/




































































































































































































































































































































































































































































Text
ебник
ГОСлГДАРСТВВННвВ НАУЧНО - ТЕХНИЧЕСКОЕ ГОРЯОВ ИЗДАТКЛЬъ1В0
И. Н. ЯКОВЛЕВ профессор л*н>^н<.рад»т»19 горном* tvtwmttmyma
алеонтомпии
1932
Г (Т) —56—5— 2
Н. И. ЯКОВЛЕВ
Проф. Ленинградского горного ин-та
1
УЧЕБНИК ПАЛЕОНТОЛОГИИ
Издание четвертое
измененное и дополненное
МОСКВА - ЛЕНИНГРАД — М ГОСУДАРСТВЕННОЕ НАУЧНО - ТЕХНИЧЕСКОЕ ГОРНОЕ ИЗДАТЕЛЬСТВО
типография ОГПЗа РСФСР „Красный Печатник*, Ленинград, Международный пр., 751
ОГЛАВЛЕНИЕ.
. ‘ 6»р.
ПРЕДИСЛОВИЕ К 4 И 1 ИЗДАНИЯМ............................ 7
ОТ РЕДАКЦИИ ..... 8
ВВЕДЕНИЕ.................................................. 13
Предмета вадами палеонтологии —13; условия сохранения остатков организмов в ископаемом состоянии—13; палеонтологическая систематика — 14; теория эволюции — 15; теория естественного отбора— 16; действие естественного отбора на организм при установившихся внешних условиях жизни и при условиях изменяющихся — 17; о причинах изменения живых существ — 18; расхождение признаков — 21; отношение палеонтологии к эмбриологии — 23; отношение палеонтологии к сравнительной анатомии — 26; отношение палеонтолмии к биологии — 27; хронология ископаемых фаун — 29; древнейшая из ископаемых фаун — 32; изменения и процесс окаменения остатков организмов в ископаемом состоянии — 33; историческое развитие палеонтологии — 34.
ПАЛЕОЗООЛОГИЯ.
I тип. ПРОСТЕЙШИЕ, PROTOZOA.................................... 37
Класс I. Корненожки, Rhizopoda........................ 38
1. Foratninifera (38) [однокамерные, Monothalamia (62), многокамерные, Polythalamia (43;]. Распространениефораминифер (49).
II. Radiolaria (51;.
Характеристика групп простейших............................... 53
Высшие животные. Metazoa.
Обшие замечания....................................... 58
II тип. ГУБКИ, PORIFERA........................................ 54
Класс I. Demospongiae................................. 57
Класс П. Hexactinellida............................... 59
Класс III. Известковые губки, Calcispongia. . . . 61
Характеристика губок.............................• ..... 62
Распространение губок....'........................ 62
III тип. КИШЕЧНОПОЛОСТНЫЕ, COELENTERATA ........ 62
Класс I. Г и д ро м е д у з ы. Н у d го m е d u s а е. 63
I. Гпдр видные полипы, Hydraria (65), II. Tabularia (66), III. Hyd-rocorallina (67).
Граптолиты, GraploTl’coidea........................... 69
Класс II. Медузы, Scyphozoa нли Acalephae............ 72
Класс III. Ко галловые полипы, A nthozoa.............. 73
Подкласс I. Zoantharia................................ 74
, I. Rugosa (Tetracoralla) (78), геологическое распространение
Rugosa (8‘) II. Hexacoralla (85).
Подкласс II. Octocoralla пли Alcyonaria (87), геоло1ичеекое странение Tabulata (91).
Распространение Anthozoa ................... 91
Характеристика групп к и ш e ч и о п о л о с т пих............. 92
IV тип. ТЕРВИ, VERMES.....................................
Мшанки, Вг у ozoa........................................ 98
I. Cydostomata (100), II. Tropnstomafa (101), 111. Cryptostomata ( 01), IV. Cheilostomata (102). . .
102 Плеченогие, Brachiopoda..............................
I. Inarticulata (109). 11. Articulata (111) [Strophomenacea (112), Pentameracea (116), Rhynchonellacea (118), Tetrabratulacea (118), Spiriferacea (119)].
Распространение плеченогих............................ 121
V тип. ЧЛЕНИСТОНОГИЕ. ARTHR0P0DA............................. 123
Подтип I. Cephalothoracica..................................... 125
Класс I. Ракообразные, Crustacea . . . . : . . . . 125
Подкласс I. Entomostraca.............................. 126
I. Phyllopoda (126), II. Ostracoda (127), III. Cirripedia (128).
Подкласс II. Malacostraca........................... 129
I. Десятиногие раки, Decapoda (130).
Подкласс III. Трилобиты, Trilobitae................... 133
Распространение трилобитов............................ 143
Характеристика групп ракообразных............................... 144
Класс II. Рако-скорпиоиы, Merostomata (Gigantostraca) 144 I. Xiphosura (145), II. Eurypterida (146).
Класс III. П ay к о о бр аз н ы e, Arae h noidea...... 148
Подтип II. Eucephalica . -...................................... 149
Класс IY. Многоножки, Myriopoda....................... 149
Класс Y. Насекомые, Insects.......................... 150
VI тип. ИГЛОКОЖИЕ, ECHINODERMATA.............................. 152
Подтип I. Pelmatozoa............................................ 154
Класс I. Морские лилии, Crinoidea..................... 154
I. Fistulala (161), II. Articulata (163), 111. Camerata (164).
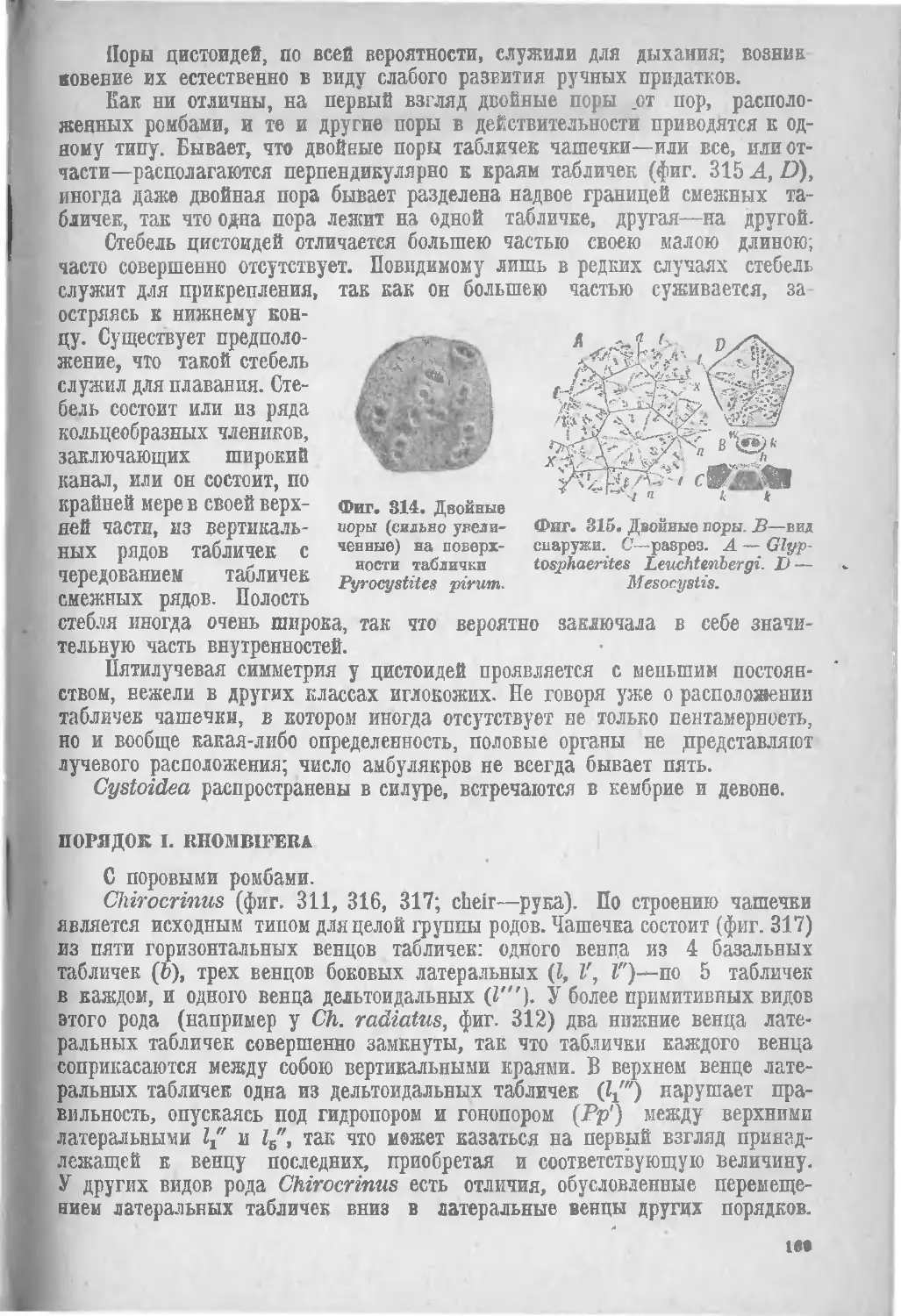
Класс II. Cystoidea................................... 164
1. Rhombifera (169), II. Diploporita (171). .
Класс III. Blastoidea................................. 172
Класс IY. Edrioasteroidea............................. 175
Геологическое развитие Pelmatozoa..................... 176
Подтип II. Eleutherozoa ......................... 176
' Класс I. Морские звезды, Stelleroidea................. 176
Подкла с I. Asteroidea............................ . 176
Подкласс II. Ophiuridea.............................. 178.
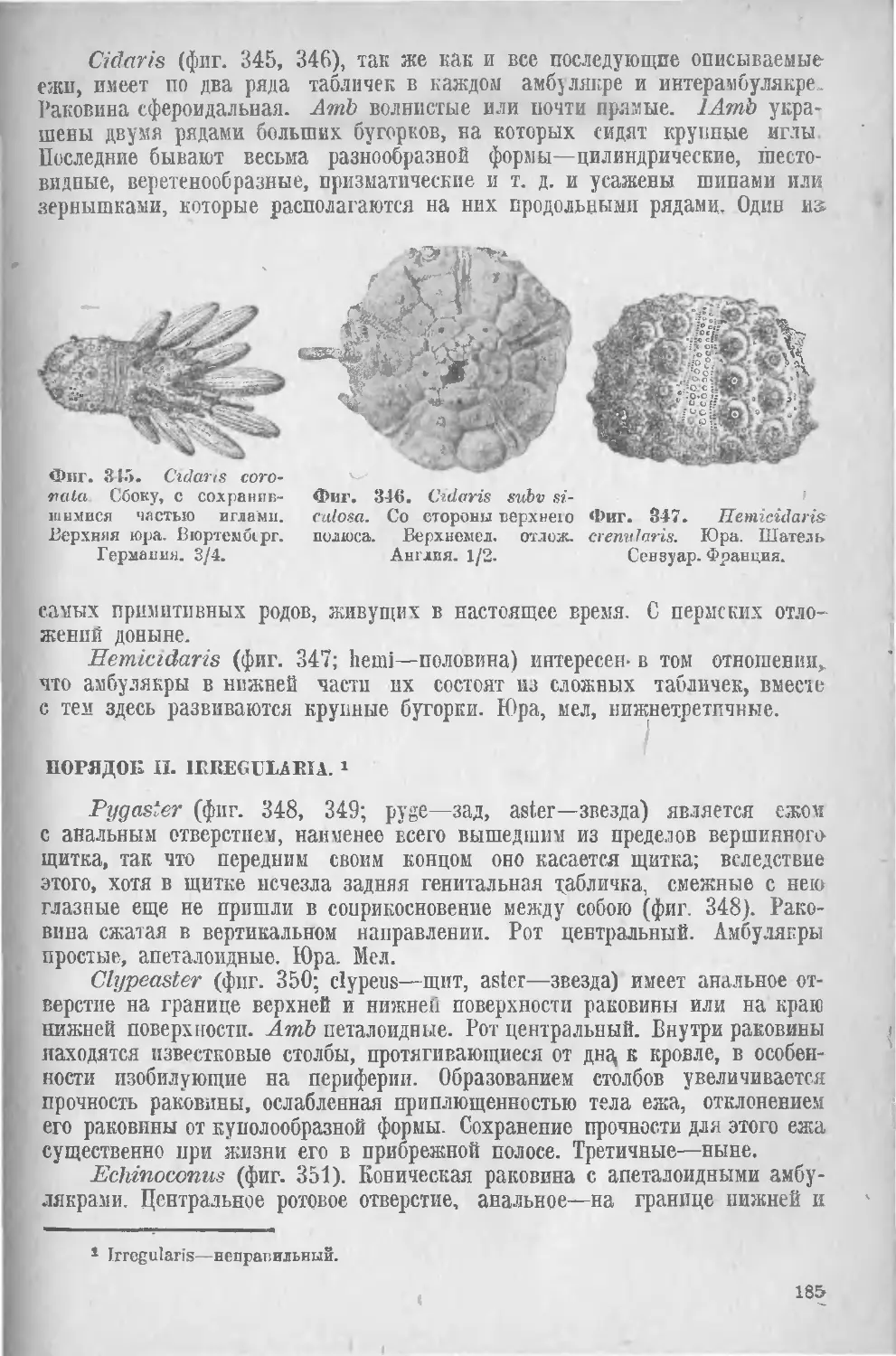
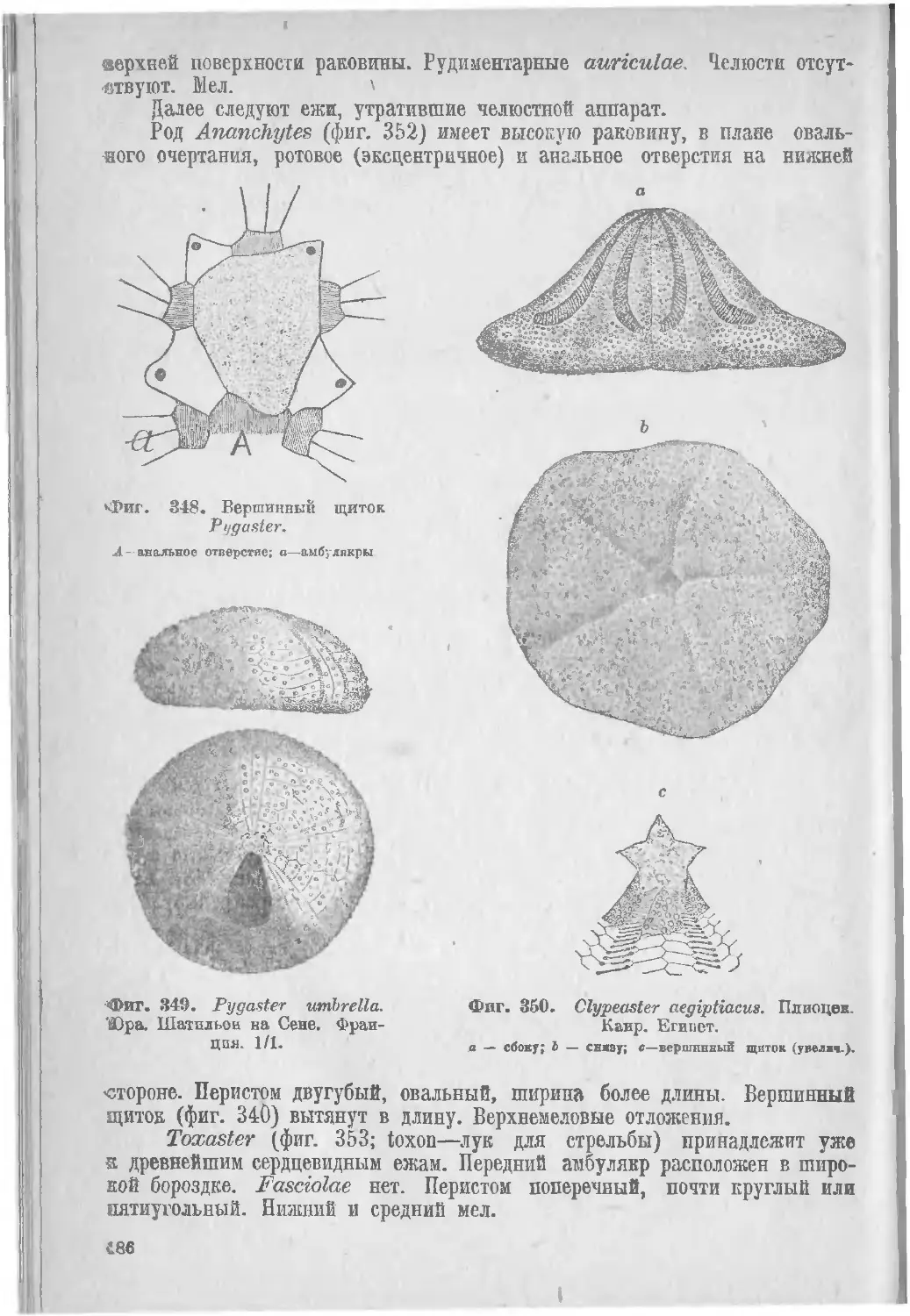
Класс II. Морские ежи, Echinoidea..................... 179
I. Regularia (183), II. Irregularia (185).......
Геологическое распространение морских ежей . .... 187
X арак т ер и с т и ка гру и п и г л о к о ж и х...'............ 187
VII тип. МЯГКОТЕЛЫЕ, MOLLUSC А.................................. 189
Класс I. Червеобразные моллюски, Amphineura 191
Класс II. Брюхоногие. Gastropoda...................... 192
Жаберные, Branichiata................................. 196
Подкласс I. Prosobranchiata........................... 196
Подкласс II. Opisthobranchiata........................ 204
Легочные, Pulmonata........................ ........ 206
Геологическое распространение брюхоногих.............. 207
Класс III. Головоногие, Cephalopoda................... 209
Подкласс I. Четырехжаберные, Tetrabranchiata.......... 210
I. Nautiloidea (210), II. Ammonoidea (217).
Подкласс П. Двужаберные, Dibranchiata................. 232
I. Десятиногие, Decapoda (232), II. Осьмнногие, Octopoda (235).
Распределение головоногих ................... 235
Класс IY. Пластинчатожаберные. Lamellihranchiata . . . 237
I. Taxodonta (245), II. Dysodonta (247), III. Hetcrodonta (250),
IV. Schizodonta (254), Y. Pachyodonta (256), YI. Desmodonta (259).
Распространение пластинчатожаберных . ... ~........... 261
Класс V. Лопатоногие, Scaphopoda...................... 262
Характеристика групп моллюсков......................................... 264
VIII тип. ПОЗВОНОЧНЫЕ, VERTEBRATA...................'. . . ............ 265
Vertebrata craniate............................................. 265
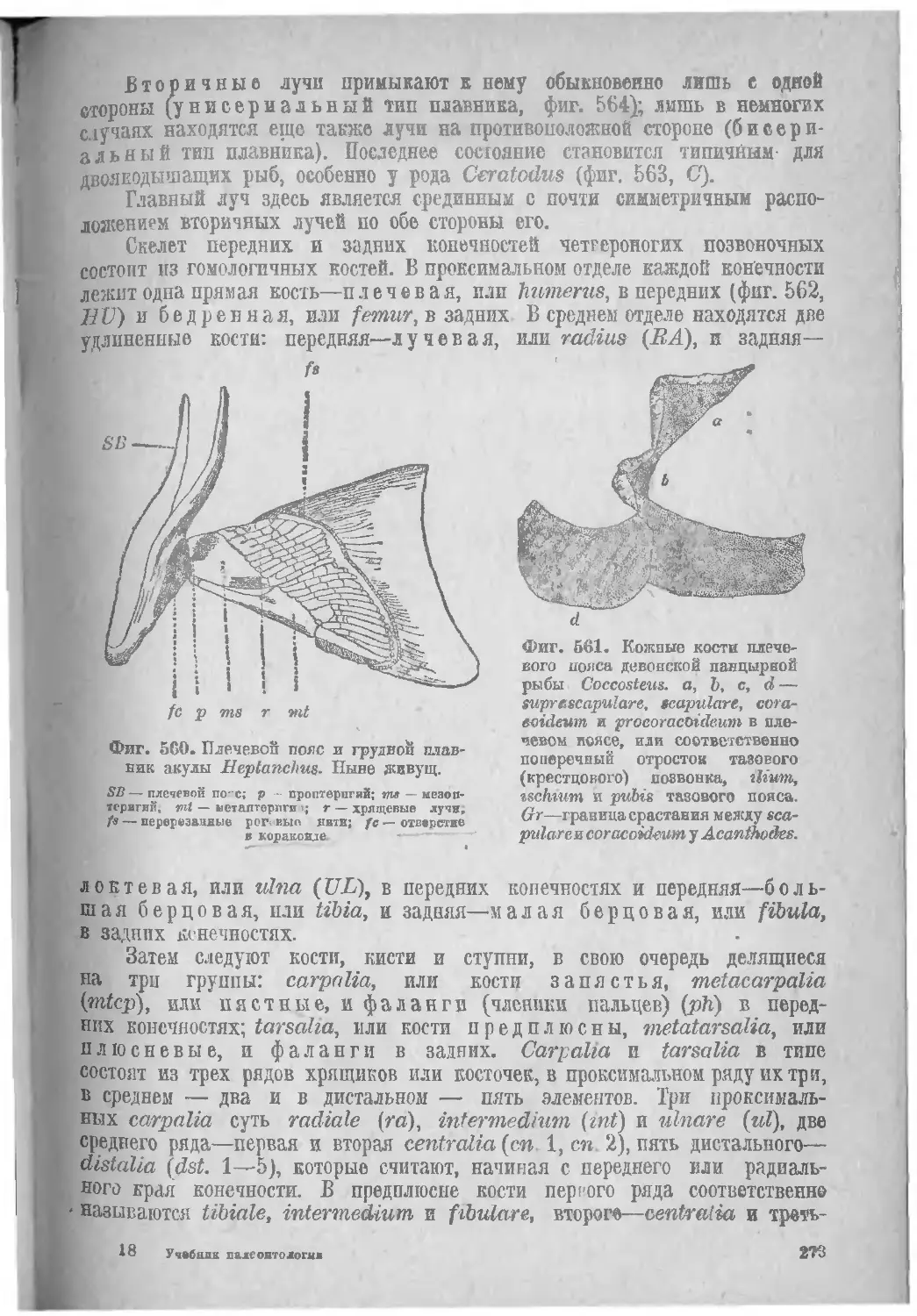
Класс I. Рыбы, Pisces....................................... 281
Подкласс I. Круглоротые, Cyclostomata...................... 284
Подкласс II. Панцирные рыбы, Placodermi..................... 284
Подкласс 111. Arthrodira.......................... . ' . . 287
Подкласс IY. Листожаберные, Elasmobranchii.................. 289
I. Selachii (290), II. Holocephali (294).
Подкласс Y. Костистые рыбы, Teleostomi...................... 295
I. Crossoptherygii (296), II. Ganoidei (299), [Heterocerci (300), Chon-drostei (300), Amioidei (301)], III. Teleostei (302), 1Y. Dipnoi (303).
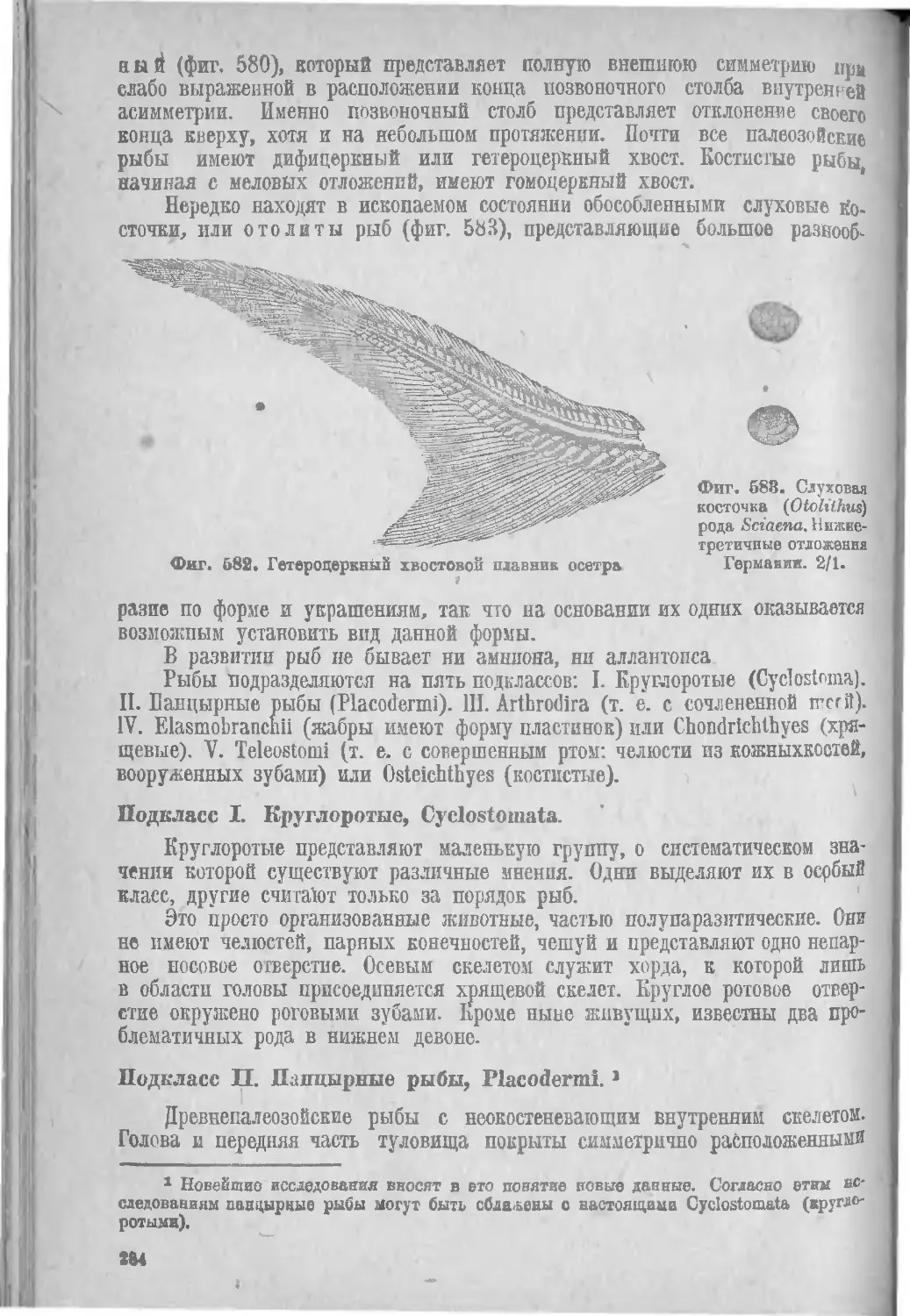
Геологическое распространение рыб.......................... ЗО'’
Характеристика групп рыб............................................... 305
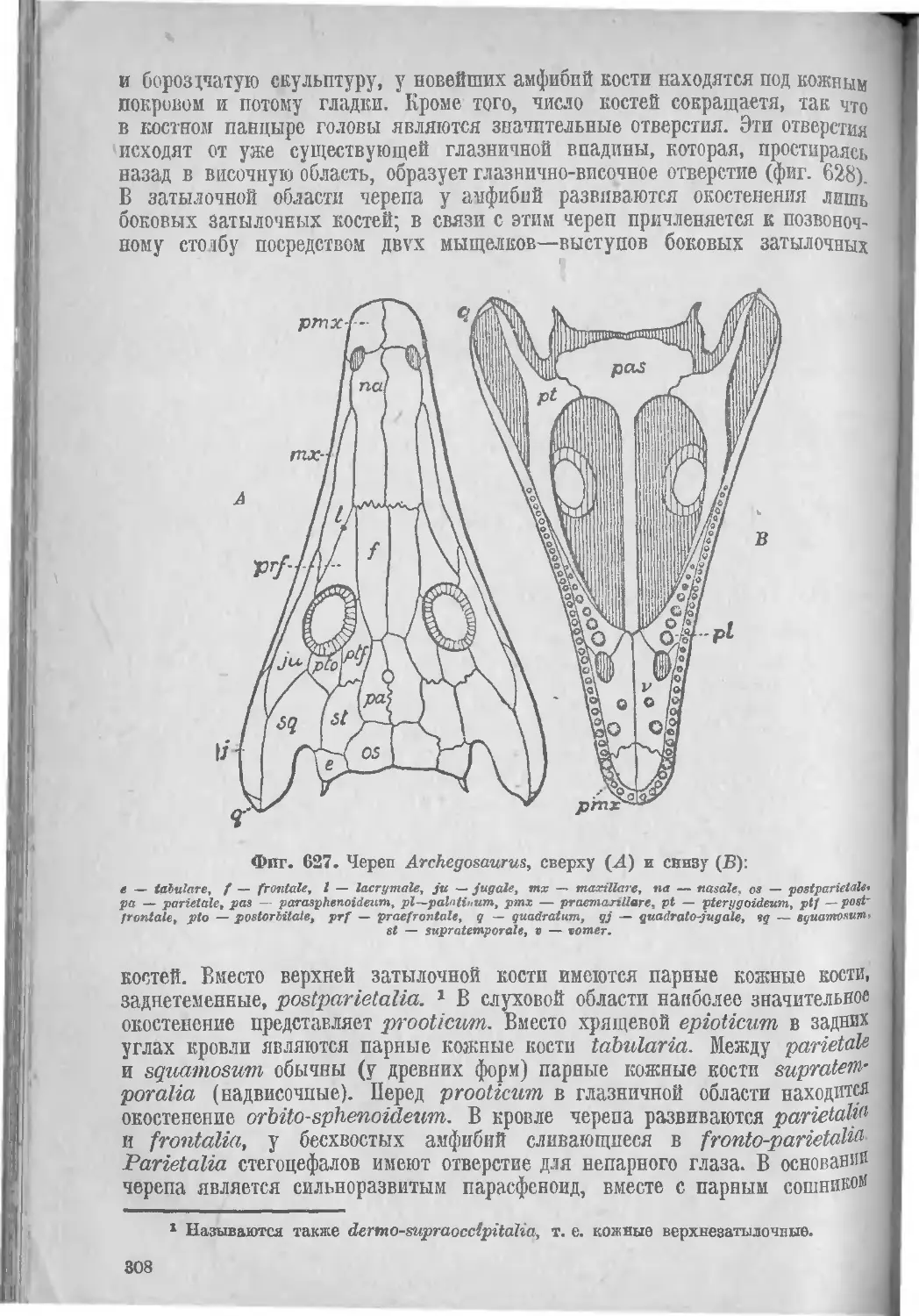
Класс II. Амфибии, Amphibia................................. 307
Подкласс I. Панцырноголовые, Stegocephali . ................ 311
I. Phyllospondyli (312), II. Lepospondyli (313), III. Temnospon-dyli (313).
Подкласс II. Амфибии собственно, Enamphibia ................ 316
I. Хостатые, Caudata (316), II. Бесхвостые, Ecaudata (317).
Геологическое распространение амфибий ............ 318
Характеристика группамфибий........................................... 318
Класе III. Пресмыкающиеся, Reptilia. . . .......... 318
Группа Anapsida............................................. 322
I. Cotylosauria (322), II. Черепахи, Chelonia (325).
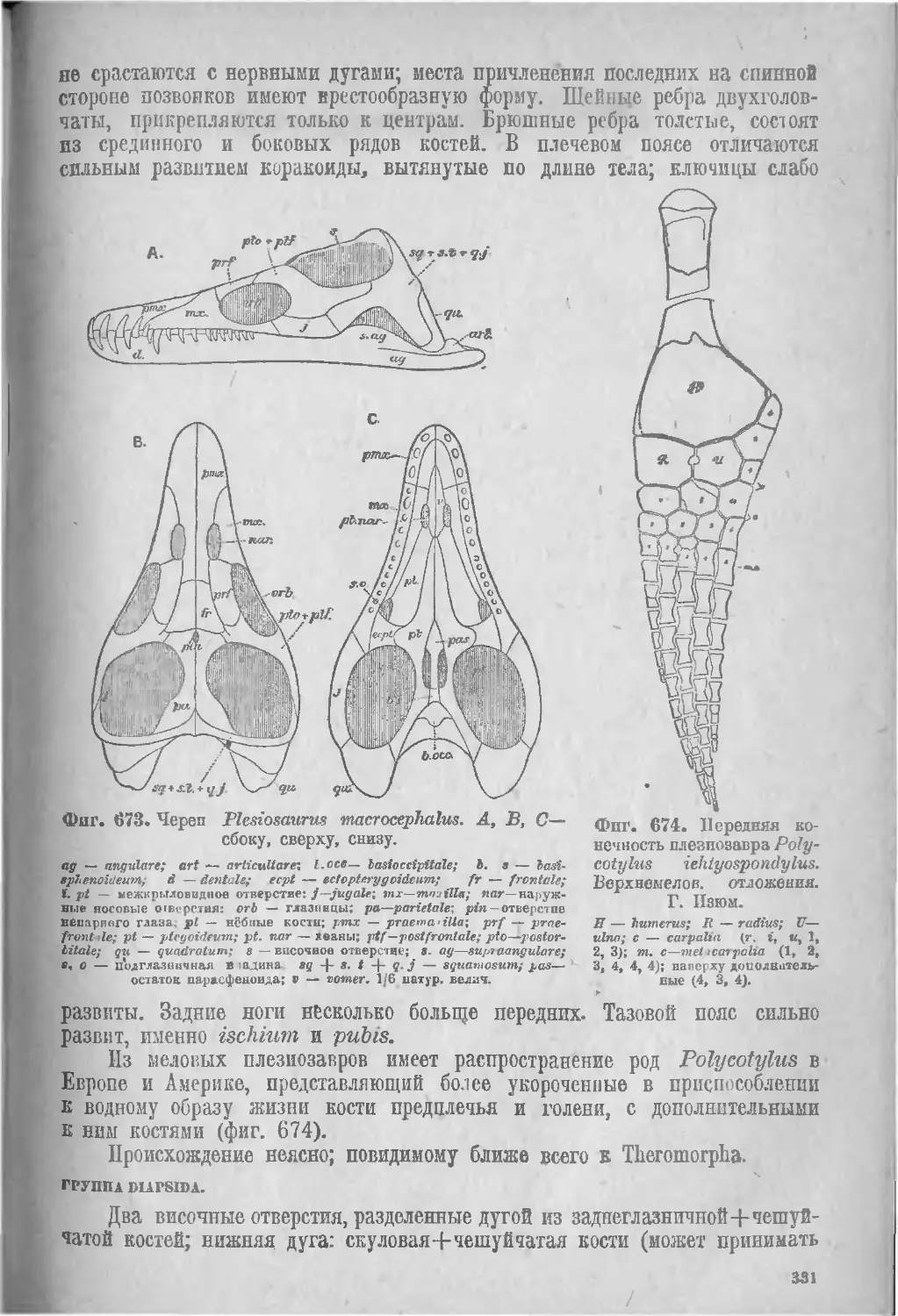
Группа Synapsida............................................ 327
III. Theromorpha (327), IY. Плезиозавры, Sauropterygia (330).
Группа Diapsida............................................. 331
V. Клювоголовые, Rhynchocephalia (322), YI. Крокодилы, Croco-dilia (322), Y1I. Динозавры, Dinosauria (334), VIII. Летающие ящеры, Pterosauria (337).
Группа Parapsida.............................................340
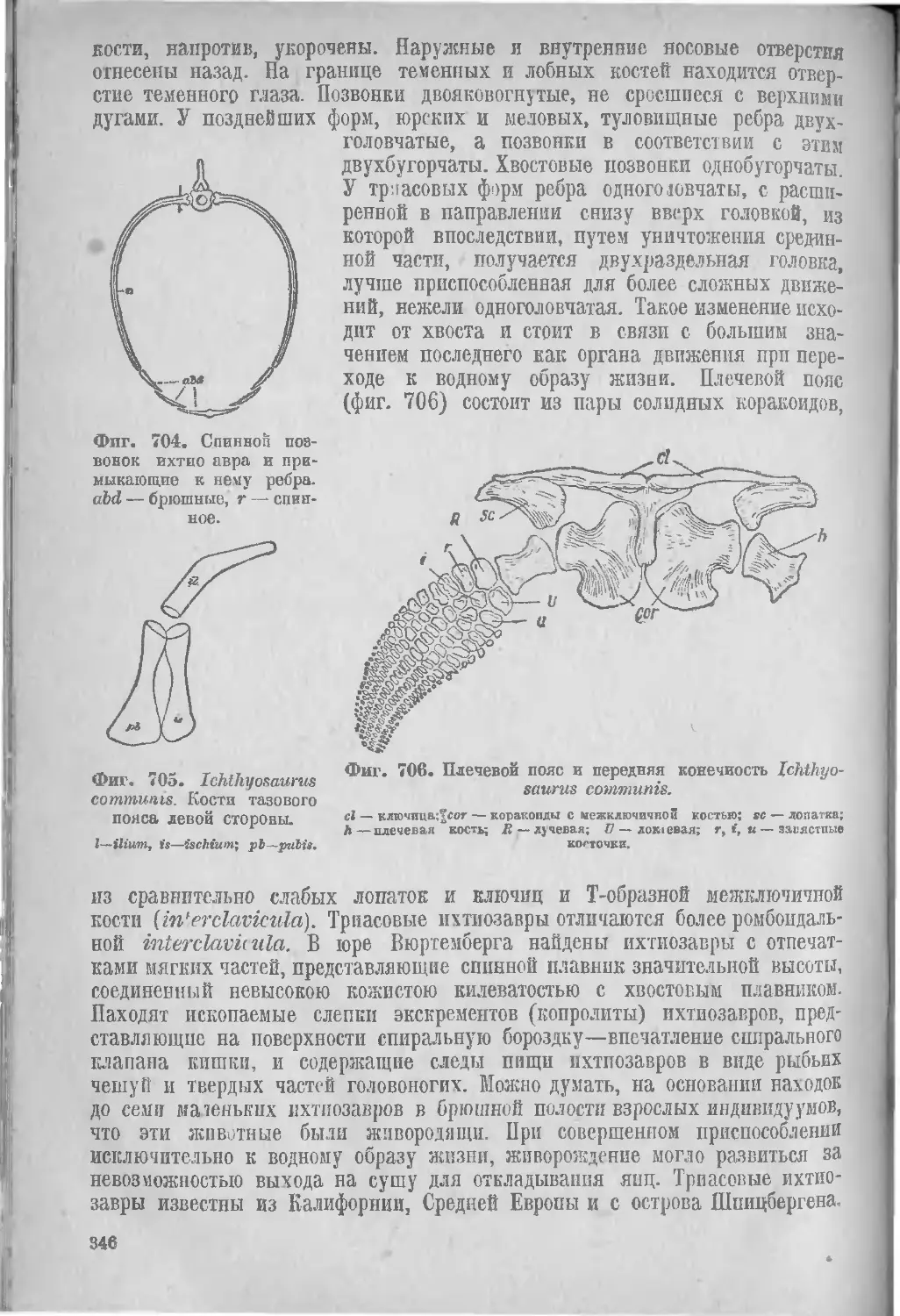
IX. Чешуйчатые, Sqnamata (840), [Ящерицы, Lacertilia (340), Mo-возавры, Mososauria (341), Змеи, Ophidia (343)], Ихтиозавры Ichthyosauria (343).
Геологическое распространение рептилий...................... 347
Характеристика групп рептилий.......................................... 347
Класс IY. Птицы, Aves....................................... 348
Подкласс I. Древние птицы, Archaeornithes................... 351
Подкласс II. Новые птицы, Neornithes........................ 353
Геологическое распространение птиц.......................... 353
Класс Y. Млекопитающие, Mammalia............................ 354
Подкласс I. Беспоследовые, Eplacentalia .................... 362
1. Однопроходные, Monotremata (362). II. Сумчатые, Marsupia-lia (363), [Multituberculata (363), Polyprotodontia (364), Dipro-todontia (365)].
Подкласс II. Последовые, Placentalia........................ 366
I. Insectivora (366), II. Chiroptera (366), III. Carnivora (366), IY. Edentata (368), V. Primates (370), [Prosimiae (370), Simiae (370], VI. Condylarthra (373), VII. Perissodactyla (375), VIII. Artiodactyla (381), [Nonruminantia (381), Ruminan-tia (381)], IX. Proboscides (385), X. Cetacea (388), XI. Sire-
nia (391). Геологическое распространение млекопитающих. 392
Характеристика групп млекопитающих.................. 392
ПАЛЕОБОТАНИКА.
I тип. СЛОЕВЦОВЫЕ, TALLOPHTTA....................... 396
Класс!. Бактерии, Schizomycetes........ . . . 396
Класс II. Син е-зелеиые водоросли, Schizophyceae . 396
Класс 111. Жгутиковые, Flagellatae...... • • 397
Класс IV. Слизевики, Myxomycetes............. 398
КлассУ. Перидине и, Peridineae. . ........... 398
Класс VI. Диатомовые водоросли. Diatomeae . . . . 398
Класс VII. Сцеплянки, Conjugatae............. 399
Класс VIII. Зеленые водоросли, С hl о гор h усе а е . . 399
Класс IX. Харовые, Charophyta........ , . . . 401
Класс X. Бурые водоросли, Phaeophyceao....... 403
Класс XI. Багряные водоросли, Rhodophyceae (F1 о г i d е а е)........................... 403
Класс XII. Грибы, Fungi...................... 405
Класс XIII. Лишайники, Lichen es............. 405
Зародышевые бессифонные, Embryophyta asiphonogama.
II тип. МОХООБРАЗНЫЕ, BRY0PHYTA............................. 410
Класс I. Мхи печеночные, Hepaticae................ 410
Класс П. Мхи лиственные, Musci frondosi. . . . . 410
III тип. ПАПОРОТНИКООБРАЗНЫЕ, PTERID0PHYTA.................... 41®
- Класс I. Псилофитовые, Psilophytales . . . . . . . . 410
Класс П. Плауновые, Lycopodiales.................. 412
Класс III. Псилотовые, Psilotaceae............... 417
Класс IV. Хвощевидные, Articulates............... 417
Подкласс I Calamophytales.....'................... 417
, Подкласс II. Pseudoborniales........................ 417
Подкласс III Sphenophyllales...................... 417
Подкласс IV. Cheirostrobales...................... 418
Подкласс V. Equisetineae.......................... 418
Класс V. Кладоксилов’ые, Cladox у dales........... 422
Класс VI. Папоротогиковые, Filicales.............. 422
Зородышевые сифоногамные, Embryophyta syphonogama.
IV тип. ГОЛОСЕМЕННЫЕ, GYMN0SPERMAE................... 429
Класс I. Папоротникосеменные, Pteridosper-males..?.............%....................... 429
Класс II. Кордаиты, Cordaitales.............. 429
Класс III. Беннетитовые, Benne titales. ..... 430
Класс IV. Саговиковыо, Cycadales............. 432
Класс V. Гинкговые, Ginkgoales.............. 435
Класс VI. Хвойные, Coniferales............... 437
Класс VII. Хвойников ые, Gnetaceac........... 439
У тип. ПОКРЫТОСЕМЕННЫЕ, ANGI0SPERMAE................- 439
Класс I. Однодольные, Monocotyledonae........ 440
Класс II. Двудольные, Dicotyledonae ......... 440
Алфавитные указатели......................... 444
в
ПРЕДИСЛОВИЕ К #-МУ ИЗДАНИЮ.
В настоящем издании даются некоторые добавления к тексту, имеющие целью облегчить усвоение учебника. Для этого названия греческого и латинского происхождения переведены на русский язык; зачастую это указывает на характерные особенности родов и групп. Затем для родов беспозвоночных, имеющих особенное геологическое и практическое значение, а также д ля растений даны таблицы-графики их распространения, и наконец в конце каждой большой систематической группы дается как бы в виде резюме краткая характеристика ее подразделений.
Некоторые добавления для позвоночных даны А. Н. Рябининым, а по палеоботанике И. В. Палибиным, а в упомянутой работе перевода латинских и греческих названий мне оказал большое содействие Е. И. Фальк.
Июль 1931 г.
Н. Яковлев.
ПРЕДИСЛОВИЕ К 1-МУ ИЗДАНИЮ.
Издаваемый курс выработан мною за время чтения лекций в Горном институте с 1898 года.
Печатая его, я прежде всего желал дать студентам Горного института учебник, соответствующий моему курсу. В этом ощущалась тем большая необходимость, что первый русский курс палеонтологии проф. И. И. Лагузена уже давно распродан.
Изложение в учебнике ведется сжато и так, что у читателя не предполагается предварительного знакомства с зоологией и ботаникой. Особенное старание было приложено к изложению общих обзоров,морфологии; в курсе введены данные относительно исторического развития групп животного и растительного царства,—указания на отйошения родства и происхождения одних групп от других, а также данные относительно причин развития тех или иных особенностей организации (например для фораминифер, иглокожих, кораллов, плеченогих). Из палеонтологической систематики приводятся лишь главнейшие подразделения; детальнее там, где это существенно для обрисовки эволюционного процесса.
Курс издан на средства, ассигнованные советом Горного иневдтута, причем дело издания было облегчено возможностью использовать клише к учебнику И. И. Лагузена. Некоторые рисунки сделаны совершенно заново по оригинальным работам авторов.
Б. Б. Ребиндеру, А. Н. Рябинину и И. А. Рейнвальду я обязан дружеским содействием по просмотру корректур и ценными указаниями, давшими возможность внести некоторые улучшения в изложение
Декабрь 1909 г.
Н. Яковлев.
ОТ РЕДАКЦИИ.
Нет надобности говорить о том, что учебники палеонтологии сейчас крайне необходимы. Правильнее, наоборот, ставить вопрос, почему их столь мало. Страна, героически созидающая новую мощную социалистическую промышленность, нуждается в больших кадрах ученых геологов, горных инженеров и техников различных разрядов и специальностей. Им знание палеонтологии необходимо. Оно нужно и для биологов. Не менее ясно, что догнать и перегнать капиталистические страны можно лишь перестроив науку на базе марксистско-ленинской методологии.
Поэтому и учебники наши в первую очередь должны коренным образом измениться в соответствующем направлении. К сожалению такого учебника палеонтологии еще нет, почему приходится переиздавать ранее существовавшие. Редакция вполне сознает все несоответствие их современным требованиям. Прежние учебники в значительной мере есть продукт отжившей эпохи,' прежней научной мысли. Отличительные особенности этой последней определяются специфическими закономерностями развития капиталистического общества. Стихийный, бесплановый, индивидуалистический характер производства и классовая борьба в капиталистическом обществе создали соответствующую организацию научной работы, придали ей определенное направление, воспитали необходимые традиции и настроения. Бесплановость, стихийность научной мысли привели к пренебрежению философией и торжеству ползучего эмпиризма.
Классовая борьба была причиной того, что великое учение Маркса, Энгельса и Ленина игнорировалось и даже оставалось по существу безвестным в обширных кругах ученых. У нас в царской России оно было буквально на нелегальном положении. Результаты налицо—едва ли не большинство наших научных работ, равно как и учебников, чужды марксистско-ленинской методологии. Советские ученые в большинстве понимают основные недостатки прежней школы, но понять еще не всегда означает выправить, перестроить на новой основе научную мысль. Для этого совершенно недостаточно бегло прочесть Энгельса, Маркса, Ленина, просмотреть некоторые статьи в марксистских журналах и т. д. Для этого нужна довольно длительная и во всяком случае настойчивая работа над собой, проверка и пересмотр буквально каждого прежнего понятия, каждого вывода старой школы, необходимо перевоспитание в новом направлении. Неудивительно поэтому, что наши учебники в большинстве случаев далеко не соответствуют тому, что требует заказ великой стройки пролетариата.
Автор предлагаемого учебника палеонтологии хорошо известен своей высокой компетенцией в палеонтологии, но недостатки прежней школы встречаются в его книге на каждом шагу.
Начать с того, что он остался верен прежним традициям описания и размещения систематического материала. Он последовательно описывает одну группу за другой с одинаковым так сказать беспартийным ко всем им вниманием. Для автора как бы не существует палеонтологии советской, призванной служить в первую очередь геолого-разведочному делу именно нашей страны, равно как и пропаганда тех идей общей биологии, в частности эволюции, которые особенно актуальны именно теперь в практике освоения природных богатств Союза, борьбы с сельскохозяйственными
вредителями, улучшения домашних пород животных и растений, наконец в практике антирелигиозной работы и т. д.
Для него существует лишь «палеонтология вообще», палеонтология внеклассовая, созерцательная. Мы не хотим этим сказать, что проф. Н. В. Яковлев не имел в виду служить практике геолого-разведочного дела. Он именно хотел этого. Но объективно получилось нечто в политическом отношении так сказать «бесскелетное». Почему бы например не указать, какие из описываемых им групп или видов имеют наибольшее значение сейчас, в нашей стране, в Донбассе, на Украине, на Урале, Кавказе и т. д., и почему именно важно их знать? Не намечает также автор и те проблемы, разрешение которых стоит на очереди соцстроительства. Его учебник превращается по существу в сухой справочник. В систематической части нет научного определения таксономических единиц-рода, вида, семейства и т. д., что служит показателем того кризисного состояния, в который загнала науку эмпирическая мысль.
Остаются по большей части совершенно обойденными общие закономерности развития. Они вынесены автором во введение. Однако, занимаясь основной частью учебника, без посторонней помощи читатель, как правило, решительно не в состоянии узнать, в чем эти общие закономерности .конкретизируются. Читатель не видит в большинстве случаев совсем развития, становления различных форм жизни. Ему самому предоставляется догадываться, какие противоречия развития вызвали к жизни ту или иную группу животных или растений, тот или иной орган какого-либо организма.
Перед читателем проходит не объединенная общей идеей, не связанная единством диалектических законов нестройная группа организмов, а зачастую даже просто пестрый конгломерат «интербрахиальных табличек», «головогрудных щитков», «створок неодинаковой величины и формы», «зубных пластинок», «заднепроходных отверстий» и т. д.
Мы очень хорошо учитываем, что в земле уцелело очень незначительное количестве живших ранее организмов, что многие соотношения и связи погибли безвозвратно. Тем не менее материал все же несомненно позволяет многое представить в движении, в развитии. Этого у автора мы, как правило, не видим. Подобные существенные недочеты устранит всякий, кто руководствуется методом марксистско-ленинской методологии.
Вводную часть мы оставили без изменений, лишь снабдив отдельные ее места примечаниями, в то время как, если бы с изданием учебника не приходилось спешить, ее следовало бы написать наново—до такой степени содержание ее страдает «пережитками прошлого», хотя бы и весьма недавнего прошлого.
Для примера: автор принимает борьбу за существование и естественный отбор односторонне, не видя противоположного начала, слитого с первым в неразрывное единство, на что указывает между прочим Энгельс. Организмы не только борются друг с другом. Приспособление в природе происходит не к чему-то «вообще», а к вполне определенной, конкретной обстановке, и в этой обстановке значительную роль играют различные организмы. В результате естественного отбора происходит взаимоприспособле-ние—нечто прямо противоположное борьбе за существование. Так сосущие насекомые (мухи, пчелы, шмели, бабочки) сплели свою жизнь в тесный «союз» с цветковыми растениями д уже не могут обходиться друг без друга: растение опыляется насекомыми, насекомые получают «за это» питание.
Растения в тенистой чаще леса борются друг с другом, отнимая корнями друг у друга воду и минеральные вещества, листьями перехватывая друг у друга свет. Однако долгая история приспособления привела к тому, что все их органы теперь приспособлены именно к такой тесноте и нехватке света. Расчистите кругом лес и тенелюбы погибнут: «враги» не могут жить друг без друга, они не только «враги», но и «друзья». Ястреб, истребляя куропаток, приносит не только вред, но и пользу: в первую очередь в его лапы попадают больные и хилые; истребляя, он в то же время оздоровляет вид куропаток. Сделано такое наблюдение: в странах, где сильно истребляются хищные птицы, наблюдаются эпизоотии среди куропаток (происходящие от паразита эймерии). Подобных примеров можно было бы привести очень много. Как видим, марксистское понимание всякого процесса как единства противоположностей находит свое подтверждение в этих процессах.
Кроме того нельзя мыслить борьбу за существование и отбор существующими с момента начала жизни в неизменном виде до наших дней. То И другое меняет свои формы, находясь, как и все, в развитии, в движении. Об этом мы здесь не можем подробно распространяться. Укажем только на следующее. Организмы какого-либо уголка жизни (стации или станции, как говорят зоо- и фитогеографы), взаммоприспособляясь, как указано выше, образуют то более, то менее сплоченное взаимосвязями, взаимо-.адвисимостями. сообщество.или биоценоз. Поспешно и неверно было бы
сравнивать его с организмом. Это—особое качество, являющееся прямым следствием процесса приспособления обычного рода (дарвиновской борьбы за существование и отбора). Подмечено, что в борьбе за существование один биоценоз сменяется другим биоценозом. Закономерности развития биоценозов' качественно иные, нежели развития отдельных видов. Об этом палеонтологу не лишне знать, так как проблема биоценозов тесно связана с проблемой фаций. Далее. Биоценозы расширяясь, видоизменяясь, образуют флоры п фауны—единицы высшего порядка. Палеонтология установила и, здесь своеобразную, опять-таки качественно иную форму борьбы за существование со своими собственными закономерностями. Фауны и флоры на протяжении истории органического мира сменяют друг друга (сплошь и рядом быстро, скачкообразно), и об этом также не лишне было бы специально упомянуть в учебнике палеонтологии, сообщив и констатированные для этого процесса занономерности.
В главе «О причинах изменения живых существ» автор различает «внешние и внутренние причины» изменений, разрывая их, считая независимыми, в то время как здесь мы имеем типичный пример единства противоположностей. «Волосяной покров животных часто находится в зависимости от климата: в Ангоре не только козы, но и кошки и пастушьи собаки имеют шерсть в виде руна»—пишет автор. Однако у кошек эта длинная шерсть все же совсем иная, нежели у коз, у собак она не похожа ни на козью, ни на кошачью. Условия жизни в Ангоре (внешняя причина) почему-то требуют длинного руна, но каждый вид в результате своих специфических закономерностей (внутренние причины) дает длинную шерсть непременно на свой образец.
В воду борьба за существование загоняла не раз многих наземных животных. При этом всегда происходит приспособление к плаванию: лапы или ноги превращаются в ласты, хвост приобретает плавательные лопасти, форма тела становится по большей части веретеновидной и т. д. Внешняя
среда дает определенный «заказ», если так можно выразиться, и организация каждого вида «выполняет» этот «заказ» каждый раз на свой образец, в зависимости от внутренних своих закономерностей,‘являющихся следствием всей предшествовавшей истории приспособления. Таким образом внутренние и внешние причины неразделимо связаны друг с другом. Отметим попутно, что автор обсуждает между прочим и этот приведенный нами пример приспособления к воде наземных животных и находит, что причины эволюции здесь «внутренние». Самая важная «причина» водная среда и необходимость приспособления к ней, как читатель видит, наоборот,—внешняя. Во «внутреннюю» же она превратилась у проф. Н. Н. Яковлева вследствие его пристрастия к метафизическим представлениям Ламарка, Копа и других псевдо-ламаркистов, которые видят причину эволюции между прочим в «сознательных стремлениях» животного приспособиться к новой среде. Нет надобности говорить, что даже и эмпирические данные ныне не позволяют признавать сознательной деятельности у животных, да еще таких низко организованных, как ихтиозавры. Эмпиризм науки капиталистической эпохи вел к механицизму, к стремлению сводить сложные процессы к более простым; например жизненные процессы механисты спешат свести на физико-химические, как будто это может нам объяснить качественное своеобразие жизни. Наоборот, такое сведение, последовательно проведенное, смазывает сущность явлений жизни. Для исследователя бывает нередко необходимо проделать и сведение, но это лишь часть работы и при этом отнюдь не самая главная. Самое же важное, наоборот, заключается в том, чтобы правильно оценить именно специфические закономерности. Так же в частности обстоит дело и с «сознанием» животных. Ламарк и его последователи, как и многие другие авторы прежней школы, награждали одним и тем же «сознанием» весь мир животных. Оно у них изменялось лишь количественно. Здесь даже не сведение сложного к простому, а хуже—сведение простого к сложному. Уже в среде эмпириков начался протест против такой «науки» о нервной деятельности животных. Скоро наметилась и обратная крайность—все стали сводить на чисто автоматические акты. Мы здесь не можем подробно обсуждать этот вопрос. Заметим лишь следующее. Качественных ступеней в развитии психики нельзя понять, если начать все сводить та к сознанию типа человеческого, то к простым рефлекторным реакциям. Каждая ступень должна быть понята в ее своеобразии. Обращаясь к вышезатронутому вопросу, мы должны подчеркнуть, что сознание человека конечно есть уже не то, что мы имеем у животных. Его качественное своеобразие не подлежит никакому сомнению. Лишь у шимпанзе в недавнее время В. Келеру удалось подметить кое-какие черты, характерные для человека. Но это и неудивительно: шимпанзе очень близок к человеку. Психика человека выросла из психики зверя, в частности человекообразной обезьяны. Но, утверждая это, мы не должны забывать, что теперь наше сознание уже приобрело свои определенные, специфические особенности, далекие от такового зверей. Вот почему невозможно видеть «сознательное стремление» у животных, какое цидит автор предлагаемого учебника. Ясно, что во-первых для качественно иного процесса должно быть и новое обозначение, иной термин. Во-вторых—и это гораздо важнее подчеркнуть—воскрешать сейчас похороненное даже капиталистической наукой «сознание» животных, точнее даже сознание в каждом органе животного (в плавниках, хвостовых лопастях и т. д. у водных животных), значит тянуть науку в, болото метафизики самого худшего сорта.
Видеть во всем мире разлитое «сознание», как это было свойственно палеонтологу Э. Копу и другим псевдо-ламаркистам, — значит проповедывать мистику.
Мы полагаем, что автор учебника этого не хотел, но так все же получилось, и это гораздо более важно, чем субъективные стремления автора. Отсутствие диалектического понимания процесса жизни видно и в том, что автор не прочь говорить о «неизменных условиях» жизни, об «отсутствии стимула для эволюции» при этих «неизменных условиях», а следовательно и о «неизменности» фауны на тот или иной срок развития. Вообще в различных учебниках палеонтологии совсем не редко говорят о дошедших до нашего времени «в неизменном» виде с палеозоя, с мезозоя, с начала третичного времени и т. д. тех или иных животных (Lingula, Limulus и др.), забывая отметить, что в действительности они дошли не в «неизменном» виде, а лишь в малоизмененном. Эволюция все же шла, но медленнее, чем у других организмов. Подобный упор на «неизменность» ведет самих же авторов к ошибкам, хотя они отлично знают, что абсолютной неизменности нет; и примером таких ошибок мы могли бы взять указания автора этого учебника на якобы отсутствие стимула для развития сумчатых в Австралии и их «неизменность» в течение третичного времени. Пользующийся диалектическим методом не может допустить подобной ошибки, так как метод не позволяет предполагать абсолютных остановок развития. ”
Мы указали на ряд ошибок как в методологии, так и в отношении фактического материала в предлагаемом учебнике. Неприменение диалектического метода влечет за собой целую систему ошибок.1
Редакция
•1 В тексте введения отмечены важнейшие случаи отдельных ошибок подстрочными примечаниями.
12
ВВЕДЕНИЕ.
Предмет и задачи палеонтологии. Слово «палеонтология», в переводе с греческого, значит учение о древних организмах. Изучая организмы как явления прошлого и процессы, лежащие в основе совокупности этих явлений, палеонтология является наукой исторической—историей органического мира. Материал, изучаемый палеонтологией, есть остатки организмов прошлых эпох истории земли. Чтобы составить себе отчетливое представление о том, что может дать палеонтология и чего не может, о пределах знания в этой области, необходимо заняться рассмотрением вопроса о том, как могли сохраниться доныне остатки организмов эпох прошлого.
Условия сохранения остатков организмов в ископаемом состоянии. Наблюдения над современным органическим миром выясняют, что трупы организмов, живущих на суше, быстро разрушаются, если они остаются значительное время на открытом воздухе. Это разрушение, даже если оно происходит только под влиянием мороза, жары и атмосферы, быстро совершаясь, ведет к полному исчезновению трупа.
Яркий пример этого явления дает вымирание бизонов в Сев. Америке. Область, населявшаяся этими крупными животными, год от году суживалась, по мере того _как местности, ими обитаемые, становились более заселенными. В местностях, где эти животные существовали не более двадцати лет тому назад, еще находили их скелеты на земле, но там, где бизоны исчезли более 20 лет,' от многочисленных остатков их, покрывавших землю, ничего не осталось.
Остатки наземных животных сохраняются только, если они находятся в защищенных местах, например в пещерах, или если они, прежде чем распадутся на части, будут занесены илом, а это случается, когда животное умерли не естественной смертью от старости или болезни, а случайно, например когда они были унесены наводнением, или если они завязли в болотах или плывучих песках и т. п.
Иногда подобным образом могут возникнуть скопления множества трупов; в настоящее время, например в степях Южной Америки, стада’ пасущихся там миллионов полудиких быков часто гибнут, испугавшись чего-либо, разбежавшись й сразбегу попадая в воду. То же бывает при давке стад на водопое, во время жары.
Вышеизложенное делает ясным, что в особенно неблагоприятных условиях сохранения находятся птицы, менее подвергаясь случайностям указанного рода вследствие своего воздушного образа жизни. Действительно палеонтологический материал по птицам еще менее значителен, нежели по четвероногим животным.
Главным хранилищем остатков прежде живших организмов является море, морское дно.
Целый ряд различных факторов, химических и механических, работает, не переставая, над разрушением выдающихся элементов рельефа земли, как бы стремясь сравнять горы с низменностями. Продукты разрушающих процессов дождем и ветром сносятся в долины и затем через посредство рек в океан.
Весь обломочный материал, накопляющийся в море, осаждается на его дно, в общем сортируясь так, что у берега отлагаются песок и галька, а вдаль от берега относятся течениями более легкие илистые частицы.
При многократно происходивших на земле изменениях в распределении суши и моря дно морское часто становилось сушей, и наоборот. В таких случаях слои осадков, образовавшихся на дне морском, могут выйти из-под уровня океана.
Трупы организмов, живущих в море или заносимых в него после смерти, падают на дно моря, мягкие части их сгнивают, а твердые — раковины, кости—сохраняются, покрываются осадками и могут быть изучены, когда дно морское станет сущей, и осадки, его составляющие, станут доступны наблюдению, будучи прорезаны оврагом, речкой, обнажены расьопками и т. п.
Палеонтологическая систематика. Изучение органического мира как современного, так и ископаемого установило, что каждый организм не есть нечто, совершенно не имеющее себе аналогии, но что организмы распределяются в системе в группы, вследствие сходства членов одной и той же группы в известных особенностях.
Эти группы опять-таки могут представлять, в большей или меньшей степени, общие их членам особенности, могут быть близки или далеки одни другим, так что их можно распределить в группы высшего порядка и т. д. Две крупнейшие группы оргайического мира, это —два царства: животных и растений. Каждое царство распадается на подчиненные группы— типы. Б царстве животных один из типов составляют позвоночные животные.
Типы разделяются на классы (например—позвоночные млекопитающие), классы на отряды или порядки (например — копытные), отряды на семейства (например— жвачные), семейства на роды ироды на виды, представляющие уже низшую систематическую единицу.
Так например к роду Equus принадлежат виды: лошадь — Equus са-ballus, зебра—Equus zebra, осел—Equus asinus\ как видно, латинское видовое название животного ставится после родового.
Двойные названия стали общепринятыми после выхода в свет труда К. Линнея «Система природы-» во второй половине XVIII века. Позднее на междунарнных конгрессах систематиков этот обычай (употреблять два названия—одно родовое—впереди и второе видовое—на втором месте) был окончательно санкционирован и считается теперь обязательным для всех. Выработан целый кодекс международных правил номенклатуры, некоторые из которых мы приводим: 1) обязательным считается то название вида (или рода), которое появилось почти первым (даже если оно неудачно, грамматически неправильно и т. п.). Все остальные названия того же организма отбрасываются как синонимы, 2) названия составляются из слов латинских или греческих, но с латинскими окончаниями. Если употребляется слово иного языка, то его окончание все же латинизируется, например Heliceprion Bessonowi Karp. Здесь от русской фамилии «Бессонов» взят латинский родительный падеж. Сзади ставится сокращенно или полностью фамилия автора, впервые описавшего вид. Напр. L означает Линней, Karp, (см. выше) Карпинский.
Теория эволюции. Палеонтология дает несомненные факты эволюции органического мира, его развития, и для нее имеет интерес объясне-ние этого развития—теория эволюции.
Для всякого биолога, является ли он палеонтологом, зоологом или ботаником, в настоящее время нет сомнений в том, что органический мир развивается. Эволюция органического мира, т. е. происхождение одних групп его от других, в настоящее время считается в биологической науке столь же несомненным началом, как например в физике взаимное притяжение тел и химическое сродство в химии.
При построении теории развития мнения все-таки еще значительно расходятся.
Ранее всего и наиболее разработанным, наиболее прочно' стоящим отделом теории развития является теория естественного отбора, или теория Дарвина; одного этого учения было довольно, чтобы с появлением его оказалось ниспровергнутым господствовавшее до того мнение о неизменяемости органического мира.
Дарвин разработал вопрос о происхождении видов вследствие естественного отбора, выживания в борьбе за жизнь индивидуумов, наиболее приспособленных к условиям жизни.
Вопрос о происхождении видов действительно имеет первостепенное значение. Вид есть систематическая единица низшего порядка; виды—наиболее близкие между собою систематические группы; если виды не изменяются и не связаны переходами, но вполне определенно отличаются друг от друга, то как же ожидать переходов лежду более разнящимися существами, принадлежащими к различным родам и другим группам высших порядков?
Между тем на практике при обособлении видов нередко представляются значительные трудности; нет критерия, который мог бы применяться и применялся бы с успехом одинаковым образом различными исследователями Изучая какую-нибудь определенную местную фауну, различные исследователи неодинаково разделяют ее на виды. Все это непонятно при определенности, неизменяемости видов, но вполне понятно, если исходить от эволюционных идей, если предполагать, что виды происходят одни от других, что вид не есть нечто существующее в природе независимо от соображений иссле-до! атзля, но есть понятие, вводимое последним для удобства, есть состояние, выхваченное из непрерывного ряда состояний текущего процесса.1 Как в геологической систематике (см. ниже) приходится употреблять хотя бы иногда совершенно искусственные подразделения, так и зоологическая систематика вводит подобные подразделения. , .
Дарвин, придя к заключению, что организмы изменяются, происходят одни от других, задался вопросом, какой фактор заставлял в мире организ
1 С та им положением, обыкновенным со времен Ч. Дарвина, совершенно нельзя соглашаться. Вид не есть только условное понятие, а есть объективно существующая реальность, имеющая свои определенность, специфичность, свою качественную характеристику, отличную от предшествовавшего состояния, равно как и от последующего. Каждый вид возникает, развивается и прекращает свое существование (превращаясь в другой вид), следуя своим собственным закономерностям. Для примера возьмем два вида—лошадь и осла. Они когда-то ранее составляли один вид. Теперь же, хотя они еще во мно.ом очень сходны, живут в сходных условиях, сходно ведут себя—тем не менее они уже специфически отличаются друг от друга хотя бы физически: продукты их. скрещивания ужо не плодущи. Значит возникло нечто, менящее соотношения внутри прежнего вида. Границы вида во времени:—от одного перерыва непрерывности до другого. P*i.
мов исчезать одни формы и возникать другие. Он нашел разрешение этого вопроса, исходя от довольно общеизвестных явлений получения человеком пород домашних животных и возделываемых растений.
Человек получает эти породы, пользуясь изменчивостью организмов, с одной стороны, наследственностью—с другой.
Вследствие изменчивости индивидуумы одной породы разнятся между собою так, что отличимы один от другого и что нет даже двух индивидуумов, абсолютно тождественных. Наследственность стремится сохранять раз возникшие особенности, передав признаки предков потомкам, и является началом консервативным, противоположным изменчивости.
Подбирая индивидуумов с наиболее сильно выраженными желательными особенностями и заставляя таких индивидуумов спариваться, давать потомство, человек получает потомство данной породы с сохраненными по наследственности (а у некоторых индивидуумов и усиленными) желательными особенностями.
Произведя подобным образом отбор в ряду поколений, человек значительно увеличивает желательные особенности породы, получает последнюю изменившуюся в том или другом направлении, — коров со способностью давать молоко почти целый год в значительном количестве, кур со способностью почти целый год нести яйца и т. п.
Когда порода установится, будет в полной мере представлять желательные особенности, отбор продолжается, имея целью уже не усиление тех или других особенностей, а их сохранение на известном уровне. При этом отбор состоит в удалении индивидуумов, отличающихся худшими качествами, чтобы они помесью не ухудшали породы.
Таким образом были получены: рогатый скот, домашняя птица, хлебные растения и т. п.,—путем искусственного отбора.
Теория естественного отбора. Дарвин находит существование в природе явления, аналогичного искусственному отбору, приходя к этому предположению путем следующих умозаключений.
Все живые существа имеют наклонность чрезвычайно размножаться, так что потомство любой пары вскоре покрыло бы всю землю, истребило бы на ней все, чем может питаться, если бы такая наклонность размножаться не встречала извне препятствий к полному осуществлению.
Способность размножаться указанным образом доказывается простыми, неоспоримыми вычислениями, основанными на данных наблюдения.
Многие обыкновенные птицы наших стран, дрозды, синицы, воробьи, откладывают пять пар яиц, выводят в лето пять пар птенцов, так что, следовательно если весною в данной местности было около миллиона птиц одной из указанных пород, то к следующему году должно быть около шести миллионов. Подобного однако совершенно не происходит; по наблюдениям всех более или менее знающих природу людей—охотников, натуралистов и-т. п.— число животных какой-либо породы остается приблизительно одинаковым из года в год, колеблясь несколько в зависимости от годичных климатических колебаний, от колебаний легкости или трудности добывания , пищи в тот или другой год.
Следовательно необходимо принять в приведенном нами примере, что каждый год умирает в пять раз больше индивидуумов указанных пород, чем остается в живых. Подобным образом пара устриц производит около 2100 000 жизнеспособных потомков, из которых в среднем выживает лишь
одна пара, заступающая место родителей на устричной отмели. Число подобных примеров можно было бы увеличить в каких угодно размерах.
* Стремление организмов чрезвычайно быстро размножаться подтверждается примерами быстрого размножения рогатого скота и лошадей, наблюдавшимися ^Австралии и в средние века в Америке, при ее открытии. В Америке до ее открытия испанцами не было ни одного из домашних животных Старого света. Лошади, коровы, свиньи, овцы и т. п. завезены в Америку европей-; цами; имея благоприятные условия прокорма и мало врагов, эти животные вскоре размножились чрезвычайно, частью одичали, образовав стада диких лошадей и быков, пасущиеся в степях Северной и Южной Америки.
Таким образом не подлежит сомнению, с одной стороны, стремление организмов быстро и в крайней степени размножиться, так же как массовая гибель организмов—с другой стороны.
Если так, если из нарождающихся лишь немногие остаются в живых, очевидно это будут индивидуумы более здоровые, более сильные, более приспособленные к условиям существования.
Происходящее таким образом в природе выживание в борьбе за жизнь цриспособленнейших есть то, что Дарвин назвал естественным отбором, находя в этом явлении многое аналогичным искусственному отбору.
Выражение «борьба за жизнь», употребленное Дарвином, требует пояснения относительно того, что под этим выражением подразумевается. Это выражение условное, употребленное для краткости. Здесь соединены явления нескольких различных категорий,—не' только прежде всего приходящая на мысль борьба активная, сознательная между индивидуумами, потребности которых приходят в столкновение, но и борьба, выражающаяся в конкуренции.
Два волка нередко дерутся из-за добычи, но те же два волка нередко являются конкурентами, борются между собою за жизнь, хотя бы они и не встречались. Так бывает в голодный год, при недостатке пищи, когда найти пищу для одного значит наверное уменьшить возможность ее получения для Другого.
Наконец под словами «борьба за жизнь» понимается борьба с неорганическими неблагоприятными условиями жизни, что испытывает например растение, когда его семена попадают в мало подходящие климатические и почвенные условия.
Действие естественного отбора на организм при установившихся внешних условиях жизни и яри условиях изменяющихся. Если внешние условия существования организмов относительно не изменяются, то естественный отбор делает то, что в каждом поколении выживают наиболее здоровые, наиболее сильные индивидуумы породы, причем отличительные особенности последней долгое время могут оставаться практически почти неизмененными. Не то, если внешние условия существования более или менее заметно^изменяются; изменяется например климат страны вследствие поднятия гор в местности, бывшей до того низменностью,—горы конденсируют атмосферные осадки, изменяют направление ветров; климат, бывший сухим, может стать влажным, и обратно. Эти изменения прежде всего отражаются на растительности, весьма чувствительной к климатическим условиям. Изменение растительности в данной стране влияет на травоядных животных, частью может быть эмигрирующих по невозможности приспособиться к новым условиям жизни, частью, наоборот, иммигрирующих именно в виду появления этих новых условий, благоприятных пришельцам.
2 Учебник палеонтологии.
17
Состав населения травоядных животных отражается на населении плотоядных, тесно связанных с первыми в своем существовании. Для них также может явиться необходимость измениться, приспособиться к новым условиям жизни, вести новый образ ее, например приобрести способность более быстрого бега, перейти к ночному образу жизни.
При изменении внешних условий существования организмы часто должны измениться или переселиться. При невозможности путем переселения найти условия, близкие к исчезнувшим: в данной местности, и при неспособности измениться виды вымирают.
В случае изменения организмов основой для этого изменения является изменчивость. Без сомнения степень изменчивости, различная у различных видов, имеет значение, определяющее, способен ли организм, изменившись, приспособиться к новым условиям жизни.
Большою изменчивостью, наклонностью к образованию разновидностей и местных видоизменений отличаются например ежевика, улитки-прудовики (Limnaeus, фиг. 1) и пр. (фиг. 2). Напротив, гусь, даже в состоянии одомашнения, в котором другие животные склонны к усиленной изменчивости, представляет поразительное однообразие. Изменившиеся условия ведут за собою усиленную изменчивость, так у воробья, водворившегося в Сев.
Америке с 1864 г., замечается значительное
усиление изменчивости в сравнении с европейским родичем, и то же самое имеет там место для нашей наземной улитки [Helix nemoralis). Вообще это обстоятельство создает лучшие условия для приспособления.
Фауна млекопитающих Австралии, как известно, является наиболее примитивной и архаичной среди современных млекопитающих:
Фиг. 1. Изменчивость Limnaeus. Озеро Щучье. Верхнеуральский округ, Оренбургская область. */s натуральной величины.
она такова, как европейская во время начала третичного периода; в Европе произошло дальнейшее развитие фауны, тогда как изоляция австралийской фауны на острове, хотя бы и столь большом, как Австралия, уменьшила для нее число стимулов для развития. 1 О причинах изменения живых существ. Различают внешние и внутренние причины, вследствие которых могут возникать изменения в органи
1 Пример автора с фауной Австралии можно назвать классическим примером того, куда ведет мысль эмпирика, не желающего считаться с диалектикой. По автору выходит, что фауна Австралии застыла на той стадии, на которой она находилась несколько десятков миллионов лет. Однако на лть уже с того, что это даже и фактически неверно Во-первых сумчатых в Европе найдено совершенно незначительное количество, тогда как различных плацентарных было очень много (в Австр 1лви, если не считать завезенных недавно человеком, одна лишь собака динго и несколько льгучих мышей относятся к плацентарным— остальная фауна млекопитающих состоит сплошь йз сумчатых и яйцеродных). Уже по одгому это ty фауна Австралии не такова как европейская „во время ретичного периода". Во-вторых найденные в Европе сумчатые совсем не таковы, как Австралийские. Некоторых, как напр. Neoplagtaulax, относят даже к иному отряду. Можно, не зная палеонтологии (даже не зная биологии вообще), категорически утверждать, что флуна Австралии кЬнеч но развилась. Наверное шло приспособление 'к вечно меняющимся условиям жизни, шла спецна’.чзаци» сумчатых. Автор противоречит самому себе, когда утверждает, что не было «стимула для развития". Выше он сам упоминал например хотя бы о законе перенаселения. Если бы перенаселение у сумчатых каким-то чудом пре» оати-лось, го остается еще перенаселение у птиц, пресмыкающихся, насекомых, у растений и т д. Эти последние вынуждены изменяться, что ведет к изменению всей жизненной обстановки и одно это должно дать „стимул для развития* чудом не перенаселяющих землю сумчатых.
зации. Прежде всего- влияние климата; климат—комплекс различных явлений, как *го: температуры, влажности воздуха, количества дождевых осадков, количества солнечных лучей и т. п., и трудно решить, какая часть в наблюдаемом результате процесса изменения выпадет на долю действия.каждого фактора в отдельности из перечисленных. Если например семена происходящие из низко расположенных долин, сеют в горах, то вырастающие растения во многом отличаются от родительского организма, давшего семя: размеры растения и его частей меньше, а окраска интенсивнее. Однако эти изменения не наследственны; при сборе семян с растения в горах и при посеве их снова в равнине взошедшие растения представляют те особенности, как все их родичи не удалявшиеся из этой местности.
Фиг. 2. Изменчивость раковины двустворчатого иеченогого животного Spvifer mosquensis. В каждом горизонтальном ряду один и тот же экземпляр в трех положениях. Каменноугольные отложения. С. Мячково, окрестности Москвы.
Волосяной покров животных часто находится в зависимости от климата, в Ангоре в Турции не только козы, но и кошки и пастушьи собаки имеют шерсть в виде руна. На различные, хотя и родственные формы животного царства климат влияет иногда весьма неодинаково, например на некоторые породы охотничьих собак, перевезенных в Индию из Европы.
У многих бабочек в году встречаются два различные поколения, зимнее, выходящее из перезимовавших куколок, и отличающееся от зимнего летнее, развивающееся из яичек зимних форм. Но из куколок, зимней формы, при естественных условиях развивающихся в летнюю, можно получать вместо последней зимнюю же, подвергая куколки действию искусственного холода; и наоборот, летние формы можно получить из куколок, дающих в природе зимнюю форму, если действовать на куколки нагреванием. Пища также оказывает влияние на окраску перьев и шерсти животных.
Изменения организации животного могут явиться также в результате стремления животного приспособиться к новым условиям жизни, например при
переходе к новому образу жизни. Это пример внутренних причин изменения. Есть несколько групп водных позвоночных животных, которые считаются происшедшими от наземных четвероногих, таковы: из пресмыкающихся—ихтиозавры, плезиозавры, мозазавры, из млекопитающих—китообразные, ластоногие. Переход от жизни на суше к плаванию в открытом море происходил с постепенностью: сначала животные должны были стать пловцами в прибрежной полосе, не теряя еще способности вылезать иа сушу, затем последняя способность в некоторых случаях утрачивалась, но всегда приобреталась способность более совершенного плавания в открытом море. При этом животное совершает более энергичные плавательные движения, главнейше хвостом, на котором зачастую образуется плавник. Энергичные движения хвоста и туловища, по крайней мере с задней части последнего, влекут за собою изменение сочленений ребер с позвонками в смысле большей легкоподвижности; сочленение так сказать выформовывается этими движениями. Отправления органов, функции их во многих случаях следовательно определяют их строение.
Вопрос о происхождении индивидуальных уклонений, о причинах изме-ения организации стал разрабатываться уже после Дарвина, причем особенно занимается им школа ламаркистов, уделяющих, подобно Ламарку, французскому предшественнику Дарвина в области эволюции, большое внимание отношениям между функцией и формой.
При изучении причины- изменения особенностей организации надо еще иметь в виду, что в организме все части тела образуют вместе единое целое; действие органов согласовано,—одни находятся в зависимости от других.
’ Тем, что передние конечности птпц преобразованы в крылья и устранены от поддерживания тела, когда птица не летает, создается целый ряд изменений организации: задние конечности одни выдерживают тяжесть тела при движении по земле,—при этом возникает укрепление места вочленения ног с туловищем. Кроме того тазовая область укрепляется срастанием крестцовых позвонков между собою и с тазом; клюв заменяет хватательные конечности, и шея очень подвижна, сочленение позвонков соответственно подвижное, вогнуто-выпуклое, число шейных позвонков увеличивается; там, где ноги длинны, например у аиста, страуса, естественно длинна и шея, причем клюв легче достигает до земли. Таким образом изменение одной части тела влечет за собою и изменение других частей. Говорят при этом, что органы тела находятся между собою в соотношении, в корреляции, и одновременные, в связи находящиеся изменения органов называют коррелятивными.
Принцип соотношения признаков, установленный Кювье, дал ему возможность произвести ряд блестящих открытий, когда он, как говорилось (между прочим' и им самим), по одной кости восстанавливал образ целого ископаемого животного. Это говорилось со значительным преувеличением и вело иногда к большим ошибкам, но принцип корреляции дает возможность, исходя от знания одного или лишь немногих органов ископаемого животного, делать заключения о форме и свойствах органов этого животного, еще остающихся неизвестными. Если например от ископаемого вида известны только передние конечности, имеющие тип конечностей для бегания, то по принципу соотношения органов мы вправе делать предположение, что неизвестные нам еще задние ноги были тоже бегающими, так как является невоз-¥
ложным, чтобы они были в этом случае например плавательными ластами или роющего типа ногами. Точно также черепу ископаемого животного, имеющему характер черепа хищных, может соответствовать нижяяя челюсть только такого же типа. Именно для палеонтолога, находящего обыкновенно не цельные скелеты, а смешение разрозненных костей, принадлежащих иногда многим индивидуумам, притом различных родов, принцип соотношения дает основания разобраться в этом смешении.
Не всегда формы производные связаны непрерывным рядом переходов с формами первичными; развитие происходит иногда скачками, сразу появляются индивидуумы, сильно отличающиеся по одному или нескольким признакам от произведших их форм. Примеры имеются как в царстве растений, так и в царстве животных. Так, в 1791 г. возникла порода овец с короткими кривыми ногами и длинной спиной, наподобие таксы,—обособилась порода так называемых анконских овец.
Расхождение признаков. Жизненное состязание должно быть наиболее жестоко между близкими формами, занимающими или стремящимися
занять одни и те же места в экономии природы. Это ПО-нятно, так как близкие формы \ / \ /
сходны по складу организа- \ / I \i/
ции, по потребностям и \ / I
привычкам. Таким образом \ / I
борьба за жизнь должна быть \\ / I /
особенно упорна между oco- <((Х V f / бями одного и того же вида N. / /
и затем между его разновид- \ /
ностями. Поэтому чем более потомки какого-либо вида &
будут различаться между со-бою строением, привычками и т. п., тем выгоднее это для \
них, тем легче они будут в состоянии завладеть более Фиг. в.
многочисленными и разнооб-
разными местами в экономии природы и следовательно тем более они будут размножаться. Другими словами, создание значительных уклонений между потомками одного и того же вида — расхождение признаков, как назвал это явление Дарвин, — есть явление, возникающее под действием естественного отбора.
Пусть вид А (фиг. 3) дал потомков, представляющих довольно резко разнящиеся личные уклонения. Расходящиеся от А линии различной длины изображают этих потомков. Только те из них, которые представляют полезные личные уклонения, выживут и дадут потомство. По всей вероятности выживут особи, личные уклонения которых наиболее различны между собою, так как таким образом известная форма может занять наибольшее количество мест в экономии природы, следовательно естественный отбор будет содействовать выживанию особей, представляющих именно крайние уклонения. ’Конечно это верно лишь вообще, иногда качественная сторона различий будет иметь перевес в значении над количественной, и, например могут выжить формы С и В вместо D\ форма D может и не выжить,
несмотря на то что она представляет наибольшие различия по сравнению с С; отличия эти могут быть значительно менее выгодны, чем представляемые формою В, так что в итоге действие естественного отбора сохранит последнюю.
Потомки формы А просуществуют почти неизменными некоторое, неодинаковое для всех время, что и выражено неодинаковою длиною линий, изображающих этих потомков, но затем без сомнения и с некоторыми из них повторится то же, что было с формою А, т. е. например формы В и С также дадут поколение потомков, личные уклонения которых представляют крайности, способные сохраниться под действием естественного отбора.
То же будет происходить и позже, так что история развития (генеалогия) потомков вида А выразится древовидно ветвящейся генеалогической схемой, и как конечный результат этого развития получатся живущие в одном геологическом периоде,—хотя бы в настоящее время,—формы Н, I, К. L, М. То, что они живут одновременно, выражается положением точек Н, I, К, L и М на одной горизонтальной линии. Что касается до формы А, она и ей подобные зачастую подвергаются вымиранию вследствие вытеснения своими потомками В и С. Это происходит, если ближайшие потомки формы А, мало отличаясь от нее, вступают с нею в конкуренцию, если они могут устроиться в экономии природы лишь на местах, уже занятых.
Бывают однако случаи, что прародительские формы могут при благоприятных условиях (изоляция от врагов в силу физико-географических причин, легкость добывания пищи) выжить, существовать далее одновременно со своими потомками.
Подобно форме А форма В будет вытеснена своими потомками N и £, форма' С—потомками F и бит. д., так что родоначальник форм Н. I, К, L и М и все формы, промежуточные между ним и этими последними, вымрут. Если во время рассмотренного процесса развития возникли значительные различия между видом-родоначальником и конечными происшедшими от него видами, то последние могут быть отнесены к иным родам, нежели вид А. Группа наиболее близких между собою форм может быть выделена в один род, другие виды в другой род и т. д.
Именно, как показывает генеалогия, родословная наших видов вероятно придется виды R и I отнести к одному роду, L и М к другому, форму же К может быть можно будет рассматривать как представителя подрода первого рода, к которому принадлежат формы Н и I. Таким образом от вида А, принадлежащего к известному роду, произошло два рода (один из них с подродом), состоящие из пяти видов.
Натуралисту, имеющему дело лишь с населением одного периода, например зоологу или ботанику, изучающим современный органический мир и в нем виды ff, I, К, L и Л£, не будут известны виды, переходные между названными формами, принадлежащие, вместе с родоначальной формой А, прошлым геологическим периодам. Новообразовавшиеся роды будут обособлены поэтому один от ’другого; эти роды ближе друг к другу, чем к другим одновременно существующим родам, происходят от общего корня и могут быть соединены в одно семейство.
Если предположить, что вид А произошел от другого, более древнего вида (положение которого на схеме будет например где-либо на прерывающейся линии, идущей От А вниз), принадлежащего к другому семей-
етву, чем виды Н, I, К, L и М, то ясно будет, что не только одни виды нроисходят от других, но такие же отношения происхождения существуют и между высшими систематическими группами.
Если предположить, что предок вида А произвел не одно семейство Н, I, К, L и И, но и другое, лежащее хотя бы вправо от Н, I, К, L и М, то эти два семейства могут составить отряд или порядок.
Таким образом, исходя из предположения, что в процессе развития действуют начала естественного отбора и расхождения признаков,, легко получить объяснение картины, представляемой систематикой,—одновременного существования подчиненных одна другой групп, не связанных переходами, так как переходные формы и группы вымерли.
Отсюда понятно значение палеонтологии как науки, призванной восстановить родословную мира организмов. Кроме того знание этой родословной дает возможность более точно устанавливать основания систематики.
Систематики распределяют в системе организмы по степени сходства. Большая или меньшая сумма сходных признаков, служащая основанием для установления различных систематических групп, соответствует близкой или отдаленной степени кровного родства организмов между собою, является результатом их происхождения от более или менее далеких предков. Таким образом следовательно систематика всегда была более или менее основана на отношениях происхождения, даже тогда, когда эти отношения не предполагались существующими, но пока систематика руководилась лишь признаками сходства и различия, она могла делать ошибки, невозможные, если систематика основывается на данных генеалогии, или, как говорят, филогении организмов.
Достоинство систематики, основанной на филогении, зависит от полноты данных последней; естественная классификация организмов будет тогда возможна, когда будет восстановлена во всех частях генеалогия мира организмов, а это никогда не будет возможно вполне, да и в настоящее время не доведено до предела возможного. Таким образом классификации соответствуют состоянию знаний в известную эпоху и изменяются, улучшаясь по мере накопления знаний. 1
При всей неполноте палеонтологического материала данные, им представляемые, незаменимы данными других отраслей науки о жизни. Вопросами развития органпчезкого мира занимаются ботаника и зоология, но они не дают прямых доказательств развития органического мира, каковые дает палеонтология, восстановляя то тут, то там полно реальные образы существ из числа предков современного мира организмов.
Отношение палеонтологии к эмбриологии. Генеалогия мира организмов может быть дана точно лишь палеонтологическими исследованиями, но не всегда,—палеонтологический материал, сохраняемый природою, отрывочен и не всегда дает точные филогенетические выводы. В таких случаях для получения хотя бы вероятных выводов, кроме палеонтологических данных, надо пользоваться и данными онтогении; данные онтогении являются не только дополнительными, но и поверочными в отношении палеонтолого-филогенетических заключений.
1 Оружие марксистско-ленинской методологии еще почти не касалось систематика. Однако уже сейчас ясно, что буржуазная систематика не дает критериев для своих подразделений (отрядов, семейств, родов и т. д.), отвечающих объективно существующим соотношениям организмов. Ред. '
Онтогения есть отдел морфологии, изучающий историю развития особей. Лишь немногие животные обладают при начале своего индивидуального существования вполне готовой, окончательной организацией; большинство животных развивается из яйца, тела сравнительно простой организации, и только мало-по-малу, путем сложных изменений формы, достигает окончательного вида. Например из яйца лягушки, представляющего зернышко лягушечьей икры, выходит личинка лягушки или так называемый головастик, состоящий из маленького, шарообразного тельца, с длинным, сжатым с боков хвбстиком. Далёе у головастика развиваются наружные, в виде мохнатых хохолков, жабры, заменяющиеся затем внутренними жабрами; появляются ноги, исчезают жабры и хвост, и головастик превращается в маленькую лягушку.
В этом примере рассмотрены изменения организма лишь после развития его из яичка, только постэмориональное развитие животного. Этому периоду развития предшествует развитие в яйце—эмбриональное развитие, во время которого из яйца, представляющего простейшее строение, какое только может иметь кусочек организованного вещества, путем постепенных изменений получается высокоорганизованная личинка.
Эмбрион значит зародыш; из эмбрионального и постэмбрионального развития слагается онтогения, история развития особи; изучением развития зародыша занимается эмбриология.
Изучение онтогении привело к заключению, что прохождение организмами различных стадий в своем развитии есть следствие наследственности, так как эти стадии соответствуют взрослому состоянию животных, которых следует считать в числе предков организма, представляющего тот или другой характер онтогении.
Таким образом одна из стадий развития лягушки потому представляет форму с жабрами и хвостом, что в числе предков лягушки хоть один был с жабрами и хвостом, был сходен с теми ныне живущими земноводными, к которым относится например протей.
На основании онтогении птиц надо думать, что они произошли от пресмыкающихся.
Рассмотрим развитие ноги птицы. Скелет ноги высшего позвоночного состоит обыкновенно (фиг. 4), идя сверху вниз, из бедра, двух костей голени (большой и малой берцовых), далее—двойного ряда предплюсны, плюсневых костей и наконец пальцев.
От этого типа йога птицы отличается тем, что ей недостает костей плюсны и предплюсневых костей, а вместо них существует одна длинная кость, так называемая цевка. Но у зародыша птицы, в яйце, строение ног еще не такое, как у взрослого животного.
Здесь существуют и плюсна и четыре предплюсневые кости, которые по своему расположению близки к костям пресмыкающихся, и только при дальнейшем развитии зародыша постепенно образуется типичная нога птицы, причем верхняя часть плюсны срастается с голенью, а цевка образуется от слития нижней части плюсны с предплюсневыми костями. Таким образом нога птицы проходит в своем развитии стадию ноги пресмыкающихся, почему и надо думать, что последние представляют родоначальную группу для птиц.
Филогенетические данные, представляемые палеонтологией относительно происхождения бесхвостых амфибий, с одной стороны, птиц—с другой стороны, вполне соответствуют данным эмбриологии.
организма в равные
Древнейшие амфибии были хвостатые и дышали жабрами.
Пресмыкающиеся появились на земле раньше птиц, и древнейшая птица (Archaeopteryx) в значительной мере отличается от ныне живущих птиц, приближаясь к пресмыкающимся.
Э. Геккель дал следующую краткую формулировку явлений индивидуального развития: индивидуальное развитие животного (онтогения) есть в главных чертах повторение исторического (генеалогического) его развития; это—эффектная, сжатая, хотя и не вполне ясная и точная формулировка. Несомненно, что развитие индивидуумов дает много указаний на характер предкоь, но не всякая стадия, не всякая особенность периоды его развития непременно представляет образы предков.
Постэчбриональное развитие бабочек например состоит в том, что вылупившаяся из яйца червеобразная личинка-гусеница переходит в стадию куколки, отличающейся неспособностью передвижения и тем, что животное в это время не в состоянии принимать пищи; из куколки вылупляется бабочка. Конечно здесь нельзя предположить, чтобы в числе предков бабочки были куколкообразные животные. Стадия куколки есть приспособление, обусловленное личинковою жизнью животного.
Одним из наиболее блестящих открытий палеонтологии является выяснение происхождения лошади, имеющей однопалые ноги, от пятипалых предков. Но эмбриология не дает указаний на происхождение лошади; у зародыша лошади ноги прямо обособляются однопалыми, так что стадии многопалости их оказываются выпущенными из эмбрионального развития.1
К категории подобных же несоответствий индивидуального развития с филогенией принадлежит у позвоночных животных неодинаково скорое развитие различных органов, причем развитие некоторых органов идет ускоренным образом по сравнению с-другими. Несомненно например, чтс chorda dorsalis (образование, из которого развивается позвоночный* столб высших позвоночных) появилась у позвоночных после нервной системы, ' а онтогенетически она появляется раньше.
Итак филогенетические указания, представляемые онтогениею, затемняются вводными явлениями (у бабочки), не имеющими отношения к филогении, некоторые филогенетические стадии подавляются в онтогении (развитие ноги лошади), одни стадии развития перестанавливаются на место других (развитие chorda dorsalis). Явления онтогении, нарушающие
Фиг. 4. 1. Скелет ноги взрослой курицы, уменьшен. 2. Скелет ноги зародыше курицы, увёличен.
f — бедро (на ряс. 2 обломлено наверх*), £-^5олгшая берцовая, Р—малая бд^1ова>я кость, tp—голень, is — предплюсна, т—плюс на, tm—цевка.
1 Это не совсем правильно. Не только в эмбриональном состоянии, но даже взрослом у лошади кроме громадного третьего пал да, на который она ступает, ствуют рудименты (остатки) второго и четгертого пальцев (под ,.щеткой”). Таким зом трехпалость есть. Речь может итти лишь об исчезновении первого и пятого пев Ред. t
и во суще-обра-паль-
•ее соответствие с филогенией, называются явлениями ценогенетическими.
Вследствие существования последних онтогения иногда может не представлять никакого сходства с тем, какие прародительские формы и в какой последовательности сменяли друг друга, и тогда следовательно из онтогении совершенно невозможно получить филогенетические указания; вообще при извлечении таких указаний из онтогении надо соблюдать большую осторожность.
Отношение палеонтологии к сравнительной анатомии. Как выяснено уже, руководствуясь лишь степенью сходства и различия признаков, можно впасть в ошибки в суждении о близости организмов, представляющих эти отличия. Сходства возникают не только вследствие общности происхождения, но и, при отсутствии этой общности, вследствие одинаковости назначения органа. В первом случае сходство обозначается названием гомологии, во втором—аналогии; соответственно называют органы гомологичными и аналогичными.
Изучением гомологий занимается сравнительная анатомия. Она прослеживает гомологию органов через различные группы животного царства, от одной группы к другой, близко родственной, а также внутри самих групп. Гомологии выясняются путем анализа формы и строения органов, а также тщательным сравнением общих отношений взаимного расположения органов. Чем более гомологичны органы, как в грубых чертах, так и в тонких деталях строения, тем более вероятно и генеалогическое родство животных, представляющих гомологии.
Например наличность одного и того же типа скелета в конечностях позвоночных, ведущих наземный, водный и воздушный образ жизни (нога четвероногого, крыло птицы, ласт кита), мы можем объяснить, лишь предполагая, что он был унаследован от общего предка. Все эти конечности гомологичны, представляя строение, производное от пятипалой конечности.
Орган движевия моллюсков, нога, обыкновенно является в виде массивного мускулистого образования, расположенного на брюшной стороне тела. Она бывает подошвообразна у улиток, топоровидна или клинообразна у двустворчатых пластинчатожаберных моллюсков и является нелегко распознаваемою в сильно измененном состоянии у головоногих моллюсков (к которым принадлежат например осьминоги и каракатицы), а именно частью образуя так называемую воронку и частью околоротовые щупальца. Нога головоногих таким образом значительно отличается от ноги других моллюсков, являясь разделенною на части, не сходные между собою и с обычною ногою, причем щупальца занимают необычное для ноги положение. Но в. молодых стадиях развития щупальца не окружают рта, как в зрелых, а сосредоточиваются с одной стороны его, между ним и воронкой; кроме того щупальца и воронка снабжаются нервами от нервного узла, который у моллюсков является ножным.
Зубы позвоночных животных представляют гомологию с чешуями некоторых рыб. Эти так называемые плакопдные чешуи являются костяными, снабжены зубовидным отростком, выдающимся над поверхностью тела. Образование плакоидных чешуй распространяется на оболочку ротовой полости, где их зубовидные отростки у рыб развиваются в еще мало отличающиеся от них настоящие зубы; у высших позвоночных зубы являются единственным остатком покрова чешуй, первоначально покрывавшего
у позвоночных всю поверхность кожи. Характерным для сравнительной анатомии является метод анализа, расположение изучаемого материала, материал же для изучения сравнительной анатомии доставляется зоологией и палеонтологией, иного сравнительная анатомия не имеет. Сравнительноанатомический метод особенно важен для палеонтологии, где мы зачастую имеем дело именно лишь с остатками, лишь с отдельными частями тела, органами прежде живших животных.
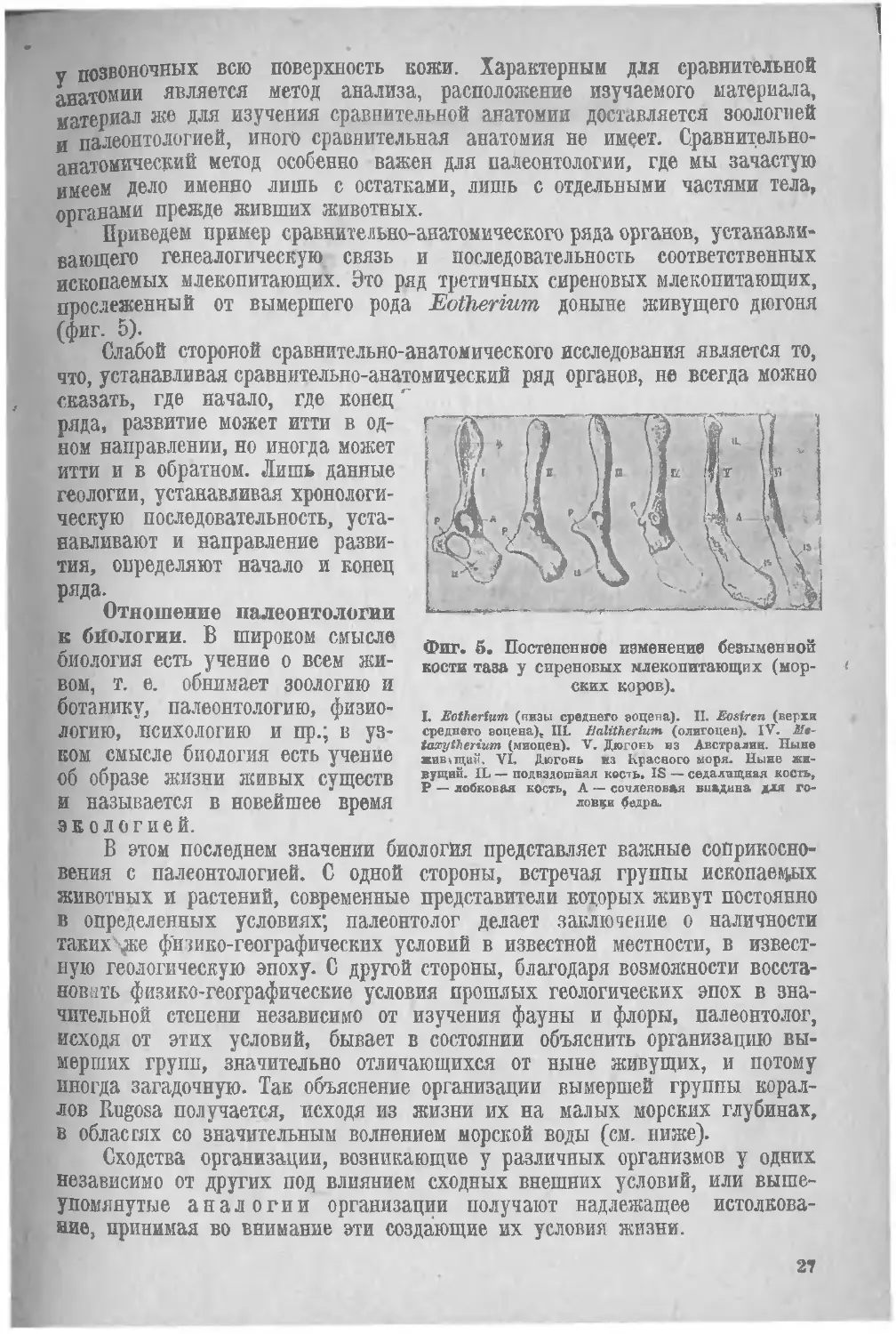
Приведем пример сравнительно-анатомического ряда органов, устанавли-
вающего генеалогическую связь и последовательность соответственных ископаемых млекопитающих. Это ряд третичных сиреновых млекопитающих, прослеженный от вымершего рода Eotlierium доныне живущего дюгоня (фиг. 5)
Слабой стороной сравнительно-анатомического исследования является то, что, устанавливая сравнительно-анатомический ряд органов, не всегда можно сказать, где начало, где конец'
ряда, развитие может итти в одном направлении, но иногда может итти и в обратном. Лишь данные геологии, устанавливая хронологическую последовательность, устанавливают и направление развития, определяют начало и конец ряда.
Отношение палеонтологии к бйологии. В широком смысле биология есть учение о всем живом, т. е. обнимает зоологию и ботанику, палеонтологию, физиологию, психологию и пр.; в узком смысле биология есть учение об образе жизни живых существ и называется в новейшее время экологией.
Фиг. S. Постепенное изменение безыменной кости таза у сиреновых млекопитающих (морских коров).
L Eothertum (низы среднего эоцена). II. Eosiren (верхи среднего эоцена), III. Halitherium (олигоцен). IV. Мв-taxytherium (миоцен). V. Дюгонь из Австралии. Ныне жив > щам. VI, Дюгонь из Красного моря. Ныне живущий. IL — подвздошная кость. IS — седалищная кость, Р — лобковая кость, А — сочленовля впадина для головки бедра.
В этом последнем значении биология представляет важные соприкосновения с палеонтологией. С одной стороны, встречая группы ископаемых животных и растений, современные представители которых живут постоянно в определенных условиях; палеонтолог делает заключение о наличности таких ^ке фишко-географических условий в известной местности, в известную геологическую эпоху. 0 другой стороны, благодаря возможности восстановить физико-географические условия прошлых геологических эпох в значительной степени независимо от изучения фауны и флоры, патеонтолог, исходя от этих условий, бывает в состоянии объяснить организацию вымерших групп, значительно отличающихся от ныне живущих, и потому иногда загадочную. Так объяснение организации вымершей группы кораллов Rugosa получается, исходя из жизни их на малых морских глубинах, в областях со значительным волнением морской воды (см. ниже).
Сходства организации, возникающие у различных организмов у одних независимо от других под влиянием сходных внешних условий, или вышеупомянутые аналогии организации получают надлежащее истолкование, принимая во внимание эти создающие их условия жизни.
Выясняя аналогию и особенно гомологию, мы выясняем степень родства, генетические отношения представляющих их организмов.
В отношении морских животных и растений, с которыми главным образом приходится иметь дело геологу, основным фактором, определяющим характер морской фауны и флоры, является глубина воды и характер морского дна в данной местности.
Континенты в прибрежной полосе уходят под воды морские, представляя под ними сначала род уступа, известного под названием континентальной платформы; этот уступ постепенно опускается до глубины около 200 метров, после чего уклон дпа усиливается, и оно нисходит круто к глубоководной, абиссальной области. Этот предел в 200 метров соответствует границе, глубже которой солнечный свет не проникает, где растения поэтому исчезают, а за ними и травоядные животные, где воды морские не подвержены волнению, и колебания температуры воздуха не чувствительны. Это весьма отчетливая биологическая граница, которая отмечает нижнюю часть прибрежной литторальной области и верхний предел глубоководной абиссальной области.
Но над этими двумя областями, характеризуемыми присутствием морского дна, на котором живет множество живых существ, находится огромная масса воды, слагающая почти весь океан, которая не соприкасается с дном и не имеет к нему отношения, это—пелагическая область, область открытого моря. Она занята множеством существ, которые никогда не живут на дне, но всегда плавают на различной глубине.
Население литторальной области вырабатывает приспособления, предохраняющие их от разрушительного действия прибоя, от сноса приливом или отливом. Эти приспособления состоят в прирастании раковин к месту их обитания, в прикреплении выделением органического вещества, в спцсоб-ности присасываться к скалам, высверливать помещение в скале, в развитии особенно прочных скелетов и раковин. На песчаном дне животные при отливе уходят глубоко в песок, имея для этого приспособления для дыхания (сифонные пластинчатожаберные). Характерными образованиями литторальной области являются рифы, сложенные колониями скелетообразующих кораллов или родственных им животных, а также известковистыми водорослями.
Абиссальная область занимает все морское дно, за вычетом литторальной области. Характерной особенностью для нее является то, что в нее никогда не проникает дневной свет, жизнь хлорбфиллосодержащих растений в ней невозможна, и основною пищею для обитателей являются трупы, падающие с поверхности. У глубоководных животных распространена, в связи с отсутствием дневного света, способность свечения, глаза их являются (например у ракообразных) или увеличенными, или, наоборот, редуцированы и даже соверщенно исчезают. Первое—у отличающихся активностью в передвижении, второе—у форм, медленно ползающих на дне. Низшие беспозвоночные, не имеющие способности передвижения, отличаются симметрией внешнего вида и хрупкостью скелетных образований (лилии, стеклянные губки). Губки и кораллы часто лежат свободно на илистом дне, причем кораллы бывают плоски, дискоидальны, что дает им возможность оставаться на поверхности рыхлого ила, не погружаясь в него.
Пелагическая область не имеет ни берега, ни дна, никакого иного предела, кроме воды и воздуха. Население ее отличается тем, что существует в посто-
но плавающем состоянии. Большая часть обитателей открытого моря в зависимости от значительного содержания воды в тканях тела являются 'вполне или отчасти прозрачными. При полной прозрачности животные так сливаются с водой, что становятся едва различимы. Например медузы, сифонофоры. Пелагические животные имеют плотность, очень близкую к плотности воды, вследствие значительного содержания ее в тканях тела; если бы животные были значительно тяжелее воды, они быстро шли бы ко дну или были бы принуждены непрерывно активно плавать, что сопряжено с большой затратой энергии. В теле мелких пелагических животных бывают делающие его легким выделения масла или воздухоносные полости. Скелеты пелагических животных малой величины отличаются сильною разветвленностью (радиолярии), а вследствие этого и большою поверхностью, при которой возрастает трение о воду, облегчающее животному возможность , держаться во взвешенном состоянии в воде. Животные более значительной величины, как некоторые ракообразные, имеют сильно разветвленные, перистые или по крайней мере нитевидно-вытянутые ноги и сяжки. Раковины пелагических моллюсков отличаются чрезвычайною тонкостью, тогда как рыбы имеют скелет, лишенный извести.
У’ крупных пелагических позвоночных, как млекопитающие китообразные, имеется ряд особенностей организации, отличающих их от родственных наземных четвероногих и служащих для облегчения плавания; этими особенностями являются: веретенообразная форма тела, гладкая, лишенная волос кожа, пропитанность костей жиром.
Кроме указанных различий в областях моря, служащих местообитанием морским животным для биологии их имеет большое значение образ жизни: активное плавание (нектонический), пассивное плавание по воле ветра, волн и течений (планктонический) или же жизнь, связанная с морским дном или с посторонними предметами, служащими субстратом (бентони ч е с к и й), причем животное или остается на субстрате неподвижно, свободно лежа или прикрепляясь, или же ползает на нем.
Нектон, планктон и бентос встречаются во всех вышеохарактеризован-ных зонах моря, литторальной, абиссальной и пелагической.
Хронология ископаемых фауп. В совокупности осадочных пластов, лежащих один над другим, каждый вышележащий пласт образовался позже нижележащих. Ести собрать «ископаемые» остатки организмов послойно, то получается представление о хронологической последовательности организмов, живших во время образования этих слоев.
Изучение ископаемых фаун приводит к заключению, что одинаковые фауны, находимые в различных местностях, одновременны, и следовательно в различных местностях слои, содержащие одних и тех же ископаемых, имеют одинаковый возраст. Обратнб — слои, содержащие различные фауны, образовались разновременно.
Последнее заключение впрочем верно с некоторыми ограничениями: и одновременно существующие фауны, например современные, бывают различны. Примером могут служить современные фауны. Эти различия вызываются между прочим климатическими условиями; для смежных областей с одинаковым климатом различия фаун вызываются существованием между ними непреодолимых для распространения организмов преград; наконец, различия морских фаун обусловливаются такими причинами, как степень глубины
моря, природа морского дна и берегов, и т.п. так называемыми фациальными причинами.
Различают фации: прибрежную, глубокого моря и др. Как изложено выше, население каждой фации обладает характерными особенностями.
Вследствие указанных причин можно иметь одновременные, но различные фауны например в мелководном песчанике и в глубоководном известняке. При определении возраста ископаемых фаун возможно сравнивать лишь фауны одинаковой фации.
Геология установила во всей массе осадочных образований большое число подразделений. Главнейшие из них следующие:
I. Ненозойская группа,.
Системы. Отделы. Ярусы.
Посдетрстичная система. Аллювий (современные послетретичные образования). Дилювий (древние послетретичные или постплиоцен).
Третичная система. Верхнетретичные. Нижнетретичные. Плиоцен. Миоцен. Олигоцен. Эоцен. Палэоцен.
II. Мезозойская группа.
Меловая система. Верхний мел. Сенон. Туроя. Сеноман.
Нижний мел. : Гольт. Неоком.
Юрская система. Верхняя юра или мальм Титон. Кимсридж. Оксфорд. Келловей.
Средняя юра или доггер. Нижняя юра или лейас.
Триасовая система. Кейпер. Раковинный известняк. Пестрый песчаник.
Верхний отдел. Нижний отдел.
III. Палеозойская группа.
Пермская система.. Каменноугольная система.
Девонская система.
Силурийская система.
Кембрийская система.
Верхний девон.
Средний девон.
Нижний девон.
Верхний силур.
Нижний силур.
Верхний кембрий. Средний кембрий. Нижний кембрий
JLV. Протерозойская группа.
V. Архейская (азойская) группа.
Группы делятся на системы, системы на отделы, ярусы и зоны.
Кроме этих подразделений слоистых пород различают еще соответственно подразделения времени.
Время, соответствующее отложению целой группы, принято называть эр ой; .время, соответствующее системе, — периодом, отделу — эпохой, ярусу — веком, а зоне—хемерой.
Что касается до основания этого подразделения, установленного в существенных чертах еще в первое время существования геологии как науки, то например в систему обособлялась совокупность осадочных образований, казавшихся составляющими одно целое как по минеральному характеру пород, так и по характеру заключающихся в них фаун.
Впоследствии различия между системами оказались иного характера, чем предполагалось при установлении систем, притом нередко меньшими. Первоначально не предполагалось связи между органическими мирами различных систем в отношении происхождения одних миров от других, установленного впоследствии, а затем сначала не было достаточно оценено значение фаунистических перерывов в осадочных образованиях исследованных тогда частей Европы.
Все системы были установлены так сказать в героический период истории геологии, сравнительно в короткий срок, при исследовании Европы. Упомянутые перерывы были следствием или того, что на границе смежных систем находились отложения, в которых остатки не находили условий благоприятных для своего сохранения, или же фаунистические перерывы в толще слоев существовали в зависимости от местного перерыва в образовании осадков. Если группа слоев может быть подразделена на две части, то эти части не всегда образовались, по времени, непосредственно одна за другой. За периодом существования моря в какой-либо местности, сопровождаемым образованием осадков на дне морском, часто следует период отступания моря, во время которого бывшее дно его становится сушей; образования же осадков с окаменелостями в данной местности на происходит, хотя оно суще-стует конечно в других местностях, покрытых морем.
С течением времени однако местность, где произошел перерыв в образовании осадков, может быть снова покрыта морем; здесь снова образуются слои осадочных пород, непосредственно налегающих на ранее образовавшиеся пласты, хотя между образованием первых и вторых протек может
быть громадный период времени. Ясно, что в этом случае фауна нижних слоев может значительно разниться от фауны верхних, но промежуточная фауна все-таки должна быть и будет находиться где-либо в других местностях.
Другими словами, если мы имеем в данной местности слои а и с, довольно резко разнящиеся между собой, то эта разница может быть вследствие отсутствия слоев Ь, находящихся где-либо в другой местности.-
Перерывы указанного происхождения в последовательности фаун, заключающихся в осадочных образованиях, дали возможность обособления систем в первое время существования геологии как науки: -при этом ошибочно полагали, что отношения, существовавшие между осадочными образованиями Европы, должны быть и повсюду. Эти отношения оказались иными; впоследствии при исследовании других частей света, при детальном переисследовании той же Европы, были найдены многие отсутствовавшие первоначально осадки и фауны; и чем более накопляются знания, тем более устанавливается постепенность смены фаун в истории органического мира, непрерывность его существования.
В настоящее время выяснилось, что границы систем, первоначально установленные так сказать наугад, во всяком случае на основании данных, иногда представляющихся скудными при современном состоянии знаний, большею-частью были намечены удачно, соответствуя например моментам усиленного изменения физико-географических условий и в соответствии с этим — моментам особенно энергичного изменения органического мира.
Что касается других систем, то, хотя в настоящее время и ясна несостоятельность и недостаточность тех оснований, которые были взяты при их установлении, эти подразделения все-таки удерживаются, потому что, изучая совокупность осадочных образований громадной толщины с' неодинаковым наслоением в разновременных отделах этой массы осадков, необходимо хоть как-нибудь, хотя бы искусственно, подразделить изучаемую толщу на отделы и дать им постоянный объем и обозначения, так чтобы с известным названием соединялось известное представление об относительной древности и о характере тех или других осадков и фаун.
В последнее время намечается возможность, даже необходимость крупных изменений в подразделении осадочных образований. Так грань между нижним и верхним силуром, составлявшими доныне два отдела одной и той же системы, в действительности невидимому имеет гораздо большее значение, соответствуя моменту сильного изменения физико-географических условий, моменту напряжения горообразовательной силы, изменению в распределении суши и моря, вымиранию крупных групп животного царства и появлению других групп.
Таким образом эта грань будет вероятно считаться за грань не только между двумя системами, но даже группами, так что лишь верхний силур может быть относим к палеозойской группе, а нижний естественно объеди-' нить хотя бы с частью археозойской группы. Наиболее естественными подразделениями ' осадочных образований, в отношении которых особенно ясна их биологическая обоснованность, являются вероятно с одной стороны все-таки группы, с другой — мельчайшие подразделения, т. е. зоны.
Древнейшая из ископаемых фаун. Древнейшая из существовавших на земле фаун, каковою является протерозойская, представляет большой интерес для теории развития.
- Эта фауна должна представлять так сказать корни мира организмов, его древнейших прародителей. Принимая теорию развития в широком смысле,— как учение, выводящее все живое из одного корня, из-простейших одноклеточных живых существ, — мы в праве ожидать, что древнейшие ископаемые фауны состоят если не из простейших животных, в большинстве не сохраняющихся в ископаемом состоянии, то из низкоорганизованных существ, не представляющих притом большого разнообразия.
К сожалению осадочные образования протерозойской группы содержат лишь редкие и проблематичные остатки организмов.
Вследствие древнего возраста этих осадочных образований они часто подвергались сильному давлению при горообразовательных процессах; давление частью непосредственно разрушало остатки организмов, частью действовало разрушающе, превращаясь в теплоту, так что не только исчезли остатки организмов, заключавшиеся в археозойских осадочных породах, но. последние приняли в некоторых случаях необычный для осадочных образований вид.
Обломочный материал — частицы глины, песку и т. п. — под действием жара превратился в кристаллический; вместо упомянутых частиц возникли кристаллические элементы кварца, роговой обманки и др.
Подобное изменение осадочных пород могло происходить кроме того в результате химического изменения осадочных пород. Древние осадочные породы отлагались на еще тонкой земной коре; эта кора, вследствие своей тонкости, подвергалась частым изломам, например вследствие извержений, и представляла выход газам, содержащимся в раскаленном ядре земли.
Газообразные, щелочные, борные, фтористые и т. п. соединения, действуя на осадочные породы, изменяли их составные части, превращая в кристаллические силикаты: полевые шпаты, слюды, амфиболы, пироксены.
Вследствие этого первая часть истории развития органического мира быть может никогда не будет написана палеонтологами; генеалогическое древо жизни скрыто от ведения палеонтологии в своей нижней части. Самое большее, чего мы можем ожидать, это то, что ветви животного и растительного царств, разошедшиеся в стороны к настоящему времени, окажутся все более сходящимися при удалении вглубь времен и менее отличными одна от другой.
Древнейшая палеозойская, кембрийская фауна содержит представителей всех типов животного царства, за исключением позвоночных, т. е. в ней находятся простейшие, кишечнополостные, черви, членистоногие, моллюски и моллюскообразные, иглокожие. Даже позвоночные могли быть в этой фауне и не сохраниться, будучи низкоорганизованными и не имея костного скелета, а лишь хрящевой.
Таким образом типы обособились от общего корня еще в докембрийское время, и надо сказать, что времени для этого обособления было достаточно, так как геологи считают возможным, что продолжительность археозойской эры превосходила все последующие, вместе взятые.
Изменения и процесс окаменения остатков организмов в ископаемом состоянии. Возможность сохранения остатков организмов в ископаемом сортоянии бывает различна, как это уже было отчасти выяснено выше. Эта возможность и род сохранения находятся в зависимости также от первоначальных особенностей строения организма и от породы, в которой заключены остатки организмов.
В редких случаях организм сохраняется вполне, даже с мягкими бес-скелетными частями, как сохранились например мамонты и носороги во льдах Сибири.
В громадном большинстве случаев органическое вещество истлевает вполне или разлагается, обугливаясь, как это бывает с так называемым хитиновым веществом некоторых животных и иногда с органическим веществом растений. Таким образом получаются обугленные ископаемые остатки, нередко хорошо сохраняя первоначальную форму органов и даже микроскопи-. ческое строение.
Даже в остатках, представляющих твердые части, — раковины, кости — в этих частях не только разрушается органическое вещество, но изменяется и неорганическое. Так, часто входящий в состав скелетов углекислый кальций растворяется в воде, содержащей углекислый газ. Если остатки заключены в водопроницаемой породе (пески, песчаники), то просачивающиеся через нее воды извлекают известковистые частицы из раковин и костей, последние становятся менее плотными по строению и, наконец, даже рассыпаются.
Бывает однако что процесс выноса минеральных частиц из твердых частей ископаемого компенсируется процессом осаждения в нем минеральных частиц, приносимых в растворенном состоянии извне. Таким образом первоначальное вещество твердых остатков организмов может совершенно заместиться другим веществом, причем первональное строение скелетного вещества может сохраниться. Так встречаются в ископаемом состоянии деревья, представляющие совершенно такие же клетки и сосуды, как живое растение, но стенки этих клеток и сосудов образованы кремнеземом вместо первоначальной клетчатки. Это является примером собственно окаменения ископаемых остатков.
Если ископаемые остатки растворяются водами, циркулирующими в породе, то, при достаточной прочности последней, в ней образуется пустота на месте раковины, кости и т. п.
Эта пустота, стенки которой например представляют отпечатки наружной и внутренней поверхности раковины, исследователем может быть заполнена гипсом, серой воском и т. п., так что получится воспроизведение тела, первоначально занимавшего пустоту. Иногда пустота заполняется осадками из минеральных растворов, проникающих в нее, так что получаются естественные слепки скелетных частей, не представляющие их первоначального микроскопического строения.
Выполнение породой внутренности раковины представляет так называемое внутреннее ядро.
Ископаемые остатки организмов, разрушившись растворением, могут совершенно исчезнуть из окружающей породы, а на месте их может образоваться минеральное выполнение пустоты, представляющее отпечаток лишь наружной поверхности ископаемых остатков; это будет так называемое внешнее ядро.
Историческое развитие палеонтологии.1 В истории развития палеонтологии можно различать три периода: древний—период более или менее фантастических вымыслов, средний — период описательного направления и новый — период эволюционных идей.
1 См. статью от редакции в начале книги. Ред.
Древний мир, с его наклонностью к спекулятивному, умозрительному рассмотрению природы, дал очень мало даже для одного только понимания происхождения ископаемых остатков организмов. Ископаемые были известны Ксенофану, Геродоту, Страбону, Аристотелю.
Арабская наука средних веков применила учение Аристотеля о происхождении низших организмов самозарождением из сырого прогретого ила к вопросу о происхождении «окаменелостей». •
Мнение это, высказанное арабским врачом Авиценной (980 — 1037), было воспринято средневековой Европой и повторяемо еще двести лет спустя, когда принимали существование в земле пластической или образовательной силы {vis plastica или virtus formativa), но допускали, что животные й растения могут окаменевать в таких местах, где есть окаменяю-щая сила. Некоторые считали окаменелости за фигурные камни {lapides figurati), подобные кристаллам, сталактитам и конкрециям, за lusus naturae, игру природы, но прихоти которой минеральные образования вылились в форму растений и животных.
Но, начиная с эпохи Возрождения наук и искусств, верное понимание ископаемых остатков животных и растений проявляется все более часто.
В этом отношении еще на границе XV и XYI столетий надо отметить знаменитого художника Леонардо да-Винчи.
Когда нелепые фантастические представления стали отступать на задний план, предстояло освободиться от возникшего под влиянием религиозных представлений мнения, что окаменелости суть остатки «допотопных» животных. Против этого мнения еще в 1517 г. привел серьезные соображения итальянец Фракасторо.
В науке мнение о происхождении окаменелостей вследствие потопа существовало еще около 100 лет тому назад. Серьезный удар потопному происхождению окаменелостей был нанесен выяснением того,' что в различные периоды на земле существовали различные животные и растения, и что нынешнее население земли принадлежит только последнему периоду и явилось на смену целому ряду вымерших растений и животных.
Ученым, установившим, что в разные времена на земле существовало различное животное население, является знаменитый Кювье (1769 —1831).
По складу своего ума, систематизирующего и классифицирующего, с успехом работающего лишь среди конкретных явлений, Кювье не был склонен 'к обобщениям, требующим полета фантазии; он был сторонником мнения о неизменяемости животных и растений. Научная палеонтология, так же как сравнительная анатомия, созданы им, причем палеонтология получила описательно-морфологическое направление.
Третий период, эволюционный, в истории палеонтологии возник после утверждения теории эволюции Дарвином (1859).
Первые применения теории эволюции к палеонтологии были даны в Европе Гильгендорфом (1866), описавшим ряд постепенно изменявшихся раковин Planortns из третичных отложений Штейнгейма в Вюртемберге. Но подобно тому как работы Кювье над позвоночными создали в публике большую славу палеонтологии, чем одновременные и весьма ценные работы Ламарка над беспозвоночными, точно так же первыми палеонтологическими работами эволюционного характера, обратившими на себя большое внимание, были работы по ископаемым позвоночным. Это были работы В. 0. Ковалевского по ископаемым
копытным, давшие образец соединения детального фактического изучения с теорией.
Кроме Кювье и Ковалевского, отмечающих этапы в историй развития науки, нельзя не упомянуть ученых, много сделавших в науке по раз уже принятому ею направлению. По позвоночным особенно значительны труды Гольдфусса, Йегера, Германа фон-Мейера, Фрааса и Иекеля в Германии, Пандера в России, Агассица в Швейцарии, Долло в Бельгии, Ричарда Оуэна в Англии, Копа, Марша и Осборна в Америке. По беспозвоночным выдающиеся труды оставили Деге и Мильн-Эдвардс во Франции, Бейрих в Германии. Иохим Барранд и Неймайр в Австрии, Давидсон в Англии, Уолькотт в Сев. Америке.
Работы по палеонтологии, часто печатаемые государственными геологическими учреждениями и в геологических журналах, с течением времени находят место для печатания в специально палеонтологических изданиях английского Палеонтографического общества и недавно возникших немецкого, американского и русского палентологических обществ.
Последняя сводка палеонтологического материала была сделана мюнхенским профессором Циттелем после семнадцати лет напряженной работы в его четырехтомном труде «Eandbuch der Palaeontologie», законченном в 1893. Сокращение этого значительно устаревшего в настоящее время труда, представляющего наиболее полный учебник палеонтологии, имеется под названием «Grundziige der Palaeontologie», перерабатывается и выходит все новыми изданиями.
По так называемой палеобиологии, изучающей главным образом организацию ископаемых форм в соответствии с внешними условиями, а следовательно и с образом жизни, поскольку он определяется этими условиями, существует обширное сочинение Dacque «Vergleichende biologische Formenkunde der fossilen niederen Here» 1921.
На русском языке существуют еще следующие учебники палеонтологии.
Лагузен, И. Краткий курс палеонтологии. 1897. Книга, хотя и старая, но вследствие подробных описаний родов могущая и до сих пор служить до некоторой степени справочником и определителем.
Борисяк, А. Курс палеонтологии.-1905, 2 ч.
Павлова, М. Палеозоология. 1927, 2 ч.
. Эти два учебника предполагают у читателя предварительные знания по зоологии, в отличие от курса Лагузена написаны на основе теории эволюции и дают хотя бы вкратце характеристики значительного числа родов.-
Штейнманн Г. Введение в палеонтологию. 1909. Представляет тоже учебник палеонтологии, переведенный с немецкого. К сожалению в последние издания этой книги автором внесены новейшие его воззрения о вымирании, о происхождении одних групп от других, встретившие решительные возражения.
Криштофович, А. Курс палеоботаники. 1928.
Все эти учебники заслуживают внимания, но, за исключением курсов Павловой и Криштофовича, представляют давно разошедшиеся издания.
Упомянем еще, что существует государственное российское палеонто-. логическое общество, в «Ежегоднике» которого рассматриваются и общие вопросы, а также даются рефераты новинок палеонтологической литературы.
JJ
ЛБОЗООЛОГИ Я.
A
I ТИП. ПРОСТЕЙШИЕ, PROTOZOA.1
Простейшие, соответственно названию типа, являются элементарными организмами. Тело простейших представляет комочек студенистого вещества — протоплазмы, состоящего главным образом из белковых веществ. В теле простейших существует ядро (фиг. 6), обособленная часть протоплазмы особого строения и свойств, являющаяся важною составною частью тела простейшаго. Ядро всегда принимает участие в процессе размножения; бывает и несколько ядер. Затем у простейших существует нередко оболочка тела, кутикула, выделяемая протоплазмою на ее поверхности или являющаяся в результате изменения поверхностного слоя протоплазмы. Частицы протоплазмы в теле простейшего находятся в движении, следствием которого может быть перемещение тела, как совокупности двигающихся частиц, с одного места на другое. При передвижении более примитивных из простейших, наприыер амёбы, протоплазма дает пальцевидные выросты тела, появляющиеся и исчезающие на любом месте, притом так, что появление выроста на одном месте тела влечет за собою убыль соответствующего объема протоплазмы на другом месте. Выросты называются ложноножками (псевдоподиями).
Ложноножки облекают своею массою посторонние тела, встречающиеся на пути животного, причем захваченные огранические тела, претерпевают химические изменения вследствие действия на них протоплазмы амёбы, а продукты, получающиеся из них в растворенном состоянии в этих процессах, усваиваются амёбою и перерабатываются в частицы протоплазматического тела.
Так происходит процесс питания.
Питание восполняет убыль вещества тела, возникающую вследствие окисления этого вещества кислородом окружающей среды. При этом процессе окисления—дыхания—получаются жидкие и газообразные продукты, являющиеся по составу более простыми сравнительно с протоплазмою. Продукты окисления организма выделяются из него во внешнюю среду или на всей поверхности тела или через посредство особых полостей, периодически возникающих в нем, расширяющихся и затем сжимающихся до исчезновения. Такой орган выделения называется вакуолей.
В процессах окисления протоплазмы выделяется теплота, затрачиваемая организмом на движение частиц протоплазмы в массе тела и в связи с этим, на передвижение последнего, на действие вакуоли.
Более высокая организация у простейших (инфузорий) выражается присутствием в оболочке более определенных отверстий для принятия пищи
1 Protos—первый, zoon—животное.
Фиг. 6. Голая Amoeba.
п — лди>» cv — вакуоля, — пищевые скопления.
и удаления неусвоенных остатков ее, развитием постоянных органов движения, в виде ресничек или жгутов, колеблющихся вследствие попеременного сжатия то одного бока реснички, то противоположного. Усложнение организации выражается также образованием раковины или скелета.
Вследствие питания происходит до известного предела рост животного и затем размножение.
При размножении делением родительский организм делится на два, приблизительно равные, дочерние организма, теряя- свое индивидуальное существование. При почковании от роди-• тельского организма отделяется небольшая его часть, путем самостоятельного роста достигающая размеров взрослого индивида, так что индивидуальность родителя при этом способе размножения не исчезает.
Наконец размножение образ )вание1. спор представляет деление родительского организма в короткий промежуток времени на значительное число мелких частей, каждая из которых дает происхождение новой особи.
Из четырех классов простейших1 лишь один, корненожек Rhizopoda (rhiza—корень, pons, podos— нога), встречается в виде ископаемых.
Rhizopoda, простейшим представителем которых является амёба, имеют лишенное кутикулы, хотя иногда снабженное раковиною тело, дающее псевдоподии, пальцевидные у амёб, нитевидные у фораминифер (фиг. 7). Из четырех порядков корненожек — амебоид-
ных, фораминифер, солнечников и радиолярий — в ископаемом состоянии сохраняются фораминиферы и радиолярии, изр°пка солнечники (в случае наличности скелета из кремнезема'
КЛАСС L КОРНЕНОЖКИ, RHIZOPODA
ИОРЯДОК I. ФОРАМИНИФЕРЫ, FORAMINTFEBA.
Существенною особенностью фораминифер (foramen — дыра, отверстие, fero — несу) является, кроме формы псевдоподий, постоянное присутствие раковины.
Последняя не всегда полностью включает протоплазматическое тело, часть протоплазмы иногда выступает на поверхность раковины, облекая ее слоем, дающим псевдоподии, так что раковина является заключенной среди протоплазмы. Иногда протоплазма, содержащаяся внутри раковины, высылает псевдоподии, наружу через поры стенок раковины (фиг. 8); в случае отсутствия пор внутренность раковины сообщается с наружной средой исключительно через устье или апертуру раковины (фиг. 7), существующее, хотя не всегда, и в случае наличности пор.
х Корненожки, жгутиковые, споровики и инфузории.
Нет сомнения, что покрытые, раковиною формы произошли от голых амёбобразных форм, стремившихся отделением наружных покровов защитить себя от неблагоприятных внешних влияний.
Такие покровы первоначально могли быть представлены просто наружным слоем протоплазмы, более плотным, чем внутренняя масса ее, как это бывает у многих амёб (фиг. 6).
Одною из дальнейших стадий усовершенствования оболочки является то состояние ее, когда она состоит из песчинок, приклеенных к телу и сцементированных более или меййе плотным видоизменением протоплазматического вещества (сем. Rhabdamminidae, фиг. 15).
При степени плотности цемента, не позволяющей первоначально шаровидной или эллипсоидальной раковине увеличиваться в размерах растяже-
Фиг. 7. Miliola, ныне живущая, с псевдоподиями, выходящими лишь из устья. В самой виутренней камере видно ядро.
Фиг. 8. Rotalia, ныне живущая, с псевдоподиями, выходящими из многочисленных пор. 14/1.
нием, это увеличение происходило наращиванием раковины при ее устье. Простейшею формою раковины, получающеюся при таком наращивании, будет трубчатая.
Таким образом возникли цилиндрические трубки, как иногда у Rhab-dammina (rhabdos—палка).
Если первоначальная раковина имела несколько устьев, то от нее исходило несколько цилиндрических трубок, образуя напр. крестовину (фиг. 9). ^Образование раковин в виде цилиндрических трубок представляет неудобства во-первых, подвергая трубки, при увеличении их длины, легкой возможности излома под действием внешней силы, во-вторых потому, что вытянутые в длину трубки сильно затрудняют движение животного. Выгоднее иметь более компактную раковину. Вследствие этого возникли формы < трубчатою раковиною свернутою неправильно, клубкообразно (фиг. 10; Ammodiscus— от некоторого сходства с рогами Аммона, египетского Зевса,
discus — диск) или в правильную плоскую спираль с прилегающими один к другому оборотами труб™.
Так шло развитие однокамерных раковин, не подразделенных внутри перегородками на отделы, камеры.
У однокамерных раковин, имеющих вытянутую трубчатую форму, бывают иногда заметны получающиеся от времени до времени перехваты, разделенные равномерными или неравномерными промежутками. Образование перехватов есть следствие периодической приостановки роста раковины.
Таким образом от первоначально простой песчанистой трубки через посредство сегментированной трубки произошли наконец раковины типа Nodosinella (nodosus—узловатый) (фиг. 13 и 14), разделенные на камеры. Раковины этого типа, однако непрочны, в особенности, благодаря существованию перехватов, вследствие чего должны были возникнуть более прочные формы раковин.
Это произошло различно. В одних случаях новые камеры становились теснее сдвинутыми, не будучи насажены на самое устье предшествующих
Фиг. 9. Rnab'la .ttnina, ныне живущая Увелич.
Фиг. 10. Ammodiscus gordialis. Каменноуг. Англия. 25/1.
Фиг. 11. Glan-dulma inflata. Юра. Гериания.
камер, но соприкасаясь с ними по окружности значительного радиуса, как например у Glandulina (фиг. 11, glandulus—имеющий ядра).
В других случаях трубчатая, с перегоро щами, раковина становилась прочнее вследствие свивания трубки в спираль плоскую или коническую— Cristellaria (фиг, 21; cristella—гребешок), навивания по меридианам эллипсоида—Quinqueloculina (фиг. 25; quinque—пять, loculus—ящичек с перегородками), иногда вследствие расположения камер в два-три ряда, примыкающие один к другому—Textularia (фиг. 16; textilis—плетеный) и ъ д.
Можно ожидать, что онтогения фораминифер будет представлять данные, свидетельствующие об указанном филогенетическом развитии. Действительно встречаются раковины фораминифер, так называемые биморф-н ы е, представляющие в ранней стадии раковину одного типа, переходящую под конец роста в раковину другого типа; встречаются также раковины триморфные. Но, обратно общему правилу, прогрессивный и филогенетически более поздний тип воспроизводится в биморфных формах в начальных стадиях раковины, а первичный тип—в конечной ее части. Мы имеем здесь 1> оптогении дело с ценогенетической перестановкой стадии развития.
Если у высших животных трудно выяснить при сложности их организма, чем объясняется например ценогенетическая перестановка в развитии chorda dorsalis и нервной системы у позвоночных (см. стр. 27), то у фораминифер указанная перестановка стадий онтогении вероятно происходит под действием естественного отбора. У фораминифер вещество раковины вырабатываемся не одной лишь поверхностью
тела, как у высших животных, во всею массою тела; у молодого, малых размеров, индивидуума отношение массы тела к его поверхности (т. е. и к поверхности отлагаемой раковины) относительно менее, чем у взрослого животного, так как при росте размеров тела объем его увеличивается пропорционально кубу, а поверхность—пропорционально лишь, квадрату этих размеров. Из этого ясно, что раковина, которую может выработать маленькое животное, будет непропорционально тонка по сравнению с толщиною ее у взрослого индивидуума. Следовательно для молодой раковины особенно ценны форма или строение, повышающие ее прочность. Животное в силу наследственности не может освободиться от воспроизведения в своей онтогении хотя бы некоторых . черт предков. Цо в то же время организм вырабатывает способность переживать прародительское состояние не соответственно ходу филогенетической истории, не в своей молодости, а позже, если1 такая перестановка для него выгодна.
