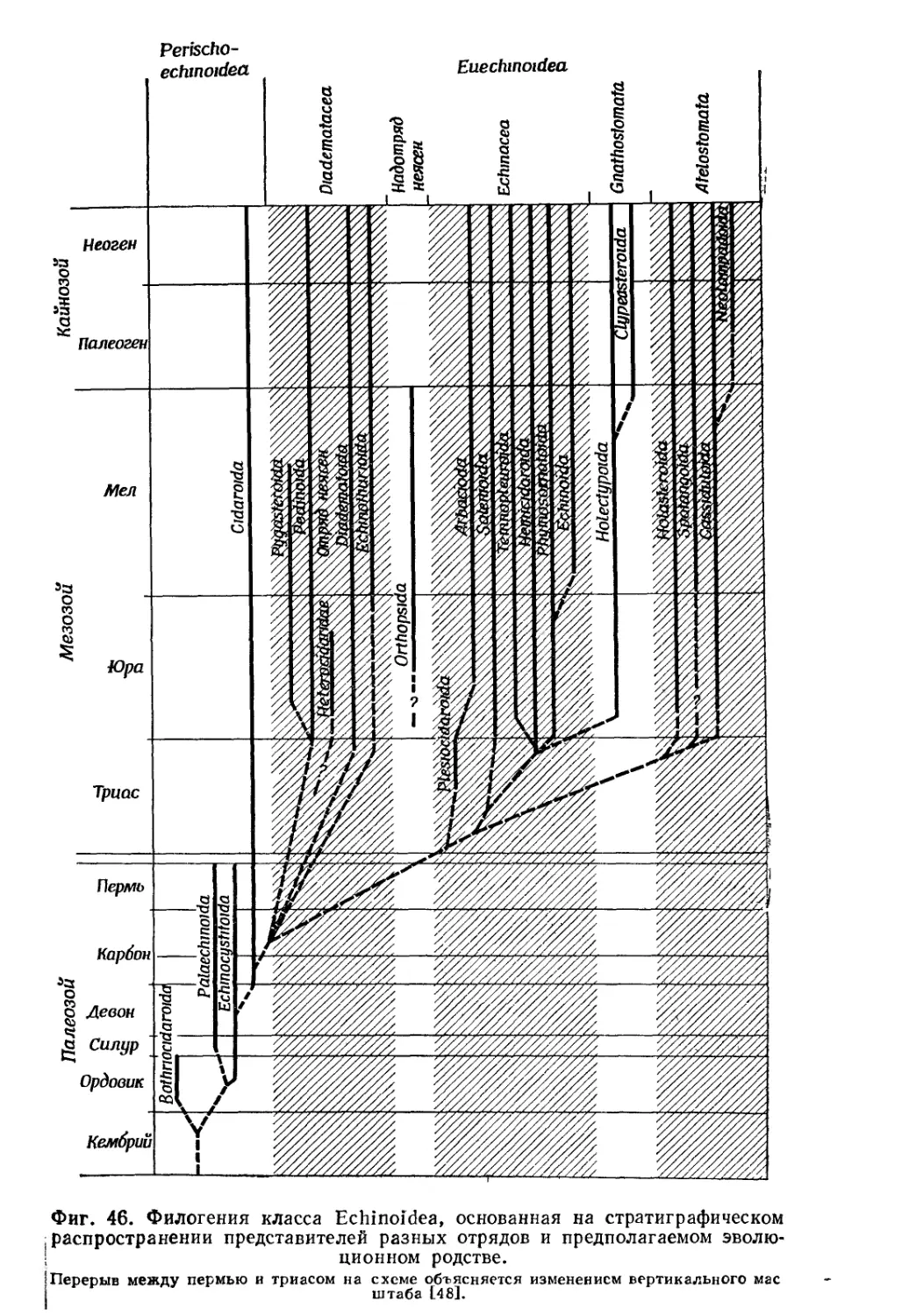
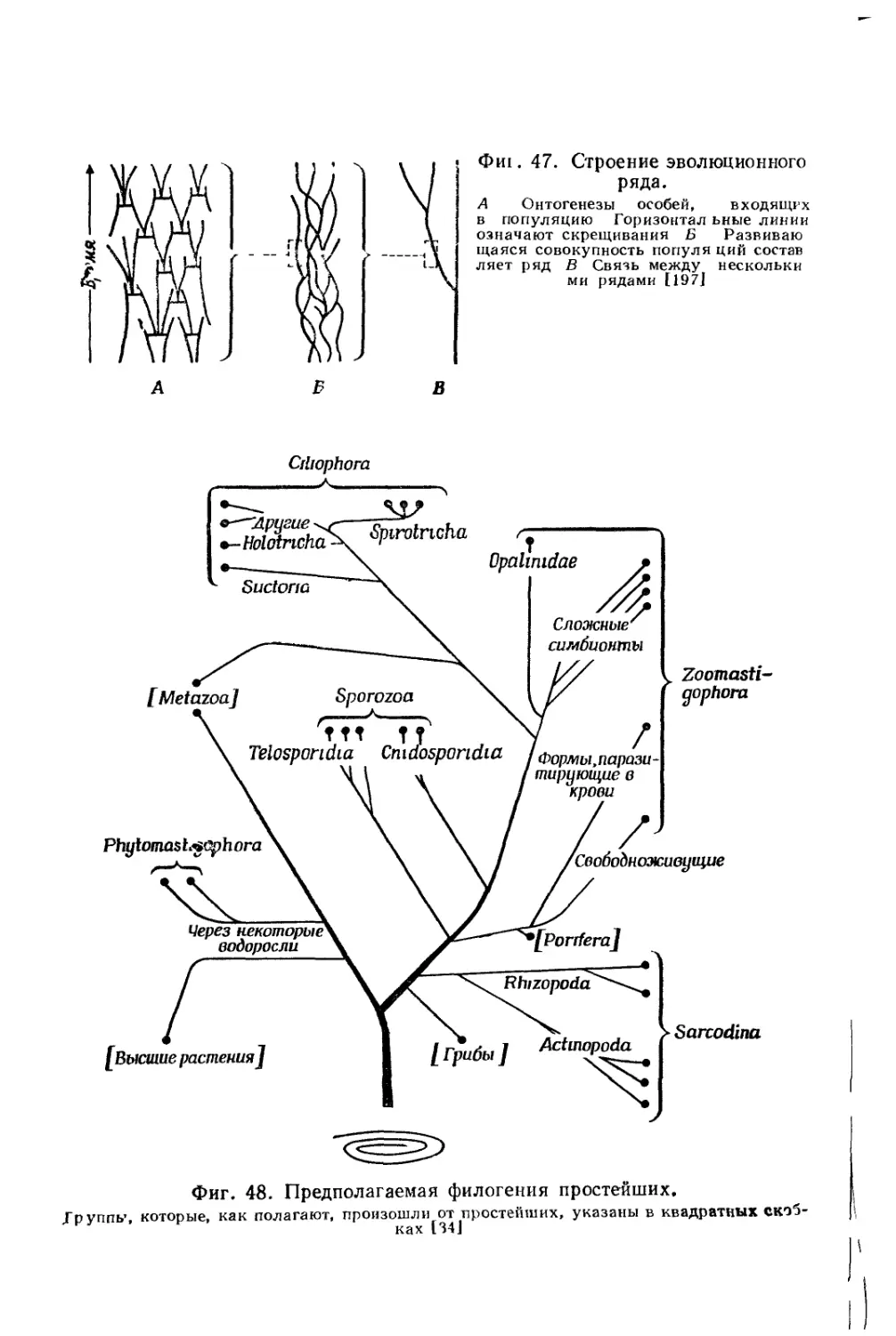
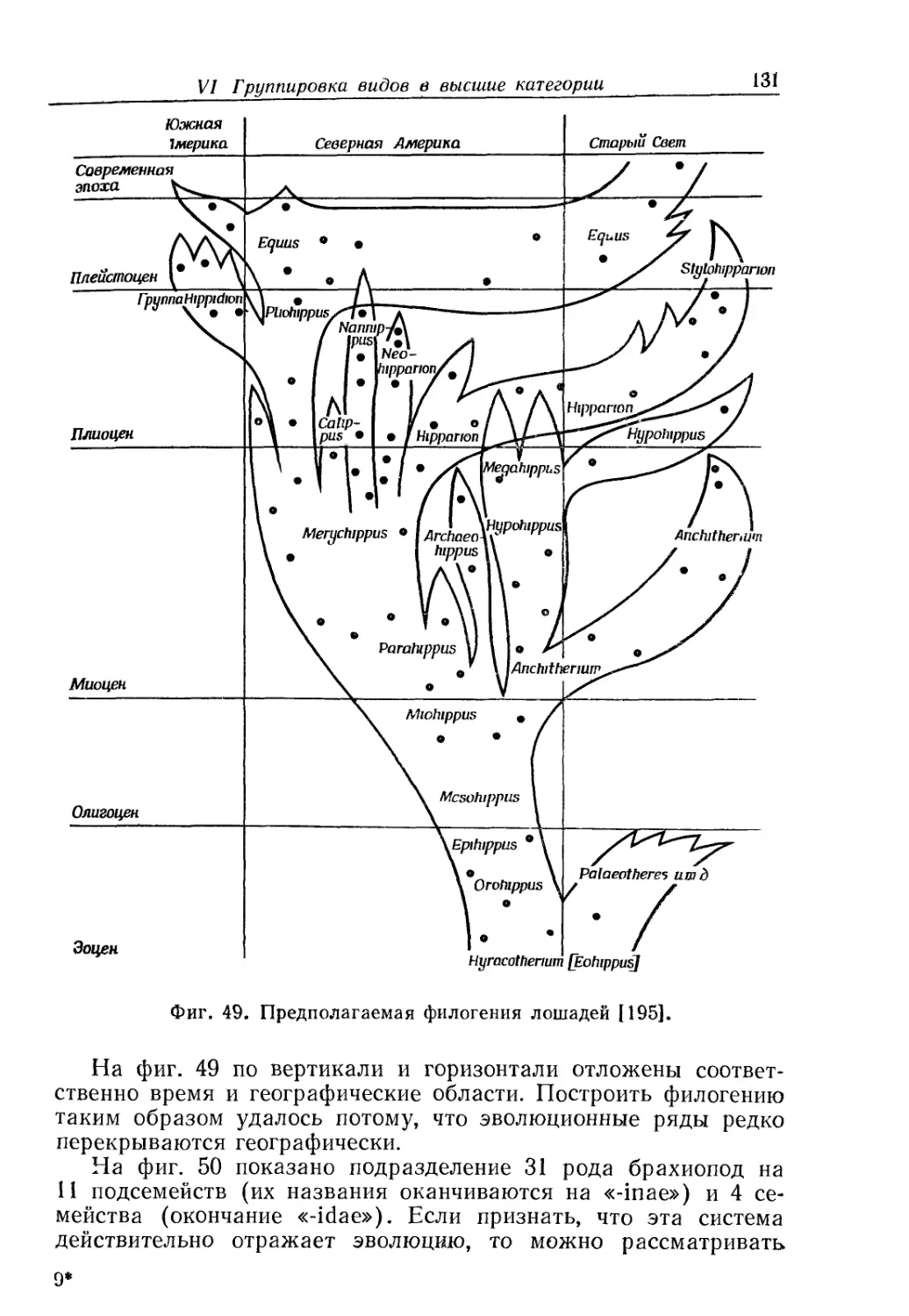
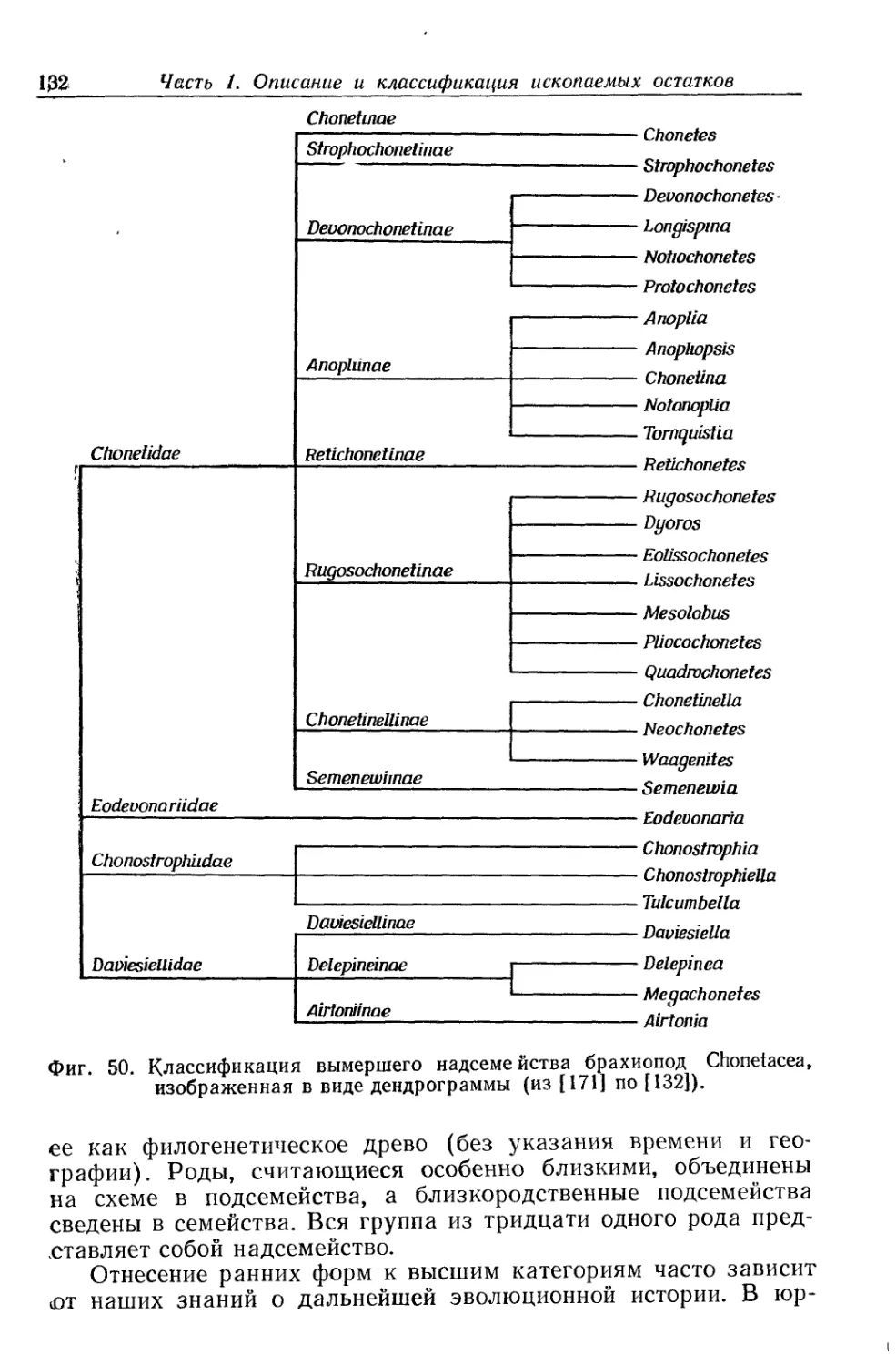
/











































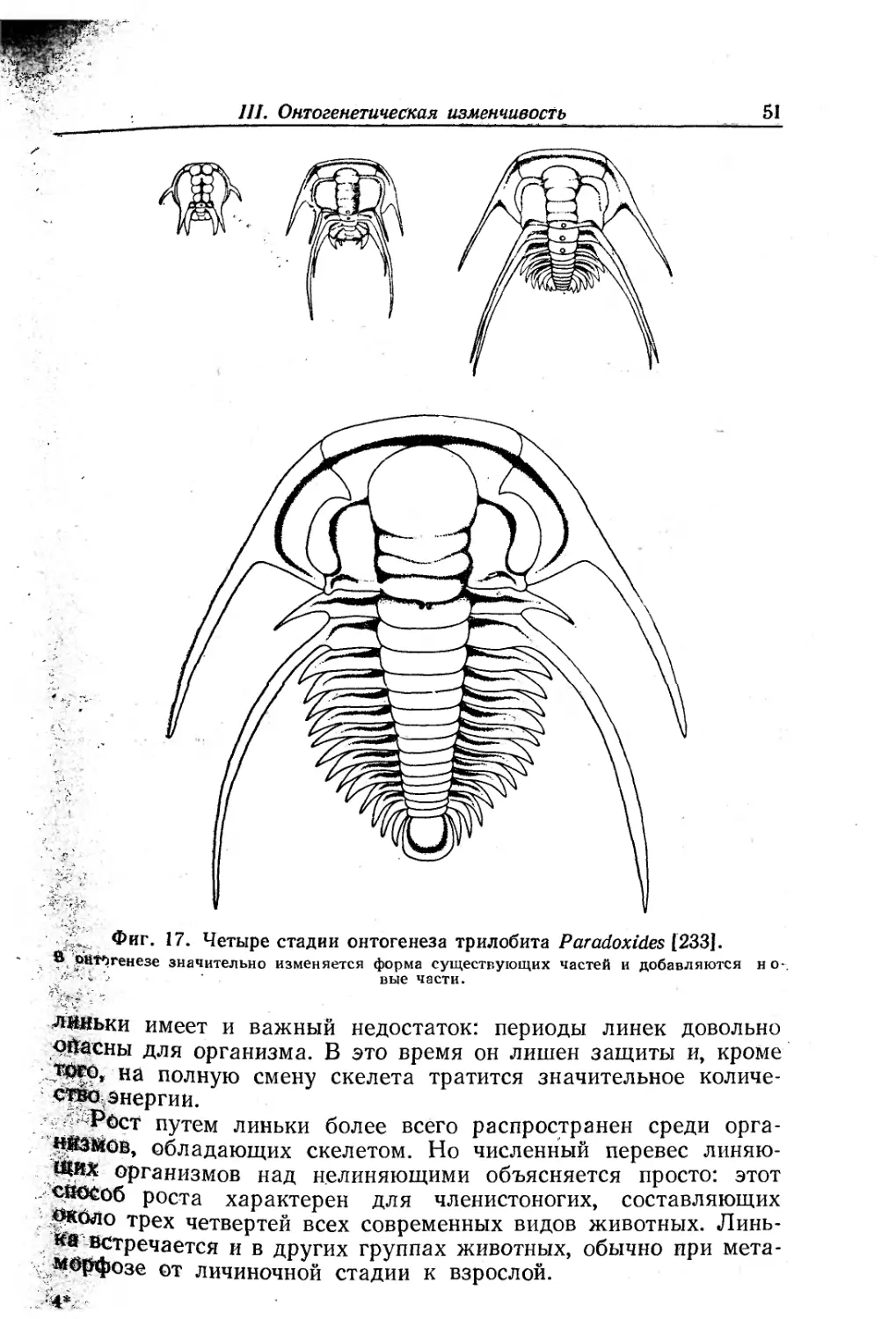
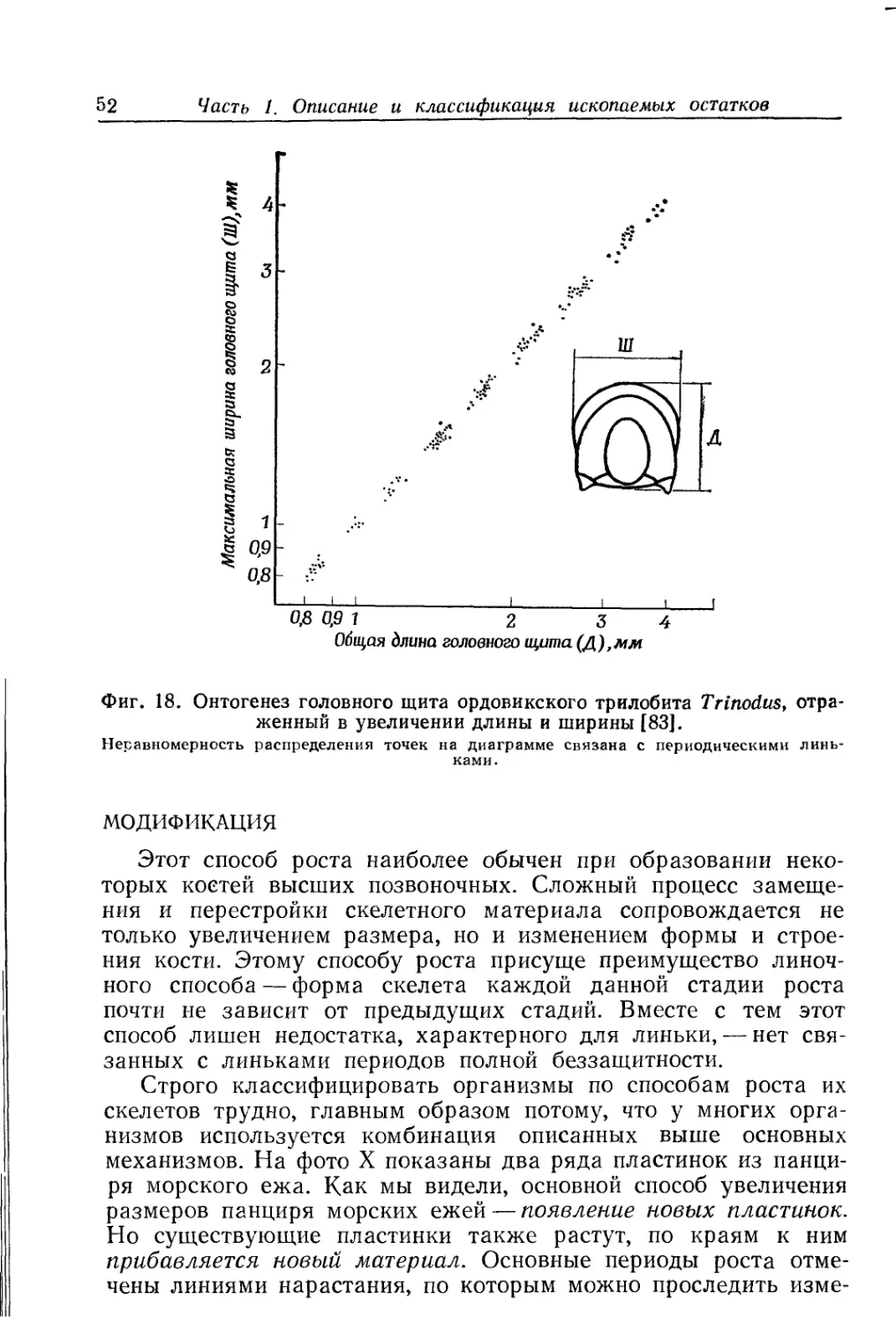

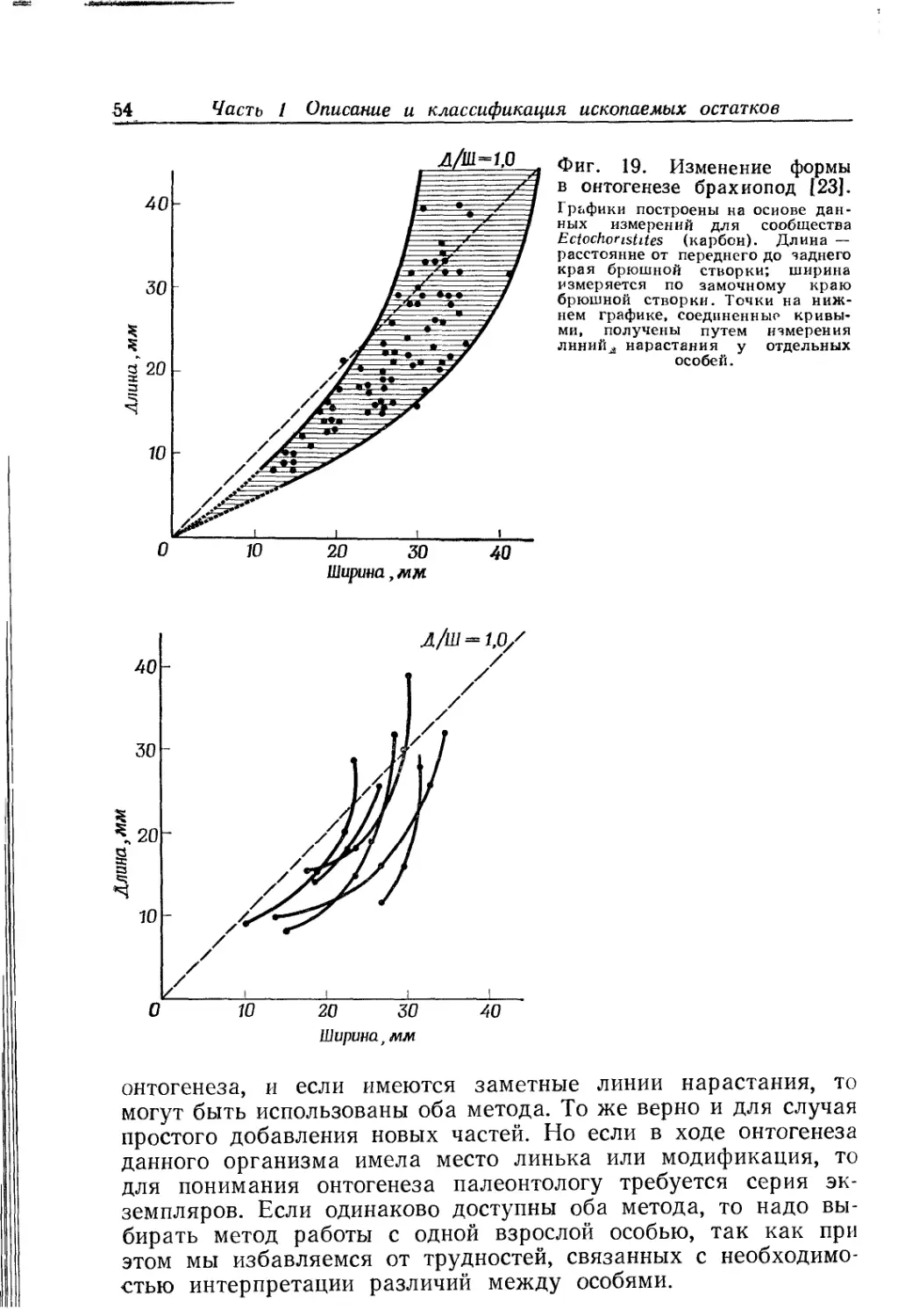

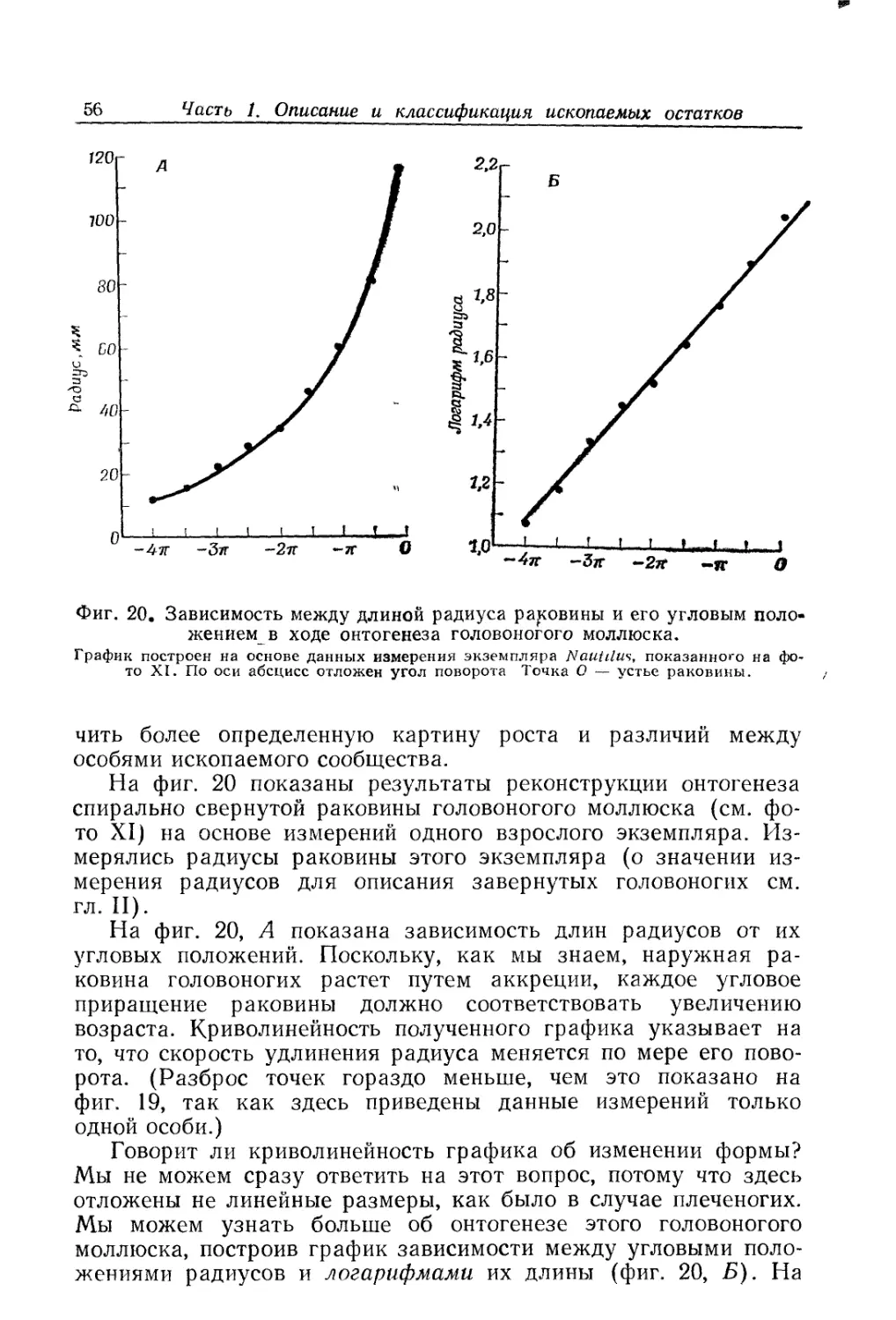




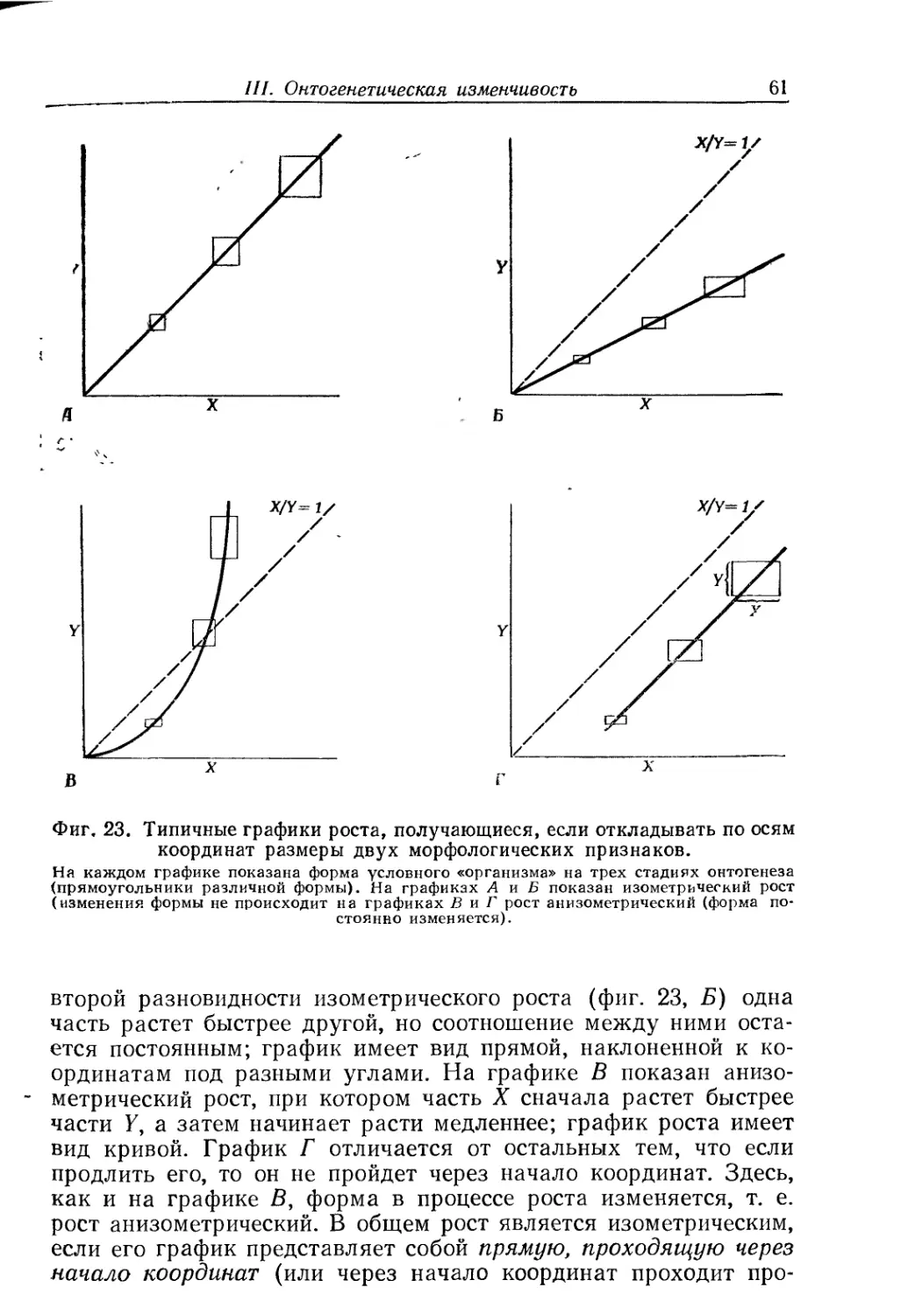







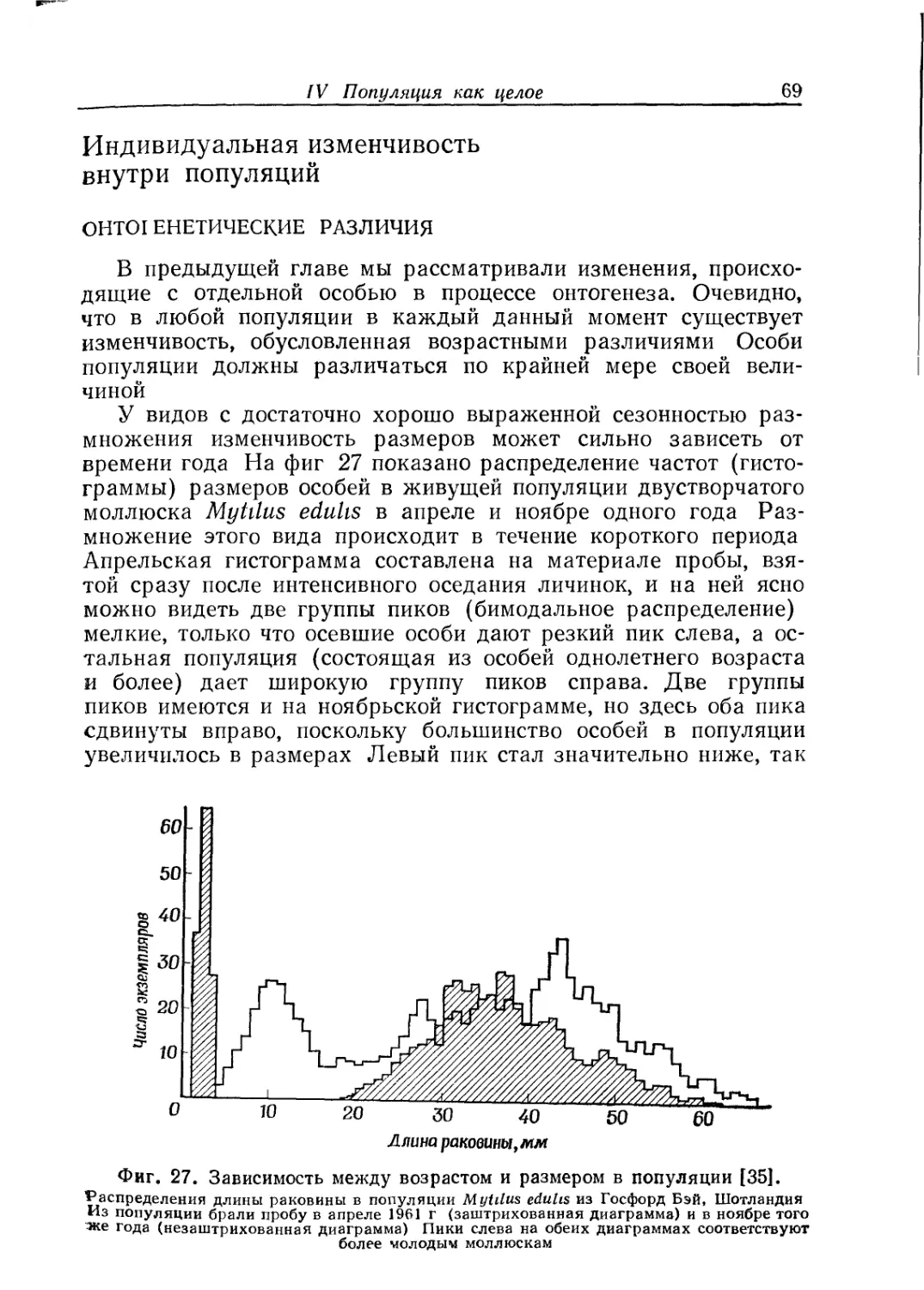

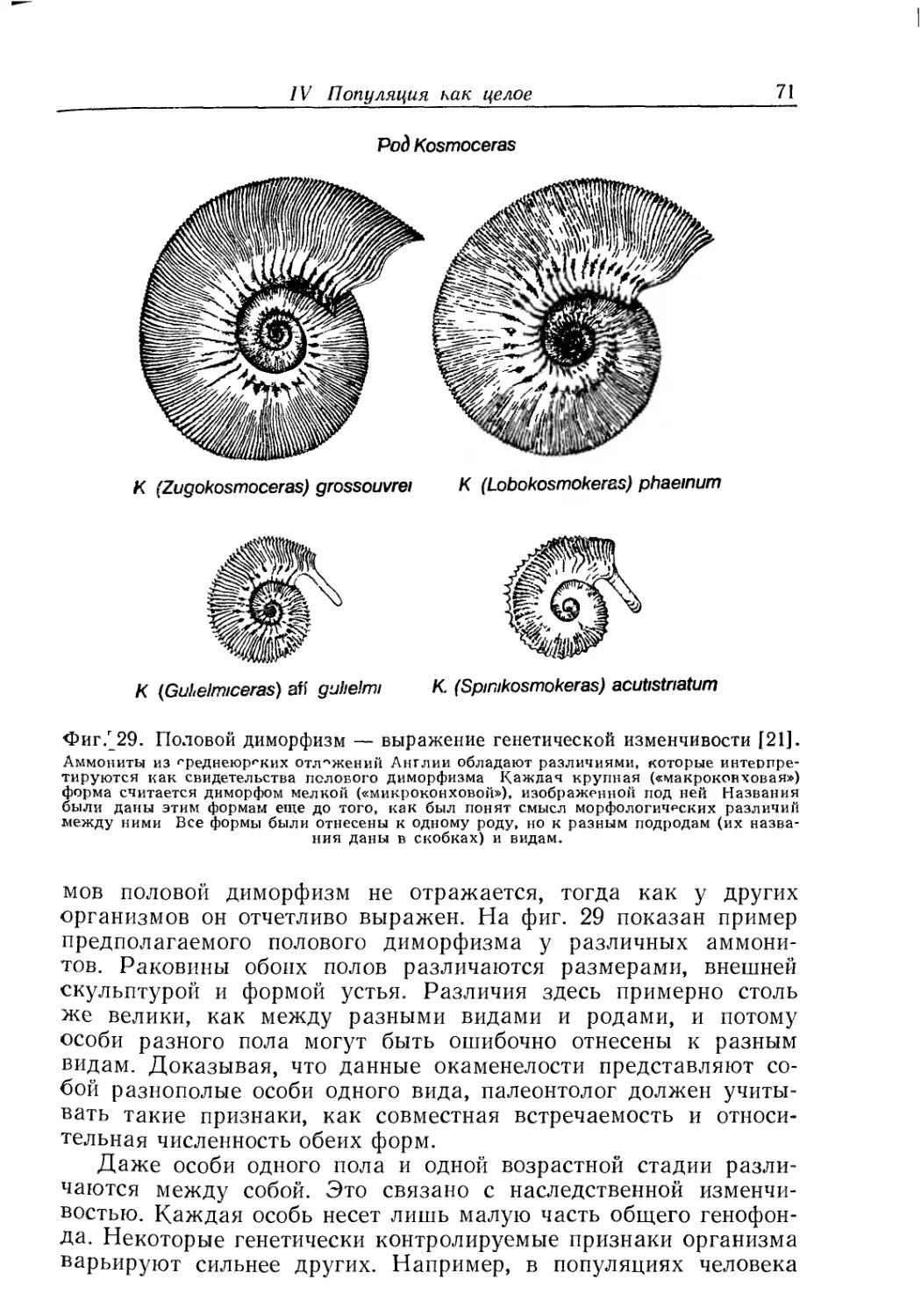










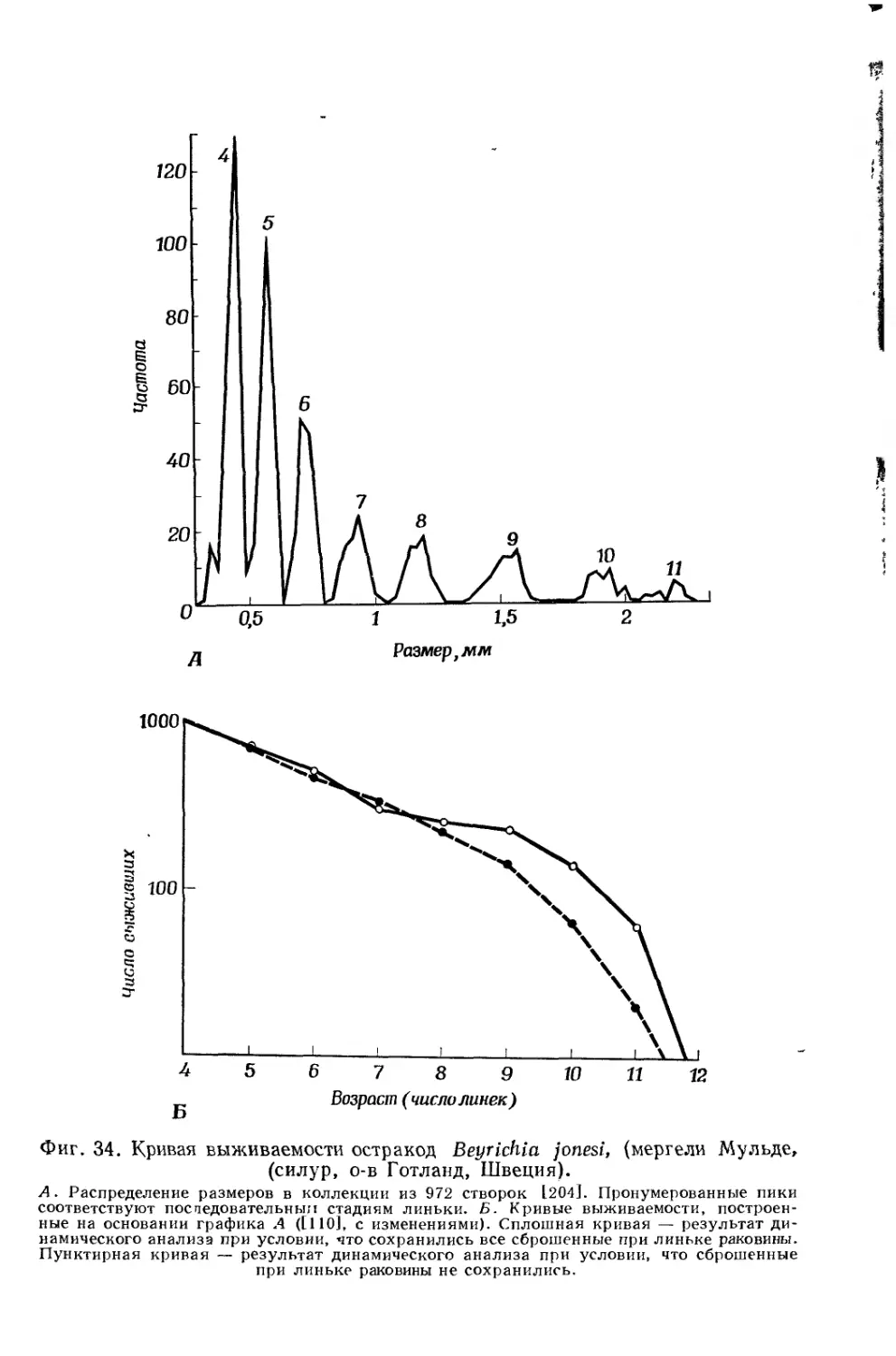








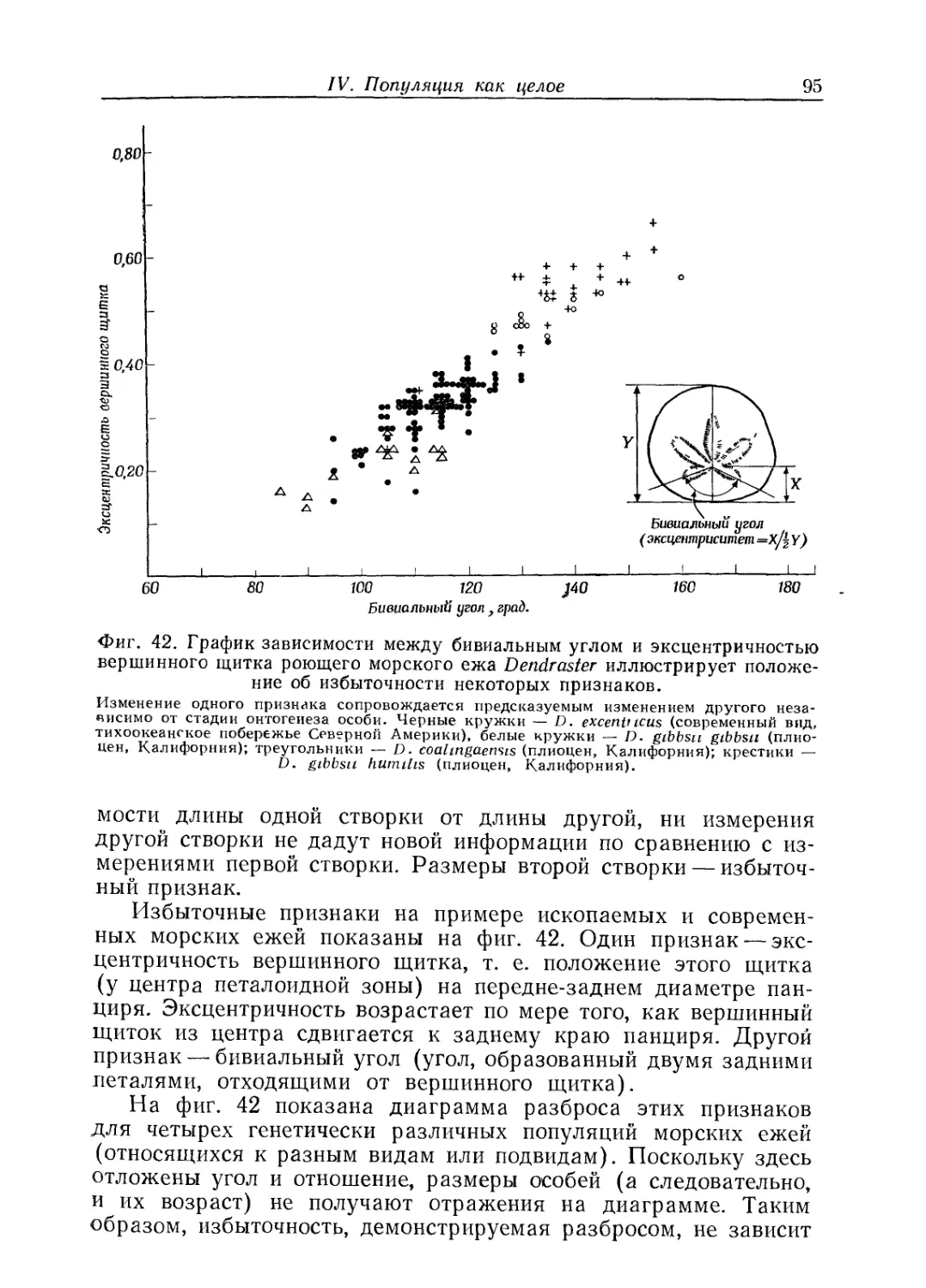












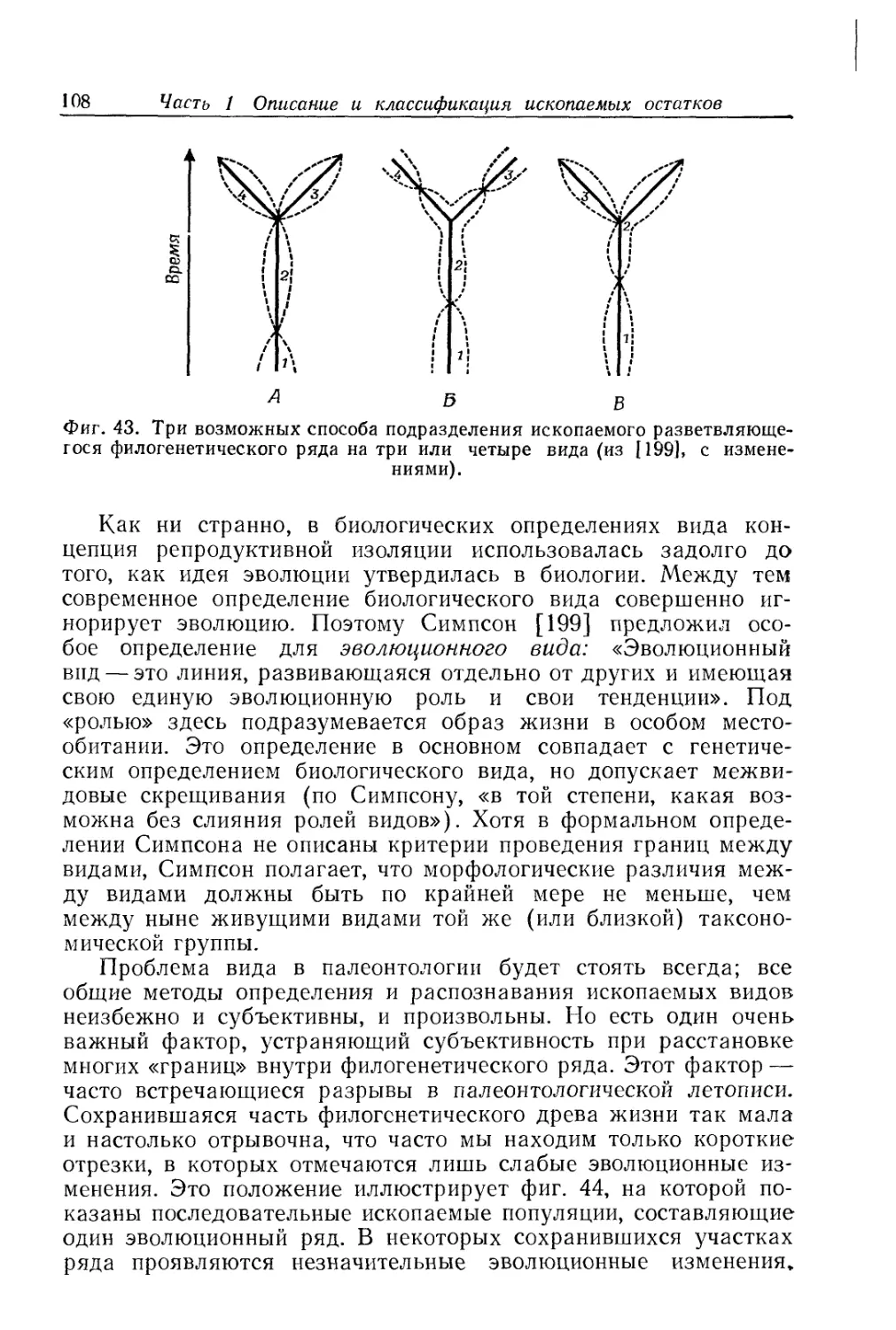
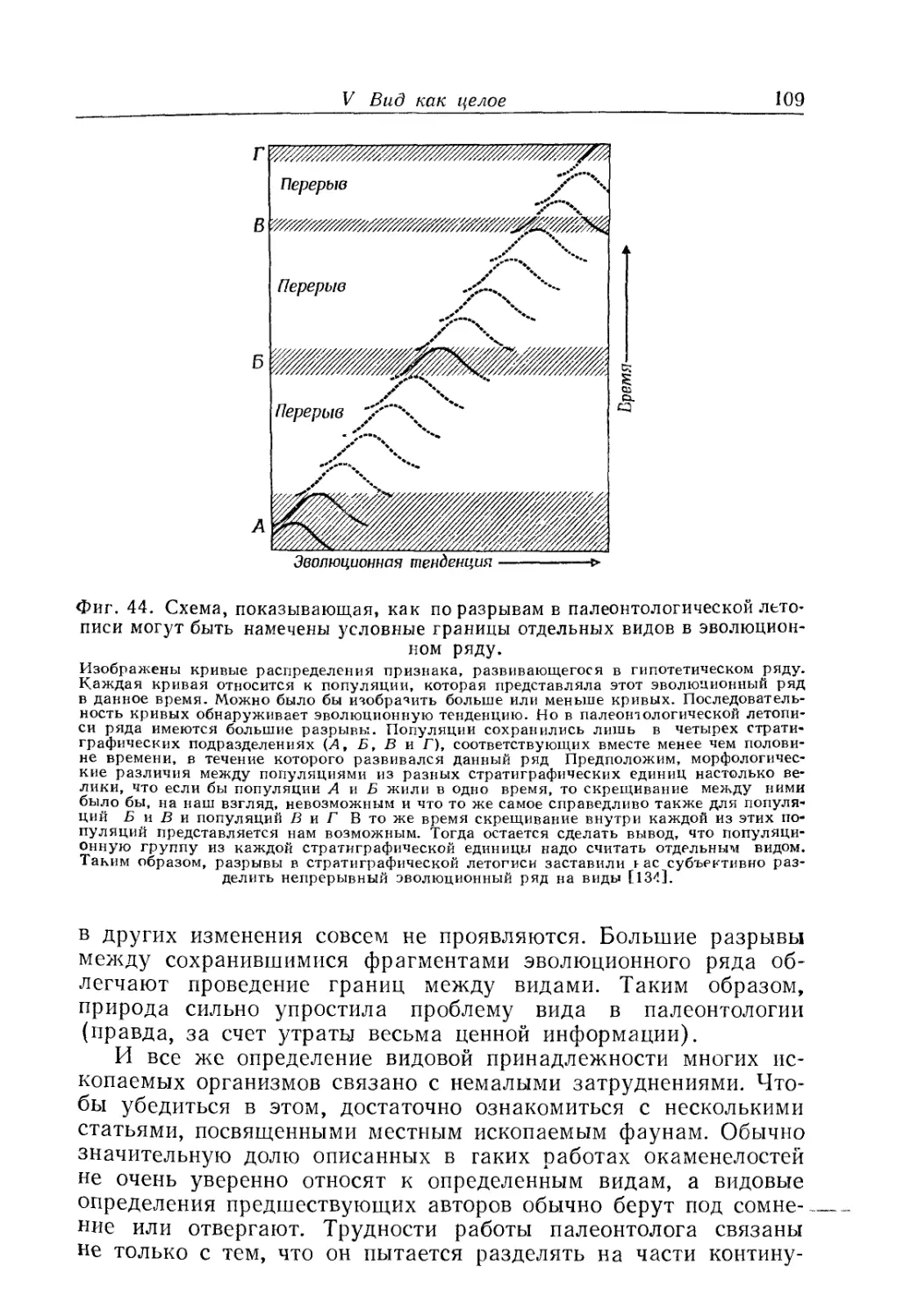


















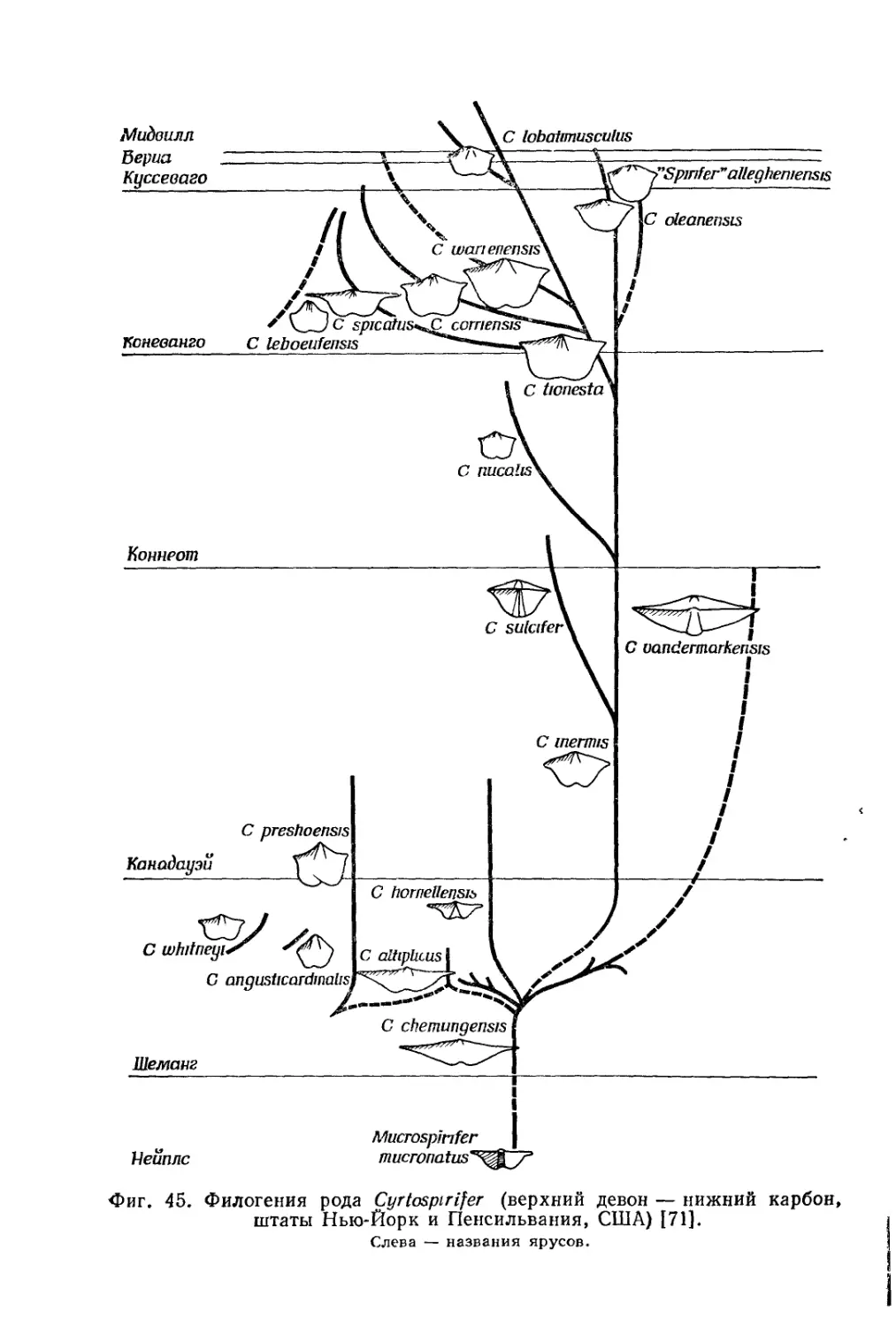



































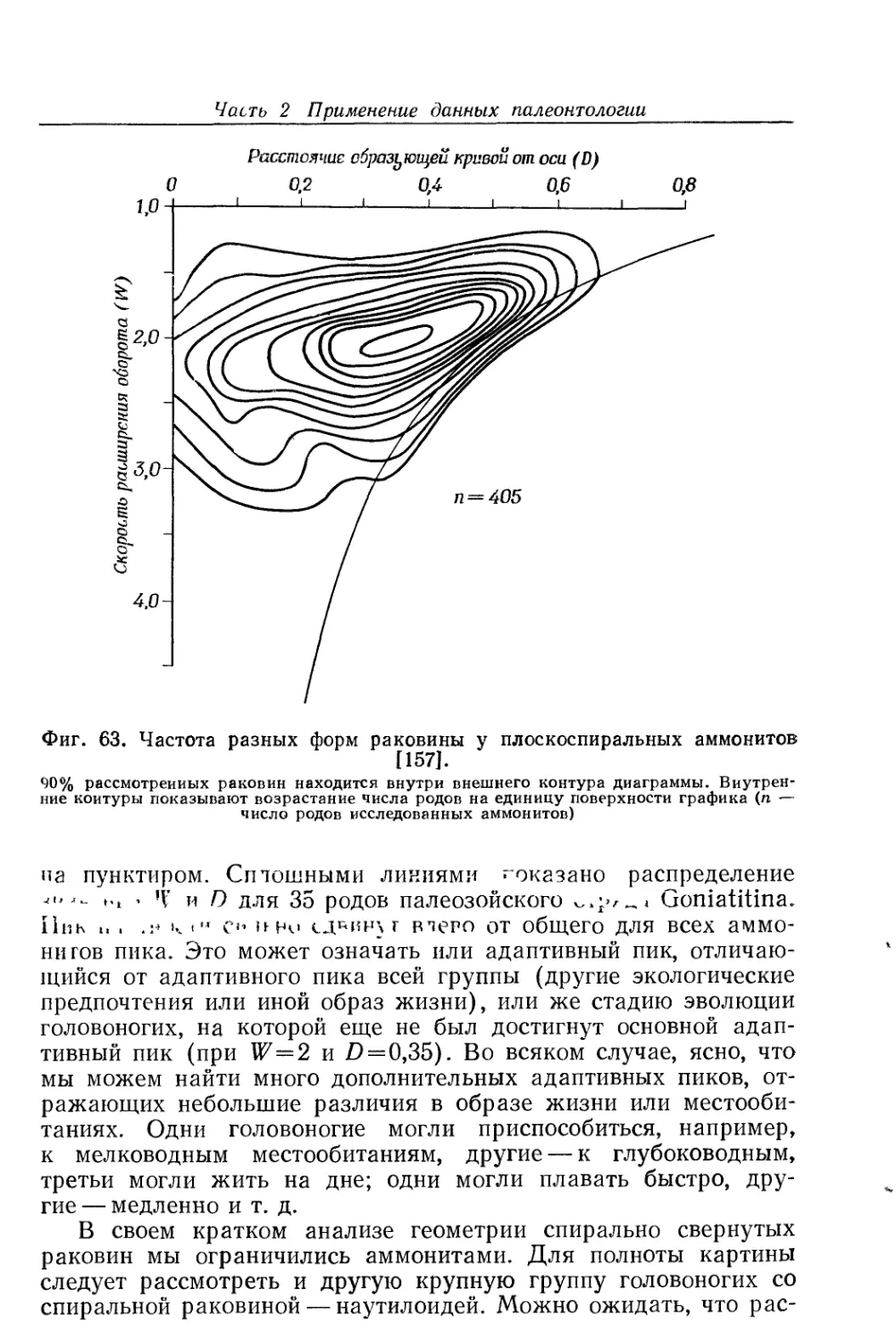



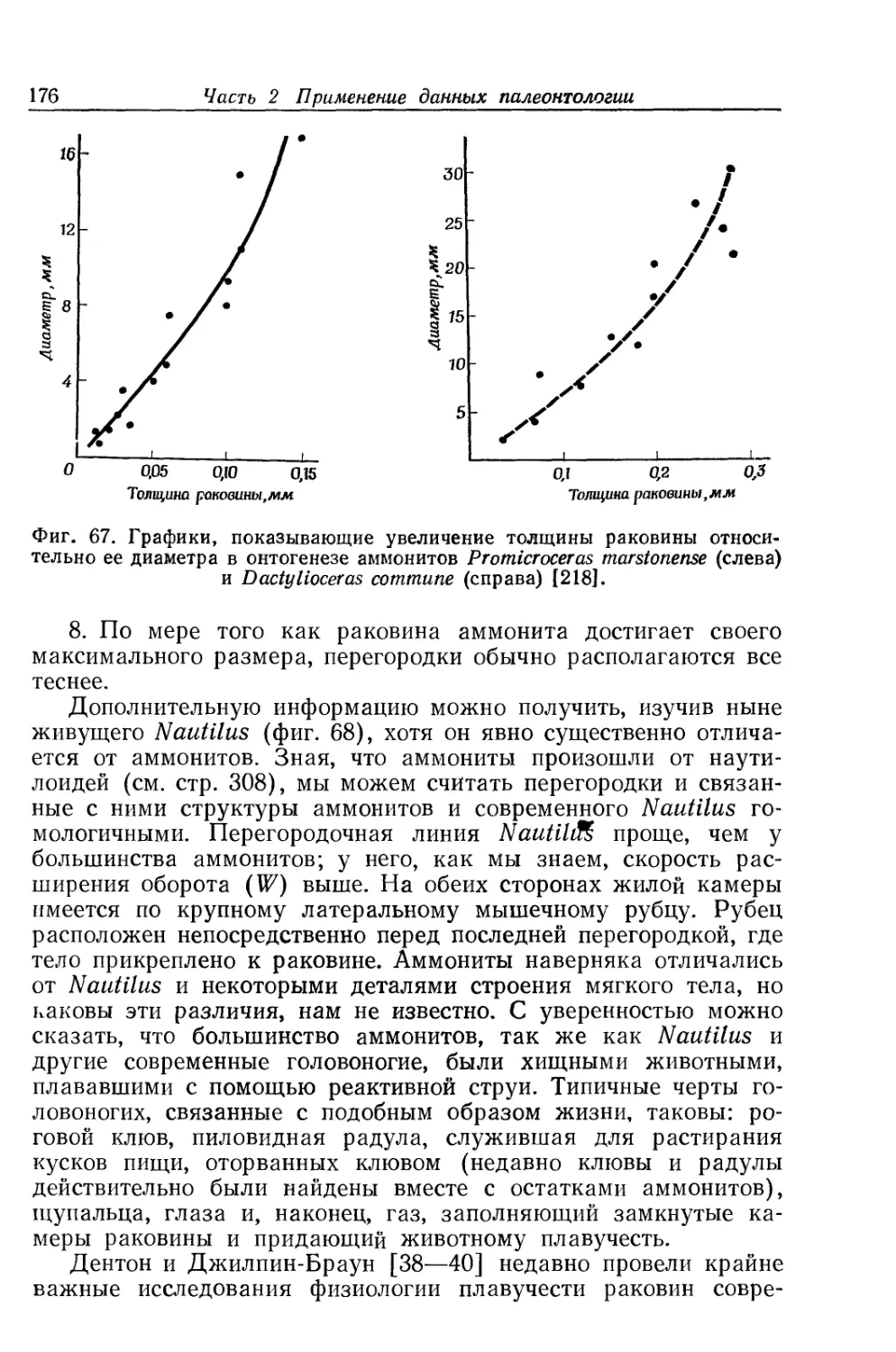

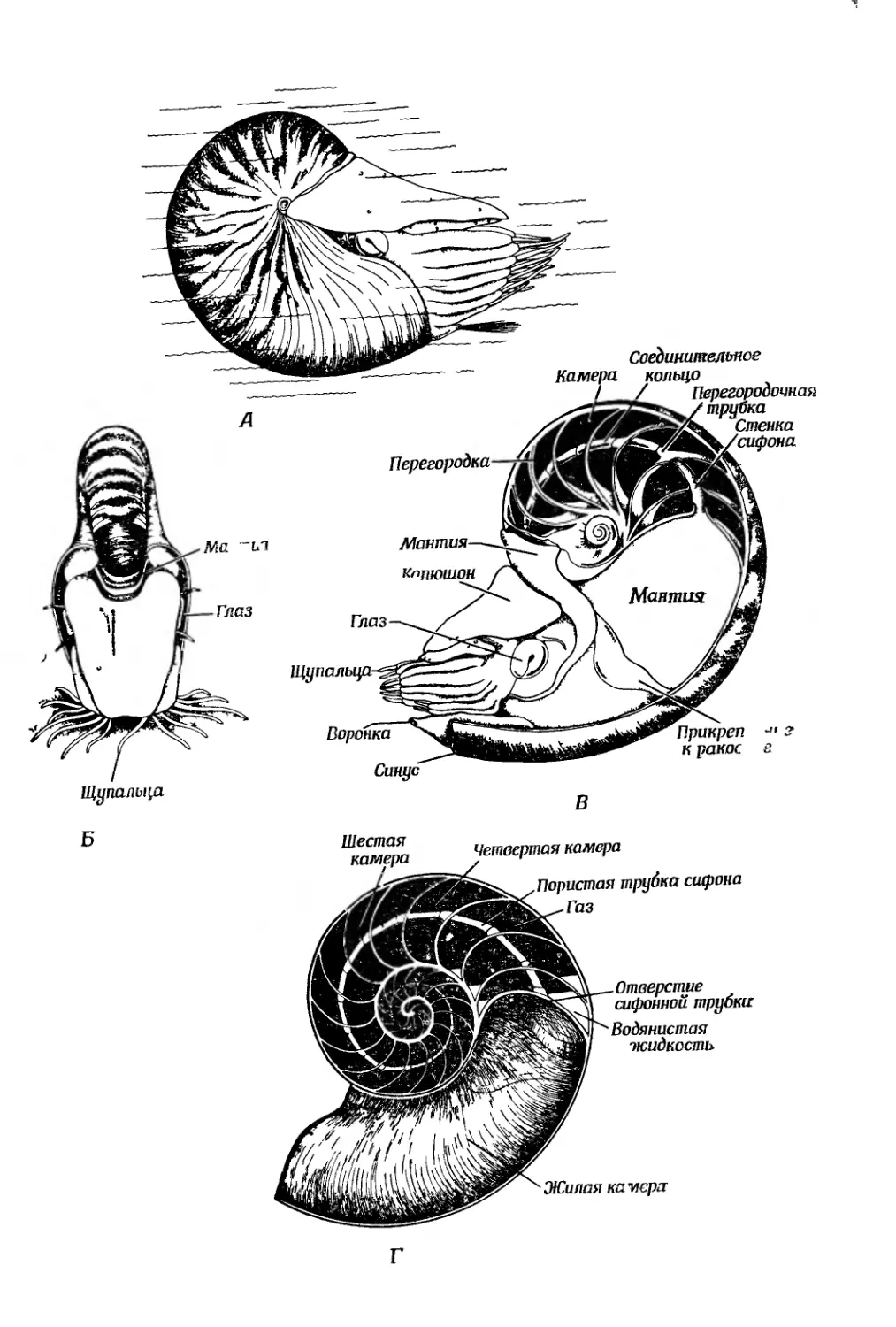



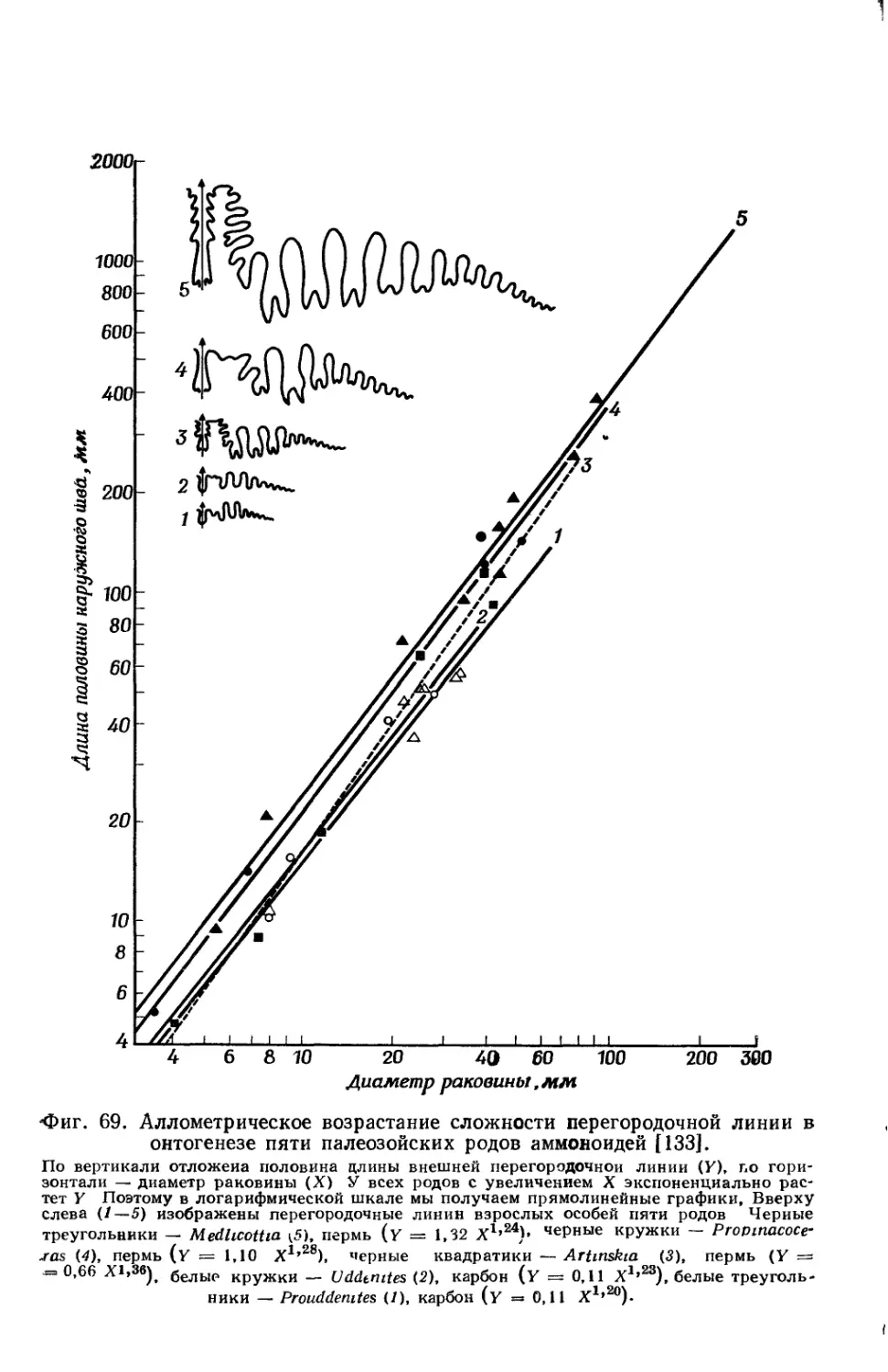




















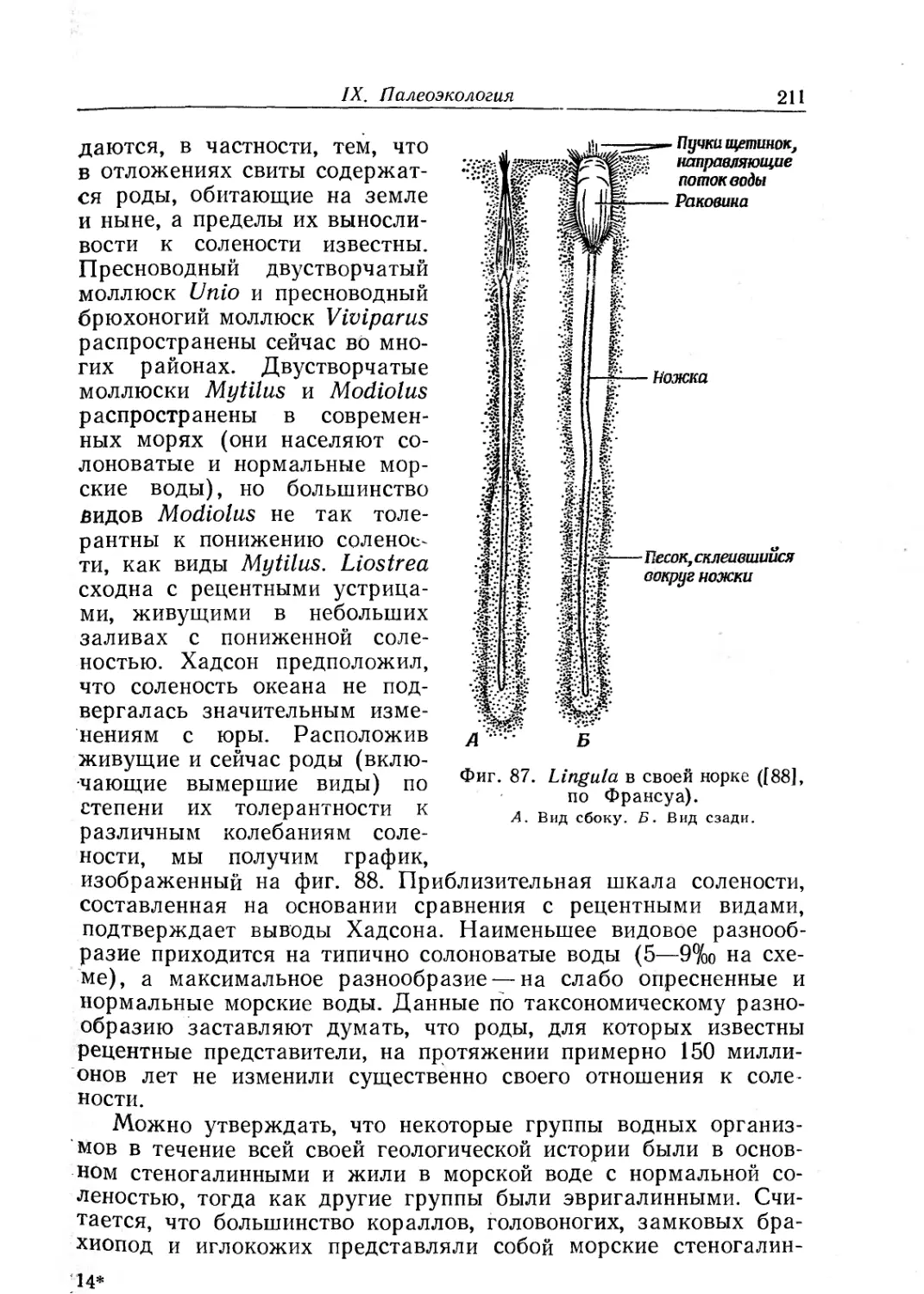
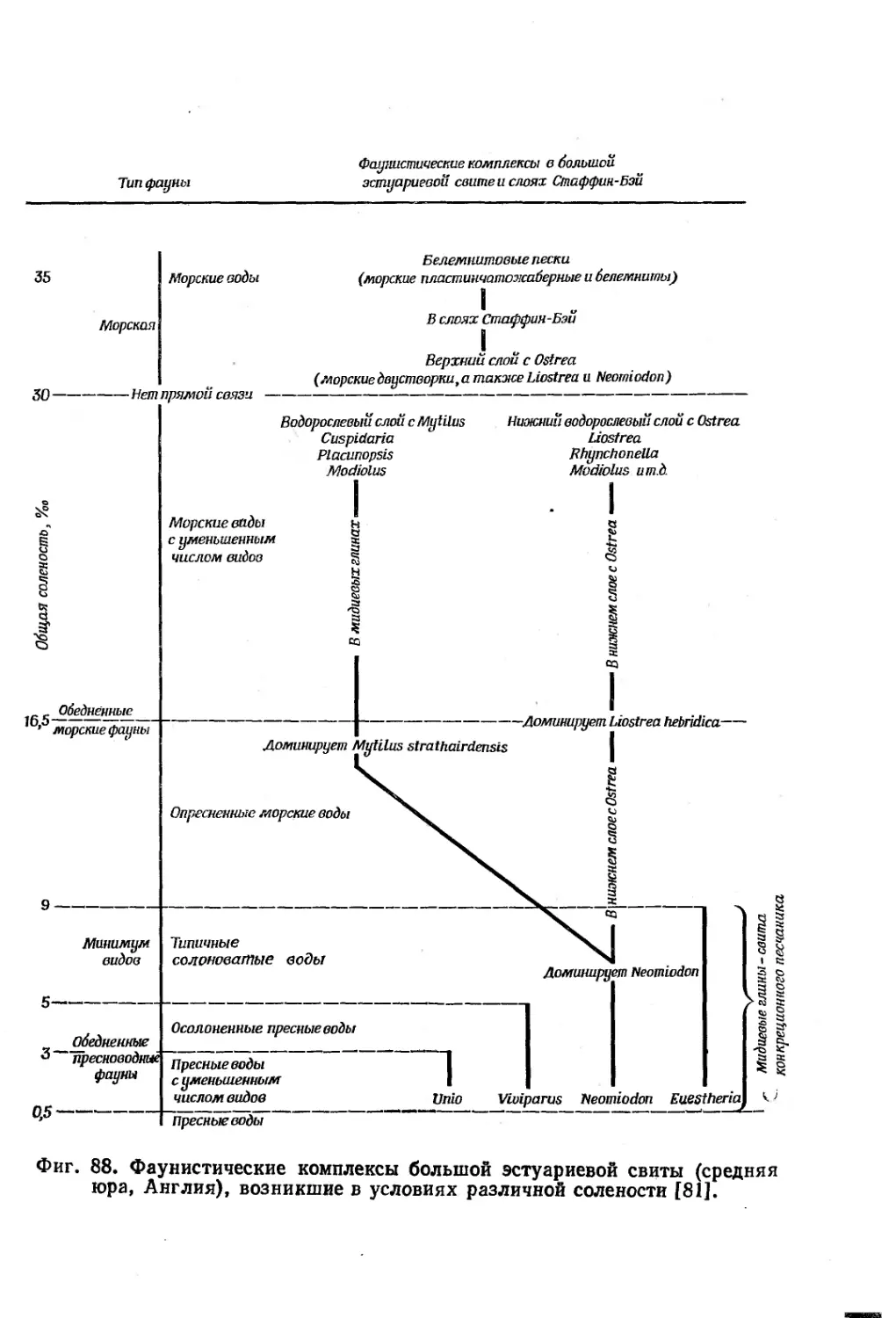



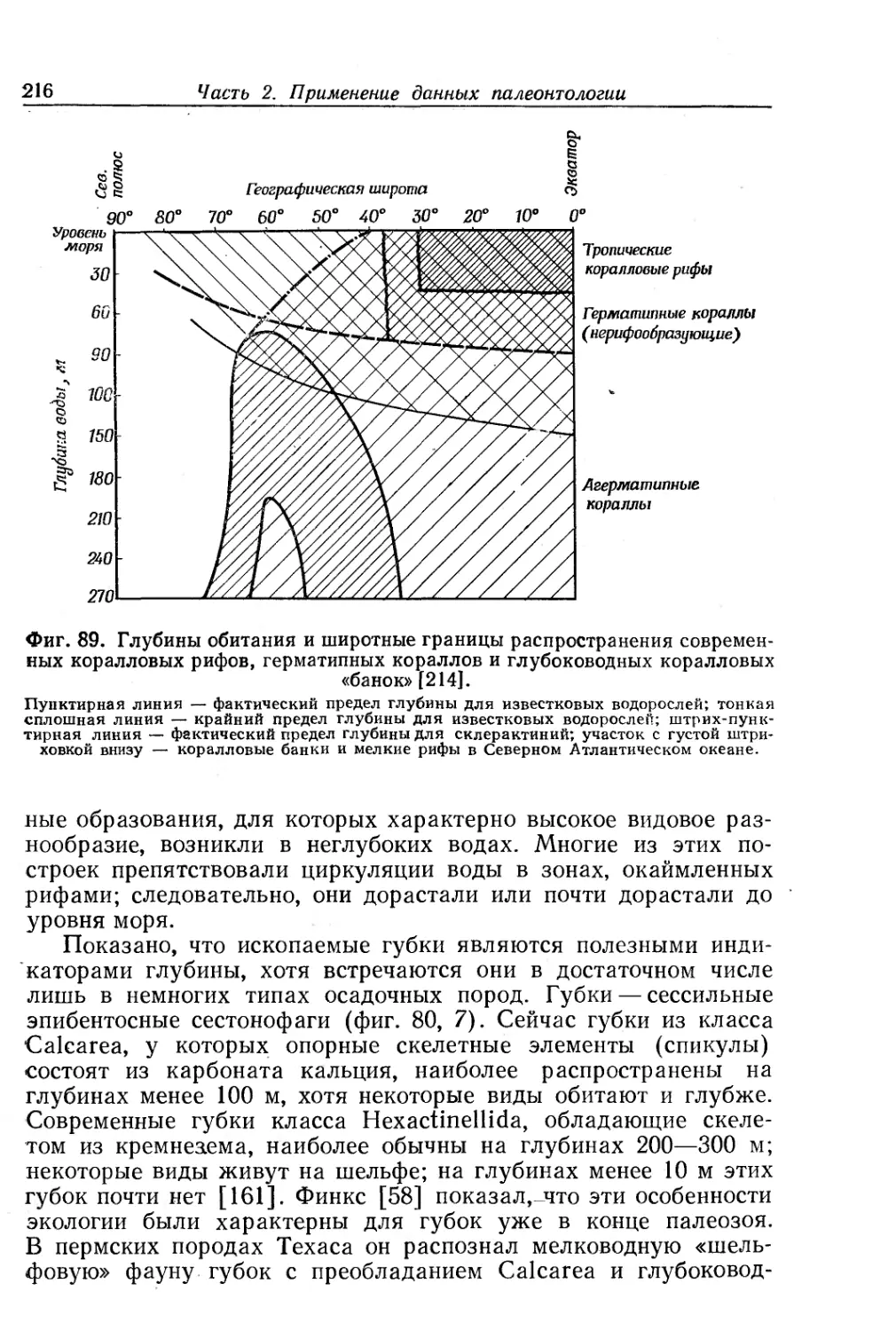






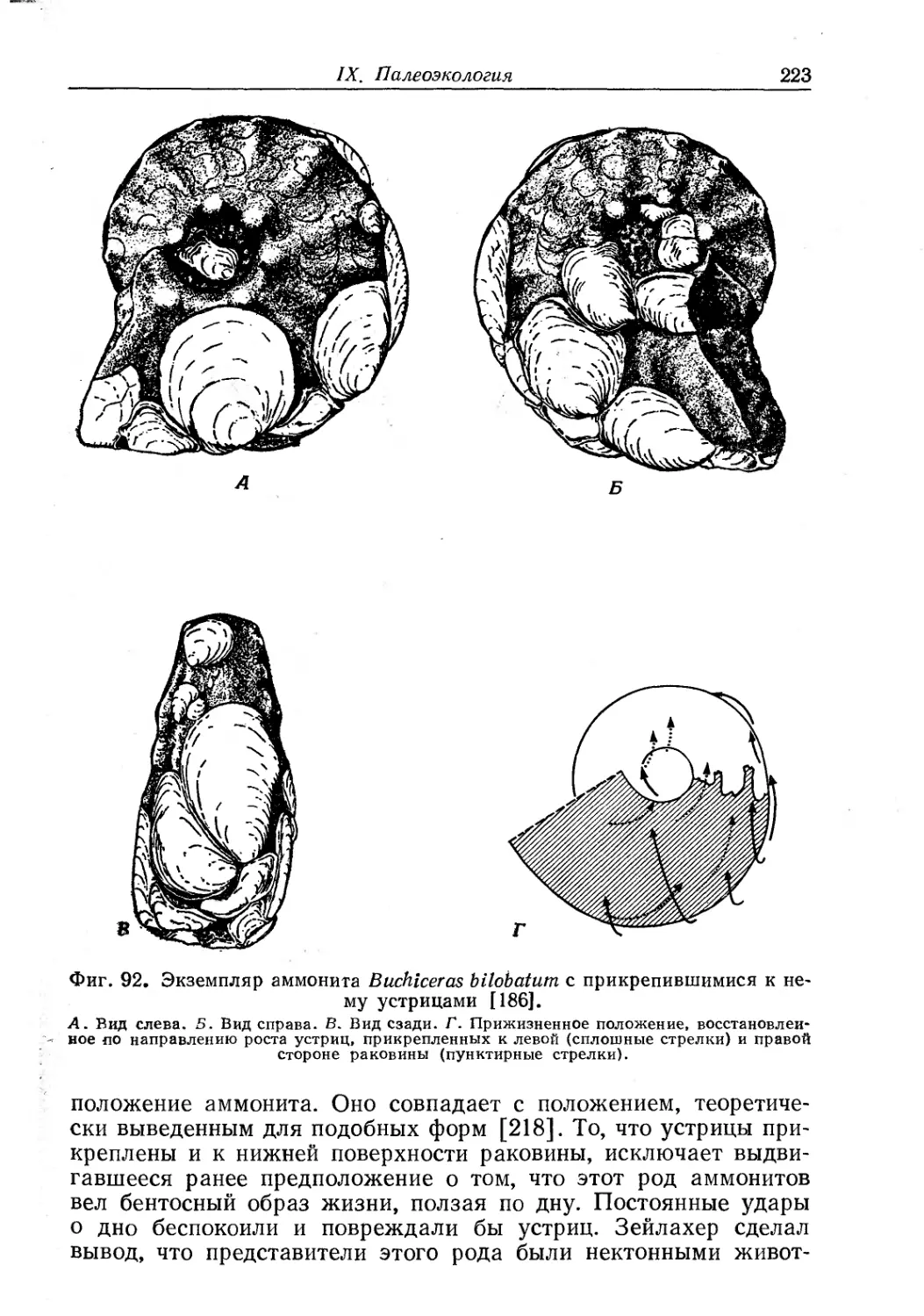
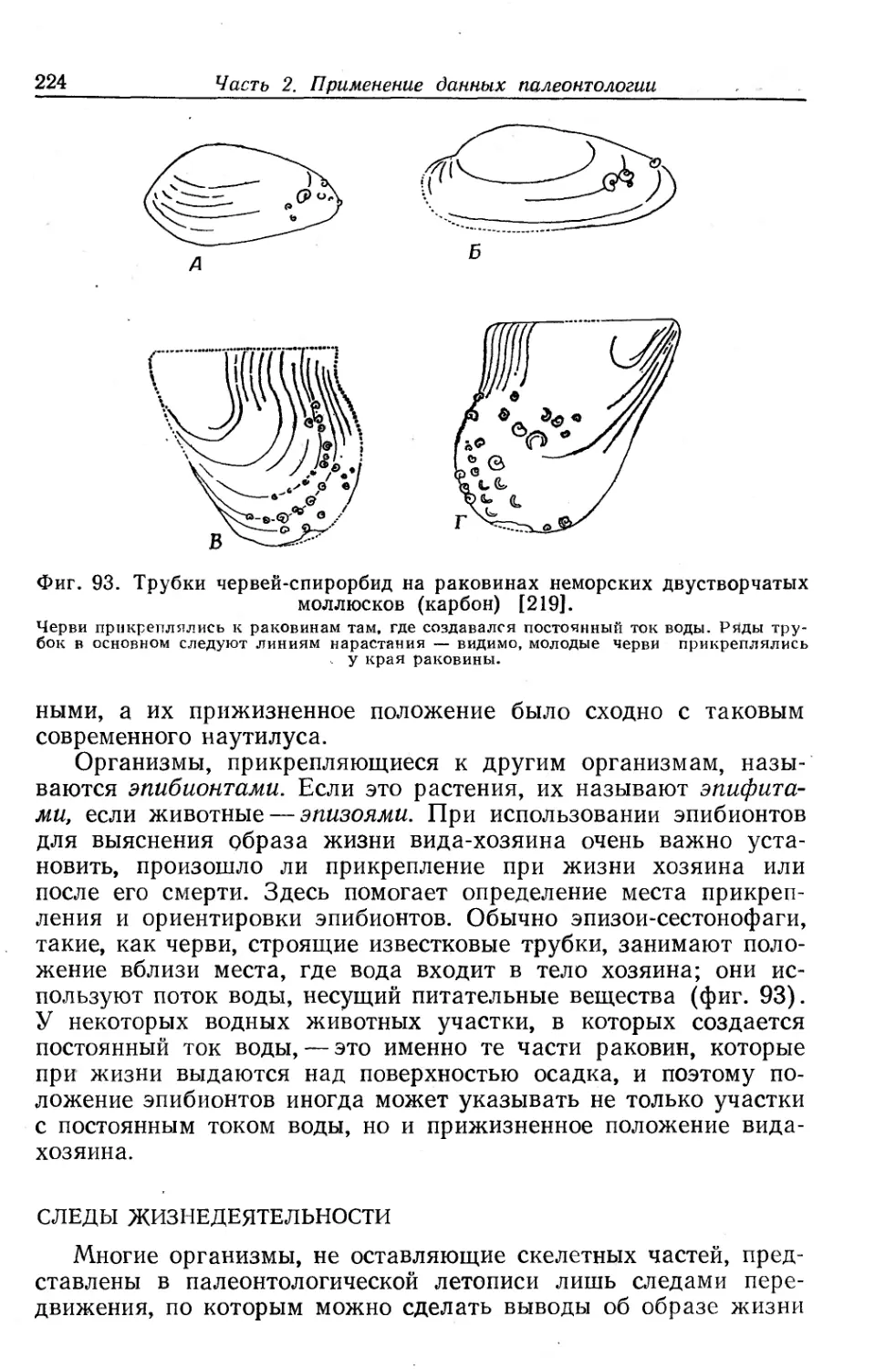



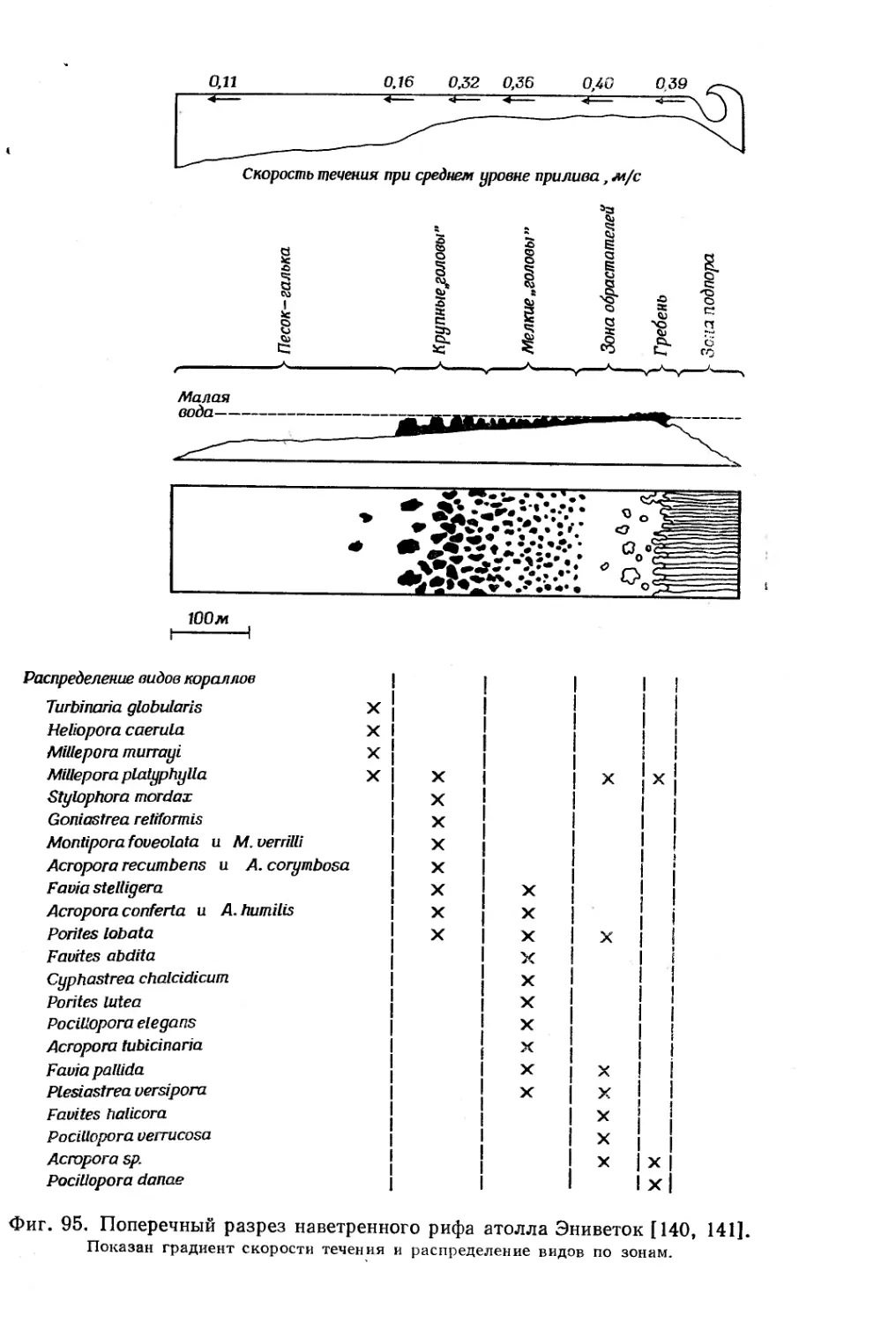
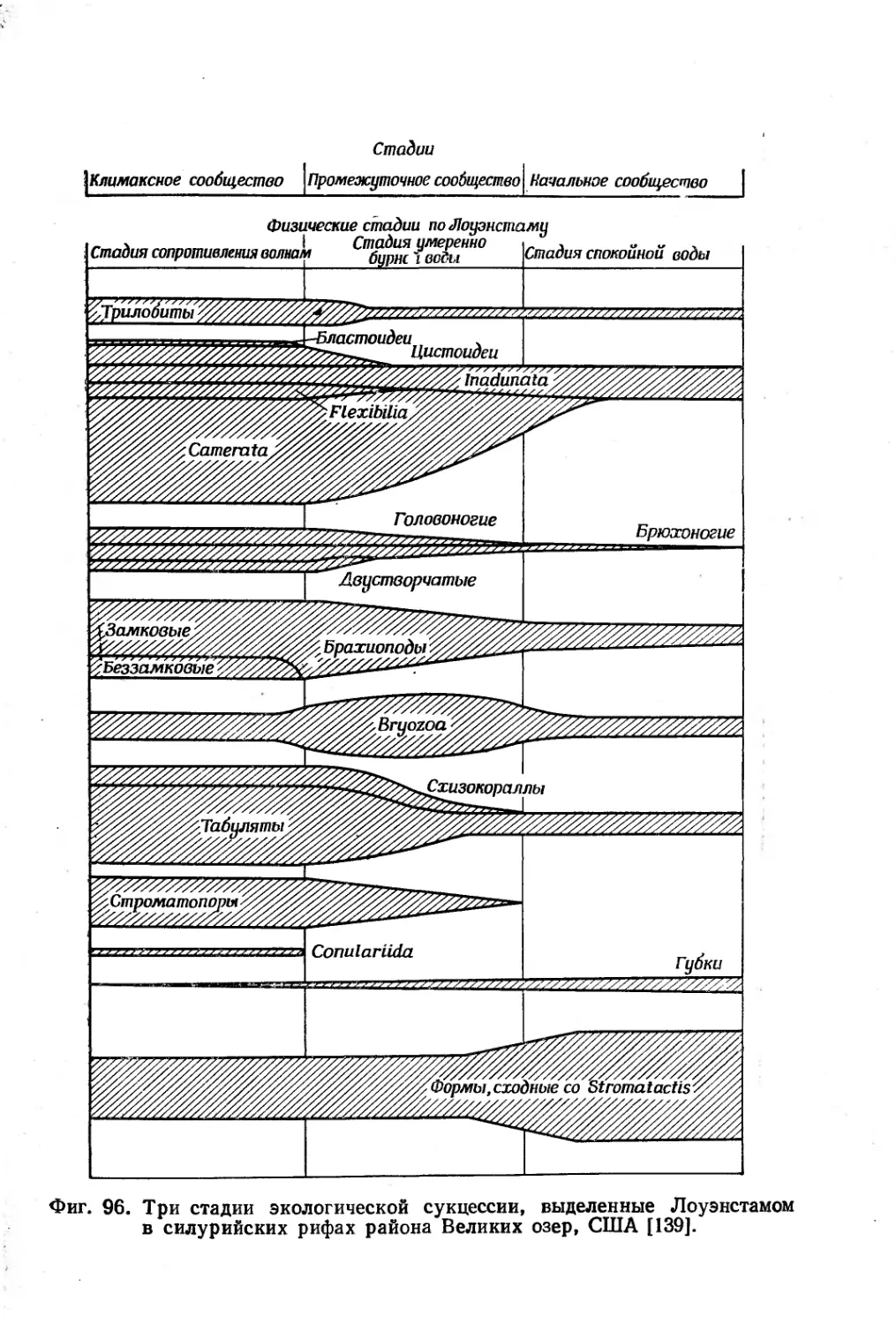





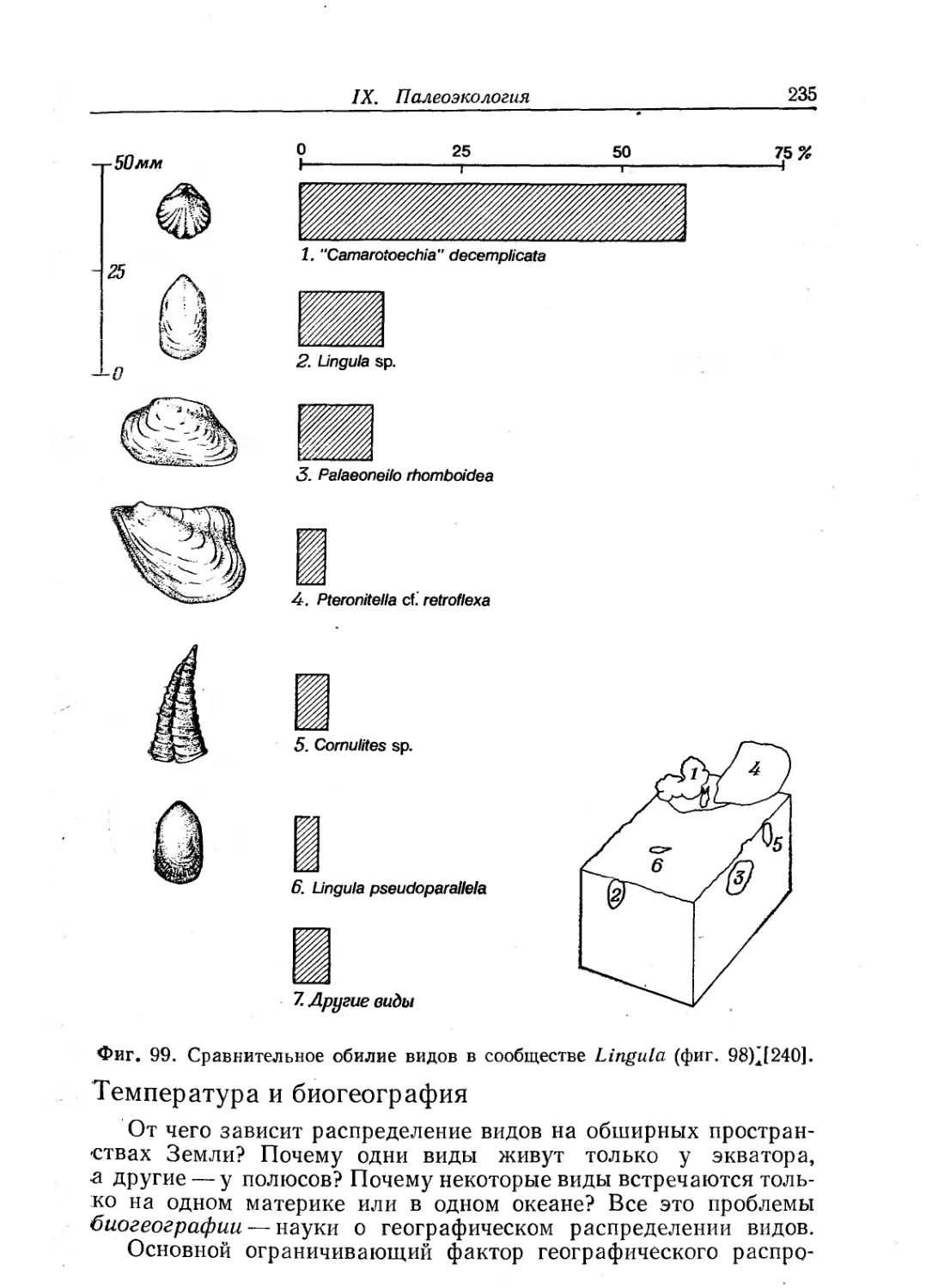


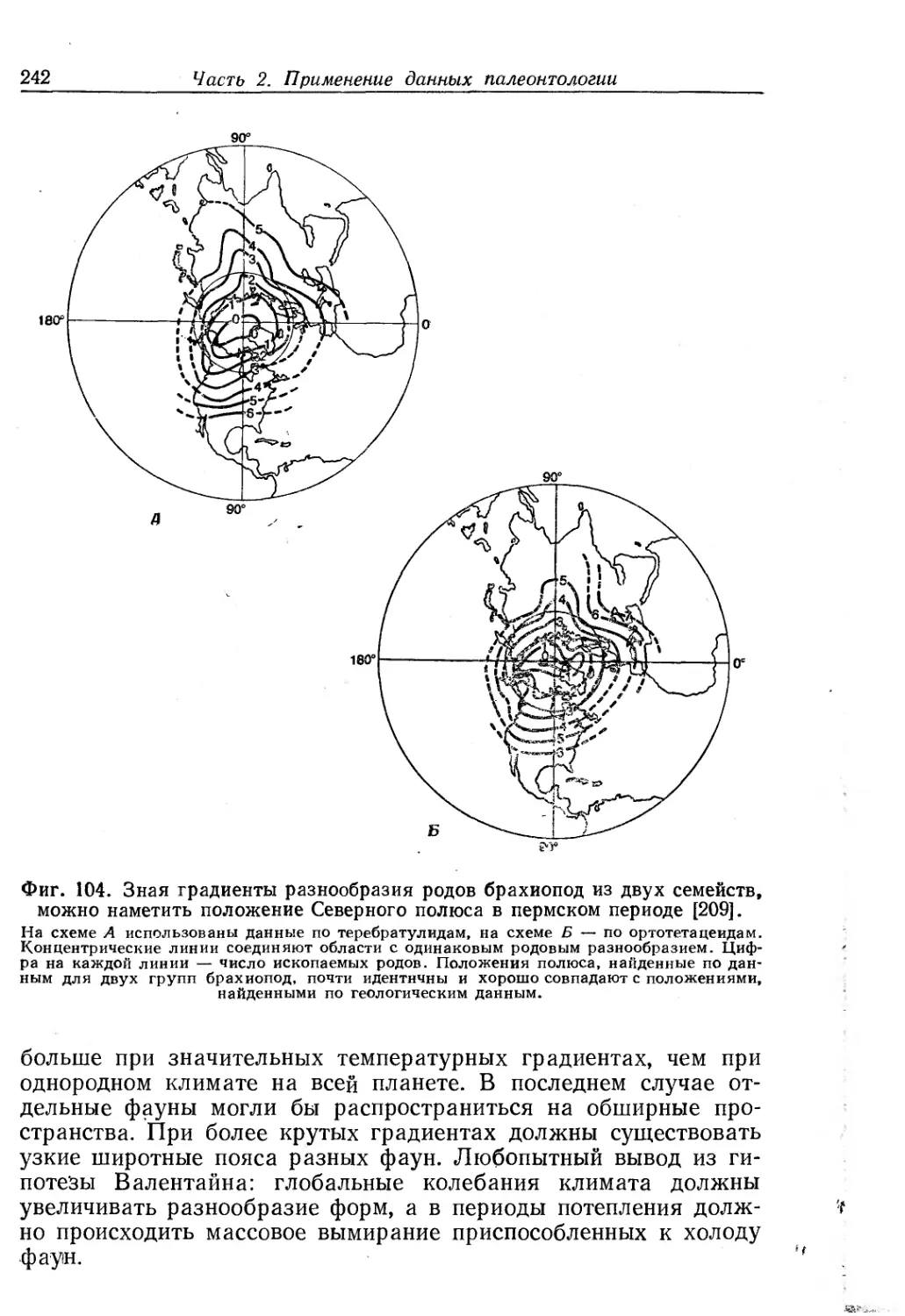



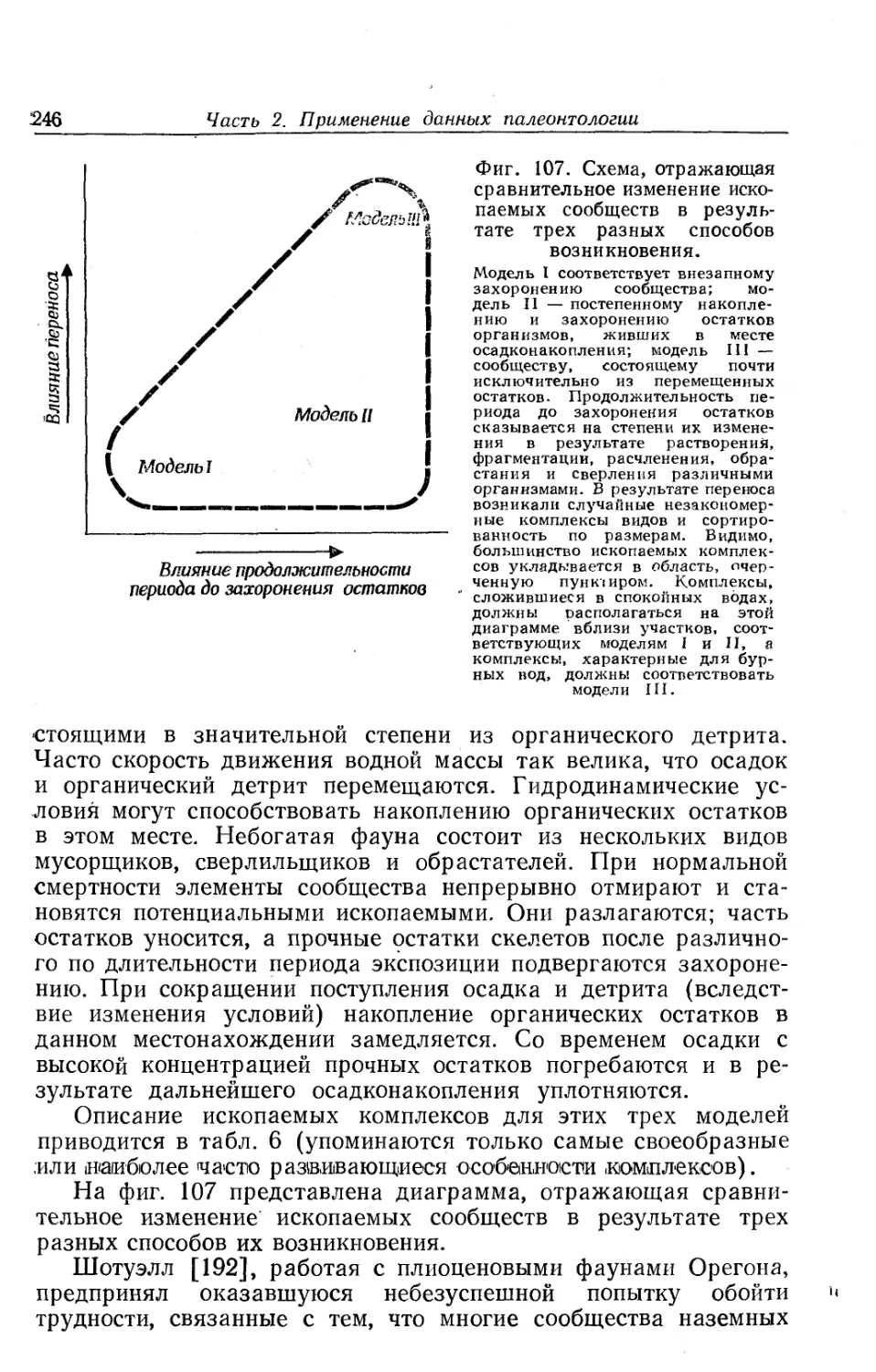
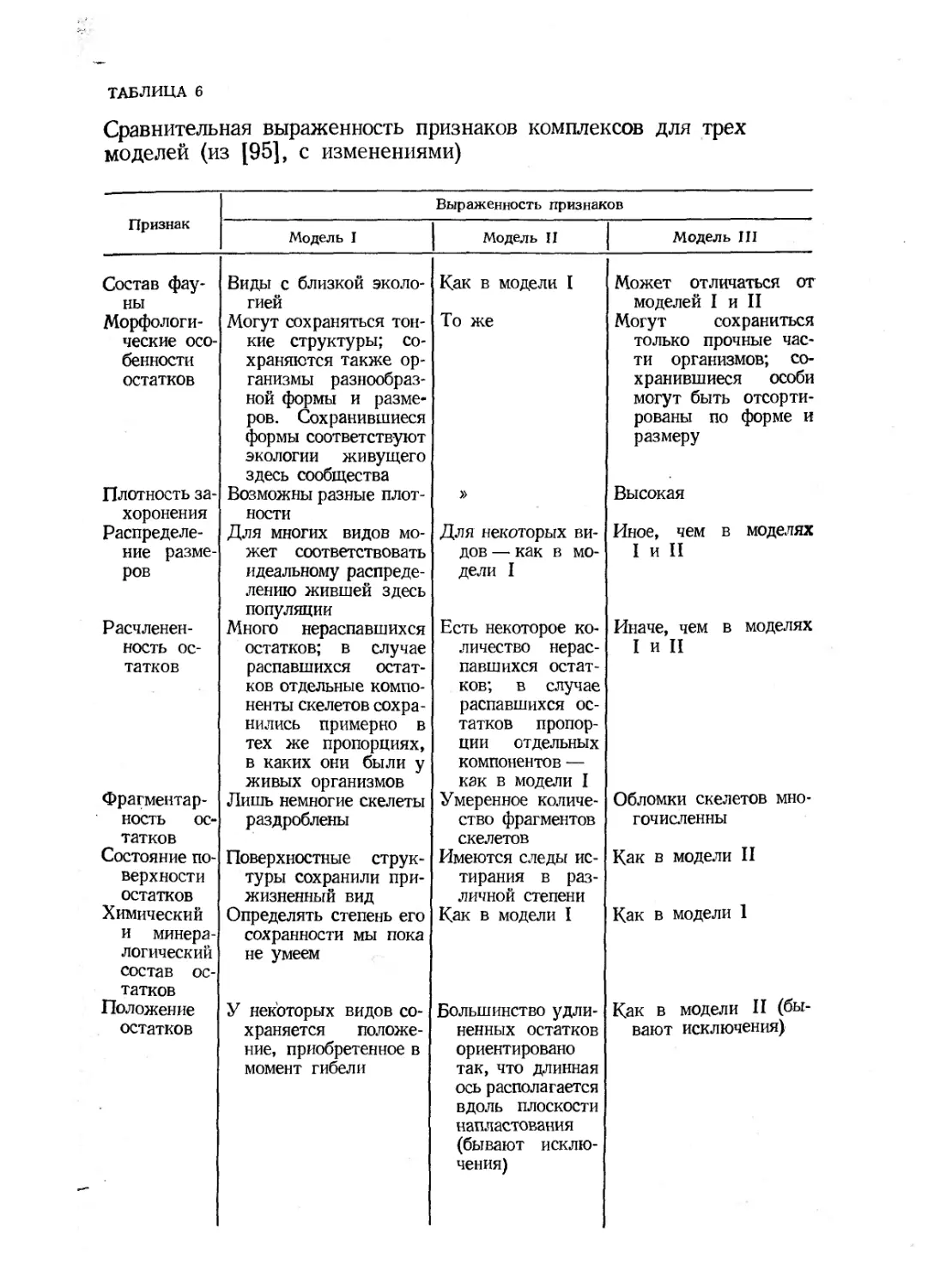

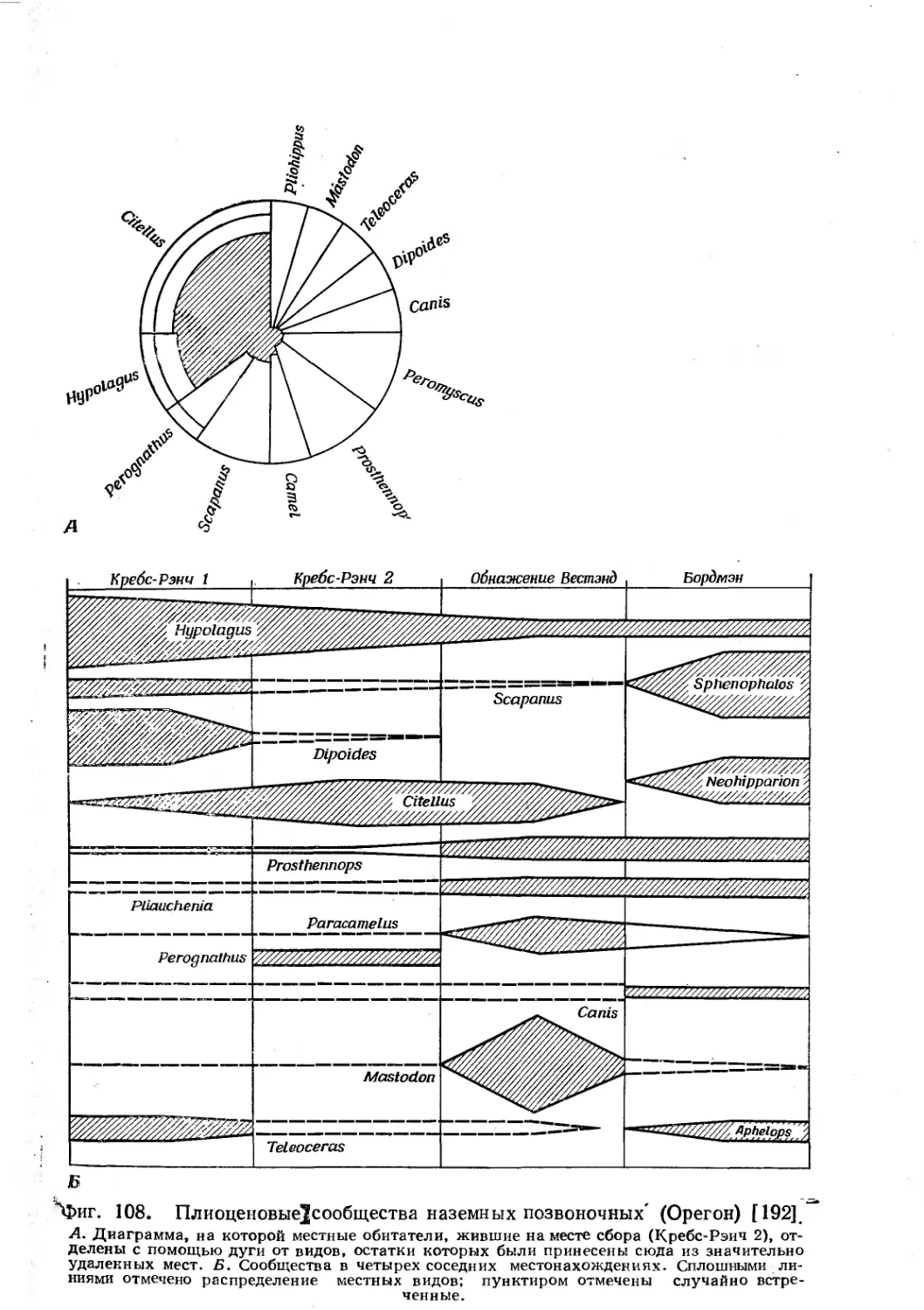







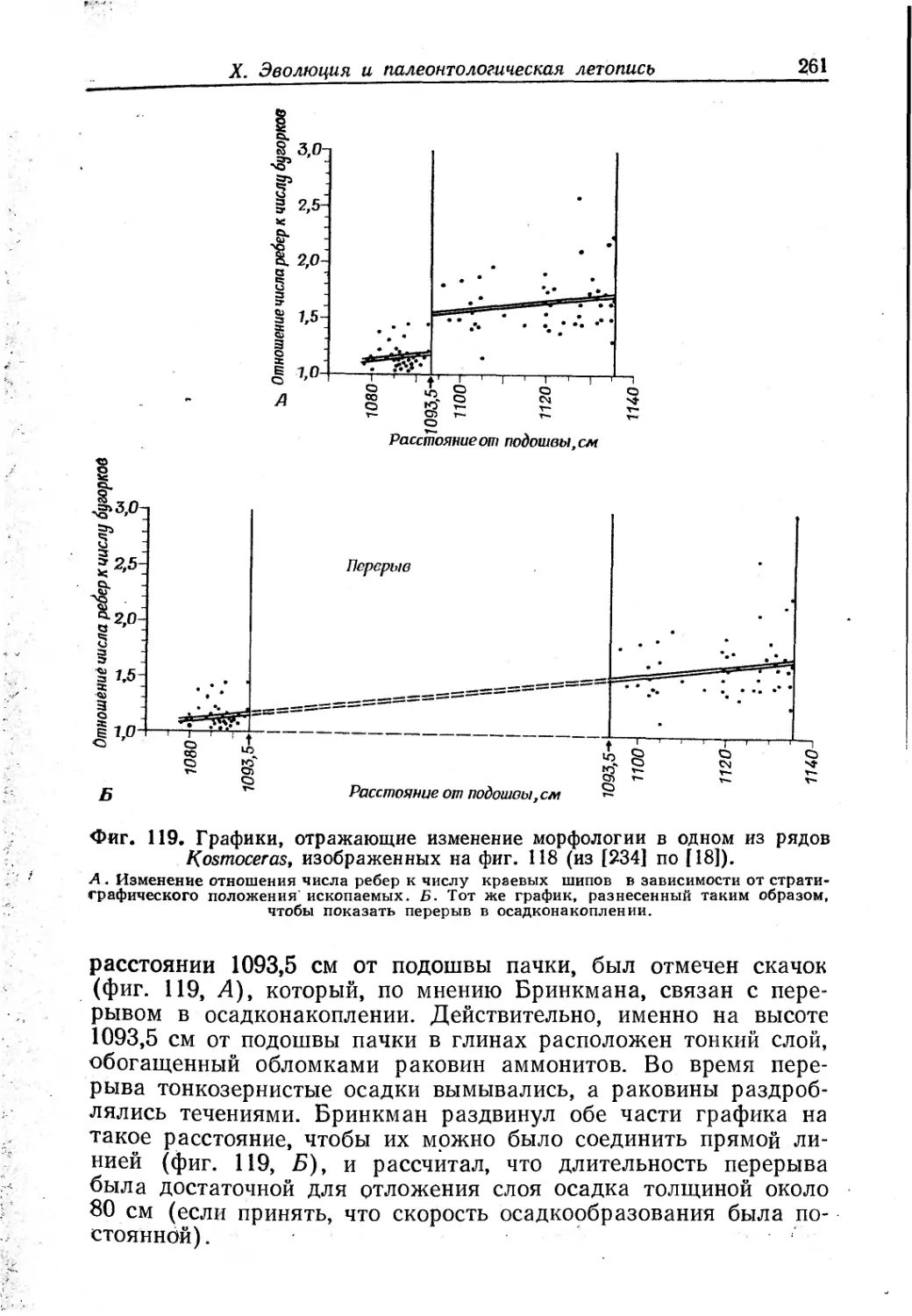


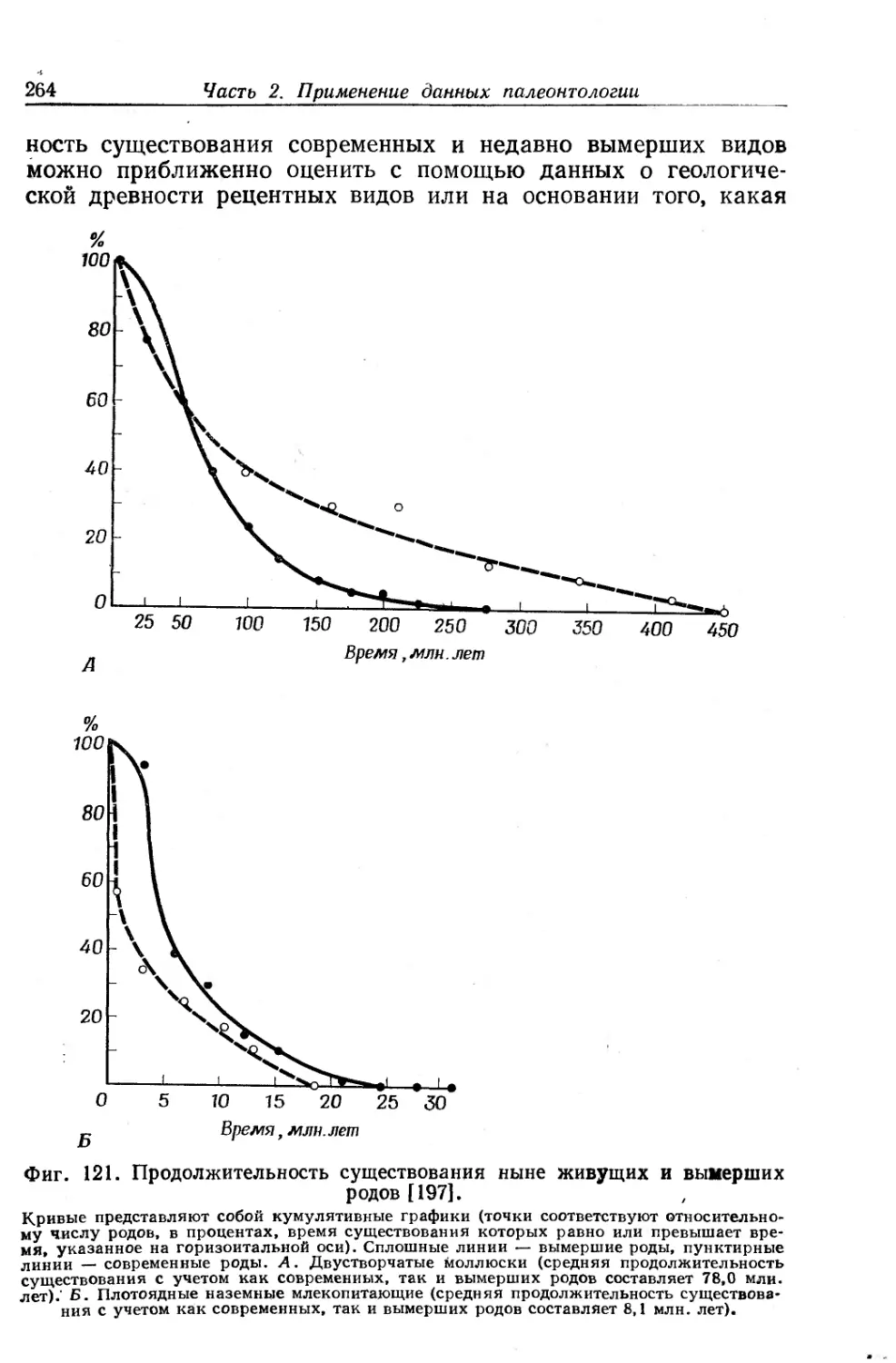


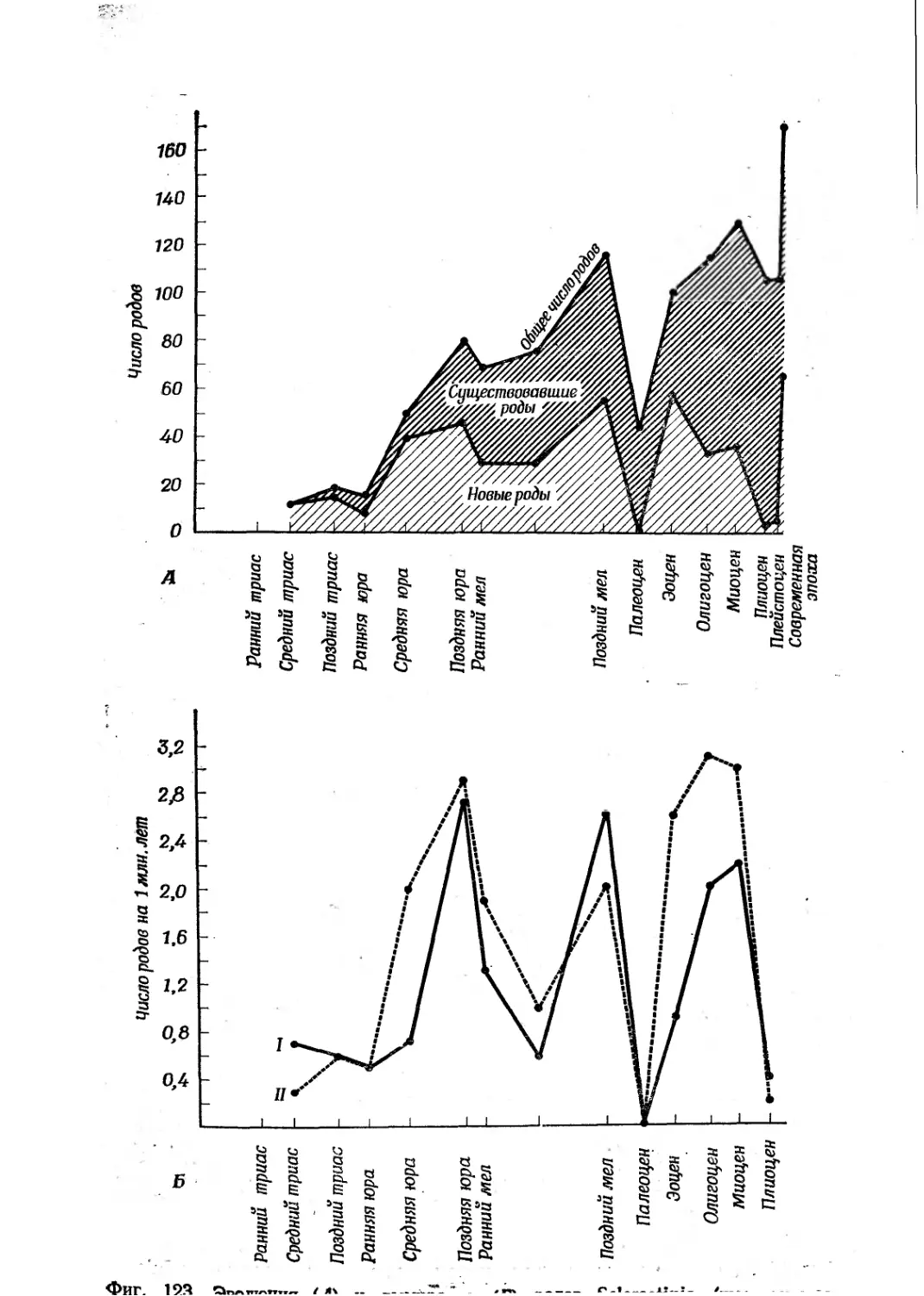











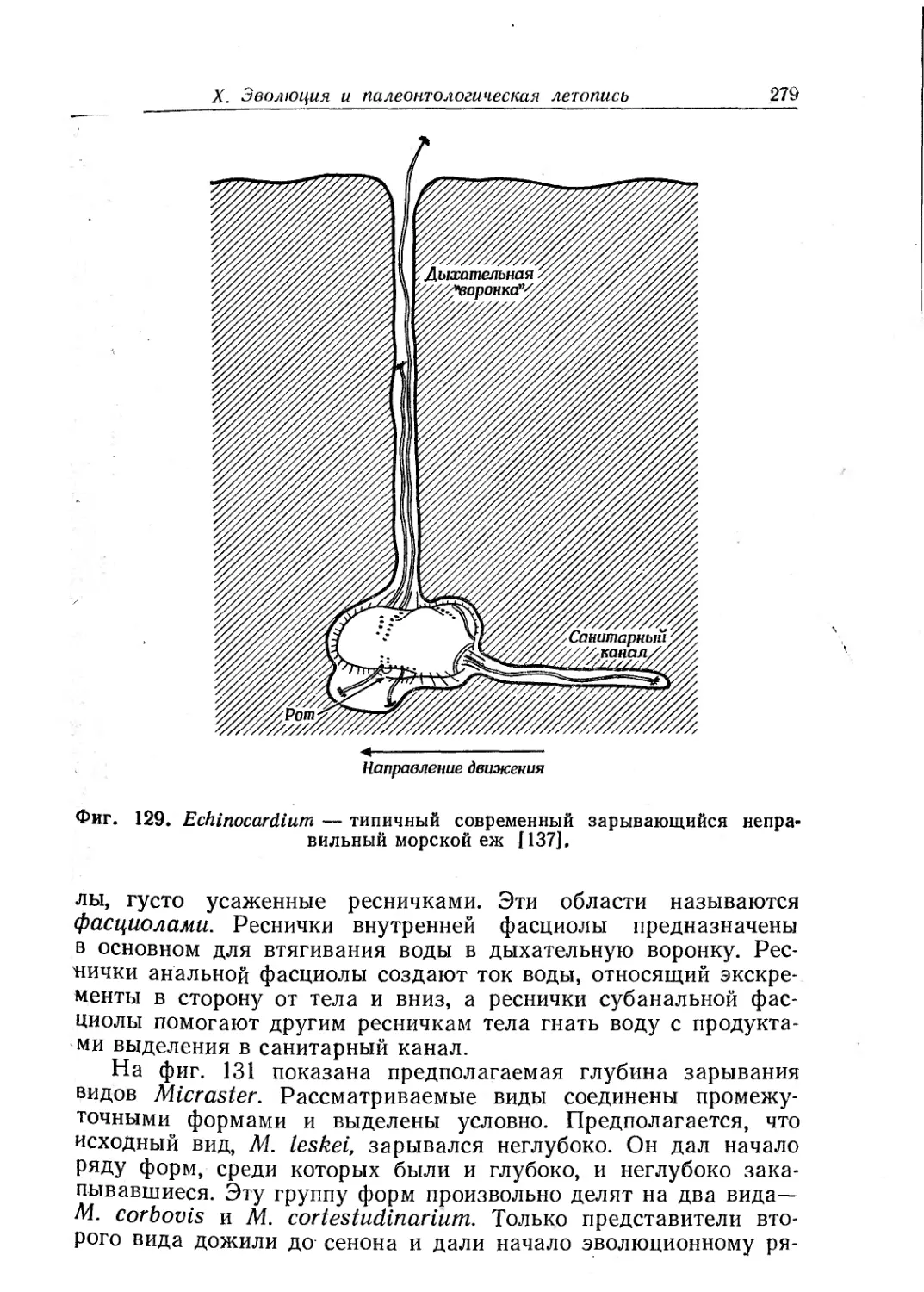





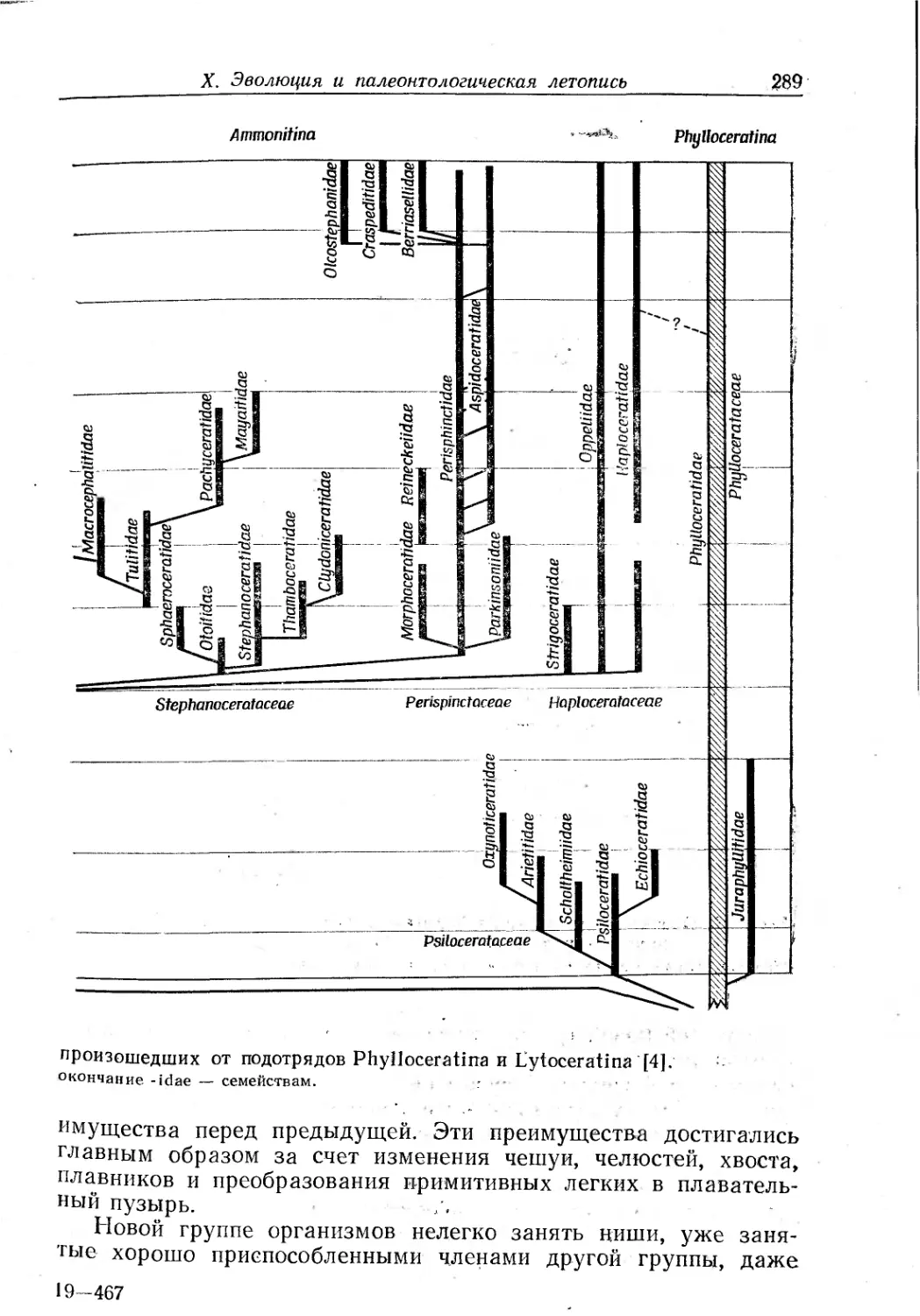
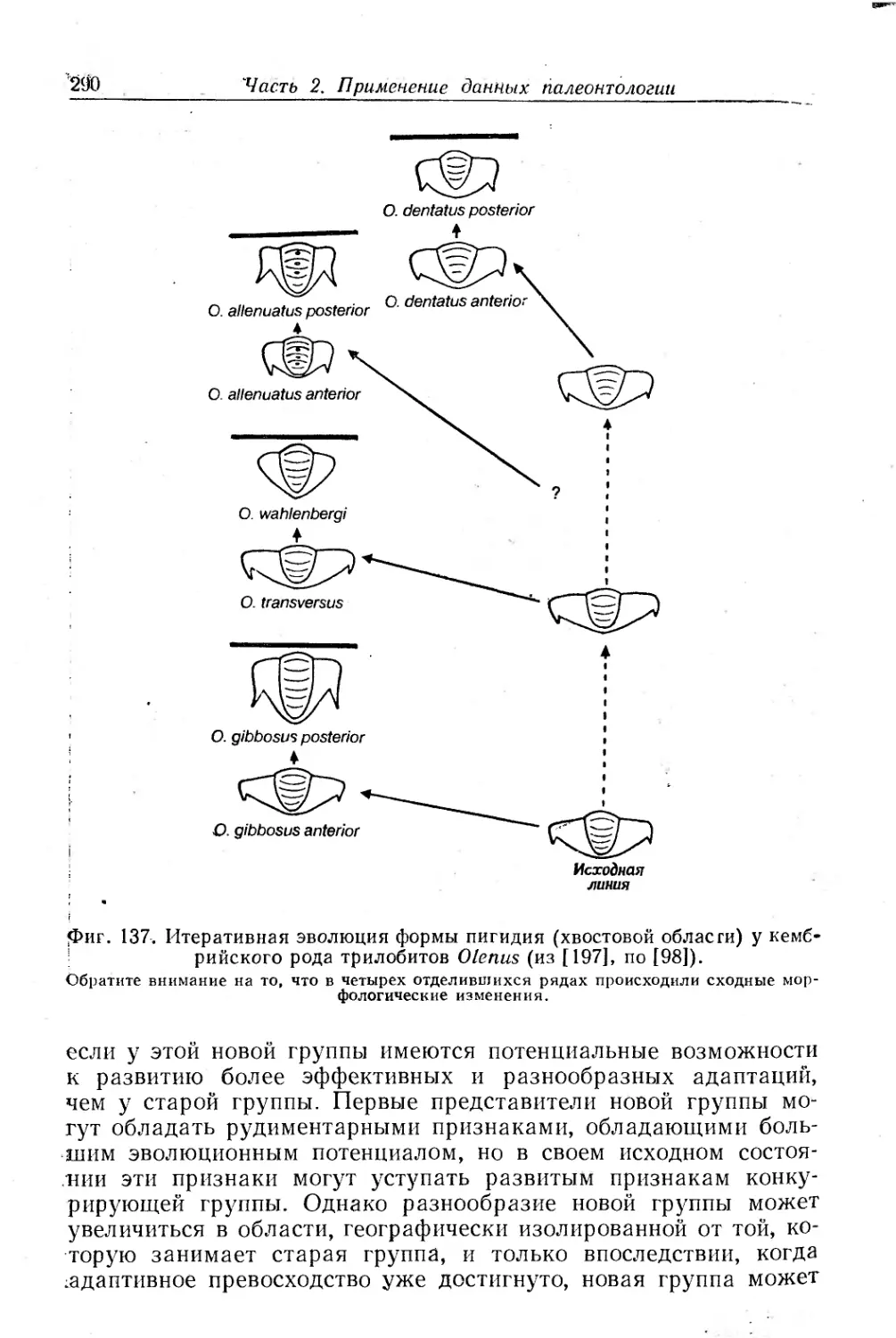

