

































































































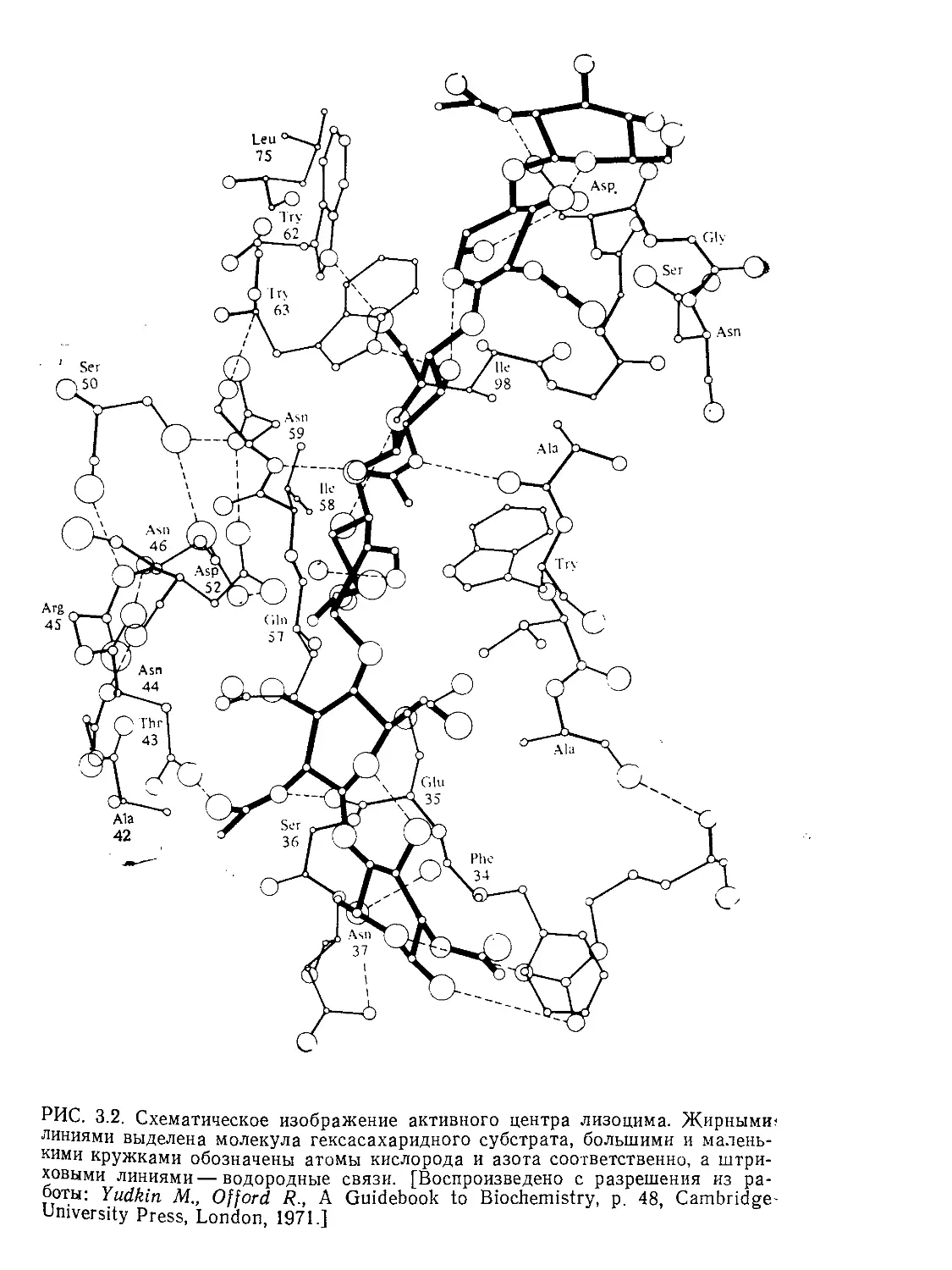
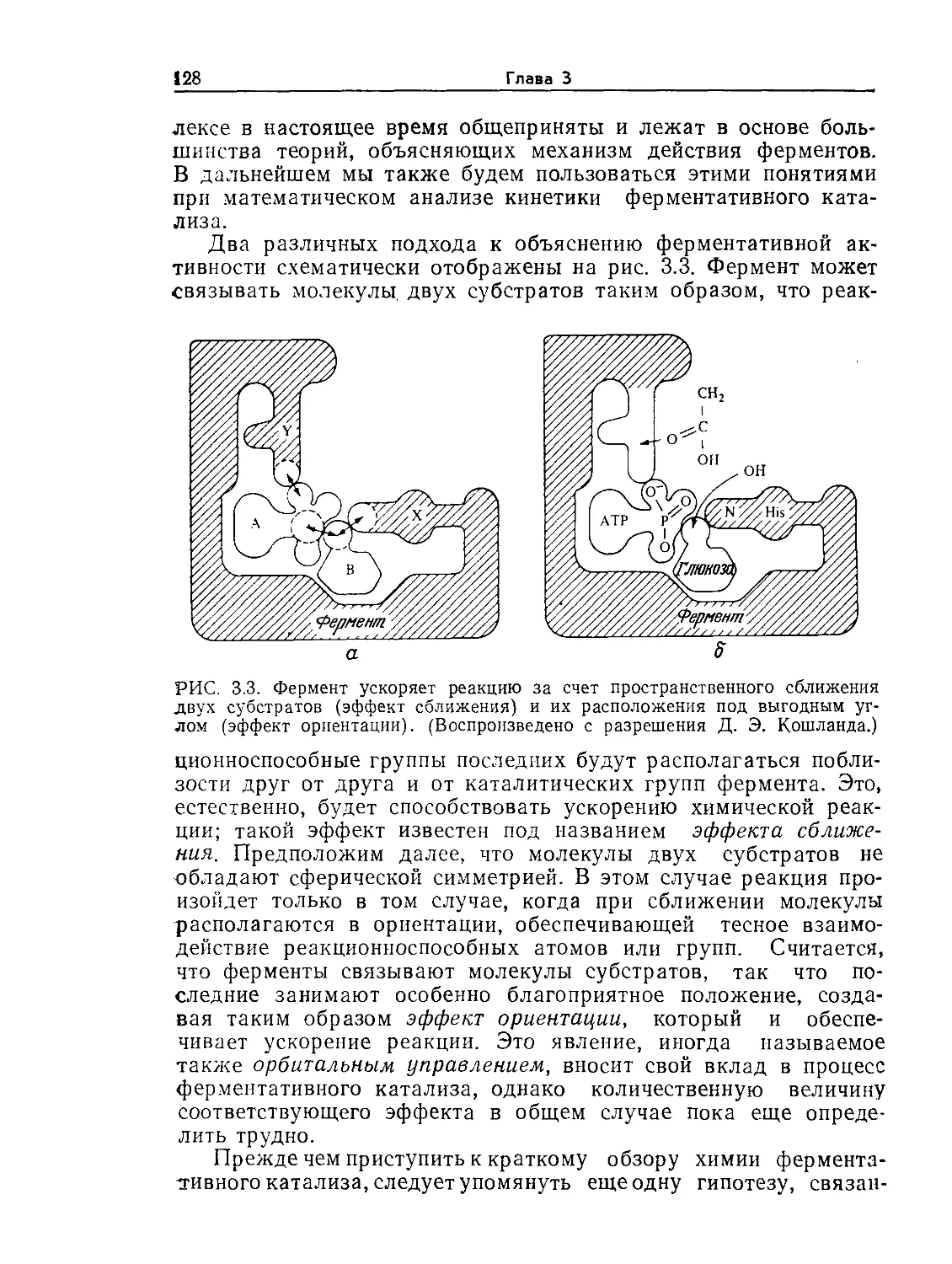

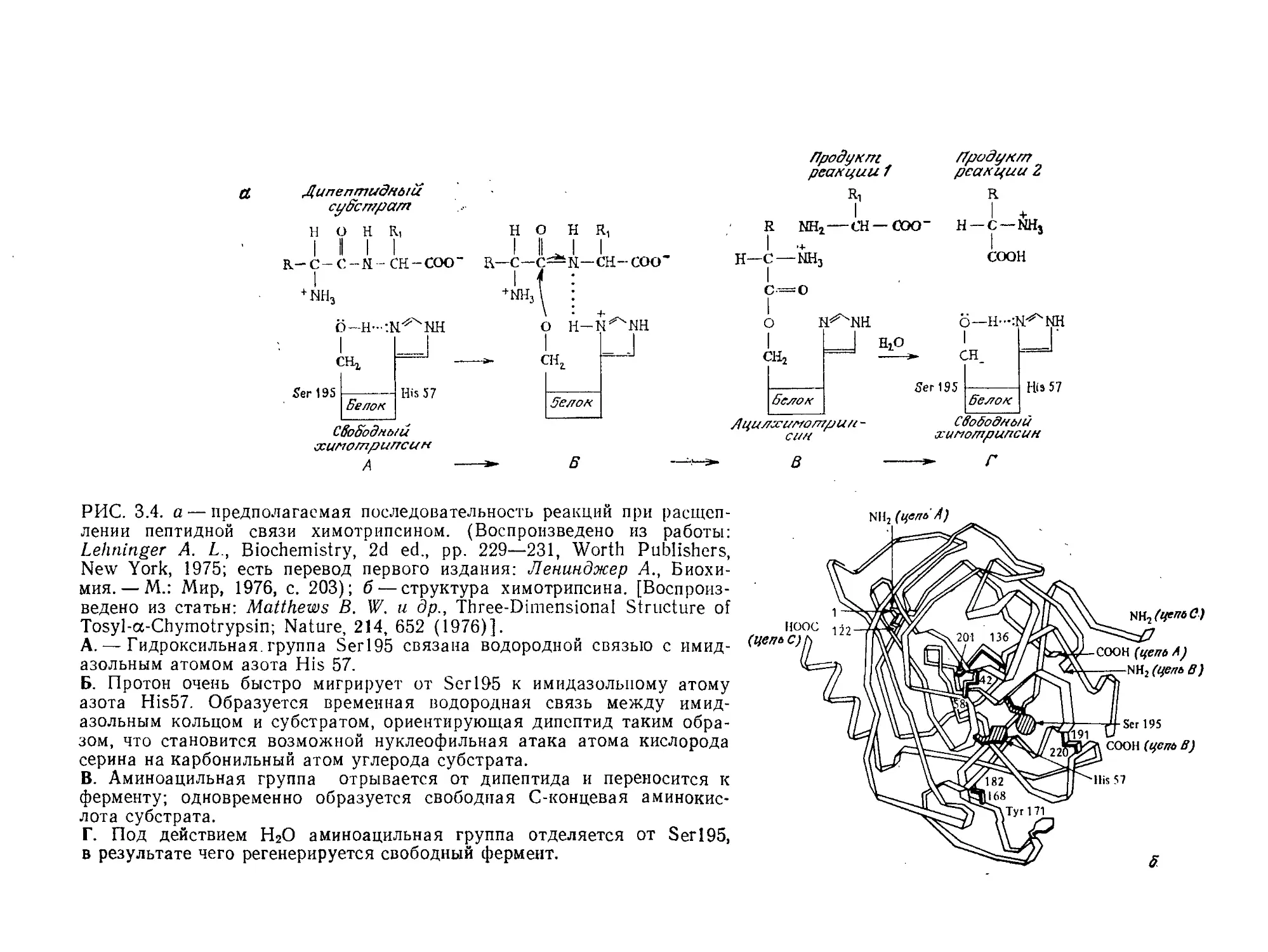


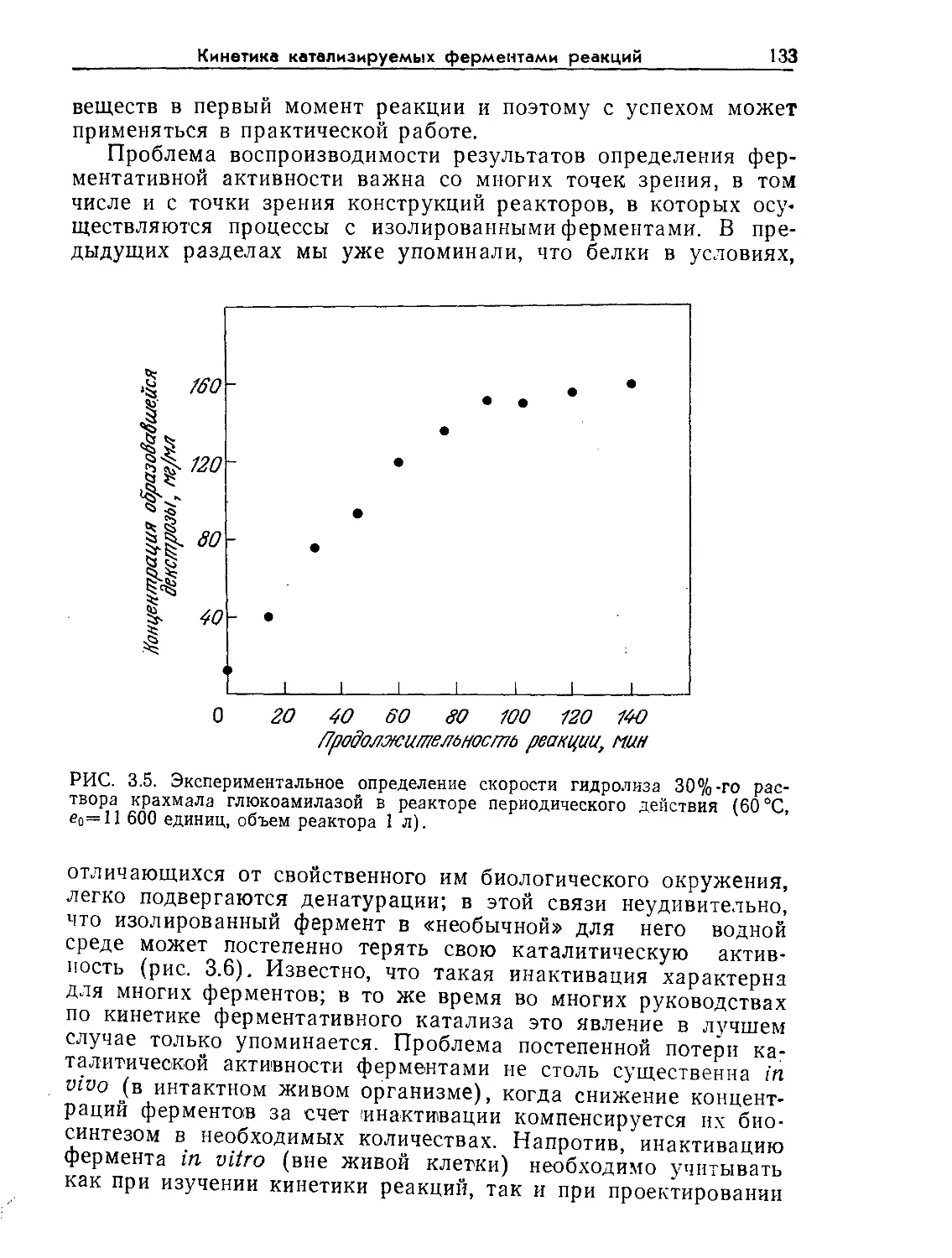














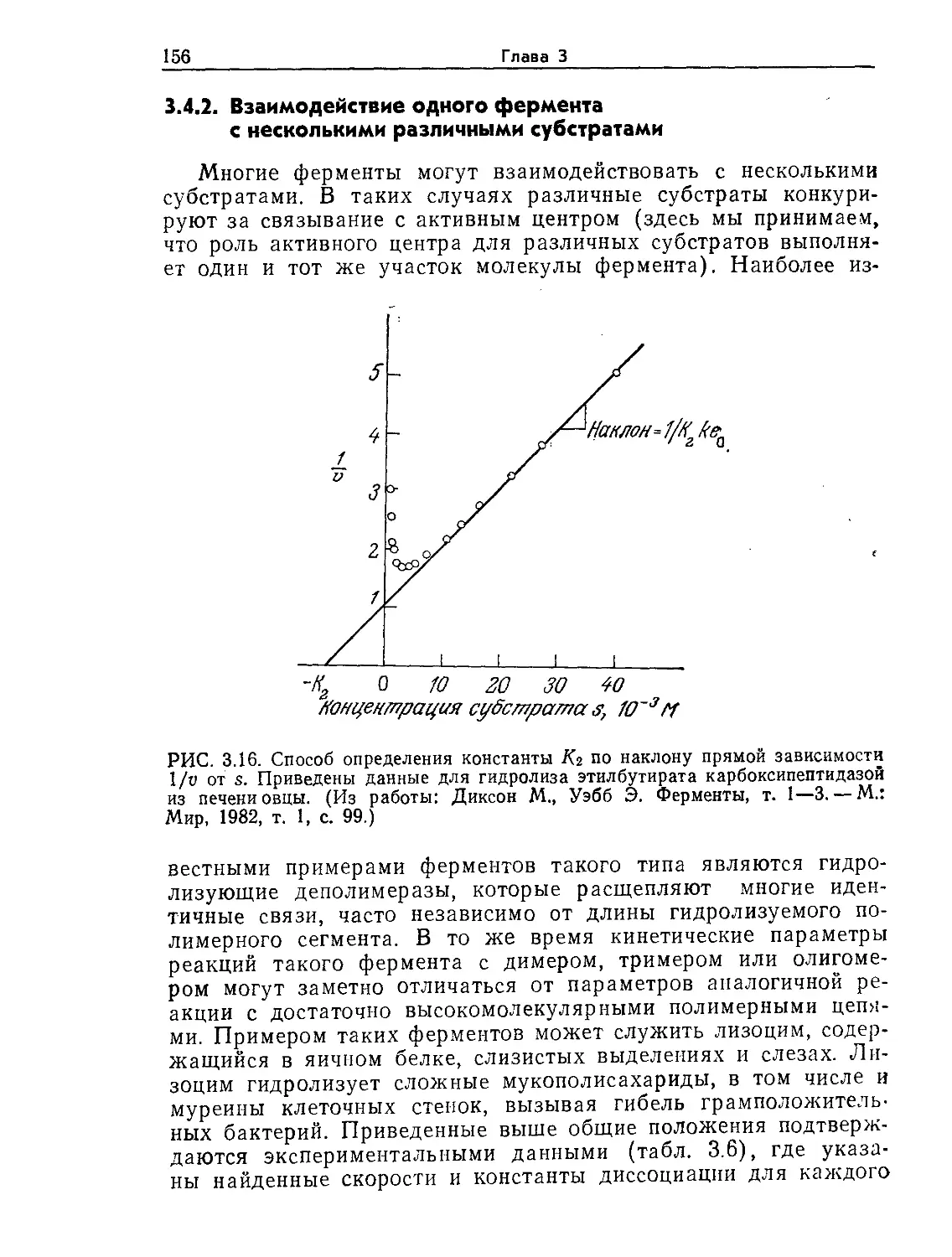

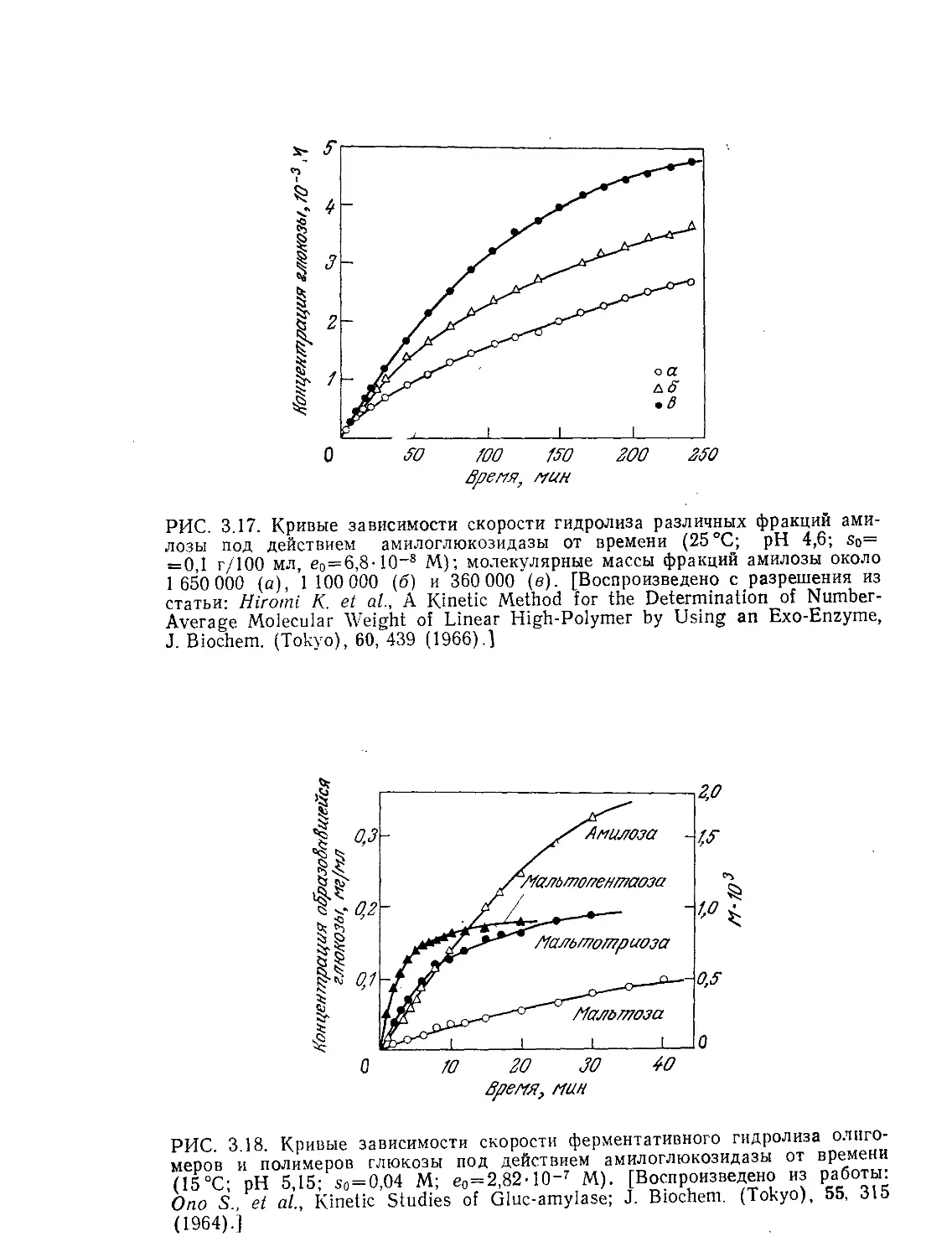










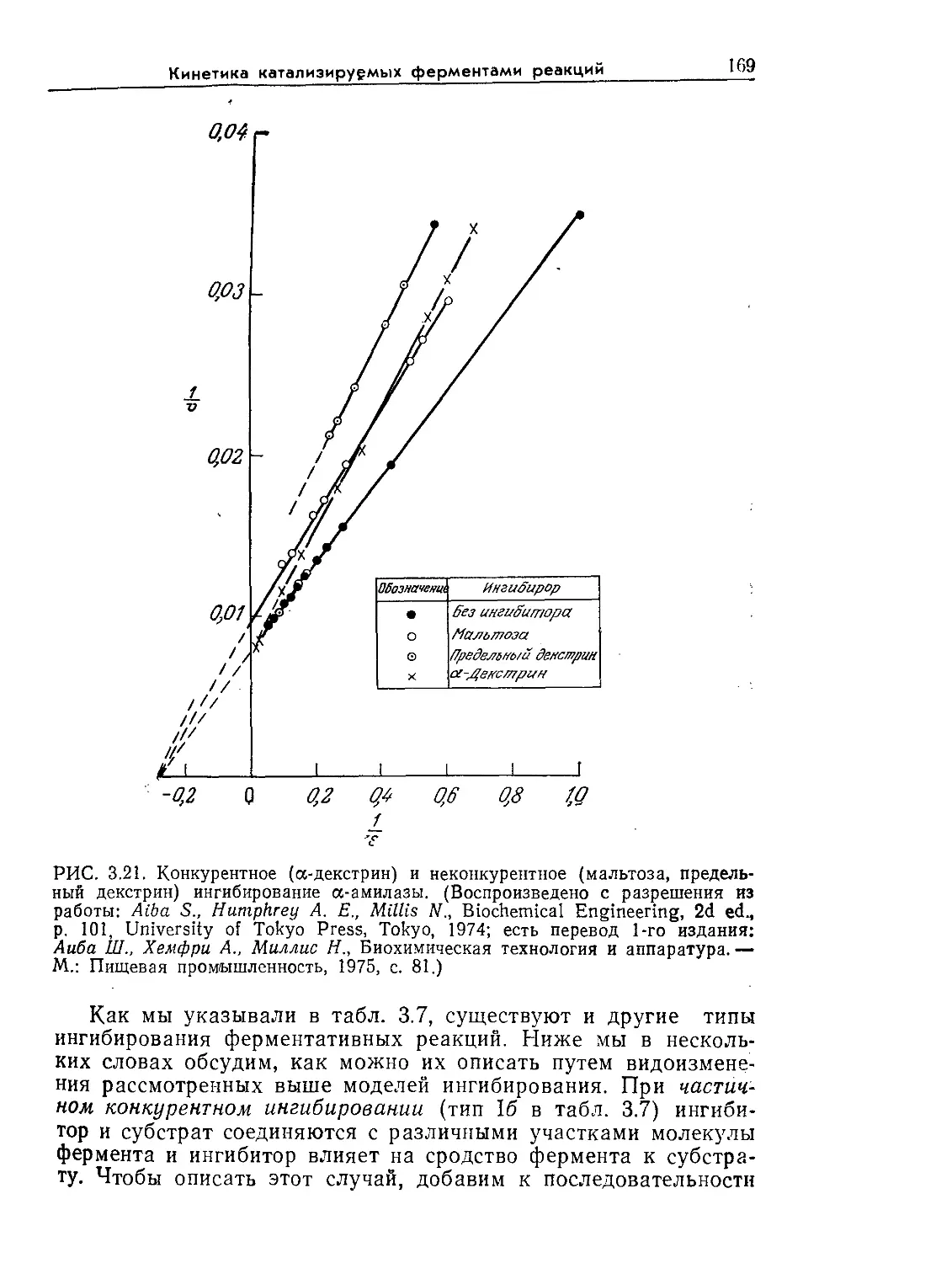


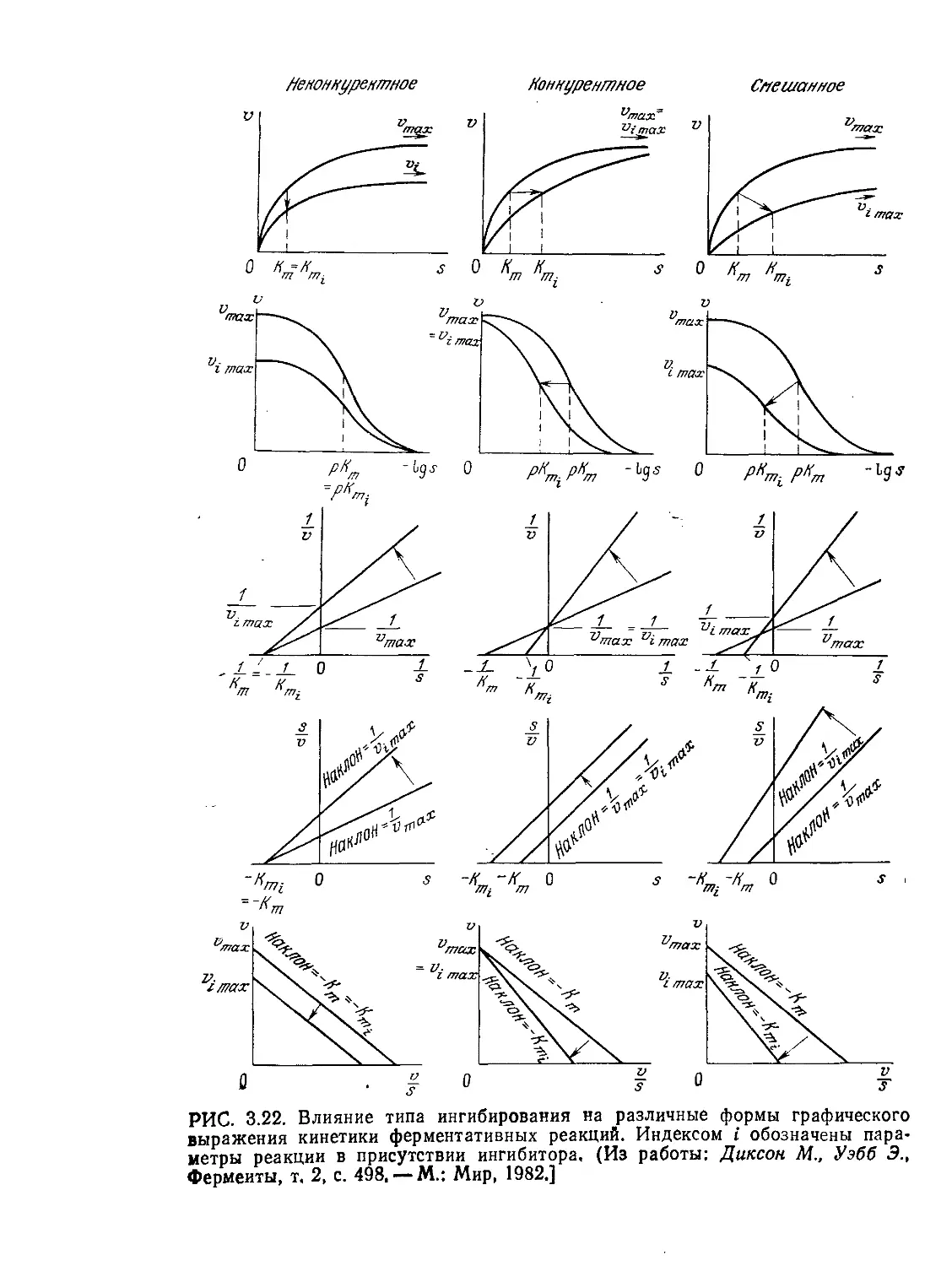

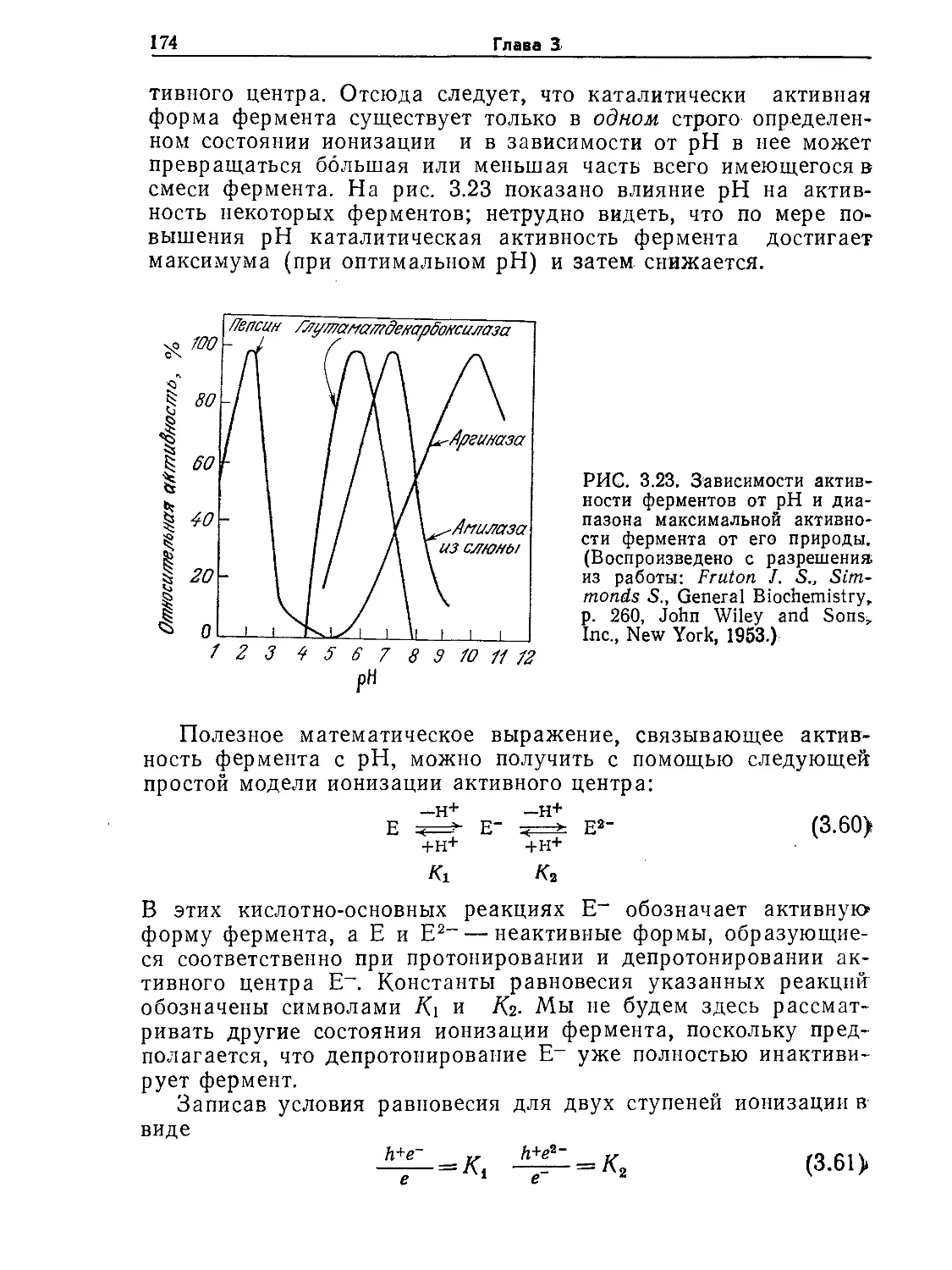


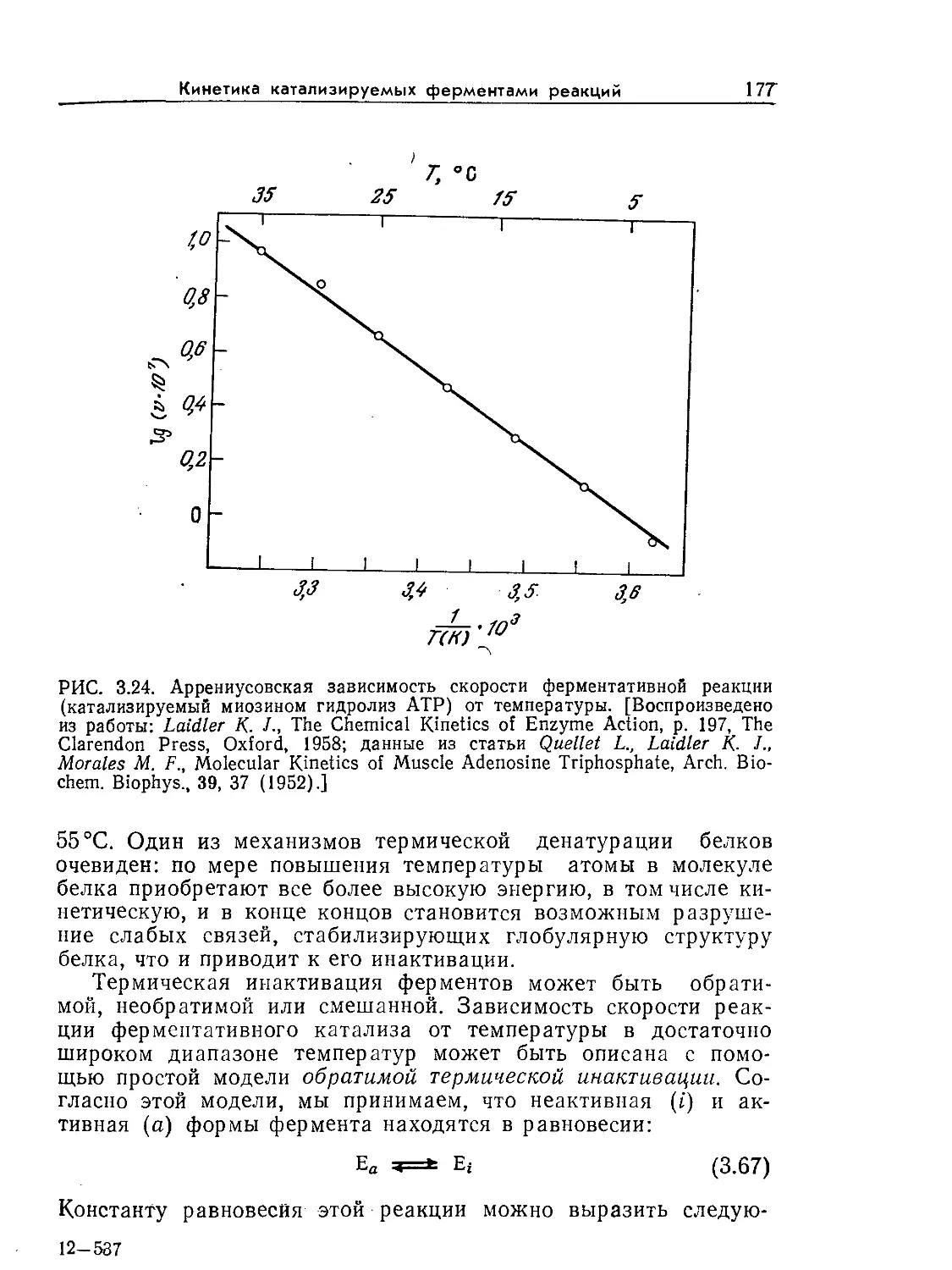
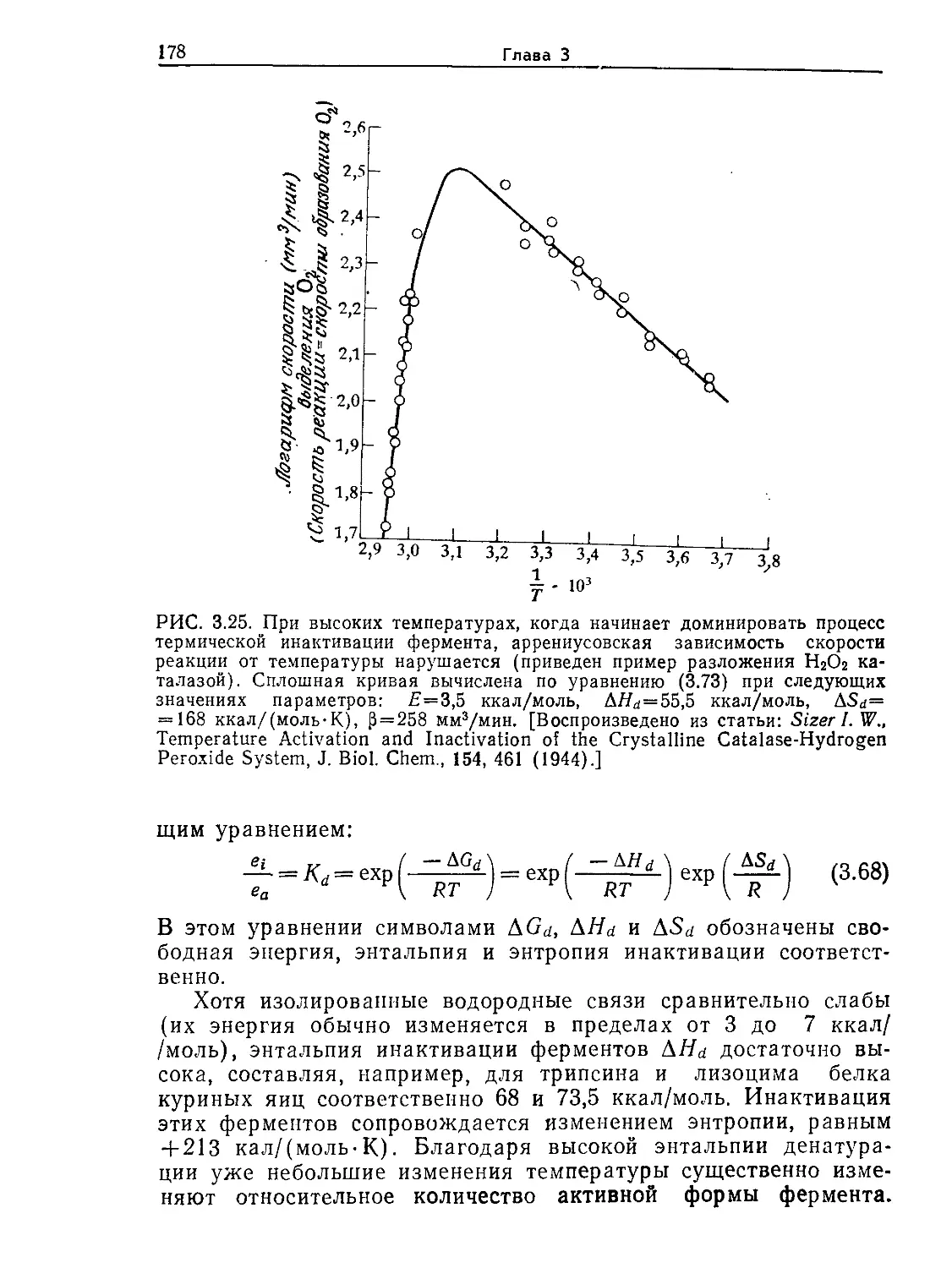



