1975).
В области функциональной и эволюционной морфологии и срав-
нительной эмбриологии в последнее время были подняты новые и
интересные проблемы. В зоологии и сравнительной анатомии бес-
позвоночных животных много новых положений и гипотез предло-
жено В. Н. Беклемишевым (1944, 1952, 1964), Н. А. Ливановым
(1945, 1955), А. В. Ивановым (1968, 1975, 1976а, 19766). В срав-
нительной эмбриологии фундаментальные работы созданы
О. М. Ивановой-Казас (1975, 1977). Наиболее спорные и сущест-
венные вопросы современной эволюционной морфологии беспозво-
ночных сосредоточены вокруг гипотез о происхождении трохофор-
ных животных, о происхождении метамерии и о месте трохофорных
и других членистых организмов в системе щивотного мира
(П. П. Иванов, 1912, 1916, 1928, 1937, 1944, 1945; Зенкевич, 1944;
В. Н. Беклемишев, 1964; Светлов, 1928, 1957, 1967, 1970; Remane;
1950; Clark, 1964, 1969; Slewing, 1969; Codreanu, 1970; Иванова-
.3
Казас, 1974; А. В. Иванов, 19766). Новые данные по индивидуаль-
ному развитию беспозвоночных занимают в этих вопросах одно из
первостепенных положений.
Методологическое значение имеет вопрос о происхождении он-
тогенеза (Северцов, 1939) и свободного пелагического личиночного
развития морских беспозвоночных. Если принять, что свободное
пелагическое развитие у многочисленных типов морских беспозво-
ночных — вторичное приспособление, то многие вопросы эволю-
ционной морфологии беспозвоночных могут оказаться вне всякой
связи с общими проблемами онтогенеза (Ежиков, 1939; Захват-
кин, 1949).
В систематике данные по индивидуальному развитию приобре-
тают все возрастающее значение. В некоторых группах беспозво-
ночных, благодаря изучению индивидуального развития, произве-
дена значительная ревизия системы (Hannerz, 1956; Наумов.,
1960; Милейковский, 1968; Ушаков, 1972).
В последние годы внимание исследователей было привлечено
к вопросам поведения животных на разных стадиях индивидуаль-
ного развития. В частности, вопросы поведения личинок беспозво-
ночных в момент посадки на дно, выбора субстрата и изменения
поведения в процессе метаморфоза получили освещение в литера-
туре (Wilson, 1954; Evans, 1971; Кутищев, 1976). Эти данные ока-
зались важными для физиологии, морфологии, систематики и по-
лезными в практике. ,
Из сказанного видно, что исследования в области индивиду-
ального развития беспозвоночных в последние годы внесли замет-
ный вклад в разрешение проблем функциональной и эволюцион-
ной морфологии, систематики, экологии и физиолюгии.
Объектом, нашего исследования послужили личинки многоще-
тинковых кольчецов. Нас интересовали закономерности индиви-
дуального и в основном личиночного развития этой группы живот-
ных. Первая задача состояла в определении видовой принадлеж-
ности личинок полихет. Для решения этой задачи потребовались
новые приемы работы с живым материалом: воспитание животных
в лабораторных условиях. Вторая задача определялась морфологи-
ческими интересами. Здесь была применена гистологическая тех-
ника, при помощи которой выявили существенные детали анато-
мического строения личицрк. Этот раздел работы дал возможность
предложить некоторые гипотезы, освещающие вопросы эволюцион-
ной морфологии аннелид. Третья задача заключалась в определе-
нии ряда экологических закономерностей, связанных со сроками
размножения и появлением личинок полихет в толще воды в за-
висимости от температуры и некоторых других условий. В работе
мы стремились вскрыть возможно больше деталей жизненного
цикла того или иного вида для более общей сравнительной оценки
явлений экологического и морфологического плана. Наконец, мы
ставили целью выявить при помощи данных по индивидуальному
развитию родственные отношения между отдельными группами по-
лихет, а также между группами трохофорных и членистых живот-
ных.
Работу проводили в прибрежной зоне Японского и Белого мо-
рей.
Основная линия настоящего исследования была определена в
незабываемом общении с моими учителями и наставниками —
/Львом Александровичем Зенкевичем и Владимиром Николаевичем
Беклемишевым. Во время работы над книгой автору очень помог-
ли советы и критические замечания профессора П. В. Ушакова,
которому приношу сердечную признательность. В подготовке мо-
нографии к печати большое участие принимали сотрудники лабо-
ратории морфологии беспозвоночных Института эволюционной
морфологии и экологии животных им. А. Н. Северцова АН СССР.
Работа не могла быть проведена без существенного содействия со
стороны академика В. Е. Соколова и чл.-корр. АН СССР А. В. Жир-
мунского.
4
ИСТОРИЯ ИССЛЕДОВАНИИ МОРФОЛОГИИ
И ЭКОЛОГИИ ЛИЧИНОК ПОЛИХЕТ
Морфологические исследования в области индивидуального
развития аннелид естественно распадаются на два этапа. Первый
этап включает работы по идентификации личинок относительно
взрослых форм. Идентификация личинок беспозвоночных осущест-
вляется чаще всего на основе внешних морфологических призна-
ков. Второй этап посвящен проблемам эволюционной морфологии,
для решения которых необходимы, кроме того, анатомические
данные.
Соответственно двум названным этапам возникли две основные
пинии исследований. Первая линия развивалась в направлении
определения градиента различий между формами трохофорных
личинок и соответствия морфологических типов личинок полихет
таксономическим подразделениям взрослых форм. Известно, что
крупным таксономическим единицам (типам и классам) свой-
ственны вполне определенные типы личинок. Например, губки име-
ют личинку амфибластулу или паренхимулу, кишечнополостные —
планулу или гаструлу, турбелляриям свойственна мюллеровская
личинка, немертинам — пилидий, моллюскам — велигер, рако-
образным — науплиус, иглокожим и кишечнодышащим — диплев-
рула. Многим представителям класса полихет свойственна трохо-
фора. Класс полихет включает около 70 семейств, взрослые формы
которых подробно изучены с систематической, анатомической и
физиологической точек зрения. Данные по эмбриональному и пост-
эмбриональному развитию этих семейств до последнего времени
оставались разрозненными и недостаточными. Сбор и анализ таких
данных позволял вскрыть вопросы о происхождении трохофорной
личинки, о единых путях развития организма на всех стадиях его
жизненного цикла и некоторые вопросы систематики полихет.
В литературе широко распространено мнение, будто все поли-
хеты имеют личинку трохофору, план строения которой обычно рас-
сматривается на примере личинки полигордия. Однако давно было
подмечено, что личинки некоторых полихет имеют весьма своеоб-
разное строение и часто совсем непохожи на типичных трохофор.
Например, И. И. Мечников (Mecznikow, 1871) описал личинку,
резко отличающуюся от обычной трохофоры, и дал ей название
митрария за характерную форму, напоминающую митру. В. Геккер
(Hacker, 1898) описал другую личинку и назвал ее рострария за
характерную форму головы в виде рострума. Эти личинки настоль-
6
ко не похожи на трохофор, что сами авторы не смогли определить
их принадлежность, которая выяснилась лишь много десятилетий
спустя (Wilson, 1932в; Милейковский, 1960а, 1961а). Оказалось,
что митрария специфична для полихет сем. Oweniidae, а ростра-
рия — для сем. Amphinomidae. Естественно, возникло предположе-
ние, что другие семейства полихет также имеют свои специфич-
ные формы личинок. Анализ обширного оригинального и взятого
из литературы материала привел нас к выводу, что специфика ли-
чинок различных семейств полихет проявляется от ранних стадий
к более поздним. Каждое семейство полихет наделено своей типич-
ной формой личинки (Свешников, 1963), однако каждую более
сложную форму можно свести к более простой форме трохофоры.
Наиболее простой формой среди трохофорных личинок, как оказа-
лось, обладают полихеты семейства Phyllodocidae. Этот факт, наря-
ду со многими другими, признаками, послужил основанием для
пересмотра положения семейства Phyllodocidae в системе полихет
(Ушаков, 1972).
Вопрос о возможных путях происхождения личинки трохофоры
не был особенно острым. Известно, что до стадии трохофоры поли-
хеты проходят стадию гаструлы (Wolterek, 1902; Wilson, 1936;
Свешников, 1971а, б, 19726). Однако в вопросе о том, как осущест-
вляются пути перехода от гаструлообразного состояния со слепым
кишечником к трохофорному состоянию со сквозным кишечником,
до сих пор нет полной ясности.
Можно наметить две позиции в теоретических построениях,
основанных на сравнительно небольшом имеющемся материале.
В. Н. Беклемишев (1944, 1964), основываясь на данных
Н. Клейненберга (Kleinenberg, 1886), Э. Мейера (Meyer, 1897,
1901) и Р. Вольтерека (Wolterek, 1902), считал трохофорных жи-
вотных плагиаксонными (поперечноосными), т. е. образовавшими-
ся в результате посадки пелагического гаструлообразного предка
ртом на грунт. По мнению В. Н. Беклемишева, первичный рот
(бластопор) вытянулся в щель, края которой посередине срослись,
а на противоположных концах щели образовались рот й анус.
Главная ось тела аннелиды и других трохофорных животных рас-
полагается, по мнению В. Н. Беклемишева, под углом 90° к глав-
ной оси тела трохофоры (или гаструлы).
Н. А. Ливанов (1955) замечает, что из работы Р. Вольтерека
(Wolterek, 1902), наиболее подробно излагающей процессы раз-
вития трохофоры полигордия, совершенно не следует, будто бы и
рот и анус трохофоры являются производными бластопора.
Н. А. Ливанов (1955) пишет: «Для категорического утверждения,
что рот и задний проход трохофоры полигордия образуются за счет
бла.стОпора путем его разделения, собственно, нет оснований» (стр.
164). Н. А. Ливанов считает, что бластопор у полигордия не дает
ануса. «Образование заднего прохода за счет бластопора можно
надумать, но факты этого не говорят» (стр. 165). Таким образом,
в противоположность В. Н. Беклемишеву, мнение Н. А. Ливанова
7
сводится к тому, что план строения высших червей принципиально
не отличается от плана строения низших червей, обе группы про-
таксонны (Светлов, 1967, 1970; Свешников, 1972а; Иванова-Казас,
1974) и представляют собой одну монофилетическую линию разви-
тия.
Гипотезу В. Н. Беклемишева о происхождении трохофорных
животных непосредственно от кишечнополостных развивали
А. Г. Шаров (1965а, б; Sharow, 1966), К. В. Беклемишев (1970), а
также Ю. С. Миничев и Я. И. Старобогатов (1972).
Вопросы происхождения трохофорных животных затрагивают
более общие проблемы происхождения группы Coelomata и вопро-
сы происхождения метамерии.
Наименее приемлемая, но развиваемая некоторыми современ-
ными исследователями (Мельников, 1971) гипотеза А. Седжвика
(Sedgwick, 1884) связывает происхождение аннелид с кишечнопо-
лостными на основе происхождения метамерии от цикломерии. По
мнению А. Седжвика и его последователей (Lameere, 1926; Bi-
nard, Jenner, 1928; Remane, 1950) , рот и анус аннелид происходят
из сифоноглиф сомкнувшегося щелевидного рта Anthozoa, пара-
подии аннелид — от щупалец полипа, а нервная цепочка анне-
лид— от нервного сплетения орального диска. Ошибочность-этих
гипотетических построений состоит в том, что аннелиды выводятся
из высокоорганизованных предковых форм, подобных Anthozoa
(Беклемишев, 1964).
Более вероятная и обоснованная гипотеза происхождения мета-
мерии путем упорядочения в расположении различных систем орга-
нов была предложена Р. Снодграссом (Snodgrass, 1928, 1938) и
развита В. Н. Беклемишевым (1944, 1952, 1964). Гипотеза мета-
мерного упорядочения систем органов хорошо объясняет процессы
утверждения метамерии у различных групп беспозвоночных.
В. Н. Беклемишев детально рассматривает многочисленные при-
меры метамерии упорядочения у турбеллярий, киноринх, немертин,
олигомерных аннелид, моллюсков, других беспозвоночных.
Фактического материала, касающегося процессов становления
метамерии в индивидуальном развитии полихет, очень мало.
П. П. Иванов (Ivanov, 1928, 1944) обосновал многочисленными фак-
тами теорию первичной гетерономной метамерии членистых живот-
ных. Сущностью этой теории является закономерная последова-
тельность возникновения в индивидуальном развитии Articulata
сначала ограниченного числа своеобразных ларвальных сегментов,
а позднее большого количества постларвальных сегментов с иными
морфофизиологическими признаками. Наиболее примитивными
среди аннелид П. П. Иванов справедливо считал Dinophilidae,
тело которых состоит из одних только ларвальных сегментов.
8
& * : *
Исследования индивидуального развития в классе полихет по-
зволяют внести большую ясность в систему этой многообразной
группы. Филогенетические отношения между семействами внутри
класса полихет остаются спорными, несмотря на большое число
/специальных исследований в этой области. До сих пор мы не име-
ем общепринятой и хорошо обоснованной системы полихет. Деле
ние на подклассы признают далеко не все специалисты, так как
оно весьма формально. П. фовель (Fauvel, 1923, 1927) делит класс
на два подкласса (Errantia и Sedentaria), к которым прибавляет
еще аберрантные дополнительные группы Myzostomaires и Archi-
annelides. Н. А. Ливанов (1940) делит полихет на три подкласса:
1) Errantia с отрядами Phyllodocemorpha, Nereimorpha и Eunice-
morpha; 2) Sedentaria с отрядами Spiomorpha, Terebellomorpha и
Serpulimorpha; 3) подкласс с одним отрядом Myzostomidae.
П. В. Ушаков (1955) в определителе многощетинковых червей
дальневосточных морей СССР приводит два подкласса — Errantia
и Sedentaria. Р. Деле (Dales, 1963) вообще не делит полихет на
подклассы. С. А. Милейковский (1968) предлагает деление класса
полихет на три надотряда — Errantia, Sedentaria и Amphinomea.
Таким образом, мы видим, что наиболее стойко во всех предлагае-
мых системах удерживаются только две группы — Errantia и Seden-
taria. Однако естественность деления на эти две группы вызывает
сомнение, так как между ними невозможно провести четкой гра-
ницы ни по признаку относительной подвижности, ни по морфоло-
гии.
Нисколько не проще обстоит дело и с делением класса поли-
хет на отряды и особенно с определением среди них наиболее при-
митивного отряда. Н. А. Ливанов (1940) выделяет среди Errantia
отряды Phyllodocemorpha, Nereimorpha и Eunicemorpha, а среди
Sedentaria — Spiomorpha, Terebellomorpha и Serpuliomorpha.
П. В. Ушаков (1955) принимает эту систему, но исключает из со-
става отряда Eunicemorpha семейства, относящиеся к архианнели-
дам. О. Гартман (Hartman, 1959) в своей системе полихет вообще
исключает ранг отряда. Р. Деле (Dales, 1962,1963) предложил но-
вую систему полихет, обосновав ее морфологическими признаками,
относящимися к строению глотки и другим производным стомодеу-
ма. Р. Деле называет 14 отрядов. Наиболее крупный отряд — Phyl-
lodocida — включает 15 семейств, в том числе HeSionidae, Pilargi-
dae и Nereidae. Построение системы по признакам строения глот-
ки, как это ни странно, совершенно размывает границы между
группами Errantia и Sedentaria. Некоторые авторы приравнивают
группу Errantia к преимущественно хищным полихетам (Rhapa-
cia), тогда как Sedentaria — преимущественно к сестофагам и дет-
ритофагам и, казалось бы, способ питания должен быть в соответ-
ствии с организацией аппарата захвата пищи (глотки). Однако по
системе' Р. Делса вторым после Phyllodocida поставлен отряд Gapi-
tellida (по системе П. В. Ушакова — настоящие Sedentaria), а да-
9
лее отряд Amphinomida (снова Errantia). Сближая эти группы по
степени примитивности глоточного аппарата, Р. Деле допускает
одностороннее суждение. Глоточный аппарат Phyllodocida, вне вся-
ких сомнений, наиболее примитивен, а глоточный аппарат Capitel-
lida и Amphinomida может быть вторично упрощен, у первых в
связи с роющим образом жизни и необходимостью заглатывать пе-
сок, а у вторых — в связи с обитанием внутри тела губок. Вместе
с тем в небольшой степени Р. Деле учитывает признаки организа-
ции выделительного аппарата. В целом его система получает ори-
гинальное выражение, так как основана на ограниченном числе
признаков. Система Р. Делса не имеет устойчивости и в некоторых
частях склоняется в сторону примитивности группы полихет
с вентрально расположенной глоткой (Eunicida, Amphinomida).
Классификация полихет, в которой впервые были учтены при-
знаки, связанные с особенностями индивидуального развития, была
предложена С. А. Милейковским (1968; Mileikovsky, 1977). В ос-
нову своей системы С. А. Милейковский положил некоторые при-
знаки строения нектохет или ювенильных стадий после заверше-
ния метаморфоза. Используя признаки главным образом внешне-
го строения личинок, С. А. Милейковский приходит к заключению,
что отряд Amphinomorpha следует повысить в ранг надотряда
Amphinomea и поставить его рядом с другими надотрядами —
Errantia и Sedentaria. Легко заметить, что при этом нарушается
принцип единообразия, взятого в качестве критерия признака,
К двум ранее известным группам, разграниченным по особенно-
стям образа жизни, добавляется третья группа по признакам внеш-
ней морфологии пелагической личинки. Существенно в системе
С. А. Милейковского то, что семейство Phyllodocidae, видимо как
наиболее примитивная группа, поставлено на первое место, как у
Н. А. Ливанова (1940)щ П. В. Ушакова (1955, 1970, 1972), а не
как у французских систематиков (Fauvel, 1923, 1927, 1959). Систе-
ма С. А. Милейковского отличается введением большого числа се-
мейств (Hartman, 1959) и новых отрядов. Некоторые семейства и
отряды сближены по признакам сходства морфологии закончивших
метаморфоз личинок.
Совершенно другие критерии кладет в основу своей системы
полихет В. Шторх (V. Storch, 1968), который, вслед за своим пред-
шественником (О. Storch, 1914, 1915), принимает тетраневрию ам-
финомид за наиболее примитивный признак и ставит эту группу в
корень филогенетического древа кольчецов. В. А. Свешников и
Цю Вень-жень (1963) показали, что в развитии амфиномиды
Notopygos первоначально закладывается только два нервных ство-
ла, подтвердив этим ранее высказанное Г. Густавсоном (Gustafson,
1930). и-Н. А. Ливановым (1940) мнение о вторичности происхож-
дения тетраневрии амфиномид.
А. Г. Шаров (1965; Sharow, 1966) считает наиболее примитив-
ной формой среди полихет род Spinther, который прежде относили
к семейству Amphinomidae. Среди примитивных признаков
10
спинтерид А. Г. Шаров называет признаки радиальной симметрии,
щелевидный рот, расположенный близко к середине брюшной сто-
роны тела, примитивность вооружения головного конца и внешнее
сходство спинтера с отпечатками пойихет, найденными М. Глес-
снером (Glaessner, 1958, 1962, 1963) в древнейших сланцах Авст-
/ралии. Концепция А. Г. Шарова была подвергнута критическому
анализу в работе С. Мэнтон (Manton, 1967). Попытка А. Г. Шаро-
ва вывести группу членистоногих из высокоспециализированной
формы кольчецов не может быть признана достаточно убедитель-
ной. Спинтериды — преимущественно комменсалы, обитающие на
губках, и специфические особенности их организации находятся
в связи с их образом жйзни.
Новейшая классификация подихет детально разработана
П. В. Ушаковым (1970, 1972). В основу этой классификации поло-
жены многочисленные и очень разносторонние критерии и приз-
наки, относящиеся к области функциональной морфологии, эколо-
гии, индивидуального развития, палеонтологии. П. В. Ушаков
предлагает деление подкласса свободноживущих полихет на от-
ряды Phyllodocemorpha, Nereimorpha и Eunicemorpha. Очень де-
тально обоснована система отряда Phyllodocemorpha, который под-
разделяется на три подотряда—Phyllodociformia, Glyceriformia и
Aphroditiformia. Двенадцать пунктов, среди которых значительное
место занимают особенности индивидуального развития семейства
Phyllodocidae, убедительно доказывают большую примитивность
организации этого семейства по сравнению с другими семействами
подотрядов Glyceriformia и Aphroditiformia. Эволютивный тип
метаморфоза, отсутствие узкоспециализированных личиночных
приспособлений, высокая плодовитость и слабо выраженная за-
бота о потомстве — вот те основные признаки примитивности, ко-
торые дополняют набор критериев, помогающих П. В. Ушакову
(1970, 1972) определить положение филлодоцид в общей системе
полихет. Предложенный П. В. Ушаковым новый и более обосно-
ванный подход к классификации полихет на примере группы
Phyllodocemorpha может и должен быть применен к остальным от-
рядам. Накопленный обширный новый материал по индивидуаль-
ному развитию полихет, по всей видимости, даст возможность, на-
конец, получить естественную систему этой филогенетически важ-
ной группы, а также и других групп беспозвоночных.
В последние годы появились отдельные морфологические рабо-
ты по изучению ультраструктур тех или иных систем органов ли-
чинок полихет и других беспозвоночных. Н. М. Ерохина-Перетол-
чина (1971а) опубликовала первые данные об ультраструктурах
нервной системы трохофоры Anaitides (Phyllodocidae). Оказалось,
что организация тонких структур зачатков дефинитивной нервной
системы трохофоры имеет большое сходство с ультраструктурами
нервной системы турбеллярии Procotyla fluviatilis (по Lentz,
1967). Исследование Н. М. Ерохиной-Перетолчиной подтверждает
мнение П. В. Ушакова о большей примитивности семейства
11
Phyllodocidae по сравнению с другими семействами полихет.
Н. М. Ерохина-Перетолчина (19716) проводит далее сравнение
организации ультраструктур нервной системы трохофоры
Phyllodocidae, турбеллярии Procotyla и взрослой полихеты Polydo-
ra (Spionidae) и приходит к выводу, что ультраструктуры нервной
системы Polydora остановились в своем развитии на уровне орга-
низации ультраструктур личинки.
* * *
Общебиологические проблемы, связанные с вопросами воспро-
изведения и продуктивности морских животных, с необходимостью
организации и ведения марикультур несколько усилили внимание
к экологическим исследованиям. В области индивидуального раз-
вития беспозвоночных, в частности полихет, были сделаны инте-
ресные работы, открывающие закономерные связи развивающихся
организмов с различными абиотическими и биотическими факто-
рами.
Более всего внимание привлекли хорошо заметные связи меж-
ду температурой воды, временем размножения животных и их
географической принадлежностью. Постоянство сроков размноже-
ния у животных определенной географической принадлежности
давно было подмечено исследователями. А. Апеллёф (Appellof,
1912) один из первых высказал мысль о том, что эмбриональные
стадии беспозвоночных протекают в более узких пределах колеба-
ния температуры, чем те, при которых существует взрослый ор-
ганизм. Поэтому температура, при которой происходит размноже-
ние того или иного животного, служит фактором, ограничивающим
его распространение. Позднее Г. Ортон (Orton, 1920) сформули-
ровал свои известные правила зависимости времени нереста мор-
ских животных от температуры воды. Серия экспериментальных
исследований С. Руннстрёма (Runnstrom, 1927, 1929, 1936) дока-
зала, что температура нормального развития яиц морских живот-
ных является величиной постоянной. Наиболее низка эта темпера-
тура для арктических видов, выше — у арктическо-бореальных
и наиболее высока у бореальных и средиземноморских. 3. Г. Пале-
ничко (1941, 1948), присоединяясь к мнению этих авторов, отме-
чает как характерную особенность биологии бореальных видов —
Pandalus borealis (Decapoda) и Mytilus edulis (Bivalvia) —их спо-
собность размножаться и в центре и на границе ареала в наиболее
теплое время года. А. Я. Базикалова (1941), основываясь на зако-
номерностях, выведенных Г. Ортоном и С. Руннстрёмом, делает по-
пытку выявить вопросы происхождения фауны бокоплавов оз. Бай-
кал. С. А. Зернов (1949) принимает положения этих авторов и
уделяет описываемой закономерности существенное место, в своей
книге. Л. А. Зенкевич (1951, 1963) в своих работах приводит ряд
фактов, подтверждающих ту же закономерность. С. Экман (Ек-
man, 1953) включает в характеристику понятия «ледниковый ре-
12
ликт» широко известную для этих форм способность размножаться
при низкой температуре. В. Л. Паули (1957), основываясь на ра-
' ботах предыдущих авторов, создал интересные построения о разви-
[ тип экологических отношений морских организмов к температуре
: и солености в процессе эволюции.
; В пользу той же закономерности свидетельствует ряд экспери-
| ментальных данных. Эксперименты В. Лузанова и Т. Дэвиса
: (Loosanoff, Davis, 1950) показали, что теплолюбивый моллюск
, Venus mercenaria может размножаться зимой, если в лаборатор-
; ных условиях его содержать при оптимальной температуре раз-
множения. По данным А. Саутвод и Д. Криспа (Southward, Crisp,
1954; Crisp, 1957), размножение Balanus balanoides и В. balanus в
лабораторных условиях происходит только при температуре, близ-
кой к той, при которой балянусы размножаются в естественных
условиях, т. е. не выше +7°. П. Корринга (Korringa, 1957) в ра-
ботах по устрицам установил, что по отношению к температуре,
обилию пищи и другим факторам наиболее чувствительным перио-
дом в жизни этих моллюсков является период раннего развития
яйцеклеток внутри мантийной полости материнского организма.
По данным П. Корринга, устрицы в различных районах европей-
ского побережья размножаются при очень близких температурах.
В ряде случаев выделяются физиологические расы, которые обна-
руживают способность размножаться при температуре или более
высокой, или более низкой, чем в основных популяциях. И. Сагара
и К. Сугавара (Sagara, Sugawara, 1957) получили интересные дан-
ные об оптимальной температуре развития морской звезды Asterias
amurensis. С. Касим (Qasim, 1956) показал, что у литоральных ры-
бок маслюков также наблюдается зависимость между сроками раз-
множения и температурой воды. В. А. Свешников (1959), иссле-
дуя размножение полихет и моллюсков Белого моря, смог опреде-
лить четыре группы стенотермных и эвритермных беспозвоночных.
Первая группа стенотермных полихет и моллюсков размно-
жается при отрицательной температуре воды — гидрологической
зимой (март — май). Вторая группа размножается гидрологиче-
ской весной или гидрологической осенью при низкой положитель-
ной температуре. Третья, малочисленная в Белом море группа
беспозвоночных размножается гидрологическим летом при высо-
кой температуре воды. Четвертая группа включает полихет и
других беспозвоночных, которым свойственна эврибионтность. Эти
животные могут размножаться большую часть года. С. А. Милей-
ковский (19606, 19616; Mileikovsky, 1968) приводит большой ма-
териал, подтверждающий правило Г. Ортона для акватории Барен-
цева и Норвежского морей. С. А. Милейковский формулирует не-
которые специфические особенности размножения беспозвоночных,
населяющих бореальные и субарктические водоемы. В субаркти-
ческих водоемах размножение беспозвоночных протекает в более
сжатые сроки, и размножение видов с различной географической
принадлежностью накладывается друг на друга в связи с тем, что
13
период прогрева поверхностных слоев воды в этих водоемах очень ।
краток. Напротив, в таких акваториях, где период прогрева поверх-1
ностных вод более длителен, границы между периодами нереста I
видов с различной географической принадлежностью хорошо ощу-
тимы и отделены большими промежутками времени.
К. А. Захваткина (1963) опубликовала интересные данные по j
фенологии личинок двустворчатых моллюсков Севастопольской I
бухты Черного моря.
Для зал. Петра Великого в Японском море некоторые данные
о сроках появления личинок полихет в планктоне опубликованы
В. А. Свешниковым (1967).
Таким образом, закономерности, открытые Г. Ортоном для ог-
раниченной акватории, оказываются действительными для водое-
мов, расположенных в различных географических широтах. Попыт-
ки некоторых, авторов (Кузнецов, 1950, 1960, 1964) отрицать эти
закономерности вряд ли можно считать убедительными.
МЕТОДЫ ИССЛЕДОВАНИЯ
Материал собирали в Японском море в зал. Петра Великого. В
i зал. Посьет—в 1965 г. в составе экспедиции Зоологического ин-
ститута АН СССР. В заливах Троица и Восток в 1969 — 1977 гг. на
; базе Института биологии моря ДВНЦ АН СССР. Проводили ка-
I явственные ловы планктона сетью Джеди из газа № 58, 61. Всего
I было собрано и обработано из Японского моря 600 проб живого
планктона. Зоологический институт АН СССР любезно предоста-
вил нам многочисленные выборки планктона из экспедиций раз-
личных годов в Охотском и Японском морях. В Белом море мате-
риал собирали на биологической станции МГУ в 1953—1959 гг.
Работу с выведением личинок вели двумя путями. В одном
случае взрослых животных добывали в момент откладки яиц или
I кладок. Отлов нерестующих животных проводили драгой, тралом
или с помощью аквалангистов. Тогда видовую принадлежность
развивающихся яиц или личинок в кладках определяли по видо-
вой принадлежности родительских форм. Собранные кладки со-
держали в небольших кристаллизаторах в лабораторных условиях,
и личинок воспитывали до возможно более поздних стадий разви-.
тия или до взрослого состояния. В другом случае видовая принад-
лежность дробящихся яиц или личинок заранее не была изве-
стна. Тогда живых личинок выбирали из свежих проб планктона
и помещали в небольшие кристаллизаторы для дальнейшего воспи-
тания в лабораторных условиях до такого ювенильного или взро-
слого состояния, когда их можно было определить по обычным
определительным таблицам. Употребляли кристаллизаторы раз-
ного диаметра. Наиболее удобными для просматривания всего
объема воды под бинокуляром оказались кристаллизаторы диа-
метром до 3 см. Каждые два-три дня сосуды доливали свежей
морской водой. Резкой смены воды не допускали, так как это,
приводило к гибели личинок. На дно кристаллизаторов добавля-
ли тонкий слой грунта, богатого органикой (детрит). Хищных
личинок полихет (Spionidae) подкармливали личинками других
видов полихет или ракообразными из микробентоса. Для умень-
шения испарения воды в маленьких кристаллизаторах их по-
мещали по несколько штук в закрытые чашки Коха и содержали
в холодном помещении при температуре около 5°. Развивающих-
ся личинок периодически извлекали из кристаллизаторов, изме-
ряли и зарисовывали под микроскопом с помощью рисовального
аппарата. Все рисунки общего вида животных сделаны автором с
15
живых личинок. Это условие мы выполняли в связи с тем, что при
фиксации личинки деформируются, а многие характерные приз-
наки (например, провизорные ресничные кольца и ресничные по-
ля, провизорные щетинки, пигментация) утрачиваются. Часть
живого материала собирали на специальных коллекторах, сделан-
ных из стеклянных, деревянных, органического стекла пластинок
или раковин мертвых моллюсков. Коллекторы, закрепленные на
якорных плотах в море, имеют высокий процент улова оседающей
молоди и приносят большой дополнительный материал.
Часть материала требовала гистологических методик. Неоце-
нимую помощь в подготовке гистологических препаратов оказала
мне М. И. Константинова. Личинок или осевшую молодь фиксиро-
вали горячим насыщенным раствором сулемы с уксусной кисло-
той. После йодирования материал хранили в 70°-ном спирте.
Тотальные препараты личинок на разных стадиях развития окра-
шивали кармином с последующей докраской пикро-индиго-карми-
ном и дифференцировали спиртом.
Для точного ориентирования серийных срезов использовали
модификацию целлоидин-парафиновой методики. Личинок зали-
вали в целлоидин. У прямоугольных целлоидиновых пластинок
размером 8X4X5 мм под острым углом отсекали одну грань, ко-
торая впоследствии служила ориентиром для определения направ-
ления среза в микротоме. В момент заливки в целлоидин личинок
укладывали под бинокуляром на пластинки относительно граней
так, чтобы по граням можно было определить положение живот-
ного. Целлоидиновую пластинку с материалом затем заливали в
парафиновый блок, грани которого совпадали с гранями целлои-
диновой пластинки. Парафиновый блок резали с помощью мик-
ротома, соблюдая точную ориентацию материала по граням цел-
лоидиновой пластинки. Личинок на разных стадиях развития ре-
зали в трех направлениях—поперечно, сагиттально и фронтально.
Наиболее ясную картину давали обычно полные серии срезов в
сагиттальном направлении. Толщина срезов 5—7 мкм. Срезы ок-
рашивали железным гематоксилином Гейденгайна, по Караччи и
Маллори. Применяли методику графических реконструкций. Ри-
сунки срезов сделаны автором при помощи рисовального аппара-
та. Этот гистологический материал почти целиком оригинален;
подобных описаний в литературе очень мало, а для большинства
видов полихет из морей СССР гистологические данные по разви-
тию в литературе совершенно отсутствуют.
МОРФО-ЭКОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА
РАЗВИТИЯ ПОЛИХЕТ
Наиболее характерны для полихет кольчатость, параподии с
пучками многочисленных щетинок и развитие, в котором, как пра-
вило, три стадии — трохофора, метатрохофора и нектохета. Стадия
пектохеты характерна только для группы полихет. В развитии дру-
' гих беспозвоночных эта стадия, как правило, отсутствует. Пол-
ный набор стадий развития характерен для морских полихет, ко-
! торые по числу видов составляют основу класса. Сравнительно
немногие полихеты приспособились к существованию в солонова-
тых водоемах, где они часто утрачивают пелагическое развитие.
Еще меньше полихет в пресных водах и в грунтовых водах пещер.
Таковы, например, Manayunkia, обитающая в реках Северной
Америки, в озерах Таймыра и в Байкале, а также Merifugia, при-
способленная к жизни в пещерах. Эти формы полностью утрати-
ли свободное пелагическое развитие. В ряде случаев утрату
свободной пелагической личинки можно поставить в связь с раз-
личными возможностями осморегуляторного аппарата на опреде-
ленных этапах онтогенеза. Наконец, известен единственный ис-
ключительный случай существования полихеты Lycastopsis catar-
ractum на плантациях бананов и какао, в пазухах листьев и среди
гниющих плодов. В исключительных случаях полихеты паразити-
руют на рыбах (Ichthyotomus sanguinaris) или на жабрах омаров
(Histriobdella homari).
। В наиболее типичном случае тело полихет построено из четы-
рех отделов: 1) головная лопасть, или простомий; 2) ларвальный
отдел, состоящий из нескольких сегментов; 3), постларвальный от-
’ дел, состоящий из большого числа сегментов; 4) анальная ло-
пасть (пигидий).
Соответственно трохофора имеет эписферу, гипосферу и зону
роста между ними. Эписфера трохофоры включает верхний купол
(полусферу), ограниченный прототрохом. На полюсе эписферы
; расположена теменная пластинка с теменным хохолком. Нередко
; на эписфере бывает, кроме того, еще и акротрох. В полость эписфе-
< ры обычно выдается значительная часть кишечника. Наиболее ин-
тересная часть эписферы — теменная пластинка. Теменная плас-
; тинка — это центр, координирующий двигательную активность ли-
; чинки. Здесь сосредоточены провизорные органы чувств: глаза,
! теменной хохолок, орган равновесий. Вместе с тем теменная пла-
стинка — это зачаток дефинитивного мозга. Теменная пластинка
17
образуется из четырех радиальных зачатков. К полюсу эписферы
сходятся меридиальные нервы. Головная лопасть взрослого живот-
ного снабжена определенным числом придатков, которые возника-
ют на стадии метатрохофоры. Так как закладка придатков головы
возникает на базе радиального зачатка (креста), невозможно го-
ворить о метамерии головного отдела. Единичные случаи сегмен-
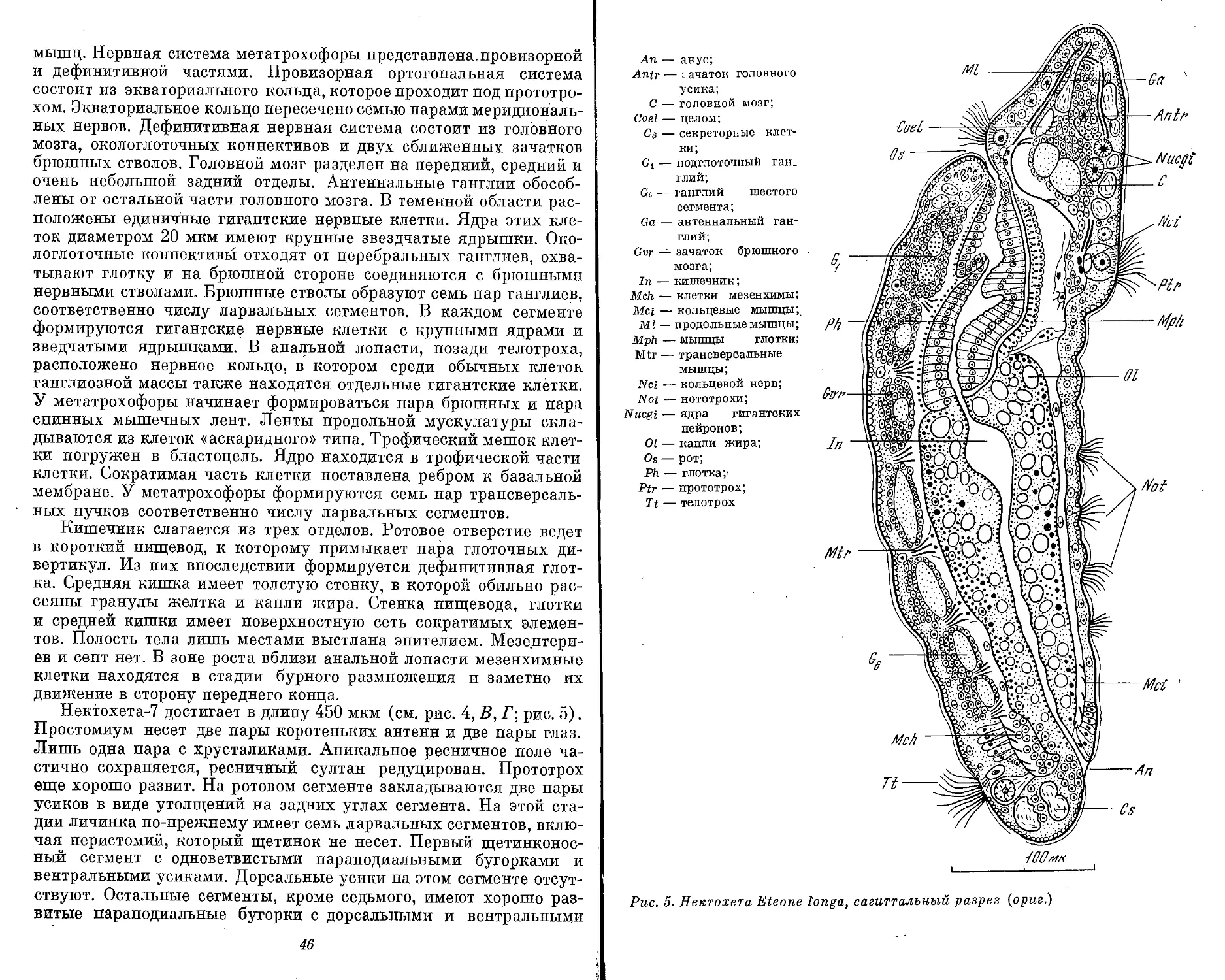
тации головного отдела у полихет (Eteone longa — Phyllodocidae;
Glycera capitata — Glyceridae) представляют собой вторичное яв-
ление. Известен случай, когда сегментация простомиума возникает,
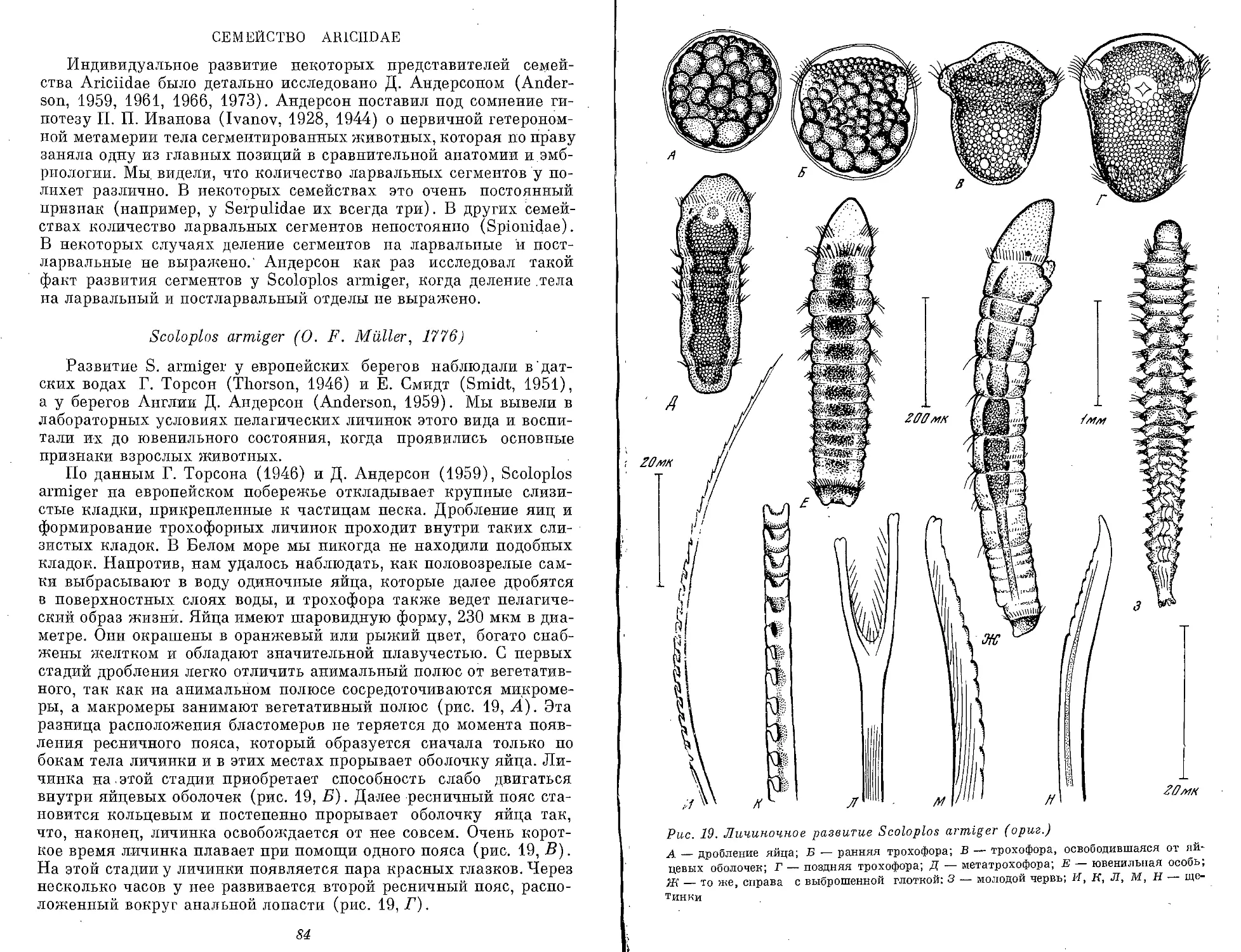
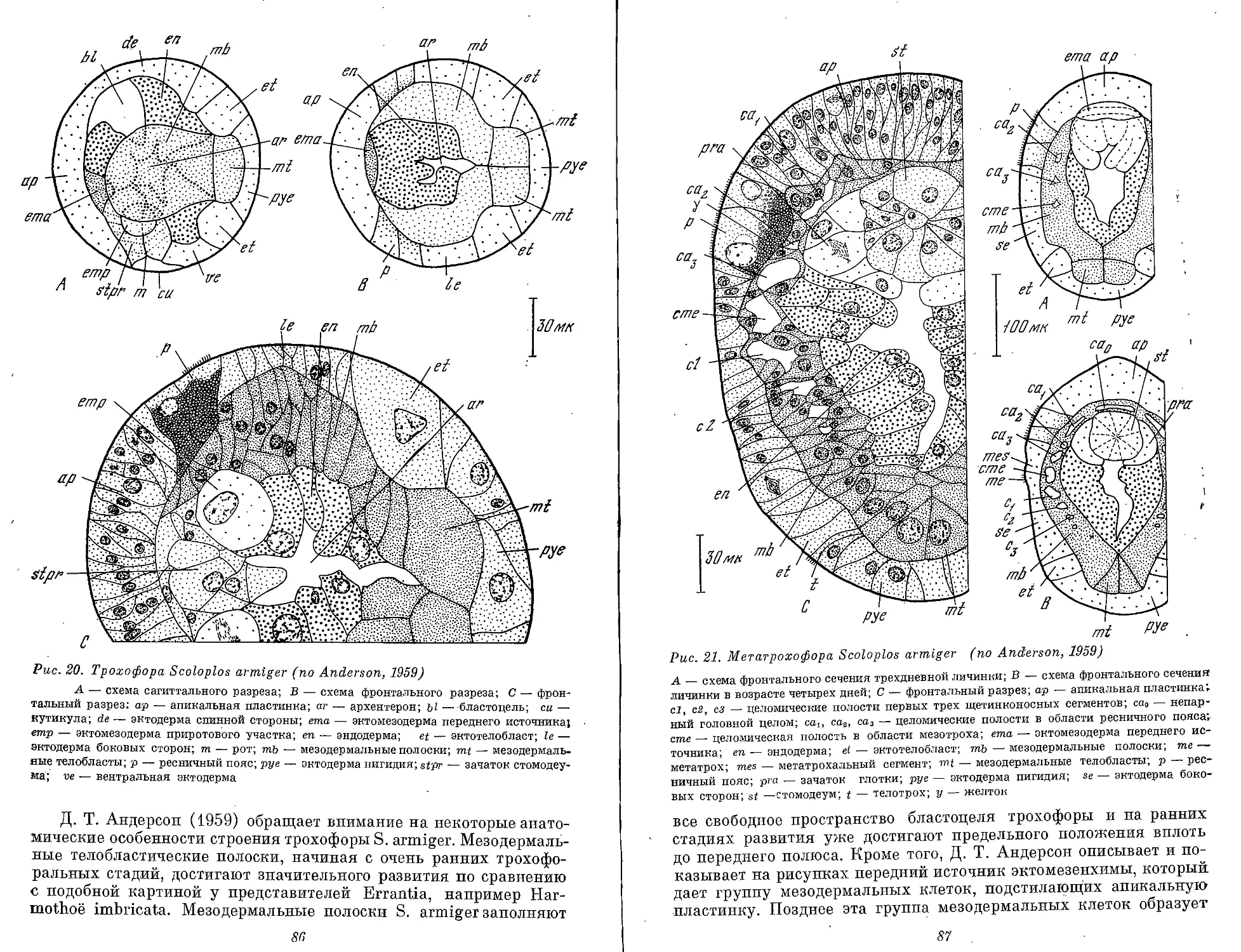
па стадии метатрохофоры Scoloplos armiger и также несет призна-
ки вторичного процесса (см. рис. 21,В).
Ларвальный отдел возникает на стадии метатрохофоры. Рото-
вое отверстие личинки располагается в первом (ротовом) сегмен-
те (перистомий) ларвального отдела. В процессе развития к рото-
вому сегменту могут примыкать несколько следующих ларвальных
и постларвальных сегментов и образовать сложный сегмент (ме-
тастомий). Ларвальный отдел состоит из определенного числа
сегментов, специфичного для отдельных семейств полихет. Коль-
чатость сегментов возникает экзогенно сначала в эктодерме, а за-
тем проникает глубже в мезодерму. Однако полость ларвально-
го тела обычно остается несегментированиОй. В ларвальных сег-
ментах не бывает половых желез, целомодуктов и хлорагогенных
клеток. После возникновения и полного развития ларвального от-
дела обычно наступает пауза, в течение которой новые сегменты
не образуются (П. П. Иванов, 1944). Число ларвальных сегментов
можно определить методом воспитания личинок в лабораторных
условиях, ориентируясь на эту паузу. В отдельных случаях лар-
вальные сегменты подвергаются частичной или полной редукции
или смешиваются с постларвальными (Anderson, 1959).
Постларвальный отдел возникает после паузы. Образование
постларвального отдела происходит путем субтерминального роста.
На переднем крае анальной лопасти образуется кольцеобразная
зона нарастания. Клетки этой зоны непрерывно размножаются,
и впереди образуется участок, состоящий из молодых, быстро ра-
стущих тканей. Кпереди от этого участка формируются новые сег-
менты тела. Каждый сегмент имеет четкие границы, пару парапо-
дий, пару брюшных ганглиев, правый и левый сомиты. Обособле-
ние мезодермальных сомитов происходит самостоятельно, а не под
влиянием метамерии эктодермы, как в ларвальном отделе. Мезо-
дермальные полоски нередко оказываются уже поделенными на
сегменты вблизи самой зоны роста, где эктодерма еще не сегмен-
тирована. В постларвальном отделе каждая система построена ме-
тамерно: В каждом сегменте повторяется определенный набор нер-
вов — невромера или невросомит; определенный набор кровенос-
ных сосудов — ангиомера или ангиосомит; определенный набор
мышц — миомера или миосомит и других систем (выделительная,
половая).
Таким образом, и в развитии, й в строении ларвальный и пост-
18
ларвальный отделы различаются весьма резко. П. II. Иванов (Iva-
nov, 1928, 1944) обозначает это различие как первичную гетеро-
номность метамерии членистых животных.
Итак, туловище полихет состоит из ларвального и постларваль-
ного отделов. Ларвальный отдел образуется на протяжении нижне-
го полушария трохофоры (на протяжении гипосферы). Постлар-
вальный отдел возникает позади ларвального.
Анальная лопасть — это, как и головная лопасть, отдел, не под-
вергающийся сегментации (кроме редких случаев вторичной сег-
ментации) . У трохофор, метатрохофор и нектохет с двумя реснич-
ными кольцами (прототрох и телотрох) анальная лопасть ограни-
чена телотрохом. Анальная лопасть у личинок несет зачатки уси-
ков, а у некоторых форм глаза.
СПОСОБЫ РАЗМНОЖЕНИЯ
И РАЗВИТИЯ
Чаще всего размножение происходит половым путем. Нередко
встречается метагенез (Syllidae, Eunicidae, Spionidae, Cirratulidae).
Бесполое размножение часто связано с эпитокией, которая сопро-
вождается значительными морфологическими изменениями той об-
ласти тела, где созревают половые продукты. Эпитокия хорошо
выражена у Glyceridae, Syllidae, Hesionidae, Nereidae, Eunicidae,
Spionidae, Cirratulidae, Chloraemidae, Scalibregmidae, Opheliidae.
В простейшем случае, когда отсутствуют специальные протоки
для выведения созревших половых клеток, эпитокия захватывает
все тело червя. Этот случай мы называем полной эпитокией. Весь
червь поднимается со дна в толщу воды, выметывает половые про-
дукты через разрывы стенки тела и после этого погибает. Полная
эпитокия встречается у представителей Glyceridae и Nereidae. Сле-
дует заметить, что осложненный способ размножения, описанный
Ф. Хемпельманом (Hempelmann, 1911) у Platynereis dumerilii
(приведен у Ливанова, 1940, стр. 103), был повторно исследован
К. Хауеншильдом (Hauenschild, 1951), который показал ошибоч-
ность данных Ф. Хемпельмана. В действительности две половые
формы Р. dumerilii следует рассматривать как два самостоятель-
ных вида: гетеронереидную, как у Р. dumerilii, и нереидную аток-
ную, как у Р. masciliensis. У некоторых Syllidae (Exogoninae)
выведение половых продуктов происходит без нарушения целост-
ности стенки тела. Тогда материнские особи после рождения по-
томства возвращаются к жизни на дне, а мужские особи гибнут.