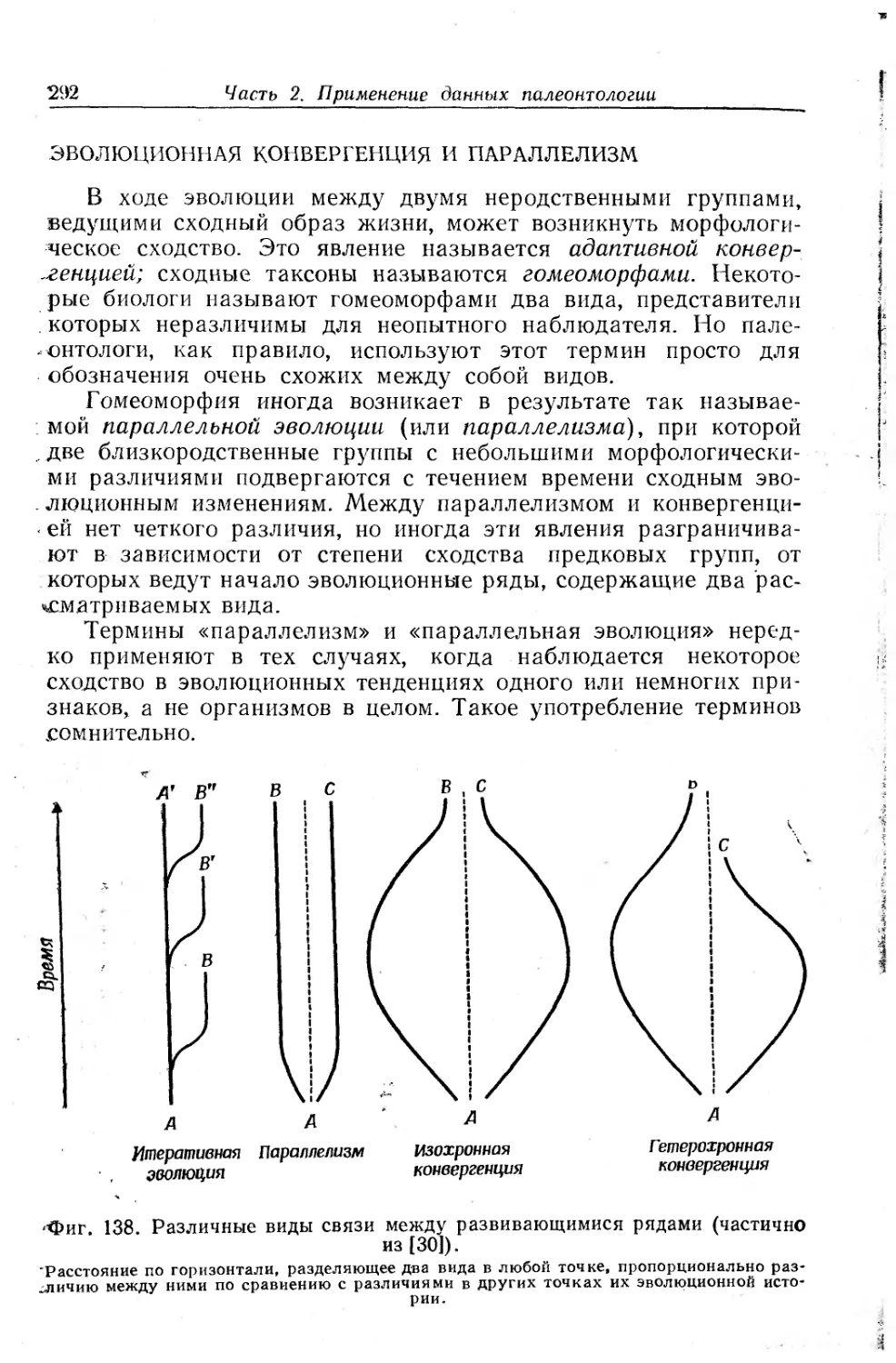
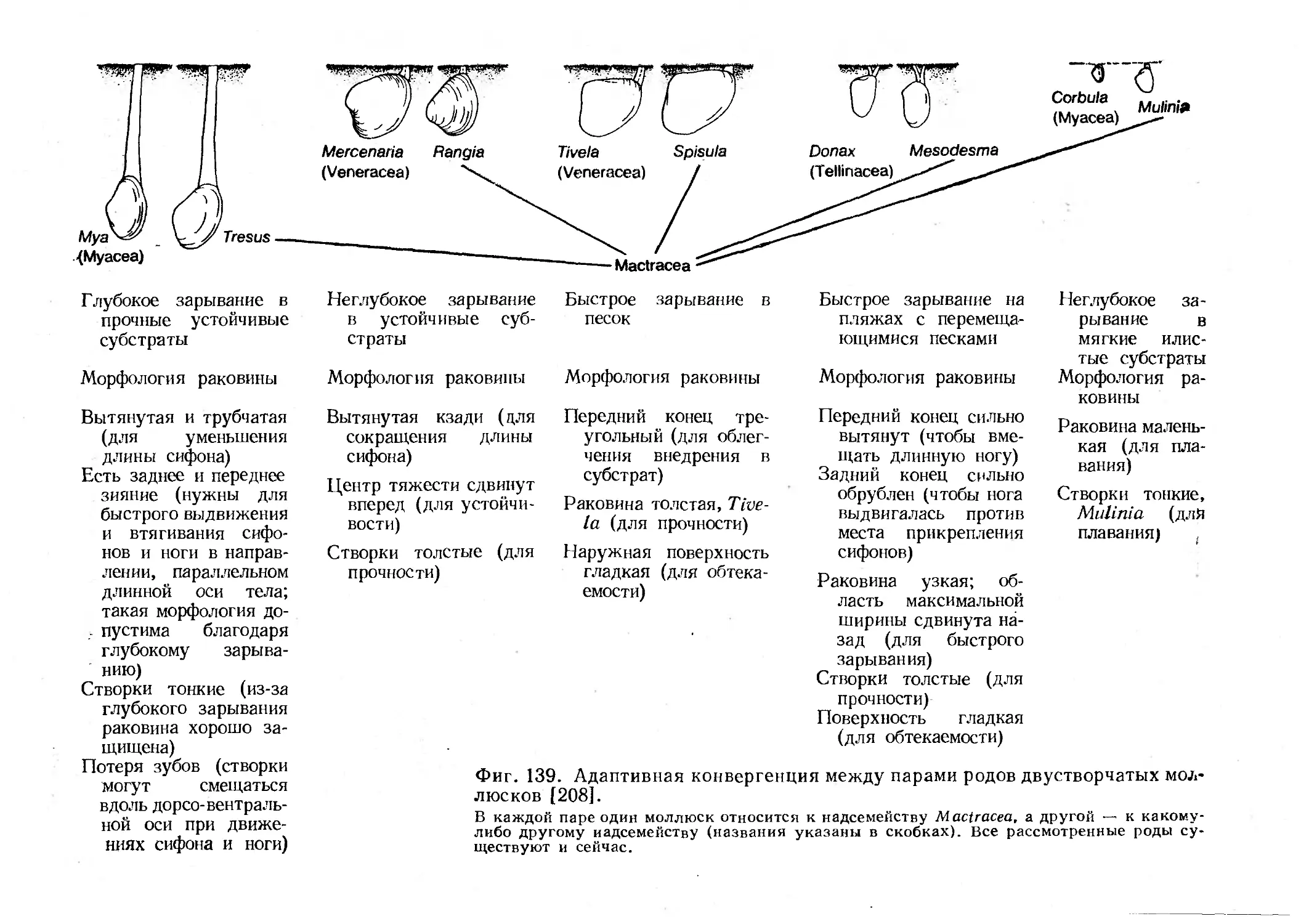












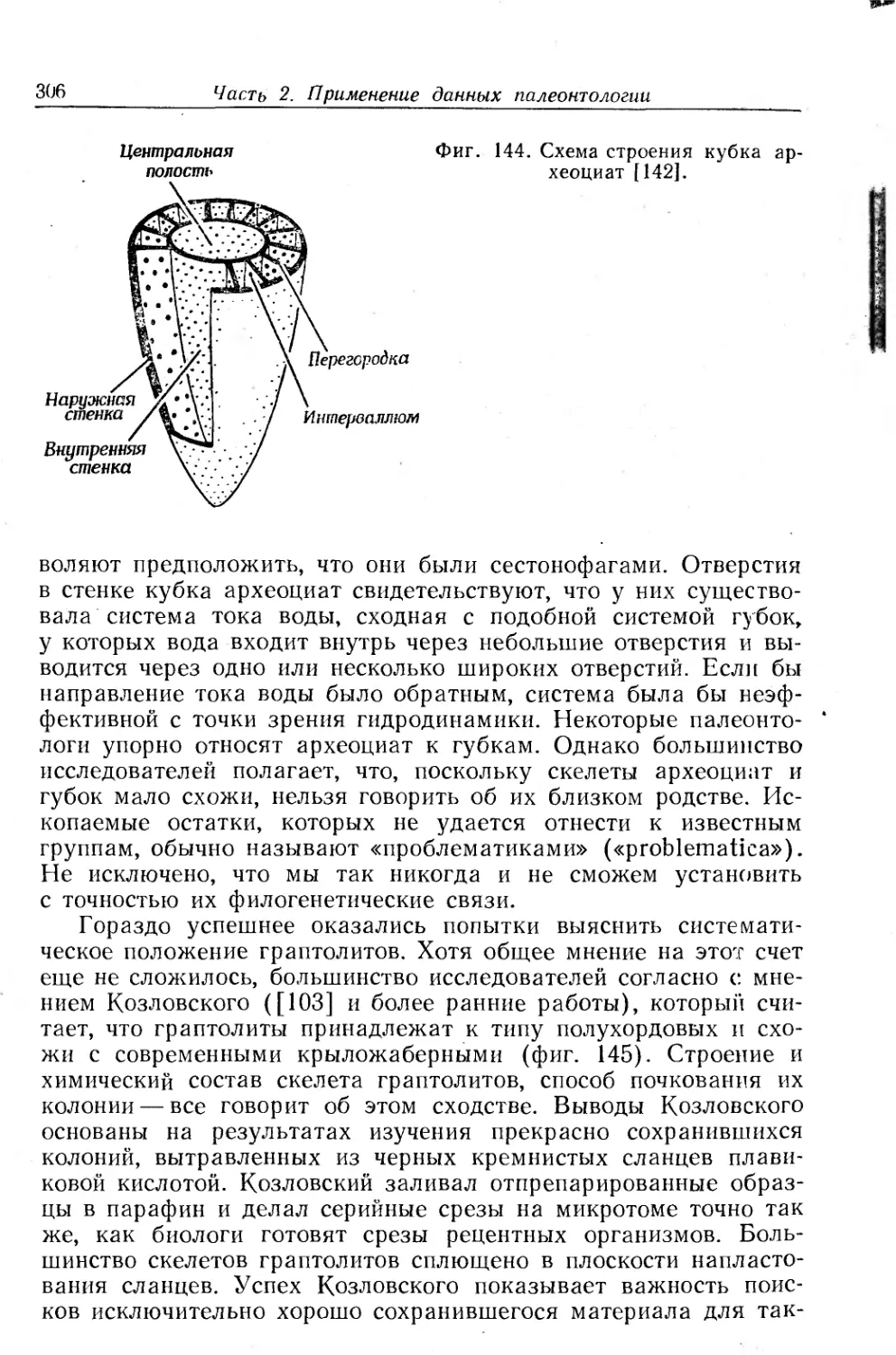
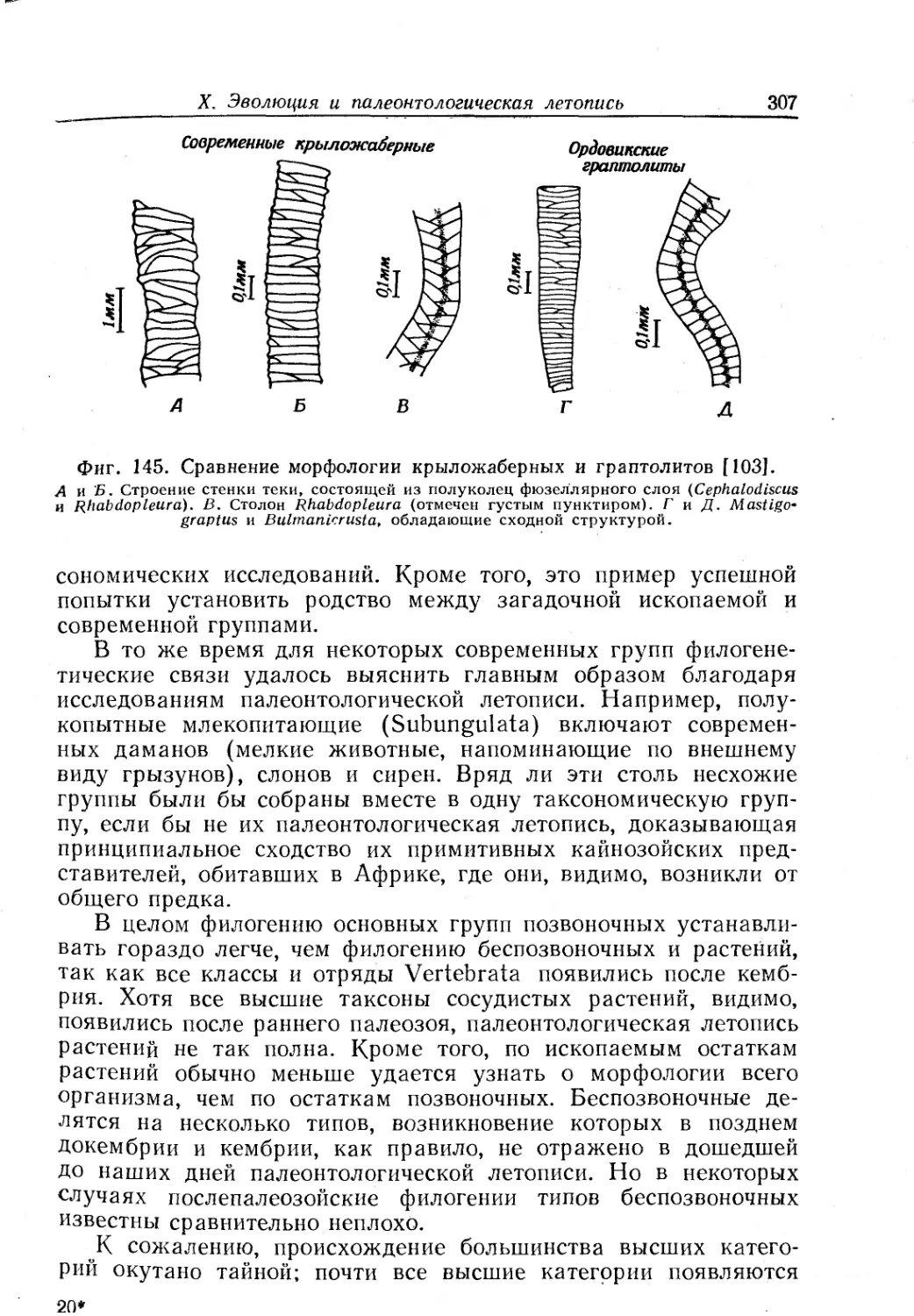

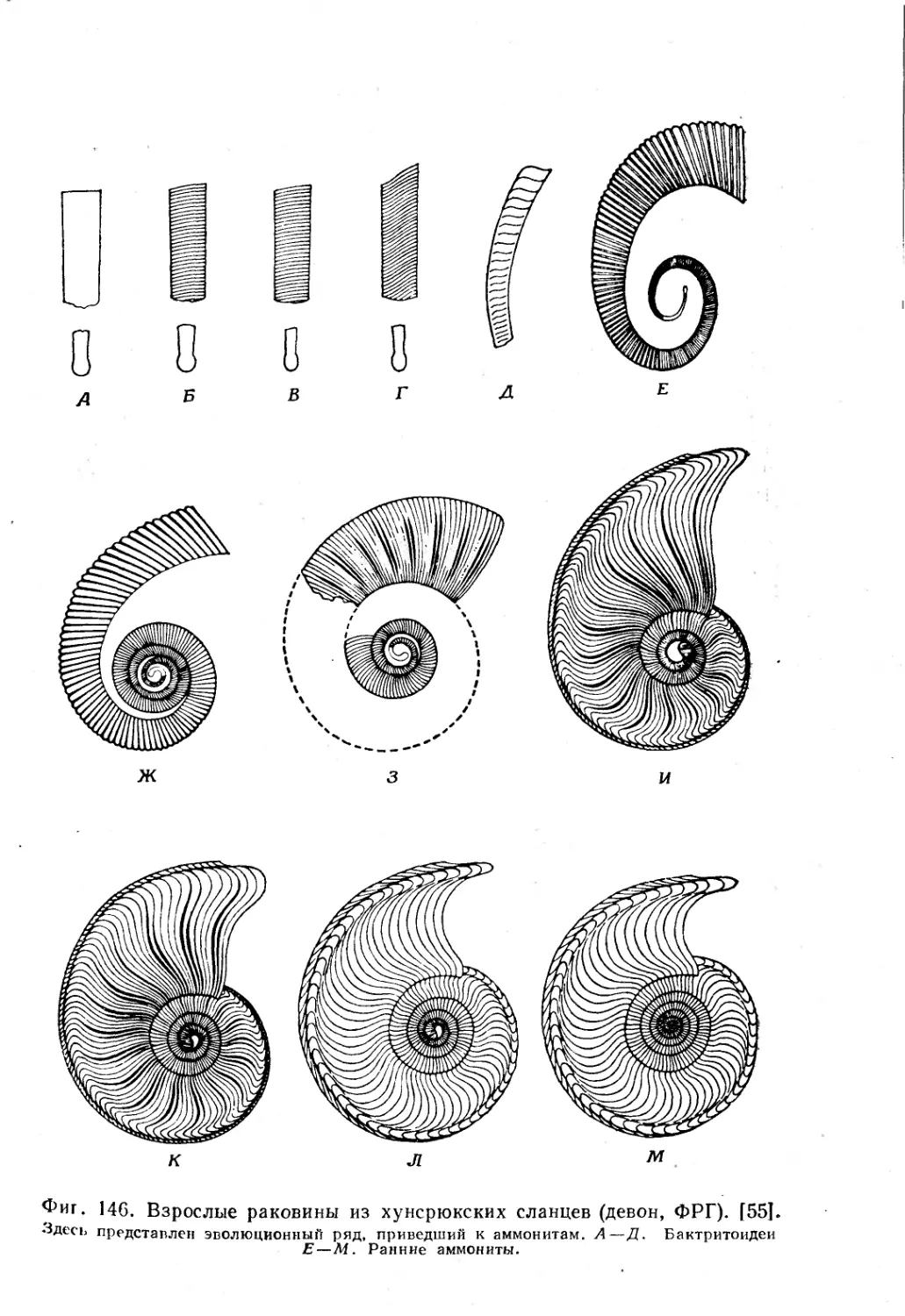

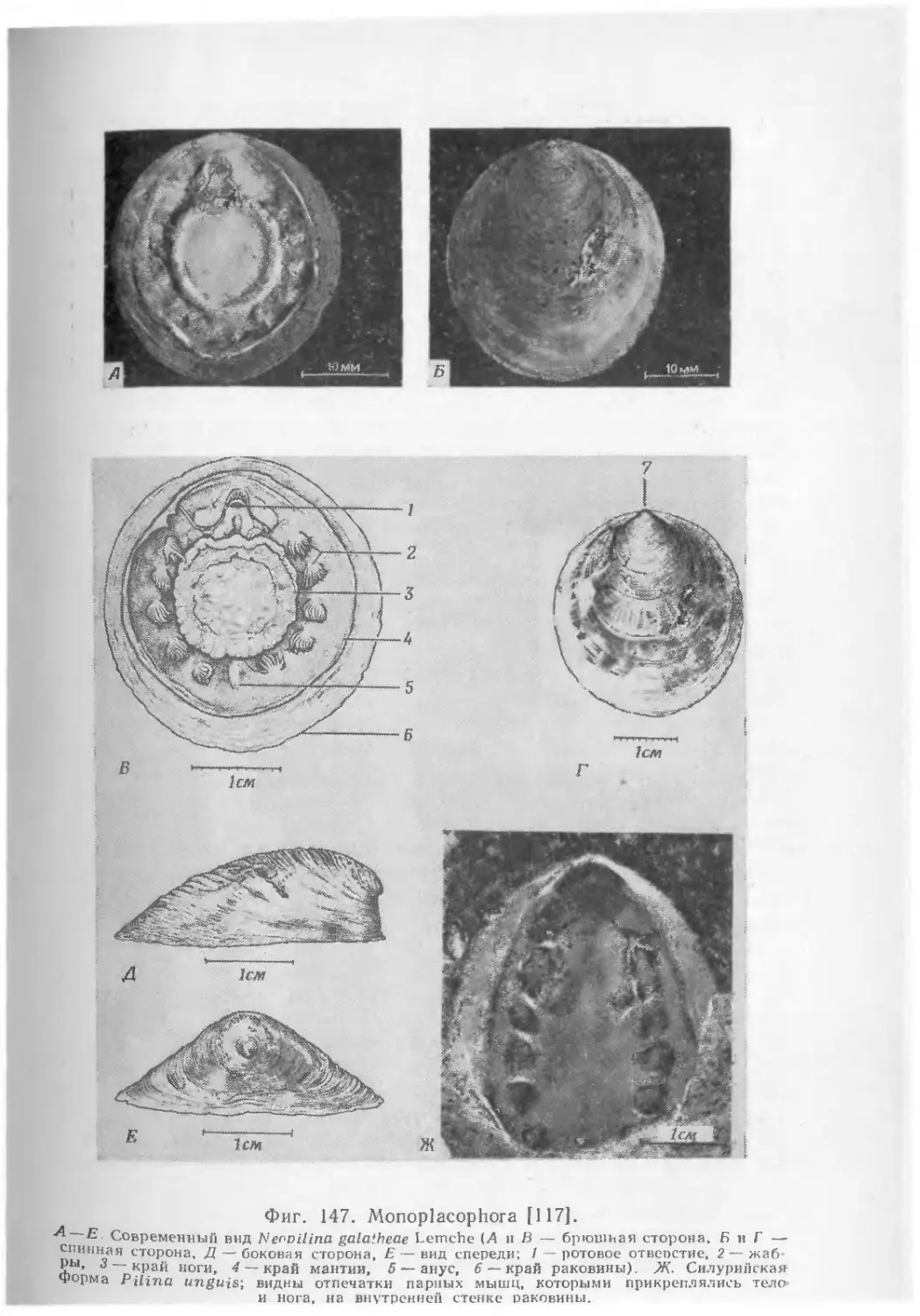
























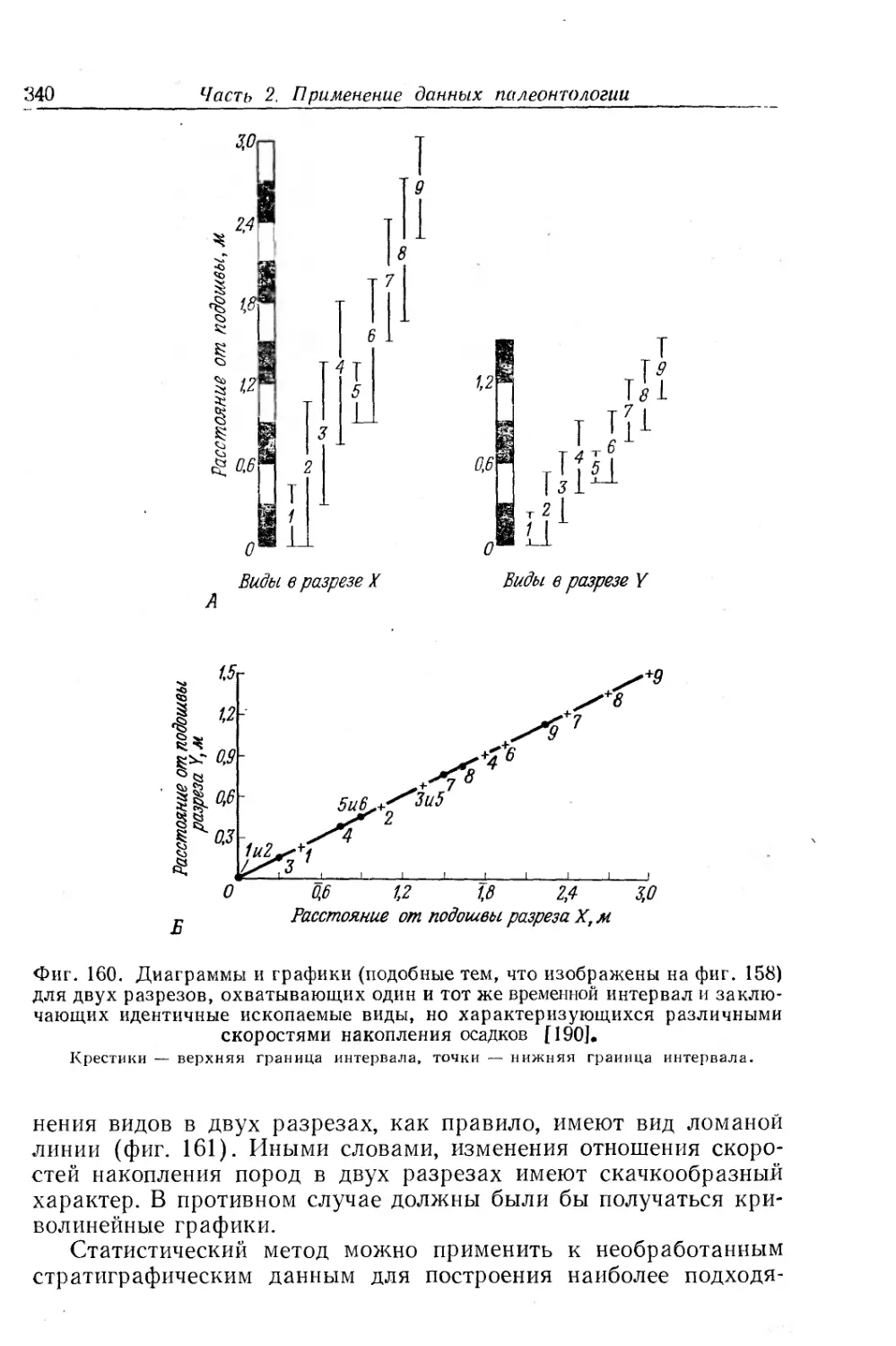
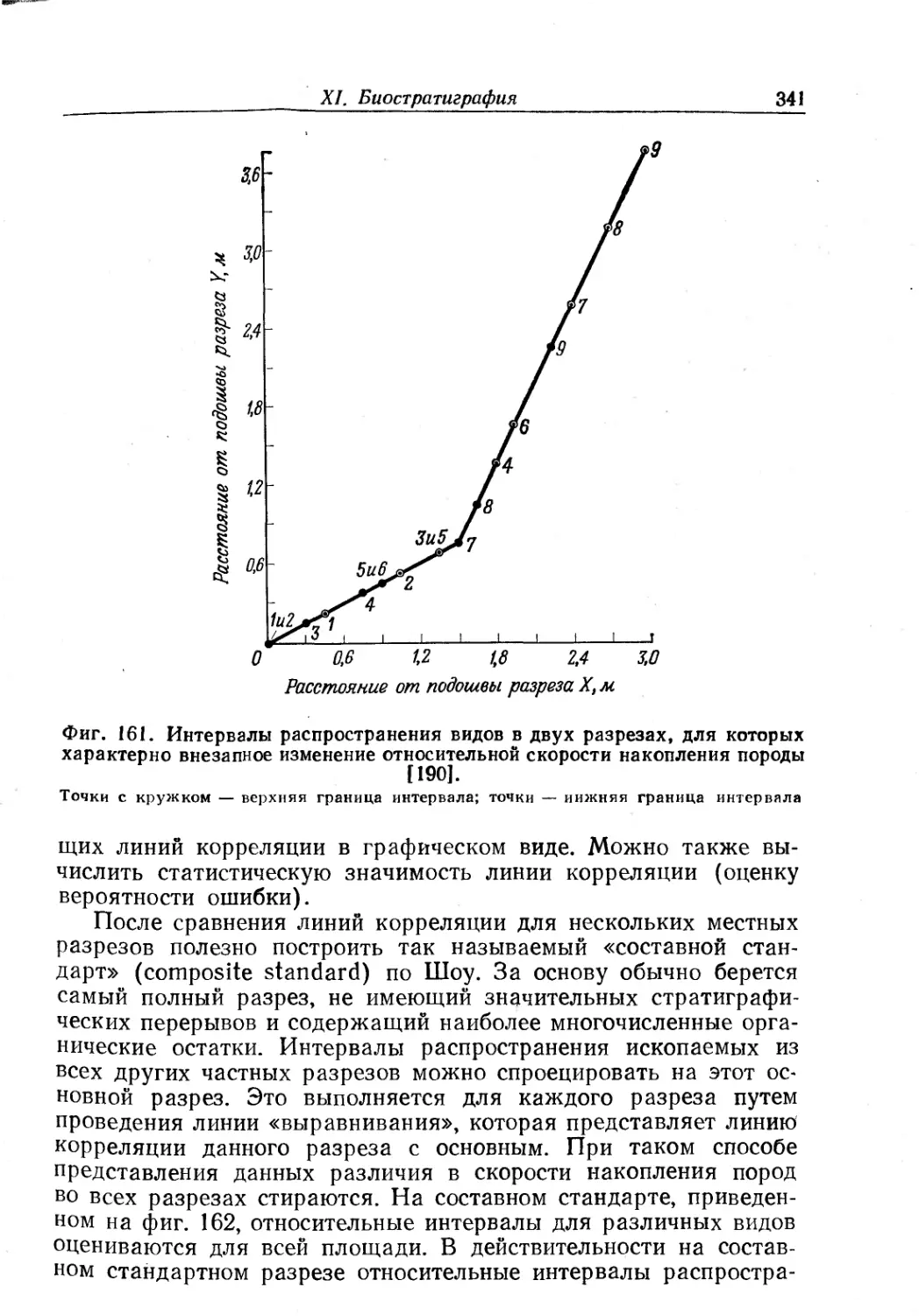
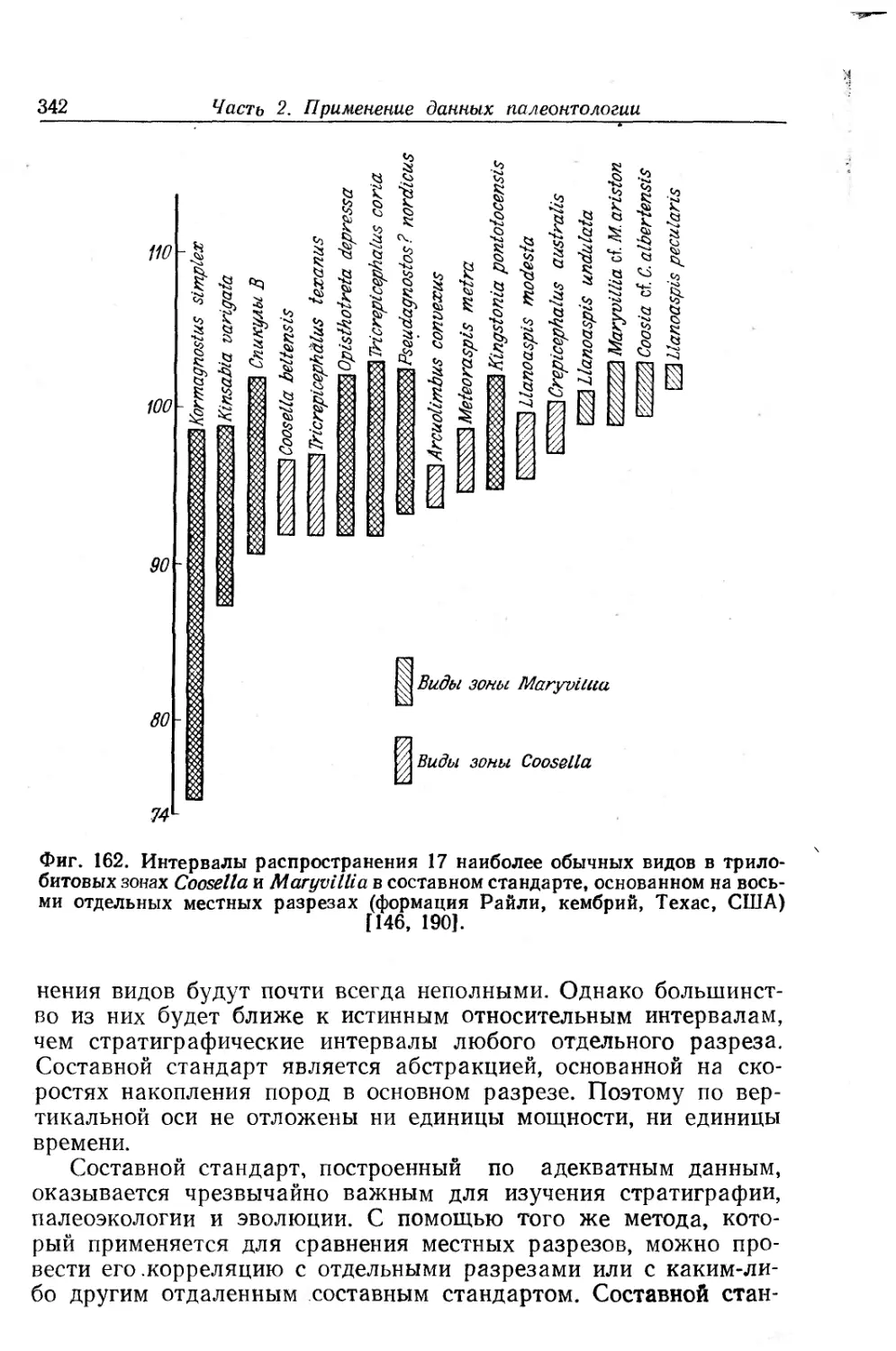


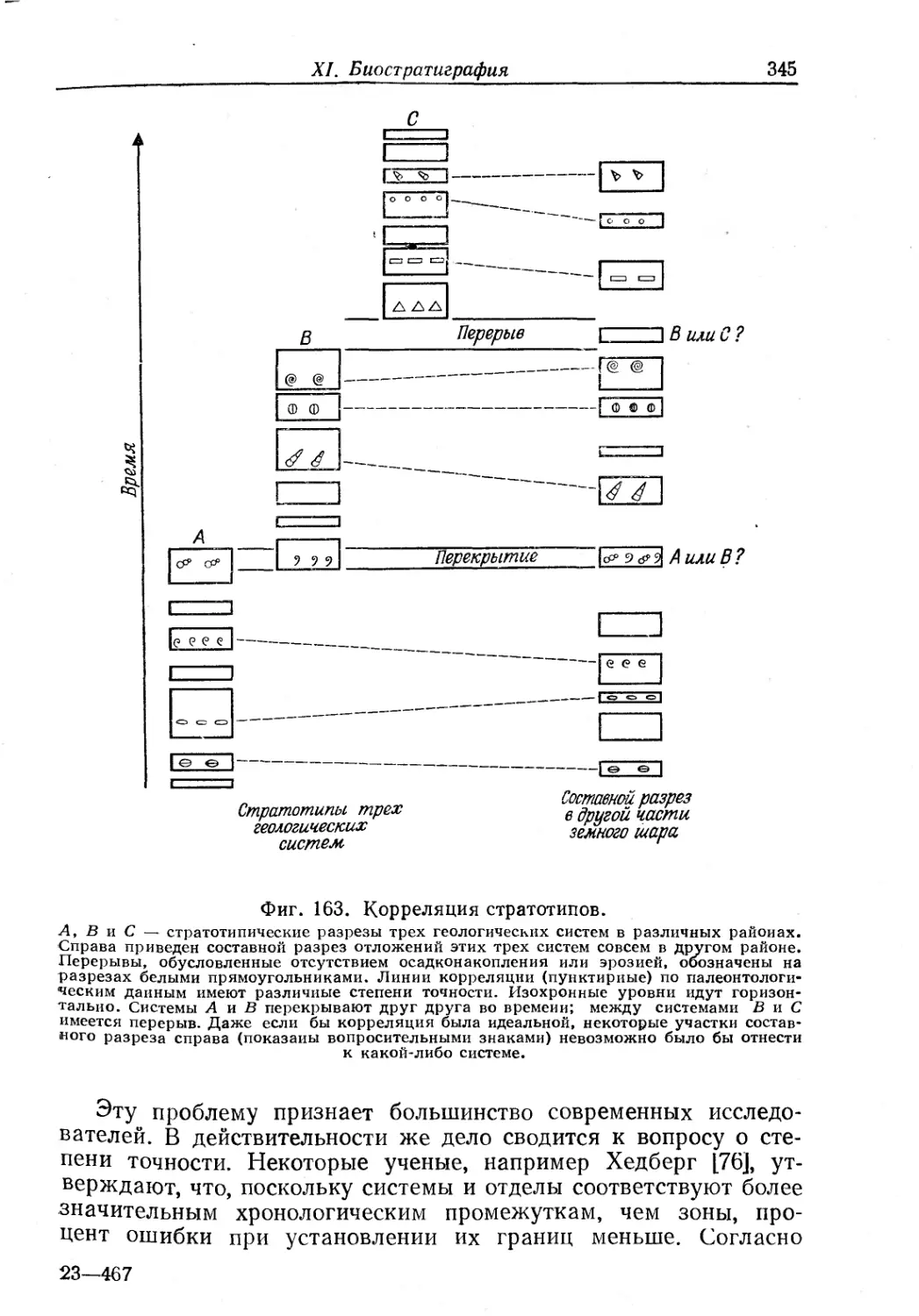


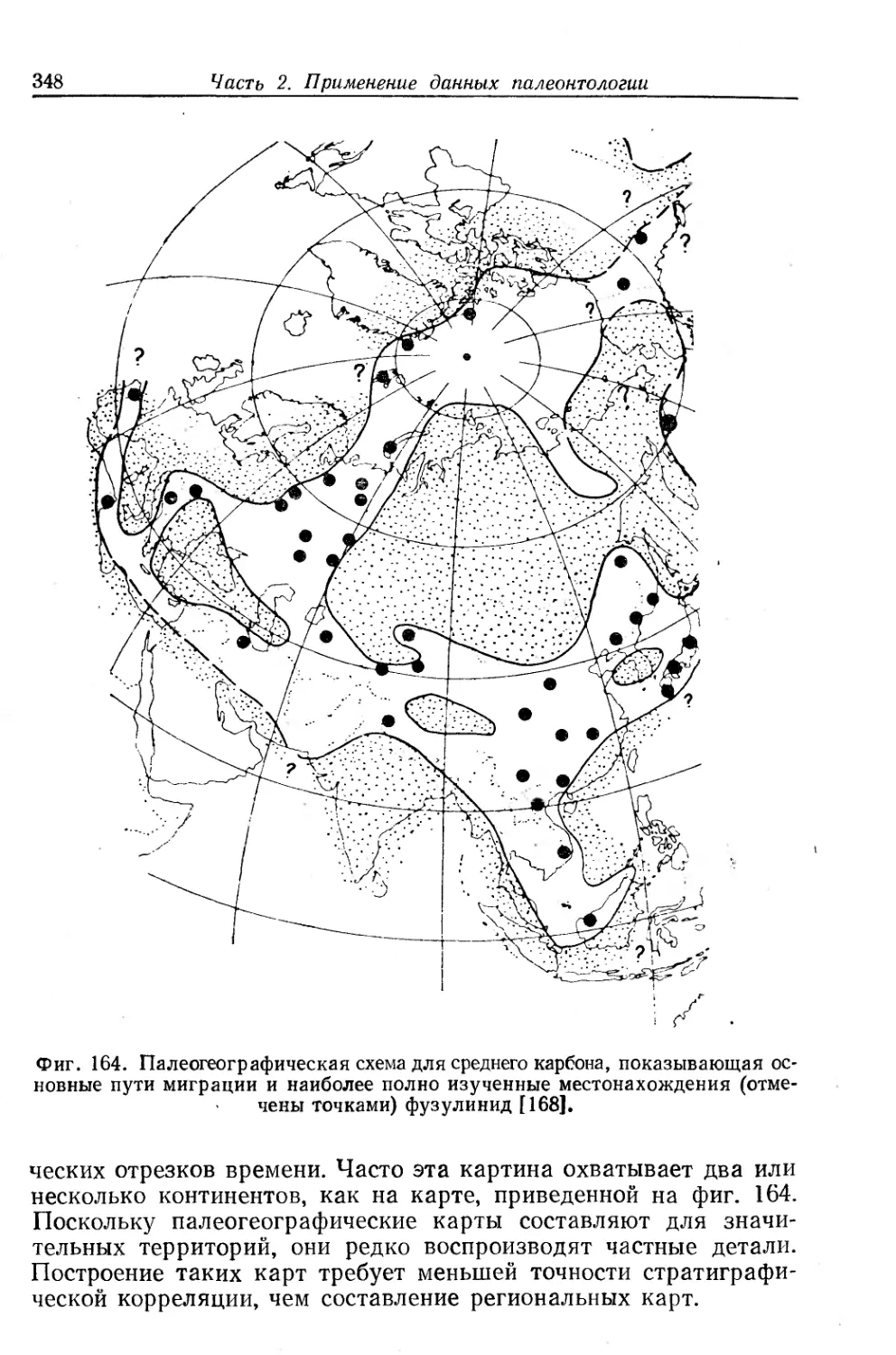











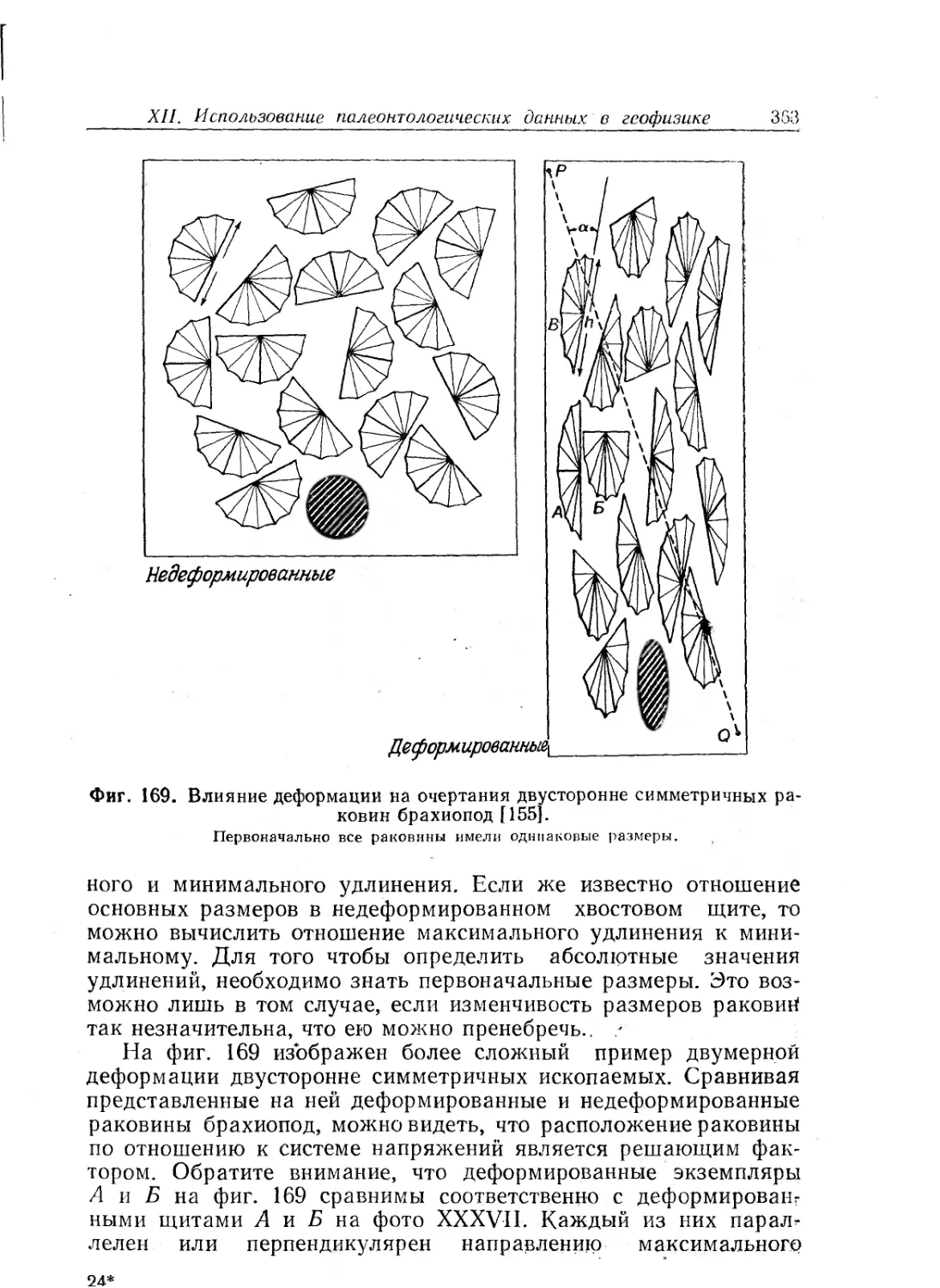
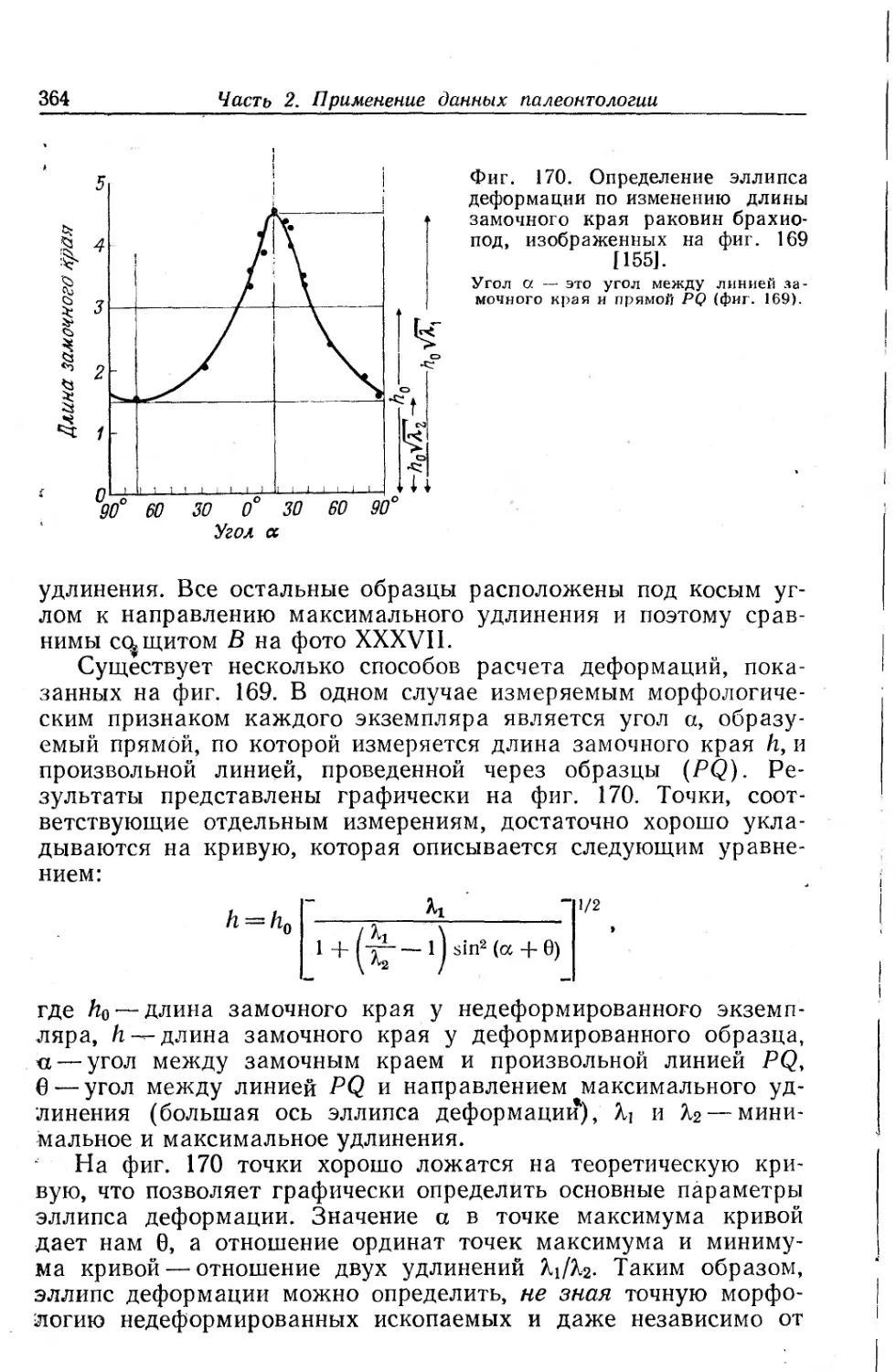