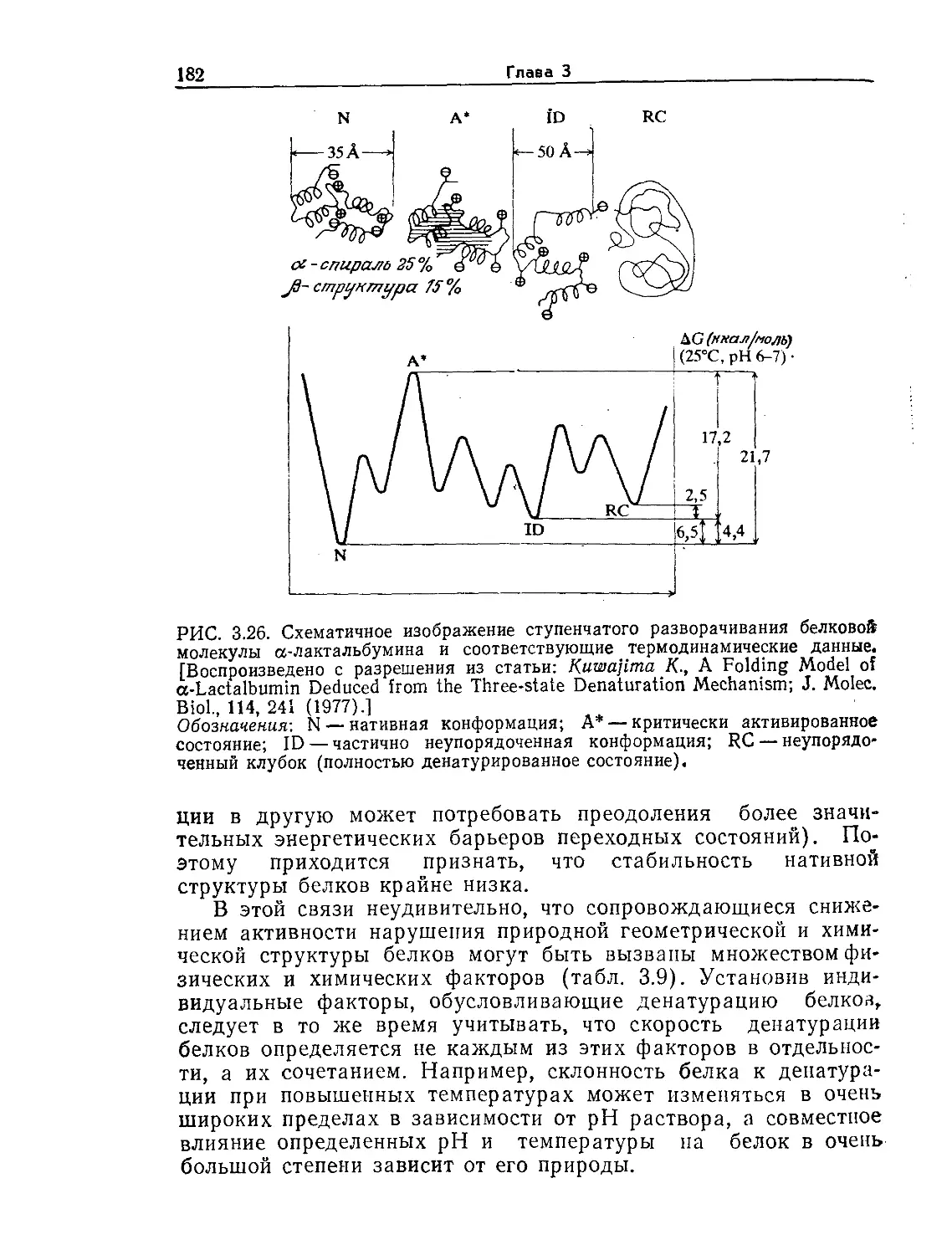











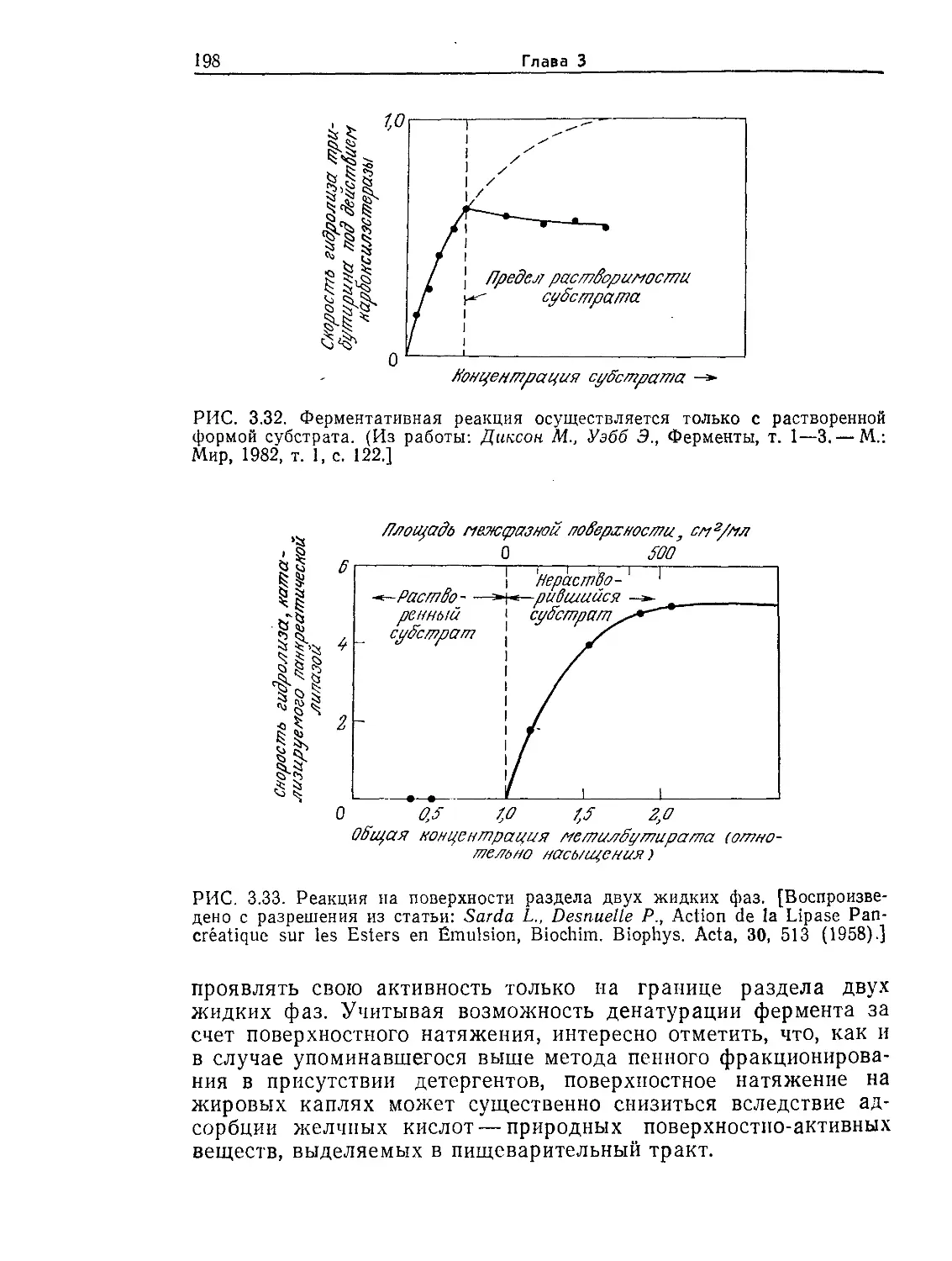







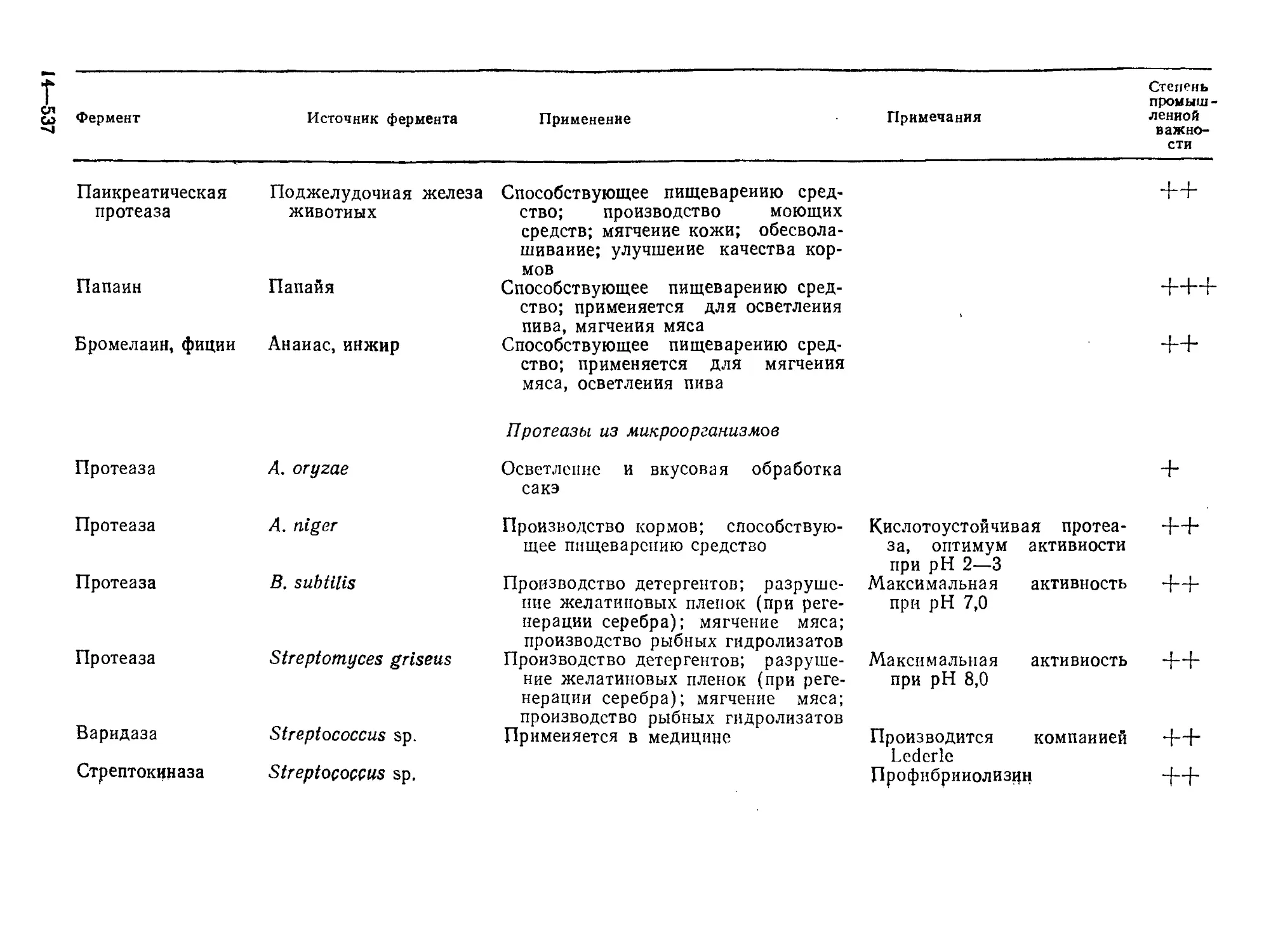

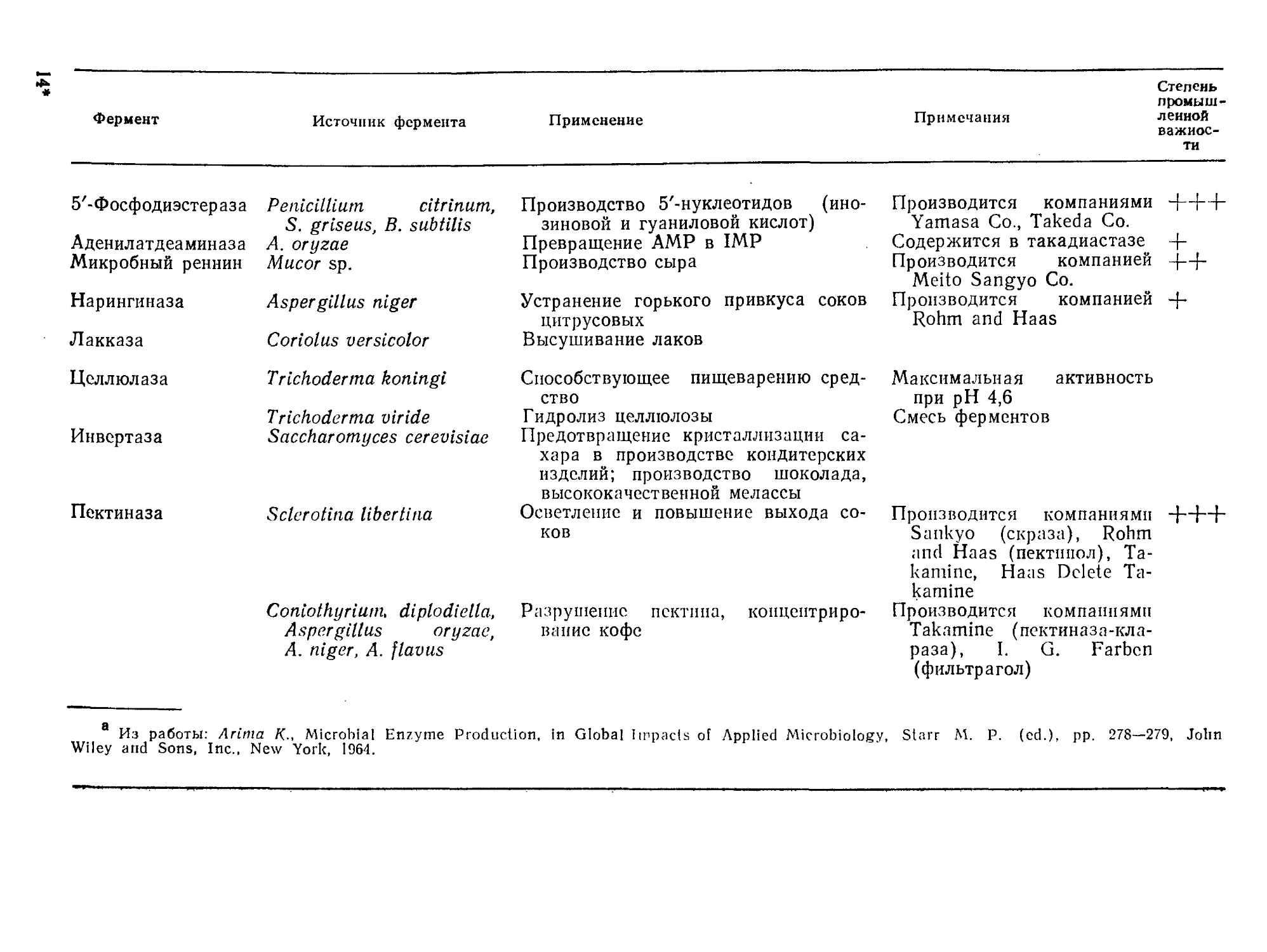









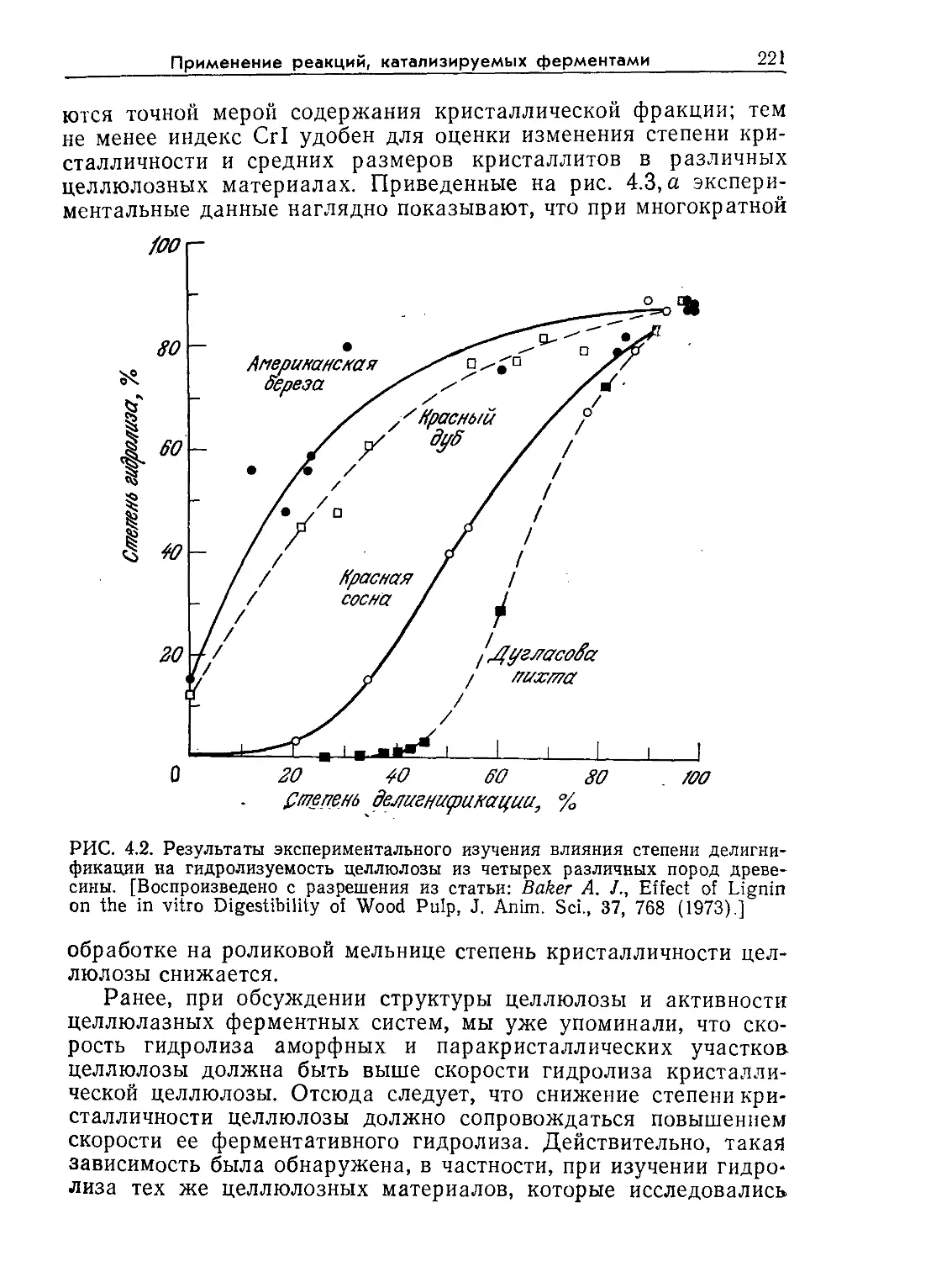
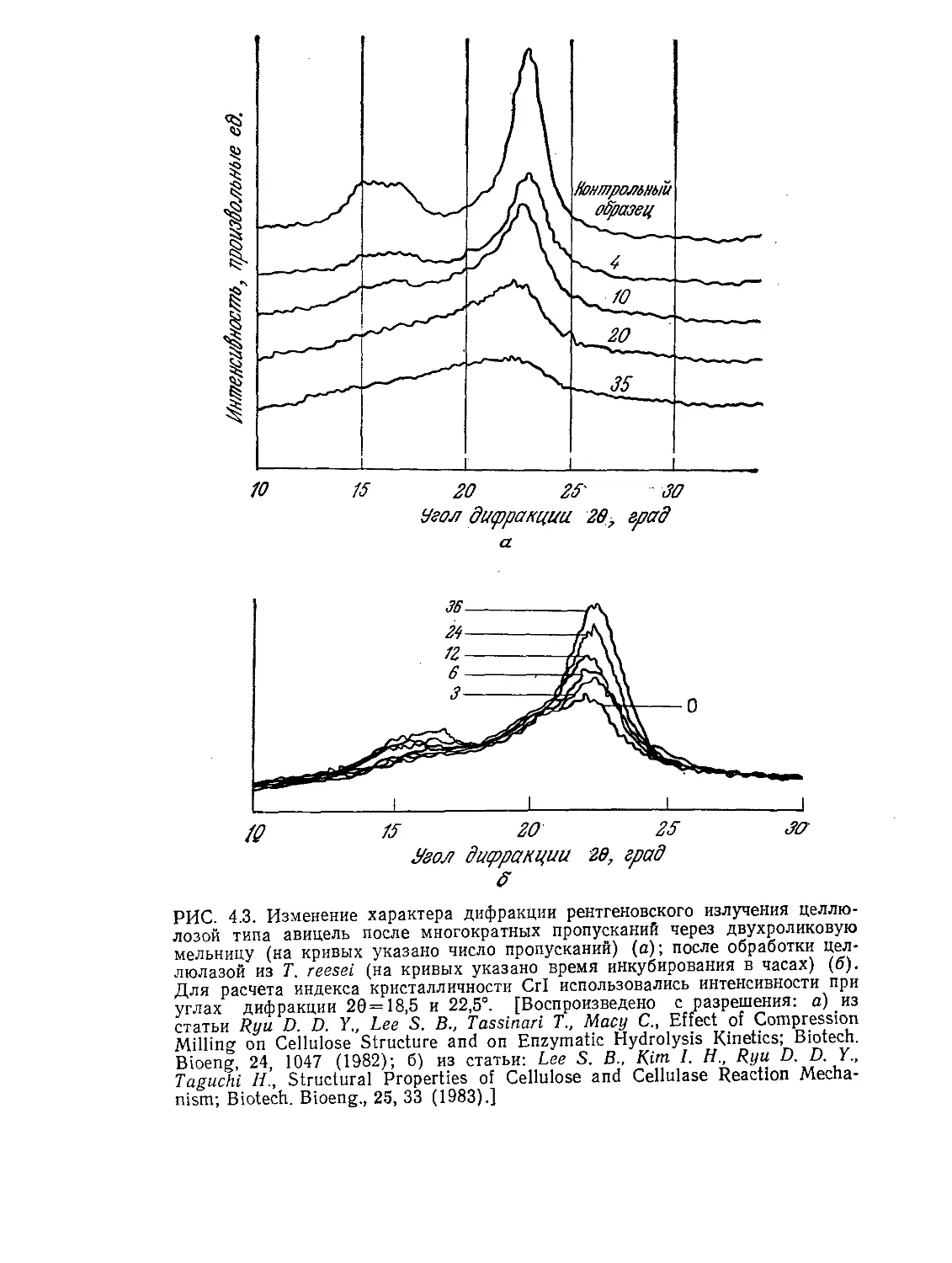


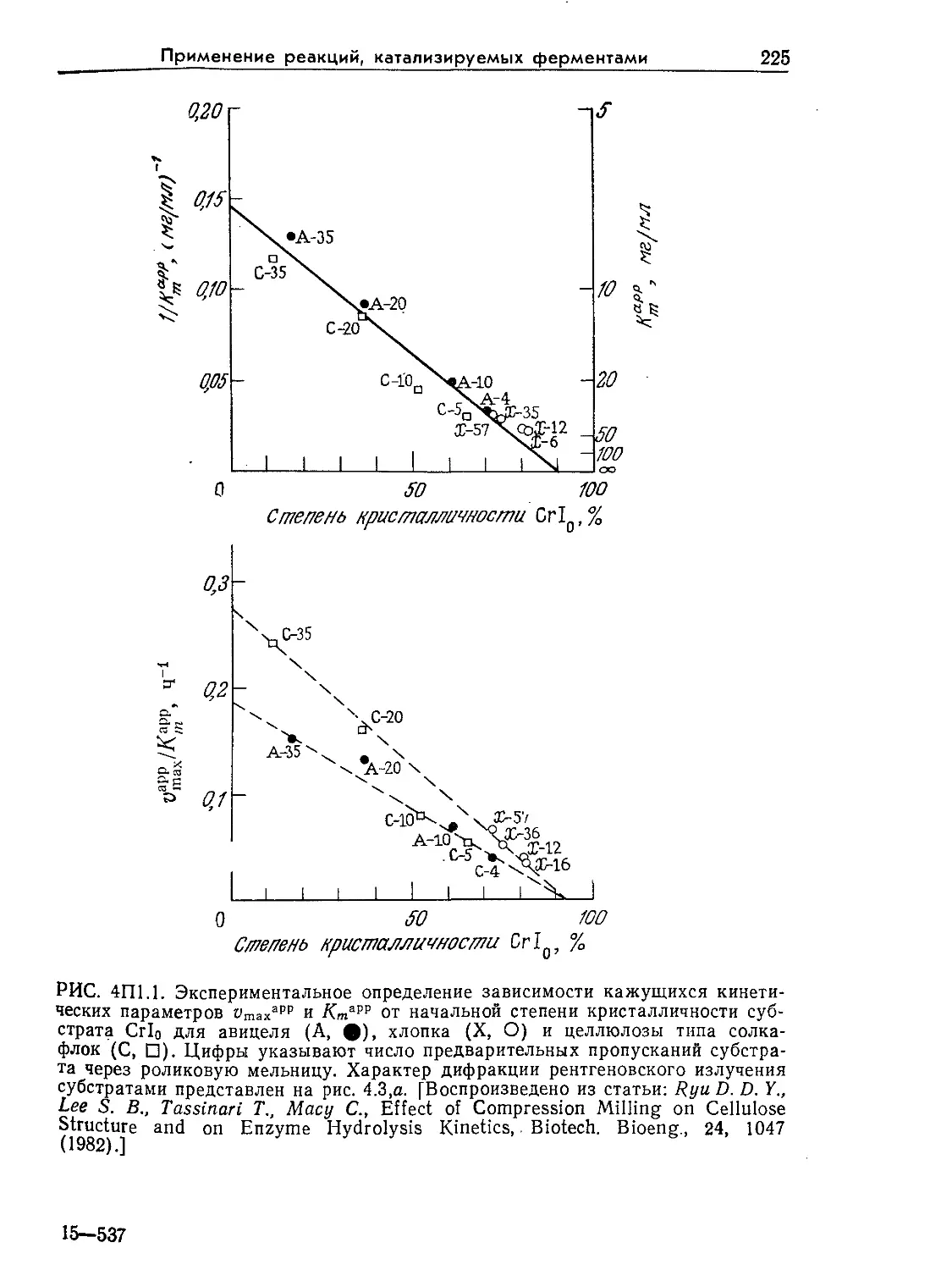













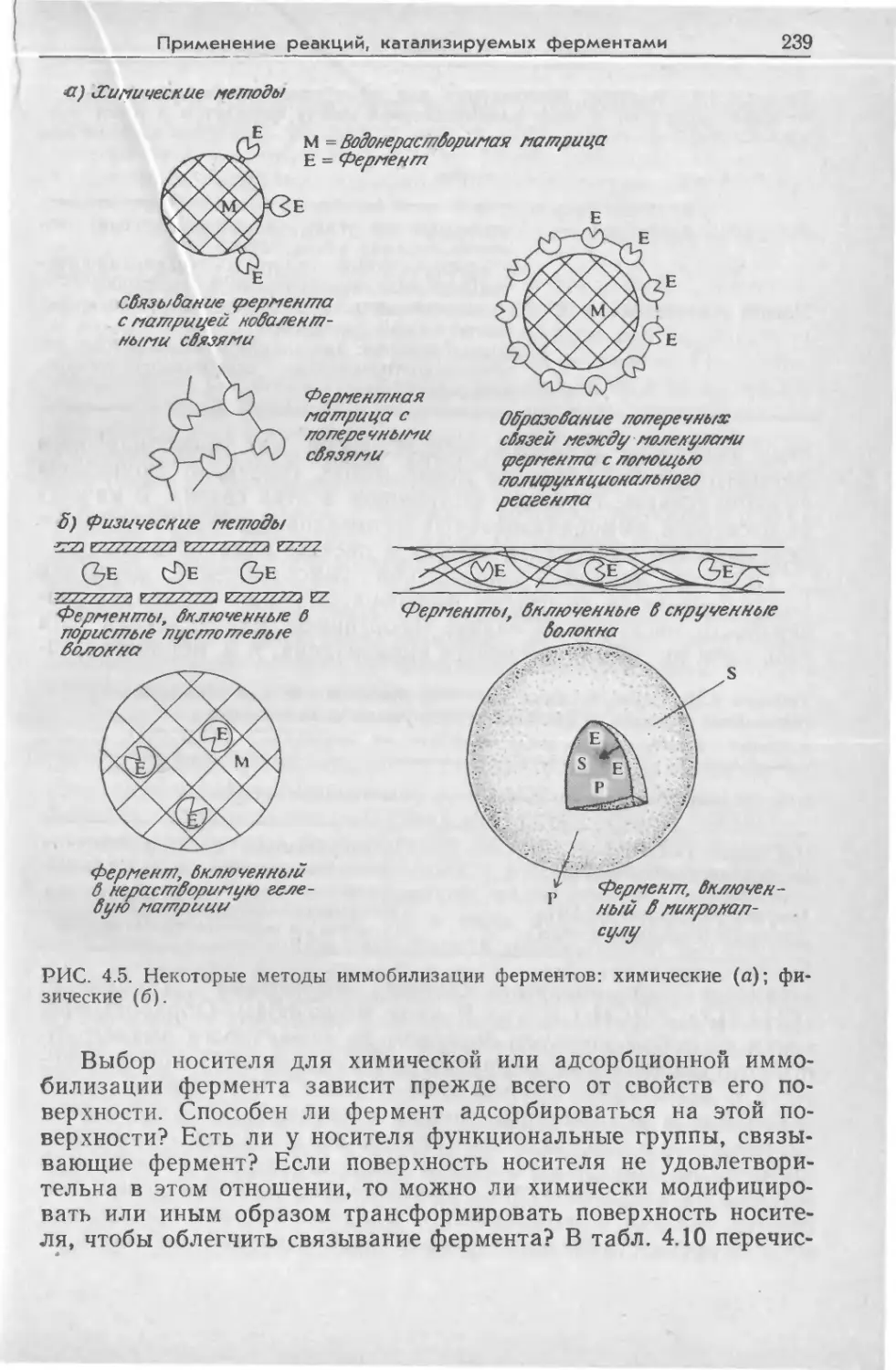


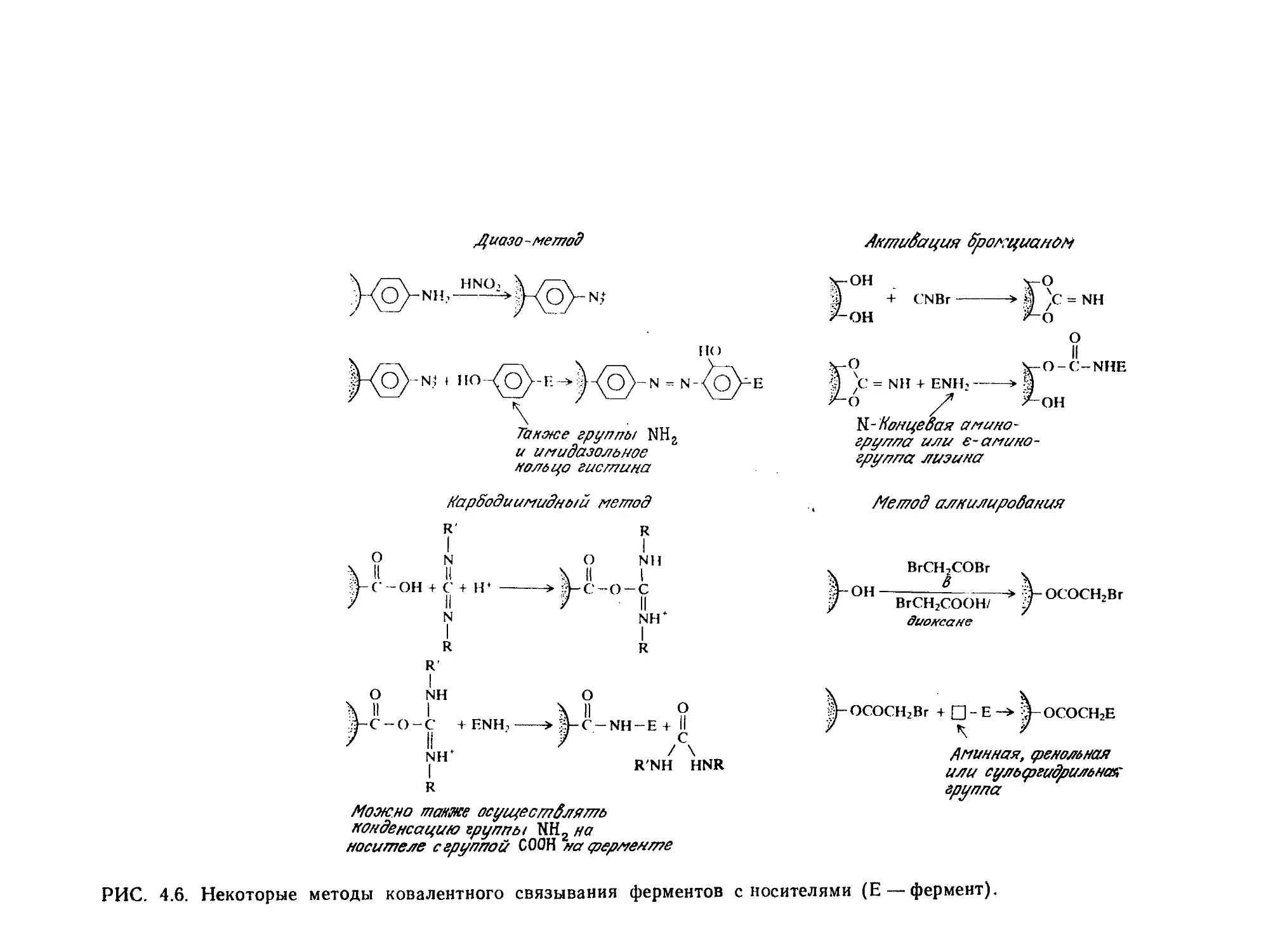
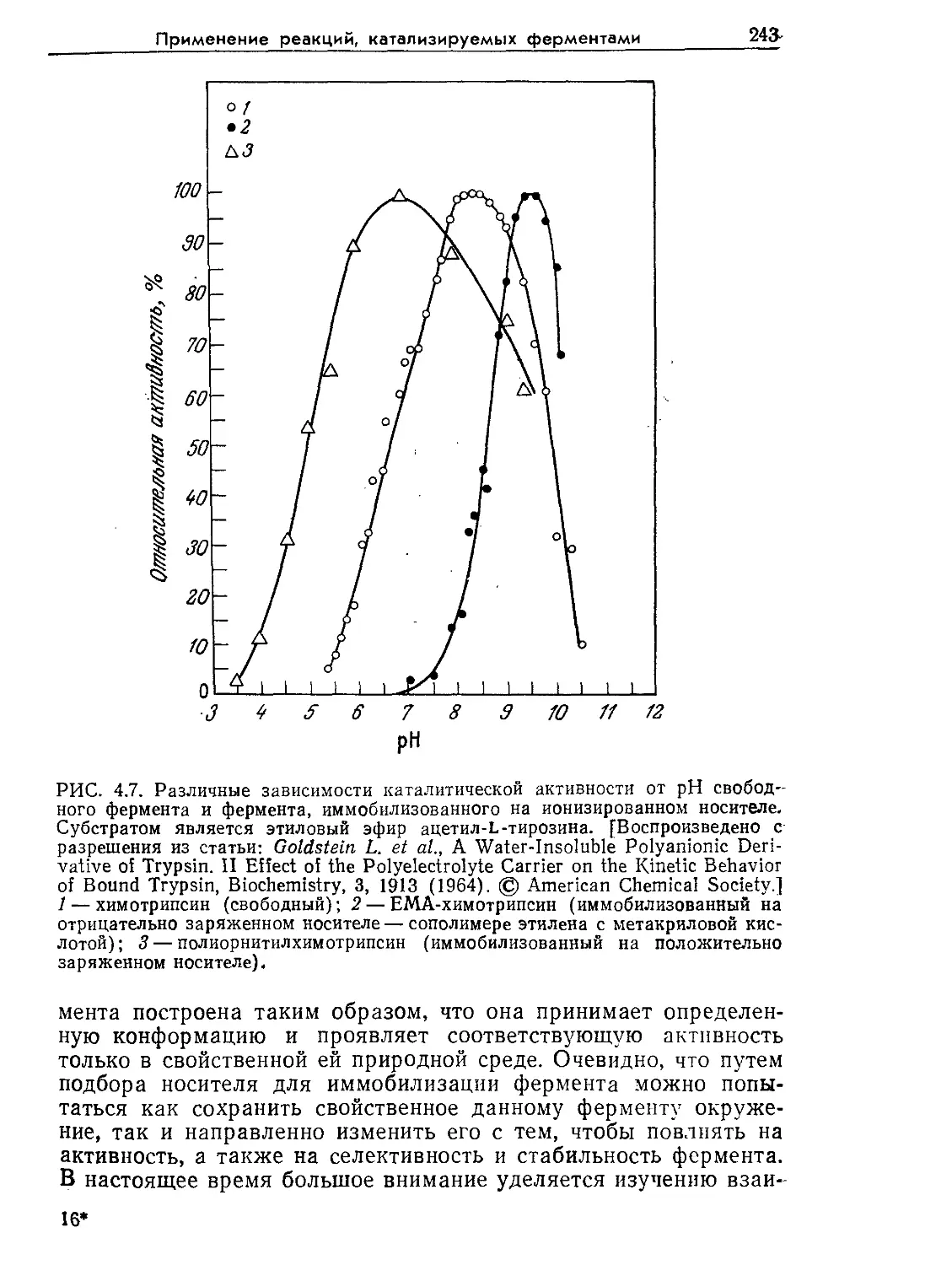






















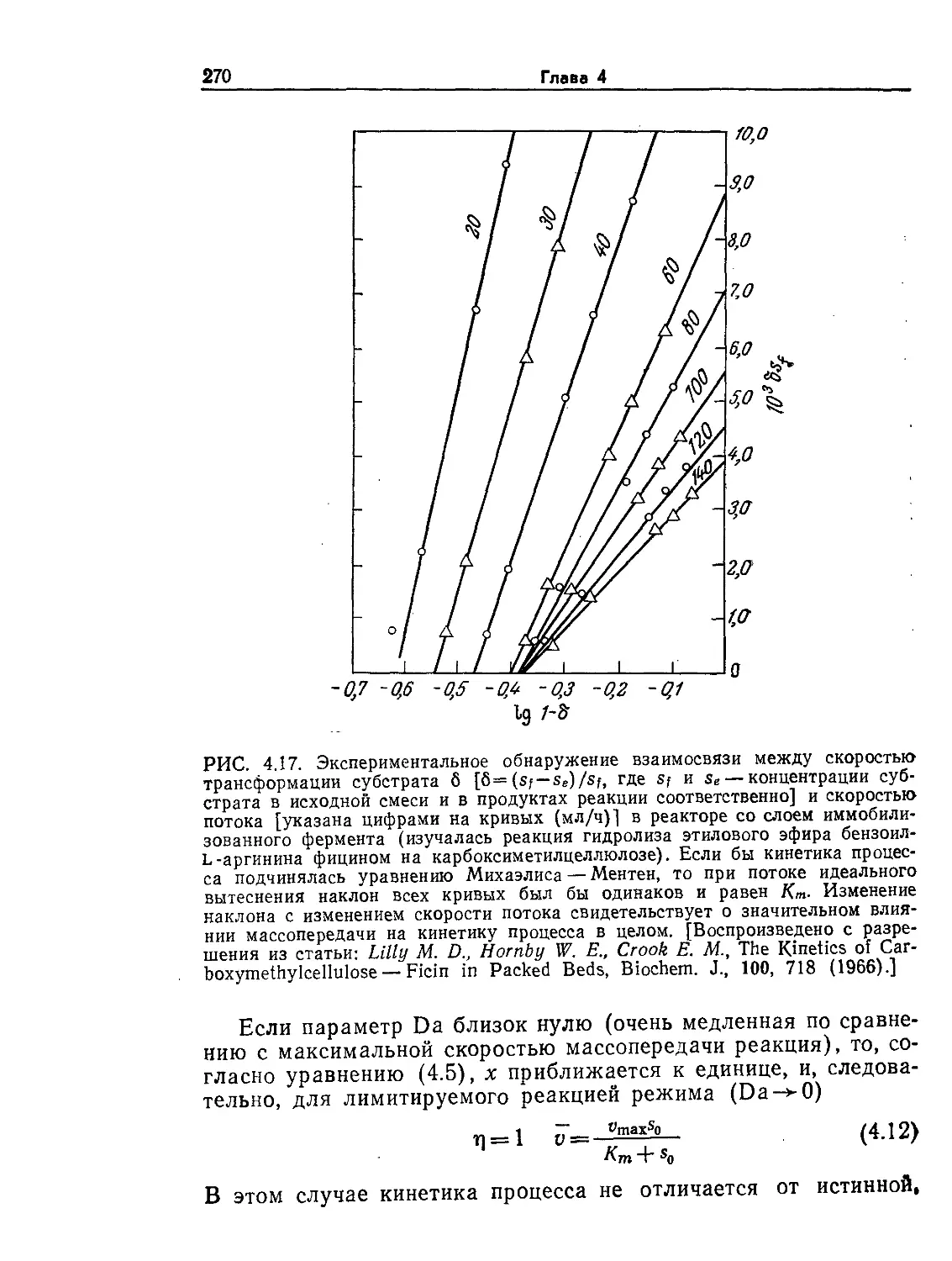
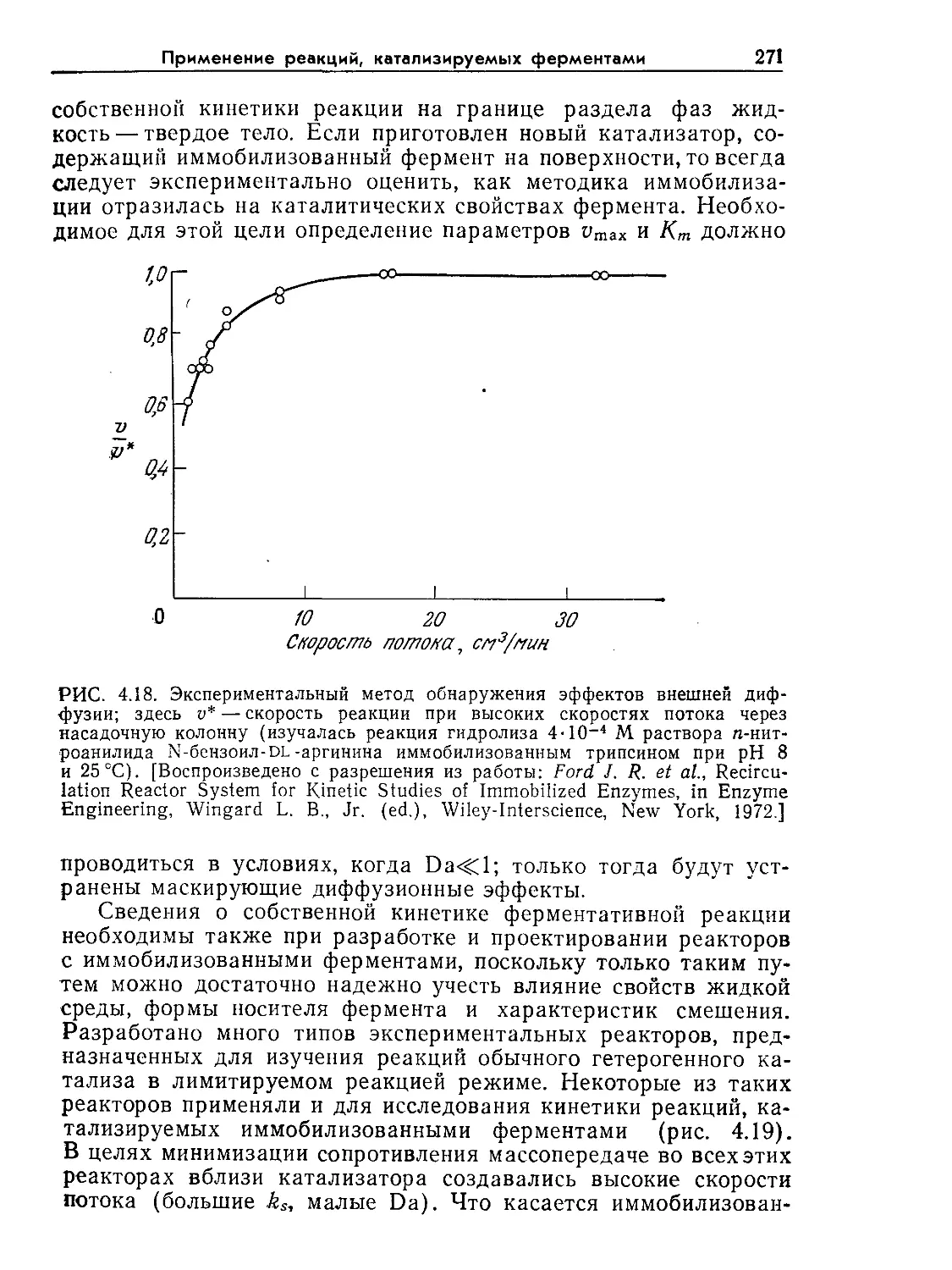