Кроме полной эпитокии у полихет может быть также и непол-
ная эпитокия. Мы различаем неполную эпитокию, которая сопро-
вождается архитомией, и неполную эпитокию, которая сопровож-
дается паратомией. В том случае, когда неполная эпитокия
сопровождается архитомией, эпитокные изменения захватывают
только заднюю часть тела (Eunice viridis — лалоло). Эта часть от-
19
рывается, всплывает в толщу воды, выметывает половые продукты
и гибнет. Атокная часть остается на дне, регенерирует недостаю-
щий задний конец и приготавливается к новому циклу.
Когда неполная эпитокия сопровождается паратомией, задолго
перед отделением эпитокной части происходит образование голов-
ного конца с придатками у эпитокной части. Эта последняя, отде-
лившись, представляет собой самца или самку, приспособленных к
пелагическому существованию и способных к сложным актам поло-
вого поведения, которое включает активный поиск самцом самки и
танец с определенной последовательностью движений (семейство
Syllidae, подсемейство Autolytinae) (Свешников, 1968). Самцы и
самки имеют вторичнополовые признаки и по своему строению зна-
чительно отличаются от атокных особей. После выбрасывания по-
ловых продуктов около самки самцы гибнут. Самки откладывают
яйца в выводковую сумку, расположенную на брюшной стороне те-
ла. Во время пелагического существования самки не питаются. Ки-
шечник у них не развит. Они удерживаются в поверхностном слое
воды, где насыщение кислородом заметно выше, и движением па-
раподий вентилируют пространство около выводковой сумки. Пос-
ле того как молодь выходит из сумки, самки погибают.
Кроме таких случаев размножения у полихет встречается раз- .
множение, не связанное с эпитокией. Половые продукты тогда вы-
водятся через нефридиальные протоки (Phyllodocidae, Aphroditi-
dae и др.), и после размножения самцы и. самки сохраняют жизне-
способность.
В редких случаях у полихет встречается гермафродитизм.
К. Хауеншильд (Hauenschild, 1954) опроверг общепринятое мнение
о принадлежности Capitomastis minimus (Capitellidae) к раз-
дельнополым животным и опытным путем доказал, что этот вид
следует относить к гермафродитам, способным в исключительных
случаях к самооплодотворению. Хауеншильд относит к гермафро-
дитам также Capitella capitata.
Живорождение встречается у Syllis vivipara (Syllidae). Парте-
ногенез известен у Dodecaceria concharum (Cirratulidae) и Grubea
pusilia (Syllidae) (Hauenschild, 1955, 1959).
Осуществление жизненного цикла полихет может происходить
при помощи свободных личинок и эпитокных стадий. Несвободные
личинки, которые проходят развитие под прикрытием оболочек и
активно не питаются в течение всего личиночного цикла, среди
полихет встречаются редко (Amphicora fabricii).
У пелагических полихет (Phyllodocidae, Alciopidae, Tomopte-
ridae, Thyphloscolecidae и некоторых Aphroditidae) весь жизнен-
ней цикл протекает в толще воды. У форм, имеющих эпитокию,
развитие может протекать двумя способами: в одном случае раз-
витие сопровождается свободной эпитокной формой и свободной
личинкой (Glyceridae, Hesionidae, Nereidae, Eunicidae), в другом
случае развитие сопровождается свободной эпитокной формой и
личинками, которые развиваются в выводковой сумке. Однако в
20
; последнем случае на очень короткое время личинки все же выхо-
дят в толщу воды. Они имеют приспособления к пелагическому
образу жизни и способны активно выбирать подходящий субстрат
для оседания. Видимо, неверно было бы считать таких личинок
несвободными.
Наконец, у многих полихет, не имеющих эпитокии, личиночное
развитие протекает свободно в толще воды и связано с активной
добычей пищи. Однако этот признак сам по себе еще не определяет
характер свободного развития. Например, пелагические личинки
Scoloplos armiger, Chone infundibuliformis во время пелагической
жизни не питаются, существуя за счет эмбрионального желтка.
Этот факт может служить примером того, что обилие желтка в яйце
не всегда приводит к подавлению свободного пелагического разви
тия.
Свободное непелагическое развитие характеризуется тем, что
личинки, освободившиеся от оболочек, ведут свободный образ жиз-
ни в придонных слоях воды или на дне (Arenicola, marina, Nicolea
zostericola, Cirratulus cirratus). Таким образом, при свободном
непелагическом способе развитйя личинки не попадают в . планк-
тон, а активно добывают себе пищу на дне. Когда и эти моменты
активной жизни по каким-либо причинам оказываются подавлен-
ными, развитие становится несвободным или прямым.
Прямое развитие у полихет встречается редко (Amphicora fab-
ricii). Частным случаем прямого развития можно считать живоро-
ждение (Syllis vivipara).
ПЕРИОДЫ, ФАЗЫ
И СТАДИИ ЖИЗНЕННОГО ЦИКЛА
Жизненный цикл вида можно разделить на эмбриональный и
постэмбриональный периоды. Изучение эмбрионального периода
в жизненных циклах полихет не входило в задачу моего Исследова-
ния. Особенность эмбрионального периода состоит в том, что с мо-
мента оплодотворения дробящееся яйцо, а затем эмбрион развива-
ются внутри оболочек. После выхода личинки из оболочек насту-
пает постэмбриональный период, который можно разделить на
фазы. В жизненном цикле полихет, как и у многих морских бес-
позвоночных, постэмбриональный период распадается на личиноч-
ную, ювенильную фазы и на фазу половозрелого состояния; Каж-
дую фазу можно разделить на стадии. ,
ОСНОВНЫЕ СТАДИИ ЛИЧИНОЧНОИ ФАЗЫ
Гаструла. Обычно у полихет постэмбриональный период и
личиночная фаза начинаются с момента выхода из яйцевых оболо-
чек трохофоры, способной к самостоятельной жизни. Однако в
редких случаях из оболочек выходит личинка на значительно более
21
I
I
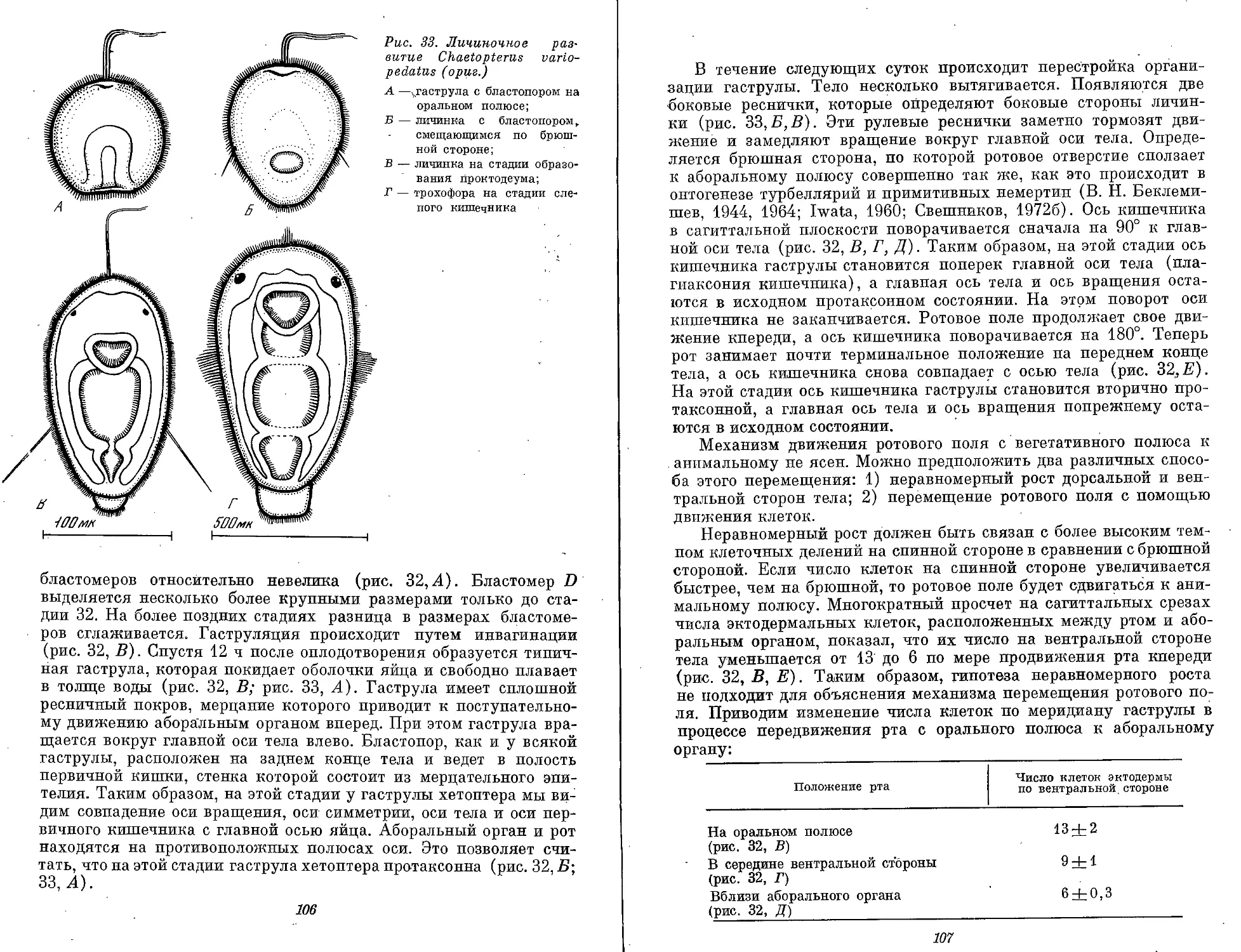
ранних стадиях развития. Например, у Chaetopterus variopedatus
из Японского моря план строения свободноплавающей гаструлы
(см. рис. 32) отвечает плану строения гаструлообразных личинок
кишечнополостных, турбеллярий и немертин. Это двуслойный ор-
ганизм с ярко выраженной гетерополярной протаксонией. Бласто-
пор находится на заднем (оральном) полюсе против аборального
органа. Движение осуществляется при помощи сплошного мерца-
тельного покрова. Полость первичного слепого кишечника также
выстлана мерцательным эпителием. Степень дифференциации и
концентрации нервной системы гаструлы полихет отвечает уровню
развития диффузной нервной системы гаструлообразных личинок
Других типов беспозвоночных. .
Протрохофора. План строения протрохофоры полихет отвечает :
плану строения высших сколецид. Сквозной кишечник. Рот сме- ;
щен вперед от орального полюса, анальное отверстие расположено
терминально. У протрохофоры накоплено ощутимое количество
мезенхимной ткани, которая еще не эпителизирована. Ресничный
покров равномерно одевает поверхность личинки. Нервная система
по степени дифференциации не отличается от таковой у гаструло-
образных личинок. Бластоцель развит слабо и выглядит как узкая
щель между кишечником и стенкой тела. Никаких дефинитивных
признаков протрохофора еще не имеет. В отдельных случаях про-
трохофора переходит в стадию трохофоры (Chaetopterus). Но
чаще протрохофора превращается в ювенильную донную форму
(Chone, Lumbriconereis).
Трохофора — личинка с мерцательным поясом по экватору.
Мерцательный пояс состоит из очень крупных клеток с длинными
ресничками, которые обеспечивают поступательное движение ли-
чинки. На анимальном полюсе — теменной хохолок и глаза. В эк-
тодерме личинки расположена система экваториальных и меридио-
нальных нервно-мышечных стволов. Шарообразное или уплощен-
ное тело разделено ресничным поясом на эписферу и гипосферу.
Кишечник слепой или сквозной. Первичная полость тела. Прото-
нефридии. Трохофора может быть с одним и двумя ресничными
поясами. Передний пояс называется прототрох. Он отделяет верх-
нее полушарие от нижнего. Рот обычно находится в нижнем
полушарии. У трохофоры Chaetopterus (мезотроха или стомария)
рот находится в верхнем полушарии. У личинок Spionidae рот
перемещается к анимальному полюсу на более поздних стадиях.
У многих трохофор свободных полихет (Errantia) прототрох — это
единственный ресничный пояс. Таких трохофор называют моно-
трохными. Монотрохное состояние может быть отдельной стадией
и затем переходит в стадию дитрохного состояния. Дитрохная тро-
хофора имеет одновременно передний и задний ресничные поясы.
Задний пояс называют телотрохом. Телотрох ограничивает аналь-
ную лопасть (пигидий). Оба ресничных пояса ограничивают ту
часть ларвального тела, которая на стадии метатрохофоры подвер-
гается ларвальной и постларвальной сегментации. У трохофоры
22
нет еще никаких дефинитивных признаков. План строения трохо-
форы полихет отвечает плану строения некоторых групп высших
сколецид, например коловраток, а также личинок некоторых низ-
ших групп моллюсков, например боконервных (Amphineura) и
двустворчатых, например Taxodonta.
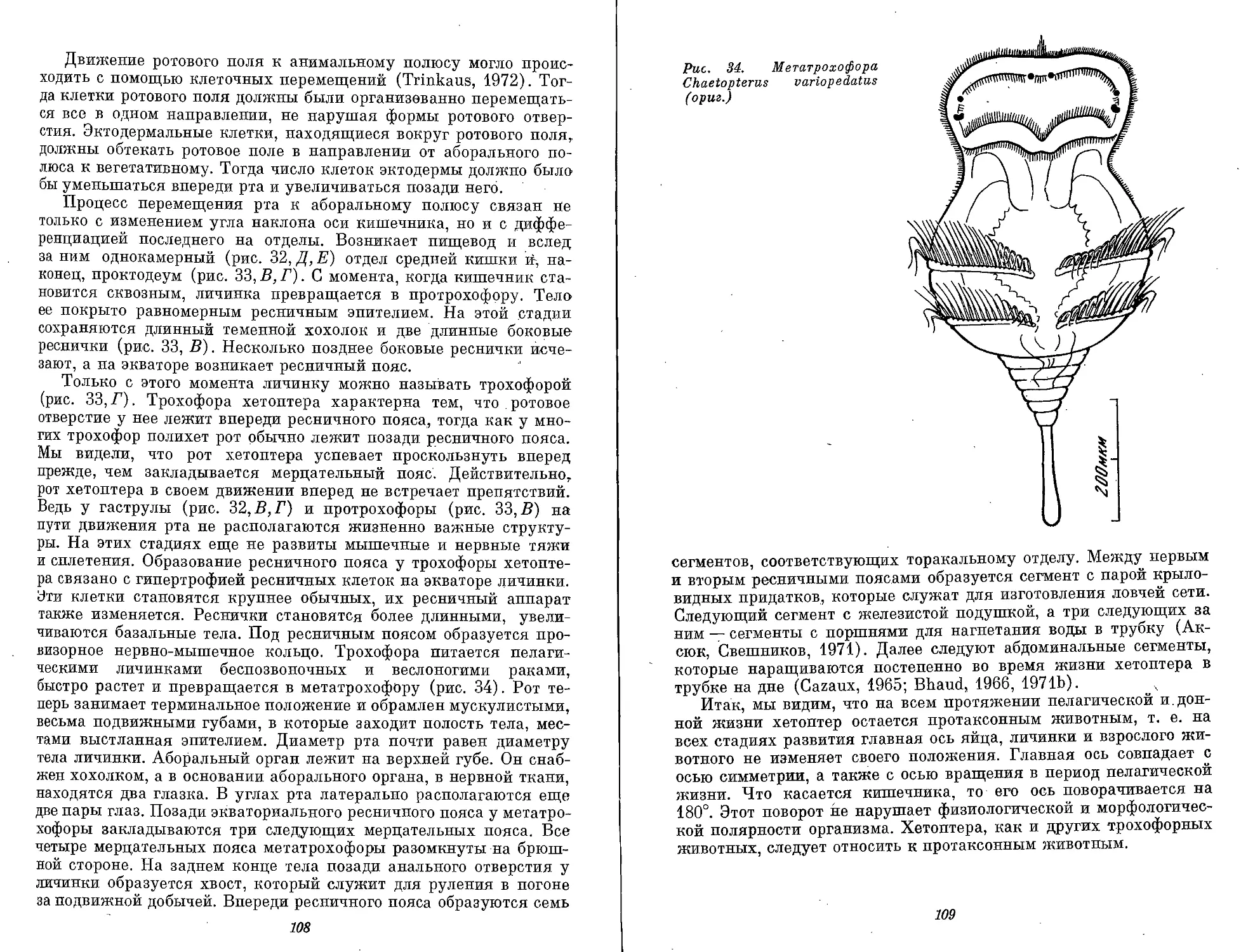
Метатрохофора может считаться наиболее типичной личинкой
полихет, потому что у нее выражена метамерия ларвального тела,
столь характерная для кольчецов. План строения метатрохофоры
весьма сложен. Метатрохофора имеет сегментированное ларваль-
ное тело. Сегментация может быть выражена в структурах экто-
дермального происхождения — ресничными кольцами, кольча-
тостью клеток эпителия, например разной их высотой. Сегменты
часто выражены также структурами смешанного экто- и мезодер-
мального происхождения, например параподиальный комплекс,
включающий нейро-, мио- и нефросомит. Число ларвальных сегмен-
тов очень различно. Наиболее часто повторяются числа 3, 7 и 13.
Деятельность метатрохофоры (движение, поиск пищи, реакция на
различные раздражители, миграции, бегство и т. д.) осуществля-
ется провизорным!! структурами, которые в сравнении с трохофо-
рой развиты у метатрохофоры сильнее. Дефинитивные структуры
функционируют у метатрохофоры в очень ограниченной степени.
План строения метатрохофоры отвечает плану строения некоторых
олигомерных организмов, например динофилид (класс Dinophili-
da) и науплиальных личинок ракообразных.
Нектохета — это личинка, у которой ларвальное тело приоб-
ретает дефинитивные признаки. Личинка удерживается в толще
воды уже не только ресничными поясами, но также или провизор-
ными, или дефинитивными щетинками. У нектохеты часто дефи-
нитивные признаки выражены ярче, чем провизорные, хотя мно-
гие из последних еще функционируют в полную силу. Из дефи-
нитивных признаков нектохеты наиболее обычны головные
придатки — пальпы, головные усики, затылочные органы. Парапо-
дии нектохет часто снабжены провизорными и одновременно дефи-
нитивными щетинками. Анальная лопасть нектохеты обычно несет
уриты (анальные усики). Очень важно подчеркнуть, что нектохе-
та претерпевает заметные преобразования ларвального отдела в
сравнении с метатрохофорой. Габитус нектохеты приближается
ко взрослому животному. Нектохету иногда удается (особенно по
щетинкам) определить по ключам для дефинитивных животных.
Число ларвальных сегментов остается неизменным с момента их
образования у метатрохофоры. Вместе с тем план строения весьма
изменяется в связи с тем, что многие провизорные структуры ме-
татрохофоры утрачивают свое значение, и у нектохеты вступают
в действие структуры ларвального отдела — параподии, мышцы,
иногда ларвальный целом. Таким образом, планы строения мета-
трохофоры и нектохеты очень близки между собой. На стадии нек-
тохеты еще острее выражен олигомерный облик организма.
23
ЮВЕНИЛЬНАЯ ФАЗА
И ФАЗА ПОЛОВОЗРЕЛОГО СОСТОЯНИЯ
Осевшая на дно или оставшаяся в толще воды нектохета, у ;
которой появились постларвальные сегменты, переходит в юве-
нильное состояние. Молодая полихета иначе движется, начинает
питаться другой пищей и другим способом, чем на личиночных
стадиях. Поэтому она иначе и по-новому реагирует на раздраже-
ния. Головной и брюшной мозг испытывают новую фазу развития.
Постларвальные сегменты постепенно нарастают, тело увеличива- i
ется в размерах. Прежние (мелкие) щетинки заменяются более ;
крупными. Появляются жабры, метанефридии, кровеносные\cocy- !
ды, хлорагогенная ткань. Во время превращения ювенильной !
формы во взрослую половозрелую форму у нее возникают поло-
вые железы или соответствующий аппарат. У ряда видов это со-
провождается явлением эпитокии, когда облик животного изме-
няется до неузнаваемости. У некоторых видов выражена забота
о потомстве. У других видов пелагические яйцеклетки обладают
защитными приспособлениями.
ОБЩАЯ МОРФОЛОГИЯ
И АНАТОМИЯ ЛИЧИНОК ПОЛИХЕТ
Для определения видовой принадлежности личинок полихет прихо-
дится принимать во внимание признаки, значительная часть кото-
рых утрачивается после фиксации. Среди комплекса признаков
особо весомы характеристики, связанные с расположением реснич-
ных полей, которые хорошо заметны только на живых объектах.
Провизорные и дефинитивные щетинки нектохет служат надеж-
ными признаками, а пигментация может варьировать в широких
пределах.
МОРФОЛОГИЯ
Ресничные поля у личинок полихет могут быть двух типов. Рес-
ничные поля, приспособленные для поступательного движения,
и ресничные поля тактильного свойства.
К ресничным полям, осуществляющим функцию осязания,
относятся апикальный султан ресничек, а также акротрох и так-
тильные реснички, расположенные на эписфере или простомиуме,
на анальной лопасти или в составе ресничных колец.
Ресничные поля, осуществляющие функцию движения, можно
разделить на собственно кинетические и гидрокинетические. Пер-
вые осуществляют движение самой личинки. Это прото-, мезо-, те-
ло, ното- и гастротрохи. Вторые служат для привлечения новых
масс воды к телу и дистанционным органам чувств, для доставки
кислорода, удаления отторгнутых отбросов и получения информа-
ции о запахах. Это реснички в области затылочных органов, в об-
ласти ротового поля, ресничной ямки и невротрохоид. Последний
иногда берет на себя двигательную функцию. Ресничные поля у ли-
чинок полихет — надежный систематический признак (Hannerz.
1956).
Глаза. У пелагических личинок всегда развита одна или не-
сколько пар глаз. Некоторые виды в развитии утрачивают глаза.
Обычно это роющие формы. У других провизорные глаза заменяют-
ся дефинитивными. Чаще всего глаза, возникшие у трохофоры, про-
должают служить и во взрослом состоянии. Количество глаз, их
строение и расположение — вполне постоянный и надежный систе-
матический признак. Глаза могут быть простыми и сложными.
Сложные глаза собраны группами на простомиуме или на жабрах.
25
Одиночные, или простые, глаза, как и сложные, могут быть с хру-
сталиками и без них.
Пигментация. У личинок полихет развита: пигментация двух
типов. Во-первых, пигменты, тонко распыленные внутри клеток
тела. Во-вторых, пигменты в виде меланофоров, хроматофоров и
зерен меланина или пигментов другого свойства в эпидермисе.
Часто на ранних стадиях пигмент развит слабее, а на более позд-
них очень интенсивно. Однако при переходе к существованию на
дне он исчезает или заменяется другим.
Личиночные провизорные щетинки. Личинки, которые ведут
длительный пелагический образ жизни, снабжены специальными
провизорными, щетинками. Они увеличивают площадь опоры и
уменьшают остаточный вес. Например, личинки семейства Oweni-
idae — митрарии — снабжены двумя длинными пучками провизор-
ных щетинок, которые сбрасываются в момент катастрофического
метаморфоза. Очень характерны провизорные щетинки у личинок
семейств Amphinomidae, Spionidae, Magelonidae, Disomidae, Sabel-
lariidae и, наконец, у личинки подкласса Myzostomida.
Личиночные дефинитивные щетинки. Наряду с провизорными
у личинок одновременно могут быть развиты и дефинитивные ще-
тинки. Они часто располагаются в ларвальных сегментах, которые
у взрослых порой не должны нести щетинок. Однако специализи-
рованные дефинитивные щетинки закладываются очень рано и мо-
гут служить хорошим признаком для идентификации вида. Напри-
мер, специализированные щетинки 5-го сегмента у Polydora или
специализированные щетинки торакального отдела Chaetopterus.
ТИПЫ ЛИЧИНОК
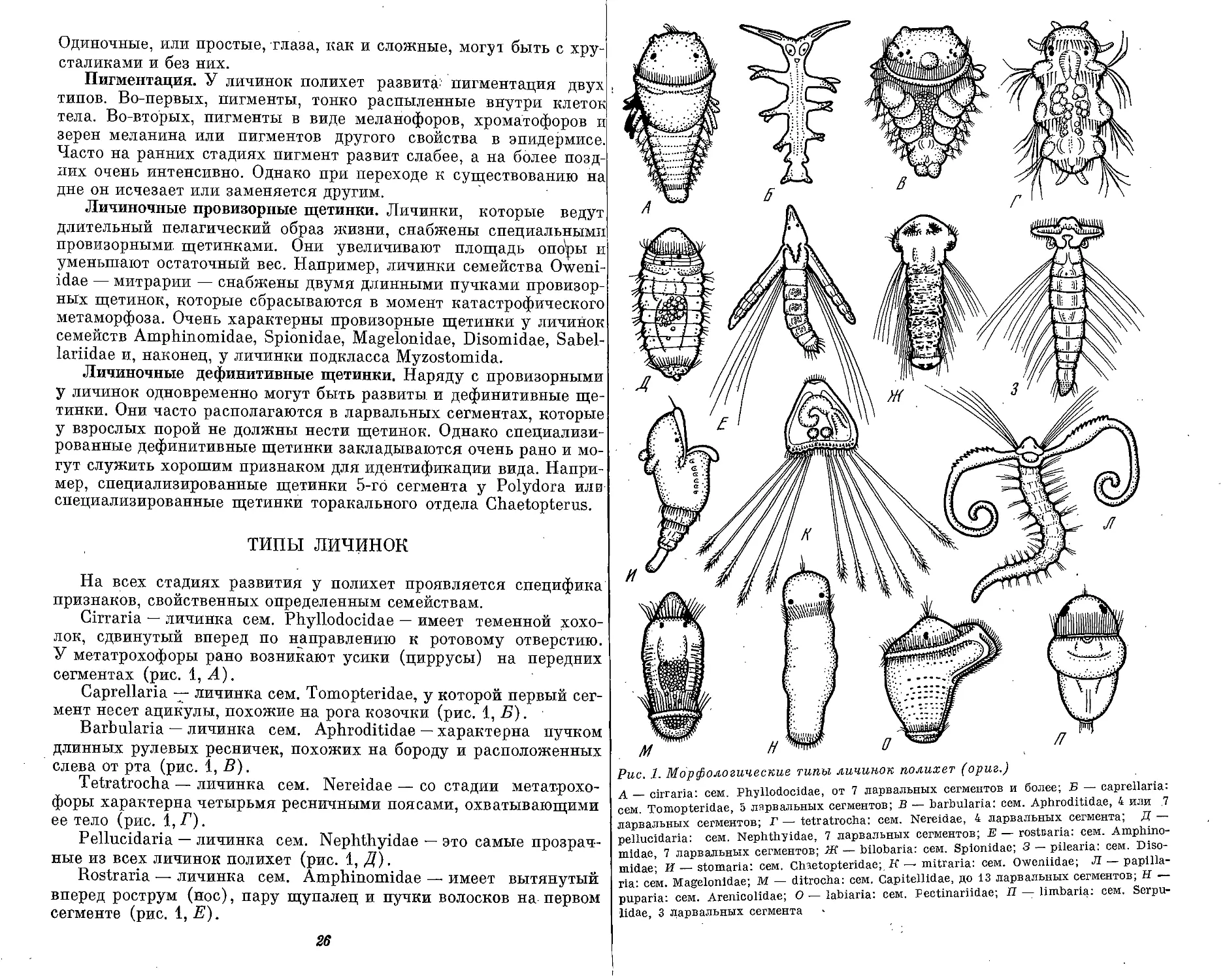
На всех стадиях развития у полихет проявляется специфика
признаков, свойственных определенным семействам.
Cirraria — личинка сем. Phyllodocidae — имеет теменной хохо-
лок, сдвинутый вперед по направлению к ротовому отверстию.
У метатрохофоры рано возникают усики (циррусы) на передних
сегментах (рис. 1, А).
Caprellaria — личинка сем. Tomopteridae, у которой первый сег-
мент несет ацикулы, похожие на рога козочки (рис. 1, Б).
Barbularia — личинка сем. Aphroditidae — характерна пучком
длинных рулевых ресничек, похожих на бороду и расположенных
слева от рта (рис. 1, В).
Tetratrocha — личинка сем. Nereidae — со стадии метатрохо-
форы характерна четырьмя ресничными поясами, охватывающими
ее тело (рис. 1, Г).
Pellucidaria — личинка сем. Nephthyidae — это самые прозрач-
ные из всех личинок полихет (рис. 1, Д).
Rostraria — личинка сем. Amphinomidae — имеет вытянутый
вперед рострум (нос), пару щупалец и пучки волосков на первом
сегменте (рис. 1, Е).
26
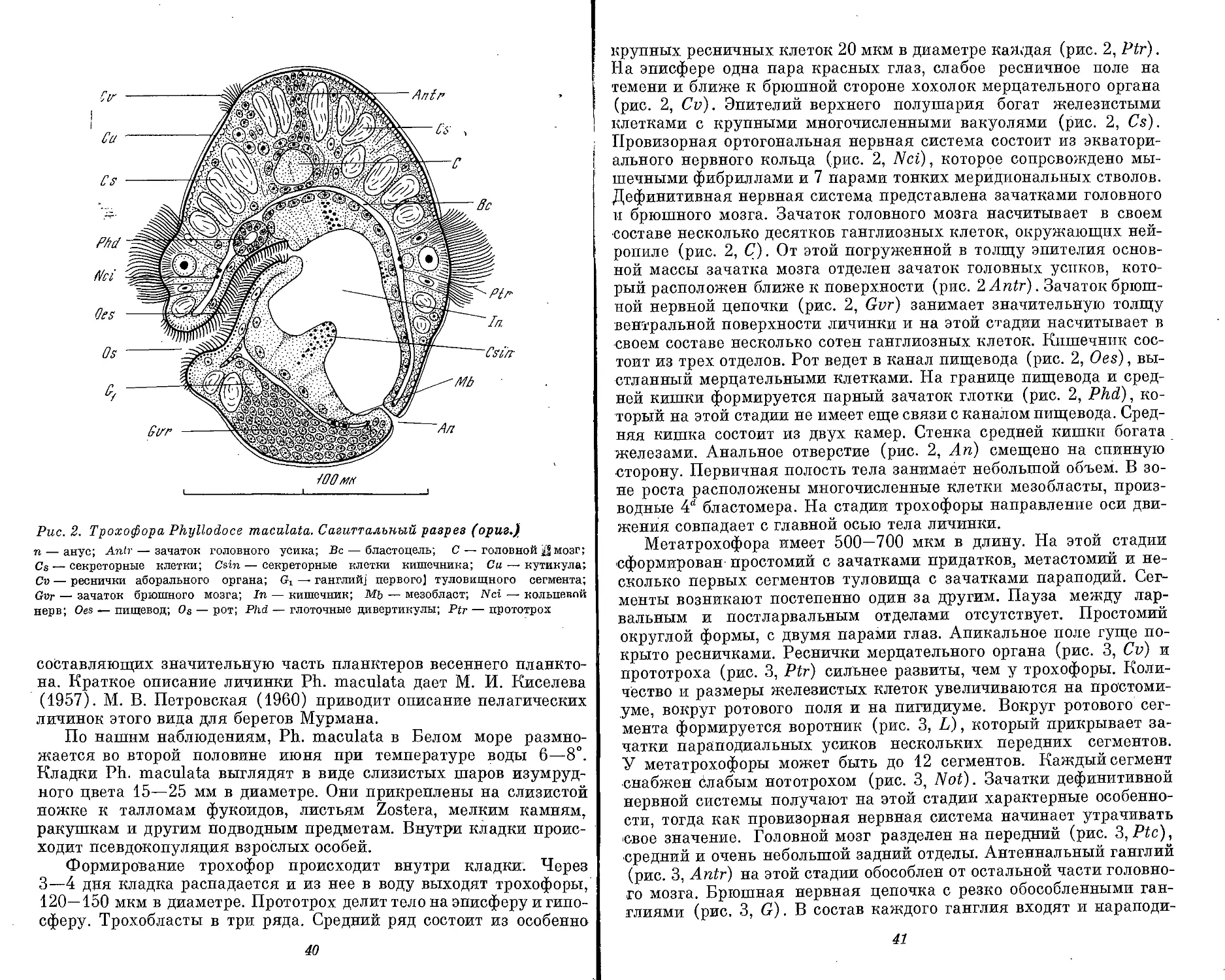
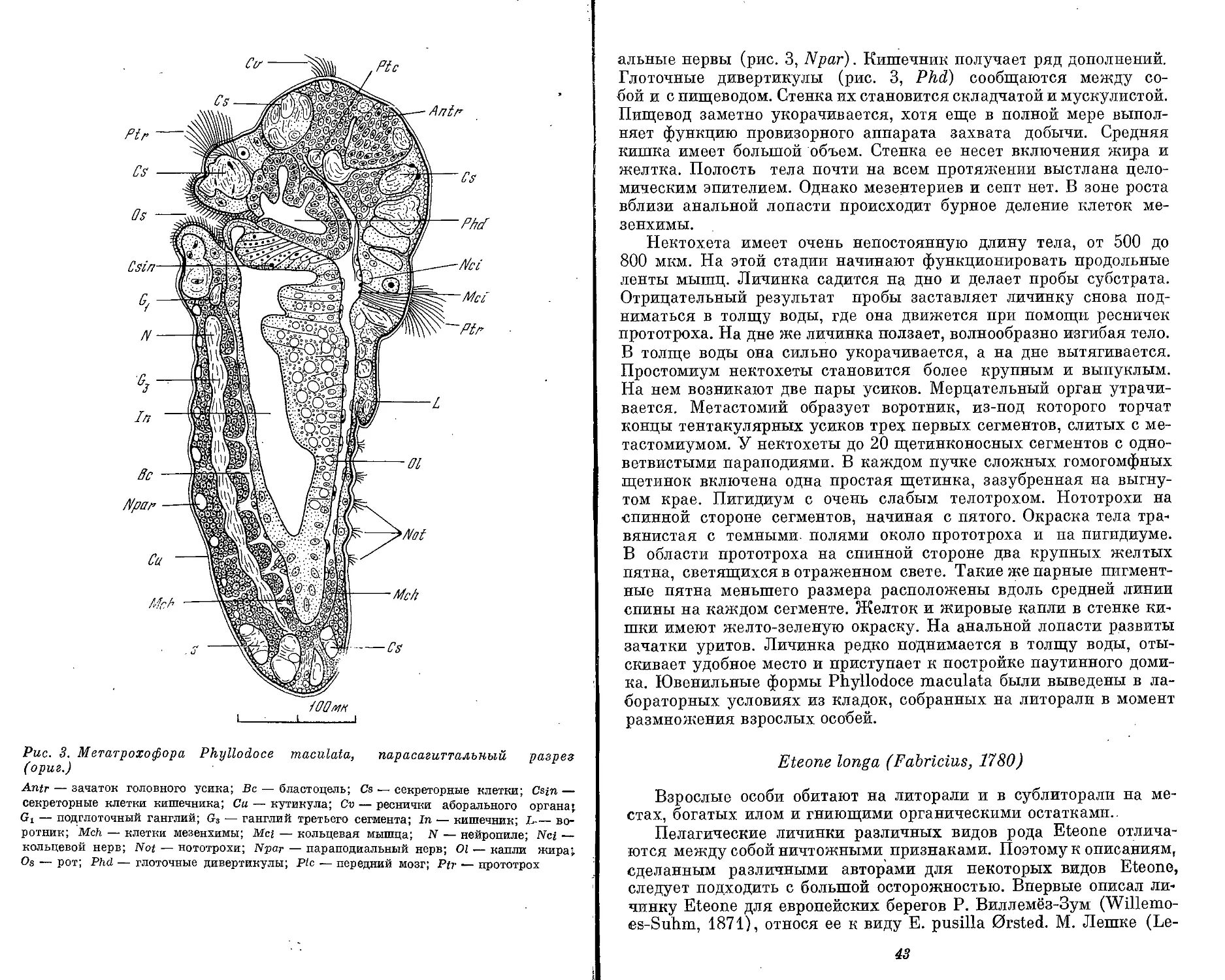
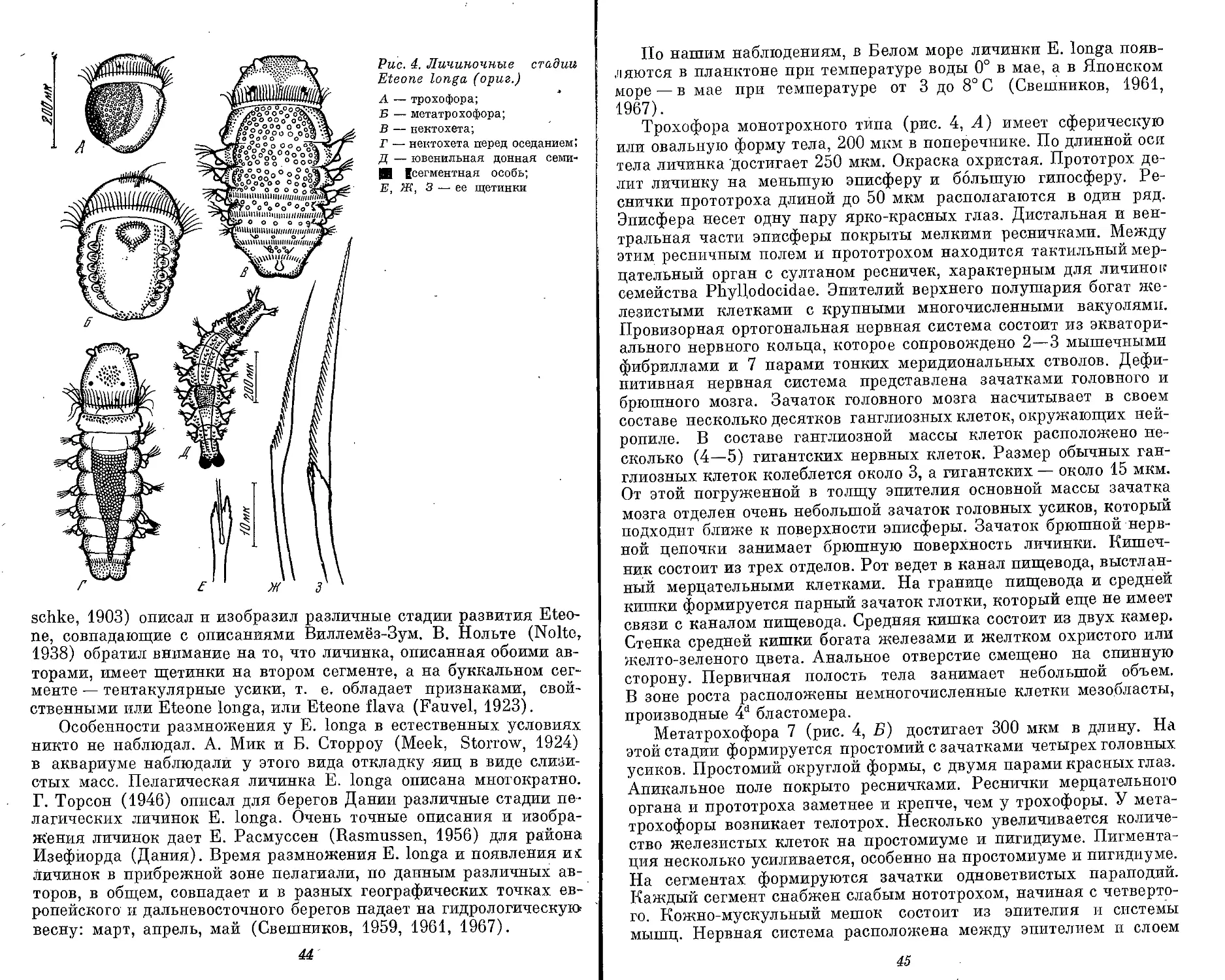
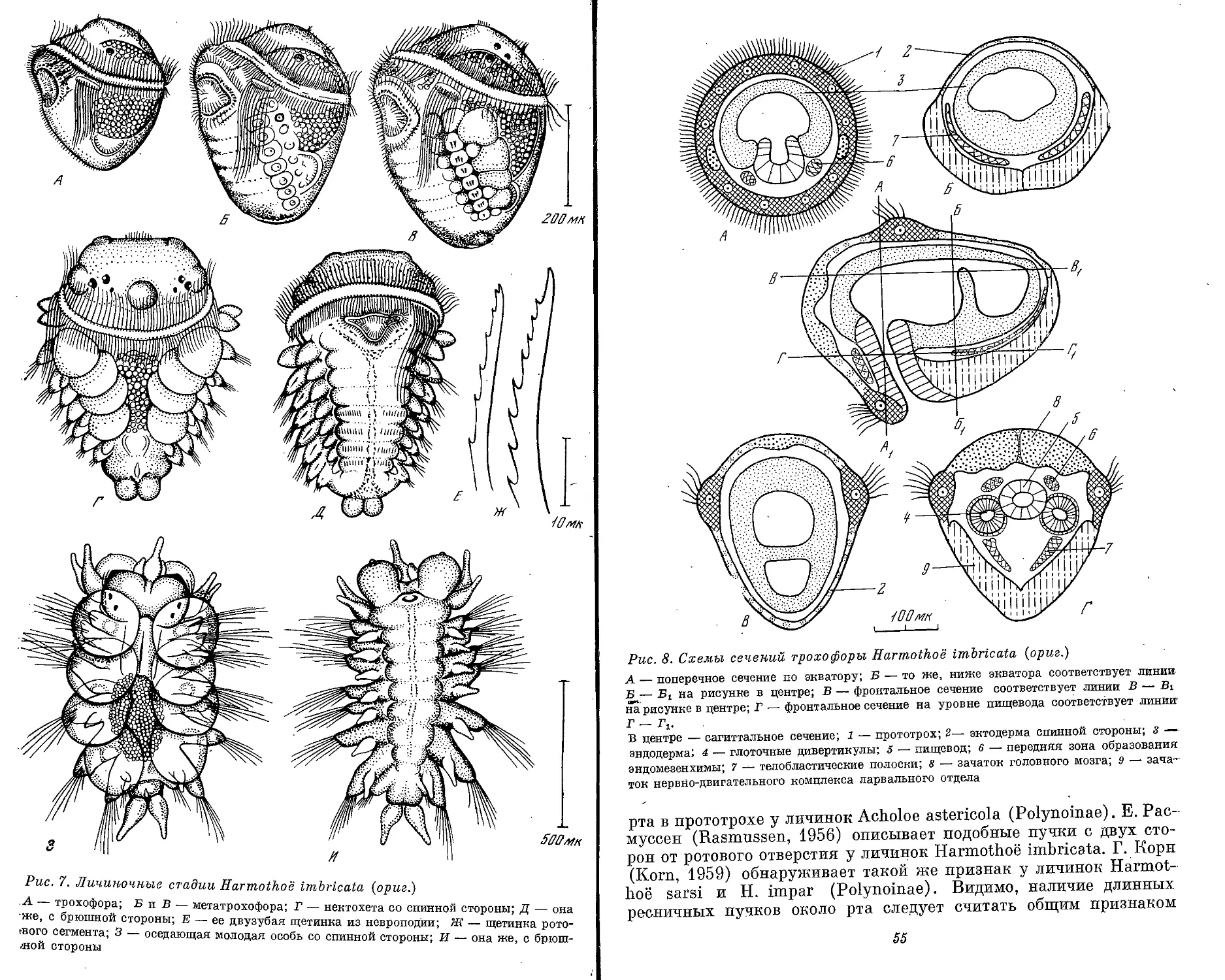
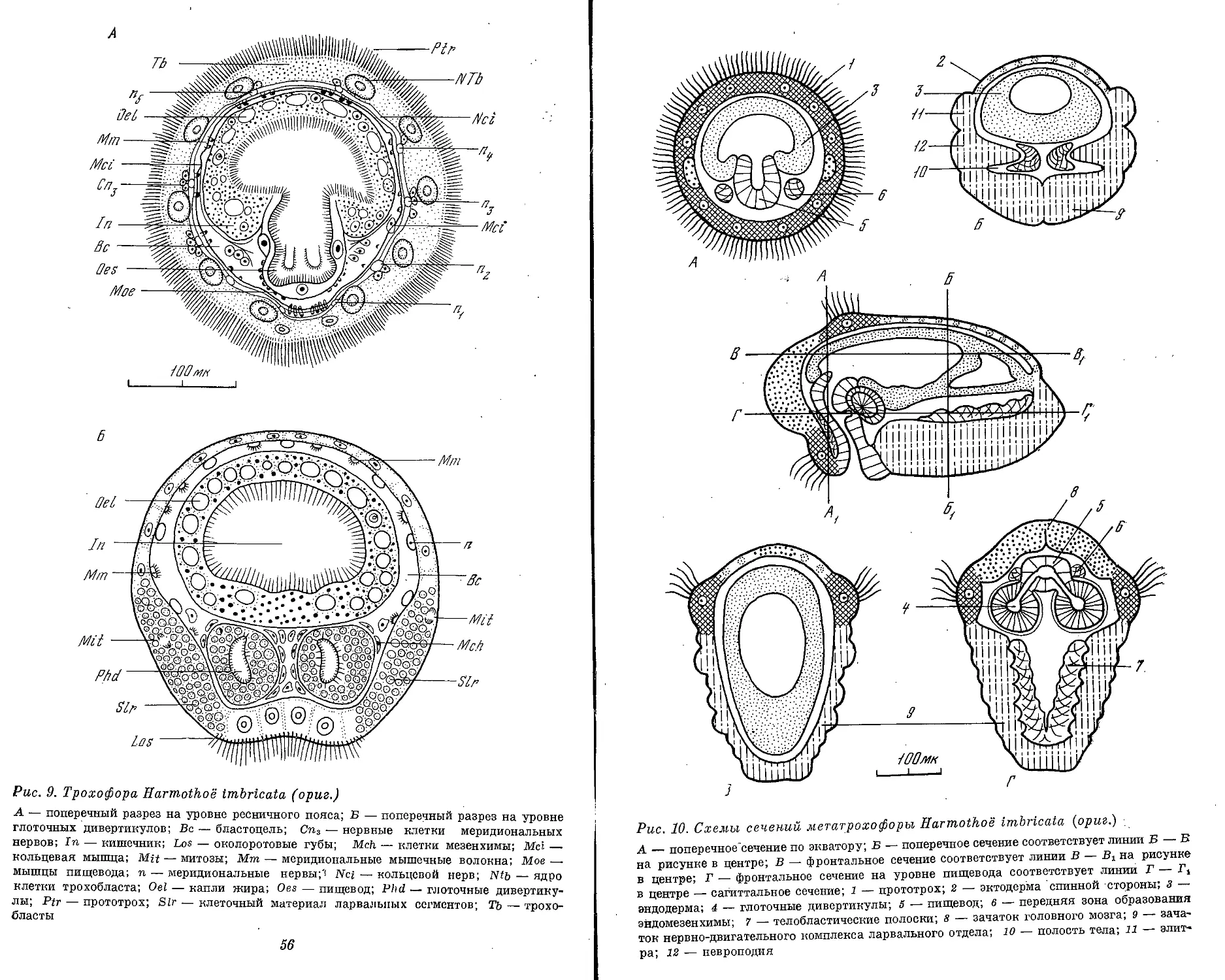
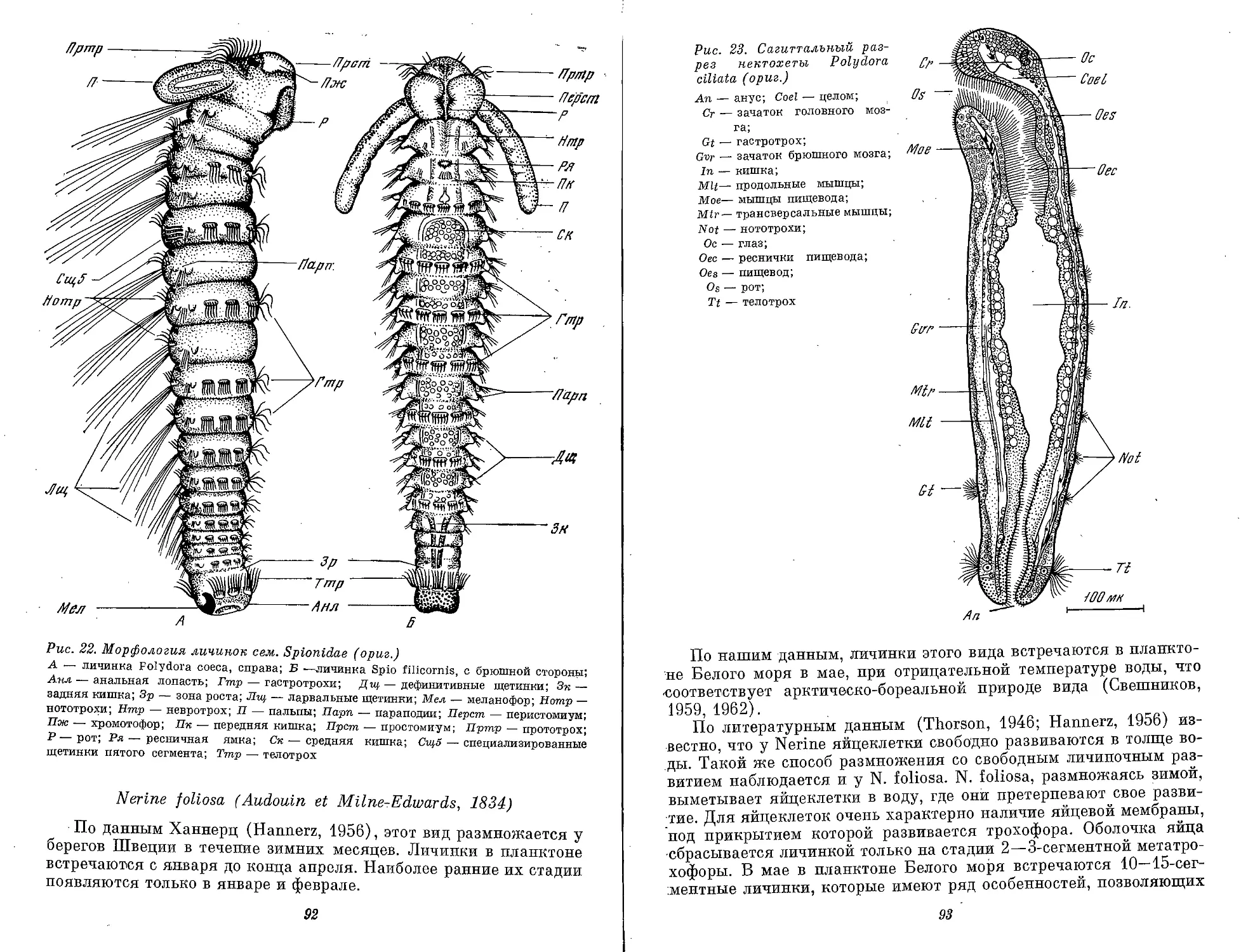
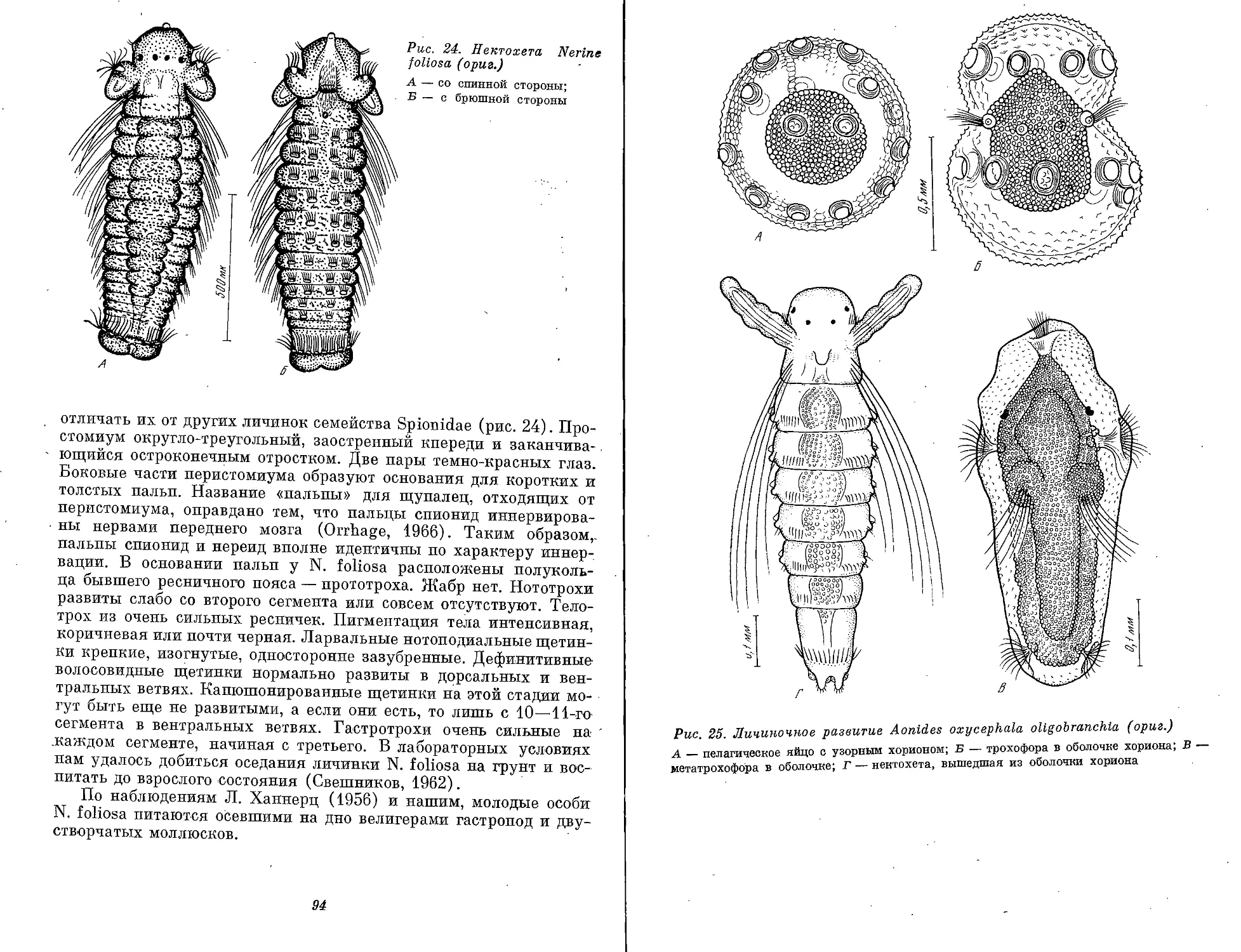
Рис. 1. Морфологические типы личинок полихет (ориг.)