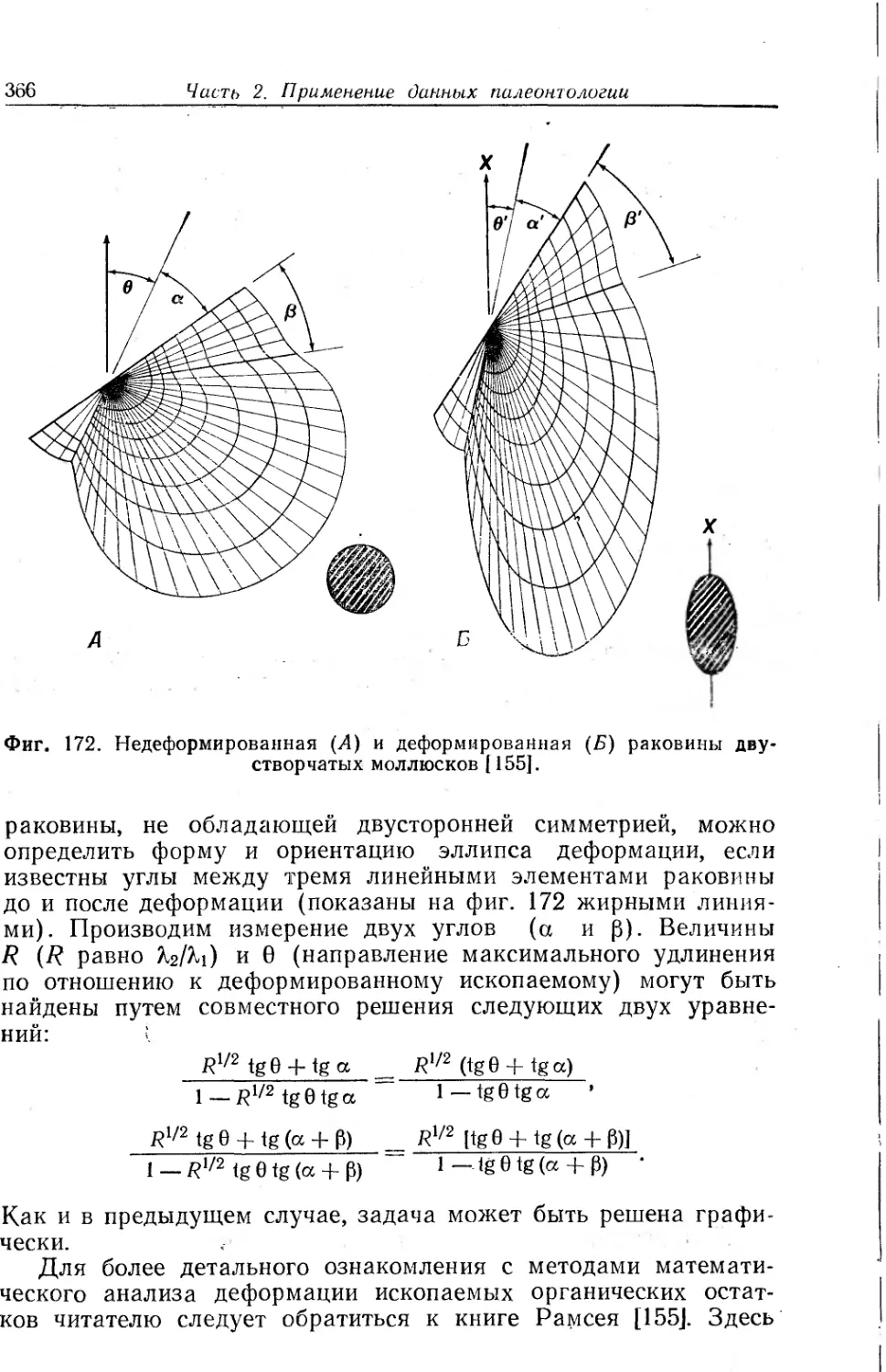

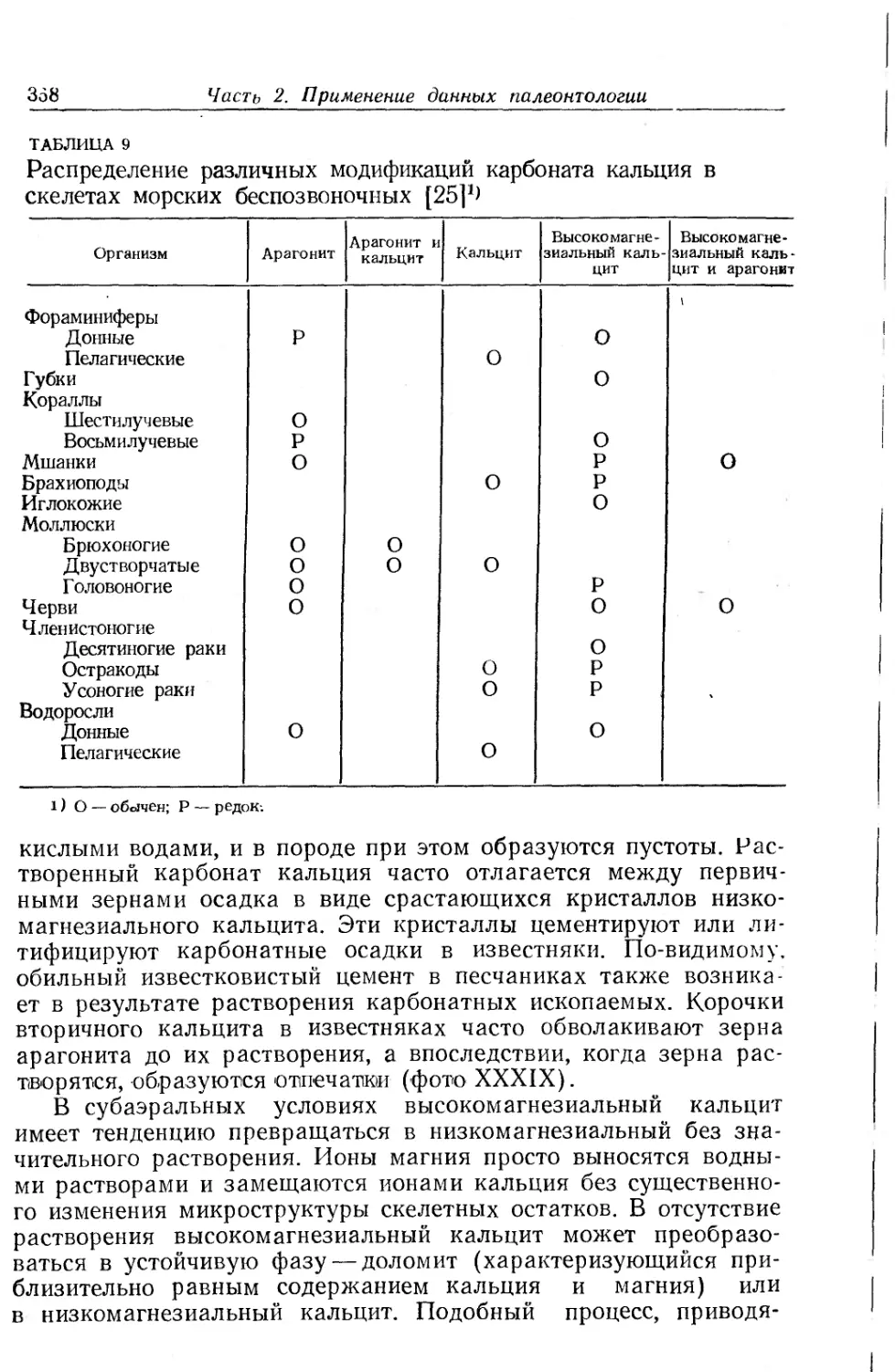
















































Text
tugia
ОСНОВЫ ПАЛЕОНТОЛОГИИ
I
PRINCIPLES OF PALEONTOLOGY
DAVID M. RAUP
University of Rochester
STEVEN M. STANLEY
The Johns Hopkins University
W. H. Freeman and Company San Francisco 1971
Д. РАУП, С. СТЭНЛИ
Р - Я.^%
ОСНОВЫ
ПАЛЕОНТОЛОГИИ
Перевод с английского
Ю. М. ФРОЛОВА и В. 3. МАХЛИНА
Под редакцией и с предисловием
д-ра геол.-мин. наук
Д. П. НАИДИНА
^246758
Омзд.ая области-вя иаучмйя библиотека вмени А.С. Йушяшт
ИЗДАТЕЛЬСТВО «МИР»
МОСКВА 1974
УДК 56
Глубокие изменения, происшедшие в палеонто-
логии за последние годы, породили потребность в
книгах, написанных по-новому. Данное издание пол-
ностью отвечает этим требованиям: в отличие от
классических учебников, в которых в систематиче-
ском порядке описывается морфология ископаемых
организмов, в ней главное внимание обращено на
общие концепции и направления современной палеон-
тологии, возникшие на основе новейших течений в
биологии. Большое внимание уделено также методам
палеонтологического исследования.
Предназначена для студентов и преподавателей
биологических и геологических факультетов, для
научных работников — палеонтологов, геологов, зо-
ологов, систематиков, океанологов, геофизиков и
геохимиков.
Редакция биологической литературы
20801—125
Р Qi) 74 125—74 (g) Перевод на русский язык, «Мир», 1974
Предисловие к русскому изданию
Книга проф. Д. Раупа (Рочестерский университет) и проф.
С. Стэнли (Университет Джона Гопкинса), перевод которой пред-
лагается вниманию читателя, задумана авторами как учебник
для студентов высших учебных заведений. Однако даже беглое
ознакомление с книгой убеждает в том, что к категории обыч-
ных вузовских учебников ее никоим образом отнести нельзя.
Прежде всего в ней нет обычного для учебника систематического
освещения фактического материала и изложения общепринятых
взглядов. Пожалуй, лишь четкое деление глав на разделы и
подразделы, многочисленные иллюстрации (их около 200) да
еще приводимые в некоторых местах действительно общеизвест-
ные элементарные сведения (образцы описания вида и ключа для
определения, примеры составления синонимики и списков видов
в геологических работах и др.) сближают ее с учебником.
Книга, как отмечают авторы в своем предисловии, написана
с перспективой на будущее, и с этой оценкой можно согласить-
ся. Основное ее назначение заключается не столько в изложе-
нии уже устоявшихся принципов и традиционных методов па-
леонтологии, сколько в формулировании новых идей и задач
этой науки, оценке новых направлений и методов исследований.
В книге отсутствует планомерное освещение вопросов морфо-
логии, систематики и развития всех (или по крайней мере ос-
новных) групп ископаемых организмов. Предполагается, что
эти сведения известны читателю (авторы упорно именуют его
«студентом»). Нет и изложения методик палеонтологических
исследований. Читатель отсылается к справочному руководству
по сбору и обработке, палеонтологического материала, в состав-
лении и редактировании которого принимал участие один из
авторов настоящей книги1.
В книге две части. Первая часть посвящена методам опи-
сания и классификации остатков вымерших организмов. Харак-
теризуется сам объект исследования — ископаемые остатки, рас-
сматриваются условия сохранения остатков организмов в иско-
1 Б. Каммел и Д. Рауп (ред.), «Методика палеонтологических исследова-
ний», изд-во «Мир», М., 1973.
6 Предисловие к русскому изданию
паемом состоянии (гл. I). Как подчеркивают авторы, палеон-
тологическая летопись, несмотря на удручающую неполноту,,
доставляет палеонтологу огромную информацию, которую нуж-
но уметь использовать. Последующие главы первой части как
раз и посвящены методам интерпретации ископаемого материа-
ла. Лейтмотив этой части: не упустить ничего из того, что запе-
чатлено в палеонтологической летописи.
Последовательно характеризуются приемы описания отдель-
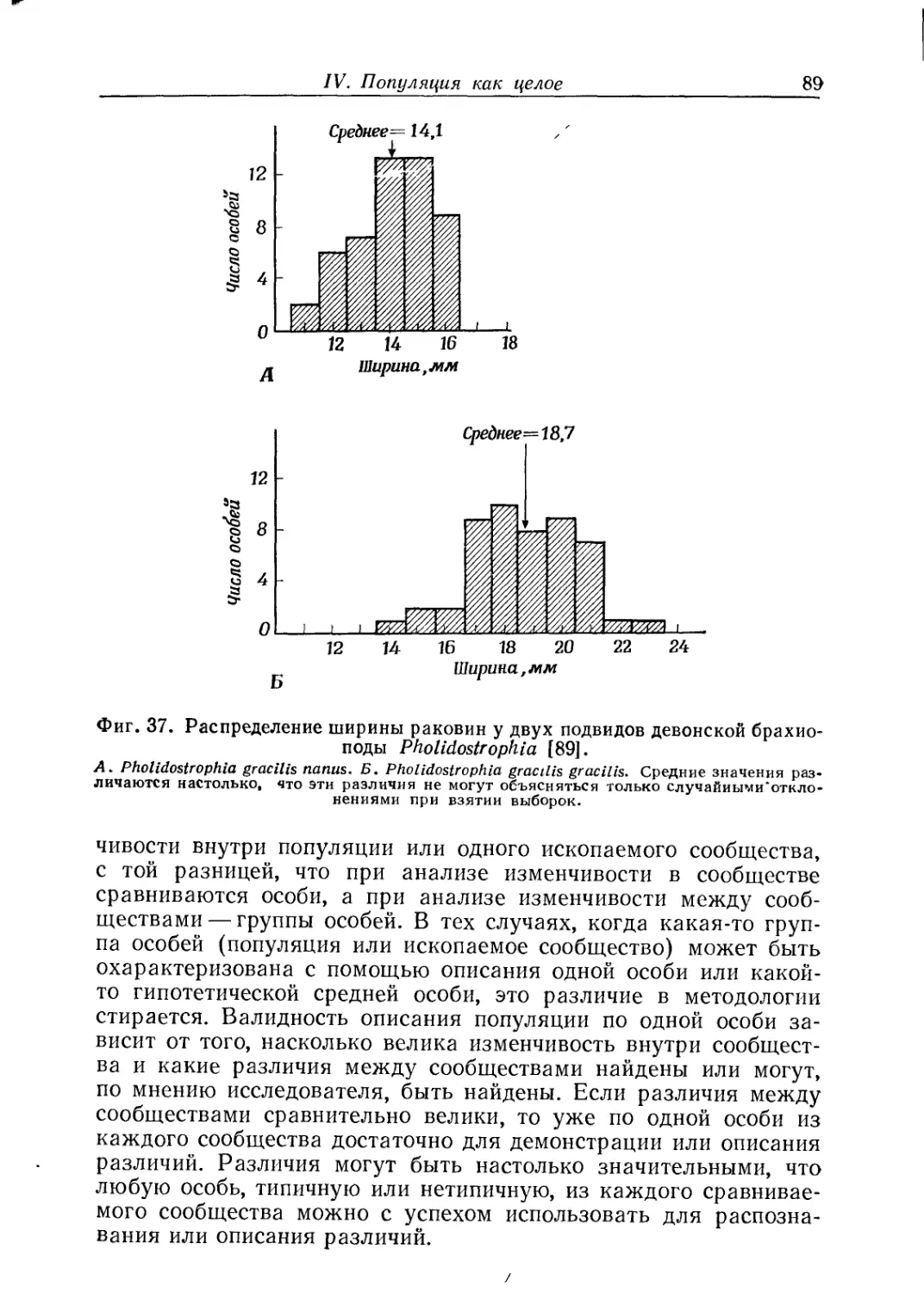
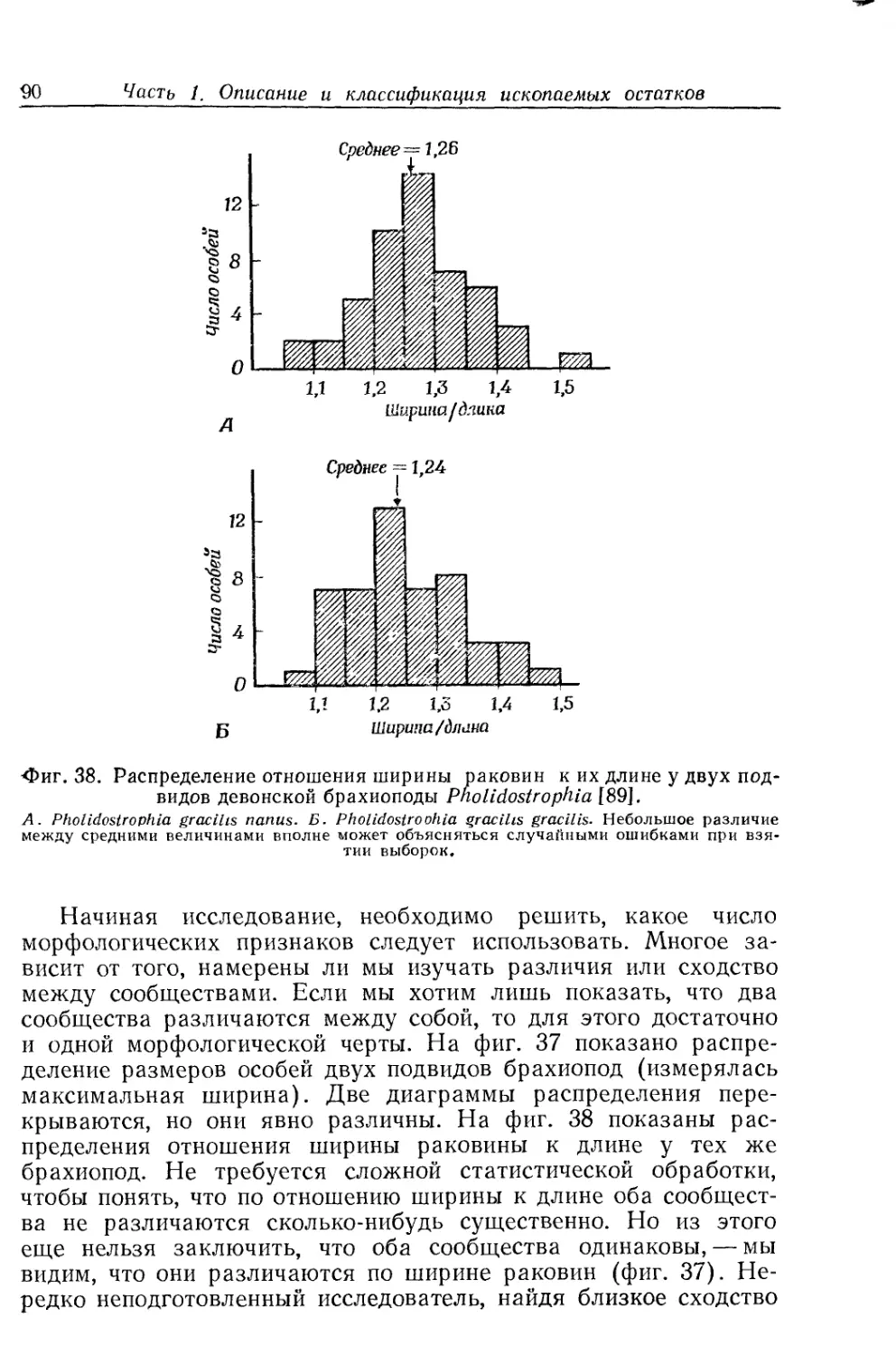
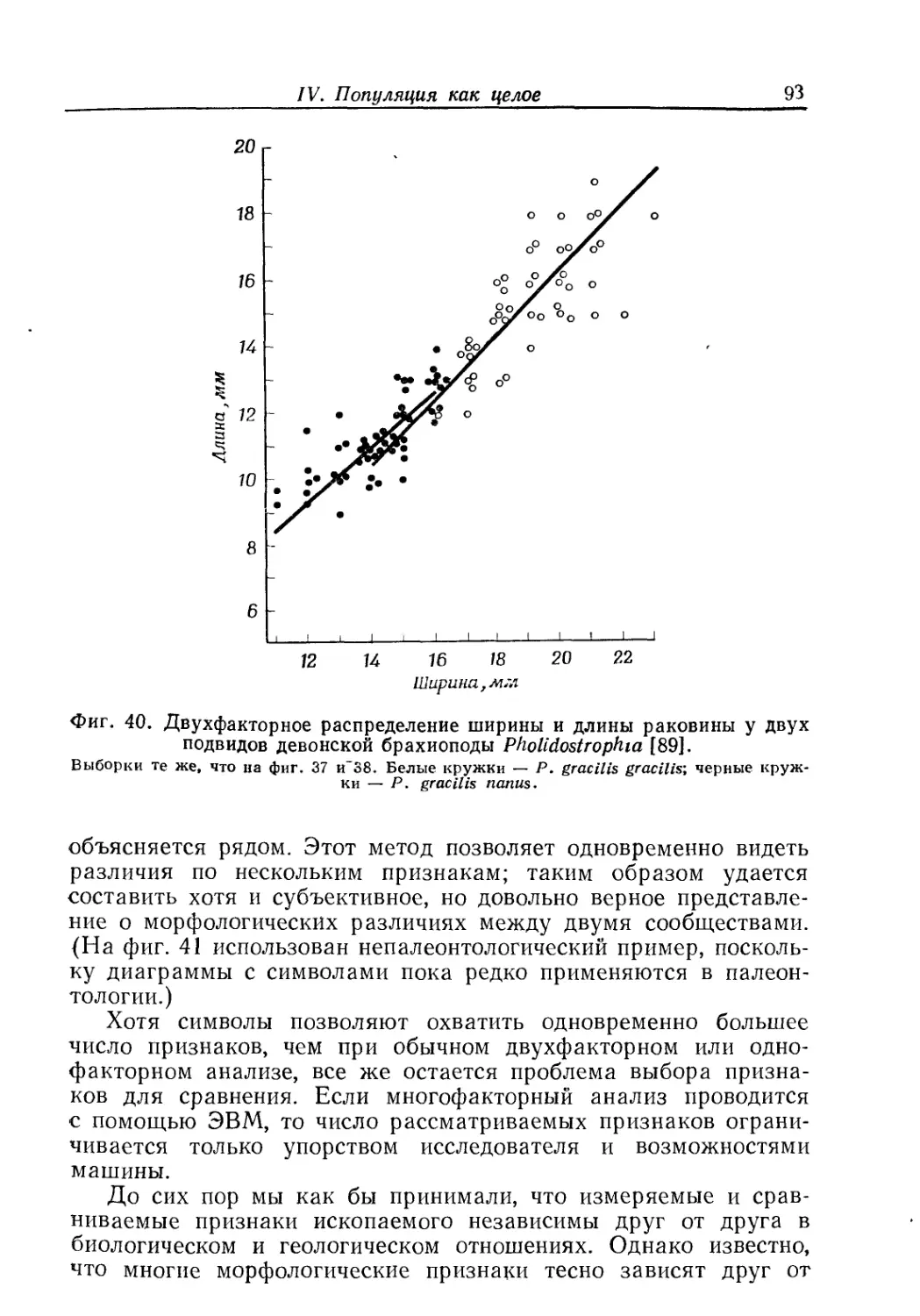
ной особи (гл. II), излагаются методы изучения онтогенетиче-
ской изменчивости (гл. III), рассматривается популяция как
целое (гл. IV).
На материале этих глав построены гл. V «Вид как целое»
и гл. VI «Группировка видов в высшие категории». Основное
внимание уделяется популяционной структуре вида: вид есть
сумма популяций, но не сумма особей; следовательно, и процесс
эволюции есть развитие популяций, а не развитие отдельных
особей. В целом авторы придерживаются концепции вида,
предложенной Э. Майром, но подчеркивают, что к ископаемому
материалу биологическое определение вида, основанное на фак-
торе репродуктивной изоляции, неприменимо. Палеонтолог име-
ет дело с трехмерной моделью вида. Поэтому разделение еди-
ного эволюционного ряда на виды — задача весьма сложная, и,
по мнению авторов, эта задача неизбежно решается субъективно
и. произвольно («проблема вида в палеонтологии будет стоять
всегда», стр. 108)
Но подобное пессимистическое заключение, как свидетель-
ствует весь труд Раупа и Стэнли, не должно обескураживать
палеонтолога. Во всех главах книги большое внимание уделено
новым направлениям в изучении палеонтологического матери-
ала: цифровому кодированию при описании ископаемых (в част-
ности, кодированию для перевода фотографической информации
в форму, пригодную для ввода в ЭВМ), математическим мето-
дам изучения различных типов изменчивости, описанию онто-
генетической изменчивости посредством метода преобразования
координат, машинным методам определения ископаемых и так
далее.
Авторы отмечают, что некоторые из этих направлений весь-
ма перспективны, но при этом подчеркивают два момента.
Во-первых, выбор того или иного метода зависит от задачи
исследования. Часто вполне можно обойтись традиционными
методами. Так, при определении родовой и видовой принадлеж-
ности некоторых групп весьма эффективны обычные ключи, а
при описании таксонов или отдельных особей — словесные их
характеристики («слово — могучее и экономное орудие», стр. 39).
Во-вторых, использовать новые методы применительно к
палеонтологическому материалу следует с осторожностью,
Предисловие к русскому изданию 7
учитывая специфику этого материала. В этой связи весьма
любопытно звучит замечание авторов, которых никак нельзя
заподозрить в консерватизме и которые, напротив, находятся
в авангарде применения нового в палеонтолог им: «Обычно счи-
тают, что только количественный подход по-настоящему объек-
тивен. Однако на самом деле измерения часто оказываются са-
мым субъективным методом описания».
Глава «Описание ископаемых» (гл. VII), завершающая пер-
вую часть книги, носит формальный характер. Именно в этой
главе, самой короткой (и наименее интересной) в книге Раупа
и Стэнли, сконцентрированы общеизвестные, элементарные све-
дения, и в целом она действительно напоминает главу учебника
п алеонтол огни.
Вторая часть книги, судя по ее названию и тому, что напи-
сано о ней в предисловии авторов, должна освещать применение
палеонтологических данных для решения различных проблем
биологии и геологии.
Несколько странным представляется понимание авторами
палеонтологических данных. «Палеонтологические данные»,
по Раупу и Стэнли, — это в сущности лишь вопросы системати-
ки, объем вида и группировка видов в крупные таксоны. А вот
вопросы адаптации и функциональной морфологии (гл. VIII),
палеоэкологии (гл. IX), эволюции (гл. X) — это уже «примене-
ние палеонтологических данных». При такой подаче материала,
например, интерпретация назначения перегородок и перегоро-
дочной линии раковин аммонитов или проблема вымирания ор-
ганизмов попадают в ту часть книги, которая называется «При-
менение палеонтологических данных» (!?).
Конечно, с подобным противопоставлением «палеонтологи-
ческих данных sensu stricto» кардинальным проблемам пале-
онтологии нельзя согласиться. Оно отражает скорее не слишком
удачную форму изложения, нежели принципиальную позицию
авторов, ибо книгу в целом отличает глубокое взаимопроникно-
вение биологии и палеонтологии.
В гл. VIII—X, составляющих немногим менее половины объ-
ема всей книги, содержится оригинальное изложение представ-
лений о путях и темпах эволюции органического мира, о причи-
нах вымирания организмов в прошлом, о главных адаптивных
приобретениях животных и растений в течение фанерозоя и
о многих других фундаментальных понятиях и проблемах. Пред-
лагаются также новые методы решения некоторых задач пале-
онтологии. В частности, большой интерес представляет попытка
анализа спирально свернутой раковины с помощью ЭВМ. Ока-
зывается, что далеко не весь спектр возможных форм, получен-
ных с помощью вычислительной машины, реализуется в приро-
де. Выяснение причин, обусловивших относительно небольшое
8
Предисловие к русскому изданию
разнообразие спирально свернутых раковин, по Раупу и Стэн-
ли, составляет задачу теоретической морфологии.
Гл. XI представляет краткий обзор применения палеонтоло-
гических данных в стратиграфии. Сопоставляются биострати-
графические и литостратиграфические подразделения, харак-
теризуются основные методы корреляции по органическим ос-
таткам, оценивается точность корреляций по ископаемым. Здесь
вызывает некоторое удивление утверждение авторов о том, что
единственным твердым принципом в стратиграфии является
«закон» последовательности напластования и что стратиграфия
относится к весьма субъективным областям исследования. С по-
добной оценкой стратиграфии, конечно, согласиться нельзя.
В гл. XII, заключительной главе книги, рассматриваются
примеры применения палеонтологических данных к решению
некоторых задач, весьма далеких от палеонтологии. Авторы ог-
раничиваются лишь тремя примерами, но число их легко можно
было бы увеличить. Глава имеет принципиальное значение, так
как убедительно свидетельствует о том, что результаты изуче-
ния остатков давно вымерших организмов могут получить самое
неожиданное применение. Что, например, может связывать ха-
рактер роста кораллов и продолжительность года? Наблюдения
различных исследователей, проведенные в последнее десятиле-
тие, позволили установить, что на эпитеке кораллов и на ракови-
не двустворчатых моллюсков запечатлены суточные следы
роста в виде линий нарастания и что, подсчитав их, можно1
определить число дней в месяце и в году. Подобные подсчеты
показали, что в палеозое в году было больше дней, чем сейчас.
Эти данные подтверждают расчеты астрономов и геофизиков
о постепенном замедлении вращении Земли и о соответственном
уменьшении числа дней в году.
Некоторые недостатки книги Раупа и Стэнли уже отмеча-
лись выше. К - ее недостаткам следует также отнести слишком
уж беглое освещение проблем палеонтологии докембрия, почти
полное отсутствие сведений о палеопатологии, а также о строе-
нии скелетных остатков, их химическом и минеральном составе.
Совершенно не затронуты и палеобиогеохимические сведения;
по мнению авторов, применение в палеонтологии изотопных ме-
тодов и различных методов изучения скелетного вещества мало
что дает вследствие существенных изменений первичных (при-
жизненных) концентраций изотопов и отдельных элементов
в процессе диагенеза. С подобным заключением трудно согла-
ситься. Во-первых, если палеонтологическую летопись, несмотря
на ее неполноту, можно тем не менее с успехом использовать,
то это точно так же справедливо и для «палеобиогеохимической
летописи». Во-вторых, несмотря на ограниченные возможности
применения в палеонтологии изотопных и многих других мето-
Предисловие к русскому изданию
9
дов, все же именно с их помощью получен ряд ценных и важ-
ных данных (например, о стадиях развития некоторых организ-
мов, о температурах роста организмов и т. д.). В-третьих, ис-
ключив чисто биогеохимические вопросы, авторы одновременно
отказались и от рассмотрения структуры скелетных образова-
ний, их минеральной формы и т. п. Между тем в изучении строе-
ния раковин моллюсков и скелетных образований других орга-
низмов в настоящее время достигнуты значительные успехи,
получены важные данные о минеральном составе скелетов
(в частности, установлено, что накопление в скелетах поли-
морфных разновидностей карбоната кальция — арагонита и
кальцита — находится под генетическим контролем и, следова-
тельно, является таксономическим признаком).
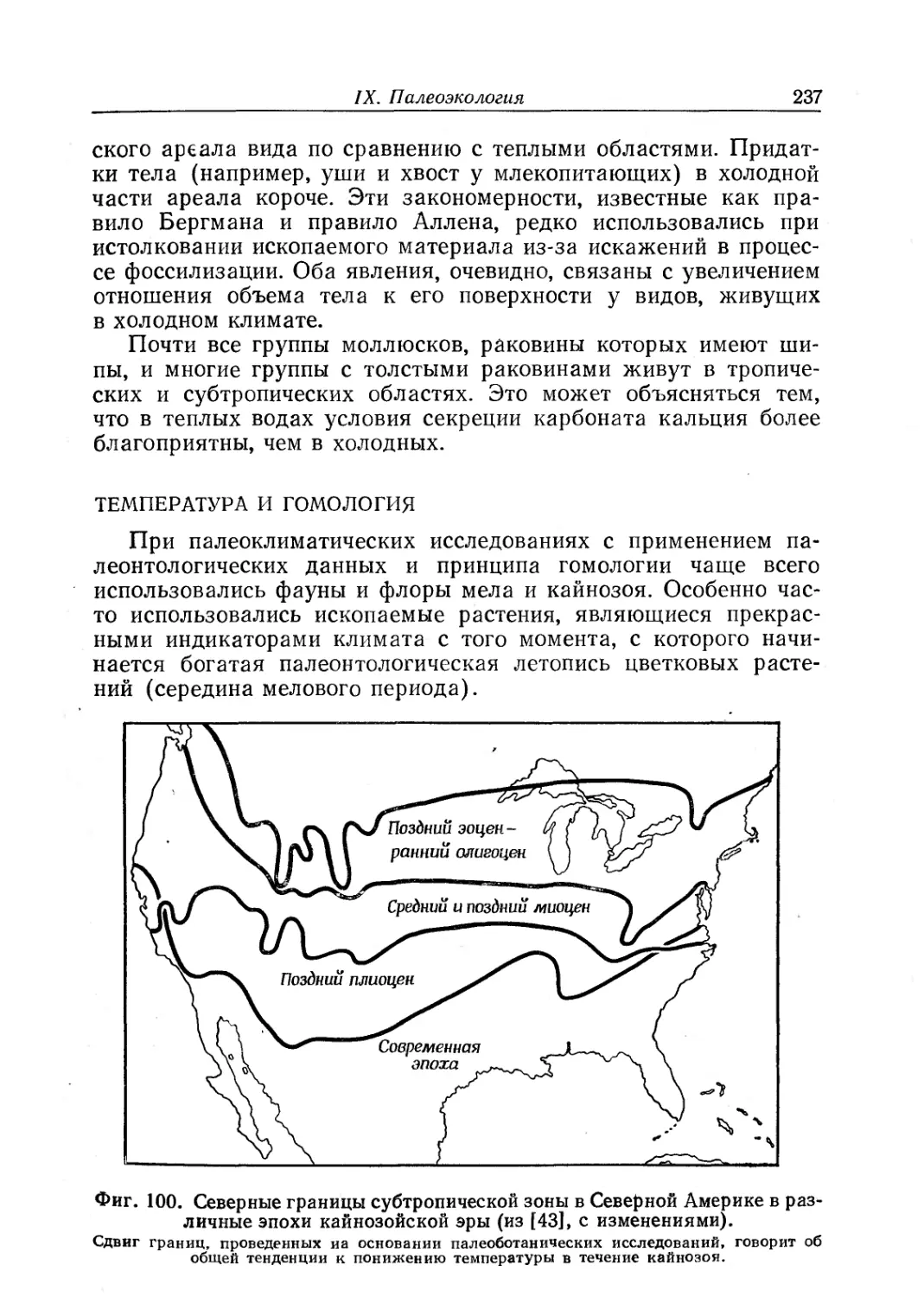
В целом несмотря на отмеченные недостатки, книга Раупа
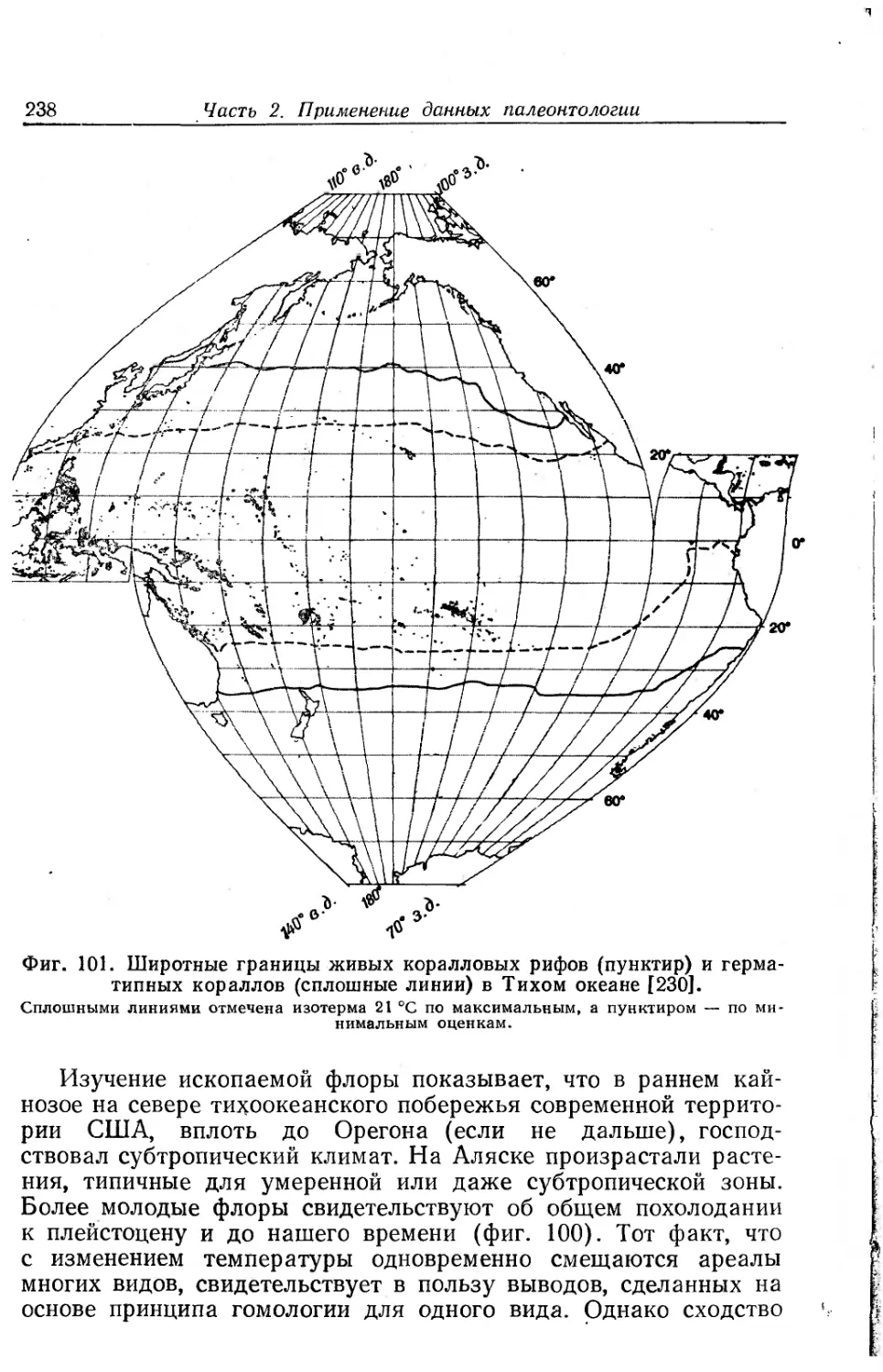
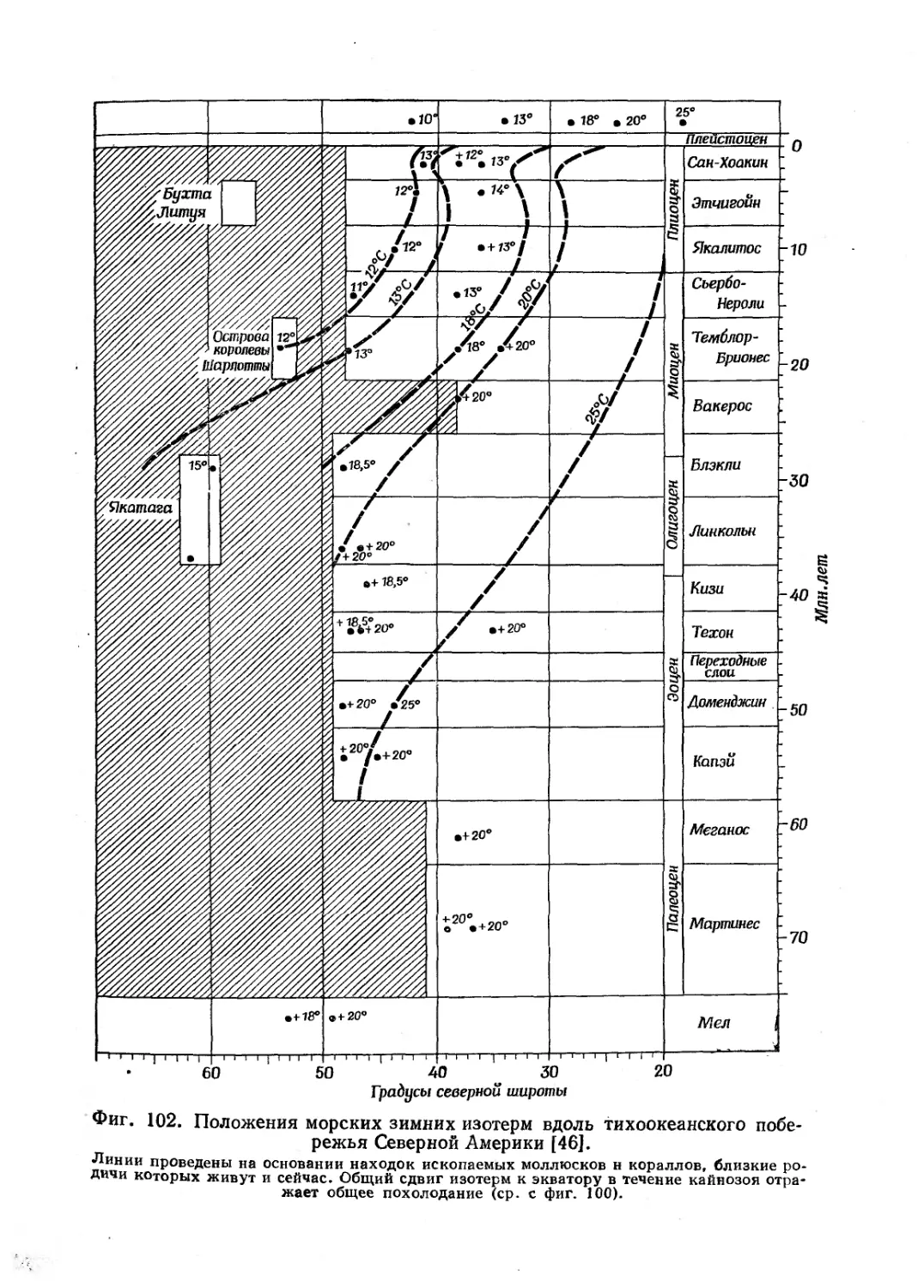
и Стэнли оставляет исключительно хорошее впечатление. Для
нее характерен новаторский подход к проблемам палеонтоло-
гии. Книга, несомненно, привлечет внимание широкого круга
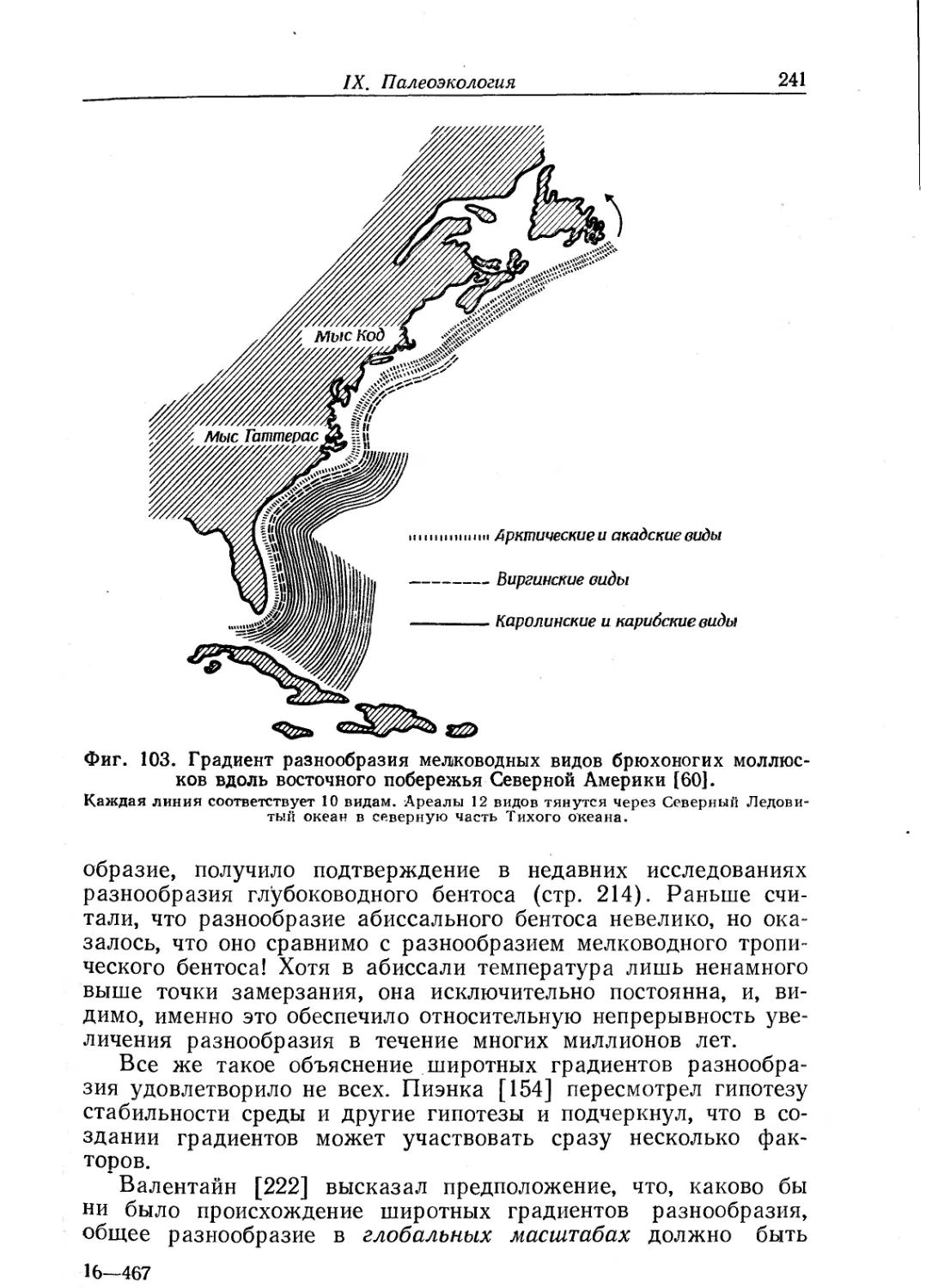
читателей еще и потому, что в ней уделено весьма большое вни-
мание новым и перспективным методам исследования. Можно
ожидать, что эта книга с большой пользой будет прочитана
нашими геологами, палеонтологами и биологами.
Перевод книги осуществлен кандидатом биологических наук
Ю. М. Фроловым (гл. I—X) и кандидатом геолого-минерало-
гических наук В. 3. Махлиным (гл. XI и XII). ,
Д. П. Найдин
Предисловие авторов
В этой книге мы стремились изложить основы палеонтологии
на уровне, приемлемом для студента высшего учебного заведе-
ния. В наши намерения не входило охватить все содержание
курса палеонтологии; книга дает учащемуся знания, необходи-
мые для понимания и усвоения этого курса, и содержит те
данные и представления, которые на лекциях и практикумах
рассматриваются лишь частично или освещаются под иным уг-
лом зрения. Обычно курс палеонтологии строится на основе
системы классификации живых существ, и учащиеся последо-
вательно переходят от одного .типа к другому. Тем яснее необ-
ходимость ознакомления студентов с общими принципами и
методами палеонтологии.
Палеонтология — увлекательная наука. Она быстро развива-
ется, и многие представления и гипотезы, изложенные здесь,
в ближайшие годы будут уточнены, а то и заменены новыми.
Мы старались охватить как можно больше новых и перспектив-
ных идей. Мы хотели посеять семена завтрашнего урожая —
пусть эта книга разбудит воображение будущих палеонтологов
и непалеонтологов.
В нашей книге двенадцать глав; если тратить на изучение
каждой главы неделю, то всего учебника как раз хватит на
средний семестр. Первые главы носят более общий характер и
содержат меньше биологических и палеонтологических приме-
ров, так что студенты должны будут уже ознакомиться с боль-
шинством таксономических групп на практикумах и лекциях,
а также при чтении справочной литературы к тому времени,
как они встретят эти группы в книге.
Первая часть учебника — «Описание и классификация иско-
паемых остатков» — начинается со вступительной главы, в ко-
торой даются общие сведения о палеонтологической летописи.
В последующих главах обсуждаются- методы изучения окамене-
лостей как отдельных представителей определенного вида, как
видов и как иерархических групп видов. Первую часть книги
можно было бы назвать «Таксономия» или «Систематика», но
она несколько отличается по содержанию от этих широких дис-
циплин.
Предисловие авторов 11
Во второй части — «Применение палеонтологических дан-
ных» — рассказывается о том, как сведения, полученные при
изучении ископаемых, применяются в различных областях гео-
логии и биологии. Кроме глав, посвященных традиционным от-
раслям науки — палеоэкологии, эволюционной теории и био-
стратиграфии,— мы включили в эту часть и такие главы, как
«Адаптация и функциональная морфология» и «Применение
палеонтологических данных в геофизике и геохимии». Функци-
ональную морфологию иногда рассматривают как раздел па-
леоэкологии, но она занимается не только экологическими
вопросами и заслуживает, как нам кажется, выделения в осо-
бую ветвь палеонтологии. Поскольку кафедры палеонтологии
по традиции входят в состав геологических факультетов, мно-
гих студентов особенно интересует вклад палеонтологии в гео-
физику и геохимию.
Возможно, вызовет критику тот факт, что мы опускаем раз-
дел, который можно было бы назвать «Биогеохимией», но нам
кажется, что многочисленные изотопные методы и методы, осно-
ванные на изучении малых элементов, за последние двадцать
лет внесли весьма незначительный вклад в палеонтологию, и
дело здесь главным образом в тех изменениях, которые проис-
ходят с органическими остатками в процессе диагенеза. Может
быть, более результативным окажется изучение некарбонатных
составляющих ископаемых органических остатков, но пока
в этой области сделано сравнительно мало.
Наконец, о названии книги. Мы раздумывали, не назвать
ли ее «Палеобиология» или «Геобиология», но решили, что
не стоит изменять название учебника, пока не изменилось на-
звание науки. Однако сами термины «палеобиология» и «гео-
биология» (последний по образцу геофизики и геохимии) до-
стойны внимания. Они подчеркивают прогрессивные перемены,
наступившие в палеонтологических изысканиях в середине XX
века.
Мы благодарны нашему постоянному помощнику Р. Итону
за выполнение фотографий и коллеге 3. Боуен за просмотр
рукописи. Мы с удовольствием примем все замечания и пред-
ложения, которые помогут улучшить книгу.
Д. Рауп
С. Стэнли
Часть первая
ОПИСАНИЕ И КЛАССИФИКАЦИЯ
ИСКОПАЕМЫХ ОСТАТКОВ
ГЛАВА I
Сохранность
органических остатков
и палеонтологическая летопись
Палеонтологическая летопись далеко не полна; в ней отра-
жена лишь небольшая часть древней жизни. К тому же то, что
сохранилось, не является представительной выборкой. В резуль-
тате действия различных биологических и геологических фак-
торов первоначальный облик органического мира оказался силь-
но искаженным в палеонтологической летописи. Изучая иско-
паемые остатки, применяя данные палеонтологии, надо ясно
представлять себе как сильные, так и слабые стороны этой
летописи. Мы должны понимать, что можно и чего нельзя
узнать по ископаемым остаткам.
Не все растения и животные имеют одинаковые шансы со-
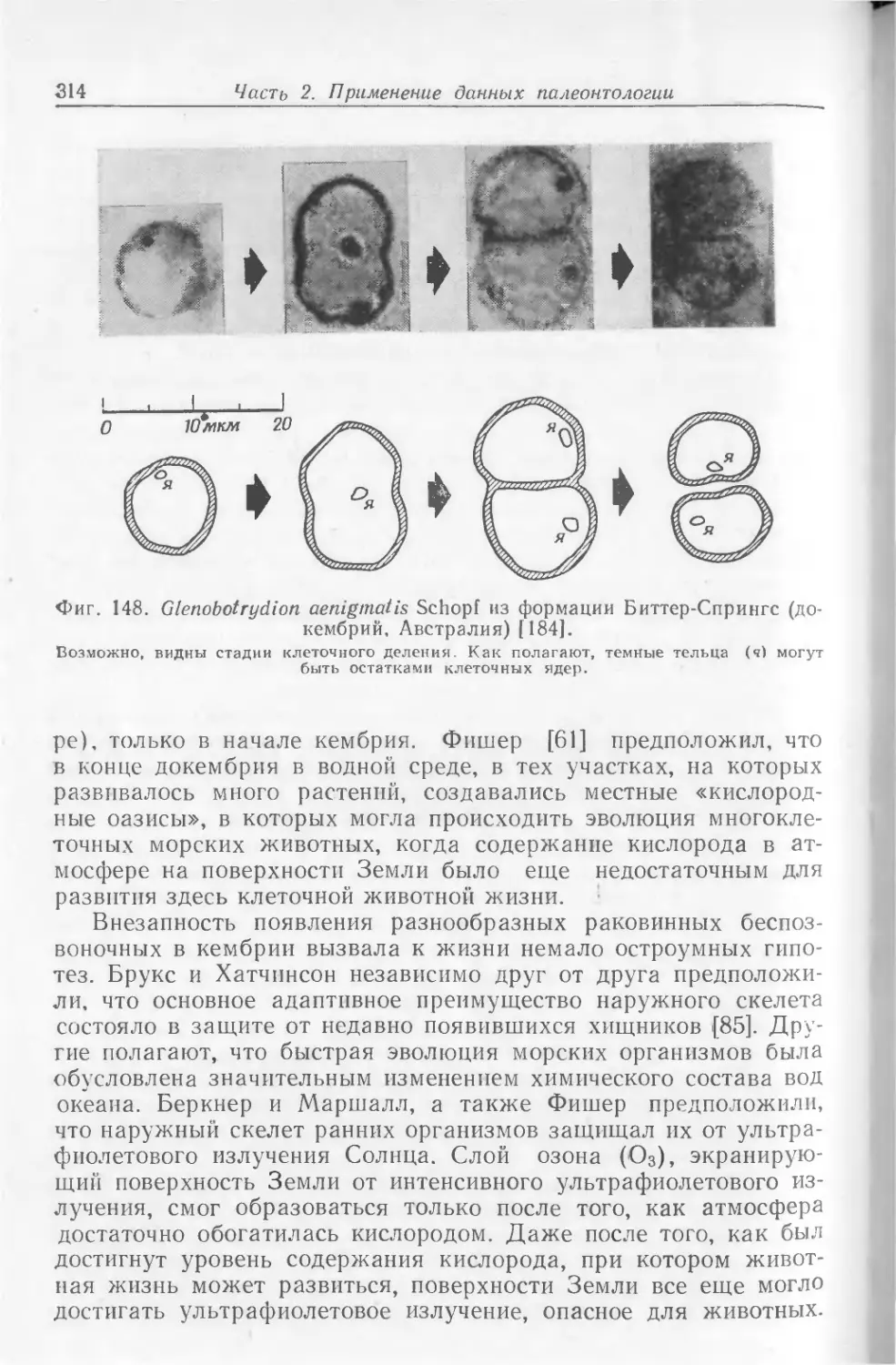
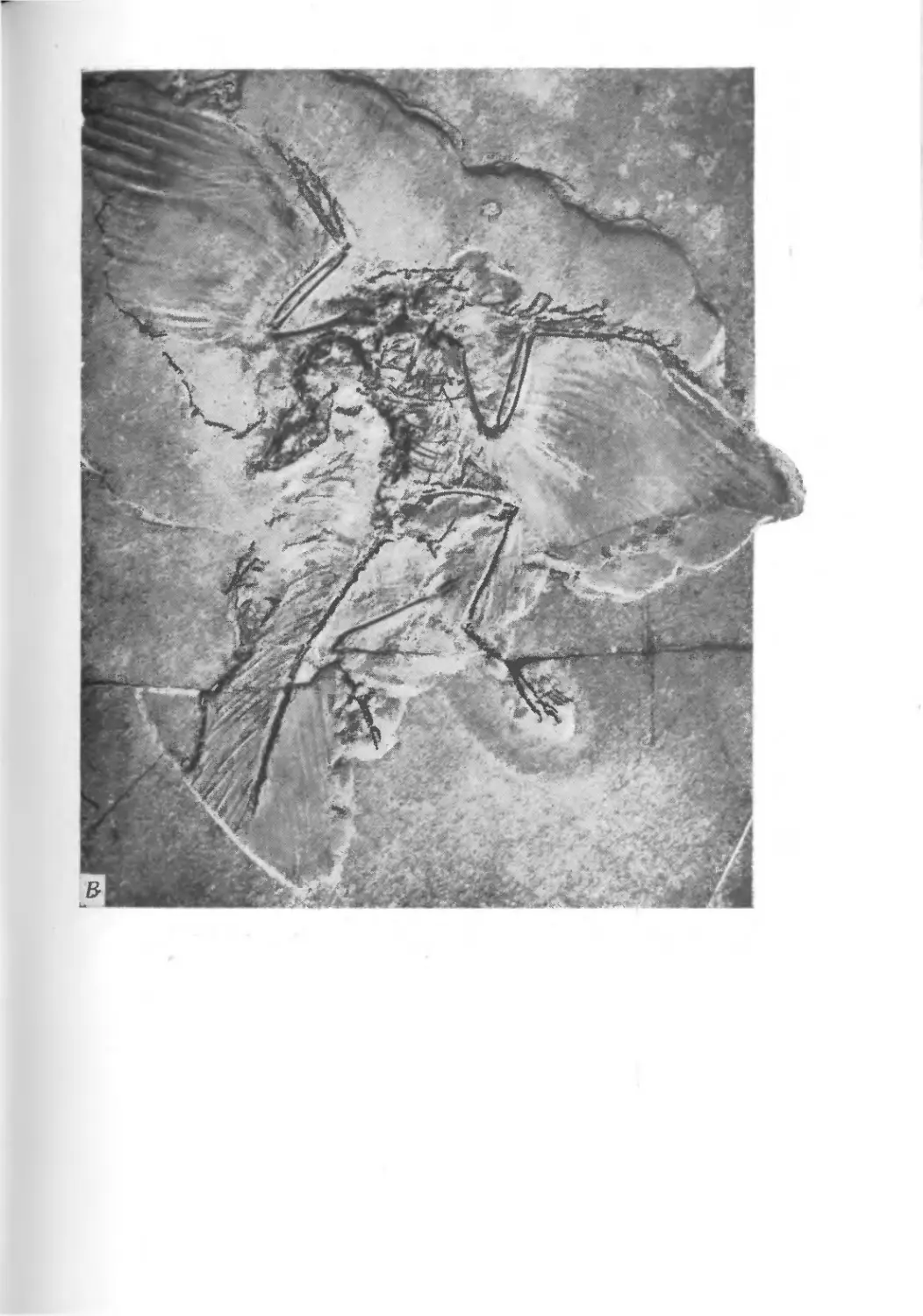
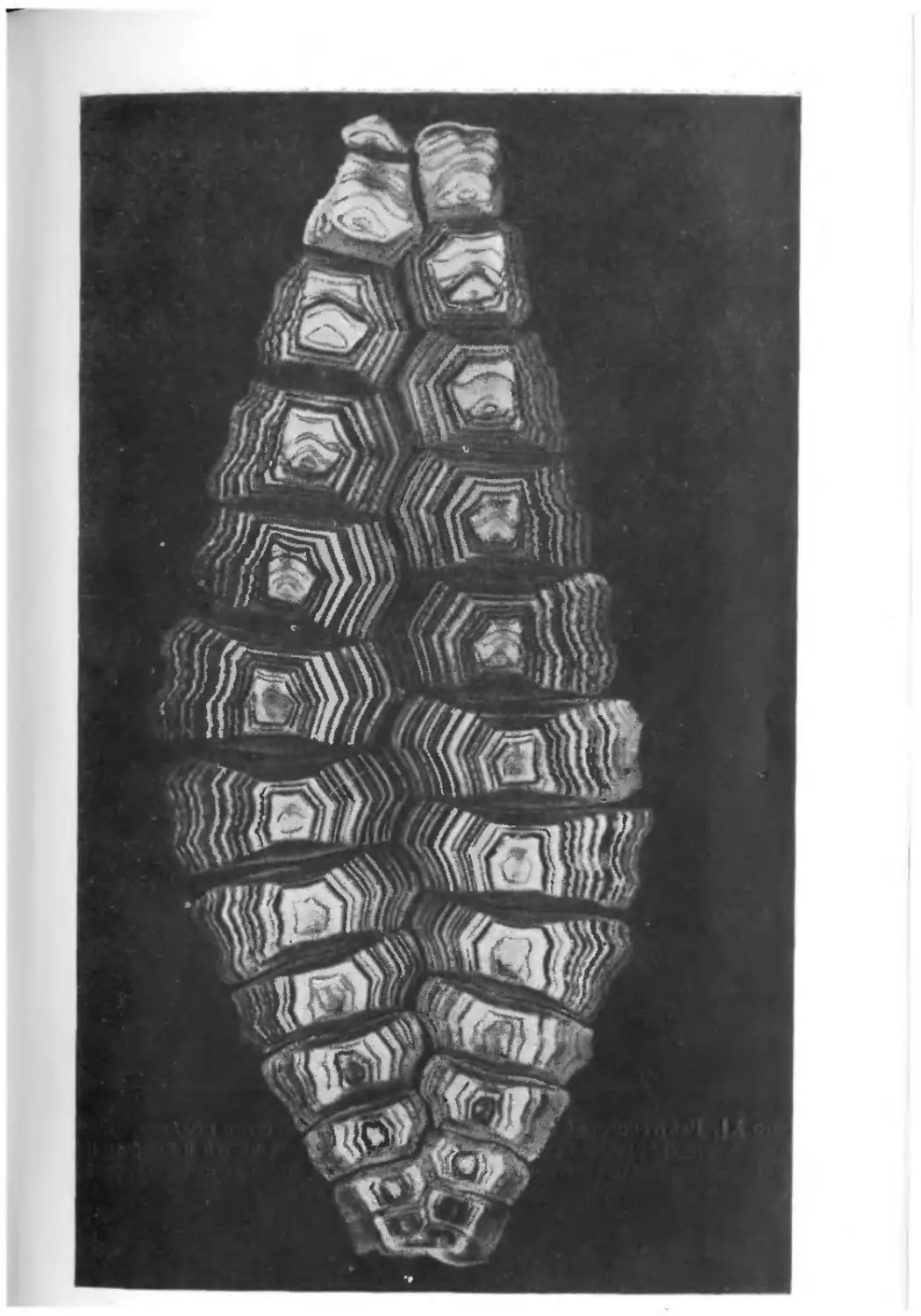
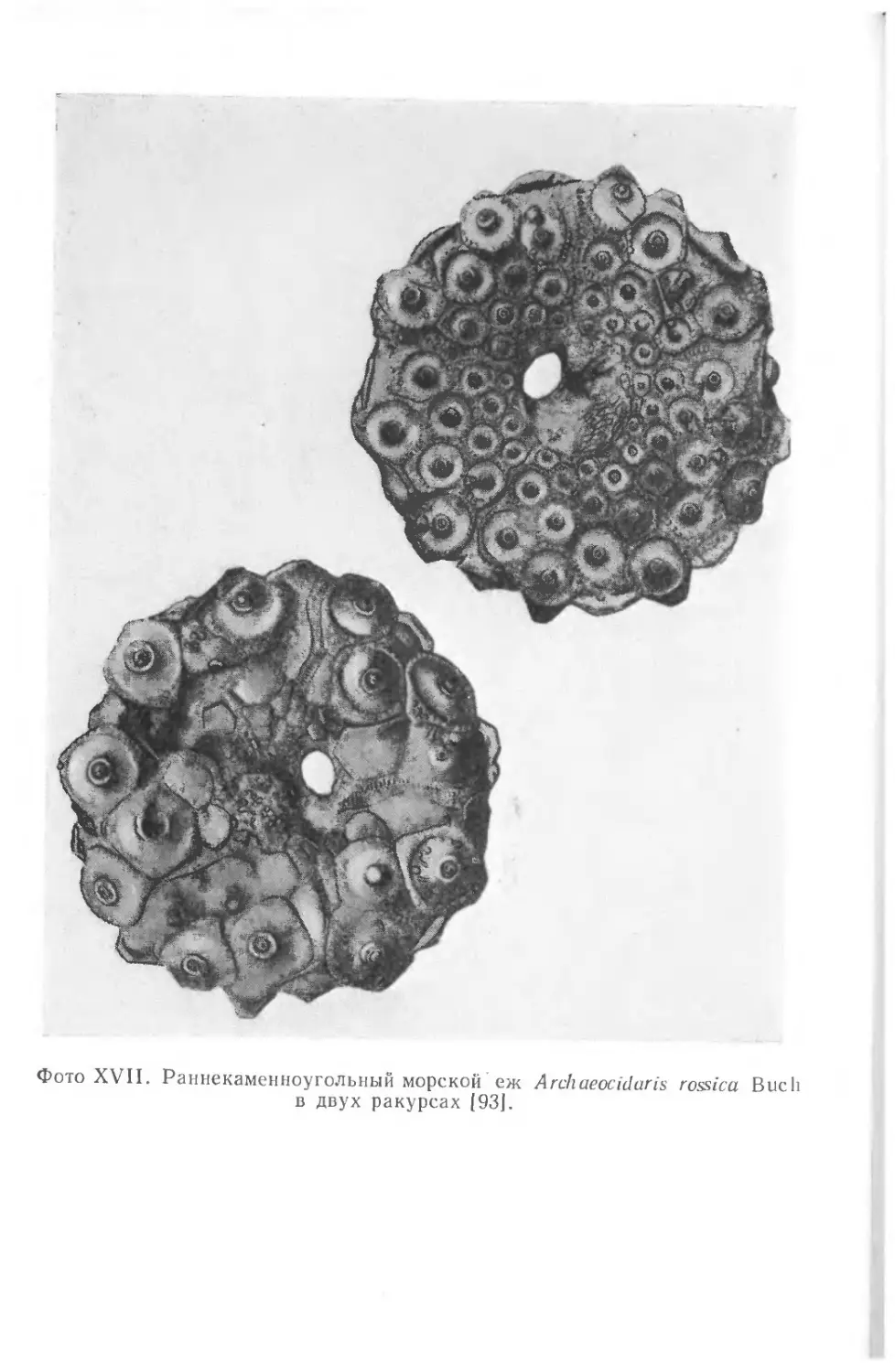
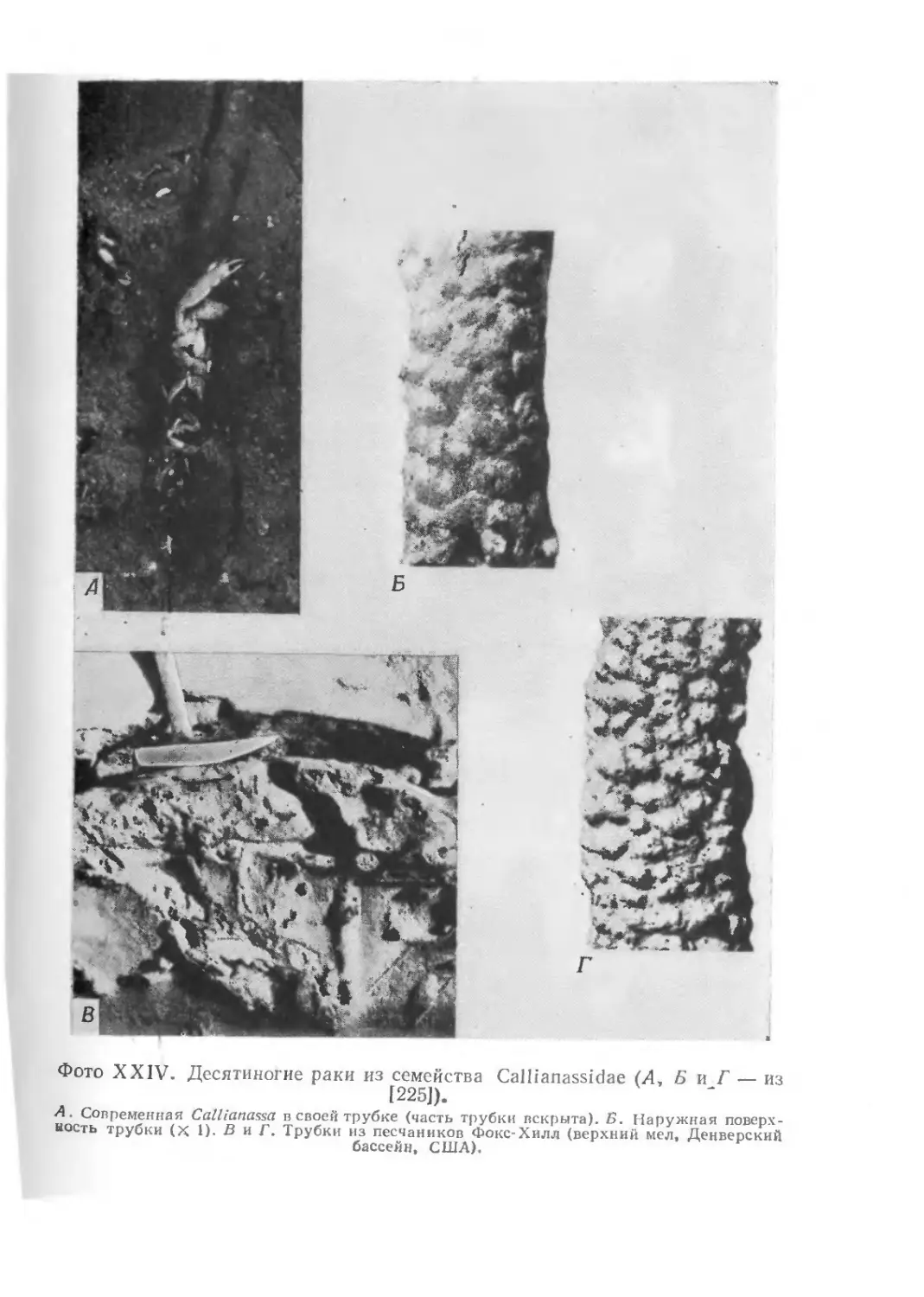
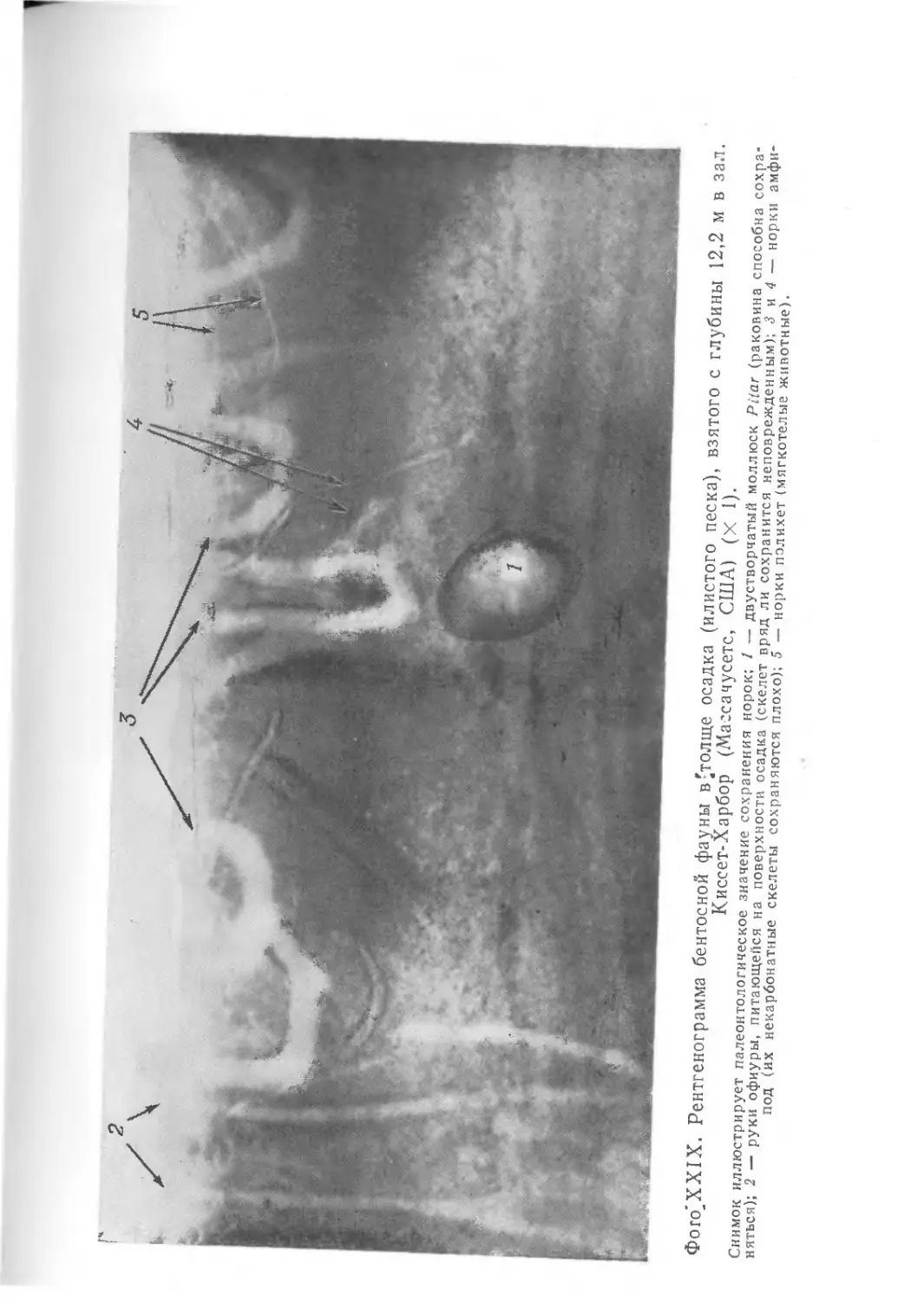
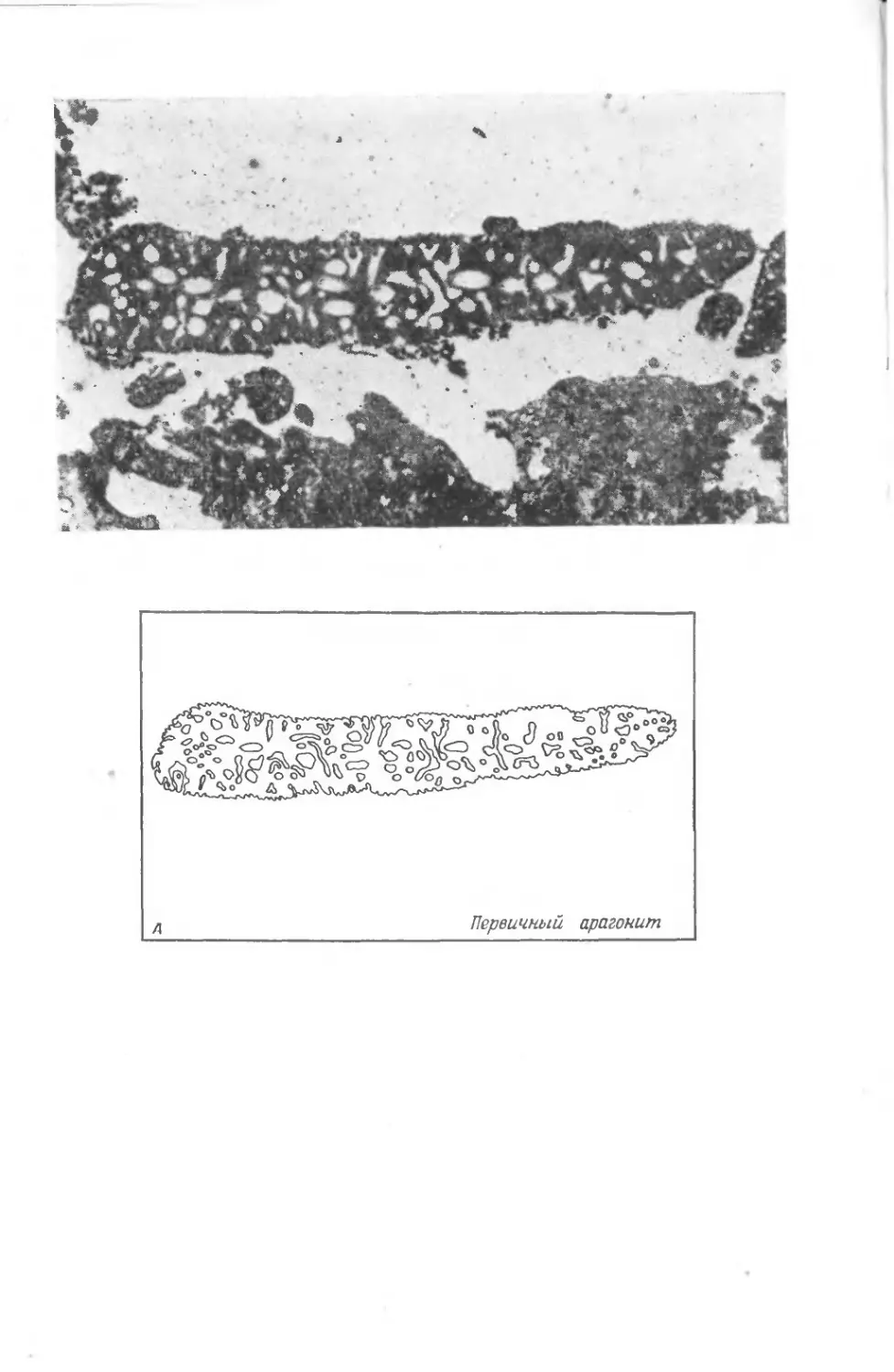
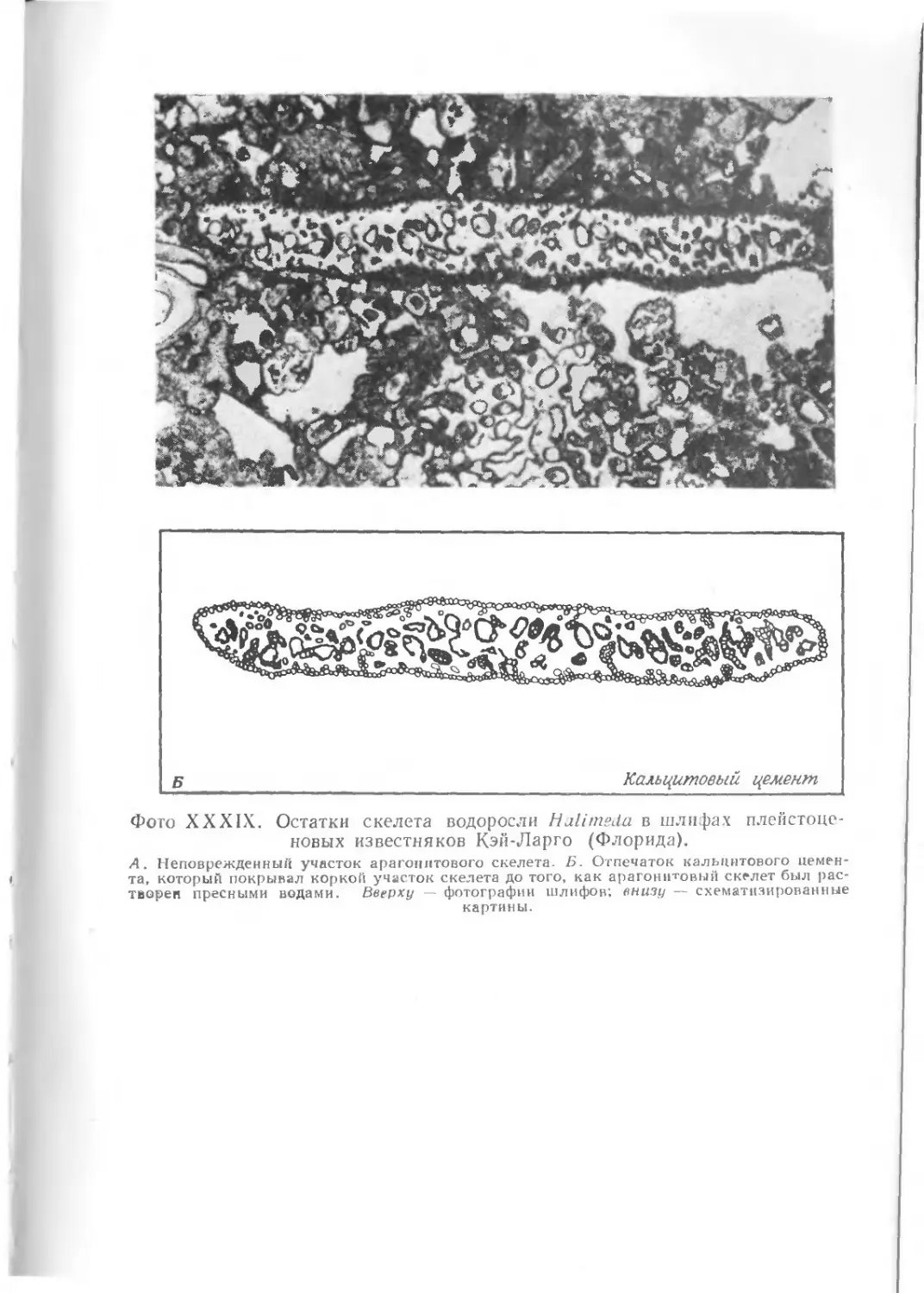
храниться в виде окаменелостей. На фото I1 показаны некото-
рые примеры необычно хорошей сохранности ископаемых остат-
ков. Все они найдены в юрских зольнхофенских известняках
(южная Бавария, ФРГ).
Скелет ракообразного Penaeus, представленный на фото I, Л,
сохранился почти в целости, причем отдельные элементы скеле-
та находятся в своем естественном положении. Само по себе
это было бы еще не так удивительно, но здесь запечатлены
некоторые интересные подробности — на фотографии они отме-
чены стрелками. Ясно видно, что тело животного упало на дно,
оставив на поверхности мягкого осадка следы падения, ныне
запечатленные в породе. При ударе о дно возникли отпечатки
спинного медианного киля (/), а также рострума (2) и глаз (3).
Затем тело перевернулось, оказавшись в том положении, в ко-
тором его ископаемые остатки и были найдены.
1 Все фотографии помещены в конце книги.— Прим. ред.
14
Часть 1. Описание и классификация ископаемых остатков
Фиг, 1. Следы пермского пресмыкающегося из Большого каньона (США).
Следы хорошо сохранились, но скелетных остатков этих животных в породах не найдено
На фото I представлен также ископаемый мечехвост Meso-
limulus. Нет ничего необычного в том, что сохранился такой
сравнительно прочный скелет, но на куске породы видны также
следы животного — последние его следы! Рабочие карьеров в
районе Зольнхофена давно уже знают, что лучший способ най-
ти ископаемое членистоногое — это идти по его следам по по-
верхности напластования известняков.
Наконец, на фото I показаны остатки небольшой птицы
Archaeopteryx. Сохранились отпечатки тонких перьев. Говорят,
первые рисунки ископаемых головоногих моллюсков из Зольн-
I. Сохранность остатков и палеонтологическая летопись
15
хофена были сделаны чернилами из сохранившихся чернильных
мешков моллюсков.
Такая поразительно хорошая сохранность — не исключение,
а скорее правило для Зольнхофена. Однако для палеонтологи-
ческой летописи в целом она не типична. По степени сохран-
ности окаменелостей в один ряд с зольнхофенскими известняка-
ми можно поставить хунсрюкские сланцы (германский девон),
сланцы Бергесс (кембрий Британской Колумбии), асфальтовые
отложения Ла-Бреа (плейстоцен южной Калифорнии), сланцы
Мэзон-Крик (карбон Иллинойса), балтийский янтарь (олиго-
цен) и многие другие породы различных стратиграфических ин-
тервалов. Каждый из перечисленных примеров отличается от
других многими биологическими и геологическими особенно-
стями.
Случаи плохой сохранности в определенном смысле сложнее.
Если в породе совершенно нет окаменелостей, то это может
означать, либо что в этом месте и вблизи него растений и жи-
вотных не было, либо что они были, но не сохранились, либо,
наконец, что они подверглись захоронению, но потом их ос-
татки были разрушены. Следы пермского пресмыкающегося, по-
казанные на фиг. 1,— прекрасный пример хорошей сохранно-
сти, однако остатки самого животного не найдены. Очевидно,
здесь преобладали условия, благоприятные для сохранения сле-
дов, но не скелетов.
Далее в этой главе мы рассмотрим условия фоссилизации
более подробно, однако прежде мы ознакомимся с общими осо-
бенностями палеонтологических данных.
Число видов
Сейчас известно около 1,5 млн. современных видов растений
и животных. Эта оценка учитывает только найденные, описан-
ные и классифицированные виды; значит, на самом деле их
гораздо больше. Ежегодно открывают новые виды, и их число
постоянно растет. Скорость, с которой растет число известных
видов, различна для разных групп животных и растений. Так,
за год описывается примерно 10 тысяч новых видов насекомых,
тогда как неизвестных видов птиц осталось, вероятно, не более
сотни. Такой контраст объясняется отчасти тем, что насекомые
отличаются гораздо большим разнообразием (сейчас их извест-
но около 850000, а птиц всего около 8600 видов), а отчасти тем,
что птицы крупнее, более заметны и издавна привлекали вни-
мание любителей и специалистов.
Число известных видов несколько завышено вследствие того,
что некоторые виды оказываются описанными в литературе по
нескольку раз под разными названиями. Но это мало влияет на
16 Часть 1. Описание и классификация ископаемых остатков
общую цифру. По мнению некоторых специалистов, после того
как работа по описанию и классификации ныне живущих видов
будет завершена, число видов растений и животных составит
примерно 4,5 млн. [70].
В то же время ископаемых видов известно всего около
130 тысяч. Это примерно 8,7% известных современных видов
и менее 3% предполагаемого числа современных видов. Такое
соотношение кажется особенно поразительным, если вспомнить,
что палеонтологическая летопись охватывает многие сотни мил-
лионов лет, тогда как современность с ее фауной и флорой —
всего лишь мгновение геологической истории. Даже при сколь-
ко-нибудь сносных шансах на сохранение число ископаемых
видов должно было бы во много раз превышать число видов,
живущих ныне.
Есть несколько возможных объяснений сравнительной мало-
численности ископаемых видов. Возможно, например, что био-
логическое разнообразие со временем возрастает. Тогда ску-
дость палеонтологической летописи просто отражает малую сте-
пень разнообразия живого мира в геологическом прошлом.
А что говорит нам сама палеонтологическая летопись об
изменениях разнообразия на протяжении геологической исто-
рии? По новейшим данным, начало этой летописи восходит к
временам, удаленным от нас не менее чем на 3,2 млрд, лет
[7, 54]. Древнейшие из известных ископаемых остатков — ока-
менелости из серии Онвервахт в Южной Африке — найдены
в породах, возраст которых не ниже этой величины. Однако
возможно, что летопись на самом деле длиннее, и не исключено,
что будут найдены еще более древние остатки.
В горных породах, образовавшихся за эти 3,2 млрд, лет, ис-
копаемые остатки распределены неравномерно. Более 99% най-
денных видов (их общее число составляет 130000) было най-
дено в фанерозойских отложениях (т. е. в породах, имеющих
возраст менее 0,6 млрд. лет). В начале кембрия обилие и разно-
образие ископаемых внезапно возрастает. Это может означать
усиленную эволюционную радиацию, т. е. увеличение разнооб-
разия. Обычно считается, что палеонтологическая летопись сви-
детельствует о постоянном повышении разнообразия, начиная
с кембрия. Верно ли такое представление? Рассмотрим факты.
Мерой разнообразия обычно служит число таксонов (видов,
родов, семейств и т. д.), существовавших в данный промежуток
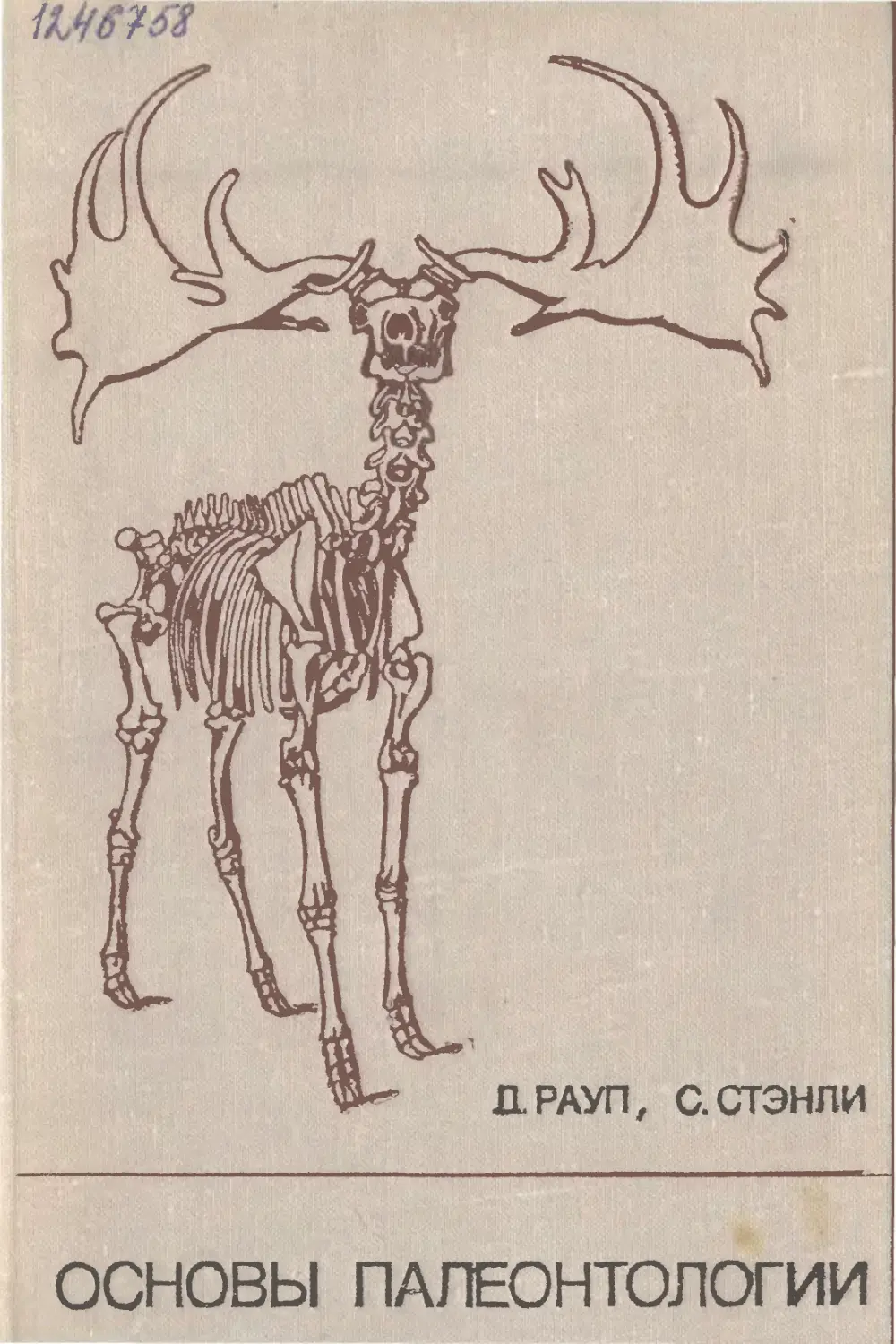
времени. На фиг. 2, А (данные взяты из [73]) показаны изме-
нения наблюдаемого разнообразия от начала кембрия до наших
дней. Ширина заштрихованных участков пропорциональна чис-
лу таксонов (как правило, семейств), известных в разные про-
межутки геологического времени. Как ясно из этой диаграммы
(и подобных диаграмм, построенных другими авторами), в па-
I. Сохранность остатков и палеонтологическая летопись
17
Мел
Юра
Плейстоцен
Плиоцен
Миоцен
Олигоцен
Эоцен
Палеоцен 4
Триас
---------<
Пермь
Карбон
Девон
Силур
Ордовик
Кембрий
А
Фиг. 2. Сравнительное число таксонов ископаемых растений и животных.
А. Известное разнообразие, по литературным данным; выражено шириной заштрихован-
ного прямоугольника [73]. Б. Мощность осадков за 1 млн- лет 1801.
леозое разнообразие неуклонно возрастало, в триасе оно умень-
шилось, а затем в течение юрского и мелового периодов и в кай-
нозое заметно увеличилось. Уменьшение разнообразия в триасе
обычно объясняют вымиранием многих форм. На более подроб-
ных диаграммах видно, что снижение разнообразия началось
еще в конце палеозоя, в середине пермского периода.
Но все обстоит далеко не так просто. На диаграмме, пред-
ставленной на фиг. 2, А, не принят во внимание тот факт, что
некоторые части стратиграфической колонки изучены лучше
других. Чем моложе горные породы, тем полнее они охаракте-
ризованы палеонтологически. Это, в частности, связано с тем,
что молодые породы, лежащие ближе к дневной поверхности,
отложены на более обширных участках по сравнению с древ-
ними. На фиг. 3 сравниваются площади обнаженных пород раз-
ного возраста на территории США. Заметьте, что площадь ме-
ловых обнажений примерно в 4 раза превышает площадь обна-
ОМСЛ .4» т
18
Часть 1. Описание и классификация ископаемых остатков
Фиг. 3. Сравнение площадей обнаженных пород разного возраста в США
[62].
Доля общей
Возраст Площадь, кмг площади
США,%
Челшершичнош период 1091940 14,17
/ Плиоцен 657860 8,54
Миоцен 594660 7.72
_ Олигоцен 121730 7,58
— Эоцен 869010 11,31
Мел 1092530 14,21
Юра 256620 3,33
Триас 126392 1,64
Пермь 295260 3,83
Карбон 1023830 13,28
Девон 21360 2,74-
Силур 205682 2j6&
Ордовик 275317 3,57
Кембрий 221470 2,94-
Докембрий 649570 8,43
жений ордовикских пород, хотя продолжительность обоих этих
периодов почти одинакова.
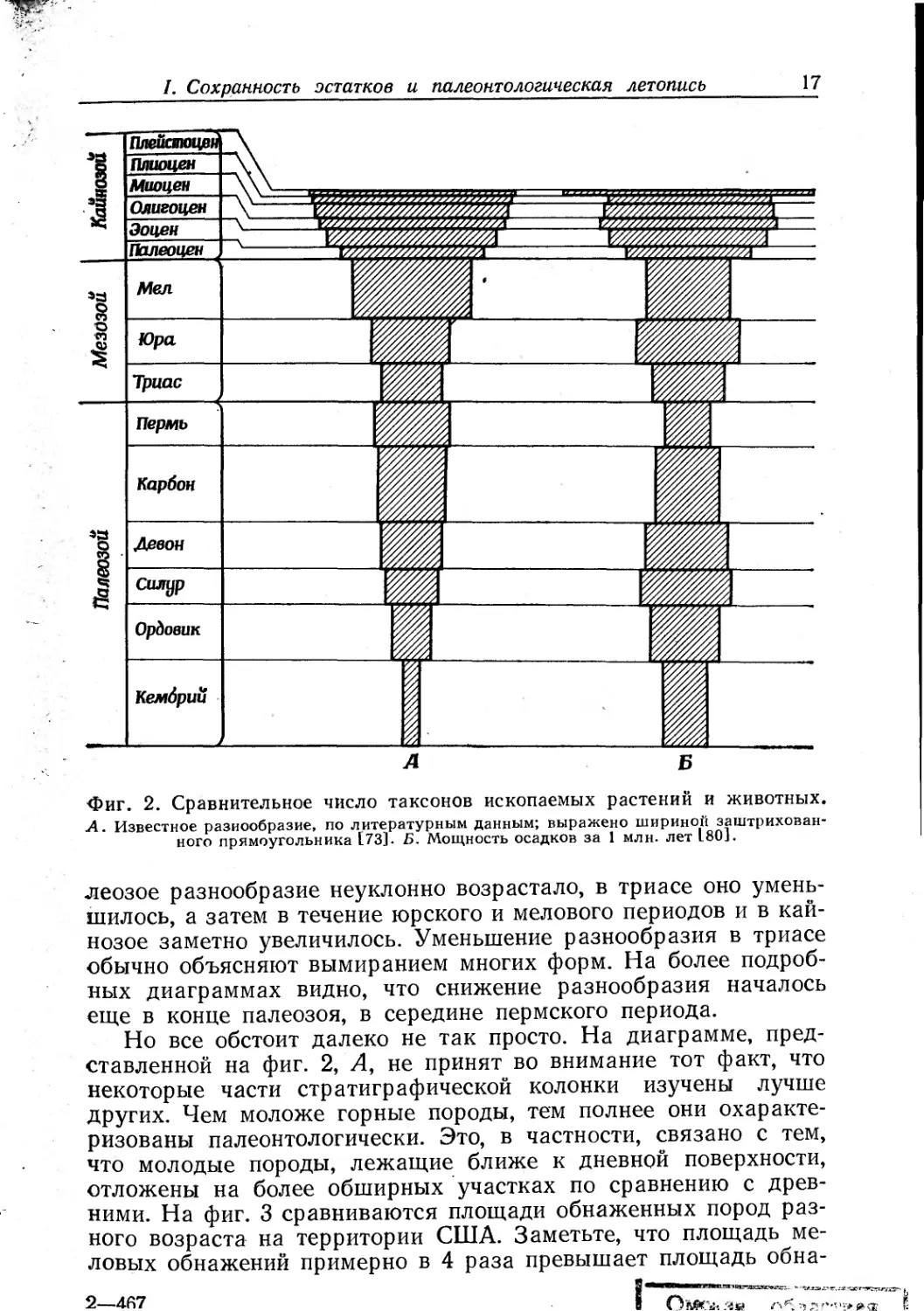
На фиг. 4 показана зависимость мощности пород от их воз-
раста. Сначала может показаться, что скорость осадконакопле-
ния, начиная с кембрия, возросла. Но это не так. Такой ход
графика объясняется тем, что более молодые породы по срав-
нению с древними полнее сохранились, лучше обнажены, и все
это создает впечатление ускорения седиментации [62].
Рассмотренные факторы и привели к тому, что мы гораздо
лучше знаем органический мир поздних периодов геологической
истории, поэтому из молодых отложений описано больше видов,,
чем из более древних. На фиг. 2 сопоставлены мощности пород
I. Сохранность остатков и палеонтологическая летопись 19
Фиг. 4. Соотношене между воз-
растом пород и мощностью их
вскрытой толщи J80].
различного возраста с данными о таксономическом разнообра-
зии. Здесь взята именно мощность, а не площадь обнажений,
так как она, очевидно, важнее для оценки относительного раз-
нообразия ископаемых. Для многих периодов существует поло-
жительная корреляция между мощностью и степенью разнооб-
разия. Поэтому можно думать, что современное разнообразие
(предполагаемые 4,5 млн. видов) не слишком сильно превышает
среднее разнообразие для геологического времени, начиная с
кембрия.
Но вернемся к вопросу о том, почему мы знаем всего 130 000
ископаемых видов. При сопоставлении диаграмм, представлен-
ных на фиг. 2, становится очевидным, что незначительность
этого числа нельзя полностью объяснить предположением о
возрастании разнообразия в процессе эволюции.
В течение геологического времени виды вымирают и сменя-
ются другими. Предприняты попытки оценить среднюю ско-
рость такой смены видов. Например, Тейхерт [213] рассчитал,
что все виды полностью сменяются новыми примерно за 12 млн.
лет. Симпсон [196] выражает скорость смены видов средней
продолжительностью существования отдельных видов. По его
мнению, между возникновением и вымиранием вида проходит
в среднем 2,75 млн. лет. Правда, он тут же замечает, что на
самом деле время существования вида может составлять от
0,5 млн. до 5,0 млн. лет. Оценки такого рода, как мы видим,
неточны.
Если принять, что сейчас на Земле имеется 4,5 млн. видов
растений и животных, что это число на протяжении фанерозоя
существенно не изменялось и что оценка Симпсона (2,75 млн.
«о*
20 Часть 1. Описание и классификация ископаемых остатков
лет) верна, то можно рассчитать, сколько видов жило на Земле
за 600 млн. лет, прошедших с начала кембрия:
(4,5-10») =982-10».
Сравнивая эту оценку с числом известных ископаемых ви-
дов (130000), мы видим, что в палеонтологической летописи
сохранились следы всего 0,013% всех видов, живших в фане-
розое.
Делались и другие оценки числа видов. Симпсон попытался
рассчитать, сколько видов обитало на Земле за всю историю
жизни, и получил цифры от 50 млн. до 4 млрд. Грант [70] по-
лагает, что с начала кембрия на Земле существовало не менее
1,6 млрд, видов. По более новой оценке Дарема [49], только
имевших шансы сохраниться морских организмов было за это
время не менее 4, а может быть и 10 млн.
Какую бы оценку мы ни сочли верной, ясно, что палеонто-
логам удалось открыть лишь крайне малую часть древнего ор-
ганического мира.
Вымершие и рецентные виды
В некоторых руководствах можно найти таблицу распреде-
ления видов по таксонам. Среди новых книг такие данные мож-
но найти у Истона [51], который перечисляет ископаемых и
рецентных животных, и у Гранта [70], рассматривающего все
известные рецентные организмы. Число видов в разных систе-
матических группах оценивают по литературным источникам.
Эти оценки часто крайне неточны. Во всяком случае ясно, что
между разными типами и классами виды распределены нерав-
номерно. Кроме того, как правило, известно гораздо больше
ныне живущих представителей какого-то типа или класса, чем
вымерших. Однако некоторые группы в прошлом имели больше
видов, чем сейчас. Так, известно около 10 000 ископаемых и
всего около 400 рецентных видов головоногих моллюсков.
При составлении таблиц числа видов в первую очередь важ-
но знать следующее: в какой мере окаменелости могут дать
представление об истинной картине органического мира далеко-
го прошлого? Как изменился состав органического мира? На-
сколько избирательность сохранения ископаемых остатков ис-
казила палеонтологическую летопись?
Около трех четвертей известных современных видов — жи-
вотные, и около трех четвертей из них (850 000) — насекомые.
Но ископаемых видов насекомых известно всего 12000 (самые
древние относятся к девону), и не потому, что раньше видов
J. Сохранность остатков и палеонтологическая летопись
21
насекомых было меньше, чем теперь, а потому, что обычные
местообитания насекомых не благоприятствуют захоронению их
остатков. В зольнхофенских известняках встречается около>
180 видов насекомых (это, кстати говоря, примерно одна чет-
верть всех найденных в Зольнхофене ископаемых видов). Мно-
го других видов ископаемых насекомых найдено в нескольких
месторождениях янтаря, который служит прекрасной средой
для захоронения насекомых. Но янтарь редок.
Важно подчеркнуть следующее. В тех случаях, когда мы
находим хорошо сохранившиеся остатки фауны насекомых, мы
видим, что они разнообразны, что фауну составляют насекомые
с широким спектром морфологических адаптаций к самым раз-
ным экологическим условиям. В своем фундаментальном обзоре
ископаемых насекомых Карпентер [24] пишет:
«Изучение фауны каменноугольного периода показывает,,
что разнообразие насекомых поразительно велико... Я уверен,,
что мы еще только начинаем знакомиться с фауной насекомых
позднего карбона... Если бы такое же число рецентных видов
было собрано из нескольких изолированных местностей, то, ко-
нечно, мы не смогли бы по этой коллекции составить верное
представление обо всем разнообразии современной фауны насе-
комых. Поэтому не исключено, что вымершие отряды насекомых
каменноугольного периода не уступали современным крупным
отрядам по разнообразию и распространенности».
Итак, можно сделать следующий вывод: скудость палеонто-
логической летописи насекомых связана в первую очередь с
тем, что они плохо сохраняются. Например, почти полное от-
сутствие остатков насекомых в пластах меловой системы скорее
всего объясняется тем, что в это время насекомые обитали в.
условиях, не благоприятствовавших сохранению их остатков.
Как уже отмечено, лишь для немногих таксономических
групп число известных ископаемых видов превышает число
рецентных. Особенно много ископаемых видов среди головоно-
гих и морских лилий. Заметен перевес вымерших форм также
и среди плеченогих, двустворчатых моллюсков и морских ежей..
Совершенно очевидно, что с течением времени в ходе эволюции,
разнообразие головоногих уменьшилось. Раковины головоно-
гих— самые обычные ископаемые во многих подразделениях
фанерозоя (особенно они многочисленны в мезозое Западной
Европы). На протяжении почти всего мезозоя головоногие про-
цветали, выделяясь как числом видов и морфологических типов,,
так и числом экземпляров. В современных морях раковинные
головоногие представлены немногими формами. Конечно, ме-
зозой можно с полным основанием назвать «эрой головоногих»..
В морях палеозойской эры процветали тысячи видов мор-
ских лилий и плеченогих. Современная бедность этих групп:
1
22 Часть 1. Описание и классификация ископаемых остатков
Фиг. 5. Изменения разнообразия четырех групп
животных со временем [73].
может быть объяснена только оскудением в ходе эволюции
(вымиранием).
История двустворчатых моллюсков и морских ежей выгля-
дит несколько иначе. Мы видим неуклонное возрастание числа
I. Сохранность остатков и палеонтологическая летопись
23
видов с ходом геологического времени. Следовательно, число
ныне живущих видов должно превышать число видов, живших
в любой момент геологического прошлого. Общее число вымер-
ших видов, относящихся к этим группам, превышает число
ныне живущих видов, но это объясняется просто тем, что остат-
ки двустворок и морских ежей накапливались в течение очень
долгого времени.
Важно отметить, что представители групп, в которых иско-
паемых видов больше, чем современных, имеют твердый скелет,
увеличивающий шансы на сохранение.
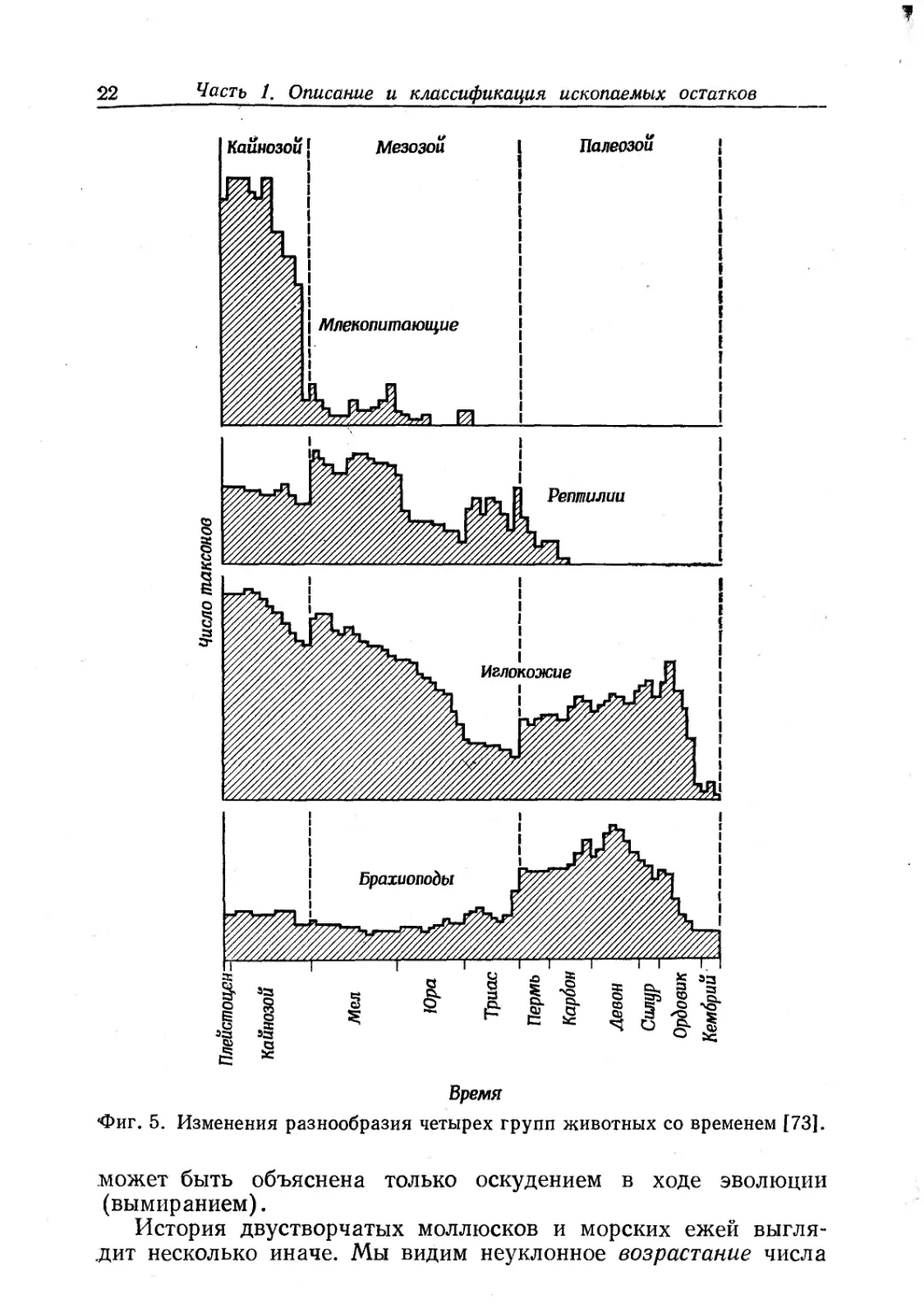
На фиг. 5 показано наблюдаемое разнообразие некоторых
групп животных в течение геологической истории. Мы уже го-
ворили об уменьшении разнообразия брахиопод, начавшемся
с середины палеозоя. Разнообразие иглокожих возросло в па-
леозое, явно уменьшилось в триасе и затем неуклонно возраста-
ло с юрского периода до наших дней. Бимодальное распреде-
ление отражает сложность рассматриваемой группы: в палеозое
дали пик стебельчатые иглокожие (особенно морские лилии),
а второй подъем, начавшийся в юрском периоде, связан с эво-
люционной радиацией неприкрепленных иглокожих, например
морских ежей и морских звезд.
Кривая разнообразия пресмыкающихся (фиг. 5) имеет два
пика — один в позднем палеозое и второй в меловом периоде.
Резкий спад в конце мелового периода соответствует вымиранию
динозавров и сопровождается впечатляющим ростом разнообра-
зия млекопитающих.
Бедность палеонтологической летописи какой-либо группы
организмов может свидетельствовать или о том, что остатки
организмов плохо сохранялись, или о том, что данная группа
отличалась небольшим разнообразием. Богатство палеонтологи-
ческой летописи, может быть обусловлено повышенной способ-
ностью данной группы сохраняться в ископаемом состоянии,
уменьшением разнообразия группы с течением времени или,
наконец, просто тем, что остатки накапливаются в течение дли-
тельного периода эволюции. Выяснение соотношения между
имеющимися палеонтологическими данными и реальной карти-
ной древней жизни — одна из важнейших задач палеонтологии.
Число особей
Наше понимание палеонтологической летописи можно допол-
нить, рассматривая современных животных и растения как по-
тенциальные ископаемые. При этом следует принимать во вни-
мание численность особей (обилие) и число видов (разнообра-
зие). Число особей одного вида может составлять от нескольких
24 Часть 1, Описание и классификация ископаемых остатков______________
Фиг. 6. Животные, собранные при драгировании с 0,25 м2 дна сублиторали
(полуостров Кии, Япония) [271].
Это двустворчатые моллюски М acoma incongrua (крупные) и Cardium hungerfordi (мелкие),
а также лопатоногий моллюск Penta Пит octangulafum^.
«сот до многих миллионов и даже миллиардов. И каждая особь—
потенциальная окаменелость.
Биологи могут помочь нам оценить порядок этих величин.
На фиг. 6 показаны макроскопические беспозвоночные орга-
низмы, которых можно найти на 0,25 м2 морского дна у полу-
острова Кии (Япония) на глубинах от 20 до 100 м. На рисун-
ке— «урожай на корню» особей трех видов: двух двустворчатых
моллюсков и одного лопатоногого моллюска. Здесь около 25 бо-
лее крупных двустворок (Масота incongrua)-, около 160 мелких
.двустворок (Cardium hungerfordi) и 12 экземпляров лолатоно-
гого моллюска (Dentalium octangulatum). И все это на одной
четверти квадратного метра! Средний возраст особей — около
двух лет. При такой продуктивности за 10 лет образуется 1000,
а за миллион лет—100 млн. потенциальных окаменелостей. Если
говорить о более обширных площадях и более длительных пе-
риодах времени, то мы получим просто ошеломляющие цифры
количества особей — тонны и тонны потенциальных ископаемых.
'Оказывается, на таком клочке морского дна за миллион лет
могло обитать больше особей моллюсков, чем было найдено
ископаемых всех видов за всю историю палеонтологии.
Конечно, приведенный здесь простой расчет страдает мно-
гими явными и неявными неточностями. Плотность жизни силь-
но варьирует в пространстве и во времени; в одних местах эта
I. Сохранность остатков и палеонтологическая летопись 25
плотность выше, чем на рассмотренном участке, а в других —
ниже. Однако этот расчет помогает нам понять, что изучаемые
нами окаменелости практически составляют бесконечно малую
долю всех существ, живших в геологическом прошлом. Если
бы палеонтологическая летопись была представительной выбор-
кой обитавших некогда на Земле растений и животных, то все
было бы значительно проще. Но представительной ее назвать
никак нельзя. Одно из осложняющих обстоятельств состоит
в том, что при прочих равных условиях богатые особями виды
имеют больше шансов сохраниться и быть обнаруженными, чем
виды малочисленные.
Условия сохранности органических
остатков
Чтобы организм попал в палеонтологическую летопись, в
отложениях должен сохраниться либо весь он целиком, либо
его части, либо какие-то следы его жизнедеятельности. Так,
следы пресмыкающегося, показанные на фиг. 1, представляют
собой часть палеонтологической летописи и могут дать нам не-
которые сведения об организме, хотя остатки мягких тканей или
скелета не сохранились. Если организм или следы его жизне-
деятельности сохранились, значит, разнообразные разрушитель-
ные процессы действовали недостаточно активно.
БИОЛОГИЧЕСКОЕ РАЗРУШЕНИЕ
Биологическое разрушение имеет место почти в любой сре-
де. Хищники и мусорщики широко распространены в мире жи-
вых существ. Одни из них крупнее тех организмов, которыми
питаются, другие — во много раз мельче своих жертв. Вряд ли
есть биологические образования, совершенно не привлекающие
мусорщиков или других разрушителей. Например, мы обычно
считаем раковину устрицы «каменной» уже при жизни моллюс-
ка. Раковина действительно очень прочна и состоит главным
образом из карбоната кальция. Но строение раковины не одно-
родно: она состоит из тонких иголочек или пластинок кальцита^
связанных органическим веществом, обычно называемым орга-
нической матрицей. Следовательно, прочность раковины частич-
но зависит от свойств этой матрицы.
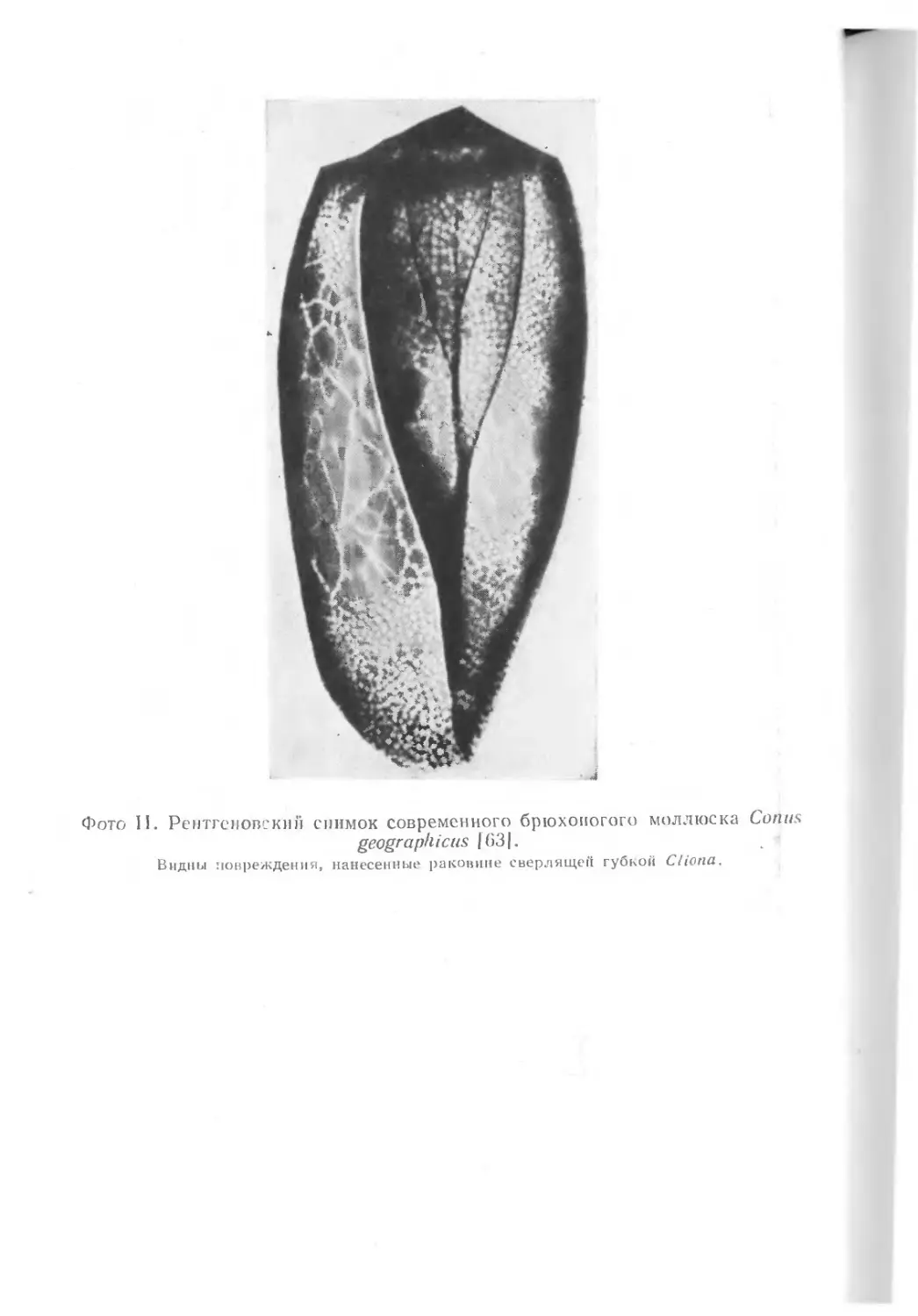
После гибели моллюска на его раковину набрасываются са-
мые разные сверлильщики — черви, губки, другие моллюски,
водоросли (фото II). Обычно там, где обитает много организ-
мов, обладающих раковинами, пустые раковины встречаются
поразительно редко, так как они уничтожаются сверлильщи-
ками.
26 Часть 1. Описание и классификация ископаемых остатков
После смерти организма его скелет подвергается захороне-
нию. Это частично предохраняет скелет от биологического раз-
рушения. Но степень такой защищенности часто преувеличива-
ют. Самые верхние слои неуплотнившегося еще осадка в обыч-
ных условиях дна никак нельзя назвать биологически инертны-
ми. В самом деле, в нескольких верхних сантиметрах осадка
интенсивно протекают процессы распада биологических тканей
под влиянием жизнедеятельности бактерий. Раковина может
сохраняться достаточно долго, чтобы осадок успел покрыть ее,
но и после этого, уже оказавшись под поверхностью раздела
осадок — вода, она может быть разрушена.
Все признают, что биологическое разрушение потенциальных
ископаемых — важный фактор, обусловливающий неполноту па-
леонтологической летописи. Но мы еще очень мало знаем об
этом процессе. Он изучался весьма слабо; особенно плохо изу-
чены деятельность бактерий и их влияние на химизм водных
растворов в толще осадков. Между тем понимание процессов
биологического разрушения существенно важно для нас, если
мы хотим глубоко понять условия захоронения и сохранения
в ископаемом состоянии остатков организмов.
МЕХАНИЧЕСКОЕ РАЗРУШЕНИЕ
Давно известно, что остатки организмов, подвергающиеся
вскоре после их смерти интенсивному воздействию ветра, волн
или потоков воды, могут быть полностью разрушены. Известно
также, что различные типы скелетов в разной степени подвер-
жены механическому разрушению, и это обстоятельство в свою
очередь искажает истинную картину органического мира прош-
лого в палеонтологической летописи.
Были проведены некоторые важные исследования процессов
механического разрушения вещества скелетов потенциальных
окаменелостей. Одно из самых важных и в то же время мето-
дически простых исследований механического разрушения было
проведено Чейвом [25]. Раковины моллюсков и скелетные об-
разования других морских беспозвоночных вместе с кремневой
галькой помещали во вращающиеся барабаны. Точно отмеча-
лось время, требовавшееся для того, чтобы разрушить раковины
до определенной степени. Некоторые результаты показаны на
фиг. 7. По оси абсцисс отложено время, по оси ординат — про-
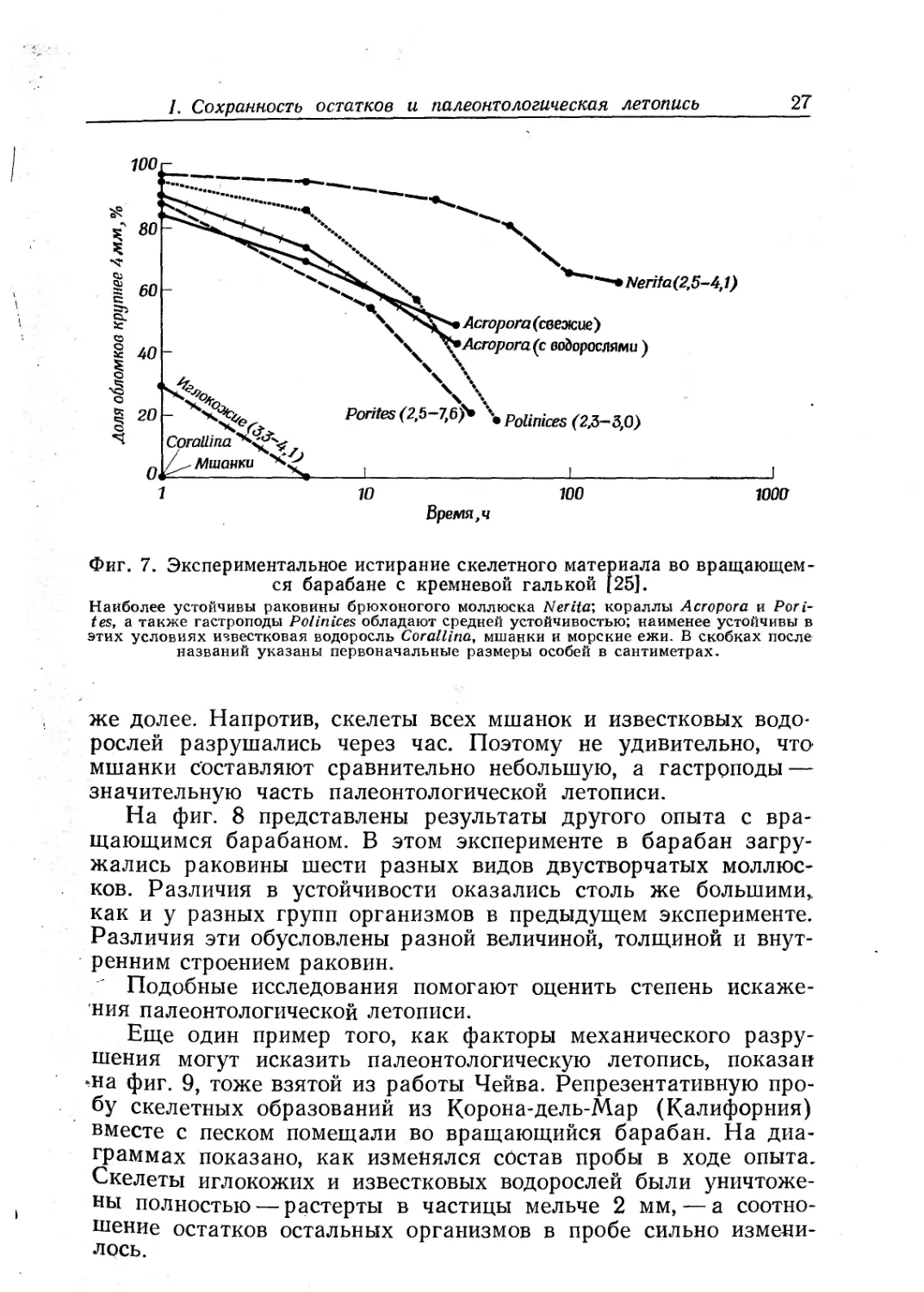
цент обломков с поперечником более 4 мм. Этот эксперимент
лроводился на скелетах гастропод, кораллов, морских ежей,
мшанок и известковых водорослей. Различия в прочности ске-
летных образований этих организмов оказались очень больши- »
ми. Более 60% скелетного вещества одного вида гастропод
остается в форме частиц крупнее 4 мм после 100 ч опыта и да-
I. Сохранность остатков и палеонтологическая летопись
27
Наиболее устойчивы раковины брюхоногого моллюска Nerita\ кораллы Асгорога и РоН-
tes, а также гастроподы Polinices обладают средней устойчивостью; наименее устойчивы в
этих условиях известковая водоросль СогаШпа, мшанки и морские ежи. В скобках после
названий указаны первоначальные размеры особей в сантиметрах.
же долее. Напротив, скелеты всех мшанок и известковых водо-
рослей разрушались через час. Поэтому не удивительно, что-
мшанки составляют сравнительно небольшую, а гастроподы —
значительную часть палеонтологической летописи.
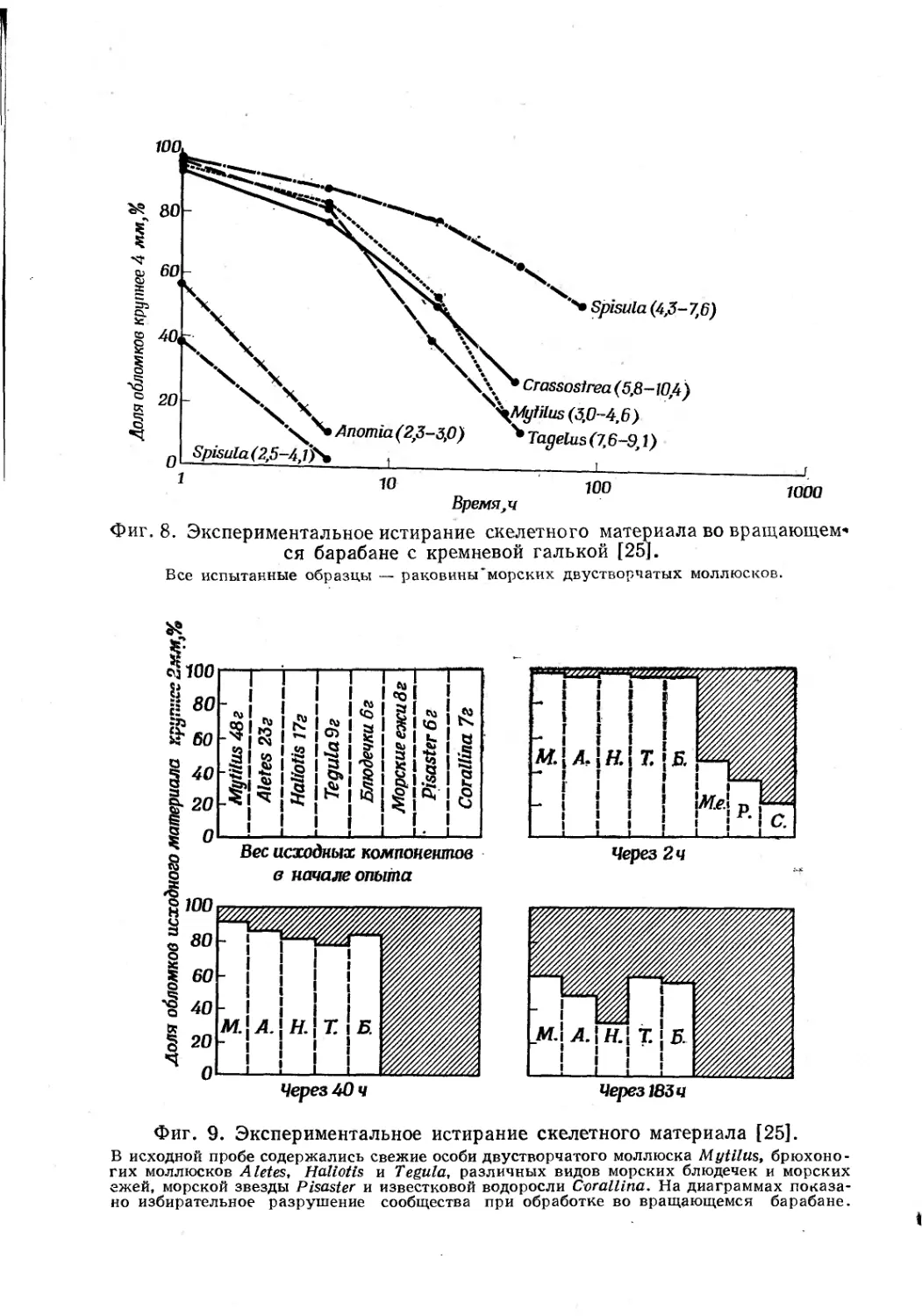
На фиг. 8 представлены результаты другого опыта с вра-
щающимся барабаном. В этом эксперименте в барабан загру-
жались раковины шести разных видов двустворчатых моллюс-
ков. Различия в устойчивости оказались столь же большими,,
как и у разных групп организмов в предыдущем эксперименте.
Различия эти обусловлены разной величиной, толщиной и внут-
ренним строением раковин.
Подобные исследования помогают оценить степень искаже-
ния палеонтологической летописи.
Еще один пример того, как факторы механического разру-
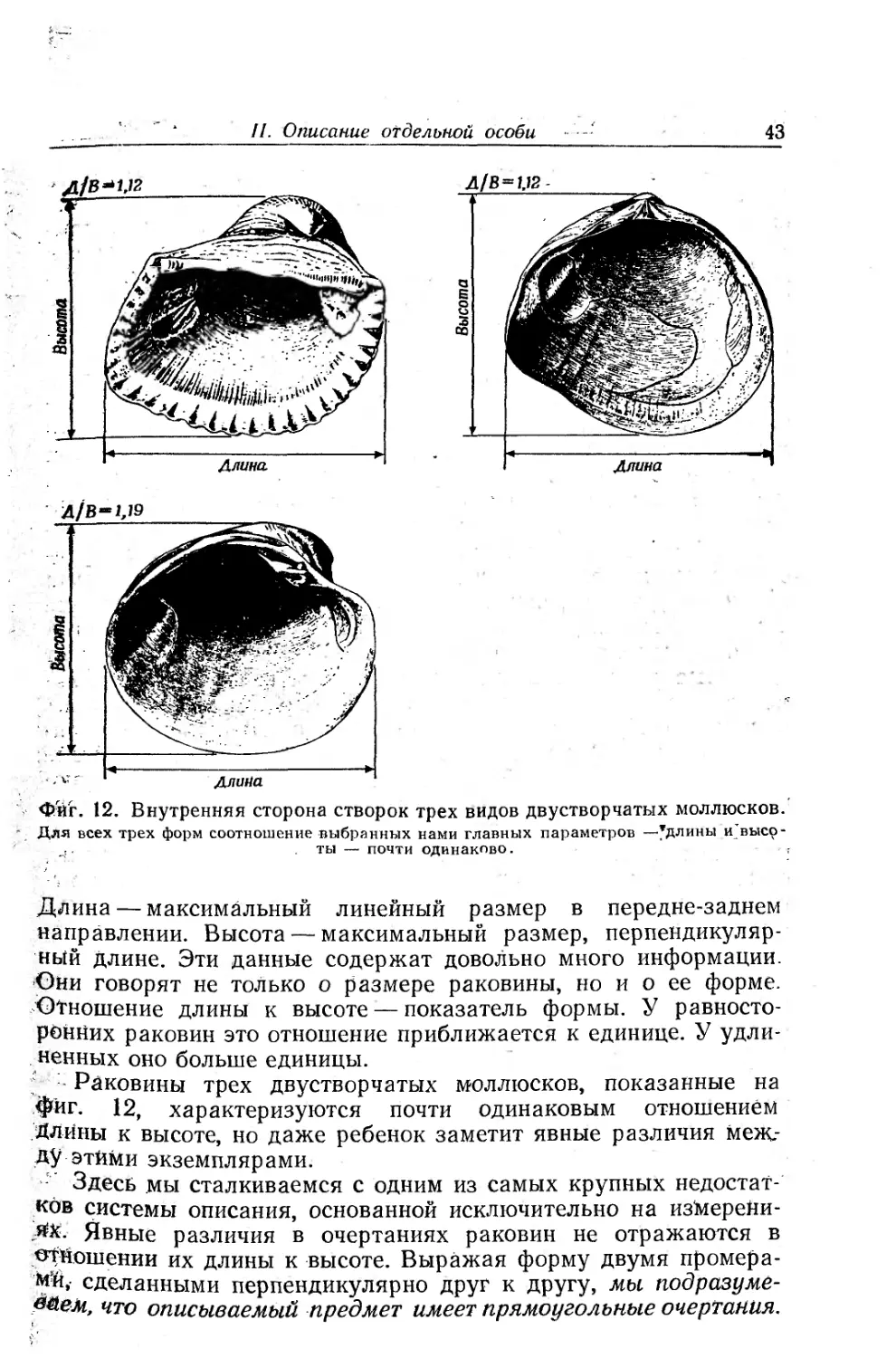
шения могут исказить палеонтологическую летопись, показан
*на фиг. 9, тоже взятой из работы Чейва. Репрезентативную про-
бу скелетных образований из Корон а-дель-Мар (Калифорния)
вместе с песком помещали во вращающийся барабан. На диа-
граммах показано, как изменялся состав пробы в ходе опыта.
Скелеты иглокожих и известковых водорослей были уничтоже-
ны полностью — растерты в частицы мельче 2 мм, — а соотно-
шение остатков остальных организмов в пробе сильно измени-
лось.
10 100
Время,ч
Фиг. 8. Экспериментальное истирание скелетного материала во вращающем*
ся барабане с кремневой галькой [25].
Все испытанные образцы — раковины*морских двустворчатых моллюсков.
___г
7000
80
60
I
1
1
и
S
I
I см
3 Й
•S1 -SJ
>1-5
20Ь^П
40
100
I .Ц
I О
• i
is
°’ I
51
I ” и tj
си I > । g
3 ч
I §,!.?!§
Вес исходных компонентов
в начале опыта
80
60
40
20
О
М.
Через 183 ч
Й100
i
о
О
I
• ।
I ।
!
I
I
Q
i
ч
Фиг. 9. Экспериментальное истирание скелетного материала [25].
В исходной пробе содержались свежие особи двустворчатого моллюска Mytilus, брюхоно-
гих моллюсков Aletes, Haliotis и Tegula, различных видов морских блюдечек и морских
ежей, морской звезды Pisaster и известковой водоросли Corallina. На диаграммах показа-
но избирательное разрушение сообщества при обработке во вращающемся барабане.
I. Сохранность остатков и палеонтологическая летопись 29
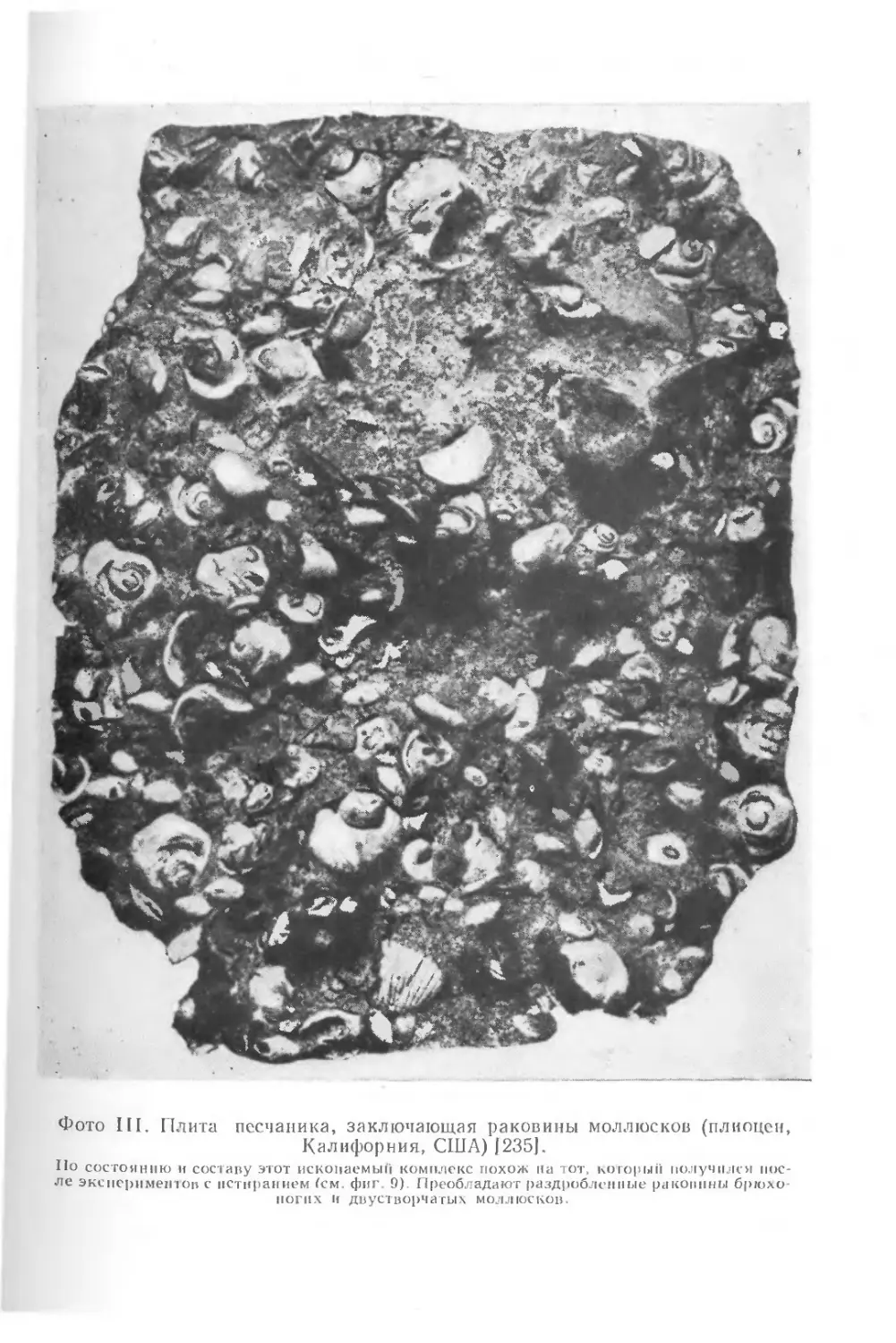
На фото III видна плита грубозернистого песчаника, заклю-
чающая раковины моллюсков. По мнению Чейва, эти раковины
подвергались примерно такому же механическому воздействию,
как и раковины в его опытах. Итак, изучая тот или иной ком-
плекс ископаемых остатков, даже, казалось бы, хорошо сохра-
нившийся, надо всегда пытаться представить себе, какие еще
организмы, впоследствии разрушенные, могли обитать в этом
месте.
ХИМИЧЕСКОЕ РАЗРУШЕНИЕ
Даже если скелет животного не подвергается биологическому
и механическому разрушению, он может не стать долговечной
частью палеонтологической летописи. Химическое раство-
рение— один из важнейших факторов, существенно ограни-
чивающих количество распознаваемых ископаемых. Химическое
растворение может происходить в любое время после смерти
животного — даже после того, как скелет давно уже фоссилизи-
ровался. Растворение может идти не только на морском дне,
но и в толще рыхлого осадка, а также уже в породе спустя
миллионы лет после гибели организма.
Способность окаменелости противостоять растворению за-
висит от ее химического состава, а также от состава и физи-
ческих свойств вод, с которыми она соприкасается. Химизм вод
в свою очередь сильно зависит от биологических факторов,
особенно от активности бактерий.
ч Если раковина была уничтожена в результате химического
растворения после ее включения в породу, то возникшая в по-
роде полость может превратиться в ядро. Подобная форма ис-
копаемых может дать некоторое представление о морфологии
организма и, следовательно, часто пригодна для палеонтологи-
ческих исследований.
; Биологические структуры,
имеющие наибольшие шансы на сохранение
Лучше всего сохраняются скелеты с большим содержанием
минеральных веществ; хуже всего сохраняются мягкие ткани,
не связанные непосредственно со скелетными образованиями.
‘Следовательно, в процессе фоссилизации происходит не только
искажающий первоначальную картину неравномерный отбор
организмов, но и отбор частей этих организмов. Так, наши
знания о древних млекопитающих часто основаны только на
результатах изучения их зубов, поскольку зубы сохраняются го-
раздо лучше других частей скелета.
Из-за разной сохраняемости так называемых твердых и мяг-
ких частей мы крайне редко можем получить сведения об ок-
30 Часть 1. Описание и классификация ископаемых остатков
раске вымерших организмов. Пигменты, за редким исключени-
ем, не сохраняются.
Эволюционные изменения могут существенно влиять на па-
леонтологическую летопись группы. Предположим, у какой-то
группы животных в ходе эволюции развились прочные скелет-
ные образования. Появление скелета резко увеличит возмож-
ность сохранения остатков этих животных в ископаемом состоя-
нии, и они займут больше места в палеонтологической летописи.
Видимо, такой эволюционный шаг сделали членистоногие. Поч-
ти у всех членистоногих есть скелет. Состав скелетов членисто-
ногих, а также их прочность, устойчивость к разложению, рас-
творению и механическому истиранию сильно варьируют. На-
пример, скелет краба /обызвествлен гораздо слабее, чем скелет
трилобита. Скелет живого трилобита был более плотным и
прочным, содержал больше чистого карбоната кальция и мень-
ше органических веществ, чем скелет краба. Поэтому скелеты
трилобитов имели больше шансов на фоссилизацию, и палеон-
тологическая летопись трилобитов гораздо более полна, чем
летопись крабов. Многие палеонтологи полагают, что относи-
тельно быстрое появление трилобитов и других ископаемых
в кембрии объясняется не внезапным возникновением многих
форм в процессе эволюции, а быстрым развитием обызвест-
вления.
Условия среды и сохранность
органических остатков
Пример прекрасной сохранности ископаемых в зольнхофен-
ских известняках подчеркивает важность физических факторов
среды в определении шансов организма на сохранение в пале-
онтологической летописи. Биологически идентичные организмы
могут превосходно сохраниться в одной среде и полностью раз-
рушиться в другой. Очевидно, для фоссилизации более благо-
приятна такая среда, в которой захоронение происходит быстро;
но имеют значение и другие факторы. Так, зольнхофенские из-
вестняки— весьма тонкозернистые, хорошо слоистые, чистые —
совершенно лишены тех признаков, которые геолог обычно
связывает с быстрой седиментацией. Происхождение этих из-
вестняков не совсем ясно, но вероятнее всего, они образовались
в результате очень медленного накопления карбоната, осаждав-
шегося без участия организмов.
Мы сможем лучше понять влияние условий среды на сохра-
нение ископаемых остатков, если будем различать области,,
в которых идет осадконакопление, и области, подвергающиеся
эрозии. Обобщая, можно сказать, что в областях, лежащих ни-
же уровня моря, осадконакопление осуществляется почти по-
I. Сохранность остатков и палеонтологическая летопись
31
всеместно, а в областях, расположенных выше уровня моря,
осадки накапливаются лишь в понижениях рельефа. Поэтому
ниже уровня моря создаются лучшие условия для сохранения
ископаемых остатков, и морская палеонтологическая летопись
несравненно полнее наземной.
В районах с постоянным интенсивным поступлением осадка
(например, в дельтах крупных рек) седиментация, конечно,
способствует сохранению окаменелостей. Однако в других об-
ластях нормальный темп седиментации часто недостаточно вы-
сок для того, чтобы обеспечить быстрое захоронение потенци-
альных ископаемых, которое предохранило бы их от действия
различных факторов разрушения.
Изучение некоторых участков земной коры, весьма необыч-
ных по своему геологическому строению, позволяет более глу-
боко познать процессы, приводящие к созданию палеонтологи-
ческой летописи. Выдающийся пример — великолепная сохран-
ность остатков млекопитающих в асфальтовых ямах Южной
Калифорнии. В данном случае интересно не только то, что ас-
фальтовые ямы — сравнительно редкое и необычное геологиче-
ское явление, но и то, что захоронение в них происходит в ре-
зультате катастроф. Кстати, роль катастроф в сохранении ока-
менелостей может быть больше, чем обычно считают. Асфаль-
товую яму никак нельзя назвать нормальным местообитанием
сохранившихся здесь организмов. Сохранение насекомых в ян-
таре во многом сходно с сохранением позвоночных в асфаль-
товых ямах.
Для всех приведенных примеров превосходной сохранности
ископаемых характерна общая черта — наличие в прошлом
биологически инертной среды, вроде асфальта (Калифорния)
или янтаря (Прибалтика). В такой среде остатки растений и
животных защищены от действия как обычных биологических
разрушающих агентов, так и физических и химических агентов.
Мы не знаем, в какой среде сформировались зольнхофенские
известняки, но, судя по степени сохранности ископаемых, среда
эта тоже скорее всего была биологически инертной. Факт сохра-
нения мягких тканей там, где седиментация была, по-видимому,
не слишком быстрой, например в среднекембрийских сланцах
Бергесс (фото IV), говорит о почти полном отсутствии бактерий
и организмов-мусорщиков. Видимо, особи некоторых видов жи-
вотных и растений сохраняются только в том случае, если их
•остатки вынесены из нормального местообитания. Пока мы не
можем сказать, насколько обычен такой перенос к месту захоро-
нения и насколько важную роль он играет в образовании пале-
онтологической летописи в целом.
Местообитание существенно влияет на сохранение остатков
•организмов. Вероятность сохранения остатков бегемота больше,
32
Часть 1. Описание и классификация ископаемых остатков
чем костей снежного барана, так как первый обитает на низ-
менных участках суши, тогда как второй обычно живет в горах,
где интенсивно действуют факторы эрозии. В самой морской
среде также существуют большие различия. Одно из самых
важных с палеонтологической точки зрения — различие между
животными, обитающими в толще осадков морского дна, и те-
ми, которые живут на его поверхности или плавают в придон-
ном слое воды. Для организмов, обитающих в толще осадка,
скорость седиментации не так уж важна, поскольку они уже
при жизни «похоронены» в толще отложений и этим до некото-
рой степени защищены от мусорщиков и почти полностью — от
механического дробления и истирания.
Посмертный перенос
Остатки разных организмов переносятся различными спо-
собами, да и остатки одинаковых организмов в разных средах
также переносятся разными способами. Одна из простейших
форм переноса — опускание остатков свободноплавающего орга-
низма на морское дно. В этом случае остатки попадают в со-
вершенно иную среду, которая может оказаться биологически
инертной. Одно из широко распространенных объяснений пре-
красной сохранности зольнхофенских окаменелостей основано
именно на такой возможности.
Почти все ископаемые, найденные в Зольнхофене, относятся
к пелагическим морским формам или наземным организмам,
остатки которых могли быть снесены в Зольнхофенское море.
Донных организмов в этом море не было — об этом говорит
не только отсутствие их остатков, но и отсутствие каких-либо
следов их жизнедеятельности. Следы волочения и касания, най-
денные в зольнхофенских известняках, — это последние, пред-
смертные следы животных, и почти все (если не все) они остав-
лены пелагическими организмами. Они не могли бы сохранить-
ся в среде с высокой биологической активностью. Это привело
к заключению, что придонные воды Зольнхофенского моря были
неблагоприятны для обитания большинства организмов и что
в палеонтологическую летопись здесь попали лишь те организ-
мы, которые были принесены из пелагиали после смерти или
неосторожно заплыли сюда и погибли от отравления.
Среди современных пелагических организмов многие, напри-
мер большинство ракообразных, держатся в толще воды благо-
даря своей двигательной активности; другие удерживаются в
толще или на поверхности воды за счет своей плавучести. Ос-
татки организмов, относящихся к первой группе, после смерти
обычно опускаются на дно. Плавучие трупы организмов второй
группы могут переноситься на большие расстояния течениями
I. Сохранность остатков и палеонтологическая летопись 33
или ветром. Хороший пример такого организма — головоногий
моллюск со спирально свернутой раковиной. При жизни*живот-
ного наполненные газом камеры раковины обеспечивают плаву-
честь. После смерти плавучесть может даже возрасти, так как
мягкое тело животного разлагается и раковина становится лег-
че. Раковина может плавать в поверхностных водах много дней,
недель и даже месяцев, пока не опустится на дно. Видимо, по-
смертный перенос раковин спирально свернутых головоногих и
других подобных животных является правилом.
Посмертный перенос скелетов донных животных обычно осу-
ществляется с помощью течений. В сравнительно неглубоких
бассейнах, где имеются сильные течения, скелет погибшего ор-
ганизма может быть перенесен на значительное расстояние до
места окончательного захоронения. Иногда трупы донных орга-
низмов всплывают, так как при распаде мягких тканей образу-
ется газ, который может задерживаться внутри скелета. К со-
жалению, неизвестно, насколько обычно это явление. Во всяком
случае, рыбы и другие животные с мягким телом при разло-
жении раздуваются и всплывают. Иногда такое явление отме-
чается и у беспозвоночных с довольно тяжелыми панцирями,
но вопрос о том, насколько оно может влиять на сохранение
окаменелостей, еще недостаточно изучен.
Пыльца и споры наземных растений, играющие важную роль
в палеонтологической летописи, могут переноситься ветром на
большие расстояния. Реки и ручьи несут массу самых разных
растительных и животных остатков. Остатки наземных орга-
низмов, особенно растений, сплошь и рядом сносятся в океан
и плывут сотни километров, пока не опустятся на дно. Инте-
ресно, что некоторые весьма глубоководные виды морских ежей,
живущие у берегов Новой Гвинеи, в своем питании полностью
зависят от растительных остатков наземного происхождения,
сносимых в океан реками.
Очень трудно сказать, насколько велика роль посмертного
переноса. О том, что перед фоссилизацией организм подвергся
переносу, могут говорить механические повреждения ископае-
мого или обнаружение его там, где организм наверняка не мог
обитать. Некоторые ископаемые не обнаруживают никаких при-
знаков посмертного переноса.
Роль посмертного переноса как фактора, искажающего па-
леонтологическую летопись, зависит от того, для каких целей
мы используем данные этой летописи. Если нас интересуют
общие тенденции эволюционного развития какой-то группы ор-
ганизмов и мы изучаем сравнительную морфологию и распреде-
ление этих организмов во времени, то перенос на несколько
километров или даже несколько сот километров не имеет зна-
чения. Несколько километров или несколько десятков километ-
3—467
34
Часть 1. Описание и классификация ископаемых остатков
ров не играют роли и при попытках реконструировать по иско-
паемым зональный климат. Но если наша задача — восстано-
вить условия среды в данном местообитании, то даже неболь-
шой перенос может оказаться очень важным.
Выводы
Мы видим, что сохранение остатков организма или следов
его жизнедеятельности — событие редкое. Чем больше мы узна-
ем о процессах фоссилизации, тем сильнее поражает нас суще-
ствование достаточно информативной палеонтологической лето-
писи. Однако растений и животных, потенциально способных
превратиться в окаменелости, так много, что даже столь мало-
вероятное событие, как их сохранение в виде ископаемых остат-
ков, становится довольно обычным. Хотя мы начинаем уже до-
вольно хорошо понимать некоторые основные закономерности
процессов захоронения и сохранения органических остатков,
здесь остались еще целые неисследованные области. Например,
в этой главе говорилось о том, что необычные геологические
явления и даже катастрофы могут способствовать сохранению
ископаемых. Но до какой степени? Имеющиеся данные не по-
зволяют ответить на этот вопрос.
Чтобы преуспеть в исследованиях, никогда не следует пол-
ностью доверять палеонтологической летописи: картина древней
жизни в ней, очевидно, сильно искажена. Следовательно, отсут-
ствие ископаемых в какой-либо толще не обязательно означает,
что в данном месте не было животных или растений.
Хотя палеонтологическая летопись во многих отношениях
ограничена, она все же содержит огромное количество инфор-
мации. Пожалуй, можно выделить две причины, по которым
довольно скудная выборка древних организмов, представленная
в палеонтологической летописи, часто удовлетворяет нуждам
палеонтологического исследования.
1. В обобщающих исследованиях темпа, тенденций и харак-
тера эволюции крупных групп растений и животных, а также
эволюционных отношений внутри этих групп палеонтологи ста-
раются использовать те группы организмов, которые сравни-
тельно хорошо отражены в палеонтологической летописи, т. е.
представлены адекватными статистическими выборками.
2t Интерпретируя ископаемые фауны и флоры отдельных
стратиграфических единиц, палеонтолог обычно выбирает по-
роды, в которых сохранилась большая доля потенциально фос-
силизируемых видов.
Итак, приступая к какому-либо исследованию, палеонтолог
выбирает определенные ископаемые группы или определенные
пласты пород..
ГЛАВА II
Описание отдельной особи
Описание ископаемых — основа почти любого палеонтологи-
ческого исследования. В каждом конкретном случае вопрос
сводится к тому, что описывать и как описывать. Если цель
работы—описать новый вид ископаемых и найти его место
в системе, то описание должно быть как можно более полным
и в то же время должно облегчать сравнение нового ископае-
мого организма с другими. Сравнение основывается обычно на
избранных признаках. Любая окаменелость имеет буквально
тысячи признаков, которые можно описать. Как же выбрать
признаки для описания? А после того как они выбраны, надо
еще разработать целесообразный и разумный способ выражения
отличий нового вида.
Проблемы описания не ограничиваются определением нового
вида и его места в системе. Предположим, палеонтолог
хочет проверить эволюционную гипотезу, согласно которой в
Ханной группе организмов в ходе эволюции происходит увели-
чение размеров особей. Это кажется несложным, но тут же
возникают некоторые вопросы. Как лучше выражать размеры?
Взять ли для этого общий вес, общий объем, максимальную
длину или ширину, площадь поверхности или какую-то комби-
нацию этих признаков? Надо ли рассматривать организм в це-
лом или можно считать, что размер каких-то его частей хорошо
отражает размеры всего организма? Выбор признаков — это
отчасти биологическая, но отчасти чисто палеонтологическая
проблема. Многие признаки, которые измерил бы биолог, пале-
онтологу недоступны — его ограничивает характер ископаемого
Материала. Такие признаки, как, скажем, вес и объем, при
фоссилизации сильно изменяются. Палеонтолог должен выби-
вать признаки, имеющие биологический смысл и в то же время
приемлемые с палеонтологической точки зрения.
' • Единой схемы описания палеонтологического материала, ко-
торая подходила бы во всех случаях и давала бы всегда пре-
восходный результат, не существует. В этой главе мы ознако-
мимся с некоторыми общими проблемами выбора признаков,
Л также метода описания. Предполагается, что у нас уже име;
Тотся собранные, отпрепарированные ’и готовые к описанию
3*
I
36 Часть 1. Описание и классификация ископаемых остатков
окаменелости. Весьма существенные вопросы сбора и препари-
рования окаменелостей здесь не затрагиваются. Эти вопросы
обстоятельно рассмотрены в руководствах, названия которых
указаны в конце книги.
Фотографический метод
В некоторых отношениях хорошая фотография окаменело-
сти— лучшее описание — и объективное, и исчерпывающее. По-
этому фотография стала неотъемлемой частью формального
описания ископаемых видов. Конечно, такие признаки как хи-
мический состав, а также некоторые мельчайшие детали строе-
ния фотографией не регистрируются.
Пример эффективного использования фотографии в палеон-
тологии— стереопары, воспроизведенные на фото V. Предвари-
тельная подготовка образцов, например опыление, и подходящее
освещение помогают выделить структуры, которые иначе оста-
лись бы невидимыми для фотокамеры, как незаметны они
глазу.
Для выявления невидимых глазу деталей можно применять
фотографирование в лучах невидимой части спектра (фото VI).
Рентгенография позволяет выявить внутренние структуры, скры-
тые от исследователя. Стереопара рентгеновских фотографий
брахиоподы (фото VI) позволяет увидеть внутреннее строение
раковины. Спиральный лофофор полностью погружен в плотную
осадочную породу, заполняющую раковину. Чтобы выявить
строение лофофора без помощи рентгенографии, понадобилась
бы кропотливая работа по изготовлению срезов и дальнейшей
реконструкции органа по серии срезов. Отсюда ясно и другое
весьма существенное преимущество фотографических методов
описания: они не требуют разрушения объекта.
Фото VI иллюстрирует также эффективность метода фото-
графии в ультрафиолетовых лучах, основанного на флуорес-
ценции малых количеств органического вещества; при помощи
этого метода выявляются скелетные структуры, невидимые в
обычном свете. Здесь же показан снимок, сделанный на пленке,
чувствительной к инфракрасным лучам. Различные структуры
по-разному поглощают тепло, и пленка улавливает эти разли-
чия, так что удается выявить структуры, неразличимые иными
способами. В последние годы большое значение в исследовании
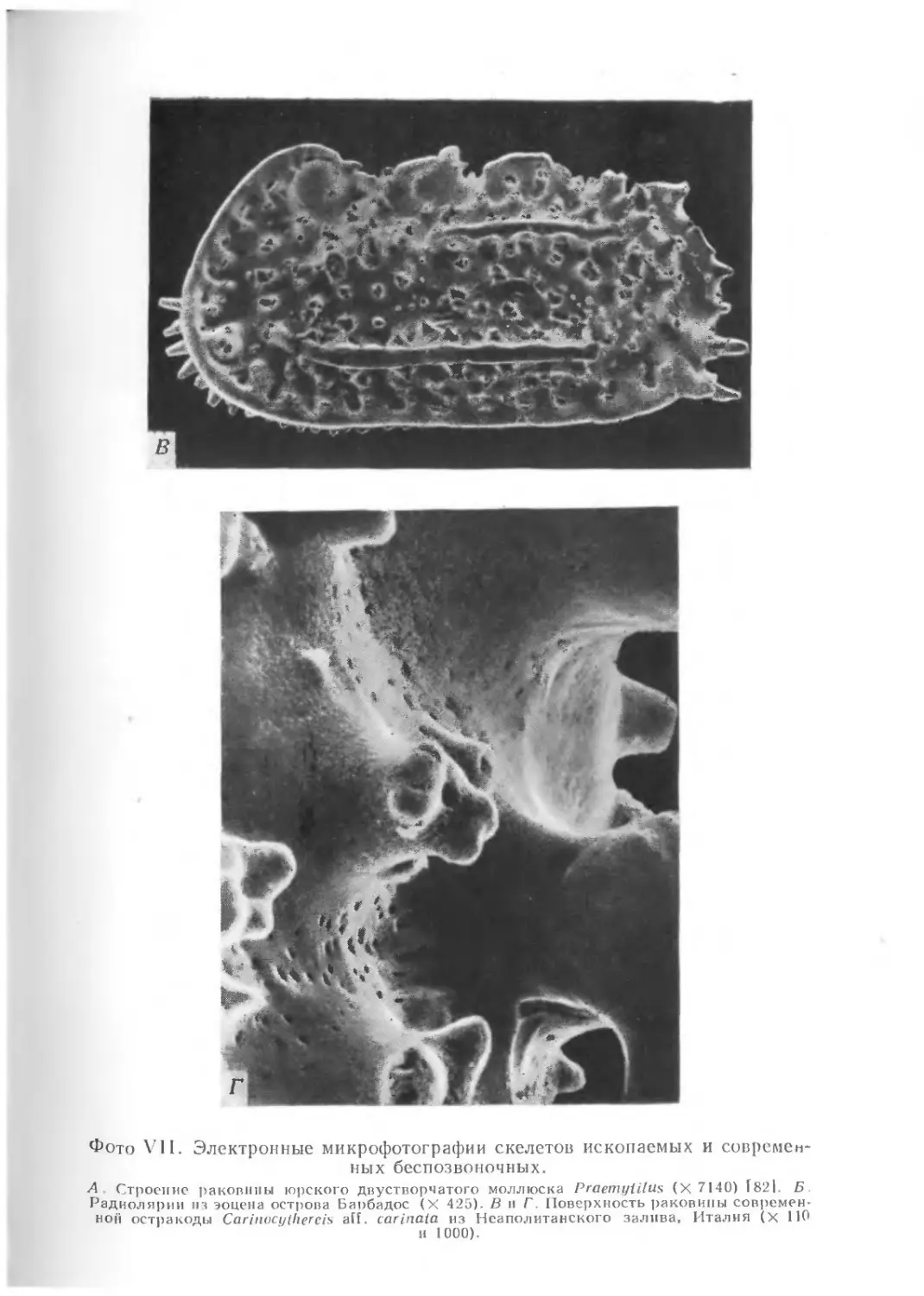
и фотографировании ультрамикроископаемых и тонких деталей
строения обычных ископаемых приобрела электронная микро-
скопия (фото VII).
Сейчас предпринимаются попытки представления фотогра-
фической информации в форме, пригодной для ввода в ЭВМ.
Этот шаг подготовлен развитием высокоскоростной вычисли-
П. Описание отдельной особи
37
тельной техники. Возможность вводить фотографическую ин-
формацию непосредственно в ЭВМ, которая автоматически и
е большой точностью оценивала бы и сравнивала признаки, вы-
глядит заманчиво.
Исследования в этой области только начались, и большей
частью они не имеют отношения к палеонтологии. Однако не-
которые из разработанных методов явно применимы и к иско-
паемым объектам.
Простейший метод перевода фотографической информации
в форму, пригодную для обработки в ЭВМ, — цифровое кодиро-
вание. Обычно это делается путем наложения на фотографию
общепринятой системы прямоугольных координат (X и У). Зна-
чения координат X и У каждой точки изображения измеряются,
регистрируются на перфокартах, бумажной перфоленте или
магнитной ленте и хранятся в памяти машины. Качество за-
писанного изображения зависит только от числа обработанных
точек.
Такой метод обработки данных годится для двумерных
структур или для структур, поддающихся изображению на плос-
кости. Но для изучения с помощью ЭВМ большинства ископае-
мых нужен метод получения цифровой информации о форме
трехмерных структур. Самый прямой способ — обработка сте-
реопар фотографий. Тогда каждую точку ископаемого задают
двумя парами координат X и У, измеренными при наблюде-
нии с двух несколько различающихся позиций; ЭВМ, триго-
, / неметрически обрабатывая эти данные, воссоздает трехмерную
картину.
Фотографии, показывающие только очертания объекта, мож-
но переработать в цифровую информацию вручную, но огром-
ная трудоемкость этой работы перевешивает все преимущества
=' хранения фотографии в форме, доступной для ЭВМ. Разработа-
, ны устройства, автоматизирующие переработку образа в цифро-
вую информацию. Одно из таких устройств показано на фиг. 10.
Фотография укрепляется на плоской поверхности, и оператор
проводит пером по тем участкам, относительно которых нужно
получить цифровую информацию. Когда перо доходит до точки,
Ч- подлежащей обработке, оператор нажимает на кнопку или пе-
даль, и координаты точки автоматически регистрируются. Бо-
лее совершенные автоматические установки производят скани-
рование изображения. Светочувствительный прибор сканирует
фотографию по нескольким разным направлениям. Отмечаю-
щиеся изменения яркости регистрируются в какой-либо подхо-
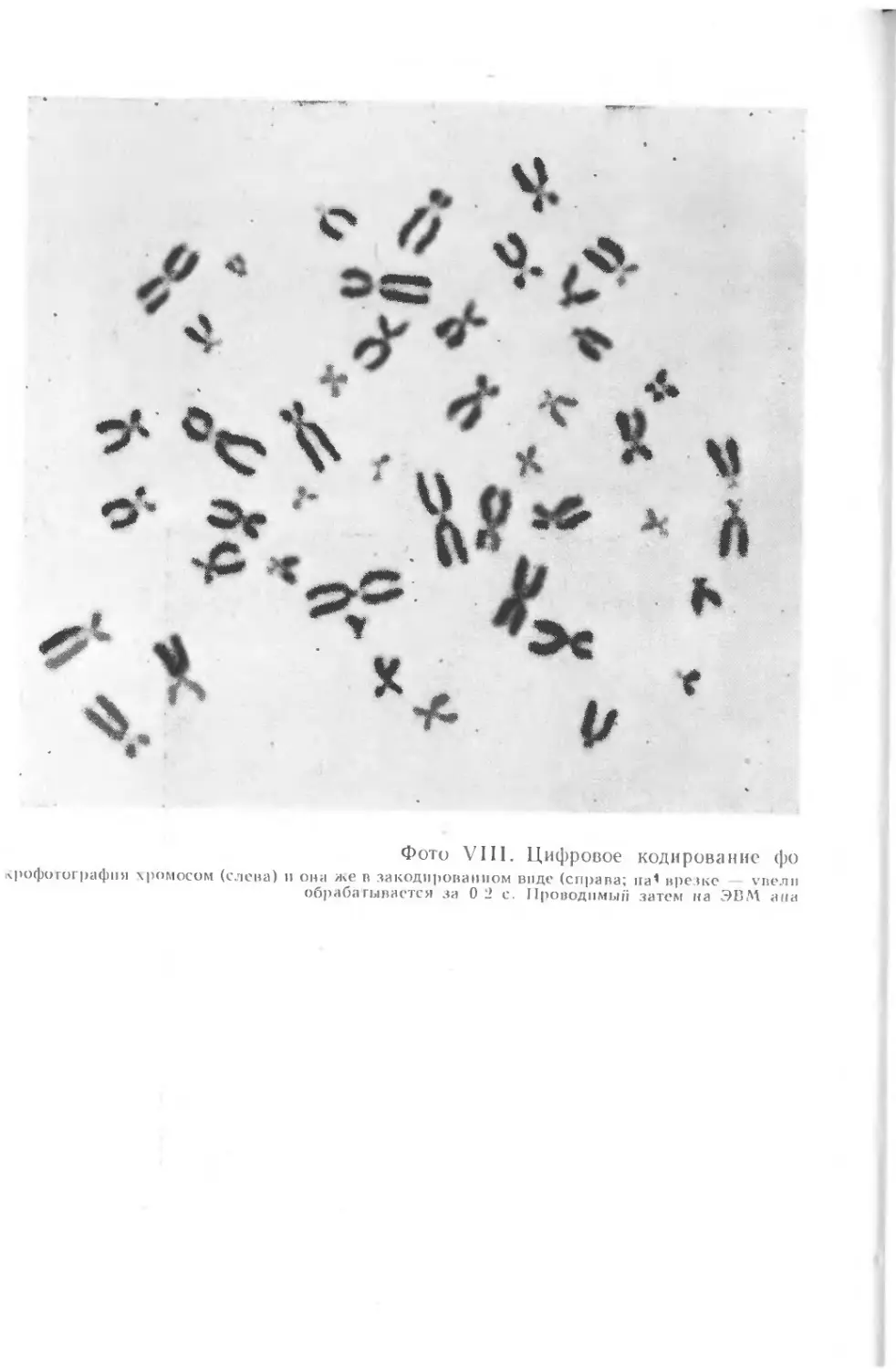
дящей для машины форме. На фото VIII слева показана исход-
ная микрофотография хромосомного набора примата, а рядом,
*4 справа, — тот же снимок в форме, пригодной для ввода в ма-
шину.
38
Часть 1. Описание и классификация ископаемых, остатков
Фиг. 10. Автоматическое кодирующее устройство.
Рисунок или фотография, подлежащая кодированию, помещается на плоскость стола. Ко-
ординаты К и Y точек, на которые указывает перо прибора, записываются на магнитной
ленте.
Ввод фотографической информации в ЭВМ сам по себе еще
не разрешает проблемы описания ископаемых, это лишь способ
задания машине информации, необходимой для такого описа-
ния. Вопрос о том, что же делать дальше с фотографией ока-
менелости, остается открытым; машина не помогает решить,
какие признаки следует выбрать для изучения данной палеон-
тологической проблемы и как выразить эти признаки. Это ос-
новной недостаток фотографического описания. Оно объективно
и потому содержит слишком много информации, в том числе
ненужной. Глядя на фотографию ископаемого, мы видим почти
все его признаки. Часть из них биологического, часть — геоло-
гического (признаки, появившиеся в процессе диагенеза) про-
исхождения. Даже если нам удастся выделить набор особенно
важных признаков, их может оказаться так много, что челове-
ческий мозг будет не в состоянии усвоить и анализировать их.
Когда студенту показывают фотографии двух окаменелостей,
он, как правило, сразу же отмечает, что это разные виды, но
не может объяснить, чем они различаются и в чем сходны меж-
ду собой. Всякое описание предполагает упрощение: надо со-
кратить число наблюдаемых и описываемых признаков до на-
бора, удобного в обращении, отбросив, в частности, те признаки,
которые кажутся неважными или излишними. Определенная
И. Описание отдельной особи
39
степень субъективности существенно важна для хорошего пале-
онтологического описания. Безупречно сделанная фотография
совершенно лишена субъективности, и именно поэтому палеон-
тологи часто предпочитают другие методы описания.
Штриховой рисунок
Самый распространенный и часто самый эффективный спо-
соб ограничить число признаков, упоминаемых в описании,—
это сделать штриховой рисунок, в котором можно отразить наи-
более важные особенности окаменелости, отметив некоторые
признаки и опустив другие.
Несколько слов о технике рисунка. Здесь нежелательна не-
аккуратность— она вносит не только ненужный «шум», но и
неконтролируемую субъективность. Аккуратный рисунок — точ-
но выполненная копия. Так как рисунок выполняется обычно
в двух измерениях, удобнее всего делать его с двумерной про-
екции объекта. Поэтому рисунки часто делают с фотографий,
калькируя контуры фотографии. Работу облегчают различные
приборы, проецирующие фотографию или оптическое изображе-
ние на плоскость рисунка. Простейший из этих приборов —
рисовальный аппарат.
Штриховой рисунок легче перевести в форму, пригодную для
ввода в ЭВМ, так как он содержит меньше информации, чем
фотография. Сканирующее устройство должно лишь отличать
белое от черного. Кроме того, машинная обработка в этом слу-
чае также облегчается, поскольку сокращается количество ин-
формации. Для некоторых палеонтологических целей штриховой
рисунок все еще слишком сложен и нуждается в дальнейшем
упрощении.
Терминология,
применяемая при описании
Наиболее обычное средство для описания какой-либо струк-
туры— это слово или сочетание слов. Слово — могучее и эко-
номное орудие, иногда оно стоит тысячи рисунков. Так, сказав,
что все вымершие лошади были одно-, дву- или четырехпалыми
(речь идет о пальцах передних ног), мы делим их этими прос-
тыми словами на валидные группы. Конечно, словам этим при-
дан весьма субъективный смысл. Используя их, мы подразуме-
ваем, что между группами лошадей существуют значительные
различия (по многим признакам) и что эти различия отражают-
ся в числе пальцев на передней конечности.
Применение для описания формы и строения специальной
терминологии значительно облегчает задачу. Главное, термине-
40 Часть 1. Описание и классификация ископаемых остатков
логия позволяет свести массу сведений к одному или немногим
словам. Правильно подобранные термины понятны сами по
себе или сравнительно легко заучиваются. Терминология быстро
становится естественной частью лексикона опытного биолога
или палеонтолога. Описательные термины очень легко кодиру-
ются, значит, их легко перевести в доступную машине форму,
и ЭВМ свободно может оперировать ими.
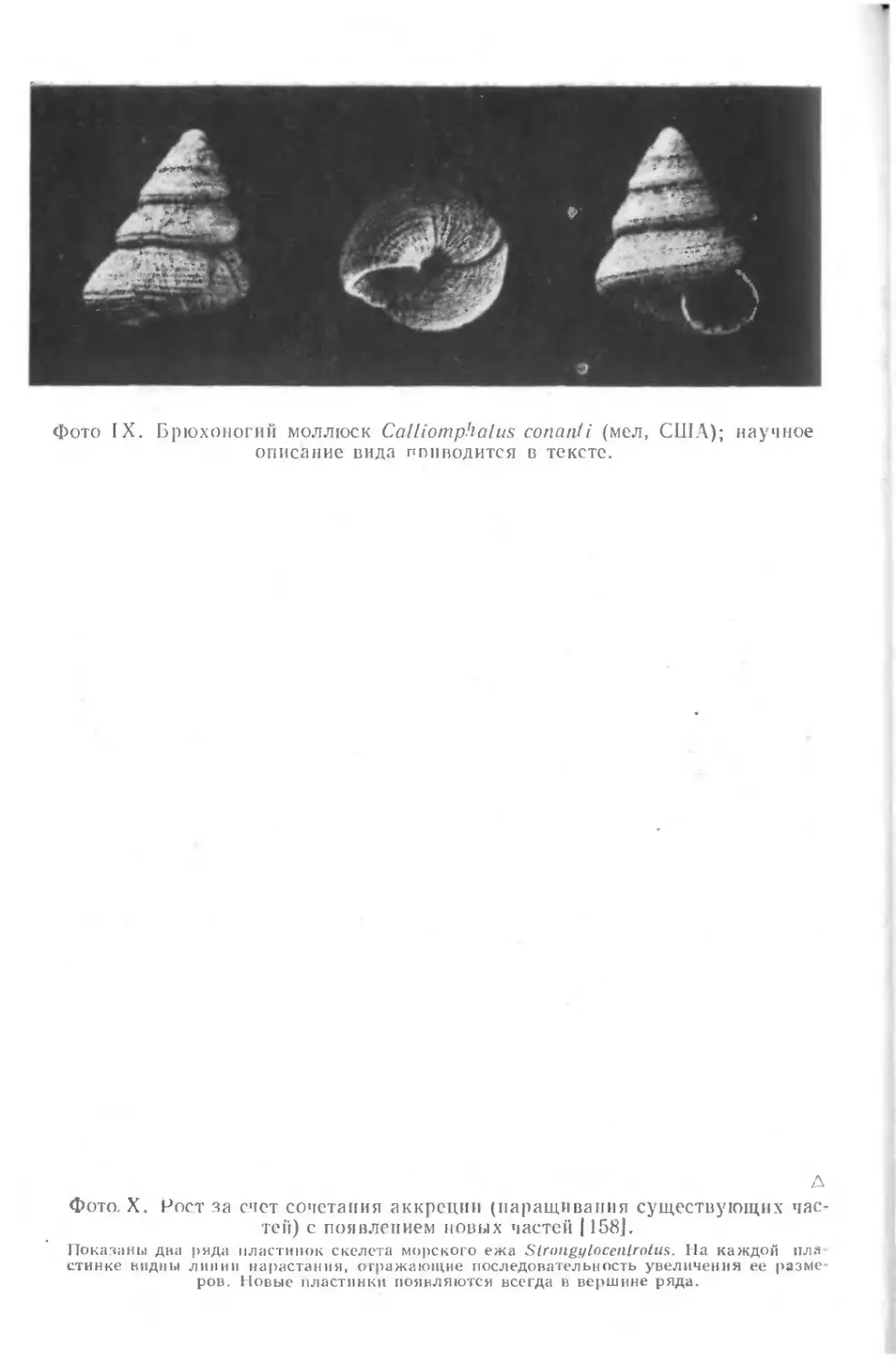
Как пример эффективного использования специальной тер-
минологии для описания ископаемых приведем формальное опи-
сание одного вида брюхоногих моллюсков (из [201]), фото-
графии которого мы также приводим (фото IX).
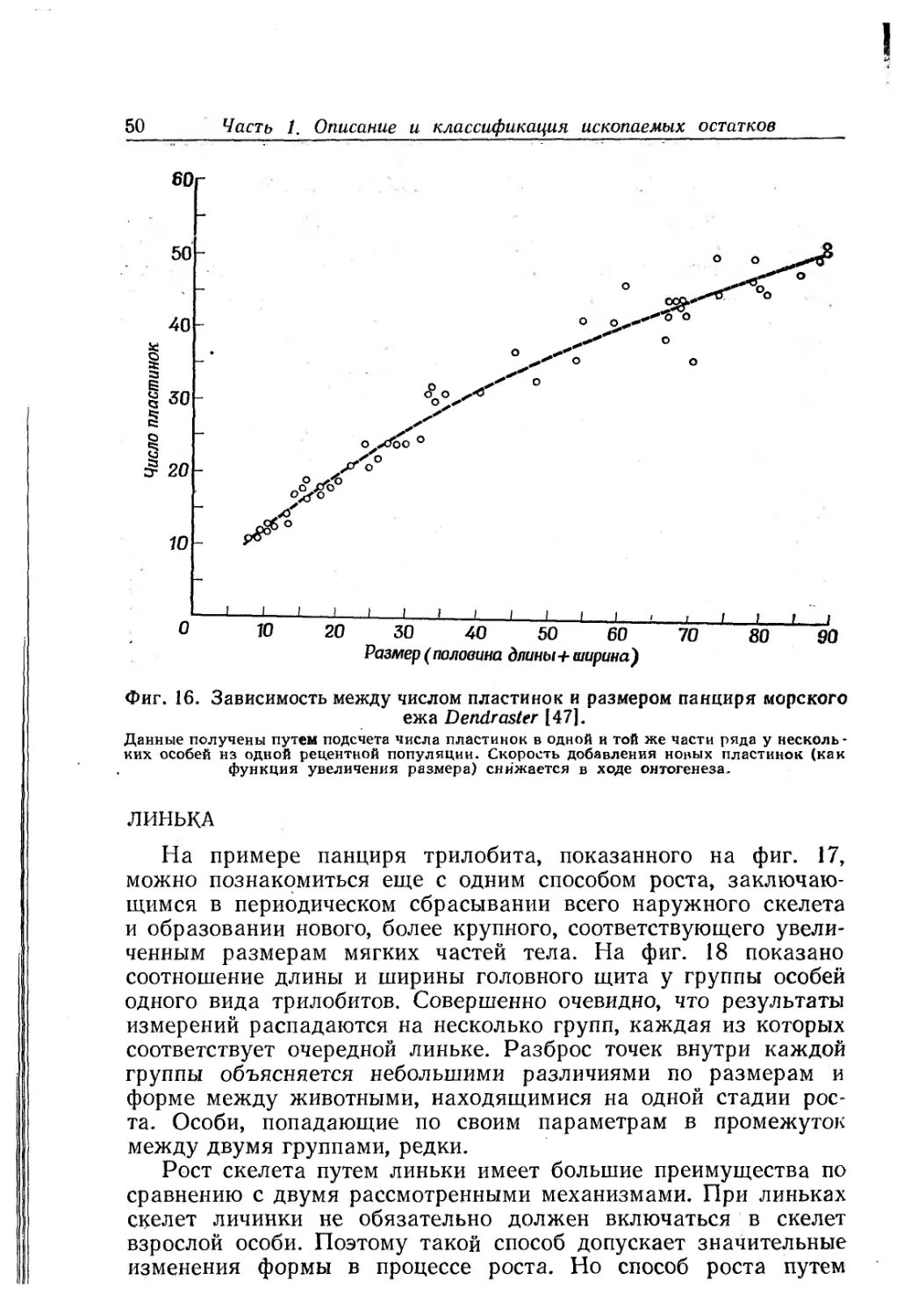
Раковина небольшая, трохусовидная, с открытым пуп-
ком, с перламутровым внутренним слоем; голотип имеет
примерно 7!/4 быстро расширяющихся оборотов. Началь-
ная часть раковины (чуть более одного оборота) гладкая,
затем появляются грубые осевые ребра и почти сразу же
вслед за ними — тонкие спиральные ребрышки; шов углуб-
ленный. Стороны оборотов имеют менее крутой наклон,
чем общий наклон поверхности завитка, и периферия каж-
дого оборота как бы нависает над следующим оборотом.
Периферия от округлой до несколько угловатой. Под пе-
риферией стороны оборота круто наклонены к широко-
округлому основанию. Скульптура из осевых и спиральных
элементов одинаково хорошо выражена; 8 спиральных
ребрышек на верхней стороне оборота в местах пересече-
ния с несколько более грубыми и более сближенными осе-
выми шнурообразными ребрами несут сглаженные бугор-
ки. Основание имеет около 10 неравномерно расположен-
ных спиральных ребрышек со слабо выраженными бугор-
ками и покрыто многочисленными осевыми ребрышками.
Пупок узкий, окаймлен краем с невысокими бугорками.
Форма устья недостаточно ясна. Оно округлое, несколько
вытянуто в ширину, в месте соединения внутренней губы
и пупкового края слегка завернуто.
Для того, кто незнаком с морфологией брюхоногих и с со-
ответствующей терминологией, это описание почти бессмыслен-
но. Но знающий человек, прочитав его, получит ясное пред-
ставление об этой группе экземпляров.
Однако любая система описательной терминологии может
привести к трудностям. Возьмем хотя бы слово «небольшая»
в первой строке описания. Оно подразумевает сравнение по
размеру с другими организмами — но с какими? Все брюхоно-
гие весьма и весьма невелики по сравнению со слонами, но
в сравнении с фораминиферами или спорами растений они —
гиганты. Слово «небольшая» означает здесь сравнение с дру-
гими брюхоногими того же типа строения или, самое большее,
II. Описание отдельной особи
41
Трохусовидная
Пупоидная
Кубаревидная
Фиг. 11. Некоторые термины, используемые при описании морфологии ра-
ковины брюхоногих моллюсков.
«равнение со всеми брюхоногими. Обычно такие термины, как
«небольшой», «большой», «широкий», «узкий» и т. п. подразуме-
вают сравнение только с близкородственными организмами.
По традиции размер раковины брюхоногого моллюска выража-
ют через высоту раковины. Значит, словом «небольшая» мы
заменяем довольно длинное описание. Ознакомившись с основ-
ным лексиконом палеонтолога, понимаешь, что этот способ опи-
сания поразительно эффективен.
Обратите внимание на термин «трохусовидная» в первой
строке приведенного выше описания. Он характеризует форму
раковины, распространенную среди брюхоногих. Раковина такой
формы вместе с раковинами других очертаний показана на
фиг. П. Такие термины (некоторым из них уже по нескольку
веков) имеют разное происхождение. Некоторые термины ука-
зывают на сходство очертаний раковины с каким-либо предме-
том: кубаревидная, например. Другие взяты из геометрии: ко-
ническая, биконическая, ортоконическая и т. д. Третьи произ-
42 Часть 1. Описание и классификация ископаемых остатков
ведены от названий отдельных организмов (родов или видов),
у которых данная форма выражена хорошо или которые так
широко распространены, что знакомы каждому биологу.
При употреблении описательных терминов возникает проб-
лема, как определить границу между двумя категориями. На-
пример, как описать раковину, по форме стоящую между шаро-
видной и кубаревидной? Один из способов решения — ввести
новые термины для промежуточных форм. В описании раковин
брюхоногих употребляются термины «высокая кубаревидная»
и «низкая кубаревидная». Но сколько бы категорий мы ни вы-
деляли, всегда найдутся окаменелости, стоящие между двумя
или несколькими категориями. Хорошая система должна содер-
жать не слишком много категорий, но все же достаточно для
разумной классификации имеющегося материала.
Одна из причин, по которым промежуточные формы обычно
причиняют не слишком много хлопот, состоит в том, что частота
разных форм в животном и растительном мире неслучайна.
Некоторые очертания встречаются чаще, другие — гораздо реже.
Если система описательной терминологии верно отражает есте-
ственные группы преобладающих форм, то промежуточные слу-
чаи будут сравнительно редкими. Таким образом, разработка
системы описательной терминологии уже сама по себе представ-
ляет значительный научный интерес, поскольку такая система
призвана дать некую основную интерпретацию органического
мира.
Конечно, есть опасность, что при разработке терминологии
могут возникнуть ошибки и появиться неверные интерпретации.
В палеонтологической литературе в прошлом неоднократно за-
креплялись многие совершенно непригодные системы описатель-
ной терминологии. Не соответствуя естественным группировкам,
они мешали верному пониманию наблюдаемых фактов.
Описание посредством измерения
Заменяя описание с помощью специальной терминологии
точными измерениями, мы избегаем некоторых трудностей, но
вместо них тут же возникают новые. При достаточно точном
измерении отпадает проблема промежуточных форм. Кроме то-
го, данные измерений пригодны для ввода в ЭВМ, т. е. для
непосредственной и эффективной обработки. Применение коли-
чественных данных пользуется хорошей репутацией среди уче-
ных. Обычно считают, что только количественный подход по-
настоящему объективен. Однако на самом деле измерения часто
оказываются самым субъективным методом описания.
Рассмотрим пример (фиг. 12). Длина и высота раковины
двустворчатого моллюска определяются линейными размерами.
11. Описание отдельной особи 43
Длина I длина
v' Длина
Ф'йг. 12. Внутренняя сторона створок трех видов двустворчатых моллюсков.
Для всех трех форм соотношение выбранных нами главных параметров —Удлини и'выср-
. ты — почти одинаково.
Длина — максимальный линейный размер в передне-заднем
направлении. Высота — максимальный размер, перпендикуляр-
ный длине. Эти данные содержат довольно много информации.
Они говорят не только о размере раковины, но и о ее форме.
Отношение длины к высоте — показатель формы. У равносто-
ронних раковин это отношение приближается к единице. У удли-
ненных оно больше единицы.
у - Раковины трех двустворчатых моллюсков, показанные на
фиг. 12, характеризуются почти одинаковым отношением
/Длины к высоте, но даже ребенок заметит явные различия меж.-
ДУ этими экземплярами.
Здесь мы сталкиваемся с одним из самых крупных недостат-
ков системы описания, основанной исключительно на измерени-
ям. Явные различия в очертаниях раковин не отражаются в
отношении их длины к высоте. Выражая форму двумя промера-
ми,- сделанными перпендикулярно друг к другу, мы подразуме-
ваем, что описываемый предмет имеет прямоугольные очертания.
44 Часть 1. Описание и классификация ископаемых остатков
Если бы все двустворчатые раковины были прямоугольными, то
все было бы просто и измерения длины и высоты или их отно-
шение исчерпывающе характеризовали бы форму. Важно пом-
нить, что, когда мы хотим описать какую-либо форму с помощью
ряда измерений, мы создаем модель этой формы, т. е. пред-
ставляем себе некий идеальный геометрический прототип и мо-
жем наблюдать или измерять отличия реального экземпляра
только по отношению к этой модельной форме.
На фиг. 13 изображены четыре гипотетические спирально
свернутые раковины головоногих моллюсков. Для каждой из
них указан наибольший диаметр d, по которому крупные фор-
мы отличаются от небольших. Но диаметр мало говорит нам
о форме раковины. Подразумеваемая модель — окружность.
В одну и ту же окружность можно вписать много разных спи-
рально закрученных головоногих; следовательно, диаметр гово-
рит нам главным образом о величине раковины. Формы, пред-
ставленные на фиг. 13, лучше описываются двумя неравными
радиусами (г4 и г2), начинающимися в морфологическом центре
спирали. Отметим, что отношение этих радиусов для формы А
больше, чем их отношение для формы Г, что, конечно, отражает
некоторые различия в очертаниях двух раковин. (Кстати, за-
метим, что сумма двух радиусов несколько меньше диаметра.)
Можно расширить использование радиусов для описания спи-
ральных форм. Вот уже полтора столетия известно, что многие
спиральные раковины беспозвоночных имеют форму, поддаю-
щуюся строгому математическому описанию. У спирально свер-
нутых раковин головоногих моллюсков отношение радиусов, из-
меренных по ходу спирали через равные угловые интервалы,
обычно сохраняется одинаковым. Это показано на фиг. 14: от-
ношение двух противоположных радиусов остается постоянным.
Помня это, вернемся к фиг. 13 и охарактеризуем каждую рако-
вину отношением радиусов, разделенных половиной оборота.
Используя этот метод описания, мы сможем различить три фор-
мы головоногих. Однако раковины Б и В нельзя различить
путем сравнения отношений радиусов, хотя на рисунке ясно
видно, что очертания этих раковин различны.
При описании посредством измерений обычно не так важны
число и точность измерений, как верный выбор подлежащих
измерению параметров. В раковине головоногих можно было
бы найти еще массу доступных для измерения признаков, но,
по-видимому, ни один другой набор измерений не будет таким
экономным или прямо достигающим цели, как отношение радиу-
сов (хотя надо помнить, что для различения раковин Б и В на
фиг. 13 потребуются и другие измерения). В идеале при измере-
нии следует производить отбор информации, т. е. использовать
лишь часть информации, отраженной на фотографии, так что
II. Описание отдельной особи
45
Фиг. 13. Четыре типа спирально свернутых раковин головоногих моллюсков
(подробности в тексте).
морфология изучаемого объекта при этом схематизируется и
упрощается. Насколько это полезно для каждого данного опи-
сания, целиком зависит от общих задач исследования и постав-
ленных вопросов. Нет какого-то одного набора измерений, ко-
торый лучше всего описывал бы данный организм. Но, как пра-
вило, для каждой группы ископаемых организмов есть основной
Набор измерений, принятый большинством палеонтологов. На-
пример, измерения, принятые для головоногих, до некоторой
Степени произвольны, но они позволяют составить весьма верное
Представление об их морфологии. Принятый набор измерений
может заменять описание с помощью терминологии.
Измерения можно применять для описания изменчивости
Особей. Располагая данными измерений большого числа особей,
Можно провести статистический анализ изменчивости.
; ' После того как мы выбрали, что следует измерять, остаются
еще две проблемы: какой метод измерения следует применить
46 . Часть 1. Описание и классификация ископаемых остатков
Фиг. 14. Обобщенная форма спи-
рально свернутой раковины голово-
ногого моллюска.
На рисунке видно, что соотношение ра*
диусов постоянно.
и к какой точности надо стремиться. До сих пор мы обсуждали
только линейные размеры — кривые поверхности для измерения
проецировались на плоскость. Проекция облегчает измерение
и, как правило, допустима. Но она всегда связана с упрощением:
кривая поверхность предполагается плоской. То же делается,
когда для измерения расстояния между двумя точками кривой
поверхности ископаемого применяется кронциркуль. Измерения,
сделанные по фотографии, достаточно точны лишь в том случае,
если плоскость фотографии совпадает с необходимой проекци-
ей. Любое оптическое проецирование предмета (включая фото-
графирование) вносит определенные искажения, которые долж-
ны быть сведены к минимуму. Допустимая степень искажения
зависит от требуемой точности и достоверности результатов.
Нет общих правил, помогающих выбрать степень точности
измерений. Она целиком зависит от поставленной задачи иссле-
дования. Например, если мы хотим отличить трилобит от дино-
завра, достаточна точность измерений до метра. Точность до
сантиметра или миллиметра в этом случае излишня. Но если
нужно различить два очень сходных организма, необходима
гораздо большая точность. Опытный работник добивается лишь
той степени точности, какая нужна в данном конкретном случае.
Для измерения ископаемых придуманы различные прибо-
ры-— сложные и простые. Многие из них сконструированы для
узких целей. Так, инструмент, показанный на фиг. 15, изобретен
в 1842 г. Альсидом д’Орбиньи для измерения так называемого
апикального угла свернутых в коническую спираль раковин
брюхоногих. Этот очень простой прибор состоит всего лишь
из транспортира и линейки, но с успехом применяется для об-
щего описания.
Для измерений часто применяется описанный ранее метод
перевода морфологической информации в цифровую. Расстоя-
II. Описание отдельной особи
47
Фиг. 15. Инструмент для измерения вершинного угла раковин брюхоногих
моллюсков, предложенный д'Орбиньи в 1842 г. [215].
ние по прямой между двумя точками легко рассчитать, если из-
вестны координаты точек. Применение метода цифрового коди-
рования для единичных или немногочисленных измерений не-
оправданно. Но если нам нужно на каждой особи выполнить
' множество измерений, то цифровое кодирование и машинная
-Обработка сильно облегчают задачу. Точность измерений, про-
водимых этим методом, целиком зависит от точности, с которой
может быть определено положение точки в системе координат.
Описание внутреннего строения
. Обсуждавшиеся выше методы, как правило, применяются
для описания внешнего строения окаменелостей. Внутреннее
утроение часто может быть не менее, а то и более важным, чем
Наружное, например строение кристаллов, слагающих раковину,
Или строение системы каналов в кости млекопитающего. Проб-
лемы, возникающие при описании внутренних структур, в ос-
новном совпадают с проблемами описания внешней формы. Но
решаются они с помощью иных методов. Как мы уже видели,
Информацию о внутренних структурах можно получить посред-
ством специальных фотографических методов, например фото-
графирования в рентгеновских лучах. Образцы, непригодные
для такого исследования, обычно распиливают или шлифуют
М Получают пришлифовку или тонкий шлиф. Пришлифовки ис-
следуются в отраженном свете, тонкие шлифы — в проходящем.
" Надо помнить, что постоянно появляются новые ме-
Мщики. Многие из них разрабатываются в областях
Науки, далеких от палеонтологии, но могут использоваться и
ври изучении внутреннего строения окаменелостей. История
^Деонтологии показывает, что иногда наибольших успехов до-
ривастся тот, кто сумел успешно применить к палеонтологиче-
ским проблемам методы, разработанные другими науками.
ГЛАВА Ш
Онтогенетическая изменчивость
Эта глава — первая в серии глав, посвященных проблемам
описания и интерпретации изменчивости ископаемых. Различия
между ископаемыми обусловлены бесчисленным множеством
биологических и геологических причин. Один из самых важных
чисто биологических факторов — различие в возрасте отдель-
ных особей в Момент их гибели. Две особи могли быть генети-
чески идентичными и жить в одинаковых условиях, и все же их
фоссилизированные остатки могут сильнейшим, образом разли-
чаться просто по той причине, что в момент гибели одна из них
была старше другой. Это различие сводится не только к раз-
мерам. Как правило, рост сопровождается изменением формы.
Особи, находящиеся на разных стадиях роста, могут настолько
различаться, что в них иногда невозможно признать предста-
вителей одного и того же вида. Особенно сильны эти различия
у организмов, у которых онтогенез (жизненный цикл) включает
метаморфоз, например у членистоногих.
Изучать онтогенез необходимо для правильного определения
видовой принадлежности данной окаменелости, для правильной
оценки и интерпретации диапазона морфологической изменчи-
вости у данного вида. Описание изменений морфологии орга-
низма в ходе онтогенеза не менее важно, чем общее описание
взрослой формы, особенно для эволюционных исследований.
Было бы недопустимым упрощением рассматривать эволюцион-
ный ряд как последовательность взрослых форм. Эволюцию
следует представлять себе как последовательность онтогенезов.
Типы роста
Рост организмов — крайне сложное явление. Оно включает
обычно несколько типов изменений, в том числе изменения раз-
меров и числа клеток, разнообразия типов клеток и относитель-
ного расположения клеток. В течение жизни форма организма
может изменяться резко (метаморфоз) или постепенно.
У всех организмов, имеющих прочный скелет для опоры,
защиты или прикрепления мышц, размеры скелетных структур
должны увеличиваться по мере роста мягких тканей. Известны
четыре типа постэмбрионального роста скелета.
III. Онтогенетическая изменчивость
49
ПРИРАЩЕНИЕ (АККРЕЦИЯ) СУЩЕСТВУЮЩИХ ЧАСТЕЙ
У большинства раковинных моллюсков размер скелета уве-
личивается просто в результате того, что к раковине добавля-
ется в течение жизни новый материал. Преимущество этого
способа состоит в том, что материал, отложенный на более ран-
них стадиях онтогенеза, продолжает использоваться и в даль-
нейшем; недостаток же его состоит в обязательном сохранении
ювенильной раковины, которая становится частью взрослой.
Хороший пример такого роста представляют раковины брюхо-
ногих (фото IX и фиг. 11). Спиральную раковину брюхоногих
можно рассматривать как конусообразную полую трубку, за-
крученную вокруг оси. По мере роста животного новый мате-
риал прибавляется к краям устья — отверстия трубки. Часто
видны линии нарастания, параллельные краям устья. Они пока-
зывают ход роста. Взрослый брюхоногий моллюск зани-
мает, как правило, всю свою раковину, и форма его мягкого
тела представляет собой фактически внутренний слепок ра-
ковины.
Рост путем аккреции встречается у многих других групп
животных, особенно у тех, которые обладают наружным скеле-
том, служащим в основном для защиты животного и прикрепле-
ния мышц.
ДОБАВЛЕНИЕ НОВЫХ ЧАСТЕЙ
У организмов, скелет которых состоит из множества одно-
родных элементов, тесно сочлененных между собой или рыхло
Лежащих в мягкой ткани, рост скелета обычно идет путем при-
бавления к нему новых частей. Так, жесткий панцирь большин-
ства морских ежей состоит из нескольких тысяч тесно сочленен-
ных кальцитовых пластинок, расположенных радиальными ря-
дами от одного полюса шаровидного скелета до другого. Рост
идет в основном за счет постоянного добавления новых пласти-
нок. График, приведенный на фиг. 16, показывает зависимость
Диела таких пластинок в участке одного из рядов от размера
скелета у разновозрастных особей одного вида морских ежей.
Отметим, что скорость увеличения числа пластинок с увеличе-
нием размера скелета изменяется. Важно, что новые пластинки
Морских ежей всегда появляются в одном месте, а именно на
^верхнем конце ряда.
; 4 У многих других организмов в процессе роста также появля-
ется новые части скелета. На фиг. 17 показаны некоторые ста-
дии онтогенеза трилобита; одно из самых заметных онтогенети-
ческих изменений — постепенное появление новых сегментов
торакса.
50 Часть 1. Описание и классификация ископаемых остатков
Фиг. 16. Зависимость между числом пластинок и размером панциря морского
ежа Dendraster [47].
Данные получены путем подсчета числа пластинок в одной и той же части ряда у несколь-
ких особей из одной рецентной популяции. Скорость добавления новых пластинок (как
функция увеличения размера) снижается в ходе онтогенеза.
ЛИНЬКА
На примере панциря трилобита, показанного на фиг. 17,
можно познакомиться еще с одним способом роста, заключаю-
щимся в периодическом сбрасывании всего наружного скелета
и образовании нового, более крупного, соответствующего увели-
ченным размерам мягких частей тела. На фиг. 18 показано
соотношение длины и ширины головного щита у группы особей
одного вида трилобитов. Совершенно очевидно, что результаты
измерений распадаются на несколько групп, каждая из которых
соответствует очередной линьке. Разброс точек внутри каждой
группы объясняется небольшими различиями по размерам и
форме между животными, находящимися на одной стадии рос-
та. Особи, попадающие по своим параметрам в промежуток
между двумя группами, редки.
Рост скелета путем линьки имеет большие преимущества по
сравнению с двумя рассмотренными механизмами. При линьках
скелет личинки не обязательно должен включаться в скелет
взрослой особи. Поэтому такой способ допускает значительные
изменения формы в процессе роста. Но способ роста путем
III. Онтогенетическая изменчивость
51
Фиг. 17. Четыре стадии онтогенеза трилобита Paradoxides [233].
“ онтогенезе значительно изменяется форма существующих частей и добавляются но-.
« вые части.
линьки имеет и важный недостаток: периоды линек довольно
опасны для организма. В это время он лишен защиты и, кроме
на полную смену скелета тратится значительное количе-
«^ЦЭнергии.
- Рост путем линьки более всего распространен среди орга-
обладающих скелетом. Но численный перевес линяю-
ШИХ организмов над нелиняющими объясняется просто: этот
. сйбСоб роста характерен для членистоногих, составляющих
ММо трех четвертей всех современных видов животных. Линь-
встречается и в других группах животных, обычно при мета-
ъ^^фозе от личиночной стадии к взрослой.
52
Часть 1. Описание и классификация ископаемых остатков
Общая длина головного щита (Д),мм
Фиг. 18. Онтогенез головного щита ордовикского трилобита Trinodus, отра-
женный в увеличении длины и ширины [83].
Неравномерность распределения точек на диаграмме связана с периодическими линь-
ками.
МОДИФИКАЦИЯ
Этот способ роста наиболее обычен при образовании неко-
торых костей высших позвоночных. Сложный процесс замеще-
ния и перестройки скелетного материала сопровождается не
только увеличением размера, но и изменением формы и строе-
ния кости. Этому способу роста присуще преимущество линоч-
ного способа — форма скелета каждой данной стадии роста
почти не зависит от предыдущих стадий. Вместе с тем этот
способ лишен недостатка, характерного для линьки, — нет свя-
занных с линьками периодов полной беззащитности.
Строго классифицировать организмы по способам роста их
скелетов трудно, главным образом потому, что у многих орга-
низмов используется комбинация описанных выше основных
механизмов. На фото X показаны два ряда пластинок из панци-
ря морского ежа. Как мы видели, основной способ увеличения
размеров панциря морских ежей — появление новых пластинок.
Но существующие пластинки также растут, по краям к ним
прибавляется новый материал. Основные периоды роста отме-
чены линиями нарастания, по которым можно проследить изме-
III. Онтогенетическая изменчивость
53
нения размера и формы пластинок. Некоторые линии нараста-
ния являются годовыми.
Второй пример комбинации этих же двух способов роста
приведен на фото XI. Это свернутая наружная раковина голо-
воногого, растущая в основном путем приращения нового ма-
териала, подобно раковинам брюхоногих. Но эти два класса
моллюсков имеют важное различие. Бюхоногий моллюск за-
полняет всю свою раковину, тогда как тело головоногого рас-
полагается в так называемой жилой камере, занимающей лишь
наружную половину или три четверти последнего оборота рако-
вины. Жилая камера отделена от остальной части раковины
(фрагмокона) обызвествленной стенкой, называемой септой.
В процессе онтогенеза тело животного увеличивается и пере-
мещается кнаружи, так что оно всегда находится вблизи устья
раковины. Время от времени, по мере того как животное дви-
жется кнаружи, образуются новые перегородки, или септы.
Возникает последовательность септ, раковина растет за счет
добавления новых частей. Различия в способе наращивания
раковины у брюхоногого и головоногого моллюсков тесно свя-
заны с физиологическими и экологическими различиями этих
моллюсков. В частности, головоногому моллюску нужна пустая
часть раковины, которая придает ему плавучесть, необходимую
для пассивного или активного перемещения.
Описание
онтогенетических изменений
При описании изменений, происходящих с организмом в хо-
де онтогенеза, биолог редко сталкивается с особыми труднос-
тями. Нередко он может непосредственно наблюдать за ростом
животного. Более того, он может точно замечать время на-
ступления онтогенетических изменений. Палеонтолог же редко
может судить по найденным остаткам о возрасте организма
в момент смерти и лишен возможности наблюдать за его рос-
том. И все же есть два способа, позволяющих получить по
ископаемому материалу важные сведения об онтогенезе. Во-
первых, можно собрать «онтогенетические ряды» особей (см.
Фиг. 16, 17 и 18) и наблюдать (или измерять) изменение фор-
мы с увеличением размера. Во-вторых, онтогенез взрослой особи
можно восстановить по некоторым признакам типа линий на-
растания (фото X), которые возникают в результате времен-
ного прекращения или замедления роста.
Выбор метода зависит от того, какой механизм (или меха-
низмы) роста используется организмом. Если рост шел за счет
простого приращения материала, без резорбции или модифи-
кации структур, возникающих в результате предшествующего
54 Часть I Описание и классификация ископаемых остатков
Ширина, лтлг
Фиг. 19. Изменение формы
в онтогенезе брахиопод [23].
Графики построены на основе дан-
ных измерений для сообщества
Ectochoristites (карбон). Длина —
расстояние от переднего до заднего
края брюшной створки; ширина
измеряется по замочному краю
брюшной створки. Точки на ниж-
нем графике, соединенные кривы-
ми, получены путем измерения
лннийл нарастания у отдельных
особей.
онтогенеза, и если имеются заметные линии нарастания, то
могут быть использованы оба метода. То же верно и для случая
простого добавления новых частей. Но если в ходе онтогенеза
данного организма имела место линька или модификация, то
для понимания онтогенеза палеонтологу требуется серия эк-
земпляров. Если одинаково доступны оба метода, то надо вы-
бирать метод работы с одной взрослой особью, так как при
этом мы избавляемся от трудностей, связанных с необходимо-
стью интерпретации различий между особями.
III. Онтогенетическая изменчивость
55
На фиг. 19 представлены графики для двух признаков бра-
хиоподы. Верхний график построен на основе измерений при-
мерно 75 ископаемых особей одного вида, найденных в одном
обнажении и, очевидно, представляющих собой одну популяцию.
Точки графика расположены веерообразно и исходят из начала
отсчета. Заметим, что этот «веер» несколько изогнут (выпук-
лостью он обращен к оси абсцисс). Из этого можно заключить,
что с увеличением размера животного скорость роста в длину
увеличивается по отношению к скорости роста в ширину. Делая
этот вывод, мы подразумевали, что размер связан со временем
некоей зависимостью. Но диаграмма не содержит никакой ин-
формации о связи между увеличением размера и абсолютным
временем. Из нее можно узнать лишь об относительной скорости
возрастания одного признака по сравнению со скоростью воз-
растания другого.
Криволинейный характер графика говорит об изменении от-
ношения между длиной и шириной, т. е. об изменении формы
в ходе роста. На фиг. 19 проведена прямая под углом 45°. Все
точки, лежащие ниже прямой, относятся к особям, у которых
ширина больше длины, а точки выше прямой — к особям, у ко-
торых длина больше ширины. В результате изменения относи-
тельных скоростей роста раковина, у которой ширина больше
длины, постепенно превращается в раковину с длиной, превы-
шающей ширину.
Этот вывод можно уточнить, проведя через разбросанные
точки кривую. Тогда мы получим обобщенную картину измене-
ния формы в ходе онтогенеза этого вида.
Почему некоторые точки не укладываются на идеальную
кривую? Это можно объяснить несколькими причинами. Одна
из них состоит в том, что данные по онтогенезу получены на
основе данных измерений большого числа особей, генетически
различающихся между собой; эти генетические различия и вно-
сят изменчивость, не имеющую отношения к онтогенезу. Кроме
того, не исключено, что скелеты некоторых особей изменены
уже после их гибели в результате диагенетических процессов
и разброс точек совсем не связан с биологическими факторами
Возможно также, что применявшийся метод измерения недоста-
точно точен и на самом деле данные могли бы гораздо лучше
укладываться на идеализированную кривую.
На фиг. 19 внизу показано несколько кривых роста раковин
гого же вида брахиопод. Точки каждой кривой получены изме-
рениями линий нарастания одной взрослой особи. График под-
тверждает, что рост особей этого вида описывается кривой; тем
самым становится ясной важность изучения онтогенеза отдель-
ной особи. Оба графика, приведенные на фиг. 19, дают в общем
одну и ту же информацию, но нижний график позволяет полу-
56 Часть 1. Описание и классификация ископаемых остатков
Фиг. 20. Зависимость между длиной радиуса раковины и его угловым поло-
жением в ходе онтогенеза головоногого моллюска.
График построен на основе данных измерения экземпляра Nautilus, показанного на фо-
то XI. По оси абсцисс отложен угол поворота Точка О — устье раковины.
чить более определенную картину роста и различий между
особями ископаемого сообщества.
На фиг. 20 показаны результаты реконструкции онтогенеза
спирально свернутой раковины головоногого моллюска (см. фо-
то XI) на основе измерений одного взрослого экземпляра. Из-
мерялись радиусы раковины этого экземпляра (о значении из-
мерения радиусов для описания завернутых головоногих см.
гл. II).
На фиг. 20, А показана зависимость длин радиусов от их
угловых положений. Поскольку, как мы знаем, наружная ра-
ковина головоногих растет путем аккреции, каждое угловое
приращение раковины должно соответствовать увеличению
возраста. Криволинейность полученного графика указывает на
то, что скорость удлинения радиуса меняется по мере его пово-
рота. (Разброс точек гораздо меньше, чем это показано на
фиг. 19, так как здесь приведены данные измерений только
одной особи.)
Говорит ли криволинейность графика об изменении формы?
Мы не можем сразу ответить на этот вопрос, потому что здесь
отложены не линейные размеры, как было в случае плеченогих.
Мы можем узнать больше об онтогенезе этого головоногого
моллюска, построив график зависимости между угловыми поло-
жениями радиусов и логарифмами их длины (фиг. 20, Б). На
111. Онтогенетическая изменчивость
57
этом новом графике все точки почти идеально укладываются на
прямую линию. Значит, угол поворота радиуса возрастает про-
порционально логарифму длины радиуса. Другими словами,
длины последовательно расположенных радиусов составляют
геометрическую прогрессию. Итак, перед нами так называемая
логарифмическая спираль. Одно из ее характерных математи-
ческих свойств — постоянство формы. Следовательно, можно
сделать вывод, что форма головоногого моллюска в ходе онто-
генеза не меняется — в процессе роста увеличивается только
число оборотов.
В общем можно сказать, что из двух методов воссоздания
онтогенеза по окаменелостям метод работы с отдельной особью
обычно проще и дает в целом более надежные результаты. Но
почти такую же информацию можно получить и при обработке
нескольких особей, если только в гетерогенной группе изменчи-
вость, не связанная с онтогенезом, не слишком велика.
В рассмотренных примерах анализа онтогенеза проводилось
измерение двух морфологических признаков и соотнесение их на
графике, что позволяет выявить относительные скорости их раз-
вития. Но такой анализ может охватывать и другие признаки.
Работая чисто графическими методами, можно ввести в анализ
третий признак, рассматривая соотношения между тремя ско-
ростями роста. Однако графический анализ не может охватить
четыре, пять или шесть признаков. Для этого употребляют бо-
лее сложные методы многофакторного статистического анализа.
Они с большим успехом используются в палеонтологии.
Совершенно другой способ описания онтогенетических изме-
нений— метод преобразования координат. Он особенно подхо-
дит для описания и истолкования сложных изменений целого
организма или его структуры. На фиг. 21 показаны очертания,
характерные для двух стадий развития трилобита. Изображе-
ния даны таким образом, чтобы величины обеих стадий были
сопоставимы. Тогда лучше заметны связанные с онтогенезом
изменения формы. На изображение первой стадии наложена ус-
ловная сетка координат. Изображение второй стадии покрыто
той же сеткой, но здесь она деформирована в соответствии с
изменением морфологии, происшедшим в онтогенезе.
Метод преобразования координат впервые предложил в
своем классическом труде «О росте и форме» д’Арси Томпсон.
Этот метод использовался редко, так как он очень трудоемок
и, кроме того, при построении деформированной решетки в ри-
сунок вносится элемент субъективности. Положение дел изме-
нилось после того, как начали использоваться быстродействую-
щие ЭВМ. Сейчас их применение в сочетании с разработанными
в других областях науки методами «анализа огибающей» позво-
ляет палеонтологу с большей точностью и быстротой строить»
58
Часть 1 Описание и классификация ископаемых остатков
Фиг. 21. Онтогенез трилобита, представленный методом преобразования ко-
ординат [147].
Условная сетка координат X — У была наложена произвольно на очертания стадии А
и были отмечены точки пересечения координатных линии Затем соответствующие точки
были найдены на рисунке стадии Б и по ним была нарисована деформированная сетка
Таким образом, деформированная сетка отражает пространственные морфогенетические
изменения, происшедшие при переходе от стадии А к стадии Б.
деформированные сетки д’Арси Томпсона, а также давать более
точное математическое описание изменений формы.
Темп роста
Некоторые растения и животные растут очень быстро, дру-
гие— очень медленно. Так, у человека рост останавливается
обычно примерно в двадцать лет. У обезьян рост завершается
примерно за два года. Онтогенез других организмов, например
насекомых, может полностью заканчиваться в течение несколь-
ких недель, а то и дней. Различия эти в основном наследствен-
ные, т. е. выработались в процессе эволюции и находятся под
генетическим контролем. Но все организмы обладают и некото-
рой изменчивостью темпа роста. Эта изменчивость не подчиня-
ется генетическому контролю, а зависит от условий, в которых
живет организм. До некоторой степени скорость роста зависит
от биологических и физических факторов среды: питания, плот-
ности популяции и т. д. Для описания возможной степени не-
наследственной изменчивости морфологического признака или
скорости его увеличения генетики используют термин «норма
реакции».
Почти у всех организмов скорость роста меняется со време-
нем, т. е. в ходе онтогенеза. Эта изменчивость в основном
контролируется генетически, но в какой-то мере зависит и от
Ill Онтогенетическая изменчивость
59
фиг. 22. Обобщенные графики роста'организма (изменение размера S со вре-
менем /) [129].
условий среды, т. е. имеет свою норму реакции. Типичная дина-
мика роста во времени показана на фиг. 22. По оси ординат
отложен размер гипотетического животного, по оси абсцисс —
время. Сначала размер увеличивается медленно, затем быстрее,
а потом рост сильно замедляется и на последних стадиях почти
прекращается. Второй график, приведенный на фиг. 22, яснее
иллюстрирует эти изменения скорости роста. Здесь на оси орди-
нат отложена скорость роста (первая производная размера по
времени), а на оси абсцисс — время. Математик сказал бы, что
вторая кривая отражает изменение крутизны первой кривой
На третьем графике фиг. 22 показана скорость изменения кру-
тизны первой кривой, и, следовательно, он представляет вторую
производную размера по времени.
Такой тип роста весьма распространен среди растений и жи-
вотных, и его можно считать самым обычным, однако рост не-
которых животных отклоняется от этого образца. Различия
касаются главным образом формы верхней части кривой, приве-
денной на первом графике. Например, у большинства млекопи-
тающих кривая роста сглаживается. Организм может жить еще
долгое время, когда его рост прекращается, то есть кривая
становится почти горизонтальной. Период после завершения рос-
та называют обычно зрелостью или взрослой стадией онтогенеза
Но у большинства растений и беспозвоночных животных рост
продолжается всю жизнь (хотя и сильно замедляется), поэтому
тут трудно строго выделить взрослую стадию. В лучшем случае
можно выделить стадию, на которой рост уже в основном за-
вершен, хотя и не прекратился.
Измерив рост конкретного организма, мы редко получаем
идеальную кривую, показанную на фиг. 22. Дело в том, что
норма реакции для скорости роста, как правило, довольно ши-
рока. Если во время развития организма вдруг происходит
улучшение питания, кривая может сместиться вверх, потеряв
плавность. Изменчивость, связанная с условиями среды, может
60 Часть 1. Описание и классификация ископаемых остатков
быть настолько значительной, что маскирует всю картину изме-
нения размера со временем.
Независимо от изменения условий среды в ходе онтогенеза
разные части организма растут обычно с разной скоростью.
Мы уже видели это на примере плеченогих (фиг. 19), у которых
длина и ширина увеличиваются с разной скоростью. Скорости
роста различных частей животного тесно координированы. Если
в результате изменения условий среды скорость роста одной
части уменьшается, то соответственно замедляется рост другой
части организма. Общеизвестно изменение в онтогенезе отно-
сительного размера головы человека: в процессе роста человека
его голова становится относительно меньше, так как она растет
не так быстро, как почти все остальное тело. Отношение раз-
мера головы к размеру других частей тела на одних и тех же
стадиях онтогенеза человека всегда одинаково, хотя факторы
среды могут усиливать или тормозить общий рост. Преимуще-
ства координированного роста с эволюционной точки зрения
ясны. Работа всего организма в значительной степени зависит
от поддержания баланса мед<ду размерами и пропорциями раз-
ных его частей. Если бы скорости роста разных частей тела
контролировались независимо, то в результате обычного изме-
нения скорости роста, вызванного средой, возникали бы нежиз-
неспособные организмы.
Явление координации роста различных частей организма
крайне важно и с палеонтологической точки зрения. Такая ко-
ординация позволяет нам пренебречь фактором абсолютного
времени и изучать развитие организма и изменение его формы
безотносительно ко времени, не пытаясь определить продолжи-
тельность развития. Это положение иллюстрируется графиками,
приведенными на фиг. 19 и 20. В обоих случаях мы получили
немало сведений об онтогенезе организмов, не имея понятия об
абсолютной скорости роста.
Почти при любом палеонтологическом исследовании онтоге-
неза измеряется изменение одного морфологического признака
по отношению к изменению другого. Можно выделить два ос-
новных типа роста: изометрический и анизометрический. При
изометрическом росте соотношение между размерами двух час-
тей организма (X и У) в ходе онтогенеза не меняется (фиг. 23,
А и Б) в отличие от того, что наблюдается при анизометриче-
ском росте (фиг. 23, В и Г). Иными словами, при изометриче-
ском росте форма организма остается неизменной, тогда как
при анизометрическом она изменяется. На фиг. 23 показаны
две разновидности изометрического роста. При изометрическом
росте, описываемом графиком А, обе части растут с совершенно
одинаковой скоростью. В этом случае график роста имеет вид
прямой, наклоненной к обеим координатам под углом 45°. При
III. Онтогенетическая изменчивость
61
Фиг. 23. Типичные графики роста, получающиеся, если откладывать по осям
координат размеры двух морфологических признаков.
На каждом графике показана форма условного «организма» на трех стадиях онтогенеза
(прямоугольники различной формы). На графиках А и Б показан изометрический рост
(изменения формы не происходит на графиках В и Г рост анизометрический (форма по-
стоянно изменяется).
второй разновидности изометрического роста (фиг. 23, Б) одна
часть растет быстрее другой, но соотношение между ними оста-
ется постоянным; график имеет вид прямой, наклоненной к ко-
ординатам под разными углами. На графике В показан анизо-
метрический рост, при котором часть X сначала растет быстрее
части У, а затем начинает расти медленнее; график роста имеет
вид кривой. График Г отличается от остальных тем, что если
продлить его, то он не пройдет через начало координат. Здесь,
как и на графике В, форма в процессе роста изменяется, т. е.
рост анизометрический. В общем рост является изометрическим,
если его график представляет собой прямую, проходящую через
начало координат (или через начало координат проходит про-
62 Часть 1 Описание и классификация ископаемых остатков
должение этой прямой). Во всех других случаях мы имеем дело-
с анизометрическим ростом.
Анизометрический рост встречается гораздо чаще изометри-
ческого. Другими словами, изменение формы в процессе онтоге-
неза не исключение, а правило. Анизометрический рост интен-
сивно изучали биологи и палеонтологи. Получены некоторые
важные выводы, главный из которых состоит в том, что очень
часто анизометрический рост двух частей организма можно при-
близительно выразить следующим уравнением:
где а и b — константы и а^1. Рост, описываемый этим уравне-
нием, называется аллометрическим. (Изометрический рост по-
лучается при о=1.) Заметим, что некоторые авторы толкуют
термин «аллометрический» шире, включая сюда всякий анизо-
метрический рост. Аллометрическому росту можно было бы
посвятить несколько глав. Отсылаем читателя к работам Хакс-
ли, Томпсона и Гулда, указанным в списке литературы [66,
67, 86, 87, 215].
Причины анизометрического роста
До сих пор мы рассматривали анизометрический рост и свя-
занные с ним онтогенетические изменения формы просто как
биологический процесс, концентрируя внимание на определении
и описании этих явлений. Но интерпретация изменений формы
в ходе онтогенеза — более интересная и гораздо более важная
задача. Современные исследователи полагают, что изменения
формы в онтогенезе не являются делом случая, а закрепились
в наследственности благодаря своей функциональной ценности
Значит, нам надо искать функциональный или приспособитель-
ный смысл наблюдаемых изменений формы или числа структур
организма. Почему, например, меняется в процессе роста форма
раковины плеченогого?
Если имеет место резкое онтогенетическое изменение, как,
например, при метаморфозе у насекомых, то оно часто может
быть связано с резким изменением образа жизни. У лягушек
развитие до взрослой стадии идет довольно постепенно. Голо-
вастик живет в водной среде и зависит от нее: он извлекает
кислород непосредственно из воды. Взрослая лягушка более
или менее связана с водой, но все же ее можно считать назем-
ным животным. Одни онтогенетические изменения в организме
лягушки связаны с появлением новых структур и исчезновением
старых, другие — с изменением относительных скоростей роста
Ill Онтогенетическая изменчивость
63
У плеченогих дело обстоит иначе. И молодая, и взрослая
особи плеченогого живут почти в одной и той же среде. Обе
являются водными формами. Обе используют принципиально
одинаковые методы добывания пищи, их жизнедеятельность
в основном одинакова. Насколько мы знаем, в ходе онтогенеза
здесь изменяется главным образом размер. Следовательно, по-
лезно было бы познакомиться с последствиями увеличения раз-
мера.
Рассмотрим кости конечности наземного позвоночного. Кость
имеет несколько функций, но одна из самых главных — служить
опорой телу. Кости должны быть достаточно прочными, чтобы
выдерживать вес тела. Прочность кости (да и любой другой
опорной структуры) приблизительно пропорциональна площади
ее поперечного сечения. Значит, независимо от длины толстая
кость должна быть прочнее тонкой. Примем во внимание, что
кость должна расти, чтобы выдерживать увеличивающийся вес
тела. Предположим, в результате роста все размеры животно-
го— общая длина тела, длина кости, ее диаметр и т. д. — удво-
ились, т. е. мы приняли, что происходит изометрический рост.
Удвоение линейных размеров животного ведет к увеличению
объема или веса тела в восемь раз (в соответствии с геометри-
ческими законами при увеличении линейных размеров любой
структуры без изменения формы ее объем возрастает как третья
степень линейных размеров). Если линейные размеры кости,
поддерживающей вес тела, удвоились, то площадь ее попереч-
ного сечения возросла только в четыре раза (площадь растет
как квадрат линейных размеров). Следовательно, объем тела
животного неизбежно возрастает быстрее, чем площадь попе-
речного сечения кости, поддерживающей тело животного. Зна-
чит, и вес животного растет быстрее, чем прочность кости. Един-
ственный способ для организма преодолеть это противоречие
состоит в следующем: площадь поперечного сечения кости
должна увеличиваться относительно быстрее, чем это было бы
в том случае, если бы все линейные размеры кости возрастали
в постоянном отношении. А это означает изменение формы кос-
ти. Чтобы выдержать возросший вес тела, кость должна стать
относительно толще. Это и происходит в процессе онтогенеза
многих наземных позвоночных.
Различия в форме опорных структур у видов, различающих-
ся размерами, часто можно бывает предсказать. Одним из пер-
вых это отметил Галилей, сформулировав принцип подобия,
который он пояснил приведенными здесь рисунками (фиг. 24).
Галилей писал:
«В качестве краткого примера только что сказанного я по-
кажу вам сейчас рисунок кости, удлиненной только в три раза,
но увеличенной в толщину в такой мере, чтобы она могла слу-
64
Часть 1. Описание и классификация ископаемых остатков
Фиг. 24. Рисунок из книги Галилея, поясняющий связь между размером и
формой костей позвоночных [16].
Размер кости растет, чтобы дать опору увеличивающейся массе животного; при этом дол-
жно возрастать отношение диаметра кости к ее длине, чтобы прочность кости была доста-
ТОЧНОЙ
жить для большого животного с той же надежностью, как мень-
шая кость служит для животного малого размера»1.
Можно расширить описанное правило, рассмотрев другие
структуры. Например, эффективность многих органов дыхания
зависит от площади поверхности газообмена. Если бы легкие
взрослого человека были просто увеличенной копией легких ре-
бенка, то эффективность дыхания взрослого была бы сильно
понижена. Требования взрослого организма к дыхательному
аппарату возрастают пропорционально третьей степени линей-
ных размеров, а поверхность легких (если не меняется их фор-
ма)— пропорционально квадрату линейных размеров. Поэтому
в онтогенезе человека весьма интенсивно развивается легочная
ткань, вследствие чего увеличивающаяся поверхность газообме-
на не отстает от роста всего тела (объема тела).
Классический пример соотношения между площадью поверх-
ности и объемом тела дают насекомые. Если бы можно было
сделать так, чтобы комар вырос до трех метров в длину, оста-
ваясь точной копией обычного комара, то он погиб бы, так как
соотношение между его поверхностью и объемом при этом сде-
лало бы невозможным терморегуляцию и дыхание. Чтобы ко-
мар таких размеров был жизнеспособным, его морфология в
онтогенезе должна была бы значительно измениться.
Вернемся теперь к структурным изменениям, которые могут
быть отражены в палеонтологической летописи. На фиг. 25 по-
казано поперечное сечение раковины двустворчатого моллюска.
Она растет путем аккреции, примерно так же, как раковины
брюхоногих и головоногих моллюсков. Как мы узнаем из после-
1 Цитируется по изданию: Галилео Галилей, Избранные труды в двух
томах, изд-во «Наука», М., 1964.— Прим, перев.
III. Онтогенетическая изменчивость
65
Фиг, 25. Поперечное сечение раковины двустворчатого моллюска иллюстри-
рует способ роста путем аккреции [149].
Раковина постоянно утолщается, так что ее прочность не отстает от увеличения общего
размера.
дующих глав, по своей геометрии раковина двустворчатых не
слишком сильно отличается от раковин других групп рассмот-
ренных нами моллюсков. Отдельную створку такой раковины
можно рассматривать как полую трубку, сужающуюся от широ-
кого открытого конца к узкому «пупку», или «макушке». Основ-
ное различие в том, что раковина двустворчатых моллюсков
сужается гораздо резче, чем раковины брюхоногих и голово-
ногих.
Как на поперечном сечении, так и на внешней поверхности
раковины двустворчатого моллюска видны линии нарастания.
Они отмечают поверхности, на которых наращивался новый ма-
териал. Новое вещество появлялось не только по внешнему
краю раковины, но и почти на всей внутренней поверхности.
Основной смысл утолщения — это упрочнение раковины, при-
способление ее к нуждам взрослого животного. У брюхоногих
и головоногих моллюсков гораздо реже происходит утолщение
имеющейся раковины. Они обходятся без этого, так как поверх-
ность их раковин искривлена гораздо сильнее, благодаря чему
раковина приобретает особую прочность в отличие от менее
искривленных раковин двустворчатых моллюсков.
Можно было бы привести много других примеров влияния
размера на онтогенетические изменения у животных, хорошо
представленных в палеонтологической летописи. Однако многое
остается неясным. Так, мы не можем предложить удовлетвори-
тельного объяснения рассмотренному ранее изменению формы
раковин брахиопод. Может быть, оно связано с размером.
Неполнота наших знаний о жизнедеятельности многих вымер-
ших групп организмов не позволяет интерпретировать наблю-
даемую онтогенетическую изменчивость. Новые сведения по-
могли бы понять не только различия между стадиями роста
одного вида, но и различия между видами.
Сказанное можно резюмировать следующим образом. Онто-
генетическое изменение формы сопровождается изменением об-
5—467
66
Часть 1 Описание и классификация ископаемых остатков
раза жизни и изменением размеров. Размеры обычно меняются
постепенно, как и форма. Образ жизни изменяется иногда по-
степенно, а иногда резко. Онтогенетическое изменение формы
брахиопод может определяться либо постепенным кумулятив-
ным изменением размера, либо каким-то (пока неизвестным)
незначительным изменением образа жизки.
Некоторые особенности онтогенеза
колониальных организмов
До сих пор мы рассматривали организмы, у которых отдель-
ные особи живут самостоятельно. Из колониальных организмов
в ископаемом состоянии чаще всего встречаются кишечнополост-
ные и мшанки.
У колониальных кораллов отдельные кораллиты часто обра-
зуют общий скелет. Влияние такого образа жизни хорошо за-
метно, если сравнить особь из центра колонии с особью с пери-
ферии. Онтогенез особей, расположенных на периферии, вслед-
ствие меньшей скученности может сильно отличаться от
развития особей, сжатых со всех сторон другими. (Иногда сход-
ное явление отмечается у неколониальных организмов, напри-
мер у устриц, живущих иногда так тесно, что рост каждой
особи влияет на форму соседей.)
У колониальных кораллов основатель колонии должен при-
крепиться к какому-то твердому субстрату, например к камню,
пустой раковине другого организма или даже к мертвому корал-
лу. При росте колонии новые особи прикрепляются к старым ее
частям. Ход онтогенеза каждой особи сильно зависит от того,
на какой стадии онтогенеза всей колонии он имеет место.
В некоторых колониях (в том числе у кораллов и мшанок)
существует морфологическая и физиологическая дифференциа-
ция особей, т. е. некоторые особи приспособлены для выполне-
ния определенных специализированных функций, скажем раз-
множения или защиты, и их деятельность идет на пользу всей
колонии. Такие особи могут генетически отличаться от других,
но могут и не иметь генетических отличий. Они могут отличать-
ся от остальных особей так сильно, что их онтогенез позволи-
тельно рассматривать отдельно от онтогенеза колонии.
ГЛАВА IV
Популяция как целое
Все виды животных и растений обладают изменчивостью;
даже между генетически идентичными близнецами имеются
небольшие различия, объясняющиеся различиями внешней сре-
ды и случайностями. Из-за существования изменчивости опи-
сание вида нельзя основывать на описании одной особи.
Популяции в биологии
Для большинства видов характерно прерывистое и неравно-
мерное географическое распространение. Разного рода случай-
ности, неравномерное распределение подходящих местообита-
ний, стадное поведение и географические преграды обусловли-
вают образование разобщенных групп. На фиг. 26 показано
прерывистое распределение одного вида наземных растений.
Для наших целей можно определить популяцию как группу
особей, живущих достаточно близко друг от друга, так что
каждая особь имеет примерно одинаковые шансы скреститься
с какой-то особью другого пола. (Сходным образом часто опре-
деляют термины локально скрещивающаяся популяция и дем.)