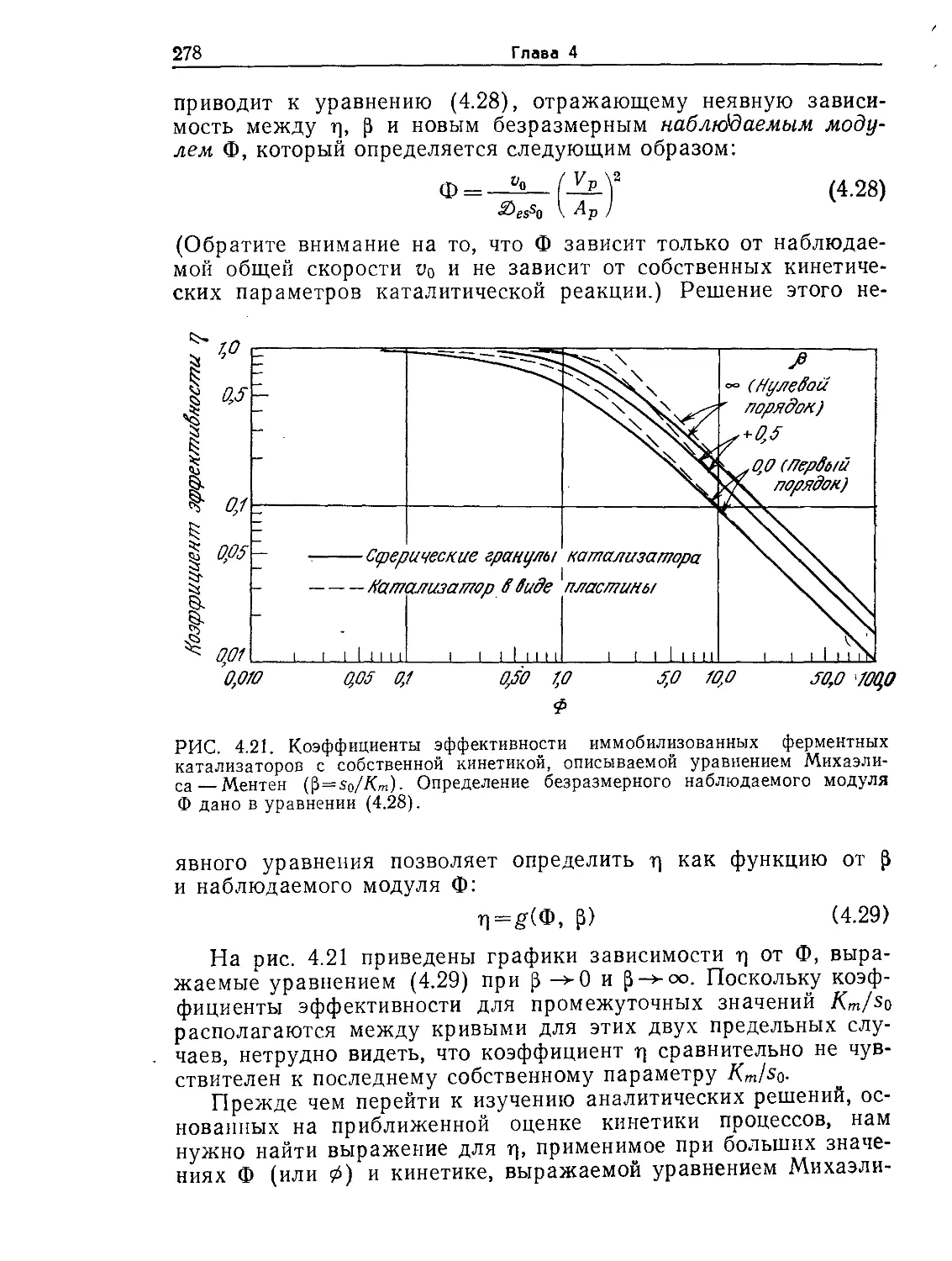





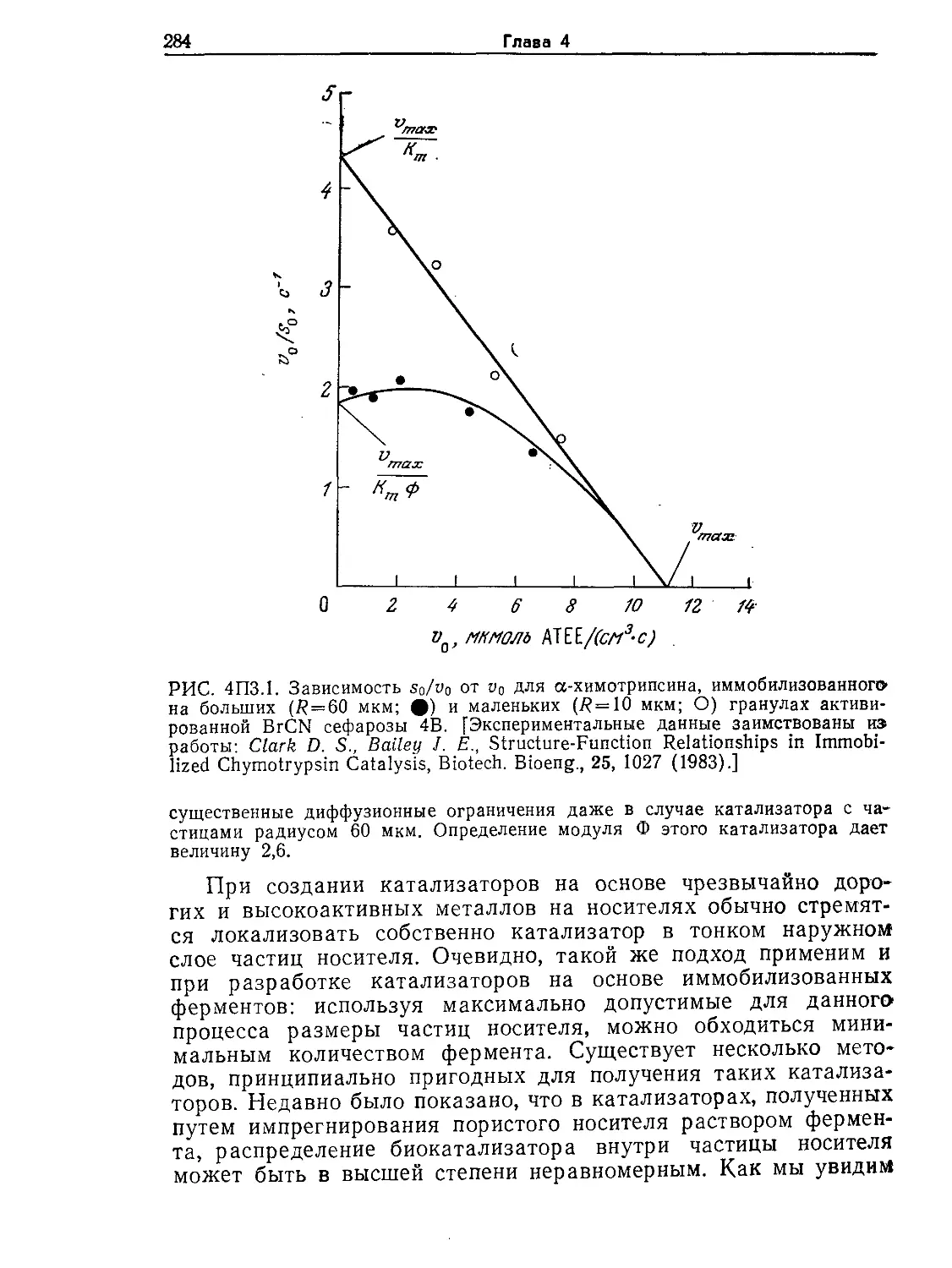














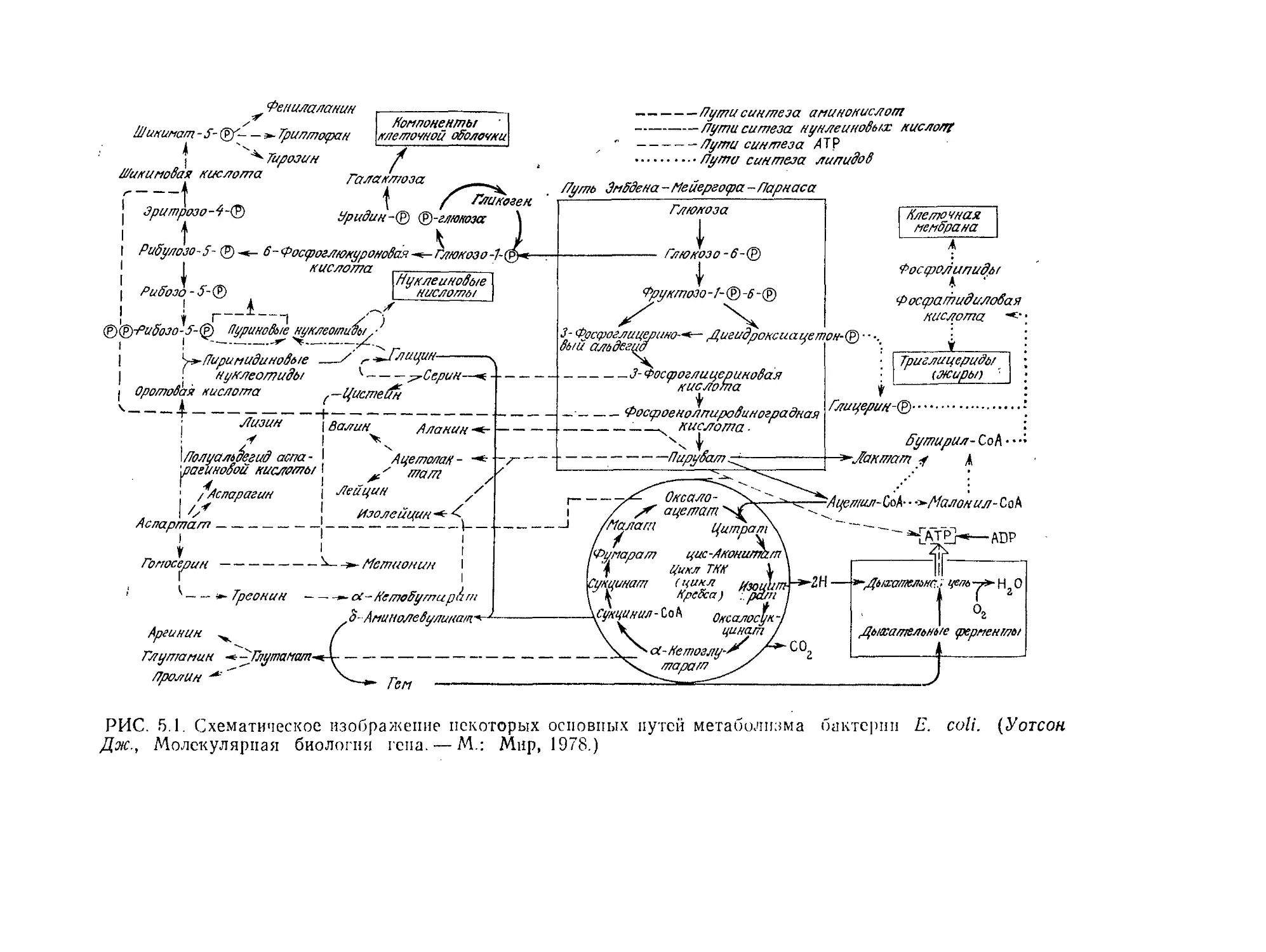













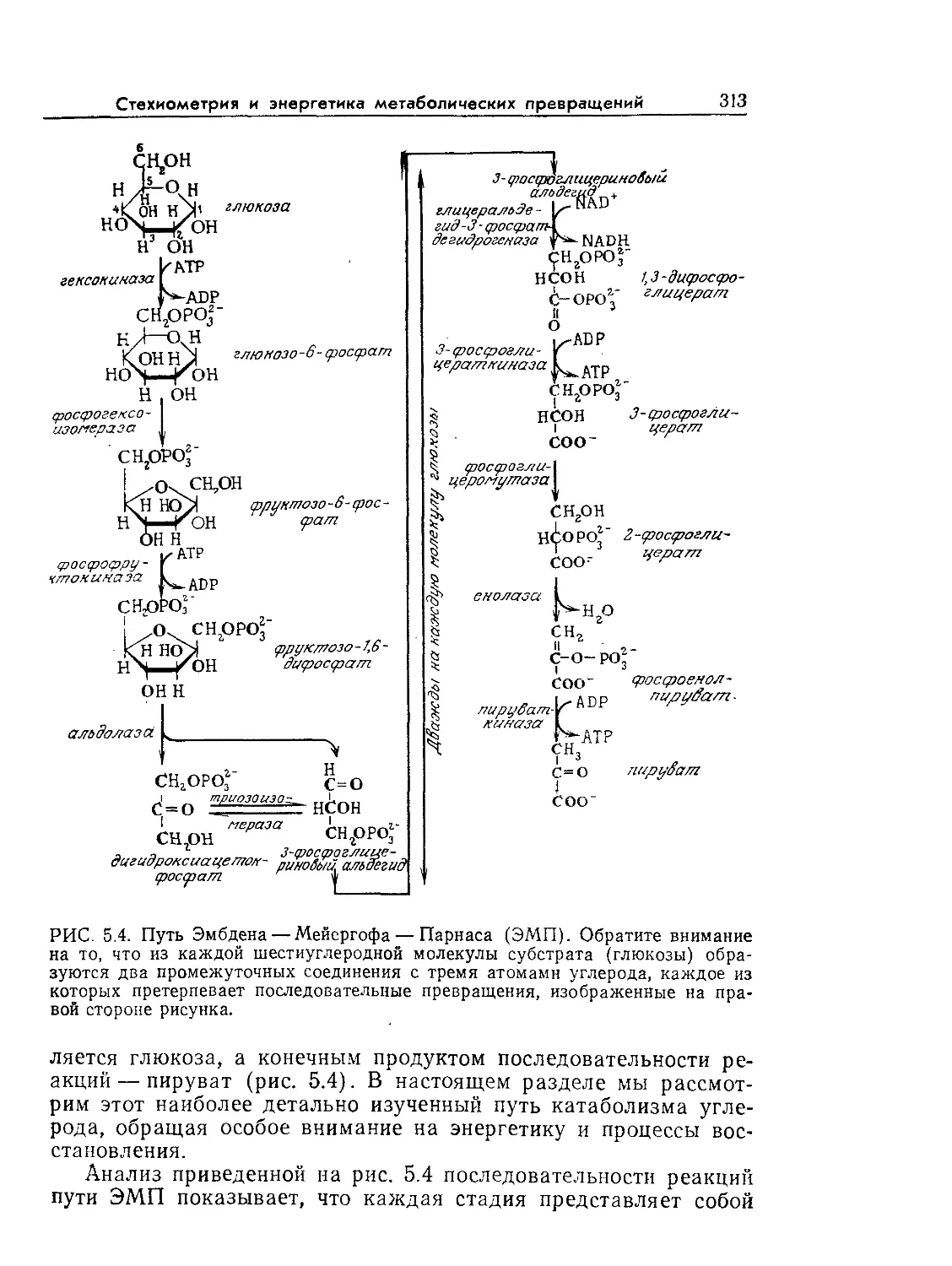




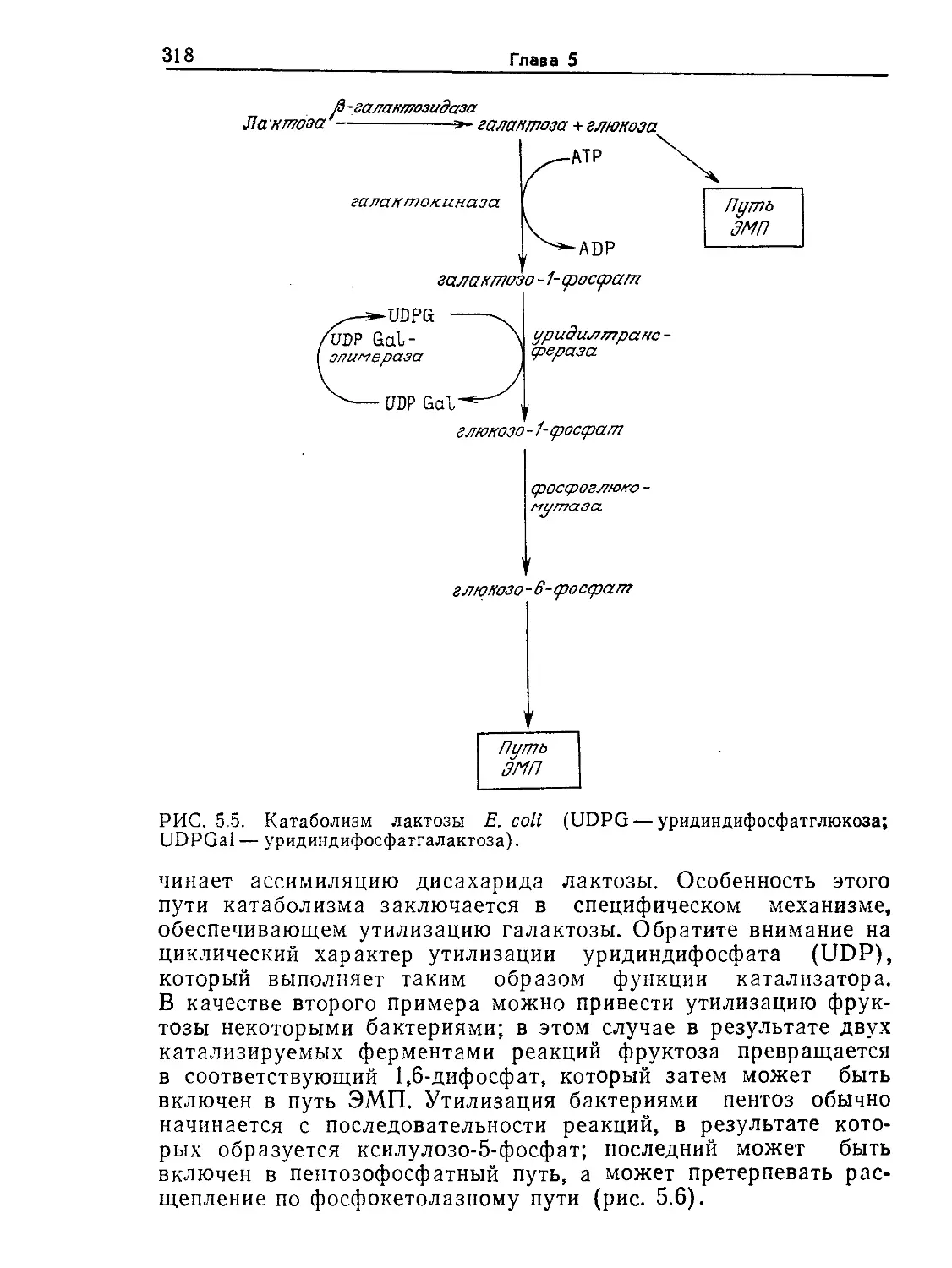



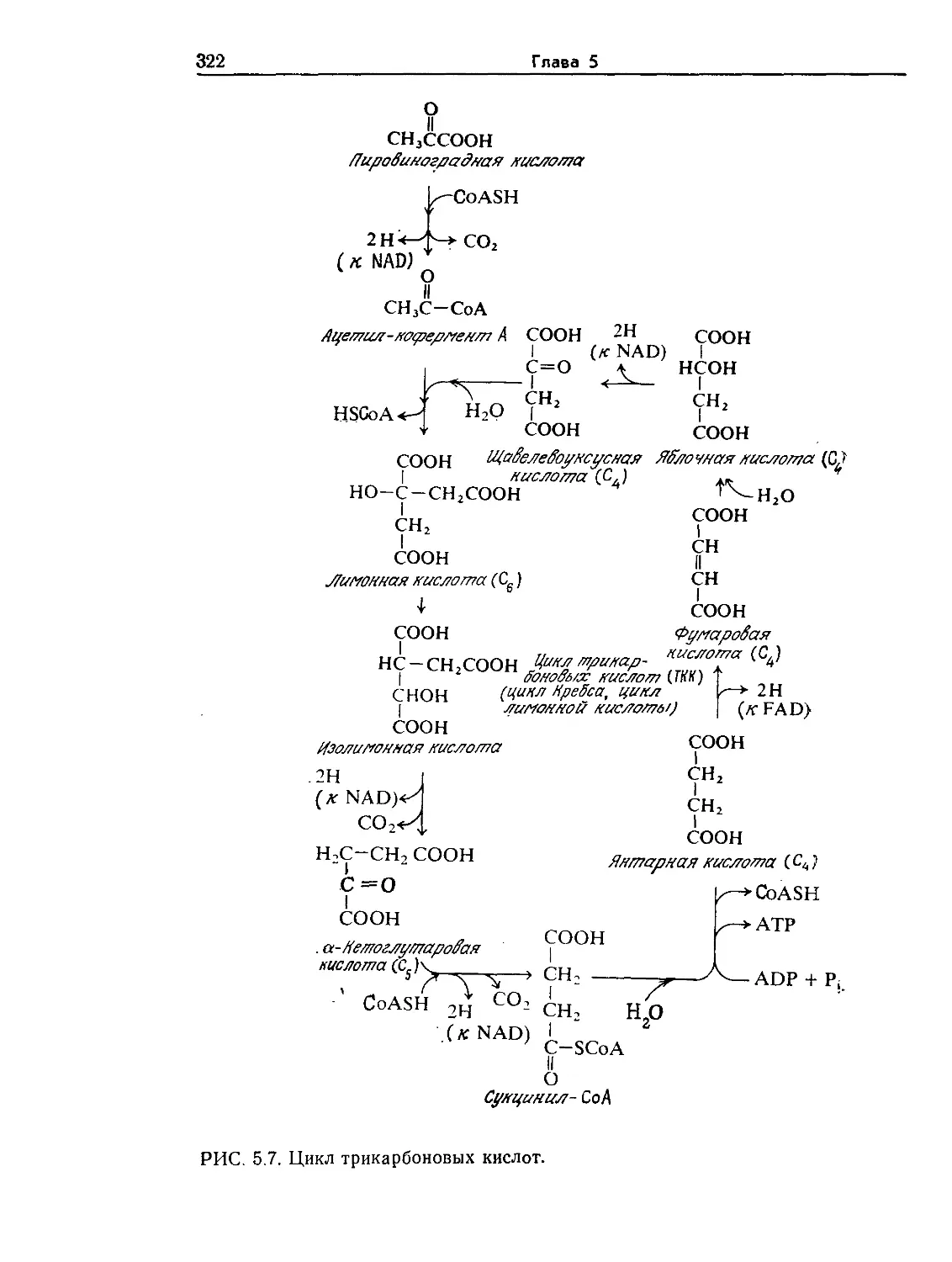






















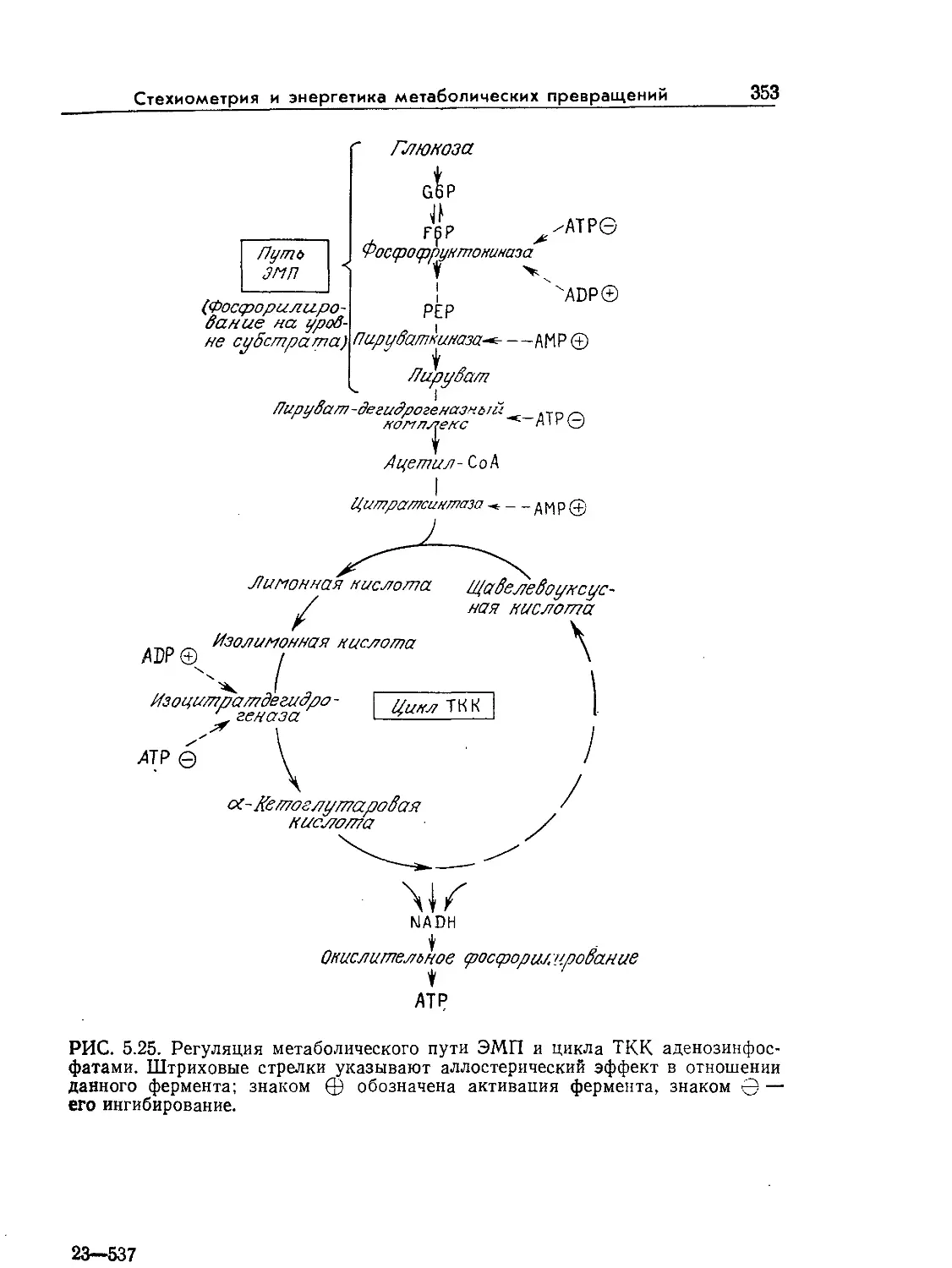



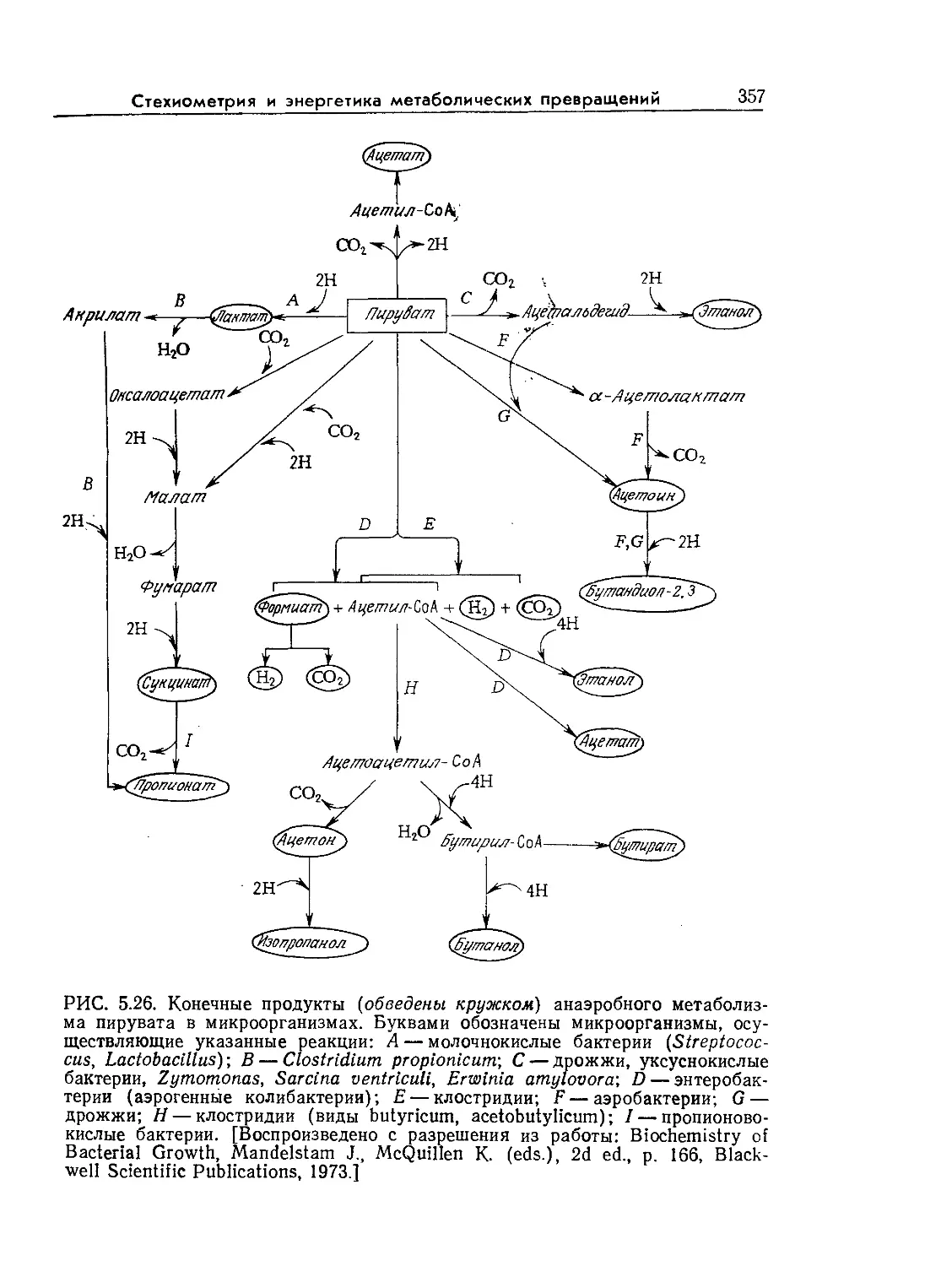


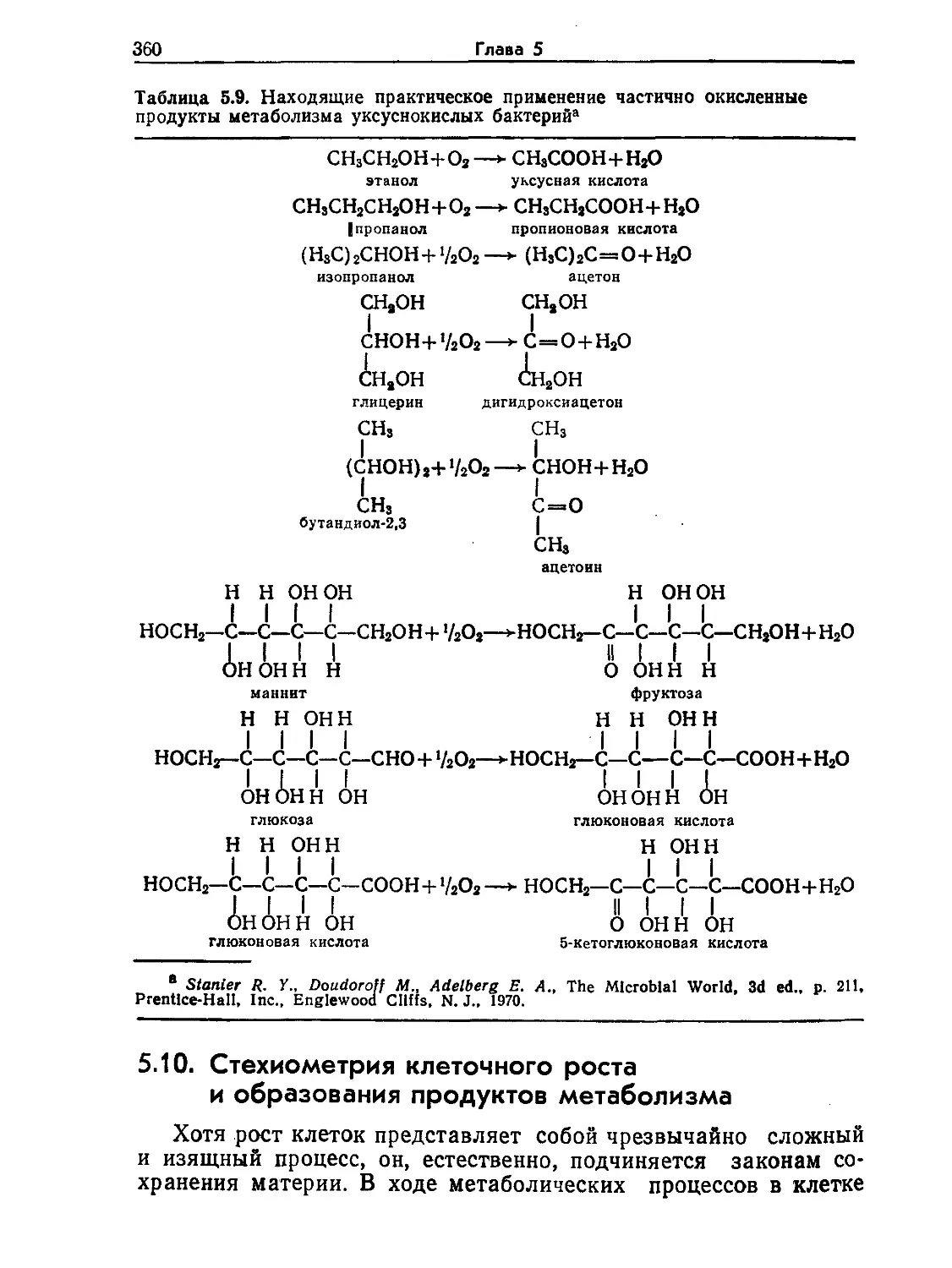
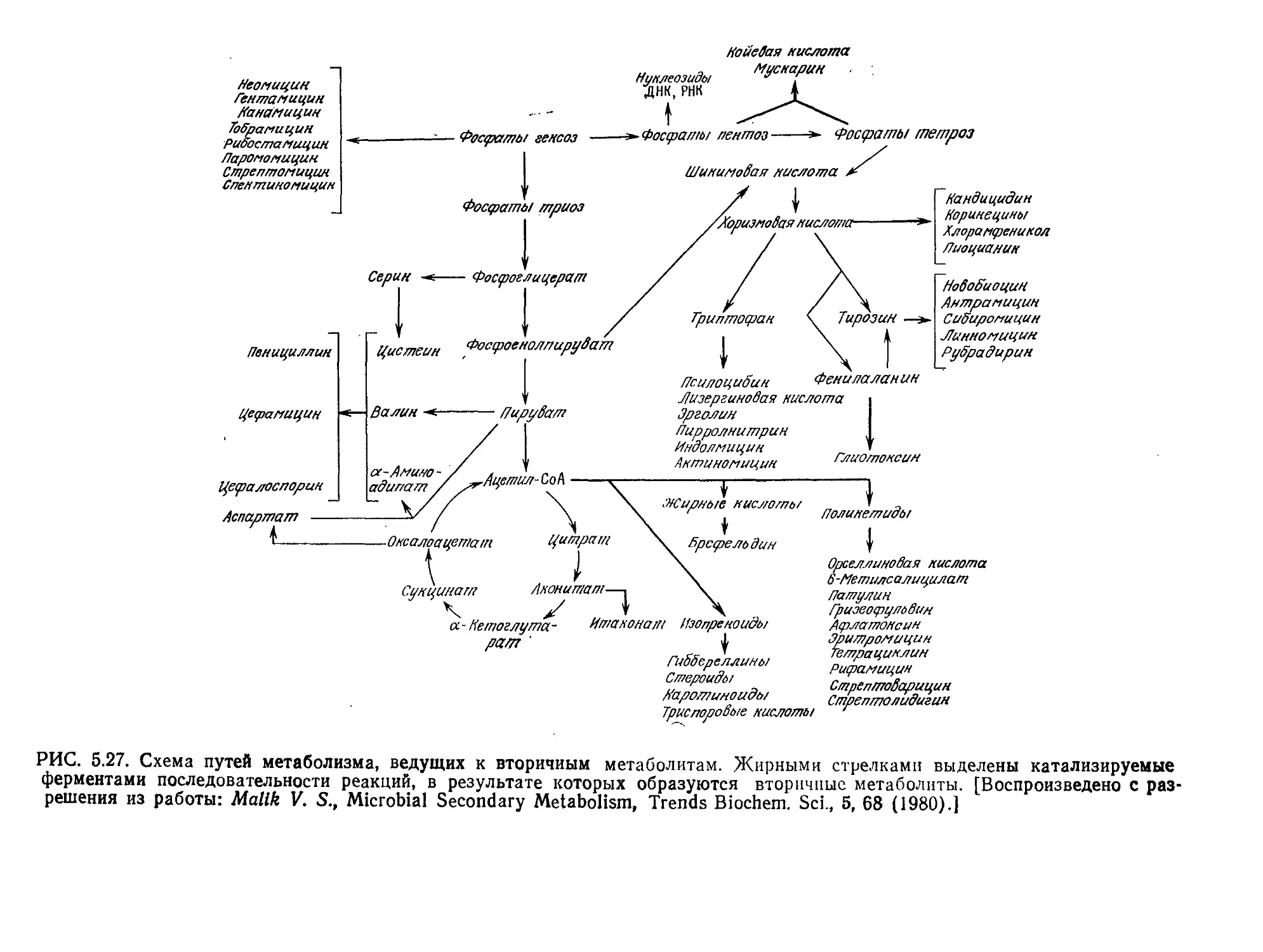
















































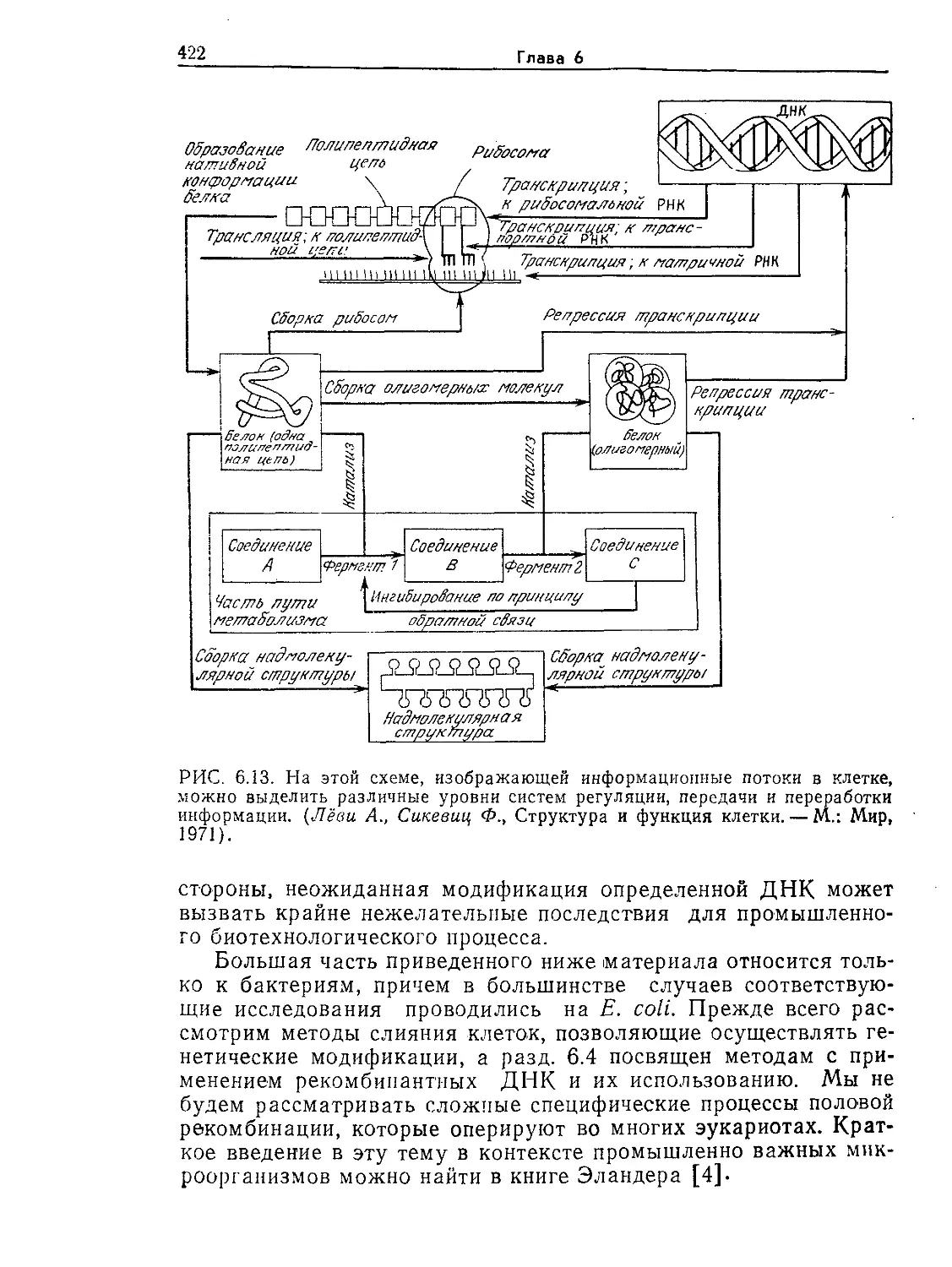




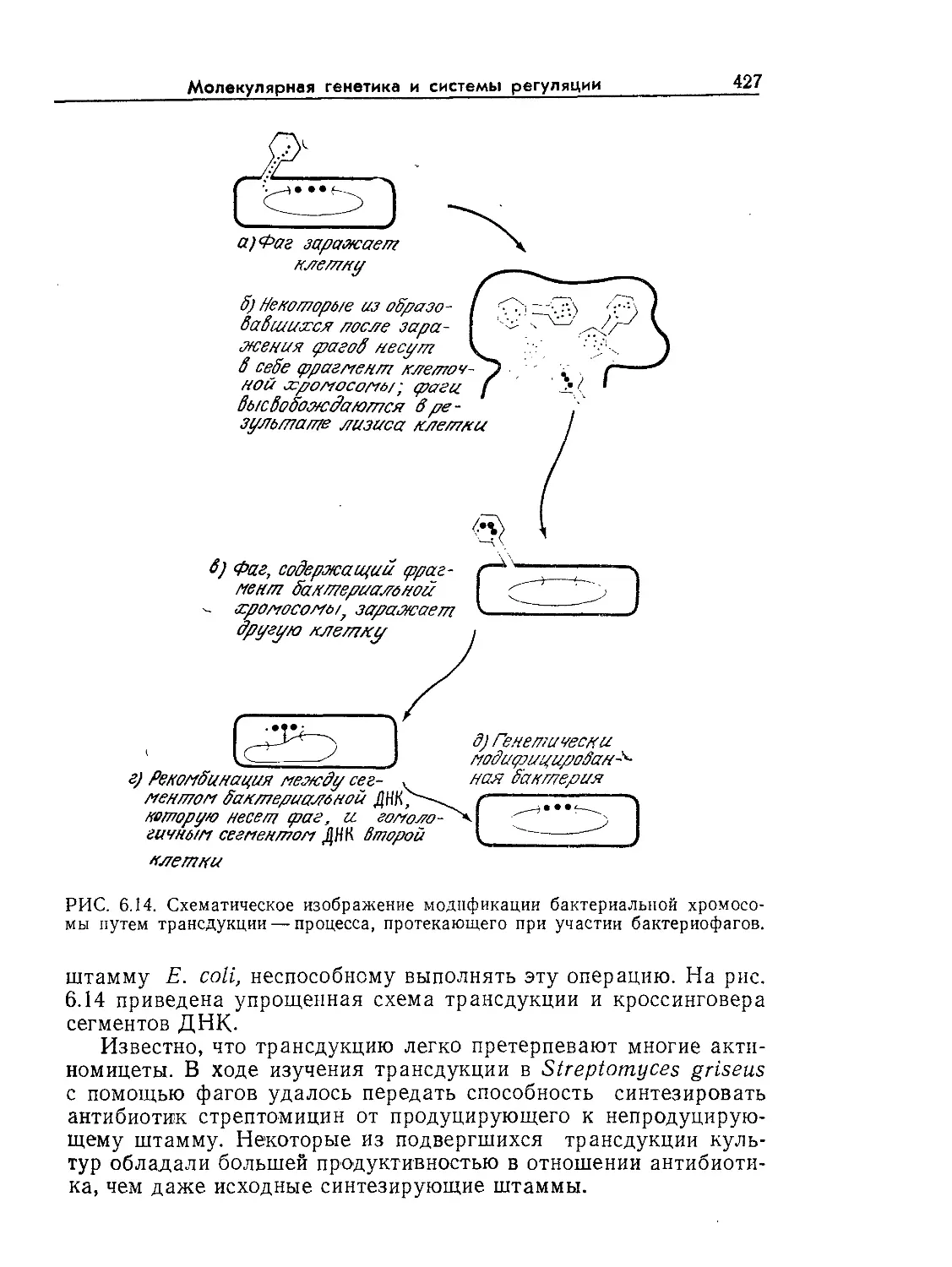

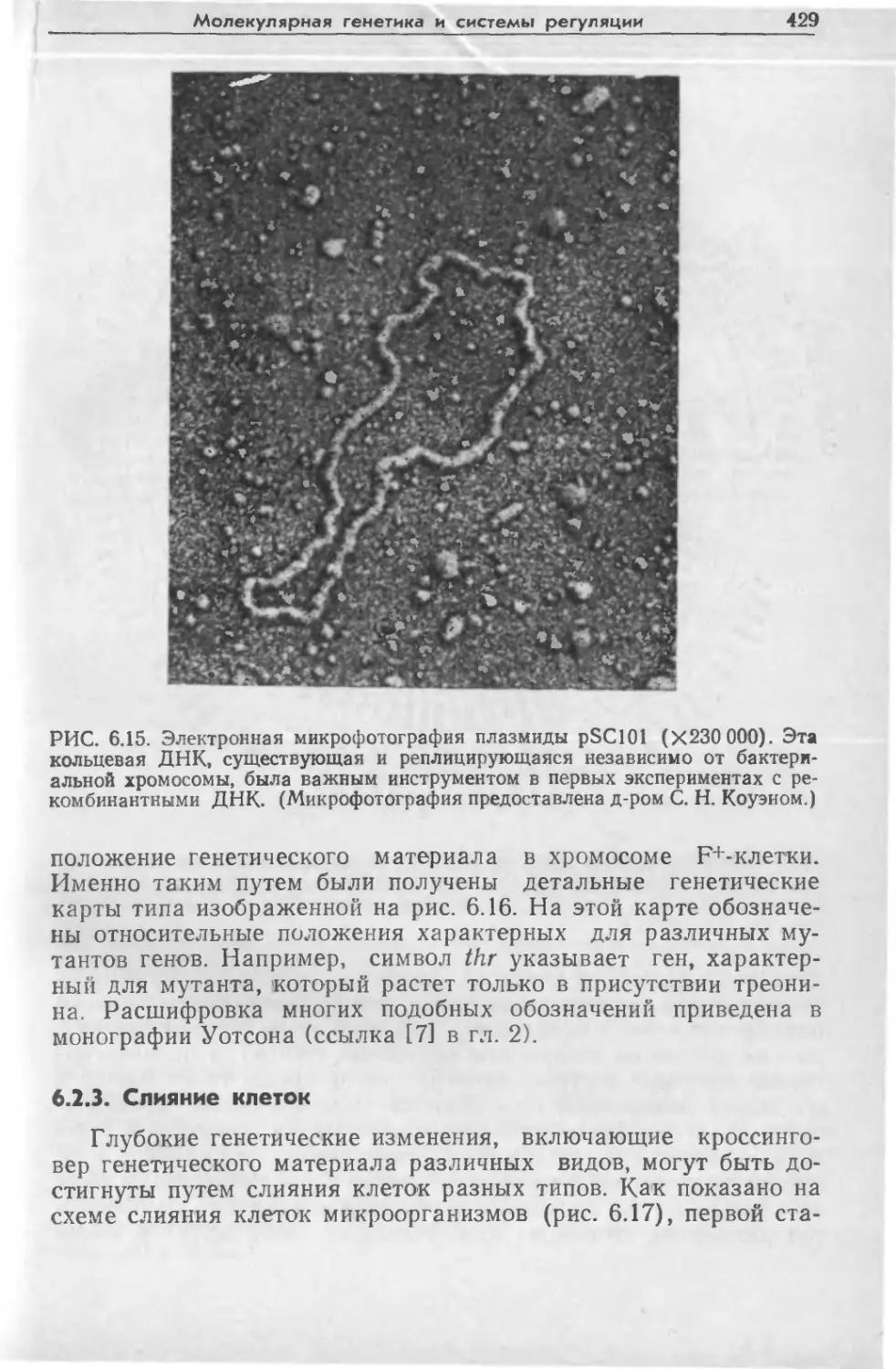

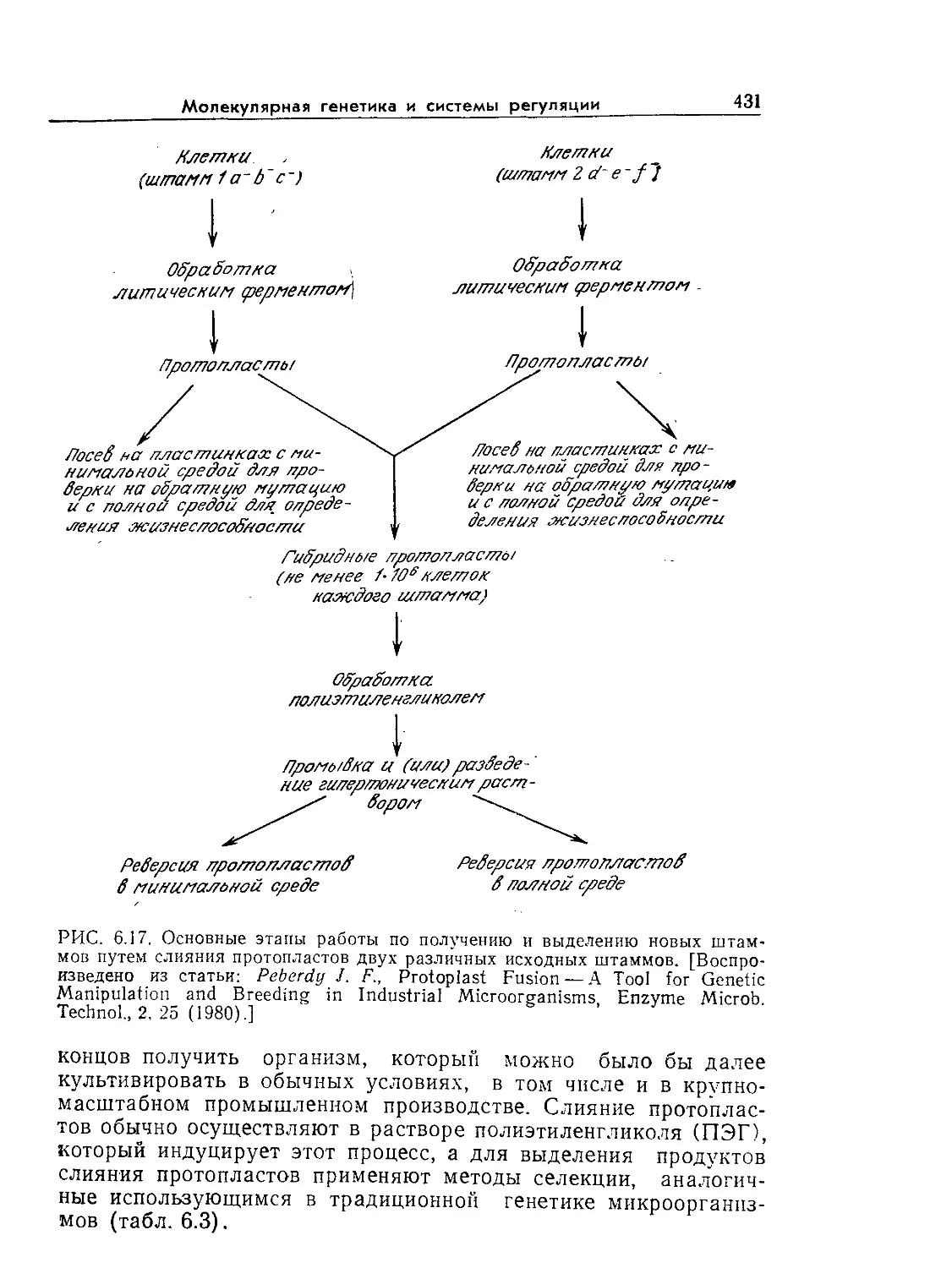

























































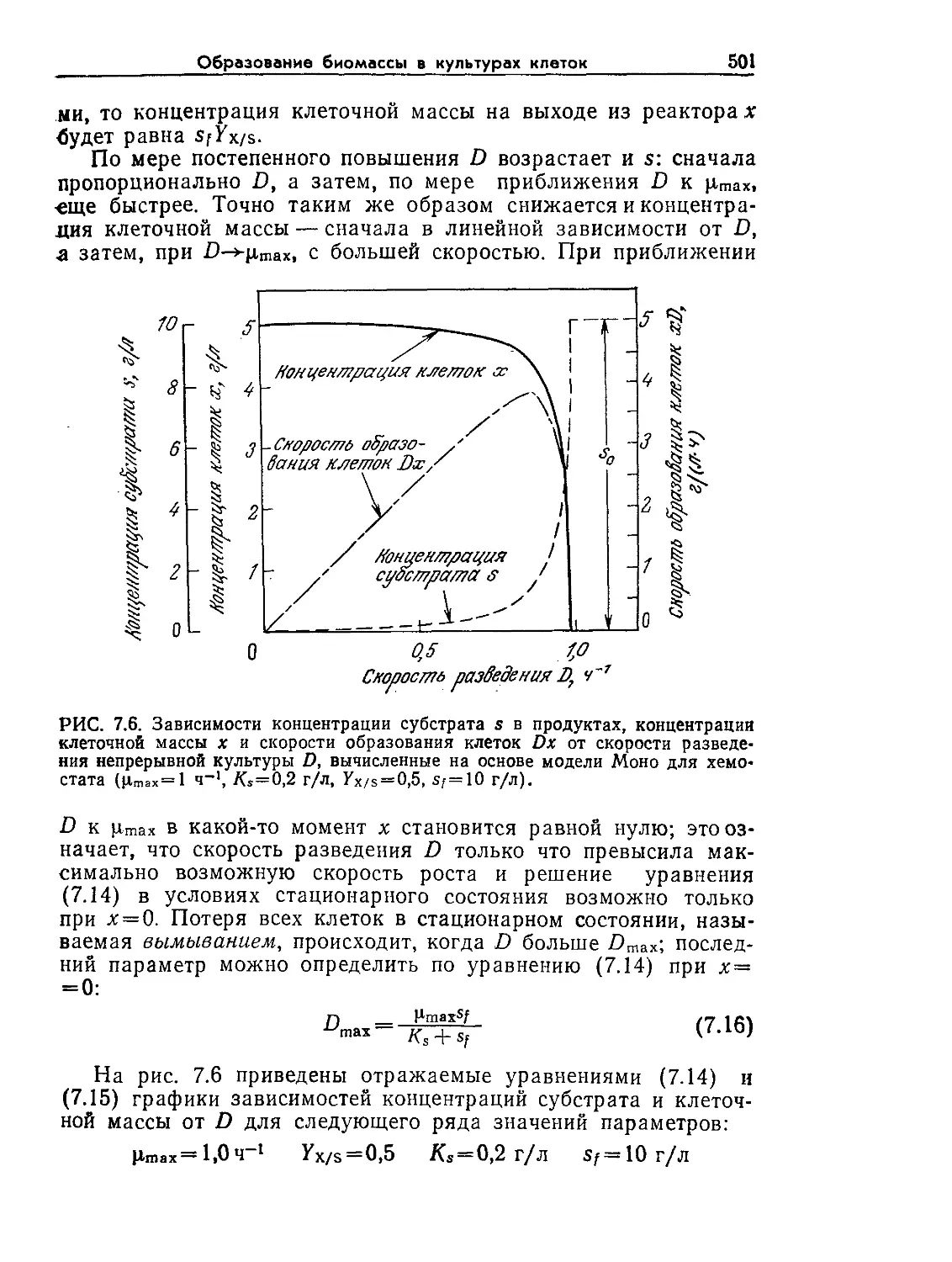

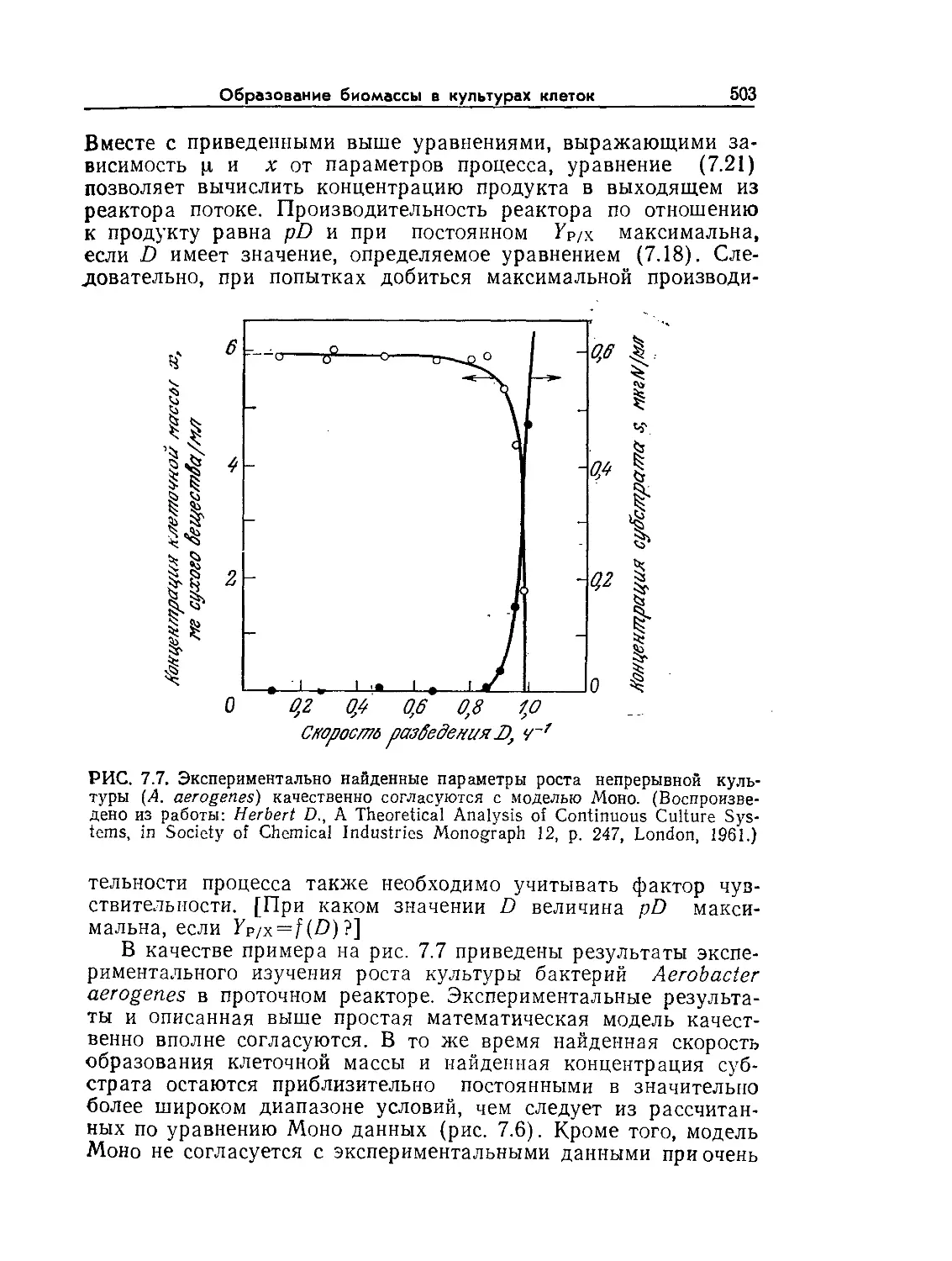
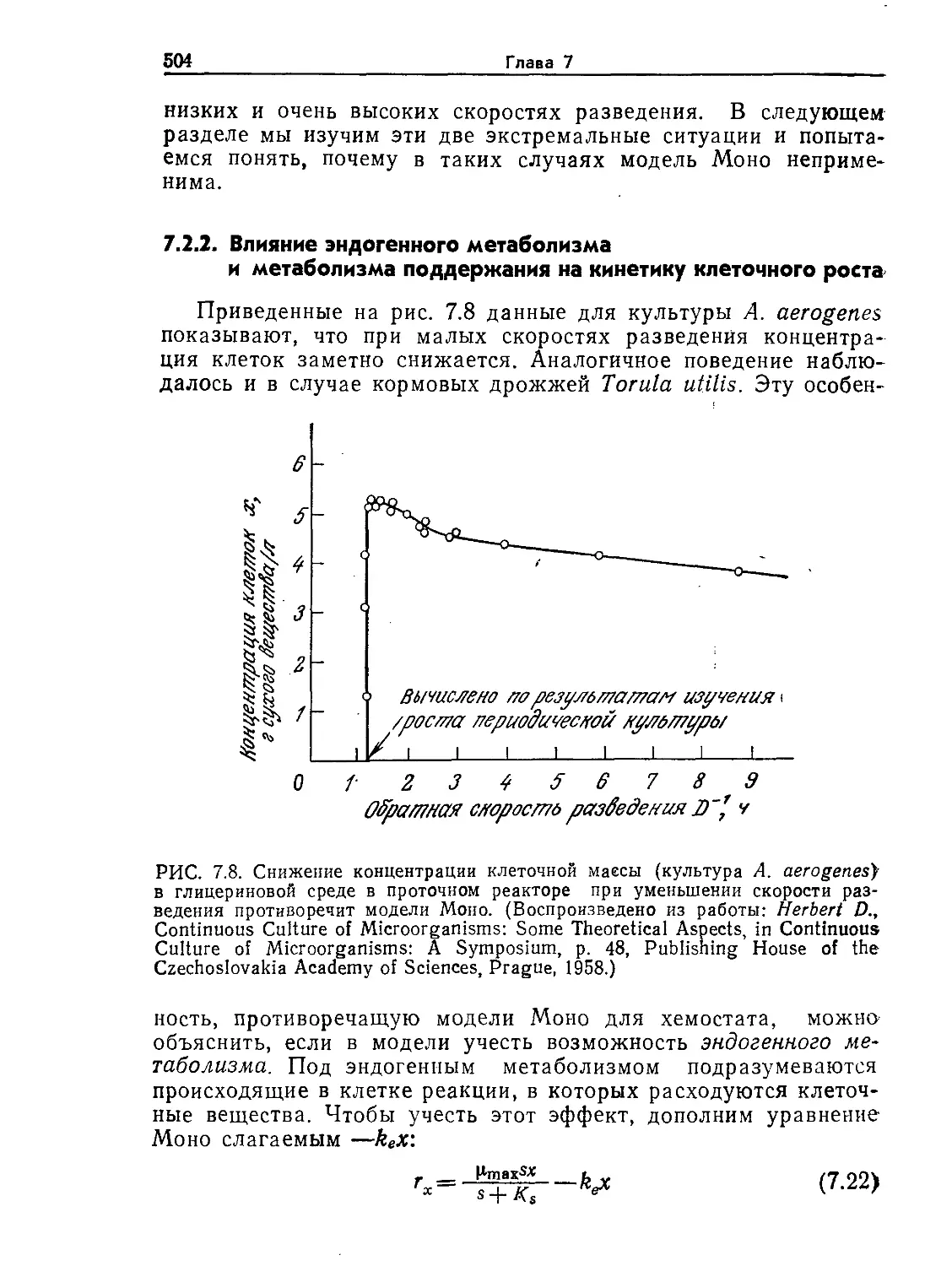
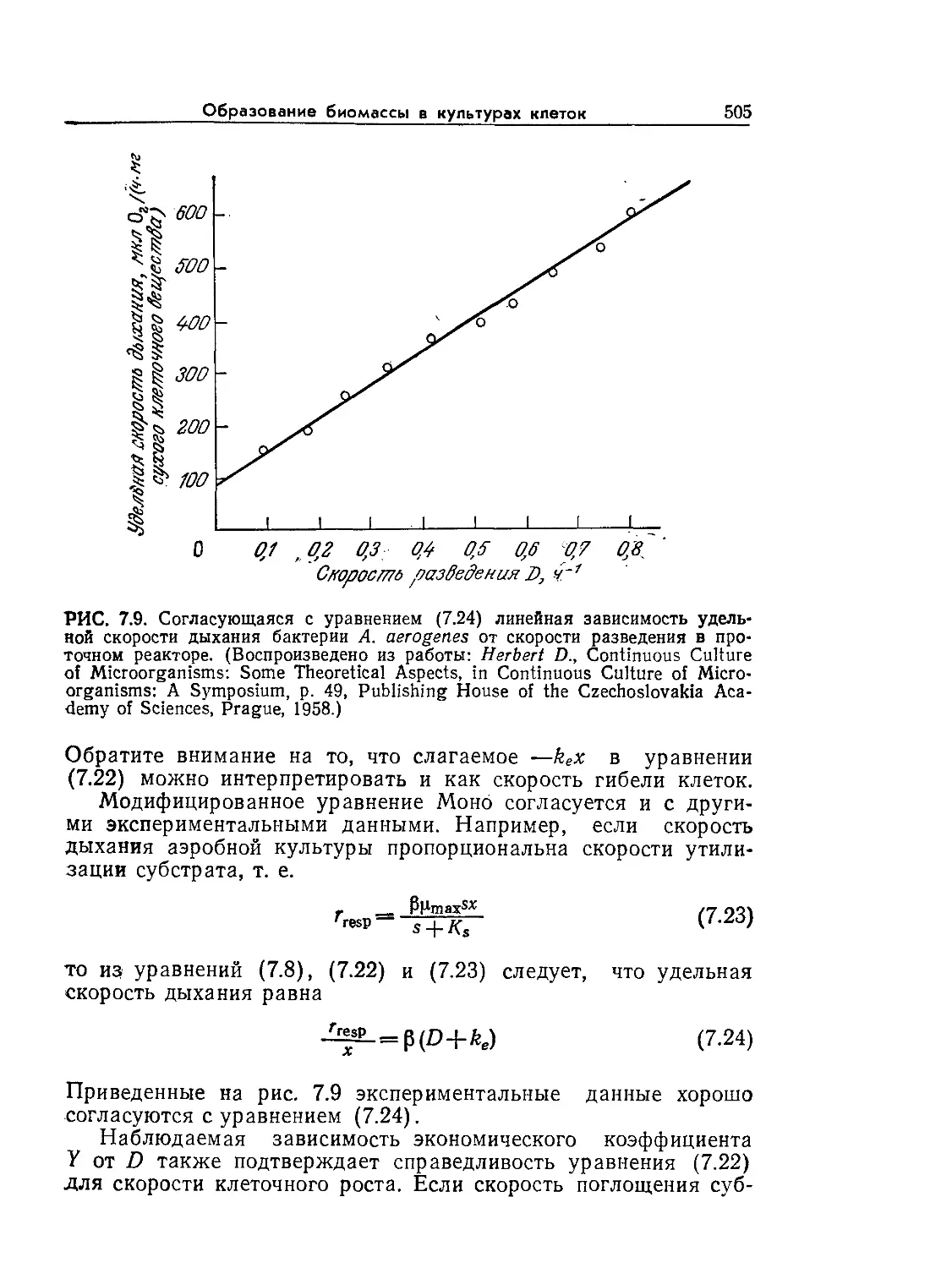