А — cirraria: сем. Phyllodocidae, от 7 ларвальных сегментов и более; Б — caprellaria:
сем. Tomopteridae, 5 ларвальных сегментов; В — barbularia: сем. Aphroditidae, 4 или .7
ларвальных сегментов; Г — tetratrocha: сем. Nereidae, 4 ларвальных сегмента; Д —
pellucidaria: сем. Nephthyidae, 7 ларвальных сегментов; В — rostoaria: сем. Amphino-
midae, 7 ларвальных сегментов; Ж — bilobaria.' сем. Spionidae; 3 — pilearia: сем. Diso-
midae; И — stomaria: сем. Chaetopteridae;. К — mitraria: сем. Oweniidae; Л — papilla-
ria: сем. Magelonidae; М — ditrocha: сем. Capitellldae, до 13 ларвальных сегментов; Н —
puparia: сем. Arenicolidae; О — labiaria: сем. Pectinariidae; П — limbaria: сем. Serpu-
lidae, 3 ларвальных сегмента
(пис 1 Ирленты, имеют строение «аскаридных» клеток с крупными трофи-
’ 'I ______ ттт ттт T/-> X<т ttttttilt r\^Y\Q □ЛТТПТ Г ТТН WFTRTTT
спе А аПа сем. Spionidae — обладают нескольким^^олов с ганглиями в каждом сегменте. Степень дифференциа-
ц фичными приспособлениями, из которых особенно характер^™ невросомита у разных представителей очень разнообразна.
обесп тастомиальные Усики (лобные доли), провизорные щетинки! Кожно-мускульный мешок личинок полихет состоит из покров-
и Р^.зветрленные меланофоры (рис. 1, Ж)РОГ°, железистого и ресничного эпителия, кольцевой и продоль-
.ной мускулатуры. Кольцевая мускулатура развита заметно слабее
-<xUJ vpxxo. л, хх7. продольной. Продольная мускулатура расположена или четырьмя
Chaetopteridae — своеобрР°Д°льными лентами (чаще у Errantia) либо сплошным тонким
юлоем (чаще у Sedentaria). На стадиях метатрохофоры и нектохе-
цы мышечные клетки, особенно клетки, составляющие продольные
----- --х- - - ж • •
ческими мешками. Параподиальные мышцы образуют сложный
^комплекс, осуществляющий тонкие движения поиска добычи, по-
стройки домика, оборонительные и другие движения.
j Первичная полость тела по объему очень различна у разных
вличинок полихет. Объем полости тела находится в обратной зави-
симости от скорости движения и в прямой зависимости от продол-
жительности пребывания личинки в планктоне.
Метамерия целома в ларвальном теле, как правило, отсутству-
ет. Парные протонефридиальные органы выделения личинок ме-
няются на органы выделения метанефридиальные или смешан-
ные.
Кишечник личинок состоит из трех отделов. Передний отдел
включает пищевод и парные карманы зачатка дефинитивной глот-
ки. Средний отдел — эндодермальная кишка — всегда состоит из
двух камер. Задний отдел — прямая кишка.
Кровеносная система развита не у всех полихет, но там, где
она есть, всегда связана с появлением постларвального тела.
Pilearia — личинка сем. Disomidae — имеет расширение голов
ной лопасти, похожее на широкополую шляпу (рис. 1,3). i
Mesotrocha (Stomaria) — личинка сем. С' . _____
разна тем, что рот ее находится в верхнем полушарии и на стадит
метатрохофоры становится необыкновенно широк. Мозг стомарш
занимает ничтожное место на верхней губе личинки (_
Mitraria — личинка сем. Owenidae — формой похожа на мит|
ру. Парит в воде при помощи двух пучков провизорных щетинок;
Катастрофический метаморфоз (рис. 1,77). f
Papillaria — личинка сем. Magelonidae — характерна своеобраз-1
ными ротовыми щупальцами, покрытыми рядами сосочко!
(рис. 1, Л).
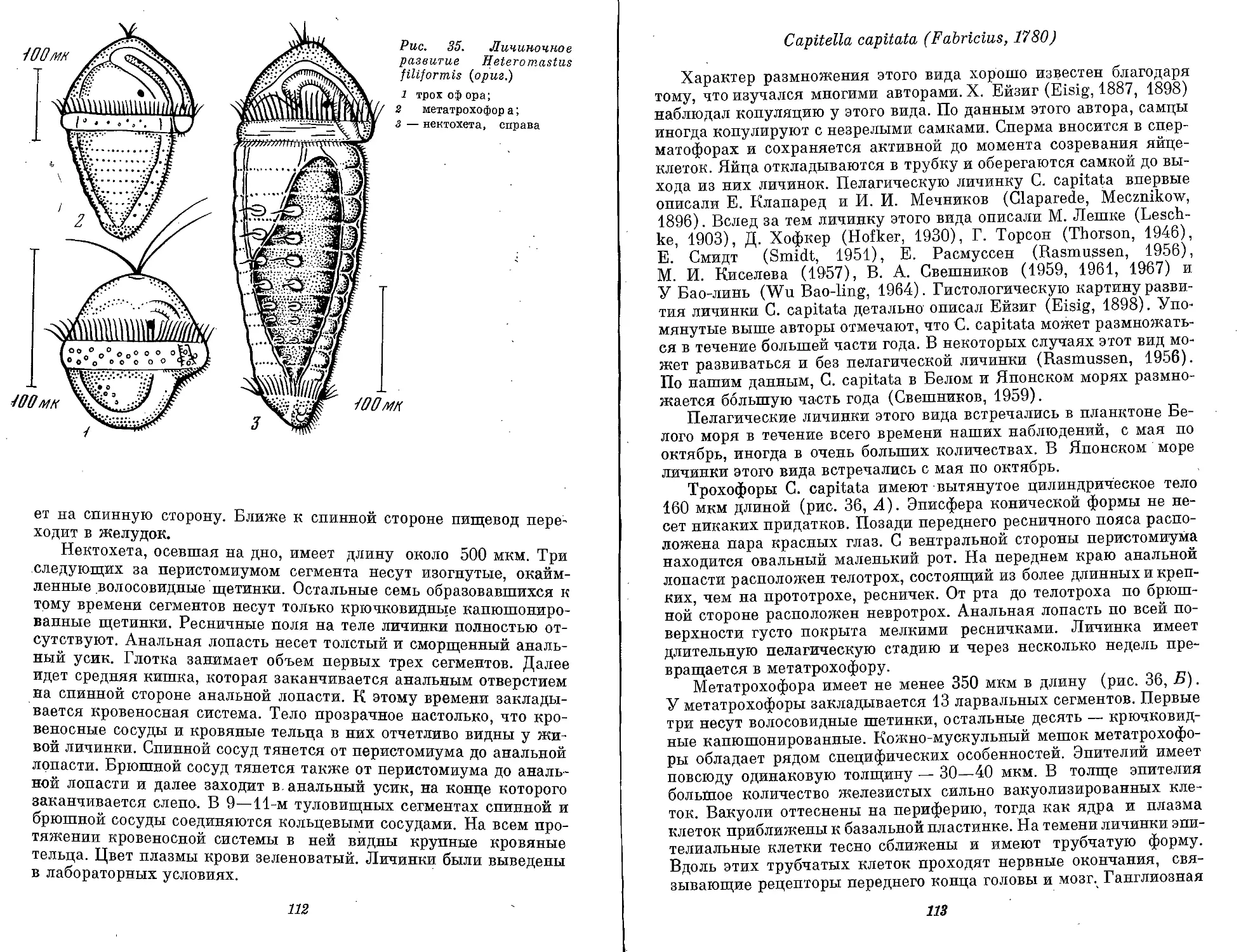
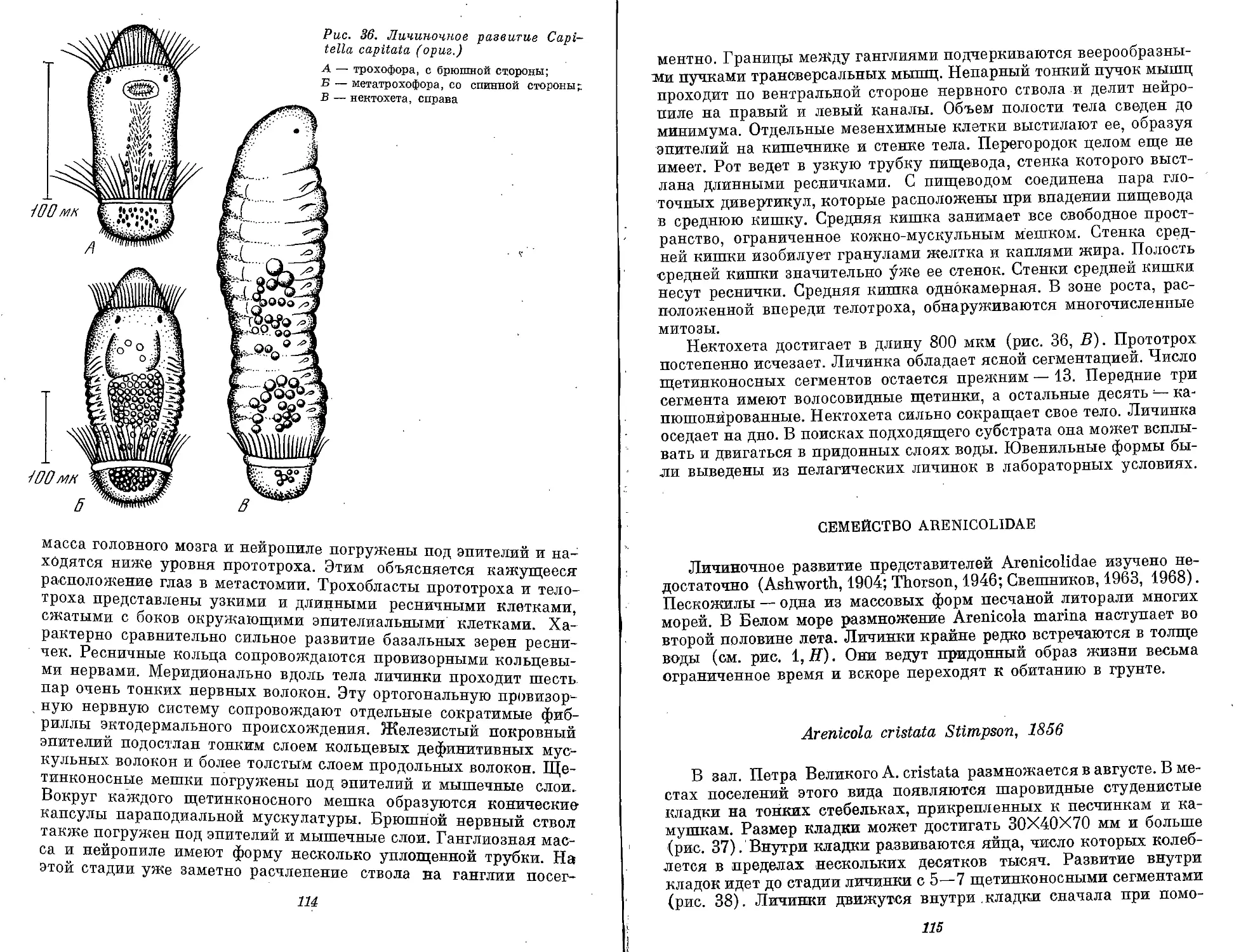
Ditrocha — личинка сем. Gapitellidae — имеет на всех стадиям
два ресничных пояса. Дорсальное тело имеет 13 сегментов
(рис. 1, М).
Puparia — личинка сем. Arenicolidae — ведет придонный медли-
тельный образ жизни и напоминает куколку (рис. 1, Н).
Labiaria — личинка сем. Pectinariidae — характерна околорото-
. выми губами, способными сильно сокращаться и вытягиваться
Пигмент расположен рядами точек (рис. 1, О).
Limbaria — личинка сем. Serpulidae — отличается железистым
валиком вокруг головы, похожим на воротник, отложенный на
вентральную сторону (рис. 1,77).
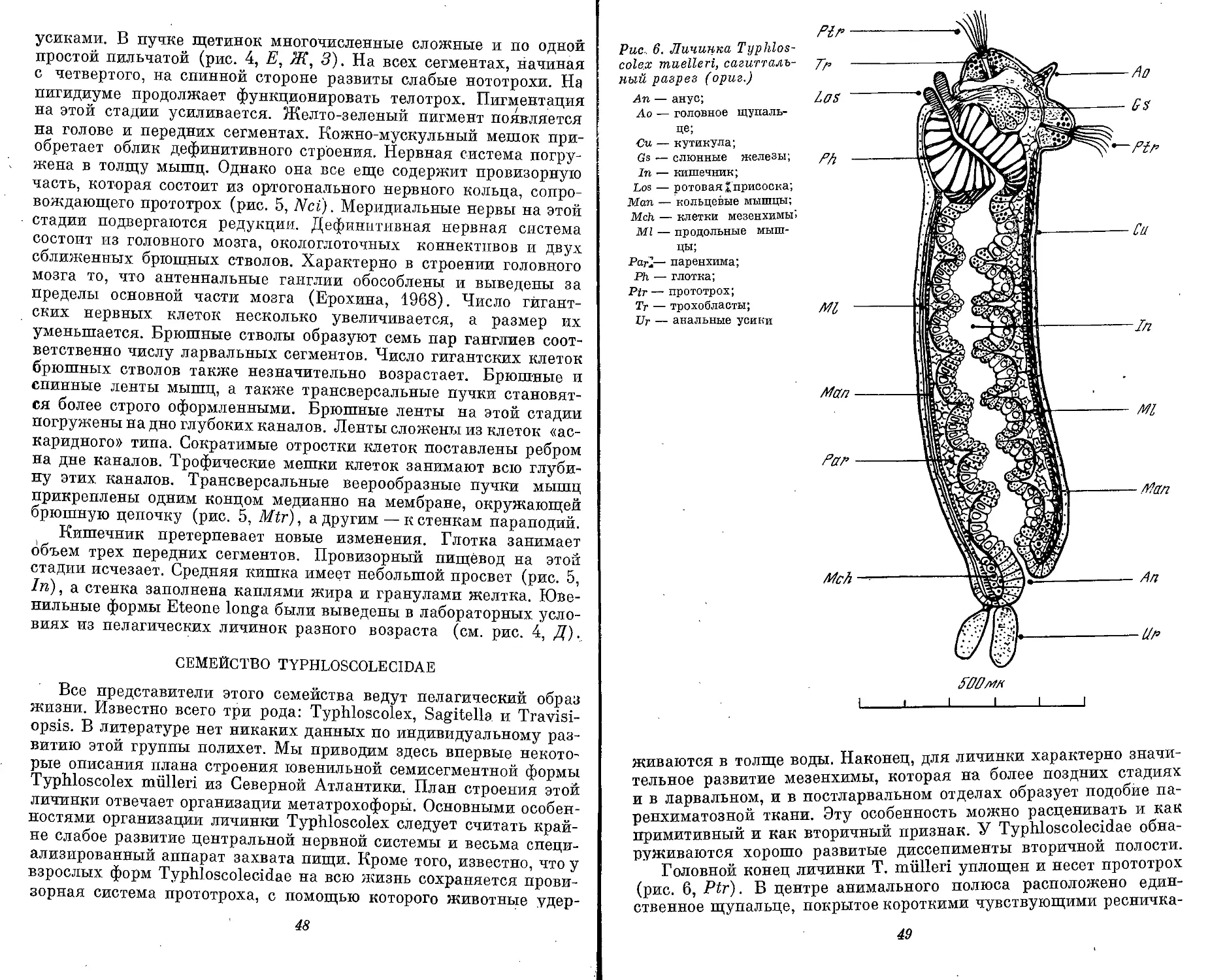
АНАТОМИЯ
В процессе метаморфоза полихет резко меняется весь план
строения. Несегментированное тело становится сегментирован-^
ным. Ортогональная нервная система трохофоры превращается в
сложно дифференцированную дефинитивную нервную систему.
Первичная полость тела замещается вторичной. Протонефридии
заменяются метанефридиями. Появляются гонады.
Полихеты имеют очень различно развитую нервную систему.
Чем сильнее дифференцирована нервная система во взрослом со-
стоянии, тем раньше появляются в процессе развития соответст-
вующие закладки. Например, стебельчатые тела головного мозга
закладываются уже у метатрохофоры Aphroditidae (Ерохина,
и др., 1970). В противоположном случае у личинок семейства Pec-
tinariidae головной мозг оказывается более дифференцированным,
чем у взрослых форм. Например, у метатрохофоры Pectinaria го-
ловной мозг поделен на три отдела, а у взрослых это деление сгла-
живается и исчезает. Брюшная нервная цепочка состоит из двух
широко расставленных (Chaetopterus) или тесно сближенных
28
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА НЕКТОХЕТ
ДО СЕМЕЙСТВА
:9(8)
; 10(H)
1(4)
2(3)
3(2)
4(1)
5(6)
6(5)
7(14)
8(9)
Прототрох и полные ресничные кольца на сегментах от-!
сутствуют. На брюшной стороне сегментов иногда развиты
слабые ресничные полукольца. Телотрох также разомкнут
на спинной стороне.
Голова оттянута в острый рострум. По бокам головы за-)Ц(Ю)
чатки гладких щупалец. Только ротовой сегмент несет' 12 (13)
пару пучков тончайших волосовидных провизорных щети-
нок, длина которых в 3—4 раза превосходит длину личин-
ки. Число сегментов до 7. Дефинитивные щетинки в Дор-
сальных и ] "
щетинки часто
вентральных ветвях. Дорсальные и вентральные 13(12)
сто разветвленные. Личинка типа fostrariaj
. . ....................... Amphinomidae (рис. 1, Е)
Голова без рострума. Зачатки щупалец по бокам головы с
рядами папилл. Ротовой и все следующие сегменты с пуч-1
ками длинных провизорных щетинок. Число сегментов 15
и больше. Дефинитивные щетинки на передних восьми
сегментах волосовидные. На 9-м сегменте щетинки шпа-
телевидные. G 10-го сегмента щетинки крючковидные,
двузубые. Личинка типа papillaria ...................
....................... Magelonidae (рис. 1, Л).
Прототрох и другие полные ресничные кольца развиты.
Личинка другого типа.
Рот расположен терминально на верхнем (анимальном)
полюсе личинки. Тело нектохеты поделено на 2 или 3 от-
дела. Передний — торакальный отдел позади ограничен
первым ресничным кольцом. В торакальном отделе 7—10
сегментов. На 4-м сегменте крупные специализированные]
щетинки. Средний и задний отделы разграничены вторым
ресничным кольцом. В этих отделах с двуветвистыми па-
раподиями брюшные щетинки в виде микроскопических
гребенчатых пластинок. На спинной стороне головного
конца два коротких придатка. Задний конец с непарным'
анальным придатком. Личинка типа stomaria ............
...............................Chaetopteridae стр. 104).
Рот расположен на перистомиуме (под прототрохом).
Личинка иного типа.
Только одно ресничное кольцо.
Личинка типа mitraria. Из-под митры вниз радиально на-
правлены 2 пучка тонких ларвальных щетинок. Также
вниз от основания пучков щетинок направлено свисаю-
щее дефинитивное тело. Дефинитивные щетинки в форме
разводного гаечного ключа. До 10 сегментов .........
.............................Oweniidae (рис. 7, К).
14(7)
15(28)
16(17)
Личинка иного типа.
В кольце прототроха, с левой стороны от рта, пучок длин-
ных рулевых ресничек. Придатки головы чаще в непарном
числе — 3 или 5. Число ларвальных сегментов от 4 до 8.
Придатки параподий в виде шаровидных, дисковидных или
кеглевидных придатков. Щетинки сложные или сабле-
видные. Личинка типа barbularia........................
.............................. Aphroditidae (стр. 51).
Личинка иного типа.
Головная лопасть плоская, с теменным хохолком. На сег-
ментах могут быть круглые, листовидные пластинки. Ще-
тинки короткие, когтевидные............................
Typhloscolecidae (стр. 48).
Головная лопасть седловидная без теменного хохолка. Ро-
товой сегмент несет направленные в стороны остроконеч-
ные придатки со стержневой щетинкой внутри. Раздвоен-
ные параподиальные лопасти выступают в стороны на
тонком теле и щетинок не несут................
............................... Tomopteridae (рис. 1, Б).
Два или больше ресничных колец.
Два ресничных кольца (прототрох и телотрох).
Тело личинки покрыто защитной оболочкой с морщинис-
той фактурой. Теменной хохолок, реснички прототроха и
телотроха продеты сквозь оболочку. Сегменты тела могут
нести провизорные пучки длинных щетинок................
.................................. Spionidae (стр. 90).
17 (16) Защитная оболочка отсутствует.
18(25) Эписфера (простомиум) с теменным хохолком.
19(20) Около рта подвижные ловчие лопасти. Вдоль ресничных
колец, на границах сегментов и на голове правильные ря-
ды черных точек. Число сегментов до 20. На спинной сто-
роне личинки в специальных, мешках. два, веерообразных
пучка больших щетинок (пали). На самых поздних стади-
ях эти специализированные щетинки выставлены наружу
впереди головы. Личинки типа labiaria........................
.............................Pectinariidae (стр. 117).
20(19) Околоротовых лопастей нет. Личинки другого типа.
21(22) Тело расчленено на головной, грудной и абдоминальный
отделы. Голова иногда с 4—6 бугорками придатков. Груд-
ной отдел вздут. На его брюшной стороне железистый во-
ротник. 3 ларвальных сегмента в составе грудного отдела.
Щетинки копьевидные. Абдоминальный отдел тонкий,
иногда из 2—3 постларвальных сегментов. Личинки часто
окрашены в оранжевые или красные тона. Личинка типа
limbaria.................................Serpulidae (стр. 122).
30
31
22(21) Тело не поделено на отделы. Личинка иного типа.
23(24) Эписфера слегка нависает над ртом. Первые 2 сегмента
без щетинок. Следующие 3 сегмента с волосовидными ще-
тинками. Крючковидные щетинки начиная с 6-го сегмен-
та................................. Capitellidae (стр. 110).
24(23) Эписфера не нависает над ртом, который мал и незаметен.
Прототрох и телотрох в виде широких поясов. Три лар-
вальных сегмента. Ротовой сегмент без щетинок. Осталь-
ные два сегмента с копьевидными щетинками. Личинка
типа puparia..........................Arenicolidae (стр. 115).
25(18) Эписфера (простомиум) без теменного хохолка.
26(27) 7 ларвальных сегментов. Дефинитивные щетинки попе-
речно-исчерченные с первого сегмента. Анальная лопасть
с непарным уритом. Личинка типа pellucidaria................
............................Nephthydae (рис. 1, Д).
27 (26) 13 ларвальных сегментов. Первые три сегмента с волосо-
видными щетинками. Капюшонированные крючковидные
щетинки с 4-го сегмента. Личинка типа ditrocha..............
. .......................... Capitellidae (стр. ИЗ).
28(15) Больше двух ресничных колец.
29(36) Эписфера (простомиум) с зачатками головных придатков
или расширена в виде широкополой шляпы.
30(31) Эписфера (простомиум) расширена в виде широкополой
шляпы. Прототрох кольцевой. 7—10 сегментов с прови-
зорными щетинками. Личинка типа pilearia....................
..................................Disomidae (рис. 1,3).
31(30) Эписфера (простомиум) не расширена. Личинка иного
типа.
32(33) Эписфера (простомиум) с двумя зачатками головных при-
датков в виде шаровидных лобных вздутий или с двумя
длинными щупальцами. Прототрох в виде двух изолиро-
ванных секций в основании щупалец. Число сегментов до '
20. Личинка типа bilobaria..................................
....Spionidae (стр. 90).
33(32) Эписфера (простомиум) с четырьмя или пятью зачатка- (
ми головных придатков.
34(35) 3—6 сегментов тела. На голове иногда 2 больших черных
пятна, экранирующих мозг. Внутри глотки челюсти. Ще-
тинки гомогомфные или гетерогомфные. 2 урита...............,
.......................................................... . . . Nereidae. (стр. 66).
35(34) 7—20 сегментов тела. На голове или на сегметах иногда
лимонно-желтые большие пятна. Челюстей внутри глотки
нет . . . ............................Phyllodocidae (стр. 39).'
36(29) Эписфера (простомиум) лишена головных придатков. Ще-
тинки с 3-го сегмента — пильчатые или лирообразные.
2—4 урита................................Ariciidae (стр. 83).
ОСОБЕННОСТИ РАЗВИТИЯ
И ЛИЧИНКИ ПОДКЛАССА ERRANTIA
Деление полихет на подклассы Errantia (бродячие) и Seden-
taria- (неподвижные, сессильные) весьма условно. В самом деле,
даже среди семейств, которых обычно признают наиболее прими-
тивными, часто можно встретить представителей, обитающих в
трубках или ходах внутри грунта (Phyllodocidae, Nereidae, Glyceri-
dae, Eunicidae). Напротив, сессильные полихеты часто совершают
миграции (Arenicolidae), а в отдельных случаях (Poecilochaetidae)
они могут совершать полный жизненный цикл в толще воды. Вмес-
те с тем нет оснований сомневаться в том, что полихетам и мно-
гим другим низшим и высшим червям вообще свойственно искать
убежище в самых разнообразных естественных укрытиях. Напри-
мер, в парагастральных полостях губок живут Aphroditidae и Syl-
lidae. Амбулакральные бороздки иглокожих, мантийные складки
моллюсков, трубки крупных полихет обычно служат постоянным
убежищем в равной мере и бродячим (Aphroditidae, Nereidae, Syl-
lidae) и сессильным (Spionidae) полихетам. Однако же многие
сессильные полихеты (Chaetopteridae, Maldanidae, Sabellariidae,
Pectinariidae, Trichobranchiidae, Terebellidae, Sabellidae, Serpuli-
dae) настолько узко адаптированы к условиям обитания в специ-
альных трубках, что по сравнению с ними так называемые бродя-
чие полихеты вполне оправдывают свое название.
В жизненных циклах Errantia и Sedentaria также можно обна-
ружить отдельные факты, противоречащие справедливости деле-
ния класса на бродячих и сессильных. Например, именно среди
форм, обитающих в трубках (Eunicidae, Chaetopteridae, Sabelli-
dae), встречаются наиболее примитивные группы гаструлообраз-
пых и протрохофорных личинок. А пелагические личинки Amphi-
nomidae — «rostraria» — и Spionidae — «bilobaria» — внешне на-
столько похожи, что можно легко прийти к ошибочному выводу о
филогенетической близости этих семейств. Однако при рассмотре-
нии большого материала по циклам развития бросается в глаза зна-
чительно большая глубина адаптивных признаков, свойственных
Sedentaria, тогда как личинки и весь цикл развития Errantia проще
и приближаются к тем нормам, которые мы считаем более прими-
тивными и общими для аннелид в целом.
2 В. А. Свешников
33
Итак, наиболее характерными признаками личинок и циклО!
развития Errantia мы считаем следующие.
1. Циклы развития Errantia чаще осуществляются свободно i
в пелагических условиях.
2. У Errantia значительно чаще встречается свободная трохо
фора без каких-либо дополнительных и специальных приспособ-
лений к обитанию и добыче пищи в пелагических условиях.
3. В циклах развития Errantia значительно реже встречено яв-
ление эмбрионизации, укорочение и выпадение каких-либо отдель-
ных стадий.
ОТРЯД PHYLLODOGEMORPHA
В отряде Phyllodocidae обособляются три подотряда: Phyllo-
dociformia (Phyllodocidae, Alciopidae, Tomopteridae, Thyphlosco-
lecidae, Lacydoniidae), Glyceriformia (Glyceridae, Goniadidae, Spae-
rodoridae, Nephthyidae), Aphroditiformia (Aphroditidae, s. lato.
Pisionidae и Chrysopetalidae) (Ушаков, 1970, 1972). Подотряд
Phyllodociformia как наиболее примитивная группа в классе по-
лихет характеризуется большим разнообразием жизненных форм
по сравнению с подотрядами Glyceriformia и Aphroditiformia.
В подотряде Phyllodociformia среди пелагических семейств мы на-
ходим и весьма древние формы, например Tomopteridae (Glaessner,
1958, 1962; Глесснер, 1963), и формы, которые на всю жизнь со-
храняют личиночные признаки (Typhloscolecidae), и такую груп-
пу (Alciopidae), которая близко примыкает к современным фил-
лодоцидам. Для каждого семейства характерны свои типы личинок.
Число ларвальных сегментов у Phyllodocidae не менее 7, у Tomop-
teridae— 5, у Aphroditidae — 4 (Sigalioninae) или 7 (Polynoinae),
у Glyceridae — 7, у Alciopidae, Typhloscolecidae и Chrysopetalidae
число ларвальных сегментов не было установлено. Мы видим не-
которое однообразие в числе ларвальных сегментов у тех семейств
Phyllodocemorpha, развитие которых было изучено. Однако этот
признак на данном этапе исследования еще не может быть признан
надежным. Единственное общее свойство личинок Phyllodocemor-
pha состоит в том, что они неизменно проходят стадию трохофоры I,
которая у них имеет общий план строения, стадию метатрохофоры
с названным выше числом ларвальных сегментов и стадию некто-
хёты. Во взрослом состоянии морфология и план строения пред-
ставителей этих семейств значительно различаются. Несомненно,
близки между собой Phyllodocidae и Alciopidae, а также Aphrodi-
tidae и Chrysopetalidae. Остальные семейства имеют специфичные
приспособления или к пелагическому образу жизни (Tomopteri-
dae, Typhloscolecidae), или, напротив, к донному образу жизни
(Aphroditidae).
У представителей Phyllodociformia известно только половое
размножение. Гаметогенез протекает сначала в целотелии, а зд)
тем в полости тела родительских особей. Оплодотворение наруж)
34
ное. Эмбриогенез протекает либо целиком в пелагиали (у пелаги-
ческих форм), либо у бентических форм только частично в бентосе.
Постэмбриональное развитие Phyllodocidae чаще всего проходит с
пелагической трохофорой типа «cirraria» (Свешников, 1963).
У Alciopidae, Tomopteridae, Typhloscolecidae и некоторых Phyl-
lodocidae (Lopadorhynchinae, lospilinae, Pontodorinae) полный
цикл развития протекает в толще воды. У остальных Phyllodoci-
dae (Phyllodocinae) и Lacydoniidae самки, как правило, делают
слизистые кладки, прикрепляя их к водорослям и подводным
предметам. Из этих кладок выходят пелагические трохофоры, ко-
торые самостоятельно питаются мелкими ракообразными и личин-
ками других беспозвоночных, реже простейшими и фитопланкто-
ном. Объекты питания определяются главным образом размером.
Однако околоротовые губы личинок Phyllodocidae очень растяжи-
мы, и часто хищница способна проглотить жертву, по размеру
превосходящую ее. Интенсивно питаясь, личинка проходит далее
стадии метатрохофоры и нектохеты. Нектохета у некоторых
Phyllodocinae с большим числом сегментов (иногда 25) оседает
на дно и у некоторых видов строит паутинный домик. Половозре-
лые донные особи обитают в толще грунта или среди водорослей,
ио для оплодотворения и произведения кладок выходят в опреде-
ленные и специфичные для каждого вида места.
Размножение и развитие отдельных представителей подотряда
изучено в незначительной степени. Развитие Lopadorhynchus
исследовано наиболее детально (Kleinenberg, 1886; Мейер, 1897).
Наблюдения за размножением и развитием Phyllodocidae у берегов
Англии проводил В. Макинтош (McIntosh, 1869, 1900), у берегов
Норвегии — О. Нордгаард (Nordgaard, 1911), у берегов Фран-
ции— Ч. Гравье (Gravier, 1923). Размножение и развитие Eula-
lia viridis описал А. Мейер (Meyer, 1938). Развитие личинки Anai-
lides groenlandica и других Phyllodocidae наблюдал, у берегов Да-
нии Г. Торсон (Thorson, 1946). Развитие личинки Phyllodoce tuber-
culata в Черном море наблюдала М. И. Киселева (1957). Личинки
Phyllodoce maculata из Баренцева моря описаны М. В. Петров-
ской (1960), а из Белого моря — В. А. Свешниковым (1961). Эмб-
риология и личиночное развитие Tomopteris helgolandica детально
описаны Б. Акессоном (Akesson, 1962; Akesson, Melander, 1967).
Описание личинки Typhloscolecidae дается здесь впервые.
Постэмбриональный период развития у многих Phyllodocemor-
pha изучен более подробно, чем у представителей других отрядов
полихет. Планы строения трохофор, относящихся к семействам
Phyllodocidae, Tomopteridae и Typhloscolecidae, различны. Это сви-
детельствует о древности каждой из упомянутых групп.
Пелагические личинки Phyllodocidae на стадиях трохофоры,
метатрохофоры, а иногда нектохеты, сохраняют общий признак —
эксцентрично расположенный хохолок (реже симметрично распо-
ложенные два хохолка), который от анимального полюса сдвинут
в сторону ротового отверстия.
35 2*
Трохофора, вышедшая из оболочек яйца или из кладки, ведет
самостоятельный и активный образ жизни. Трохофоры Phyllodoci-
dae обладают только одним ресничным кольцом, и мы условно бу-
дем обозначать эту стадию — трохофора I. Форма тела шарообраз-
ная. Ресничное кольцо проходит но экватору. Пегментация у раз-
ных трохофор различна, но преобладают зеленые, желтые и оран-
жевые тона. У некоторых личинок Phyllodocinae на эписфере, a j
метатрохофор на простомиуме, воротнике и пигидиуме развиты
крупные, лимонного цвета парные пятна, отражающие падающий
свет. Глаза обычно красные, с хрусталиками или без них. Тело тро-
хофоры и метатрохофоры разделено экваториальным ресничным
поясом. Ресничный пояс составлен из крупных клеток, располо-
женных в три ряда. Средний ряд состоит из наиболее крупных кле-
ток и ресничек. Экваториальный пояс сопровождается нервно-мы-
шечным кольцом, которое пересечено меридиональными нервно-
мышечными тяжами (от 5 до 7 пар). Число меридиональных тяжей
неодинаково в верхнем и нижнем полушариях. В эписфере их от
U до 15. По брюшной стороне проходит непарный тяж, а осталь-
ные расположены симметричными парами. Все тяжи около ани-
мального полюса ветвятся и истончаются. Непарный нервно-мы-
шечный тяж раздваивается в нижнем полушарии, охватывая рото-
вое отверстие. Пара спинных мышечных тяжей в нижнем полуша-
рии охватывает анальное отверстие. Кроме экваториального
нервно-мышечного кольца у трохофоры могут быть два или три
полных или неполных кольца в верхнем и одно кольцо в нижнем
полушариях. Эти кольца расположены в плоскостях, параллельных
экватору (Мейер, 1897; Akesson, 1967а). Вся эта нервно-мышеч-
ная система ранней трохофоры имеет провизорное значение, она
развивается и усиливается у метатрохофоры, а на стадии нектохе-
ты постепенно перестает функционировать и исчезает. В толще
эктодермы анимального полюса у трохофоры происходит диффе-
ренциация клеток и образуется зачаток головного мозга. На
вегетативном полюсе формируется зачаток брюшной нервной
цепочки.
Эпителий трохофоры и метатрохофоры состоит из покровных и
многочисленных железистых клеток.
Значительный объем трохофоры занимает кишечник. Ротовое
поле переходит в узкую трубку пищевода, который состоит из кле-
ток с сильными ресничками. Вздутая часть кишки всегда состоит
из двух камер, которые сообщаются между собой щелью в перего-
родке. Обе камеры имеют ресничную выстилку. Задняя камера
открыта наружу анальным отверстием в центре гипосферы. В тол-
ще стенки кишки расположены многочисленные гранулы желтка.
У трохофор, как правило, возникает парный зачаток глотки.
Стадия метатрохофоры начинается с момента появления лар-
вальной сегментации.
У личинок рода Eteone число ларвальных сегментов равно 7.
Ротовой сегмент щетинконосных мешков не несет. Пауза выраже-
86
па продолжительной задержкой в появлении новых постларваль-
ных сегментов. Метатрохофора и нектохета претерпевают значи-
тельные морфологические изменения, а число сегментов при этом
остается постоянным.
У метатрохофор рода Phyllodoce очень характерен железистый
воротник вокруг ротового сегмента. Эпителий личинок Phyllodoce
очень богат железами (см. рис. 2, 3, Cs). С помощью секрета этих
желез осевшая нектохета строит между песчинками паутинный
домик и живет в нем первое время. Оседающая на дно нектохета
может иметь 15—20 сегментов. Сегментация метатрохофоры разви-
вается сначала в эктодерме, а затем охватывает и мезодермальные
образования. Бластоцель на этой стадии не сегментирован (Вс),
и в нем свободно расползаются мезенхимные клетки (Meh) в нап-
равлении от анального поля к анимальному полюсу.
Стадия нектохеты характерна тем, что параподии ларвального
отдела сформированы настолько, что получают подвижность, а де-
финитивные щетинки вырастают из мешков наружу и способству-
ют парению в толще воды. Личинка на этой стадии имеет габитус
взрослого животного и готова к посадке на дно. Однако у большин-
ства личинок Phyllodocinae некоторые провизорные системы все
еще сохраняют свое значение, и благодаря им нектохета ведет пе-
лагический образ жизни. Например, у нектохеты Eteone longa со-
храняются полные ресничные кольца и спинные полукольца, ор-
тогональное нервно-мышечное кольцо, в значительной части бла-
стоцель и другие провизорные приспособления. Вместе с тем у не-
ктохеты Eteone longa развиты все приспособления, необходимые
для донной жизни. Например, олигомерное, семисегментное тело,
способное к изгибанию при помощи подвижных параподий со ще-
тинками. Дефинитивная нервная система сформирована и функ-
ционирует. Так, головной мозг разделен на отделы, и каждый от-
дел дифференцирован; имеются обычные ганглиозные и гигант-
ские нейроны. Вблизи переднего и среднего отделов мозга некто-
хеты сформированы одна непарная и две парные целомические
полости (см. рис. 5, Coel). У взрослой Eteone longa головной мозг
окружен одной непарной и четырьмя парными целомически-
ми полостями (Ерохина, 1968). Брюшной мозг нектохеты раз-
делен на семь ганглиев, соответственно числу ларвальных сегмен-
тов.
Таким образом, мы видим, что у нектохеты дефинитивные сис-
темы в составе ларвального тела развиты сильнее, чем провизор-
ные, и организм, обитающий на этой стадии в толще воды, полно-
стью приготовлен к жизни на дне.
Переход к обитанию на дне у личинок Phyllodocidae сопровож-
дается многократными пробными посадками и всплытиями в поис-
ках наиболее благоприятного места. У личинок родов Phyllodoce и
Eulalia эти пробные посадки особенно многочисленны, а поиски
продолжительны. Поэтому осевшая на дно личинка часто имеет
очень большое число сегментов. Первое время ювенильная форма
37
живет в паутинном домике, а потом переходит к свободному образу
жизни или обитанию в илу (или илистом песке).
Постэмбриональное развитие некоторых пелагических Phyllo-
docidae детально изучено Н. Клейненбергом (Kleinenberg, 1886),
Э. Мейером (1897; Meyer, 1901) и В. Акессоном (Akesson, 1967а) J
Существенных различий в процессах развития и в планах строе-
ния трохофор, метатрохофор и нектохет донных и пелагических
Phyllodocidae обнаружить не удается. Более того, наблюдается оп-
ределенное сходство. Оно выражается в том, что, например, у тро-
хофор Phyllodoce и Lopadorhynchus провизорные системы удовлет-
воряют одному плану. Так называемый теменной орган — cirrus —
у личинок обоих родов, а также у трохофоры Eulalia (Лебский,
1970а) закладывается в виде парного образования. Однако потом
он развивается преимущественно только с правой стороны (у Eula-
lia развиваются обе части). Очень близка архитектоника провизор-
ных эктодермальных мышц и ортогональной нервной системы.
К этому выводу мы приходим, учитывая новые данные, получен-
ные Акессоном (Akesson, 1967а) по нефридиальной системе тро-
хофор Lopadorhynchus.
Процессы образования дефинитивных систем протекают у на-
званных представителей сходным образом. Дефинитивная нервная
и мышечная системы у Lopadorhynchus, Phyllodoce и Eteone форми-
руются весьма близкими путями. Головной мозг закладывается в
гиде парного образования вблизи анимального полюса. Зачаток
брюшного мозга также парный. Характерно для трохофоры Lopa-
dorhynchus то, что поверхность гипосферы уплощена и расположе-
на под углом к главной оси личинки. В эктодерме формируются
щетинконосные мешки, сопровождаемые параподиальными нерва-
ми и мышечными лентами. Раньше других образуются парные
продольные брюшные и спинные ленты. Несколько позднее появ-
ляются тангенциальные и параподиальные пучки мышц. Число
ларвальных сегментов и пауза после их образования у Lopador-
hynchus не установлены.
Большое сходство можно наблюдать и в процессах формирова-
ния парных глоточных дивертикул, а затем и глотки. Это сходство
выражается в том, что парные глоточные дивертикулы у Phyllodo-
cidae, для которых известно индивидуальное развитие, возникают
как слепые карманы стомодеума или пищевода. Морфологические
особенности и расположение парных глоточных дивертикул личи-
нок Phyllodocidae весьма напоминают подобные структуры и рас-
положение глоточных карманов у некоторых Archiannelida, на-
пример Dinophilus (Schimkewitsch, 1895) и Polygordius (Salensky,
1907). Наконец, закладка постларвальных сегментов происходит
одинаково у всех Phyllodocidae. Однако в этом процессе, видимо,
нельзя ждать большого разнообразия у всех тех животных, вто-
ричная полость тела которых образуется телобластически.
Индивидуальное развитие пелагических полихет, относящихся
к семействам Alciopidae и Typhloscolecidae, изучено недостаточно.
38
семейство phyllodocidae
Развитие пелагических видов протекает в толще воды. Разви-
тие донных форм, как правило, на ранних этапах (дробление, фор-
мирование трохофоры) протекает внутри слизистых кладок, при-
крепленных к водорослям и камням.
Трохофоры I формируются внутри кладок и через 2—3 дня вы-
ходят в толщу воды. Трохофоры питаются другими личинками по-
лихет, велигерами моллюсков, охотно поедают науплиусов и взрос-
лых Copepoda. Очень прожорливы, иногда с усилием натягивают
себя на крупную жертву и на время принимают очертания съеден-
ной добычи. Быстро растут и совершают метаморфоз. Форма тела
шаровидная. Размеры трохофоры меняются в пределах от 150 до
350 мкм.
Метатрохофора характерна большим числом сегментов (10—15).
Пауза, которая обычно у личинок наступает после закладки лар-
вальных сегментов, здесь незаметна или отсутствует. На голове
обычны зачатки четырех головных усиков. Непарный усик появля-
ется уже после оседания личинки на дно. Две пары глаз. Одна па-
ра в виде небольших скоплений пигмента. Вторая пара глаз с хру-
сталиками. Характерен железистый воротник на спинной стороне
ротового сегмента. Основания усиков трех передних сегментов
прикрыты этим воротником. На голове и спинной стороне воротни-
ка иногда парные крупные лимонно-желтые хроматофоры отра-
жают падающий свет.
Нектохеты крупные, многосегментные. Тело сильно растягива-
ется и сокращается. Параподиальные придатки в виде спинных и
брюшных усиков. Параподии одноветвистые. Щетинки сложные.
Эпителий переднего конца тела богат железами, которые выделяют
паутинный секрет. Личинки, осевшие на дно, строят из паутины
домики, маскированные песчинками, частицами детрита и водо-
рослями. Способы размножения и различные типы кладок описа-
ны для берегов Дании Торсоном (Thorson, 1946), для Черного моря
К. А. Виноградовым (1949), для берегов Мурмана М. В. Петров-
ской (1960). Морфология пелагических личинок описана многими
авторами (Agassiz, 1867; Claparede, Mecznikow, 1869; Thorson, 1946;
Киселева, 1957; Петровская, 1960; Свешников, 1961).
Phyllodoce maculata (Linne, 1767)
Личинки Ph. maculata на стадии метатрохофоры (10 сегментов)
были впервые описаны и изображены А. Агассицем (Agassiz,
1867) для берегов Северной Америки. А. Агассиц отмечает в ка-
честве черты, общей для метатрохофор рода. Phyllodoce, воротник
вокруг пёристомиума или, как он неудачно его называет, «кара-
пакс». Г. Торсон (1946) в датских проливах обнаружил нектохет,
39
Рис. 2. Трохофора Phyllodoce maculata. Сагиттальный разрез (ориг.)
п — анус; Antr— зачаток головного усика; Вс — бластоцель; С — головной й мозг;
Cs — секреторные клетки; Csin — секреторные клетки кишечника; Си — кутикула;
Cv — реснички аборального органа; G, — ганглий] первого) туловищного сегмента;
Gvr — зачаток брюшного мозга; In — кишечник; Мь — мезобласт; Nd — кольпевой
нерв; Oes — пищевод; Os — рот; Phd — глоточные дивертикулы; Ptr — прототрох
составляющих значительную часть планктеров весеннего планкто-
на. Краткое описание личинки Ph. maculata дает М. И. Киселева
(1957). М. В. Петровская (1960) приводит описание пелагических
личинок этого вида для берегов Мурмана.
По нашим наблюдениям, Ph. maculata в Белом море размно-
жается во второй половине июня при температуре воды 6—8°.
Кладки Ph. maculata выглядят в виде слизистых шаров изумруд-
ного цвета 15—25 мм в диаметре. Они прикреплены на слизистой
ножке к талломам фукоидов, листьям Zostera, мелким камням,
ракушкам и другим подводным предметам. Внутри кладки проис-
ходит псевдокопуляция взрослых особей.
Формирование трохофор происходит внутри кладки; Через
3—4 дня кладка распадается и из нее в воду выходят трохофоры,
120—150 мкм в диаметре. Прототрох делит тело на эписферу и гипо-
сферу. Трохобласты в три ряда. Средний ряд состоит из особенно
40
крупных ресничных клеток 20 мкм в диаметре каждая (рис. 2, Ptr).
На эписфере одна пара красных глаз, слабое ресничное поле на
темени и ближе к брюшной стороне хохолок мерцательного органа
(рис. 2, Cv). Эпителий верхнего полушария богат железистыми
клетками с крупными многочисленными вакуолями (рис. 2, Cs).
Провизорная ортогональная нервная система состоит из экватори-
ального нервного кольца (рис. 2, Net), которое сопровождено мы-
шечными фибриллами и 7 парами тонких меридиональных стволов.
Дефинитивная нервная система представлена зачатками головного
и брюшного мозга. Зачаток головного мозга насчитывает в своем
составе несколько десятков ганглиозных клеток, окружающих ней-
ропиле (рис. 2, С). От этой погруженной в толщу эпителия основ-
ной массы зачатка мозга отделен зачаток головных усиков, кото-
рый расположен ближе к поверхности (рис. 2 Antr). Зачаток брюш-
ной нервной цепочки (рис. 2, Gvr) занимает значительную толщу
вентральной поверхности личинки и на этой стадии насчитывает в
своем составе несколько сотен ганглиозных клеток. Кишечник сос-
тоит из трех отделов. Рот ведет в канал пищевода (рис. 2, Oes), вы-
стланный мерцательными клетками. На границе пищевода и сред-
ней кишки формируется парный зачаток глотки (рис. 2, Phd), ко-
торый на этой стадии не имеет еще связи с каналом пищевода. Сред-
няя кишка состоит из двух камер. Стенка средней кишки богата
железами. Анальное отверстие (рис. 2, Ап) смещено на спинную
сторону. Первичная полость тела занимает небольшой объем. В зо-
не роста расположены многочисленные клетки мезобласты, произ-
водные 4d бластомера. На стадии трохофоры направление оси дви-
жения совпадает с главной осью тела личинки.