Изложенная теория соотношений онтогении н филогении у фораминифер, принадлежащая германскому зоологу Румблеру, подверглась критике со стороны австрийского палеонтолога Шуберта. Шуберт указывает, что прежде всего нельзя принимать для фораминифер ценогенетическую перевернутость биогенетического закона; если есть например представители Frondicularia, (см. стр. 47) с А'ос/озда-га-образными конечными камерами (род Amphimorphina), то бывают и Nodosaria с /^-ои-Угс'к/лгга-образиыми конечными камерами (Staffа). Затем, спрашивает Шуберт .почему Nodosaria с его невыгодной с механической точки зрения раковиной сохранился от силура доныне и даже имеет особенно значительное распространение в позднейших отложениях? Наконец роды фораминифер в широкой степени являются полнфилетичнымн, так что например судя по начальной части двуформенной раковины род Textularia может происходить н от Textula-ridae с трехрядным расположением камер (Gaudryina'), и от плоско-спнральных форами-ннфер (Spiroplecta}, и от конически-спиральных роталоидных (Pseudotextula'ia). Шуберт приводят факты, указывающие, что конечная часть Nodosaria-об-развой формы отличается по своему внутреннему устройству (положение устий в последовательных камерах относительно друг друга) от так сказать нормальных нодозарий. Надо сказать, что при простоте организации мягкого тела простейших возможно -появление сходных форм в разное время, как новообразований, независимо одни от других развивавшихся.
Тип Nodosaria был достигнут, исходя по крайней мере восемь раз от различных предков. Раковины с неправильным, в беспорядке, расположением камер встречаются при прирастании фораминифер к субстрату, как у Planorbulina, Carpenteria, Polymorphina. Спирально свернутые раковины получаются при расположении устья не по оси трубки. Большая или малая длина камер зависит от степени вязкости протоплазмы, камера бывает мала при слабо вязкой протоплазме (например у Glandulina).
Смежные камеры раковин фораминифер сообщаются через отверстия^ существующие в перегородках и бывшие в свое время устьями раковины. Мягкое тело животного выполняет всю внутренность раковины.
Кроме формы раковины важные признаки отличия—состав вещества стенок и микроструктура раковины фораминифер.
Бывают хитиновые (из вещества, подобного роговому), обыкновенно однокамерные раковины, с плотными стенками, представляющие одна лишь отверстие—устье. Песчаные раковины состоят из песчинок, взятых извне, кварцевых, известковистых, песчинок вулканического происхождения, раковинок фораминифер и т. п.; эти песчинки связаны протоплазматическим или по преимуществу неорганическим—кремнеземистым, известковистым—цементом, выделяемым организмом. Стенки таких раковин или совершенно плотны, и сообщение животного с наружной средой происходит через устье, или пронизаны каналами, рассеянными на поверхности раковины, через которые выходят наружу псевдоподии.
Наконец известковистые раковины встречаются у большинства фораминифер, образованы углекислым кальцием, вырабатываемым организмом и отлагаемым на его поверхности. Эти раковины также бывают пористыми или лишенными пор в стенках, в зависимости от существования или отсутствия вышеупомянутых каналов для выхода псевдоподий; поры являются
отверстиями этих каналов. Пористые известковые раковины отличаются стекловатым характером—стекловатым по блеску, более или менее полной бесцветности и прозрачности. Известковистые раковины без пор мелоподобно или фарфоровидно белы и непрозрачны.
Некоторые части известковистых пористых раковин у известковых родов бывают все-таки лишены пор. Эта особенность встречается лишь в высших типах раковин фораминифер и главнейше в таких, у которых перегородки являются двойными (фиг 12, В), т. е. у которых камеры имеют совершенно самостоятельные стенки.
Части, лишенные поровых каналов, образованы вторичными отложениями известковистого вещества. Эти отложения находятся на наружной поверхности раковины, заполняя бороздки, соответствующие внутренним перегородкам, и серединные наружные углубления, лежащие в спиральносвернутых формах с обеих сторон раковины по оси навивания (такое углубление называется пупком—umbo). Вторичные отложения извести иногда
покрывают раковину толстым слоем снаружи или образуют в раковине внут-
Фиг. 12. 4 — сечение фораминиферы, у которой каждая перегородка состоит из одной пластинки. В — у которой перегородка состоит из двух пластинок.
У велич.
•а— соединительные отверстия между камерами; Ь — перегородки; с — передняя стенка последней камеры; d — дополнительный скелет.
Фиг. 13. Ныне жив \ тая Saccam-тгпа Стенка раковины взломана одного бока. 6/1.
Фиг. 14. No. dosinella index Нчж-некаменноуг. Тульский округ. 12/1.
Фиг. 15. No~ dosinella La-husem. Нчж-некаменноуг. Тульский округ. Вертик. разрез. 30/1.
с
ренние столбики, увеличивая таким образом ее прочность. Эти вторичные отложения представляют так называемый дополнительный или промежуточный скелет и обыкновенно пролизаны системой пнтерсеп-тальных каналов; последние иногда сообщаются с системой обыкновенных поровых каналов.
Хитиновые раковины характерны для пресноводных фораминифер. Это является особенно очевидным при бывающем иногда переходе фораминифер от жизни в морской воде в солоноватую или пресную, когда, известковистые раковины лишаются извести, заменяясь хитиновыми, например у Polysto-mella. Песчаные раковины встречаются исключительно в придонной фауне, а в планктоне поверхности моря исключительно пористые известковистые, У рода Miliola в прибрежной фации раковпны известковистые, в солоноватых водах—хитиновые, на морском дне на больших глубинах—песчанистые. Наличность дополнительного скелета свойственна бентонным формам.
Подпорядок I. Однокамерные. Monothalamia Сем. Rhabdammi-n i d а е. Раковина хитиновая или состоящая из песчинок обыкновенно значительной величины; однокамерная, часто разветвляющаяся раковина иногда сегментирована пережимами, но никогда не разделена на камеры; она покрыта нечастыми и неправильно расположенными порами. Роды: Вассаттгла
!
12
(saccos—мешок) с шаровидной раковиной (фиг. 13); с силура до настоящего времени, Rhabdammina (фиг. 9), с раковиной или в. виде прямолинейной цилиндрической трубки или лучисто или неправильно разветвляющейся. Юра—ныне.
Подпорядок II. Многокамерные. Polythalamia. Сем. Nodosinel-lidae. Раковина песчанистая-или более или менее известковистая. Пористая или без пор; многокамерная, состоящая из прямолинейного или лишь немного согнутого ряда камер.
Вероятные предки всех высших фораминифер.
Nodosinella (фиг. 14 и 15). Каменноугольные отложения — мел.
Сем. Textularidae. Раковины песчанистые, известковисто-песчанистые или чисто известковистые, большею частью пористые, редко без пор. Камеры расположены, чередуясь, в двух или трех рядах.
Textularia. Камеры расположены, чередуясь, в двух рядах; апертура или представляет щель при основании конечной стенки последней камеры (фиг. 16), или заменена несколькими небольшими отверстиями ситовидно продырявленной конечной стенки. От кембрия доныне.
Bigenerina (фиг. 17; bi — два, genus — род). Раковина двуформенная (биморфная); ранние камеры располо-
жены как у текстуларий, позднейшие — в одном прямолинейном ряду, как у Nodosinella.
С каменноугольной системы доныне.
Фиг. 16. Textularia Фиг. 17. Вгде-textulariformis. Верх- mrina gracilis. некамеяноуг. Капний- Ннжнекаменноуг. окне Альпы. Виды Слобода. Туль-сверху и сбоку. 25/1. ский округ. 11/1.
Строение раковины у Textularia и Bigenerina представляет некоторые оригинальные особенности; оиа состоит нз наружного грубопористого слоя, состоящего из песчинок, связанных цементом, известковистого или кремнеземистого, и прозрачного внутреннего известковистого слоя, прорезанного многочисленными' тонкими поровыми каналами. Наружный слой постоянно существует, внутренний же. очевидно образующийся после первого, не представляет такого поетоянстга
Сем. Nodosaridae. Раковина всегда известковистая
и тонко-пористая; камеры четковидно примыкают одна к другой в прямолинейном, дугообразно согну-
Фиг. 186 Frondicula-ria Zfoelleri. Юрская глина. Рязанский округ. 30/1.
Фиг. 18. No-rfose*-?- bacil-1ищ. Эоцен (синяя глина). Киев. 4/1.
том или свернутом в плоскую спираль ряду.
Nodosaria (фиг. 18). Раковина прямая или согнутая, круглая в поперечном сечении; с апертурой, обыкновенно расположенной центрально. От кембрия доныне.
Frondicularia (фиг. 19 и 20; fron-diculus — листик). Раковина сжатая, плоская, камеры вполне или в значительной мере имеют форму обратного V, обхватывают одна другую; первичная камера обособлена, выдаваясь среди последующих. С пермских отложений доныне, преимущественно с триаса до третичных.
Oristellaria (фиг. 21). Раковина спирально-свернутая, сжатая, чечевицеобразная, иногда представляет пуговкообразное утолщение в пупке или тонкий гребень на периферии, связывающей камеры. Круглое или щелевидное устье приближено к наружной стороне оборотов раковины. С трйаса до-
ныне, распространены с юры.
Фиг. 20. Frondicularia Cordai с раскрытыми камерами. Верхний йсл. Симбирский округ (Ульяновский). 30/1.
Фиг. 21. Oristellaria roiulata var. Roemeri. Юрская глина. Рязанский округ. 20/1.
При крайней тонкопористости раковины Nodosaridae (поперечник пор у фораминифер вообще измеряется долями миллиметра) возможно, что эти поры служат но для выхода пседвоподий, как вообще у фораминифер, а лишь для доступа воды к мягкому телу, для усиления процесса дыхания.
Сем. Miliolldae. 1 Раковина не имеет пор, большею частью известко-виста и различным образом свита.
Из этого семейства рассмотрим лишь представителей подсемейств i Miliolinae, характеризующегося трубчатой раковиной, навитой на концы удлиненной Оси, так что в каждом обороте находятся лишь две камеры, лежащие или в одной плоскости, или в различных, пересекающихся по оси. Вследствие такого расположения камер, устье.раковины во время роста ее попеременно находится со стороны то одного, то другого конца оси и обра
щезо попеременно в противоположные стороны (см. стр. 43,
Фиг. 22. Spiroloculina badensis. Миоцен. Бадей около Вены, Австрия. 30/1.
фиг. 7).
Фиг. 23. Thloculina, siiuplex. Миоцен. Вена-
Австрия 15/1.
Spiroloculina (фиг. 22; splra—спираль, loculosus — разделенный перегородками). Все камеры расположены в одной плоскости и все видны снаружи. От перми доныне.
Biloculina(фиг. 20 и 23; bis — дважды). Камеры расположены водной плоскости, последующие камеры охватывают предыдущие, так что снаружи видны лишь две последние камеры. Триас — ныне.
1 Milium—> просо.
Triloculina (фиг. 24). Навивание камер происходит в трех плоскостях, так что снаружи видны три последние камеры. Юра — ныне.
Quinqueloculina (фиг. 25). Навивание происходит в пяти плоскостях. Мел — ныне.
Фиг. 26. Biloeulina Bradyi. Ныне живущая.
А — исшоиорф’пая; В—триморфная форма.
Фиг. 24. Triloculina •nflata. Миоцен. Вена. 15/1.
Фиг. 25. Quinqueloculina saxorum. Эоцен. Гриньон. Франция. 12/1.
Встречаются экземпляры Biloeulina и Triloculina, внутренние обороты которых представляют иное навивание, нежели наружные. Так средн билокулин попадаются экземпляры, внутренние обороты которых навиты как у Quinqueloculina-, бывает и так, что за первыми оборотами, навитыми по типу Quinqueloculina, следуют навитые как у Triloculina, и затем уже наконец идет билокулиновое навивание (фиг. 26, В). Также встречаются экземпляры Triloculina с первыми оборотами, навитыми по типу Quinqueloculina. Таким образом среди Miliolinae встречаются би- и триморфные формы, так что напр. случай, представленный на фиг. 23, В, вадо истолковывать так, что Biloeulina переходит в Quinqueloculina, проходя как промежуточную стадию тип Triloculina. Указанное превращение прогрессивно с механической точки зрения: увеличение числа облекающих раковину снаружи конечных камер повышает
прочность раковины, потому что всякое внешнее давление, сопровождаемое опасностью излома раковины, распределяется на все большее число камерных стенок, так что отдельная стенка тем менее в опасности, чем более
Фиг. 28. Endothyra Фиг. 29. Haploph-Pandiri. Верхнека- ragmium globulis. меня. Тульский ок- Нижнекамск и. руг. 22/1. Тульский округ.8/1.
Фиг. 27. Alveolina Bosci. Эоцен. Париж.
Alveolina (alveolus — небольшая полость). Раковина спирально-свернутая, с оборотами большею частью вытянутыми в направлении, параллельном оси навивания; при такой вытянутости форма раковины является яйцевидной, веретеновидной (фиг. 27), цилиндрической, но бывает и шаровидной. Обороты делятся на камеры перегородками, параллельными оси навивания. Каждая камера по бокам распространяется далее предшествующих камер, увеличивая этим длину раковины по оси. Камеры коротки в направлении, перпендикулярном оси навивания, и подразделены на отделения вторичными вертикальными перегородками, перпендикулярными этой оси. На поверхности раковины имеются бороздки, параллельные оси навивания, соответствующие границам между камерами. С .деловых отложений доныне, особенно часто
в третичных, где иногда являются породообразующими (альвеолиновые известняки).
Сем. Endothyridae. Трубчатая раковина свернута в плоскую или в низко коническую спираль. Иногда камеры разделены на отделения. Устье раковины всегда приближено к стезке предпоследнего оборота. Пористые или без пор. Песчанистые, известково-песчанистые или вполне известковые.
Подсем. Endothyrinae. Раковина песчаниста, большею частью со
значительным содержанием извести; иногда пориста.
Endothyra (фиг. 28; endos — внутренний, thyra — дверь). Раковина обыкновенно образует два или три-оборота по несимметричной слабоконической спирали. Устье в виде одной щели или заменено группой грубых пор,
как у Textularia. Встречается от каменноугольных отложений до триаса.
Haplophragmium (фиг. 29; haplus — простой, phragmos — разделение). Раковина биморфная, в более старой части — типа Endothyra, к устью — типа Nodosinella. С каменноугольных отложений доныне.
Подсем. Fusulininae. Обороты с наклонностью сильно вытягиваться
в направлении оси завивания.
Фиг. 80. Поперечное сечение • Fusulina pusilia. ВерхЬека-менн. Карпинские Альпы.
Каждый оборот вполне объемлет предыдущие. Раковина всегда известковистая и пористая.
Эту группу надо считать за происшедшую от Endothyridae путем роста раковины главным образом в направлении оси завивания, с сохранением слабой асимметрии.
Фиг. 31. Fusulina longissima. Верх-некаменн. Царев курган, блгз Жигулей. Средневолжский край. 8/1.
Fusulina (фиг. 30 и 31; fusus — веретено). Раковина веретенообразная или цилиндрическая с закругленными концами. Перегородки на всем протяжении от одного конца до другого гофрированы, складчаты в своей нижней ближайшей к оси завивания части. Противолежащие складки смежных перегородок одновременно или выпуклы, или вогнуты в противоположном направлении, так что складки смежных перегородок соприкасаются и срастаются, и вследствие этого камеры в нижней части, примыкающей к предыдущему обороту, являются подразделенными на отделения, сообщающиеся в верхней части камер. Устье в виде срединной щели. Каменноугольные отложения и пермские.
Schwagerina (фиг. 32; по имени исследователя фораминифер—Швагера). Перегородки в раковинах представляют ровную плоскость на большей части своей длины, не обнаруживают складчатости "там, где последняя развита у фузулин, но, приближаясь к оси завивания, становятся почти внезапно волнистыми, разветвляются, сплетаясь этими разветвлениями в сложную перегородчатую сеть. Устье раковины в виде срединной щели, как у фузулин, или же в виде ряда круглых отверстий. Швагерины обыкновенно отли
чаются от фузулин и по наружной форме, будучи шаровидными, но есть виды, раковина которых удлинена по оси завивания, приближаясь в то же время к раковинам фузулин и по другим особенностям. Каменноуг: и пермокарбон.
Сем. Rotalidae (rota—колесо). Раковина всегда известковистая и пористая, свернутая спирально; вследствие асимметрии спирали иногда с одной ^гуроны видимы все обороты, а с другой—только последний. Высшие формы имеют дополнительный скелет и интерсептальные каналы. Устье раковины примы-
кает к поверхности предыдущего оборота. Группа вероятно вышла из Endothyridae.
Globigerina (фиг. 33; globus— шар, gero—несу). Раковина грубо-
Фиг. 32. Scnwagerma princeps. Верхнекаменн. Серегово. Обл. Комн. 5/1. Левый рисунок представляет разрез по продольной оси, обломанный снизу
и справа.
пористая. Камеры расположены спирально, иногда не вполне правильно, или вздуты и составляют неправильной формы скопление. С кембрия доныне, чаще в кенозойских отложениях.
Глобигерины — животные пелагические, живущие в открытом море; раковины их
в настоящее время находятся в громадных количествах в глубоководных отложениях. Пелагической! жизнью объясняется оригинальный характер раковины в виде неправильного скопления шарообразных камер; при втом раковина представляет оольшую поверхность, чем если бы ранние камеры были скрыты позднейшими. При значительности поверхности раковина испытывает большое трение в воде, благодаря чему облегчается возможность плавать во взвешенном состоянии. Пелагические виды имеют раковину, покрытую чрезвычайно тонкими известковистыми
шинами, очень легко отпадающими по смерти живот-
ного и не сохраняющимися у ископаемых форм. Фиг. 33. Globigerina bulloides. Мио-Существование шипов еще более увели- пен. Вена. 50/1.
чивает поверхность раковины; последняя, при
пелагическом образе жизни животного, не должна необходимо отличаться прочностью так как у большинства фораминифер, ползающих по дну морскому, чем и объясняется мало прочное между собою соединение камер.
Nummulites (фиг. 34—38; nummulus — монетка). Раковина чечевицеобразная или дискообразная, состоящая из многочисленных спиральных оборотов, разделенных перегородками на камеры. Возрастание раковины происходит главным образом по периферии ее, отчего она и приобретает Дислоидальную форму. Последующие обороты охватывают предыдущие вполне, так что наружная поверхность раковины есть поверхность последнего обо
рота. Существуют отложения дополнительного скелета в виде цилиндрических участков в стенках раковины; оконечности этих участков выдаются на поверхности стенок в виде бугорков (фиг. 34}. Рели эти отложения в смежных оборотах приходятся одни над дру-
Фиг. 34. Nummuli-tes dis'ans. Эоцен. Крым. 1/1.
гими по прямой линии, то сливаются в столбики, проходящие через несколько оборотов нормально к поверх-
Фиг. 36. Nummulites, разрезанный поперек и частью вдоль.
Фиг. 35. Поперечный развез Nummuhtes laevigatuS. Видны радиально направленные столбики дополнительного скелета. У велич.
Фиг. 37. Nummulites laevigatus. Часть раковины. Круговой разрез, обнаруживающий камеры двух наружных оборотов и наружную поверхность третьего. 1/1.
Фиг. 38. Nummulites Grt-zehensis. Круговой разрез. Видны камеры четырех наружных оборотов и наружная поверхность следующего. 1/1.
Фиг. 39. Orbitmdes expanset. Миоцен -Чамахаттн Ар пения. 1/1.
ности их (фиг. 33). Кроме
л
Фиг. 40. Orbitoides раругасеа. Эоцен. Кресренберг. Бавария. Германия. Увелич.
« — центральная камера; а—первичные; b — вторичные камеры.
перечника в 60 мм.
ности их (фиг. 33). Кроме того отложения дополнительного скелета сплошь образуют периферическую часть раковины, место перегиба от одной плоской стороны раковпны к другой. Обороты раковины наиболее отстоят один от другого при спинном стержне, на периферии раковины (фиг. 33), в середине же ее весьма сближены, перегородки, в которых соединительные отверстия между каперами лежат в средней плоскости близ спинного стержня предыдущего оборота, имеют продолжения и в указанные узкие пространства между оборотами, причем являются здесь иногда неправильными по своему ходу (фиг. 36), иногда сплетаются между собою, образуя сеть (фиг. 35). Наиболее кр^пнле нуммулиты достигают поперечника в 60 мм. Каменноугольные отложения (?) — ныне; особенно часто, массами, в нижнетретичных отложениях (эоцен).
Orbitoides (фиг. 39, 40; orblta — круговая линия). Раковина или совершенно такая же чечевицеобразная, как у Nummilites, или с утолщениями на поверхности, пуговицеобразным центральный, а иногда и радиальными, в связи с которыми поверхность приобретает звездчатый вид, и очертание раковины становится многоугольным. Камеры сначала располо-
Фпг. 41. Orbitoides apiculata. Меловые отло... эиия. Мастрихт. Бельгия. Часть разреза ири значительном увеличении. J3/1.
Фиг. 42. Orbitoides minor. Меловые отложения. Гора Дибрар. Юго-вост. Кавказ. Азербайджанская ССР. Разрез. 6/1.
Фпг. 43. Orbitoides (Orthoph-гадтгпа) Pratti. Эоцен. Далмация. Разрез.
Фиг. 44. Orbitoides (Orthophragmi/iu) итЫН-cat а. Эоцен, Новая Каледония. Часть разреза при значительном увеличении. С — центр раковины.
жены в плоскую спираль с оборотами лишь касающимися, а затем позднейшие камеры располагается в той же плоскости циклически (по концентри-
ческим кругам). На плоских боках диска, таким образом получающегося, находятся слои мелких вторичных камер. Через эти слои, перпендикулярно к ним, проходят столбики дополнительного скеле га. В слое главных первичных камер сообщаются не смежные камеры оборота, как обыкновенно бывает, но камеры смежных оборотов. В стенках между оборотами и в перегородках имеется система интерсептальных каналов (фиг. 41, 42, 43, 44). С верхнемеловых отложений до миоцена, особенно часто в эоцене и олигоцене.
Распространение фораминифер.
Громадное большинство фораминифер живет в море, именно в прибрежной полосе, ползая по морскому дну, по растениям, здесь находящимся, и т. п. Фораминиферы наиболее крупной величины встречаются в мелко-
водной полосе тропиков. Некоторые однако, как например Gtobigerina, являются животными пелагическими, обитателями открытого моря, и в связи
4 I. Учебшп палеонтологах.
49
с этим остатки их могут быть и на больших глубинах. В древних палеозойских отложениях фораминиферы сравнительно редки и плохо сохранены, так что трудно составить себе представление о всей фауне фораминифер, тогда существовавшей. В кембрии указаны Textularia, Nodosaria, Lagena
Геологическое распространение фораминифер.
Отделы или ярусы Роды Кембрий Силур Девон Карбон Пермь Триас Юра \ Мел Эоцен Олиго цен Миоцен Плиоцен Современные
Nodosinella ....
1
Textularia .... Bigenerina .... Nodosaria .... Frondieularia . . . Cristellaria ....
0 1
1
Spiroloculina . . . Biloculina .... Triloeulina .... Quinqueloeidina . . Alveolina ?
Endothyra . . . Haplophragmium . Fusulina . . * . . Schzvagerina ....
Globigerina .... Nummulites .... Orbitoides 1
?
и Cristellaria. Из силура известны Rhabdammiiiidae (Sacpammina), Textu-laridae, Rotalidae (?), Nodosaridae и глобигерины.
В каменноугольных отложениях большое разнообразие родов форамини-? фер, причем раковины последних, скопляясь иногда массами, являются
породообразующими. Так Saccammina в значительной мере образует известняки в Великобритании, Бельгии.* «
В верхнекаменноугольных отложениях СССР, Китая, Японии и Сев. Америки распространены фузулиновые известняки. В каменноугольных отложениях встречаются также Nodosinella, Haplophragmium, Endothyra, Textularia, Bigenerina, Schwagerina, Nummulites (?). Лишь здесь типы спирально-свернутых фораминифер становятся распространенными.
В пермских отложениях фораминиферы не так обильны, как в каменноугольных, по большей части те же роды, что и в каменноугольных, можно отметить появление Frondicularia.
Триас также не богат фораминиферами. Endothyra, Haplophragmium, Nodosaria, Nodosinella, Textularidae, появляются представители Miliolldae {Bilocidina).
В юрских отложениях представители фораминифер не играют значительной роли, это по большей части мелкие и рассеянно встречающиеся формы.
Породообразующими являются фораминиферы в верхнемеловых отложениях: так называемый белый - мел состоит из раковинок текстуларий, кристелларий, глобигерин, милиолид и т. д.
В третичных отложениях многие роды, например Nummulites, также играют видную роль в образовании пород: третичные известняки с Milio-lidae в окрестностях Парижа, нуммулитовые известняки Сев. Африки, Крыма и т. д., орбитоидные породы Кавказа.
Наиболее специализированные роды фораминифер (фузулины, нуммулиты) появляются после долгого существования более простых типов.
ПОРЯДОК II. РАДИОЛЯРИИ RADIOLARIA.1
Тело радиолярий разделено хитиновой перепонкой (фиг. 45) на две части: внутреннюю, содержащую ядро, шаровидную или яйцевидную центральную к а п с у л у и наружную, внекапсулярную часть; обе части протоплазмы сообщаются через поры капсулы. Внекапсулярная протоплазма дает нитевидные псевдоподии. Образуются скелеты, редко отсутствующие, из кремнезема, реже сернокислого стронция, из кальдиево-алюминиевого силиката; скелеты последнего рода растворяются в воде по смерти животного, почему и неизвестны в ископаемом состоянии. Скелеты в высшей степени разнообразны по форме; они проникают через всю массу протоплазмы, или находятся лишь во внекапсулярпой части, состоят из отдельных игол (спику-лей) или имеют решетчатое строение, часто с выдающимися шипами, иногда представляют несколько концентрически окружающих друг друга и соединенных между собою радиальными столбиками шаров (фиг. 45), иногда дискоидальны (фиг. 48). Радиолярии часто размножаются делением, обыкновенно же путем образования спор в центральной капсуле. Радиолярии находятся в морях, на всяких глубинах, во всех климатах, местами образуют в настоящее время радиоляриевый ил. Радиолярии в большинстве случаев микроскопически малы, почему и не имеют большого геологического значения. .Во многих случаях все-таки радиолярии принимали существенное участие в образовании кремнеземистых и известково-кремнистых пород (кремнистых
1 Radioing — лучик, от radius — луч.
сланцев, кремней, яшм и т. п.). Известны даже из более древних отяоже-ний, нежели другие представители животного царства, — из докембрийских. Ископаемые представители тадиолярий невидимому не уступают в разно-
Фиг. 45. Actinomma asteracanthibn. Все животное в оптическом сечении.
о—ядро; Ъ — стенка центральной капсулы; е—самая внутренняя кремнеземистая раковтна заключена в ядре; с1 — средняя раковина, заключенная в центральной капсуле; с3 — наружная раковина, лежащая б экстракац-сулярнсй протоплазме; видны четыре радиальных кремнеземистых шипа, связывающие три сферические кремнеземистые раковины. Бедно тонколучнстое строение протоплазмы и тонкие экстракапсулярные псевдоподии.
Фиг. 46. Eucyritditim cranioides. Ныне живущая. С псевдоподиями. У велич.
образии ныне живущим и принадлежат к одним с последними семейства!» и даже родам. Найдены между прочим в девонских кремнистых сланцаv и яшмах на Урале. Верхнемеловые (белого мела) кремни вероятно обязаны
Фиг. 47. Ltthoca-тре Tschernyschewi. Девон. Урал. Уве-лич в 100 раз.
Фиг. 4S. Stilorlietya tnultispina. Ныне живущая. Мессина. Сицилия. Италия. Увелич.
своим происхождением переотложенному веществу скелетов радиолярий; особенно известны третичные радиодяриевые мергеля на острове Барбадосе, в Сицилии и т. д.
j Характеристики групп простейших.
I 1 класс: Rhizopoda, корненожки. Одноклеточные животные, снабженные ложнонож-
I’ ками; большею частью водные, делятся на 4 порядка.
Порядок: Amoebina. С изменяющейся формой тела и толстолопастными ложноножками. Бесскелетны и в ископаемом состоянии неизвестны.
Порядок: Foraminifera. 0 тонкими, нитевидными, образующими сеть ложноножками, с раковиной, лишенной пор или пористой, известковистой, песчанисто или только хитиновой, однокамерной или многокамерной. Большею частью морские бентонныё формы, встречаются с нижнего кембрия.
1 подпорядок: Monothalamia, однокамерные, хитиновые или песчанистые.
2 подпорядок: Polythalamia, многокамерные, песчанистые или известковистые.
Порядок: Heliozoa. Шаровидные, с обособлением в протоплазме коркового и сердцевинного вещества, с тонкими довольно неизменными лучистыми ложноножками, с более плотною осевой нитью в них, иногда с кремнеземистым скелетом, в виде решетчато-продырявленных раковин. Послетретичные. *
Порядок: Radiolaria. Шаровидны, с центральной капсулой, заключенной в перепонку и внекапсулярной протоплазмой с тонкими лучистыми ложноножками. Большею частью с решетчатым скелетом, а реже в виде игол. Морские планктонные; встречаются с докембрия.
II класс: Mastigophora (Flagellata), жгутиковые. Одноклеточные животные с одним или двумя жгутиками, органами движения. Иногда снабжены хлорофиллом, зеленым красящим веществом растений, иногда выделяют известковый или кремнеземистый скелет, с кембрия. Палеоботаники относят Flagellata к водорослям, описание их см. в отделе палеоботаники,
III класс: Infusoria. С многочисленными ресничками неизменной формы, с отверстиями, соответствующими ротовому и заднепроходному. С хитиновыми оболочками известны с древних послетретичных.
IV класс: Sporozoa. Паразитические формы, производные от Amoebina и Mast:--gophora, известны только ныне живущие.
ВЫСШИЕ ЖИВОТНЫЕ.
ОБЩИЕ ЗАМЕЧАНИЯ.
Элементарными составными частями тела высших животных являются к л е т к и, — образования,' но форме своей сходные с организмом тех или других простейших животных. Каждая отдельная клетка тела высших животных состоит из основного протоплазматического вещества и ядра или ядер, клетки могут образовать псевдоподии и реснички, выделять жидкие, кутикулярные и скелетные вещества, могут принимать пищу, усваивать и т. д., словом—сходны с организмом. простейших не только по форме, морфологически, но в значительной степени и по отправлениям, физиологически.
У более низко организованных Metazoa1 (например личинки губок) клетки тела группируются в два слоя—наружный, эктодерму, и заключенный в ней внутренний слой, энтодерму. У других животных является еще третий, промежуточный слой—м е з о д е р м а. При таких различиях в положении слоев клетки их конечно не могут иметь совершенно одинаковых отправлений, последние определяются соответственно указанному различию в положении слоев, так что например развитие двигательных ресничек составляет принадлежность наружного слоя, а внутренние клетки играют главную роль в процессе обмена веществ и т. д. В связи
1 Meta—после, zoon—жиьотное.
с различиями в отправлениях клеток первичных слоев существует и различие в форме их, в способе соединения между собою.
Первичный слой может быть соединительнотканный или эпителиальный. Эпителием называется комплекс плотно примыкающих одна к другой клеток, выстилающий свободные поверхности, будет ли это наружная поверхность тела, или поверхность его внутренних полостей:
Соединительная ткань, в отличие от эпителия, никогда не находится на открытых поверхностях, но всегда в подэпителиальных замкнутых пространствах животного тела и обыкновенно состоит. не из плотно сомкнутых клеток, но из таких, которые или вовсе не . связаны между собою, или связаны без примыкания вплотную и заключены в жидком или твердом основном межклеточном веществе.
Эпителий и соединительная ткань собственно не всегда являются тем, что принято называть тканью, т. е. комплексом клеток одинаковой формы и одинакового отправления. И в эпителии и в соединительной ткани могут находиться различные клеточные элементы: нервные, кроме формы характеризующиеся высокою степенью чувствительности; мускульные, характеризующиеся способностью сильно сокращаться в одном направлении—в направлении длины с одновременным расширением в поперечном направлении; железистые клетки, выделяющие известные вещества, и т. п.
В ранних стадиях индивидуального развития Metazoa эпителий и соединительная ткань состоят из совершенно одинаковых клеток и могут быть названы тканями, притом основными, так как из них развиваются ткани: нерв ная, мускульная и др. Для Metazoa характерен еще встречающийся исключительно у них процесс полового размножения. В основе этого процесса лежит слияние двух половых клеток, яйцевой и семенной; яйцевая клетка при этом, как говорят, оплодотворяется и приобретает способность развиться в зрелый организм путем роста, деления и диференциацип образующихся клеток в различные ткани.
II ТИП. ГУБКИ, PORIFEKA. 1
Губки—животные, лишенные способности передвижения с одного места на другое, за немногими исключениями прикрепленные в зрелом возрасте неподвижно к посторонним предметам. Губки—животные водные и в подавляющем большинстве морские. Тело одних губок имеет постоянную форму, обыкновенно бокаловидную или мешкообразную, у других губок форма тела непостоянна, находясь в зависимости например от предметов, к которым губка прирастает, в значительной степени облекая их.
Тело губки покрыто очень тонким эпителием на своей гладкой или бугристой наружной поверхности. Из многочисленных отверстий тела микроскопически малые поры служат для входа воды вовнутрь тела, для -выхода же воды наружу служат большие отверстия (psculum). Мягкое тело губки пронизано системой полостей, представляющих водоносные пути. Большею частью эти полости являются каналами; каналы местами прерываются расширениями, тан называемыми жгутиковыми камерами (фиг. 49, 50). Стенки камер покрыты жгутиковым эпителием, удлиненные клетки которого на краю свободной поверхности снабжены кольцевым кутикулярным ворот-
1 Рогоз—пора, скважина, (его—несу. По-гречески губка — spongia, ср. названия многих групп губок.
ничком, выходящим в полость камеры, а в центре этой поверхности жгутиком; колебания жгутиков вызывают ток воды в известном направлении Тон воды приносит твердые частицы, из которых пищевые удерживаются
и перевариваются жгутиковыми клетками, дыхание же и выделение проис-
ходит вероятно через посредство всех эпителиальных клеток.
Жгутиковые воротничковые клетки эпителия свойственны исключительно губкам, отсутствуя у всех других Metazoa. Главная масса мягкого
тела губки образована мезодермической соединительной тканью, лежащей под эпителием. В соединительной ткани там и сям образуются половые клетки, яйцевые и семенные, наконец она служит как опорное вещество для эпителиальных полостей. Яйцевые и семенные клетки развиваются из так называемых амебоидных, способствующих своими движениями распределению продуктов пищеварения. Лишь редко, у бесскелетных мясистых губок, для опоры достаточно хрящеподобного основного вещества соединительной ткани; большая часть губок обладает кремнеземистым, известковистым или рого-
Фиг. 49. Разрез тела ныне живущей губки (Cortictum) с известковым скелетом. Рицны две жгутиковые камеры. Увелич.
вым скелетом, который возникает в соединительнотканных клетках в виде отдельных маленьких скелетных элементов—иго лочек (spicula). Каждый скелетный элемент образован первоначально одной клеткой, но впоследствии в его наращивании принимают участие и другие клетки. Форма ске-
летных элементов имеет большое систематическое значение, так как оказы-
вается, что у каждого рода губок скелет состоит из одного или из немногих сортов игол. Бак ни разнообразны скелетные элементы губок, они приводятся к немногим основным типам, причем состоят из того или ЛРУГОГО числа лучей, исходящих радиально от / центра спикули, т. е. от начального пункта, от которого совер-
I /ШК-р' шается ее образование.
ЬЛмШ ^Учн располагаются п0 определенному числу осей, про-I ходящих через центр, так что число лучей самое большее
Фиг- 50. является вдвое большим числа осей., жгутиковая Различают следующие основные типы спикулей губок:
''d 1) Одноосный тип, построенный по одной оси и по-
тому имеющий форму просто палочки или иглы (фиг. 51, « и Ъ). Одноосная спикуля может быть или однолучевая (фиг. 51, Ъ), или двухлучевая (фиг. 51, о); в последнем случае оба лежат на одной оси. Ось может быть прямая или согиггая.
2) Трехосный тип (фиг. 51, е). Спикуля образована отложением по трем осям, пересекающимся в общем центре под прямыми углами, так что спикуля самое большее шестилучевая.
3) Четырехосный тип (фиг. 51, с, сГ), который можно рассматривать как происшедший отложением скелетного вещества по четырем осям, пересекающимся под равными углами так, как пересекаются перпендикуляры, опущенные из центра тетраэдра на середины его граней.1
1 Луч по-лат. — actis, actinis, по-гречески один--monos, четыре—tetra, шесть— Пеха, отсюда названия групп губок.
Многочисленные уклонения от этих типов, часто представляемые спи-кулями, происходят различными путями. Спикули покрываются мелкими
Фиг. 61. Скелетные элементы:
« — прямой и согнутый двухлучевые одноосные; b — одаол) чевой одноосный; с и d—четырехосный трехлучевой и четырехлучевой; е — трехосный шестилучевой.
шипами, шишковатыми или корневидными выростами (фиг. 52); лучи раздваиваются или неправильно разветвляются на концах (фиг. 53); при неразвитии некоторых лучей получаются скелетные элементы (фиг. 54), принадлежность которых к одному из вышеперечисленных основных типов нелегко устанавливается; между прочим, вследствие недоразвития числа лучей, получаются элементы, подобные одноосным, в действительности же принадлежащие к одному из остальных типов.
По оси каждой спикули расположен осевой к а и а л, который при жизни губки наполнен органи ческим веществом. При определении типа спикули осевые каналы дают ценные указания, так как они во-первых не имеют продолжений в различные шиповидные или корневь ;ные выросты, с другой стороны напри sep
5( Ч,
истинная природа кажущегося одноосного элемента, происшедшего из трехосного, познается по маленькому овтатку реей исчезнувших лучей (фиг. 54), кажущийся одноосный элемент является в действительности трехосным с лучами, не развившимися по двум осям (фиг. 54). Кроме собственно скелетных элементов в теле губкп существуют еще мясные иглы, малой ве^лчиьы, не связанные между собою и потому редки сохраняющиеся в ископаемых губках.
Скелетные элементы одних губок остаются не-
связанными Между собою ИЛИ СВЯЗаНЫ ЛИШЬ Орга- Фиг. 52. < пикули, ничееким веществом, спонгином, близким по
составу в шелку; у других губок скелетные элементы прочно соединяются
разветвлениями своих концов или же отлагающимся между
концами неор-
Фиг. 53. Четырехдучевые скелетные элементы Callopegma acaule. соединенные между со-
бою разветв. енными концами лучей. Рерхнемелов. Сенои. Ганновер. Германия. 40/1.
ганическим веществом. При прочном "П соединении скелетных элементов губки способны в ископаемом состоянии хорошо сохранять наружную форму тела.
* Скелет губок состоит из спонгина, или из аморфного кремнезема, или содержит в различной пропорции эле-j менты из того и другого вещества,
[ или наконец состоит из углекислого
J кальция.
ф .. У ископаемых губок вследствие Неразвитые вторичного переотложения вещества лучи, кремнеземистые элементы часто ока-
зываются превращенными в известковистые и наоборот, так что при определении приходится руководствоваться не столько минералогическим характером, сколько формой скелетных эле-
ментов.
Размножение губок происходит бесполым и половым путем. Бесполое размножение почкованием приводит к образованию колоний губок,
Фиг. 55. Sttilis-pongia glomerate,. Виднц три oseula. Верхняя юра. Нат-гейм. Вюртемберг.
Германия. 1/1.
(состоящих из соединенных гтесте индивидуумов, число которых бывав? иногда трудно определить. Наиболее легко определяется индивидуум но существованию отдельного osculum (фиг. 55); но иногда ьоги1а малы и неотличимы от отверстий отдельных каналов, иногда стенки тела губки склад-чаты так, что складки соприкасаются и срастается, образуя более или менее замкнутые полости, с отверстиями, которые не являются первый взгляд и могут быть приняты за них.
При половом размножении из оплодотворенной яйпевой клетки развивается личинка, сначала свободно плавающая действием ресничек, покрывающих тело, а затем прикрепляющаяся и развивающаяся в зрелое животное.
Губки разделяются на классы таким образом, что во-первых обособляются губки с известковистым скелетом (Calcispongia). Из остальных губок резко обособляется группа с трехосными спикулями (Hexactinellida). По выделении этих двух групп остается большое количество губок, хотя и крайне разнообразных, но связанных постепенными переходами, так что они почти все могут |быть соединены в класс, получивший название Demo-spongiae; это именно губки, скелет которых или состоит
из кремнеземистых спикулей различных типов, но никогда не трехосных, пли из волокон спонгина, или из комбинации названных спикулей и волокон, пли наконец губки совершенно бесскелетные.
КЛАСС I. DEMOSPONGIAE.
Из этих губок в ископаемом состоянии сохраняются лишь губки с кремнеземистым скелетом, состоящим из четырехлучевых спикулей (Tetract,inel-lida) или однолучевых (Monactinellida).
У Tetractmellida основная четырехлучевая форма спикулей представляет разнообразные видоизменения, получающиеся вследствие изменения угла между лучами, вследствие ветвления или атрофии некоторые из них, искривления лучей и покрытия их выростами, так что в конце концов скелетные элементы могут быть совершенно неправильной формы (фиг. 52), называются тогда десмами1 и характеризуют группу так называемых каменных губок (Lithistida), скелет которых отличается большою прочностью вслед-.ствпе сцепления смежных скелетных элементов сильно разветвленными на концах лучами, благодаря чему общая форма тела губки хорошо сохраняется в ископаемом состоянии.
Aulocoptum (фиг. 56, 57; aulos — трубка, copeion — кусок). Тело губки яблоковидно, коротко-стебельчато; на основании губки плотный концентрически морщинистый покров. Глубокая воронкообразная центральная полость, в которую открываются широкие дугообразные каналы, параллельные боковой поверхности тела. Более узкие каналы лучисто расходятся от основания губки к боковой поверхности. Скелетные элементы явственно четырехлучевые. В силуре.
1 В последнее время для скелетных элементов губок и для частей одного элемента выработана обширная терминология.
Jerea (фиг. 58). Грушевидное тело губки снабжено стеблем с корневидными разветвлениями на нижнем конце. Центральная полость заменена
Фиг. 56. Атксоргит aurantium. Ив силурийских ведунов Силезии. 1/2.
Фиг. 57. То же. Разрез, о — центральная полость; г - радиальные; с — концентрические каналы. 1/2.
Фиг. 58. Jetea piriformis Сеноман. Влекдоун. Англия. 1/2.
широкими продольными, у оси тела вертикальными, к периферии дугообразными каналами. Имеется система узких радиальных каналов. Скелетные элементы, правильно четырехлучевые, разветвляются. Средний и верхний мел.
Pemmatites (фиг. 59). Бесстебельча-тое тело формы шарообразной, хлебообраз-но-сплющенной, пронизано системой кана-
Фиг. 59. Pemmatites macroporus. Ядро, состоя щее лишь из выполнении каналов кремнеземом. В промежутках между петлями сети каналов порода. Верхнекаменноугольн. Р. Аша. Урал. 3/2.
А— вертикальный разрез; В — вид сверху; В, — радиальные стволы каналов; Р, Pti Рл — поперечные соединения между н 1ии, 74 — разрезанное по длине, — Рл разрезанное поперек.
лов, радиально расходящихся к поверхности от средины нижней стороны тела. Радиальные каналы соединены поперечными перемычками. Скелетные элементы в виде осевого бугорчатого стержня, от которого отходят боковые короткие отростки. В верхнекаменноугольных и пермских, артинских отложениях.
Astylospongia (фиг. 60; а—без, stylos—стержень). Бесстебельчатое тело губки шарообразно с небольшим углублением на верхней стороне.
Фиг. 60. Astylospongia praemorsa. Вид сверху и сбоку. Нижний силур. Таллин. Эстония. 1/1. S—отдельный скелетный элемент. Из валунов Силезии. 44/1.
Радиальные и меридиональные дуговые концентрические каналы; отверстия первых—на боках, последних—в углублении верхней стороны. Наружная поверхность кроме того имеет мёридианальные желобки, которые представляют зачатки концентрических каналов. Скелет из четырехлучевых элементов, лучи которых разветвляются уже близ утолщенного центра. В верхнем и нижнем силуре.
КЛАСС II. HEXACTINELLIDA.
При большом разнообразии во внешней форме эти губки отличаются неплотным сложением и сравнительно тонкими стенками скелета. В связи с этим отсутствует сложная система каналов, развивающаяся в солидной массе тела Lithistidae. В стенках тела губок бокаловидного, грибовидного, цилиндрического, реже неправильно комковатой формы, проходят нормально к его поверхности снаружи и совнутри простые короткие, обыкновевно слепо ! оканчивающиеся, реже сквозные каналы, часто размещенные правильными рядами, иногда же распределяющиеся неправильно. Иногда каналы вовсе не обособлены. Спикули или соединены концами лучей при посредстве кремнеземистого цемента, или лежат свободно. Палеозойские Hexactinellida принадлежат исключительно к более примитивной группе со spiculae, не соединенными между собою.
Ventriculites (фиг. 61; venter—брюхо). Форма тела бокаловидная или цилиндрическая, иногда с верхними краями, отогнутыми, как у тарелки. Центральная полость большая и глубокая. Стенка тонкая, мелкоскладчатая, так что представляет вертикальные ребра-и разделяющие их борозды, а в поперечном разрезе волниста.
В бороздах наружной поверхности имеются перемычки, так что промежутки между перемычками представляют отверстия, расположенные вертикальными рядами. Короткие прямые каналы перпендикулярны к стенкам; отходят от наружной и внутренней стенок, оканчиваясь слепо. Шестилуче- < вые спикули соединены лучами, образуя скелет с кубическими промежут-। ками между соединенными лучами (фиг. 62). Узловая часть спикули полая,
так что соединение лучей ее производится перемычками, расположенными по ребрам октаэдра. Губка была снабжена, разветвленными корневидными отростками, состоящими из кремнеземистых волокон. Верхний мел.
Astraeospongia (фиг. 63; aster—звезда). Неприкрепленное, вогнуто-
OSO.
Фиг. 61. Ventriculites. Реставрация 1/1. osc. — csculum, г — корневидные отростки.
выпуклое, блюдцеобразной фермы тело. Спикули большие, в несколько миллиметров размерами, не соединены между собою; в спикуле два луча, перпендикулярные к поверхности тела, укорочены, тогда как в плоскости, параллельной поверхности тела, вследствие разветвления лучей их бывает не 4, а 6. Силур, девон.
Фиг. 63. Astraeospongia meniscus. Верхний силур. Тенесси. Сев. Америка. 1/1.
Фиг. 62. Ventriculites striatus. Сенов. Ганвовер. Германия. Скелет. 12/1.
)
К Hexactinellida, притом со скелетными элементами, на связанными между собою, многие относят группу Receptaculidae (фиг. 64; receptaculum— вместилище), встоечающихся в силуре и девоне. Это все свободные, шаровидные, конические или чашкообразные тела, имеющие центральную полость и покрытые на наружной, а иногда и на, внутренней поверхности ромбическими табличками, расположенными косыми рядами. Эти таблички пред
ставляют расширенные концы двух лучей, лежащих нс одной оси, нормальной к стенке тела; четыре остальных луча, лежащие в одной плоскости,
Фиг. 64. А. Нижняя часть Receptaculites orbis. Нижний силур (глауконитовый известняк). Швеция. В. Часть стенки В. Neptuni в меридиональном разрезе. С. Часть выветрелой наружной (верхней) поверхности. В. В eptuni. Средний девон Эйфель. Рейнская Пруссия. Германия. 5/1. В. Polygonosphaerites tesselatus. Средний девой. Нассау. Германия. Вид тангенциальных разветвлений лучей совнутри, с нанесенными жирным черным очертаниями снаружи прилегающих табличек. 8/1.
d — луч, обращенный к нижнему полюсу; рг— противоположный ему луч; I—концевые части боковых лучен соседних щитмков • spl — осевое тельце бокового луча близ центра щитика; ы — налегание наружных щитиков двух соседних скелетных элементов. 1/1.
отходят от этой оси под прямыми углами близ наружной поверхности тела. Самая принадлежность рецептакулитов е губкам спорна.
КЛАСС Ш. ИЗВЕСТКОВЫЕ ГУБКИ, CALCISPONGIA 1
Calcispongia представляют скелет из элементов, - связанных между собою лишь в исключительных случаях, типично же свободных. Скелетные элементы состоят из углекислого кальция, бывают одноосные, трех- или четырехлучевые. Спонгина никогда не бывает. К этой группе принадлежит род Stellispongia (фиг. 55; Stella - звезда).
1 Calx—известь.
Характеристики групп губок.
I класс: Demospongia. Разнообразные по наружной форме, большею частью толстостенные формы с четырех-или одноосевыми кремнеземистыми иглами, или со скелетом из кремнезема и спонгина, или только спонгина, н наконец бесскелетные. От кембрия доныне.
1 порядок: Tetractinellida. Разнообразны пб форме; от кембрия доныне.
2 порядок: Monactinellida. Скелет из кремнезема или и из спонгина. Большею частью обитатели морского мелководья, встречаются и в пресной воде. От кембрия доныне.
3 порядок: Lithistida. Скелетные элементы неправильной формы с разветвлениями на концах лучей, сцепление которых сообщает особенную прочность форме н способность хорошо сохраняться в ископаемом, состоянии. С докембрия доныне.
4 порядок: Ceraospongia. Со скелетом из спонгина. Морские, большею частью мелководные. В ископаемом состоянии достоверно неизвестны.
5 порядок: Myxospongia. Бесскелетны. В ископаемом состоянии неизвестны.
II класс: Hexactinellida. Большею частью тонкостенные, часто крупные и весьма правильной формы обитатели спокойных вод, с правильными трехосными иглами из кремнезема; иногда в скелете имеются н волокна спонгина. С кембрия доныне.
III класс: Calcispongia. Известковые губки. Большею частью небольшой величины; с девона доныне.
Распространение губок.
Порядки, семейства и даже, как правило, роды губок отличаются космополитизмом в своем географическом распространении. Классы губок более отличаются их батиметрическим (по степени глубины моря) распределением, чем географическим распространением. Так Calcispongia живут в прибрежной полосе, в границах отлива и прилива, губки из класса Demospongiae с прочным скелетом из разветвленных и связанных скелетных элементов характеристичны для глубин оТ 200 до 300 м тогда как Hexactinellida в настоящее время встречаются в глубоких водах, притом имеющие прочный скелет—близ берега, на меньшей глубине, a Hexactinellida со свободными скелетными элементами—на больших глубинах, вдали от ' берега. Древние формы Hexactinellida были мелководны. Глубоководные губки отличаются правильностью формы тела. В ископаемом состоянии, конечно преимущественно встречаются губки с прочным скелетом. В палеозое, до каменноугольных отложений, губки сравнительно редки, хотя известны уже из кембрия (Hexactinellida, Demospongiae). В силуре также встречаются Hexactinellida, — группы последних, отличающиеся оригинальностью, к которым принадлежат Astraeospongia, рецептакулиты; Demospongiae (Aulo-copium, Astylospongia). С девона изредка попадаются Calcispongia. В каменноугольных и перми можно указать Pemmatites. Губки становятся весьма многочисленны в юре и мелу (Jerea, Ventriculites').
Ill ТИП. КИШЕЧНОПОЛОСТНЫЕ, COELENTERATA
Животные с лучистой симметрией тела, имеющие одну внутреннюю полость (coelenteron), наподобие мешка открывающуюся наружу одним отверстием, служащим для принятия пищи и для выведения наружу ее непереваренных остатков. Стенка тела состоит из эктодермы и энтодермы, между которыми находится слой бесструктурного вещества, в виде тонкой кутикулярной пластинки, или же студенистый слой значительной толщины (у медуз). В эктодерме находятся особые стрекательные клетки, при своем сокращении выбрасывающие полую заостренную нить, через которую выли-62
вается из клетки жгучая жидкость,- парализующая добычу или прогоняющая неприятеля. Кишечнополостные разделяются на четыре класса: Hydrozoa* Scyphozoa, Anthozoa и Cumophora.
Последние, стоящие особняком, по своей организации—планктонные животные, в ископаемом состоянии не встречаются.
КЛАСС I. ГИДРОМЕДУЗЫ HYDROZOA ИЛИ HYDROMEDUSAE.
Основною формою тела животных этого класса является форма тела полипа пли гидранта (фиг. 65, A; hydra—полип). Полип прикрепляется одним концом своего мешкообразного тела, тогда как на другом, верхнем конце, находится ротовое отверстие, окруженное рядом отростков, называемых щупальцами. Полость тела продолжается в эти отростки. Все тело очень сократимо и постоянно изменяет форму. Здесь нет еще однако специальных мускульных клеток, являющихся у высших животных; здесь цилиндрические клетки эктодермального и энтодермального эпителиев в своих частях, обращенных к промежуточной опорной пластинке, дают корневидные отростки, обладающие сильною сократимостью. Отростки клеток одного слоя расположены под прямым углом к самым клеткам и параллельно продольной оси тела, отростки клеток другого слоя — в поперечном направлении. При сокращении первых уменьшается высота тела, но оно расширяется; при сокращении последних происходит обратное. Отличием от высших животных является еще то, что слой мускулистых отростков распространен по всему телу, не имея местного характера — характера органа, причем этот слой не служит для выполнения лишь совершенно определенных движений, как это-происходит при передвижении других животных этого класса — свободноплавающих медуз; у полипов слой мускульных отростков, сокращаясь то в одних, то в других местах, производит различные движения тела.
У полипов появляются уже особые разветвленные чувствительные клетки—нервные. Дыхание происходит через посредство обоих эпителиев.
Животные класса гидромедуз почти всегда колониальны, и в отношении колоний говорят о составляющих их полипоидных и медузоидных особях, в отличие от свободно живущих полипов и медуз классов Anthozoa и Scyphozoa.
Медузопд по внешему своему виду представляет полушаровидное тело с трубчатым хоботком, находящимся, подобно языку колокола, в центре вогнутой стороны тела (фиг. 65, С, D). Отверстие этого хоботка является ротовым.
Как ни отличается на первый взгляд медузоид от полипоида, первый является по своему строению производным от последнего (фиг. 65) и даже легко приводится к его типу. Надо представить себе лишь, что поверхность полипоида между ртом и щупальцами впячивается вовнутрь, приближаясь к стенке противоположной части тела, что в последний, главным образом при выпуклой поверхности, утолщается опорная пластинка, превращаясь в массивный студенистый слой. Медузы описанного строения по крайней мере часть жизни проводят свободно плавая. Плавание происходит толчками, вследствие выжимания из колокола воды. Понятно, что при движении медуза обращена выпуклой поверхностью в сторону движения.
Медуза представляет различные новообразования сравнительно с полипом. В связи с указанным образом движения медузы мускулатура лбкализирова-
на: на выпуклой поверхности тела мускулы вполне отсутствуют, на вогнутой же находится широкое мускульное кольцо, при сокращении которого уменьшается полость колокола под его вогнутой поверхностью, а при уменьшении этой полости часть воды, в ней находившейся, выбрасывается наружу, вследствие чего животное получает толчок и двигается. Подобным образом.
Оиг. 65. Схема происхождения медузоида от гидранта.
ect.—эктодерма, обозначена точками; end—энтодерма, штрихи ми* ei>t. cat.—пищеварительная полость; ms — студенистая опорная пластинка, черная; Л — гидра«та в вертикальном разрезе; С—медузоид в вертикальном разрезе и D — в горизонтальном по линии аЪ; В — промежуточная между А и В форма; mth— poi; I — щу-пальцы; hyp — околоротовая площадка; тппЬ — хоботок; rad — радиальные^ каналы;
cir—кольцевой капал; г—velum; nv, nv'—двойное нервное кольцо.
нервные клетки сосредоточиваются на краю колокола, и отсюда присходит иннервация, во-первых, мускулов, а кроме того и особых органов чувств, помещающихся на краю колокола. Это или органы зрения — глаза, или статоцисты — органы, при посредстве которых животное ориентируется относительно положения своего тела в пространстве, органы чувства равновесия, Статоцисты представляют открытые или закрытые сумки с заключенными в них известковистыми конкрециями, производящими различное
давление на стенки сумки при различных положениях тела животного. Глаза состоят из чувствительной сетчатой оболочки, к которой примыкает прозрачная чечевица, представляющая местное утолщение кутикулы.
Наконец, для медуз, принадлежащих к классу гидромедуз, характерною особенностью является кольцевая кайма (velum—фиг. 65, С), расположенная на краю колокола, совнутри его. Размножение гидромедуз также представляет особенности, характеризующие этот класс животных: из двух главных форм индивидуумов—полип размножается бесполым путем, дает путем почкованья медуз, медузы же развивают половые продукты, и из оплодотворенной яйцевой клетки медузы получается полип. Это — так называемое размножение чередующимися поколениями.
Порядки гидромедуз следующие: Hydraria, Hydrocoralllna, Tabularia, Саш-panularia. К гидромедузам относя? также и Graptolithoidea.
ПОРЯДОК I. ГИДРОИДНЫЕ ПОЛИПЫ, HYDRARIA.
Одиночные животные или колонии с размножением чередующимися поколениями. В состав колонии входят или только полипоиды и медузоиды,
или еще другие индивидуумы, развивающиеся из полипоидов, выполняющие специально функции схватывания добычи или обладающие высокою чувствительностью и т. п.
Колониальные формы чаще всего растениевидны, кустообразны (фиг. 66). Стеблеобразные разветвления колонии имеют по оси каналы, посредством которых устанавливается сообщение, передача питательных соков между различными частями колонии. Колония прикрепляется к посторонним предметам горизонтальным сплетением ветвей в основании колонии. Стенки ветвей колонии состоят из тех же слоев клеток, как и тело отдельных особей. Одиночные формы Hydraria не имеют твердых частей, а колониальные выделяют наружный скелет из хитина или углекислого кальция. У кустообразновет-вящихся колоний скелет образует тонкую хитиновую трубчатую оболочку снаружи ветвей и стволов; эта оболочка у одних гидроидов оканчивается при основании отдельных индивидуумов, у других же образует чашечкообразное вместилище для полипов, теку. Оболочка стеблей называется
Фиг. 66. Часть колонии Obeha geJatinosa
р — полип с сжа!ЫМ1 щупальцами; рх — с рас простерты я и, w — медузоидная ссобь в го потеке gi th — тека.
65
5 Учебник иалевптологии.
перидерм, а оболочка особей-—гидротека; в случае существования гидротек медузоиды также защищены хитиновыми капсулами, так называемыми гопотеками (половые особи называются гонофорами). Различные гидроиды представляют различные градации редуцирования медузоид-ных особей.
В кустообразных колониях полипоиды прикреплены посредством стебельков или без них, сидят рядами, вплотную один к другому или с промежутками, с верхней стороны ветвей или с нцжней.
Кроме кустообразной разветвленности, колонии гидроидов,представляют и другие формы.
ПОРЯДОК II. TABULAR! А
Так род Tlydractinia (фиг. 67) образует скелет в виде корки на раковинах улиткообразных моллюсков. Эта корка бывает хитиновою или известко
вистою и состоит из пластинок, параллельных поверхности обрастаемой раковины, лежащих одна над другой; пластинки соединены между
Фиг. 67. Ilydraclinia.
Ныне живущая.
Фиг. 68. Millepora nodosa. Ныне живущая.
А — наружная поверхность скелета: В—перпендикулярный разрез; к — гастропоры; с —дактилопоры
собою перемычками, перпендикулярными к пх поверхности, так что между пластинками существует свободное пространство. На поверхности поднимаются полые шипы или бугорки различной величины, и к ней же примыкает отлагающее скелет мягкое основание колонии, от которого поднимаются вверх на стебельках различные индивидуумы, не имеющие твердой оболочки. Пластинки пронизаны каналами, приводящими в сообщение межпластиночные пространства Род Ilydr actinia встречается в ископаемом состоянии в меловых и Третичных отложениях; живет и в настоящее время.
Скелет гидрокораллов известковист и имеет форму масс, облекающих посторонние предметы или ветвистых. Он состоит из сплетения палочек (фиг. 68), в которых находятся открывающиеся на. поверхности трубчатые углубления двух родов: более крупные называются гастропорами и имеют горизонтальные перегородки, мелкие называются д а к т и л о п о ра м и. Скелет отлагается сплетением ветвей мягкого тела, последнее обособляется еще в поверхностный слой. Полипы возвышаются над этим слоем и бывают двух родов: питающие колонию индивидуумы, или гастрозооиды, поме-
дающиеся в гастропорах, и дактилозооиды, занимающиеся ловлей добычи, снабженные в изобилии стрекательными органами, помещающиеся в дакти шпорах.
ПОРЯДОК III. HYDROCO!!AL!JNA.
Millepora1 является ныне живущим представителем гидрокораллов; принимает большое участие в образовании коралловых рифов. Б ископаемом состоянии встречается, хотя и редко, в третичных отложениях. Есть вымершие
Фиг. 69. Actinostroma intertexlum. Уинлок. Англия. 12/1.
4—«пФ параллельно верхней поверхности, представляет вертикальные столбики и радиально расходящиеся от них гориэентальпые ветви; В— вертикальный разрез.
Фиг. 70. Stromatopora tuber-culata. Девон. Онтарио. Канада. 1/1.
Фиг. 71. Stromatopora centrotum. Вертикальное и тангенциальное сечеиие. Кижн девон. Сев. Америка. Увелич. в 10 раз.
Фиг. 70-А. Stromatopora. Поперечный (вертикальный) излом. Девон. Р. Ухта. Обл. Коми.
ископаемые группы, которые с известной вероятностью могут быть сопоставлены с гидрактиниями и гидрокораллами.
С гидрактиниями сходна полеозойская группа Stromatoporoldea. Известковистые скелеты строматопороид весьма различны по форме; они могут быть полушаровидны или шарообразны, древовидно разветвлены, могут неправильно облекать в виде корки различные предметы; часто представляют значительного размера массы. Скелет состоит из ряда параллельных между собою пластинок, разделенных промежуточными пространствами; пластинки пересечены под прямым углом столбиками, дающими мутовчато, через известные промежутки, ветки; эти ветви, соединяясь между собою, с^ствен-
*'Mills— тысяча, poros—пора.
но и образ) ют пластинки. Нижняя поверхность скелета часто покрыта тонким,- концентрически морщинисты я, лишенным пор покровом. Поверхность пластинок неровная, и на верхней стороне их у некоторых форм находятся неглубокие бороздки, образующие звездчатые фигуры.
Actinostroma (фиг. 69; actis — луч, stroma — слой). Вертикальные (или радиальные — при шаровидных колониях) столбики расположены через довольно равномерные промежутки один от другого, пересекаю?
все или часть пластинок, и потому в вертикальном разрезе представляют сеть с четырехугольными петлями. Пластинки пористы, состоя из сплетения ветвей столбиков, и на их поверхности концы вертикальных столбиков выдаются в виде бугорков. Очень часто в девоне, редко в верхнем силуре.
Stromatopora (фиг. 69, 70, 70-А, 71; stroma—слой, рогов—пора). В разрезе столбики и пластинки трудно различаются, сливаясь в сетку с мелкими
Фиг. 72. ilalacoslroma timanicum. Каменноугольные С. Серегово. Р. Вымь.
А Обл. Коми.
а — выветрелая поверхность в поперечном изломе Натуральная величина. Ъ~- горизонтальный разрез, увел, в 5 раз. с — вертикальный разрез, увелич. в 5 раз
петлями. В этой основной массе имеются трубчатые цилиндрические углубления, с днищами, вероятно, вмещавшие полипов; эти углубления все одинаковой величины.
Часто в девоне, реже в верхнем силуре. Actinostroma более сходна с Hydractinia, a btromatopora с Millepora. Stromatoporoidea иногда почти сплошь образуют слои древнепалеозоиских известняков.
Неясно систематическое положение следующих форм, близких к гидроидным полипам. В нижнекаменноугольных отложениях Урала и Зап. Европы встречаются так называемые Spongiosiromidae, об организации которых дает представление род Malacostroma (фиг. 72,; malacos—мягкий, ostracon—рако-68
винарСкелет его состоит из вертикально чередующихся более плотных (темных) и менее плотных слоев, без вертикальных столбиков. Сложение тех
и других слоев зернистое и с каналами большего и меньшего размера (каналы колониальные и каналы ткани).
Еще более примыкает к гидрак-тпниям род Umlotimania, слагающий целые рифы на Урале и Тимане в верхнекаменноугольных отложениях. Полипияки (фиг. 73) его образуют корковидные пластинки, до 2/3 см толщиною. Эти корки состоят из толстых вертикальных пластин, местами соединенных между собою перемычками (фиг. 73, &). Корка плотна у нижней поверхности, наиболее ячеиста у верхней поверхности, закрытой тонким покровным слоем с бугорчатой поверхностью и с отверстиями между бугорками, ведущими во внутренние пустоты корки.
а
Фиг. 73. Vuilotimanla reticulata. Верхнекаменноугольные отложения. Дыроватый Камень. Луньевка. Сев. Храл.
а — вертикальный рнзреа породы с заключенными в ней пласт ивкамн UralotTtnanta. Натуральная величина. Ъ—горизонтальный разрез, увел, в 2*/а раза
ГР А ПТОЛИТЫ, GRAPIOLITHOIDEA.* 2
Вымершая нижнепалеозойская группа. Встречаются обыкновенно обособленные ветви колоний. Ветви всегда представляют трубчатый канал, соединительный для ячеек отдельных полипов, примыкающих к нему, располагаясь в ряд вдоль него. Независимо от канала бывает полая или сплошная прутообразная ось (virgula), служащая опорою ветви. У простейших форм ось находится с одной стороны канала, а ряд ячеек с противоположной стороны его. Ячейки открываются не только вовнутрь, в канал, по и наружу, устьями на дистальном конце.3 Ячейки расположены или в непрерывном ряду (Monograptus, фиг. 74; monos—один), непрерывно примыкая одни к другим, или разделены промежутками (Rastrites, фиг. 75; rastrites—скребок).
У более сложных форм полипняк (polyparium) представляет расположение ячеек в два ряда, с противоположных стофон канала (Diplograptus, фиг. 78; diplous — двойной) или даже в четыре ряда (Phyllvgrapttis, фиг 76; phyllon — лист); в таких случаях virgula занимает центральное между рядами положение. Скелет граптолитов состоял из органического, хитинового вещества, подобного роговому, зачастую обугливающегося при сохранении в ископаемом состоянии.
В индивидуальном развитии граптолитов наиболее раннею известною стадиею является так называемая ножка (sicula). Это образование (фиг. 77, А) it
1 Существует мнение, что скелет Malacoslroma образуется отложением извести микроско-ьическими водорослями.
2 Graptos—писаный, lithos—камень.
2 Дистальный — находящийся наиболее далеко от центра тела или шеста прикрепления, противоположное понятие— проксимальный.
. ь?
(
представляет полый конус, открытый во всем основании. Simula уже имеет virgula. Первые ячейки образуются от sicula с одной стороны ее или попеременно с двух противоположных сторон. Обыкновенно рост sicula прекращается, как только образовались первые ячейки, и иногда она со
вершенно исчезает вслед за тем.
Не так давно были найдены экземпляры хорошо сохраненных колоний
Diplograptus (фиг. 78). Ветви,
Фиг. 74. JUonograptus priodon. Силур. Окр. Праги. Чехо-Словакия.
в— в натуральную величину; b •— увеличенный яродольный разрез; с —канал; д — стержень; th — ячейка (thiea); а? —наружное отверстие ячейки; с — вид со стороны, протинопсложнои ячейкам; d — Mon&graptus lohe-micus.
попадавшиеся до сих пор отдельно, образуют розеткообразные колонии, сходясь своими virgulae в центральной части розетки; здесь virgulae соединяются, и в месте их соединения находится маленькое, почти четырехугольное тело (соединение в розетку с указанным телом в пен-
Фиг. 75. Rastrites Linnei. Верхний силур. Окрестности Праги. Чехо-Словакия.
Фиг. 76. vhyllograptus. Поллпняв разрезан поперек на две части и верхняя часть приподнята, чтобы видеть четыре ряда ячеек. Нижний силур. Англия. 1/1.
тре, скрепляющим virgulae, было ранее известно для некоторых граптоли-тов); это тело окружено группой яйцевидных хитиновых капсул, считаемых за гонофоры, так как они содержат siculae-, siculae иногда выходят из капсул наружу и развиваются в отдельные колонии, но в других случаях они остаются прикрепленными к родительской колонии, развивая в ней отдельные новые ветви. Над центральным телом и гонофорами находится большое пузыреобразное тело, вероятно, наполненное воздухом при жизни животного и служившее для удержания колонии в определенном положении при плавании, подобно тему как это бывает у щзыреносок (сифонофор), одного из порядков ныне живущих гидромедуз.
Некоторые граптолпты были животными прираставшими
Например, в верхнекембрийских, силурийских и девонских отложениях встречаются кустообразно разветвленные колонии, вс'ви которых снабжены ячейками для полипов и соединены поперечными перемычками.
Фиг. 77. А - Diplograptus gracilis, sicula В—sicula с 5-ю первыми гидро-теками, (t1—I6); С -Monogruptus dhbius, sicula и 3 первые гидротеки, сильно увеличено.
Фиг. 78. Diplograptus pristis. Нижний силур. Нью-Йорк. CACHL 2/3.
Dictijonema (dictyon—сеть, пета—нить) образует корзинкоподобную или воронкообразную колонию, прикрепленную, внизу снабженную острием (фиг. 79). Устья ячеек для
полипов находятся на внутренней стороне ветвей. Кембрий, силур, девон.
От гидроидов грапто-литы отличаютсясущество-ванием siculae и virgulae-, хотя они представляют сходство с теми или другими гидроидами по расположению ячеек полипов, по присутствию плавательного пузыря и гонофор, эти сходства могут быть только внешними; в гонофорах, например, были siculae, может быть, не имевшие никакой аналогии с медузами, находящимися в гонофорах ги-дроидсв. Во всяком слу-
Фгг. 79. а — Dictijonema retiformis. Верхний силур. Нью-Йорк. САСШ. Ъ — Dictyonema sp. Веточка с ячейками из валунов Сев. Германии.
Фиг. 30. ВИаЪ-dopleura. Ветка колонии с трубчатым концом (£/.); S'—аналог virgula', th— тека. Увелич. Ныне живущ.
чае, если даже грапто-
литы принадлежали к Coelenterata, приближаясь к гидроидам, они представляют достаточно отличий для выделения в особый вымерший порядок.
Граптолиты встречаются чрезвычайно часто в глинистых сланцах, реже в известняках верхнекембрийских и силурийских отложений. Вымирают в нижнем девоне. Граптолиты имеют широкое географическое распространение, объясняющееся тем, что они передвигались плаванием, и потому имеют большое геологическое значение, представляя хорошие руководящие формы.
Monograptus часто в среднем и в верхнем силуре. Rastrites в верхнем силуре. Diplograptus и Phyllograptus в нижнем силуре.
Граптолиты подразделяются на три группы: прикрепленных Dendroidea и Axonolipa и свободно плавающих планктонных Axonophora. К Dendroidea принадлежит Dictyonema, к Axonolipa—Phyllograptus, к Axonophora—Monograptus, Diplograptus, Rastrites. Примитивнейшими считаются Dendroidea, встречающиеся уже в верхнем кембрии. Из них могли выйти Axonolipa, лишенные оси и встречающиеся главным образом в нижнем силуре. Из Axonolipa вышли имеющие распространение главным образом в верхнем силуре Axonophora, снабженные осью.
За последнее же время найдены точки близкого соприкосновения граптолитов с прирастающими колониями так называемых кишечно-жаберных червей (род Rhabdopleura). У Rhabdopleura индивидуумы колонии малы и, так же как у Monograptus, расположены в прямолинейный ряд, одной стенкой прирастающий; в этой стенке находится трубчатое образование, совершенно аналогичное virgula (фиг. 80).
Строение стенок ствола колонии также аналогично, наконец дистальный конец колонии представляет трубчатое образование, несколько сходное с sie.ula и содержащее зародыши новых особей, обособляющихся почкованием.
В отношении строения мягкого тела кишечио-жаберные черви принадлежат к высшим беспозвоночным животным и совершенно отличны от кишечнополостных. Таким образом граптолиты являются одною из вымерших групп, об организации которых мы можем лишь догадываться с некоторою вероятностью, исходя от организации сколько-нибудь сходных ныне живущих животных.
КЛАСС II. МЕДУЗЫ SCYPHOZOA или ACALEPHAE. 1
Животные, достигающие значительной величины, одиночные (они-то и известны в общежитии под названием медуз). По сравнению с медузоид-ными особями колоний полипомедуз, организация собственно медуз является более сложною. Край зонтикообразного тела рассечен на лопасти, в связи с чем отсутствует velum. Кроме того край колокола иногда снабжен щупальцами. На нижней поверхности колокола находится толстый слой концентрических мускулов, своим сокращением выгоняющих воду из желудочной полости, причем животное получает толчок и передвигается. Края ротового отверстия также рассечены на лопасти, иногда удлиненные, служащие для схватывания добычи. Медузы размножаются чередующимися поколениями. Половое животное—медуза производит полипа; полип существует кратковременно,.уничтожаясь в процессе деления, которое происходит поперек тела полипа и дает медуз.
Тело медуз состоит из бесскелетной студенистой или хрящеподобной массы, содержит в своем составе много воды. Тем не менее, при благоприятных условиях, медузы дают отпечатки в тонкозернистых породах, например в кембрийских сланцах Британской Колумбии в Канаде, в юрском литографском камне Баварии (фиг. 81), в мелу. В кембрийских породах различных стран находятся четырехлучевые (реже пятилучевые) 2 тела, принимаемые за слепки (ядра) желудочной полости медуз (фиг. 82).
1 Scyphos—чаша, zoon—животное, acalephe - крапива.
3 Изменчивость числа лучей существует и у нцце живущих медуз.
КЛАСС Ш. КОРАЛЛОВЫЕ ПОЛИПЫ, ANTHOZOA.1
Коралловые полипы отличаются от гидроидных более сложной) организацией); края ротового отверстия у них имеют продолжение вовнутрь полипа, в виде глоточной трубки, открывающейся нижним концом в coelenteron.
Сильно развиты радиальные выросты
Фиг. 81. Hhizostomites admirandus. Отпечаток. Вад свиву, со стороны четырехлопастно! о хоботка. Пунктиром дополнены недостающие части. Верхняя юра (литографский сланец). Золенгофен. Бавария. Германия. 1/8.
стенки тела—мезентериальные пе- .« регородки, также вдающиеся вовнутрь на некоторое расстояние. Этими перегородками coelenteron разделяется на мезентериальные камеры, сообщающиеся между собою в окол®
Фиг. 82. Medusites Lindstromi. Кембрий. Окрестности Таллина. Эстония. 2/3.
осевой части coelenteron, куда перегородки не достигают. В верхней части полипа перегородки доходят до глоточной трубки, примыкая к ней. Свободные края мезентериальных перегородок извилисты, снабжены стрекательными
органами, железистыми клетками; здесь же развиваются половые клетки. Anthozoa характеризуются отсутствием пер-
вичной радиально-лучевой симметрии, имеющейся у низших Coelenterata; в ранних стадиях развития Anthozoa во всех органах ясно выражена двусторонняя симметрия, в зрелом состоян”и часто маскируемая расположением перегородок, когда она становится радиальнолучевой.
Двусторонняя симметрия прежде всего выражена в ротовом отверстии, постоянно вытянутом
Фиг. 83. Схематический поперечный рагрез полила Zoan-thar<a. Стрелки означают плоскость симметрии. Известковый .скелет показан жирной черной линией.
м — меаситер п; s — серегоролкв; 1 — шесть перегородок, 1 цикла, 2 — шесть перегородок II цикла.
в одном направлении, определяющем плоскость симметрии.
Затем по плану двусторонней симметрии построены мезентериальные складки. Мезентериальные складки покрыты на обеих сторонах мускульными волокнами, на одной стороне складки волокна большею частью образуют утолщения (фиг. 83).
Как видно, даже при строго лучевом расположении перегородок в зрелой стадии коралла мускульные
утолщения перегородок, по своему расположению,
вредставляют двустороннюю симметрию: правая и левая половины фиг. 83 симметричны.
Крупнейшими подразделениями Anthozoa являются Zoanthariaи Alcyonaria.
1 Anthos — цветок.
ПОДКЛАСС I. Zoantharia. *
Zoantharia имеют перегородки в переменном числе, кратном двух. Щупальца простые^ трубочкообразные. У некоторых групп существует скелет, состоящий из игольчатых кристаллов (фибр) аррагонита.
Скелет отлагается наружным слоем мягкой стенки тела, эктодермой, и представляет образование, сходное по форме с нижнею частью мягкого тела животного; скелетная ячейка открыта сверху, и здесь из нее часто выдается верхний конец мягкого мешкообразного тела; в таком случае мягкое тело может вполне втягиваться в ячейку, в углубление ее верхнего конца, называемое чашечкой. Главными составными частями скелета ячейки являются стенка (theca) и перегородки (septae). На фиг. 84 видно соотношение скелетной стенки к мягкой стенке тела; первая образуется в
«Риг. 84. Схематический разрез коралла Mussa. Жирной черной линией показан разрез мягкого тела полипе, В левых двух кораллитах разрез сделан через перегородку, в правом—через межперегородочное пространство.
S — перегородка; isl — интерсептальиая камера; с — costae; id — интеркосталь-ное пространство; ps. th—pseudotheca; ер— эпптека; d— днссепиыенты.
кольцевой складке второй (фиг. 83); в подобных же складках, идущих от стенки тела радиально вовнутрь,' образуются радиальные перегородки; складки, их образующие, лежат посредине мезентериальных камер; пространство между двумя соседними перегородками представляет интерсептальную камеру. Перегородки на краях бывают зубчаты; иногда на внутреннем краю их наверху резко обособляется глубокой выемкой крупный зубец. Такой зубец, если смотреть на чашечку сверху, кажется самостоятельным образованием, независимым от перегородки, хотя и лежит в одной плоскости с нею, и называется свайкой (palus). Перегородки бывают сплошные и пористые. Поверхность перегородок бывает гладкая или неровная—в последнем случае она покрыта струйками, ребрышками, бугорками соответственно характеру эктодермальной поверхности радиальных складок мягко тела. Отложение перегородки происходит слоями, параллельными ее боковым поверхностям, на обеих этих поверхностях. Бугорки на обращенных одна к другой боковых
1 Zoon—животное, anthos—цвето!.
поверхностях смежных перегородок при своем росте и утолщении могут прийти в соприкосновение, образуя при этом п е р е м ыч к и (synapticulae). Иногда бугорки в вертикальных рядах тоже сливаются, образуя вертикальные перемычки. Все эти перемычки скрепляют коралл, так что при сильном развитии их иногда стенка отсутствует. Иногда стенка образована концами перегородок, расширенными на периферии до соприкосновения (ложная стенка, pseudotheca).
В некоторых случаях стенка бывает покрыта снаружи особым концентрически морщинистым известковым покровом, эпитекой. Эпитека образуется так называемой краевой зоной тела полипа (фиг. 84), краем стенки тела, свешивающимся сверху вниз, за скелетную стенку. Эпитека существует и на основании ячейки, и здесь эпитекальный покров представляет даже большее постоянство, чем на боках, иногда он только на основании и имеется, вообще развитие эпитеки идет от основания вверх.
На наружной поверхности стенки находятся иногда ребра, представляющие части перегородок, выдающиеся наружу; такие ребра называются costae. У некоторых кораллов наружные ребра соответствуют промежуткам между перегородками и называются тогда pseudocostae или rugae.
Коралл надстраивает свою ячейку все выше, наращивая стенку и перегородки, причем мягкое тело перемещается снизу вверх. Части ячейки, становящиеся свободными при этом перемещении мягкого тела, заполняются известковистыми образованиями, отлагаемыми основанием мягкого тела полипа' Уже вышеупомянутые перемычки (synapticulae) могут образоваться из противолежащих бугорков лишь тогда, когда мягкое тело будет находиться над бугорками, удалившись из пространства между ними, так что, хотя бугорки представляют части перегородок, образующиеся в радиальных складках мягкого тела, окончательное соединение бугорков в перемычку производится основанием полипа.
Основание полипа отлагает также известковистые листочки, плоские или выпуклые, горизонтальные или косо расположенные по отношению к стенке коралла (фиг. 84). Между этими листочками существуют промежутки, так что совокупность листочков представляет подобие пузырчатой ткани. Эти листочки (disseplmenta) обыкновенно расположены лишь на периферии полипняка, реже заполняют все поперечное сечение (род Cystiphyllum). Основание полипа отлагает также так называемые днища (tabulae), горизонтальные пластинки, плоские, вогнутые или выпуклые, занимающие все поперечное сечение коралла или находящиеся лишь в срединной его части, свободной от перегородок. Последнее скелетное образование, отлагаемое основанием мягкого тела полипа, есть столбик (columella), столбчатый или пластинчатый, расположенный центрально, совпадая с осью коралла. Столбик отлагается центральною частью основания полипа, в таком случае впяченною вовнутрь, так что во впяченности отлагается столбик.
Столбик бывает образован иногда внутренними концами перегородок, соединяющимися по оси коралла. Такой столбик называется ложным, и его надо отличить от настоящего столбика, образованного основанием мягкого тела полипа.
Настоящий столбик бывает или плотный, состоящий из пластинчатых концентрических слоев, или губчатого сложения, или состоит из пучка грифелевидных палочек.
Все скелетные образования, возникающие внутри ячейки коралла, называются эндотекальиыми, так что говорят, например, о пузырчатой эндотеке.
Экзот скальным образованием является цененхима—отложение скелетного вещества вне ячейки, соединяющее иногда ячейки у колониальных кораллов. Цененхима отлагается ценосарком—частями мягкого тела полипов, разрастающимися наружу, за пределы отдельных чашечек.
Микроструктура скелета. Микроскопические исследования тончайшей структуры скелета кораллов, ставшие более частыми за последнее время, с одной стороны, лучше объясняют происхождение различных элементов скелета, а с другой—дают возможность устанавливать наличность различий тогда, когда их трудно найти при исследовании лишь макроскопическом.
Скелет кораллов состоит из последовательно отлагавшихся известковистых слоев или пластинок. Пластинин состоят из фибр аррагонита, направ
Фиг. 85. Перегородка ныне живучего коралла.
i — трабекулы? If ~ ливня соприкосновения тра беку л; Id—линия расхождения трабекул; ер — эпитека, справа нижний обломленный край. 4/1.
Фиг. 86. Поперешое течение верегородок и ложной стенки Hexacoralla. Ныне живущ. *%.
ленных перпендикулярно к отлагавшей их поверхности. Известковистые фибры в перегородках и стенке коралла группируются в элементы высших порядков—в пучки фибр, а пучки—в балочки или трабекулы.
Пучки фибр представляют лучистые и в то же время концентрически слоистые аггрегаты, округленной или неправильной формы (фиг. 88), в несколько десятых долей миллиметра в поперечнике.
Трабекулы имеют вид столбиков, состоящих из ряда пучков фибр (фиг. 85). В перегородках различных кораллов трабекулы расположены различно. Иногда они расходятся веерообразно от некоторой линии, занимающей более или менее срединное положение в перегородке (фиг. 85); в этом случае трабекулы идут вверх от указанной линии, отгибаясь затем с одной стороны ее кнаружи, с другой стороны вовнутрь. Зона, лежащая наружу от линии расхождения, стоит в связи с развитием costae и отсутствует при неразвитии последние. Иногда трабекулы в перегородке бывают горизонтальны, так что верхний край перегородки образован всего одной трабекулой, а внутренний—оконечностями всех их; бывают и иные положения трабекул.
Окончания трабекул на краю перегородки иногда являются в виде зубчиков.
Сплошные и пористые перегородки возникают в зависимости от того, сомкнуты ди соседние пучки фибр совершенно плотно или с промежутками
в перегородках дает возможность точно
)
Фиг. 87. Поперечное сечение перегородок (3-х) и настоящей стенки Hexacoralla. Ныне живущ. 16/1.
Ребрышки и струйки, находящиеся на боковых поверхностях перегородок, представляют, так сказать, выдающиеся на поверхности перегородки бока трабекул. Бугорки на боковых поверхностях перегородок представляют выдающиеся части пучков фибр, так что обыкновенно бугорки группируются в ряды соответственно трабекулам, слагаемым этими пучками.
Центры пучков фибр часто имеют значительно более темный цвет, чем остальные части (центры находятся в несколько иных условиях отложения известковистого вещества, соответствуя началу процесса отложения); поэтому и осп трабекул, состоя из этих центров, лежащих близко один от другого, являются темноокрашенными; также темно окрашены и средние линии перегородок в поперечном разрезе коралла. В некоторых случаях радиальное расположение фибр относительно центров стушевывается, сохраняется лишь перпендикулярное их положение по отношению к боковым поверхностям перегородок; тогда темные линии перегородок являются совершенно непрерывными (фиг. 87). .
Фиг. 88. Поперечное сечение перегородки НехасогаПа. Нынеживущ. Увелич. ffl— линия нарастания; сс—центры пучков фибр.
Существование темных линий
различить их в разрезах кораллов, отличать от днищ, лежащих иногда почти диаметрально, различать разрезы диссепиментов, примыкающих иногда к концам перегородок, представляя как бы непрерывное продолжение последних. Диссепименты, днища—вообще скелетные образования, отлагаемые основанием мягкого тела полипа и утолщающиеся отложением вещества лишь с одной стороны,—состоят просто из слоев фибр, не представляя пучков их и трабекул, а следовательно и темных линий (фиг. 89).
Подобно перегородкам построена и скелетная стенка кораллов. Понятно, что ложная стенка (фиг. 86), состоя из расширенных периферических концов перегородок, является сложенною из трабекул. Настоящая стенка, в которой между периферическими концами перегородок введены независимые от
перегородок участки, отделенные от перегородок швами и представляющие самостоятельные центры отложения известковистого вещества, тоже состоит из трабекул (фиг. 87). Швы и расположение центров дают возможность раз-
личать под микроскопом ложную стенку от настоящей.
Размножение коралловых полипов. Полипы в своем развитии исходят от свободно плавающей личинки, развивающейся половым путем гермафродитно. Личинка прикрепляется и
развивается в зрелое животное, причем полип или остается одиночным и размножается далее лишь половым путем, или путем почкования (иногда деления) развивается целая колония. При почковании дочерние полипы обособляются иногда с боковых стенок родительского организма, от эктодермы, причем возникает древовидно ветвящаяся колония (фиг. 90, Л); такой способ почкования называется боковым. В других случаях почки развиваются на
Фиг. 89. Сечение диссепимента Galaxea. д — пластинки нарастания. Увелич.
Г ?
Фиг. 90. Cyaihophyllvm arliculatu.n. Почкование верхушечное (краевое) в Б, боковое в >1. Верхний силур. Канада.
верху полипа, в его чашечке; такое почкование называется верхушечным (фиг. 90, В). Иногда почки развиваются на трубчатых выростах основания полипа, хотя такой способ в рассматриваемой группе встречается редко, являясь, напротив, очень частым у Alcyonaria. Встречается и размножение делением, причем новообразующиеся полипы или совершенно отделяются от родительского организма, или остаются в соединении с ним, образуя колонии. Если отдельные индивидуумы колонии (кораллиты) обособлены друг от друга на значительном протяжении, то колония получается древовидная; если же они соединены между собою на всем протяжении стенок, то полипняк колонии является массивным. Иногда, при неполном делении, индивидуумы колоний образуют ряды с сообщающимися чашечками, число которых не может быть определено (Leptoria) вследствие недостаточной их обособленности.
Zoantharia разделяются на несколько порядков, из которых порядки актиний не образуют скедета; палеонтологический интерес представляют образующие скелет кораллы Rugosa и Hexacoralla.
ПОРЯДОК I. RUGOSA (TETRACORAL1A).
Вымершая палеозойская группа. Симметрия коралла определяется четырьмя первичными перегородками (protosepta), по отношению к которым последующие перегородки расположены характеристичным образом. Первичные перегородки доходят до нижнего, заостренного конца полипа. Вторичные перегородки (metasepta) развиваются последовательно с ростом коралла, начинаясь на все бьлее вышележащем уровне в полипняке (фиг. 91). Эти перегородки возникают парами, члены которых лежат по обе стороны плоскости
симметрии. Боковая поверхность покрыта rugae (ruga—морщина, от этого rugosus—морщинистый); часто развита эпитека. Из первичных перегородок две, неодинаковые, лежат в плоскссти симметрии, а две другие, одинаковые, расположены по обе стороны этой плоскости. Двусторонняя симметрия иногда от"етлпво выражена в отверстии чашечки зрелого коралла, иногда же перегородки здесь расположены радиально; это расположение называется псевдорадиальным, так как получается лишь вследствие известного роста в устье, в остальной части ячейки расположение остается строго двустороннесимметричным. Во многих случаях, при слабом развитии эпитеки, расположение перегородок отчетливо выражено на боках полипняка; именно о нем дают представление rugae, как известно, чередующиеся с перегородками, расположенными внутри ячейки. Как видно на фигуре 91, в двух из четырех систем вторичных перегородок они расположены перисто по отношению к симметрично разделяющей их первичной перегородке, называемой главной Расположение перегородок в двух остальных квадра нт ах
Фиг. 91. Полипняк Streptelasma сбоку; слева—главная, справа—боковая перегородка.
Фиг. 92. Вид полипняка ео стороны нижнего заостренного-конца.
видно лучше на фиг. 92, представляющей вид коралла с его заостренного конца. Здесь видно, что в главных квадрантах, т. е. прилегающих к главной первичной перегородке (/г), вторичные перегородки расположены перисто относительно нее и—смежные, по крайней мере,—параллельно относительно первичных перегородок, называемых боковыми (S).
В двух остальных квадрантах, разделенных так называемой противоположной первичной перегородкой, смежные с ней вторичные перегородки расположены параллельно по отношению к ней; в этих квадрантах, кроме того, вторичные перегородки * отходят перисто от боковых первичных. Это расположение вторичных перепородок Rugosa по отношению к первичным чрезвычайно характерно, дает возможность всегда отличить представителей Rugosa от кораллов других групп. В случае, если бока коралла закрыты эпитекой, расположение перегородок можно обнаружить, удалив эпитеку шлифованием или растворением в кислоте.
Кроме указанных первичных и вторичных перегородок у Rugosa. в самой верхней части полипняка бывают еще перегородки третьего порядка,
короткие и разом появляющиеся во всех интерсептальных камерах, образованных первичными и вторичными перегородками. Что касается до главной первичной перегородки, она бывает иногда очень коротка и лежит в так называемом септальном углублении (fossula), резко выделяясь в отвер-
стии чашечки (фиг. 93). Главная перегородка располагается обыкновенно на выпуклой стороне коралла, изредка на вогнутой стороне
Бывают fossulae и при боковых перегородках, но более редко, нел.ели ш при главной и еще реже при противоположной. Fossulae. образуются,
Фиг. 94. Схема развития перегорел--', у Rugosa. 81—боковая перегородка; аа, cd — устьевые края.
Фиг. 93. Zaphrentis еогпжоргае. Нижнекаменн. Турне. Бельгия. Увелич. Fossula с главной перегородкой вертикальны.
"4
Фиг. %. Четыре стадии развития перегородок у кораллов Rugosa (схема). Первая слева, лишь с первичными перегородками.
5—боковая перегородка; Slt Ss — вторичные neperoj едки
вероятно, потому, что при первичных перегородках интерсептальные камеры бывают иногда шире, чем смежные (фиг. 94;. Мягкое тело, опускаясь в эти уширенные пространства, мешает развитию первичной перегородки до нормального размера. Возможно, что fossulae. служили направляющими желобками для движения воды ко рту и от него
Рассмотрим вопрос о причинах развития характеристичного для Rugosa расположения mctascpta в отношении prolosepta.
Эти причины определяются следующим образом. Определяющими моментами в развитии типичных Rucosa можно считать согнутое состояние их полипняка и механические условия развития, создаваемые этой вогнутостью.
Устье конического полипняка Rugosa обыкновенно перпендикулярно к вогнутой стороне полипняка, как это изображено на фиг. 94.
Характеристичное для Rugosa расположение вторичных перегородок (перистое в огно-шеяии главной перегородки и параллельные в отношении противоположной) необходимо является потому, что вторичные перегородки имеют наклонность расти, как и вообще пе
регородки у Anthozoa верпендик' ляряо к плоскости устья чашечки. При наличности в со путем водипняке перегородок вторичные перегородки могут расти указанным обра аом беспрепятственно лишь в двух квадрантах из четырех, разграниченных первичными перегородками. Именно на фиг 94— 95 это будет в квадранте, расположенном со стороны вогнутой поверхности первичной боковой перегородки (8). В квадрантах, расположенных со стороны выпуклой поверхности боковых перегородок, вторичные перегородки не могут расти перпендикулярно к устью чашечки вследствие того, что но крайней мере в части, близлежащей к выпуклой поверхности боковой первичной перегородки, она своею выпуклостью будет препятствовать вторичным перегородкам расти указанным образом. Поэтому в квадрантах, лежащих с выпуклой стороны боковых первичных перегородок, вторичные перегородки развиваются не перпендикулярно к краю чашечки,а иначе— именно, как наблюдается параллельно этой выпуклой стороне.
При таком характере развития перегородок они располагаются на вогнутой стороне кораллу параллельно находящейся здесь первичной перегородке, так как она тоже пер-
пендикулярна к устью чашечки; эта первичная перегородка является, следовательно, так называемою противоположною
На выпуклой стороне коралла новые вторичные перегородки, смежные с находящейся здесь первичной, отходят от нее под углом вверх. Это будет главная первичная перегородка.
Фиг. 9в. Strepti last.ria europaewm из валунов Силезии. 1/2.
а — экземпляр 9 выдох лен пою частью края чашечки; Ъ — экземпляр, разрезанный вдоль посередине.
Фйг. 97. Strepl 'lasma robustum Половина поперечного сечения. Силур. Канада.
Что касается до боковой первичной перегородки ($), то смежные с нею вторичные перегородки расположены параллельно по отношению к ней в главном квадранте, а в противоположном отхидят под углом вверх.
Характерными эндотекальными образованиями являются днища и пузырчатая эндотека. Rugosa размножаются половым путем и почкованием, но не делением. Для них именно характерно вну гричашечное верхушечное почкование.
Первичными Rugosa надо считать одиночные кораллы с согнутым по-липнякич, с двусторонней симметрией как в молодых стадиях, так и в зрелом состоянии.
К роду Streptelasma (силур; streptos— скрученный, elasma—пластинка' принадлежат (фиг. 96—97) одиночные, несколько согнутые, конические кораллы с весьма многочисленными (82—130), постоянно двусторонне-симметрично расположенными перегородками, из которых более длинные изгн баются, переплетаясь около оси. Имеются пузырчатая эндотека и многочисленные сближенные горизонтальные днища.
Характеристичное расположение перегородок отчетливо выражено на боковой поверхности полипняка.
Роды Zaplirentis и Amplexus (силур — каменноугольные отложения, особенно последние; za—очень, plirentis—перегородка, amplexus—окруженный)
характеризуются присутствием fossula у главной перегородки. У обоих развиты днища; у Zaphrentis хотя и слабо—пузырчатая эндотек?. У Zaph-
rentis полипняк конический, с длинными перегородками (фиг 93, 99),
Фиг. 98. Amplexus coralloides. Два эк-вемпляра, из Авглии (верт. разрез) и Бельгии. Нижвецамев-воуг. 1/2.
Фиг. 99. Zaphrentis Phillipsi. Нижвека-меввоуг. Бельгия.
1/1-
у Amplexus, вследствие малого угла конусности,—почти цилиндрический, с короткими перегородками (фиг. 98)
У следующей группы кораллов, примыкающей к рассмотренной, в центральной части полипняка развиты днища, к периферии переходящие в пузырчатую эндотеку.
Из них—CyaHiophylium (cyat-hos — чаша, phyllon — лист), к которому принадлежат кораллы одиночные и колониальные, иногда с ложным столбиком (фиг. 100 — 102). У оди-
Фиг. 100. Cyathophyllum-hexagonwm. С] еднвй левов. Геролыптейв. Эйфель. Германия 1/2.
ночных кораллов с хорошо развитыми перегородками почти всегда имеется более или менее развитая fossula, у колониальных она отсутствует. Силур— каменноугольные; особенно в девоне.
Фиг. 101. Cyat-hophyllwn ceratites. Средний девон. С. Ку-; иново. Ливенского у., Орловской губ. 1/1.
ствуют в начальных
Фиг. 102. Cyathophy tam caespitosum Разрезы Средний девон. Эйфель. Германия. 2/1.
Род Сапг'пга (фиг. 109) отличается тем, что имеет утолщзния перегородок в главных квадрантах. Эти утолщения стереоплазмой—плотным веществом, находятся в средней части полипняка, не достигая периферии и не доходя также до центра.
и
Имеется /ossuia и днища, выпуклые вверх. Утолщения перегородок отсут-в конечных стадиях полипняка. Каменноугольные
отложения.
Lithostrotion (фиг. 105; litlios—камень, strotion—балка), сложный полипняк; ячейки имеют настоящий грифелевидный столбик. Каменноуголь-
ные отложения.
Роды Clisiophyllum (фиг. 107; elision—палатка), Lonsdaleia (фиг. 103— 104; по имени ученого Лонсделя) и Dibunophgllum (фиг. 108; dis—два, buEOS—холм, phyllon—лист) принадлежат к одной группе, отличающейся присутствием ложного столбика, образованного не только закручивающимися в центре полипняка концами перегородок, но и выпяченными в центре
кверху днищами.
Фиг. 103. Lonsdaleia floriformis. Нижнёкаменвоуг. Подмосковный бассейн. 3/1.
Фиг. 104. Lonsdaleia Bi onni. Верхнекаменноуг. Подольск, Московская обл.
1/1.
Фиг. 105. Lilhosi-rotion basal titorme. Вид сверху Нижнекаменн. Бристоль. Англия.
У Lonsdaleia происходит сокращение перегородок, отсутствующих у наружной стенки; пузырьки s
Фиг. 10G. Cystiphyllum vesieulosum Разрезанный вертикально Средний девон. Хермания. 1/1.
эндотеки располагаются так, что образуется как бы внутренняя стенка; у Dibunophyllum (фиг. 1С8) столбик во всю свою
толщину пересекается про-
Фиг. 108. Dibunophyllum tarbinatum. Разрез. Нижие-каменноугольвые. Южн. Урал.
2/1.
Фиг. 107. CliswphyPwm densi amellatum. Разрез. Нчжнекгмеиноугольные.
Аганово. Южн. Урал. 2/1.
должением одной из первичных перегородок. Все три рода встречаются только в каменноугольных отложениях.
Род Gystipliyllum (фиг. 106)—одиночные кораллы—отличается заполнением чашечки одной лишь пузырчатой эндотекой; перегородки или вполне отсутствуют, или являются в виде ребер, едва возвышающихся над поверхностью пузырчатой эндотеки на дне чашечки. Силур, девон.
6*
вэ
С другой стороны к Cystiphyllum примыкает группа Rugosa, снабжен-пых крышкой, закрывающей отверстие чашечки; из них Rhlzophyllwin (rhiza—корень, phyllon—лист) и Calceola (туфля)—в виде согнутого полуконуса.
' Bhizophyllum (фиг. 110) представляет некоторую неправильность в форме полипняка, с поверхности которого отходят корневидные отростки.
Фпг. 10'3. Caninia cylin Irica.
Гориво сгальный раз рез. Турнэ. Бельгия. 1/2. ,
Фпг. 111. Calceo'a san palina. Средний девой Эйфель. Германия. 1/1.
Фиг. 112.
Фиг. 110. Rkizophyllwm gotla псИсит.'Вертсняа силур Готланд. Швеция. 1/1.
Goniophylhim pyramidale. Верхний силур. Готланд.
Швеция. 1/1.
Здесь эти отростки служили для почкования и образования колоний. Имеется пузырчатая эндотека и слабо развитые в высоту перегородки. Силур—нижний девон.
У Calceola свободно лежавший на дне морском полипняк имеет правильную полуконическую форму, лишен корневидных отростков (фиг. 111). Средний девон.
Правильность формы полипняка Calceola п прпнлющенаость его с одной стороны невидимому объясняется тем, что это был коралл не прирастающий, но свободно лежащий, именно приплющенной стороной на дне.
(?omop7&yZZim(gonia—угол) имеет прирастающий полипняк в виде четырехгранной пирамиды с крышкой из четырех частей, также пирамидальной (фиг. 112). Силур.
f"'
Геологическое распространение Rug-osa.
Роды
Отделы или ярусы
Streplelasma .... Г
Zaphrentis..........:
Amplexus............
Cyathophyllum . . .
Caninia . .
Lithostretion ...
Clisiophyllum . ...
Lonsdaleia..........
Dibunophyllum ....
Cystiphylluni ....
IlhtsopkyUum ....
Calceola............
Goniophyllum ....
НОРЯДОК II. HEXACORALEA.
Перегородки образуют несколько циклов, т. е. групп, члены каждой йз которых имеют одинаковые размеры, хотя и не всегда развиваются одновременно. Число первичных перегородок шесть (по гречески—hexa): и так как последующие перегородки развиваются во всех интерсептальных камерах, то числа перегородок в последовательных циклах будут 6, 12, 24, 48, и т. д.; число перегородок в каждом цикле равняется, следовательно, еумме перегородок во всех предшествующих циклах, и полное число перегородок является кратным шести.
Cyclolites (фиг. ИЗ; cyclos—окружность, lithos—камень) представляет пример Hexacoralla с пористой стенкой и перегородками. Полипняк простой, свободный, дискоидальной или эллиптической формы. Наружная стенка его развита лишь в виде плоского основания. От нее поднимаются чрезвычайно многочисленные, тонкие, радиальные перегородки, простирающиеся до центра полипняка. Между перегородками synapticulae и dissepimenta. Нижняя поверхность стенки покрыта концентрически морщинистой эпитекой. Меловые, реже третичные отложения.
Leptoria (фиг. 114; leptos—тонкий). Полипняк массивный. Стенка и перегородки имеют., плотное строение. Ячейки соединены между собою
в извилистые ряды" с неразвитыми в промежутках между отдельными ячейками стенками. Вследствие этого перегородки почти параллельны между собою, и столбики сливаются в пластинчатое образование, протягивающееся
посредине ряда ячеек. Юра—третичные.
Сравнение групп Rugosa и Hexacoralla.
До самого последнего времени принимались такие отличительные особенности Rugosa. и Hexacoralla: Puigosa представляют числи перегородок, кратное четырем (отсюда довольно распространенное название Telraco-
Фиг. 113. Cyclolites undulata. Виды снизу и сбоку. Верхнемедовые отложения.
Гозау. Австрия. 1/1.
ralla), обладают двусторонней симметрией; Hexacoralla имеют перегородки в числе кратном шести, обладают радиально-лучевой симметрией.
Так как Rugosa являются
единственными скелет образую- r *-!® * г
шими палеозойскими Zoantharia, то представляется возможным вывести позднейших скелет образующих Zoantharia, т е. Hexacoralla, филогенетически только из Rugosa. В настоящее время, в результате новейших исследований, оказывается, что Rugosa имели не четыре первичных перегородки, а шесть, подобно Hexacaralla, и лишь дальнейшее развитие перегородок у этих групп, Zoantharia шло раз-
Фиг. 114. Leptoria tenella Верхн. юра Судак. Крым. 1/1.
Фиг. 115. Поперечное сечение кЬЛама из группы Rugosa (Lophophyllum proliferum. Верхчекаменн. Сев. Америки).
I— первичные перетородкЯ — главпая, G — противоиол жвал, 8—боковы« ; а — d, at — 2>t — вторичные перегородки, последовательность развития коюрых соответствует по ледова ель-вости букв алфавита; а— перегородки третьего порядка. Противоположная перегородка соединена со столбиком.
лично. Именно, у Hexacoralla последующие перегородки развиваются во всех первичных интерсептальных камерах, а у Rugosa лишь в четырех из шести, так что число первичных и вюричных перегородок у взрослых Rugosa будет 2п + 2щ + 2 (фиг. 104) или, при равенстве шип, что часто
бывает, 4n + 2; с перегородками 3-го порядка общее число перегородок будет 2 (2n + 2m + 2), т е. кратное 4-м (фиг 115).
Первичные интерсептальные камеры Rugosa, в которых не развивается вторичных перегородок, являются всегда прилежащими к противоположной первичной перегородке.
Возможно, что Rugosa перешли в Hexacoralla вследствие перехода от бокового прикрепления к осевому, причем согнутое состояние полипняка сменилось прямостоячим, при этом утратилась и побудительная причина к характерному для Rugosa расположению перегородок и двусторонняя симметрия сменились радиальной, свойственной организмам прикрепленным,
у которых осевая линия прикрепления (губки, кораллы, грибы) и которым свойственна радиальная симметрия.
Резкое различие между Rugosa и Hexacoralla существует в способе их прикрепления: у Rugosa оно боковое (фиг 116), у Hexacoralla осевое (фиг. 117), причем у первых
тела пряма и перпендикулярна к поверхности
Фиг. 116. Боковое прирастание Cyathophyllum ceratites. Дна конических О1ИНОЧНЫХ полипняка, приросшие к цилиндрическому колониальному. Девон. Ухта. Обл. Коми. 3/2.
Фиг. 117. Paras-milia centralis. Осевое прирастание. Верхнемелов. отлож. Англия. Натур, ве-лич.
необходимости отги-
ось коралла не пересекает субстрата (объекта прирастания), а у последних, наоборот, ось упирается в субстрат. Боковое прирастание Rugosa является при-
чиной согнутого состояния их полипняка вследствие бания устья от почвы, а согнутое состояние, как изложено выше, является причиною характерного для Rugosa двустороннесимметричного расположения
перегородок.
Подкласс II. Octocoralla, или Д1суоиаг1а.1
Полипы этой группы отличаются тем, что всегда имеют одно и то же число щупалец и мезентериальных складок, именно 8; щупальца всегда полые и каждое в отдельности перистой формы. Скелет Alcyonaria бывает 'известковый или роговой; известковый состоит из спикулей, подобных спи-кулям литистидных губок, .или из фибр, подобно тому как у Zoantharia. Бывают скелеты, состоящие из перемежающихся роговых и известковистых участков, или могут быть известковые спикули, рассеянные в основной массе рогового вещества или мягкого тела колонии. Одиночные полипы редки, обыкновенно Alcyonaria животные колониальные.
Колонии развиваются из материнского организма почкованием, причем характерно то, что почки развиваются не прямо от материнского орга-Н-----—---------------,
1 Octo—ВОС“1»*>.
низма или от дочерних, ни образуются на трубчатых выростах тела полипов. Таким образом полости тела полипов колонии Alcyonaria никогда не сообщаются между собою непосредственно, но всегда посредством трубчатых выростов (столоны или солении); эти выросты могут быть длинны, сильно разветвлены или могут быть рудиментарны, так что на первый взгляд полипы будут казаться
сообщающимися непосредственно.
Фиг. 119. Heliopura covulea. Один иолип и прилегающие мягкие части.
в—выдающаяся часть полвпа с восемью пери" стымк щупальцами; Ъ — нижняя часть полипа» с — эктодерма, d — слой с каналами; е — быкол-неннн цененхим аль них трубочек, ныне жинущ»
Фиг 118. Часть лолип-няка Tubipora musu.^,. Ныне живущ. 1/1.
так называемый осевой скелет,
Этот
или он
Колонии Alcyonaria отсутствует.
Фиг. 120. Chaetet.es radians.
Нижвекаменн. отлож. 1/1.
или имеют скелет представляет твердую ось колонии, находящуюся вне тела полипов; полипы примыкают к оси проксимальным концом тела, которое остается мягким; осевой скелет следует раз-
ветвлениям колонии.
У Alcyonaria, не имеющих осевого ске-й» лета, тело отдельных полипов иногда отлагает скелет, подобно тому как у Zoantharia. Ячейки этого скелета всегда снабжены днищами, что же касается до перегородок, то они зачаточны, иногда представлены лишь вертикальными рядами мелких шипов или же совершенно отсутствуют; притом если существуют, то по числу часто не соответствуют щупальцам и мезентериальным складкам, бывая, например, ’в числе 12; поэтому вообще перегородки Alcyonaria называются ложными перегородками (pseudosepta). Вымершие представители Alcyonaria без осевого скелета, распространенные в палеозое и затем почти сразу вымершие, обыкновенно объединяются в группу Tabulata, которою мы и будем исключительно заниматься.
Из ныне живущих Alcyonaria близки к Tabulata—и даже могут быть отнесены к ним—лишь два рода: Heliopora и Tubipora. Род ТиЫрота (фиг. 118; tubus—трубка, poros—пора) представляет скелет, состоящий из цилиндрических трубок, разделенных небольшими промежутками и соеди-88
ненных горизонтальными пластинами. .Пластины, через которые проходят трубки, расположены поперек колонии на известных расстояниях, так что из них торчат трубчатые ячейки. Эти пластины пронизаны каналами, лежа-
щими в плоскости их; через посредство этих каналов полости ячеек находятся между собою в сообщении, так что
С
У?*'
И».
I»
s’?4
Фиг. 12Т. а, Ъ, с—Heliolites рцгоеа. Общий вид, поперечный и продольный разрез. Средней девон. Эйфель. Германия, d — Heliopora Partschi. Верхвемелов. Гозау. Австрия
каналы аналогичны столонам. Внутри ячеек ТиЫрота бывают воронкообразные днища. Перегородки отсутствуют.
Heliopora (фиг. 121, d; hellos—солнце, рогоз—пора) представляет ячейки с радиальными перегородками, рассеянные в цененхиме, состоящей
из многочисленных, плотно прилегающих одна к другой вертикальных трубочек; как ячейки, так и "рубочки имеют горизонтальные днища. Ценосарк, образующий цененхиму, имеет вид тонкой оболочки снаружи колонии (фиг. 119) и дает вниз полые слепые придатки, отлагающие трубочки цё-ненхимы.
К Heliopora, в ископаемом состоянии встречающейся с меловых отложений, примыкают палеозойские роды Heliolites и Chae-tetes.
Chaetetes, подобно Heliopora, имеет стенки ячеек, состоящие из
Фиг. 122. Схема строения коралла Heliopora и Helio-liteo ci — пёнгихимальные трубочки, у которых стенка ксядой состоит из двух пластин I—XIX. трубочки, па месте которых воави-кает ячейка полипа.
Фиг. 123. Halysites catenularia. Верхний силур. Готланд.
Швеция. 1/1.
пучкоз фибр (фиг. 120), так что стенка состоит из одного слоя таких пучков, так сказать, однослойна, и принадлежит сразу двум ячейкам; позднейшая из них пристраивается к готовой стене, не представляя в соприкосновении с последнею собственной, самостоятельно развитой стенки. У Chaetetes все ячейки приблизительно одинакового по размерам поперечного сечения и неправильно многоугольной формы. Имеются днища, перегородок нет. Размножаются делением, причем ячейки подразделяются продольными выростами внутренней поверхности стенок, растущими вовнутрь ячейки до взаимного соприкосновения. В начальной стадии развития эти выросты ь. по-
перечном разрезе ячеек могут быть приняты за перегородки. Девон, каменноугольные.
Heliolites (фиг 121, а, Ъ, с; hellos—солнце) представляет большое сходство с Heliopora. Полипняк также состоит из цилиндрических ячеек, связанных цененхимой, состоящей из тонких призматических трубочек. Стенки ячеек и трубочек плотного строения (отличие от Heliopora). Ячейки полипов снабжены 12 ясно развитыми перегородками. Силур, девон.
Фиг. 124. bi/ringoporaramulosa. Каменноуг. Фихтельгебирге. i < р-цания Общий вид и разрез двух ячеек. 1/1.
Фиг. 125. Favosites ро’утогрна. Средний девон Германия. Левый экземпляр увеличен.
Фигура 122 дает понятие о том, как из полип-Нлка с однородными ячейками типа Chuctetes могли возникнуть полииняки типа Heliolites, Heliopora,— именно путем остановки роста, полного или частичного, 19 ячеек. При этом понятно и возникновение 12 перегородок.
Halysites (фиг. 123; halysis — цепь). Полипняк состоит из трубчатых ячеек эллиптического поперечного сечения. Ячейки срастаются между собою своими узкими сторонами и располагаются цепочкообразными рямми, которые пересекаются между собою под различными углами. Такое расположение
получается вследствие почкования только с одной стороны. Днища многочисленны и горизонтальны. 12 ложных перегородок представлены рядами шипиков. Ячейки покрыты на боках морщинистой эпитекой. Иногда между большими ячейками находятся группы малых (соотношение с Heliolites). Силур.
Sgringopora (syrinx — дудка, poros — пора) является представителем другой группы Tabulata, именно тех, у которых каждая ячейка самостоятельно развивает свою соб -твенную стенку (фиг. 124). Цилиндрические ячей и на различных горизонтах соединены короткими поперечными трубочками, перегородки в виде вертикальных 5ядов известковых шипов, нища хорошо развиты, обыкновенно воронкообразны, но ьндоте и или являются просто согнутыми. Силур — каменноугольные отложения.
Favosites (favus—пчелиные соты) имеет массивный, реже ветвистый полипняк. Кораллиты призматические, многоугольного поперечного сечения, плотно прилегающие друг к другу стенкамп, которые просверлены крупными порами (фиг. 125), приводящими в сообщение полости ячеек. Перегородки отсутствуют или зачаточны, в виде вертикальных рядов шипов. Днища обыкновенно хорошо развиты и горизонтальны. Силур — каменноугольные.
иногда образуют подобие пузырчатой
эо
Favosites легко выводится из типа Syringopora, если представить себе коралл» ты последнего пришедшими в соприкосновение, как это иногда и бывает у Syringopora, между некоторыми из смежных кораллитов колонии и на некотором протяжении длины кораллитов.
Распространение Anthozoa.
Современные коралловые полипы по своему распространению могут быть разделены на две группы: одиночных и колониальных рифовых. Одиночные кораллы главным образом живут в спокойной воде, на больших глубинах и находятся почти во всех широтах. Рифообразующие кораллы ограничены в своем распространении глубиною и температурою морской воды, встречаясь лишь на небольших глубинах, в мелких морях или в прибрежной полосе глубоких морей, и только в постоянно теплой воде, так что по распространению сосредоточиваются почти исключительно в поясе тропиков. Кроме того, рифовые кораллы могут жить лишь в чистой, свободной от мути воде. В соответствии с этим, ископаемые кораллы встречаются главным образом в известняках, реже в глинистых сланцах и особенно редко в песчаниках. Известны ископаемые рифы.
В палеозое распространены группы, затем вымершие—Tetracoralla и Tabulata. В нижнепалеозойских отложениях Cyathophyllum, Streptelasma, Calceola, Cystiphyllum, Favosites, Heliolites, Halysites\ в верхнепалеозойских (каменноугольных и пермских) характеристичны Zaphrentis, Amplexus, Caninia, Llthostrotion, Clisiophyllum, Lonsdaleia, Dibuno-phyllum. С мезозоя получают распространение Hexacoralla, невидимому занявшие место Tetracoralla так же, как современные Alcyonaria заняли место Tabulata. Палеозойские рифы образованы, главным образом, не кораллами Zoantharia, преобладающими в современных рифах, а строматопороидами, гидрактиниями и Tabulata. Малое значение Tetracoralla при образовании рифов, вероятно, стояло в связи с неспособностью их размножаться делением а давать массивные полипняки.
Далее описывается группа археоциат (Archaeocyathidа; arehaios—древний, cyathos — чаша) (фпг. 126), систематическое положение которых не установлено. Это воронкообразные тела со срединною центральною полостью, достигающею до узкого основания. Стенки пористые, промежуток между наруж-
пою стенкой и стенкой центральной полости заполнен радиальными перегородками (А), иногда к ним присоединяются горизонтальные днища (С) или пузырчатая ткань; наконец, последняя может заполнять все пространство между
Фиг. 126. Археоциаты. A. Archaeoeyathus patulus. Бей-Булук, Сибирь.
5/1. В. и D. Cyclocyatus Yakov! evi. Камешки. Минусинск. Н. Кембрий.
<*. Coscinocyalhus. Кембрий. Сардиния. Е. Archaeoeyathus Vassiltevskii. Камешки. Н. Кембрий F.Spirocyathus subradiutus. Бей-Булук. Н. Кембрий. (?. Archaea yathus aculus. Камешки. Н. Кембрий. Н. Archaeoeyathus Sibin-cus. Камешки
стенками Систематическое положение археоциат не установлено. Их причисляют или к кораллам, или к губкам, даже к известковым водорослям. Весьма распространены в кембрийских отложениях, главным образом в нижнем кембрпе Южной Европы, Сибири, Северной Америки, Австралии, где как бы занимают место Zoantharia (Rugosa), являющихся с силура.
Характеристи/ги групп кишечнополосп.ных.
I класс: Hydrozoa. Имеют гидрополипов и гидрометуз. Те и другие часто связаны чередованием поколений, н.п иль те или другие могут отсутствовать. Колониальные формы большею частью с хитиновым реже известковые, скелетом ко/опии. С кембрия доныне.
1 порядок: Hydrana. Большею частью пресновоАные формы, без перидерма. В иско-..аемош состоянии неизвестны.
2 порядок: Tubulana. Большей частью колониальны; полипы с хитиновым перидермой, который у гидракгиний при основании иногда становится известковистым и состоит из концентрических пластинок и радиальных столбиков. От силура доныне.
3 порядок: Hydrocorallina. Морские, колониальные, с обизвестковлевным перидермой. С триаса доиыве.
4 порядок: Campanularia. Перидерм хитиновый, отлагается не только при основании нолипов, но и около стебля с чашечкой. С кембрия доныне, хотя и редки в ископаемом состоянии.
Graptolitha С хитиновым перидермом; разветвленные зли палочкообразные колонии, часто с осью. Начальные стадии воронкообразны Верхний кембрий —нижний карбон
П класс: Scyphozoa. Медузы с толстым студенистым слоем, с краем, рассеченным на лопасти и потому без плавательной каймы. В ископаемом состоянии хотя и не часто, во дают отпечатки, известные уже с кембрия.
Ill класс: Anthozoa. Полипы, часто колониальные, с глоточной трубкой и радиаль-нымр мезентериями, 'часто с известковым скелетам. От нижнего силура доныне.
I подкласс: Zoantharia. Перегородки в переменном числе. Щупальца просты з Скелет ив волокон аррагоната.
1 порядок: Rugosa (Tetracoralla). Полипняк конический, с боковым прикреплением В расположении перш сродэк двусторонняя симметрия. Число перегородок кратное четырех Нижний силур—пермь.
• 2 порядок: Hexacoralla. Полипняк с осевым прикреплением. Расположение перего-
родок радиально-симметричное. Число их кратное шести. Триас — ныне-
I
И подкласс: Octocoralla (Aieyonaria) Восемь перегородок, щупальца перистые. Силур - ныне.
Приба зление: Archaeocyathida. Воронкообразные тела, с двумя стенками, наружной л внутренней, в промежутке между ними перегородки и пузырчатая ткань. Стенки пористы. Кембрии.
Ill класс: Ctenophora. Со студенистым телом, с 8 меридиональными рядами плава-гельныл. придатков. Планктонные животные. В ископаемом состоянии неизвестны.
IV ТИП. ЧЕРВИ, VERKES.
Черви, так же как и все еще более высшие животные, отличаются от уже рассмотренных низших Metazoa присутствием в теле, по крайней мере, двоякого рода полостей: 1) пищеварительного канала, протягивающегося спереди, от рта, назад до анального отверстия, выводящего »з тела непереваренные остатки пищи, и 2) полости, помещающейся между пищеварительным каналом и стенкой. тела, так называемой общей полости Teia, целома (coelom). По присутствию целома черви и все еще . более высшие животные объединяются в одну группу под названием Coelo-
А
th- В
Фиг. 127 А а В. Схима строения тело Polygordius.
А — поперечный разрезаем — кутикула; ср— эпидермис; тр — мускульные плче1ипки, ее, ее — цЁломическиЙ эпителии; de—кпшечкый эпителий; dm— срединная перегорэдка (мезентерии); dvr vt> — спиниой и брюшной кривенйсныо сосуды; п—органы выделения; ТУ — брюшной нервный ствол В — продольный разрез; о—)<*?, ph — глотка; 9*и—кишка; г—прямая кишка; ап—анальное отверстие; sp — посегментные перегородкй {дпссекнкодгы ; д— мозг; яр —нерв, отходящий от него в щупальца.
mata, противополагаемую остальным M.etazoa—Coelenterata (в широком смысле, со включением губок), у которых существует всего одна полость в теле, как бы она ни была разветвлена. В ископаемом состоянии до последнего времени были известны лишь остатки кольчатыхчервей (Annelida), обладающих сегментацией тела, наружной и внутренней.
Целом кольчатых червей состоит из двух отделов (фиг. 127/ А), протягивающихся во всю длину тела справа и слева от кишки, между нею и стенкою тела. Целомические полости покрыты изнутри слоем эпителия, с одной стороны входящего в состав стенки тела, а с другой—в состав стенки кишки. Над кишкою и под нею целомические полости соприкасаются, и эпителий их образует вертикальную срединную перегородку (мезентерий) в теле, состоящую, понятно, из двух слоев эпителия. Эта перегородка удерживает пищеварительный канал в срединном положении. Каждый из двух названных отделов общей полости тела делится поперечными перегородками на камеры соответственно наружному подразделению тела на
сегменты. У простейших кольчатых червей все сегменты тела одинаковы, за исключением переднего—головного и заднего—анального, с выводным отверстием кишки.
Стенка тела состоит из наружного эпителия/ или эпидермиса, отделяющего снаружи кутикулу, из лежащего под эпидермисом слоя мускулов, поперечных и продольных, и, как указано выше, целомического эпителия. В состав стенки кишки, кроме целомического эпителия, входит еще внутренний, кишечный эпителий. Камеры общей полости тела наполнены бесцветной прозрачной целомической жидкостью, которая является посредствующей средой, служащей для распределения по всему организму питательные соков, получающихся при пищеварении.
При восхождении от более простых типов животного царства к более сложным, у червей впервые встречается кровеносная система, со-
стоящая из двух продольных сосудов,
Фш’. 127 С. Поперечный разрез тела высшего кольчатого червя:
at а' —спинной и брюшной cap подои; с — жабры; d— ще нитки параподнй.
расположенных в срединной вертикальной перегородке тела, один над кишкою, другой под нею. Эти сосуды сообщаются в каждом сег менте боковыми сосудами.
Продольные сосуды не имеют самостоятельных стенок, а являются просто промежутками между двумя слоями эпителия вертикальной перегородки. На пути этих < осу-дов имеются отделы, отличающиеся сильной сократимостью; при сокращении их кровь проталкивается по системе сосудов и приводится в движение.
Кровь может быть рассматриваема как видоизменение сиеди-никльной ткани; основное вещество ее представляет жидкость,
в которой плавают клеточные элементы. Кровь, так же как и цёломическаг
жидкость, распределяет по телу питательные соки, кроме того Она поглощает
продукты разложения вещества тела и выделяет их в выделительных органах (почках). Затем кровь играет роль в процессе дыхания. У низших червей этот процесс совершается на всей поверхности тела, у высших — в особых органах дыхания, жабрах; поглощаемый в этих органах извне кислород распределяется по телу кровью. Жабры представляют обыкновенно сильно разветвленные придатки по бокам тела или в головной части (фиг. 127, С).
Органы выделения имеются в каждом сегменте в виде трубочек, на одном конце открывающихся воронкообразным расширение и в камеру полосги тела; на другом конце эти трубочки открываются наружу отверстием в стенке тела, на брюшной стороне его.
На брюшной стороне тела находится брюшной нервный ствол, у низших форм примыкающий к эпидермису (фиг. 127, А и В), у высших лежащий несколько глубже внутрь (127, С) и представляющий ряд посег-ментных утолщений—нервных узлов. В передней части этот нервный ствол соединен с головные мозгом., лежащим над пищеводом, в головном
сегменте; соединение устанавливается боковыми спайками с каждой стороны пищеварительного канала.
Головной мозг иннервирует органы чувств, находящиеся на голове: щупальца, глаза и статоцисты (органы чувства равновесия).
В каждом сегменте имеются органы движения. Сбоку и вентрально развиваются обособления кутикулы в виде щетинок, расположенные в сегментах пучками. Нижние части щетинок углублены в тело, располагаясь в сумках кожи (фиг. 127, С}. Дно сумки отдатяется или приближается к поверхности тела действием мускулистых тяжей, прикрепленных к нему и к стенке тела; вместе с тем втягиваются вовнутрь или выдвигаются наружу щетинки. С увеличением числа и размеров щетинок у величивался и мускуль
Фиг. 128. Serpula gordialis. Турон. Дрезден. Саксония.
Фиг. 129. Spirorbis mnpkhalodes, приросшие к раковине брахиоподы Orlholheles umbraculum. Средний девон. Эйфель. Германия.
Фиг. 130. Serpula I mix. Доггер. Германия. 1/1.
ный аппарат, и все это вело к выпячиванию всего органа с кожей наружу; так возникли ножные бугорки, или параподии. Щетинки, будучи* подвижны указанным образом, способствуют движению животного, происходящему также просто вследствие извивов мускулистой стенки тела.
В сегментах тела из целомического эпителия развиваются половые продукты, семенные и яйцевые клетки, выходящие наружу через трубочки вы длительных органов. Кроме полового размножения, некоторые кольчатые черви размножаются и бесполым путем, делением, что возможно вследствие однородного сегментирования тела этих червей.
В ископаемом состоянии чаше всего встречаются известковые трубки, отлагаемые поверхностью тела морских кольчатых червей трубкожилов (Tubicolae) и заключающие в себе тело червя при его жизни.
К роду Serpula (serpo—ползу; от силура доныне; главнейше с юры), из трубкожилов, относят круглые или килеватые на поверхности трубки (фиг. 128, 130), неправильно извивающиеся или спирально свернутые,
замкнутые на одном конце, обыкновенно прирастающие к посторонним телам.
К роду Spirorbis (spirorbis—спиральный завиток; от силура доныне) принадлежат маленькие, улиткообразные трубки, прирастающие к раковинам, растениям и т. п. (фиг. 129).
в
Фиг. 132. Слева Machairodus incurvus, справа Prioniodus ele-даг^. Нижний силур. Ленинградская обл. 15/1.
Фиг. 131. A. Canadia spinosa (из Chaetopoda). Вид сбоку. 4/3. (редкий кембрий Бричал<уи»я Колумбия. Канада. В. Oftoia prolific!. 2/3 (из Gephyrea). Оттуда же С. AnMcwia sagittiformes. 2/1 (из Caetognatha). Оттуда же.
Кольчатые щетинконогие (ChaetopoOa) морские черви, живущие свободно, крупные, хищные, имеют хитиновые зубчатые челюсти, длиною в несколько мм, напоминающие по своей форме зубы акуловых рыб, за какозые даже были приняты первоначально, • но совершенно отличные от этих зубов по своему строению. Такие челюсти, конодонты (Prioniodus и hlachairodus, фиг. 123), у нас известны из силурийских отложений.
Наши познания относительно ископаемых червей разом сильно увеличились в 1910 г., вследствие находок, сделанных в кембрийских сланцах Сев. Америки, в Британской Колумбии, где была найдена чрезвычайно богатая фауна разнообразных червей. Найдены представители пелагических червей класса Chaetognatha (фиг. 131, D), червей подкласса Annelida Gephyrea, (фиг. 131, С).
В отличие от ныне живущих Gephyrea, кембрийские представители их имеют следы сегментации тела. Annelida Chaetopoda также имеют в этой фауне любопытных представителей, отличающихся, подобно ныне живущему роду Aphrodite, присутствием чешуевидных роговых образований в коже, у ископаемой формы сосредоточенных на брюшной и спинной стероне параподий (фиг. 131, Л).
Фгг. 133. Ne-'cites loomisi Следы двух червей. Средний девон Кровельный „ланей. Хазельбах. Тюрппген. Германии.
В ископаемом состоянии сохраняются разнообразные следы жизнедеятельности червей,—следы ползания на поверхности ила, или ходы вглубь толщи ила. Следы ползания являются конечно, углубленными, желобовид-
г
Фиг. 134. Helminthoides labyrinMca. Следы нескольких червей. Эоден. Флиш. Брегенц-ский лес Германия 1/2
ними с гладкою поверхностью или с рельефом наподобие заплетенной косы волос (фиг. 133, Nereites)', последнее стоит, вероятно, в свяпт с присутствием парных параподий. Гладкие следы иногда правильно изгибаются.
*7 Учебник палеонтологии.
97
Фиг. [3 ? RliizocoraHium Девсн. U-образиые ходы.
так чт( в последовательных изгиоах движение черви происходило в противоположные стороны, а самые изгибы весьма сближены между собою у Helmin-Иго/des (фиг. 134), вследствие чего ил, поглощаемый червем при движении для питания, наиболее полно использовывался. Ходы, углубленные в толщу ила, являются U-образ-пыли, двухколенчатосог-чутыми {RhizocoroUium, фиг. 135), или кустовидно разветвленными, в связи с чем они получают название фукоидов (фиг. 136, Fucus—название весе ма распространенной водоросли). В трубках Rhizoco-rallium, открывавшихся наружу 'вумя отверстиями, через эти отверстия выдавались наружу оба конца животного. Дугообразная нижняя часть хода, соединявшая оба вертикальные колена, перемещается с течением времени все ниже,
так что старые дуги животное заполняет землистым материалом, причем в расколе породы сохраняются сл^ды этих дуг. Fucoides (фиг. 137) имеют одно выводное наружу отверстие, но р; возможность помещения тела двумя концами в двух соседних разветвлениях,— изобилие последних давало возможность более, полно использовать толщу ила в отношении содержащихся в ней питательных веществ. Указанные следы деятельности червей хорошо характеризуют некоторые геологические горизонты, иногда кроме них ничего не содержащее. Встречаются с кембрия доныне.
I ПРИБАВЛЕНИЕ К ТИПУ Ч Е Ч В Е Й.
Мииышм, Bryozoa. »
По организации примыкают к червям.
Животные, прикрепленные к месту обитания, почти исключительно коло
ниальные. Отдельные индивидуумы колонии сидят в ячейках, отлагаемых стенкой тела. Ячейки бывают кожистые, роговые или известковистые.
Из ячейки, через имеющееся в ней отверстие, устье, высовывается наружу и втягивается обратно часть мягкого тела животного с ротовым
Фиг. 136. Fucus. Ходы червей Polidora просверленные в породе. А— схема U-обравного хода в вертикальном разрезе. _В--отверстия U-образных ходов.
’ Вгуоп—мох, zoon—животное.
отверстием, окруженным щупальцами и находящимся около него анальным отверстием. Иногда имеется подвижная крышечка (фиг. 138), прикрывающая устье ячейки. Пищеварительный канал петлеобразно изогнут, свешивается петлей в полость ячейки, состоит из пищевода, желудка и кишки. В области передней части петли находится нервный узел.
Полость тела, наполненная жидкостью, находится в прямом сообщении с внутренностью щупалец, которые полы и исполняют роль органов дыхания, приводя жидкость полости тела в ближайшее соседство с морской, видрй. Щупальца иногда помещаются на подковообразной площадке (л о ф о-фор), обособленно поднимающейся над околоротовым полем. Щупальца покрыты ресничками, действием которых производится ток воды, доставляющий ко рту пищевые частицы. В полости тела развиваются яйцевые и
семенные клетки.
Яйца йногда помещаются в особых капсулах, овицеллях (фиг. 139) расположенных на вершине ячеек. Тела отдельных индивидуумов находятся в некотором .
сообщении между собою посредством отверстий !
Фиг. 137. Fucoides. Типичнее строевве фукоидов. Ходы, разветвляющиеся вниз.
Фиг. 138. Мшанка с высунувшейся из ячейки околоротовой частью тела.
а — щуцальца; с — ротовое отверстие; fанальное; I —крышечка. Ныне жи-„ вущ. Увелич.
в стенках ячеек или посредством каналов столонов, от которых иногда отходят индивидуумы. Это сообщение объясняет происхождение почкованием целой колонии из одного индивидуума, получающегося из личинки. Личинка же развивается из оплодотворенного яйца. Мшанки гермафродитны.
Сообщение между индивидуумами колонии делает возможным разделение труда между ними, дифференциацию индивидуумов. Кроме индивидуумов уже шисанпого типа, существуют еще так называемые авикулярии и в и брак у л и. Те и другие прикрепляются к верхней части нормальных ячеек. У некоторых новейших мшанок авпкулярни существуют при каждой ячейке, в виде придатка, напоминающего по форме птичью голову, сидящую па коротком стебле (фиг. 140). В авикулярии-имеются сильные мускулы, приводящие в движение, наподобие нижней челюсти, нижнюю меньшую часть авикулярии. При жизни животного авикулярия качается на своем стебле, с нижнею постоянно двигающеюся «челюстью», причем схватываются и удерживаются сравнительно большие животные, приближающиеся к колонии. Авикулярии возникают видоизменением нормальных индивидуумов колонии, причем мускулы развиваются за счет дегенерировавшего мягкого Тела животного, ячейки становятся значительно меньше, представляют от
личную от нормальной форму и выдаются наружу среди нормальных ячеек; мешшая, подвижная часть авикулярии, представляет видоизмененную крышечку нормальной ячейки. Вибракули, представляющие гибкие, приходящие в колебательное движение хлысты, также соответствуют крышечке нормаль-
ных ячеек.
Колонии мшанок представляют большое разнообразие в отношении
I
общей формы, являются кустообразными, в виде корок на камнях, коралловидными и т. и. Отдельные ячейки едва различимы простым глазом.
История индивидуального развития которую они проходят, дает основание
Фпг. 139. Fhistra _ а — авпкудярпп; о — овицелли; жирным черным закрашены \стья открытые в различной степени. Ныне жи-вущ. Увелич.
мшанок, особенно стадия личинки, предполагать ближайшее родство мшанок с червями. Личинка их, подобно личинке червей, имеет дугообразно согнутый пищеварительный канал, сравнительно боль-
Фиг. 110. Авикулярия.
ек — ячейка, с? крышечка, у — руди мент кишки; — мускулы крышечки
шой головной мозг и органы зрения, глазки. У червей организация развивается далее, личинка же мшанок, вследствие прикрепления, испытывает задержку в развитии, не только не приобретает усложняющейся в дальнейшем развитии организации, как у червей, но далее регрессирует. Именно, в связи с прикреплением, совершенно атрофируются органы зрения и головной мозг, остается лишь брюшной нервный узел, сегментация тела также не развивается.
Большая часть современных мшанок живет в море, лишь немногие в пресной воде (постедние имеют исключительно кожистые ячейки). Они находятся большею частью скоплениями на различной глубине. В ископаемом состоянии часто встречаются во всех системах, начиная с нижнего силура, и иногда образуют рифоподобные массивы в миниатюре.
ПОРЯДОК I. CYCLOSl'OMA'IA.
Cyclostomata характеризуется простыми известковистыми трубчатыми ячейками, иногда с поперечными днищами, с конечными, во все поперечное сечение ячейки, без крышечек, устьями. В< гречаются с нижнего силура доныне, но распространены лишь В палео юе и мезозое.
Из этой группы род Ceriopora (cerion—пчелиные соты, рогов — пора) (фиг 141) образует корковидные, массивные, ветвящиеся или лопает-
ICO
I
яые нолипняки, иногда состоящие из двух или более слоев ячеек, расположенных один на другом. Ячейки почти одинаковы, плотно примыкают друг к другу, и их круглые или многоугольные отверстия покрывают всю верхнюю поверхность колонии. С силура доныне.
ПОРЯДОК II. TREPOSTOMATA.
Trepostomata характеризуется известковистыми ячейками, в началь-
ных стадиях роста вплотную прилегающими друг к другу и имеющими многоугольное строение. У большинства форм этого порядка зрелая стадия
развития колонии отличается от начальной тем, что в зрелой стадии сечение ячеек округляется, и пространство между соседними ячейками заполняется добавочными образо-
в
' Фиг. 141а. Ceriopwa irregularis. фпг*Ceriopora spongites. А—Вия, Туроа. Марсель. Франция. сбоку. Мед. Эссен Германия. В— То же, вид сверху. 1/1,
V
ваниями. сходными по форме с начальными стадиями развития ячеек, называемыми мезопорами. Как в ячейках, так и в мезопорах зачастую , имеются днища в некоторых формах неполные или центрально про-
дырявленные (род Tabulipora). По внутреннему строению порядок Trepostomata разделяется на две группы: Amalgamata и Integrata. У Amal-gamata стенки соседних ячеек сливаются в одно целое, без ясно видимого разделения, у Integrata между стенками соседних ячеек проходит прослой иного строения, легко отличимый в сечениях по более темной окраске. В обеих группах кроме мезопор встречаются нередко акантопоры, представляющие тонкие короткие трубчатые полости в стенках ячеек у наружной поверхности колонии.
Форма колоний Trepostomata весьма разнообразная, от тонких корковых обрастаний на посторонних предметах до массивных бесформенных рифоподобных тел; часты ветвистые разности, иногда дающие сложные кусто-видные или даже сетчатые колонии.
Некоторые представители Trepostomata имеют большое внешнее сходство - с кораллами Tabulata. Известны с нижнего силура до конца мезозоя, имея наибольшее значение в нижнем палеозое.
ПОРЯДОК III. С PYPTOSTOMATA.
Устье у Cryptostomata круглое, как у Cyclostomata, но ведет не в самую ячейку, обыкновенно короткую, а в переднее пространство, отделенное от собственно ячейки неполной перегородкой. Наиболее часты в палеозое.
Особенно часты и важны такие представители их, ветвистая или листообразная колония которых несет ячейки только йа одной стороне, как веефо-
видные или воронкообразные колонии рода F&nestella (фит. 142; fenestella— оконце). У Fenestella колония состоит из восстающих ветвей, соединенных поперечными перемычками; первые несут по два ряда, разделенные киле-ватостью, устий ячеек. Силур—пермь-
ПОРЯДОК IV. CHEILOSTOMATA.