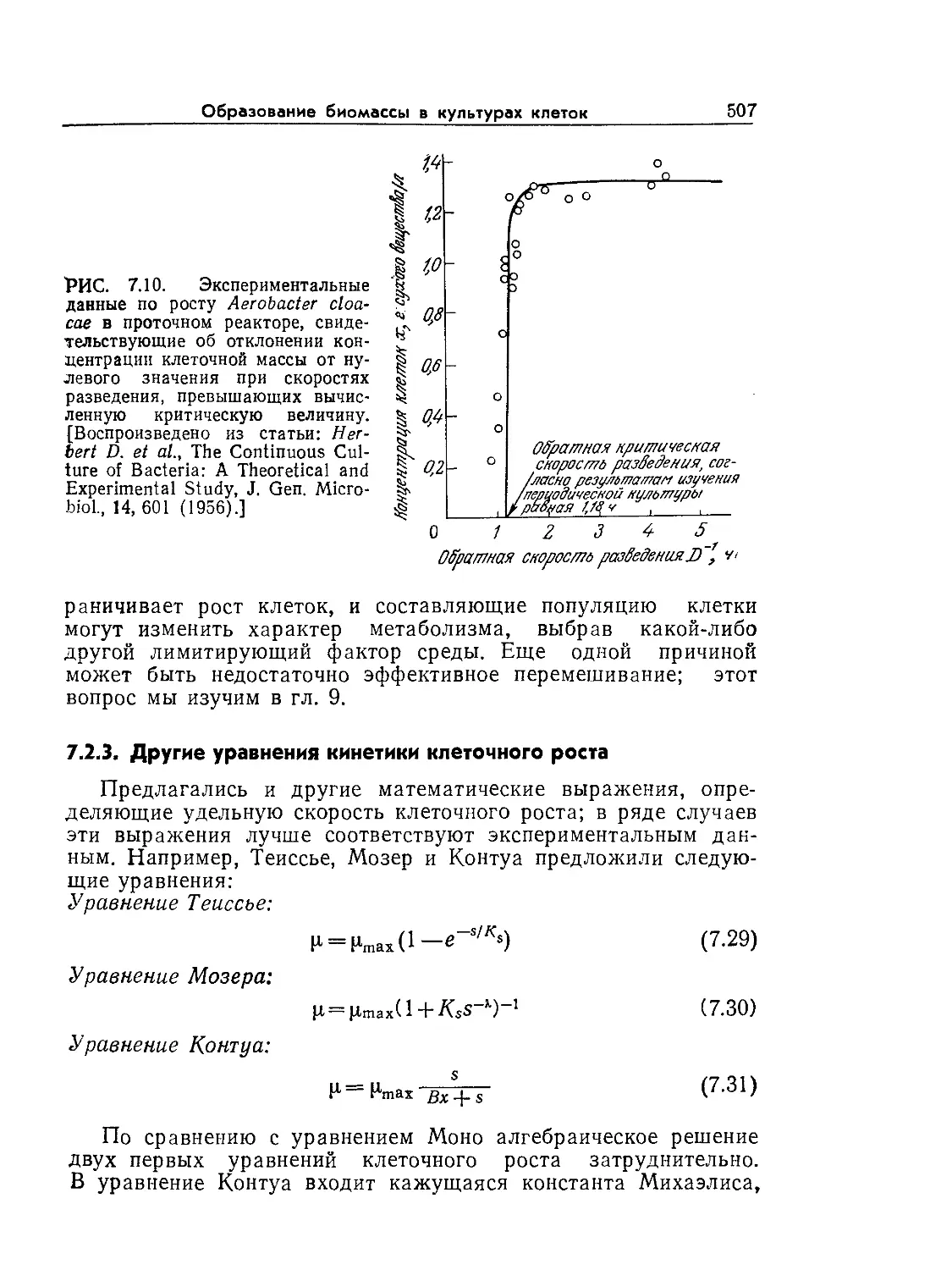


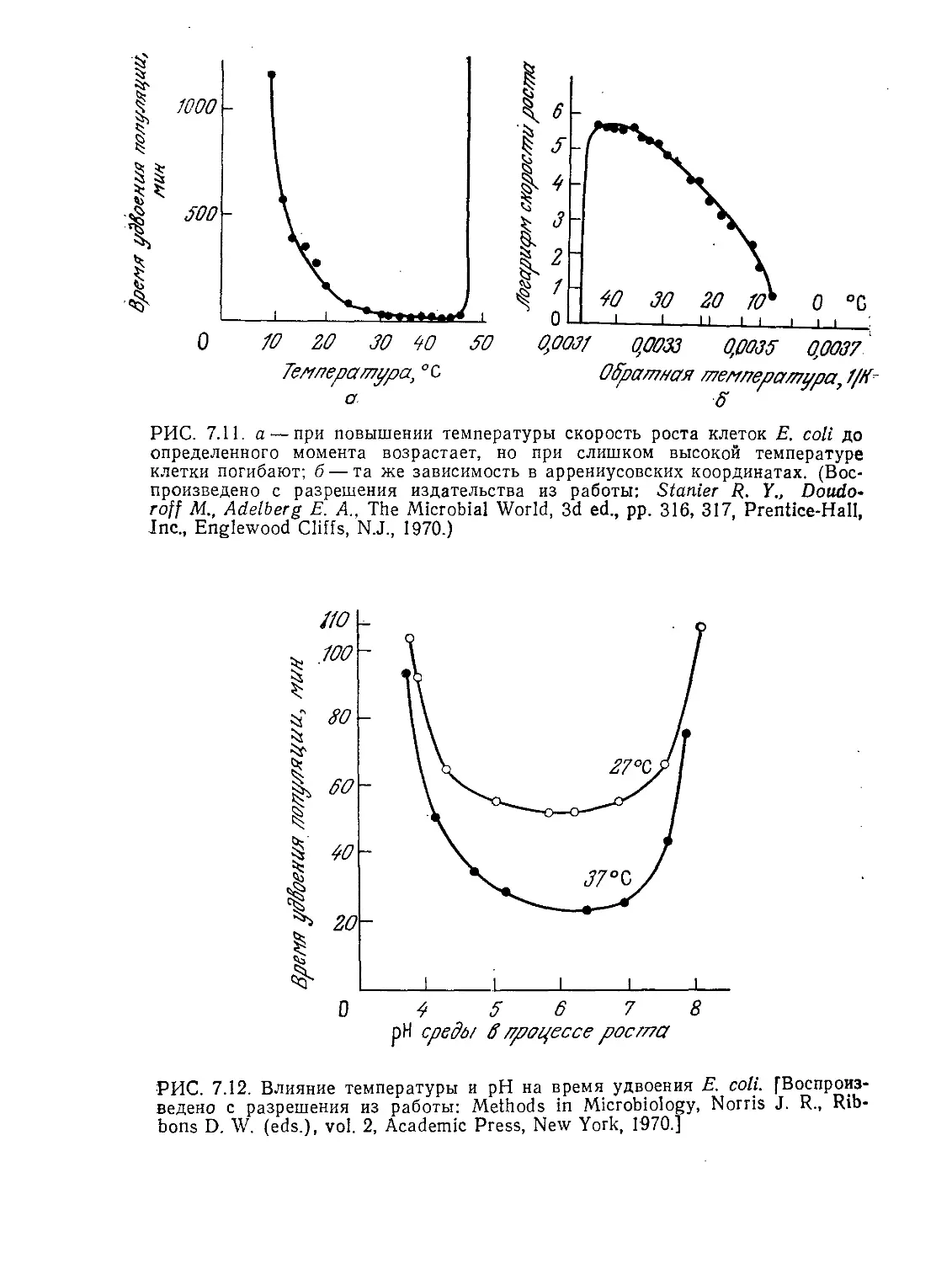

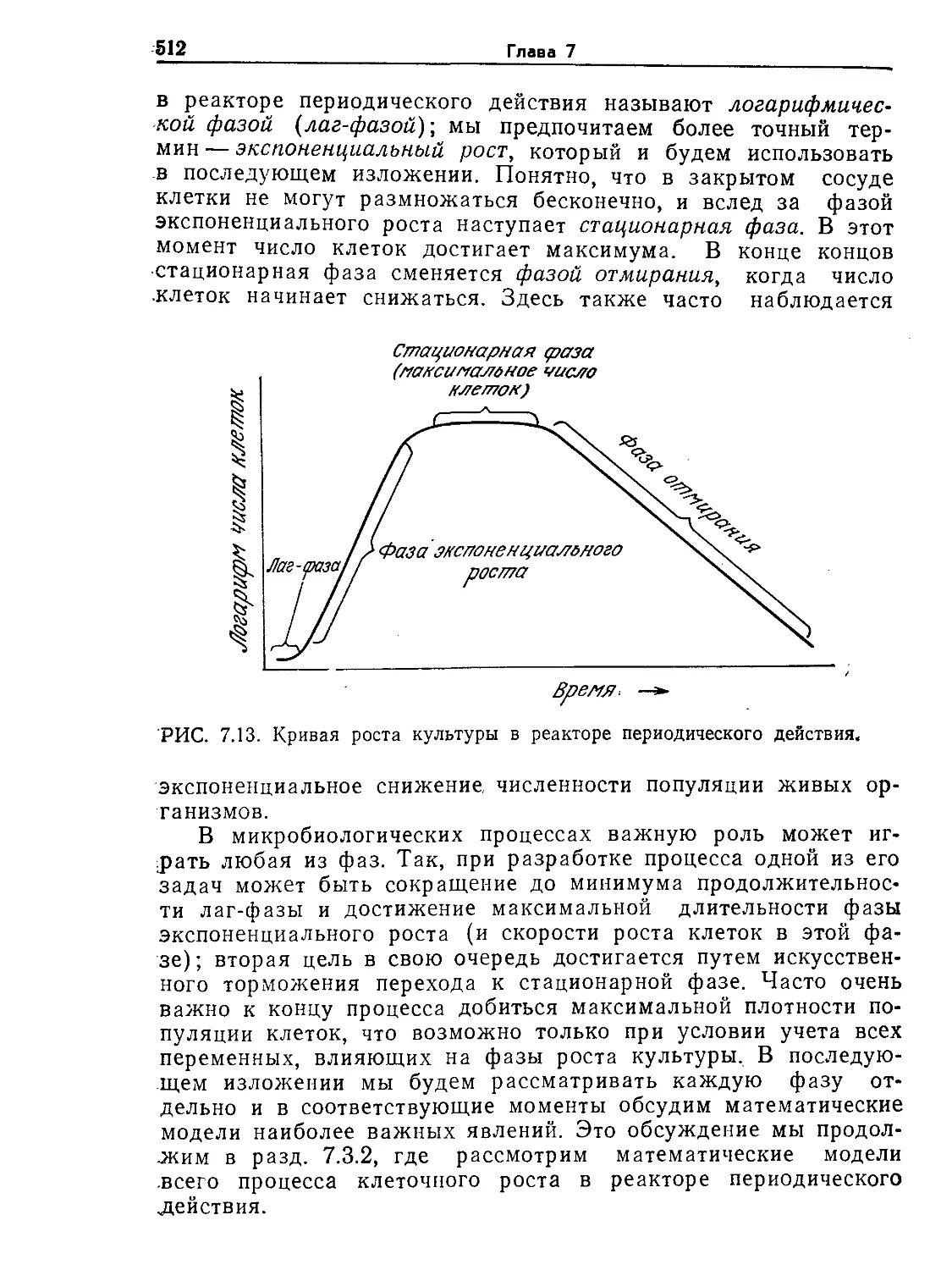

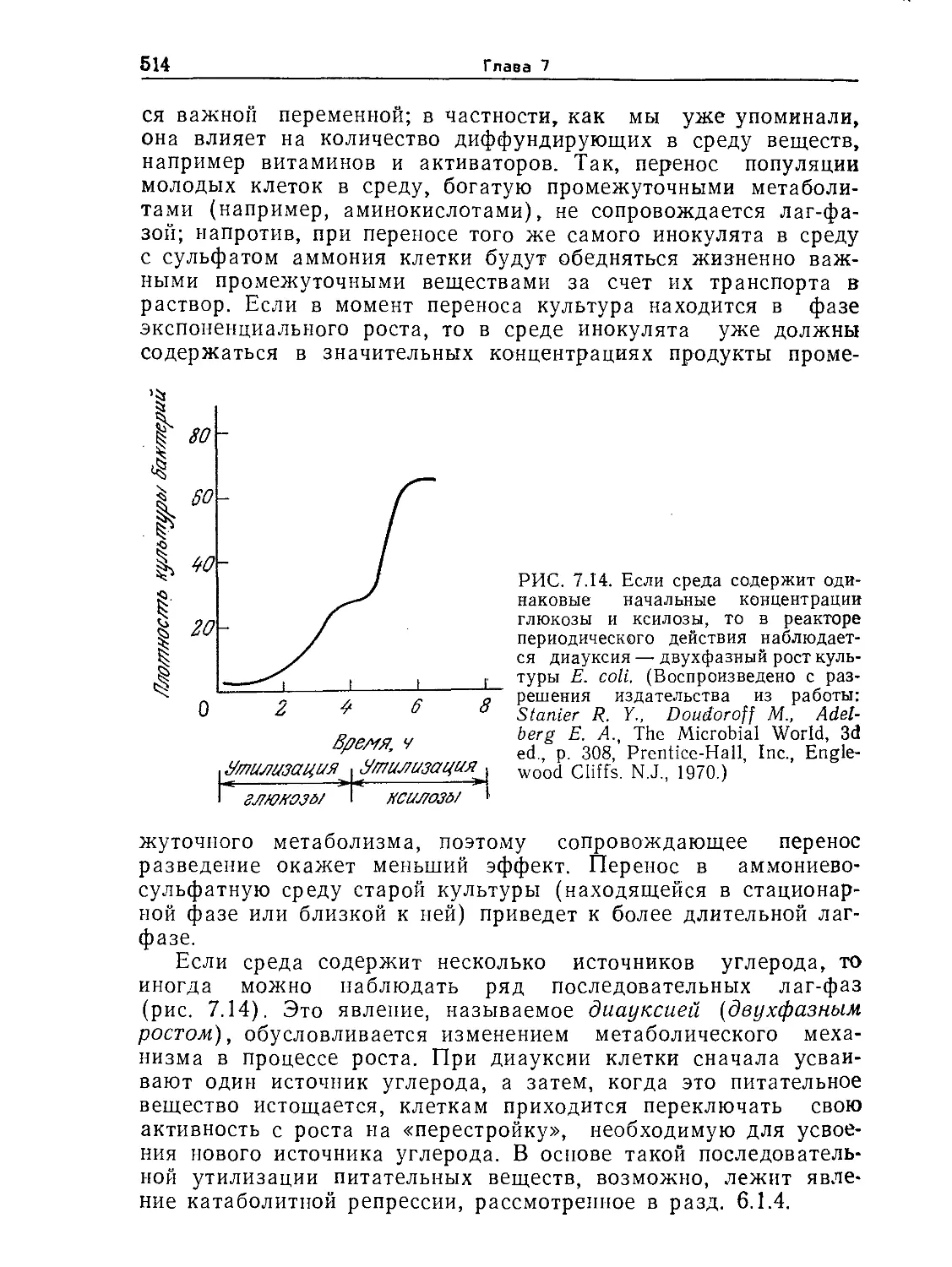



















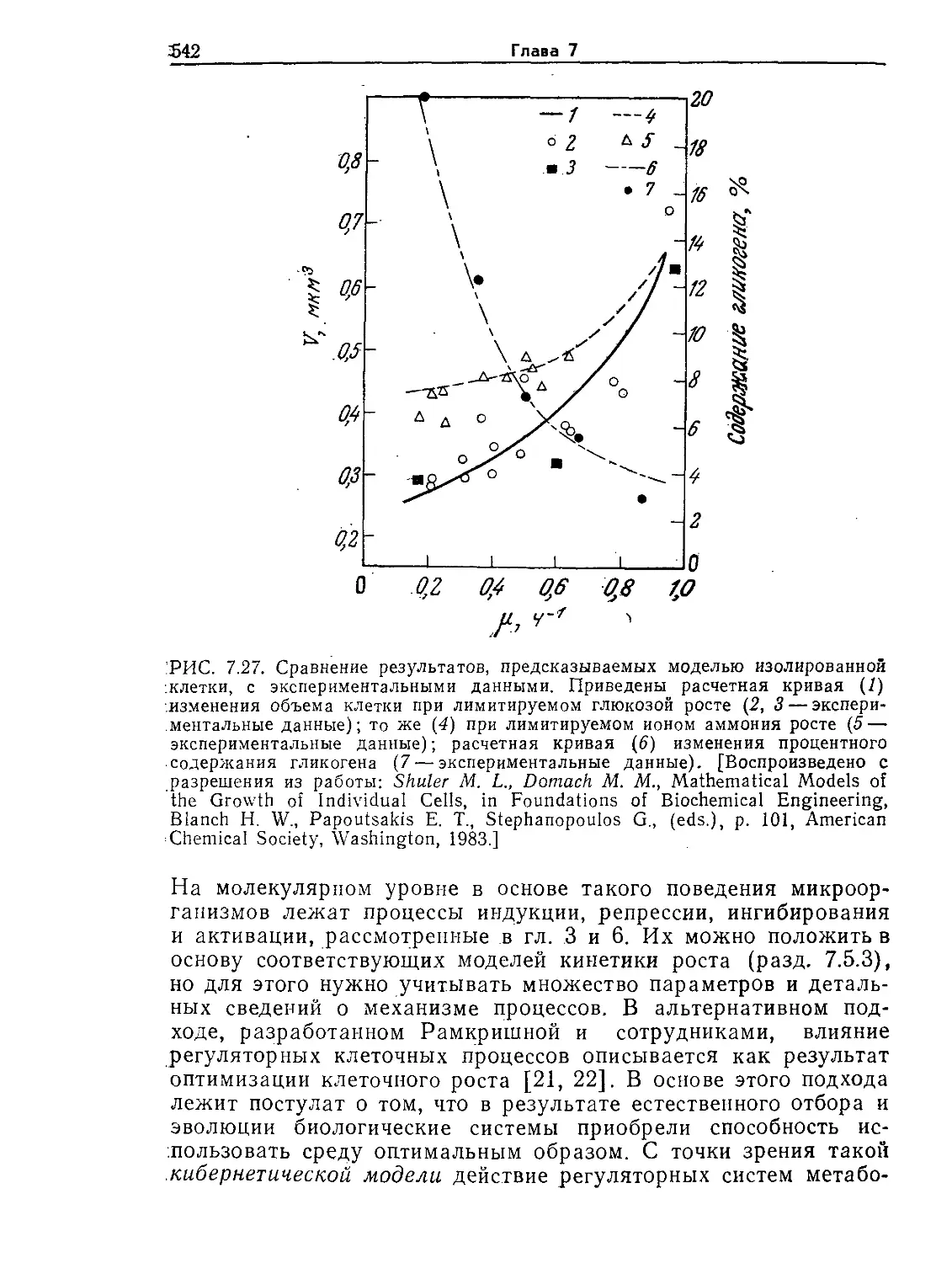

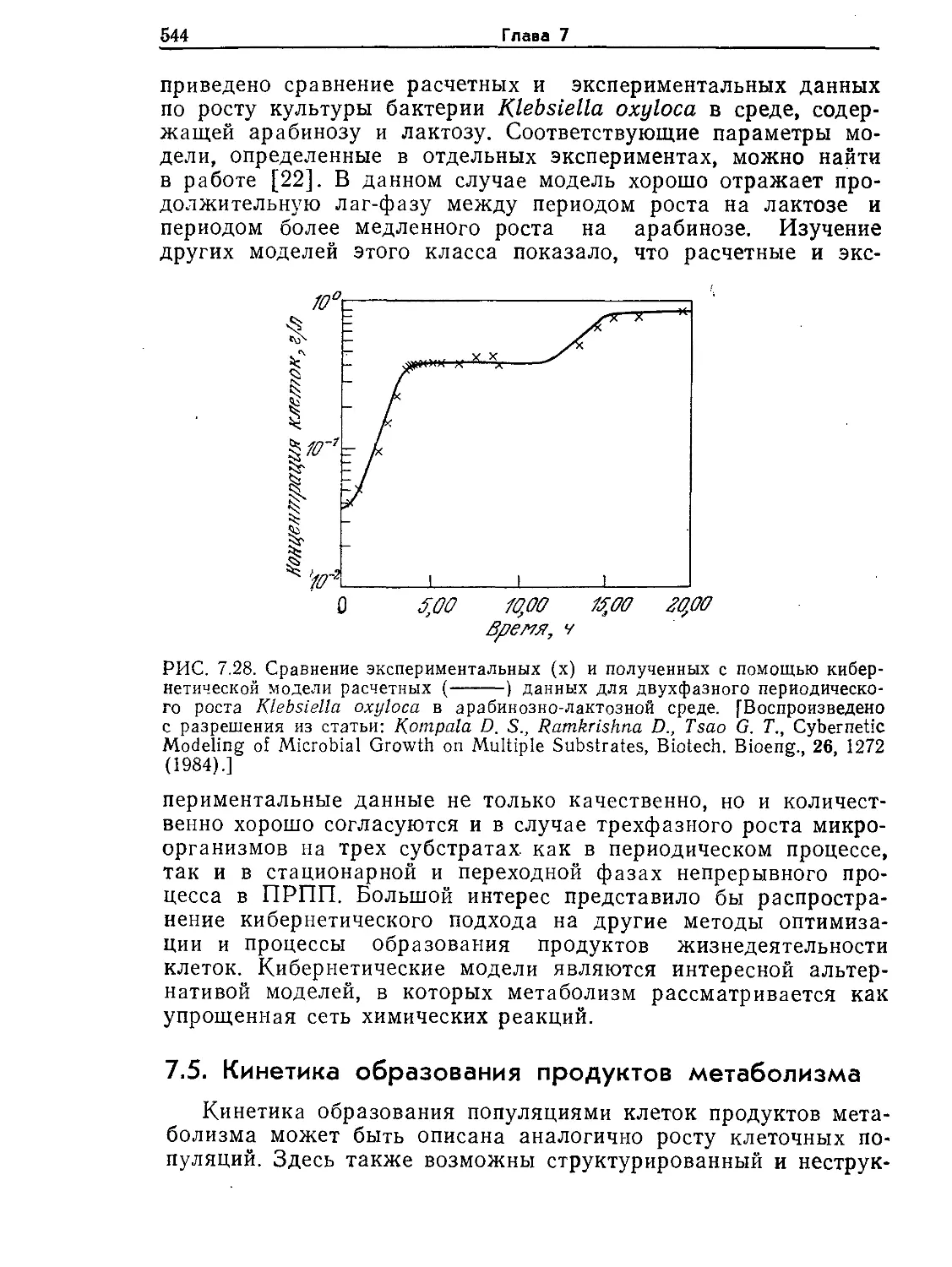


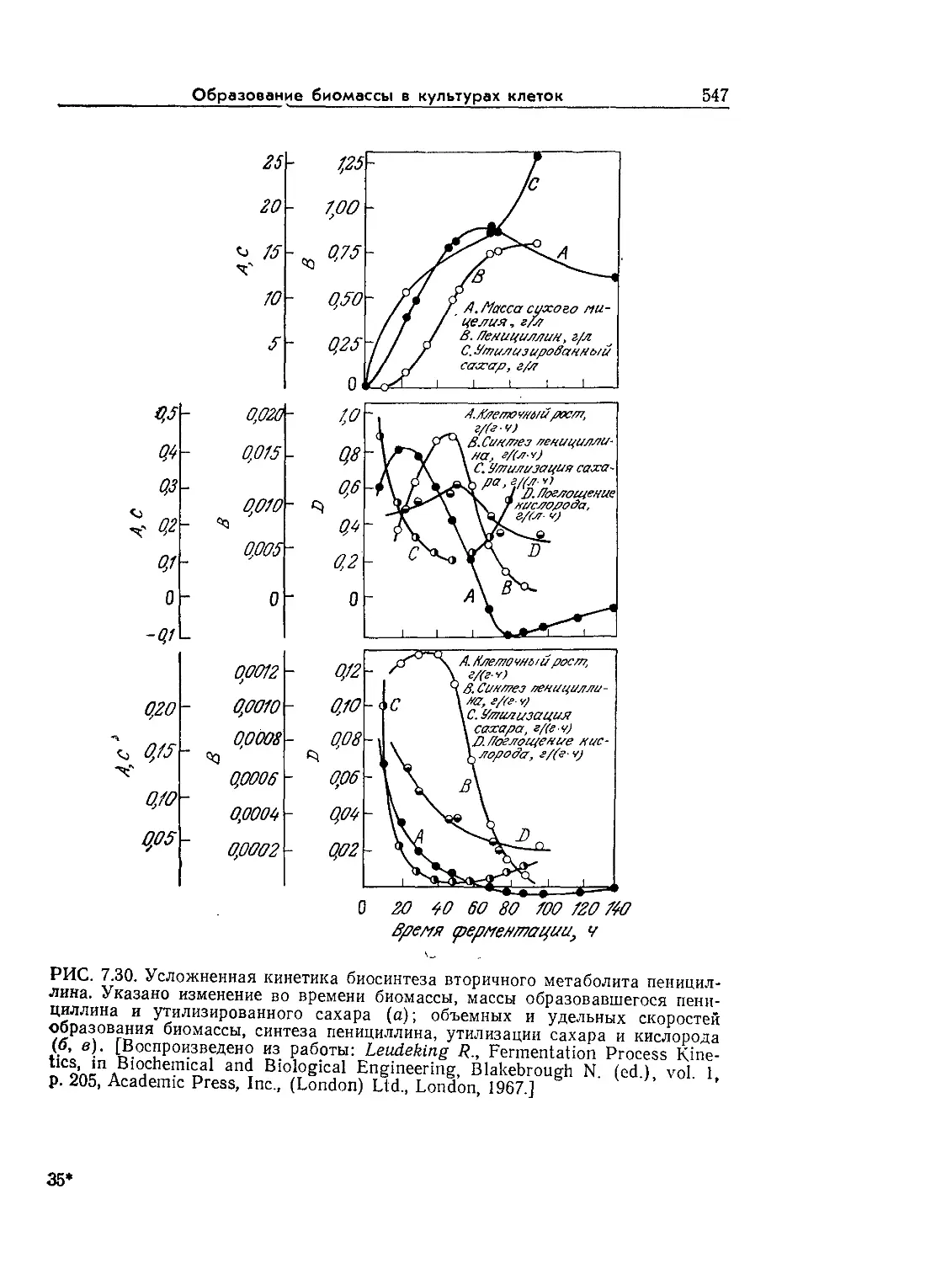

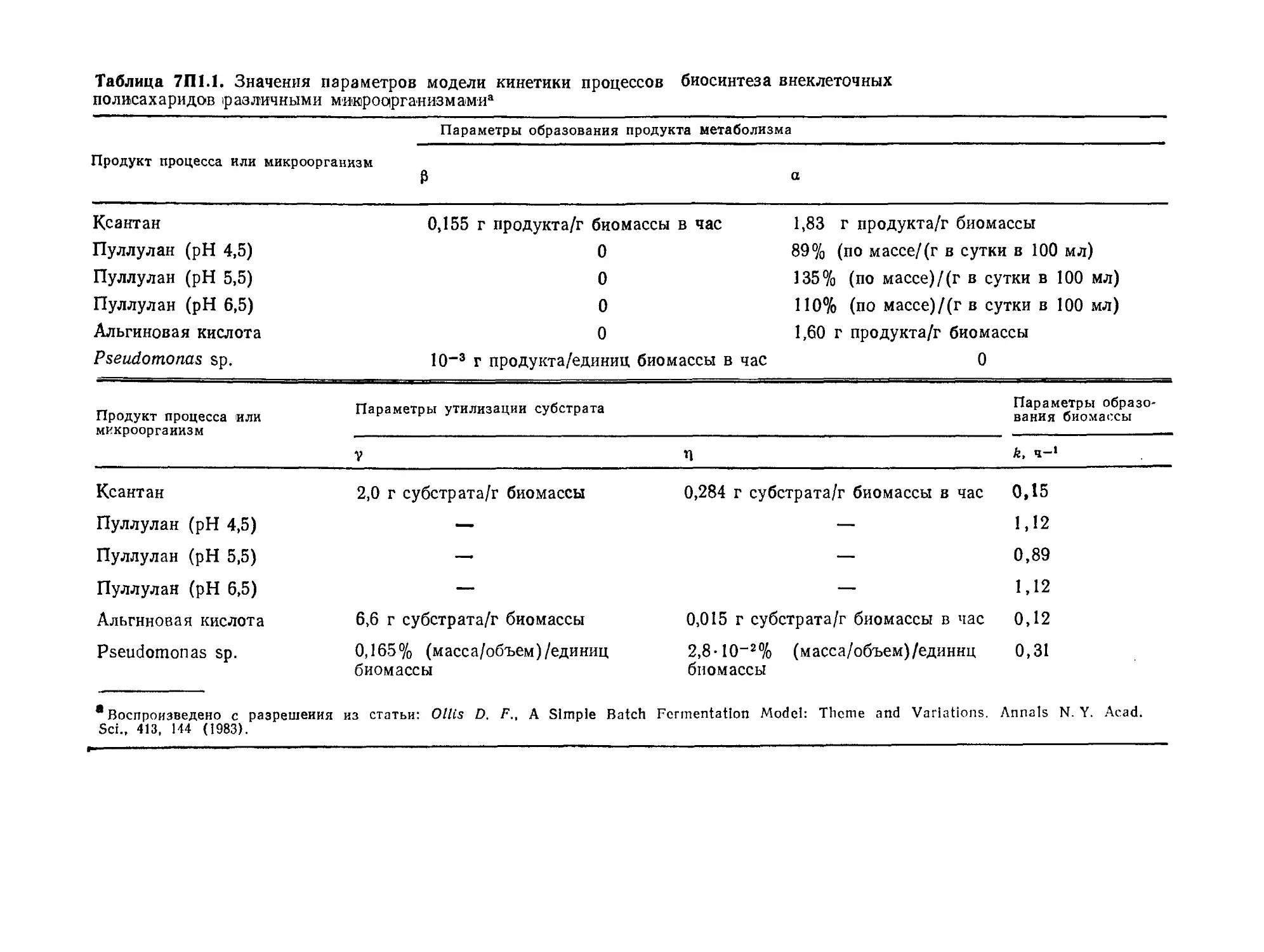

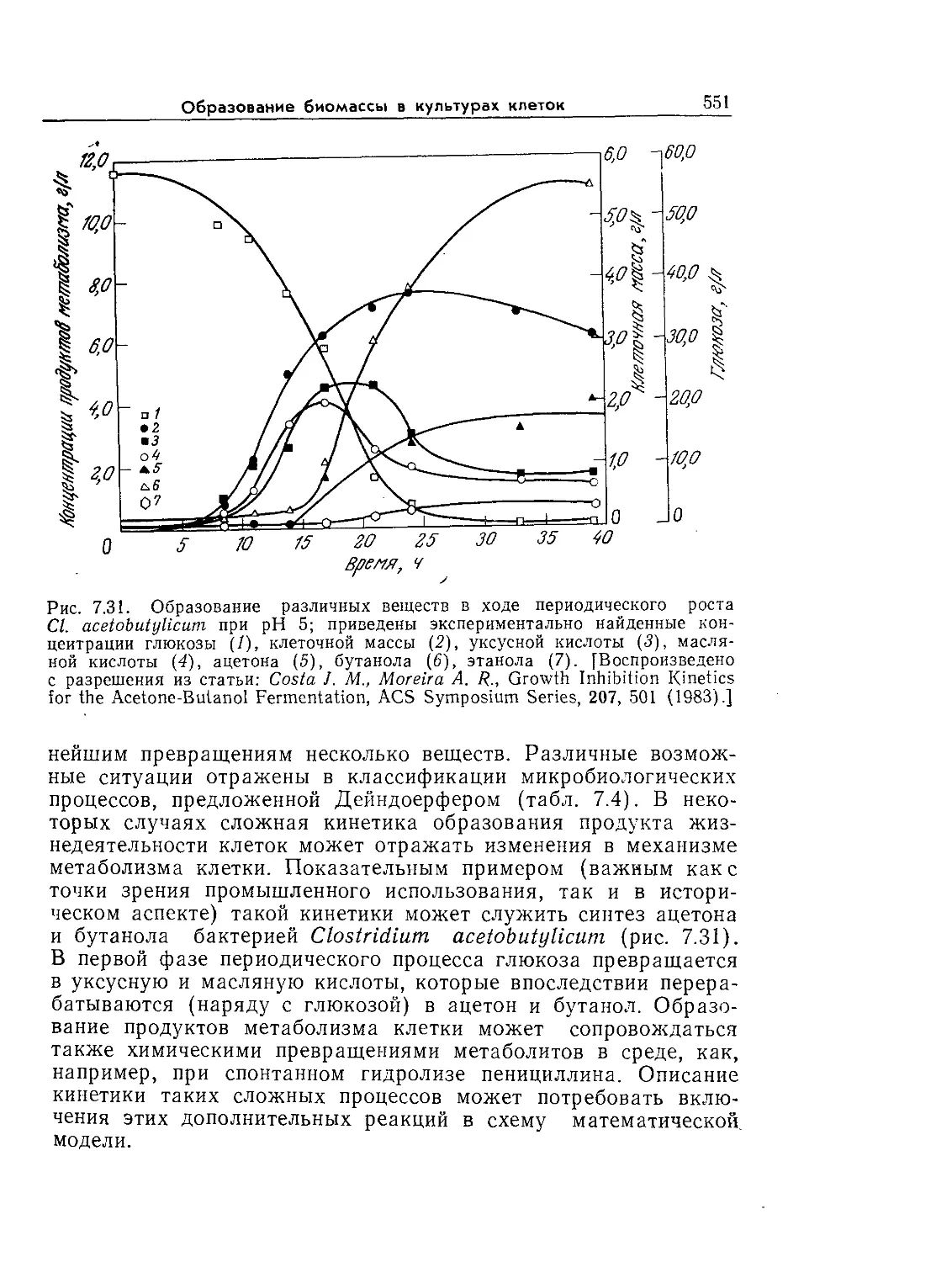

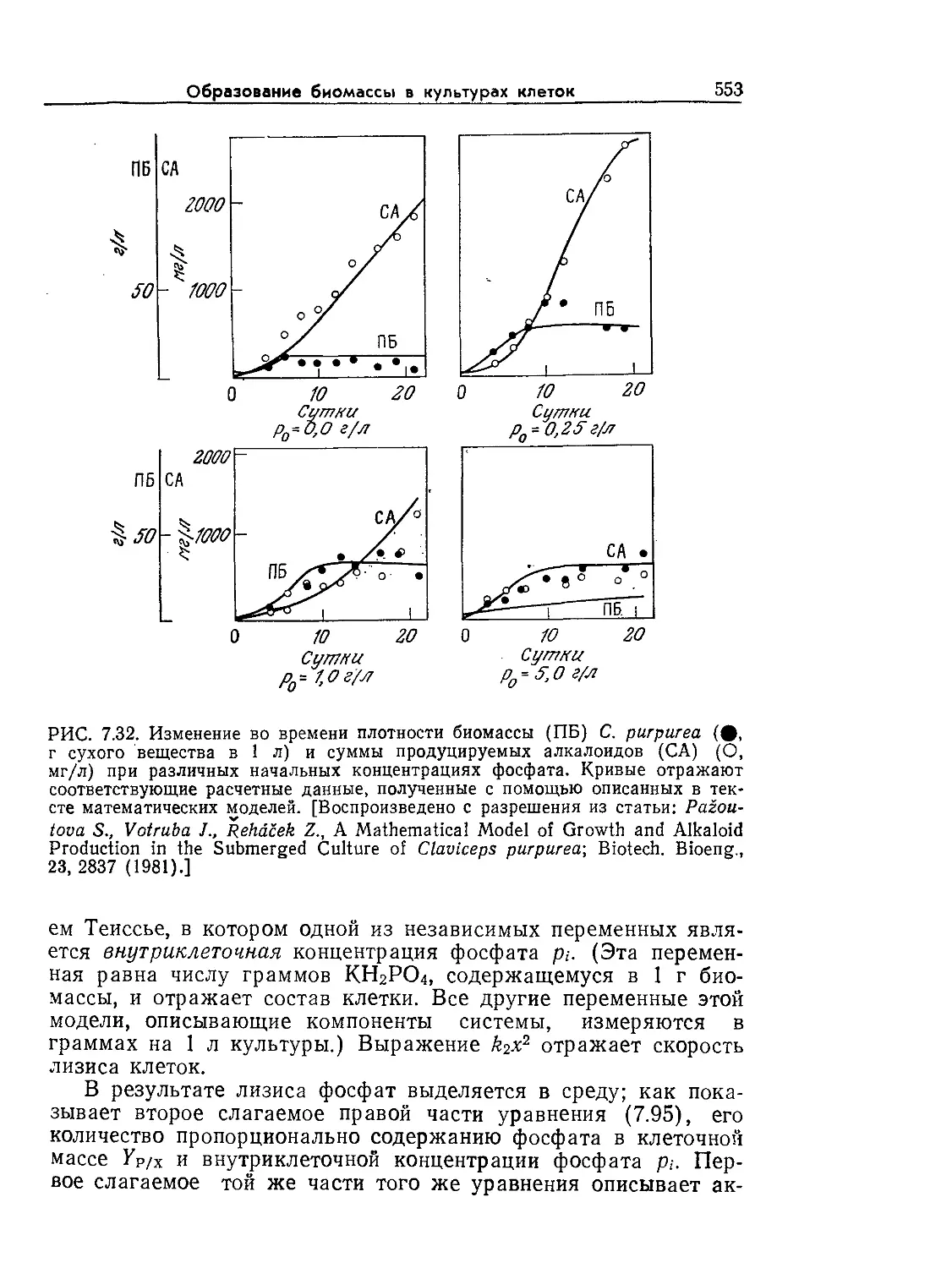

















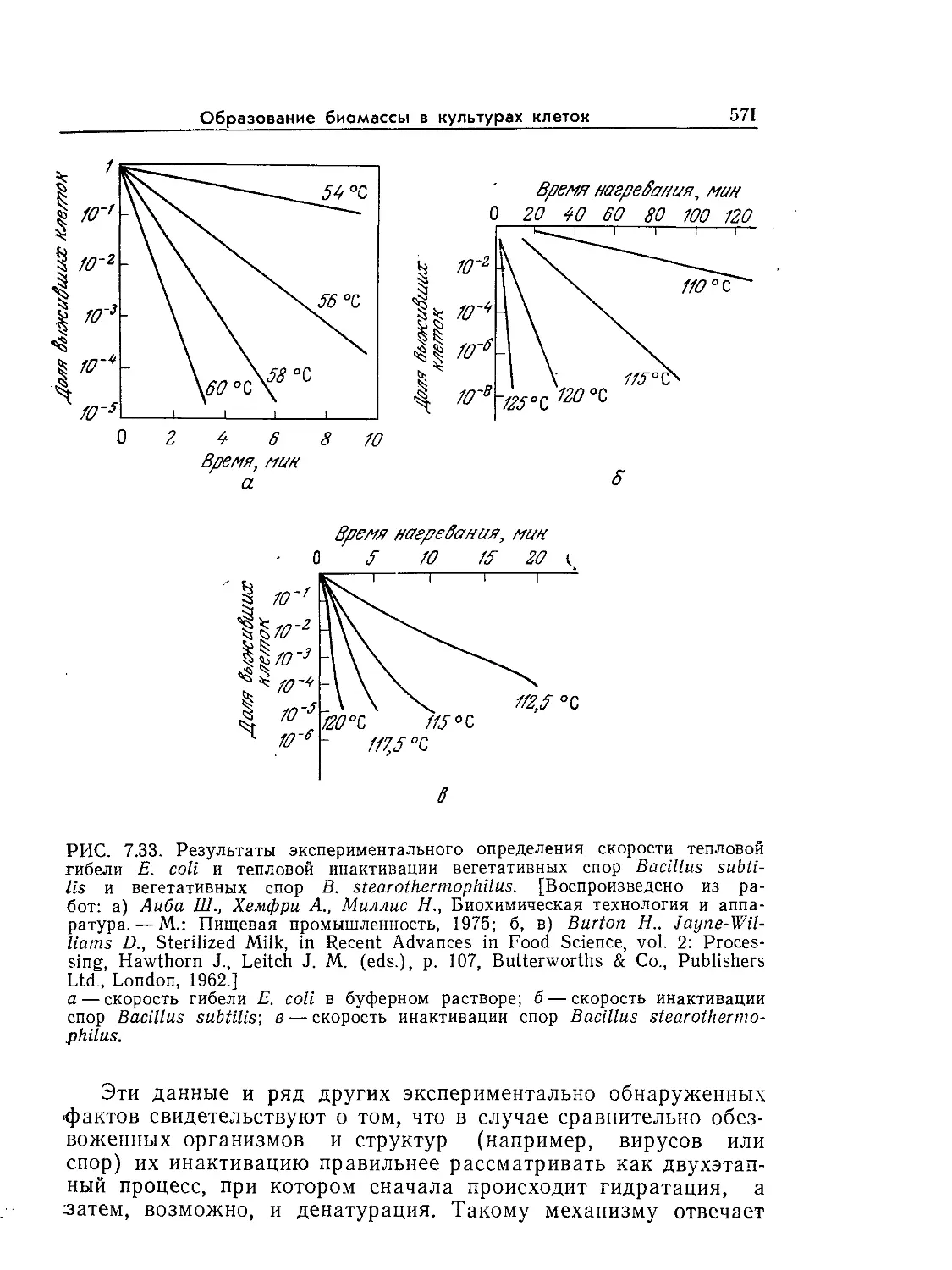



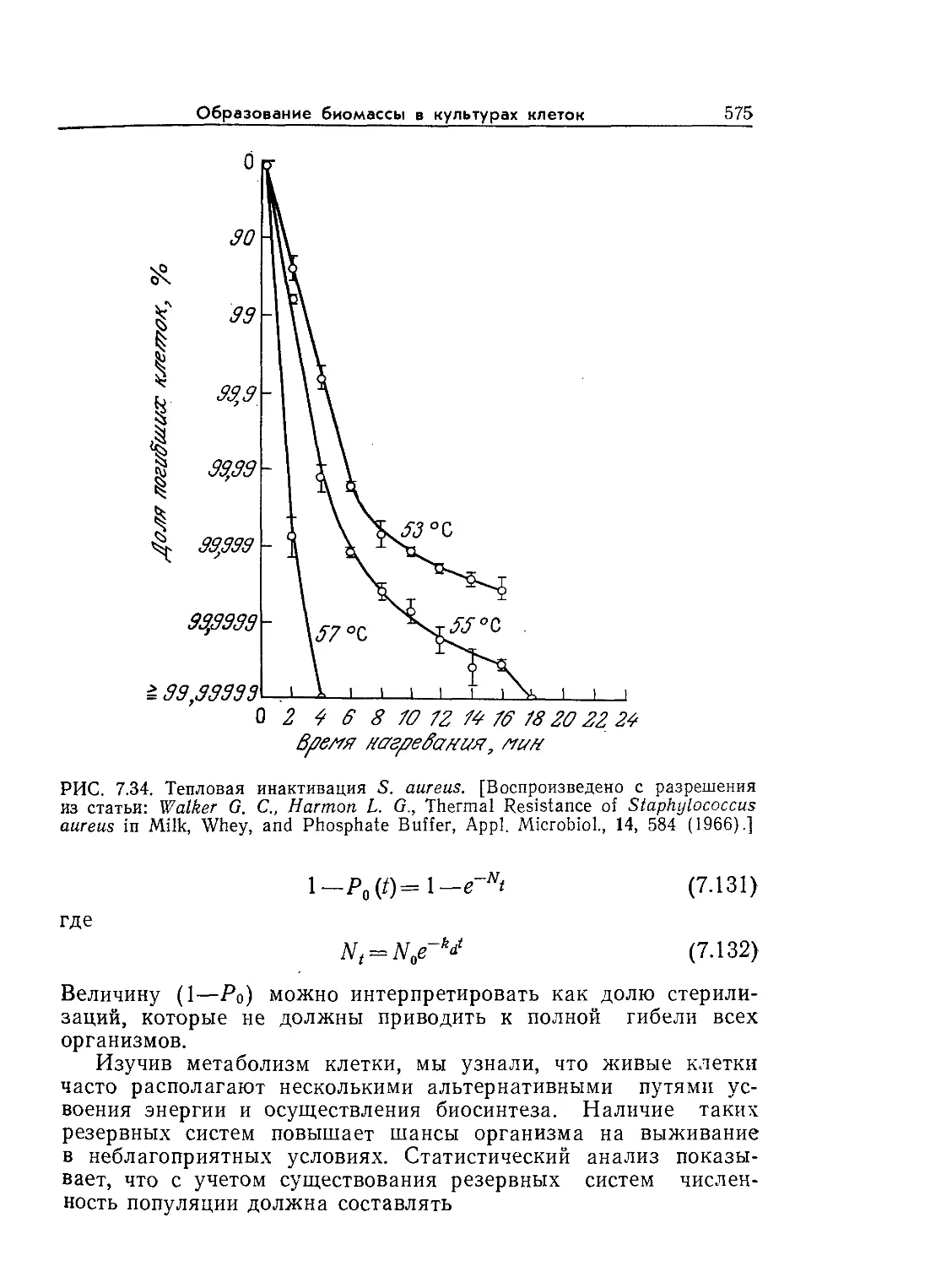






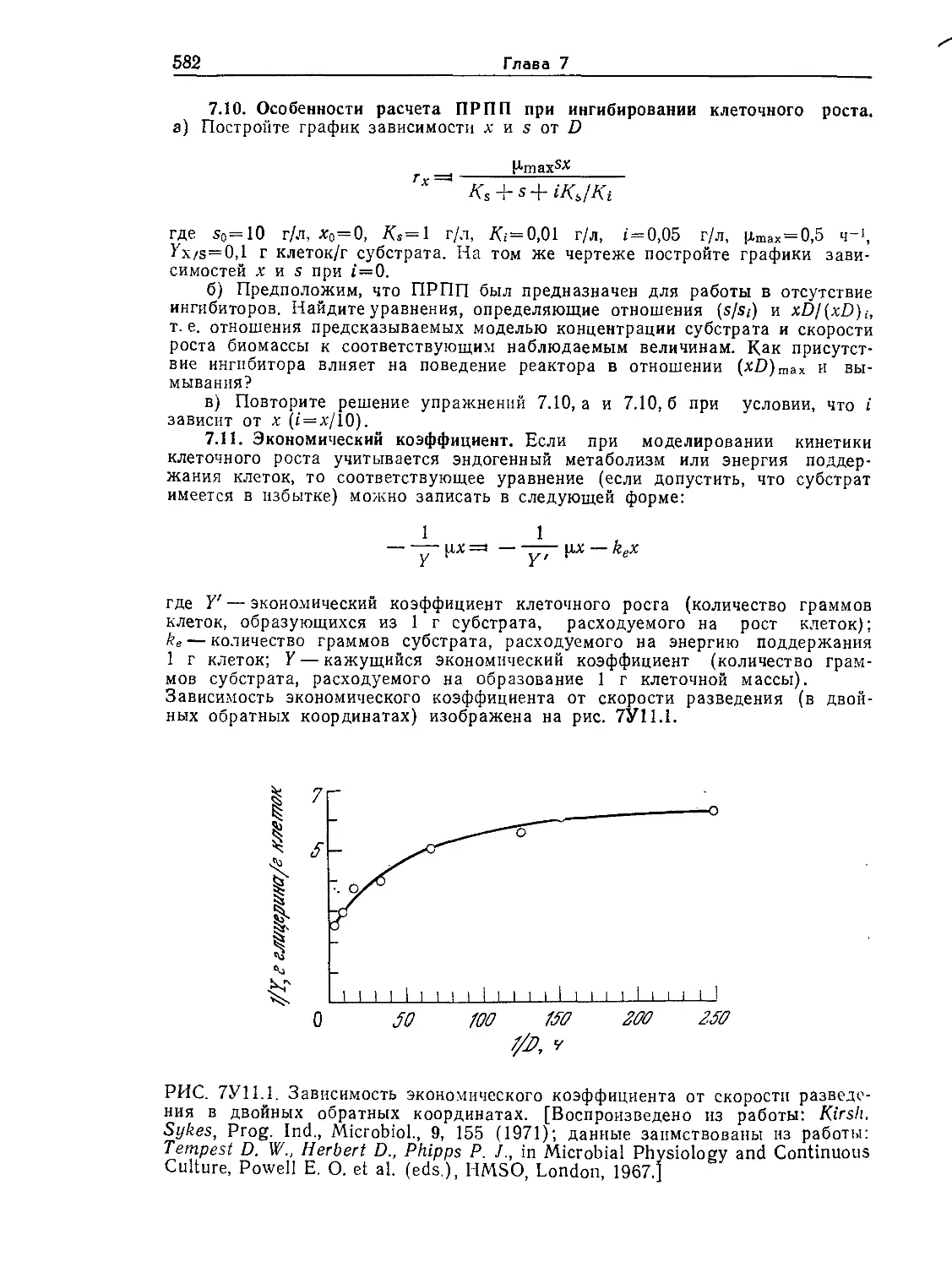
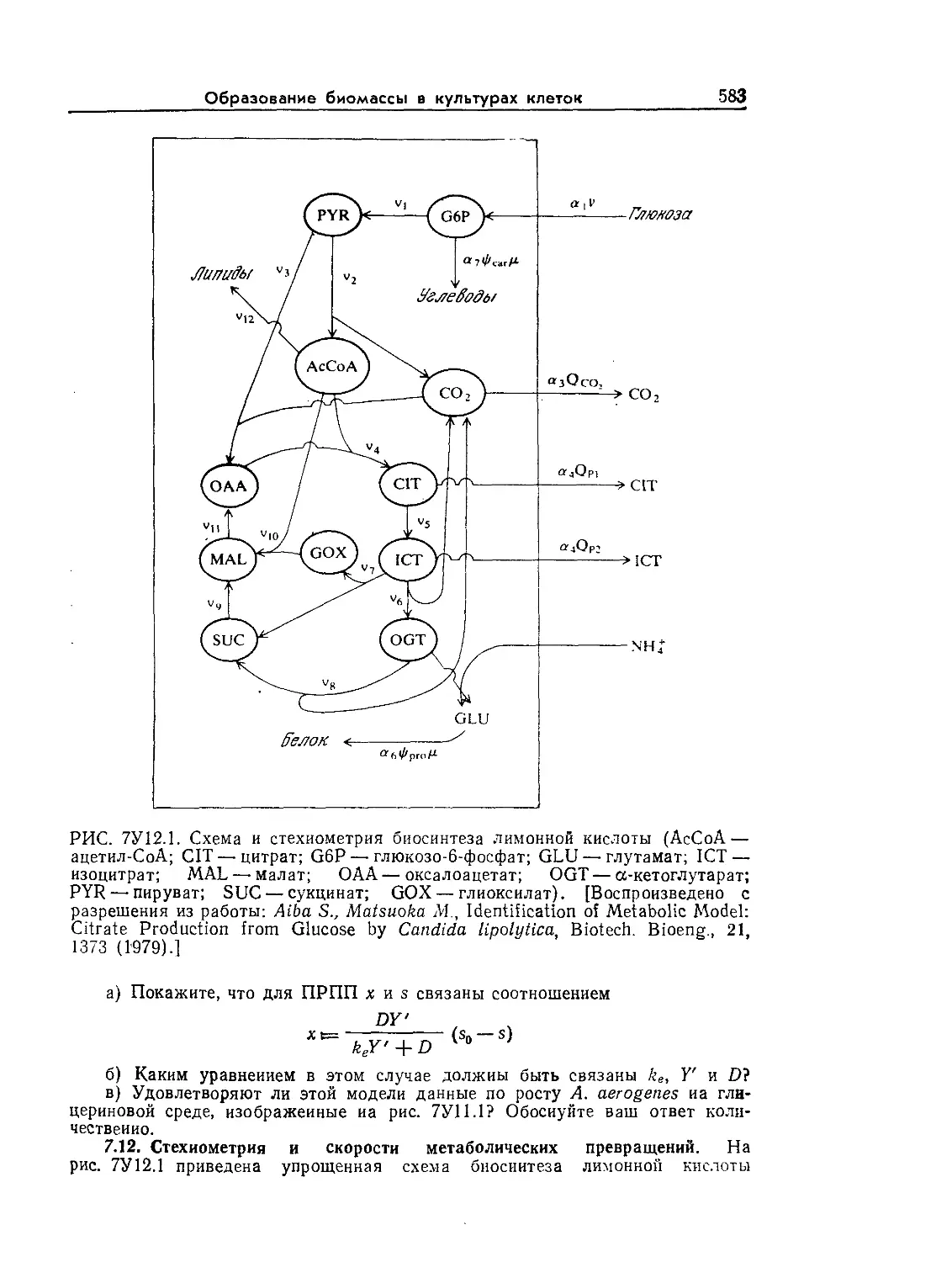