Метатрохофора имеет 500—700 мкм в длину. На этой стадии
сформирован простомий с зачатками придатков, метастомий и не-
сколько первых сегментов туловища с зачатками параподий. Сег-
менты возникают постепенно один за другим. Пауза между лар-
вальным и постларвальным отделами отсутствует. Простомий
округлой формы, с двумя парами глаз. Апикальное поле гуще по-
крыто ресничками. Реснички мерцательного органа (рис. 3, Cv) и
прототроха (рис. 3, Ptr) сильнее развиты, чем у трохофоры. Коли-
чество и размеры железистых клеток увеличиваются на простоми-
уме, вокруг ротового поля и на пигидиуме. Вокруг ротового сег-
мента формируется воротник (рис. 3, L), который прикрывает за-
чатки параподиальных усиков нескольких передних сегментов.
У метатрохофоры может быть до 12 сегментов. Каждый сегмент
снабжен слабым нототрохом (рис. 3, Not). Зачатки дефинитивной
нервной системы получают на этой стадии характерные особенно-
сти, тогда как провизорная нервная система начинает утрачивать
свое значение. Головной мозг разделен на передний (рис. 3,Ptc),
средний и очень небольшой задний отделы. Антеннальный ганглий
(рис. 3, Antr) на этой стадии обособлен от остальной части головно-
го мозга. Брюшная нервная цепочка с резко обособленными ган-
глиями (рис, 3, G). В состав каждого ганглия входят и иараподи-
41
Рис. 3. Метатрохофора Phyllodoce maculata, парасагиттальный разрез
(ориг.)
Antr— зачаток головного усика; Вс — бластоцель; Cs — секреторные клетки; Cstn —
секреторные клетки кишечника; Си — кутикула; Cv — реснички аборального органа*
Gi — подглоточный ганглий; G3 — ганглий третьего сегмента; In — кишечник; L— во-
ротник; Meh — клетки мезенхимы; Met — кольцевая мышца; N — нейропиле; Nci —
кольцевой нерв; Not — нототрохи; Npar — параподиальный нерв; 01 — капли жира;
Os — рот; Phd — глоточные дивертикулы; Pic — передний мозг; Ptr — прототрох
альные нервы (рис. 3, Npar). Кишечник получает ряд дополнений.
Глоточные дивертикулы (рис. 3, Phd) сообщаются между со-
бой и с пищеводом. Стенка их становится складчатой и мускулистой.
Пищевод заметно укорачивается, хотя еще в полной мере выпол-
няет функцию провизорного аппарата захвата добычи. Средняя
кишка имеет большой объем. Стенка ее несет включения жира и
желтка. Полость тела почти на всем протяжении выстлана цело-
мическим эпителием. Однако мезентериев и септ нет. В зоне роста
вблизи анальной лопасти происходит бурное деление клеток ме-
зенхимы.
Нектохета имеет очень непостоянную длину тела, от 500 до
800 мкм. На этой стадии начинают функционировать продольные
ленты мышц. Личинка садится на дно и делает пробы субстрата.
Отрицательный результат пробы заставляет личинку снова под-
ниматься в толщу воды, где она движется при помощи ресничек
прототроха. На дне же личинка ползает, волнообразно изгибая тело.
В толще воды она сильно укорачивается, а на дне вытягивается.
Простомиум нектохеты становится более крупным и выпуклым.
На нем возникают две пары усиков. Мерцательный орган утрачи-
вается. Метастомий образует воротник, из-под которого торчат
концы тентакулярных усиков трех первых сегментов, слитых с ме-
тастомиумом. У нектохеты до 20 щетинконосных сегментов с одно-
ветвистыми параподиями. В каждом пучке сложных гомогомфных
щетинок включена одна простая щетинка, зазубренная на выгну-
том крае. Пигидиум с очень слабым телотрохом. Нототрохи на
спинной стороне сегментов, начиная с пятого. Окраска тела тра-
вянистая с темными, полями около прототроха и па пигидиуме.
В области прототроха на спинной стороне два крупных желтых
пятна, светящихся в отраженном свете. Такие же парные пигмент-
ные пятна меньшего размера расположены вдоль средней линии
спины на каждом сегменте. Желток и жировые капли в стенке ки-
шки имеют желто-зеленую окраску. На анальной лопасти развиты
зачатки уритов. Личинка редко поднимается в толщу воды, оты-
скивает удобное место и приступает к постройке паутинного доми-
ка. Ювенильные формы Phyllodoce maculata были выведены в ла-
бораторных условиях из кладок, собранных на литорали в момент
размножения взрослых особей.
Eteone longa (Fabricius, 1780)
Взрослые особи обитают на литорали и в сублиторали на ме-
стах, богатых илом и гниющими органическими остатками..
Пелагические личинки различных видов рода Eteone отлича-
ются между собой ничтожными признаками. Поэтому к описаниям,
сделанным различными авторами для некоторых видов Eteone,
следует подходить с большой осторожностью. Впервые описал ли-
чинку Eteone для европейских берегов Р. Виллемёз-Зум (Willemo-
es-Suhm, 1871), относя ее к виду Е. pusilia 0rsted. М. Лешке (Le-
43
Рис. 4. Личиночные стадии
Eteone longa (ориг.)
А — трохофора;
Б — метатрохофора;
В — нектохета;
Г — нектохета перед оседанием;
Д — ювенильная донная семи-
Н Всегментная особь;
Е, ЯС, 3 — ее щетинки
schke, 1903) описал и изобразил различные стадии развития Eteo-
ne, совпадающие с описаниями Виллемёз-Зум. В. Нольте (Nolte,
1938) обратил внимание на то, что личинка, описанная обоими ав-
торами, имеет щетинки на втором сегменте, а на буккальном сег-
менте — тентакулярные усики, т. е. обладает признаками, свой-
ственными или Eteone longa, или Eteone flava (Fauvel, 1923).
Особенности размножения у Е. longa в естественных условиях
никто не наблюдал. А. Мик и Б. Сторроу (Meek, Storrow, 1924)
в аквариуме наблюдали у этого вида откладку яиц в виде слизи-
стых масс. Пелагическая личинка Е. longa описана многократно.
Г. Торсон (1946) описал для берегов Дании различные стадии пе-
лагических личинок Е. longa. Очень точные описания и изобра-
жения личинок дает Е. Расмуссен (Rasmussen, 1956) для района
Изефиорда (Дания). Время размножения Е. longa и появления их
личинок в прибрежной зоне пелагиали, по данным различных ав-
торов, в общем, совпадает и в разных географических точках ев-
ропейского и дальневосточного берегов падает на гидрологическую
весну: март, апрель, май (Свешников, 1959, 1961, 1967).
44
По нашим наблюдениям, в Белом море личинки Е. longa появ-
ляются в планктоне при температуре воды 0° в мае, а в Японском
море — в мае при температуре от 3 до 8° С (Свешников, 1961,
1967).
Трохофора монотрохного типа (рис. 4, А) имеет сферическую
или овальную форму тела, 200 мкм в поперечнике. По длинной оси
тела личинка достигает 250 мкм. Окраска охристая. Прототрох де-
лит личинку на меньшую эписферу и большую гипосферу. Ре-
снички прототроха длиной до 50 мкм располагаются в один ряд.
Эписфера несет одну пару ярко-красных глаз. Дистальная и вен-
тральная части эписферы покрыты мелкими ресничками. Между
этим ресничным полем и прототрохом находится тактильный мер-
цательный орган с султаном ресничек, характерным для личинок
семейства Phyllodocidae. Эпителий верхнего полушария богат же-
лезистыми клетками с крупными многочисленными вакуолями.
Провизорная ортогональная нервная система состоит из экватори-
ального нервного кольца, которое сопровождено 2—3 мышечными
фибриллами и 7 парами тонких меридиональных стволов. Дефи-
нитивная нервная система представлена зачатками головного и
брюшного мозга. Зачаток головного мозга насчитывает в своем
составе несколько десятков ганглиозных клеток, окружающих ней-
ропиле. В составе ганглиозной массы клеток расположено не-
сколько (4—5) гигантских нервных клеток. Размер обычных ган-
глиозных клеток колеблется около 3, а гигантских — около 15 мкм.
От этой погруженной в толщу эпителия основной массы зачатка
мозга отделен очень небольшой зачаток головных усиков, который
подходит ближе к поверхности эписферы. Зачаток брюшной нерв-
ной цепочки занимает брюшную поверхность личинки. Кишеч-
ник состоит из трех отделов. Рот ведет в канал пищевода, выстлан-
ный мерцательными клетками. На границе пищевода и средней
кишки формируется парный зачаток глотки, который еще не имеет
связи с каналом пищевода. Средняя кишка состоит из двух камер.
Стенка средней кишки богата железами и желтком охристого или
желто-зеленого цвета. Анальное отверстие смещено на спинную
сторону. Первичная полость тела занимает небольшой объем.
В зоне роста расположены немногочисленные клетки мезобласты,
производные 4d бластомера.
Метатрохофора 7 (рис. 4, Б) достигает 300 мкм в длину. На
этой стадии формируется простомий с зачатками четырех головных
усиков. Простомий округлой формы, с двумя парами красных глаз.
Апикальное поле покрыто ресничками. Реснички мерцательного
органа и прототроха заметнее и крепче, чем у трохофоры. У мета-
трохофоры возникает телотрох. Несколько увеличивается количе-
ство железистых клеток на простомиуме и пигидиуме. Пигмента-
ция несколько усиливается, особенно на простомиуме и пигидиуме.
На сегментах формируются зачатки одноветвистых параподий.
Каждый сегмент снабжен слабым нототрохом, начиная с четверто-
го. Кожно-мускульный мешок состоит из эпителия и системы
мышц. Нервная система расположена между эпителием и слоем
45
мышц. Нервная система метатрохофоры представлена.провизорной
и дефинитивной частями. Провизорная ортогональная система
состоит из экваториального кольца, которое проходит под прототро-
хом. Экваториальное кольцо пересечено семью парами меридиональ-
ных нервов. Дефинитивная нервная система состоит из головного
мозга, окологлоточных коннективов и двух сближенных зачатков
брюшных стволов. Головной мозг разделен на передний, средний и
очень небольшой задний отделы. Антеннальные ганглии обособ-
лены от остальной части головного мозга. В теменной области рас-
положены единичные гигантские нервные клетки. Ядра этих кле-
ток диаметром 20 мкм имеют крупные звездчатые ядрышки. Око-
логлоточные коннективы отходят от церебральных ганглиев, охва-
тывают глотку и на брюшной стороне соединяются с брюшными
нервными стволами. Брюшные стволы образуют семь пар ганглиев,
соответственно числу ларвальных сегментов. В каждом сегменте
формируются гигантские нервные клетки с крупными ядрами и
зведчатыми ядрышками. В анальной лопасти, позади телотроха,
расположено нервное кольцо, в котором среди обычных клеток
ганглиозной массы также находятся отдельные гигантские клетки.
У метатрохофоры начинает формироваться пара брюшных и пара
спинных мышечных лент. Ленты продольной мускулатуры скла-
дываются из клеток «аскаридного» типа. Трофический мешок клет-
ки погружен в бластоцель. Ядро находится в трофической части
клетки. Сократимая часть клетки поставлена ребром к базальной
мембране. У метатрохофоры формируются семь пар трансверсаль-
ных пучков соответственно числу ларвальных сегментов.
Кишечник слагается из трех отделов. Ротовое отверстие ведет
в короткий пищевод, к которому примыкает пара глоточных ди-
вертикул. Из них впоследствии формируется дефинитивная глот-
ка. Средняя кишка имеет толстую стенку, в которой обильно рас-
сеяны гранулы желтка и капли жира. Стенка пищевода, глотки
и средней кишки имеет поверхностную сеть сократимых элемен-
тов. Полость тела лишь местами выстлана эпителием. Мезентери-
ев и септ нет. В зоне роста вблизи анальной лопасти мезенхимные
клетки находятся в стадии бурного размножения и заметно их
движение в сторону переднего конца.
Нектохета-7 достигает в длину 450 мкм (см. рис. 4, В, Г-, рис. 5).
Простомиум несет две пары коротеньких антенн и две пары глаз.
Лишь одна пара с хрусталиками. Апикальное ресничное поле ча-
стично сохраняется, ресничный султан редуцирован. Прототрох
еще хорошо развит. На ротовом сегменте закладываются две пары
усиков в виде утолщений на задних углах сегмента. На этой ста-
дии личинка по-прежнему имеет семь ларвальных сегментов, вклю-
чая перистомий, который щетинок не несет. Первый щетинконос-
ный сегмент с одноветвистыми параподиалытыми бугорками и
вентральными усиками. Дорсальные усики па этом сегменте отсут-
ствуют. Остальные сегменты, кроме седьмого, имеют хорошо раз-
витые параподиальные бугорки с дорсальными и вентральными
46
An — анус;
Antr — i ачаток головного
усика;
С — головной мозг;
Coel — целом;
Cs — секреторные клет-
ки;
Gj — подглоточный ган-
глий;
Gs — ганглий шестого
сегмента;
Ga — антеннальный ган-
глий;
Gvr — зачаток брюшного
мозга;
In — кишечник;
Meh — клетки мезенхимы;
Met — кольцевые мышцы;
Ml — продольные мышцы;
Mph — мышцы глотки;
Mtr — трансверсальные
мышцы;
Net — кольцевой нерв;
Not — нототрохи;
Nucgt — ядра гигантских
нейронов;
01 — капли жира;
Os — рот;
Ph — глотка?
Ptr — прототрох;
Tt — телотрох
Рис. 5. Нектохета Eteone longa, сагиттальный разрез (ориг.}
усиками. В пучке щетинок многочисленные сложные и по одной
простой пильчатой (рис. 4, Е, Ж, 3). На всех сегментах, начиная
с четвертого, на спинной стороне развиты слабые нототрохи. На
пигидиуме продолжает функционировать телотрох. Пигментация
на этой стадии усиливается. Желто-зеленый пигмент появляется
на голове и передних сегментах. Кожно-мускульный мешок при-
обретает облик дефинитивного строения. Нервная система погру-
жена в толщу мышц. Однако она все еще содержит провизорную
часть, которая состоит из ортогонального нервного кольца, сопро-
вождающего прототрох (рис. 5, Net'). Меридиальные нервы на этой
стадии подвергаются редукции. Дефинитивная нервная система
состоит из головного мозга, окологлоточных коннектнвов и двух
сближенных брюшных стволов. Характерно в строении головного
мозга то, что антеннальные ганглии обособлены и выведены за
пределы основной части мозга (Ерохина, 1968). Число гигант-
ских нервных клеток несколько увеличивается, а размер их
уменьшается. Брюшные стволы образуют семь пар ганглиев соот-
ветственно числу ларвальных сегментов. Число гигантских клеток
брюшных стволов также незначительно возрастает. Брюшные и
спинные ленты мышц, а также трансверсальные пучки становят-
ся более строго оформленными. Брюшные ленты на этой стадии
погружены на дно глубоких каналов. Ленты сложены из клеток «ас-
каридного» типа. Сократимые отростки клеток поставлены ребром
на дне каналов. Трофические мешки клеток занимают всю глуби-
ну этих каналов. Трансверсальные веерообразные пучки мышц
прикреплены одним концом медианно на мембране, окружающей
брюшную цепочку (рис. 5, Mtr), а другим — к стенкам параподий.
Кишечник претерпевает новые изменения. Глотка занимает
объем трех передних сегментов. Провизорный пищевод на этой
стадии исчезает. Средняя кишка имеет небольшой просвет (рис. 5,
In), а стенка заполнена каплями жира и гранулами желтка. Юве-
нильные формы Eteone longa были выведены в лабораторных усло-
виях из пелагических личинок разного возраста (см. рис. 4, Д).
СЕМЕЙСТВО TYPHLOSCOLECIDAE
Все представители этого семейства ведут пелагический образ
жизни. Известно всего три рода: Typhloscolex, Sagitella и Travisi-
opsis. В литературе нет никаких данных по индивидуальному раз-
витию этой группы полихет. Мы приводим здесь впервые некото-
рые описания плана строения ювенильной семисегментной формы
Typhloscolex mulleri из Северной Атлантики. План строения этой
личинки отвечает организации метатрохофоры. Основными особен-
ностями организации личинки Typhloscolex следует считать край-
не слабое развитие центральной нервной системы и весьма специ-
ализированный аппарат захвата пищи. Кроме того, известно, что у
взрослых форм Typhloscolecidae на всю жизнь сохраняется прови-
зорная система прототроха, с помощью которого животные удер-
48
Рис. 6. Личинка Typhlos-
colex muelleri, сагитталь-
ный разрез (ориг.)
Ап — анус;
Ао — головное щупаль-
це;
Си — кутикула;
Gs — слюнные железы;
In — кишечник;
Los — ротовая X присоска;
Man — кольцевые мышцы;
Meh — клетки мезенхимы;
Ml — продольные мыш-
цы;
Раг2— паренхима;
Ph — глотка;
Ptr — прототрох;
Тт — трохобласты;
Ur — анальные усики
живаются в толще воды. Наконец, для личинки характерно значи-
тельное развитие мезенхимы, которая на более поздних стадиях
и в ларвальном, и в постларвальном отделах образует подобие па-
ренхиматозной ткани. Эту особенность можно расценивать и как
примитивный и как вторичный признак. У Typhloscolecidae обна-
руживаются хорошо развитые диссепименты вторичной полости.
Головной конец личинки Т. mulleri уплощен и несет прототрох
(рис. 6, Ptr). В центре анимального полюса расположено един-
ственное щупальце, покрытое короткими чувствующими ресничка-
49
ми. Большую часть анимального полюса занимают трохобласты
(рис. 6, Тг), тогда как на долю элементов нервной системы прихо-
дится ничтожная часть объема эпителия. Эта характерная морфо-
логическая особенность строения личинки (и взрослого животного)
должна совпадать с относительно пассивным образом жизни. Об'
этом свидетельствует не только слабое развитие и ничтожная диф-
ференциация нервной системы, а также отсутствие органов зрения.
Второй характерной особенностью личинки является строение
аппарата захвата пищи — околоротовых губ и глотки со слюнными
железами. Морфология аппарата захвата пищи такова, что глотка
не может выворачиваться, как у хищных полихет. По своей струк-
туре глотка Т. miilleri представляет собой типичный глотательный
аппарат типа насоса, сильно вздута, почти шарообразна.
Мышечные тяжи расположены радиально и на расстоянии друг
от друга, что позволяет предполагать возможность значительного'
изменения общего объема глотки (рис. 6, Ph). Околоротовые губы
также снабжены сильной мускулатурой и вытянуты трубочкой:
под некоторым углом вверх. У основания околоротовых губ с дор-
сальной стороны открывается канал слюнных желез (рис. 6, Gs)..
Вслед за глоткой следует кишечник с глубокими перетяжками в.
каждом сегменте. Создается впечатление, что кишечник в ларваль-
ном теле поделен на семь камер. Клетки в стенке каждой камеры
распределены так, что группы железистых клеток сосредоточены
главным образом на границах между камерами, тогда как группы
клеток, всасывающих пищу,— в средних участках камеры. Такое-
поочередное расположение клеток эпителия кишечника функцио-
нально оправдано тем, что благодаря этому создаются дифферен-
цированные участки для переваривания и всасывания пищи (рис.
6, In). Подобная картина известна и для других полихет (Догель,.
1938). Однако у Т. miilleri строение кишечника носит явные чер-
- ты примитивности, и, по нашему мнению, это связано с питанием
полужидкой пищей.
В самом деле, изложенные особенности пищеварительной си-
стемы наталкивают на мысль о том, что Т. mulleri представляет
собой эктопаразитическую форму, способную питаться тканями
пелагических кишечнополостных (сифонофор, медуз, гребневи-
ков). На основании анализа морфологических данных я полагаю,,
что Т. miilleri вертикально висит в толще воды. Движение сильно
развитых ресничек прототроха вызывает перемещение животного-
вертикально вверх. Прикасаясь апикальным щупальцем к пленке
натяжения, Т. miilleri выключает прототрох и медленно тонет.
Опустившись на некоторую глубину, животное вновь включает
прототрох и поднимается к пленке натяжения. Такое поведение-
очень типично для велигеров моллюсков и некоторых других пе-
лагических организмов, например щупальцевых гребневиков. Од-
нако у названных животных функцию аппарата захвата пищи в
значительной мере выполняют у велигеров волюм, а у гребневиков,
щупальцы. У Т. mulleri кажется возможным предположить следу-
50
ющий способ питания. Поднимаясь вертикально к поверхности,
Т. miilleri может случайно натолкнуться на медузу или сифонофо-
ру. Околоротовые губы направлены вверх, сложены трубочкой и
могут присасываться к поверхности жертвы при помощи сосущих
движений глотки. Слюнные железы выделяют пищеварительный
сок, растворяющий покровы жертвы в точке ее соприкосновения
с околоротовыми губами. Дальнейшие сосательные движения обес-
печивают поступление полужидких тканей в кишечник Т. mulle-
ri. Учитывая значительные различия в размерах Typhloscolecidae
и пелагических кишечнополостных (сифонофор и сцифоидных ме-
дуз), можно думать, что разовое всасывание не приводит к гибели
жертвы. Если это так, то предполагаемый способ питания Typhlo-
scolex mulleri следует относить к эктопаразитическому, а не к хищ-
ническому способу. Typhloscolecidae в целом обнаруживают яв-
ные черты примитивной организации, а также специализации в
отношении способа питания. К примитивным чертам плана строе-
ния относится прежде всего сохранение во взрослом состоянии
провизорных систем, например ресничного пояса на головной ло-
пасти. Черты специализации аппарата питания были разобраны
выше. К этому надо прибавить некоторые особенности строения
кожно-мускульного мешка. Например, поперечное сечение часто
имеет форму квадрата или прямоугольника; преимущественное
развитие получают сначала четыре, а позднее шесть продольных
мышечных лент; параподиальные мышцы развиты очень слабо.
Листовидные придатки не имеют специальной мускулатуры и, ви-
димо, слабо управляемы; они увеличивают общую поверхность те-
ла и способствуют парению в толще воды. Наконец, частичное за-
полнение полости тела паренхимой, возможно, связано с ослаб-
лением или полным отсутствием изгибательного способа движения
и с факультативным эктопаразитизмом.
Итак, мы видели, что особенности индивидуального развития
Phyllodociformia укладываются в границы представлений о фило-
генетических связях внутри этой группы (Ушаков, 1970, 1972).
Большая примитивность всего отряда отражается в значительном
морфологическом разнообразии составляющих групп. В этом от-
ношении, например, план строения Aphroditidae или некоторых се-
мейств Nereimorpha более стабилен. Изложенные выше данные по
строению личиночных форм позволяют заключить, что в семействе
Phyllodocidae выражены наиболее примитивные особенности пла-
на строения трохофор, метатрохофор и нектохет.
СЕМЕЙСТВО APHRODITIDAE
У Aphroditiformia известно только половое размножение. Оп-
лодотворение наружное или внутреннее (Daly, 1972). Эмбриоге-
нез протекает либо целиком в пелагиали (у пелагических форм,
например Nectochaeta Grimaldii), либо у бентосных форм частич-
51
но в толще воды, а частично в бентосе. Например, Harmothoe вы-
нашивает потомство под элитрами до -полного формирования тро-
хофор, способных самостоятельно питаться (Izuka, 1912; Daly,
1972). Постэмбриональное развитие Aphroditiformia обычно про-
ходит с пелагической трохофорой типа «barbularia» (Свешников,
1963). Пелагические трохофоры питаются диатомовыми водоросля-
ми типа Coscinodiscus. Личинки далее проходят стадию метатро-
хофоры и нектохеты. Число ларвальных сегментов на этих стадиях
у различных видов чаще всего постоянно. Для Sigalioninae — 4;
для Polynoinae — 7 или 8; для Pisionidae — не больше 8.
Размножение и развитие отдельных представителей этого под-
отряда изучено в незначительной степени. Наиболее подробно
изучено развитие одного из массовых видов у берегов Европы —
Harmothoe imbricate (Sars, 1845; Muller, 1851; Kallenbach, 1883;
Hacker, 1894; McIntosh, 1900-1923; Leschke, 1903; Izuka, 1912;
Nolte, 1936; Smidt, 1944; Thorson, 1946; Rasmussen, 1956; Киселе-
ва, 1957; Korn, 1959; Милейковский, 1959; Свешников, 1959, 1961,
1963, 1967, 1968; Константинова, 1961; Ерохина и др., 1970). По
размножению и развитию других групп этого подотряда сведений
в литературе немного (Thorson, 1946; Киселева, 1957; Свешни-
ков, 1960). Некоторые особенности развития и морфологии пела-
гических личинок комменсала морских звезд — Acholoe asterico-
la — известны по работе Д. Давенпорт (Davenport, 1954).
О размножении и развитии представителей семейства Chryso-
petalidae в литературе имеются весьма ограниченные сведения
(Милейковский, 1962). Последние данные о развитии Pisionidae
сведены в работе Б. Акессон (Akesson, 1961).
Aphroditidae — обитатели донных и пелагических биоценозов.
По способу питания это — активные хищники или полифаги
(Стрельцов, 1966а, б, 1968). Хорошо развитая глотка с четырьмя
челюстями занимает не менее У3 объема тела животного. Тело,
как правило, короткое. Параподии выдаются далеко в стороны.
Поступательное движение своеобразное. В нем сочетаются шага-
тельные и ундулярные движения (Lawry, 1970). Рецепторные при-
датки головы хорошо развиты. Хеморецепция осуществляется
крупными пальпами. Тактильная рецепция осуществляется тре-
мя антеннами. Две или три пары глаз. Характерны щетинки на
ротовом сегменте. У многих полихет других семейств щетинки па
ротовом сегменте отсутствуют. Гетерономная метамерия выраже-
на чередованием сегментов, вооруженных элитрами, с сегментами,
вооруженными спинными усиками. Элитры образуют сплошной
ряд круглых щитков, ограничивающих выводковую камеру для вы-
нашивания потомства. У пелагических представителей (Nectocha-
eta Grimaldii и др. ) элитры не развиты. Параподии двуветвистые.
Нотоподии часто выполняют защитную роль, и поэтому пучки но-
топодиальных щетинок направлены вверх.
Развитие пелагических видов протекает в толще воды. Разви-
тие донных видов, как правило, на ранних этапах (дробление, фор-
52
мирование трохофоры) протекает внутри выводковой камеры на
спине материнской особи. Трохофоры или метатрохофоры покида-
ют выводковую камеру, и дальнейшее развитие проходит в тол-
ще воды.
В высокой Арктике у тех же видов обычно все развитие и ме-
таморфоз происходят в выводковой сумке материнского организ-
ма (Thorson, 1952),
Harmothoe imbricata (L.)
Первое описание развития этого вида сделал М. Cape (Sars,
1845). По его данным, размножение Н. imbricata у берегов Норве-
гии происходит с февраля по март. Яйца диаметром 50 мкм розо-
вого цвета, находятся под элитрами материнской особи. В. Мак-
Интош (McIntosh, 1900) у берегов Англии наблюдал размноже-
ние и развитие этого вида в марте. А. Ицука (Izuka, 1912)
у берегов Исландии наблюдал размножение Н. imbricata в апре-
ле — мае. Е. Смидт (Smidt, 1944) отмечает появление личинок
в планктоне у берегов Дании только в холодное время года (с нояб-
ря по март). Г. Торсон (Thorson, 1946), напротив, в прол. Зунд на-
ходил личинок, похожих на Н. imbricata, все лето. Однако точную
идентификацию путем выведения в лаборатории Г. Торсон не де-
лал. Е. Расмуссен (Rasmussen, 1956) также для берегов Дании
описывает размножение Н. imbricata с января по март. Наиболь-
шее обилие личинок в планктоне около 250 экз/М3 падает на ян-
варь, когда береговая полоса покрыта льдом. М. И. Киселева
(1957) наблюдала в Черном море пелагических личинок Н. im-
bricata с января по март, когда они, по ее словам, являются поч-
ти единственными представителями личинок полихет в планктоне.
Г. Корн (Korn, 1958, 1959) в Кильской бухте собирал личинок
этого вида в марте.
По нашим данным, в Белом море (Свешников, 1959) Н. imbri-
cata размножается в апреле — мае. В это время море покрыто
льдом, и температура воды не поднимается выше 0°. В начале мая
в планктоне Белого моря появляются трохофоры. В Японском мо-
ре (Свешников, 1967) размножение этого вида происходит также
весной, в мае.
Пелагические трохофоры имеют шарообразную форму от 250
до 400 мкм в диаметре (рис. 7, А). Ресничный пояс проходит не-
много выше экватора и надо ртом прогнут вверх. Создается впе-
чатление, будто рот в своем движении к анимальному полюсу
смещает и прогибает ресничный пояс (Свешников, 1971а, 1972а).
Ресничный пояс состоит из трех рядов трохобластов. Верхний и
нижний ряды состоят из более мелких клеток, а средний состав-
лен из крупных клеток, реснички которых достигают длины
120 мкм (рис. 9, A, Tb, Ntb). С левой стороны от рта в нижнем ряду
имеется пучок длинных, сильных рулевых ресничек, длина кото-
рых достигает 200 мкм (рис. 7, А — Д'). Д. Давенпорт (Daven-
port, 1954) отмечает наличие такого же пучка с левой стороны от
53
Рис. 7. Личиночные стадии Harmothoe imbricata (ориг.)
же /бптптлй’п Б И В метатР°хо*°Ра; Г — нектохета со спинной стороны; Д — она
®ого сегмента 3 ™Р™Ы; Е ~ ее двузубая щетинка из невроподии; Ж - щетинка рото-
рной стороны ’ оседающая молодая особь со спинной стороны; И — она же, с брюш-
Рис. 8. Схемы сечений трохофоры Harmothoe imbricata (рриг.)
А — поперечное сечение по экватору; Б — то же, ниже экватора соответствует линии
Б — на рисунке в центре; В — фронтальное сечение соответствует линии В — Bi
на рисунке в центре; Г — фронтальное сечение на уровне пищевода соответствует линии-
Г - Л-
В центре — сагиттальное сечение; 1 — прототрох; 2— эктодерма спинной стороны; з —
эндодерма; 4 — глоточные дивертикулы; б — пищевод; 6 — передняя зона образования
эндомезенхимы; 7 — тепобластические полоски; 8 — зачаток головного мозга; .9 — зача-
ток нервно-двигательного комплекса ларвального отдела
рта в прототрохе у личинок Acholoe astericola (Polynoinae). Е. Рас-
муссен (Rasmussen, 1956) описывает подобные пучки с двух сто-
рон от ротового отверстия у личинок Harmothoe imbricata. Г. Корн
(Korn, 1959) обнаруживает такой же признак у личинок Harmot-
hoe sarsi и Н. impar (Polynoinae). Видимо, наличие длинных
ресничных пучков около рта следует считать общим признаком
55
Рис. 9. Трохофора Harmothoe imbricata (opus.)
A — поперечный разрез на уровне ресничного пояса; Б — поперечный разрез на уровне
глоточных дивертикулов; Вс — бпастоцель; Сп3 — нервные клетки меридиональных
нервов; In — кишечник; Los — околоротовые губы; Meh — клетки мезенхимы; Mel —
кольцевая мышца; Ми — митозы; Мт — меридиональные мышечные волокна; Мое —
мышцы пищевода; п — меридиональные нервы;'; Net — кольцевой нерв; Шь — ядро
клетки трохобласта; Ое1 — капли жира; Oes — пищевод; Phd — глоточные дивертику-
лы; Ptr — прототрох; Sir — клеточный материал ларвальных сегментов; Тъ — трохо-
бпасты
56
Рис. 10. Схемы сечений метатрохофоры Harmothoe imbricata (рриг.)
А — поперечное сечение по экватору; Б — поперечное сечение соответствует линии Б — В
на рисунке в центре; В — фронтальное сечение соответствует линии В — Вц на рисунке
в центре; Г — фронтальное сечение на уровне пищевода соответствует линии Г — 1\
в центре — сагиттальное сечение; 1 — прототрох; 2 — эктодерма спинной стороны; з —
эндодерма; Л — глоточные дивертикулы; 5 — пищевод; в — передняя зона образования
эндомезенхимы; 7 — телобпастические полоски; 8 — зачаток головного мозга; 9 — зача-
ток нервно-двигательного комплекса ларвального отдела; 10 — полость тела; 11 — элит-
ра; 12 — невроподия
личинок (на стадии трохофоры и метатрохофоры), принадлежащих
к семейству Aphroditidae. На эписфере Н. imbricata находится сла-
бый акротрох. От рта до анального отверстия по брюшной стороне
узкой полоской тянется невротрох. Трохофора имеет одну пару
глаз, снабженных линзами (рис. 7, А). Под ресничным поясом про-
ходит экваториальное нервное кольцо (рис. 9, A, Net), сопровож-
даемое экваториальным мышечным кольцом (рис. 9, A, Mei). В ме-
ридиональном направлении проходят пять пар нервных тяжей, со-
ставляющих вместе с экваториальным и параэкваториальными
кольцами провизорную ортогональную нервную систему личинки
(рис. 9, А и Б, ni-5). Ортогональная нервная система сопровож-
дается соответствующими мышечными меридиональными и эква-
ториальными и параэкваториальными фибриллами (рис. 9, А и В,
Мт). Провизорная мышечная система трохофоры является произ-
водной эктомезенхимы личинки и тесно примыкает к эктодерме.
Подобная организация ортогона описана Э. Мейером (1897) и
Б. Акессоном (Akesson, 1967) для трохофоры Lopadorhynchus.
Кишечник у трохофоры Н. imbricata сквозной. Рот снабжен
верхней губой (рис. 7, А). Пищевод (рис. 9, A, Oes), выстланный
мелкими ресничками, поднимается ото рта вверх и впадает в сред-
нюю кишку как бы с анимального полюса (рис. 8). Средняя киш-
ка трохофоры (и метатрохофоры) двухкамерная. Передняя каме-
ра отделена от задней камеры перегородкой (рис. 8). Короткая
задняя кишка трохофоры Н. imbricata открывается анусом, сме-
щенным к спинной стороне (рис. 7, В и Г). На границе пищево-
да и средней кишки у трохофоры инвагинацией образуется пара
глоточных дивертикулов, которые представляют собой зачаток бу-
дущей глотки (рис. 8 Г; 9, Phd). В их образовании принимают
участие многочисленные клетки эндомезенхимы, производные 4d
бластомера (рис. 9, Meh). Стенка средней кишки (рис. 9, А, Б, In,
Oel) заполнена желточными гранулами темно-карминного или си-
реневого цвета, который определяет общий тон окраски личинки
и может служить отличительным признаком для личинок это--
го вида.
Метатрохофора (рис. 7, Б и Б; рис. 10) несколько вытягивает-
ся в длину и приобретает яйцевидную форму. Длина ее увели-
чивается до 600 мкм. Диаметр наиболее широкой части тела
доходит до 800 мкм. Ресничный покров сохраняется в первоначаль-
ном виде, и только в центре эписферы возникает слабый апикаль-
ный пучок ресничек. К первой паре глаз прибавляются еще две
пары новых глаз, также с преломляющими линзами (рис. 7, Б, В,
Г). У метатрохофоры образуются восемь пар щетинконосных меш-
ков. Это число ларвальных сегментов характерно для личинок
Н. imbricata и для личинок Н. sarsi, тогда как у Н. impar закла-
дываются семь ларвальных сегментов (Korn, 1958). На этой ста-
дии зачатки частей параподий расположены в три ряда. Первый
ряд с дорсальной стороны представляет собой зачатки спинных
усиков и элитр. Первый в этом ряду зачаток в дальнейшем разо-
58
вьется в спинной усик ротового сегмента. Второй бугорок круп-
нее первого и является будущей элитрой. Третий маленький за-
чаток — снова дорсальный усик. Крупные четвертый и пятый —
зачатки элитр. Шестой зачаток — будущий усик, седьмой —
элитра, восьмой — усик. Второй ряд бугорков почти одинакового^
размера. Бугорки второго ряда разовьются впоследствии в невро-
подиальные ветви параподий. В глубине их уже имеются заклад-
ки ацикул и хет. Третий ряд одинаковых по размеру бугорков
представляет собой будущие брюшные усики.
На стадии метатрохофоры одновременно существуют провизор-
ные и дефинитивные системы. Зачатки дефинитивной нервной си-
стемы у метатрохофоры заметно развиты и дифференцированы
(Ерохина, и др., 1970). Головной мозг метатрохофоры включает
зачатки грибовидных тел (ассоциативные центры), ганглиозную'
массу и нейропиле (рис. 11, Ср, С). Брюшной мозг метатрохофо-
ры состоит из ганглиозной массы и нейропиле. Ортогональная
нервная система метатрохофоры медленно дегенерирует. Движе-
ние осуществляется все еще только с помощью ресничного пояса.
Однако относительная нагрузка на ресничный пояс увеличивается
вместе с увеличением размеров тела, а скорость движения умень-
шается.
Нектохета увеличивается до 850 мкм (рис. 7, Г, Д, 3, И). Одна-
ко число сегментов остается равным восьми. На простомиуме по-
являются зачатки трех антенн и двух пальп. Формируется пара-
подиальный комплекс. Элитры становятся плоскими, а невропо-
диальные ветви со щетинками сильно выступают в стороны. Пара-
подии начинают функционировать и помогают работе ресничного
пояса. Щетинки невроподиальной ветви уплощены и несколько
изогнуты на конце. По выпуклой стороне изгиба имеются шипы.
Щетинка двузубая на конце (рис. 7, £). На этой стадии на всех
сегментах, кроме первого, нотоподиальных бугорков еще нет.
Но каждый сегмент внутри нотоподиальных хетоносных мешков
уже имеет ацикулы и щетинки. Щетинки ротового сегмента спе-
циализированы, серповидно изогнуты и сильно зазубрены
(рис. 7, Ж).
Ортогональная провизорная нервная система нектохеты пол-
ностью утрачивается. Напротив, зачатки дефинитивной нервной
системы продолжают развиваться. Число клеток значительно воз-
растает и может достигать порядка 10 000. Оформляются грибо-
видные тела, ганглиозный отдел дифференцируется на определен-
ные центры. Особенно четко определены зрительные и пальпар-
ные центры (рис. И, Ср, С) (Ерохина и др., 1970).
Парные уплотненные полоски мезодермы, вытянутые вдоль
главной оси от анального поля до анимального полюса, распада-
ются на парные мешочки, число которых совпадает с числом лар-
вальных сегментов (рис. 12, Coel.). Каждый мешочек образован
примерно 200 мезенхимными клетками. Мешочки, которые соот-
ветствуют сегментам с элитрами, несколько крупнее, чем те, ко-
59
торые прилежат к сегментам со спинными усиками. Эти мешочки
в процессе развития увеличиваются в объеме, и стенки их сопри-
касаются друг с другом, со стенкой кишечника и со стенкой тела.
Таким образом формируются диссепименты и мезентерии, а так-
же спланхно- и соматоплевра в ларвальном теле нектохеты
(рис. 12, Sep.). Обращает на себя внимание непарная целомиче-
ская полость в головной части личинки, образующаяся, по всей
видимости, из производных источника мезодермы в области гло-
точных дивертикулов. Непарная целомическая полость обнаруже-
на ранее также в головной части архианнелид Protodrilus и Sac-
cocirrus, а позднее у полихет Scoloplos armiger (Anderson, 1959)
и Eteone longa (Ерохина, 1968).
Кроме указанных туловищных целомов у нектохеты в области
пищевода формируются еще три пары целомических камер.
По своему положению они находятся ковнутри (медианно) от
туловищных целомов и непосредственно примыкают к пищеводу
(рис. 12). Ранее эти дополнительные целомические полости у по-
лихет описаны не были.
Все развитие, от момента оплодотворения до оседания некто-
хеты на дно, в Белом море длится около полутора месяцев. Осев-
шая молодь задерживается в поясе ламинарий, на порогах и бы-
стринах, под камнями, где их можно обнаружить в большом ко-
личестве, начиная с первой половины июня.
Ювенильные формы Harmothoe imbricata были выведены нами
в лабораторных условиях из пелагических личинок различного
возраста.
ОТРЯД NEREIMORPHA
В состав отряда Nereimorpha авторы вводят разные семейства.
Н. А. Ливанов (1940) называет шесть семейств — Syllidae, Не-
sionidae, Nereidae, Nephthydae, Sphaerodoridae и Amphinomidae.
П. В. Ушаков (1955, 1972) относит семейства Nephthydae и
Sphaerodoridae к отряду Phyllodocemorpha, семейства Syllidae,
Hesionidae, Pilargidae (Otopsidae) и Nereidae, имеющих настоя-
щие метанефридии,— к отряду Nereimorpha, сем. Amphinomidae
возводит в ранг отряда Amphinomorpha. С. А. Милейковский
(1968; Mileikovsky, 1977) предлагает повысить Amphinomorpha
в ранг надотряда, равный по значимости Errantia и Sedentaria.
Признавая обособленность Amphinomorpha, мне кажется, преж-
девременным повышение этой группы в ранг надотряда. Своеоб-
разие личинки рострарии, свойственной этой группе полихет, не
представляет исключительного случая. Есть и другие аберрант-
ные типы личинок, например митрария, стомария и др. Это, мне
кажется, не дает основания возводить в ранг отряда или надотряда
соответствующие группы полихет до тех пор, пока для этого не
будет избран достаточно убедительный критерий.
У представителей отряда Nereimorpha известны разные спо-
собы размножения, развития и разные типы личинок. Наряду с по-
60
Рис. 11. Метатрохофора Harmothoe imricata на стадии формирования лар-
вальных сегментов, парасагиттальный разрез (рриг.)
Вс — бластоцель; С — головной мозг; Chs — щетинконосные мешки; Ср — зачаток гри-
бовидных тел мозга; Gvr — зачаток брюшного мозга; 1са — передняя камера кишки;
1ср — задняя камера кишки; Is — перегородка, разделяющая камеры кишки; Los —
верхняя губа; Meh — клетки мезенхимы; Ми — митозы; N — нейропиле; Phd — гло-
точные дивертикулы; Ptr — прототрох; Tib — телобласт
ловым размножением встречается и бесполое. Например, у предста-
вителей сем. Syllidae известно правильное чередование бесполого
и полового размножения (метагенез). У некоторых представителей
Hesionidae, и особенно часто у Nereidae, наблюдается явление
эпитокии. У некоторых видов Nereis плавающие эпитокные особи
выводят половые продукты наружу сквозь разрывы стенки тела.
61
Рис. 12. Нектохета Harmothoe imbricata, фронтальный разрез
Слева — на уровне элитр; справа — на уровне щетинконосных мешков невро-
подий (ориг.)
С — головной мозг; Ch — щетинки; Ch$ — щетинконосный мешок; Сое! — целомическая
камера; Ср — грибовидное тело головного мозга; Е1 — элитра; Meh. — клетки мезенхи-
мы; Md — мезодерма; Phd — глоточные дивертикулы; Ptr — прототрох; Sep — септы;
Ur — анальные усики
После этого вся размножающаяся эпитокная часть популяции по-
гибает.
Личиночное развитие в отряде Nereimorpha изучено неравно-
мерно. Процессы личиночного развития Syllidae изучены недоста-
точно. Личинки Hesionidae описаны достаточно детально (Casano-
va, 1954; Banse, 1956; Davenport, Hickok, 1957; Свешников, 1961).
Личинки Pilargidae неизвестны. Личиночное развитие Nereidae
изучено наиболее подробно.
62
СЕМЕЙСТВО HESIONIDAE
Личинку Ancistrosyllis sp. (Harpochaeta cingulata) описал
E. Коршельт (Korschelt, 1893). Личинку Magalia perarmata Marion
et Bodretzky описала Л. Казанова (Casanova, 1954). Развитие Cas-
talia punctata описано К. Банзе (Banse, 1956). Д. Давенпорт и
Д. Хайкок (Davenport, Hickok, 1957) впервые описали личиночное
развитие факультативного комменсала Podarke puggetensis John-
son, живущего среди амбулакральных ножек морских звезд. Ли-
чинки Ancistrosyllis sp. и Podarke puggetensis значительно отлича-
ются от других личинок Hesionidae. По данным О. Хартман (Hart-
man, 1947), Ancistrosyllis следует относить к семейству Pilargiidae,
Castalia punctata (О. F. Muller)
О размножении и развитии этого вида имеются короткие наблю-
дения В. Макинтоша (McIntosh, 1908), Г. Аугенера (Augener,
1939) и Г. Торсона (Thorson, 1946). Более детально личиночное
развитие Castalia punctata описано К. Банзе (Banse, 1956). Все
эти авторы отмечают, что размножение С. punctata происходит у
европейских берегов или весной, или осенью.
По нашим данным, размножение этого вида в Белом море про-
исходит в первой половине июня при температуре воды 5—6° С.
Половозрелые (гетеронереидные) самцы и самки всплывают на по-
верхность воды и выметывают здесь половые продукты. Яйцеклет-
ки розового цвета около 200 мкм в диаметре. Между временем не-
реста и появлением личинок проходит 2—3 недели. После этого
перерыва в планктоне появляются трохофорные гаструлы. Они
имеют яйцевидную форму около 240 мкм длиной (рис. 13, 7). Ли-
чинки прозрачные, водянисто-изумрудного цвета. Прототрох про-
ходит по экватору личинки и делит ее приблизительно на равные
по объему половины. На эписфере имеется небольшой акротрох и
одна пара латерально расположенных маленьких красных глаз. От
ротового отверстия, густо усаженного ресничками, по брюшной
стороне узкой полосой тянется невротрох. Стомодеум узким про-
ходом, снабженным ресничным эпителием, поднимается вверх и
впадает в слепой двухкамерный кишечник. Анальное отверстие
отсутствует. Строение кишечника личинки вызывает интерес. По
существу такая личинка отвечает признакам гаструлы, ибо облада-
ет мешкообразным слепым кишечником. К организации гаструлы
прибавляется несколько новых существенных признаков. Это,
прежде всего, разделение кишечника на отделы: пищевод и двух-
камерная средняя кишка. Во-вторых, экваториальный ресничный
пояс. Совокупность перечисленных признаков позволяет нам назы-
вать эту личинку — трохофорной гаструлой. Создается
впечатление, что у личинки Castalia punctata бластопор, образовав-
шийся на оральном полюсе гаструлы, смещается вперед и упира-
ется в край ресничного пояса. По пути движения бластопора
63
Рис. 13. Личиночные стадии Castalia punctata (ориг.)
1 — трохофора со слепым кишечником, справа;’^’— метатрохофора, справа; з — пелаги-
ческая нектохета, с вентральной стороны; 4 — она же, со спинной стороны; 5 — пяти-
сегментная донная особь, со спинной стороны; в — девятисегмептпая донная особь; Т —
сложная двузубая щетинка-из вентральной части невроподиалыюго пучка
вперед образуется невротрох. Подобную картину миграции бласто-
пора с орального полюса вперед мы наблюдали в развитии Nereis,
Chaetopterus variopedatus и Pectinaria. Этот момент мы подчерки-
ваем здесь потому, что в развитии полихет, как это будет видно
дальше, не увидим однообразной картины в процессах формиро-
вания сквозного кишечника.
Метатрохофора (рис. 13, 2) имеет в длину около 300 мкм. Ги-
посфера вытягивается и несколько заостряется. На эписфере пер-
воначальная пара глаз приобретает преломляющие хрусталики, а
кроме того, появляются еще две пары глаз. Акротрох приобретает
вид полукольца. Вдоль нижнего края прототроха появляется узкая
полоска коричневой пигментации. У личинки закладывается до 6
пар щетинконосных клеток, соответствующих передним щетинко-
носным сегментам. Нам никогда не приходилось наблюдать сколь-
ко-нибудь заметной паузы в развитии сегментов тела личинки. Это
не позволяет говорить о ларвальном или постларвальном отделе
тела у Castalia punctata. Анальная лопасть на этой стадии несет
пару небольших бугорков — будущих анальных усиков. Невротрох
еще хорошо развит и тянется от рта до анальной лопасти. Стомо-
деум от вестибулюма поднимается почти до верхнего полюса эпи-
сферы и впадает в кишечник. На этой стадии кишечник становит-
ся сквозным.
Нектохета имеет размер 400 мкм (рис. 13, 3, 4). Эписфера при-
обретает вид простомиума. На нем ближе к вентральной стороне в
виде полушарий возникает пара пальп. В передней части эписферы
(простомиума) возникают зачатки парных антенн. На спинной сто-
роне эписферы образуется непарная антенна, которая у взрослых
Castalia punctata, как известно, отсутствует. Перистомий несет рот,
покрытый нежными ресничками и две пары щупальцевидных уси-
ков, из которых вентральные вдвое меньше дорсальных. Ротовой
сегмент щетинок не несет. Следующий сегмент является первым
щетинконосным. Спинные усики этого сегмента превосходят по
длине все остальные придатки. Они плоские, изогнуты назад и на-
поминают по форме и расположению ласты морской черепахи (Che-
lone imbricata). Видимо, они играют роль рулей при плаванье и
препятствуют вращению личинки вокруг продольной оси. Следую-
щий, третий сегмент лишен брюшного усика, что является для
нектохеты С. punctata очень характерным признаком, который
сохраняется и после метаморфоза до стадии в 10—20 сегментов.
На остальных сегментах дорсальные и вентральные усики разви-
ты нормально. Вентральные ветви параподий несут сложные ще-
тинки. Дорсальные ветви развиты слабо и несут по 1—2 волосо-
видных щетинки с зубчиками по наружному краю. Анальная ло-
пасть с вентральной стороны несет на конце непарный выступ и
пару двучленистых анальных усиков, прикрепляющихся ближе к
дорсальной стороне. На этой стадии начинает формироваться глот-
ка. Через некоторое время прототрох начинает исчезать, и личин-
ка опускается на дно, имея 6—7 щетинконосных сегментов.
3 В. А. Свешников
65
Осевшая на дно и развившаяся до 9 сегментов молодая поли-
хета имеет длину около 1 мм (рис. 13, 5, 6). У нее сохраняется
непарная антенна. Парные антенны и пальпы значительно увели-
чиваются. Спинные усики сегментов становятся очень длинными,
анальные усики — многочленистыми, а непарный вырост анальной
лопасти выглядит в виде длинного шипа. Сложные щетинки стано-
вятся двузубыми (рис. 13, 7) в вентральной части невроподиаль-
ных пучков и остаются по-прежнему однозубыми в дорсальных
частях. Глотка занимает объем 3—4 передних сегментов. По брюш-
ной стороне тела появляются зеленовато-коричневые зерна пиг-
мента. Весь ряд развития был получен в лабораторных условиях
после искусственного выведения из яиц, взятых от эпитокных ро-
дительских особей.
СЕМЕЙСТВО NEREIDAE
В семействе нереид мы встречаемся с большим разнообразием
способов размножения и развития. Возможно, это следует, поста-
вить в связь и с широкой способностью нереид приспосабливаться
к различным условиям существования. Они обитают в морях, эсту-
арных участках, входят в пресные воды, а отдельные виды (Lycas-
topsis) приспособились даже к наземному существованию на бана-
новых плантациях в сотнях километров от морского берега.
Нереиды вызывают значительный интерес у исследователей.