Популяция обладает общим генофондом. Генофонды соседних
популяций вида могут быть частично или полностью изолиро-
ваны. Если две популяции скрещиваются, то говорят, что между
ними существует поток генов.
Разделение видов на отдельные популяции наглядно выра-
жено на островах Вест-Индии и других группах островов. При-
уроченность к островам неизбежно делит наземные виды на
генетически изолированные популяции. Подобным же образом
многие морские организмы связаны с мелководьем и потому
изолированы от своих сородичей, живущих вокруг других остро-
вов. Это крайние случаи, но сходную картину распределения
организмов можно обнаружить во всей биосфере. Кроме того,
Даже если географические преграды отсутствуют, удаленность
между разными частями ареала прерывает поток генов.
5*
Фиг. 26. Пятнистое распределение растений Clematis fremontii.
Точки \внизу слева) — отдельные растения Заметьте, что пятнистость распределения
проявляется в разных масштабах (крупные скопления состоят из мелких).
IV Популяция как целое
69
Индивидуальная изменчивость
внутри популяций
OHTOIЕНЕТИЧЕСКИЕ РАЗЛИЧИЯ
В предыдущей главе мы рассматривали изменения, происхо-
дящие с отдельной особью в процессе онтогенеза. Очевидно,
что в любой популяции в каждый данный момент существует
изменчивость, обусловленная возрастными различиями Особи
популяции должны различаться по крайней мере своей вели-
чиной
У видов с достаточно хорошо выраженной сезонностью раз-
множения изменчивость размеров может сильно зависеть от
времени года На фиг 27 показано распределение частот (гисто-
граммы) размеров особей в живущей популяции двустворчатого
моллюска Mytilus eduhs в апреле и ноябре одного года Раз-
множение этого вида происходит в течение короткого периода
Апрельская гистограмма составлена на материале пробы, взя-
той сразу после интенсивного оседания личинок, и на ней ясно
можно видеть две группы пиков (бимодальное распределение)
мелкие, только что осевшие особи дают резкий пик слева, а ос-
тальная популяция (состоящая из особей однолетнего возраста
и более) дает широкую группу пиков справа. Две группы
пиков имеются и на ноябрьской гистограмме, но здесь оба пика
сдвинуты вправо, поскольку большинство особей в популяции
увеличилось в размерах Левый пик стал значительно ниже, так
Фиг. 27. Зависимость между возрастом и размером в популяции [35].
Распределения длины раковины в популяции Mytilus eduhs из Госфорд Бэй, Шотландия
Из популяции брали пробу в апреле 1961 г (заштрихованная диаграмма) и в ноябре того
же года (незаштрихованная диаграмма) Пики слева на обеих диаграммах соответствуют
более молодым моллюскам
70
Часть 1 Описание и классификация ископаемых остатков
Фиг. 28. Pyrgo ftscheri (рецептный вид) [51].
А . Вид со стороны апертуры Б Вид сбоку В Экваториальное сечение мегасфериЧеской
(возникающей при бесполом размножении) раковины Г Экваториальное сечение микро-
сферической раковгны (половое поколение) На В и Г некоторые камеры пронумерованы.
как некоторые из молодых особей погибли, и шире, так как
скорость роста особей различна.
Средний возраст популяции может влиять не только на из-
менчивость размеров. В предыдущей главе мы говорили о пле-
ченогом, у которого форма раковины систематически изменялась
в течение жизни (фиг. 19). Всякое описание формы особей в по-
пуляции будет сильно зависеть от соотношения особей разного
возраста в данной выборке.
Некоторые таксономические группы характеризуются слож-
ным жизненным циклом, дающим разные типы онтогенетической
изменчивости. Например, большинство видов фораминифер име-
ет жизненный цикл, в котором половое размножение сменяется
бесполым. Поэтому многие виды диморфны. Формы, появившие-
ся в результате полового размножения, обычно крупнее, хотя
они вырастают из очень маленькой зародышевой камеры. Осо-
би, появившиеся путем бесполого размножения, мельче, но раз-
виваются из более крупной зародышевой камеры. На фиг. 28
показаны обе формы распространенного кайнозойского рода
Pyrgo. Некоторые камеры пронумерованы, чтобы облегчить
сравнение. Диморфизм особенно сильно выражен у крупных
видов фораминифер. У них форма, появившаяся в результате
полового размножения, может быть крупнее другой в пять раз.
НАСЛЕДСТВЕННЫЕ РАЗЛИЧИЯ
У большинства раздельнополых организмов особи противо-
положного пола различаются морфологически. Это — следствие
основных генетических различий. На скелетах одних организ-
IV Популяция как целое
71
Род Kosmoceras
К (Zugokosmoceras) grossouvrei
К (Lobokosmokeras) phaeinum
К (Guhelmiceras) at) guhelmi
К. (Spinikosmokeras) acutistnatum
Фиг/29. Половой диморфизм — выражение генетической изменчивости [21].
Аммониты из ('реднеюр''ких отложений Англии обладают различиями, которые интерпре-
тируются как свидетельства полового диморфизма Каждая крупная («макроконховая»)
форма считается диморфом мелкой («микроконховой»), изображенной под ней Названия
были даны этим формам еще до того, как был понят смысл морфологических различий
между ними Все формы были отнесены к одному роду, но к разным подродам (их назва-
ния даны в скобках) и видам.
мов половой диморфизм не отражается, тогда как у других
организмов он отчетливо выражен. На фиг. 29 показан пример
предполагаемого полового диморфизма у различных аммони-
тов. Раковины обоих полов различаются размерами, внешней
скульптурой и формой устья. Различия здесь примерно столь
же велики, как между разными видами и родами, и потому
особи разного пола могут быть ошибочно отнесены к разным
видам. Доказывая, что данные окаменелости представляют со-
бой разнополые особи одного вида, палеонтолог должен учиты-
вать такие признаки, как совместная встречаемость и относи-
тельная численность обеих форм.
Даже особи одного пола и одной возрастной стадии разли-
чаются между собой. Это связано с наследственной изменчи-
востью. Каждая особь несет лишь малую часть общего генофон-
да. Некоторые генетически контролируемые признаки организма
варьируют сильнее других. Например, в популяциях человека
72
Часть 1 Описание и классификация ископаемых остатков
число пальцев на руках и ногах практически постоянно, тогда
как другие наследственные признаки — цвет волос, размеры
тела, отпечатки пальцев и другие — отличаются сильной измен-
чивостью.
Наследственная изменчивость в популяции может быть не-
прерывной или прерывистой, т. е. она может выражаться в пол-
ном спектре промежуточных перекрывающихся форм или же
в дискретных и резких различиях. Пример прерывистой измен-
чивости— направление навивания раковины у некоторых мол-
люсков и фораминифер. Раковина может быть левозавитой (си-
нистральной) или правозавитой (декстральной). Особи с разным
направлением навивания, принадлежащие к одному виду, обыч-
но идентичны, с той разницей, что каждая из них представляет
собой зеркальное изображение другой. Оба направления нави-
вания могут встречаться в одной скрещивающейся популяции.
На фото XII показан пример прерывистой изменчивости,
взятый из палеонтологической летописи. У силурийских трило-
битов, считающихся одним видом, варьирует расстояние между
бугорками на хвостовом щите (пигидии). Бугорки находятся
на выпуклых кольцах, составляющих осевую часть пигидия.
В материале, изученном Бестом [13], первый и третий бугорки
(считая от переднего края пигидия) пигидия трилобитов раз-
делены шестью, семью или восемью кольцами. Бест счел эту
изменчивость наследственной (возможно, этим признаком управ-
лял один ген, имевший два аллеля). На основании соотношения
особей с разным расположением бугорков он также вывел за-
ключение, что изученные им трилобиты являются случайной
выборкой из одной популяции.
Генетически контролируемая изменчивость гораздо чаще бы-
вает непрерывной. Особенно распространена непрерывная из-
менчивость размеров тела особей одной возрастной стадии и
одного местообитания. Однако причиной ее могут быть не только
изменения наследственного материала, но также случайные
события, влияющие на рост, и неявные различия условий среды.
Насколько широким может быть диапазон наследственной
изменчивости внутри одной популяции? Ясный ответ на этот
вопрос был бы очень полезен, особенно для палеонтолога, кото-
рый знал бы, какую изменчивость он может встретить в иско-
паемой популяции; более широкий диапазон изменчивости сви-
детельствовал бы, что мы имеем дело с остатками не одной
популяции. К сожалению, различия в размере и сложности
генофондов делают строгое предсказание пределов изменчиво-
сти трудным, а то и невозможным. Кроме того, проблема услож-
няется еще и тем, что о генетических различиях между иско-
паемыми популяциями можно судить только по их морфологи-
ческому проявлению.
IV. Популяция как целое
73
Диапазон изменчивости в популяции варьирует не только
от вида к виду, но и внутри вида в зависимости от времени
и места. Ниже мы приводим некоторые из факторов, влияющих
на диапазон изменчивости.
Давление отбора. Наследственная изменчивость популяции
всегда подвержена естественному отбору. Всегда какие-то вари-
анты лучше других приспособлены к определенной среде, и они
дадут более жизнеспособное потомство; поэтому состав генофон-
да отражает сравнительную приспособленность разных вариан-
тов. Там, где среда «терпит» лишь немного вариантов, существу-
ет, как мы говорим, высокое давление отбора.
Мутационное давление. Первичный источник почти всякой
наследственной изменчивости — генная мутация. Частота мута-
ций различна у разных видов и в разных популяциях. При про-
чих равных условиях изменчивость выше в той популяции, в ко-
торой выше частота мутаций.
Способ размножения. У организмов, всегда размножающих-
ся бесполым путем (простым делением или почкованием), сме-
шение генетического материала внутри популяций невелико и
низок потенциал морфологической изменчивости. Как мы виде-
ли, половое размножение может чередоваться с бесполым. Та-
кая смена способов размножения широко распространена у рас-
тений. Она приводит к групповой изменчивости, при которой
особи одной группы очень схожи друг с другом, но явно отлича-
ются от особей других групп.
Мы упомянули лишь некоторые факторы, влияющие на на-
следственную изменчивость в популяциях. Предсказать диапа-
зон изменчивости в популяции нельзя, но палеонтолог или био-
лог, работающий с какой-то определенной группой организмов,
привыкает к диапазону изменчивости, свойственной популяциям
этой группы, и научается сразу распознавать отклонения от
обычной изменчивости.
НЕНАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ
В гл. III говорилось о «норме реакции». Мы видели, что ге-
нетические механизмы не полностью подчиняют себе морфоло-
гию организма. Условия среды и случайности также могут вы-
зывать изменения.
Ненаследственная изменчивость связана главным образом
о экологическими различиями. Крайние примеры прерывистой
ненаследственной изменчивости встречаются у общественных
насекомых, например у пчел и термитов. У этих насекомых раз-
личия между рабочей особью и царицей могут возникать просто
из-за того, что одна особь на ранней стадии развития получала
больше пищи (или иную пищу), чем другая.
74
Часть 1 Описание и классификация ископаемых остатков
Для раковин многих беспозвоночных, обитающих в расще-
линах или полостях твердого субстрата, характерна непрерыв-
ная ненаследственная изменчивость. Раковины таких видов при-
нимают форму, близкую к форме соответствующих пустот.
Некоторые ненаследственные изменения, возникшие под
влиянием среды, имеют функциональное значение и могут быть
полезными для организма, тогда как другие, по-видимому, со-
вершенно не связаны ни с какой функцией. Пример первого
случая — развитие мозолей на руках и ногах приматов. Мо-
золь— приобретенный признак, возникающий в результате взаи-
модействия между живой тканью и внешней средой. Часто мо-
золь полезна организму, защищая кожу от истирания и
ушибов.
Наследственные признаки организма (или популяции орга-
низмов) редко можно наблюдать в чистом виде. Отделить на-
следственные признаки от ненаследственных особенно трудно1
потому, что иногда одни и те же морфологические варианты
могут появляться как под действием условий среды, так и под
действием генетических факторов. Например, у разных видов
иглокожих содержание в скелете изотопов 16О и ,8О может
сильно различаться. Эти различия могут быть эволюционными
(наследственными), но не меньшие различия обусловлены влия-
нием разной температуры воды.
Ископаемая популяция
В процессе фоссилизации органических остатков картина
изменчивости популяции искажается. Основной геологический
процесс, увеличивающий видимую изменчивость, — это уплотне-
ние осадков или деформация осадочных пород. Обычно гораздо
чаще в результате фоссилизации происходит уменьшение из-
менчивости В основном это связано с тем, что при фоссилиза-
ции многие признаки организма уничтожаются и различия,
бывшие очень резкими в живой популяции, сглаживаются. По-
этому редко ископаемая популяция обладает такой же сильной
изменчивостью, как близкая к ней ныне живущая.
Палеонтолог не надеется найти сохранившиеся мягкие ткани
организма; он знает, что информация о них отсутствует и что
с таким положением дел просто приходится мириться. Поэтому
важнее обратить внимание на искажающее влияние фоссилиза-
ции Процесс фоссилизации как бы фильтрует изменчивость:
хорошо сохраняющиеся организмы составляют непропорцио-
нально большую часть палеонтологической летописи. В общем
отдельные особи с прочным и твердым скелетом сохраняются
лучше, чем особи того же вида, скелеты которых оказались бо-
лее легкими и хрупкими. Если забыть об этом, то можно при-
IV Популяция как целое
75
Станция Общее число Доля правых створок,%
1 45 11
2 154 13
3 11 18
4 51 Около 50
5 73 Около 50
6 284 46
7 27 Около 50
8 450 87
Фиг. 30. Сортировка створок пелеципод при посмертном переносе [124].
Соотношение правых и левых створок в разных местах на восточном берегу Тринидада
различно, однако здесь наблюдается определенная система Обе створки раковины Pitar
dione одинаковы, за исключением того, что одна из них — зеркальное изображение
другой
писать обнаруженному виду скелет более мощный, чем было
в действительности. Вполне обычно также избирательное раз-
рушение некоторых стадий онтогенеза. Так, в палеонтологиче-
ской летописи четвертичного периода (за исключением погре-
бений) встречается мало детских черепов, так как они отлича-
ются хрупкостью. Явление избирательного сохранения чрезвы-
чайно сложно и до сих пор до конца еще не изучено. Это одна
из самых многообещающих областей палеонтологического ис-
следования. Шансы на сохранение малы, и сохранение именно
данной особи может зависеть от незначительного морфологиче-
ского отличия.
Сортировка во время посмертного переноса также сильно
влияет на состав ископаемого сообщества. Поразительный при-
мер этого влияния, особенно ценный с палеонтологической точки
зрения, показан на фиг. 30. П. Мартин-Кэй [124] изучал рако-
вины двустворчатых моллюсков, собранные на пляже Тринида-
да. Эти организмы ведут роющий образ жизни в прибрежных
биотопах. К своему удивлению, исследователь обнаружил, что
на каждом ограниченном участке пляжа число левых и правых
створок не совпадает. Далее, оказалось, что соотношение между
числом левых и правых створок довольно систематически меня-
76
Часть 1. Описание и классификация ископаемых остатков
ется на протяжении одного пляжа. Все виды, изучавшиеся
Мартин-Кэем, имеют практически неразличимые створки, яв-
ляющиеся просто зеркальным изображением одна другой. Ти-
пичные створки одной особи показаны на фото XIII. Обнару-
женный характер сортировки легко объяснить, если допустить,
что действие волн не единственный фактор, влияющий на пере-
нос раковин. Так как волны обычно подходят к пляжу под
углом и создают направленное вдоль берега течение, раковины
чаще отлагаются на берегу не прямо напротив местообитания
живого организма, а подхватываются течением, несутся вдоль
берега и наконец либо разрушаются в результате истирания,
либо выбрасываются на берег в некотором отдалении от место-
обитания. Поскольку показанные на фото XIII створки пред-
ставляют собой зеркальные изображения друг друга, волны
переносят их по-разному.
Эффект дифференцированного переноса тринидадских рако-
вин поразителен, и его интерпретация довольно ясна, так как
мы знаем, что в исходной популяции число левых и правых
створок было одинаковым. Если бы мы сравнили раковины,
собранные на пляже, с раковинами исходных популяций, то мы,
очевидно, нашли бы и другие проявления связи между морфо-
логией и переносом.
Искажения, внесенные в палеонтологическую летопись диф-
ференцированным переносом, трудно оценить. У некоторых
групп, например у нектонных головоногих моллюсков, посмерт-
ный перенос является скорее правилом, чем исключением. Но
мы не знаем, насколько важную роль играет такой перенос
среди разных типов растений и животных, нормально обитаю-
щих на твердом субстрате. Недавние работы океанографов и
морских геологов доказали существование крупных течений,
способных переносить на значительные расстояния некоторые
донные организмы.
Для оценки дальности переноса раковин донных морских
организмов, обладающих двумя створками, обычно применяют
метод, основанный на подсчете процента разделившихся на
створки раковин. Например, у некоторых плеченогих после
смерти раковины разделяются легче, чем у других. По морфо-
логическим признакам часто удается оценить характер и ин-
тенсивность волнения и переноса, в результате действия кото-
рых раковины данного вида распались на створки. По степени
разрушения и износа створок также можно оценить действие
этих факторов.
Так, Буко, Брас и Демер [17] изучали влияние переноса на
сохранность раковины у трех родов нижнедевонских брахиопод.
Два рода были представлены почти равным числом брюш-
ных и спинных створок (Leptocoelia — 879 брюшных и 893 спин-
IV. Популяция как целое
77
ных створки, Platyorthis— 561 брюшная створка и 548 спин-
ных). Третий род, Leptostrophis, был представлен 378 брюш-
ными створками и только 35 спинными. Авторы сделали вывод,
что раковины первых двух родов не переносились далеко от
места обитания животных, тогда как особи, относящиеся к
третьему роду, очевидно, жили где-то далеко и после смерти
их раковины переносились донными течениями. Из двух родов,
захороненных вблизи их местообитания, раковины Leptocoelia
гораздо чаще бывают неповрежденными, чем раковины Platyor-
this. Авторы заключили из этого, что раковины Leptocoelia
расчленялись не так легко, как раковины Platyorthis.
Динамика популяций
Изучение динамики популяций — это изучение изменений
размера и состава популяций во времени. В основном учитыва-
ются рождаемость, скорость роста и смертность. У большинства
групп животных размножение обычно связано с определенной
стадией онтогенеза, начинаясь после наступления половой зре-
лости и оканчиваясь со смертью или незадолго до смерти. Пло-
довитость (скорость размножения) индивидуума может менять-
ся в течение репродуктивной стадии или оставаться постоянной.
Рост обычно сначала идет быстро, потом замедляется; затем
он или резко прекращается (у человека), или постепенно за-
медляется, продолжаясь до самой смерти (у большинства бес-
позвоночных). Смертность часто бывает высока в начале жизни
(у человека); позднее она часто опять возрастает — резко (у че-
ловека) или постепенно (у многих беспозвоночных).
Биологи, изучающие популяции, часто используют так назы-
ваемую кривую выживаемости — график зависимости числа осо-
бей от их возраста. График может относиться к одной или
нескольким популяциям вида. Обычно данные откладываются
в логарифмической шкале, поскольку тогда смертность дает
прямую линию (фиг. 31, построенная для гипотетической попу-
ляции, в которой ежегодно умирает 90% особей).
Палеонтолог, пытающийся изучать динамику популяций,
прежде всего должен удостовериться в том, что найденное иско-
паемое сообщество действительно представляет собой бывшую
популяцию. Как показано в предыдущем разделе, проблема
эта нелегка. Дело не только в том, что процессы разрушения
и сортировки изменяют картину. Часто бывает трудно опреде-
лить, одновременно ли жили все особи данного сообщества.
Раньше многие палеонтологи полагали, что простой график
«размер—частота» позволяет отличить измененное ископаемое
сообщество раковин или скелетов от неизмененного. Считалось,
что высокая смертность на очень ранних стадиях онтогенеза
78 Часть 1. Описание и классификация ископаемых остатков
Фиг 31. График выживаемости при
постоянной смертности 90% в год.
В полулогарифмическом масштабе можно
получить график в виде прямой линии.
характерна для большинства животных и что накопление ра-
ковин или скелетов в условиях, когда отсутствует избиратель-
ный перенос и разрушение (или когда действие этих факторов
незначительно), должно создавать ископаемые сообщества, в
которых мелких особей значительно больше, чем крупных. Сле-
довательно, зависимость между размером и частотой для такого
сообщества должна быть сходна с графиком, приведенным на
фиг. 32, А, а колоколообразная кривая нормального распреде-
ления размеров (фиг. 32, Б) должна свидетельствовать о том,
что при переносе шла сортировка, причем средний размер опре-
делялся такими факторами, как скорость транспортирующих
течений.
Фиг. 32. Сдвинутая влево (Л) и нормальная (Б) кривые распределения раз-
меров в ископаемом комплексе, состоящем из одного вида.
Раньше считалось, что первая кривая говорит о наличии незначительного переноса или
сортировки, а вторая — о сортировке течениями.
IV. Популяция как целое
79
Фиг. 33. Получение сдвинутых влево и нормальных кривых распределения
размеров при наложении разных величин смертности на одну кривую
роста [35].
А. Типичная кривая роста Cardium edule (сердцевидки съедобной) Б. Четыре возможных
кривых выживаемости В. Кривая распределения размеров раковин мертвых сердцеви-
док, основанная на кривой роста А и кривоп выживаемости // (постоянная смертность) Г.
Кривая распределения размеров раковин, основанная на кривой роста А и кривой выжи-
та эмости 1IJ (смертность постоянно возрастает).
Но Крейг и Халлам [35] указали, что у большинства мор*
ских донных беспозвоночных ранняя смертность падает в ос-
новном на планктонные личиночные стадии, захороняемые от-
дельно от взрослых. Значит, у них постларвальные ископаемые
сообщества не отражают высокую раннюю смертность. У боль-
шинства видов смертность возрастает в постларвальный период
жизни. Крейг и Халлам показали, что, рассматривая дан-
ные измерений скорости роста Cardium edule (сердцевидки
съедобной) и различные возможные кривые выживаемости, па-
леонтолог может теоретически рассчитать возможные ископае-
80
Часть 1. Описание и классификация ископаемых остатков
мне сообщества с разными кривыми распределения размеров.
На фиг. 33 показана нормальная кривая роста Cardium (Д),
четыре возможные кривые выживаемости (Б) и кривые распре-
деления размеров (В и Г), полученные путем комбинирования
кривой роста с различными кривыми выживаемости — при по-
стоянной смертности (кривая выживаемости II) и при постоян-
но возрастающей смертности (кривая выживаемости III).
Другими словами, для отдельной популяции или отдельного
вида в зависимости от скорости роста и выживаемости можно
получить несколько возможных распределений размера. Распре-
деления на фиг. 33, В и Г сходны с кривыми, приведенными
на фиг. 32; ясно, что колоколообразная кривая нормального
распределения не обязательно говорит о сортировке течениями,
а асимметричная кривая не обязательно указывает на высокую
смертность на ранних стадиях.
Важно помнить, что кривые роста и выживаемости варьиру-
ют в разных популяциях одного вида и с течением времени
внутри одной популяции. Изменения кривой выживаемости вида
обусловлены главным образом воздействием факторов внешней
среды. Однако наличие множества неизвестных переменных,
участвующих в процессе образования ископаемого сообщества,
нередко очень затрудняет анализ динамики ископаемой попу-
ляции.
Для изучения динамики популяций ископаемых таксонов
самое главное — суметь выделить внутри вида возрастные груп-
пы, основываясь на таких признаках, как стертость зубов
(у млекопитающих), кольца роста (на чешуйках рыб, пластин-
ках иглокожих и раковинах моллюсков) и стадии линьки (чле-
нистоногие). По некоторым из этих признаков организмы четко
делятся на годичные классы. Так, кольца роста обычно появля-
ются в результате очень медленного зимнего роста, а стадии
линьки иногда связаны с ежегодным сбрасыванием наружного
скелета. Если нет четких структур, отмечающих ход роста,
выделить возрастные группы иногда удается по графикам рас-
пределения размеров (фиг. 27).
Куртен [109] предложил два подхода к изучению динамики
популяций применительно к ископаемым сообществам. Первый
подход применим к таким ископаемым сообществам, которые
состоят из остатков организмов, погибших от естественных при-
чин (естественная смертность) за какой-то период. Куртен изу-
чал остатки плейстоценовых европейских пещерных медведей,
популяциям которых была свойственна повышенная смертность
в период зимней спячки. Поэтому скелеты скапливались в пе-
щерах дискретными годичными группами. Важно, что при ис-
пользуемом здесь динамическом подходе не имеет значения,
оставлена ли группа сохранившихся скелетов животными, ро-
IV. Популяция как целое
81
дившимися в один год или за несколько лет. Главное, чтобы
в популяции скорость роста и выживаемость мало изменялись
во времени.
Другой подход применим к сообществам, образовавшимся
в результате внезапной гибели всей популяции (массовая
смертность). Это, по Куртену, временной подход. Катастрофи-
ческие события (например, бури) могли привести к гибели
целых популяций; иногда в палеонтологической летописи уда-
ется распознать ископаемые сообщества, возникавшие подобным
образом.
Анализируя ископаемое сообщество, важно установить, ка-
кой из двух подходов — динамический или временной — приго-
ден в данном случае. Попробуем приложить оба подхода к
данным по силурийской остракоде Beyrichia jonesi, опублико-
ванным Спьельднесом [204].
На фиг. 34, А, показано распределение размеров в коллек-
ции 972 створок из мергелей Мульде с острова Готланд. Рако-
вина остракод имеет две створки. Найдя целую раковину,
Спьельднес считал ее за две створки. К сожалению, он не со-
общает о том, сколько было левых и сколько правых створок,
но нераспавшихся раковин было 41. Как и другие членистоно-
гие, остракоды растут путем линьки, поэтому их скелеты должны
группироваться по признаку размера. Первые три линочные ста-
дии Beyrichia jonesi очень малы; на фиг. 34 они не отражены
вследствие их плохой сохранности. Как правило, сброшенную
при линьке створку или раковину невозможно отличить от
створок и раковин, опустевших после смерти рачков. Сообщество
из мергелей Мульде отличается от других ископаемых сооб-
ществ этого вида гораздо лучшей сохранностью. Заметим, что
на фиг. 34, А каждый следующий пик ниже предыдущего. Ви-
димо, это сообщество образовалось из локальной популяции
при постепенном накоплении сброшенных и опустевших из-за
смерти рачка раковин. Значит, здесь пригоден динамический
подход.
В табл. 1 в двух левых столбцах представлены данные, на
основе которых построен график 34, А. Если мы примем, что
сохранились почти все створки, то полных раковин было вдвое
меньше; их число в каждой размерной группе указано в треть-
ем столбце. Прежде всего встает вопрос, какая доля раковин
сброшена при линьке, а какая опустела после смерти. Рассмот-
рим возрастной класс 4, в котором 155 раковин (столбец 3).
Можно считать, что каждый рачок, переживший возрастную
стадию 4, оставил раковину возрастного класса 5, либо перели-
няв (если он перешел в класс 6), либо погибнув. Значит,
105 раковин возрастной стадии 5 представляют всех сохранив-
шихся особей, прошедших линьку на стадии 4. На стадии 5
Фиг. 34. Кривая выживаемости остракод Beyrichia jonesi, (мергели Мульде,
(силур, о-в Готланд, Швеция).
А. Распределение размеров в коллекции из 972 створок 1204]. Пронумерованные пики
соответствуют поспедовательныи стадиям линьки. Б. Кривые выживаемости, построен-
ные на основании графика А ([110], с изменениями). Сплошная кривая — результат ди-
намического анализа при условии, что сохранились все сброшенные при линьке раковины.
Пунктирная кривая — результат динамического анализа при условии, что сброшенные
при линьке раковины не сохранились.
ТАБЛИЦА 1
Таблицы жизненного цикла силурийской остракоды Beyrichia jonesi
Подход Возраст (число линек) Число створок Соответст- вующее число це- лых рако- вин Число перели- нявших особей Число по- гибших за возрастную стадию Число вы- живших к началу воз- растной стадии Число вы- живших к началу ста- дии из 1000 живых осо- бей на пер- вой стадии
Динамический; 4 309 155 105 50 155 1000
принимается, что все сбро- 5 210 105 73 32 105 677
шенные при 6 146 73 46 27 73 471
линьке ракови- ны сохрани- 7 92 46 38 8 46 297
лись 8 75 38 35 3 38 245
9 70 35 21 14 35 226
10 42 21 10 11 21 135
11 19 10 0 10 10 65
Временной 4 309 155 0 50 155 1000
5 210 105 0 32 105 677
6 146 73 0 27 73 471
7 92 46 0 8 46 297
8 75 38 0 3 38 245
9 70 35 0 14 35 226
10 42 21 0 11 21 135
11 19 10 0 10 10 65
Д инамический; 4 309 155 0 155 483 1000
принимается, что сброшен- 5 210 105 0 105 328 679
ные при линь- 6 146 73 0 73 223 462
ке раковины не сохранились 7 92 46 0 46 150 311
8 75 38 0 38 104 215
9 70 35 0 35 66 136
10 42 21 0 21 31 64
11 19 10 0 10 10 21
-
6*
84
Часть 1 Описание и классификация ископаемых остатков
перелиняли 73 особи. Так можно заполнить весь столбец 4.
Число погибших в каждой возрастной стадии (столбец 5) по-
лучаем, вычитая число перелинявших особей (столбец 4) из
общего числа раковин данной стадии (столбец 3). Общее число
мертвых особей в пробе составляет 155; это число двустворча-
тых раковин. Вычитая из него 50 — число особей, погибших на
стадии 4, — получаем 105 — число особей, доживших до ста-
дии 5. Так можно рассчитать все цифры столбца 6. Для построе-
ния кривой выживаемости удобно рассчитать число выживших
на 1000 особей, существовавших в начале первой рассматри-
ваемой возрастной стадии, как это сделано на графике, при-
веденном на фиг. 31. Данные такого пересчета представлены
в столбце 7. Построенная на их основе кривая выживаемости
показана на фиг. 34, Б сплошной линией.
Геологические свидетельства того, что рассмотренное сооб-
щество Beyrichia возникло в результате массовой смертности,
отсутствуют.
Но для сравнения построим теперь кривую выживаемости
по данным фиг. 34, А, пользуясь временным подходом. Предпо-
ложим, что все сохранившиеся раковины принадлежат внезапно
погибшим животным, а сброшенных при линьке раковин нет.
Следовательно, мы принимаем, что в данном сообществе отра-
жен состав популяции в какой-то момент времени. Значит, все
особи одного возрастного класса появились на свет одновремен-
но. Рассмотрим вторую группу цифр в табл. 1. Первые три
колонки не изменились. Мы примем, что смертность во всех
возрастных группах была одинаковой и что рождаемость во
всей популяции не изменялась (оба предположения не всегда
верны). Тогда если в 4-й возрастной группе 155 особей, то
столько же их должно было бы быть в каждой сохранившейся
возрастной группе. Меньшее число раковин в каждой старшей
группе (столбец 3) — это число особей, доживших до начала
соответствующей возрастной стадии (столбец 6). Число особей,
погибших на той или иной возрастной стадии (столбец 5), рав-
но числу выживших к началу этой стадии минус число выжив-
ших к началу следующей стадии.
Оказывается, что столбцы 5—7 и соответствующие кривые
выживаемости, рассчитанные с использованием разных подхо-
дов, идентичны. Причина этого — вычитание линочных стадий
при динамическом подходе. Поэтому для сообщества, включаю-
щего все сброшенные при линьке наружные скелеты, мы при
правильном анализе должны получить те же результаты, что
и для такого же сообщества, возникшего при массовой гибели
организмов. Использование динамического подхода дало бы
иные результаты, если бы мы применили его для анализа сооб-
щества животных, не линяющих в ходе роста, например сооб-
IV. Популяция как целое
85
щества двустворчатых моллюсков или плеченогих. Посмотрим,
к каким выводам мы придем, если примем, что сообщество
остракод состоит только из постепенно накапливавшихся рако-
вин мертвых животных, а сброшенные при линьке раковины не
сохранялись. В этом случае число особей, погибших в течение
каждой возрастной стадии (столбец 5), просто равнялось бы
числу раковин в столбце 3; вычитать перелинявших особей не
надо. Число особей, доживших до начала очередной возрастной
стадии (столбец 6), было бы равно сумме числа погибших в
течение этой стадии и числа погибших в течение всех после-
дующих стадий. Цифры в столбце 7 рассчитываются, как и ра-
нее. Кривая выживаемости, построенная по данным столбца 6,
показана на фиг. 34, Б пунктиром. Рассчитанная на основе
«безлиночного» подхода, она почти не отличается от кривой,,
рассчитанной с помощью других подходов.
Судя по данным о сохранности, приводимым Спьельднесомг
верен, очевидно, первый динамический подход. Но другой под-
ход дал практически тот же результат. Если сохранность изу-
ченного сообщества настолько полна, как думает Спьельднес
и как это следует из графиков, приведенных на фиг. 34, А, то
мы получили точное представление о кривой выживаемости
этого вида, по крайней мере в данном местообитании. Для
нелиняющих видов кривая выживаемости, полученная путем
динамического анализа, значительно отличалась бы от кривой,
получившейся в результате временного анализа. Поэтому очень
важно тщательно изучить, как образовалось исследуемое сооб-
щество.
Хотя рождаемость и в особенности смертность могут сильно
варьировать в различных популяциях данного вида, а также
в одной популяции с течением времени, для некоторых ископае-
мых видов можно вывести характерные кривые выживаемости.
Затем можно попытаться связать такие кривые с жизненным
циклом, образом жизни и способом размножения вида.
Методы описания изменчивости
Мы рассмотрим биологические и геологические причины из-
менчивости в отдельном ископаемом сообществе и в разных со-
обществах. Важно уметь описать изменчивость ясным и разум-
ным образом.
ИЗМЕНЧИВОСТЬ ВНУТРИ СООБЩЕСТВА
Один из самых простых и иногда самых эффективных мето-
дов описания изменчивости — применение разновидности фото-
графического метода, рассмотренного в гл. II. На фиг. 35 изо-
86
Часть 1 Описание и классификация ископаемых остатков
Фиг. 35. Пиктографический метод выражения морфологической изменчиво-
сти [50].
Очертания выбранных особей двустворчатого моллюска Carbontcola (карбон, Англия) рас-
положены так, чтобы продемонстрировать основные тенденции отклонения от «нормы»
(в центре) Подобные диаграммы используются для оценки и описания изменчивости в
видовых сообществах, содержащих одни и те же варианты
Сражено множество близкородственных двустворчатых моллюс-
ков из каменноугольных пластов Англии. Изображенные особи
специально подобраны таким образом, чтобы продемонстриро-
вать основные тенденции изменчивости. В результате получи-
лась пиктограмма. Экземпляры расположены в соответствии
с их морфологическими отношениями. Так, тенденция к удли-
нению раковины (возрастание отношения длины к высоте) от-
ражена в рядах, идущих на пиктограмме от центра.
На фиг. 36 показаны так называемые диаграммы изменчи-
вости, построенные на основании анализа пиктограммы. Черные
кружки — отдельные формы из данного ископаемого сообщест-
ва Диаграмма изменчивости в сочетании с аккуратно построен-
ной пиктограммой позволяет в общем оценить морфологическую
изменчивость. Однако этот метод нельзя назвать строгим: пик-
тограмма очень субъективна и два исследователя, обрабаты-
IV Популяция как целое
87
Фиг. 36. Диаграммы изменчивости [50].
Три ископаемых сообщества Carbonicola проанализированы по образцу, показанному на
фиг 35 Каждый кружок символизирует одну особь Кружки размещены на диаграмме
в соответствии с морфологическим сходством между особями Тве диаграммы справа по-
строены для сообществ, отстоящих друг от друга в стратиграфическом разрезе на 15 см
Они находились примерно на 152 см ниже сообщества, отображенного на левой диа-
грамме
вающие независимо друг от друга один и тот же материал,
получат скорее всего разные пиктограммы. Для более эффек-
тивного анализа изменчивости необходимо обратиться к более
формальным статистическим методам.
Простейший статистический метод, позволяющий охаракте-
ризовать изменчивость, — построение гистограмм или диаграмм,
показывающих частоту каждого варианта. Это пример одно-
факторного анализа. С его помощью мы можем многое узнать
об изменчивости одного признака — степень отклонения от сред-
него значения, симметрична ли изменчивость и т п. Для анали-
за однофакторного распределения разработано немало тонких
методов. Палеонтолог должен быть знаком с ними, чтобы уметь
эффективно использовать и интерпретировать однофакторное
распределение.
Как уже говорилось при рассмотрении онтогенетической
изменчивости (гл. III), многие морфологические соотношения
лучше описывать, рассматривая одновременно два признака.
Диаграмма разброса — простейшая графическая форма такого
двухфакторного анализа. Экспериментальные точки можно ап-
проксимировать прямой, проходящей через начало координат
(как сделано, например, на графике идеального изометрическо-
го роста). Усредненную зависимость между двумя переменными
обычно выражают наклоном прямой, лучше всего «подходящей»
к данным экспериментам. Этот наклон можно определить раз-
ными способами. Очень часто разброс точек соответствует пря-
мой, не проходящей через начало координат. Чтобы получить
представление об усредненной зависимости между рассматри-
ваемыми признаками, надо знать не только наклон прямой, но
и ее смещение по вертикали.
88 Часть 1, Описание и классификация ископаемых остатков
Что делать, если разброс точек не удается аппроксимировать
прямой? Мы встречались с такой ситуацией, рассматривая он-
тогенетическую изменчивость. Иногда график аллометрического
роста становится близким к прямой, если применить логариф-
мическую шкалу. Если и это не помогает, приходится прибегать
к другим, более тонким методам «подгонки кривыми». В случае
двухфакторного распределения диапазон и тип изменчивости
можно выразить только после того, как к полученным данным
подобрана какая-либо кривая (прямая или нет). Отклонения
от «нормы», или средней величины, соответствуют отклонениям
от этой кривой.
Описывать группу особей на основе изменчивости лишь од-
ного или двух признаков, — это значит упростить ситуацию.
Мы видели, что некоторые признаки в популяции почти не ме-
няются, тогда как другие значительно варьируют. Более полную
картину изменчивости можно получить, включая в анализ боль-
шее число признаков и применяя последовательно или одно-
временно однофакторный и двухфакторный методы. Это так
называемый многофакторный анализ — статистический метод,
позволяющий измерить тип и диапазон изменчивости в выборке.
Сбор и обработка данных для многофакторного анализа отни-
мают много времени, и потому здесь очень полезны быстродей-
ствующие ЭВМ, которые можно применять не только для на-
копления данных путем их цифрового кодирования, но и для
статистической обработки этих данных. Редкая область науки
имеет дело с большим количеством сырых данных, чем пале-
онтология, и поэтому не удивительно, что статистическая оценка
изменчивости с помощью ЭВМ заняла важное место в палеон-
тологических исследованиях.
Однако применение даже самых современных и сложных си-
стем кодирования и машинной обработки не снимает проблем,
связанных с количественным описанием изменчивости (гл. II).
Поскольку невозможно описать все аспекты изменчивости (да
это и не нужно), приходится делать выбор. Выбор предполага-
ет субъективную, часто априорную, оценку того, какие призна-
ки популяции важны, интересны или имеют непосредственное
значение для решения какой-то более общей проблемы. Измен-
чивость можно описывать почти бесконечным числом способов;
какого-то единственного или безупречного способа здесь нет.
МЕЖПОПУЛЯЦИОННАЯ ИЗМЕНЧИВОСТЬ
Для описания различий между живыми популяциями или
ископаемыми сообществами с успехом применяются многие
изобразительные, графические и численные методы. Некоторые
из них сходны с методами, используемыми для описания измен-
IV. Популяция как целое
89
Фиг. 37. Распределение ширины раковин у двух подвидов девонской брахио-
поды Pholidostrophia [89].
A. Pholidostrophia gracilis nanus. Б. Pholidostrophia gracilis gracilis. Средние значения раз-
личаются настолько, что эти различия не могут объясняться только случайиыми'откло-
нениями при взятии выборок.
чивости внутри популяции или одного ископаемого сообщества,
с той разницей, что при анализе изменчивости в сообществе
сравниваются особи, а при анализе изменчивости между сооб-
ществами— группы особей. В тех случаях, когда какая-то груп-
па особей (популяция или ископаемое сообщество) может быть
охарактеризована с помощью описания одной особи или какой-
то гипотетической средней особи, это различие в методологии
стирается. Валидность описания популяции по одной особи за-
висит от того, насколько велика изменчивость внутри сообщест-
ва и какие различия между сообществами найдены или могут,
по мнению исследователя, быть найдены. Если различия между
сообществами сравнительно велики, то уже по одной особи из
каждого сообщества достаточно для демонстрации или описания
различий. Различия могут быть настолько значительными, что
любую особь, типичную или нетипичную, из каждого сравнивае-
мого сообщества можно с успехом использовать для распозна-
вания или описания различий.
90 Часть 1. Описание и классификация ископаемых остатков
4>иг. 38. Распределение отношения ширины раковин к их длине у двух под-
видов девонской брахиоподы Pholidostrophia [89].
A. Pholidostrophia gracilis nanus. Б. Pholidostroohia gracilis gracilis- Небольшое различие
между средними величинами вполне может объясняться случайными ошибками при взя-
тии выборок.
Начиная исследование, необходимо решить, какое число
морфологических признаков следует использовать. Многое за-
висит от того, намерены ли мы изучать различия или сходство
между сообществами. Если мы хотим лишь показать, что два
сообщества различаются между собой, то для этого достаточно
и одной морфологической черты. На фиг. 37 показано распре-
деление размеров особей двух подвидов брахиопод (измерялась
максимальная ширина). Две диаграммы распределения пере-
крываются, но они явно различны. На фиг. 38 показаны рас-
пределения отношения ширины раковины к длине у тех же
брахиопод. Не требуется сложной статистической обработки,
чтобы понять, что по отношению ширины к длине оба сообщест-
ва не различаются сколько-нибудь существенно. Но из этого
еще нельзя заключить, что оба сообщества одинаковы, — мы
видим, что они различаются по ширине раковин (фиг. 37). Не-
редко неподготовленный исследователь, найдя близкое сходство
IV. Популяция как целое
91
Фиг. 39. Пары гипотетических кри-
вых распределения иллюстрируют
важность изменчивости в определе-
нии смысла наблюдаемых различий
между средними.
В случае Б гораздо более вероятно, чем
в случае iA, что найденные различия
объясняются случайностью.
в
£
о
Е
CJ
В
Разность между средними
для двух проб
Измеряемый параметр
в распределении каких-либо морфологических признаков у двух
сообществ, делает вывод об идентичности этих сообществ, хотя
между ними могут существовать важные различия, которые
просто не отражены в использованных параметрах.
На фиг. 37 и 38 показаны крайние случаи, а между ними
существует бесчисленное множество промежуточных. Для раз-
личения промежуточных случаев необходимы статистические
методы. Прежде всего важно уметь определять, соответствует
ли найденная изменчивость истинному положению вещей (т. е.
можно ли ее интерпретировать с геологической или биологиче-
ской точки зрения) или же она объясняется просто случайно-
стью (в пробу случайно попали особи с такой изменчивостью).
Для того чтобы выяснить, не объясняется ли обнаруженная при
однофакторном анализе разница между средними простой слу-
чайностью, можно применить метод статистической проверки.
Для этого нужно оценить не только разность между средними,
но и диапазон изменчивости в каждом из двух сообществ. На
фиг. 39 представлены две пары гипотетических распределений,
иллюстрирующие важность оценки диапазона изменчивости.
Здесь различия между средними одинаковы, но изменчивость
значительно больше в выборке 5, чем в выборке А. Очевидно,
в случае выборки Б более вероятно, что отмеченная разница
между средними объясняется простой случайностью. Следова-
тельно, вполне разумно будет включить выражение для измен-
чивости в оценку достоверности разности средних. Статистиче-
ская проверка не дает однозначного ответа типа «да» или «нет»;
мы получаем лишь вероятность того, что найденная разность
средних не случайна.
Всегда остается вопрос, какой должна быть эта вероятность,
чтобы с уверенностью можно было сказать, что разность сред-
92 Часть 1. Описание и классификация ископаемых остатков
них достоверна. Обычно за порог принимается вероятность 0,05.
Иными словами, выборки можно считать различающимися, если
мы на 95% уверены в том, что разность не объясняется случай-
ностью. На первый взгляд это довольно жесткое требование.
Но оно хорошо обосновано. Одно из главных соображений в
пользу этой строгости — возможность ошибок даже при таком
ограничении: если исследователь получил двадцать результатов
и вероятность каждого из них 0,95, то не исключено, что один
из результатов окажется неверным! Еще важнее, что если ис-
следователь делает заключение, основанное на последователь-
ности результатов, полученных ранее с таким же ограничением,
то вероятность справедливости заключения оказывается значи-
тельно ниже 0,95. Если рассчитанная вероятность ошибки 0,05,
то разность средних называют «статистически достоверной» при
пятипроцентном пороге. Статистическая проверка сама по себе
не дает никакой геологической или биологической информации,
она лишь говорит нам, не могут ли найденные различия объ-
ясняться случайностью. Если полученная разность статистически
достоверна, то остается проблема интерпретации происхождения
обнаруженных различий.
Количественное изучение морфологических различий между
ископаемыми сообществами обычно основывается на двухфак-
торном анализе. На фиг. 40 показано типичное двухфакторное
сравнение данных по двум сообществам. Пользуясь математи-
ческим методом, через точки данных для каждой выборки про-
вели прямую. Полученные прямые различаются по тому, в какой
точке они пересекают ось ординат, и по наклону. Значит, два
рассматриваемых подвида различаются по форме и по развитию
формы в онтогенезе. Но достоверны ли эти различия стати-
стически? Здесь необходимо применить критерии достоверности.
Если окажется, что различия между прямыми достоверны, то
можно будет заняться биологической или геологической интер-
претацией этого факта.
Заметим, что на фиг. 40 приведены те же данные, что и на
фиг. 37 и 38. Представленный на фиг. 40 график, показывающий
связь двух параметров, объясняет, почему данные на фиг. 37
в отличие от фиг. 38 свидетельствуют о значительных различиях
между исследуемыми выборками.
Для описания межпопуляционной изменчивости с учетом
более чем двух факторов вместо формального многофакторного
анализа используются также графические методы. Один из них,
с применением символов, иллюстрируется на фиг. 41. Его основу
составляет двухфакторный анализ. Измерялись некоторые при-
знаки организмов, выбранных из двух популяций. Из них два
параметра отложены по осям координат. Остальные признаки
показаны на графике различными символами, значение которых
IV. Популяция как целое
93
Фиг. 40. Двухфакторное распределение ширины и длины раковины у двух
подвидов девонской брахиоподы Pholidostrophia [89].
Выборки те же, что на фиг. 37 и~38. Белые кружки — Р. gracilis gracilis', черные круж-
ки — Р. gracilis nanus.
объясняется рядом. Этот метод позволяет одновременно видеть
различия по нескольким признакам; таким образом удается
составить хотя и субъективное, но довольно верное представле-
ние о морфологических различиях между двумя сообществами.
(На фиг. 41 использован непалеонтологический пример, посколь-
ку диаграммы с символами пока редко применяются в палеон-
тологии.)
Хотя символы позволяют охватить одновременно большее
число признаков, чем при обычном двухфакторном или одно-
факторном анализе, все же остается проблема выбора призна-
ков для сравнения. Если многофакторный анализ проводится
с помощью ЭВМ, то число рассматриваемых признаков ограни-
чивается только упорством исследователя и возможностями
машины.
До сих пор мы как бы принимали, что измеряемые и срав-
ниваемые признаки ископаемого независимы друг от друга в
биологическом и геологическом отношениях. Однако известно,
что многие морфологические признаки тесно зависят друг от
Часть 1. Описание и классификация ископаемых остатков
Число остей на нижних
цветковых чешуях:
О 0-1
0 2-3
<Ь 4
Число остей на
колосковых чешуях:
О 4-5
0 6-7
С<8-9
Форма клина
(цифры в скобках-
отношение длины к ширине)
О Короткий и широкий
(0,5-1,0)
Aegilops variabilis
О
Длина верхнего .междоузлия
а
2$
О Промежуточный
(1,1-1,5)
Ъ Узкий и длинный
(1,6 и более)
Число рудиментарных
колосков:
О 3-4
О 2
<У1
Фиг. 41. Диаграмма с условными значками, использованная для анализа
морфологических различий между двумя видами Aegilops, ближайшего ро-
дича пшеницы Д241].
друга. Связь эта может быть чисто механической. Например,
тюбая структура, содержащая какие-либо внутренние структу-
ры, всегда будет крупнее этих последних. Если построить гра-
фик зависимости между размером одной структуры и размером
другой, находящейся внутри нее, то станет ясно, что при увели-
чении первой будет увеличиваться и вторая.
Более распространены функциональные связи между призна-
<ами. Например, створки раковины брахиоподы сходятся по
чинии комиссуры. Поскольку очертания створок имеют одина-
<овую форму и одинаковый размер, ни высокий коэффициент
сорреляции, получающийся при построении графика зависи-
IV. Популяция как целое
95
Фиг. 42. График зависимости между бивиальным углом и эксцентричностью
вершинного щитка роющего морского ежа Dendraster иллюстрирует положе-
ние об избыточности некоторых признаков.
Изменение одного признака сопровождается предсказуемым изменением другого неза-
висимо от стадии онтогенеза особи. Черные кружки — D. excenti icus (современный вид,
тихоокеанское побережье Северной Америки), белые кружки — D. gtbbsu gtbbsu (плио-
цен, Калифорния); треугольники — D. coaltngaensts (плиоцен, Калифорния); крестики —
D. gtbbsu humtlis (плиоцен, Калифорния).
мости длины одной створки от длины другой, ни измерения
другой створки не дадут новой информации по сравнению с из-
мерениями первой створки. Размеры второй створки — избыточ-
ный признак.
Избыточные признаки на примере ископаемых и современ-
ных морских ежей показаны на фиг. 42. Один признак — экс-
центричность вершинного щитка, т. е. положение этого щитка
(у центра петалоидной зоны) на передне-заднем диаметре пан-
циря. Эксцентричность возрастает по мере того, как вершинный
щиток из центра сдвигается к заднему краю панциря. Другой
признак — бивиальный угол (угол, образованный двумя задними
кеталями, отходящими от вершинного щитка).
На фиг. 42 показана диаграмма разброса этих признаков
для четырех генетически различных популяций морских ежей
(относящихся к разным видам или подвидам). Поскольку здесь
отложены угол и отношение, размеры особей (а следовательно,
и их возраст) не получают отражения на диаграмме. Таким
образом, избыточность, демонстрируемая разбросом, не зависит
96
Часть 1. Описание и классификация ископаемых остатков
от стадии онтогенеза. Эта избыточность довольно просто объ-
ясняется, если вспомнить, что точка окончания петалей на пе-
риметре скелета ежа анатомически жестче фиксирована, чем
положение вершинного щитка. Величина бивиального угла не
дает для сравнения популяций новой информации, которая уже
не содержалась бы в данных о положении вершинного щитка.
Какой из этих двух признаков использовать в том или ином
исследовании, зависит от того, какое измерение легче выполня-
ется. В этом примере, видимо, лучше отказаться от измерения
бивиального угла, так как его трудно провести с достаточной
точностью.
В распоряжении палеонтолога имеется целый арсенал опи-
сательных методов для сравнения ископаемых сообществ. Мы
коснулись лишь отдельных методов. Поскольку разные методы
предназначены для решения совершенно разных задач, очень
важно правильно выбрать метод. Выбор можно сделать только
после того, как ясно поставлена задача исследования. Поэтому
обычно в первую очередь необходимо сформулировать именно
задачу и уже затем выбрать методы для ее решения.
ГЛАВА V
Вид как целое
Огромное разнообразие формы, физиологии и поведения
организмов не беспорядочно. Если бы наше воображение было
способно создать картину всего спектра возможных биологиче-
ских форм, то мы обнаружили бы, что все живущие и вымер-
шие организмы, вместе взятые, составляют лишь малую часть
этого воображаемого целого* Рецентные виды образуют внутри
этого спектра дискретные группы, так что существует сравни-
тельно немного основных типов, и каждый из них использует
особенности и преимущества своего положения в отдельных
местообитаниях. Другими словами, для большинства видов
характерна приспособленность к одному или немногим типам
среды.
Для сохранения биологического разнообразия виды должны
оставаться обособленными. На языке биологов это означает,
что межвидовая гибридизация должна быть редким явлением.
Конечно, она существует, особенно среди растений. Иногда в
результате естественной межвидовой гибридизации появляются
новые структуры или новые комбинации структур, каких не бы-
ло ни у одного из родительских видов. В конечном итоге успеш-
ная гибридизация «смешивает» родительские признаки, и в ре-
зультате вместо двух видов образуется один. Тот факт, что
ярко выраженное видовое разнообразие неизменно существует
на протяжении миллионов лет, свидетельствует о сравнительной
редкости межвидовой гибридизации.
Разнообразие сохраняется потому, что виды репродуктивно
изолированы один от другого (хотя они могут обитать в одной
географической области и занимать сходные биотопы). Таким
образом, в основе происхождения видов лежит возникновение
репродуктивной изоляции. Чтобы понять биологическую кон-
цепцию вида, необходимо зрать, как возникает репродуктивная
изоляция. Палеонтолог обычно имеет дело только с результа-
том, а не с процессом эволюции вида, но знание процесса так
же важно для палеонтолога, как знание гидродинамики — для
литолога.
98 Часть 1. Описание и классификация ископаемых остатков
Биологическое определение вида
Наиболее общепринятое биологическое определение вида
сформулировал Э. Майр: вид — группа действительно или по-
тенциально скрещивающихся популяций, репродуктивно изоли-
рованных в естественных условиях от других таких же групп.
Оно неприменимо в качестве служебного определения в палеон-
тологии, так как скрещивание ископаемых видов невозможно
наблюдать. Но даже и большинству биологов редко приходится
применять это определение непосредственно. Биологи разрабо-
тали некоторые непрямые методы разграничения видов. Многие
из этих методов применимы и к ископаемым остаткам.
Приведенное выше определение состоит из нескольких су-
щественных элементов. Вид определяется как группа популя-
ций, чем подчеркивается тот факт, что большинство видов гео-
графически распадается на субъединицы или скрещивающиеся
внутри себя популяции. Определение указывает, что такие по-
пуляции действительно или потенциально скрещиваются между
собой. Можно выделить два типа взаимоотношений между по-
пуляциями. Аллопатрическими называют популяции, занимаю-
щие изолированные географические области. Симпатрические
популяции занимают одну и ту же область или же области их
обитания частично перекрываются. Аллопатрические популяции
могут быть разделены географическими преградами, препят-
ствующими обмену генами. В таких случаях говорят о геогра-
фической, но не о репродуктивной изоляции (хотя на самом
деле географические преграды могут исключать возможность
скрещивания). Две популяции считаются репродуктивно изоли-
рованными только в том случае, если между ними не происходит
скрещивания, хотя живут они в одной области. Значит, слово
«потенциально» в определении вида особенно важно.
Суть определения вида состоит в том, что группа популяций,
являющаяся видом, репродуктивно изолирована от других по-
добных групп: два вида могут быть симпатрическими, но они
не скрещиваются между собой. Должна ли репродуктивная
изоляция видов быть абсолютной? Если нет, то какой поток
генов можно считать допустимым? Если две популяции успешно
скрещиваются (т. е. дают плодовитое потомство), то обяза-
тельно ли их надо считать принадлежащими к одному виду?
Четкого ответа на эти вопросы мы дать не можем; отметим
лишь — поскольку нас интересует сохранение индивидуально-
сти вида, — что такой обмен генами, который не приводит к
уничтожению различий между видами, не мешает считать виды
репродуктивно изолированными.
Важно, что в определении вида подчеркивается репродук-
тивная изоляция видов «в естественных условиях». Отмечено
V. Вид как целое
99
немало примеров скрещивания видов в неволе или одомашнен-
ных видов. Объясняется это тем, что часто репродуктивная
изоляция поддерживается довольно незначительными экологи-
ческими или этологическими преградами, которые не действуют
в условиях неволи или одомашнивания. Способность организмов
скрещиваться в неволе обычно не считается признаком того,
что они принадлежат к одному виду.
Рассмотрим пример, показывающий трудность практическо-
го применения определения вида. Предположим, мы подозре-
ваем, что две аллопатрические популяции принадлежат к одно-
му виду. В согласии с определением мы можем проверить наше
предположение, только перенеся особей из одной популяции
в местообитание другой. Но при этом мы неизбежно нарушим
естественные условия обитания одной или обеих популяций и,
следовательно, можем устранить какие-то экологические или
этологические основы репродуктивной изоляции.
Видообразование
Видообразование — эволюционный процесс возникновения
вида. Известно два типа видообразования: филетическое и гео-
графическое. Видообразование первого типа представляет собой
эволюцию одного вида в другой с течением времени. Поскольку
в результате этого процесса один вид заменяет в «эволюцион-
ном ряду» другой вид, общее число видов, живущих в каждый
данный момент, не возрастает. Географическое, или аллопатри-
ческое, видообразование заключается в том, что один вид пре-
вращается в два или более сосуществующих видов; это приво-
дит к возрастанию общего числа видов.
ФИЛЕТИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
Филетическое видообразование является результатом дейст-
вия естественного отбора на генетический состав одной или
нескольких популяций. Изменения в генофонде вызывают изме-
нения в фенотипе (форме, строении, физиологии и поведении)
организмов, составляющих популяцию. Воздействуя на фено-
типическую изменчивость, естественный отбор вызывает со вре-
менем изменения в генетическом составе популяции, а любое
такое изменение, пусть даже небольшое, и есть эволюция.
Эволюционные изменения генетического состава популяции
могут означать или прогрессивную адаптацию к постоянным
условиям среды, или приспособление к меняющимся внешним
условиям. С каждым последующим поколением увеличивается
разрыв между генофондом поколения, которое мы условно при-
мем за исходное, и генофондом современного поколения. По-
скольку популяция изменяется, почти неизбежно (статистиче-
7*
100 Часть 1 Описание и классификация ископаемых остатков
ски) должны появиться различия, мешающие особям с изменив-
шимся фенотипом скрещиваться с особями, обладающими ис-
ходным предковым фенотипом.
Если представить себе хронологический ряд популяций и
выбрать одну из них в качестве исходной, то в этом ряду будет
точка, в которой накопившиеся отличия уже таковы, что все
последующие популяции были бы репродуктивно изолированы
ют исходной, если бы жили в одно время с ней. Эта точка и
•соответствует возникновению нового вида.
Такое представление об эволюции популяции во времени
можно распространить на вид, состоящий из большого числа по-
пуляций. Если между этими популяциями существует обмен
генами, то процесс филетического видообразования захватывает
целый вид. Но по иронии судьбы при наличии генетического
контакта между всеми популяциями процесс этот неизбежно
приводит к вымиранию предкового вида. Такой тип вымирания
называется филетическим вымиранием.
ГЕОГРАФИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
Рассмотрим теперь популяции одного вида, живущие в одно
время, но географически изолированные одна от другой. В этом
случае две или несколько групп организмов одного вида эволю-
ционируют и претерпевают филетическое видообразование не-
зависимо. Если географические преграды между популяциями
остаются непреодолимыми достаточно долго, то может развить-
ся репродуктивная изоляция. Когда она возникает, мы говорим,
что произошло географическое видообразование: из одного вида
возникло два или несколько одновременно живущих видов.
Разграничение между филетическим и географическим видо-
образованием несколько искусственно, поскольку оба процесса
основаны на естественном отборе. Главное различие состоит в
том, что филетическое видообразование идет в отсутствие гео-
графической изоляции, а географическое видообразование тре-
бует такой изоляции.
При географическом видообразовании новые виды могут
появиться в обоих эволюционных рядах или только в одном.
В любом случае число видов увеличивается. Может показаться,
что процесс географического видообразования должен неизбеж-
но вести к постоянному увеличению общего числа видов, но на
деле этому противодействует вымирание. Эволюционные линии
время от времени обрываются вследствие быстрых изменений
условий среды, к которым вид не может приспособиться путем
филетического видообразования. Таким образом, число видов,
живущих на Земле в любой данный момент, представляет собой
результирующую позитивного процесса географического видо-
образования и негативного процесса вымирания.
V. Вид как целое
101
ПОДВИДЫ
Мы можем рассматривать подвиды как зарождающиеся ви-
ды. Подвидами обычно называют географически изолированные
популяции или группы популяций, генетически различные, но не
настолько, чтобы между ними существовала репродуктивная
изоляция. С палеонтологической точки зрения подвидами можно
назвать и промежуточные стадии замещения одного вида дру-
гим в процессе филетического видообразования.
КЛИНЫ И КОЛЬЦЕВЫЕ ПЕРЕКРЫВАНИЯ
Некоторые аллопатрические популяции разделены участками
среды, неблагоприятными для их обитания. Так, популяции
птиц на отдельных островах какого-либо архипелага разделены
значительными водными пространствами, а равнинные популя-
ции наземных растений разделены горными цепями. Другие
аллопатрические популяции не разделены такими резко выра-
женными преградами.
Обмен генами между популяциями, расположенными на
краях географического ареала вида, может быть сильно ослаб-
лен уже вследствие большого расстояния между ними. В ре-
зультате может возникнуть градиент генетического состава
популяций, выражающийся в так называемой клине. Обнару-
жено, .что у некоторых видов популяции, живущие на противо-
положных краях ареала, репродуктивно изолированы друг от
друга, но как бы соединены цепью скрещивающихся популяций.
Если такие репродуктивно изолированные популяции с гра-
ниц ареала мигрируют так, что их ареалы перекрываются, то
образуется так называемое кольцевое перекрывание. Популя-
ции, некогда разделенные значительным расстоянием, живут
симпатрически, оставаясь четко выраженными видами, но меж-
ду ними существует непрерывный ряд скрещивающихся попу-
ляций.
К счастью для биолога-практика, клины и кольцевые пере-
крывания встречаются сравнительно редко. Палеонтологу же
обычно вообще не удается выделить клины и кольцевые пере-
крывания вследствие того, что палеонтологическая летопись
недостаточно полна, а определения возраста недостаточно
точны.
Возникновение видов в отсутствие
изоляции
Было много споров о том, могут ли из одного вида возник-
нуть два или несколько живущих одновременно, если часть по-
пуляций вида не подверглась географической изоляции. Впрочем,
102
Часть 1 Описание и классификация ископаемых остатков
здесь мы можем не касаться этого вопроса, так как и биологи,
и палеонтологи согласны в том, что если это и случается, то
чрезвычайно редко.
Скорость видообразования
Определить скорость, с которой происходит видообразова-
ние,— задача скорее для палеонтолога, чем для биолога. Био-
лог видит только популяции, живущие в данный момент, и для
реконструкции истории изучаемого вида может опираться толь-
ко на догадки. Палеонтолог же работает со стратиграфически-
ми последовательностями ископаемых, по которым можно вос-
становить хронологический порядок популяций по их сохранив-
шимся остаткам; следовательно, положение палеонтолога более
выгодно и у него гораздо больше возможностей расширить на-
ши знания о том, как в процессе эволюции появляются виды.
Рассматривая вопрос о скорости видообразования, мы долж-
ны помнить разницу между филетическим и географическим
видообразованием. Нас интересует скорость изменений в одном
эволюционном ряду, с одной стороны, и влияние физической
изолированности популяций, необходимой для географического
видообразования, — с другой. Самыми важными факторами,
определяющими скорость филетического видообразования, явля-
ются: 1) время генерации, 2) давление отбора, 3) мутационное
давление и 4) размер популяции.
Время генерации — основная единица измерения времени
в случае естественного отбора. Успех или неуспех вариантов
из одной скрещивающейся популяции выражается в том влия-
нии, которое эти варианты оказывают на генетический состав
последующего поколения. Ясно, что вид с малым временем ге-
нерации будет давать больший прирост генетических изменений
за тысячу лет, чем вид с большим временем генерации. Следо-
вательно, малое время генерации (этот признак контролируется
генетически) можно рассматривать как приспособление, спо-
собствующее быстрой эволюции. Поэтому можно ожидать, что
группы организмов с малым временем генерации должны обла-
дать большим разнообразием, чем группы с большим временем
генерации.
Если популяция обитает в очень благоприятных условиях,
то большое число форм может иметь одинаковый успех и ге-
нетический состав будет медленно изменяться от поколения
к поколению. Но там, где биологические или физические фак-
торы среды обусловливают усиление конкуренции между осо-
бями, «непригодные» особи быстрее элиминируются из популя-
ции и степень генетических различий между популяциями воз-
V. Вид как целое
103
растает. Поэтому так называемое давление отбора сильно влия-
ет на относительные скорости эволюции в данном ряду.
Для эволюционных изменений существенно важно наличие
генетической изменчивости в популяции. Без такой изменчивости
популяция или остается неизменной из поколения в поколение,
или начинает вымирать, в зависимости от степени «жесткости»
условий среды. Поскольку в конечном счете источником генети-
ческой изменчивости являются генные мутации, частота мута-
ций прямо влияет на скорость эволюции. Частоту мутаций на-
зывают мутационным давлением. При прочих равных условиях
возрастание мутационного давления вызовет увеличение ско-
рости эволюции.
Размер популяции играет важную роль в определении ско-
рости эволюции, так как от него зависит абсолютная частота
нового варианта, появление которого вызвано мутацией, а также
скорость, с которой благоприятная мутация может распростра-
ниться в популяции. Здесь действует несколько противоположно
направленных процессов, но в целом совершенно правильно
будет считать, что скорость эволюции мелких популяций выше,
чем крупных.
Рассмотренные факторы влияют на скорость изменений в
ряду поколений и, следовательно, на скорость географического
и филетического видообразования. Но для того чтобы происхо-
дило географическое видообразование, нужна географическая
изоляция между популяциями. Она зависит от нескольких фи-
зических и биологических факторов. Организмы, живущие на
- мелководье на окраине материка, имеют не много шансов по-
пасть в условия географической изоляции, так как пригодные
для них биотопы могут составлять непрерывную цепь. Но по-
пуляции того же вида, обитающие у побережий островов, ско-
рее всего будут претерпевать географическую изоляцию, так
как пригодные для них биотопы могут быть разделены рас-
стоянием в десятки и даже сотни километров. Не удивительно
поэтому, что на мелководье у островов Вест-Индии встречается
гораздо больше видов на единицу площади дна, чем, например,
вдоль восточного побережья Африки.
Поразительный пример географического видообразования
представляют фауны морских беспозвоночных, обитающих на
мелководье у атлантического и тихоокеанского берегов Панам-
ского перешейка. Условия среды здесь практически одинаковы,
но по обе стороны перешейка живут лишь некоторые виды.
Перешеек уже несколько миллионов лет находится выше уровня
моря и со времени своего последнего поднятия представляет
собой барьер, разделяющий популяции морских видов. Посте-
пенно в результате географического видообразования фауны
двух побережий стали совершенно различными.
104
Часть 1. Описание и классификация ископаемых остатков
Иногда на основании изучения группы ископаемых видов,
распределенных в соответствии с географическими условиями
мест их обитания, удается сделать определенные палеогеогра-
фические выводы. Таким способом удалось очень точно просле-
дить историю поднятий и опусканий Панамского перешейка за
последние 120 млн. лет. Очевидно, периоды быстрого геогра-
фического видообразования связаны с периодами тектонической
активности.
Один и тот же географический элемент может представлять
собой преграду для обмена генами между популяциями одного
вида и не служить такой преградой для другого вида. Потенциал
расселения организма — важный биологический фактор, влияю-
щий на географическое видообразование. Некоторые виды на
протяжении всего жизненного цикла очень ограничены в воз-
можностях передвижения. В таком случае такой фактор, как
расстояние, приводит к полной изоляции отдельных популяций
вида. Например, у брюхоногого моллюска Purpura lapillus
оплодотворенные яйца в течение всего периода своего развития
остаются на дне моря, а взрослая особь за всю свою жизнь мо-
жет переместиться всего на несколько метров. Расселение идет
буквально «черепашьим шагом», ускоряясь только за счет
случайных факторов — штормов или прикрепления к дрейфую-
щему плавнику.
У других видов того же рода совсем иной способ размноже-
ния: из оплодотворенных яиц выходят свободноплавающие ли-
чинки, ведущие планктонный образ жизни целых 6 недель.
У этих видов потенциал расселения чрезвычайно высок, и не
удивительно, что для них в меньшей степени характерно гео-
графическое видообразование, чем для Purpura lapillus.
Расселительный потенциал может сильно различаться даже
у близкородственных видов, и поэтому палеонтологу важно
знать расселительные возможности организмов. По раковине
брюхоногого или двустворчатого моллюска часто можно узнать,
обладал ли организм свободноплавающей стадией, — для этого
надо изучить личиночную раковину, сохраняющуюся у вершины
взрослой. Следовательно, по морфологии палеонтолог может
узнать кое-что о потенциале расселения.
Биологические методы разграничения
видов
На практике проблема различения видов обычно сводится
к следующему вопросу: считать ли две морфологически разли-
чающиеся популяции разными видами или же это таксоны низ-
шего ранга (например, подвиды)? Здесь биологу больше всего
помогает общепринятое положение, согласно которому морфо-
V. Вид как целое
105
логически различные популяции могут быть симпатрическими
только в том случае, если они принадлежат к разным видам.
Конечно, при этом подразумевается, что разделение одного вида
на два или большее число видов без репродуктивной изоля-
ции— настолько редкое событие, что его не стоит принимать
во внимание. Рассуждая таким образом, биолог, когда ему
доводится встретить два морфологически различающихся орга-
низма с перекрывающимися ареалами, довольно уверенно делает
вывод, что эти организмы относятся к двум разным видам.
Почти все аллопатрические популяции обладают некоторыми
различиями-—морфологическими, физиологическими, этологи-
ческими или экологическими. Оценивая вероятность репродук-
тивной изоляции в случае двух подобных популяций, пале-
онтолог основывается главным образом на сравнении со сте-
пенью различий между популяциями, встречающимися симпат-
рически и потому с уверенностью относимыми к разным видам.
Такой способ разграничения видов не всегда надежен. Все со-
гласны, что есть очень слабо различающиеся виды (так назы-
ваемые «виды-двойники»), а есть виды, различающиеся настоль-
ко, что сомнения в обоснованности придания им статуса вида
не возникает.
В общем степень различий между видами внутри каждой
эволюционной группы довольно постоянна, так что специалист
постепенно приобретает опыт, помогающий ему интерпретиро-
вать видовые различия между аллопатрическими популяциями.
Кроме того, большинство современных видов обладает доста-
точно четкими отличиями, так как эти виды давно находятся
в репродуктивной изоляции и успели приобрести множество
заметных отличительных признаков.
Тот факт, что выделение видов в значительной степени ос-
новано на опыте исследователя, нашел выражение в неофици-
альном, но поразительно действенном определении вида: «вид
является видом, если это признает опытный специалист».
В выделении видов может помочь также географическое
распределение аллопатрических популяций. Если популяции
разделены уже давно, то шансы на завершение процесса видо-
образования у них выше, чем у популяций, живущих в тесной
близости и не испытавших продолжительной географической
изоляции. Поэтому таксономисты часто обращаются к биогео-
графии. С определенным основанием можно сказать, что попу-
ляции, разделенные большим расстоянием, относятся к разным
видам, даже если морфологические различия между ними не
очень велики. Такой подход может быть весьма полезным в ка-
честве вспомогательного, но если выделять виды только на ос-
новании географической разделенности, то это может привести
к значительным затруднениям.
106
Часть 1. Описание и классификация ископаемых остатков
Подводя итог, можно сказать, что при определении границ
между видами биолог опирается в основном на фенотипические
различия между популяциями. Иногда он использует данные
биогеографии, иногда ему помогает симпатрическое распростра-
нение исследуемых популяций. Биологическое разграничение
видов прежде всего является интерпретацией эволюционной ис-
тории изучаемого организма.
В разное время появлялись предложения расстаться с си-
стемой, основанной на определении вида, которое редко может
применяться объективно; говорили, что это определение надо
заменить другим, основанным на морфологии, в котором гра-
ницы между видами будут установлены произвольно, с помощью
статистических методов. Для этой цели предлагались различные
определения «морфологического вида». Так, предлагалось счи-
тать разными видами две популяции, статистически достоверно
различающиеся по одному или нескольким морфологическим
признакам. Такая система привлекательна тем, что позволяет
избавиться от многих практических проблем.
Подобные предложения никогда не получали всеобщей под-
держки, в основном потому, что результаты эволюционного
процесса не соответствуют условным границам между видами.
Во многих случаях было показано, что различие между популя-
циями, достаточное для репродуктивной изоляции, лишь весьма
приближенно коррелирует с абсолютной степенью различий
между популяциями. Однако совершенно несомненно, что ре-
продуктивная изоляция — основное условие разделения массы
организмов на дискретные адаптивные единицы. Поэтому так-
сономисты считают, что лучше описывать результаты эволюции
теоретически безупречным, хотя и с трудом приложимым на
практике методом, чем методом, теоретически неверным, осно-
ванным на соглашении.
Проблема вида в палеонтологии
Если бы мы могли построить трехмерную модель эволюции
живого, так чтобы время было отложено по вертикальной оси,
а эволюционные различия между группами выражались бы в
расстоянии между ними по горизонтали, то получилось бы не-
что вроде сложного разветвляющегося кверху дерева. Изучая
живые формы, биолог видит только горизонтальную, двумерную
верхнюю плоскость дерева — верхушки самых высоких ветвей.
Другими словами, он не имеет дела с эволюционными рядами
(кроме очень коротких, в которых происходят незначительные
эволюционные изменения). Биологические виды явственно раз-
личаются между собой потому, что эволюционные ряды, к кото-
рым они принадлежат, уже долгое время репродуктивно изоли-
V. Вид как целое
107
рованы один от другого. Если какой-то ряд расщепляется в
настоящее время, то, как мы видели, определить, имеется ли
репродуктивная изоляция между зарождающимися видами или
ее нет, оказывается делом весьма нелегким.
Палеонтолог должен учитывать еще одно измерение — время,
и потому в палеонтологии невозможно применять общеприня-
тое биологическое определение вида (основанное на репродук-
тивной изоляции) и в то же время рассматривать виды как
дискретные реально существующие единицы. Отсюда возникла
так называемая «проблема вида в палеонтологии». В биологии
аналогичная проблема возникает из-за существования клин и
кольцевых перекрываний, но эти явления встречаются не на-
столько часто, чтобы создавать большие затруднения.
Предложено множество различных подходов к определению
и распознаванию ископаемых видов. Некоторые исследователи
полагают, что биологическая концепция вида должна быть
отброшена в основном потому, что ее невозможно применить
к эволюционному континууму; кроме того, невозможно судить
о репродуктивной изоляции ископаемых видов. По-видимому,
большинство палеонтологов если и стоит за применение био-
логической концепции вида, то лишь в непрямой и неполной
форме. На практике это можно осуществить, относя к ископае-
мому виду лишь те экземпляры, которые, по мнению палеонто-
лога, принадлежали бы к одному «биологическому» виду, если
бы жили в одно время. Мы подчеркивали, что, хотя биолог
определяет виды, основываясь на генетических соображениях,
распознает он их чаще всего с помощью морфологических
критериев. Палеонтолог может поступать так же, опираясь на
степень морфологических различий между биологическими ви-
дами (желательно — видами рецентной группы, близкородствен-
ной изучаемой ископаемой группе).
Но остается одна существенная проблема: коль скоро мы
определили количественно морфологическую изменчивость
внутри вида, где провести границы между видами одного эво-
люционного ряда? На фиг. 43 показаны три возможных решения
Для случая простого эволюционного разветвления. Некоторые
исследователи предпочитают проводить границы между видами
в точках ветвления, с тем чтобы избежать ситуации, изобра-
женной на фиг. 43, Б. Поскольку добавляется еще одно изме-
рение — время, — некоторые исследователи используют для ви-
дов, представляющих условно отграниченный отрезок эволюци-
онного ствола, специальный термин, например «последователь-
ный вид» (successional species) или «палеонтологический вид»
(paleospecies)1.
1 В. Г. Гептнером предложен термин «фратрия».— Прим,, перев.
108
Часть 1 Описание и классификация ископаемых остатков
Фиг. 43. Три возможных способа подразделения ископаемого разветвляюще-
гося филогенетического ряда на три или четыре вида (из [199], с измене-
ниями).
Как ни странно, в биологических определениях вида кон-
цепция репродуктивной изоляции использовалась задолго до
того, как идея эволюции утвердилась в биологии. Между тем
современное определение биологического вида совершенно иг-
норирует эволюцию. Поэтому Симпсон [199] предложил осо-
бое определение для эволюционного вида: «Эволюционный
вид — это линия, развивающаяся отдельно от других и имеющая
свою единую эволюционную роль и свои тенденции». Под
«ролью» здесь подразумевается образ жизни в особом место-
обитании. Это определение в основном совпадает с генетиче-
ским определением биологического вида, но допускает межви-
довые скрещивания (по Симпсону, «в той степени, какая воз-
можна без слияния ролей видов»). Хотя в формальном опреде-
лении Симпсона не описаны критерии проведения границ между
видами, Симпсон полагает, что морфологические различия меж-
ду видами должны быть по крайней мере не меньше, чем
между ныне живущими видами той же (или близкой) таксоно-
мической группы.
Проблема вида в палеонтологии будет стоять всегда; все
общие методы определения и распознавания ископаемых видов
неизбежно и субъективны, и произвольны. Но есть один очень
важный фактор, устраняющий субъективность при расстановке
многих «границ» внутри филогенетического ряда. Этот фактор —
часто встречающиеся разрывы в палеонтологической летописи.
Сохранившаяся часть филогенетического древа жизни так мала
и настолько отрывочна, что часто мы находим только короткие
отрезки, в которых отмечаются лишь слабые эволюционные из-
менения. Это положение иллюстрирует фиг. 44, на которой по-
казаны последовательные ископаемые популяции, составляющие
один эволюционный ряд. В некоторых сохранившихся участках
ряда проявляются незначительные эволюционные изменения.
V Вид как целое
109
Перерыв
В
Перерыв
Б
Перерыв
А
Эволюционная тенденция
Фиг. 44. Схема, показывающая, как по разрывам в палеонтологической лето-
писи могут быть намечены условные границы отдельных видов в эволюцион-
ном ряду.
Изображены кривые распределения признака, развивающегося в гипотетическом ряду.
Каждая кривая относится к популяции, которая представляла этот эволюционный ряд
в данное время. Можно было бы изобразить больше или меньше кривых. Последователь-
ность кривых обнаруживает эволюционную тенденцию. Но в палеонтологической летопи-
си ряда имеются большие разрывы. Популяции сохранились лишь в четырех страти-
графических подразделениях (А, Б, В и Г), соответствующих вместе менее чем полови-
не времени, в течение которого развивался данный ряд Предположим, морфологичес-
кие различия между популяциями из разных стратиграфических единиц настолько ве-
лики, что если бы популяции А и Б жили в одно время, то скрещивание между ними
было бы, на наш взгляд, невозможным и что то же самое справедливо также для популя-
ций Б и В и популяций В и Г В то же время скрещивание внутри каждой из этих по-
пуляций представляется нам возможным. Тогда остается сделать вывод, что популяци-
онную группу из каждой стратиграфической единицы надо считать отдельным видом.
Таким образом, разрывы в стратиграфической летогиси заставили гас субъективно раз-
делить непрерывный эволюционный ряд на виды [134].
в других изменения совсем не проявляются. Большие разрывы
между сохранившимися фрагментами эволюционного ряда об-
легчают проведение границ между видами. Таким образом,
природа сильно упростила проблему вида в палеонтологии
(правда, за счет утраты весьма ценной информации).
И все же определение видовой принадлежности многих ис-
копаемых организмов связано с немалыми затруднениями. Что-
бы убедиться в этом, достаточно ознакомиться с несколькими
статьями, посвященными местным ископаемым фаунам. Обычно
значительную долю описанных в таких работах окаменелостей
не очень уверенно относят к определенным видам, а видовые
определения предшествующих авторов обычно берут под сомне-
ние или отвергают. Трудности работы палеонтолога связаны
не только с тем, что он пытается разделять на части контину-
по
Часть 1 Описание и классификация ископаемых остатков
умы (эволюционные линии), но и с тем, что на ископаемых
остатках не удается изучить многие признаки, обычно доступ-
ные биологу (как морфологические, так и этологические, эколо-
гические и географические).
Мы уже видели, что для описания вида недостаточно одной
особи, и рассмотрели примеры изменчивости внутри популяции
и между популяциями. Раньше, даже после того как теория
эволюции получила широкое признание, многие таксономисты
считали, что существует идеальная форма вида и что можно
выбрать отдельную особь — типовой экземпляр, или голотип,—
которая представляла бы эту идеальную форму; идея внутри-
видовой изменчивости при этом игнорировалась. В XX веке, ког-
да мы лучше стали понимать генетику популяций, такой «ти-
пологический подход» был почти вытеснен. Сейчас всеми при-
знано, что морфология вида лучше всего описывается стати-
стически, что позволяет учитывать значительную изменчивость.
Представление исследователя о живом или ископаемом виде
отражено в тех известных ему особях, которых он относит к
этому виду. Симпсон [194] предложил для такой группы особей
полезный термин гиподигм (hypodigm). Все особи гиподигма,
включая описанную ранее в качестве типового экземпляра, име-
ют одинаковый вес при описании вида. На практике возникает
проблема передачи представления о виде другим исследовате-
лям. Конечно, в большинстве случаев невозможно описать в
таксономических статьях все особи гиподигма. Этих особей мо-
жет быть несколько тысяч или даже миллионов. Проблема
частично разрешается помещением в начале описания вида так
называемой синонимики. Это перечисление особей или популя-
ций, описанных предыдущими авторами и включаемых автором
статьи в свой гиподигм. Многим из этих особей или популяций
предыдущие авторы могли давать другие видовые наименова-
ния, но эти наименования отвергаются составителем синонимики
и перечисляются только для исторической справки. Синонимика
редко отражает весь гиподигм, тем не менее она представляет
собой краткий обзор гиподигма автора, и ее можно использовать
для сравнения.
Формальное именование и описание
вида
Биологу или палеонтологу приходится выделять новый вид
в тех случаях, когда обнаруживаются особи, до того никем не
названные, или когда, по мнению исследователя, известный ра-
нее вид на самом деле состоит из двух или нескольких разных
видов. В палеонтологии обычно действуют одновременно обе
причины: исследователь может выделять новый вид, основы-
V Вид как целое
111
ваясь на новом ископаемом материале, но включая в него особи
или популяции, ранее относившиеся к другим видам. Как мы
видели, индивидуальное представление исследователя о виде,
его мнение о делении родов на виды — дело субъективное.
Именование видов, напротив, регламентировано определенными
правилами. В противном случае возник бы номенклатур-
ный хаос.
МЕЖДУНАРОДНЫЙ кодекс зоологической
НОМЕНКЛАТУРЫ
В разное время предлагалось множество разных систем
именования и описания видов. Одни продержались недолго,
другие дольше. Часто в разных странах использовали разные
системы. Системы варьировали от таксона к таксону. Все это
приводило к серьезной путанице. Но за последние полвека ис-
следователи, работавшие в разных странах с различными
группами растений и животных, сумели постепенно прийти к
соглашению. Теперь большинство вопросов, связанных с видо-
выми названиями, находится под контролем международных
организаций. Одна из самых важных — Международная комис-
сия по зоологической номенклатуре. Она работает под эгидой
Постоянного комитета международных зоологических конгрес-
сов и занимается проведением в жизнь и усовершенствованием
Международного кодекса зоологической номенклатуры.
Кодекс был принят V Международным зоологическим конг-
рессом, состоявшимся в Берлине в 1901 г. Это довольно про-
странный официальный документ, говорящий о правилах уста-
новления названий видов и других таксономических групп, а
также о проблемах, связанных с названиями, предложенными
в рамках старых систем. Комиссия действует как судейская
коллегия и законодательный орган, рассматривающий вопросы
таксономической процедуры.
Международный кодекс действителен и для современных,
и для ископаемых организмов, и это, конечно, удобно, так как
мало кто из таксономистов работает и с теми и с другими. Если
бы не унифицирующее действие Международного кодекса, мог-
ли бы развиться две совершенно независимые и, возможно,
взаимоисключающие системы.
Международный кодекс касается таксономических категорий
от подвида до надсемейства, но в этой главе мы будем говорить
в основном о его приложении к категории вида.
Международный кодекс применим почти ко всем вопросам
таксономии животных. Ботаники используют Международные
правила ботанической номенклатуры. Кроме того, для некото-
рых особых групп, например для бактерий, существуют еще
112
Часть 1. Описание и классификация ископаемых остатков
один-два свода правил. Но небольшие различия между этими
кодексами нас не касаются, и для общего ознакомления с пра-
вилами номенклатуры мы рассмотрим зоологический кодекс.
Что касается установления новых видовых названий, кодекс
дает указания по выбору названия, его опубликованию, описа-
нию нового вида и обозначению одного или нескольких типовых
экземпляров.
Чтобы вид был признан официально, ему надо дать название.
Выбор названия регламентируется некоторыми правилами.
Главное из них — название должно быть биноминальным, т. е.
состоять из двух слов. Официальное название вида, к которому
принадлежит человек, — Homo sapiens; sapiens — видовое на-
звание, а Ното — название рода, к которому относится этот
вид. В соответствии с Международным кодексом видовое на-
звание (называемое иногда тривиальным) — в данном случае
sapiens — не имеет смысла без родового. На практике большин-
ство заново открываемых видов легко может быть отнесено к
уже существующим родам, так что при описании нового вида,
как правило, приходится придумывать только одно название.
Но если новый вид явно отличается от всех установленных ро-
дов, то одновременно дается и новое родовое название.
Исключая только что упомянутое отнесение к какому-либо
роду, кодекс не требует полной таксономической идентифика-
ции нового вида, признавая, что полная классификация часто
затруднительна или невозможна, особенно если новый вид резко
отличается от всех других известных видов.
Самое главное требование к новому названию — это чтобы
оно не было уже занято («преоккупировано»). Это ограничение,
препятствующее возникновению гомонимов, относится к соче-
танию родового и видового названий, т. е. можно поставить во
вторую часть биномена название, использованное уже для ви-
дов в другом роде, но оно не должно совпадать с видовыми на-
званиями в данном роде. Договорились также не применять
одинаковых названий в близких родах, так как с углублением
наших знаний об эволюционных взаимоотношениях родов родо-
вая принадлежность видов часто меняется.
В качестве названий для видов и родов употребляются ла-
тинские или латинизированные слова. В выборе слов допускает-
ся значительная свобода — в качестве названия используются ла-
тинизированные названия местностей, имена и фамилии людей,
различные прилагательные. Впрочем, выбор ограничивается не-
которыми практическими и эстетическими соображениями, пере-
численными в кодексе.
. Чтобы название нового вида стало официально признанным,
его надо опубликовать в каком-либо распространенном издании,
т. е. оно должно быть напечатано типографским способом, до-
V. Вид как целое
113
статочно большим тиражом и должно распространяться по обыч-
ным каналам. Точные правила, регламентирующие форму пуб-
ликации, сложны. Их назначение — обеспечить всем таксоно-
мистам мира возможность публикации новых названий. Новое
название не признается официально, если оно просто занесено
в этикетку музейного экземпляра или если новый вид описан
устно перед ученым собранием.
Виды нельзя описывать анонимно. Поэтому авторство ново-
го вида — существенная часть описания. Чаще всего виды опи-
сываются одним автором, но вполне допустимо участие двух или
нескольких лиц.
Необходимость перечисленных требований к публикации яс-
на. Ежегодно описываются тысячи новых видов на многих язы-
ках, во множестве разных изданий, от серийных монографий,
издаваемых музеями, до крупных международных журналов.
Конечно, возникают сложные библиографические проблемы.
Международная комиссия попыталась их упростить, предложив,
чтобы публикации делались на нескольких рекомендованных
языках. Обычно рекомендуются французский, немецкий, анг-
лийский и русский, хотя кодекс не придает этой рекомендации
силы закона.
Международный кодекс не предписывает сопровождать пуб-
ликацию нового вида фотографией или другой иллюстрацией
с изображением описываемого экземпляра, но в приложении
к кодексу иллюстрирование настоятельно рекомендуется, и оно
вошло в традицию. Исходя из практических соображений, ил-
люстрирование можно считать обязательным.
О форме описания Международный кодекс говорит порази-
тельно мало. Основное требование — чтобы название сопровож-
далось «описанием», цель которого — выделить признаки, отли-
чающие данный таксон. Некоторые общие рекомендации Меж-
дународной комиссии, опубликованные в виде приложения к
кодексу, раскрывают это требование подробнее. Кроме того, сло-
жились уже традиционные формы описания.
Кодекс указывает, что каждое описание нового вида должно
сопровождаться обозначением типового экземпляра или серии
экземпляров. Это единственные экземпляры, к которым офици-
ально относится название нового вида. Кодекс требует из прак-
тических соображений, чтобы эти типовые экземпляры были
четко этикетированы, чтобы были приняты необходимые меры
для их сохранения и чтобы они были доступны заинтересован-
ным в их изучении исследователям. Обычно типовые экземпляры
сдаются в большой музей, где хранение хорошо поставлено.
Кроме рассмотренных правил именования видов, кодекс со-
держит многочисленные указания для казусов, связанных с ис-
торическим развитием системы именования и необходимостью
8—467
114
Часть 1. Описание и классификация ископаемых остатков
включить в систему виды, описанные не по правилам. Бывает,
что новое название предлагается для уже описанного и назван-
ного кем-то вида. В этом случае применяется так называемый
закон приоритета. За исключением некоторых особых случаев,
название, предложенное первым, предпочитается всем предло-
женным позже. Международный кодекс содержит указания по
разъяснению случаев такой путаницы и по улаживанию споров,
которые могут возникнуть.
В номенклатуре видов могут возникать и возникают самые
разнообразные затруднения. Все сказанное здесь —только вве-
дение к проблеме именования видов.
ФОРМА ОПИСАНИЯ
Майр, Линсли и Юзингер [128] предложили следующую
форму описания нового вида.
Научное название
Синонимика (если есть синонимы)
Голотип (включая указания о стратотипической мест-
ности и месте хранения)
Диагноз
Описание
Измерения и другие численные данные
Сравнительные замечания
Ареал (географический)
Местообитание (экологические замечания) и гори-
зонт (для ископаемых)
Перечень просмотренного материала
Научное название должно быть дано в биноминальной фор-
ме, согласно правилам Международной комиссии. Диагноз —
перечисление признаков нового вида, которыми он отличается
от всех других видов. Описание — полное перечисление призна-
ков без указания на сходство с другими признанными видами
или на отличие от них. Число измерений и других количествен-
ных данных, включаемых в описание, может быть различным
у разных авторов и меняется в зависимости от описываемой
группы организмов. Как минимум надо включать основные из-
мерения типового экземпляра или экземпляров. В разделе «За-
мечания» может содержаться информация о негенетической из-
менчивости, стадиях онтогенеза, эволюционном родстве с дру-
гими видами, а для ископаемых форм, кроме того, и сведения
о степени сохранности. Здесь также может разъясняться смысл
названия вида.
В разделе «Ареал» обычно перечисляются, кроме типового
местонахождения, другие места, где найден новый вид. Что
касается местообитания, палеонтолога особенно интересуют
V. Вид как целое
115
сведения о геологическом распространении. «Перечень просмот-
ренного материала» обычно включает ссылки на коллекции
музеев, где хранятся особи, не считающиеся официально ти-
повыми.
Чтобы лучше ознакомить читателя с формами описания, мы
здесь приводим три оригинальных описания вида, взятые из ли-
тературы, с фотографиями типовых экземпляров (фото XIV—
XVI). Эти примеры иллюстрируют однотипность формы описа-
ния и показывают допустимые отклонения от стандарта.
Девонская брахиопода Chonetes pachyactis Imbrie (фото XIV).
Отличительные признаки: Отношение длины к шири-
не невелико; резко выраженные широкие округлые ребра; уме-
ренная и одинаковая глубина створок; крупные размеры. Cho-
netes pachyactis наиболее близок к С. macltirea Norwood et
Pratten из известняков Иллинойса, но последний отличается
менее толстой спинной створкой и большей вздутостью ма-
кушки.
Описание. Крупная раковина; ширина взрослых особей
обычно равна 30 мм. Очертания от поперечно-полукруглых до
почти четырехугольных. Ребра отчетливые, широкие, округлые,
почти одинаковые по ширине и форме, со струйками; увеличение
числа ребер происходит в результате бифуркации (ветвления),
а иногда путем вставления; у переднего края насчитывается
около 7 ребер в расчете на отрезок 5 мм.
Брюшная створка. 6—8 шипов по каждую сторону макушки
вдоль замочного края.
Спинная створка. Боковой и передний профили умеренно
и равномерно вогнуты; толщина примерно равна толщине
брюшной створки. Задне-боковые области вогнуты несколько
менее, чем вся поверхность створки.
Размеры голотипа. Длина 20 мм, ширина 32 мм, тол-
щина 5,7 мм.
Распространение. Формация Норвэй-Пойнт (местона-
хождения 41, 46, 47), Мичиган.
Голотип. Экземпляр U.S.N.M.1 № 124397; формация Нор-
вэй-Пойнт, местонахождение 47.
Замечания. Анализ стадий роста этого вида показывает,
что основные пропорции в ходе роста регулярно меняются, так
что более крупные особи относительно шире и толще мелких.
Нижнетриасовый аммонит Xenoceltites youngi Kummel et Steele
(фото XV) [108].
Раковина с объемлющими оборотами, поперечное сечение
которых сжатое, дискоидальное. Брюшная сторона узкоокруг-
лая, боковые стороны выпуклые, пупковый край закругленный,
1 Национальный музей США.— Прим, перев.
8*
116
Часть 1. Описание и классификация ископаемых остатков
но хорошо выражен, пупковая стенка почти плоская и подходит
к пупковому шву под углом примерно 45°. К этому виду отне-
сены 3 экземпляра из коллекции. Их размеры:
Измеряемая величина Голотип MCZ1)5266 Паратип MCZ 5268 Паратип MCZ 5267
д 56,4 38,7 28,0
ш 13,7? 8,7? 6,1
в 22,9 16,8 12,5
Ду 18,3 11,5 7,6
ш/д 24,3? 22,5? 21,8
в/д 40,6 43,4 44,6
Ду/Д 32,4 29,7 27,1
1) Музей сравнительной зоологии, США.
Боковые стороны внутренних оборотов несут невысокие сла-
бо выраженные ребра. Эти невысокие ребра имеются на голоти-
пе и на крупном паратипе, но на меньшем паратипе внутренние
обороты совершенно гладкие, хотя в остальном он полностью
сходен с голотипом и с другим паратипом. На наружных обо-
ротах видны грубые линии нарастания, периодически сближаю-
щиеся, так что образуется нечто вроде извилистых ребер, не
заметных на внутреннем ядре. Грубые линии нарастания пере-
секают брюшную сторону, образуя иногда слабые пережимы
на брюшной стороне.
Перегородочная линия сохранилась плохо; она состоит из
двух тонко рассеченных боковых лопастей и низких широких
седел.
Замечания. Отнесение этого нового вида к роду Xenocel-
tites основано на наличии неясных невысоких ребер на внутрен-
них оборотах, на рисунке перегородочной линии и общем внеш-
нем сходстве с Xenoceltites subevolutus Spath (Frebold, 1930,
p. 14, pl. 3, figs. 1—3; Spath, 1934, p. 130, 131, pl. 2, fig. 2; pl. 8,
fig. 2; pl. 9, fig. 4; pl. 11, fig. 2). Большинство видов, отнесенных
Спэтом (1934) к роду Xenoceltites, имеет довольно ясно выра-
женные пережимы на внешних оборотах. Эта морфологическая
черта выражена у X. youngi слабо. Периферические пережимы
особенно выражены у X. spitsbergensis Spath и X. gregoryi Spath
(это формы с более эволютной раковиной, чем X. subevolutus).
Спэт (1934) пишет, что он имел 100 экземпляров Xenoceltites
со Шпицбергена; по Спэту, все три вида составляют последо-
вательный ряд переходных форм. Нельзя с уверенностью гово-
рить о диапазоне изменчивости шпицбергенских видов, но, судя
V. Вид как целое
117
по имеющимся данным, X. subevolutus мог иметь сильно реду-
цированные периферические пережимы на внешних оборотах,
в такой же степени, как у X. youngi.
В составе рода Xenoceltites: X. cordilleranus (Smith) и X. in-
termontanus (Smith) (известняки c Meekoceras, юго-восток Айда-
хо); X. hannai, X. douglasensis и X. matheri Mathews, 1929
(зона Anasibirites, Форт-Дуглас, Юта); X. robertsoni и X. war-
reni McLearn, 1945 (зона Anasibirites, Британская Колумбия);
X. russkiensis Spath, 1934 (Ceratites minutus Diener 1895, non
Waagen) (скифский ярус, район Владивостока, Сибирь); X. evo-
tutus Waagen, 1895 (цератитовые известняки нижнего триаса,
Соляной кряж, Пакистан).
Местонахождение. Пласт «а», зона Meekoceras graci-
litatus, Криттенден-Спринг, графство Элко, Невада.
Место хранения. Голотип MCZ 5266, паратипы
MCZ 5267 и не изображенный паратип MCZ 5268.
Верхнекембрийский трилобит Pterocephalops tuberculineata Clark
(фото XVI) [26].
Диагноз. Вид Pterocephalops со слегка суживающейся
глабелью, несущей глабелярные борозды и лишенной срединно-
го глабелярного бугорка.
Описание. Ширина кранидия примерно равна его длине.
Глабель почти прямоугольная, слегка сужающаяся к лобовому
краю. Наибольшая выпуклость глабели расположена поперечно,
кпереди от средней точки глабели. Две пары направленных
назад боковых глабелярных борозд видны на отпрепарирован-
ных экземплярах; на поверхности панциря они видны хуже.
Окципитальная борозда выражена хорошо. Окципитальное
кольцо шире всего в своей средней части и несет окципитальный
бугорок.
Выпуклая дублюра резко загнута вниз до уровня, ко-
торый значительно ниже глабели. Кайма той же ширины, что
и дублюра; она плоская, но край загнут. Лохман-Балк (1959)
рассматривает это как широкую краевую борозду при узкой
кайме; возможно, мы неверно толкуем эту структуру. Непод-
вижные щеки поднимаются круто вверх от спинных борозд поч-
ти до уровня вершины глабели. Пальпебральные лопасти не
сохранились. Ширина неподвижных щек —примерно одна треть
ширины глабели. Неподвижные щеки, начиная от точки, лежа-
щей слегка кпереди от центра глабели, резко загнуты вниз
спереди и сзади. Задняя часть неподвижных щек широка у ос-
нования и четко обозначена задней краевой бороздой. Лицевые
швы перед глазом и за глазом расширены слабо.
Свободные щеки, туловище и хвостовой щит не известны.
На поверхности разбросаны точечные бугорки, на кайме
поперечный ряд равномерно расставленных бугорков.
118
Часть 1. Описание и классификация ископаемых остатков
Размеры. U.S.N.M. 158449а: At: 8,4; А2 : 6,8; В : 4,7; Bi: 5,7;
D: около 3,6; J, : 10,1; J2 : 7,6; J3: 7,4; К: 4,1; Ki: 3,9; Li : 3,6.
U.S.N.M. 158449b: : 6,9; A2 : 5,5; В : 3,7; Bj : 4,6; Ji: 9,4;
J2 : 6,8; K:3,l; Ki : 2,9; Li: 3,0. Заметим, что A2 — сагиттальное
расстояние от задней границы окципитального кольца до осно-
вания крутого предглабелярного поля; оно может быть негомо-
логично применяемой обычно окципитальной интрамаргиналь-
ной длине головного щита (Шоу, 1957, стр. 194—195), но имен-
но такое измерение кажется логичным принять для данного
головного щита.
Сравнение. Этот вид очень сходен с Р. acrophthalma
Raselti, 1944 , но оба имеющихся образца отличаются от обоих
экземпляров Разетти, во-первых, несколько более сильной су-
женностью к глабели, во-вторых, довольно многочисленными
бугорками (на рисунках Разетти их видно немного), в-третьих,
наличием двух пар глабелярных борозд, которых нет даже на
очищенных частях Р. acrophthalma, и, в-четвертых, наличием
окципитального бугорка. Эти небольшие различия кажутся по-
стоянными. Очевидно, оба вида очень сходны.
Замечания. Видовое название tuberculineata напоминает
•о наличии ряда бугорков на кайме.
Голотип. U.S.N.M. 158449а (1 головной щит).
Паратип. U.S.N.M. 158449b (1 головной щит).
ДИАГНОЗ И ОПИСАНИЕ
Форма и содержание диагноза и описания в значительной
мере варьируют в зависимости от времени и места опубликова-
ния. Усилившийся интерес к внутривидовой изменчивости су-
щественно повлиял на стиль и содержание диагнозов и описа-
ний. Поскольку одна из главных целей в данном случае — пе-
редача информации, понятно, что здесь высоко ценится унифи-
кация. Например, везде, где только возможно, используется
стандартизованная терминология. Если бы каждый таксономист
изобретал новые термины для описания окаменелостей, возник
бы полнейший хаос.
Дать абсолютно полный диагноз или описание невозможно.
Как мы знаем из гл. II, для описания окаменелости можно ис-
пользовать бесконечное число признаков. Описать исчерпываю-
ще один экземпляр или группу экземпляров невозможно, да
и не нужно. В диагноз следует включать столько признаков,
сколько необходимо для того, чтобы отличить особи нового ви-
да от особей близкородственных видов. Обычно особое внима-
ние уделяют признакам, наиболее заметным при любой степени
сохранности. Если сообщается слишком много признаков, то
система становится громоздкой и внимание отвлекается от са-
V. Вид как целое
119
мых важных признаков. Поэтому в диагнозе могут отсутство-
вать некоторые признаки, которые являются тем не менее
диагностическими. Вот в этом случае особенно важными ста-
новятся хранящиеся экземпляры, которые служат «поддержкой»
диагнозу.
Описание в отличие от диагноза служит нескольким целям,,
и не последняя из них — перечисление признаков, которые в
будущем могут оказаться существенными для диагноза. Если
вид входит в хорошо изученную группу и во многих отношениях
сходен с другими ее членами, то описание может быть кратким,,
так как существуют описания близкородственных видов. При
этом можно ограничиться простым диагнозом. Виды, относя-
щиеся к сравнительно мало изученным группам, надо описывать
подробнее, чтобы в случае последующего обнаружения близких
видов можно было провести необходимое сравнение.
Желательно, чтобы описание содержало сведения об онто-
генезе, особенно если он сопровождается изменениями формы.
Важно описать изменчивость, обнаруженную внутри популяций
и между популяциями вида. Если новый вид устанавливается
на основании лишь нескольких экземпляров или их фрагментов,
то описать изменчивость невозможно.
Довольно много спорили о том, сколько экземпляров необ-
ходимо для установления нового вида. Некоторые утверждали,
что, поскольку внутривидовая изменчивость очень важна, нель-
зя устанавливать новый вид на основании одного экземпляра
или фрагмента. Но тут не может быть универсального правила.
В каждом конкретном случае решение зависит от степени от-
личия от родственных видов. Часто и на единственном экземп-
ляре видно, что организм отличается от всех известных, и нет
смысла ожидать, пока накопится достаточно много экземпля-
ров. Многие важные палеонтологические открытия основаны на
одиночных экземплярах или даже на фрагментарном материале.
С другой стороны, если новый вид принадлежит к хорошо изу-
ченной группе, в которой различия между видами невелики, то
надо накопить обширный материал для того, чтобы описание
было полным и эффективным.
Многие требования к объему и содержанию описания про-
диктованы практическими соображениями. Описание не должно
быть слишком длинным, иначе его трудно будет прочитать и
усйоить. Общий объем диагноза и описания нового вида вместе
с замечаниями обычно не превышает 2000 слов. Ввиду быстрого
развития автоматических методов хранения, переработки и из-
влечения информации многие таксономисты с надеждой
смотрят на открывающиеся возможности расширения емкости
описания. Во многих случаях было бы сравнительно легко за-
кодировать описание нового вида, чтобы оно могло храниться.
20
Часть 1. Описание и классификация ископаемых остатков
е занимая много места, и быть извлечено в любой момент для
равнения с другими видами. В последнее время разработаны
екоторые принципы такой обработки и проведены предвари-
ельные исследования. Конечно, это заманчивая возможность —
ократить информацию, содержащуюся в двух тысячах слов
писания ископаемого вида, до перечисления 30—40 морфоло-
ических признаков, которое займет всего несколько сантимет-
ов магнитной ленты. Можно будет даже отказаться от диагно-
а в его обычной форме — описания признаков разных видов
югут сравниваться автоматически, а различия, отмеченные ма-
циной, и будут представлять собой диагноз.
Особенность строения организма, использованная в диагно-
е, называется таксономическим признаком. Чем таксономиче-
кий признак отличается от других признаков? Здесь мы по-
ддаем в заколдованный круг: таксономический признак — это
[ризнак, оказавшийся полезным для таксономического разгра-
[ичения. Это признак, пригодный для выявления различий меж-
(,у таксономическими группами.
Чтобы быть пригодным для различения видов, признак дол-
кен удовлетворять нескольким условиям. Поскольку различе-
[ие видов в конечном счете сводится к оценке степени генети-
[еских различий между популяциями, таксономическим может
>ыть признак, проявляющий минимум негенетической изменчи-
юсти. Этот признак должен быть хорошо заметным, что особен-
to относится к ископаемому материалу, так как неполнота
охранное™ часто приводит к неполноте общего представления
>б организме. В идеале таксономический признак должен при-
утствовать и быть различимым на протяжении всего онтогене-
а, а не только на некоторых его стадиях.
Многие морфологические признаки, отвечая перечисленным
ребованиям, все же не могут служить для целей таксономии.
Это обычно признаки, хотя и подчиненные генетическому контро-
но, но подверженные сильной генетической внутрипопуляцион-
юй изменчивости. Пример — цвет волос у человека.
Насколько эффективен данный таксономический признак,
дожет зависеть от того, какую таксономическую категорию мы
шссматриваем. Многие признаки слишком изменчивы для того,
[тобы пользоваться ими при различении видов, но могут быть
срайне важными при описании подвидов. Другие признаки не
вменяются от вида к виду, но важны при различении родов
I семейств.
Биологи часто выбирают таксономические признаки, связан-
1ые с анатомией мягких частей тела или с поведением, но сви-
щтельства о них редко сохраняются в палеонтологической ле-
юписи. Из-за этого возникают трудности в таксономии тех
фупп, которые имеют и живых, и вымерших представителей.
V. Вид как целое
121
Очень часто палеонтолог в таких случаях переисследует живые
виды и находит набор вторичных таксономических признаков,
основываясь на хорошо сохраняющихся частях организма. Но
некоторые живые виды не удается различать по подобным при-
знакам. В результате обычно под одним названием объединя-
ется несколько ископаемых видов, хотя если бы мы имели пол-
ное представление об их морфологии, то мы бы разделили их.
Подведем итог. Таксономические признаки выбираются в
первую очередь по их полезности для целей таксономического
различения. Их выбор в значительной мере основан на опыте.
В целом таксономические признаки являются, вероятно, наибо-
лее важными с биологической точки зрения.
ТИПОВЫЕ ЭКЗЕМПЛЯРЫ
Выбор типовых экземпляров связан с некоторыми трудностя-
ми. Если обозначается один экземпляр, его называют голоти-
пом. Если выбрано несколько экземпляров, они называются
синтипами. Правила допускают оба варианта, хотя Междуна-
родная комиссия настоятельно рекомендует использовать не
серию синтипов, а голотип, так как всегда остается возмож-
ность, что последующие исследователи разделят серию на два
или несколько видов.
Раньше типовые экземпляры широко использовались для
описания видов и как специальные эталоны для сравнения. Но
теперь (частично благодаря принятию популяционного подхода
в таксономии) мы знаем, что многие из этих «эталонов» не
являются самыми представительными экземплярами своих ви-
дов. Даже в большой выборке может не найтись особи, которая
являлась бы некой средней особью по всем наблюдаемым мор-
фологическим признакам. Кроме того, средняя величина измен-
чивого признака ничего не говорит о размахе изменчивости.
В современной биологии и палеонтологии типовой экземпляр
не служит более основой для диагноза вида (кроме тех случа-
ев, когда других особей нет). Вместо этого он стал «носителем
названия». Когда вид получает название, оно формально да-
ется только одному или нескольким типовым экземплярам. На
практике типовые экземпляры ископаемых видов не являются
типичными представителями гиподигма; обычно они уклоняются
в сторону большего размера и лучшей сохранности.
На разных стадиях таксономической работы важную роль
играют и некоторые другие виды типовых экземпляров. Пара-
тип— всякий экземпляр, кроме голотипа, который формально
обозначен автором вида как использованный при описании ви-
да. Таким образом, обозначение одного голотипа и серии пара-
типов позволяет сохранить некоторые преимущества и системы
122
Часть /. Описание и классификация ископаемых остатков
голотипа, и системы синтипа. Голотип остается носителем на-
звания, а паратипы, которых может быть много, служат для
более полного выражения авторской концепции данного вида.
Лектотип — экземпляр, исходно обозначенный как синтип, но
затем выделенный в качестве окончательного типового экзем-
пляра вида. В этом не было бы необходимости, если бы автор
сразу обозначил голотип и серию паратипов.
Случается, что типовые экземпляры уничтожены или потеря-
ны. Международная комиссия установила порядок обозначения
новых типовых экземпляров-—неотипов — взамен утраченных.
Для видов, описанных много лет назад, часто важно бывает
заново описать экземпляры и дать их новые изображения, осо-
бенно если оригинальное описание составлено по правилам,
которые теперь не применяются. Плезиотипы — типовые экзем-
пляры, используемые для переописания существующего вида.
Выделяются также другие категории типовых экземпляров.
Например, топотип — экземпляр, не входящий в оригинальный
типовой материал, но собранный в типичном местообитании.
Изменение видовых названий
Исследователь может изменить название таксона, если его
использование нарушает какое-то правило номенклатуры или
если он считает, что таксон был классифицирован неверно. Мы
рассмотрим только случаи изменения видовых названий.
Одинаковые видовые названия, относящиеся к разным ви-
довым группам, называются гомонимами. Первичные гомони-
мы— это идентичные названия, установленные для разных так-
сонов (с разными голотипами), относящихся к одному роду.
Автор гомонима, появившегося позднее, заблуждался, не зная,
что использованное им видовое название уже занято. После
обнаружения такой ошибки оставаться в силе может только
опубликованный раньше, старший гомоним. Кодекс предписы-
вает всегда отвергать младший гомоним. Вторичные гомонимы
возникают, когда вид переносится в другой род, в котором уже
есть вид с тем же видовым названием. Согласно кодексу, ни
один из авторов этих двух видов не совершил ошибки, посколь-
ку видам, принадлежащим к разным родам, можно давать одно
и то же видовое название. Проблема возникает в том случае,
если впоследствии оба вида относят к одному роду. Исследова-
тель, обнаруживший вторичные гомонимы, по правилам дол-
жен отвергнуть младшее наименование. Отвергнутый первич-
ный или вторичный гомоним должен быть замещен самым ста-
рым из пригодных названий, а если ранее опубликованные
названия непригодны, то новым.
Два разных названия, данные одному таксону, называют
V. Вид как целое
123
синонимами. Объективные синонимы — это различные названия,
предложенные для одного типового экземпляра (или экземпля-
ров) . Здесь никакой проблемы не возникает; старший (т. е. ра-
нее опубликованный) синоним должен быть сохранен, младший
же синоним всегда отвергается. Субъективные синонимы — это
названия, установленные для разных типовых экземпляров, ко-
торые впоследствии были отнесены каким-либо автором к одно-
му виду. Этот автор должен применить к обоим типовым эк-
земплярам и ко всем экземплярам гиподигма, к которому он
их отнес, старший синоним. Но другой исследователь может
полагать, что эти типовые экземпляры относятся к разным ви-
дам; он не сочтет названия синонимами и сохранит оба. Итак,
кодекс автоматически устраняет младший объективный сино-
ним, тогда как младший субъективный синоним остается при-
годным названием, и его использование целиком зависит от
того, на какой позиции стоит исследователь.
Иногда на основании приоритета приходится заменять на-
звания, ставшие привычными. Многие исследователи были не-
довольны изменением родового названия всем знакомого эоце-
нового предка лошади Eohippus на Hyracotherium. Кодекс
предоставляет право Комиссии по зоологической номенклатуре
отменять в некоторых случаях действие закона приоритета.
Благодаря этому сохранились многие привычные названия, ока-
завшиеся младшими гомонимами или синонимами.
Перспективы
Для палеонтолога таксономическая процедура — очень слож-
ный и важный предмет. В действительности она значительно
сложнее, чем описано здесь. Сложность этой процедуры и много-
численность, казалось бы, устаревших и неэффективных правил
часто поражают и озадачивают студентов. Появление возмож-
ности изучать биологию и палеонтологию организмов с других
планет дало толчок многим реформаторским идеям. Часто
утверждают, что исследование космоса дает нам повод испра-
вить ошибки и ввести новую, современную систему.
Нельзя отрицать возможность улучшения применяющихся
ныне методов. Самым большим препятствием к широкой рефор-
ме служит значительность связанных с нею затрат. Изменить
систему именования и классификации 1 500 000 современных и
130 000 ископаемых видов — задача гигантская. Масштаб проб-
лемы тормозит прогресс, но он же оказывает стабилизирующее
действие и предотвращает путаницу, которая, несомненно, воз-
никла бы в результате проведения частичных реформ. Вероят-
но, следующее поколение ученых займется усовершенствованием
системы, но вряд ли она будет изменена существенно.
ГЛАВА VI
Группировка видов
в высшие категории
Именование видов обычно идет рука об руку с их организа-
цией в систему. Виды с общими признаками группируют вместе
и отделяют от других групп, что облегчает сообщение инфор-
мации о них. Например, можно сказать: «Я видел, как птица
ела червяка». Но сейчас известно около 8600 видов птиц и око-
ло 25 000 видов червей, и если слова «птица» и «червяк» заме-
нить в этой фразе на видовые названия, то фраза окажется не
понятной ни для кого, кроме специалистов. Используя слово
«птица», мы описываем пожирателя червяка как члена четко
определенной группы сходных существ, тем самым исключая
из рассмотрения всех ныне живущих животных, кроме 0,1%.
Такая формулировка допустима и в тех случаях, когда уточнять
вид нет необходимости или если это затруднительно.
Возможно, главная причина, заставляющая объединять виды
в категории более высокого ранга, состоит в том, что человек не
может выучить и запомнить названия и отличительные признаки
полутора миллионов разных организмов. В примере с птицей
и червем хорошему орнитологу, возможно, удалось бы опреде-
лить вид птицы (особенно если он знаком с фауной данного
района), но вряд ли он смог бы определить червя. В этом от-
ношении объединение видов в более крупные категории ком-
пенсирует недостаточный объем человеческой памяти. Обычный
порядок определения экземпляра состоит в том, чтобы опреде-
лить сначала тип, а потом постепенно идти от общего к част-
ному.
Как лучше классифицировать растения и животных? Группу
видов можно подразделять различными способами. Скажем,
можно разбить все виды на 26 категорий в алфавитном порядке
в зависимости от того, с какой буквы латинского алфавита на-
чинается название вида. Несколько более логично было бы рас-
положить виды по годам их открытия. Но такие системы не
помогут наблюдателю, поскольку организмы не имеют видимых
признаков, которые позволили бы классифицировать их в соот-
ветствии с такими системами.
Другие системы классификации основаны на легко наблю-
даемых признаках: на внешнем виде, типе местообитания, осо-
VI. Группировка видов в высшие категории
125
бенностях поведения, географическом или стратиграфическом
распространении. ?4ногие из ранних систем классификации
позвоночных были основаны на типе местообитания. Водные
позвоночные (рыбы) отделялись от наземных (пресмыкающие-
ся, птицы и млекопитающие) и т. д. Хотя некоторые группы
(например, водные млекопитающие) не укладывались в такую
систему, она была полезна во многих отношениях.
Самые удачные системы классификации основаны на морфо-
логии. Это слово мы применяем в широком смысле, включаю-
щем не только внешнюю форму, но и внутреннюю анатомию
и даже некоторые детали физиологии, биохимии и поведения.
Большинство групп организмов классифицируется сейчас
в соответствии с системами, возникшими задолго до того, как
теория органической эволюции стала общепринятой. Системати-
ки додарвиновской формации обычно считали, что виды, с кото-
рыми они работают, были созданы независимо друг от друга.
И все же некоторые ранние системы, особенно в том, что ка-
сается деления на высшие таксоны, изменились после распрост-
ранения эволюционной теории лишь весьма незначительно.
Классификация и эволюция
Если классификация должна служить в первую очередь це-
лям описания и определения, то главным критерием для выбора
системы является ее полезность. С повышением интереса к эво-
люции от классификации стали также требовать, чтобы она
выражала эволюционные отношения.
В классификации большинства групп растений и животных
повышение интереса к эволюции не вызвало особых переворо-
тов главным образом потому, что морфологическое сходство
тесно связано с эволюционной близостью. Поэтому система
классификации, призванная отражать эволюционные взаимоот-
ношения, в основном должна быть построена на тех же чертах
сходства и различия, что и система, облегчающая определение
организмов.
Классификации постоянно изменяются. Частично это явля-
ется результатом расширения наших знаний о богатстве живого
мира. Часто приходится создавать новые высшие категории,
чтобы отразить различия между новыми и давно известными
видами. Кроме того, изменения в классификации обусловлены
накоплением теоретических знаний о механизмах эволюции. По-
этому можно сказать, что классификация всегда отражает со-
временный ей уровень эволюционного мышления. Системы клас-
сификации, используемые разными одинаково высококвалифи-
цированными учеными, обычно различаются просто потому, что
по-разному интерпретируется эволюция.
126
Часть 1. Описание и классификация ископаемых остатков
Таксономические категории
Линней в 1759 г. использовал всего шесть таксономических
категорий: царство, класс, отряд, род, вид и разновидность. Из
них высшими категориями мы считаем царство, класс, отряд
и род. Поскольку таксономическую категорию царство Линней
употребил только для отделения животных от растений, в своей
зоологической работе он применял только три высшие катего-
рии: класс, отряд и род.
Во времена Линнея число известных видов было сравни-
тельно невелико, и этих категорий было достаточно для под-
разделения всего множества форм на удобные в обращении
группы. Громадный рост числа известных видов потребовал
выделения добавочных категорий, ниже царства и выше вида.
Главные добавления — тип и семейство, так что теперь, как
правило, используется следующая иерархия: царство, тип,
класс, отряд, семейство, род. Для некоторых групп употребля-
ют также категории подтип, подкласс, надотряд, подотряд, над-
семейство, подсемейство и подрод.
Каковы определения этих категорий? Проще всего опреде-
лить каждую из них как часть ближайшей высшей категории.
Международный кодекс зоологической номенклатуры содер-
жит немало правил пользования высшими таксономическими
категориями. Сам кодекс включает правила образования кате-
горий до уровня надсемейства. Но в рекомендациях Междуна-
родной комиссии, приложенных к кодексу, содержатся указания
по всем высшим категориям (если не непосредственные, то под-
разумеваемые). Правила почти параллельны тем, что относят-
ся к видам. Например, мы уже видели, что новый вид должен
быть отнесен хотя бы к ближайшей высшей категории (роду).
Подразумевается, что каждый род может быть отнесен к се-
мейству, отряду и так далее, хотя допускается, что эволюцион-
ные связи рода могут быть неясными. В таком случае род
может оставаться изолированным («incertae sedis»)1. Сущест-
вуют правила образования названий высших категорий, что
позволяет определять таксономический ранг по окончанию на-
звания.
Концепция типа распространяется и на высшие категории.
Когда предлагается новый род, оригинальное описание должно
сопровождаться обозначением типового вида. Типовой вид, та-