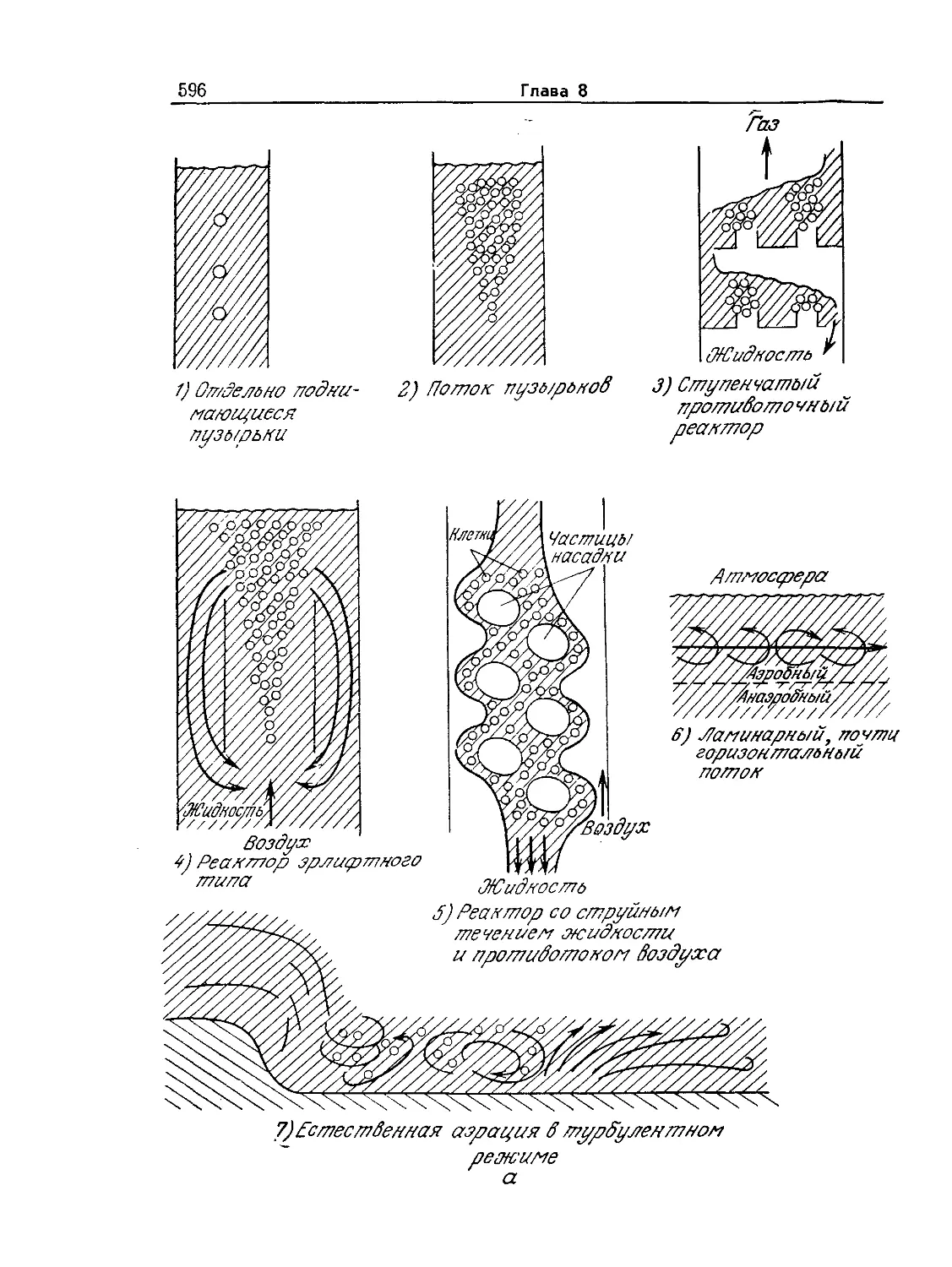
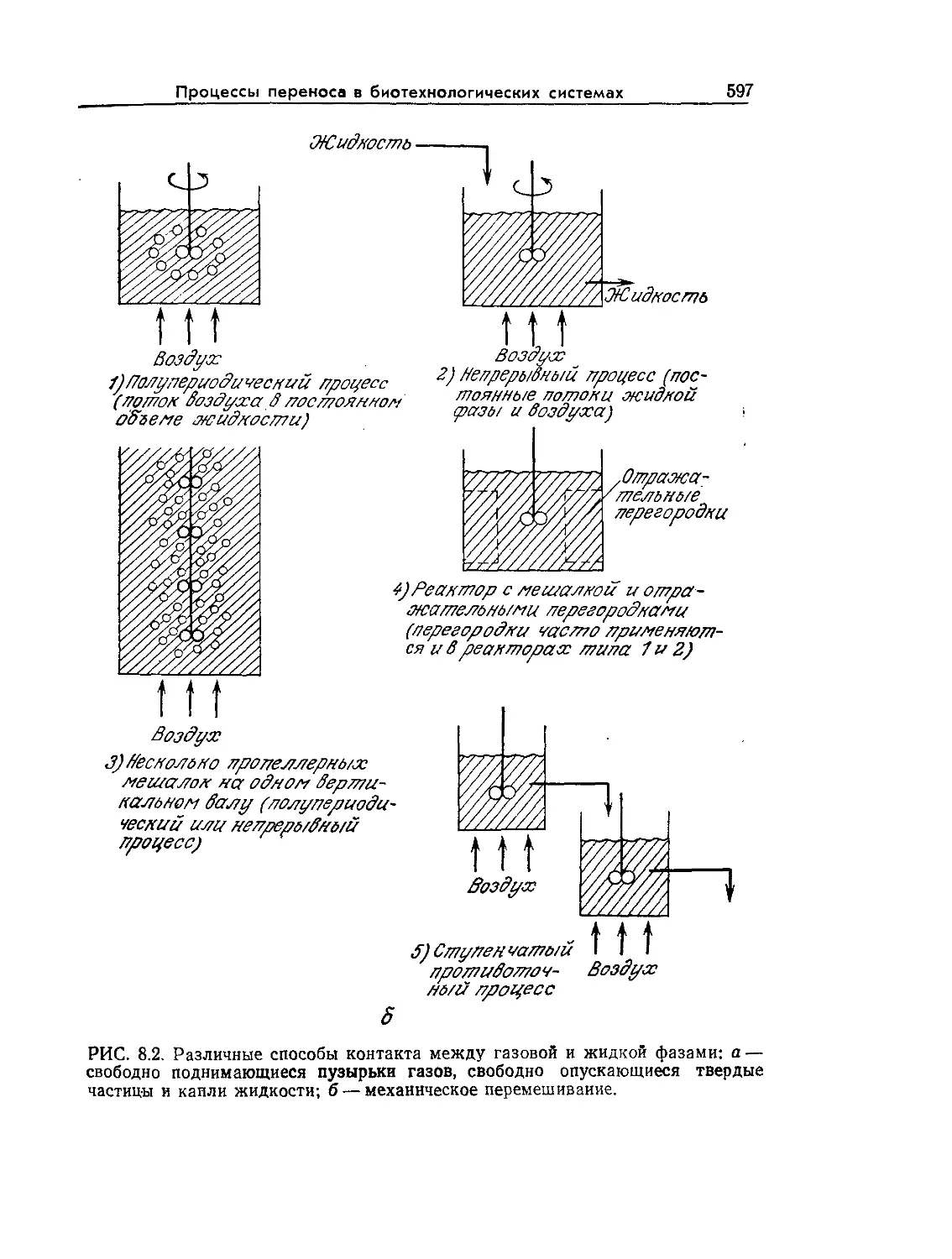



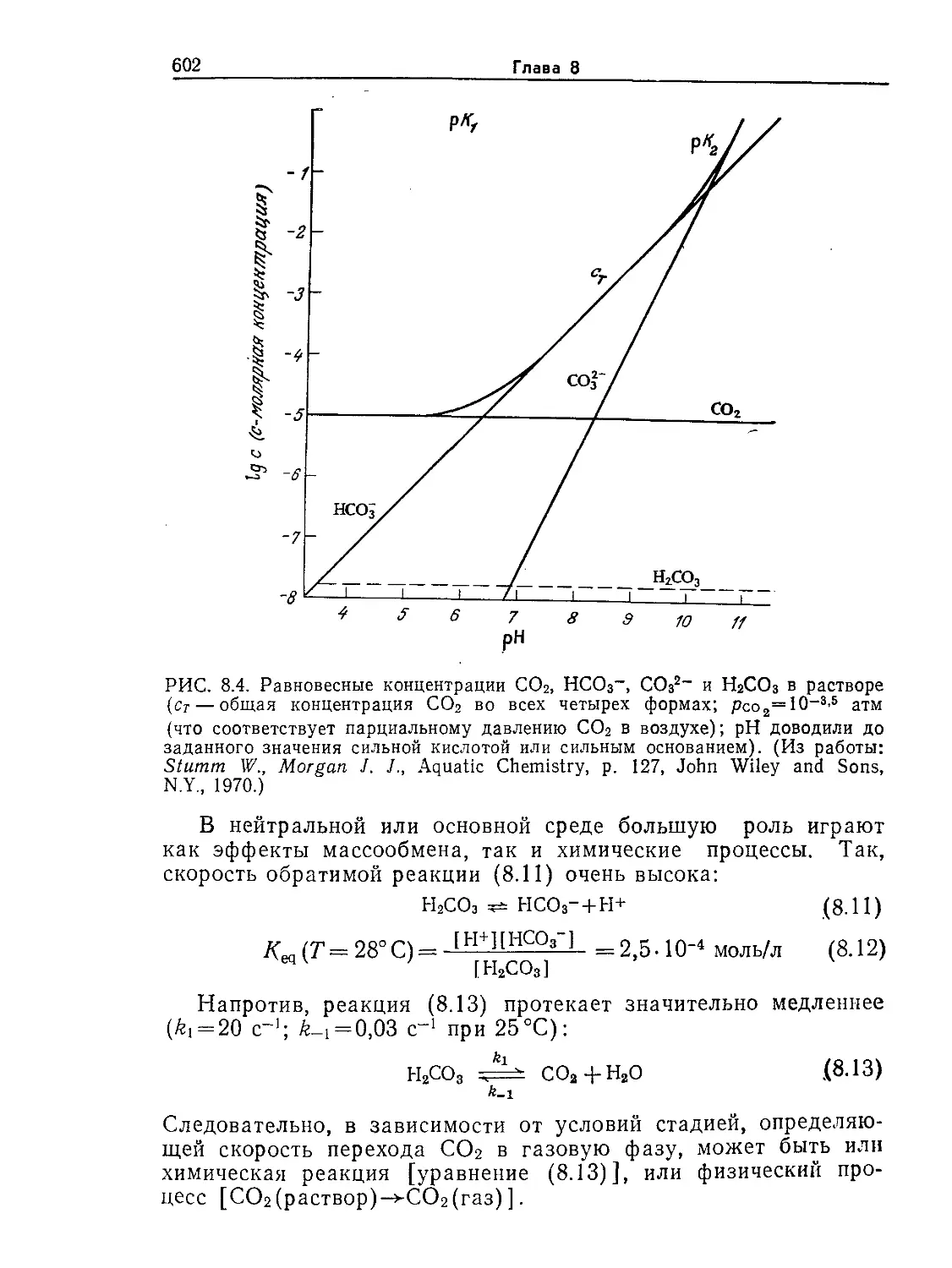


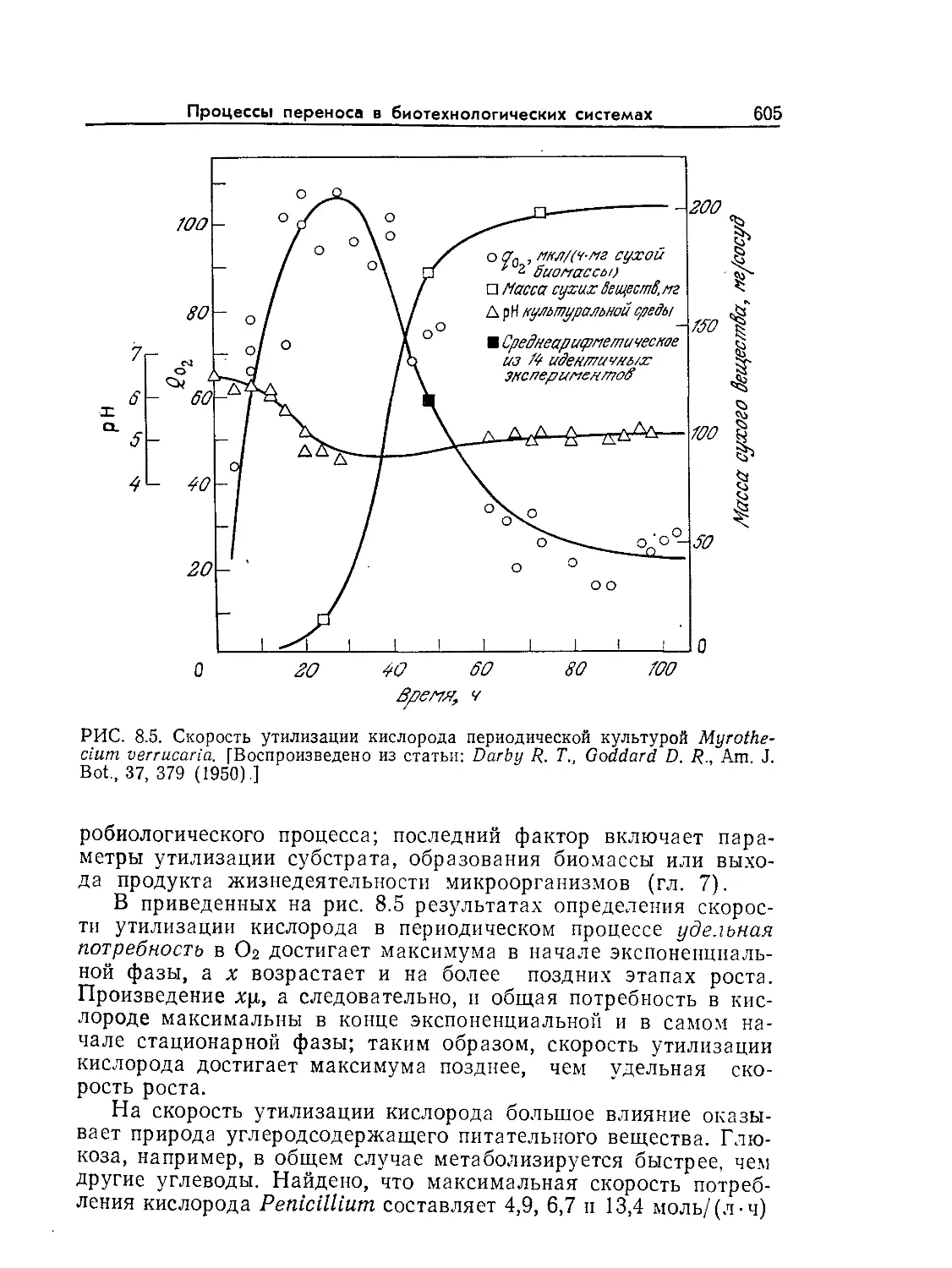




























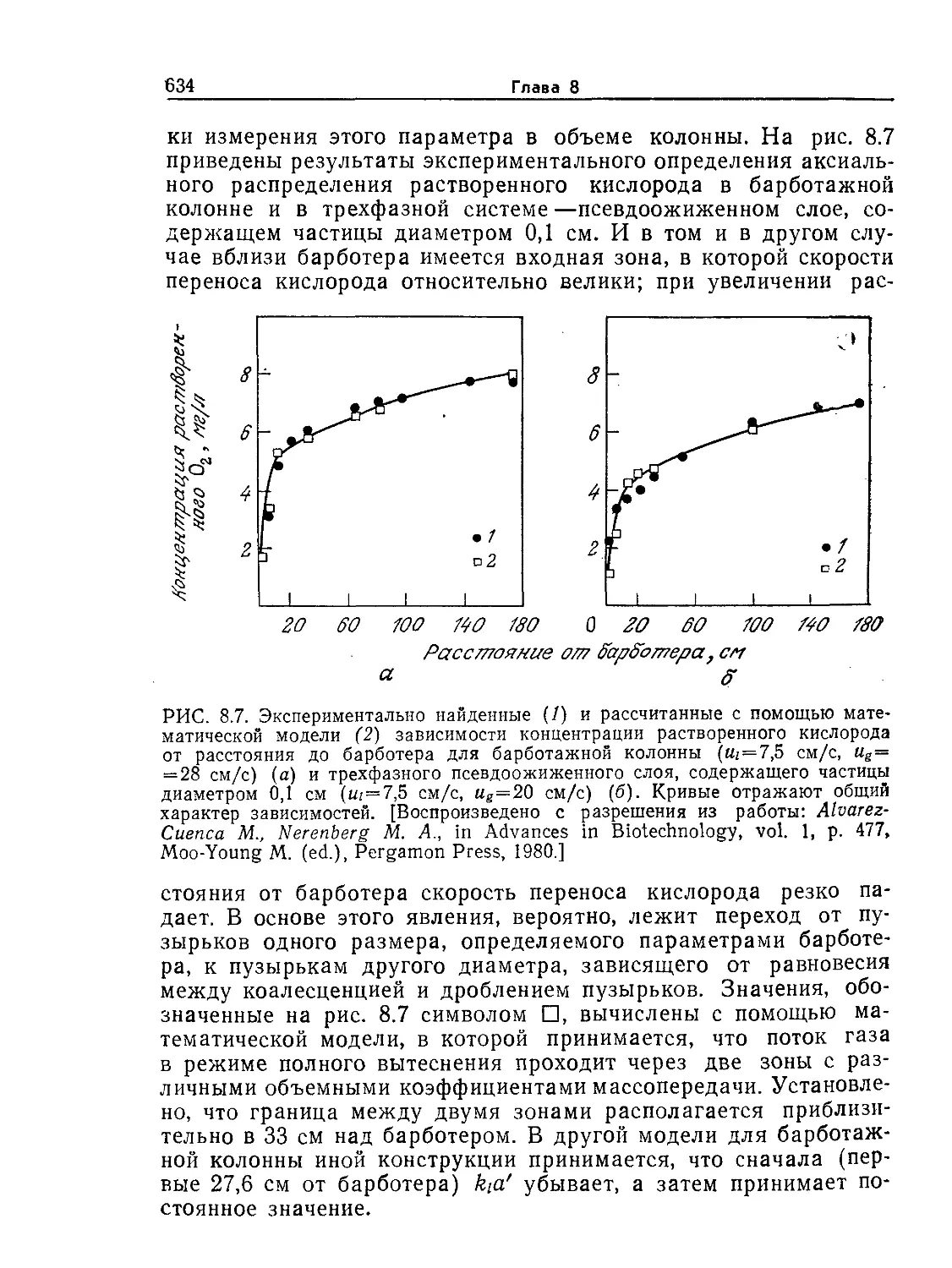

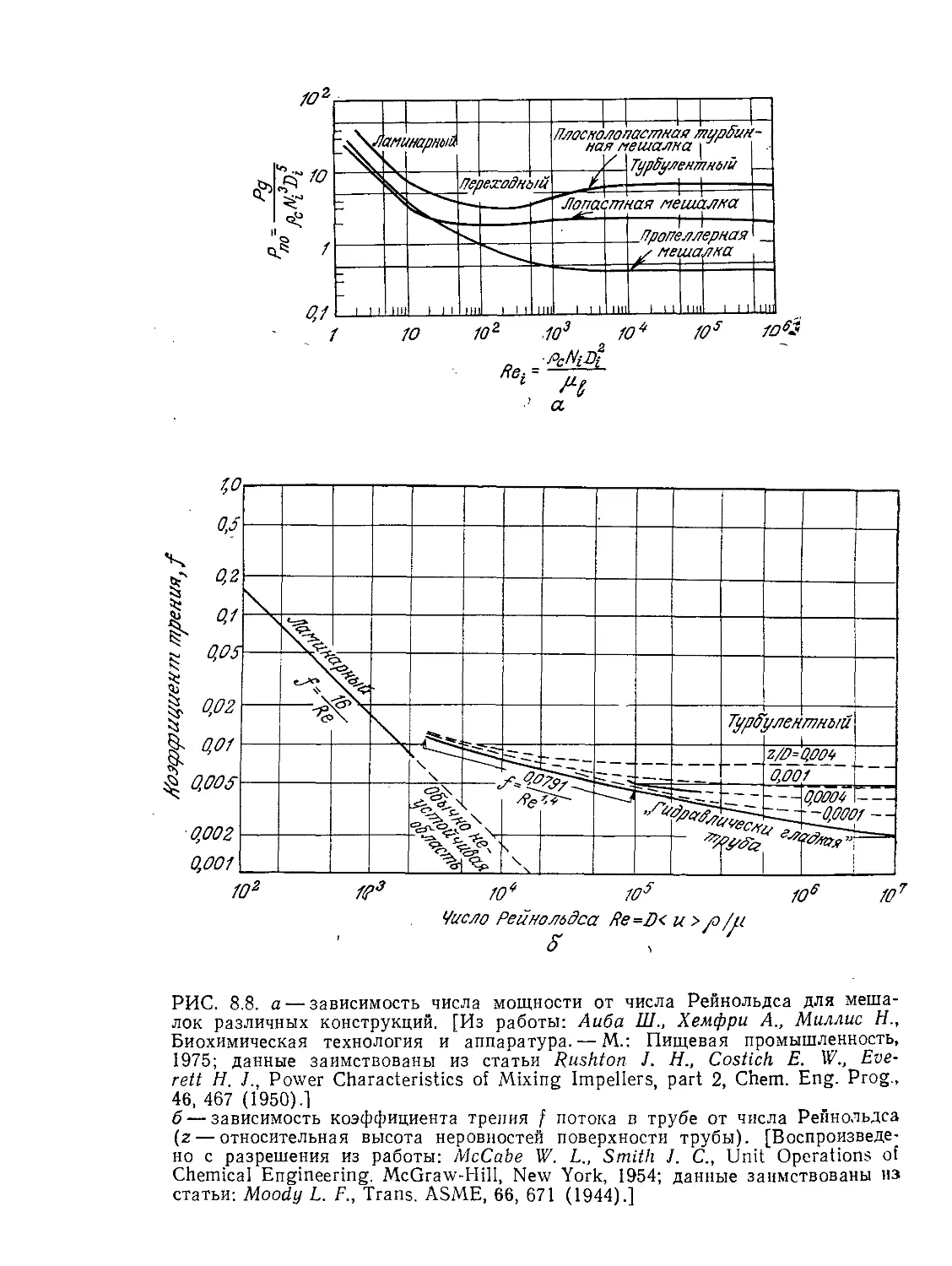
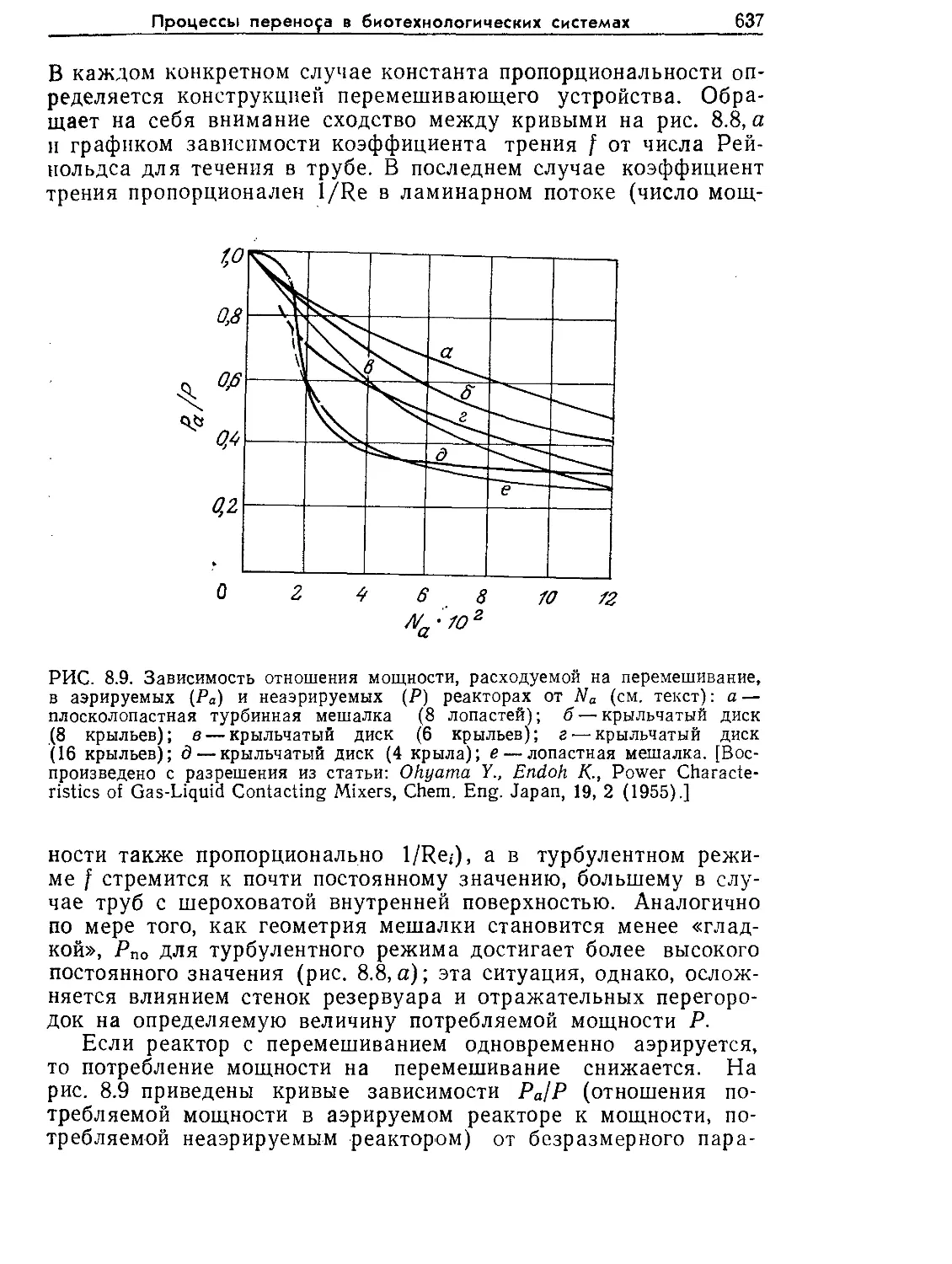

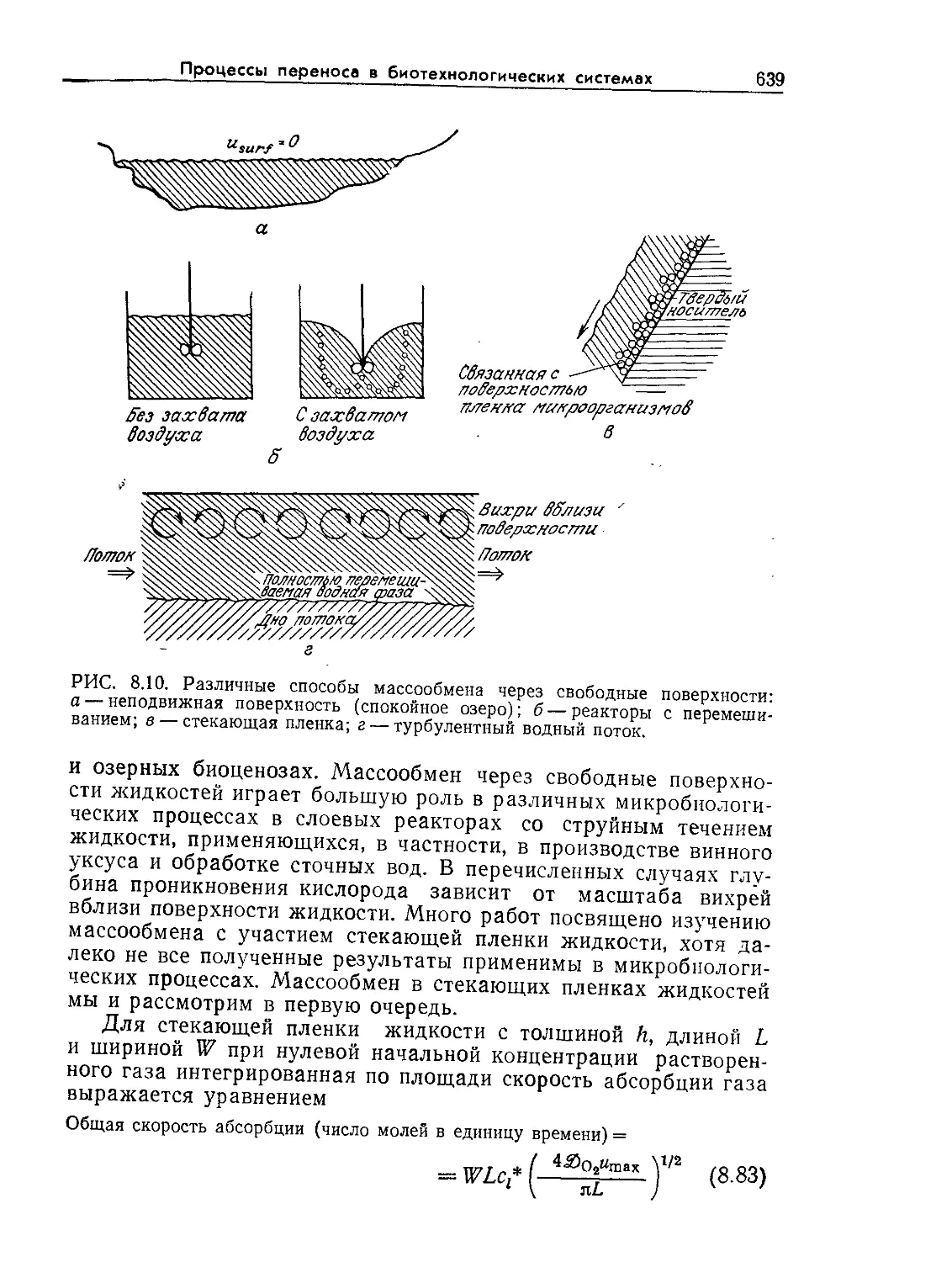





















































Tags: биологические науки в целом молекулярная биология общая биофизика, общая биохимия и общая физиология химия биология биохимия биофизика издательство мир биохимическая инженерия
ISBN: 5-03-001028-9
Year: 1989