По данным Д. Рейш (Reisch, 1957), известно, что из 20 видов не-
реид 10 имеют свободное пелагическое развитие с хорошо выра-
женной эпитокией у взрослых. Это Nereis succinea (Frey, Leuckart),
развитие которого описано К. Банзе (Banse, 1954) и О. Кинне
(Kinne, 1954). Далее, свободное пелагическое развитие имеют
N. grubei (Kinberg) — по данным Д. Рейш (Reisch, 1954), N. irro-
rata (Malmgren), Perinereis marione (Audouin, Milne-Edwards) и
P. cultrifera (Grube)—по данным P. Герпин (Herpin, 1925) и
К. Казо (Cazaux, 1969). Однако у Р. cultrifera, по наблюдениям
В. Заленского (Salensky, 1882), Д. Пэр и П. Ранкурель (Peres,
Rancurel, 1948), происходит и прямое развитие без пелагической
стадии. Кроме того, свободное пелагическое развитие имеется у
Nereis pelagica — по данным Д. Вильсон (Wilson, 1932а), N. рго-
cera Ehlers и Platynereis bicanaliculata (Baird) — по наблюдениям
Д. Губерле (Guberlet, 1934), а также Р. dumerilii (Audouin, Mil-
ne-Edwards) — по данным К. Хауэншильд (Hauenschild, 1951) и
К. Казо (Cazaux, 1969). Nereis vexillosa Grube имеет эпитокию, но
развитие яиц у этого вида проходит внутри слизистых кладок
(Johnson, 1943).
Роение без эпитокии с последующим свободным развитием
наблюдается у Nereis japonica (Izuka) — по данным А. Ицука
(Izuka, 1908, 1912). Роение без эпитокии с последующим разви-
тием яиц внутри слизистых кладок описано Е. и К. Беркли (Вег-
66
keley, Berkeley, 1953) у Micronereis nanaimoensis Berkeley, Ber-
keley и, по данным Ф. Рюлье (Rullier, 1954), у N. variegata Cla-
parede. У двух последних видов наблюдается забота о потомстве,
выраженная у самцов.
Размножение без роения и эпитокии с последующим развити-
ем яиц в слизистых кладках под защитой самки или самца идет у
Ceratonereis costae (Grube) — по наблюдениям М. Дюрпюн (Dur-
chon, 1956) и у Neanthes caudata (delle Chiaje) — по данным
Д. Рейш (Reisch, 1957).
Прямое развитие имеется у Nereis diversicolor Muller — по
наблюдениям М. Богуцкого и Р. Дейле (Bogucki, 1954; Dales,
1950). Однако Г. Торсон (Thorson, 1946) указывает, что под влия-
нием солоноватых вод N. diversicolor может становиться живоро-
дящим.
Живорождение отмечено Р. Смитом (Smith, 1950) у Neanthes
lighti Hartman. У этого вида полное развитие яиц и личинок до
30-сегментной стадии происходит в течение 3—4 недель внутри
целома родительской особи.
Гермафродитизм наблюдается у трех видов: Hamanereis ran-
euensis — по данным X. Фаерборн (Feuerborn, 1932), N. indica
(Southern)— по наблюдениям Р. Айяр (Aiyar, 1935) и Platynereis
massiliensis (Moquin-Tandon) — по данным К. Хауэншильд
(Hauenschild, 1951). У двух последних видов это протандриче-
ский гермафродитизм.
Не остается сомнений в том, что характер развития у нереид
обладает значительной пластичностью и изменяется под действи-
ем внешних условий гораздо легче, чем у других полихет. Это до
некоторой степени и послужило основанием для выбора представи-
телей рода Nereis как объектов для акклиматизационных меро-
приятий в Каспийском море (Зенкевич, 1953).
Nereis virens Sars, 1846
Наряду с тем, что развитие одних нереид изучено с большими
подробностями, развитие других нереид почти совсем неизвестно.
В частности, данные о размножении и развитии Nereis virens очень
ограниченны.
В. Макинтош (Macintosh, 1910) наблюдал у берегов Англии в
мае нерест N. virens, которые выбрасывали яйцеклетки зеленова-
того цвета 20 мкм в диаметре. В октябре Макинтош обнаружил в
планктоне трохофор 228 мкк в диаметре и, не смутившись десяти-
кратным различием поперечников, отнес их к виду N. virens.
Г. Торсон (Thorson, 1946) выражает сомнение в том, что яйцеклет-
ки и трохофоры, описанные Макинтошем, принадлежат к одному и
тому же виду. Сам Торсон не наблюдал у берегов Дании размноже-
ния N. virens, но он приводит устные данные Густафсона, кото-
рый будто бы наблюдал непелагическое развитие этого вида у
берегов Швеции.
67
3*
Мы проследили размножение и развитие этого вида в планкто-
не Белого моря и в лабораторных условиях (Свешников, 1960).
N. virens ведет скрытый, закапывающийся образ жизни, а период
нереста и связанный с ним выход в толщу воды очень кратковре-
менны. Вероятно, поэтому червь долгое время не был обнаружен
в Белом море (Сластников, 1939). Теперь N. virens обнаружен поч-
ти на всей акватории Белого моря (Кудерский, 1959). Л. А. Зен-
кевич (1947) считает, что N. virens вселился в Белое море в связи
с современным потеплением Арктического бассейна и ранее там
не обитал. П. П. Анненкова и 3. Г. Паленичко (1947), напротив,
предполагают, что N. virens в Белом море представляет собой ре-
ликтовую форму. На основании данных по размножению и разви-
тию N. virens, изложенных ниже, мы склоняемся в сторону этого
последнего предположения.
В Белом море размножение N. virens неизменно из года в год
происходит при температуре воды 9° (Свешников, 1959) и. длит-
ся всего 2—3 дня. Особенно большое количество гетеронереидных
особей появляется в поверхностных слоях воды в том случае, если
оптимальная температура размножения совпадает с фазой ново-
луния или полнолуния. Эпитокные пелагические самцы и самки
длиной от 30 до 45 см всплывают на поверхность в штилевые ночи
и плавают, выставив из воды голову и сильно изгибая тело. В мо-
мент нереста самки и самцы N. virens вылавливались в эмалиро-
ванные тазы, в которых они выметывали половые продукты. Че-
рез некоторое время небольшие порции оплодотворенных яиц пе-
реносились в садки или кристаллизаторы, в которых происходило
дальнейшее их развитие.
Сперматозоиды имеют шаровидную головку, небольшой кони-
ческий перфораторий, вздутую шейку и обладают очень длинной
хвостовой нитью, до 50 мкм, тогда как головка имеет размер около
5 мкм. Общий габитус сперматозоида N. virens несколько напоми-
нает таковой у Micronereis variegata, описанный Ф. Рулье (Rul-
lier, 1954), хотя между ними имеются существенные различия.
'Яйцеклетки N. virens, выметанные в воду, имеют в диаметре
200 мкм. Большие скопления яиц имеют розоватую окраску. После
оплодотворения оболочка яйца сильно взбухает, и ее диаметр уве-
личивается до 500 мкм. Ядро расположено в центре яйца; ядро ок-
ружено жировыми и белковыми включениями. Как указывает
Р. Дейле (Dales, 1950), наличие в зрелых яйцах крупных жировых
капель, по-видимому, характерно для видов нереис, имеющих пе-
лагическую личинку. У видов Nereis, развивающихся без пелаги-
ческой личинки, например N. diversicolor (Беляев, 1953), жировые
капли в яйцеклетках отсутствуют. Содержимое яйца N. virens, по
нашим наблюдениям, имеет нежную желто-зеленую окраску.
Взбухшая оболочка абсолютно прозрачна.
Дробление неравномерное, гетероквадрантпое. Бластомер D
много крупнее остальных бластомеров. Гаструляция происходит
путем одновременно и параллельно идущих процессов эпиболии и
68
инвагинации. Бластопор возникает на оральном полюсе в виде от-
верстия с небольшим диаметром. В это же время определяется эк-
ваториальное расположение ресничного кольца. Другие эктодер-
мальные клетки, кроме трохобластов, ресничек не несут.
Личинку на этой стадии мы предлагаем называть трохофорной
гаструлой, так как она обладает одновременно признаками гаст-
рулы и трохофоры. В результате гаструляции образуется слепой
кишечник, ось которого на этой стадии совпадает с главной осью
тела. Поперек этой оси располагается экваториальный ресничный
пояс (рис. 14, Б). На следующей стадии бластопор трохофорной га-
струлы начинает смещаться вперед и останавливается в непосредст-
венной близости от ресничного пояса. В результате этого процесса
ось кишечника наклоняется и становится под прямым углом к
главной оси тела. Во время смещения бластопор сохраняет малый
диаметр, который имел в момент возникновения. Поэтому у живой
личинки бластопоральное отверстие почти невозможно заметить
(рис. 14, Б и 15, 1). На пути перемещения бластопора возникает
нейротрох (рис. 14, В, Г и рис. 15, 1). Следует особо отметить, что
бластопор не имеет щелевидной формы. Дальнейшие преобразова-
ния бластопора превращают его в ротовое отверстие, которое ведет
в слепой кишечник. Анальное отверстие возникает значительно
позднее и вне связи с бластопором. Подобная картина строения
стомодеума и положения бластопора известна у других нереид
(Costello, 1945).
Метатрохофора I (рис. 15, 2) имеет почти те же размеры, но
более вытянутую яйцевидную форму. На эписфере появляется па-
ра маленьких красных глазков. На теле метатрохофоры возникают
дополнительно три ресничных кольца — паратрохи. Все три пара-
троха смещены ближе к нижнему полюсу. Под последним паратро-
хом возникает такой же коричневый пигмент, как и под прототро-
хом. На этой стадии по бокам от кишечника образуются шесть пар
хетоносных мешков — три пары дорсальных и три пары вентраль-
ных. Передние пары хетоносных мешков находятся почти на уров-
не прототроха.
Метатрохофора II (рис. 15, 3) еще больше вытягивается. На
этой стадии оформляется ресничный пищевод. Паратрохи оказы-
ваются расположенными на равном расстоянии от прототроха и
друг от друга. В то же время хетоносные мешки отодвигаются
назад и благодаря этому две первые пары оказываются расположен-
ными между прототрохом и первым паратрохом, две вторые па-
ры — между первым и вторым паратрохами и, наконец, две третьи
пары оказываются расположенными между вторым и третьим
паратрохами. Создается впечатление, будто бы ротовой сегмент
несет щетинки. Однако Дальнейший сдвиг хетоносных мешков
назад, как мы увидим, меняет картину.
Ранняя нектохета (рис. 15, 4) имеет тело 250 мкм длиной. На
эписфере появляются еще две пары более крупных глаз. С боков
на каждом сегменте образуются сначала сглаженные, а затем
69
Рис'. 14. Процессы образования и перемещения бластопора у Nereis virens (ориг).
А — дробление яйца на стадии 8 бластомеров; Б — начало гаструляции, сагиттальный разрез; В — начальная стадия перемещения бпас-
топора у трохофорной гаструлы, сагиттальный разрез; Г -продолжение процесса перемещения бластопора, сагиттальный разрез; Вр-блас-
топор; Ch — хромосомы; L — желток; Nir — невротрохоид; Ptr —- прототрох
Рис. 15. Личиночные стадии Nereis virens (ориг.)
1 — трохофорная гаструла; 2 — ранняя метатрохофора; з — поздняя метатрохофора;
4 — ранняя нектохета; 3 — поздняя нектохета; в — челюсти нектохеты; 7 — семисег-
ментная донная ювенильная особь; 8 — челюсти на этой стадии; 9 — гетерогомфная
(слева) и гомогомфная (справа) щетинки из невроподиального пучка параподий личинки
более выпуклые бугорки параподий. Пучки сложных щетинок вы-
ставляются далеко за пределы тела. В момент плаванья у N. virens
щетинки плотно прикладываются к телу, которое может слегка
изгибаться. На каждом щетинконосном сегменте имеется паратрох.
Анальная лопасть окрашена коричневатым пигментом и несет
пару маленьких анальных усиков. Щетинки в щетинконосных
мешках закладываются, начиная с дистальной части концевого чле-
ника, затем появляется весь членик, затем образуется дистальный
конец рукоятки и в последнюю очередь основная часть щетинки.
Окончательно сформировавшаяся нектохета имеет размер
430 мкм (рис. 15, 5). Голова приобретает округлую, почти шаровид-
ную форму и резко граничит с первым хетоиосным сегментом. С бо-
ков головы появляются очень характерные на этой стадии крупные
пигментные пятна, занимающие значительную часть головы. Эти
пятна исчезают сразу после естественной смерти или после фикса-
ции личинки. Простомиальиая часть головы несет пару антенн и
пару шаровидных пальп. Прототрох, значительно уменьшающийся
к этому времени, проходит позади пальп и по верхней гра-
нице крупных глазных пятен, разграничивая простомиум от пе-
ристомия. На брюшной стороне по этой границе несколько ниже
прототроха лежит ротовое отверстие. С боков перистомия появля-
ется сначала только одна пара тентакулярных усиков. Только те-
перь становится ясно, что перистомий формируется за счет участ-
ков, расположенных между прототрохом личинки и первым хето-
носным мешком и никаких щетинок не несет. Границы сегментов
тела становятся очень ясными благодаря необыкновенно сильно
развитым на этой стадии хетоносным мешкам. Таким образом, в
процессе развития еще у метатрохофоры первые две пары хетонос-
ных мешков возникают на уровне прототроха и постепенно пере-
мещаются назад, оказываясь, наконец, позади перистомия. За пер-
выми парами хетоносных мешков соответственно располагаются и
остальные — вторая и третья пары. Тогда, считая перистомий пер-
вым сегментом туловища, у Nereis virens насчитывается не три, а
четыре ларвальных сегмента. У сформировавшейся нектохеты на
первом щетинконосном сегменте щупальцевидные усики развиты
гораздо слабее, чем на двух других сегментах. Параподиальные
бугорки на этом сегменте тоже выражены гораздо слабее. Пучки
щетинок второго и третьего сегментов опираются на коричневые
ацикулы. Кишечник на этой стадии состоит из трех отделом.
В глотке, занимающей объем всего первого хетоиосиого сегмента,
закладываются челюсти. Первоначально челюсти состоят из двух
отделов — большого концевого зубца и небольшого придаточного от-
дела округлой формы, снабженного двумя зубчиками (рис. 15, 6).
Этот придаточный отдел впоследствии обламывается, так что его
можно считать личиночным. Подобное сбрасывание личиночных
зубцов на челюстях Micronereis variegata в процессе метаморфоза
наблюдал Ф. Рюлье (Rullier, 1954). В среднем отделе кишечника
еще имеются остатки зеленоватого желтка и несколько крупных
73
прозрачных вакуолей. Задний отдел кишечника имеет ресничную
выстилку. Анальная лопасть несет пару анальных усиков, опушен-
ных на самых кончиках чувствительными ресничками. На этой ста-
дии личинка садится на дно, и некоторое время прироста новых се-
гментов на ее теле не наблюдается. После закладки четырех пер-
вых сегментов у N. virens протекает длительная пауза, в течение
которой новые сегменты не образуются. Уже в бентосе через не-
которое время у личинки появляются постларвальные сегменты.
Личинка на стадии семи сегментов (рис. 15, 7) имеет округлую
и четко отделенную от туловища голову. Простомий несет спереди
пару опушенных редкими ресничками антенн, а снизу пару корот-
ких почти шаровидных пальп. С боков средней части головы еще со-
храняются красно-коричневые большие глазные пятна. В затылоч-
ной части головы имеются две пары собранных в группы дефинитив-
ных глаз. Перистомий несет две пары щупальцевидных усиков, из
которых дорсальная пара вдвое крупнее вентральной. Параподии
первого щетинконосного сегмента оказываются совсем неразвиты-
ми. На этом сегменте имеется пока одна пара дорсальных усиков, а
вентральные находятся в зачаточном состоянии. Пучки щетинок
на этом сегменте редкие. Щетинки в них держатся слабо и легко
выпадают. Параподии остальных сегментов развиты нормально.
В них появляются ното- и невроподиальные ветви с развитыми вен-
тральными и дорсальными усиками. В нотоподиальиых пучках, ко-
торые закладываются с опозданием, щетинки чаще гомогомфные.
В невроподиальных пучках щетинки в одинаковой мере могут
быть и гомо- и гетерогомфные (рис. 15, 9). Анальные усики за-
метно удлиняются. Кишечник дифференцируется на отделы.
Глотка становится объемистой, и в ней хорошо видны многозубые
челюсти. Между зубчиками челюстей как бы натянута прозрач-
ная мембрана (рис. 15, 8). Позади глотки обособляется малень-
кий желудок. Глотка и желудок занимают объем двух первых
сегментов тела. Далее идет средняя кишка, наполненная остат-
ками зеленоватых зерен желтка. Задний отдел кишечника зани-
мает объем двух последних сегментов и открывается на спинной
стороне пигидиума анальным отверстием.
На стадии восьми сегментов происходят следующие изменения
в строении, молодой полихеты. Пальпы становятся двучленистыми
и хорошо видны сверху. Первый хетоносный сегмент окончатель-
но теряет щетинки и передвигается ближе к голове. На нем появ-
ляется до сих пор отсутствующий вентральный усик, а дорсальный
усик этого сегмента оказывается длиннее обоих щупальцевидных
усиков перистомия. Окончательно потеряв щетинки и продвигаясь
далее вперед, этот сегмент образует при слиянии с перистомием
метастомий. Из сказанного видно, что в состав головы у Nereis вхо-
дят, помимо простомия и ротового сегмента, еще и первый щетин-
коносный сегмент червя, т. е. только два из четырех ларвальных
сегментов, а не все ларвальные сегменты, как это считает
В. Н. Беклемишев (1952).
74
Таким образом, развитие N. virens в Белом море происходит с
типичной пелагической личинкой и длится не больше недели. До
сих пор во взрослом состоянии N. virens не найден в проме-
жутке между Бергеном (Западная Норвегия) и Белым морем.
Короткий период пелагической стадии N. virens исключает возмож-
ность проникновения личинки червя со струями теплого течения
от берегов Норвегии в Белое море, так как этот путь должен был
бы продолжаться несколько месяцев. Поэтому N. virens следует
считать тепловодным реликтом в фауне Белого моря (Свешни-
ков, 1955). В случае дальнейшего потепления Арктики ареалы
Nereis virens в Белом и Норвежском морях могут сомкнуться.
ОТРЯД EUNICEMORPHA
Размножение и развитие различных представителей отряда
Eunicemorpha изучено в некотором отношении лучше, чем у дру-
гих отрядов полихет. Здесь известны многие детали раннего эмбри-
огенеза (Akesson, 1967b), личиночное развитие (Claparede, Meczni-
kow, 1869; М. И. Киселева, 1957; Свешников, 1967), эпитокия
(Woodworth, 1907; Mayer, 1908; Caspers, 1961) и различные спосо-
бы защиты потомства (Thorson, 1946, 1952). Сопоставление дан-
ных, заимствованных из литературы, приводит к убеждению о
том, что Eunicemorpha имеют значительное и интересное разно-
образие способов размножения и развития. Разнообразны способы
эмбриогенеза, удивляют некоторые факты морфогенеза, например
необычная форма образования стомодеума у Eunice kobiensis.
Разнообразны формы личинок. Интересны олигомерные формы
Eunicemorpha, напоминающие метатрохофор (Korschelt, 1893;
Akesson, 1967b). Наконец, обращает на себя внимание то, что
именно в отряде Eunicemorpha встречаются формы, приспособ-
ленные к паразитированию на червях, иглокожих, ракообразных и
рыбах (семейства Lumbriconereidae, Histriobdellidae и Ichthyoto-
jnidae).
СЕМЕЙСТВО EUNICIDAE
Представители этого семейства размножаются различными спо-
собами. У некоторых видов наблюдается эпитокия (Eunice viridis
и Е. fucata). Е. viridis у островов Фиджи и Самоа образует эпи-
токную часть на заднем конце тела. Передний атокный конец тела
остается на дне до следующего года, а задний эпитокный конец
всплывает на поверхность для размножения половым путем не-
пременно в октябре — ноябре. Атлантический палоло (Е. fucata) у
Антильских островов таким же способом размножается в июле.
Эмбриогенез изучен только у отдельных видов, а особенно под-
робно у Eunice kobiensis. Мы воспользуемся данными Б. Акессона
75
(Akesson, 1967b) по эмбриогенезу Eunice kobiensis, которые им
были полуиены в лаборатории Харбор на архипелаге Сан-Жуан
(Атлантика). В наших водах Японского моря Eunice kobiensis так-
же встречается. Однако для япономорской популяции этого вида
данных по эмбриогенезу в литературе нет.
Eunice kobiensis McIntosh, 1885
Б. Акессон (1967b) пользовался осмированным материалом,
залитым в глютаральдегид. Личинок после фиксации и заливки ре-
зали на ультратоме стеклянным ножом (1 мкм). Окраска желез-
ным гематоксилином по Гейденгайну.
Шаровидно дробящиеся яйца имели диаметр около 310 мкм.
Самой ранней стадией в материале была бластула, сформировав-
шаяся спустя 24 часа после оплодотворения. Это целобласту'ла с
небольшим бластоцелем. Вегетативный полюс занят клетками,
производными основного квартета, которые загружены желтком
(рис. 16, А и В).
Гаструляция происходит частично путем инвагинации, частич-
но путем эпиболии. В результате этого образуется гаструла с объ-
емистым, глубоким архентероном (рис. 16, В и Г). Гаструляция
заканчивается 30 ч спустя после оплодотворения. Трохобласты рас-
полагаются по экватору гаструлы. Такая личинка заслуживает на-
звания — трохофорная гаструла. На границе бластопора с правой и
левой сторон архентерона образуются телобластические полоски,
вытянутые вдоль главной оси тела. Никаких признаков плагиак-
сонии не наблюдается.
Через 38 ч после оплодотворения по середине вентральной сто-
роны под ресничным поясом возникает новая зона образования
стомодеума (рис. 17, А и Б). Бластопор в это время несколько
уменьшается в диаметре, а объем полости архентерона сокращает-
ся. Размеры парных телобластических полосок увеличиваются. На
протяжении третьего, четвертого и пятого дней стомодеум все
глубже выпячивается внутрь, диаметр бластопора и объем поло-
сти архентерона уменьшаются. Однако все это время бластопор
и вторично возникший рот существуют вместе, можно сказать на
противоположных полюсах животного (рис. 17, А — Е).
Б. Акессон, по работе которого мы излагаем материал, подчер-
кивает, что такое соотношение между положением бластопора и
независимо от него возникающим (вторичным) ртом очень харак-
терно для вторичноротых животных. Близкую картину образова-
ния стомодеума независимо от бластопора можно наблюдать сре-
ди первичноротых в развитии немертин и волосатиков (Gordi-
асеа).
На вторые сутки развития Eunice kobiensis на вершине архен-
терона (рис. 17, В), почти по главной оси тела, возникает непар-
ный зачаток головной мезодермы. По описаниям и рисункам
Б. Акессона этот зачаток возникает вне связи с телобластическими
7в
Рис. 16. Ранние стадии эмбрионального развития Eunice ko-
biensis (по Akesson, 1967; с изменениями)
А -а Б — продольное сечение бластулы; Ви Г — продольное сечение гаструлы: 1 — бпа-
стоцепь; 2 — архентерон; а — анимальный полюс; V — вегетативный полюс
полосками, которые к этому времени еще не достигают области, где
закладывается мезодерма головы (рис. 17, В). Таким образом,
Б. Акессон описывает два источника возникновения мезодермы.
Подобную картину мы видели в развитии Harmothoe imbricata
(см. рис. 8, 6). Дальнейшее развитие мезодермального зачатка го-
ловы приводит к образованию непарного головного целома. Непар-
ный головной целом описан для других полихет различными авто-
рами (Anderson, 1959; Ерохина, 1968). У форонид непарный голов-
ной целом трохофорной личинки актинотрохи, образующейся сход-
ным способом, носит название аксоцель.
Дальнейшие процессы развития Eunice kobiensis приводят к
формированию личинки, у которой последовательно развиваются
сначала щетинконосные мешки, а затем характерный челюстной
аппарат внутри глоточного мешка.
77
Рис. 17. Образование сто-
модеума вне связи с бла-
стопором у Eunice ko-
biensis (по Akesson,
1967)
А — сагиттальный разрез
личинки в возрасте
38 ч;
В — то же, двух дней;
В — трех дней;
Г — четырех дней;
Д — пяти дней;
Е — семи дней;
1 — эктодерма переднего
конца;
2 — зачаток прототроха;
з — эктодерма заднего кон-
ца;
4 — эндодерма;
S — архентерон;
е — зачаток брюшной нерв-
ной цепочки;
7 — стомодеум;
в — граница мезодермаль-
ной полоски;
9 — мезодерма головного
конца
Lumbriconereis japonica Marenzeller, 1879
Для Японского моря развитие Lumbriconereis было впервые
описано С. Окуда (Okuda, 1946). С. Окуда считал, что имел дело
с L.latreilli And. et M.-Edw., но M. Имаджима и О. Хартман (Ima-
jima Hartman, 1946) относили этот вид к L. japonica Marenzeller.
Детальные описания развития этого вида касались только внеш-
ней морфологии. Мы впервые даем для личинок L. japonica анато-
мическое строение.
В зал. Посьет кладки этого вида появились в массовом количе-
стве в первой половине июня при температуре 16° на глубине 0,7—
2 м. Кладки были прикреплены к листьям Zostera marina, на мерт-
вой ракушке и на раковинах живых брюхоногих и двустворчатых
моллюсков. При кладках находились и взрослые особи Lumbricone-
reis. Чаще других носили на себе эти кладки Tegula rustica и Но-
75
melopoma sangerense *. Слизистые сферические кладки яркого оран-
жевого цвета имели в диаметре от 1,5 до 3 см. Внутри слизистой
массы шнур яиц уложен зигзагами или по спирали. Яйцо овальной
формы имеет 300 мкм в ширину и 400 мкм по длинной оси. Яйцо
нагружено желтком оранжевого цвета. Спустя несколько часов под
яйцевыми оболочками проходит стадия трохофоры (рис. 18, Л).
Трохофора несет следы процесса эмбрионизац'ии. Многие спе-
циальные провизорные приспособления имеют примитивную форму
или не развиваются совсем. Прототрох в обычном значении этого
слова отсутствует. Личинка имеет кольцевое ресничное поле до
250 мкм шириной при общей длине тела 350 мкм. Это ресничное
поле опоясывает личинку по экватору. Реснички слабые, не более
5 мкм длиной, обеспечивают лишь вращение личинки вокруг глав-
ной оси и аэрацию. На апикальном поле личинки — небольшой хо-
холок. Маленькое ресничное поле есть и на вегетативном полюсе.
Таким образом, большая часть поверхности тела личинки песет
ресничный покров. На этой стадии ресничный эпителий представ-
лен плоскими клетками не более 3 мкм толщиной. Под эпителием
залегает синцитиальная масса, наполненная гранулами желтка.
Никакой более глубокой дифференциации клеточный материал
трохофоры Lumbriconereis не испытывает. Весь ход развития про-
текает у Lumbriconereis внутри слизистой кладки. Отсутствие сво-
бодного развития приводит к сглаживанию границ между ста-
диями. Здесь невозможно отделить стадии метатрохофоры и нек-
тохеты, так как они утрачивают свою специфику. Дальнейшие
преобразования идут в следующем порядке. Ресничный пояс при-
обретает более интенсивную оранжевую окраску по сравнению с
остальной частью тела. В задней трети тела образуется зона, в ко-
торой эктодермальные клетки делятся и формируют1 пару щетин-
коносных мешков (рис. 18, В, В). В каждом мешке формируется
по три щетинки разного строения. Через двое суток тело личинки
достигает 550 мкм в длину. Теперь можно различить два отдела,
разделенные перетяжкой. Передний отдел состоит-из части, кото-
рая соответствует эписфере личинки и простомиуму взрослого жи-
вотного.
На более светлом фоне клеток эктодермы выступают симмет-
ричные оранжевые пятна. Позади эписферы располагается реснич-
ный пояс прототроха до 120 мкм шипиной. Пояс состоит из птэа-
вой и левой половин, между которыми проходит узкая щель, ли-
шенная ресничек. Перетяжка между головой и туловищем окра-
шена значительно слабее, чем прототрох и туловище. Последнее
разделено на две равные части. Передняя половина туловища
представляет собой первый щетинконосный сегмент. Он ограничен
спереди и сзади правильными рядами желтых пигментных. пятен
удлиненной формы. Сегмент несет пару весьма подвижных щетин-
коносных мешков со щетинками. В каждом мешке развиты ще-
1 Моллюски, на которых находились эти кладки, любезно были определены
А. Н. Голиковым.
79
Рис, 18. Личиночное раз-
витие Lumbriconereis jap'o-'
nica (ориг.)
А — протрохофора в оболоч-
ке яйца;
Б — личинка на стадии одного
щетинконосного сегмента;
В — личинка трехдневного воз-
раста;
I — личинка с пятью сегмен-
тами, вышедшая из клад-
ки (с брюшной стороны)
тинки трех видов. Самая крупная щетинка внешне напоминает
в этом возрасте щетинки взрослых серпулид. Изогнутый конец ру-
коятки имеет два зазубренных ребра, которые соединяются в тон-
кое остро оттянутое лезвие. Вторая щетинка, круглая в сечении,
окаймлена с двух сторон и кончается вытянутым острием. Третья
щетинка копьевидная. Щетинки обнаруживают такую подвиж-
ность, которая позволяет личинке выбраться из слизистой клад-
ки и перейти к самостоятельной жизни в грунте.
Головной и брюшной мозг имеют сравнительно небольшие раз-
меры. Индифферентной ткани в теле личинки на этой стадии
немного. Личинка малоподвижна. Основные ее усилия тратятся на
медленное движение внутри слизистой массы кладки.
Личинка, благополучно покинувшая оболочку кладки, далее
формируется следующим образом. У личинки последовательно,
один за другим, образуются щетинкоиосные сегменты (рис. 18, Г).
Под ресничным кольцом прототроха возникает ротовое отверстие.
Кишечник становится сквозным на стадии двух или трех щетинко-
носных сегментов. Однако и на этой стадии эндодерма содержит
еще большое количество недоиспользованного запаса энергии в
виде гранул желтка. Челюстной аппарат в глотке формируется на
стадии пяти щетинконосных сегментов. Только в это время
Lumbriconereis приступает к активному питанию. Ювенильные
формы были получены из кладок в лабораторных условиях.
80
* * *
Анализ литературных данных по развитию других Eunicemor-
pha показывает следующую общую картину. Наиболее примитив-
ную форму онтогенеза имеют представители Ophryotrocha (семей-
ство Dorvilleidae — Akesson, 1967b). Свободная личинка снабжена
минимальным количеством желтка, опоясана несколькими реснич-
ными кольцами, активно питается и имеет метаморфоз эволютив-
ного типа. В результате ряд личиночных адаптаций сохраняется
на всю жизнь. Например, ресничные кольца у Ophryotrocha суще-
ствуют наряду с пучками щетинок. Олигомерное тело и небольшие
размеры подчеркивают характеристику.
У представителей Lumbriconereis (Lumbriconereinae) онтоге-
нез представлен в двух вариантах. В Средиземном море (Clapare-
de und Mecznikow, 1869) имеет место пелагический тип развития.
В Японском море (Okuda, 1946; Свешников, 1967) перегрузка яиц
желтком и формирование кладок приводят к полной утрате сво-
бодного развития.
У Lysidice (подсемейство Eunicinae) яйца имеют большой за-
пас желтка (М. И. Киселева, 1957). Это изменяет облик личинки.
Тонкий ресничный эпителий личинки составлен из очень мелких
клеток, несущих нежные реснички по всей поверхности тела. Хо-
рошо развиты глаза. Личинка ведет свободный образ жизни на
всех стадиях. Однако она*не питается активно. Обычные морфоло-
гические границы между стадиями сглаживаются. Различий меж-
ду ларвальным и постларвальиым телом не образуется.
Эмбрионизация процессов в этом ряду достигает, наконец, выс-
шего предела у Eunice kobiensis. Здесь процессы перемещения
бластопора идут с такой скоростью, что получается эффект вто-
ричноротости, как это замечает и автор работы по развитию Е. ko-
biensis (Akesson,1967b).
В. Н. Беклемишев (1964), разбирая вопрос о происхождении
двусторонней симметрии, пишет, что ее возникновение связано у
всех Bilateria с перемещением рта в одну из антимер. «Scolecida
и Deiiterostomia начинают ползать, лежа на одной из антимер,
первичный рот сдвигается со своего места на вегетативном полюсе
и располагается на ползательной стороне, в оральной плоскости,
которая в силу этого становится единственной плоскостью симмет-
рии— сагиттальной плоскостью» (том I, с. 339). Трохофорных
животных В. Н. Беклемишев исключал из той группы Bilateria,
которой свойственны процессы перемещения бластопора. Теперь,
когда стали известны работы Акессон (Akesson, 1967b) и наши
(Свешников, 1971, 1972), мы можем с полным правом и трохофор-
ных животных относить к таким Bilateria, которым свойственны
процессы перемещения бластопора.
ОСОБЕННОСТИ РАЗВИТИЯ
И ЛИЧИНКИ ПОДКЛАССА SEDENTARIA
Деление полихет на подлассы Errantia (бродячие) и Sedenta-
ria (неподвижные) можно принять лишь условно. Условность та-
кого деления подтверждается, например, тем, что сидячая поли-
хета Poecilochaetus serpens может совершать весь свой жизненный
цикл в толще воды. Многие пелагические личинки Sedentaria на-
чинают строить основы своего домика еще во время пелагической
жизни (Chaetopteridae, Pectinariidae, Terebellidae), а иные с этим
домиком навсегда остаются в составе пелагических организмов
(Terebellidae).
Вместе с тем при рассмотрении материала по развитию Seden-
taria бросается в глаза значительно большая глубина адаптивных
признаков личинок сидячих полихет.
Наиболее характерными признаками личинок и циклов разви-
тия Sedentaria мы считаем следующие.
1. Циклы развития Sedentaria чаще протекают внутри трубок,
капсул, кладок или под прикрытием материнского организма.
2. Яйцеклетки Sedentaria богаче, чем у Errantia, снабжены пи-
тательными веществами и соответственно у Sedentaria чаще прояв-
ляется эмбрионизация, укорочение или выпадение каких-либо от-
дельных стадий, составляющих обычную последовательность ряда
(гаструла, протрохофора, трохофора, метатрохофора, нектохета,
ювенильная донная форма). Часто сглаживается или исчезает гра-
ница между ларвальным и постларвальным отделами.
3. Среди Sedentaria встречаются семейства, где личиночные
приспособления уводят организм далеко от нормальных линий
развития, в сторону катастрофического метаморфоза (Oweniidae).
Сумма особенностей онтогенеза Sedentaria позволяет сказать,
что этот подкласс более специализирован в сторону вторичных
приспособлений по сравнению с Errantia. Однако в отдельных слу-
чаях, когда количество желтка в яйцеклетках падает до минимума
(например, у некоторых Chaetopteridae), неожиданно проявляют-
ся архаичные признаки развития. У Chaetopterus variopedatus яй-
цевые оболочки покидает настоящая гаструла, бластопор которой
мигрирует к переднему полюсу и обнаруживается полная идентич-
ность с подобными процессами у турбеллярий и немертин (Свеш-
ников, 19726).
Деление подкласса на отряды достаточно стабильно удержива-
ется десятилетиями. Н. А. Ливанов (1940) называет всего три от-
ряда — Spiomorpha, Terebellomorpha и Serpulimorpha. П. В. Уша-
52
ков (1955) делит подкласс Sedentaria на четыре отряда — Spiomor-
pha, Drilomorpha, Terebellomorphli и Serpulimorpha. Исследование
индивидуального развития ряда семейств, входящих в состав этих
отрядов, показывает, что нет достаточных оснований для значи-
тельно более дробного деления подкласса на отряды, как это дела-
ет Р. Дейле (Dales, 1962, 1963), который называет 14 отрядов по-
лихет. Некоторые современные специалисты по аннелидам вообще
отказываются от необходимости деления класса полихет на отря-
ды (Hartman, 1959; Bellan, 1964). Мы принимаем деление класса
полихет на отряды, предложенное П. В. Ушаковым (1955, 1972).
ОТРЯД SPIOMORPHA
Способы размножения и развития в отряде Spiomorpha весьма
разнообразны. Некоторые виды размножаются бесполым способом.
Например, Pygospio elegans на литорали датского побережья раз-
множается путем фрагментации. Фрагменты из 2—7 сегментов ре-
генерируют недостающие части тела. По мнению Е. Расмуссена
(Rasmussen, 1953), бесполое размножение Р. elegans на датском
побережье — весьма обычное явление в течение летнего времени.
В зимнее время наступает период полового размножения. Беспо-
лое размножение наблюдается также у Cirratulidae. У Dodecaceria
caulleryi отшнуровываются задние сегменты, из которых каждый
образует четыре новых червя. В этом случае отделившиеся сегмен-
ты в свою очередь размножаются, а не только регенерируют (De-
horne, 1933; Martin, 1933, 1936; Herlant-Meewis, 1958). У той же
Dodecaceria наблюдается партеногенетическое развитие яиц в по-
лости тела матери до стадии трехсегментной личинки, которая за-
тем переходит к самостоятельному образу жизни.
Развитие у различных Spiomorpha протекает либо в толще во-
ды (некоторые Ariciidae, Spionidae, Chaetopteridae), начиная с пе-
лагического яйца и кончая поздней нектохетой, либо дробящиеся
яйца, трохофоры и метатрохофоры формируются внутри слизи-
стых кладок, внутри материнской трубки, а более поздние ста-
дии — нектохеты и многосегментные ювенильные особи выходят
в толщу воды (некоторые Spionidae, Magelonidae, Poecilochaetidae).
Наконец, Cirratulidae проявляют заботу о потомстве, которое за-
щищают и выкармливают в собственных щупальцах. У некоторых
Chaetopteridae личинки на ранних стадиях имеют примитивную
гаструлообразную форму, а на поздних стадиях весьма специали-
зированы (Stomaria, см. рис. 1, И). Однако глубина специализа-
ции личинок Chaetopteridae не достигает таких уровней, которые в
других отрядах приводят к катастрофическому метаморфозу (Owe-
niidae, Drilomorpha). Глубина специализации личинок Chaetopte-
ridae соответствует глубине специализации взрослых форм, и раз-
витие протекает путем постепенных (эволютивных) изменений.
83
СЕМЕЙСТВО AR1CIIDAE
Индивидуальное развитие некоторых представителей семей-
ства Ariciidae было детально исследовано Д. Андерсоном (Ander-
son, 1959, 1961, 1966, 1973). Андерсон поставил под сомнение ги-
потезу П. П. Иванова (Ivanov, 1928, 1944) о первичной гетероном-
ной метамерии тела сегментированных животных, которая по праву
заняла одну из главных позиций в сравнительной анатомии и эмб-
риологии. Мы. видели, что количество ларвальных сегментов у по-
лихет различно. В некоторых семействах это очень постоянный
признак (например, у Serpulidae их всегда три). В других семей-
ствах количество ларвальных сегментов непостоянно (Spionidae).
В некоторых случаях деление сегментов на ларвальные й пост-
ларвальные не выражено.' Андерсон как раз исследовал такой
факт развития сегментов у Scoloplos armiger, когда деление .тела
на ларвальный и постларвальный отделы не выражено.
Scoloplos armiger (О. F. Miiller, 1776)
Развитие S. armiger у европейских берегов наблюдали в дат-
ских водах Г. Торсон (Thorson, 1946) и Е. Смидт (Smidt, 1951),
а у берегов Англии Д. Андерсон (Anderson, 1959). Мы вывели в
лабораторных условиях пелагических личинок этого вида и воспи-
тали их до ювенильного состояния, когда проявились основные
признаки взрослых животных.
По данным Г. Торсона (1946) и Д. Андерсон (1959), Scoloplos
armiger на европейском побережье откладывает крупные слизи-
стые кладки, прикрепленные к частицам песка. Дробление яиц и
формирование трохофорных личинок проходит внутри таких сли-
зистых кладок. В Белом море мы никогда не находили подобных
кладок. Напротив, нам удалось наблюдать, как половозрелые сам-
ки выбрасывают в воду одиночные яйца, которые далее дробятся
в поверхностных слоях воды, и трохофора также ведет пелагиче-
ский образ жизни. Яйца имеют шаровидную форму, 230 мкм в диа-
метре. Они окрашены в оранжевый или рыжий цвет, богато снаб-
жены желтком и обладают значительной плавучестью. С первых
стадий дробления легко отличить анимальный полюс от вегетатив-
ного, так как на анимальном полюсе сосредоточиваются микроме-
ры, а макромеры занимают вегетативный полюс (рис. 19, Л). Эта
разница расположения бластомеров не теряется до момента появ-
ления ресничного пояса, который образуется сначала только по
бокам тела личинки и в этих местах прорывает оболочку яйца. Ли-
чинка на этой стадии приобретает способность слабо двигаться
внутри яйцевых оболочек (рис. 19, В). Далее ресничный пояс ста-
новится кольцевым и постепенно прорывает оболочку яйца так,
что, наконец, личинка освобождается от нее совсем. Очень корот-
кое время личинка плавает при помощи одного пояса (рис. 19, В).
На этой стадии у личинки появляется пара красных глазков. Через
несколько часов у нее развивается второй ресничный пояс, распо-
ложенный вокруг анальной лопасти (рис. 19, Г).
84
Рис. 19. Личиночное развитие Scoloplos armiger (ориг.)
А — дробление яйца; Б — ранняя трохофора; В — трохофора, освободившаяся от яй-
цевых оболочек; Г — поздняя трохофора; Д — метатрохофора; Е ювенильная особь,
Ж___то же, справа с выброшенной глоткой: 3 — молодой червь; И, К, Л, М, Н ще-
тинки
de ™
Рис. 20. Трохофора Scoloplos armiger (no Anderson, 1959)
A — схема сагиттального разреза; В — схема фронтального разреза; С — фрон-
тальный разрез; ар — апикальная пластинка; аг — архентерон; Ъ1 — бластоцель; си —
кутикула; de — эктодерма спинной стороны; ета — эктомезодерма переднего источника]
етр — эктомезодерма приротового участка; еп — эндодерма; et — эктотелобласт; 1е —
эктодерма боковых сторон; т — рот; mb — мезодермальные полоски; mt — мезодермаль-
ные телобласты; р — ресничный пояс; руе — эктодерма пигидия; st'pr — зачаток стомодеу-
ма; ve — вентральная эктодерма
Д. Т. Андерсон (1959) обращает внимание на некоторые анато-
мические особенности строения трохофоры S. armiger. Мезодермаль-
ные телобластические полоски, начиная с очень ранних трохофо-
ральных стадий, достигают значительного развития по сравнению
с подобной картиной у представителей Errantia, например Наг-
пюЬЬоё imbricata. Мезодермальные полоски S. armiger заполняют
86
mt PW
Рис. 21. Метатрохофора Scoloplos armiger (no Anderson, 1959)
A — схема фронтального сечения трехдневной личинки; В — схема фронтального сечения
личинки в возрасте четырех дней; С — фронтальный разрез; ар — апикальная пластинка;
cl, с2, сз — целомические полости первых трех щетинконосных сегментов; сао — непар-
ный головной целом; са1} са2, са3 — целомические полости в области ресничного пояса;
сте — целомическая полость в области мезотроха; ema — эктомезодерма переднего ис-
точника; еп — эндодерма; et — эктотелобласт; mb — мезодермальные полоски; те —
метатрох; тез — метатрохальный сегмент; mt — мезодермальные телобласты; р — рес-
ничный пояс; pro. — зачаток глотки; руе — эктодерма пигидия; se — эктодерма боко-
вых сторон; st —стомодеум; t — телотрох; у — желток
все свободное пространство бластоцеля трохофоры и на ранних
стадиях развития уже достигают предельного положения вплоть
до переднего полюса. Кроме того, Д. Т. Андерсон описывает и по-
казывает на рисунках передний источник эктомезенхимы, который
дает группу мезодермальных клеток, подстилающих апикальную
пластинку. Позднее эта группа мезодермальных клеток образует
87
стенку непарного головного целома (рис. 20, А, В, С). Подобный
источник, образующий непарный головной целом, мы наблюдали
и у Harmothoe imbricata (см. рис. 8, Г; рис. 10, Г; рис. 12). Таким
образом, в строении трохофоры S. airmiger и Н. imbricata обнару-
живаются некоторые черты сходства, которые заключаются в том,
что у обоих видов передний источник эктомезенхимы дает начало
стенке непарного головного целома.
Дальнейшее развитие приводит к формированию метатрохофо-
ры (рис. 19, Д), на удлиненном теле которой один за другим по-
являются новые ресничные пояски — паратрохи. Это проявление
метамерии в эктодерме соответствует развивающейся метамерии
в мезодерме.
По мнению Д. Т. Андерсон (1959), мезодерма в развитии мета-
трохофоры S. armiger ведет себя иначе, чем в развитии других по-
лихет. Особенности развития мезодермальных полосок выражены
в том, что внутри групп клеток, их составляющих, последователь-
но образуются парные целомические полости (рис. 21, А, В., С).
Д. Т. Андерсон различает помимо непарного переднего целома
(Са0) три пары передних целомов (Са15 Са2, Са3), расположенных
в области ресничного кольца. Следующая за ними пара цело-
мов (сте) располагается у метатрохофоры на уровне метатро-
ха (те). Все целомические полости последовательно возникают
сзади наперед. В последнюю очередь возникают пара передних
целомов (caj и непарный целом вблизи анимального полюса
(рис. 21, А, В). Вслед за тем образуются целомические полости бу-
дущих щетинконосных сегментов (Cl, С2, СЗ и т. д.) (рис. 21,
В, С). Характерно, что никакой паузы в образовании последова-
тельного ряда целомических полостей у метатрохофоры S. armiger
наблюдать не удается. Полости этих сегментов Андерсон обозна-
чает как полости щетинконосных сегментов (рис. 21, В, С — С1,
С2, СЗ и т. д.), таким образом отделяя их от передних ларвальных
сегментов. Многие различия между ларвальным и пост ларвальным
отделами тела y.S. armiger скрыты и затушеваны процессами эмб-
рионизации, которые обычно сильнее проявляются при гипертро-
фии объема питательных веществ в яйце. Яйца S. armiger следу-
ет относить именно к такому типу, так как количество желтка в
них достаточно обильно. Вместе с тем одно из наиболее суще-
ственных различий между характером образования ларвальных и
постларвальных сегментов остается. Ларвальные сегменты Scolop-.
los armiger, как это следует по теории П. П. Иванова (1944), за-
кладываются сзади наперед и почти одновременно, а постларваль-
ные — спереди назад и последовательно в течение длительного
времени.
12-сегментная нектохета имеет длину около -600 мкм
(рис. 19, Е). Ясно выделяется конический простомиум, на кото-
ром образуется дополнительный венчик ресничек — акротрох, рас-
положенный выше глаз двумя разорванными участками. Прото-
трох расположен на границе простомиума и перистомиума. Пер-
88
вые два сегмента, следующие за головой, имеют очень слабые
паратрохи и лишены щетинок. Остальные сегменты, начиная с
третьего, несут более развитые паратрохи и снабжены пучками ще-
тинок. На этой стадии еще нет никаких признаков бугорков пара-
подий. Сначала у личинки появляются только волосовидные ще-
тинки (рис. 19, IT). Они искривлены и по внешнему краю имеют
зубчатость. Первоначально эта зубчатость имеет у основания ще-
тинки форму сложных тупых узоров, а на конце щетинка остро
зазубрена. Только на более поздних стадиях зубчатость волосовид-
ных щетинок приобретает характерную для S. armiger ступенча-
тую форму (рис. 19, К). На стадии 12-сегментной нектохеты ли-
чинка имеет анальное отверстие, расположенное между двумя бу-
горками на конце пигидиума. Телотрох на этой стадии развит еще
очень хорошо. Личинка может иногда выходить из грунта и пла-
вать в придонных слоях воды. Вскоре после этой стадии у личинки
исчезает в акротрох и паратрохи на первых двух сегментах. Ки-
шечник начинает дифференцироваться на отделы. Образуется глот-
ка, способная выбрасываться наружу (рис. 19, Ж). Обособляется
передний, средний и задний отделы кишечника. Личинка оконча-
тельно переходит к донному образу жизни и уже никогда не вы-
ходит из грунта.
Окончательно сформировавшееся молодое животное имеет в
длину 3—4 мм (рис. 19, 3). Голова округляется и полностью теря-
ет ресничный покров, как и все тело. Глаза погружаются очень
глубоко под покровы и становятся плохо заметными. На сегмен-
тах тела появляются бугорки ното- и невроподий. Помимо волосо-
- видных щетинок в пучках появляются щетинки другой формы.
Первый сегмент (перистомий) совсем не несет щетинок. В нотопо-
диях второго сегмента имеются только волосовидные щетинки с
характерной ступенчатой зубчатостью. В невроподиях этого сег-
мента имеются кроме волосовидных еще и толстые короткие ще-
тинки (рис. 19,' М), слабо изогнутые и округло зазубренные на вы-
пуклом краю. Эти щетинки на конце тупо двузубые. Такие же
щетинки имеются в невроподиях третьего, четвертого и пятого сег-
ментов. В нотоподиях этих сегментов, в пучках среди обычных
волосовидных щетинок, находится по одной вилообразной щетинке
(рис. 19, Л). Один конец вилки несколько длиннее другого. Каж-
дый кончик слегка раздвоенный. По внутреннему краю вилки на-
ходятся волоски, направленные концами к средней линии вилки.
Начиная с шестого сегмента, в невроподиях появляются короткие
ацикуловидные щетинки с однозубым кончиком (рис. 19, Н). На
спинной стороне каждого сегмента, начиная с шестого, образуются
небольшие бугорки будущих жабр. Пигидий заканчивается
придатками, между которыми открывается анальное отверстие.
По данным Торсона (Thorson, 1946), S. armiger в датских водах
размножается в апреле — мае. По данным Смидта (Smidt, 1951),
этот вид, также у берегов Дании, размножается с мая по октябрь
с максимумом в весенне-летнее время. По нашим данным (Свеш-
89
ников, 1959, 1960), S. armiger в Белом море размножается в сере-
дине июня при температуре воды 5—7°. Пелагическая стадия
длится не более трех недель.
СЕМЕЙСТВО SPIONIDAE
Личинки семейства Spionidae изучены полнее, чем личинки
многих других семейств полихет. Для берегов Дании личинки спи-
онид описаны Торсоном (1946). Для берегов Швеции полные и
детально иллюстрированные описания личинок спионид приводит
Л. ХаннерцДНаппегг, 1956). Для Черного моря спиониды описа-
ны М. И. Киселевой (1957), а интересные экспериментальные ис-
следования по выбору субстрата личинками спионид проделала
Г. А. Киселева (1967). Пелагические личинки Белого и Японского
морей описаны мною (Свешников, 1962, 1967). Личинок спионид
Средиземного моря детально описали М. Бо (Bhaud, 1967а( Ь,
1971b) и К. Казо (Cazaux, 1970, 1972). Другие работы, касающие-
ся отдельных видов личинок спионид, приведены мною ниже.
Личинки семейства Spionidae обладают рядом специфических
приспособлений, которые отличают их от других личинок полихет.