ким образом, служит носителем названия рода. Подобным же
образом семейство должно иметь типовой род, и т. д.
При описании и именовании нового вида необходимо прило-
жить все усилия, чтобы найти ему подходящее место в сущест-
1 Неопределенного положения (лат.) — Прим, перев.
VI. Группировка видов в высшие категории
127
вующих высших таксонах. Если это оказывается невозможным,
то иногда предлагают новые категории; может быть предложено
не только название типа, но и новые названия класса, отряда,
семейства и так далее. Это иллюстрирует важную черту обыч-
ной системы классификации: высшая категория может быть
монотипической. Тип может иметь всего один класс; в классе
может быть всего один отряд, и т. д.
Система классификации
как интерпретация эволюции
В большинстве случаев системы классификации подразуме-
вают, что эволюция следует по определенному пути. В гл. IV
мы говорили о двух способах видообразования: филетическом
и географическом. Первый представляет собой постепенный пере-
ход одного вида в другой по одной эволюционной линии, а вто-
рой связан с разветвлением одной линии на две, что часто
выражают графически в виде филогенетического древа. На
фиг. 45 и 46 показаны два различных филогенетических древа.
На фиг. 45 показано возникновение нескольких близкородст-
венных видов за короткое время; каждый участок древа явля-
ется частью развивающейся линии.
Филогенетическое древо в лучшем случае является упроще-
нием действительности. В каждый момент времени вид обычно
представлен многочисленными географически разделенными
популяциями. Элементы эволюционного ряда могут дивергиро-
вать с появлением географической изоляции, а позднее, если
географические преграды исчезают, они конвергируют. «Внут-
реннее строение» эволюционной линии показано на фиг. 47.
На фиг. 46 показана эволюция большой группы видов, объ-
единяемых в отряды, в течение длительного промежутка време-
ни. Следовательно, одна линия на фиг. 46 представляет собой
комплекс многих развивающихся линий, каждая из которых со-
стоит из развивающихся популяций.
Какая система координат используется при построении фи-
логенетического древа? Применяется несколько систем; важно
понять разницу между ними. На фиг. 45 и 46 по вертикали
отложено время, по горизонтали — степень родства между ли-
ниями. На таких филогенетических древесах окончание ветви
означает вымирание эволюционной линии.
На фиг. 48 показано «безразмерное» филогенетическое дре-
во. Здесь конец ветви не означает вымирания эволюционной
линии, хотя п может совпадать с ним, а расстояние между вет-
вями не говорит о географическом или эволюционном расстоя-
нии. Время отражено на диаграмме только в указании хроноло-
гической последовательности эволюционных событий.
I
I
Mucrospinfer |
mucronatus'^^^JT
Нейплс
Фиг. 45. Филогения рода Cyrtospirifer (верхний девон — нижний карбон
штаты Нью-Йорк и Пенсильвания, США) [71].
Слева — названия ярусов.
Perischo-
echinoidea
Euechinoidea
Фиг. 46. Филогения класса Echinoidea, основанная на стратиграфическом
;распространении представителей разных отрядов и предполагаемом эволю-
! ционном родстве.
IПерерыв между пермью и триасом на схеме объясняется изменением вертикального мае
штаба [48].
Cdiophora
Фш.47. Строение эволюционного
ряда.
А Онтогенезы особей, входящих
в популяцию Горизонтал ьные линии
означают скрещивания Б Развиваю
щаяся совокупность популя ций состав
ляет ряд В Связь между нескольки
ми рядами [197J
о—'Другие
•-Holotricha -
Phytomastsgcphora
Spirotncha
Rhizopoda
Сложные'
симбионты
Через некоторые
водоросли
Т
Opalinidae
Suctona
[Metazoa]
Sporozoa
Zoomasti-
yophora
Telosporidia
Cnidosporidia
Формы,парази-
тирующие в
крови
Свободноживущие
[Porrfera]
[ Грибы] Actinopoda
> Sarcodina
Фиг. 48. Предполагаемая филогения простейших.
Группы, которые, как полагают, произошли от простейших, указаны в квадратных скоб-
ках (34J
Фиг. 49. Предполагаемая филогения лошадей 1195].
На фиг. 49 по вертикали и горизонтали отложены соответ-
ственно время и географические области. Построить филогению
таким образом удалось потому, что эволюционные ряды редко
перекрываются географически.
На фиг. 50 показано подразделение 31 рода брахиопод на
11 подсемейств (их названия оканчиваются на «-inae») и 4 се-
мейства (окончание «-idae»). Если признать, что эта система
действительно отражает эволюцию, то можно рассматривать
9*
132
Часть 1. Описание и классификация ископаемых остатков
Chonehnae
——————————— — Chonetes Strophochonetinae
OUUpilUCllUlieibC)
Deoonochonetinae - Devonochonetes —— I Zl/P Oin/V
l on gispina — . Агл At rt rt 9~r> c*
lyullULilunElSd
гою cnoneies
Anophinae Д / lOpllCl
П OpHupolo Г'ЬлпоИпп
hllUIKUllU in
MUlUHUpUCL Tnrnmristi л
Chonetidae Retlchonetlnae 1 XJI I
г lit tXCz/1 Ul Di
Rugosochonetinae
L-/IJUJ CAj Т*П 1 ЮС П П nrt fd/ic
bOLL^ocnuncies
. Л//С
• “ 1 iVittbUlULJub — DIinmr hniw foe
i llUJilZsLt-x)
yuauivuHoiieieb ("'.hrtTiPtinpll п
Chonetinellinae xsl L\Ji IZsLLJ
_ ТЛ/zr/TZTO Ш I z>c
Semenewiinae у r LLUUCt
Eodevonariidae
LOueUOnUHU
Chonostrophiidae Choiiostivphid
" ” ^nunosiiupnisiiu Tnlf'nfn
1 Uli* lAlll tCv DaviesieUinae
Daoiesiellidae Delepineinae
— - veiepineu
Airtoniinae ' iweuucrionEicif Airtonia
Фиг. 50. Классификация вымершего надсемейства брахиопод Chonetacea,
изображенная в виде дендрограммы (из [171] по [132]).
ее как филогенетическое древо (без указания времени и гео-
графии) . Роды, считающиеся особенно близкими, объединены
на схеме в подсемейства, а близкородственные подсемейства
сведены в семейства. Вся группа из тридцати одного рода пред-
ставляет собой надсемейство.
Отнесение ранних форм к высшим категориям часто зависит
ют наших знаний о дальнейшей эволюционной истории. В юр-
VI. Группировка видов в высшие категории
133
ских зольнхофенских известняках были найдены ископаемые,
которые считают остатками первой настоящей птицы. По своей
морфологии это пресмыкающиеся (у них есть зубы, длинные
хвосты, да и общий план строения напоминает рептилий), но
у них имеется нечто вроде функциональных крыльев с перьями.
Зная, что в позднем мезозое эта модификация в анатомии
пресмыкающихся привела к сильной адаптивной радиации
(класс Aves), мы придаем зольнхофенским экземплярам особое
эволюционное значение. Мы определяем и классифицируем их
как птиц (класс Aves), хотя они ближе к мезозойским пресмы-
кающимся, чем к современным птицам. Если бы не было из-
вестно, что зольнхофенская птица дала начало такому широко-
му разнообразию организмов, сильно отличающихся от репти-
лий, то мы скорее всего сочли бы ее просто аберрантной
рептилией.
Тут можно задать вопрос, имеют ли высшие таксономические
категории объективное содержание. Имеют и не имеют в одно
и то же время. В определении биологического вида использует-
ся концепция репродуктивной изоляции; ее рассматривают как
важное биологическое явление, позволяющее развивающимся
линиям оставаться генетически различными. В этом смысле
данное Симпсоном определение эволюционного вида имеет не-
которое объективное основание, хотя подразделение эволюци-
онных линий на виды условно. Но семейство уже не имеет тако-
го объективного содержания. Высшие таксономические катего-
рии используются для выражения эволюционных соотношений,
но сейчас, на современном уровне наших знаний, мы не имеем
специальных правил для выделения этих категорий. Другими
словами, между родом и семейством нет такого порога, как
между подвидом и видом (в каждый данный момент времени).
И все же разделение на высшие категории в какой-то мере
объективно. Так, классификация в целом основана на предпо-
ложении, что все виды одного рода произошли от одного пред-
кового ряда. Это следствие нашего представления о том, что на
надвпдовом уровне филогения всегда выражается в постоянном
ветвлении. Если можно доказать, что два вида имеют общего
предка, не являющегося предком третьего вида, то первые два
вида можно объективно поместить в высший таксон, к которо-
му не относится третий. Но это не поможет объективно решить,
где кончается уровень семейства и начинается уровень отряда,
так как различие между этими уровнями только количествен-
ное. Не удивительно, что системы классификации, разработан-
ные разными таксономистами, часто различаются по составу
высших категорий. Однако системы, основанные на одинаковом
понимании филогении, должны быть взаимно коррелируемы.
Биолог или палеонтолог редко имеет точное представление о
134
Часть 1. Описание и классификация ископаемых остатков
филогении группы, поэтому система отражает современное ей
состояние знаний.
Подходы биологов и палеонтологов к проблемам, связанным
с высшими таксономическими категориями, несколько различа-
ются, так как палеонтолог может прямо наблюдать изменения
во времени, а биологу приходится делать косвенные выводы
о хронологии развития в эволюционных рядах. Палеонтолог
иногда способен доказать, что один вид произошел от другого,
так как он находит их в последовательно залегающих пластах.
Критерии выделения высших
категорий
Если классификация должна отражать филогению, то мы
должны уметь оценивать «филогенетическое расстояние», или
степень ’ генетического различия между видами. Генетическое
различие нельзя прямо наблюдать ни у ископаемых, ни у боль-
шинства современных форм. Однако морфологические разли-
чия можно использовать как приблизительную меру генетиче-
ских различий (так же мы поступаем, работая с видами). Два
вида из группы, имеющие максимум общих морфологических
признаков, скорее всего произошли от общего предка и, значит,
должны быть отнесены к одной высшей категории.
Большое число видов, обладающих множеством таксономиче-
ских признаков, можно распределить по группам многими очень
различными способами. Какую классификацию из нескольких
имеющихся следует применить? Какая из них верно отражает
филогению? Иногда кажется, что ответов на эти вопросы столь-
ко же, сколько таксономистов. Говорили даже, что\ таксономия
на этом уровне не наука, а искусство и что ее методы не под-
даются четкому и логичному объяснению. Рассмотрим некото-
рые часто употребляемые методы. \
Очень часто a priori выделяют несколько признаков как
имеющие большее эволюционное значение, чем остальные, и
подразделение основывается на этих избранных признаках. Та-
кое взвешивание признаков проводится различны^ образом.
В приведенном выше случае с остатками из Зольнхофена кри-
терием для отнесения их к классу Aves стало наличие перьев.
Основано такое суждение главным образом на тбм, что перья
послужили важнейшим признаком, обеспечившим организму
возможность занять ряд местообитаний, в которых не могли
жить его предки, и благодаря этому стала ^бзможной его ог-
ромная эволюционная экспансия.
В других случаях выбор признаков основан на оценке влия-
ния взвешивания на создаваемую классификацию. На фиг. 51
показаны чисто произвольные данные, с помощью которых мы
VI Группировка видов в высшие категории
135
иллюстрируем этот способ взвешивания. Каждый из десяти ги-
потетических видов имеет по десять морфологических признаков.
Каждый из признаков представлен у вида в одном из двух воз-
можных состояний, отмеченных плюсом или минусом (напри-
мер, «большой» и «маленький», «красный» и «белый», «с зуба-
ми» и «без зубов» и т. д.).
Попробуем разделить эти виды на две группы (любого раз-
мера). Примем, что эти две группы соответствуют разным вет-
вям филогенетического древа; для выделения этих групп мы
должны использовать морфологические данные. Разделить де-
сять видов на две группы можно 637 разными способами. Нам
надо найти способ, дающий наиболее близкий к истине ре-
зультат.
Некоторые из возможных подразделений с морфологической
точки зрения более оправданны. На фиг. 52 показаны три спо-
соба подразделения. С помощью первого из них мы получаем
две группы, гетерогенные по всем морфологическим признакам.
Второй дает группы, гомогенные по одному из признаков (чет-
вертому). Третий дает гомогенность по трем из десяти призна-
ков (1, 7 и 8). Можно сказать, что последний способ «под-
тверждается» 30% признаков. Заметим, что эту систему класси-
фикации «почти поддерживает» еще один признак (6-й): одна
группа полностью гомогенна по признаку 6 (все плюсы), а дру-
гая—почти гомогенна (у всех видов, кроме одного, минусы).
Три признака, безупречно поддерживающие такую группировку,
можно назвать «превосходными» таксономическими признака-
ми, потому что все они подтверждают одну и ту же систему.
Признак 6 можно назвать «пригодным» таксономическим при-
Признаки
1 2 3 4 5 6 7 8 9 10
А -— + —_ + — + + — + —
В — — — + + + — — +
С + + + + — — — + — +
D + + —— — — — — + + +
Виды Е + — + — 4- 1 + — + — —
F + — —_ + + — — + — +
G — — “Г —_ + + + — + —
Н — + — + — + + — + +
I + — + — + — — + + +
J — + — — — + + — —- +
Фиг. 51. Закодированная морфология 10 гипотетических видов.
Система 1 Признаки
1 2 3 4 5 6 7 8 9 10
А — 4- — + — 4- 4- — 4- —
В — — — — 4- 4- 4- — — 4- Род 1
С 4- 4- 4- 4- — — — 4- — 4-
Виды D 4- 4- " — — — — 4- 4- 4-
Е 4- 4- — 4- 4- — 4- — —
F 4- — — + 4- — — 4- — 4-
G — — 4- — 4- 4- 4- —.— 4- — Род 2
Н — 4- — 4- —— 4- 4- — 4- 4-
I 4- — 4- — 4- — — 4- 4- 4-
J .— 4- — .— —— 4- 4- . 4-
Система 2
Признаки
1 2 3 4 5 6 7 8 9 10
А — 4- — 4- — 4- 4- — 4- —
С 4- + 4- 4- — — — 4- — 4- Род 1
F 4- — — 4- 4- •— — 4- — +
Н — 4- — 4- 4- 4- — 4- 4-
Виды В .—. — -— — 4- 4- 4- — — 4-
D 4- 4- — — — — — 4- 4- 4-
Е 4- — 4- — 4- 4- — 4- — — Род 2
G — — + — 4- 4- 4- — 4- —
I 4- — — — 4~ — — 4- 4- 4-
J — 4- 4- — ——•— 4- 4- — •— 4-
Система 3 Признаки
1 2 3 4 5 6 7 8 9 10
А — 4- — 4- — 4- 4- — 4- •
В — — — — 4- 4- 4- — — 4-
G — — 4- — 4- 4- 4- — 4- — Род 1
Н — 4- — 4- — 4- 4- — 4- 4-
Виды J — 4- — — — 4- 4- — — 4-
С 4- 4- 4- 4- — — — 4- — 4-
D + 4- — — — — — 4- 4- 4-
Е + — + — 4- 4- — 4- — — Род 2
F 4- — — 4- 4- — — 4- — 4-
I 4- — 4- — 4- — — 4- 4“
Фиг. 52. Три возможные классификации 10 гипотетических видов на 2 рода.
VI Группировка видов в высшие категории
137
знаком, так как он указывает на систему, очень близкую к той,
которую подтверждают первые три.
Если надо разделить на группы большее число видов (на-
пример, несколько сот), то сложность проблемы сильно воз-
растает— число возможных систем классификации увеличива-
ется почти до бесконечности. Все их рассмотреть невозможно,
но на основании предварительного изучения десяти видов мож-
но заранее сказать, что признаки 1, 7 и 8 будут полезны также
для разделения многих видов. Можно добавить и признак 6,
потому что его распределение на 90% согласуется с распреде-
лением первых трех. Признаки 1, 7, 8 и 6 можно проверить на
большей группе видов. Если по каждому из них можно разде-
лить большую группу одинаковым (или почти одинаковым)
образом и если получающиеся подразделения обоснованы с гео-
графической, экологической или стратиграфической точек зре-
ния, то можно сделать вывод, что эти 4 признака играют ключе-
вую роль в определении главных эволюционных делений. Испы-
тание признаков можно было бы продолжить; постепенно мы
выяснили бы их сравнительную важность во всех последующих
таксономических решениях, т. е. провели бы взвешивание таксо-
номических признаков.
В нашем вымышленном примере классификацию можно раз-
вить, разбив группы на более мелкие подразделения. Назовем
нашу исходную группу из десяти видов семейством. Первона-
чальное деление даст два рода, дальнейшее деление — подроды
и так далее. Конечно, признаки, используемые для выделения
родов, не могут быть использованы для выделения подродов,
так как роды гомогенны по этим признакам. Следовательно,
проводя процедуру взвешивания признаков, мы исключаем воз-
можность их использования на более низких таксономических
уровнях.
В большинстве случаев взвешивание признаков проводится
способом, сходным с приведенным в нашем примере. Например,
присутствие или отсутствие перьев у позвоночных считается
признаком с большим весом, так как перья всегда имеются
у группы организмов, объединяемых и по другим признакам
(у птиц), тогда как у других позвоночных перьев нет. Следо-
вательно, присутствие или отсутствие перьев аналогично призна-
кам 1, 7 и 8 в нашем примере. Отсутствие зубов характерно
почти для всех птиц, но Archaeopteryx имел зубы. Следователь-
но, наличие или отсутствие зубов — признак, соответствующий
признаку 6 в нашем примере, и может быть использован в ка-
честве таксономического признака.
Таксономист редко работает по определенному плану, он
может сам не осознавать логических шагов, ведущих к конеч-
ному результату. Не удивительно, что процесс классификации
138
Часть 1. Описание и классификация ископаемых остатков
иногда называют интуитивным и даже считают его «искус-
ством».
Очевидно, самым опасным заблуждением в истории создания
высших таксономических группировок было естественное жела-
ние найти какой-то один решающий признак для группировки
видов. При установлении отрядов или подклассов беспозвоноч-
ных возникло множество таксономических проблем. Так, неуда-
чей окончились попытки объединить семейства брахиопод в
отряды, основываясь только на микроструктуре раковины или
морфологии брахидиума. Не удались и попытки разделить три-
лобитов на отряды только на основании очертаний лицевых
швов. Для Bivalvia предлагались две схемы, каждая была ос-
нована на одном морфологическом признаке: многие биологи
ратовали за использование строения жабр для группировки
семейств в отряды, а другие биологи и большинство палеон-
тологов стояли за использование строения замка раковины.
Появились две несовместимые системы классификации. Сейчас
хорошо известно, что эуламеллибранхиатный тип строения жабр
развился и в группах зарывающихся двустворчатых моллюсков
с гетеродонтным замком, и у устриц, совсем не имеющих замко-
вых зубов и произошедших от гребешков. Некоторые формы
замка также возникали независимо в двух или нескольких так-
сономических группах. Современные исследователи не пытаются
больше распределить по отрядам семейства двустворчатых мол-
люсков на основании только одного признака. Обычно исполь-
зуется несколько признаков, включая строение замка и жабр,
причем все эти признаки различным образом взвешиваются.
Сейчас выделение высших категорий по одному признаку
проводится редко. Впрочем, для некоторых групп этот способ
оказался в общем верным. Как мы видели, перья являются
решающим таксономическим признаком для класса птиц. По-
добным же образом семейства динозавров делят на два боль-
ших отряда по строению таза (фиг. 53). Но объединение этих
отрядов в один подкласс можно назвать искусственным; родство
между ними не ближе, чем родство каждого из этих отрядов
с отрядом крокодилов, произошедшим от той же предковой
группы.
Численная таксономия
Нельзя ли передать работу по классификации машине?
Нельзя ли с помощью информации, приведенной на фиг. 51,
запрограммировать ЭВМ так, чтобы она рассмотрела все 637
возможных комбинаций и выбрала способ группировки по прин-
ципу наибольшего числа признаков? Современные ЭВМ, очевид-
но, могли бы делать это за несколько долей секунды1 Кроме
Триас | Юра | Мел
Фиг. 53. Два отряда динозавров, имеющих общее происхождение, но разли-
чающихся отроением таза.
4. Птицетазые динозавры (Ornithischia). Б. Ящеротазые динозавры (Saurischia) 132]
Подвздошная
140
Часть 1. Описание и классификация ископаемых остатков
того, можно так составить программу, что при выборе «лучшей»
классификации ЭВМ будет руководствоваться большим числом
различных критериев. Выбор системы классификации по прин-
ципу наибольшего числа признаков — только один из многих
возможных критериев.
Численная таксономия — наука о классификации организмов
при помощи машинных или математических методов. Если бы
мы разработали численный метод оценки 637 возможных клас-
сификаций для десяти воображаемых видов на фиг. 51, то это
было бы переходом к численной таксономии. Численная так-
сономия немногим моложе обычной, но почти все главные ее
успехи достигнуты во второй половине 50-х годов, что связано
с развитием вычислительной техники.
До сих пор все системы классификации, разработанные с по-
мощью численных методов, основывались только на наблюдае-
мых признаках организмов. Такие факторы, как экология, био-
географическое или стратиграфическое распределение, не учи-
тывались. Поэтому созданные таким образом системы класси-
фикации называют фонетическими — они основаны на фенотипе
(в широком смысле слова, включающем физиологию, биохимию,
поведение и то, что мы обычно понимаем под морфологией).
Много спорили о том, насколько «естественны» такие системы
классификации, отражают ли они филогению. Ответ на этот
вопрос в какой-то мере зависит от того, какое значение в клас-
сификации придается нефенетическим данным. В действитель-
ности большинство обычных систем классификации во многом
или даже полностью основано на морфологии, и методы числен-
ной таксономии мало меняют дело.
Большинство методов численной таксономии можно исполь-
зовать на любом таксономическом уровне: основными единица-
ми, подлежащими классификации, могут быть отдельные особи,
виды и даже роды и высшие категории. Единицы, подлежащие
классификации, называются операционными таксономическими
единицами (ОТЕ).
Обычно работа методами численной систематики распадает-
ся на следующие этапы: 1) выбор ОТЕ, подлежащих класси-
фикации; 2) выбор группы фенетических признаков (обычно
50—100) для описания ОТЕ; 3) сравнение каждой ОТЕ с каж-
дой другой; 4) выявление групп, или «кустов», ОТЕ на основа-
нии рассчитанных степеней подобия. Так, 10 гипотетических
видов на фиг. 52 являются ОТЕ, каждая из которых характери-
зуется десятью признаками.
Для оценки подобия между ОТЕ применяется множество
приемов. Многие из них представляют собой сложные статисти-
ческие методы, которые невозможно рассмотреть в этой книге.
Но можно привести простой пример использования методов
VI. Группировка видов в высшие категории
141
численной таксономии, когда подобие определяется процентом
признаков, по которым две ОТЕ совпадают или сочетаются.
Мера подобия, таким образом, может изменяться от 0 (совпа-
дений нет) до 100 (полное совпадение по всем признакам).
В табл. 2 показаны степени подобия для десяти гипотетических
видов.
ТАБЛИЦА 2 Матрица подобия для 10 гипотетических видов J
А (по данным фиг. 51) I
В С D Е F G Н
А 50 30 40 20 20 60 90 10 70
В 20 30 50 50 70 60 40 80
С 70 50 70 10 40 60 30
D 40 60 20 50 70 50
Е 60 60 10 70 30
F 20 30 70 30
G 50 50 50
Н 20 80
I 20
J
Табл. 2 показывает, что некоторые пары ОТЕ обладают
гораздо более высокой степенью подобия, чем другие. Значения
меры подобия довольно четко распадаются на две группы.
В табл. 3 матрица подобия перестроена так, чтобы резче выра-
ТАБЛИЦА 3
Матрица подобия видов из табл. 2 с выделением групп
Н J
А В G С D Е F I
А 50 60 90 70 30 40 20 20 10
В 70 60 80 20 30 50 50 40
G 50 50 10 20 60 20 50
Н 80 40 50 10 30 20
J 30 50 30 30 20
С 70 50 70 60
D 40 60 70
Е 60 70
F I 70
142 Часть 1. Описание и классификация ископаемых остатков
Фиг. 54. Дендрограмма, основанная на табл. 3.
зить эти выявившиеся группировки (ABGHJ и CDEFI). Подобие
велико при сравнении членов одной группы и незначительно
между членами разных групп.
Дальнейшее изучение исходных данных по подобию позво-
ляет подразделить полученные группировки на более мелкие
единицы и выявить полную картину фенетического распреде-
ления ОТЕ. Результаты полного анализа обычно выражаются
в форме так называемой фенограммы или дендрограммы
(фиг. 54). Это обычный способ отображения численных так-
сономических исследований. Шкала слева указывает подобие
между ОТЕ или группами ОТЕ, связанными горизонтальными
линиями (подобие между группами ОТЕ обычно рассчитыва-
ется на основании усреднения отдельных значений подобия).
Заметим, что две группировки третьего варианта системы клас-
сификации, предложенной на фиг. 52, на фиг. 54 разделяются
на самом низшем уровне шкалы подобия.
Рассмотренный способ группировки пригоден в том случае,
когда число ОТЕ невелико. Если, напротив, их число велико,
то применяются другие методы, позволяющие использовать
ЭВМ. Эти методы различаются по критериям выделения групп,
и потому дендрограммы, построенные разными методами по
одинаковым данным, могут различаться. Важно хорошо пони-
мать суть метода группировки, применяемого в данном иссле-
довании (так же как метода измерения подобия).
Есть ли соответствие между дендрограммой и филогенети-
ческим древом, между фенетической классификацией и обычной?
И если есть, то каково оно? Многие сторонники численной таксо-
номии заявляют, что фенетическая классификация столь же
реальна, как и обычная. Они считают, что группы на фенограм-
ме соответствуют таксонам высшего ранга, причем ранг таксо-
на определяется по расстоянию на шкале подобия (например,
Фиг. 55. Дендрограмма, полученная в результате численно-таксономическо-
го анализа брахиопод надсемейства Chonetacea (ср. с классификацией, раз-
работанной обычными методами,— фиг. 50) [171].
144
Часть 1. Описание и классификация ископаемых остатков
на фиг. 54 ветвление, намечающееся между делениями 60 и 70,
может иметь ранг родовой границы, ветвление ниже деления
60 может соответствовать границе подсемейства и т. д.). Эти
границы условны, но ведь так же обстоит дело и в обычной
таксономии.
Эффективным способом проверки численной таксономии слу-
жит сравнение ее результатов с обычной системой классифика-
ции хорошо изученных групп организмов. Проведенные провер-
ки говорят в пользу численной таксономии. В очень многих от-
ношениях оба подхода дают одинаковые результаты. Многим
сторонникам обычной таксономии, утверждавшим, что построе-
ние системы — скорее искусство, чем наука, пришлось пересмот-
реть свою методологию. Предметом спора является, в частности,
взвешивание признаков. Большинство приемов численной так-
сономии не предусматривает проведения этой процедуры, одна-
ко получаемые результаты часто оказываются сравнимыми с
результатами тех методов, существенной частью которых яв-
ляется взвешивание. Возможно, это означает, что взвешива-
ние— это всего лишь способ ускорить работу, а если операции
перепоручаются ЭВМ, то оно не требуется. С другой стороны,
возможно, что взвешивание имеет биологический смысл, а успех
численной таксономии, несмотря на исключение этой процеду-
ры, объясняется просто-напросто тем, что группы организмов
хорошо отграничены одна от другой и поэтому даже несовер-
шенный метод позволяет их выделить.
На фиг. 55 показана дендрограмма почти того же набора
родов брахиопод, который был уже классифицирован обычными
методами на фиг. 50; дендрограмма получена методами числен-
ной систематики. Хотя между этими двумя классификациями
имеются явные различия, в основном они поразительно хорошо
совпадают. Так, род Eodevonaria на фиг. 50 отделен от всех
других родов на уровне семейства; на фиг. 55 он отделяется от
всех других на одном из самых низких уровней сходства (фене-
тического). Роды Dyoros и Eolissochonetes очень тесно связаны
на фиг. 55 и входят в одно подсемейство на фиг. 50. Подобное
же совпадение обнаружено для родов Chonostrophia, Chonostro-
phiella, Tulcumbella.
Численная таксономия находится еще в колыбели. Пока
она чаще применяется к рецентным видам. Однако ее значение
в биологии и палеонтологии, несомненно, должно возрасти. Ме-
тоды и приемы будут изменяться, но основная цель — ускорить
процесс классификации, сделать классификацию более строгой —
останется.
ГЛАВА VII
Определение ископаемых
Определить ископаемое — значит отнести его к одному из
таксонов какой-либо существующей классификации. При этом
специалист по данной таксономической группе всегда форму-
лирует свою собственную концепцию избранного таксона, ко-
торая может отличаться от концепции других специалистов.
Таким образом, определение и классификация тесно связаны
в сознании исследователя. Но в этой главе описывается подход
неспециалиста, который охотно принимает классификации, раз-
работанные ранее.
Конечно, есть классификации более или менее общеприня-
тые. Сложности, связанные с проблемой вида в палеонтологии,
приводят к тому, что результаты определения видов неспеци-
алистом оказываются ненадежными.
Но в разных случаях требуется различная точность опреде-
ления, и обычно определение до вида не является необходимым.
Так, для палеоэкологического анализа при условии хорошо раз-
работанной стратиграфии изучаемого района определения до ви-
да обычно не требуется. Например, морское происхождение
осадочных пород можно установить просто по наличию ископа-
емых остатков Cephalopoda (определение до класса) или Echi-
nodermata (определение до типа). Подобным же образом при
изучении стратиграфии или геологическом картировании иногда
удается различить почти идентичные местные стратиграфические
единицы разного возраста по ископаемым, определенным до
типа, класса или отряда.
ОБЫЧНАЯ МЕТОДИКА
При разработке классификации обычно начинают с низших
таксономических категорий, постепенно переходя к высшим.
Определение, как правило, проводится в обратном порядке.
Определив вначале тип или класс, мы сразу сильно сужаем
круг видов, к которому может принадлежать исследуемый эк-
земпляр.
Итак, сначала определяют тип, и потому важно хорошо
представлять себе таксономические признаки, используемые для
146
Часть 1. Описание и классификация ископаемых остатков
распознавания типов. Запомнить их нетрудно. В палеонтологи-
ческой летописи отражен с десяток главных типов, и каждый
характеризуется в среднем 6—10 признаками.
Определить класс, к которому принадлежит ископаемое,
значительно труднее. Палеонтологи имеют дело примерно с
30 классами. Мало кто умеет верно распознавать представите-
лей всех классов с первого взгляда, но отличительные признаки
большинства классов достаточно хорошо описаны в современ-
ных монографиях, что позволяет проводить определение и не-
специалисту.
Спускаясь ниже по лестнице таксономических категорий,
приходится все глубже погружаться в специальную литературу.
Поэтому для неспециалиста часто оказывается проще послать
свои экземпляры окаменелостей для определения признанному
специалисту.
Познакомимся с обычным ходом определения на примере
ископаемого, показанного на фото XVII. Экземпляр был найден
в пластах нижнего карбона под Москвой. Он несколько повреж-
ден, поэтому здесь приведены также фотографии родственного
современного организма (фото XVIII).
Окаменелость покрыта многоугольными пластинчатыми обра-
зованиями, располагающимися радиальными рядами. Ряды
пластинок прослеживаются от одного «полюса» почти сфери-
ческого скелета до другого. Многие пластинки имеют один или
несколько шишковидных выступов, называемых бугорками, ко-
торые, как можно понять по строению живых родичей ископае-
мого организма (фото XVIII), служили местами прикрепления
подвижных игл. Судя по этой комбинации признаков, наш эк-
земпляр относится к типу Echinodermata, к классу Echinoidea.
Такие признаки отсутствуют у всех остальных классов этого
типа и у всех остальных типов. Значит, все классы,.кроме од-
ного, сразу отпадают, и нам остается выбрать из примерно
8000 видов (если экземпляр не принадлежит к новому, ранее
не известному виду). Следовательно, нас не интересуют более
120 000 других ископаемых видов и миллион с лишним совре-
менных.
Класс в этом случае можно было бы определить и по дру-
гим признакам, однако у данного экземпляра именно описан-
ные признаки сразу же бросаются в глаза и сохранились луч-
ше других.
В классе Echinoidea, как принято считать, около 75 семейств.
Примерно две трети из них обладают наряду с радиальной яв-
ной билатеральной симметрией. Поэтому морских ежей, хотя
это и не вполне верно с теоретической точки зрения, делят на
две группы: правильных (радиальных) и неправильных (била-
теральных). Показанная здесь окаменелость не имеет билате-
VII. Определение ископаемых
147
ральной симметрии, так что в поле нашего зрения остается все-
го около 25 семейств.
Посмотрев внимательнее, замечаешь, что пластинки, из ко-
торых состоят выступающие ряды, делятся на две группы. Одни
(амбулакральные) несут узкие углубления, или поры (две на
пластинке), а на других (интерамбулакральных) пор нет. Да-
лее, амбулакральные пластинки значительно мельче интерам-
булакральных (из-за посмертного смещения пластинок видны
не все амбулакральные ряды). У современного вида имеется
пять двойных рядов интерамбулакральных пластинок, а иско-
паемый экземпляр имеет более десяти интерамбулакральных
рядов. Виды с числом интерамбулакральных рядов, превышаю-
щим 10, встречаются только в шести семействах морских ежей.
Представители всех этих семейств найдены в палеозойских
отложениях. Из них только у представителей семейства Archae-
ocidaridae имеются бугорки того же типа, что и у экземпляра,
представленного на фото XVII.
Заметим, что по строению и расположению бугорков совре-
менный вид (фото XVIII) в общем не отличается от ископаемого
(если не входить в подробности, которые нас сейчас не интере-
суют) . Но современный вид относится к другому семейству
(Cidaridae), которое мы отбросили, основываясь на числе ин-
терамбулакральных рядов. Строение бугорков позволило вы-
брать одно семейство только после того, как все семейства,
кроме шести, отпали по другим причинам.
Подчеркнем еще раз, что для определения можно было бы
выбрать и не те признаки, которые мы здесь использовали. Наш
анализ был ограничен минимальным числом признаков, и при-
том таких, которые легче всего распознаются. Но из осторожно-
сти обычно пользуются большим числом признаков; это нужно
для того, чтобы результаты определения можно было под-
твердить.
Таким образом, определение семейства, к которому относит-
ся окаменелость, велось по следующим этапам: 1) выбор типа
и класса; 2) констатация того, что наш экземпляр относится
к правильным морским ежам; 3) выбор шести семейств пра-
вильных морских ежей; 4) выбор одного семейства из шести.
Можно было бы идти и другими путями. Мы обошли уровни
подкласса и отряда, применив не совсем строгое деление на
правильных и неправильных морских ежей, не признанное в
таксономии официально, поскольку эти две группы не считают-
ся истинно эволюционными. Все же такое деление удобно в
практике определения.
Мы не будем далее спускаться ниже уровня семейства. Род
и вид определяют подобным же образом, просто число исполь-
зуемых признаков больше. При определении вида важно иметь
1П*
148
Часть 1 Описание и классификация ископаемых остатков
возможность сравнивать определяемый экземпляр с экземпля-
рами, принадлежащими к разным «подозреваемым» видам, или
с их фотографиями.
Ключи
Предпринимались многочисленные попытки упорядочить
процедуру определения. Лучшим инструментом для этого ока-
зался ключ. На стр. 150 воспроизведен ключ, обычно употреб-
ляемый при определении морских ежей. Ключ состоит из ряда
парных альтернативных утверждений, касающихся отдельных
морфологических признаков. Из каждой пары выбирается то
утверждение, которое точнее описывает морфологию экземпля-
ра, что приводит к другой паре утверждений, и т. д. В конце
концов мы приходим к названию таксономической группы. Это
может быть вид (как в ключе на стр. 150), семейство и т. д.,
смотря по таксономическому уровню ключа. Использование
данного ключа предполагает, что высшая таксономическая ка-
тегория, к которой относится определяемый экземпляр, извест-
на. Ключ помогает определять низшие таксоны. Таким образом,
у каждого ключа есть верхний и нижний таксономический
предел.
Альтернативные утверждения (теза и антитеза) ключа мо-
гут касаться одного признака или сочетания признаков. Выби-
рать признаки надо с осторожностью. Если на каком-то этапе
исследователь не сможет сделать выбор между тезой и анти-
тезой, он собьется или остановится. Чтобы ключ был пригодным
и широко применимым, надо выбрать для него такие признаки,
которые хорошо сохраняются и могут быть диагностированы
однозначно. Выбор признаков в значительной мере зависит от
материала, для которого предназначен ключ. Так, ботаники
часто составляют совершенно разные ключи для одной и той
же группы растений в зависимости от того, когда надо опреде-
лять эти растения: во время цветения или нет, в облиственном
состоянии или без листьев. Живые деревья обычно определяют
по так называемым «габитусным ключам», которые могут быть
целиком основаны на таких признаках, как вид коры, общая
форма дерева, морфология ветвей и побегов. Такие ключи мож-
но применять в любое время года, независимо от того, покрыты
деревья листвой или нет. Разработаны параллельные ключи,
основанные на форме листа или же на признаках цветков и
плодов. Ключ, разработанный биологом, может не иметь ничего
общего с ключом, составленным палеонтологом. Все эти раз-
личия подчеркивают тот факт, что таксономические признаки,
обычно используемые для определения видов, выбраны из
множества признаков, которые можно было бы применить. Сту-
VII Определение ископаемых
149
Pod Histocidans
magnrfica
cobosi
sharreri
uariabilis
misakiensis
elegans
crassispma
formosa
acutispma
denticulata
recuruata
cannata
australiae
nuitingi
Фиг. 56. Дендрограмма рода морских ежей Histocidaris, построенная на^осно-
вании ключа, воспроизведенного на стр. 150.
' Цифры соответствуют номерам тез и антитез ключа
денты должны хорошо усвоить, что признаки, обычно исполь-
зуемые при определении вида, не единственные, отличающие его
от других. К сожалению, часто случается, что биолог или па-
леонтолог, обнаружив невозможность различить два вида по
какому-то одному ключу, делает вывод, что они идентичны,
хотя в действительности их можно различить по многим призна-
кам, перечисленным в оригинальных описаниях видов или типо-
вых экземпляров.
На фиг. 56 показана в виде дендрограммы структура ключа
для определения одного рода морских ежей. Каждое развет-
вление соответствует паре утверждений, номер которой постав-
лен в кружке. Как соотносится дендрограмма такого типа с
дендрограммой, отражающей филогению? Другими словами,
насколько определение связано с филогенией? Можно построить
ключ, целиком основанный на филогении, но поскольку главное
назначение ключа — способствовать определению, многие, и
притом наиболее эффективные, ключи следуют филогении лишь
в небольшой степени. Например, определяя в начале этой
главы морского ежа, мы отбросили около двух третей семейств
на том основании, что наш экземпляр не обладал билатераль-
ной симметрией. Такая симметрия появлялась у морских ежей
независимо несколько раз, так что на филогенетическом древе
нет разветвления, отделяющего всех радиальных ежей от всех
билатеральных. И все же сортировка по этому признаку четко
делит почти всех морских ежей на две группы и очень полезна
при создании ключа.
150
Часть 1. Описание и классификация ископаемых остатков
В работе с одними таксономическими группами ключи упо-
требляются чаще, с другими — реже. В значительной мере это
связано с традицией, но дело еще и в том, что при определении
организмов разных групп возникают разные проблемы. Чтобы
построить эффективный ключ, мы должны достаточно хорошо
знать таксономию группы. Группа должна «поддаваться» по-
строению ключа, т. е. классификация должна быть основана на
сравнительно четких признаках, по возможности выражающихся
в присутствии или отсутствии какой-то морфологической харак-
теристики. Ниже мы приводим ключ для рода Histocidaris.
Ключ к видам рода Histocidaris
1. Первичные иглы гладкие, самое большее — с несколькими незазуб-
ренными продольными ребрами. В зоне амбитуса изогнуты
вниз ...............................................Н. magnifica
Первичные иглы с более или менее тонкими зазубринами или шипа-
ми; иглы амбитуса не изогнуты вниз............................ 2
2. Первичные иглы однообразно зазубрены......................., 3
Первичные иглы аборальной стороны с разбросанными длинными
шипами, главным образом у основания...........................11
3. Зазубрины первичных игл очень мелкие, микроскопические ... 4
Зазубрины первичных игл грубее, ясно различимы невооруженным
глазом........................................................ 9
4. Маргинальный ряд амбулакральных туберкул очень нерегуляр-
ный .................................................Н. cobosi
Маргинальный ряд амбулакральных туберкул регулярен или иног-
да слегка нерегулярен у крупных экземпляров................... 5
5. Межпоровая зона с четко выраженной, глубоко вдавленной сред-
ней линией..........................................Н. sharreri
Межпоровая зона без четко выраженной, глубоко вдавленной сред-
ней линии .................................................... 6
6. Створки крупных тридентных педицеллярий более или менее лож-
кообразные, внутренняя поверхность вогнута .................. 7
Створки крупных тридентных педицеллярий более или менее уз-
кие, не ложкообразные, обычно с небольшой вогнутостью над апо-
физом ........................................................ 8
7. Первичные иглы цилиндрические....................Н. variabilis
Первичные иглы веретенообразные..................Н. misakiensis
8. Первичные иглы тонкие цилиндрические..............Н. elegans
Первичные иглы довольно толстые, веретенообразные Н. crassispina
О. Первичные иглы в зоне амбитуса изогнуты вверх, несколько упло-
щены к концу; зазубрины расположены в основном так, что обра-
зуют пару латеральных килей..........................Н. formosa
Первичные иглы в зоне амбитуса не изогнуты вверх и не уплощены
к концу.......................................................10
10. Диаметр вершинного щитка составляет около 50% горизонтального
диаметра...........................................Н. acutispina
Диаметр вершинного щитка составляет около 36% горизонтального
диаметра.........................................Н. denticulata
11. Иглы амбитальной зоны несколько уплощены и изогнуты вверх в на-
ружной части........................................Н. recurvata
Иглы амбитальной зоны не уплощены и не изогнуты вверх в наруж-
ной части.....................................................12
VII. Определение ископаемых
151
12. Створки крупных тридентных педицеллярий очень широкие ....
.............................................Н. carinata
Створки крупных тридентных педицеллярий узкие, тонкие ... 13
13. Длина первичных игл чуть превышает горизонтальный диаметр . .
.............................................Н. australiae
Длина первичных игл приблизительно в 2,5 раза превышает горизон-
тальный диаметр................................И. nuttingi
Машинные методы определения
Процедуру определения начинают с высших таксонов из
соображений удобства и простоты. Противоположный подход
практически недоступен, так как пришлось бы систематически
сравнивать определяемый экземпляр с экземплярами всех из-
вестных видов, пока не обнаружится сильное сходство. Однако,
если бы время нас не ограничивало, такой подход мог бы ока-
заться полезным, поскольку применяемый обычно способ не
лишен недостатков. Взять хотя бы трудности, возникающие в том
случае, если при определении одного из высших таксонов была
сделана ошибка. Если бы при определении морского ежа мы
ошиблись в выборе семейства, то это привело бы к ошибочному
установлению нового вида.
ЭВМ освободила науку от некоторых процедур, к которым
ранее прибегали с целью экономии времени. ЭВМ делает выбор
в миллионы раз быстрее человека. Это позволяет разработать
метод определения низшего таксона для экземпляров, система-
тическое положение которых совершенно неизвестно, т. е. соз-
дать программу, сравнивающую морфологические признаки не-
известного экземпляра с признаками всех известных видов даже
очень большой таксономической группы.
В гл. VI мы рассмотрели некоторые основные методы коди-
рования морфологического описания видов и отметили, что
диагностическое описание вида может быть записано на одной-
двух перфокартах или на коротком отрезке магнитной ленты.
Если мы располагаем такими записями отличительных призна-
ков большого числа видов, то весьма нетрудно запрограммиро-
вать ЭВМ на сравнение определяемого экземпляра с этими
записями.
Сейчас определение ископаемых с помощью ЭВМ применя-
ется редко, ведь методология такого определения появилась
еще совсем недавно (всего около 10 лет назад). Но эти методы
неизбежно будут применяться все шире. Важным фактором,
тормозящим развитие методов машинного определения, явля-
ется также сложность перевода морфологической информации
в форму, пригодную для ввода в ЭВМ. Пока существует эта
трудность, «человеческий элемент» будет оставаться важным
в процедуре таксономического определения. Поэтому сейчас
152 Часть 1. Описание и классификация ископаемых остатков
главная проблема — подбор таких признаков, которые были бы
пригодны для машинной обработки, однако без риска умень-
шить'научную ценность результатов.
Представление
результатов определения
Определялся ли вид для публикации или просто для состав-
ления музейной этикетки, результаты определения должны быть
представлены в удобной форме. Выработаны некоторые тради-
ционные формы изложения результатов.
Принято набирать видовые и родовые названия курсивом.
Родовое название пишется с прописной буквы, видовое — со
строчной. Сразу после видового названия приводится имя автора
вида. Эти традиционные правила соблюдаются во избежание
путаницы.
Ниже мы приводим список ископаемых форм, найденных в
одном третичном сообществе. Отметим, что фамилии некоторых
авторов видов заключены в скобки. Это означает, что со времени
установления вида род был изменен. Обратите внимание на то,
что название рода иногда сокращают до его первой буквы,
если вслед за одним видом упоминается еще один или несколь-
ко видов того же рода. Если сокращение может привести к пу-
танице, то его не делают. В палеонтологических работах родо-
вые названия часто сокращают, а фамилии авторов видов
опускают.
Список видов, найденных в ископаемом сообществе
Rhabdocidaris sp. cf. R. zitteh Loriol
Porocidaris schmidelii (Miinster)
Pedinopsis? melo, n. sp.
Porosoma lamberti Checchia-Rispoli
Ambipleurus rotundatus, n. sp.
A. douvillei (Lambert)
Echinolampas fraasi Loriol
Plesiolampas curriae, n. sp.
P. aut aduensis, n. sp.
Conoclypus delanouei Loriol
Echinocyamus polymorph a (Duncan et Sladen)
Brightonia macfadyeni, n. sp.
Leviechinus gregoryi (Currie)
Pharaonaster sp. cf. P. ammon (Desor)
Opissaster farquharsoni Currie
0. auraduensis, n. sp.
0. somaliensis Currie
0. derasmoi Checchia-Rispoli
O. derasmoi var. angulatus n. var.
H emi aster (Trachyaster) sp.
Schizaster africanus Loriol
S. (Paraster) hunti, n. sp.
VII. Определение ископаемых
15а
S. (Paraster) sp. cf. S. (P.) meslei Peron et Gauthier
S. (Paraster) karkarensis, n. sp.
S. (Paraster) beloutchistanensis (Archiac)
S. (Paraster) duroensis, n. sp.
Linthia somaliensis Currie
L. caternosa Loriol
Lutetiasier maccagnoi Checchia-Rispoli
Arcaechinus auraduensis, n. sp.
Migliorinia migiurtina Checchia-Rispoli
Eupatagus cairensis i,Loriol
E. dainelli (Checchia-Rispoli)
E. jecundus (Checchia-Rispoli)
E. sp. cf. E. cordiformis Duncan et Sladen
Brissopsis sp. cf. B. raulini Cotteau
Для нескольких видов списка указан также подрод (Para-
ster). Название подрода принято заключать в скобки и ставить
вслед за родовым. Сокращение «sp.», помещенное вместо видо-
вого названия после названия рода или подрода, указывает, что-
вид определить не удалось. В нескольких случаях после «sp.»
стоит «cf.» и видовое название, что указывает на неточное или
сомнительное определение вида. Стоящие после названия вида
буквы «и. sp.» означают, что автор списка впервые дал этому
виду название.
Если мы занимаемся такой группой, в которой связи между
видами и родами ясны, а классификация и номенклатура отно-
сительно устойчивы, то простой комбинации родового и видово-
го названий с фамилией автора вида вполне достаточно для
однозначного обозначения вида. Но названия многих организ-
мов во избежание недоразумений не могут быть просто приве-
дены в такой форме, как в этом списке. Рассмотрим для при-
мера морского ежа, показанного на фото XVII.
Он определен как Archaeocidaris rossica (Buch). Ниже при-
водится так называемая синонимика этого вида. Синонимика —
краткая история таксономической трактовки вида с библиогра-
фическими ссылками на важные работы.
Archaeocidaris rossica (Buch)
(?) Cidaris deucalionis Eichwald, 1841, стр. 88. [Невозможно установить, како-
му виду соответствует описание, поэтому название нельзя сохранить.J
Cidaris rossicus Buch, 1842, стр. 323.
Cidarites rossicus Murchison, Verneuil et Keyserling, 1845, стр. 17, табл. 1,
фиг. 2,a—2,e.
Palaeocidaris rossica L. Agassiz et Desor, 1846—1847, стр. 367.
Echinocrinus rossica d'Orbigny, 1850, стр. 154.
Palaeocidaris (Echinocrinus) rossica Vogt, 1854, стр. 314.
Eocidaris rossica Desor, 1858, стр. 156, табл. 21, фиг. 3—6.
Echinocrinus deucalionis Eichwald, 1860, стр. 652.
Eocidaris rossicus Geinitz, 1866, стр. 61.
Archaeocidaris rossicus Trautschold, 1868, табл. 9, фиг. 1 —10,b; 1879, стр. 6,
табл. 2, фиг. },а—l,f, 1, h,\, iA, k, \,l; Quenstedt, 1875, стр. 373,.
табл. 75, фиг. 12; Klem, 1904, стр. 55.
154
Часть 1. Описание и классификация ископаемых остатков
Archaeocidarts rossica Loven, 1874, стр. 43; Tornquist, 1896, фиг. на стр. 27,
табл. 4, фиг. 1—5, 7, 8.
Archaeocidaris rossicavar. schellwieni Tornquist, 1897, стр. 781 табл. 22, фиг. 12.
Cidarotropus rossica Lambert et Thiery, 1910, стр. 125.
Данный вид, по-видимому, был впервые описан в 1841 г.
Эйхвальдом под названием Cidaris deucalionis, но автор сино-
нимики [93] отвергает это название, так как описание Эйхваль-
да было слишком нечетким. Второй пункт синонимики — ссылка
на описанный Бухом вид Cidaris rossicus. Название rossicus,
использованное при первом валидном описании вида, имеет при-
оритет перед всеми последующими названиями этого вида (на-
писание это в дальнейшем было изменено на rossica лишь для
того, чтобы устранить грамматическое несоответствие, возник-
шее в связи с переносом вида в другой род).
Третий пункт синонимики — упоминание о том, что Мурчи-
сон, Вернейль и Кейзерлинг отнесли этот вид к роду Cidarites
(что значит «ископаемый Cidaris»). Затем следуют пункты,
отражающие последующие перемещения вида, связанные глав-
ным образом с изменениями представлений об эволюционном
родстве вида или с разногласиями по этому вопросу. Один
пункт синонимики выделяется среди других: речь идет о на-
звании Echinocrinus deucalionis, которое употребил в 1860 г.
Эйхвальд, очевидно считавший свою публикацию 1841 г. валид-
ной.
В связи с родовым названием Echinocrinus возникает дру-
гая номенклатурная проблема. Это название было предложено
(вполне валидно) в 1841 г. Л. Агассицем. Через три года Мак-
кой совершенно независимо предложил для той же группы
морских ежей название Archaeocidaris. Согласно правилам,
следует употреблять название Echinocrinus, поскольку оно было
предложено первым. Но в 1955 г. Международная комиссия по
зоологической номенклатуре сделала специальное исключение
из правил, отчасти потому, что название Echinocrinus редко
употреблялось специалистами, а отчасти потому, что оно могло
ввести в заблуждение, будучи очень схожим с родовыми на-
званиями, обычными в других классах Echinodermata, особенно
среди морских лилий.
В списке синонимов могут присутствовать видовые названия,
совершенно не связанные в номенклатурном отношении с опи-
сываемым видом. Это случается тогда, когда, по мнению автора
синонимики, два или более видов, рассматривавшиеся ранее
как отдельные систематические группы, являются на самом де-
ле единым видом. В таком случае название, предложенное пер-
вым, используется в качестве видового названия для объеди-
ненного вида, если на этот счет нет официальной оговорки.
Поскольку синонимика носит отчасти исторический характер,
VII. Определение ископаемых
155
отчасти же отражает субъективную интерпретацию таксономи-
ческой проблемы, весьма часто у разных авторов синонимики
одного и того же вида не совпадают.
БИБЛИОГРАФИЯ
По палеонтологии (особенно по таксономии) имеется об-
ширная литература, восходящая к восемнадцатому столетию,
на многих языках и в самых разных изданиях — от регулярных
периодических изданий до нерегулярно появлявшихся моногра-
фий, издаваемых музеями и правительственными учреждения-
ми, и даже частных изданий. Палеонтолог в большей мере
зависит от литературы, чем, скажем, физик или инженер.
Для определения очень полезны монографии по данной так-
сономической группе или по ископаемым, найденным в данном
интервале стратиграфической колонки. В хороших монографиях
можно найти ссылки на всю важную литературу. Статьи, по-
явившиеся после выхода в свет крупной монографии, можно
найти в других библиографических источниках. Работая с мо-
нографией, надо знать, насколько подробной намеревался ее
сделать автор. Например, авторы «Treatise on Invertebrate
Paleontology» старались охватить все роды, но не пытались пе-
речислить все известные виды.
Если нет современного обзора, палеонтолог должен обра-
титься к таким изданиям, как, например, «Zoological Record»,
публикуемый Лондонским зоологическим обществом. В каждом
томе дается достаточно полный обзор зоологической и палео-
зоологической литературы за минувший год. «Zoological Record»
делится по таксономическому принципу на 18 выпусков; каж-
дый из них содержит список работ, авторов, библиографические
ссылки и подробный предметный указатель. В указателе
приводится полный список всех таксономических названий, ис-
пользованных в цитированных работах. «Zoological Record» —
ценное пособие, позволяющее проследить за упоминаниями в
литературе рода или вида на протяжении многих лет.
Но «Zoological Record» неполон. Для составления полной
библиографии понадобилось бы слишком много времени. По-
этому данные «Zoological Record» обычно приходится допол-
нять, просматривая реферативные издания, такие, как «Biologi-
cal Abstracts» и «Bibliography of North American Geology».
Часть вторая
ПРИМЕНЕНИЕ
ДАННЫХ ПАЛЕОНТОЛОГИИ
ГЛАВА VIII
Адаптация
и функциональная морфология
Мы рассматривали эволюцию с разных точек зрения и в раз-
ных контекстах. Основным был тезис, что виды (или популя-
ции) появляются в результате естественного отбора и потому
приспособлены к среде. Адаптация позволяет организму про-
тивостоять изменениям условий среды, захватывать новые мес-
тообитания и более эффективно использовать каждое место-
обитание.
Поскольку приспособление шло в бесчисленных отдельных
эволюционных рядах на протяжении сотен миллионов лет, мы
наблюдаем сейчас в природе поистине поразительное разнооб-
разие форм. Отчасти оно может отражать разнообразие воз-
можных типов образа жизни. Однако нельзя утверждать, что
типов образа жизни столько же, сколько биологических видов.
Многие организмы, развивавшиеся независимо друг от друга,
ведут теперь очень сходный образ жизни. У таких организмов
встречается иногда явление конвергенции, т. е. независимого
появления сходных морфологических черт со сходными функ-
циями. В то же время часто организмы приспосабливаются к
одинаковой среде совершенно разными способами. Поэтому чис-
ло морфологических типов, возникших в результате адаптивной
эволюции, значительно выше числа основных типов образа
жизни.
В этой главе мы подробнее рассмотрим результаты адапта-
ции и попытаемся измерить успех организмов во взаимодейст-
вии со средой и в ее использовании. Мы примем, что морфология
в основном адаптивна, т. е. что наблюдаемые морфологиче-
ские черты полезны для жизнедеятельности организмов. Теоре-
тически эволюция может привести и к нефункциональной мор-
фологии (хотя это маловероятно). Современные палеонтологи
VIII. Адаптация и функциональная морфология
157
и биологи считают, что это относительно редкое явление, возни-
кающее в необычных в генетическом или экологическом отно-
шении обстоятельствах.
Неслучайность
результатов адаптации
Если бы мы могли описать спектр биологически возможных
планов строения и сравнить его с действительно появившимися
формами, то обнаружили бы, что адаптация не является неопре-
деленной, т. е. эволюция дала далеко не все возможные формы,
а существующие планы строения используются не одинаково
часто. Здесь можно провести аналогию с работой конструктора
автомобилей: сравните разнообразие возможных конструкций
автомобилей с теми, которые выбираются для запуска в про-
изводство. Если бы мы могли как-то выразить все разнообразие
возможных форм автомобилей, а потом сравнить нашу оценку
с тем, какие модели существуют в действительности, то оказа-
лось бы, что распределение реально существующих автомобилей
(и «живых», и «вымерших») чрезвычайно «пятнисто» и сущест-
вует сравнительно немного основных моделей, оказавшихся
эффективными.
Неслучайность адаптации привлекала внимание многих био-
логов-эволюционистов. Добжанский [41] так писал об этой
проблеме:
«Каждый организм обладает определенным сочетанием органов
или признаков и сочетанием генов, обеспечивающих развитие
этих признаков. Разные организмы имеют разные сочетания
генов и различаются также некоторыми генами. Число мысли-
мых сочетаний генов, имеющихся у разных организмов, разу-
меется, огромно. Действительно существующие сочетания сос-
тавляют лишь бесконечно малую часть числа потенциально воз-
можных или хотя бы мыслимых. Можно представить себе, что
все эти сочетания образуют многомерное пространство, в кото-
ром каждый существующий или возможный организм имеет
свое место.
Существующие и возможные сочетания можно рассортиро-
вать по их приспособленности к существующим на земном шаре
местообитаниям. Многие из мыслимых сочетаний (большинство)
непригодны для выживания в какой-либо среде. Другие позво-
ляют занять некоторые местообитания и экологические ниши».
Другой биолог-эволюционист, Сьюэл Райт, предложил для
выражения этих принципов графический метод. Вертикальная
и горизонтальная оси на фиг. 57 соответствуют двум генети-
чески контролируемым признакам. Площадь, ограниченная дву-
мя осями, включает, таким образом, все возможные сочетания
158
Часть 2 Применение данных палеонтологии
Фиг. 57. Графическая интерпретация, по Сьюэлу Райту, адаптивных пиков
и долин для случая двух генетически определяемых признаков (отложены по
горизонтальной и вертикальной осям [41]).
Пики отмечены знаком «плюс»
двух признаков. Например, на горизонтальной оси может быть 1
отложено число ребер на раковине двустворчатого моллюска,
а на вертикальной — сравнительная высота этих ребер.
В любой среде приспособительное значение некоторых соче-
таний морфологических признаков окажется большим по срав-
нению с другими сочетаниями. Вариации адаптивности схема-
тически отражены на фиг. 57 в виде «адаптивных пиков», раз-
деленных «адаптивными долинами». На схеме имеется несколько
адаптивных пиков. Это означает, что существует несколько
адаптивных сочетаний двух морфологических признаков (хотя
природа адаптивной ценности, очевидно, различна для каждого
сочетания). В промежутках между адаптивными пиками на-
ходятся формы, плохо приспособленные к любой среде, доступ-
ной этой группе организмов. Развивающиеся линии обычно
«взбираются» на адаптивные пики и избегают адаптивных до-
лин (йли быстро их пересекают). Если бы мы действительно
измерили какой-либо морфологический признак у множества
разных организмов (например, ребристость у двустворчатых *
моллюсков), то нашли бы, что выраженность признака распада-
ется на адаптивные пики.
VIII Адаптация и функциональная морфология
159
Теоретическая морфология.
Спиральная раковина
Лишь немногие ископаемые группы изучены настолько хоро-
шо, что для них удается провести анализ, представление о ко-
тором мы получили в предыдущем разделе (фиг. 57), и опре-
делить спектр возможных форм Конечно, при этом необходимо
описывать как появившиеся в ходе эволюции, так и не реали-
зовавшиеся формы. Такой анализ относят к теоретической мор-
фологии. По-видимому, лучше всего подходит для подобного
анализа наружная форма спирально свернутой раковины бес-
позвоночного. Эту модель мы и используем как пример, ил-
люстрирующий возможности теоретической морфологии и ана-
лиза адаптивных пиков у ископаемых.
Многие беспозвоночные обладают спирально свернутыми
раковинами. Примеры можно найти во многих группах — от
одноклеточных фораминифер до высокоразвитых организмов —
моллюсков и брахиопод. Спиральная свернутость развилась
независимо во многих эволюционных линиях, и ее функциональ-
ная роль различна.
Основы геометрии спиральной раковины можно разобрать
на примере обычного брюхоногого моллюска. У большинства
представителей этого класса моллюсков раковина имеет форму
полой конусовидной трубки, открытой на широком конце
(устье). По мере роста животного трубка удлиняется в резуль-
тате приращения нового материала к устью. При этом на одной
стороне устья приращение материала идет быстрее, чем на
другой. В результате возникает спиральность, трубка как бы
закручивается вокруг фиксированной оси, называемой осью
навивания.
Некоторые параметры спирали в ходе роста сохраняют пора-
зительное постоянство, так что на них можно основывать эф-
фективную систему описания. Особенно полезны четыре пара-
метра, показанные на фиг. 58. Первый из них — форма попе-
речного сечения оборота; ее называют обычно формой образую-
щей кривой. На фиг. 58 образующая кривая — окружность.
(Более строго образующая кривая определяется как линия пе-
ресечения расширяющейся трубки с плоскостью, в которой ле-
жит ось навивания. У большинства гастропод она совпадает
с формой устья.) Второй параметр — скорость расширения обра-
зующей кривой по мере обращения ее вокруг оси. Она часто
называется скоростью расширения оборота-, это — отношение
любого линейного размера (например, диаметра) двух обра-
зующих кривых, разделенных полным оборотом спирали. На
фиг. 58 скорость расширения оборота равна 2, т. е. при каждом
обороте вокруг оси линейный размер образующей кривой удваи-
160
Часть 2. Применение данных палеонтологии
Фиг. 58. Схема части раковины брюхоногого моллюска [156].
вается. Третий параметр — положение и ориентировка образую-
щей кривой по отношению к оси. На фиг. 58 образующая окруж-
ность отстоит от оси на расстоянии, равном половине своего
диаметра. Ориентация по отношению к оси важна и в тех случа-
ях, когда образующая кривая не является окружностью. На-
конец, четвертый параметр — перемещение образующей кривой
вдоль оси — так называемый перенос оборота. Он лучше всего
выражается отношением перемещения вдоль оси к перемещению
от оси на протяжении любой части оборота вокруг оси. Изме-
ряется перемещение геометрического центра образующей кри-
вой. У некоторых форм перенос отсутствует и трубка обраща-
ется вокруг оси в одной плоскости, образуя плоскоспиральную
раковину.
Форма образующей кривой, скорость расширения оборота,
положение образующей кривой по отношению к оси и перенос
оборота обычно (хотя и не всегда) в ходе роста не изменяются.
Но при сравнении различных видов брюхоногих моллюсков
обнаруживаются существенные различия в этих параметрах.
У одних образующие кривые близки к окружности, у других
они очень сложны по форме и с трудом поддаются математиче-
скому описанию. Однако вариации формы образующей кривой
у гастропод ни в коем случае не являются случайными.. Если
бы мы рассмотрели большое число видов брюхоногих моллюс-
ков, живых и вымерших, то мы нашли бы, что одни формы
образующей кривой встречаются часто, другие — редко, а третьи,
ло-видимому, вообще не встречаются.
VIII. Адаптация и функциональная морфология
161
Фиг. 59. Раковина брюхоногого моллюска,
у которого величина переноса оборота в ходе
онтогенеза уменьшается [156].
Среди брюхоногих отмечается и значительная изменчивость
скорости расширения оборота — от немногим более 1,0 (теоре-
тически возможный минимум) до 4 или 5. Положение образую-
щей кривой может быть различным; крайние случаи — сопри-
косновение с осью или отделение от нее так называемым
пупком (конусообразной полостью, тянущейся от основания до
вершины раковины).
Очень сильной изменчивости подвержен перенос оборота;
он варьирует от нуля (плоскоспиральные раковины) до очень
высоких значений (башенковидные раковины). Среди живых
современных видов брюхоногих нет настоящих гь.оскоспираль-
ных форм, но у брюхоногих раннего палеозоя такие раковины
были довольно распространены.
Рассмотренные четыре параметра хорошо описывают морфо-
логию спирально свернутой раковины, но не исчерпывает ее.
Можно было бы назвать и другие параметры, например направ-
ление навивания оборотов. Некоторые гастроподы имеют э-
завитую (синистральную) раковину, другие — правозавг ю
(декстральную). Интересно, что огромное большинство бр< -.о-
ногих обладает декстральной раковиной.
Для того чтобы полнее описать форму брюхоногих, ам
нужны также приемы описания отклонений от только что опи-
санной модели. Пример отклонения — раковина, изображенная
на фиг. 59, у которой величина переноса оборота уменьшается
в онтогенезе, так что раковина имеет вогнутый боковой про-
филь (вместо обычного прямого профиля спирали). Такое онто-
генетическое отклонение от нормальной геометрии обычно спе-
цифично для вида и контролируется генетически; по-видимому,
оно имеет адаптивноегзначение.
Кроме упомянутых черт морфологии, в нашу общую спи-
ральную модель не вошли многие мелкие признаки, например
отпечатки мышц на внутренней стенке раковины, окраска рако-
вины, скульптура ее поверхности, форма крышечки и т. д.
Проверить математическую модель можно, попытавшись
восстановить форму раковины с помощью ЭВМ. Выбрав форму
11—467
162
Часть 2 Применение данных палеонтологии
образующей кривой, ее положение, скорость расширения и ве-
личину переноса оборота, мы можем получить контур раковины,
который возник бы в процессе ее роста при заданных парамет-
рах. На фото XIX показана смоделированная на ЭВМ раковина
гастроподного типа. Образующая кривая — окружность, сопри-
касающаяся с осью навивания. Скорость расширения оборота
равна 2,0. Перенос оборота довольно высок. Эти характе-
ристики были введены в виде электрических сигналов в анало-
говую ЭВМ. На экране осциллографа можно было наблюдать
«рост» образующей кривой. На фото XIX приведена осцилло-
грамма, представляющая собой обобщенную форму поверхности
раковины. Результат подтверждает, что наша модель, основан-
ная на четырех параметрах, верно отражает главные аспекты
процесса образования раковины. Но, что еще важнее, моделиро-
вание на ЭВМ позволяет создать контуры раковин гастропод,
которые теоретически возможны, но в природе не появились.
На фото XX показано 25 форм, созданных с помощью той
же аналоговой ЭВМ. Для простоты изменяли только величину
переноса оборота и скорость расширения оборота. Поэтому по-
лученная группа форм представляет собой лишь небольшую
часть спектра типов раковин, которые могли бы образоваться
в ходе эволюции брюхоногих. Некоторые из этих форм обычны
для брюхоногих моллюсков, другие (особенно те, что находятся
в левой нижней части) редки или вовсе отсутствуют. Таким
образом, на фото XX имеется один или несколько адаптивных
пиков (собранных в верхней части таблицы) и адаптивные до-
лины (особенно в левом нижнем углу).
Модель можно расширить так, чтобы она включала спираль-
но свернутые формы, существующие во многих других группах
животных. Спирально свернутые раковины головоногих по своей
форме мало чем отличаются от раковин многих брюхоногих.
У большинства головоногих раковины плоскоспиральные (ну-
левой перенос). Образующие кривые — эллипсоиды или окруж-
ности— симметричны по отношению к плоскости навивания.
У многих форм соседние обороты перекрываются, так что вся
образующая кривая не видна. Если увеличить скорость расши-
рения оборота или отодвинуть образующую кривую от оси, то
обороты не будут перекрываться и образующая кривая будет
полностью видна. Большинство раковин головоногих по своей
геометрии относится к крайним формам, показанным на фото XX
в правом верхнем углу.
Конечно, головоногие и брюхоногие моллюски сильно разли-
чаются и раковины их имеют разные функции. Раковина голо-
воногого разделена внутренними перегородками на ряд запол-
ненных газом камер, создающих плавучесть. Но на наружной
форме и общей геометрии раковины наличие внутренних камер
VIII. Адаптация и функциональная морфология
163
никак не отражается. Поэтому с чисто описательной точки зре-
ния можно рассматривать раковину головоногих как частный
случай основной модели, описывающей раковину брюхоногих.
У двустворчатых моллюсков скорость расширения оборота,
как правило, значительно выше, чем у брюхоногих и головоно-
гих со спирально завернутой раковиной. Скорость расширения
оборота у брюхоногих редко превышает 5, а у двустворчатых
моллюсков она нередко превышает 1000 000. Такие скорости
расширения оборота сопровождаются у двустворчатых еще и
смещением оборота —у таких форм, как гребешки, оно мини-
мально, но имеется всегда и вносит в строение раковины асим-
метрию. Среди форм, смоделированных на ЭВМ (фото XX),
некоторые (особенно в правом нижнем углу) приближаются по
морфологии к раковине двустворчатых моллюсков, но ракови-
ны большинства двустворчатых не входят в эту группу мо-
делей.
Сравнение между двустворчатыми моллюсками и другими
организмами со спирально свернутыми раковинами имеет ско-
рее геометрическую, нежели физиологическую основу. Скелет
двустворчатого моллюска состоит на самом деле из двух сочле-
ненных спирально свернутых раковин (одной декстральной и
одной синистральной). Как мы увидим далее, геометрические
отличия раковин двустворчатых моллюсков от раковин всех
других организмов со спиральной раковиной могут быть связа-
ны с функцией раковины.
Рассмотрим теперь еще одну группу животных со спирально
свернутой раковиной — брахиопод Каждая створка раковин
брахиопод представляет собой плоскоспиральную раковину. Как
и у двустворчатых моллюсков, целый скелет брахиопод состоит
из двух сочлененных спирально свернутых раковин. Скорости
расширения оборота у них всегда велики, что практически не
позволяет спутать геометрию раковины брахиопод с геометрией
раковин плоскоспиральных головоногих. У брахиопод варьирует
в основном форма образующей кривой. Для других рассмот-
ренных групп удовлетворительную модель раковины можно по-
строить, приняв, что ведущий (растущий) край раковины соот-
ветствует форме поперечного сечения расширяющейся раковины
(в плоскости, в которой лежит ось навивания). У брахиопод
ведущий край раковины часто неплоскостной и не совпадает по
форме с теоретической образующей кривой. Особенно ясно это
видно на формах с хорошо развитыми на переднем крае рако-
вины седлом и синусом. Неплоскостной край образуется, когда
некоторые части раковины растут быстрее других. Это не озна-
чает, что у раковины нет образующей кривой в геометрическом
смысле. Просто рост ведущего края во времени отклоняется
от простой геометрической модели. Таким образом, для того
11*
164
Часть 2 Применение данных палеонтологии
чтобы работать со множеством брахиоподных форм, нам приш-
лось бы ввести добавочный параметр, отражающий неравномер-
ность роста.
Раковины многих фораминифер имеют спиральную форму,
по-видимому, укладывающуюся в модель, разработанную для
брахиопод и моллюсков. Раковина имеет камеры, характер
строения которых значительно сильнее отражается на внешней
морфологии, чем это наблюдается у головоногих. Поэтому гео-
метрия раковин фораминифер не кажется столь регулярной.
Однако проведенные измерения показывают, что по основной
геометрии раковины фораминифер не отличаются от раковин
других беспозвоночных. В этой области остается еще многое
сделать: необходимо подтвердить полученные результаты и раз-
работать модель, применимую к фораминиферам.
На фото XXI показана другая группа форм раковин, смоде-
лированных на аналоговой ЭВМ. В центре — обобщенная форма
с образующей кривой, имеющей форму окружности и соприка-
сающейся с осью, с нулевым переносом и сравнительно медлен-
ным расширением оборота. Морфологический эффект изменения
каждого из этих параметров показан отходящими от центра
рядами форм. Возрастание только скорости расширения дает
форму брахиоподного типа. Удаление образующей кривой от
оси приводит к появлению формы, очень типичной для спираль-
но свернутых головоногих. Увеличение переноса при сохранении
сравнительно медленного расширения оборота дает гастропод-
ные типы.
На фиг. 60 эти соотношения для форм с образующей кривой
в виде окружности отражены на трехмерной блок-диаграмме.
Перенос оборота возрастает от нуля (справа) до 4 (слева).
Скорость расширения оборота возрастает от 1 (теоретически
возможный минимум, вверху) до 1 000 000 (внизу). Расстояние
от оси до образующей кривой (в долях ее диаметра) возрастает
в глубь диаграммы. Значит, этот «блок» содержит все геометри-
ческие формы, возможные в рамках модели.
Заштрихованные области блок-диаграммы (фиг. 60) соот-
ветствуют формам, наиболее распространенным среди четырех
таксономических групп. Гастроподы, как правило, занимают
область медленного расширения оборота, но характеризуются
большим разнообразием по величине переноса оборота и рас-
стоянию образующей кривой от оси. У двустворчатых перенос
обычно невелик, но высока скорость расширения оборота. Бра-
хиоподы почти не отличаются от них по скорости расширения,
но геометрически отделены отсутствием переноса оборота. Спи-
рально завернутые раковины головоногих в большинстве своем
плоскоспиральны, но скорость расширения оборота этих рако-
вин всегда ниже, чем у брахиопод, так что области, соответ-
1
Фиг. 60. Упрощенная^блок-диаграмма распределения форм раковины среди четырех основных
групп беспозвоночных, обладающих спирально свернутой раковиной [156J.
166
Часть 2 Применение данных палеонтологии
ствующие формам раковин этих двух групп, не перекрываются.
На трехмерной блок-диаграмме, приведенной на фиг. 60,
четыре эволюционные группы занимают практически неперекры-
вающиеся области в доступном для них геометрическом про-
странстве. Это и не удивительно: все эти группы сильно разли-
чаются по функциям раковины и по своей экологии, поэтому они
неизбежно занимают разные адаптивные пики.
Больше всего поражает тот факт, что основная часть блок-
диаграммы пуста. Этому может быть несколько причин. Воз-
можно, пустой объем соответствует адаптивным долинам. Не
исключено, что формы, которые заняли бы этот объем, с биоло-
гической точки зрения невозможны. Наконец, возможно, что
различные формы, которые заняли бы всю доступную геометри-
ческую область, просто еще не успели развиться. Наиболее
правдоподобным кажется первое предположение.
Методы функционального
морфологического анализа
i
Нет единого метода расшифровки функций по морфологи-
ческим признакам. Здесь возможны различные подходы. В ка-
кой-то м£ре подход зависит от имеющейся информации. При
изучении вымерших организмов неполная сохранность материа-
ла и отрывочное знание образа жизни и местообитаний ограни-
чивают набор возможных приемов анализа. При исследовании
живых организмов обычно удается непосредственно изучить
анатомию мягких частей, наблюдать образ жизни, выявить
предпочтения по отношению к местообитаниям, что в общем
повышает точность интерпретации.
Наиболее обычный подход к интерпретации фоссильных
структур — сравнение с рецентными видами. Рассмотрим сна-
чала случай, когда ископаемый и рецентный таксоны, обладаю-
щие какими-либо сходными структурами, близкородственны, и
сами структуры, как полагают, имеют общее происхождение. *
Тогда по гомологии (одинаковому происхождению) можно счи-
тать, что фоссильная структура выполняла те же функции, что
и рецентная. В других случаях сходство структур может быть
более поверхностным, возникшим независимо в разных таксо-
номических группах. Но и тогда можно считать, что фоссильная
структура выполняла ту же функцию, что и рецентная. Тогда
говорят о гомоплазии (одинаковой форме). Когда установлено,
что гомопластические структуры служат для одинаковой или
сходной функции, их называют аналогичными. Гомология и го-
моплазия— это скорее основа для сравнения, а не для рас-
шифровки функции.
VIII. Адаптация и функциональная морфология
167
В последние годы признано, что в конечном счете интерпре-
тация функций часто основана на анализе механических свойств
структуры, даже когда этот анализ применялся к ископаемым
таксонам на основе принципов гомологии или гомоплазии. По-
скольку окаменелости — это обычно остатки скелетов, а боль-
шинство скелетов — жесткие поддерживающие структуры, фос-
сильные структуры в основном несли при жизни механические
функции. Часто мы можем наблюдать механическую функцию
структур в современных таксонах и по гомологии переносить
наши выводы на родственные ископаемые таксоны. Так, мы
можем наблюдать, что осенью самцы оленей сражаются между
собой, используя при этом рога. Поскольку рога имеются толь-
ко у самцов, подрастают и сменяются ежегодно и полного раз-
вития достигают к осеннему сезону размножения, можно сде-
лать вывод, что первичная их функция — служить оружием во
внутривидовой борьбе за самок (что не исключает их вторичных
функций). По гомологии можно сделать вывод, что сходно
устроенные рога ископаемых оленей имели ту же первичную
функцию.
Но функция фоссильных структур редко бывает так ясна,
как в рассмотренном случае. Даже если известны рецентные
аналогичные или гомологичные структуры, функции их часто
недостаточно изучены. Обычно анализ проводят следующим
образом. Сначала для подобной структуры у рецентной группы
постулируют гипотетические функции. Затем проверяют это
предположение: насколько полезной могла быть такая функция
для организма с данным образом жизни и данным местооби-
танием, а также насколько предполагаемая функция вероятна
с точки зрения морфологии организма, его образа жизни и эко-
логических предпочтений.
В рамках этого основного плана возможны разные вариан-
ты. Если вымершая группа тесно связана с рецентной и иссле-
дуемые структуры считаются гомологичными, то для проверки
гипотезы используют данные о живущей и вымершей группах.
Если фоссильные и рецентные структуры негомологичны, то
бывает необходимо доказать справедливость выводов, получен-
ных на рецентной группе. Например, при изучении ископаемых
сообществ удается доказать, что вымершие таксоны по образу
жизни и по экологическим предпочтениям действительно были
сходны с современными.
Если данных о живых организмах недостаточно или они во-
обще отсутствуют, то с успехом применяется вариант вышеопи-
санной процедуры, называемый парадигматическим подходом.
Применение парадигм строго описано Рудвиком [172]. Соглас-
но схеме Рудвика, для данной структуры постулируется одна
или несколько функций, а затем на основании этих предполо-
168
Часть 2 Применение данных палеонтологии
жений строятся абстрактные механические модели, называемые
парадигмами, — одна парадигма для каждой вероятной функ-
ции. Каждая парадигма показывает, как должна была бы вы-
глядеть структура, чтобы наилучшим образом соответствовать
данной функции. Возникает набор чисто гипотетических струк-
тур (иногда полезно построить трехмерные изображения или
машинные модели этих структур). Парадигма, наиболее сход-
ная с реальной структурой, имеет в своей основе наиболее
вероятную функцию этой структуры.
Важное правило, которое надо соблюдать при парадигмати-
ческом подходе, состоит в том, что гипотетические идеальные
структуры должны быть определены в соответствии с генетикой
и физиологией организма. Так, можно принять, что головоногий
моллюск функционирует как подводная лодка, но постройка
судна вовсе не аналогична росту моллюска, и это различие
(наряду с другими) исключает подводную лодку в качестве па-
радигмы.
При любом подходе к функциональной морфологии важно
учитывать то, что можно назвать факторами множественного
действия. Здесь возможны три основных варианта: 1) структура
может иметь несколько функций; 2) развитие и функционирова-
ние одной структуры может зависеть от нескольких генов;
3) один ген может влиять на несколько структур. Третий случай
генетики называют плейотропией.
Благодаря этим факторам многие структуры подвержены
давлению отбора сразу в нескольких направлениях. Можно
сравнить такую структуру с механической системой, в которой
на объект действуют две или более противоположно направлен-
ные силы. В этом случае направление и величина каждой силы
могут быть изображены в виде вектора, а направление и вели-
чина результирующей силы (которая при достаточной вели-
чине сдвинет предмет) определяются взаимодействием отдель-
ных сил.
Точно так же и направление эволюции большинства морфо-
логических признаков является некой «равнодействующей». На-
пример, большинство наружных раковин моллюсков выполняет
защитную функцию и служит для поддержания мышечной си-
стемы. Очень толстые стенки раковин были бы полезны для
защиты от многих хищников. Но зато большая подвижность,
свойственная чаще всего моллюскам с тонкими, легкими рако-
винами, позволяет организму успешно избегать хищников. По-
этому толщина раковины у большинства моллюсков есть резуль-
тирующая двух противоположных давлений отбора. Надо также
учитывать, что вымерший вид мог представлять собой стадию
эволюции, на которой конечный «компромисс» еще не был до-
стигнут.
VIII Адаптация и функциональная морфология
169
Ниже приведены некоторые примеры функционального ана-
лиза, иллюстрирующие разнообразие существующих методов и
подходов. Некоторые из них основаны главным образом на не-
онтологической информации; другие сравнительно независимы
от данных о рецентных организмах.
Функциональная морфология
спиральной раковины
Вернемся к спирально свернутым раковинам. При взгляде
на блок-диаграмму, приведенную на фиг. 60, возникает важный
вопрос: почему различные эволюционные группы занимают бо-
лее или менее неперекрывающиеся области? Ответов может
быть много, а некоторые детали проблемы выходят за рамки
нашей книги. Но мы можем коснуться некоторых главных фак-
торов. Заметьте, что на двух обращенных к нам стенках блок-
диаграммы изображена пунктирная кривая. Это вычисленная
кривая; области, лежащей выше этой линии, соответствуют ра-
ковины, обладающие объемлющими оборотами, а ниже ее — ра-
ковины с открытой спиралью. Раковины, лежащие на этой кри-
вой, имеют соприкасающиеся обороты. Две пунктирные кривые
на блок-диаграмме (фиг. 60) на самом деле представляют собой
край кривой поверхности, идущей по направлению к нижнему
правому переднему углу. На фиг. 61 показаны две гипотетические
формы, находящиеся выше и ниже кривой. Одна раковина со
скоростью расширения оборота (W), равной 10, имеет спираль
с несоприкасающимися оборотами; другая, со скоростью рас-
ширения оборота, равной 3,5, имеет объемлющие обороты.
Важное следствие описанных закономерностей состоит в том,
что раковина, у которой обороты не накладываются друг на
друга, характеризуется свободным внутренним краем образую-
щей кривой. У двустворчатых форм (брахиопод и двустворча-
тых моллюсков) этот внутренний край образует замочный край
раковины. Вдоль него обычно располагаются зубы и зубные ям-
ки для сочленения створок; они увеличиваются по мере роста
образующей кривой. Развитие замочного края было бы крайне
затруднено у форм с перекрыванием последующих оборотов:
у них замочный край пришелся бы на внешний край предыду-
щего оборота и не осталось бы места для развития связочной
площадки, или интерареи. Видимо, именно поэтому раковины
двустворчатых и брахиопод соответствуют области, располо-
женной на блок-диаграмме (фиг. 60) ниже пунктирно?! линии.
Необходимо также объяснить сравнительную редкость одно-
створчатых раковин (брюхоногих и головоногих) в нижней час-
ти f блок-диаграммы. Функции такой раковины — поддержание
мягкого тела моллюска и его защита. Если устье раковины
170
Часть 2 Применение данных палеонтологии
Фиг. 61. Влияние изменения скорости рас-
ширения оборота (IF) на степень перекрыва-
ния между последовательными оборотами
[156].
Т — величина переноса ^оборота; D — расстояние
образующей кривой от оси
слишком широко, то страдает защитная функция; кроме того,,
раковина с широким устьем имеет относительно меньшую внут-
реннюю поверхность для поддержания и прикрепления мягкого
тела. Неудобство слишком широкого устья для брюхоногого
моллюска можно понять, если представить себе, что он зани-
мает одну створку раковины брахиоподы.
Некоторые одностворчатые моллюски, например блюдечко,
имеют, конечно, очень большие устья по отношению к общим
размерам, но такие формы живут, прикрепившись мускулистой
ногой к твердому субстрату. Широкое устье дает большую пло-
щадь прикрипления ноги к субстрату, и поскольку животное
остается прикрепленным, раковина достаточно хорошо защища-
ет его.
Теперь займемся несколько более подробно анализом адап-
тивных пиков и рассмотрим геометрию раковин спирально свер-
нутых головоногих, размещающихся в верхнем левом углу пра-
вой стенки блок-диаграммы на фиг. 60. На фиг. 62 показана
группа гипотетических форм, соответствующих этой области.
Скорости расширения оборота (W) варьируют от 1 до 4, а рас-
стояние между образующей кривой и осью (D) — от 0 до 0,6.
Показана также линия, разделяющая раковины с объемлющи-
ми оборотами и раковины с несоприкасающимися оборотами.
Именно эту область занимает и занимало в геологическом
прошлом большинство головоногих со спиральной раковиной.
Та же система координат применена на графике, отражающем
различную частоту разных геометрических типов среди 405 ро-
дов аммонитов позднего палеозоя и мезозоя (фиг. 63). Были
измерены скорость расширения оборота (W) и расстояние обра-
зующей кривой от оси (D) у типичных видов из 405 родов.
VIII Адаптация и функциональная морфология
171
о
ip-f-
1,5-
4,0-1
0,5
0,6
Расстояние образующей кривой от оси (D)
0,1 0,2 0,3 0,4
—I____________i_____________i____________i___
Фиг. 62. Группа теоретически возможных плоскоспиральных раковин, по-
лученных при варьировании скорости расширения оборота и положения обра-
зующей кривой [157].
Каждый вид был отмечен точкой, а затем очерчены области
плотного размещения этих точек.
Наиболее обычная для рассмотренных аммонитов геометрия
соответствует области с W=2 и Z) = 0,35. Можно сказать, что
она была самой «популярной». Эта область повышенной плот-
ности — общий адаптивный пик всей группы.
Функциональная морфология головоногих со спирально свер-
нутой раковиной не так проста, как может показаться, если
судить по фиг. 63. Внутри класса Ammonoidea есть более мел-
кие группы, приспособившиеся к своим дополнительным адап-
тивным пикам. Пример показан на фиг. 64. Масштаб здесь тот
же, что и на фиг. 63; область, занимаемая аммонитами, очерче-
Часть 2 Применение данных палеонтологии
Расстояние образующей кривой от оси (D)
О 0,2 0,4 0,6 0,8
1,0 -|-----1-------1------1-------------1-------1------1------1
Фиг. 63. Частота разных форм раковины у плоскоспиральных аммонитов
[157].
90% рассмотренных раковин находится внутри внешнего контура диаграммы. Внутрен-
ние контуры показывают возрастание числа родов на единицу поверхности графика (п —
число родов исследованных аммонитов)
на пунктиром. Сплошными линиями - оказано распределение
н» ’ Т и Г) для 35 родов палеозойского Goniatitina.
Инь и . .р ч ।,Л с»1 нни сдвинут влево от общего для всех аммо-
нитов пика. Это может означать или адаптивный пик, отличаю-
щийся от адаптивного пика всей группы (другие экологические
предпочтения или иной образ жизни), или же стадию эволюции
головоногих, на которой еще не был достигнут основной адап-
тивный пик (при W=2 и £) = 0,35). Во всяком случае, ясно, что
мы можем найти много дополнительных адаптивных пиков, от-
ражающих небольшие различия в образе жизни или местооби-
таниях. Одни головоногие могли приспособиться, например,
к мелководным местообитаниям, другие — к глубоководным,
третьи могли жить на дне; одни могли плавать быстро, дру-
гие — медленно и т. д.
В своем кратком анализе геометрии спирально свернутых
раковин мы ограничились аммонитами. Для полноты картины
следует рассмотреть и другую крупную группу головоногих со
спиральной раковиной — наутилоидей. Можно ожидать, что рас-
VIII Адаптация и функциональная морфология
Фиг. 64. Частота различных форм раковины у гониатитов [157].
Пунктирной линией очерчена «аммонитовая область» — линия внешнего контура и
фиг. 63 (и — число исследованных родов).
пределение плотностей у наутилоидей несколько отличается от
распределения у аммонитов, оставаясь, однако, в той же части
спектра спиральных форм. Интересно, что единственный ныне
живущий род наутилоидей — Nautilus — выпадает из «аммони-
товой области» (фиг. 63). Раковина Nautilus имеет скорость
расширения оборота W около 3,5, т. е. значительно большую,
чем это характерно для аммонитов. Можно только гадать о том,
случайно ли это различие или же оно отражает тот факт, что
Nautilus (а может быть, и все наутилоидеи) в ходе эволюции
приспособился к другому адаптивному пику.
Проблема перегородочной линии
головоногих
Как иллюстрацию к парадигматическому подходу рассмот-
рим часто обсуждавшийся вопрос о функции сложных по ри-
сунку перегородочных линий раковин аммонитов. Конечно, мы
не разрешим здесь эту проблему окончательно, но по крайней
мере можем пролить на нее некоторый свет.
Наружная
оценка раковины
Перегородочные линии
Перегородочные линии
не сохранились
Наружная стенка раковины
частично удалена
Фиг. 65. Морфология перегородок у аммоноидей [57].
A Goniatites kentuckiensis Miller Видны перегородочные линии гониатитового типа и по-
верхность одной перегородки (нижний карбон Кентукки, США) Б Xenaspis skinneri Mil-
lei et Furnish с цератитовым типом перегородочных линий (пермь штата Техас, США, и
Мексики) В Phylloceras (Hypophylloceras) onoense Stanton Там где наружный слой
раковины снят, видны перегородочные линии аммонитового типа (ннжний мел Калифор-
нии США) Г Поверхность перегородки Baculites — оаспространенного мелового рода
с прямой раковиной
VIII Адаптация и функциональная морфология
175
Сначала рассмотрим некоторые основные черты морфологии
раковины аммонитов (фиг. 65—68):
1. Типичная раковина аммонита тоньше и потому более
хрупка, чем типичная наутилоидная раковина.
2. Перегородки (септы), делящие раковину аммонита на
камеры, имеют характерную складчатую структуру края, кото-
рым они примыкают к наружной трубке раковины (фиг. 65)
Перегородки, как и завернутая раковина, секретировались мя-
систой мантией, выстилавшей при жизни жилую камеру.
3. Рисунок перегородочной линии (линии соединения пере-
городки и внешней трубки раковины), как правило, усложняет-
ся в ходе онтогенеза (фиг. 66).
4. Рисунок перегородочной линии у разных видов сильно
различается и служит важным для классификации аммонитов
признаком.
5. Камеры соединены только сифоном — мясистым тяжом,
идущим от жилой камеры ко всем остальным.
6. Толщина наружной стенки раковины возрастает в ходе
онтогенеза не так быстро, как диаметр оборота (фиг 67).
7. Аммоноидные перегородки обычно выпуклы к устью, а на-
утилоидные перегородки вогнуты
Фиг. 66. Усложнение рисунка пере-
городочной линии юрского аммонита
Cremceras renggen (Oppel) в ходе
i онтогенеза [145].
Показаны только части перегородочных
линий, в правой половине раковины
Стрелка у казьвает направление к устью
176
Часть 2 Применение данных палеонтологии
Фиг. 67. Графики, показывающие увеличение толщины раковины относи-
тельно ее диаметра в онтогенезе аммонитов Promicroceras marstonense (слева)
и Dactylioceras commune (справа) [218].
Толщина раковины, мм
8. По мере того как раковина аммонита достигает своего
максимального размера, перегородки обычно располагаются все
теснее.
Дополнительную информацию можно получить, изучив ныне
живущего Nautilus (фиг. 68), хотя он явно существенно отлича-
ется от аммонитов. Зная, что аммониты произошли от наути-
лоидей (см. стр. 308), мы можем считать перегородки и связан-
ные с ними структуры аммонитов и современного Nautilus го-
мологичными. Перегородочная линия Nautili проще, чем у
большинства аммонитов; у него, как мы знаем, скорость рас-
ширения оборота (W) выше. На обеих сторонах жилой камеры
имеется по крупному латеральному мышечному рубцу. Рубец
расположен непосредственно перед последней перегородкой, где
тело прикреплено к раковине. Аммониты наверняка отличались
от Nautilus и некоторыми деталями строения мягкого тела, но
каковы эти различия, нам не известно. С уверенностью можно
сказать, что большинство аммонитов, так же как Nautilus и
другие современные головоногие, были хищными животными,
плававшими с помощью реактивной струи. Типичные черты го-
ловоногих, связанные с подобным образом жизни, таковы: ро-
говой клюв, пиловидная радула, служившая для растирания
кусков пищи, оторванных клювом (недавно клювы и радулы
действительно были найдены вместе с остатками аммонитов),
щупальца, глаза и, наконец, газ, заполняющий замкнутые ка-
меры раковины и придающий животному плавучесть.
Дентон и Джилпин-Браун [38—40] недавно провели крайне
важные исследования физиологии плавучести раковин совре-
VIII Адаптация и функциональная морфология
177
менных головоногих. Они изучали раковины Nautilus и двух
сильно отличающихся родов сепиид— Sepia и Spirula. Рако-
вина Nautilus наружная, как и у аммонитов; раковины Sepia
и Spirula внутренние. У представителей всех трех родов рако-
вины разделены на камеры, а удельный вес всех животных
почти равен удельному весу морской воды и остается неизмен-
ным независимо от глубины погружения, так что плавучесть
животных почти нейтральна. В замкнутых камерах содержится
смесь газов, близкая по составу к воздуху; в недавно образо-
вавшихся камерах имеется некоторое количество водянистой
жидкости. Эти детали строения Nautilus показаны на фиг. 68.
Жидкости больше всего в последней камере; по составу она
напоминает морскую воду, но концентрация солей в ней ниже.
Давление газа в камерах — ниже 1 атм, причем оно меньше все-
го в недавно образованных камерах. В более старых камерах
давление газа около 0,9 атм. Моллюски всех изученных групп
используют соотношение газа и жидкости в камерах для управ-
ления плавучестью.
Nautilus растет очень быстро: примерно каждые две недели
появляется новая перегородка. В промежуток между телом и
последней перегородкой животное секретирует жидкость, кото-
рая помогает ему противостоять внешнему давлению, когда
тело перемещается вперед от последней перегородки, а также
когда образуется новая перегородка. Когда новая перегородка
кальцифицируется, жидкость откачивается через сифон, в ко-
тором пористые роговые и известковые слои окружают цент-
ральный мясистый стебель. Пространство, освобожденное от
жидкости, наполняется газом путем диффузии через сифон.
Такая последовательность событий объясняет, почему в камере
т;ем больше жидкости и тем ниже давление газа, чем позже она
образовалась.
Давление газа в замкнутых камерах всегда ниже 1 атм,
независимо от глубины погружения, и стенки раковины должны
выдерживать все давление окружающей воды. Nautilus погру-
жается до глубины 600 м и ниже. При погружении на 600 м
раковина должна выдерживать давление около 60 атм. Дентон
и Джилпин-Браун [39], а также Рауп и Такахаси [160] под
вергали в лаборатории раковины Nautilus (с закупоренными
сифонами) воздействию высоких давлений воды. Раковины
трескались при давлениях, соответствующих несколько большим
глубинам океана, чем предел погружения Nautilus. Таким обра-
зом, максимальная глубина погружения этого моллюска, види-
, мо, определяется прочностью раковины.
Дентон и Джилпин-Браун пришли к выводу, что поразитель-
ное сходство строения сифона и разделенной на камеры спи-
ральной раковины у ископаемых и живых групп головоногих
12—467
Щупальца
Б
Шестая
камера
Жилая камера
Отверстие
сифонной трубка
Водянистая
жидкость
Четвертая камера
Пористая трубка сифона
Газ
Г
VIII Адаптация и функциональная морфология
179
указывает на общность способов управления плавучестью у всех
этих групп, включая аммонитов Их аргументы неотразимы, так
как физиология разных рецентных групп оказалась сходной
Вернемся к проблеме функции перегородочной линии у ам
монитов Существует три основные гипотезы, объясняющие
сложный рисунок перегородочных линий
Согласно одной гипотезе, образование складок на перего-
родке необходимо для повышения устойчивости раковины к
разности давлений внутри (и снаружи Очевидно, многие ам-
мониты жили на значительных глубинах и подвергались как
значительным внешним давлениям, так и быстрой смене давле-
ний при подъемах и погружениях
По другой гипотезе сложный рисунок перегородочной линии
связан со способностью животного регулировать свой общий
удельный вес Часто высказывалось предположение, что мыш
цы, закрепляющие животное в его жилой камере, возможно,
располагались так, что позволяли моллюску изменять объем
заполненного газом пространства, которое, как считали, су
ществовало между телом и последней* перегородкой Согласно
этой гипотезе, тело животного представляло собой как бы пор
тень, который мог двигаться взад или вперед, и в результате
менялся удельный вес всего животного, а следовательно, и его
плавучесть, что позволяло ему перемещаться по вертикали в
столбе воды Мышцы, прикреплявшиеся к седлам перегородки,
толкали тело вперед, а те, что прикреплялись к лопастям, тяну
ли назад
Третья гипотеза состоит в том, что складчатость края пере
городок объясняется складчатостью мантии Складчатая мантия
увеличивала относительную поверхность выделения газа или
жидкости в пространство между мантией и последней перего
редкой
Для каждой гипотезы можно построить парадигму Если
принять, что складчатость перегородок служила прежде всего
Для укрепления наружной стенки раковины, то в этом случае
парадигмой будет механическая структура, способная выдержи-
вать большие давления при минимуме затраченного материала
(для экономии энергии организма) Такая структура должна
одинаково прочно поддерживать наружную раковину во всех
точках Чем больше расстояние данной точки раковины от внут-
ренней опорной структуры, тем меньшей поддержкой для нее
"Служит эта структура Инженер, которому поручили бы создать
Фиг 68 Биология современного наутилуса
и* Наутилус плывущий назад по механизму реактивного движения [57] Б Наутилус
цепляющийся щупальцами за дно [57] В Внутреннее строение наутилуса (часть ра
’ ковины удалена) [57] Г Разрез раковины [39]
J2*
180
Часть 2 Применение данных палеонтологии
такую конструкцию, разработал бы систему двутавровых балок,
отходящих под соответствующими углами от стенки раковины
и сходящихся внутри раковины, в центре. Он мог бы применить
решетчатую опорную систему. Но моллюск должен выделять
вещество раковины своей мясистой мантией, представляющей
поверхность для секреции углекислого кальция. Кроме того,
у раковинных головоногих, по-видимому, ранее образованные ка-
меры раковины обязательно должны быть изолированы от жи-
лой камеры. Даже сифон, поскольку он заполнен тканями тела,
не обеспечивает свободного обмена жидкостями между жилой
камерой и остальными камерами.
Следовательно, парадигмой для гипотезы прочности должна
быть система сплошных поперечных перегородок, выделяемых
мантией по мере роста наружной раковины. Задача состоит в
том, чтобы обеспечить эффективную поддержку стенок при ми-
нимальной затрате материала. Реальная прочность раковины
как целого будет зависеть от самого ее непрочного участка. Ин-
женерные принципы говорят нам, что гофрированность края
опорных перегородок помогает создать лучшую опору для меж-
перегородочных участков раковины. Амплитуда этой складча-
тости должна быть велика по сравнению с шириной межперего-
родочных промежутков, иначе значительные участки раковины
останутся без опоры. Поскольку мы предположили, что гофри-
рованность края перегородок нужна для увеличения прочности
наружной стенки, а не самих перегородок, достаточно, чтобы
они были волнистыми только по краям — там, где они соеди-
няются со стенками. Можно предположить, что в ходе онтогене-
за число перегородок должно увеличиваться или должен услож-
няться рисунок складчатости, так как толщина наружной стенки
аммонитов в онтогенезе уменьшалась.
Можно назвать по крайней мере три важных фактора, кото-
рые могли создавать значительную межвидовую изменчивость
рисунка складчатости, описанной в парадигме: 1) разные му-
тации в разных эволюционных рядах должны были привести к
различным конструктивным решениям проблемы увеличения
прочности; 2) на разных глубинах и при разных режимах смены
давления различные таксоны приспособились к действию давле-
ния по-разному; 3) у моллюсков с разной формой спиральной
раковины должны были появиться разные системы, повышаю-
щие прочность.
Теперь сравним рассмотренную парадигму с известной мор-
фологией перегородок аммонитов. Оказывается, сильно измен-
чивый рисунок периферической складчатости перегородок аммо-
нитов очень хорошо укладывается в парадигму. Усложнение
рисунка перегородочной линии в ходе онтогенеза у аммонитов —
хорошо известное явление; оно было предсказано парадигмой
VIII Адаптация и функциональная морфология
181
как механизм, компенсирующий уменьшение относительной тол-
щины завернутой наружной раковины. Ньюэлл [133] собрал
данные, показывающие, что у группы позднепалеозойских аммо-
нитов длина перегородочной линии возрастала в ходе онтогене-
за гораздо быстрее, чем диаметр раковины (фиг. 69). Тенден-
ция к сближению перегородок в ходе онтогенеза также играет
компенсаторную роль, хотя некоторые авторы предполагают,
что такое сближение — побочное следствие замедления роста
по мере приближения раковины к максимальному размеру.
Следует отметить одно слабое место этой гипотезы: давление
воды передается на последнюю перегородку телом моллюска и
неупругой жидкостью. Следовательно, кроме увеличения проч-
ности наружной раковины, складчатость перегородки должна
была служить и для увеличения прочности самой перегородки.
Мы отмечали, что перегородки аммонитов обычно выпуклы в
сторону жилой камеры, а перегородки наутилоидей вогнуты.
«Аммонитовая» конструкция лучше противостоит давлению жид-
кости в жилой камере (вспомните вогнутое дно многих аэро-
зольных баллончиков).
Мы не будем подробно разрабатывать парадигму для вто-
рой гипотезы, объясняющей складчатость края перегородки.
Эта гипотеза предполагает, что складчатость обеспечивала при-
крепление мышц, с помощью которых животное перемещалось
в жилой камере в глубь нее или к устью. Достаточно сказать,
что любая парадигма, построенная для иллюстрации этой гипо-
тезы, предусматривает следы прикрепления мышц на перего-
родке или на внутренней стенке раковины. Животное подверга-
лось большим давлениям, и для продвижения к устью раковины
ему потребовались бы очень сильные мышцы, а следовательно,
и очень глубокие рубцы для прикрепления этих мышц. Крупные
мышцы, закрепляющие современного Nautilus в его камере, сла-
бо прикреплены к внутренней стенке раковины. Им приходится
очень быстро (до 2—3 см в месяц) перемещаться вперед, чтобы
успевать за ростом спирально свернутой раковины. У аммони-
тов не найдено глубоких или обширных отпечатков мышц.
Третья гипотеза говорит, что складчатость края перегоро-
док— это побочное следствие увеличения площади поверхности
мантии, необходимого для усиления секреции газа или жидко-
сти в пространство между мантией и последней перегородкой.
Парадигма к этой гипотезе должна предусматривать складча-
тость или волнистость всей перегородки, а не только ее краев.
Значит, эта парадигма не подходит к наблюдаемой конфигура-
ции перегородок у аммонитов.
В общем из рассмотренных парадигм к известным особенно-
стям морфологии перегородок лучше всех подходит первая, ос-
нованная на предположении, что перегородка с гофрированным
2000
Диаметр раковины,мм
Фиг. 69. Аллометрическое возрастание сложности перегородочной линии в
онтогенезе пяти палеозойских родов аммоноидей [133].
По вертикали отложена половина длины внешней перегородочной линии (У), по гори-
зонтали — диаметр раковины (X) У всех родов с увеличением X экспоненциально рас-
тет У Поэтому в логарифмической шкале мы получаем прямолинейные графики, Вверху
слева (/—5) изображены перегородочные линии взрослых особей пяти родов Черные
треугольники — Medlicottia <5), пермь (у = 1,32 X1’24)’ чеРные кружки — Ргортасосе-
-г as (4), пермь (у = 1,10 X1’28), черные квадратики — Artinskta (3), пермь (У =
~ -X1’36), белые кружки — Uddtnttes (2), карбон (у = 0,11 X1’23), белые треуголь-
ники — Prouddetiites (1), карбон (у = 0,11 X1’20)-
VIII Адаптация и функциональная морфология
183
краем должна была поддерживать внешнюю стенку, сопротив-
лявшуюся гидростатическому давлению. Это еще не значит, что
здесь все ясно. Мы не рассмотрели всех возможных функций;
вполне возможно, что перегородки со складчатым краем выпол-
няли у аммонитов не одну функцию. Во всяком случае, пара-
дигма к «гипотезе поддержки» должна быть разработана гораз-
до подробнее и строже, чем это сделали мы.
Механические свойства челюстей
динозавра
Подвижные скелетные элементы челюстей позвоночных дей-
ствуют обычно как рычаги, приводимые в движение челюстными
мышцами. Чтобы понять функционирование челюстей ископае-
мых форм, приходится восстанавливать морфологию мышц по
скелету или на основании гомологии с ныне живущими родст-
венными формами. Проведенный недавно анализ механики че-
люстей меловых рогатых динозавров пролил свет на способы их
питания [144]. Эти рогатые четвероногие (фиг. 70) обычно
считались травоядными, но в отличие от большинства травояд-
ных, имеющих дробящий и размалывающий зубной аппарат,
у цератопсов были режущие зубы.
Рассмотрим здесь анализ механических свойств челюстей
рода Triceratops, проведенный Остромом (фиг. 71). Точка опо-
ры, относительно которой вращалась нижняя челюсть Tricera-
tops, расположена вблизи заднего конца челюсти. Предполага-
ется, что главная мышца, поднимавшая один конец челюсти,
прикреплялась к выдающемуся вверх отростку челюсти, так
называемому венечному отростку. Другой конец мышцы был
прикреплен к мощному костному воротнику черепа. Остальные
мышцы, очевидно, играли незначительную роль в движении че-
люсти, и сейчас ими можно пренебречь. Направление тяги
главной мышцы показано на фиг. 71, Г жирной пунктирной
стрелкой. Нижняя челюсть большинства позвоночных действу-
ет как рычаг третьего рода: мышечная сила, подйимающая че-
люсть, прилагается к ней в точке между опорой и точкой при-
ложения противодействующей силы (причиной ее возникновения
служит пища, находящаяся между зубами). Простой рычаг
третьего рода показан на фиг. 71, Б. Действие каждой из двух
сил определяется длиной плеча (расстоянием от точки прило-
жения силы до точки опоры). Момент мышечной силы равен
произведению силы на длину плеча (Ь на фиг. 71, В). Если
поднимающая сила действует под острым углом к рычагу (как
на фиг. 71, В), то ее момент уменьшается; он равен произведе-
Часть 2 Применение данных палеонтологии
Фиг. 70. Реконструкция рогатых динозавров, близких к Triceratops [5].
Л Chasmosaurus в позе ползания, обычной для музейных и книжных реконструкций це-
ратопсов Б. Torosaurus в «приподнятой» позе, на котооую указывают новые исследова-
ния механических свойств конечностей динозавров
нию обычного момента на синус острого угла. Следовательно,
момент силы, действующей на рычаг под углом 45°, равен
Поднимающая сила х b х sin 45° = Поднимающая сила х Ь'.
Чем меньше угол между силой и рычагом, тем меньше момент.
Рассмотрев устройство такого рычага, мы можем предска-
зать, что эффективность челюсти позвоночного максимальна,
когда мышечная сила действует на челюсть под прямым углом
и приложена как можно ближе к переднему концу челюсти. Но
такое устройство вносит затруднения: рот будет еле приоткры-
ваться и в лицевой области не окажется места для прикреп-
ления мощных мышц. К тому же при удалении мышц от точки
опоры скорость закрывания рта будет уменьшаться.
VIII Адаптация и функциональная морфология
185
Поднимающая
сига
a t b W
Сила
сопротивления
Сопротивления
В
А
Фиг. 71. Механические свойства челюсти Triceratops [144].
А. Поперечное сечение челюсти, показывающее стригущие поверхности зубов Б Прос
той рычаг третьего рода с параллельно направленными силами В. Простой рычаг третье-
го рода с непараллельно направленными силами Г Череп, на котором показано располо-
жение главной мышцы-аддуктора Д Нижняя челюсть Е Схема действия сил на ниж-
нюю челюсть
Но есть способ увеличить длину плеча и усилить действие
рычага, даже если действие мышцы направлено под острым уг-
лом к челюсти: надо поднять точку приложения силы над точ-
кой опоры, удлинив плечо, на которое действует сила. У Trice-
ratops использовался именно этот способ благодаря венечному
отростку. На фиг. 71, Е показано, как венечный отросток удли-
няет плечо главного аддуктора.
Остром [144] рассчитал для нескольких видов цератопсов,
какая доля силы главного аддуктора действует на пищу при
разном ее положении на челюсти. Чтобы провести расчет, он
186
Часть 2 Применение данных палеонтологии
просто сравнивал длину плеча мышечной силы с длиной плеча
сопротивляющейся пищи при разных положениях (это также
длина плеча «равной и противоположно направленной» силы,
приложенной к пище). На фиг. 71, Е показано, что главная
мышца-аддуктор действует под углом 80° к линии, соединяю-
щей место ее прикрепления с точкой опоры. Поэтому действи-
тельным плечом становится Ь'у а доля силы главного аддуктора,
действующая на пищу, рассчитывается таким образом:
Сила, приложенная к пище _ Ь’
Сила, развиваемая главным аддуктором аг 4- а2 ‘
Остром рассчитал, что у большинства видов на пищу у клю-
ва действовало 30%, у передних зубов — 50%, а у задних зу-
бов, очень близко к точке опоры, — 250—300% силы главного
аддуктора. Судя по предполагаемым размерам главных мышц,
их сила была весьма велика.
Особенности челюсти цератопсов, в частности появление вы-
сокого, мощного венечного отростка, способствовали передаче
на зубы мощной срезающей силы. Остром считает, что, если
цератопсы действительно были вегетарианцами, то они специ-
ализировались на поедании жесткой растительной пищи, напри-
мер пальм и цикадовых, которых было много в конце мелового
периода.
Недавние исследования механических особенностей конеч-
ностей динозавров [5] показывают, что многие виды динозав-
ров, передвигавшихся на четырех ногах, были совсем не так
неуклюжи, как предполагалось ранее. В реконструкциях перед-
ние ноги часто помещали в позу ползания, сильно расставляя
их в стороны (фиг. 70). Сейчас полагают, что у большинства
этих форм, в том числе у Triceratops, передние ноги были рас-
положены ближе к средней линии тела и могли поэтому слу-
жить эффективными бегательными конечностями. Возможно,
рогатый Triceratops был похож на носорога. Судя по проведен-
ному Остромом анализу его срезающих челюстей, не исключено
даже, что Triceratops был хищником!
Парус пеликозавров
Интерпретация функции бывает особенно убедительна, когда
она поддерживается теоретическими математическими сообра-
жениями (см. принцип подобия, стр. 63). Пример — наиболее
широко принятая интерпретация функции спинного «паруса»
звероподобных пресмыкающихся пермского рода Dimetrodon.
Парус был образован кожей, натянутой на сильно вытянутые
остистые отростки позвонков (фиг. 72). Было выдвинуто пред-
положение, что парус служил для регуляции температуры тела.
VIII Адаптация и функциональная морфология
187
Фиг. 72. Скелет Dimetrodon [167].
Максимальная длина около 3,5 м
По этой гипотезе, Dimetrodon мог сильно повышать или пони-
жать температуру своего тела, изменяя положение паруса по
отношению к направлению ветра или к солнечному свету. Ско-
рость поглощения или потери тепла могла также до какой-то
степени регулироваться изнутри, посредством изменения тока
крови в парус и из него. Ромер [166] показал, что площадь
паруса была непропорционально велика по сравнению с пло-
щадью поверхности тела у более крупных видов Dimetrodon.
Соотношение можно описать простым уравнением, сходным с
уравнением для анизометрического роста (стр. 62):
Площадь паруса — (Площадь поверхности тела)1-6 (1)
Если парус был терморегулирующим органом, то, исходя из
принципа подобия, можно ожидать, что в процессе эволюции
увеличение размеров тела должно было сопровождаться увели-
чением площади паруса пропорционально объему тела (т. е.
тому объему, который надо нагревать и охлаждать), так как
лишь при этом условии эффективность терморегуляции могла
сохраняться на прежнем уровне. Примем, что форма тела не из-
менялась (хотя это не совсем верно); тогда объем тела должен
был возрастать как куб линейных размеров, а площадь поверх-
ности тела — как квадрат линейных размеров. Другими слова-
ми, (объем тела) з должен был оставаться пропорциональным
(площади поверхности тела)“ независимо от размера тела.
Если площадь и объем измеряются в одних и тех же единицах,
то пропорция превращается в равенство:
Объем тела = (Площадь поверхности тела)2 .
Если площадь паруса пропорциональна объему тела, то:
Площадь паруса = (Площадь поверхности тела) 2
(2)
188
Часть 2. Применение данных палеонтологии
Хорошее совпадение между наблюдаемым соотношением
[уравнение (1)] и теоретическим расчетом [уравнение (2)] го-,
ворит в пользу гипотезы о терморегулирующей функции паруса.
Специальные адаптации у брахиопод
» Рудвик [173] провел анализ функций различных структур
•брахиопод и предложил интерпретацию функции морфологии
раковины у двух поразительно сходных мезозойских таксонов,
принадлежащих к разным надсемействам (фиг. 73). Необычная
деталь строения их раковин — четыре узких выроста по краю
раковины. Мантия, секретировавшая раковину, очевидно, при
жизни организма выстилала изнутри выросты обеих створок.
Рудвик отклонил гипотезы, согласно которым эти выросты слу-
жили для повышения прочности раковины или увеличения по-
верхности мантии, через которую происходил газообмен, по-
скольку нет явных причин для упрочнения раковины, да и вы-
росты лишь ненамного увеличили бы площадь мантии. Рудвик
высказал предположение, что выросты служили для предупреж-
дения об опасности, действуя подобно ряду антенн. Чувстви-
тельная мантия, доходившая до кончиков выростов, восприни-
мала прикосновения, и животное таким образом чувствовало
приближение постороннего объекта к открытой мантийной по-
лости. Створки быстро закрывались, избавляя животное от
опасности. Предположение Рудвика подтверждается также тем,
в
Фиг. 73. Конвергентная морфо-
логия видов брахиопод [173].
A. Tetractinella trigonella (средний
триас). Б. Cheirothyrie fleuriausa (верх-
няя юра). В. Предполагаемые контуры
поля чувствительности края мантии.
Рисунок показывает, как краевые
выросты раковин у обоих видов могли
служить «антеннами».
VIII. Адаптация и функциональная морфология
189
что длина отростков почти одинакова, они равномерно распреде-
лены по краю мантии и очень тонкие (для экономии материала
раковины).
Зрение трилобитов
Кларксон [29] применил необычный подход к функциональ-
ной морфологии, изучая зрение силурийских трилобитов под-
отряда Phacopina, в частности вида Acaste downingiae (Salter)
(фиг. 74 и фото XXII). Факопины обладали сложным глазом,
состоявшим из многочисленных фасеток, собранных в ряды.
Фасетки имели угловатые очертания, что обеспечивало наиболее
плотную упаковку (впрочем, между ними существовали узкие
промежутки, как видно на фото XXII, Д). При жизни каждая
фасетка венчала собой наружный конец цилиндра, передавав-
шего свет к нижележащим светочувствительным клеткам.
Чтобы определить ориентацию осей отдельных фасеток гла-
за, Кларксон использовал бинокулярный микроскоп с поворот-
ным предметным столиком, на котором был укреплен трилобит.
Фиг. 74. Зрение Acaste downingiae 129].
Поля зрения левого и правого глаз, показанные в стереографической проекции.
Вид головного щита спереди в предполагаемом прижизненном положении. Показаны
«Широтные» пределы зрения по мере приближения объекта, имеющего высоту h, от точки
а к точке с.
190
Часть 2 Применение данных палеонтологии
Кларксон построил стереографические проекции осей ориента-
ции отдельных фасеток по отношению к плоскости билатераль-
ной симметрии животного Собранные вместе, эти проекции
составляют зрительное поле глаза (фиг 74, А, Б) Фасетки
были расположены рядами, поэтому зрительное поле Acaste
состояло из отдельных полос, и трилобиты воспринимали свет
как ряд светлых и темных полос
Проведя сравнение со сложными глазами современных чле-
нистоногих, Кларксон сделал вывод, что сложные глаза трило
битов были недостаточно высоко организованными, чтобы вос-
принимать форму объектов Но расчленение поля зрения на
зрительные полосы, вероятно, позволяло трилобитам воспри
нимать горизонтальные перемещения объекта, его размер, ско-
рость и направление движения Биологические исследования
показали, что при наличии глаз такого типа, как у факопин, для
восприятия движений на фоторецепторы должна была проеци-
роваться прерывистая картина Зрительные полосы у Acaste
как раз и образуют такую прерывистую картину Движение
тени по горизонтали через поле зрения будет поочередно вос-
приниматься зрительными полосами Как показано на фиг 74, В,
животное могло также воспринимать приближение объекта,
отмечая, что объект регистрируется все более высоко располо
женными фасетками глаза
На основании выводов Кларксона о зрении трилобитов мож
но убедительно интерпретировать прижизненное положение це-
фалона (головного щита) Acaste Поля зрения обоих глаз пол-
зающего трилобита почти наверняка были ориентированы так,
что животное обладало 360-градусным боковым зрением Это
могло быть только в том случае, если передний край головного
щита быт дугообразно приподнят над субстратом, а не приле-
гал к нему плотно Действительно, головной щит экземпляра,
изображенного на фото XXII, А—В, ориентирован именно так,
хотя эти трилобиты сохранились в свернутом положении, а не
в нормальной позе питания или движения Животные, очевидно,
свернулись для защиты, и многие в этом положении и погибли.
Форма раковины у двустворчатых
моллюсков
Как пример большого значения наблюдений над живыми ви-
дами для интерпретации функциональной морфологии ископае-
мых форм рассмотрим два слабо связанных надсемейства дву-
створчатых моллюсков — Агсасеа и Carditacea Морфологиче
ские тенденции их эволюции можно связать с изменениями
образа жизни В обоих надсемействах имеются представители
двух экологических групп (фиг. 75). Одна группа состоит из
VIII Адаптация и функциональная морфология
191
Фиг *75 Конвергенция между роющими и прикрепленными при помощи
биссуса двустворчатыми моллюсками Агсасеа и Carditacea (из [208], с изме-
нениями).
А и Б Anadara otalis и A antiquata (Агсасеа) В и Г Venencardta borealis и Cardita floridana
(Carditacea) На верхних рисунках четыре вида показаны в их прижизненных положе
пнях На нижних рисунках два вида моллюсков с равносторонними раковинами зарыва
юи I в осадок два других вида с вытянутыми раковинами показанные тут же при про
икновении в осадок встречали бы гораздо большее сопротивление
видов, прочно прикрепленных к субстрату биссусом (пучком
роговых нитей, секретируемых ногой моллюска) В другую вхо-
дят закапывающиеся животные, живущие неприкрепленными
(или очень слабо прикрепленными тонким биссусом) в мягких
осадках. Большинство современных Агсасеа прикреплено к по-
верхности субстрата прочным биссусом Эти виды обычно обла-
дают гораздо более удлиненной раковиной, чем закапывающие-
ся Виды Carditacea, прикрепляющиеся прочным биссусом, так
же более вытянуты, чем их неприкрепленные родичи Некото-
рые из них прикрепляются к твердым предметам в толще осад-
ка, а не на его поверхности
Прижизненное положение типичных зарывающихся видов
этих двух надсемейств показано на фиг 75 Компактная равно-
сторонняя форма раковин обеспечивает малую площадь брюш-
ной стороны раковины и соответственно малое сопротивление
при закапывании. Брюшной край к тому же закруглен для об
легчения проникновения в субстрат Удлиненные раковины ви-
дов, прикрепленных прочным биссусом, гораздо хуже прони-
192
Часть 2 Применение данных палеонтологии
кали бы в субстрат. Неглубоко закапывающиеся животные мо-
гут быть вымыты из субстрата волнами и течением, и им важно
уметь быстро закапываться.
Видам, имеющим толстый биссус, для прочного прикрепле-
ния нужно, чтобы при сокращении ретракторов биссуса брюшной
край раковины плотно прилегал к поверхности прикрепления
Компактная форма и закругленный брюшной край зарывающих-
ся видов Агсасеа и Carditacea позволял бы им контактировать
с плоской поверхностью только в одной точке, и прикрепление
было бы непрочным.
Зная роль формы раковины у современных представителей
Агсасеа и Carditacea, можно сделать выводы об образе жизни
вымерших видов. Палеонтологическая летопись Агсасеа прости-
рается по меньшей мере до девона, и, рассматривая ее, мы
узнаем, что в палеозое эта группа была почти исключительно
представлена формами с удлиненными раковинами, которые
при жизни животного, очевидно, были прикреплены к субстрату
прочным биссусом. Зарывающиеся Агсасеа с компактной ра
ковиной появились, по-видимому, только в мезозое Обе эколо-
гические группы не вымерли и распространены в современных
морях.
Ранняя палеонтологическая летопись Carditacea недостаточ-
но известна, но, вероятно, ход эволюции группы был подобен
только что рассмотренному. Большинство ранних Carditacea
(пермь и триас) имели вытянутые раковины, очевидно прочно
прикреплявшиеся к субстрату. Более компактные, свободно жи-
вущие роющие формы стали распространенными только в конце
мезозоя и в кайнозое. Если верно мнение Хислипа [74], то
некоторые кайнозойские линии компактных свободно роющих
форм дали начало биссусным формам типа Venertcardia (Clat-
bornicardta) nasuta, которые вернулись к образу жизни своих
эволюционных предшественников (фиг. 76).
Функциональная морфология формы раковины Агсасеа и
Carditacea особенно интересна тем, что в обоих надсемействах
отмечаются одинаковые способы эволюционной дивергенции на
экологические группы Эволюционная конвергенция формы ра-
ковины и образа жизни — одно из важнейших доказательств
того, что морфология скелета в основном адаптивна По-види-
мому, выраженное сходство между рассмотренными таксонами
брахиопод, изученными Рудвиком (фиг. 73), также является
следствием адаптивной конвергенции.
Заключение
Рассмотренные нами случаи — лишь отдельные примеры ин-
терпретации различных функций ископаемых организмов Эти
примеры показывают сложность возникающих задач, для реше-
VIII Адаптация и функциональная морфология
193
Фиг. 76. Кайнозойская фйлогения Venericardia (Claibornicardid) с'берегов
Мексиканского залива и восточного побережья Северной Америки [74].
ния которых необходимо привлекать различные аналитические
методы. Функциональная морфология — крайне важная область
исследования. Это объясняется двумя причинами во первых,
в применении к отдельным ископаемым видам она может су-
щественно способствовать палеоэкологическому анализу—ис-
следованию взаимодействия видов с их средой (их образа жиз-
ни, экологических предпочтений, связи с другими видами и
т. п.); во-вторых, в применении к таксономическим группам
она может показать, в какой степени морфологические тенден-
ции эволюции, наблюдаемые в палеонтологической летописи,
являются адаптивными (фиг. 76).
ГЛАВА IX
Палеоэкология
Экология — наука о взаимодействии организмов с их средой.
Палеоэкология занимается теми же вопросами, но на ископае-
мых видах.
Существует много побудительных причин к изучению палео-
экологии. Главная задача геологии — исследование истории
Земли и ее обитателей. Хотя большинство детальных палео-
экологических анализов ограничено краткими промежутками
геологического времени и географическими областями, изме-
ряющимися метрами, километрами или десятками километров,
результаты множества таких работ позволяют представить об-
щую историю жизни и условий среды на Земле. Поскольку в
стратиграфической летописи имеются пробелы, а ископаемые
остатки дают лишь неполную биологическую информацию, нам
никогда не удастся полностью восстановить эту историю. Все
же палеоэкология может дать немало сведений об истории жиз-
ни на Земле, и сейчас мы только начинаем понимать возмож-
ности этой науки.
При палеонтологическом исследовании важно не попасть в
логический порочный круг. В любом палеонтологическом иссле-
довании необходимо четко установить, собираемся ли мы ре-
конструировать условия среды по непалеонтологическим данным
и затем, зная условия, интерпретировать данные о вымерших
видах или же мы будем реконструировать условия среды по
организмам. В одной работе можно использовать оба подхода,
но они не должны перемешиваться, что создало бы порочный
круг и свело к нулю ценность окончательных выводов.
Первый шаг в палеоэкологическом исследовании — определе-
ние того, произошло ли захоронение изучаемых органических
остатков в том же месте, где обитали организмы, а если нет,
то как далеко остатки были перенесены. Этот вопрос мы рас-
смотрим ниже. Сначала сделаем несколько общих замечаний.
Как указывалось в гл. I, морские животные (в основном это
беспозвоночные) обитают в обширном осадочном бассейне, где
вероятность сохранения остатков организмов в общем выше,
чем на суше. Среди наземных организмов преобладают насеко-
мые, но их остатки сохраняются редко. Чтобы остатки наземных
IX. Палеоэкология
195
организмов, которые могут сохраняться (это главным образом