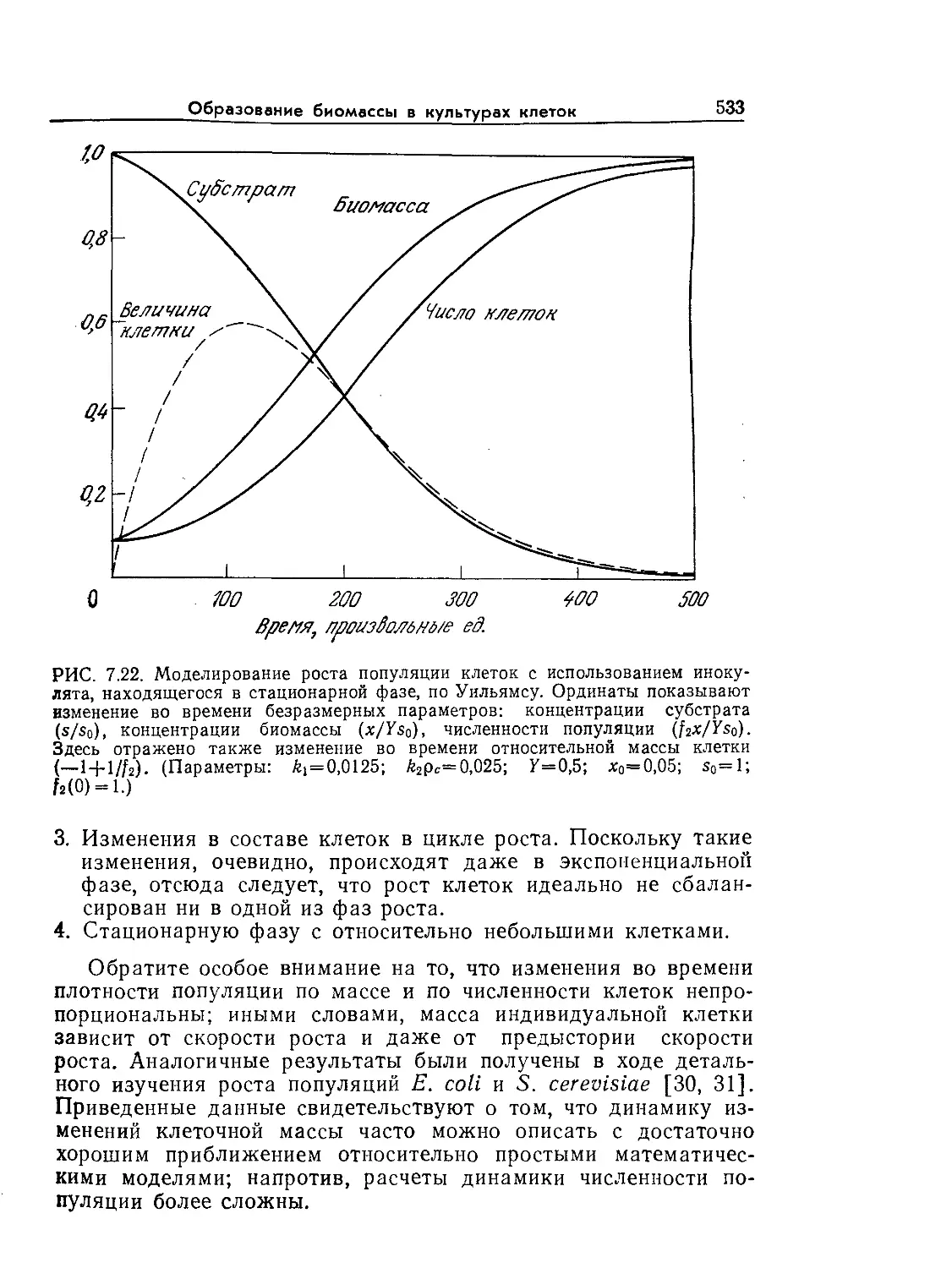
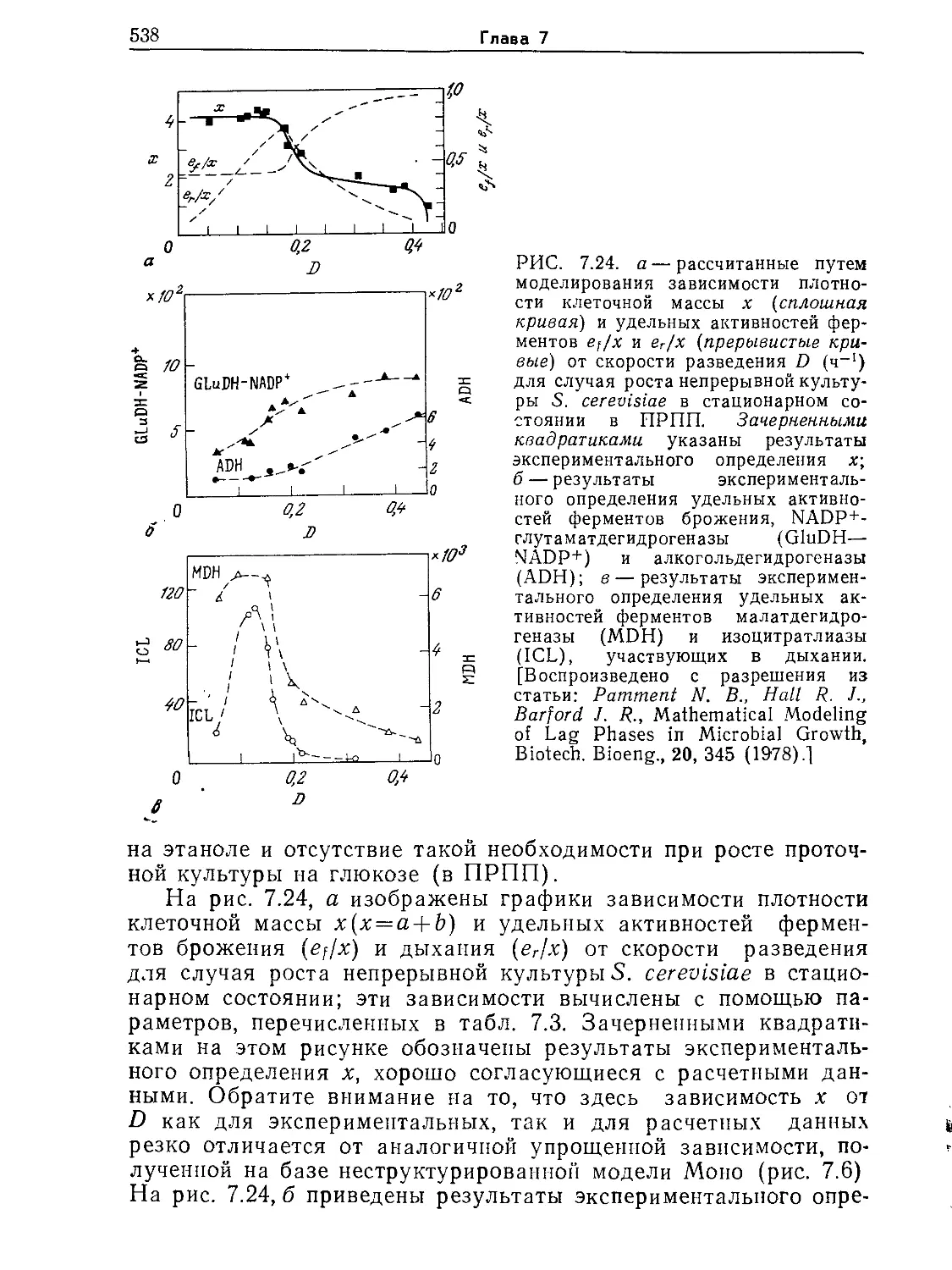
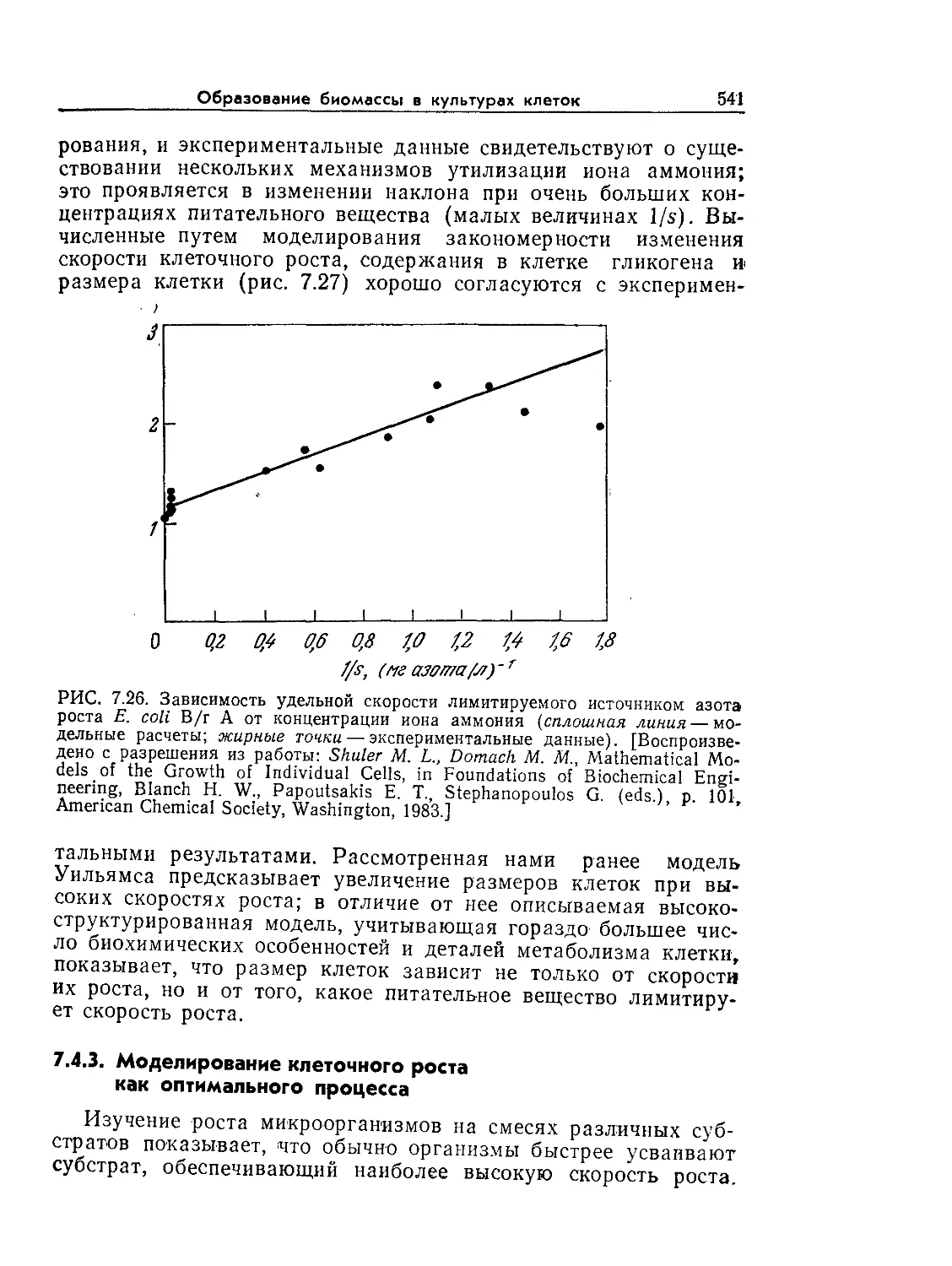
Основы биохимической инженерии
BIOCHEMICAL
ENGINEERING
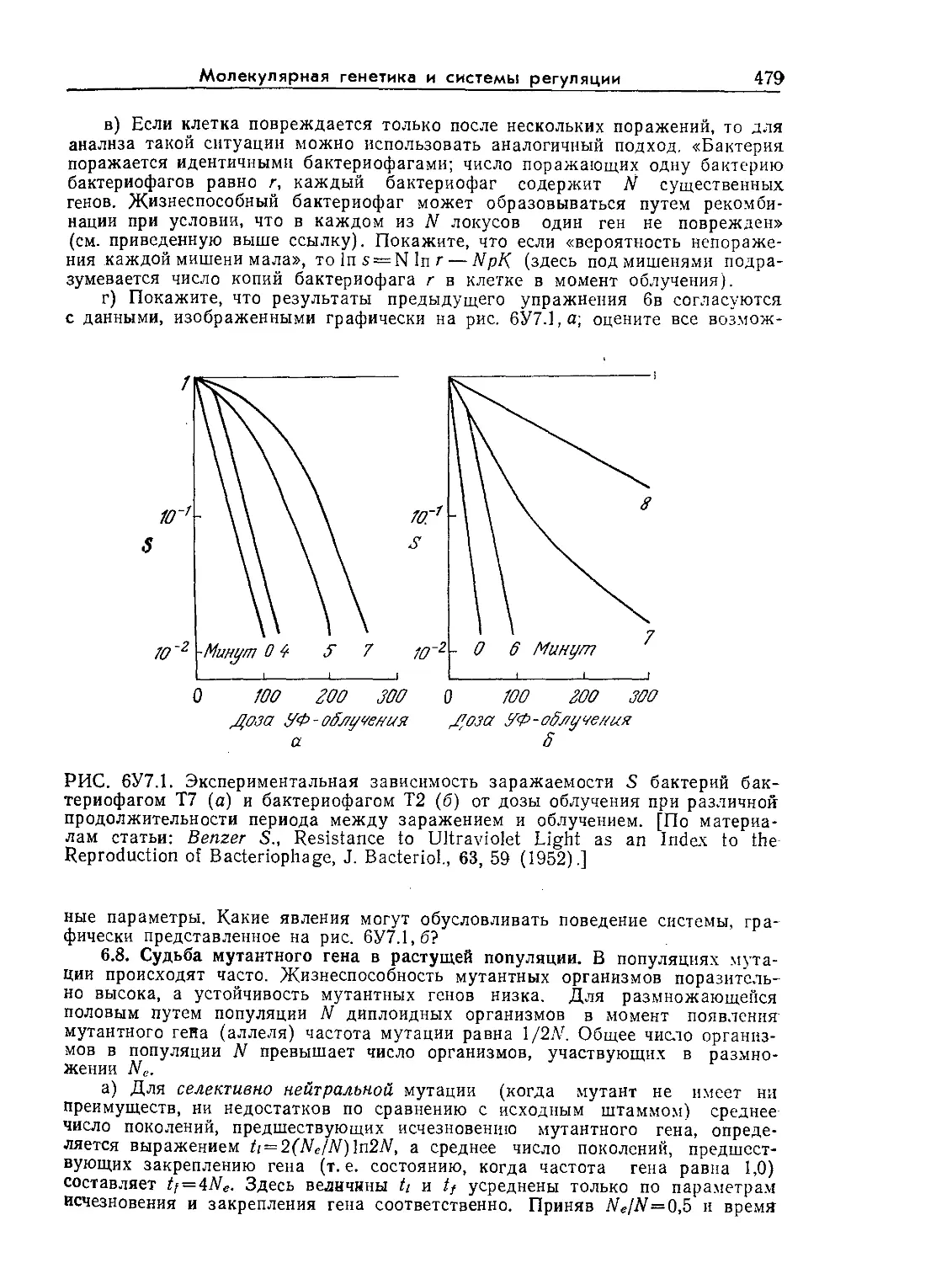
FUNDAMENTALS
Second Edition
James E. Bailey
California Institute of Technology
David F. Ollis
North Carolina State University
McGraw-Hill Book Company
New York St. Louis San Francisco Auckland Bogota
Hamburg Johannesburg London Madrid Mexico Montreal
New Delhi Panama Paris Sao Paulo Singapore Sydney
Tokyo Toronto
Дж. Бейли, Д.Оллис
ОСНОВЫ
биохимической
инженерии
В 2-х частях
1
Перевод с английского
А. А. Кирюшкина
Москва «Мир» 1989
ББК 28.07
Б 40
УДК 57.04
Бейли Дж., Оллис Д.
Б 40 Основы биохимической инженерии. Пер. с англ, в 2-х
частях. Ч. 1. — М.: Мир, 1989. — 692 с., ил.
ISBN 5-03-001028-9
Фундаментальный труд, написанный известными американскими специали-
стами в области проектирования, разработки и применения процессов, связанных
с переработкой биологических материалов и использованием биологических аген-
тов, в первую очередь ферментов и клеток. Он написан с целью заполнить про-
бел, часто существующий между биологией и технологией на их пути к биотех-
нологии.
В русском переводе книга выходит в двух частях.
В первой части рассматриваются теоретические основы микробиологии, био-
химии и биологии клетки, ферментативного катализа и молекулярной генетики.
Отдельные главы посвящены стехиометрии и энергетике клеточных процессов, ки-
нетике роста популяций клеток, транспортным явлениям в процессах с участием
биокатализаторов и микроорганизмов.
Предназначена для студентов и преподавателей биологических н технологи-
ческих специальностей, научных работников, инженеров, работающих в микро-
биологической и медицинской промышленности.
Б
1901000000—196
041(01)—89
93—89, ч.
1
ББК 28.07
Редакция литературы по химии
ISBN 5-03-001028-9 (русск.)
ISBN 5-03-001027-0
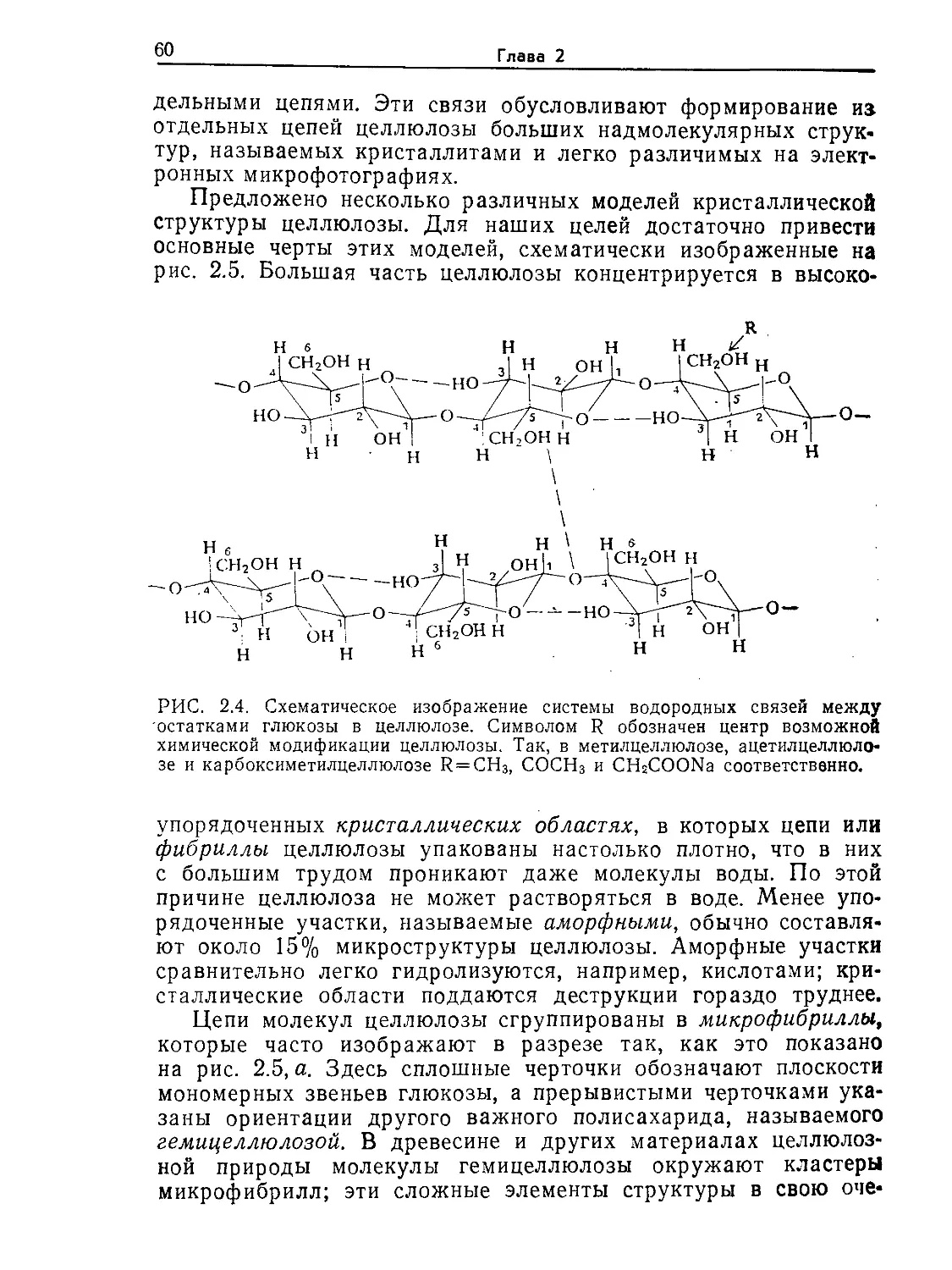
ISBN 0-07-003212-2 (англ.)
© 1986, 1987 by McGraw-Hill, Inc.
© перевод на русский язык, «Мир», 1989
От переводчика
Биохимическая технология (биотехнология) —одна из самых
старых и одновременно одна из самых молодых наук и отраслей
промышленности, если под биотехнологией понимать использо-
вание живых организмов или биологических процессов в про-
изводстве,— ведь такие процессы, как хлебопечение, сыроделие,
виноделие и др., известны человечеству с незапамятных времен.
Однако термин «биотехнология» привился только с середины
70-х годов, когда биотехнология пережила свое второе рожде-
ние в связи с появлением генетической инженерии.
В настоящее время процессы биохимической технологии ши-
роко используются при производстве ценных биологически ак-
тивных веществ (антибиотиков, ферментов, гормонов и др.),
для предотвращения загрязнения окружающей среды, защиты
растений от болезней и вредителей, в крупномасштабном про-
изводстве белков и аминокислот, предназначенных в качестве
добавок к кормам в животноводстве. Развитие генетической и
клеточной инженерии позволило получать ранее недоступные
вещества — в первую очередь лекарственные препараты (ин-
терфероны, гормоны роста, инсулин человека и др.). Другое
перспективное направление биотехнологии связано с получени-
ем гибридом и продуцируемых ими антител, последние уже
сейчас широко используются в качестве лекарственных и диа-
гностических препаратов и специфических реагентов.
Для того чтобы все возможности генетической инженерии
стали реальностью, необходимо перенести ее методы из стек-
лянной лабораторной пробирки в стальные промышленные
реакторы*. Биохимическая технология в настоящее время пе-
реживает период чрезвычайно бурного роста, который был сти-
мулирован успехами в фундаментальных и теоретических ис-
следованиях в области наук о жизни, заметно опередивших
практическое использование результатов этих исследований. До-
стижениям фундаментальных наук посвящено немало изданий,
опубликованных как в СССР, так и за рубежом; в то же время
* Промышленная микробиология и успехи генетической инженерии: Пер,
с англ. — М.: Мир, 1984, с. 6.
6
От переводчика
чрезвычайно важные проблемы, связанные с использованием
этих достижений в производстве, освещались главным образом
в специальной периодической литературе.
В какой-то мере эту диспропорцию поможет исправить пред-
лагаемый читателю перевод книги известных американских
специалистов в области биохимической и химической техноло-
гии Дж. Бейли и Д. Оллиса, которые попытались охватить тео-
ретические и практические аспекты всех основных проблем био-
химической технологии*.
Основное внимание авторы уделяют математическому мо-
делированию и методам расчета и проектирования аппаратов
и процессов с участием биокатализаторов (клеток или фермен-
тов). Эти проблемы рассматриваются ими главным образом
с точки зрения взаимного влияния биологических и биохимиче-
ских процессов, с одной стороны, и явлений переноса и кон-
струкций биореакторов — с другой. Они сравнительно подробно
останавливаются на особенностях иммобилизованных фермен-
тов и клеток и их использовании в биотехнологии, вопросах
стехиометрии и энергетики биотехнологических процессов, яв-
лениях массо- и теплопередачи, конструкциях реакторов, типах
и принципах работы контрольно-измерительной и регулирую-
щей аппаратуры, методах выделения и очистки продуктов био-
процессов, экономических аспектах биотехнологии, проблемах
биологической очистки сточных вод.
Поставленная авторами задача настолько широка и много-
гранна, что, естественно, ряд проблем и тем освещены ими
в какой-то степени фрагментарно. Этот недостаток частично
компенсируют включенные в текст примеры конкретных про-
цессов или расчетов, а также многочисленные упражнения и
задачи, приведенные в конце каждой главы. Упражнения по-
добраны таким образом, чтобы читатель мог не только прове-
рить степень усвоения материала, но и расширить приобре-
тенные знания, поскольку решение задач часто требует озна-
комления с дополнительной литературой, выполнения расчетов,
а иногда и экспериментальных работ.
* Что касается литературы, изданной в СССР, то по широте и глубине
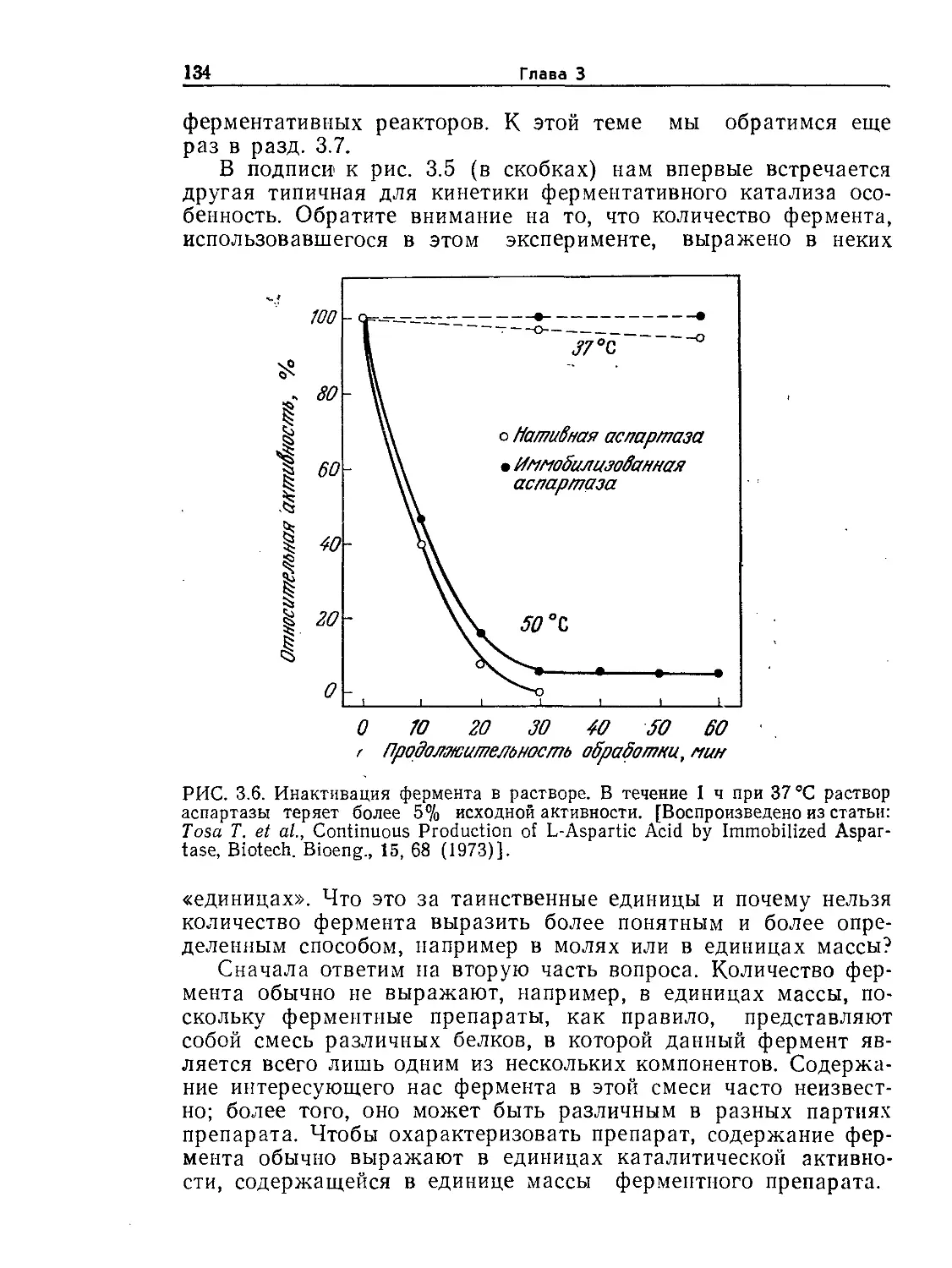
охвата основных проблем биотехнологии (особенно ее практических аспектов)
с настоящей книгой может конкурировать, пожалуй, только одна монография:
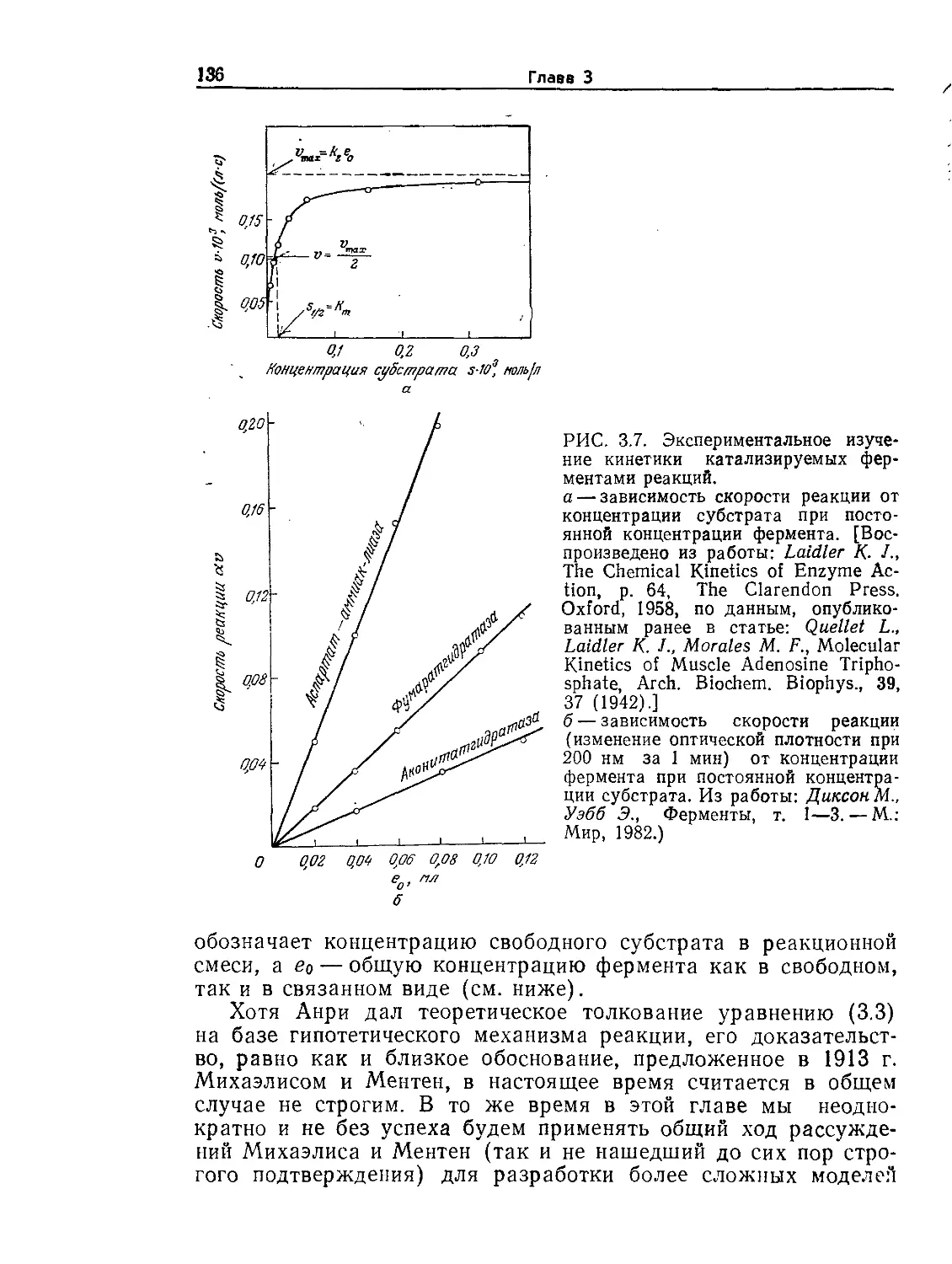
Аиба Ш., Хемфри А., Миллис Н., Биохимическая технология и аппаратура. —
М.: Пищевая промышленность, 1975. Последняя, однако, уже в известной сте-
пени устарела и, будучи издана небольшим тиражом, давно стала библио-
графической редкостью.
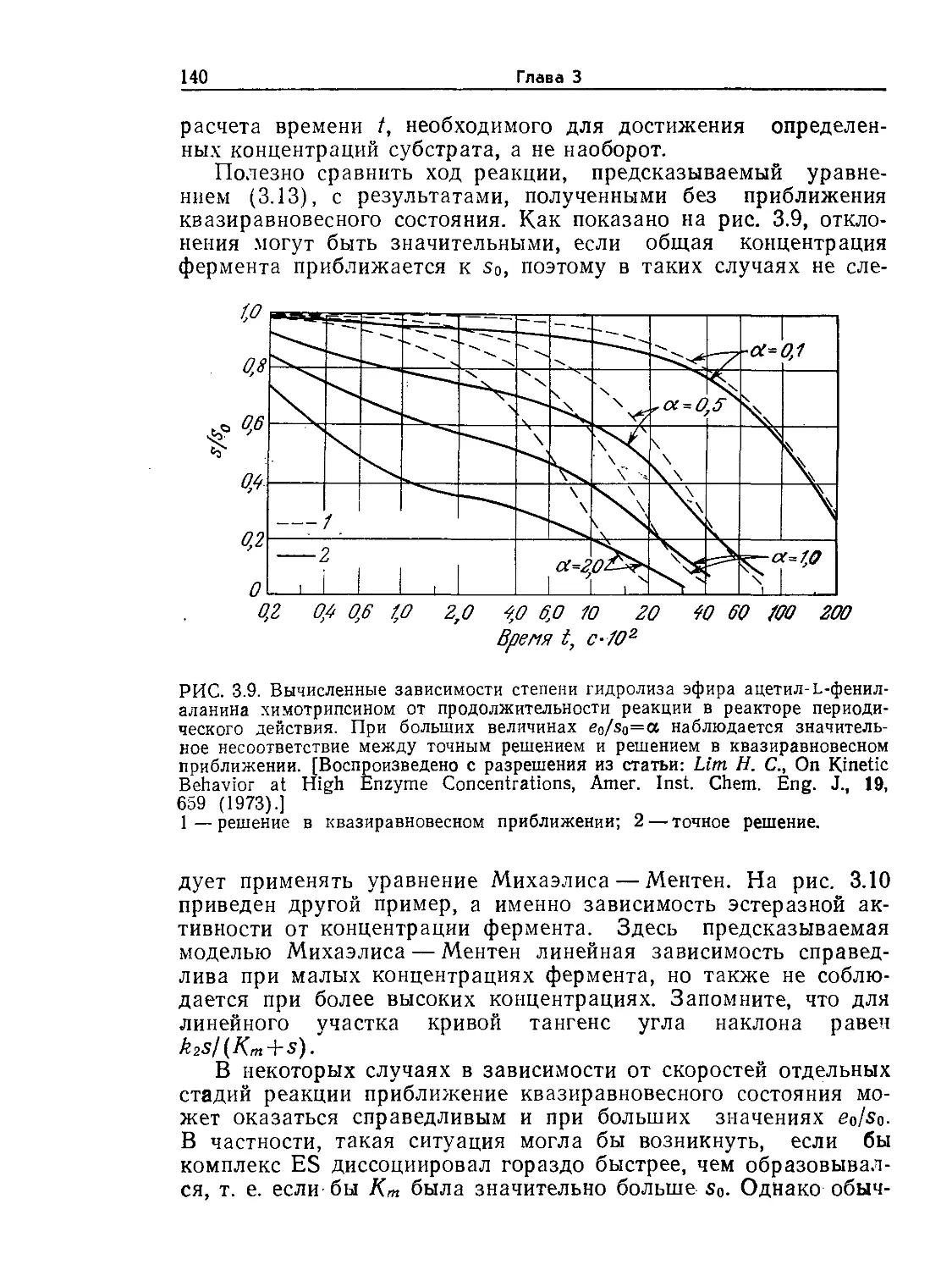
В 1988 г. выпущена еще одна книга, написанная коллективом авторов
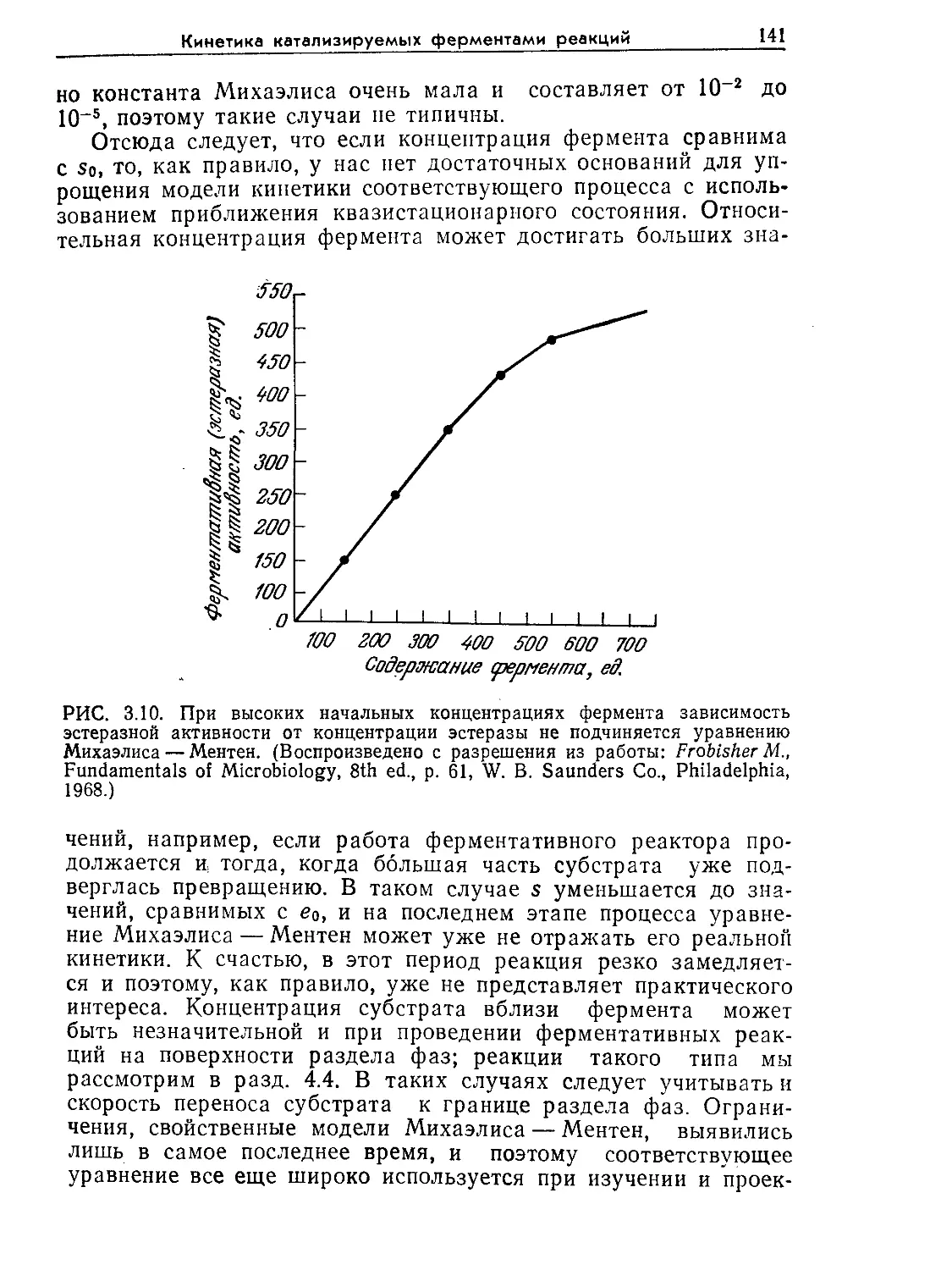
(Англия, США, Швейцария), представляющая собой учебник по биотехноло-
гии, где особое внимание уделено связи биотехнологии и химической техноло-
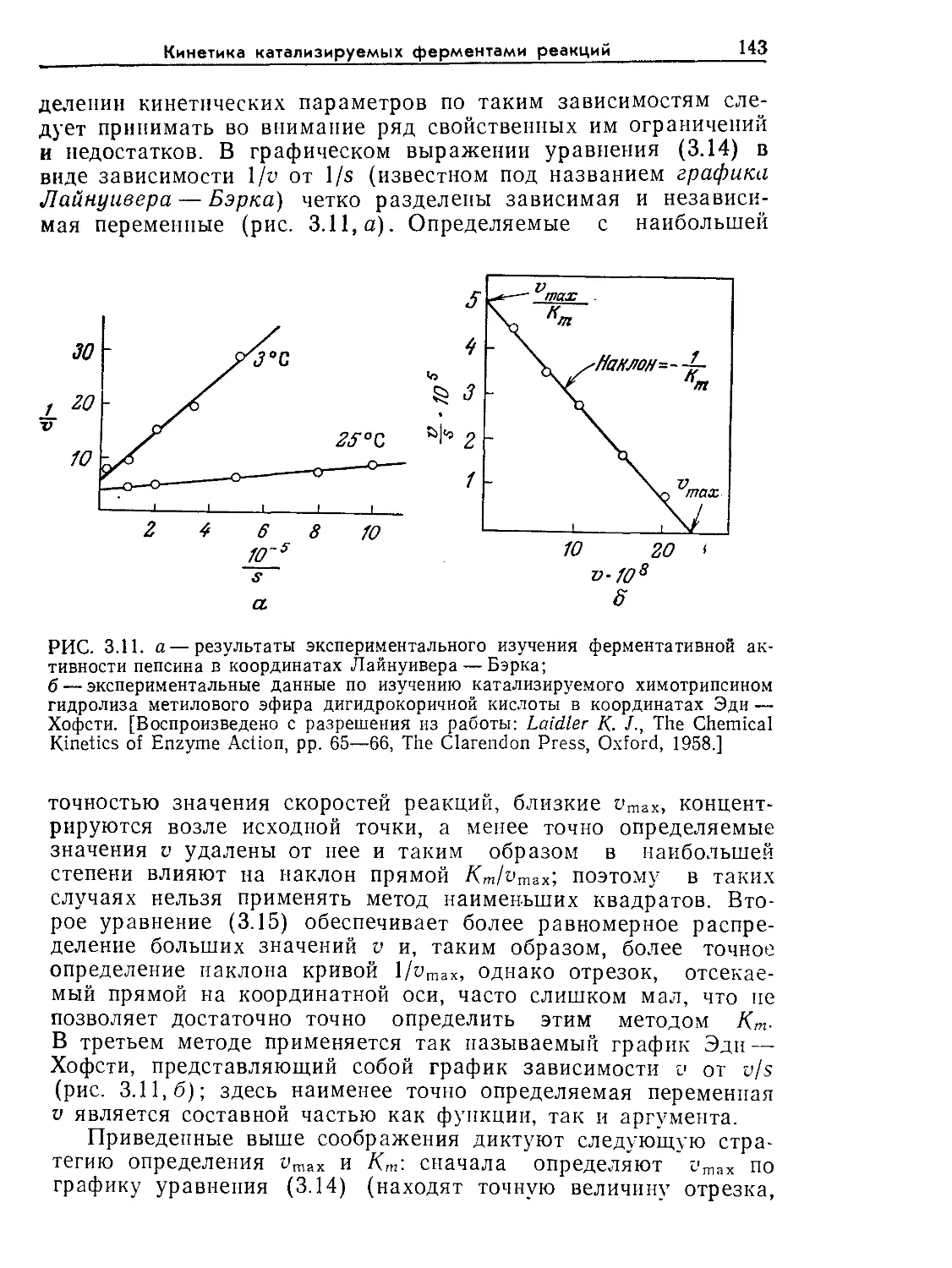
гии: Биотехнология. Принципы и применение: Пер. с англ./Под ред. И. Хиг-
гинса, Д. Беста, Дж. Джонса. — М.: Мир, 1988.
От переводчика
7
Книга Бейли и Оллиса представит большой интерес для
очень широкого круга читателей. В первую очередь она пред-
назначена для студентов и аспирантов, специализирующихся
в области биохимической технологии. Она полезна и для ин-
женерно-технических работников предприятий химической, мик-
робиологической, медицинской и фармацевтической промыш-
ленности, для конструкторов и технологов проектных организа-
ций соответствующего профиля. И наконец, этой книгой можно
пользоваться и при самостоятельном изучении предмета, по-
скольку анализу любых проблем собственно биотехнологии
в ней всегда предшествуют главы или разделы, в которых на
очень доступном и в то же время вполне современном уровне
изложены основы микробиологии, биологии и биохимии клетки,
ферментативного катализа и молекулярной кинетики.
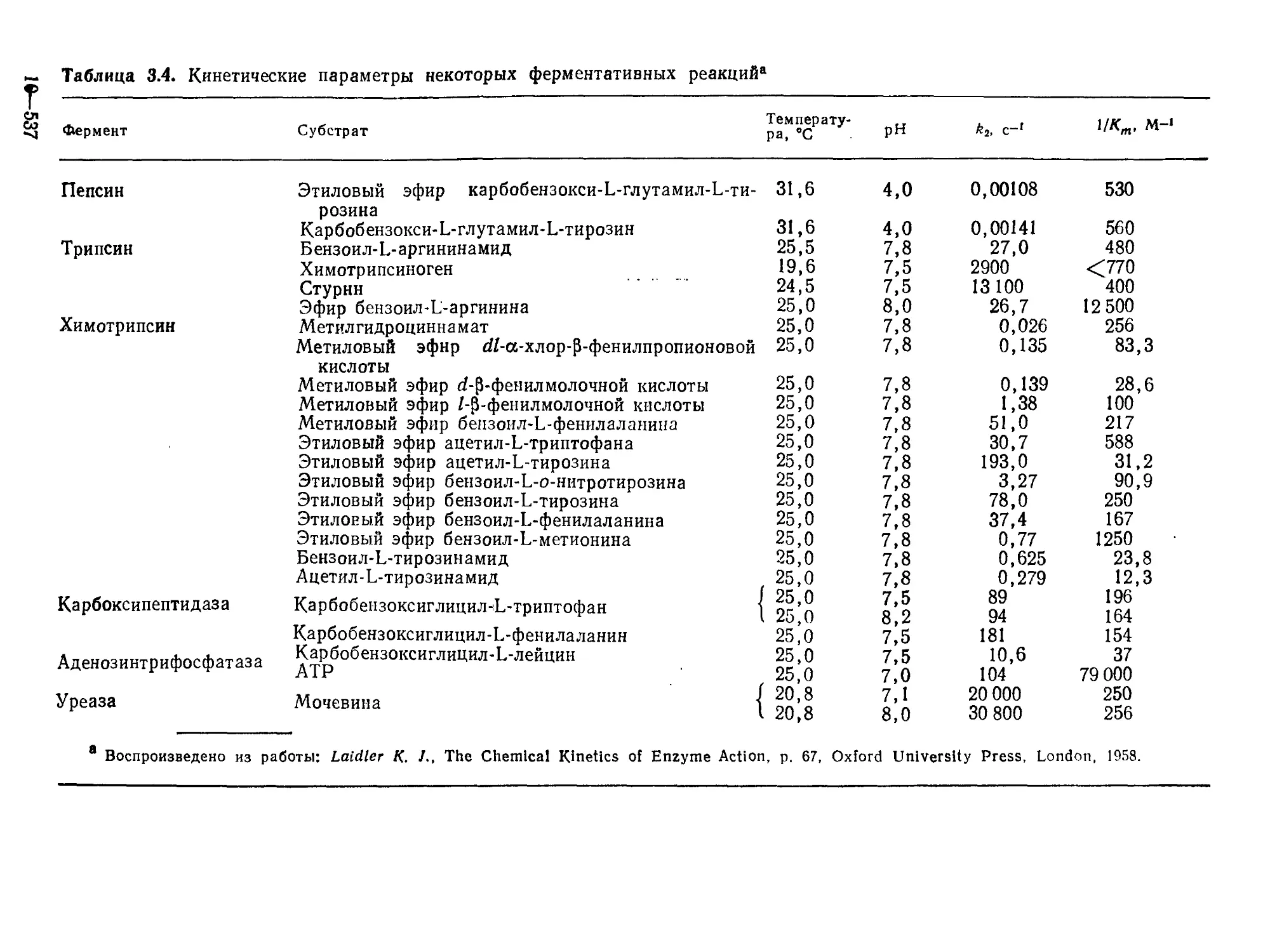
А. Кирюшкин
Предисловие авторов
Биохимическая технология имеет дело в основном с процес-
сами, связанными с переработкой биологических материалов
и с использованием биологических агентов, например клеток,
ферментов или антител. Для успешного решения задач био-
химической технологии необходимо владеть знаниями как основ
биологии, так и методологией и стратегией химической техно-
логии. Решение современных проблем биотехнологии базирует-
ся на использовании новейших данных и самой передовой тех-
нологии из обеих областей, и на этой основе разрабатываются
новые биотехнологические процессы, способы их контроля и
управления и намечаются пути их дальнейшего развития и оп-
тимизации. Для того чтобы стать хорошим специалистом в этой
области, нужны многолетние тщательное теоретическое изуче-
ние предмета и практическая работа.
Главная задача настоящего учебного пособия заключается
в том, чтобы помочь читателю сделать первые шаги на этом
чрезвычайно интересном и трудном пути. При этом все основ-
ные теоретические концепции рассматриваются с точки зрения
их практического применения в биохимической технологии.
В книге излагаются также основы способов проведения реак-
ций и процессов разделения, используемых в настоящее время
в биотехнологии. Особое внимание уделяется той центральной
роли, которую играют биологические свойства систем в дости-
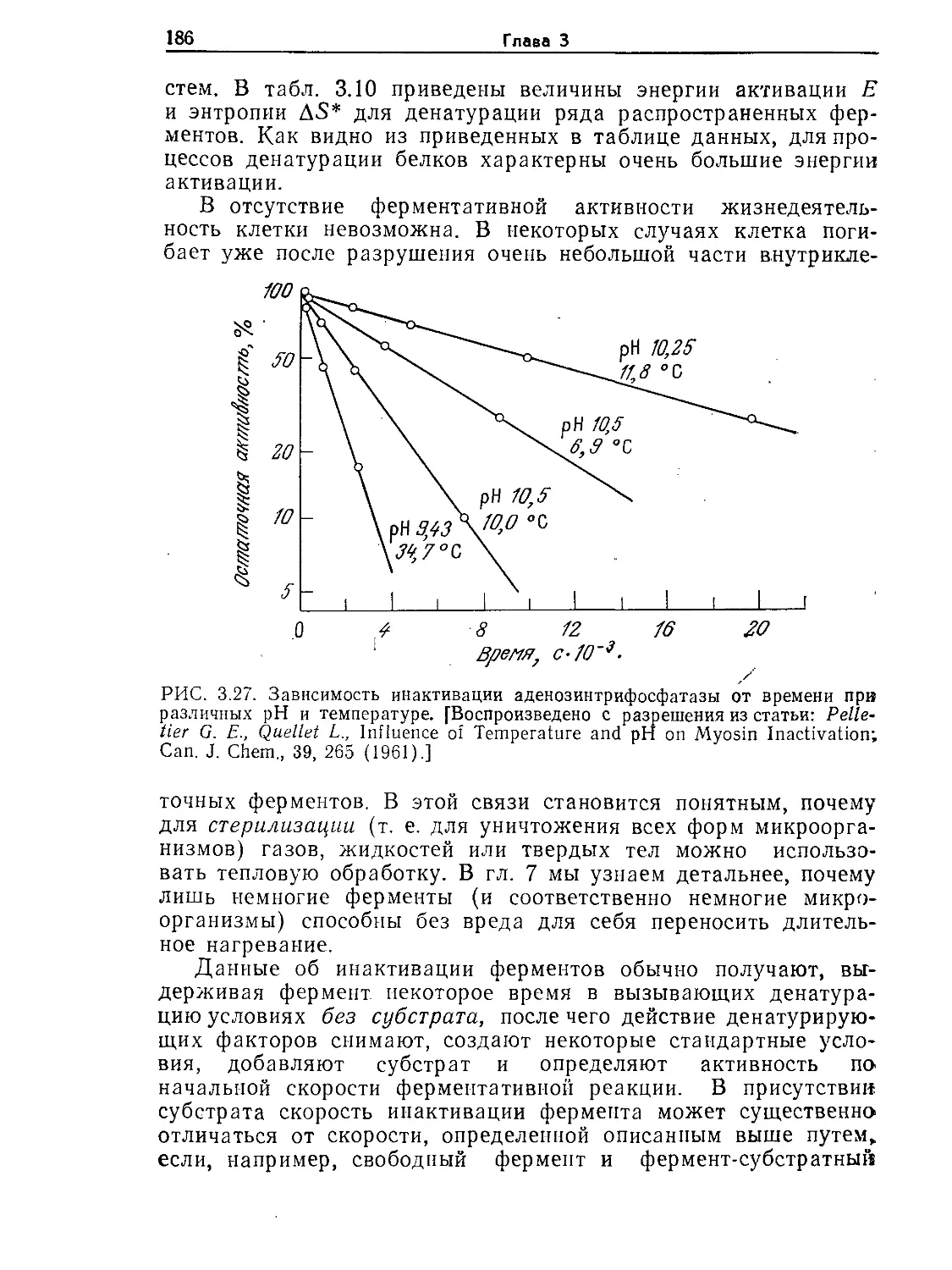
жении цели данного процесса оптимальным путем. Мы также
остановимся на различных осложнениях и ограничениях, обу-
словленных чувствительностью и неустойчивостью биологиче-
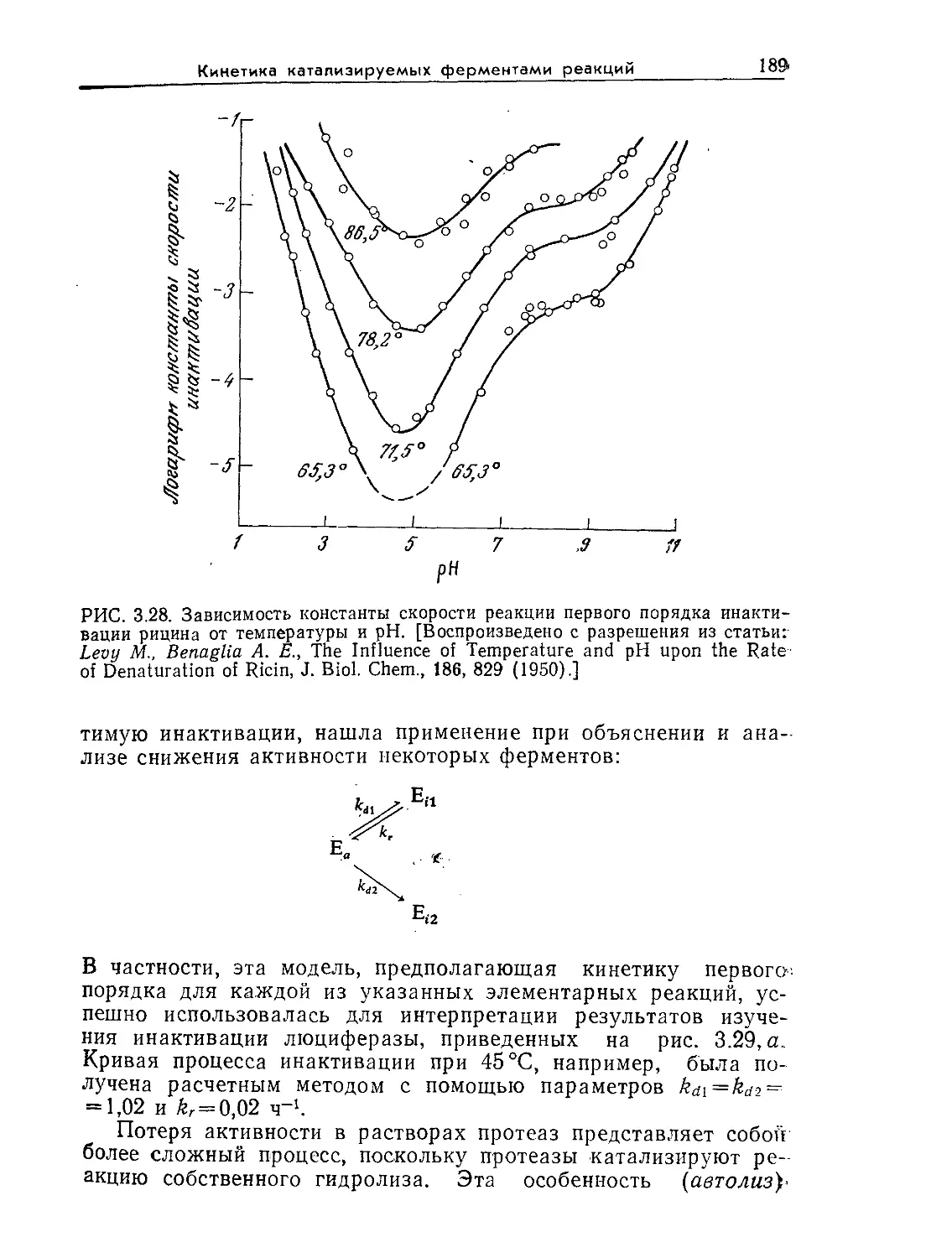
ских компонентов систем. Концентрируя внимание на фунда-
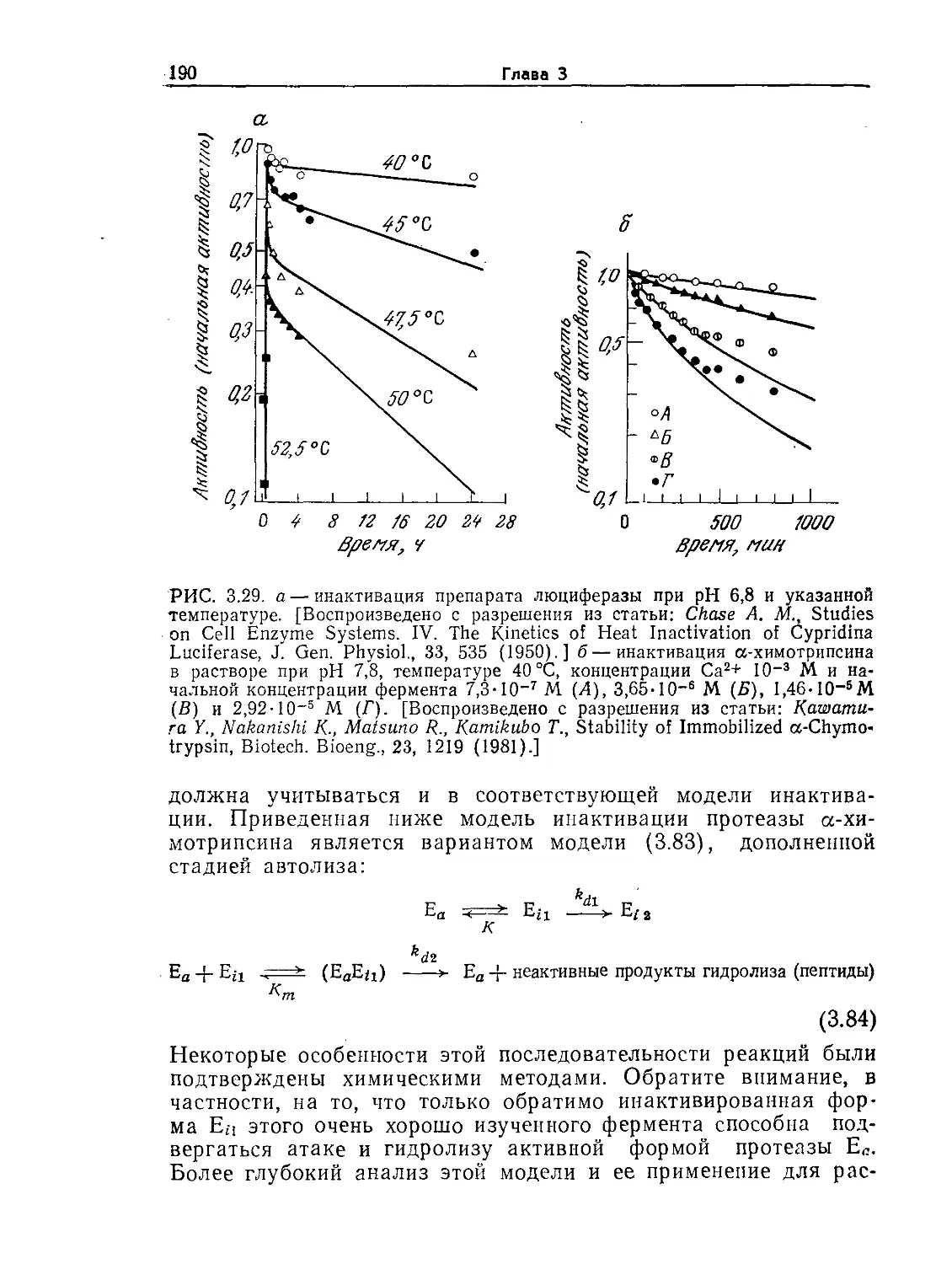
ментальных принципах биологических наук и технологии и
постоянно подчеркивая необходимость их комплексного при-
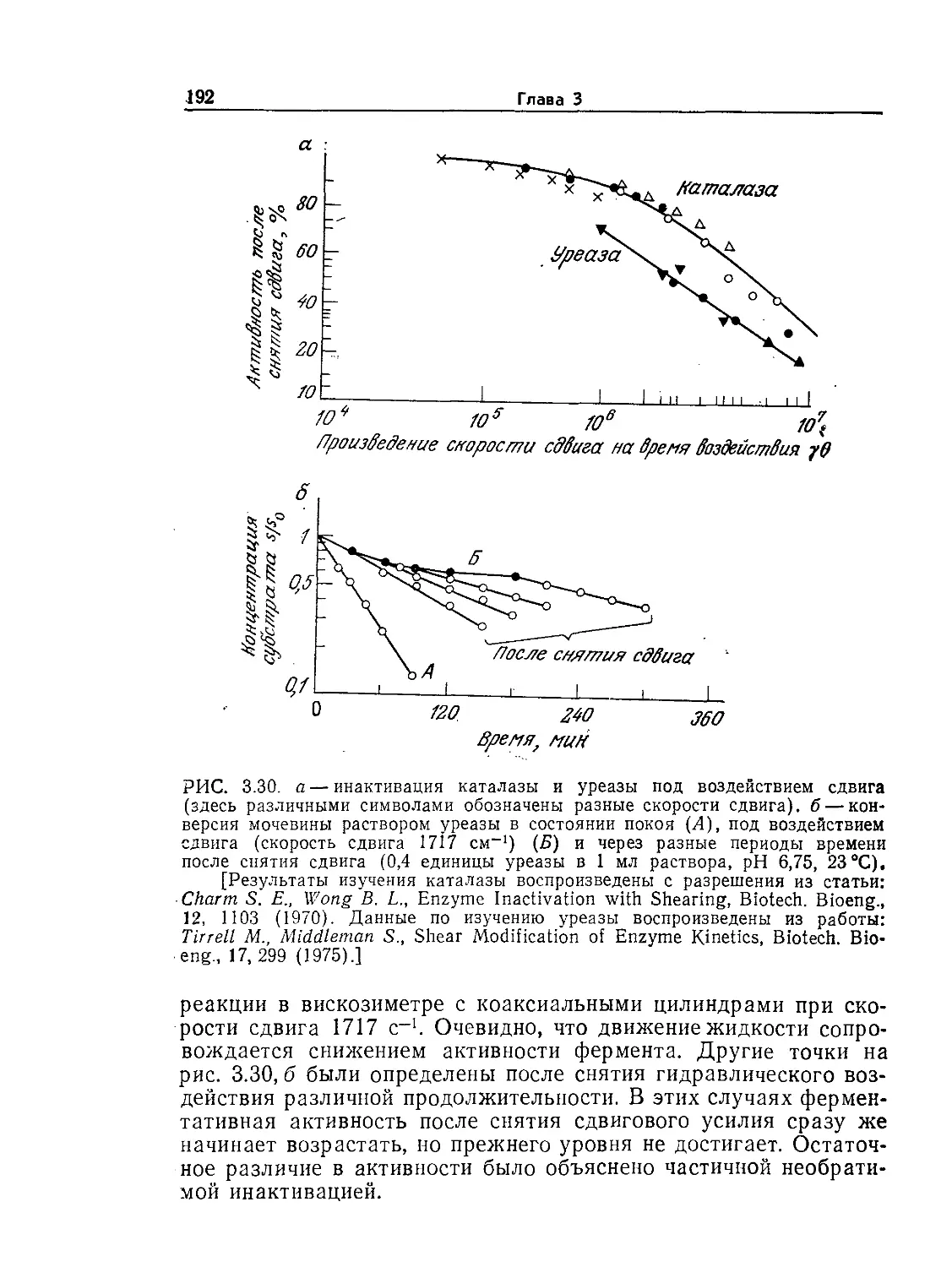
менения, мы стремились в первую очередь помочь читателю
создать достаточно прочную основу для дальнейшего более
тщательного их изучения и практической работы. Однако мно-
голетняя успешная деятельность в такой чрезвычайно быстро
развивающейся области, как биотехнология, возможна только
в том случае, если специалист следит за всеми новейшими ре-
зультатами фундаментальных исследований в биологии.
Предисловие авторов
9
Настоящая книга представляет собой вводный курс по био-
химической технологии, изучаемой студентами старших курсов
и аспирантами, специализирующимися в области химической
технологии. В то же время отдельные разделы и части книги
могут послужить основой для других курсов по химической
технологии, технологии пищевых продуктов и ряду других от-
раслей промышленности, в том числе связанных с охраной
окружающей среды. Как и в первом издании, весь материал
изложен систематично, в строгой логической последовательно-
сти, начиная с самых общих принципов биологии. Поэтому
книгой можно пользоваться и при самостоятельном изучении
предмета, например работникам промышленности.
Чтобы сделать ее более доступно?! для самостоятельного
изучения и создать необходимую базу для одно- или полуго-
дичного факультативного или вводного курса, в текст в качестве
самостоятельных разделов включены основные положения био-
химии, биологии клетки, кинетики ферментативного катализа
и молекулярной генетики. Само собой разумеется, что в таких
разделах дается только введение в эти отрасли науки и даже
в совокупности они ни в коей мере не претендуют на полноту
изложения основ наук о жизни в той мере, в какой это необ-
ходимо для тех, кто будет глубоко изучать биохимическую
технологию или работать в этой области. Последним, очевидно,
необходимо более глубокое изучение основ биологии и методов
практической работы в индивидуальном порядке или по соот-
ветствующим программам высших учебных заведений.
В ходе изложения основ биологических наук мы старались
постоянно иллюстрировать их примерами технологических про-
цессов и методов анализа. На связь основ биологических наук
с биохимической технологией часто указывается уже в разде-
лах, посвященных основным концепциям биологии, а разработ-
ка конкретных инженерных решений рассматривается только
после изучения соответствующего теоретического материала.
Так, вопросы кинетики ферментативного катализа и технологии
соответствующих процессов изложены непосредственно после
описания белков и других биологически важных соединений,
а кинетика роста клеток рассматривается вслед за описанием
стехиометрии и регуляции путей метаболизма.
^Примеры, приведенные в тексте, и упражнения в конце каж-
дой главы дают читателю возможность самому проверить прак-
тическую применимость основных концепций и расширить пред-
ставление об изучаемом предмете. Решение этих 150 упражне-
ний различной степени сложности потребует от читателя
необходимых обсуждений, самостоятельной работы с литера-
турой или расчетов.
Основной причиной выпуска в свет второго издания настоя-
ю
Предисловие авторов
щей книги послужил скачкообразный прогресс в биологических
науках, революционизировавший наши представления о воз-
можностях трансформации организмов и материалов и созда-
нии на этой основе новых веществ и процессов. Технология
рекомбинантных ДНК и гибридом послужила базой для созда-
ния совершенно новой биотехнологической промышленности.
В текст книги включены новые данные о методах клонирования
и экспрессии генов, слияния клеток, а также об основных путях