Вследствие атрофии переднего пространства и обусловленного этим выступания внутреннего, суженного устья, из Cryptostomata, вероятно,
o«iieat jL>t
Фиг. 142. Fenestella retiformis. Слева общий вид колонии в натуральную величину; посредине— увеличенная часть колонки со стороны устий; справа увеличенная часть колонии с противоположной стороны. Пермские отложения. Пёснек. Тюрингия. Германия.
возникли Cheilostomata, в ископаемом состоянии известные с юры и с третичных отложений, приобретающие перевес над Cyclostomata.
У Cheilostomata ячейки разнообразной формы, обыкновенно коротки,
овального очертания, с устьями меньшего диаметра, чем ячейки, расположенными сбоку, прикрывающимися подвижной крышечкой. Только в этой группе имеются вибракули, авикулярии и овицелли; конечно, в ископаемом состоянии вибракули и авикулярии не сохраняются, но о былом присутствии их свидетельствуют особые поры, через посредство которых происходило причлененг.е названных органов к
Фиг. 148. Membranvpora iupidosa Слева общий вид колонии в натуральную величину. Справа — увеличенная часть колонии, внизу ячейки с тыльной стороны, вверху ячейки вывалились, и виден отпеча-ок на юроде их устьевых сторон. Миоцен.
Сарматский ярус. Подол ия.
телу колонии.
Не все Cheilostomata имеют вполне обызвестленные стенки ячеек, у некоторых стенки являются роговыми и гибкими, у других передняя стенка ячеек перепончата, а остальные известковистые
Так у рода Memhrampora (фиг. 143; membrana—перепонка), у которого коло-
ния состоит из одного слоя ячеек, причем все имеют роговые стенки с одной и той же стороны колонии. С меловых отложений доныне.
II ПРИБАВЛЕНИЕ К ТИПУ ЧЕРВЕЙ
Ллеченогил, Brachiopodc.
Мягкое тело плеченогих, подобно тому как у мшанок, представляет пищеварительный канал, изогнутый дугообразно так, что анальное отверстие находится близ ротового на переднем конце тела. Кроме того, опять-таки подобно мшанкам, а также многим кольчатым червям, около ротового отвер-
стия находится два кожистых бахромчатых выроста (фиг. 144, 145), соответствующие жабрам, но здесь называемые руками. Руки особенно сходны с лофофором мшанок Рот помещается между основаниями рук. Тело всегда покрыто раковиной, состоящей из двлх обособленных частей — створок. Створки отлагаются поверхностью мягкой кожистой двухлопастной мантии, представляющей два перепончатых выроста стенки тела. Лопасти мантии соответствуют по своей форме створкам, внутреннюю поверхность которых они выстилают. *
В подавляющем большинстве плеченогие—животные, прикрепленные к месту обитания. Прикрепление происходит или посредством мускулистой
ножки, суженного заднего продолжения тела, или раковина прикрепляется прирастанием одной створки; ножка покрыта тонким роговым чех ликом, оконечность которого разветвлейа в корневидные отростки; этими отростками животное прикрепляется к посторонним предметам, находящимся на морском дне. »
Спирально свернутые руки помещаются в большей передней части мантайной полости, задняя же
Фиг. 144. Ныне живущая Lingtda.
Мягкое тело животного, удаленное из раковины. Ближняя лопасть мантии отворочена вниз, так что наверху гидны спирально закрученные руки; а — ротовое отверстие. На обнаруженных внутренних частях поверхности мантии видны впечатления кровеносных сосудов.
Фиг. 145. Вертикальный разрез иыне жищщей IValdheimia, жирной червой краской boj азава ман-тийная полость.
» — раковина (брюшная створка); тп — мантия; а — руки; Ьг — бахромчатый край их; с — аппарат ручных поддержек; d — пищеварительный канал; п — органы выделения; т — ыускулы-закрывателн; mf— мускулы открыватели; st— ножка.
занята собственно телом. Кишка иногда оканчивается слепо, если же имеется анальное отверстие, то оно расположено асимметрично. Нервная система представляет два нервных узла — надглоточный (головной мозг,) меньший, и подглоточный (брюшной), больший. Эпителий кожи обладает чувствительностью, осязанием, органы слуха и зрения отсутствуют, без сомнения, в связи с сидячим образом жизни и наличностью раковины, прикрывающей тело. Органы выделения имеются в виде двух пар воронкообразных трубочек; только эти органы и представляют, хотя и слабо, первичную, здесь почти утраченную сегментацию тела.
Есть сердце, представляющее орган кровеносной системы, обладающий большой сократимостью, причем он приводит в движение кровь. Полость тела представляет продолжение в мантию в виде сосудов, выдающихся на поверхности мантии (фиг. 144); соответственно этому внутренняя поверхность раковины представляет васкулярные впечатления, (фиг. 147).
Плеченогие размножаются исключительно половым путем, причем почти
всегда раздельнополы.
Створки раковины называются брюшною и спинною, плоскость симметрии дельт каждую из них пополам. Прикрепление происходит посредством брюшной створки, котовая обыкновенно больше спинной. Каждая створка в своей наиболее старой
части» представляет макушку, обыкновенно. более значительною в брюшной створке. Край створок около макушки называется з а д-ним, в противоположной частп— передним, остальные два края называются боковыми. Расстояние ст макушки до переднего края представляет длину створки, перпендикулярный размер между боковыми краями — ш и-
Фиг. 146. Прикрепление ножками ныне живущих Waldheimtae.
Фиг. 147. С.магор-horia Humbletoneksis. Пермь. Англия. Внутреннее ядро с васкулярными впечатлениями.
рину створки, толщина раковины представляет расстояние между центрами створок. Ножка выходит или
в промежутке между задними краями створок, или через отверстие в макушке
Фиг. 14S. Створки ныне живущей Waldheimia совнутри; слева спинная, справа брюшная створка.
с—впечатление передних замыкателей; д'—впечатление задних замыкателей; ЪЪГЪГ*— впечатление ножных мускулов; сс' — замочный отросток; I — ручной аппарат $ — зубные ямки; f— foramen, d — дельтидиальные пластинки, t — замочный зуб, с — впечатление передних отмыкателеи; — впечатление задних отмыкателеи
брюшной створки, или под нею. Если отверстие в макушке, то оно круглое (foramen); если под макушкой, то треугольное (delthyrium, delta—дельта, iliyra—дверь). Оба могут существовать одновременно. Delthyrium иногда закрыт отчасти или вполне; он может быть закрыт или двумя табличками, дель-тидиальнымй1, составляющими deltidium (фиг. 148), или одной (pseu-dodeltidium (фиг. 149), покрывающей delthyrium сводообразно от одного бока к другому. Дельтидиальные таблички или могут быть ’разделены проме-
чутком по плоскости симметрии раковины, или, при более значительном развитии их, притти в соприкосновение в этой плоскости, ограничивая foramen снизу (фиг. 148) или оставляя для ножки- круглое отверстие между собою.
pseudQdeltidium или ограничивает отверстие для ножки сверху, или замыкает его со всех сторон. Deltidium и pseudodeltidiiim, невидимому, возникают различно, отлагаясь различными частями мягкого тела животного. Треугольное отверстие, соответствующее deltliynum, находится иногда и на спинной створке, под макушкой, здесь оно называется chilidium.
Задний край створок иногда бывает прямым и в этом случае обыкновенно является основанием треугольной площадки, вершиною которой является макушка створки, в боковые стороны обособляются резким ребристым перегибом между заднею и боковыми частями поверхности створки. Такая треугольная площадка — area может быть на брюшной или на обеих створках: в последнем случае она все-таки сильнее развивается на брюшной створке.
Створьи раковины соединены между собою или посредством мускулов, проходяших через тело от внутренней поверхности одной створки к другой, или. кроме того, шарнирно сочленены на заднем краю.
Шарнир, пли замок, состоит из двух выступов на краю брюшной створки—з у б о в, по одному с каждой стороны delthyrium, входящих в зубные ямки спинной створки. Кроме того в сочленении принимает участие замочный отросток спинной створки, расположенный под* макушкой *ее и входящий между зубами
брюшной створки так, что вследствие расширенности оконечности замочного отростка створки можно разнять обыкновенно лишь с повреждением зубов
Фиг. 140. Orthwian squmnata. Брюшная створка совнутрн. Нижний силур. Кук-керс. Эстония.
или отростка.
Замочные ямки с внутренней стороны ограничены замочными пластинками (фиг. 148), которые часто поддерживаются вертикальными ил в наклонными септальными пластинками, простирающимися до дна раковины. Зубы брюпгной створки подобным образом поддерживаются иногда зубными пластинками.
Кроме зубных пластинок, достигающих иногда значительных размеров, внутри створок бывают еще другие пластинчатые образования, служащие большею частью для прикрепления мускулов пли ручного аппарата. Чаще всего бывает срединная перегородка, различной высоты п Длины, которая начинается под макушкой и иногда доходит до переднего края. Иногда срединная перегородка, или ребро, развита и в спинной Кворке, начинаясь обыкновенно от замочного отростка Зубные пластинки в брюшной створке иногда сходятся посредине, ограничивая между собою Углубление и продолжаясь впереди от него в перегородку, идущую к переднему краю. Это образование называется spondylium (фиг. 149).
Аналогичное образование, но реже, бывает в спинной створке.
Раковина закрывается смыканием краев створок пли открывается их Удалением друг от друга. Открывание и закрывание раковины совершается посредством мускулов. Му ску лы закрыватели (diductorcs) пересекают тело, прикрепляясь на внутренней поверхности обеих створок на некотором расстоянии от макушек (фиг. 145, 181). При их сокращении створки смы-
таюгсд, притягиваясь одна к другой. Мускулы открыватели (adducto res) прикрепляются в брюшной створке на некотором расстоянии от макушш а в спинной створке к замочному отростку. При сокращении их замочный отросток, перемещаясь в глубину дельтириального отверстия, приближается я внутренней поверхности брюшной створки, спинная створка поворачивается около линии, соединяющей зубы, как около оси, и отходит на переднем и боковых краях от брюшной створки; раковина приоткрывается.
Кроме открывателей и закрывателей есть еще ножные мускулы Kadjustores), посредством которых может происходить некоторое движение тела на ножке.
На внутренней поверхности створок места прикрепления мускулрв являются в виде более или менее ясных впечатлений и располагаются при плоскости симметрии раковины (фиг. 148).
У беззамочных плеченогих мускулы более многочисленны, они " производят некоторое боковое сдвигание или вращение одной створки относительно другой, закрывание и открывание раковины (последнее достигается
Фиг 1^0. Спинная створка современной IlhynchonMa с простым ручным аппаратом в вида двух изогнутых отростков.
Фиг; 151. Спинная створка TereLralvla с петлевидным ручным аппаратом, сг — crura.
Фцг. 15S. Спинная створка Tet Лг alula.
частью разбуханием мягкого тела, происходящим по крайней мере в передней его части).
У многих брахиопод существует и имеет весьма важное значение для систематики особенный известковый аппарат, который служит опорой для спирально свернутых рук. Этот ручной аппарат (bracJiidium] прикрепляется всегда к замочному краю спинной створки.
В простейшем случае он состоит из двух более или менее коротких изогнутых отростков, так называемых с^ига, расположенных симметрично (тносительно средней линии створки (фиг. 150). У других форм эти отростки продолжаются в тонкие лентообразные известковые пластинки, которые образуют род петли, то короткой (фиг. 151), то длинной (фиг. 152), свободно висящей в полости раковины, или прикрепленной еще посредством двух поперечных пластинок к продольной средней перегородке, проходящей на” дне спинной створки (фиг. 152), или Же петли, всецело прирастающей к этой створке. Наконец, в некоторых му чаях ручные поддержки, образовав первоначально петлю, далее продолжаются в две симметрично расположенные известковые конические спирали, выполняющие большую часть раковины. Эти конические спирал^ обращены основаниями друг к другу, а вершинами противоположны, обращены к боковым краям (Spirifer, фиг. 153), или же основаниями обращены к брюшной створке, а вершинами тогда к спинной (Atrypa, фиг. 154, 155). Для систематики, для установления ближайшего родства важна прежде всего ферма рук и их положение относительно друг 106
яруга и створок. Наличность более или менее далеко идущих известковых ручных поддержек имеет второстепенное значение. Так, существуют ближайшие соотношения между Atrypa и 'EtiynchAn^la, живущей ныне и представляющей прикрепленные е коротким crura руки, также-* спирал! но свернутые и с таким же расположением спиралей, как в Atrypa с ее аппаратом ручных поддержек, простирающихся на все протяжение *рук. у Terebratula и ей подобных имеется аппарат ручных поддержек, в виде петли, а затем следуют неимеющие твердых • поддержек, конически свернутые части рук. Е этой группе примыкают спприфериды, имеющие не только начальную ручных поддержек.
Фиг. 153. что видны
Раковина Spirifer, взломанная так, известковые конические спирали ручного аппарата.
петлю, но
и следующие за нею известковые конусы
Фиг. 154. Atrypa aspera со спинной створкой, взломанной так, что видны об-' ращенные к ней вершинами известковые конические спирали ручного аппарата. Средний девой. Эйфель.
Германия. _
различных групп бра-Strophomenacea spon-
В случае развития в брюшной, а изредка и в спинной Створке spondyliwm открыватели и заьрыватели прикрепляются к ним вместо того, чтобы прикрепляться ко дну-створок Spondylium развивается чаще всего при значительной глубине створки; такая глубина является обыкновенно следствием прикрепления животного к месту обитания и следствием необходимости удлинять створку в случае быстрого накопления осадков на дне морском В указанном случае раковина удлиняется у индивидуумов вида, обыкновенно при условиях медленного накопления осадков, представляющего низкую раковину; при переходе животного к глубокой, удлиненной раковине мускулы должны йыли бы ненормально растягиваться; возникновение spondyl'um, на который переносятся места прикрепления мускулов, дает возы )жвость компенсировать возрастание глубины Створки, . так что длина мускулов остается твкая же, как у индивидуумов с дчзкой створкой.
Так как возникновение spondylwm связано с возрастанием в высоту (в длину) брюшной створки, то ясно, что spondylium может возникать у представителей совершенно <аопод, приобретаясь одними независимо от других. Так, среди dylium приобретает Orthisina, среди Spinferacea—Spi-nfer, Spinferina (превращаясь в Cyrtrna). Таким образом, У представителей различных групп происходит схождение признаков, конвергентноеть. Конвергентное развитие есть процесс, противоположный процессу развития С расхождением признаков. Вследствие конвергентного развития сближаются в своих особенностях роды, ироисхо-опцие от различных корней. Конвергентноеть, однако, не может итти столь далеко, чтобы ат различных корней полечилось одинаковое потомство, чхобы совершенно одинаковая организация получилась различными путями. Вследствие конвергентнбети возникают сходства лишь в некоторых особенностях, ио не абсолютное тождество организации. Конвергентноеть возникает вследствие действия одинаковых внешних условий на различные Организмы.
| Приведенные примеры конвергентного развития являются также и примерами °Ррел ятивного развития признаков. Возрастание глубины брюшной створки вле-ет за собою развитие spondylium, влечет за собою обыкновенно возрастание area в.
Фиг. 155. Atrypa reticularis, спинная створка. Средний девон. Эйфель. Германия.
в связи с увеличением дельтириального отверстия, развитие на нем pseudodeltitltum (Afeekella, Cyrtnia) таким образом коррелятивно с возрастанием глубины створки развивается целый ряд особенностей организации во многих случаях неосновательно принятых в основу обособления не только видов, но даже родов "(фиг. 156); в действительности такие отличия могут быть просто лишь индивидуальными.
Фиг. 156. Ряды форм, связанных между собою, отличающихся лишь удлинением брюшной створки и коррелятивно с удлинением возникающими особенностями А—F—виды Terebratellc Мел. Англия. А — TerMatella р еШа, В— то же, С—Terebratella (Trigonosemus) incertus, D — Ter. ^Tngonoseimis eltgans), E, F—Terebratella {lerebrirostro) lyr, (т - К —Bpirifor. it—К—Cyrtia eyrtaena. Силур
. Гопанд. Швеция.
У тех брахиопод, у которых зубы атрофировались (Proddcfus, фиг. 183) г зубгые пластинки отсутствуют, при возрастании в глубину брюшной створки spondylium не может развиться, и действие углубленности створки на мускулы компенсируется значительным удлинением замочного отростка (Л uloste-ges, фиг. 186); здесь н° место прикрепления мускулов в брюшной створке приближается к замочному отростку, перемещаясь ва spondylium, а наоборот: оконечность замочного отростка, сл] жащая местом прикрепления мускулов, приближается ко дил брюшной створки.
Наружная форма и украшения раковины представляют важные отличительные признаки. Большею частью створки неодинаковой величины, брюшная . больше спинной и более выпукла; спинная иногда плоска или даже вогнута. Углублению, синусу, находящемуся посредине и впереди брюшной створки, соответствует возвышение или седло спинной створки. Прикрепленная брюшная створка более варьирует по форме, у индивидуумов одного и того же вида, чем спинная. Украшения раковины состоят из простых или дихотомически раздваивающихся, радиально расходяпи хся от макушки складок (у Articulata), ребрышек, иногда линейных, иногда таюйе из шипов пли тонких полых трубчатых выростов раковины. Area бывает лишена украшений
По своему строению раковина брахиопод Спивает совершенно известковой или состоит из перемежающихся слоев органическою, рогового и известкового вещества. Известке вые раковины состоят из призматических фибр,
образующих с поверхностью раковины тупой угол (фиг. 157, с). Часто такие раковины проникнуты каналами, проходящими через всю толщу раковины, так что на наружной поверхности раковины отверстия этих каналов являются в виде пор, иногда различимых простым глазом. Каналы заключают в себе трубчатые выросты мантии, не имеют сообщения с наружной средой, так как вся поверхность раковины покрыта наружным перепончатым хитиновым слоем. Назначение каналов не выяснено, возможно, что они доставляют органическое вещество, частью рассеянное даже между призматическими фибрами. Вышеупомянутые тонкие трубчатые вы-
росты поверхности раковины находятся в связи с каналами, пронизывающими толщу ракойины; полость каналов, __ __
иногда в тагом случае сравнительно широки?, переходит в полость выростов.
Личинки брахиопод подвижны, передвигаются посреди вом ресничек, после прикрепления испытывают атрофию органа зрения и редукцию других органов чувств. Животное производит реенпчками рук движенье воды, приносящей при этом пищевые частицы, микроскопические водоросли и т.п.
Брахиоподы—животные исключительно морские, живут преимущественно на глубинах до 640 м, чаще всего до 200 м и все-таки не у самого побережья. Если брахпоподы уже имеются в какой-либо местности, то обыкновенно в большом коли-
Фиг. 1м. j
а—наружная точечная поверхность ТетеГта-tula (слабо увелич.)! Ъ — вертикальный рас рез через раковину (100/1); с—
внутренняя поверхность раковины Waldketmia с отверстиями каналов (100/1) •
честве, или же, -напротив, совершенно отсутствуют. Что касается до систематики брахиопод, то прежде всего они подразделяются на беззамочных (Inarticulata) и замочно-створчатых (Articulata).
Плеченогпе и мшанки имеют столько же общего между собою, как
и с червями, так что в систематике то соединяют плеченогих и Ешанок в один тип, то обособляют каждую группу как самостоятельный тип, то считают за подотделы типа червей.
ПОРЯДОК I. PIA.R1TICLLAT4
Замок и известковые ручные поддержки у Inarticulata отсутствуют. Раковина известково-роговая пли известковая. На краю мантии имеются длинные щетинкп, служащие для недопущения посторонних предметов вовнутрь раковины. '
Наиболее примитивные из известных в настоящее время брахиопод являются в то же время и наиболее древними. Они отличаются дельтири-альныи отверстием для ножки, не закрытым pseudodeltidium, и слабо выраженными мышечными впечатлениями. Такова Ruslella, широкоовального очертания, из нижнего кембрия.
Следующую стадию развития представляют роды с отверстием для ножки, более или менее прикрытым pseudodeltidium. Такова кембрийская 1 Pater ma и отличающаяся от нее более глубокой брюшной створкой, с высокой area, Volborthia (по имени русского ученого, Фольборта) из русского силура.
Из Rustellacea выходят, с одной стороны, Kutorgmacea с зачаточной
area, большим дельтириальным отверстием и мускульными впечатлениями, предобразующими • мускульные впечатления iStrophomenacea (низших Articu-laia). С другой стороны, из Bustellacea выходят Obolacea.
а
Ъ
Фчг. 158. Obolus.
а — брюшная створка Obolus Apolhnis. Верхний кем бри а (унвулитовыи песчаник), Кен-гисепп, Ленинградская обл; Ъ — спинная створка О. Quen-stedti. Детское село.
Род Obolus (фиг. 158, 159; название древнегреческой монеты). Раковина круглого или овального очертания, умеренно выпуклая,. гладкая, известковороговая, блестящая. В обеих створках имеется area, более значительная в брюшной створке. На area брюшной створки находится желобок для ножки, идущий от макушки к замочному краю. От последнего по дну створок, на некотором протяжении, идет по направлению к лобному краю срединная перегородка {septum). Наружная поверхность створок покрыта
ленинградская обе и. тонкими продольными реорышками и концентрическими линиями нарастания раковины. Верхний кембрий.
Lingula (фиг. 160; lingula—язычок). Раковина тонкая, известково роговая, блестящая, почти равностворчатая, приплющенная, удлиненно-четырех-
угольная, ложечковидная или овальная, заостряющаяся к макушке, усеченная напереди, обыкновенно гладкая или с тонкими концентрическими, реже с концентрическими и радиальными струйками. Ножка преходит между створками, не имеющими даже желобка для ножки. От силура доныне, главным образом в силуре и девоне.
Фиг. 159. Брюшные створки Obolus Apollinis совнутри.
Оорма и тонкостенность раковины Lingula связаны с приспособление" животного к совершенно особому, необычному для брахиолод образу жизни именно: Lingula при посредстве необыкновенно длинной гибкой ножки зарывается в песок морскою дна. Lingulacea могут считаться производными от Obolacea.
Известково-роговые раковины с отверстием для ножки в брюшной створке имеются у родов Siphonotreta, Discina.
Фиг. 160. LingiUahicarinata. Средний д< в >н. Сиворицы, Ленинградской обд. 1/1.
Siphonotreta (фиг. 161, 162; siphon-сифон, Пегоз-просверленныЙ). Раковина удлиненно-овальная, умеренно-выпуклая, неравностворчатая. Макушка
Фиг. 161. Siphonolreta
verrucosa. Нижний силур (вагикатовый известняк). Слуцк, Ленинградской обл. 1/1.
Фиг. 162. bijjlwnotieta unguiculata. Нижний силур
брюшной створки просверлена отверстием для ножки; поверхность раковины украшена концентрическими линиями и ребрышками и покрыта полыми шипами. Силур.
Discina (фиг. 163, 164;
discus—диск). Раковина почти круглого очертания, с коническими створками, украшенными концентрическими, реже радиальными струйками. При почти центральной макушке брюшной
(эхиносферит. известняк). Р. Волхов; Ленннгр. оол. 1/1. Раковина обломлена с одного бока.
створки находится щелевпдное отв( рстие для ножки, протягивающееся
ио
отсюда к замочному краю, но все-таки замкнутое перед ним. Третичные—ныне.
Ножка у Огеста коротка, перпендикулярна к плоскости соприкосновения створок, поэтому брюшная створка приплющена, менее выпукла, нежели спинная, и раковина приобретает круглое очертание. Discinidae известны с нижнего силура.
Crania (фиг. 165;cranion—череп) имеет раковину совершенно известковистую, лишена ножки, прирастает к посторонним предметам макушкой менее выпуклой брюшной створки.
Обе створки конические, совнутри имеют широкую приплющенную, и
Фаг. 164. Discina \ficmzotteta} elliptica. Нижний силур. Р. Поповка, около Слуцка, Ленвчгр. Нбл. а—Спинная створка, Ъ— вся рако-' вина сбоку; с—брюшная створка.
Фиг. 163. Discin a (Discinisca) lamel-losa. а—Брюшная створка совнутри; Ъ—вся раковина сбоку; с—брюшная створка снаружи. Ныне живущ. 1/1.
иногда с зернистою поверхностью, полосу соприкосновения створок. Каждая створка с сильно вдавленными мышечными впечатлениями; из них два перед
замочным краем, два
Фпг. 165. Crania (Ancistrocrania) parisiensis var volgensis. Белый ме_ С. Языкове, около Ульяновска. Средне-Волжский край. Спинная и брюшная створки—совнутри; вся раковина снаружи.—сбоку. 1/1.
близ середины створки, между последними в нижней створке находится треугольный вы сгуп. Мышечные впечатления с этим носо
видным выступом сообщают створке вид мертвой головы. От макушек створок по наружной поверхности их
расходятся радиальные ребрышки. Нижняя створка, прирастая приобретает рубец на месте прикрепления. От силура доныне.
Округленная форма; радпально лучевая симметрия створок приобретается как следствие совершенного прикрепления. Ножка утрачивается, и отверстие для нее, находившееся между мускульными впечатлениями, зарастает. Вероятно происхождение Crania от Discinidae
ПОРЯДОК II. ARTICULATA.
Обе створки соединены посредством замка, постоянно известковисты, с аппаратом ручных поддержек или без него. Край мантии не имеет щети-/Нок; как у Inarticulata, но часто является волнистым в связи с присут ствием складок на поверхности раковины. В складчатой раковине створки могут быть менее приоткрыты, чем в нескладчатой, и потому нет такой нгдобности в присутствии щетинок. Кишка оканчивается слепо, так чтъ анального отверстия нет.
Подпорядок I. Stropliomonacea. Простейшие замковые плеченогие без аппарата ручных поддержек. Среди поздней пих из них, более чем в какой-либо дру. ”ой группе, происходит зарастание отверстия для ножки и атрофия последней, При этом развиваются особые приспособления, отчасти или вполне исполняющие функции ножки. Именно происходит прирастание брюшной створки, иногда посредством полых шипов, развивающихся в особенном изобилии, а
<Фнг. 166. Producing gigantms. Спинная створке совнутри.
«I — впечатления замыкателей; Ъ — возвышения, служившие* опорою для мягких pjK. v — почковидные впечатления.
Фпг. 167. Oi this
[Sc&zoph&ria) iesu- Фвг.1(^8. Orlln^(()i /ham-pinata. Нижне-ка- bonitrP) calligramma менвоуг. P Кадужка, Нижний силур (ва-c. Рождество, около гинатовый известняк). Калуги, Московской Пулково, около Левнн-области. 3/4. града. 1/1.
иногда исключительно в околомакушечной части поверхности створки, т. е в той части, где проходит нормально ножка. На спинной створке шипы или вовсе не развиваются, или во всяком случае развиты менее, нежели на брюшной.-
Вместе с тем брюшная створка становится более глубокой и выпуклой, макушка ее более загнутой, и отверстие для ножки совершенно исчезает.
Фиг. 169. Orthis striat&la. Средний дрюн Эйфель Германия.
а — брюшная створка; Ь — енннняя створка-
Фиг. 170. Oithisina anomala Нижний cnflvp (невский яр)с). Эстония. 1/1
Раковина в таком случае не обращена к почве замочным краем, как в случае присутствия ножки, но лежит на выпуклой поверхности брюшной створки.
Прикрытие дельтирпума осуществляется посредством псевдоде льтиди$ма.
Иногда имеется рудиментарный ручной аппарат в впде согнутых пластинок, приросших к внутренней поверхности спинной створки и ограничивающих так называемые «почковидные впечатления» (Chonetes, фиг. 177, 179; Productus, фиг. 166).
Orthis (фиг. 167, 168, 169; orthis—прямой, по характеру замочного края). Раковина изменяется по очертанию от четырехугольного до овального,
еоВерхность ее покрыта радиальными струйками или ребрышками. Створки более пли менее выпуклы; спинная иногда почти плоская или слегка вогнутая. Area хорошо развита в обеих створках и обыкновенно представляет открытый дельтириум. Зубы имеют значительную величину и поддер живаются хорошо развитыми зубными пластинками. Зубные ямки спинной, створки глубокие.
а Ъ
Фиг. 171. Strophomena Stephani. Нижний девой. Богемия. Чехо-Словакия.
а — брюшная створка со стороны замочного края; Ь — со стороны лобного края. 1/1.
Часто имеются в обеих створках внутренние средние ребра. От силура до каменноугольных, особенно часто в силуре. Подразделяется на многочисленные подроды. Представители Orthidae известны уже в кембрие, напр. Nisusia.
Orthisina (фиг. 149, 170; inns—подобный) отличается высокою area яа брюшной створке, закрытой псевдодельтидиумом; в последнем отверстие для ножки. Зубные пластинки срастаются в spondtjhim. Нижний силур.
Strophomena (фиг. 171—173; strophes—согнутый, тепе—луна). Рако-
вина изменяется по очертанию от четырехугольного до полукруглого, радиально струйчатая. Длинный, прямой замочный край соответствует наибольшей ширине раковины. Брюшная створка выпуклая, спинная
Фиг.172. Strophomena гтЬгех. Нижний силур. Пулково, около Ленинграда. Со стороны брюшной створки. 1/1.
Фиг. 173. Strophoi.iero, planumbona. Нижний силур. Оксфорд. Огайо. САСШ.
а — брюшная створка совнутри в околомакушечиои частя; b — спинная створка в таком же положении 2/1.
вогнутая, поэтому створки сближены так, что для мягкого тела имеется лишь узкое пространство.
Обе створки с area-, макушка большой створки в молодости просверлена, затем замкнута. Имеется псевдодельтидиум: зубы расходящиеся и поддерживаемые зубными пластинками, два больших мышечных впечатления разделены срединной перегородкой. Силур.
Ortlwthetes (фиг. 174—176) отличается от предыдущего раковиной тоско-выпуклой или двояко-выпуклой, иногда делающейся с возрастом вогнуто-выпуклой. Всегда выпукла спинная створка. Более существенным отличием является отсутствие отверстия на макушке брюшной створки. Orthothetes
® I Учебник палеонтологии. 113
появляется как бы на смену Strophome па, имея распространение от верх-пего силура до перми.
Роды Chonetes и Productus имеют рудиментарный аппарат ручных поддержек, ограничивающий «почковидные впечатления».
Фиг. 174. Orthotheies crenistna. Нижнекамеввоуг. Уиксфирд. Англия.
а — Енутреваяя поверхность брюшной створки около замочного края; Ъ — спинная створка с внутренней стороны (А, А' — замыкатели; R—отмыкатели, J— замочной отюосток; d— зубные ямки). 1/1.
Chonetes (фиг. 177—179; chone — чашка) имеет раковину поперечновытянутую, полукруглую, вогнуто-выпуклую. Обе створки имеют area и pseudodeltidum. Брюшная створка с сильно развитыми зубами; края area примыкающие
Фиг. 175. Orthotheies arach-
noidea. Нижнекамеввоуг. Валдай, "пниигр. обл. 1/1.
Фиг. 176. Orthotheies (Streptorhynchus) pelargona» tus. Пермские отложения. Тёнстолль. Англия 1/1.
к макушке, усажены рядом полых шипов. В спиной створке имеется раздвоенный замочный отросток. Силур—пермь.
Фиг. 177. Chonetes striatella. Верхний силур. Готланд. Швеция. 1/1.
в — со стороны спинной створки; Ъ—спинная створка совнутри.
Фиг. 178. Chonetes carbonifera-Верхнекаменноуг. С. Мячково-около Москвы. 3/1.
Productus (фиг. 166, 180—183; productus—продолженный). Area отсутствует или рудиментарна. Брюшная створка сильно выпуклая, с большой загнутой макушкой. На поверхности створок, главным образом в околома-кушечной части брюшной створки, имеются трубчатые иглы, служивши®
прикрепления раковины к посторонним предметам, причем иглы охватывали этот предмет, обрастали вокруг него. Поверхность раковины покрыта - радиальными ребрышками, а иногда и концентрическими л складками. Зубов нет, замочный отросток развит. Нерх-Z । ний девон, каменноугольные и пермские отложеш;..
w J Л
Фпг. 180. Productus se-mireliculatus. Нижнека-менн. вг^естпяк Визе. Бельгия. 1/1.
Фиг* 179. Choneles sarctnulata. Нижи девой. Кобленц. Германия. Наверху спинная створка совнутри, со срединной перегородкой и пластинками, ограничивающими зубвые ямки. 1/1.
Strophalosia (фиг. 184—184; strophes— согнутый, halos круг) отличается от Рго-ductus достигающей значительной высоты area на брюшной створке, с pseudodel-tidzum\ покров из трубчатых игол развит в высокой степени; иглы имеются даже и tidium. Прирастание происходило иногда прямо стенкой раковины, особенно макушкой, отчего на раковине оставался рубец. Каменноугольные и пермские отложения.
Различные виды Strophalosia повидимому выходят из различных видов Productus; тот или другой представитель рода Productus, прикрепленный к морскому дну, в местности с быстрым накоплением осадков был поставлен в необходимость быстро наращивать брюшную створку, особенно в околомакушечной части, посредством которой происходит прикрепление; таким образом возникала area значительной высоты, и усиливался покров трубчатых игол, служащих для прикрепления. Крайнюю степень раз-
Фпг. 181. Pr'-dudus giganteus. Разрез по длине раковины нормально к обеим створкам.
отмыкатеяи.
мадьно к обеим
а — замыкатели; Ъ —
на поверхности pseudodel-
Фиг. *82. Prodwcfws korti-dus. Пермские отложения (цех-штейн). Гера. Тюрингия. Германия. 1/2.
Фиг. 183. Productus 'horridus. Виутрен-яя сторона спинной створки. Пермские тложения. 1ёснек Тюрингия. Германия.
Фиг. 184. Strophalosia Goldfussi: вид сбоку, со стороны спинной створки, вид спинной створки совнутри. Пермские отложения. Гера. Тюрингия. Германия. 1/1.
вития area в высоту с значительным удлинением замочного отростка и изредка развитие днищ представляет Aulosteges (фиг. 186).
Подпорядок И. Pentameracea. Pentameracea имеют раковины, створки которых большею частью выпуклы. Area и pseudodeltidium слабо развиты или отсутствуют. Брюшная створка с сильно развитыми зубными пластинками, образующими spon-
dylium. В спинной имеются замочные пластины, иногда с отходящими от них crura. Короткий замочный край.
Представители Pentameracea известны в кембрие. наир. Syntrophia", отличается длинным замочным краем. Поверхность гладкая, delthyrium открытый; у Hvenella поверхность складчатая. '1о же в кембрие.
Porambonites (фиг. 187—189: pores—птпеп-
Фиг. 185. Strophalosia horres-
cens. Пермские отложения.
Усть-Bara. Северный Край. 1/1.
Фиг. 186. Aulosteges Wangenheimi. Пермский известняк Гребени, Оренбургский край. 1/1.
а — спинная створка совнутрн; Ъ — брюшная снаружи; с— то же, со стороны area; d — спинная створка со стороны замочного края
Фиг. 187. Porambonites aequirostris. Нижний силур Эхиносферитовый известняк. Пулково, около Ленинграда,
стие, ambon—возвышение). Раковина шаровидно вздутая, со створками, мало разнящимися по величине, поверхность покрыта радиальными рядами точечных углублений. Обе створки с низкою area, без псевдодельтидиума Длинные зубные и замочные пластины, иногда сходящиеся своими концами на дне створки и в брюшной створке даже сливающиеся в срединную перегородку. Нижний силур.
Pentamerus (фиг. 190—194; penta-пять, meros — часть). Раковина часто достигает зна пгтельной величины, имеет вздутую брюшную створку с большой и сильно загнутой макушкой. Сильно развиты зуб
ные и замочные пластины. Первые образуют spondylitim. Псевдодельтн-диум обыкиоЕЗНно отсутствует, а если имеется, то в связи с сильной
загнутостыо макушки является вогнутым. Поверхность раковины гладкая или радиально-ребристая. Силур, девон.
а - Ъ
Фиг. 188. Рогат-bonttes reiiculalus. Нижний силур. Глауконитовый извест ник. Пулково, около Ленинграда.
Фиг. 189. Porambonites aequirostris. Нижний силур (невский ярус). Свитам. Эстония.
а — брюшная створка, b — спнпная.
®пг. 190. Pentamerus voguhcas. нижний девон Пнжний Туринск., Уральской области. Урал. 1/1.
Фиг. 191. Р. Knighti, разбитый
Фиг. 192. Р. esthonus Верхний силур, Эстония 1/1.
вдоль по снондилнуму; вид со стороны поверхности разлома. Верхний силур. Англия. 1/1,
Фиг. 193. Р. ga.le.a-tus. Средний девон. Эйфель. Германия. 1/1.
Фиг. 194. Р. acuto-lobatus.
Пнжний девон. Богословск.
Уральская обл. Урал. 1/1.
Camaroplioria (фиг. 147, 195, 196; camara — сводчатое помещение, phoieo—несу), отличается от Pentamerus меньшими размерами и наличностью spondylium в обеих яворках. Имеются длинные, тонкие crura Каменноугольные, пермские.
Подпорядок 1П. Rliynclionellacea У Rhyndio-nella (фиг. 150, 197 — 199; rhynchos — клюв, rhynchonella — клювик) имеются дельтидиаль-ные пластинки в отверстии дельтириума, иногда соприкасающиеся в плоскости симметрии раковины, ограничивая снизу отверстие для ножки. В спинной створке имеются crura. От силура доныне. Подпорядок IV. Terebratulacea. Отличаются присутствием петлеобразного аппарата ручных
Фиг. 195. Cmnarophoria supertes. Нижнепермский известняк. Кириллов, Ленинградской обл. 1/1.
поддержек. Прикрытие дельтириума осуществляется посредством дельтлдиаль-ных пластинок. Terqaratula (фиг. 151, 200: уменьшительное от terebratus про-
Фпг. 196. Camarophgi ш Schloiheium . Перм ские отложения. Гера. Тюрингия. Германия. Слева и наверху раковина, внизу ядро. Справа— внутренность створок в околомакушечной части рт—замочный отросток; с—crura", x—sptndylinm брюшноя створки, д—спинной створки; s и s'—средние перегородки
Фиг. 197. Rhynchonella Меуеп-dorffr. Верхний девон. Псков. 1/1.
сверленный) имеет гладкую, редко ребристую раковину, на лобном крае спинной створки часто с выемкой, ограниченной по бокам одной или двумя складками. Ручной аппарат короткий. От триаса доныне, особенно в мезозое.
Фиг. 199. В. loxia. Верхневолж- Фиг. 200. Tere^ratnla ские слои. Хорошово, около сагпса. Белый мел-Москвы. 1/1. Ульяновск. 1/1.
Фиг. 198. В. cuboides. Верхний девон Котлубан. Урал. 1/1.
Палеозойские формы этого типа принадлежат главным образом к роду Pielasma (фиг. 201) или сходные с ним. Dielasma (di—два, elasma—пластинка) отличается сильно развитыми зубными пластинами. Девон—пермь.
Фиг. 202. WaUL-heimia Fijche-riana. Верхние волжские слои. Хорошово, около Москвы.
Waldheimia (фиг. 145, 146, 148, 202; по имени ученого Вальдгейма) отличается весьма длинным ручным аппаратом. 0 мезозоя доныне.
Stringocephalus (риг. 203; strmgo — натягиваю, cephale — голова) представляет весьма специализированный род Terebratu-lacea. Раковина большая, гладкая. Брюшная створка с острой, большой и сильно загнутой макушкой, под которой находится ограниченный дельти диальны ми пластинками foramen.
Фиг. 201. Dielasma dong ata. - Цех-штейн. Гёмблетон. Англия. Справа внутренность раковины в околомакушечной части, увелич.
Брюшная створка с
сильно развитой срединной перегородкой, спинная с необыкновенно длинным замочным отростком. BracJiiduim длинный, следующий краю створки при
Фиг. 203. Stringocephahis Burtini. Средний девон. Пафрат, около Кельна, Германия.
л—наружный вид раковины; Ъ —экземпляр с ручным аппаратом: с —молодой экземпляр с большим макушечным отверстием; сггяняая creepка ® ручвым аппаратом (Ъ — замочный отросток; d — зубные ямки; с — стига; I— ручной аппарат; вт — septum; а — замыкатели).
бллзительно на всем его протяжении, с радиальными шиповидными отростками в лобной части. Девон.
Подпорядок V. Spiri-
feraeea. К brachidtum
типа Terebratulacea у Spiri-feracea присоединяются известковистые ленты, образующие спирально свер-иутые конусы.
Spirifer (фиг. 153, 204—207; spiro — спираль, fero—несу) имеет
обращенными внутрь
Фиг. 204. В. Spirifer Verneuili (Archiaci). Верхний девон. Елен, Ц. Черноземной обд.3/4.
Фиг. 204 A. SpiriferVetneuili (disjunctus). Верхний девон. Р. Девица, на Ю. от Воронежа, Ц. Черноземной обе 1/1.
конусы ручных поддержек, направленные основаниями друг к другу и к плоскости симметрии раковины, вершинами в противоположные стороны к боковым краям. Замочный край прямой, с area и треугольным отверстием
Фиг. 205. Spirifer musqutnsis. Верхнекакен-ноуг. Мячковс, около Москвы. 2 2.
Фиг. 206. Spirifer fasciger Вер* векаменноуг. Куткан'.ау ва Юрс-зани. Урал. 3 4.
дельтириума. От силура до триаса. Представляет многочисленные подроды. Путем удлинения area с возникновением псевдодельтидиума возникает Cyrtia (фиг. 156 Н—К, cyrtos — согнутый), при развитии кроме того споидилиума в брюшной створке получается Cyrtina (фиг. 208).
Фиг. 208. Cyrtina carbonaria. Нижне-камевноуг. Кендалль.
ния. 1/2.
Ирландия. 1/1.
Фиг. 207. Spirifer undulaius. Пермские отложения. Цехштеин. Герма-
©иг. 209. Retzia tulensis. Девонско-каменноуг. отложения (малевка-мурае-внпнский ярус). Ма-левка, к Ю. от Тулы,
Московск. обл. 1/1.
замочным краем; ра•
коротким изогнутым
Retzia (фиг. 209) отличается
ребристая. Брюшная створка с выдающейся, усеченной отверстием форамена макушкой, под ним находится дельтидиум. От силура до триаса.
Uncites (фиг. 210; uncus — крючок) вероятно возникает из Retzia путем удлинения околомакушеч-ной
ковина радиально
части, сильно изгибающейся. Девон.
Фиг. 211. A Tigris concent-rica. Верхний девон. Спинная створка с ручным аппаратом. 1/1.
Фиг. 210. Uveites gryphus_ Средний девон. Бенсберг, около Кельна. Германия. 1/2.
Фиг. 212. Athyns pectini-fera. Пермский известняк. Баитуган, Самарский край. 2/1-
Aihyris (фиг. 211—213; a—без, thyris—маленькая дверь; намек на отсутствие delthyrium) имеет раковину гладкую или с концентрическими украшениями. От силура до триаса, главным образом в -девоне и каменно-
угольных.
фиг. 213. AtJiyris Pus-ehiana. Малевка-мураев-нинский ярус. Мура-евня, к С. от Ранен-бурга, Московской оба.
1/1.
Фиг. 214. Atrypa as-рега. Средний девон. Эйфель. Германия. 1/1.
Фиг. 215. Atrypa reticularis.
Средний девон. Геролыптейи.
Эйфель. Германия. 1/1.
Atrypa (фиг. 154, 155, 214—215; а—без, trypa—отверстие; сначала ошибочно предполагалось отсутствие форамена) отличается от вышеописанных Spiriferace расположением конусов ручных поддержек; конусы обращены основаниями к брюшной створке, в связи с этим менее выпуклой, чем спинная. Раковина радиально ребристая, редко гладкая. Распространен в си-
луре и девоне-
Распространение пле^еног^лх.
Плеченогие дают большое количество важнейших геологических руководящих форм, отличаясь значительной распространенностью во времени и в пространстве и способностью к хорошему сохранению в ископаемом состоянии Их раковины состоят главным образом из известкового шпата и менее разрушаются при окаменении, чем состоящие из аррагонита раковины моллюсков. Но определения ископаемых брахиопод в отношении родов и видов затрудняются необходимостью установления внутренних особенностей раковины и сильною изменчивостью в связи с прикреплением к месту обитания.
Ез двух главнейших подразделенийбрахиопод беззамочные являются конечно, древнейшими, они особенно распространены и разнообразны в палеозое. Из них простейшие, не имеющие форамена более всего распространены в кембрии и нижнем силуре (Obolus, Lingula и др.), менее—в верхнем силуре и девоне и почти вымирают в каменноугольных, когда остается лишь род Lingula.
Беззамочные, имеющие форамен, распространены главным образом в нижнем силуре (Siphonotreta, Crania и др.), после чего выживают лишь Discinidae и Crania. Strophomenacea и Pentameracea встречают* я с кембрия, наибольшее количество родов их в силуре: Orthidae — главным образом в нижнем силуре, Strophomenidae—в верхнем; Productidae в каменноугольных. Strophomenacea имеют и ныне живущих представителей, f fcfe как Pentameracea вымирают в конце палеозоя, за исключением Rhyncho-Bcllidae. Sporiferacea встречаются от силура до юры, a Terebratulacea от си-лУра доныне.
Геологическое распространение брахиопод.
Роды Отделы или ярусы
Кембрий Силур Девой Карбои Пермь Триас Юра । Мел Третнч-! ные i Совре- 1 менные 1
Paterina . . . Volborthia . . Rustella . . . Obolus .... Lingula . . . Siphonotreta . Discina . . . Crania .... Orthis .... Orthisina . . Stropkomena . Orthotheies . . Chonetes . . . Productus . . Strophalosia . Aulosteges . . Porambonites Pentamerus . Camarophoria Rhynchonella . Terebratula Waldheimia . Dielasma . . Stringocephalus Spirifer . . . Cyrtina . . . Retzia .... Uncites . . . Athyris . . . Atrypa . . . —
—
—
W
—
—
F ТИП. ЧЛЕНИСТОНОГИЕ, ARTHROPODA. 1
Членистоногие (раки, пауки, многоножки и насекомые) во многих отношениях близки к кольчатым червям и прежде даже были соединены с последними в один тип.
Тело членистоногих представляет сегментацию внешнюю и внутреннюю Внешняя сегментация, членистость тела членистоногих выражена даже более резко, чем у аннелид, вследствие образования твердого наружного кожного панцыря, хитинового или известкового. Такой твердый панцырь делал бы тело животного неподвижным, если бы он не распадался на отдельные части —сегменты; границы частей совпадают с границами внутренних сегментов.
С другой стороны, членистоногие отличаются от аннелид в общем большей диференциацией, специализацией различных отделов тела, слабой у низших членистоногих (многоножки), значительной у высших. Голова всегда состоит из нескольких слитых сегментов, следующая группа сегментов образует грудь {thorax)- и наконец третья группа сегментов составляет брюшко {abdomen).
Наиболее характерным для артропод отличием являются их конечности—членистые, что и отразилось в названии типа.
В то время как аннелиды обладают самое большее конечностями в виде параподцй, нерасчлененных выступов брюшной стороны тела, не двигающихся самостоятельно, конечности артропод сочленены с телом посредством суставов и способны к самостоятельному движению как в целом, так и в отдельных частях. Здесь по сравнению с аннелидами происходит переме'ще-ние двигательной деятельности из стенки тела в конечности. Первично каждый сегмент имеет одну пару конечностей, как и у аннелид, но затем происходит видоизменение конечностей соответственно отделу тела, на котором находятся данные конечности.
Прежде всего находящиеся на голове конечности частью делаются носителями органов чувств (сяжки, антенны), частью же превращаются в органы для схватывания и пережевывания пищи, т. е. в челюсти. Это происхождение сяжков и челюстей отчасти доказывается непосредственно тем, что у зародышей и личинок многих раков (стадия науплиус), а иногда даже и у взрослых животных названные органы являются в виде настоящих, притом действующих ног.
Но обыкновенно локомоторная деятельность этих, самых передних конечностей у артропод исчезает, и происходит сближение и слияние соответствующих сегментов тела, образование из них головного отдела, состоящего У ракообразных, соответственно числу на нем сяжков и челюстей, из пяти сегментов. Происхождение головы слиянием сегментов доказывается также непосредственно тем, что границы между сегментами, впоследствии слившимися, еще сохраняются у зародыша. Изредка они сохраняются и у взрослых животных.
Локомоторные конечности всегда существуют на среднем отделе тела— г Р у д и, тогда как брюшные ноги могут отсутствовать совершенно или быть приспособлены для целей дыхания или размножения.
Дыхание у артропод происходит или через поверхность тела—у живот-
1 Arthros—член, pens, podos—нога
ных малых размеров, с мягкими покровными частями, или посредством специальных органов: жабер—у водных животных, легких и трахей—у наземных.
Жабры являются в виде выростов кожи, листоватых или кустоватых, с нитевидными разветвлениями, прикрепляются на конечностях, своими движениями постоянно доставляющих приток воды к жабрам; иногда сами конечности превращены в жабры.
Трахеями называются длинные разветвленные трубочки, наполненные воздухом, открывающиеся наружу и проникающие все части тела; легкие являются складчатыми органами, в полостях с ограниченным сообщением со внешней средой.
При наличии специальных локализованных органов дыхания получает значительное развитие кровеносная система (сердце, кровеносные сосуды); при дыхании же путем диффузии, через кожу поверхности тела, кровеносная система развита слабо или вовсе не развита. Понятно, что если кислород окружающей среды поступает в тело лишь в немногих местах тела, в специальных органах дыхания, то развитие кровеносной системы является необходимым, ибо только при этом кровь может достигать органов дыхания даже из отдаленных частей тела.
Нервная система членистоногих тесно примыкает к нервной системе аннелид по присутствию брюшных нервных стволов, с нервными узлами па них, и по присутствию головного мозга, соединенного по сторонам глотки с брюшным.
Срастание или тесное соединение смежных сегментов в отделы тела влечет за собою слияние соответствующих этим сегментам нервных узлов.
Органы чувств являются в виде расположенных на некоторых местах осязательных волосков, подобных же обонятельных и вкусовых органов, а также глаз, находящихся на голове.
Глаза представляют прозрачные светопреломляющие тела: чечевицеобразный хрусталик (являющийся местным утолщением хитинового покрова тела) и лежащее под ним стекловидное тело (видоизменение эпидермиса); под стекловидным телом находится сетчатая оболочка с примыкающими к ней нервными клетками. Глаза "бывают простые и сложные (фасеточные), представляющие скопление—до нескольких тысяч—простых глаз.
Соответственно высокой организации и специализации отделов тела артропод бесполое размножение у них не встречается.
Что касается классификация артропод, то надо сказать, что еще недавно объединяли вместе пауков, многоножек и насекомых в группу Tracheata, т. е. животных, дышащих внутренними воздухоносными полостями, обособляя их от дышащих жабрами Branchiata, раков.
Несомненно однако, что пауки на основании многих других особенностей организации много ближе к ракам, чем к остальным Tracheata, и что вовсе нет необходимости считать наличие внутренних воздухоносных дыхательных полостей за свидетельство тесной степени родства. Такие полости могли быть приобретены различными группами независимо одними от других. Как раз внутренние воздухоносные дыхательные полости древнейших паукообразных, именно «легкие» скорпионов, нетрудно вывести из жабер известных раков—мечехвостов (см. ниже); и это тем более имеет значение, что характер расчлененности тела и число конечностей этих обеих групп замечательным образом одинаковы.
Затем ракообразные и паукообразные представляют слияние головы и груди в головогрудь (cephalothorax), тогда как многоножки и насекомые отличаются, наоборот, обособленною головою. .
На основании вышеизложенного членистоногих можно подразделить на два подтипа: Cephalothoracica и Eucephalica.
Подтип I. Cephalothoracica.
КЛАСС I. РАКООБРАЗНЫЕ. CRUSTACEA. 1
Фиг. 216. Важнейшие формы конечностей речного рака:
1—первая аатениа; 2—мандибула; 3 и 4 —максиллы: 5—7— ногочелюсти; 8—хорная нога;
9» «брюшная ножка.
Хитиновый панцырь ракообразных часто делается более прочным вследствие отложения в нем известкового вещества.
Членистость тела никогда не выражена полно, но проявляется присутствием посегментных конечностей. Сегменты головы обыкновенно не выражены; сегменты груди также могут в различной степени сливаться между собою и с головными сегментами (головогрудь). Во всяком случае нет резко обособленной головы. Брюшко всегда расчленено.
Часто со спинной стороны головогруди разрастается на бока обособленная от них складка кожи, отлагающая своею поверхностью раковину или в виде нераздельного спинного щита, или двустворчатую, покрывающую все тело или некоторые части его.
Ракообразные, за немногими исключениями, типичные водные животные; этим определяется характер органов дыхания (жабры) и форма двигательных конечностей —п лава тельных-ножек.
Ножки ракообразных первично являются расщепленными, состоя — или во время всей жизни, или в зародышевом состоянии — из двух ветвей, сидящих на короткой основной части (фиг. 216).
Головной отдел типичных ракообразных несет пять пар конечностей. Первые две пары, которые У личинок и у низших раков еще часто служат
для гребли, обыкновенно становятся исключительно носителями органов чувств осязания, обоняния и других (1 и 2 антенны), три следующие пары головных конечностей окружают рот и действуют как челюсти, посредством превращенной в жевательную пластинку основной части, на которой сидят более или менее незначительные по размерам ветви; далее всего идет это превращение у передних челюстей (мандибул), менее далеко у остальных двух пар челюстей (1 и 2 максиллы).
Кроме того, часто еще принятию пищи содействуют передние грудные ноги (ногочелюсти), не будучи однако превращены в настоящие челюсти.
1 Crusta — корка.
Двигательные ноги могут служить для передвижения различным образом, даже у одного и того же животного в различных частях его тела. ¥вк, туловищные конечности у высших раков, утрачивая одну ветвь, превращаются в длинные ходные ноги, которые несут тело, опираясь на почву поэтому некоторые раки, прнобревшие такие ноги, сделались животными наземными вполне или отчасти (мокрицы, крабы). Одновременно с ходкими грудными ногами могут быть и плавательные брюшные, сохраняющие рас
Фиг. 217. а—nauplius; Ь—zoea.
щепленное состояние.
У многих ракообразных переход от конечностей одной категории к конечностям смежного отдела выражен постепенностью изменения характера конечностей при переходе от одного конца тела к другому. Таков переход от челюстей через ногочелюсти к ходным ногам обыкновенного речного рака (фиг. 216).
У некоторых Crustacea индивидуальное развитие является прямым, без метаморфоза, так что молодое животное отличается от взрослого лишь размерами, но не формой и организацией, но обыкновенно развитие совершается с метаморфозом, прпчед одни раки проходят личинковую стадию, называемую nauplius, а другие zoea (фиг. 217).
У nauplius тело несегментиро-вано п имеет три пары конечностей, представляющих две пары антенн и мандибулы. У zoea имеются и некоторые из средних конечностей, а брюшко сьгментиросано, но без конечностей.
Ракообразные делятся на четыре подкласса: 1) Entomostraca, 2) Mala-costraca, 3) Trilobitae, 4) Gigantos-traca, или Merostomata.
Возможно, что новые успехи палеонтологии и эмбриологии дадут основание для выделения обеих последних групп в качестве самостоятельных классов, равноценных Crustacea, Arachnoidea. Myriopoda и Insecta.
Подкласс I. Entomostraca.1
Наиболее примитивные, большею частью небольшой величины ракообразные, с переменным количеством сегментов и конечностей (от менее десяти пар конечностей до нескольких десятков), с панцирем или без него, в индивидуальном развитии проходящие стадию nauplius
ПОРЯДОК I. ЛИСТОНОГИЕ, РНУLLOPODA. 1 2
Называются так по характеру своих ног, являющихся пластинчато* сплющенными и едва членистыми. Ногочелюстей нет, брюшко без конечностей. По внешнему виду весьма разнообразны. У одних имеется спинной щит (Apus, фиг. 218), у других двустворчатая раковина (Estheria, фиг. 219),
1 Entomos—разделенный, ostracon—черепок.
2 Piiytton—лист, pons, podos—нога
бывает и отсутствие панцыря. Древнейшие (кембрий)—морские животные, позднейшие—обитатели пресных или солоноватых вод.
Estheria (фиг 219; от собственного имени). Раковина равностворчатая тонкая, роговая, овальная или четырехугольная, с прямым замочный
Фаг. 218. Apus cancriformis. Слева вид со спинной, справа—с брюшной стороны.
краем; макушки створок находятся впереди или посредине. Наружная поверхность концентрически ребристая или струйчатая, а между ребрышками пересекающиеся или ветвящиеся струйки. Девон—ныне.
Род Apus, характеризующийся непарным спинным щитом, встречается с триаса доныне.
ПОРЯДОК II. РАКОВИНЧАТЫЕ, OSTRACODA.
Маленькие ракообразные, вполне заключенные в двустворчатую раковину; с немногими неясными сегментами (фиг. 220) и рудиментарными нелистоватыми ногами. Две пары антенн служат для схватывания добычи и для плавания, три пары челюстей, две пары ног.
Створки раковичнатых соединены посредством эластичной связки, находящейся на спинном краю, открывающей их, и мускула, идущего поперек ют одной створки к другой и закрывающего их. Раковина известковая или роговая, иногда несет украшения на поверхности в виде бугров,
струек и т. п.
Живут стаями
Фиг. 210. Estheria
rninuta. Триас. Зинг-сгейм. Баден. Гер-• мания. 6/1.
Beyrichia (фиг. 222; по имени ученого Бейриха). Раковина маленькая,
в мелкой воде и питаются трупами животных.
Lepev&tia (фиг. 221; lepis—чешуя; dittos—двойной). Раковина продолговатая, расширяющаяся назади, спинной край прямой, брюшной закругленный. Створки не одинаковы, правая шире и охватывает брюшной край левой створки. Поверхность раковины гладкая, в исключительных случаях (L. Moelleri) покрыта бугорками; напереди и близ спинного края находятся глазные бугорки. На внутренней поверхности створок находится приблизительно центральное большое мышечное впечатление. От силура до каменноугольных.
краем; поверхность ее вздута в не-
равностворчатая, с прямым замочным
Фнг. 220. Cypridina mediierranea.
£1—сердце; SSI — мускулы: О — парный сложный глаз; 0'—непарный глаз; G—головной мозг; Sts—лобный орган; F—орган оплодотворения; Т— семенные железы; Fu— вильчатое раздвоение заднего конца тела.
Фиг. 221. Leperdi- Фиг.222. Beyrichia На Hisingeri. Верх- tubercuiata. Из си-иий силур. Гот (анд. лурийских валунов Швеция. 1/1. Восточной Пруссии.
Фиг. 223. Сурп з faba. Множен.
Энинген. Швейцария. 15/1.
многочисленные крупные бугры, разделенные бороздками; поверхность бугров бугорчата. От силура до каменноугольных.
Cypris (фиг. 223). Раковина почковидная или овальная, тонкая, гладкая или шиповатая, или с точечною псверхностью. Брюшной край иногда вогнутый. В пресной воде. От третичных доныне.
ПОРЯДОК III. УСОНОГИЕ, CIHE1PEDIA 1
Животные по внешнему виду настолько Непохожие на ракообразных, что их долго причисляли к моллюскам. Такое уклонение от типа обусловливается сидячим образом жизни усоногих в зрелом состоянии. Свободно пе-
1 Cirri—усики. Pes, pedis—нога.
передвигающиеся личинки их близки к типичным ракообра чым, представляют остракодовую, так называемую стадию cipris Ци ирисовые личинки
прикрепляются тонною частью тела и зачем теряют двустворчат )ю раковину, а в cuticula остаю'щейся мягкой мантии часто развиваются изиежовые таблички (фиг. 224—226). Членистость тела, челюсти, глаза ис.е^ают,
жабры ичсердце отсутствуют, 4—6 пар дву-ветвнетых, закрученных и высовывающихся из полости мантии «усоножек» производят своими движениями движение воды, пригоняю-
Фиг. 224. Разрез Lulanus.
Ти—раковина, Ос—яичник; Od~яйцеировод; Ов— отверстие после диего; 4 aductor; Sc— scuiuwt Тв—te gum.
Фиг. 225. Lepas ananjeza.
S—scutum; T—lergum; C—carmu; /‘—стебел* Ныне живую..
щее пищу. Гермафродиты. Большей частью морские животные. С каменно-
угольных отложений.
Lepas развивает из прикрепленной передне-головной части длинный мягкий стебель и в мантии пять известковых табличек, по бокам jiapnue terga (находящиеся наверху), scuta (находящиеся у стебля) и непарную спинную carina (фиг. 225). Третичные—ныне.
У Balanus (фиг. 224 и 226; balanos — жолудь) стебель заменен пластинкой, которой животное прирастает к камням, раковинам и другим предметам прибрежной полосы; на периферии этой пластинки от нее поднимаются особые известковые пластинки, образуя бокаловидную раковину, в отверстии которой лежа'? terga и scuta, образуя крышечку отверстия. Третичные — ныне.
и
Фиг. 226. Схема раковины баланса.
С — carina; R — rostrum; В — baste, CL—carino-laterale; L—lateralc; RL— roslro—iaterule; a—alae, r—racii, p—parietes.
Подкласс П. Malacostraca. 1
Из Malacostraca rpjnna Leptostraca представляет некоторые особенности, общие с Entomostraca. Именно разделенные сегменты груди, дв) ветвистые пластинчатые грудные ноги, вильчато-раздвоенный хвостовой сегмент и дву
1 Malacos—мягкий, ostracon—черепок, раковнвь
® Учбик аале«жт»догж«
129
створчатую раковину (фиг. 227) Но по числу сегментов тела—5 в голове 8 в груди, 7 в брюшке—Leptostraca являются близкими к Malacostraca.
В настоящее время Leptostraca немногочисленны, но есть много ископаемых палеозойских форм, принадлежаших, вернее всего, к Leptostraca. Конечности их мало известны, и различение родов производится главным образом на основании формы панцыря и хвостовой части. Большие сомнения вызывает отнесение к этому подклассу рода Spathiocans (фиг. 228), самая принадлежность которого к ракообразным вообще является спорною (см. ниже об анаптихах в главе о головоногих моллюсках).
Фиг. 227. Nebalia Geoffгоуг.
А! и А"—антенны; О—стебельчатый глаз; Л—желудок; D—кишка.
паемом состоянии неизвестны.
Фиг. 228. Spalhiocaris. Верхний девон. Тиман. Обл. Коми. 1/1.
Подкласс Alai icostraca делится на три группы: Leptostraca, Arthrostraca и Thoracostraca. Первая ив них уже охарактеризована; Arthrostraca отличаются отсутствием или слабым развитием головогрудного панцыря, Thoracostraca — псгтоя! но сильным развитием его. Arthrostraca—так же, как и Leptostraca -вследствие ела! ого развития панцыря или слабой связи его с сегментами плохо сохраняются в ископаемом состоянии. Отчасти поэтому — еще вероятно и вследствие слабого развития, малой распространенности во время палеозоя— Arthrostraca редки в отложениях последнего
К Arthrostraca принадлежат порядки Isopoda (равноногие) и Amphipoda (бокоплавы).
Isopoda, характерными представителями которых являются мокрицы, имеют тело, сжатое в сппнно-брюшном направлении, с коротким брюшком, часть сегментов которого срастается. Плавают и ползают яри помощи грудных ног. Палеозой (?), триас—ныне.
Amphipoda—маленькие морские и пресноводные рачки, плавающие и прыгающие; имеют сжатое с боков тело, три пары передних плавательных и три пары задних прыгател> ных ног. Достоверно известны ляль из третичных отложений, имеютс.. проблематичные находки из палеозоя.
К Thoracostraca принадлежат подпорядки Schizopoda, Stomatopoda, Decapoda и Cumacea; у Schizopoda (расщеплеиноногих) хитиновый спинной павцырь прикрывает все туловищные сегменты, срастаясь лишь с тремя лоредними сегментами груди. Расщепленные ноги па головогруди. Палеозой (?) триас—ныне.
У Stomatopoda (ротоногих) панцирем прикрыты тахже лишь передние (о) сегменты туловища. Отличительно преоблазание развития брюшка над зередней частью. Задние туловищные ноги расщепленные. Туловищные ноги второй пары имеют большие размер»* I служат для схватывания до ычи. Пал озон (?), юра—ныне.
Cumacea в состав гоювогр,ди входят лишь той передние сегмента груди; в иско-
ПОРЯДОК I. DECAPOD А, ДЕС ЛИНОГИЕ РАКИ.
Голова и, почти всегда, все сегменты туловища покрыты большим и обыкновенно прочным головогрудным щитом (а—с; фиг. 229); он часто под-
разделен на передний, .оловной и задний, грудной, отделы затылочной бороздкой (фиг. 2z9,&, 232). Щит напереди продолжается в остроконечно,
С
Фпг. 229. Glyphea regleiana. Верхняя юра.
а-с—головогрудь; Ъ—затылочная бороздка, a— rostrum, д—глаза, h-o—копеч-
косчп головогрудного отдела, - абдомен; e-f—хвостовой плавник
4 а, а**—антенны (сяжки)- тд—тпх—челюсти; 8—последвяя иогочелюсть; 9—18—родные ноги; 14—19—плава-ельные коги; <?«—яички; 19—20— хвостовою илаввик, Ad'—Afa'—детали челюстей; Гр2—вторая ходная нога;
Ар— третья плавательная нога; Л/р3—вторая иогочелюсть.
rostrum (а, фиг. 229). Жабры помешаются в полостях под боковыми, вниз °т спины идущими продолжениями головогрудного щита. Головогрудь несет
13 пар придатков — пять принадлежащих голове и восемь груди. На голове находятся две пары сяжков (а, а", фиг. 230, Л) и три пары челюстей. Три передние пары конечностей груди служат для движения и для принятия
Фиг. 231. Eryon propinquus.верхняя юра (литографский сланец). Ззлеигофе Бавария. Германия 1/1.
пиши, почему очи называются ногочелюстями, задние пять пар суть одио-ветвистые хотные ноги; некоторые из них оканчиваются клешнями Abdomen несет шесть4или менее) пар двувеiвистых плавательных hoi (фиг 230, Л), служащих у самок также для прикрепления икры (яичек); самая задняя
пара этих ног сплющена в спиннобрюшном направлении я образует с конечным сегментом юла хвостовой плавник.
Decapod 1 подразлеляется на две группы- долгохвостых Масгвга и короткохвостых. и та крабов, Bracbytira. Первые встречаются с триаса (Рет-pjiir, фиг. 232), изобилуют в юре (Glyphea, Егуоп, фиг. 229, 231). у Brachyura, встречающихся с пермских отложений и достигших наибольшего распространения в настоящее время, abdomen короток, без хвостового плавника, загнут на головогрудь, помещаясь в продольной впадине ее брюшной поверхности.
Подкласс Ш. Трилобиты, Trilobitae.
Вымершая палеозойская группа, получившая свое название (trilobos— трехчленный) от разделения тела на три части двумя спинными бороздками, протягивающимися от переднего конца до заднего (фиг. 233). Срединная непарная продольная часть тела, заключенная между упомянутыми бо
Фиг. 233. Phacops (T)alma-nia) cauilalus. В ‘рхний силур.
Дедлей. Англия.
Л—голов ой щит; г—туловище; з— хвостоьыи шит. s/j—s/j—поперечные бороздки глабели; iZ—щечные остро-ковеч,1я; оу осевая лопасть туло-винта: I—краевая каемка головного Щита, I'—хвостового щита; st1 шиловидное продолжение последнего»
Фиг. 234. Головной щит Phacops (Palmania) Haus-manui. Верхний силур. Богемия.
pZ—лимб, sin—краевая бороздка; а— щечные остроконечня; гла-бель 1—3—поперечные бороздки глабели; х /2— пере идея, средняя И за идея боковые лопасти Гибели; 80—затылочная боровгка, а— затылочное кольцо; а—лицевой шов; ос—зрительная поверхность глаз; d— пальпебральная лопасть.
роздками, называется осевою (rhachis), она выпукла; две боковые части {pleurae) понижены относительно средней части (фиг. 236). Тело покрыто панцирем, отличающимся значительным содержанием хитинового вещества,. В передне задаем напра 'Ленин тело также подразделено на три всегда явственно обособленные части: головной пиг, туловище, состоящее из Переменного числа подвижных сегментов, и хвостовой щит (фиг. 233).
Головной шит обыкновенно полукруглого очертания и примыкает прямым задним краем к твловищу. Осевая часть головного щита называется гла-белью (glabella), боковые щеками (genae). Краевая часть головного Щита обыкновенно отделена от остальной его поверхности бороздками: у заднего края затылочною (фиг. 234), обособляющей от щита за th-
Фиг. 2м5. Р acops latifrons.
ql — гл а бель; sf — глаз; pf — продолжение головного щита над глазам, з — хвостовой щит
X
л очное ко и.цо, на остальном протяжении краевою бороздкою, отделяю-щей от щита выпуклое краевое вздутие или плоскую краевую кайму (limbus). "Щит представляет заворот набрюшную поверхность. Задние углы щек иногда бывают выткнуты в щечные остроконечна.
Перед затылочной бороздкой глабель представляет обыкновенно парные так называемые поперечные бороздки (sulci laterales) в числе 1—3 пар, разделяющие глабель на боковые лопасти. Вся часть глабели, лежащая перед передними боковыми бороздками, .называется лобною;
иногда боковые бороздки одной и той же пары соединяются посредине глабели. Надо думать, что боковые лопасти глабели соответствуют конечностям, расположенным на нижней стороне -тела около рта. 1
Щеки подразделяются на чатси так ’называемыми швами, проходящими по поверхности головного щита в виде линейных бороздок. Предполагается, ч,о в соединении по этим швам существовала некоторая подвижность соединенных частей щита После смерти животного часто происходит распадение головного щита на части по этим швам (фиг. 242, 245).
Главным из них является лицевой шов (sutura facialis), отсутствующий лишь у немногих родов трилобитов. Он начинается двумя ветвями (по ветви на каждой из щек) на заднем краю щита, или в задних углах щек, или впереди этих углов на наружном краю, идет оттуда к глазам, шибает их с внутренней стороны и затем направляется наперед; при этом обе ветви его или соединяются перед глабелыо у лобнотс крал щита, или переходят несоединенными на нижнюю поверхность щита в лобной его части (например Calymmene), будучи часто соединены на завороченной вниз части щита особым параллельным лобному краю ры лицевым швом (sutura ros'ralis). В. этом случае лицевые и рыльцевой шов отделяют на завороте щиток, также называемый рыльце-вым (rostrate). Лицевой шов делит щеки на наружную подвижную ча(ть и на внутреннюю неподвижную, прилегающее к глабели, часть. В лобной
части головного щита на брюшной поверхности его к краю заворота примыкает пластинка своеобразной фирмы различной у различных трилобитов, называющаяся г и постимой (hypostoma) и гомологичная верхней губе ракообразных (фпг. 237, В?/р).
Глаза отсутствуют у немногих сравнительно трилобитов; чаще имеются по одному на каждой щеке, обыкновенно являясь сложными, фасеточ-н ы м и. Линзы (фасетки) в различи >м числе, о ыкновенно в большом, доходящем до 15 000. Обыкнивензо весь глаз покрыт сплошной роговой оболочкой, иногда прозрачной, реже общая оболочка непрозрачна и тогда снабж два отверстиями соответственно каждому отдельному глазку.
Фиг. 236. Схематический разрез через туловищный сегмент трилобита.
Ър—основные частя ног; еп—внутренние- ег—наружные ветви конечно'той; Ъг—жабры (под вопросом) аг -железы (под вопросом); d-кишка, п —нервные стволы; А—кровеносный сосуд: ер—* осевая часть спинной поверхности сегмента, pl—плевры.
Туловище (thorax) состоит из подвижно сочлененных между собою сегменте в, число которых варьирует между 2—29 у различных родов. Осевые части сегментов имеют напереди продолжение, подходящее под впереди лежащий смежный сегмент. Благодаря этому трилобиты большею частью способны свертываться (фиг. 235) так, что головной и хвостовой щиты соприкасаются, а вся мягкая брюшная поверхность будет закрыта и в безопасности от нападения; только при этом упомянутые передние продолжения осевой части сегментов становятся
видимы.
Боковые части (pleurae) сегментов иногда представляют на своей наружной поверхности продольную бороздку или ребро. Каждая плевра на некотором расстоянии от спинной бороздки представляет перегиб, так что наружная часть от внутренней отогнута вниз и обыкновенно в то же время назад, так что при свертывании трилобита эти отогнутые части совершенно замыкают' его тело с боков.
Наружная от перегиба часть плевры или сохраняет одну и ту же ширину до конца, закругляясь тогда на нем, или суживается к конпу и даже вытягивается в длинный шип. Наружный конец плевры представляет заворот панциря на брюшную поверхность.
Хвостовой щит (pygidium) состоит из слитых в одно целое
Фиг. 287. Реставрация нижней стороны три лобита.
Нур—гвпостоыа: At— антеввы: Л/d — мандибулярные ко вечности; максиллярные конечности; Л/р—
вогочелюсти затылочного кол»ца; Edp— внутренние, ходкие; Ехр—наружные плекачельные ветви чудовищных конечностей Ър— основные части ослодних; Н—трубочка (просвечивают); А— анальное отверстие.
сегментов, границы которых иногда
намечаются бороздками на всей поверхности щита, иногда ясны лишь на осевой части и наконец могут совершенно отсутствовать. Осевая лопасть простирается до заднего края щита, или является укороченной, сосредоточиваясь в передней части его (фиг. 259) Край хвостового щита часто представляет, подобно головному щиту, лимб, цельный или зазубренный выдающимися концами сегментов. Число сегментов хвосювого щита изменяется от 2 до 28.
Конечности у трилобитов имеются в большом количестве, находясь на каждом сегменте (фиг. 237) туловища и хвостового шита (за исключением последнего, анального сегмента) у тех немногих трилобитов, у которых вообще были открыты сохранившимися ноги. Конечности являются двувет-вистычи (фиг. 236, 237), состоят из основной трехчленистой части и отхо-
адтцтгх вт нее под спинными бороздками двух ветвей: наружной — плавательной, покрытий волосками, с листоватыми жабрами (фиг. 236, Вг), к ней при реиляющикися, и внутренней — ходной, снабженной на концах коготками.
Под головным щитом также имеются конечности, прежде всего четыре пары ногочелюстей и челюстей, расположен аых около рта с прикреплением
Фиг. 238. Схематический продольный разрез трилобита-.
Лу—гяпоЛома; ss—рыльцевой щеток; к—головной щит; гг—затылочное кольцо- г—ттловищттые сегменты; «—хвостовой щит; Ьр — перепончатая брюшна поверхность тела; о—рот; ое—пищевод; dj—передний желудок» ds—желудок; da—кишка; а—анальное отверстие.
их под задними лопастями глабели и под затылочным кольцом. Впереди челюсчей находятся антенны, пли сяжки, соответствующие следующей паре лопастей глабели.
Головной щит трилобитов состоит из постоянного числа сегментов, пяти, по мнению некоторых (но, может быть, более). Это можно думать на основании сегментации
глабели (фиг. 234), а также на основании числа конечностей под головным щитом. Лицевой шов, предполагается, соответствует границе я ° ° между первым спереди сегментом и следующим.
Фиг. 239. Три ио'-лвд.)вагельн1 ie ран-
в и. стадии разни ня трллобита.
Нлииолее развитый отдел пищеварительной системы, жзлу.’ок, пометов под головным щитом (фиг. 253), с чем св <заны раздвинутость глаз и наблюла мое нередко расширение глабели в лобной ча< ти.
Пнливидуальиое развитие трнлобитоь, судя по молодым ®кз мн ярам их (размерами от0,4 мм).
находимым совместно со взрослыми, в общих чертах протекает следующим образом Первона-
чально тело овально и дискоидатьно в очертании и состоит лишь из головною и хвостового (всегда меньшего по размерам) щитов, граница между которыми намечена поперечной бороздкой, сначала развитой в околоосевой части тега и затем ра про.траняю-щеися до его боковых краев (фиг 2)9). Сегм нтацпя первоначально едва намечена, затем усили, ается. Швы и глаза отсутствуют, затеи появляются со сто, оны наружных краев и постепенно подвигаются к глабели. На фиг 239, бис, подвижные части щ| к являются в виде узких полосок, глаз еще нег. Так как годовио щит все। да представляет отчетливую пятичленистоеть, то можно предполагать присутствие на брюшной поверхности ei о пяти пар конечно, тей, а под хвостовым щитом чи до конечностей, соответствующее числу сегм< нтов.
Описанные личинки трилобитов можно считать близкими к личинке науплиус,хотя более примитивными (prvionaupliitt).
Расщепленные ноги трилобитов свидетельствуют о принадлежности их к ракообразным. Большая изменчивость числа сегментов сближает их
Фиг. 240. Trinu-cleus Goldiussi. Нижний силур. Веселя. Богемия. Че-хо-Слоиакия. 1/1.
трилобитов, СЧИ-
c Entomosti мз, из которых особенно близки к трилобитам листорогие по характеру жабер и по изобилию конечностей
Трилобиты все-таки примитивнее Phyllopoda, это явствует из следующего: 1) все они морские животные, 2) все сегменты их тела, за исключе-ЕИем анального, несут пару придатков, 3) все придатки, за исключением щупалец, двуветвисты; 4) типично голова несет следы происхождения из пяти сегментов; 5) личиночная форма трилобитов более примитивна, чем личиночная форма (nauplius) Phyllopoda.
Отличия трилобитов от Entomostraca настолько значительны, что трилобиты MorjT быть обособлены как группа, равноценная Entomostraca или Malacostraca, а может-быть и всему классу Crustacea.
В систематике трилобитов одним из руководящих оснований является положение лицевого шва, — начало его на заднем краю головного щита пли на переднем. Соответственно различают группы Opisthoparia и Prop aria, возможно, независимо одна от другой, вышедшие из оощего корня. К первой группе близки трилобиты с лицешм швом, начинающимся в щечных углах. Наконец есть трилобиты без лицевого шва, отсутствующего вероятно вследствие совершенного срастания подвижной части щек с неподвижной, причем обыкновенно атрофируются и глаза Кроме этого, вторичного отсутствия лицевого шва, считается возможным и первичное его отсутствие у примитивных форм.
Примитивность развития у трилобитов выражается в весьма слабом развитии хвостового щита сравнительно с туловищем, причем хвостовой щит состоит из 3—4 сегментов. Удлиненные глаза и так называемые глазные линии, — валикообразные возвышения, идущие от глаз к глабели соответственно ходу глазных нервов ((Menus, Pty-choparia),—также свойственны примитивным формам. Приближенность глаз к заднему краю головного щита и в то же время к глабели, на -основании данных онтогении
тается признаком далеко ушедшего вперед развития. Признаком такого же ра (вития является способность свертываться.
При классификации трилобитов надо иметь в виду, что у родственных родов бывают значительные различия, а у неродственных значительные сходства, возникающие вследствие приспособления в 1-м случае — к различному образу жизни, а во 2-м, по аналогии, — к сходному образу жизни
Приплющенный, слабо выпуклый головной щит, с центральными глазами, и хвостовой щит, оканчивающийся острием (у Olenelhis, Dalmania), иог\т считаться возникающими при бентоническом, роющем образе жизни ® полосе мелководья. Вздутая напереди гладкая глабель {Phacops) при округленном хвосте принимается за указание на нектонпческий образ жизни.
Phv Атрофированные глаза (Aeglina) вероятно связаны с активным передвижением в зоне, куда слабо проникает дневной свет. Головной щит, ^сплющенный на краю в плоскую каВму, и отсутствие глаз (Harpes, ™nmic..eus) могут быть связаны с бентоническим образом жизни в глубоко-лежащей, лишенной света зоне моря. Слепота может возникать у трилоби-гов и при роющем образе жизни на малой глубине.
ГРУППА OPISTHOPAEIA
Из Opisthopana род Olenus (фиг. 241) отличается маленькими глазами, соединенными с глабелью глазными линиями. Свободные щеки и хвостовой щит малы. Бока глабели параллельны. Вечхний кембрий
Proelus (мифологическое имя; фиг. 242) отличается краевым возвы-
шением головного щита, рудиментарными, а то боковыми бороздками гла-
и вовсе отсутствующими
Фиг. 241. Olenus trunca tus. Верхний кембрий. Аидрарум. Швеция. 1/1.
бели и присутствием на глабели задних основных лопастей, примыкающих к затылочной бороздке. Верхний силур, девон.
К Proetus примыкает род Phtllipsia (фиг. 243, по имени ученого, Филлит а), имеющей гла-сель более i ыдающуюся, ограниченную параллельными бороздками, с более
Фиг. 242. Pi oelus erenuta. Девон. Богемия. Чехо-Сло-вакия. Наверху части головного щита. Ввпзу хвостовой щит.
резко выраженными основ
ними лопастями глабели. Туловище состоит из 9 сегментов, осевая лопасть хвостового щита представляет до 18 колец. От верхнего девона до перми
включительно.
Ptychopana (фиг. 244; ptyche—складка, pareia —щека) имеет лицевые швы, начинающиеся в щечных углах, глабель, суженную напереди, с 3 парами боковых бороздок; имеет глазные дрнии. Средний кембрий.
Фиг. 243. Plnllipsia дет-mPlifera (реставрация). Нихшекаменноуг. 1/1.
Фиг. 244. Phychopnr а striata. Средний кембрий. Гинец. Богемия. Чехо-Сло-вакия. 1/2.
Фиг. 245 Peltura scara baeoides. Верхний кембрий. Норве'ия.
Peltura (фиг. 245, pelte — полулунный щит, ига — х <ост) отличается закругленными щечными углами, глабель достигает до лобного края; хвостовой щит с зубчатый свободным краем. Верхний ьембрий,
Dicellocephalus (фиг. 246; dicella — инструмент для копания, ке-phale — голова). Глабель с параллельными, непрерывно во всю ширив) глабели протягивающим! ся подеречными бороздками. Верхний кембрий.
CaU/mme^e (фиг. 247; kekalymenos — скрытый) имеет лицевые швы, начинающиеся в щечных угяах. Боковые бороздки отчетливо обособляют на суживающейся кпереди глабе и боковые лопасги округленного очертания. 13 сегментов туловища. Са’уттепе способна свертываться совершенно. Силур.
Homalonotus (фиг. 248; homalos—на том же самом уровне, notos — зад). Удлиненное, суживающееся кзади тело неясно разделяется на три части продольными бороздками, глабель гладкая, приплющенная и почти квадратная. Хвостовой щит удлиненный, трехугольный. Силур, девон.
Olenellus (фиг. 250, 251) имеет большие узкие полулунные глаза, приближенные глабели обоими своими концами. Щеки и
к
Фиг. 246. Dicellocephalus mtnimeso-tensis. Кембрий. Висконсин. САСШ.
Фи. 248. ffomolonatus delphtnocepfialus. Верхний силур. Локпорт. Нью-Йорк. САСШ. 1/1.
Фиг. 247. Caiyinmene conspicua.
Верхний силур. Эзель. Эстония. 1/1.
плевры продолжаются в остроконечия. Конечный сегмент хвостового щита иногда шиповидно удлинен. Нижний кембрий.
Paradoxides (фиг. 249; paradoxes — удивительный) имеет большое число (от 16 до 20) туловищных сегментов, широкий головной и узкий маленький хво'товые щи гы. Щечные остроконечия сильно развиты; плевры оканчиваются на свободном конце шипами, направленными назад. Глабель расширена напереди, широкая с 3 поперечными бороздками. Лицевые швы,
в отличие от Olenellvs, явственно выражены, они несколько расходятся от глаз взад и наперед. Достигает 0,6 м длины. Средний кембрий.
Harpes (фиг. 252; Ьагре — крючок), под бно Olenus, имеет глаза, соединенные с глабелью поперечными ребрышками, и маленький хвостовой щит. Вздутыми щеками глабелыо и широким плоским лимбом головногв
щита, покрытым точечными сквозными отверстиями, напоминает Tr’nucleus. Число туловищных сегментов доходит до 29. Силур, редко девон.
Trmucleus (фиг 240 tri—три, nucleus-ядро) имеет туловищные сегменты в малиМ количестве, именно, 6 сегментов. Хвостовой щщ
Фиг. 249. Parade i ides bohe-.nteus. Средний кембрий. Гинеи. Богемия. Чехо-Слоьакия.
1/£.
Фиг. 250. OlenMus Gilbert Нижнги кембрай. Вермонт. САСШ. 1/1.
Фиг. 251. Olenethis {Rleso-nans} Rlickvitzi. Рестав-раци.< Нижний кембрий.
Эстония. 1/1.
Хвостовой щит явственно
значительно меньших размеров, нежели головной.
сегментирован; головной иногда представляет слабую сегментацию, имеет щечные остроконечия У взрослых индивидуумов глаз нет. Свободный плоский край (лимб) головноцо щита широк и покрыт ямками. Нижний силур.
ГРУППА PItOFAEIA.
К Proparia относят, следующие роды за исключением разве Illaenus.
Asanhus (фиг. 253—255; asaplius — неясный) Головной и хвостовой щиты равновелики, со слабо вырал.^нной сегментацией, на хвостовом щите явственно членисга срединная лопасть, членистость боковых лишь намечена слабыми линиями. Ветви лицевого шва соединяются перед расширенною переднею оконечностью глабели. Гл, бель не сегментирована. Глаза приближены к заднему краю и сидят на возвышениях или даже на длинных стебельках. 8 туловищных сегментов. Гипостома вилообразно раздвоенная на заднем косце. Нижний силур.
Подрод Megalasms (фиг. 256; megale — большой, aspis — щит) отличается от Asaphus более резко обюобледной и цилиндрической глабель», а также заострением головного и хвостового щитов. НчжниЧ силур
Illaenus (фиг. 257, ^58; illaino — восоглазить) представляет полное отсутствие сегментации не только на глабели, но и на осевой лопасти хвосто вого щита; и гибель, и осевая лопасть хвостового щита кроме того отграничиваются лишь пс бокам от остал! ной поверхности щишв. Ветви тацевого шва соединены на завороте головного щпта рыльцевым швом. Число туловищах сегментов 10 Нижний силур.
Bronteus (фиг. 259, 260; мифологическое имя) отличается расширенною напереди глабелью со слабыми следами боковых лопастей. Большой
Фи.’. 252. Harpes ungula. Верхний свл'р. Прага. Богемия. Чехо-Словакия. 1/J.
хвостовой щит представляет крайне укороченную осевую лопасть, от которой радиально расходятся бороздки, отграничивая плоские участки хвостового щита, соответствующие вошедшим в состав его сегментам. Силур, девой.
Фиг. 253. Asapbus expanses. Нижний сил)р. Р. Поповка около Сл’ ика Ленинградской обл. 1/1.
Фиг. 254. Asaphus Wetsst Нижний гилур. С Пулково около Ленинграда. 1/1.
Фиг. 255. Asaphus Kama leuAkyi. Нижний силур. С. Пулково около Ленинграда. 1/1.
Фиг. 250. Meyaiaspis exte-nuatus. Нижний силур.
Швеция. 1/1.
Agnostus (фиг. 261; agnostns — неизвестный) имеет только два туловищных сегмента. Головной шит подобен хвостовому; последний представляет следы трех-четырех сегментов, его составляющих. Следы сегментации есть и на глаеди.
I Адп <• us мыяетея специ шпигованным в некоторых от ошениях, .имени) п cnw(K)!in<Tii гве,iMidTbcr и по слаоий си’менТироьапностм осевЛ части тела. Кембрий Пил. ий силур.
Phacops (фиг. 2b2, 263; phakos — линза; ops — глаз) имеет взду. тую, расширяющуюся напереди глабель; впереди ее соединяются ветви лицевого шва, начинающиеся на боковых краях головного щита. Боковые бороздки выражены в различной степени отчетливо или отсутствуют; щеч-ные утлы закруглены или продолжаются в щешые остроконечия. Глаза
Фиг. 867. Шаепи
Фиг. 259. Bronteus palifer. Нижний девон. Коньепрусс. Boienna Чсхо-Сло-вакия. 1/1.
Флг. 262. PhacojA Sternbergi. Девон. Гостии. Богемия. Чехо-Словакпя.
mam. Нижней <• С. Пулковооколо 1< града. 1/1.
Фиг. 260. BronteuK vmbe[life.r. Нпжний девон. Сливеиец. Богемия Чехо-Сло-вакия. 1/1.
Фиг 258 Illaenuscras-sicauda. Нижний силур. Далекарлия. Швеция. Свернувшийся индивидуум. 1/1.
Фиг. 263. Fliaeops lah-frons. Средний девой. Эйфель. Германия. 1/1-
Фиг. 261. Agrostus pis1-jormis. Кембрий. 2/1.
большие, выдающиеся, с крупными фасетками. 11 туловищных сегментов. Подрод Dalmania отличается менее вздутою глабелыо и явственными бороздками глабели (фиг. 233, 234). Силур, девон.
Chasmops (фиг. 264; hasma — пасть, ops — глаз) отличается от Dalmania сходящимися к плоскости симметрии тела двумя передними боковыми бороздками глабели, так что ими занвчпваются с обеих сторон глабели большие треугольные передние боя ..ые лопасти; эти лопасти весьма характерны и легко распознаются. Следующие кзади боковые лопасти, напротив, стеснены в развитии, зачаточны и имеют вид короткою бокового острия, но
Фиг. 264. Chasmops Odieti. Нижний силур. Таллин. Эстония. 3/4
Гтлгтигающего вершиной до спинных бороздок тела. Задние боковые лоласги образуют как бы ножку чашевидного абриса глабели Нижний силур.
* Encnnurus (фпг. 265, 266; сппо — разделять, ига — хвост). Головной щит f бугорчатый; глабель, расширяющаяся, напереди, выпуклая; свободные части щек разделены в лобной части узким рыльце-вым щитком; глаза маленькие, на конических возвышениях; 11 туловищных сегментов; хвостовой щит треугольной формы, с большим числом сегментов на срединной лопастп и с меньшим на боках. Бороздки глабели слабо развиты. Силур.
распространение трилобитов.
Уже нижнекембрийские трилобиты свидетельствуют о глубокой древности этой группы,—они уже настолько разнообразны, что возникновение группы трилобитов надо отнести к далекому докембрийскому времени Наибольшее распространение трилобиты имеют в кембрии и нижнем силуре, после чего быстро уменьшаются в количестве и разнообразии.
Фиг. 265. Encrinu- Фпг. 266. Епсгг-rus puneiatus. Си- nurus mvltiseg-лур. Готланд. Шве- mentatus. Силур.
ция. 1/1. Эстония.
Геологическое распространение трилобитов.
Роды Отделы или ярусы
Кембрий Силур Девой Карбон Пермь
Olenus 1‘lyclioparia . . . Peltura Dicellocephalus . . Cal ,mmene . . . homalonolus . . . Olenellus .... Paradoxides . . . Harpes ... Trinucleus .... Asaphus Megalaspis .... Illenus Praoetus Philhpsia .... Bronteus Agnostus .... Phacops . ... . Chasmops .... Encrinurus . . . j™.
—
Для кембрия характерны Agnostus, Olenellus, Paradoxides, Olenus Ptychoparia, Peltura, Dicelloceghalus.
Для сил1"’-' Trinucleus, Harpes, Cdlymmene, Asaphus, lllaemis. Для девона Bron. rroetus, Phacops. Для каменноугольных отложений—Phil, lipsia. В в ..чих вымирают последние трилобиты сем. Proetidae, род Phyllipsia.
Некоторые роды трилобитов космополитичны в своем распространении, но большею частью распространение их ограниченное^
Характеристика групп ракообразных.
I подкласс: Entomostraca. Преимущественно маленькие ракообразные с телом, чрезвычайно различным по виду, из различного числа сегментов. Задний последний сегмент часто оканчивается вильчатым раздвоенном. Складки кожи образуют раковину, которая бывает нередко двустворчата. Обыкновенно развитие со стадией науплиус. От кембрия доныне.
1 порядок. Phyllopoda. Маленькие, с листообразными плавательными ногами, с небольшим или со значительным числом сегментов, часто 6 двухстворчатой раковиной. Современные большую частью в пресной воде. С кембрия доныне.
2 порядок: Ostracoda. Имеют двухстворчатую раковину, покрывающую голову и семь пар конечностей, задние из которых служат для ползания и плавания. С кембрия доныне.
3 порядок: Cirripedia. Во взрослом состоянии все ведут прикрепленный образ жизни или паразитический. Большей частью морские животные. Кембрий? Нижний силур—ныне.
4 порядок: Copepoda. Увеличенные сяжки служат для плавания. Не имеют раковины, не сохраняются в ископаемом состоянии.
II подктасс: Malacostraca, нормально имеют двадцать сегментов.
1 группа: Leptostraca. Имеют двухстворчатую раковину. Конечности листовидные. Имеют постоянно 21 сегмент. Вильчаторазвегвденный хвостовой плавник. С кембрия доныне.
2 группа: Arthrostraca. Только передний грудной сегмент сливается с головой. Отсутствует головогрудный паипырь. С девона доныне.
1 порядок: Amphipoda. Тело сжатое с боков, на вытянутом брюшке позади находятся прыгательные ноги. Палеозой. Третичные — ныне.
2 порядок: Isopoda. Тело сжатое в спинно-брюшном направлении, с широким брюшком. В море, пресной воде п на суше. Девон (?). Карбон. Трнас — ныне.
3 группа: Thoraeostraca. Вместе с головой несколько сегментов (иногда все) груди входят в (©став головогруди, покрытой общим панцирем.
1 порядок: Gumacea. С головогрудью, в состав которой входят три грудных сегмента. Маленькие, морские. Известны лишь ныне живущие.
2 порядок: Schizopoda, Головогрудь содержит в себе все сегменты груди и все ноги последней являются расщепленными. Девон (?). Карбон. Триас—ныне.
3 порядок: Stomatopoda. В головогрудь входят пять сегментов груди. На ней имеются хватательные ноги, расщепленные ноги. Карбон (?). Юра—иыне.
4 порядок: Decapoda. Головогрудь заключает в'себе все сегменты груди и совершенно не имеет расщепленных ног. Пермь—ныне.
III подкласс: Trilobiti. Величиною большею частью в несколько см с непостоянным числом сегментов. Спинной панцыръ трехраздельный и вдоль и поперек. Имеются состоящие из нескольких сросшихся сегментов головвой и хвостовой щиты, и туловище из подвижных сегментов. На головном щите обыкновенно два глаза, так называемый лицевой шов, и с нижней стороны головного щита пластиика верхней губы, так называемая гипостома. Конечности расщепленные. Кембрий — пермь.
КЛАСС П. РАКОСКОРПИОНЫ, MEROSTOMATA ИЛИ GIGANTOSTRACA. 1
Представители его отличаются от остальных ракообразных тем, что имеют 6 пар конечностей головогруди, причем 1-я пара, подобно последующим, оканчивается клешнями. Многочленистый абдомен. снабжен 6 па-
1 Meros — часть, stoma — рот.
/
рамп двJ ветвистых листоватых ног и хвостовыым сегментом (telson), большею часгыо шиповидным; имеют два боковых сложных глаза в два срединных простых.
ПОРЯДОК I. МЕЧЕХВОСТЫ, X1PHOSURA.
Тело явственно подразделено вдоль на три части, подобно тому как у трилобитов. Abdomen представляет от семи до десяти сегментов. Хвостовой сегмент вытянут в длину, шиповиден, подвижен. Единственным ныне ЖВ1 щим представителем Xiphosura является Limulus(§m. 267—269; limns—
Фиг. Kt .Limulus polyphemus. Ныие живущий. 1 — головогрудь; 2 — а' Ло-теп; 3—хвостовой шив; 4—глазки; 5—глаза. 1/3.
Фиг. 268. Limulus palypTumus. — Ныне живущий.
Л—головогрудь. В—abdomen, С— часть хвостового шипа. 1—6—конечности головогруди; 7—ГУ—конечвостн абдомена; 7- operculum-®—12— извечности, несущие т—рот
Уменьшено.
ил)—рак-мечехвост, живущий на тихоокеанском побережья Азии, Сев. Америки и на Малайском архипелаге. Тело представляет широкий сильно выпуклый головогрудный щит и подвижно сочлененный с ним abdomen, юсго-ящий из сегментов, слитых так, что границы их остаются лишь слабо выраженными. Конечности являются в виде ходных ног, окружающих рот, снабжены клешнями, а основные членики их представляют жевательные лопасти. Конечности абдомена пластинчаты, первая’ служит крышечкой (operculum) для остальных, на задней поверхности которых находятся жабры. Абдомен личинок восьмичленист (фиг. 269), тельсон листовиден.
Трехраздельность в продольном направлении, широко расставленные сложные, глаза, помещающиеся при килеватых возвышениях головогрудного Щпта, напоминающих лицевой шов трилобиюв, и абдомен личинки (фиг. 269) в «трилобитовой» стадии развития Limulus — сближают Limulus с трилобитами. Встречается от перми доныне. Пермские и триасовые представители малы величиною, юрские достигают более значительных размеров, а третичные
10 Учебник валеонто fогяи
145
и современные—наибольших. Интересны палеозойские представители Xiphd sura (известны с девона), ио своей организации в зрелом возрасте соответ, ствующие трилобитовой стадии LtmuLus и трилобитам.
Таков например Belinuru-s (фиг. 270, belos — копье, ига — хвост). Головогрудный щит представляет широкую краевую каемку, продолжающуюся на задних углах щита г длинные остроконечия. Абдомен из восьми сегментов, не считая длинного тонкого шипа; по vU следние три (6 — 8)
Фиг. 271
Рогциккшь,
Ч» . л&й&'й .£>’
Euryptcrus Fischen. Верхняя сидур. о. Эзель. Эстония.
II— VZ—конечности, расположенные под головогрудным щитом; 1—О—вдомв-ватьные cei енгы, под которыми расположены ждбры; В— разрез тела ио срединной оси н лорю-вгатрально, с—третий абдоминальный сегмент в поперечном разрезе, d—поперечный разрез абдоминального сегмента из задан*» не несущих конечностей.
Фпг. £69. Трило-битовая стадия личинки Limulus. Уве-лич.
Фиг. 270. Belmunis гедгпае Кшеиво-угольн. соложения. Ирландия. 1/1.
срастаются, остальные свободны. Девон и каменноугольные отложения Европы и Сев. Америки. *
ПОРЯДОК II. EURYPTERIDA.
Распространены исключительно в палеозое, размером достигают до 2 и в длину.
Под головогрудью находятся шесть пар одноветвистых ног; ноги передней пары малы, находятся впереди рта и снабжены клешнями; абдомев
стоит из 13 сегментов; хвостовой сегмент иногда лопатообразной формы; т°‘ рест'1 передним сегментам абдомена на брюшной поверхности соответствуют ' 1а"ные пластинчатые образования, видимо гомологичные ншам Limulus, ccynniM жабры. ।
Eurypterus (фиг. 271, 272: eurus — широкий, pteron — крыло). На ерхней стороне головогрудного щита, кроме больших почковидных боковых.
глаз, находится пара маленьких точечных глазков, приближенных к срединной линии, а на брюшной стороне его расположены конечности. Из них 2—4-я пары покрыты шипами, задняя пара наибольшая по величине с незначительными основными члениками, представляющими жевательные лопасти, и широкими дистальными концами, превращенными в плавательные лопасти. Между
метастома, I—F—сегменты с пластинчатыми конечаостями. 7— 12—сегмент -л без кояечяостеа.
Фиг. 273. Ptervgotus onilien-sis со сиинвоб стороны. В рх-иий силур. 1’отцикюЛ, о. Эзель. Эстония. Уиеш.ш.
основными члениками задней пары ног расположена большая яйцевидная пластинка {metastoma). Шес(и передним сегментам абдомена, снабженным лишь коротким внутренним заворотом, соответствуют на брюшной стороне пять пар пластинчатых образований, которые череиицеобразно налегают на смежные сзато пластинки. Из них передняя первая пара, аналогичная крышечке Limulus’^, представляет еще непарный средний членистый отрссток, кроме того подразделена поперечной бороздкой на две части и соответствует двум сегментам спинной стороны. От верхнесилурийских до каменноугольных отложения.
Pterygotus (фиг. 273; pteron — крыло, ous ухо) отличается от Eurypt&riis боковыми глазами, расположенными на краю головогрудного щита, сильным развитием передней пары конечностей и хвостовым сегментом овальной формы. Верхний силур
Этот порядок распространен от кембрийских до пермских отложений наиболее в верхнем силуре и девоне.
КЛАСС Ш. ПАУКООБРАЗНЫЕ, AKACHNOIDEA.
Членистоногие, дышащие внутренними воздухоносными полостями, с головогрудью, несущею шесть пар конечностей; абдомен лишен конечностей (имеет их лишь в состоянии зародыша), глаза простые. Преимущественно наземные животные.
Конечности (фиг. 274) сходны между собою; подобно тому как у Енгур-
Фиг. 275. Palaeophonus vuntvus. Со спинной стороны. Верхний силур Готланд. Швеция.
имя caledomcus.C6"iorn-ной стороны. Верхний силур. Ленаркшаир.лнг-лия.
Фиг. 274. Scorpio europaeus. Ныне живущ. С брюшной стороны.
й — гребни {pti Яка); J — етжгмы.
иногда легкие, возникающие набрюшной стороне абдомена из мешкообразных впяченностей, на передней стенке которых развиваются листообразные выросты с находящимися в них кровеносными сосудами. Эти легочные мешки открываются наружу парными отверстиями на брюшной поверхности сегментов — стигмами. Иногда вместо легких имеется система трахей, открывающихся наружу также стигмами.
В палеозое есть скорйионы, столь же высоко организованные, как и современные, и существенно от последних не отличающиеся, но есть и низко организованные. Таков Palaeophonus (фиг. 275, 276). Ходные ноги его состоят из члеников почти одинаковой величины и невидимому не имеют когтей. Позади шести нар передних конечностей есть пара видоизмененных конечностей, вероятно соответствующих так называемым ресипа 1 скорпионов. В отличие от других позднейших скорпионов Palaeophonus, повидИ'
1 Органы оеизания, как предполагают.
мому является водным животным, так как он найден вместе с окаменелостями, принадлежащими к морской фауне. В соответствии с этим он повидиммому йе имеет стигм и дышал вероятно жаберными ножками, подобно Еигур-terida. Встречается в силуре.
Интересно, что верхнепалеозойские представители Eurypterida находятся Е отложениях пресных вод, тогда как древнейшие — в морских отложениях Эти особенности распространения Eurypterida
и скорпионов, а также соприкосновение представителей обеих групп в анатомической организации делают несомненной связь скорпионов с Eurypterida.
Фиг. 276 J'uphrtjnus
Prestivitchii. Верхнее i-менноуг. Англия.
Фиг. 277. Eup!ioberia ormtgera. Каменноугольные отложеидя. Илли нойс. САСШ (Из Diplopoda).
В палеозое уже встречаются паукообразные жгутоногие, или скорпионовые пауки (Pedipalpi), существующие и ныне, в некоторых отношениях промежуточные между скорпионами и настоящими пауками. Именно Pedi-palpi имеют явственно членистый абдомен и в конечностях представляют сходство со скорпионами, а по отсутствию pectina и присутствию паутинных желез сходны с пауками. На фиг. 276 В изображен паук из числа имеющих членистый абдомен.
Из палеозоя известны уже и настоящие пауки, отличающиеся нерасчле-ненным абдоменом.
Подтип II. Eucephalica.
КЛАСС IV. МНОГОНОЖКИ, MYRIOPODA.
Животные с удлиненным телом, содержащим не менее 12 (а иногда значительно более) одинаковых, за исключением головы, сегментов.
Голова, как у насекомых, несет сяжки и три пары челюстей, затем глазки, обыкновенно простые. Дышат сегментально расположенными трахеями. Сегменты тела, за исключением конечного, анального, снабжены конечностями.
По общей фбрме тела и однородности сегментов его напоминают анне-гид. И Современные многоножки распределяются в два подкласса: Chilopoda и biplopoda.
Первая группа может считаться более примитивною; на голове развито полное число конечностей (4 пары). За ними следует пара ногочелюстей первого туловищного сегмента. Каждому сегменту туловища соответствует пара конечностей. Хищники.
Diplopoda (фиг. 277) имеют редуцированное число конечностей на голове, так как во время эмбрионального развития исчезает задняя пара челюстей.
Растительноядные, в связи с этим лишенные ногочелюстей. Сегменты тела слиты попарно, так что каждый имеет две пары конечностей.
Палеозойские многоножки распределяются также в эти две группы и известны с девона.
КЛАСС V. НАСЕКОМЫЕ, IXSECTA.
Тело насекомого подразделяется на голову, thorax и abdomen. Голова состоит из слитых сегментов; она- несет четыре пары придатков—пару сяжков и три пары челюстей. В туловище три -сегмента, каждый с парою ног; кроме того thorax обыкновенно несет две пары крыльев на. своей спинной поверхности. Abdomen состоит из нескольких (обыкновенно 10) сегментов, которые лишены конечпостей. Насекомые дышат трахеями. В индивидуальном развитии насекомые испытывают так называемый метаморфоз: из яичка вылупляйся животн ю в состоянии значительно разнящемся от состояния зрелого животного, к которому подходит, постепенно изменяя свой вид. Иногда, как например у чешуекрылых (бабочки), метаморфоз весьма значителен, резок, из яйца выходит первоначально червеобразная личинка, в других случаях собственно личинки не бьиает или вовсе, или личинковая стадия отличается от зрелого состояния' только отсутствием крыльев (тараканы, саранча и пр.).
Приобретение насекомыми лпчипковых стадий, коренным образом разнящихся от образа взрослого животного, есть явление ценогенетическое, представляющее нарушение филогении, воспроизводимой в онтогении. Чем далее ушла какая-либо группа насекомых от их прототипа, тем более было в ее истории так сказать вводных элементов, тем резче могут разниться стадии метаморфоза; напротив, примитивные типы в своей онтогении представляют постепенное развитие, отдельные стадии которого мало разнятся между собою, так же как и от состояния зрелого животного; в общем можно сказать, что насекомые с неполным метаморфозом примитивнее насекомых с полным метаморфозом. Этому положению соответствует и геологическая последовательность появления порядков насекомых: ранее существуют порядки с неполным метаморфозом,, позже появляются порядки с полным метаморфозом.
Другим показателем примитивного состояния или ушедшего значительно вперед развития является состояние челюстного аппарата; жевательные челюсти, лежащие симметрично по бокам ротового отверстия, представляют первичное состояние, а сосательный ротовой аппарат, образуемый сращением челюстей с изменением их, есть производное об азовапие.
Насекомые известны в ископаемом состоянии .со среднекаменноугольных отложений.
В палеозое известны представители Orfhoptera (прямокрылые, с жевательными челюстями, с неполным метаморфозом; тарзкаповые в палеозое; кузнечики, саранча—с юры), Hem’ptera (полужесткокрылые, с сосущими челюстями, с неполным метаморфозом; э о—группа, к которой принадлежат клопы, вши, тли), и с перми Neu-optera (сетчатокрылые, с жевательными челюстями, с полным или неполным метаморфозом, напр. муравьиные львы).
С триаса к этим группам присоединяются Coleoptera (жуки, хотя и с первичными жевательными челюстями, но- с полным метаморфозом) *
фиг. 278. Stennciictya lobata. Комментри? Алье. Франция. Каыенноугольн. 2/9 Представитель Palaeodictyoptera.
с юры Hynienoptera (перепончатокрылые, с полным метаморфозом, р которым принадлежат муравьи с жевательными челюстями, пчелы—с сосательными челюстями), Diptdra (двукрылые, напр.-мухи, комары,— с колющими и сосущими челюстями и полным метаморфозом), Lepidoptera (чешуекрылые, бабочки—с сосущими челюстями и полным метаморфозом).
Три последние порядка, к которым исключительно или отчасти принадлежат насекомые, питающиеся соками, высасываемыми цз цветов растений, естественно могли развиться лишь тогда, когда возникли и сколько-нибудь значительно распространились цветковые расте-
ния, т. е. с мезозоя.
Кроме представителей порядков, живущих в на-
стоящее время, в ископае-
всрхнекамензоугольных отложениях, ветре-
ном состоянии, а именно в верхнекаменноугольных отложениях, встречаются Palaeodictyoptera (фиг. 278), мало специализированные большие насекомые, личинки которых вероятно жили в воде и постепенно развивались в крылатых взрослых насекомых, имевших, как и личинки, жевательные челюсти. Palaeodictyoptera отличаются тем, что их thorax еще сохраняет следы происхождения из трех слитых сегментов, так как промежуточные между этими сегментами бороздки еще не изгладились. Кроме того, у них имеется третья пара крыльев небольшой величины на переднем сегменте торакса а боковые крыловидные выросты на всех сегментах брюшда и так называемые cerci на последнем сегменте. Эти последние членистые придатки являются видоизмененными брюшными конечностями. Присутствие их и членистость торакса представляют весьма примитив-
Фпг. 279. Т исаеття ovalis. Мезон Крик.
Иллинойс САСШ. Каменноугольн. 4/9.
Иые особенности. Наличность крыловидных придатков на всех сегментах торакса и абдомена дает некоторое основание для сопоставления Palaeo-dictj optera с трилобитами. Крылья двигались лишь вертикально.
Другие верхнекаменноугольные насекомые хотя и примыкают к современным порядкам (Orthoptera, Hemiptera и др.), но отличаются от них более примитивными особенностями, так что их собственно выделяют в особые порядки. Верхиекаыенноугольцда цадедоыые достигают по сравнению с со-
временными необыкновенно большой величины, до 75 сантиметров между концами раскрытых крыльев. Такая величина, а также то обстоятельство, что все палеозойские насекомые обладали неполным метаморфозом, ставится в связь с теплым климатом того времени на том основании, что в настоящее 1ремя насекомые особенно большой величины и с неполным метаморфозом являются по преимуществу обитателями теплых стран, без резко выраженной зимы, тогда как насекомые с полным метаморфозом большею частью живут в местностях с холодным климатом. Вымирание палеозойских насекомых и появление в триасе порядков с полным метаморфозом ставится в связь с переменою теплого климата на холодный (пермская ледниковая эпоха). Развитие насекомых в прошлые геологические периоды таким образом представляет яркий пример связи с изменением окружающих неорганических и органических условий жизни (климат и растения).
ч
VI 711-1. ИГЛОКОЖИЕ, ЕСШКОИЕВМАТЛ.
Один из наилучше охарактеризованных и наиболее отличимых типов жг.ввеого царства. Почти все животные этого тппа представляют пятилу-
Фиг. 280. Разр-з иаралл -льно поверх-вости панциря современного морского ежа (SphaerechirLiis). Сильно увеличено.
Фиг. 281. Схема амбу-якральной системы мор ско8 звезды.
чевую симметрию, имеют подэпидермический скелет, состоящий из углекислого кальция, с характерной решетчатой микроструктурой (фиг. 280), и обладают системой мешочков и трубок (амбулякральная система), несущих году по всему телу. Главнейшими частями амбулакральной системы являются пять радиальных сосудов, даюших наружу с обеих сторон маленькие ветви, известные под названием ножек (фиг. 261).
Радиальные сосуды отходят от кольцевого око-лоротового сосуда, в который вода, поступая извне, проходит через каменистый канал (8Yc канал с отложениями извести в стенках), снаружи оканчивающийся ситовидно-продырявленной м а д р е,п о-ровой пластинкой (Af). Амбулакральные ножки (р) представляют мешочки с мускулистыми сократимыми стенками; при наполнении водою, вгоняемой сократимыми пузырьками — ампулламп (Л), ножка вытягивается, при сокращении стенок — укорачивается. При движении животное вытягивает ножки, присасывается ими к предмету, по которому ползет, и затем сокращением ножек передвигает тело.
Внешним образом, по существованию лучистой симметрии, иглокожие сближаются — и еще не так давно были соединяемы — с кишечнополостными, но достаточно присутствия обособленных кишки и общей полости тела (coelom), чтобы считать иглокожих более высшими животными, чем кашечш полостные.
Подобно амбулакральной системе, различные органы животного представляют большею частью пятилучевое расположение.
Амбулякральпая система образуется, обособляясь в молодых стадиях животного от его общей полости тела; последняя, кро_ме того, у взрослого животного состоит из нескольких отделов.
Кровеносная система не представляет сосудов с самостоятельными стенками, но образована промежутками между органами.
Дыхание происходит при посредстве амбулякральной системы и через стенку тела, если она достаточно тонка, причем в стенке иногда развиваются специальные выросты или складки; наконец дыхание, происходит также при поглощении воды через ротовое пчи анатгное отверстие^ в последнем случае вода выбрасывается обратно наружу.
У иглокожих нет особой выделительной системы; гомологом ее по происхождению может быть является амбулакральная система
В одном из более значительных отделов общей полости тела—осевой полости — находится осевой орган. В осевом органе происходит развитие половых клеток. Осевая поесть окружена кишкой, и поэтому можно думать, что из кишки питательная жидкость осмотически проходит в осевой орган, который таким образом аналоги’ часги кровеносной системы, помимо отп вления функций размножения.
Органы чувств — осязания, зрения слуха—развиты слабо. Органы осязания щ ставлены амбулакральными придатками, : шипами, покрывающими снаружи стейку тс
Фиг. 283. Схема личинки иглокожего при переходе к лучистой симметрии.
Фпг. 282. Схема личинки иглокожего на дгу-сторонне-симметричнон стадии развития.
Индивидуальное развитие иглокожих идет от двусторонне-симметричной червеобразной личинки (фиг. 282), у которой рот (О) лежит напереди и вентрально, анус (As)—назади или задневентрально. От одного конца тела к другому по длинной оси тела проходит кишка с желудочным расширением. Слева ir справа от кишки симметрично расположены отделы общей полости тела (rpc, Ipc, rhe, llic). Личинка реснитчата, причем реснички располагаются лишь в некоторых областях, образуя обыкновенна полосу, замкнутую кольцом.-
Проплавав некоторое время, личинка прикрепляется спинною частью к поверхности (фиг.283), лежащею позади ротового отверстия, так что рот (О) переходит на задний конец личинки, теперь обращенный кверху; рот располагается справа от анального отверстия’ (Ля), чем объясняется происходящее возрастание органов левой стороны на счет органов правой стороны, причем некоторые органы правой стороны совершенно не развиваются, а кишка изгибается не в простую плоскую петлю, но несколько винтообразно, свиваясь по направлению часовой стрелки.
Следствием прикрепления животного было не только усиленное развитие органов одной стороны тела, ослабление органов другой стороны и развитие лучистой симметрии; в связи с прикреплением развиваются бороздки, идущие от рта по направлению лучей, и несущие реснички, производящие приток воды ко рту; эти бороздки снабжены радиальными ответвлениями амбуляк-ральной системы. Анус и мадрепорит, находящиеся на боковой поверхности тела, представляя препятствие распространению амбулякров в направлении их местонахождения, оказываются лежащими между амбулякрами, интеррадиально, что служит вообще исходным пунктом для разнообразных уклонений от лучистой симметрии, наблюдаемых у различных иглокожих.
Органы пищеварения и мадрепорит не подчиняются пятилучевой симметрии, нарушают ее, причем два луча, задние, заключающие анус и мадрепорит, составляют bivium, а три остальные (передний, правый передний и левый передний) известны под названием trivium. Половые продукты у иглокожих образуются в осевой полости и первоначально невидимому выводились через отверстие в стенке тела между ртом и анусом, как у голотурий и некоторых Cystoidea. Впоследствии, у позднейших иглокожих, и половые органы становятся в более тесную связь с лучистой симметрией.
Иглокожих разделяют на прикрепленных в зрелом возрасте (Pelmatozoa) и на свободно живущих (Eleutherozoa), каковы например морские звезды и ежи. Eleutherozoa оказывается в своем индивидуальном развитии проходят прикрепленную стадию, частью сохраняя связанные с нею особенности и в зрелом состоянии, что заставляет считать Pelmatozoa более примитивною группою, давшею происхождение Eleutherozoa. Иглокожие суть исключительно морские животные.
*
Подтип I. Pelmatozoa.1
КЛАСС I. МОРСКИЕ ЛИЛИИ. CRINOIDEA.
Нормальная лилия (фиг. 284) состоит из кроны (К, corona), прикрепленной ее спинной (т. е. противоротовой) оконечностью к стеблю (S, columna), который прикрепляется к морскому дну или к какому-либо твердому телу посредством корня (Д, radix).
Крона состоит из теки (си), содержащей внутренности, и из пяти рук 'ar, brachia), которые могут более или менее ветвиться. Часть теки, ниже места отхода от нее свободных рук, называется дорзальною (т. е. спинною) чашечкою или просто чашечкою; брюшная часть, лежащая выше мест отхода рук, т. е. околоротовая поверхность, называется tegmen или иногда диском.
Скелетная и некоторые другие системы имеют радиальное расположение, в котором преимущественно выражены пять лучей. Вследствие этого все животное может быть разделено на пять почти одинаковых отделов пятью воображаемыми радиальными плоскостями, проходящими через вертикальную спинно-брюшную ось и через места отхода рук.
Элементы скелета по своему положению являются радиальными или интеррадиальными.
’ Pelm», atos—стебель, zoon -животжое.
Дорзальная чашечка в ев простейшей форме составлена двумя иля тремя .горизонтальными рядами табличек, венцами, каждый из пяти табшчек, так что таблички одного венца чередуются' с табличками соседях венцов (фиг. 285). Из них наиболее важными являются таблички, поддерживающие руки, так называемые радиальные таблички (.R); таблички, под ними интеррадиально лежащие, называются базальными (В), потому что у многих лилий они образуют основание чашечки, покоясь на стебле.
У некоторых лилий имеется венец радиально расположенных infrabasalia
фиг. 284. Bofryoc'-inus decadactylus. OOmu i Над лилЯИ. Силур.
©ООО© ©О’
яопшй».
Фпг. 285. Анализ двух типов устройства чашечки: вверху—с одвопикловым основанием, внизу—с дву цикловым. Справа и слева разные прпцхы изображения одной и той же разложенной чашечки.
^.R
Фпг. 286. Две формы диска морской лилии: 1 — простейшая (одни oralia)', 2 — более сложная (между oraita образуются ambttlacralia).
(IB), нчхо^яшихся под базальными, которые иные авторы называют парабазальными. Первый тип основания называется одноцикловым, последний—д вуцикловым.
Подобным образом tegmen в его' простейшей форме (Лиг. 286) состоит из пяти оральных табличек—oralia (О), чередующихся с радиаль-
ними (ft), но здесь всегда почти находятся также ambulacralta (Amb), покрывающие желобки, которые радиально идут к рукам яад соприкасающимися краями oral1 а или между ними. Рот—или под oralia, или между ними; в последнем случае он прикрыт амбулякральными табличками. Задняя оральная табличка во многих примитивных формах бывает просверлена отверстием, представляющим сообщение амбулякральной системы с наружною средою (Му, иногда это отверстие занято мадрепоровой пластинкой. Анус (Ля) лежит между заднею оральною и прилегающею (R) и закрыт табличками.'
Руки в их простейшей форме состоят из ияда члеников, называемых bracluaha (Вт), идущих прямо от радиальных табличек (фиг. 287). Каждый членик руки округлен на наружной или спинной поверхности и выем чат на внутренней или брюшной поверхности.
Фиг. 287. Простая аеразветвленная оу-ка (Hybocrinus).
Фиг. 288. Расположение мягких частей в желобке руки морской лилии; сильно увеличено.
Брюшная ручная бороздка, таким образом получающаяся, содержит следующие мягкие части, следуя от брюшной стороны к спинной (фиг 288).
Желобки (fg), приводящие ко рту пищу (инфузорий, микроскопические диатомовые водоросли и ракообразных), выстланные на поверхности реснитчатым эпителием (et), который производит и направляет струю воды кровеносный сосуд (Ь); водоносный сосуд (w), от которого отходят трубочки (р) к серии щупалец (/) — амбулякральных ножек, лишенных присосков, окаймляющих амбулякральные желобки и служащих для осязания и Дыхания; (stc) и (сс) — продолжения отделов цёлома и (де)— канал, содержащий генитальный (несущий половые продукты) ствол. Кроме того имеются нервы, 156
показанные жирна! черной краевой ОР, п2, ws), и чувствительные сосочки на щупальцах (я).
Все эти мягкие ч«,сти защищены в орюшной бороздке кроющими пластинками (с.р. фиг. 287, 288), также называемыми амбулякраль-ными (АтЬ), которые могут закрывать и открывать амбулакральные желобки, смотря по надобности.
От рук можно проследить ход различных органов з текальную полость.
Пищеприводные желобки проходят поверх tegmen ко рту, в который они вливают приносимую ими воду. Рот ведет во внутренние пищеварительные органы, делающие оборот спирали; у некоторых ископаемых лилий в средине текальной полости находится свернутая пластинка, представляющая остаток объизвестненной стенки пищеварительного пути (фиг. 295).
Водоносные сосуды (радиальные амбулакральные каналы) соединяются с околопищеводным водоносным кольцом (hydrocircusy, эти органы снабжены мускулами; здесь не имеется а м п у л л, существующих (фиг. 281. А)
Фиг. 289. Схемы различного ветвления рук морских лилий.
1 2 3. 4 5
Фиг. 290. Схема эволюции от гдиоряд-ной руки к ^вурядной.
у тех иглокожих, у которых амбулякральная система выполняет функцию движения. У более просто организованных лилий водоносное кольцо сообщается с полостью тела посредством одного реснитчатого канала в заднем интеррадиусе, а полость тела сообщается с наружною средою одним отверстием в задней оральной табличке, иногда сливающимся с анальным отверстием. У некоторых форм эта система приведена в соответствие с радиальной симметрией развитием в каждом интеррадиусе пор, проводящих воду в полость тела.
Скелетные элементы теки распределяются в две категории; 1) первичные элементы, или элементы первичной чашечки, развивающиеся первыми как в онтогении, так и в филогении, иннервирующиеся от камерного органа; это именно, членики стебля, таблички дорзальной чашечки (IB, В, В), членики рук (Br, brachialia) и их разветвлений, затем таблипки tegmen—оральные и амбулякральные; 2) вторичные, дополнительные элементы, или элементы вторичной чашечки, для понимания происхождения которой необходимо сначала изучить руки, или brachia.
Руки редко являются простыми. Первою ступенью в изменении простых рук (фиг. 289) является ветвление раздвоением, постоянно повторяемое определенным образом (I, правильная дихотомия, или из о то-' мия). Но в каждой половине руки первая ветвь справа, затем первая слева и т. д. могут быть менее, нежели другие ветви (2, 3). Таким образом возникает главный ствол, дающий маленькие ветви справа и слева попеременно.
. При *енденции к уничтожению разветвлений этих ветвей (фиг. 289, 4, 5) получается наконец главный ствол руки с маленькими ветвями, правильно размещенными по противоположном сторонам его последовательных brachialia (фиг. 289, 5); эти ветви тогда называются pinnulae, и рука является снабженной pinnulae (фиг. 284, р). Pi-inula представляется совершенно гомологичной с маленькими конечными ветвями простой дихо-томирующей руки (фиг. 289, 7), но в руке с pinnulae последние, служа целям размножения и сосредоточивая в себе зрелые половые клетки, приобретают назначение, отсутствующее у разветвлений первоначальной дихото-мирующей руки.
В простой дихотомирующэй руке каждое brachiate, пчд.,ерживаюш.ее две ветви, делает ото двумя верхними покатыми сторонами, или плечами, равной величины (фиг 290, Д), каждая с вентральной выемкой. Такой члеиик называется плечевым (axillare). Если размеры части руки, помещающейся на одном плече, уменьшаются, уменьшается и это плечо. При дальнейшем ходе этого процесса широкое плечо становится близким к положению, параллельному с нижнего сочленовною поверхностью плечевой таблички (фиг. 290, 3). Таким образом рука с pinnulae состоит из серии плечевых табличек, в которых попеременно правое и левое плечи широки а почти параллельны, между тем как другие плечи стали малы и несут pinnulae.
Рука с pinnu7ae может быть простой или ветвистой, хотя и никогда не в столь значительной степени, как в последней; в соответствии с ветвлением различают ручные и плечевые таблички различных порядков.
Вследствие атрофии некоторых рук, число их может быть не 5, а 3, 1, и дажи все руки могут уничтожиться, как это бывает у некоторых верхне-иалеозойских лилий Indocrinus, Monobrachiocrinus, Embryocrinidae.
Ряды амбулякральных табличек (АтЬ) необходимо ветвятся вместе с brachialia, также образуя различные серии или порядки.
Руки и их разветвления, в которых членики расположены в огни ряд, представляя более или менее параллельные поверхности соприкосновения, называются о д н о р я д-н нм и. Простые руки у Crinoidea всегда одворядвы. Руки с pinnulae претерпевают изменение. Так как в таких руках плоскос ти сочленений попеременно покаты направо и налево, brachialia имеют наклонность приобрести клиновидную форму, приобретаемую онтогенетически- и филогенетически в позднейших стадиях (фиг. 290, 3). Наконец bra chtalia располагаются в два чередующихся ряда, и тогда рука называется двурядной (фиг. 290, 4, 5). Такое строение, удваивая число pinnulae в данной длине руки, дает выгоды в отношении сбора пищи и в отношении функций размножения.
Рука, таким образом построенная, может претерпеть еще дальнейшие изменения. Прилегающие правый и левый членики двурядвой руки могут сливаться, образуя сложное brachia е (в противность простому первичному brachiate), несущее непременно две pinnulae, по одной с каждой стороны.
Первичная чашечка, расширяясь, переходит во вторичную. Расширение дорзалыщй чашечки происходит таким образом, что в нее входят нижние части рук, а брюшная сторона передвигается вверх по отношению этих частей рук. Расширение дорзальной чашечки может произойти, как у Icthyo-crinus (фиг. 291), вследствие непосредственного бокового соединения проксимальных 1 brachialia, причем проксимальные ambulacralia входят в tegmen.
У некоторых современных лилий проксимальные brachialia соединены гибким покровом, содержащим маленькие дополнительные таблички; таким образом текальная полость замкнута не только первичными, но и вторич-
1 Прокси м а л ь на—часть, ближайшая к месту прикрепления, листель на— часть, находящаяся наиболее далеко от места прикрепления.
' I
ними элементами. Маленькие таблички могут возрастать в размерах и прочно связывать вместе руки и их ветви (фиг 292); такие таблички называются interbrachialia различных порядков. Подобным образом есть mterambulacralia различных порядков. Interbrachialia иногда,- х,отя редко, опускаются, раздвигая радиальные таблички. За исключением примитивных форм, interbrachialia располагаются в определенном порядке и сходны в различных интеррадиусах. Эта правильность часто нарушается в заднем интеррадиусе, который расширяется введением анальных табличек, называемых так потому,
что они непосредственно примыкают к таблкШм, поддер-
Фиг. 291. IcJthyo-crinus ptrifoi mis.
Фиг. 292. Sagenocnnus etrpansus. В состав чашечки кроме brachialia (Вг) входят interbrachialia (iBr) различных порядков.
Фиг. 298. Диск tegmen лилии Cyatho-crinus та vaceus 1 — амбулакральные таблички на месте; 3-амб. табл, сняты, видны края oralia. 1/1.
живающим анальную трубку (proboscis, хоботок), если этот орган имеется (фпг. 295, 298).
Итак первичными элементами чашечки являются IB, В, В, Вг, АтЪ, а вторичными IBr, lamb.
• Видоизменения чашечки не ограничиваются введением в нее brachialia а. interbrachialia, но выражаются и в изменениях первичной д< рзальной чашечки.
Данные палеонтологии, сравнительной анатомии и эмбриологии свидетельствуют, что некоторые’ лилии с одноцикловым основанием произошли от лилий с двуцикловым основанием путем атрофии IB.
Затем надо отметить наклонность к слиянию табличек проксимального венца В или IB. В этом венце бывают 5, 4, 3, 2 таблички и наконец иногда все сливаются в одну.
Рассмотрим видоизменения tegmen. Мы уже упоминали о часто находящихся в нем интеррадиальных oralia, имеющихся у некоторых ныне йшвущих лилий в зрелом возрасте, временно существующих у некоторых лилий в молодых стадиях развития, наконец встречающихся и у ископаемых форм. Вообще можнб считать примитивным tegmen, образованный указанными оразьными табзичками.
От этого примитивного патеозойского типа выходят три линии развития:
1) Амбушкры проходят над краями oralia (фиг. 293, Cyathocrinus, Cupressocrinus) — Ш'ряюк Fistulata.
2) Амбулякры проходят между oralia, оставляя открытым рот, между тем как oralia (фиг. 294, О) постепенно атрофируются (Taxocrinus}— порядок Articulata.
3) Ачбулякры проходят под оральными табличками и под дпупми тегминальными, которые развились одновременно со включением brachialia в чашечку — порядок Camerata (фиг. 295).
Б группе Camerata таблички твгмепа утолщаются и, плотно примыкая одна к дру-юй, образуют прочную сводообразную кровлю чашечки, не изменяющуюся в форме; амбулякры у них следуют внутренней поверхности тегмена (фпг. 295), заключаясь в трубках, из мелких табличек; происхождение такою тегмена и амбуликров ставится в связь с удобствами, обусловленными постоянством формы тегмена. При гибкости теглена брюшная поверхность приплющивается, когда
Фиг. 294 Диск Taxocrinus mtermed us. А—анальный инторрадичс; гАтЪ, illAmb, MAmb — mterambulacra-lia I, II, III порядка.
Фиг. 295. Actinocrinus probosctdialis. Диск вскрыт; видны: трубчатые амбулакральные каналы и остаток объизвестненной стенки пищеварительного пути (в середине). 1/1.
руки раскрыты, и выпячивается вверх, когда руки сомкнуты; эфи изменения сопровождаются перерешениями внутренностей—неудобством, исчезающим при развитии негибкого тегмена. Амбулякрадьные табличку, может быть, были сомкнуты вначале, но так как, вследствие выпячивания тегмена, борозд..л стали более открыты, потребовалась защита их иным образом. В связи с этим они опустились ниже поверхности, а тегминальные таблички сомкнулись иад ними. Опустившиеся сосуды были заключены в трубку, частью (сверху) состоящую из ambulacralia.
Стебель достигает у некоторых лилий длины в несколько метров; у других же он укорочен или даже атрофирован, так что чашечка прикрепляется непосредственно основанием или является совершенно свободной, неприкрепленной. Стебель состоит из коротких члеников, в примитивном стебле располагающихся пентамерно, так что стебель является сложенным из пяти вертикальных рядов шестиугольных табличек, причем таблички смежных рядов чередуются (фиг. 296).
Следующей стадией в развитии стебля были та, в которой таблички уже не чередуются, но рас-
1 2
Фиг. 297. Сочленовные поверхности радиальных таблп-чск (/—Bisocnnus olhda.2— Bath'jcrioMS aldrickianus').
Фиг. 296. Строение пои-митиввого стебля: 1— сечение; 2—вид сбоку.
положены рядами, как вертикально, так и горизонтально. Наконец пеп-тамеры каждого горизонтального ряда слива * тся, образуя членики стебля c&lumnalia, еще с широким центральным отверстием.
Наружная нентамерность стоит в соответствии и вероятно в связи с лентамерностыо внутреннею; от каждою из пяти отделов камерного органа в стебель идут кровеносные сосуды и не, вные стволы. Полость стебля часто подразделяется в желобки выростами твердой наружной с енки его; эти желобки содержат в себе нервные стволы, а иногда даже могут образовать замкнутые каналы, заключающие стволы. Осевой канал стебля некоторых ископаемых форм отличался значительным поперечником и, возможно, содержал некоторые мягкие части све х тех, что у ныне живущих лилий. Членики стебля бывают круглыми, эллиптическими, многоугольными (пятиугольными). Боковые членимые придатки стебля называются cirri, усики (фиг. ЗОЬ).
Различные таблички или соединены неподвижно между собою, плотно примыкая одни к другим, или подвижно. *
Подвижное сочленение существует например часто в таком виде, что на сочленовных поверхностях табличек развиваются валикообразные возвышения (фиг. 297, f), рядом с возвышениями между члениками существуют прлости; полости эти заполнены мускулами (фиг. 297, mf) или эластичными связками (dl, И). Таблички чашечки всегда соединены между собою неподвижно; членики стебля у одних лилий—подвижно, у других — неподвижно;, членики рук—всегда подвижно. При неподвижном соединении примыкающие поверхности могут быть неровны, например радиально струй-чаты у члеников стебля (фиг. 296, 2).
Мы рассмотрим несколько морских лилий, принадлежащих к трем порядкам: Fistulata, Articulate и Camerata, характеризующимся тремя вышеуказанными типами tegmen.
Другое подразделение морских лилий основано ие на характере тегмена, а на характере дорзальной чашечки, причем руководящим является состав ее основания из одного или двух венцов базальных табличек. Таким образом морские лилии разделяются на Mqnocyclica и Dicyclica. Надо не упускать из виду, что кроме Monocyclica, ряд предков которых всегда представлял одвоцикловое основание, есть Monocyclica, приобрев-шие одноцикловый базис вторично, происшедшие от Dicyclica, у которых один венеп основания подвергся уменьшению в размерах и наконец исчез. За исключением этого группы Monocyclica и Dicyclica существуют обособленно с древнейших отложений, невидимому будучи в главной массе не связаны между собою отношением происхождения одной от другой (см. ниже, в главе о Cystoidea), а происходят независимо одна от другой из общего корня, от класса Cystoidea. Группы Monocyclica и Dicyclica обнаруживают в своем развитии параллелизм, причем у некоторых представителей их независимо развивается например proboscis и tegmen типа Camerata; таким образом группа Camerata не является единою в своем происхождении, ио сборною, содержа представителей как Monocyclica, так и Dicyclica.
ПОРЯДОК I. FISTULATA. 1
Руки без pinnulae или с ними. Чашечка первичная: членики рук не входят в состав чашечки; амбулякры расположены над табличками последней. Для морских лилий этой группы характерно также присутствие анальной трубки {proboscis), .конической, цилиндрической или ретортообразной (фиг. 298); таблички этой трубки часто снабжены маленькими щелевидными или круглыми порами. В анальном интеррадиусе, кроме интерраднальной анальной таблички, большею частью находится еще одна табличка (бывает и более одной), расположенная между радиальными, а внизу вдвигающаяся между базальными табличками. Это так называемая радиа-нальная табличка (га, фиг. 299), некоторыми исследователями считающаяся табличкой, отчленившейся от одной из радианальных (г, фиг. 299).
Типичным представителем этой группы являетея род Cyathocrinus (фиг. 293 и 300; cyathos—чаша, crinon—лилия).
Чашечка кубкообразная. 5 1В\ В большие; R с широкой выемкой для причленения рук. Имеется только одна интеррадиальная анальная табличка (фиг. 300, х). Руки дихотомически ветвящиеся, без pinnulae. Стебель круглый, с пятилопастным каналом и с радиальною струйчатостью на соприкасающихся поверхностях члеников. От силура до пермских отложений.
1 Fistula—канал.
I «1
11 Т.абяжж й« можтааarm.
Е роду Cyathocrinus примыкают палеозойские роды Poteriocrinus (фи1. 301; poterion—кубок) и Cromyocrinus (фиг. 299, 302) с руками, снабженными ptnnulae, с 2—3 аналогичными табличками.
Cromyocrinus огличается от Poteriocrinus менее разветвленшлми руками и низкой ча печкой с венцом 1В, маленьких, но все таки выдающихся из-под стебля на нгружную поверхность чашечки.
Cyathocrinus и Poteriocrinus радиальные таблички выемчаты наверху, в месте сочленения с руками, у Cromyocrinus этот край прямой. Руки состоят из одного ряда члеников, каждый с одной pinnulat; pmnulae смежных члеников расположены поочередно, то с одного, то с другого бока руки.
Poteriocrinus в девоне и каменноугольных. Cromyocrinus в каменноугольных.
С рассмотренной группой родов, типом которой является Cyathocrintis, пред -ставляет некоторые соотношения
В то время как
У
род Cupressocrinus
R—
о^сгсго
Фиг, ЗлО. Cyathocrinus longimanus. Верхний силур. Готланд. Швеция. Наверху сочленовная поверхность членика стебля; внизу анализ чашечки.
Фиг. 299. Анализ чашечки Cromyocrinus.
lb -infrabatalia; Ъ—lasana; r—radiaUa; гц—radianale; a't e”, a'"—interradialia aualia.
•Фиг. 298. Лидия (Cyathocrinus) е вш нугой анальной трубкой.
(собственно Cypressocrinus, по сходству с кипарисом—Cypressus) являющийся весьма оригинальным, специализированным. Чашечка (фиг. 303) отличается совершенным отсутствием анальных табличек IB сливаются в одну; массивные руки не ветвятся, состав чены из одного ряда малочисленных широких Вг, опираются на радиальные таблички во всю их ширину; tegmen не представляет совершенно замкнутого покпова из табличек, как у Cyathocrinus, в таком покрове здесь нет надобности, так как tegmen защищают плотно смыкающиеся руки, но 5 oralia характерных для Cyathocrinus, сохранены в виде пятилопастной розетки («консолидационный аппарат» фиг. 303, 3); анальная О более остальных и просверлена отверстием, к кото
рому примыкает анальный хоботок. Стебель четырехугольного сечения с осевым и четырьмя периферическими каналами.
ПОРЯДОК II. ARTICULATA.1
Руки бывают довольно сильно разветвлены, хотя имеют pinvulae Первичная чашечка существует лишь в ранних стадиях индивидуального развития, а затем tegmen поднимается вверх между нижними частями рук.
Фиг. 301. Poteriocrinus multiplex. Верхний каменноугольный известняк. Мячково, около Москвы. Сарана анализ чашечки, а—таодички анальною интеррадиуса.
Анальные таблички исчезают в связи с уменьшением размеров proboscis-, одна анальная табличка существует у зародыша. Оральные таблички существуют у зародыша, редко в зрелом состоянии: обыкновенно в этом состоянии tegmen кожистый, с заключенными в нем известковыми частица «и, или состоит из мелких табличек, неправильно расположенных; амбулякральные желобки и рот открыты. IB отсутствуют или малы и сливаются с верхним члеником стебля. Редкие представители этой грузны (Sternmatocr inns'} находятся уже в верхнепалеозойских отложениях; большое распространение с мезозоя доныне.