Одна из основных особенностей заключается в том, что ранние ли-
чиночные стадии спионид часто проходят под прикрытием специ-
альных оболочек (слизь, капсулы и пр.), и в планктон выходят
личинки, имеющие уже тело из нескольких сегментов. Свободная
трохофора имеется лишь у Nerine, Aonides, Spiophanes и Prionospio,
но во всех этих случаях трохофора закрыта специальной оболоч-
кой, под покровом которой происходит ее развитие в планктоне.
Личинки очень рано формируют на сегментах парные пучки длин-
ных волосовидных щетинок. Длина этих ларвальных щетинок
очень различна у разных видов, но чаще всего равна или несколь-
ко превосходит длину тела. Особенно длинными обычно бывают
щетинки первого сегмента. Ввиду того что личинки спионид очень
рано развивают ларвальные щетинки, к ним невозможно приме-
нить термины «метатрохофора» и «нектохета» в обычном смысле.
Поэтому мы условимся различать стадии личинок спионид по чис-
лу сегментов их тела. Характерно для личинок спионид их дли-
тельное пребывание в планктоне, где они могут формировать длин-
ное, многосегментное тело. Для поддержания этого длинного тела,
иногда состоящего из двух десятков сегментов, служат очень мощ-
ные ресничные поля, возникающие на сегментах.
Простомиум образуется из верхнего полушария (эписферы)
трохофоры. У большинства представителей Spionidae сформировав-
шиеся личинки имеют простомиум округлой формы. В ряде случа-
ев простомиум имеет лобные рога, например у личинок Spiophanes
bombyx. У Nerine на стадии трех сегментов впереди рта на грани-
це простомиума и перистомиума возникает конусообразный вы-
рост, который позднее вырастает в остроконечный придаток про-
стомиума.
90
Обычно у трохофоры первоначально формируется одна пара ли-
чиночных глаз. При дальнейшем развитии число глаз увеличива-
ется. По числу глаз сформировавшихся личинок всех Spionidae
можно разделить на две большие группы. В первую группу попа-
дают личинки, имеющие две пары глаз — Nerine, Laonice, Prionos-
pio, Nerinides, Aonides и Spiophanes. В другую группу, характери-
зующуюся тремя парами глаз, попадают остальные представители
подсемейства Spioninae и виды рода Scolelepis. Личинки в обеих
группах различаются по цвету глаз. У одних глаза красные, у дру-
гих — черные.
У пелагических личинок спионид рано формируются пальпы,
которые иннервируются от надглоточного ганглия.
Первый сегмент у личинок спионид несет наиболее короткие
ларвальные щетинки. Следующие сегменты имеют все более корот-
кие провизорные щетинки, которые выпадают при переходе к дон-
ному образу жизни. Жабры у пелагических личинок развиваются
чаще всего перед оседанием.
Ресничные поля имеют специфичное расположение у каждого
вида. Прототрох обычно состоит из двух изолированных участков
в основании пальпофоров. Нототрохи располагаются правильными
полукольцами на спинной стороне сегментов. Гастротрохи распо-
лагаются полукольцами на брюшной стороне сегментов (рис. 22, В).
Расположение гастротрохов может служить видовым признаком.
Например, у Spio гастротрохи выражены на нечетных сегментах,
начиная с третьего. У Polydora соеса на 3, 5, 7 и 9, а далее на каж-
дом сегменте (рис. 22, А). У Polydora ciliata — на 7, 10, 13, 15.
Подобно другим ресничным полям, гастротрохи исчезают при пе-
реходе личинок к донному существованию.
Ресничная ямка — это углубление на вентральной стороне од-
ного из трех первых сегментов. Углубление ведет в сложно развет-
вленной железистый аппарат, который характерен только для пред-
ставителей семейств Spionidae, Disomidae и Poecilochaetidae.
Ресничная ямка — личиночный орган, который редуцируется пос-
ле оседания личинки на дно.
Нухальные органы несут функцию хеморецепторов й распола-
гаются у личинок и взрослых спионид на спинной стороне просто-
миума, заходя иногда на первые сегменты тела. Пигментация из-
менчива. Характерны разветвленные, меняющие свою форму ме-
ланофоры.
Дефинитивные щетинки почти у всех личинок спионид возни-
кают во время пелагической жизни. Волосовидные щетинки сохра-
няются или заменяются более крупными. Крючковидные капюшо-
нированные щетинки у некоторых видов также сохраняются (Po-
lydora), а у других (Nerine) выпадают из передних сегментов и
остаются только в сегментах второй половины тела.
Многие личинки спионид — хищники. Рот ведет в обширный,
покрытый ресничками пищевод, который переходит в объемистый
кишечник, способный вместить обильную пищу. Полость тела срав-
нительно невелика и выстлана целомическим эпителием (рис. 23).
91
Рис. 22. Морфология личинок сем. Spionidae (ориг.)
А — личинка Polydora соеса, справа; Б —личинка Spio filicornis, с брюшной стороны;
Аня — анальная лопасть; Гтр — гастротрохи; Дщ — дефинитивные детинки; Зп —
задняя кишка; Зр — зона роста; Лщ — ларвальные щетинки; Мел — меланофор; Натр —
нототрохи; Нтр — невротрох; П — пальцы; Парп — параподии; Перст — перистомиум;
Пж — хромотофор; Пи — передняя кишка; Прст — простомиум; Пртр — прототрох;
Р — рот; Ря — ресничная ямка; Ск — средняя кишка; Сщ5 — специализированные
щетинки пятого сегмента; Ттр — телотрох
Nerine foliosa (Audouin et Milne-Edwards, 1834)
По данным Ханнерц (Hannerz, 1956), этот вид размножается у
берегов Швеции в течение зимних месяцев. Личинки в планктоне
встречаются с января до конца апреля. Наиболее ранние их стадии
появляются только в январе и феврале.
92
Рис. 23. Сагиттальный раз-
рез нектохеты Polydora
ciliata (ориг.)
Ап — анус; Coel — целом;
Ст — зачаток головного моз-
га;
Gt — гастротрох;
Gvr — зачаток брюшного мозга;
In — кишка;
МП— продольные мышцы;
Мое— мышцы пищевода;
Mir — трансверсальные мышцы;
Not — нототрохи;
Ос — глаз;
Оес — реснички пищевода;
Oes — пищевод;
Os — рот;
Tt — телотрох
По нашим данным, личинки этого вида встречаются в планкто-
не Белого моря в мае, при отрицательной температуре воды, что
соответствует арктическо-бореальной природе вида (Свешников,
1959,1962).
По литературным данным (Thorson, 1946; Hannerz, 1956) из-
вестно, что у Nerine яйцеклетки свободно развиваются в толще во-
ды. Такой же способ размножения со свободным личиночным раз-
витием наблюдается и у N. foliosa. N. foliosa, размножаясь зимой,
выметывает яйцеклетки в воду, где они претерпевают свое разви-
тие. Для яйцеклеток очень характерно наличие яйцевой мембраны,
под прикрытием которой развивается трохофора. Оболочка яйца
•сбрасывается личинкой только на стадии 2—3-сегментной метатро-
хофоры. В мае в планктоне Белого моря встречаются 10—15-сег-
:ментные личинки, которые имеют ряд особенностей, позволяющих
93
отличать их от других личинок семейства Spionidae (рис. 24). Про-
стомиум округло-треугольный, заостренный кпереди и заканчива-
ющийся остроконечным отростком. Две пары темно-красных глаз.
Боковые части перистомиума образуют основания для коротких и
толстых пальп. Название «пальпы» для щупалец, отходящих от
перистомиума, оправдано тем, что пальцы спионид иннервирова-
ны нервами переднего мозга (Orrhage, 1966). Таким образом,,
пальпы спионид и нереид вполне идентичны по характеру иннер-
вации. В основании пальп у N. foliosa расположены полуколь-
ца бывшего ресничного пояса — прототроха. Жабр нет. Нототрохи
развиты слабо со второго сегмента или совсем отсутствуют. Тело-
трох из очень сильных ресничек. Пигментация тела интенсивная,
коричневая или почти черная. Ларвальные нотоподиальные щетин-
ки крепкие, изогнутые, односторонне зазубренные. Дефинитивные
волосовидные щетинки нормально развиты в дорсальных и вен-
тральных ветвях. Капюшонированные щетинки на этой стадии мо-
гут быть еще не развитыми, а если они есть, то лишь с 10—11-го
сегмента в вентральных ветвях. Гастротрохи очень сильные на
.каждом сегменте, начиная с третьего. В лабораторных условиях
нам удалось добиться оседания личинки N. foliosa на грунт и вос-
питать до взрослого состояния (Свешников, 1962).
По наблюдениям Л. Ханнерц (1956) и нашим, молодые особи
N. foliosa питаются осевшими на дно велигерами гастропод и дву-
створчатых моллюсков.
94
Рис. 25. Личиночное развитие Aonides oxycephala oligobranchia (ориг.)
А — пелагическое яйцо с узорным хорионом; Б — трохофора в оболочке хориона; В —
метатрохофора в оболочке; Г — нектохета, вышедшая из оболочки хориона
Aonides oxycephala oligobranchia Wu.
В зал. Посьет удавалось проследить развитие этого вида в пер-
вой декаде июня при температуре 14—15° (Свешников, 1967).
Пелагические яйцеклетки имеют характерную оболочку диаметром
1050 мк. Шестигранная фактура оболочки с короткими шипиками на
углах. На поверхности оболочка несет два круга полусферических
лунок до 100 мк в диаметре каждая. Больший круг содержит 11 лу-
нок, меньший — 7 (рис. 25,4). Дробящееся яйцо 500 мк в диаметре
богато снабжено желтком лимонного цвета. Анимальный полюс
зародыша обращен в сторону малого круга лунок. На стадии тро-
хофоры оболочка принимает грушевидную форму (рис. 25, Б).
Между меньшим и большим кругами лунок образуется прогиб обо-
лочки на уровне прототроха. Трохобласты соприкасаются с этим
прогибом изнутри, сквозь оболочку снаружи просовываются рес-
нички прототроха. Таким образом, трохофора оказывается одетой
защитной рубашкой, в рукава которой продеты реснички
(рис. 25, 5).
Метатрохофора в рубашке достигает 500 мк в длину и 200 мк в
ширину (рис. 25, В). Сквозь проемы в защитной оболочке у ме-
татрохофоры продеты реснички теменного хохолка, прототроха и
телотроха. Только провизорные щетинки ротового сегмента уже
сформированы. На эписфере появляется пара алых глаз.
На стадии восьми сегментов пелагическая личинка достигает
800 мк в длину и все еще находится в оболочке яйца. Сквозь эту
оболочку выставлены телотрох, прототрох, гастротрохи, а также
длинные провизорные и маленькие дефинитивные щетинки. Фор-
мирование щетинконбсных мешков, нервной системы, пищевари-
тельного тракта происходит за счет эмбрионального желтка, так
как активное питание все еще отсутствует.
Характерно также отсутствие ресничной ямки (провизорный
хеморецептор) -и невротрохоида. Гастротрохи на всех сегментах,
начиная со второго. Провизорные волосовидные щетинки на всех
сегментах. Провизорные щетинки первого сегмента немного превы-
шают длину тела, и щетинконосные мешки этого сегмента снабже-
ны более сильной мускулатурой. На этой стадии личинка сбрасы-
вает наконец яйцевую оболочку. После этого у личинки формиру-
ются вздутия простомиальных усиков и нухальные органы между
ними (рис. 25, Г). Капюшонированные щетинки двузубые. На ста-
дии восьми сегментов, пройдя весь путь эмбрионального и пост-
эмбрионального развития в пелагиали и под покровом яйцевых обо-
лочек, личинка садится на'дно. Ювенильные формы были выве-
дены из пелагических личинок в лаборатории.
Развитие личинок Aonides paucibranchiata Southern в Белом
море детально описано В. К. Лебским (19706).
96
Рис. 26. Пелагическая ли-
чинка Prionospio malmgre-
ni (ориг.)
А — со спинной стороны;
В — справа
Prionospio malmgreni Claparede, 1869
Л. Ханнерц (Hannerz, 1956) наблюдал в планктоне появление
пелагических яиц этого вида у берегов Швеции в июле — августе.
В Белом море 5—7 сегментные личинки появляются в конце мая..
В зал. Петра Великого многосегментные личинки встречаются в
толще воды в начале сентября. Длина личинок колебалась от
500 мкм до 2 мм. Простомиум округлый, с двумя парами красных
глаз. Пальпы в виде двух небольших вздутий по бокам головы. Про-
тотрох расположен двумя полями у оснований пальп. Нототрохи
отсутствуют. Невротрох развит на коротком расстоянии от нижней
губы рта до ресничной ямки, расположенной на первом сегменте.
Гастротрохи на всех сегментах, начиная с первого. Ларвальные
щетинки у молодых личинок тонкие, волосовидные, длинные.
У многосегментных личинок они выпадают задолго до момента осе-
дания ювенильной особи на дно. На стадии десяти сегментов аналь-
ная лопасть принимает коническую форму и заканчивается аналь-
ными усиками, число которых нарастает от 4 до 6. Дефинитивные
щетинки — капюшонированные крючковидные — появляются в вен-
тральных ветвях параподий после оседания личинки на дно и имеют
на концах пять зубцов, посаженных в виде гребня. Нижний зуб
крупнее других. Личинки почти лишены пигмента. Тело тонкое и
длинное. Мало подвижны (рис.'26). Личинки были выведены в ла-
бораторных условиях.
4 в. А. Свешников
97
ЯЮмкм
Рис. 27. Пелагическая ли-
чинка Spio filicornis (ориг.)
А — со спинной стороны;
Б — с брюшной стороны
Spio filicornis (О. F. Muller, 1776)
Размножение и развитие этого вида описано Торсоном (Thor-
son, 1946), Окуда (Okuda, 1946) и очень детально Ханнерцем
(Hannerz, 1956). По данным Окуда, S. filicornis откладывает студ-
невидную кладку, прикрепляя ее вблизи своей жилой трубки.
Эмбрионы развиваются внутри кладки до стадии 4 сегментов, пос-
ле чего переходят к пелагическому существованию. У европейских
берегов этот вид размножается с февраля по май. В Белом море —
в апреле — мае (Свешников, 1959, 1962). У берегов Японии — в
апреле — июне. В зал. Петра Великого личинки этого вида обиль-
ны в мае и сентябре. В это время они находятся на стадии 15—
20 сегментов. Простомиум округлой формы с тремя парами черных
глаз. Пальпы длинные, достигают третьего щетинконосного сег-
мента. Прототрох расположен двумя полями на уровне глаз с бо-
ковых сторон головы. Нухальный орган в виде гребня на спинной
стороне головы продолжается до первого щетинконосного сегмен-
та. Ресничная ямка развита на вентральной стороне второго сег-
мента. Нототрохи — на всех сегментах, начиная со второго. Гастро-
трохи развиты на 3, 5, 7, 9, 11, 13, т. е. на всех нечетных сегментах.
Небольшие жабры — начиная с первого сегмента. На дорсальной
стороне каждого сегмента имеется по паре желез, которые проду-
цируют слизистый секрет. Провизорные щетинки тонкие, длин-
PS
ные, легко выпадают. Меланофоры на спинной стороне каждого
сегмента расположены в форме прямоугольников. Хроматофоры
в виде двух ярко-желтых пятен в основании пальп. Пищевод зани-
мает длину первых пяти сегментов. Стенка кишечника наполнена
желто-зеленым желтком. Дефинитивные волосовидные щетинки
представлены в дорсальных и вентральных ветвях каждого сегмен-
та. Дефинитивные капюшонированные, крючковидные, двузубые
щетинки развиты в невроподиях с десятого сегмента (рис. 27). Ли-
чинка, перешедшая к обитанию на дце в илистом песке, немедлен-
но в течение нескольких минут строит домик. Личинки были выве-
дены в лабораторных условиях.
Pygospio elegans Claparede, 1863
У Р. elegans известны бесполый и половой способы размноже-
ния. Бесполое размножение путем аутотомии описано Расмуссе-
ном (Rasmussen, 1953). Самцы и самки отделяют (обрывают)
задние концы или фрагменты тела, которые регенерируют. На
переднем конце фрагментов отрастает постоянное число сегмен-
тов, а на заднем конце фрагментов — непостоянное (Иванов, 1908,
1912, 1916; Маликова, 1975). По наблюдениям Расмуссена, беспо-
лый процесс идет в течение лета и быстро восстанавливает числен-
ность популяции, сократившуюся под влиянием зимних неблаго-
приятных условий. Весной происходит половое размножение. Ли-
чиночное развитие этого массового литорального вида описано для
разных точек европейского побережья разными авторами (Leschke,
1903; S6derstro,m, 1920; Thorson, 1946; Smidt, 1951; Hannerz, 1956).
В Белом море размножение этого вида происходит в мае, а в
зал. Петра Великого — дважды в год — в мае и августе.
Пелагические личинки живут в толще воды до стадии 15—
20 сегментов. Простомиум округло-треугольный с тремя парами
черных глаз. Пальпы могут доходить до 5 или 7-го сегмента тела.
Прототрох расположен двумя секциями у оснований пальп на
уровне глаз. Нухальный орган в виде гребня расположен на спин-
ной стороне головы. Ресничная ямка отсутствует. Нототрохи раз-
виты на всех сегментах, начиная со второго. Гастротрохи имеются
только на 5 и 7-м сегментах. Жабры отсутствуют. На спинной сто-
роне сегментов, начиная с 3-го, развиты железы в форме веретен,
по три на каждом сегменте. Провизорные щетинки тонкие,
длинные. По бокам сегментов попарно расположены разветвлен-
ные меланофоры. Пара меланофоров — по сторонам от ротового
отверстия. Хроматофоры расположены по бокам от рта, а иногда
на пальпах и на пигидиуме. Пищевод тянется на протяжении трех
передних сегментов. Стенка кишки наполнена желтком. Дефини-
тивные волосовидные окаймленные щетинки развиты в вентраль-
ных и дорсальных ветвях параподии. Капюшонированные крюч-
ковидные щетинки с двузубым кончиком — с 8-го сегмента
(рис., 28). Личинки были выведены в лабораторных условиях.
99
4*
Рис. 28. Пелагическая ли-
чинка Pygospio elegans
(ориг.)
А — со спинной стороны;
Б — с брюшной стороны
Polydora ciliata ciliata (Johnston, 1838)
Личинки рода Polydora отличаются от всех других спионид
тем, что у них еще во время пелагической жизни развиваются круп-
ные специализированные щетинки на пятом сегменте тела. Эти
щетинки служат для высверливания ходов в известковом субстра-
те. Представители этого рода факультативно или постоянно ведут
образ жизни перфораторов, высверливая ходы в раковинах брю-
хоногих и двустворчатых моллюсков, а также в створках баляну-
сов. Наиболее обычный перфоратор — Polydora ciliata поселяется
в Белом море в стенках раковин Littorina littorea, поражая швы,
устье и столбик. На раковине одного моллюска можно обнаружить
несколько десятков взрослых Р. ciliata (Свешников, 1963).
В Японском море этот вид перфорирует створки Crenomytilus
grayanus и других моллюсков с толстой раковиной. Личинку Р. ci-
liata очень детально на всех стадиях развития изображает и описы-
вает Вильсон (Wilson, 1928). Точное описание личинок Р. ciliata
приводит Ханнерц (Hannerz, 1956). Черноморских личинок Р. ci-
liata описывает М. И. Киселева (1957). У берегов Швеции размно-
жение этого вида происходит дважды: в марте и сентябре. Прибли-
зительно в это же время размножается Р. ciliata в Черном море.
В Белом море Р. ciliata успевает размножаться один раз в год, в
июне—июле. В это время температура воды в прибрежных участ-
ках моря достигает 8—9° С. Такие сроки размножения соответству-
ют бореальной природе вида. В ходах Р. ciliata в конце июня мы
обнаруживали яйцевые капсулы в виде цепочек, содержащих по
7—9 звеньев. Внутри каждого звена находилось по 30—40 личинок
100
Рис. 29. Пелагическая
личинка Polydora cili-
ata ciliata (ориг.)
А — со спинной стороны;
.Б — с брюшной стороны;
В — специализированная
щетинка пятого сег-
мента;
Г — шиловидная щетинка
пятого сегмента;
Д — двузубчатая щетинка
пятого сегмента
и большое количество клеток-кормилок. Личинки, готовые поки-
нуть свои капсулы, имеют три сегмента и общую длину до 250 мкм.
Сформировавшаяся пелагическая личинка на стадии 13—15 сег-
ментов (рис. 29) имеет длину несколько более 1 мм.
Простомиум округлой формы с тремя парами черных глаз.
Пальпы достигают длины 3—5 сегментов. Прототрох расположен
двумя полукольцами по краям головы. Нухальный орган в виде
гребня на спинной стороне головы продолжается на первом щетин-
коносном сегменте. Ресничная ямка отсутствует. Нототрохи на всех
сегментах, начиная с третьего. Гастротрохи — на 7, 10, 13 и 15-м
сегментах. Жабры отсутствуют. Провизорные щетинки тонкие, не
слишком длинные. Меланофоры по бокам от ротового отверстия с
вентральной стороны парные и крупные, а на спинной стороне
каждого сегмента — разветвленные с изменяющейся формой. Ме-
ланофоры могут быть также и на пигидиуме. Хроматофоры раз-
виты в основании пальп и на пигидиуме. Пищевод тянется до чет-
вертого, а у более крупных личинок — до седьмого сегмента. Стенка
кишечника заполнена зеленым желтком. Дефинитивные волосовид-
ные щетинки развиты в дорсальных и вентральных ветвях парапо-
дий. Капюшонированные крючковидные двузубые щетинки начи-
наются с седьмого сегмента. Специализированные щетинки пятого
сегмента различны. Первая из крупных щетинок — толстая и одно-
зубая (рис. 29, В). Вторая — шилообразная, изогнутая (рис. 29, Г).
Третья — толстая, двузубая (рис. 29, Д'). Все следующие щетин-
ки — толстые и двузубые. Между ними располагаются окаймлен-
ные копьевидные щетинки. Личинки были выведены в лаборатор-
ных условиях.
101
Рис. 30. Пелагическая,
личинка Polydora соеса
( ориг.)
А — со спинной стороны;
Б — с брюшной стороны;
В — специализированная
щетинка пятого сег-
мента;
Г — саблевидная щетинка
пятого сегмента;
Д — капюшонированная
щетинка невроподии
десятого сегмента
Polydora соеса (Oersted, 1843)
Личинки Р. соеса описаны Торсоном (1946) для берегов Дании
и Ханнерцем (Hannerz, 1956) для берегов Швеции, где они встре-
чаются в планктоне с начала марта до конца апреля.
В Белом море личинки этого вида встречаются с конца мая до
середины июня, когда температура воды колеблется от 1,5 до 4,5°.
Такие сроки размножения и последующего появления личинок в
планктоне при низкой положительной температуре соответствуют
арктическо-бореальной природе вида (Свешников, 1959). Личинка
с окончательно сформировавшимися признаками имеет длину не
более 1 мм и 15—18 сегментов тела (рис. 30, А, К). Простомиум с
тремя парами черных глаз. Пальпы небольшие, достигают середи-
ны первого щетинконосного сегмента. Прототрох расположен двумя
полукольцами по бокам головы. Нухальный орган уплощен и захо-
дит на спинную сторону первого сегмента. Ресничная ямка отсут-
ствует. Нототрохи на всех сегментах, начиная со второго. Гастро-
трохи на 3, 5, 7, 9 и далее на всех следующих сегментах. Жабры
отсутствуют. Провизорные щетинки тонкие, длинные. Меланофоры
на голове отсутствуют. Меланофоры на спинной стороне каждого
сегмента расположены округлыми пятнами по медианной линии те-
ла, а в виде длинных пятен окаймляют передний край сегментов.
Два больших хроматофора расположены на пигидиуме. Пищевод
занимает три первых сегмента. Стенка кишечника заполнена
желтком. Дефинитивные волосовидные щетинки развиты в дор-
сальных и вентральных ветвях парацодий. Капюшонированные
двузубые крючковидные щетинки — с 7-го сегмента (рис. 30, Д').
Специализированные толстые щетинки 5-го сегмента имеют форму
102
Рис. 31. Личинка Polydora caulleryi (ориг.)
А — пелагическая личинка с брюшной стороны; Б — ювенильная донная особь; В —
капюшонированная щетинка восьмого сегмента; Г — специализированная щетинка пя-
того сегмента
ложки (рис. 30, В). Между ними располагаются тонкие, окаймлен-
ные на конце щетинки (рис. 30,Г). Личинки были выведены в ла-
бораторных условиях и охотно оседали на раковинах моллюсков.
Polydora caulleryi Mesnil, 1893
Личинки этого вида редко встречаются в планктоне Белого мо-
ря. Взрослые формы в Белом море до сих пор не найдены. По дан-
ным Ханнерц (Hannerz, 1956), личинки Р. caulleryi встречаются у
берегов Швеции с конца февраля до конца апреля:. По нашим на-
блюдениям, Р. caulleryi в планктоне Белого моря встречается с кон-
ца мая до середины июня. Сформировавшаяся пелагическая личин-
ка на стадии 13—15 сегментов (рис. 31, Л) имеет длину около 1 мм.
Простомиум округлой формы с тремя парами черных глаз. Пальпы
короткие. Прототрох расположен двумя полукольцами по краям го-
ловы. Нухальный орган в виде плоского гребня на спинной стороне
головы продолжается на первом щетинконосном сегменте. Реснич-
ная ямка развита на брюшной стороне второго сегмента. Нототрохи
развиты на всех сегментах, начиная со второго. Гастротрохи распо-
лагаются на 3, 5, 7, 10 и 12-м сегментах. Жабры появляются после
оседания на дно на 7, 8 и 9-м сегментах (рис. 31, В). Провизорные
щетинки тонкие, длинные. Меланофоры мелкими зернами распола-
гаются поперечными рядами на спинной стороне каждого сегмента,
начиная с 7-го. Глотка, задняя кишка и пигидиум интенсивно пиг-
103
ментированы. Пищевод занимает два передних сегмента. Стенка
кишечника заполнена желтком. Дефинитивные волосовидные ще-
тинки развиты в дорсальных и вентральных ветвях параподий. Ка-
пюшонированные двузубые крючковидные щетинки начинаются с
7-го сегмента (рис. 31,5). Специализированные щетинки 5-го сег-
мента характерны гребневидным окончанием с одним крупным
заостренным зубцом (рис. 31,5). Кроме того, в составе пучка ще-
тинок 5-го сегмента развиты тонкие окаймленные копьевидные ще-
тинки и окаймленные волосовидные. Личинки были выведены в ла-
бораторных условиях.
СЕМЕЙСТВО CHAETOPTERIDAE
Всесторонний интерес исследователей по отношению к пред-
ставителям этого замечательного семейства полихет объясняется
многими удивительными особенностями биологии, экологии, физио-
логии и эмбриологии хетоптерид. Из многочисленных работ, сделан-
ных на различных видах хетоптера, можно упомянуть некоторые,
освещающие разнообразие подходов в исследовании одного и того
же объекта. Биология и поведение Chaetopterus variopedatus осве-
щались несколькими авторами (Wells, Dales, 1951). Свечение хе-
топтера описал Дж. Никол (Nicol, 1952). Поршневой способ нагне-
тания воды в жилую трубку у хетоптера открыт Т. С. Аксюк и
В. А. Свешниковым (1971). Морфология нервной системы Spiochae-
topterus typicus детально исследована Л. Оражем (Orrhage, 1966).
Индивидуальное развитие хетоптерид исследовано к настояще-
му времени детальнее, чем у других полихет. Исследования процес-
сов созревания яиц, дробления, гаструляции, процессов миграции
бластопора, метаморфоза, дыхательной активности и различные
физиологические эксперименты, связанные с развитием хетоптерид,
дают многогранное представление об особенностях онтогенеза этой
интересной группы (Lillie, 1906; Enders, 1909; Thorson, 1946; Wer-
ner, 1953; McNair-Scott, 1957; Allen, 1961; Cazaux, 1965; Bhaud,
1966, 1971b; Свешников, 1971, 1972).
Chaetopterus variopedatus (Renier, 1848)
Крупные экземпляры хетоптера, собранные в зал. Петра Вели-
кого, содержали зрелые Половые продукты. Искусственное оплодо-
творение в лабораторных условиях дало сотни тысяч нормально
развивающихся яиц. Процент яиц, отклоняющихся от нормального
развития, колебался (около 20). В результате удалось проследить
развитие Ch. variopedatus.
Наблюдения за развитием Chaetopterus variopedatus показали,
что эти крупные полихеты (некоторые экземпляры достигали чет-
верти метра) откладывают яйца диаметром всего 100 мкм и весьма
бедные желтком. При температуре 20° первая борозда дробления
появляется спустя 1 ч после оплодотворения. Каждое следующее
дробление происходит примерно через 40 мин. Разница в размере
104
Рис. 32. Дробление и процессы перемещения бластопора у Chaetopterus
variopedatus (ориг.)
А — стадия четырех бластомеров; Б — стадия восьми бластомеров; В — гаструла с бла-
стопором на оральном полюсе; Г — начало процесса перемещения бластопора; Д — ста-
дия дифференциации слепого кишечника; Е — начало образования проктодеума
Рис. 33. Личиночное раз-
витие Chaetopterus vario-
pedatus (ориг.)
А —^гаструла с бластопором на
оральном полюсе;
Б — личинка с бластопором t
смещающимся по брюш-
ной стороне;
Б — личинка на стадии образо-
вания проктодеума;
Г — трохофора на стадии сле-
пого кишечника
бластомеров относительно невелика (рис, 32,4). Бластомер D
выделяется несколько более крупными размерами только до ста-
дии 32. На более поздних стадиях разница в размерах бластоме-
ров сглаживается. Гаструляция происходит путем инвагинации
(рис. 32, В). Спустя 12 ч после оплодотворения образуется типич-
ная гаструла, которая покидает оболочки яйца и свободно плавает
в толще воды (рис. 32, В; рис. 33, А). Гаструла имеет сплошной
ресничный покров, мерцание которого приводит к поступательно-
му движению аборальным органом вперед. При этом гаструла вра-
щается вокруг главной оси тела влево. Бластопор, как и у всякой
гаструлы, расположен на заднем конце тела и ведет в полость
первичной кишки, стенка которой состоит из мерцательного эпи-
телия. Таким образом, на этой стадии у гаструлы хетоптера мы ви-
дим совпадение оси вращения, оси симметрии, оси тела и оси пер-
вичного кишечника с главной осью яйца. Аборальный орган и рот
находятся на противоположных полюсах оси. Это позволяет счи-
тать, что на этой стадии гаструла хетоптера протаксонна (рис. 32, В;
33, А).
106
В течение следующих суток происходит перестройка органи-
зации гаструлы. Тело несколько вытягивается. Появляются две
•боковые реснички, которые определяют боковые стороны личин-
ки (рис. 33,В,В). Эти рулевые реснички заметно тормозят дви-
жение и замедляют вращение вокруг главной оси тела. Опреде-
ляется брюшная сторона, по которой ротовое отверстие сползает
к аборальному полюсу совершенно так же, как это происходит в
онтогенезе турбеллярий и примитивных немертин (В. Н. Беклеми-
шев, 1944, 1964; Iwata, 1960; Свешников, 19726). Ось кишечника
в сагиттальной плоскости поворачивается сначала на 90° к глав-
ной оси тела (рис. 32, В, Г, Д'). Таким образом, на этой стадии ось
кишечника гаструлы становится поперек главной оси тела (пла-
гиаксония кишечника), а главная ось тела и ось вращения оста-
ются в исходном протаксонном состоянии. На этом поворот оси
кишечника не заканчивается. Ротовое поле продолжает свое дви-
жение кпереди, а ось кишечника поворачивается на 180°. Теперь
рот занимает почти терминальное положение на переднем конце
тела, а ось кишечника снова совпадает с осью тела (рис. 32, Е).
На этой стадии ось кишечника гаструлы становится вторично про-
таксонной, а главная ось тела и ось вращения попрежнему оста-
ются в исходном состоянии.
Механизм движения ротового поля с вегетативного полюса к
анпмальному не ясен. Можно предположить два различных спосо-
ба этого перемещения: 1) неравномерный рост дорсальной и вен-
тральной сторон тела; 2) перемещение ротового поля с помощью
движения клеток.
Неравномерный рост должен быть связан с более высоким тем-
пом клеточных делений на спинной стороне в сравнении с брюшной
стороной. Если число клеток на спинной стороне увеличивается
быстрее, чем на брюшной, то ротовое поле будет сдвигаться к ани-
мальному полюсу. Многократный просчет на сагиттальных срезах
числа эктодермальных клеток, расположенных между ртом и або-
ральным органом, показал, что их число на вентральной стороне
тела уменьшается от 13 до 6 по мере продвижения рта кпереди
(рис. 32, В, Е). Таким образом, гипотеза неравномерного роста
не подходит для объяснения механизма перемещения ротового по-
ля. Приводим изменение числа клеток по меридиану гаструлы в
процессе передвижения рта с орального полюса к аборальному
органу:
Положение рта
Число клеток эктодермы
по вентральной, стороне
На оральном полюсе
(рис. 32, В)
В середине вентральной стороны
(рис. 32, Г)
Вблизи аборального органа
(рис. 32, Д)__________________
13 + 2
9 + 1
6 + 0,3
107
Движение ротового поля к анимальному полюсу могло проис-
ходить с помощью клеточных перемещений (Trinkaus, 1972). Тог-
да клетки ротового поля должны были организованно перемещать-
ся все в одном направлении, не нарушая формы ротового отвер-
стия. Эктодермальные клетки, находящиеся вокруг ротового поля,
должны обтекать ротовое поле в направлении от аборального по-
люса к вегетативному. Тогда число клеток эктодермы должно было
бы уменьшаться впереди рта и увеличиваться позади него.
Процесс перемещения рта к аборальному полюсу связан Не
только с изменением угла наклона оси кишечника, но и с диффе-
ренциацией последнего на отделы. Возникает пищевод и вслед
за ним однокамерный (рис. 32, Д, Е) отдел средней кишки й, на-
конец, проктодеум (рис. 33, В, Г). С момента, когда кишечник ста-
новится сквозным, личинка превращается в протрохофору. Тело
ее покрыто равномерным ресничным эпителием. На этой стадии
сохраняются длинный теменной хохолок и две длинные боковые
реснички (рис. 33, В). Несколько позднее боковые реснички исче-
зают, а на экваторе возникает ресничный пояс.
Только с этого момента личинку можно называть трохофорой
(рис. 33,Г). Трохофора хетоптера характерна тем, что ротовое
отверстие у нее лежит впереди ресничного пояса, тогда как у мно-
гих трохофор полихет рот обычно лежит позади ресничного пояса.
Мы видели, что рот хетоптера успевает проскользнуть вперед
прежде, чем закладывается мерцательный пояс. Действительно,
рот хетоптера в своем движении вперед не встречает препятствий.
Ведь у гаструлы (рис. 32, В, Г) и протрохофоры (рис. 33,5) на
пути движения рта не располагаются жизненно важные структу-
ры. На этих стадиях еще не развиты мышечные и нервные тяжи
и сплетения. Образование ресничного пояса у трохофоры хетопте-
ра связано с гипертрофией ресничных клеток на экваторе личинки.
Эти клетки становятся крупнее обычных, их ресничный аппарат
также изменяется. Реснички становятся более длинными, увели-
чиваются базальные тела. Под ресничным поясом образуется про-
визорное нервно-мышечное кольцо. Трохофора питается пелаги-
ческими личинками беспозвоночных и веслоногими раками,
быстро растет и превращается в метатрохофОру (рис. 34). Рот те-
перь занимает терминальное положение и обрамлен мускулистыми,
весьма подвижными губами, в которые заходит полость тела, мес-
тами выстланная эпителием. Диаметр рта почти равен диаметру
тела личинки. Аборальный орган лежит на верхней губе. Он снаб-
жен хохолком, а в основании аборального органа, в нервной ткани,
находятся два глазка. В углах рта латерально располагаются еще
две пары глаз. Позади экваториального ресничного пояса у метатро-
хофоры закладываются три следующих мерцательных пояса. Все
четыре мерцательных пояса метатрохофоры разомкнуты на брюш-
ной стороне. На заднем конце тела позади анального отверстия у
личинки образуется хвост, который служит для руления в погоне
за подвижной добычей. Впереди ресничного пояса образуются семь
108
сегментов, соответствующих торакальному отделу. Между первым
и вторым ресничными поясами образуется сегмент с парой крыло-
видных придатков, которые служат для изготовления ловчей сети.
Следующий сегмент с железистой подушкой, а три следующих за
ним — сегменты с поршнями для нагнетания воды в трубку (Ак-
сюк, Свешников, 1971). Далее следуют абдоминальные сегменты,
которые наращиваются постепенно во время жизни хетоптера в
трубке на дне (Cazaux, 1965; Bhaud, 1966, 1971b). ч
Итак, мы видим, что на всем протяжении пелагической и.дон-
ной жизни хетоптер остается протаксонным животным, т. е. на
всех стадиях развития главная ось яйца, личинки и взрослого жи-
вотного не изменяет своего положения. Главная ось совпадает с
осью симметрии, а также с осью вращения в период пелагической
жизни. Что касается кишечника, то его ось поворачивается на
180°. Этот поворот не нарушает физиологической и морфологичес-
кой полярности организма. Хетоптера, как и других трохофорных
животных, следует относить к протаксонным животным.
109
ОТРЯД DRILOMORPHA
Индивидуальное развитие в разных семействах отряда Drilo-
morpha изучено неравномерно. Например, до сих пор никто не на-
блюдал пелагических личинок в семействе Chloraemidae (Flabel-
ligeridae). Это может служить некоторым основанием для причи-
сления жизненного цикла Chloraemidae к голобентическому типу.
Б семействе Opheliidae не только изучено личиночное развитие, но
проведено большое число опытов, доказывающих способность ли-
чинок Opheliidae определять тонкие различия в свойствах грунта
и выбирать наиболее подходящий субстрат для оседания (Wilson,
1954, 1955). Развитие пелагических личинок семейства Capitelli-
dae изучено достаточно детально.
СЕМЕЙСТВО CAPITELLIDAE
Представители семейства Capitellidae относятся к широко рас-
пространенным, эврибионтным формам. Capitella capitata, напри-
мер, имеет всесветное распространение и обитает иногда в чрез-
вычайно неблагоприятных условиях. По данным Д. Рейш (Reish,
1955), в некоторых американских гаванях с сильным загрязнени-
ем нефтяными отходами не может обитать ни одно беспозвоноч-
ное, кроме Capitella capitata. Этот вид может обитать не только
свободно, но легко переходит к обитанию на других животных
(Clark, 1956). Явления эпитокии у представителей этого семейст-
ва нет. Бесполое размножение также неизвестно. Многие Capitel-
lidae имеют сложный копулятивный аппарат. Копуляция происхо-
дит внутри трубки, где в это время находятся самец и самка. Сам-
ка откладывает яйца или внутри своей трубки, или около трубки в
виде слизистых кладок различной формы. Личинки часто лецито-
трофного типа. Наряду с обычной раздельнополостью присущ так-
же гермафродитизм. Хауеншильд (Hauenschild, 1954) опроверг об-
щепринятое мнение о принадлежности Capitomastus minimus к раз-
дельнополым животным и опытным путем доказал, что, этот вид
следует относить к гермафродитам, способным в отдельных случа-
ях к самооплодотворению. Для перекрестного оплодотворения Ca-
pitomastus minimus сохраняет копулятивный аппарат. Хауен-
шильд относит к гермафродитам также Capitella capitata. Видимо,
у этих форм обычно половой процесс происходит путем перекрест-
ной копуляции, но при длительной изоляции появляется способ-
ность к самооплодотворению.
Heteromastus filiformis (Claparede, 1864)
По данным П. Фовеля (Fauvel, 1927), Н. filiformis размножа-
ется у берегов Франции с сентября по апрель, т. е. в холодное вре-
мя года. О. Линке (Linke, 1939), напротив, наблюдал размножение
110
♦
этого вида в районе о-ва Гельголанд исключительно весной.
Е. Расмуссен (Rasmussen, 1956) впервые дал очень точные и под-
робные описания различных стадий развития Heteromastus filifor-
mis. При этом Расмуссен обнаружил несоответствие в числе тора-
кальных сегментов у Н. filiformis, по описаниям Фовеля, и у тех
особей, которых он сам наблюдал у берегов Дании. По описаниям
Фовеля (1927), род Heteromastus характеризуется тем, что имеет
длинные волосовидные щетинки на первых пяти сегментах. По
данным Расмуссена (1956), в районе Изефиорда (Дания) только
что осевшие молодые особи и половозрелые самки и самцы всегда
имели только четыре торакальных сегмента с волосовидными ще-
тинками. На основании этих данных, Расмуссен предполагает, что
вид, развитие которого он наблюдал, является или разновидностью,
или новым видом рода Heteromastus. По данным Расмуссена, эта
форма размножается в Изефиорде в начале апреля при темпера-
туре воды 6—7°.
По нашим данным, Н. filiformis размножается в Белом море
в середине июня при температуре 5—7°, а в Японском в конце
мая — около 12° (Свешников, 1959, 1967). В нижних горизонтах
литорали на песчаном грунте половозрелые самки откладывают
шарообразные слизистые кладки, прикрепленные к грунту длин-
ными ножками. Диаметр кладки 4—5 мм. На кладку сверху нали-
пают песчинки и детрит. Яйцеклетки желто-зеленого цвета имеют
в диаметре 110 мк..Кладка содержит несколько сотен яиц. Через
3—4 дня внутри кладки формируются молодые трохофоры, кото-
рые вращаются и, постепенно освобождаясь от оболочек, выходят
наружу.
Сформировавшаяся трохофора имеет в Белом море 115 мкм в
диаметре (рис. 35, 7), а в Японском — 200 мкм. Эписфера в области
ротового поля заметно нависает над гипосферой. От апикальной
пластинки отходит длинный (около 100 мкм) султанчик, играю-
щий роль осязательного органа. На эписфере имеется пара черных
и сравнительно крупных глаз. Прототрох, расположенный по эк-
ватору, имеет один ряд ресничек. Гипосфера меньше эписферы.
Ротовое отверстие густо усажено мелкими ресничками. От рта к
нижнему краю гипосферы тянется поле невротроха. Пищевод под-
нимается от рта вверх, где впадает в слепой кишечник, содержа-
щий желто-зеленый желток.
Метатрохофора увеличивается в длину до 350 мкм (рис. 35, 2).
Кишечник остается слепым. Метатрохофора имеет девять ларваль-
ных сегментов.
Нектохета имеет размер около 400 мкм (рис. 35, 3). Только три
передних торакальных сегмента несут волосовидные щетинки. Ос-
тальные — капюшонированные крючковидные. От ротового поля
по брюшной стороне тела до анального поля тянется невротрох.
Пигидиум несет хорошо развитый телотрох. Рот, расположенный
под нависшей эписферой, ведет в тонкий и длинный пищевод. Пи-
щевод поднимается в верхнюю часть эписферы, откуда поворачива-
111
/00мк
Рис. 35. Личиночное
развитие Ueteromastus
fillformis (ориг.)
1 трок оф ора;
2 метатрохофор а;
3 — нектохета, справа
ет на спинную сторону. Ближе к спинной стороне пищевод пере'
ходит в желудок.
Нектохета, осевшая на дно, имеет длину около 500 мкм. Три
следующих за перистомиумом сегмента несут изогнутые, окайм-
ленные волосовидные щетинки. Остальные семь образовавшихся к
тому времени сегментов несут только крючковидные капюшониро-
ванные щетинки. Ресничные поля на теле личинки полностью от-
сутствуют. Анальная лопасть несет толстый и сморщенный аналь-
ный усик. Глотка занимает объем первых трех сегментов. Далее
идет средняя кишка, которая заканчивается анальным отверстием
на спинной стороне анальной лопасти. К этому времени заклады-
вается кровеносная система. Тело прозрачное настолько, что кро-
веносные сосуды и кровяные тельца в них отчетливо видны у жи-
вой личинки. Спинной сосуд тянется от перистомиума до анальной
лопасти. Брюшной сосуд тянется также от перистомиума до аналь-
ной лопасти и далее заходит в. анальный усик, на конце которого
заканчивается слепо. В 9—11-м туловищных сегментах спинной и
брюшной сосуды соединяются кольцевыми сосудами. На всем про-
тяжении кровеносной системы в ней видны крупные кровяные
тельца. Цвет плазмы крови зеленоватый. Личинки были выведены
в лабораторных условиях.
112
Capitella capitata (Fabricius, 1780)
Характер размножения этого вида хорошо известен благодаря
тому, что изучался многими авторами. X. Ейзиг (Eisig, 1887, 1898)
наблюдал копуляцию у этого вида. По данным этого автора, самцы
иногда копулируют с незрелыми самками. Сперма вносится в сцер-
матофорах и сохраняется активной до момента созревания яйце-
клеток. Яйца откладываются в трубку и оберегаются самкой до вы-
хода из них личинок. Пелагическую личинку С. capitata впервые
описали Е. Клапаред и И. И. Мечников (Claparede, Mecznikow,
1896). Вслед за тем личинку этого вида описали М. Лешке (Lesch-
ke, 1903), Д. Хофкер (Hofker, 1930), Г. Торсон (Thorson, 1946),
Е. Смидт (Smidt, 1951), Е. Расмуссен (Rasmussen, 1956),
М. И. Киселева (1957), В. А. Свешников (1959, 1961, 1967) и
У Бао-линь (Wu Bao-ling, 1964). Гистологическую картину разви-
тия личинки С. capitata детально описал Ейзиг (Eisig, 1898). Упо-
мянутые выше авторы отмечают, что С. capitata может размножать-
ся в течение большей части года. В некоторых случаях этот вид мо-
жет развиваться и без пелагической личинки (Rasmussen, 1956).
По нашим данным, С. capitata в Белом и Японском морях размно-
жается большую часть года (Свешников, 1959).
Пелагические личинки этого вида встречались в планктоне Бе-
лого моря в течение всего времени наших наблюдений, с мая по
октябрь, иногда в очень больших количествах. В Японском море
личинки этого вида встречались с мая по октябрь.
Трохофоры С. capitata имеют вытянутое цилиндрическое тело
160 мкм длиной (рис. 36, А). Эписфера конической формы не не-
сет никаких придатков. Позади переднего ресничного пояса распо-
ложена пара красных глаз. С вентральной стороны перистомиума
находится овальный маленький рот. На переднем краю анальной
лопасти расположен телотрох, состоящий из более длинных и креп-
ких, чем на прототрохе, ресничек. От рта до телотроха по брюш-
ной стороне расположен невротрох. Анальная лопасть по всей по-
верхности густо покрыта мелкими ресничками. Личинка имеет
длительную пелагическую стадию и через несколько недель пре-
вращается в метатрохофору.
Метатрохофора имеет не менее 350 мкм в длину (рис. 36, Б).
У метатрохофоры закладывается 13 ларвальных сегментов. Первые
три несут волосовидные шетинки, остальные десять — крючковид-
ные капюшонированные. Кожно-мускульный мешок метатрохофо-
ры обладает рядом специфических особенностей. Эпителий имеет
повсюду одинаковую толщину — 30—40 мкм. В толще эпителия
большое количество железистых сильно вакуолизированных кле-
ток. Вакуоли оттеснены на периферию, тогда как ядра и плазма
клеток приближены к базальной пластинке. На темени личинки эпи-
телиальные клетки тесно сближены и имеют трубчатую форму.
Вдоль этих трубчатых клеток проходят нервные окончания, свя-
зывающие рецепторы переднего конца головы и мозг. Ганглиозная
113
Рис. 36. Личиночное развитие Capi-
tella capitata (ориг.)
А — трохофора, с брюшной стороны;
Б — метатрохофора, со спинной стороны ‘
В — нектохета, справа
масса головного мозга и нейропиле погружены под эпителий и на-
ходятся ниже уровня прототроха. Этим объясняется кажущееся
расположение глаз в метастомии. Трохобласты прототроха и тело-
троха представлены узкими и длинными ресничными клетками,
сжатыми с боков окружающими эпителиальными клетками. Ха-
рактерно сравнительно сильное развитие базальных зерен ресни-
чек. Ресничные кольца сопровождаются провизорными кольцевы-
ми нервами. Меридионально вдоль тела личинки проходит шесть
пар очень тонких нервных волокон. Эту ортогональную провизор-
ную нервную систему сопровождают отдельные сократимые фиб-
риллы эктодермального происхождения. Железистый покровный
эпителий подостлан тонким слоем кольцевых дефинитивных мус-
кульных волокон и более толстым слоем продольных волокон. Ще-
тинконосные мешки погружены под эпителий и мышечные слои..
Вокруг каждого щетинконосного мешка образуются конические
капсулы параподиальной мускулатуры. Брюшной нервный ствол
также погружен под эпителий и мышечные слои. Ганглиозная мас-
са и нейропиле имеют форму несколько уплощенной трубки. На
этой стадии уже заметно расчленение ствола на ганглии посег-
ментно. Границы между ганглиями подчеркиваются веерообразны-
ми пучками трансверсальных мышц. Непарный тонкий пучок мышц
проходит по вентральной стороне нервного ствола и делит нейро-
пиле на правый и левый каналы. Объем полости тела сведен до
минимума. Отдельные мезенхимные клетки выстилают ее, образуя
эпителий на кишечнике и стенке тела. Перегородок целом еще не
имеет. Рот ведет в узкую трубку пищевода, стенка которого выст-
лана длинными ресничками. С пищеводом соединена пара гло-
точных дивертикул, которые расположены при впадении пищевода
в среднюю кишку. Средняя кишка занимает все свободное прост-
ранство, ограниченное кожно-мускульным мешком. Стенка сред-
ней кишки изобилует гранулами желтка и каплями жира. Полость
средней кишки значительно уже ее стенок. Стенки средней кишки
несут реснички. Средняя кишка однокамерная. В зоне роста, рас-
положенной впереди телотроха, обнаруживаются многочисленные
митозы.
Нектохета достигает в длину 800 мкм (рис. 36, В). Прототрох
постепенно исчезает. Личинка обладает ясной сегментацией. Число
щетинконосных сегментов остается прежним — 13. Передние три
сегмента имеют волосовидные щетинки, а остальные десять — ка-
пюшонйрованные. Нектохета сильно сокращает свое тело. Личинка
оседает на дно. В поисках подходящего субстрата она может всплы-
вать и двигаться в придонных слоях воды. Ювенильные формы бы-
ли выведены из пелагических личинок в лабораторных условиях.
СЕМЕЙСТВО ARENICOLIDAE
Личиночное развитие представителей Arenicolidae изучено не-
достаточно (Ashworth, 1904; Thorson, 1946; Свешников, 1963, 1968).
Пескожилы — одна из массовых форм песчаной литорали многих
морей. В Белом море размножение Arenicola marina наступает во
второй половине лета. Личинки крайне редко встречаются в толще
воды (см. рис. 1, 77). Они ведут придонный образ жизни весьма
ограниченное время и вскоре переходят к обитанию в грунте.
Arenicola cristata Stimpson, 1856
В зал. Петра Великого A. cristata размножается в августе. В ме-
стах поселений этого вида появляются шаровидные студенистые
кладки на тонких стебельках, прикрепленных к песчинкам и ка-
мушкам. Размер кладки может достигать 30X40X70 мм и больше
(рис. 37). Внутри кладки развиваются яйца, число которых колеб-
лется в пределах нескольких десятков тысяч. Развитие внутри
кладок идет до стадии личинки с 5—7 щетинконосными сегментами
(рис. 38). Личинки движутся внутри . кладки сначала при помо-
115
J0 мкм /00 мкм
Рис. 37. Кладка Arenicola cristata в береговой зоне Японского моря (ориг.)