растения и позвоночные), действительно сохранились, они долж-
ны быть перенесены из исходных местообитаний организмов в
районы осадконакопления, например в озера или реки. Боль-
шинство ископаемых сообществ состоит из остатков видов, при-
несенных из нескольких местообитаний. Перенос и смешение
мешают проведению палеоэкологических исследований ископае-
мых сообществ наземных организмов. Зато такие сообщества
широко использовались для региональных фаунистических и
флористических исследований, много давших для понимания
древних климатов и палеобиогеографии. Морские организмы не
только реже подвергаются переносу, но и лучше сохраняются.
Кроме того, в геологическом прошлом численность раковинных
морских беспозвоночных превышала численность сохраняемых
наземных растений и животных. Озерные биоты по своей при-
годности для палеоэкологических исследований стоят между
морскими и наземными. Они обычно не подвергаются значи-
тельному посмертному переносу, но озера занимают незначи-
тельную. часть поверхности Земли, а озерные отложения часто
разрушаются эрозией. Поскольку морские раковинные беспоз-
воночные были объектом большинства палеоэкологических ис-
следований, мы будем говорить в основном об этих животных.
Основные принципы экологии
Самая крупная единица, рассматриваемая экологами, — эко-
система— произвольно выбранный участок среды вместе со все-
ми живущими здесь организмами. В понятие экосистемы входят
также все взаимодействия — физические, химические и биологи-
ческие,— установившиеся в границах выбранного участка. Эко-
системой мы с одинаковым правом можем назвать и всю био-
сферу, и лужицу дождевой воды, в которой водятся два-три
вида простейших.
Местообитание — среда, в которой живет организм. Это мо-
жет быть скалистый морской берег, степь или — для паразита
(блохи, например) — вид-хозяин (например, собака). В экоси-
стеме может быть несколько местообитаний. Экологическую ни-
шу часто определяют как положение организма в местообита-
нии, включая сюда его образ жизни и роль в экосистеме. Но
некоторые ученые предпочитают определять экологическую ни-
шу как комплекс условий среды, позволяющих виду существо-
вать. В любом случае, нас интересует взаимодействие вида
с его окружением. Обд определения различаются не по своей
сути, а по тому, что подчеркивается в каждом из них. В первом
упор делается на физиологические и поведенческие адаптации
вида к среде, а во втором — на экологические границы приспо-
13*
196
Часть 2 Применение данных палеонтологии
собленности вида. Первый способ определения обычно облегча-
ет описание отдельной ниши, а второй способствует сравнению
между двумя или несколькими сходными нишами. Большинство
местообитаний занято несколькими видами, и каждый из них
имеет свою экологическую нишу. Обычно каждый вид пред-
ставлен несколькими особями (не менее двух), составляющими
популяцию (гл. IV). Популяции двух или нескольких видов,
занимающие местообитание, обычно называют сообществом.
Как правило, сообщество называют по одному-двум самым за-
метным и многочисленным его компонентам. Всем известные
примеры — пихтово-еловый лес северных широт и балянусо-ми-
диевое сообщество скалистых океанских побережий. Точно так
же, как в экосистеме может быть несколько местообитаний, так
и местообитание может содержать несколько сообществ. Тер-
мин «сообщество»» пока не определен в экологии достаточно
строго; о нем мы поговорим подробнее в дальнейшем.
В каждой экосистеме взаимодействует множество живых и
неживых компонентов. Основные взаимодействия связаны с пере-
носом энергии и вещества. Схема движения вещества в простой,
но типичной экосистеме показана на фиг. 77. В большинстве
сообществ органические соединения синтезируются из окружаю-
щих веществ продуцентами — фотосинтезирующими растения-
ми. Они потребляются травоядными организмами, часть из ко-
торых идет в пищу хищникам. Паразиты, питающиеся живыми
организмами, не обязательно убивая их, и мусорщики, питаю-
щиеся мертвыми организмами, также различным образом вклю-
чаются в экосистему. Последовательность видов от продуцентов
до хищников, на которых никто не охотится, называется пище-
вой цепью или пищевой сетью. Органическое вещество проду-
центов, травоядных и хищников, не потребленное видами, обра-
зующими последующие звенья пищевой цепи, разлагается орга-
низмами-редуцентами, в основном бактериями. Наконец, другие
организмы, называемые преобразователями (это также в основ-
ном бактерии), химически изменяют некоторые продукты разло-
жения, делая их снова пригодными для продуцентов. Таким
образом, бактерии чрезвычайно важны для успешного замыка-
ния цикла; без них экосистемы не могли бы существовать в том
виде, в каком они нам известны.
Следует подчеркнуть, что в круговороте, показанном на
фиг. 77, участвует вещество. Энергия также движется в круго-
вороте по часовой стрелке, но она не восстанавливается. Общее
количество живого вещества (включая накопленные питатель-
ные вещества), имеющегося в любой данный момент в экосис-
теме, называется биомассой.
Поток вещества или энергии через пищевую цепь можно
изобразить с помощью простой диаграммы. На фиг 78 показана
IX Палеоэкология
197
Второстепенные компоненты
Основные компоненты
>► Консументы
Живые компоненты
Неживые
компоненты
Фиг. 77. Схема потоков вещества через типичную экосистему (из [28],
с изменениями).
так называемая «энергетическая пирамида», отражающая пе-
ренос энергии. Первичный источник энергии для большинства
экосистем — Солнце, поскольку большинство продуцентов ис-
пользует для синтеза органических соединений солнечную энер-
гию. Продуцентов поедают травоядные, тех поедают хищники,
а они в свою очередь идут в пищу «верховным» хищникам.
Крутизна пирамиды отражает потерю энергии на каждом уров-
не, объясняющуюся несовершенством механизмов обмена и раз-
ложением никем не съеденных особей. Подобную пирамиду
можно построить и для биомассы (в данный момент времени)
компонентов экосистемы. Хищники, как правило (но не всегда),
крупнее своих жертв. Поэтому к вершине пищевой пирамиды
уменьшается не только биомасса, но и численность организмов.
Обычно пищевые пирамиды имеют три-четыре уровня. Чем
меньше уровней в пирамиде, тем меньше потери энергии.
Палеоэкологи практически лишены возможности определять
поток энергии и вещества через экосистемы. Но для некоторых
ископаемых сообществ удается реконструировать хотя бы не-
198
Часть 2. Применение данных палеонтологии
Фиг. 78. Энергетическая пирамида для экосистемы в Силвер-Спринге, Фло-
рида [140].
Трофические уровни: П — продуценты, Т — травоядные животные, X — хищники,
ВХ — «верховные» хищники. Р — редуценты. Каждый прямоугольник соответствует об-
щему потоку энергии через данный трофический уровень. Темные полосы соответствуют
энергии, накопленной биомассой в изученной области; светлые полосы соответствуют по-
тере энергии при дыхании и в результате перемещения организмов за пределы исследуе-
. мой области.
полную картину пищевых цепей. Например, палеоэколог может,
как правило, распознать хищные виды и их жертвы.
Морская экосистема
Современная морская экосистема используется большинст-
вом палеоэкологов как общая модель для интерпретации древ-
них морских отложений и фаун. Один из недостатков такого
подхода состоит в том, что на протяжении примерно двух третей
времени, прошедшего с начала кембрия, климаты Земли были
значительно теплее современных [43]. Кроме того, если сейчас
континенты сравнительно сильно выдаются над уровнем моря,
то в течение большей части палеозоя, мезозоя и кайнозоя об-
ширные участки континентальной коры были покрыты неглу-
бокими эпиконтинентальными морями. Примеры современных
эпиконтинентальных морей — Балтийское море и Гудзонов за-
лив, но оба эти моря расположены в холодных северных ши-
ротах.
На фиг. 79 представлена упрощенная блок-диаграмма, опи-
сывающая окраины современного материка. Покрытый водой
край материка называют шельфом. На глубинах около 200 м
проходит край шельфа. За ним до глубин около 5000 м тянется
континентальный склон, переходящий затем в ложе океана.
Окраинная зона моря, периодически то заливаемая прили-
вом, то обнажающаяся при отливе, называется приливно-отлив-
ной или литоральной зоной. Поверхность шельфа называется
сублиторальной зоной, а поверхность континентального склона
образует батиальную зону. Ложе океана—абиссальная равнина,
прерываемая глубоководными впадинами, подводными горными
IX. Палеоэкология
199
Фиг. 79. Окраина современного материка (схема).
цепями й другими менее значительными топографическими эле-
ментами. Фотической зоной называют толщу воды, пронизы-
ваемую светом. Глубина расположения ее нижней границы в
разных местах различна в зависимости от прозрачности воды,
но в общем более или менее совпадает с границей шельфа.
Только верхняя половина фотической зоны (верхние 100 м)
освещена достаточно сильно для того, чтобы в ней мог проте-
кать эффективный фотосинтез. На схеме (фиг. 79) океан вда-
ется в сушу, образуя мелководное эпиконтинентальное море.
Так же как и шельф, находящийся дальше от материка, дно
этого моря — так называемый сублиторальный субстрат.
Большинство осадочных пород морского происхождения,
которые мы находим ныне на материках, отлагалось именно
в эпиконтинентальных морях подобного типа, и лишь немногие
возникли на абиссальных глубинах.
Экологическая классификация морских организмов основана
на том, где они живут и способны ли они к самостоятельному
передвижению (фиг. 80). Совокупность обитателей дна называ-
ют бентосом (прилагательное от этого термина — бентонный,
или бентосный}. Бентосные животные называются эпифауной,
если живут на поверхности субстрата, и инфауной, если живут
в субстрате. Способные к передвижению бентосные формы на-
зываются вагилъными, а неподвижные — сессилъными. Организ-
мы, обитающие в толще воды над дном, называются пелагиче-
скими. Пелагические организмы, способные активно передви-
гаться, называются нектонными, а передвигающиеся в основном
за счет волн и течения — планктонными. Растительный планктон
называют фитопланктоном, а животный — зоопланктоном. На
фиг. 81 приведена упрощенная схема круговорота пищи в океа-
IX. Палеоэкология
201
не. Пищевые цепи морских экосистем обычно сравнительно не-
сложны. В современных морях основными продуцентами явля-
ются одноклеточные фитопланктонные организмы, главным об-
разом диатомовые и динофлагелляты. В водах открытого моря
единственные фотосинтезирующие продуценты — организмы фи-
топланктона, но на шельфе, где фотическая зона достигает дна,
в производстве пищи участвуют и бентосные растения, в том
числе покрытосеменные.
В открытом море первичными консументами являются мик-
роскопические организмы зоопланктона. В современных морях
большинство взрослых форм в планктоне относится к двум
\ группам ракообразных. Кроме того, в зоопланктоне многих об-
ластей массами встречаются мельчайшие личинки беспозвоноч-
ных, на взрослой стадии входящих в состав бентоса. На океан-
ском дне первичными консументами являются бентосные бес-
позвоночные. Продукты метаболизма и разложения растений,
по-видимому, составляют основную пищу и зоопланктона и пер-
5 вичных бентосных консументов. Отсутствие растений на абис-
[ сальных равнинах свидетельствует о том, что животные пита-
ются здесь главным образом неживым органическим веществом.
# В большинстве морских местообитаний пирамида биомассы,
как правило, перевернута (фиг. 82). Перевес зоопланктона над
фитопланктоном обычно объясняют тем, что жизненный цикл
; организмов фитопланктона короче (высокая скорость размно-
жения). Однако некоторые исследователи полагают, что про-
- дукты метаболизма фитопланктона, постоянно высвобождаю-
щиеся в воду, в значительной мере служат пищей для зоопланк-
; тона и бентоса.
Хищники, потребляющие зоопланктон,— это в основном нек-
тонные рыбы. В мелководных районах к зоопланктону добав-
ляется множество бентосных беспозвоночных, также питающих-
ся фитопланктоном. Бентосные беспозвоночные живут, кроме
Фиг. 80. Образ жизни различных морских беспозвоночных. ,
1 — морская улитка (брюхоногий моллюск); 2 — морской еж (иглокожие, Echinoidea);
3 — медуза (кишечнополостные, Scyphozoa); 4 — кальмары (головоногие моллюски); 5 —
морская звезда (иглокожие, Asteroidea); 6 — краб (ракообразное); 7 — губка (Porifera);
3 — мидии (двустворчатые моллюски); 9 — сердцевидка (двустворчатый моллюск);
Ю — червь-хетоптерида (многощетинковый кольчец); 11 — макома (двустворчатый мол-
люск); 12 — пектинария (многощетинковый кольчец); 13 — морской огурец (иглокожие,
Holothurioidea). Животные 3 и 4 ведут пелагический образ жизни (3 — планктонный, а
4 -— нектонный). Остальные животные относятся к бентосу, населяют скалистый (спра-
,ва) или мягкий субстрат. Из бентосных животных 9, 10, 11 и 12 относятся к инфауне,
остальные — к эпифауне. Только губка (7) относится к типичным сессильным орга-
низмам; хетоптернда (10) и мидии (8) способны к очень незначительному перемещению и
могут считаться сессильными; остальные изображенные здесь бентосные животные яв-
ляются вагильными формами, хотя степень их активности может различаться; 1 и 2 —
соскребатели, питающиеся бентосными водорослями; 4, 5 и 6 — хищники, но могут пи-
таться и трупами животных; 7, 8, 9 и 10 питаются взвешенной в воде органикой; 11, 12 и
13 — грунтоеды. Стрелками, показаны токи воды, фильтруемой организмами.
202
Часть 2. Применение данных палеонтологии
Фиг. 81 Пищевой цикл в океане [28].
того, на батиальных и абиссальных глубинах, но там их чис-
ленность ниже, так как там меньше пищи.
Морские палеоэкологи занимаются в основном бентосными
организмами, так как виды, живущие на субстрате или в его
толще, гораздо реже пелагических видов подвергаются значи-
тельному посмертному переносу. Перед тем как подвергнуться
захоронению, остатки пелагических организмов непременно
должны были упасть на дно, и субстрат, на который они попа-
ли, мог не иметь совершенно никакого отношения к среде, в ко-
торой жили сами организмы.
Хант [84] предложил широко распространенную классифи-
кацию бентосных консументов (фиг. 80). Он выделил среди
обитателей мягких грунтов три группы по способу питания:
хищники — «животные, питающиеся главным образом другими
животными, в живом виде или в виде трупов» (трупоедство
присуще многим хищникам, а многие мусорщики поедают жи-
вых животных, поэтому строгое разделение мусорщиков и хищ-
ников неоправданно); сестонофаги (пожиратели взвеси)—«жи-
вотные, питающиеся взвешенными в окружающей воде микро-
организмами и частицами детрита»; грунтоеды— «питаются
детритом, накопленным на дне, и микроорганизмами, связанны-
ми с детритом». К трем группам Ханта мы можем добавить
IX. Палеоэкология
203
Пролив
Лонг-Айленд
Пролив
Ла-Манш
Зоопланктон а
зообентос
Фитопланктон
Фиг. 82. Перевернутые пирамиды
биомассы в двух неглубоких мор-
ских бассейнах [140].
Биомасса (в г/м2) указана для столба
воды с поперечным сечением 1 м2
и для участка дна под ним пло-
щадью 1 м2.
четвертую — соскребателей. Они избирательно снимают с по-
верхности субстрата органическую пленку (состоящую в основ-
ном из микроскопических водорослей). Соскребатели живут
главным образом на твердых субстратах, и их не было в фауне,
для которой Хант разработал свою классификацию.
Ограничивающие факторы
и местные условия среды
До сих пор мы говорили в основном об экологии современ-
ных организмов. В этом и в следующем разделе будут приведе-
ны примеры приложения данных и принципов экологии к пале-
онтологической летописи.
Главная цель палеоэкологии — точно реконструировать усло-
вия среды обитания животных, остатки которых сохранились.
Физические, химические и биологические свойства среды, огра-
ничивающие распространение какого-то вида, называются обыч-
но ограничивающими факторами.
Большинство бентосных беспозвоночных размножается, вы-
пуская гаметы в воду, где и происходит оплодотворение. Из
оплодотворенного яйца появляется планктонная личинка; су-
ществование планктонных личинок способствует расселению
вида на большие расстояния.
Торсон [216] подчекнул, что стадия планктонной личинки
чувствительнее других стадий жизненного цикла к ограничи-
вающим факторам. Личинка обычно менее устойчива, чем взрос-
лый организм.
Запас пищи должен служить ограничивающим фактором
для всех потребителей. Но пока мы плохо представляем себе
аппетиты большинства сестонофагов, грунтоедов и соскребате-
лей. Поэтому мы не в состоянии судить о значении запаса пищи
как ограничивающего фактора для большинства нехищных мор-
ских организмов. Доказано, что тип субстрата и соленость —
главные факторы, управляющие распределением отдельных
бентосных популяций.
204
Часть 2. Применение данных палеонтологии
Тип субстрата
Твердые, скалистые субстраты редко становятся частью па-
леонтологической летописи. Осадки обычно смываются с таких
субстратов, на них идет не осадконакопление, а эрозия. Средой
захоронения для большинства органических остатков являются
мягкие субстраты. Они особенно важны для палеоэкологии —
ведь, если не считать самих окаменелостей, характер и рас-
пределение осадочных пород служат единственным источником
сведений для реконструкции древних местообитаний.
Осадочные породы запечатлевают процессы осадкообразо-
вания; в частности, по форме и размеру частиц, слагающих тер-
ригенную породу, можно определить условия, в которых шло
осадкообразование. Размеры частиц осадка сильно варьируют.
Например, размер частиц глины менее 0,004 мм, частиц ила
0,004—0,0625 мм и зерен песка 0,0625—2 мм; далее идут гравий,
галька и валуны. Ил составляют частицы размером от глинис-
тых до илистых. Размер зерен обычно отражает степень под-
вижности воды (силу волн или течений) в том участке бассейна,
где происходило осадконакопление. Тонкозернистые осадки от-
лагаются обычно в спокойных водах; хорошо отсортированные,
грубозернистые осадки более типичны для районов с сильными
течениями и волнением. Большинство морских терригенных
осадков относится к одному из двух типов: 1) частицы, слагаю-
щие осадки, имеют наземное происхождение и состоят из сили-
катных обломков, особенно из кварца, полевого шпата и гли-
нистых частиц; 2) частицы, слагающие осадки, представлены
карбонатом кальция — скелетными остатками и игольчатыми
кристаллами арагонита, накопленными водорослями или осаж-
денными в результате неорганических процессов. Насколько
мы знаем, расселение бентосных организмов не зависит от рас-
пределения этих типов осадков, но оно зависит от размера зерен
осадка.
Одно из главных проявлений роли субстрата в экологии бен-
тосных организмов — распределение механизмов питания. Сэн-
дерс [176] показал, что процент грунтоедов выше всего на или-
стых осадках, а процент сестонофагов — на песчаных (фиг. 83).
Сэндерс объясняет эту связь главным образом пищевыми запа-
сами. Мельчайшие взвешенные частицы органического вещества,
необходимого грунтоедам, оседают на дно в спокойных водах,
где накапливается также ил, но из грубозернистых песков, под-
верженных действию более сильного волнения, это органическое
вещество быстро вымывается. Сэндерс подчеркнул также, что
запасы взвешенной пищи в спокойных водах возобновляются
медленно и неспособны поддерживать большое число организ-
мов. Однако, как сравнительно недавно показали Д. Родс и
IX. Палеоэкология
205
Фиг. 83. Обилие (выраженное общим сухим весом тканей) животных с раз-
личными типами питания на 8 станциях в проливе Лонг-Айленд [176].
Станции перечисляются слева направо в порядке возрастания процентного содержания
илистых и глинистых частиц.
Д. Янг, на самом деле запасы взвешенного органического ве-
щества достаточно велики и в спокойных водах, но илистые
частицы забивают улавливающие аппараты многих сестонофа-
гов, что препятствует распространению этих животных с песков
на илистые осадки.
206
Часть 2. Применение данных палеонтологии
Фиг. 84. Моллюски из надсемей-
ства Nuculacea.
1 — Nucula и 2 — Void'd; зти организмы
питаются с помощью отростков, распо-
ложенных в околоротовой области, —
так называемых придатков ротовых
щупалец.
Связь между типом осадка и механизмом питания хорошо
прослеживается в палеонтологической летописи даже в палео-
зое. Например, во многих палеозойских аргиллитах и сланцах
встречаются двустворчатые моллюски из надсемейства Nucula-
cea (фиг. 84, фото XXIII). Современные их представители в ос-
новном относятся к грунтоедам и живут на илистых осадках.
Поскольку основные черты морфологии Nuculacea связаны со
свойственным группе способом питания, можно по гомологии
и по положению в стратиграфической летописи предположить,
что большинство палеозойских Nuculacea также были грунто-
едами.
От типа субстрата во многом зависят также способы при- -
крепления и передвижения бентосных животных. Рассмотрим
четыре распространенных типа субстрата. На скалистом дне
обычно расселяется прикрепленная эпифауна. Здесь можно
встретить лишь небольшое число видов, относящихся к инфауне,
они вбуравливаются в твердый субстрат или гнездятся в естест-
венных трещинах. Скалистые субстраты редко имеют строго
горизонтальные поверхности. Кроме того, они подвержены ак-
тивному действию течений и волн. Поэтому большинству видов,
населяющих скалистые субстраты, необходимо прикрепление.
Из мягких субстратов наиболее трудны для освоения бентосом
подвижные пески. Их обычно населяют отдельные высокопод-
вижные виды инфауны, способные быстро закапываться, когда
создается угроза их вымывания течением или волнами. Отсут-
ствие или скудость ископаемых остатков во многих прибрежных
отложениях обусловлены не только тем, что условия оказались
неподходящими для захоронения, но и непригодностью этих
условий для жизни. Дальше от берегов, в спокойных зонах моря
на дне накапливаются мягкие и вязкие илы. Раковинная ин-
фауна здесь в основном представлена мелкими животными, час-
то обладающими тонкостенными раковинами. Крупное животное
с тяжелой раковиной погрузилось бы в мягкий ил; в таких
условиях механизмы питания и дыхания многих организмов не
смогли бы нормально функционировать. На фиг. 85 приведен
график зависимости между весом животного и процентным со-
IX. Палеоэкология
207
0 s----1---i—J------1----1---1----i---i___i ।
o 20 40 60 80 100
Содержание ила, %
Фиг. 85. Максимальный вес особей разных видов двустворчатых моллюсков
с различных субстратов [208].
Крупные особи приурочены к осадкам с низким содержанием ила.
держанием ила в субстрате. Данные относятся к двустворчатым
моллюскам из разных местообитаний. Крупные виды с тяжелой
раковиной приурочены к песчаным осадкам. На мягких, илис-
тых субстратах эпифауна представлена немногими видами, так
как здесь трудно прикрепляться и бороться с погружением.
Субстраты, сложенные смесью ила и песка, обычно отлича-
ются от движущихся песков и защищенных от волнения илистых
грунтов, будучи устойчивыми и плотными. Обычно на таких суб-
стратах видовое разнообразие бентоса весьма велико. Живот-
ные, строящие постоянные жилые норы (трубки или туннели),
как правило, приурочены к илистому песку — они используют
его клейкость.
СОЛЕНОСТЬ
Соленость — мера содержаний растворенных солей в природ-
ных водах — выражается обычно в промилле (%о). Соленость
нормальной океанической воды составляет около 35°/оо- В табл. 4
приводится общепринятая классификация природных вод по их
солености.
208
Часть 2. Применение данных палеонтологии
ТАБЛИЦА 4
Классификация природных вод по
солености [101]
Тип вод Соленость, %0
Пресные воды 0—0,5
Солоноватые воды 0,5—30
Морские воды 30—40
Осолоненные воды 40—80
Рассолы 80
Отношение разных организмов к солености может быть весь-
ма различным как по средней предпочитаемой солености, так
и по диапазону переносимых значений солености. Виды с узким
диапазоном выносливости называются стеногалинными, с широ-
ким — эвригалинными.
Наиболее наглядно экологическая роль солености выступает
при сравнении видового разнообразия фауны в местообитаниях
с различной соленостью. Большинство водных организмов —
морские формы. Многие из них стеногалинны; они обитают в
водах открытого моря, где заметные колебания солености от-
сутствуют. Пресноводных стеногалинных форм среди водных
организмов меньше. Сравнительно немногие виды обитают в со-
лоноватых или осолоненных бассейнах. Однако многие формы,
которые выносят солоноватую или осолоненную воду, способны
жить и в той и в другой. Осолоненные и солоноватые, массы
развиваются в прибрежных зонах (например, в заливах и лагу-
нах) в тех случаях, когда нарушается связь с водами откры-
того океана (хороший пример — Балтийское море, фиг. 86). Со-
леность колеблется с приливом и отливом, в зависимости от
дождей, штормов и сезонных изменений климата. Обитатели
такой нестабильной среды должны быть эвригалинными. По-
видимому, в любой период геологической истории эвригалинных
видов было сравнительно немного. Немногие виды, живущие
в условиях ненормальной солености, обычно отличаются высо-
кой численностью, так как здесь межвидовая конкуренция за
экологические ниши ослаблена. Большинство солоноватоводных
видов произошло не от пресноводных, а от морских форм. Лишь
немногие из них способны жить в пресной воде, но многие спо-
собны переносить нормальную соленость вод океана.
Случаи сниженного видового разнообразия, связанные с осо-
лонением или опреснением, распознаются обычно в палеонтоло-
е Гоглсмд
Фиг. 86. Связь между разнообразием морских моллюсков и соленостью
в Балтийском море.
А. Схематический разрез через Балтийское море от Финского залива до Готландской впа-
дины (июль, 1933 г.). Сплошными линиями показана соленость в промилле, пунктиром —
температура в градусах Цельсия [96] Б. Разнообразие моллюсков и градиент солености
примерно по тому же разрезу [203].
14—467
210
Часть 2. Применение данных палеонтологии
гической летописи по стратиграфическим признакам, свиде-
тельствующим о том, что осадки накапливались в прибрежной
зоне. Особенно веским доказательством накопления осадков
в прибрежной зоне является их связь с расположенными рядом
континентальными отложениями (озерными, речными и т. п.),
которые хорошо распознаются по многим морфологическим и
палеонтологическим признакам.
Осолоненные бассейны встречаются главным образом в жар-
ких, аридных климатах. Осадки подобных бассейнов обычно
содержат минералы, формирующиеся при химическом осажде-
нии в условиях выпаривания (галит, гипс, ангидрит), и, кроме
того, они иногда связаны с эоловыми отложениями. Если толща
содержит растительные остатки, а рядом расположены дельто-
вые или озерные отложения, то все это свидетельствует о ее
накоплении в солоноватом, а не в осолоненном бассейне.
Иногда форма раковины морского животного явно коррели-
рует с соленостью воды в его местообитании. Так, чем меньше
соленость, тем меньше ребер на раковине и тоньше сама рако-
вина сердцевидки съедобной (Cardium edule). Возможно, дело
здесь в затрудненной секреции углекислого кальция, однако
вопрос этот недостаточно изучен, и переносить этот вывод на
ископаемые виды было бы рискованно. У вымершего вида кор-
реляция могла быть и обратной. Кроме того, отношение к соле-
ности— не такой уж важный эволюционный признак по сравне-
нию, например, с основными морфологическими адаптациями,
связанными с питанием или передвижением.
Рискованно также переносить выводы об отношении рецепт-
ного вида к солености на родственный вымерший вид. Мы зна-
ем один род, который, по-видимому, остается эвригалинным уже
многие миллионы лет. Это беззамковая брахиопода Lingula
(фиг. 87), ныне обитающая в прибрежных, часто несколько
опресненных водах. Палеонтологическая летопись этого «живого
ископаемого» простирается в глубь времен по меньшей мере до
ордовика. Множество фактов свидетельствует о том, что на про-
тяжении всего времени своего существования Lingula входила
главным образом в состав прибрежных фаун с низким видовым
разнообразием.
Хадсон [81] ' предпринял попытку выяснить по характеру
распределения фауны в юрской большой эстуариевой свите
(Англия) и по ее разнообразию соленость воды бассейна, в ко-
тором формировались отложения свиты. Он выделил несколько
фаунистических сообществ, каждое из которых представлено
группой видов, обычно встречающихся вместе в определенных
породах. Сообщества не образуют простой стратиграфической
последовательности, и их горизонтальное распространение про-
слежено. Выводы Хадсона иллюстрирует фиг. 88. Они подтверж-
IX. Палеоэкология
211
даются, в частности, тем, что
в отложениях свиты содержат-
ся роды, обитающие на земле
и ныне, а пределы их выносли-
вости к солености известны.
Пресноводный двустворчатый
моллюск Unio и пресноводный
брюхоногий моллюск Viviparus
распространены сейчас во мно-
гих районах. Двустворчатые
моллюски Mytilus и Modiolus
распространены в современ-
ных морях (они населяют со-
лоноватые и нормальные мор-
ские воды), но большинство
видов Modiolus не так толе-
рантны к понижению соленос-
ти, как виды Mytilus. Liostrea
сходна с рецептными устрица-
ми, живущими в небольших
заливах с пониженной соле-
ностью. Хадсон предположил,
что соленость океана не под-
вергалась значительным изме-
нениям с юры. Расположив
живущие и сейчас роды (вклю-
чающие вымершие виды) по
степени их толерантности к
различным колебаниям соле-
ножка
Пучки щетинок,
направляющие
поток воды
Раковина
Песок, склеившийся
вокруг ножки
£
ё-
В
д-
'•йГ
В’
Фиг. 87. Lingula в своей норке ([88],
по Франсуа).
А. Вид сбоку. Б. Вид сзади.
ности, мы получим график,
изображенный на фиг. 88. Приблизительная шкала солености,
30
А •
Ж
Б
составленная на основании сравнения с рецентными видами,
подтверждает выводы Хадсона. Наименьшее видовое разнооб-
разие приходится на типично солоноватые воды (5—9°/оо на схе-
ме), а максимальное разнообразие — на слабо опресненные и
нормальные морские воды. Данные по таксономическому разно-
образию заставляют думать, что роды, для которых известны
рецентные представители, на протяжении примерно 150 милли-
онов лет не изменили существенно своего отношения к соле-
ности.
Можно утверждать, что некоторые группы водных организ-
мов в течение всей своей геологической истории были в основ-
ном стеногалинными и жили в морской воде с нормальной со-
леностью, тогда как другие группы были эвригалинными. Счи-
тается, что большинство кораллов, головоногих, замковых бра-
хиопод и иглокожих представляли собой морские стеногалин-
14*
35
30
Тип фауны
Фаунистические комплексы в большой
эстуариевой свите и слоях Стаффин-Бэй
Морские воды
Морская
Нет прямой связи
_ Обеднённые
16,5--------—-----
’ морские фауны
Белемнитовые пески
(морские пластинчатожаберные и белемниты)
В слоях Стаффин-Бэй
Верхний слой с Ostrea
(морские двустворки, а также Liostrea и Neomiodon)
Водорослевый слой с Mytilus
Cuspidaria
Placunopsis
Modiolus
Нижнии водорослевый слой с Ostrea
Liostrea
Rhynchonella
Modiolus ит.д.
Морские вады
с уменьшенным
числом видов
------------------------------------Доминирует Liostrea hebridica-
Доминирует Mytilus strathairdensis I
Осолоненные пресные воды
__Обедненные__________________________
3 пресноводные Пресныеводы
фауны с уменьшенным
числом видов
-------------------------------------
• пресныеводы
Vnio Viuiparus Neomiodon Euestheria*
Мидиевые глины - свита
конкреционного песчаника
Фиг. 88. Фаунистические комплексы большой эстуариевой свиты (средняя
юра, Англия), возникшие в условиях различной солености [81].
IX. Палеоэкология
213
ные формы. Остракоды, двустворчатые и брюхоногие моллюски
обычно населяли и пресные и морские воды, и многие виды
этих групп были эвригалинными. Фораминиферы с агглютини-
рованной раковиной сейчас особенно распространены в при-
брежных местообитаниях с низкой соленостью, и их используют
как палеоэкологические индикаторы соответствующих условий.
Познакомившись с экологией Lingula и родов, рассмотрен-
ных Хадсоном, мы можем сделать вывод, что все виды (как
современные, так и вымершие) одного рода обладают примерно
одинаковым отношением к солености. Но надо помнить, что это
упрощение. Многие современные роды включают виды с разной
толерантностью к изменениям солености. На уровне семейства
отношение к солености отличается еще большим разнообразием.
Так, многие семейства моллюсков включают как стеногалинные,
так и эвригалинные формы. Некоторые семейства, например
Ostreidae, Mytilidae и Neretidae, содержат в основном солоно-
ватоводные формы, но среди них есть и некоторые стеногалин-
ные морские виды. Чтобы определить соленость бассейна, в ко-
тором накапливались осадочные толщи, необходимо сочетать
строгий таксономический анализ с данными по видовому разно-
образию, а также с седиментологическими и стратиграфически-
ми сведениями. И все же выводы, как правило, получаются
Очень приблизительными. Мы можем лишь различать пресно-
водные, солоноватоводные, морские и гипергалинные ископае-
мые сообщества. Чем дальше мы уходим от современности, тем
меньше точность наших методов.
ГЛУБИНА
Это одна из самых важных характеристик местообитания, но
по составу морского ископаемого сообщества ее часто очень
трудно определить даже приблизительно. Зная глубину древних
морей, мы могли бы представить себе их береговую линию,
уточнить наши знания о палеогеографии. Глубина самым не-
посредственным образом воздействует на организмы через гид-
ростатическое давление, но ее непосредственное влияние на
/распределение жизни в море сравнительно невелико и сказыва-
ется только в случае больших перепадов глубины, чем те, кото-
рые обычно интересуют палеоэкологов. Кроме того, глубина
мало влияет на твердый скелет, и приспособленность к большим
глубинам не связана прямо с прочностью скелета. Быстрое из-
менение давления сильно сказывается на мягких тканях. Со-
временные животные, быстро поднятые тралом или дночерпа-
телем с абиссальной равнины на поверхность, страдают от рез-
кого спада давления. Хотя во многих таксономических группах
в ходе эволюции возникли формы, способные жить на разных
214
Часть 2. Применение данных палеонтологии
глубинах, в распределении организмов по глубинам редко уда-
ется найти какие-либо таксономические или функционально-
морфологические закономерности.
Глубина может влиять на распределение видов и опосредо-
ванно, через такие связанные с глубиной факторы, как осве-
щенность, соленость, температура, количество растворенного
кислорода, запас пищи. Например, многие современные бентос-
ные виды опускаются глубже в той части своего ареала, которая
ближе к экватору, а у полюсов живут в мелководных районах.
Это связано с тем, что неглубокие воды вблизи экватора слиш-
ком теплы для них.
Синтез питательных веществ путем фотосинтеза невозможен
за пределами освещенной зоны, и количество пищи на глубинах
более 200 м значительно снижается. Давно известно, что чис-
ленность организмов падает на протяжении континентального
склона к абиссальной равнине. Недавно Сэндерс, Хесслер и
сотр. [177] провели широкое исследование с применением со-
временных приборов для взятия проб и показали, что, хотя чис-
ленность глубоководных бентосных организмов более высока,
чем считалось до сих пор, описанная тенденция действительно
существует. Оказалось, кроме того, что таксономическое разно-
образие глубоководного бентоса гораздо выше, чем полагали
ранее. Следствия этого открытия будут рассмотрены ниже
(стр. 240—243). Показано также, что в глубоководных осадках
явно преобладают грунтоеды. Из-за низкой концентрации взве-
шенных частиц пищи питание ими оказывается сравнительно
невыгодным.
Освещенность определяет распределение растений в море.
Это ясно уже хотя бы из самого факта наличия фотической зо-
ны, в которой освещенность с увеличением глубины постепенно
падает до нуля. Свет красного участка видимого спектра погло-
щается быстрее, чем синие лучи. Большинство растений способ-
но в той или иной степени использовать весь видимый спектр,
но оптимальная длина волны для разных видов различна. Из
морских водорослей зеленые водоросли поглощают в основном
красные лучи и приурочены поэтому главным образом к лито-
рали и неглубоким зонам сублиторали. Красные водоросли
спускаются в сублитораль значительно глубже. В обеих груп-
пах имеются виды, способные секретировать скелеты из карбо-
ната кальция. Именно поэтому красные и зеленые водоросли
представлены в палеонтологической летописи. Присутствие ис-
копаемых зеленых водорослей, не подвергшихся переносу, обыч-
но говорит о том, что в данном древнем местообитании глубина
воды была не более 60—70 м.
Один из важнейших экологических эффектов света с точки
зрения морской палеоэкологии — влияние освещенности на рас-
IX. Палеоэкология
215
пределение рифообразующих кораллов на разных глубинах.
Среди современных кораллов (Scleractinia, шестилучевые ко-
раллы) выделяют две экологические группы. Экологические раз-
личия между ними связаны с тем, что в мягких тканях корал-
лов одной группы живут определенные водоросли, зооксантел-
лы. Физиология их связи с кораллом не до конца изучена, но,
по-видимому, водоросли помогают кораллу, усваивая отходы
его метаболизма или способствуя построению известкового ске-
лета. Во всяком случае, крупные, массивные колонии образуют
только те шестилучевые кораллы, которые содержат зооксан-
теллы. Это так называемые герматипные (от греч. herma —
подводная скала) кораллы, основные строители тропических
коралловых рифов. Виды кораллов, не содержащие зооксантелл,
называются агерматипными. Зооксантеллы нуждаются в свете,
и потому герматипные кораллы приурочены к неглубоким во-
дам. На фиг. 89 показано, что рифы герматипных кораллов
редко могут процветать на глубинах более 20 м, хотя встреча-
ются иногда и на глубинах несколько более 50 м (а отдельные
герматипные кораллы иногда встречаются на глубинах, превы-
шающих 100 м). Следовательно, настоящие ископаемые корал-
ловые рифы могут служить хорошим индикатором мелководных
условий осадконакопления. Современные рифы строятся много-
численными видами кораллов совместно с известковыми во-
дорослями и богатой фауной беспозвоночных. Глубоководные
коралловые «банки», отмеченные на фиг. 89, строятся немноги-
ми агерматипными видами в сообществе с малоразнообразной
фауной беспозвоночных; известковые водоросли там отсутству-
ют. Исходя из этого, Тейхерт [214] сделал вывод, что извест-
ные из верхнего триаса Южной Аляски коралловые рифы (ске-
леты, из которых они состоят, относятся всего к восьми видам
кораллов) на самом деле были, вероятно, сходны с современ-
ными глубоководными и холодноводными коралловыми банками
у берегов Норвегии.
Переносить выводы об ограничивающих факторах, сущест-
венных для современных коралловых рифов, на ископаемые
рифы следует с большой осторожностью. Достаточно много
позднетриасовых и раннеюрских таксонов живет и сегодня, так
что можно утверждать, что более ранние (среднетриасовые) ри-
фообразующие шестилучевые кораллы также содержали зоо-
ксантеллы и обитали примерно на тех же глубинах, что и со-
временные формы. Основные палеозойские группы рифообра-
зующих кораллов, например табуляты и строматопоры, таксо-
номически отличались от современных шестилучевых кораллов.
Возможно, мы так никогда и не узнаем, были ли эти организмы
связаны с зооксантеллами, но многие независимо полученные
факты указывают на то, что палеозойские рифы и рифоподоб-
216
Часть 2. Применение данных палеонтологии
Географическая широта
Фиг. 89. Глубины обитания и широтные границы распространения современ-
ных коралловых рифов, герматипных кораллов и глубоководных коралловых
«банок» [214].
Пунктирная линия — фактический предел глубины для известковых водорослей; тонкая
сплошная линия — крайний предел глубины для известковых водорослей; штрих-пунк-
тирная линия — фактический предел глубины для склерактиний; участок с густой штри-
ховкой внизу — коралловые банки и мелкие рифы в Северном Атлантическом океане.
ные образования, для которых характерно высокое видовое раз-
нообразие, возникли в неглубоких водах. Многие из этих по-
строек препятствовали циркуляции воды в зонах, окаймленных
рифами; следовательно, они дорастали или почти дорастали до
уровня моря.
Показано, что ископаемые губки являются полезными инди-
каторами глубины, хотя встречаются они в достаточном числе
лишь в немногих типах осадочных пород. Губки — сессильные
эпибентосные сестонофаги (фиг. 80, 7). Сейчас губки из класса
Calcarea, у которых опорные скелетные элементы (спикулы)
состоят из карбоната кальция, наиболее распространены на
глубинах менее 100 м, хотя некоторые виды обитают и глубже.
Современные губки класса Hexactinellida, обладающие скеле-
том из кремнезема, наиболее обычны на глубинах 200—300 м;
некоторые виды живут на шельфе; на глубинах менее 10 м этих
губок почти нет [161]. Финке [58] показал, что эти особенности
экологии были характерны для губок уже в конце палеозоя.
В пермских породах Техаса он распознал мелководную «шель-
фовую» фауну губок с преобладанием Calcarea и глубоковод-
IX. Палеоэкология
217
ную «океанскую» фауну, лишенную Calcarea, но богатую стек-
лянными губками.
Одна из самых резких экологических границ в море — грани-
ца между литоралью и сублиторалью. Обитатели литорали под-
вержены быстрым и сильным изменениям физико-химических
условий среды, особенно температуры и солености. На мягких
грунтах многие литоральные формы, в частности черви (анне-
лиды), ракообразные и двустворчатые моллюски, закапываются
в субстрат, защищаясь от этих изменений. Существование та-
ких форм может отмечаться в палеонтологической летописи со-
хранившимися ходами, пробуренными когда-то в осадке. На
фото XXIV показан глубокий ход .рачка Callianassa, живущего
сейчас на песчаной литорали, а также ход, оставленный иско-
паемым представителем Callianassa в породах верхнего мела.
Благодаря столь широкому стратиграфическому распростране-
нию этот рачок — прекрасный индикатор литоральных отложе-
ний. Зейлахер [188] и Родс [164] сравнивают глубокие норки
литоральных животных с незаглубленными горизонтальными
ходами глубоководных видов, которым не нужно глубоко зака-
пываться. Зейлахер предполагает, что глубоководные закапы-
вающиеся виды в основном относятся к грунтоедам (это не
удивительно — см. стр. 214), которые прокладывают свои ходы
так, чтобы наиболее полно охватить слои осадка, содержащие
пищу (фото XXIV).
Строматолиты представляют собой характерно слоистые, по-
душковидные образования, сложенные последовательно нарас-
тавшими слоями водорослей (фото XXV). Их присутствие обыч-
но указывает на литоральное происхождение осадочных пород.
Каждый слой, как правило, создан одним или несколькими ви-
дами сине-зеленых водорослей. Эти нитчатые водоросли при
жизни покрыты клейкой слизью, к которой прилипают обломоч-
ные частицы карбоната кальция (сами водоросли не секрети-
руют скелета). Когда поверхность водорослевого слоя перекры-
вается осадком, нити водорослей прорастают через осадок, со-
здавая новый слой, который затем также покрывается осадком.
Строматолиты — очень устойчивые растительные сообщества. Их
история прослеживается даже в докембрий, что делает их осо-
бенно ценными для палеоэкологии. Их внешняя форма обычно
отражает такие факторы внешней среды, как местные течения
и волнения.
К определению глубины бассейна седиментации и местооби-
тания ископаемого сообщества можно подходить различными
путями. Часто окончательный вывод говорит только о расстоя-
нии от берега («прибрежный», «удаленный от берега»), а не
о глубине. Осадки, накопившиеся в зоне действия прилива и от-
лива, часто распознаются по физическим и химическим особен-
218
Часть 2. Применение данных палеонтологии
костям, таким, как трещины усыхания и тонкозернистый доло-
мит, а также по палеонтологическим данным. Изучение регио-
нальной стратиграфии и других геологических данных обычно
позволяет распознать мелководные лагунные отложения. Отло-
жения песчаных пляжей распознаются по многим признакам —
размеру зерен, особенностям залегания, малому видовому раз-
нообразию и другим. Глубоководные отложения также облада-
ют характерными седиментологическими и биологическими
признаками. Детальное разграничение, однако, удается редко.
Особенно трудно оценить глубину образования сублиторальных
отложений и фаун отложений эпиконтинентальных морей и
шельфа, а они, к сожалению, составляют большинство извест-
ных нам осадочных пластов.
Пространственное распределение
популяций
Мы отмечали уже, что популяции обычно распределяются
в пространстве характерным образом.
Многие бентосные морские формы живут группами. Такое
распределение объясняется обычно двумя причинами: неравно-
мерностью условий среды и видовыми особенностями репродук-
тивного и общественного поведения. Видимо, вторая причина
более важна. Прекрасный тому пример — виды морской травы,
населяющие мелководные области многих частей света
(фиг. 90). Они способны размножаться отростками, распростра-
няющимися под поверхностью осадка. Если семя зостеры по-
падает на свободный от растений участок и прорастает здесь,
то благодаря отросткам может создаться пятнистый узор рас-
пределения травы. Пятна травы разделены свободными участ-
ками дна. Трава изменяет условия среды на дне, укрепляя суб-
страт и действуя как ловушка для тонкозернистых осадков. Она,
кроме того, предоставляет новые субстраты (свои стебли и кор-
ни) для множества организмов. Некоторые виды двустворчатых
и брюхоногих моллюсков, морских ежей и червей приурочены
к таким «подводным лугам».
Другой фактор, создающий пятнистое распределение бентос-
ных беспозвоночных,— образование локальных популяций, кото-
рое происходит различными способами. Многие бентосные виды
производят большое число личинок, живущих какое-то время
(в среднем три недели) в составе планктона, а затем оседаю-
щих на дно и проходящих метаморфоз. Много спорят о том, спо-
собны ли личинки выбирать подходящий субстрат для оседания
и могут ли неподходящие условия на дне замедлять оседание
личинок. Во всяком случае, личинки одного вида обычно пере-
носятся течениями вместе как одна популяция и оседают на од-
IX. Палеоэкология
219
Фиг. 90. Морская трава зостера в Средиземном море у берегов Испании.
Обратите внимание на корни, заметные там, где течения размыли дно. На таких размы-
тых участках обычно встречается меньше видов бентосных животных, чем на соседних, за-
росших травой.
ном участке. Часто «стадность» проявляется у них при оседа-
нии — их привлекают взрослые особи своего вида, и в этом есть
биологический смысл: ведь в том месте, где уже прижились
другие, очевидно, сможет жить и молодая особь. Поэтому удач-
ное распределение часто оказывается пятнистым, и эта пят-
нистость, по-видимому, не связана с неравномерностью условий
среды на дне.
Распределение бентосных видов, не обладающих стадией
планктонной личинки, еще чаще бывает пятнистым. Для многих
бентосных видов характерна забота о яйцах и личинках, поэтому
молодые особи остаются вблизи взрослых.
Наконец, особи некоторых вагильных видов собираются
вместе уже на взрослой стадии. Видимо, они скапливаются там,
где больше пищи или условия среды по каким-то другим причи-
нам особенно благоприятны.
Иногда скопления бентосных форм обусловлены неравномер-
ностью физических условий среды. Здесь особенно важны свой-
ства субстрата, а не температура или соленость. Мидии (Myti-
220 Часть 2. Применение данных палеонтологии
las) прикрепляются к твердым предметам нитями биссуса. Да-
же на песчаном пляже несколько мидий-«пионеров» могут за-
крепиться на случайных обломках раковин или на гальке, а их
раковины в свою очередь служат местом прикрепления для дру-
гих мидий. В результате на больших пространствах может со-
здаться пятнистое распределение мидий (фото XXVI). Видимо,
подобным же образом возникли и скопления палеозойских бра-
хиопод; это доказано Циглером и сотрудниками для некоторых
силурийских групп. Раковины мертвых брахиопод становились
центрами колоний, а некоторые особи прикреплялись к живым
соседям. Палеозойский род брахиопод Pentamerus оставил мно-
гочисленные колонии тесно скученных особей, сохранившихся в
прижизненном положении (фото XXVII). Неясно, прикрепля-
лись ли взрослые особи ножками (мясистыми стебельками,
которые есть у многих родов брахиопод) или свободно лежали
на субстрате. Во всяком случае, область ножки каждой ракови-
ны направлена вниз, и можно думать, что особи не были при-
креплены одна к другой, а просто скучились в результате сов-
местного оседания личинок (взрослые брахиоподы неспособны
передвигаться).
Там, где морские популяции с пятнистым распределением
подверглись захоронению в месте обитания, ископаемые сооб-
щества могли сохранить оригинальный рисунок распределения.
Как правило, время является главным фактором, изменяющим
картину распределения. В районах, в которых распределение
планктона (а значит, и личинок бентосных организмов) из года
в год менялось, а. скорость осадконакопления была невелика,
палеонтологическая летопись содержит усредненную, сглажен-
ную запись распределения бентоса.
Важно уяснить себе, что, хотя процветание какого-то вида
в определенном месте всегда объясняется факторами внешней
среды, отсутствие вида в других сходных местообитаниях мо-
жет быть случайным, связанным с превратностями переноса ли-
чинок и кратковременностью существования многих локальных
популяций. Равномерность условий в месте осадкообразования
не обязательно приводит к равномерности распределения попу-
ляции или к образованию равномерно распределенного ископае-
мого сообщества.
Образ жизни
Мы рассмотрели факторы, контролирующие расселение ны-
не живущих и вымерших видов. Рассмотрим теперь образ жизни
вида в свойственном ему местообитании. В случае ископаемых
видов мы можем получить лишь отрывочные сведения. Напри-
мер, какова была ориентация организма при жизни? Переме-
IX. Палеоэкология
221
щался ли он, а если перемещался, то как? Была ли это при-
крепленная или свободноплавающая форма? Как организм до-
бывал пищу? Как он размножался?
СВЕДЕНИЯ, СОХРАНЯЮЩИЕСЯ
В ПАЛЕОНТОЛОГИЧЕСКОЙ ЛЕТОПИСИ
Самый простой способ ознакомления с образом жизни вы-
мерших видов заключается в непосредственном наблюдении от-
дельных особей, захороненных в прижизненном положении (фо-
то XXVIII). Иногда удается наблюдать уникальные условия
захоронения, например в результате катастрофы.
гомология
Менее прямой способ изучения образа жизни — применение
принципа гомологии (стр. 166). Говорят, например, что если
бы не было живых черепах, то мы не могли бы представить себе
образ жизни черепах вымерших. Можно было бы предполагать,
что черепаший панцирь служил защитой какому-то странному
созданию, но мы наверняка не смогли бы выявить по ископае-
мым остаткам различие в образе жизни между сухопутной ко-
робчатой и пресноводной каймановой черепахой. Конечно, из-
ложенные в гл. VIII выводы об образе жизни аммонитов сдела-
ны в значительной мере благодаря тому, что существует и изу-
чен современный Nautilus. Не будь этого, много лет могло бы
уйти только на то, чтобы доказать, что аммониты относились
к головоногим. Лишь недавно удалось найти вместе с аммони-
тами характерные для головоногих челюсти («клювы»).
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ
Функциональная морфология (более подробно см. в
гл. VIII) — главный источник информации для реконструкции
образа жизни, особенно если она соединяется с седиментологи-
ческим исследованием, дающим сведения об условиях в место-
обитании. Пример — работа Гранта [68] о брахиоподах рода
Waagenoconcha из пермских пород Пакистана. Неполовозрелые
особи несли вблизи макушки шипы, использовавшиеся, вероят-
но, для того, чтобы удержать организм на бентосных растениях,
губках или мшанках, на которых, очевидно, оседали личинки
(фиг. 91). Взрослые особи жили на тонкозернистом осадке, ни-
как не прикрепляясь. Положение, в котором захоронены живот-
ные, и большой вес брюшной створки позволяют заключить, что
эта шиповатая створка была обращена к субстрату. Видимо,
шипы в первую очередь служили для поддержки питающегося
222
Часть 2. Применение данных палеонтологии
Фиг.^91. Реконструкция образа жизни брахиоподы W aagenoconcha abichi
из пермских отложений Пакистана [68].
Молодые особи изображены прикрепленными к водорослям.
взвесью животного на вязком илистом дне, хотя Грант предпо-
лагает, что они, кроме того, служили также якорями. Шипы,
словно лыжи, не давали животному погрузиться в мягкий суб-
страт. Легко понять, какова была бы судьба молодых брахио-
под, если бы они не прикреплялись к растениям. Из-за их ма-
лого размера линия комиссуры еле выдавалась бы из ила, и
малейшее взмучивание субстрата привело бы к забиванию пи-
щевого аппарата, прекращению нормальной жизнедеятельности
и гибели большинства особей.
\
ТЕСНЫЕ МЕЖВИДОВЫЕ СВЯЗИ
Нередко выводы об образе жизни ископаемого организма
можно сделать по находкам, показывающим физическую связь
между видами. Пример — раковина аммонита Buchiceras, опи-
санная Зейлахером из меловых пород Перу (фиг. 92). Раковина
усажена устрицами. Расположение устриц доказывает, что они
прикреплялись не к пустой раковине, лежавшей на дне, так как
они равномерно распределены по обеим боковым сторонам. По
характерной ориентации устриц в зависимости от наклона по-
верхности прикрепления удается восстановить прижизненное
IX. Палеоэкология
223
Фиг. 92. Экземпляр аммонита Buchiceras bilobatum с прикрепившимися к не-
му устрицами [186].
А. Вид слева. Б. Вид справа. В. Вид сзади. Г. Прижизненное положение, восстановлен-
ное по направлению роста устриц, прикрепленных к левой (сплошные стрелки) и правой
стороне раковины (пунктирные стрелки).
положение аммонита. Оно совпадает с положением, теоретиче-
ски выведенным для подобных форм [218]. То, что устрицы при-
креплены и к нижней поверхности раковины, исключает выдви-
гавшееся ранее предположение о том, что этот род аммонитов
вел бентосный образ жизни, ползая по дну. Постоянные удары
о дно беспокоили и повреждали бы устриц. Зейлахер сделал
вывод, что представители этого рода были нектонными живот-
224
Часть 2. Применение данных палеонтологии
Фиг. 93. Трубки червей-спирорбид на раковинах неморских двустворчатых
моллюсков (карбон) [219].
Черви прикреплялись к раковинам там, где создавался постоянный ток воды. Ряды тру-
бок в основном следуют линиям нарастания — видимо, молодые черви прикреплялись
. у края раковины.
ними, а их прижизненное положение было сходно с таковым
современного наутилуса.
Организмы, прикрепляющиеся к другим организмам, назы-
ваются эпибионтами. Если это растения, их называют эпифита-
ми, если животные — эпизоями. При использовании эпибионтов
для выяснения образа жизни вида-хозяина очень важно уста-
новить, произошло ли прикрепление при жизни хозяина или
после его смерти. Здесь помогает определение места прикреп-
ления и ориентировки эпибионтов. Обычно эпизои-сестонофаги,
такие, как черви, строящие известковые трубки, занимают поло-
жение вблизи места, где вода входит в тело хозяина; они ис-
пользуют поток воды, несущий питательные вещества (фиг. 93).
У некоторых водных животных участки, в которых создается
постоянный ток воды, — это именно те части раковин, которые
при жизни выдаются над поверхностью осадка, и поэтому по-
ложение эпибионтов иногда может указывать не только участки
с постоянным током воды, но и прижизненное положение вида-
хозяина.
СЛЕДЫ ЖИЗНЕДЕЯТЕЛЬНОСТИ
Многие организмы, не оставляющие скелетных частей, пред-
ставлены в палеонтологической летописи лишь следами пере-
движения, по которым можно сделать выводы об образе жизни
IX. Палеоэкология
225
Фиг. 94. Ископаемые следы, так назы-
ваемые Helminthoidea labyrinthica (из
[189], по Рихтеру).
Подобные следы найдены в меловых и эоце-
новых отложениях Альп и Аляски.
и поведении организма. Немецкие исследователи, много сделав-
шие для изучения этих следов, называют их «Lebensspuren»
(«следы жизни»). Образование разнообразных «следов жизни»
можно наблюдать ныне как в лаборатории, так и в природе.
Один из плодотворных методов — рентгенография животных,
закапывающихся в грунт в аквариуме. Передвижение животных
регистрируется с помощью цейтраферной съемки или кино-
съемки (фото XXIX).
Пионер в области исследования ископаемых «следов жизни»
Рудольф Рихтер анализировал образование, показанное на
фиг. 94. Оно оставлено червеобразным грунтоедом, двигав-
шимся в мелкозернистом осадке так, чтобы систематически по-
глощать этот осадок, не пропуская через себя один и тот же
материал дважды. Для того чтобы мог возникнуть подобный
след, были необходимы четыре условия: 1) животное должно
было двигаться горизонтально в пределах одного слоя осадка;
2) после прохождения определенного расстояния делать U-об-
разный поворот; 3) двигаться в непосредственной близости к
уже пройденным участкам туннеля, но так, чтобы 4) сохраня-
лось некоторое минимальное расстояние от ранее пройденных
участков. При соблюдении двух последних «условий» возникает
узор, в котором отрезки туннеля разделены одинаковыми про-
межутками. Эти особенности поведения должны были быть за-
писаны в форме генетических инструкций, в соответствии с ко-
торыми животное реагировало на химические и тактильные
раздражители в окружающей среде. Подобные инструкции мож-
но ввести в ЭВМ и получить рисунок, сходный с наблюдаемым.
Одни «следы жизни» соответствуют местам отдыха живот-
ных, другие — более или менее постоянным укрытиям (фо-
то XXIV), а некоторые являются просто следами, оставляемыми
при ползании.
Много интересного могут рассказать и следы наземных поз-
воночных. Большая часть мощной триасовой красноцветной
226 Часть 2. Применение данных палеонтологии
толщи в долине Коннектикут (северо-восток США) накопилась
на болотистых низменных пространствах. На поверхностях на-
пластования песчаников, слагающих толщу, видны следы не-
скольких видов динозавров (фото XXX).
Следы животных особенно важны в тех случаях, когда не
сохранились скелетные остатки. В триасовых породах долины
Коннектикута костей динозавров практически не обнаружено,
но многочисленные их следы свидетельствуют о разнообразной
фауне динозавров. Теплый, влажный климат области, в кото-
рой сформировались красноцветные толщи, очевидно, был не-
благоприятен для сохранения скелетов, несмотря на то что эта
область была осадочным бассейном, в котором за несколько
миллионов лет накопилась мощная толща осадков. Нередко
и в «Немых» морских осадочных породах сохранившиеся «следы
жизни» говорят о том, что бентос был обильным.
Ископаемые сообщества
Мы говорили об образе жизни и локальном распределении
популяций одного вида. Обычно эти вопросы объединяют под
названием аутэкологии — изучения взаимодействий вида с его
средой. Отрасль экологии, изучающая взаимодействие двух и
большего числа видов между собой и со средой, называется
синэкологией.
Изучение видов, живущих в ассоциации, привело к концеп-
ции биологического сообщества. Современные исследователи,
как правило, называют сообществом всякое естественное скоп-
ление видов, живущих в данной области. Мы будем использо-
вать именно это определение.
Уже давно известно, что определенные группы растений и
животных часто встречаются вместе. Поэтому некоторые иссле-
дователи считают, что сообществом можно назвать только та-
кую совокупность организмов, которая почти без изменений
встречаются во многих областях. Мы будем считать такую ассо-
циацию особым типом сообщества — повторяющимся сообщест-
вом (recurrent community).
Наконец, некоторые экологи полагали, что понятие сооб-
щества должно подразумевать взаимодействие и взаимную за-
висимость видов. Иногда такое активное взаимодействие очевид-
но, в других случаях связи между некоторыми видами неясны.
Сейчас считается, что указание на взаимодействие не должно
входить в определение сообщества.
В наземных биотопах и вдоль границ между сушей и водой
иногда имеет место так называемая экологическая сукцессия.
Это длительное последовательное изменение сообществ, живу-
щих в каком-то районе, в идеале — от начального сообщества
IX. Палеоэкология
227
к окончательному, климаксному сообществу. С установлением
климаксного сообщества достигается приблизительное равнове-
сие, и в дальнейшем в нем происходят, лишь незначительные
биологические изменения, если не -вмешиваются крупномасштаб-
ные изменения внешней среды. Обычно сукцессию можно на-
блюдать там, где недавно произошло крупномасштабное и рез-
кое изменение условий среды (например, лесной пожар или от-
ступание ледника), освобождающее место для колонизации
начальным сообществом. Основная причина сукцессии состоит,
видимо, в том, что некоторые сообщества постепенно так
изменяют свою среду, что она становится более пригодной для
других сообществ. Это явление особенно характерно для высших
растений, которые изменяют почву и обычно затеняют друг
Друга.
В море явление экологической сукцессии по разным причи-
нам не имеет широкого распространения. Прежде всего, сукцес-
сия должна идти в определенной области. Пелагические виды
активно плавают или пассивно перемещаются в толще воды. На
морском дне местообитания стационарны, но продолжительность
жизни большинства бентосных видов невелика, и во многих
районах популяции из года в год появляются и исчезают, что
зависит от многих факторов, в том числе от превратностей пе-
реноса личинок. В большинстве случаев главными продуцен-
тами здесь являются не высшие растения, а одноклеточные ор-
ганизмы фитопланктона, которые мало изменяют физически
донные местообитания. Очевидно, экологическая сукцессия в
бентосных сообществах имеет место только там, где организмы,
подобно морской траве (стр. 218), заметно изменяют физические
условия среды на дне. Пожалуй, самый наглядный пример эко-
логической сукцессии в море представляют органогенные рифы.
Рассмотрим этот случай подробнее.
ОРГАНОГЕННЫЕ. РИФЫ
Морская палеоэкология обособилась в качестве ветви пале-
онтологии в 50-х годах. Многие из ранних морских палеоэко-
логических исследований посвящены изучению органогенных
рифов, отчасти потому, что древние рифы служат ловушками
нефти, отчасти же вследствие большого научного интереса к
самим рифам.
Геологи давали термину «риф» множество разных определе-
ний. Важнейший признак рифа, входящий в большинство опре-
делений, — наличие прочного остова, образованного организма-
ми и стоящего (или стоявшего некогда) на морском дне. В наше
время органогенные рифы строятся в основном скелетообра-
зующими герматипными кораллами вместе с корковыми извест-
ковыми водорослями и многими другими не столь важными
15*
Малая
100м
--------1
Распределение видов кораллов
Turbinaria globularis X
Heliopora caerula X
Millepora murrayi X
Millepora plaiyphylla X
Stylophora mordax
Goniastrea retiformls
Montiporafoveolata u M. verrilli
Acropora recumbens u A. corymbosa
Favia stelligera
Acropora conferta u A.hmnilis
Porites lobata
Fauites abdita
Cyphastrea chalcidicum
Porites lutea
Pocillopora elegans
Acropora tubicinaria
Fauia pallida
Plesiastrea uersipora
Favites halicora
Pocillopora uemicosa
Acropora sp. [
Pocillopora danae ।
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
Фиг. 95. Поперечный разрез наветренного рифа атолла Эниветок [140, 141].
Показан градиент скорости течения и распределение видов по зонам.
Стадии
^Клцмаксное сообщество ^Промежуточное сообщество^ Начальное сообщество
Физические стадии
Стадия сопротивления волнам
Стадия умеренно _ , ,
бурнс 1 воды Стадия спокойной вооы
Двустворчатые
Фиг. 96. Три стадии экологической сукцессии, выделенные Лоуэнстамом
в силурийских рифах района Великих озер, США [139].
230
Часть 2. Применение данных палеонтологии
организмами. По красоте и разнообразию видов, составляющих
рифы, они принадлежат к числу самых живописных биологиче-
ских сообществ (фото XXXI).
Древние рифы особенно удобны для палеоэкологического
анализа по трем причинам: 1) большинство рифообразующих
организмов обладает прочными, хорошо сохраняющимися ске-
летами; 2) скелеты, образуя прочное сплетение, скрепляя его
или включаясь в него, редко подвергаются посмертному пере-
носу и часто сохраняются в прижизненном положении; 3) рифы
представляют собой непрерывную запись состава сообщества,
изменявшегося со временем. Кроме того, с геологической точки
зрения рифы важны и потому, что изменяют условия среды как
непосредственно вокруг себя, так и в соседних областях. Рифы
часто разрастаются вверх до уровня моря (или почти до этого
уровня; см. стр. 215) и заслоняют прибрежную зону от действия
волн и течений, делая возможным накопление мелкозернистых
осадков. Обычно можно наблюдать выраженную зональность
в распределении организмов от наветренной к подветренной
стороне рифа, так как противодействие рифа волнам и течениям
создает «градиент энергии» (фиг. 95).
Многие рифы растут тысячи лет, образуя свою собственную
палеонтологическую летопись. Часто рифовое сообщество изме-
няется по мере приближения к уровню прилива. Лоуэнстам
[120] изучал силурийские рифы области Великих озер в США.
Эти рифы образованы не существующими ныне группами орга-
низмов и представляют собой структуры в виде холмов, в кото-
рых в ходе роста происходила экологическая сукцессия
(фиг. 96). Начальное сообщество, поселившееся на глубинах,
на которых волнение не проявлялось, состояло из сравнительно
немногих групп организмов. По мере своего роста риф подни-
мался в перемешиваемые волнением слои воды, фаунистический
состав рифа изменялся и обогащался; перевес получили виды,
обладавшие прочным скелетом. Наконец, вблизи уровня моря
образовалось .устойчивое к волнению климаксное сообщество.
В нерифообразующих сообществах, сохранившихся в палеонто-
логической летописи, экологическую сукцессию наблюдать прак-
тически не удается.
СООБЩЕСТВА МЯГКИХ ГРУНТОВ
Представление о повторяющихся бентосных сообществах
ввел Петерсон [153], основываясь на обширных исследованиях,
проведенных в Северном море. Петерсон обнаружил, что виды
обычно собраны в группы, причем эти группы повторяются,
т. е. встречаются на морском дне в разных изолированных друг
от друга районах. Петерсон подчеркивал, что найденные им по-
вторяющиеся сообщества — всего лишь статистические группы.
IX. Палеоэкология
231
Он не допускал, что повторяющиеся сообщества могут объеди-
няться вследствие экологической взаимозависимости составляю-
щих их видов. Петерсон полагал, что виды, входящие в состав
повторяющихся сообществ, связаны только общими требования-
ми к внешней среде. Некоторые исследователи считали, что ста-
тистические данные не доказывают существования повторяю-
щихся сообществ, и указывали, что, поскольку нет двух совер-
шенно идентичных бентосных биотопов, не может быть и двух
одинаковых сообществ. Сейчас большинство биологов полагают,
что концепция повторяющихся сообществ верна по крайней ме-
ре для некоторых биотопов мягких грунтов, однако для оконча-
тельного суждения данных еще недостаточно.
Можно думать, что концепция повторяющихся сообществ
лучше всего подходит для сообществ с тесной взаимозависимо-
стью видов, например для рифовых сообществ,- в которых одни
виды служат субстратом для других. Многие экологи вслед за
Петерсоном признают, что консументы сообществ мягких грун-
тов сравнительно слабо связаны между собой и что большинст-
во бентосных консументов сосуществуют лишь потому, что
предъявляют сходные требования к среде. Отсутствие побочных
продуцентов, малая длина пищевых цепей, колебания числен-
ности видов из года в год — все это позволяет думать, что меж-
ду видами сообщества мягких грунтов существует лишь слабая
взаимозависимость (если отвлечься от взаимоотношений типа
симбиоза и «хищник — жертва»). Советский биолог Е. П. Тур-
лаева [220] изучала взаимодействие бентосных организмов при
конкуренции за пищу. Она обнаружила, что среди видов одного
местообитания, питающихся одинаковым способом, обычно вы-
деляется большой биомассой какой-то один вид. Эта ситуация
иллюстрирует постулат экологии, часто называемый принципом
конкурентного исключения1, согласно которому из двух сосу-
ществующих видов, нуждающихся в одном и том же факторе,
количество которого в среде ограничено, численность более
сильного конкурента всегда значительно выше численности вто-
рого вида. Со временем слабый конкурент может быть вытес-
нен из биотопа. В реальном сообществе в любой момент вре-
мени может существовать небольшое число еще не вытесненных
слабых конкурентов. В сообществах, описанных Турпаевой, по-
видимому, количество пищи ограничено. В табл. 5 перечислены
виды, входящие в сообщество, и указаны их способы питания.
Многие из видов, составляющих небольшую долю биомассы
каждой пищевой группы, могут быть слабыми конкурентами в
борьбе за пищу. Во многих бентосных сообществах мягких
1 Другое название этого закона — принцип Гаузе, по имени советского
биолога Г. Ф. Гаузе, обосновавшего его экспериментально.— Прим. ред.
^32 Часть 2. Применение данных палеонтологии
грунтов 90% биомассы или более обычно составляют лишь не-
сколько видов. По-видимому, это объясняется тем, что в боль-
шинстве местообитаний возможно лишь ограниченное число
способов добывания пищи. В каждой пищевой группе обычно
доминирует один вид и в общем будет процветать небольшое
число видов. Во многих сообществах мягких грунтов нет при-
крепленных растений, не устанавливаются сложные пищевые
цепи и условия среды непостоянны. Поэтому здесь не достига-
ется полного устранения слабых конкурентов.
ТАБЛИЦА 5
Доминирующая группа в биоценозе Astarte crenata
в Баренцевом море
(по данным с 49 станций)
Виды Тип питания Биомасса, г/м2 Число осо- бей на 1 м2
Astarte crenata Фильтраторы-А1^ 60,80 23
Spirochaetopterus typicus Собиратели 5,08 106
Phascolosoma margaritaceum Заглатыватели-А 5,00 3
Cardium ciliatum Фильтраторы 3,94 0
Ctenodiscus crispatus Заглатыватели 2,49 3
Macoma calcarea Собиратели 1,84 13
Мшанки Подстерегающие 1,54 —
Saxicava arctica Фильтраторы-А 1,47 1
Nephthys sp. Заглатыватели 1,45 6
Ophiocantha bidentata Собиратели 0,66 2
1) Фильтраторы-А — сестонофаги (пожиратели взвеси), фильтрующие непосредственно
прилегающий к дну слой воды. Собиратели — избирательные грунтоеды, собирающие детрит с
поверхности осадка. Заглатыватели — неизбирательные грунтоеды, перерабатывающие осадок.
Подстерегающие — пожиратели взвеси, выхватывающие пищу из потоков воды; у них нет соб-
ственной системы создания тока воды.
Начало палеоэкологическим исследованиям сообществ поло-
жила работа Циглера и сотр. [240]. Изучая нижнесилурийские
отложения Англии, они выделили пять сообществ, которые, оче-
видно, повторялись как в пространстве, так и во времени. Эти
сообщества были выделены на основании видового состава и
географического распределения (фиг. 97). Пять частично пере-
крывающихся сообществ, располагавшихся от прибрежной зоны
в сторону открытого моря, пронумерованы цифрами от 1 до 5.
Соседние сообщества (например, 1 и 2 или 4 и 5) могут иметь
несколько общих видов, а разобщенные сообщества (например,
1 и 4 или 2 и 5) совершенно различны по видовому составу.
Ближайшее к берегу сообщество называется сообществом
Lingula (фиг. 98 и 99). Lingula упоминалась на стр. 210 как
IX. Палеоэкология
233
Край шельфа
Зал.Кардиган
ЬМ Направление течении
Современная береговая линия
Области, в пределах которых
обнажены нижнесилурииские
породы
кт
Фиг. 97. Палеогеография Уэльса в начале позднего лландовери (ранний
силур [238]).
Показано распределение пяти бентосных сообществ в зонах, параллельных древним бере-
говым линиям,
/ — сообщество Lingula-, 2 — сообщество Eocoelia-, 3 — сообщество Penfamerus; 4 — со-
общество Stricklandia-, 5 — сообщество Clorinda-, Т — турбидиты (осадки, образованные
мутьевыми потоками); Г — граптолитовые илы.
род, обитавший уже с раннего палеозоя в прибрежных районах.
Разделяя это силурийское сообщество Lingula, по примеру
Турпаевой, на пищевые группы, мы видим, что самый многочис-
ленный вид —замковая брахиопода «Camarotoechia» decempli-
cata — был сестонофагом, обитавшим на поверхности осадка.
Lingula — второй по численности вид — также принадлежал к
сестонофагам, собиравшим пищу с поверхности осадка, но вел
роющий образ жизни. Palaeonetlo, представитель нукулид (Bi-
valvia), был, судя по сходству с некоторыми современными фор-
мами, грунтоедом. Таким образом, эти три вида относились к
разным пищевым группам.
Даже если есть все основания полагать, что обнаруженный
фаунистический комплекс отражает состав ископаемого сооб-
щества, результаты анализа могут быть обесценены двумя важ-
ными источниками ошибок: во-первых, разделение на пищевые
группы может быть неверным (например, мы можем не знать,
934
Часть 2. Применение данных палеонтологии
Фиг. 98. Реконструкция сооб-
щества Lingula с фиг. 97 [240].
на какой глубине питались некие брахиоподы) и, во-вторых, со-
хранившееся «сообщество» может не включать остатков мягко-
телых видов, относившихся к доминирующим формам. Сообще-
ство Lingula, очевидно, хорошо укладывается в модель Турлае-
вой. Распределение трех видов по пищевым группам, видимо,
правильно; численность же этих видов настолько высока, что
предположение об их доминировании также скорее всего верно.
И все же, учитывая превратности сохранения, мы должны при-
знать наши выводы весьма шаткими. Подобный анализ силурий-
ских сообществ открытого моря представил бы гораздо большие
трудности, поскольку для них характерно большее видовое раз-
нообразие, а образ жизни составляющих их видов известен
хуже.
Исследование силурийского сообщества Lingula позволяет
сделать еще один вывод. По гомологии мы можем предполо-
жить, что силурийские виды Lingula жили в прибрежных место-
обитаниях с неустойчивой соленостью (стр. 210). Низкое видо-
вое разнообразие сообщества Lingula по сравнению с четырьмя
сообществами открытого моря также говорит о неблагоприят-
ных изменениях солености (стр. 210). Эти выводы подтвержда-
ются и стратиграфическими данными. На палеогеографических
картах сообщество Lingula приурочено к прибрежным областям,
где соленость должна была колебаться.
IX. Палеоэкология
235
2. Ungula sp.
3. Palaeoneilo rhomboidea
4. Pteronitella cf. retroflexa
5. Cornulites sp.
6. Ungula pseudoparallela
7. Другие виды
Фиг. 99. Сравнительное обилие видов в сообществе Lingula (фиг. 98)2(240].
Температура и биогеография
От чего зависит распределение видов на обширных простран-
ствах Земли? Почему одни виды живут только у экватора,
•а другие — у полюсов? Почему некоторые виды встречаются толь-
ко на одном материке или в одном океане? Все это проблемы
биогеографии — науки о географическом распределении видов.
Основной ограничивающий фактор географического распро-
236 Часть 2. Применение данных палеонтологии
странения организмов — температура. Однако она мало влияет
на местное распределение (за исключением тех зон, где су-
ществуют быстрые и контрастные колебания температуры, как,
например, на литорали). Температура определяет главным об-
разом широтное распределение видов. Одно из немногих место-
обитаний, в которых температура мало влияет на биогеогра-
фию,— глубоководные зоны, всегда характеризующиеся низки-
ми температурами.
Другие важные биогеографические факторы будут рассмот-
рены ниже, так как они особенно существенны для понимания
органической эволюции и важны для биостратиграфии. Сейчас
мы займемся только проблемами, связанными с температурой.
Палеоклимэтические выводы чаще всего основываются на
небиологических данных. Полезную информацию о климатах
прошлого можно получить, изучая ледниковые отложения, эоло-
вые осадки, известняки, несущие достоверные признаки накоп-
ления в тропиках, й многие другие типы пород.
Млекопитающие и птицы способны к поддержанию постоян-
ной температуры тела, но почти все остальные организмы при-
нимают температуру окружающей среды. И теплокровные и хо-
лоднокровные животные обычно приурочены к определенным
диапазонам температур, пригодных для выживания или успеш-
ного размножения. Виды, способные переносить широкий диапа-
зон температур, называются эвритермными, а приуроченные к
узкому интервалу — стенотермными.
При проведении палеоклиматических и палеобиогеографиче-
ских реконструкций следует помнить о нескольких источниках
возможных ошибок. Исследования палеомагнетизма убедили
многих геологов, что в ходе истории Земли материки дрейфова-
ли. О путях дрейфа материков пока мало что известно, и прак-
тически невозможно наметить единую глобальную схему клима-
тической зональности для домелового и, может быть, даже для
докайнозойского времени. Все же местные климатические усло-
вия любого периода можно восстановить по соответствующим
породам и окаменелостям.
Такие факторы, как крупные океанские течения, обычно на-
рушают простую картину климатической зональности. Темпера-
тура воды вдоль юго-восточного побережья США гораздо выше,
чем на тех же широтах вдоль западного побережья Северной
Америки. Силы Кориолиса несут на север вдоль атлантического
побережья теплый Гольфстрим, а вдоль тихоокеанского побе-
режья на юг направляют холодное Калифорнийское течение.
ТЕМПЕРАТУРА И МОРФОЛОГИЯ
Температура, как и соленость, слабо отражается на морфо-
логии скелета. Все же удается отметить тенденцию к возраста-
нию среднего размера особей в холодных областях географиче-
IX. Палеоэкология
237
ского ареала вида по сравнению с теплыми областями. Придат-
ки тела (например, уши и хвост у млекопитающих) в холодной
части ареала короче. Эти закономерности, известные как пра-
вило Бергмана и правило Аллена, редко использовались при
истолковании ископаемого материала из-за искажений в процес-
се фоссилизации. Оба явления, очевидно, связаны с увеличением
отношения объема тела к его поверхности у видов, живущих
в холодном климате.
Почти все группы моллюсков, раковины которых имеют ши-
пы, и многие группы с толстыми раковинами живут в тропиче-
ских и субтропических областях. Это может объясняться тем,
что в теплых водах условия секреции карбоната кальция более
благоприятны, чем в холодных.
ТЕМПЕРАТУРА И ГОМОЛОГИЯ
При палеоклиматических исследованиях с применением па-
леонтологических данных и принципа гомологии чаще всего
использовались фауны и флоры мела и кайнозоя. Особенно час-
то использовались ископаемые растения, являющиеся прекрас-
ными индикаторами климата с того момента, с которого начи-
нается богатая палеонтологическая летопись цветковых расте-
ний (середина мелового периода).
Фиг. 100. Северные границы субтропической зоны в Северной Америке в раз-
личные эпохи кайнозойской эры (из [43], с изменениями).
Сдвиг границ, проведенных иа основании палеоботанических исследований, говорит об
общей тенденции к понижению температуры в течение кайнозоя.
238
Часть 2. Применение данных палеонтологии
Фиг. 101. Широтные границы живых коралловых рифов (пунктир) и герма-
типных кораллов (сплошные линии) в Тихом океане [230].
Сплошными линиями отмечена изотерма 21 °C по максимальным, а пунктиром — по ми-
нимальным оценкам.
Изучение ископаемой флоры показывает, что в раннем кай-
нозое на севере тихоокеанского побережья современной террито-
рии США, вплоть до Орегона (если не дальше), господ-
ствовал субтропический климат. На Аляске произрастали расте-
ния, типичные для умеренной или даже субтропической зоны.
Более молодые флоры свидетельствуют об общем похолодании
к плейстоцену и до нашего времени (фиг. 100). Тот факт, что
с изменением температуры одновременно смещаются ареалы
многих видов, свидетельствует в пользу выводов, сделанных на
основе принципа гомологии для одного вида. Однако сходство
t-ю
1-20
ИЗО
-60
к-70
20
60
50 40 30
Градусы северной широты
Фиг. 102. Положения морских зимних изотерм вдоль тихоокеанского побе-
режья Северной Америки [46].
Линии проведены на основании находок ископаемых моллюсков н кораллов, близкие ро-
дичи которых живут и сейчас. Общий сдвиг изотерм к экватору в течение кайнозоя отра-
жает общее похолодание (ср. с фиг. 100).
Млн.лет
1-40
H50
240 Часть 2. Применение данных палеонтологии
высокогорной флоры средних широт с обычной флорой высоких
широт может вводить в заблуждение. Судя по местной иско-
паемой флоре, можно решить, что данная область когда-то име-
ла умеренный климат, тогда как на самом деле изучаемая фло-
ра была высокогорной, а вокруг нее в долинах процветали тро-
пические растения, по которым и надо было судить о климате
области. К счастью, флоры долин и низменностей, самые на-
дежные индикаторы климата, сохраняются чаще благодаря
рельефу своих местообитаний.
Коралловые рифы растут только там, где температура воды
редко опускается ниже 18 °C. Почти все современные рифы на-
ходятся в пределах полосы, ограниченной примерно 30° широты
по обе стороны от экватора (фиг. 101). Ископаемые рифы широ-
ко используются как индикаторы древних тропических или
близких к тропическим климатов. Палеозойские рифы, созда-
вавшиеся различными сообществами, также обычно считаются
свидетельством существования теплых морей. Дарем [46] со-
брал данные по географическому распределению кайнозойских
рифообразующих кораллов и моллюсков тихоокеанского побе-
режья Северной Америки (эти данные составили огромный
том). Его выводы (фиг. 102), основанные на соображениях го-
мологии (главным образом на уровне родов), подтверждают
только что рассмотренные данные палеоботаники.
ГРАДИЕНТЫ ТАКСОНОМИЧЕСКОГО РАЗНООБРАЗИЯ
Градиенты таксономического разнообразия — возможно, са-
мый поразительный пример влияния температуры на глобальное
распределение организмов. Видовое разнообразие большинства
высших таксономических категорий заметно возрастает от по-
люсов к экватору (фиг. 103).
Фишер [60] показал, что абиотические условия среды предо-
ставляют в умеренных областях не меньшее число экологиче-
ских ниш, чем в тропических, хотя видообразование в тропиках
может идти несколько быстрее. Фишер и другие исследователи
подчеркивают, что разнообразие — это продукт эволюции и что
паузы в процессе эволюционного возрастания разнообразия пре-
пятствуют возникновению максимального, «насыщающего» раз-
нообразия в биотопе. Такие паузы, связанные, как правило,
с климатическими изменениями, гораздо более часты и опасны
в холодных зонах, чем в тропиках. Возможно, что массовые вы-
мирания чаще происходили именно в высоких широтах. Тропи-
ческие сообщества, находясь в условиях непрерывной, постоян-
ной эволюции, ближе подходят к зрелости и в них сохраняется
высокое видовое разнообразие. Представление о том, что не тем-
пература сама по себе, а степень стабильности условий среды
является главным фактором, контролирующим видовое разно-
IX. Палеоэкология
241
Фиг. 103. Градиент разнообразия мелководных видов брюхоногих моллюс-
ков вдоль восточного побережья Северной Америки [60].
Каждая линия соответствует 10 видам. Ареалы 12 видов тянутся через Северный Ледови-
тый океан в северную часть Тихого океана.
образие, получило подтверждение в недавних исследованиях
разнообразия глубоководного бентоса (стр. 214). Раньше счи-
тали, что разнообразие абиссального бентоса невелико, но ока-
залось, что оно сравнимо с разнообразием мелководного тропи-
ческого бентоса! Хотя в абиссали температура лишь ненамного
выше точки замерзания, она исключительно постоянна, и, ви-
димо, именно это обеспечило относительную непрерывность уве-
личения разнообразия в течение многих миллионов лет.
Все же такое объяснение широтных градиентов разнообра-
зия удовлетворило не всех. Пиэнка [154] пересмотрел гипотезу
стабильности среды и другие гипотезы и подчеркнул, что в со-
здании градиентов может участвовать сразу несколько фак-
торов.
Валентайн [222] высказал предположение, что, каково бы
ни было происхождение широтных градиентов разнообразия,
общее разнообразие в глобальных масштабах должно быть
16—467
242
Часть 2. Применение данных палеонтологии
90°
Фиг. 104. Зная градиенты разнообразия родов брахиопод из двух семейств,
можно наметить положение Северного полюса в пермском периоде [209].
На схеме А использованы данные по теребратулидам, на схеме Б — по ортотетацеидам.
Концентрические линии соединяют области с одинаковым родовым разнообразием. Циф-
ра на каждой линии — число ископаемых родов. Положения полюса, найденные по дан-
ным для двух групп брахиопод, почти идентичны и хорошо совпадают с положениями,
найденными по геологическим данным.
больше при значительных температурных градиентах, чем при
однородном климате на всей планете. В последнем случае от-
дельные фауны могли бы распространиться на обширные про-
странства. При более крутых градиентах должны существовать
узкие широтные пояса разных фаун. Любопытный вывод из ги-
потезы Валентайна: глобальные колебания климата должны
увеличивать разнообразие форм, а в периоды потепления долж- :г
но происходить массовое вымирание приспособленных к холоду
фаун. 4
IX. Палеоэкология
243
Стели попытался использовать градиенты таксономического
разнообразия как индикаторы местонахождения полюсов в древ-
ности (фиг. 104). Используя данные по разнообразию для от-
дельных таксонов и для определенных геологических периодов,
можно с помощью математических методов определить древние
положения полюсов, если считать, что широтные градиенты бы-
ли тогда такими же, как и теперь. Данные по разнообразию
следует оценивать с осторожностью. Число видов, найденных
в данной стратиграфической единице, может определяться не
столько географической широтой местообитания, сколько усло-
виями в нем. Оно зависит также от тщательности сбора и от
того, преобладает ли в систематике данной группы тенденция
к «объединению» или «дроблению». Кроме того, следует учи-
тывать, что полученные недавно геофизиками данные, свиде-
тельствующие о значительном дрейфе материков в геологиче-
ском прошлом, также могут усложнить использование градиен-
тов разнообразия ископаемых таксонов.
Посмертная утрата информации
Историю отдельной особи от ее рождения до обнаружения
ее фоссилизированных остатков удобно разделить на этапы
(фиг. 105). Палеоэкология в узком смысле слова занимается
только этапом от рождения до смерти организма. Хотя приве-
денные на фиг. 105 названия ветвей палеонтологии, изучающих
последующие этапы, не являются общепринятыми, они позволя-
ют создать полезную классификацию. Данный раздел вместо
«Посмертная утрата информации» можно было бы назвать
«Тафономия».
Термин «ископаемое сообщество», или «ископаемый фауни-
стический комплекс», мы использовали для обозначения группы
ископаемых остатков (одного или нескольких видов), найден-
ных вместе в стратиграфической летописи. Необходимо уточнить
это понятие. Чтобы комплекс считался единым, он должен быть
почти гомогенным или однотипно гетерогенным по видовому
составу. По вертикали фаунистические комплексы обычно огра-
ничены одним стратиграфическим слоем, но могут распростра-
няться и на несколько слоев. По горизонтали они могут прости-
раться на сантиметры, метры или (реже) километры. По вер-
тикали и по горизонтали комплекс может сменяться (постепенно
или резко) или «немыми» слоями, или другими комплексами.
Гомогенный или однотипно гетерогенный состав комплекса от-
ражает его экологию и историю сохранения.
Ископаемые комплексы классифицировали разными спосо-
бами. Одна система показана на фиг. 106. Ископаемый комп-
лекс, целиком состоящий из остатков видов, принадлежащих
к одному сообществу и захороненных в том же месте, где они
244
Часть 2. Применение данных палеонтологии
Этапы формирования
ископаемого
Ветви палеонтологии
Обнаружение —- Окончательное захоро- нение Исследование диагенеза — Тафономия
Биостратономия
—
1 иоель Рождение Палеоэкология
Фиг. 105. Ветви палеонтологии, изучающие ископаемые организмы от момен-
та рождения или выхода из яйца и до обнаружения их ископаемых остат-
ков [П4].
жили, мы назовем прижизненным комплексом. Если ископаемый
комплекс целиком состоит из остатков видов, перенесенных от-
туда, где они жили (даже если они жили вместе), мы будем
называть его посмертным комплексом. Комплексы из остатков
видов, живших ранее в двух или нескольких местообитаниях,
мы назовем смешанными посмертными комплексами.
При изучении таксономического состава ископаемого комп-
лекса исследователь ищет ответ на два основных вопроса: ка-
кие здесь имеются виды и каково было их численное соотноше-
ние при жизни? Исследования на беспозвоночных, позвоночных
и растениях показали, что на второй вопрос редко удается по-
лучить хотя бы приблизительный ответ.
Как уже говорилось в гл. IV, для определения того, захоро-
нены ли организмы в том месте, где они жили, применяются
различные критерии. Сохранение в прижизненном положении
Живое сообщество
Ископаемое (ые)
сообщество (а)
Захоронение in situ Перенос
I
Прижизненный комплекс Посмертный комплекс
(ископаемое сообщество) (танатоценоз)
I
Автохтонное ископаемое Постороннее сообщест-
сообщество, находящееся во, перенесенное из
в той же среде, что и другой современной
исходное живое сообщество среды
I
Переотложенное из
других, более древ-
них пород
Фиг. 106. Классификация ископаемых сообществ по способу их возникнове
ния [35].
ц
IX. Палеоэкология
245
с уверенностью позволяет исключить посмертный перенос. Ко-
ралловые рифы и следы животных особенно ценны для палео-
экологии в том отношении, что они не подвергаются переносу.
Иногда сообщества или популяции могут внезапно покрываться
слоем осадка, причем множество особей подвергается захороне-
нию в прижизненном положении.
О посмертном переносе остатков бентосных организмов часто
можно судить по самим остаткам. При переносе остатки орга-
низмов расчленяются, ломаются, истираются и сортируются по
размеру. Джонсон [95] проанализировал условия, при которых
возникают захоронения бентосных организмов, дав три модели
процесса сохранения. Конечно, три модели Джонсона-—лишь
отдельные точки непрерывной последовательности. Обычно ис-
копаемые комплексы не согласуются полностью ни с одной из
трех моделей, хотя о многих комплексах можно сказать, что
они в большей мере соответствуют какой-либо одной из них.
Вот описание этих трех моделей.
Модель I. Сообщество обитает в ограниченной зоне мор-
ского дна ниже уровня малой воды, но выше максимального
уровня действия волнения. Водная масса движется над дном с
небольшой скоростью, которая иногда может повышаться до
такой степени, что начинается размыв осадков. Субстрат — тер-
ригенные осадки, местами с примесью обломков скелетов.
Обильная и разнообразная фауна состоит из мягкотелых орга-
низмов и организмов с твердым скелетом. При нормальном
уровне смертности элементы сообщества постоянно отмирают
и становятся потенциальными ископаемыми. Они разлагаются;
некоторые скелетные остатки, пролежав на дне какое-то время,
подвергаются захоронению. Затем внезапно происходит гибель
и захоронение всего сообщества: оно быстро покрывается тол-
стым слоем осадка, сходного с материалом субстрата. Осадки,,
заключающие ископаемый комплекс, в процессе дальнейшего’
осадконакопления постепенно уплотняются.
Модель II. Местообитание сходно с описанным в модели I.
Элементы сообщества в условиях нормальной смертности не-
прерывно отмирают, становясь потенциальными ископаемыми.
Они разлагаются; часть остатков уносится, другая часть через
какое-то время подвергается захоронению. Местные условия
среды меняются со временем, и сообщество постепенно заме-
няется другим. Ископаемый комплекс погребается все глубже;
заключающие его осадки в результате дальнейшего осадкона-
копления все более уплотняются.
Модель III. Сообщество обитает на ограниченной зоне мор-
ского дна ниже уровня малой воды, но выше максимального-
уровня действия волнения. Водная масса движется над дном с
умеренной скоростью. Дно покрыто терригенными осадками, со-
246
Часть 2. Применение данных палеонтологии
Влияние продолжительности
периода до захоронения остатков
Фиг. 107. Схема, отражающая
сравнительное изменение иско-
паемых сообществ в резуль-
тате трех разных способов
возникновения.
Модель I соответствует внезапному
захоронению сообщества; мо-
дель II — постепенному накопле-
нию и захоронению остатков
организмов, живших в месте
осадконакопления; модель III —
сообществу, состоящему почти
исключительно из перемещенных
остатков. Продолжительность пе-
риода до захоронения остатков
сказывается на степени их измене-
ния в результате растворения,
фрагментации, расчленения, обра-
стания и сверления различными
организмами. В результате переноса
возникали случайные незакономер-
ные комплексы видов и сортиро-
ванность по размерам. Видимо,
большинство ископаемых комплек-
сов укладывается в область, очер-
ченную пункчиром. Комплексы,
сложившиеся в спокойных водах,
должны располагаться на этой
диаграмме вблизи участков, соот-
ветствующих моделям I и II, а
комплексы, характерные для бур-
ных вод, должны соответствовать
модели III.
стоящими в значительной степени из органического детрита.
Часто скорость движения водной массы так велика, что осадок
и органический детрит перемещаются. Гидродинамические ус-
ловия могут способствовать накоплению органических остатков
в этом месте. Небогатая фауна состоит из нескольких видов
мусорщиков, сверлильщиков и обрастателей. При нормальной
смертности элементы сообщества непрерывно отмирают и ста-
новятся потенциальными ископаемыми. Они разлагаются; часть
остатков уносится, а прочные остатки скелетов после различно-
го по длительности периода экспозиции подвергаются захороне-
нию. При сокращении поступления осадка и детрита (вследст-
вие изменения условий) накопление органических остатков в
данном местонахождении замедляется. Со временем осадки с
высокой концентрацией прочных остатков погребаются и в ре-
зультате дальнейшего осадконакопления уплотняются.
Описание ископаемых комплексов для этих трех моделей
приводится в табл. 6 (упоминаются только самые своеобразные
.или 1иа1ибюлее часто развивающиеся особенности комплексов).
На фиг. 107 представлена диаграмма, отражающая сравни-
тельное изменение ископаемых сообществ в результате трех
разных способов их возникновения.
Шотуэлл [192], работая с плиоценовыми фаунами Орегона,
предпринял оказавшуюся небезуспешной попытку обойти
трудности, связанные с тем, что многие сообщества наземных
ТАБЛИЦА 6
Сравнительная выраженность признаков комплексов для трех
моделей (из [95], с изменениями)
Признак Выраженность признаков
Модель I Модель II Модель III
Состав фау- ны Виды с близкой эколо- гией Как в модели I Может отличаться от моделей I и II
Морфологи- ческие осо- бенности остатков Могут сохраняться тон- кие структуры; со- храняются также ор- ганизмы разнообраз- ной формы и разме- ров. Сохранившиеся формы соответствуют экологии живущего здесь сообщества То же Могут сохраниться только прочные час- ти организмов; со- хранившиеся особи могут быть отсорти- рованы по форме и размеру
Плотность за- хоронения Возможны разные плот- ности » Высокая
Распределе- Для многих ВИДОВ МО- Для некоторых ви- Иное, чем в моделях
ние разме- ров жет соответствовать идеальному распреде- лению жившей здесь дов — как в мо- дели I I и II
Расчленен- популяции Много нераспавшихся Есть некоторое ко- Иначе, чем в моделях
ность ос- татков остатков; в случае распавшихся остат- ков отдельные компо- ненты скелетов сохра- нились примерно в тех же пропорциях, в каких они были у живых организмов личество нерас- павшихся остат- ков; в случае распавшихся ос- татков пропор- ции отдельных компонентов — как в модели I I и II
Фрагментар- Лишь немногие скелеты Умеренное количе- Обломки скелетов мно-
ность ос- татков раздроблены ство фрагментов скелетов гочисленны
Состояние по- верхности остатков Поверхностные струк- туры сохранили при- жизненный вид Имеются следы ис- тирания в раз- личной степени Как в модели II
Химический и минера- логический состав ос- татков Определять степень его сохранности мы пока не умеем Как в модели I Как в модели 1
Положение У некоторых видов со- Большинство удли- Как в модели II (бы-
остатков храняется положе- ние, приобретенное в момент гибели ненных остатков ориентировано так, что длинная ось располагается вдоль плоскости напластования (бывают исклю- чения) вают исключения)
Часть 2. Применение данных палеонтологии
Продолжение табл. 6
Признак Выраженность признаков
Модель I Модель II Модель III
Распределе- ние остат- ков в про- странстве Структура и текстура осадка Распределение нерас- павшихся остатков некоторых видов мо- жет сохраняться тем же, что и при жизни Соответствует предпо- лагаемым предпочте- ниям фауны; харак- терна для сравни- тельно спокойных вод Иное, чем при жиз- ни Как в модели I Как в модели II Не обязательно соответ- ствует предполага- емым предпочтениям фауны; характерна для сравнительно не- спокойных вод
позвоночных представляют собой смешанные посмертные сооб-
щества. Шотуэлл изучал сообщества из разных местонахож-
дений, которые относились примерно к одному стратиграфиче-
скому интервалу. Он принял, что все виды, найденные в данном
месте сбора, жили в этой области, но некоторые сохранились
вблизи места, где жили, а другие переносились на значительные
расстояния. Дальность переноса каждого вида оценивалась по
степени сохранности скелета. По методу Шотуэлла, степень со-
хранности вида в данном месте рассчитывается как отношение
Общее число скелетных элементов
Минимальное число сохранившихся особей "
В этом выражении минимальное число особей — это наи-
меньшее число организмов, от которых могло остаться найден-
ное количество скелетных элементов. Например, если найдено
9 скелетных элементов одного вида, а одинаковых среди них
только два, то минимальное число особей равно 2. Чтобы сде-
лать полученные для разных видов цифры сравнимыми, их со-
относят с числом потенциально способных сохраниться скелет-
ных элементов каждого вида. Тогда ископаемый комплекс мож-
но отобразить с помощью диаграммы, приведенной на фиг. 108, А.
Каждый вид представлен сектором, угол которого соответствует
относительному обилию вида. Отношение радиуса затемненной
части к радиусу всего сектора отражает полноту сохранения.
Разделение видов на живших вблизи места сбора и живших
в отдалении сделано на основании того, выше или ниже число
скелетных элементов на особь данного вида по сравнению со
средним числом скелетных элементов для всей группы. Конеч-
но, этот критерий несколько спорен. Виды, отнесенные к мест-
ным формам, отмечены дугами.
На фиг. 108, Б показана общая картина распределения мест-
ных видов в четырех районах сбора. Почти все эти вымершие
Кребс-Рэнч 2
Обнажение Вестэнд
Бордмэн
>Рэнч 1
Hypolagus Dipoides Scapanus
Neohipparion /
Prosthennops
PUauchenia Perognathus Х////////////////УУ//////////У
Mastodon PeLeoceras Cants ~~ '' "'/////////, flphelop4
Б
’"'фиг. 108. Плиоценовые^сообщества наземных позвоночных' (Орегон) [192].
А. Диаграмма, на которой местные обитатели, жившие на месте сбора (Кребс-Рэич 2), от-
делены с помощью дуги от видов, остатки которых были принесены сюда из значительно
удаленных мест. Б. Сообщества в четырех соседних местонахождениях. Сплошными ли-
ниями отмечено распределение местных видов; пунктиром отмечены случайно встре-
ченные.
[>иг. 109. Основные местообитания, занимавшиеся фаунами, приведенными
на фиг. 108 [192].
IX. Палеоэкология
251
Хищники
Фиг. ПО. Количественное соотношение хищников и травоядных в двух пли-
оценовых местонахождениях (Маккэй-Резервер, Орегон, и Гемпхилл, Техас,
США) и в плейстоценовой асфальтовой яме Ла-Бреа (Калифорния, США) [191].
Последняя диаграмма сильно напоминает перевернутую пищевую пирамиду.
таксоны имеют близких рецентных родичей. Таким образом,
соображения, основанные на принципе гомологии, позволяют
с высокой степенью достоверности определить образ жизни и
характер местообитания плиоценовых родов.
Такая реконструкция подтверждает результаты анализа, по-
казанные на фиг. 108, Л; здесь выявляется, что разные локаль-
ные сообщества занимали различные местообитания, причем
имел место переход от пруда и берега пруда к лесу и, наконец,
клугу (фиг. 109).
Сейчас трудно наблюдать процесс образования ископаемых
сообществ наземных позвоночных. Представление о нем могут
дать знаменитые плейстоценовые асфальтовые ямы в Ла-Бреа
(Калифорния). Животные гибли в этих ловушках. Некоторые
хищники приходили сюда в поисках мертвых или умирающих
животных и часто также вязли в асфальте. Поэтому в ископае-
мом комплексе несообразно много остатков некоторых живот-
ных, питавшихся падалью. Среди них — гигантский волк Canis
dirus и смилодоны (саблезубые кошки), которые в других мес-
тообитаниях были, как известно, активными хищниками. Ске-
леты многих смилодонов несут следы прижизненных поврежде-
ний: по-видимому, к асфальтовым ямам приходили питаться
падалью больные и раненые хищники. В комплексе из Ла-Бреа
численное соотношение организмов сильно искажено. Поэтому
комплекс в целом создает ложное впечатление о перевернутой
пищевой пирамиде (фиг. НО).
Однако местные плиоценовые фауны Орегона, изученные
Шотуэллом, дают картину нормальных пищевых пирамид, в ко-
торых численность травоядных значительно превышает числен-
ность хищников. Эта общая картина может примерно отражать
сравнительную численность пищевых группировок при жизни,
но сравнительная численность отдельных ископаемых видов
вполне может оказаться сильно искаженной. Как и в случае
беспозвоночных, сохранность видов зависит от образа жизни
и прочности скелета.
ГЛАВА X
Эволюция
и палеонтологическая летопись
Со времени опубликования дарвинского труда «Происхож-
дение видов» (1859) идея органической эволюции стала одной
из наиболее важных идей, лежащих в основе современных пред-
ставлений о месте и роли человека во Вселенной.
Механизм эволюционных изменений рассматривался выше,
особенно в гл. IV—VI. Наше представление об этом механиз-
ме, в котором главное — это генные мутации и естественный
отбор, основано на неонтологических науках, изучающих орга-
низмы на протяжении очень короткого периода, составляющего
всего лишь недолгий момент в геологической истории. Палеон-
тологическая летопись мало что добавляет к нашему пониманию
основного механизма эволюции, однако она служит единствен-
ным источником данных о макроэволюции органического мира.
Без палеонтологической летописи макроэволюция живого так
и не стала бы общепризнанным фактом и оставалась бы всего
лишь гипотезой.
Палеонтологическая летопись дает информацию по пяти
вопросам, существенным для понимания эволюции: 1) филоге-
нетическая связь между крупными таксономическими группа-
ми; 2) время появления главных адаптаций; 3) скорость; 4) на-
правление и 5) пути эволюции. Нашему пониманию последних
трех пунктов особенно способствовали работы Дж. Симпсона.
Для измерения эволюционных изменений используют и мор-
фологический и таксономический критерии (мы применяем тер-
мин «морфологический» в широком смысле слова, включая сю-
да генетические, биохимические и структурные признаки). Са-
мый главный признак — структура генотипа, но даже биолог не
знает точно, что за код заложен в хромосомной ДНК- Изучая
палеонтологическую летопись, мы вынуждены исследовать не-
полный набор анатомических признаков и ничтожные, часто
сильно измененные по химическому составу образцы организ-
мов. При изучении таксономических изменений ископаемых ор-
ганизмов нам приходится использовать некоторые категории
искусственной системы классификации, которую мы наклады-
ваем на эволюционный континуум. В этой классификации меж-
ду установленными таксонами обычно существуют большие
F
X. Эволюция и палеонтологическая летопись 253
филогенетические разрывы. Однако, несмотря на свою непол-
ноту, палеонтологическая летопись позволяет сделать важные
выводы об эволюции, вымирании и филогении.
Географическое видообразование
В палеонтологической летописи очень мало хорошо докумен-
тированных примеров географического видообразования. Это
объясняется стратиграфическими перерывами и плохой сохран-
ностью ископаемых. Известные примеры обычно ограничивают-
ся кайнозойскими ископаемыми. Рассмотрим работу Уоллера
[224] по филогении гребешка рода Argopecten из неогена Се-
верной Америки (фиг. 111).
Считают, что позднемиоценовый Argopecten comparilis жил
как в биотопах открытого моря, так и в замкнутых заливах —
г от штата Виргиния на севере до Карибского моря на юге. Он
проник морским путем и в Тихий океан. Когда в плиоцене по-
L явился Панамский перешеек, произошло географическое раз-
*’ деление генофонда A. comparilis на атлантический и тихоокеан-
ский. Географическая изоляция привела к эволюционной дивер-
| генции. Тихоокеанские потомки — A. comparilis, A. circularis и
| A. purpuratus — остались более эврибионтными, а атлантиче-
fc ские экологически специализировались: одни виды приспособи-
лись к биотопам открытого моря, а другие — к изолированным
В? бухтам. Разумно предположить, что более интенсивное видо-
образование среди атлантических потомков объясняется тем, что
С? шельф у побережья Атлантики и Мексиканского залива зна-
j чительно шире, чем у тихоокеанских берегов. Кроме того, здесь
% больше лагун, лежащих за барьерными островами вдоль при-
брежных равнин. Тихоокеанское же побережье Северной и Юж-
Й ной Америки во многих местах прилегает к горным цепям. По-
I этому во многих районах шельф здесь очень узкий и быстро
Г сменяется крутым континентальным склоном.
й' Чтобы разработать подробную филогению наподобие той,
« что показана на фиг. 111, необходимы детальные морфологиче-
ские и стратиграфические исследования. Такие реконструкции
Л не только показывают роль географического видообразования,
но и оправдывают использование понятия вида в палеонтологии
р даже при неизбежно произвольной расстановке границ внутри
| эволюционных рядов.
I Скорость эволюции
Скорость эволюции — просто мера биологических изменений,
gZ соотнесенная со временем. В идеале скорость эволюции следует
измерять в абсолютных единицах времени (например, в мил-
I
г
254
Часть 2. Применение данных палеонтологии
Атлантический
A. irradians
A. nucleus
A. gibbus
A. ptrpuratus
A. circulans
A anteamplicostatus
A v'ivenarius
А. Вид а
A comparHis
A. oboreus
A charltonius
A choctawhatcheensis
A nicholsi
А. Вид b
Фиг. 111. Филогения гребешков рода Argopeden в кайнозое Северной Амери-
ки 1224].
Цифрами 1—8 обозначены изохронные поверхности, размещенные в соответствии с дан-
ными по абсолютному возрасту (7 — 19 млн. лет; 2 —16 млн. лет; 3—13 млн. лет; 4—8 млн.
лет; 5—6 млн. лет; 6—1,8 млн. лет; 7—0,25 млн. лет; 8— настоящее время, 0 млн. лет);
каждой поверхности соответствует стратиграфический горизонт, послуживший источ-
ником ископаемых, а также источником других данных для реконструкции филогении.
Сплошные линии на поверхностях 5, 6 и 8 символизируют географический барьер, отде-
ливший Атлантический океан, Мексиканский залив и Карибское море от Тихого океана
после миоцена. Пунктирные линии на поверхностях 3, 5, 6 и 8 символизируют экологи-
ческий барьер, отделяющий биотопы замкнутого залива (вверху справа) от открытых
морских (внизу слева) на Атлантическом побережье.
лионах лет). Если интервал времени точно не известен (а это
наиболее обычный случай), то степень эволюционных измене-
ний соотносят, как правило, с мощностью осадочных пластов
или с относительным геологическим временем. Например, мож-
но говорить о степени эволюционных изменений на тысячу мет-
ров отложений в непрерывной осадочной последовательности
или на эпоху данного геологического периода.
X. Эволюция и палеонтологическая летопись
255
Рассматривая темп эволюции, мы будем использовать два
сходных по звучанию, но различных по смыслу прилагатель-
ных— филогенетический, что значит относящийся к филогении
(причем рассматриваться может как один эволюционный ряд,
так и два или несколько родственных рядов), и филетический —
более узкий термин, означающий принадлежность к одному
эволюционному ряду.
Мы рассмотрим три основных подхода к измерению скоро-
сти эволюции на основании палеонтологических данных: 1) фи-
логенетическую скорость, измеряемую по морфологическим
критериям; 2) филогенетическую скорость, измеряемую по так-
сономическим критериям; 3) скорость, измеряемую по числу
таксонов.
Симпсон [197] подчеркивает, что важно различать филоге-
нетическую скорость и скорость изменения числа таксонов. Фи-
логенетическая скорость характеризует изменения в одном ряду
(филетические изменения) или в нескольких близкородственных
рядах. Например, в мезозойском семействе кораллов Rhipidogy-
fidae (фиг. 112) мы можем измерить скорость морфологических
изменений в эволюционном ряду от Codonosmilia до Placogyra
или от Codonosmilia до Acanthogyra, основываясь на измене-
ниях разных структур. При другом подходе, основанном на из-
мерении таксономических изменений, можно учитывать сред-
нюю продолжительность существования родов (в миллионах
лет на род) в каждом ряду. Скорость изменения числа таксонов
говорит о темпе возникновения новых таксономических групп
внутри высшего таксона. Возьмем, например, подотряд Сагуо-
Щ| Agariclicae
lllllll Fungiicae
Poriticae
BBSS Styfophyllicae |Ю§| Caryoph ynifcae
НИ Faviicae
Flabellicas
Фиг. 113. Филогенетическое древо Scleractinia (шестилучевые кораллы) [229].
Показаны подотряды (их 5), надсемейства (различаются штриховкой) и семейства.
X. Эволюция и палеонтологическая летопись
257
Фиг. 114. Скорости эволюции новых семейств и родов Scleractinia (шестилу-
чевые кораллы) [229].
I — новые роды, Н — новые семейства.
phylliina (к которому относятся Rhipidogyridae) или весь от-
ряд кораллов Scleractinia (фиг. 113). Можно, например, изо-
бразить на графике число новых родов или семейств, возник-
ших в мезозое и кайнозое. На фиг. 114 показан такой график
для всего отряда. Далее в этой главе мы проанализируем этот
график.
ФИЛОГЕНЕТИЧЕСКАЯ СКОРОСТЬ,
ИЗМЕРЯЕМАЯ ПО МОРФОЛОГИЧЕСКИМ КРИТЕРИЯМ
В палеонтологической летописи очень трудно выделить на-
стоящий эволюционный ряд, включающий три или более таксо-
нов. Большинство схем, предложенных для описания таких ря-
дов, основано на неполных данных и представляет собой силь-
ное упрощение сложных филогений. Кроме того, большинство
морфологических тенденций, показанных в более простых из
предложенных схем, охватывает очень короткие интервалы вре-
мени. Их абсолютную продолжительность трудно измерить, так
что невозможно оценить и скорость на основе абсолютного вре-
мени. Все это не позволяет установить темп эволюции.
Один из простейших типов морфологических эволюционных
изменений — изменение размеров. На фиг. 115, Б показаны раз-
меры тела в эволюционном ряду рогатых динозавров. В этом
ряду длина животных заметно возрастает, как и в большинстве
других рядов, показанных на фиг. 115, А. Изменение наклона
кривой на графике служит мерой скорости возрастания разме-
ра. В нашем примере скорость, очевидно, уменьшается, однако:
17—467
258
Часть 2. Применение данных палеонтологии
Ланс
Хеллкрик
Эдмонтон
Киртланд
Белли-Ривер
Меса-Берде
Proioceratops
$,жадокта
Фиг. 115. Возрастание длины
тела в процессе эволюции ро-
дов рогатых динозавров (верх-
ний мел, США) [31].
А. Филогения североамериканских
рогатых динозавров, Б. Увеличение
Длины тела в ходе эволюции от
Proioceratops к Triceratops.
следует учитывать, что простая кривая, проведенная через три
точки, конечно, может быть слишком большим упрощением.
На трех родах цератопсов (фиг. 115, Б) можно наблюдать
еще один важный тип морфологических изменений. При увели-
чении длины тела в ряду меняются и пропорции тела. На
фиг. 116 за постоянную принята длина спины и сравниваются
относительные размеры других частей тела. Хорошо заметно
уменьшение относительной длины конечностей и хвоста, тогда
Как относительный размер черепа остается почти неизменным.
Хотя подобных примеров различных скоростей морфологи-
ческой эволюции известно много, такие графики построены
Для небольшого числа случаев. Один пример показан
Triceratops
Monoclonius
Proioceratops
Фиг. 116. Пропорции скелет ay трех родов рогатых динозавров[31].
Ланс
Белли-Ривер
Джадокгпа
60
50
40
borealis
J_______________I_______I_____________I________L
Эоцен Олигоцен Миоцен Плиоцен
30
25
---------------------->
Время
Фиг. 117. График изменения со временем двух параметров зубов в филоге-
нетическом древе лошадей [197].
В ряду, приведшем к роду Neohipparion, представители которого питались травой,"отно-
шение высоты паракона (пунктирная кривая) к длине эктолофа (сплошная кривая) рас-
тет быстрее, чем в ряду, приведшем к роду Hypohippus, представители|которого питались
листьями.
17*
260
Часть 2. Применение данных палеонтологии
1
К. phaeinum
К. compressum
3 -
4
< <-<>
К. aculeatum К- ornatum
-1300
-900
К. grossouvrei
К. castor
К. grossouvrei
К. obductum
К obductum
К. castor
К. gulielmii
К. gulielmii
К. jason
К. medea
К nodosum
800
-700
-100
-0
Расстояние от подошвы, см
Фиг. 118.'Почти’непрерывные эволюционные ряды аммонитов рода Kosmoce-
ras (оксфордские глины, верхняя юра, Питерборо, Англия) (из [234] по ] 18]).
на фиг. 117; здесь отложены два параметра зубов лошадей.
Ясно, что два эволюционных ряда различались по темпу
эволюции.
Бринкман [18] изучал изменение двух признаков у аммо-
нитов рода Kosmoceras (юрские отложения Англии). Он собрал
окаменелости из пачки глин мощностью 13 м, строго фиксируя
положение каждого экземпляра от подошвы пачки. Он выделил
четыре основные филетические последовательности (или четыре
эволюционных ряда). Их деление на виды несколько спорно
(фиг. 118). Теперь считают, что боковые ушки (выросты около
устья раковин в рядах 3 и 4) — это половые признаки, присущие,
видимо, самкам. Поэтому можно думать, что вместо четырех
здесь имеется на самом деле два эволюционных ряда — живот-
ные рядов 3 и 4 являются просто самками форм 1 и 2.
Бринкман обнаружил филетическое увеличение отношения
числа ребер к числу краевых шипов. При построении кривых,
описывающих это отношение для экземпляров одного ряда на
X. Эволюция и палеонтологическая летопись
261
Фиг. 119. Графики, отражающие изменение морфологии в одном из рядов
Kosmoceras, изображенных на фиг. 118 (из [234] по [18]).
А. Изменение отношения числа ребер к числу краевых шипов в зависимости от страти-
графического положения ископаемых. Б. Тот же график, разнесенный таким образом,
чтобы показать перерыв в осадконакоплении.
расстоянии 1093,5 см от подошвы пачки, был отмечен скачок
(фиг. 119, Л), который, по мнению Бринкмана, связан с пере-
рывом в осадконакоплении. Действительно, именно на высоте
1093,5 см от подошвы пачки в глинах расположен тонкий слой,
обогащенный обломками раковин аммонитов. Во время пере-
рыва тонкозернистые осадки вымывались, а раковины раздроб-
лялись течениями. Бринкман раздвинул обе части графика на
такое расстояние, чтобы их можно было соединить прямой ли-
нией (фиг. 119, Б), и рассчитал, что длительность перерыва
была достаточной для отложения слоя осадка толщиной около
80 см (если принять, что скорость осадкообразования была по-
стоянной) .
262
Часть 2. П рименение данных палеонтологии
Возраст, млн. лет
Фиг. 120.
А. Степень развития новых признаков у ископаемых родов двоякодышащих рыб, выра-
женная условными средними баллами степени развития многих (26) признаков (крести-
ки — баллы Уэстолла; кружки — отклоняющиеся роды) (из [197], данные из [232]).
Б. Скорость эволюции ископаемых двоякодышащих рыб. Показана зависимость между
первой производной (наклон кривой графика А) каждой точки кривой и временем.
Дальнейшие исследования подтвердили выводы Бринкмана,
показав, что найденные им филетические изменения имели ме-
сто на большом пространстве. Нектонный образ жизни большин-
ства видов аммонитов способствовал их широкому расселению,
и их захоронение в осадках (после смерти их раковины опу-
скались на дно) не зависело от местных условий на дне (кроме
тех условий, которые могли влиять на сохранение). Работа
Бринкмана важна в том отношении, что в ней документально
показаны морфологические и таксономические филетические
изменения на видовом уровне.
Лерман [119] полагает, что тринадцатиметровая пачка глин,
изученная Бринкманом, накапливалась в течение примерно
X. Эволюция и палеонтологическая летопись „, 263
1,5 млн. лет. Однако его заключение основано на сравнении с
Известными мощностями юрских аммонитовых зон и обычными
оценками продолжительности их образований, а эти Оценкй
могут быть ошибочными.
Часто бывает желательно сравнить скорости изменения раз-
ных признаков в одном эволюционном ряду, что позволяет оце-
нить среднюю скорость изменения всего морфологического об-
лика организма. Уэстолл [232] применил интересный метод
(хотя и субъективный, но весьма полезный) для исследования
ископаемой двоякодышащей рыбы. Он выбрал 26 признаков
скелета ископаемых родов и, сравнив их с соответствующими
признаками самой древней и примитивной двоякодышащей ры-
бы и с признаками самых молодых специализированных форм,
обозначил степень развития каждого признака условным бал-
лом. Общий балл рода — сумма баллов отдельных признаков.
Симпсон [197] модифицировал систему баллов и построил гра-
фики, показанные на фиг. 120. Ясно, что в ранний период своей
истории двоякодышащие развивались быстро, но с позднего па-
леозоя изменялись довольно слабо.
ФИЛОГЕНЕТИЧЕСКАЯ СКОРОСТЬ,
ИЗМЕРЯЕМАЯ ПО ТАКСОНОМИЧЕСКИМ КРИТЕРИЯМ
Используя для измерения филогенетической скорости эво-
люции таксономические критерии, мы снова встречаемся с не-
которыми затруднениями, связанными с использованием мор-
фологических критериев; дело в том, что часто бывает нелегко
распознать настоящие эволюционные ряды и трудно определить
абсолютную продолжительность эволюции.
Филогенетические изменения в рядах аммонитов Kosmoceras,
изученных Бринкманом, можно измерить не только по морфо-
логическим, но и по таксономическим критериям. Работая на
высших таксономических уровнях со сложными рядами близко-
родственных таксонов, обычно удается сравнительно точно оце-
нить темп эволюции в абсолютных единицах времени. На
фиг. 121 показаны построенные Симпсоном графики продолжи-
тельности существования родов двустворчатых моллюсков и
плотоядных наземных млекопитающих. Симпсон полагает, что
различия, выявляемые на этих графиках, могут быть несколько
преувеличены, но все же средняя продолжительность существо-
вания родов хищников (около 8 млн. лет) гораздо меньше про-
должительности существования родов двустворчатых моллюсков
(около 78 млн. лет). Это различие говорит о более быстрой
(в среднем) эволюции хищников.
Один из самых интересных моментов в изучении эволю-
ции— выяснение скорости видообразования. Продолжитель-
264
Часть 2. Применение данных палеонтологии
ность существования современных и недавно вымерших видов
можно приближенно оценить с помощью данных о геологиче-
ской древности рецентных видов или на основании того, какая
Фиг. 121. Продолжительность существования ныне живущих и вымерших
родов [197].
Кривые представляют собой кумулятивные графики (точки соответствуют относительно-
му числу родов, в процентах, время существования которых равно или превышает вре-
мя, указанное на горизонтальной оси). Сплошные линии — вымершие роды, пунктирные
линии — современные роды. А. Двустворчатые моллюски (средняя продолжительность
существования с учетом как современных, так и вымерших родов составляет 78,0 мли.
лет).’ Б. Плотоядные наземные млекопитающие (средняя продолжительность существова-
ния с учетом как современных, так и вымерших родов составляет 8,1 млн. лет).
X. Эволюция и палеонтологическая летопись
265
Фир. 122. Процентное содержание современных видов моллюсков в олиго-
ценовых и неогеновых отложениях Венесуэлы [123, 226, 227].
доля вымерших видов, живших в течение какого-то геологиче-
ского периода, уцелела до нашего времени. В обоих случаях
мы имеем дело с продолжительностью существования отдель-
ных видов (тех, которые дожили до наших дней). Многие виды
растений и беспозвоночных миоцена существуют и сейчас, но
из миоценовых или плиоценовых видов млекопитающих лишь
немногие дожили до современной эпохи. Данные Симпсона по
наземным хищникам и двустворчатым моллюскам (фиг. 121)
приводят к заключению, что в кайнозое млекопитающие разви-
вались быстрее беспозвоночных. Как указано в гл. V, скорости
географического и филетического видообразования зависят
от факторов среды, а также от репродуктивных и генетических
особенностей, присущих развивающимся популяциям. Посколь-
ку вымирание той или иной группы должно освобождать эколо-
гические ниши, которые она занимала, можно полагать, что
после крупных вымираний видообразование должно было уско-
ряться. Наступание ледника в плейстоцене и быстрое изменение
условий среды могли повлиять на скорости видообразования
многих таксономических групп кайнозоя; поэтому интерпретиро-
вать данные о скоростях эволюции таксонов, существовавших
в плейстоцене, следует с осторожностью.
Как сказано в гл. I, Симпсон [196] рассчитал, что средняя
продолжительность существования вида животных составляет
примерно 2,75 млн. лет. Эта величина, по мнению Симпсона,
может колебаться от 0,5 до 5 млн. лет. Маркс [123] и Вейсборд
[226, 227] показали, что многие современные виды моллюсков
266 Часть 2. Применение данных палеонтологии
из Венесуэлы существуют 10 млн. лет и более, а некоторые
виды — даже по 20—30 млн. лет (фиг. 122). Продолжитель-
ность существования многих ископаемых видов, как полагают,
значительно превышает симпсоновскую среднюю оценку.
ЧАСТОТА ПОЯВЛЕНИЯ НОВЫХ ТАКСОНОВ
Скорость эволюции очень часто измеряется по частоте по-
явления новых таксонов. Мы уже приводили график числа но-
вых родов и семейств за 1 млн. лет для склерактиний (шести-
лучевые кораллы) (фиг. 114). На фиг. 123, А показан график
числа родов склерактиний в разные эпохи мезозоя и кайнозоя;
на нем выделены новые роды. По этому графику можно рас-
считать скорость эволюции. Та же информация представлена
в другом виде на фиг. 123, Б, где отдельно показаны появив-
шиеся и вымершие роды. Эти две кривые очень сходны; они
имеют по три пика, означающие в каждом случае три поворота
в эволюции склерактиний. Возможны графики других типов.
Выбор типа графика зависит от того, на что надо обратить осо-
бое внимание — на скорость увеличения разнообразия или на
скорость вымирания.
Склерактинии делятся на две экологические группы — гер-
матипные и агерматипные формы (стр. 215); поэтому графики,
приведенные на фиг. 114 и 123, представляют собой упрощение.
Многие семейства состоят только из герматипных или только
агерматипных родов, но есть и смешанные семейства. По-ви-
димому, эволюция герматипных родов должна отражать изме-
нения климата или других условий среды, влиявших на рост
рифов, тогда как эволюция агерматипных кораллов, менее ог-
раниченных экологически, должна быть в целом менее зависи-
ма от изменений среды.
На фиг. 124 приведены графики, характеризующие скорости
эволюции родов герматипных и агерматипных кораллов. Гер-
матипные кораллы появились раньше агерматипных, и число
герматипных родов сильно увеличилось задолго до того, как
стало повышаться разнообразие агерматипных групп. Действи-
тельно, пик в позднемеловую эпоху на фиг. 123 обусловлен поч-
ти исключительно развитием герматипных родов.
Если отвлечься от кажущегося замедления эволюции в па-
леоцене, то число агерматипных родов с момента их появления
и до наших дней постоянно возрастало (фиг. 124, Л). График
для герматипных кораллов далеко не столь монотонен, здесь
имеются три пика разнообразия, совпадающие с пиками на
фиг. 123. Стратиграфическая летопись свидетельствует о том,
что первые два пика совпадают с периодами интенсивного рифо-
образования, от которых осталось много окаменелостей и кото-
Фиг. 193 о
Число родов
S, _ М 4s Ст> со о <\э 4s о»
’“OOCjOOOOcsQ
I I I I I I I I I I I I I I [“т *г
Ранний триас
Средний триас
Поздний триас
Ранняя юра
Средняя юра
Поздняя юра
Ранний мел
Поздний мел
Палеоцен
Эоцен
Олигоцен
Миоцен
Плиоцен
Плейстоцен
Современная
эпоха
Фиг. 124. Развитие родов внутри двух крупных экологических групп Sclera-
/'tinio Г99О1
X. Эволюция и палеонтологическая летопись
269
рые за короткий срок привели к увеличению разнообразия ко-
раллов.
На фиг. 123 доминирующие тенденции эволюции герматип-
ных Scleractinia сильно затемняют ход развития агерматипных
родов, хотя число герматипных родов превосходит число агер-
матипных менее чем в два раза. Конечно, для интерпретации
частот появления новых таксонов в крупных группах животных
очень важен выбор включаемых в эти группы таксономических
подразделений.
При интерпретации графиков следует учитывать также не-
полноту и искаженность палеонтологической летописи. Практи-
ческое отсутствие данных о сроках появления и вымирания ро-
дов в палеоцене, очевидно, связано со скудостью морской оса-
дочной летописи этой очень непродолжительной эпохи. Вероят-
но, некоторые вымирания, отнесенные к позднему мелу, и
появления новых таксонов, отнесенные к эоцену, на самом деле
имели место в палеоцене. Другое любопытное явление — кажу-
щееся увеличение числа родов кораллов после плейстоцена
(фиг. 124), а дело здесь главным образом в том, что рецентные
фауны известны нам лучше, чем плейстоценовые. Многие из
вымерших герматипных кораллов плейстоцена, найденных в ри-
фах, подвергавшихся действию прибоя, очень плохо сохранились
и не поддаются определению. Многие агерматипные роды при-
урочены к глубоководным районам, и потому большая доля
плейстоценовых родов может находиться в отложениях, недо-
ступных палеонтологу. Наконец, не исключено, что немалое чис-
ло родов, относящихся к обеим экологическим группам, хорошо
сохранилось, и притом в доступных для исследования областях,