Encrinus (фиг. 304). Низкая чашечка с дицикловым базисом. IB 5 очень маленьких, скрытых под верхним члеником стебля. В 5 больших. В состав чашечки входят два порядка Вг.
Руки состоят из двух ветвей, внизу из одного, но немного выше из двух рядов члеников. Стебель круглый, с круглым центральным каналом, с радиально струйчатыми соприкасающимися поверхностями члеников.
Articula*"'»—членистый.
Примыкает к Cromyocrinus. Триас.
Pentacrinus (Лиг. 305; penta—пять). IB существует лишь у различных юрских видов. Руки ветвятся вполне или почти дихотомически, состоят из одного ряда члеников. Стебель
Фиг. 302. Cromyocrinus geminatus. Верх* ний каменноугольный известняк. Мячково, около Москвы.
снабжен усиками, расположенными мутовчато.
Сечение стебля пятиугольное, соприкасающиеся поверхности члеников стебля представляют углубления, составляющие пятилопастную розетку; в этих углублениях помещаются связки. С триаса доныне, всего чаще в юре.
ПОРЯДОК III. CAMERATA.
Тегмен, лишенный гибкости, состоящий из довольно больших толстых табличек, руки нижнею частью иногда входят в состав чашечки при участии интербрахиальных табличек. Ротовое отверстие под поверхностью тегмена. Анальное отверстие часто расположено на конце хоботкообразного выроста. Руки однорядно- или дву-ьядноч тенистые, снабжены pinnulae. Исключительно палеозойская группа.
Platy crinus (фиг. 306; pla-tys—плоский). Чашечка состоит из одного венца базальных табличек (3 таблички) и одного радиальных. Руки ветвятся 2—3 раза, внизу однорядно-, выше двурядночленистые. Стебель близ чашечки состоит из круглых, а далее из эллиптических члеников. Послед-
ние сочленены между собой так, что стебель является скрученным. Каменноугольные отложения, редко девон.
Actinocriniis (фиг. 295, 307, 308; aktin—луч) Коническая чашечка, представляет на свои£ табличках укра пения в виде радиальных складок 3 В.В в анальном интеррадиусе раздвинуты анальными табличками (а1), доходящими до В. гВг во всех интеррадиусах. Каменноугольные отложения.
КЛАСС П. CYSTOIDEA.1
Группа примитивных Pelmatozoa, скелетированная часть стенки тела которых (тека) состоит из многоугольных табличек, снабжена стеблем или утрачивает его.
1 Kustts- яолссть »•*
Тека замкнута до рта, оставляя лишь у последнего выход для амбу-лякральных радиалгных сосудов; таблички ее пронизаны особыми порами. Амбулякральные сосуды, по выходе из теки, распространяются по ее повер!
ности или прямо от рта поднимаются над этой поверхностью; в том и в другом случаях они переходят на своих концах или на концах своих разветвлений на ручные придатки (brachi-olae), поднимающиеся над поверхностью теки.
Brachiolae не ветвятся, состоят из двойного ряда, объизвест-ненных члеников, на брюшной стороне снабжены лишь краевыми пластин;:; ми, неимеяуишмгАяе.
Кишка свернута по направлению часовой стрелки: рот лежит обыкновенно в центре верхней стороны тела; anus лежи, интеррадиально, в боковой стенке теки, большею частью закрывается клапаном, состоящим из треугольных створок, в сомкнутом состоянии образующих пирамиду, выдаю-
Фиг. 303. Cupressocnnus crassus. Средн, девон, Эйфель. Германия.
1—сотяеховвая поверхность членика стебля; Я—центральная пластинка в основанпн чашечки; 3—консолидациейвый аппарат; 4—-поперечный разрез рук со спирально свернутыми pinnulat. 1/1.
щуюся на поверхности теки.
Имеются иногда сливающиеся (фиг. 309) отверстие первичного каменистого канала (гидропор) и отверстие канала, выводящего половые продукты (г о н о п о р).
Фиг. 304. Enrmns. А и В—Е. moniliformis. С—Е. lilitormis. ’гриас.
Число brachiolae изменяется в обратном соотношении с их величиною, бывая минимум—два (при двух амбулякрах животного).
Число табличек, входящих в состав теки, изменяется в больших пределах у различных родов—от десятков до сотен. Уменьшение числа табличек сопровождается приобретением ими больших размеров - и более правильной формы, обыкновенно шести-
угольной, вместе с тем становится более определенным относительное положение табличек, они располагаются в горизонтальные венцы с чередованием элементов соседних венцов.
Фиг. 305. Pentacrinus. briar aides. Лейас.
1—стебель Р. lugalttformir, 3—стебель Р. eubangulartt.
Определенность (пентамерность) в расположении табличек теки возникает в значительной мере благодаря влиянию амбулакральных сосудов и стебля. Последний и brachio-lae, двигаемые мускулами, создавая напряжение в теке, дают импульс к определенной группировке табличек соответственно с этими напряжениями; так ириамбулякральные таблички группируются в ряды соответственно амбулякрам, в то время
Фиг. 306. Ilatyc.vnus synvnetricus.
Я—общев вид; D—сочленовная поверхность члеников винтообразно изгибающегося стебля; ас—осевой канал; А—сочленовное ребро.
как интеррадиальные таблички расположены еще без порядка. С другой стороны, на нижнем конце чашечки, опирающемся на стебель, замечается по крайней мере один венец пяти табличек, даже тогда когда на остальной поверхности теки таблички расположены беэ порядка.
Текальные поры имеют правильную форму и расположение и приводят в сообщение пространство внутри теки с наружной средой. По внутреннему строению пор и их отношению к табличкам теки различают два типа пор: ромбовые поры и двсйные.
1) Ромбовые поры располагаются ромбами, причем половина ромба находится на сдвой табличке, другая половина на другой, смежней табличка,
1б< « >
форма отдельных пор, входящих в состав ромбов, весьма различна у различных цистоидей.
Простейшие поры существуют у некоторых представителей древпего и во всех отношениях примитивного рода Chirocrinus (фиг. 311); отверстие
поры представляет (фиг. 312, а) удлиненную щель, ведущую в такой же длины внутреннюю складку (Jnir. 313, a, at), выдающуюся на внутренней
Фиг. 307. Actinocrinus steiiin <6. Нижний каменноугольный известняк. Турин. Бельгия.
поверхности теки (самое большее на несколько миллиметров). Наиболее
распространенными являются поры с щелью, «нем ее стенок {Ь и с, фиг. ЗГ2; Ь, фиг. 303) Щель у типай (фиг. 312) смыкается с промежутками, оставляя между последними дырчатые отверстия, иногда же имеются лишь два
Фиг. 309. Относительное расположение отверстий на теле цистоидей: О—рот; а—анус; Рг — гидропор; Рр — гонопор; I— Г—радиусы амбулякральпой си-стомы.
замкнутой в средние сраста-
Фиг. 311. Chirocrinus atavus. Слева—чашечка со стеблем, справа— часть чашечки (видны 2 амбул. желобка и поровый ромб между ними)
Фиг. 310. Расположе-' иве краевых складок на табличках цн<тоидеи (Lichenoides prtscus).
крайние отверстия, наиболее удаленные одно от другого, разделенные значительною заросшею частью щели (фиг. 312, е), причем эти два отверстия иногда прикрыты снаружи ситовидно продырявленной кнопкой.
У Ь chinosphaeriies поры наиболее оригинальны, белее стоят особняком, представляя, кроме поровых каналов, пронизывающих таблички, теки перпендикулярно к ее пов^хности, еще каналы, параллельные поверхности
стенки, идущие под наружным слоем ее; эти последние каналы соединяют поровые каналы (f, фиг. 312; А, В фиг. 308).
Как сказано выше, кроме собственно пор, находящихся в самых табличках теки, у цистоидей иногда бывают еще так называемые поровые складки, лежащие, выдаваясь на внутренней поверхности теки. Такие складки, чрезвычайно тонкостенные, были найдены у некоторых форм (фиг. 313, а, Ь), но у других, даже часто встречающихся, их не было найдено.
Объяснить последнее, в виду родственности форм той и другой группы, будет всего естественнее, предположив, что эти складки были и у форм, у которых их не найдено, но были мягкими необъизвествеиными, так что не могли со~.ранитьси на окаменелостях
Наконец что касается до самых простых пор типа, изображенного на фнг. 313 а и Ь, то надо
Фиг. 812. Различные типь ромС'>вых пор цистоидей, вид снаружи.
Фиг. 313. Ромбовые поры цистоидей в вертикальных разрезах.
а, а„ Ъ—Chiroerinus (соответствуют ей» фпг. 312): в, 8—продол иые, а,—поперечный разрез Л, E—Echinosphaerites (соот-ветств. f4>nr. 312), Л-норовые каналы; f— поровые складки; п~ tps вица между см.жвыми табличками.
I
сказать, что оии, а следовательно и поры вообще, выводятся из еще более простых образований другой категории, из складок, покрывающих поверхность табличек теки многих древних иглокожих, складок, направленных перпендикулярно к краям табличек и находящих собе продолжение в подобных же складках смежных табличек (фиг. 312 а и 310); надо только предположить, что этн складки стали глубже, уже и, в части выдающейся вовнутрь теки, тонкостениы,—и мы получим поровые складки.
Складки, идущие на поверхости табл и тек теки древних иглокожих перпендикулярно к их сторонам, расположены соответственно напряжениям, возникающим в соприкасающихся табличках при их росте; складки эти, делая поверхность таблички волнистою, сообщают последней таким образом большую прочность.
2) Двойные поры располагаются на одной и той же табличке, а не на двух смежных как ромбовые.
Вти поры—парные, так что поры каждой пары всегда находятся на определенном расстоянии одна от другой на наружной поверхности теки (фиг. 314, 315), хотя внутренние концы поровых каналов могут этой определенности не представлять. Наружные парные отверстия пор расположены в овальном углублении, окруженном выдающимся на поверхности теки ва ликом (подобные валики существуют иногда и вокруг поровых ромбов)
Доры цистоидей, по всей вероятности, служили для дыхания; возник вовение их естественно в виду слабого развития ручных придатков.
Как ни отличны, на первый взгляд двойные поры от пор, расположенных ромбами, и те и другие поры в действительности приводятся к одному типу. Бывает, что двойные поры табличек чашечки—или все, или отчасти—располагаются перпендикулярно к краям табличек (фиг. 315 А, О), иногда даже двойная пора бывает разделена надвое границей смежных табличек, так что одна пора лежит на одной табличке, другая—на другой.
Стебель цистоидей отличается большею частью своею малою длиною; часто совершенно отсутствует. Невидимому лишь в редких случаях стебель служит для прикрепления, так как он большею частью суживается, за
остряясь к нижнему концу. Существует предположение, что такой стебель служил для плавания. Стебель состоит или из ряда кольцеобразных члеников, заключающих широкий канал, или он состоит, по крайней мере в своей верх-
Фиг. 314. Двойные
поры (сильно увеличенные) на поверхности таблички Pyrocystvtex pirum.
Фиг. 315. Двойные норы. В—вид снаружи. С—разрез. А - - GJgp-tosphaerites Leuchtenbergi. I)— Mesocystis.
ней части, ез вертикальных рядов табличек с чередованием табличек смежных рядов. Полость стебля иногда очень широка, так что вероятно заключала в себе значи-
тельную часть внутренностей.
Пятилучевая симметрия у цистоидей проявляется с меньшим постоянством, нежели в других классах иглокожих. Не говоря уже о расположении табличек чашечки, в котором иногда отсутствует не только пентамерность, но и вообще какая-либо определенность, половые органы не цредставляют лучевого расположения; число амбулякров не всегда бывает пять.
Cystoidea распространены в силуре, встречаются в кембрие и девоне.
ПОРЯДОК I. КПОМВП'ЕКА
С поровыми ромбами.
Chirocrinus (фиг. 311, 316, 317; cheir—рука). По строению чашечки является исходным типом для целой группы родов. Чашечка состоит (фиг. 317) из пяти горизонтальных венцов табличек: одного венца из 4 базальных табличек (б), трех венцов боковых латеральных (I, I', Г)—по 5 табличек в каждом, и одного венца дельтоидальных (I'"). У более примитивных видов этого рода (например у Ch. radiatus, фиг. 312) два нижние венца латеральных табличек совершенно замкнуты, так что таблички каждого венца соприкасаются между собою вертикальными краями. В верхнем венце латеральных табличек одна из дельтоидальных табличек (Z/") нарушает правильность, опускаясь под гидропором и гонопором (Рр') между верхними латеральными Z/' и Z6", так что может казаться на первый взгляд принадлежащей к венцу последних, приобретая и соответствующую величину. У других видов рода Chirocrinus есть отличия, обусловленные перемещением латеральных табличек вниз в латеральные венцы других порядков.
Поры у некоторых древних видов имеют вид щелей, открытых во всю длину поры, у большинства же—с частичным срастанием продольных стенок щели. Из базальных ромбов находится постоянно, по крайней мере, оба нижние, идущие на 1г и 12, вместе с ними часто еще полуромбы между Ъа и bj или Ья. Амбулякры пентамерны, дают по 3—5 ветвей; ручные придатки не длиннее высоты теки. Большое геогрфиче-сьое распространение: окрестности Ленинграда, Богемия, Канада, Нижний силур.
Echinoencrinus (фиг. 318,319, echinos—морской еж)
,а гз И 1‘ и fi
Фиг. 316. Ckirocrinus рсппгдег. Нижний
силур. Эстония. а—увел, амбулакральное Фш. 317. Анализ чашечки Ckirocrinus поле. radiatus.
отличается малым числом поровых ромбов. Поровые щели со стенками, срастающимися в средней части. В основании чашечки обыкновенно два поровых ромба между табличками Ъй и /2, Ъ2 и/х), реже четыре, как на
Фиг. 318. Л чал из чашечки Echinoen-erbius Lahuseni.
Фиг. 313. Околоро-тсчая площадка
Eckinoencrinus. а— места отхода Ьга-chiolae, G—гонопор, Н—гидропор, .1/— ротовое отверстие. 3/1.
Фиг. 320. Расположение поровых ромбов на табличках примитивной пистопдеи. в местах прилегания кишки к теке (см. пунктир) ромбы исчезли.
Аз—анус.
фиг. 318; anus окружен 3—4 табличками. 5—12 brachiolae, расположенных на маленькой околоротовой площадке, горизонтально притупляющей вершину чашечки. Первичные поры—гидропор (Н) и гонопор (6/)—рудиментарны, заключены в упомянутой площадке. Все таблички с сетчатой скульптурой.
Вследствие уменьшения размеров чашечки число поровых ромбов уменьшилось.
Изчезновенпе поровых ромбов прежде всего произошло в частях стенки чашечки, лежа щих вблизи кишки (фиг. 320) прп уменьшении размеров чашечки кишка прилегала к стенкам и, стесняя дыхательную деятельность прилежащих пор, была причиною их неразвитая.
Так было у Chirocrinus-, у Eckinoencrinus кроме того поры ве развиваются между табличками /" вследствие малых размеров этих табличек.
Echinoencrimts встречается в нижнем силуре, в окрестностях Ленинграда, в Швеции.
Eclwnosphaerttes (фиг. 321; echinos—морской еж, sphalra—wap). Тека— шаровидная или грушевидная—образована многочисленными, неправильно многоугольными и без порядка расположенными табличками; без стебла дли с коротким многорядночленистым полым стеблем. Амбулякры сосредоточены около рта, каждый с одной брахиолей. Околородовое поле, несущее brachiolae, большею частью хоботкообразно приподнято над окружающею поверхностью теки. Число brachiolae 2—4, реже 5—8. Anus в верхней трети теки. Над ним и несколько влево находится гидропор. Поры оригинальны по своему строению и расположены ромбами, с несколькими рядом лежащими тангенциальными поровыми ходами, лежащими в стенке теки; имеются два, реже три тангенциальных хода; они находятся в средней части стенки теки (фиг. 313, А и 2>), отходя под очень острыми углами от поровых каналов, и идут далее параллельно между собою. Поры закрыты наружным слоем стенки теки, что указывает на ослабленность функционирования пор. Нижний силур. Прибалтийский край, Богемия и Англия.
ПОРЯДОК II. DIKLOPORITA.1
Таблички теки с двойными порами.
Glyptosphaerites (фиг. 315 — 322; glyptos— скульпт) рный). Тека л блоковндная, образована многочисленными, довольно большими, неправильно многоугольными и без по
рядка расположенными тонкими табличками; снабжена
коротким многорядно-члени-стым стеблем. Пять амбуля-рров распространяются на всей верхней половине поверхности теки, где им соот-ветств) ют слабо врезанные бороздки, прикрывавшиеся краевыми табличками; по
следние не сохраняются, но на краях бороздок заметны следы их прикрепления. Амбулякры с малым числом ветвей и brachiolae (Вг)\ последние малы, судя по соответствующим им сочленовным площадкам, находящимся на концах боковых ветвей амбулякров.
Фиг. 321. 1 — Echinosphaerilts aurantium;
2 — JEchinosphaeriles infaustus.
1 — нижний силур, Богемия; 2~ p. Волхов, Ленинградская обл.; 8, 4t б—околорОтовые поля индивидуумов с ДЕумя (4), тремя (5) ж четырьмя (5) trachfalae.
Diploos—двойной, рогоз—пор*
Разветвления амбулакров яе отличаются определенностью, желобки разветвлений пересекают таблички теки без определенного отношения к их границам или центрам, и даже brachiolae не всегда приходятся в центре несущих их табличек, притом иногде две brachiolae помещаются на одной табличке. Ветвление амбулякров большею частью происходит справа и слева попеременно, но часто также несколько ветвей расположены подряд на одной стороне, а иногда и все. Таблички, несущие brachiolae, по своей форме и расположению еще мало отличаются от остальных табличек теки, еще не обособились среди них. Поры окружены овальными валиками.
Ротовое отверстие (О) пятиугольного очертания, с большими покрывающими табличками. Имеется большая мадрепоровая пластинка (V)
Фиг. 322. Gtyptosphae,ttes Leucbtenbergi Нижний силур. Окресв Ленинграда.
Фиг. 323. Детали строения амбулакрального желобка Cod. trilobutvs
пс — нервный канал ланцетовидной пла стянкж Li h— щель гпдроеппры; Л, р— крав таблички.
треугольного очертания, губчатого строения; блчз нее, * несколько, ниже, находится гонопор (G), а еще ниже—большое анальное отверстие (Лз).
Встречаются в нлжне.ч силуре Богемии, Швеции и в окрестностях Ленинграда.
КЛАСС Щ BLASTOIDEA.1
У Blastoidea текальные таблички распределяются в определенном положении и числе в три венца (фиг. 324. 326), а именно: три базальных таблички (Д) — две большие (образованные слиянием йо две четырех из пяти первичных базальных табличек) и одна маленькая; пять радиаль яых (2?), часто вильчатой формы, образующих замкнутый венец; пять дель-тоидальных йнтеррадиальных по положению, опирающихся на отростки радиальных и часто окружающих ротовое поле своими верхними концами.
Пять аыбулякральных радиальных сосудов идут от рта, располагаясь на поверхности ланцетовидной пластинки, окаймленной краевыми табличками несущими brachiolae.
Радиальные и дельтоидалгные таблички с каждой стороны амбулякров образуют складки, расположенные вкрест шва между этими табличками; эти
1 Biastoe—бутон, eidos—вад
Фиг. 324. Codaster t ilobatus.
1—околородовая поверхность; 3—то же, с обнаженными ланце х о видным и пластинками; 3—анализ чашечки; 4—чашечка сбоку; б—чашечка снизу; 6—поперечиое сечение через радиальную пластинку.
складки свешиваются в текальную полость, образуя гидроспиры, служащие для дыхания.
Codaster является наименее специализированным из blastoidea (фиг. 323, 324; codon—колокол, aster — звезда), 3 В образуют конь тесную чашечку (фиг. 324, 3 — 5), за которой следует более цилиндрический венец из пяти й, отростки которых наверху отгибаются почти под прямым углом, образуя часть усеченной вершины чашечки. Примыкая к верхним краям радиальных табличек, идут пять дельтоидалькых, окружающих пятиугольное ротовое отверстие (фиг. 3z4, 1—3, А). Эти таблички снабжены срединным возвышением.
Между прилегающими дельто-идальными табличками и проходя вник в выемку каждой радиальной, находится длинная пластинка, соответственно форме ее называемая ланцетовидной (фиг. 324", 2, 6—L); края ее частью прикрывают радиальные и дель-тоидальные таблички; ее верхняя поверхность представляет бороздку, проходящую ко рту между смежными дельтоидальными.
По бокам ланцетовидной пластинки (фиг. 323 L; 324, 6) лежат маленькие боковые таблички (sp и osp), но не в простом ряду, а зигзагом, так что каждая пара образует ромбоэдр, разделенный швом между табличками. На этих
швах помещаются места отхода brachiolae (Ьг). Амбулякральные бороздки были прикрыты подвижными маленькими краевыми табличками (fgr фиг. 323), впечатления которых (рубчики) видны по бокам бороздок. Рот был прикрыт подобным же образом. Прилегающие края дельтоидальных табличек образуют ряд резко выраженных складок (гидроспир), идущих вкрест шва между этими табличками (фиг. 324, I, 2—п\ 6). Складки эти, очевидно одинакового происхождения и назначения с поровыми складками в поровых ромбах цистоидей. Ясно выраженная пятилучевая симметрия в строении чашечки нарушается только анальным отверстием (Лв), расположенным в одном из интеррадиусов на границе между делыгоидальной табличкой и радиальными. В этом интеррадиусе складки гидроспир обыкновенно отсутствуют (одна или две существуют на некоторых экземплярах). Анальное отверстие было закрыло маленькими табличками.
Codaster представляет настолько много общего с цистомдеями, что мог бы быть отнесен к ним, если бы не было промежуточных форм между ними и типичными Blastoidea. Встречается от силура до каменноугольных отложений в Европе и Северной Америке.
Род Pentremites (pente —пять, remos — табличка), встречающийся в изобилии в каменноугольных отложениях Северной Америки (фиг. 325—328) является уже сильно специализированным представителем Blastoidea Складки
гидроспир (фиг. 325) здесь сосредоточиваются на более тесном пространстве и над ними распространяются, скрывая их, боковые таблички (sp, osp) и частью ланцетовидная (L). Соединением ближайших складок гидроспир правой и левой стороны образуется вторая, внутренняя ланцетовидная пластинка, лежащая под наружною (si, фиг. 325, 5).
Боковые таблички также двоякого рода: большие внутренние (?р) и маленькие наружные (osp); между последними находятся попы Ср). Через эти поры вода входит в гидроспиры, омывает их, выходит же через отверстия у ротового поля—спи раку ли (фиг. 321, Ь; фиг. 322); спиракули сообщаются с каналом, лежащим м ищу ланцетовидной пластинкой и гид-роспирами с каждой стороны амбулякра, так что в каждое spiraculum одновременно открываются два канала, принадлежащие смежным ачбулякрам. В одном из тпиракулей помещается анальное отверстие.
Прикрытие гидроспир боковыми табличками, возникновение spiracula это черты, типичные для Blastoidea; развитие же пор между наружными боко
Фмг. 325. Петли амбулакрального поля Pentremites (сильно увелич.).
S—вид сверху; 4—одна сторона между ланцетовидной пластинкой и радиальной табличкой; увеличено еще более; б—сеченпе поперек амбулакрального поля.
Фиг. 326. Pentremites f odoni. Камевяоугольн. Иллинойс. САСШ. а— сбоку, Ъ—сверху, с—со стороны основания.
выми табличками, развитие каналов между гидроспирами и ланцетовидной пластинкой—явления, характеризующие Pentremites.
Blastoidea встречаются от силура до пермских отложений.
Эта группа несомненно должна считаться за происшедшую от Cystoidea общее между ними указано частью при описании рода Codaster; что касается до табличек ланцетовидных и боковых, то подобные приамбулякральные таблички, независимо от табличек собственно теки, развиваются у довольно многих Cystoidea, выше при описании этой группы оставленных без рассмотрения.
Все предыдущие классы Pelmatozoa объемлют прикрепленных животных, живущих со ртом, обращенным кверху, и получающих пищу посредством тока морской воды, направляемого ко рту вдоль реснитчатых пищеприводных желобков. У этих Pelmatozoa пищеприводные желобки помещаются на особых образованиях, brachia или bracniolae, обыкновенно поднимающихся от тела, и амбулякральные сосуды хотя могут приходить в эти придатки, ио не проходят через табличатую стенку чашечки.
КЛАСС IV. EDEIOASTEEOIDEA. 1
Иглокожие, выделяемые в класс Edrioasteroidea, вет образ жизн„ подобно Pelmatozoa и причисляются к последним, но имеют пищеприводные желобки, расположенные непосредственно на поверхности табличек чашечки,
и —по крайней мере в некоторых случаях — эти таблички чашечки пронизаны отверстиями для прохода амбулакральных ножек. Некоторые из Edrioasteroidea (как Аде-lacrinus, agele — стадо; Cyaikocystis} вероятно были животными, прикрепленными во время всей своей жизни. У них не было найдено пор между табличками чашечки, образующими дно пищеприводных желобков. Другие представители Edr.oa-steroidea (как Edr toaster, фиг. 329; edrlon — сидение) вероятно обладали способностью передвигаться е одного места на другое. Пищеприводные желобки Edrioaster весьма сходны с таковыми
я
Фиг. 327. Pentremit.es Godoni. Каменноу.ольные.
Иллинойс. САСШ.
С—ЪтасЫо1 е покрывают амбулл-крпльное поле; о и у—brach. сн>ты; р— лницетоЕП ная пластинка без к, аевых, а—си <та ланцетовидная пластинка^ видны щели глдроспир.
Фиг. 328. Pentremites Godoni. Вскрытое послойно амбулякральное поле, внизу боковые края вырезаны, видна полость чашечки,pi—pinnula (остальные удалены); 5 пар верхних боковых разветвлений аыбулякр. желобков представлены с сохран. двурядными табличками, прикрывающими желобки, на иижиих 5 парах эти таблички сняты.
Л.8—краевые таблички с ворами (р) между ними; з—наружные таблички, примыкающие к ланцето-ввдк*и пластинке (I); hy—гидроспиры. Увеличено.
Фиг. 329. 1' di ioaster Его, sbyi с ротовой стороны-атЪ — к, оющие таблички двух амбулякров; в остальных эти таблички удалены и видны снабженные амбу-лякральными порами таблички чашечки (ad/, ps— рот; As—anus; р— пиры. Нижний силур. Оттава.
Канада,
желобками морских звезд, за исключением тсто, что у Edrioasteroidea они покрыты амбулакральными кроющими табличками, как у морских лилий. Таким образом намечается путь развития Eleutherozoa из Pelmatozoa. Edrioasteroidea встречаются от силура до ваменноу-ольных.
Edraios—сидящий.
Геологическое развитие Pelmatozoa.
Близость различных групп Pelmatozoa видна уже из того, что нередко Blastoidea и Edrioasteroidea считаются за порядки класса Cystoidea. С другой стороны, несомненна близость Crinoidea и Cystoidea, именно Cystoidea Iiiiom-bifera, приближающихся к морским лилиям по правильности строения чашечки и по числу венцов ее. Главное отличие цистоидей от лилий в конце концов составляет присутствие у первых пор на табличках чашечки. Но путь к превращению Cystoidea в Crinoidea намечается ясно выраженной тенденцией к уменьшению числа поровых ромбов у Cystoidea Rhombifera. Есть основание думать, что превращение цистоидей в морских лилий произошло путем уничтожения двух венцов, на которых всего упорнее удерживались последние ромбы, а именно базального и верхнего латерального (базальный венец лилий затем образовался заново, свободным от пор). Уничтожение венцов одних табличек при сохранении табличками других венцов своей величины могло происходить только путем скачкового изменения, так что постепенных переходов в этом случае быть не может. Уничтожение происходило сразу, в одном поколении, в молодых стадиях индивидуума. Встречаются изредка аномальные экземпляры, у которых вследствие задержки в индивидуальном развитии сохраняются уничтожающиеся таблички, уменьшенной величины, со следами ромбов, соответствующие по числу уничтожающимся венцам, причем становится очевидным, что уничтожаются или два венца, или три; в первом случае из Cystoidea Rhombifera получаются Crinoidea' Dicyclica, а во втором — Crinoidea Monocyclica.
Что касается Blastoidea, Edrioasteroidea и вымершей группы Carpoidea, небольшой, недолго существовавшей и отличающейся резко выраженной двусторонней симметрией,—нет сомнения, эти группы вышли из Cystoidea.
Подтип II. Eleutlierozoa.1
КЛАСС I. МОРСКИЕ ЗВЕЗДЫ, STELLEROIDEA. ’
Подкласс I. Asteroidea.
В теле настоящих морских звезд, Asteroidea 8, можно различать цен тральный кружок и отходящие от него—большею частью в числе пяти—л учи (фиг. 330). У одних морских звезд преобладающе развиваются лучи, у других преобладает центральный кружок, так что лучи являются лишь в виде пяти углов его. Бывают и промежуточные состояния. Рот и амбулакральные ножки расположены на нижней, брюшной стороне тела животного; рот в центре тела, амбулякральные ножки вдоль лучей, в амбулякральных желобках, идущих посредине лучей. На спинной стороне, почти в центре ее, располагается анальное отверстие (если оно не атрофировано), а эксцентрически, в одном из интеррадиусов—мадрепоровая пластинка.
Кожа морских звезд отлагает скелет из известковых пластинок, соединенных между собою подвижно, так что лучи звезд при жизни животного гибки.
Скелет лучей в их поперечном сечении представляет: амбулякраль-
1 Eleutheros—свободный, soon—жжвотаов-’ Stella—звезда (лат.).
• Aster—звезда (грея }
ные пластинки, находящиеся на брюшной стороне (фиг. 331), образующие скелет дна амбулякральных жечобков; менее постоянны бывают адамбулакральные пластинки, лежащие по краям амбулякральных желобков, и краевые, образующие боковые стенки лучей. Спинная поверхность лучей покрыта мягкой кожей, с заключенными в ней различными более или менее мелкими табличками. Органы морских звезд располагаются частью в полости тела, частью в амбуля- Д
кра’льном желобке. В полости тела /жй
помещается пищеварительный ка-нал, половые железы, центральные д
части амбулакральной системы, ‘ каменистый канал, опускающийся гЖЗЦ
от мадрепоровой пластинки к коль-цевому околоротовому водоносному сосу; у. ^4/5
На дне амбулякральных же-лобков проходит радиальный водо-носный сосуд с ампул лам и, отходящими от него вовнутрь, Фиг. 330. Pa,aeaster eucharis. Девон. Пью„ МСЖДУ ачбулякральными таоличка- Иорк. СЛСШ. С нижней стороны два луча МИ, И с ОТХОДЯЩИМИ наружу амбу- оборваны. 1/1.
лякральяыми ножками.
Концы амбулякральных ножек могут присасываться к посторонним предметам, к которым затем притягивается тело животного сокращением
1 2 3 4
Фиг. 331. Л—поперечное сечение луча ныне живущей морской звезды Astropecten aurantiacus.
а— аябс тякрз тъные таблички; ad—адамбулакральные; m’, т2—краевые; й—амбулякральпьте ножки; с- целом 4- -слепые отростки пищеварптетького канала; и—нерв; д—кровеносный; w—водоносный сосуд
3—расположение амбулякральных табличек ныне живущей морской звезды.
3—поперечное сечение луча современной офиуры.
а— „позвонок11 (сросшиеся амбулякральные таблички); т—боковые таблички; г—спинная, Й—брюшная табличка, «, д и n—сосуды в верт
4—расположение амбулякральных табличек ископаемой звезды Aspidosoma petaloides Нижний девон. Рейн. Германия.
ножек. Таким образом животное передвигается. В амбулякральных желобках I помещаются также радиальный кровеносный сосуд и радиальный нерв. Лучи г оканчиваются дистально глазками. На спинной поверхности тела, а иногда также и на других частях его, находятся трубчатые* выросты кожи, служа-i щие для дыхания, — кожные жабры.
12 Учебник цалеоптотогш. 177
После смерти морских звезд скелет их тела обыкновенно разваливается та образующие его таблички.
Значительная часть палеозойских морских звезд противополагается остальным в том отношении, что первые имеют амбулакральные таблички расположенные чередуясь справа и слева (фиг. 331, 4), тогда как последние (к которым относятся все современные) отличаются расположением амбулакральных табличек поперечными рядами (фиг. 331, 2).
Первая группа вымерла в конце палеозоя, представляя лучи менее гибкие вследствие чередования табличек, чем лучи современных звезд, у которых каждая пара амбулякральных пластинок, лежащих поперечно одна против другой, образует так называемый «позвонок».
Морские звезды являются распространенной в настоящее время группой „гивотн”х; они наиболее часто живут на умеренной глубине, но встречаются также и на большой. В ископаемом состоянии встречаются с силура.
Подкласс II. Офпуры, Ophiuridea.
Подкласс Ophiuridea, или змеехвостки, отличается от вышеописанных настоящих морских звезд многими особенностями.
Лучи офиурид тонки и резко отграничены от центрального кружка (фиг. ?32, 333). Главная масса луча состоит из амбулякральных пластинок,
Фиг. 332. Onhiocten ferrugineum. Юра. Баден. Германия. С ротовой стороны.
Фиг. 333. Ophiothnx cchtnata. Ныне живут. С ротовой стороны; дистальные части лучей опущены.
о/—хмбулякральвые ножки; b—выводные отверстия для половых продуктов; Jp—брюшные пластинки рук, sp—боковые пластинки, з—центральный кружок; тп— мздрепоровад пластивка с гвдропорох; розовы» хцнткы; г—зубы.
срастающихся между собою в каждом «позвонк°», так что общая полость тела является сдавленною, сохраняется лищь в виде щелевидных пространств (фиг. 331, 3). Вследствие этого, в отличие от настоящих морских звезд, в общей полости тела лучей офиур нет внутренностей—печеночных мешков, половых желез. С другой стороны, боковые края первоначально существующих амбулякральных желобков срастаются снизу, превращая амбулякральные желобки *в совершенно замкнутые каналы.
Кроме непарных (вследствие срастания в каждом позвонке) амбуляк-ральных пластинок скелет рук представляет еще при каждом позвонке одну спинную, две боковые и одну брюшную табличку; последняя развивается во вновь возникшем снизу покрове. Амбулякральные ножки оттеснены с брюшной стороны лучей на бока широкими брюшными пластинками, а так как ножки притом еще утрачивают присоски, то они перестают служить органами движения, превращаются восязательныеидыхате льны е органы, а движение животного на морском дне происходит вследствие гребли пуками.
Б ископаемом состоянии известны с силура. Палеозойские офиуры не представляют всех вышесказанных отличий от мэрских звезд; у некоторых из них бороздки рук не вполне замкнуты, а амбулякральные таблички еще расположены чередуясь и не срослись в позвонки.
КЛАСС П. МОРСКИЕ ЕЖИ, ECHINOIDEA.
так называемых правильных, противоположных, нижнем и
Фиг. 334. Раковина правильного ежа с аборального (анального) полюса. R—амбулякральное; J—• интерамбулякральное поле. В правом переднем поле расположена мадрепоровая пластинка.
Шорские ежи имеют тело шаровидное, сердцевидное или диекоидальное, покрытое шипами. Наружный скелет представляет раковину, образованную многочисленными известковыми табличками, в большинстве случаев неподвижно между собою соединенными.
У ежей более древнего, исходного типа, •. рот и анальное отверстие расположены, на ] верхнем, полюсах тела. Большая часть раковины образована меридианальными рядами табличек, радиальными амбулакральными и мнтерамбулякральными (фиг. 334).
Амбулякральные таблички снабжены отверстиями для прохода изнутри раковины амбулакральных ножек, каждой ножке соответствуют два рядом лежащие отверстия, так что ножка проходит через пару пор двумя корнями.
В каждом радпусе расположены по крайней мере два меркдианальных ряда амбулакральных табличек, в интеррадиусах може* быть иногда только по одному ряду. Таблички околоротогого поля (перистома) залегают в перепончатой основе, подвижно соединены между собою и представляются видоизмененными амбулякральными табличками, так что таблички, лежащие радиально, против концов амбулякров, даж( сохраняют отверстия для ножек.
В околоанальном поле наиболее важны л пять больших, лежат иьтеррадигльно, снабжены одним или несколькими большими отверстиями, выводными для продуктов половых желез,--это так называемые генитальные таблички (фиг 334); чередуясь с ними и приходясь против концов амбулякральных полей, лежат глазные таблички, меньшей величины, снабженные маленьким отверстием для щупика. Одна из генитальных табличек ситовидно продырявлена и является мадре-
постоянны десять табличек*
норитом. Совокупность глазных и генитальных табличек называется в е*р пт и и и ы м щитком.
Кишка спирально свернута (фиг. 335). Начальная, околоротовая часть ее окружена жевательным аппаратом, в виде куполообразной основы («ари-
Фиг. 335. Схематический вертика 1ьный разрез морского ела (Echinus).
и—анус; ар—ампуллы; й—лгабра; соет.—окол&пищеБОдное кольцо возоносной системы; do—сынявод орган; #—гонада; гл- мадрепорит, mt—рот; яг—нервное кольцо; уо, рг—отростки края околоротового now; pd—ножки; ph—глотка; рр—околоаналыюе поле; ру—пирамиды (секторы) жевательного аппарата; 8, г—пнщеварй тельный канал, г, w, е—нервный радиальный ствол; se.—Каменистый канал, sp— шип, tb—бугорок; ъ- Болят в пузырь на околопищево ном кольце. <'лена сечение проходит на стенке раковины через амбух.кральныс таблички, справа через ннтераыбулякральные.
стотелев фонарь»), состоящей пз пяти секторов, челюстей; каждая челюсть снабжена твердым зубом, челюсти соединены подвижно и приводятся в движение мускулами, так что зубы смыкаются и размыкаются.
Кольцевой канал водоносной системы лежит над жевательным аппа-
ые каналы, идущие вдоль амбулякральных полей по внутренней поверхности раковины.
Амбулякральные таблички ежей бывают простые и сложные. Простые имеют только одну пару, сложные представляют несколько пар, состоя из соответственного числа сросшихся простых табличек. Иногда на поверхности сложной таблички еще сохраняются в виде бороздок границы между простыми сложной (фиг. 336, а).
ратом, от которого отходят радиал
Фиг. 336. Сложные таблички.
n—Pseudodiadema hnnisphaerfeum (юра) образована из трех простых табличек. />—габ дичка JF.rhznomeira, состоящая из Ь простых табличен.
табличками, вошедшими в сост;
Срастание простых табличек в сложную происходит вследствие взаимного давления, возникающего при увеличении с возрастом числа табличек в меридианальном ряду и при увеличении размеров табличек.
Не все ежи проявляют способность к образованию сложных табличек. Эта способность наблюдается у тех ежей, у которых амбулякральные поля при границе их с о-.оло-
рото! ым имеют так Bast веемые auricula'’, известковистые выступы вовнутрь раковины, к которым прикрепляются иускуш двигатели аристотелева фонаря Иногда auriculae, как например у рода Cularus, рЖюлагант интерамбулякральио и тогда не возникает сложных амбулакральных табличек, иногда же auriculae располагаются при нижнем конце амбулакров и тогда развиваются сложные таблички. Вероятно потому, что находящиеся внизу auru ulae представляют так сказать устои, при которых взаимное давление табличек становится особенно сильным.
Последствием взаимного давления прости': табличек при срастании их в сложную является приобретаемая некоторыми из них клиновидность формы, причем они не доходят до бокового края сложной таблички, к которому обращены клиновидные концы. Такие укороченные в ширину простые пластинки называются полу и ластинками (фиг. 336, Ь).
Если поверхность раковины в направлении снизу вверх представляет более или менее равномерную кривизну, то амбулякральные ножки в разных
частях одного и того же амбулакра находятся в сходных условиях действия; не то у новейших ежей, у которых раковина резко делится на верхнюю и нижнюю половины. При таком разделении амбулякральные ножки, расположенные близ верхнего полюса тела, не могут служить для движения животного ислу-- жат как жабры (фиг.
337). В связи с этим возникают так назы-
Фиг. 337. Brissopsis lyrifera, ползущий.
af— амбулякральные ножки; к—амбутякральные кожки, превращенные в жабры, /фасциоли или семиты.
ваемые петалоид-ные амбулякры, названные так по сходству с венчиком цветка. Именно, в то время как обыкновенно ряды пор амбулякрои идут равномерно и прямолинейно сверху до низу, в петалоидных амбуляк-рах обе поровые полоски каждого амбулякрального пиля при макушке очень сближены между собою, затем расходятся и далее, при совершенном образовании петалоидов (фиг. 350), снова сбшжаются еще на верхней поверх» ности раковины, сообщая таким образом здесь амбулякральному пространству форму лепестка. Петалоиды у некоторых родов совершенно замкнуты внизу, у других же обе ветви петалоида не соединяются, так что лепесток остается открытым. Во внепеталоидпой части амбулякра, при совершенном развитии петалоидов, амбулякральные поры становятся редко рассеянными и неясными, и могут даже совершенно исчезнуть.
При вышеупомянутом действии верхних амбулякральных ножек, исключительно как жабр, выбрасывание экскрементов посреди них становится неудобным для животного; поэтому у таких форм анальное отверстие перемещается к задней части тела животного.
Это перемещение назад анального‘отверстия обыкновенно балансируется перемещением вперед ротового отверстия.
Анальное отверстие последовательно перемещается из вершинного положения на бок раковины, на границу боковой и нижней стероны, на нижнюю сторону. Ротовое отверстие получает более ограниченное перемещение, в пределах лишь нижней стороны, становясь из центрального эксцентричным.
Таким образом еж лишается пятилучевой симметрии, приобретая двустороннюю. В этом отношении, коррелятивно с перемещением анального и ротового отверстий, возникает целый ряд изменении организации.
Рот становится сжатым, двугубым (фиг. 338), с переднею и заднею губою. Первая помещается на краю впадины, отступая таким образом, впячиваясь вовнутрь тела; при задней губе поверхность раковины, наоборот, несколько выпячивается наружу так, что губа приобретает вид черпака; при движении животного по морскому дну вперед нижг нею губою загребается ил и песок с них мелкими животными, идущими в нищу ежу. Челюстной
Фиг. 339. Вершенный щигик I'chino-bnssus sculalus (увели ч.).
.Ag> г ..^ -Ц®
Фиг. 33S. Дву-1убый рот МлСГОг-ster corangui-нит.
мзме-
Фпг. 340. Вершинный щиток Ananchytes ovata.
заключенными в
же аппарат атрофируется вследствие затруднительности работы челюстями при сжатом, двугубом рте. Образ жизни ежа в отношении питания изменяется. С выходом анального отверстия из вершинного щитка последний няется. Пере вижение анального отверстия происходит в интеррадиусе, обращенном назад при движении животного. Поэтому при выходе анального отьерстия из вершинного щитка, генитальная табчичка последнего, соответствующая названному интеррадиусу, сначала получает от анального отверстия выемку, становится вильчатой, а затем прорывается анальным отверстием при дальнейшем его движении и, будучи разрушена таким образом, атрофируется.
У неправильных ежей, двусторонне симметричных, вершинный щиток становится состоящим лишь из девяти табличек (фиг. 339), причем иногда является вытянутым в направлении спереди назад, как у рода Ananchytes (фиг. 340). У Ananchytes глазные и генитальные таблички становятся попарно соприкасающимися. Далее тот же процесс удлинения вершинного щитка спереди назад ведет к обособлению двух задних глазных табличек,от остальных. Между теми и другими вдвигаются интеррадиальные таблички боковой поверхности раковины; так у Collyrites (фиг. 341). Три передние амбулякра, обособленные таким образом от задних, получают название trivium, два задние—bivium. Обособление trivium's, ot bivium’a, менее резкое, чем у Collyrites, существует у всех двусторонне-симметричных ежей. У двусторонне-симметричных, сердцевидных ежей амбулякры помещаются в углублениях поверхности раковины В этих углублениях у родителей помещаются, пока достаточно не вырастут, молодые
341.
Фиг.
Вершинный щиток Со!1д-riteselliptica. (увелич.)
ежи. Название иглокожих в наибольшей степени приложимо к ежам благодаря их наружному покрову из известковых игол Этот покров иногда является тонким, состоя из тонких коротких щетинковидных иголочек (фиг. 337), в других случаях иглы хотя коротки, но довольно толсты (фиг. 345), наконец бывает покров из игол сравнительно малочисленных, но длинных, длина которых в несколько раз превышает поперечник тела. Все иглы на нижнем конце имеют утолщенную головку с углублением; этим углублением головка иглы сидит на бугорке поверхности раковины; за головкой следует сжатая часть иглы, шейка и затем тело иглы. Иглы служат для защиты животного от неприятелей, употребляются иногда и для ходьбы по морскому дну.
Бугорки поверхности раковины, к которым причленяются иглы, бывают мелкие и крупные. В амбулякрах последние бывают лишь при наличности сложных амбулякральных табличек.
Фиг. 342. Bothriocidaris PtMeni. Нижний силур. Неммис. Эстония.
а—сбоку (в натуральную велвчлпу); Ъ—вершинный щиток; с—рот с 5 челюстями (увелич.).
Фасциолями, или семитами (fasciolae, semitae), называются гладкие пространства поверхности раковины (фиг. 337, у; 354),’ покрытые исключительно тонкими щетинками. Эти пространства обыкновенно имеют вид узких полосок с сомкнутыми концами. Фасциоли по положению их называются: перипетальная (околопеталоидная), анальная и пр.
Морские ежи, в отличие от морских лилий, представляют группу, усилившуюся в распространении к настоящему времени.
Морские ежи, разделяются на два порядка: правильных ежей (Regularia) и неправильных (Irregularia). Первые отличаются центральным положением рта и анального отверстия, пятилучевой симметрией и равномерным развитием амбулякров; неправильные ежи имеют по крайней мере эксцентрично расположенное анальное отверстие, а также двустороннюю симметрию с более или менее ясно выраженным обособлением bivium’s, и trivium's,. Regularia являются древнейшей, исходной для развития Irregularia группой, существовавшей с древнепалеозойских отложений. Irregularia существуют с юрских отложений. 1
ПОРЯДОК I. REGULARIA.
Палеозойские представители Regularia (regularis—правильный) существенно отличаются от позднейших, представляя число меридиапальных рядов табличек или меньшее, или большее двадцати, тогда как послепалео-зойские представители Regularia имеет 20 меридиапальных рядов табличек, по два ряда в каждом радиусе и интеррадиусе.
Bothriocidaris (фиг. 342). Небольшая шаровидная раковина. В каждом интерамбулякре лишь один ряд табличек, в амбулякре два ряда. Ни глаз-
Фиг. BiB.Archaeocidm s Worth eni. Нпжне-иаменноуг. С.-Луи, Миссури. САСШ. А— околоротовая поверхность. Амбулякральные и интерамбулякральные таблички продолжаются на перистом, но неплотно, чешуевидно налегая. В центре видны раздвинувшиеся части челюстного аппарата. В,С— Archaeovidaris rossica. Верхнекамевн. известняк. Мячково, Московской обл.
В~с аборальной стороны; С—сбоку; р—сектор с зубом; г—вводная часть.
Чэлюстиой аппарат.
личек. Бугорки очень мелкие,
ные, ни генитальные таблички не имеют пор (вопреки фпг. 342). Иглы в албулякрах, а иногда и в интерам-булякрах, мелкие. Нижний силур.
Archaeocidaris (фиг. 343; archalos—древний, cidaris—тюрбан) Раковина обладает некоторой гибкостью вследствие несколько черепитчатого налегания одних табтичек на другие. В интерамбулякрах 4 ме-ридианальных ряда табличек. В амбу лякрах по- два ряда. Длинные веретенообразные иглы усажены спирально расположенными шипами. Каменноугольные, пермские.
Melonites (фиг. 344; melon— дыня). Раковина весьма значительных размеров, сферическая, с мери-дианальными складками. АтЬ широкие, вогнутые с обоих боков срединного ребра, с 6—16 рядами табличек. Таблички несколько черепитчато налегающие, средние ряды наиболее широкие. 1ЛтЪ с 4—11 рядами таб-иглы маленькие. Камен-
многочисленные,
но угольные отложения. а
Ъ
Фиг. ”44. Melonites mullipora: а — со стороны анального полюса; Ъ — сбоку (в половину натуральной величины); с — часть амбуляьрального веля ^увелпч.). Каменноуг.
Сев. Америка.
Cidaris (фиг. 345, 346), так же как и все последующие описываемые ежи, имеет по два ряда табличек в каждом амбрякре и интерамбулякре. Раковина сфероидальная. АтЬ волнистые или почти прямые. 1АтЪ украшены двумя рядами больших бугорков, на которых сидят крупные иглы Последние бывают весьма разнообразной формы—цилиндрические, шестовидные, веретенообразные, призматические и т. д. и усажены шипами или зернышками, которые располагаются на них продольными рядами. Один иа
Фиг. 315. Cidaris сого-vata Сбоку, с сохранившимися частью иглами. Верхняя юра. Вюртеыбгрг. Германия. 3/4.
Фиг. 3h>. Cidaris subv si-culosa. Co стороны верхнею полюса. Верхнемел. отлож. Англия. 1/2.
Фиг. 347. Hemicidaris crenvlaris. Юра. Шатель Севзуар. Франция.
самых примитивных родов, живущих в настоящее время. С пермских отложений доныне.
Hemicidaris (фиг. 347; hemi—половина) интересен- в том отношении,, что амбулакры в нижней части их состоят из сложных табличек, вместе с тем здесь развиваются крупные бугорки. Юра, мел, нижнетретпчные.
ПОРЯДОК П. IRREGULABIA. 1
Рудaster (фиг. 348, 349; pyge—зад, aster—звезда) является ежом с анальным отверстием, наименее всего вышедшим из пределов вершинного-щитка, так что передним своим концом оно касается щитка; вследствие этого, хотя в щитке исчезла задняя генитальная табличка, смежные с нею глазные еще не пришли в соприкосновение между собою (фиг. 348). Раковина сжатая в вертикальном направлении. Рот центральный. Амбулакры простые, апеталоидные. Юра. Мел.
Clypeaster (фиг. 350; clypeus—щит, aster—звезда) имеет анальное отверстие на границе верхней и нижней поверхности раковины или на краю нижней поверхности. АтЪ петалоидные. Рот центральный. Внутри раковйьы находятся известковые столбы, протягивающиеся от дна к кровле, в особенности изобилующие на периферии. Образованием столбов увеличивается прочность раковины, ослабленная приплющенностью тела ежа, отклонением его раковины от куполообразной формы. Сохранение прочности для этого ежа существенно при жизни его в прибрежной полосе. Третичные—ныне.
Echinoconus (фиг. 351). Коническая раковина с апеталоидными амбу-лякрами. Центральное ротовое отверстие, анальное—на границе нижней и
1 Irregularis—неправильный.
верхней поверхности раковины. Рудиментарные auriculae. Челюсти отсутствуют. Мел. \
Далее следуют ежи, утратившие челюстной аппарат.
Род Ananchytes (фиг. 352) имеет высокую раковину, в плане овального очертания, ротовое (эксцентричное) и анальное отверстия на нижней
<Риг. 348. Вершинный щиток Piigaster.
Л.—анальное отверстие; о—амбулякры
Фит. 349. Pygaster umbrella. Юра. Шатплъон на Сене. Франция. 1/1.
Фиг. 350. Clypeailer aegiptiacus. Плиоцен. Каир. Египет.
а — сбоку; Ъ — снизу; с—вершинный щиток (увелжч.).
•стороне. Перистом двугубый, овальный, ширина более длины. Вершинный щиток (фиг. ЗфО) вытянут в длину. Верхнемеловые отложения.
Toxaster (фиг. 353; toxon—лук для стрельбы) принадлежит уже я древнейшим сердцевидным ежам. Передний амбулякр расположен в широкой бороздке. Fasciolae нет. Перистом поперечный, почти круглый или пятиугольный. Нижний и средний мел.
Micraster (фиг. 354). Имеет все амбулакры в бороздках (передний апеталоидный), субанальную фасциолю. Перистом явственно двугубый, с выдающеюся нижнею губою. Анальное отверстие на усеченней задней стороге. В средне- и верхнемеловых отложениях, в третичных.
Фиг. 351. Echinoconus atbogalerus. Белый мел. Англия 1/1.
Фиг 353. Toxaster сот planatus. Веском. Оксер. Франция. 3/4
Фиг. 352. Ananchytes ovata. Белый мел Вест" фалия. Германия. Сверху увеличено.
>Фиг. 354. jylicraster corangunium. Сеион. Франция. 3/4.
Геологическое распространение морских ежей.
Ископаемые ежи известны с силурийской системы (Bothriocidaris). В каменноугольных отложениях и девоне встречаются ежи с большим числом рядов АтЪ и 1АтЪ, как Melonites. Когда со временем таблички раковины становятся толще, гибкость раковины утрачивается, ежи с числом меридиональных рядов большим двадцати исчезают.
В мезозое появляются Irregularia, начинаясь в юре родом Pygaster
От форм, подобных Pygaster, исходят: линия развития, приводящая к Clypeaster, и другая линия, ведущая к утрате челюстного аппарата,— к сердцевидным ежам.
Характеристики, групп иглокожих.
1 полтин. Pelmatozoa. Прикрепленные к морскому дну стеблем, или непосредстве! но аборальной стороной чашечки, или неподвижно, хотя и свободно лежащие. Тело мешковидное, шарообравное, или сплющенное от одного бока к другому, покрытое на поверхности известковыми табличками, проявляющими наклонность группироваться по пяти
в горизонтальные венцы. На ротовой стороне пиша приводится ко рту но большею частью разветвленным амбулакральным желобкам действием находящихся на них мерцательных ресничек. Желобки, так же как и почти постоянно центральный рот, защищены кроющими табличками и продолжаются в членистые подвижные ручные придатки. Заднепроходное отверстие иитерамбулякралыю и почти всегда на верхней стороне. Кембрий—ныне.
1 класс: Crinoidea, морские лилии. Редко бесстебельчаты и свободно двигаются. Чашечка со стороны, противоположной рту, и на боках представляет наружный скелет из горизонтальных венцов табличек, по пяти (или н кратном от пяти), а на ротовой стороне (крышечка чашечки) реже представляет пенамерность и пятикратность в расположении табличек, чем неправильное расположение. На периферии крышки отходят от чашечки пять обыкновенно разветвленных рук, часто снабженных pinnulae. С кембрия доныне.
1 порядок: Larviformia. Большею частью маленькие формы с круглым стеблем, чашечка представляет по одному венцу В, В и О. Руки простые, большею частью неразветвленные, редко двурядиочленистые. Силур—ныне.
2 порядок: Fistulata. Чашечка часто с IB и постоянно с IR.A, крышечка из тонких табличек со значительным анальным хоботком. Руки большего частью разветвленные, часто двурядночленистые. Силур—пермь.
3 порядок: Articulata. Нередко бесстебедьчатые, большею частью с толстыми табличками чашечки. IB большею частью в редукции. IR редко встречаются; крышечка чашечки, покрытая тонкими табличками или кожистая, заключающая в себе основания рук. Руки почти постоянно разветвленные, очень редко двурядночленистые. Рукп с pinnulae. С триаса доныне.
4 порядок: Camerata. Часто большой величины, постоянно стебельчатые. Большею частью с многочисленными IR. Крышечка прочно сложенная табличками. Нижняя часть разветвленных, большею частью двурядночденпстых рук неподвижна, будучи заключена и стейку чашечки. Силур—пермь.
2 класс: Cystoidea. Большею частью короткостебельчаты. Чашечка со всех сторон прочно образована табличками, имея их в пятикратном числе и группировке строго радиальной и интерраидиальной, пли в неправильном расположении п в изменчиво большом количестве; чашечка пронизана порами; рукп короткие, не ветвятся, не имеют pinnulae, находятся или непосредственно около рта, пли при удлиненных амбулякральных желобках, в числе от двух до пяти идущих от рта по поверхности тела. Силур—девон.
1 порядок: Rhombifera. С поровыми ромбами. Силур, девон.
2 порядок: Diploporita. С двойными порами. Силур, девон.
3 класс: Blastoidea. Чашечка правильно пятилучевая; основание из трех табличек; затем следуют пять радиальных со значительными, а иногда и очень большими выемками для амбулякральных полей и с гидроспирами, возникшими из ромбовых пор цистоидей, наверху еще пять околоротовых табличек. Многочисленные мелкие ручные придатки без pinnulae. Анальное отверстие поблизости от рогового. Силур—пермь.
4 класс: Edrioasteroidea. Бесстебельчатые, часто свободно лежащие или прирастающие поверхностью тела, шаровидного, мешкообразного или дискообразного; амбулякральные желобки ие ветвящиеся и без ручных придатков.
П подтип: Eleutherozoa. Неприкрепленные, свободно двигающиеся иглокожпе.
1 класс: Asterozoa. Обитатели морского дна, Тело из нейтрального диска и пяти лучей, от него отходящих. Рот и пять амбулякральных бороздок, от него отходящих на иижней, брюшной стороне тела. Постоянно имеется особенный внутренний ротовой скелет и пять радиальных двойных рядов табличек ambulaeralia и adambulacralia. С срегнего кембрия доныне.
1 подкласс: Asteroidea, морские звезды. Кружок переходит в нередко обособленные, иногда очень короткие лучн. Амбулякральные бороздки на них открыты и в них находятся амбулякральные ножки. С кембрия доныне.
2 подкласс: Ophiuridea. змеехвостки. С цилиндрическими лучами, резко обособленными от диска. Внутри их имеются только ambulaeralia, слитые в ряд ручных позвонков, и амбулякральная бороздка закрыта кожею. Вместо ножек имеются щупальца. Нет анального отверстия. С силура доныне.
2 класс: Echinoidea, морские ежи. Свободные обитатели морского дна. Шаровидные, конические, дискообразные или сердцевидные с плотным панцирем из табличек (изредка черепитчато налегающих), расположенных в пятикратном число меридианальными рядами. От рта, расположенного на брюшной стороне, идут пять амбулякральных нолей (снабженных двойными пдрами) к верхнему полюсу, где находится вершинный щиток аз особенных табличек. Заднепроходное отверстие находится в нем или в заднем между-188
амбтлякре. Амбулакральные ножки частью нередко Превращены в органы дыхания. От силура доныне.
1 порядок: Rcgularia, правильные ежи; шаровидны, рот и анальное отверстие иа противоположных полюсах; развит челюстной аппарат; силур—ныне.
2 порядок: irregularia, неправильные ежи. Различной формы; заднепроходное отверстие в заднем интеррадиусе, рот большею частью без челюстного аппарата, иногда впереди, таким образом с ясной двусторонней симметрией. Амбулакральные поля часто петалопдпые или углубленные. С триаса доныне.
3 класс: flolofh urioidea. Морские кубышки. Червеобразные свободные обитатели морского дна, большею частью лишь с рассеянными маленькими кожными скелетными телятами. Рот на переднем конце. Изредка пелагические формы. Кембрий—ныне.
VII ТИП. МЯГКОТЕЛЫЕ, MOLLUSCA.
Самые древние мягкотелые, подобно более примитивным из ныне живущих, являются животными с двусторонней симметрией, без первичной продольной сегментировки.
В развитии всей организации мягкотелых определяющим моментом явилась концентрация, стяжение кожномускульного мешка 1 на брюшной стороне тела, где затем развился особый мускулистый орган передвижения, служащий для ползания, плавания, зарывания и т. п.,—нога. Вследствие возникновения ноги стала невозможною членистость тела, как она выражена у Arthropoda и Annelida, со всеми ее последствиями, как то: расчлененным кожным скелетом, членистыми придатками туловища, сегментальными органами и т. п.; кроме того мягкая, богатая слизеотделительными железами и во многих местах богатая ресничками кожа могла выделить защищающую тело раковину только со спинной стороны.
Затем нарастающий край раковины оттянул от тела кожную складку (мантия или епанча), огибающую спину.
В этом типичном наружном строении мягкотелых у различных групп произошли во многих случаях изменения, так что не только утратился первоначальный червеобразный вид тела, но у значительной части мягкотелых развилась и асимметрия всего организма. При всех изменениях организма моллюски все-таки представляют известные общие особенности внутренней организации, которые свидетельствуют о тесном родстве этих животных между собою, как бы ни разнились они по наружному виду. Раковина состоит в основе своей из рогового или хитиноподобного вещества, конхиолина, отделяемого кожей спины и позднее проникнутого минеральными солями, главным образом углекислым кальцием (невидимому аррагони-том). Типично раковина состоит из следующих слоев: 1) рогового наружного слоя, который однако иногда отсутствует и вообще тоньше остальных слоев, 2) срединного слоя, состоящего из тонких известковых призм, расположенных под углом к поверхности слоя (призматический илй фарфоровидный слой), 3} внутреннего перламутрового слоя из пластинок, параллельных поверхности слоя.
Роговой и призматический слои раковины наращиваются утолщенным краем мантии, так что при этом возрастает поверхность раковины, перламутровый слой, напротив, отлагается всею поверхностью мантии, так что от него по преимуществу увеличивается толщина раковины.
1 Т. е. кожа с подлежащим, прилегающим к ней мускульным слоем.
Пищеварительный канал нормально разделяется на три отдела: передний—ротовую полость и пищевод, средний—желудок, задний—кишку.
. В ротовой полости у большинства мягкотелых находятся двоякого рода роговые образования: челюсти и radula. Челюсти бывают обыкновенно парными: боковыми (у большинства Gastropoda) или спинною и брюшною (у Cephalopoda).
Radula представляет хитиновый покров языка, покрыта хитиновыми же зубчиками различной формы, расположенными симметрично продольными и поперечными рядами. Radula служит для измельчения’ пищи, существует как у наиболее примитивных, так и у обладающих усложненной организацией мягкотелых, и отсутствует, очевидно будучи утрачена, у представителей наиболее специализированных' типов, напр. у пластинчатожаберных, пища которых не требует измельчения, причем у них отсутствует обособленная ротовая полость. В ротовую полость открываются слюнные железы, в средний отдел пищеварительного канала доставляет свои выделения пищеварительная железа—«печень».
Общая полость тела мягкотелых сильно сокращена, редуцирована по своим размерам, разделяется на отделы: околосердечный, гонадиальный, содержащий половые железы, и почечный. Эти отделы иногда соединены между собою, что представляет более примитивное состояние. Почки в виде парных трубочек.
В связи с ограниченными размерами целома пространство между внутренностями в значительной степени заполнено соединительной тканью, в которой остаются промежутки, служащие путями кровообращения. Из них кровь собирается по близости от жабер в настоящие сосуды^ имеющие свои собственные стенки, — жаберные артерии, несущие кровь к жабрам (реже легким), и затем из жабер кровь поступает в непарное или парные (соответственно числу жабер) предсердия, а из них в единственный желудочек сердца.
Нервная система мягкотелых в простейшей форме (Amphineura) подобна нервной системе некоторых червей, состоя пз головного мозга (окологлоточное кольцо), двух брюшных и двух боковых стволов с многочисленными поперечными перемычками; у остальных моллюсков брюшные стволы редуцируются, так что остается на них всего лишь одна пара узлов (ножные), а на боковых стволах возникают три пары нервных узлов.
Кроме слабо развитых органов чувств, находящихся в коже (осязания — обыкновенно в щупальцах, вкуса и обоняния), моллюски имеют статоцисты, расположенные на ноге, и пару глазе сетчатой оболочкой, выстилающей кубкообразную поверхность, полость которой или остается открытой, или замкнута и заполнена светопреломляющим стекловидным телом.
Моллюски размножаются исключительно половым путем, раздельнополы или гермафродитны. В своем развитии проходят стадию личинки, которая сходна с личинкой червей по присутствию предротового реснитчатого кольца; при дальнейшем развитии это кольцо большею частью превращается в лопастной орган (velum), служащий для плавания животного.
Моллюски являются животными по преимуществу водными, но даже члены одного и того же класса ведут самый различный образ жизни. Большинство живет в море, немногие в пресной воде, один порядок гастропод и еще немногие отдельные их представители приспособлены к наземному 190
образу жизни. Моллюски представлены во всех трех больших группах водных организмов, именно: в бентосе, состоящем из ползающих или прикрепленных животных, населяющих глубины моря; нектоне, состоящем из активно плавающих животных, способных передвигаться против течений; планктоне, содержащем животных, плавающих пассивно по течению.
Первая группа заключает моллюсков прибрежных и моллюсков больших глубин, дающих также представителей нектона. Другие две группы состоят из пелагических моллюсков; цефалоподы принадлежат исключительно к нектону, тогда как свободно плавающие гастроподы и личинки различных групп моллюсков принадлежат к планктону.
Современные моллюски распределены по всей поверхности земли и под всеми широтами. Наземные найдены на высочайших горах, пелагические не только распространены по поверхности моря, но могут нисходить до глубины в 750 м, не достигая дна морского. Моллюски больших глубин найдены во всех океанах, с распространением до глубины в 850 м от поверхности.
Обыкновенно моллюски—животные, живущие свободно, и плавают, зарываются в почву или сверлят твердые предметы, в которых помещаются, но некоторые являются прикрепленными в зрелом состоянии, именно некоторые гастроподы и пластинчатожаберные.
Современная прибрежная (литторальная) фауна моллюсков разделяется, в зависимости от климата, на три области: северную и южную полярные области и тропическую.
Фауна больших глубин, или абиссальная, не распределяется в отдельные области, и некоторые из ее видов обладают универсальным распространением как в северной, так и в южной части одного и того же океана пли в нескольких различных океанах.
Пелагическая фауна состоит из видов, весьма широко распространяющихся, но все-таки распределяющихся в две полярные и одну тропическую области. Лишь небольшое число групп моллюсков покинуло море, перейдя к пресноводному образу жизни, именно некоторые пластинчатожаберные и гастроподы. Наземный образ жизни ведут лишь некоторые гастроподы, главным образом легочные.
Морские группы проникли в пресные воды особенно в странах с теплым климатом и чаще всего там, где море является опресненным.
КЛАСС I. ЧЕРВЕОБРАЗНЫЕ МОЛЛЮСКИ, AMPHINEURA.
Двусторонне-симметричные моллюски с удлиненным червеобразным телом, с подошвообразной ногой на брюшной стороне. Главные представители Amphineura, Chitonidae (фиг. 355), животные с широким, сщатым дорзо-вен-трально телом, с приплющенной брюшной и умеренно выпуклой спинной стороной. Спинная сторона одета мантией и покрыта раковиной, состоящей из 8 пластинок, поперечно вытянутых и соединенных между собою подвижно, с черепитчатым налеганием каждой пластинки на передню часть пластинки, позади лежащей. Таким образом части раковины соединены между собою подвижно, что дает возможность животному свертываться с загибанием переднего и заднего концов на брюшную сторону.
Двусторонняя симметрия существует не только во внешнем виде животного, но и во внутреннем его строении. Рот и анальное отверстие находятся
шинство их живет в приорежнои полосе,
а
Фиг. 355. а—Chiton sqvamosus. Ныне живущий. Ь, с—Chiton pns' iis. Каменноугольные отложения. Бельгия.
на противоположных концах тела. Голова слабо обособлена, в ротовой полости есть radula, но нет челюстей. Симметрично с обоих боьоз тела, в желобке между ногой и мантией, расположены многочисленные жабры. Вдоль тела в нем тянутся четыре нервных ствола. Обособления нервных узлов нет. Гонада (половая железа) одна, обособлена так, что открывается своими выводными отверстиями прямо наружу. Парным жабрам соответствуют парные предсердия Силур, ныне.
Chitonidae— морские животные, передвигающиеся ползанием, медленно и лишь на небольшие расстояния. Они обыкновенно травоядны, и боль-присасываясь подошвой ноги к скалам, но некоторые живут и на больших глубинах. Распространены во всех морях.
Основные отличия всех остальных классов моллюсков (Gastropoda, Lamelllbranchiata, Scaphopoda и Cephalopoda) от Amphineura в значительной степени вызваны сокращением брюшной стороны тела животного так, что концы пищеварительного канала становятся сближенными, а самый канал, вместо того чтобы итти прямолинейно от одного конца тела к другому, становится дугооб-
разно изогнутым ввеох. Внутренности выпячиваются вообще вверх, образуя внутренностный мешок; как изложено ниже, вследствие этого возникают отклонения в организации от двусторонней симметрии
У всех этих моллюсков развивается между мантией и телом особая мантийная полость, содержащая жабры. Нервная система представляет обо-обленные нервные узлы.
К IACC II. БРЮХОНОГИЕ, GASTROPODA.
Асимметричные моллюски с более или менее ясно обособленной головой, на которой находятся щупальца, глаза и radula; с хорошо обособленной ползательной пли плавательной ногой, с мантийной полостью, находящейся на спине и открывающейся наперед. Все мягкое тело заключено в раковину, не представляющую частей, обособленных совершенно, имеющую вид конуса, свернутого спирально или не свернутого. Признак, которым брюхоногие уже внешне отличаются от всех остальных моллюсков, это—асимметрия их организации, обусловленная определенным поворотом задней части тела по отношению к передней.
Асимметрия возникает прежде всего на формах, которые отличаются уже от Amphineura с их червеобразно вытянутым телом — развитием внутренностного мешка; эти формы обладают, вместо первоначально задней жаберной полости (с анальным отверстием; устьями почек и гонад, жабрами), полостью, расположенною под отвисающим назад (при движении животного вперед) внутренностным мешком, как это видно на ли-
чинках гастропод (фиг. 356, I). Такое брюшное положение жаберной полости о вышеперечисленными расположенными в ней органами не наносит ущерба действию их лгшь у животных плавающих, у которых тело находится в воде во взвешенном состоянии, так что внутренностный мешок не производит цавления на жаберную полость. Таковы плавающие личинки гастропод и цефалоподы. У животных, ползающих на ноге, каковы все типичные гастроподы, внутренностный мешок давит сверху на мантийную полость с заключенными в ней органами, делая невозможным или затруднительным их действие (фиг. 356,2), этого давления при ползавии не будет лишь
Фиг. 356. Две стадии еародиша лужанкп (Ра1шлпа vivvpora)
а — анальное отверстие; с — околосердечная сумка (целом); со—предсердие, d — кишка; dk — крышечка. в — глаз, f — нога (в I показано пунктиром^ расположение ноги зрелого индивидуума, чтобы показать ее несовместимость с брюшным положением мантийной полости1 w); д — гонада; h — мозг; » — висцеральный узел; к — первоначально правая жабра (в II вторая жабра н в I обе жабры дополнены пунктиром); I — левый париетальный уз<л; т — мантийная полость; п — почка; п' — отве стие почечного капала, о — рот; р — ножной узел; г — правый париетальный узел; в — внутренностный мешок, покрытый мантией в раковиной; I — щупальце; ® — velum.
в том случае, если внутренностный мешок при движении будет наклонен не назад, а вбок. Последнее происходит вследствие асимметричного роста, при котором внутренностный мешок так поворачивается, что примыкающая к нему мантийная полость отворачивается от брюшной стороны вбок и в сторону спины, устьем своим наперед, совершенно освобождаясь этим от давления внутренностного мешка (фпг. 356, 11).
Вращение может бытьили паправо1(у большинства гастропод, фиг. 356,72), или налево (у меньшинства гастропод). Это вращение и возникающая йслед-ствпе него асимметрия многих внутренних органов были у древнейших гастропод необходимым последствием усвоения передвижения ползанием.
К этому присоединилось дальнейшее изменение первоначально симметричной организации. Растущий внутренностный мешок мог уже у симметричных предков гастропод удлиняться не только в прямом направлении, но также спирально свертываться в срединной плоскости вверх и вперед, на спинную сторону (эктогастрически), не нарушая двусторонней симметрии, как это бывает в особенности у головоногих.
У гастропод же, гри асимметричном строении их мягкого тела, и раковина получает асимметричный вид конической спирали (фиг. 357), ко-
1 От наблюдателя, если стать лицом к головной части тела улитки
13 Учебник пале онтологии
)
19S
торая представляет то преимущество, что при одинаковой длине является более компактною, чем спираль плоская.
Действие поворота задней части тела на организацию проявляется в органах кровеобращения и дыхания, на нервной системе и почках. Головной мозг и близлежащие ножные нервные узлы не подвержены действию поворота тела вследствие своего расположения напереди, по поворот те/а действует на задние нервные узлы, изменяя их положение.
Именно так называемые париетальные нервные узлы и лежащие под кишкой висцеральные (внутренностные) узлы перемещаются с поворотом задней части тела, правые поднимаются вверх и перемещаются над кишкой налево, левые опускаются под кишку и перемещаются затем направо, так что нервные стволы, несущие эти узлы, перекрещиваются, вместо того чтобы итти параллельно по бокам тела, как было первоначально.
Первоначально у гастропод жаберная полость находилась на заднем конце тела, так что пара жабер в ней была направлена назад, и сердце, помещавшееся под кишкой, получало кровь из жабер через свои предсердия сзади. Вследствие развития внутренностного мешка, мантийная полость переместилась на спинную сторону, и положение жабер и сердца сделалось обратным—жабры напереди, сердце позади; таким образом есть возможность различать з а д н е ж а б е р н ых (Opisthobranchiata) и переднежаберных (Prosobranchiata) гастропод.
Но жабры и сердце не просто принимают обратное положение.
То обстоятельство, что животное стало носить спирально свернутую коническую раковину на одном боку, чаще всего на правом, повлияло на органы, расположенные на этом боку; эти органы от давления раковины испытывают затруднение в своем развитии и развиваются слабее таких же органов левого бока или совершенно атрофируются. Та жабра и то предсердие, которые должны были бы лежать на правом боку, теперь не развиваются, так что различают более примитивных прозобранхиат с двумя жабрами и двумя предсердиями (Diotocardia) и производных от них с одною жаброю и одним предсердием (Monotocardia).
Подобно жабрам, на правой стороне стесняется развитие почки, так что у Monotocardia одна почка.
Раковина бывает двух типов, симметричная и спиральная. Первая бывает слюдцеобразная или низко-коническая и встречается у немногих сравнительно гастропод. Спиральная раковина состоит из свернутой в спираль конической трубки. В исключительных случаях спиральная раковина бывает неправильно свернутою (у Vermetus) или дискоидальной, свернутой в плоскую спираль (например Bellerophon). Обыкновенно раковина представляет коническую винтовую спираль.
Держа ваковину вертикально, так чтобы макушка была наверху, а устье, пли апертура, внизу и обращено к наблюдателю, называют раковину завитой направо, когда устье ее приходится с правого боку, и завитой налево, когда оно на левом боку.
Спирально свернутая раковина состоит из завитков, или оборотов; линия, разграничивающая на раковине смежные завитки, называется шовною (su, фиг. 357). Все обороты раковины, за исключением последнего, образуют вместе спираль раковины (5). Нижняя часть последнего завитка представляет основание раковины. Спираль представляет различие у различных видов и подов; иногда она образована значительным числом
оборотов, иногда небольшим, и может быть длинна, коротка или сжата. В соответствии с этим изменяется вершинный угол спирали, т. е. угол между линиями, проведенными от макушки к основанию раковины по противоположным сторонам наружной поверхности оборотов (угол между образующими конуса спирали, лежа! (ими в одной плоскости, проходящей через ось конуса). Часто внутренние части оборотов соприкасаются, сливаясь между собою в ось ракшины, идущую от макушки к середине основания раковины (фиг. 357) и известную под названием столбика [columella). В других случаях внутренние части не сливаются, и вместо столбика получается трубчатое пространство, суживающееся в поперечнике от основания раковины кверху; это пространство называется пупком [umbilicus). Отверстие пупка иногда является отчасти заполненным отложением вещества раковины, образующим мозолистость [callus). Животное прикреплено к столбичной части раковины посредством мускула, сокращение которого втягивает тело животного в раковину; когда же животное не втянуто, раковина вмещает в себе лишь внутренностный мешок.
Обыкновенно Полость раковины гастропод непрерывно тянется от макушки до устья, но в некоторых случаях в более старых частях раковины образуются поперечные перегородки, совершенно обособляющие пространство позади них от пространства, впереди них лежащего (фиг. 357). Форма апертуры значительно изменяется в различных родах и имеет большое значение в классификации; апертура по форме может быть круглою, овальною, продолговатою и т. д. Устьевой край раковины состоит из наружной губы [L) и лежащей в около-столбпчной части внутренней губы.
При ползании гастропод раковина помешается на спинной поверхности тела, с макушкой, обращенной назад и вверх и с апертурой
вниз; вследствие этого дальнейшая от маку гаки часть апертуры будет передней, а ближайшая к макушке задней. Иногда, как у Naltca, край апертуры непрерывен и тогда называется цельным, или голостомным; в других случаях передний край выемчат, или выемчатость переходит в удлиненный раструб [de), в котором помещается приводящий воду в жаберную полость трубчатый сифон; такие формы называются сифоностомными; иногда имеется также другой канал на заднем краю апертуры с помещающимся в нем ВЫВ01ЯЩИМ или анальным сифоном.
Наружная губа может быть тонкою и острою или утолщенною. Внутренняя губа бывает образована или стенкой предпоследнего обороту раковины, или налегающим на него мозолищым утолпцнием.
Наружные украшения обыкновенно состоят или из врезанных на поверхности раковины бороздок, или из ребрышек, складок, бугорков, шипов и т. п. Различают продольные или спиральные украшения, идущие
Фиг. 357. Продольный разрез Triton. Верхняя часть раковины многократно обособлялась перегородками.
а — макушка; «м — шов; S — спираль, L — наружпая губа апертуры; ас — передний канал; рс — ваднкн канал.
параллельно швам, и поперечные, направленные более или менее поперечно по направлению к швам.
Многие гастроподы имеют крышечку, закрывающую устье раковины, когда животное скрыто в ней. Эта крышечка прикреплена к задней части ноги, бывает известковая или роговая и нарастает или концентрически круговыми линиями, или спирально.
Все гастроподы, снабженные жабрами, являются животными водными. Есть проснои дные и наземные, с легочною полостью, приспособленною для воздушного дыхания. Большею частью гастроподы—обитатели мелководья, прибрежной полосы моря; распространены преимущественно до глубины в 100 метров.
Большая часть гастропод питается растениями; но некоторые плотоядны. Есть и хищники (Natica, Buccinum, Мигех), которые пробуравливают посредством radula раковины других моллюсков и поедают их мягкое тело
Жаберные, Branchiata.
Подкласс I. Prosobranchiata.1
Роды Prosobranchiata описаны ниже, начиная с принадлежащих к группе liiotocardia, как наиболее примитивных, сохранивших еще все парные
органы в полном числе.
Pleurotomaria ( pleura—бок, tome—разрез) проходит через все геологические периоды, из которых известны ископаемые фауны, т. е. от кембрия доныне. Коническая, умеренно высокая раковина (фиг. 358) представляет на наружной губе вырезку (синус) с параллельными между собою и со швом краями. Эта выемка при росте раковины зарастает в более старых частях ее, простираясь всегда лишь на небольшое сравнительно протяжение назад от апертуры.2 Заросшая синусовая выемка является в виде полоски на боках завитков, етграничен-
Фпг. 358. Pleurotomaria ell-piica. Нижний си- НО® ОТ Остальной НОЬСрХНОСТИ лур. Справа часть синусовой полосни, увели- раКОВИНЫ ПО крайней Мере ПО
’еиная. тому, что синусовая полоска
представляет на своей поверхности лишь линии нарастания, дугообразные и обращенные выпуклостью дуги назад, к вершине раковины.
Форча раковины весьма разнообразна, с боками завитков круглыми (фпг. Зэ8) или угловатыми вследствие присутствия продольных килеватосгей (фиг. 359), гладкими или покрытыми спиральными и поперечными ребрыш-
1 Proso—кпереду, branchia—жабры.
8 Синусовая выемка соответствует разрезу мантийного края, а этот разрез по своем) положению соответствует оконечности кишки и дает возможность более скорого удал«нш вкскрементов и воды из жаберной полости
ками или рядами бугорков, раковина с пупком или без него, с оборотами в разной степени объемлющими, так что вйделяют значительное число подродов, проходящих иногда через несколько геологических- систем. В настоящее время представители Pleurotomaria редки, в фаунах палеозоя и мезозоя многочисленны и разнообразны.
Фпг. 359. Pleurotomaria Buchi. Юра. Макарьев на р. iaaco. Ивановской сбл.
Фиг. 360. Wiaphistoma qualtenaium.
Нижний силур. 'Галлин. Эстония.
от Pleurotomaria своею удлиненною, башенковою раковиною. От кембрия до триаса включительно.
У Ehaphistoma (фиг. 360) синус не является закругленным на конце своем, обращенном назад, но угловатым, причем вершина этого острого угла лежит на киле, отграничивающем верхнюю, плоскую часть поверхности завитков от нижней округленной. Раковина с пупком, с низкою спиралью. Кембрий—силур.
Фпг. 361. а — Bellerophon (Bwania) decussatus. Нижпе-каменпиуг. С. Слобода, около Одоевска, Ц. промышленной обл.; (6 — Бе lerophon (Euphhnus) Urii, о.туда же; с — Bellerovhon tuberculatus. Средний девон. Нассау. Германия.
Оиг. 362. Fissu-reUaitahca.tAno-цен. Грунд около Вены. Австрия.
Bellerophon (фиг. 361; мифологическое название) имеет вполне симметричную завитую в плоскую спираль раковину, с синусовыми выемкой и полоской, находящимися в области плоскости симметрии раковпны. Поперечная или, кроме того, и продольная скульптура. Силур—пермь.
Возможно, что Bellerophop сохраняет симметрию благодаря передвижению плаванием, а не пелзанием.
Fissurella (fissus—отверстие) имеет коническую колпачковидную раковину с отверстием на вершине конуса (фиг. 362). С юры доныне.
Коническая колпачковидная форма раковины возникает у представителей различных групп гастрспод как приспособление при жизни в полосе прибоя, дающее возможность раковине примыкать, присасываться к поверхности скал. Индивидуальное развитие ныне живущих Fissureha представляет в первых стадиях раковину спирально свернутую, слабо
асимметричную, с синусом па наружной губе наподобие раковины Bellerophon. Затем при росте раковины синус превращается в отверстие, сначала лежащее на боку раковины а затем иа ее вершине.
Euomphalus (фиг. 364; еп—широкий; omphalus—пупок) представляет
раковину с широким пупком, с низкою, инона почти плоскою или даже углубленною спипалью, обороты продольно-киловатые или иногда усажены буграми. Наружная губа выемчатая на уровне верхнего киля. От силура до триаса.
Фиг. 363. Strapnrollus Dio-
nysui. Иижнекахен ноугольяые. Визе. Бельгия.
Фч". 364. Euomphalus catillus Ка-менноуг. Кильдар. Ирландия.
Strapctrollus (фиг. 363; strabos—завороченный) отличается от Еиот-halus круглыми боками оборотов. От силура до юры.
Флг. 365» Platyschisma uchten-sts. Средний девон. Р. Ухта.
Фиг. 366. Haliotis volhynica. Мионен. Венский бассейн.
Фиг. 367. Turbo chersonensis. Третичные. Сарматский ярус. Д. Анновка на 11нгуле, около Зиновьевска. УССР
Platyschisma (фиг. 365; pla*ys—широкий, schisma—разрез) отличается
от двух предыдущих родов узких пупком. Наружная гуоа с широкий выем
Фиг. 36S. Turbo
Puscln Юра. Оренбургский край.
кой, чем этот род приолнжается к Pleurotomaria. От силура до пермских.
Haliotis (фиг. 366; hals — море, otis—ухо). Раковина плоская, ушкооб-разная, с весьма значительным расширенным последним оборотом и устьем, с рядом отверстий на наружном левом краю; отверстия эти соответствуют синусовой вырезке Pleu-
Фиг. 369. Trochus rotomaria и, подобно этой вырезке, podoticus. Миоцен, впоследствии зарастают в верхней Кишинев. Бессарабия. > части раковины. С верХНвМеЛОВЫХ
отложений доныне.
Turbo (фиг. 357, 368; turbo—вершина). Раковина конусовидная или волчкообразная; бока оборотов округленные и устье почти круглое. Украше
ния в виде бугорков и ребрышек, имеется известковая спиральпо-парастаю щая крышечка. От силура доныне.
Trochus (фиг. 369; trochus—коле :о) отличается приплющенными бо
ками и оснозанием, с угловатым перегибом между ними; в связи с этим
апертура имеет угловатое очертание. Роговая концентрически нарастающая крышечн?. От силура доныне.
Patella (фиг 370; patella—чашка) по форме раковины сходна с Fissu-rella, отличается лишь отсутствием отверстия. От силура доныне.
Отличается от Diotocardia, имея лишь одно предсердие и одну жабру. Примыкает к 1 Diotocard а и отличается от Monotocardia по наличию двух почек.
Фиг. 370. Patella rugosa. Юра. Франция.
Фиг. 371. Naticopsi s carbonarius. Нижнека-менноуг. Из окрестн. Скопина, Московской обл.
Древнейшие предста-гптели Monotocardia вероятно обособляются в палеозое от Trochidae.
371; nato—
вероятно род Naticopsis (фиг.
К таковым принадлежит
Фиг. 372. Platyceras parasiticus. На чвшечке Cromyocrinus. Верхне-каменноуг. С. Мячкево, около Москвы.
плавать). Спираль короткая, слабо выдающаяся над значительным последним оборотом. Раковина гладкая или с линиями нарастания, утолщающимися в ребрышки в верхней части оборотов у шва, или с бороздками, расположенными подобно ребрышкам. Внутренняя губа с утолще-яиями наверху и с продольными ребрышками. Девон—триас.
Platyceras (фиг. 372; platys—широкий, keras— рог) представляет раковину, по форме
Фпг. 373. Natiea millepun-data. Раковина и крышечка. Плиоцен. Монте Марио, около Рина. Италия.
напоминающую головной колпак, образованную главным образом последним оборотом; макушка еще довольно развита, представляя несколько оборотов спирали, тогда как она редуцирована у послепалеозойских форм, принадлежащих к роду Capulus. Platyceras встречается от кембриа до триаса. Capulus от триаса доныне. Формы мало передвигающиеся, почти всю жизнь остающиеся на одном месте; так Platyceras живет па морских лилиях, вероятно питаясь их экскрементами и прикрепляясь над их анальным отверстием, или на камнях, припем устье раковины принимает неправильную форму в зависимости от субстрата.
Natica (фиг. 373J имеет раковину шаровидную или полушаровидпую, обыкновенно гладкую, без пупка или с пупком, иногда прикрытым отчасти
мозолистым утолщением. Роговая или известковая крышечка. С триаса доныне.
Paludina (фиг. 374; paludis—болото). Коническая или башенковая раковина; обороты выпуклые, гладкие, бугорчатые, или с продольными ребрами. Без пупка или с узким пупком. Роговая крышечка с концентрическими линиями нарастания и эксцентрическою начальною частью. Апертура ограничена непрерывным краем вследствие отложения известкового вещества на внутренней губе. Эпидермический слой раковины толстый, как это вообще часто бывает у пресноводных. Живет в пресной или слабо соленой воде. С юры доныне
Фиг. 374. Ряд фор1»» постепенно нзменялдепся Paludina из нижпеплиопеновых отложений Славонии:
а Paludina Keumayri из самых нижних слоев; к — Paludina ВаегмъЪ яз самых верхних слоев, Ъ—i—промежуточные формы иа промежуточных слоев.
I
Из вер’гнетретпчпых отложений юго-востсчной Европы Не ймайром списаны знаменитые ряды палюдин, постепенно изменяющихся от нижних горизонтов к высшим. Развитие начинается от гладких форм и приводит к килееатым; происходило оно в морях ввутрегнпх, 01 реснепных, находящихся в слабом сообщении с океаном. При таких условиях 8миг| шин были затруднительны: предки и потомки находятся в одном местона-холадвнии.
Turitella (фиг. 375, 376; furls—башня). Раковина высокая, башен-когая. Устье кру>лое или четырехугольное, цельное. Наружная губа тонкая. Раковина покрыта продольными спиральными реорышкаыи. Триас— ныне.
Vermetus (фиг. 377; vermis—червь). Раковина прирастающая, развернутая трубчатая, с макушечной частью все-таки спирально свернутой, снабженной поперечными перегородками. Напоминает Serpula, а иногда ffairorbis из червей. Тркас —яыкэ.
!
Loxonema (фиг. 380; loxos—косой; пета—нить). Раковина башенко-вая; обороты выпуклые с S-образно изогнутыми линиями нарастания. Высота устья раковины более ширины. От силура до триаса.
Zygopleura (фиг. 381; zygon—иго, pleura—бок). Отличается от предыдущего рода поперечными ребрышками, образующимися ния. От девона до нижнемеловых отложений.
Prouiathildia отличается от предыдущей распадением поперечных ребрышек па бугорки, а иногда еще и развитием спиральных ребрышек. От перми доныне.
Через посредств» Promathildia и некоторых других родов на границе палеозоя из голостомиых Monotocardia выходят си-фоностомные, про-стейшпи представителем которых является род Centluum (фиг. 378, 379), представляющий башенковую } каналом напереди устья. Соединение поперечной скульптуры с продольною. От триаса доныне.
К Cerithium примыкает своеобразный род Nerinea (фиг. 382, 383). При той же общей форме раковины, как у Cerithium, и нередко 8-образных
Фиг. 375. Turi-telle, turret. Миоцен. Ульм. Вюртемберг. Германия.
Фиг. 376. Тип tella imbricata-гга. Эоцен. Гри-ньон. Франция.
по линии нараста-
Фиг. 377. Гете-м' intovtus. Плиоцен. Флоренция. Италия.
раковину с копотким сифонным
Фиг. 378. Cerithium ’ nssiensc Верхняя юра. Каменка на Донце. УССР.
Фиг. 379. СегМ'гит тагдапмееит Олигоцен. Гессен-Дарм-штадт. Германия.
Фиг. 380. Loxc пета sinuosum. Верхний силур.
Фиг. 381. Zygop-leura hybnda. Триас.
линиях нарастания, как бывает у всего комплекса перечисленных родов, начиная с Loxonema, Nerinea отличается сильно развитыми продольными килями, находящимися внутри оборотов на трех или на всех четырех сторонах поперечного сечения оборотов. Такие складки иногда бывают и у представителей рода Cerithium, но лишь на столбике. Юра—мел.
Складчатость внутри раковины является следствием складчатости мантии, а иослед-ияя складчато ггь—следствием несоответствия ыыкду размерами поверхности мантии и поперечником полости обер тон, который непропорционально мал, почему мантия может прилегать к внутренней поверхности оборотов, лишь образовав складки.
Aporrhats (фиг. 384) отличается спиралью несколько укороченною сравнительно с ближайшими предыдущими родами, апертурой, представляю-
Фпг. 382. Nerinea Ег hwaldi. Верхняя юра. Каменка на р. Донце, УССР.
Фпг. 383. Nerinea astrakhanica. Меловые отложения. Оз. Баскунчак, Иижне-Волжскои области. Вид сбоку н разрез по вертикальной оси.
щей дза канала—передний и задний, и пальцевидные отростки на наружной губе. От юры доныне.
Strombus (фиг. 385; strombos—сосновая шишка) имеет короткую спираль с весьма значительным последним оборотом. Узкая, длинная апертура,
Фиг. 384. А рот/ hais pespehcan'. Мпицен (сарматский яр\с). Виш-невец. Польша.
с коротким согнутым каналом напереди. Наружная губа
Фиг. 385. Str >mbus coro-
крыловидво расширенная, нагереди с выем ь ой. От мела доныне.
Сургаеа (фиг. 386)иред-ставляет крайне укороченную спираль, в зрелом возрасте объемлемую более или менее совершенно весьма большим последним оборотом. Длинное щелевидное устье на обоих концах с коротким каналом. Губы зазу >рены совнутри.
natus. Ыисден. ВенскийОт Юры Д0НЫН6.
бассейн. Австрия. Buccinum (фиг. 387;
римская военная труба). Раковина вздутая, брюшистая; спираль сравнительно короткая. Обороты гладкие или с ребрами попереоными. Апертура широкая с коротким, но широким и согнутым 1 аналом, наружная губа тонкая, внутренняя губа мозолиста и со складкой. Троичные—ныне.
Tref.ospi.ra (фпг. 388) с короткою спиралью и весьма значительным последним Очоротом. Линии нарастания представляют в верхней части завитков синусовый выгио назад. Канал короткий. Поверхность покрыта спиральными ребрышками или бороздками. Наклонность к периодическому возникновению утолщений вдоль линий нарастания. Пермь—триас.
Fusus (фиг. 389; fusus—веретено). Веретенообразная раковина с длинного спиралью и длинным прямым сифонным каналом Апертура овальная, наружная губа острая, внутренняя—гладкая. Мел—ныне.
r Pleurotoma (фиг. 390; pleura — бок,
tome—разрез). Веретенообразная раковина с прямым сифонным каналом имеет на, наруж-Л ной губе под швом довольно глубокую сифон-ную выемку. Мел—ныне.
Conus (фиг 391) отличается от преды-
Фиг. 387. Вис-cinum baccatwm.
Дармштадт. Германия.
Сарматский «рус. Таганрог.
Фиг. 386. Сур raea subexcisa. Олигоцен. Вейн-гейм- Гессен-
Фиг. 388. Tretospura dives-ошЫ са. Пермские отложения. Поволжье.
дущего рода весьма укороченною спиралью. Апертура длинная, узкая, с параллельными краями, с сифонной выемкой на переднем конце. Наружная губа острая. Внутренний сл *й раковины впоследствии утончается (рассасываясь) до предпоследнего оборота включительно. Мел—ныне.
Фиг. 389. Fusus longirostris Миоцен. Баден, около Вены. Австрия.
Фиг. 890. Pleurotoma asperulata. Миоцен. Венгрия.
Фиг. 391. Conus роп-derosus. Миоцен. Вен-
грия.
От Monotocardia отделяют как равноценный им подторядок Heteropoda, киленогих, отличающихся, впрочем, только ногой, большею частью превращенной в вертикальный плавник, на котором еще сохраняется рудимент ползательной ноги. Раковина иногда отсутствует, а если имеется, то симметрична, тонка и легка. Особенности раковины и ноги возникают как при сиособление к пелагическому образу жизни, который ведут киленогие, передвигающиеся плаванием (фиг. 392), причем брюшная сторона тела животного с плавником обращена кверху, а раковина вниз.
Подкласс II. Opisthohrtn chiata.1
Эта группа совмещает гастропод весьма различных анатомически. Есть и первично-примитивные типы, есть типы, приобретшие примитивность вторично, вследствие деградации, есть наконец и формы, столь ate высоко организованные. как в среднем Prosobranchiata.
Сиг. 392. Са, папа. Ныне живущл
р — хоботок; t — щупальце; f — нога; d — рудимент ползательпои ноги. • — жабры; s ~ раковина. Животное изображ. ш в но на льном положения, в состоянии плавания на поверхности моря.
Деградация проявляется в яеразвитни жабр, мантийной полости, рако-
вины, проявляется в более высокой организации личинок сравнительно с зрелыми инд! в щуумами. Раковина Opisthobranchlata не отличается никакою
Фиг. 398. Actaeonetla mgantea. Турон Австрия.
существенною особенностью от раковины Prosobranchiata. Ископаемые формы могут быть относимы к этой группе, лишь исходя
Фиг. 394.
Actaeonella саи-caitca. Мел. За-. кавказье.
от родового сходства с современными ее представителями. Животные морские. Девон—ныне.
Ac',aeonella (фиг. 393—39-1; от мифологического имени). Толстостенная раковина, вздутая. гладил. Обороты объемлощи так, что спираль коротка или даже углублена. Столбик напереди утолщенный, с тремя острыми складками. В средне- и верхнемеловых отло-
жениях.
Возможно, что к Opisthobranchiata принадлежит род Nerinea.
Крылоногие, Pteropoda, находятся в таком же отношении к Opisthobranchiata как Heteropoda к Prosobranchiata, представляя группу, отличительные особенности Китирой обязаны своим возникновением приспособлению
Opisthen—сзади, branchia—жабр»
к пелаги гескому образу жизни. Йога их превращена в две крыловидных плавательных лопасти (фиг. 395); тело вытянуто или свернуто в спираль, иногда покрыто прозрачной, тонкой раковиной, но чаще,-голое. Животные малых размеров, живущие массами в открытом море па некоторой глубине и только на ночь поднимающиеся к его поверхности. Раковинки их часто скопляются в огромном количестве на морском дне, образуя известковистый птероподовыя ил.
Ископаемые крылоногие известны из верхнемеловых и третичных отложений; в палеозое есть ископаемые, скорее всего принадлежащие к птеро-подам, хотя большею частью и отличающиеся необычно большими для последних размерами.
В палеозое и позже вшречаются крылоногие еще со спирально свернутой рчковииой, как например низко-коническая раковигка Spirialis (от третичных доныне).
Conularia Ci иг,. 396. 397). Раковина тонкостенная, прямая, пирамидальная, с квадратным или ромбическим поперечным сечением, гриострез-
Фпг. 395. styKeHa гес ta. Ныне живущая.
Фиг. 896. Conulana cancellcta. Верхний силур. Хотим на р. Дне тре. Бессарабия. Справа часть noeei хнозтн в увеличенном виде.
ная пли усегечная на заднем конце. В молодости прирастает, позже свободная. Каждая из поперечно струйчатых или ребристых боковых поверхностей разделена вдоль пополам наружной бороздкой, соответствующей внутреннему ребру. Задняя часть раковины разделена поперечными перегородками Апертура сужена четырьмя треугольными или языкообразными лопастями переднего края, загнутыми к продольной осп раковины. Достигает длины в 20 см. Часто в силуре и девоне, доходит до пнжнеюрских отложений.
Tentaculites (фиг. 398, 399; tentaculum—щупальце, lithos—камень). Раковина толстостенная, удлиненно-коническая с круглым поперечным сечением, на заднем конце приостренная плп оканчивающаяся шарообразным расширением. Поверхность украшена выдающимися поперечными naj аллель ними кольцами. Макушечная часть раковины часто разделена поперечными перегородками. Иногда наполняет целые слои в силурийских и девонских отложениях.
Myliolina, (фиг. 400; stylos—столбик). Раковина маленькая, игловидная или удлиненно-коническая, прямая или слегка согнутая, с неравномерным углом возрастания. Поперечное сечение круглое. Задний конец раковины
тупоокругленный пли снабженный шаровидным раздутием первичной раковины. Поверхность гладкая или покрыта продольными струйками. Ниж..ий девон. >
Styliolina интересна в том стношепии, что является одним ив палеозойских родов ваиболее определенно относимых к Pteropoda на основании сходства с ныне живущими представителями этой группы. Именно Styholina весьма сходна с ныне живущею Styholat последняя отличается главным обрезом отсутствием вздутой первичной раковины и продольных струек.
Фиг 3?7. Conulana quad) isuicata. Верх-иекаменноуг. Шотландия.
Фиг. 398. Tentacu-lites ormolus. Верхний силур. Англия.
Фпг. 399. Tentocu-lites acuarius. Киж-ниа девон. Покровское, восточный склон Урала, Уральская оол. 10/1.
Фиг. 400. Styho-1гпа nucleata. Девон. Урал. Покровское. Уральская обл.
Легочные, Pulmonata.
Мантийная полость видоизменена так, что представляет легкое. Жабер нет. Дыхание производится посредством сплетенкя кровеносных сосудов, находящегося на внутренней поверхности легкого. Главным образом наземные и пресноводные формы. Анатомически имеют некоторые общие с Opistho-branchiata особенности, например неперекрещивающиеся нервные стволы, но вообще вероятие обособлялись в различные периоды от разных групп морских брюхоногих. Дышат воздухом, и поэтому водные формы должны периодически подниматься на поверхность воды для возобновления запаса воздуха. Древнейшие Puimonata изредка встречаются в девоне и каменноугольной системе, немногочисленные в юре и мелу, чаще встречаются в третичных отложениях и достигают максимума распространения в настоящее время, i в пресной воде.
Limnaeus (фиг. 401; limus—ил) имеет острую, умеренно выс- кл’ю спираль. Апертура широкая, овальная; наружный край острый. Верхняя юра—пчне.
Hercynella (фиг. 402; от географического названия) является девонским представителем Pulmonata, притом из живущих в прибрежной полосе моря или в сол «и «атых лишь водах эстуариев. Раковина подобная тому, что у Patella, отличается складкой, от вершины доходящей почти до края, внутри существует соответствующая впадина.
Helix (фпг. 403; helix—завиток)—род наземных Pulmonata, имеет шаровидную коническую пли дискоидальную раковину. Апертура косая, полу
Фиг. 401. Limnaewpachy-jasier. Миоцен. Ульм. Вюртемберг. Германия.
Фиг. 402. Не) cynella bohe-mica. Девон. Ло иков. Богемия. Чехо-Словакия.
Фпг. 403. Hehx Brocchvi
Плиоцен. Италия.
лунная или округленная, с разъединенными поверхностью последнего оборота краями. Третичные—ныне. Родственные роды известны с каменноугольных отложений.
Геологическое распространенгге брюхоногих.
Среди всех моллюсков брюхоногие представляют наибольшее количество видов. Встречаясь уже в кембрии, к настоящему времепи они достигают максимума своего развития. Интересно, что кембрийские гастроподы зачастую колпачковидны. Из Prosobranchiata, вообще составляющих большинство гастропод, в палеозое встречаются лишь голостомиые, в собенности Pleurotomariidae, Turbinidae и Trochidae, Turritellidae, Loxonematidae. Последние дают начало в триасе сифоностомным гастроподам, которые становятся преобладающей группой в третичном периоде. Opisthobranchiata известны с каменноугольных отложений. Pulmonata с девона в виде водных форм и с камепноуготьных отложений в виде наземных. Heteropoda известны лишь с миоцена; вообще редко сохраняются в ископаемом состоянии. Pte-ropoda известны с палеозоя, в последнем частью в виде условно относимых к этой группе форм (Conularia, TentacuUtes). Крупнейших размеров достигают позднейшие представители гастропод (Triton, Fusus и др.), у позднейших же и более сложная скульптура раковины. Древнейшая скульптура в виде поперечных ребер (Zygopleura'), соответствующих линиям нарастания раковины; затем эти ребра распадаются на бугорки, группирующиеся поперечными рядами; затем к поперечной скульптуре присоединяется и продольная (Promathildia}. Изредка продольная скульптура, хотя и слабо выраженная, бывает первичною уже у рекоторых палеозойских гастропод.
Геологическое распространение брюхоногих.
Роды Отделы или ярусы
Кембрий] i ! Силур Девон Карбон Пермь i Триас Юра Мед Третий- I ные ! Ill J II 1 и 1 ] 01ЧНИЭЯ -adar)
Pleurotomaria Murchisonia . Tth.opkistoma . Bellerophon . Eissurella . . Euomphalus . Straparollus . Platyschisma . Haliotis . . . Turbo . . . Trochus . . . Patella . . . Nalicopsis . . Platyceras . . Capulus . . . Natica . . Palulina . . Turritella . . Vermetus . . Loxonema . . Zygopleura . . Promathildia . Cerithium . . Nerinea . . Aporrhais . . Strombus . . . Cypraea . . . Buccinum . Tretospira . , Fusus .... Plcurotoma Conus .... Actaeonella Spirialis . . . Conularia . . 'lentaculites . Styliolina . . Sercynella . . Limnaeus . . Helix ....
?
г
•
—
КЛАСС Ш. ГОЛОВОНОГИЕ, CEPHALOPODA.
Головоногие имеют целый ряд особенностей организации, общих им с брюхоногими, а именно: обособленную ст туловища голову, сходно устроен ный внутренностный мешок, нераздельную раковину и т. д. Йри всем том головоногие резко отличаются от брюхоногих вполне двусторонней симметрией своего строения и расположением мантийной полости на брюшной стороне тела животного (фиг. 404). Такое положение мантийной полости у головоногих возможно благодаря тому, что головоногие—животные водные и притом передвигающиеся главным образом плаванием, в отличие от пол зающих брюхоногих.
Позади головы на брюшной стороне тела находится трубчатый муску листый орган—воронка (фиг. 404 tr), открывающийся передним узким концом наружу, а основанием в полость мантии. В мантийную полость,
в щель вокруг шейной части тела животного, извне входит вода, выходящая через воронку из мантийной полости при сокращении ее мускулистых стенок. Сокращение это происходит быстро, вода с силою выбрасывается из воронки, благодаря
Фиг. 404. Боковой вид головоного (каракатицы, Sepia) с нанесенными ор. анаыи.
i—слепой придаток желудка, с—сердце, d—кишкв, dr — анальное отверстие, f — хватательная рука, д — го и ад а, к — жабра, I— печень, tn—желудок, ♦nr — край мантии, tnt — мантии нал полость, п — почка, р — околосердечная сумка (coelom), si — сифонная полость, st — раковина с перегородками, t—чернильный мешок (закрещен жирным черным), tr — воровка.
узкости ее наружного конца, и животное перемещается толчками в на-
правлении, противоположном истечению, т. е. назад. Воронка гомологична ноге моллюсков, возникая из двух боковых лопастей брюшной стенки, срастающихся или только
соприкасающихся; кроме того, во-
ронка иннервируется от ножного нервного узла. Но воронка гомологична
лишь части ноги моллюсков; другая часть ноги представлена околорото-выми придатками—руками; руки у всех ныне живущих представителей головоногих за исключением вообще особняком стоящего Nautilus, пред
ставляют вытянутые от проксимального конца к дистальному придатки, снабженные щупальцами, присосками или крючочками.
Число рук у головоногих доходит до 10; они или сильно вытянуты и снабжены присосками (Sepia), или укорочены по сравнению с предыдущим случаем, но имеют удлиненное основание и покрыты рядом щупалец (Nautilus).
Аналогичные рукам головоногих, усаженные чувствительными осязательными сосочками, щупальца имеются у личинок более архаичных гастропод Trochidae, в числе 4-х пар на задней части ноги. Есть аналогия также со Scaphopoda (см. ниже).
В индивидуальном развитии головоногих руки развиваются попарно, сначала ближайшее к брюшной стороне тела, а затем более далекие, так что при развитии последних рот становится окружен руками, чего не бывает сначала. Ход развития рук и иннервация их от ножного нервного узла заставляют считать руки гомологичными части ноги моллюсков. Перемещение их на голову является вторичным.
После вышеизложенного ясно, что название класса — «головоногие» — является естественным.
14 Учебаик иа...о оологии.
Головоногие являются моллюсками наиболее высоко организованными по строению нервной системы, кровеносной, органов чувств и т. д.
Нервная система характеризуется скоплением нервных узлов в голове
' Кровеносная система состоит из замкнутой сети кровеносных сосудов и из сердца. Органами дыхания являются перистого вида жабры. Существуют печки, печень, слюнные железы. В мантийную полость открывается, между прочим, выводное отверстие так называемого чернильного мешка, железы, выделяющей сепию — жидкость черного цвета. Выделенная животным сепия, выброшенная через воронку, делает мутною окружающую воду, облегчая этим бегство от преследователей.
В ротовом отверстии находятся две челюсти, верхняя и нижняя, совместно напоминающие клюв попугая, роговые, иногда, у позднейших форм, начиная с триаса, с известковыми оконечностями (фиг. 405, 406). В ротовой полости находится radula
Фиг, 405. Челюсти вине аив)щ. Nautilus pompilius.
а — нзжняя челюсть сбоку, Ъ — верхняя челюсть сбоку, с — та же челжкть снизу.
На верхней конечности головы помещаются два большие глаза, которые у ныне живущих головоногих (за исключением Nautilus} почти так же высоко организованы, как глаза позвоночных животных.
Головоногие всегда ра-дельнополы. Некоторые формы не имеют раковины; если же она существует, то бывает наружною (фиг. 407) или внутреннею (фиг. 404); в последнем случае она заключена под мантией в епгнной части тела животного.
Головоногие — животные исключительно морские; передвигаются плаванием, но иногда и ползают при посредстве рук. Разделяются на два подкласса: четырехжаберных и двужаберных.
Подкласс I. Четырехжаберные, Tetrabranchiata.
Четырехжаберные головоногие имеют четыре жабры и наружную многокамерную раковину; разделяются на два порядка Nautiloidea и' Ammonoidea; последние — животные совершенно вымершие; из первых сохранился доныне род Nautilus, исходя от организации которого можно составить себе представление об организации вымерших форм.
ПОРЯДОК I. NAUTILOIDEA.
Судя по ныне живущему Nautilus, животные этой группы помещались в передней, всего позже образующейся камере раковины; эта камера и называется жилою в отличие от остальных, так называемых воздушных камер (фиг. 407). Руки Nautilus являются в виде шести дугообраз-
Фиг. 406. Nautilus pompilius, ныне живущий, без раковины, вид спереди. •
а — руки, t — щупальца, tr — воронка, с — капюшои, тп, m — чедюст*. Капюшои сдвинут к един кой стороне.
ных пластинчатых образований, расположенных симметрично по обе стороны рта (фиг. 406). Руки снабжены многочисленными щупальцами. Есть основание думать (см. ниже), что у вымерших палеозойских Nautiloidea числе рук колебалось от 2 до 8.
Воронка, состоящая из несрастающихся двух лопастей, находится на наружной, периферической стороне раковины; эта поверхность раковины называется брюшною. На противоположной, спинной части головы находится плотная кожистая верстие, апертуру раковины, кргда тело животного втянуто в нее. Основание челюстей роговое, но вершинные части их известковисты. Мягкое тело животного прикреплено двумя мускулами по бокам к стенкам жилой камеры. Мантия в промежутке между мускулами также прирастает Е стенке раковины на протяжении кольцеобразно идущей вокруг и поперек тела полоски (annulus)-, таким образом раковина с заднею частью животного герметически отделена от передней части. Как мускулы, так и annulus оставляют след прикрепления на внутренней поверхности раковины. Благодаря мускулам животное может несколько высовываться из раковины и втягиваться в нее. От задней стенки тела через все камеры се до самой старой тянется ПОЛЫЙ КОЖИСТЫЙ ТЯЖ—СИфОН.
Оифон многих более древних палеозойских Nantiloidea имел значительный поперечник и вероятно заключал в себе некоторые внутренности. Вообще можно считать, что сифон представляет задний отдел тела, суженный образованием перегородок.
У Nautilus нет чернильного мешка.
Раковина Nantiloidea представляет трубку с поперечным сечением различной формы, округленным или многоугольным, суживающуюся наподобие конуса к наиболее старой части. Эта трубка бывает прямою, дугообразно согнутою или спирально свернутою; в спиральных формах обороты могут быть обособлены один от другого, или сипряктсаться на всем протяжении, или лишь н известной части ддины раковины; последующие обороты могут значительно охватывать предыдущие, закрывая бока их (и н в о л ю т н ы е раковины).
СчИТСЗТСЯ СО8ИИКШИМ ИЗ СЛИ..ГИЯ двух спинных ицпатец.
Полость раковины не непрерывна во всю длину, как у брюхоногих, но разделена, на камеры перегородками (septa) \ величина камер возрастает к апертуре раковины. Сифон проходит через отверстия в перегородках; перегородки часто представляют трубчатое продолжение, охватывающее сифон снаружи; эти сифонные дудки (фиг. 415; обыкновенно направлены от перегородок назад. Длина их составляет часть расстояния между смежными перегородками, равна ему или даже несколько больше, так что свободные задние концы дудок входят в предыдущие дудки. Вовнутри сифон часто представляет различные известковые образования определенной формы и расположения.
Апертура раковины имеет в некоторых случаях края простого очертания, лежащие в одной плоскости или немного от нее отклоняющиеся; в других случаях от краев апертуры отходят отростки, иногда полые, трубчатые (фиг. 421). У некоторых форм: Gomplio-сегаз(фиг. 417), Не-xameroceras (фиг. 408) апертура сильно сужена сравнительно с поперечным сечением жилой камеры и является разделенною на лопа сти. В апертуре по следнего типа непарная часть ее отвер стия приближена к брюшной стороне и, предполагается, соответствует воронке, а симметрично расположенные парные части—рукам Линия, по которой край
перегородки примыкает к наружной стенке раковины, называется су тур-ной, или шовной (suture —шов); она хорошо видима на внутренних ядрах раковины, где наружная стенка отсутствует.
Отличительною особенностью раковины Nautiloidea является простая форма сутурлой линии; она бывает, как говорят, прятал (лежащая в одной плоскости) или изгибающаяся (так наз. л о п а стная). Изгибы сутуркой линии расположены симметрично относительно плоскости симметри; кривая сутурной линии попеременно уклоняется своими изгибами те вперед, то назад; части кривой, обращенные выпуклостью вперед, называются с е д-лам и, а промежуточные между ними части, обращенные выпуклостью назад, называются лопастями.
Первичная раковина Nautiloidea редко сохраняется, вероятно вследствие слабого объизвестнения или его полного отсутствия; если раковину
1 Наполним, что и у некоторых брюхоногих (папр. Euomphalus, Trifon, Vermetvs бывают перегородки в вериинной части раковины.
Ins разрезать по плоскости симметрии, то в центре ее видно небольшое свободное пространство (фиг. 409), в котором у ископаемых форм находится отпечаток первичной раковины.
У Nautilus .первичная раковина может быть развивается в виде не сохраняющегося кожистого пузырька, оставляющего на задней стенке первой воздушной камеры рубец (фиг. 410).
У рода Orthoceras известна яйцевидная известковая первичная раковина, впоследствии обыкновенно не сохраняющаяся, отламывающаяся от позднейшей части раковины. Сифон у Orthoceras свешивался своим задним концом в полость первичной раковины.
Каждая из воздушных камер в свое время была жилою; затем животное наращивало раковину, перемещалось в ней вперед, а задним концом
тела отлагало новую перегородку.
Камеры называются воздушными, так как они наполнены воздухом у пойманных близ морской поверхности животных. Есть однако предполо-
Фиг. 408. Phragmo-ceres (Hexameroceras) Pander Силур. Eoi e-мия Чехо-Словакия.
Фиг. 409. Первый оборот ракови-вы Nautilius роы-pilius, разрезанный продольно.
в —« сифон, с — слепой
Фиг. 410. Первая воздушная камера с рубцом о" прикрепление первичной раковины.
конец его» х — свобод-
ное пространство.
жение, что камеры не всегда наполнены воздухом, что на значительной глубине они содержат воду с растворенными в ней в изобилии под значительным давлением газами. Когда животное быстро поднято на поверхность, газы, расширяясь, вытесняют из камер воду. Возможно, что совокупность воздушных камер играет роль гидростатического аппарата, изменяя вес животного при втягивании его в раковину и при выдвигании из нее и способствуя этим его вертикальному перемещению.
Поверхность раковины Nautiloidea бывает гладкая или представляет различную скульптуру — продольные кили, поперечные ребра; бугорки, различным образом группирующиеся в ряды.
Раковины Nautiloidea бывают прямыми, согнутыми и спирально свернутыми Нельзя утверждать, что в каждой в отдельности из трех этих групп ее представители находятся между собою в ближайших отношениях Родства, происхождения. Возможно, что различные представители Nautiloidea с прямой раковиной приобретают согнутую раковину независимо одни от Других. Роды, ближайшие к какому-либо представителю Nautiloidea с согну
той раковиной, могут быть не среди родов с такою же раковиной, а среди родов с прямой.
Присутствие апертур, сжатых выростами с боков, также не составляет принадлежности какой-либо одной группы. Такие апертуры бывают среди Nautiloidea с прямой раковиной, так ясе как и среди имеющих согнутую или свернутую ракогину.
Род Orthoceras (фиг. 411, 412; orthos —• прямой, keras — рог) принадлежит к числу наиболее распространенных. Имеет прямую или слабо согнутую, коническую или почти цилиндрическую раковину, круглого или
Фчг. 411. Orthoceras timi-йип’.Герхний :илур. Боге-<ил. Чехо -Словакия.
Фиг. 41а. Orthoceras aniMlatum. Верхний силур. Богемия. Чехо-Словакпя.
।
Фиг. 413. Endoceras duplex (слева)—с раскрытым внизу ядром сифона. Киниекуле. Швеция. Нижний Силур. Endoceras со'т-липе (справа;—в си-фоиальиой полости коническое острие одной из цоронок. Нижний си: ур. Ораниенбаум. Ленинградская обл.
Фиг. 414. Endoeeras pro-tec forme. Продольное сечение, обнаруживают.! о воронки сифональиой полости и эндисифон. Нижний силур. Нью-Йорк. САСШ.
овального поперечного сечения, с сифоном малого попер “чника, центральным или эксцентричным. Сифонные дудки короткие; шовная линия прямая-Апертура иногда гжата с боков, представляя три лопасти. Раковины с различной продольной и поперечной скульптурой. От кембрия до триаса включительно.
В нижнекембрийских отложениях встречается род Volborthella, являющийся древнейшим представителем цефалопод. Морфологически он может быть не отличается от Orthoceras, но отличается малыми размерами раковины (всего несколпко мм) и тем, что раковина повишмому состояла исключительно из органического вещества. По крайней мере думать это дает основание то, что зо-первых сама раковина никогда не сохраняется, а лишь
ядра и затем то, что жилая камера всегда сплюшена—вероятно вследствие относительно малой прочности.
Endoceras (фиг. 413—416; endos—внутри, keras — рог). Раковина прямая, достигающая дз}х метров в длину, с сифо-ном необыкновенно большого диаметра, приближение м к краю раковины. Длина сифонных дудок
Фиг. 415. Схема продольного сечения Endo-ceras commune, пока-зывакийяя сифонные
ДУ ки.
Фиг. 417. Gomphocercs bdhemicum. Верхний силур р. Богемия. Чехо-Словакия.
Фиг. 416. Endoceris vaginatum. Нижи, силур. Вайвара. Ленинградская обл.
Фиг. 418. Phragmoceras Broderipi. Верхний силур. Богемия. Чехо-Словакия.
превосходит расстояние между смежными перегородками. Внутри сифона имеются извеЬтковистые отложения в виде вложенных друг в друга полых, конических воронок, обращенных вершиной к начальной части раковины. 1ерез отверстия вершин воронок в сифоне проходит узкой внутренний канал—э н д о с и ф о н, имеющий собственную стенку. Нижний силур.
Gomphoceras (фиг. 417; gompbos — палица). Короткие, прямые раковины; поперечник круглый, наибольших размеров в средней части рако-
215
I
вины, так что она нередко имеет грушевидную форму. Апертура суженная Т-образной формы. Перегородки сближены. Сифон центральный или эксцен-
тричный, цилиндрический или представляет цеждудамерные разду гия. Силур—каменноугольные.
Phraginoceras (фиг. 408, 418; pbragmo — разделение) отличается от Gomphoceras главным образом, согнутостыо раковины. Размеры последней быстро возрастают к апертуре.
Опт. 420. Estonwceras imperfe ctum. Нижний силур. Таллин. Эстония. 1/2.
1>иг. 419.
Lituites lituus. Из чижве-
Фиг. 421. Nautilus (Pteronautilus) cornutus. Пермь. Красповидово на
Волге, близ Казани. 1/4
силурийских
валунов Вос- Фиг. 422. Nautilus (Metacoceras) tr^gonotuberculatus. Справа изобра-
то'.иой Прус- жена отделено жилая камера. Пермокарбон. С Никитовна Донецкий сии. 1/2. бассейн. 1/2
Овальное поперечное сечение. Сифон обыкновенно приближен к вогнутой стороне раковины, почти цилиндрический, иногда содержит радиальные, листоватые, известковые внутреобрайования. Силур.
Estonioceras (фиг. 420) имеет раковину с обособляющеюся от спирали конечною частью. Внутренние обороты лишь касаются друг друга, наружные слабо объемлющи, так что раковина дискоидальна. Поперечное сечение внутренних оборотов подобно сечению линзы, с приострениями по бокам соответственно килю, проходящему по бокам оборотов. Бока наружного оборота закруглены. Сифон приближен к наружной стороне оборотов, причем на внутренних оборотах лежит вплоть у нее, а на наружном на некотором расстоянии. Нижний силур.
Lituites (фиг. 419; lituus — музыкальная труба) отличается длинною, прямолинейно вытянутою конечною частью раковины и лопастной апертурой. Нижний силур.
Nautilus 421,422; nautilus—мореплаватель). Раковина с оборотами весьма различно объемлющими, так что форма ее изменяется* от дискоид-альной до эллипсоидальной, с широким или узким пупком или совершенно-без него.
Поперечное сечение оборотов весьма разнообразно, овальное или. многоугольное.. Сифон центральный или приближен к наружной, реже ко внутренней стороне оборотов, узкий. Поверхность раковины гладкая или представляющая скульптуру в виде поперечных ребер, струек, бугорков, продольных килей и т. п. Формы с широким пупком и украшениями встречаются в палеозое и триасе; в позднейших отложениях, лишь гладкие формы с узким пупком. Род Nautilus разделен на значительное число групп, считаемых подродами и даже самостоятельными родами (Metacoceras, Dis-cites, Pteronautilus и др.).
ПОРЯДОК II. AMMONOIDEA.
Различия между Nautiloidea и Ammonoidea (по крайней мере, древнейшими палеозойскими представителями последних) не могут считаться резко обособляющими обе группы. Все особенности организации являются присущими представителям обеих групп, но со временем накопляются значительные различия в степени развития тех или других особенностей.
Так в отношении сифона для Ammonoidea является характерным признаком малый диаметр его, отсутствие в нем внутренних известковистых отложений и положение сифона-обыкновенно вплотную у брюшной стороны раковины. Характерна постоянно известковистая яйцевидная или шаровидная первичная раковина вместо состоящей из органического вещества первичной раковины Nautiloidea, быстро развивающаяся во всех группах сложность, сильная расчлененность лопастной линии. Только у древнейших Ammonoidea лопастная линия такого же характера, как у большинства Nautiloidea, а затем очень скоро, уже в девоне, возрастает число лопастей и седел и их глубина или длина. Во всех случаях возникает наружная или сифонная лопасть (фиг. 423, eZ), находящаяся при плоскости симметрии раковины на брюшной, наружной стороне ее, затем возникают верхняя и нижняя, или первая и вторая, боковые лопасти (фиг. 423, sZj, sZ2) и далее, к линии, разграничивающей смежные обороты, другие, так
личением высоты поперечного сечения т es lss п is
si
•фиг. 423. Лопастная линия аммонита, представленная в одной плоскости.
т—средняя плоскость оборота; п—шов; «Z—наружная допаять; si, н sZa — первая и вторая боковые лопасти; ее — наружное седло; lst н 2sa ~~ первое и второе боковые седла; is—внутреннее седло; t*Z—внутренняя лопасть.
называемые вспомогательные лопасти; приходящаяся на эту линию называется шовной, а находящиеся на внутренней стороне оборотов — внутренними, в том числе при плоскости симметрии —антиси-ф он н а я лопасть.
Седла имеют аналогичные названия: наружные — по одному с каждой стороны наружной лопасти и т. д. Не только число, но также и форма лопастей и седел разнообразны уже у девонских аммоноидей.
В конце палеозойской эры у большинства Ammonoidea лопастная линия испытывает дальнейшее усложнение: становится более сложным очертание лопастей и седел; края седел и лопастей, бывшие прежде цельными, при дальнейшем развитии получают мелкие зазубрины или более крупные зубцы. Далее лопасти и седла становятся в своем очертании рассеченными, типичными для отдела аммонитов (мезозой).
Появление вспомогательных лопастей и число их стоятв связи с уве-оборотов; у аммонитов с оборотами низкими и только соприкасающимися, но не объемлющими, число . лопастей не бывает более шести.
Увеличение числа лопастей и седел и большая разветвленность их делают более прочным соединение перегородок с наружною стенкою раковины, а потому прочность всей раковины возрастает.
В развитии лопастных линий отдельного индивидуума выражается филогения рода, последовательные перегородки являются все более сложными. Так Medli-
cottia в своем развитии представляет перегородки, соответствующие зрелому состоянию родов: Ibergiceras, Prodaraelites, Subpronorites и Prosicanites (фиг. 424, 1 — ff).
Кроме лопастных линий, важные указания на филогению дает онтогенетическое развитие скульптуры раковины.
Сифонные дудки перегородок у древнейших форм отдела г он и ат и-тов направлены назад, как у Nautilus; у новейших форм перед перегородкой возникает короткое воротникообразное продолжение дудки, и- наконец обращенная назад часть дудки уничтожается, так что дудка является вполне обращенною наперед. Что каеается до формы раковины, то в различное время (триас, юра, мел) и у представителей различных генетических линий проявляется наклонность к переходу от раковины с сомкнутыми оборотами к различным образом развернутой раковине (фиг. 447 — 449, 467 — 469).
Этот процесс в большинстве случаев происходит так, что прежде всего •обособляется от предшествующего оборота жилая камера, а затем последовательно всё более внутренние обороты. Обороты остаются при этом в одной плоскости — Grioceras (фиг. 467) или делаются извитыми по виню: ой конической спирали, как у гастропод — Turrilites (фиг. 449). Бывают также раковины со срединною прямою частью, к которой примыкают на ее концах свернутая начальная и согнутая конечная части раковины — Scaphites (фиг. 469). Наконец возникают совершенно 218
прямые (за исключением небольшой начальной части), как у Orthuceras, раковины — Baculites (фиг. 448).
Жилая камера вообще значительно длиннее, чем у Nautiloidea. Она со
фит. 424.1—9 онтогения Medlieo tia artiensis. Пермокарбон. Урал. Развитие лопастной линии: »—стадия J'^ergiceras. 3—стадия Prodarieiites. я—стадия Subpronorites. 5—стадия Prosica-oites. В 9 лопастная линия изображена незаконченно справа. 10—лопастная линия Med-licottia primas. Пермокарбон. Индия.
•—Плоскость симметрии раковины; и—шовная линия.
Фиг. 425. а—Slephcmoceras Braiken ridgi. Апертура с боковыми ушками До гер. Франция 6 — Aptychu? laevis. Верхняя юра, Золенгофен. Бавария. Германия, с—аммонит, устье которого прикрыто аптихом. d — вертикальный разрез аптиха.
ставляет (т г/я до 1х/а оборотов. Край жилой камеры редко является прямым; обыкновенно у большей части палеозойских форм на наружной стороне раковины существует выемка для воронки, как у Nautiloidea, а у новейших
форм, напротив, во шикает большею частью прямой, но иногда согнутый придаток, вероятно образующийся отложением известкового вещества по-
верхностью воронки.
По бокам апертуры иногда имеются боковые ушки (фиг. 425,о). Во многих случаях ротовой край апертуры ограничен позади пережимом (фиг. 427) или, наоборот, ребровидным утолщением раковины (фиг. 453). Такие пережимы и ребровидные утолщения раксвины нередко наблюдаются и на более старых частях раковины, разделяясь равномерными промежут-
ками и соответствуя прежним устьям раковины.
В жилой камере некоторых аммонитов находят иногда известковые или ооговые образования, служившие крышкою дтя устья раковины, подобно крышке раковины гастропод. Это или две пластинки (фиг. 425,6 и с), примыкающие одна к другой прямыми краями в плоскости симметрии раковины, или одна, соответствующая совокупности упомянутых двух. Эти образования иногда находили замыкающими устье; известковые пластинки (ап тихи) все-таки лишь отчасти соответствуют крышке гастропод: судя по форме и строению 1 аптихов, предполагают, что они представляют образование, аналогичное капюшону Nautilus, но объизвестненное. Тогкие, по преимуществу роговые, отиноч-ные пластинки называются анаптиха-м и; такие пластинки некоторыми исследова-
Фиг. 428. Bact-ritts elegans.
Верхний девон. Эйфель. Рейнская Пруссия. Германия.
Фиг. 427. Prolobrtes delphinulus. Верхний девон. Верхнеуральск
телями считаются за панцыри ракообразных (см. фиг. 228).
Древнейших представителей аммоноидей соединяют под названием го-ниатитов, имея в виду, как характерные общие признаки цельно-крайную лопастную линию, слабое развитие скульптуры раковины и др. Число лопастей бывает часто менее шести, нормального для позднейших аммоноидей — аммонитов. г
Обе группы — гониатиты и аммониты — связаны переходами и не могут быть резко отграничены одна от другой.
Bactrites (фиг. 426; bactron— палка) имеет раковину такой же формы, как у Orthoceras, и вообще настолько близок к последнему, что считается звязующим звеном между Nautiloidea и Ammonoidea. Лопасти в сутурной линии едва намечены, и едва ли не одно лишь краевое положение сифона заставляет отнести этот род к Ammonoidea. Силур, девон.
Существуют роды, близкие к Bactrites по простоте сутурной линии, с раковиною в разной стддени свернутости, подобно древнейшим Nautiloidea.
1 Позволяет предполагать в срединном слое пластинки значительное присутствие органического вещества.
У Prolobites (фиг. 427) раковина шарообразная с объемлющими оборотами. Стенка раковины перед устьем имеет широкий поперечный пережим, 'акие пережимы имеются и на внутренних оборотах. Поверхность раковины
• 4
Фиг, 42S. Agoniatites lohemicus. Деьон. Хлу- Фиг. 429. 2 .посЛев emaciatus. Девон
бочеп. Богемия. Чехо-Словакия. Хлубочеп. Богемия. Чехо-Словакия.
гладкая или покрытая поперечными струйками. Лопастная линия представляет глубокую наружную лопасть и по одной, слабо выраженной боковой лопасти Верхний девон.
simplex. Верхний девон. Фиг. 431. Manticoceros mtumescens. Верхний девон. Тиман.
Симан. Устье р. Чути. Р. Лыа ’оль. Обл. Коми. 1/2.
Agmuatites (фиг. 428; а —без, gonia — угол) имеет раковину с оборотами, приплющенными на брюшной стороне, с пупком умеренно широким ила узким. Седла и отчетливо выраженная боковая лопасть округлены. Девон.
Pinacites (фиг. 429; pinax — планка). Раковина инволютная, сжатая с боков. Седла првострены. Девон.
Tornoceras (ф ir. 430) имеет округленную раковину с узким пупком и глубокую боковую лопасть. Верхний и средний девон.
.лМд/ШМ-ч
Фиг. 433. Prolecanites lunu-Фиг. 432. Manticoceras ВаМипЛг. Р. Чуть. Ти лап. licosta. Верхний девон. Нас-
Верхпий девой. Обл. Комн. 1/1. са;. Германия.
/
Следующие роды являются большею частью уже усложненными по строению раковины, представляя наружную лопасть, подразделенную вторичным седлом, и нередко скульптуру в вию более или менее крупных поперечных ребер.
< Фиг. 434. Tvnjmtes acutus. Верхний девон. Тиман. Р. Ярега-иоль. Обл Коми. 1/2.
Manticoceras (фиг. 431, 432) имеет глубокую наружную лопасть, разделенную широким вторичным седлом, боковая лопасть ограничена широким выдающимся наружн im седлом. Раковина инволютная или со слабо объемлющими оборотами. В первом случае — собственно Manticoceras, в последнем Gephyrocetas. Скульптура иногда в виде поперечных ребер. Девон.
Фиг. 43а. Gastrloceras Jossae. Пермокарбон. Артинский saaoi на Урале.
hmanites (фиг. 434; по нахождению в Тиманском кряже). Сжатая с боков раковина с приостренной наружной стороной. Лопастная линия имеет две боковых лопасти и два седла Верхний девон
Prolecamtes (фиг. 433, lekane—широкая чашка). Раковина с широким пупком, дискоидальная, гладкая или поперечноструйчатая. Седла просты©
узкие, напереди округленные, булавовидные вследствие некоторой суженности при основании Лопасти приостренные. Верхний девон, каменноугольные.
। Gastrioceras (фиг. 435) с оборотами мало объемлющими, трапецои-, дальнего сечения. Поверхность раковины представляет тонкие продольные (спиральные) ребрышки, поперечные струйки (иногда корикие поперечные ребра на боках оборотов) и поперечные пережимы. Сифонная лопасть разделена срединным седлом. Основания лопастей иногда приострены. Каменноугольные отложения, пермокарбон.
Фиг.
Р. Шартымка на
436. Pronorites cyclolobus var. uralensis. Верхнекаменноуг. восточной склоне Южного Урала.
а, b — внутренние обороты; d, е — наружный оборот; с — лопастная линия.
Далее следуют роды с зазубренными, кроме сифонной, лопастями.
Pronorites (фиг. 430; от географического названия). Бока оборотов раковины параллельны между собою, гладкие, лишь со струйками нарастания.
Сифонная лопасть в основании трехзубчатая или разделенная седлом с вырезкой на вершине, первая боковая лопасть двузубчатая, остальные лопасти простые заостренные. Каменноугольные отложения и пермокарбон.
<
J
Фиг. 43S. Medli-cottia artiensts.
Пермокарбон. Хол-динская гора на р.' Косьве. 1/1.
Medlicottia (фиг. 424, 437 и 438; по имени ученого Медликотта). Раковина плоская, подобно тому как у Pronontes. На узкой наружной стороне проходит срединная бороздка, ограниченная с обеих сторон килями, иногда б}горчатыми. Обороты сильно объемлющие, так что пупок очень мал. Сифонная лопасть глубокая и ограниченная высокими узкими наружными седлами с вырезками на боках седел. Все лопасти в основании двузубчаты, седла на боках иногда зазубрены. Пермокарбон.
Cyclolobus (фиг. 439; kuklos — круг, lobus—лопасть). Шарообразная раковина с узким пупком. Имеет
Фиг. 437. Medlicottia ОгЫдпуапа. Пермокарбон. Гора Каткобаш в окрестностях Артинсксю аавода.
поперечные пережимы или ребровидные утолщения и покрыта тонкими поперечными струйками. Седла с боковыми зубцами, а на вершине округленные. Лопасти в основании дву- и трехзубчатые. Пермокарбон.
с
Г
F
Фиг. 439. Cyclolobus stachei. Пермокарбон. Сицилия. Италия.
Фиг. 440. Arcesles -intuslcbiatus. Триас Галыптадт. А львы. Австрия.
Д1
в: л:
G
Arcestes (фиг. 440) отличается от Cyclolobus главным образом сильно рассеченными ачмонитовыми лопастями и седлами. Триас.
В то время как девонские и каменноугольные гониатиты большею частью развиваются в пермском и триасовом периодах в аммонитов, в девоне гониатиты дают две специализированные боковые ветви — цл и м е н и й характерных для верхнего девона, и цератитов — для триаса.
Clymenia (фиг. 441, 442; kllmax— лестница) отличается малообъемлющими ступенчатыми оборотами, положением сифона на спинной стороне их. с сифонными дудками, обращенными назад. Лопастная линия гониатитового характера, но наружная лопасть обыкновенно отсутствует, а на ее месте развивается широкое наружное седло. Верхний девон.
Г
Цератиты отличаются мелко зазубренным основанием лопастей сутурлой линии при гладких седлах
а
Si
фиг. 441. Лопастные линии различных кли-мении (Oxyclymenia, Cymaclymet..a, Gomo-clymmia).
Фпг. 442. Clymewia (Oxycly-menta) undulata. Верхний девон Эльберсреит. Фихтельге-бирге. Германия.
Фиг. 44 J. Нипдатг-tes Raddet. Пермские. Джульфа. Закавказье.
Hungarites (Otoceras) (фпг. 443; Hungaria — Венгрия) имеет заостренный киль на брюшной стороне и стрельчатое поперечное сечение. II. Rad-dei имеет лопастную линию, еще мало отличающуюся от гониатитовой. Пермокарбон—триас.
Ceratites (фиг. 444). Умеренно-навитые обороты, широкий пупок. Поперечное сечение почти квадратное, обыкновенно высота больше ширины. Скульптура состоит из поперечных ребер на боках оборотов, начинающихся бугорками у пупка. Триас.
Нижеследующие аммониты юры и мела перечислены начиная с простейших.
Phylloceras (фиг. 445; phyllon — лист) имеет более или менее глубоко, отчасти тонко рассеченные лопасти; оконечности седел и их разветвлений округленно листовидны. Лопасти и седла миогочис •пенны. Без пупка или с узким пупком. Раковина округленная и обыкновенно гладкая или струйчатая. Юра, нижнемеловые отложения.
Фиг. 444. Ceratites nodosus. Раковинный известняк. Триас. Вюрцбург. Бавария. Германия.
Lytoceras (фиг. 446). Обороты округлены и мало объемлющп, гладки или покрыты простыми, неветвящимпся ребрами. Вспомогательные лопасти отсутствуют или слабо развиты. Концы разветвлений седел неявственно
15
Учебник палеонтологии.
225-
листоваты. Лопасти и седла обыкновенно являются двусторонне-симметрично и двукратно разветвленными. Юра, нижнемеловые отложения.
К Lytoceratidae принадлежат многие, преимущественно меловые роды, представляющие наклонность к образованию развернутой крюкообразной {Hamites, фиг. 447; hamus — рыболовный крючок), прямолинейно вытянутой (К acuities, фиг. 448; baculum — палка) или улиткообразной, башенковой (Тиг-r Hites, фиг. 449; turris — башня) раковины.
Arietites (фиг. 450;aries—
Фиг. 445. Phylloceras heterophyllum. Ee;J ний лгйас. баран). Плоская ДИСКОИдаль-Поркшайр. Англия. * ная, с широким пупком ра-
ковпна, с крупными про-гчыми изгибающимися вперед ребрами. Наружная сторона с килем, ограниченным двумя бороздами. Число лопастей и сегел близко к нормальному.
Фиг. 447. _Ня-mites rolwida-
/ш. Гольт. Фольк-стон. Англия.
Нижний лейас.
В верхнем лейасе отсюда выходит род Harpoceras (фиг. 451; harpe—серп) с серпообразно изогнутыми линиями нарастания, или ребрами, с килем, с умеренношироким или узким пупком. Лопастная линия с большим числом лопастей и седел. Верхний лейас и доггер.
Amaltheus (фиг 452)болыпею частью с узким пупком; снаружи ариостренпый или узловатый киль; бока гладкие струйчатые или с простыми ребрами. Лопасти
Фиг.446. Lytoceras Liebigi. Титов. Шграм-берг. Моравия. Чехо-Словакия.
и седла сильно раз-ветвгены, в увеличенном числе. Лейас
Далее следует значительная группа родов с поперечными реорами, раз-
Д{ляюшимися у округленной в большинстве случаев наружной поверхности раковины. Лопастная линия сильно рассеченная, кроме двух боковых лопа-г1ей имеется еще шовная 1 лопасть, образуемая несколькими неглубокими и в совокупности своей отступающими назад вспомогательными лопастями. .Апертура нередко с боковыми ушками; при вей обыкновенно пережим на
j аковпне.