развития перспективных технологических процессов крупно-
масштабного производства сверхчистых веществ белковой при-
роды.
Во втором издании большее внимание уделено ряду проблем
технологии: например, включены новые главы, посвященные
процессам разделения, приборного обеспечения и контроля био-
технологических процессов и их экономической эффективности,
а также впервые освещены также другие важные темы, в том
числе стехиометрия метаболических превращений, конструкция
многофазных реакторов, технология процессов с участием жи-
вотных и растительных клеток.
Кроме того, в процессе подготовки второго издания в текст
были внесены многочисленные изменения, позволившие улуч-
шить систематизацию и изложение материала. В частности,
был сокращен, но тем не менее стал более информативным
основной описательный материал, а также более системно рас-
смотрены вопросы стехиометрии, кинетики и конструкции био-
реакторов. Во втором издании также более подробно изложены
важные проблемы коалесценции и диспергирования в много-
фазных реакторах.
Много ценных улучшений во второе издание было внесено
благодаря обоснованным критическим замечаниям М. Шюлера,
Д. Лауффенбергера, П. Рейлли, Ф. Арнолд, Д. Кирвана и
Э. Гэйдена. В настоящем издании учтены также отдельные
предложения, связанные с проблемами освещения фундамен-
тальных исследований и (или) новыми упражнениями, которые
внесены нашими коллегами, студентами и уже закончившими
обучение специалистами.
Мы, наконец, хотели бы выразить нашу искреннюю призна-
тельность многочисленным друзьям, коллегам, студентам и ру-
ководителям, стимулировавшим наше становление как специа-
листов в биохимической технологии за годы, прошедшие с пер-
вого издания. Во многих отношениях они также являются
соавторами этой книги.
Джеймс Э. Бейли
Дэвид Ф. Оллис
Глава 1
Введение в микробиологию
Жизнь человека связана многочисленными нитями с кро-
хотными живыми существами, называемыми микроорганизма-
ми. В масштабе биосферы в целом, включающей все регионы
планеты, где существует жизнь, микроорганизмы играют важ-
нейшую роль в усвоении солнечной энергии. Биологическая
деятельность микроорганизмов является одним из основных
этапов в кругооборотах углерода, кислорода, азота и других
необходимых для жизни элементов. С другой стороны, микро-
организмы вызывают различные заболевания человека, живот-
ных и растений.
В настоящей книге основное внимание будет уделено ис-
пользованию микроорганизмов человеком. Эти универсальные
биологические катализаторы служат человечеству уже многие
тысячи лет. Древние греки приписывали богу Дионису изобре-
тение процесса брожения в виноделии, а на «Голубом мону-
менте», датируемом седьмым тысячелетием до нашей эры, изо-
бражен процесс пивоварения в Вавилоне. В питании человека
уже давно большую роль играют процессы брожения, с по-
мощью которых получают, например, сыр, хлеб, йогурт и сое-
вый соус. В конце прошлого века Пастер и Тиндалл показали,
что любые процессы брожения инициируются микроорганизма-
ми; тем самым было положено начало микробиологии как нау-
ки. Эти работы стимулировали дальнейшие исследования, и уже
в начале двадцатого века Бухнер, Нейберг и Вайцманн разра-
ботали технологические схемы производства этанола, глицерина
и других химикатов.
В сороковых годах нашего столетия успехи биологии, гене-
тики микроорганизмов и технологии ознаменовали начало эры
антибиотиков, позволивших избавить человечество от ряда за-
болеваний и существенно повысить среднюю продолжитель-
ность жизни. В этот период и родилась биохимическая техноло-
гия, т. е. технология, в которой используются катализаторы,
исходные вещества и (или) сорбенты биологического проис-
хождения. Началось превращение биотехнологии из эмпириче-
ского искусства в область предсказуемой, поддающейся плани-
рованию и оптимизации технологии.
12
Глава 1
На следующем этапе развития биохимической технологии
были разработаны способы получения стероидов, предназна-
ченных для контроля рождаемости, а также для лечения артри-
та и воспалительных процессов. Применение методов культиви-
рования растительных и животных клеток позволило наладить
массовое производство вакцин и других ценных биологических
препаратов. Не вызывает сомнения тот факт, что успешное
использование и целенаправленное изменение клеточных про-
цессов человеком оказало огромное воздействие на различные
стороны жизни прошлых и живущих ныне поколений, в том
числе на проблемы здравоохранения, экономики, охраны окру-
жающей среды и на социальные вопросы.
Результатом комплексных работ в области молекулярной
биологии и генетики микроорганизмов явилось понимание основ
различных способов контроля и катализа, лежащих в основе
биосинтетических процессов в живых клетках. Эти теоретиче-
ские работы послужили основой для разработки методов тех-
нологии рекомбинантных ДНК, возможности применения кото-
рых настолько велики, что с трудом поддаются воображению.
Таким путем уже производятся новые вакцины и лекарственные
препараты, но это только начало будущих поистине революци-
онных преобразований.
В изучении биохимической инженерии наша задача будет
заключаться в том, чтобы читатель смог понять суть биотехно-
логических процессов, научился их анализировать и на этой
основе разрабатывать и рационально использовать новые про-
цессы. Для этой цели, однако, необходимо овладеть минимумом
знаний о росте и функциях клетки. Эти факторы и ряд других
особенностей биологического характера обычно определяют весь
биотехнологический процесс. Представим себе, что живой мик-
роорганизм в самом первом приближении можно рассматривать
как саморасширяющийся химический реактор, который потреб-
ляет из среды определенные соединения, называемые питатель-
ными веществами, растет, самовоспроизводится и выделяет про-
дукты жизнедеятельности в окружающую среду. В таких слу-
чаях, как очистка сточных вод, цель технологического процесса
состоит в усвоении питательных веществ (в данном случае за-
грязняющих органических веществ). Если же микроорганизмы
выращивают в качестве источника пищи или компонента пи-
щевой смеси, то в результате процесса получают микробную
массу. Напротив, в процессе очистки сточных вод эта микроб-
ная масса, образующаяся в процессе усвоения питательных
веществ, оказывается нежелательным побочным продуктом, и ее
количество должно быть сведено к минимуму. Наконец, во мно-
гих естественных процессах и в промышленном производстве,
например при получении пенициллина или этанола, основной
Введение в микробиологию
13
интерес представляют продукты, образующиеся в результате
жизнедеятельности клеток и выделяемые ими в среду. На отно-
сительные скорости усвоения питательных веществ, роста клеток
и выделения продуктов жизнедеятельности в очень большой
степени влияет тип клеток, а также температура, состав и спо-
соб перемешивания среды. Для понимания этих взаимосвязей
необходимо знать основы биохимии, биофизики и биологии
клетки. Значительная часть настоящей книги будет посвящена
этим дисциплинам, поскольку их изучение обычно не включают
в программы подготовки инженеров и технологов.
Во всех случаях, когда это только представляется возмож-
ным, мы будем давать не только качественную оценку биоло-
гических процессов, но и приводить соответствующие матема-
тические выражения, характеризующие количественную сторону
этих процессов. Часто эти математические модели будут чрез-
мерно упрощенными и идеализированными, поскольку даже
единичный микроорганизм представляет собой очень сложную
систему. Тем не менее основные концепции микробиологии бу-
дут полезными для создания математических моделей и про-
верки их корректности так же, как, например, основные поло-
жения гидродинамики позволяют понять корреляцию между
коэффициентом трения и числом Рейнольдса.
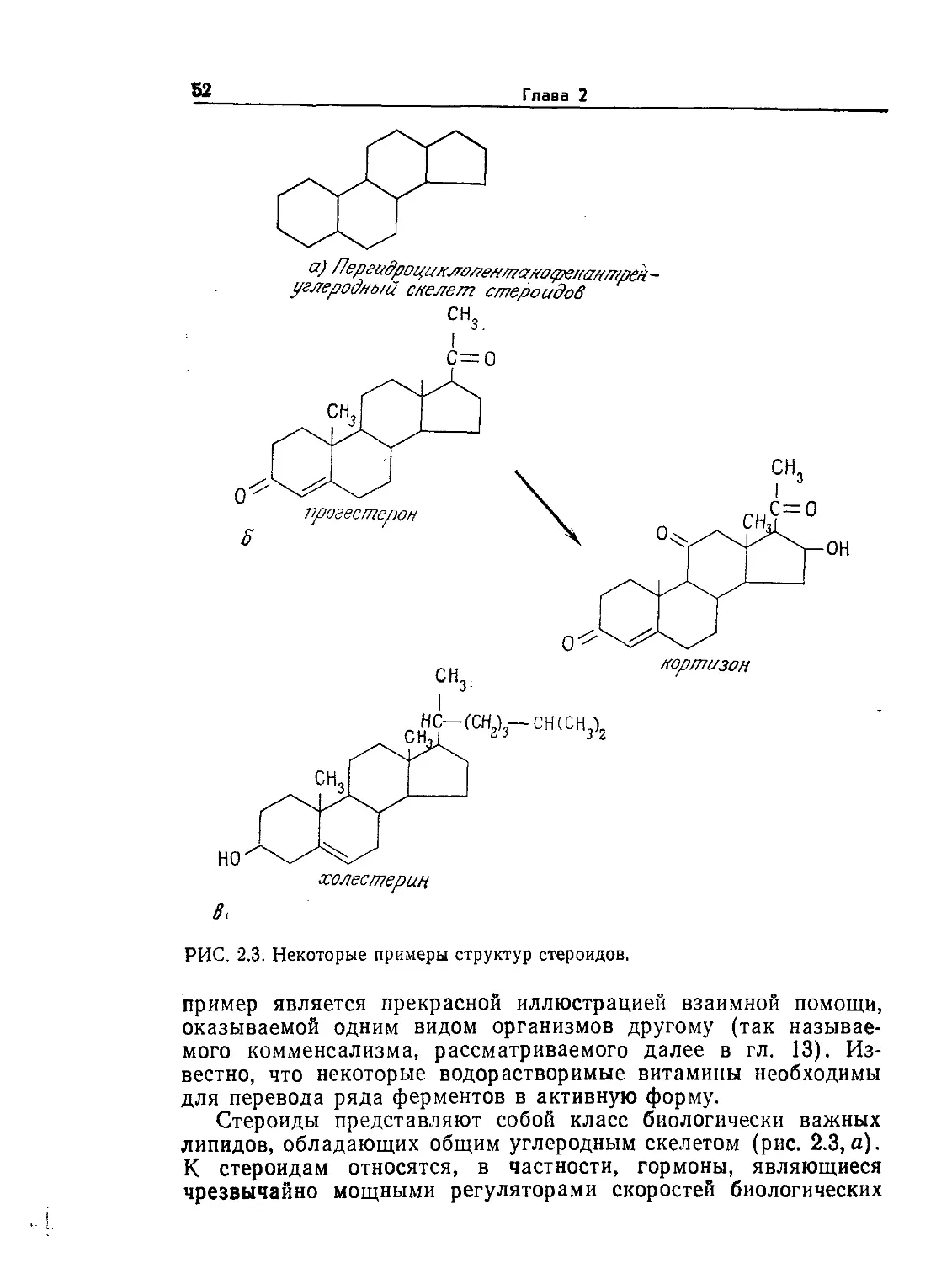
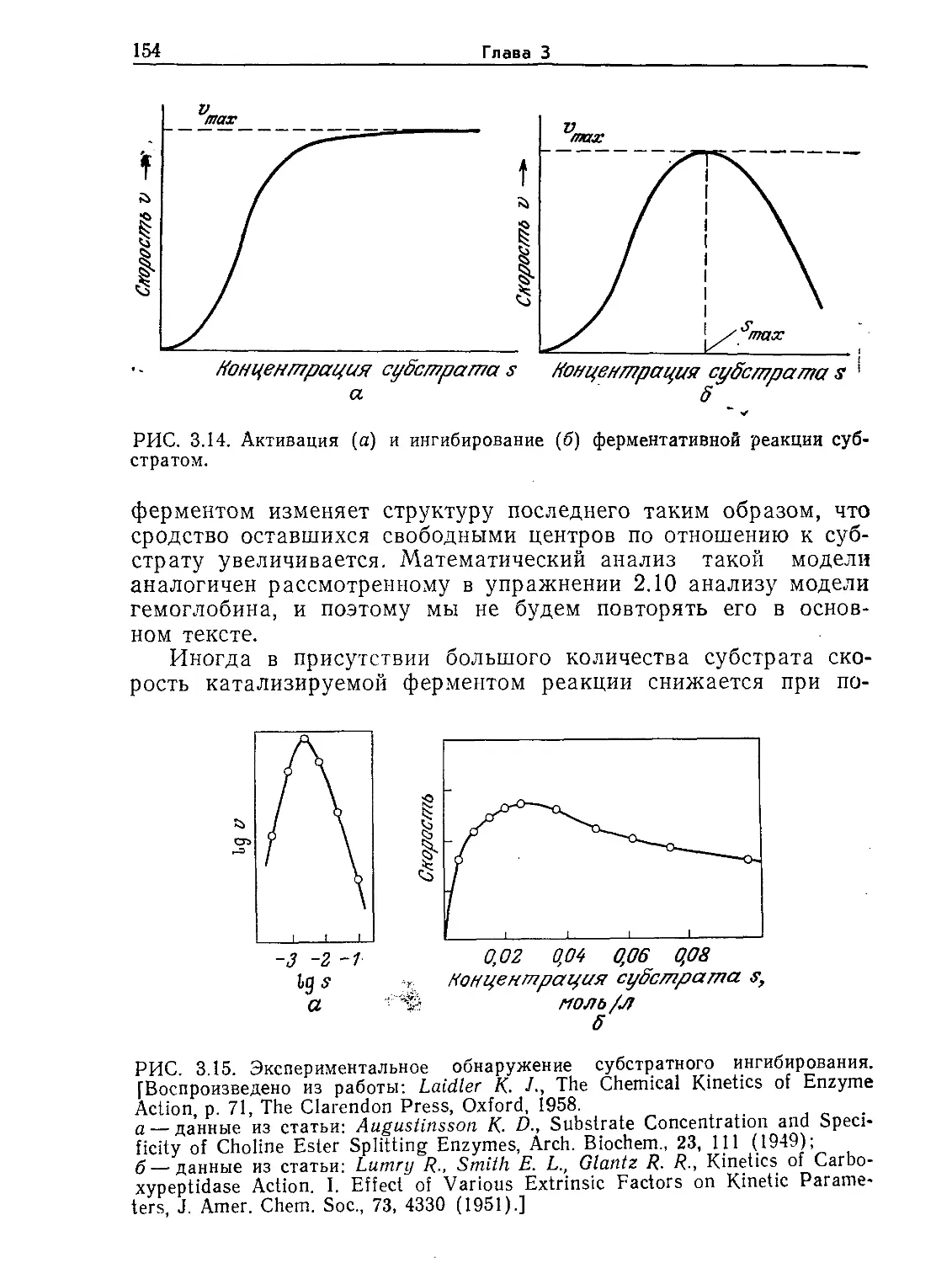
1.1. Биофизика и клеточная теория
Микробиология — это наука, изучающая живые организмы,
не видимые невооруженным взглядом. Как правило, диаметр
микроорганизмов не превышает 0,1 мм. В настоящее время мы
знаем, что даже простейшие микроорганизмы вмещают слож-
нейший комплекс удивительно эффективных и в высшей степе-
ни координированных химических реакторов, систем информа-
ции, контроля, управления и массопередачи. Эти данные были
получены в результате многочисленных экспериментальных ис-
следований с использованием методов, заимствованных из фи-
зических и химических наук. Поскольку такой подход оказался
чрезвычайно плодотворным, применимость основных положений
химии и физики к биологическим системам является в настоя-
щее время широко распространенной в комплексе наук о жизни
рабочей гипотезой. Для более четкого ограничения области
науки, в которой биологические проблемы решаются физиче-
скими методами, иногда употребляют термин «биофизика».
Поворотным пунктом в понимании живых систем можно
считать 1838 г., когда Шлейден и Шванн впервые сформули-
ровали клеточную теорию, постулирующую, что все живые си-
стемы состоят из клеток и продуктов их жизнедеятельности.
Таким образом была создана концепция основного модуля или
14
Глава 1
основного строительного элемента жизни. Эта концепция об
общем для всех живых организмов элементе структуры позво-
ляет разделить изучение живых систем на два этапа, на первом
из которых исследуют составляющие систему клетки, а затем
на этой основе стараются понять структуру всего организма.
Целесообразность такого разделения исследований базиру-
ется на том факте, что выделенные из самых разнообразных
организмов клетки имеют много общего как в отношении их
строения, так и в отношении выполняемых ими функций. По-
этому данные, полученные при экспериментах на клетках од-
ного организма, во многих случаях удается успешно перенести
на клетки других типов. Наличие общих для всех клеток осо-
бенностей упрощает и нашу задачу изучения поведения микро-
организмов. Сконцентрировав внимание на самых универсаль-
ных функциях клеток, можно создать основу для понимания
функций всех живых систем.
Нам не хотелось бы, однако, чтобы у читателя создалось
впечатление, будто бы все клетки одинаковы. Напротив, клетки
мышц, например, резко отличаются от клеток глаза или мозга.
Точно так же существует множество типов одноклеточных ор-
ганизмов, которые в соответствии с описанными ниже особен-
ностями их организации могут быть разделены на две основные
группы.
1.2. Строение клеток
С помощью электронного микроскопа было установлено, что
существуют два существенно различающихся типа клеток. Хотя
им свойственны и некоторые общие черты, все же клетки одного
типа настолько отличаются от клеток другого типа по структуре
и функциям, что целесообразно рассматривать их раздельно.
Все известные в настоящее время клетки относятся к первому
или второму типу.
1.2.1. Прокариотические клетки
Прокариотические клетки не имеют заключенного в мембра-
ну ядра. Прокариоты отличаются относительно небольшими
размерами и простотой строения. Обычно они существуют изо-
лированно, вне связи с другими клетками. Линейные размеры
этих клеток, которые могут иметь сферическую, палочкообраз-
ную или спиральную форму, как правило, составляют от 0,5
до 3 мкм*. Чтобы представить себе эти размеры, полезно срав-
* 1 м (метр) = 103 мм (миллиметров) = 106 мкм (микрометров; ранее так-
же употреблялся термин микрон) = 109 нм (нанометров) = 1010 А (ангстремов).
В в едение в микробиологию
15
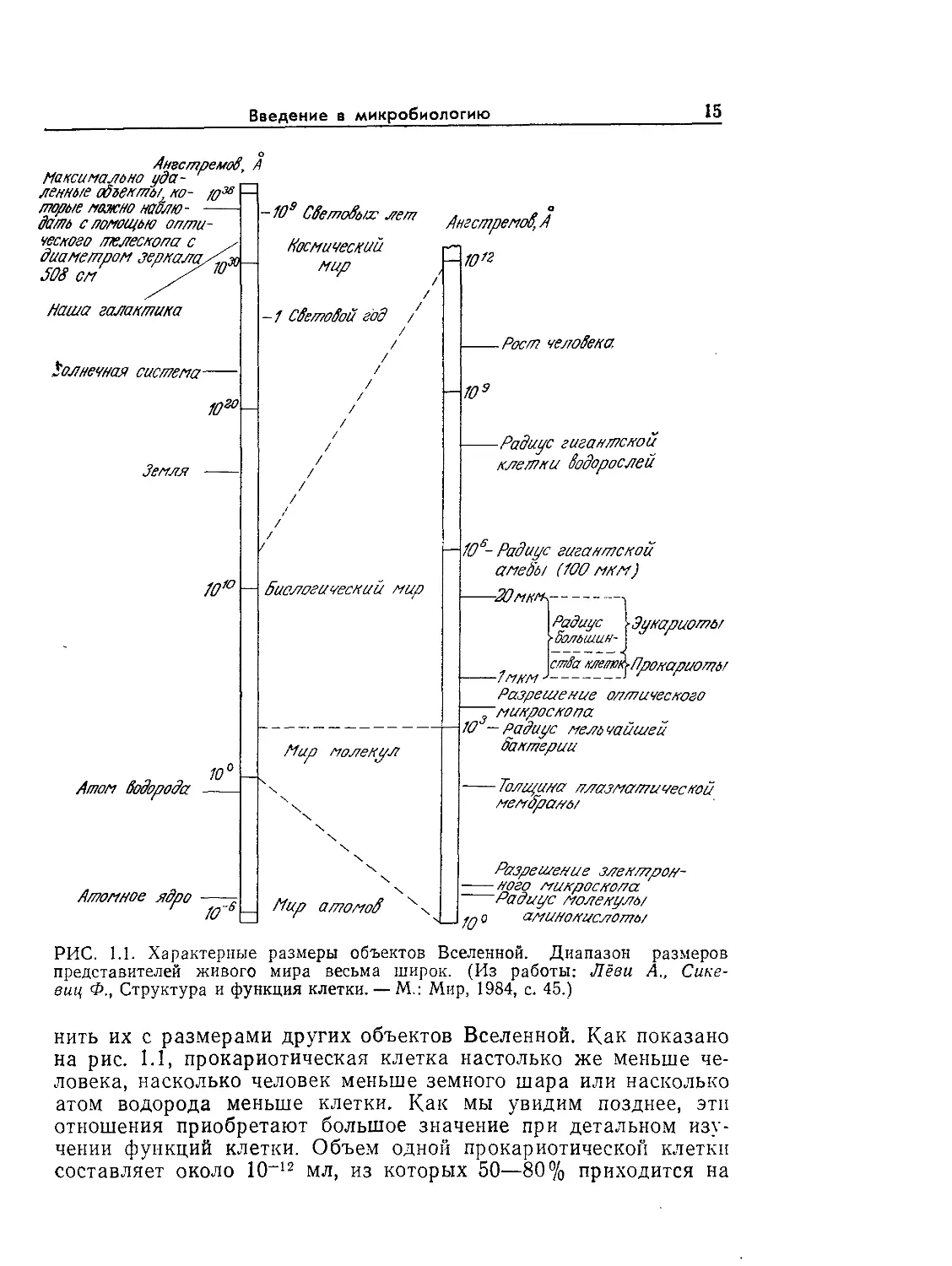
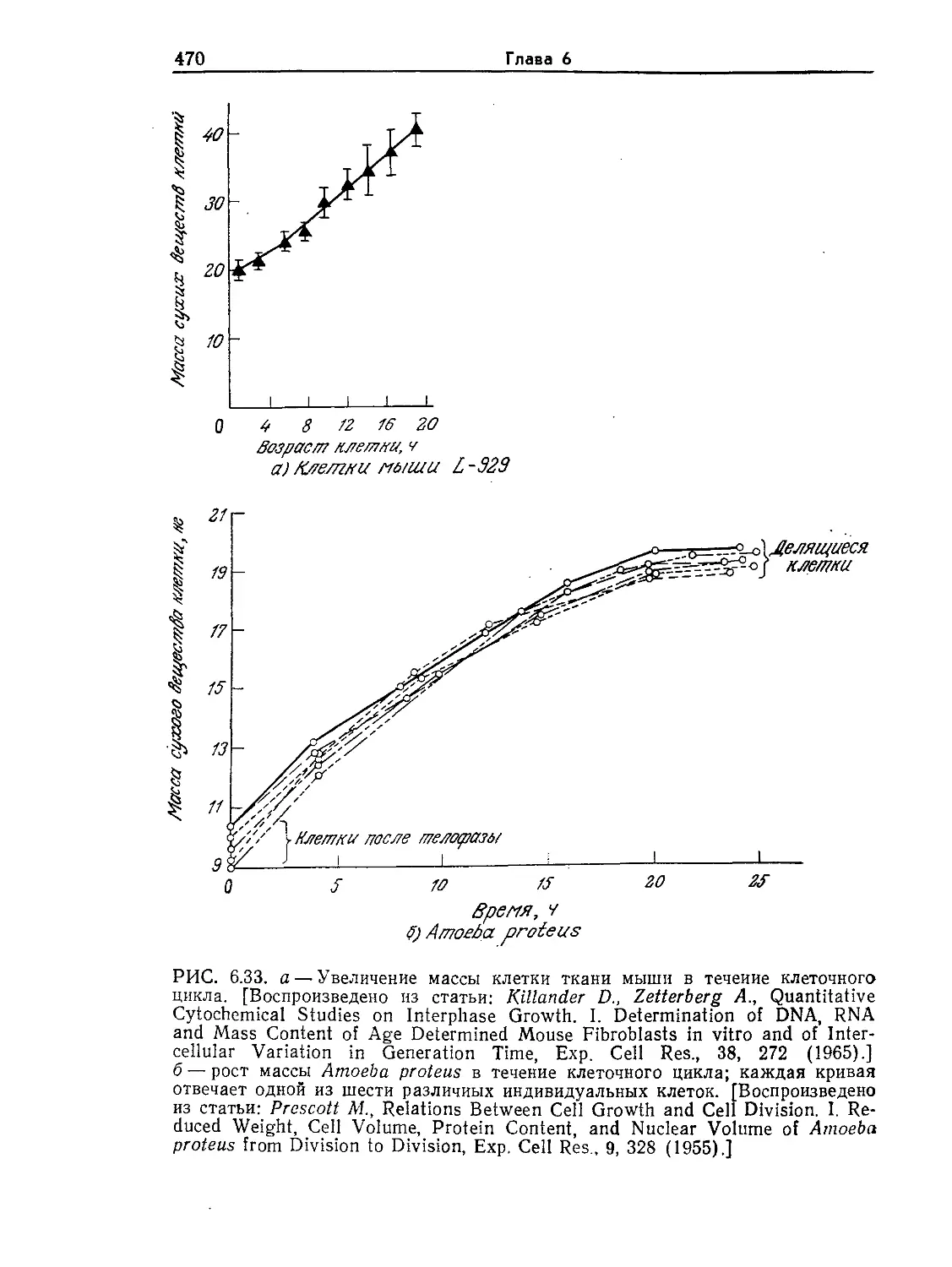
о
Ангстремов, Л
Максимально уда-
ленные объекты, ко- ю36
торые можно наилю- —
дать с помощью опти-
-109 Световых лет
веского телескопа с
диаметром зеркалалйкзо—
603 см / /z? II
Маша галактика
Солнечная система-----
10го
Земля ——
/0ю
10° -
Атом водорода ------
Атомное ядро------
г 10"в
Космический
мир
-7 Световой год
биологический мир
Мир молекрл
Мир атомов
Ангстремов, А
10™
----Рост человека.
Ю9
----Радиус гигантской
клетки водорослей
10 s- Радиус гигантской
амевы (100 мкм)
----20мкм.---------
Радиус
> большин-
ства теток
----1мкм ----------
Эукариоты
Прокариоты
Разрешение оптического
~микроскопа
-10 —радиус мельчайшей
бактерии
---Толщина плазматической
мембраны
Разрешение злектрон-
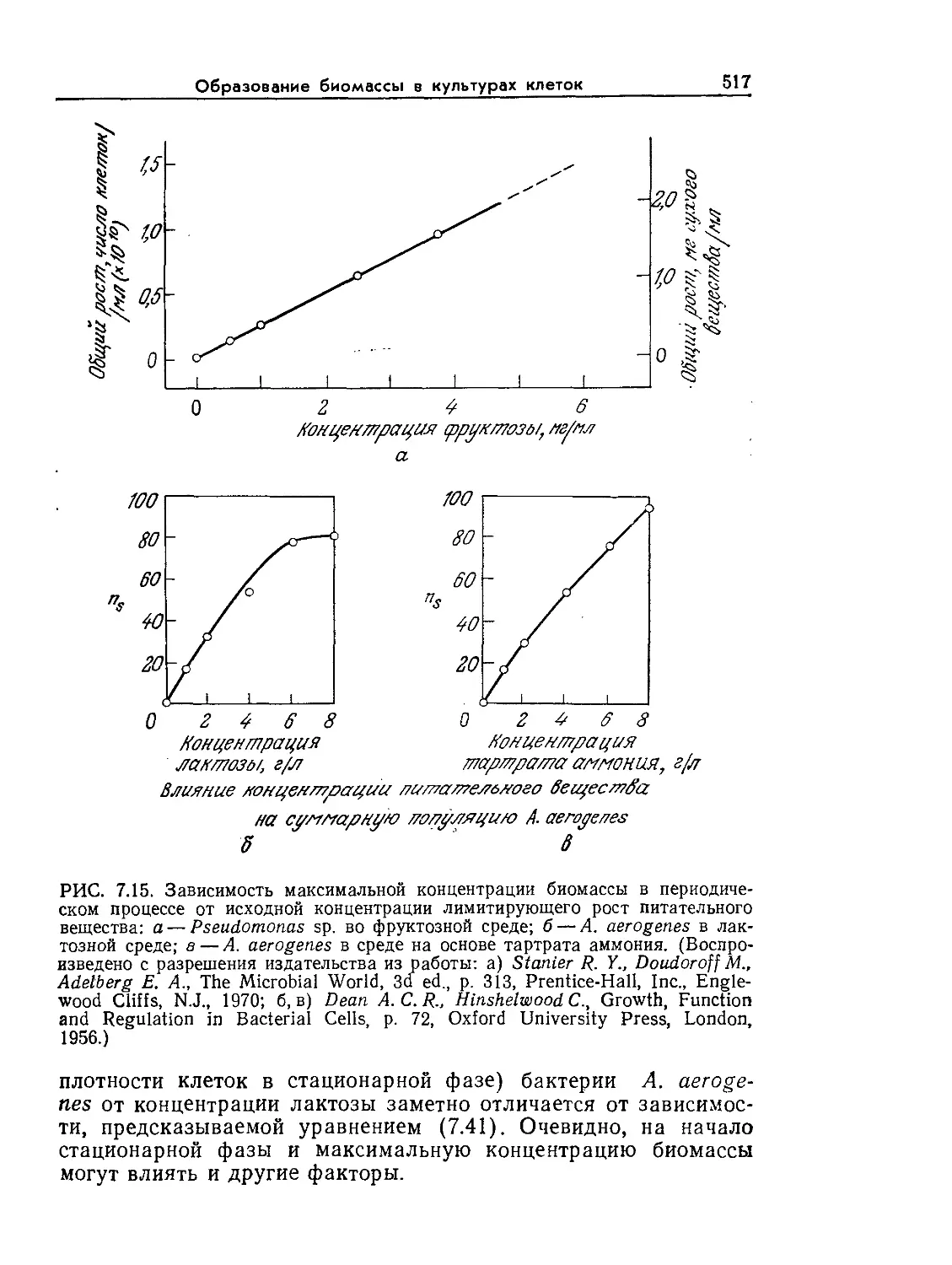
---кого микроскопа
Радиус молекулы
J_I jq о аминокислоты
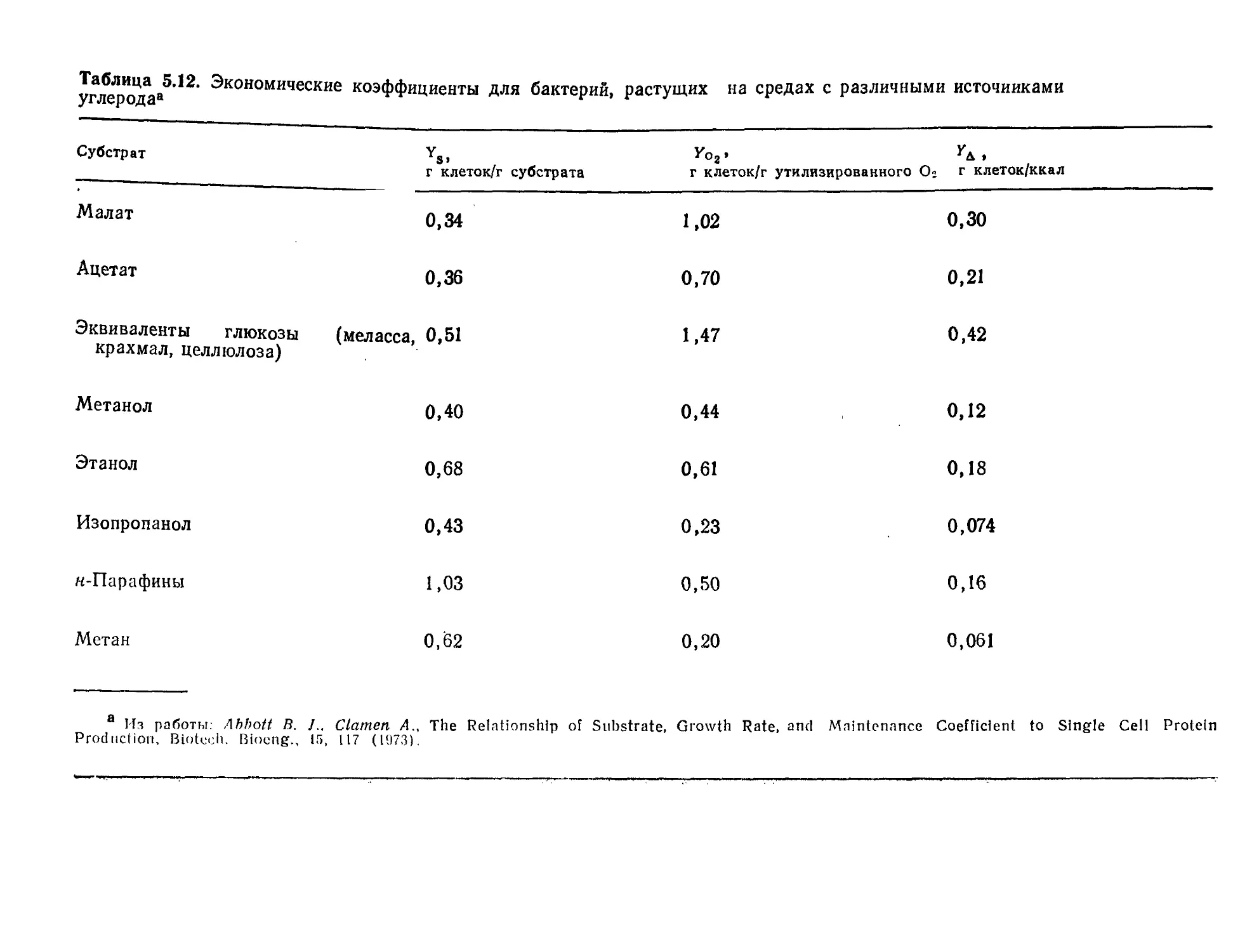
РИС. I.I. Характерные размеры объектов Вселенной. Диапазон размеров
представителей живого мира весьма широк. (Из работы: Лёви А., Сике-
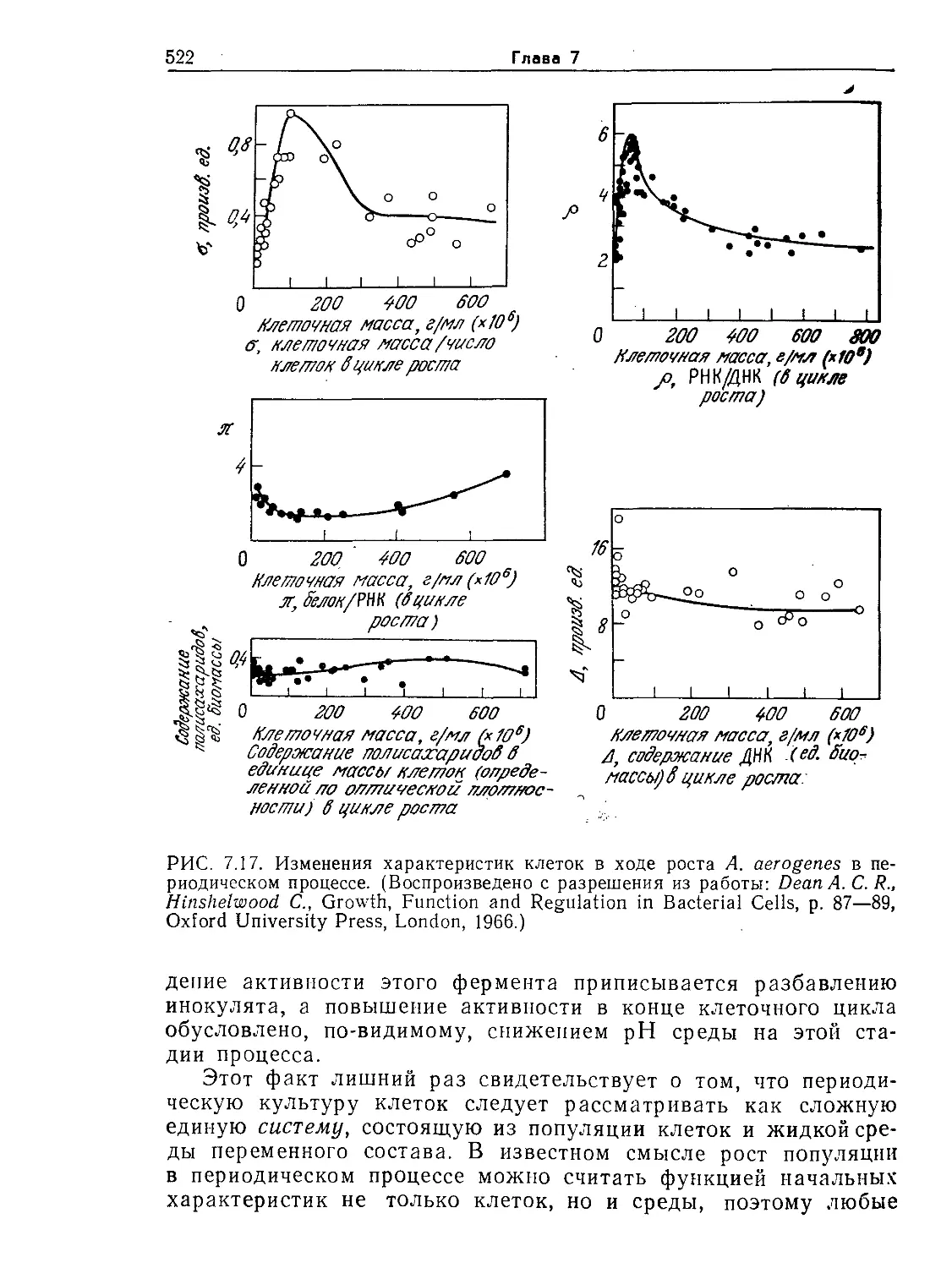
виц Ф., Структура и функция клетки. — М.: Мир, 1984, с. 45.)
нить их с размерами других объектов Вселенной. Как показано
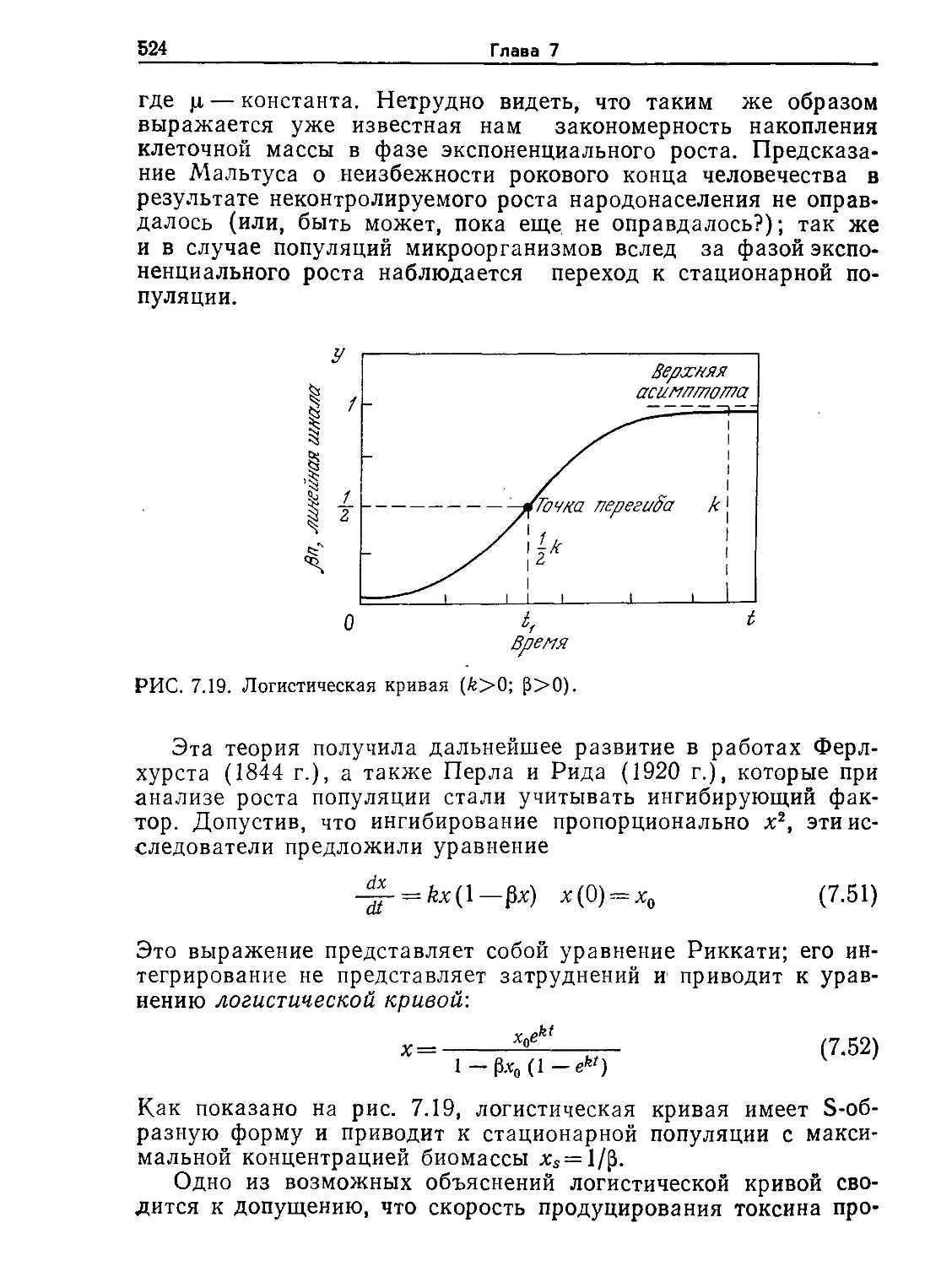
на рис. l.l, прокариотическая клетка настолько же меньше че-
ловека, насколько человек меньше земного шара или насколько
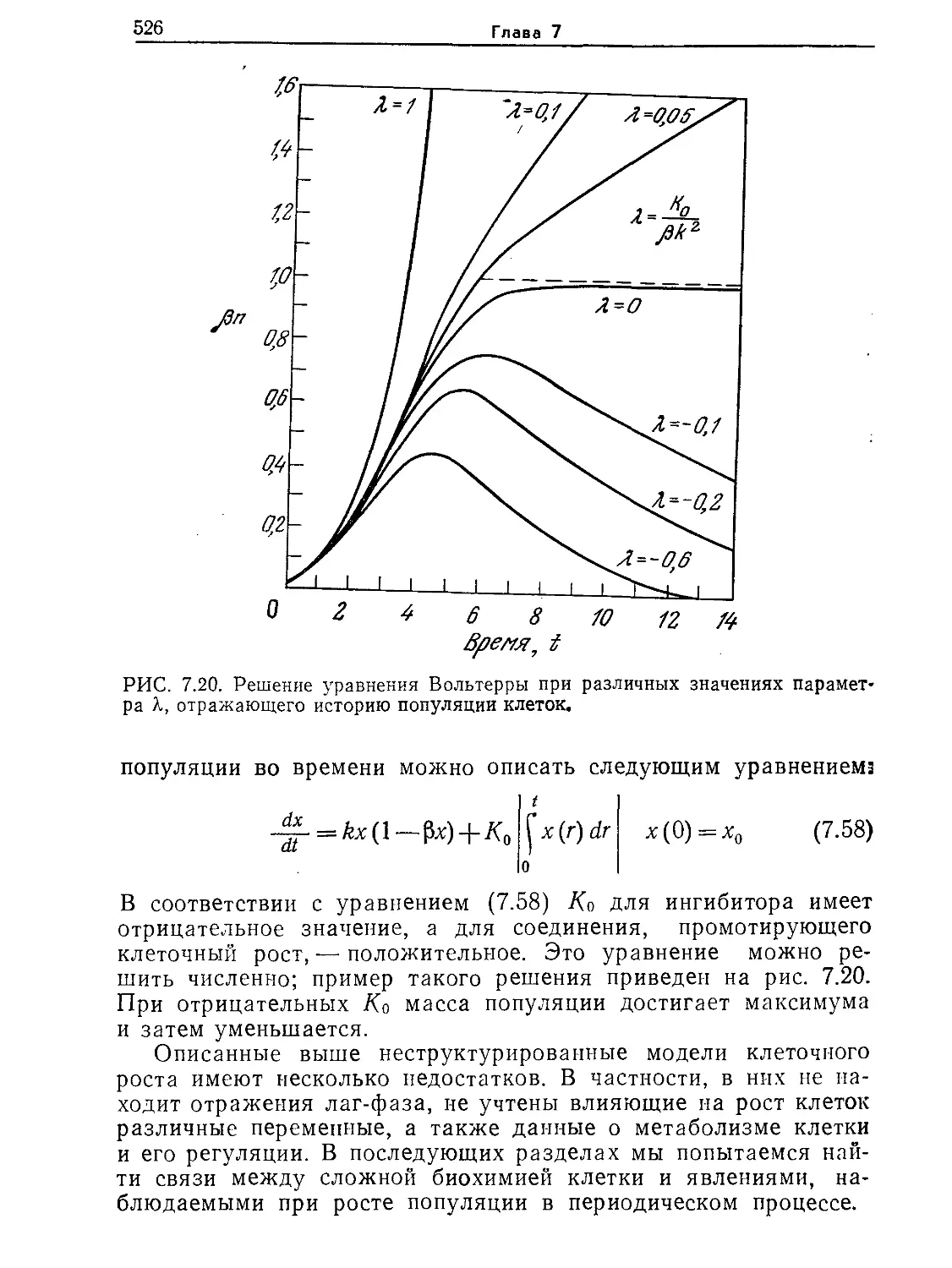
атом водорода меньше клетки. Как мы увидим позднее, эти
отношения приобретают большое значение при детальном изу-
чении функций клетки. Объем одной прокариотической клетки
составляет около 10~12 мл, из которых 50—80% приходится на
16
Глава 1
воду. В первом приближении можно считать, что масса одного
прокариота составляет 10~12 г.
Микроорганизмы этого типа растут очень быстро и широко
распространены в природе. Некоторые прокариоты могут удваи-