но они пока не открыты.
Филетические тенденции
Тенденцией (trend) мы назовем направление адаптивных
измененйй в эволюционном ряду. При изучении тенденций проб-
лема выделения истинных. эволюционных рядов стоит не так
остро, как при изучении скоростей эволюции, поскольку во
многих случаях тенденции можно определить даже по довольно
отрывочным палеонтологическим данным.
ОРТОГЕНЕЗ
В прошлом некоторые исследователи выдвигали концепцию
ортогенеза, т. е. филетической эволюции, идущей в некоем по-
стоянном направлении. Цногда говорят, что ортогенез—.это
.270
Часть 2. Применение данных палеонтологии
прямолинейная эволюция. Примером может служить постоян-
ное увеличение размеров в ходе эволюции. Некоторые исследо-
ватели считали, что ортогенетическая эволюция, однажды на-
чавшись, продолжает идти дальше в одном и том же направле-
нии, даже если соответствующее изменение перестало быть
приспособительным и может в конце концов привести к выми-
ранию ряда. Эта идея несовместима с концепцией приспособи-
тельных изменений под действием естественного отбора.
Классический пример, часто цитировавшийся сторонниками
теории ортогенеза, — эволюция рода Megaloceras, к которому
относился гигантский олень, живший в плиоцене и плейстоцене.
Рога этого животного были огромны даже для его громадного
тела (фиг. 125). Рога ежегодно сбрасывались и вырастали
снова, что требовало больших затрат энергии. В ходе эволюции
рода размер тела возрастал, но размер рогов возрастал гораздо
’ИГ/ 125. Megaloceras—гигантский ископаемый олень плейстоцена; размах
рогов около 2,5 м [167].
X. Эволюция и палеонтологическая летопись
271
быстрее. Ранее предполагали, что тенденция к увеличению рогов
снижала приспособленность и привела к вымиранию рода. Джу-
лиан Гексли [86] показал, что увеличение относительного раз-
мера рогов в процессе онтогенеза характерно и для современ-
ного благородного оленя. Поскольку это увеличение в общем
аллометрично (фиг. 126), Гексли предположил, что увеличение
размера рогов в эволюции гигантского оленя могло происхо-
дить подобным же образом. Если размер тела и размер рогов
были в эволюции генетически связаны, то давление отбора в на-
правлении увеличения размера тела одновременно могло при-
водить и к увеличению размера рогов, которое могло быть либо
полезным, либо вредным. Если в какой-то момент увеличение
размера рогов стало вредным, то эволюция в направлении уве-
личения рогов могла прекратиться лишь после того, как давле-
ние отбора, направленное на увеличение размеров тела, урав-
новесилось давлением отбора против увеличения рогов. Нет
никаких данных, которые заставили бы предположить, что под
влиянием некоего таинственного «ортогенетического механиз-
ма» комплекс тело — рога перешагнул за это равновесие и даль-
нейшее увеличение размеров рогов стало отрицательно сказы-
ваться на приспособленности. Но Гексли подчеркивает следую-
щее: если размер организма, например Megaloceras, увеличи-
вается до того, что оказывается на грани допустимого, то это
может привести к слишком сильной специализации и любое
272
Часть 2. Применение данных палеонтологии
изменение условий среды нарушит баланс и приведет к выми-
ранию таксона. В быстро сменявшихся климатических зонах и
биотических районах плейстоцена род Megaloceras мог погиб-
нуть именно таким образом.
Предполагают также, что аллометрический рост рогов у Ме-
yaloceras приводил к их увеличению до гигантских размеров
только у старых животных; таким образом, размеры рогов мог-
1И оказаться вредными у старых самцов, которые уже не уча-
ствовали в размножении из-за физиологического старения или
юражений в боях с молодыми самцами. Тогда отрицательное
гриспособительное значение рогов могло никак не сказываться
та эволюции.
Некоторые исследователи предполагали, что завернутость
»аковины грифеидных устриц возрастала до тех пор, пока это
те стало препятствием для их открывания. Это и привело к вы-
тиранию грифеид. Огромные размеры клыков саблезубых кошек
айнозоя также выдвигались в качестве причины вымирания
тих животных. Против этих гипотез можно привести те же воз-
ажения, что и против гипотезы об «антиприспособительном»
арактере рогов гигантского оленя.
Сторонники «смягченной версии теории ортогенеза» просто
читают «прямолинейную эволюцию» общим механизмом эво-
юционных изменений, не упоминая о внешних движущих силах
золюции и об «антиприспособительных» тенденциях. Многие
сследователи (самый выдающийся из них — Дж. Симпсон)
чровергли эту форму концецции ортогенеза. В начале XX века
ногие считали, что развитие Современного рода Equus из эоце-
эвой «ранней лошади» Hyracotherium (бывший Eohippus) шло
эямым путем. Однако Симпсон показал, что такие процессы,
1к увеличение размера тела и высоты зубов (фиг. 117), умень-
ение числа пальцев, действительно имевшие место в общей
юлюции лошадей, не составляли единой филетической тенден-
ш. Действительно, изменения происходили спорадически и
разными скоростями в разных рядах сложного разветвляю-
егося древа. Даже древо, показанное на фиг. 49, упрощено,
>тя все множество рядов действительно делится на две боль-
ие группы, различающиеся по эволюции зубов. Ранние лошади
[тались мягкими листьями, и строение их зубов было приспо-
блено к такому способу питания. Но затем, по-видимому, в ре-
льтате распространения жесткой, богатой кремнеземом тра-
нистой растительности (в начале кайнозоя) в некоторых ря-
х развились более сложные коренные зубы, пригодные для
ретирания таких растений. Увеличилась сложность рисунка
розд на коронках коренных зубов, увеличилась высота коро-
к, появился цемент. Лошадь, питающаяся травой, была од-
м из многих новых травоядных родов, развивавшихся в се-
X. Эволюция и палеонтологическая летопись
273
Ьедине и конце кайнозоя; консервативная, менее удачная фило-
генетическая группа лошадей, питавшихся листьями, существо-
вала до плиоцена.
* Как подчеркнул Симпсон, в палеонтологической летописи
«практически нет примеров прямолинейной эволюции. В самом
деле, любая новая адаптация по определению означает измене-
ние направления хода эволюции.
Рекапитуляция и гетерохрония
В начале XIX столетия Бэр отметил, что некоторые стадии
эмбрионального развития высших животных напоминают стадии
эмбрионального развития низших животных. Так, зародыш че-
ловека последовательно проходит стадии, на которых он внеш-
не напоминает эмбрион рыбы, амфибии, рептилии и низших
Млекопитающих. Эта закономерность стала известна под на-
званием биогенетического закона. Позднее Геккель и другие
ранние эволюционисты расширили эту концепцию, создав так
Называемый закон рекапитуляции. Согласно этому закону, выс-
шие животные проходят стадии, сходные с последовательными
взрослыми стадиями их предков в филогенезе. Эта концепция,
обычно формулируемая так: «Онтогенез есть повторение фило-
генеза», в основном ошибочна. Первоначальное обобщение
Вэра ближе к истине: развитие высших групп часто напоминает
развитие предковых форм. Но при этом обычно в эволюции
.сильно изменяются скорости развития и последовательность его
'Этапов. Для понимания таких изменений полезно представить
себе эволюцию как последовательность онтогенезов (или их от-
резков, в зависимости от того, когда в онтогенезе начинается
размножение).
> На любой стадии онтогенеза могут происходить разного ро-
да эволюционные изменения, не влияющие непосредственно на
доследующие стадии. Например, у личинок морских беспозво-
ночных обычно имеются сложные признаки, развившиеся в зна-
чительной мере независимо от изменений взрослой формы.
В процессе эволюции нередко возникают также глубокие из-
менения взрослых форм, обусловленные изменением в онтоге-
незе предков этих форм. Например, может меняться очередность
развития двух органов или же развитие одного из них может
запаздывать настолько, что он становится рудиментом или сов-
сем исчезает. Эволюционное изменение последовательности раз-
вития органов называется гетерохронией. Обнаружено множест-
во различных случаев гетерохронии, и для каждого есть назва-
ние. Мы не будем рассматривать здесь этот обширный и гро-
моздкий словарь; всех, кто заинтересуется этим вопросом,
отсылаем к прекрасному обзору де Бера [37]. Здесь мы рас-
смотрим лишь несколько примеров.
18—467
274
Часть 2. Применение данных палеонтологии
Фиг. 127. Граптолит Monograptus argentus [20].
У этой формы только образовавшиеся первыми ячейки, или теки, обладают
новой (крючкообразной) формой (внизу).
Стрелки указывают, куда направлено устье теки. В ходе эволюции были
утрачены поздние стадии роста колонии и у некоторых видов крючкооб-
разная форма утвердилась на всех стадиях.
В XVIII и в начале XIX века многие палеонтологи, занимав-
шиеся мезозойскими аммонитами, пытались применять идеи,
близкие геккелевскому принципу рекапитуляции, ко всем видам
аммонитов, полагая, что ход эволюции аммонитов можно вос-
становить по изменениям орнаментации раковины и рисунка
перегородочной линии в онтогенезе. В 1901 г. А. П. Павлов раз-
венчал ортодоксальную формулировку закона рекапитуляции,
показав, что в некоторых юрских рядах аммонитов новые эво-
люционные признаки появлялись на ранних стадиях онтогенеза;
лишь позже в соответствующих рядах эти признаки удержива-
лись у взрослых форм. Другими словами, развитие новых приз-
наков в онтогенезе запаздывало по отношению ко времени на-
ступления половой зрелости.
В некоторых эволюционных рядах граптолитов наблюдается
гетерохрония в образовании колонии (правда, образование ко-
лонии— это не настоящая онтогенетическая последовательность:
ведь колония состоит из множества, особей, каждая из которых
имеет свой онтогенез, начинающийся с очередного почкования).
Булмен [20] отметил, что многие структурные изменения, воз-
никшие в эволюции граптолитов, появились на ранних стадиях
образования колонии. В результате дальнейшей эволюции они
закрепились на поздних стадиях развития колонии (фиг. 127).
X. Эволюция и палеонтологическая летопись
275
III
IV
П
Фиг. 128. Схема, показывающая возможность перехода от Rugosa (четырех-
лучевые кораллы) к Scleractinia (шестилучевые кораллы) [131].
;Прказан вид ячеек сверху. Изображено развитие септ в онтогенезе типичного четырехлу-
.'чёвого коралла (/), типичного шестилучевого коралла с почти идеальной радиальной сим-
Метрией (IV) и кораллов промежуточных форм (II — представитель’Rugosa с радиаль-
ным расположением септ у взрослой формы; III — раннемезозойский представитель
‘Scleractinia с двумя малыми «секстантами»). Шесть первичных септ всюду изображены
Жирными линиями. К — кардинальная септа; А — алярные септы; Пр — противополож-
ная септа; Д — диалярные септы’ (Rugosa); П — первичные септы (Scleractinia). Септы
Второго, третьего и т. д. порядков, расположенные между септами первого порядка, изо-
бражены тонкими линиями. Предполагается, что эволюционный переход от четырехлу-
Чевых к шестилучевым кораллам произошел путем внедрения "септ высших порядков в об-
v.. ластях, которые на рисунке покрыты точками.
Гетерохрония могла играть важную роль в возникновении
некоторых высших таксонов. Шиндевольф и Клауд предполага-
ют, что именно так из палеозойских четырехлучевых кораллов
; (Rugosa), которые, очевидно, вымерли в конце пермского перио-
да, возникли современные шестилучевые кораллы (Scleractinia).
Самые древние шестилучевые кораллы известны из среднего
триаса. Как показано на фиг. 128, септы (вертикальные плас-
тинчатые перегородки) шестилучевых кораллов расположены
почти радиально кругами по 6 или в числе, кратном шести.
У четырехлучевых кораллов септы были расположены
в четырех квадрантах (у взрослых форм). Но такое располо-
жение септ по четыре у взрослых четырехлучевых кораллов
развивалось в онтогенезе из шести септ первого порядка. Воз-
можно, в результате гетерохронии у взрослой формы сохрани-
лись все шесть септ первого порядка, и, таким образом, Rugosa
Дали ^начало шестилучевым кораллам. У некоторых позднепа-
деозойских четырехлучевых и раннемезозойских шестилучевых
48*
276
Часть 2. Применение данных палеонтологии
кораллов наблюдаются промежуточные типы морфологии. И все
же нельзя с уверенностью утверждать, что шестилучевые корал-
лы произошли от четырехлучевых, поскольку их скелеты раз-
личаются не только по морфологии, но и по минералогическому
составу (у шестилучевых скелет построен не из кальцита, а из
арагонита). Возможно, шестилучевые кораллы развились неза-
висимо из бесскелетных актиниеподобных предков.
ПРАВИЛО КОПА
Знаменитый палеонтолог XIX века Э. Коп, работавший пре-
имущественно в области палеонтологии позвоночных, отметил
общую тенденцию к увеличению размеров в ходе филогенеза
в различных таксономических группах. Эту тенденцию нельзя
считать ни ортогенетической, ни всеобщей, но тем не менее она
широко распространена.
Мы уже говорили о тенденции к увеличению размеров, ха-
рактерной для эволюции рогатых динозавров (фиг. 115). В об-
щей картине эволюции лошадей наблюдается та же тенденция,
как явствует из сравнения современного рода Equus и его чет-
вертичных родичей с эоценовой ранней лошадью Hyracotherium,
бывшей величиной примерно с фокстерьера. Увеличение разме-
ров началось только после эоцена, а в некоторых эволюционных
рядах вместо увеличения шло уменьшение. Несколько приме-
ров явного увеличения размеров в филогенезе беспозвоночных
рассмотрел Ньюэлл [133].
Причины увеличения размеров различны в разных эволю-
ционных рядах. Некоторые животные благодаря увеличению
размеров, возможно, становились сильнее и могли нападать на
других животных или успешно защищаться. Теплопродукция
у теплокровных животных пропорциональна объему их тела
(т. е. кубу линейных размеров), а потеря тепла пропорциональ-
на поверхности тела (т. е. квадрату линейных размеров). В ре-
зультате крупное животное лучше сохраняет тепло в холодном
климате, чем мелкое животное. Другое преимущество больших
размеров состоит в том, что, хотя крупное животное бегает
лишь ненамного быстрее мелкого, оно гораздо выносливее и
может дольше бежать с большой скоростью. Мышцы крупного
животного сокращаются медленнее, но зато размах каждого
движения шире (например, шире шаг при беге). Поэтому при-
мерно при одинаковой скорости передвижения крупное живот-
ное на том же расстоянии сделает меньше движений, чем мел-
кое, т. е. его выносливость опять-таки будет больше. Этот фак-
тор мог иметь большое значение в эволюционном увеличении
размеров лошадей, питавшихся травой.
Важно помнить, что с увеличением размеров связаны, и не-
которые затруднения (это вытекает из принципа подобия). На-
X. Эволюция и палеонтологическая летопись
277
пример, увеличение размера травоядных должно было вызвать
затруднения в добывании пищи, так как жевательная поверх-
ность зубов увеличивалась пропорционально квадрату линейных
размеров, а объем живых тканей, для поддержания которых
Нужна пища,— пропорционально кубу линейных размеров. Симп-
сон интерпретирует сложные эволюционные изменения коренных
зубов у лошади (фиг. 117) как адаптации, способствующие ре-
шению этой проблемы. В целом увеличение размеров — само-
ограничивающийся процесс. Человек тридцатиметрового роста
не мог бы ходить и с трудом выдерживал бы собственный вес,
поскольку вес увеличивается пропорционально кубу линейных
размеров, а мышечная сила (которая пропорциональна площа-
ди сечения мышцы) — пропорционально их квадрату. Многие
функции, связанные с различными поверхностями организма, при
увеличении общих размеров также становятся менее эффектив-
ными. Например, потребность в пище растет вместе с объемом
тканей, который пропорционален кубу линейных размеров, тогда
как добывание пищи у таких организмов, как брахиоподы, мшан-
ки, двустворчатые моллюски, зависит от снабженных ресничками
Поверхностей, а их эффективность пропорциональна лишь квад-
рату размеров. Потребность в кислороде тоже возрастает
пропорционально кубу линейных размеров, тогда как погло-
щение кислорода идет на двумерных поверхностях жабр или
легких.
Можно взглянуть на правило Копа и по-другому: ведь здесь
можно говорить не о тенденции к увеличению размеров в про-
цессе эволюции, а об эволюции от малых размеров. Хорошо
известно, что многие высшие таксоны развились из групп орга-
низмов, обладавших сравнительно малыми размерами. Во мно-
гом это объясняется тем, что предковые группы обычно бывают
относительно неспециализированными, а мелкие особи гораздо
чаще бывают неспециализированными, чем крупные. В боль-
шинстве случаев у видов, для которых характерны крупные
размеры, возникло слишком много специальных адаптаций, и
они вряд ли смогут дать начало большой новой группе с на-
столько важными отличиями, чтобы ее пришлось выделить в
новый высший таксон. Например, очень маловероятно, чтобы
слоны могли дать начало совершенно новому отряду четверо-
ногих. Присущие слонам особые признаки, в том числе сильно
утолщенные конечности, поддерживающие массивное тело, уко-
роченная шея, несущая голову, и хобот, заменяющий длинную
подвижную шею [162], значительно ограничивают их эволюци-
онный потенциал. Новые группы обычно возникают не от круп-
ных специализированных предков, а от мелких неспециализиро-
ванных, и, может быть, именно поэтому правило Копа адекват-
но отражает действительность.
278
Часть 2. Применение данных палеонтологии
ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ МОРФОЛОГИЧЕСКИХ
ТЕНДЕНЦИЙ
Приспособительное значение многих морфологических тен-
денций, отмечаемых в палеонтологической летописи, остается
загадкой. В качестве примера можно привести изменение
скульптуры раковины, обнаруженное Бринкманом в эволюци-
онном ряду Kosmoceras (стр. 260). Беспозвоночные очень мно-
гочисленны в морских местообитаниях, и поэтому они чаще
позвоночных оставляют в палеонтологической летописи почти
непрерывную запись филетических изменений. Один из наибо-
лее подробно исследованных ископаемых рядов — род морских
ежей Micraster из меловых пластов «мела» Англии. Первым
филетические тенденции Micraster выявил палеонтолог-люби-
тель Роув [170]. Позже Кермак [100] подтвердил с помощью
биометрических исследований основные наблюдения Роува, но
отверг некоторые детали его работы. Совсем недавно Никольс
[137, 138] проанализировал приспособительное значение най-
денных у Micraster морфологических тенденций. Совокупность
работ этих палеонтологов — классический пример исследования
эволюции.
Echinocardiutn — неправильный морской еж из группы мор-
ских ежей, сменивших исходную радиальную симметрию на би-
латеральную. Это изменение связано с однонаправленным пере-
движением животного по осадку или в его толще. Типичный
современный роющий вид показан на фиг. 129. Он находится
в незамкнутой норке и движется вперед, прорывая ход в осадке
своими подвижными иглами, находящимися главным образом
на брюшной поверхности. Связь с придонной водой поддержи-
вается через вертикальную «воронку», проделанную особыми
дыхательными амбулакральными ножками. Животное питается
осадком, богатым органическим веществом, собирая этот оса-
док специальными амбулакральными ножками, окружающими
рот. Некоторое количество пищи попадает в рот и с дорсальной
поверхности, по передней борозде (фиг. 130). Иглы и амбула-
кральные ножки, окружающие анус, образуют задний санитар-
ный канал, куда животное по мере своего продвижения выделя-
ет экскременты.
На фиг. 130 показана морфология трех видов позднемелово-
го рода Micraster и трех современных неправильных морских
ежей. Очень редко встречаются ископаемые морские ежи, у ко-
торых сохранились иглы, но о морфологии и расположении игл
можно многое узнать, изучая шишковидные бугорки в местах
прикрепления игл, так называемые туберкулы. Почти у всех
видов сердцевидных морских ежей некоторые участки панциря
несут ряды туберкул, к которым при жизни прикрепляются иг-
X. Эволюция и палеонтологическая летопись
279
◄-------------------
Направление движения
Фиг. 129. Еchinocardium — типичный современный зарывающийся непра-
вильный морской еж [137].
лы, густо усаженные ресничками. Эти области называются
фасциолами. Реснички внутренней фасциолы предназначены
в основном для втягивания воды в дыхательную воронку. Рес-
нички анальной фасциолы создают ток воды, относящий экскре-
менты в сторону от тела и вниз, а реснички субанальной фас-
циолы помогают другим ресничкам тела гнать воду с продукта-
ми выделения в санитарный канал.
На фиг. 131 показана предполагаемая глубина зарывания
видов Micraster. Рассматриваемые виды соединены промежу-
точными формами и выделены условно. Предполагается, что
исходный вид, М. leskei, зарывался неглубоко. Он дал начало
ряду форм, среди которых были и глубоко, и неглубоко зака-
пывавшиеся. Эту группу форм произвольно делят на два вида—
М. corbovis и М. cortestudinarium. Только представители вто-
рого вида дожили до сенона и дали начало эволюционному ря-
Позднемеловые неправильные морские ежи
Micraster (Isomicraster)
senonensis
Micraster corbovis Micraster coranguinum
Неглубокая
Глубокая
’’Гладкая”
„Разделенный ’
\ / петалоид
Гомогенные
петалоиды
Нет губы
Неглубокий петалоид
Глубокий петалоид
Желобок
широкая
фасциопа
Узкая'фасциола
Высшая
точка
Нет желобков
Выражен-
7 Гладкая
зона
Гпанулированная
зона Г "
Hem фасциолы
Нет заднего
возвышения
Высшая
точка
Заднее
возвышение
Панцирь
высокий
Анус расположен
высоко
Анус расположен
низко
Фиг. 130. Морфология и образ жизни современны
Рецептные неправильные морские ежи
it
Echinocardfum cordatum
Spatangus purpureas
Spatangus raschi
3***!
Амбитальные
чувствительные амбу-
лакральные ножки
Центрально
Оральные амбулакраль-
ные ножки рот
Перипластрональ^^. .
ные амбулакраль- Суоанальные
ные ножки ножки
Пластрон
ножки
Анус
Губа
ЛаМерально
Анрльная
фасциола
ископаемых неправильных морских ежей [138]
Субанальная
фасциола
282
Часть 2. Применение данных палеонтологии
ду, составляющему вид М. coranguinum. Считается, что неглу-
боко закапывающийся вид М. (Isomicraster) senonensis по-
явился в районе исследования в раннем сеноне, развился же
он в географически изолированной области и поэтому долгое
время не имел связи с основной популяцией. Он, видимо, не
достиг той степени генетической изоляции, которая препятство-
вала бы скрещиванию с М. coranguinum (хотя палеонтологи
рассматривают их как два отдельных вида); найдены промежу-
точные формы, свидетельствующие о скрещивании между этими
двумя группами морских ежей.
По гомологии с рецентными формами и на основании адап-
тивной морфологии ископаемых Micraster Никольс сделал выво-
ды об их образе жизни. Предполагаемая глубина зарывания
трех видов Micraster и известная глубина зарывания трех со-
временных ежей показаны на фиг. 130. Изучив некоторые при-
способительные особенности, развившиеся в ряду М. cortestudi-
narium — М. coranguinum, Никольс выявил тенденцию к уве-
личению глубины зарывания. Ниже мы перечислим эти особен-
ности (фиг. 130).
1. Расширение панциря, при котором самая широкая и вы-
сокая область сдвигается назад. Это изменение, очевидно, свя-
зано с приобретением обтекаемой формы и с отсутствием необ-
ходимости сбрасывать осадок с дорсальной поверхности (по-
следнее важно для животных, которые лишь частично зарыва-
ются в донный осадок).
2. Усложнение скульптуры в области дыхательных амбула-
кральных ножек. Видимо, это должно увеличивать площадь рес-
ничной зоны, что способствует созданию сильного тока воды
через длинную дыхательную «воронку».
Фиг. 131. Изменение образа жизни в процессе эволюции Micraster (верхний
мел, Англия) [138].
X. Эволюция и палеонтологическая летопись 283
3. Увеличение числа туберкул на брюшной поверхности, что,
?-очевидно, приводит к увеличению площади ресничной зоны.
При движении животного реснички гонят тонкозернистую фрак-
. цию осадка назад.
4. Расширение субанальной фасциолы, благодаря чему улуч-
шается очистка глубокой норки от отходов жизнедеятельности.
-На панцире М. (Isomicraster) senonensis, который зарывался
неглубоко и как бы «пахал» дно, нет субанальной фасциолы.
В большинстве случаев эволюционные тенденции, отмечае-
. мые в палеонтологической летописи и интерпретируемые с точ-
\ ки зрения функциональной морфологии, выражены слабее и до-
• кументированы хуже, чем только что рассмотренные тенденции
у Mier aster. Например, главные тенденции в эволюции лошадей
“выделены на основании разорванной палеонтологической лето-
писи. Выделение и интерпретация непрерывных или почти не-
j прерывных ископаемых рядов — главная цель палеонтологиче-
ского исследования. Однако эта цель редко достигается.
Пути эволюции
Обсуждая филетические тенденции, мы занимались отдель-
• ными рядами или филогенией этих неполных рядов. Существо-
вание определенных путей эволюции можно установить, срав-
нивая темпы эволюции или ее тенденции в двух или нескольких
рядах.
Эволюционная дивергенция
’ Для эволюционных изменений характерен не ортогенез, а по-
вышение разнообразия и дивергенция. Разнообразие растет
в первую очередь благодаря географическому видообразованию,
в результате которого один ряд разделяется на два или на
несколько рядов. Дивергенция — это увеличение различий меж-
Ду двумя или несколькими рядами; о дивергенции можно гово-
рить в том случае, если эти ряды имеют общее происхождение.
Для анализа характера ветвления можно выбрать любой таксо-
номический уровень, любую таксономическую единицу. Чем
^мельче единица, тем сложнее характер ветвления, а самое вы-
сокое таксономическое разрешение достигается при анализе на
Уровне вида. Любое самое сложное филогенетическое древо
-Можно в конце концов свести к последовательности отдельных
юнтогенезов (фиг. 47).
Крупномасштабное увеличение разнообразия и дивергенцию
эволюционных рядов называют адаптивной радиацией. Адаптив-
ная радиация обычно происходит непосредственно вслед за
Эволюционным «прорывом» — приобретением новых адапта-
284
Часть 2. Применение данных палеонтологии
Время, млн.лет
Фиг. 132. Скорости эволюции надсемейств и родов двустворчатых моллюс-
ков, питающихся через сифон (сравните с фиг. 114) [207].
I — надсемейства, II — роды.
ций — и приводит к образованию новой, высшей категории.
В качестве примера приведем некоторых двустворчатых мол-
люсков [207]. Огромное большинство палеозойских родов от-
носилось к семействам — представителям эпифауны и примитив-
ной инфауны. В мезозое и кайнозое появляется много таксонов,
представители которых вели роющий образ жизни. В резуль-
тате их дифференциации широко распространились зарываю-
щиеся двустворчатые моллюски, столь обильные в современных
морях. Сходство по таким фундаментальным признакам, как
микроструктура раковин, механизм замка, строение жабр и си-
фонов, свидетельствует о том, что большинство послепалеозой-
ских роющих таксонов вполне могло возникнуть от общих пред-
ков путем радиации. Большинство этих таксонов входит в под-
класс Heterodonta. Причина адаптивной радиации разнозубых
двустворчатых моллюсков состоит в том, что они совершили
крупный адаптивный прорыв: края мясистой мантии, выстилаю-
щей створки раковины, у них слились, образовав сифон; появи-
лись другие признаки, связанные с сифоном. Многие из новых
двустворчатых моллюсков роют значительно быстрее, чем при-
митивные группы. Многие способны жить в более глубоких сло-
ях осадка и потому лучше защищены (что особенно важно на
литорали). По сравнению со всеми остальными современными
морскими таксонами разнозубые Bivalvia, питающиеся с по-
X. Эволюция и палеонтологическая летопись
285
Современная
эпоха
Кайнозой
Мея
Юра
Триас
Пермь
Карбон
Девон
Силур
Ордовик
Фиг. 133. Схема геохронологического распространения основных групп
рыб [32].
мощью сифона, сильно преобладают в таких нишах, где можно
вести роющий образ жизни и питаться взвешенным веществом.
В возникновении и дифференциации основных высших таксо-
нов большую роль сыграли также многие другие изменения,
возникшие в процессе эволюции. Наиболее замечательные из
них — покрытое скорлупой амниотическое яйцо рептилий, бла-
годаря которому ранние рептилии перестали быть связанными
в своем размножении с водой, что позволило им занять в позд-
нем палеозое сухопутные местообитания, непригодные для их
предшественников — амфибий; размножение голосеменных (сос-
на, ель и т. д.) и покрытосеменных (цветковых) растений с по-
мощью пыльцы и семян, что избавило эти организмы от зави-
симости при оплодотворении от водной среды и позволило им
заселить многие наземные местообитания, недоступные для их
бессемейных предшественников; противопоставление большого
пальца и стереоскопическое зрение, давшие ранним приматам
возможность производить разнообразные действия руками и из-
готовлять орудия, что в свою очередь способствовало развитию
крупного, сложного мозга.
Адаптивная радиация обычно идет определенными путями.
Высшие таксономические категории, входящие в новую группу,
286
Часть 2. Применение данных палеонтологии
Фиг. 134, Представители трех крупных групп костных рыб, перечисленных
на фиг. 133 [32].
Эволюция шла в направлении развития все. более, тонкой чешуи и более симметричного
хвоста. Тазовый плавник сдвигался вперед, челюсти укорачивались.
А. Рalaeoniscus (палеониски, пермь). Б. Pholidophorus (костные ганоиды, юра). В. Ыиреа
(костистые рыбы, кайнозой).
появляются сравнительно рано, а скорость их появления -в про-
цессе радиации уменьшается. Низшие таксономические катего-
рии в ходе радиации, напротив, появляются все быстрее. Это
положение для группы питающихся с помощью сифона высоко-
развитых двустворчатых моллюсков иллюстрируется на фиг. 132.
То же наблюдалось у шестилучевых кораллов, относящихся
к Scleractinia (фиг. 114).
Такая закономерность отражает тот факт, что новые приспо-
собительные возможности, появляющиеся у группы вместе с но-
выми признаками, используются группой очень быстро. После
того как образ жизни группы в основном определится, проис-
ходит все более дробная дифференциация низших таксонов.
В любое время ветвление может изменить свой характер или
же — в случае вымирания — совершенно прекратиться.
Известны случаи, когда в геологическом прошлом одна
группа получала столь важное эволюционное преимущество,
X. Эволюция и палеонтологическая летопись
287
Фиг. 135. Скорости появления (число соответствующих таксонов за 1 млн.
лет) отрядов, семейств и родов млекопитающих [197].
/ — отряды; II — семейства; III — роды.
что побеждала другую группу со сходным образом жизни и
близкими экологическими предпочтениями, в результате чего
эта вторая группа вымирала. Это явление называется экологи-
ческим вытеснением. О таком вытеснении можно судить только
по сосуществованию двух групп во времени и последующему
постепенному исчезновению той группы, которую можно считать
менее приспособленной. Примеры эволюционного вытеснения
известны в геологической истории рыб.
Примитивные щитковые, бывшие главным образом мусорщи-
ками или грунтоедами, в девоне, во время развития пластино-
кожих, постепенно исчезли (фиг. 133). Пластинокожие были
первыми челюстными позвоночными. Их челюсти послужили тем
новым адаптивным приобретением, которое и обусловило их ра-
диацию в девоне и, возможно, привело к тому, что они вытес-
нили многих щитковых. Пластинокожие и сохранившиеся
бесчелюстные в позднем палеозое уступили место акулоподоб-
ным рыбам с хрящевым скелетом и костным рыбам. По-видимо-
му, основное преимущество этих двух новых групп состоит в
288
Часть 2. Применение данных палеонтологии
Нижняя юра I Средняя юра | ' Верхняя юра
Батский
Байосский.
Тоарский
Ярусы Lytoceratina
Ammonitina
Кимериджский
Оксфордский
Келловейский
Плинсбахский
Синемюрский
Геттангский
Пурбекский
3
( Ьс
-----------
j
Портлендский^
Фиг. 136.^Итеративная эволюция семейств юрских аммонитов,
Окончание -асеае присваивается надсемействам
наличии хорошо развитых плавников, благодаря которым эти
рыбы плавали гораздо лучше своих предшественников.
Последующая адаптивная радиация костных рыб прошла
через три главных этапа; на фиг. 134 показаны особи, относя-
щиеся к трем таксономическим группам, поочередно домини-
ровавшим в прошлом. Каждая группа имела адаптивные пре-
X. Эволюция и палеонтологическая летопись
289
произошедших от подотрядов Phylloceratina и Eytoceratina [4].
окончание, -idae — семействам. -
имущества перед предыдущей. Эти преимущества достигались
главным образом за счет изменения чешуи, челюстей, хвоста,
плавников и преобразования примитивных легких в плаватель-
ный пузырь.
Новой группе организмов нелегко занять ниши, уже заня-
тые хорошо приспособленными членами другой группы, даже
19—467
'290
Часть 2. Применение данных палеонтологии
О. dentatus posterior
♦
О. wahlenbergi
4
О. gibbosus posterior
4
Исходная
линия
фиг. 137. Итеративная эволюция формы пигидия (хвостовой области) у кемб-
рийского рода трилобитов Olenus (из [197], по [98]).
Обратите внимание на то, что в четырех отделившихся рядах происходили сходные мор-
фологические изменения.
если у этой новой группы имеются потенциальные возможности
к развитию более эффективных и разнообразных адаптаций,
чем у старой группы. Первые представители новой группы мо-
гут обладать рудиментарными признаками, обладающими боль-
шим эволюционным потенциалом, но в своем исходном состоя-
нии эти признаки могут уступать развитым признакам конку-
рирующей группы. Однако разнообразие новой группы может
увеличиться в области, географически изолированной от той, ко-
торую занимает старая группа, и только впоследствии, когда
^адаптивное превосходство уже достигнуто, новая группа может
X. Эволюция и палеонтологическая летопись
291
распространиться в ареале старой группы. Правда, если старая
группа широко распространена, то такой поворот событий ма-
ловероятен. В мезозое дифференциации ранних млекопитаю-
щих — это были мелкие и неспециализированные животные —
мешало господство рептилий, особенно динозавров, в наземных
местообитаниях по всему земному шару. Как известно, радиа-
ция млекопитающих, начавшаяся вслед за массовым вымирани-
ем динозавров в позднемеловую эпоху, привела к тому, что по
своему адаптивному разнообразию они превзошли мезозойских
рептилий. Дифференциация млекопитающих в кайнозое
(фиг. 135) — прекрасный пример типичной адаптивной радиа-
ции. Как экологические «наследники» динозавров млекопитаю-
щие представляют пример так называемого экологического за-
мещения (оно не обязательно подразумевает вытеснение в ре-
зультате конкуренции, но большинство исследователей считают
вытеснение частным случаем замещения).
В особом случае последовательного экологического замеще-
ния исходная родительская группа дает одну за другой группы
высших таксонов, каждая из которых замещает предыдущую.
Этот процесс называется итеративной эволюцией. Видимо, та-
кой тип эволюции был свойствен мезозойским группам аммони-
тов, хотя здесь трудно точно установить отношения между
предковой группой и группами, произошедшими от нее. Заль-
фельд [174] сделал вывод, что два медленно развивавшихся,
подотряда, Phylloceratina и Lytoceratina, жили главным обра-
зом в глубоководных районах и дали начало быстро сменявшим
друг друга группам мелководных таксонов, объединяемых в под-
отряд Ammonitina. Таким образом, этот подотряд имеет поли-
филетическое происхождение: (фиг. 136).
Известно, что трилобиты кембрия также претерпели итера-
тивную эволюцию. Лучше всего документирован пример, при-
водимый Кауфманом [98, 99] и обсуждаемый Симпсоном [197].
Кауфман нашел в последовательности пластов верхнего кемб-
рия в Швеции четыре сменявших друг друга эволюционных ряда
рода (Menus. Каждый ряд он разделил на три вида! или под-
вида (фиг. 137). В трех рядах наблюдаются непрерывные мор-
фологические переходы. Во всех рядах выявляются опре-
деленные морфологические тенденции, из которых самая за!-
метная — удлинение пигидия (хвостового щита). Ранние пред-
ставители первых двух рядов почти идентичны. Полагают, что
эти четыре ряда последовательно отделялись от медленнее эво-
люционировавшей географически удаленной исходной группы.
Каждый ряд вселялся в изученную область, развивался здесь,
следуя отмеченным нами тенденциям, а затем вымирал, осво-
бождая место для новых таксонов, произошедших от исходной
группы.
19*
292
Часть 2. Применение данных палеонтологии
ЭВОЛЮЦИОННАЯ КОНВЕРГЕНЦИЯ И ПАРАЛЛЕЛИЗМ
В ходе эволюции между двумя неродственными группами,
ведущими сходный образ жизни, может возникнуть морфологи-
ческое сходство. Это явление называется адаптивной конвер-
генцией; сходные таксоны называются гомеоморфами. Некото-
рые биологи называют гомеоморфами два вида, представители
которых неразличимы для неопытного наблюдателя. Но пале-
онтологи, как правило, используют этот термин просто для
обозначения очень схожих между собой видов.
Гомеоморфия иногда возникает в результате так называе-
мой параллельной эволюции (или параллелизма), при которой
две близкородственные группы с небольшими морфологически-
ми различиями подвергаются с течением времени сходным эво-
люционным изменениям. Между параллелизмом и конвергенци-
ей нет четкого различия, но иногда эти явления разграничива-
ют в зависимости от степени сходства предковых групп, от
которых ведут начало эволюционные ряды, содержащие два рас-
сматриваемых вида.
Термины «параллелизм» и «параллельная эволюция» неред-
ко применяют в тех случаях, когда наблюдается некоторое
сходство в эволюционных тенденциях одного или немногих при-
знаков, а не организмов в целом. Такое употребление терминов
сомнительно.
Время
-Фиг 138 Различные виды связи между развивающимися рядами (частично
из [30]).
"Расстояние по горизонтали, разделяющее два вида в любой точке, пропорционально раз-
личию между ними по сравнению с различиями в других точках их эволюционной исто-
рии.
Гетерохромная
конвергенция
Глубокое зарывание в
прочные устойчивые
субстраты
Морфология раковины
Вытянутая и трубчатая
(для уменьшения
длины сифона)
Есть заднее и переднее
зияние (нужны для
быстрого выдвижения
и втягивания сифо-
нов и ноги в направ-
лении, параллельном
длинной оси тела;
такая морфология до-
- пустима благодаря
глубокому зарыва-
нию)
Створки тонкие (из-за
глубокого зарывания
раковина хорошо за-
щищена)
Потеря зубов (створки
могут смещаться
вдоль дорсо-вентраль-
ной оси при движе-
ниях сифона и ноги)
Неглубокое зарывание
в устойчивые суб-
страты
Морфология раковины
Вытянутая кзади (для
сокращения длины
сифона)
Центр тяжести сдвинут
вперед (для устойчи-
вости)
Створки толстые (для
прочности)
Быстрое зарывание в
песок
Морфология раковины
Передний конец тре-
угольный (для облег-
чения внедрения в
субстрат)
Раковина толстая, Т[ve-
la (для прочности)
Наружная поверхность
гладкая (для обтека-
емости)
Фиг. 139. Адаптивная конвергенция между парами родов двустворчатых мол-
люсков [208].
В каждой паре один моллюск относится к надсемейству Mactracea, а другой — к какому-
либо другому надсемейству (названия указаны в скобках). Все рассмотренные роды су-
ществуют и сейчас.
Быстрое зарывание на
пляжах с перемеща-
ющимися песками
Морфология раковины
Передний конец сильно
вытянут (чтобы вме-
щать длинную ногу)
Задний конец сильно
обрублен (чтобы нога
выдвигалась против
места прикрепления
сифонов)
Раковина узкая; об-
ласть максимальной
ширины сдвинута на-
зад (для быстрого
зарывания)
Створки толстые (для
прочности)
Поверхность гладкая
(для обтекаемости)
Неглубокое за-
рывание в
мягкие илис-
тые субстраты
Морфология ра-
ковины
Раковина малень-
кая (для пла-
вания)
Створки тонкие,
Mulinia (длй
плавания) ,
294
Часть 2. Применение данных палеонтологии
Параллелизм сходен с итеративной эволюцией, но он встре-
чается не в сменяющих друг друга, а в одновременно сущест-
вующих рядах. Два конвергентных ряда могут претерпевать
одинаковые изменения одновременно или в разное время. Клауд
[30] выделил поэтому два типа адаптивной конвергенции: изо-
хронную и гетерохронную конвергенцию. Различия между ите-
ративной эволюцией,' параллелизмом, изохронной конвергенци-
ей и гетерохронной конвергенцией показаны на фиг. 138. На-
стоящий параллелизм редко отражается в палеонтологической
летописи, хотя, по-видимому, это явление и было распростра-
нено. Напротив, примеры конвергенции весьма многочисленны;
ведь для того, чтобы найти конвергенцию, достаточно обнару-
жить две сходные формы, относящиеся к неродственным высшим
таксонам. Для выявления же параллелизма нужны по меньшей
мере четыре таксона (по два из каждого ряда).
При изучении палеозойских брахиопод найдено много при-
меров гомеоморфии, связанной с адаптивной конвергенцией.
Один из случаев гомеоморфии рассматривался выше с точки
зрения функциональной морфологии (фиг. 73). Хорошо извест-
ный пример адаптивной конвергенции в широком масштабе
представляют сумчатые и плацентарные млекопитающие.
В меньшем масштабе это явление можно наблюдать у родов
двустворчатых моллюсков надсемейства Mactracea (фиг. 139).
Вымирание
Вымирание сильно влияет на пути эволюции, но не всегда
является частью самой эволюции.
Чтобы скрещивающаяся популяция достигла успеха, необ-
ходимо хорошее адаптивное соответствие между ее генофондом
и средой. Поскольку генетический состав популяций обычно
изменяется главным образом под действием естественного от-
бора, нарушение адаптивного баланса, как правило, бывает
связано только с изменениями условий среды. Когда среда ста-
новится неблагоприятной, то популяция может приспособить-
ся к новым условиям путем эволюции, мигрировать в район
с более благоприятными условиями или вымереть. Если ни од-
ной популяции вида не удастся справиться с неблагоприятными
условиями путем эволюции или миграции, то вид вымрет. Вы-
мирание вида происходит и в том случае, когда накапливается
слишком много филетических изменений и старый вид стано-
вится новым видом; это так называемое филетическое выми-
рание.
Изменения среды, приводящие к вымиранию, могут быть
физическими, химическими или биологическими. Мы уже виде-
X. Эволюция и палеонтологическая' летопись
295
ли, как одна таксономическая группа может вытеснить другую
благодаря своему адаптивному превосходству. Чрезмерное хищ-
ническое истребление одной группы другой также может послу-
жить причиной вымирания первой группы.
Вследствие перерывов в стратиграфической летописи редко
удается точно определить время вымирания ископаемого вида.
Значительная часть ископаемых видов известна из пластов, ко-
торые образовались за интервал времени, составляющий лишь
часть времени существования этих видов. Следует также пом-
нить, что даже если бы палеонтологическая летопись была пол-
ной, то и тогда деление рядов на отдельные виды все равно
было бы субъективным и спорным. Есть и еще одна сложность,
связанная с тем, что не обязательно вымирают сразу все попу-
ляции широко распространенного вида.
ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ
Самое наглядное доказательство важной роли географиче-
ских факторов в вымирании дают реликтовые группы — таксо-
ны, уцелевшие в ограниченных областях, в то время как повсю-
ду их родичи давно вымерли. Один из самых замечательных
примеров — существование бластоидей в позднем палеозое в
Южном полушарии. Древнейшие представители этих прикреп-
ленных иглокожих, обладавших стеблем и питавшихся сесто-
ном, найдены в силуре. Расцвета они достигли, по-видимому,
в раннем карбоне. Уже в отложениях среднего и верхнего карбо-
на найдены остатки только трех или четырех родов. Но в перм-
ском периоде, видимо, произошла новая экспансия группы,
с центром в районе острова Тимор (Зондские острова), где об-
наружены остатки 16 родов. Небольшая часть тиморских родов
и еще несколько других обнаружены также в пермских породах
Австралии и Азии. В Западном полушарии найден только один
пермский вид — в арктической области Канады.
Сумчатые млекопитающие Австралии — важная реликтовая
группа, живущая в наши дни. В конце мезозоя сумчатые были
распространены по всей Земле. Но в раннем кайнозое, когда
началась широкая адаптивная радиация млекопитающих, пла-
центарные почти во всем мире заместили сумчатых. Два мате-
рика— Австралия и Южная Америка — на протяжении всего
кайнозоя оставались изолированными. Поэтому здесь сумчатые
смогли существовать дольше, не вступая в конкуренцию с пла-
центарными и не страдая от хищнического истребления с их
стороны. Но незадолго до конца плиоцена между Северной и
Южной Америкой возник узкий сухопутный мост — Панамский
перешеек. Северные виды плацентарных мигрировали к югу,
вытесняя сумчатых и уничтожая их. Это привело к вымиранию
295
Часть 2. Применение данных палеонтологии
сумчатых; выжили только опоссумы — примитивные формы, ра-
нее, в период изоляции, давшие начало различным южноамери-
канским группам сумчатых. Один вид опоссума добился необы-
чайного успеха и расширил свой ареал к северу, заняв значи-
тельные пространства Северной Америки. Остальные сумчатые
процветали по-прежнему только в изолированной Австралии.
Некоторые острова также послужили убежищами для важ-
ных реликтовых групп. Так, лемуры, предшественники высших
приматов, хорошо представлены на Мадагаскаре, тогда как в
Африке осталось лишь несколько видов лемуров. Единственный
ныне живущий представитель примитивной группы рептилий
Rhynchocephalia (круглоголовые)—род Sphenodon — живет
только на нескольких мелких островах у берегов Новой Зелан-
дии; он возник в триасе и близок к предкам настоящих ящериц
и динозавров. Интересно, что в районе Австралии и Новой Зе-
ландии существуют и многие другие реликтовые группы, в том
числе различные замковые брахиоподы, Nautilus, примитивный
двустворчатый моллюск Neotrigonia и, наконец, сумчатые. Оче-
видно, и суша и море в этом районе уже многие миллионы лет
сохраняют в какой-то мере биогеографическую изолирован-
ность.
Географическая изоляция может спасти некоторых предста-
вителей таксономической группы, которые сохраняются в виде
реликтовых форм. Однако группа в целом может при этом по-
страдать. Очень ограниченная изолированная географическая
область предоставляет мало возможностей для развития боль-
шого числа полуизолированных популяций, в которых могли бы
проявиться новые адаптации. Кроме того, чем меньше эта об-
ласть, тем меньше в ней разнообразие биотопов, поэтому силь-
ные изменения условий среды могут вредно сказаться на всей
фауне.
Прекрасный пример уменьшения разнообразия форм в ре-
зультате изоляции — современные рифообразующие кораллы
района Карибского моря [223]. В раннем кайнозое в море Тетис
сосуществовали коралловые фауны Средиземноморья, Индий-
ского океана и Восточноиндийской области. В позднем олиго-
цене большая часть европейских компонентов карибской фауны
вымерла. Остались почти исключительно те роды, которые жи-
вут в Карибском регионе и сейчас. В миоцене морская связь
между Средиземным морем и Индийским океаном прервалась.
Появление сухопутного барьера и последующее общее похоло-
дание привели практически к прекращению роста рифов в Сре-
диземном море и к образованию двух фаун кораллов, которые
остаются раздельными и поныне: вест-индской и индо-тихо-
океанской фаун. До появления в позднем плиоцене Панамского
перешейка эти фауны и без того были связаны слабо, а после
X. Эволюция и палеонтологическая летопись
297
его появления связь была полностью утрачена. Изолированная
карибская фауна, занимающая небольшую географическую об-
ласть, после миоцена сократилась примерно до 36 видов, а ши-
роко распространенная индо-тихоокеанская фауна дифференци-
ровалась до более чем 500 видов.
МАССОВОЕ ВЫМИРАНИЕ
Одна из величайших загадок палеонтологической летопи-
си — тенденция высших таксономических групп к вымиранию за
сравнительно короткое время. За массовыми вымираниями, как
мы видели, обычно следует интенсивная адаптивная радиация,
сопровождающаяся экологическим замещением. Многие крупные
вымирания приблизительно совпадают по времени с рубежами
периодов и эпох. Причина этого проста: часто условные грани-
цы помещали там, где в палеонтологической летописи были от-
мечены значительные переломы. Наиболее известные крупные
вымирания животных произошли в конце палеозоя и в конце
мезозоя.
Массовые вымирания морских организмов происходили в
конце кембрия, ордовика и девона. Но гораздо более драмати-
ческим, если говорить о числе исчезнувших таксонов, оказался
кризис морской фауны в конце пермского периода. Среди самых
важных вымерших групп были фузулиниды из простейших, про-
дуктиды из брахиопод, многие таксоны мшанок и аммонитов.
Высшие таксоны, вымершие, как полагают, в конце пермского
периода, перечислены на фиг. 140. В отличие от морских бес-
позвоночных наземные позвоночные на границе пермь — триас
потеряли сравнительно мало высших таксонов. Разнообразные
фауны земноводных начала пермского периода в значительной
мере уступили место в середине перми фаунам пресмыкающих-
ся. Таким образом, в середине пермского периода начался
расцвет рептилий, продолжившийся и в триасе, т. е. уже в ме-
зозое.
В конце триаса произошло первое крупное вымирание по-
звоночных, затронувшее как наземные, так и морские группы.
На фиг. 141 показано, что из девятнадцати верхнетриасовых
отрядов и подотрядов рептилий вымерло восемь. Одновременно
из состава фауны земноводных исчезли многочисленные ранее
представители лабиринтодонтов. Из беспозвоночных существен-
но сократились аммониты, достигшие в триасе огромного разно-
образия. До юрского периода дожила только небольшая группа
филлоцератин, давшая в этом периоде начало литоцератинам.
В юрском и меловом периодах в группах Phylloceratina и Lyto-
ceratina возникло много надсемейств и семейств (см. стр. 291
Карбон Пермь Триас Юра ' Л
FUOUtUHUU Conulariida Tabulate Streptelasmatina Columnariina Astrocoeniina Fungiina
► Кишечнополостные
Faoiina J Trilobita л Eurypterida Beyrichiida Leperditiida Thysanura Palaeodictyoptera Megasecoptera Protohemiptera > Orthoptera
-
> Членистоногие
f 1
Carditacea Cardiacea Myacea
’ Двустворчатые
Bellerophontacea > PLatyceratacea Subulitacea
i
Patellacea Trochacea ' Брюхоногие
uiwnmcea
C&i ithiumu V
Naticacea d Solenochilida Goniatitina Ceratitina Phylloceratina > Trepostomata ? Cryptostomata J Dalmanellacea Productacea J Blastoidea ? Inadunata Flexibilia. Camerata 7
• Головоногие
а
Мшанки 1
- Брахиоподы
Стебельчатые
иглокожие
Articulate J Rhechitomi Stereospondyli J tchthyosaurie > Sauropterygie Rhynchocephalia | Sguamate
Амфибии
> Рептилии
Arctiosaurie Pelycosaurie Ictidosauria >
•j
Фиг. 140. Позднепермское вымирание и появление в раннем триасе высших |
таксонов животных [182]. 1
.5
X. Эволюция и палеонтологическая летопись
299
Верхний триас Нижняя юра
Амфибии
----------------- Стереоспондили
Рептилии
----------------- Диадектоморфы
----------------- Проторозавры
----------------- Нотозавры
----------------- Плакодонты
----------------- Псевдозухии
----------------- Фитозавры
----------------- Дицинодонты
----------------- Териодонты
—------------------------------------- Черепахи
--------------------------------------Ихтиозавры
-------------------------------------- Плезиозавры
--------------------------------------Эозухии *
---------------------------------------- Хоботноголовые •
--------------------------------------Ящерицы ’
-------------------------------------— Крокодилы
-------------------------------------- Звероногие динозавры
-------------------------------------- Птиценогие динозавры*
--------------------------------------- Иктидозавры
---------------------- Птеродактили
-------------------- Зауроподы
-------------------- Стегозавры
Фиг. 141. Крупные группы амфибий и рептилий на рубеже триас — юра [33].
Звездочками отмечены группы, остатки которых пока не найдены в нижнеюрских поро-
дах, но поскольку они обнаружены в отложениях верхнего триаса и верхней юры, ясно,
что эти животные существовали и в ранней юре.
и фиг. 136). В юрском и меловом периодах большого разно-
образия достигают также морские и наземные позвоночные.
Затем, в конце мелового периода, волна вымирания уничтожи-
ла динозавров. На этом же рубеже исчезают аммониты и дру-
гие, в основном морские группы (в том числе белемниты и не-
которые важные таксоны брюхоногих и двустворчатых моллюс-
ков). Таксономический эффект мелового кризиса фауны показан
на фиг. 142. По своему масштабу он соперничает с пермо-триа-
совым кризисом.
В кайнозое произошло только одно крупное массовое выми-
рание, которое относится к плейстоцену, когда исчезли многие
группы наземных млекопитающих. Но это вымирание, очевид-
но, не было связано с изменениями климата в плейстоцене, об-
условившими, как известно, развитие в Северном полушарии
четырех крупных оледенений. Ведь среди других экологических
групп, например наземных растений и мелководных морских
беспозвоночных, крупных вымираний не произошло. Сейчас
9S
Юра Мел Кайнозой Четвертичны период
-
— '
_ ... 1
Isoptera
Aphaniptera
Strepsiptera
Drdssenacea
Rudistae
Euomphalacea
Troctionematacea
Nerineacea
Pyramidellacea
Atlantacea
Lamellariacea
SpirateUacea
Phylloceralina
Lyfoceratina
Ammonitma
Belemnoidea
Pycnodontiformes
Aspidorhynchiforrnes
Urodela
tchthyosauria
Sauropterygia
Mesosuchia
Pterosauria
Theropoda
Sauropoda
Ornithischia
Odonfhognaihae
Palaeognathae
Neognathae
Caenolestoidea
Inseciiuora
Chiroptera
Taeniodonta
Primates
Carnivora
Sirenia
Condylarthra
Litopterna
Notoungulata
Perissodactyla
Arfiodaclyla
Edentata
Rodentia
Насекомые
Двуствор-
чатые
^Брюхоногие
' Головоногие
Рыбы
Амфибии
• Рептилии
Птицы
л
Моллюски
Млекопитающие
Фиг. 142. Вымирание высших таксонов животных в позднемеловую эпоху и
появление новых таксонов в начале кайнозоя (182].
X. Эволюция и палеонтологическая летопись ЗОТ
многие исследователи полагают, что большинство исчезнувших
наземных животных было доведено до вымирания новым хищни-
ком — человеком.
Массовые вымирания групп высших растений, как правило,
не совпадают с вымираниями животных. Поражает, что два
крупных изменения наземных флор предшествовали значитель-
ным изменениям наземных фаун. В эволюционной истории выс-
ших растений можно выделить три основные флоры, доминиро-
вавшие последовательно в неморских местообитаниях: палеозой-
ская флора споровых растений, мезозойская флора голосемен-
ных и кайнозойская флора покрытосеменных. Переходы между
ними не совпадают с границами геологических эр, а предшест-
вуют им примерно на половину геологического периода. Воз-
можно, каждый переход был связан с экологическим вытесне-
нием. Флора голосеменных стала господствующей в поздней
перми, а флора покрытосеменных — в позднемеловую эпоху.
Для эволюции высших растений крупномасштабные, массовые
вымирания не так характерны, как для эволюции животных.
Просто по мере расцвета замещающей группы старая постепен-
но исчезала.
Для объяснения причин массовых вымираний, отмеченных
в геологической летописи, выдвигалось множество гипотез,
иногда совершенно несостоятельных. Некоторые гипотезы пы-
тались объяснить какой-то отдельный случай массового выми-
рания, другие — все случаи. Невозможно рассмотреть здесь все
эти гипотезы — упомянем лишь некоторые. Многие из них прак-
тически нельзя проверить. Например, такие факторы, как изме-
нения солнечной радиации или эпидемии микробных заболева-
ний, вполне могли привести к вымираниям, но, видимо, не могли,
оставить прямых геологических свидетельств.
Поэтому вполне естественно, что внимание геологов привле-
кают те несколько гипотез, которые объясняют массовые вы-
мирания разрушительными силами, следы действия которых
могли запечатлеться в стратиграфической летописи. Такими
силами могли быть инверсии магнитного поля Земли, изменения
климатических условий и колебания относительного уровня мо-
ря. Так, предполагалось, что периодические обращения магнит-
ной полярности Земли в геологическом прошлом могли временно
снимать защиту от космического излучения, которую обеспечи-
вает магнитное поле. В результате поверхность Земли подвер-
галась воздействию более интенсивной радиации, а это могло
значительно повысить частоту мутаций и соответственно уско-
рить темп эволюции и вымирание. Исследования остаточной на-
магниченности пород различного возраста подтверждают, что
магнитная полярность менялась, но пока не доказано, что эти
инверсии совпадают во времени с периодами быстрой эволюции
302
Часть 2. Применение данных палеонтологии
или массовыми вымираниями. Кроме того, массовые вымирания
обычно происходили за отрезки времени, по-видимому, превы-
шающие время, необходимое для инверсии магнитного поля.
Изменения климатических условий и колебания относитель-
ного уровня моря — явления, не связанные друг с другом. И те
и другие вместе и по отдельности привлекались для объяснения
массовых вымираний. Геологическая летопись показывает, что
в пермском периоде во многих регионах на континентах сущест-
вовали обширные, в высшей степени аридные области, сменив-
шие болота, преобладавшие в каменноугольном периоде. Кроме
того, пермский период, по-видимому, был периодом резких кли-
матических контрастов, а возможно, и колебаний климата.
В пермских толщах сохранились свидетельства и оледенения,
и аридности. Но летопись плейстоцена содержит данные, ме-
шающие признать изменения климата основной причиной массо-
вого вымирания животных различных групп. В плейстоцене ис-
чезли многие группы млекопитающих, но морские беспозвоноч-
ные не претерпели существенных изменений. Пермо-триасовый
кризис фауны, напротив, затронул в основном морских беспо-
звоночных.
Ньюэлл [136] подчеркнул роль колебаний относительного
уровня моря в вымираниях. Поскольку многие области конти-
нентов обладали сравнительно сглаженным рельефом, даже не-
значительные колебания уровня моря вызывали быстрые транс-
грессии или, наоборот, приводили к осушению обширных пло-
щадей. При этом резко изменялись условия среды. Основное
доказательство важной роли колебаний уровня моря в выми-
раниях— явная приуроченность многих ископаемых видов к вре-
менным интервалам, отмеченным крупными регрессиями древ-
них морей. Многие важнейшие стратиграфические границы сов-
падают как с несогласиями в залегании пластов, так и с мас-
совыми вымираниями ископаемых групп. Но колебаниями уров-
ня- моря трудно объяснить полное вымирание таких широко
распространенных и богатых формами групп, как динозавры,
трилобиты или аммониты. Попытки связать скорость эволюции
и вымирания с периодами горообразования в основном не
удались.
Изменения климата или колебания уровня моря могли со-
провождаться изменениями солености древних океанов. Бейрлен
[14] предположил, что, в частности, пермо-триасовый кризис
был следствием понижения солености воды в океанах. При этом
в наибольшей степени были, затронуты в основном стеногалин-
ные морские группы (мшанки, кораллы, брахиоподы, морские
лилии, аммониты, трилобиты); наименее пострадавшие группы
имеют сегодня многих представителей в солоноватых водах
(двустворчатые и брюхоногие моллюски, рыбы). Бейрлен пола-
X. Эволюция и палеонтологическая летопись
303
гает, что наиболее вероятной причиной уменьшения общей со-
лености океанов явилось замедление поступления солей. Фишер
[59] выдвинул другую гипотезу: по его мнению, в окраинных
морских бассейнах в результате испарения формировались креп-
кие рассолы, которые вследствие своей большой плотности пере-
текали из этих окраинных бассейнов в глубоководные зоны
океана. Там рассолы и накапливались, лишь слабо смешиваясь
с остальной водной массой, которая постепенно становилась бо-
лее соленой. Прекращение процесса образования рассолов или
уменьшение его интенсивности, происшедшее в конце пермского
периода, привело, по мнению Фишера, к полному перемешива-
нию глубинных рассолов с океанской водой и восстановлению
нормальной солености воды в океане. В пользу гипотезы и ее
вариантов говорит все то, что мы знаем о существовании эва-
поритовых бассейнов пермского возраста, особенно в бассейне
Делавэр в Техасе. И все же это только гипотеза.
Вряд ли можно объяснить все вымирания действием какого-
либо одного фактора. Разные массовые вымирания, вероятно,
имели разные причины, и, кроме того, здесь могли действовать
многочисленные факторы, один из которых — взаимосвязанность
видов, не только через пищевые цепи, но и через такие меха-
низмы, как образование субстратов (вспомним коралловые ри-
фы) и симбиоз.
В свете этих рассуждений попробуем привлечь данные по
морскому фитопланктону к решению проблемы крупных выми-
раний. Эволюционная история морского фитопланктона, как и
история позвоночных, беспозвоночных и наземных растений, де-
лится на три основных этапа, приблизительно совпадающих
с тремя геологическими эрами. История основных групп мор-
ского фитопланктона показана на фиг. 143. Акритархи1 и другие
группы были разнообразно и обильно представлены в раннем
палеозое. В позднем девоне начался упадок раннепалеозойских
фитопланктонных флор, продолжавшийся в триасе. В интервале
от каменноугольного периода до конца триаса фитопланктон-
ные сообщества были очень редкими — ни одна из крупных
таксономических групп не является обильной или широко рас-
пространенной.
В ранней юре произошла адаптивная радиация новых групп,
включая новые таксоны акритарх, кокколитофорид и динофла-
геллят. Эта радиация достигла максимума в конце мелового
1 Группа акритарх (Acritarcha Evitt, 1963) предложена в качестве нео-
пределенной систематической категории, объединяющей микрофоссилии раз-
личного происхождения, но сходные по внешним морфологическим призна-
кам. В советской литературе описаны акритархи из докембрия и нижнего па-
леозоя (в основном, очевидно, это оболочки одноклеточных водорослей).—
Прим. ред.
301
Часть 2. Применение данных палеонтологии
Кайнозой
Мел
Карбон
Кембрий
Докембрий
Девон
Силур
Ордовик
Юра
Триас
Пермь
t
сз
Е
&
S
е
Q
Е
<з
3
Фиг. 143. Схема геохронологического распространения основных групп
морского фитопланктона (из [44], с изменениями).
периода, но затем внезапно оборвалась в результате крупных
вымираний, охвативших новые группы.
В пластах нижнего палеоцена наблюдается та же картина,
что и в породах карбона — триаса: остатки фитопланктона
очень скудны. В конце палеоцена вновь началась адаптивная
радиация, давшая современную флору фитопланктона. Выми-
рания, особенно в олигоцене, сократили относительное обилие
некоторых групп, процветавших в раннем кайнозое, и в совре-
менных морях преобладают диатомовые и динофлагелляты.
Толкование палеонтологической летописи фитопланктона в
какой-то степени спорно. Не исключено, что в эпохи кажуще-
гося уменьшения разнообразия существовало множество групп
организмов, остатки которых просто не сохранились. Но сокра-
щение акритарх в позднем палеозое и их новая адаптивная
радиация в юре (фиг. 143) — почти бесспорные факты. Бесспор-
X. Эволюция и палеонтологическая летопись
305
но и то, что в позднем мелу происходили крупные вымирания
фитопланктона.
Таппан [212] предположила, что на протяжении фанерозоя
фитопланктон сильно влиял на фауну. Это объяснялось тем,
что фитопланктон — первичный продуцент пищи и производитель
кислорода. Таппан считает, что крупные вымирания и умень-
шение продуктивности фитопланктона могли следовать сразу
же за эпохами выравнивания рельефа суши, когда замедлялось
поступление в океаны питательных веществ с продуктами эро-
зии материков. Резкое сокращение фитопланктона могло вы-
звать крупные вымирания организмов-консументов, а также
групп, нуждающихся в кислороде.
Тот факт, что крупные изменения наземной флоры несколь-
ко опережали массовые вымирания наземных животных, может
объясняться подобным же образом. Вероятно, достаточно круп-
ные изменения флоры в море и на суше могли способствовать
обширным вымираниям животных, уже столкнувшихся с други-
ми неблагоприятными условиями среды.
Любая гипотеза, претендующая на объяснение массовых вы-
мираний, должна объяснить загадочную тенденцию, выражаю-
щуюся в том, что вымирания затрагивают не экологические,
а таксономические группы. Фузулиниды, продуктиды, аммони-
' ты и динозавры обладали широким спектром экологических
адаптаций, и тем не менее эти группы внезапно исчезли как
таксономические единицы. Это заставляет думать, что факторы,
обусловившие массовые вымирания, действовали на фундамен-
тальные генетические и физиологические признаки, а не на
менее существенные экологические адаптации внутри групп. Не-
зависимо от того, разрешима ли проблема массовых вымираний,
она будет и в дальнейшем волновать воображение ученых. Во
всяком случае, в последнее десятилетие появилось немало но-
вых данных, проливающих свет на эту проблему.
Выяснение филогенетических связей
Определить филогенетические связи таксономических групп
можно на основании биологических и палеонтологических дан-
ных, используя их раздельно или вместе. При изучении вымер
ших групп с неясными филогенетическими связями обычно ста-
раются установить родство с типами, имеющими рецентных
представителей. Это стремление, часто заставляющее выдавать
желаемое за действительное, привело ко многим неверным вы-
водам. Интересный пример — кембрийские археоциаты
(фиг. 144). Бокаловидные скелеты археоциат создавали рифо-
подобные постройки, особенно многочисленные в Австралии.
Морфология и прикрепленный образ жизни этих животных поз-
20—467
306
Часть 2. Применение данных палеонтологии
Фиг. 144. Схема строения кубка ар-
хеоциат [142].
воляют предположить, что они были сестонофагами. Отверстия
в стенке кубка археоциат свидетельствуют, что у них существо-
вала система тока воды, сходная с подобной системой губок,
у которых вода входит внутрь через небольшие отверстия и вы-
водится через одно или несколько широких отверстий. Если бы
направление тока воды было обратным, система была бы неэф-
фективной с точки зрения гидродинамики. Некоторые палеонто-
логи упорно относят археоциат к губкам. Однако большинство
исследователей полагает, что, поскольку скелеты археоциат и
губок мало схожи, нельзя говорить об их близком родстве. Ис-
копаемые остатки, которых не удается отнести к известным
группам, обычно называют «проблематиками» («problematica»).
Не исключено, что мы так никогда и не сможем установить
с точностью их филогенетические связи.
Гораздо успешнее оказались попытки выяснить системати-
ческое положение граптолитов. Хотя общее мнение на этот счет
еще не сложилось, большинство исследователей согласно с мне-
нием Козловского ([ЮЗ] и более ранние работы), который счи-
тает, что граптолиты принадлежат к типу полухордовых и схо-
жи с современными крыложаберными (фиг. 145). Строение и
химический состав скелета граптолитов, способ почкования их
колонии — все говорит об этом сходстве. Выводы Козловского
основаны на результатах изучения прекрасно сохранившихся
колоний, вытравленных из черных кремнистых сланцев плави-
ковой кислотой. Козловский заливал отпрепарированные образ-
цы в парафин и делал серийные срезы на микротоме точно так
же, как биологи готовят срезы рецентных организмов. Боль-
шинство скелетов граптолитов сплющено в плоскости напласто-
вания сланцев. Успех Козловского показывает важность поис-
ков исключительно хорошо сохранившегося материала для так-
Фиг. 145. Сравнение морфологии крыложаберных и граптолитов [103].
А и Б. Строение стенки теки, состоящей из полуколец фюзеллярного слоя {Cephalodiscus
и Rhabdopleura). В. Столон Rhabdopleura (отмечен густым пунктиром). Г и Д. Mastigo-
graptus и Bulmanicrusta, обладающие сходной структурой.
сономических исследований. Кроме того, это пример успешной
попытки установить родство между загадочной ископаемой и
современной группами.
В то же время для некоторых современных групп филогене-
тические связи удалось выяснить главным образом благодаря
исследованиям палеонтологической летописи. Например, полу-
копытные млекопитающие (Subungulata) включают современ-
ных даманов (мелкие животные, напоминающие по внешнему
виду грызунов), слонов и сирен. Вряд ли эти столь несхожие
группы были бы собраны вместе в одну таксономическую груп-
пу, если бы не их палеонтологическая летопись, доказывающая
принципиальное сходство их примитивных кайнозойских пред-
ставителей, обитавших в Африке, где они, видимо, возникли от
общего предка.
В целом филогению основных групп позвоночных устанавли-
вать гораздо легче, чем филогению беспозвоночных и растений,
так как все классы и отряды Vertebrata появились после кемб-
рия. Хотя все высшие таксоны сосудистых растений, видимо,
появились после раннего палеозоя, палеонтологическая летопись
растений не так полна. Кроме того, по ископаемым остаткам
растений обычно меньше удается узнать о морфологии всего
организма, чем по остаткам позвоночных. Беспозвоночные де-
лятся на несколько типов, возникновение которых в позднем
докембрии и кембрии, как правило, не отражено в дошедшей
до наших дней палеонтологической летописи. Но в некоторых
случаях послепалеозойские филогении типов беспозвоночных
известны сравнительно неплохо.
К сожалению, происхождение большинства высших катего-
рий окутано тайной; почти все высшие категории появляются
20*
308 Часть 2. Применение данных палеонтологии
в палеонтологической летописи внезапно, без промежуточных
предковых форм. Симпсон [197] перечисляет несколько причин
такой внезапности. Вот некоторые из них:
1. Появление новой высшей категории обычно отмечает
крупный адаптивный прорыв, который часто сопровождался за-
нятием ранее свободных ниш. В таких условиях эволюция мо-
жет происходить очень быстро.
2. Все ряды предковой группы, которые были достаточно
сходны с новой группой, чтобы вступить с ней в конкуренцию,
должны были быстро вытесняться.
3. Часто появление высшей категории приходится на пере-
рыв в стратиграфической летописи (иногда быстрый темп эво-
люции и стратиграфические несогласия имеют одну и ту же
причину — изменение физико-географических условий на обшир-
ных площадях).
4. Обнаружению некоторых переходных форм могла поме-
шать смена местообитания во время адаптивного прорыва.
5. Крупные адаптивные прорывы случались обычно в срав-
нительно небольших популяциях или таксономических группах.
6. Переходы происходили обычно в таксонах, члены ко-
торых отличались от предковой и потомковой высших категорий
сравнительно небольшими размерами.
7. Переходы обычно происходили в географически ограни-
ченных районах; возможно, одинаковые переходные процессы
происходили в разное время в разных районах.
Иногда в палеонтологической летописи все же сохраняется
так называемое «недостающее звено» — вид, вероятно, пред-
ставляющий собой переходную стадию между высшими таксо-
нами. Одна из таких форм — позднеюрская птица Archaeopterix,
о которой уже говорилось в другой связи (фото I). Археопте-
рикс обладал чертами рептилий и птиц. Наличие оперения го-
ворит о теплокровности (теплокровны и современные птицы),
а крупные зубы, массивные кости и другие особенности скелета
напоминают пресмыкающихся.
Наглядный пример перехода можно наблюдать в описанном
Эрбеном [55] последовательном морфологическом ряду голово-
ногих из нижнедевонских хунсрюкских сланцев (ФРГ). В 30-х
годах Шиндевольф выдвинул мысль о происхождении аммонитов
от бактритоидей. При этом он подчеркнул сходство начальных
камер бактритов и ранних аммонитов. Сифон (стр. 175) бактри-
тов занимал краевое положение, как и у большинства аммони-
тов. Морфологический ряд, описанный Эрбеном, видимо, под-
тверждает гипотезу Шиндевольфа. Ряд составляют бактриты,
аммониты и промежуточные формы (фиг. 146). Относительное
стратиграфическое положение таксонов, входящих в ряд, не изу-
чалось. Но то, что все эти таксоны встречены в одной местной
о
ши
Фиг. 146. Взрослые раковины из хунсрюкских сланцев (девон, ФРГ). [55].
•Здесь представлен эволюционный ряд, приведший к аммонитам. А—Д. Бактритоидеи
Е—М. Ранние аммониты.
310
Часть 2. Применение данных палеонтологии
последовательности пластов, говорит в пользу гипотезы о про-
исхождении аммонитов от бактритов.
Недостающее звено иногда удается выявить также благода-
ря открытию неизвестных прежде рецентных организмов. В ка-
честве примера приведем глубоководного моллюска Neopilitia.
В 1952 г. участники датской экспедиции на «Галатее» подняли
представителей этого необычного рода с океанского дна у Тихо-
океанского побережья Центральной Америки, с глубины около
3500 м. Лемхе и Вингстранд [117] описали анатомию этого
моллюска, которому присущи многие примитивные признаки.
Наиболее необычен общий план строения: моллюск имеет ряды
парных мышц ноги, органов, напоминающих почки, жабр
(фиг. 147). У других групп моллюсков имеется только по одной
паре этих органов, а если их по нескольку пар, то это вторичное
(не примитивное) явление.
За два десятилетия, предшествовавших открытию Neopilina,
некоторые палеонтологи, в том числе Венц и Найт, предполага-
ли, что раннепалеозойские ископаемые, принадлежащие к клас-
су Monoplacophora’ (фиг. 147), были блюдцеобразными предка-
ми брюхоногих, а может быть, и других высших моллюсков.
Обнаруженная в 1952 г. Neopilina была отнесена к моноплако-
форам, хотя ископаемые представители группы были известны
только из отложений нижнего и среднего палеозоя. Neopilina—
«живое ископаемое», представляющее группу, которая, видимо,
на много миллионов лет нашла убежище на дне глубоководной
части океана.
Эволюционное положение моноплакофор, анатомия которых
известна только по Neopilina, все еще является спорным. Во-
первых, неясно, можно ли считать полусегментированное строе-
ние тела примитивных моноплакофор доказательством того,
что моллюски произошли от сегментированных кольчатых чер-
вей. Уже давно аннелиды и моллюски считаются родичами из-
за сходства ранних стадий их онтогенеза. Во-вторых, возник
вопрос, действительно ли моноплакофоры, вроде Neopilina, да-
ли начало высшим классам моллюсков. На большинстве схем,
реконструирующих облик «первого моллюска», имеется только
по одной паре каждого органа. Если Neopilina находится на
главной линии ранней эволюции моллюсков, то возникновению
высших классов моллюсков должно было предшествовать ис-
чезновение сегментации. Нет веских доказательств того, что
группа, к которой относится Neopilina, дала начало какому-
либо другому классу моллюсков. Несмотря на свое неясное фи-
логенетическое положение, Neopilina — один из самых важных
живых таксонов, открытых в последние десятилетия.
Наиболее плодотворным для выяснения филогенезов оказы-
вается совместное рассмотрение палеонтологических и неонто-
Фиг. 147. Monoplacophora [117].
Е Современный вид NeWtlina galatheae Lemche (А п В — брюшная сторона. Б и Г —
спинная сторона, Д — боковая сторона, £ — вид спереди; 1 — ротовое отверстие, 2 — жаб-
?Ь1> *“1Край ноги, 4 — край мантии, 5 — анус, 6 — край раковины). Ж. Силурийская
<рорма РIlina unguis; видны отпечатки парных мышц, которыми прикреплялись тело-
и нога, на внутренней стенке раковины.
312 Часть 2. Применение данных палеонтологии
логических данных. Использование только палеонтологических
или только неонтологических данных может привести к ошиб-
кам. Мы не будем приводить конкретных примеров, но несостоя-
тельность большинства гипотез палеонтологов и неонтологов
о связях между таксономическими группами обусловлена имен-
но использованием односторонней информации.
Эволюция экосистем
В истории жизни большую роль сыграли различные новше-
ства, имевшие большое адаптивное значение. Многие из них
имели лишь весьма небольшое значение для общей экологиче-
ской истории Земли, тогда как другие были очень важны, по-
скольку прямо или косвенно приводили к новому образу жизни
и к освоению новых местообитаний. Так появлялись новые эко-
системы, и биосфера сильно изменилась с тех пор, как впервые
появилась жизнь.
Видимо, первым важным событием в истории жизни было
возникновение механизма самокопирования с помощью нукле-
иновых кислот. К сожалению, по палеонтологической летописи
трудно датировать это событие. С тех пор прошли сотни мил-
лионов лет, на протяжении которых осадки в значительной мере
изменялись.
Вторым важным событием в органической эволюции было
возникновение клетки. Древнейшие остатки организмов, обла-
давших, по-видимому, клеточным строением, найдены в форма-
ции Онвервах,т в Южной Африке (3,2 млрд, лет)- Эти остатки
представляют собой мельчайшие палочковидные и сферические
структуры, сохранившиеся в кремнистых сланцах и других мел-
козернистых осадочных породах. Баргхурном и Шопфом [7J опи-
саны лучше сохранившиеся палочковидные образования, напо-
минающие бактерий, и водорослевые сферы из формации Фиг-
Три в Южной Африке (минимальный возраст 3,1 млрд. лет).
Из древних докембрийских пород известны также строматоли-
ты (однако широкое распространение они, видимо, получили
только в среднем докембрии).
Наиболее полно изученные докембрийские отложения, содер-
жащие остатки микроорганизмов — кремнистые сланцы форма-
ции Ганфлинт (Онтарио, Канада) (около 1,9 млрд. лет). Все
микроорганизмы, описанные из этой формации, делятся на два
основных типа: структуры, сходные с сине-зелеными водоросля-
ми (иногда эти структуры образуют строматолиты), и напоми-
нающие бактерий структуры (фото XXXII). Сине-зеленые водо-
росли и бактерии отличаются от всех других современных кле-
точных организмов некоторыми примитивными чертами. Самая
важная из них состоит в том, что генетический материал таких
X. Эволюция и палеонтологическая летопись
313
клеток не упакован в хромосомы в специальной клеточной струк-
туре— ядре. Такие клетки, не имеющие хромосом и ядра, на-
зываются прокариотическими. Прокариотическая клетка может
содержать большой набор генов, и одна мутация вряд ли может
сильно изменить клетку, а поскольку прокариоты размножают-
ся бесполым путем, мутации медленно распространяются в по-
пуляции. Этим может объясняться необычайно низкая скорость
эволюции бактерий и сине-зеленых водорослей и сходство их
докембрийских представителей с современными [184].
Хромосомы и оформленное ядро характерны для клеточных
эукариот, к которым относятся все высшие организмы (в том
числе высшие водоросли и простейшие). Возникновение эука-
риотической организации было одним из важнейших шагов ор-
ганической эволюции. Видимо, оно открыло путь быстрой эво-
люции и дифференциации таксонов одноклеточных и многокле-
точных. Шопф [184] описал несколько остатков эукариотических
форм (возможно, это были высшие водоросли и грибы) из фор-
мации Биттер-Спрингс в Австралии (около 1 млрд, лет; фиг-
148). Отсутствие подобных форм в сходных породах формации
Гаифлинт и других изученных разрезах среднего докембрия при-
вело Шопфа к мысли, что переход от прокариотов к эукариотам
мог произойти не раньше позднего докембрия.
О том, как появились высшие (эукариотические) водоросли
и примитивные многоклеточные животные, палеонтологическая
летопись пока дает очень мало сведений. Древнейшие известные
фауны многоклеточных животных — фауна Эдиакары в Австра-
лии (фото XXXIV) и сходные фауны с других континентов.
Фауна Эдиакары состояла, по-видимому, только из мягкотелых
животных, часть которых можно отнести к кишечнополостным,
аннелидам и иглокожим (или сравнить с ними). Возраст фауны
Эдиакары известен недостаточно точно, но породы, вмещающие
ее, древнее тех, в которых найдены первые известные трилоби-
ты, и моложе пород, возраст которых, определенный радиомет-
рическим методом, составляет 700 млн. лет. Практическое от-
сутствие следов передвижения организмов, ходов или других
следов жизнедеятельности в докембрийских породах заставляет
предполагать, что до позднего докембрия многоклеточная жизнь
не была распространена- Видимо, она возникла вскоре после
перехода от прокариотов к эукариотам и быстро достигла боль-
шого разнообразия.
Предполагается, что в первичной атмосфере Земли не было
кислорода, а его накопление — главным образом результат фо-
тосинтезирующей деятельности растений. Беркнер и Маршалл
[11] предположили, что содержание кислорода в атмосфере до-
стигло уровня, при котором возможна жизнь клеточных живот-
ных (около 1 % содержания кислорода в современной атмосфе-
314
Часть 2. Применение данных палеонтологии
Фиг. 148. Glenobotrydion aenigtnatis Schopf из формации Биттер-Спрингс (до-
кембрий, Австралия) [184].
Возможно, видны стадии клеточного деления. Как полагают, темные тельца (я) могут
быть остатками клеточных ядер.
ре), только в начале кембрия. Фишер [61] предположил, что
в конце докембрия в водной среде, в тех участках, на которых
развивалось много растений, создавались местные «кислород-
ные оазисы», в которых могла происходить эволюция многокле-
точных морских животных, когда содержание кислорода в ат-
мосфере на поверхности Земли было еще недостаточным для
развития здесь клеточной животной жизни.
Внезапность появления разнообразных раковинных беспоз-
воночных в кембрии вызвала к жизни немало остроумных гипо-
тез. Брукс и Хатчинсон независимо друг от друга предположи-
ли, что основное адаптивное преимущество наружного скелета
состояло в защите от недавно появившихся хищников [85]. Дру-
гие полагают, что быстрая эволюция морских организмов была
обусловлена значительным изменением химического состава вод
океана. Беркнер и Маршалл, а также Фишер предположили,
что наружный скелет ранних организмов защищал их от ультра-
фиолетового излучения Солнца. Слой озона (О3), экранирую-
щий поверхность Земли от интенсивного ультрафиолетового из-
лучения, смог образоваться только после того, как атмосфера
достаточно обогатилась кислородом. Даже после того, как был
достигнут уровень содержания кислорода, при котором живот-
ная жизнь может развиться, поверхности Земли все еще могло
достигать ультрафиолетовое излучение, опасное для животных.
X. Эволюция и палеонтологическая летопись
315
Каковы бы ни были причины внезапного появления различ-
ных раковинных морских животных в кембрии, приобретение
наружного скелета открыло множество новых путей для эволю-
ции. Наружные скелеты служили и для защиты от хищников, и
в качестве опоры для мышц в тех группах, которые не могли бы
существовать, оставаясь мягкотелыми.
Мы здесь перечислим адаптивные приобретения фанерозоя,
имевшие широкое экологическое значение.
1. Возникновение скелета позвоночных (ордовик?).
2. Развитие челюстей и плавников у рыб, приведшее к ши-
рокому использованию нектонных морских и пресноводных ме-
стообитаний (силур и девон).
3. Возникновение наземных растений, обеспечившее выход
позвоночных на сушу (силур—девон).
4. Возникновение ног и легких у предков амфибий, обусло-
вившее завоевание суши позвоночными (девон).
5. Появление амниотического яйца рептилий. Благодаря ему
рептилии освободились от необходимости размножаться только
в водной среде и смогли приобрести разнообразные приспособ-
ления, позволившие им занять различные ниши в наземных
местообитаниях (карбон).
6. Развитие дыхательного аппарата и крыльев у насекомых,
способствовавшее быстрому увеличению разнообразия этих жи-
вотных (карбон).
7. Появление у растений способа размножения с помощью
пыльцы и семян, позволившее голосеменным растениям размно-
жаться не только во влажных средах и занять множество на-
земных местообитаний (поздний палеозой).
8. Широкая дифференциация раковинных морских беспозво-
ночных, приобретение ими специальных органов, обеспечивших
возможность обитания в субстратах (сюда входят сифоны дву-
створчатых и брюхоногих моллюсков и петалоидные амбула-
кральные ножки морских ежей; все эти группы в мезозое до-
стигли громадного разнообразия).
9. Появление у рептилий таза, характерного для динозавров.
Это обеспечило динозаврам замечательные способности к пере-
движению и позволило им занять огромное множество наземных
местообитаний (триас).
10. Возникновение покрытых перьями теплокровных птиц,
которые значительно превзошли своих предков — летающих
рептилий — по диапазону адаптации (юра).
11. Появление цветка и защищенного семени у покрытосе-
менных. Это приобретение открыло путь интенсивной адаптив-
н°й радиации покрытосеменных в ранее не занятые ниши (ран-
ний мел).
316
Часть 2. Применение данных палеонтологии
12. Появление основных признаков млекопитающих, особен-
но плацентарное развитие плода, за чем последовали многие
сложные адаптации млекопитающих кайнозойской эры (триас—
.мел).
13. Появление травянистых покрытосеменных растений. Бла-
годаря им возникло множество новых луговых и степных ниш
для млекопитающих и ниш в зарослях морской травы для бен-
тосных беспозвоночных (ранний кайнозой).
14. Возникновение хватательных рук и стереоскопического
зрения у приматов, обеспечившее огромное увеличение возмож-
ностей их мозга (ранний кайнозой). •'
Точных датировок большинства крупных адаптивных прио-
бретений все еще не существует. Но поскольку эти приобрете-
ния означали большой экологический успех и обусловливали
появление новых высших таксонов, можно думать, что они поя-
вились незадолго до интенсивных адаптивных радиаций. Таким
образом, палеонтологическая летопись может служить довольно
надежным источником данных о приблизительном времени воз-
никновения главных адаптивных новшеств и о хронологии за-
селения основных местообитаний на Земле.
ГЛАВА XI
Биостратиграфия
Стратиграфия изучает распространение, состав и хронологи-
ческую последовательность образования осадочных горных по-
род. Особое внимание уделяется истории этих пород и содержа-
щихся в них ископаемых остатков. Биостратиграфия является
ветвью стратиграфии, изучающей главным образом ископаемые
остатки. Биостратиграфия в широком смысле включает в себя
большинство тем, затронутых в этой книге. Биостратиграфия
в узком смысле, как она рассматривается в данной главе, изу-
чает пространственное распространение и временные связи ис-
копаемых остатков и вмещающих их пород.
В 1961 г. Американская комиссия по стратиграфической но-
менклатуре опубликовала в Бюллетене Американской ассоциа-
ции геологов-нефтяников Кодекс стратиграфической номенкла-
туры. Этот стратиграфический Кодекс — не просто свод правил
номенклатуры; он в большей мере, чем Международный ко-
декс зоологической номенклатуры, освещает общие научные
концепции, и поэтому в нем содержится больше противоречивых
утверждений, чем в зоологическом кодексе. Тем не менее это
ценная сводка господствующих воззрений в стратиграфии, спо-
собствовавшая созданию единообразия в классификации и на-
званиях стратиграфических подразделений. Она, несомненно,
потребует пересмотра в соответствии с изменениями общих
принципов и методов в будущем.
В общем стратиграфия имеет лишь очень немногочисленные
однозначные основополагающие принципы и характеризуется в
значительной мере субъективностью подхода. Чуть ли не един-
ственным непреложным стратиграфическим принципом является
«закон» суперпозиции, гласящий, что в любой ненарушенной
осадочной толще наиболее древний пласт подстилает толщу, а
наиболее молодой располагается в ее кровле. При решении стра-
тиграфических проблем обычно применяются многие принципы
палеоэкологии, эволюции, седиментологии и геоморфологии. На-
пример, так называемый «закон биологической сукцессии», ча-
сто обсуждавшийся на первых, ставших теперь историческими
Дискуссиях по биостратиграфии, утверждает, что биоты в тече-
ние геологического времени сменяли друг друга в определенной
318
Часть 2. Применение данных палеонтологии
последовательности. Эта последовательность является просто
результатом органической эволюции и могла бы соответственно
рассматриваться как часть эволюционной палеонтологии, заим-
ствованная биостратиграфией.
Литостратиграфические единицы
Стратиграфия имеет дело, во-первых, с подразделениями
толщ пород (для расчленения стратиграфических разрезов) и,
во-вторых, с единицами геологического времени (для деления
геологической истории). На основании литологического состава
и физических свойств пород в стратиграфической летописи вы-
деляются объемные толщи. Подразделения горных пород, часто
называемые литостратиграфическими единицами, выделяются
вне их временных отношений или содержания в них ископаемых
остатков, за исключением тех случаев, когда последние влияют
на физический или химический состав пород. Подразделения
устанавливаются на основе литологического однообразия, оцен-
ка которого неизбежно связана с большой долей субъективно-
сти. Основной литостратиграфической единицей является свита
(formation). Свиты объединяются в единицы более высокого
ранга, называемые сериями (groups), и подразделяются на еди-
ницы более низкого ранга, называемые пачками (members).
Считается, что необходимым условием установления свиты яв-
ляется возможность ее картирования. Однако вследствие боль-
шого разнообразия масштабов геологических карт это условие
имеет сомнительную ценность. Подобно таким во многом про-
извольным таксономическим подразделениям, как «род» или
«семейство», свита является категорией, не поддающейся коли-
чественному описанию. Как и в случае высших таксономических
подразделений в биологии, в геологии многие исследователи
также пытаются развить общий принцип установления объема
литогенетических единиц. Вместе с тем в стратиграфии, как
и в таксономии, не существует единого мнения об объеме того
или иного подразделения. Как правило, с увеличением объема
информации возрастает степень детализации подразделений.
Хорошо известно, что стратиграфические данные далеко не
полны. Наличие поверхностей размыва и несогласного залега-
ния свидетельствует о существовании больших перерывов в
осадконакоплении.
Большинство литогенетических подразделений, которые, ка-
залось бы, являются результатом непрерывного процесса осад-
конакопления, на самом деле формировалось с перерывами. При
определенных условиях, например в замкнутых эвапоритовых
бассейнах, химическое осаждение может приводить к почти
непрерывному осадконакоплению. Однако в большинстве других
XI. Биостратиграфия 31 &
бассейнов накопление осадков происходит спорадически. Почти
все морские обломочные осадки подвергаются в той или иной
степени переносу, прежде чем достигнут места окончательного
осаждения. Разумеется, многие из них переносятся в морские
бассейны осадконакопления с суши. Если осадочный материал
доставляется в бассейн осадконакопления почти непрерывно
(обычно скорость поступления непостоянна), то окончательное
накопление осадочных слоев в каждый данный момент проис-
ходит только на определенных участках дна бассейна. В других
участках бассейна в это время осадки могут не накапливаться
или может даже происходить размыв ранее накопившихся сло-
ев- Периодические кратковременно действующие факторы, на-
пример штормы, могут порождать волны и течения, размываю-
щие мощные осадочные толщи, которые накапливались в тече-
ние многих лет.
Отдельный слой или прослой отражает единичное событие
в процессе накопления. Поверхности между слоями или прослой-
ками в таком случае указывают на перерывы в процессе осад-
конакопления так же, как несогласия свидетельствуют о подоб-
ных же перерывах на обширных территориях. Еще в начале
XX века Баррел предложил различать непрерывное накопление
и накопление с перерывами. Перерывы, соответствующие крат-
ковременному прекращению осадконакопления или размыву
осадков, он предложил называть диастемами.
Один из наиболее поразительных примеров прерывистости
процесса накопления осадочных пород выявляется при изуче-
нии сохранившихся следов роющих животных- На фиг. 149 и фо-
то XXXIII, взятых из работы Роудса [163], представлен штуф
силурийской породы, вырезанный перпендикулярно напластова-
нию. В породе видны три отчетливых типа ходов, оставленных
роющими животными. Некоторые ходы нарушают слоистость.
Ходы животных проникают вниз от границы раздела осадок —
вода на глубину нескольких сантиметров. Если накопление было
непрерывным и деятельность роющих организмов происходила
одновременно с ним, то можно предполагать, что роющие жи-
вотные в период накопления осадков оставались в нескольких
верхних сантиметрах и сильно разрушали первичное напласто-
вание.
На разрезе штуфа видны четыре слоя (А—D), в которых
слоистость не была целиком разрушена. Верхние прослойки
в этих слоях (особенно во втором слое снизу) частично разру-
шены роющими животными. Роудс пришел к заключению, что в
этом штуфе запечатлены четыре отчетливых периода осадкона-
копления. Тщательные наблюдения показывают, что четыре от-
четливо слоистых слоя, мощность каждого из которых составля-
ет несколько сантиметров, последовательно отлагались друг за
320
Часть 2. Применение данных палеонтологии
*st
Граница. 4
— Граница 3
Граница 2
10 см
Фиг. 149. Схематический разрез штуфа породы из формации Стонхауз (си-
лур, Новая Шотландия) с ходами животных (см. фото XXXIV),.
-----Граница 1
другом. Каждый новый слой формировался над подстилающим
слоем очень быстро. Граница осадок—вода, на которой отлагал-
ся каждый слой, показана на фото XXXIII. Во многих случаях
нижняя поверхность слоев отличается хорошей сохранностью
вследствие того, что роющие организмы локализовались лишь
в верхней части слоя, у границы осадок—вода.
Перемещение организмов в вертикальном направлении пока-
зано на фиг. 149 жирными стрелками, пересекающими обычные
ходы, большинство которых ориентировано горизонтально. На-
личие этих вертикальных ходов подтверждает гипотезу о вне-
запном осаждении, поскольку, очевидно, они проделывались
беспозвоночными для возврата вверх, к границе раздела вода —
осадок. Некоторые животные оказались не столь «удачливыми»,
вероятно потому, что были неспособны проложить себе путь
наверх через вновь сформированный слой осадка. Гипотеза
о «мгновенном» осаждении подтверждается еще и тем, что слой
В, нижние прослойки которого в наименьшей степени наруше-
XI, Биостратиграфия
321
ны роющими организмами, был наиболее мощным из всех че-
тырех.
Сходный анализ, проведенный Зейлахером [187], показал,
что многие известные флишевые толщи Европы формировались
путем последовательного осаждения новых слоев, отлагавшихся
в короткие периоды времени и с перерывами. Большинство сле-
дов зарывания и ползания во флишевых отложениях сохраняет-
ся в подошвах песчаников, подстилаемых глинистым сланцем.
В течение многих лет не существовало единого мнения о том,
происходило ли зарывание только на границе между глинами
и песком или же организмы проникали через всю толщу одно-
родного песчаника, причем следы их жизнедеятельности сохра-
нялись лишь на нижней поверхности пласта. Используя различ-
ные доказательства, Зейлахер пришел к выводу, что одни орга-
низмы оставляли следы только на поверхности осадка, тогда
как другие проникали в осадок на значительную глубину. Он
далее ограничился рассмотрением последней группы организ-
мов и показал, что каждый вид проникал до определенной, ха-
рактерной для него глубины. Поэтому если каждый слой обра-
зовывался за короткий период времени, то должна была суще-
ствовать какая-то максимальная мощность для всех песчаных
пластов, в которых встречаются различные типы подповерхно-
стных зарываний. На фиг. 150 представлены данные по различ-
ным типам зарываний, подтверждающие такое предположение
и гипотезу о быстром осаждении.
Приведенные примеры представляют благоприятную воз-
можность для демонстрации прерывистого накопления. Есть
основания полагать, что большинство осадочных пород осажда-
лось подобным же образом независимо от того, происходило ли
накопление отдельных слоев с перерывами продолжительностью
в дни, годы или в тысячи лет.
Осадки, отлагающиеся одновременно в разных местах, раз-
личаются по своим химическим и физическим свойствам. Обра-
зовавшееся на большой площади в течение определенного про-
межутка времени осадочное тело может отражать изменения,
I происходившие на земной поверхности. Части осадочного тела,
] отражающие изменения по простиранию, называются фациями.
I При распознавании фаций основное внимание обращают на
литофации, выделяемые по характерным литологическим свой-
ствам, и биофации, выделяемые по содержащимся в них орга-
ническим остаткам. Фации можно выделять внутри литогенети-
ческих подразделений, которые, согласно принятым правилам,
делятся на серии, свиты и пачки. Фации используются для оп-
ределения боковых границ между литогенетическими единица-
ми, хотя эти границы обычно бывают не резкими, а постепен-
ными.
21—4R7
Постседиментационные следы
жизнедеятельности
Председиментационные следы
жизнедеятельности
Й> _ д» § и § я .а | 2 II 3 дЗ § ’й -S S 3 S <в "§. G S? 3 и о .о Й §§. 5-2 § “ °- ° Palaeodictvon й <3 g Я* лат «Я» rm гтч жкч S. 8.» Ш vi Ш И 8&- Я
л Х'ЧЬ1 • -VZ'l (МИ 11 1
4
0-1 21-3 6 - - 1-2 41 12 16 19 3 5 2-3 33 14 18 18 7 8 3-4 19 17 9 15 9 7 -5 18 14 7 15 7 4 -6 17 15 8 10 14 4 4-1 10-2 3-- 13 28 1276 512 12 17 451 423 1-- 941 -4- 212 1-- 934 131 1-1 446 1-1 1 - - 3-- 2 - - 4 - - 2 - - 1 - - 9-- 3-- 415 31- 1- 121- 5-1 1-- 726 12- 52- 8 - - 52- 2 - - 237 2 - - 21- 43- 3 - - 1-- 416 33- 13- 12- 441 1-- 123 221 41- 25-
-7 12 11 10 11119 -8 10 11 8 7114 5 16 ЦКот — 6-2 1 -1 liQRff 415 л. 5 2- 3 - - -1- 2-6 1-- 12- 23-
-9 12 14 6 10134 -10 12 5 2 12 4 2 -12 13 17 5 11 16 3 -14 7 16 4 7 16 4 — 2 -2 1 - - 6 - - «Кв 3 13 Д 1 - - 1 - - 1-2 1-3 11- 41- 32- 2 - - 1-- 21- 21- - 1 - 51- 673 21- 44- 11- 1-- 273 -3- 13- -3-
-16 5 5 5 6 5 5 -18 7 6 1 7 6 1 -1 - 2 - - 1-- 2 - - 2 - - 312 1 - - 21- 1-- -2- 31- 2 - -
-20 8 8 - 6 8 - -25 14 10 — 12 10 - -30 9 10 1 791 -35 10 6 - 10 6 - -40 4 3 - 4 3 - -45 7 7 — 7 7 - -50 4 7 - 3 7 - -60 8 3 - 8 3 - -л -л 1 —а | w СП Ю 1 -* 1 | —а. —а | | 1 1 1 1 1 1 1 1 Ml 1 । । . —* —а —а —а | —*> | | 1 1 i 1 1 1 1 1 — 1 1 -* 1 1 1 > | | | -4. -А J | 1 1 1 1 1 1 1 1 Mi i -* 1 1 М 1 | | -4. -Л | _Л | | 1 1 1 1 1 1 1 1 СО —• 1 ГО М СО СП ГО 1 -Л j | _* -Л _4. | 1 1 1 1 1 1 1 1 СО 1 1 1 1 — СО I —а । —а «и. —а —а | _а 1 1 1 1 1 1 1 1
-80 7 5 - 7 5 - -100 2 3 - 2 3 - -150 12 8 - 12 7 - -200 4 7 - 3 7 - —* ГО I 1 I 1 1 1 1 1 1 1 -‘111 1 — 1 1 till 1 i I -* -* 1 1 Illi 1 Ю 1 -* Mill 1 1 1 1 -*111 1111 1 J ’ 1 1
Сумма 655 493 6 130 60 28 72 24 121 49 >9 75
Фиг. 150. Следы зарывания и
ползания, сохранившиеся на
нижних поверхностях пластов
в трех разрезах (А, В и С) фли-
ша северной Испании [187].
Постседиментационные типы следов
жизнедеятельности приурочены к
пластам с небольшой мощностью;
председиментационные следы жизне-
деятельности не связаны с мощно-
стью слоев.
XI. Биостратиграфия
323
Биостр атигр афические подр аз деления.
Биозона
На систему литогенетических подразделений накладывается
система подразделений, установленных на основании присутст-
вия тех или иных ископаемых. Единицы этой системы подразде-
лений, называемые биостратиграфическими единицами, пред-
ставляют собой вполне реальные осадочные толщи, но их гра-
ницы определяются на основании различных палеонтологиче-
ских критериев, таких, как появление, максимальное распро-
странение и исчезновение ископаемых видов или родов в раз-
личных местных разрезах.
Основное биостратиграфическое подразделение — зона. Из-
вестны зоны разного типа. Существует понятие биозоны — это
зона, охватывающая все породы земного шара, образовавшиеся
за время существования соответствующего вида. Биозона — это,
в сущности, абстрактное понятие, так как выделить ее на самом
деле никогда не удается.
Ни один вид не бывает представлен во всех породах соот-
ветствующей ему биозоны.
Почему же виды не заполнили свои биозоны? Можно пред-
положить, что многие виды возникли в результате эволюцион-
ной дивергенции географически изолированных популяций ранее
существовавшего вида. Заполнение возникшим таким образом
видом соответствующей биозоны частично зависело от того, на-
сколько быстро этот вид распространялся в разных районах зем-
ного шара, в которых он потенциально мог обитать. В некото-
ром смысле оказывается, что мы сравниваем скорости эволюции
и географического расселения.
Если бы не наличие эффективных преград, то большинство
видов заняло бы обширные географические площади довольно
быстро по сравнению со скоростью их эволюции. Многочислен-
ные примеры быстрого расселения видов при условии доступно-
сти новых ареалов приведены в превосходной книге Элтона [53]
по экологии нашествий. Однако в действительности расселению
видов мешают различные географические преграды. Биогеогра-
фические границы, особенно их протяженность по широте, оп-
ределяются условиями среды, в первую очередь температурой.
Кроме того, океаны служат преградами для расселения назем-
ных животных, а суша—для расселения обитателей моря. Эф-
фективными преградами могут быть также резкие изменения
высоты над уровнем моря или океанские глубины.
Трудно сформулировать общие критерии для определения
эффективности тех или иных преград в распространении вида.
Иногда преграды преодолеваются, причем отчасти это дело слу-
чая. Несомненно, расселение некоторых видов, несмотря на, ка-
21*
324
Часть 2. Применение данных палеонтологии
залось бы, непреодолимые преграды, произошло за счет дейст-
вия совершенно случайных факторов, таких, как сильные ветры,
дрейфующий плавник и т. д. Симпсон [193] назвал случайные
агенты распространения «счастливыми путями». Например, цепи
островов часто оказываются «счастливыми ступеньками» при
миграции как наземных, так и мелководных морских видов с од-
ного континента или континентального склона на другой.
Более непрерывные, но узкие пути распространения иногда
фигурируют под наименованием «коридоров миграции». Вслед-
ствие своей узости такие коридоры в ходе геологической исто-
рии часто то закрывались, то вновь открывались. Например,
Панамский перешеек выступал в роли континентального кори-
дора, ведущего в Южную Америку, для североамериканских
плацентарных, что позволило им вытеснить в конце плиоцена
ранее изолированные южноамериканские группы сумчатых.
У многих групп растений и животных выработались специ-
альные приспособления для преодоления преград. Например,
личиночные формы, ведущие планктонный образ жизни, можно
считать расселительными стадиями жизненного цикла некото-
рых бентосных морских беспозвоночных. Выяснилось, что у
большинства планктонных личинок на какой-то период задержи-
вается метаморфоз, если им не удается обнаружить подходя-
щий субстрат. Долгое время считалось, что перенос личинок
мелководных беспозвоночных через Атлантику невозможен.
Однако Шелтема [180] выяснил, что личинки мелководных бен-
тосных видов, обнаруживаемые на обоих побережьях Атланти-
ческого океана, могут переноситься через Атлантику в направ-
лении с запада на восток и с востока на запад с помощью ос-
новных морских течений (фиг. 151). Распространение многих
групп наземных растений обусловлено прежде всего ветровым
переносом спор и семян. При неблагоприятных условиях среды
многие водные и наземные одноклеточные организмы перехо-
дят в пассивную, покоящуюся стадию (цисты) и в такой фор-
ме, подобно спорам, могут переноситься ветром; так вид может
преодолевать преграды, непреодолимые иным способом.
Довольно часто бывает так, что вид исчезает из большей ча-
сти своего ареала, но продолжает сохраняться в пределах ог-
раниченной области, образуя здесь одну или более так называ-
емых реликтовых популяций.
Отсутствие ископаемых остатков вида во многих частях его
биозоны может объясняться, в частности, тем, что даже в пре-
делах своего географического ареала виды, как правило, при-
урочены к определенным благоприятным для них условиям
среды. Соответственно отмечается приуроченность многих иско-
паемых видов к определенным породам. Такие формы можно
назвать «фациальными ископаемыми». В большинстве случаев
XI. Биостратиграфия
325
Фиг. 151. Распределение планктонных личинок гастропод сем. Architectoni-
cidae в Атлантическом океане [180].
Точки с кружками отмечают станции, в которых были обнаружены личинки Philippia
krcbsii. Взрослые особи, принадлежащие к сем. Architectonicidae, обитают на мелководье
в тропиках. Стрелки показывают главные поверхностные морские течения.
их локальное распространение отражает первоначальную эко-
логию. Кроме того, ни один вид не сохраняется во всех литоге-
нетических толщах, формировавшихся в тех условиях, в кото-
рых он обитал. И наконец, возможно, что тот или иной вид на
самом деле присутствует в литогенетической толще, но из-за
неправильного определения, обусловленного плохой сохранно-
стью или ошибкой исследователя, он там не найден.
Во многих частях земного шара преобладающие ветры и
морские течения нарушают широтные температурные градиенты,
Резкие колебания температур и горные преграды обусловлива-
ют тенденцию к ограничению распространения групп видов
пределами определенных регионов, которые именуются биогео-
графическими провинциями. Виды, обитающие только в данном
районе, называются эндемичными. В большинстве ныне выделяе-
мых биогеографических провинций имеется некоторое число эн-
демичных видов, но в них встречаются также виды, представ-
22—467
326
Часть 2. Применение данных палеонтологии
фиг. 152. Биогеографические провинции, выделенные различными авторами
для Атлантического побережья США и побережья Мексиканского залива
/221].
ленные в других провинциях. Поэтому границы провинций не
всегда оказываются четко очерченными; расхождения во мнени-
ях относительно точного разграничения провинций и наиболее
удобной степени подразделения обусловили разнообразие си-
стем деления на провинции для некоторых частей света.
Например, для прибрежных вод восточной части Северной
Америки предложено несколько различных подразделений на
провинции (фиг. 152). Восточная и западная границы выделяе-
мых биотических провинций — это континентальный склон и бе-
реговая линия. Гольфстрим поворачивает от берега у мыса Гат-
XL Биостратиграфия
327
терас, что вызывает резкий перепад температуры. Лабрадорское
течение, направляющееся на юг вдоль северо-восточного побе-
режья, отклоняется в сторону океана у мыса Код.. Северная и-
южная границы провинций в других частях света не обязатель-
но совпадают с границами, представленными на фиг. 152 (как
по широте, так и по среднегодовой температуре).
В главе, посвященной палеоэкологии, обсуждалось соотно-
шение между температурными градиентами и градиентами так-
сономического разнообразия. Можно было бы ожидать, что ши-
роко распространенные теплые климаты прошлого должны бы-
ли обусловить возникновение обширных биогеографических
провинций, а резкие температурные градиенты приводили к су-
жению таких провинций. Лучше всего изучен в этом отношении
юрский период. Аркелл [3] и другие авторы отметили, что мно-
гие раннеюрские таксоны (особенно головоногие моллюски)
были космополитами; например, для каждого установленного
североамериканского вида аммонитов имеется европейский
двойник. Наоборот, фауны средней и поздней юры четко распре-
делены по провинциям. Аркелл различал три крупные поздне-
юрские фаунистические провинции, которые он называл
областями (realms)1: Бореальная область (северные широты);
Тихоокеанская область и область Тетиса (вся остальная часть
земного шара, но особенно характерны для этой области иско-
паемые фауны Средиземноморья). Области эти возникли не
сразу, а ареалы их фаун время от времени смещались и ча-
стично перекрывались. Однако в целом эти три области сохра-
нялись, и их можно распознать также в меловом периоде.
Большинство исследователей считали, что отчетливая обособ-
ленность этих областей была обусловлена развитием резких тем-
пературных градиентов и физиографических преград, препятст-
вующих расселению животных. Однако Халлам [72] высказал
предположение, что Тихоокеанскую область Аркелла следует
считать частью области Тетиса. Он предположил также, что
причиной появления и сохранения Бореальной области послу-
жило вовсе не воздействие низких температур или наличие фи-
зиографических преград; он считал, что эта область представ-
ляла собой огромное внутреннее море с несколько пониженной
соленостью. Халлам сравнивал фауны этого моря, характери-
зовавшиеся незначительным таксономическим разнообразием,
с фаунами современного Балтийского моря. Но что бы ни яви-
лось причиной фаунистической дифференциации в юре, не-
1 В советской литературе биогеографической провинции (province) авто-
ров настоящей книги соответствует палеобиогеографическая область, разде-
ляемая на палеобиогеографические провинции. Единицу палеобиогеографи-
ческого районирования крупнее области некоторые советские исследователи
именуют поясом [соответствует области (realm) Аркелла!. —Прим. ред.
22*
328
Часть 2. Применение данных палеонтологии
трудно понять, почему межконтинентальная корреляция, как
правило, оказывается более простой и четкой для нижнеюрских
отложений, чем для верхнеюрских. <
Валентайн [221] отметил, что биогеографическим подразде-
лениям определенного ранга должны соответствовать биострати-
графические единицы определенного ранга.
Корреляция по ископаемым
Поскольку ископаемые виды не полностью занимают интер-
валы своих биозон, возникает вопрос: в какой мере стратигра-
фическое распределение ископаемых можно использовать для
установления относительного геологического возраста? Ясно, что
при помощи биостратиграфических подразделений временные
соотношения удается определять лишь весьма приближенно.
Установление временной эквивалентности двух пространствен-
но разобщенных стратиграфических единиц называется корре-
ляцией. К сожалению, некоторые исследователи используют
термин «корреляция» также в значении эквивалентности типов
пород или обнаруженных ископаемых независимо от фактора
времени. Для описания связей такого рода лучше использовать
термины «литологическая эквивалентность» и «биостратиграфи-
ческая эквивалентность».
Хронологическую корреляцию можно проводить различны-
ми методами; лишь некоторые из этих методов основаны на
изучении ископаемых остатков, и тем не менее ископаемые ока-
зываются наиболее важным инструментом, используемым при
хронологической корреляции. Прежде всего, как правило, уста-
навливают биостратиграфическую эквивалентность, а затем уже
эту эквивалентность можно интерпретировать как приблизи-
тельно соответствующую хронологической. В последующих раз-
делах речь пойдет о нескольких типах биостратиграфической
эквивалентности.
ИНТЕРВАЛЫ СТРАТИГРАФИЧЕСКОГО
РАСПРОСТРАНЕНИЯ И ЗОНЫ
Большинство стратиграфических исследований основано на
изучении вертикального распространения ископаемых остатков
в местных стратиграфических разрезах, вскрывающих последо-
вательность пластов и доступных для изучения. Стратиграфиче-
ский разрез представляет собой трехмерное обнажение пластов
горных пород, часто располагающееся на склоне холма или
вдоль русла ручья. При корреляции обычно считается, что раз-
рез имеет одно измерение- Двумерные колонки, используемые
для изображения стратиграфических разрезов в геологической
XI. Биостратиграфия
329
Описание
Серые, мягкие мергелистые известняки
с фукоидами
Серые, плотные известняки с многочисленными
Idiohamltes comanchensis и I. spp.
Серые карбонатные аргиллиты, содержащие
немногочисленные прослои известняков и желези-
стые конкреции прослои мергелистых известняков
Серые^лергелистые известняки, мергели и карбо-
натные аргиллиты с прослоями песчаников и
многочисленными Pholadomya sanctisabae,
Proiocardia texana, Tapes ^.,Cryphaca navia,
а также редкими остатками Oxytropidoceras
belknapi
Желтые и желтовато-коричневые мергели и
мергелистые известняки с многочисленными
Gryphaea navta
Черные, тонкослоистые восковые аргиллиты
(вверху-известковые, внизу—песчанистые, с
прослоями известняков, содержащими Cuprime-
ria texana и Oxytropidoceras belknapi)
-^^Желтовато-коричневые мергелистые известняки
с многочисленными Cyprimeria texana и
^^Oxytropidoceras belknapi
ТКелтовато-коричневые известковые слабопесча-
^\нистые аргиллиты
Светлые плотные известняки
Известняки
Прослои
песчаников
Мергели
Мергелистые
известняки
Карбонатные
аргиллиты
Аргиллиты
Фиг. 153. Стратиграфическая колонка формации Киамичи (нижний мел,
Мичем-Филд, штат Техас, США) [152].
литературе, служат только для того, чтобы показать типы по-
род на различных уровнях (фиг. 153). Выбор методики изуче-
ния распределения фауны в стратиграфическом разрезе опре-
деляется условиями местности, структурными и текстурными
особеннсстями пород, а также характером самих ископаемых.
Обычно при стратиграфических исследованиях мощности пла-
стов измеряются в футах или метрах. Ископаемые тщательно
Часть 2. Применение данных палеонтологии
Сводный
разрез Отдел Зона Подзона
р-^ирджилиен
зазрили Цискоан
^^рМиссурий
i
$
сз
§
h-M Бендиан
ррили Морроу
§
I
§
ts
Формация
Парадис
Trittcttes
cullomensls
В этой работе
не выделена
Wedekindelllna
ultimata
Fusulina
eximia
Fusulina
girtyi
Wedekindelllna
euthysepta
Fusulina
hayensts
Fusulinella
whitensls
Fusulinella
prospectensis
Fusulinella
devexa
Зона Profusulinella
Зона Millerella
Фиг. 154. Сводный разрез нижней и средней части известняков
собираются по всему разрезу, их положение точно фиксируется
и отмечается интервал их распространения (фиг. 154).
В обычном методе корреляции учитываются нижняя и верх-
няя границы стратиграфического интервала распространения
одного вида в двух или более стратиграфических разрезах. При
определенных обстоятельствах эти «самое раннее» или «самое
позднее» появления указывают на приблизительный временной
эквивалент. Однако они могут соответствовать разным момен-
XI. Биостратиграфия
331
Стратиграфическое распространение видов фузулинид
Хорквилла (карбон, штат Аризона, США) [169].
там времени — вследствие миграции и различий во времени вы-
мирания вида в разных районах. Палмер [148] установил, что
позднекембрийские трилобиты в пределах различных участков
Большого бассейна (США) появлялись и исчезали в разное вре-
мя. Каждая из трех фаун, показанных на фиг. 155, по-видимому,
представляла ветвь основного ствола трилобитов, развивавше-
гося в глубоководных зонах моря. Детальная корреляция пока-
зывает, что после своего появления вторая фауна мигрировала
332
Часть 2. Применение данных палеонтологии
Запад
Восток
'X
Берег
у Фауна 3
-v Исчезновение
> Фауна 2
х Исчезновение
Сохранившиеся данные
Океан
Зона Cedaria
Зона Bolasptdella
Исчезновение
Зона Bathyuriscus-Elrathina
Фиг, 155. Схема, показывающая последовательные трансгрессии мелковод
ных позднекембрийских морей в западной части США [148].
По мере развития трансгрессий на восток распространялись трилобиты, главный ствол
которых находился в глубоководных зонах на западе. Горизонтальные штриховые линии
представляют изохронные уровни.
из глубоководных зон к береговой линии, по-видимому, вытес-
няя прежнюю фауну по мере своего продвижения к востоку.
После того как эта вторая фауна претерпела значительную эво-
люцию, она оказалась вытесненной третьей фауной, также миг-
рировавшей в восточном направлении. Таким образом, границы1
биостратиграфического подразделения, определяемые по каждой
из трех фаун (например, подразделения, заключающего птеро-
цефалиид), не совпадают. Миграция не была «мгновенным» со-
бытием. Объясняется это, вероятно, тем, что расселение живот-
ных происходило постепенно и являлось результатом экологи-
ческой конкуренции. Нет никаких данных, которые указывали
бы на то, что изменение фаун было связано с изменениями ок-
ружающей среды, поскольку основные фаунистические сдвиги
XI. Биостратиграфия
333
не совпадают с основными изменениями в характере отложений.
В свое время применялся также метод корреляции, основан-
ный на прослеживании тех интервалов разрезов, в которых ка-
кой-либо вид был представлен особенно обильно. Однако на
обилие вида при жизни, равно как и на посмертные изменения
его остатков, сильно влияют местные условия среды; поэтому
этот метод непригоден для корреляции разрезов, удаленных .
друг от друга на значительные расстояния. В то же время в ка-
ком-то определенном районе, в котором преобладали определен-
ные условия среды, корреляции, основанные на разумном ис-
пользовании данных о максимальном обилии, могут быть до-
статочно точными.
Корреляция, основанная не на одном, а на нескольких ви-
дах, более точна. Большинство признанных биостратиграфиче-
ских зон выделено на основании совместной встречаемости ос-
татков двух или более видов. Такая зона, обычно называемая
комплексной зоной (assemblage zone), как правило, получает
наименование по одному характерному роду или виду (причем
совсем не обязательно, чтобы этот род или вид был представлен
в зоне как по всей площади, так и в разрезе).
ПРОЦЕНТНОЕ СОДЕРЖАНИЕ ОБЩИХ ТАКСОНОВ
Некоторые корреляции были установлены путем сравнения
местных ископаемых комплексов со «стандартными» комплекса-
ми из тех районов, в которых биостратиграфические связи вы-
ражены более четко. Сходство в таких случаях, как правило,
выражают отношением числа таксонов (обычно видов), найден-
ных в исследуемом и в стандартном (типовом) комплексе.
Например, можно вычислить (в процентах) отношение числа
таксонов, найденных в комплексе, к числу таксонов, найденных
в другом комплексе, и использовать это отношение как коэф-
фициент для сравнения. Можно также использовать отношение
числа общих таксонов к общему числу уникальных таксонов в
обоих комплексах.
Симпсон [198] предложил для сравнения двух ископаемых
фаунистических комплексов следующий коэффициент:
Число общих таксонов х 100
Число таксонов в менее богатом комплексе
Этот коэффициент наиболее удобен в тех случаях, когда один
комплекс намного богаче другого. (Более бедная фауна обычно
дает менее полное представление об истинном комплексе.) Сим-
псон рассмотрел также методы сравнения комплексов по от-
носительным количественным показателям. Эти методы особен-
но важны в случае очень схожих фаун.
334
Часть 2. Применение данных палеонтологии
Корреляции, основанные на процентном отношении общих ви-
дов, можно проводить без учета стратиграфического распростра-
нения таксонов, однако точность корреляций при этом в значи-
тельной степени снижается в связи с влиянием экологических
факторов. Поэтому этот чисто статистический метод следует
применять с учетом наших знаний о границах биозон, а также
об экологии видов.
РУКОВОДЯЩИЕ ИСКОПАЕМЫЕ
Таксоны, на которых основывается корреляция, обычно на-
зывают руководящими ископаемыми. Идеальные руководящие
ископаемые должны удовлетворять следующим требованиям:
они должны иметь широкое географическое распространение,
обладать экологической толерантностью и высокими темпами
эволюции, иметь четко выраженные морфологические особенно-
сти и быть многочисленными. Руководящие ископаемые особен-
но важны для целей межрегионально?! и межконтинентальной
корреляции.
Лучшими морскими руководящими ископаемыми обычно яв-
ляются планктонные и нектонные виды, поскольку они широко
распространены и в меньшей степени зависят от местных усло-
вий, чем бентосные формы. Примером могут служить многие
ордовикские и силурийские граптолиты и мезозойские аммони-
ты. Некоторые бентосные группы, такие, как кембрийские три-
лобиты и позднепалеозойские фузулиниды, также являются
важными руководящими ископаемыми.
К сожалению, геологи часто не совсем верно используют тер-
мин «руководящие ископаемые». Часто при геологическом кар-
тировании таксон рассматривается как «руководящее ископае-
мое» и в том случае, когда он характерен лишь для данной ли-
тостратиграфической единицы и поэтому может использовать-
ся для сопоставления разрезов изолированных обнажений, ког-
да одних литологических данных для этого недостаточно. Упо-
требление термина «руководящие ископаемые» в этом значении
вносит путаницу, и его следует избегать.
МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
Хорошие результаты при корреляции (особенно при сопо-
ставлении разрезов кайнозойских отложений) можно получить,
используя внезапные морфологические изменения, появляющие-
ся у какого-либо вида или рода. Особенно характерен пример
с изменением направления навивания раковины у планктонных
фораминифер. Так, изменение направления навивания у Globo-
rotalia menardii использовалось для установления границы меж-
ду плиоценом и плейстоценом по колонкам донных осадков
Фиг. 156. Изменения в направлении навивания раковин у ископаемых фора-
минифер (отнесены к виду Globorotalia menardii, но, возможно, принадлежат к
нескольким видам) [56].
впрУ^аггТКИ отбоРа колонок донных осадков. Б. Направление навивания раковин (Л —
®°: 11 впРаво). Граница между плиоценом и плейстоценом, установленная по изме-
- ию направления навивания раковин форамииифер. показана пунктирной линией.
336
Часть 2. Применение данных палеонтологии
океанов (фиг. 156). Направление навивания раковин планктон-
ных фораминифер использовалось также для сопоставления раз-
резов более древних кайнозойских отложений. По-видимому,
направление навивания изменялось под влиянием температуры,
и широко распространенное обращение направления навивания,
очевидно, было вызвано значительным изменением климата.
Вероятно, само по себе направление навивания не имеет при-
способительного значения. Скорее всего это неадаптивный при-
знак, определяемый одним геном или группой генов, контроли-
рующих также адаптацию к температуре ([126], стр. 236).
Возможность использовать изменения направления навива-
ния для целей корреляции обусловлена тем фактом, что это из-
менение, видимо, наступает внезапно и затрагивает данную
группу на обширных пространствах.
ЭКОЛОГИЧЕСКИЕ МОДЕЛИ
В некоторых случаях для различных фаций и характерных
для них ископаемых удается выявить признаки, имеющие стра-
тиграфическое значение и позволяющие сопоставлять два или
более разрезов. На модельной схёме, представленной на
фиг. 157, показано последовательное развитие трансгрессии на
запад, которая затем сменилась регрессией в обратном направ-
лении. Во время отложения гипотетических литогенетических
Фиг. 157. Гипотетический разрез последовательности осадочных пород, воз-
никшей во время трансгрессии, сменившейся затем регрессией [92].
«Временная» линия от А до G, соединяющая точки наиболее глубоководных отложений
каждой колонки, представляет время максимальной трансгрессии в западном направ-
лении.
XI. Биостратиграфия
337
подразделений на западе была суша, а на востоке — море. Бере-
говая линия первоначально, по мере того как море наступало
на сушу, передвинулась к западу, а потом, по мере отступания
моря, сместилась к востоку. Каждое из подразделений морских
отложений представляет отчетливую фацию с характерными ис-
копаемыми и литологическими особенностями. Морские фации
представляют собой смежные зоны, простиравшиеся параллель-
но береговой линии. Точка, отмечающая самую западную око-
нечность каждой фации, соответствует времени максимальной
трансгрессии к западу. Линия, соединяющая эти точки для всех
фаций, соответствует одному и тому же периоду истории раз-
вития последовательности отложений. На практике для изуче-
ния доступны лишь изолированные выходы пород или разрезы
скважин. Все же, определив положение глубоководных фаций
для всех разрезов, можно установить приблизительное положе-
ние изохронной поверхности.
КОЛИЧЕСТВЕННЫЕ МЕТОДЫ КОРРЕЛЯЦИИ
Шоу [190] предложил статистический метод корреляции, ос-
нованный на детальном знании местных разрезов, содержащих
многочисленные общие виды ископаемых.
Отрезки графика представляют расстояния от подошвы раз-
резов до их кровли, обычно измеряемые в метрах. Каждому
виду ископаемых, встречающемуся в двух разрезах, соответст-
вуют на графике точки, отмечающие нижнюю и верхнюю грани-
цы интервала распространения вида (фиг. 158). Если интервалы
распространения видов в двух разрезах (X и У) идентичны, то
линия, соединяющая все точки, будет иметь вид прямой и обра-
зует с каждой осью координат угол 45°.
Условимся, что время появления и исчезновения в двух ме-
стонахождениях каждого из девяти видов, общих для обоих
рассматриваемых разрезов (фиг. 158), было одинаковым. Тогда
линия, соединяющая точки, будет линией приблизительной кор-
реляции.. То, что эта линия не обязательно является линией точ-
ной корреляции, даже если разрезы идентичны, обусловлено на-
шей гипотетической ситуацией; ведь мы приняли допущение, что
только каждая точка является точкой корреляции. Если скоро-
сти накопления породы в течение интервала времени, соответ-
ствующего расстоянию между двумя точками, различны, то
истинная линия корреляции не совпадает с прямой между этими
двумя точками. Крайними случаями при подобного рода анали-
зе являются такие, в которых накопление породы в интервале
между двумя точками в одном разрезе произошло до начала
накопления породы, в другом разрезе. Такие случаи (фиг. 159)
позволяют определить максимальную возможную ошибку, просто
Виды в разрезе Y
4>мг. 158. График, показывающий идентичность интервалов распространения
девяти ископаемых видов в двух гипотетических разрезах [190].
А. Вертикальные интервалы распространения видов в двух разрезах. Б. На график нане*
сены нижняя (точки) и верхняя (крестики) границы интервала распространения каждого
вида в двух разрезах. При соединении всех точек получается прямая линия, идущая под
углом 45° к осям координат.
XI. Биостратиграфия
339
Фиг. 159. Часть графика, подобного тому, что изображен на фиг. 158J190J.
Показывает случаи абсолютно несинхронного осадконакопления в двух разрезах в тече-
ние некоторого интервала времени. А и В — два различных события, такие, как появле-
ние или исчезновение видов, отмечающие начало и конец временного интервала в разре-
зах X и У. Интерполяция прямой АВ является точной только в том случае, если скоро-
сти осадконакопления в течение всего интервала были идентичны в обоих разрезах. От-
резки ВМ и BN показывают максимальную возможную ошибку, которая возникает в том
случае, если все осадки, накопившиеся в течение определенного интервала либо в разрезе