Stephanoceras (фиг. 425,453, 454; stephanos — корона). Обороты развиваются в ширину больше, чем в высоту. Пупок умеренно широкий (до широкого) и глубокий. Бока у пупка с короткими, относительно толстыми, прямымиреб-рами, образующими бугорки на перегибе к наружной стороне _. . оборотов. От бугор-
! Еа ребро разделяется
на 2—3 более тонких ребра. Пережи-L мов нет. Доггер — ^4-4 Оксфорд.
Фиг. 450. Artetit“s ВисЫапЯл. Нижний лейас . Англия.
Фиг. 448. Баеи-htes anceps. Обломок. Верхние «еловые. Маст-рихт. Лимбург.
Фиг. 451. Harpoccras serpentinum. Верхний лейас. Англия.
Cadoceras (фиг. 460). С узким и глубоким пупком. Наружные обороты низкие и широкие с исчезнувшей скульптурой или ослабевшей, так что на наружной стороне ребра слабо развиты, исчезая на небольшом расстоянии от перегиба к пупку. По направлению наза’Д от жилой камеры скульптура (поперечные ребра) все усиливается, а поперечное сечение оборотов изменяет
1 Швом называется линия, разграничивающая смежные обороты раковины в лупке.
15* 227
i
форму, становясь постепенно высоким и менее широким. Позади апертуры пережим. Келловей и Оксфорд.
Macrocep aides (фиг. 455; macros — длинный, cephak — голову Обороты округленные, довольно высокие и умеренно широкие, сильно объем-
Фиг.
Step} tmewras Humphntsi.
Англия.
Фиг. 452. Amaltkeus mafgarita-/its. Средний лейас. Вюртемберг. Германия.
Фиг. 454. Stephanocera» cwonalum.
Келловей. Франгня. 1/3.
Фиг. 455 Macrocephahles mactoce-phalus. Келловей. Энинген. Вюртемберг. Германия. 1/2.
лющие, с узким пупком. Нет резкого перегиба от пупочной части к наружной, нет бугров и пережимов. Ребра раздваиваются сравнительно недалеко от пупка. Скульптура и форма оборотов изменяется, как у Cadoceras, по в менее резкой степени. Верхний доггер, Оксфорд.
Cardioceras (фиг. 456, 457; сот—сердце). Довольно ипволютные формы с килеватой наружной стороной, покрытые редкими ребрами, которые кнаружи
делятся на две или более ветвей, делающих киль зубчатым; часто еще имеются многочисленные короткие наружные вводные ребра. Оксфорд и ки-
- '
меридж.
Pensphinctes (фиг 458, sphincter— кольцевой мускул). Раковина плоская, большею частью с широким пупком; скульптура состоит из прямых, расщепляющихся (на 2 и более) безбугор-чагых ребер, место деления их близко к наружной стороне. Длина жилой камеры около одного оборота. Бывают ушки и пережимы. Средний доггер—неоком.
Olcostephanus (фиг. 459) отличается от Perisphinctes более короткой жилой камерой, ребрами, расщепляющимися пучкообразно или повторно, с одного боку. Нередко бугры на месте первоначального деления ребер. Верхняя юра и нижний мел.
Далее следуют аммониты, производные от только что рассмотренной группы, отличающиеся возникновением бугров на поперечных ребрах и иногда слабым развитием частей ребер между буграми. Именно ребра несут большею частью околопупочные и краевые наружные бугры и прерываются на гладкой, бескилевой наружной поверхности более или менее широкой бороздой. Число вспомогательных лопастей 1—2.
Фиг. 456. Cardioceras alternans. Оксфорд. М«р.ввики, около Москвы. 1/1.
Фиг. 457. Cardoceras cordatam. Оксфорд. Д. Никитина на р. Оке. Рязанский район. 1/2.
Фиг. 458. Perisphincte.4 Nikttiiu. Нижневолжский ярус. Кашпур на р, Волге, Средне-1 илжскиь район. 1/3.
Cosmoceras (фиг. 462, 463). Пупок невелик, пережимов нет. Ребра многочисленные, тесно сближенные, вилообразно расщепляющиеся; у наружной борозды, а часто и у пупка п в месте раздвоения ребер находятся бугры или шипы. Доггер—неоком.
Schloenbaehia (фиг. 461; по имени ученого Шленбаха). Раковина с довольно широким пупком. Наружная сторона с острым, редко расчленяющимся в бугры килем, который при устье образует выдающийся отросток, иногда отгибающийся назад. Сильно выраженные простые или раздваивающиеся ребра, часто снабжен
ные бугорками. Лопасти мало разветвленные, иногда почти цератнтового
характера. Меловые отложения.
I Hoplites (фиг. 464, 465; hoplites—тяжело вооруженный греческий воин). Ребра, изгибающиеся наперед, образующие бугры или бугровидные утолщения у
пупка или на краю наружной стороны, последняя иногда с бороздой. Пупок умеренной величины, пережимов большею частью нет, место деления ребер
Фиг. 461. Cosmoceras ornatum. Кел-ловеи Вюртемберг. Германия.
Фиг. 462. Cosmoceras Dunvam. Верхний келловей. Свистово, ь районе Рязани
Фиг. 459. Olcottephanus virgatus. Ilua: неволжсъ.иа яр,с. Мневники около Москвы J/3
Фиг. 460. Cadoceras Elatmae. Нижеий «елловей. Елатьма на р. Оке, в paBoi о Тамбова 2/3.
Фиг. 463. ScMoeribachta cri 'ata. Го4ьт Франция.
ближе к пупку, чем у PerispMnctes. Шовная лопасть не отступает назад, 2-я боковая очень мала Титон нижний мел.
rAcanthoceras (фиг. 466; akantha—шип). Ребра простые или раздваивающиеся, прямые, утолщающиеся кнаружи, с боковыми и краевыми буграми. Наружная сторопа широкая, со срединным рядом бугров .Мел.
К Cosmoceratidae примыкают, подобно тому как к Lytoceratidae, некото-
иг. 464. Hoplites kirghisensis.
Сакмарскин городок близ Оренбурга. 3/4.
Кимеридж.
Фиг. 465. Hoplites eudoxus. Кимеридж.
Городище на Волге, врайоне Ульяновска.3/4.
Фиг. 466. Acanthoceias Rhowmagense. Сеноман. Франция.
Фпг. 46". Cnoceras sAnbirs-kensis. Нижнемелов. Ульяновск. 2/3.
рые побочные формы с более или менее развернутыми оборотами раковины, распространенные преимущественно в нижнемеловых отложениях.
Cnoceras (фиг. 467; crion—баран, keras—рог) имеет раковину, завивающуюся в одной плоскости, состоящую из немногих не соприкасающихся оборотов. Поверхность с поперечными ребрами, редко расщепленными, с буграми и шипами, обособляющимися на ребрах. К собственно Crioceras относят формы со спирально свернутой раковиной, к Ancyloccras (фиг. 468)—
с последний оборотом, сначала прямолинейно удлиненным, а затем крюко-образно изогнутым: Scaphites (фиг. 469; scaphe—лодка) отличается раковиною с сильно объемлющими оборотами и, вследствие этого, узким пупком.
Подкласс II. Двужаберлые, Bibranchiata. •
К двужаберным принадлежат наиболее общеизвестные ныне живущие головоногие, как осьминоги и каракатицы. Двужаберные имеют две жабры, раковину, скрытую под мантией, редуцированную, рудиментарную, или они
Фиг. 468. Aacylo-ceras Malhooma-пит. Неоком. Франция.
Фиг. 469. Scaplntes equalis. Сеноман. Франция.
бычи; лишь вздутые концы их имеют
совершенно лишены раковины; в случае существования раковины она служит не органом внешней защиты тела животного, как у Tetra-branchiata, но внутренней опорой для мантии и плавникообразных придатков. Руки в числе восьми или десяти, воронка сплошная, без разоеза на боку, и чернильный мешок. Челюсти роговые.
ИОРЯДОК I. ДЕСЯТИ НОГПЕ, DECAF 3DA.
Из десяти рук две, д тинные и сократимые, служат для ловли до-восемь остальных ру к
коротки, снабжены присоскам*! или крючочками по всей длине с внутренней стороны. Всегда имеется внутренняя раковина.
У древнейших представителей десятиногих, распространенных в мезозое и вымерших к концу его, раковина отличается сравнительно большими размерами, отчасти соответствует раковине четырехжаберных и, подобно тому как у последних, в молодости, должно быть, заключала в себе тело животного, в зрелом состоянии, обратно,—раковина заключалась в теле.
У Belemnites (фиг. 470—473, 475; belemnon—копье) часть раковины, первоначально возникающую в индивидуальном развитии, представляетphrag-moconus, сходный с раковиной Orttioceras, за каковую иногда может быть принят по первому взгляду. Phragmoconus представляет полый прямой конус, разделенный вогнутыми перегородками на камеры с краевые сифоном; имеет шарообразную известковую первичную раковину. Стенка фрагмоконуса представляет на спинной стороне пластинчатое продолжение, значительно выдающееся вперед, известное под названием proostracum. В последней, наиболее значительной камере фрагмокону са, должно быть, помещалось молодое животное; затем развивалась вторая часть раковины—rostrum, облекавший напереди фрагмоконус. который помещался в коническом углублении ростра—а л ь в е о л е. Образование ростра начиналось на заднем конце фрагмоконуса, отсюда ростр возрастал главным образом назад, но нарастал и
вперед, постепенно облекая фрагмоконус так, что наконец последний становился почти весь заключенным в альвеоле ростра. В эту пору животное переставало быть заключенным во фрагмоконусе, мантия распространялась на бока и назад, отлагая ростр, на поверхности которого замечаются впечатления кровеносных
Ph
сосудов мантии. Ростр представляет плотное известковое образование и обыкновенно только он и сохраняется & ископаемом состоянии, реже фрагмоконус и еще реже proostracum. Ростр бывает цилиндрической, конической, веретенообразной формы. Задний конец его при-острен. Длина доходит до 40 см, диаметр альвеоли также достигает размера в несколько дюймов. Ростр нарастает
Фиг. 471. Belem-mtes Panderi. Оксфорд. Ниже Костромы на Волге.
Фиг. 473. Реставрация -животного с нанесенной раковиной белемнита. Вид с брюшной стороны.
Фиг. 470. Продольный разрез раковины белемнита.
ростр, Ph — фрагыо-ковус; и—первичная »-а-мера; st—сифон; с—камерная, лишенная стенки часть фрагмоконуса.
Фиг. 478. Belem-mtes bipartitus. Неоком. Франция.
0®
елоями, отлагаемыми концентрически первичной раковины фрагмоконуса к
вокруг осевой линии, проходящей от приостренному концу ростра; ось эта расположена не центрально, но ближе к нижней поверхности ростра. В связи с ростом ростра преимущественно назад, слои нарастания его более тонки впереди и утолщены назади. Слои состоят из фибр, призм кальцита, направленных перпендикулярно к поверхности слоя и, в общей сложности, расходящихся радиально от осевой линии. Поверхность ростра гладкая или зернисто-бугорчатая, иногда имеет продольную бороздку на нижней (реже на верхней) поверхности, а также боковые бороздки.
Собственно Belemnites (фиг. 471, 472) встречается в юре и мелу.
Триасовые представители белемнитов отнесены к особым родам, более примитивным, более близким по раковине к Tetrabranchiata. Именно раковина триасовых белемнитов отличается от раковины позднейших слабым
р швитием ростра сравнительно с фрагмоконусом.
Верхнемеловой род Belemnitella (фиг. 474) отличается от Belemmtes щелевидным отверстием на передней части брюшной поверхности стенки альвеоли. Со'1тветст-
Фиг. 474. Belemnitella muc^nala. Се-вон. Вестфалия. Гер-
Фнг. 475. Продольные разрезы раков ш: a—Belemnites, Ъ—Belosepta, с—Sepia. Во всех разрезах р. о.—proostracum, si—сифон и д—ростр. Стенка камерной части и псре-
венно фрагмокон;с имеет на стенке выдающееся продольное ребро. Кроме того, на спинной
*
Фиг. 476. Раковина ныне живущей Sepia officinalis с брюшной
стороны.
мания.
гооодкн показаны жирными линиями.
«стороне фрагмоконуса также замечается обыкновенно ребро, которому соответствует бороздка в альвеоле. Цилиндрический ростр оттянут на заднем конце в тонкое острие.
Род Belosepia (фиг. 475, &), из эоцена очевидно, является произвсд-йым от мезозойских белемнитов. Здесь ростр имеет уменьшенные размеры, перегородки фрагмокон; са протягиваются в нем боод вперед от расширенной полости сифона к спинной стороне. Они многочисленны и сближены. Этот род, как и выражено его названием, от белемнитов приводит к каракатицам (Sepia), живущим ныне и встречающимся в ископаемом состоянии в третичных отложениях.
Раковина каракатицы (фиг 475, с. 476), так называемая os sepiae, заключена в мантии животного на его спинной стороне (фиг. 404). На зад
нем конце ее находится острие, представляющее совершенно рудиментарный ) остр Proostracum 2 ваяется в виде широкой пластинки, находящейся на спинной стороне; с брюшной стороны он i покрыта рыхлым известковистым отложением, в основе состоящим из тонких пластинок, соответствующих перегородкам, еще явственно различимым у Belesepia. 1
ПОРЯДОК II. ОСЬКИНО ГИК, OCTOPODA.
Имеют лишь восемь одинаковых длинных рук, не имеют раковины—за исключением Argonauta, у которого раковина наружная Отпечатки формы, близкой к ныне живущим осьминогам, известны из верхнемеловых отложений Ливанских гор.
Argonauta (фиг. 477, 478; аргонавты—мифологические мореплаватели) имеют раковину лишь у самок. Она спирально свернута, как у Tetrabran-
Фиг. 477. Животное Argonauta (ныне живущее) с раковиной, А — расширенная лопасть задней рука, В — раковина, С—воровка, D—передние руки.
Фиг. 47&. Раковина Argonauta.
chiata, но не имеет внутренних перегородок и отлагается не столько мантией, сколько расширенными концами двух задних рук; в ископаемом состоянии известен из плиоцена.
^аспросгпранемие головоногих.
Nautiloidea. Встречаются, хотя редко, уже в кембрийских отложениях,— Volborthella, Orthoceras и др. До окончания эпохи нижнего силура прямые и югнутые формы преобладают над спирально-свернутыми. В верхiем силуре Nautiloidea достигают максимума своего распространения. В силуре встречаются особенно часто Orthoceras, Endoceras, Lituites. В каменно-угольн > х—триасе в изобилии встречаются виды и подроды Nautilus, с юры остается лишь собственно Nautilus, притом лишь виды с гладкой, без круп ных ребер и бугров, и инволютной раковиной. Формы с кольчатыми поперечными украшениями известны в нижнем силуре, несколько позже - с продольными ребрами, а также ребристо- и бугорчгтобокие формы.
Наибольшее разнообразие в отношении строения сифена—в нижнем силуре, в отношегпи обшей формы—в силуре и сутурной линии—в триасе.
Ammonoidea. Го татиты известны с верхнего силура, распространены главны!" образом в девоне—Tornoceras, Manticoceras, Timanites, Bact-rites -и каменноугольных—Gastrioceras.
Кишении ограничены в распространении девоном, цератиты встречаются в триасе, редко в пермских.
Геологическое распространение головоногих.
Роды Отделы или ярусы
Кембрии Силур Девон Карбон Пермь Триас Юра Мел Третичные 1 эиннэк -odaoQ
Orthoceras . . FoZiorZ/ieZZe . Endoceras . . Gomphoceras . Phragmoceras Estonioceras . L Unites . . . Nautilus . . . Eactrites . . . Prolobites . . Agoniatites Pinacites . . Tornocerasi . . Prolecanites . Gasirioceras . Pronorites . . Medlicottia Cyclolobus . . Areestes ... Clymenia . . Hungarites . . Philloceras . . Lytoceras . . Ariectites . . Harpoceras . . Amalthens . . Stephana eeras Cadoceras . . Macro cepha lites Cardioceras Perisphinctes . Olcostephanus Cosmoceras Schloenbachia Hoplites . . . Aeanthoceras Crioeeras < .
—
ч
—
—
Аммониты известны с каменноугольных. Триасовые семейства, как Агсе-stidae, Ceratitidae и др., за исключением Phylloceratidae, не переходят в юру и заменяются в последней новым.! формами. Исключительно в нижнем лейасе встречается например Arietites, в среднем и верхнем лейасе являются Наг-poceras, Amaltheus, Lytoceras и древнейшие формы Stephanoceratidae. Эти семейства проходят и в остальные отделы юры. Прибавляются Cosmoceratidae. На границе юры и мела снова происходит значительное изменение фауны . аммонитов. Наиболее стойкими являются Phylloceratidae и Lytoceratidae, из Stephanoceratidae вымирают уже в нижнем мелу Perisphinctes и Olcoste-ptianus, на место Cosmoceratidae яв !яются Soplites, Acanthoceras. Характерным для мела является значите пятое развитие более или менее развернутых форм, как Hamites, Baculites, Crioceras, Ancyloceras. Представляя максимум распространения в лейасе, аммониты разом вымирают в конце мелового периода. Отличаются большим горизонтальным распространением, распространены преимущественно в отложениях глубоководных, глинистых и । звштковистых, реже встречаются в образованиях прибрежной фации.
Dibranchiata более многочисленны и разнообразны в современной фауне, чем в прошлые геологические периоды. Некоторые формы принадлежат к фауне пелагической, другие—к»абиссальной.
Типичны для береговой фации осьминоги и каракатицы. Dibranchiata не звестны в палеозое, древнейшие представители их появляются в триасе. Белемниты распространены главным образом в юре и в мелу, особенно часто в глинистых породах.
Двужаберные сравнительно редки в эоцене.
КЛАСС IY. ПЛАСТИНЧАТОЖАБЕРНЫЕ, LAMELLIBRANCHIATA.
(Acephala, Bivalvia, Pelecypoda).
Пластинчатожаберные моллюски имеют тело с внутренней и внешней симметрией. Головная область тела зачаточна, и мантия, разделяющая я на правую и левую лопасти, выделяет двустворчатую раковину, которая покрывает и заключает в се >е все тело (фиг. 479).
Головная область снабжена с каждого бока парой губных щупалец. Нога, без плоской подошвы, находится на брюшной стороне, обыкновенно /приспособлена к тому, чтобы зарываться в почву. Мантийные лопасти соединены одним или двумя мускулами, расположенными поперек тела и смыкающими створки, к коюрым они прикреплены. Под мантией находятся две боковые симметричные гребенчатые жабры. Пластинчатожаберные лишены radula и питаются микроскопическими организмами; последние приносятся током воды, возникающим от действия ресничек, расположенных на некоторых мягких частях тела. Ротовое отверстие находится на переднем конце тела, анальное—на заднем. Раковина состоит из двух выпуклых створок одинаковой или неодинаковой величины (равностворчатая, неравностворчатая). Относительное положение частей раковины определяется, рассматривая животцое сзади и сверху. Вправо от плоскости соприкосновения створок (обыкновенно являющеюся и плоскостью симметрии) при этом будет правая створка, влево—л е в а я. Раковипа покрыта всегда концентрическими линиями нарастания ее. При верхнем крае выдаются исходные точки роста раковины — макушки. Длиною раковины называется
наибольшая прс^женность ее спереди назад, высотою — сверху вниз, тол-щ и н о ю—наибольшее расстояное между наружными поверхностями створок. Края раковины называются: передним, задним, нижним (или брюшным), верхним (спинным или замочным).
о
t
Фиг. 479. Муа arenaria. Ныло живущая. Обращена головною часты» вниз, брюшною направо. Левая створка, мантия и половина сифонов удалены.
а — передний, аг — задний замыкающий мускул; Ъ — внутренностный мешок; с-клоакальная камера, и которую открывается анальное отвер стие; f — йога; д — жабры; h—сердце; т — разрез края мантии; о — рот; р—бахромчатость края маитпн; s — жаберный сифон; "s' — аиальвын сифон; t—губные щупики; «—анальное отверстие. Наверху стрелками показаны входнщий н выходящий ток воды.
myaria). Передний мускул г (одномускульные, Monomyaria),
Открывание и закрывание раковины совершается посредством органов, действие которые на раковину противоположно: таковыми являются связка и замыкающие мускулы.
Замыкающие мускулы первично являются в числе двух: переднего, находящегося около рта, и заднего—около анальниго отверстия. Сжимаясь, укорачиваясь, мускулы закрывают створки. Их места прикрепления являются на створках в виде углубленных впечатлений.
Лонасти мантии соединены между собою всегда на спинной стороне тела и иногда в других частях их периферии. Иногда края лопастей мантии назади сложены или срастаются так, что образуют раструбы, служащие для входа и выхода воды, называемые сифонами. Мантия плотно прилегает ко внутренней поверхности створок и на периферии их обыкновенно прирастает к ним по некоторой линии. Эта линия оставляет впечатление на внутренней поверхности створок в виде непрерывной линии, называемой мантийной. Вне мантийной линии край епанчи свободен и иногда снабжен примитивными органами зреиия. Концы мантийной линии обыкновенно примыкают к отпечаткам закрывающих раковину мускулов (фиг. 480), расположенных: один—на переднем, другой на заднем конце тела. В случае сколько нибудь значительного развития сифонов (верхний—анальный, нижний—жаберный) они втягиваются вовнутрь раковины мускулами, берущими начало в мантии; чем больше сифоны, тем более и эти мускулы и тем более они вдаются вовнутрь, делая мантийную линию выемчатою, с синусом (фиг. 480). Синус отсутствует лишь у очень немногих из имеющих сифоны форм, которые лишены способности втягивать сифоны в ракогмну.
Кроме впечатлении замыкателей, на раковине имеются еще над ними или между ними меньшие впечатления ножных мускулов.
Замыкателей бывает два или один- Если два, то они бывают или одинаковой величины, или различной (равно- и разномускульные пластинчатожаберные, Isomyaria и Hetero- или Anise -изменяется в размерах до полного исчезновения причем эти изменения обусловливаются
механическими условиями его действия в связи с относительным положе-
нием мягкого тела животного
При равномускульности линия, соединяющая впечатления замыкателей в одной и той же створке, параллельна краям замочному и брюшному; при разномускульности Ва линия пересекает замочный край под углом, причем меньший передний мускул всегда более близок к замочному краю, чем больший задний, и величина переднего мускула тем меньше, чем он ближе к замочному краю (фиг. 481, В, D, F).
Задней мускул, напротив, удаляется от замочного края и увеличивается в размерах в то время, как передний уменьшается.
Указанное изменение положения мускулов связано с поворотом мягкого тела в раковине, происходящем так, что рот, подобно переднему замыкателю, приближается к замочному краю, а анальное отверстие, подобно заднему замыкателю, удаляется от замочного края. Мускулы действуют, всегда преодолевая препятствие со стороны эластичной, как пружина, связки, находящейся у замочного края, подобно тому как это имеет место у Ostracoda. Чем дальше расположен мускул от замочного края, тем более, так сказать, плечо рычага, на который мускул действует, тем сильнее действие мускула; и обратно, чем ближе мускул к замочному краю, тем менее его действие. В послед-
в раковине.
Фиг. 480. Meretrix chwne. Ныне живущая. А—вид обеих створок со спины. В — внутренность левой
CTBOpi.H.
а — передний; р — задний край; d — спинной; v — брюшной 1и—луночка; и — макушка: 2— связка; аа —впечатление перед него замыкателя; ра — заднего; pl—мантийная линия s—ее-синус; ж, у — кардинальные зубы; я — передний боковой зуб
Фиг. 481. Правые створки четырех различных пластинчатожаберных, с различно развитыми мускулами-замыкателями. А — Pectuncuhis, В — Муппаг D — Modiola, F — Pecten.
а — передний замыкатель; р — задний; ar, рт, рт* — пожные мускулы; Ъ — маСушка; И—связка; pi — маитинная линия t — вубы замка.
нем случае полезное действие мускула невелико, и он уменьшается в размерах, а затем атрофируется, как находящийся в невыгодных условиях действия. У равномускульных форм мускулы расположены симметрично
в отношении замка, действуют одинаково и равны, у одномускульных форм конечным этапом в развитии единственного мускула является занятие им в створке центрального или почти центрального положения, при котором он действует равномерно на всю окружность створок.
Связка (фиг. 480) состоит из двух частей: наружной—темной, подобно наружному слою раковины состоящей из конхиолина, и внутренней— светлой, упругой как хрящ, содержащей кроме органического вещества известь. Связка, сокращаясь, стремится сблизить замочные края обеих створок, заставляя их вращаться, как на шарнире, около оси обыкновенно цилиндрического тела связки. Мускулы, закрывая створки, противодействуют сокращению связки, растягивают ее поперек. При толстой связке, впрочем, лишь наружные (т. е. наиболее далекие от тела животного) слои ее растягиваются при действии мускулов, а внутренние при этом сжимаются, так что когда связка открывает раковину, то наружные слои связки действуют сжимаясь, .а внутренние расширяясь. Между сжимающимися и расширяющимися слоями связки будет граница, по которой не происходит ни сжатия, ни расширения. По этой-то границе и происходит обособление обеих частей связки.
Первоначально связка не представляла обособленной части раковины; раковина на первых стадиях своего развития, так же как вероятно и у древнейших пластинчатожаберных, состоит лишь из рогоподобного конхиолина и имеет блюдцеобразную форму. Затем эта раковина под действием возникающих мускулов надламывается посредине, так что образуются две полукруглые створки с прямым замочным краем, объизвестняющиеся за исключением соединительной части, которая представляет первичную связку. В этом состоянии связка
не является ни внутреннею, ни наружною, каковые типы связки развиваются в дальнейшем историческом развитии пластинчатожаберных (фиг. 480, 486). Бывает в связке одновременно наружная и внутренняя части, как у Mactra (фиг. 482), и вообще даже наружная связка является внутреннею в части своей, которой она примыкает к мягкому телу и наращивается.
Первичная связка находилась под макушками как впереди, так и позади их. При дальнейшем развитии она сосредоточивается позади макушки.
Наиболее совершенным типом связки является такой, когда она, располагаясь позади макушек, представляет полуэллипсоид, длинными краями прикрепленный к створкам или к особым выступам их, называемым связочными нимфами. Другая форма связки, это — сплющенный тяж, прот гивающийся от одной макушки к другой [Lm, Spondylus, фиг. 484), с длинною осью, перпендикулярною к плоскости разделения створок; этот тип сосуществует с area, находящейся под макушками, простирающейся по обе стороны их. Наконец третий тип представляет связку, состоящую из нескольких отдельных частей, расположенных в отдельных углублениях area, находящейся под макушками; этот тип развивается из предыдущего— сложная связка (фиг. 503), называема! так в отличие от простой вы неописанных типов.
Фиг. 482. Околозамочная часть левой гтворки ныне живущей Mactra. •а — внутренняя, Ь — наружная часть связки.
В индивидуальном развитии сложная связка исходит из простой связки.
У молодых Perna связочные ямки постепенно увеличиваются в числе. Сначала имеетсг одна ямка, как у Avicula, зчтвм выделяющий связку участок спинного гребня мягкого тела разделяется на две обос<б юнные части, отлагающие каждая отдельную часть сложной связки, и т. д. Нечто подобное происходит у Arcidae, у которых связка помыкается в нескольких коленчато-согнутых бороздках. Каждая из последних начинается одной ямкой, cootbi т етвующей вершине колена, затем, вследствие раздвоения отлагающего связку участка мягкого тола и все растущего удаления полученных раздвоением частей, возникают все бодее расходящиеся колена.
Связка с area, простирающейся как вперед, так и назад от макушек является типом более древним, чем связка, сосредоточивающаяся исключи
тельно позади макушек.
Когда связка находится вполне позади макушек, часть area, находя щаяся впереди их, обособляется в так называемую луночку (фиг. 480 483), позади же макушек связка занимает тогда лишь часть щитка
подобного луночке, но более значительных размеров (фиг. 483).
Замок. Подобно тому как у брахиопод замочные края створок раковины пластинчатожаберных утолщаются, расширяются в замочную площадку, и на них развиваются пластинчатые пли бугровидные выступы —• з у-б ы. У пластинчатожаберных зубы и зубные впадины развиваются в обеих створках; в каждой створке зубы расположены, чередуясь
ка и связочные нифмы Astarte Duboi-si. Нижневолж-скпе слон. Хо-рошово, около Мобквы.
Фиг. 484. Связка Span-dylus. Справа прикреплев-вая створка, посредине area которой от макушки протягивается желобок, имеющий ввязку.
с зубными впадинами.
У наиболее примитивных пластинчатожаберных не существует замочной площадки, и замочный край подобен остальным краям, между прочим и в том отношении,
чту к нему, как и к ним, от ма-
ку'шки поверхности area, под макушкой находящейся, доходят радиальные ребрышки, часто покрывающие поверхность раковины у пластинчатожаберных, рти ребра делают раковины зубчатыми, что способствует более прочному соедьнеяню етж рок при закрытии их. Эти зубцы под действием естественного отбора усиленно развиваются там, где действие их бо^ее постоянно, т. е. на том краю, где створки и при открывании не вполне отходят одна от другой,—на замочном краю. Раз начавшись, развитие зубов шло неуклонно, они остались на замочном краю и тогда, когда ре'ра, первоначально имевшиеся между макушкой и замоным краем, исчезли с этого пространства.
Изучение индивидуального развития раковины пластинчатожаберных свидетельствует, что зубы — если и не у всех, то у большей части пластинчатожаберных — онтогенетически развиваются из образований, представляющие ребра внутренней поверхности раковины.
Ребра на внутренней поверхности раковины вообще бывают у пластинчатожаберных, хотя много реже, чем на наружной поверхности Эти вну
16
У «ебни аямоктологаа
2*1
тренние ребра правильно чередуются с наружными, так что те и другие представляют как бы складки гофрированной раковины. В случае присутствия внутренних ребер ясно, как те из них, которые расположены при замочном крае, при развитии раковины дают начало зубам. Таким образом онтогения пластинчатожаберных, так же как и геологически древнейшая дчауна их, согласно свидетельствуют об указанном происхождении зубов замка.
Обыкновенно внутренние ребра отсутствуют, а первичные зубные пла-
стинки в индивидуальном развитии часто утрачивают первоначальную пластинчатую форму. Так (см. фиг. 485) из первичных зубных пластинок обособляются кардинальные зубы, лежащие поперек замочной площади и радиально расходящиеся от макушки; эти зубы обособляются именно из части зубных пластинок под макушками, представляющей оюгну-тость вверх или вниз; из частей первичных зубных пластинок, отдаленных от макушек, в этом типе обособляются латеральные зубы.
Фиг. 485. А и D — развитие замка Lutina neglecta. Стадия D не непосредственно следует за А. Налево — левая, направо — правая створка. В А-первичные зубные пластинка I(LA)II, Л1\ъ D обособившиеся из нпх зубы гетеродонтного замка, кардинальные 2, За, 36, 45 и латеральные, передние — LAI, LAII, и задние—LPI, LPI1 (LAI в приводимом примере не дает каршиадьных зубов, целиком образуя латеральный). L — связка.
Различают следующие типы замка:
А. Дизодонтный (или плагибдонтный, т.-е. несовершенно- иля косозубый), состоящий из внутренних ребер или первичных зубных пластинок, иногда укороченных (вплоть до п чезновения) в зрелом возрасте ио сравнению с молодыми стадиями индивидуума (фпг. 495, 5иЗ); замочная площадка (расширение замочного края) часто не развита.
В. Таксодонтный, состоящий из многочисленных одинаковых зубов (фиг. 486). Название указывает на «стройный» ряд зубов.
С. Гетеродонтный (разнозубый) — из небольшого числа средних кардинальных зубов (расходящихся ит-под макушки поперек замочного края), а иногда и боковых, латеральных (параллельных краям раковины). Этот тип замка представляет подразделения на:
1) . собственно гетеродонтный тип (фиг. 485),
2) шизодонтный (расщепленнозубый)'—срединный зуб одной створки разделен на две ветви, направленные одна вперед, другая назад;
в другой створке нет зуба, соответствующего промежутку между этими ветвями (4b7),
3) пахиодонгный (толстозубый), состоящий из немногих толстых согнутых или шиповидных, главным образом кардинальных зубов (фиг. 535, 537). н наконец.
D Десмодонтный (связочнозубый), отличцощийся слабым развитием зубов, вследствие зияния раковины назади, а часто и напереди. Животное, помещаясь в сделанном им углублении почвы, не имеет надобности плотно закрывать раковину для безопасности. О другой стороны, герметическое закрывание является и невозможным, по крайней мере назади, где находятся сифоны. При зиянии раковины являются бесполезными и не развиваются латеральные з)бы, служащие направляющими при движении створок во время закрывания. Связка же развивае1ся хорошо, как это вообще бывает прп слабом развитии зубов, так как в этом случае лишь при таком развитии связки створки будут достаточно прочно соединены.
Фиг. 486. Таксодонт-вый замок и внутренняя связка. Nucula. 2/1.
Фиг. 487. Шизодонтный замок. Trigoiaa.
Существуют воззрения двоякого рода относительно происхождения замка десмодонт-ноготпла (фиг. 541—543).
По мнению одних, десмодонтный замок представляет специализированную разновидность замка гетеродонтного. Связка десмодонтвых пластинчатожаберных часто помещается на ложковидном отростке под макушкой одной створки, подходящем под углубление макушечной части другой створки. Ложковидный отросток считается за видоизмененный зуб створки. Согласно другому воззрению, выступы и впадины замочного края створок Desmodonta не чередуются в одной створке и не соотвеютв'ют друг друге в противоположных створках, как они чередуются и соответствуют у Heterodonta По атому воззрению, зубовидные выступы у впадины замочною края у Desmodonta являются образованиями вроде первичных зубдов края раковины пластинчат жаберных, а не развиваются из первичных зубных пластинок. Вышеописанное расположение связки аа зубовидном отростке, считающееся характерным для Desmodonta, являете! вторичным, возникает у форм например Муа\, пок >ящихся на почве в боковом положении, на одной створке причем вообще створки развиваются неодинаково, а на верхи й развивается зубовидны i отросток, как бы свешивающийся вниз и представляющий видоизменение лигаментной евнзрчвои нимфы.
С каждой стороны внутренностного мешка, над ногою и обыковенно па протяжении от околоротовых щупалец до задних замыкающихся мускулов, находятся жабры.
В обшей отдельная жабра состоит из продольного ствола, от *отор< го с обоих боков отходят листоватые или нитевидные ответвления. В более
архаических формах (Nucula) эти ответвления пластинчаты, соединены между собой лишь посредством несущего их ствола.
Следующими этапами на пути усложнения строения жабер являются жабры с нитевидными ответвлениями главного ствола и жабры сетчатого сложения, возникающего вследствие соединения нитей поперечными перемычками
Все пластинчатожаберные — животные водные. Большинство из них живут в море, но некоторые семейства проникли в пресные воды. Питаются микроскопическими организмами, главным образом водорослями. В общем пластинчатожаберные являются животными, зарывающимися в ил дна, на котором они живут. В зависимости от свойства дна изменяется положение животного на нем, изменяется строение мягкого тела животного и форма его раковины.
Первично древнейшие пластинчатожаберные, происшедшие, подобно первичным гастроподам, от Amphineura-ао^обпых моллюсков, должны были иметь плоскость соприкосновения створок перпендикулярную, к почве (орто-конхные формы). Тело их необходимо симметрично в отношении этой плоскости. Наиболее примитивны покоящиеся на брюшном краю (Nucula), Таковые лишены сифонов, створки их вполне закрываются, организация мягкого тела очень примитивна; хотя они уже не передвигаются ползанием, подобно гастроподам и амфиневрам, но нога их по форме еще приближается (Nucula) к ноге гастропод более, чем у других пластинчатожаберных. Сюда принадлежат свободные формы (Nucula) или прикрепленные к месту пребывания (Агсл). Прикрепление происходило у пластинчатожаберных, живущих в полосе прилива и отлива и смежно с ней, там, где вследствие приб >я, приносившего изобильную пищу, животному не было надобности передвигаться в поисках пищи.
Животное прикрепляется к скалам, к обломкам раковин и т. п. посредством byssus— пучка нитей, представляющих продукт выделения желез ноги (может быть, аналогичных слизеотделительным железам брюхоногих); это выделение твердеет и аггломерируется с посторонними предметами.
Производными от описанной группы с горизонтальной продольной осью тела являются пластинчатожаберные, живущие зарывшись головной частью вниз, с вер шкальной продольной осью. Одни из них — бесспфонные или иногда представляют зачатки сифонов (Unionidae, Astartidae, Cyrenidae, Cyprinidae); другие имеют развитые сифоны.
Сифоны развиваются у форм, зарывающихся глубоко, вероятно в зависимости от рыхлости почвы, причем животное естественно уходит вертикально в почву передним концом, на котором находится нога. Погружение в почву, в ил может происходить и просто вследствие собственного веса животного.
Невидимому всегда вторичным является положение животного на боку, на одной из створок, плоскость соприкосновения которых тогда бывает параллельна поверхности дна (плевроконхные формы). Это происходит, м<>жет быть, при переходе от мягкого морского дна к твердому, в которое нельзя углубиться, тогда животное естественно ложится на тот или другой бол. При этом пластинчатожаберные бывают с уплощенной верхней створкой, приобретающей форму крышечки, и углубленной нижней створкой; плевро* конхные пластинчатожаберные бывают свободны или прикреплены. Обыкновенно прикрепляются прй высокой температуре морской воды, мало*
глубине я волнении: при высокой температуре животное производит в изобилии известь, малая глубина связана с волнением, а последнее обусловливает прикрепление (Chama, Diceras, Rudistae), отчасти происходящее и потому, что вследствие значительного веса раковины животное испытывает затруднения в передвижении, остается жить все на одном месте. Прикрепление увеличивает прочность, почему прикрепленные и выживают. Прикрепление имеет последствием образование формы раковины, округленной в плане, вследствие одинаковости окружающих условий со всех сторон, и углубление прикрепленной створки.
Животное, подвешенное к крышечкообразной верхней створке, представляет отвисающую вниз массу тела, соответственно которой внутренняя поверхность нижней створки формуется вогнутой. Нижняя створка иногда становится коралловидно-кони ческой и в нижней части своей внутри представляет ряд вогнутых днищ, лежащих одно над другим и развивающихся по мере удлинения створки, отделяя нижнюю часть створки от верхней, занятой мягким телом (рудисты, Hippurites).
Все вышеизложенное относится к Isomyaria.
Anisomyaria— ортоконхны; плоскость соприкосновения створок является плоскостью симметрии. Все ныне живущие — и, вероятно, и ископаемые — Anisomyaria снабжены биссусом, по форме клиновидны, подобно Mytilus— митилусообразяы.
Если представить себе Isomyaria, снабженных биссусом, вроде Area, прикрепленными в прибрежной полосе, где существует значительное волнение воды, то, вследствие выхода биссуса ближе к переднему концу тела, он будет оказывать давление на передний замыкающий раковину мускул, затрудняя его развитие, оттесняя его всё более' к макушке. В связи с этим уменьшится напереди и мантия, а потому раковина получит суженную напереди, мптилусообразную форму, характерную для Anisomyaria.
Mouomyaria все плевроконхны, сифоны у них конечно отсутствуют. Те из них, которые менее удалились от исходных для них Anisomyaria (Avicula и некоторые Pecten), имеют еще отличительный для последних byssus. Теряя его, они становятся свободными (Pecten) или прикрепленными раковиной (Ostrea).
Пластинчатожаберные передвигаются посредством своей ноги, зарывая ее в ил и затем подтягивая на ней, как на якоре, свое тело, или передвигаются довольно быстро попеременным открыванием и захлопыванием створок (Lima, Pecten)-, некоторые передвигаются, с силою выбрасывая воду через анальное сифонное отверстие.
В общем пластинчатожаберные преобладают на небольших глубинах, но некоторые морские формы и сзоем распространении доходят до глубины почти 850 м.
ПОРЯДОК I. ТАХОВОЛТА
Равномускульные пластинчатожаберные, с цельной, редко слабо выед-чатой мантийной линией. Силур — ныне. Исключительно морские
Уже с силура подразделяются на две группы: Агсасеа и Nuculacea. Первые имеют между макушкой и замочным краем треугольную area с наружной связкой, а зубы’часто пластинчатые и параллельные замочному краю. Nuculacea имеют короткие зубы, расположенные поперек замочной
площадки, связку наружную или внутреннюю, сосредоточивающуюся непо-
Macrodon (488; macros— длинный, odous — зуб) имеет зубы удлиненные и горизонтальные назади, короткие и косые впереди. Раковина с прямым замочным краем, удлиненная впе-
ср дегвенно под макушками.
Фиг. 488. Macrodon Keyserluigt. Оксфорд. Д. Новоселки на р. Оке. Рязанский край.
редн^-заднем направлении, с макушками в передней части. От девона до третичных, главным образом в каменноугольных.
Cyrtodontarci (фиг. 489) имеет area с наружной связкой, характерную примитивного типа, именно: напереди путем пережима подразделенные на
для Агсасеа, и замок из зубов весьма находятся коленчато-согнутые, иногда
Фиг. 489. Cyrtodu datса ba\eu:dlioides. Замок правей створки; левая створка п area правой. Пермь. Донецкий бассейн.
Фиг. 490. Д ylihis as-ре> Ср Юра.
Ка ьвад ic, Франция.
Флг. 491. Mytrhis sublaevis. Ср. Юра. Минчингсм стон. Англия.
Фиг. 493. Си-cullaea Golow-kiHskii. Пижне-мелов. Саратов
Фиг. 492. А гса dilu-vit. Плиоцен. Сиена. Италия.
части зубы, назади пластинчатые, вытянутые параллельно замочному краю. Верхрекаменноугольные — пермские
Этот рот связывает Arcidae с Cyrtodonfidae, типом которых является род Cyrtodonta, весьма примитивный, силурийский и девонский. Cyrto-don'а имеет такие же зубы, как у Cyrtodon area, но свяжу простую, распо-ложе ную сзади макушек. Cyrtodontarca представляет тип, имеющий точки соприкосновения не только с Arcidae, но и с, Aviculidae ц даже с Heteiodonta.
Mijtilus (фиг. 490, 491) также относится то к Taxodonta то к Dy-sodonta (Aviculidae). Имеет равностворчатую, но весьма неравностороннюю, разномускульную раковину, с макушками напереди замочного края. Раковина несколько зияет напереди благодаря бпссусу. Сзади макушек протягивается наружная связка. Раковина представляет толстый эпидермический и тонкий перламутровый слой; замок в зрелом возрасте отсутствует или рудиментарен, в виде мелких коленчато согнутых зубов. Раковина гладкая или е радиальными ребрышками. Триас — ныне.
Area (фиг. 492) отличается радиальной скульптурой, многочисленными короткими зубами, расположенными поперек замочной площадки. Третичные — ныне.
Cucullaea (фиг. 493) отличается короткими поперечными зубами посредине замочной площадки и длинными косыми по концам ее. Юра — ныне.
Nucula (фиг. 486; nucula — маленький орешек). Раковина овальная или трехугольная, с концентрической скульптурой: многочисленные короткие зубы расположены поперек замочного края в два ряда, расходящиеся от макушки и разделенные под нею внутренней связкой. От силура доныне.
Главным образом в палеозое встречается род Palaeonei.o (от силура до юры), отличающийся наружной связкой, так что зубы расположены непрерывным рядом.
ПОРЯДОК IT. DTSODONTA.i .
Разномускульные и одномускульные пластинчатожаберные, в большинстве случаев прикрепленные биссусом, которому иногда соответствует выемка под передним ушком одной из створок, дающая возможность створкам плотно замыкаться, несмотря на проходящий между ними наружу биссус. Силур—ныне.
Avicula iPteria) (фиг. 494; avicula—маленькая птица, pteron- крыло). Раковина косая, расширенная задневентрально, с ушками при концах длинного прямого замочного края, перавностворчатая; более плоская правая створка снабжена выемкой для биссуса. Треугольная наружная связка расположена под макушками, в средней части треугольника area. У современных видов Avicula зубы отсутствуют или редуцированы (до одного) в зрелом возрасте, более развиты в ранних стадиях. У палеозойских Avicula ‘зубы развиты в виде косых первичных зубных пластинок, расходящихся наперед и назад от макушек. Украшения в виде концентрических или радиальных ребрышек. Силур — ныне.
Pterinea (фиг 495), близкий к Avicula палеозойский род; отличается от Avicula постоянно имеющимися зубами, косо направленными спереди назад, напереди короткими, назади длинными; переднее мышечное впечатление углубленное; связка на площадке, имеющей форму прямоугольника. От силура до каменноугольных.
Особенности area со связкой не являются резким отличием Pterinea от Avicula; как показывает изучение онтогении ныне живущих форм, узкая, треугольная, сосредоточивающаяся под макушками связка в индивидуальном развитии может превращаться в связку, более распространенную на поверхности area.
Существуют Aviculae с менее косой раковиной и редуцированными ушками (Oxytoma, пермь — ныне), намечающие переход от Avicula к Pseudomonotis (фиг. 496), отличающемуся выпуклой левой и плоской
1 Dys — без; odous, odontos — зуб.
правой створкой; переднее ушко последней крайне слабо развито. Скульптура радиальная. Девон—юра.
Aucella (фиг. 497) является производною от PseudomonOtis', отличается крайнею редукцией обоих ушков. Вздутая левая створка имеет
Фиг. 494. Aviculaden-sicostata. Меловые. Ульяновск.
Фпг. 495. Ptennea lacvis. Девон. Нассау. Германия.
большую, спчьяо загнутую макушку. Украшения преобладающе концентрические. Верхняя юра, нижний мел.
Posidonomya (фиг. 498). Тонкая, сжатая с боков раковина, равностворчатая, концентрически складчатая Замочный край прямой, без ушков, беззубый; макушка почти посредине замочного края, рдва выдающаяся. Силур — юра.
Своеобразное развитие в отношении связки представляют примыкающие к Avicula роды Inoceramus и Perna, именно: связка этих и некоторых других родов является в виде нескольких обособленных одна от другой частей, лежащих каждая в отдельной ямке замочного края (сложная связка).
Древнейшие, более примитивные предста-
Фиг. J96. Pseud omonotts eehinaia. ВИТели ЭТОЙ группы Доггер. Сёттон. Англия. (принадтежащейк одномускульным пластинчатожаберным), как Gervillea (фиг. 503; по имени ученого Герсилля), еще имеют зубы в виде немногочисленных косо идущих шастинок и связку, помещающуюся иногда лишь в небольшом числе (3 — 4) ямок. Триас — эоцен.
Perna (фиг. 499). Равпостворчатая раковина по форме изменяется от овальной до четырехсторонней. Макушка острая, на переднем конце; замочный край широкий, беззубый, с многочисленными ямками для связки Напереди выемка для биссуса. Триас — ныне.
Inoceramus (фиг. 500—502; inus—подобный, keramis— черепитчато налегающий). Округленно-яйцевидная, бо.ее или менее неравностворчатая раковина, с концентрическими, редко с рад на юными украшениями. Макушка выдающаяся, находящаяся напереди. Сильно развит призматический слой раковины. Юра и мел, главным образом средний и верхний.
Простую связку имеют Pectinidae—одномускульные пластипчатожаЗер ные, производные от Avicn’.idae. У Pectinidae,, цдагиодонтный замок пред-
Фчг. 498. Postdonomya Bechert. Нижнекамевноуголь-ные. Нассау. Термания.
ставляет видоизменение, состоя из немногочисленных зубов, расположенных симметрично по обе стороны связки, — изодонтный замок.
Pecten (фиг. 481, 504, 505, pecten—гребень). Раковина почти равнобокая, иногда в высокой степени неравностворчатая, с хорошо развитыми,
Фиг, 499. Puna mytiloi-des Оксфорд. Р. Печорг Сев. край. Сбоку и со спвнеой стороны.
Фиг. 502. Inoceramus suliatus. Гольт. Франция.
Фиг. 500. Pnoceramus Cripsit. Се-нон. Гозау. Австрия.
Фиг. 601. Inocetamus ret-rorsus. Верхняя юра. Р. Оленек. Якутс.;ая область.
почти равными ушками; выемка для биссуса более или менее слабо развита ' вязка внутренняя, срединная, треугольная; по обе стороны связки немногочисленные зубы в виде косых первичных зубных пластинок. Распадается на многочисленные подроды (Entolium, Vola и др.). Девон — ныне.
Lima (фиг. 506). Косоовальная раковина, часто несколько вытянута, наперед, равностворчата, обыкновенно несколько зияет напереди в связи с наличностью биссуса; замочный край беззубый, с ушками. Макушки острые, удаленные одна от другой. Скульптура в виде радиальных ребрышек. От каменноугольных доныне, главным образом в мезозое.
Spondylus (фиг. 484, 507; spondylos— замок). Отличается короткими немногочисленными зубами, симметрично, как вообще у Pectinidae, расположенными по обе стормы связки. Раковина или свободная—и тогда почти Равностворчатая, или прирастающая правой створкой, тогда увеличиваю-
щейся в размерах, приобретающей высокую area-, прирастание происходит околомакушечной частью, которая вследствие этого сильно варьирует по форме. На радиальных ребрышках поверхности раковины часто находятся шиповатые или пластинчатые выросты. Триас—ныне.
Фиг. 503. Gerwllea awculmdes Верхняя юра. Оксфордская ”лпна. Франция.
Фиг. 504. Pecten' asper. Сеноман. Франция. 1/2.
«лиг. 505. Pec еп impenah^
Неоком. Ганновер. Германия. 1/3.
Фиг. 506. Limn еопмЪппа.
Волжский ярус Хорошово, около Москвы.
Ostrea (фиг. 508—510; ostracon—раковина). Также сильно прирастает одной, вследствие этого сильно варьирующей по форме створкой. Раковина неправильного очертания, неравностворчатая, иногда весьма толстая, листоватого сложения, с сильно развитым призматическим слоем. В зрелом возрасте зубы отсутствуют или рудиментарны, в виде зубчиков. Иногда со скульптурой в виде радиальных ребер или складок. Каменноугольные—ныне, особенно в третичных
Gryphaea (фиг. 511 — 513; сходство с загнутым клювом грифа) отличается сильно выпуклой левой створкой с загнутой макушкой. В молодости прирастает, в зрелом состоянии свободна; правая створка плоская, крышечковидная. Лейас до третичных, главным образом мезозой.
ПОРЯДОК III. HETERODONTA.1
Heterodonta представляет группу, специализированную в отношении развития замка; в этой же группе развивается наиболее совершенный тип связки, цилиндрической, наружной, лежащей позади макушек. Наконец } Heterodonta почти всегда имеются сифоны, хотя иногда столь слабо развитые,
1 Heteros — равличный.
Фиг. 507. Spondyhis spinosus. Турон.' Дрезден. Саксония., Герма-
что мантийная линия не имеет синуса (Integrlpalliata, т е. с ненарушенной линией».
Integripalliata. Первичными являются формы палеозойские, снабженные зубами всех категорий — передними и задними боковыми и кардинальными; к таковым принадлежат мало еще отличающиеся друг от друга в палеозое Cyprinidae и Astartidae
Род Сурггпа (фиг. 514; от географического названия), установленный на основании современных его представителей, имеет раковину овальной или округленной формы, вздутую, концентрически струйчатую; макушки выдающиеся, загнутые наперед Передние боковые зубы уже подверглись некоторой редукции, именно: в левой створке передний боковой зуб уменьшение размерах, вместе с тем он смещен на замочной площадке под передний кардинальный. В правой створке передний боковой вовсе отсутствует. Мезозойские представители рода Cyprina представляют более первичное состояние переднего бокового зуба, который расположен нормально, впереди замочных Юра — ныне.
Сугепа (фпг. 515; от географического названия) отличается от Cyprina хорошо развитыми передними боковыми зубами. Боковые зубы вообще являются довольно удаленными от кардинальных и иногда могут быть
Фиг. 508. Osh еа callifera. Олигоцен. Майнцский бассейн. 1/2.
покрыты тонкими струйками на своих боковых поверхностях. В пресных водах или опресненных морских. Юра — ныне.
Может быть, к Cyrenidae примыкает род Dreissensia (фиг. 517 — 518), возникающий из Heterodonta подобно тому, как Mytilus из Taxodonta, т. е. вследствие прикрепления биссусом и скученной жизни большого лисла особей.
Dreissensia при митилусоооразиой общей форме раковины имеет под ма ’ ушками внутри створок перегородку, служащую для прикрепления переднею
замыкателя и ножного мускула. В пресных и солоноватых водах. С эоцена доныне.
Родственная Congeria (фиг. 516) отличается почти квадратным очей-тапием сильно вздутых, больших, толстых и тяжелых створок. Связка внутренняя, удлинена в направлении замочного края Впечатление ножного мус-
Фиг* 509. Ostr. a digi tali/га. Верхне-третичные. Штейнабрунна. Венски и бассейн. Австрия.
Фиг. 510. Ostrea Khoroschovensis. Волжский ярус. Мневники, около Москвы.
Фиг. 511. Qryphaea vesicularis. Белый мел. Остр. Рюген.
Фиг. 512. Gryphaea 'Kauf-таппг. Верхнемелов. Ри-штана в Ферм некой области. 1/3.
Фиг. 513. Gryphaea arcuata. Нижний лейас Вюртемберг. Германия-
куда обособлено от впечатления переднего замыкателя, лежит на перегородке кзади и несколько вовнутрь от последнего. С миоцена доныне.
Astarte (фиг. 483, 519, 520). Округл -нно-трехугольная иди овальная, толстая, гладкая или концентрически струйчатая; боровые зубы у мезозойских и кенозойских форм иногда отсутствуют (фиг. 519); кардинальные зубы иногда с наклонностью к уменьшению их до 2 в каждой створке; существует хорошо выраженная луночка. С девона доныне.
Pleurophorus (фиг. 5 21,522; pleuron—ребро, phoros—ношение) отличается поперечно удлиненной, четырехугольной раковиной. Макушка отодвинута наперед. Поверхность раковины с концентрическими струйками и немногими радиальными ребрышками, идущими косо назад. Переднее мышечное впечатление сильно углублено. По два кардинальных зуба в каждой створке и, кроме того, задние боковые, удлиненные, пластинчатые. Девон—триас.
Cardium (фиг 523, 524; сот— сердце) имеет кардинальные (по 2 в каждой створке) и латеральные зубы (по одному напереди и назади). Первые повернуты так, что располагаются не рядом, а почти что один над другим на замочной площадке. Раковина сердцевидной формы, ради
ально ребристая, с зубцами, соответствующими ребрам, на краях. С триаса доныне.
Cardiola (фиг. 525) вероятно является палеозойским представителем Cardildae, подобно тому как Posidonomya по отношению к Pernidae. Отли-
Фиг. 515. Cyrena fluminalis. Замок нравов и левой створок. Постплиоцеи. С. Александров-Гай, Нижневолжский район.
чается вздутой тонкой раковиной, наличностью area под макушкой, отсутствием зубов. Кроме радиальной скульптуры, еще концентрическая. Силур, девон.
Sinupalliata. 1 Мантийная линия с синусом. Формы с кардинальными и боковыми зубами, или с одними лишь боковыми.
К первым принадлежит например род Mactra (фиг. 482, 527) со связкой главным образом внутренней, с передними и задними боковыми зубами, кардинальные зубы находятся перед ямкой для связки, в левой створке дву
1 Sinus—выемка, pallium—мантия
ветвистый, коленчато согнутый зуб, входящий во впадину между двумя зубами правой створки. С меловых отложений доныне.
Род Venus (фиг. 52t>; название мифологическое) представляет по три кардинальных зуба в s 1
каждой створке; иногда рудимент заднего бокового. Кардинальные зубы надрезаны слабою борозд- / кою на две части. Скульптура концентрическая, № края зазубренные С юры доныне
Фиг. 517. Dreissensia Braidi Миоцен ВеиСенау, <юдо Майнца.
Фйг. 51(>. Congeria cong lobata. Мпоцен. Иннерсдорф, окол° Вены.
Фиг. 518. Dreissensia ros-triformis. Постплиоцен Черный Яр, Астраханский край.
Фиг. 519. Astarte porrecta. Нижнемеловые отложения. Ульяновск.
ПОРЯДОК IV. SCHIZODONTA. 1
С цельной мантийной линией, с наружной связкой.
Myophoria (фиг. 528) овальной или треугольной формы, с простым замком, подобно тому как у Astarte, состоящим из одних кардинальных зубов; один крупный зуб в левой створке и два охватывающих его зуба п >авой створки. Поверхность раковины покрыта концентрическими струйками или (у триасовых форм) радиальными ребрышками; есть киле-ватый перегиб от задней и дорзальной части к передней и вентральной. Девон—триас.
1 Scbizo—расщепляю, odous, odonts—зуб.
Пермские формы с толстым треугольным, подразделенным на две ветви зубом левой створки относятся к роду Schizodus (фиг. 529, 530; .schizo — ртсщепляю, odous—зуб).
Фиг. 521. Pleurophorus oblonijus. Наружное и внутреннее ядро. Пер мокарбон. Донецкий бассейн.
Фиг. 52й. Pleviophorw-costatvs. Пермь, Англия.
фиг. 520. Astarte Voltsvi. Средняя юра. Эльзас.
Фиг. 525. А—Cardiol interrupter. Верхний силур. Прага. В — Саг-diola alata. Верхний силур. Богемия. Чехо-Сло-вакия. Вид со стороны агса.Макушкаобломана.
Фиг. 523. Carthum Fittoni. Миоцен (сарматский ярус). Таганрог.
Фиг. 524. Cardiwn absoletum. Сарматский ярус. Коше-умоика, близ г. Александровска, Екате-оцнославсклй край.
Фиг. 526. Venus cutcta. Миоцен. Гейнфарн, около Вены. Австрия.
Фиг. 527. Mactra podolica. Сарматский ярус. Вена. Австрия.
Юрские и меловые представители этого типа—род Trigonia (фиг. 487, 531, 532) имеют поперечно-зазубреьные на боковых поверхностях зубы, разнообразную концентрическую и радиальную скульптуру, _ обыкновенно различную впереди и позади от указанного (у Myophoria) перегиба.
В мезозое род Trigonia представлен большем количеством видов, в настоящее время—немногими.
Unionidae (фиг.533) считаются пресноводными представителями Schizodonta, в раздое время переходивших к жизни в пресной воде, с наклонношью зубов и скульптуры становиться неправильными и наконец исчезать.
Фиг. 528. А— Myophoria Keferstemi. Триас (кейпер) Рапб1ь, Каринтия. В — Myophora Goldfussi. Раковинный известняк Вюртемберг. Германия. С и D —ламок левой и правой створок М. Whate leuae. Кейпер. Юлвые Альпы.
К и К* 1 — перегиб, т — переднее мышечное впечатление;" ггяр —
пдастиикообразные зубы, тгя — нераздельный треугольный зуб левей створки, шя/ — пластяиковые зубы правой стаоркя-
Фиг. 529. Schlzothti Wheelen. Верхнева-ыенноуг. Миссури. САСШ.
Unionidae отличаются значительным развитием перламутрового слоя раковины. Последняя обыкновенно вытянута в направлении длины; это при макушках, расположенных напереди, влечет за собою вытянутость назад элементов задней части замка. Палеозойские представители, например Carbo-nicola (каменноугольные, пермские), отличаются от позднейших всегда простой скульптурой в виде концентрических струек.
Фиг. 539. Schizodus obscurus.
А — ядро правой створки. Пермь Ганау. В и С— замок левой и правой створок; обозначения зубов те же, что на фиг. 528 С х В.
Фиг. 531. Trigonia cla-vellata. Оксфорд. Попе-1ян1, Ковенщина.
Фиг. 532. Trigonia со-stata. Средняя юра. Вюртемберг. Германия
ПОРЯДОК V. PACHYODONTA. »
Толстостенные, сильно выпуклые, с круглыми массивными зубами. Раковины до-юрских представителей свободны и равностворчаты, позднейшие прирастают и становятся неравностворчатыми. Макушки сильно завиты. Мышечные впечатления, резко выраженные, часто находятся на выдающихся площадках. У крайних из прирастающих форм, наиболее отличных по форме
—.
1 Pachys — тогсга*
раковины от нормального тпьа пластинчатожаберных, развиваются сифоны, связка изчрзает, раковина становится коническою и приобретает необычное строение, а именно — в чей развивается призматический слой, состоящий из призм, полых и расположенных параллельно поверхности стенки. Силур — мел.
Megalodon (фиг. 534; megas— большой, odous-—зуб) отличается выдающимися, сильно занятыми наперед макушками. На высокой замочной площадке 2 — 3 валикообразных зуба, большею частью изогнутых, приблизительно параллельны задней части замочного края. Девон — юра. Может быть сближен с некото рыми Heterodonta более обычного типа, например с Astartidae.
Dice,-as (фиг. 535; di — два, keras — рог)имеет выпуклую, толстую, гладкую, неравностворчатую раковину, прирастающую макушкой большей (то правой, то левой) створки; макушки выдающиеся, сильно закручены спирально наружу и наперед. В связи с такой закрученностыо связка расщепленна надвое в более старых частях, помещающихся в бороздках, идущих cnri-рально по наружной поверхности раковины
В одной створке дугообразно изогнутый зуб, помещающийся между двумя зубами другой створки. Заднее мышечное впечатление на выдающейся с внутренней поверхности створки площадке. Верхняя юра.
От Diceratinae исходят две линии развития: одна, которой принадлежит жнвушмй и в ^стоящее время род Chama, и другая, представленная вымершею группою рудистов.
Фиг. 533. L'nionidae flabrllatus. Плиоцен (конгериевые слои) Комори. Венгрия. Левая створка совнутри и снаружи.
т, п — мышечные впечатления; ftn — впечатление ножного мускула; mzi9 — кардинальные зубы; hzo — задние боковые, mt, — соответствует передней, hzu ~ задней ветви треугольного зуба Trigoniat hz— зубная ямка.
от мак) шек к замочному краю.
Фиг. 534. Megalodon cucullatus. Средний девон. Пафрат, около Кельна. Германия.
Chama (фиг. 536) прирастает и имеет связку подобно Dicer as. Замок 5 ранних стадиях развития представляет кардинальные и редуцированные боковые зубы. Из н, х получаются в зрелом возрасте два зуба на прикрепит Учгбввк нале 'Втологпв 257
ленной и один на свободней створке; зубы развиваются вследствие отложе ния известного вещества при первоначальных кардинальных, зубах; это отложение связывает, изменяет, частью закрывает цервоначальные зубы Строение раковины отличается от обычного* строения раковины пластинчатожаберных и прмближштся к строению вымерших руд.чстов по своей труб-чатости и сетчатости. Мел—ныне
Крайнюю степень специализации, обусловленную прирастанием, пред-ставляют так называемые руднсты, к которым принадлежит например род Hippurites (фиг. 537). Имеет коническую нижнюю, прирастающую, и зрышечковидную верхнюю створки. Макушки совершенно не закручены. Нижняя часть глубокой конической нижней створки содержит поперечные несколько вогнутые днища, обособляющие наверху пространство, занятое мягким телом животного. Крышечксвидная створка снабжена отростками, служившими для прикрепления мускулов. Наружная поверхность прпкреплен-
Фиг. bbu. Левая и правая створки Vicerus aw'et&ium (Оксфорд, Франция) и D. Ziileh (титон, Моравия):
я — переднее, а* — заднее мускульное впечатление; с — большом замочный зуб; d — зубная ямка; I — бороздка связке; в — площадка с задним мускульным впечатлением.
Фиг. 536. Chama squatnv/a. Эоцен. Гампшайр. Англия.
ной створки несет с одного боку три бороздки, идущие продольно от макушки до верхнего края; одна из этих бороздок соответствует исчезнувшей связке, дзе остальные, рядом лежащие позади первой, соответствуют глубоким складкам поверхности раковины, так называемым столбикам, вдающимся вовнутрь и служившим опорою дл^ сифонов; последние выходили через два отверстия крышечки, расположенные соответственно столбикам. Открывание раковины могло происходить например вследствие набухания ноги, обычного у свободных пластинчатожаберных при выходе ноги из раковины для движения животного. Кроме того и при закрытой раковине, должно быть, существовало некоторое сообщение внутренности раковины с наружной средой через пористую на поверхности и снабженную внутри радиальными каналами крышечку. Подобно всем прикрепленным животным, рудисты при-надлежит прибрежной фации; для защйты от прибоя возникала значительная толщина раковины, а необходимость значительного выделения извести де нала возможным нахождение рудистов лишь в теплых водах, где именно происходит такое выделение. Меловые отложения
ПОРЯДОК VI. DFSMODONTA.
Палеозойские Desmodonta отличаются тонкостью раковины и цельною, без синуса, мантийной линией. Связка наружная, зубов нет, макушки н передней половине длины раковины. Раковина гладкая или с украшениями, лишь редко не концентрическими. К числу их принадлежит, например, род
Фиг. 537. Bippurites radiosus. Верхнемеловые отложения. Франция.
1 — верхняя створка с зубовидвымя отростками с, с' и сгг; а — мышечное впечатление;
2—внутренняя поверхность нижней створ! и; А—связочная бороздка В передний, С— звдвнй столбик; d, d' в d” ~~ зубные впадины: аа1 — мышечные впечатления, х — жилое пространство; V—вид Н ppuriles сбоку; 4— со стороны К| ышкв в последнем внизу сохранилась пористая наружная поверхность; наверху, где видны два отве- стия для сифона, поверхность стерта, так что видны радиальные каналы.
Edmondia (фиг, 538), с раковиной косоовальпого очертания. Каменноугольные, пермские.
В палеозое же появляются первые Desmodonta с синусом мантийной лини", Таков род Allorisma (фиг. 539; alios — изменчивый, ereisma — связка) с равностворчатой, удлиненной, выпуклой, слабо зияющей назади
1 Desmos—свллжп.
I
• t
раковиной. Передняя сторона короткая, иногда с луночкой. Зубов нет. Девон— пермские.
В мезозое появляются Desmodonta, снабженные замком. В простейшей форме замок является у рода Pleuromya (фиг. 540, 541). Именно здесь под макушками створки имеют по язычковидному выступу с вогнутою поверх-
ностью; выступ правой створки налегает на выступ левой. Раковина неколько зияет назади. Связка наружная, позади макушек. Триас — нижнемеловые.
У рода Рапораеа (фиг. 542; мифологическое название)
фйг. 53S. Ed-mcndia nibras-ceivas. P Коде-ма, Донецкий бассейн. Пермокарбон.
Фиг. 539. Allonsma iegularis. Слобода, Тульский грай. Нижнекамеивоуг.
1л
Фиг. 541. Замок Pleuromya margmata. Юра. Франция.
D, G— правая и левая створки, L связка.
выступы замочного края являются уже зубовидными. Раковина зияет спереди и сзади. Сифоны своею длиною в несколько раз превосходят длину раковины. Последняя у ныне живущих представителей достигает в длину до 0,5 м. Триас-ныне.
Род Муа (фиг. 479, 543) представляет довольно резкую внутреннюю асимметрию при наличии внешней симметрии: связка, здесь внутренняя, помещается на видоизмененном зубе—ложковидном отростке левой створки, подходящем под макушку правой. Триас—ныне.
Pholas (фиг 545; pholeos—нора, скважина) является родом, представители которого помещаются в высверленных ими в дереве или в углублениях камней.
Раковина сильно зияет как напереди, так и назади: напереди имеет она сетчатую и шиповатую скульптуру. Связка отсутствует, открывание раковины производится мускулами, прикрепленными снаружи к отвороту около макушечной части и идущими поперек от одной створки к другой. Эти мускулы бывают прикрыты снаружи дополнительными известковыми пластинками. Под макушками находятся известковые отростки, служащие для прикрепления ножного мускула. Юра—ныне.
7 eredina (фиг. 544; teredon—сверлильщик) имеет речно от макушек к брюшному краю, но створки соединены неподвижно известковым отложением на месте зияния спереди и назади; назади это отложение докрывает и сифоны, которые заключены таким образом в из вестковую трубку. Юра —ьыне.
Фиг. 540. Pleuromya регедппа. Хорошово, у Москвы. Верхняя юра.
раковину формы Pholas, вытянутого попе-
Распространение пластинчатожаоерных.
Из кембрийских отложений пока известны лишь немногочисленные и недостаточно ясные остатки единичных пластинчатожаберных. Из. силура известна уже богатая фауна, содержащая представителей Heteromyaria,
Фиг. 544. Teted-na psi sonata. Эоцен Франция.
а - раковина; Ь, с — отложеаия извести около сифонов и напереди створок.
Taxodonta, Desmodonta и, может быть, также Heterodonta. Возможно, что обособление этих групп произошло в докембрийское время.
Во всяком случае, как этого и следо- - rfgri- -вало ожидать, палеозойские представи- »->>.
тели вышеперечисленных групп нередко являются более примитивными и более -7?*.. ->.
сближают эти группы, чем послепале-
ОЗОЙСКИе формы. Достаточно напОМНИТЬ на- фиг* 645. Pholas Levesquei Эоцен, пример, палеозойских Aviculidae, Astartidae, фРан«ия (макушкой сравне' род Cyrtoaontarca. Во всяком случае,
| наиболее специализированные типы: по замку—Trigoniidae, Spondylidae, по способности прирастания—Ostreidae, Spondyeidae Rudistae, развиваются пиль к мезозою, и во время него сильнее распространяю!ся отномускульные и снабженные сифонами формы. Taxodonta существуют довольно неизменно
ж различные яры как в отношении численности, так и в отношении однообразия. Немногочисленные, притом бессифонные в палеозое (Heterodonta Astartidae, Megalodontidab и др.), в мезозое представляют большое число семейств, специализированную ьетвь Ра< hyodonta и типичные формы с синусом мантийной линии В девоне начинается ветвь перешедших к жизни в пресной воде Unionidae.
Desmodonta представляют в палеозое, уже с девона, формы с синусом мйнтпйчой линии (AHorisnia). Периодами наибольшего разнообразия Aniso-myaria являются верхпеиалеозойскпй и нижнемезозойский. В кенозое характерны преобладание Heterodonta и крупнейшие размеры некоторых пластинчатожаберных.
КЛАСС V. ЛО1ГАТОНОГИЕ, SCAPIIOPODA.
Морские двусторонне-симметричные моллюски; тело и раковина конически (почти цилиндрически) вытянуты в передне-заднем направлении. Мантия двулопастная, как у пластинчатожаберных, со сращением краев мантии на брюшной стороне, но с отверстиями напереди н назади. Широкое отверстие раковины является передним, имеет тонкий нарастающий край; заднее отверстие узко, край его соответствует более старой начальной части раковины и потому являеся утолщенным.
Голова рудиментарна, лишена глаз, имеет парные придатки, снабженные нитевидными щ-пальцами, со вздутиями на концах. Эти придатки могут считаться соответствующими пластинчатым рукам Nautilus. Имеется radula, жабр нет. Нервная система симметрична в своем строении, подобно тому как у пластинчатожаберных.
Цилиндрическая нога служит для зарывания в песок, причем в воду выдается лишь заднее отверстие раковины, служащее для входа воды вовнутрь и для выведения ее наружу. Более всего сходны с пластин«ато-
Фпг. 646. Dentalium, продольный разреа.
а — анальное отверстие, d — кишка, f — нога, д — половая железа, Л — головной узел, к —ок< лоротоиые придатки, I — иечень, т — ножяой мускул, wit —мантийная полость, pd — ложной увел, • — раковина, i — щупальца, v — висцеральный узел.
жаберными, отличаясь от них присутствием radula. Животные хищные, питаются фораминиферами, диатомовыми водорослями и т. п., живут от прибрежной полосы до глубины 800 м.
Род Dentalium (риг. 54G; dens—зуб) в ископаемом сос^ошии ветре* чается с силура, живет и ныне.
Геологическое распространение пластинчатожаберных
Роды Отделы или ярусы
। Кембрий 1 Силур Я . о Кагбон Пермь Триас Св о Мел -hn.Ls) 1Д4 ©ТЧННтМ 1 -э<коЭ 1 1
Macrodon . . Cyrtodontarca Cyrlodonia Al у Ulus . . . Area .... CucuUaea . . Nueula . . . Palaeoneilo A ci’cu a . . . Ptcrinea . . . Or io ma . . . Pseudo nionotis Aucella . . . Posidonom ya Geruillea . . . Perna . . . Inoceramus Pecten . . . Lima .... Spondylus . . Ostrea .... Gryphaea . . Cyprina . . . Cyrena . . . Dreissensia Congeria . . Astarte . . . Pleurophorus Cardium . . . Cardioia . . . Mactra . . . I’eirus .... Myophoria . . Pehizoilus . . Trigonia . . Carbonicola Vnio .... Mega'odon . . Diceras . . . Chama .... Lippuritcs . . Edmondia . . A'torisma . . Pleuromya . . Punopaea . . Муа I bolas . . Teredina . .
г
-1
X
1
Характеристики групп моллюсков.
1 класс. Amphineura. Обитатели морского дна. С полною двустороннею симметриеи тела. Тело округленное червеобразное или сплющенное в спинно-брюшном направлении, покрыто известковыми шипами или раковиной из нескольких табличек. Силур—ныне.
2 класс. Gastropoda. Во всех местах обитания кроме воздуха. Обыкновенно с головой и ползательной ногой, жабрами или легкими. Маития непараан и обыкновенно ненарнан асимметричная известковистая раковина, свернутая в коническую спираль и снабженная нередко роговой или известковистой крышечкой. Кембрий—ныне.
1 подкласс: Prosobranchiata. Большей частью обитатели морского дна с перекрещенными соединениями нервных узлов, сердце с одним или двумя предсердиями; обыкновенно одна или две жабры. Кембрий—ныне.
1 порядок: Diotocardia. Два предсердия, две жабры. Раковина часто колпачковндная или ушкообразная. Почти исключительно морские. Кембрий—ныне.
2 порядок: Monotocardia. С одним предсердием и одной жаброй. Большею частью морские, редко пресноводные, часто снабжены сифоном. Кембрий—ныне.
3 порядок: Heteropoda. Пелагические Monotocardia с плавательной ногой. Раковина ечень тонкая, двусторонне симметричная или отсутствует. Третичпые~ныне-
2 подкласс: Opisthobranchiata; только в море, частью бонтонные, частью планктонные формы. Предсердие и жабра позади желудочка сердца. Только частью с спиральной или прямой раковиной. Девон—доныне.
Порядок Pteropoda. Пелагические Opisthobranchiata. Верхний мел, третичные, ныне.
3 класс: Pulmonata. Морские, наземные, пресноводные. Легкое и предсердие перед желудочком, раковина большею частью спиральная или атрофирована. С девона доныне.
4 класс: Cephalopoda. Исключительно морские, двусторонне симметричные, снабженные рогорыми челюстями в виде клюва. Хищные животные, плавающие или ползающие.
1 подкласс: Telrabranchiata. Постоянно с наружной раковиной, содержащей воздушные камеры п сифон и свернутой обыкновенно в плоскую спираль; устье раковины иногда сужено. Ныне живущий род с очень многочисленными щупальцами и четырьмя жабрами. Кембрий—ныне.
1 порядок: Nautiloidea. Раковина в виде плоской спирали, согнутая или прямая. Перегородки вогнутые наперед, шовная линия простая; сифон часто широкий, с обращенными назад сифонными дудками; начинается на задней стенке первой чашечкообразной камеры, которая имеет назади рубец. У прямых раковин существует мешкообразный эмбриональный пузырь. Кембрий—ныие.
2 порядок: Ammonoidea. Раковина обыкновенно плоскоспиральная, редко совершенно развернутая. Перегородки выпуклые наперед, швы большею частью рассеченные. Сифон постоянно узкий и прилежащий к стенке раковины, почти постоянно к наружной, и большею частью с обращенными вперед епфонными дудками, начинается на конце почти всегда спиральной эмбриональной камеры. Часто существует крышечка. Девон—мел.
2 подкласс: Dibranchiata. С двумя жабрами и 8—10 руками, снабженными присосками или хитиновыми крючками, большею частью с чернильным мешком. Раковина почти постоянно внутренняя ми отсутствует. С триаса доныне.
1 порядок: D capoda. С 10 руками часто покрытыми крючками и большей частью с внутренней раковиной; последняя обыкновенно представляет спинной щит, а кроме того еще часто большею частью прямую и коническую раковину с воздушными камерами и сифоном и позади массивный ростр. Триас —ныне.
2 порядок: Octopoda. С 8 руками без крючков, без раковины, только самки Argonauta со вторичной, наружной плоскоспиральной раковиной. От верхнего мела доныне
5 класс: Lamellibranchiata. Морские и пресноводные обитатели. С двухсторонней симметрией, без головы, с топорообразной роющей ногой и большею частью с пластинчатыми жабрами, одеты парной мантцей, которая выделяет раковину, состоящую из двух створок, расположенных по бокам тела, закрываемую двумя или одним мускулами и открываемую эластичной связкой. От кембрия (?) доныне.
1 порядок. Taxodonta. Равномускульные пластинчатожаберные, с цельной, редко слабо выемчатой мантийной линией. Исключительно морские. Силур—ныне.
2 порядок: Dysodonta. Равномускульные и одномускульные, большею частью прикрепленные бпссусом. Морские. Силур-ныне.
3 порядок Heterodonta. Двумускульные, почти всегда с сифонами. Силур-ныне.
4 порядок: Schizodonta. С цельной мантийной ’•инией с наружной связкой. Морские, пресноводные. Левой—ныне.
5 порядок: Pacbyodonta. Двумускульные, с цельной мантийной линией. Голстостен-иые раковины. Морские. Девон—ныне.
6 порядок: Desmodonta. Двумускульные, тонкая раковина, зубов нет. Связка наружная. Морские. Силур —ныне.
6 класс: Scaphopoda. Морские, двусторонне симметричные, с роющей ногой, без жабер. Мантия и раковина трубообразны. Силур—ныне.
VIII тип. ПОЗВОНОЧНЫЕ, VERTEBRATA.
Отличительные особенности позвоночных следующие: двусторонняя симметрия, продольная сегментация стенки тела; кишка, открывающаяся наружу ртом напереди и анальным отверстием назади и снабженная—если не у взрослого животного, то по крайней мере в ранних стадиях жизни,— парными боковыми жаберными щелями; спинной нервный ствол; расположенная между ним и нижележащей кишкой спинная струна; продолжение тела кзади от анального отверстия в форме сегментированного хвоста; хорошо развитый целом и обособленная сосудистая кровеносная система.
Позвоночные разделяются на два подтипа: бесчерепных—Vertebrata Acrania и собственно позвоночных, снабженных черепом*—Vertebrata Сга-niata. К первым относятся: так называемый ланцетник с немногими родственными ему- родами, дегенерировавшая группа оболочников, или туникат, и еще одна группа животных—Enteropneusta (кишечнодышащие), действительная принадлежность которых к типу не является установленной.
Vertebrata Craniata.
Собственно позвоночные делятся на классы: круглоротых—Cyclostomata (миноги и миксины), рыб—Pisces, амфибий—Amphibia, пресмыкающихся— Reptilia, птиц—Aves и млекопитающих—Mammalia.
Тело Vertebrata явственно подразделяется на три части: голову, туловище и хвост.
Первичное состояние органов наиболее стойко сохраняется лишь в средней из этих трех частей—туловища. Сегментация выражена резче всего у низших представителей позвоночных, рыб, в гораздо меньшей .степени и притом только во время эмбрионального периода их жизни—у высших, птиц и млекопитающих. Например первичная сегментация мускулов стенки тела исчезает у высших позвоночных вследствие слияния первичных сегментов и возникновения таким путем новых мускулов. Мускульные сегменты головы утрачивают у позвоночных свое первично-сегментальное расположение, приспособляясь, в соседстве со ртом, главным образом к отправлению жевательных функций. В дорзальной части головы происходит усиленное развитие головного мозга, занимающего значительное пространство. В вентральной половине головы находятся передние чаоти пищеварительной системы. Между вышеупомянутыми жаберными щелями здесь лежат жаберные дуги, носители жабер у рыб и амфибий, тогда как у высших животных весь этот аппарат является лишь преходящим образованием зародышевого состояния. Хвост, являющийся у низших форм органом движения при плавании, у высших—с развитием двух пар конечностей как органов движения—теряет значение и наконец становится рудиментарным. Наружный кутикулярный скелет—такой, как у членистоногих,—здесь не развивается; кожа позвоночных мягка, и кожный скелет их представляет скорее орган защиты мягкого тела, чем опоры для мускулов. Взамен этого по продольной
оси /ела образуется внутренний скелет из плотной гкани, являю щейся в форме хряща или кости и образующей позвоночник и череп.
Хрящ и кость могут считаться специализированными видоизменениями соединительной ткани, которая распространяется во все части тела позвоночного, соединяя их и доставляя для них опору.
Внутренний скелет позвоночных есть собственно лишь местное видоизмените соединительнсй ткани, пред с iявляющее большую прочность, луч шую опору и лучшие механические условия для действия мускулов.
Хрящ онтогенетически и филогенетически предшествует кости. Хрящ есть ткань, в которой клетки выделяют прочную эластичную основу, более или менее однородную.
У высших позвоночных костная ткань усиливается в значении как преобладающая скелетная ткань, в конце концов вытесняющая хрящевую основу скелета. Происходит так сказать борьба мея.ду хрящем и костью, в которой кость побеждает.
Кость развивается пли на месте хряща, разрушающегося тогда при ее развитии (преформированная хрящем юють), или независимо от хряща—в соединительной ткани. В случае костей, префпрмированных хрящем, наружные ?лои их могут быть отложены под надкостницей, оболочкой кости, без предшествующ го возникновения хряща, это—перихондральная кость; кость, развивающаяся на месте хряща с изменением его в кость, называется эндохондральной костью. Кости, префор-мированные соединительной тканью, называются кожными. Кость возникает как результат деятельности особых костеобразовательных клеток— остеобластов, которые могут сохраниться в готовой костной ткани, так что вмещающие их маленькие полости (костные тельца) дают во все стороны тончайшие канальцы (фиг. 548, 549).
ош
С -А
'2^1 ч
-.1 >•
Фиг. 549. Продольный разрез трубчатой кости. Гаверсовы каналы и костные тельца.
виг. 548. Поперечный разрез черев трубчатую кость:
G — Гаверсовы каналы, К — коствое тельце, I—слои кости.
Фиг. 547. Стекловидный хрящ.
• Более значительные части скелета позвоночных представляют еще сравнительно большие каналы, которые содержат в себе кровеносные сосуды (Гаверсовы каналы). При образовании зубов костная масса отлагается образующими ее клетками только с одной стороны, так что она никогда не включает костных телец.
Пногда хрящ уплотняется вследствие отложения в нем углекислой извести, без участия костеобразовательных клеток и без пр. об(етения характерного строения кости. Таким обиазом пол$ чается так называе-8в«
st
sec-
$г
в
sp
с
Фиг. 650. Развитие пера птипы. А, В'- первичного, С, D— вторичного.
р — сосочек (жирным показан евпдермнс); Ъ — влагалище пера; — душка; зр — очин, расходящийся кверху в пучок лучей или бородок al; sec—сидящие на последних вторичные лучи; II— стержень пера, F—оиахало.
иый известковый хрящ, именно у низших рыб, каковы Elasniobraiichla (акулы),
ГТоврЬвы тела. У позвоночных покровным образованием является кожа, состоящая из эпидермиса, наружного многослойного эпителия, и соединительнотканной собственно кожи, cutis И та и другая часть кожи облашют способностью производить твердые образования. Эпидермис дает происхождение рогоьым образованиям, волосам, чешуям, перьям, когтям, рогам и т. п. В cutis могут развиться кости, принадлежащие к категории вышеупомянутых кожных костей.
Волосы, перья и роговые чешуи—образования гомологичные. При об азованип ч>шуй в эпидермисе возникают местные рогогые утолщения то в виде бугорчатых, то че-репитчато налегающих чешуй.
Развитие пера птицы указывает на его родство с чешуями пресмыкающихся. Первоначальный зачаток пера является в виде сосочка кожи, приподнимающего эпидермис, причем эти утолщения подобно чешуям направляются потом своими вершинами к заднему концу тела животного (фиг. 5Ь0, А).
Кожа вокруг основания сосочка углубляется и образует кругом него влагалище (фвд. 550, В). Затем эпидермический покров сосочка расщепляется от вершины вниз, образуя нитевидные лучи*, которыеввнзу кисточко-видно сливаются в о ч п-н е пера. По мере роста пера сосочек отстает от вершины, причем покрывающий его эпидермический слой следует за нам, отделяя на своем пути роговые чехлнкп, образующие душку первичного пера.
Впоследствии первичное, предварительное перо сменяется вторичным, окончательным.
Вторичное перо отличается от первичного тем, что вместо равномерно развитых, расположенных веером
лучей последние расположены в нем перисто, по бокам стержня, образован.
него двумя срединными лучами, срастающимися и усиленно сравнительно с остальными личами развивающимися (фиг. 550, С и D).
Из всех твердых образований собственно кожи наиболее распространенными и основными являются чешу и рыб—разумеется, совершенно отличные от эпидермических роговых чешуек змей, птиц и т. п.
Формою чешуи, исходною для других чешуи и даже вообще для кожных костей, являются плакоидные чешуи, или кожное зубы (.фиг. 551), существующие вообще у всех селахпй (акулы, скаты)
Плакоидные чешуи возникают при конических, сосочковидных, проникающих в эпидермис возвышениях cutis, причем 1) выпяченный этим воз-
вышением
эпидермис отлагает над сосочком чехчик из твердого однородного вещества (э м а л ь); 2) сосочек отлагает на своей поверхности и непосредственно под эмалью слой кости без клеток, прони-р кнутый радиальными канальцами (д е н т и н); ден-«ь тин облекает центральную > сосочка [pulpa), содержащую в себе кровеносные сосуды; 3) при дентинном основании зуба
~ . У „ часть
с содеР;
к нему примыкает плоская основная пла-
Фиг. 5&1. Профильный разрез плакоидной чешуйки еб-временной ai улы Scymnus.
К — .мал»* Э — дентиновые канальцы, Р — пульпа; О — одо!тобла сты; h — Гаверсов канал, В — основная пластянка, С — кожа с волокнами соединительной гканн
с т и н к а, состоящая и t костного вещества обыч ного строения. Вырос-
ший зуб прорывает эпидермис и свободно выступает наружу. Собственно выдающаяся часть и называется кожным зубом.
Плакоидные чешуи является образованиями, исходными для двоякого рода образований кожи. Переходя на оболочку -ротовой полости, предста-
вляющую непосредственное продолжение кожи, плакоидные чешуи превра щаются в орган пищеварительной системы—зуоы полости рта. При этом вместо основной пластинки плакоидной чешуи развивается покров корневой столбчатой части зуба, состоящий, подобно основной пластинке, из костного вещества с костными тельцами (цемент). У низших позвоночных зубы находятся не тол >ко на краях челюстей, но и на небе и на жаберных дугах, у выси их—только на краях челюстей.
В коже, покрывающей тело снаружи, плакоидные чешуи представляю” весьма значительные и разнообразные видоизменения. У так называемых ганоидных и костистых рыб исчезает верхняя коническая часть пл?войди ых чешуй, и наружный чешуйчатый покров состоит из основных пластинок. При этом каждая чешуя или вовсе не приподымает кожи, когда чешуи (именно у ганоидных рыб) -лежат в одной плоскости, одни возле других, пли кожа слабо приподнята—когда чешуи расположены черепитчато, с налеганием заднего края каждой чешуи на передний край смежной, назади тежащей чешуи. Путем слияния основных пластинок плакоидных чешуй возникают значительного размера кожные кости
; при этом
Первичный скелет древнейших рыи иы i наружным. Затем возник внутренний костный, прафорыированный хрящом скелет, или образованный известковым хрящом, или наконец возникший частью вследствие распространения процесса окостенения снаружи в участки соединительной ткани, лежащие внутри тела. Кроме того, кожные кости, возникшие на поверхности тела, могли войти в тесную связь с подлежащими, преформированными хрящем костями черепа и плечевого пояса, как это будет изложено ниже.
Древнейшею частью скелета туловища позвоночных является спинная струна (chorda dorsalis), онтогенетически развивающаяся из срединного продольного выроста спинной стенки кишечника.
Это образование, состоящее из клеток, покрыто двойной прочной эластичной кутикулярной оболочкой. Уже у многих рыб в связи с хордой развивается позвоночный, столб. Передвижение рыб совершается при посредстве изгибов вбок хвоста и задней части туловища мускульных ^сегментов отражается на хорде, давая импульс к возникновению вокруг нее скелета, сначала хрящевого, а затем костного.
Развитие хряща и кости происходит от периферии хорды к центру, начинаясь в окружающей хорду соединительной ткани. Кроме того, каждый позвонок первоначально состоит из отдельных хрящей или косточек, образующих затем, по мере развития животного, своим слиянием целый позвонок.
4 Позвонки развиваются соответственно числу сегментов мускулатуры, которая является определяющей в их развитии.
Сначала возникают две пары хрящей — верхние и нижние дуги позвонков, — из которых одна, исходя от хорды вверх, охватывает ствол спинного мозга; другая, исходя от хорды вниз, охватывает целом. Затем развивается трубчатый покров вокруг самой хорды, причем, соответственно каждому позвонку, первоначально развивается несколько отдельных окостенений. Так, в туловищной части тела древнейших четвероногих, например пермского Arche-gosaurus, кроме верхних, так называемых нервных дуг (neurdlia или neurapophyses, фиг. 552), различают
лежащий напереди срединный брюшной hypocentrum и задние, парные, боко-вые preurocentra\ это — так называемые рахитомные позвонки. С соприкасающимися частями всех этих троякого рода элементов сочленены ребра. В дальнейшем развитии позвонка в теле его преимущественное развитие получает hypocentrum, растущий своими боковыми концами вверх, до соприкосновения этих концов между собою на спинной стороне от хорды, так что образуется костяное кольцо вокруг нее. Pleuroccntra в общем развиваются менее, чем hypocentrum, иногда они невидимому вовсе не окостеневают, оставаясь в хрящевом состоянии, или имеют малые размеры и вытисняются наверх преобладающим в своем развитии hypocenttum, так что окончательно тело позвонка может быть образовано только последним. Примитивные и древнейшие каменноугольные амфибии имеют так называемые эмболомерные, или диплоспондиловые позвонки (фиг. 554), в которых плеври-
Фиг. 552. Передний туловищный позвонок. Arche gosawrus Deche-пг сбоку; налево—перед, направо—зад позвонка.
1Гс—hypocenirwm. Pc—pleur госепЪ art Jfe—нервные дуги. Df— остистый отросток, Z, — сочленовые отростки W — высота иозволка.
центры и гипоцентры являются в виде двух просверленных дисков почти
равной величины; уменьшение верхней части гипоцентров и нижней части плевроцентров приводит к рахитомному состоянию пермских форм;
Фиг. 553. Туловищные позвонки гановдпой рыбы Calloptcnis Agas-sizt. Верхняя юра. Германия.
hyc — гипоцентры; pic — пл евро центры; е — ребра; п—верхние дуги; df—остистые рт-росгкн.
дальнейшее уменьшение плев-юцентров при одновременном увеличении гипоцентров ведет к стереоспонд и левому позвоночному столбу триасовых амфибий, где каждый позвонок является двояковогнутым. Хорда, сначала (в индивидуал! ном развитии и у древнейших типов) покрытая лишь сравнительно тонкой костяной обложкой, затем, при дальнейшем надвигании костного вещества к центру хорды, уменьшается в поперечнике, сжимается внутри тела позвонка (вертебрально) и расширяется между позвонками (интер-вертебрально), так что интервертебральное раздутие ее выфпрмовываег обращенные друг к другу поверхности смежных позвонков в виде коничеш их углублений.
Таким образом получается наиболее примитивный тип—двояко-вогнутого, амфицельного позвонка, свойственный низшим позвоночным. Иногда, у более низших типов, такой позвонок просверлен в центре
отверстием для сохраняющейся хорды, а у высших тиков хорда совершенно
исчезает, п позвонок не имеет отверстия в центре.
У высших животных обращенные друг к другу оконечности позвонков
представляют вогнутые и выпуклые поверхности, так что является подвижное сочленение позвонков, при котором позвоночный столб может совершать различные и значительно изгибы, а вместе с гем тело — различные движения. Такие позвонки по форме сочленовных поверхностей бывают описго-цёльными (спереди выпуклы, сзади вогнуты) или процёльными (спереди вогнуты, сзади выпуклы), могут представлять седлообразную сочпеновиую поверхность (выпуклую в одном направлении и вогнутую в другом, перпендикулярном), или
Фиг. 554. Зинне хвостовые возвонги ганоидной рыбы Euryeuimus. Верхняя юра. Германия.
Нс — гипоцентры; Р с — плевроцрнтры оР — верхние и кЛ — нижние дуги. На конце хвоста и евро центры исчезают, ьытес-жяась наверх.
наконец эта поверхность
является плоской. Последние дсе формы свойственны наиболее высшим позвоночным. Кроме сочленений между телами позвонков, у четвероногих развиваются еще сочленения между верхними дугами их. При втом верх-
I
ниедуги напереди и назади несут парные косые сочленовные отростки (фиг. 552).
Дуги позвонков, верхние и нижние, иногда и у выс.пчх позвоночных,
когда тело позвонка является уже сплошным и непросверлепным, остаются еще несросшимися с телами позвонков и соединены с последними посредством хрящевых прослоев; у наиболее же высоко стоящих позвоночных дуги
срастаются с телом в одно нераздельное целое.
Нижпие или гемальные дуги позвонков в хвостовой области, там куда уже не достигает общая полость тела, также соединяются между собою внизу, подобно невральным дугам; в свободном пространстве, остающемся между ними и телом позвонка, проходят кровеносные сосуды. Далее наперед гемальные дуги часто имеют наклонность исчезнуть, будучи представлены могут продолжаться
лпйь короткими отростками тел позвонков; или как обособленные от тел позвонков ребра, охватывающие брюшную полость, так называемые плевральные или нижние ребра. Кроме того различают настоящие или верхние ребра, лежа цие горизонтально, на границе между спинною и брюшною частью мускулатуры (фиг. 555). У большинства позвоночных имеются пли только нижние (большинство рыб), пли только верхние ребра (прочие классы позвоночных). У некоторых рыб (Crossopterygii) одновременно существуют и верхние и нижние ребра.
Одни считают ребра за выросты позвонков, отчленившиеся от последних; другие считают ребра образованиями, соответствующими жаберным ду1ам. У многочисленных весьма древних
они
ск
и примитивных четвероногих поперечные
отростки позвонков, служащие для прнчленения ребер, самостоятельны, лежат между позвонком и собптвенно ребром. Последнее состоит из двух участков, спинного и брюшного; брюшной при
Фиг. боб. Схематический поперечный разрез туловища рыбы:
тт — спинные куску лы; Ът — брюш ные мускулы; ch— хорда, ob— верхняя дуга d — остистый отро чок; юк — нервный кв в ал, pi — нижние ребра г — верхние ребра; с — coelom; ft — лучсвосцы» fs — плавна» новые лучи
мыкает к грудной кости, связывающей ребра левой и правой половины тела. Таким образом между позвонком и грудной костью является
дуга, совершенно так же трехчленистая, как
жаберные дуги. Грудная кость соответствует совокупности copulae у жаберных дуг (фиг. 5(17).
Происхождение, одинаковое с ребрами и жаберными дугами, может быть иьеют так называемые пояса конечностей: плечевой, или
гояс передних конечностей, и тазовой, или пояс задних конечностей. Каждый пояс представляет ряд костей, промежуточный между скелетом туловища и скелетом конечностей; к этому ряду костей, а не прямо к скелету туловища примыкает скелет конечностей.
В каждом поясе различают спинной отдел, кверху от сочленовной лля конечности впадины, и брюшной—книзу от этой впадины. Первичный скелет плечевого пояса во всех крупнейших группах позвоночных представляет более или менее ясно четыре части: suprascapulare и scapulare (или scapula, лопатка) в спинном отделе, coracoideum и procoracovkeum
в рюшном (фиг 556—558). Такое же расчленение представляет первичное устройство тазового йояса у низших позвоночных (Фш 559). В тазе четвероногих различают в спинном отделе подвздошную кость (ilium), гомо логичную scapula, переднюю кость в брюшном отделе—лобковую (pubis) и заднюю—седалищную (ischium)-, как в плечевом поясе, так и в тазовом третья часть его, coracoideum или соответственно ischium,
отклоняется назад, так что для восприятия давления конечности получается простейший механизм в виде треноги, с сочленовной впадиной в центре. Это отклонение обусловливает и отклонение пояса от типа жаберной дуги. Впрочем, при слабо действующих конечностях, когда от них не передается давление на пояса, как у рыб, пояса являются снова в форме, близкой к первичным жаберным дугам. У рыб отсутствует спинной отдел в тазовом поясе. Кроме названных костей, развивающихся на месте первоначальных хрящей, в плечевом поясе напереди развиваются еще кожные кости: ключица, clavicula, налегающая иа коракоидную часть и сходящаяся посредине с ключицей другого бока, и кнаружи от нее вторая кость—задняя ключица,
cleithrum (фиг. 633, 638, 655).
Ч
-,U
Фиг. 558. Плечевой пояс примитивной рыбы Acantlodesws пермских отложений Германии
Фпг. 559. Тавовой иояс пермской амфибии >1/-ehegosavrut
Фиг. 556. Плечевой пояс примитивного ныне живущего пресмыкающегося Sphenodon.
Фиг. 557. Плечевой пояс примитивного ныне живущего млекопитающего Огт-thovhynchiis
Значеиие букв о, Ъ, е, d—см. фиг.
561.
Пояса поддерживают парные конт чности, передние и задние. Конечности бывают двоякого рода: плавники у водных позвоночных, именно у рыб, и пятипалые ноги четвероногих
У селахий, принадлежащих к низшим рыбам, передние вли грудные плавники построены так (фиг 560), что соединены с плечевым поясом тремя основными хрящевыми члениками, которые различаются как Piopteiyyium, Meso- и Metapterygium К ним примыкает, в мозаикообразном расположении, комплекс маленьких хрящевых члеников, связанных между собою тонкими, прочными прослойками < оединительной ткани. Па периферии находится кайма вз роговых нитей.
Что касается до задмих или брюшных плавников 'елахий, то они подобны передним, но стоят на низшей ступени развития, что выражается прежде всего в уменьшении числа основных члеников,—не развивается мезоптеригий, а иногда также и проптеригий
Данные эмбриологии и палеонтологии заставляют считать главным лучом всего Плавника метаптеригиальный луч, состоящий из метаптеригия (фиг. 560) с лежащими по направлению его оси члениками. Именно этот луч закладывается ранее других у зародыша и, с другой стороны, сильно развит у древнейших ископаемых селахий.
Вторичные лучи примыкают к нему обыкновенно лишь е одной стор !НЬ1 (у ни сериальный тип плавника, фиг. 564); лишь в немногих случаях находятся еще также лучи на противоположной стороне (б и сериальный тип плавника). Последнее состояние становится типичным- для двоякодышащих рыб, особенно у рода Ceratodus (фиг. 563, С).
Главный луч здесь является срединным с почти симметричным расположением вторичных лучей по обе стороны его.
Скелет передних и задних конечностей четгероногих позвоночных состоит из гомологичных костей. В проксимальном отделе каждой конечности лежит одна прямая кость—плечевая, или humerus, в передних (фиг. 562, HV) и бедренная, или femur, в задних В среднем отделе находятся две удлиненные кости: передняя—л у ч е в а я, или radius (RA), и задняя— fs ~
Фиг. 5С0. Плечевой пояс и грудной плавник акулы Heptanchus. Ныне живущ.
SB — плечевой по' с; р проптеригий; те — мезоп-тернгип, nil — «етаптерпги i; т — хрящевые лучн, Л — перерезанные рог< вью пяти; fc — отверстие в коракоиде
Фиг. 531. Кожпые кости плечевого пояса девонской панцирной рыбы Coccosteus. а, Ъ, с, d — suprsscapulare, tcapulare, ста-eoideum и procoracbaleum в плечевом поясе, ми соответг .зенно поперечный отросток тазового ^крестцового) ц эввонка, ilium, ischium и pubis тазового пояса. Сгг—гра 1ица срастания между sca-pularen ccracoidewn у Acanihodes.
локтевая, или ulna (UL), в передних конечностях и передняя—большая берцовая, или tibia, и задняя—малая берцовая, или fibula, в задних конечностях.
Затем следуют кости, кисти и ступни, в свою очередь делящиеся на три группы: carpalia, или кости запястья, mel'icarpalia (mtcp), или пястные, и фаланги (членики пальцев) (ph) в перед них конечностях; tarsalia, или кости предплюсны, metatarsalia, или плюсневые, и фаланги в задних. Carpalia и tarsalia в типе состоят из трех рядов хрящиков или косточек, в проксимальном ряду их три, в среднем — два и в дистальном — пять элементов. Три проксимальных carpalia суть radiale (га), intermedium (mt) и ulnare (ul), две среднего ряда—первая и вторая centralia (сп 1, сп 2), пять дистального- -distalia (dst. 1—5), которые считают, начиная с переднего или радиального края конечности. В предплюсне кости первого ряда соответственно называются tibiale, intermedium и fibulare, второго—centralia и треть-
18
Учебник палеонтологи*
2?3
Qvo—distalia. Metacarpalia и metatarsalia представляют собою по пяти палочковидных костей, сочленяющихся каждая с отдельной distdle-, за ними следуют фаланги, от одной до пяти в каждом пальце. Первый палец передней и задней конечности (I) называется большим; пятый палец каждой конечности - м и з и н ц е м.
II ipn .ie плавники рыб и пятипалые конечности четвероногих, должно быть, связаны между собою отношением нрэиехожде ня. По что из них является первичным, что производным,— вопрос, решаемый до сих пор в противоположном смысле двумя группами ученых в связи с тем, что одни предполагают происхождение четвероногих от рыб, а другие обратное. Затем самый переход от конечностей одного рода к конечностям другого не выяснен еще детально, промежуточные формы не найдены нн зоологией, ни палеонтологией. По вопросу о происхожде-
нзи парных плавников существуют три весьма различные теории.
1. По одной из них (Гегеибаура) парные плавники являются производными от жаберного аппарата; жаберные дуги образуют пояса, а сидящие па них лучи, постепенно изменяя свое относительное расположение, превращаются в основные и вторичные лучи симметричного бисериаль-ного плавника, который таким образом представляет первичный тип плавника (фиг. 563). Положение Плавников назади, в особенности брюшных плавников, объяснялось вторичным смещением назад задних жаберных дуг, которые утратили свои первоначальные жаберные функции.
2,- Вторая теория (Бальфура, Т а-чераи Майварта) считает,что парные плавники по своей природе таковы же, как
Фиг. 5G2. Схема строения передней конечности с плечевым поясом.
gl — сочленовная ямка, SCP — scapulare; COR — coracoideum; р. cor — procora-cotdeum. I — V — пальцы* CL — c'avicula; HU — bumerus; RA — radium- UL — ulna; та — radiale, ul — ulnare; ini — intermedium, cn. 1, cn. 2 — centralia; dst 1-5 — distaha; mtep. 1 - 5 — metacarpalia;
ph — фаланга.
Фиг. 563. Гипотетическое происхождение парных плавников (С) из жаберных дуг (Л). Три последовательных стадии.
и непарные, и развились из парных продольных боковых плавниковых складок (фиг. 564); лучи возникают здесь для уплотнения плавника, подобно тому как в непарных плавниках, а пояса развиваются распространением вовнутрь тела базальных частей лучей, обособлением, а затем срастанием их так, что получается твердая опора в основании плавника Авторы обеих этих теорий считают пятипалую конечность производною от плавника.
3. Третья теория, созданная Екелем, считает на основании палеонтологических данных (плавники девонского и каменноугольного рода селахий Cladoselache) исходным типом плавника унисериальиый тип с двумя основными лучами, из которых задний (метаптеригий) является главнейшим, а передний (проптеригий) добавочным. Как в плавнике, большая часть лучей (пальцев) пятипалой конечности примыкают к заднему основному лучу, обозначенному ulna в передней конечности и fibula в задней. Такое относительное расположение лучей особенно очевидно сказывается в их иннервации У современных позвоночных (фиг. 565), а, кроме того, заметно и у некоторых примитивных ископаемых форм, например у некоторых представителей рода Plesiosaurus (фиг. 566). Таким образом гомология парных плавников и йог четвероногих несомненна. Е к е л ъ
считает конечности позвоночных не возникшими у них заново, независимо от предков позвоночных, но унаследованными от этой прародительской группы, каковою Е к е л ь (и некоторые др.) считает ракообразных с их двуьетвистыми расщепленными ногами.
В связи с позвоночным столбом находится скелет непарных плавников рыб. Таковыми являются: спинной, часто поделенный на дг.а плавника — передний и задний, хвостовой и расположенный позади заднего прохода—анальный (фиг. 564). Непарные плавники получаются в результате расчленения одной сплошной кожной складки, которая тянулась по спиле, загибалась на месте образования хвостового плавника и доходила по брюшной стороне до анального отверстия. Эта складка волнообразно колебалась, усиливая движения животного, происходящие от такого же колеба-
тельного движения туловища. Такой непрерывный плавник встречается у
—у"" .......у ............— i зародышевом
колебавшпеся
некоторых рыб в состоянии их зрелости и у многих рыб в или личиночном состоянии, а также у некоторых амфибий — тритонов, личинок лягушек (головастиков) и др. При колебательном движении плавника в нем должны были возникнуть, так сказать, узловые участки, менее всего
gw
Фиг. 566. Проксимальная часть скелета передней: конечности плезиозавра Crs:ptoclidus oxoniensis.
Фиг. 564. Гипотетическое происхождение парных и непарных плавников из плавниковых складок. Наверху рыба с плавниковыми складками н намеченными плавниками, вппзу — рыба с плавниками:
г—спинной, «—хвостовой, в— анальный, bf — брюшной н Ът — грудной плаввикн.
Фиг. 565. Пи-иер’алия ноги Ranodon sibi-ricus.
Н—humerus, Т— Н~ humerus, R— tibia, i'--iibu'a. radius, 8—unify
или вовсе не испытывавшие колебательного движения и потому атрофировавшиеся. Плавники рыб, вскоре после их возникновения, должны были приобрести дающий им стойкость остов. Этот остов состоит из палочковидных, хрящевых, костных пли роговых лучей; лучи имеют опору в лученосцах, примыкающих к остистым отросткам позвонков (фиг. 555).
В связи с передним концом спинпой струны, где нет позвонков, развиваются некоторые части головного скелета, пли черепа. В черепе различают две части: верхнюю — краниальную, являющуюся вместилищем органов чувств и головного мозга, и нижнюю — висцерал ьную, охватывающую собою переднюю часть кишечника и состоящую из нескольких пар скелетных дуг, возникших в простенках между жаберными щелями и называемых висцеральными. Череп развивается у зародыша из двух пар хрящевых пластинок, симметрично примыкающих к переднему концу хорды по бокам и следовательно, продолжающих с^бою ряд центров позвонков. По мере того как идет развитие, эти пластинки срастаются между собою и обрастают передай конец хорды, образуя дно черепа,
№.
подстилающее зачаток головного мозга. При разрастании этого пласти» чгтого дна вверх получается мозговая хрящевая коробка. К этой коробке с боков прирастают хрящевые капсулы, содержащие органы чувств: назади слуховые, напереди носовые. Лежащие между ними зрительные капсулы, в связи с подвижностью глаз, остаются свободными от соединения с черепом. Зрительные капсулы представляют склеротику, наружную оболочку глаза.
Таким образом возникает первичный хрящевой череп, например акул (фпг. 567).
Скелет висцеральной части черепа является в виде некоторого количества хрящевых стержней, которые расположены во всех жаберных дугах сверх) вниз и представляют скелет, развивающийся независимо от краниальной части и охватывающий переднюю часть пищеварительного канала в виде параллельных между собою цуг. Самая передняя из них называется челюстной дугой, так как она образует остов челюстей, верхней и нижней.
Фиг. 567. Головной скелет акулы Squattna vulgaris.
Etn—bocoeoS втмгмдальныЙ отдел; Кг — хрящ брызгалъца, Ро — за^веглазнвчный, поствр-бетальвый отросток; Осс — затылочный отдел; PQ—нёбно-квадратный хрящ; Z, Z', L*'— губные хрящи; Ed — мандибулярный хрящ; НцЕ—гио-мавдибуляриый хрящ; hy — гиоидный (подъязычный хрящ. Со — copula гиоидной дуги я жаберных дуг; HI — VII — жаберные Дуге; £Вг, С£г, НВг — части жаберной дуги. BR — жаберные лучи гио-кандибуляряого хряща; Вг — жаберные лучн гиоидной дуги.
Она постоянно представляет два отдела; верхний из них — квадратный (Quad: alum) — соединяется сочленением или прирастает неподвижно к боковой стенке собственно черепа непосредственно перед слуховой капсулой и высылает вперед и вверх к черепу крыловидно-нёбную (Pterygopa-latmum) ветсь, а также несет на своем нижнем конце нижнюю половину скелета челюстей — нижнечелюстной Меккелев хрящ (Mandibulare). Крыловидно-нёбный хрящ образует верхний край рта, нижнечелюстной хрящ постоянно сбраз] ет нижний край рта и соединяется в срединной плоскости тела с таким же хрящем другого бока. Следующие жаберные дуги в большей степени расчленены и соединяются на брюшной стороне миленькими соединительными чаиячи (copulae), образ я корзинкоподобный скелет, который только через посредство самой передней из этих дуг, подъязычной пли гиоидной (H'joideum), прикрепляется с одной стороны к скелету челюстей, а с другой—к черепу (фиг. 567), Ilyvideumявляется
единственною частью скелета жаберных дуг, которая удерживаете^ всеми позвоночными, тогда как остальные жаберные дуги у всех живот«ых, у которых отсутствуют жабры, отчасти также исчезают или примыкают в рудиментарном состоянии к Hyoideum. Между челюстной и гиоидной дугой лежит первая жаберная щеть, которая или является рудиментарной (кругло-ротые, миноги), или видоизмененной (брызгальце, или spiraculutti, рыб и среднее ухо прочих позвоночных).
У высших позвоночных череп не остается первичным, в виде сплошной хрящевой коробки; у них развивается вторичный череп, возникающий вследствие развития в хрящевых стенках первичного черепа костей
Фиг. 568. А — схема строения вторичного черепа позвоночного в вертикал ином разрезе. Хрящевые копти обозначены прямым шрифтом, кожные — курсивом; В— поперечный разрез черепа в затылочной облаетг; С—в теменной области; D—в лобной области;
Е -в носовой области.
Ifск. С—ниягпечелгечной или Меккелев, хрящ. Хрящевые кости: AL, SPH — alt-3phenofdeum; ART — articu>are: В. ОС — basiocciptlale; В. SPLi — basis-phenci-deum-, EC. ET— setoethmoideumt OP. ОТ—epsoticum EX. ОС — ‘Exoccipilals, BY. SI — hgo-mandtbularc', HE. ET ~ metoettonoideum, OP. ОТ epist? oticum; OR. SPB— orbaosphenotdeum', PAL — paUUnum: PR. ОТ — prooticum; IB. SPH—prues-phenoideufn; PTG — pterygofdeum; QU — quadratum; S. ОС — карта occipital#'- В. BR, C. BR, PH. BR — костя жаберной дуга. Кожные госта: DNT~- dentale: ER — frontale* ИХ — ma‘ilia; HA — nasals', PA —parietale: PA, SPB — paraepkencideum; PMX — praemaxilla\ SQ — squamosum to — сошник. Выдающийся хрящевой передали конец черепкой коробки образует rostrum. ,,
в определенном количестве. Кроме этих костей, развивающихся на месте хряща, в состав черепа входят еще кожные кости, образовавшиеся в соединительной ткани вне хряща. Эти кожные кости пли прилегают и черепу и тогда сравнительно легко отделяются от него, или же (у высших позвоночных) срастаются с остальными костями черепа таким же образом, как последние между собою, становятся подобно последний неотделимы от черепа м неотличимы от костей, возникших в хряще.
В затылочной области черепа развиваются четыре кости вокруг затылочного отверстия (фиг. 568); непарная нижняя —основная затылочная, basioccipitale, парные —боковые затылочные, exoccipiiaHa, и непарная верхняя затылочная — supraoccipitale. В стенках слуховой капсулы развивается ряд костей oii-м (каменистая кость, petrosum четвероногих позвоночных), обыкновенно три: prooticum— спереди, opisth-Шсит — сзади и epio .cum ~ наверху. В этой же области развивается еще
вожпая кость, лежащая выше и впереди otica, и именно позади и сбоку лобных костей, pteroticum или squamosum (чешуйчатая, или височная кость). Спереди от basioccipilale в дне черепа развивается кость, называемая основной клиновидной — basisphenoideum. С каждой стороны от has sphenoideum лежат парные крыло-клиновидные кости, ali-sphenoid?» (AL. Si’ll), принимающие участие в (бразозапии боковых стенок межглазничной области. Впереди basisj.hcnoideum находится передняя клиновидная кость—praesphenoideum, с которой связаны парные г л а з н и ч но-клиновидные — orbitosphenoidea (OR. SP II), дополняющие боковые стенки межглазничной области. Basioccipilale, basisphenoideum и praesphenoideum образуют вместе основание костного черепа. В задней части хряща, лежащего между обонятельными капсулами, появляется вертикальная костная пластинка, средняя решетчатая к о с т ь — mesoethmoideum (ME. ЕТ), а наружные стенки обонятельных капсул окостеневают парными боковыми решетчатыми окостенениями — ectoethmoiclea (ЕС. ЕТ). Наверху полость черепа замыкается двумя парами кожных костей: теменных—parietalia (РА), развивающихся непосредственно впереди supraoccipitale и снизу обыкновенно сходящихся с ali-sphenoidea, и лобных — frontalia (FB), находящихся перед parielalia и пшзу часто соединяющихся с orbitosphenoidea. Пара носовых— nasalia (NA) развивается над обонятельными капсулами, непосре (ственно перед trontalia-, наконец под основанием черепа развиваются две важные кожные кости; впереди сошн и к — vomer (vo), который может быть парным, и paraspheiioideum (РА. SPH) сзади. Под глазной орбитой рыб протягивается подглазничное кольцо кожного происхождения, соответствующее скуловой дуге четвероногих. Одна из костей его (infraorbitalia), лежащая у переднего края, достигает весьма значительной величины и называется слезной, lacrimae
Первичные хрящевые челюсти, окостеневая, также получают сложное строение, состоя из нескольких костей.
Первичная верхняя челюсть или palato-quadratum окостеневает тремя главными хрящевыми костями с каждой стороны: нёбной — palatinwm (PAL- спереди, затем крыловидной (у рыб — нескольких крыловидных) pteryqoideum (PTG) и квадратной—quadratum (QU) сзади. Е ним присоединяются кожные кости: межчелюстные — praemaxilla (РМХ) и верхнечелюстные — maxilla (MX), которые в костном черепе и образуют настоящую переднюю границу рта. Проксимальный конец первичной ппжпей челюсти окостеневает в хрящевую кость — articulare (ART), с помощью которой но («егаена нижняя челюсть; остальная ее часть остается в виде тонкого пеокостеневающего хряща, который может совершенно исчезнуть в - взрослом состоянии. Вторичная нижняя челюсть образована изменяющимся числом кожных костей, из которых самая важная передняя — з у б н а я, dentate (DNT).
У современных позвоночных нёбно-квадратный и мандибулярйын хрящи являются спло.' ними । бр-is ьанпмчи и зоологи считают их за первичные элементы челюстной д\'1и, ничеп' нэ говоря о том, куда же девались остальные элементы ее как жаберной дуги. И-, иалеоиго.югпческис данные, хотя и немногочисленные пока, указывают, что первоначально верхняя и нижняя челюсти имели более сложный состав, чем впоследствии. Именно, у и.тлезойско;о представителя селахий, рода Acanthodes, верхняя челюсть состоит л.ч трех частей (фиг. 569), причем две пз вид, Ь3. п а3, должны принадлежать одной жаберной дуге, которая и является собственно челюстью, а часть а3 принадлежит
дугв, лежащей еще далее впереди. Также и hyo-manthbv’are акул представляет здесь двух-члеиистость, соответственно нормальному подразделению прочих висцеральных дуг. Нижняя челюсть состоит из 2 первичных частей, так что вся челюстная дуга состоит из четырех, как вообще жаберные дуй. Возможно, что впереди са в верхней челюсти находилась 4-я составная часть ее <.«,), хрящевая, не сохранившаяся, дополнявшая верхнюю челюсть в длину соответственно длине нижней челюсти У современных акул все эти элементы версией челюсти слиты в одно целое, чем ей сообщаемся бол шля прочность, яеобходп ая при хватанпи добычи. Но и у со- j еменаых акул существ) ют части, которые должны считаться за рудименты висцеральных дуг Это так называемые губные хрящи (фиг. 567), которые принадлежат двум дугам, частью вошедшим (как можно думать, судя по Acanlhodes) в состав передней чаыи верхней челюсти. Схема фиг. 570 поясняет состав околоротового скелета в первоначальном его виде, причем не делается такого различия между хрящевыми и кожными стями челюстей, как в схеме фиг. 568.
Фиг. 570. Наверху — схематическое изображение первЩаною состояния пяти жаберных луг, вошедших в состав головы. Внизу схема расположения частей этих дуг но образовании челюстей.
АГ — нос, А — глаз, О — ухо, Gr — зад-♦ над граница головы.
ко-
Фиг. 560. Схема строения челюстей на-еозойской акулы Acant-hodes.
1~4—жабррные дуги, < оптедппте в состав чслюс1вй, 3— собств* sno челюстная, -1— подъязычная дуга; a-d — четыре части каждой не жаберных д)г;Ап ЪЛ1 ^—губные хрящи.
Е заключение коснемся еще некоторых особенностей' мягких частей тела позвоночных. Пр.жде всего — строения сердца. Сердце высших позвоночных четырехкамерное, с совершенным обособлением правой половины от левой. Каждая половина в верхнем (или переднем) конце представляет предсердие, в нижнем (или заднем) желудочек, сообщающиеся между собой отверст ем с клапаном в разделяющей их перегородке. Левая половина сердца содержит кровь, освободившуюся в легких от углекислого газа и насыщенную кислородом (артериальную), она распределяется отсюда по телу. Правая половина сердца содержит кровь, поступившую из всего тела по выполнении процессов окисления, насыщенную углекислым газом (венозную), идущую отсюда в легкие. Таково строение млекопитающих и птиц. У рептилий и амфибий желудочки не вполне отделены, поэтому происходит смешение артериальной и венозной криви; дых'одяпая из сердца х' и производящая в теле процесс окисления крсвь не бывает с самого начала вполве артериальной, т. е. свободной от углекислого газа, и процесс окисления происходит не так энергично, как v млекопитаюпшх и птиц; у рыб сердце состоит из одного предсердия и одного желудочка, является следовательно венозным.
Фиг. 571. Череп амфибии Alasto-doiisav us (hganteus с верхней стороны. Слизевые каналы показаны жирныцж линиями.
Kptuie того у млекопитающих и птиц кровообращение происходит по двум кругам, соприкасающимся в сердце, один — от левого желудочка до правого предсердия, большой круг, в котором кровь совершает процесс окисления, и другой—от правого желудочка до левого предсердия, малый круг, в котором кровь в легких восстановляет свою окисли юльную способность. Таким образом каждая капля крови проходит дважды через сердце, преж е чем сделать весь свой пут^ по телу дважды получает в сердце толчок, застаьтяющий ее двигаться. У рыб кровь проходит путь в одном кругу и только один раз получает импульс к движению. Здесь кровь Двигается медленнее. Наконец дыхание жабрами в воде со слабым содержанием растворенного кислорода происходит не с такой энергией, как в воздухе. Поэтому млекопитающие и птицы развивают наибольшую энергию в процессе окисления, рыбы — наименьшую, рептилии и амфибии занимают промежуточное положение; первые две группы представляют животных теплокровных, с постоянною температурою тела, не зависящих поэтому в своей жизнедеятельности от температуры окружающей среды; последние же три группы — животные холоднокровные, с переменной температурой тела, зависящей от температуры окружающей среды.
Кроме хорошо развитой, присущей всем позвоночным, функционирующей пары глаз, у многих из них существует еще пара более или менее рудиментарных органов, стоящих в связи с чувством зрения. Именно, верхняя стенка головного мозга образует два один за другим лежащих выступа: передний — теменной (или париетатьный)) орган и задний — эпифиз. Иногда эти органы бывают глазоподобны, представляют сетчатую оболочку и светопреломляющее чечевицеобразное тели. Таг у пресмыкающихся бывает
значительно развит париетальный глаз, нерв которого проходит через особое теменное отверстие черепа, а у миног, наоборот, более развит эпифиз. Предполагают, что это — два глаза одной пары, смещенные из своего первоначального положения так, что лежат не по обе стороны плоскости симметрии тела, но в этой плоскости один за другим.
Кроме органов тех чувств, которые свойственны наьемным четвероногим, рыбы, водные амфибии и личинки наземных имеют органы так называемой боковой линии, кожного ми 6-го чувства. Это чувство предполагается до известной степени средним между чувствами слуха и осязания, во лишь более томим, чем каждое из последних в нек< торых отношениях. Оно воспринимает слабое колебательное движение водной среды, не ощутимое ни слухом, ни осязанием. Органы кожного чувства расположены по известным линиям на голове и в виде одной, двух или более продольных линий по бокам Туловища. В простейшем случае по этим линиям находятся так называемые слизевые каналы (фиг. 571). В других случаях эти каналы опус каются с поверхности тела на некоторую глубину, сообщаясь
тогда с наружной средой отверстиями, расположенными черва извесиые промежутки.
Позвоночные представляют еще увакительвые различия в ошогаеиип яекоторых особенностей эмбрионального развития, В этом развитии у позвоночных яфипотся известные преходящие образование питательный желток, амнион и аллантоис. ПитательиыЛ желток есть часть яйца, служащая для питания еще неспособного к совершеиио самостоятельной жизни животною. Он скопляется в брюшной стейке кишки, растягивая ее в форме мешка (фиг. 572); именно у большей части рыб и амфибий, причем при из-рас хо ювании желтке, этот _ешок исчезает. Но уже у селахий п затем у так на; ываемых амниот желточный мешок так отшнуровывается от тела, что между ними остается только трубчатое соединение, стебел нчтообрвзньй пупочный тяж.
Амнион есть оболочка зародыша, которая существует лишь у высшпх позвоночных. рептилий, птиц и млекопитающих, объединяемых поэтому под названием амниот. Амнион возникает таким, образом, что зародыш впадает в желточный мешок, а складка—
края впадины—стягивается и наконец срастается вад за-। Лишем Внут]вннпй лист складки, пузыреобраано окружающий зародыш, есть собственно амнион, который под зародышем продолжа тся в иаруж ын слой пупка и далее в стенку тела вародыша. Наружные лист складкн, который отделяется от амниона, а
Фиг. 572.7—желточный мешок рыб и ааврюнй. Ли III —желточный мешок, аллантоис и амнион амниот.
е — тело аз-родышз., из кишка которого (d) выпячивается желточный мешок ds; al — аллантово, а— амнион, з — s гола, вя —воренвки ее.
также от желточного мешка, покрывает собою все остальные части сародыша и назы-ва тся серозной оболочкой (serosa). Ал лаптопе амниот
есть во что иное, как брюшной мочевой пузырь амфибий и отчасти более высших позвоночных. У большинства млекопитающих последовых (Placentalia) мочевой пузырь есть новооб азовавие, хотя" и стоящее в непосредственной связи со стеблем аллантоиса. У Placentalia аллантоис разрастается, выполняя пространство между желточным мешком и зародышем, срастается с serosa н затем, через ее посредство, с маткой материнского тел., образуя послед (placenta) и служит также целям питания плода вместо рудимен-
тарного желточного мешка.
КЛА.СС I. РЫБЫ, PISCES.
Рыбы в течение всей жизни дышат жабрами, расположенными в виде мясистых бахромок на жаберных дугах. Первичным числом жаберных щелей (считая и брызгальце) должно быть по крайней мере восемь. Первичные челюсти, происшедшие главным образом из челюстной жаберной дуги, бывают различно прикреплены к черепу. Квадратная часть верхней челюсти или прямо сливается со слуховой областью черепа, или поддерживается промежуточным между нею и черепом элементом, hyo-mandibulare, спинным элементом гиоидной дуги; эти два типа черепа и сочленение с гим челюстей называются автостилическим и гиостилическим.
В связи с жабрами развивается иногда аппарат жаберных крышек, подвижных кожных костей, прикрывающих жаберные щели.
Чешуя, покрывающая тело рыб, бывает:
1) плакоидного типа, т. е. типа кожных зубов (фиг. 57S), причем она часто сменяется, подобно зубам низших 'позвоночных. Плакоидная чешуя обыкновенно является в виде маленьких ромбических табличек, звездчатых, листовидных (фиг. 574) заостренных, лопатовидных или даже ко-
нических образований, которые лежат плотно вместе и образуют шероховатую поверхность (шагрень);
2) ганоидного типа (фиг. 575); такая чешуя достигает большей величины, чем плаковдная. Она ромбической или округленной формы, и
в последнем случае смежные
чешуи налегают черечитчато од а на другую. Ромбические обы гноьеньо расположены пра-
Фпг. 578. Чешуйка современного ската (JtnJa) в натуральную величину.
Фиг. 57-1. Че-шуГдси современной акулы (Саг-chanas) в увеличенном виде.
Фиг. 575. Ганоидная чешуйка Elomohthys stratus, Камен-яоуг. а — снар .л:и, Ъ — со-впутри. 3/1.
Фиг. 577. Циклоидная че-
шуйка соврем иной костистой рыбы (Naucrates), сольно увеличена, сбоку показана в на-
фиг. 578. Ктеноидная чешуя современной костистой рыбы (Solee). Слева
Фиг. 576. Две ганоидные чешуи современной кв-стеиерой рыбы (Г'0'ypterusi с внутренней стороны.
турельную величину.
в натуральи. величину.
Фпг. 579. Ихтиодорулит Cteticcanthus. Нижнекаг
ВИЛЬПЫМИ рядами И ПОДВИЖНО сочленены зубовид- ыенвоугольиые оиоженпя НЫМ отростком верхнего края, ВХОДЯЩИМ В соответ- реД“еИ части ствующее углубление внутренней стороны приле-
гающей чешуи (фиг. 57-6). Открытая наружная поверхность покрыта слоем эм 1ли. Он о дыкновенно толст и блестящ, иногда темно окрашен; эмаль гладкая, шероховатая или ребристая;
3) типа циклоидных п ктеноидных чешуй. Это—тонкие гибкие чешуи округленной, эллиптической, четырех, пяти или шест»)гельвой формы (Фпг. 577 и 578). Ктенондные чешуи происходят вследствие образованы выступов лини? нарастания, так что возникают зубцы, расположенные по радиусам па свободном заднем краю (фиг 578).
Иногда у рыб в коже, на теле и на голове развиваются большие костяные пластинки, щитки и шипы.
Большие шипы, лхтиодорулиты (фиг. 579), обыкновенно встречаются на переднем краю непарных, более редко парных плавников, ши па голове акуловых рыб (Sulachil). Часто достигают весьма значительной величины и ктрсчаютея обособленными в ископаемом состоянии, причем палеозойские экземпляры их иногда характеризуются разнообразной, сильно развитой ску тьптурой. Лучи плавников у высших рыб также являются костными; бывают цельными или подразделенными па многочисленные мелкие членики, причем часто разв 'твляются к дистальному концу.
3j6w бывают на всех хрящах или костях ротовой и жаберной полостей; но форме бывают коническими (хватающие) или сжатыми с двух противоположных боков, так что являются подобными наконечнику стрелы, с режущими краями.
На одном основании могут находиться несколько зубцов. Для раздробления пчщи, иногда состоящей из снабженных раковинами моллюсков, служат птоские, полусф рические или булыжникообразные зубы. В молодости почти у всех рыб зубы прикрепляются к хрящу или кости челюстей волокнистой соеди-
Фиг. 580. Гояоцеркяып хвостовой плавник фиг. 581.Дифпц<'ркнЫи хвостовой плав-лосося, ник современного Polypterus.
нптетьпой тданыо, а у акул и многих костистых рыб такое прикрепление существ ет во время всей ® :зяп. У костистых рыб и многих ганоидных корни зубов срастаются, сливаются с несущею их костью, и обыкновенно вокруг н 1жней части зуба вырастает чашечковидное возвышение. В исключительных сл\чаях зубы сидят в обособленных*** ячейках. Идущие на смену зубы обыкновенно развиваются сбоку сменяемых и выталкивают послезние.
В позвоночном столбе различают туловищные и хвостовые позвонки, отличающиеся; п -рвые — подвижными ребрами, последние — неподвижными гемапо^изами (юмальными дугами).
Задн й конец позв ночного столба входит в хвостовой плавник, фирма котор* г > различна, представляя филогенетическое и системашческое значение Первичным, исходя от низших । ыб, надо считать состояние д и-фицеркного плавника (фиг. 581). В нем позвоночный столб протягивается по прямой линии до конца туловища и симметрично обрамлен сверху и снизу хвостовым плавником. Гетероцеркный плавник (фиг. 582) представляет позвоночный столб отклонившимся в верхнюю лопасть хвостовою плавника, которая обыкновенно щ евосходит по своим размерам нияснюю лопасть. Производным от гетероцеркного плавника является гомоцерк-
ный (фиг.. 580), который представляет полную внешнюю симметрию при слабо выраженной в расположении конца позвоночного столба внутренней асимметрии. Именно позвоночный столб представляет отклонение своего конца кверху, хотя и на небольшом протяжении. Почти все палеозойские рыбы имеют дифицеркный или гетероцеркный хвост. Костисые рыбы начиная с меловых отложений, имеют гомоцеркный хвост.
Нередко находят в ископаемом состоянии обособленными слуховые йо-сточку, или отолиты рыб (фиг. 583), представляющие большое разйооб-
Фиг. 682. Гетероцеркный хвостовой плавник осетра
Фиг. 583. Слуховая косточка (Otolithus) рода Sciaena. Нижке-третичпые отложения Гермаиии. 2/1.
разив по форме и украшениям, так что на основании их одних оказыва лея возможным установить вид данной формы.
В развитии рыб не бывает ни амниона, ни аллантоиса
Рыбы подразделяются на пять подклассов: I. Круглоротые (Cyclostoma). II. Панцирные рыбы (Placodermi). Iff. Artbrodira (т. е. с сочлененной ггсгй). IV. Elasmobrancnii (жабры имеют форму пластинок) или Chondrichthyes (хрящевые). V. Teleostomi (т. е. с совершенным ртом: челюсти из кожныхк лтей, вооруженных зубами) или Osteichthyes (костистые).
Подкласс I. Круглоротые, Cyclostomata.
Круглоротые представляют маленькую группу, о систематическом значении которой существуют различные мнения. Одни выделяют их в оербый класс, другие считайт только за порядок рыб.
Это просто организованные животные, частью полупаразитические. Они не имеют челюстей, парных конечностей, чешуй и представляют одно непарное носовое отверстие. Осевым скелетом служит хорда, к которой лишь в области головы присоединяется хрящевой скелет. Круглое ротовое отверстие окружено роговыми зубами. Кроме ныне живущих, известны два проблематичных рода в нижнем девоне.
Подкласс П. П..нцырные рыбы, Placodermi.1
Древнепалеозойские рыбы с неокостеневающим внутренним скелетом. Голова и передняя часть туловища покрыты симметрично расположенными
1 Новейшее исследования вносят в его понятие повив данные. Согласно етим в®' следованиям панцирные рыбы могут быть сблажевы о ^стоящими Cyclostomata (жругдо-ротымн).
костяными пластинками. Парные плавники отсутствуют; напротив, часто существуют покрытые панцирем органы гребли или хватания, подвижно сочлененные с телом, На голове находятся слизевые каналы и теменной глаз Зубы, если имеются, не обособлены от челюстей; последние не всегда имеются.
У рода Pleraspis (фиг. 584; pteron—крыло, aspis— щит) костяной панцирь, находящийся на передней части тела, состоит из спинной, брюшной и боковых частей. Граница между головой и туловищем не выражена подразделением костяного панциря, но намечается лишь положением внутренних органов. Спинная часть панциря выдается клювообразно вперед, а
Фиг. 584. Реставрация aspis rostrata. Нижний де юн. Англия.
назади представляет шип, находящийся в срединной плоскость тела. По бокам находились парные глаза, а лежащий несколько позади их непарный глаз не выходил наружу, оставляя лишь углубление совнутри спинного щита. На внутренней же поверхности последнего с каждой стороны существуют впечатления 6 жаберных дуг. Жаберною щелью вероятно было отверстие на боковой части панциря, позади глаз, с каждого бока. Сзади панцыря тело было покрыто наверху ромбическими, на боках удлиненными чешуямп. Передние конечности вероятно отсутствовали, подобно тому как у личинок амфибий.
Фиг. 586. Головной щит Ceph-alaspts Murchisoni с отпечатком жаберных щелей (жирным) и жаберных дуг, вид снизу.
Нижний девон. Англия
Вероятно эти животные имели способное выпячиваться ротовое отверстие) Верхний силур и нижний девон.
У Cepluilaspis (фпг. 585, 586; kephale — голова, aspis — щит) панцырь в приспособлении к жизни на дне, сплюснутый дорзовентрально, с глазами, переместившпмися на средину верхней стороны. Панцырь без подразделения на отдельные кости, глапшй, округленный напереди, усеченный назади, с остроконечными продола ениями задних углов. Имеется шесть жаберных дуг и соответственно им семь пар жаберных отверстий, симметрично расположеньях на нижней стороне головного щита (фиг. 586). Тут же находилось ротовое отверстие? Тело в поаеречном сечении треугольное, с каждого бока Покрыто тремя продольными рядами чешуй. На животе мелкие чешуи. У основания гстероцеркного хвоста находился треугольный спинной плавник. Верхний ецлур, девон.
Далее следуют панцырные рыбы, по своей организации приближающиеся к обычному типу организации рыб.
У Pterichthys (фиг. 587; pteron — крыло, ichthys— рыба) голова укорочена по сравнению с туловищною частью тет; череп состоит из костей, могущих быть поставленными в параллель костям нормального черепа позвоночных. Парные глаза с кольцевыми окостенениями склеротики; третий глаз помещен в четырехугольной пластинке между парными глаз!мн На нижней сдороне головы, на переднем краю ее, находятся две поперечно вытянутые
Фпт. 587. Реставрация Plerichthyg hltUeri. Девон. Шотландия. А—[вид со спинной стороны, В — с (брюшной, С—сбоку. Пунктиром показано распространение слизевых каналов.
С — ключицы, Е — epioticum, Fr— лобные кости, J —jugale, Ic — wterclartcula, межклгочвца, т — верхние челюсти, Os — верхняя затылочная кость, Ра — теменные кости, Ра — задние ключицы, Prf — предлобные кости, Ptf— заднелобные костя, R — плавнпко бразпыд орган, St—supralemporalc, надвксочиая кость. 1/2.
пластинки (т, фнг. Б), правая и левая, которые должны приходиться впереди ротового отверстия и поэтому, по крайней мере условно, могут быть сочтены за «верхние челюсти». 1 Выемки у задне-боковых углов этих пластинок могли находиться в связи с присутствием тут носовых отверстий.
Панцырь, покрывающий туловище, является почти плоским па нижней стороне и крытевиднэ-вздутым наверху. Он состоит из одиннадцати таблиц из которых три являются непарными, срединными, а восемь — парными, и они соединены между собою с налеганием краев одних па подходящие п°Д них края других таблиц
1 Существует мнение, что ротовые части у Ptenckthys не содержали твердых частей, к рот в лпячивал :я наружу.
аде
Из этих таблиц парные брюшные могут считаться гомологами передних и задних ключиц, а непарная срединная соответствует interclavicula. К передним ключицам примыкают оригинальные листообразные придатки, сочлененные с ключицами пбдвижно. Эти придатки считаются одними исследователями за соответствующие грудным плавникам, а другими — за задние шины головного щита, соответствующие шипам у Cephalaspis. Придатки покрыты снаружи костными табличками, по длине подразделяются на две части, соединенные между собою с некоторою подвижностью. В виду того, что эти придатки часто представляют зубцы на краю, который будет внутренним, когда придатки обращены свободными концами вперед, возможно, что они не столько были плавательными органами, сколько служили для схватывания добычи, зажимаемой между зазубренными краями. На спине находятся две крышеобразно перегибающиеся в плоскости симметрии пластинки, на боках— по две боковые. ✓
Хвост покрыт маленькими, округленными, слегка черепитчато налегающими костяными чешуйками. На спинной стороне позади панцыря находится маленький плавник. Вдоль спинного гребня находятся чешуи в форме маленьких узких продольных срединных табличек; позади спинного плавника они удлинены и подобны фулькрам ганоидных. Задняя конечность рыбы образована совершенно гетероцеркный хвостовым плавником, треугольным по форме п выемчатым назади, не не двулопастным.
На теле и голове существует хорошо выраженная система слизевых каналов, намеченная бороздками на наружной поверхности некоторых из костяных таблиц; бороздки эти нередко смешивают со швами.
К Pterichthys близки роды Asterolepis (aster — звезда, lepis — чешуя) и Bothriolepis (botrion— бороздка).
Asterolepis отличается от Pterichthys большею величиной и тем, что передняя непарная спинная пластинка у первого налегает на край обеих боковых пластинок (фиг. 588), тогда как у Pterichthys она налегает лишь на переднюю из них и перекрывается заднею. Затем у Asterolepis несколько большее число костей в составе плавникообразных органов.
Bothriolepis (фиг. 589) отличается длинными плавникообразными органами, превосходящими длину панцыря, а также V-образной формой участка слизевых каналов, находящегося на голове назади глаз. Верхний девон.
Подкласс III. Artbrodira.
Выделяется из группы папцырных по наличию нижней челюсти, плечевого пояса и сочленения между головой и жаберной областью: отсюда имя— с сочлененной шеей, т. е. с отграниченной головой.