Рис. 38. Личинка Arenicola cristata (ориг.)
А — пятисегментная личинка из кладки; Б — нотоподиальиые и невроподиальная
(сверху) щетинки третьего сегмента
щи ресничных колец, а позднее при помощи пучков щетинок, раз-
витых к моменту выхода из кладки. Личинки короткое время жи-
вут в придонных слоях воды, а затем закапываются в грунт.
Простомиум с двумя парами красных глаз, из которых перед-
няя — с хрусталиками. Переднее ресничное кольцо широкое, распо-
ложено на границе простомиума и ротового сегмента. Второе рес-
ничное кольцо расположено на границе между анальной лопастью
и зоной роста. Невротрох широкой полосой тянется вдоль вент-
ральной стороны тела от рта до анальной лопасти. У пятисегмент-
ной личинки параподиальные бугорки не выражены. Щетинки со
второго сегмента. В нотоподиальных пучках развиты волосовидные
окаймленные и копьевидные окаймленные щетинки. В невроподи-
альных пучках — крючковидные щетинки с двумя мелкими и одним
крупным зубцами (рис. 38). Пигментация в виде желтовато-ко-
ричневых гранул развита на головной и анальной лопастях. Глотка
занимает первый сегмент и на этой стадии не выворачивается.
Стенка кишки заполнена гранулами желтка. Личинки были выве-
дены из кладок в лабораторных условиях.
116
ОТРЯД TEREBELLOMORPHA
Представители различных семейств отряда Terebelomorpha раз-
виваются или с пелагической личинкой, или их развитие проходит
в придонных слоях воды с бентосной личинкой, или внутри сли-
зистых кладок, прикрепленных к материнской трубке. Организа-
ция личинок'этого отряда весьма различна. Одни из них (Pectina-
riidae) имеют типичные приспособления к пелагическому образу
жизни, а другие (Terebellidae) обильно обеспечены питательными
веществами, питание лецитотрофное, и все развитие их протекает
в условиях дна.
СЕМЕЙСТВО PECTINARIIDAE
Пелагические личинки рода Pectinaria обладают характерны-
ми особенностями, отличающими их от личинок других полихет.
На ранних стадиях развития у них развиваются околоротовые ло-
пасти и появляется характерный точечный пигмент на эписфере
и на границе между возникающими сегментами.
Общий облик личинок этого семейства описан многими авто-
рами. Н. В. Бобрецкий (1873) кратко описал пелагическую ли-
чинку Pectinaria neapolitana. Ф. Гревли (Gravely, 1909а, Ь) наб-
людал и описал личинок Pectinaria у берегов Англии. Д. Вильсон
(Wilson, 1936) проследил развитие личинок Р. koreni от трохофоры
до осевшей нектохеты. Г. Торсон (1946) описал последовательное
развитие трех видов Pectinaria у берегов Дании. М. И. Киселева
(1957) детально описала развитие черноморской Р. neapolitana.
В Белом море встречаются два вида этого рода — Pectinaria
koreni и Р. hyperborea.
Pectinaria koreni (Malmgren, 1865)
Д. Нильсон (Nilsson, 1925) наблюдал размножение этого вида
в районе Кристинеберга (Швеция) в мае — июле. Животные выб-
расывали половые продукты в воду, где проходило дробление яиц.
и развитие личинок. Д. Вильсон (Wilson,1936) проследил развитие
Р. koreni от трохофорной гаструлы до осевшей нектохеты. Г. Тор-
сон (1946) проследил стадии развития этого вида у датских бере-
гов. По нашим наблюдениям, Р. koreni размножаются в Белом мо-
ре в конце июля. В это время или в начале августа в планктоне
появляются трохофоры.
Трохофора (рис. 39, А) имеет размер около 130 мкм. Эписфера
несет одну пару красных глаз и апикальный пучок ресничек дли-
ной около 50 мкм. Прототрох состоит из одного ряда ресничек, рас-
положенных па поясе, который окрашен в оранжевый цвет. Впере-
ди рта образуются лопасти, способные сокращаться и вытягивать-
ся. От рта до анального отверстия тянется невротрох. Под
околоротовыми лопастями, на уровне ротового сегмента, в поле рес-
ничек невротрохп находится ресничная ямка. Анальная лопасть
117
Ри-С. 39. Личиночное развитие Pectinaria koreni (ориг.)
А — трохофора; Б — метатрохофора; В — ранняя нектохета- г
оседанием на дно Ранняя нектохета, Г - нектохета перед
несет сильный телотрох. На эписфере, а также вдоль прото- и те-
лотроха располагаются округлые или кольцеобразные меланофо-
ры темно-зеленого, почти черного цвета.
Метатрохофора имеет размеры от 360 до 500 мкм в длину (рис.
39, Б). Пропорции тела метатрохофоры изменяются по сравнению
с таковыми у трохофоры. Ротовые лопасти могут расходиться и сбли-
жаться, подниматься и опускаться и, наконец, подтягиваться так,
что тело становится совершенно гладким со всех сторон. Послед-
нее наблюдается в тех случаях, когда личинка стремительно дви-
жется вперед, вращаясь вокруг оси тела. Тело метатрохофоры сег-
ментировано. Границы сегментов отмечены рядами пигментных
пятен, так же хорошо развитыми здесь, как на головной лопасти
и на пигидиуме. Число сегментов в начале образования доходит
всего до 8—10, а позднее непрерывно увеличивается.
Ранняя нектохета имеет длину до 700 мкм (рис. 39, В). Тело-
становится массивным, так что околоротовые лопасти уже не ка-
жутся столь крупными. Эписфера теряет апикальный пучок ресни-
чек. Меланофоры густо покрывают головную лопасть, так что она
напоминает окраску пантеры. Ротовой сегмент на спинной стороне
развивает одну пару щупальцевидных усиков. Сегменты тела ста-
новятся многочисленными. Число их может превышать 20. Вдоль
каждого сегмента правильными рядами располагаются меланофо-
ры. Только сегменты, находящиеся непосредственно вблизи зоны
нарастания, лишены пигмента. Зона роста располагается, как и у
всех полихет, впереди пигидиума. Никаких признаков скафы у
ранней нектохеты не было замечено. На уровне ротового сегмента
образуется шарообразный по форме нефридий зеленоватого цвета.
Ниже этой железы, на уровне второго — седьмого сегментов тела,,
формируется пара мешков с шестью щетинками опахала в каждом.
Сегменты тела также имеют хетоносные мешки, в которых разви-
ваются волосовидные и крючковидные щетинки.
Дальнейший метаморфоз происходит внезапно, в очень корот-
кие сроки, и совершенно меняет облик личинки. Все это время она
держится в толще воды, хотя полностью теряет ресничный покров.
Видимо, ткани ее тела имеют удельный вес, соответствующий мор-
ской воде, так что она не тонет. Такая пелагическая личинка име-
ет размер несколько более 1 мм (рис. 39, Г). Голова небольшая,
втянутая в передние сегменты. Голова по спинному краю несет
на этой стадии три выроста, напоминающих антенны. Эти выросты
позднее превращаются в зазубренный край мембраны. С вентраль-
ной стороны голова снабжена парой крупных, опушенных реснич-
ками, хватательных щупалец, похожих на пальпы. Между хвата-
тельными щупальцами располагается щелевидный рот, усажен-
ный мелкими ресничками. Ротовой сегмент несет две пары
щупальцевидных усиков. Со спинной стороны голову обрамляет-
опахало, состоящее из золотистых гребневидных щетинок, направ-
ленных вверх. Опахало (пали) очень характерно для представите-
лей семейства Amphictenidae. Голова отграничена от тела заметной
119
Рис. 40. Пелагическая ли-
чинка Pectinaria hyperbo-
rea (opus.)
перетяжкой, похожей на шейный отдел. Этот отдел состоит мини-
мум из трех-четырех сегментов и соединяет голову с торакальным
отделом. Торакс состоит из 15—17 щетинконосных сегментов. Ще-
тинки волосовидные и крючковидные. На этой стадии параподии
несут точечные меланофоры. Остальной пигмент на голове и теле
к этому времени почти совсем пропадает, образуется задний отдел
тела — скафа. Скафа образуется благодаря некоторому смещению
вперед зоны нарастания. Новые сегменты тела закладываются те-
перь между скафой и торакальным отделом. Скафа имеет ромби-
ческую форму и заканчивается маленьким анальным усиком. Ска-
фа сегментирована. Сквозь прозрачное тело животного виден пет-
леобразный кишечник. Пищевод очень длинный, проходит через
все тело и выстлан внутри ресничным эпителием. В задней части
тела пищевод переходит в объемистый петлеобразный желудок,
который соединяется с задней кишкой. Личинка в планктоне стро-
ит тонкую, прозрачную органическую трубку и вместе с ней опус-
кается на дно. Ювенильные формы были получены из пелагиче-
ских личинок в лабораторных условиях.
Pectinaria hyperborea (Malmgren, 1865)
X
Личинки Р. hyperborea появляются в планктоне Белого моря
в конце июня — начале июля. Сформировавшаяся нектохета (рис.
40) имеет размер около 560 мкм. Околоротовые лопасти имеют
меньший размер по сравнению с лопастями у личинки Р. koreni.
Эписфера несет одну пару красных глаз. Прототрох заходит на око-
120
пороговые лопасти, которые покрыты сплошным ресничным покро-
вом. От ротового отверстия, которое помещается между лопастями,
до анального отверстия тянется невротрох. Непосредственно под
око'лоротовыми лопастями находится ресничная ямка. На границе
анальной лопасти проходит хорошо развитый телотрох. Эписфера
покрыта точечными меланофорами. Вдоль верхней и нижней гра-
ницы прототроха проходят правильные ряды точечных меланофо-
ров. Такие же правильные ряды меланофоров проходят по границе
каждого сегмента. На этой стадии личинка имеет 15—18 сегментов.
В ното- и невроподиальных щетинконосных мешках закладывают-
ся щетинки. В нотоподиальных мешках — волосовидные, а в невро-
подиальных — клювовидные. Анальная лопасть густо усажена то-
чечными меланофорами и несет анальное отверстие, расположен-
ное ближе к спинной стороне. На уровне первого — пятого
сегментов тела, ближе к спинной стороне, находится пара больших
мешков, в которых закладываются щетинки опахала (пали). На
более поздней стадии щетинки опахала резким движением выво-
рачиваются наружу. Такая нектохета строит прозрачную органи-
ческую трубку в планктоне и садится с ней на дно. Ювенильные
формы были получены в лабораторных условиях из пелагических
личинок.
ОТРЯД SERPULIMORPHA
Представители отряда Serpulimorpha обычно обеспечивают свое
потомство значительным количеством питательных веществ. Ли-
чинки представителей этого отряда питаются лецитотррфно, т. е.
за счет желтка, независимо от того, ведут ли они пелагический об-
раз жизни или развиваются под прикрытием оболочек. Эта особен-
ность позволяет представителям серпулиморфа осуществлять рас-
селение на значительное расстояние за короткое время, так как
они обладают высокой скоростью движения и не питаются в это
время планктотрофно. Например, пелагическая личинка Chone in-
fundibuliformis по скорости движения превосходит многих личи-
нок полихет и других беспозвоночных (Константинова, 1969).
Serpulimorpha, которые развиваются без пелагической личинки,
часто образуют популяции с высокой плотностью поселения осо-
бей. Непелагическое развитие позволяет некоторым представите-
лям этого отряда обитать в пресной воде. Например, недавно от-
крытая тропическая Caobangia, перфорирующая раковины пре-
сноводных моллюсков (Jones, 1974).
Размножение и личиночное развитие представителей серпули-
морфа изучено неравномерно (Claparede, Mecznikow, 1869; Lo Bi-
anco, 1899; Zenkewitsch, 1935; Segrove, 1941; Иванов, 1944; Okuda,
1946; Свешников, 1961, 1963, 1967).
121
Рис. 41. Пелагические ли-
чинки Spirorbis (ориг.)
А — трохофора Sp. spirillum,
с брюшной стороны;
Б — нектохета того же вида,
со спинной стороны;
В — трохофора Sp. spirorbis,
с брюшной стороны;
Г — нектохета того же вида
с брюшной стороны
СЕМЕЙСТВО SERPULIDAE
Представители этого семейства размножаются только половым
путем. У некоторых видов Spirorbis встречается гермафродитизм
(Willemoes-Suhm, 1871; Leschke, 1903). В толщу воды в прибреж-
ной зоне выходят характерные личинки типа limbaria (Свешников,
1963). Несколько видов личинок Черного моря детально описала
М. И. Киселева (1957). Личинки серпул Японского моря описаны
Окуда (Okuda, 1946) и мною (Свешников, 1967).
Spirorbis spirillum (L., 1767)
Размножение и развитие этого вида описано несколькими авто-
рами (Claparede, Mecznikow, 1869; Thorson, 1946). В Белом море этот
вид размножается в июле — августе. В это время в трубках Sp. spi-
rillum находятся цепочки яиц и эмбрионов, которые располагают-
ся между телом материнского организма и стенкой трубки. В це-
почках насчитывается от 12 до 36 развивающихся яиц. Яйца име-
ют оранжево-коричневую или красную окраску.
122
На стадии трохофоры личинка достигает 210 мкм в длину (рис.
41, А). Простомиум несет теменной хохолок и три пары глаз. На
ротовом сегменте развит свисающий на брюшную сторону ворот-
ник, в котором развита железа, продуцирующая секрет для строи-
тельства трубки. Анальная лопасть несет слабый телотрох и две
длинные хвостовые нити. По брюшной стороне тянется невротрох.
На стадии нектохеты (рис. 41, Б) личинка имеет по длинной оси
около 250 мкм. Три первых торакальных сегмента несут щетинки.
На этой стадии появляются абдоминальные сегменты. В это вре-
мя эмбрион активно освобождается от оболочки капсулы и начи-
чает оживленно плавать. Движение стремительное, прямолиней-
ное. Пелагическая жизнь такой личинки длится не более суток.
Личинка живет в береговой полосе среди зарослей Zostera. Несколь-
ко позднее на простомиуме появляются парные зачатки жабер.
Личинка начинает плавать у дна, прикрепляется на соответствую-
щий субстрат и немедленно строит известковую трубку. Ювениль-
ные особи были выведены из яиц в лабораторных условиях.
Spirorbis spirorbis L '.
В Белом море этот вид размножается также в июле — августе
(Свешников, 1961). Цепочки эмбрионов располагаются между
стенкой трубки и материнским телом. В цепочке насчитывается
от 14 до 52 яиц темно-коричневого или красного цвета.
На стадии трохофоры личинка имеет размер около 270 мкм
(рис. 41, В). Простомиум несет теменной хохолок и две пары глаз.
Перистомий снабжен свисающим на брюшную сторону воротни-
ком, по боковым сторонам которого развиты мерцательные поля.
Личинки освобождаются из-под оболочек капсулы и уходят из-под
опеки материнского организма на стадии нектохеты (рис. 41, Г).
Ювенильные особи были выведены в лабораторных условиях.
ПРОИСХОЖДЕНИЕ
ТРОХОФОРНЫХ ЖИВОТНЫХ
Группа трохофорных животных (Trochozoa) включает типы
моллюсков, сипункулид, эхиурид, аннелид, тихоходок, онихофор
и членистоногих. Для аннелид, эхиурид, сипункулид и моллюсков
характерна пелагическая личинка — трохофора, которая обладает
слепым или сквозным кишечником, первичной полостью тела и
протонефридиями. Трохофора аннелид приобретает первичную сег-
ментацию и превращается в метатрохофору, план строения кото-
рой сравнивается с планом строения пелагической личинки низ-
ших членистоногих — науплиуса. Единство происхождения группы
трохофорных животных не вызывает сомнения.
Существует несколько гипотез происхождения трохофорных жи-
вотных. Одни авторы считают, что трохофорныо животные про-
изошли от кишечнополостных, близких к актиниям. А. Седжвик
(Sedgwick, 1884), а вслед за ним А. Ламеер (Lameere, 1926),
А. Бинар и Р. Жане (Binard, Jenner, 1928) и, с некоторыми допу-
щениями, А. Ремане (Remane, 1950) считают, что аннелиды про-
исходят от предков, похожих на Anthozoa. Рот и анус аннелид,
по мнению этих авторов, происходят из сифоноглиф сомкнувшего-
ся щелевидного рта актинии; брюшная нервная цепочка — от нер-
вного сплетения орального диска; параподии — от щупалец по-
липа. Эти авторы считают, наконец, что брюшная сторона аннелид
гомологична оральной стороне тела Anthozoa. О. А. Мельников
(1971) делает попытку вывести гетерономную метамерию Articu-
late из цикломерии Anthozoa. В. Н. Беклемишев (1944—1964)
справедливо называет точку зрения Седжвика и его последовате-
лей парадоксальной, потому что невозможно себе представить
происхождение аннелид из высокоспециализированных форм —
Anthozoa.
В. Н. Беклемишев (1944, 1964) предложил оригинальную гипо-
тезу происхождения трохофорных животных от таких пелагиче-
ских гаструлообразных предков кишечнополостных, которые опу-
стились на дно ртом вниз. После оседания гаструлы на дно рот
стал щелевидно растягиваться, а тело соответственно вытянулось
вдоль так называемой бластопоральной поверхности; щелевид-
ный рот сомкнулся посередине, кишечник стал сквозным, а на
концах его образовались рот и анус. Беклемишев считает, что в наи-
более примитивных случаях взрослая аннелида представляет со-
бой спинной вырост трохофоры, образовавшийся под прямым углом
124
к главной оси тела личинки. Поэтому Беклемишев относит анне-
лид, а вслед за ними и других Trochozoa к так называемым плаги-
аксонным животным. В противоположность трохофорным живот-
ным сколециды отнесены к протаксонным животным, потому что
главная ось тела у них совпадает на всех стадиях развития с глав-
ной осью яйца. В основу гипотезы о плагиаксонии аннелид Бекле-
мишев кладет толкования фактов о развитии некоторых архиан-
нелид и полихет, взятых из работ Р. Вольтерека (Wolterek, 1904),
Н. Клейненберга (Kleinenberg, 1886), Э. А. Мейера (1897) и
Д. Вильсона (Wilson, 1932а). Беклемишев следующим образом
истолковывает факты, приведенные в классической работе Вольте-
река (1904) по развитию полигордия: бластопор вытягивается в
длинную щель и, замыкаясь посередине, дает начало рту и анусу.
Таким образом, у трохофоры возникает «бластопоральная по-
верхность», которую Беклемишев сравнивает с оральной поверх-
ностью актиний и гребневиков. Брюшная сторона взрослого червя
представляет собой разросшуюся в длину «бластопоральиую по-
верхность» трохофоры. Задний конец аннелиды с расположенным
на нем анусом соответствует, по мнению Беклемишева, заднему
краю «бластопоральной поверхности» трохофоры, а не оральному
полюсу личинки или, соответственно, не заднему концу тела тур-
беллярии. В результате Беклемишев считает, что процессы разви-
тия низших червей (турбеллярий) кардинально отличаются от про-
цессов развития высших червей и что сколециды и трохофорные
животные независимо друг от друга происходят от предков ки-
шечнополостных. Сам Вольтерек описывает судьбу бластопора по-
лигордия следующим образом: бластопор несколько вытягивается,
а затем замыкается сзади наперед (об этом упоминает и Беклеми-
шев). В результате образуется слепой (без ануса) кишечник с ро-
товым отверстием, смещенным вперед вследствие процесса замы-
кания бластопора. Позднее анальное отверстие прорывается близ-
ко к месту первичного расположения бластопора. Таким образом,
на основании описаний Вольтерека можно утверждать, что бла-
стопор, замыкаясь, смещается вперед, а анальное отверстие обра-
зуется позднее, поблизости к исходному положению бластопора.
Факты, описанные Вольтереком, не дают полного основания для
утверждения, будто бы анальное отверстие полигордия образуется
за счет бластопора. Тогда в значительной степени лишается осно-
вания вывод Беклемишева, что планы строения плоских червей и
аннелид радикально различны.
Следующее положение Беклемишева состоит в том, что тело
взрослой аннелиды представляет собой вырост спинной стороны
трохофоры, и поэтому главная ось трохофоры расположена перпен-
дикулярно главной оси взрослой аннелиды. Это положение основы-
вается па фактах, описанных Н. Клейненбергом (Kleinenberg,
1886) и Э. А. Мейером (1897), по развитию трохофоры лопадорин-
ха (Phyllodocidao), который, по мнению Беклемишева, обладает
примитивным для аннелид плагиаксонным расположением осей.
J25
Положение осей у всех остальных аннелид Беклемишев выводит
из этого наиболее, по его мнению, примитивного положения осей,
свойственного Lopadorhynchus. Однако сам Э. А. Мейер (1897)
указывал на то, что и трохофора и взрослый лопадоринх по срав-
нению с другими видами семейства Phyllodocidae являются вто-
рично измененными формами в связи с переходом к голопелагиче-
скому существованию. Итак, мы видим, что литературные источ-
ники, которыми Беклемишев пользовался для аргументации
гипотезы независимого происхождения плоских и кольчатых чер-
вей от кишечнополостных, не дают достаточно убедительного фак-
тического материала, безукоризненно подтверждающего кардиналь-
ные различия проморфологии турбеллярий и аннелид. Принимая
положения Беклемишева, Ю. С. Миничев и Я. И. Старобогатов
(1972), без достаточных оснований, пытаются сравнивать схема-
тизированные ими «проморфологические перестройки» у личинок
аннелид с процессами регуляции осей у моллюсков, проходящими
под действием силы тяжести.
Гипотеза Беклемишева о первичной плагиаксонии аннелид, ко-
торая была положена им в основу представлений о происхождении
трохофорных животных, была проанализирована с позиций срав-
нительной эмбриологии и личиночного развития (Светлов, 1967;
Свешников, 1971а, 1972а, б; Иванова-Казас, 1974, 1977; А. Ива-
нов, 1975, 1976а, б). Этот анализ показал, что гипотеза Беклеми-
шева представляет собой вариант цикломерной гипотезы, против
которой он сам возражал.
Фактический материал, приведенный в настоящей книге, не да-
ет подтверждений в пользу гипотезы Беклемишева и его последо-
вателей. Среди исследованных видов не найдено ни одного такого,
в развитии которого ротовое и анальное отверстия одновременно
были бы производными бластопорального отверстия. Напротив, по-
лучено несколько убедительных фактов, которые показывают, что
анальное отверстие не имеет непосредственного отношения к бла-
стопору, если он в процессе личиночного развития перемещается от
орального полюса в сторону аборального. У ряда исследованных
видов, в процессе перемещения бластопора в сторону аборального
полюса, кишечник остается слепым, т. е. анальное отверстие долгое
время отсутствует. Такую картину мы наблюдаем у Castalia punc-
tata, Nereis virens, Chaetopterus variopedatus, Heteromastus filifor-
mis (см. рис. 13, 14, 32, 33, 35). Д. Вильсон (Wilson, 1936) описал в
развитии Pectinaria koreni раннюю трохофору, имеющую слепой
кишечник. Трохофоры, у которых слепой кишечник сохраняет это
свое состояние в процессе перемещения бластопора, известны у
некоторых моллюсков. Например, у Chiton polii, по данным
А. О. Ковалевского (1883), главная ось тела личинки сохраняет
исходное состояние, а ось кишечника меняет угол наклона. П. Пел-
зенеер (Pelseneer, 1906) иллюстрирует неизменность главной оси
личинки и изменение угла наклона оси кишечника в развитии
устриц. Вне связи с бластопором образуется анальное отверстие у
126
пластинчатожаберных моллюсков Unio и Dreissena (Lillie, 1895;
Meisenheimer, 1901). Главная ось тела личинок полихет и
некоторых моллюсков совпадает с осью их вращения во время
поступательного движения. Ось вращения и главная ось тела
остаются неизменными у рассмотренных мною личинок полихет на
всем протяжении их индивидуального развития. Вдоль главной оси
тела личинок полихет располагаются у трохофор меридиональные
провизорные нервные стволы, связанные с аборальным органом, а
у метатрохофор и нектохет — зачаток брюшной нервной цепочки и
продольные ленты мышц. Ось кишечника может изменять угол
своего наклона относительно главной оси тела. Величина этого угла
зависит от степени перемещения бластопора в сторону аборально-
го полюса. Угол этот может у разных организмов иметь разную ве-
личину от нуля до 180°. О. М. Иванова-Казас (1974) дает следую-
щее определение главной оси у животных, относящихся к группе
Spiralia: «Хотя морфологическая ось есть некая воображаемая ли-
ния, но она соединяет две конкретные материальные точки: у Spi-
ralia анимальный полюс представлен на стадии бластулы, гаструлы
и личинки клетками розетки и развивающимся из них апикальным
органом чувств, а вегетативный — макромерами и бластопором»
(стр. 7). Если безоговорочно принять, что главная ось животного
проходит через анимальный полюс и противолежащий ему бласто-
пор, то вопрос о значении в сравнительной морфологии понятия
«морфологическая ось» становится не -столь существенным по срав-
нению с тем первостепенным значением, которое ему придавал
В. Н. Беклемишев (1964).
У плоских, круглых червей и немертин первичный рот (бласто-
пор) также меняет свое положение, перемещаясь к переднему кон-
цу тела. В развитии различных турбеллярий описаны факты пере-
мещения рта к анимальному полюсу. Степень перемещения раз-
лична в разных группах турбеллярий. У бескишечных турбеллярий
рот или остается в первоначальном положении на заднем полюсе
либо немного перемещается вперед (Lang, 1894; Богута, 1970). У
многоветвистых турбеллярий рот перемещается до середины брюш-
ной стороны (рис. 42, Л) (Surface, 1907). У некоторых прямоки-
шечных рот перемещается в переднее положение.
В развитии свободноживущих нематод недавно были открыты
процессы образования щелевидного бластопора. Бластопор Апо-
plostoma vivipara и некоторых других нематод вытягивается в длин-
ную щель и, замыкаясь посередине, дает начало рту и анусу (Ма-
лахов, Черданцев, 1974, 1975; Малахов, Акимушкина, 1976).
В развитии немертин известны процессы миграции бластопора
(Dawydoff, 1928; Сое, 1943; Iwata,1960). Процесс перемещения рта
в переднее положение показан для палеонемертины Procephaloth-
fix simulus (Iwata, 1960) (рис. 42, Б). Мы видим достаточно полное
сходство в процессах перемещения бластопора в развитии турбел-
лярий, немертин и полихет (рис. 42, В). Сходство наблюдается так-
же и в изменении наклона оси кишечника относительно главной
127
оси тела. Если это так, то отпадают основания считать, что у сколе-
цид и трохофорных животных одни и те же процессы, связанные с
изменением наклона оси кишечника, приводят первых в состояние
протаксонии, а вторых — дейтераксонии. Ясно, что процессы, свя-
занные с изменением угла оси кишечника относительно главной
оси тела, не имеют значения для определения кардинальных расхо-
ждений в плане строения названных групп животных. Один приз-
нак не может служить основанием для суждения о положении
крупной группы организмов в общей системе животного мира.
Итак, анализ приведенных данных показывает, что нет доста-
точных оснований для суждений о кардинальных различиях в пла-
нах строения сколецид и трохофорных животных. Остается выяс-
нить степень близости этих двух групп.
В. А. Догель (1938,1940) сближает аннелид с предковыми форма-
ми низших несегментированных червей. Признаки становления ме-
тамерии, наблюдающиеся у ряда турбеллярий и немертин, а также
сходство трохофорных личинок аннелид с мюллеровской личинкой
турбеллярий и пилидием немертин позволяют Догелю предполо-
жить, что низшие и высшие черви родственны между собой. Наи-
более примитивными среди кольчецов Догель считает олигомерных
архианнелид. Б. Гатчек (Hatschek, 1876, 1888, 1893) считал пред-
ковой формой аннелид гипотетический трохофорообразный орга-
низм— трохозоон, план строения которого сближал с пла-
нами строения гребневиков и коловраток. Н. А, Ливанов (1945,
1955) защищает гипотезу происхождения аннелид от немертино-
образных предковых форм, основываясь на строении невросомита
аннелид, развивающегося функционально на основе нервных спле-
тений и двух продольных стволов нервной системы немертинообраз-
ных червей. А. Ремане (Remane, 1967) и ряд других крупных за-
рубежных зоологов (Ulrich, 1951; Jagersten, 1955, 1959; Marcus,
1958; Ах, 1960; Brien, 1969) развивают теорию Spiral i а. Сущ-
ность этой теории основана на признании родства между аннели-
дами и турбелляриями. Последователи теории Spiralia находят в
организации турбеллярий признаки глубокого упрощения, которое
плоские черви будто бы претерпели в процессе регрессивной эво-
люции на пути от целомических сегментированных предков, общих
с аннелидами.
Филогенетические отношения между сколецидами, трохофор-
ными животными и другими группами многоклеточных проясни-
лись благодаря анализу новейших зоологических данных, прове-
денному А. В. Ивановым (1968, 1973, 1975, 1976а, б) и его учени-
ками (Иванов, Мамкаев, 1973). Открытие в современной морской
фауне фагоцителлообразного организма — Trichoplax adhaerens
(Grell, 1971; Иванов, 1973) позволило заполнить то недостающее
звено между одноклеточными и многоклеточными, которое
прежде искусственно заполнялось организмами растительного про-
128
Рис. 42. Последовательные
стадии перемещения бласто-
пора (bl)
А — у турбеллярий (по Surfa-
ce, 1907);
Б — у немертин (по Iwata,
1960);
В — у полихет
исхождения, подобными Volvox. За этим открытием последовало ут-
верждение нового надраздела животных — Phagocitellozoa (Ива-
нов, 1973, 1976), от предковых форм которого произошли три ветви
многоклеточных: Mesozoa, Enantiozoa и Enterozoa. Mesozoa и
Enantiozoa оказались слепыми ветвями, а первичные Enterozoa'.
дали начало двум новым ветвям — радиальносимметричным Coelen--
terata и первичным билатеральным организмам — Archibilateria..
Эти последние дивергировали и дали начало двум ветвям. Одна ИВ;
них развилась в группу Scolecida. Другая ветвь, происшедшая от Ar-
chibilateria, легла в основу первичных целомических организмов -
Procoelomata. Потомки Procoelomata дали начало нескольким груп-
пам, среди которых находятся .истинные Protostomia и Deuterosto-
mia, а также группы, не принадлежащие ни к первично-, ни ко
1/25 В- А. Свешников 72Р
ARTHROPODA
CHORDATA
ENANTIOZOA ENTEROZOA MESOZOA
phagocytellozoa
PROTOZOA
Первичные
EUKARYOTA
Puc. 43. Положение трохо-
форных животных в общей
системе животного мира
вторичноротым (Иванов, 1968, 1975,1976; Codreanu, 1970; Salvini-
Plawen, 1972; Свешников 19726; Иванов, Мамкаев, 1973). А, В. Ива-
нов приходит к выводу, что понятие Protostomia не может теперь
использоваться в качестве названия таксономической группы, про-
тивопоставляемой названию Deuterostomia. В самом деле, среди
так называемых первичноротых известны организмы, формально
не отвечающие этому названию. Например, у волосатика Paragor-
dius рот не имеет отношения к бластопору и прорывается вторич-
но, как у настоящих вторичноротых (Montgomery, 1904). У поли-
хеты Eunice kobiensis (см. рис. 17) рот также прорывается вторич-
но вне всякой связи с бластопором. Что касается вторичноротых, то
они сохраняют свое таксономическое значение и остаются такой си-
стематической группой, которая сравнима по рангу с группой тро-
хофорных животных, а также с группами щупальцевых, погонофор
и щетинкочелюстных (рис. 43). Отсюда следует, что современные
целомические организмы представлены несколькими крупными
группами, равными по своему таксономическому рангу. Появление
вторичной полости тела у предков современных целомических жи-
вотных это ароморфоз, который открыл новые пути для
адаптивной радиации многообразной группы Coelomata. Известно,
что целом животных относится к числу мультифункциональных ор-
ганов. Одна из первостепенных функций целома направлена на
поддержание гомеостатического режима полостной жидкости, т. е.
внутренней среды, организма (Флоркэн, 1947). Кроме того, це-
1
130
лом выполняет транспортную, выделительную, половую, опорную
и, до некоторой степени, двигательную функции (Зенкевич, 1944;
Беклемишев, 1964; Clark, 1964; Иванов, 1976а). Возникновение це-
лома в эволюции животных несомненно связано с развитием всего
комплекса перечисленных функций, а не какой-либо одной из них,
например опорной. Именно благодаря мультифункциональности
целом стойко удерживается в многочисленных ветвях Coelomata.
Функция поддержания гомеостатического режима внутренней сре-
ды организма всегда остается главенствующей. Остальные функ-
ции могут быть развиты или подавляться в разной степени в связи
с разнообразием экологических условий.
Эволюция трохофорных животных была направлена по пути
развития метамерии. Целом в разных группах Trochozoa то при-
обретал, то терял метамерное строение. Исходное нерасчлененное
состояние целома не связано с метамерией тела (Ливанов, 1955;
Иванов, 1975, 1976а). Схизоцельный целом у животных появляет-
ся раньше, чем развивается истинная метамерия.
Надтип Trochozoa эволюирует в трех направлениях. Каждое на-
правление характеризуется функциональными преобразованиями
целома и степенью развития метамерии. Соответственно намечают-
ся три группы трохофорных животных. В первую группу можно
отнести эхиурид, сипункулид и моллюсков, у которых признаки ме-
тамерии целома или отсутствуют, или выражены очень слабо. Эхи-
уриды имеют исходно нерасчлененный целом. У сипункулид перас-
члененный личиночный целом превращается у взрослых в двураз-
дельную полость тела (Clark, 1964, 1969). У моллюсков одна часть
целома представлена изолированной околосердечной сумкой — пе-
рикардом, а другая — целомической полостью, функционально свя-
занной с открытой кровеносной системой. Ко второй группе отно-
сятся аннелиды. Аннелиды имеют разные степени и формы мета-
мерного расчленения целома. В ряду трохофорных животных
развитие метамерного целома аннелид более цсего связано с раз-
витием метамерии тела, физиологией, экологией и особенностями
способов движения. В состав типа Annelida входят классы Dino-
philida, Archiannelida, Myzostomida, Polychaeta, Oligochaeta
и Hirudinea. У динофилид и мизостомид признаки метамерии выра-
жены в значительной степени системами органов эктодермального
происхождения, а целом не сегментирован. Трудно согласиться с
мнением А. В. Иванова (19766), что целом Trochozoa возник как
гидроскелет, т. е. в связи с развитием одной только опорной функ-
ции. Нет также прямых доказательств, что динофилиды — это уп-
ростившиеся потомки полимерных кольчецов. Движение динофилид
при помощи ресничных колец никак не отнимает возможности воз-
никновения у них целомической полости в связи с многочисленны-
ми функциями, которые выполняет целом. Таким образом, вопрос
о положении динофилид в типе кольчецов пока остается открытым.
У представителей других классов кольчецов целом принимает раз-
ную форму в зависимости от экологии, физиологии, степени упоря-
131
5*
дочённости метамерии и способа движения. У полихет, олигохет и
пиявок можно наблюдать несколько разных способов движения и
соответственно разное строение целома. Развитие многочисленных
Парных целомических камер у свободноживущих полихет функцио-
нально связано с изгибанием их тела в горизонтальной плоскости
й сопровождается развитием параподиальных выростов. Преобла-
дание перистальтического способа движения у роющих полихет и
олигохет сопряжено с частичной утратой поперечных перегородок
полости тела и в ряде случаев параподиальных выростов. У пия-
вок преобладает изгибание тела в вертикальной плоскости. Парные
целомические камеры при таком способе движения теряют свое
функциональное значение. Опорная функция переходит к паренхи-
ме, а гомеостатическая, транспортная и выделительная функции
сохраняются за целомической полостью, которая приобретает фор-
му лакунарной системы. Развитие полимерного типа строения —
одна из характерных особенностей аннелид. В. А. Догель (1954)
считает, что полимерия вносит элемент двойственности — извест-
ный диссонанс — в процесс эволюции числа метамерных органов.
Далеко идущая полимеризация органов ведет к дезинтеграции ор-
ганизма как целого. Напротив, тенденция к интеграции органов ве-
дет к олигомеризации систем и к гетерономности сегментов в раз-
ных частях тела. Взаимодействие двух встречных путей эволю-
ции — полимеризации и олигомеризации — приводит к интересным
комбинациям планов строения метамерных организмов. Третья
группа трохофорных животных включает типы Tardigrada, Onycho-
phora и Arthropoda. В этой группе взаимодействие двух встречных
путей эволюции — полимеризации и олигомеризации — приводит,
особенно у Arthropoda, к удивительному богатству и разнообразию
адаптивной радиации в направлении гетерономной метамерии.
Tardigrada — своеобразная и вполне самостоятельная группа. Оли-
гомерный план строения и гомономность сегментации стойко со-
храняется у представителей этого небольшого типа. Энтероцельное
образование парных целомических полостей, хотя и носит вторич-
ный характер, но ставит тихоходок в особое положение среди дру-
гих типов этой группы. Onychophora генетически связаны с пред-
ками Annelida и сохраняют многие черты организации кольчецов.
Наземный образ жизни Onychophora вырабатывает у них некото-
рые признаки, свойственные членистоногим. Например, дыхание
при помощи первичных трахей, смешанная с кровеносным руслом
вторичная полость тела (миксоцель), движение при помощи конеч-
ностей. Arthropoda — наиболее разнообразная и богатая видами
группа, генетически связанная с предками аннелид. Одно из глав-
ных направлений эволюции членистоногих — переход от водного
обитания к жизни на суше — блестяще обосновано М. С. Гиляро-
вым (1970). Выход на сушу сопряжен с коренной перестройкой ме-
ханизмов водообмена. Членистоногие обладают удивительными
приспособлениями, которые позволяют им переходить из водной
среды на сушу в процессе индивидуального развития. Гомеостати-
132
ческий режим их внутренней среды, малая проницаемость покро-
вов, олигомеризация и многие, другие кардинальные направления
эволюции членистоногих приводят эту группу трохофорных живот-
ных к необычайному расцвету. Наконец, приспособления к полету
позволили насекомым завоевать воздушный океан раньше, чем это-
го достигли позвоночные.
Итак, мы видим, что трохофорные животные — это одна из
самых крупных и разветвленных частей филогенетического древа
(рис. 43). Trochozoa — прогрессивная, наиболее богатая типами
группа, завоевавшая все возможные среды обитания и генетически
связанная с исходной группой Procoelomata. Организация трохо-
форной личинки, метаморфоз, в результате которого формируется
первичная гетерономная метамерия сегментированного тела,
наконец, функциональное многообразие структуры целомической
полости — все эти признаки до некоторой степени роднят трохо-
форных животных с другими надтипами Coelomata.
Надтип Tentaculata включает типы Phoronida, Bryozoa и
Brachiopoda. Формы личинок у представителей всех трех типов
надтипа Tentaculata позволяют предполагать их родственные
связи с первичными Coelomata и Trochozoa.
Надтип Pogonophora — вполне самостоятельная группа цело-
мических организмов, которую нельзя относить ни к вторичноротым,
ни к так называемым первичноротым. Некоторые признаки
поверхностного сходства с Annelida, несомненно, относятся к числу
параллелизмов и конвергенций (Иванов, 1975, 1976).
Надтип Chaetognatha — одна из наиболее обособленных групп
Coelomata. Целом закладывается энтероцельно, и в процессе инди-
видуального развития становится сначала двух-, а затем трехраз-
дельным. Подавляющая часть представителей Chaetognatha обита-
ет в толще воды.
Последний надтип группы Coelomata составляют Deuterostomia,
которые включают три генетически родственных типа — Hemi-
chordata, Echinodermata и Chordata.
ЗАКЛЮЧЕНИЕ
Приведенные выше данные по морфологии личиночного развития
одной из узловых групп трохофорных животных показывают, что'
план строения Trochozoa кардинально не отличается от такового'
у Scolecida. Организация трохофоры на определенных стадиях
развития весьма близка к организации Scolecida, и эту близость
невозможно объяснить конвергенцией. Первичная полость тела,
слепой кишечник, протонефридии — весь этот комплекс органиче-
ски связанных между собой систем органов обнаруживает призна-
ки общего плана строения низших и высших червей. Соотношение
морфологических осей, которым прежде придавали большое значе-
ние в зоологии и филогенетических построениях (Беклемишев,
1964), потеряло свой вес после того, как обнаружилось, что. мор-
фологические оси у всех билатеральных организмов изменяют по-
ложение в значительной степени произвольно и не могут служить
положительным критерием для определения места той или иной
группы организмов в общей системе животного мира.
Поиск нового критерия для оценки генетических отношений
между крупными таксономическими группами животных привел к
пересмотру значения морфофизиологических признаков. Среди
узловых систем органов вновь привлекло внимание исследователей
сравнительно-морфологическое и функциональное значение вто-
ричной полости тела. Критический анализ многочисленных теорий
происхождения целома, проделанный А. В. Ивановым (1976а),
привел к заключению, что схозоцельный путь возникновения по-
лости тела, обоснованный Н. А. Ливановым (1955), остается наи-
более правдоподобным и подтверждается новым фактическим
материалом. Например, в индивидуальном развитии полихеты
Harmothoe imbricata (см. рис. 9, И, 12) обнаружены стадии схизо-
цельного образования исходно нерасчлененного целома у метатро-
хофоры и дальнейшей сегментации вторичной полости ларвального
тела у нектохеты. Подобные факты, подтверждающие схизоцельный
путь образования целома в индивидуальном развитии полихет, не
единичны и их число может быть легко умножено (Anderson, 1959).
Отсутствие принципиальных различий между телобластическим и
энтероцельным способами образования целома, обнаруженное
В. Н. Беклемишевым (1964), позволило устранить прежде ка-
завшийся существенным барьер между группами первично- и вто-
ричноротых организмов. А. В. Иванов (19766) справедливо остав-
134
ляет значение таксономического ранга только за надтипом
вторичноротых, которых наряду с другими равными по рангу груп-
пами целомических животных объединяет в подраздел Coelomata.
Положение надтипа трохофорных животных в составе под-
раздела Coelomata было оценено с позиций морфологии личиноч-
ного развития полихет (Свешников, 19726) и с точки зрения но-
вейших морфофизиологических и анатомических данных (Иванов,
1976а, б). Трохофорные животные — это одна из самых крупных
ветвей Coelomata, эволюция которых развивалась по пути сочета-
ния многообразных форм метамерии и функциональных преобразо-
ваний вторичной полости тела. Узловое положение среди трохофор-
ных животных занимают аннелиды, в составе которых полихеты
представляют наиболее многообразную и во многих отношениях
интересную группу. Индивидуальное и личиночное развитие поли-
хет изучено все еще неполно. Первые шаги в направлении исследо-
вания личиночного развития уже дали некоторые новые основания
для разрешения вопросов систематики полихет (Ушаков, 1972;
Mileikovsky, 1977). Однако многие положения в системе полихет
остаются спорными. Анализ общего плана строения личинок поз-
волил определить среди них ряд характерных морфологических
типов (см. рис. 1).
Пока еще остается совершенно незатронутым вопрос о со-
отношениях между морфологическими типами и жизненными фор-
мами у личинок полихет. С полным основанием можно говорить о
делении морфологических типов личинок полихет по крайней мере
на две жизненные формы. Личинки, приспособленные к длитель-
ному существованию в толще воды, составляют одну жизненную
форму (Amphinomidae, Spionidae, Magelonidae Poecilochaetidae, Sa-
bellariidae). Личинки, приспособленные к кратковременному
существованию в толще воды, относятся к другой жизненной фор-
ме (Arenicolidae, Serpulidae). Специальное исследование в этой
области даст возможность создать классификацию жизненных
форм личинок полихет. Эти исследования дали бы возможность
определить широту адаптивной радиации в индивидуальном раз-
витии полихет. Некоторые общие положения в этой области были
разработаны для личинок беспозвоночных Г. А. Шмидтом (1951,
1953, 1968). Однако многие вопросы морфологической экологии
индивидуального развития полихет еще остаются открытыми.
Одно из важных и перспективных направлений морфо-
логической экологии составляет проблема жизненных циклов ви-
дов. В. Н. Беклемишев (1964) впервые придал экологическую
окраску понятию жизненного цикла вида, предложив этот термин
в качестве нового морфоэкологического критерия в биологии. С тех
пор в разработке этой проблемы сделаны лишь первые шаги
(Jagersten, 1972; Свешников, 1977). Сложность проблемы жизнен-
ных циклов заключается в необходимости обобщения многообраз-
ного спектра данных, полученных на основе изучения популяцион-
ных закономерностей развития видов. Таковы некоторые перспек-
135
тивы, которые открываются на основе исследования, проделанного
автором настоящей книги.
Разрешение современных вопросов динамики онто- и филогене-
за невозможно без всесторонних исследований в области морфоло-
гии, физиологии, биохимии, экологии и других разделов биологии и
палеонтологии. Еще не проведено сопоставление нового филогене-
тического древа беспозвоночных животных с данными палеонто-
логии и геологической хронологии. Прежние оригинальные постро-
ения, проведенные Д. М. Федотовым (1966), требуют пересмотра
в свете новых данных. Обширный поток современных данных по
индивидуальному развитию аннелид и других беспозвоночных
существенно изменил представления об отношениях между глав-
ными стволами животного мира. В результате закрепляется мето-
дологически важное положение о монофилетическом развитии ор-
ганического мира.
ЛИТЕРАТУРА
Аксюк Т. С., Свешников В. А. Поршневой способ нагнетания воды у Cha-
etopterus variopedatus (Polychaeta, Annelida)ДАН СССР, 1971, 197,
№ 3, 724 — 727.
Анненкова В. II., Паленичко 3. Г. Нахождение бореальной полихеты в Бе-
лом море.— Природа, 1947, № 5, 64 — 67.
Базикалова А. Я. Материалы по изучению размножения Байкальских ам-
фипод.— Изв. АН СССР, 1941, № 3, 407 — 426.
Беклемишев В. В. Основы сравнительной анатомии беспозвоночных. М.,
«Сов. наука», 1944.
Беклемишев В. В. Основы сравнительной анатомии беспозвоночных.
Изд. 2-е. М., «Сов. наука», 1952.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных.
Т. 1: проморфология. Т. 2: органология. Изд. 3-е. М., Изд-во «Наука»,
1964.
Беклемишев К. В. Соотношение морфологических осей и таксономическая
близость основных групп первичноротых животных.— Журн. общ. биол.,
1970, 31, № 3, 302-315.
Беляев Г. №. Биология Nereis succinea в Северном Каспии. Сборник работ
об ' акклиматизации Nereis в Каспийском море.— Материалы к позна-
нию фауны и флоры.— Зоология, 1953, вып. 33(48), 243 — 284.
Изд-во МОИП.
Бобрецкий В. В. К истории развития аннелид.— Зап. Киевск. об-ва естесТв.,
1873, 3, вып. 3, 333—345.
Богута К. К. Новая турбеллярия. Anaperus biaculeatus sp. nov. из сублито-
рали Белого моря.— Зоол. журн. 1970, 49, вып. 2, 198—209.
Виноградов К. А. К фауне кольчатых червей (Polychaeta) Черного моря,—
Труды Карадагск. биол. ст., 1949, № 8, 3—84.
Гиляров №. С. Закономерности приспособлений членистоногих к жизни
на суше. М., «Наука», 1970.
Глесснер М. [Glaessner №. А]. Древнейшие бесскелетные организмы.— При-
рода, 1963, № И, 73 — 78.
Догель В. А. Сравнительная анатомия беспозвоночных, т. I. Л, Учпедгиз,
1938.
Догель В. А. Сравнительная анатомия беспозвоночных, т. 2. Л., Учпедгиз,
1940.
Догель В. А. Олигомеризация гомологичных органов как один из главных
путей эволюции животных. Изд-во ЛГУ, 1954.
Ежиков И. И. О типах развития многоклеточных из яйца.— Сб. «Памяти
академика А. Н. Северцова», т. I. М., Изд-во АН СССР, 1939.
Ерохина В. №. О плане строения мозга полихет.— ДАН СССР, 1968, 179,
К» 4, 986 — 989.
Ерохииа-Перетолчина Н. №. Ультраструктура ' дефинитивного зачатка
головного мозга трохофоры Anaitides maculata (Phyllodocidae, Poly-
chaeta).— Вести. МГУ, Биология, 1971а, № 3, 17—21.
Ерохина-Веретолчина Н. №. Ультраструктура нейронов головного мозга
Polydora ciliata (Polychaeta, Sedentaria).— Там же, 19716, № 5, 3—7.
Ерохина В. №., Константинова №. И., Свешников В. А. Развитие головного
6 В, А. Свешников
137
мозга Harmothoe imbricata (L.).— Сб. «Вопросы эволюционной морфо-
логии и биоценологии». Изд-во Каз. ун-та, 1970, 46—60.
Захваткин А. А. Сравнительная эмбриология низших беспозвоночных. М.,.
«Сов. наука», 1949.
Захваткин Ю. А. Проморфология яйца насекомых.— Журн. общ. биол.,
1969, 30, № 4, 426—441.
Захваткин 10. А. Эмбриология насекомых. М., «Высшая школа», 1975.
Захваткина К. А. Фенология личинок двустворчатых моллюсков Севасто-
польской бухты.— Труды Севаст. биол. ст., 1963, 16, 173—175. ,
Зенкевич Л. А. Очерки по эволюции двигательного аппарата животных.—
Журн. общ. биол., 1944, 5, № 3, 129—171.
Зенкевич Л. А. Фауна и биологическая продуктивность моря, т. 2. Моря.
СССР. Л., «Сов. наука», 1947.
Зенкевич Л. А. Фауна и биологическая продуктивность моря, т. 1. Мировой
океан. М., «Сов. наука», 1951.
Зенкевич Л. А. Теоретические обоснования (акклиматизации Nereis в Кас-
пийском море).— В кн.: Сборник работ об акклиматизации Nereis в.
Каспийском море. М., МОИП, 1953, 10—34.
Зенкевич Л. А. Биология морей СССР. М., Изд-во АН СССР, 1963.
Зернов С. А. Общая гидробиология. М.— Л., Изд-во АН СССР, 1949.
Иванов А. В. Происхождение многоклеточных животных. Филогенетические
очерки. Л., «Наука», 1968.
Иванов А. В. Trichoplax adhaerens — фагоцителлообразное животное.— Зоол.
журн., 1973, 52, вып. 8, 1117—1131.
Иванов .4. В. О происхождении Coelomata.— Журн. общ. биол., 1975, 36,
№ 5, 643 — 653.
Иванов А. В. О происхождении целома.— Зоол. журн., 1976а, 55, вып. 6,
805—814.
Иванов А. В. О соотношении между Protostomia и Deuterostomia и система
животного мира.— Зоол. журн., 19766, 55, вып. 8, 1125—1137. .
Иванов А. В., Мамкаев Ю. В. Ресничные черви (Turbellaria), их происхожде-
ние и эволюция.VI., «Наука», 1973.
Иванов И. И. Регенеративные процессы у многощетинковых червей и от-
ношение их к онтогенезу и морфологии аннелид. СПб., 1912.
Иванов И. И. Регенерация и онтогенез у Polychaeta.— Зоол. вести., 1916, 1,
319—450.