X, либо в разрезе У, отложены до начала осадконакопления в другом разрезе. В любом
случае корреляционная ошибка будет составлять 15 м.
соединив две точки прямой линией. Определение действительной
ошибки является результатом оценки, основанной на геологи-
ческой информации. Шоу считает, что метод прямой линии при-
меним для большинства разрезов, сложенных морскими отло-
жениями, накопившимися в центральных регионах континентов.
Выражение «накопление породы» (rock accumulation) часто
употребляют вместо «накопление осадка» (sediment accumula-
tion) в связи с тем, что различные типы осадков после отложе-
ния уплотняются в разной степени. Поэтому мощность пород
в «стратиграфической записи» не всегда пропорциональна пер-
воначальной мощности осадков.
Рассмотрим теперь случай, когда скорость накопления по-
роды в каждом из двух разрезов почти постоянна, но различ-
на в разных разрезах. (Допущение, что стратиграфический диа-
пазон каждого из девяти видов в двух разрезах одинаков, оста-
ется в силе.) Тогда вертикальные отрезки, соответствующие ин-
тервалу распространения каждого вида в двух разрезах, будут
различными, и линия, соединяющая точки на графике, уже не
будет образовывать с осями координат угол в 45° (фиг. 160).
Для любого данного отрезка времени существует определен-
ное отношение скоростей накопления породы в двух изучаемых
разрезах. Это отношение может меняться. Шоу обнаружил, что
графики, используемые для сравнения интервалов распростра-
340
Часть 2. Применение данных палеонтологии
Фиг. 160. Диаграммы и графики (подобные тем, что изображены на фиг. 158)
для двух разрезов, охватывающих один и тот же временной интервал и заклю-
чающих идентичные ископаемые виды, но характеризующихся различными
скоростями накопления осадков [190].
Крестики — верхняя граница интервала, точки — нижняя граница интервала.
нения видов в двух разрезах, как правило, имеют вид ломаной
линии (фиг. 161). Иными словами, изменения отношения скоро-
стей накопления пород в двух разрезах имеют скачкообразный
характер. В противном случае должны были бы получаться кри-
волинейные графики.
Статистический метод можно применить к необработанным
стратиграфическим данным для построения наиболее подходя-
XI. Биостратиграфия
341
Фиг. 161. Интервалы распространения видов в двух разрезах, для которых
характерно внезапное изменение относительной скорости накопления породы
[190].
Точки с кружком — верхняя граница интервала; точки — иижняя граница интервала
щих линий корреляции в графическом виде. Можно также вы-
числить статистическую значимость линии корреляции (оценку
вероятности ошибки).
После сравнения линий корреляции для нескольких местных
разрезов полезно построить так называемый «составной стан-
дарт» (composite standard) по Шоу. За основу обычно берется
самый полный разрез, не имеющий значительных стратиграфи-
ческих перерывов и содержащий наиболее многочисленные орга-
нические остатки. Интервалы распространения ископаемых из
всех других частных разрезов можно спроецировать на этот ос-
новной разрез. Это выполняется для каждого разреза путем
проведения линии «выравнивания», которая представляет линию
корреляции данного разреза с основным. При таком способе
представления данных различия в скорости накопления пород
во всех разрезах стираются. На составном стандарте, приведен-
ном на фиг. 162, относительные интервалы для различных видов
оцениваются для всей площади. В действительности на состав-
ном стандартном разрезе относительные интервалы распростра-
;4
342
Часть 2. Применение данных палеонтологии
Фиг. 162. Интервалы распространения 17 наиболее обычных видов в трило-
битовых зонах Coosella и Maryvillia в составном стандарте, основанном на вось-
ми отдельных местных разрезах (формация Райли, кембрий, Техас, США)
[146, 190[.
нения видов будут почти всегда неполными. Однако большинст-
во из них будет ближе к истинным относительным интервалам,
чем стратиграфические интервалы любого отдельного разреза.
Составной стандарт является абстракцией, основанной на ско-
ростях накопления пород в основном разрезе. Поэтому по вер-
тикальной оси не отложены ни единицы мощности, ни единицы
времени.
Составной стандарт, построенный по адекватным данным,
оказывается чрезвычайно важным для изучения стратиграфии,
палеоэкологии и эволюции. С помощью того же метода, кото-
рый применяется для сравнения местных разрезов, можно про-
вести его .корреляцию с отдельными разрезами или с каким-ли-
бо другим отдаленным составным стандартом. Составной стан-
XI. Биостратиграфия
343
дартный разрез делает возможным применение количественно-
го метода корреляции значительно удаленных друг от друга
районов. По-видимому, его можно применить также для меж-
континентальной корреляции, причем в этом случае следует ис-
пользовать интервалы стратиграфического распространения ро-
дов, а не видов. Путем сравнения серий разрезов с составным
стандартом в принципе возможно" продемонстрировать геогра-
фическую миграцию видов. Подобным же образом в некоторых
районах могут быть распознаны реликтовые популяции- Кроме
того, в составном стандартном разрезе почти всегда гораздо бо-
лее точно отражена картина относительного времени появления
и исчезновения видов, чем в любом единичном разрезе.. Поэто-
му применение составного стандарта обеспечивает большую до-
стоверность распознавания эволюционных линий и филогенети-
ческих связей.
Хронологические
и хроностратиграфические
подразделения
Региональные стратиграфические разрезы расчленены на
крупные подразделения, отличающиеся одно от другого по со-
держащимся в них ископаемым. Именно по этому принципу бы-
ли установлены в течение нескольких десятилетий XIX века все
системы, от кембрийской до четвертичной; при этом исследова-
тели действовали без какого-либо заранее хорошо продуманного
плана. Они следовали естественной тенденции связывать гра-
ницы систем с основными фаунистическими и флористическими
перерывами. Даже в наши дни многие установленные в то вре-
мя границы приходится признать очень удачно установленны-
ми, поскольку, несомненно, некоторые из использованных в ка-
честве критериев перерывов соответствовали крупным эволю-
ционным изменениям или массовым вымираниям, происходив-
шим в течение коротких промежутков времени в глобальном
масштабе. Однако приходится признать, что весьма распростра-
ненная в наши дни тенденция рассматривать границы между
геологическими системами как естественные перерывы, хорошо
приуроченные к наиболее подходящим стратиграфическим го-
ризонтам, неверна. В действительности часто столь же или да-
же более целесообразно было бы использовать другие границы.
В процессе становления современной геохронологической шка-
лы был велик элемент произвольности.
Геохронологическая шкала, разумеется, долго оставалась
шкалой относительного времени. Только после того, как были
разработаны радиометрические методы датирования (в начале
344
Часть 2. Применениеданных палеонтологии
XX века), эту шкалу удалось довольно точно связать с абсо-
лютным временем, измеряемым годами.
Объем геологических систем все еще определяется на осно-
вании сравнения с типовыми разрезами, или стратотипами, пред-
ставляющими собой местные стратиграфические разрезы в стра-
тотипических районах, где впервые были установлены эти систе-
мы. Большинство стратотипических местностей находится в Ев-
ропе. Одна из проблем, связанных с этой традицией, заключает-
ся в том, что не все стратотипы достаточно полны, чтобы пред-
ставлять объем систем, принятых в настоящее время во всем
мире. В настоящее время многие исследователи считают необ-
ходимым пересмотр систем, с тем чтобы установить их границы
в новых районах, в пределах которых существуют непрерывные
или почти непрерывные разрезы, содержащие ископаемые.
Схема, представленная на фиг- 163, показывает обычную
процедуру, применяемую при прослеживании границ системы
от ее стратотипа в других биогеографических провинциях или
на других континентах. Когда одна система выделяется в од-
ном районе, а примыкающая к ней — в другом, может оказать-
ся, что в системы вошли породы, возраст которых частично сов-
падает. Столь же вероятно, что между двумя системами, уста-
новленными в двух различных районах, может существовать пе-
рерыв во времени. Иными словами, даже стратотипические раз-
резы геологических систем, согласно их современному опреде-
лению, не дают четкого расчленения стратиграфической колон-
ки. Корреляция пород стратотипа с отложениями других райо-
нов по ископаемым остаткам не является, как мы видели,
достаточно точной; к тому же этот метод не позволяет уста-
новить точную синхронность границ систем в глобальном мас-
штабе.
Системы разделяются на отделы, а отделы — на ярусы. Все
эти единицы чаще всего устанавливаются по палеонтологиче-
ским данным. И системы, и отделы, и ярусы относятся к катего-
рии хроностратиграфических подразделений. Хроностратиграфи-
ческие подразделения должны быть изохронными, т. е. они
должны отграничиваться поверхностями, соответствующими по-
всюду одному и тому же возрасту. Именно так определяются эти
подразделения в Кодексе стратиграфической номенклатуры,
упомянутом в начале главы. Однако термин «хроностратигра-
фическое подразделение», используемый в этом смысле, может
ввести в заблуждение. Поскольку системы, отделы и ярусы вы-
деляются прежде всего на основе биостратиграфической эквива-
лентности (за исключением их типовых районов), установлен-
ные границы подразделений никоим образом не могут быть
в большей мере изохронными, чем биостратиграфические гра-
ницы, используемые для их определения.
XI. Биостратиграфия
345
Фиг. 163. Корреляция стратотипов.
А, В и С — стратотипические разрезы трех геологических систем в различных районах.
Справа приведен составной разрез отложений этих трех систем совсем в другом районе.
Перерывы, обусловленные отсутствием осадконакопления или эрозией, обозначены на
разрезах белыми прямоугольниками. Линии корреляции (пунктирные) по палеонтологи-
ческим данным имеют различные степени точности. Изохронные уровни идут горизон-
тально. Системы А и В перекрывают друг друга во времени; между системами В и С
имеется перерыв. Даже если бы корреляция была идеальной, некоторые участки состав-
ного разреза справа (показаны вопросительными знаками) невозможно было бы отнести
к какой-либо системе.
Эту проблему признает большинство современных исследо-
вателей. В действительности же дело сводится к вопросу о сте-
пени точности. Некоторые ученые, например Хедберг [76], ут-
верждают, что, поскольку системы и отделы соответствуют более
значительным хронологическим промежуткам, чем зоны, про-
цент ошибки при установлении их границ меньше. Согласно
23—467
346
Часть 2. Применение данных палеонтологии
этому мнению, в большинстве случаев границы систем, просле-
женные ;в глобальном масштабе, настолько близки к синхрон-
ным, что термином «хроностратиграфическое подразделение»
вполне можно пользоваться. Уэллер [228] предлагал сохра-
нить термин «хроностратиграфическое подразделение» при
описании систем и отделов, но с оговоркой, что в большин-
стве случаев эти подразделения являются лишь приближенно
изохронными. Однако Данбар и Роджерс ([45], стр. 293) высту-
пают против использования термина «хроностратиграфическое-
подразделение»- Они пишут: «Исторически отделы и системы
всегда ставились стратиграфами в зависимость от состава иско-
паемых, как и зоны, и если зона является биостратиграфическим
термином, то и эти подразделения надо считать биостратигра-
фическими». Вопрос о том, допустимо ли рассматривать систе-
мы и отделы как хроностратиграфические подразделения, оста-
ется предметом споров.
Сама по себе шкала геологического времени есть не что
иное, как абстракция. Геохронологические подразделения этой
шкалы определяются границами между системами в стратоти-
пах. Как видно при рассмотрении фиг. 163, эти геохронологиче-
ские подразделения могут частично совпадать или разделяться
перерывами. Таким образом, хотя время само по себе непрерыв-
но, геохронологическая шкала в том виде, в каком она прини-
мается теперь, не дает возможности четко подразделять этот
континуум.
Нетрудно понять, почему многие исследователи в настоящее
время стремятся к установлению геологических систем заново
в других районах, нежели первоначальные типовые, причем с ис-
пользованием границ, проходящих в породах с ископаемыми. 1
При этом нижняя граница системы была бы выделена в одном
месте, а верхняя — в другом. Типовой разрез для каждой грани-
цы можно было бы выбрать так, чтобы граница была установ-
лена по надежным руководящим формам, заключенным в по-
родах, образовавшихся в процессе почти непрерывного осадко-
накопления. Новые границы систем можно было бы установить
таким образом, чтобы они как можно ближе совпадали с тра-
диционными. Некоторые из старых стратотипов можно было бы
сохранить, но их лучше использовать для установления границ
между системами, а не для определения границ какой-то одной
системы. Отделы, для которых также имеются стратотипы, мож-
но было бы подвергнуть такой же ревизии, хотя многие из них
в настоящее время выделены в районах, где они представлены
хорошо обнаженными напластованиями, богатыми ископаемы-
ми остатками.
Другое предложение было внесено Беллом и его сотрудника-
ми [10]. В этом предложении, идущем в разрез с некоторыми
XI. Биостратиграфия
347
идеями и правилами, закрепленными в Кодексе стратиграфиче-
ской номенклатуры, и отражающем точку зрения меньшинства
геологов, указывается, что радиометрические определения аб-
солютного возраста дали временную шкалу в миллионах лет.
Эта шкала независима от так называемой «относительной гео-
хронологической шкалы», основанной на стратиграфических и
палеонтологических данных. Они предлагают использовать тер-
мин «хронологический» лишь применительно к радиометрически
установленной временной шкале. По их мнению, термин геохро-
нологический следует использовать применительно к традицион-
ной временной шкале эр, периодов и эпох. Белл предлагает на-
зывать хроностратиграфическими подразделениями толщи пород
(системы и отделы), соответствующие современным геохроно-
логическим подразделениям. В таком случае хроностратиграфи-
ческие подразделения можно было бы определить, используя
биостратиграфические подразделения (зоны), а также любые
другие подходящие критерии. Границы хроностратиграфических
подразделений не обязательно совпадали бы с границами био-
стратиграфических единиц, но обычно они совпадали бы. Вре-
менной протяженности каждого геохронологического подразде-
ления соответствовали бы широко распространенные толщи по-
род (хроностратиграфические подразделения) вместе со всеми
перерывами в этих толщах, возникшими в периоды отсутствия
осадконакопления или эрозии. Стратотипический разрез при
таком построении принимается во внимание, но с учетом при-
сущих ему ограничений. По аналогии с таксономическими кон-
цепциями (см. стр. ПО и 122) стратотип, согласно этой схеме,
можно до некоторой степени уподобить типовому экземпляру
вида, хроностратиграфическое подразделение — гиподигму
(стр. ПО), а геохронологическое подразделение — виду.
Точность корреляции
Противоречивость в определении групп, систем и ярусов в
качестве хроностратиграфических единиц обусловливает значи-
тельные разногласия в вопросе о точности корреляции страти-
графических подразделений по ископаемым остаткам.
Известно, что определенные фауны и флоры прошлых эпох,
как и в наши дни, сосредоточивались в отдельных, отчасти изо-
лированных географических провинциях. Поскольку биоты в
пределах одной провинции отличаются значительным единооб-
разием, корреляция стратиграфических подразделений внутри
провинции гораздо точнее, чем корреляция между различными
провинциями. Стратиграфическая корреляция между провин-
циями обычно используется для установления обширных при-
близительно синхронных «поверхностей» при реконструкции
палеогеографической картины определенных коротких геологи-
23*
348
Часть 2. Применение данных палеонтологии
Фиг. 164. Палеогеографическая схема для среднего карбона, показывающая ос-
новные пути миграции и наиболее полно изученные местонахождения (отме-
чены точками) фузулинид [168].
ческих отрезков времени. Часто эта картина охватывает два или
несколько континентов, как на карте, приведенной на фиг. 164.
Поскольку палеогеографические карты составляют для значи-
тельных территорий, они редко воспроизводят частные детали.
Построение таких карт требует меньшей точности стратиграфи-
ческой корреляции, чем составление региональных карт.
XI. Биостратиграфия
349
Большинство межконтинентальных корреляций основано на
методе руководящих ископаемых, а не на сравнении обширных
ископаемых фаун и флор. Обычно руководящие ископаемые,
взятые из двух сравниваемых провинций, принадлежат к двум
видам одного и того же рода; иногда они относятся к одному и
тому же виду. Если принять (ввиду отсутствия других данных
по этому вопросу), что оценка средней продолжительности су-
ществования одного вида, составляющая, по Симпсону (197],
0,5—5 млн. лет, верна, то этот интервал устанавливает прибли-
зительную точность корреляций, основанных на отдельных иско-
паемых видах. В действительности продолжительность сущест-
вования многих видов морских беспозвоночных превышала
5 млн. лет (фиг. 121). Точность корреляции, основанная на срав-
нении родов, а не видов, определяется интервалом от 8 млн. лет
(продолжительность существования родов кайнозойских плото-
ядных) до 78 млн. лет (продолжительность существования родов
кайнозойских двустворчатых моллюсков) [197]. Продолжитель-
ность существования отдельных родов и видов могла значитель-
но изменяться. Далее, в различных семействах и отрядах сред-
няя продолжительность жизни составляющих их родов и видов
может сильно различаться. Совершенно очевидно, почему одно
из важнейших требований, предъявляемых к надежному руко-
водящему ископаемому,— относительно короткий стратиграфи-
ческий интервал распространения. Ясно также, что использова-
ние нескольких таксонов для проведения одной корреляции
очень ценно.
Многие межконтинентальные корреляции проводятся после-
довательно в два или несколько этапов. При этом принимается,
что одна биота существовала одновременно со второй, вторая—
с третьей и т. д- Таким образом, третья биота примерно корре-
лируется с первой, хотя они могут иметь мало (или совсем не
иметь) общих видов или родов. Подобные многоэтапные корре-
ляции приводят к значительным ошибкам. В большинстве кор-
реляций значительная доля субъективности вносится при выбо-
ре таксонов, особенно при определении их первого появления
и исчезновения.
Многие исследователи считают," что границы стратиграфиче-
ских подразделений, прослеживаемые по всему земному шару,
представляют собой почти синхронные уровни. Другие, напри-
мер Уэллер ([228], стр. 562), придерживаются иной точки зре-
ния. Уэллер пишет: «Вероятно, в силу более или менее значи-
тельной обособленности сходство между различными фаунисти-
ческими и флористическими провинциями выражено меньше,
чем между разными частями одной провинции. Следовательно,
сравнения и корреляции должны проводиться на основе более
общих эволюционных изменений, что обусловливает меньшую
350 Часть 2. Применение данных палеонтологии
точность результатов. Неясно, какую степень неточности можно
допустить при рассмотрении этих менее прямых связей... В це-
лом ошибка межпровинциальных корреляций, основанных на
палеонтологических данных, составляет, вероятно, четверть гео-
логического периода».
В большинстве случаев при проведении4 корреляций в преде-
лах одной биогеографической провинции разрезы бывают удале-
ны друг от друга всего на несколько километров (редко более
100 км), а общая протяженность изучаемой области достигает
десятков или сотен километров (редко более 2000 км). Корре-
ляция такого масштаба базируется на биотах, обладающих
многими общими элементами флоры и фауны. Однако здесь и
точность корреляции должна быть значительно выше, чем при
корреляции для разных провинций. Для проведения местных
палеонтологических исследований в пределах одной провинции
часто необходима исключительно точная корреляция. Скорость
эволюции видов слишком мала, чтобы ее можно было исполь-
зовать при определении изменений ископаемых. Очень полезны-
ми для целей корреляции могут оказаться почти изохронные
литогенетические единицы. Например, особенно большую цен-
ность представляют с этой точки зрения прослойки вулканиче-
ского пепла или другие прослойки, образовавшиеся в результа-
те очень кратковременных процессов осадконакопления, широко
распространившихся на большой площади; обычно их называют
«маркирующими слоями».
При определенных условиях присутствие сходных биот в от-
ложениях различных районов может указывать на различный
возраст пород. Предположим, например, что нужно определить ।
какую-то ископаемую фауну как принадлежащую комплексу,
обитавшему в пределах узкой береговой полосы, заливавшейся
приливом. Обнаружение такой биоты в двух обособленных обна-
жениях, находящихся в 300 км друг от друга, в направлении,
перпендикулярном данной береговой линии, дает возможность
с уверенностью предполагать, что возраст слоев этих двух обна-
жений различен.
Некоторые ученые выражают надежду на то, что усовершен-
ствование радиоактивных методов датирования и расширение
возможностей определения абсолютного возраста пород обес-
печат значительно более высокую точность корреляции по срав-
нению с тем, что могло дать использование ископаемых. К со-
жалению, это оптимистическое предположение оправдывается
только для очень молодых пород.
Точность радиометрического метода определения абсолютно-
го возраста обычно оценивается стандартной ошибкой. Неточ-
ность является результатом статистической изменчивости для
образцов одинакового происхождения и неизбежных ошибок из-
XI. Биостратиграфия
351
мерений. Кроме того, значительные ошибки возникают в ре-
зультате утери измеряемых дочерних продуктов радиоактивного
распада вследствие, например, выветривания или нагревания.
Шкала абсолютного геологического времени (фиг. 165), со-
ставленная Калпом [106], основана на измерениях возраста от-
дельных образцов, которые, как предполагается, оставались
фактически закрытыми системами. Поэтому стандартные ошиб-
ки его определений являются общими для большинства подоб-
ных определений. На фиг. 166 на оси ординат отложены значе-
ния абсолютного возраста, использованные для построения пос-
лекембрийской части шкалы геологического времени, а по оси
абсцисс — стандартные ошибки. Видно, что с возрастом пород
сильно возрастает стандартная ошибка. Можно предположить,
что точность корреляции по ископаемым для более древних
толщ пород также оказывается более низкой вследствие худшей
сохранности ископаемых и меньшего числа обнажений таких
пород. Вообще говоря, достоверность корреляций, проведенных
при помощи палеонтологических данных или при помощи радио-
метрических методов определения абсолютного возраста, может
быть большей или меньшей, в зависимости от данного конкрет-
ного исследования. 4
Попытки соотнести данные радиометрических исследований
с установленными биостратиграфическими подразделениями
также связаны с определенными трудностями. Одна из них со-
стоит в том, что с помощью радиометрических методов редко
удается установить непосредственно абсолютный возраст иско-
паемых. Объясняется это тем, что лишь часть измерений абсо-
лютного возраста проведена для минералов, образование кото-
рых совпало по времени с накоплением осадков, заключающих
ископаемые. Многие определения абсолютного возраста прове-
дены по вулканическим толщам, стратиграфическое положение
которых можно установить лишь приближенно, на основе соот-
ношения с осадочными породами, содержащими ископаемые.
Свою геохронологическую шкалу (фиг. 165) Калп основывал
только на определениях по минералам, время образования ко-
торых укладывается в биостратиграфическую схему.
Дополнительные трудности возникают вследствие недоста-
точной биостратиграфической корреляции тех ископаемых, кото-
рые тесно связаны с минералами, возраст которых определен ра-
диометрическими методами. Идеальным было бы установление
абсолютного возраста границ всех систем и ярусов в стратоти-
пических разрезах. Однако эта цель практически недостижима,
поскольку определение абсолютного геологического возраста
возможно лишь по определенным минералам и в определенной
геологической обстановке. Данные для границ систем и ярусов
в геохронологической шкале Калпа установлены на основе опре-
Приблизитель- м ное начало Эра Период Эпоха интервала Четвертичный Плейстоцен f __ Плиоцен § Миоцен а; Олигоцен 36 3 ал“с^1лл-лл-лл-ллл-«1г - Палеоцен $ лет tad 0 50 100 150 200 250 300 350 400 450 500 550 600
Маастрихт „ , Кампан _ _ Поздним Синтон ----------л-лг-ВЛ ., Коньяк / 90 МвЛ Турон , — Сеноман Раннии - 120 Неоком Кб - § Поздняя _ Средняя --™ 166 Ранняя Баиос - ,. ]& J —-— ~ Поздний Триас Средний 200 ~ ______ Ранний
Поздняя Пермь 26О “ Ранняя
Карбон "" Пенсильвания J
_ Визе 320 Миссисипии Турне
8 i Поздний '™ — п ' 365 Девон Средний п 390 - Раннии __ — 405—: Силур — 425— Поздний Трентон 445 _ Ордовик Средний Ранний _
1 1 иии ” Поздний 530 Кембрий Средний ~ Ранний ~ >"" ? ? ? ? ? _ ? ? ? ? Ч
Фиг. 165. Абсолютная геохронологическая шкала по Калпу [106].
XI. Биостратиграфия
353-
200
400
О
500
Фиг. 166. Величины абсолютного возраста, использованные Калпом [106J
для построения послекембрийской части абсолютной геохронологической
шкалы (фиг. 165).
На оси абсцисс — вычисленная погрешность.
делений абсолютного возраста, проведенных в районах, сильно
удаленных от местонахождений стратотипов этих систем и яру-
сов.
Проблемы, возникающие при установлении абсолютного воз-
раста биостратиграфических границ системы, рассмотрены на
примере силура. Согласно геохронологической шкале Калпа
[106] , силурийский период приходится на интервал 405—425 млн.
лет назад. По геохронологической шкале Холмса [80] границы
силура датируются соответственно в 400 и 440 млн. лет. Други-
ми словами, по Калпу продолжительность силурийского перио-
да составляет 20, а по Холмсу — 40 млн. лет. Абсолютный воз-
раст нижней границы силура, по данным этих исследователей,
354 Часть 2. Применение данных палеонтологии
колеблется в пределах 15 млн. лет. Правда, Калп отмечает, что
возраст нижней границы силурийской системы сомнителен.
Радиометрические методы определения возраста едва ли
пригодны для локальных корреляций, особенно для разрезов
древних пород. Большинство местных фациальных и палеоэко-
логических исследований требует значительно большей точно-
сти, чем это могут обеспечить радиометрические измерения, да-
же при изучении кайнозойских отложений, абсолютный возраст
которых определяется с ошибкой порядка 0,5 млн. лет.
Можно надеяться, что применение количественных биостра-
тиграфических методов, подобных тому, который был предло-
жен Шоу [190], позволит в будущем увеличить точность корре-
ляций. Однако использование этого метода связано с необходи-
мостью чрезвычайной скрупулезности при проведении страти-
графических исследований и с высокими требованиями, предъ-
являемыми к коллекции ископаемых. Поистине астрономическое
число человеко-часов потребовалось бы, если бы метод Шоу
начали широко применять в стратиграфической практике, осо-
бенно при установлении составных стандартных разрезов.
Некоторые ученые вообще отрицают пригодность количест-
венного подхода для целей корреляции. Их аргументы сходны с
аргументами тех таксономистов, которые возражают против
использования численной таксономии (стр. 144). Возникает
вопрос, можно ли с помощью этого метода дать при корреляции
равную статистическую оценку для всех ископаемых таксонов
(или, в таксономии, всем измеряемым морфологическим харак-
теристикам). Так же как некоторые таксономисты отстаивают
•особую ценность определенных «ключевых характеристик» при
классификации таксономических групп, так и некоторые страти-
графы отстаивают особую пригодность некоторых руководящих
ископаемых для корреляции (см., например, [94]. Существует
мнение, что если использовать достаточное число измерений, то
малозначительные данные просто растворяются в общей массе
данных и ценность статистических выводов не уменьшается.
Очевидно, проблема сводится к отысканию экономичных путей,
для получения необходимого объема данных с тем, чтобы оправ-
дать затраты, связанные с широким внедрением статистических
методов. j
Биостратиграфы постоянно стремятся совершенствовать ме-
тоды корреляции. Наш разговор о погрешностях корреляции ни
.в коей мере не умаляет значения и ценности применяемой в на-
стоящее время стратиграфической классификации. Цель его
состояла в том, чтобы показать «ловушки», подстерегающие ис-
следователя в его работе по биостратиграфической корреляции,
и по возможности стимулировать совершенствование существую-
щих корреляционных схем и методов корреляции.
ГЛАВА XII
Использование
палеонтологических данных
в геофизике и геохимии
Данные об ископаемых организмах служат непосредствен-
ной основой для изучения биологической истории Земли. Одна-
ко, как мы уже видели, эти данные могут быть использованы и
в других областях исследования, далеких от чистой биологии.
Так, в биостратиграфии палеонтологические данные помогают
восстановить хронологию событий в истории Земли. В главе, по-
священной палеоэкологии, мы видели, как можно использовать
данные об ископаемых для реконструкции физико-географиче-
ских условий прошлого (очертания массивов суши и морей, рас-
пределение возвышенностей и глубоководных впадин, характер
седиментационных процессов и т. п.). Подобную информацию
можно непосредственно применять при решении чисто геологи-
ческих проблем, например при изучении дрейфа континентов,
скорости погружения отдельных участков земной коры, развития
бассейнов осадконакопления.
В этой главе мы рассмотрим примеры прямого использова-
ния данных об ископаемых для решения вопросов геофизики и
геохимии.
Линии нарастания
у морских беспозвоночных
В скелетах, образующихся путем последовательного нараста-
ния, часто можно ясно видеть последовательные линии или коль-
ца нарастания. Мы уже отмечали это, рассматривая онтогене-
тическую изменчивость у морских ежей (гл. III). Линии нара-
стания отчетливо выражены у кораллов и моллюсков. На
фото XXXV показан обычный характер роста эпитеки кораллов.
Линии нарастания используются при изучении как измене-
ний, происходящих в процессе онтогенеза, так и функциональ-
ного значения этих изменений. Они особенно ценны тем, что
дают возможность изучать изменение в онтогенезе внешней фор-
мы при наличии лишь одного взрослого экземпляра. Кроме того,
сравнение скорости нарастания у одного и того же или у близ-
кородственных видов позволяет судить об условиях среды, в ко-
торых они развивались. Так, например, известно, что современ-
356
Часть 2. Применение данных палеонтологии
ные виды рифообразующих кораллов растут быстрее в теплой
воде. Эти данные помогают судить о том, каким был климат
геологического прошлого.
До недавнего времени почти не удавалось интерпретировать
линии нарастания в единицах абсолютного времени. Не раз вы-
сказывались предположения, что каждая линия соответствует
годичному циклу роста. Делались попытки определить с по-
мощью линий нарастания возраст данного ископаемого экземп-
ляра в годах по аналогии с определением возраста дерева по
числу годичных колец. Это, однако, не дало существенных ре-
зультатов, потому что возраст организма в момент гибели очень
редко имеет значение для видового определения ископаемых ос-
татков, а также потому, что до сих пор остается неясным, дей-
ствительно ли линии нарастания являются годичными. Линии
нарастания у беспозвоночных нередко наводят на мысль о ка-
кой-то незакономерной цикличности или же о наличии более
сложных циклов, чем простые годичные циклы.
В 1963 г. Дж. Уэллс [231], специалист по ископаемым и ны-
не живущим кораллам, занялся этой проблемой более глубоко,
чем любой из предшествующих исследователей. Он установил,
что пояски нарастания у некоторых современных кораллов рас-
положены гораздо теснее, чем если бы они соответствовали го-
дичным приращениям. Уэллс обнаружил примерно 360 тончай-
ших линий в скелетной массе, образовавшейся, по-видимому, за
один год, и высказал предположение, что каждая из этих линий
соответствует нарастанию за одни сутки. С физиологической
точки зрения подобное объяснение не лишено оснований, так
как кальцификация у рифообразующих кораллов, по-видимому, 1
зависит от фотосинтетической активности их симбионтов — во-
дорослей, которая заметно различается в разное время суток.
Далее Уэллс обратился к изучению нескольких хорошо со-
хранившихся экземпляров кораллов из девонских и каменно-
угольных отложений, у которых наличие годичных поясков на-
растания считалось хорошо установленным. При этом он обна-
ружил также более тонкие пояски нарастания, число которых
было гораздо больше, чем следовало ожидать. У девонских эк-
земпляров это число в среднем составляло 400 (от 385 до 410),
а у экземпляров каменноугольного возраста колебалось от 385
до 390. Многие исследователи сочли бы это доказательством
того, что тонкие пояски не являются суточными или что толстые
пояски ошибочно считались годичными. Но Уэллсу были извест-
ны предположения, основанные на геофизических исследовани-
ях, что в девоне год состоял более чем из 365 дней.
Исходя из чисто теоретических рассуждений, геофизики и
астрономы доказали, что скорость вращения Земли вокруг ее
оси уменьшается вследствие приливного трения, что, безусловно,.
XII. Использование палеонтологических данных в геофизике
357
Время, млн. лепг
Фиг. 167. Изменение числа дней в году с течением геологического времени
[231].
Подсчет основан на предположении об уменьшении скорости вращения Земли вокруг оси
в результате приливного трения.
должно приводить к уменьшению числа дней в году. Геофизи-
ческие методы позволяют рассчитать это замедление вращения
Земли, что в свою очередь позволяет рассчитать уменьшение
числа дней в году (фиг. 167). Длина суток, пр-видимому, увели-
чивается примерно на 0,0016 с за столетие. Благодаря открыти-
ям археологов, давшим нам возможность познакомиться с астро-
номическими наблюдениями древних египтян, эта оценка полу-
чила некоторое подтверждение; однако время, прошедшее от
египетской цивилизации до наших дней, так мало, что это нельзя
считать достаточно убедительным доводом. А вот данные Уэллса
по девонским и каменноугольным кораллам удивительно хорошо
подтверждают теоретические предсказания. '
Эти данные по кораллам служат хотя и не окончательным,
но весьма знаменательным подтверждением теоретических рас-
четов. Необходимы дальнейшие исследования для проверки и
определения соотношения между геологическим временем и ско-
ростью вращения Земли. Если бы можно было точно установить
изменение числа дней в году, представилась бы возможность
подойти к этой проблеме с другой стороны. Так, например, если
известно число дней в году, наблюдаемое в линиях нарастания
У данного ископаемого животного, можно попытаться опреде-
лить его геологический возраст. Этот метод геохронологии име-
358
Часть 2. Применение данных палеонтологии
ет явные преимущества перед методами, основанными на радио-
активном распаде. Он не зависит от сингенетичных минералов
или от сопутствующих изверженных пород. Геохронология, осно-
ванная на изучении линий нарастания, дает возможность более
точно установить соответствие между «относительной» шкалой
времени, применяемой в биостратиграфии, и «абсолютной» шка-
лой времени, основанной на определении радиоактивности. Кроме
того, линии нарастания не подвержены химическим изменениям
(обусловленным диагенезом и метаморфизмом), которые иска-
жают данные, полученные на основе определения радиоактивно-
сти. Конечно, датирование по линиям нарастания в большей ме-
ре зависит от степени сохранности материала, но это скорее
морфологическая сохранность, чем химическая. Главный огра-
ничивающий фактор геохронологии, основанной на изучении ли-
ний нарастания, состоит в том, что число дней в году изменя-
ется со скоростью примерно один день за 10 млн. лет.
После первой работы Уэллса подобные исследования прово-
дились как на кораллах, так и на других организмах, скелеты
которых увеличиваются путем нарастания. В 1965 г., например,
Скраттон опубликовал детальные исследования циклического
нарастания скелета у некоторых девонских кораллов. Хотя он
и не обнаружил убедительных доказательств годичной перио-
дичности роста, он все-таки нашел небольшие приращения
(морфологически схожие с суточными поясками роста, обнару-
женными Уэллсом); эти пояски были сгруппированы в более
крупные полосы, содержащие в среднем по 30,6 мелких поясков.
Скраттон высказал предположение, что продолжительность
лунного месяца в девоне составляла 30,6 суток. С биологиче-
ской точки зрения это вполне правдоподобно, поскольку извест-
но, что у многих морских организмов, в том числе и у кораллов
ритмы физиологических процессов связаны с лунными или при-
ливными циклами.
Скраттоновский месяц, равный 30,6 дня, лишь незначитель-
но превышает продолжительность современного приливного
цикла (29,5 дня), но эта разница оказывается существенной.
Некоторые данные Скраттона приведены в табл. 7. Самое мень-
шее среднее число поясков в каждой полосе нарастания для
одного образца коралла составляет 29,9. Определенная по это-
му образцу продолжительность девонского месяца почти на по-
ловину суток длиннее, чем нынешний месяц. (Следует отметить,
что общепринятая длина лунного месяца, равная 28 дням,— все-
го лишь условность, введенная для удобства и не имеющая под
собой твердой научной основы.)
Заключения Скраттона носят предварительный характер, но
они открывают возможности для дальнейшего использования
данных о линиях нарастания в геофизике при изучении эволю-
XII. Использование палеонтологических данных в геофизике
359
ТАБЛИЦА 7
Установление продолжительности лунного месяца в
девоне по числу поясков нарастания на эпитеке
ископаемых кораллов [185]
Номер образца Число анализиро- ванных полос на- растания Среднее число поясков на полосу
ВМ R44851 12 30,9
OUM ДТ2 13 30,85
оим дтз 10 30,7
OUM ДТ4 16 30,7
OUM ДТ5 12 30,4
OUM ДТ6 14 30,8
OUM ДТ7 12 31
OUM ДТ8 6 30,6
OUM Д232 9 30
оим дгзз 8 29,9
Среднее по 10 образцам 30,59
ции системы Земля—Луна в масштабах геологического време-
ни.
Некоторые из наиболее детальных исследований, связанных
с изучением линий нарастания, были проведены на раковинах
двустворчатых моллюсков. Баркер [9] выявил гораздо более
сложные циклы, чем все известные до этого. Баркер изучил шли-
фы раковин четырех современных видов двустворчатых, собран-
ных на морских отмелях от острова Принца Эдуарда (Канада)
до Флориды. Различия в текстуре, составе и толщине линий на-
растания позволили выделить пять циклов разной периодично-
сти: годичный, полугодовой, 14-дневный, суточный и шестичасо-
вой. В табл. 8 приведены эти циклы и соотношение каждого из
них с ростом раковины.
Результаты, полученные Баркером, были подтверждены Пан-
неллой и Макклинтоком [149] при изучении других современных
двустворчатых моллюсков (фото XXXVI). Работа Скраттона
по изучению длины лунного месяца также была продолжена
Паннеллой, Макклинтоком и Томпсоном [150]. Их данные позво-
ляют предположить (фиг. 168), что в течение геологического
времени изменение числа дней в лунном месяце, возможно, про-
исходило незакономерно.
ТАБЛИЦА 8
Периодические линий нарастания у двустворчатых моллюсков [9]
Линия нарастания Толщина Сублинии Условия роста Вероятный период роста
5-го порядка 1—16 мкм • Карбонат кальция Раковина открыта или закрыта при высоком или низком приливе 6 ч
Конхиолин Раковина в процессе открывания при смене при- лива отливом
4-го порядка 5—60 мкм Толстый 5-го порядка . Тонкий 5-го порядка День (быстрый рост) Ночь (медленный рост) 24 ч
3-го порядка 0,1—0,9 мм Толстый 4-го порядка Тонкий 4-го порядка Максимальная приливная амплитуда Минимальная приливная амплитуда 15 дней
2-го порядка 0,9—11,0 мм Богатый конхиолином . Бедный конхиолином Нормальные условия Условия равноденствий 1/2 года
1-го порядка 2—22 мм Толстый 2-го порядка, бога- тый конхиолином Тонкий 2-го порядка, бедный конхиолином Лето Зима 1 год
XII. Использование палеонтологических данных в геофизике 361
Фиг. 168. Изменение числа дней в лунном месяце с течением геологического
времени, установленное по линиям нарастания скелетов различных ископае-
мых [150].
Все это новое направление по изучению линий нарастания
развивается чрезвычайно быстро и, несомненно, может способ-
ствовать значительному пополнению наших знаний в тех разде-
лах геологической истории, которые зависят от динамики систе-
мы Земля—Луна—Солнце.
Использование ископаемых остатков
при изучении деформации горных пород
Вторым примером использования палеонтологического мате-
риала для решения вопросов, не относящихся к биологии, слу-
жит использование их в структурных и геофизических исследо-
ваниях механики и деформации горных пород. Особый интерес
представляет установление основных направлений напряжений,,
вызывающих деформации. Для определения величины и харак-
тера деформации необходимо, чтобы порода имела какие-либо
особенности структуры и включала частицы, первоначальные
размеры и форма которых были бы известны. Лишь немногие
абиогенные структуры отвечают этому требованию. Во многих
исследованиях используются образцы, первичная форма кото-
рых предполагается шарообразной (оолиты, конкреции и т. п.);
однако нередко такое предположение сделать трудно. Недефор-
мированные оолиты, например, варьируют по форме, хотя в
среднем они имеют шарообразную форму. Только при работе
с крупными образцами имеется возможность допустить перво-
начальную сферичность. Кроме того, сферическая форма слиш-
ком проста, что ограничивает возможности определения давле-
ния и напряжений в образце по его деформации. Редко бывают
известны и первоначальные размеры образцов.
24—467
362
Часть 2. Применение данных палеонтологии
Ископаемые организмы в этом отношении гораздо более пер-
спективны. Во-первых, они более широко распространены (по
крайней мере в осадочных породах). Во-вторых, их первона-
чальная форма, как правило, лучше известна и может быть
определена с большей точностью (путем сравнения с недефор-
мированными экземплярами тех же видов). В-третьих, первона-
чальная форма органических остатков обычно более сложна, и
это обеспечивает получение более полных данных об имевших
место деформациях. Достаточно просто могут быть определены
первоначальный объем или размеры таких животных, как три-
лобиты, развитие которых сопровождается линькой и последова-
тельные стадии роста которых морфологически различны. Если
ископаемые сохраняются в породе в виде отпечатков, они из-
меняются во время деформации так же, как и сама вмещающая
порода. Очень часто какая-либо неорганическая частица, на-
пример оолит, выдерживает большее напряжение, чем вмещаю-
щая порода; поэтому по деформации такой частицы не удается
точно определить напряжение, приложенное ко всей породе.
Здесь мы будем рассматривать только двумерную дефор-
мацию, т. е. деформацию ископаемых, залегающих в плоскостях
напластования. При этом мы ограничимся рассмотрением того,
что геофизики называют «однородным напряжением», т. е. из-
менения объема и формы, при котором прямые линии остаются
прямыми, параллельные — параллельными, а удлинения или
укорочения первоначально одинакового направления имеют при-
близительно одну и ту же величину.
В каждом примере мы будем сталкиваться с определением
«эллипса деформации» — фигуры, которая получилась бы при
действии тех же самых напряжений на круг. Наиболее важны-
ми характеристиками эллипса являются: направление большой
оси (направление максимального удлинения), большая ось (ве-
личина максимального удлинения) и малая ось (величина ми-
нимального удлинения). Эти данные ничего не говорят
о давлениях, вызывающих деформацию, потому что идентичные
картины напряжения можно получить при весьма разнообраз-
ных системах давления. Например, данный эллипс деформации
может быть получен растяжением, параллельным направлению
максимального удлинения, или сжатием в перпендикулярном на-
правлении.
На фото XXXVII показаны три деформированных хвостовых
щита трилобитов. На фото XXXVII, А и Б деформация сим-
метрична по отношению к первоначальной двусторонней симмет-
рии трилобита; направление максимального удлинения парал-
лельно оси двусторонней симметрии (Л) или перпендикулярно
этой оси (5). Если известна первоначальная морфология пан-
диря трилобита, то можно определить направления максималь-
XII. Использование палеонтологических данных в геофизике 363
Фиг. 169. Влияние деформации на очертания двусторонне симметричных ра-
ковин брахиопод [155].
Первоначально все раковины имели одинаковые размеры.
кого и минимального удлинения. Если же известно отношение
основных размеров в недеформированном хвостовом щите, то
можно вычислить отношение максимального удлинения к мини-
мальному. Для того чтобы определить абсолютные значения
удлинений, необходимо знать первоначальные размеры. Это воз-
можно лишь в том случае, если изменчивость размеров раковий
так незначительна, что ею можно пренебречь., /
На фиг. 169 изображен более сложный пример двумерной
деформации двусторонне симметричных ископаемых. Сравнивая
представленные на ней деформированные и недеформированные
раковины брахиопод, можно видеть, что расположение раковины
по отношению к системе напряжений является решающим фак-
тором. Обратите внимание, что деформированные экземпляры
А и Б на фиг. 169 сравнимы соответственно с деформирован-
ными щитами А и Б на фото XXXVII. Каждый из них парал?-
лелен или перпендикулярен направлению максимального
24*
364
Часть 2. Применение данных палеонтологии
Фиг. 170. Определение эллипса
деформации по изменению длины
замочного края раковин брахио-
под, изображенных на фиг. 169
Ц55].
Угол а — это угол между линией за-
мочного края и прямой PQ (фиг. 169).
удлинения. Все остальные образцы расположены под косым уг-
лом к направлению максимального удлинения и поэтому срав-
нимы с(^ щитом В на фото XXXVII.
Существует несколько способов расчета деформаций, пока-
занных на фиг. 169. В одном случае измеряемым морфологиче-
ским признаком каждого экземпляра является угол а, образу-
емый прямой, по которой измеряется длина замочного края h, и
произвольной линией, проведенной через образцы (PQ). Ре-
зультаты представлены графически на фиг. 170. Точки, соот-
ветствующие отдельным измерениям, достаточно хорошо укла-
дываются на кривую, которая описывается следующим уравне-
нием:
где h0~ длина замочного края у недеформированного экземп-
ляра, /г —длина замочного края у деформированного образца,
а— угол между замочным краем и произвольной линией PQ,
6 — угол между линией PQ и направлением максимального уд-
линения (большая ось эллипса деформации), Xj и 2.2— мини-
мальное и максимальное удлинения.
На фиг. 170 точки хорошо ложатся на теоретическую кри-
вую, что позволяет графически определить основные параметры
эллипса деформации. Значение а в точке максимума кривой
дает нам 0, а отношение ординат точек максимума и миниму-
ма кривой — отношение двух удлинений Х1/Л2- Таким образом,
эллипс деформации можно определить, не зная точную морфо-
логию недеформированных ископаемых и даже независимо от
XII. Использование палеонтологических данных в геофизике 365
Фиг. Г 171. Деформированные раковины
брахиопод, иллюстрирующие влияние раз-
личной первоначальной ориентировки на
деформацию [155].
Объяснения см. в тексте.
того, обладали ли ископаемые первоначально двусторонней
симметрией. Важно, однако, иметь возможность допустить, что
первоначально все ископаемые имели одинаковые размеры!
Любая изменчивость образцов по размерам приводит к увели-
чению разброса точек, как это видно на фиг. 170. Если измен-
чивость слишком велика и если число экземпляров мало, то
следует применять другие методы анализа.
На фиг. 171 изображены две деформированные раковины
брахиопод разных видов. Для построения эллипса деформации
нам достаточно знать, что обе раковины до деформации обла-
дали двусторонней симметрией. Необходимо измерить три угла:
угол (ф) между первоначальной морфологически срединной ли-
нией и перпендикуляром к замочному краю (на каждом экземп-
ляре) и угол (а) между замочными краями обоих экземпляров.
Для отношения Х2/М (задающего форму эллипса деформации)
и угла 6 (ориентация длинной оси эллипса по отношению к од-
ному из ископаемых) получаем следующие два уравнения:
1/17 tge
1/ ^2 _ tg (6 + а)
V Xi tg(e + a + VB)’
Для решения этих уравнений можно воспользоваться гра-
фическим методом (с помощью круга Мора).
На фиг. 172 схематически изображена створка раковины
двустворчатого моллюска до и после деформации. Даже для
366
Часть 2. Применение данных палеонтологии
Фиг. 172. Недеформированная (Л) и деформированная (5) раковины дву-
створчатых моллюсков [155].
раковины, не обладающей двусторонней симметрией, можно
определить форму и ориентацию эллипса деформации, если
известны углы между тремя линейными элементами раковины
до и после деформации (показаны на фиг. 172 жирными линия-
ми). Производим измерение двух углов (а и 0). Величины
R (R равно WM) и 6 (направление максимального удлинения
по отношению к деформированному ископаемому) могут быть
найдены путем совместного решения следующих двух уравне-
ний: I
Я1/2 tg 6 + tg а _ Rl/2 (tg 6 + tg а)
1—jR^tgOtga 1 tg6tgq ’
Rl/2 tg 6 + tg (а + P) __ Rl/2 [tg 6 + tg (a + p)J
1 _ tg e tg (cc + p) 1—tgetg(a + p) •
Как и в предыдущем случае, задача может быть решена графи-
чески.
Для более детального ознакомления с методами математи-
ческого анализа деформации ископаемых органических остат-
ков читателю следует обратиться к книге Рамсея [155J. Здесь
XII. Использование палеонтологических данных в геофизике 2>&1
мы ограничились лишь кратким и далеко не полным введением
в эту область.
Рассмотренные примеры иллюстрируют важную роль иско-
паемых остатков в изучении деформаций осадочных пород.
Эти примеры показывают также, что геофизик должен быть
знаком с палеонтологией, а палеонтолог должен владеть мето-
дами геофизических исследований. Влияние деформации горных
пород на форму ископаемых означает, что морфологическую^
изменчивость того или иного вида нельзя понять без учета воз-
можных деформаций.
Реконструкция
диагенетических обстановок
Изучение химического диагенеза требует знания минерало-
гии, химического состава и структуры первоначального осадка.
Без этих сведений геохимик может видеть лишь конечные про-
дукты диагенеза. Многие известняки образуются главным обра-
зом из остатков скелетов морских беспозвоночных. Для иско-
паемых остатков организмов, у которых имеются близкие роди-
чи среди ныне живущих видов, обычно удается довольно точно
определить первоначальный скелетный материал. Таким обра-
зом, окаменелости часто служат ключом для восстановления
процессов диагенеза.
Большинство морских беспозвоночных, имеющих скелет, вы-
деляют карбонат кальция в одной или нескольких следующих
модификациях: низкомагнезиальный кальцит, высокомагнези-
альный кальцит и арагонит. Разграничение двух первых моди-
фикаций достаточно произвольно (при содержании изоморфно-
го магния ниже 4% кальцит считается низкомагнезиальным, а
при содержании его выше 4% — высокомагнезиальным).
В решетке арагонита содержание магния никогда не бывает
значительным. Распределение трех модификаций карбоната
кальция в скелетах ныне живущих беспозвоночных показано
в табл. 9.
Как высокомагнезиальный кальцит, так и арагонит неустой-
чивы по сравнению с низкомагнезиальным кальцитом при тем-
пературах и давлениях, существующих на поверхности Земли.
В морской воде, однако, они обычно не изменяются, оставаясь
в состоянии, получившем название «метастабильного». Если эти
минералы подвергаются действию пресной воды в субаэраль-
ных условиях, то они преобразуются в низкомагнезиальный
кальцит и другие минералы.
Арагонит, содержащийся в скелетах, в ряде случаев превра-
щается в низкомагнезиальный кальцит без видимого изменения
микроструктуры. Однако обычно он полностью растворяется
358
Часть 2. Применение данных палеонтологии
ТАБЛИЦА 9
Распределение различных модификаций карбоната кальция в
скелетах морских беспозвоночных [25]1)
Организм Арагонит Арагонит и кальцит Кальцит Высокомагне- зиальный каль- цит Высокомагне- зиальный каль- цит и арагонит
Фораминиферы
Донные Пелагические Р 0 О
Губки Кораллы 0
Шестилучевые О
Восьмилучевые р о
Мшанки о р 0
Брахиоподы о р
Иглокожие Моллюски 0
Брюхоногие о о
Двустворчатые 0 0 о
Головоногие 0 р
Черви Членистоногие 0 0 0
Десятиногие раки о
Остракоды 0 р
Усоногие раки о р
Водоросли
Донные Пелагические 0 о о
1)0 — обычен; Р — редок-.
кислыми водами, и в породе при этом образуются пустоты. Рас-
творенный карбонат кальция часто отлагается между первич-
ными зернами осадка в виде срастающихся кристаллов низко-
магнезиального кальцита. Эти кристаллы цементируют или ли-
тифицируют карбонатные осадки в известняки. По-видимому,
обильный известковистый цемент в песчаниках также возника-
ет в результате растворения карбонатных ископаемых. Корочки
вторичного кальцита в известняках часто обволакивают зерна
арагонита до их растворения, а впоследствии, когда зерна рас-
творятся, образуются отпечатки (фото XXXIX).
В субаэральных условиях высокомагнезиальный кальцит
имеет тенденцию превращаться в низкомагнезиальный без зна-
чительного растворения. Ионы магния просто выносятся водны-
ми растворами и замещаются ионами кальция без существенно-
го изменения микроструктуры скелетных остатков. В отсутствие
растворения высокомагнезиальный кальцит может преобразо-
ваться в устойчивую фазу — доломит (характеризующийся при-
близительно равным содержанием кальция и магния) или
в низкомагнезиальный кальцит. Подобный процесс, приводя-
XII. Использование палеонтологических данных в геофизике
369
щий к избирательной доломитизации высокомагнезиального
кальцита окаменелостей, наблюдается в некоторых породах
(фото XXXVIII).
Использование геохимиками карбоната ископаемых для объ-
яснения процессов диагенеза аналогично использованию в струк-
турной геологии деформированных окаменелостей для расшиф-
ровки геологических процессов. Однако очень часто ископаемые
скелетные остатки в породе настолько обильны, что оказывают
влияние на ход диагенеза. В таких случаях геохимик, изучаю-
щий осадочные породы, не может дать полное и точное объясне-
ние процесса диагенеза, не обратившись к рассмотрению биоло-
гических факторов. Весьма вероятно, что эволюционные изме-
нения в типе скелетного материала, вырабатывавшегося
организмами, соответствующим образом сказывались на диа-
генезе. Например, процент таксонов беспозвоночных, выраба-
тывающих арагонит, в палеозое был, очевидно, гораздо ниже,
чем сейчас. Поэтому преобладание кальцитовых скелетов в па-
леозое, возможно, отразилось на характере диагенеза палеозой-
ских известняков.
Выводы и перспективы
дальнейших исследований
При подготовке палеонтологов приходится все в большей ме-
ре обращаться к данным и методам других наук; в связи с этим
рассмотренные в этой главе исследования привлекают особое
внимание. Подобное взаимопроникновение наук вовсе не озна-
чает, что чисто описательные палеонтологические исследования
утратят свое значение. Напротив, все упомянутые здесь специ-
альные исследования возможны лишь благодаря накоплению
огромной массы описательных данных. Мы можем, таким обра-
зом, считать исследования, рассмотренные в этой главе, «побоч-
ным продуктом» биологического изучения ископаемых. Несом-
ненно, большинство предстоящих в будущем важных открытий
потребует как глубокого понимания исследуемой проблемы, так
и использования сопредельных н5ук. Пример такого сочетания
продемонстрировал Уэллс, когда он понял, что наличие 400 по-
ясков в годичном приращении скелетов девонских кораллов не
было ни отклонением, ни ошибкой исследования, а, напротив,
представляло собой научные данные, которые можно было ис-
пользовать для подтверждения результатов астрономического
процесса.
СПИСОК ЛИТЕРАТУРЫ
1. Ager D. V., Principles of Paleoecology, New York, McGraw-Hill, 1963,
371 p.
2. American Commission on Stratigraphic Nomenclature, Code of stratigrap-
hic nomenclature, Bull. Amer. Assoc. Petrol. Geol., 45, 645—665 (1961).
3. Arkell W. J., Jurassic Geology of the World, London, Oliver and Boyd,
1956, 806 p.
4. Arkell W. J., Introduction to Mesozoic Ammonoidea. In: Moore R. C.,
ed., Treatise on Invertebrate Paleontology, Boulder, Colo., Geological Socie-
ty of America and University Press of Kansas, Part. L, Mollusca, 4, 81 —
129 (1957).
5. Bakker R. T., The superiority of dinosaurs, Discovery (Peabody Mus. Nat.
Hist.), 3, 11—22 (1968).
6. Barghoorn E. S., Schopf J. W., Microorganisms from the Late Precambrian
of central Australia, Science, 150, 337—339 (1965).
7. Barghoorn E. S., Schopf J. W., Microorganisms three billion years old
from the Precambrian of South Africa, Science, 152, 758—763 (1966).
8. Barghoorn E. S., Tyler S. A., Microorganisms from the Gunflint Chert,
Science, 147, 563—577 (1965).
9. Barker R. M., Microtextural variation in pelecypod shells, Malacologia,
2, 69—86 (1964).
10. Bell W. C., Kay M., Murray G. E., Wheeler H. E., Wilson J. A., Note
25 — Geochronologic and chronostratigraphic units, Bull. Amer. Assoc.
Petrol. Geol., 45, 666—670 (1961).
11. Berkner L. V., Marshall L. C., The history of growth of oxygen in the earth's
atmosphere. In: Brancazio P. J. and Cameron A. G. W., eds., The Origin
and Evolution of Atmospheres and Oceans, New York, Wiley, 1964, 102—
126.
12. Berry W. B. N., Growth of a Prehistoric Time Scale, San Francisco,
W. H. Freeman and Company, 1968, 158 p.
13. Best R. V., Intraspecific variation in Encrinurus ornatus, Jour. Paleont.,
35, 1029—1040 (1961).
14. Beurlen K-, Der Faunenschnitt an der Perm-Trias-Grenze, Z. Deut. Geol.
Ges., 108, 88—99 (1956). 9
15. Blackwelder R. E.. Taxonomy: A Text and Reference Book, New York,
Wiley, 1967, 698 p.
16. Bonner J. T., Morphogenesis, Princeton, Princeton University Press, 1952,
296 p.
17. Boucot A. J., Brace W., deMar R., Distribution of brachiopod and pele-
cypod shells by currents, Jour. Sedim. Petrology, 28, 321—332 (1958).
18. Brinkmann R., Statistisch-biostratigraphische Untersuchungen an mittel-
jurassischen Ammoniten fiber Artbegriff und Stammesentwicklung, Got-
tingen, Abhandlung Gesell. Wiss., Vol. 13, No. 3 (1929).
19. Brown C. A., Palynological Techniques, Baton Rouge, La., C. A. Brown,
1960, 188 p.
20. Bulman О. M. B., Programme-evolution in the graptolites, Biol. Rev., 8,
311—334 (1933).
Список, литературы
371
21. -Callomon J. Н., Sexual dimorphism in Jurassic ammonites, Trans. Leices-
ter Lit. Philos. Soc., 57, 21—56 (1963).
22. Camp C. L., HannaG-. D., Methods in Paleontology, Berkeley, University
of California Press, 1937, 153 p.
23. Campbell K. S. IF., A Lower Carboniferous brachiopod-coral fauna from
New South Wales, Jour. Paleont., 31, 34—98 (1957).
24. Carpenter F. M. The geological history and evolution of insects, Amer.
Scientist., 41, 256—270 (1953).
25. Chave К. E., Skeletal durability and preservation. In: Imbrie J. and Ne-
well N. D.,eds., Apporoaches to Paleoecology, New York, Wiley, 1964,
p. 377—387.
26. Clark M. G., Shaw A. B., Paleontology of northwestern Vermont. XV,
Jour. Paleont., 42, 382—396 (1968).
27. Clark W. E., Le Gros, ed., Essays on Growth and Form Presented to
D'Arcy Wentworth Thompson, Oxford, The Clarendon Press, 1945, 408 p.
28. Clarke G. L., Elements of Ecology, New York, Wiley, 1965, 560 p.
29. Clarkson E. N. K., Schizochroal eyes and vision of some Silurian acastid
trilobites, Paleontology, 9, 1—29 (1966).
30. Cloud P. E., Some problems and patterns of evolution exemplified by fos-
sil invertebrates, Evolution, 2, 322—350 (1949).
31. Colbert E. H., Evolution of the horned dinosaurs, Evolution, 2, 145- 163
(1948).
32. Colbert E. H., Evolution of the Vertebrates, New York, Wiley, 1955,
479 p.
33. Colbert E. H., The Age of Reptiles, New York, Norton, 1965, 228 p.
34. Corliss J. O., Comments on the systematics and phylogeny of the Protozoa,
Syst. ZooL, 8, 169—190 (1959).
35. Craig G. Y., Hallam A., Size-frequency and growth-ring analyses of My-
tilus edulis and Cardium edule, and their paleoecological significance, Pa-
leontology, 6, 731—750 (1963).
36. Dacque E., Vergleichende biologische Formenkunde der Fossilen niederen
Tiere, Berlin, Gebriider Borntrager, 1921, 777 p.
37. DeBeer G. R., Embryos and Ancestors, London, Oxford University Press,
1958, 197 p.
38. Denton E. J., Gilpin-Brown J. B., The buoyancy of the cuttlefish Sepia
officinalis (L.), Jour. Marine Biol. Assoc. United Kingdom, 41, 319—342
(1961).
39. Denton E. J., Gilpin-Brown J. B., On the buoyancy of the pearly nautilus,
Jour. Marine Biol. Assoc. United Kingdom, 46, 723—759 (1966).
40. Denton E. J., Gilpin-Brown J. B., On the buoyancy of Spirula spirula,
Jour. Marine Biol. Assoc. United Kingdom, 47, 181 —191 (1967).
41. Dobzhansky T., Genetics and the Origin of Species, New York, Columbia
University Press, 1951, 364 p.
42. Donovan D. T., Stratigraphy, Chicago, Rand McNally, 1966, 199 p.
43. Dorf E., Climatic changes of the past and present, Amer. Scientist, 48,
341—364 (1960).
44. Downie C., The geological history of the microplankton, Rev. Paleobotany
and Palynology, 1, 269—281 (1967).
45. Dunbar C. 0., Rodgers J., Principles of Stratigraphy, New York, Wiley.
1957, 356 p. (К. Данбар, Дж. Роджерс, Основы стратиграфии, ИЛ, М..
1963.)
46. Durham J. IF., Cenozoic marine climates of the Pacific Coast. Geol. Soc.
Amer. Bull., 61, 1243—1264 (1950).
47. Durham J. IF., Classification of clypeasteroid echinoids, Univ. Calif.
Publ. Geol. Sci., 31, 73—198 (1955).
48. Durham J. W., Classification. In: Moore R. C., ed., Treatise on Invertebra-
te Paleontology. Boulder, Colo., Geological Society of. America and Uni-
versity Press of Kansas, Part U, Echinodermata, 3, 270—295 (1966).
372
Список, литературы
49. Durham J. W., The incompleteness of our knowledge of the fossil record.
Jour. Paleont., 41, 559—565 (1967).
50. Eagar R. M. C., A study of a nonmarine lamellibranch succession in the
Athraconaia lenisulcata zone of the Yorkshire Coal Measures, Philos. Trans.
Roy. Soc. London, ser. B, 233, 1—54 (1947).
51. Easton W. H., Invertebrate Paleontology, New York, Harper, 1960, 701 p.
52. Eicher D. L., Geologic Time, Englewood Cliffs, Prentice-Hall, 1968,
150 p.
53. Elton C. S., The Ecology of Invasions by Animals and Plants, London,
Methuen, 1958, 181 p.
54. Engel A. E. J. et al., Alga-like forms in Onverwacht Series. South Africa:
oldest recognized lifelike forms on earth, Science, 161, 1005—1008 (1968).
55. Erben H. K., Ober den Ursprung der Ammonoidea, Biol. Rev., 41, 641 —
658 (1966).
56. EricsonD. B., EwingM., WollinG., Pliocene-Pleistocene boundary in deep-
sea sediments, Science, 139, 727—737 (1963).
57. Fenton C. L., Fenton M. A., The Fossil Book, Garden City, Doub eday,
1958, 482 p.
58. Finks R. AL, Late Paleozoic sponge faunas of the Texas region, Bull.
Amer. Mus. Nat. Hist., 120, 1—160 (1960).
59. Fischer A. G., Brackish oceans as the cause of the Permo-Triassic marine
faunal crisis. In: Nairn A. E. M., ed., Problems In Palaeoclimatology, Lon-
don, Interscience, p. 566—580 (I960).
60. Fischer A. G., Latitudinal variations in organic diversity, Evolution, 14,
64—81 (1960).
61. Fischer A. G., Fossils, early life, and atmospheric history, Proc. National
Acad. Sci., 53, 1205—1215 (1965).
62. Gilluly J., Distribution of mountain building in geologic time, Geol. Soc.
Amer. Bull., 60, 561—590 (1949).
63. Ginsburg R. N., Early diagenesis and lithification of shallow-water carbo-
nate sediments in south Florida. In: LeBlanc R. J. and Breeding J. G.,
eds., Regional Aspects of Carbonate Deposition, Soc. Econ. Paleont. Mi-
neral. Spec. Publ., 5, 80—100 (1957).
64. Ginsburg R. N., Lowenstam H. A., The influence of marine bottom com-
munities on the depositional environment of sediments, Jour. Geol., 66,
310—318 (1958).
65. Glaessner M. F., Pre-Cambrian fossils, Biol. Rev., 37, 467—494 (1962).
66. Gould S. J., Allometry and size in ontogeny and phylogeny, Biol. Rev.,
41, 587—640 (1962).
67. Gould S. J., Evolutionary paleontology and the science of form, Earth-
Science Rev., 6, 77—119 (1970).
68. Grant R. E., Spine arrangement and life habits of the productoid brachio-
pod Waagenoconcha, Jour. Paleont., 40, 1063—1069 (1966).
69. Grant R. E., Structural adaptation in two Permian brachiopod genera, Salt
Range, West Pakistan, Jour. Paleont., 42, 1—32 (1968).
70. Grant V., The Origin of Adaptations, New York, Columbia University
Press, 1963, 606 p.
71. Greiner H., «Spirifer dixjunctus»: Its evolution and paleoecology in the Cat-
skill Delta, Bull. Peabody Mus. Nat. Hist., Yale Univ., II, 75 (1957).
72. Hallam A., Faunal realms and facies in the Jurassic, Paleontology, 12,
1—18 (1969).
73. Harland IF. B. et al., The Fossil Record, London, Geol. Soc. London, 1967,
827 p.
74. Heaslip W. G., Cenozoic evolution of the alticostate venericards in Gulf
and East Coastal North America, Palaeontographica Americana, 6, 65—135
(1968).
75. Hecker R. F., Introduction to Paleoecology, New York, Elsevier, 1965,
166 p.
Список литературы
373
76. Hedberg Н. D., Nature of time-stratigraphic units and geologic-time units,
Bull. Amer. Assoc. Petrol. Geol., 35, 1077—1081 (1951).
77. Hedgpeth J. W., ed., Treatise on Marine Ecology and Paleoecology, 1, Eco-
logy, Geol. Soc. Amer. Mem., 67, 1296 (1957).
78. Hessler R. R., Sanders H. L., Faunal diversity in the deep-sea, Deep-Sea
Research, 14, 65—78 (1967).
79. Heywood V. H., McNeil J., eds., Phenetic and Phylogenetic Classification,
Systematics Assoc. Publ., 6, 164 (1964).
80. Holmes A., A revised geological time-scale, Edinb. Geol. Soc. Trans., 17,
183—216 (1960).
81. Hudson J. D., The recognition of salinity-controlled mollusc accemblages
in the Great Estuarine Series (Middle Jurassic) of the Inner Hebrides, Pa-
laeontology, 6, 318—326 (1963).
82. Hudson J. D., The microstructure and mineralogy of the shell of a Jurassic
mytilid (Bivalvia), Palaeontology, 11, 163—182 (1968).
83. Hunt A. S., Growth, variation and instar development of an agnostid tri-
lobite, Jour. Paleont., 41, 203—208 (1967).
84. Hunt O. D., The food of the bottom fauna of the Plymouth fishing grounds,
Jour. Marine Biol. United Kingdom, 13, 560—599 (1925).
85. Hutchinson G. E., The biologist poses some problems, Amer. Assoc. Adv.
Sci. Publ., 67, 85—94 (1958).
86. Huxley J. S., The relative size of antlers in deer, Proc. Zool. Soc., London,
1931, 819—864 (1931).
87. Huxley J. S., Problems of Relative Growth, New York, Dial Press, 1932,
276 p.
88. Hyman, The Invertebrates. IV. Smaller Coelomate Groups, New York,
McGraw-Hill, 1959, 738 p.
89. Jmbrie J., Biometrical methods in the study of invertebrate fossils, Bull.
Amer. Mus. Nat. Hist., 108, 211—252 (1956).
90. Imbrie J., Brachiopoda of the Traverse Group (Devonian) of Michigan,
Part I, Bull. Amer. Mus. Nat. Hist., 116, 345—410 (1959).
91. Imbrie J., Newell N. D., Approaches to Paleoecology, New York, Wiley,
1964, 432 p.
92. Israelsky M., Oscillation chart, Bull. Amer. Assoc. Petrol. Geol., 33, 92—
98 (1949).
93. Jackson R. T., Phylogeny of the Echini with a revision of Paleozoic species
Boston Soc. Nat. Hist., Mem., 7, 443 (1912).
94. Jeletsky J. A., Is it possible to quantify biochronological correlation? Jour.
Paleont., 39, 135—140 (1965).
95. Johnson R. G., Models and methods for the analysis of the mode of forma-
tion of fossil assemblages, Geol. Soc. Amer. Bull., 71, 1075—1086 (1960).
96. Jurva R., Seas, In A General Handbook on the Geography of Finland,
Fennia, 72, 136—160 (1952).
97. Kaesler R. L., Numerical taxonomy in invertebrate paleontology. In: Teic-
hert C. and Yochelson E. L., eds., Essays in Paleontology and Stratigrap-
hy, Lawrence, University Press of Kansas, 63—81 (1967).
98. Raufmann R., Variationsstatistische Untersuchungen fiber die «Artabwan-
dlung» und «Artumbildung» an der oberkambrischen Trilobitengattng
Olenus Dalm, Abh. Geol. Pal. Inst. Univ. Greifswald, 10, 1—54 (1933).
99. Kaufmann R., Exakt-statistische Biostratigraphie der Olenus-Arten von
Sudoland, Geol. Foren. Stokholm ForhandL, 1935, 19—28 (1935).
100. Kermack K. A., A biometrical study of Micraster coranguinum and M.
(Isomicraster) senonensis, Philos. Trans. Roy. Soc. London, B, 237, 375—
428 (1954).
101. Kinne O., The effects of temperature and salinity on marine and brackish
water animals. II. Salinity and temperature-salinity combinations, Ocea-
nogr. Mar. Biol. Ann. Rev., 2, 281—339 (1964).
102. Koch D. L., Strimple H. L., A new Upper Devonian cystoid attached to a
374
Список литературы
discontinuity surface, Iowa Geological Survey Report of Investigations,
*5,49 (1968).
103. ’ Kozlowski R., On the structure and relationship of graptolites, Jour. Pa-
leont., 40, 489—501 (1966).
104. Kraft P., Die Mikrophotographie mit infraroten Strahlen, Internal. Cong,
of Photog. 8, Dresden, 1931, Berichte, 341—345 (1932).
105. -Krumbein W. C., Sloss L. L., Stratigraphy and Sedimentation, 2nd ed.,
San Francisco, W. H. Freeman and Company, 1963, 660 p.
106. Kulp J. L., Geologic time scale, Science, 133, 1105—1114 (1961).
107. ' Kummel B., Raup D. M., eds., Handbook of Paleontological Techniques,
San Francisco, W. H. Freeman and Company, 1965, 852 p. (Б. Каммел,
Д. Рауп (ред.), Методика палеонтологических исследований, изд-во
«Мир», М., 1973.)
108. Kummel' В., Steele G., Ammonites from the Meekoceras gracilitatus zone
at Crittenden Spring, Elko County, Nevada, Jour. Paleont., 36, 638—703
(1962).
109. Kurten B., On the variation and population dynamics of fossil and recent
< mammal populations, Acta Zool. Fennica, 76, 1—22 (1953).
110. Kurten B., Population structure in paleoecology. In Imbrie J. and New-
ell N. D., eds., Approaches to Paleoecology, New York, Wiley, 1964, 91 —
106.
111. Ladd H. S., ed., Treatise on Marine Ecology and Paleoecology, 2, Paleoe-
geology, Geol. Soc. Amer. Mem., 67, 1077 (1957).
112. Land L. S., Diagenesis of skeletal carbonates, Jour. Sedim. Petrology, 37,
914—930 (1967).
113. Laporte L. F., Ancient Environments, Englewood Cliffs, Prentice-Hall,
1968, 116 p.
114. Lawrence D. R., Taphonomy and information losses in fossil communities,
Geol. Soc. Amer. Bull., 79, 1315—1330 (1968).
115. Ledley R. S., High-speed automatic analysis of biomedical pictures, Sci-
ence, 146, 216—223 (1964).
116. Leich H., Eine neue Lebensspur von Mesolimulus walchi und ihre Deutung,
Aufschluss, 1, 5—7 (1965).
117. LemcheH., Wingstrand K- G., The anatomy of Neopilina galatheae Lemche,
1957 (Mollusca, Tryblidiacea), Galathea Rept. (Copenhagen), 3, 1—71
(1959).
118. Leon R., Ultraviolettes Licht endeckt Versteinerungen auf «leeren» Plat-
ten. Ein Pantpod im Jura-Kalk, Senckenberg Naturf. Gesell., Natur. Mus.,
63, 361—364 (1933).
119. Lerman A., On rates of evolution of unit characters and character comple-
xes, Evolution, 19, 16—25 (1965).
120. Lowenstam H. A., Niagaran reefs of the Great Lakes area, Jour. Geol., 58,
430—487 (1950).
121. McLean J. D., Jr., Manual of Micropaleontological Techniques, Alexan-
dria, Va., McLean Paleont. Lab. (1959-date).
122. MacurdaD. B., ed., Paleobiological Aspects of Growth and Development,
a Symposium, Paleont. Soc. Mem., 2, 119 (1968).
123. Marks J. G., Especies vivientes de moluscos que se encuentran en las for-
maciones Terciarias de Venezuela, Acta Cientifica Venezolana, 3, 135—
136 (1952).
124. Martin-Kaye P., Sorting of lamellibranch valves on beaches in Trinidad,
B. W. I. Geol. Magazine, 88, 432—434 (1951).
125. Mayr E., Systematics and the Origin of Species, New York, Columbia
University Press, 1942, 334 p.
126. Mayr E., Animal Species and Evolution, Cambridge, Harvard University
Press, 1963, 797 p. (Э. Майр, Зоологический вид и эволюция, изд-во
«Мир», М., 1968.)
127. Mayr Е., Principles of Systematic Zoology, New York, McGraw-Hill,
Список литературы
375
1969, 428 р. (Э. Майр, Принципы зоологической систематики, изд-во
«Мир», М., 1971.)
128. Mayr Е., Linsley Е. G., IJ singer R. L., Methods and Principles of Systema-
tic Zoology, New York, McGraw-Hill, 1953, 328 p. (Э. Майр, Э. Линсли,
P. Юзингер, Методы и принципы зоологической систематики, ИЛ, М.,
1956.)
129. Medawar Р. В., Size, shape, and age. In: Clark W. E., Le Gros, ed., Essays
on Growth and Form presented to D'Arcy Wentworth Thompson, Oxford,
The Clarendon Press, 157—187 (1945).
130. Moore H. B., Marine Ecology, New York, V/iley, 1958, 493 p.
131. Moore R. C., LalickerC. G., Fischer A. G., Invertebrate Fossils, New York,
McGraw-Hill, 1952, 766 p.
132. Muir-Wood H. M., Chonetidina. In: Moore R. C., ed., Treatise on Inverte-
brate Paleontology, Boulder, Colo., Geological Society of America and
University Press of Kansas, Part H, Brachiopoda, 412—439, 1965.
133. Newell N. D., Phyletic size increase —an important trend illustrated
by fossil invertebrates, Evolution, 3, 103—124 (1949).
134. Newell N. D., Fossil populations, System. Assoc. Publ., 2, 63—82 (1956).
135. Newell N. D., Adequacy of the fossil record, Jour. Paleont., 33, 488—499
(1959).
136. Newell N. D., Paleontological gaps and geochronology, Jour. Paleont.,
36, 592—610 (1962).
137. Nichols D., Changes in the Chalk heart-urchin Micraster interpreted in re-
lation to living forms, Philos. Trans. Roy. Soc. London, B, 242, 347—437
(1959).
138. Nichols D., Mode of life and taxonomy in irregular sea-urchins, System.
Assoc., 3, 61—80 (1959).
139. Nicol D., The biotic development of some Niagaran reefs— an example of
an ecological succession or sere, Jour. Paleont., 36, 172—176 (19(52).
140. Odum E. P., Fundamentals of Ecology, Philadelphia, Saunders, 1959,
546 p.
141. Odum H. T., OdurnE. P., Trophic structure and productivity of a windward
coral reef community on Eniwetok Atoll, Ecol. Monogr., 25, 291—320
(1955).
142. Okulitch V. J., Archaeocyatha. In: Moore R. C., ed., Treatise on Inverte-
brate Paleontology, Boulder, Colo., Geological Society of America and
University Press of Kansas, Part E, Archaeocyatha and Porifera, 1—20,
1955.
143. Olson E. C., The Evolution of Life, London, Weidenfeld and Nicolson,
1965, 300 p.
144. Ostrom J. H., A functional analysis of jaw mechanics in the dinosaur Tri-
ceratops, Postilia (Peabody Mus. Nat. Hist., Yale Univ.), 88, 1—35 (1964).
145. PalframanD. F. B., Variation and ontogeny of some Oxfordian ammonites:
Taramelllceras richei (de Loriol) and Creniceras renggeri (Oppel), from
Woodham, Buckinghamshire, Palaeontology, 9, 290—311 (1966).
146. Palmer A. R., The faunas of the Riley Formation in central Texas, Jour.
Paleont., 28, 709—786 (1955).
147. Palmer A. R., Ontogenetic development of two olenellid trilobites, Jour.
Paleont., 31, 105—128 (1957).
148. Palmer A. R., Biomere — a new kind of stratigraphic unit, Jour. Paleont.,
39, 149—153 (1965).
149. Pannella G., MacClintock C., Biological and environmental rhythms reflec-
ted in molluscan shell growth. Paleont. Soc. Mem., 2, 64—80 (1968).
150. Pannella G., MacClintock C., Thompson M. N., Paleontological evidence
of variations in length of synodic month since Late Cambrian, Science, 162,
792—796 (1968).
151. Павлов А. П., Le Cretace enfcrieur de la Russie et sa fauna. Nouv. Soc.
Imp. Nat. Moscou, livr., 21, 87 p. (1901).
376
Список литературы
152. Perkins В. F., Biostratigraphic Studies in the Comanche (Cretaceous) Se-
ries of Northern Mexico and Texas, Geol. Soc. Amer Mem., 83, 138 (1960).
153. Peterson C. G. J., Valuation of the sea. II. The animal communities of the
sea bottom and their importance for marine zoogeography, Rep. Danish
Biol. Sta., 21, 1—44 (1913).
154. Pianka E. R., Latitudinal gradients in species diversity: a review of con-
cepts, Amer. Nat., 100, 33—46 (1966).
155. Ramsay J. G., Folding and Fracturing of Rocks, New York, McGraw-Hill,
1967, 568 p.
156. Raup D. M., Geometric analysis of shell coiling: general problems, Jour.
Paleont., 40, 1178—1190 (1966).
157. Raup D. M., Geometric analysis of shell coiling: coiling in ammonoids,
Jour. Paleont., 41, 43—65 (1967).
158. Raup D. M., Theoretical morphology of echinoid growth, Paleont. Soc.
Mem., 2, 50—63 (1968).
159. Raup D. M., Michelson A., Theoretical morphology of the coiled shell,
Science, 147, 1294—1295 (1965).
Г60. Raup D. M., Takahashi T., Experiments in strengthin cephalopod
shells (abs.),‘Geol. Soc. Amer. Paper 101, 172—173 (1968).
161. Reid R. E. H., Bathymetric distribution of Calcarea and Hexactinellida
in the past and present, Geol. Magazine, 105, 546—559 (1968).
162. Rensch B., Evolution Above the Species Level, New York, Columbia Uni-
versity Press, 1960, 419 p.
163. Rhoads D. C., Missing fossils and paleoecology: Discovery (Peabody Mus.
Nat. Hist., Yale Univ.), 2, 19—22 (1966).
164. Rhoads D. C., Biogenic reworking of intertidal and subtidal sediments in
Barnstable Harbor and Buzzards Bay, Massachusetts, Jour. Geol., 75,
461—476 (1967).
165. Richter R., Psychische Reaktionen fossiler Tiere, Palaeobiologica, 1, 226—
244 (1928).
166. Romer A. S., Relative growth in pelycosaurian reptiles. Robert Broom
Commem. Vol. Special Publ. Roy. Soc. South Africa, 45—55 (1948).
167. Romer A. S., Vertebrate Paleontology, Chicago, University of Chicago
Press, 1966, 468 p.
168. Ross C. A., Development of fusulinid (Foraminiferida) faunal realms, Jour.
Paleont., 41, 1341—1354 (1967).
169. Ross C. A., Sabins F. F., Early and Middle Pennsylvanian fusulinids from
southeast Arizona, Jour. Paleont., 39, 173—209 (1965).
170. Rowe A. W., An analysis of the genus Micraster, as determined by rigid
zonal collecting from the zone of Rhynchonella cuvieri to that of Micraster
coranguinum, Quart. Jour. Geol. Soc. London, 55, 494—547 (1899).
171. Rowell A. J., A numerical taxonomic study of the chonetacean brachio-
pods. In: Teichert C. and Yochelson E. L., eds., Essays In Paleontology
and Stratigraphy, Lawrence, University Press of Kansas, 113—140 (1967).
172. Rudwick M. J. S., The inference of function from structure in fossils, Brit.
Jour. Philos. Sci., 15, 27—40 (1964).
173. Rudwick M. J. S., Adaptive homeomorphy in the brachiopods Tetractinel-
la Bittner and Cheirothysis Rollier, Palaont. Z., 39, 134—146 (1965).
174. Salfeld H., Uber Artbildung bei Ammoniten, Zeitschr. Deutsch. Geol.
GeselL, Monatsber., 65, 437—440 (1913).
175. Sandberg P. A., Hay W. W., Study of microfossils by means of the scan-
ning electron microscope, Jour. Paleont., 41, 999—1001 (1967).
176. Sanders H. L., Oceanography of Long Island Sound, 1952—1954. X. The
biology of marine bottom communities, Yale Bingham Oceanographic
Coll., 56, 345—414 (1956).
177. Sanders H. L.f Hessler R. R., Hampson G. R., An introduction to the study
of deep-sea benthic faunal assemblages along the Gay Head-Bermuda tran-
sect, Deep-Sea Research, 12, 845—867 (1965).
Список литературы
377
178. Savory Т., Naming the Living World, London, English University Press
Ltd., 1962, 128 p.
179. Schafer W., Aktuo-Paleontologie, nach Studien in der Nordsee, Frankfurt
Waldemar Kramer, 1965, 666 p.
180. Scheltema R. S., Dispersal of larvae by equatorial ocean currents and its
importance to the zoogeography of shoal-water tropical species, Nature
• 217, 1159—1162 (1968).
181. Schenk E. T., McMasters J. H., Procedure in Taxonomy, 3rd ed., Revised
by A. M. Keen and S. W. Muller, Stanford, Calif., Stanford University
Press, 1956, 119 p.
182. Schindewolf О. H., Neokatastrophismus? Deutsche Geol. Gesell. Zeitschr.,
114, 430—445 (1962).
183. Schlanger S. 0., Subsurface geology of Eniwetok Atoll. U. S. Geol. Surv.
Prof. Paper 260-BB, 991- 1066 (1963).
184. Schopf J. W., Microflora of the Bitter Springs Formation, Late Precambri-
an, central Australia, Jour. Paleont., 42, 651—688 (1968).
185. Scrutton С. T., Periodicity in Devonian coral growth, Palaeontology, 7,
552—558 (1965).
186. Seilacher A., Epizoans as a key to ammonoid ecology, Jour. Paleont., 34,
189—193 (1960).
187. Seilacher A., Paleontological studies on turbidite sedimentation and erosi-
on, Jour. Geol., 70, 227—234 (1962).
188. Seilacher A., Biogenic sedimentary structures. In: Imbrie J. and Ne-
well N. D., eds., Approaches to Paleoecology, New York, Wiley, 296—
316, 1964.
189. Seilacher A., Fossil behavior, Sci. Amer., 217, 72—80 (1967).
190. Shaw A. B., Time in Stratigraphy, New York, McGraw-Hill, 1964, 365 p.
191. Shotwell J. A., An approach to the paleoecology of mammals, Ecology, 36,
327—337 (1955).
192. Shotwell J. A., Intercommunity relationships in Hemphillian (mid-Plio-
cene) mammals, Ecology, 39, 271—282 (1958).
193. Simpson G. G., Mammals and land bridges, Jour. Washington Acad. Sci.,
30, 137—163 (1940).
194. Simpson G. G., Types in modern taxonomy, Amer. Jour. Sci., 238, 413—
431 (1940).
195. Simpson G. G., Horses, New York, Oxford University Press, 1951, 247 p.
196. Simpson G. G., How many species? Evolution, 6, 342 (1952).
197. Simpson G. G., The Major Features of Evolution, New York, Columbia
University Press, 1953, 434 p.
198. Simpson G. G., Notes on the measurement of faunal resemblance, Amer.
Jour. Sci., 258-A, 300—311 (1960).
199. Simpson G. G., Principles of Animal Taxonomy, New York, Columbia
University Press, 1961, 247 p.
200. Simpson G. G., Rowe A., Lewontin R. C., Quantitative Zoology, New York,
Harcourt, Brace, 1960, 440 p.
201. Sohl N. F., Archeogastropoda, Mesogastropoda and Stratigraphy of the
Ripley, Owl Creek and Prairie Bluff Formations, U. S. Geol. Surv. Prof.
Paper No. 331-A, 151 (1960).
202. Sokal R. R., Sneath P. H. A., Principles of Numerical Taxonomy, San
Francisco, W. H. Freeman and Company, 1963, 359 p.
203. Sorgenfrei T., Molluscan Assemblages from Marine Middle Miocene of
South Jutland and their Environments, Geol. Surv. Denmark, II Ser., No.
79, 503 (1958).
204. Spjeldnaes N., Ontogeny of Beyrichia jonesi Boll., Jour. Paleont., 25, 745—
755 (1951).
205. Srb A. M., Owen R. D., Edgar R. S., General Genetics, 2nd ed., San
Francisco, W. H. Freeman and Company, 1965, 557 p.
378
Список литературы
206. Stanley S. AL, Paleoecology and diagenesis of Key Largo Limestone, Flo-
rida, Bull. Amer. Assoc. Petrol. Geol., 50, 1927—1947 (1966).
207. Stanley S. M., Post-Paleozoic adaptive radiation of infaunal bivalve mol-
luscs — a consequence of mantle fusion and siphon formation, Jour. Pa-
leont., 42, 214—229 (1968).
208. Stanley S. M., Relation of Shell Form to Life Habits in the Bivalvia (Mol-
lusca), Geol. Soc. Amer. Mem., 125 (1970).
209. Stehli F. G., Permian zoogeography and its bearing on climate. In: Na-
irn A. E. M., ed., Problems in Palaeoclimatology, London, Interscience,
537—549, 1964.
210. Stoll N. R. et al., eds., International Code of Zoological Nomenclature,
London, Internal. Trust for Zool. Nomen., 1961, 176 p.
211. Sylvester-Bradley P. C., ed., The Species Concept in Paleontology System.
Assoc. Publ. 2, 145 (1956).
212. Tappan H., Primary production, isotopes, extinctions and the atmosphere,
Palaeogeogr., Palaeoclimat., Palaeoecol., 4, 187—210 (1968).
213. Teichert C., How many fossil species? Jour. Paleont., 30, 967—969 (1956).
214. Teichert C., Cold- and deep-water coral banks, Bull. Amer. Assoc. Petrol.
Geol., 42, 1064—1082 (1958).
215. Thompson D’A. W., On Growth and Form, New York, Cambridge Universi-
ty Press, 1942, 1116 p.
216. Thorson G., Reproductive and larval ecology of marine bottom invertebra-
tes, Biol. Rev., 25, 1—45 (1950).
217. Thorson G., Bottom communities (sublittoral or shallow shelf), Geol. Soc.
Amer. Mem. 67, 1, 461—534 (1957).
218. Trueman A. E., The ammonite body-chamber, with special reference to
the buoyancy and mode of life of the living ammonite Quart. Jour. Geol.
Soc. London, 96, 339—383 (1941).
219. Trueman A. E., Supposed commensalism of Carboniferous spirorbids and
certain non-marine lamellibranchs, Geol. Magazine, 79, 312—320 (1942).
220. Турпаева E. П., Food interrelationships of dominant species in marine
benthic biocoenoses, Trans. Inst. Oceanol. and Oceanogr., 11, 198—211
(1957).
221. Valentine J. IF., Biogeographic units as biostratigraphic units, Bull. Amer.
Assoc. Petrol. Geol., 47, 457—466 (1963).
222. Valentine J. IF., Climatic regulation of species diversification and extin-
ction, Geol. Soc. Amer. Bull., 79, 273—276 (1968).
223. Vaughan T. IF., Wells J. IF., Revision of the Suborders, Families and Ge-
nera of the Scleractinia, Geol. Soc. Amer. Spec. Paper 44, 363 (1943).
224. Waller T. R., The Evolution of the Argopecten gibbus Stock \Mollusca:
Bivalvia), with Emphasis on the Tertiary and Quaternary Species ofEas-
tern North America. Paleont. Soc. Mem., 3, 125 (1969).
225. Weimer R. JHoyt J. H., Burrows of Callianassa major Say, geologic indi-
cators of lithoral and shallow neritic environments, Jour. Paleont., 38.
761-767 (1964).
226. Weisbord N. E., Late Cenozoic gastropods from northern Venezuela, Bull.
Amer. Paleont., 42, 1—672 (1962).
227. Weisbord N. E., Late Cenozoic pelecypods from northern Venezuela, Bull.
Amer. Paleont., 45, 1—564 (1964).
228. Weller J. M., Stratigraphic Principles and Practice, New York, Harper,
1960, 725 p.
229. Wells J. IF., Scleractinia. In: Moore R. C., ed., Treatise on Invertebrate
Paleontology, Boulder, Colo., Geological Society of America and Universi-
ty Press of Kansas, Part F, Coelenterata, 328—444, 1956.
230. Wells J. W., Coral reefs, Geol. Soc. Amer. Mem., 67, 2, 609—631 (1957).
231. Wells J. IF., Coral growth and geochronometry, Nature, 197, 948—950
(1963).
Список литературы
379
232. Westoil Т. S., On the evolution of the Dipnoi. In: Jepsen G. L., Simp-
son G. G. and Mayr E. (eds.), Genetics, Paleontology, and Evolution,
Princeton, Princeton University Press, 121 —184, 1949.
233. Whittington H. B., The ontogeny of trilobites, Biol. Rev., 32, 421—469
(1957).
234. Woodford A. 0., Historical Geology, San Francisco, W. H. Freeman and
Company, 1965, 512 p.
235. Woodring IF. P., Stewart R., Richards R. W., Geology of the Kettleman
Hills Oil Field, California, U. S. Geol.Surv. Prof. Paper 195, 170 (1940).
236. Wright S., The roles of mutation, inbreeding, crossbreeding, and selection
in evolution. Proc. Sixth Intern. Cong. Genetics, 1, 356—366 (1932).
237. Zangerl R., Radiographic techniques. In: Kummel B. and Raup D. M.,
eds., Handbook of Paleontological Techniques, San Francisco, W. H. Fre-
eman and Company, 305—320, 1965.
(Ь. Каммел, Д. Рауп (ред.), методика палеонтологических исследований,
изд-во «Мир». М., 1973.)
238. Ziegler А. М., Silurian marine communities and their environmental signi-
ficance, Nature, 207, 270—272 (1965).
239. Ziegler A. M., Boucot A. J., Sheldon R. P., Silurian pentameroid brachio-
pods preserved in position of growth, Jour. Paleont., 40, 1032—1036
(1966).
240. Ziegler A. M., Cocks L. R. M., Bambach R- K., The composition and struc-
ture of Lower Silurian marine communities, Lethaia, 1, 1—27 (1968).
241. Zohary D., Feldman M., Hybridization between amphidiploids and the
evolution of polyploids in the wheat (Aegilops-Triticum) group., Evolution,
16, 44—61 (1962).
25*
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Абиссальная равнина 198, 214
Абсолютный возраст, определение ра-
диометрическое 350, 351, 358
Агерматипные кораллы 215, 266
Адаптационная конвергенция 155,
192 292___294
Адаптация 99, 156, 157, 278—283; см.
также Функциональная морфоло-
гия
— к температуре 336
Адаптивная конвергенция 292
— радиация 283—292
Адаптивные долины 158
— пики 158, 171
Аллена правило 237
Американская комиссия по страти-
графической номенклатуре 317
Аммониты, жилая камера 176 '
— вымирание 297
— половой диморфизм 71, 72, 260
— челюсти 221
— эволюция 260, 261, 263
Аналогичные структуры 166
Археоциаты 305
Аутэкология 226
Амфибии, эволюция 315
Бактерии, генетика и эволюция 313
— докембрийские 312
— в пищевой цепи 196
— в посмертном разрушении живот-
ных 26
Балтийский янтарь 15, 31
Батиальная зона 198, 199
Бентос 199—201, 214
— инфауна 199
— пищевые взаимоотношения 202, 203
— субстрат 204—207
— таксономическое разнообразие 241
— эпифауна 199
Бергесс сланцы 31
Бергмана правило 237
Библиография источников палеонто-
логической информации 155
Биогенетический закон 273
Биогеографические провинции 325—
328
Биогеография 335—337
Биозоны 323
Биомасса 196
Биостратиграфическая корреляция
см. Стратиграфическая корреляция
— эквивалентность 328
Биостратиграфические единицы см.
Стратиграфические единицы
Биостратиграфия 317—354; см. также
Стратиграфия
Биофации 321
Биттер-Спрингс формация 313
Брахиоподы см. Плеченогие
Брюхоногие, личинки, расселение 104
— механическое разрушение 26, 27
— описательная морфология 40—42,
46
— рост 49
— устойчивость к солености 211, 212
— функциональная морфология 169
Вагильные организмы 199
Взвешивание признаков 134
Вид см. также Видообразование,
Разнообразие
— гиподигм ПО
— именование формальное 110, 111
— клины 101
Предметный указатель
381
— количество особей 23—25
— кольцевые перекрывания 101
— описание ПО, 114—118
— определение 98, 145—148
— «палеонтологический» 107
— «последовательный» 107
— проблема в палеонтологии 106—
НО
— продолжительность существования
265—269
— синонимика ПО, 153—155
— смена 19, 20
— типовой 126
— эволюционный 108
Видообразование 99, 100
— и вымирание 267
— географическое 99, 100, 253
— скорость 102—104, 263—265
— филетическое 99
«Виды-двойники» 105
Водоросли докембрийские 312—314
— зеленые 214
— зооксантеллы 215
— известковые 215, 227
— карбонат кальция, модификация
368
— красные 214
— механическое разрушение 26
— и рифы 227
— сине-зеленые 312
— строматолиты 217, 218, 312
— экология 214—217
Выживаемость 77, 78, 80
Вымирание 19—23, 100
— и видообразование 265
----географические факторы 295—
297
— кораллов 266—269
— массовое 297—305
— пермо-триасовое 298
— в позднемеловую эпоху 300
— триас — юра 299
— филетическое 100,294
Высшие категории 126, 287
Вытеснение экологическое 287
Ганфлинт формация 312
Генерация, время 102, 103
Генофонд 67
Геологическое время 346, 351, 357
Геофизика и палеонтология 355—
367
Геохимия и палеонтология 355, 367—
369
Гетерохрония 273—277
Гибридизация (скрещивание) 97, 282
Гиподигм НО
Глубина водоема 213—218
Головоногие см. также Аммониты
— белемниты 299
— геометрия спиральной раковины
164, 170—173
— описание 44
— перегородочная линия 173—183
— посмертный перенос 33
— разнообразие 20, 21
— рост 53—57
— устойчивость к солености 211, 212
Голотип ПО, 121
Гомеоморфия 292, 294
Гомология 166
— использование в палеонтологии
237—240
Гомонимы 122
Гомоплазия 166
Граница эволюционного ряда 107
Граптолиты, систематическое положе-
ние 306, 307
— эволюция 274
Грибы 313
Грунтоеды 202
Губки, индикаторы глубины 216
— карбонаты кальция, модификации
368
Давление отбора 73, 102, 103
«Двойники» (слабо различающиеся
виды) 105
Двустворчатые моллюски, деформа-
ция ископаемых 365, 366
----классификация 134
— — линии нарастания 65, 355
----личиночное расселение 104
----образ жизни 190—192. 206, 207
----описание 42, 43
— — разнообразие 21, 209
----функциональная морфология
169
----эволюция 192, 207, 253, 265, 284,
285
----экология 226
Двухфакторный анализ 87, 90
Дендрограммы 142—144, 149
Деформация ископаемых двустворча-
тых моллюсков 365, 366
— горных пород 74, 361—367
— плеченогих (ископаемых) 363—365
— трилобитов 362, 363
Джонсона модели сохранения 245
Диагноз в описании видов 114, 118—
121
Диастемы 319
382
Предметный указатель
Дивергенция эволюционная 283—291
Диморфизм половой 70, 71, 260
Динозавры, классификация 138
— конечности, механические особен-
ности 186
— скорость эволюции 258
— «следы жизни» 226
— филогения 139, 315
— челюсти, механические свойства
183—186
Докембрийские отложения 312—314
Естественный отбор 73, 99, 102, 270
Закон биологической сукцессии 317
— приоритета 114
Замещение экологическое 291
Зарывающиеся организмы 217, 225,
293, 319
— двустворчатые моллюски 291, 292
— иглокожие 279—283
— ракообразные 217
Захоронение в прижизненном поло-
жении 221
Зоны 323
Зооксантеллы 215
Зоологическая номенклатура, между-
народный кодекс 111—114, 126, 317
Зоопланктон 199, 200
Иглокожие см. также Морские ежи
— изотопный состав 74
— карбонаты кальция, модификация
368
— разнообразие 23
— филогенез 129
— эволюционная радиация 23
Изменчивость, диаграммы 89
— межпопуляционная 88—96
— внутри сообщества 84—88
Измерение ископаемых 42—46
----приборы 46, 47
Изоляция географическая 98
Изотопный состав скелетов ископае-
мых 74
Инфауна 199, 206
Инфракрасные лучи в фотографиро-
вании ископаемых 36
Итеративная эволюция 291
Карбонат кальция, модификации в
составе скелетов ископаемых 367—
369
Категории Линнея 126
Кислород, изотопы 74
Кишечнополостные см. Кораллы и
Медузы
Классификация 124, 125
— дендрограммы 142—144
—критерии 134—138
— применение ЭВМ 138—144
— фенетическая 140
— фенограммы 142—144
— и численная таксономия 138—144
----эволюция 125, 127
Климаксное сообщество 227
Климат см. Температура
Клины 101
Ключи для определения видов 148—
151
Кодекс стратиграфической номенкла-
туры 317
Количество видов см. Разнообра-
зие
Колониальные организмы, дифферен-
циация особей 66
---- рост 66
Кольцевые перекрывания 101
Комплексная зона 333
Конвергенция адаптивная 292
— эволюционная 156, 188, 189, 292—
295
Консументы 197
Континентальный склон 198, 214
Копа правило 276, 277
Коралловые рифы см. Рифы
Кораллы агерматипные 215, 216, 266
— герматипные 215, 266
— карбонат кальция, модификации
368
— механическое разрушение 26, 27
— пояски нарастания 356
— скорость эволюции 259, 266, 267
—устойчивость к солености *210, 212
— эволюция 255, 266—269
— экология 215, 216, 227
Корреляция см. Стратиграфическая
корреляция
Кошки саблезубые 251
Ла-Бреа асфальтовые отложения 15,
251
Лектотип 122
Линия нарастания у двустворчатых
65, 355
------кораллов 356
— ----морских беспозвоночных и
геохронология 355—337
Литоральная зона 198, 217, 350
Л итостр атигра фические единицы
318—323
Литофации 321
Личинки, расселение 104, 203, 324
Лошади, филогения 131, 272, 273
Предметный указатель
383
«Маркирующие слои» 350
Медузы 200, 201
Международный кодекс зоологиче-
ской номенклатуры 11—114, 317
Местообитание 195
Миграция трилобитов 331
Микроископаемые см. Бактерии,
Пыльца, Споры, Фораминиферы
Миграция млекопитающих 295
Минерализация ископаемых 367—369
Млекопитающие, биогеография 295
— плиоценовые сообщества 249, 250
— полукопытные 307
— разнообразие 23
— стертость зубов 80, 272
— эволюция 265, 291, 315
Многофакторный статистический ана-
лиз 57, 88
Модели морфологические 159—166
— сохранения по Джонсону 245, 347
Моллюски см. также Брюхоногие,
Головоногие, Двустворчатые
— геометрия,- раковины 159—166
— карбонат кальция, модификации
368
— линии нарастания 355
— функциональная морфология 169—
183
Моноплакофоры 310, 311
Морская трава 219
Морские ежи 26
----зарывающиеся виды 278—283
----морфология 278—283
----механическое разрушение 26—
29
----определение видов 148, 149
----рост 9, 49, 52, 53
— лилии, разнообразие 21
Мульде мергели 81
Мусорщики 196, 197
Мшанки, вымирание 297
— карбонаты кальция, модификации
367
— разрушение механическое 26
— рост 66
Мэзон-Крик сланцы 15
Мягкие ткани, сохранение 29
Направление навивания у моллюсков
72, 161
------фораминифер 72, 334—336
Насекомые ископаемые 21
— сохранность 21
— эволюция 315
Нектон 199
Неотип 122
Норма реакции 58, 59, 73
Ограничивающие факторы в экологии
203
---------глубина 213—218
---------освещенность 214, 215
---------соленость 207—213, 302.
303
--------- типы субстрата 204—207
Однофакторный анализ 87
Онвервахт формация 312
Онтогенез 48; см. также Рост
— возрастные группы популяций 80
Онтогенетическая изменчивость 48—
66
— — метод преобразования коорди-
нат 57
— — многофакторный анализ 57
Операционные таксономические еди-
ницы 140—142
Описание ископаемых 35—47
— посредством измерения 42—47
Описательная морфология брюхоно-
гих 40—42
----использование ЭВМ 36—38
---- внутреннего строения 36, 47
---- измерение 42—46
----нового вида ПО, 114—118
----приборы 46, 47
----терминология 39—42
Определение видов 145—148
— — ключи 148—152
----машинные методы 151, 152
Ортогенез 269—273
Осадки, размеры частиц 204
Освещенность 214, 215
Остракоды, динамика популяции 81
— устойчивость к солености 213
Отделы стратиграфические 344
Отряд 126
Палеогеография и фоссилизации 104,
347, 348
Палеоклимат см. Температура
Палеонтологический вид 107
Палеонтология и геофизика 355—367
----геохимия 355, 367—369
Палеоэкология 194, 243
Парадигма 168
Парадигматический подход 167, 180—
183
Паразиты 196, 197
Параллельная эволюция 292—294
Параллелизм 292—295
Паратип 121
Пачки'стратиграфические 318
Пеликозавры, парус 186—188
----функциональная морфология
186, 187
384
Предметный указатель
Первичные консументы 201
Пигменты, сохранность 14, 30
Пиктограмма 86
Пищевая сеть 196, 197
Планктон 199, 200; см. также Зоо-
планктон, Фитопланктон
— ракообразные 201
Плезиотип 122
Плейотропия 168
Плеченогие 90
— вымирание 297
— деформация 363—365
— карбонат кальция, модификации
368
— классификация 133, 144
— онтогенез 55, 56
— разнообразие 23
— рентгенография 36
— рост 54, 55
— сортировка створок 76
— устойчивость к солености 210
— филогения 128
— функциональная морфология 188,
189, 221, 222
— экология 210, 220, 221, 222, 232,
233
Подвиды 101
Подготовка образцов для фотографи-
рования 36
Позвоночные, возникновение скелета
315
— кости, рост и функции 63
— плиоценовые сообщества 249, 250
Популяции 67—84, 196
— аллопатрические 98
— возрастное распределение особей
69, 70, 80
— возрастные группы 80
— географическое распределение 67,
218—220
—давление отбора 73, 102, 103
— дем 67
— динамика 77—84
— ископаемая 74—78, 85—96, 241—
243
— кривая выживаемости 77, 78, 81
— массовая смертность 81
— мутационное давление 73, 102, 103
— ненаследственные изменения 74
— наследственные различия 70—73
— онтогенетические различия 69, 70
— остракод 81
— поток генов 67
— размер 103
— симпатрические 98
— трилобитов 72
— эволюция 100
«Последовательный» вид 107
Посмертное разрушение 243—254
---- биологическое 25, 26
----механическое 26—29
----химическое 29
Посмертный комплекс 244
Посмертный перенос 32—34, 195,
243—254
----сортировка 75, 76
Поток вещества, схема 197
— генов 67
Потенциал расселения 104
Преобразователи 196
Пресмыкающиеся, следы 15, 226
— разнообразие 23
— сохранность 15
Приборы для измерения ископаемых
46, 47
Прижизненный комплекс 244
Признаки, взвешивание 134
Приливно-отливная зона 198; см.
также Литоральная зона
Принцип конкурентного исключения
231
Принцип подобия 63, 276, 277
Продуценты 196, 197, 201
Продолжительность существования
видов у млекопитающих 265
— —-----моллюсков 265
Прокариоты, эволюция 313
Простейшие см. также Фора мини -
феры
— филогения 130
Прочность раковины брюхоногих 65
----головоногих 177—181
----двустворчатых моллюсков 65
— скелета позвоночных 63
Птицы 14, 137, 308
— эволюция 133, 315
Пыльца, перенос 33
— эволюция 315
Радиация эволюционная см. Адап-
тивная радиация
Размер, определение 42, 43
— и функциональная морфология 187
— увеличение в ходе эволюции 181.
270—272, 276—278
Разнообразие ископаемых видов 21,
23, 97, 208—211, 240—243, 283
— головоногих 20, 21
— градиенты 240—243
— двустворчатых 22, 23
— иглокожих 23
—и соленость 208—211
— млекопитающих 23
•— морских лилий 21
Предметный указатель
385.
--плеченогих 21, 22
— пресмыкающихся 23
----эволюция 97
Раковина, геометрия у брюхоногих
163
------головоногих 44, 5з—57, 162,
163
------двустворчатых 163
------фораминифер 164
Ракообразные, карбонат кальция, мо-
дификации 368
— планктонные 201
— сохранность 13
Расселение 323—327
— случайные агенты 324
— «счастливые пути» 324
Растения см. также Морская трава,
Пыльца, Споры, Фитопланктон
— гибридизация 97
Регрессия моря 336
Редуценты 196
Рекапитуляция 273—277
Реконструкция диагенетических об-
становок 367—369
Реликтовые популяции 295, 343
Рентгенография см. Фотографирова-
ние ископаемых
Репродуктивная изоляция 97
— система 70, 73, 316
Рептилии см. Пресмыкающиеся, Дино-
завры
Рифы 227—230
— из археоциат 305, 306
Род 126
— изолированный 126
— типовой 126
Рост аллометрический 62
— анизометрический 60—62
— брюхоногих 49
— годичные линии нарастания 356
— головоногих 53—57
— двустворчатых 64, 355
— и линька 51, 81
— — метаморфоз 48, 62
— изометрический 60—62
— кораллов 355—358
— костей у позвоночных 63
— моллюсков 49, 65, 355
— морских ежей 9, 49, 52, 53
— плеченогих 54, 55
— путем аккреции 49, 64, 355—361
----добавления новых частей 49, 53
----модификации 52, 53
-соотношение частей тела 61
— суточный 356
— трилобитов 49—51, 58
— темп 58—62
— членистоногих 51, 81
Руководящие ископаемые 334, 349
Рыбы, возраст, определение 80
— двоякодышащие 263
— эволюция 287—289, 315
Свиты стратиграфические 318
Семейство 126
Серии стратиграфические 318
Сессильные организмы 199
Сестонофаги 202, 306
Символы в иллюстрации
изменчивости 92, 93
Синонимика НО, 123, 153—155
Синтип 121
Синэкология 226
Скелет ископаемых, изотопный со-
став 74
— — минерализация 367—369
Следы жизнедеятельности ископае-
мых 224—226
------сохранность 13—16, 224—226,
249
«Следы жизни» 224—226
Смертность массовая 81
Смешанный посмертный комплекс
244
Соленость 207—213, 302, 303
Сообщества ископаемые 226—237
— повторяющиеся 226, 230
Сообщество климаксное 227
Соскребатели 202
Сохранность 13—34, 243—251
— модели Джонсона 245
— мягких тканей 226
— насекомых 21
— пигментов 14, 30
---- в янтаре 15, 21
Спиральная раковина, геометрия 159,
160—164
------ моделирование при помощи
ЭВМ 161 — 164
----функциональная морфология
169—173
Споры, перенос 33
Статистический анализ 88
---- критерии достоверности 92
Стеногалинные организмы 208—211,
302
Стенотермные организмы 236
Стратиграфическая корреляция 328—
354
----использование радиометрическо-
го метода 350
----количественные методы 337—-
343
386
Предметный указатель
----по морфологически измененным
ископаемым 334—336
----по руководящим ископаемым
334
— — точность 347—354
----по экологическим признакам 336
— номенклатура, кодекс 317
— система 344
Стратиграфические единицы 34, 323,
328
— ранги 328
Стратиграфия см. Геологическое вре-
мя, Стратиграфическая корреляция
Строматолиты 217, 218, 312
Сублиторальная зона 198
Сублиторальный субстрат 199
Субстраты 204—207
Сукцессия экологическая 226, 227
Таксономический признак 120, 134—
138
Таксономическое разнообразие 240
Таксономия см. Классификация
Таксоны 16
— новые, частота появления 266—269
Тафономия 243
Температура и градиенты разнообра-
зия 242, 243
----ископаемая флора 238—240
----морфология 236, 237
Тенденции филетические 269
Теоретическая морфология 159—166
Тип 126
Типовой род 126
Типовые разрезы 344
— экземпляры НО, 121, 122
Топотип 117
Травоядные 196, 197
Трансгрессия моря 336
Трилобиты, генетическая изменчи-
вость 72, 73
— деформация 362, 363
— зрение и функциональная морфо-
логия 189, 190
— классификация 138
— рост 49—51, 58
— сохранность 30
— эволюция 290, 331
Ультрамикроископаемые, фотографи-
рование 36
Устрицы, посмертное разрушение 25
—эпизои 223, 224
Фации 321
Фенетическая классификация 140
Фенограммы 142—144
Фенотип 99, 100
Филетическое вымирание 100
Филогенетическая скорость эволюции
255—266
Филогенетические связи 305—312
Филогенетическое древо 127
Фитопланктон 199—201, 303, 304
Фораминиферы, карбонат кальция,
модификации 368
— направление навивания 72, 334—
336
—раковина, геометрия 164
— репродуктивные особенности 70
•—устойчивость к солености 213
Фотическая зона 199—201
Фотографирование ископаемых в ин-
фракрасных лучах 36
----в ультрафиолетовых лучах 36
---- подготовка образцов 36
— — рентгенографическое 36, 225
----стереопары 36
----электронная микроскопия 31
Фотографический метод 36—39
Фотосинтез 198
Фузулиниды, вымирание 297
Функциональная морфология голово-
ногих 173, 176—178
---- двустворчатых 190—192
----динозавров, механические осо-
бенности конечностей 183—186
---------свойства челюстей 183—186
---- методы анализа 166—169
----в палеоэкологии 221, 222
----паруса пеликозавров 186—188
----перегородочной линии 183
----плеченогих 188, 189
----спиральной раковины 169—173
----трилобитов 189, 190
----факторы множественного дейст-
вия 168
Хищники 196, 197, 202
Хроностратиграфические подразделе-
ния 343—347
Хунсрюкские сланцы 15
«Царство» (категория Линнея) 126
Цератопсы, морфология и размер 258
Цифровое кодирование 37—39, 47
Черви, карбонат кальция, модифика-
ции 367
— экология 224
Численная таксономия 138—144
Членистоногие, рост 49—51
— сохранность 13—15
Прейметный указатель
387
Шельф 198
ЭВМ, использование для целей клас-
сификации 138—144
— в моделировании формы раковины
161 — 164
---описании ископаемых 36, 37, 39
--- определении видов 151, 152
— цифровая информация 37—39, 47
Эволюционный вид 108, 133
Эволюционная радиация иглокожих
23
Эволюционные линии ПО
Эволюция см. тщоке Видообразова-
, ние, Вымирание
— и адаптация 156, 278—291
— двустворчатых моллюсков 192, 253
— иглокожих 129
— итеративная 291
— и классификация 125, 127
— кораллов 257, 258
— лошадей 131, 272, 273
— ортогенез 269—273
— параллелизм 292
— плеченогих 132
— подвидов 101
— простейших 130
— скорость 253—266
— и увеличение размера 181, 270—
272, 276—278
— филогенетическое древо 127
— частота появления новых таксонов
266—269, 285, 286
— экосисте м 312—316
Эвригалинные организмы 208—211
Эвритермные организмы 236
Экологические ниши 195
Экологическая сукцессия 226, 227
Экологическое замещение 291
Экосистемы 195—203
— морская 198—203
— эволюция 312—316
«Эллипс деформации» 362
Эндемичные виды 325
Энергетическая пирамида 197, 198-
Эпибионты 224
Эпизои 224
Эпиконтинентальные моря 198
Эпифауна 199, 236
Эпифиты 224
Эукариоты, эволюция 313
Янтарь балтийский 15, 31
— ископаемый 15, 31
Ярусы стратиграфические 344
ОГЛАВЛЕНИЕ
Предисловие к русскому изданию.......................... . 5
Предисловие авторов ..................................... ... Ю
ЧАСТЬ ПЕРВАЯ
ОПИСАНИЕ И КЛАССИФИКАЦИЯ ИСКОПАЕМЫХ
ОСТАТКОВ
Глава I. Сохранность органических остатков и палеонто-
логическая летопись...................................... 13
Число видов................................................ 15
Вымершие и рецентные виды...................................20
Число особей.............................................. 23
Условия сохранности органических остатков ..................25
Биологические структуры, имеющие наибольшие шансы на сохра-
нение ......................................................29
Условия среды и сохранность органических остатков .... 30
Посмертный перенос..........................................32
Выводы.................................................... 34
Глава II. Описание отдельной особи............................ 35
Фотографический метод.......................................36
Штриховой рисунок...........................................39
Терминология, применяемая при описании......................39
Описание посредством измерения..............................42
Описание внутреннего строения ............................. 47
Глава III. Онтогенетическая изменчивость...................... 48
Типы роста..................................................48
Описание онтогенетических изменений.........................53
Темп роста................................................ 58
Причины анизометрического роста.............................62
s Некоторые особенности онтогенеза колониальных организмов 66
Глава IV. Популяция как целое................................. 67
Популяции в биологии ...................................... 67
Индивидуальная изменчивость внутри популяций .............. 69
Ископаемая популяция ...................................... 74
Динамика популяций ........................................ 77
Методы описания изменчивости................................85
Глава V. Вид как целое........................................ 97
Биологическое определение вида..............................98
Видообразование ........................................... 99
Возникновение видов в отсутствие изоляции..................101
Оглавление 3b9
Скорость видообразования ................................... 102
Биологические методы разграничения видов.....................104
Проблема вида в палеонтологии.................. . 106
Формальное именование и описание вида .... ПО
Изменение видовых названий . . . 122
Перспективы . . • 123
Глава VI. Группировка видов в высшие категории ... 124
Классификация и эволюция.....................................125
Таксономические категории....................................126
Система классификации как интерпретация эволюции . . 127
Критерии выделения высших категорий......................... 134
Численная таксономия................................... . 138
Глава VII. Определение ископаемых...............................145
Ключи...................................................... 148
Машинные методы определения..................................151
Представление результатов определения ...................... 152
ЧАСТЬ ВТОРАЯ
ПРИМЕНЕНИЕ ДАННЫХ ПАЛЕОНТОЛОГИИ
Глава VIII. Адаптация и функциональная морфология 156
Неслучайность результатов адаптации............. . 157
Теоретическая морфология. Спиральная раковина . . 159
Методы функционального морфологического анализа . . . 166
Функциональная морфология спиральной раковины . . 169
Проблема перегородочной линии головоногих ... 173
Механические свойства челюстей динозавра .... . . 183
Парус пеликозавров...........................................186
Специальные адаптации у брахиопод ..... .188
Зрение трилобитов.......................... . . . 189
Форма раковины у двустворчатых моллюсков . . 190
Заключение...................................................192
Глава IX. Палеоэкология.........................................194
Основные принципы экологии...................... . . 195
Морская экосистема ..........................................198
Ограничивающие факторы и местные условия среды .... 203
Тип субстрата ......................................... . . 204
Пространственное распределение популяций.....................218
Образ жизни..................................................220
Ископаемые сообщества....................................... 226
Температура и биогеография...................................235
Посмертная утрата информации.................................243
Глава X. Эволюция и палеонтологическая летопись ... 252
Географическое видообразование........................ ... 253
Скорость эволюции............................... . . 253
Филетические тенденции.......................................269
Пути эволюции............................................... 283
Вымирание....................................................294
Выяснение филогенетических связей............................305
Эволюция экосистем...........................................312
Глава XI. Биостратиграфия.......................................317
Литостратиграфические единицы................................318
Биостратиграфические подразделения. Биозона..................323
390 Оглавление
Корреляция по ископаемым......................................328
Хронологические и хроностратиграфические подразделения . . . 343
Точность корреляции...........................................347
Глава XII. Использование палеонтологических данных в
геофизике и геохимии........................................355
Линии нарастания у морских беспозвоночных.....................355
Использование ископаемых остатков при изучении деформации
горных пород.................................................361
Реконструкция диагенетических обстановок......................367
Выводы и перспективы дальнейших исследований..................369
Список литературы ............................................... 370
Предметный указатель..............................................380
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении,
качестве адресу: перевода и другие просим присылать по 129820, Москва, И-НО, ГСП 1-й Рижский пер., д. 2, Издательство «Мир»
Д. РАУП, С. СТЭНЛИ
ОСНОВЫ ПАЛЕОНТОЛОГИИ
Редактор Е. Э. Казакевич Художник А. Ь. Шипов
Художественный редактор Ю. Л. Максимов Технический редактор В. П. Сизова
Корректоры В. И. Киселева и О. Ф. Туркова
Сдано в набор 26/IV 1974 г. Подписано к печати 6/IX 1974 г. Бумага тип. № 2.
60X90’/ie= 13,75 бум. л. Усл. печ. л. 27,50. В т/ч 3 п. л. вкл. Уч.-изд. л. 27,63.
Изд. № 4/6730. Цена 2 р. 72 к. Зак. 467.
ИЗДАТЕЛЬСТВО «МИР»
Москва, 1-й Рижский пер., 2
Московская типография № 11 Союзполиграфпрома при Государственном комитете Совета
Министров СССР по делам издательств, полиграфии и книжной торговли. Москва, 113105.
Нагатинская ул., !.
Фото I. Окаменелости иззольнхофенских известняков (юра, Бавария, ФРГ).
А . PenaeUs spcciosus ( 1 2). Стрелкой / отмечен след падения карапакса; 2 - - след росгру-
ма; 3 — след глаз. Н. Л1 esolimulus walchi (>' 1 Сохранились последние следы его пере-
движения (Л): ниже - искривленные бороздки, оставленные телом животного, когда оно
скользило по поверхности осадка к месту окончательного .захоронения. И. Archaeopteryx
lithogiaphica (\ 1/2). Сохранились отпечатки перьев.
Фото II. Рентгеновский снимок современного брюхоногого моллюска Conus
geographic us [63|.
Видны повреждения, нанесенные раковине сверлящей губкой СНопа.
Фото III. Плита песчаника, заключающая раковины моллюсков (плиоцен,
Калифорния, США) [235].
По состоянию и составу этот ископаемый комплекс похож на тот, который получился пос-
ле экспериментов с истиранием (см. фиг. 9). Преобладают раздробленные раковины брюхо-
ногих и двустворчатых моллюсков.
Фото IV. Окаменелости из сланцев Бергесс (среД
A. Ayshcaia pea'iz/icuZa/a, относящаяся к Onychophora, животным, стоящим по морфологии
Г. Marella splendens и Burgessia bella, трилобитообразные членистоногие. Д и Е. Декстро
известных трилобитообразных найдены только в сланцах Бергесс. На фотографиях В и Д
жения, кроме Д, даны с увеличением
нии кембрий, Британская Колумбия, Канада).
между кольчатыми червями и членистоногими. Б. Canadia sctidera, кольчатый червь. Б и
латеральный н дорсальный вид U aptia fieldcnsis. другого трилобптообразпого. .
в виде темных силуэтов заметны сохранившиеся остатки внутренних органов. Исе пзоо!
в 2 — 3 раза; Д увеличено в 6 раз.
ФотоУ. Стереофотографии брахиоподы Echinauris opuntia (пермь, Пакистан)
[69|.
Рассматривая стереолары через карманный стереоскоп, можно видеть обьемные
изображения.
Фото VI. Использование фотографии для исследования ископаемых.
Роденовские снимки (стереопары) брахноиоды Spittocyrlia euruteines (девон, Огайо,
ША); видны структуры брахидпума [2371. Б. Поверхность напластования зольпхофеп-
ких известняков в видимом свете (слева; видны только дендриты) и в ультрафиолетовых
лучах (справа; видна лич.<а десятиногого ракообразного) I11XJ. И. Грантолиг Diplo-
graptus gracilis в видимом свете (слева) и в инфракрасном свете (справа) I.104J.
Фото VII. Электронные микрофотографии скелетов ископаемых и современ-
ных беспозвоночных.
А Строение раковины юрского двустворчатого моллюска Praemylilus (X 7140) f82l. Б.
Радиолярии пч эоцена острова Баобадос (X 425). В и Г. Поверхность раковины современ-
ной остракоды Carinocythercis all. carinata из Неаполитанского залива, Италия (X 110
и 1000).
iniiiiimii; < -
ftfcgi.
W”1
И
IW/
П ГП fl fl II n II I I
II111 fl ‘ ‘ * * * *
IM I fl
IM I I
-w.
А
mi®
^iiik
:
.diri rtUflr
•n
»fir
MH-
w
#№•
Uh
M
I
1
I
I
III
I I I
11 I
I 1 I
I I I I I
I I I I I
W
I
I
I
..1,1 • 9,1
вчит
fir
i4?«L
’ЙГ
"bffillj
fi'JWW'
цЕЦнНЫ’ «’
!i W«u
V-
ш.
.ж»
!*-h'7 K' , ,
flit-
чи-М^Чий^
ilFih fliir*
•rll1*»*?. ,^1,.
irtil.ififtft.j-'tsS
.“> 'We
Д1»1 m*
’ЙЧ"Х
WiJr «<
I I I
I
I 11
I I I
I I I I I I I I
I I I I I 1 1 1 I I I I
III 11
III 11
III
III
III
111
I
I 'I 4111.(1
««К
Ip fl
w
,чв,
tf .
»V> 5»
i“ W":
w
W'"l
111
5i'!'
iuiiittiiTHviii
....--- , j
I I
|'Жи
rHi 'fo
V
rtl..
IK-4’
тографической информации
l&r I1
irfriwi
I ltllllllllll,llliilllllllll.I
! '1,1.1г . ......
inn
11!
Ill»
[115].
ченный фрагмент). Процесс кодирования полностью
лиз позволяет выявить хромосомные нарушения.
автоматизирован, одна фотография
Фото VIII. Цифровое кодирование фо
крофотографня хромосом (слепа) и она же и закодированном виде (справа; па’ врезке свели
обрабатывается за 0 2 с. Проводимый затем на ЭВМ ана
фото IX. Брюхоногий моллюск Calliotnphalus conanli (мел, США); научное
описание сида приводится в тексте.
А
Фото. X. Рост за счет сочетания аккреции (наращивания существующих час-
тей) с появлением новых частей 1158].
Показаны два ряда пластинок скелета морского ежа Strongylocetilrotus. Па каждой ила
стинке видны линии нарастания, отражающие последовательность увеличения ее разме-
ров. Новые пластинки появляются всегда в вершине ряда.
Фото XI. Рентгеновский снимок современного головоногого моллюска Nau.'
tilus, иллюстрирующий рост за счет добавления новых частей и аккреции.
В онтогенезе увеличивается длина раковины (аккреция) и число камер (появляются
новые септы).
1
Фото XII. Генетическая изменчивость в ископаемой популяции [13].
Хвостовые части (пигидии) силурийского трилобита Encritiurus с разным расположением
первых (антерпорных) трех осевых бугорков. Между вторым и первым бугорком имеется
3 или 4 кольца, между третьим и вторым — также 3 или 4 кольца. Соотношение экземпля
ров с разным расположением осевых бугорков, как полагают, свидетельствует о том, что
этот ископаемый комплекс — представительная выборка из менделевской популяции,
находившейся в состоянии равновесия (/ — передний коней пигидия; 2 — первый буго-
рок; 3 — второй бугорок; 4 — третий бугорок; 5 — задний конец пигидия).
Фото XIII. Две створки Pitar dione (/ — правая; 2 — левая).
Створки совершенно одинаковы, только одна представляет собой зеркальное изображе-
ние другой (см. фиг. 30)
Фото XIV. Девонская брахиопода Chonetes pachyactis Imbrie (Xl).
Фото XV. Аммонит Xenoceltit es youngi Kummel et Steele (X 1).
Фото XVI. Трилобит Pterocephalops tuberculineata Clark (X 4).
Фото XVII. Раннекаменноугольный морской еж Archaeocidaris rossica Buch
в двух ракурсах [93].
Фото XVI11. СовременньпУморской еж Eucidaris tribuloides (Lamark) (пример
морфологии цидароидных морских ежей).
Фото XIX. Контур раковины брюхоногого моллюска, построенный на анало-
говой вычислительной машине [159].
Фото XX. Группа из225 смоделированных на аналоговой ЭВМ раковин [159].
Велич Ина переноса оборота возрастает справа налево, а скорость расширения оборота —
сверху вниз.
Фото XXL Серия контуров раковины, построенных на вычислительной ма-
шине, демонстрирует изменение формы раковины с изменсние1М геометричес-
ких параметров [156].
Фото XXII. Зрение Acaste downingiae [29].
A — В Трилобиты в свернутом положении. Г и Д. Правый глаз; видны линзы
V-»
Фото XXIV. Десятиногие раки из семейства Callianassidae (А, Б и Г — из
[225]).
А. Современная Callianassa в своей трубке (часть трубки вскрыта). В. Наружная поверх-
ность трубки (X 1). В и Г. Трубки из песчаников Фокс-Хилл (верхний мел, Денверский
бассейн, США).
Фого XXI11. Моллюски Clinopisthia и Polidevcia (вид сзади и со стороны
правой створки).
CliiwpitUiia (Д) — каме1ноугольный род, по образу жизни, видимо, сходный с Nuctila\
Polide'Jcia (Б) — каменноугольный род, по образу жизни, очевидно, сходный с Yoldia.
Фото XXV. Современные и ископаемые строматолиты.
А. Современные строматолиты (залив Шарк, Западная Австралия). Строматолиты растут
в литоральной зоне осолоненной лагуны, их уплощенные вершины достигают уровня вы-
сокой воды. На переднем плане справа — мелкие, активно растущие незрелые формы.
Внутри строматолитов имеются четко выраженные слои, состоящие из известковых ооли-
тов и обломков раковин фораминифер (песок со знаками ряби между строматолитами —
это и есть раковины фораминифер, принесенные из сублиторальной части шельфа). Песок
переносится волнами к берегу и здесь захватывается и удерживается липкой слизью
нитчатых сине-зеленых (бесскелетных) водорослей. Строматолиты быстро цементируются
неорганически осаждающимся арагонитом. Подобные строматолиты встречаются на от-
крытом прибою мысе, где они подвержены сильному действию волн. Длина линейки, раз-
личимой в верхней части снимка, 1,5 м. Б. Строматолиты из того же района, но живу-
щие на несколько более защищенном участке берега и ориентированные параллельно на-
правлению набегания волн (перпендикулярно береговой линии). Лопата на среднем пла-
не дает представление о масштабе. В. Плоскости наслоения столбчатых строматолитов.
Отдельные строматолиты вытянуты по направлению древних течений, перпендикулярно
древней береговой линии (протерозой, район Большого Невольничьего озера, Канада;
ср. с Б). Г. Поперечный разрез ветвящихся столбчатых строматолитов; показана часть
двадцатиметрового пласта столбчатых строматолитов, сложенных доломитом. Вертикаль-
ная часть линейки — 30,5 см. Д. Поперечный разрез сросшихся куполообразных строма-
толитов, сложенных доломитом и кремнистым сланцем. Одно деление линейки - пример-
но 3 см (протерозой, район Коппермайн-Ривер, Канада).
Фото XXVI. Скопления раковин мидии Mytilus edulis на широком песчаном
приливно-отливном пляже у Барнстейбл-Харбор (Массачусетс, США).
-Фото XXVII. Нижняя поверхность скопления раковин брахиопод Pentame-
rus oblongiis, сохранившихся в прижизненном положении ( 0,45) [239].
Формация Ред Маунтин (силур, Алабама, США)
Фото XXVIII. Цистоидеи Adocet ocyst is williamsi, сохранившиеся в прижиз-
ненном положении прикрепленными к древней поверхности эрозии (х 1,3)
[102].
Формация Шелл-Рок (девон, Айова, СШ В работе Г1021, откуда взята фотография, впер-
вые сообщалось о нахождении ископаемых цистоидеи, явно прикрепленных к субстрату
стеблями.
Фого’ХХ1Х. Рентгенограмма бентосной фауны в^толще осадка (илистого песка), взятого с глубины 12,2 м в зал.
Киссет-Харбор (Массачусетс, США) (X 1).
Снимок иллюстрирует палеонтологическое значение сохранения норок; 1 — двустворчатый моллюск Pitar (раковина способна сохра-
няться); 2 — руки офиуры, питающейся на поверхности осадка (скелет вряд ли сохранится неповрежденным); 3 и 4 — норки амфи-
под (их некарбонатные скелеты сохраняются плохо); 5 — норки пэлихет (мягкотелые животные).
Фото XXX. Следы динозавра из Роки-Хилл (триас, Коннектикут, США).
Животное сошло с твердого глинистого б< рега, на котором видны отпечатки капель дож
дя, в пруд пли ручеП. На его мягком илистом дне видны знаки ряби. Стрелка указывает
на не1лубокнн след. Обратите внимание на два более глубоких следа.
Фото XXXI. Верхняя часть кораллового рифа на глубине нескольких мет-
ров вблизи Норт-Рок, Бермудские острова.
Масштабная липепка имеет длину 50 см. Крупные кораллы-мо.зговики внизу справа и в
Hein ре слева относятся к разным видам рода />iploria. Листовидный коркообразукицпй
коралл ниже лнненкн относится к роду ,Иontastrea. Ветвящиеся древовидные формы —
восьмилученые кораллы
Фото XXXIII. Разрез штуфа породы из формации Стонхауз (силур, Новая
Шотландия, Канада) с сохранившимися ходами животных [163].
Фото XXXII. Остатки микрофлоры из кремнистых сланцев Ганфлинт
(докембрий, район оз. Верхнего, Канада) [8].
В- Kakabekia umbellate.. Г и Д. Eosphaera tyleri. Е и Ж.. Eoastrion simplex. Системати-
ческая принадлежность этих трех видов неясна. 3 и И. Gunf lintia grand is (водорослеподоэ-
ные нити).
Фото XXXIV. Представители фауны Эдиакары (Австралия) |Ь5].
A. Dickensonia, загадочная сегментированная форма ( !).£>. Spriggina, форма, наномн
нающая кольчатого червя ( J.7). В. Parvincorina, род неясного систематического положе
пня ( 1,33). Г. Belfane/la - очевидно, медуза (< 0,7). Д. Ran%ea, напоминающая неко-
торых сидячих колониальных кишечнополостных ( 0,6). Е rribruchidium, возможно,
относится к иглокожим ( 1 2) Ж- Arborea, род неясного систематического положения
IX 1,1',.
Фото XXXV. Пояски нарастания на эпитеке кораллов Holophragma calceo-
loides из девонских мергелей Висби, о. Готланд (Швеция) [231].
Фото XXXVI. Суточные линии нарастания на поперечном сечении створки раковины современного двустворчатого мол-
люска тридакны [149].
Цифры указывают число дней. Изменение формы и толщины слоев позволяет предполагать их зависимость от 14-дневного приливного
цикла.
Фото XXXVII. Хвостовые щиты трилобитов, деформированные в результате
тектонических движении |155|.
Фото XXXVIII. Избирательная доломитизация]высокомагнезиального каль
цита скелета водорослей Corallina в верхнекайнозойских известняках атолла-
Эниветок [183].
А. Светлоокрашенные ромбы доломита приурочены к темным нитям Corallina (X 30).
Б. Отдельная нить водоросли, видная в нижней правой части фото А, при большем
увеличении.
Фото XXXIX. Остатки скелета водоросли Halimsda в шлифах плейстоце-
новых известняков Кэй-Ларго (Флорида).
Л. Неповрежденный участок арагонитового скелета. Б. Отпечаток кальцитового цемен-
та, который покрывал коркой участок скелета до того, как арагоннтовый скелет был рас-
творен пресными водами. Вверху — фотографии шлифов; внизу — схематизированные
картины.