Черты высшей организации и образование внутреннего скелета делают неясным систематическое положение этой девонской группы. Указывается отношение к высшим отрядам костистых (двоякодышащим) и осетровым.
Представителем подкласса, наиболее изученным, является девонский род Coccosteus.
Coccosteus (фиг. 590) (kokkos—зерно, osteon—-кость) является единственною панцирною рыбою с окостеневшим, хотя лишь отчасти, позвоночным столбом, —окостеневают дуги позвонков. Существуют две парь; боковых табличек на туловище, которые могут считаться соответствующими: передние—
костям крышечки, а задние — костям спинного отдела плечевого пояса (см также фиг. 556)
Задняя половина тела голая На границе между головой и жаберной
Фиг. 588. Asterolepis о nat is. Верхний девов. А — сверху В—снизу, С — плавниь?-образиый придаток.
областью находится сочленение. Края челюстей зубчаты, но обособленных зубов нет. Глаза боковые, окруженные склеротикальным кольцом, состоящим из 4 пластинок. Сравнительно короткий ластообразный орган (st). Положение его на границе между головой и жаберной областью не позволяет считать его (также у Pte-ricMhys и др.) грудным плавником. Но на существование грудных плавников, может быть, указывает округленный вырез (pf), а таз ( ?)—на наличность брюшных. Вертикальная костная пластинка (fc) вероятно служила основанием переднего спинного плавника. З.'дний спинной плавник (d) хорошо
у,. развит. Так же и заднепроходной плавник,
Фиг. 589. Панцырь Botnnolr. is r * г гь
canadensis. Реставрация. Девиц. ПОВИДИМОМу ИМеВШИЙ ОПОрОЮ ПЛаСТПНКу а. На Каы_да. 1/2. головных и туловищных щитах имеются слизе-
вые каналы Девон Европы и Сев. Америки. Coccosteus соединяет признаки Placodermi, как образование панцыря. отсутствие настоящих зубов, с признаками химер (распространение слизевых каналов, спинной шип) и ганоидных (аппарат жаберных крышек и скульп-
гура щитков) и даже четвероногих (строен/. плечевого пояса, характер брюшного панциря—как у амфибий-стегоцефалов).
Подкласс IV. Ли^тожаберные, ElasmoLranchii (Chondrichthyes).
Вообще говоря, низко организованы и представляют различные примитивные особенности Настоящие ьос’н не развиваются ни в позвоночнике, ни в виде кожнь х костей черепа В связи с этим череп и челюсти являются здесь в простейшей форме, представил сплошные хрящевые образования. 1 Кожный скелет состоит исключительно из плакоидных чешуй. Иногда основания соседних чешуй могут сливаться, и, зероятно, таким путем образовались кожные таблицы, известные у ископаемых форм. Ихтиодорулиты
фпг. 530. Coccosteus decipiens. Нижний девон. Шотландия. А — реставрация. Пунктиром показаны слизевые каналы. В — верхняя жаберная крышечка (ор') с сочленовной головкой.
К— головяая область:
я — носовые ямки, si — непарный глаз: wtd — ввжияя челюсть; р — теменная кость; ее — яя-тыдочная; вр — epfottcum 6 - непарная шиннэя пластинка; ор. ср*—жаберные крышки, at — подвижный шип; 1 —4— аде кенты плетеного иояса, Л — основная пластинка переднего спинного плавника; 4 — задний спинной плавник;
а — опора анальиеге плавника; t — брюшной плавник, pf — место грудного плавника.
могут считаться за большие специализированные кожные зубы,, часто имея коническую полость, вдающуюся в зуб от его основания. Внутренний скелет остается хрящевым; но он часто приобретает большую прочность, особенно на поверхности, отложением известковых солей, обыкновенно в форме призматических табличек. У всех ныне живущих хрящевых рыб, за исключением сильно специализированных Holocephali, жаберные щели (6—7) открываются непосредственно наружу. Подобным образом это было и у ископаемых рыб. У одних число жаберных щелей более, нежели у других, и это является примитивной особенностью. Гиостплический череп должен считаться более примитивным, чем автостилический. У сампов в связи с задними лучами брюшных плавников развиваются парные придатки с идущими вдоль них бороздами, это — так называемые pterygopodia, играющие роль органов совокупления, так что через них в тело самки вводятся сперматозоиды (семя) и яйца оплодотворяются в материнском организме в отличие от прочих рыб. Это совершенство организации и специальные приспособления для развития рода и обеспечили процветание группы, вообще примитивной, от палеозоя Доныне.
Elasmobranchif делятся на два порядка: Selachli (акулы и скаты) и Но-locepbali (химеры).
1 Отсюда название Chondrichthyes. Другое имя указывает иа своеобразное строение жабор не параллельных пластинок в ли листков, прирастающих основанием кхрящезым духам.
19 Гч.бяж»
26°
Ко^’ндо:; i. selachii (plaommtomi, иоиеречноротые).
Жаберные щели всегда открываются непосредственно наружу. У более примитивных палеозойских форм ротовая щель находится на конце рыла, как вообще у позвоночных, а у позднейших, более специализированных селахий, она перемещается па брюшную сторону головы, причем перетняя оконечность последней образует приостренный хрящевой rostrum. Череп селахий обыкновенно является гиостилическим, т. е. гио-мандибулярный и нёбно-кватратный хрящи образуют свободно двигающийся подвесок для сочленения нежней челюсти с черепом.
Позвонки становятся прочными вследствие отложений извести в хряще, которые сначала принимают форму сжатого посредине цилгндра, развива-
Фиг. 591. Схематический поперечный разрез посредине:
а — цшгтоспонд илового, астероспопдплевого, текстпсцондитозого лозвопков.
С — снцнцяя струна; D—центральное известковое кольцо; Е—наружная эластичная оболочка спинной струны; N— жеврапофизы; Н—гемапофизы.
ющегося во внутреннем слое хряща (фиг. 591 а), вокруг хорды. Дополнительно к нему могут появиться радиально расположенные продольные пластинки между основаниями дуг (фиг. 591 б), или концентрические цилиндры, развивающиеся снаружи первоначального (фиг. 591 с). Таким образом являются три типа позвонков: цик лоспон диловые, астероспондиловые и тектиспондиловые, и соответственно три группы селахий.
В ископаемом состоянии, кроме позвонков, часто находят зубы, ихтио* дорулиты и иногда кожные таблички. Зубы акул принадлежат к числу, окаменелостей, которые в средние века описывались под названием glosso-petrae, окаменелых птичьих и змеиных языков.
Селахии являются большею частью хищными рыбами, живущими в на-ътоящее время на различной морской глубине, от поверхности до глубины около 2000 м. Хотя типично морские животные, иногда они заходят сравнительно далеко в реки и немногие являются постоянными обитателями пресной воды. Между ними совершенно нет небольших по величине, а некоторые акулы являются наибольшими из всех существующих рыб. Пища их состоит из рыб, моллюсков, ракообразных и других беспозвоночных и даже из жи отных планктона.
Selachii разделяются на акул (Squaloidei) и скатов (Batoidei); первые отличаются цилиндрическим, редко сплюснутым .телом, боковыми жаберными щелями и свободными грудными плавниками. Зубы большею частью приострены.
Cladoselache (Cladodus, klados— ветвь, odous —зуб) (фиг. 592—595) является древнейшей и примитивнейшей из известных акул. Хесстовой плавник гетероцеркный, по наружному очертанию равнодольчатый, с круто загибающимся кверху концом позвоночного столба, с большою нижнею лопастью. Вдоль хьоста протягиваю ся горизонтальные кожистые ей hi. Имеются два спинных плавника, передний иногда с ихтиодорулнтом. Нет анального плавника. Парные грудные и брюшные плавники имеют длинную расчлененную главную ось, к наружному краю которой впереди прнчпенены параллельные лучи (фиг. 593). Rostrum отсутствует, и ротовая щель находится на конце рыла; орбиты окружены двойным кольцом из кожных табличек, вероятно образованных слитыми чешуйками. Кожа покрыта маленькими чешуйками ромбической формы. Зубы с широким основанием (фаг. 594),
Фпг. 592. Cladoselache Fyleri. Верхний девон. Огайо. САСШ. с большим срединным и меньшими боковыми зубцами, тесно сидят на челюстях поперечными рядами. Жаберных дуг пять, даже, может-быть, шесть или семь. Спинная струна должна была находиться в первичном неразделенном состоянии, но невральные и гемальные дуги существовали. Достигает 1,5 —1,8 м. Верхний девон — пермские.
Pleuracanthus (фиг. 596, 597; plenron — бок, akantha — шип) также имеет почти конечный рот: в затылочной части головы в плоскости симметрии прикреплен большой шпп. Непарные плавники окружают почти все тело в срединной плоскости, за исключением брюшной поверхности. Именно плавниковая кайма на спине тянется от затылка до оконечности хвоста. Единственная дифференциация,, испытываемая ею, это — слабая выемка приблизительно посредине ее длины, отделяющая длинный спинной плавник от столь же однообразного хвостового Последний — почти дифпцеркный, испытывает дальнейшую специализацию в том, что на передам корце своей вентральной части сильнее скелетирован и вследствие этого представляет выступ из каймы Далее на брюшной поверхности слезет анальный плавник, подразделенный на две части. Грудные плавники с бисерпальным расположением вторичных лучей, в большем количестве находящихся на передней, чем на задней стороне оси. Брюшные плавники почти моносе-
риалъны. В роговом сколете большой нёбно-квадратный хрящ причленяется своею заднею частью к посторбитальной части черепа, тогда как его передний челюстной отдел, как у всех более новых акул, примыкает к носовой области впереди глаза. Нюлняя челюсть подвешена к нёбно-квадратному хрящу (palato-quadratum) в hyo-mandibulare. В то время как на заднем краю palato-qwdratum еще существуют рудиментарные жаберные лучи челюстной дуги в виде нескольких маленьких хрящей, гиоидная дуга назади усажена весьма большим числом чрезвычайно длинных жаберных лучей, которые образуют широкую жаберную крышечку над следующими 5 жаберными дугами. Эiа жаберная крышечка, аналогичная крышечке Holotepliali, есть признак специализации.
Позвонки представляют чисто хрящевые тела, но верхние и впжнпе дуги пх обгизвестнепы.' Ребра представляют еще необычайно большое для селахпй развитие. Кожный скелет будучи представлен маленькими чешуйками около рта, вдоль спины и на птеригоподгях самца. Челисти усажены большими острыми трехзубчатыми зубами. От девона до верхнего триаса, глагным образом в ьаменноу гол! ных и пермских.
Acanthodes i фиг. 559,598, 599;
Фиг. 593. Cladcs-elache Fy-leri с брюшной -стиролы, реставрация.
Фпг. 594. Зубы Cladosclacke (подобные отдельные bj6.j описывались под названием Cladudus).
мало
развит,
Фиг. 505. Зуб Cladoselache lamntrides. Со стороны узкого бика. Верди®
каменное голь-иые. Мичково, около Москвы.
akantha—шип, odous—зуб). Чешуйки шагрени утолщены, имеют квадратное очертание, их плоские и покрытые эмалью поверхности тесно примыкают. Существуют кожные кости на черепе и плечевом поясе. Челюсти соединены с черепом гиостплачески. Орбиты окружены кольцом из четырех костяных пластинок. Хвостовой плавник гетероцеркный, остальные имеют каждый в передней части по пхтподору литу. Сзади лежащая часть плавника представляет мягкую перепонку, прикрепленную к телу и к ихтиодо-рулиту. Лишь в грудных плавниках в перепонке имеются лучи. Челюсти снабжены зубами. Жаберные щели открываются прямо наружу. В позвонках скелетированы лишь дуги. По присутствию кожных костей отличаются от типичных хрящевых. Нижний девон — пермские.
Вышеприведенными тремя родами исчерпывается йсе, что имеется из более или менее полно известных палеозойских акул. Кроме этого, известны пз палеозоя в высшей степени разнообразные зубы и ихтиодорулиты, так что палеозой можно считать эрой наибольшего разнообразия и распростра-
яенпя акул. Довольно примитивным представителем акул, дожившим от иезозоя доныне, является Cestracion (фиг. 600) с пятью жаберными ще-
ФиС. 596. Pleuracanthus De^heni. Голова с ихтиодо-рулитом и передняя часть позвоночного столбе с грудными плавниками. Пермские отложения Наверху в Ъ В/бы. Брауиау. Богемия. Чехо-Словакия. 1/2.
лями, с хорошо развитыми центрами астероспондиловых -позвонков, со ртом, расположенным почти на конце рыла. Зубы Cestracion двоякого рода: напереди маленькие, заостренные, назади тупые, сплющенные в дробящие цластинки. Юра —ныне.
Плоские, подобно тому как у Cestracion, с точечной поверхностью, четырехугольные зубы встречаются у рода Psamino-dus (фиг. 601; psammos— песок, odous —зуб). Каменноугольные.
Срединный, находящийся в месте симфиза (сращения) двух ветвей нижней (и верхней) челюсти ряд зубов, подобный тому, что у Cestracion, но лишь отличающийся большею величиною. свернутый спирально, со сплющенными с боков зубами, существует у рода Hclicoprion (фиг. 602; helix — завиток, prion — пила) из пермокарбона и верхнекаменноугольных. I *Скаты представляют ветвь селахий, специализированную в приспособлении к жизни на морском дне. Скаты имеют тело сплющенное дорзовен-трально, жаберные щели на нижней стороне и большие, прирастающие к голове грудные плавники. Зубы большие частью с плоскою верхнею поверхностью. Они известны с мезозоя; в литографском
юрском с шнце найденодин
из примитивнейших и более близких к акулам, представителей — РМ-nobatus (фиг. 603; rhine — напилок, bates — колючий куст), доживший
доныне. Туловище хотя и сжатое, но более вытянутое в длину, чем у позднейших <катов Грудные плавники тоже сохраняют примитивный ха-
рактер, не достигая своими передними лучами до оконечности морды, как бывает у позднейших скатов. Форма переходная между акулами и скатами,
Фиг. 597. Plcuracanihus sessilis II, рмскйе отл'же-вия. Лебах, у СаарбрЮхСва. Реставра jin. 1/4.
хотя и недостаточно выясненная по организации, известна из девона или нижнекаменноугольных.
ПОРЯДОК П. ХИМЕРЫ, HOLOCEPHALI
(ЦЕЛЬНО 1ОЛОВЫЕ).
Химеры представляют группу морских рыб, примыкающих по скелету и по жабрам к селахиям, хотя и уклоняющихся от последних во многих отноше
Фиг. 598. Реставрация Aeanthodes Wardi. Каменноугольные отложения. Англия.
ниях. Голова с весьма большими глазами, сжата с боков, передняя часть черепа превращена в межглазничную перегородку, череп автостилический; нижняя челюсть (МСК, фиг. би4) снабжена с каждой стороны одной весьма значительной зу ной пластинкой (£>"), в верхней челюсти соответственно находится одна или две пары зубных пластинок (1), D'}. Жаберные щели закрыты складкою кожи,
Фиг. 599. Чешуи Aeanthodes. а — снаружи, Ъ — совнутри, с — отдельная чешуйка.
так что сообщение жаберной полости с наружной средой происходит посредством одной лишь щели. Позвонки не обособлены, спинная струна окружена лишь объизвестненными хрящевыми кольцами, имеющимися в числе большем, нежели дуги. Передний спинной плавник снабжен большим ихтио-дирулитом. У современных форм кожа голая, у некоторых ископаемых отчасти покрыта шагренями и пластин! ами.
Современные Holocepliali представляют лишь незначительный остаток группы, более распространенной в прошлые геологические периоды, и появившейся с девона.
Ptyctodus (ptyktos— несок, odous—зуб) известен только но зубным пластинкам, сжатым с боков и представляющим каждая перетирающие пищу участки (триторы). Зубных пластинок всего
четыре, по две в каждой челюсти; на каждой пластинке по одному тритору (фиг. 606), состоящему из очень твердых, с точечной поверхностью, налегающих одна на другую шастин, направленных
Фиг. 600. Челю ь Cestracion PJulippt, ныне живуль
косо к функционирую-
щей поверхности тритора. Девон Европы и Сев. Америки.
Подкласс V. Костистые рыбы (Teleostomi или Ostoichthyes).
Отличаются
а
Фиг. 601. Зуб Fsammodus ап-oustus. Верхне-гамевноугольн. Мяуково, около Москвы.
развитием: кости на месте хряща во внутреннем скелете; чешуй ганоидных или циклоидных вместо плако-идных, в покровах, вторичного, костного черепа на месте’ первичного хрящевого; развитием нижних ребер вместо или вдо-
Фиг. 602. Спиральный срсДйиный ряд зубов. Не-Исорпол Eesic ioivi. Пер-окарсоя. Красноуфимск.
бавок к верх-
ним ребрам хрящевых рыб; аппарата костяных жаберных крышек, прикре-
пленного к гиоидной дуге. Вроме того, у расматряваемых рыб развивается.
Фэт. 603. Tthinobalus mirabihs. Верхняя юра. Литографский сланец. Эйхштедт. Бавария. Герма! ня
го — rostrum; ев вововые капгулы ft — ротогчя щ₽лъ; Ът — же. ы& ехелет; р— трудно, пл.ввчк.^ V — брюшвов плавник; i -плвввпковыв Туча; с th, — плечевой поле, ft.о — тжзовов пояс; v pt — ироптчр, д’; m. pt — мезоаторвггй ml pt — истмперпгй.
как вырост стенки пищевода, плавательный пузырь, интересный мо-жет-быть ие стол „ко как гидростатический аппарат (так как иногда он отсутствует), сколько по своему происхождению, аналогичному происхождению жаберных -мешков (может-быть, является видоизменением задней пары последних) и по способности превращаться в орган воздушного дыхания (у двоякодышащих).
Костистые рыбы делятся на порядки: кистеперые (Gressopierygii), ганоидные (Ganoidei), двоякодышащие (Dipnoi) и собственно костистые рыбы (Teleostei).
ПОРЯДОК I. КПСТЕПЕ-
РЫЕ, CBOSSOPTEKVGH.
Crossopterygii, или кистеперые, названы такса основании строения их парных плавников, имеющих выдающуюся из тела осевую л спасть значительной величины, покрытую чешуями. Скелет парных илавяиков, редко известный у ископаемых кистеперых, бисермального тина. Чешуи ромбические или круглые. В черепе существует наклонность к слиянию напереди лежащих костей (praemaxilla, nasalia, ethmoidea и даже frontalia и ргае-frontalia) в сплошной щит, покрывающий рыло (фиг. 609).
Область щек покрыта несколькими костями жаберной крышки. Снизу
Фиг. 605. Раскрытая пасть ныне живущей химеры Callorhynchus pt—еёбвые зубные пластнн-
кн, si — eyot ыв njaeififii.H нижнеи челюсти, t—триторы ti — сешяакоЕЫе »убы
фиг. 604, Череп выве живущей Chimaera. ОС и OR — хрящи крышечки.
головы, между ветвями нижней челюсти, развивается из чешуек но крайней мере пара костяных горловых пластинок; кроме того, бывают иногда чаленькие боковые горловые пластинки или передняя.
Зубы двоякого рода- наружные — мелкие и внутренние—крупные, хватающие; последние, с сильно складчатой стенкой. Существуют окостенелые позвонки, или более или менее сохраняется спинная струпа.
Хвостовой плавник дифицеркный или более или менее гетс-роцеркный. Но в последнем случае верхняя лопасть только немного меньше, чем нижняя (гетеро-ди-фицеркный ник).
Часто встречаются в девоне и камен-
плав-
ноугольных, затем
уменьшаю гея в числе; в настоящее время два Polypterus).
Holoptychius (фиг. 607, 608; holos — целый, ptyx — ижладка) Грудные плавники прпостренные, брюшные, менее остролопастные. Хвост гетероцеркиый. Пет отверстия теменного глаза. Позвонки не окостеневают.
рода в Африке (один из них
Фкг. 606. Трнтор Ptyctodus obliquus. Верхний девон. С. Чу-дово. Леиияградская обд.
Чешуи круглые и сильно налегающие: свободная поверхность их представляет украшения вви в л горковилн более или менее радиальных рей ыш к. Зубы приобретают весьма сложную складчатость стенок, особенно около основания. Девон Европы и Сев. Америки
Фиг. 607. Реставрация Holoptychius nobilissimus.
Osteolepis (фиг. 609, 610; os г—кость, lepis — чешуя). Чешуи падкие, ромбические. Есть средняя горловая пластинка. Кости черепа часто
сливаются между собою, особенно напереди. В лобных костях отверстие те-
Фиг. €08. а, Ь — хватающие зубы Holoptychius (Deni.ojus' Ырт talus', — спьио увеличенный поперечный разрез 3j6a: d—чешуя.
Фпг. 603. Реставрация Osteolepis n.acrolejndotvs.
а/— анальный плавппк, cf—хвостовой, df — второй сппннои, pcf — грудной, pf — брюшной плавник, ор — жаберная крышечка.
Фиг. 610. Реставрация черепа Osteo7epis, разложенного на составные кости. А — вид черед! сверху. Б — Be "ви нил ней челюсти с горловыми пластинками между ними.
Fr — лобные поста, Ра — теяённъзе, ос — затылочные, md — япжнял челюсть, тх — верх_ няя; op, pop, sop — жаберные крышки; ju— скуловая кость; пунктиром показано распро странепне слизевых каналов
мерного глаза. Зубы круглого сечения. Полые позволен. Гетероцеркный хвосг. Девол. 1
ПОРЯДОК II. ГАНОИДНЫЕ, GANOIDE1.
Туловище и хвост обыкновенно покрыты ганоидными чешуями, редко голые или с костяными та личками. Черен — покрытый кожными костями или совершенно окостенелый, нёбно-квадрагная дуга подвижно сочлененная с черепом (гиостилическая); жаберные щели прикрыты костяной крышечкой. состоящей из нескольких костей. Часто имеется брызгальце. Позвоночный столб хряшевой или в разной степени окостенения. Плавниковые лу ч?и членистые, и передние края плавников, особенно непарных, обыкновенно покрыты рядом двускатных табличек — фулькр.
Парные плавники хорошо развиты, хвостовой — дифицеркный или ге-терон.еркный. Наиболее приметною, если и не вполне отличительною особенностью ганоидных рыб, является их кожный покров. Никакая другая группа рыб не обладает чешуями, которые состояли бы из толстого костяного внутреннего слоя и из наружного эмалевого слоя. Ни в какой другой группе рыб нет сочленения чешуй посредством выступа и впадины.
Фиг. 611. Реставрация Palaeoniscus macropomus. Верхнепермские отхоже ния. Германия. 1 2-
столь характерного для ганоидов с ромбической чешуей. Бывают, все-таки, ганоидные чешуи округленной формы, налегающие друг на друга совершенно подобно циклоидным и ктенопдным чешуям, причем толщина их не бывает заметно больше, чем у последних; но эти округленные чешуи (JMegalurus, Amici) под покровом эмали имеют внутренний слой с костными клетками, хотя иногда весьма уменьшенной толщины.
Ганоидные рыбы представляют наибольшее развитие в палеозое, триасе и юре; с начала мелового периода они все более и более замещаются собственно костистыми рыбами (Teleostei). Немногие еще существующие пыггс ганоиды живут или исключительно, или по крайней мере отчасти в пресной воде, тогда как огромное большинство ископаемых форм встречается в чисто морских отложениях. Как правило, только тоетичные ганоидные встречаются* в пресноводных отложениях.
I Ганоидные могут быть разделены на три группы: I. Heterocerci П. Chondrostei;1 111. Amioidei.
1 См. примечание на стр. 284 о новейшей классификации рыб в связи с новым взглядом °’, хряшевых ганоидов как на примитивную, а не дегенеративную группу, близкую к селахиям.
Фиг. 612. Реставрация черепа Palaeoniscus.
qp, уор, жаберные крышки; Ът—жаСерные лучи
гнондной дуги. Пунктиром по казаны слваевые каналы.
Подпорядок I. Heteroeerci. Хвостовой плавник гетероцеркный. Один спинной и один анальный плавник, с членистыми лучами, более многочисленными, чем лученосцы. Спинная струна сохраняется, но дуги, остистые отростки и скелет плавников более или менее окостенелые; голова покрыта кожными ко-стяяг ми пластинками. Непарные и обыкновенно парные плавники окаймлены фулькрами. Покров чепуй с тела продолжается на хвостовой плавник, особенно на его верхнюю лопасть. Чешуи ромбические или ромбоидальные, редко круглые. Palaeonis'us (фиг. 611,
Ы 2; palaios—древний, oniscus— мокрица). С тонким телом ма-
Фиг. 613. Реставраци* Plaly^.junus stnatus. Пермские отложения. Англия.
лых или средних размеров. Зубы прностренные, небольшие. Плавники неболипие, лучи членистые и ветвящиеся дистально. Чешуи ромбические, с неправильными косыми бороздками. Пермские.
Platysomus (фиг. 613; platys — широкий, soma — тело). Тгловище, вытянутое в высоту, ромбическое или дискоидальное. Парные плавники малы. Спинной и анальный велики, расположены один против другого. Чешуи ромбические, вытянутые сверху вниз, каждая с внутренним вертикальным килем, образующим выдающуюся оконечность на верхнем краю. Кам енио угольные, пермские.
Подпорядок II. Cliondrostei (хрящевые ганопды). Одними считаются за группу регрессивного характера, вышедшую из Palaeo-niscidae, а другими за промежуточную ступень на пути прогрессивного развития рыб от хрящевых (акуловых)к костистым. Тело остается веретенообразным с типичным гетероцеркный хвостом (фиг. 5с2,614).
Морда продолжается вперед рта и обонятельных капсул. Рот на нижней стороне, с связи с жизнью на дне. Примитивную особенность пр 'дставляет присутствие sairaculum. Особенностями дегенеративного характера могут быть утрата чегауйного покрова из туловище (сохраняется на хвосте), жаб-рных лучей гиоидной дуги, средпцяой горловой пластинки, редукция костей жабсрлой
ЗОО
крышечки, зубов, относительный недостаток окостенения во внутреннем скелете, в котором позвоночный столб остается хрящевым В связи с жизнью на дне тело приплющено на брюшной стороне. Вместо чешуйвого покрова, сохраняющегося лишь на хвосте, развивается пять продольных рядов костяных щитков, одо* спинной и четыре боковых. В пресных водах и внутренних
Фиг. 614. Acipenser, осетр, ныне живущий о — рфТ.
иорях Европы, Азии и Сев. Америки. В ископаемом состоянии с мезозоя В личинковых стадиях имеют конические зубы и всю серию кожных кост
ных образований относительно более развитую, чем в зрелом возрасте. Интересно отметить,
что Palaeoniscidae иногда выказывают наклонность к особенному развитию пяти продольных рядов чешу й, одно: о спинного и оста тьных — парных боковых, намечая таким образом первые стадии в развитии пяти рядов костяных табличек осетровых.
Лейасовый род С'юп-drosteus (фиг. 615; cbon-drcs— хрящ,' os — кость) является переходным между Palaeoniscidae и современными Acipense-ridae (осетровыми), представляя более короткий
Фиг. 615. Реставрация головы Chondrosteus acipei se-roides.
7—Р—кости жаберной крышки. £0—hyo-manditilart. IS, 17, 18— кости нижней челюстк. 13 — ж^бервыв лучи.
ростр, чем у последних, более развитой и менее вытягивающийся наружу челюстной аппарат, жаберные лучи гиоидной дуги, расположение костей верхней поверхности черепа, более близкое к Palaeoniscidae. Челюсти беззубые. Тело голое. Нижний лейас.
Подпорядок III. AinioiJei. Позвонки в разной степени окостенения, иногда с сохраняющеюся спинною струною. Имеется одна горловая пластинка, иногда фулькры. Хвостовой плавник внешне гомоцеркный, внутренне гетеро-Церкный (геми-гетероцеркный) Чешуи очень тонкие, черепитчато налегающие, круглые или ромбические; несмотря на их тонкость, они все-таки представляют типичные особенности настоящих ганоидных чешуй; основание содержит костные тельца, поверхность покрыта эмалью. Только один род
(Amia) существует в реках Центральной Америки и смежной части Северной Америки, он известен из третичных отложений. Ископаемые формы ветре таются с верхнего лейаса.
Megalurus (фиг. 616, 617: megale—большой, онга—хвост). Позвоночный столб загибается в верхнюю часть округленного хвостового плавника; в хвостовой части состоит из нолунозвонков (плеврбцентры и гипоцентры),
Фиг. 616. Megalurus elegantissiwus. Верхняя юра. Золенгофен.
Германия. 1/2.
1ерез один несущих верхние и нижн ie дуги. Передняя часть позвоночного столба состоит из полных позвонков. Чешуя очень тонкая, круглая, есть фулькры. Верхняя юра, мел.
ПОРЯДОК Ш. СОБСТВЕННО КОСТИСТЫЕ РЫБЫ, TELEOSTEI.
Фиг. 617. Megalurus potyspowtylus. а — часть позвоночного столба из хвостовой области; Ъ—чешуи. Верхняя юра. Кельгейм.
Почти все современные Osteichthyes принадлежат к этому высшему порядку рыб. Для Teleostei характерны утрата эмалевого покрова на костях наружного скелета, округленные, тонкие, черепитчато-налегающие чешуи, простые амфицельные костяные тела позвонков, гомоцеркный хвост, отсутствие горловых пластинок и брыз-гальца.
Чешуи костистых рыб бывают циклоидные и ктеноидные; иногда вместо них развиваются более или менее значительные костяные пластинки. Во внутреннем ухе именно этих рыб развиваются отолиты сложйой формы (фиг. 583), с особенной микроструктурой. Более примитивные из костистых рыб сохраняют соединение плавательного пузыря с пищеводом (как у ганоидных),
оно исчезает у других, составляющих большинство среди третичных и современных рыб.
Костистые рыбы вероятно развивались из амиоидпых в триасе и вскоре превзошли своих предков по разнообразию форм. Они являются значительно распространенными уже в верхнеюрскпх и нижнемеловых отложениях (напр. Leptolepis\ leptos—маленький, lepis—чешуя, фпг. 618), но достигают наибольшего распространения в третичных отложениях и в настоящее время. Живут частью в море, частью в пресной воде.
Костистые являются наиболее высоко стоящими рыбами по совершенству внутреннего скелета; последний представляет хорошую опору для действия мускулов, а это, в связи с гибкостью чешуи, делает костистых лучшими пловцами, чем предшествовавшие им и уступившие им место ганоидные.
ПОРЯДОК IV. DIPNOI, ДВОЯКОДЫШАЩИЕ.
Покрытые круглой, черепитчато налегающей чешуей рыбы, дышат жабрами и легкими. Череп автостилический, без spiraculwm и теменного глаза, с неразвитыми кожными костями челюстей и, в связи с этим, отсутствием на ротовом краю зубов.
На крыловидно нёбных костях существуют гребенчатые зубные пластинки, гребни которых иногда зубчаты, свидетельствуя о происхождении из слившиеся конических зубов обычной для рыб формы. О том же свидетельствуют и данные эмбриологии относительно индивидуального развития ныне живущих родов. Имеется жаберная крышка. Окостеневают дуги позвонков и ребра, сохраняется спинная струна. Парные плавники бисериального типа, в осевой
Фиг. 618. Реставрация Lrpto1 pis dztbius. Верхняя юра. Бавария. Германия.
части покрытые чей уею, подобно тому как у Crossopterygii, из которых Dipnoi вероятно развились. У древнейших есть горловые пластинки и слизевые каналы на поверхности черепа. Хвостовой плавник гетероцеркный или двфи-церкный, иногда сливающийся в одно целое с остальными непарными плавниками. В связи с жизнью в периодически высыхающих водных бассейнах развивается дыхание не только жабрами, но и действующим тогда как легкие видоизмененным плавательным пузырем, внутренние стенки которого приобретают ячеистый вид. Зате^ обонятельные ямки сообщаются отверстиями на нёбе (внутренние носовые отверстия, хоаны) с полостью рта, так что дыхание легкими возможно при закрытом рте, подобно тому как у четвероногих позвоночных. У остальных рыб хоан нет.
Группа, представляющая наибольшее разнообразие в палеозое, в настоящее время представленная лишь тремя родами в пресных водах в Африке, Америке и Австралии (Pro top ter us, Lepidosiren, Ceratodus).
Среди палеозойских Dipnoi есть формы co строением черепа, более приближающимся к нормальному типу черепа костистых; таков каменноугольный Ctenodus (фиг. 620; ktenos—гребень, odous—зуб). От таковых исходит, во-первых, линия, ведущая к современным Dipnoi (фиг. 625), а затем линия специализированных Dipnoi (фиг. 621), вымирающих в палеозое.
Phaneropleuron (фиг. 619; phaneros—открытый, pleuron—бок) является представителем таких наиболее примитивных Dipnoi-, имеет дифицеркный
хвост, сливающийся в одно цеЛде со спинным плавником, анальный плавинв обособлен. Парные теменные и лобные кости. Кроме зубных пластинок имеются конические зубм на краю челюстей Две горловые пластинки. Девон Канада и ШотланДия.
Фиг. 619. Реставрация Phaneropleuron. Девой. I/O.
&
р-ра.
ptrr
Ф-'
Фиг. г -
620. Череп Ctenodus. лобкые; ра — томе иные жостж.
Dipterus (фиг. 621—624; dipteros—двукрылый) с гетероцеркным хвостом двумя спинными п одним анаДьныи плавником. Кости черепа многочисленны, развита серия крупных срединных костей, а теменные и лобные малы Девон.
Ceratodus (фиг. 625, 626; ceros — рог, odous—зуб) покрыт тонкой эластичной циклоидной чешуей без костной основы. Непарные плавники слиты в «дно целое с дифицеркным хвосторыз. Приплющенная всрлняя поверхность черс па образована двумя большими срединными костями, расположенными одна за другой по длине черепа, и двумя парами боковых костей (фиг. 625, В). На нижней поверхности черепа (фиг. 625, С) находится длинный парасфеноид (PSpti), с ромбическим передним расширением, ограниченный с боков нёбно-крыловидными костями (Pt)". Последние соприкасаются впереди и каждая несет большую треугольную зубную пластинку (d) с радиапьными ребрами.
Нт сошнике (Го) находится пара острых долотовидных зубов (d')- На каждой ветви
нижш й челюсти тоже по одной зубной пластинке. Число гребней на зубных пластинках изменяется у различных видов от 4 до 6. Триас—ныне. В настоящее время живет в реках Австралии, питается растениями.
Геологическое распространение рыб.
Древнейшие рыбы известны в настоящее время в виде отдельных чешуй-табличек и других отрывочных и плою определимых остатков из нижнего силура Сев. Америки. Из верхнего силура Прибалтики, Шотландии и Сев. Америки известны простейшие Placodeimi (Pteraspidae. Cepbalaspidae) и Sela-chli (Acantbodidae). В девоне рыбы являются весьма распространенными, именно Placoderml, Artbrodira, Crossopterygn (максимум распространения), Gano-idei (Heterocerci), Dipnoi. Selachii (Cladoselache. Acanthodes) и Holocephali
Каменноугольная фауна представляет максимум развития селахий: С1а-dodvs (Cladcselache}. Psammodus, PPUccpr/nn.
В пермских отложениях достигают наибольшего распространения Heterocerci.
Фиг. 621. Dipterus Valenciennesiz. Девок. Шотландия, а—часть нёбньй поверхности черепа с двумя зубами; б—нижняя челюсть.
Фиг. 622. Зуб Dipterus platycephalus. Девон.
Р. Сясь. Ленинградская область. Зуб на обломке челйстной костиА_
adf.
•Фиг. 623. Реставрация jDijpfents FaZenciennesit. Девон. 2/3.
В первой половине мезозоя распространены ганоидные Amioidei, весьма разнообразные, и Chondrostei; с второй половины начинают преобладать собственно костистые рыбы.
Наиболее древние типы кости
стых, как Leptolepis (фиг. 618), еще имеют эмаль на чешуях; хвост близок к форме хвоста у Amioidei, но центры позвонков цельные, нет горловой пластинки. Триас—мел.
В третичных отложениях очень часто встречается Ыиреа, сельдь.
tmepucmuKU групп рыб.
1 класс. Рыбы, Pisces. . Дышащие жабрами водные позвоночные с хрящевым или костным скелетом, в большинстве случаев с парными плавниками, и всегда с непарными. За исключением Cyclostomata имеют в коже чешуи.
Фпг. 624. Реставрация ’-оловы Dipterus. Пунктиром показано распространение слизевых каналов.
20
Учебник палеонгологии.
305
1 нодклалз: Cyclostomata. Хрящевой скелет. Не имеют чешуи. Не имеют челюстей в парных конечностей. Носовое отверстие непарное. Осевой скелет в виде хорды. Ныт живущие, в нижнем девоне два проблематичные рода.
2 подкласс: Placodermi. Не окостеневающей внутренний скелет. Нет типичных парных конечностей позвоночных, челюстей и зубов. Голова и передняя часть тела покрыты костяными пластинками, возникающими из слияния костяных чешуй. Гетероцеркный хвост. Силур, девон.
3 подкласс: A”‘hrodire Сходны с Placodermi по присутствию костяного панцыря
Фиг. 626. А—Ceratodus Forsteri. Ныне живущий. Австралия. В — вид черепа сверху; С — снизу. В В хрящевые части покрыты точками.
ВС — d, d1 — зубы: Vo — сошник; Pi — нёбко-кры говядине кости; Psph — парасфеноид; па — носовые отверстия; Qu — квадратный хрящ; Вг — жаберная волость, г — самое переднее ребро.
в передней части тела, но отличаются наличием нижней челюсти, плечевого пояса и со-членения между головой и туловищем. Окостеневают части дуг позвонков. Девон.
4 подкласс: Elasmobranchii Рыбы с хрящевым или объизвественным, но не окостенелым внутренним скелетом и с кожными зубами-чешуями. Челюсти первичного xapai гера, из palatoquadratum и mandibulare. Имеются парные грудные и брюшные плавник- Большею частью 6 — 7 жаберных щелей. Морские животные.
1 порядок: Selachii, Нёбноквадратный хрящ обособлен от черепа. Все жабе ; вые щели открытые. Силур—ныне.
2 порядок: Holocephali. Нёбнрквадратный хрящ срастается с черепом. Жаберные щели прикрыты. Девон—ныне.
5 подкласс: Teleostomi. С костями во внутреннем и наружном скелете. Большею частью, пичлоидные и хтеноидные чешуи. Зубы не только на челюстях но и на костях
Фиг. 626. Зуб. Ceratodus Каиррг. Триас. Вюртемберг. Германия.
полости рте и на жаберных дугах. Жабры прикрыты костяными крышечками. Большею частью имеют плавательный пузырь. Девон—ныне.
1 порядок: Crossopterygii. Парные плавники имеют осевую лопасть, покрытую чешуей Скелет плавников бисериальнл о типа. Хвост дифицеркный или (у древнейших форм) чаще гете-родеркный. Позвонки более или мене окостеневают. Девон—ныне.
2 порядок;- Ganoidei. Ганоидные чешуи. Внутренний скелет в разной степени окостенения. Вообще довольно различны, так что делятся на несколько подпо-рядкоп (Heterocerci, Chondrostei, Amioidei). Девон—ныие.
3 порядок: Dipnoi. Обитатели пресной воды,/внутренний скелет главным образом хрящевой. Palato-quadratum слито с
черепом. Вместо зубов немного-
численные зубные пластины. Парные плавники с осевою лопастью, как у Crossopterygii. Дышат жабрами и (воздушное дыхание) видоизмененным плавательным пузырем, в связи с чем имеют хоаны. Хорда отчасти сохраняется. Девон—ныне.
4 порядок: Teleostei. С сильным окостенением внутреннего скелета. Циклоидные чешуи. Парные плавники моносериальные, с окостенелыми лучами. Хвост гомоцеркный. Юра—ныне.
КЛАСС П. АМФИБИИ, AMPHIBIA.
Древнейшие амфибии, или земноводные, подобно ганоидным и многим костистым рыбам, имели сплошной костяной панцырь поверхности черепа. Таковы панцырноголовые, или стегоцефалы (фиг. 627), у которых головной панцирь оставляет свободны ии лишь глазные впадины и носовые отверстия. Здесь вошли в тесное соединение с покровными костями собственно черепа такие кости, которые у рыб, более или менее подвижно, принадлежали боковой области головы, именно кости подглазничного кольца, дающие происхождение слезной (lacrymale), скуловой (jugale) и квадратно-скуловой (quadrato-jugale) костям.' Palato-quadratum здесь непосредственно примыкает к черепу в двух пунктах: перед слуховой областью черепа и в предглазничной части; кроме того, передние концы palato-quadratum, соединенные напереди у селахий, здесь являются раздельными.
При прямом примыкании palato-quadratum к черепу, hyo-mandibulare теряет свое значение, представляемое мм у рыб, — подвеска к черепу челюстного аппарата, — и становится все более и более рудиментарным и получает иное назначение, а именно в связи с развитием у четвероногих среднего уха, соответствующего брызгальцу рыб, hyo-mandibulare образует косточки: operculum, закрывающую отверстие (слуховое окно), и столбчатую косточку columella (или stapes высших классов), примыкающую одним концом к operculum, а другим к squamosum и quadratum. Редукция hyo-mandibulare происходит в связи с изменением способа дыхания. С утратою жабер утрачивается аппарат жаберных крышек, а с утратою последнего hyo-mandibulare, примыкающий к нему, лишается значительной части св< ей функции У новейших амфибий, производных от стегоцефалов, происходят сильные изменения в черепе. В то время как у стегоцефалов кости черепа находятся в покровах и потому представляют на своей поверхности ямчатую
и бороздчатую скульптуру, у новейших амфибий кости находятся под кожным покровом и потому гладки. Кроме того, число костей сокращаетя, так что в костном панцыре головы являются значительные отверстия. Эти отверстия исходят от уже существующей глазничной впадины, которая, простираясь назад в височную область, образует глазнично-височное отверстие (фиг. 628). В затылочной области черепа у амфибий развиваются окостенения лишь боковых затылочных костей; в связи с этим череп причленяется к позвоночному столбу посредством двух мыщелков—выступов боковых затылочных
Фиг. 627. Череп ArcheguMurus, сверху (4) и снизу (В):
« — tabufare, f — frontale, I — lacrymale, ju — jugale, mx — mattfUare, na — nasale, os — postparietale* pa — parietale, pas — parasphenoideum, pl—palatinum, pmx — praema-nllare, pt — pterygoideum, ptf—post' frontale, pto — postorbitale, prf — praefrontale, g — quadrat um, gj — quadrate-jug ale, sg — squamosum f st — supratemporale, v — vomer.
костей. Вместо верхней затылочной кости имеются парные кожные кости, заднетеменные, postpariefalia. 1 В слуховой области наиболее значительное окостенение представляет prooticum. Вместо хрящевой epioticum в задних углах кровли являются парные кожные кости tabularia. Между parietale и squamosum обычны (у древних форм) парные кожные кости supratein' poralia (надвисочпые). Перед prooticum в глазничной области находится окостенение orbito-sphenoideum. В кровле черепа развиваются parietalif1 и frontatia, у бесхвостых амфибий сливающиеся в fronto-parietalid Parietalia стегоцефалов имеют отверстие для непарного глаза. В оснований черепа является сильноразвитым парасфеноид, вместе с парным сошником
1 Называются также dertno-supraoccipitaha, т. е. кожные верхнезатылочаые.
к несущим их костям иногда при
Фиг. 628. Череп лягушки Discoglossus pictus, ныне живущ. А — сверху, Б — снизу, С — сбоку.
апд — angulare, аг — articular с, са — columella, d — dentale, 9 — jugate, m — тахШаге, ps — parasphenoi deum, pt — ptery-goidewn, v — vomer, q — quadratum, e — ethmoideum, eo — exoccipiiale, fp — Jronto-parietale, n — nasale, p — palatinum, pro prooticum, pm — praemaxillare.
(vomer) образующий кровлю ротовой полости в главной ее части. С боков в состав этой кровли входят крыловидные кости (pten/goidea). из переднего отдела palatoquadratum возникают нёбные кости (palatwa). В связи с тем, что носовые полости служат здесь и для прохождения воздуха в органы дыхания, существуют внутренние носовые отверстия, хоаны. Зубы находятся не только на краях челюстей, но и на костях кровли ротовой полости: на vomer a, palatina, transversa (поперечные, или ectopterygoidea рыб) и parasphenoideum. Зубы п] посредстве неглубоких зубных впадин; у некоторых позднейших стегоцефалов зубы представляют сложную складчатость дентина стенок, подобно тому как у кистеперых рыб (лабиринтовые зубы, фиг. 642). Нижняя челюсть состоит из многочисленных (до 10) костей, несущих крупные зубы, а кроме того мелкие «шагреневые».
Позвоночный столб более или менее окостенелый и в общем может быть подразделен на шейную, туловищную, крестцовую и хвостовую области. У палеозойских амфибий спинная струна иногда покрыта лишь тонкой костяной обложкой,
или позвонки состоят из отдельных частей, т. е. являются эмболомерными и рахитомными. При более совершенном окостенении они бывают амфицельны, про- или опистоцёльны; два последние видоизменения развиваются при возникновении межпозвоночных хрящей и при прирастании их сзади или спереди к амфицельному позвонку.
Туловищные позвонки, за исключением первого шейного, снабжены короткими прямыми ребрами, не соединенными с грудной костью. Один позвонок (редко два или три) поддерживает таз. У бесхвостых амфибий хвостовые позвонки срастаются в одну кость, кобчиковую.
Индивидуальное развитие амфибий происходит с метаморфозом, в связи с чем у зародыша не развивается ни амниона, ни аллантоиса, причем, за немногими исключениями, дыхание первоначально происходит жабрами, иногда остающимися и в течение всей жизни.
Ископаемые, вымершие формы, объединяемые в подкласс стегоцефалов (Stegocephali, т. е. панцырноголовые), появляются в нижнекцменноугольных отложениях уже резко диференцированными на 3 группы высшего порядка, развивающиеся параллельно. При этом позднейшие формы ближе к типу амфибий, чем древние, каменноугольные или пермские. Эмболомерная палеозойская ветвь ведет к пресмыкающимся (Cotylosauria).
Из трех порядков, на которые подразделяются современные амфибии, рассмотрим два главнейшие, представители которых встречаются в ископае
мом состоянии: хвостатых амфибий (Urodela или Caudata) и бесхвостых (Acura или Ecaudata).
Третий порядок Gymnophiona (т. е. голые змеи, или Apoda, безногие,, обязанный своим названием голому змееобразному телу, представлен лишь
в современной тропической фауне немногими слепы «и формами. Роющий, подземный образ жизни своеобразными упрощениями организации еще более затемняет их положение в • системе, Сплошной покров черепа-костяные чешуйки в коже и двойной ряд зубов у некоторых родов дают основание
Фиг. 629. Череп стегоцефала Diceratosaurus рт с-tohneatvs. Снизу. Каменноугольн. отложения Сев. Америки, Огайо.
Ер — tabular^, Jg — jugate, — marillare, Ра — palatinum. Pm — praemaxillare, Ps -^parasptienoideum, Pt — p*er goideum, Qu — quadratum, Psq — squamosum, Tr — transcersvm, to— vomer. Ос— затылочные мыщелки.
Фиг. 631.
Фпг. 680. Чешуи брюшной по верхвости стегоцефал.
а — Arche gosauruSy Ъ — Sclerocephalu» е — Petrobates (последний — из Micro-sauna, см. ввэке).
tt 3 — позвонок Eranchiosaurus, c6okv и с конца, 3, 4, 5 — позвонок представителя Aficrosauna {Byte plesion) снизу, сбоку и с конца; с — канал гг иеной струны, he — п поцентр; п — нервным канал; р» поперечный отросток.
видеть в них дегенерировавших потомков или непосредственно стегоцефалов, или же ветвь хвостатых.
Anura и Urodela имеют голую кожу; Stegocephali, напротив, защищенную, особенно на жизоте покровом чешуй (фпг. *30, 632, В), округленных, жальных, ромбоидальных—подобно чешуе ганоидных рыб, или палочко зидных.
Подкласс I. Паицырноголовые, Stegocephali.
Получили свое название потому, что череп их сверху обычно представляет сплошной костяной покров, прерываемый лишь глазными и носовыми отверстиями. Нет имеющихся у высших позвоночных височных отверстий и скуловых дуг. Кости кровли черепа представляют на наружной поверх-I пости ямчатую и бороздчатую скульптуру (фиг. 641). От современных амфибий стегоцефалы отличаются большим числом костей в составе черепа; подобно тому как у рыб имеются заднелобные и заднеглазничные кости; всегда существует, подобно тому как у некоторых кистеперых рыб, отверстие для теменного глаза. Развиты слизевые каналы на голове; плечевой пояс отличается сильным развитием, состоит из ключиц, передних и задних (cleithra) лопаток и межключичной кости (interclavicula, фиг. 633). Строение позвоночного столба у стегоцефалов представляет большое разнообразие, позвонки бывают: 1) темноспондилового типа. (т. е. расчлененные на части): а) эмболомерные или диплоспондиловые (из двух дисков), б) рахитомные (из сегментов) и в) стереоспондиловые сплошные, слабо амфицельные с небольшим отверстием в центре для хорды (у триасовых форм); 2) у Microsauria позвонки лепоспондиловые (тип оболочки, гильзы), имеют форму цилиндров, окружающих хорду, со сжатием посредине (фиг. 631, 3—5); 3) у Branchiosauria позвонки филлоспондиловые (тип листочков кости), бочкообразные, покрыты тонкой костной обложкой из четырех частей; две верхние представляют нервные дуги, разросшиеся вниз, две нижние могут считаться соответствующими гипоцентру (фиг. 631, 1); плевроцентры считаются неразвившимися. Запястье и предплюсна остаются в хрящевом состоянии у многих стегоцефалов, представляют от 6 до 10 элементов.
Древнейшие четвероногие—стегоцефалы представляют в своей организации много особенностей, общих им с известными группами рыб—с кистеперыми ганоидами и с панцырными рыбами. С первыми они весьма сходны в расположении и числе костей черепа, в отношении лабиринтового строения зубов, в присутствии кольца окостенений склеротики, непарного глаза. С Placodermi у стегоцефалов является общим строение плечевого пояса, иногда существующая вытянутость заднебоковых углов черепа (Ceraterpe-ton, Cephalaspis, Pterichtys, Coccosteus). В виду этого, а также большей близости между амфибиями и рыбами, нежели между другими классами четвероногих и рыбами, иногда объединяют рыб и амфибий под названием Ichthyopsida,—предполагают (это делается большинством палеонтологов), что амфибии произошли от рыб, именно от кистеперых ганоидов. Но существует и противоположное предположение, имецно, что рыбы представляют четвероногих, приспособившихся к жизни в воде, подобно тому как приспособились к ней вышедшие из наземных четвероногих китообразные, ластоногие и многие группы пресмыкающихся—ихтиозавры, плезиозавры, мозазавры и др. Такой . переход от жизни наземной к водной происходил у позвоночных в истории земли многократно, и приспособление к водному образу жизни достигало высокой степени совершенства. Обратный же переход, отводной жизни к наземной,наблюдается (у некоторых рыб) редко и не бывает полными совершенным. При этом воззрении сходства между Stegocephali и Crossopterygii могут быть следствием происхождения их, не одних от других, но от одной исходной прародительской группы. В будущем палеонтология, может быть, прольет свет на вопрос ь о генетических соотношениях рыб и амфибий путем выяснения того, К какая конечность является первичной: плавник или пятипалая конечность.
ПОРЯДОК I. PHYLLOSPOKDTLI.
Brancliiosauria. Маленькие саламандрообразные формы с плоской, широкой, закругленной наререди головой, слабым окостенением позвоночника, чешуйчатой кожей. Вероятные предки Urodela и Anura (возможно и Gymnophiona).
Branchiosaurus (Protriton) (фиг. 632—635;branchia—жабры, saurus— ищер). Голова короткая и широкая со сравнительно большими глазничными впадинами. Имеется окостенение склеротики в виде кольца костяных пласта
Фиг. 633. Плечевой пояс Branchiosaurus.
с—scapula, cl— clar^ula, e-intercla-cicula, sc — chithrum. 3/1.
Фиг. 635. Тазовый пояс Branchiosaurus. i—ilium, is—ischium.
Фиг. 634. Обломок челюсти Branchiosaurus с sy-бами.
нок и, кроме того, окружающее его дополнительное окостенение в виде мозаикообразного скопления более мелких табличек. Зубы полые, конические (фиг. 634). Все туловищные и передние хвостовые позвонки снабжены ко
роткими прямыми ребрами. Затем идут задние хвостовые позвонки без ребер и, должно быть, еще значительное число совершенно неокостеневающих позвонков, так как существуют отпечатки Braneluosaurus-c. мягкими частями, показывающие, что хвост по длине равнялся туловищу с головой. Крестцовые ребра не срастаются, но сочленены с поперечными отростками крестцового позвонка, подобно тому как остальные позвонки. Задние конечности имеют по 5 пальцев, передние по 4. Плечевой пояс состоит из interclavi-cula, clavicula, cleithrum и scapula. Коракоиды остаются в хрящевом состоянии. Не окостеневают кости запястья и предплюсны и концы длинных костей конечностей. В тазе (фиг. 635) не окостеневает pubis. Брюшная сторона тела покрыта чешуями, черепитчато налегающими, ромбоидально округленной формы, расположенными в поперечные ряды, как показано на фиг. 632, В. Молодые экземпляры имеют жаберные дуги. Пермские отложения.
ПОРЯДОК II. LEPOSPONDYLI (MICROSAURIA). t
Microsauria стоят на высшей ступени развития, нежели Branchiosauria. Имеют амфицельные позвонки, хотя и с сохра
няющеюся и представляющей еще большой поперечник хордой (фиг. 631, 3—5), ребра иногда двухголовчатые, притом длиннее и тоньше, чем обыкновенно у стегоцефалов. Лобковые кости всегда окостеневают (фиг. 636). Кости запястья п предплюсны тоже нередко окостеневают. Зубы простые с большой пульпой. Каменноугольные — пермские. Пример Dicerato-saurus (фиг. 629; dis — двойной; Keras — рог). Верхнекаменноугольные.
Фпг. 636. Тазовый пояс DiscoqauruA i — 'dium, is — ischium, p —pub-s.
ПОРЯДОК III. TEMNOSPONDYLI (LABYEINTHOBONTIA).
ГРУПП* I. EMBObOMEBI.
Примитивная группа, исходный тип позвонков, двудисковый. Затылочный мыщелок 1 или 3-членный. Нижняя поверхность черепа близка к первичному типу пресмыкающихся: basioccipitale и basisphenoideum развиты за счет parasphenoideum, нёбо закрыто широкими птеригоидами. У Anthra-cosaurus не обособляется крестцовый позвонок. Типичные эмболомерные позвонки Cricotus (пермь Техаса). Нижнекаменноугольные до нижнепермскпх.
ГРУППА П. HHACBITOMI.
Позвонкп расчлененные на части; полулунные гипоцентры и малые парные плевроцентры. Небольшие нёбные отверстия, постепенно увеличивающиеся—от палеозоя к триасу. Зубы с радиальными складками дентина. Верхнекаменноугольные до нижнего триаса.
Archegosaurus (фиг. 552, 559, 627, 637—640; archegos—предок). Череп более длинный, нежели широкий, причем лицевая часть все более Удлиняется с возрастом. Глаза окружены кольцевым окостенением склеротики. Наружные кости представляют ямчатую скульптуру, но слизевые каналы не выражены явственно. Кроме зубов, находящихся на краю челюстей, есть внутренние зубы, некоторые из них большие, на сошнике и нёбных костях. Стенка зубов представляет простые радиальные складки.
Спинная струна сохраняется, окружена гипоцентрами и плевроцентрлми (фиг. 552 и 637). Позвонки рахитомные, окостеневают неодинаково в различных частях тела, в хвостовой области плевроцентры представляют два
отдельных окостенения, верхнее и нижнее.
Фиг. 637. Archegosaurus Decheni. Нижнепермские отложения. Саарбрюккен. Германия. 1/1.
Межключичная кость, ромбоидальной формы (фиг. 638), к ней примыкают напереди трехугольной формы ключицы (подобно ей представляющие радиально ямчатую скульптуру), а назади также трехугольные лопатки,покрытые сбоку, на наружном краю, ложкообразными cleithra. Коракоиды не окостеневают. Передние ноги по размерам значительно меньше задних. Кости запястья и предплюсны окостеневают. Существует брюшной панДырь из налегающих, поперечновытянутых чешуй; кроме того, маленькие, неправильные кожные окостенения рассеяны по туловищу и ногам (фиг. 639). Молодые индивидуумы имеют жаберные дуги. Достигают полутора метров в длину \
ГРУППА III. STEBEOSPONDHJ
Группа, отличающаяся строением своих зубов, сложных радиально-складчатых, подобно тому, как у кистеперых рыб. Позвонки стереоспондиловые, массивные; преобладают гипоцентры. Двойной затылочный мыщелок. Разви-
той парасфеноид, недоразвитые основные кости черепа; большие нёбные отверстия. Триас.
1 У нас в пермских пресноводных отложениях на С. Двине найдены остатки интересного представителя рахитомных — Dvinosaurus. Животное имело около 1 м длины, представляло смешение примитивных черт со специализированными и сохранило жаберный аппарат во взрослом состоянии. Возможно, что это форма вторичного приспособления к постоянно-водному регнму в виду неблагоприятных для амфибий условий жизни в пермском сухом и холодном климате.
571, 641, 642; mastos— сосок, odon— зуб)
Mastodonsaurus (фиг.
Фиг. 638. Плечевой пояс Аг-chegosaurus. Ншкнепермские. Саарбрюккен. 01— clavicula, Clt— cleithrum, It — interelavicula. Sc — scapula, humerus показан пунктиром, coracoideutn не окостеневает.
является крупвейшим из известных стегоцефалов: длина черепа достигает 1,25 и*. Череп сжат дорзовентрально, длина немного более ширины. Наружные кости покрыты ямчатой скульптурой. Имеются слизевые каналы, не на всем протяжении выраженные вполне отчетливо. Зубы отличаются весьма сложными извивами радиальных складок (лабиринтовое строение). Позвонки короткие, амфицельные (тело их соответствует гипоцентрам), с отверстием в центре для прохода хорды; каждый, за исключением
Фиг. 639. Чешуи Ai chegosaurus De cheia, удлиненные— брюшные, округленные — ножные.
Фиг. 640. Зубы Archc-gosaurus, цельный и с и„ -ломом. Увеличены.
первого шейного, несет пару ребер и несрастающиеся с телом позвонка нервные дуги. Неизвестно, существовал ли покров из чешуй. Триас.
I Фиг. 641. Череп Mastodonsaurus дгдап-teus. Триас. Гаильсдорф Вюртемберг. Германия. 1/8.
Фиг. 642. Поперечный разрез зуба Masto^onsaw Jaegeri, 3/1.
Подкласс II. Амфибии собственно (Euampliibla).
ПОРЯДОК 1. ХВОСТАТЫЕ, UBODELA ИЛИ CATDATA.
Хвост сохраняется в течение всей жизни; лобные и теменнные кости разделены; конечности являются примитивными, но своему строению приближаясь к конечностям стегоцефалов. Метаморфоз не выражен резко.
Фиг. 643. Череп современной формы Megalobalrach s japomcus. А—сверху, В — снизу, С — часть нижней челюсти.
апд — angulare, art — articulare, d — зубы, Exo — exoccipltale, Er — frontale, Mx — maxtllare, N — носовое отверстие, Ea — nasale, Oiph—orbfto-suhenoideum; Pa — parietale, Pmx — praemaxillare, Prf—praefrontale, Pt — pterygoideum, Qu — quadratum, Sq — squamosum
Древнейший представитель Urodela описан из пермских отложений Сев. Америки (Lysoropfyiis). Он представляет примитивные двояко-
Фиг. 644. Megalobatrachus Scheuchzeri. Миоцен. Энинген.
Баден. Германия. 1/8.
вогнутые позвонки с сохраняющеюся спинной струной, тогда как у позднейших хвостатых позвонки бывают вогнуто -выпуклые. Череп отличается от черепа позднейших хвостовых присутствием supraocci-pitale (фиг. 645). Эта особенность, гак
же как в сочлене
ние черепа с позвоночником, в котором принимают участие три затылочные кости, имеют рептильный характер, тогда как окостеневшие жаберные дуги связывают Lysorophus с амфибиями. Это — форма, указывающая на связь
двух классов. Следующий ископаемый представитель хвбЬтатых известен лишь из отложений переходных от юрских к меловым (Hylaeobatrachus'). Известен и верхкемечовой представитель Caudata. В третичных отложениях встречаются уже и принадлежащие к ныне существующим родам хвостатые, например Megalobatrachus из миоцена Бадена, описанный первоначально, в качестве ископаемого человека, погибшего во время потопа (Homo diluvii testis, фиг. 644). Современные представители рода Megalobatrachus, живущие в Японии и Китае, являются крупнейшими из ныне живущих амфибий (фиг. 643).
Caudata считаются деградировавшими потомками Branchiosauria, представляя с ними следующие общие особенности: короткие п )ямые, ребра; круп-
Фиг. 64а. Lysorophus trica-rinatus. Пермские отложения. Сев. Америка, f—frpnfa'e, п — nasal-,, р—parietale, pf— praBiontale, so — supraocci-pitale, q — quadratum, sq — squamosum, pa — части перво: о шейного позвонка.
сос
Фиг. 646. Fa’aeobatracfeus qrandtpes. Миоцен. Зи-бенбюрген. Австрия. 2/3.
сос — копчиковая кость, f — femur, h — humerus, fl — ilium, tu — предплечье, Ifi — голень.
ные поперечные отростки, отходящие от всех позвонков; одно и то же число предтазовых позвонков; сходство в строении плечевого пояса, отличающегося дегенеративным характером (окостеневает только scapula} у Caudata; сходство в тазовом поясе, в котором у Caudata так же как у Branchiosaurus не окостеневает pubis, одинаковое число пальцев—4 на передних и 5 на задних конечностях.
ПОРЯДОК И. БЕСХВОСТЫЕ, АЛЕКА ИЛИ ECAUDATA.
Тело широкое н короткое, лишенное хвоста, две пары конечностей, неравных по длине. Задние конечности более длинные, подразделены на четыре отдела, каждый значительной длины (один отдел состоит из проксимального ряда предплюсны/; служат для прыгания. Теменные кости срастаются с лобными (фиг. 628 А, С)-, метаморфоз резко выраженный.
Древнейшие представители ископаемых Anura известны из верхнеюрских отложений САСШ и Испании (Eobatrachus) и в третичных отложениях средней и южной Европы (Palaeobatrachus) (фиг. 646).
Геологическое распространение амфибий.
Branchiosauria встречается в каменноугольных и пермских отложениях, также и Microsauria; лабиринтодонты встречаются, начиная с каменноугольных отложений и кончая триасом; каменноугольные лабиринтодонты мало известны.
Стегоцефалы являются главным образом наземными и пресноводными животными; некоторые повидимому жили в дуплах деревьев, судя по нахождению их в полостях лепидодендронов и сигиллярий. В последнее время стали известны морские лабиринтодонты, с дельфинообразно-удлиненным рылом, из триаса Шпицбергена и Индии.
В различных отложениях, начиная с девона и до триаса включительно, встречаются следы ступней четвероногих животных, в большинстве случаев приписываемые стегоцефалам, например считаемые за следы лабиринтодонтов отпечатки в триасе, известные под названием Chirotherium (cheir — рука, therium—зверь).
Собственно амфибии (Urodela') встречаются, начиная с пермских отложений (Lykorophus из Сев. Америки), в отложениях переходных от юры к мелу (Hplaeobatrachus), в верхнем мелу, в миоцене (Megalobatrachus} и доныне. Второй порядок амфибий, Anura, встречается с верхней юры и доныне. Третий, Gymnophiona, в ископаемом состоянии не найден.
Характеристики групп амфибий.
Класс Amphibia. Почти все дышат в молодых стадиях жабрами, снабжены пятипалыми конечностями. Голые или покрытые чешуей, внутренний скелет обыкновенно окостеневает неполно, череп низкий и большею частью широкий, с парными затылочными мыщелками и большим парасфеноидом. В настоящее- время пресноводные и наземные формы. С девона (?), нижнекаменноугольных доныне.
1 подкласс: Stegocephali, панцырноголовые. Череп представляет наибольшее число костей, покров которых прерывается лишь носовыми и глазничными отверстиями. Зубы часто радиальноскладчатые. Плечевой пояс постоянно с тремя кожными костями. Позвонки большею частью не совсем окостенелые, хвост длинный. Почти только в пресноводных отложениях. От нижнего карбона до триаса включительно.
1 порядок: Phyllospondyli. Позвонки представляют на периферии тонкие, не сросшиеся между собой пластинчатые окостенения. Верхний карбон и нпжняя пермь.
2 порядок: Lepospondyli. Амфицельные позвонки, но с хордой, занимающей еще большую часть поперечника позвонка. Каменноугольные—пермские.
3 порядок: Temnospondyli. В массе позвонка преобладает костное вещество, позвонки вмболомериые, рахитомные или стереоспондилоные. Карбон—триас.
2 подкласс: Euamphibia. С уменьшенным числом костей в черепе.
1 порядок: Candata. Хвостатые, е голой кожей. Верхний мел—иыне.
2 порядок: Anura. Бесхвостые, лягушки. С нрыгательными ногами. Верхняя юра, третичные, ныне.
3 порядок: Gymnophiona. Роющие, подземного образа жизни, покрытые чешуями. Ныне живущие.
КЛАСС III. ПРЕСМЫКАЮЩИЕСЯ, REPTIL1A.
Название подсказано обликом и походкой ныне живущих их представителей, коротконогих и волочащих при движении живот по земле, но есть вымершие группы с телом, поставленным высоко на ногах. Пресмыкающиеся 318
Фиг. 647. Череп Spnenodon pvnrtatum А — глазница; В — верхнее, 8' — ннжчее височное отверстие.
откладывают яйца или живородящи, развиваются онтогенетически без метаморфоза, с амнионом и алантоисом, имеют тело голое или покрытое роговыми чешуями, костяными щитками, дышат исключительно легкими, холоднокровны, имея не вполне разделенное сердце и кровообращение
В черепе рептилий отличительно присутствие одной или двух пар височных отверстий, верхних и нижних (фиг. 647); последние иногда отсутствуют вледствие уничтожения перемычки', ограничивающей их снизу.
J низших и филогенетически более древних рептилий височные отверстия отсутствуют, и череп в этом отношении сходен с черепом стегоцефалов; кроме того, для них характерно первичное нёбо, на котором внутренние носовые отверстия, или хоаны, представляют более или менее длинные щели; разделенные между собою сошником (фиг. 648).
У высших и филогенетически позднейших форм развивается вторичное нёбо (фпг. 649); оно образуется соединением по средней линии лежащих в одной плоскости выростов верхнечелюстных и нёбных костей. При этом образуется полость между вторичным нёбом и параллельным с ним первичным; сошники также дают продольный вырост по средней линии, перпен-
днкулярно К ИХ
Pinx _
Ch
Vo
Jv
Во Оро 4 *
648. Череп прими-ныне
foil
Фиг.
тивного ныне живущего пресмыкающегося Spheno-don с первичным нёбом.
СЛ — яоавы, лежащие напереди, разделенные сошником Го Р1 — йёбиые к осте. Pt—крыловидные, Qu — квадратная кость.
плоскости, так что этот вырост разделяет пространство между первичным и вторичным нёбом на два рядом лежащие продольные каналы. Внутренние носовые отверстия при этом являются отнесенными все далее назад и притом не находятся в кровле ротовой полости—вторичном нёбе, которое таким образом является совершенно твердым и более прочным по сравнению с первичным. Вторичное нёбо распространяется все далее спереди назад, так что первичное принимает все меньше участия в ограничении сверху ротовой полости, rarasphenoideum недоразвит; есть только передний киль (ростр).
Соединение черепа с позвоночным столбом происходит посредством одного затылочного мыщелка, образованного основною затылочною костью, иногда при участии и боковых затылочных. В сочленении с нижнею челюстью находится лишь квадратная кость, иногда подвижная. Нижняя челюсть (фиг. 650) покрыта двумя передними (dentale и spleniale) и двумя задними (angulare, articulare) кожными костями и несет еще наверху при венечном отростке, одну, две или даже три дополнительные кости, coronoideumn supraangulare. Зубы у более
примитивных форм, кроме края челюстей, находятся на mmera, transvei sa и pterygoidea, у высших — только на краю челюстей; зубы прирастают своим основанием к челюстям, бывая плевродонтного типа (фиг. 651, 7; прирастание боком, со внутреннего края челюсти) или акродонтного
типа (фиг. II; зубы сидят на верхнем краю челюсти), наконец, зубы сидят в альвеолах (фиг. Ill) — текодонтные зубы
Фиг. 649. Вторичное небо пресмыкающихся.
Z — нёбо ныне живущей черепахи Tesitido; II — представителя Theromorpha (Udenodo npusillus) из пермо-риаса Юк ной Африки; III — ныне живущего крокодпта. В I вторичное нёбо зачаточно; сошник г, видимы । во всю длину, разделяет своим передним концом внутренние носовые отверстия, лишь немного прикрытые с одного края выростами верхнечелюстных костей (тп). В II сошники скрыты под вторичным нёбом, образованным межчелюстными (P»i) и нёбными (Z7) костями. В III вторичное нёбо образовано меж елюстными (Рш), верхнечелюстными нёбными (Р1) я крыловигиымн (Pt) костями; между последними находятся хоаны (£Л).
Позвоночный столб является укрепленным вследствие вытеснения хорды; позвонки углубленные с одного пли с обоих концов.
В противоположность тому, что у амфибий, ребра рептилий (также как и высших амниот) длинны и изогнуты, огибая брюшную полость и соединяясь отчасти на брюшной стороне, причем в результате этого соединения
Рга
Фиг. 650. Нижняя челюсть мозазавра Clidastes tortor из верхнемеловых отложений Сев. Америки, с внутренней стороны. 1/8.
Апд — angulare, Art — articulare, Cor — coronoideum, Sur—supraongulare, Cor и Sur в месте соприкосновения образуют венечный отросток.
образуется грудная кость. Вследствие укорочения и неразвитая ребер в областях, лежащих впереди и позади этой области грудных ребер, обособляются шейные и поясничные (последние между грудными и крестцовыми) ребра. Ребра одно- или двухголовчаты. В задней части хвоста
имеются парные нижние, дуги, которые расположены межпозвоночно, иногда имея опору в особых пластинках (так называемые интерцентры, см. у „хтпозавров, фиг. 703, D).
Таз первично прикреплен в двум крестцовым позвонкам, но часто -еще а и.,;, аольким добавочным. Имеется отверстие (foramen obturatorium) в os pubis.
У некоторых, обыкновенно более древних, групп существуют так называемые брюшные ребра (фиг. 671, 678, 702, 704), состоящие из палочковидных частей, возникшие из брюшного покрова чешуй стегоцефалов; у позднейших групп они исчезают.
В ископаем!м состоянии редкие представители встречаются уже с каменноугольных отложений.
Систематика рептилий находится в состоянии значительной неустойчивости. Существует довольно большое, но меняющееся число порядков и подпорядков, но объедивеиие их в более значительные группы не удается в такой степени, чтобы это было достаточно прочно и пользовалось широким признанием. Одну из более удачных группировок представляет подразделение по числу и расположению височных отверстий черепа на группы:
I II III
Фиг. 651. Зубы: I — плевродонтного типа, II — акродонтного, Ill — текодонтного.
а — зуб, Ь — челюсть.
Aaapsida1 (порядки Cotylosauria и Chelonia) без височных отверстий, подобно тому как у амфибий стегоцефалов, Parapsida — с височным отверстием наверху черепа (например Sqnamata, Icbthyo-auria), Synapsida—с отверстием внизу (например Theromorpha, Plesio-sauria) и выводимые из них Diapsida с двумя височными отверстиями (Crocodilia, Dino-sanria и др.). Определяющим является положение отверстия относительно костей: squamosum и poslorbitale. Так, у Parapsida височное отверстие располагается над комплексом костей squamosum + poslorbitaee, а у Synapsida — под ним (или перед ннм). У Diapsida два отверстия разделены перемычкой, состоящей нз упомянутых костей, jugale и quadrato-jugale (иногда) и отросток squamosum образуют нижнюю дугу.
Возможные генетические отношения этих групп можно представить схемой:
/Stegocephain AnaT)sida/ParapSida
(EmbolomerJ ^Synapsida — DiapsMa (-> Aves)
I
(Mammalia).
Если фация красного песчаника девона говорит о широком развитии материков, в связи с чем появляется первая наземная фауна амфибий, достигающая расцвета в каменноугольный период, богатый заболоченными водоемами жаркого климата, то инжияя
1 Названия составляются из слова apsis, что обозначает дугу, перемычку, свод (ср. абсиды в архитектуре). Поэтому Anapsida значит — без дуги, Synapsida — имеющие составную дугу, Diapsida — две дуги, Parapsida — особую дугу, параллельное образование. Подробнее объясняется в тексте в соответственных местах.
пермь — представляет новую перемену лика земли и климата, приводящую к засушливому я холодному климату пермского периода. Фауна амфнбнн, связанная биологически с водоемами, клонится к упадку, а получают распространение и развитие рептилии, уже раньше евшиеся от земноводных. Пермь — время усиленного формообразования пресмыкаю-
?. -..к, достигающих расцвета в товасе. Уже в пермский период мы имеем главные ветви наземных четвероногих, представленные и в современной фауне. Наследием стегоод-фа юв являются наши саламандры и тритоны, группа Anapsida выжила в лице черепах, Parapsida—ящериц, Diapsida — крокодилов и новозеландского Sphenodon’z (Halleria), а также с мела — птиц; потомство Synapsida представлено господствующими с кенозоя млекопитающими. Смена этих классов обменяется вымиранием прежних владык земли при резкой перемене климата и быстрым развитием новых форм, как бы дожидавшихся своего времени на. втором плане в предшествующий период — по своим этологическим данным. Возможные черты переходной организации вероятно скрыты в эмбриональной жизни, а остатки взрослых организмов нэ дают указаний иа прямое происхождение высших систематических единиц; все являются с чертами специализации и определенных приспособлений. Такое противоречие между требованиями теории медленной и постепенной эволюции и фактами многочисленных богатых находок и успехов изучения и методов исследования ископаемых форм может получить разрешение только в теории скачковых изменений.
ГРУППА ANAPSIDA.
Височные дуги отсутствуют. В сплошном покрове черепа нет височных отверстий. Зияние лишь в затылочной области, откуда могут возникать, развиваясь кпереди, вырезки (слуховое отверстие), впадины для мощной жевательной мускулатуры, иногда закрывающиеся вторично сплошным покровом, отверстым назади. Сюда относятся: родоначальная группа древнейших форм — порядок Cotylosauria и невидимому давно вышедшие из нее и дожившие п доныне, многочисленные и разнообразные представители порядка черепах (Chelonia или Testndinata).
ПОРЯДОК I. COTTLOSAURIA.1
Наиболее низко стоящие рептилии, с двояковогнутыми позвонками, с черепом стегоцефалоподобным, представляющим или сплошной щит, или одно широкое височное отверстие. Есть отверстие теменного глаза. Кроме зубов на краях челюстей, есть зубы на нёбе.
Большое разнообразие форм, от мелких ящерицеобразвых до грузных звероподобных. Можно отметить связь с низшими стегоцефалами (эмболомерными и рахитомными) и эволюцию в направлении к черепахам и тероморфам (группа Synapsida, дающая начало млекопитающим!.
Примитивнейшими и замечательными по мозаике признаков амфибий и рептилий в каждом органе являются Seymouriidae — Seymouria из перми Техаса н Kotlhssia (выеденная в подрод} с С. Двины, — изученные исключительно полно и совершенно, особенно первая, за последние 10—15 лет. Недостает только последних позвонков хвоста и нескольких фаланг конечностей.
Это были небольшие ()/а—1 метра) насекомоядные животные, с коротким хвостом, неуклюжие, скорее похожие на амфибий, приспособленные к роющему образу жизни.
Череп — низкпй, совершенно стегоцефаловцй с поверхности, с многочисленными скульптурными костями черепа и нижней челюсти, с височными вырезками для наружного слухового отверстия. Нёбо первичное, как у эмболомерных, ио сближенные хоаны и нпчюжный киль (ростр) парасфеноида — черты рептилий. В затылке ие окостеневает supraoccipita!e; затылочный мыщелок —трехчленный Зубы в один ряд по краям челюстей, лабиринтового строения, но кроме того на нёбных костях по 2 с каждой стороны (черта стегоцефалов), а в нижней челюсти даже на 3-й венечной (coronoideum), что встреча
1 Название обозначает ящеров с отверстием, впадиной (cotyle), так как первые черепа были найдены без костей основания черепа, легко выпадающих за отсутствие» связи швом с мозговой коробкой.
юсь только у кистеперых рыб (Osteolepis). Кроме этих теко^донтных bjOob—нёбо и челюсти усыпаны мельчайшими „шагреиеными" зубчиками.
Решающим признаком класса репгилгй являются позвонки, с мощными невральными дугами и горизонтальными сочленовными отрожками Но ’''-роение позвонков —
Фиг. 652. Pareiasaurus Karpinskis (по имени проф. Карпинского). Сев. Двина. Пермотриас.
явно эмболомерного происхождения, с крупными дпсвгми гипоцентров (интерцеитров) во всех отделах вплоть до 5 хвостовых, которые несут отросткп для головки ребер. Ребра двухюловчатые. Крестцовый позвонок 1 (2 у Kutlassial). Пояс плечевой низшего типа, но без с^ег’Йгииг^пласгпнчатый таз. Общий вид конечностей — как у древних стегоцефалов, но окостенение полное, и число фаланг и когти — как у рептилий. Плечевая кость имеет /огаигеп ulnare рептилий, в стопе замечательно отдельное окостененье intermedium.
Фиг. 653. (Череп Pareiasaurus Karpmskii. Сев. Двина. Пермотриас.
Pareiasaurus (фиг. 652—655).1 Представляет массивное животное, около 3 метров длиною, с коротким хвостом и весьма солидно построенными ногами. Голова широкая, короткая и сжатая сверху вниз. Череп в виде сплошного щита, со скульптурой лаби-ринтодонтового типа, представляющей соединение ямчатости с радиальною бороздчатостыо. Напереди отверстия двух ноздрей, разделенные, но сближенные между собою; маленькие орбиты помещаются сбоку, и имеется значительное отверстие теменного ' глаза
(фиг. 653). Весьма значительный единственный затылочный мыщелок является раздвоенным. Нёбо (фиг. 654) представляет ряды зубов на vomera, palatina и ptenjooidea. Края челюстей снабжены рядом более крупных
> Название указывает на замечательное развитие костей в области щек (pavela).
зубов, крона которых сжата в направлении снаружи вовнутрь, имеет наверху при остренный ребристый и зазубренный край округленного очертания. Ретвинлж-ней челюсти снизу представляют пс выросту, направленному вниз (фиг. 652).
Фиг. 654. Схема нёба Pareiasawus Baini. Пермотриае. Южно-Афр. Союз.
fat — осяопвая затылочная; Ъ» — основная клиновидная; ecpt — область вктоптервгоила (наружная ветвь кры ювидний ко<ту): ipt — иитерптеригондная (между ветвями крыдоввдвой вост) ролость, открывающаяся назад; те — верхняя челюсть; р. pi — задние боковые отрос тки крыловидной костя, т ротягнвающисся к квадратной кости; pl — обласгь нёбных костей: pt — пгрчдвяя середввная часть крыловидной сбл;сти; pi, nd — трапы; qu — квадратная кость, v — обтасгь сошкака. Маленькие кружки означают основания конических зубов. Ij7.
Фиг. 655. Плечевой иоле. Pareiasaurus. Пермо-трпас. Южн. Африка.
С1 __ clavfcvla, CUh — cleithrum; Cr — coracoidevm, F. gl — сочтеновпая впадина длн плечевой костя,
It — interciavicula, Per — procoruemdevm, Sc - scapula Плечевой пояс представлен развернутым на ндос-кости, вад с брюшной яоверлностн.
Позвонки двояковогнутые, со сраввптельло маленькими клиповидными интерцентрами в шейной и спинной облвстп. Шейные ребра двухголовчатые, туловищные одноголовчатые. Крестец составляют два первичных позвонка, но кроме того в него входят обыкновенно задний туловищный и
передний хвостовой. Плечевой пояс (фиг. 655), по составляющим его элементам, сходен с плечевым поясом лабиринтодонтов, представляя claviculae, tnterclavicula, scapulae, coracaidea, cleithra', кроме того здесь окостеневает procorai oideum. Таз является уже несколько снециатзированным по сравнению с тазом стегоцефалов, в котором pubis и ischium пластинообразны, лежат в одной плоскости и связаны между собою хрящей. Здесь же все кости таза сращены между собою. Ulna отличается своею массивностью. В предплюсне tibiale, fibulare и intermeaiiim слиты в одну кость.
Конечности имеют по пяти пальцев, снабженных ногтевыми конечными
Фиг. 656. Скелет нын& живущей черепахи (En-ys еигораеа); часть брюшнсге щита видна влево от скелета.
Г — позвонковые щитки; С — реберные йиткя; ЛГ — краевые щптки; JT — затылочный щиток; Р — хвостовые щитки, BIJS — брюшной щит, С1 — ключичный щиток; 1с1 — ыежключичиый щеток; Sc — лопптка; -4с — ее акромиальный отросток; Со — коракоид; РЬ — лобковая кость; Is — седалищная кость; II — подвздошная кость! Н — плечевая кость; R — лучевая Kocib; U — локтевая кость; Re — бедреыая кость; 21 — большая берцовая. R — мал „я берцовая кость.
члениками.
Вдоль
о&ные
на р.
оюстенения,
спины расположены
С. Двине и в
в виде щитков Южной Африке.
в небольшом числе рядов небольшие начатой поверхностью. Пермо-триас
с
ПОРЯДОК П. ЧЕРЕПАХИ, CHEIONIA.
Черепахи отличаются коротким и широким телом, с охватывающим его
костяным
имеют
пащырем как
роговую
обложку.
на свине, так и на животе; челюсти
без зубов
Панцырь состоит из более или иенее выпуклого спинного щита и плоского брюшного шита (фиг. 656), соединенны:; между собой по бокам тела. В этом костном ящике остаются большей частью лишь два отвер. стия: переднее — для головы, сидящей на довольно длинной и гибкой шее, и для передних конечностей, заднее отвер; тие—для задних конечностей и хвоста.
Костный панцырь черепах состоит из широких костных пластинок, которые спаяны между собою посредством швов. В состав спинного панциря входят, главным образом, спинные позвонки и ребра: позвонки — своими расширенными наверху и приплющенными остистыми отростками, ребра— пластинчато расширившись взад и вперед до взаимного соприкосновения; кроме того, на всем краю возникшего таким образом спинного щита развивается кольцо неболошпх кожных окостенений.
Брюшной щит состоит из непарной кости впереди и нескольких пар парных костей позади. Хотя эти кости не находятся в прямой связи с внутренним скелетом, так как плечевой пояс и таз обыкновенно прикреплены к панцырю лишь посредством связок, но все-таки непарная пластинка.
Фиг. 657. Ei.rystervum Wagleri со спинной стороны. Верхи, юра. Эйхштедт. Германия. 1/3.
быть может, соответствует interclavicula низших четвероногих, как кости кожной, а ближайшая пара также соответствует claviculae.
У большинства черепах эпидермический слой кожи над панцирем образует большие роговые щитки, которые отделены друг от друга бороздками и срастаются с костными тастинкачп панцыря. Роговые пластинки в общем группируются соответственно рядам костяных, но границы тех и других не совпадают ме кду собой—и число их не одинаково. Череп с одной парой височных впадин пли совершенно без вех. В плечевом поясе, под панцирем, существует coracoideun п scapula-, так называемый ргосога-coideum есть акромиальный отросток лопатки.
Иногда ребра срастаются так, что между дистальными концами их остается свободное пространств-). Таково, между прочим, состояние панцыря у черепахи, известной из верхнеюрских отложений и принадлежащей к роду Eurysternum (фпг. 657; eurys — широкий, sternum — грудная кость). Брюшной панцырь в подобных случаях также представляет отверстия.
Черепахи невидимому выходят из котилозавров, из группы Diadectes (dia—поперек), представляющих значительное пластинчатое расширение ребер. Еще ближе к предкам черепах Eunotosaurus (си —широкий, notes— спина, — пермь Ю. Африки) и наконец древнейшая черепаха—в верхнем триасе Прусской Саксонии Triassochelys — типичная примитивная черепаха, с уменьшенным числом костей в первично-сплошном сводчатом покрове черепа. Имеется suprtdemporale и 2 задних отверстия. Зубы рудиментарные в челюстях и в виде гребчя на сошнике, крыловидных и парасфеноиде. В плечевом поясе cleithrum, коракоид, зачаток акромиального отростка на лопатке. В боюшном панцыре принимают участие расширенные брюшные ребра.
ГРУППА STHA1 81BA.
Одно височное отверстие (первично — под дугой заднеглазничной + чешуйчатой костей; образуется вследствие отхождения этой дуги от скуловой)
Фиг. 65Ь. Реставрация Deatcrosanrus. Пермские отложмия (медистый песчаник). Ключевский рудник, Башкирская АССР. 1/12.
Снизу ограничено скуловой + чешуйчатой костью. Обычно к ней примыкают сверху части верхней дуги, откуда и название — со сложной дугой.
В эту группу входят порядки: Theromorpha (зверообразные) и Sauropte-rygia или Plesiosauria (ящеро-ластые или близкие к ящерам).
ПОРЯДОК III. THEROMORPHA.1
Представители его имеют двояковогнутые позвонки, височные отверстия, вубы разделены на резцы, клыки и коренные, подвергающиеся смене (Dia-
1 Ther—животное, шогрЬе — вид.
Фиг. 659. Реставрация Bhopalodon. Пермские отложения (медистый песчаник). Башкирская АССР. 1/12.
demotion), как у- млекопитающих; зубы па нёбе редки и мало развиты; существует наклонность к образованию вторичного нёба (фиг. 649, II): затылочный мыщелок трехраздельный (фиг. 661), а иногда имеются два мыщелка, вследствие выведения basioccipitale из образования мыщелка" подобно тому как у млекопитающих, нижняя челюсть образована, главным образом, зубной костью (фиг. 663—664); в плечевом поясе scapula некоторых родов несет так называемый акромиальный отростбк, к которому примыкает ключица (фиг. 666), и в то же время пояс утрачивает cleithrum, так что становится таким, как плечевой пояс низших млекопитающих— Monotremata. Крестец образован 4 — 5 позвонками; в тазе ischium и pubis становятся почти такими,
как у млекопитающих, благодаря присутствию широкого foramen obtura-lorium (фиг. 670). Число фаланг на пальцах 2 3. 3. 3. 3, как у млекопитающих.
У примитивнейших представителей этой группы лопатка без акромиального отростка, нёбо первичное, зубы, хотя и представляют дифферен-
Фиг. 660. Череп Inostrancevia Alexandra Пер-мотриас. Сев. Двина Сев Край.
Фиг. 661. Вид черепа Lystrosaurus dcclivis со стороны затылка. Пермо-трпас. Капская колония. Африка.
sq — чешуйчатая кость; Ъ. осс — основная яатылошая кость; ех. осс — боковые затылочные (верхняя часть — ta bulare); f. т — затылочное мозговое отвср«тие; ра, — теменные кости; в. осс — вгрхияя затылочная (верхняя часть — inter parielale, ср стр. 308 vostyartetdlia).
циацию на три категории, как например у Deuterosaunis (фиг. 658), но коренные простые однозубчатые. У рода Rhopalodon (фиг. 659) имеется склеротикальное кольцо, череп более удлиненный, нежели у Deuterosaurus. Более далеко ушедшие в развитии представители Theromorpha, как Dicy-notion, представляют вторичное нёбо, хотя и недалеко еще распространявшееся спереди назад (фиг. 549, II). Значительную дифференциацию зубов представляет род Inostrancevia (фиг. 660), хищник, достигающий 3 метров длины, с необыкновенно мошно развитыми клыками верхней челюсти и с обособлением нескольких зубцов на задних коренных зубах. Inostrancevia
(по имени профессора Иностраяцева) имеет хорошо развитые височные отверстия.
Роды Dicynodvn и Udenodon (фиг. 649, 666, 664) являются весьма сиециалмзгриьаяными представителями этой группы. Они довольно далеко
Фиг. 662. Нижняя челюсть Procolophon, представителя Cotylosauria.
Фит. 663. Нижняя челг сть Lycosuchus с дифференциацией зубов и с возрастанием deniale, так что ею образован венечный отросток.
Фиг. 664. Нижняя челюсть Cynognalhus, -рода, близкого к Inostrancei га. Длина den-talc почти равна дд^не челюсти.
Изменение нижней челюсти Theromorpha в направлении характера нижней челюсти млекопитающих.
Фиг. 665. Примитив- nodon с акромиальным носа, примитивного иый пояс Procolophon. отростком лопатки (&). млекопитающего.
Изменение плечевого пояса Theromorpha в направлении характера пгечевого ' пояса млекопитающих.
Фиг. 668. Таз Procolophon, представителя Cotylosauria.
Фиг. 669. Таз Ude-nodon.
Фиг. 670. Таз Diadcmodon срас-ширенньм дистально ilium (il) и с большим отверстием (pozctnen obtu-ratomum) между ischium и pubis.
Изменение тазового пояса Theromorpha в направлении к тазу м. екодитающих.
ушли в своем развитии, представляя вторичное нёбо (фиг. 649, TI), акромиальный отросток лопатки (фиг. 666) и расширенный в месте причленения к крестцу Шит (фиг. 669). Очевидно, они произошли от форм с высокой диференциацией зубной системы, от которой у Dicynodon сохранились лишь пара клыкообразных зубов в верхней челюсти, а у Udenodon, ыолет быть являющегося просто самкою Dicy-riodon, атрофированы все зубы На
месте резцов можно предполагать присутствие роговой обложки челюстей Все Theromorpha отличаются, сравнительно с Cotylosauria, гладкостью костей черепа. Описанные здесь роды встречаются в пермо-триасе (пермских? отложениях) у нас, в Ост-Индии, Южной Африке и в отложениях Шотландии, которые могут принадлежать триасу.
ПОРЯДОК IV ПЛЕЗИОЗАВРЫ, SAUROPTERTGIAP
Примитивные водные пресмыкающиеся, с длинной шеей и умеренно коротким хвостом. Череп маленький, с теменным отверстием и парий больших
Фиг. 371. Plesiosaurus dohchodeirus. Нижний лейас. Англия. С брюшной стороны 1/18
височных отверстий (фиг. 673). Снизу последние ограничены пирокой дугой,
повидимому сложного состава—из скуловой, чешуйчатой и заднеглазничной
костей, т. е. как бы слившейся из верхней и нижней дгг, причем замкнулось бывшее между н ши нижнее височ-
ное отверстие. Нёбо первичное. Зубы конические, сидящие в ячейках, в одном ряду на краю челюстей. Позвонки слабо двояковогнутые, почти с плоскими поверхностями, обращенными друг к другу.
Спинные ребра одно головчатые; имеются брюшные ребра; конечности более или менее листообразные, пятипалые, с укорочением костей пред-
Фиг. 672. Plesiosaurus macrocephalus. плечья и голени и с увеличением числа Нижний лейас. Реставрация. ЧЛ6НИКОВ пальцев, ИНО1ДЯ развито КОЛ1 ДО окостенений склеротики.
Род Plesiosaurus (фиг. 671—673; plesion—близкий; sauros—ящерица) отличается коническими, продольно бороздчатыми зубами. Центры позвонков
1 Sauros — ящерица; ptc-ygion -крылышко.
не срастаются с нервными дугами; места применения последних на спинной стороне позвонков имеют крестообразную форму. Шейные ребра двухголовчаты, прикрепляются только к центрам. Брюшные ребра толстые, состоят из срединного и боковых рядов костей. В плечевом поясе отличаются сильный развитием коракоиды, вытянутые по длине тела; ключицы слабо
Фпг. 674. Передняя ко вечность плезиозавра Poly-cotylus iehiyospondylus. Верхнемелов. от. ожепия.
Г. Пзюм.
И — humerus; R — radius; U— ulna; с — carpalia (r, i, «, 1, 2, 3); m. c—met icarpalia (1, 2t 3, 4, 4, 4); наверху дополнительные (4, 3, 4).
Фиг. 673. Череп Plesiosaurus macrocepJialus. А, Р, С— сбоку, сверху, снизу.
од — angulare; art — articultare; I.осс— lasioccipttale; Ъ. s — Ъаз$-splenoideum, d — dentate; ecpt — ectopte.ryg aide am; fr — frontale;
J. pt — межкрыловидное отверстие: j—jugate; mr—таз ilia; nar— наружные носовые о «верстая: orb — глазницы; pa—parietale; pin — отверстие непарного глаза, pl — нёбные кости; ртх — ргаепю>Ша; prf — ргае-front de; pt — ptegoidevm; pt. nar — хоаны; ptf—postfrontale; pto—poster* bitale; qu — quadratum; 8 — височное отверстие; s. ад—supiaangulare; s, о — подглазничная в адина eg -f- s. / + / — squamosum; pas—
остаток парасфеноида; » — vomer. 1/6 натур, велич.
развиты. Задние ноги несколько болыце передних. Тазовой пояс сильно развит, именно ischium и pubis.
Из меловых плезиозавров имеет распространение род Polycotvlus в Европе и Америке, представляющий более укороченные в приспособлении к водному образу жизни кости предплечья и голени, с дополнительными к ним костями (фиг. 674).
Происхождение неясно; невидимому ближе всего к Theromorpha.
ТРУПЕ* DIAPSIBA.
Два височные отверстия, разделенные дугой из заднеглазничной+чешуй-татой костей; нижняя дуга: скуловая+чешуйчатая кости (может принимать
участие и квадратно-скуловая). Происходит из группы Synapsida (с первично-нижним отверстием) вследствие отхождения теменной кости от верхней дуги. Порядки: Ehynchocephalia (клювоголовые, с ныне живущей новозеландской гаттерией Sphenoclon), Crocodilia (крокодилы), Dinosauiia (ужасные ящеры), Pterosauria (летающие ящеры).
ПОРЯДОК V. КЛЮВОГОЛОВЫЕ, RHPNCHOCEPHA1U.
Череп с двумя парами височных отверстий. Первичное нёбо; зуба иногда не только на краях челюстей, но и на других костях нёба; амфи-цёльные, иногда просверленные хордой позвонки, с интерцентрами. Ребра
Фиг. 675. Шечевов пояс молодого Spkenodon ^хрящевые участки затушеваны точками).
cl — ключиц*; t. cl— ыетйключичная кость; ее—лопатка; ». sc — надлопвтка; сог—коракоид.
Фиг. 676. Плечевой пояс взрослого Sphenodon.
Cl — ключица; F gl — сочленовная впадина для плечевой кости, Est — ыежключица; St — грудина; Сг—коракоид; Sc—лопатка.
с так называемыми крючковатыми отростками, отходящими от середины ребра назад с налеганием на соседнее ребро. Имеется покров брюшных ребер. Inter clavicula с грудным отростком. Humerus с 2 отверстиями: для прохода кровеносного сосуда—foramen ulnarе и для нерва—/. radiale (фиг. 677). Две-три проксимальных carpalia и tarsalia. Pubis и ischium разделены вторичным лобково-седалищным окошком. 2 или 3 тазовых позвонка. Триас — ныне.
К этому порядку принадлежит примитивнейшее из ныне живущих пресмыкающихся — новозеландский Sphenodon (Hatteria) (фиг. 647, 648, 675 — 678) с амфицельными, просверленными хордою позвонками; тазовых позвонков два. Параллельные ряды зубов на челюстных и нёбных костях. Брюшные ребра составлены угловатым непарным срединным и двумя боковыми элементами (фиг. 678). Humerus (фиг. 677) с foramen ulnare и foramen radiale. Проксимальный ряд carpalia представляет три косточки. Имеются две centralia. В плечевом поясе (фиг. 6/5, 676) коракоид с отверстием, имеются ключицы и узкая Т-образная interclavicula. Тазовой пояс с широким лобково-седалищным отверстием.
ПОРЯДОК TI. КРОКОДИЛЫ, CROCODILIA.
Животные большею частью крупные, с длинным, служащим для плавания хвостом, покрытые роговыми и костяными щитками, с зубами лишь на краях челюстей, сидящими в альвеолах. Позвонки у древнейших форм двояковогнутые. позднее большею частью процётьные. Существует грудная кость и слабо развитые брюшные ребра, спинные ребра с крючковатыми отроет-
вами (process, uncinati). В предплюсне проксимально находятся две большие косточки (фиг. 682), astragalus (=tibiale + intermedium + centmle) и calcaneiim (fibulare), последний, как у многих млекопитающих, опирается ва почву трлько заточным бугром. Триасовые представители крокодилов (fie-lodon) имеют двояковогнутые позвонки и первичное нёбо, со внутренними носовыми отверстиями, примыкающими к сошникам сзади (фиг. 679). У позднеСших крокодилов существует вторичное небо, у юрских — с хоанами, примыкающими сзади к нёбным костям (фиг. 681); позвонки вогнуты Начиная
с
А
Pmx---
пе-
Мх—
Ъа,— Prf
Мл
Фиг. 6/‘J.
Рог
Sq-£
Фиг. 677. А — левая рсдняя конечность bpheno-don punctatum (ныне живут.); В — НИЖНИЙ конец плечевой кости с обратной стороны.
Л—плечевая жесть; est—foremen radial*; еп — foramen uhiare; J?—-лучевая; If—локтевая кость; r, i, « — костбчки проксимального ряда запястья; с eentrale; (1—5) — косточки дистального ряда.
Рт.<
юрских крокодилов еще двоякомеловых отложений, крокодилы
Р1 ...
PI ..
Ju
Qu
Exo
То -Ch..
Во
Belodon Kapffi. Трнас. Штутгарт, Вюртемберг. Германия. А — череп сверху, В — снизу.
Л—гтазпнцы; Во—о новпая затылочиая кость; Ch—хоаны; D—предглаз-
нвчнов отверстие; Ею- боковая затылочная кость; Fr—лобная, Ju—скуловая кость; La— слезная; //а;-верхнечелюстная; Ра—носовые кости; JV—носовое отверстие; Ра—теменная кость; Р1- нёбная кость; Рт—межчелюстная; Prf предлобная; 11—крыловидная, Qu- квадратная, в 8— височные отверстия, верхнее нныжнее; 8д—чешуйчатая кость; 1о—сошнкк.
Фиг. 678. Схема брюшных ребер (6, I/} и собственно ребер (г, г') i'phenodon.
U — крючковатый отросток.
(Crocodilus') имеют процельные позвонки и вторичное нёбо (фиг. 649,111), в строении которого принимают участие крыловидные кости, и в них, Далеко назади, открываются хоаны. Отверстие теменного глаза отсутствует. Таз крокодилов (два крестцовых позвонка) представляет оригинальную особенность в том, что pubis не входит в состав сочленовной впадины для ведра, прикрепляясь к передней части ischium.
Костя черепа и щитки туловищного панцыря покрыты ямчатыми vrjv-бленпями.
Современные крокодилы являются обитателями пресной воды, мезозойские были большею частью животными морскими; переход к жизни в пресной воде, в устьях рек, произошел с эпохи нижнегс мела.
ПОРЯДОК VII. ДИНОЗАВРЫ, DINOSAUEIA. 1
Рептилии обыкновенно значительных, а иногда и гигантских размеров, с длинной шеей и длинным хвостом, с конечностями, приспособленными к наземному образу жизни. Задние конечности обыкновенно длиннее передних иногда хождение только на них и происходит, подобно тому как у птиц
Фиг. 680. Teleosaurus cadomenns. Юра. Франция. Околотазовая часть скелета с панцирем. а — со спинной, Ъ — с брюшной стороны.
Череп крайне мал сравнительно с размерами всего тела. Также относительно весьма мала вместимость мозговой полости. Нет отверстия теменного глаза. Зубы текодонтные, сосредоточеннЕте на краях челюстей. Позвонки опистоцёль-ные пли почти плоские на обоих концах, редко амфицельные; крестец содержит от двух до десяти сросшихся позвонков. Ребра двухголовчатые, грудная кость не вполне окостеневающая, ключицы и прокоракоиды отсутствуют. Лопатка весьма длинная, иногда срастающаяся с коракоидом. В состав сочленовной для бедра впадины входят все гри элемента газа. Ilium — широкая, вытянутая в переднезаднем направлении кость.
1 Deinos — страшный, cauros - ящерица
Ischia — обыкновенно тонкие и вытянутые, часто соединяющиеся напереди в плоскости симметрии. Pulis направлен вниз и вперед, иногда развивает задний отросток (post-pubis), направляющийся назад параллельно ischium (фиг 680), подобно тому как направлен pubis у птиц.
Femur расположен вер-
тикально, а его головка направляется, как у птиц, в поперечном направлении, входя в сочленовную впадину таза. Astragalus тесно примыкает к tibia, с наклонностью образовать tibio-tarsus, как у птиц.
Череп является типичным черепом рептилий по присутствию двух пар височных отверстий, по характеру квадратной кости, совершенно неподвижной противоположно тому, что у птиц. Иногда имеется покров из кожных костных табличек, также брюшные ребра. Сходства с птицами возникают отчасти конвергентно, вследствие сходного образа жизни — хождения только на задних ногах. Есть хищные динозавры, со сжатыми с боков коническими зубами, и есть травоядные—с плоскими лопатообразными зубами.
К динозаврам принадлежат крупнейшие из всех когда-либо существовавших наземных четвероногих.
Все древнейшие (триасовые) динозавры были животными хищными, с зубами режущими и распределенными более или менее равномерно по длине челюстей; в связи с хищным Образом жизни они имели, в облегчение скелета,
Фиг. 681. Teleosaurus cadomensis. Юра. Франция. Череп сверху (4), снизу (В) и сбоку (С).
А — глазницы, апд — угловая кость; art — сочленовная кость. Во — основная ватылочнзя кость; Bsph — основная клиновидная, Ch — хоаны; В — отверстие иижнеи челюсти; d — зубная кость; Fr — лебная; Ju — скуловая; La — слезная; Л/ж — верхнечелюстная; Ва — носовая; Ра — теменная; Р1 — нёбная; Ртх — межчелюстная; Prf — предлобная; Qu — квадратная. Quj — квалрат-носкуловая; S к S' — височные отверстия, верхнее и нижнее;
«Sg — чешуйчатая кость.
полые длинные кости конечностей и полые, с весьма тонкой стенкой, позвонки. Туловищные позвонки платидёльные или амфицельные. Эти животные со слабо развитыми передними конечностями и хождением на задних ногах образуют группу Theropoda (триас — мел).
Группа Sauropoda представляет более или менее массивных, по преиму-
ществу травоядных, динозавров с четырьмя • почти равными по величине
ногами, так что, предполагается, животное опиралось при ходьбе на все ноги. Позвонке с полостями по б<-кам. Еости конечностей солидные. Ноги
пятипалые.
Седа принадлежит например род Diplodo-cus (фиг. 683, 684), представители которого достигают в длину до 23 м. 1 Череп Diplodocus сравнительно весьма мал, прыгоднят в задней части и сжат с боков. Носовые отверстия, слившиеся в одно, помещаются на макушке черепа, как это бывает у животных, дышащих воздухом и в то же время остающихся подолгу под вододо. На боковой поверхности черепа находятся, кроме глазниц, треугольные предглазничные отверстия и перед ними еще небольшие отверстия. Зубы тонкие, цилиндрические, находятся лишь на переднем краю челюстей и употреблялись вероятно лишь для схватывания рыб, служивших пищей. Плечевой пояс состоит из удлиненных лопаток, коракоидов и пары окостенений, считаемых за окостенения грудной кости. Шея и хвост весьма длинны, первая давала возможность дышать даже стоя на дне, в при-
г и ш IV
Фиг. 682. Предал осна правой задней ноги крокодила.
F — fibula; f — ftbulare (calca-neum)\ I — IV* — плюсневые кости; T-rtHta; t, i, c ~ tibtale, fate* medium, entrale, сросшиеся в алгола из; N? — tarsale и vne-tatarsile 5; 1, 2, 3 — первое — третье tarsale, сросшиеся в одну косточку, 4 — четвертое tarsale.
брежной полисе моря, последний являлся органом равновесия и плавания. В хвосте цижнже дуги представляют на свободном конце отростки, направленные взад и вперед. Спинномозговой канал сильно расширяется в крестце. Юра Северной Америки.
Orthopoda, или Praedentata, представляют группу травоядных динозавров, опиравшихся при ходьбе на все четыре ноги или только на задние. Характеризуются тазом, в котором pubis представляет часть, направленную вперед, п часть, направ
ленна ю назад, притом лобковые кости правой и левой стороны, подобно тому как у птиц, не срастаются в срединной плоскости тела. Кроме того, челюсти спереди беззубы, может быть были покрыты роговым покровом, а нижняя челюсть спереди образована особой предзубной (praedentale) костью. Юра и
Фиг. 683. Череп Diplodocus longus. Сбоку и сверху. Верхняя юра, каньон Сити. Колорадо. САСЩ-
мел. Сюда принадлежит напри-
мер из европейских родов, хорошо изученный, благодаря большим раскопкам
1 Таков экземпляр, гипсовая копия которого находится в Академии Наук, в ее Па-Нозологическом Институте.
произведенным в Бельгии, род Igua-nodon (фиг. 685 — 688), с лопатообразными, зазубренными по краям зубами, с тремя пальцами на задних ногах и пятью на передних. Внутренний палец последних представляет лишь один, превращенный в шпорец, членик. Позвоночник скреплен приросшими к остистым отросткам око-’ стеневшими сухожилиями. До 10 метров длиною. Мел.
Род Trachodon (trachys—неровный, odous — зуб), из верхнего мела Сев. Америки, имеет удлиненно-низкий череп с ложкообразно расширенной мордой наподобие утиного клюва. Зубы в несколько рядов, запасные находятся в бороздках зубной кости, общим числом до 2000. Ischium с длинным узким концом. Конечные фаланги в форме расширенных или удлиненных копытец. Общая длина скелета до 10 метров. Близкий род Mandschurosaurtis найден недавно у нас на р. Амуре в верхнемеловых отложениях. Реставрированный скелет находится в Центральном Геологическом Музее (бывш. Геологического Комитета) в Ленинграде.
ПОРЯДОК VIII. ЛЕТАЮЩИЕ ЯЩЕРЫ, | PTEROSAURIA
Главные особенности в строении Ptcrosauiia, или птеродактилей, как их нередко называют, были связаны с их способностью летать, причем органами полета, как у птиц и ле-тучил мышей, служили передние конечности; летание происходило посредством летательной перепонки, как У летучих мышей п белок-летяг. По способу прикрепления летательной перепонки едва ли не наибольшая аналогия представляется именно с белками-летягами.
У птеродактилей последний палец локтевой стороны кисти удлинен (.подобно пальцам летучей мыши) и
Фиг. 685. Тазовой пояс Iguanodon bermssarten-sis. Нижнемеловые отложения. Бельгия.
а — сочленовная для бздра впадина; гг — ilium; is — ischium, р —pucic; р' ~ pcstpubis.
поддерживал складку кожи (фиг. 690 — 691), которая шла назад к задним конечностям и хвосту. Кости скелета отличаются легкостью, будучи точки, но в то же время, как у птиц, прочны, состоя из плотного костного вещества. Позвонки и длинные кости конечностей заключают в себе воздухоносные полости. Череп сидел на шее под прямым углом, как у птиц, был относительно больших размеров и по общему виду походил на птичий, особенно по удлиненному и заостренному на конце клюву. Костя черепа срастаются с исчезновением швов, как у птиц, особ нно в лицевой части. Глазницы большие; имеется кольцо окостенений склеротики. Челюсти снабжены зубами (фиг. 689) или, у позднейших, меловых форм, беззубы (фиг. 691) и вероятно имели роговой покров.
Туловищные позвонки процёль^ы, хвостовые ачфппёльны; имеется от четырех до семи крестцовых позвонков, обыкновенно слитых между собою. Пальцы передних конечностей малы, а иногда и рудиментарны, с утоняющимися проксимальными концами. Один из пальцев представляет рудимент лишь в-виде направленной к плечу пястной
кости. Плечевая
кость сильно
Iguanodon
Фиг. 687. Зуб
Mantelli. Нижнемеловые отложения. Остров Уайт. Англия.
1/2.
Фиг. 686. Iguanodon bernissartensts. Нижне-мел. отложения. Берниссар. Бельгия. 1/80.
расширена проксимально. Грудная кость широкая, иногда со срединным килем. Ребра тонки. Из костей плечевого пояса развиты лишь удлиненные ключицы и коракоиды. Задние конечности слабы. Pubis, как и в тазу кро
кодилов. не участвует в образовании сочленовной впадины для бедра. Имеются брюшные ребра. Хвост был или длинный, пнегда с расширением на конце (Rampliorhynclius., фиг. 690; rhamphos—изогнутый клюв, rhyuchos—
Фиг. 688. Череп Iguanodon berm-sartensis,
л — глазница; art — сочлейовяая кость; d — зубная; Fr — лобная кость; Ju — екуловая; La — слез тая; Л/ж — верхнечелюстная; N — носовое отверстие; /а — темечяая кость; Ртх — межчелюстная; Ptf — заднелобная; Qu — квадратная, Quj — квадратно-скуловая;
5 — височное отверстие; Sq — чеш йчатая кость; 8ут — предзубиая (prAedentuh);
х, у — надглазничные косточки.
клюв) или короткий (Pterodactylus, Nyctoda tylus, nyctos — ночь, dactylus — палец, фиг 691). По величине некоторые птерода тити были не больше воробья, другие достигали 6 метров в размахе крыльев. Встречаются в юре и меловых отложениях, достигая крупнейших размеров в позднейших, меловых формах. Сходства в организации с птицами считаются возникшими
Фиг. 689. Череп Rharnphorliynchus.
Не — носовое отверстие; Ле — предглазничиая внадниа; О — глазница; Т— верхнее височное отверстие.
исключительно вследствие способности летания: одна группа не находится в отношении происхождения к другой. Возможно происхождение Pierosauria от примитивных Dinosauria или, вернее, из общей родоначальной группы с динозаврами и крокодилами.
группа pabapsida.
Верхняя височная дуга из чешуйчатой и заднегчазнпч гой костей, огра ничивающая снизу верхнее височное отверстие. Верхняя граница дана те-
Фиг. ‘ЙО. Реставрация RhamphorhynclMs 1/2.
ленной костью, и сзади часто остаток tabulare. Образуется из Anapsida отступанием верхней дуги от теменной кости и утончением снизу чешуйчатой, вплоть до полной потери дуги в некоторых случаях.
Одновременный рост в вышину квадратной кости, обычно приобретающей подвижное сочленение с чешуйчатой.
Порядки: Squamata (чешуйчатые—ящерицы, мозазавры и змеи) и Icbtbyosauria (рыбоящеры).
ПОРЯДОК IX. ЧЕШУЙЧАТЫЕ. SQUAMATA.
11одпорядок I. Ящерицы, Lacertilia. Животные с первичным плп зачаточным вторичным небом, с позвонками большею частью пропёльными, редко амфицельными; височная дуга, ограничивающая верхнее височное от-
Фиг. ЫН. Syctodaetyl'^s gracilis. Верхкемеловыв отложеинл. плпзас. САСШ. I/O.
верстие снизу, часто отсутствует, подвижная высокая квадратная кость Часто имеется отверстие теменного глаза (фиг. 692).
Примитивное строение черепа, содержащего остатки tabulare, transversum, первичное нёбо— все говорит о древности группы. Некоторый свет на происхожденье от Anapsida проливает находка в нижней перми Техаса Araeoscelis, маленькой (Va л«) ящерицеобразной форты с параноидным черепом, высокой Квадратной костью, неподвижно закрепленной широкой пластинкой чешуйчатой кости. Конечности, позвонки, ребра — все напоминает ящерицу. Такого Hie типа остатки Кadaliosaurus из перми Саксонии.
Настоящие ящерицы скудно встречаются от триаса Капской колонии (Pallgiiana) до мела Южной и Средней Европы, а г :авным образом с нижнетретичных отложений доныне.
Фиг. 692. Череп ныне живущей ящерицы Monitor niloticus.
Col— columella l, столбик; Его—боковая ватылочиая кость; Fr—лобная кость; Ju—скуловая кость; La—слезная; ЛЛг—верхнечелюстная; N—носовое отверстие; На—носовая кость; Ра—теменная ко^ть; Р1— вёбная; Pwx—межчелюстная; Prf— предлабная; Рто—переавеслу-овая; Pt — крыловидная; Ptf — заднелобная; — квадратная;
Qvj — квадратно-скуловая; 5* — верхнее височное отверстие; Sb—верхняя затылочная кость; —чешуйчатая; Г— поперечная
кость; F—надг лазничн ал.
Дают начало двум значительным ветвям: своеобразно специализированным мозазаврам, которые быстро развились, широко распространи-лись и вымерли в мелсвых морях, и утратившим конечности змеям, процветающим в современной фауне рядом с ящерицами. Интересно отметить тенденцию к змеевидному развитию и утрате конечностей и различной степени среди представителей современных ящериц.
Подпорядок II. Мозазавры, Mosasauria (Ящеры с Мааса). Большие морские пресмыкающие с двумя парами листообразных конеч-
1 Columella (Col) не должно смешиватв с слуховыми косточкаки (стр 307 и 355) тою же названия: его окостенение вёб<о квадратного хряща, особенно ха-ракт< расе для ящериц, носвт еще имя cpipterygoideu/m, указывающее на связь с крыловидной костью.
Г
Фпг. 693. Скелет мозазавра P'aleeat pus co.yp'iaeus. Верхвемелов!ie одоления.
Канзас. САСШ. 1/30.
Фиг. 694. Позвонки мозазавров (вид сзади).
Л — шейным Gw^astes tortor сгадн, представ тяющий знгантруы В — епмнмоп иэзвинокг / lutecarpus coryphaeus. без зигантрум;
С двостоври позвонок Tylosam us dtspelor.
Фпг. 695. Череп Clidastes velox. А сбоку, Б — сверху, С — снизу.
Л — глязяяды; art — articular?'. hs — basisp^-enoideum; co — condylus occipitalis-, t or — corouofdeum; d — rent ile; f—frontale-, (a — praefron-iale; fop — poxtfruwta e; J jugale; ini i-ruemt illare; I — lacry-niule; mx - m jillue; N — нковы'1 (имрстя: n — nascle, os —-8vpr oe ijitale: p — pariiiale\ pa — palatinum; p4 — petrosum — пере ве< луковая коеть (у rooticuiM; pit ptg — pterygoideum\ q — quad-raiuni; s — височные отверстия; за — supraangulare; sp — angulare; spa — spleniale sgr — squamosum f наружный элемент; а внутреннем, примыкающий к теаевной кости, pari etale, представляет собою tabu-are); V — vomer; я соч^еиеаво посредине ветвей важней «влюетж.
ностей (фиг. 693). Череп типа черепа ящериц; отличается просверленною слуховым отверстием квадратною костью (фпг. 695).
• Большие конические зубы сидят в альвеолах; кроме края челюстей, вводятся на крыло-видных костях. Doe ветви нижней челюсти соединены между собою подвижно посредщвом связок, кроме того посредине каждой ветви нахо 1ится подвижное сочленение между передней и задней половиной ветви । фпг. 650 и 695, -4), что давало возможность широко ра зевать пасть и заглатывать крупную добычу. Позвонки процельные, почковидного или • треугольного поперечного сечения (фиг. 694), иногда с сочленением зигосфен - зигантрум, как у змей (фиг. 696), или без него (l/osa-saurus, род, встречающийся также и в меловых отложениях СССР). Кресца нет. Проксимальные части конечностей укорочены; существует гиперфалан-тпя — увеличение числа члеников пальцев. Развит покров из че-шуй. В своем распространении, географически весьма широком— Сев. и Южная Америка, вся Европа, Новая Зеландия, — мозазавры ограничены верхнемеловой эпохой.
Подпорядок III. Змеи, Ophidia. Без ног. Позвонки процельные, со сложным сочленением (зигосфен-зигаитрум) между верхними дугами (фиг. 696). В черепе отсутствуют височные дуги (фиг. 697).
Подобно ящерицам, достоверно известны лишь с третичных отложений
Фпг. 696. Позвонок ныне живущей змеи.
А — спереди, В — сзади; а, Ь — зигапофизы, обыкновенные сочленовные отростки; с — площадки для прнчленения ребер; za, zs— зигантрум, знгосфеи.
Фиг. 697. Черен ныне живущей змеи Сго-talus horn us.
Art — сочленовая кость нижней челюсти; Bs—основная клпиогид чая* D — зубп >я; Et — атмоидаль ая; J1’—лобная; Лпг—межчелк сть; ЛЬг—верхняя челюсть; Е — носовая; ОсЬ — основная затылочная, Oct — боковая затылочная; Осз— верхняя затылочная; Р—теменная Pf—заднелобная; Р1— »ёбная; t— пе-редиеслуховая, Prf - ппедлобная; Pt — крыловид-бья; Qu — квадратная; Sq — чешуйчатая; Тг — поперечная»
ПОРЯДОК X. ИХТИОЗАВРЫ, IPHTHYOSAURIA.
Также группа примитивных пресмыкающихся (фиг. 698—708) с первичным нёбом, с двояковогнутыми позвонками, с одною парою височных
Фиг. 698. Туловищный позвонок Ichthuiisaw us. Лейас, Вюртемберг, Германия. 2/3.
отверстий. Зубы радиально складчаты, помещаются или в слабо выраженных просто сидят в желобке вдоль краев
подобно тому как у лабиринтодонтов; альвеолах, или последних нет, и зубы челюстей, удерживаясь в нормальном
положении лишь мясом. Ихтиоза еры встречаются с триаса до меловых «тллжений включительно. У древн пших, триасовых форм ноги более приближаются к типу конечностей наземных четвероногих, представляя более
удлиненные и разделенные промежутком кости предплечья и голени нормальную пятипалую ми даже с числом пальцев менее пяти конечность (фиг. 699, Л, Б). У позднейших увеличено не только число члеников в пальцах, но и число самых пальцев (фиг. 699, С). 1 У древнейших форм задние конечности больше передних, подобно тому как это вообще часто бывает у наземных четвероногих, например у амфибий; у позднейших, обратно, передние ноги больше задних, так как при водном образе жизни
Фиг. 699. Эволюция передньй т-о-ечности ихтпозатфа-
Л—передняя конечность Ifixosaurus ffornt lianas. Трпас. Ломбардия Италия. В—передняя коиечноггь Merrtan ia Yitteli Трмае. Калифорния. С~передняя конечность Ichthyosaurus. Conybeari Юра Англия
вообще передние конечности играют ботее значительную роль, нежели задние. У позднейших в движении принимает участие сильно развитой вертикальный хвостовой плавник с отгибающимся в нем вниз позвоночником.
Фиг. 700. Скелет Ichtyosaurus quaHnscissus с отпечатком мя.ких частей плавников. Герхний лейас. Гольцмаден, Вюр-темберй Германия.
фпг. 701. Тае ихтио-вавра foretocnemus califor mens, Tptac. Ka лпфорния. tl — ilium, si —ischium’, of—fwa-men obtw ai orium; pb— pubis
Таз древнейших ихтиозавров обычного типа таза древнейших четвероногих (фиг. 701) с пластинчатыми ischium и pubis’, у позднейших форм таз, в соответствии с уменьшением задних конечностей, сильно редуцирован, состоит из тонких костей и вероятно не был связан с позвоночным столбом. Глазничные отверстия с окостенениями склеротики имеют относительно большие размеры у позднейших форм в приспособлении к пребыванию на
* Таким образом—гииерфалангця вместе с гипердактилией
большой глубине. Череп удлиненный, особенно у позднейших форм, образуется прпостренный rostrum, составленный главным образом удлиненными межчелюстными и носовыми костями (фиг. 702); верхнечелюстные и лобяыо
Фиг. 702. Реставрация Ichthyosaurus quadriscissus Верхний лейас. Вюртемберг. Германия.
-В — Череп. Вид сверху.
С — Черег Вид снизу
F—Зуб и его поперечное сечение.
Фпг. 703. А, В, С, D, Е, F—Ichtyosaurus longifrons. Верхний лейас. Нормандия. Франция.
а — anffulare; art — articulare; Ъо — I asioccipitale; be — basispltenmdeum; pas — остаток парасфеноида; d — &evta e, ex o.—eroccipitale; t—intercentra ipt— межкрыловидиое отверстие; j—jugate; la—lacrymale; ma— *na ilia- na — vasal#. « — наружные носовые отверстия; nf — нервные дуги; Con— затылочный мыщелок; ®JP—opfaf1 oticum. pa—parietal#; у tn—теменное отверстие; pl—jalatmmn; pmx—ptacmaxilla, prf~praefrontale; Pt^-pterygoideum; ch—хоаны; ptf—posifrontale; pto—postorbitale., q. j—quadratojugale; qu— quadratum, * — ребро; s_. a — supraangulare; 9. e—supraoccipitalei st—squamosum; sq—suprasquamoswn (supratempeirait);
9 — vomer.
кости, напротив, укорочены. Наружные и внутренние носовые отверстия отнесены назад. На границе теменных и лобных костей находится отверстие теменного глаза. Позвонки двояковогнутые, не сросшиеся с верхними пугам". У позднейших форм, юрских и медовых, туловищные ребра двух-
Фиг. 704. Спинной поз-
вонок ихтио авра и примыкающие к нему ребра. abd — брюшные, г — спинное.
Фиг. 705. Ichthyosaurus communis Кости тазового поя за левой стороны.
I—ilium, is—ischium; ph—pubis.
головчатые, а позвонки в соответствии с этим двухбугорчаты. Хвостовые позвонки однобугорчаты. У триасовых форм ребра одного тончаты, с расширенной в направлении снизу вверх головкой, из которой впоследствии, путем уничтожения срединной части, получается двухраздельная головка, лучше приспособленная для более сложных движений, нежели одноголовчатая. Такое изменение исходит от хвоста и стоит в связи с большим значением последнего как органа движения при переходе к водному образу жизни. Плечевой пояс (фиг. 706) состоит из пары солидных коракоидов,
Фиг. 706. Плечевой пояс и передняя конечность Ichthyosauri's communis.
cl — ъхачткы&от — коракоиды с межключичной костью; sc — лопатка; Л —плечевая кость; R — л)чевая; V — дешевая; г, £, и — засястпые корточки.
из сравнительно слабых лопаток и ключиц и Т-образной межключичной кости (in^erclaricula). Триасовые ихтиозавры отличаются более ромбоидальной interclavii ula. В юре Вюртемберга найдены ихтиозавры с отпечатками мягких частей, представляющие спинной плавт пк значительной высоты, соединенный невысокою кожистою килеватостью с хвостовым плавником. Находят ископаемые слепки экскрементов (копролиты) ихтиозавров, представляющие на поверхности спиральную бороздку—впечатление спирального клапана кишки, и содержащие следы пищи ихтиозавров в виде рыбьих чешуй и твердых частей головоногих. Можно думать, на основании нахи док до семи маленьких ихтиозавров в брюшной полости взрослых индивидуумов, что эти животные были живородящи. При совершенном приспособлении исключительно к водному образу жизни, живорождение могло развиться за невозможностью выхода на сушу для откладывания яиц. Триасовые ихтиозавры известны из Калифорнии, Средней Европы и с острова Шпицбергена.
4
Позднейшие представители встречаются почти во всех частях света, причем некоторые верхнеюрские и меловые отличаются совершенною беззубостью. Можно думать о происхождении ихтиозавров непосредственно от лабиринто
Фиг. 707. Эволюция межключичиой кости ихтиозавров.
а — JUixosaurus atacus Триас. Ъ ~ Hlxosaurus Cornalianus. Триас. с — Ichthyosaurus quauriscissus. Юра.
Фиг. 708- иопролит ихтиозавра.
донтов, на основании характера зубов, позвонков, interclavicula, на основании находок в п следнее время ихтиозавров и довольно сходных с ними морских лабиринтодонтов в триасе Шпицбергена.
Теологическое распространение рептилий. «
Древнейшими пресмыкающимися из Anapsida являются Cotylosauria, встречающиеся с карбона до триаса. С триаса же начинаются и Clielonia.
Группа Synapsida появляется в верхнем карбоне в виде примитивных Tberoinorpha, которые вымирают в конце триа’са. Sauropterygia распространены с триаса до мела включительно.
В группе Diapsida Rliychocephalia и Crocodilia в широком смысле этого последнего порядка известны с триаса и до ыне; Dinosauria и Pterosauria, появляясь в триасе, вымирают в конце мелового периода.
Fpjnna Anapsida начинается в нижней перми (Araeoscelis} и в виде ящериц (Lacertilia) и змей (Ophidia) продолжается доныне (Squamata); Mosasauria встречаются исключите, ьно в верхнемеловых отложениях.
£ *
Характеристики групп пресмыкающихся.
Класс. Пресмыкающиеся: Replilia.
Группа Anapsida. Подобно стегоцефалам без височных отверстий.
1 порядок: Cotylosauria. С двояковогнутыми позвонками. С теменным отверстием. Дна таз вых позвонка. Пермь. Триас.
2 порядок: Chelonia. Черепахи, короткие и широкие, беззубые. Покров черепа редуцировал сзади с нижнего края. Почти постоянно со спинным и брюшным панцирем, костяным и роговым. Пояса находятся внутри панцыря. Триас—ныне.
Грхппа Synapsida. Одно височное отверстие под дугой, образованной squamosum и postorbitale.
•> порядок: Theromorplia. С двояковогнутыми позвонками. Зубы дчфференцированы; из трехраздельного затылочного мыщелка иногда образуются два; 4—5 тазовых позвонков. Пояса сташвятся, как у млекопитающих. Наземные и пресноводные. Пермь. Триас.
4 поряюк: Sauropterygia, плезиозавры. Маленький череп, длинная шея. Теменное отверстие. Позвонки плати- иди амфицельные. Короткий хвост, сильно развитые пояса. Брюшвые ребра. Триас—мел.
Группа Diapsida. Два височные отверстая, разделенные дугой из postorbllale и squamosum.
5 порядок: Rhynchocephalia. Первичное небо, зубы на челюстях, а иногда и на других костях нёба Амфицельные позвонки, иногда просверленные хордой Два тазовых
о(вовка. Брюшные ребра. Большею частью теменное отверстие. Наземные н плаваю щие. Пермь—ныне.
6 порядок: Crocodilia. Вторичное нёбо у позднейших крокодилов. Процельные, пла-тицёльные или амфицельные позвонки. Брюшные ребра. Обыкновенно также панцырь из щитков, кожных костей. Триас—ныне.
7 порядок: Dinosauria. Наземные, достигающие гигантских размеров. Небольшой череп, без теменного отверстия. Шея и хвост длинные; от трех до десяти крестцовых еозвонков. В плечевом поясе только большие лопатки и коракоид. В тазу большая под. вздошная кость, вытянутые седалищная и лобковая. Триас—мел.
8 порядок: Pterosauria. Шейные и туловищные позвонки процельные. Брюшные ребра. Большая грудная кость. Пятый палец передних конечвостей удлинен и между ннм. н туловищем помещается летательная перепонка. Череп вытянут и клювообразен как у птиц. Триас—мел.
Группа Parapsida. Височное отверстие располагается над перемычкой из squamosum и postorbitale.
9 порядок: Squamata. Подпорядок I: Lacertilia, ящерицы. С первичным или зачаточным вторичным нёбом, с позвонками большею частью процельными, редко амфицельными; височная дуга, ограничивающая верхнее височке отверстие снизу, часто отсутствует. Часто имеется отверстие теменного глаза. Триас—ныне.
Подпорядок И: Mosasanria. Квадратная кость просверлена слуховым отверстием. Нижняя челюсть состоит из четырех подвижно между собою сочлененных частей. Позвонки процельные., В связи, с морским образом жизни нет крестца и существует гиперфалангия. Верхний мел.
Подпорядок III: Ophidia. Вез ног. Позвонки продельные с сочленением знгосфен-зигантрум. В черепе отсутствуют височные дуги. Третичные—иыне.
10 порядок: Ichtbyosauria. С заостренным рылом и короткой шеей. С первичным нёбом, с двояковогнутыми позвонками. Зубы радиально складчатые С ластами, с гипер-фалангией и гипердактилией конечностей. Брюшные ребра. Имеется спинной плавник и хвостовой плавник с продолжающимся в иижнюю лопасть позвоночником. Таз редуцирован. Триас—мел.
КЛАСС IV ПТИЦЫ. AYES.
Кроме оперения, характерными особенностями птиц являются передние конечности, приспособленные исключительно для полета (крылья), и задние конечности, на долю которых выпадает нести тело на твердой почве (ноги).
Класс птиц сравнительно со всеми другими отличается однородностью их организации, соответствующей однородности отдельных порядков других классов позвоночных. Это объясняется односторонним приспособлением всех птиц в одном направлении к совершенно определенному образу жизни. По своей организации даже современные птицы (и еще более ископаемые) весьма близки к пресмыкающимся, так что были объединяемы вместе с последними в группу Sauropsida, и, нет сомнения, птицы связаны с пресмыкающимися отношением происхождения. При этом переходе от пресмыкающихся к птицам главным шагом было развитие покрова, перьев, который имеет назначение двоякого рода: создание органа летания и защиту от потерь животной теплоты, особенно значительных у животных, перемещающихся летанием в воздухе. В связи с последним находится и теплокровность птиц с их высокой и малоколеблющейся температурой тела. Как было указано в общем обзоре позвоночных, перья являются образованиями ороговевшего эпидермиса, подобно чешуям рептилий. Затем, скелет передних конечностей путем недоразвития снабженной когтями кисти превратился в скелет крыла (фиг. 709), непригодного для иного рода движения, кроме полета. В связи с этим произошло значительное усиление мускулов, действующих при полете, прикрепляющихся к брюшной части плечевого пояса
(грудные мускулы), и увеличение поверхности их прикрепления, именно грудной кости и ее срединного продольного гребня. Наконец тем, что передние конечности птиц прсоб) азозаны в крылья и устранены от поддне-Екпванвя тела, когда птица не летает, создается целый ряд измерений организации. Вследствие того, что задние конечности одни выдерживают тяжесть тела при движении по земле, произошло образование крестца из значительного числа позвонков; кроме двух первично - крестцовых позвонков рептилий, особенно отчетливо выделяющихся у птиц в зародышевом состоянии, в состав крестца в следующих стадиях индивидуального развития входит значительное число поясничных и хвостовых позвонков,
причем все позвонки длинного крестца срастаются между собою и с вытянутою спереди назад подвздошною костью. Таким образом задние конечности получают весьма солидную опору в тазе
Происходит образование клюва. Древнейшие, мезозойские птицы еще имели зубы, но так как принятие пищи, за исключением схватывания
добычи на-лету, ’ происходило только склевыванием, толчками, с почвы, то зубы потеряли значение и исчезли, тогда как
Фиг. 709, Скелет ныне живущеЗ птицы.
ccf — запястье; С1— ключица; Со ~~ коракоид; Ви —ннжвви отростки грудных позвонков, F — малая бердовая кость, Ре — бедро, Н — плечевая кость; Л — подвздошная кость; Js — седалищная кость; Мс—пястье; pft р", р"'—фаланги трех пальцев; РЬ—лобковая кость; Ри —крючковатые отростки; R — лучевая кость; Rh — шейные ребра; 8с—лопатка; St — грудная кость; Stc — грудинный отд* л ребер; Т—боль таая берцовая кость; Тт — цевка {tarsometatarsu^}* V
локтевая кость; Z — пальцы.
твердые челюсти с их роговым покровом, вообще возникающим
у различных позвоночных при отсутствии зубов, развились в птичий клюв. Клюв заменяет хватательные руки, и для увеличения его подвижности шея очень подвижна, сочленение позвонков вместо первоначально амфицельного делается описто-
цельным, притом с особенш й, седлообразной формой весьма подвиж-йых сочленовных поверхностей (фиг. 710). Число шейных позвонков
Увеличивается.
В связи с часто существующим зерноядением развился соответствующий ему, служащий для подготовки к пищеварению, зоб; в связи с усовершенствованием полета — система воздухоносных полостей, проницающих многие кости конечностей, затылочную область черепа и пр.
Скелет является легким, но, вследствие прочного соединения срастанием многих костей, и весьма прочным; у взрослой птицы срастаются, с исчезновением границ, кости черепной коробки, тазовые позвонки, хвостовые по-
звонки, тела грущых между собой, срастаются с телами позвонков шейные ребра, срастаются ребра при посредстве окостенелых здесь крючковатых отростков, срастаются в дужку ключицы; в особенно высокой степени срастание происходит в костях конечностей. Срастается редуцированная малая берцовая кость с большою
Фиг. 710. Грудной позвонок Hesperomis берцовою, предплюсна срастается ча-regalis. Верхнсысловые отложения. Канзас. СТЬЮ С ГОленыО, частью С ПЛЮСНв-САСШ. а —вид сбоку; 6 —спереди. 1/2. ВЫМИ КОСТЯМИ, В ЦбВКу (ф11Г. 4).
Таким образом в строении скелета птиц в высокой степени наблюдается закон соотношения,
корреляции признаков.
Череп П1Иц является весьма близким к черепу рептилий, нёбо остается первичным (фиг. 711); сочленение черепа с позвоночником совершается при посредстве одного затылочного мыщелка; имеется подвижная, подобно тому как например у ящериц, квадратная кость.
Имеются коракоиды, кольцо окостенений склеротики.
Фиг. 711. Череп ныне живущей птицы, а — сбоку, Ъ — снизу.
AZs — алисфеноид; Аид — угловая кость; Art — сочленовная; Bt — basttemporale (остаток парасфеноид»)/ С — мыщелок; 1) — зубная кость; Et — этмоидальная; Аг — лобная; J — скуловая; Jmx — межчелюст ’ L—слезная; Л/г — верхне елюсгная; N— носовая; 0b —основная затылочная юсть; 01— боковая 3aTblj. вая; Оз— верхнезатылочная; Ра — теменная; Pal — нёбная; Pt — крыловидная; Q — квадрitii 1я;„ .
квадратно-скуловая; 8т — межглазничная перегородка; Spb — основная клиновидная; Sq — чеш)Нча
Уо — сошник.
Крыло птиц содержит в своем дистальном отделе части трех пальцев с наиболее полно развитым и длинным срединным; последний, вместе с проксимально следующими отделами конечности, служит для прикреплено ряда маховых перьев (фиг. 713), образуя край крыла.
Птицы вероятно происходят от примитивных динозавров группы Theropoda. Существует, однако и другое мнение, что птицы могли произойти от ящериц (Lac rtiha).
Птицы делятся на два подкласса, — Archaeornithes и Neornithes. Е пер
вому относятся два рода: древнейшие юрские птицы — Archaeopteryx и Archaeornis (фиг. 712, 713).
Подкласс I. Древние птицы. Archaeornithes.
Archaeopteryx и Archaeornis оба происходят из юрского литографского сланца Золепгофена и Эйхштедта в Баварии. Первый из них находится в Берлинском, а второй в Лондонском музее естественной истории.
Archaeopteryx отличается от Archaeornis коракоидом, сходным с коракоидом Ratitae, посредине более узким, чем вверху и внизу, III metacarpal?, расширенная сверху и снизу, а не цилиндрическая, как у Archaeornis. Лобковые кости с симфизом. В отношении происхождения Archaeopteryx является древнейшей формой, чем Archaeornis.
Череп Archaeornis имеет такую форму, как у типичных птиц; мозговая коробка сравнительно велика, составляющие
Фиг. 712. Archaeornis Simensii. Литографский сланец. Юра. Эсхштедт. Бавария. Германия.
с — заплел е; cl — ключица; со — коракоид; Л — плечевая кость; т — лучевая; и — локтевая; X—/II—пальцы.
ее кости слиты между собою, и квадратная кость невидимому свободна. Имеется кольцо окостенений склеротики, а также конические зубы, равномерно
расположенные в ряд и невидимому сидящие в альвеолах. Тела позвонков, насколько известно, амфицёльны или, может быть, платицельпы, т. е. с плоскими сочленовными поверхностями. Ребра очень тонкие, шейные ребра свободны, грудные без крючковатых отростков.
Грудная кость неизвестна, может-быть. вследствие предполагаемого некоторыми слабого развития и потому большой возможности затеряться. П-образная дужка подобна имеющейся у современных птиц. Коракоид сходен с коракоидом Cariuatae.
Крылья относительно малы и представляют три пальца, каждый окан-
Фиг. 713. Реставрация крыла A’-cLaeornts Маховые перья прикрепляются лишь к среднему из трех пальцев, который по преимуществу и сохраняется у позднейших птиц
чивается когтем.1 В тазе лобковые кости без симфиза, ilium значительно вытянута спереди назад; сочленовая внадина для бедра просверлена; швы между косьями таза не сглажены Задние конечности такого же характера, как у ныне живущих птиц, с четырьмя пальцами, снабженными когтями; кости предплюсны слиты между собою. Имелись тонкие брюшные ребра. В хвосте перья располагались парами, по бокам каж .ого из многочисленных (20) хвостовых по-
звонков, не слитых в хвостовую косточку современных птиц. Эта птица была величиною с грача. Кости Archaeornis были поводимому не пневматичны, и летательная способность невелика.
Фиг. 714. Hesptrornis regalis. Верхне-еловые отлохемия. Канзас. САСШ. 1/8.
1 У довольно многих ныне хйвущих птиц, притом из числа более примитивных, в крыльях имеются, как рудимент, св (бодрые пальцы с ко гтчми, иногда даже служащие при лазании по деревьям
Подкласс II. Новые птицы, Neornithes.
Тип этого подкласса выражен во всех ссыльных птицах, в особенности живущих ныне. Подкласс подразделяется на две группы/ Ratitae, с грудной костью без киля и недоразвитыми крыльями, и Carinatae с килеватою грудною костью, с хорошо развитыми крыльями. Ratitae, без сомнения, происходят от Carinatae, но, вероятно, от (флее низко стоящих, чем современные Carinatae.