Иванов И. И. Общая и сравнительная эмбриология. М.— Л., ОГИЗ — Био-
медгиз, 1937.
Иванов И. И. Первичная и вторичная метамерия тела,— Журн. общ. биол.,
1944, 5, № 2, 61—95.
Иванов И. И. Руководство по общей и сравнительной эмбриологии. Л., Учпед-
гиз, 1945.
Иванова-Казас О. М. К вопросу о соотношении морфологических осей у
Spiralia.— Зоол. журн., 1974, 53, вып. 1, 5—18.
Иванова-Казас О. М. Сравнительная эмбриология беспозвоночных животных.
Новосибирск, «Наука», 1975.
Иванова-Казас О. М. Сравнительная эмбриология беспозвоночных животных.
М., «Наука», 1977.
Киселева Г. А. Оседание личинок Polydora ciliata (Johnston) на различные
субстраты.— Республ. сб. «Донные биоценозы и биология бентосных
организмов Черного моря». Серия «Биология моря». Киев, 1967, 85—90.
Киселева М. И. Пелагические личинки многощетинковых червей Черного
моря.— Труды Севаст. биол. ст., 1957, 9, 58—112.
Ковалевский А. О. Embryologische Studien an Wurmern und Arthropoden.—
•Mem. A. s. SPb, Ser. VII, 1871, 16, 70. (Эмбриологические исследования
членистоногих. Избр. работы, 123—266.)
Ковалевский А. О. Embryogenie du Chiton polii avec quelques remarques sur
le developpement des autres chitons.— Ann. Mus. hist. nat. Marseille, 1883.
N 7, 7—46.
138
Константинова М. И. Формирование мускулатуры в процессе морфогенеза
Harmothoe imbricata (Polychaeta).— ДАН СССР, 1961, 137, № 1, 232—235.
Константинова М. И, Характеристика движения пелагических личинок мор-
ских беспозвоночных.— ДАН СССР, 1966, 170, № 3, 726—729.
Константинова М. И. Движение личинок полихет.— ДАН СССР, 1969, 188,
№ 4, 942—945.
Кудерский Л. А. Распространение топловодной полихеты Nereis virens S.
в Белом море.— Природа, 1959, № 3, 106—107.
Кузнецов В. В. Периоды размножения ракообразных в прибрежных зонах
восточного Мурмана.— ДАН СССР, 1950, 75, № 2, 315—319.
Кузнецов В. В. Белое море и биологические особенности его флоры и фауны.
М.— Л., Изд-во АН СССР, 1960.
Кузнецов В. В. Биология массовых и наиболее обычных видов ракообразных
Баренцева и Белого морей. М.— Л., «Наука», 1964.
Кутищев А. А. Избирательная способность личинок дальневосточной мидии
Crenomytilus gray anus (Dunker) при оседании на субстрат.— ДАН СССР,
1976, 230, № 3, 737—740.
Лебский В. К. Строение нервной системы Eulalia viridis (L.) (Polychaeta,
Phyllodocidae).—ДАН СССР, 1970a, 190, № 6, 1486—1489.
Лебский В. К. Развитие Glycera capitata 0rsted и Aonides paucibranchiata
Southern (Annelides, Polychaeta). Биология Белого моря.— Труды Бе-
ломорск. биол. ст. МГУ, 19706, 3, 91—97.
Ливанов Н. А. Класс полихет. Руководство по зоологии, т. 2. М,—Л., Изд-во
АН СССР, 1940, 10—136.
Ливанов И. А. Пути эволюции животного мира.— Уч. зап. Казанск. ун-та,
1945, 105, № 3, 5-199.
Ливанов И. А. Пути эволюции животного мира. М., «Сов. наука», 1955.
Малахов В. В., Акимушкина М. И. Эмбриональное развитие свободноживу-
щей морской нематоды Enopleus brevis.— Зоол. журн., 1976, 55, вып. 12,
1—12.
Малахов В. В., Черданцев В. Г. Ранние стадии эмбрионального развития
свободноживущей морской нематоды Anoplostoma vivipara.— Труды
Беломорск, биол. ст. МГУ, 1974, 4, 154—161.
Малахов В. В., Черданцев В. Г. Эмбриональное развитие свободноживущей
морской нематоды Pontonema vulgare.— Зоол. журн., 1975, 54, вып. 2,
165—174.
Маликова И. Г. Регенерация у Pygospio elegans на разных стадиях онтоге-
неза.— Вести. Ленингр. ун-та, 1975, № 9, 35—40.
Мейер Э. Исследования над развитием кольчатых червей.— Труды Об-ва
естоствоисп. Казанск. ун-та, 1897, 31, вып. 4, 1—356.
Мельников О. А. О первичной гетерономии сегментов тела Articulata.—
Журн. общ. биол., 1971, 32, № 5, 597—612.
Милейковский С. А. Размножение и личиночное развитие полихеты Наппо-
thoe imbricata L. в Баренцевом и других морях,— ДАН СССР, 1959, 128,
№2,418—421.
Милейковский С. А. О принадлежности личинки полихеты типа ростра-
рия из планктона Норвежского и Варенцова морей к виду Euphrosyne
borealis и всего данного типа личинок к семействам Euphrosynidae и
Amphinomidae.— ДАН СССР, 1960а, 134, № 3, 731—734.
Милейковский С. А. О связи между температурными границами нереста вида
и его зоогеографической принадлежностью у морских беспозвоночных.—
Зоол. журн., 19606, 39, вып. 5, 666—669.
Милейковский С. А. О принадлежности двух личинок полихет типа ростра-
рия из планктона северо-западной Атлантики к видам Amphinome pal-
lasi и Chloenea atlantica (Amphinomorpha).— ДАН СССР, 1961a, 141, № 3,
754—757.
Милейковский С. А. Некоторые проблемы экологии размножения морских
донных беспозвоночных с пелагическим развитием.— Труды Мурм.
морск. биол. ин-та, 19616, вып. 3(7), 147—169.
139
6
Милейковский С. А. О морфологии и систематике полихет семейства Chrysope-
talidae Ehlers, 1864.— Зоол. жури., 1962, 41, вып. 5, 648—659.
Милейковский С. А. Морфология личинок и систематика Polychaeta.—Зоол.
журн., 1968, 47, вып. 1, 49—51.
Миничев Ю. С., Старобогатов Я. И. Проблема торсионного процесса и про-
морфологические перестройки у личинок трохофорных животных,—
Зоол. журн., 1972, 51, вып. 10, 1437—1449.
Наумов Д. В. Гидроиды и гидромедузы морских, солоноватоводных и пре-
сноводных бассейнов СССР. Определители по фауне СССР. Изд. ЗИН
АН СССР, 1960, 70.
Паленичко 3. Г. Распространение и биология креветки Pandalus borealis в
Баренцевом море.— Зоол. журн., 1941, 20, вып. 3, 398—415.
Паленичко 3. Г. Особенности биологии беломорской мидии.— Зоол. журн.,
1948, 27, вып. 5, 411—420.
Паули В. Л. К истории развития экологических отношений морских орга-
низмов к температуре и солености,— Труды Севаст. биол. ст., 1957, 9,
282—290.
Петровская М. В. К экологии многощетинковых червей Восточного Мурмана
и некоторые данные о периодах их размножения и личиночных фор-
мах.— Труды Мурм. морск. биол. института, 1960, 5, 28—67.
Светлов П. Г. Исследование над развитием кольчатых червей.— Труды зоол.
лабор. и Севастоп. биол. ст., 1928, № 11—13, 95—329.
Светлов П. Г. О первичной гетерономии состава тела позвоночных.— Арх.
анат., гистол., эмбриол., 1957, 34, № 2, 3—22.
Светлов П. Г. Соотношение морфологических осей в онто- и филогенезе раз-
ных групп животных.— Журн. общ. биол., 1967, 28, Я» 5, 567—579.
Светлов П. Г. Морфологические оси кольчатых червей и других животных.—
Сб.: Вопросы эволюционной морфологии и биоценологии, Изд-во Казанск.
ун-та, 1970, 125—146.
Свешников В. А. Размножение и развитие Nereis virens Sars.— ДАН СССР,
1955, 103, № 1, 165—167.
Свешников В. А. Типы размножения и развития многощетинковых червей
в связи с их географическим распространением.— Зоол. журн., 1959, 38,
вып. 6, 829—841.
Свешников В. А. Пелагические личинки некоторых полихет Белого моря.— .
Зоол. журн., 1960, 39, вып. 3, 343—355.
Свешников В. А. Пелагические личинки полихет Белого моря.— Зоол. журн.,
1961, 40, вып. 2, 164—177.
Свешников В. А. Пелагические личинки семейства Spionidae (Polychaeta)
Белого моря. Биология Белого моря.— Труды Беломорск, биол. ст. МГУ,
1962, 1, 146—170.
Свешников В. А. О типах личинок полихет (Polychaeta).— ДАН СССР, 1963,
150, № 6, 1393—1396.
Свешников В. А. Личинки архианнелид и полихет залива Посьет Японско-
го моря.— Исследования фауны морей, 1967, 5(13), 125—159.
Свешников В. А. Тип кольчатые черви (Annelida). Класс многощетинковые
кольчецы (Polychaeta). В кн. «Жизнь животных», т. 1. М., «Просвеще-
ние», 1968, 465—490.
Свешников В. А. Проморфология полихеты Chaetopterus variopedatus (Reni-
er).— Научные сообщения Ин-та биологии моря, Владивосток, ДВНЦ,
1971а, вып. 2, 196—199.
Свешников В. А. Протаксония полихеты Chaetopterus variopedatus (Renier)
(Polychaeta, Annelides).—Вести. МГУ, Биология — почвоведение, 19716,
№ 3, 91—93.
Свешников В. А. Соотношение морфологических осей в онтогенезе Аннелид.
Значение проморфологического критерия в сравнительной анатомии.—
Журн. общ. биол., 1972а, 33, № 2, 157—165.
140
Свешников В. А. К вопросу о происхождении трохофорных животных.—
Вести. МГУ, Биология — почвоведение, 19726, № 6, 63—70.
Свешников В. А. Структура жизненного цикла Crenomytilus grayanus.—ДАН
СССР 1977, 236, № 4.
Свешников В. А., Дю Венъ-жень. Рострария — личинка Notopygos sp. (Amphi-
nomidae, Polychaeta) из Южно-Китайского моря.— ДАН СССР, 1963,
149, № 3, 738—742.
Северцов А. Н. Морфологические закономерности эволюции. М.— Л., Изд-во
АН СССР, 1939.
Сластников Г. С. Новый вид Nereis в Белом море.— Природа, 1939, № 1,
80—81. '
Стрельцов В. Е. Биология питания плотоядного многощетинкового червя
Harmothoe imbricata (L.) в Дальнезеленецкой губе Баренцева моря,—
Труды Мурм. морск. биол. ин-та АН СССР, 1966а, вып. 11(15), 115—121.
Стрельцов В. Е. Закономерности постэмбрионального роста многощетинко-
вого червя Harmothoe imbricata L. (Polychaeta, Errantia) на литорали
южной части Баренцева моря.— ДАН СССР, 19666, 169, № 6, 1442—1445.
Стрельцов В. Е. Количественная характеристика питания Harmothoe imbri-
cata (L.) (Polychaeta, Errantia).— Труды Мурм. морск. биол. пн-та АН
СССР, 1968, вып. 17(21), 96—104.
Тринкаус Дж. [Trinkaus J. Р.]. От клеток к органам. Перевод с англ. М.,
«Мир», 1972.
. Ушаков П. В. Многощетинковые черви дальневосточных морей СССР. Опре-
делители по фауне СССР, № 56. М., Изд-во АН СССР, 1955.
Ушаков П. В. О положении филлодоцид в общей системе многощетинковых
червей. Отчетная научная сессия по итогам работ 1969 г. Тезисы докл.
Л., «Наука», 1970, 3—4.
Ушаков П. В. Многощетинковые черви, 1. Фауна СССР. Нов. серия, № 102.
Л., «Наука», 1972.
Федотов Д. М. Эволюция и филогения беспозвоночных животных. М., «Нау-
ка», 1966.
Флоркэн М. [Florkin Ж] Биохимическая эволюция. Перев. с англ. М., Гос-
издат иностр, лит., 1947.
Шаров А. Г. Происхождение и основные этапы эволюции членистоногих
(Arthropoda). I. От аннелид к членистоногим.— Зоол. журн., 1965а, 44,
вып. 6, 803—817.
Шаров А- Г. Происхождение и основные этапы эволюции членистоногих. 2.
Происхождение и филогенетические взаимоотношения основных групп
членистоногих.— Зоол. журн., 19656, 44, вып. 7, 963—979.
Шмидт Г. А. Эмбриология животных, ч. 1. Общая эмбриология. М., «Сов.
наука», 1951.
Шмидт Г. А. Эмбриология животных, ч. 2. М., «Сов. nayKa»j 1953.
Шмидт Г. А.. Типы эмбриогенеза и их приспособительное значение. М.,
«Наука», 1968.
Agassiz A. On the young stages of a few annelids.— Ann. Mag. Natur. History,
Ser. 3, 1867, 19, 203—218, 242, 257.
Aiyar B. G. Hermaphroditism in Lycastis indica (Southern).—Current Sci.
(India), 1935, N 3, 367—368.
Akesson B. On the histological differentiation of the Larvae of Pisione remota
(Pisionidae, Polychaeta).— Acta zool., 1961, 42, 177—225.
Akesson B. The embryology of Tomopteris helgolandica (Polychaeta).—Acta
zool., 1962, 43, 135—200.
Akesson B. On the nervous system of the Lopadorhynchus larvae (Polycha-
eta).— Arkiv zool., Ser. 2, 1967a, 20, N 2, 55—78.
Akesson B. The embriology of the Polychaete Eunice kobiensis.— Acta zool.,
1967b, 48, 141—192.
Akesson B., MelaMer Y. A preliminary report on the early development of the
polychaete Tomopteris gelgolandica.— Arkiv zool., Ser. 2, 1967, 20, N 6, 141—
146.
141
Allen M. T. Cytochemical aberrations of oxydative activity in the developmen-
tal stages of Chaetopterus pergamentaceus, using succinate and DPNH as
substrates.- Biol. Bull. Woods Hole, 1961, 121, 379—380.
Anderson D. T. The embriology of the Polychaete Scoloplos armiger.— Quart.
J. Mier. Sci., 1959, 100, pt. I, 89—166.
Anderson D. T. The Development of the Polychaete Haploscoloplos fragUis.—
Quart. J. Mier. Sci., 1961, 102, pt. 2, 257—272.
Anderson D. T. The comparative embryology of the Polychaeta.— Acta zool.,
1966,47,1—42..
Anderson D. T. Embryology and phylogeny in Annelids and Arthropods.-r-
Pergamon press, 1973.
Appelloj A. Uber die Beziehungen zwischen Fortpflanzung und Verbreitung
mariner Tierformen.— Verhandl. 8, Intern. Zool. Kongr. Graz, 1910. Jena,
1912, 303—311.
Ashworth J. II. Reproduction and development in Arenicola.— Liverpool Biol.
Soc., Proc., 1905, 19, 15.
Augener H. Beitrag zur Polychaetenfauna der Ostsee.— Kieler Meeresforschung,
1939, 3, H. 1, 133-147.
Ax P. Die Entdeckung neuer Organisationstypen im Tierreich.— Die nene
Brehm-Biicherei, 1960, 258, 1—116.
Banse K. Uber Morphologie und Larvalentwicklung von Nereis (Neanthes)
succinea (Leuckart) 1847.— Zool. Jahrb. Abt. Anat. Ontogen. Tiere, 1954,
74, H. 1, 160-171.
Banse K. Uber die Entwicklung von Castalia punctata.— Veroffentl. Inst.
Meeresforsch. Bremenhaven, 1956, 4, 17—24.
Bellan G. Contribution a I’Etude systematique bionomique et ecologique des
Annelides Polychaetes de la Mediterranee. Theses Fac. Sci. Univ. Marseille,
1964.
Berkeley E., Berkeley C. Micronereis nanaimoensis sp. n.: with some notes on
its life-history.— J. Fish. Res. Board Canada, 1953, 10(2).
Bhaud M. Etude des development et le 1’ecologie de quelques larves de Chaeto-
pteridae.— Vie et Milieu, 1966, 17, (ЗА), 1087—1120.
Bhaud M. Contribution a 1’ecologie des larves pelagiques de 1’Annelides 'Poly-
chetes a Banyuls-sur-Mer. Comparaison avec les regions septentrionales.—
Vie et Milieu, 1967a, 18 (2B), 273—315.
Bhaud M. Etude du developpement de quelques larves d’Annelides Polychetes
a Banyuls-sur-Mer.— Vie et Milieu, 1967b, 18 (ЗА), 531—558.
Bhaud M. La phase larvaire d’Oxydromus propinques (Hesionidae).—Vie et
Milieu, 1971a, 22 (1A), 153—161.
Bhaud M. Aspects systematiques et biogeographiques de 1’etude des larves
Planctoniques d’Annelides Polychetes. These de doctorat d’etat et sciences
naturelies. Paris, Arago, 1971b.
Binard A., Jenner B. Sur 1’existence de cavites coelomiques dans le segment
palpaire des Polychetes.— Rec. Inst. Torley-Rousseau, Bruxelles, 1928, 1,
173—189.
Bogucki M. Adaptacja Nereis diversicolor (O. F. M.) do rozcienczoney wody
morskiej i wody slodkiej.—Polskie arch, hydrobiol., 1954, 2 (15), 237—
251.
Brien P. Principes de Phylogenese animale appliques a 1’interpretation des
Metazoaires diploblastiques et a leur evolution en Metazoaires triploblas-
tiques.— Mem. Acad. roy. Belgique. Cl. Sco., 1969, 38, 3—110.
Casanova L. Note sur le developpement de Magalia perarmata.— Rec. Tran.
Station Marine d’Endoume Fasc., 1954, 13, 155—162.
Caspers H. Beobachtungen fiber Lebensraum und Schwarmperioda des Palolo-
wurmes Eunice viridis.— Rev. ges. Hydrobiol., 1961, 46, 2, 175—183.
Cazaux C. Developpement larvaire de Chaetopterus variopedatus (Renier).—
Actes Soc. Linneenne Bordeaux, 1965, 102, N 1, Ser. A, 1—31.
Cazaux C. Etude morphologique du developpement larvaire d’Annelides Poly-
chetes (Bassin d’Arcachon). II. Phylodocidae, Syllidae, Nereidae.— Arch.
Zool. Exp. Gen., 1969, 110, 2, 145—202.
142
•Cazaux C. Recherches sur 1’ecologie et le developpement larvaires des poly-
chetes de la region d’Arcachon. These Fac. Sc. Univ. Bordeaux, 1970, 477—
543.
Cazaux C. Developpement larvaire d’Annelides Polychetes (Bassin d’Arca-
chon).— Arch. Zool. Exp. Gen., 1972, ИЗ, 1, 71—108.
Claparede E., Mecznikow E. Beitrage zur Kenntnis der Entwicklungsgeschichte
der Chaetopoden.— Z. wiss. Zool., Leipzig, 1869, 19, 163—205.
Clark R. B. Capitella capitata as a commensal, with a bibliography of parasi-
tism and commensalism in the polychaetes.— Ann. Mag. Natur. History,
1956, 9, N 102, 433—448.
Clark R. B. Dynamics in metazoan evolution. The origin of the coelom and
segments. Oxford, Clarendon Press, 1964.
Clark R. B. Systematics and phylogeny: Annelida, Echiura, Sipuncula.— Chem.
Zool, 1969, 4, 1-68.
Clark R. B. Reproduction, Speciation and Polychaete Taxonomy. In «Essays
on Polychaetous Annelids». In Memory of Dr. Olga Hartman, edited by
D. J. Reish, K. Fauchald. A special publication of Allan Hankock Founda-
tion, Los Angeles, Univ. South Calif. Press, 1977, 477—502.
Codreanu R. Grands Problemes controverses de 1’Evolution Phylogenetique des
Metazoaires.— Ann. Biol, 1970, 9, F. S. 11—12, 671—709.
Coe W. R. Biology of the nemerteans of the atlantic North America.— Reans.
Connecticut Acad. Artsand. Sci, 1943, N 35, 129—328.
Costello D. P. Experimental studies of germinal localization in Nereis, I. The
development of isolated blastomeres.— J. exp. Zool, 1945, 100, 19—66.
Crisp D. J. Effect of low temperature on the breeding of marine animals.—
Nature, 1957, 179, N 4570, 1138—1139.
Dales R. Ph. The reproduction and larval development of Nereis diversicolor
0. F. Muller.— J. Marine Biol. Ass. U.'K, 1950, 129, 321—360.
Dales R. Ph. The Polychaete stomodeum and the interrelationships of the
families of Polychaeta.— Proc. Zool. Soc. London, 1962, 139, pt. 3, 389—428.
Dales R. Ph. Annelids. London, Hutchins. Univ. Libr, 1963.
Daly J. M. The maturation and breeding biology of Harmothoe imbricata
(Polychaeta: Polynoidae).— Marine Biol, 1972, 12, N 1, 53—66.
Davenport D. Notes on the early stages of the commensal Polynoid Acholod
aistericola (Delle Chiaje).—J. Marine Bion. Ass U. K, 1954, N 33, 123—127.
Davenport D., Hickok J. Notes on the early stages of the facultative commensal
Podarke pugettensis Johnson (Polychaeta, Hesionidae).— Ann. Mag. Nat.
History, 1957, Ser. 12, 10, 625—631.
Dawydoff K. N. [Давыдов К. H.]. Sur 1’embryologie des Protonemertes.— Compt.
rendus. Acad. Sci, 1928, N 186, 531—533.
Dehome A. La Schizometameria et les segments tetragemmes de Dodecaceria
caulleryi n. sp.— Bull. biol. France et Belgique, 1933, 67, 298—326.
Drew G. The anatomy, habits and embryology of Yoldia limatula Say.— Mem.
Biol. Lab. Hopkins Univ, 1899, 4, 1—37.
Durchon M. Mode de reproduction et developpement de Nereis costae Grube.—
Arch. Zool. exp. gen, 1956, 93, 57—69.
Etsig H. Die Capitelliden des Golfes von Neapel. Fauna und Flora des Golfes
von Neapel, Monogr, 16, Berlin, 1887.
Eisig H. Zur Entwicklungsgeschichte der Capitelliden.— Mitt. zool. Stat. Neapel,
1898, 13, 1—292.
Ekman S. Zoogeography of the Sea. London, 1953.
Enders H. E. Study of the life history and habits of Chaetopterus variopedatus
(Renier).— J. Morphol, 1909, 20, 479—531.
Evans S. M. Behaviour in polychaetes.— Quart. Rev. Biol.. 1971, 46, N 4, 379—405.
Fauchald K. The polychaete worms. Definitions and Keys to the Orders, Fami-
lies and Genera.— Nat. Hist. Mus. Los Angeles Country. Sci, Ser, 1977, 28,
1—190.
Fauvel P. Polychetes errantes.— Faune France, 1923, 5.
Fauvel P. Polychetes sedentaires.— Faune France, 1927, 16.
Fauvel P. Classe des Annelides Polychetes.— Traite zool, 1959, 5, f. 1, 13—196.
143
Feuerborn H. Eine Rhizocephale und zwei Polychaeten aus dem Siisswasser von
Java und Sumatra.— Intern. Vereinig. theoret. Limnol. Stuttgart Verh.,
1932, 5, 618—660.
Florkin M. Aspects moleculaires de 1’adaptation et de la phylogenie. Paris,
Masson, 1966.
Glaessner M. F. New fossils from the base of the Cambrian in South Australia.—
Trans. Roy. Soc. S. Austr., 1958, 81, 185—188.
Glaessner M. F. Рте-cambrian fossils.— Biol. Rev., 1962, 37 (4), 467—494.
Gravely F. H. Polychaeta larvae of Port Erin.— Proc. Trans. Liverpool .Biol.
Soc., 1909a, 23, 575—653.
Gravely F. H. Studies on Polychaete larvae.— Quart. J. Microsc. Sci., 1909b,
53, 597—627.
Gravier Ch. La ponte et 1’incubation chez les Annelides Polychetes.— Ann. Sci.
natur. Zool., 1923, Ser. 10, 6, 153—247.
Grell K. G. Embryonalentwicklung bei Trichoplax adhaerens К. E. Schulze.—
Naturwiss., 1971, 58, 11, 570. .
Guberlet I. E. Life history studies on some Pacific Annelides.— Anat. Rec.,
1934, 60, 58.
Gustafson G. Anatomische Studien liber die Polychaeten Familien Amphinomi-
dae und Euphrosynidae.— Zool. Bidrag. Uppsala, 1930, 12, 305—471.
Hannerz L. Larval development of the Polychaete families Spionidae Sars,
Disomidae Mesnil and Poecilochaetide n. fam. in the Gullmar'Fjord (Swe-
den) .— Zool. Bidrag, Uppsala, 1956, 31, 1—204.
Hartman O. Polychaetous annelide, pt. 8, Pilargiidae.— Allan Hancock Pacific
Exped., 1947, 10, N 5, 482—523.
Hartman O. Catalogue of the Polychaetous Annelids of the world.— Allan
Hancock Found. Pubis. Occasion, Paper, 1959, N 23, pt I, II, 1—628.
Hatschek B. Beitrage zur Morphologic und Entwicklung der Annelids.— Sit-
zungsber. Akad. Wiss. Wien, 1876, 50, 26.
Hatschek B. Lehrbuch der Zoologie, eine morphologische Ubersicht des Tier-
reichs. Jena, 1888—1891, 1—3.
Hatschek B. System der Anneliden, ein vorlaufiger Bericht.— Lotos, Prag,
1893, 13, 123—126.
Hauenschild C. Nachweis der sogenannten atoken Geschlechtsform des Poly-
chaeten Platynereis dumerillii als eigene Art auf Grund von Zuchtver-
suchen.— Zool. Jahrb., 1951, 63, H. 1, 105—128.
Hauenschild C. Uber Sexualitat und Entwicklungszyklus von Capitomastus
minimus (Annel. Polych. sedent).— Zool. Jahrb., 1954, 65, H 1, 54—58.
Hauenschild C. Obligatorische Parthenogenese bei dem Polychaeten Grubea
pusilia.—Naturwissenschaften, 1955, 42, N 22, 611—612.
Hauenschild C. Weitere Kreuzungsversuche zur Frage der Geschlechtsbestim-
mung bei dem Polychaeten Grubea clavata.— Z. Naturforsch., 1959, 14b,
N 2, 89—92.
Hacker V. Uber die Metamorphose der Polynoinen.— Ber. Naturforsch. Ges.
Freiburg, 1894, 9, 131—136.
Hacker V. Die pelagischen Polychaeten und Achaetenlarven der Planktonexpe-
dition der Humboldt-Stiftung 1889.— Ergebn. Plankt. Exp. Kiel und
Leipzig, 1898, 2, 1—50.
Hempelmann F. Zur Naturgeschichte von Nereis dumerilii.— Zoologica Stutt-
gart, 1911, 62, H. 1, 1—135.
Herlant-Meewis H. La reproduction asexuee chez les annelides.— Annee biol.,
1958, 34, N 3—4, 133-166.
Herpin R. Recherches biologiques sur la reproduction et le development de
quelques Annelides Polychetes.—Bull. Soc. Sci. Nat. ouest France, 1925,
Ser. 4, 5, 1—250.
Hofker J. Faunistische Beobachtungen in der Zuidersee wahrend der Trocken-
legung.— Z. Morphol. Oekol. Tiere, 1930, 18, 189—216.
Imajima M., Hartman O. The Polychaetous Annelids of Japan.— Allan Hancock
Found. Pubis. Occasion. Paper, 1964, N 26, pt I, pt II, 1—237, 238—452.
144
Ivanov P. P. [Иванов П. Д'.]. Die Entwicklung der Larvalsegmente bei den
Anneliden.— Z. Morphol. Oekol. Tiere, 1928, 10, 62—161.
Iwata. Fum.io. Studies on the comparative embryology of Nemerteans with
special reference to their interrelationship.— Puhi. Akkeshi Marine BioL
Stat., Sapporo, 1960, N 10, 1—51.
Izuka A. On the breeding habit and development of Nereis japonica n. sp.—
Annot. zool. japan, 1908, 6, 295—305.
Izuka A. The errantiale Polychaeta of Japan.— J. Coll. Sci. Imper. Univ. Tokyo,
1912, 30, art. 2, 1—262.
Jagersten G. Studies on the morphology, larval' development and biology of
Protodrilus.— Zool. Bidrag. Uppsala, 1952, 29, 425—512.
Jagersten G. On the early phylogeny of the Metazoa. The Bilaterogastrea-theo-
ry.— Zool. Bidrag. Uppsala, 1955, 30, 221—354.
Jagersten G. Further remarks on the early phylogeny of the Metazoa.— Zool.
Bidrag. Uppsala, 1959, 33, 79—108.
Jagersten G. Evolution of the Metazoan life cycle. A comprehensive theory.
London — New York, Acad. Press, 1972.
Johnson M. W. Studies on the life history of the marine annelid Nereis vexil-
losa.—Biol. Bull. Woods Hole, 1943, 84, 106—114.
Jones M. L. On the Caobangiidae, a new family of the Polychaeta, with a Re-
description of Caobangia billeti Giard.— Smiths, contrib. Zool., 1974, N 175,
1—55.
Kallenbach E. Ueber Polynoe cirrata O. F. M. Ein Beitrag zur Kenntniss der
Fauna der Kieler Bucht. Diss. Jena, Eisenach., 1883.
Kinne O. Uber das Schwarmen und die Larvalentwicklung von Nereis siccuniea
Leuckart (Polychaeta).— Zool. Anz., 1954, 153, N 5—6, 114—126.
Kleinenberg N. Die Entstehung des Annelids aus der Larvae von Lopadorhyn-
chus.— Z. wiss. Zool., 1886, 44, 1—227.
Korn H. Zur Unterschiedung der Larven von Harmothoe Kinberg 1857.— Kieler
Meeresforsch., 1958, 14, H. 2, 177—186.
Korn H. Vergleichend embryologische Untersuchungen an Harmothoe Kinberg
1857 (Polychaeta, Annelida). Organogenese und Neurosekretion.—Z. wiss.
Zool., 1959, 161, N 3—4, 346—443.
Korringa P. Water temperature and breeding throughout the geographical range
of Ostrea edulis.— Ann. biol., 1957, 33, N 1—2.
Korschelt E. Ueber Ophryotrocha puerilis Clap-Metsch. und die polytrochen
larven eines anderen Anneliden (Harpochaeta cingulata, n. g. n. sp.). —
Z. wiss. Zool. Leipzig, 1893, 57, 224—289.
Lameere A. Les organes segmentaires des Polychetes.— Ann. Soc. roy. zool.
Belgique, 1926, 56, 113—119.
Lang A. Lehrbuch der vergleichenden Anatomie der wirbellosen Thiere, Jena,
1894.
Lawry J. V. Mechanisms of locomotion in the polychaete Harmothoe.— Compar.
Biochem. Physiol., 1970, 37, N 2, 167—179.
Lentz T. L. Fine structure of nerve cells in planarian.— J. Morphol., 1967, 121,
N 4, 323—331.
Leschke M. Beitrage zur Kenntnis der pelagischen Polychaetenlarven der Kieler
Fohrde.— Wiss. Meeresunters., N. f. Abt. Kiel, 1903, 7, 113—134.
Lillie Fr. R. The embryology of the Unionidae.— J. Morphol., 1895, 10, 1—100.
Lillie Fr. R. Observations and experiments concerning the elementary phenome-
na of embryonic development in Chaetopterus.— J. Exp. Zool., 1906, N 3,
153—268.
Linke O. Die Biota des Jadebusenwatters.— Helgolander Wiss. Meeresunters.,
1939, 1, H. 3, 201—348.
Lo Bianco S. Notizie biologische riguardanti specialmenta il periodo di matu-
rita sessuale degli animali del golfo di Napoli.— Zool. Stat. Neapel, Mitt.,
1899, 13, 448—573.
Loosanoff V. L., Davis H. C. Conditioning Venus mercenaria for spawning in
winter and breeding its larvae in the laboratory.— Biol. Bull. Woods Hole,
1950, 98, 60—65.
145
Manton S. M. The polychaete Spinther and the origin of the Arthropoda.—
J. Natur. History, 1967, 1, N 1, 1—22.
Marcus E. On the evolution of the animal phyla.— Quart. Rev. Biol., 1958, 33,
24—58.
Martin E. A. Polymorphism and methods of asexual reproduction in the annelid
Dodecaceria of Vineyard Sound.— Biol. Bull. Woods Hole, 1933, 65, 99—105.
Martin E. A. Asexual reproduction in Dodecaceria fimbriatus.— Biol. Bull. Woods
Hole, 1936, 71, 396.
Mayer A. G. The annual breeding swarm of the Atlantic palolo.— Par. Tortagas
Lab. Garnegie Inst. Wash. D. C„ 1908, 1, 102—112.
McIntosh W. C. Early stages in the development of Phyllodoce maculata.— Ann.
Mag. Natur. History, Ser. 4, 1869, 4, 104—108.
McIntosh W. G. A monograph of the British Annelids, vol. 1, pt. 2. Polychaeta.
Amphinomidae to Sigalionidae.— London Roy. Soc., 1900, 215—442.
McIntosh W. C. A monograph of the British Annelids, vol. 2, pt. 1. Polychaeta.
Nephthydidae to Syllidae.— London Roy. Soc., 1908, 1—232.
McIntosh W. C. A monograph of the British annelids, vol. 2, pt 2. Polychaeta.
Syllidae to Ariciidae.— London Roy. Soc., 1910, 233—524.
McIntosh W. C. A monograph of the British marine annelids, vol. 3, pt. 1. Poly-
chaeta. Opheliidae to Amphictenidae.— London Roy. Soc., 1914, 1—368.
McIntosh W. C. A monograph of the British marine annelids, vol. 3. pt. 1. Poly-
chaeta. Hermellidae to Sabellidae.— London Roy. Soc., 1922, 1—250.
McIntosh W. C. A monograph of the British marine annelids, vol. 4, pt 2.
Polychaeta. Sabellidae to Serpulidae.— London Roy. Soc., 1923, .251—538.
Me Nair-Scott D. B. The effect of sugars on gastrulation of Chaetopterus
embyos.— Biol. Bull. Woods Hole, 1957, 113, 332—333.
Mecznikow E. [Мечников И. И.]. Uber die Metamorphose einiger Seethiere.—
Z. wiss. Zool., 1871, 21, 233—244.
Meek A., Storrow B. On a pelagic phase of Arenicola marine and Eteone arcti-
ca.— Ann. Mag. Natur. History London, ser. 9, 1924, 14, 453—544.
Meisenheimer I. Entwicklungsgeschichte von Dreissena polymorpha.—Z. wiss.
Zool., 1901, 69, 1—137.
Meyer A. Der Rogen und die Entwicklung der Trochophora von Eulalia viridis.—
Biol. Generalis, 1938, 14, 2, 334—389.
Meyer E. [Мейер 3.]. Studien fiber den Korperbau der Anneliden.— Mitt. Zool.
Stat. Neapel, 1901, N 14, 247—585.
Mileikovsky S. А. [Милейковский C. A.]. Some common features in the drift
of pelagic larvae and juvenile stages of bottom invertebrates with marine
currents in temperature regions.— Sarsia, 1968, N 34, 209—216.
Mileikovsky S. А. [Милейковский C. A.]. On the Systematic Interrelationships
within the Polychaeta and Annelida. An attempt to Create an Integrated
System Based on their larval Morphology. In «Essays on Polychaetous An-
nelids». In Memory of Dr. Olga Hartman, edited by D. J. Reish, K. Fauchald.
A special publication of Allan Hancock Foundation. Univ. South. Calif.
Press, Los Angeles, 1977, 503—524.
Montgomery Th. H. The development and structure of the larva of Paragor-
dius.— Proc. Acad. Natur. Sci., 1904, N 56, 738—755.
Muller M. Uber die Entwicklung und Metamorphose der Polynoen.— Miillers
Arch. Anat. Physiol. Berlin, 1851.
Muller M: Uber die weitere Entwicklung von Mesotrocha sexoculata.— Mullers
Arch. Anat. Physiol. Berlin, 1885.
Nicol J. A. Studies on Chaetopterus. III. Factors affecting the light response.—
J. Mar. Biol. Ass. U. K., 1952, N 31, 113—144.
Nilsson D. Ein Beitrag zur Kenntnis des Lebensdauer einiger Polychaeten, nebsl
Bemerkungen fiber den Rohrenbau der Amphicteniden.— Arkiv. zool.,
Stockholm, 1925, 17A, N 11, 1—34.
Nolte W. Annelidenlarven.— Nordisches Plankton. Kiel — Leipzig, Lipsius und
Tischer, 1936, Lief. 23, 59—169.
Nolte W. Annelidenlarven.—Nordisches Plankton. Kiel — Leipzig, Lipsius und
Tischer, 1938, Lief. 24, 171—282.
146
Rordgaard 0. Faunistiske og biologiske Jakttagelser veg den Biologiske Station
i Bergen.— Kgl. Norske Vidensk. Selsk. Skrifter, 1911, N 6, 1—58.
Okuda S. Studies on the development of Annelida Polychaeta.— J. Fac. Sci.
Hokkaido Imper. Univ., Ser. 6, 1946, 9, N 2, 115—219.
Orrhage L. Uber die Anatomie des centralen Nervensystemes der sedentaren
Polychaeten.— Arkiv zool., 1966, Ser. 2, 19, N 3, 99—133.
Orton Y. H. Sea temperature, breeding and distribution of marine animals.—
J. Marine Biol. Ass. U. K., 1920, N 12, 339—366.
Pelseneer P. Mollusca. A treatise on zoology. London, 1906, 5.
Peres J. M., Rancurel P. Observations sur la ponte de Perinereis cultrifera
Grube dans le Golfe de Marseille.— Bull. Soc. zool France, 1948, 73, N 2,
97—100.
Qasim S. Z. Time and duration of the spawning season in some marine teosts
in relation to their distribution.— J. conceil perman. internat. explorat.,
1956, 21, N 2, 144—155.
Rasmussen E. Asexual reproduction in Pygospio elegans Claparede (Polychae-
ta, Sedentaria).— Nature, 1953, 171, 1161.
Rasmussen E. Faunistic and biological notes on marine invertebrates, III.—
Biol. Medd. Dan. Vid. Selsk., 1956, 23, N 1, 1—84
Rasmussen E. Systematics and Ecology of the Isefjord Marine Fauna (Den-
mark) .— Ophelia, 1973, 11, 1—495.
Reish D. J. The life history and ecology of the Polychaetous Annelid Nereis
grubei (Kinberg).— Allan Hancock Foundation Publications, 1954, 14.
Reish D. J. The relation of Polychaetous Annelids to Harbor Pollution.— Pub-
lic Health Repts, 1955, 70, N 12, 1168—1174.
Reish D. J. The life history of the Polychaetous Annelid Neanthes caudata
(delle Chiaje). Including a Summary of development in the family Nerei-
dae.— Pacific Sci., 1957, 11, N 2, 216-228.
Reish D. J., Fauchald K. (ed). Essays on Polychaetous Annelids. In Memory
of Dr. Olga Hartman. A special publication of Allan Hancock Foundation.
Univ. South Calif. Press., Los Angeles, 1977.
Reisinger E. Die Evolution des Orthogons der Spiralier und das Archicolomaten
problem.— Z. Zool. Syst. Evolutionsforsch., 1972, 10, N 1, 1—43.
Remane A. Die Entstehung der Metamerie der Wirbellosen.— Zool. Anz., Suppl.,
1950, 14, 16—23.
Remane A. Die Geschichte der Tiere. In «Die Evolution der Organismen»,
I. Hrsg. G. Heberer, 1967, 589—677.
Rullier F. Recherches sur la morphologie et la reproduction du nereidien Micro-
nereis variegata Claparede.— Arch. zool. exp., gen., 1954, 91, fsc. 2,
00—00.
Runnstrom S. Uber die Thermopathie der Fortpflanzung und Entwicklung
marinen Tiere in Beziehung zu ihrer geographischen Verbreitung.— Ber-
gens Mus. Arbok. Naturv. Rekke, 1927, N 2, 1—67.
Runnstrom S. Weitere Studien liber die Temperaturanpassung der Fortpflan-
zung und Entwicklung maraner Tiere.— Bergens Mus. Arbok., 1929, N 10,
1—45.
Runnstrom S. Die Anpassung der Fortpflanzung und Entwicklung mariner Tiere
an die Temperaturverhaltnisse verschiedener Verbreitungsgebiete.—Ber-
gens Mus. Arbok., 1936, N 3, 1—36.
Sachs M. Observations on the embryology of an aquatic gastrotrich, Lepido-
dermella squammata (Dujardin, 1841).— J. Morphol., 1955, 96, N 3,
473—496.
Sagara J., Sugawara K. On the optimum temperature for the development of the
starfish, Asterias amurensis.— Bull. Tokai Reg. Fish. Res. Lab., 1957, N 16,
49—52.
Salensky W. [Заленский В.]. Studies sur le developpement des Annelides.
P. I, IL— Arch. Biol., 1882, 3, 345—378.
Salensky W. [Заленский В.]. Etudes sur le developpement des Annelides.
Deuxieme partie. Developpement de Branchiobdella.— Conclusions et ref-
lexions.— Arch. Biol., 1887, 6, 1—65, 589—654.
147
Salensky W. [Заленский В.]. Morphogenetische Studien an Wiirmer.— Mem.
Acad. St. Petersbourg, 1907, 19, Nil.
Salvini-Plawen L. Zur Morphologie und Phylogenie der Mollusken.— Z. wiss..
Zool., 1972, 184, 205—394.
Sars M. Zur Entwicklung der Anneliden.— Arch. Naturgesch. Jahrg., 1845, 11,
1, 11—19.
Schimkewitsch V. M. [Шимкевич В. M.]. Zur Kenntnis des Baues und der
Entwicklung des Dinophilus vom Weissen Meere.— Z. wiss. Zool., 1895, 59,
46—79.
Sedgwick A. On the origin of metameric segmentation and some other morpho-
logical questions.— Quart. J. micr. Sci., N. S., London, 1884, 24, 43—82.
Segrove F. The development of the Serpulid Pomatoceros triqueter L.—
. Quart. J. micr. Sci., 1941, 82, 467—540.
Sharow A. G. [Шаров А. Г.]. Basic Arthropoda stock. International Series of
Monographs in Pure and Applied Biology. Oxford, Zoology Division,
1966, 30.
Slewing R. Lehrbuch der vergleichenden Entwicklungsgeschichte der Tiere. Ham-
burg — Berlin, 1969.
Smidt E. L. B. Biological studies of the invertebrate fauna of the harbour of
Copenhagen.— Vid. medd. Dansk naturh. foren., 1944, 107, 235—316.
Smidt E. L. B. Animal production in the Danish Waddensea.— Meddr. Damn.
Fisk.-og Havunders., Fiskeri, 1951, 11, N 6, 1—151.
Smith R. I. Embryonic development in the viviparous nereid polychaete Nean-
thes lighti Hartman.— J. Morph., 1950, 87, 417—455.
Snodgrass В. E. Morphology and evolution of the insect head and its appenda-
ges. Smithson. Misc. Coll., 1928, 80, N 1.
Snodgrass R. E. Evolution of the Annelida, Onychophora and Arthropoda.
Smithson. Misc. Coll., 1938, 97, N 6.
Southward A., Crisp D. Becent Changes in the Distribution of the Intertidal
Barnacles Chthamalus stellatums Poli and Balanus balanoides L. in the
British Isles.— J. Anim. Ecol., 1954, 23, N 1, 163—177.
Soderstrom A. Studien fiber die Polychaeten familie Spionidae.— Dissertation..
Uppsala. Almquist, Wiksell, 1920, 1—286.
Storch O. Vergleichend-anatomische Polychaetenstudien, II.—Verhandl. Ges.
Nat. Arzte, Berlin, 1914, 85, 709—711.
Storch O. Ein Beitrag zur Anatomie von Hermodice carunculata.— Zool. Anz.r
1915, N 45, 35—44.
Storch V. Zur vergleichenden Anatomie der segmentalen Muskelsysteme und
zur Verwandtschaft der Polychaeten-Familien.— Z. Morphol. Tiere, 1968,
63, 251—342.
Strathmann R. R. Toward understanding complex life cycles of benthic inverteb-
rates.— Ecology of Fouling Communities U.S.— U.S.S.R. Cooperative Pro-
gram, Part I, 1975, Beaufort, North Carolina, 1—20.
Surface F. M. The early development of a polyclad Planocera unquilina Wh.—
Proc. Acad. Natur. Sci. Philadelphia, 1907, N 54, 514—557.
Thorson G. Reproduction and Larval development of Danish marine bottom
invertebrates, with special reference to the Planktonic larvae in the Sound
(0resund).— Meddz. Danm. Fisk.-og Havunders., Plankton, 1946, 4, N 1,
1—523.
Thorson G. Zur jetzigen Lage der marinen Bodentier-Ukologie.— Zool. Ges.
Wilhelmshaven, 1952, N 34, 270—327.
Ulrich W. Vorschlage zu einer Revision der Grosseinteilung des Tierreichs.—
Verh. Deutsch. Zool. Ges. Marburg., 1951, 215—271.
Wells G., Dales R. Spontaneous activity patterns in animal behaviour. The
irrigations of the burrow in the Polychaetes Chaetopterus variopedatus and
Nereis diversicolor.— J. Marine Biol. Ass. U. K., 1951, N 29, 661—680.
Werner B. Beobachtungen fiber den Nahrungserwerb und die Metamorphose
der Metatrochophora von Chaetopterus variopedatus.— Helgolander Wiss.
Meer, 1953, 4, N 3, 225—238.
148
Willemoes-Suhm R. Biologische Beobachtungen liber niedere Meeresthiere.—
Z. wiss. Zool. Leipzig, 1871, 21, 380—396.
Wilson D. P. Die larvae of Polydora ciliata Johnston and Polydora hoplura
Claparede.— J. Marine Biol. Ass. U. K., 1928, 15,129—149.
Wilson D. P. The development of Nereis pelagica L.— J. Marine Biol. Ass. U. K.,
1932a, 18, 203—217.
Wilson D. P. On the Mitraria larva of Owenia fusiformis delle Chia]‘e.— Philos.
Trans. London, set. B, 1932b, 221, 231—334.
Wilson D. P. Notes on the early stages of two Polychaetes, Nephthys hombergi
Lamarck and Pectinaria koreni Malmgren.— J. Marine Biol. Ass. U. K.,
1936, 21, 305-310.
Wilson D. P. The attractive factor in the settlement of Ophelia bicornis Savi-
gny.— J. Marine Biol. Ass. U. K., 1954, 33, 361—380.
Wilson D. P. The role of microorganisms in the settlement of Ophelia bicornis
Savigny.— J. Marine Biol. Ass. U. K., 1955, 34, 531—543.
Wilson E. B. Experiments on the cleavage mosaic in Patella and Dentalium.—
J. exp. zool. Inst. Philadelphia, 1904, N 1, 197—268.
Woltereck R. Trochophora-Studien. Uber die Histologie der Larvae und dio
Entstehung des Annelids bei den Polygordius-Arten der Nordsee.— Zoologi-
ca, 1902, 34, 1—71.
Woltereck R. Wurmkopf, Wurmrumpf und Trochophora.— Zool. Anz., 1904, 28,
273—322.
Woodworth W. The palolo worm Eunice viridis (Gray).— Bull. Mus. Compar.
Zool. Harward, 1907, 51, 1—21.
Wu Bao-ling. Subspecific differentiation and ecological characteristics of Capi-
tella capitata (Fabricius).— Oceanol. Limnol. Sinica, 1964, 6, N 3, 260—271.
Zenkewitsch L. А. [Зенкевич Л. 4.]. Ueber das Vorkommen der Brackwasser
Polychaete Manayunkia polaris (n. sp.) an der Murmankiiste.— Zool. Anz.,
Leipzig, 1935, 109, 7—8, 195-203.
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ.................................... 3
ИСТОРИЯ ИССЛЕДОВАНИЙ МОРФОЛОГИИ И ЭКОЛО-
ГИИ ЛИЧИНОК ПОЛИХЕТ ......................... 6
МЕТОДЫ ИССЛЕДОВАНИЯ......................... 15
МОРФО-ЭКОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА РАЗВИ-
ТИЯ ПОЛИХЕТ................................. 17
Способы размножения и развития.............. 19
Периоды, фазы и стадии жизненного цикла..... 21
ОБЩАЯ МОРФОЛОГИЯ И АНАТОМИЯ ЛИЧИНОК .... 25
Морфология , . , ........................... 25
Типы личинок................................ 26
Анатомия ................................... 28
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА НЕКТОХЕТ ДО СЕ-
МЕЙСТВА .........................,.......... 30
ОСОБЕННОСТИ РАЗВИТИЯ И ЛИЧИНКИ ПОДКЛАССА
ERRANTIA............................... 33
Отряд Phyllodocemorpha.................................... 34
Семейство Phyllodocidae............................... 39
Семейство Typhloscolecidae........................... 48
Семейство Aphroditidae................................ 51
Отряд Nereimorpha......................................... 60
Семейство Hesionidae................................. 63-
Семейство Nereidae.................................... 66
Отряд Eunicemorpha........................................ 75
Семейство Eunicidae................................... 75
ОСОБЕННОСТИ РАЗВИТИЯ И ЛИЧИНКИ ПОДКЛАССА
SEDENTARIA............................... о2
Отряд Spiomorpha.......................................... 83
Семейство Ariciidae . . . .......................... 84
Семейство Spionidae................................... 90
Семейство Chaetopteridae............................. 104
150
Отряд Drilomorpha...........;....................... 110
Семейство Capitellidae........................... 110
Семейство Arenicolidae........................... 115
Отряд Terebellomorpha............................... 117
Семейство Pectinariidae.......................... 117
Отряд Serpulimorpha................................ 121
Семейство Serpulidae............................. 122
ПРОИСХОЖДЕНИЕ ТРОХОФОРНЫХ ЖИВОТНЫХ................. 124
ЗАКЛЮЧЕНИЕ.......................................... 134
ЛИТЕРАТУРА.......................................... 137
Владимир Александрович Свешников
МОРФОЛОГИЯ ЛИЧИНОК ПОЛИХЕТ
Утверждено к печати
Институтом эволюционной морфологии и экологии
животных им. А. Н. Северцова
Академии наук СССР
Редакторы издательства А. М. Гидалевич, Т. Н. Маркова
Художник А. П. Николаев
Художественный редактор Н. Н. Власик
Технический редактор Н. Н. Плохова
Корректор М. К, Запрудская
ИБ № 7495
Сдано в набор 2.9.77. Подписано к печати 10.3.78.
Т-00175. Формат бОхЭО1/^. Бумага типографская № 2
Гарнитура обыкновенная. Печать высокая
Усл. печ. л. 9,5. Уч.-изд. л. 10,1. Тираж 1000 экз. Тип. зак 3027
Цена 1 руб.
Издательство «Наука»
117485, Москва, в-485. Профсоюзная ул., 94а
2-я типография издательства «Наука»
121099, Москва, Г-99, Шубинский пер., 10
ИСПРАВЛЕНИЕ
Страница Строка Напечатано • Должно быть
34 12 св. Phyllodocidae Phyllodocemorpha
Зак. 3027. Ъ. А. Свешников