Древнейший представитель Ratitae известен из верхнемеловых отложений штата Канзас ₽ Северной Америке. Именно, это род Hesperornis (фиг. 714). Он отличается от всех кенозойских птиц присутствием зубов в челюстях. Имея недоразвитые крылья и отсутствие киля на грудине, Hesperornis в подробностях строения скелета не столько примыкает к ныне живущим Ratitae, животным наземным, подобно страусу, бегающим, сколько к некоторым из ныне живущих 'Carinatae, именно гагарам. Hesperornis (hesperos—вечер) имел на HOiax все четыре пальца обращенными вперед и соединенными перепонкою и был птицею водною, ныряющею, подобно пингвинам. Таким образоы уже в вепх-немеловых отложениях встречаются птицы, приспособленные к специализированному образу жизни, и, следовагезьно, в эту эпоху должно было существовать разнообразие среди птиц, между тем найдены лишь единичные роды. Из них, например, Ichtliyomis (фиг 715), также из верхнемеловых отложений Канзаса, с зубами, каковыми особенностями он г менных Carinata.
Фиг. 715. Ichthyoni :s victor. Верхне-меловые отложенья. Кспзас. САСШ. 1/4.
с двояковогнуты ИИ позвонк ШИ и отличается существенно от совре-
Геологичесаое распространение птиц.
Древнейшими являются юрские птицы Archaeornis и Archaeopteryx.
Большое зияние, которое существует между ними и верхнемеловым Hesperornis, еще не заполнено и доныне палеонтологическими находками.
Другой представитель зубастых верхнемеловых птиц Ichtyornis, по новейшим данным, невидимому уже близок к современным птицам из Carina-tae, представители которых, как-то: утки, курицы и болотные птицы, найдены в верхнем мелу Америки и были, быть может, частично зубастыми. Большинство порядков ныне живущих птиц встречается в третичных отложеньях.
В верхнем эоцене известны роды современных птиц, настолько умножающиеся в миоцене, что в нем встречаются уже представители почти каждого из ныне существующих порядков.
23
Учебник палеонтологии
353
КЛАСС V. МЛЕКОПИТАЮЩИЕ, MAMMALIA.
Млекопитающие являются животными с телом, обыкновенно покрытым волосами, и вскармливающими своих детенышей молоком.
Замечательное явление в этом классе представляет крайнее разнообразие приспособлений к различному образу жизни и в связи с этим соответствующее разнообразие в развитии органов, при посредстве которых животное ближайшим образом становится в соотношение с окружающей средой, именно органов движения и принятия пищи, т. е. конечностей и зубной системы. *
Как было выяснено в общем обзоре позвоночных, волосы образуются, подобно чешуям и перьям, роговым слоем эпидермиса. У млеко титающих сравнительно редко встречаются роговые
чешуи и щитки кожных костей (ящеры, броненосцы). Роговое вещество образует рога на лбу жвачных животных, собственно лишь роговую оболочку костяных выростов, выходящих из лобных костей. Наконец рога оленей не имеют ничего общего с кожей, представляя большею частью костяные выросты черепа, лишь сначала покрытые кожей, затем стирающейся. Эти рога ежегодно сбрасываются с пеньков, а затем снова вырастают на последних, благодаря деятельности сосудов кожи. Весьма распространенными, сосредоточивающимися лишь на концах пальцев, ороговениями являются когти, ногти и ко-
А — у Theromorpha (C'ynognathys), приближающихся к млекопитающим; В — у зародыша млекопитающего (Tatusia).
Апд — angulare; Ar — articular#; D — dental#; Inc — incus; Alai — malleus; memb. ty — барабанная перепонка; Qu — qua-dratum; St — stapes; Ty — tympanicum.
Кожа млекопитающих отличается от кожи других позвоночных изобилием различных желез; наиболее замечательны из них млечные, у высших млекопитающих приуроченные к сосцам, тогда как у низших сосцы не обособляются на гладкой поверхности живота.
Развитие черепа млекопитающих находится совершенно под влиянием сильно разрастающегося головного мозга. В связи с последним межглазничная перегородка, развитая у других классов позвоночных между областями носовою и клиновидных костей, чрезвычайно укорачивается, а иногда и совершенно не развивается, тогда как глазницы отходят от срединной линии и сдвигаются на бока черепа. Обонятельная кость (ethmoideum), называемая у млекопитающих решетчатою (в связи с сильною проды-рявленностью обонятельными нервами), образует стенку между передней частью мозговой полости и заднею носовой. Мозговая голость получает развитие, преобладающее над лицевою частью. Скуловая дуга'назади отходит от squamosum, и в месте отхода ее на последней н ходится сочленение для нижней челюсти. Таким образом quadratum у млекопитающих не
принимает участия в сочленении черепа с нижней челюстью. Quadratum, подобно articulare в нижней че^ости, превращается в одну из слуховых костичек, перемещаясь в полость среднего уха, причем /фиг.” 716) quadra-turn — наковальне {incus), a articulare = молоточку {malleus). Наковальня причленяется к стремени {stapes), представляющему собой, подобно columella предшествующих классов, видоизменение верхней части гиоидной дуги и несущему отверстие для артерии.
Нижняя челюсть млекопитающих состоит только из одной зубной кости. Эти- изменения, как изложено выше, намечаются у некоторых рептилий Theromorpha из группы Theriodontia. 1
Череп сочленяется с позвоночным столбом посредством двух мыщелков, принадлежащих главным образом затылочным костям, но отчасти и основной затылочной,
В черепе млекопитающих многие кости являются сложными, слившимися из костей, существующих отдельно у низших классов позвоночных. Так, затылочная часть черепа лишь у молодых особей, у зародыша, состоит из отдельных костей. У взрослых животных- она представляет одну затылочную кость, возникшую от слияния боковых, верхней и основной затылочной. Сбоку череп ограничен тоже сложною височною костью {temporale)-, она представляет: 'чешуйчатую часть {squamosum), каменистую {petrosum) и барабанную {tympanicum). Чешуйчатая кость, в отличие от низших классов позвоночных,, принимает участие в образовании стенки черепа, очевидно, в связи с развитием головного мозга и расширением черепной полости. Каменистая кость {petrosum) млекопитающих происходит на счет сливающихся друг с другом pro-, epi- и opisthoticum и заключает в своей толще слуховой лабиринт. Она принимает значительное участие в образовании черепной коробки, а именно ее дна, отчасти прикрываясь снаружи другими костями, например кожной барабанной костью—tympanicum. В простейшем 1 * * * * * * В
1 У некоторых Theriodontia, наиболее напоминающих млекопитающих и по диффе-
ренцировке и даже смене зубов, наблюдается роет зубной кости навстречу чешуйчатой
и редукция костей сочленовного отдела нижней челюсти, которые явно теряют свою
функцию и не могут служить для прикрепления мощной жевательной мускулатуры. 1? то же время роет венечного отростка и соответственное усиление чешуйчатой кости, подавляющей квадратную до ничтожного остатка, характерен для всей группы Theriodontia. Наконец, недавно обнаружено (у Diademodon) параллельное существование двух сочле-
нений нижней челюсти с черепом: наружное, вновь образованное чешуйчатой костью
с зубной'позади венечного отростка, а совнутри, рядом с ним, прежнее—квадратной
с сочленовной. С другой стороны, эмбриология указывает на гомологию слуховых и прилежащих к ним косточек зародыша млекопитающих со многими элементами заднего отдела нижней челюстп, из которых особенно интересно отметить гомологию барабанной (tympanicum) кожной кости с угловой (angulare).
В подтверждение того же взгляда о пэремеиэ функции говорит сравнительная анатомия мускулов и нервов: 1) отсутствие у млекопитающих мускулатуры, имевшей назначение оттягивать назад нижнюю челюсть и получавшей начало иа шее пли затылке, а прикрепление в заднем отделе нижней челюсти (muse, depressor mandibulae), 2) образование мускула, напрягающего барабанную перепонку, начинающегося иа молоточке и снабженного нервом от ветви тройничного нерва, заведующей жевательной мускулатурой (muse, tensor tympani). Кроме того, у многих Theromorpha имеется характерное для млек .питающих отверстие для артерии в stapes (стремя), а общий вид черепа, с мощной височной дугой и соответствующим высоким венечным отростком нижней, челюсти, говорит о сильно развитой мускулатуре жевательного аппарата с перекреплением мышц на новые места—и освобождение, ослабление и легкий переход на службу слуховому аппарату костей иижней челюсти вокруг прежнего сочленения puadrafo-articulare рептилий.
случае, .у млекопитающих низших порядков, tympunicum является < виде неполного кольца, на котором натянута-*барабанная перепонка. Иногда это кольцо, разрастаясь, образует при основании черепа особый пузырь (bulla tympanica), играющий роль резонатора. Некоторые авторы считают tympani сит гомологом angular е низших четвероногих.
Скуловая дуга образована соединением скуловой кости (jugale) со скуловым отростком squamosum. В основании черепа иногда срастаются все находящиеся здесь клиновидные кости (basi- и praesphenoideum, а1г-и orbitopshenoiden) и примыкающие к ним небольшие крыловидные (pterygoided).
(Виг. 717. Череп собаки (Cams famihans) сверху и снизу
ар — иёбные отверстия; Cav gl — сочленовная для нвжнеи челюсти впадина; С. осс— затылочные мыщелки;
Fr— frontale; Jp — Interparietale; Ju—jugate; L — lacrymale; SI, Six— maxSUare; S'a _I nasak; Occ. bas— basioccipitale; P — palatinum; Pa — panetale; Pit — скуловой отросток височной кости; Рюкв, PSlx — praemaxiUare; Pt— pterygoideum; SO — supraoctpitale; Sph— alisphenoideum; Sphf — basisphenoideum; Sph"— praesphenoideum; Sq — squamosum; T—tympanzcwn; Vo — vomer.
Существует хорошо развитое вторичное нёбо. Ни praefrontalia, ни postfronidlia у млекопитающих не развиваются.
Зубы текодонтны, расположены в один ряд на краю челюстей. Образование солидного вторичного нёба, образование более прочной, состоящей из одной кости, нижней челюсти, укорочение челюстей, при котором достигается лучшее действие их как рычагов, — все это явления одного и того же порядка, сопровождаемые кроме того развитием высших форм отдельных зубов и редукцией смены их. Зубы в разных частях челюстей приспособляются для разных функций, отсутствующих иногда у низших классов четвероногих, как например пережевывание, и зубная система из гомодонтной низших классов становится гетеродонтной, разделяясь на резцы, клыки и корен -
•нъье .зубы Нормально у млекопитающих два поколения зубов: мощное и окончательное. Из них первое является исторически более древним. Однако у некоторых низших млекопитающих существуют зачатки зубов еще одного поколения, а иногда и двух. Некоторые млекопитающие совершенно не имеют смены поколений зубов. Вторично зубная система может стать упрощенной, гомодонтдий, как у китообразных, или зубы могут совершенно исчезнуть
Первичною формою зуба млекопитающих является коническая, как у рептилий Первое осложнение, которое испытывает зуб гладкой формы (гапло-донтный), состоит в приобретении дополнительных зубцов, одного с передней стороны первоначального конуса, другого задней; в то же время корень зуба получает продольную борозду, вследствие наклонности раздвоиться. При более сильном развитии дополнительных зубцов получается триконодо&т-ный зуб (фиг. 718,' Т, а, &). Триконодонтное состояние зубов удерживается до верхней юры. Со времени последней появляется третья ступень в развитии зуба. Предполагается, что допол-нительные зубцы сдвигаются в верхней челюсти наружу, в ниж-— внутрь. Получающаяся таким образом тргтуберку-яярная форма зуба (фиг. 718, I, с) является господствующей до эпохи верхнего эоцена и исходной для развития зубов современных млекопитающих. Трехзубчатые жевательные поверхности, чередующиеся в той и дру-[ гой челюстях, приспособлены для разрезывания мелкой добычи вроде насекомых. Такое чередование зубов в челюстях сохраняется у нынб живущих хищных и, несколько стушевываясь, сохраняется у жвачных.
Тритуберкулярный зуб испытывает дальнейшее усложнение вследствие развития придаточного у'еппа сза.ти (пятка'; таким образом получается бугорчато-режущий зуб, особенно типично выраженный в так называемом плотоядном зубе хищных (фиг. 718, I, d). Пятка увеличивается и получает три добавочных бугорка^ Получающийся таким образом шёстибугорчатый
л о О о
X? О-Ч )—О )mcJ
i”1 f>a m
f роса mob
Фиг. 718 II. Схема равви-тин и расположения верхние и нижних вубов у различных млекопитающих. Зубы верхней челюсти показаны жирным.
А — гаплодоитныв Зубы; В — трико-водоишые; С — триту беркуляриые; D — наверху тритуберку лярные, внизу бугорчато-релущне; Е — то же с увеличением числа бугорков ш, пятке; F — то же, с придаточным бугорком наверху; G — зубы шестибугорчатые внизу и наверху.
Фиг. 7181. Разни гие основной формы зубов млекопитающих
а — трикоподонтпый (начальная стадия) JJr»malherTvtn\ триас; b — Micro сопо-dtm, триас; с—гря-туберк "оный, Тгг isodon и з Creodon-tia\oov.ev.\d~ бугорчато - реж\ щнп яуб, Viterra (ныне жв-вущ); л, b, d — сбоку; с — сверху.
।
зуб представляет форму зуба, основную для коренных зубов копытных, хищных и приматов. Из него развиваются несколько типов. При приспособлении жевательной поверхности к перемалыванию растительной пищи по-
Фиг. 719. Зубы млекопитающих.
а — бунодоптЕыи зуб 'Uuracotherfum; b — лофодонтгый зуб Dinolhetium; с — отдельный бугорок, с наклонностью стать полулунным, flvopotamus; d — буио селенодонтный зуб Palaeoeyopsy е — селенодонтный, Proto сегав, с несколько стерший дребняып; f — селенодонтный, Prodremothenutn с нестертыми грсбпгми
верхность пятки приходит иа один уровень с поверхностью остальной (первоначальной) части зуба, и вся поверхность становится четырехбугорчатой вследствие редукции остальных бугорков. Самые бугорки становятся менее заострьннымй. ТакЙ4Г^«оЗон«гны^ зубя (фиг. 719, а) существуют еще и в настоящее время например у свиньи. Если бугорки вытягиваются в направлении поперек челюсти до взаимного прикосновения в поперечных рядах, то возникают два поперечных гребня, оунодонтный зуб — основная форма для зубов всеядных животных — превращается в^лофодоыпный (фиг. 719, Ь). Большею частью при этом оба наружные бугорка" также соединяются между собою продольным гребнем, так называемой' наружной стенкой.
С другой стороны бугорки бунодонтного з^ба могут принять V-образную форму вследствие появления на каждом двух расходящихся гребней (фиг. 719 с), и затем из таких бугорков, округлением их формы могут получиться снабженные бугорками полулунной формы — селенодонтные ' (фиг. 719, е и /') коренные зубы жвачных. Дальнейшее усложнение зубов этого типа происходит путем возникновения складчатости эмали гребней ши полулун.
Если крина бунодонтного коренного зуба испытывает лишь ньбольщое измененье, то она остается в общем низкою а покрывается толстым
Фиг. 720. Переход ст бра-хиодовтного вуба Ancluth -rium (а) к •-ипсодов гному лошади (о); b — промежуточная стадия Higpanon.
слоем эмали, тогда как корень удлиняется, прекращая свой рост за известным пределом. Только корень бывает тогда покрыт цементом. Эти так называемые брахиодонтные зубы ведут путем постепенного изменения в^гипссдонткым зубам многих копытных. Именно последнего рода зубы
встречаются у жвачных, непарнокопытных и хоботных. Они лишь поздно развивают корни, притом (фиг. 720) короткие, -тогда как крона становится весьма высокой вследствие продолжающегося роста, которым компенсируется постоянное стирание жевательной поверхности при растительной пище, ссдер-
жапей обыкновенно частицы земли, песку. В соответствии с таким ростом зуоа бугорки и гребни превращаются в столбики или вертикально-^удлинен-
ные пластины, которые покрыты лишь тонким слоем-эмади. В промежуточных между буграми и гребнями углублениях отлагается цемент.
Состав зубной системы млекопитающих выражают формулой в виде дроби, fra месте числителя которой обозначены числа зубов разных категорий в одной половине верхней челюсти, на месте знаменателя — числа зубов в половине нижней челюсти. При этом первая цифра как над чертой, так и под нею, представляет число резцов, вторая — клыков, третья — ложнокоренных и четвертая— настоящих коренных.
Так например зубная формула человека и обезьян Старого света будет следующая: =—а всего 32.
3-1 . 3 . !
Для многочисленных сумчатых формула g ~—g---, а для
3 • 1 - 4 3 последовых - - - - -—g .
Формулою, исходною для млекопитающих, гипотетически 5 • 1 • 4 • 5
считается ————g (впрочем число настоящих коренных доходит да,.;е до семи); она не получает полного развития ни у одного мле".опптающ'го хотя вероятно, могла бы быть установлена в онтогении.
Позвоночный столб млекопитающих состоит из позвонков платицёльных, т. е. с плоскими примыкающими поверхностями. Только шейные позвонки копытных, отличающиеся особенною подвижностью, являются опи-стоцёльными. Число позвонков колеблется сравнительно
Фиг. 721. Разные стадии гипсоддитии и отложения цемента (доказан пунктиром) на верхних коренных зубах, в виде сверху и в вертикальном разрезе. Разрезы эмали показаны жирным
4 — Aceratheriwn} Б — Rhinoceros pachygnatl.va; С — Rhinoceros апИдю-tatis; D — Elasmotherutm
И
А
с другими классами позвоночных лишь в узких пределах;
в шее их бывает за немногими исключениями, всего лишь семь. В шейной
области позвонки срастаются с рудиментарными вильчатыми ребрами (про-
Фе г. 723, Шейный позвонок собаки.
свет развилка называется foramen transversariumf, грудные ребра частью соединены с узкой, большей частью членистой, грудной костью, частью свободны, имеют хрящевые нижние половины.
Крестец первоначально состоит только из двух позвонков, как у рептилий, позже с ними срастаются еще несколько хвостовых позвонков, остальные хвостовые позвонки часто, как и самый хвост, рудиментарны. Плечевой пояс низших, так называемых однопроходных, млекопитающих состоит еще из тех же частей, что и у рептилий; но у всех остальных млекопитающих он является значительно редуцированным. У однопроходных имеется и коракоид, упираю-
щийся своим нижним концом в грудную кость, и про-коракрид (фиг. 724); лопатки имеют акромиальный выступ, к которому примыкаю? ключицы. У остальных млекопитающих proeoracotdeum отсут-
Фиг. 724. Плечевой пояс (с правого бока) ехидны (Echidna hjstrij.).
sc — лопатка (s — гребень; а — акромиальный отросток); сот — коракоид; рс — прокоракоид; дс — сочленовная ямка для плечевой кости; fc — inter claticula; cl — ключица, ps — трудная кость; г — ребро
а — акромиальный отроет ок: с — корачонднын отросток; д—- сочленовная впадина; <8 — гребень лопатки.
ствует, a coracoideum является лишь в виде отростка лопатки, садостоя-тельного у зародыша, но затем сливающегося с лопаткой.
Ключицы у млекопитающих также йспытывают редукцию (напр. у хищных), а иногда и совершенно атрофируются. Таким образом лопатка получает более свободное положение и постепенно становится единственным носителем конечности; в соответствии с последним, под влиянием сильно увеличивающихся мускулов, прикрепляющихся к ней со стороны ее спинного края, лопатка возрастает в размерах и развивает гребень, оканчивающийся в акромиальном отростке.
Таз состоит из трех типичных костей, и обе половины соединены в лобковом сращении.
Наибольшие различия у млекопитающих представляются в дистальном отделе конечностей, .типов стопоходящих и пальцеходящих.
Первоначальною формою была стопоходящая (или полу-стопоходящая) конечность с концевыми члениками пальцев, средними по форме между когтями и копытами; лучевая кость предплечья вращалась около локтевой, сообщая вра-' щение кисти. Локтевая кость представляет локтевое сочленение с плечевой (/laments); это сочленение дает возможность угловых движений предплечья относительно плеча. Задние конечности отличаются проксимальным рядом ко
стей предплюсны, состоящим из двух костей: таранной (astragalus) и пяточной (calcaneum). От хождения на стопе многие млекопитающие перешли к хождению на пальцах. Последнее происходит главным образом при посредстве средних пальцев, боковые пальцы в меньшей степени служат для опоры при ходьбе. В некоторых случаях животное при движении опирается о почву исключительно концевыми члениками пальцев (копытные), причем боковые пальцы не достигают до почвы, не действуют и потому атрофируются, тогда как работающие срединные, наоборот, увеличиваются в размерах. При этом пястные и плюсневые кости удлиняются, и концевые фаланги превращаются в расширенный остов копыта. В то яге время radius, первоначально располагавшийся рядом с ulna, становится впереди ее; нижняя часть ее часто атрофируется и ulna срастается с radais, так что он утрачивает способность вращательного движения около оси предплечья
и перенимает от ulna ее угловое движение. Также атрофируется fibula в задних конечностях Таким образом возникает колоннообразная, упрощенная, стойкая, сообщающая быстроту бегу, ограниченная -в своих функциях лишь качательным движением взад и вперед нога копытных.
Запястье у млекопитающих обыкновенно со’стоит из двух рядов косточек У примитивных форм имеется, подобно тому что у рептилий, одна из косточек третьего ряда, centrale (йшг. 729). В примитивном запястьи все косточки обособлены, и расположение их в основе является сериальным, т е. каждая косточка первого ряда покоится на одной косточке второго ряда, косточки расположены не только горизонтальными, но и вертикальными рядами. У позднейших млекопитающих, у которых по их образу жизни является надобность в особенной солидности и стойкости ног (копытные), элементы двух рядов косточек запястья чередуется между собою, причем это чередование развилось постепенно вместо расположения косточек как горизонтальными, так и вертикальными рядами. Такое чередующееся расположение косточек, подобно тому что в кирпичной кладке, сообщает запястью большую прочность, сопротивляемость весу тела.
первоначальной
большую прочность, сопротивляемость
Ь
Фиг. 726. Ступ ня ноги пальцеходящего, собаки ।кости покрыты мясом).
mt — metatarsale, b, Ъ', b,f — мягкие do душечки.
Фиг. 725. Ступня девой ноги стопоходящегб Coryphodon апах
1 — tibia, а — astragalus, с — caleanewn, сЪ — cuboidcs (4 -j- 5 tarsale), m ну — my - metalarsalta.
В настоящее время может считаться достаточно установленным происхождение млекопитающих от пресмыкающихся. Выше, при рассмотрени1 среди рептилий порядка Theromorpha, были отмечены переходные от этого порядка к млекопитающим формы поясов, плечевого и тазового, зубной системы, а также наличность у некоторых Theromorpha, свойственного всем млекопитающим вторичного нёба. Был намечен также переход от нижней челюсти рептилий к нижней челюсти, млекопитающих. В связи с последним находится судьба quadratum, не принимающей у млекопитающих участия в сочленении черепа с нижней челюстью; принимается, на основании данных эмбриологии, что у млекопитающих две слуховые косточки из трех, имеющихся в среднем ухе, именно наковальня {incus) и молоточек {malleus? возникают,’соответственно, из quadratum и articulate (стр. 355).
Млекопитающие подразделяются на подклассы: Eplacentalia (эмбриональное развитие без последа, плацентц) и Placentalia (с последом).
Подкласс I. Беспоследовые, EplaceiitaHa.
К этому подклассу причисляются: 1) так называемые однопроходные, Monotremata, к которым принадлежат низшие из ныне живущих млекопитающих (утвднос и ехидна), и 2) сумчатые Marsupialia.
ПОРЯДОК 1 ОДНОПРОХОДНЫЕ, МОгГОТРЕМАТД.
Monotremata являются животными, у которых существует, как и у тизших классов позвоночных, клоака — полость, открывающаяся в нижний
*Фиг. 727, Ступвя ноги копытоходящего, лошади.
ръ — фаланги; я?2, ти8 — metacarpalta
Фиг. 72S. Предплюсна левой задней ноги Phena-codus primaevus. Эоцен.
Сев. Америка
а — таранная (astragaliis), с — > пяточная (caleantum) кость; п — natncidare (centrale), cb — cuboi-des (44-5 tarsale), I — T — плюсневые кости
Фиг. 729. Ступня левой передней ноги.
С
Фпг. 730. Запястные и пястные кости правой ноги лошади.
Фиг. 731. Кости таза с прилежащей частью позвоночного столба ныне живущего сум‘ атого (Macro-pus).
Jl — подвздошная; РЪ — лобковая; Js—седалищная; Л?—су-ночные кости; А — вертлужная впадина, £ — два крестцовых позвонка.
конец прямой кишки; в клоаку открываются отверстия мочевых и половых путей и анальное (отсюда название). Отверстия млечных желез открываются на поверхности живота, лишенной сосцов. Шейпые ребра не срастаются с позвонками. Коракоид велик и примыкает к грудине; есть прокоракоид, "Т-образная interclavictda. Имеется пара сумочных костей, отходящих
вперед от лобковых костей и поддерживающих стенки сумки (складка кожи на животе), которая иногда имеется (у ехидны) и служит для ношения деюнышей. Размножаются, откладывая яйца. Зубы если и имеются, то лишь в молодости, заменяясь потом роговыми пластинками.
В настоящее время Monotremata живут исключительно в Австралии, Но-в й Гвинее и Тасмании. Из Австралии же из послетретичных отложенпЙ известны и ископаемые остатки, принадлежащие двум ныне живущим родам: ехидна (Echidna) и утконос (Ornithorhynchus).
ЫОРЯДОК И. СУМЧАТЫЕ, MA3SUPIALIA.
Сумчатые (Marsupialiaj характеризуются постоянным присутствием сумочных костей (фиг. 731) и сумки и не имеют клоаки. Коракоид и interclavicula редуцированы. Рождают детенышей. В развитии зародыша послед (placenta) ,шляется в виде исключения, поэтому аллантоис, за отсутствием соединения зародыша с материнским организмом при посредстве последа, превращается в мочевой пузырь. Зубы чрезвычайно разнообразны, так что по зубной системе можно различать не только плотоядных и травоядных сумчатых, но и типы хищных, насекомоядных, грызунов и копытных.
У сумчатых, смена зубов редуцирована, типично сменяется лишь задний молочный зуб. Редукция смены зубов может стоять в связи с*гем, что детеныши Marsupialia рождаются недоразвитыми в сравнении с детенышами последовые (Placentalia)', помещаясь в сумке, они висят, присосавшись к сосцам. Такое состояние могло препятствовать развитию зубов, быть причиною позднего развития молочного поколения зубов и неразвития на смену им зубов окончательных.
В настоящее время сумчатые населяют Австралию и (двуутробки) жив] т в Южной Америке. Ископаемые формы, кроме этих частей света, встречаются и в Евразии. Сумчатые разделяются на три подпорядка: Multltuberculata, Polyprotodontia, Diprotodontia.
Подпорядок I. Многобугорчатые, Multitubercolata. Multituberculata характеризуется своими многобугорчатыми коренными зубами (фиг. 732).
Фиг. 733. Neoplagiaulax еоеае-nus. Цижний еоцен. Реймс. Германия. Левая вв!вь нижней челюсти совнутри. 2/1.
т1 — ю»2 — коренные зубы; р — ложпо-кореппои; i — резец.
Фиг. 732. Многобугорчатые коренные зубы группы Multituberculata (род Cimolomys}.
В. мел САСШ. Увеличены.
Miiltitvberculata встречаются в отложениях от триаса до эоцена. Из триаса известны лишь отдельные зубы их (Microlestets).
Многобугорчатые зубы являются настолько специализированными, что представляется затруднительным вывести их, подобно зубам обычного типа, млекопитающих, из триконодонтных и тритуберкулярных зубсв. Возможне, чтс они принадлежат особой линии развития, шедшей особым путем. Заелу-
живают быть отмечеными и особенные, иногда сплющенные с боков и бороздчатые на них, режущие ложнокоренные зубы, встречающиеся в нижней челюсти у Multituberculata (фиг. 733), а также у некоторых современных сумчатых Diprotodontia. Наконец резцы у Multituberculata имеются всего в числе двух в каждой челюсти, подобно тому как у грызунов из последовых, а также удлинены, подобно резцом грызунов, опять-таки подобные резцы имеются у современных Diprotodontia.
Многобугорчатые зубы Multituberculata представляют некоторое сходстве с первоначальными зубами утконоса из ныне живущих Monotremata, почему можно предполагать также принадлежность Multituberculata к Monotremata а не к Marsupialia.
Иодпорадок И. Многорезцовые, Polyprotodontia. Polyprotodontia отличаются от рептилий раздвоенностью корней у коренных зубов и несколько большим продолжением назад сочленовного выступа зубной кости. Они отличаются и от высших млекопитающих слабым развитием выемки под венечным отростком
1>иг. <34. Di omaiherium silvestre. Верхний триас. Сев. Америка.
Увеличено почти вдвое.
Фиг. Г35. Phaseolotherv'm ’Bucklamdi. Лева., ветвь нижней челюсти, совнутри. Нижняя юра
Апгли". 3 1.
т — место коренных зубов; pvt — ложноко ровных; с — клы ка; i — резцов.
нижней челюсти, а также расположением резцов в виде ряда, понижающегося наперед к симфизу (сращению) ветвей нижней челюсти. Число резцов до 5 в верхней и до 4 в нижней челюсти. Триас—ныне.
Dromatherium1 (фиг. 718, 734). Нижняя челюсть малой величины б зубной формулой 3 • 1 • 4 • 6. Резцы стержневидные, прямые, расположенные уменьшаясь по величине кпереди и нисходя к симфизу, так что верхние резцы вероятно также представляли отгибающийся книзу ряд. Клыки выдающиеся, согнутые. Три передних ложнокоренйых зуоа прямые, грифелевидные, остальные коренные зубы триконодонтны. Имеется зачаток пятки на коренных зубах. Верхний триас Сев. Америки.
Сюда же относятся и мелкие мезозойские (юрские) млекопитающие Triconodontidae.
Таков например Phascolotherium (фиг. 735), известный по нескольким нижним челюстям из среднеюрских отложений Англии. Зубная формула 4 • 1 • 2 • 5 Настоящие коренные зубы несут три зубца: средний большой и по бокам его меньшие. Большой венечный отросток.
Мезозойские Polyprotodontia хотя не являются прочно установленными по своему систематическому положению по отношению к Monotremata и Marsupialia, но представляют интерес как группы, промежуточные между рептилиями и типичными млекопитающими; промежуточны они по характеру зубной системы, представляющей иногда большее число зубов, нежели
1 dromas— бег
у позднейших млекопитающих: по характеру венечного отростка нижней челюстщ по присутствию на внутренней стороне ее бороздки в связи с Меккелевым хрящем (например у Dromcdherinm).
После отхода BeiBH Monotremata ствол класса млекопитающих выделил две ветви — сумчатых и последовых. Принимается, что предки сумчатых были все животными насекомоядными, лазающими по деревьям, в связи с чем, с одной стороны, имели упрощенный (по сравнению с Reptilia и Monotremata) и дающий возможность более свободных движений плечевой пояс, с другой стороны — приобрели приспособление стопы для схватывания за ветви, связанное с лазанием.
Исходя от этой стадии, вышли различные линии развития современных сумчатых, перешедшие в отношении пищи от насекомоядности к плото
Фпг. 7S3. Diprotodon australis. Постплиоцен. АвсТ; алия. Реставрация.
Фиг. 737. Задние ноги сумчатых: А — ныне живущего Trichosurus vul-pinus. Австралия. В — вымершею Diprotodon australis, a—astragalus с—calcaneum.
ядности, всеядности и травоядностм, а в отношении способа движения — от древо л азалия к наземному, роющему, прыгающему и бегающему типам движения В- эти линии развития распределяются именно ископаемые кено-зойские сумчатые. Некоторые группы их, вымершие, стоят более обособленно, представляя, как это часто бывает с вымершими группами и формами, признаки специализации.
Подпорядок ILL Двурезцовые, Diprotodontia. Увеличенная в размерах пара средних резцов. Третичные —ныне. К числу резко специализированных форм принадлежит Diprotodon (фиг. 736, 737, В) — животное, величиною с носорога, достигавшее следовательно не существующих у ныне живущих сумчатых больших размеров. Это животное имеет два пальца, из средних, редуцированных, как у сумчатых древолазов; в связи же с совершившимся переходом от древолазанил к наземному образу жизни и необходимостью нести массивное тело происходит необыкновенное увеличение костей предплюсны и голени и соответствующих костей передних конечностей, как компенсация уменьшения пальцев, унаследованного от древолазающих предков. В большом черепе пара средних резцов увеличены в размерах, а боковые резцы напротив уменьшены. Коренные зубы являются типичными лофодон-тными, двухгребенчатыми. Послетретичные отложения Австралии.
Подкласс II. Лоследорые, Placentalia.
Сумка и сумочные кости отсутствуют, обыкновенно отсуетвует и клоака. Детеныши развиваются в матке, питаясь при посредстве последа. Еак правило, резцы, клыки и ложяокоренные зубы испытывают смену. Первичною зубною х 3-1-4-3
формулою является •
ПОРЯДОК I. НАСЕКОМОЯДНЫЕ, INSECTIVORA.
Животные малой величины, являющиеся наиболее низко стоящими по организации из последовых. Представляют иногда некоторые особенности, свойственные сумчатым, например отогнутый вовнутрь задний угол нижней челюсти и отверстия на нёбе. Передняя часть головы обыкновенно вытянута в рыло, что находится в соответствии с образом жизни, преимущественно насекомоядным. .Межчелюстные кости соответственно удлинены. Зубная формула I ^-| . Животные длиннохвостые, с зубами остробугорчатыми, триконодонтными, тритуберкулярными, с наклонностью стать четырехбугорчатыми. Пальцеходящи или стопоходящи, почти всегда пятипалы.
Пальцы снабжены когтями.
Уже из эоцена известны представители различных семейств, так что формы более обобщенного характера должны были существовать в мезозое. II действительно некоторые роды мезозойских млекопитающих, известные лишь по их нижним челюстям, могут быть отнесены и относятся различными исследователями то к сумчатым, то к насекомоядным.
Весьма вероятно, что древнейшие насекомоядные были зачастую животными со способностью древолазания и насекомоядно-всеядными. Из ныне живущих сюда относятся, например землеройки (Sorex), крот (Talpa) и еж (Erinaecus'), появляющиеся в Европе с миоцена.
ПОРЯДОК И. РУКОКРЫЛЫЕ, CHIROPTERA.
Маленькие животные, питающиеся насекомыми или реже плодами, с сильно удлиненными передними конечностями, соединенными перепонкой и образующими летательный орган. Сюда относятся летучие мыши; из них нетопырь (Vespertilio) появляется с миоцена.
Весьма близки к насекомоядным, составляя, по всей вероятности, спе-циализованную ветвь, отошедшую в мезозое от сумчатых Polyprotodontia.
ПОт>ЯгтОК Ш. ХИЩНЫЕ, CARNIVOR/
Отличительные особенности животных этого порядка прежде всего в их зубной системе. Зубы всех трех категорий. Клыки обыкновенно большие, конические, изогнутые и острые. Коренные зубы обыкновенно сжатые с боков и режущие, особенно передние. Из этих режущих зубов большое развитие приобретают так называемые плотоядные зубы, по одному с каждого бока в обеих челюстях. В верхней челюсти плотоядным зубом бывает последний (спереди) ложнокоренной, в нижней — первый из настоящих коренных.
Посредине крыши черепа находится часто высокий гребень, разделяющийся впереди на две расходящиеся ветви п служащий для прикрепления
сильных жевательных мышц. Конечности иногда у хищных, ведущих водный образ жизни {Pinnipedia—тюлени и пр.), превращены в ласты.
Древнейшие хищные Creodontia представляются весьма сходными с насекомоядными, отличаются от последних глав дым образом большей величиной, отсутствием удлинения рыла и мея.челюствых костей, большими клыками, большими в не имеющими столь-острых бугорков, как у насекомоядных, коренными вубам *, и образом жизни более наземным, нежели древесным.
От позднейших, настоящих хищных Creodontia отличаются постоянно полною зубною формулою, почти всегда трехбугорчатыми коренными зубами, едва намеченным плотоядным зубом. Срастание косточек проксимального ряда запястья, существующее у позднейших хищных, здесь еще не произошло
Примитивнейшие из Creodontia эоценовые Creodontia Primitive, может быть, даже принадлежат к Insec'jvora. Вторая группа, Creodontia Adapnva, являются предками настоящих хищных. Третья группа; Creodontia Inadaptiva, является группою вымершею, параллельною собственно хищным. Inadaptiva отличаются плотоядными зубами, развивающимися только из коренных зубов, r противность настоящим хищиыл Эта группа представляет семейства, п траллетьные кошачьим, собачьим и гиенсвмм.
Сем. Camdae (собаки) является с верхнего эоцена, зубная фор-3-1 • 4 • 2 мула полная у древнейших представителей, у новейших д—1—ъ
(фиг. 738); в ныне живущей фауне хищных являются наименее уклонившимися от древнего прототипа Представителями.
Фиг. 738. ?убы ныне жглчщ rj волка (ranis lupus). Фиг. 739. Черв! Ursus spe-А — верхние: Ъ — нижние. 3/; - • laeus 1/5.
с — клыки; pj — pi~ ложпоко) енныс; w> — w3 — паст ящяе коренные.
Медведи (Ursidae) являются с миоцена, представляют тогда ту же зубную формулу, что и новеййше собаки, отличаясь тем, что, в связи со всеядностью их, плотоядные зубы не выражены, а коренные велики и имеют большое число небольших бугорков. Более древние формы примыкают по не столь большим коренным зубам к Amphicyon, миоценовому роду Canidae.
Пещерный медведь (Ursus spelaeus, фиг. 739, 740), современник пещерного человека, павнялся по величине крупнейшим ныне живущим медведям, но превосходил их толщиной. Передние ‘ лапы были ужасающей величины. Отличался крутым подъемом от лицевой части черепа к лобной, отсутствием двух передних ложнокорелных зубов (фпг. 739, 740; у ныне живущих медведей они существуют); последнее, вместе со слабым развитием фаланг, несущих когти, и с тупо-бугорчатостью зубов, делает очевидным, что он не являлся весьма страшным неприятелем для пещерного человека. Распространен в после-третичных отделениях Европы (между прочим Европейской, но не-Азиатской России).
Кошки (Felidae) в позднейших своих представителях отличаются ’крайнею среди хищных редукцией числа коренных зубов, сильным развитием плотоядных и редукцией бугорчатых зубов. У ныне живу-
3 • 1 щих Felidae число коренных зубов , у ископаемого рода Machat-
2 1
rodus (фиг. 741) (т—.—у . Machairodus* 1 — животное большой вели-Z — х ±
айны, с необыкновенно длинными, сжаты ди с боков, саблевидными клыками
Фиг. 740. 3} бы Ursus spela&ns. А —. верхние В—нижние. 1/2.
Фпг. 741. Череп Machairodus neogaeus. Постплиоцен. Южная Америка. 1/5
верхней челюсти, с зазубренной кялеватостыо на их вогнутой стороне. Клыки таковы, что вероятно выдавались за нижнюю челюсть даже и при состоянии наибольшей разинутости пасти, при этом препятствуя поглощению больших кусков добычи, а так как коренные зубы в то же время отличаются слабым развитием, то является предположение, что Machair edits представляет специализированный тип — животное, питавшееся только кровью своей добычи, причем клыки могли служить для прокусывания шеи. Оли гоцен — послетретичные.
Felidae встречаются с миоцена, Felts — с плиоцена.
Ластоногие Pinnipedia известны с миоцена, но есть уже эоценовые формы хищных, представляющие изменения в направлении ластоногих.
ПОРЯДОК IV. НЕПОЛНОЗУБЫЕ, EDENTATA.
Животные, отличающиеся редукцией зубной системы, или совершенно беззубые, или по крайней мере без резцов. Зубы не сменяются, без эмали, без корней, состоят из дентина и цемента. Нередко существует нисходящий отросток скуловой дуги, представляющий отличительную особенность. Кожа часто с роговыми чешуями, у семейства броненосцев — с панцырем из костяных щитков. У некоторых коракоид более обособлен (отверстием) от лопатки, чем у остальных последовых. Неполнозубые могут считаться за сильно специализированную ветвь некоторых, весьма низко стоящих и древних — вероятно мезозойских — млекопитающих.
Ископаемые формы, с нижнетретичных отложений до конца плиоцена, ограничены в своем распространении Южной Америкой; затем распространяются и в ближнюю часть Сев. Ам рики.
_____________
1 Machaira — меч, odons — зуб.
Среди ископаемых заслуживают быть отмеченными отличающиеся исполинскими размерами Megatherium (исполинский ленивец) и Glyptodon (испопинский броненосец).
Megatherium, (фиг. 742), животное в 7. м длиною, отличается массивностью задней части скелета; задние ноги, вместе с хвостом, должно быть, служили для поддержания туловища в стоячем положении, при котором
Фиг. "42. Mogafhorium americomum. Постпдиоцен. Южная Америка. А — коренной зуб (сбо! у).
животное могло доставать листья с деревьев, служившие пищей. Кости конечнистей Megatherium несут значительные гребни, указывающие на силу мускулов этого животного. При ходьбе животное опиралось главным образом на наружную сторону ног, вследствие чего последние являются как бы скр) ченныши. Средние пальцы хорошо развиты и вооружены огромными
А
Б С
Фиг. 743. А — Glyptodon reticulatus. Постплиоцен. (Южная Америка.
А—Общий вид Glyptodon repticukerlaius. Б—три панцирных щитка (1/3) Glyptodon lae-vis. С — три зуба нижней челюсти (1/2) Glyptodon reticulakaus.
когтями, боковые редуцированы. Передняя часть нижней челюсти ложковидно вытянута, очевидно в приспособлении к длинному, сильно вытягивающемуся языку, который мог применяться для схватывания листьев с деревьев (подобно тому как у ныне живущего жирафа). Коренные зубы в небольшом числе, перетирающие, росли по мере истирания и, состоя из перемежающихся мягких и твердых пластинок зубного вещества, представляла поперечные ребра. Новейшие послетретичные отложениия Южной Америки.
Glyptodon1 (фиг. 743) встречается тоже в Южной Америке, отличается
1 Glyptos — скульптурный, odon—зуб.
24 Учебяжк палеонтолога» 369
2 , J .2__3*3
-к——к-----5—д-. Коренные зхбы большею частью тупобугорчаты.
Z • 1 • 2 — d • 3
отделены перемычкой или стенкой от височных отверстий. В w
от современных броненосцев тем, что костяные щитки его панцыря были расположены не поперечными рядами, как у ныне живущих броненосцев, благодаря чему последние могут свертываться, а как ячейки в пчелиных сотах, вследствие чего панцырь отличался неподвижностью, как у черепах. Вероятно при опасности ноги прижимались к туловищу. Хвост и верх головы покрыты панцырем. Спинные и поясничные позвонки сращены в неподвижное целое. Зубы, состоя из вещества различной твердости, при истирании представляли своеобразный рисунок. Длина тела около 4 м. Послетретичные.
Все вышерассмотренные порядки отличаются пальцами, снабженными когтями; от одного из этих порядков вышла группа приматов {Primates) с пальпами. оканчивающимися ногтями, реже когтями.
ПОРЯДОК V. приматы? "primates.
Приматы стопоходящи. Всегда пятипалы и имеют зубы всех трех категорий, мало специализированную зубную систему. Зубная формула
Глазницы
щем приматы по характеру конечностей и зубной системы—признакам, наиболее важным систематически,—являются животными мало специализированными и менее удалившимися от прототипа млекопитающих, чем большая часть других порядков. Зато они достигают исключительной специализации в совершенно новой для животных области — необычного развития головного мозга. Могут считаться вышедшими в мезозое из насекомоядных (Insectivora).
Подпорядок I. Полуобезьяпы, Prosimiae (Lemuroidea). Низшие приматы с хватательными передними и задними конечностями, большею частью снабженными ногтями. Коренные зубы иногда остробугорчаты; существует лишь перемычка между глазничной и височной впадиной. Ветви нижней челюсти несросшиеся в симфизе. Зубная формула Г 2 • 1 • 3 3 п
большею частью-,.—,—п—В зацястьи большею частью существует ы • 1 • О • О*
отдельная centrale.
Полуобезьяны ниже обезьян по развитию головного мозга, по выдающейся лицевой части черепа, по присутствию кроме пары грудных сосцов часто еще пары брюшных. Известны с эоцена {Adapis, фиг. 744). Современные лемуры ограничены в распространений Мадагаскаром, Южной Африкой и Ост-Индией.
Подпорядок II. Обезьяны, Simiae. Обезъяны имеют тупобугорчатые коренные зубы, совершенно замкнутую назади глазничную впадину и слитые в симфизе ветви нижней челюсти.
Обезьяны делятся на обезьян Нового Света — Platyrrhlul и обезьян Старого света — Catarrhini. Первые более примитивны, имеют большое
3-3 #
число коренных зубов g-.g, широкую хрящевую носовую перегородку н
1 Primates—первые по достоинству.
потому ноздри, направленные наружу, хвост длинный, содержит не менее 14 позвонков, иногда хвост бывает хватательный. Вторые имеют зубну ю фор-2 - 3
мулу <5—q > узкую носовую перегородку и ноздри, опущенные вниз; хвост
никогда не бывает хватательным. Ископаемые обезьяны известны с олигоцена (Mesopithecus, фиг. 745).
Из узконосых обезьян наиболее близко к человеку сем. Anthropomor-phae, человекообразных обезьян (ныне живущие представители: гиббон,
в
Фиг. 744. Череп и зубы ископаемой полуобезьяны Adapts parisiensis. Верхний эоцен. Керси. Франция. Л — черед; Б — верхний, С — нижний ряд зубов.
Vi т,* т? fi* р*р?р с р
оранг-утан, горилла и шимпанзе). Человекообразные лишены хвоста; лазающие животные с сильно искривленными пальцами, поэтому по земле ходят на наружном крае ног, опираются и на передние конечности, более длинные, нежели задние.
Еще более, нежели ныне живущие человекообразные обезьяны, близок к человеку Pithecanthropus из верхнеплиоценовых или нижнепостплиоценовых отложений Явы. Его череп является промежуточным по своей вместимости
Фиг. 745. Череп и зубы верхней челюсти ископаемой обезьяна Старого Света, Mesopithecus pentehcus. Верхний миоцен. Пикорми, около Афин. Гиеция.
между черепом человека и ныне живущих человекообразных обезьян. Возраст слоев с Pithecanthropus оценивается в 400 или 500 тысяч лет. За последнее время открыты близкие к Pithecanthropus роды: Eanthropus из плиоцена (?) Англии и Sinanthropus из плейстоцена Китая.
Что касается ископаемого человека, выделяемого в группу Anthropidae
Фиг. 746. Череп неандертальского человека из Спи в Бельгии.
(Hominidae),, то остатки его достоверно известны лишь из послетретичных отложений, причем принадлежат к двум типам. Один {Homo sapiens) тст же, что и представляемый современными расами человечества; другой— так называемый неандертальский человек (Homo пеа* derthalensts), названный так по местонахождению в Неандертале у Дюссельдорфа в Германии. Неандертальский человек имел череп низшего типа, меньшей вместимости, с покатым лбом (фиг. 746), с сильно выдающимися, подобно тому как у некоторых человекообразных обезьян, вздутиями кости на верхнем краю глазных впадин.
Кроме того, неандертальский человек отличается от современного человека по характеру нижней челюсти — отсутствием подбородка,- т. е. выступа челюсти вперед на нижнем краю.
В связи с присутствием подбородка передняя поверхность нижней челюсти от ряда резцов бывает поката вперед и вниз, тогда как при отсутствии подбородка, у обезьян, она поката назад и вниз (фиг. 745). У неандертальского человека, при соответствующем положении черепа, нижняя челюсть очерчи
вается напереди линией, близкой к вертикальной, т. е. челюсть в указанном отношении имеет промежуточный характер.
Одновременное существование в прошлом Homo sapiens и Ното neandertka^nsis является препятствием для того, чтобы считать последнего прародителем первого.
Кроме того, есть некоторые особенности Homo neanderthalensis в строении зубов, например, слияние корней, препятствущие считать его прародителем современного человека.
Возможно, что неандертальский человек представляет боковую ветвь человеческого рода, вымершую расу, причем яв”яется интересным ее широкое распространение (по всей Европе, у нас найден в Крыму). *
Homo neanderthalensis, как и pithecanthropus, интересны тем, что если они и не принадлежат к числу прямых предков современного человека, то все-таки предсгазляют отдельные особенности организации, промежуточные между особенностями организации ныне живущих человека и человекообразных обезьян.
Последние в свою очередь, выходя в прошлом из общего с человеком корня, могут быть ветвью, пошедшею в другом направлении, нежели родственная ей ветвь человеческого рода.
Известен еще, хотя только по нижней челюсти, Homo heidelbergensis, еще более дреьний и более примитивный, чем неандертальский (с более покатой назад нижней челюстью).
Подобно тому как есть ископаемые, вымершие виды человека, есть и вымершие так сказать подвиды или разновидности Homo sapiens. В Европе таковою является так называемый Cro-Magnon, (по имени одной местности во Франции), отличавшийся высоким ростом (1,94 м у взрослого мужчины), существовавший около 25 тыс. лет назад. Коо-Каньон имел черти азиатского характера.
Вообще же считается возможным возникновение человека полифилите-372
* 'Г
Череп современ-европейца.
Фиг. 747. ного
чески в различных местах земного шара, независимо в одних местах от других
Во всяком сличав нахождение промежуточных ступеней от своевременного человека (Ното sapiens)—к низшим приметам наталкивается на значительные затруднения во-первых вследствие трудности установления относительного хронологического возраста отдельных находимых остатков, а затем и вследствие неполноты и особых трудностей нахождения самих остатков первобытного человека. Трудность состоит в том, что человек жил во время последнего геологического периода, в течение которого не успели еще произойти значительные изменения в перемещении суши и воды, так что иско-I паемые остатки, лучше всего сохраняющиеся в водных отложениях еще не успели обнажиться и стать доступными наблюдениям.
Некоторыми исследователями предполагается существование человека в третичном периоде, по крайней мере с олигоцена, но это предположение строится на шатком основании находок подобия кремневых орудий. Древнейшие орудия этого рода представляют просто оббитые куски кремня; подобные обломки могут возникать однако, в природе просто под действием перемен температуры и под действием перекатывания кремней движущейся водой.
Достоверно известно, что человек существовал во время дилювия, одновременно с мамонтом, шерстистым носорогом, пещерным медведем и другими вымершими—отчасти, несомненно, вследствие истребления человеком—крупными млекопитающими.
На обломках костей, на стенах пещер найдены рисунки этих животных, сделанные их современником, доисторическим человеком (фиг. 747).
Скудость имеющихся в настоящее время данных относительно ископаемых: предков человека не мешает, на основании сравнительно-анатомического изучения, ставить человека в порядок приматов, причем они выделяют человечество в группу, эквивалентную Prosimiae и Simiae, т. е. в нашем изложении—подпорядку, другие считают Hominidae просто за семейство группы Catantini.
Далее следуют группы млекопитающих, снабженных копытами; эти группы вышли из форм, для которых характерно присутствие когтей на пальцах, именно из Creodontia. Для копытных характерны высокие конечности, построенные исключительно для быстрого перемещения по почве и исключающие возможность лазания, рытья и схватывания.
ПОРЯДОК VI. ПЕРВИЧНЫЕ КОПЫТНЫЕ, CONDVLARTHRA.
Древнейшие (нижнеэоценовые) копытные малой величины—Condylarthra имели, подобно Creodontia, маленький низкий череп с низким и длинным [Теменным гребнем и с широко открытыми назад глазницами. Существуют также точки соприкосновения с Insectivora, например: удлиненные межчелюст-
1 В южной Африке открыт череп Homo Rhodesianus ив четвертичных отложений, 'надлежащий также к разновидности Homo sapiens.
ные кости, короткие прямые клыки. Зубная формула
3 • 1 • 4 • 3.
3 -1 - 4 • 3. ’
зубы рас
положены непрерывным рядом, ктыки маленькие, коренные зубы брахиодон-тны, бунодсниы или селенодонтны. Па ногах по пяти пальцев; концевые
Фиг. 748. Phenacodus pr’maevus. Нжжвьй воден. Уайоминг. Сев. Америка.
членики пальцев, всех или некоторых, утрачивают сжатую с боков, когтевидную форму и приобретают приплюшенность спереди назад — копытообразность (фиг. 749). Косточки запястья сернальны или чередуются. Иногда уцелевает самостоятельное centrale. Имеется длинный хвост. Condy larthra были животными стопоходящими или пальцеходящими с редукцией боковых пальцев.
Знаменит род Phenacodus (фиг. 748), объединяющий в себе те особенности, которые теоретически были приписаны американским палеонтологом Копом прототипу копытных. Через несколько лет после создания образа этого прототипа он получил
Фпг. 749. Кисть TTuracops sccialit-Иидвий аоцев.
реальное осуществление в виде открытого п описанного тогда* Копом Phenacodus. Это — животное, величиною с собаку, очевидно быстронсгое, цальцеходяшее; о почву опирались главный образом три средние пальца, из которых средний был значительно удлинен. Они имели расширенные концевые членики, в то время к<*к более короткие боковые были боле®
сжа-ы. У Euprotogonia, представителя этого семейства из еще более древних слоев эоцена, все пальцы были сжаты с боков и все „дирались о почву.
Фиг. 750. Ihenacodus jpimaevus. Зубы, верхние и нижние, начиная с клыюв.
ПОРЯДОК VII. НЕПАРНОКОПЫТНЫЕ, PERISSODACTVLA.
В настоящее время представлены тапиром, лошадьми и носорогами.
Животные с редуцированным числом пальцев, нечетным по крайней мере на задних ногах (3 или 1). На передних ногах атрофия боковых пальцев происходит менее скоро, чем
циональною специализированностью передней конечности. Вес тела покоится
на задних, в связи с меньшею
Фиг. 751. С чет ныне живущего тапира (Tapirus indicus).
Фиг. 752. Зубы Тарггиз priscus. Миоцен. Эппельсгейм, близ Бориса. Гессен.
А — коренные зубы верхней челюсти: В — коренной зуб пинг*челами.
исключительно прежде всего на III пальце, через который проходит ось конечности; в предплюсне astragalus представляет/блоковидную сочленовную с tibia поверхность Коренные зубы лофодонт-ны илц селено-лофо-донтны; ложнокоренные квадр^ного сечения, подобно коренным.
Из ныне живущих непарнокопытных при-
мити вней шим является тапир (Tapirus), живущий в южной Америке и юго-восточной Азии
_ - ~ й , 3 • 1 4 • 3. „
Тапир имеет зубы всех категорий, зубную формулу --g- g . Ко-
ренные зубы брахисц )нты, без цемента, лофодонтны, с двумя поперечными
375 /
среонямп (фиг. 752). Глазная орбита и височная впадина находятся в широком сообщении между собой. Четыре пальца на них
ногах. С верхнего миоцена в Евразии и Сев.
'•
Ж
' < • у'
Фиг. 753l Череп Acerath~riwm megafothts. Верхний миоцен.
Колорадо. САСШ.
передних и три на зад-Америке.
Сем. носорогов, Rhinocerottdae. Грузные травояд иле, с толстой ко'жей и обыкновенно скудным волосяным покровом. Позднейшие формы отличаются одним или двумя рогами на носовых костях, представляю-щимиэпидермическое образование как бы из склеенных волос.
Широкое соединение между орбитой и височной впадиной. По четыре пальца на передних но
гах и по три на задних. Резцы и клыки у позднейших форм в различной степени редуцированы. Коронные зубы лофодонты (фиг. 729); на верхней челюсти гребни соединены наружной стенкой, кроме того в пространство между гребнями отходят складки (шпорцы) от наружной стенки и от обоих гребней. На нижних зубах гребни дугообразно изогнуты (фиг. 755). С развитием рогов развивается дающая опору ка между ноздрями.
Aceratherium (фиг. 753) не имеет рогов (или имеет лишь зачаточн. рог) и носовой перегородки, имеет резцы и клыки, четыре пальца на передних и три на за дних ногах .Олигоцен и миоцен Европы, плиоцен Азии.
Рсд Rhinoceros встречается с миоцена. Имеет по три пальца на ногах. Rhinoceros tichorhivus{ — R.antiquitatis, фиг. 754), вымершая форма по-слетретичного периода, был распространен в ледниковую эпоху в Европе и Сибири, отличался отсутствием передних зубов, густым волосяным покровом и присутствием двух рогов на носовых костях, нередкий рог был очень велик; между ноздрями существовала солидная костяная перегородка R. tichorhinus, как и R. Merckii (фиг 156), наиболее близки к почти вымер-
костям костяная перегород-
носовым
Фиг. 754. Rhinoceros antiqu fatis. Послетретичные. КраВбург Верхняя Бавария. Германия.
эте
части Еьропейской Росии и ь Гер-
Фиг 755. Верхний (Л) и нижний (В) корен ные зубы носорога.
аст\ ст — шжорцы
тему в настоящее время двурогому африканскому носорогу.1
Elasmotherium 1 2 * * s (фиг. 757)— вымерший послетретичный носорог, живший в южной Сибири, смежной мании. Этот весьма специализированный носорог отличался присутствием огромного рога на лобных, а не на носовых костях; лобные кости имеют соответственное основанию рога вздутие. Из зубов имелись лишь коренные, в высокой степени гипсодонтные, с сильным развитием цемента и с сильно выраженною мелкосклад-чатостыо нмали. Зубы его являются приспособленными к жизни в полосе степей, с питанием жесткими, иссти-рающими зубы травами.
Baluciiitherium, открытый в Белуджистане, Монголии, является представителем группы носорогов, оригинальных по своему внешнему виду и отличающихся гигантским ростом (вдвое более современного индейского носорога). Из СССР известен близкий к нему род Indricotherium из олигоцена Тургайской степи. Это безрогие, подобно Aceratherium, носороги, встречаю
Фиг. 756. Голова 'Rhinoceros Мегскй. В. Яна, Верхоянский округ, в северо-восточной Сибири.
щиеся в миоцене и верхнем олигоцене (пределы распространения еще точно не остановлены). Они отличаются клыкообразным развитием вторых верхних резцов и вероятно таким же соответствующих нижних (фиг. 758).
1 Двурогие носорог? объединяются в особую группу Ricerorhinus Животное теплого
климата, Ъ. Mt.rckn находился в Европе в гижн. постплиоцене, вместе с ранне-неандор-
тальскими расами, а уже с 3-го оледенения (средний постплиоцен) появляется в Европе
и приспособившийся к северному климату шерстистый носорог (D. aniigwtatis или tick or fanus'i. '
s jEZasma — пластична
Вторая важная особенность, это—лошадинообразное удлинение передней части черепа. Шея удлинена более, чем это обыкновенно бывает у носорогов, и вместе со сравнительно малой величиной головы это ставится предположительно в связь с питанием листвой деревьев 1 Палеонтологическая история развития семейства лошадиных Equidae, как известно, изучена весьма хорошо, представляя довольно заезженный образцовый пример
Фиг. 757. JSlasmofherium sibiriciim. Черги с вывалившимися зубами и обломанными с наружного ьрая зуСвыки впадивами; нижняя челюсть и коренные зубы.
в — ауб верхней челюсти, С — ауб нижсей.
последовательных постепенных филогенетических изменений. Древнейшие представители, известные из эоцена, представляют маленьких животных, немного более 0,3 м высотою, с ногами, подобно тому как у тапира, имеющими 4—3 пальца, с брахиодонтными зубами, снабженными округленными бугорками, с глазной орбитой, широко соединенной назади с височной впадиной. Таково подсемейство Hyracotheriinae и в нем род Ryracotherium (фиг. 719, 759, 760). Развитие этих животных в следующие геологические эпохи приводит к однопалой лошади с рудиментами боковых пальцев в виде так называе-
1 Реставрированный скелет fniricotherium выставлен • в Палвозоологичепко" Ии титуле Академии Наук.
мых «грифельных» (пястных и плюсневых) костей. В связи с возрастанием длины ног удлиняется шея и голова, давая возможность удобного срывания травы с почвы; удлинение черепа происходит главным образом за счет удлинения лицевой части так, что с одной стороны расширяется пространство для помещения возрастающих в размерах зубов, с другой— глаза к выгоде для животного являют
ся ПОДНЯТЫМИ над гори- Фиг. 758. Череп Baluchitheriwm Grangers. Миоцен. Мон-зонтом. Глазницы отде- голая
ляются от височных от-
верстий костяной перемычкой. Коренные зубы приобретают лунчатые бугорки; извивы эмали услож-
няются; в промежутках между последними отлагается цемент; в связи с переходом от жизни на болотистой почве и питания чистою, сочною мякотью растущих там крупных, отчасти кустарниковых, растений, к жизни на травянистых, покрытых злаками равнинах, зубы быстрее стираются от более твердой и с примесью песка пищи, и в связи с этим становятся гипсодонтными со сложною поверхностью, приспособленною для перетирания жестких злаков.
За подсемейством Нуга-cotheriinae следует Anchith-eriinae. У них число функционирующих пальцев 3—3, боковые пальцы еще достигают земли. Орбита не замкнута. В зубах имеются уже лунки, но еще нет цемента, и зубы брахиодонтны.. Представителями этого семейства
Фиг. 759. А, Б, С — черепа Ilyracatfkeriwn, Meso- ЯВЛЯЮТСЯ например ОЛИГОЦ0-fcippws, Шрынпоп. новый род Mesohtppus, ле-
жащий на прямой линии развития ныне живущих лошадей, и близкий к нему Anchitherium, лежащий на боковой линии. Последний существует и в миоцене, а в плиоцене эта
соковая линия развития представляет конечный этап в роде Hippanon, уже с гипсодонтными (фиг. 720) зубами, боковыми пальмами, не достигающими земли, с замкнутыми назади глазницами. В зубах есть цемент. Собственно ло-
С
Фиг. 760. А, Б, С— коренные зубы ily. acot'ieriwin, Anchithcriuw1, Equus.
Соответственные бугорки обозначены соответственными буквами. Разрезы емали обозначены Жирным, разрезы дентина — точками, цемента — вертикальной штриховкой. Зуб А стерт наименее, В — более и С — еще более.
шадь, Equus, впервые появляется в в°рхнеь плиоцене Евразии и Сев. Америки. Ископаемые виды этого рода имели весьма широкое распространение, но ныне живущие находятся исключительно в Азии и -Африке. Ряд предков лошади представлен более полно в Америке, и там именно происходило развитие линии непосредственных предков лошади; европейские ископаемые предста-
Фиг. 761. Левая нога игереди и совнури Ancliitherium (Л, A'), Lipparion (В, Б'), Equus (С, С').
вители Etpndae Hyracotherium, Anchitherium, Sipparion, за исключением одного Hyrctcotherium, являются невидимому лишь эмигрировавшими в разное время в Европу. Род Equus, развившись в Америке и отсюда населив Европу, затем вымер в Америке, оставшись лгшь в Евразии.
НОРЯДОК УШ. ПАРНОКОПЫТНЫЕ. ABTIOBACTYbA.
Животные, опирающиеся при ходьбе главным образом на III и IV пальцы, которые поэтому сильно развиты, тогда как остальные редуцированы иногда до полного исчезновения. Парнокопытные отличаются тем, что astragalus нх имеет блоковидную суставную поверхность не только проксимально, как у непарнокопытных, но и дистально (фиг. 762). Первоначальная зубная формула g ' g-. Верхние резцы и клыки имеют наклонность к исчезновению. Нижние клыки сохраняется, но большею частью примыкают к резцам, становясь и по форме подобными им. Парнокопытные резделяются на жвачных и нежвачных. Первые отрыгивают полу-пережеванную и затем полупередаренную пищу в рот для вторичного пережевывания, после которого она, поступая вторично в желудок, идет в иное его отделение, нежели первоначально. Приспособление, дающее возможность набирать наско| о запас пищи в желудок, для того чтобы затем занят] ся ее переработкой в условиях безопасности.
га.
Фиг. 762. Правая таранная кость (astragalus) вымершего рода оленей (Ра-laeomeryx),
Фиг. 763. Sis erymanthius. Верхний миоцен. Пи-•'еоми около Аф ш. Верхние коренные зубы. 1/2.
Подпорядок I. Нежвачные, Nonruminantia. Броме устройства желудка, отличаются полной зубной системой, бунодонтны; имеют несросшиеся пястные и плюсневые кости III и IV пальцев, разьитой лобно-теменной гребень.
У рода Sus, свиньи, клыки увеличены по сравнению с первичным их состоянием, иногда значительно. Так же и коренные зубы представляют увеличенное, против первичного, число бугорков (фиг. 763), хотя увеличение происходит путем развития гспомогзтельных бугорков; число главных бугорков 4. Орбита назади незамкнута. Существуют, хотя- укороченные, два боковые пальца. В ископаемом состоянии с плиоцена.
Подпорядок II. Жвачные, Rnminantia. Селенодонтны; пястные я плюсневые кости III и IV срастаются в «дудку». Орбита назади замкнут» лобно-теменной 1ребень обыкновенно отсутствует.
Сем Cervidae, олени (Cervicornia, оленерогие). Самцы большею частью с ро-гамг: если рога отсутствуют, то верхние клыки, большею частью хорошо выраженные, увеличены. Сохраняются фаланги боковых пальцев (фиг. 76" С).
Олени представляют группу, расцвет которой произошел в послетре-тичный период и достиг максимума в настоящее время. Рога оленей (фиг. 764) развивались постепенно с течением времени, таг что у древнейших фоцм они очень просты, представляют мало разветвлений, у позднейших ови
Фиг. 764. Рога различных оленей
А — Cervulus thcranoceros, нижний плиоцен; В — Cei vulus pardinensts, верхний плиоцен; С, В — Corvus elap 1л (ныне живущий и в постплчоцене), на втором году и взрослый; Е—ныне живущий мунтжак, Cervulus mun'ijac; t—Вата, ныне живущая лань.
становятся все более разветвленными. Филогенеткческое развитие рогов воспроизводится в их онтогении. Так например у ныне живущего благородного оленя. Cervus elaphus, рога на пери и году жизни являются в виде сравнительно
Фиг. 765. Megaceroe hibernicus (= Cervut giganteus).
Постллиоцен. Ирландия.
невысоких и неразвст-вленных конических образований. На втором году жизни эти рога сбрасываются, а заново развивающиеся на месте сброшенных являются уже вильчато-разветвленными (фиг. 764, С); на вырастающих год спустя прибавляется еще одна ветвь и т. д., так что у взрослого оленя рога являются с большим числом ветвей (фиг. 754, D).
Е оленям с простейшими рогами, не-разветзленными или лишь виль шп раздвоенными, принадлежит род Сарг&Лгм косуля (нижний плиоцен — ныне).
Из после третичных отложений Европы известен живший одновременно с доисторическим человеком во время ледникового периодаMegaceros (группа лани, Вата) hihernicus (фиг. 765), рога которого в ширину захватывают более трех метров и отличаются широколапчатой формой. Безрогие предшественники оленей известны с олигоцена.
Сем. Cavicornia—полорогие (антилопы, быки, козы, овцы); как известно, отличаются ротовой оболочкой центрального костяного стержня рогов; верхние клыки рудиментарны или отсутствуют;
рудиментарные боковые
Фиг. 736. Cervus elaphus. Ныне живущий благородный олень.
А—верхний коренной, В—нижний коренной зуб, С—передняя, D—задняя конечность.
пальцы уже утратили фаланги, плюсневые и пястные кости функционирующих пальцев срастаются в дудку. Род Bos с плиоцена. Из видов, существовавших еще в историческое время, заслуживает упоминания Bos primigenius ' (фиг. 767, В и 768), тур древних славян, от которого произошли вероятно некоторые породы домашнего скота; в общем однако он был истреблен, а ие одомашнен; вымирание его закончилось около трехсот лет тому назад, когда он водился лишь в особых парках-зверинцах около Варшавы.
А В
Фиг. 767. А — Bos (Bison) pristus, В — Bos primigenius. Черепа. Постплиоцен.
Bos primigenius отличается длинным узким лбом и S-образно согну тыми рогами.
Bos (Bison) europaeus, зубр, и Bos (Bison) arnei icanus, бизон, вымирают в настоящее время; вымершую форму, родственную зубру, представляет В. prisons (фиг. 767, А).
НОГЯД&К IX. ХОБОТНЫЕ, PROBOSCIDEA
Фиг. 768. Коренные зубы Bos primige-mus. С—геряний, В — нижний.
Нине живущие хоботные являются оригинальными животными, благодаря присутствию хобота представляющего вытянутый нос, и пары больших бивней, в верхней челюсти (у вымерших, более древних форм и в нижней) — гипертрофчр ванных резцов верхней челюсти, растущих в течение всей жи.ши. Хоботные пятипалы, с вторично-сериальным расположением костей ва-пястья и предплюсны. Зубная система вообще является специализированной, клыки отсутствуют, а коренные зубы приобретают огромную величину и оригинальное строение. Зубная формула 1 • 0 • 3 • 3 .
тс—тг—з—к, но коренные зубы у позд-V • V • и • о
нейших представителей хоботных не существуют одновременно, как обыкновенно у млекопитающих, а появляются один за другим в течение всей жизни животного:* каждый последующий выдвигается сзади челюсти на место износившегося функционировавшего, и являются в каждый данный момент в числе не более одного вполне образованного и функционирующего зуба с каждого бока челюстей. Что касается оригинально о строения коренных зубов крайних представителей слоновых, то оно все-таки приводится к лофодонтному типу, так что число гребней является сильно увеличенным сравнительно с тем, что обыкновенно доходит до 27 у ныне живущего индийского слона. Зуб становится гипсодонтным, растущим в течение долгого периода, так что гребни становятся вытянутыми в направлении высоты, с почти параллельными боковыми поверхностями (фиг. 770, В), а ложбины между ними заполняются цементом, отсутствующим у более древних хоботных. В связи с необходимостью
л®
Фиг. 769. Череп мидийского слома (Е1е-phas indicus) в вертикальном разрезе:
So — верхняя ватыяочная ?ость; со — затылочный мыщелок; Ра — теменная кость; Рг — лобная; Их— верхнечелюстная; — межчелюстная; НЕ — решетчатая; ее — мозговая целость; а — носовая полость; I — бивень; «*„ тпл — первый в второй коренные зубы.
поддерживать голову с тяжеловесными бивнями, череп представляет большую высоту и большую затылочную поверхность, к которой прикрепляются шейные мускулы, поддерживающие голову на везу Увеличение размеров черепа
являются воздухоносными,
Фиг. 770. Коренной зуб (схематически) слона в в де сверху (А) и в вертикалью м прлдоль иом разрезе (В). Разрезы эмали показан:! жирным, дентин i—косыми штрихами, цемент докрыт точками.
достигается за счет утолщения стенок его, верхняя стенка мозговой полости например имеет толщину большую, чем высота мозговой полости (фиг. 769). В то же время утолщение стенок таково, что не ВЛ1 чет - за собою значительного увеличения веса черепа; стена и его имея крупноячеистое сложение. В связи с присутствием хобота и бивней носовые отверстия на черепе приближены к лобной части. Слоны пальцехидящи, но под пальцами имеется эластичная подушка; но’и колоннообразны, различные отделы их расположены не под углом один относительно другого, как у других’ млекопитающих, но по npiMofl линии. Кости предплечья и голени не испытывают той редукции, что у копытных.
Роды Elephas, Mastodon и Dinothe-rium только и были извешны из слоновых до XX века, как группа, обособленная от млекопитающих обычного типа. Находки последнего времени, сделанные в нижнетретичных отложениях Ливийской пустыни, восполняют этот пробел в отношении переходов.
Род Moeritherium* 1 (фиг. 761, Л), из верхнего эоцена и нижнего олигоцена Египта, представляет небольшое тапирообразное животное. Moeritherium вероятно жил в болотистых местностях и питался сочной, полуводяной растительностью этого времени. В Moeritherium мало что напоминает собственно слоновых позднейшего времени и если бы не сj ществование переходных форм, можно
было бы вообще сомневаться в принадлежности Moeritherium к слоновым. Все-таки задняя часть черепа уже представляет зачатки развития ячеистой ткани, носовые отшретт'я и носовые кости уже начинают отступать, указывая на присутствие хватательной верхней губы, и началась редукция зубов, причем в каждой челюсти вторая пара резцов начала развиваться в бивни.
• 3 1 • 3 • 3
Зубная формула . Зубы бунодонтны, четырехбугерчаты,
А) * v * О • О 1
с наклонностью бугорков к соединению в гребни. Moeritherium должно быть имел около 1 м в высоту 2
Palaeomaitodon (фиг. 771 В), из нижнего олигоцена Египта, болеь слонообразен, чем его пре1шеств(ннчк Aioeritherinm, и большей величины. Череп возрос в высоту. Маленькие носовые кости с носовыми отверстиями отступили так, что лежат непосредственно перед орбитами. Это должно было быть связано с присутствием довольно значительного хобота, как у тапира.
1 От названия озера Мерн.
1 Находки в Ливийской пустыне установили еще связь слоновых с морскими коровами (Siren-c —представлены ныне живущими дюгонями в Красном море и Иидийскоа. океане и вымершей недавно Стеллеровой коровой в Беринговом море), сблизив Mcsrtthe rium с Eolheriwn, и выяснили эволюцию морских коров (см. фиг. 5, стр. 10).
25 Учебявх палеонтологах.
385
За исключением довольно сильно развитых бивней, все передние зубы исчезли.
Бивни верхней челюсти велики, загнуты вниз и с полосой эмали лишь на части поверхности, обращенной наружу. Нижняя челюсть предста-
Фиг. 771. Л, В, С, I) — боковой вид чеу зпа слоновых и в"д их коренных зубов сверху. Л—Moeritherium. Верхний эоцен. В — Palaeomastodon. Нижний олч roue л С—Mastodon (Bunolophodon) angustiders. Миоцен. Савсан. Фра ция D— Mastodon агпепсл-nus. Постилиицен. Сев. Америка.
вляет значительно удлиненный симфиз, и бивни ее направлены прямо вперед, как у Moeritherium. Зубная формула . Коренныо зубы
шестпбугорчаты.
Год Mastodon (фиг. 771, (7, D) с миоцена до поелетретичных отложений включительно; более древние формы (Bunolophodon) имели четыре
бивня в обеих челюстях с полосою эмали на наружной поверхности Коренные з>бы настолько возросли в размерах, что только два зуба развиты сдн временно с каждого бока челюсти. Носовое отверстие отнесено еще далее назад, чем у Palaeomasiodon, свидетельствуя о значительном развитии хобота.
Позднейшие формы Udastodon несут лгшь верхние бивпи, нижние атрофированы, хотя нижняя челюсть еще сохранила длинный симфиз;
Фиг. 772. А — скелет мамонта, Elephas primigenius, иа мерзлой почвы Сибири.
В — коренной втб мамонта.
буно-лофодонтные коренные зубы представляют большое число гребней. Собственно слон. Elephas (фиг. 772) является с плиоцена. Нз ископаемых слонов необходимо отметить мамонта (Elephas primigenius}, жившего во время ледникового периода. Мамонт был современником доисторического человека. Мамонт имел распространение от Западной Европы через Сибирь и Аляску в САсШ. Мамонт, в приспособлении к холодному климату местностей, где он обитал, был покрыт длинною шерстью буро-рыжего цвета. Бивни мамонта отличаются большою величиною—до 3 метров длины. Мамонт питался травами, хвоей и шишками хвойных, и, в соответствии с этой грубой пищей, коренные зубы его имели большое число тонких гребней и действовали перетирающе. Трупы мамонта, сохранившиеся с мясом, кожей и волосами, находят во льдах севера Сибири. В Зоологическом музее Академии Наук в Ленинграде находятся остатки такого мамонта, вывезен-
пого с устьев Лены в 1806 г., а также чучело, скелет и мягкие части ма-
монта, привезенного в первые годы XX века с Березовки в северо-восточной Сибири.
В то время как вышеог ис шные роды слоновых могут находиться на
Фиг. 773. Пзображ ние иамонта яа стене ецеры в Комбар лль, во Франции, (деланное дои.то-рич,ским человеком. Отчетливо представлен покров из длинных волос.
Фиг. 774. Dinotliei lwn giqnnteum. Верхний - touch. Энпельс.сим. Гс«.сен. Германия. 1/20.
прямой линии развития, род Linotherium (фиг. 774, 775) представляется специализированным, лежащим на ветви, которая является боковою от главной линии развития слоновых. Именно он является специализированным в отношении бивней нижней челюсти, которые отгибаются круто вниз.
В верхней челюсти (недостаточно сохранившейся у единственного известного до сих пор экземпляра черепа) бивни невидимому отсутствовали. Имелось по 5 лофодонтых двух- и трехгребенчатых коренных зубов. Несомненно Dmotherium
должен был обособиться доволь-
но рано от главного ствола слсновых, от рода не более древнего и не более специализированного, нежели Palaeomastodon. Dinotherium встречается в миоцене и в нижием плиоцене Евразии.
ПОРЯДОК X КИТООБРАЗНЫЕ, CETACEA.
Голые, с гладкой кожей, рыбообразные водные животные с большой необособленш й от туловища головой и с носовыми отверстиями, лежащими на верхней стороне рыла, близ лобной части (фпг. 77 b). Передние конечности превращены в ласты, задние — отсутствуют.
Имеется горизонтальный хвостовой плавник, а иногда и вертикальный, спинной, без
твердого плавникового скелета.
Челюсти весьма длинны, так что образуется длинное рыло за счет межчелюстных и верхнечелюстных костей. Верхнечелюстные кости у позднейших форм о >ъемлют межцй постные, образуя весь край верхней челюсти, снабженный зубами. В задней части черепа кости, у позднейших форд,
образом состагляющая кровлю
иаоборот, стеснены в развитии. Носовые кости очень малы, теменные смещены со средней линии на бока черепа, так что верхнезатылочная кость, главным ) черепа, приходит в соприкосновение с лоб-
л
Фиг. 775. Dinofherium bavc.. гейт. Верхний ми цеч. Бреитен-брои. Бавария. Германия. А—коренные зубы верхней, В—коренные зубы нижней челюсти. V*-
м
Фиг. 776. Череп ныне живущего дельфина. А—сверху, В—сбо су.
occipftale; Ft — frontale^ Ju —jugate^ JHd — нижняя челюсть; ME — mesafJimtrilntm; кость; N—носовые отверстия; Na - nasaia; Pa —pane аИл\ PMx—praetnaiii-tare; SO — eupraoccipiiale- Sq — equamoaum.
-condylus; Er О — «с -г- верхнечелюстная
ными костями. Соединенные носовые отверстия (брызгальце) ведут в носовую полость, протягивающуюся почти прямо вниз. Носовая полость не содержит но овых раковин (разветвленных образований обонятельной кости) и является исключительно органом дыхани% но пе обоняния.
У новейших китообразных зубы многочисленны, однородны, гапло-донтны, у более древних они иногда дифференцированы, так что являются коренные зубы с двумя-тремя корнями и зубчатой1 кроной. Беззубые киты представляют зубы лишь у зародышей, а в поста я бри овальном состоянии животное имеет китовый ус,—роговые пластины, свешивающиеся с верхних челюстей, бахромчатые на краю и служащие для отцеживания воды от до-
Фиг. 777. Череп Profccetus aiavus. Снпяу и сбоку. Верхний годен. Моккатам бита lijfipa Ei и нет.
бычч, состоящей из мелких ракообразных. Шейпые позвонки укорочены и большею частью срастаются. Сочленения позвонков между собой и ребер с позвонками редуцированы. Передние конечности представляют укороченное плечо и предплечье и удлиненную кисть. Кости предплечья неподвижны в локте Существует гиперфалантия. Существ йот рудиментарные кости таза, не еоетиненные с позвоночным столбом. Это. рудимент—так же как например ушных мускулов, носовых раковин, волосяного покрова и к жных желез—свидетельствует, что китообразные происходят от наземных четвероногих млекопитающих, перешедших к водному образу жизни. Всего вероятнее исходною для китообразных была насекомоядно-креодонтная группа млекопитающих.
В верхнем эоцене Старого и Нового света встречаются примитивные роды зубишх китообразных — Protocetif» (фиг. 777) в Египте, имеющий здо
дифференцированную формулой уТрТЧТз
Фиг. 778. Реставрация че епа Zcuglodon Osiris. В рхний воден. Египет.
Ар — место носового бтверствя; С — клыки; Fr— лобная кость; J — резцы; «/« — скуловая ь с<ь; И — коренные з>бы; йя — верхнечелюс гная кость; Na — но совая кость; Oct — con iylus occipita F — ложнокорен-Ные зубы; Fa — теменная Кость; Pm — межчелюстная кость; Sq — чешуйчатая кость.
зубную систему с нормальной для млекопитающих , с коренными зубами режущими, уже несколько упрощенными по сравнению с зубами Creodontia вследствие редукции внутренних бугорков (передние зубы изменены, как вообще у четвероногих, перешедших к водному образу жизни, служа для схватывания добычи). Носовое отверстие находится впереди, сзади него длинные носовые кости.
К Protocetus примыкает Zaugloclon (фигД778; zeugl—иго), являющийся несколько специализированным в отношении большой величины и характера коренных зубов, представляющих своеобразную зубчатую крону и совершенное исчезновение внутренних бугорш У него так же, как у Protocetus, имеется на черепе лобно-теменной гребень. Шейные позвонки не короче спинных и не
сливаются между собою. В приспособлении к жизни в прибрежной полосе имелся панцырь из многоугольных костяных табличек, покрывавших с обоих боков весь спинной плавник и вероятно прилегающую часть туловища.
Ископаемые китообразные, близкие к современным дельфинам (фиг. 776), кита.1 и кашалотам, встречаются нередко с миоцена.
Взаимные отношения и происхождение этих 4 групп недавно выяснены довольно удачно. Zeuglodontidae рассматриваются как боковая ветвь, отчленившаяся еще в эоцене от Creodontia, достигшая расцвета в верхнем эоцене и вымершая в миоцене. Современные китообразные берут начало в общей группе Microzeuglodo'ntidae. В нижнем олигоцене «тчленяются зубатые киты 1 (Agoropliiiis), достигающие расцвета уже в миоцене На границе олигоцена с йиоценом возникают предки беззубых китов (Patriocetus), развивающихся в плиоцене. Неясно происхождение ветви дельфинов, расцвет которой падает на современную эпоху.
ПОРЯДОК XI. МОРСКИЕ КОРОВЫ, SIRENIA.
Сиреновые или морские коровы рассматриваются в настоящее время ; как ветвь копытных животных, приспособившаяся к морскому образу жизни. Полагают, что морские коровы из всех копытных всего ближе стоят к хоботным (Proboscidea), имея с ними общих предков. Первичные свреновые, зубная система которых в настоящее время редуцирована, обладали полным комплектом зубов, так что имеются верхние клыки и многочисленные коренные зубы, снабженные двумя поперечными гребнями. Из сиреновых доныне сохранились в тропических морях дюгонь (Ilalicore) и ламантин (Manatus), и лишь jb конце XVIII века окончательно была истреблена мор
I
ская корова (Rhytina), жившая у берегов Берингова моря. Скелет ее (Rhytina Stelleri) хранится в Зоологическом музее Академии Наук.
Нынешние сиреневые обладают ластами, вполне приспособленными для плавания.
Раскопки в эоцене Европы показали однакэ, что у предков сиреновых имелась правильно функционировавшая пятипалая конечность.
Постепенная редукция костей заднего пояса прекрасно прослеживается у сиреновых, начиная со среднего эоцена доныне. '1аьов ряд Eothenum и Eostren из среднего эоцена, Halitherium из олигоцена, Metaxytherium из миоцена и современные Halicore dugong из Аветрагаи и Halicore tdbernaculi из Красного моря.
Хорошо известен и сохранился скелет Halitlierwm Schinzi (фиг. 779) из среднего олигоцена Германии.
Изучение ископаемых сиреновых служит одним из примеров возмож-
Фиг. 779, Halitherium Schinzi.
ности построения генетического ряда предков современных животных на основании палеонтологических данных.
Геологическое распространение млекопитающих.
Древнейшими представителями млекопитающих являются их триасовые формы из Multituberculata Microlestes) и Polyprotodontia (Droma-therium).
Нельзя однако сказать, что до сих пор существ1,'ет полное единогласие в признании их млекопитающими. Существует мнение, что их все еще слезет относить к рептилиям.
Несомненно все-таги, что Marsupicdia произошли от первичных Synapsida. Большим пробелом в познании геологического распространения млекопитающих является незначительность находок их в меловых отложениях, между тем как уже в эоцене наблюдается большое их разнообразие.
Однако, лишь в плиоцене, а не в миоцене, как у птпц, появляется большинство родов ныне живущих млекопитающих. Можно сказать, таким образом, не вдаваясь в подробности, что временем наибольшего развития млекопитающих являются третичный и четвертичный периоды.
Характеристики групп млекэпитагогцих.
1 подкласс: EplacentaVa, бесчослотовне.
1 порядок: Monotremata, однопроходные. Беззубые, со свободными шейными ребрами. Имеются coracoidea, mterclavicula и сумочные кости. Послетре.ичные отложения н ныне.
2 порядок: Marsupialia, сумчатые. Постоянно сумочные кости и сумка. Меняется лишь задний ложнокоренной зуб. Триас — ныне.
1 лодпорядок: Multituberculata. Многобугорчатые коренные зубы. 2 резда. Триас— воден.
2 подпорядок: Polyprotodontia. Число резцов до 5 в одной ветви челюсти. Коренные зубы остробугорчаты. Триас — иыие.
3 подпорядок: Diprotodontia. Увеличенная в размерах пара средних резцов. Третнч-иые — ныне.
2 подкласс: Placentalia, последовые. Сумка и сумочные кости отсутствуют; обыкновенно отсутствует и клоака. Смена резцов, клыков и ложнокоревных зубов.
1 порядок: Insectivora. Зубы триконодонтные, тритуберкулярные с наклонностью стать четырехбугорчатыми. Юра? Мел? Третичные—ныне.
2 порядфс: Chiroptera. Летательная перепонка, натягиваемая удлиненными пальцами передних конечностей и соединяющая их с туловищем. Третичные —ныне.
3 порядок: Carnivora. Сильные клыки, задние коренные иногда с режущими ребрами, к,. Нижняя челюсть двигается только вертикально. Третачные—ныие.
В 4 порядок Edentata. Обыкновенно только коренные зубы; без эмали. Покров волосы к или роговые чешуи или костяные щитки. Третичные—ныне.
Е; 5 порядок: Primates. Глазницы отделены от височных отверстий. Уменьшенное число зубов. Третичные—ныие.
1 подиорядок: Prosimiae. Хватательные конечнести, служащие для лазания. Лишь
Иперомычка отделяет глазницы от височных впадин. Симфнз ветвей нижней челюсти. ИТретичные—ныне.
В 2 подпорядок: Simiae. Имеют тупобугорчатые коренные зубы, совершенно замкнутую назадн глазничную впадину и сросшиеся с исчезновением шва в симфизе ветви нижней
| челюсти. Третичные (с олигоцена)—ныне."
6 порядок: Condylarthra. Маленькие, крайне примитивные копытные с низкими зубами в полном числе. Пятипалы, с сериальным расположением. Стопоходящн. Эоцен.
i 7 порядок: Perissodactyla. От четырех до одного пальца, сильнейший третий. Пальцеходящи. Зубная система большею частью полная. Коренные зубы лофодонтны, редко бунодонтиы. Третичные, иыпе.
8 порядок: Artiodactyla. Сильно развиты третий и четвертый пальцы. Astragalus блоковиден сверху и снизу. В зубной часто отсутствуют резцы и клыки. Зубы буно-донтны, селенодонтны. Третичные—ныне.
1 подиорядок: Nonruminantia. Полная зубная система; бунодонтиы; несросшиеся f пястные и плюсневые кости III и IV пальцев. Третичные, ныне.
Г 2 подпорядок: Ruminantia. Желудок в соответствии жвачке; селенодонтны; пястиые к и плюсневые кости III и IV срастаются. Верхнетретичные, ныне.
I 9 порядок: , Proboscidea. Имеют бивни, видоизмененные резцы, а все остальные Ь передние зубы, часто даже вообще все нижние резцы бывают атрофированы. Коренные зубы имеют от двух и почти до тридцати поперечных рядов бугров или гребней; В последние часто высокие. Череп почти постоянно очень высокий и короткий. Высокие В пятипалые ноги, почти без редукции костей. Ноги с сериальным расположением костей, В пальцеходящи с копытообразными ногтями. С верхнего эоцена доныне.
В 10 порядок: Cetacea. Рыбообразные, с голой кожей, достигающие огромной величины В водные млекопитающие. Горизонтальный хвостовой плавник. Задние конечности атрофи-В рованы. Передние конечности имеют внешне вид плавников.
В Череп с очень длинными верхнечелюстными, межчелюстными и верхнезатылочной В костями. Носовые кости редуцированы, наружные носовые отверстия постоянно позади В конца, вадние позади основания часто длинного рыла. Шейные позвонки короткие. В Хвост длинный. С верхнего эоцена доныне.
, 11 порядок: Sirenia. Большие, неуклюжие водные млекопитающие. Травоядные,
с зубной системой, находящейся в редукции. Толстые ребра. Передние конечности ластообразны, задние редуцированы. Горизонтальный хвостовой плавник. В настоящее время у берегов тропических морей и в реках. Ископаемые с эоцена.
ggBS3
4
Как ни резко обособлены царства растений и животных в высших представителях, бывает затруднительно отнести их низшие формы определенно к одному из царств. Это удается легче, если руководствоваться не ' одними морфологическими, но и физиологическими особенностями организма.
Клетки большинства растений снабжены оболочкой из клетчатки (целлулозЬт), по химическому составу представляющей углевод.
Растения питаются главным образом простыми по составу неорганическими соединениями. Растения извлекают из земли перерабатываемые ими в вещество своего тела неорганические соли й воду и получают из воздуха главную составную часть своего тела — углерод. Углекислый газ воздуха разлагается в зеленых частях растений с выделением $ атмосферу кислорода.
Растения претворяют названные пищевые элементы в органические составные части их тела: крахмал, клетчатку, жиры, белки и др, служащие затем пищею для животных. Последние не могут питаться простыми неорганическими веществами, требуют готовых органических соединений и зависят в своем питании от растений, вырабатывающих эти органические соединения.
Указанное физиологическое отличие существует для громадного большинства растений, но все-таки не для всех: грибы например не имеют _ активно действующего красящего вещества (хлорофилла) и живут как паразиты других организмов, или питаются разлагающимися органическими , веществами.
Затруднения, встречаемые при отграничении животного царства от растительного, являются следствием развития обоих из общего корня. При этол развитие шло по двум различным направлениям, по которым создались крупные различия, характерные особенности высших животных и растений. Мы еще далеки от построения идеальной картины развития животного и растительного мира, так как громадное число связующих типов не сохранилось до нашего времени, и связь между группами пока еще далеко не выяснена.-
В то время как принятые в прежнее время системы растительного царства ставили своей задачей главным образом разбить существующие растения на группы ясно отличимые одна от другой, современная систематика старается выразить главным образом генеалогические отношения отдельных групп, отказываясь’ связывать между собою отделы, обладающие лишь внешним схо ством. Так приходится отказаться от деления на «слоевищные» и «стебельные», на «споровые» и «семенные», а тем более на
«тайнобрачные» и «явнобрачные», как на рубрики,, не имеющие строго генеалогического содержания. В настоящем учебнике при изложении при i-ята в основе наилучше разработа пая из современных систем—именно система А. Зиглера с некоторым дополнени м, обуслов шваемым большими достижениями в области па еоботаники в тече"ие последних лет. С другой стороны в интересах единства системы расположения палсонтоло гического и па ^оботани еского материала в учебнике были приняты общие приемы класси 1икации. Так были приняты наименования для крупных подразделений на типы и классы.
»
I тип. Слоевцовые, Thallophyta.
Класс I Бактерии, Schizomycetes.
, П Сииезеленые водоросли, Sckizophyceae.
, III Жгу тиковое, Hagellatae.
„ IV Слизевики, Myxomycetes.
, V Перидииеи, Peritiineae.
, VI Диатомовые водоросли, Biatomaceae.
, VII Сц!'плянки, Con;ugatae.
, VIII Зеленые водоросли,, Chlorophyceae.
, IX Харовые, Charopkyta.
» X Бурые водоросли, /InSeophyeeae.
ж XI Б/иряные водоросли, liiiodophyeeae.
, XII Грибы, Pungi.
, XIII Пиилаиники, Lichenes. '
II тип. Мохообразные, Bryophyta.
Класс I Мхи печеночные, Ilepaticae.
, II Мхи лиственичные, Musci frondosi.
Ill тип. Папоротникообразные, Pteridophyta.
Класс I Пеилофитовые, Psiloplylales.
„ II Блауновые, Lyconodiales.
„ III Псилофиты, Psilotaceae.
, IV Хвощевидные, Articulates.
„ V Бладоксиловые, Cla-lopyitales.
, VI Папоротниковые, Filicales.
IV тип. Голосеменные, Gymnospermae.
Класс I Папоотникосеменные, Pleridospermales.
„ II Бордантовые, Cordailes.
„ III БеннетитовЫе, Bennetitales.
„ IV Саговниковые, Cycadales.
’ . ¥ Гин кговые, Ginkgoales.
„ VI Хвойные, Coniferales.
„ VII Хвойниковые, Gnelales.
I
V тип. Покрытосеменные. Angiospermae.
Класс I Однодольные, Monocofyledonae.
„ II Двудольные, Dicotyledonae.
Недостатки палеоботанических данных лишают пока возможности выяснить в целях эволюции роль каждой отдельной естественной группы (типа или-класса) в течение ее исп рнческого развития или угасания, но во всяком случае все, что дает современная палеоботаника, многое выясняет в отношении понимания современного нам растительного мира в значительно бол! шей степени, чем может нам дать изучение морфологии современных растений. Физиоло
гически конечно растения могут быть далее разгруппированы по различным признакам. До известной степени впрочем можно наметить, как выражение хода эволюции, и известное объединение групп на основании некоторых морфологических признаков, однако без универсального значения. Так, у всех групп растений, до мохообразных, тело растений не показывает расчленения на стебель, лист и корень, а представляет слоевище или таллус, могущий быть и расчлененным, но никогда не дающий образований, гомологичных образованиям побега стеблевых растений. В группе мхов наблюдается развитие как таллуса, так и стебля, в то время как высшие члены все имеют стебель, впрочем иногда биологически снова редуцирующийся до видимой внешности таллуса.
I ТИП. СЛОЕВЦОВЫЕ, THALLOPHYTA.
КЛАСС I. БАКТЕРИИ, SCHIZOMYCETES.
Одноклеточные растения, живущие одиночно или в виде колоний, составленных из отдельных клеток, образующих нити, цепочки и группы комочков; ядра незаметны или слабо намечены, пигмента не имеют; размножаются простым делением. В ископаемом состоянии можно установить наличность следующих типов бактерий:
1) Железные бактерии, в виде бацилл, известны из верхнеальгонкских рудных месторождений С. Америки, мезозойских железосодержащих оолитов 3. Европы и наконец описанная Эренбергом миоценовая форма юга европейской части СССР, под именем Gallionella.
2) Серные бактерии — уже известны из медистых сланцев Германии. Известны также из миоценовых отложений Баварии (Beggiatoa}, в живом и полуископаемом состоянии такие бактерии встречаются у нас на дне Черного моря уже от глубины 200 м в глубинах, заполненных сероводородом.
3) Известковые бактерии образуют микрококки в альгонкских отложениях. 1
Мы совершенно не коснемся вопроса о сапрофитных 2 и патогенных бактериях, как не имеющих для нас реального значения. Несомненно, что бактерии существовали на земле с древнейших времен как возбудители всякого рода гниения, но все. же роль их в процессе образования углей прежними авторами была сильно преувеличена. Большинство опубликованных находок ископаемых бактерий вряд ли можно считать относящимися к этим организмам.3
КЛАСС II. СИНЕЗЕЛЕНЫЕ ВОДОРОСЛИ, SCHIZOPHYCEAE ИЛИ CYANOPHYCEAE.
Водоросли характеризуются присутствием одной или нескольких оболочек, причем внешняя часто превращена в слизистый панцырь; характерной чертой является их окраска от оливковозеленого до голубого, коричневого и даже почти черного цвета. Эта окраска обусловливается присутствием нескольких пигментов, кроме хлорофилла встречается здесь каротин
1 Альгонкским называется верхний отдел археозойской группы.
8 Живущих на разложившихся трупах животных и растений.
8 Многие ор.аиншы, считавшиеся раньше принадлежащими к бактериям, ныне от-яосят к разным другим группам водорослей.
Фиг. 780. - A. Gloeocapsomorpha prisca. Ор.юввций. Эррас—Салив. Эстовия 360/1. В. Cladiscotlwllus Keppeni. Нижве-камен-ноугольиые отложении. . Донецкий бас-
и особые пигменты—различные фикоцианы. Размножаются посредством образования гормогоний—распадения нитей или путем образования из одной нити нескольких спор.
Древнейшими типами синезеленых водорослей являются открытые в адь-гопкских слоях С. Америки, Camasia, Collenia, Cryptozoon и др. Синезеленые водоросли — весьма древняя группа, как видно из следующих примеров, известных в* данное время.
Сем. Cliroococcaceae. Сюда относятся описанные Рено: Gleioconis Borneti из пермских отложений Франции и Subtetrapedia russiana из русских нижнекаменноугольных отложений (Реш).
Сем. Protophyceae. Сюда отно-] сятся два рода—один, описанный! Залесским, представляющий грозде-1 впдьо-соединенные клетки со слизистой, легко разбухающей оболочкой—
Gloeocapsomorpha prisca, из балтийского силура, образующая скопления, известные под именем горючего сланца или куккерсита (Эстония, Ленинградская обл.). К ней близка водоросль Cladiscothadus Keppeni—из ваших нижнекаменноугольных отложений (фиг. 780Б), (Рено).
Сем. Oscillatoriaceae. Сюда относится описанная Уолкоттом из среднекембрийских отложений Британской Колумбии Marpolia spissa. Форма близкая была описана недавно Вологдиным из Казакстанских палезойских отложений под именем М. (?) tscliingisi.
Сем. Spongiostromata. Сюда относятся образования, напоминающие наслоенные пластинки известняка, как Collenia, Camasia, Cryptozoon из альгонкских отложений С. Америки и Gymnosolen из нижнего силупа СССР.
КЛАСС 14. ЖГУТИКОВЫЕ, FLAGLLL АТАЕ.
Одноклеточные подвижные организмы с пульсирующей вакуолею и снабженные обычно в передней части одним или несколькими жгутиками размножаются дел ждем; многие обладают способностью инцистирования (покрытия особыми временными оболочками). Питание по растительному и по животному типу. Сюда относятся кокколитофоры (сем. Coccolitliophoraceae) и кремневые жгутиковые (Dictyochaceae или Sillcoflagellatae).
Ж первым принадлежат морские планктонные водоросли южных морей размером менее 1/100 мм, большею частью передвигающиеся при помощи двух жгутиков; их оболочка имеет характерный панцирь, состоящий из большего или меньшего числа известковых пластинок (кокколитов) цельных или прободенных, имеющих вид игол, колоколов или бокалов. В ископаемом состоянии коккольтофочы известны уже с кембрия, а более сложные формы лишь с мела; они играют значительную роль в строении писчего мела, их находили в эоценовых глинах Крессенберга (Бавария). Современные формы отлагают известь в глубоководных (4000 м) глобигериновых осадках в Атлантическом океане. Архангельский описал две кокколитофоры из сено
манских отложений саратовского Поволжья—Cribrosphaera Murrayi я С. Ehrenber(;i„ (фиг. 781).
Кремнистые жгутиковые. Строение их мягкого тела еще мало известно; имеют больше*» частью одно ядро и один жгутик, у некоторых видов имеются мелаие хроматофоры. Скелеты кремнистых жгутиковых у одйих имеют форму кольца, спабкенпого игольчатыми выростами (род Mesocena), перегородками (род Dtc'.yocha), из которых возникают формы с дв\ мя кольцами, соединенными перекладинами (род Distephanas) и еще более сложные скелеты. Эти жгутиковые известны не древнее сенона или даже юры, обычны в третичных отложениях, особенно в сармате (диатомовые породы Конки у Орехова и в Подэлии); ныне обычны в морях и океанах
КЛАСС IV. СЛИЗЕВИКИ, MYXOMYCETES. ,
Организмы представляют голые протоплазматические массы, лишенные хлорофилла, с многочисленными ядрами, размножающиеся спорами, образующимися в особых вместилищах. Крайняя нежность этих организмов, лишен
Фиг. 781. а—Cribmsnhacra Murrayi. В— Cn'jrospJiaera Ehrenbergii. Кокколиты. Сеноман. С ратов. 2250/1.
Фиг. 782. Feridinium ^дго-phorttm. Мел. Де ич. Пров. Саксония. Германия. 300/1.
ных оболочки в их амебоидной стадии, является причиной редких случаев сохранения их в ископаемом состоянии.
Два представителя слизевиков известны из палеозойских отложений Франции Myxomycetes Mangini (пермокарбон) и Bretonia Hardingheni (из верхне-карбона и перми)—оба сомнительны. Однако древность этой примитивной группы очевидна.
КЛАСС V. ЛЕРИДПИЕИ, PEKIDINEAE (DIN0F1AGELLATAE).
Одноклеточные водоросли (фпг. 782), облеченные панцырем из целлу-лозных щитков, движущиеся при помощи жгутиков, лежащих в желобках. Достоверно известны из верхнеюрскнх отложений [Peridmium cf. cinctum) и меловых отдожений Европы (Эрепберг). Перидннеп обычны ныне в морском планктоне и отчасти в солоноватых бассейнах (Аральское море).
КЛАСС VI. ДИАТОМОВЫЕ ВОДОРОСЛИ, DIATOMEAE.
Одноклеточные, микроскопически малые 0,105 — 0,25 л/м водоросли; особи-клетки иногда соединяются в колонии например посредством студенистого выделения Наружная форма клеток весьма разнообразна и в то же время представляет симметрию: дгустороннюю или' радиалч юлучистую.
nt
•up
(T
I
tp(i
dtLtf да.
Клеточная оболочка проникнута аморфным, прозрачным, как стекло кремнеземом, является как бы в виде двух створок, из которых одна немясго больше и надета на другую, как крышка на коробь у -Обе створки соеди-нейы в поверхности соприкосновечпя, в пояске (фиг. 783). Оболочка бывает снабжена на нару<кной поверхности различной скульптурой в виде ребрышек, бугорков- и т. п ; иногда имеет тонкие щели, через которые соприкасаются с водой протоплазматические тяжи, сообщающие водоросли движение. Размножаются продольным делением, причем отдельные створки отходят к дочерним клеткам; последние образуют заново по одной недостающей створке. Диатомеи встречаются всегда скопления ли в пресной и соленой воде; иногда, и особенно в неглубоких озерах, массами, так что скелеты их могут образовать целые слои тонкозернистых осадков (трепел, инфузорная земля). В тысячах видов диатомеи распространены по всему земному шару; отличаются вообще широким распространением в пространстве и во времени. Одни и те же виды встречаются в настоящее время живущими как в жарком, так и в холодном поясах.
Многие ископаемые виды тождественны с современными и очень немного ископаемых родов, которые не пмелп бы современных представителей.
Когда появились первые диатомовые—неизвестно. Есть сомнительные указания*на их нахождение в силуре и девоне; дос северные находки диатомовых относятся к лейасу. Их отсутствие в палеозое вероятно имеет причиной сравнительно легкую растворимость кремнезема. Hei оторые плотные кремнистые породы древнейших времен (кремнистые сланцы, роговики, кремни) могут состоять из инфузорной земли, в которой скелеты диатомей были растворены, и кремнезем вторично осажден из раствора, так что не осталось следа диатомей.
Г
UP
ф-
SOO
Фиг. 783. Navi-’ula т-ridis. Ныне живущая (и верхнетреичаая).
Л — клетка с плоской стороны, В — со сто; они пояска, С — в поперечном сечении (s' — s' в At; .up — евжняя, меньшая; ор—верхняя, большая створка; gi, да — поясковые стороны ствопок; I — щели; тк — бугорки; nl, дг — насечки
КЛАСС VII. СЦЕПЛЯНКИ, CONJUGATAE.
Одноклеточные или нитчатые водоросли зеленого цвета. Бесполое и половое размножение (деление и копуляция апланогамет, т. е. неподвижных элементов на произведшей их особи). Ископаемые типы сцеплянок относятся к двум семействам:
1) Семейство Desmidiaceae. Ископаемые десмидиевые водоросли известны из девона, юры и мела — род Xanthidium.
2) Семейство Zygnemaceae. Ископаемые зигнемовые известны лишь из эоценовых сланцев, где найдена нитчатка &pi:~ogyra и ныне живущая.
КЛАСС УШ. ЗЕЛЕНЫЕ ВОДОРОСЛИ, CHLOROPHYCEAE.
Одно- или многоклеточные зеленые растения; клетки с одним или многими ядрами, несущие хроматофоры с зеленым пигментом—хлорофиллом. Размножаются вегетатигным путем, делением клеток или зооспорами, половое
размножение—копуляция изогамет до образования оогамет, оплодотворяемых сперматозоидами. Зеленые водоросли играют огромную роль между ископаемыми слоевцовыми, особенно те из них, которые имеют известковый скелет.
Из числа семейств, относящихся к зеленым водорослям, упомянем:
1. Сем. Volvocineae—подвижные формы, близкие к жгутиковым. Наиболее важньтё Reinschia, составляющая массу сапропеля образовавшаго пермские угли.
2. Сем. Chlamydomonadaceae—сюда относится Gloeocystis oxfordiensis из верхнеюрских отложений.
3. Сем. Phacotaceae. Род. Phacotus — подвижная форма с двумя жгутиками, чечевицеобразной формы с кальцинированной оболочкой. Phacotnis lenticularis образует в миоценовых отложениях Швейцарии толщи известняков; встречается в ледниковых отложениях Зап. Европы и у нас.
4. Сем. Botryococcaceae. Род Botryococcus встречается в солоноватых водах Евразии и образует сапропель на дне озер (напр. В. Втаип1 в оз. Балхаша в Средней Азии). Сюда же относится род Pila из каменноугольных и юрских отложений Евразии; она составляет главную массу богхедов в Зап. Европе и у нас.
5. Сем. Кодиевые, Codiaceae. Слоевище водоросли состоит из трубочек, являющихся каждая крупными клетками, сильно ветвящимися на местах отхождения. Центр клетки занят крупной вакуолью, а в стенкоположной протоплазме расположены мелкие хроматофоры и многочисленные ядра. Размножение гаметами, образующимися в особых гаметангиях; мужские гаметы значительно меньше женских. Сюда относятся Halimeda—водоросль, тело которой пропитано известью и состоит из члеников, разделенных перетяжкой, обычно слабо инкрустированной; известна с эоцена (Н. Saportae); Halimeda ныне весьма обычна для теплых морей, где обитает по коралловым рифам. Воиета Hochstetteri в пермских отложениях Сербии, равно как близкая к ней Gymncordium bellerophontis из тех же отложений, в Тироле и на Балканском полуострове. Древнейшим типом Codiaceae являются Ра-laeoporella variabilis и Р. grandis из нижнего силура.
6. Мутовочные сифонеи, Dasycladaceae (Siphoneae verticillatae). Слоевище таких водорослей представляет сильно увеличенную разветвленную многоядерную клетку, несущую закономерно расположенные разветвления главной оси; эти разветвления представляют густые мутовки по 10—15 в кольце. Первичные ветви в свою очередь трижды ветвятся, образуя по четыре членика. На вершинах ветвей образуются гаметангии мужские и женские, копулирующие друг с другом; после копуляции гаметы прорастают. Все растения обычно сильно покрыты известью, которая откладывается вокруг главной оси растения во внешнем ослизняющемся слое ее оболочки, а затем захватывает и боковые ростки, причем все растение погружается в известковую оболочку почти целиком; растение отмирает, а известковый скелет сохраняет структуру растения.
Dasycladaceae были всегда широко распространены в теплых морях с силурийского времени; число ископаемых родов почти в шесть раз превышает число форм, обитающих в современных морях. Ископаемые формы значительно превосходят формы современные по размерам и красоте форм.
Наиболее характерными являются следующие роды.
Primocoralha—ствол несет редко сидящие ветви, суженные к основанию, слабо покрыт известью. Нижний силур, Сев. Америка.
Rhabdoporella—ствол представляет толстый неразветвлепный цилиндр с булавовидными ветвями, сильно облеченный известью Верхний силур Готланда; из палеозоя Казакстана Вологдин описал Rhabdoporella dispar-rimosa, близкую шведскому виду — R. pachyderma (фиг, 784).
Cyclocriniis (фиг. 785)—ствол на верхушке булавовидный со сферически расходящимися тонкими ножками, несущими грибовидные расширения, в общем смыкающимися между собою путем соприкосновения своих шляпок, образуя сферическое тело, выделяющее оболочки из извести. Нижний силу р.
Diplopora—ствол образует толстый известковый панцырь с конусовидными влагалищами у выхода ответвлений оси из этого панцыря (фиг 786). Средний триас Альп.
Фиг. <84. Rliab- Фиг. 78И.Cycloertmts Фиг. 786. 1. Поперечные разревы Diplo-dopordla pachy- porosus. Ордовиций. рога. Триас. Тироль. 2/1. 2. Diplopora, derma. Верчнчй Германия Схематиче- вид совнутри. Триас. Эвино. Натур, ве/чч. сипур. Гот анд. ский разрез. 3/i
Швеция. 90/1.
Triploporella— ствол удлиненный, булавовидный, густо покрытый ветвями. Верхняя юра и средний мел Европы.
Cymopolia—ствол остоит из четковидных члеников, связанных участками, лишенными твердого скелета. Встречался в морях с мелового периода до настоящего времени.
Acicularia—тело водоросли грибообразное, с звездчато-расположенными члениками клиновидной формы, прикрепленное к длинной, тонкой ножке. Чокракские отложения Крыма и Кавказа и в третичных отложениях всей Европы; доныне один вид сохранился в водах Антильских островов.
Dasycladaceae—но своему древнему происхождению и разнообразию налеозойскид и мезозойских форм могут служить руководящими ископаемыми для определения геологического возраста отложений.
КЛАСС IX. ХАРОВЫЕ, CHARj)PHYTA.
Прямостоящие, многоклеточные, богатые хлорофиллом водоросли пресной и солоноватой воды, с членистыми стеблем и ветвями и мутовчатым разветвлением. Стебель состоит из чередующихся длинных и коротких клеток;
26 Учебник палеонтологии.
401
последние образуют узлы, из ксиоры» выходят ветви второго Порядка. Бесполое размножение совершенно отсутствует. Женские органы—яйцепочки (так же, как и производящие сперматозоиды мужские)—помещаются на узлах ветвей и видимы простым глазом. Оболочки плодов, а иногда и стебля, в известный период объизвестняются и тогда сохраняются в ископаемом состоянии Почти космополитическое семейство, известное с карбона, а может быть и девопа.
По устройству яйцепочки разделяются на три группы:
I. Собственно Characeae, имеющие яйцепочку, состоящую из 5 покровных клеток, часто скрученных спиралью и увенчанных коронкой. Сюда относятся роды: Nbtella, известная уже с олигоцена, и собственно Cham. Хары (фиг. 787) часто встречаются в пресноводных отложениях; иногда встречаются целые скопления инкрустированных стеблей; клетки стеблей (Д гг) большею частью окружены оболочкой из тонких корковых клеток (rz). Яйцевидные или шаровидные плоды (4—В) окружены оболочкой из .5 спирально и налево накрученных трубчатых покровных клеток. Покровные клетки образуют на вершине яйцепочки коронку Ifc), состоящую из пяти клеток, отделившихся от концов покровных меток. Такне плоды современных
Фиг. 787. Chara. А — Gy года ites vulgar is. Ныне живущая. П/од. В — E—G. medico ginula. Bepi нгЧ олигоцен. Монморанси. Парижский бассейн. В—сбоку, С—сверху, D—снизу, к— коронка; а место нрикреплени." коронки; е — нижнее отверстие плода; Е—кусок стебля: клетка междоузлия (гг); корковые клет <и («)..
Фиг. 788. Troche fiseus bulbifomus, сверху п сбоку. Девон. Марьвно, около
Па злавска
Фиг, 789. Вусгд.ыт melo. Девон. Урал А—сбоку, В—сверху, С — снизу.
хар встречаются в пресноводных отложениях от юры и доныне. Хары обычпы в третичных отложениях и в дилювии.
В виду того, что в ископаемом состоянии чаете находят стебли и яйце-почкн отдельно и трудно установить связь междз ним:;, ныне предпочитают описывать, отдельно в известных случаях, то и другое. Стебли хар описываются под названием Characeites (с палеоцена), а4 яйцеклетки под названием Gyrogonites (например Gyrogonites medicaiimila, фиг. 787, Б—Е). Яйцеклетки, относимые к этому искусственному роду, известны с лейаса до настоящего времени. В палеоцене Европы встречался род Lagynophora с яйцепочками бутыльчато-укороченной формы, затем род Kosmogym, тоже вымерший, с одной спиралью клеток и с выдающимися буграми на их поверхности; представители рода изваяны с верхней юры.
II. Palacocharaceae—древние типы хар имели более пяти покровных клеток—сюда относится род Palaeochara из верхнего карбона.
III. Clavatoraceae-—покровная клетка одиночная, образованная листочками,—сюда относится род Clavator из верхней юры.
Сюда ясе относите/ невидимому плохо связанная с новейшими типами
особая древняя группа форм Characeae из палеозоя. Морские отложения (или отложения солоноватых вод) девона (у нас, Сев. Америка) доставляют особые остатки, по существу одинаковые с плодами хар, отличающиеся покровными клетками, закрученными вправо, притом имеющимися в большом числе—9 (8, 10) или 18 (16, 20). Таков род Trociiiliscus (фиг. 788). Из девона нашего и Рейнской Пруссии описан род Sycidiiim (фиг. 789) с плодами яйцевидной пли грушевидной формы и, большею частью, с 18 меридионально проходящими ребрышками. Так как покровные клетки Chara в первых стадиях индивидуального развития также идут прямо от одного полюса плода к другому, то очевидно Sycidium представляет первичную особенность в характере ребрышек.
КЛАСС X. БУРЫЕ ВОДОРОСЛИ, PHAEOPHYCEAE.
Высокоразвитые многоклеточные водоросли бурого цвета, зависящего от примеси к хлорофиллу бурого пигмента фикофепна.
Размножаются изогаметами и гетерогаиетаыи; кроме полового известно и вегетативное размножение.
Живут почти исключительно в морях, часто достигая колоссальной величины (Macrccystis в Саргассовом море имеет до 300 метров длины). Некоторые антарктические формы (Lessonia) по массивности напоминают подводные деревья.
За водоросли раньше принимали многие ископаемые,. относимые к другим группам, и особенно такие, которые называли фукоидами, из которых многие не относятся к этой группе организмов (см. черви).
Не имея скелета, бурые водоросли плохо сохранились со времени своего появления в силурийский период. Сведения об ископаемых водорослях весьма скудны. Представители семейств Desmarestiaceae (Desmarestia) и Laminariaceae ‘ (Lessonia) известны из миоцена Калифорнии. Zoopliycus— в эоцене Италии, Cliordaceae—ископаемые, представлены пока родом Chor-dites из эоцена Франции.
Сем. Fucaceae достоверно известно с палеозоя, Hymanthaliopsis и Orestovia (пермокарбон Кузбасса), -принимавшие участие в образовании углей (томит): Lasiophycus из нижнего лейаса Франции; Palaeohalidrys из миоцена Калифорнии; Ascophyllum из неогена Сев. Кавказа (Грозный);, Cystopliyllum п Cystoseira из третичных отложений Европы, Сев. Кавказа и Майкопских сарматских отложений; род Sargassum известен уже в эоцене Европы..
В девонских п силурийских отложениях нередко встречается крупный представитель бурых водорослей; стволы его, имеющие от 20 см до 1 м в диаметре, состоят из рыхлого сплетения гиф—это Prototaxites (или Ne-matophycus) палеозойских морей.
КЛАСС XI. БАГРЯНЫЕ ВОДОРОСЛИ, RHODOPHYCEAE (FIORIDEAE).
Чрезвычайно разнообразные по внешнему виду водоросли, имеющие пурпуровую или багряную окраску, в которой хлорофилл замаскирован особым пигментом — фикоэритрином. Слоевище часто сильно разветвленное пли лопастное.
Размножаются бесполым или половым путем, вплоть до сложного процесса оплодотворения с образованием карпогония, соответствующего зародышу высших растений и помещающегося в цистокарпии. Половые органы мужские и женские (антергдии и карпогонии) развиваются на одном и тем
же индивидууме однодомно.
Из карпогониев развиваются особые образования—цистокарпии в виде полостей полулунной формы в сечении с центральным возвышением на дне и выходящим наружу отверстием на вершине (фиг. 790, В).
В цистокарпиях развиваются споры
Из числа многих сомнительных указаний на нахождение багрянок следует отметить немногие: из сем. Bhodymeniaceae—Lomentarites из нижнего
лейаса и из сем. Ltelesseriaceae—представителей рода Delesseria, начиная с верхнего мела.
В палеоботанике имеют значение по преимуществу формы, обладающие известковым скелетом. К числу их относятся:
9
Фпг. 790. Ldhothamiaum polymurphum. Ныне живущая (берег Бретани).
4 — гертикалышй разрез через
желвакообразное тело бодо}ослп; х — углубления, вызванные постороння ми телами; с— цистокарпии. В — разрез через цистокарпий (с) с шипом («) и конечным отверстием о)', С — вид с поверхности, представляющий
Фиг. 791. Ldhothamnwm gosavunst;. Се-нон. Франция.
4 — наружный вид (ж — место прикрепления); В — продольный, С — поперечный разрез через клеточную ткань г; t — тетраснорангии, расположенные слоями.
углубленные отверстия цистокарпиев ($г).
1. Сем. Solenoporaceae, род известковых водорослей из силура и девона, который некоторые авторы (например Пчелинцев) склонны относить к Tabulata (трубчатым кораллам).
Sole.iopora—слоевище имеет форму шара величиной с яблоко, которое на вертикальном разрезе обнаруживает многочисленные клетки, располагающиеся часто в концентрическом порядке. Встречались от нижнего силура до юры.
2. Сем. Corallinaceae имеют слоевище весьма разнообразной формы, желвакообразное с шишковатыми выступами на поверхности, кустисто-разветвленное с короткими толстыми разветвлениями, прирастающее к субстрату или корковидное, облекающее субстрат. Слоевище состоит из плотно- сомкнутой ткапи многочисленных кубических или параллелепипеда иных клеток от 0,005 до 0,016 мм величиной,, стенки которых большею частью твердеют
от отложения извести (составляющей 90% веса всего растения), так что образуются камневидные, кажущиеся неорганическими тела. Сюда относятся различные типы багрянок, способных отлагать углекислый кальций, образуя в одних случаях пленки на камнях, в других—кустики известковатых стеблей, в третьих — известковые скопления, обычно шаровидной формы; сюда относятся Arclieolithothamnium, LitJiothamnium (фиг. 791), Derma-lithon и Lithophyllum, последние из них являются нуллипоровыми водорослями, участвуя в образовании коралловых рифов в южных морях. В ископаемом состоянии известны уже с нижнего мела.
Особенно большую роль в образовании нуллипоровых известняков в мелководных частях тропических морей играют представители рода Litho-thamnium, образующие иногда огромные скопления рифов. Род известен уже с нижнего мела; ныне известно более 50 видов рода из меловых и третичных отложений и до 100 видов ныне обитающих в холодных и теплых морях всего света.
КЛАСС XII. ГРИБЫ, FUNGI.
Одно- или многоклеточные растения паразитного или сапрофитного образа жизни, никогда не имевшие хлорофилла. Размножение бесполое или половое, чаще конидиями или другими типами спор. Остатки грибов в ископаемом виде нередки, но часто остаются незамеченными. Известно до сорока семейств ископаемых грибов со среднедевонского времени. Регопо-sporites antiquarius из каменноугольных растений. Весьма многочисленны остатки грибов в третичных отложениях.
Остатки грибов обычно плохо сохраняются и имеют мало значения для геологических целей.
КЛАСС XIII. ЛИШАЙНИКИ, LICHENES.
Лишайники являются организмами, представляющими симбиоз между грибами .и зелеными водорослями. Известны со времени каменноугольного периода; род Opegrapha известен уже с верхнемелового времени. В третичных (янтароносных и буроугольных) отложениях Германии остатки лишайников встречаются нередко.
Зародышевые бессифонные, Embryophyta asiphonogama.
Редко слоевищеобразные, обычно расчлененные на стебель и листья растения, с ясно выраженным чередованием поколений. Оплодотворенная сперматозоидом из антеридия (носитель мужских половых клеток) яйцевая клетка архегония (носитель женских половых клеток) дает зародыш, который впоследствии развивается в бесполое поколение (спорофит), производящее споры, из которых в свою очередь прорастает новое половое поколение, или гаметофит. Производящий оплодотворение сперматозоид обладает произвольным движением п, вышедши из антеридия, попадает к устью архегония через посредство жидкой среды, например—капли росы.
К ним принадлежат подотделы Bryophyta (мохообразных) и Pterido-phyta (папоротникообразных). У первых основная генерация растения, соб
ственно «мох», представлена гаметофитом (половым поколением), слоевчще-образпа или расчленена на стебель и листья, но не имеет проводящих пучков или таковые самого простого устройства. Наоборот, гаметофит папоротникообразных проявляется лишь в виде мелкого, часто едва заметного глазу заростка, или проталлия, и все функции жизнедеятельности растения, кроме полового процесса, осуществляются спорофитом (бесполым поколением), представляющим расчлененное на стебель, листья и корень растение. Кроме того, у них развита система сложно устроенных проводящих пучков. Так как, в противоположность мхам и другим ниже организованным группам растений, проводящая пли сосудистоволокнистая система всех высших растений, начиная с папоротникообразных, имеет Много общего, то здесь именно будет уместно коснуться анатомии этих растений.
Черты анатомии папоротникообразных и высших зародышевых растений. Водоросли, грибы, лишай и мхи (представители последних 3 групп почти не имеют палеонтологического значения) являются организмами без дифференциации тканей, состоят из более пли менее однородных клеток. Остальная часть царства растений представляет организмы с обособившимися в теле тканями, состоящими из клеток определенной формы и функции. Лишь две ткани обособлены без отношения к функции слагающих их клеток: прозенхимой называют ткань, состоящую из вытянутых в одном направлении клеток с заостренными концами (фиг. 792, Е), тогда как остальные типы тканей называют паренхимой; клетки типичной паренхимы имеют приблизительно одинаковые размеры по всем направлениям (фиг. 792, D],
Ирозенхимные клетки дают разнообразные видоизменения (фиг. 792); стенки их могут нарастать совнутри, местами, так что на стенке являются тонкие и утолщенные места; при этом на внутренней поверхности оболочки получаются узоры, просвечивающие через стенку, видимые снаружи. Иногда утолщение клетки представляет спиральную полоску (фпг. 792, G) пли отдельные кольца, расположенные в отдельных поперечных сечениях клеток. Часто встречается сетчатое утолщение стенки (фиг. 792 А), с неодинаковыми петлями сетки. Просветы петель иногда, представляются снаружи точечными или щелевпднымп («поры»). Бывают клетки с особыми, сравнительно сложного строения окаймленными порами, очерчивающимися двумя концентрическими кружками.
Прозенхпмные клетки при расположении их в ряды по длине стебля, с прилеганием заостренных концов, нередко дают происхождение длинным трубкам; прилегающие участки стенки уничтожаются или продырявливаются таким образом, что устанавливается непрерывное сообщение полостей целого ряда клеток. Такие новообразования, составленные из вышеперечисленных типов прозенхимных клеток, называются сосудами; соответственно типам клеток различают спиральные, коленчатые, сетчатые сосуды. Растения, имеющие сосуды, противоположно слоевищу бессосудистых, представляют корень, стебель и листья. К сложным образованиям, возникающим подобно сосудам из клеток, принадлежат ситовидные трубочки. Степки этих трубок (фиг. 792, F) представляют участки, пронизанные действительными, а не кажущимися лишь, как у сосудов, отверстиями. Протоплазматическое содержимое соседних трубок сообщается сквозь отверстия сит. Ситовидные трубки постоянно находятся в коре растений: назначение их—передача по телу растения органических веществ, вырабатываемых листьями.
В коре не бывает сосудов, составляющих главным образом древесину. Полости сосудов лишены протоплазматического содержимого, утратили его посте слияния слагающих сосуд клеток; сосуды состоят лишь из мертвых оболочек клеток ~------------- ------ тт
и
наполнены водою. Назначение сосудов—передача
по
Д—часть лестничного сосуда с удаленной отчасти стенкой.
D и Е — клетки паренхимы и прозенхимы. F—часть ситовидной трубки с ситовидными участками Se. pl. G—часть сп"оальииго сосуда со спиральным волокном. И— часть клетки скелетного элемента.
ар с
С—схематизированное сечение концевой части стебля по длине, через точку роста, представляющее макушечную клетку (ар. с), клетки, от нее отщепляющиеся (Seg е.). ар тег—верхушечная образовательная ткань; ер —кожица; hyp—скелетные элементы; sei—то же; par—паренхима; Ъ. sh—оболочка сосудистого пучка; sv. t—ситовидная трубка; sp. v—спиральные сосуды;
sc. е—лестничные сосуды. Фпг. 792. Анатомические элементы высших растений.
ИЗ
растению воды с растворенными в ней солями, извлекаемыми корнями почвы. Сосуды и ситовидные трубки служат органами питания растения
В растении существуют и скелетные элементы, дающие оперу, эти элементы бывают прозенхим цыми и паренхимными. Скелетные элементы
толстостенны: утолщение стенки доходит до того, что полость клеток крайне сокращается. Это большею частью отмершие, не содержащие протоплазмы и наполненные воздухом клетки (фиг. 792, Н), стенка которых часто приобретает особенную прочность, благодаря изменению целлулозы в древесное вещество. В коре находятся прозенхимные скелетные элементы л у о а.
Как элементы, проводящие воду, так и элементы, проводящие органические вещества, не разбросаны по растению без всякого порядка, а собраны в особые пучки, и при этом так, что часть пучка занята элементами, проводящими воду, а остальная часть пучка—элементами, проводящими органические вещества. Эти пучки называются проводящими, сосудистыми или, вследствие присутствия в них и скелетных элементов, — сосудистоволокнистыми. Часть сосудистого пучка, занятая элементами, проводящими воду, называется кселемой, первичной древесиною частью. Часть пучка, занятая элементами, проводящими органические вещества, называется флоэмой. Различают несколько типов сосудистых пучков на основании расположения в них ксилемы .и флоэмы.
Так называемые коллатеральные сосудистые пучки получили свое название от того, что в них ксилема и флоэма примыкают друг к другу только одной своей стороной, располагаясь по направлению одного и того же радиуса, так что вообще ксилема пучка обращена внутрь, а флоэма—наружу (фиг. 793, A, D).
В одних коллатеральных сосудистых пучках ксилема непосредственно , примыкает к флоэме. Такие пучки называются закрытыми (фиг. 793, 2). В других же коллатеральных сосудистых пучках между флоэмой и ксилемой лежит особый слой, состоящий пз нескольких рядов клеток, вытянутых в тангенциальном направлении, с тонкими оболочками из чистой клетчатки, с ядром и богатых протоплазмой. Этот слой клеток называется» камбием. Сосудистые пучки, содержащие камбий, называются открытыми (фиг. 793, В, D). Камбиальные клетки способны делиться. Клетки, отделяемые наружу, в конце концов превращаются в элементы флоэмы: клетки же, отделяемые внутрь, превращаются в элементы ксилемы.
Второй тип сосудистых пучков представляют концентрические сосудистые пучки. Концентрические пучки характеризуются тем, что в них одна часть пучка, состоящая из элементов одной категории (обыкновенно ксилемы), окружена со всех сторон элементами другой части, другой категории. По типу концентрических пучков построены также и стеллы папоротников, раньше разсматривавшиеся как концентрические пучки (фиг. 793, С).
Первоначальная древесина (например в одногодовом стволе древесного растения) происходит не из камбия, а из первичной образовательной ткани и называется первичной древесиной, в противоположность вторичной, возникающей вследствие деятельности камбия. В одногодичном стволе хвойных или двудольных имеется в основной ткани лишь одно кольцо сосудистых пучков коллатерального типа, открытых (фиг. 793, В). Основная ткань состоит из обыкновенных паренхимных клеток и по своему положению относительно сосудистых пучков распадается на три части. Часть, лежащая внутри, называется сердцевиной. Часть, лежащая наружу от сосудистых пучков, называется первичной корой. Наконец участки основной ткани, лежащие между пучками, называются первичными сердцевинными лучами. Различают пучковый камбий (камбий сосудистых пучков) и межпучковый. Оба названные камбия составляют одно сплошное камбиальное
кольцо, отлагающее внутрь вторичную древесину, а наружу—вторичную кору. В деятельности камбия наших древесных растений сказывается периодичность в зависимости от климатических условий Она выражается в образовании так называемых жодичных колец. Вообще весной камбий отлагает сосуды с более ширгеим просветом поперечного сечения и с тонкими стенками; к осени отлагаются сосуды, сплюснутые в радиальном
Фиг. 7S3. Поперечные разрезы главных типов сосудистых пучков.
А — коллатеральный закрытый пучок злака, сахарного тростника (Saccharum offictnarum). PG и RG — сосуды. Cbf — элементы флоэмы; пучок окружен скслетЕшга элементами (темные стенки) И совсем снаружи — основной паренхимой. В — коллатеральный oi крытый пучок двудольного лютика (Ranunculus repensy, с — камбий, s — сосуды, ® — ситовидные трубки, vg — оболочка пучка С — концентрический пучок папоротника (Pieris aquRina)\ b. sZi — оболочка сосудистого пучка, «и. t — ситовидная трубка, sc. v в sp v — сосуды 1) — схематический поперечный разрез через одногодичный стебель, нарастающий впоследствии в толщину, с 8-ю сосудистыми пучками; с — кольцо камбия; ш — сердцевина; тк — первичные сердце винные лучи; h — первичная древесина; р — первичная кора.
направлении, меньшего поперечного течения и покрытые толстыми оболоч ками. Образование древесины прекращается на зиму и начинается потоп снова весной с образования широкопросветных элементов. Соответственно этому между весенними и осенними слоями существует резкая граница, видимая простым глазом и могущая служить для определения возраста соответствующей части растения на основании числа годичных колец. У пальм между корой и древесиной нет кольцевидного слоя камбия, поэтому ствол их растет в толщину лишь в молодом состоянии, когда еще все клетки способны делиться, затем ствол растет лишь в вышину.
В листьях сосудието-волокнистые пучки являются в виде жилок, называемых нервами.
Тип II. Мохообразные, Bryopliyta.
К этому типу относятся два. весьма характерных класса, представители которых легко различимы по внешнему виду.
Определенно выявлена смена поколений, одно несет половые органы, антеридии и архегонии, из которых вырастает другое, бесполое поколение, _ несущее споры.
КЛАСС I. МХИ ПЕЧЕНОЧНЫЕ, HEPATICAE.
Стебель имеют в виде зеленого слоевища, не имеющего дифференциации на стебель и листья. Печеночники известны с каменноугольного времени—Hepalicites Willsi (продуктивный ярус Стафордшира); Palaeohepatica (в Галицийском кейпере), Marchantites [юрские отложения Англии, юга европейской части СССР (Каменка) и в вельде Уссурийского края].
КЛАСС II. ЛИСТВЕННЫЕ МХИ, MUSCI FRONDOSI.
Лиственные мхи имеют определенно дифференцированные стебель и'листья; сюда относятся: общераспространенные виды, как кукушкин лен (Polytrichum) или торфяной мох (Sphagnum), имеющие типично выраженный стебель со спирально расположенными на нем листьями.
Мхи лиственные известны уже с верхнекаменноугольного времени (Musettes polytrichaceus)-, формы близкие современным встречались уже в третичное время.
Хорошо отличимые формы лиственных мхов известны из Балтийского янтаря (олигоцен), отложений Энингена (верхний миоцен); играющие большую роль в растительном ландшафте нашего времени торфяные мхи появились в конце третичного времени около ледникового периода и являются ныне активными торфообразователями. В ледниковых и послеледниковых отложениях Евразии мхи довольно обычны.
Тип. III. Папоротникообразные, Pteridopliyta.
Сюда относятся высокоорганизованные сосудистые споровые растения, имеющие смену поколений и сложное анатомическое строение. Существует одно- или разноспоровость.
В настоящее время известно шесть основных классов папоротникообразных: 1) Псилофитовые—Psilophytales, 2) Плауновые — Lycopodiales, 3) Псилотовые-т-Psilotales (ископаемые виды неизвестны), 4) Хвощевидные — Articulatales, 5) Кладокснловые — Cladoxylales (только вымершие виды) и 6) Папоротники—Fllicales.
КЛАСС L ПСИЛОФИТОВЫЕ, PSILOPHYTALES.
Растения мелкие или средней величины, дихотомически ветвящиеся, по облику напоминающие то плауны, то водоросли, часто с щетинистыми листьями или без таковых, несущие споровместилища, расположенные на
концах разветвлений побегов в форме коробочки, напоминающей спорогонии мхов. Анатомически характеризуются присутствием центрального ствола, состоящего нз трахеид, облеченных флоэ"ой.
Сюда относятся древнейшие типы наземных растений, сохранивших мнопе черты сходства с водорослями, свойственные верхнесилурийским и девонским отложениям Евразии и Сев. Америки (В. Канада).
Rhynia
Сшу го »е -V nighani.
фдг. 794. Rhyma. Де-юн. Реставрация. Слева более ветвистое растение, чем справа. На верхушках ветвей споранх'ии. Уменьшено.
Сюда относятся следующие роды: Rhyma (фиг. 794) (девон нижней и средней Шотландии и Германии) HickUngia (девон Шотландии), Lo-дапеИа (девон В. Канады), haliserites (нижний девон Германии), Нотпеа (девон нижней и средней Шотландии), Sporogonites (нижний девон Норвегии), Pseudosp&rochnus (средний девон Чехо-Словакии), Psilophyton (нижний девон В. Канады, Европы и СССР—Воронеж, губ.), Dawsonites (нижний девон В. Канады, Норвегии и С. Франции), Asteroxylon (нижний и средний девон Шотландии, Средней и С. Европы и южной Сибири—Минусинский край).
Тип И. Мохообразные, Bryophyta.
К этому типу относятся два. весьма характерных класса, представители которых легко различимы по внешнему виду.
Определенно выявлена смена поколений, одно несет половые органы, антеридии и архегонии, из которых вырастает другое, бесполое поколевие,. несущее споры.
КЛАСС I. МХИ ПЕЧЕНОЧНЫЕ, HEPATICAE.
Стебель имеют в виде зеленого слоевища, не имеющего дифференциации на стебель и листья. Печеночники известны с каменноугольного времени—Hepaticites Willsi (продуктивный ярус Стафордшира); Palaeohepatica (в Галицийском кейпере), Marchantites [юрские отложения Англии, юга европейской части СССР (Каменка) и в вельде Уссурийского края].
КЛАСС II. ЛИСТВЕННЫЕ МХИ, MLSCI FRONDOSI.
Лиственные мхи имеют определенно дифференцированные стебель и'листья; сюда относятся: общераспространенные виды, как кукушкин лен (Polytrichum) или торфяной мох (Sphagnum), имеющие типично выраженный стебель со спирально расположенными на нем листьями.
Мхи лиственные известны уже с верхнекаменноугольного времени (Musettes polytrichaceus)\ формы близкие современным встречались уже в третичное время.
Хорошо отличимые формы лиственных мхов известны из Балтийского янтаря (олигоцеп), отложений Энингена (верхний миоцен); играющие большую роль в растительном ландшафте нашего времени торфяные мхи появились в конце третичного времени около ледникового периода и являются ныне активными торфообразователямп. В ледниковых и послеледниковых отложениях Евразии мхи довольно обычны.
Тип III. Папоротникообразные, Pteridopliyta.
Сюда относятся высокоорганизованные сосудистые споровые растения, имеющие смену поколений и сложное анатомическое строение. Существует одно- или разноспоровость.
В настоящее время известно шесть основных классов папоротникообразных: 1) Псилофитовые—Psilophytales, 2) Плауновые — Lycopodiales, 3) Псилотовые-т-Psilotales (ископаемые виды неизвестны), 4) Хвощевидные — Articulatales, 5) Кладокснловые — Cladoxylales (только вымершие виды) и 6) Папоротники—Filicales.
КЛАСС L ПСИЛОФИТОВЫЕ, PSILOPHYTALES.
Растения мелкие или средней величины, дихотомически ветвящиеся, по облику напоминающие то плауны, то водоросли, часто с щетинистыми листьями или без таковых, несущие споровместилища, расположенные на
концах разветвлений побегов в форме коробочки, напоминающей спорогонии мхов. Анатомически характеризуются присутствием центрального ствола, состоящего из трахеид, обтеченных флоэмой.
Сюда относятся древнейшие типы наземных растений, сохранивших многие черты схотства с водорослями, свойственные верхнесилурийским и девонским отложениям Евразии и Сев. Америки (В. Канада).
Фог. 704. Rliynia. Девон. Реставрация. Слева более ветвистое растение, чем справа. На верхушках ветвей спорангии. Уменьшено.
Сюда относятся следующие роды: Rhynia (фиг. 794) (девон нижней и средней Шотландии и Германии) Hicklingia (девон Шотландии), Lo-ganella (девон В. Канады), Haliserites (нижний девон Германии), Ногпеа (девон нижней и средней Шотландии), Sporogonites (нижний девон Норвегии), Pseudosporochnus (средний девон Чехо-Словакии), Psilophyton (нижний девон В. Канады, Европы и СССР—Воронеж, губ.), Dawsonites (нижний девон В. Канады, Норвегии и С. Франции), Asteroxylon (нижний и средний девон Шот ландии, Средней и С. Европы и южной Сибири—Минусинский край).
КЛАСС И. ПЛАУНОВЫЕ, LYCOPODIALES.
В современной флоре, так же как и в новейших ископаемых, эта группа представлена только травянистыми плаунами, тогда как в палеозойское время преобладали древовидные представители, обособляемые в группу Lepidophyta.
Им присуща большая часть признаков организации плауновых. Дихотомически ветвящиеся стеблевые части с тесно примыкающими, сравнительно небольшими игловидными или чешуевидными, однонервными листьями, расположенными спирально; большие спорангии, сидящие поодиночке в пазухах листьев или на верхней поверхности, у их основания. К этому присоединяются чуждые для новейших плауновых признаки, как-то: вторичное нарастание в толщину древесинного цилиндра, покров ствола большими листовыми подушками и рубцами и особый вид корневых органов.
Из современных плаунов наиболее обыкновенные являются равноспоровыми и без «язычка» (ligula) на поверхности нормальных листьев, это будут Lycopodiales eligulatae — представители семейства ликоподиевых (Lycopodiaceae), куда относятся роды Lycopodium (плауны)—около 180 видов из тропических и умеренных стран всего света, и род Phylloglossum (Австралия, Тасмания и Нов. Зеландия).
Из числа современных Lycopodiales ligulatae известно до 700 представителей рода Selaginella.-
Ископаемые формы ликоподиевых Lycopodiales подразделяются на два основных типа, представителями которых являются хорошо изученные роды Lepidodendron и Sigillaria.
Lepidophyta примыкают по своей разноспоровости и по присутствию ligula к менее изобилующей своими представителями в современной флоре умеренного пояса группе плаунов (Selaginellaceae).
У обыкновенных плаунов (Lycopodium') заростки хорошо развиты, а у селагвнелловых являются' упрощенными, редуцированными, оставаясь в пределах оболочки споры, после того как эта оболочка лопнула вследствие развития заростка.
Lepidodendron (фиг. 795, lepis — чешуя, dendron — дерево) имеет прямой ствол, ветвящийся в верхней части и достигающий в высоту до 30 м. Корни ветвятся, подобно стволу, дихотомически, покрыты многочисленными корешками, росли в горизонтальном направлении, вероятно, в болотистой почве или, возможно, частично под водой. Некоторые из более тонких разветвлений ствола оканчивались коническими или овальными спороносными колосками. Игловидные листья, значительно варьирующие в отношении длины у различных видов, покрывают поверхность молодых побегов по тесно прилегающим спиралям, а их расширенные основания или листовые подушки образуют сплошной покров ствола, налегающий на кору. Вследствие деятельности зоны образовательной ткани, окружающей цилиндр древесины, главный ствол и ветки нарастают из года в год в толщину; годичные кольца отсутствуют, так что нет указаний на резкую смену времен года.
Хорошо сохраненная листовая подушка Lepidodendron—наиболее заметно отличительная особенность рода—ромбоидальна или веретенообразна, достигая в исключительных случаях длины до 8 см. Оставляя в стороне детальные отличия, типичная листовая подушка Lepidodendron характера ...а
ьияуися ясно выраженной гладкой площад :ой, часто расположенной в средней части (фиг. 795, С). Эта площадка представляет листовой рубец, или исто прикрепления основания листа. На листовом рубце находятся три мапых рубчика, из которых наибольший, срединный, отмечает положение
Фиг. 795. Lepidodendron.
А — реставрация растения, г — спороносный колосок. В — поверхность ствола Z. obovatum, показлваюгцая положение и форму листовых подушек. С — одна отд лызая лясювая по-душка, схематически: I— вход в камеру при основании язычка; ш — срединная килсватость н ixcueu части подушки; п — листовой рубец; о — дыхательные отверст я; s — бо; овые рубчики; х — трсхугобьное возвышение. В— часть продольного разреза спороносного волоска. а — ось; 8b, sb' — споролистик; sjp — спорангий. Е— макроспор». E — i микроспоры. G — спороносный колосок.
единственного сосудистого пучка, проходящего в лист из ствола; боковые рубчики представляют сечения дв\х пучков паренхиматозной ткани. Над листовым рубцом находится рубчик, отвечающий месту входа в камеру (Z) при основании язычка. Листовая подушка в целом образует возвышение с килеватостыо в нижней части. По сторонам згой килеватостп под листовым рубцом находятся два больших отверстия (о), парихны — дыхательные или выделительные отверстия.
Иногда имеются отпечатки экземпляров с облезшими наружными слоями покровов ствола. На таких отпечатках вышеописанные подробности поверхности листовых подушек не выражены. Тогда сохраняются обыкновенно лишь части листовой подушки, примыкающие, как к основе, в сосугистому пучку; в связи с этим листовая подушечка является в виде сосочковидного образования, направленного косо кверху, на коническом округленном конце которого находится листовой рубец, в виде широкого углубления. Таковые отпечатки описывались под названием Knorria Формы с поперечно-ромбическими листовыми подушками (фиг. 796), которые притом налегают чере-питчато и представляют вершину с листовым рубцом внизу, выделяются в род Lepidophlolos. Бугорчатые побеги, несущие шишки, обычно описываются под именем Hcdonia. Плодоношения oi ык-новенно помещаются .на концах крайних стеблевых разветвлений в виде более или менее значительных шишек (фиг. 795, А, г) и описываются под названием Lepidostrobus (strobus—конус). У другого близкого рода такие шишки являлись сидячими на боковой поверхности стволов, производя тогда на ней крупные конические углубления, расположенные в два противолежащих продольных ряда. Ствол у него гладкий, без парихи и язычков, — этот род называется Ulodendron (ule — древесина), свойственный верхнему карбону. Сюда же приближается род Bothrodendron, характеризуемый гладкой корой и отсутствием листовых подушечек, заме
ненных спирально расположенными маленькими листовыми рубцами, имеющий язычок и такое же расположение шишек, как и Ulodendron.
Спороносные колоски, находимые иногда изолированными и в окремне-лом состоянии (д), представляют многочисленные, большей частью спирально расположенные споролистики (фиг. 795, D, s';—sb'), которые отличаются от простых листьев значительной длиной и шириной, а также почти горизонтальным положением своей нижней части, несущей единственный большой спорангий (sb'). Спорангии, достигающие до 2 см длиной (фпг. 795, D, sp), содержат многочисленные споры—микроспоры или макроспоры. Таким образом лепидодендроны несомненно были растениями разпоспоровыми.
Известньт макроспорангии, в которых преобладающе развивалась за счет других одна макроспора (Lepklocarpon), притом поверх оболочки споранпы здесь развивается еще оболочка, исходящая из споролистика, снабженная вдоль спорангия щелевидным отверстием на верхней стороне. Благодаря присутствию этой оболочки с отверстием спорангий приближается к семени сифоногамных растений. Остается впрочем неизвестным, идет ли сходство далее, происходило ли оплодотворение семени пыльцой; присутствие зародыша в макроспоре открыто не было. Во всяком случае, получается
Фиг. 796. Листовая подушка Leyiclopkloios тас-rolepidotus.
д — рубчик прилистника; I — рубчик сосудистого пучка листа; п листовой рубец; « — боковые рубчики.
i
указание на возможность связи плауновых через посредство Lepidophyta с голосеменными саговиковыми или хвойными.
Строение ствола. Отличительными особенностями леппдофптов являются: 1) малая ветчина центрального цилиндра древесины (фиг.797,L,h) по сравнению с диаметром ствола, 2) присутствие слоя вторичной древесины, окружающей первичную, 3) образование вторичной коры цилиндром образовательной ткани, расположенным вплоть у вых подушек.
5 Р*
строения ствола
В 18,
области листо-
распространенным во всех каменноу голь-
30 метров, в виде обык-
Фпг. 799. Реставрация сигиллярии со стигыа-рией.
новенно неразветвлен-ной колонны, с куполообразной вершиной, образованной линейны-
Фпг. 798. Части поверхности ствола сигиллярий.
а — Sigillaria Brardi. Каменноугольные. Вештин. Германия. Ъ — Sig. ВоЫауг. Каменпоуголън. Рурский бассейн. Германия, с — Sigillaria, elegantula. Рурский бассейн. Германия.
Фиг. 797. Поперечные сечения стволов, Le-ptdodendron.
А — Lepid. irerifolizimz т, — сердцевина; р — зона первичной древесины; s—вторичной (на границе обеих имеется место отхода боковой ветви); ж — зона внутренней коры, здесь исчезнувшей; pd — наружная кора; 1>Ъ—основания листьев; между ж и pd лежит наружная кора. В—tepid. sslagtnotdes. Молодой ствол перед образованием вторичной древесины: h — первичный древесинный цилиндр; b — зона дуба со следами листьев (£6); ir —внут^епжяя кора; иг и pd— наружная кора; Ър —• осиовапжя листьев с язычком (1д).
Род Lepidodendron появляется в девоне, становится весьма обычным в ни®-некаменно -угольных отложениях, остается
ных, за исключением самых верхних, и только в редких представителях проникает в пермь.
Род Sigillaria ((риг. 798, 799; sigilium—печать) появляется уже в верхнем девоне, изобилует в средней и нижней части верхнего карбона и исчезает в пермп, представители его имеют прямой ствол высотой до
а ,Ь с
ми травовидными ди-
стьями, реже главный ствол дихотомически ветвится. Более молодые части ствола и его разветвлений у некоторых видов покрыты листьями.
Листовые ругцы—шестиугольного очертания, расположены вертикаль
ными и в то же время спиральными рядами, без листовых подутчек, иногда
на Гладкой поверхности ствола, а иногда на поверхности вертикально-ребристой (фиг. 798). Рубцы или прилегают друг к другу или разделены промежутками. Ствоты сигиллярий имели вторичное нарастание в толщину
Плодоношения в виде колосков или шишек, сидящих на стебельках, располагались наверху ствола, рядами-мутовками, возникавшими на стебле через некоторые промежутки по его высоте, так что старые мутовки распознаются по рубцам прикрепления их колосков.
Спорангии находились на верхней стороне споролистиков и заключали в себе по нескольку спор. Сигиллярии были растения разноспоровые. Встреч ,ются от нижнекаменноугольных отложений до пермских включительно.
Подземные части сигиллярий (и лепидодендронов) известны под названием стигмарий (Stigmaria) (фиг. 800). Стигмарии представляют Дихотоми-
Фиг. 800. Stiff ма ia ficoides.
A. — общий вид сверху. Каменноугольные Клег тон. Иоркшогр. В — то же самое. Вогезы. Кусок с рубцами (п) и с сохранившимися корешками. С — поперечное сеч-ине цвландра древесины, т — сердцевинная полость. I) — поперечное сечение корешка; ат — наружная, шт — средняя, 1т — внутренняя часть коры; Л — древесина
чески ветвящиеся, покрытые корешками остатки стволов. Корешки легко спадают, оставляя круглые рубцы (фиг. 800, В).
К классу плауновых принадлежат еще семейства; Isoetaceae, представленные одним родом Isoetes, довольно широко распространенным в настоящее время (до 60 видов) и появившимся еще в нижнем мелу. Ископаемые формы мало распространены или не сохранялись. Растения с клубневидными, растущими в ширину стволами, несущими у основания многочисленные листья. У основания их имеется язычок (лигула) с ямкой, в которой расположен спорангий, где заключены макро- или микроспоры.
Pleuromejaceae—неветвистые растения, высотой до 1 м, несущие четыре отходящих в стороны лопасти с придатками на верхушке; кора напоминает кору сигиллярий, покрытую листозыми рубцами, на конце побега—крупная конечная шишка.
Известны два вида: Р. Sternbergii и Р. oculma, встречающиеся в триасе в пестрых песчаниках Германии, среднем триасе Франции и нижнем триасе Русского острова у Владивостока.
КЛАСС' Ш. ПСИЛОТОВЫЕ, PSILOTALES
В ископаемом виде неизвестны до сих нор, но несомненно существуют. Род Tmesipteris распространен в Австралии и соседних с ней областях; род Psilotum — ь тропиках и субтропиках обоих полушарий.
КЛАСС IV. ХВОЩЕВИДНЫЕ, ARTICULATALES.
. Отличительной особенностью хвощевых -является мутовчатое расположение ветвей и листьев; мутовки прикрепляются в узлах, между которыми находятся лишенные ветвей и листьев междоузлия. Листья бесчерешчатые, сидячие, маленькие, шиловидные, однонервные, реже клиновидные и вильчатонервные, иногда удлиненные и образующие влагалище срастанием своих оснований. Спорангии прикрепляются к видоизмененным листьям, спорофиллам, располагающимся также мутовчато, так что сближенные мутовки их образуют колоски на концах ветвей. Спорофиллы иногда щитовидны (фиг. 805, А), и спорангии прикрепляются на стороне щитка, обращенной к стеблю. Споры бывают или все одинаковой величины (равноспоровые хвощевые), или двоякого рода (разноспоровые хвощевые); в последнем случае большие споры — макроспоры, развивающиеся в макробпорангпях, дают женские заростки; меньшие споры, микроспоры, развиваются в микроспорангиях и дают при прорастании мужские заростки.
Подкласс I. Каламофитовые. Calamophytales.
Растения с членистыми стеблями и ветвями и ветвистыми спорофил лами. Сюда относятся роды Calamophyton (calamus—камыш) из среднего девона Германии и Hyenia из верхнего девона той же страны, а также Норвегии. Очень редкая, мало изученная группа.
Подкласс II. Псевдоборниевые. Pseudoborniales.
Напоминают хвощи, несущие четыре дланевидно-рассеченных листа-Известен лишь один вид—Pseudobornia из верхнего девона о-ва Медвежьего-
Подкласс III. Клинолистовые. Sphenopliyllales.
Характеризуются мутовчатым расположением листьев в числе равном трем на членистом стебле. Листья клиновидные, цельные или рассеченные, мутовки нечередующпеся. Стебель обладает треугольным пучком, лишен сердцевины, вторичное нарастание. Спорангии в шишкообразных плодоношениях на ножках, также налагающиеся.
Sphenophyllum (фиг. 801, sphen — клин). Тонкие, нежные, по' общему виду схожие с каламитами, растения палеозойской эры, отличающиеся от остальных хвощевых по преимуществу клиновидными листьями, числом листьев в мутовках, кратным трех с трехкрыльчатым строением древесинного цилиндра, а также отсутствием сердцевинной пол'Ости.
Нижний стебель клинолиста покрыт бороздками, которые непрерывно продолжаются через узлы, как у Asterocalamites. Разветвление слабое, пазушное. Спороносные колоски на конце ветвей (фиг. 801). Листья в числе 6, 9,12 или большем, кратном трех, без чередования в смежных мутовках, большей частью ширококлпновидные и зазубренные на усеченном конце (-21, Е} с нервами, ветвящимися повторно вилчато; концы разветвлений -нервов окан
27
Учебник палеонтологии.
417
чиваются каждый в одном из зубцов края (Е), реже листья глубоко рассечены подобно тому, как у Asterocalamites (А). Иногда оба рода листьев встречаются на одпом и том же растении. Trizygia называют такие формы, у которых 6 листов группируются в 2 больших и 1 маленькую тру или наоборот. Троекратность числа листьев стоит в связи с трех членистым расположением сосудистых пучков ствола. В спороносных ко-
Фяг. Sphenophyllum. А—8 angustifolia. Верадекамепкоуг. Германия. А—ветвь с двумя спороносными ко лесками. В — часть колоска; а—ось;з—спорангия; го- споролистики. С поперечное сечение через узловую линию. S.sfe-phanense. Верхнекаменноушльные. Сент-Эттен. Кора отсутствует. Листовые пучки (Ъд) ответвляются от трехкрыльчктого первичного стержня древесины и проходят через вторичною (здесь четырехстопную) древесину. D —- 8 cmei fo гит. Верхнекаменноугольные. Мутовка листьев сверху. К — нервация. 8. Shlotheimi.
F — 8 (Trizigia) speciosum. Пермские.
лосках (SphenopliyUostachijs), которые найдены на листоиос-ных побегах (А), и не имеют Мутовок бесплодных листьев, узкие споролистики несут ло одному (Ь), по 2 итп по 3 спорангия, сидячих или стебельчатых. Разрез через узловую часть стебля (С), кроме центрального трехкрыльчатого первичного древесинного щг^индра, утолщенного вторичной древесиной из крупных клеток, представляет еще листовые тяжи (Ьд), отходящие от первичной древесины и в числе 3x2 обособленно проходящие через вторичную древесину (и кору). Едаио листы проявились в верхнем девопе; обычны в каменноугольных и перм--ских отложениях сев. полушария. Природа клянолпстов мало изучена: одни авторы считают их водяными растениями, другие— наземными, с зыощимися стеблями.
Подкласс IV. Хейростробовие, CEeirostrobales.
Сохранился один род с одним видом Cheirostrobzis, известным из кульма Шотландии, по шишке весьма сложного строения, без вегетативных частей. Систематическое положение этой группы еще не выяснено.
Подкласс V. Хвотцеобразные, Equisetineae.
Разделяются на трп семейстра: на каламиты. 1) Asterocalamitaceae, 2) C’alaniitaceae и настоящие хвощи, 3) Equisetaceae. Растеши, достигавшие древесного роста, часто с низким, но сильным ростом стеблей и мутовочным расположением ветвей и побегов. Спорофиллы колоскообразные, несущие одпо-пли разносторонне побеги.
1. Сем. Астерокаламлты, Asterocalamita с еае (4stero-calamites, фиг. 802). Древовидное растение с разветвленным стеблем. В ископаемом состоянии встречаются часто ядра центральной полости с проходящими бороздками и ребрами, реже—куски корней и листья. Последние в мутовкаV длинны (до 10 см) и разветвлены многократно дихотомически, подобно том - как ветвятся листья у первобытных хвощеобразных (Cala-
Asterocalamites scro6; cuiatus Лижвекаменяоугольвые.
Л—ядро части ствола. Моравия. В—ядро части корня.
Тюрин гея. покрытая лкстьяхв ветвь. отдельной
лист. Альтеидврф.
Фиг. 802.-
mophyton и Hyenia). Спороносные органы, еще не вполне хорошо известные, невидимому являются колосками, образованными (подобно тому как у хвоща) исключительно споронесущими листиками Строение стволов отличается существенно от каламитов, ребрами и листьями, не чередующимися в соседних междоузлиях, %о противопоставленными последним.
На основании редко встречающихся рубцов—следов прикрепления ветвей—можно думать, что стволы этих растений отличаются колоннообразным видом. Встречаются в девоне и особенно в нижнем карбоне.
2. Сем. Каламиты, Са-lamitaceae. Каламиты (фиг. 803) отличаются от хвощей своим древовидным ростом и сильно развитым цилиндром древесины. Остатки представителей этрй группы встречаются очень часто в палеозое. Особенно часто встречаются выполнения породой широкого сердцевинного цилиндра, дающие возможность различить ход сосудистых пучков, ибо последние реброобразно выдаются, как у хвоща, в разрушающуюся паренхиму сердцевины и, вследствие своей плотности, оставляют на ядрах сердцевинной полости
отпечатки в Виде продольных борозд (А, В, которые при узлах (п) перекрещиваются с поперечными бороздами. У большинства стволов каламитов (Catamites— фиг.803) сосудистые пучкирасположеиывсмежныхмежд0узлиях,чередуясь(С),
как у хвоща; у Asterbcakimites (фиг. 80?) сосудистые пучки смежных междоузлий не чередуются, а соответствуют друг другу, образуя поэтому на ядрах сердцевинной полости прокодящие через узлы линии (фиг. А, В).
Род Calamodendron (dendron—дерево)—(верхний девон и пермь) харак-
теризуется более широкими и сложнее построенными лучами.
Обугленные части стволов каламитов и выполнения породой централь-
ной полости чрезвычайно частые в каменноугольных отложениях, представляют различного рода ветвление; рубцы от прикрепления ветвей заметны на узловых линиях как более или менее большие чашечкообразные углубления (фиг. 803 A, zn), в направлении к которым сходятся сверху и снизу (Д средина) сосудистые пучки. Стволы развиваются, начинаясь
коническими основаниями на ползущих корневищах с мутовчато - расположенными цилиндрическими корнями (Е).
Строение стволов (D). Широкая центральная полость возникает на месте первоначально имеющейся, а затем разрушающейся паренхимы. Центральная полость бывает окружена толстым цилиндром древесины, разделенной на клиновидньГе участки промежуточными сердцевинными лучами. Внутренние концы этих клиньев вдаются реброобразно в центральную полость и производят бороздки на поверхности ядер выполнений этой полости.
Листья (F, G) постоянно простые, однинервные, линейные или ланцетовидные. Когда они приподняты и не срастаются основаниями, они называются Aster opivy Hites (F), а когда распространяются в плоскости, причем часто, особенно в молодости, срастаются наподобие^ влагалища при основании, то называются Annularia (G).
Вальцевидные спороносные колоски Caiamostachys (фиг. 804) подобны по общей форме колоскам хвоща, но достигают более значительной величины. Неплодущие чешуйчатые листочки колосков собраны в мутовки, чередующиеся с мутовками плодущих-спорангиофоров. Есть виды, образую-
Фиг. S03. Calmmtrs.
А—Ci pauctramts Черхяевамвжнеугольные Нижплл Силезия Часть ствола с ветвями и рубцами от ветвей (Zn). Меаынче, тесно сближекиые рубцы узловой линии представляют места прикрепления листьев. В -С cruciatns. Всрхйскамеииеугольпые Донецкий бассейн Узел с одним большим рубцом от ветви и многими точками схождения бороздок. С — схема расположения сосудистых пучков при узле; i'c — подузловые каналы. D — кусок цилиндра древесины {— Arlhropitys). Пермские Хемниц. Впереди сердцевинная полость с узловыми линиями (п); наверху, в пиперечиом сечеппн, обнаружены клиновидные участки (А) древесинного ци шндра, обособтонные лучами сердцевины. Е — корневище с корнями и конически суженными начальными ча тямн стеблей С. Suckowi Верхнекаменноугольные Луареной бассейн F — покрытая листьями ветвь Acte? ^phyllites equisetiformie Верхвекаменвоуглльвые Коммевтри. G — ветвь с листьями Annularia stellate
* Верхнекаменноуг. Цввкау. Германия.
щпе одного рода споры, и есть виды, споры которых двоякого рода, т. е. макроспоры и микроспоры.
Геологическое распространение каламитов простирается на каменноугольные и нижнепермские отложения (красный лежень сев. полушария; они отсутствуют в так называемой глоссонтериевой флоре). Наибольшее распространение в верхнекаменноугольных.
3. Сем. Настоящие хвощи, Eqnisetaceae. Группа, богатая вымершими родами, которые несомненно относились к различным семействам в конце палеозоя и мезозоя.
В современной флоре к хвощевым принадлежит род Equisetum, к которому относятся травянистые растения, ллшь в исключительных случаях достигающие значительных размеров (Е. giganteum в Средней и Южной Америке со стеблями до 12 м высотой и до 2 см диамегром). Растение имеет далеко распространяющиеся подземные побеги, корневища. Полый, покрытый продольными полосками, стебель разделен на междоузлия (фиг.805), причем его срединная полость прерывается днищами, диафрагмами, расположенными в узлах. Каждое междоузлие переходит наконец
в мутовку листьев, которые срастаются, образуя влагалище, так что у отдельных листочков, вошедших в состав его, остаются свободными лишь вершинки в виде зубцов на краю влагалища
Центральная полость окружена паренхимой, в которой лежит круг сосудистых пучков с водоносными полостями. Сосудистым пучкам соответствуют на наружной поверхности стеблей ребра, продольно идущие, тогда как промежуточные между ребрами бороздки лежат в одних радиусах с широкими воздухоносными бороздковыми полостями коры, которые
Фиг. 804. Спороносные колоски Са-lamostachys typ<ca, каламита. Верхне-каменноугольные. . Радниц. Богемия.
Фиг. 805. ЕдгизеГищ,
А—спорофилл. В—узловая часть стебля Ныне живущ. С—междоузлие. D—егс диафрагма Триас. Штутгарт. Германия. sch—щиток; st—ножка; «тэ—спорангии; s'—влагалище; I — бахромка коры; г — свободные концы листьев.
расположены попеременно с сосудистыми пучками в паренхиме коры (р). При каждом узле сосудистые пучки соединяются здесь в сосудистый пучок прилегающего междоузлия, так что пучки (а вместе с ними ребра и бороздки наружной поверхности) в смежных междоузлиях расположены попеременно.
В настоящее время -хвощи в количестве свыше 20 видов распространены во всех частях света, за исключением Австралии. Ископаемые остатки (Equisetit.es), по всем различным особенностям сходные с ныне живущими, определенно могут быть прослежены отныне до триаса, даже, может быть, до каменноугольных.
Представители триасовых видов достигают огромных размеров (20 см толщины ствола), юрские представляют уже уменьшение размеров, третичные не превосходят по размерам современных.
Ископаемые остаткй хвощей находят часто в виде песчаниковых выполнений центральной полости, на которых часто различимы также листовые влагалища (С), также встречаются обособленные отпечатки диафрагм (D).
Ближе к настоящим хвощам, чем к каламитам, стоит характерный род Schizoneura с листовыми влагалищами, разорванными на две пластинки, напоминающие два супротивных ланцетных листа. С пермотрпаса до нижней юры
КЛАСС V. КЛАДОКСИЛОВЫЕ, CLADOXYLALES,
Вымершая группа папоротникообразных, представленная одним родом Cladoxylon из кульмских отложений Германии, со стволом до 4 см и перисто-рассеченными листьями, покрывающими верхушки побегов.
КЛАСС VI. ПАПОРОТНИКОВЫЕ, FILICALES.
Растения, обычно имеющие развитые листья. Стебель у травянистых видов подземный,.у древовидных — высоко поднимается над землей и бывает покрыт рубцами от опавших листьев и остатками листовых черешков. Ствол некоторых папоротников покрыт плотным покровом воздушных корней. Сосудистые пучки округленного или линейного сечения различно разбросаны в паренхиме. Вторичное нарастание древесины существует лишь иногда, притом бывает лишь слабо выражено. Листья папоротников называются ваяли или перышками у травянистых папоротников большие сравнительно со стеблем; у многих папоротников вап даже чрезвычайно крупных размеров, обыкновенно многократно перисторазделенные (ветвление в несколько порядков). Иногда при основании листового черешка (так называемые рахисы) или его разветвлений находятся особенной формы дополнительные перышки или афлебии, являющиеся аналогами прилистников высших растений. Афлебии служат для прикрытия и для защиты листьев, пока они молоды и находятся еще в спирально свернутом (от вершины листа к основанию) состоянии. У взрослого растения афлебии отпадают. Споровместилища, спорангии — в, виде маленьких округленных скоплений — возникают в большом числе на нижней стороне листьев, разбросанно по всей поверхности листьев или только по краям их, иногда собраны в кучки (Sori) и снабженные выростком кожицы, называемым покрывальцем. Заростки однодомны, т. е. один и тот же заросток производит и женские и мужские половые клетки.
Современные папоротники различаются главным образом на основании особенностей строения оболочки их спорангиев: 1. Eusporangiatae, 2. Proto-sporangiatae и 3. Leptosporangiatae, причем у первой группы спорангии имеют от двух и несколько слоев клеток в оболочке, вторая группа промежуточная между первой и второй, и наконец у третьей спорангии состоят из однослойной оболочки. Эта группа во многих отношениях архаичного характера; стенка спорангия (фиг. 806, а) толста, состоит из нескольких слоев клеток и не имеет так называемого кольца (annulus), имеющегося у остальныхбольшинства современных папоротников. У этих последних спорангии имеют тонкую, однослойную стенку и снабжены кольцом, развитым в различной степени: полным периферическим (фиг. 806, Ъ — косое) или неполным периферическим (фиг. 806,с), замкнутым (фиг. 806,d), или недоразвитым, в виде группы утолщенных клеток на боковой стороне под вершиной (фиг. 806, е). Типичное кольцо состоит из одного ряда несколько выдающихся клеток (фиг. 806, с) с утолщенными радиальными и внутренними стенками. По созревании спорангия, высыхая, внешний ряд стенок сокращается, и кольцо распрямляется, обусловливая тем вернее разрыв стенки спорангия и высевание созревших спор.
Стремясь объединить все, как современные, так и ископаемые формы, в одной системе филогенетического происхождения, палеоботаника по возможности размещает ископаемые папоротники, спороношение которых доста-422
точно известно, по естественным группам, соответствующим ныне живущим семействам и родам, или по крайней мере устанавливает возможное к ним приближение. _
Главнейшие папоротники из типа Eusporangiatae. Особняком стоят древние типы папоротников, возникших еще в девоне и получивших развитие в каменноугольных и пермских отложениях. Они известны большей частью по остаткам стволов и стеблей, представляющих тип первобытного устройства стелы и листовых следов. Роды Stauropteris п Mctaclepsydropsis имеют листовой след в форме двойного якоря; у Clepsydropsis и Zygopteris— в форме песочных часов; у Botryopteris листовой след имеет форму близкую к омеге — о> (греч. буква), у Asterochlaena (пермь) он имеет звездчатую разветвленную форму. Листевые побеги и органы размножения у многих т. н. «первобытных папоротников» совсем’ неизвестны до сих пор.
Фиг. 806. Типы спорангиев.
а—Angiopteru, Ъ—Cyahed (г- кольцо, s—споры); с—Polyslichum', d—Schizaea', е—Os-mwnda.
Clepsydropsis найден в кульмскпх отложениях Киргизских степей (С. Kirgisica).
Затем следует отметить другие главные роды из сем. Marattiaceae, к числу которых относится Pearonizis (фиг. 807) — окремнелые стволы последнего, часто встречающиеся в верхнекаменноугольных п особенно в ниж-непермскпх отложениях, представляют в поперечном разрезе осевой части ствола сосудистые пучки пластиночного вида, в окружающей зоне коры проходят многочисленные корешки, выходившие наружу. Отпечатки, Соответствующие этим окремнелым стволам, описываются под названием Саи-loptens\ окаменелые стволы древовидных папоротников' из рода Protoptei is найдены в верхнеюрскпх отложениях Волги и мелевых отложениях Приуралья.
Из мараттпевых типичны Marattiopsis со спорангиями, сросшимися в синангий с оберткой, открывающейся в виде лодочки. Ваи церистые с линейными долями типа Taepiopteris. С рэта через всю юру.
Следующие роды относятся к типу Protoleptosporangiatae:
Tiianuioptens (фпг. 810; tliamnus — куст). Толстые цилиндрические протостельпые стволы, покрытые снаружи спирально расположенными основаниями листьев; посредине черешка листьев в поперечном разрезе обнаруживается сосудистый пучок подкововидного сечения. Основания вай образуют большую часть поперечного сечения ствола, собственно ствол — значительно меньшую. Весьма близки сюда роды Zalessky a, Chasmatopteris, Petcheropteris. Эти вымершие роды относятся к доныне живущей группе Osmundaceae. Пермские отложения Приуралья.
Osmundites — род, близкий по анатомическому строению к современному (Jsmunda от юры до третичных в обоих полушариях.
Todites — род по строению вай близкий к Todea, от кейпера до рэта.
Cladophlebis—характеризуется косопоставленными к стержню языковидными сегментами вай и несколько раз дихотомпрую^ими вторичными нервами Характеризуют флору мезозойского типа, начиная от кейпера до верхней юры.
Следующие роды относятся к типу Leptosporangiatae-.
Klukia (фиг. 811). Ветки с трижды рассеченными ваями, из кото
eV aw© ©©
Фиг. 807. Psaromus Gutbieri. Нижнепермские. Хемниц, аги — сечение воздушных корешков.
Фиг. 808. PecoptensPlufikenetti. Часть плодоносной ветви с семенами, прикрепленными близ концов перышек. 2/1
Фиг. 809. Sphevoptens, А—Sph. affinvs. В—перышко Sph. optu-siloba. Камсввоугольв. Англия. С—то же, целое перо. Каменноугольв. Авглия.
рых каждая несет на концах группы спорангиев, вскрывающихся в виде колпачка. От лейаса до рэта.
Gleichenites. Ваи вильчатые или вееровидные. Спорангии расположены группами от двух до четырех, с полным горизонтальным или круглым концом. От кейпера до верхнего мела.
Lacvopteris Им.еет перистые ваи со спорангиями, расположенными в виде черепитчатых розеток по бокам главного нерва. От рэта до вельда.
Hymenophyllites — с»нежными ваями из одного слоя клеток, со спорангиями, имеющими экваториальное кольцо, расположенными па окончаниях жилок, выходящих из основания пластинки листа. В каменноугольных и юрских отложениях (Краков, С. Донец).
Sfachypteris. Ваи трижды рассеченные, несущие продолговатые спорангии па разветвлениях вюрого порядка Юра Англии и средн. Азии.
Gonwpteris—с сегментами вай сфеношероидной или пекоптероидной формы, спорангии по краю редуцированной пластинки, часто до стержня несущего лишь булавообразные дву-створч тые спорангии. Близок к современным Dicksonia. Типичен для нижней и средней юры.
Замечательны представители обширной, почти вымершей ныне, группы Dipteridaceae; особенно из сем. Camptoptendaceae, именно' роды
Фиг. 810. Thamnoptcns Schlechtendaht. Перлские.
а — разрезы листовых черешков, Ъ — разрез ствола с—стелла.
Clathropteris (фпг. 812), Dictyo-phyllum и Cmnptopteris, с сетчатым жилкованием п дланевидной основной формой пластинки ван Спорангии голые, на ипягней сти
Фиг. 811. К1икга exilis Пижв. юра. Галиция. 10/1.
4—часть ваи, В— то же самое, более увелич. и отдельно спорангий
роне листа, кольцо неполное. Из Dipteridaceae весьма интересен род Haus-mannia; от перми до нижнего мела.
Сюда же относятся также Salvinia (фиг. 813).—водяные папоротники, имеющие горизонтально плавающий стебель с двумя парами зеленых бес плодных и одним подводным, плодущим листом рассеченным на нитевидные дольки. Разноспоровые растения от верхнего мела до настоящего времени.
На основании остатков листьев устанавливаются некоторые роды ископаемых папоротников, например: . 1
Archaeopteris (archaios— древний, pteris—крыло); (фиг. 814)с большими, двукратно перистыми листьями. Перья первого порядка почти противолежащие, косо восходящие, усажены тесно примыкающими перышками, каковые нахотятся также даже на главном стержне (фиг. 814, А). Перышки удлиненно-яйцевидной формы, в основании сильно суженные, несколько нисходящие по стеблю, на краю иногда зазубренные, имеют многочисленные нервы, которые параллельно входят в перышко, лучисто расходятся и дихотомически ветвятся. Плодоносные перышки являются весьма редуцированными в отношении пластинки, которая несет многочисленные веретенообразные спорангии, сидячие или стебельчатые, одиночные или группами по два-три, не имеющие кольца (annulus).
В девсле, особенно в верхнем, имеет широкое географическое распространение (Канада, Медвежий остров, Бельгия, Германия, у вас), редко в нижнем девоне.
Pecopteris (фиг 808). Перышки, прикрепленные широким основанием и почти под прямым углом, большей частью цельнокрайние и параллельнобокие, с протягивающимся главным нервом, от которого также почти под
Фиг. 812. Clathroptens, из средвего кейпера. ЛунЦ Альпы Австрия. 1/3. Вправо, часть листа 3/1
Фиг. 813. Saltinia Reussi. Два листика. Миоцен. Видии. Чехо-Словакия.
Фиг. 814. Archaeopte/ris hibwnica, со спорофиллами. Верхний девон. Ирландия.
прямым углом отходят побочные, иногда вильчатораздвоенные нервы. Главным образом в каменноугольных и пермских отложс ниях
Sphenopteris (фиг. 809 A, В, C\ sphen — клин) имеет двух, трех или четырехкратноперистые перышки; небольшие перышки, суженные в основании или даже клиновидные, с краями более или менее глубоко- и часто перисто-лопастными или рассеченными. Дихотомически ветвящиеся нервы расходятся радиально от основания перышек или отходят под тупым углом от среднего нерва. Анормальные перышки (прилистники—афлебии) не-
Фиг. 816. Alethoptaris Sent. Камевно-угольн. Саарский бассейн. Германия.
Фиг. 815. Cardioptens polymorpha, , Кульм.
Фиг. 817. а — Neuropteris hetcrophylla. Каме.:ио)год!.й. Саарский бассейн. Германия Ъ — N. ScM.ch.ani. Каменноугольн. Верхняя Силезия.
редки. Не резко отграничены от типа листьев Archaecp+eris. Формы со сфе-ноптерондной нервацией встречаются от девона до перми; главное распространение — в верхнекаменноугоЗшных отложениях.
Ископаемые папоротники — именно палеозойские, отчасти нижнемезозойские— во многих отношениях отличаются от позднейших и современных. Важнейшими отличиями представляются следующие:
Они были разнообразнее и среди них более преобладали древовидные формы, нежели теперь; листья, нередко мощно развитые, часто располагались на стволах продольными рядами. Часто встречается вильчатое ветвление вай, также и разнообразнейшие переходы к перистому расположению. Листья без срединного нерва, как у Archaeopteris, в среднем пелеозое очень частые, позже редки. Сетчатонервные листья до перми весьма редки, тогда как в настоящее время они чрезвычайно распространены. Спорангии палеозойских папоротников архаичны, большей частью типа редких в настоящее время Ма-гаШасеае.
Уже давно обращало внимание то обстоятельство, что для многих ро-
дов ископаемых папоротнцков, как Alethopteris (фиг. 816, alethos — настоящий), Ncuropteris (фиг. 817) и некоторых Pecoptens, никогда не было обнаружено существование обычных для папоротников кучек спорангиев
Плодоношения стали известны лишь в последнее десятилетие, причем оказалось, что они являются в виде так называемых микро- и макроспорангиев; последние представляют черты сходства с семенем низших семенных, именно саговиковых. Таким образом представилась возможность выделить их в особую группу Pteridospermales.
Зародышевые сифоногамные, Embryophyta siphonogama.
Этот отдел растений, называвшихся ранее «явнобрачными», «семенными» и пр., характеризуется тем, что для достижения яйцеклетки микроспора вырастает в трубку (сифон), заключающую генеративные ядра, откуда и название сифоногамных.
Возникновение разноспоровости составило важный шаг вперед в смысле более рапнего разделения полов. Разделение полов, происходящее у разноспоровых лишь на половом поколении, заростке, начало определяться уже на бесполовом поколении, которое и стало давать мужские спорангии (или микроспорангии) и женские спорангии (или лакроснорангии). Вместе с тем развитие половых индивидуумов сводится все более собственно к развитию одних лишь половых органов, тело же полового индивидуума, или заросток, несущий эти органы, все более редуцируется. У плаунового Selaginella заростки по своему объему выходят из пределов споровой оболочки. У низших явнобрачных редукция полового поколения идет еще дальше. Макроспора, называемая здесь зародышевым мешком, вполне погружена в макроспорангий или семяпочку, у покрытосеменных обычно заключенную еще в двойной покров завязи. Микроспоры не производят сперматозоидов, так как здесь оплодотворение лишь в исключительных случаях происходит при посредстве воды; как правило, оно производится при помощи ветра или насекомых. Микроспоры являются в виде цветковой пыльцы; стенка микроспоры (пыльцы) выпячивается или вырастает в впде трубки (сифона), по которому генеративные ядра и достигают яйцеклетки. Мужской заросток крайне редуцирован в пыльцевом зерне, где развиваются на его счет генеративные ядра пыльцевой трубки. В зародыше различимы уже части, соответствующие корешку и стеблю будущего растения, но он останавливается в своем развитии на известной стадии развития и заключен в остальной, преобладающей по размерам части — семени, представляющей заросток с сильно развитым запасом питательных веществ. В семени зародыш находится в покоящемся состоянии, в состоянии приостановки роста. Это состояние скрытой в семени жизни может продолжаться чрезвычайно долго и прекращается при условии обильного доступа влаги; тогда зародыш развивается в зрелое растение.
Сифоногамные растения распадаются на два типа, весьма различные по своему строению: 1) Gymnospermae пли голосеменные растения и 2) Angio-spermae или покрытосеменные растения. У последних семяпочка заключена в замкнутой полости — завязи, образующейся путем смыкания вокруг нее плодолистиков. Таким образом у покрытосеменных семяпочка защищена от вредных внешних воздействий покровом, отсутствующим у голосеменных. Типичные цветки, ярко окрашенные в периферической части — околоцветнике, цветки с выделением соков, приманивающих насекомых, существуют лишь у покрытосеменных.
Голосеменные появляются в каменноугольном .периоде, покрытосеменные достоверно начинаются с мела и с тех пор преобладают над голосеменными.
IV ТИП. ГОЛОСЕМЕННЫЕ, GYMNOSPERMAE.
К голосеменным принадлежат классы папоротникосеменных (Ptendo-spei males), кордаитов (Cordaitales), беннетитов (Bennetitales), #саговпков (Су-cadales), гинкговых (Ginkgoalet), хвойных (Coaiferalcs) а хвийлпковых (Gnctales).
КЛАСС I. ПАПОРОТНИКОСЕМЕННЫЕ, PTERIDOSPERMALES.
Pteridospermales представляют своеобразный характер, имея листву папоротникового типа, органы размножения, уклоняющиеся в сторону сифо-
ногамных растений; кроме того, они представляют вторичное нарастание древесины (фиг. 818), почему и должны быть относимы к голосеменным растениям.
Более детально изучен род Lyginopteris-, известно, что семена этого растения, описанные под именем Lagenostonu (фиг. 819), принадлежат растениям, стволы которых, имеющие вторичную древесину, известную под именем Lyginodend-tor., черешки (ваи) — под именем Rhachi oplens, а листва Splienopteris Hoeninghausi. Возможно, что отпечатки, известные как Tdanghim, являются микроспорофиллами того же растения.
С листьями типа Neuropteris и стволами Ме-dullosa, имеющими вторичное утолщение черешками Mgeloxylon, объединяются семена Trigono-carpus, достигавшие величины голубиного яйца.
К этой группе «ееменных папоротников» несомненно относится целый ряд растений, листва которых была раньше описываема из каменноугольных отложений, как папоротниковая, под названием Manopteris, Alethopteris (фиг. 816), Neuropteris (фпг. 817) и некоторые из Spheno-pteris (фиг. 809), а также Pecopteris (фиг. 808), но сейчас еще не представляется возможности точно провести границу между настоящими папоротниками и голосеменными, имевшими внешность папоротников.
Фиг. 818. I yginoplcris Old-hamia Верхнекаменвоугольв. Англия.
ат, ir — наружная в внутрснпяя часть коры: — Ьа — сосудистые
пучки, проходящие в листья; pd — слой пробковой ткани, к центру от которого находятся древесина и сердцевина
КЛАСС II. КОРДАИТЫ, CORDAITALES.
Эта группа, распространенная от девона до перми, представляет всего лишь один род (с подродами):
Cordaites, по имени палеонтолога Корда (фиг. 820). Кордаиты были высокие (30—40 Л1) сравнительно тонкоствольные деревья, со стволами, достигающими значительной высоты без разветвления и несущими на вершине густую крону. Последняя состояла из ветвей различных порядков, на которых находились в изобилии простые листья большой величины. Листья
варьировали по форме у различных подродов; они были различной формы от лопатчатой с тупыми концами или ланцетовидной, приостренные на концах (фиг. 820, В, С), до линейной, весьма узкой и длинной, подобной листьям злаков. Листья всегда параллельнонервные с коллатеральными сосудистыми пучка ни, почему (и по наружной Форме) сходны с отдельными перышками листьев саговиковых.
Стволы представляют сердцевину большого поперечника, которая при дальнейшем росте расщепляется поперек, образуя сближенные горизонтальные пластины (дна Прагмы,фиг. 820, F). Цилиндр древесины проникнут многочисленными сердцевинными лучами, представляет первичную и вторичную древесину; последняя сходна с древесиной хвойных. Ствол покрыт толстой корой
Цветы кордаитов сережковидны, однополы. Мужские и женские цветки не всегда различаются по наружному виду; в некоторых формах все-таки (фиг. 820, (?) ось женского соцветия несет одиночные семяпочки, сопровождаемые лишь немногими кроющими листками (прицветниками, фиг. 820, Г).
Мужские цветки (Я, К) представляют солидную ось (а), с многочисленными чешуевидны hi листочками и цилиндрическими пыльцевыми мешочками, сидящими на длинных ножках, так что цветы представляют тычинки (рЪ). Внутри пыльцевых зерен (Я) перед оплодотворением образуется группа клеток (?),• которая может считаться за заросток. На окремнелых экземплярах можно наблюдать пыльцевые зерна (Я) в удлиненной вершине пыльцевой камеры (sp) и в ней самой (pty.
Семена кордаитов часто встречаются обособленными в верхнекаменноугольных и пермских отложениях. Они овальны или сердцевидны (О, Р), имеют толстую, снаружи мя-
систую (/), внутри деревянистую (Л) оболочку, подобно семенам саговиковых и папоротникообразных.
Из палеозойских родов кордаитов необходимо отметить Neoggeiatiopsis из нижней Гондваны и тунгусского яруса Сибири, имевшего важное значение в образовании угольных толщ Кузнецкого бассейна. Кордаиты вообще играли значительную роль в образовании каменного угля.
Этот класс, имевший пышное развитие в палеозое, угас без прямых потомков в мезозое.
Фиг. 819. Реставрация семени Lyginodendron (Lage nostoma Lo-tnaxt). Семя представлено окруженным открытым чашечковидным покровом, снабженным головчатыми железками.
КЛАСС III. БЕННЕТИТОВЕ1Е, BENNETITALES.
Растения весьма сходные с саговпками и лишь весьма недавно получили строго установленное место среди зародышевых сифонояишых растений. Будучи сходны с саговиками по строению ствола и листве, беннетито-вые сильно отличаются своими органами размножения, напоминающими устройство цветка покрытосеменных; их женские цветки в виде ортотрой-ных семяпочек в центральной части, мужские цветки представляют спирально расположенные микроспорофпллы папоротникового вида, найоминаю-щие собрание тычинок покрытосеменных.
В отличие от ныне живущих саговиковых стволм бенпетитов (фиг. 823) низкие, приземистые, разветвленные, имеют цветы не верхушечные, а расположенные на боках ствола, благодаря чему при значительном числе их производилось и значительное количество сей ян В останьиом стволы почти что обычного строения стволов саговиковых.
Фиг. 820. Cordaites.
*
А — общий впд растения (ствол разрезан для укорочения), В, С — листья I) — поперечное сечение листа, показывающее коллатеральный характер сосудистого пучка; Ъ — скелетные элементы коры; р — соединитесь пая ткань сосу цист ого пучка; з», tn,' — механическая ткань; в — эпидермис. Е — кусок ствола с листовыми рубцами (Ъп); h — древесина; т — цилиндр древесины (Artisia). F — кусок ствола в продольном и поЧРречном разрезе: т — цплевдр сердцевины; d — его диафрагма; А — цилиндр д швеелны, г — кора.
— ветвь с мужскими (справа — В) и женскими (слева — I) цветами; Ът — прицветники; Ъп — листовые рубцы. К — продольный разрез через мужской цветок: а — ось; Ъг — прицветник; Ъз — ножка; pb — пыльцевой мешочек (L). А1 — зерно пыльцы (= JV, рк): а — оболочка; i — внутренние группы клеток. N— продольный разрез через семяпочку; а — иар^жцая оболочка; п — ядрышко; рк — „пыльцевая камера" с 2 зернышками пыльцы; зр — ее вершина. О — семя {СогЛагзрегтичп Gutbieri). В — семя в продольном разрезе (С. angustodunensey f — наружная, мясистая, Л — внутренняя, деревянистая оболочка; рк — выль целая камера; е — внутренняя часть семени.
Цветы бенпетитов являются в виде кругловатых почкообразных тел, не выдающихся за пределы покрова ствола из листовых черешков (фиг. 824). Цветы могут считаться сидящими на редуцированных боковых ветвях ствола и, таким образом, положение их не представляет ничего исключительно оригинального. Цветы их более сложны, чем у ныне живущих саговиков, представляют совмещение пыльцевых мешочков с семяпочками в одном цветке, так что цветок является обоеполым
У рода Cycadoidea (фиг 821 сти конического осевого выступа, 1
Фиг. 821. Cycadoidea marylandica. Ствол с почти тридцатью незрелыми цветками а, рассеянными между старыми основаниям!: листьев. Ниллеагдов. Сев.
Америка. 1/8.
824) семяпочки помещаются на поверхпо-орый является в середине цветка как бы окончанием его оси. Микроспорофиллы, несущие пыльцевые мешочки, являются у данного рода построенными сложно, сложнее, чем у высших растений Именно они перисто-разветвлены, так что каждый микроспорофилл напоминает листовое образование папоротника. Эти органы размножения на периферии цветка объемлются покровом околоцветника из высоких волосистых листочков. Сходство в устройстве цветка у беннетитов и у покрытосеменных позволяет предполагать, что первые дали начало последним (фиг. 822)
Близкими родами являются Веп-netites и Wilhamsonia, и замечательны по строению разветвленных стволов, несущих на разветвлениях цветы, были роды Wielandiella и TFz7-liamsoniella.
Беннетпты были широко распространены от верхнего триаса до начала мела, преимущественно в сев. полушарии.
КЛАСС IV. САГОЕЛКОВЫЕ, CYCADALES.
В настоящее время саговикивые ограничены в своем распространении тропическими и субтропическими областями. По своему большей частью не-разветвленному стволу и по большим перистым листьям (фиг. 833) саговиковые по общему виду сходны с древовидными папоротниками и пальмами. Перистые листья образуют венец на вершине ствола: основания отпавших листьев большей частью сохраняются и вместе с чешуевидными низовыми листьями одевают ствол плотным чешуйчатым покровом. Перистые листья кожисты, однонервны или многонервны (редко с пе-рпетым жилкованием). Саговиковые — растения двудомные. Цветы образуются на вершине ствола, имеют форму шишек. Чешуйки таких шишек <микро п мешочки или завязи (фиг. 825). Опло-)м сперматозоидов, развивающихся из
Фиг. 822. Неразвериувшийся цветок Cycadoidea dacotensis, в продольном сечении.
с — осевой выступ с семяпочками Ъ; с — еще закрученные микроспорофиллы; d — ио-кровные листики цветка., Пижвемелов. Сев. Америка. 112.
макроспорофиллы) несут пыльцевые дотворение совершается посредств
микроспор. Эту особенность представляют среди сифоногамных еще лишь гинкговые.
Ствол представляет сердцевину большого диаметра с .полостями в ней. Древесинный цилиндр представляет вторичное нарастание (фиг. 826).
В ископаемом состоянии от саговиковых чаще всего находят листья, отчасти сходные с листьями ныне живущих представителей, отчасти уклоняющейся от последней формы. Цветы и плоды сравнительно редки, напротив, стволы встречаются часто, особенно в юре и мелу.
Многие остатки, могушие принадлежать к Bennetitales и Cycadales и также некоторым близким к ним классам, пока еще мало изучены.
Фиг. 823. Совершенно раскрывшийся цветок Cycadoide.a ingens. 1/t.
Некоторые из таких отпечатков листьев имеют нередко важное стратиграфическое значение, но остаются не связанными с определенными родами и пока описываются самостоятельно, как особые роды.
К числу их относятся:
Pterophyllum (фиг. 827). Мелкие или крупные вайи с удлиненными узкими лопастями, сидящими по сторонам главного нерва. От среднего карбона до вельда.
Taeniopteris (фиг. 828, А, В, tainia -- лента). Листья большие, лентовидные, параллельнокрайние, с ясно выраженным срединным нервом. Побочные нервы перпендикулярны к срединному, простые или дихотомически ветвящиеся. От каменноугольных отложений до мела включительно.
Glossopteris (фиг. 829, А, В, С, glosse— язык) имеет простые языковидные или лопатчатые листья, суживающиеся к основанию (Л), непосредственно тесными скоплениями прикрепляющиеся в стволу. Нервация сетчатая. Подобные нервы, под острым углом отходящие от срединного, слива
28
Учебник палеонтологии.
ются в сетку с удлиненными петлями (В). Плодоношения недостаточно еще изучены.
Корневища, принадлежащие этому роду, были описываемы под названием Vertebraria (фиг. 829, С), продолыюскладчаш и снабжены попереч-
Фпг. 824. Половина поперечного сечения ствола Су-cadoidea Saxbyana.
d — сердцевикная полость; я? — древесина; ph — флоэма, вместе образующие кольцо коллатеральных сосудистых пучков; It — сосудистые пучки, проходящие наружу через кору и повторно разделяющиеся, распределяясь по листовым черешкам; 1Ь — основания листьев, образующие пок* ров вокруг стола. Нижнемелов.
Англия
Фиг. 826. Четверть поперечного сечения ствола Cycas revolutcr. sp — сердцевинно-паренхимная часть.
ha,1, йв3 и т. д. суть последовательно один за других? образованные слои древесины; рй*, р№, ph3 н т. д. также последовательно образовавшиеся слои флоэмы; между ph1 п Ля’ особенно широкая зона основной паренхимы; х — последний возникший, еще деятельный камбий; г — паренхима коры с радиально и тангенциально проходящими пучками; р — периферическая часть коры (черные места ее лежат ниже плоскости сечения).
Фиг. 827. Pterophyllum Brauni. Триас. Верхняя Силезия. 1/1.
А
Фиг. 825. Макро- (А) и микро-спорофилл (В) Су-саз (ныне живущ.). В нижней части первого находится 5 завязей, одна сильно возросшая в размерах (справа). Микр^спорофилл представлен снизу, покрыт пыльцевыми мешочками.
ными ребрами, на которых прикрепляются нижние листья. Весьма распространенная в перми и триасе южного полушария «глоссоптерисовая флора», найден? также на Сев.' Двине. Пермокарбон до рэта.
Nilssonia. Листва, напоминающая Taeniopteris, если пластинка не является разделенной на сегменты. От верхов триаса до рэта (фиг. 830).
Zamites Вайи крупные, несущие сидящие на мозолистых утолщениях кинжаловидные сегменты. Юра (фиг. 831).
Podozamites (фиг. 832). На нижних стержнях сидят двуряднорасположенные ланцетовидной формы вайи с параллельными частыми жилками Обычны в мезозойских отложениях.
Фпг. 829. А, В, С — G-lossopteris. А — Gl. Browniaina. Пермь. Иоганвесбург. Трансвааль. В — Gl indico. Пермь. Ост-Индия. С — vertebraria. Пермь. Индия.
КЛАСС V. ГИНКГОВЫЕ, GINKGOALES.
Эта группа представлена в современной флоре лишь видом Ginkgo-biloba деревом, сохранившимся в восточной Азии, замечательным по при сутствию у него в процессе оплодотворения сперматозоидов с произвольным движением. Будучи несколько несходны по устройству плодоношения с тисовыми, гинкговые характеризуются оригинальной вееровидной пластинкой листа цельной или рассеченной на узкие доли. Ginkgo появляются уже с девона, будучи широко распространены в перми и мезозое.
Род Ginkgo-—листья вееровидные, трехугольноромбические (фиг. 834) от почти цельных до сильнорассеченных. Нервация вильчатая. Мужские цветки сережкообразны, тычинки с 2 или несколькими пыльниками, женские
обычно несут две семяпочки. Процветали в юре (G. digitata, G. sibvrica) существовали в мелу и третичном периоде до плиоцена.
Фиг. 830. Nilssoma orientalis. Юра. Ка-ленка. Донбасс. 1/2.
Фиг. 831. ZamitesFe-neonis. Верхняя юра. Франция.
Род Багега, известный с красного лежня до вельда, отличался большой узкостью долей листа и большим числом пыльников и семяпочек в цветках. Более сомнительны:
Фиг. R32. Podozami-tes distaus. Рэт-Лейас.
Франция. 2/3.
Фиг. 833. Cycas circinalis. Переднее дерево с плодовыми листьями в кроне п молодыми Wedtln.
мокарбона. Б настоящее время Psygmophyllum
Czekcmowskia с узколинейными или почти ните • видными долями (от юры до мела), Ginkgodium со слабо намеченным расщеплением удлиненной пластинки (юра); Phoenicopsis с выходящими пучками и сидящими на укороченных побегах узкими листьями.
Другой цикл родов, отличающихся крупностью сильно рассеченных пластинок листа, характерен для палеозоя: Psygmo-pliyllum (фиг. 835) с дла-невидными листьями, характерен от девона до пер-определенно относят к па-норобпикам. Rhipidopsis — виды с дланевидно-рассеченными обратно-клиновидными листьями, известен из нижне-гондванских слоев Индии, Урала
и Аргентины.
КЛАСС V. ХВОЙНЫЕ, CONIFERALES.
Хвойные отличаются прежде всего своими листьями, маленькими, неразделенными, игольчатыми (хгэи) или чешуевидными, однонервными многолетними. По листьям хвойные внешне сходны с плаунами, по ветвлению ствола — обильному, большей частью с мутовчатым расположением ветвей— с хвощами.
Цветки однополые, голые, без околоцветника, растения однодомные1 или реже двудомные. Мужские цветки сережчатые, микроспорофиллы более
или менее щитовидной формы. Женские цветки обыкнотенно образуют шишки, состоящие из деревенеющих со временем макроспорофиллов, тесно сближенных и спирально расположенных на оси. На верхней стороне спорофиллов находятся макроспорангии (семяпочки). В анатомическом отношении хвойные характеризуются развитием в древесине трахеид, отличающихся от собственно сосудов тем, что поперечные стенки в примыкающих по длине концах клеток не уничтожаются; трахеиды снабжены окаймленными порами.
Хвойные известны в ископаемом состоянии с каменноугольных отложений и, вместе с саговиками, они преобладают в растительности мезозоя до появления в мелу покрытосеменных. В то время как многие третичные,
1 Т. е. с мужскими и геенскими цветками иа одном и том же индивидууме растения.
437
даже меловые, формы могут быть отождествлены в отношении признаков родового и даже видового характера с ныне живущими, большинство древнейших, известных часто только по бесплодным ветвям и по стволам, сохранившим структуру, могут быть лишь приблизительно размещены в системе. Древнейшие хвойные часто не- имеют годичных колец древесины.
Род Walchia (фиг. 836—837; по имени ученого Бальха), встречающийся в верхнекаменноугольных отложений и особенно отличительный для пермских, представляется близким к ныне живущей Araucaria, известной в ископаемом состоянии с юры. Walchia — деревья с мутовчатых расположением ветвей. Последние разветвляются двурядно, перисто. Иглы короткие. Окаменелые стволы Walchia, известные под названием араукаритов или Агаи-carioxylon, встречаются например в изобилии в верхнекаменноугольных отложениях Донецкого бассейна.
Фиг. 836. Walchia piniformis. Нижнепермские. 1/1.
Фиг. 837. А — шишка с семенами Walchia рг-niformis. В — W. fer-tilis. Мужской цветок. Нижнепермские. Отэи. Франция.
Б пермокарбоне и пермских отложениях встречается род Ulmannia, отличающийся маленькими толстыми листьями языковидной формы (фиг. 838). В юре и меле был обычен род Brachyphyllum.
Представителем группы, особенно частой в нижнем мезозое, отличающейся глубокими зубцами на краю чешуек шишек, является род Voltzia (фиг. 839; по имени ученого Вольца). Большие деревья, с мутовчаторасположенными ветвям к последние разветвляются попеременно. Листья концов ветвей длинны и плоски, листья проксимальных частей ветвей коротки и киловаты (4). Пермь, особенно триас. На основании характера шишек считаются предками ныне живущих тиссовых.
Болотный кипарис Taxodium distichum, растущий теперь в области побережья Мексиканского залива, в миоцене и плиоцене принадлежал к распространениейшим хвойным сев. полушария, включая полярные страны. То же положение, начиная с юры, занимал род Sequoia, ныне сохранившийся только на западе в Сев. Америке.
Лиственницы, ели, сосны принадлежат к семействам вероятно существовавшим уже в триасе и во всяком случае в юре. Эта группа изобилует
в мелу и в третичных; ее представители во время олигоцена образовали залежи янтаря в Б. Пруссии и Прибалтийском крае.
Большая часть обычных хвойных лесных пород существует с третичного времени и мало изменилась со средне- и верхнетретичного периода, изменив лишь ареалы своего распространения. Находимые в ледниковых отложениях остатки лиственницы {Larix-europed), сосны {Pinus sylvestris) и ели {Picea excelsa) почти ничем не отличаютея от форм современных.
Фиг. 838. Ulmannia Bronni. а — медистый сланец Франкенберга. 6 — увеличена. Средний красный лежень. Фридрихсрода. Тюрингия Германия
Фиг. 839. Voitzia. А — Т> — V. heterophylla. Триас. Зульцбад. Эльзас.
Л — ветвь с длинными и короткими листьями. В — ветвь с раскрывшейся шишкой. С — мужской цветок. D — ветвь с рубцами от листьев Е — V. Ltebei. Верхненермские. Требниц Тюрингия. Гермавин. Чешуя шишки с двумя зачатками семян (s).
КЛАСС VII ХВОКНИКОВЫЕ, GNETACEAE.
Вымирающая древняя группа, сохранившая доныне три резко различных рода: Хвойник {Ephedra), Вельвичия {Tumboa) и Гнетум {Gnetum). В ископаемом состоянии с постоверностыо неизвестна.
V ТИП. ПОКРЫТОСЕМЕННЫЕ, Angiospermae.
Плодолистики образуют завязь, в которой скрыты семяпочки; завязь превращается в плод, с заключенными внутри него семенами. Цветки однополы; деревья, кустарники и травы (последние редко сохраняются в ископаемом состоянии или в виде семян и плодов).
Сюда принадлежит большинство ныне живущих цветковых растений. Покрытосеменные появляются в мелу (находки, не представляющиеся достоверными, известны в юре), где были уже обособлены обе подгруппы их — однодольные (Monocotyleaonae) и двудольные (Dicotyledonae).
Ископаемые покрытосеменные представляют мало существенных отличий от современных форм; несовершенство сохранения остатков часто мешает отнести ряд родов с уверенностью к определенным семействам, несмотря на постоянство их присутствия в тех или иных отложениях (меловые Credneria, которые ныне сближают с платанами) (фиг. 840), эопеновые Dewalquea и др.
Древнейшие покрытосеменные известных из отложений, одновременных гольту, из потомакских слоев Америки, валанжинского яруса Португалии, из Англии, с Мадагаскара, Уссурийского края и др.
Фиг. 840. Credneria triacuminata. Сенов. Гарц. Германия. 2/3 натур, велич.
КЛАСС!. ОДНОДОЛЬНЫЕ, MOXOCOTYLEDONAE.
Кроме особенностей семени, характеризуются еще простой нервацией листьев (параллельнонервны) и древесиной с закрытыми сосудисто-волокнистыми пучками, лишенной поэтому способности втори-июго нарастания в толщину; пучки лежат разбросанно в основной паренхиме; камбия нет, стволы расту т в толщину лишь в молодости.
В ископаемом состоянии довольно часто встречаются пальмы, появляющиеся с верхнемеловых и распространенные в третичных отложениях. С эоцена известны как перистолистные, так и вееролистные пальмы. Таковы в олигоцене Phoe-nicites (фиг. 841), родственный ныне живущей финиковой пальме Chamaerops (фиг. 842) (в сармате и плиоцене 3. Закавказья).
Широко распространеп был в эоцене и олигоцене род Sabal, известный и у нас из Могильно
на Волыни и из окрестностей Аджамки. Присутствие пальм является особенно ценным критерием для суждения о бывшем климате стран, где они были находимы.
Злаки также появляются с мелового периода, род Bambusium известен в эоцене Украины; там же указаны остатки бананов (Musa).
КЛАСС II. ДВУДОЛЬНЫЕ, DHOTY LEDO ХАЕ.
Представляют кольцевое расположение сосудистых пучков и камбия. Нервация листьев сложная, сетчатая. 5же в сеномане получают решительное преобладание над голосеменными и споровыми.
Начиная с мела, известно много сережкоцветных, причем из третичных отложений часто в формах, близких к современным, как то: тополи (с гольта Гренландии), ивы, березы, ольхи (Salix, Betul'i, Alnus), каштаны (Castanea), буки (Fagtus) и дубы (Quercus).
Фиг. 841. Перистолистная пальма (Fhoenicites borealis). 1/4. Из олигоцена Вейссенфельского
буроугольного бассейна Германии.
Фиг. 842. Вееролистная пальма (Chamaer ops helvetica). Олигоцен. Германия.
0з сеномана известен Dryophyl-lum, родственный дубам и каштанам.
Вообще двудольные оережкоцвет-ные, как тополя (в гольте Гренландии), ивы, березы, дубы, являются древними.
Широко распространены были виды грецкого ореха, например Jug-lans cinerea, достигавший р. Алдана; Хлебное дерево (Artocarpus) было находимо до Гренландии. В меловых и третичных отложениях из- . вестны многочисленные виды фикусов (напр. Ficus tiliaefolia, фиг. 843). Тюльпанное дерево (Liriodendron) было в мелу и третичном периоде широко распространено по всему северному полушарию, произрастая теперь в двух разрозненных ареалах (Китай и атлантические штаты САСШ).
Магнолиевые и в настоящее время имеют примитивные особенности строения, сближающие их с голосеменными, именно — отсутствие сосудов в древесине; они имеют лишь рами.
Фиг. 843. Ficus tiliaefolia. Лист (1) и плодовая ветьа (J2). Третичные. Энниген
Германия,
трахеиды с окаймленными по-
"Фиг. 844. Platanus aceroides. Миоцен. Шротцбург. Швейцария.
Лавры, камфарные и коричные деревья являлись важными членами третичной флоры Европы, Америки, до арктических стран, но например в век сармата были еще многочисленны на Кавказе. Род платанов (Platanus, фиг. 844) процветал в мелу и в третичном периоде (на склонах Урала,), причем может быть к ним должны быть отнесены и характерные, но довольно загадочные по систематическому положению креднерии (Credneria, фиг. 840), так типичные для сеномана и сенона (у нас в Уральской областии на Сахалине). Ныне китайский род айлант (Ailant/ius) широко распространен был по третичной Европе (у нас в Донбассе на р. Крынке). Один из наилучше изученных из ископаемых род Acer (клены) достоверно встречается с низов третичной системы. Из сем. виноградных род Vitis (виноград) известен с эоцена, a Gissites является широко распространенным растением сеноманской эпохи, в том числе у нас с Эм-бы. Интересно бывшее широкое распространение миртовых—теперь преимущественно жителей тропиков. Евкалип-ты, характерные ныне для Австралии, росли с мелового периода в Европе, Чехо - Словакии, Азии (Сибирь) и Америке, что доказывается находками не только характерных листьев, но и плодов. Плоды ав-
стралийского рода£ер-tospermum известны между прочим из олигоцена Волыни.
Аралии пользовались обширным распространением еще в меле (у нас в Уссурийском крае). Может быть, к тому же семейству относятся и De-walquea с дланевидными листьями, типичные для эоцена, но до сих пор загадочные по своим родственным отношениям (найдены у нас на Волге у Камышина). Зонтичные по плодам достоверно установлены лишь с третичного периода. Сложноцветные достоверно известны из олигоцена.
Наиболее достоверно нахождение таких родов, которые дали, кроме отпечатков листьев, остатки плодов или цветов, как например отпечатки из балтийского янтаря. Хотя большинство ископаемых древесин семенных
растений принадлежит голосеменным, в частности хвойным, в кайнозойских отложениях находятся нередко и остатки покрытосеменных; из однодольных — пальм (Palmoxylon — с мела), а из двудольных.— многих родов, чаще других березы {Betlinum), дуба (Quercinium), бука (Fegoniiim), лавра и др. Ряд ископаемых древесин пока не мог быть выяснен и помещен в то или другое из ныне существующих семейств.
Флора покрытосеменных мелового периода характеризуется тем, что, при отсутствии известных у них плодов, многие настолько еще далеки от современных нам форм, что с трудом укладываются в наши семейства.
Растения начала третичного периода узнаются легче, но находимые в средних широтах, например в средней Европе, формы или их близкие родичи теперь произрастают лишь в субтропиках или тропиках.
Лишь во флоре постплиоцена мы находим формы, живущие и теперь на тех же местах, или даже более северные, и весьма редко — вымершие в ближайших районах или странах (как напр., Trapa, Brasenia), имеющие ныне разорванные ареалы распространения.
АЛФАВИТНЫЕ УКАЗАТЕЛИ
Латинский
А.
abdomen 123, 150.
Acalepbae 72.
Acanthoceras 231, 236, 237.
Acanthodes 279, 292, 304.
Acanthodidae 304.
Acephala 237.
Acer 442.
Acerafherium 359, 376, 377.
Acicularia 401.
Acipenser 301.
Acipenseridae 301.
Acrania 265.
Actaeonella 204, 208.
Actin ocrinus 160, 164.
Actinomma 52.
Actinostroma 68.
Adapis 370. adductores 106. adjustores 106.
Aeglina 137.
Agelaciinus 175.
Agnostus 141, 143.
Agoniatites 221, 236. '
Agoropliius 391.
Ailanthus 442.
Alcyonaria 73, 87. Alethopteris 427, 429. alisphenoideum 278, 356. Allorlsma 259, 262, 263. Alnus 441.
Alveolina 45, 50.
Amaltheus 226, 236, 237. ambulacralia 156, 158.
Amia 299. 301, 302.
Amioidei 299, 301, 305, 307.
Ammodiscus 39.
Ammonoidea 215, 235, 264.
Amoeba 38.
Amphibia 265, 307, 318.
Amphicyon 367.
Amphimorphina 41. Amphineura 191, 244, 264.
Amphipoda 144.
Ancistocrania 111.
An plexus 81, 85. Ananchytes 182, 186. Anapsida 321, 322, 347.
Anehitherimae 379.
Anehitherium 358, 379, 380.
Ancyloceras 237;
Angiospermae 395, 439. angulare 319.
Anisomyaria 238, 245.
Annelida 96, 189.
Annnlaria 420.
Annulus 211, 422. 425.
Anthozoa 63, 73.
Anthracosaurus 313.
Ai.thracosia 256, 263.
Anthropidae 371.
Anthropomorphae 371.
Anura 310, 317, 318.
Apoda 310.
Aporrhais 202, 208.
Apthychus 219.
Apus 126.
Arachnoidea 126, 148.
Araeoscelis 341, 347.
Araucaria 438.
Araucarioxylon 438.
Area 244, 247, 263.
Arcacea 245.
Arcestes 224, 236.
Arcestidae 237.
Archaeocidaris.
Archaeocyathida 91.
Archaeocyathus 92.
Archaeolithothamnium 405.
Archaeopteris 425, 427.
Archaeopteryx 351, 353.
Archaeoridaris 184.
Archaeornis 351, 353.
Archaeornitbes 351.
Archegosaurus 269, 313.
Arcidae 246.
area 105, 240, 250.
Argonauta 235.
Arietites 226, 236, 237.
Arthrodira 284, 287, 304, 306.
Arthropoda 123, 189.
Artbrostraca 144. articulare 278, 319, 355.
Articulata 1(8,109,111, 159, 161, 163, 188.
Articulates 395, 410, 417.
Artiodactyla 381, 393.
Artocarpus 441.
Asaphus 140, 143.
Ascophyllum 403.
Aspidosoma 177.
Astacus 131.
Astarte 241, 253, 254, 263.
Asfarlidae 244, 261, 262.
Asterocalamitaceae 418.
Asterocalamites 417, 4:8.
Asterochlaena 423.
Asteroidea 176, 188.
Asterolepis 287.
Asfropecten 177.
Asterophyllites 420.
Asteroxylon 411.
Asterozoa 188.
Astraeospongia 60. astragalus 333, 360.
Astylospongia 59.
Athyris 121, 122.
Atrypa 106, 121, 122.
Aucella 248, 263.
Aulocopium 57.
Aulosteges 108, 116, 122.
Auriculae 181.
Aves 265, 321, 348.
Aricula 241, 247, 263.
Aviculidae 246, 248, 261. axillare 158.
Axonolipa 72.
Axonophora 72.
B.
Bactrites 230, 235, 236.
Baculites 219, 226, 237.
Baiera 436.
Balanus 129.
Baluchiterium 377.
Bambusium 440. basioccipitale 277, 278, 313. basisphenoideum 278,313,356.
Bathycrinus 160.
Batoidei 290.
Beggiatoa 396.
Belemnitella 234.
Belemnites 232, 233.
Belinurus 146.
Bellerophon 194, 197, 208.
Belodon 333.
Belosepia 234, 235.
Bennenites 432.
Bennettitales' 395, 429, 430, 433.
Betula 441.
Betulinum 443.
Beyrichia 128.
Bigenerina 43, 50.
Biloculina 44, 50.
Bison 383.
Biyalvia 237.
Bivium 154, 182.
Blastoidea 172, 176, 188.
Bos 383.
Bothriocidaris 184, 187.
Bothriolepis 287.
Bothrodendron 414.
Bothryococcaceae 400.
Bothryococcus 400.
Bothryocrinus 155.
Bothryopteris 423.
Boueina 400.
brachia 154, 157.
Brachial ia 157.
Brachiata 196.
brachidium 106.
brachiolae 165.
Bracliiopoda 102.
Brachyphylum 438.
Brachyura 133.
Branchiata 196.
Branchiosauria 311, 312, 317.
Branchiosaurus 312, 317.
Brasenia 443.
Bretonia 398.
Brissophis 181.
Bronteus 141, 143.
Bryophyta 395, 406, 410.
Bryozoa 98.
Bucemum 195, 202, 208. bulla tympanica 356.
Bunolophodon 386. byssus 244.
C.
Cadoceras 227, 236.
Calaminaceae 418.
Calamites 419.
Calamodendron 419.
Cala-nophytales 417. Calamophyton 417, 418.
Calamostachys 414, 420.
Calcaneum 333, 360.
Calceola 84, 85.
Callopegma 55.
Callorhynchus 297.
Callus 195.
Calymmene 134, 139, 143.
Camarophoria 118, 122.
Camasia 397.
Camerata 159, 160, 161, 164, 188.
Camptopteridaceae 425.
Camptopteris 425.
Canidae 367.
Cams 358-
Capreolus 382.
Capulus 199, 208. .
Carcharias 272.
Cardioceras 228, 236.
Cardiola 253, 263.
Cardium 253, 263.
Carina 129.
Carinaria 204.
Carinatae 351, 353.
Carnivora 366, 393. carpalia 273.
Carpentaria 41.
Carpoidea 176.
Castanea 441.
Catarrhini 370-
Caudata 310, 316, 318.
Caulopteris 423.
Cavicornia 383. centrale 361, 273. Cephalaspis 285, 311. Cephalopoda 209, 264, 304.
Cephaiothoracica 122.
Cephalothorax 125.
Ceraterpeton 311.
Ceratites 225.
Ceratitidae 237.
Ceratodus 273, 304.
Ceriopora 100.
Cerithium 201, 208.
Cervicornia 381.
Cervidae 381.
Cervas 382.
Cestracion 293.
Cetacea 388, 393.
Chaetetes 89, 91.
Chaetognatlia 96.
Chaetopoda 96.
Chama 245, 257, 263.
Chamaerops 440.
Chara 402.
Charophyta 395, 401.
Chasmatopteris 423.
Chasmops 137, 142, 143.
Cheilostomata 102.
Cheirostrobus 418.
Cheirostrobales 418.
Chelonia 321, 322, 325, 347.
Chilidium 105.
Chilopoda 149.
Chirocrinus 167, 169.
Chiroptera 366, 393.
Chirotherium 318.
Chitonidae 191.
Chlorophyceae 395, 399.
Chondrichthyes 284, 289.
Chondrostei 299, 300, 305, 307.
Chondrosteus 301.
Chonetes 112, 114, 122 Choraceae 402.
Chordaceae 403. chorda dorsalis 25, 40, 269. Chordites 403.
Chrooeoccaceae 397. ' Cimotomjs 336.
cirri 160.
Cirripedia 128, 144.
Cladiscothailus 397.
Cladodus 305.
Cladophlebis 424.
Cladoselache 274, 291, 304.
Cladoxylales 410, 422.
Cladoxylon 422.
Clamydo monadoceae 400.
Clatbropteris 425.
C avator 402.
Clavatoraceae 402. clavicula 272, 313.
cleithrum 272, 311, 313, 314 323.
Clepsydropsis 423.
Chdastes 342.
Clisiophyllum 83, 85.
Clupea 305.
Clymenia 224, 236.
Clypeaster 185, 187.
Coccolithophoraceae 397.
Coccosteus 287, 311.
Codaster 173, 320.
Codioceae 400.
Coelenterata 62. coelenteron 62, 73. coelom 87, 93.
Coleoptera 150.
Collennia 397.
Collyrites 182. columel a 75, 195, 307, 341. columna 154.
Columnalia 160.
Condylarthra 373, 393.
Congeria 252, 263.
Con ferales 395, 429, 437.
Coniopteris 425.
Conjugatae 395, 399.
Cpnuiaria 205, 207, 208.
Conus 203, 208.
Copepods 144-copalae 276. coracoideum 271, 325, 360. Corallinaceae 404.
Cordaitales 429.
Cordaites 395, 429, 430. corona 154.
coronoideum 319. eorfichim 55.
Coscinocyathus P3.
Cosmoceras 229, 236.
Cosmoceratidae 231, 237.
Cotylosauria 309, 321, 322.
347.
Crania 111, 122.
Craniata 265.
Credneria 440, 442.
Creodontia 357, 367, 391.
Cricotus 313.
Cribrospherae 397.
Crinoidea 154, 158, 176, 188.
Crioceras 218, 231, 236; 237.
Cristellaria 40, 44, 50.
Crocodilia 332, 333, 347.
Cromyocrinus 162, 164.
Crossopterygii 296, 304, 307, 311.
Crura 106.
Crustacea 125, 126.
Cryptocludus 275.
Cryptostomata 101.
Cryptozoon 397.
Ctenodus 303.
Ctenacanthus 282.
Ctenophora 63.
Cucculaea 247, 263.
Cumucea 144,
Cupressocrinus 159, 162. cutis 267.
Cyanophyceae 396.
Cyathocrinus 159, 161.
Cyathocystis 175.
Cyathophyllum 82, 85.
Cycadales 395, 429, 432.
Cycadof dices 417.
Cycadoidea 432, 434.
Cyclocrinus 401.
Cyclolitbs 85.
Cyclolobus 224, 236.
Cyclostoma 284.
Cyclostomata 100, 265, 306.
Cymopalia 401.
Cynognathus 329.
Cypraea 207, 208.
Cypress us 162.
Cyprina 2j1, 263.
Cyprinidae 244.
Cypris 128.
Cyrena 251, 263.
Cyrenidae 244, 251.
Cyrtina 107, 120, 122.
Cyrtodonta 246, 263.
Cyrtodontarca, 246, 261, 263.
Cyrtodontidae 246.
Cystiphvllum 75, 83, 85.
Cystoidea 161, 164, 174, 176, 188.
Cystoiseira 403.
Cystopbyllena 403.
Czekanowskia 436.
D.
Dalmania 137, 142.
Dama 383.
Dasycladaceae 400.
Dawsonites 411.
Decapbda 144, 232, 264.
Dellesseria 404.
Dellesseriaceae 404. delthyrium 104. deltidium 104.
Demospongiae 57.
Dendroidea 72. dentale 278, 319. dentalium 262, 319. Dermalithon 405.
Desmarestioceae 403. Desmodonta 259, 261, 265. Deuterosaurus 327, 328. Dewalquea 440, 442.
Diadectes 326.
Diademodon 327, 329, 355.
Diapsida 321, 331, 347.
Diatomeae 395, 398.
Dictiocha 397.
Dictiochaeae 397.
Dictyophyllum 425.
Dibranc iata 232, 237, 264.
Dibunophyllum 83, 85.
Dicellocephalus 138.
Diceras 245, 257, 263.
Diceratinae 257.
Diceratosaurus 313.
Dicksonia 425.
Dicotyledonae 395, 439, 440.
Dictyonema 71.
Dicyclica 161.
Dicynodon 328, 329.
Diductores 105.
Dielasma 118, 122.
Dinoflagellatae 398. Dinosauria 332, 334, 347. Dinotherium 358, 385. Diotocardia 194, 196, 264.
Diplodocus 336.
Diplograptus 69, 70.
Diplopora 401.
Diploporita 149, 171, 188.
Dipnoi 296, 303, 304, 307.
Diprodonla 365.
Diprotodontia 363, 365, 393.
Diptera 144, 150.
Dipteridaceae 425.
Dipterus 304.
Discina 110, 122.
Discites 215.
Discoglassus 309. dissepimenta 75. distalia 273, 274. Distephanos 395.
Dorypyge 143.
Dreissensia 251, 263.
Dromatherium 357, 364, 365, 392,
Dryophyllum 441.
Dvinosaurus 314.
Dysodonta 247, 264.
E.
Ecaudata 310, 317-
Echidna 360, 363.
Echinoconus 185.
Echinoencrinus 170.
Echinoidea 179, 188.
Echinosphaerites 167, 171.
Echinus 180.
Ectopterygoideum 309.
Edentata 368, 393.
Edmondia 259, 263.
Edrioaster 175.
Edrioasteroidea 175, 176, 188.
Elasmobranchii 267, 284, 289,
306.
Elasmotherium 359, 376.
Elephas 385, 387.
Eleutherozoa 154, 176, 188.
Embolomeri 313, 321.
Embryocrinidae 158.
Embryophyta asiphonogama 405.
Embryophyta si phonogama 428, Encrinurus 143.
Encrinus 163.
Endoceras 214, 235, 236.
Endothyra 46, 50.
Endothyridae 46.
Enteropneusta 265.
Entolium 249.
Entomostraca 126, 129, 144.
Eoanthropus 371.
Eobatrachus 317.
Eophrynus 149.
Eosirea 392.
Eotherium 27, 392.
Ephedra 439.
epioticum 308.
Epipterygoideum 341.
Eplacentalia 361, 362, 392.
Equidae 380.
Equisetaceae 420.
Equisetineae 418.
Equisetites 421.
Equisetum 420.
Erinaceus 366.
Eryon 133.
Estheria 121, 126.
Estonioceras 215, 236.
ethmoideum 296, 354.
Euamphibia 316, 318.
Eudothyrinae 46.
Eucephalica 149.
Eunotosaurus 326.
Euomphalus 198, 208, 212.
Euphoboria 149.
Euprotogonia 375.
Elonichtys 282.
Emyis 325.
Eurycormus 270.
Eurypterida 146.
Eurypterus 147.
Eurysternum 328.
Eusporangiatae 422.
Exoccipitale 277.
F.
fasciolae 183.
Favosites 90, 91.
Fegonium 441, 443.
Felidae 368.
Felis 368.
femur 273.
Fenestella 102.
fibula 273. fibulare 273, 325. Ficus 441.
Filicalee 395, 410, 422.
Fissurella 197, 208.
Fistulata 159, 161, 188.
Flagellatae 395, 397.
Florideae 403.
Flustra 100. foramen 104. „ obturatorium 321, 328. „ transversarium 359.
„ ulnare 323.
Foraminifera 38. fossula 80.
Frondicularia 41, 43. frontalia 278, 296, 308. frontoparietalia 308.
Fucaceae 403.
Fungi 395, 405.
Fusulina 46.
Fusulininae 46.
Fusus 203, 208.
G.
Gallionella 396.
Ganoidei 296, 299, 304, 307.
Gastrioceras 223, 235, 236.
Gastropoda 192, 264.
Genae 133
Gephyrea 96.
Gephyroceras 222.
Gervillea 248, 263.
Gidaris 181, 185.
Gigantostraea 126, 144.
Ginkgo 435.
Ginkgoales 395, 429, 435.
Ginkgobiloba 435.
Ginkgodium 436.
glabella 133.
Glandulina 40, 41.
Gleich enites 424.
Gleiocomis 397.
Gleocapsomorpba 397.
Globigerina 47, 50.
Gloecestis 400. glossopterae 290. Glossopteris 433, 435.
Glyphea 133.
Glyptodon 369.
Glyptosphaerites 171.
Gnetacea 439.
Gnetales 395, 429.
Gnetnm 439.
Gomphoceras 212, 214, 236.
Goniophyllum 84, 85.
Graptolithoidea 65, 69.
Gryphaea 250, 263.
Gymnocordium 400.
Gymnophiona 310, 313.
Gymnospermae 395, 429.
Gyrogonites 402.,
H
Halicore 391, 392.
Halimeda 400.
Haliotis 198, 208.
Haliserites 411.
Halitherium 392.
Halonia 414.
Halysiles 90, 91.
Hamites 226, 237.
Haplophragmium 50.
Harpes 137, 140, 143.
Harpoeeras 226, 236, 237.
Hatteria 322.
Hausmannia 425.
Helicoprion 293, 295, 305.
Heliolites 89, 91.
Heliopora 88.
Helix 207, 208.
Hemicidaris 185.
Hemiptera 150.
Hepaticae 395, 410.
Hepaticites 410.
Heptanchus 273.
Hercynella 207, 208.
Hesperornis 350, 352, 353.
Heterocerci 293, 300, 304, 306, 307.
Heterodonta 246,250,261, 262, 264.
Heteromyaria 238, 261.
Heteropoda 203, 204, 264.
Hexacoralla 76; 85.
Hexactinellida 57, 59.
Hexameroceras 212.
Hicklingia 411.
Himanthaliopsis 403.
Hipparion 358, 380.
Hippurites 245, 258, 263.
Holocephali 289, 292, 294, 304, 306.
Holoptychius 297.
Holothyrioidea 189.
Homalonotus 139, 143.
Hominidae 371, 373.
Homo 372.
Homo Cro-Magnon 372.
Homo diluvii 317.
Hoplites 229, 236, 237.
Horea 411.
humerus 273.
Hungarites 225, 236.
Hydractinia 66.
Hydraria 65. . .
Hydrocircus 157.
Hydromedusae 63.
Hydrozoa 63.
Hyenia 417, 419.
Hylaeohatrachus 317, 318-
Hyloplesion 310.
Hymenophillites 424.
Hymenoptera 150. hyoideum 276. hyomandibulars 292, 307. Hyopotamus 358. hypocentrum 269. hypostoma 134.
Hyracotheriinae 378.
Hyraeotherium 358, 378, 380»
I.
Ibergiceras 218. Ichthyoerinus 158, 159.
Ichthyornie 353.
Ichthyosauria 321, 340, 343-jerea 58.
Iguanodon 337, 338. ilium 272.
Illaenus 141, 143.
Inarticulata 109. incus 355.
Indocrinae 158.
Indricotherium 377. infrabasalia 155. infraorbitalia 278. Inoceramus 248, 263. Inostrancevia 328.
Insecta 126, 150.
Insectivora 366, 373, 393.
Integripalliata 251. interbrachialia 159. interclavicula 311, 362. intermedium 273, 323, 325. , irregularia 183, 189.
ischium 272, 325.
Isoetaceae 416.
Isoetes 416.
Isomyaria 238, 245.
Isoj.oda 144. jugale 307, 356. luglans 441.
K.
Kadaliosaurus 341.
Klukia 424.
Knorria 414.
Kosmogyra 402. Kotlassia 322, 323. Kutorginacea 110.
ь.
Labyrinthodontia 313.
Laccopteris 424.
Lacertilia 340, 347. iacrymale 278, 307.
Lagenostoma 429.
Lagynophora 402.
Lamellibranchiata 237, 264.
Laminareaceae 403.
Larix 439.
Larviformia 188.
Lasiophycus 403. Lemuroidea 370.
Lepas 129.
Leperditia 128.
Lepidocarpon 414.
Lepidodendron 412.
Lepidophloios 414.
Lepidophyta 412, 415.
Lepidoptera 150.
Lepidostrobus 414.
Lepospondyli 313, 318.
Leptjlepis 302, 305.
Leptoria 78, 85.
Leptosphermum 442. Leptostraca 129, 144.
Lichenes 395, 405.
Lima 240, 245, 249, 263.
Limbus 134.
Limnaeus 18, 207, 208.
Limulus 145.
Lingula 110, 122.
Liriodendron 441.
Lithistida 57, 59.
Lithophyllum 405. Lithostrotion 82, 85. Lithothamnium 405.
Lituites 215, 235, 236.
Loganella 411.
Lonsdaleia 83, 85. Loxonema 201, 208. Loxonematidae 207.
Lucina 242.
Lycosuchus 329.
Lycopodiales 395, 410, 412.
Lycopodium 412.
Lycopodiaceae 412.
Lyginodendron 429. Lysorophus 316, 318.
Lytoceras 225, 236, 237.
Lytoceratidae 226, 231, 237.
11
Machairodus 96, 368.
Macrocephalites 228, 236.
Macrocystis 403.
Macrcdon 246, 263.
Macropus 362.
Macrura 133.
Mactra 240, 253, 263.
Malacostraca 126, 129, 144.
Malacostroma 68-malleus 355.
Mammalia 265, 321, 354.
Manatus 391. mandibula 125. mandibulare 276. Mandschurosaurus 337. Manticoceras 221, 222, 235. Marattiaceae 423, 427. Marcbantites 410.
Mariopteris 429.
Marpolia 397.
Marsupialia 363, 392. Mastodon 385, 386. Mastodonsaurus 280, 315. maxilla 125, 278.
Medlicottia 217, 224, 236.
Medullosa 429.
Megaceros 382, 383. Megalaspis 140, 143. Megalobatrachus 317, 318. Megalodon 257, 263. Megalodontidae 261. Megalurus 299, 302.
Megatherium 369. Melonites 184, 187. Membranipora 102. Moretrix 239.
Merostomata 126, 144.
Mesoeena 397.
Mesocystis 169. mescethnuiideum 278.
Mesohippus 379, Mesopithecus 371. metacarpalia 273. Metaclepsy dropsis 423. metacoceras 215. metapterygium 272.
Metasepta 78.
Metastoma 147. Metatarsalia 273. Metaxytherium 392. Metazoa 53.
Micraster 187.
Microlestes 363, 392.
Microsauria 311, 313, 318. Microzeuglodontidae 391. Miliola 42.
Miliolidae 44.
Miliolinae 44.
Millepora 67.
Mixosaurus 347.
Modiola 239.
Moeritherium 385. Monactinellida 57. Monitor 341.
Monobrachiocrinus 158. Monocotelodonae 395. Monocoty ledonao 439, 440. Monocyclica 161.
Monograptus 69. Monomyaria 245.
Monothalamia 42
Monotocardia 194, 199, 264.
Monotremata 362, 392.
Morattiopsis 423.
Mosasauria 341, 347.
Multituberculata 363, 392.
Murchisonia 197, 208.
Murex 196.
Musa 440.
Muscites 410.
Musci frondosi 395, 410.
Mya 238, 260, 263.
Myeloxylon 429.
Mvophoria 254, 255, 263.
Myriopoda 126, 149.
Myrina 239.
Mytilus 246, 247, 251, 263.
Myxomycetes 395, 398.
N.
Naricula 399. nasalia 278, 296.
Natica 195$ 196, 199, 208.
Naticopsis 199, 208.
Naucrates 282.
Nauplius 126.
Nautiloidea 210, 235, 264.
Nautilus 209, 210, 215, 235, 236.
Nebalia 130.
Nem .tophycus 403.
Neoggeratiopsis 430.
Neoplagiaulax 363.
Neopteris 429.
Neornithes 351, 353.
Nerinea 201, 204, 208. neuralia 269.
neurapophyses 269.
Neuroptera 150.
Neuropteris 427, 429.
Nillsonia 435.
Nitella 402.
Nodosaria 41, 43.
Nodosaridae 43.
Nodosinella 40, 43, 50 Nodosinellidae 43.
Nonruminantia 381, 393. Nucula 244, 247, 263.
Nuculacea 245.
Nummulites 47, 50. Nyctodactylus 339.
0.
Obelia 65.
Obolus 110, 122.
Octocoralla 87.
Octopoda 235, 264.
Olcost phanus 229, 236, 237.
Olenellus 137, 139, 143.
Olenus 137, 143. operculum 145, 307.
Ophidia 343, 347.
Ophiuridea 178, 188.
Opisthobranchiata 194, 294,264.
Opisthoparia 137, 138, opisthoticum 276. oralia 149, 155, 159, 162. Orbitoides 49, 50. orbitosphenoideum 278, 308, 356.
Orestovia 403.
Ornithorhynchus 363.
Orthidae 113.
Orthis 112, 122.
Orthisina 107, 113, 122.
Orthoceras 213, 214, 219, 235, 236.
Orthopoda 336.
Orthoptera 150.
Orthothetes 113, 122.
i Oscillatoriceae 397.
; Osmunda 424.
t. Osmundia 424.
. os sepiae 234.
\ osculum 54.
, Osmundaceae 223.
’ Ospubis 321.
Osteichthyes 284 295, 302.
Osteolepis 297, 323.
Ostracoda 121, 127, 144.
Ostrea 250, 263.
Ostreidae 261.
otica 277.
Otoceras 225.
Otolithus 284.
Oxytoma 247, 263. .
P.
Pachyodonta 256, 262, 265.
Palacosyops 358.
Palaeaster 177.
Palaeobatrachus 318.
Palaeocharaceae 402.
Palaeodiehoptera 151.
Palaeohalidrys 403.
Palaeohepatica 410.
Palaeomastodou 385.
Palaeomeryx 381.
Palaeoneilo 247, 263.
Palaeoniscidae 301.
Palaeoniscus 300.
Palaeophonus 148.
Palaeoporella 400. palatinum 278, 309, 323. palatoquadratum 278, 292, 307, 309.
Paliguana341.
Palmoxylon 443.
Paludina 193, 200, 208.
Palus 74.
Panopaea 260, 263.
Paradoxides 139, 143.
Parapsida 321, 340.
parasphenoideum 298, 309, 313 319.
Pareiasaurus 323. parietale 278, 308. Patella 199, 207, 208. Paterina 109, 122.
Patriocetus 391.
Pecten 239, 245, 249, 263.
Pectina 148.
Pectinidae 249.
Pectrunculus 239.
Pedipalpi 149.
Pelecypoda 237.
Pelmatozoa 154,187.
Peltura 138, 143.
Pemphix 133.
Pentacrinus 164.
Pentameracea 116.
Pentamerus 116, 122.
Pentremites 173, 175.
Peridineae 395, 398.
Perisphinctes 229, 236, 237.
Perissodactyla 374, 393. Perna 241, 248, 263.
Petrosum 276.
Petscheropteris 423.
Phacops 137, 142, 143.
Phacotaceae 400.
Phacotns 400.
Phaeophyceae 403.
Phaneropleuron 303.
Phascolotherium 364. Phenacodus 362, 374. Phenicites 440.
Phillipsia 138, 143.
Pholas 260, 263.
Phragmoceras 214, 236.
Phragmoconus 232.
Phylloceras 225, 236.
Phylloceratidae 237.
Phylloglossum 412.
Phyllograptus 69.
Phyllopoda 126, 144.
Phyllospondyli 312, 318.
Picea 439.
Pila 400.
Pinacites 221, 236.
Pinnipedia 367, 368. pinnulae 157, 161.
Pinus 439.
Pisces 265, 305.
Pisocrinus 160.
Pithecanthropus 371, 372. placenta 281, 366.
Placentalia 281, 366, 393.
Placodermi 284, 288, 304, 306.
Plagiostomi 290.
Planorbis 35.
Planorbulina 41.
PIaronin= 423.
Platanus 442.
Platecarpus 341.
Platyceras 199, 208.
Platycrinus 164.
Platyrrhini 370.
Platyschisma 198, 208.
Platysomus 300.
Plesiosauria 321, 327.
Plesiosaurus 274, 330.
Pleuracanthus 291. pleurae 133, 135. pleurocentrum 269. Pleuromejaceae 416. Pleuromya 260, 263. Pleurophorus 253, 263. Pleurotoma 203, 208.
Pleurotomaria 196, 208.
Pleurotomariidae 207.
Podozamites 435.
Polycotylus 331.
Polygonosphaerites 61.
Polymorphina 41.
Polyparium 69.
Polyprotodontia 363, 364, 366, 392, 393.
Polypterus 282, 297.
Polystomella 42.
Polythalamia 43.
Polytrichum 410. Porambonites 116, 182.
Porifera, 54.
Posidonomya 248, 263. postfrontale 356.
Procolophon 329. postparietale 308. Poteriocrinus 162.
Praedentata 336.
Praefrontale 296, 356. praemaxilla 279, 296. praesphenoideum 278, 356.
Preridinium 398.
Primates 370, 393.
Primocorallia 401.
Prioniodus 96.
Proboscidea 384, 391, 393.
Proboscis 159, 161. processus uncinati 333. procoiacoi leum 271, 326, 359. Prodai ael tes 218.
Prodremotherium 358.
Productus 108, 112, 114, 115, 122.
Proetus 138, 143.
Prolecanites 223, 236.
Prolobites 221, 236.
Promathildia 201, 207, 208.
Pronorites 223, 236.
Proostracum 232, 235.
Prooticum 277, 308.
Proparia 137, 140.
propterygium 272.
Protopteris 423.
Protosporangiata 422.
Prosicanites 218.
Prosimiae 370, 373, 393.
Prosobranchiata 194, 196, 204, 207, 264.
Protocetus 390, 391.
Protoleptosporangiatae 423.
Protophyceae 397.
Protonauplius 136.
Protosepta 78.
Prototaxites 403.
Protozoa 37.
Protriton 312.
Psammodus 2, 293, 295, 305.
Pseudoborniales 417.
Pseudocosta 75. pseudodeltidium 97, 104. Pseudomonotis 247, 263. pseudosepta 88.
Pseudosporochnus 411. Pseudorbia 417.
Pseudotextularia 41. pseudotheca 75.
Psilophytales 395, 410.
Psilophyton 411.
Psilotaceae 395.
Psilotales 410, 417.
Psilotum 417.
Psygmophyllum 436.
Pteraspidae 304.
Pteraspis 285.
Pteria 247.
Pterichthys 286, 311.
Pteridophyta 395, 406.
Pteridospermae 429.
Pterinea 247, 263.
Pterodactylus 339.
Pteronautilus 215. Pterophyllum 433, 434. Pteropoda 204, 264.
Pterosauria 332, 337. pteroticum 278.
pterygoideum 278, 308, 319, 323, 356.
pterygo-palatinum 276. pterygopodia 289.
Pterygospermales 395.
Pterygotus 147.
Ptychoparia 137, 138, 143.
Ptyctodus 295. pubis 272, 313, 317. Pulmonata 206, 264. pulpa 268.
Pygaster 185, 187.
Pygidium 135.
Pyrocy estites 169.
Q quadrato-jugale 307. quadratum 276, 307, 355.
Quercinium 443.
Quercus 441.
Quinqueloculina 40, 45, 50.
ft.
radiale 273.
Radius 273, 360.
Radix 154.
Radula 190, 192, 195, 210, 237, 262.
Raja 282
Ranodon 275.
Rastrites 69.
Ratitae 353.
Receptaculidae 60.
Receptaculites 61.
Regularia 183, 189.
Reinschia 400.
Reptilia 265, 318, 347.
Retzia 120, 122.7
Rhabdammina 39, 43.
Rhabdamminidae 39, 42.
Rhabdopleura 72.
Rhabdoporella 401.
Rhachiqpteris 429.
Rhachis 133.
Rhachitomi 197, 208.
Rhaeopbyceae 395.
Rhamphorhynchus 339.
Rhenikopsis 436.
Rhinobatus 293.
Rhinoceros 359, 376.
Rhinocerotidae 376.
Rbipidopsis 436.
Rhizophyllum 84, 85.
Rhizopoda 38.
Rhodophyceae 395, 403-Rhodymeniaceae 404.
Rhombitera 169, 171, 188.
Rhopalodon 328.
Rhynchocephalia 332, 347.
Rhynchonella 107, 118, 122.
Rhynia 411.
Rhytina 392.
rostrum 131,232,291,319,345.
Rotalia 43.
Rotalidae 46.
Rudistae 245, 261.
Rugae 75.
Rugosa 81, 78, 85.
Ruminantia 381, 393.
Rustellacea 110, 122.
S.
Sabal 440.
Saccamina 42, 50.
Sagenocrinus 159.
Salix 441.
Sargassum 403.
Sauropoda 335.
Sauropsida 348.
Sauropterygia 327, 330, 347.
Scaphites 218, 232.
Scaphopoda 209, 262, 265.
Scapula 271, 313.
Scapularo 271.
Schizodonta 254, 264.
Schizodus 255, 263.
Schizomycetes 395, 396.
Schizoneura 421.
Schizophyceae 395, 396.
Schizopoda 144.
Schloenbachia 229, 236.
Schwagerina 46, 50.
Sciaena 284.
Scorpio 148. scuta 129. Scyphozoa 63, 72.
Selachii 283, 289, 290, 304, 306.
Selaginella 412.
Selaginellaceae 412.
Semitae 183.
Sepia 209, 234.
Septa 74, 212.
Sequoia 438.
Serpula 95.
Seymouria 322.
Sicula 69.
Sigillaria 412, 415.
Silicof'lagellatae 397.
Simiae 370, 373, 393.
Sinantropoi 371.
Sinupalliata 253.
Siphoneae 400.
Siphonotreta 110, 122.
Sirenia 391, 393.
Solenopora 404.
Solenoporaceae 404.
Solvinia 425.
Sorex 366.
Sori 422.
Spathiocaris 130, Sphagnum 410. Sphenodou 322, 332. Spbenophyllales 417.
Sphenophyllostachys 418.
Sphenophyllum 417. Sphenopteris 427, 429. Sphaere- hnius 152. spicula 65.
spiracultfm 277, 300, 303.
Spirialis 205, 208.
Spirifer 19, 106, 119, 122.
Spiriferacea 107, 119.
Spiriferina 107.
Spirogira 399.
Spiroloculina 44, 50.
Spiroplecta 41.
Spirorbis 96.
Spleniale 319.
Spondylidae 261.
spondylium 105.
Spondylus 240, 249, 263.
Spongiostromata 397. Spongiostromidae 68. Sporogonites 411.
Squaloidei 290.
Squamata 321, 340, 347. squamosum 278, 307, 308,354. Stachypteris 425.
Stapes 355.
Stauropteris 423.
Stegocephali 309, Sil, 321.
Stenodutya 151.
Squatina 276.
Solea 282.
Stelleroidea 176.
Stellispongia 57, 61. Stemmatocrinus 163. Stephanoceras 219, 227, 236. Stephanoceratidae 237. Stereospondyli 314.
Stigmaria 416.
Stomatopoda 144.
Straparollus 198, 208. Streptelasma 81, 85. Stringocephahis 119, 122, 318. Stromatopora 68.
Stromatoporoidea 67. Strombus 202, 208. Strophalosia 115, 122. Strophomena 113, 122. Strophomenaceae 107,110,112. Styliolina 205, 208. Subpronorites 218.
Subterapedia 397 sulci laterales 134. supraangulare 319. supraoccipitale 277, 316. suprascapulare 271. supratemporale 308.
Sus 381. sutura 134, 212. Sycidium 403.
Synapsida 321, 327, 347, 392.
Synapticula 75.
Syringopora 90, 91.
T. tabulae 75. tabulare 308.
Taeniopteris 423, 433, 435.
Talpa 366.
Tapirus 375.
Tarsalia 273.
Taxocrinus 159.
Taxodium 438.
Taxodonta 245, 261, 264. tegmen 154, 157.
Telangium 429.
Teleosaurus 334.
Teleostei 296, 299, 301, 302.
Teleostomi 284, 295, 306.
Temnospondyli 313, 318. temporale 355.
Tentaculites 205, 207, 208.
Terebratula 107, 118, 122.
Terebratulacea 118.
Teredina 260, 263.
Terga 122, 129.
Testudinata 322.
Tetrabranchiata 210, 264.
Tetracoralla 78-Tetractinellida 57.
Textularia 40, 43, 50.
Textularidae 43.
Thallophyta 395, 396.
Thamnopteris 423. theca 65, 74, 154. Theriodontia 355.
Theromorpha 320, 821, 327, 347, 355.
Theropoda 335.
Thoracostraca 144. thorax 123, 135, 150. tibia 273.
tibiale 273, 325.
Timanites 223, 235.
Tissites 442.
Tmesipteris 417.
Todea 424.
Todites 424.
Toretocnemus 344.
Tornoeeras 222, 235, 236.
To .vaster 186.
Trachodon 337.
Transversum 309, 319.
Trapa 443.
Tretospira 202, 208.
Triassochelys 327.
Trigonia 243, 255, 256, 263.
Trigoniidae 261.
Trigonocarpus 429.
Trilobitae 126, 133.
Triloculina 45, 50.
Trinucleus 140, 143.
Tripl oporella 401.
Triton 195, 212.
Trivium 154, 182.
Trizygia 418.
Trochidae 199, 207, 209.
Trochiliscus 403.
Trochus 199, 208.
Tubicolae 95.
Tumboa 439.
Tubipora 88.
Turbo 198, 208.
Turbinidae 207.
Turrilites 218.
Turritella 200, 208.
Turritellidae 207. tympanicum 355.
U.
Udenodon 328.
Ullmannia 438.
Ulna 273, 325, 360.
Ulnare 273.
Ulodendron 414;
Umbilicus 195.
Umbo 42.
Uncites 120, 122.
Unio 257, 263.
Unionidae 244, 256, 262, Uralotimania 69.
Urodela 310, 316, 318.
Ursidae 367, Ursus 367, 368.
V.
velum 65, 72, 190.
Ventriculites 59.
Venus 254, 263.
Vermes 93.
Vermetus 194, 200, 208, 212.
Vertebraria 434.
Vertebrata 265.
Vespertilio 366.
virgula 69.
Vitis 442.
Vola 249.
Volborthella 214, 235, 236.
Volborthia 109, 122.
Voltzia 438.
vomer 278, 309, 319, 323.
W.
Walchia 438.
Waldheimia 119, 122.
Wiellandiela 432.
Williamsonia 432.
Williamsoniella 432.
X.
Xiphosura 145.
Xauchidium 399.
Z.
Zalesskya 423
Zamites 435.
Zaphrentis 81, 85.
Zeuglodon 391.
Zeuglodontidae 391.
Zoantharia 73, 74.
Zoea 126.
Zoophycas 403, Zygopleura 201, 207, 208.
Zygopteris 423.
Русский.
А.
Абиссальная область 28.
Авикулярии 99.
Автостилический череп 281. Адамбулякральн. пласт. 177. Айлаит 442.
Акрододтные зубы 319.
Акромиальн. отроет. 359. Акулы 289, 290.
Аллантоис 281.
Альвеола 232, 320.
Альвеолии. известняк 40.
Амбулякральн. ножка 152.
„ пластинка 157,
177.
„ система 152.
Дм ба 37.
Ам бовидные 38.
Аммониты 217, 237.
Амнион 281.
Амииоты 281.
Ампуллы 152, 157, 177.
Амфибии 307, 316.
Амфииевры 100, 244.
А.мфицельи. позвои. 281. Аналогия 26.
Анальи. табл. 159.
„ трубка 159.
Анальное отверстие 93, 182. Анальи. плавн. 275.
Анаптих 220.
Аннелиды 96, 123, 189.
Антенны 123.
Антеридий 405.
Антилопы 383.
Антисифоин. лоп. 217.
Апертура 38, 194.
Аптих 22о.
Аралии 442.
Ареа 105, 240.
Аристот. фонарь 180.
Аррагониты 74.
Артериальн кровь 279.
Архегоний 405.
Археоциатиды 91. Асимметрия молл. 193. Астерокаламиты 418. Астероспои ил. позв. 290. Афлебии 422, 427.
Б.
Багрянки 403.
Базальные таблички 155.
Бактерии 396.
Балочки 76.
Бананы 440.
Барабанная кость 356.
Бедренаи кость 273.
Беззамочн. брахиоподы 106, 121.
Беззубые киты 391.
Безногие 310.
Белемниты 234.
Бениетиты 429, 430.
Бентос 29.
Береза 441, 443.
Берцовые кости 273.
Беспоследовые 362.
Бесеифонные 405.
Бесхвостые амфибии 310,317.
Бесчерепные 265.
Бивни 387.
Биморфные раковины 44.
Биология 27.
Биссериальиый плавник 272.
Биссус 244.
Бластоидеи 172.
Боковая линия 280.
Боковые затылочн. к. 277.
„ лопасти 217.
„ перегородки 80.
„ решетчатые 278.
» таблички 173.
Бокоплавы 130.
Большой круг кров. 280. Бороздки трилоб 133—134 Борьба за жизнь 17.
Брахиодонтные зубы 358.
Брахиоподы 109, 121.
Броненосцы 368.
Брызгальпе 277.
Брюхоногие 196.
Брюшиые плавники 273.
Бугорчато - режущий зуб 357.
Бугорчатые зубы 357.
Бук 441.
Буиодонтный зуб 358.
Бурые водоросли 403-Быки 383.
В.
Баи 422, Вакуоля 37. Васкулярное впечатление 103. Век 31.
Веиечи. отросток 319.
Венозная кровь 279.
Венцы крииоидей 155.
» морских лилий 155.
Верхнечелюсти. кость 278.
Верхние ребра 271.
Верхняя затылочн. к. 277.
Верхушечное почкование 78.
Вершинный угол 195.
„ щиток 180. о Внбракуди 99.
Вид 16.
Виноград 442.
Височная кость 278.
Височные отверстия 322.
Висцеральные дуги 275.
Влагалище 267.
Внутренний скелет 266.
Виутреииие лопади 217.
Внутренностный мешок 192.
Виутренияя связка 240.
Водоносное кольцо 157.
Водоносные сосуды 157.
Водоросли 396.
Воздушные камеры 210, 213.
Волосы 267.
Воронка 209.
Воротничковые клетки 54, 55.
Вспомогательные лопасти 217. Вторичная древесина 408, 409.
„ кора 409.
, чашечка 157.
Вторичное нёбо 319.
„ перо 267.
Вторичные лучи 272.
, перегородки 78.
Вторичный черен 277.
Выделительи. органы 94.
Высшие животные 53.
Высшие зародышевые 406.
Г.
Гаверсовы каналы 266.
Гагары 353.
Гаметофит 405.
Ганоидные рыбы 296, 299.
Ганоидные че.иуи 282.
Гаплодонтиые зубы 357.
Гастрозооид 66.
Гастроподы 192.
Гастропор 66.
Геккель 25.
Гемальиые дуги 271.
Генеративные ядра 428.
Генитальные таблички 179.
Гетеродонтная зуб. сист. 356.
Гетеродонтиый замок 242.
Гетероцеркный плавник 283.
Гиббон 371.
Гидрактинии 66.
Гидрант 63.
Гидроидный полип (J5-
Гидрокораллы 66.
Гидромедузы 63.
Гидропор 165.
Гидроспира 173.
Гидротека 66.
Гиоидная дуга 276, 355.
Гиостилический череп 281, 290, 292.
Гипердактилия 344.
Гиперфалангия 344.
Гипостома 134.
Гипоцентр 302.
Гипсодонтный зуб 358.
Глабель 133, 136.
Главная перегородка 79.
Глаза 64, 124.
Глазиично-в.: сочное отв. 308.
Глазничио - клипов. кость 278.
Глазные таблички 179.
Глоссоптерис. флора 434.
Годичные кольца 409.
Голова 126, 265.
Головной мозг 9,123,125, 854.
Головогрудь 125.
Головоногие 209, 285.
Голосеменные 428, 429.
Голостомные молл. 195, 201.
Гомодонтная зубн. сист. 356.
Гомология 26.
Гомоцеркный плави. 283.
Гонада 192.
Гониатиты 217, 220, 224.
Гоиопор 165.
Гонотека 66.
Гонофор 66, 70.
Горилла 371.
Горловые пластинки 297.
Граптолиты 69.
Грибы 405.
Грудная кость 271, 320, 351.
Грудной плавник 272.
Грудь членистоногих 123.
Губки 54.
Губиые хрящи 279.
Губы морских ежей 182.
„ моллюск. 195.
Д.
Дактилозооид 66.
Дактилопор 66.
Дарвин 20.
Двойн. поры 166, 168.
Двояковвгнутый позв. 270.
Двоякодышащие 296, 303.
Двугубый рот м. ежей 182.
Двудольные 439.
Двужа1бериые 232.
Двукрылые 151.
Двурезцовые 365.
Двурядные руки 158.
Двустворч. раковина 237.
Двусторови. симметрия 73.
Дв. цикл, диск 155.
Деление 38.
Дельтидиальи. таблички ЮЗ.
Дельтоидальи. табл. 172.
Дельфии 391.
Дея тин 268.
Десмодонтный замок 243.
Десмы 57.
Десятиногие 130, 232.
Диатомовые 398.
Диафрагмы 420.
Дизодоитный замок 242.
Динозавры 334.
Диплоспоидил. позв. 26», 311.
Диск мор. лилий 154.
Диссепимеиты 77.
Дистальный 69.
Дифицеркный плав. 283.
Дихотомия 157.
Днища 75.
Древесина 407.
Древние птипы 351.
Древовиди. папор. 422, 432.
„ плауны 412.
„ хвощи 418.
Дубы 441, 443.
Дуги жаберные 268, 271.
„ позвонков 269.
Дудки сифонные 212. Душка пера 267.
Дюгонь 391.
Е.
Евкалипт 442.
Еж 366.
Ежи морские 179.
Ель 438.
Епанча молл 189.
Ехидна 365.
Ж.
Жаберная дуга 268. „ крышка 281.
„ полость молл 192.
, щель 277, 281.
Жаберные 196.
Жабры 94, 124.
„ моллюск 194,196, 290.
„ позвои. 265, 274.
Жвачные 381.
Жгутиковые 397.
Жгутиков, камеры 54. Желобки амбул. 155. Желток 281.
Желточный мешок 281.
Жилая камера 210.
Жираф 369.
Жуки 150.
3.
Завитки раковины 194.
Заднежаберные 194. *
Задняя ключица 272.
Закрытые сосуд, пучки 408.
Замок моллюсков 241.
„ плеченог. 105.
Замочная площадка 241.
Замочио-створчатые 109. Замочные пластинки 105.
Замочный отросток 105.
Замыкатели 238.
Запястье 273.
Зародышев. бессиф 405.
, сифоии. 42§.
Затылочная бороздка 130, 133.
Затылочное кольцо 134.
Затылочные кости 277.
Звезды морские 176.
Зеленые водор. 399.
Землеройка 366.
Земноводные 307.
Зигосфен-зигантрум 342.
Змеевиди. звезды 168.
Змеехвостки 188.
Змеи 310, 343.
Зрительн. капсула 276.
Зубатые киты 391.
Зубная кость 278.
„ формула 359.
Зубные пластинки 105.
„ ямки 105.
Зубр 383.
Зубы млекопит 356.
„ моллюсков 241.
„ морск. ежей 180.
, плечеиогих 105.
, позвоночных 268.
И.
Ивы 441.
Иглокожие 152, 187.
Иглы губок 55.
Иглы морск. ежей 183.
Известк. губки 61.
я раковины 41.
„ хрящ. 267.
Изменчивость 17.
Изодоитный замок 249.
Изотомия 157.
Инволютиая раков. 211.
Интербрахнальиые таблички
159.
Интеррадиальн. таблички 155.
Иитерсептальн. камера 74.
„ канал 48.
Интерцентры 321.
Инфрабазальн. табл. 155.
Инфузории 37.
Ихтиодорулиты 283.
Ихтиозавры 343.
К.
Каламиты 417, 418.
Каламофитовые 407.
Камбий 408.
Каменистая кость 277.
Каменные губки 57.
Каменистый канал 152.
Камеры прост. 44. головоног. 210.
Камфарные 442.
Капсула 56.
Капюшон 211.
Каракатицы 232.
Кардинальные зубы 242.
Кашалоты 391.
Каштаны 441.
Квадратная кость 278.
Квадратно-скулов. кость 307.
Квадратный хрящ 276.
Киленогие 203.
Кнстеперые 296, 311.
Кисть позвоночных 273.
Китовый ус 390.
Китообразные 388.
Киты 391.
Кншечнодышащне 265.
Кишечножаберн. черви 72.
Кишечнополостные 62, 92.
Кладокснловые 410, 422.
Клен 442.
Клетки 53.
Клетчатка 394.
Клименви 224, 2У5.
Клиновидн. кости 278.
Клниолистовые 417.
Клоака 362, 366.
Клюв 349.
Клювоголовые 332.
Ключпца 272.
Клыкн 356.
Кобчнковая кость 309.
Ковалевский 38.
Кожа позвоночных 267.
Кожномуск. мешок 189.
Кожное чувство 280.
Кожные жабры 177.
• „ зубы 268.
, кости 266.
Кожный скелет 265.
Козы 383.
Кокколитофоры 401.
Коллатеральн. пучки 408.
Колокол граптолитов 64.
„ медуз 63.
Колонии 56.
Колоски 417, 421, 423.
Кольцевая кайма 65.
Кольцо папор. 422.
Кольчатые черви 193.
Конвергенция 107.
Конечности позвои. 273.
, членнст. 123.
Конодонты 96.
Коисолндац. аппарат 162.
Конхиолин 189.
Концентр, сосуд, пучки 408.
Копролиты 346.
Копытные 373.
Кора 407.
Кораллов полнпы 73.
Корданты 429.
Коренной зуб 357.
Корень раст. 406.
Корень иглок. 154.
Коричневое дерево 442.
Корненожки 38, 53.
Корреляция 20, 107, 350.
Костистые рыбы 295, 302.
Костная ткань 266.
Костные тельца 266.
Кость 266.
Кошки 368.
Крабы 126.
Краевые пластинки 177.
„ влем. глабели 134.
Краниальный отдел 275.
Креднерня 442.
Кремнистые жгутиковые 397.
Крестец 300.
Кровообращение 265, 280.
Кровь 94, 279.
Крокодилы 322, 332.
Крона 154.
Крот 365.
Кроющие пластинки 157.
Круглоротые 284.
Крыло 150, 350, 355.
Крыловидная кость 278.
Крыло-кли ’овидная кость 278.
Крыло-небная дуга 276.
Крылоногие 204.
Крышечка молл. 196.
„ мшанок 89.
„ ракообр. 129.
, рыб 281.
Ксилема 408.
Ктенондная чешуя 282.
Куккерсит 397.
Кукушкин лен 410.
Кутикула 37.
Кучки напор 442.
Кювье 20.
Л.
Лабиринтовые зубы 309, 343.
Лабиринтодонты 313, 347.
Лавровые 442.
Ламарк 20.
Лани 383.
Ланцетник 265.
Ландетовнди. пластинка 172, 173.
Латеральные зубы 242.
Легкие 124. 148.
Легочиые 206.
Леннвцы 369.
Лепидодендрон 416.
Лепоспондиловый позв. 311.
Летающие ящеры 337.
Летучие мыши 337, 366.
Лилин морские 154.
Лнмб 134.
Линзы трилоб. 134.
Лист 406.
Лиственница 438.
Лнствеин. мхи. 410.
Лнстожаберные 289.
Листоногне 126.
Лнсточкн коралл. 75.
Литторальнаи фауна 28.
Лицевой шов 134.
Личинка иглокож 15g.
» мечехв. 145.
„ молл. 190.
„ насек. 150.
„ ракообр. 126.
„ трилоб. 136.
Лишайники 405.
Лобковые кости 272.
Лобная кость 278.
Лобн. части глаб 134.
Ложная стенка 75.
Ложные перегородки 8 .
„ ребра 75.
Ложный столбик 75.
Ложноножки 37.
Локтевая кость 273.
Лопасти головой. 213.
Лопастной орган 190.
Лопатка 271.
Лопатоногие 262.
Лофодонтные зубы 358.
Лофофор 99.
Лошадь 379.
Луб 408.
Лужаикн 193.
Луночка моллюск. 241.
Лучевая кость 273.
, симметрия 152.
Лучеиосцы 275.
Лучи иглокож 152.
, пера 267.
, плавниковые 275.
М.
Магнолиевые 441.
Мадрепоров. пластинка 152, 176, 180.
Макроспора 414, 416, 417,428.
Макроспорофнлл 432.
Макушка плеченог. 104.
„ моллюск. 194.
Малый круг кров. 280.
Мамоит 387.
Мавднбулярный хрящ 278.
Мантийная лнння 238.
Мантийная полость 193, 209.
Мантия плеченог. 103.
„ моллюск. 189.
Маховые перья 351.
Медведи 367.
Медузоид 63.
Медузы 63, 72.
Межглазничи. перегородка 294.
Междоузлия 417, 419.
Межключичная кость 311,326. Межпучковый камбий 409. Межчелюстная кость 278.
Мезентериальв. перегородка 73. Мозентериальн. складки 73. Мезентерий 93.
Мезодерма 53.
Me: оптеригии 272.
Меккелев хрящ 276. Метаморфоз 150. Метаптеригий 272. Метастома 147.
Мечехвосты 145.
Микроспора 414,416,417,428. Микроспорангии 417.
Микроспорофилл 432. Микроструктура и гл ок. 152. Микроструктура корал. 76. Миксины 265.
Миноги 265.
Миртовые 442. Млекопитающие 326, 354. Млечные железы 354. Многобугорчатые 363. Многобугорчат, зубы 363. Многокамерные 43. Многоножки 149.
Многорезцовые 364. Мозазавры 341.
Мозолистость раковины 195. Моллюски 264.
Молоточек 355.
Молочн. зубы 357.
Морские ежи 179. 187, 188.
„ звезды 176, 188.
„ коровы 27, 391.
„ кубышки 189.
в лилии ?54, 188. Мохообразные 410. Мочевой пузырь 281. Мускульные впечатл. 106, 238, , клетки 54, 63.
„ утолщения 73
Мхи 410.
Мшанки 98.
Мыщелки затыл. 308, 314, 328.
Мягкотелые 189. Мясные иглы 56.
Н.
Надвисочяая кость 308. Надкостница 266. Наковальня 355.
Наружная связка 240.
„ стенка зуба 358. Насекомоядные 366, 370. Насекомые 150. Наследственность 17. Науплиус 136. Настоящий столбик 75. Настоящие хвощи 416. Неандертальск. раса 372-
Нёбная кость 278. Нёбно-квадратн. хрящ 278, _ 292.
Нёбо вторичное 319.
„ первичное 319. Неврапофизы 269. Нежвачные 381. Нектон 29. Непарнокопытные 375. ^парный глаз 280.
„ плавник 275. Неподвижн. щеки 134. Неполнозубые 3'18. Неправильные ежн 189. Нарвация листьев. 409. Нервная слет. 57.
„ » молл 190.
Нервные дуги 271.
„ клетки 63-
„ узлы 94, 192. Нетопырь 366. Нижнечелюстный хрящ. 276. Нижние ребра 271. Нижняя челюсть 278. Нимфы 240.
Новые птицы 353.
Нога молл. 189.
, позвоночн. 272.
„ член ист. 122, 124. Ногочелюсти 125.
Ножка граптол 69.
„ иглокож. 152.
„ морск. ежей 176.
, плеченог. 103. Ножные муск. 106. Носовая капсула 273.
„ кость 278. Носороги 376.
Нуллипоры. 405.
Нуммулит, известняк .51-Нуммулиты 48.
О.
Обезьяны 370.
Ободочки зародыша 281. Оболочники 265.
Обонятельные кости 354. Общая полость тела 93, 152. Овицелли 99.
Овцы 383.
Однодольные 439, 440. $
Однокамерные 40. 42.
Однопроходные 362. Однорядные руки 158. Одноцикловый диск 155. Околоротовый скелет 278. Оленерогие 381.
Олени 381.
Ольха 441.
Онтогения 23, 40. Опистоцёльн. позв. 270.
Опорная пластинка 63.
Оральные табл. 155. Оранг-утан 371.
Орех- -111.
Ортоконхн. раковина 244. Осевой канал игол 56, „ оргаи 153.
Осетровые 301.
Основная затылочн. кость 277.
„ кливовидн. 278.
„ пластинка зуба 268.
Остеобласты 226.
Остистый отросток 269. Остроконечия щечные 134. Осьминоги 232, 235. Отбор естественный 17.
„ искусственный 17. Отверстия черепа 308. Открыватели 106-Открытые сосуд, пучки 408. Отолиты 283.
Офиуры 178. Очин пера 267.
П.
Палеоботаника 394.
Пальмы 432, 440, 443. Пальцеходящие 361. Пальцы 273.
Панцырноголовые 309, 311. Панцырные рыбы 284, 311. Панцырь броиеносц. 368.
„ динозавров 334.
, крокодил 334.
в черепах 326.
Папоротники 410, 422, 429. Папоротниковые 422. Папоротникообразные 406,410. Папоротникосемевяые 429. Парабазальн. табл. 155. Параподни 95, 97. Парасфеноид 278. Париетальный орган 280. Парнокопытные 381, Парные плавн. 272. Паукообразные 148. Пахиодонтный замок 243. Пелагическая фауна 28. Первичная древесина407,408. Первичная кора 409.
„ раковяна 212, 217. сегментация 265.
„ чашечка 157.
Первичное нёбо 319, 843.
, перо 267.
Первичные копытные 373.
я перегородки 78. Первичный череп 276-Перегородки головоногих 212. „ коралл. 74.
межглазные 349, 351.
Перегородки носорогов 376. , плеченогих 105.
, простейших 34.
Переднежаберные 144.
Передн. клиновидная 278.
Перемычки 75.
Перепончатокрылые 151.
Перерывы фаун 23.
Передннеи 398.
Перидерм 66.
Период 31.
Перистом 179.
Перихондральная кость 266.
Перламутров слой 189.
Перо 367.
Перышки папортн. 422.410.
Песчаные раковины 41.
Петалоидн. амбулякры 181.
Печеночники 410.
Пещерный медведь 367.
Пищевар. канал 93.
Плават. пузырь 295.
Плавники 273.
Плагиодонтный замок 242.
Плакоидные чешуи 26,268,281.
Планктон 29.
Пластинчатожаберные 237, 261.
Пластическая сила 35.
Платан 440, 442.
Платицельные позв. 335, 359.
Плауновые 410, 412.
Плауны 410, 412.
Плевральные ребра 271.
Плевродонтные зубы 319.
Плевроконхные формы 244.
Плевроцентры 302.
Плезиозавры 330.
Плечевая кость 273.
„ табл. 152.
Плечевой пояс 271. Плеченогие 102, 121.
Плечо мор. лилий 158. Плодолистики 439, 428, 437.
Плоские позвонки 270.
Плотоядный зуб 366.
Плюсневые кости 273.
Подвздошная кость 272.
Подвижные щеки 134.
Подглазничное кольцо 278.
Подъязычная дуга 276.
Позвонки морск. звезд 178.
, позвоночных 269.
Позвоночные 265.
Позвоночный столб 269, 352.
Покрытосеменные 439.
Полип гидроидн. 65.
, коралл. 73.
Полипияк граптолитов 69.
Полидоид 65.
Полорогие 383.
Полость тела кишечноп. 62.
, , общая 93.
Полужесткокрылые 150.
Полуобезьяны 370.
Полупластинки 181.
Полупозвонки 302.
Поперечноротые 290.
Поперечные отростки 271.
Поры иглок. 166.
Послед 281.
Последовые 361, 366.
Почкование 38, 56.
Пояса конечностей 271.
Правильные ежи 189.
Предплюсна 273.
Пресмыкающиеся 318.
Преформированн. кость 266.
Призматическ. слой 189.
Прикрепление иглокож. 153.
Прилистники 427.
Приматы 370.
Проводящие пучки 408.
Прозенхима 406.
Происхождение видов 7.
Проксимальный 69.
Проптеригий 272.
Простейшие 37.
Противопол. перегородка 79.
Протонауплиус 136.
Протоплазма 37.
Протоплазматическое тело 38.
Процельные позв. 270.
Прямокрылые 150.
Псевдоборниевые 417.
Псевдоподии 37.
Псилотовые 410, 417.
Псилофитовые 410.
Птеригоподий 289.
Птеродактили 339.
Птероподовый ил 205.
Птицы 348.
Пузырчатая эндотека 76.
Пупок 281.
» раковины 195.
Пучки кораллов 76.
Пучки сосудистые 408.
Пучковый камбий 409.
Пыльца 428.
Пястные костн 273.
Пятилуч. симметрия 152.
Пятипал, конечность 272.
Пятка зуба 357.
Пяточная кость 333, 360.
Р.
Равномускульные 238.
Равноногие 130.
Равноспоровые 412, 417.
Радиальная симметрия 73, 87. Радиальные табл. 155, 161. Радиолярии 38, 51.
Радуля 190, 210.
Развитие лопасти, линии 217.
„ особи 22.
» позвоночн. 281.
Развитие скачками 21.
Размножение высш. 54, 65.
» прост. В1'.
Разномускульные 238. Разиоспоровые 412, 417. Раковина ежей 179.
» молл 189.
„ плеч 103.
, прост. 42.
„ ракообр. 125.
Раковинчатые 127.
Ракообразные 125, 144.
Рако-скорпионы 144.
Расхождение признаков 21. Расщепленные ножки 125. Рахитомные позв. 269, 311. Ребра 105, 241, 269, 271, 320. Резец 356.
Рептилия 318, 347.
Решетчатая кость 354.
Рога млекоп. 354, 376,377, 381.
Роговые нити 2 2.
„ образов, позв. 267.
Ромбовые поры 166.
Ростр 131, 232, 319, 345.
Рот 63.
Рудисты 245, 258.
Руки 103, 154.
Рукокрылые 366.
Ручной аппарат 105. Рыбы 281, 281.
Рыльце трилобита 134.
С.
Саговиковые 429, 432. Свайка 74.
Свертывание трилоб. 135.
Свиньи 381.
Связка 240.
Связочные ннмфы 240.
Сегментация 93, 123, 265.
„ трилобитов 135.
Седалищная кость 272.
Седло головоног. 212.
„ плеченогих 108.
Седлообразные позв. 270.
Селахии 268.
Селенодонтн. зубы 358.
Сельдь 305.
Семенная клетка 54.
Семенные 394.
Семенные папоротники 429.
Семиты 183.
Семя 54.
Семяпочка 428, 431, 437, 439.
Сепия 210.
Сеитал’ н. пластинки 105.
Септальное углубление 80. Сердце 124, 190, 279.
Сердцевидные ежи 179.
Сердцевина 408.
Сердцевинные лучи 409.
Сережкоцветиые 441.
Сериальное запястье 361.
Серозная оболочка 281.
Сетчатокрылые 150.
Сигиллярии 416.
Синезеленые водор. 396.
Синофоры 70.
Синус моллюсков 196, 238.
„ плеченогих 107.
Сиреневые 392.
Систематика геолог. 30.
„ зоолог. 23.
„ палеонтол. 23.
„ палеофит. 394.
Ситовидные трубки 406.
Сифои 238, 428.
Сифонные дудки 212, 215.
Сифонные лопасти 217. Сифоностомные молл. 195,201. Скаты 289, 290, 293.
Скелет гидрокоралл, 67.
„ граптолитов 62.
„ губок 55, 61. , иглокож. 152, 154. „ коралл. 74, 87.
„ позвоночн. 265.
„ прост. 42.
Скелетн. элем, растений 408.
Склеротика 276.
Скорпионов, пауки 149.
Скорпионы 149.
Скуловая дуга 278, 356.
„ кость 356.
Скульптура раковины 220.
Слезная кость 278, 307.
Слизевики 398.
Слизевые каналы 280.
Слоевище 406, 410.
Слоевищные 394.
Слоевцовые 396.
Сложная связка 240-
Сложи, табл, ежей 180.
Слоны 387.
Слуховая капсула 276.
Слуховые кости 361, 277.
Смена зубов 283, 357.
Собаки 367.
Соединительная ткань 54, 662.
Солении 88.
Солнечники 38.
Сосна 438.
Сосочек 268.
Сосудпсто-волокн. пучки 408.
Сосудистые растения 408.
Сосуды раст. 408.
Сохранение остатков 15.
Сочленовные отростки 271.
Сошник 278.
Сперматозоид 402,405,432,435.
Спикули 51, 55.
Спинная струна 269.
Спинной плавник 275.
Спиракули 174.
Спираль молл. 193. » плечен. 103.
Спонгин 56.
Спорангии 422.
Споровые 394.
Спорофит 405.
Споры 38, 428. .
Сравнит, анатомия 26.
Среднее ухо 277.
Средняя решетчатая кость 278.
Статоцисты 64, 95, 190.
Стволы некой, раст. 418, 415.
Створки водор 399.
„ молл. 237.
, плечеи. 103.
Стебель 154, КО.
„ раст. 406.
Стегоцефалы 309.
Стенка кораллов 74.
Стереоепондил. позв. 2’10, 311.
Стигмарии 416.
Стигмы 148.
Столбик коралл. 75. „ мол. 195, 258.
„ позвон. 307, 341, 355.
Столон 88.
Стрекательн. клетки 62.
Стремя 355.
Строматопориды 67.
Ступня 273.
Сумочвые кости 363.
Сумчатые 363.
Сутурная линия 212.
Схождение признаков 107.
Сцеплянки 399.
Сяжки 123.
Т.
Таблички иглокож. 179.
Тазовой пояс 272.
Тайнобрачные 395.
Таксодонтный замок 242.
Тапиры 375.
Таранная кость 360.
Текальные поры 166.
Текодонтные зубы 320, 3^3, 356.
Тектиспондилов. позв. 290.
Тельсон 145.
Теменная кость 278.
Теменной орган 280. Темноспондилов. позв, ЗЦ. Теплокровные 280.
Тетраспорангии 403.
Ткани 54, 406.
Тополя 441.
Торфяные мхи 410.
Трабекулы 76.
Трахеи 124, 148.
Трахеиды 441.
Трепел 399.
Триконодонтн. зубы 357.
Трилобитовая стадия 145 Триторы 295.
„Тритуберкуляри. зубы 357.
Трубкожилы 95.
Туловище 265. .
Туннкаты 265-
Тур 383.
Тюлени 367.
Тюльпанное дерево 441.
У.
Узлы 401.
Узлы гервн. 94, 192.
Унисериальнын плавник 272.
Усики 160.
Усоногие 128.
Устье 39.
Утконос 364.
Ухо 274.
Ушки боковые 220.
Ушные кости 284.
Ф.
Фаланги 273.
Фарфоровидн. слой 189.
Фасеточн. глаза 124, 134.
Фасциолн 183.
Фация 30.
Фибры коралл. 74.
Фикус 441.
Филлоспондил. позв. 311.
Филогения 23.
Флоэма 408.
Фораминиферы 38, 49.
Фрагмоконус 233.
Фукоиды 403.
Фулььры 299.
X.
Харовые 9, 401
Хвойниковые 429, 439.
Хвойные 429, 435.
Хвост 265.
Хвостатые амфибии 310, 316.
Хвостовой плавник 275, 283.
„ сегмент 132.
Хвощевидные 410, 417.
Хвощеобразные 418.
Хвощи 408.
Хемера 31.
Хепростребовые 418.
Химеры 289, 294.
Хитин 41.
Хитоны 191.
Хищные 366.
Хлебной Царево 441.
Хлорофилл 394.
Хоаны 303, 309.
Хоботные 384.
Хоботок медуз 63.
„ морск. лилий 159.
Холоднокровные 280.
Хронология 29.
Хрящ 266.
„ известковый 269.
Хряп евой череп 277.
Хрящевые ганоиды 300.
„ рыбьи 272.
ц-
Целлулоза 394.
Цёломич. полость 93.
Целыюголовые 294.
Цемент зубов 268.
Цененхима 76.
Ценогения 26.
Цёнбсарк 76.
Центральная капсула 51.
Центральный кружок 176.
Цератиты 224, 235.
Цефалоподы 209.
Циклоидные чешуи 282, 295. Циклоспондилов. позв. 290. Циклы морск. лилий 155.
Циприс 129.
Цистоидеи 164.
Цистокарпий 404.
Ч.
Чашечка коралл. 74.
„ морск. лилий 154. Человекообразные 371. Челюсти молл. 190.
„ мшанок 99.
„ червей 96.
„ члени.стон 123.
Челюстная дуга 276.
Челюстные кости 278.
Червеобразные молл 191.
Черви 93.
Чередование поколений 65.
Череп 275.
Черепахи 325.
Чернильный мешок 210.
Четвероногие 270.
Четырехжаберные 210.
Чешуекрылые 151.
Чешуи рыб 268.
„ четвероног. 267.
Чешуйчатая кость 278.
Чешуйчатые 340.
Членистоногие 123.
Членистость жаб. дуг. .279.
Чувство равновесия 64, 95.
III.
Шагрень 282, 309.
Швы головой 218.
„ трилоб. 134.
Шизодонтный замок 242 Шимпанзе 371.
Шип хвостовый 139.
Шипы плеченогих 110.
щ
Щеки трилобитов 133.
Щетинки плеченогих 110.
Щетинки червей 95.
Щетинконогие 96.
Щит трилобитов 135.
„ черепах 326.
Щиток вершинный 180.
„ пластинчатожаб. 241.
„ рыльцевой 134.
Щупальца 155, 262.
Э.
Эволюция 20.
Экзотекальн. образован. 76.
Экология 27.
Эктодерма 53.
Эмаль 268.
Эмболомерн. позвонок269, 311.
Эмбриология 24.
Эндосифоа 214.
Эидотека 76.
Эндохондральн. кость 266.
Энтодерма 53.
Эпидермис 94, 267.
Эпитека 75.
Эпителий 54.
Эпифиз мозга 280.
Эпоха 31.
Эра 31.
Я.
Явнобрачные 395.
Ядро внешнее 34.
„ внутреннее 34.
„ клеточное 37.
Яйцевая клетка 54.
Яйцеклетка 428. •
Ячейка 74.
Ящерицы 340.
Ящеры летающие 337.
Сдано в набор 27 ноября 1931 г.
Поступило к печати 13 марта—23 мая 1932 г.
Формат бумаги 62 X 94.
Количество бумажных листов 14б/1в.
Количество печ. знаков в бум. листе 106506.
Инд. Г(Р)-56-5-2.
Ответственный редактор Е. Фальк.
Технический редактор А. И. У с о в а.
Горгиз № 2723-28.
Ленгорлит № 35196.
Заказ № 1079. Тираж 18500 зкз.