ваться в размере, массе и числе за 20 мин. Прокариоты, как
правило, биохимически универсальны в том смысле, что они
могут усваивать самые разнообразные питательные вещества
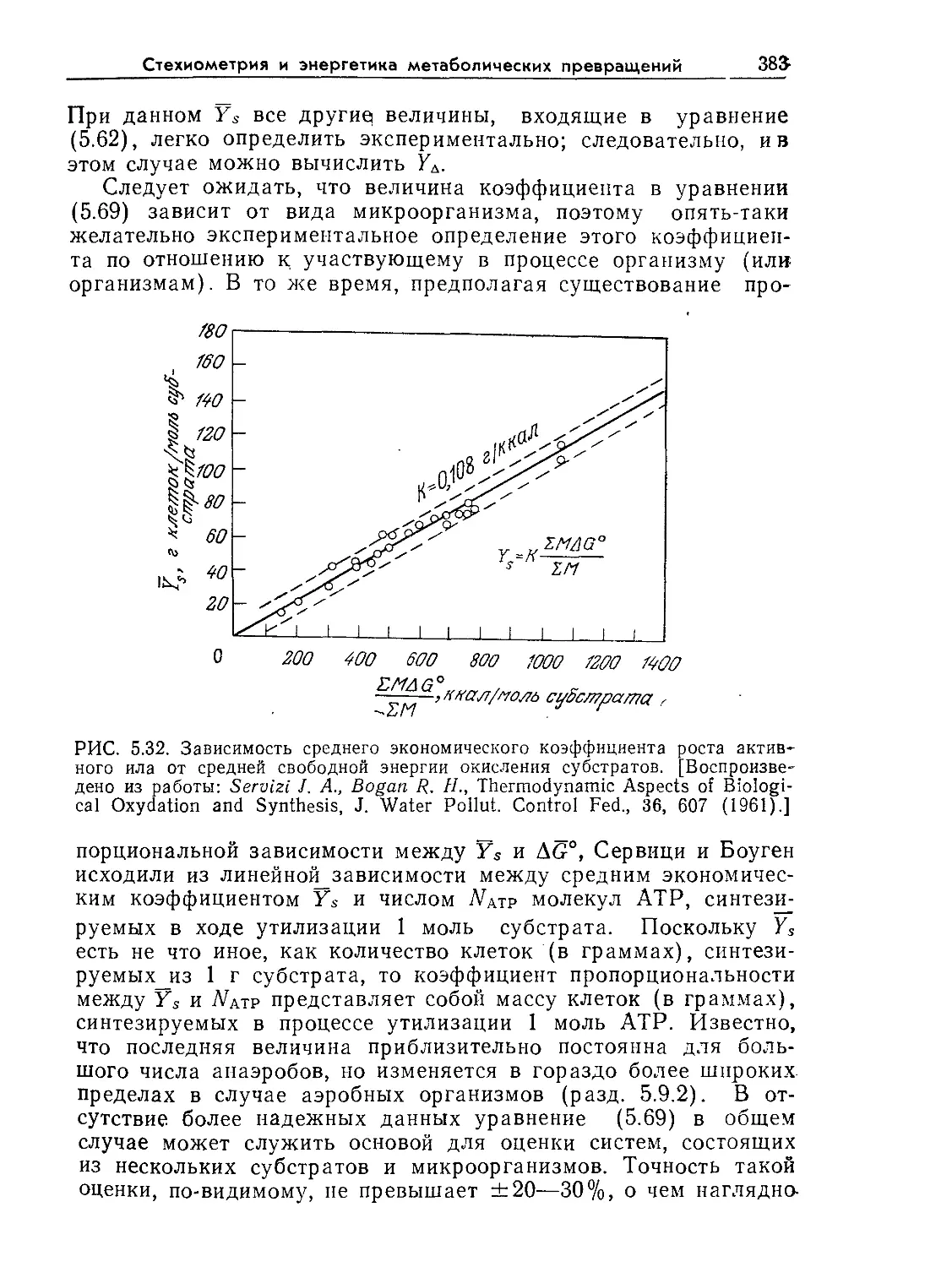
и, более того, способны выбирать наилучшие питательные ве-
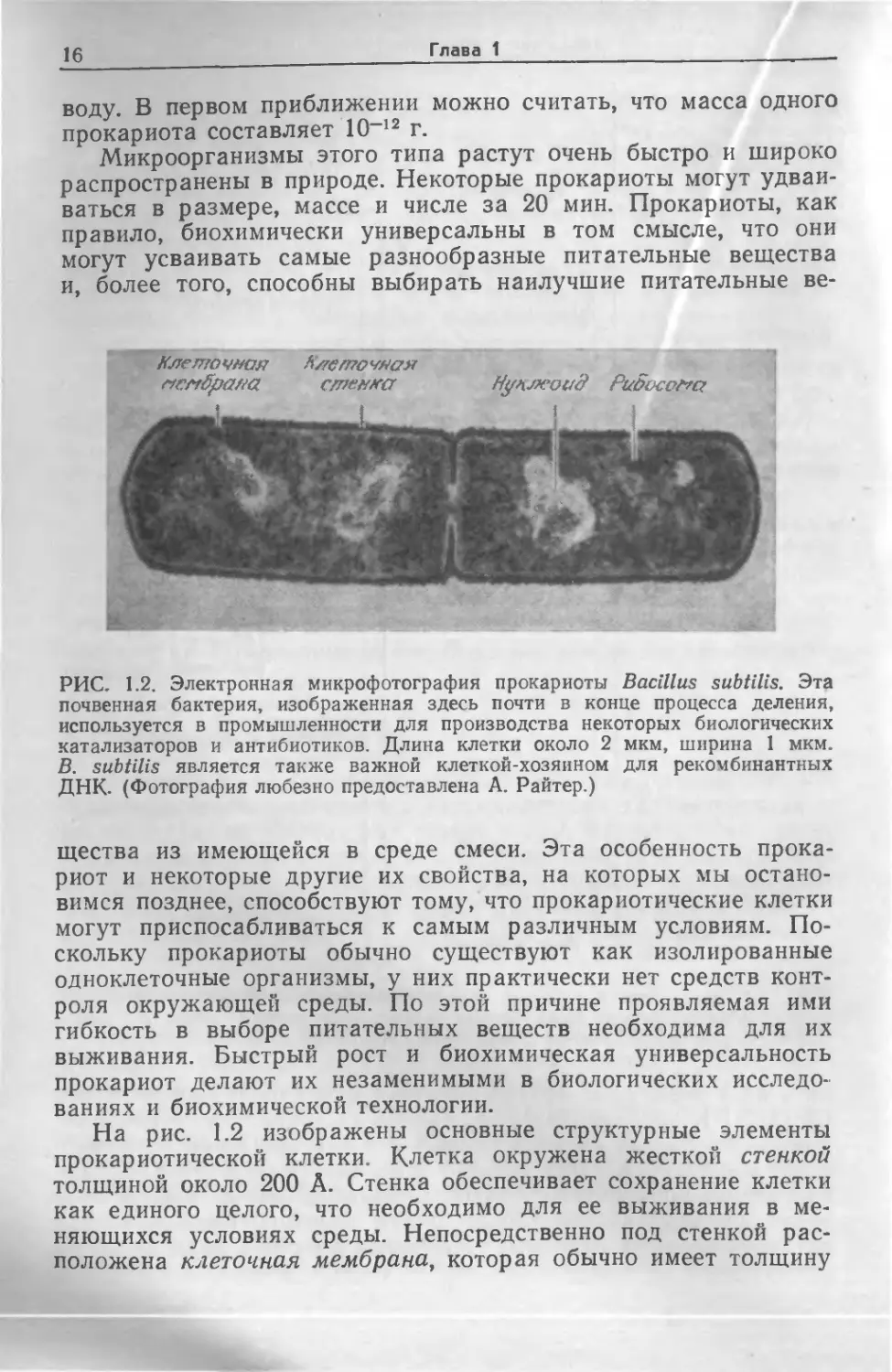
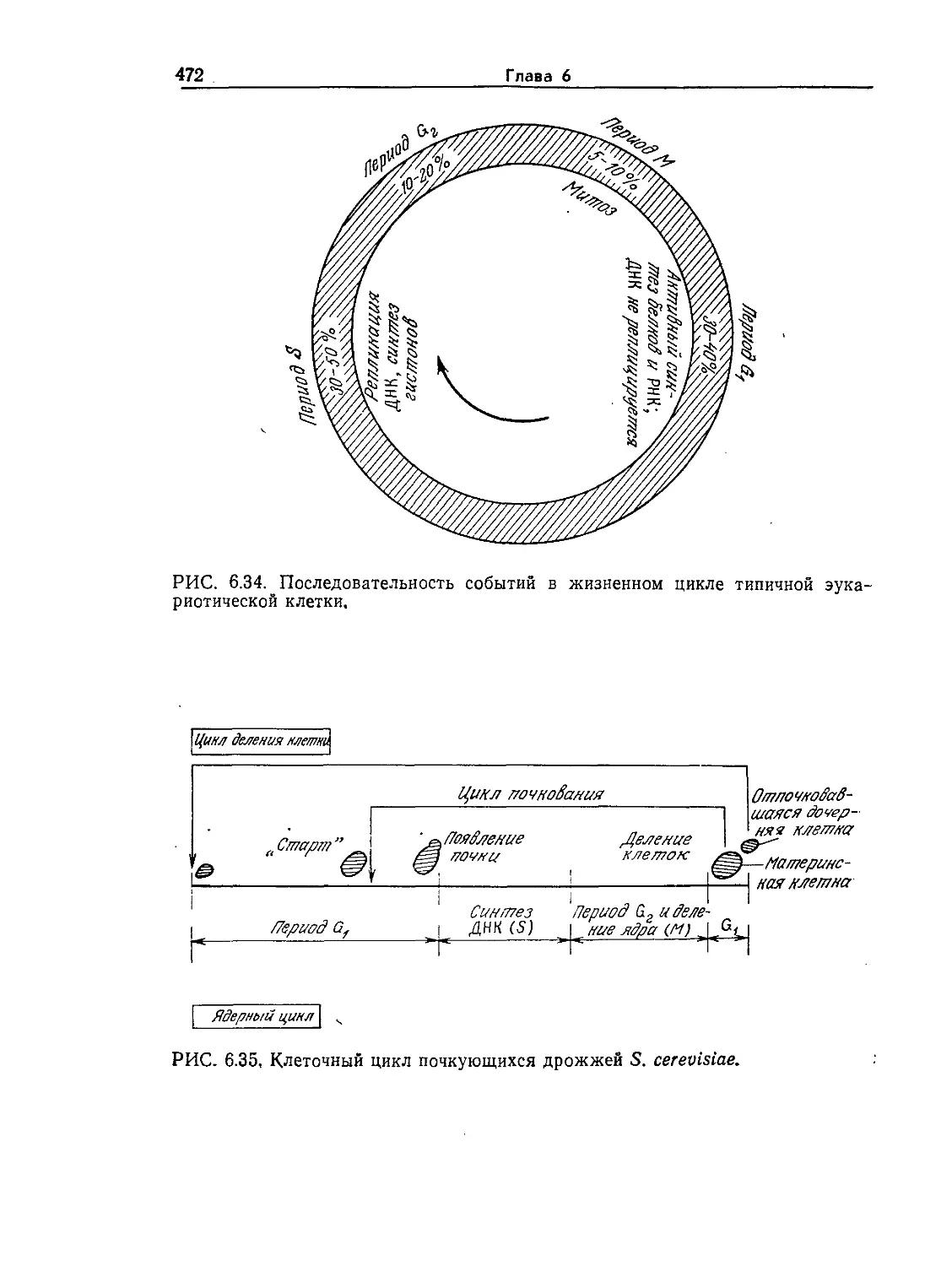
РИС. 1.2. Электронная микрофотография прокариоты Bacillus subtilis. Эта
почвенная бактерия, изображенная здесь почти в конце процесса деления,
используется в промышленности для производства некоторых биологических
катализаторов и антибиотиков. Длина клетки около 2 мкм, ширина 1 мкм.
В. subtilis является также важной клеткой-хозяином для рекомбинантных
ДНК. (Фотография любезно предоставлена А Райтер.)
щества из имеющейся в среде смеси. Эта особенность прока-
риот и некоторые другие их свойства, на которых мы остано-
вимся позднее, способствуют тому, что прокариотические клетки
могут приспосабливаться к самым различным условиям. По-
скольку прокариоты обычно существуют как изолированные
одноклеточные организмы, у них практически нет средств конт-
роля окружающей среды. По этой причине проявляемая ими
гибкость в выборе питательных веществ необходима для их
выживания. Быстрый рост и биохимическая универсальность
прокариот делают их незаменимыми в биологических исследо-
ваниях и биохимической технологии.
На рис. 1.2 изображены основные структурные элементы
прокариотической клетки. Клетка окружена жесткой стенкой
толщиной около 200 А. Стенка обеспечивает сохранение клетки
как единого целого, что необходимо для ее выживания в ме-
няющихся условиях среды. Непосредственно под стенкой рас-
положена клеточная мембрана, которая обычно имеет толщину
Введение в микробиологию
17
около 70 А и по строению не отличается от мембран других
клеток. Иногда ее называют плазматической мембраной. Важ-
нейшая функция мембраны заключается в транспорте веществ
из клетки в среду и наоборот, причем от мембраны зависит,
какие вещества и с какой скоростью будут транспортироваться
в клетку и из клетки. Внутри клетки имеется довольно большая,
четко не ограниченная область, называемая нуклеоидом, кото-
рая играет основную роль в контроле жизненно важных функ-
ций клетки. Темные пятнышки неправильной формы внутри
клетки изображают рибосомы — центры важнейшего биохими-
ческого процесса — белкового синтеза. Цитоплазмой называется
жидкость, занимающая весь остальной объем клетки. В про-
кариотической клетке имеются также светлые, напоминающие
пузырьки области, называемые резервными гранулами; их не
видно на приведенном рисунке, но можно различить на неко-
торых других микрофотографиях. Строение и функции перечис-
ленных структурных элементов прокариотической клетки мы
рассмотрим более детально позднее, после того как будут из-
ложены необходимые основные положения и объяснены соот-
ветствующие термины.
Обладая многими общими структурными и функциональны-
ми элементами, различные прокариоты могут в то же время
существенно отличаться друг от друга. У сине-зеленых водо-
рослей, например, имеются мембраны, способные улавливать
энергию света и использовать ее для фотосинтеза. В этом слож-
ном процессе утилизации солнечной энергии клетки обеспечи-
ваются необходимым для их жизнедеятельности органическим
веществом и выделяют в атмосферу кислород.
1.2.2. Эукариотические клетки
Второй основной тип клеток составляют эукариотические
клетки. Эукариотическими называют клетки, ядро которых за-
ключено в мембрану. Как правило, эукариотическая клетка по
объему в 1000—10 000 раз больше прокариотической. К этому
типу клеток принадлежат все клетки высших организмов. Эука-
риотические клетки отличаются большим разнообразием форм,
что необходимо, в частности, для обеспечения различных спе-
циализированных функций. В составе высших организмов эти
клетки сосуществуют и взаимодействуют друг с другом различ-
ными путями и поэтому не нуждаются в биохимической гибко-
сти и приспособляемости, столь необходимых для прокариот.
К эукариотам относятся и многие важные виды микроорганиз-
мов. В следующем разделе мы приведем ряд примеров одно-
клеточных эукариот.
2—537
18
Глава 1
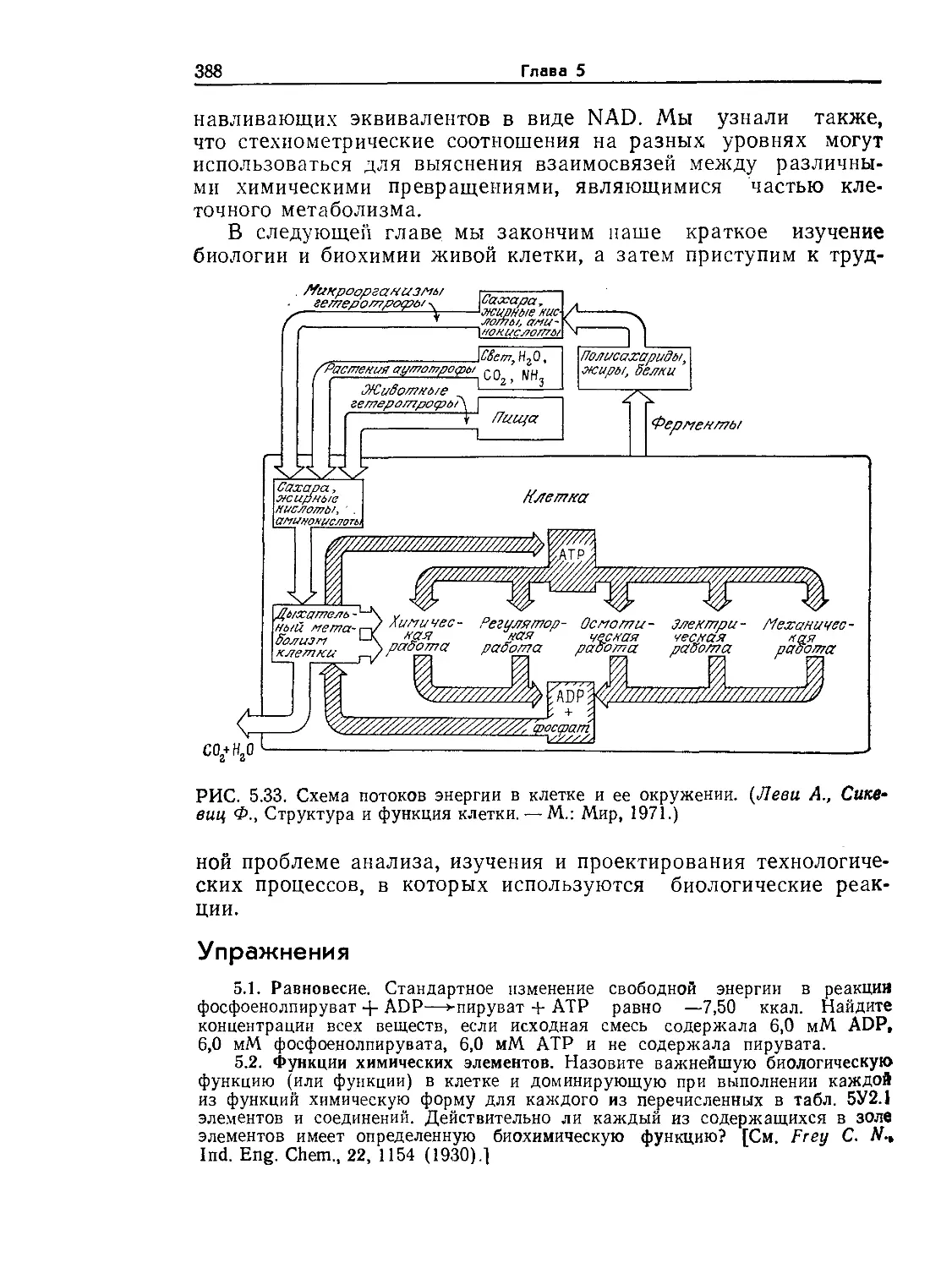
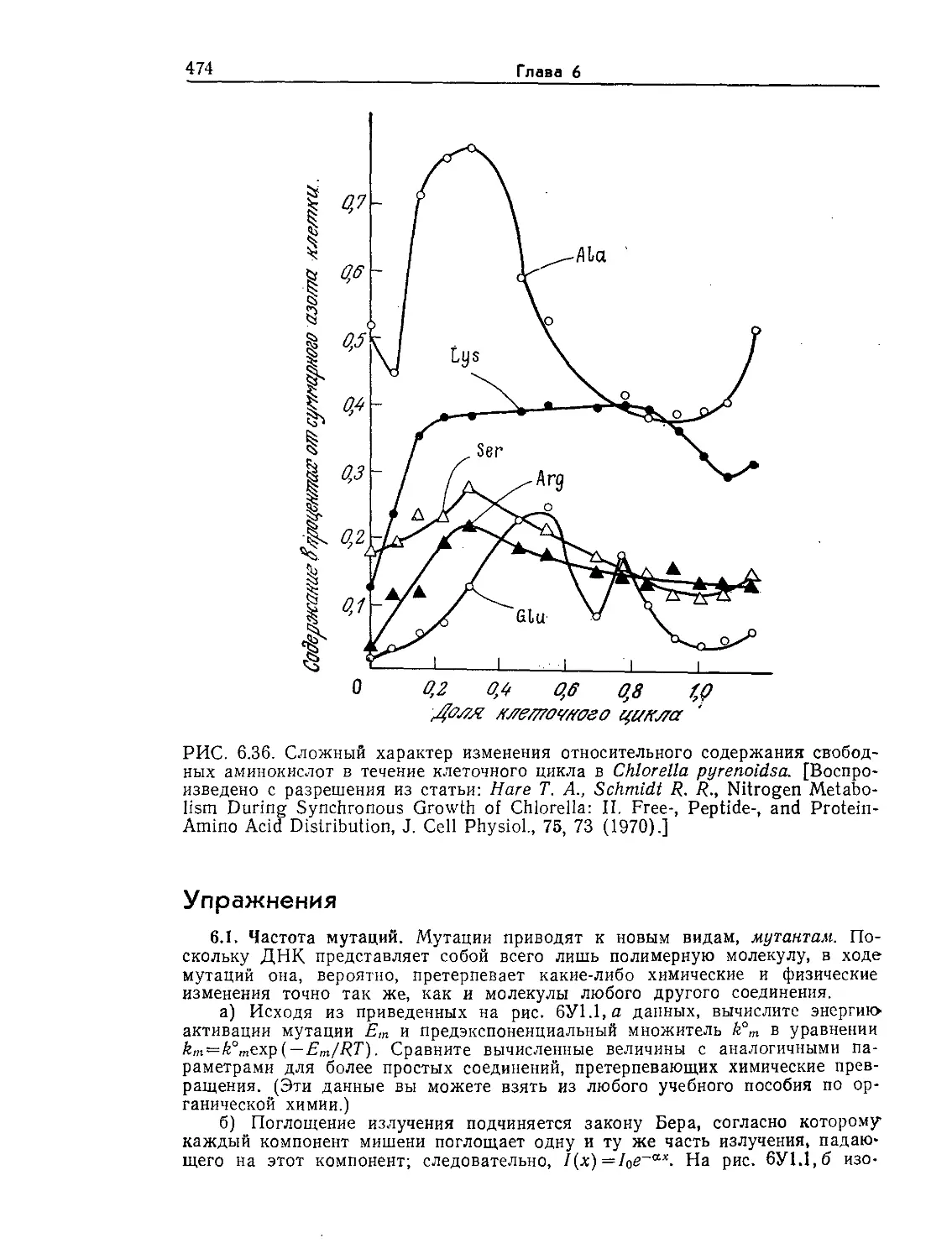
РИС. 1.3. Типичная эукариотическая клетка. Такой идеализированной клетки
с указанной обобщенной структурой в природе не существует; на самом деле
эукариоты существенно различаются по своей организации. Тем не менее не-
которые общие черты и структурные элементы характерны для многих эука-
риот, поэтому концепция типичной эукариоты в ряде случаев оказывается
удобной и полезной.
Как показано на рис. 1.3 и 1.4, по степени сложности внут-
ренней структуры эукариоты намного превосходят прокариоти-
ческие клетки. Для эукариот характерна высокая степень про-
странственной организации и дифференциации отдельных эле-
ментов клеточной структуры. Внутренний объем клетки разделен
на ряд четко ограниченных структурных компонентов, которые
подробнее мы рассмотрим позднее; каждый из этих компонентов
имеет свою структуру и функцию, необходимую для нормальной
жизнедеятельности всей клетки. Здесь мы обсудим только самые
основные детали строения эукариотических клеток.
Клетка окружена плазматической мембраной, аналогичной
мембране прокариот. Снаружи эта мембрана может быть за-
Введение в микробиологию
19
ХЛА
шт*.
Пга
' Л . it
,у Комплекс
ЧЗчЁЯШМЯ| ГЬллджа
dpnutfio
{яраяео. kj
W.nfl jfaGeeOMCr
>Л
.^fjffopa
'^*Дд м&мёране
Ядро
w<?
Х^ммал^а ~
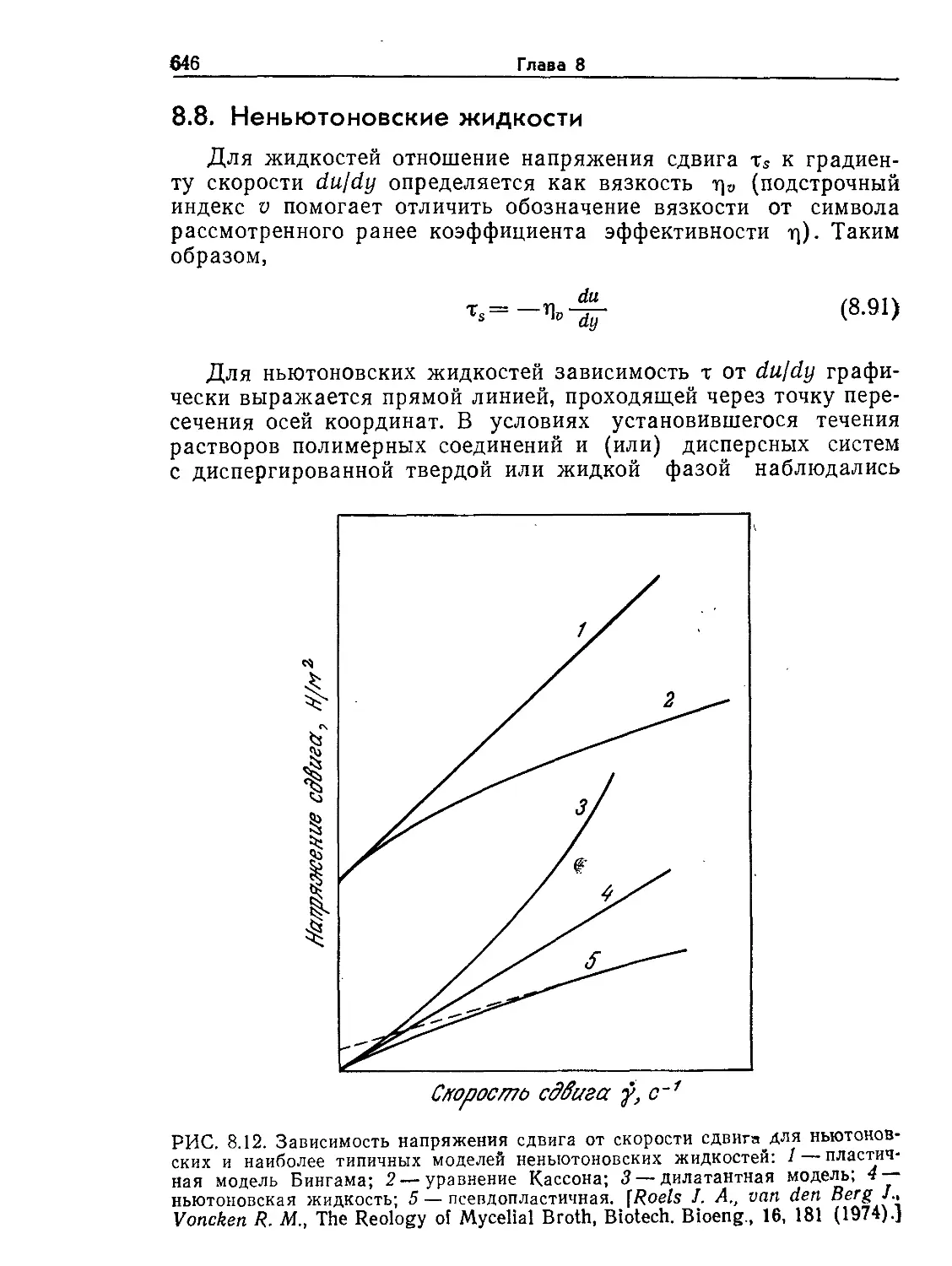
\яшчес#(ш
.ре/т/мрлун
НдКЮЗЮНдрия
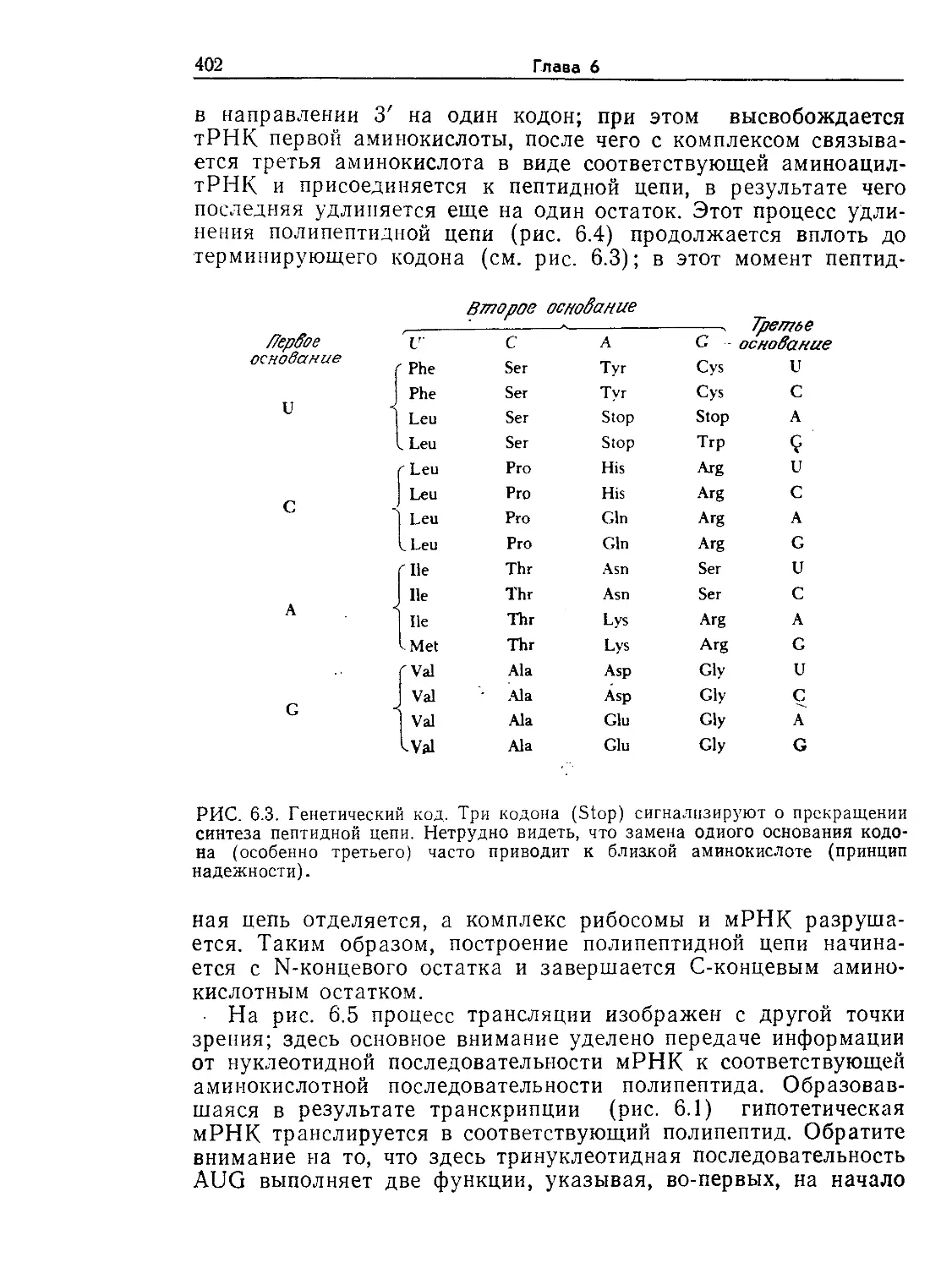
РИС. 1.4. Электронная микрофотография клетки печени крысы (X11 000).
(Фотография любезно предоставлена Дж. Э. Пэйладом, Йельский универси-
тет.)
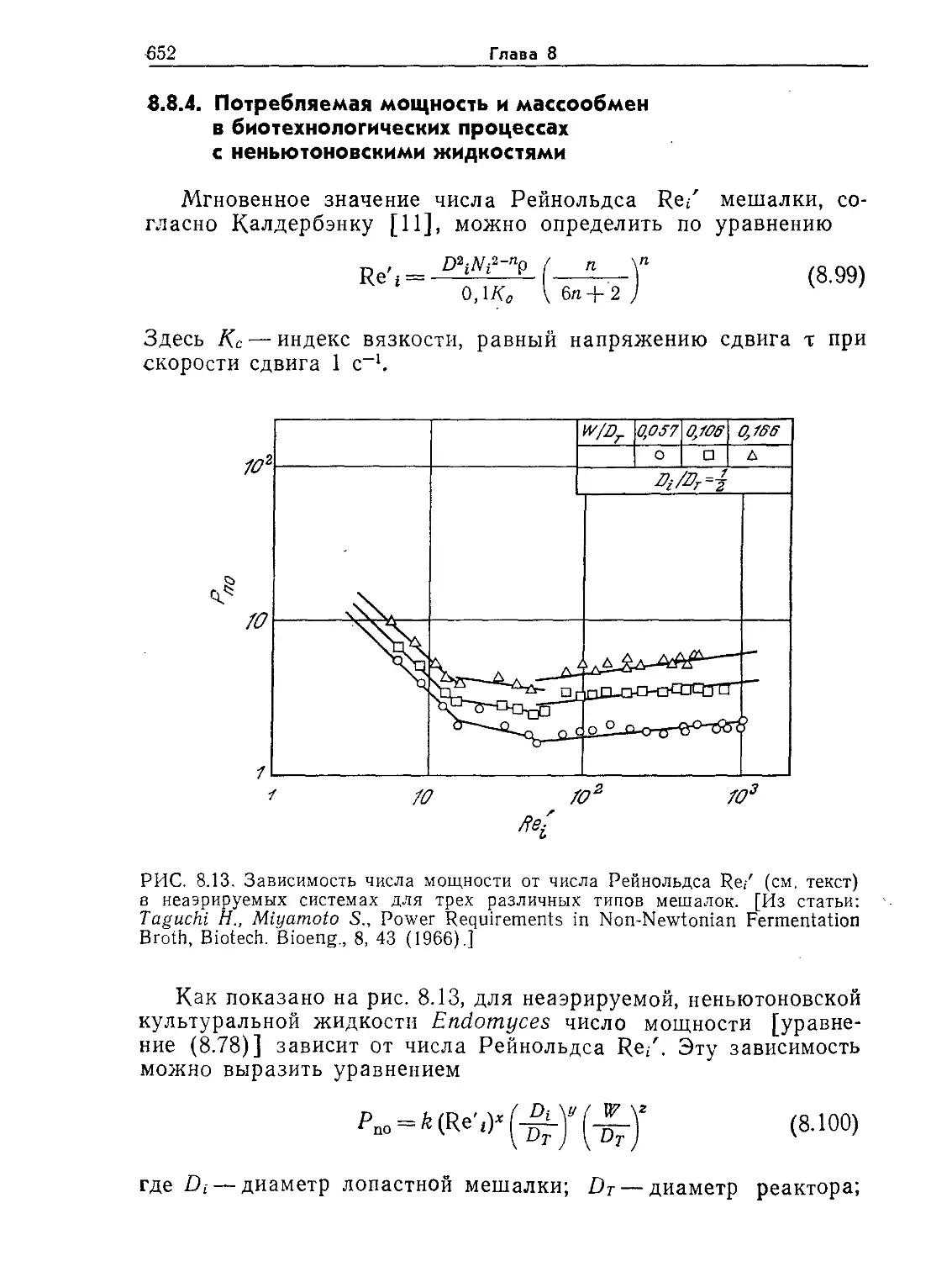
щищена клеточной стенкой, или оболочкой. Природа других
покровных структур клетки зависит от ее типа. Так, клетки
высших животных обычно окружены тонкой оболочкой, особые
адгезивные свойства которой существенны для связывания кле-
ток друг с другом и последующего образования специализиро-
ванных тканей и органов (например, печени). Клетки растений,
напротив, обычно окружены очень толстой и прочной стенкой.
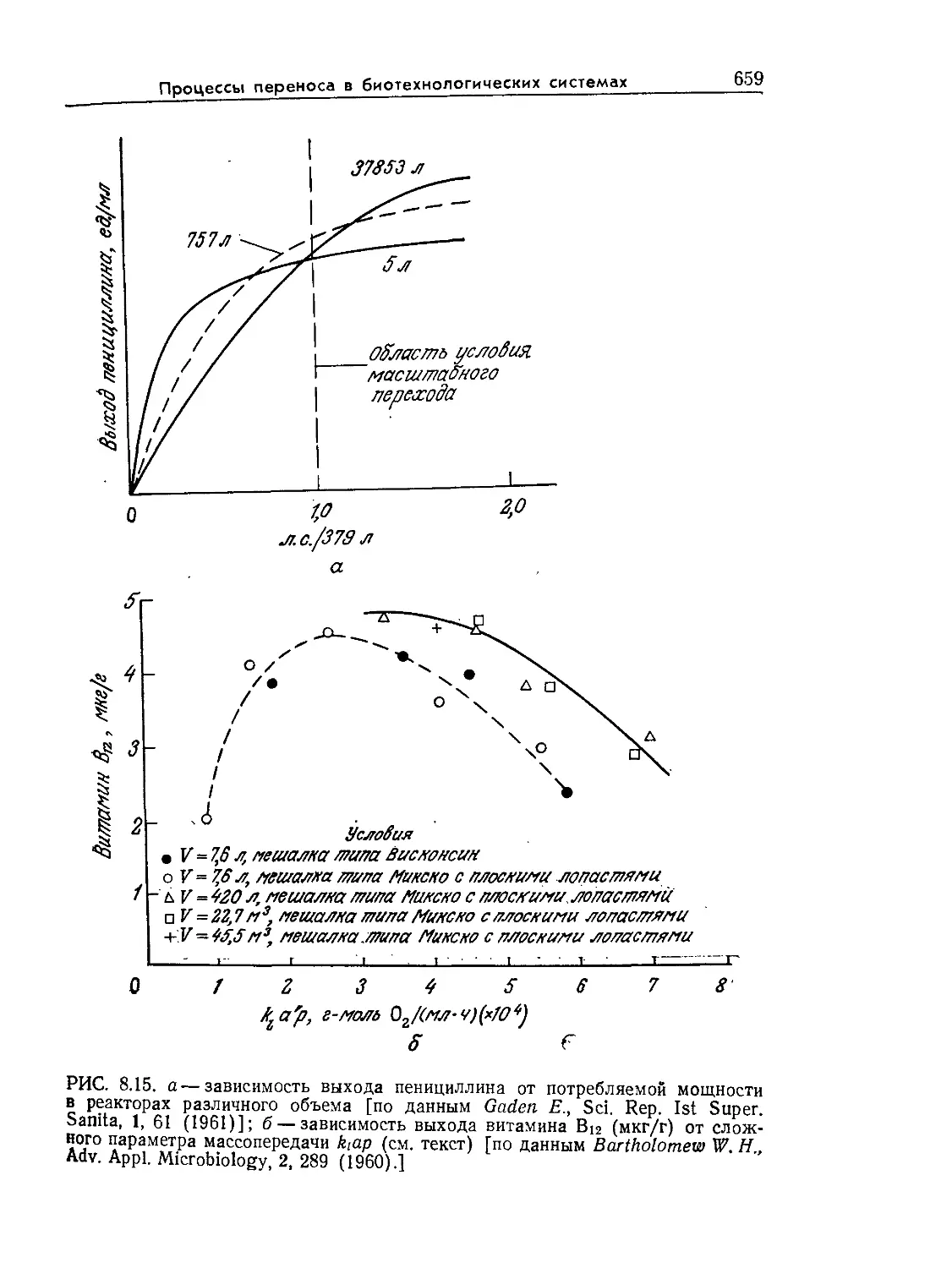
Стенки отмерших клеток деревьев представляют собой основ-
ную составную часть древесины.
В специализации различных структурных элементов эука-
риот большую роль играют внутриклеточные мембраны. От
клеточной мембраны внутрь клетки отходит сложная мембран-
ная система, называемая эндоплазматическим ретикулумом или
эндоплазматической сетью. Ядра эукариот окружены пористыми
мембранами. К поверхности большинства элементов эндоплаз-
матического ретикулума прикреплены рибосомы — центры бел-
кового синтеза, о чем уже упоминалось в разделе, посвященном
2*
20
Глава 1
прокариотам. Рибосомы последних, однако, несколько меньше
рибосом эукариот.
Основной функцией ядра эукариот являются контроль и ре-
гулирование каталитической активности рибосом, причем вы-
деляемые ядром химические посредники (информационные
и др. РНК) не только регулируют скорость реакций, но и опре-
деляют последовательность присоединения аминокислот при
синтезе белка.
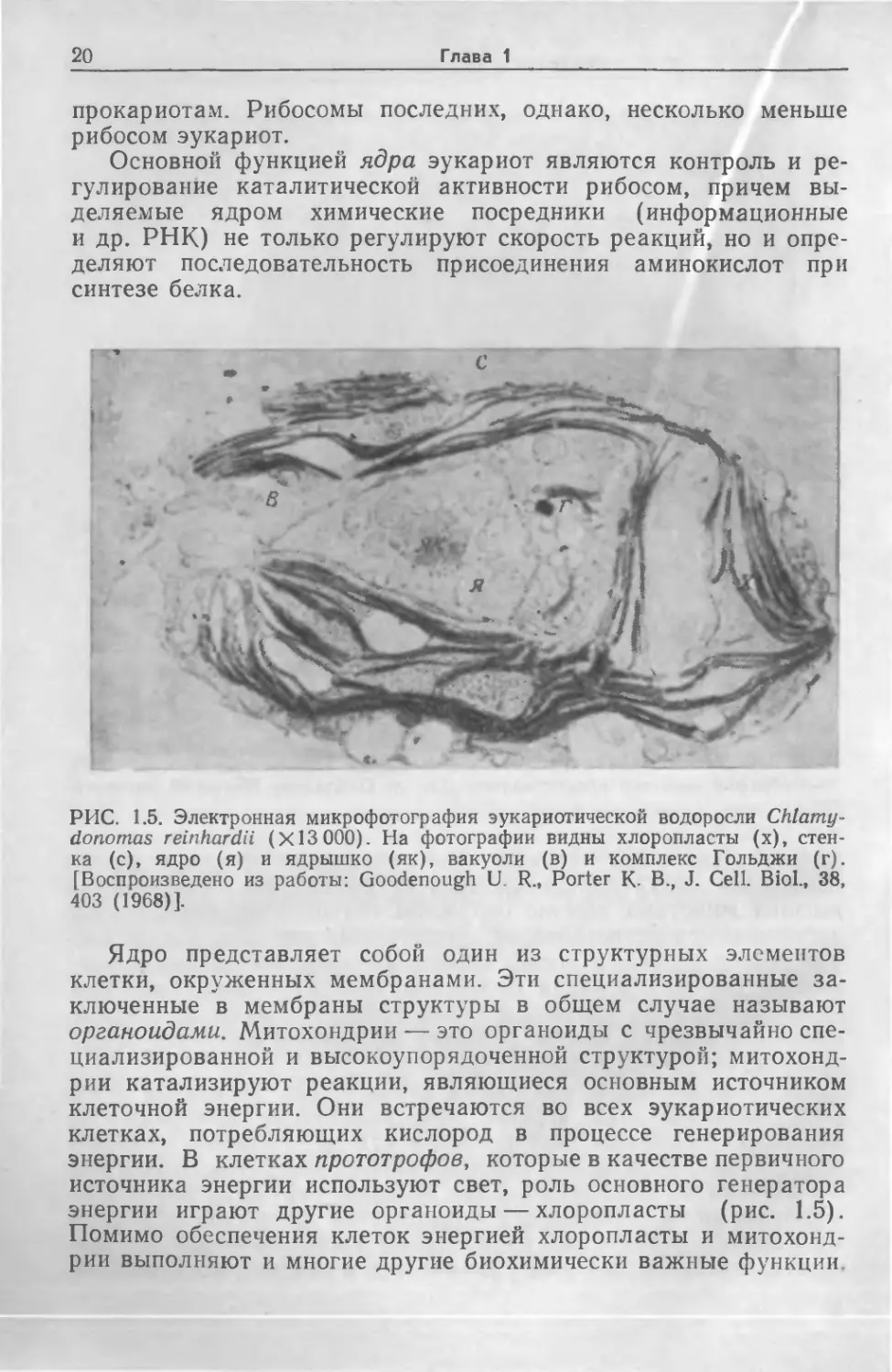
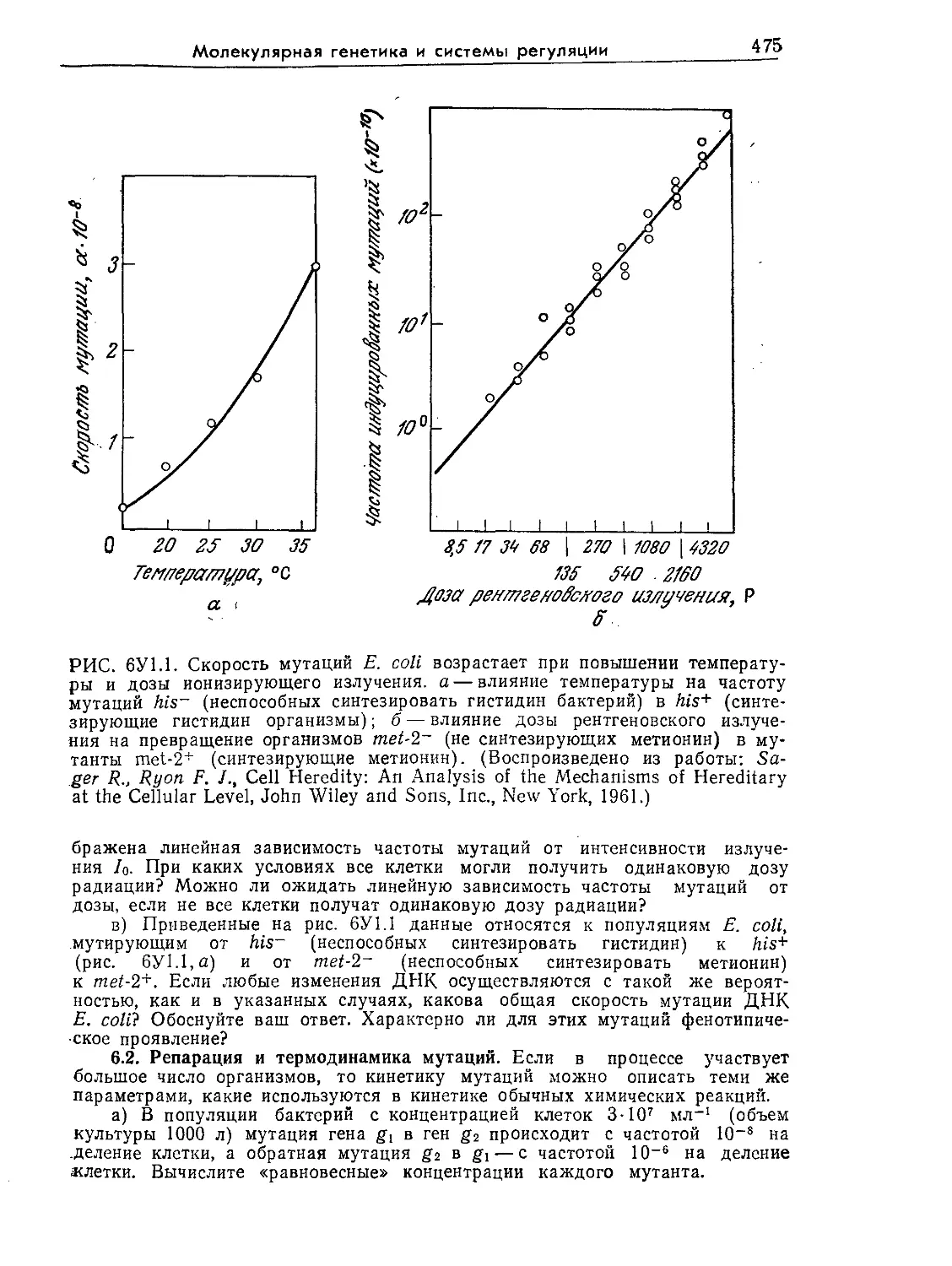
РИС. 1.5. Электронная микрофотография эукариотической водоросли Chlamy-
donomas reinhardii (Х13 000). На фотографии видны хлоропласты (х), стен-
ка (с), ядро (я) и ядрышко (як), вакуоли (в) и комплекс Гольджи (г).
[Воспроизведено из работы: Goodenough U. R„ Porter К- В., J. Cell. Biol., 38,
403 (1968)].
Ядро представляет собой один из структурных элементов
клетки, окруженных мембранами. Эти специализированные за-
ключенные в мембраны структуры в общем случае называют
органоидами. Митохондрии — это органоиды с чрезвычайно спе-
циализированной и высокоупорядоченной структурой; митохонд-
рии катализируют реакции, являющиеся основным источником
клеточной энергии. Они встречаются во всех эукариотических
клетках, потребляющих кислород в процессе генерирования
энергии. В клетках прототрофов, которые в качестве первичного
источника энергии используют свет, роль основного генератора
энергии играют другие органоиды — хлоропласты (рис. 1.5).
Помимо обеспечения клеток энергией хлоропласты и митохонд-
рии выполняют и многие другие биохимически важные функции.
Введение в микробиологию
21
На рис. 1.3—1.5 изображены и другие органоиды — комплекс
Гольджи (аппарат Гольджи, пластинчатый комплекс), лизосомы
и вакуоли. В самых общих чертах их функции сводятся к осу-
ществлению некоторых химических реакций и к компартмента-
лизации (т. е. к приуроченности к определенным участкам клет-
ки) ряда соединений, обеспечивающей изоляцию последних от
остальной цитоплазмы. Процессы компартментализации важны
как с точки зрения эффективности реакций, так и с точки зре-
ния предотвращения нежелательных взаимодействий между со-
держимым органоидов и другими компонентами клетки.
Обнаружение описанных выше типов органоидов в самых раз-
личных эукариотах позволило по-новому оценить основные пре-
имущества клеточной теории. Теперь различные стороны жизне-
деятельности клеток можно рассматривать как сумму происхо-
дящих в органоидах процессов, каждый из которых в свою
очередь можно изучать отдельно. Считается, что органоиды
одного типа выполняют аналогичные операции и функции не-
зависимо от природы клеток, к которым они принадлежат; пока
что не было обнаружено исключений из этого правила.
Таким образом, основной путь изучения клетки заключается
в определении химического состава, строения и биохимической
активности органоидов. Большая часть имеющихся в настоящее
время данных о биохимии клетки получена именно таким путем.
Поэтому в следующем разделе мы вкратце рассмотрим методы
центрифугирования, широко применяющиеся для выделения
составных частей клеток.
1.2.3. Фракционирование клеток
Основная проблема в изучении свойств определенных орга-
ноидов из данного типа клеток заключается в получении доста-
точного для последующего биохимического анализа количества
этих органоидов. Обычно для этой цели выделяют большое число
органоидов из большого числа клеток (из так называемой по-
пуляции клеток). Как правило, стандартная методика выделе-
ния органоидов включает в себя в качестве первой стадии гомо-
генизацию суспензии клеток в специальном растворе с помощью
трубки с вращающимся пестиком или ультразвука. Таким путем
пытаются разрушить клетки, не затрагивая содержащиеся в них
органоиды и не нарушая их структуру. Следующая стадия за-
ключается во фракционировании полученной суспензии, которая
в идеальном варианте представляет собой смесь выделенных
из клеток целых органоидов.
Как инженеры-технологи, мы знаем, что любой процесс раз-
деления основан на различиях в физических п (или) химиче-
ских свойствах разделяемых компонентов. Обычный метод
22
Глава 1
фракционирования органоидов клетки базируется на различиях
в их физических характеристиках: размере частиц, их форме
и плотности. Упрощенно процесс центрифугирования рассмот-
рен в приведенном ниже примере.
Пример 1.1. Изучение движения частицы в условиях центрифугирования.
Предположим, что сферическая частица радиусом и плотностью рр поме-
щена в центрифужную пробирку, содержащую жидкую среду плотностью
Центрифужная
пробирка
РИС. 1П1.1. При вращении центрифуги с высокой скоростью суспендирован-
ные в центрифужных пробирках частицы движутся в направлении от оси
вращения. Поскольку скорость движения этих частиц зависит от их размеров,
формы и плотности, метод центрифугирования позволяет разделять частицы,
различающиеся по этим параметрам.
р, и вязкостью Цс. Если затем эту пробирку укрепить в центрифуге и вра-
щать с угловой скоростью со (рис. 1П1.1), то можно вычислить параметры
движения частицы, используя следующее уравнение (какие допущения при-
няты в этом уравнении?) :
Сила сопротивления среды = выталкивающей силе
6л|лс/?ц,. = ^^-С(рр — pf) (1П1.1)
где иг — скорость частицы в направлении г:
</,=-£- (1П1.2)
и G — центробежное ускорение:
G = co2r (Ш1.3)
Введение в микробиологию
23
Для определения действующей на частицу силы сопротивления среды в урав-
нении (1П1.1) использован закон Стокса, так как в условиях центрифугиро-
вания скорости частиц (и, следовательно, их числа Рейнольдса) обычно
очень малы. Коэффициент силы тяжести в уравнении (1П1.1) отсутствует,
поскольку направление г перпендикулярно направлению силы тяжести (см.
рис. 1П1.1). В то же время при вращении ротора центрифуги с большой уг-
ловой скоростью развивается ускорение G, которое обычно во много раз пре-
восходит силу тяжести и может достигать величин 600—600 000 g.
Гомогенат
Центри/ругиро- Супернатант, содержа-
бание супернатанта щий растворимые пом-
ири 15000у б течение 5 нин поненты цитоплазмы
Ядра и ин та кт- Митохондрии, Фрагменты эндо-
ные клетки лизосоме/ плазматического
ретикулума, ридосомы
РИС. 1П1.2. Основные стадии процесса разделения структурных элементов
клеток методом центрифугирования. На более поздних стадиях процесса вы-
деляют все более и более мелкие компоненты клеток.
Интегрированием указанного выражения можно определить время, необ-
ходимое для движения частицы из положения rj в положение г2:
Нс
t = ---------------------1п —
2 со2Я2 (рр - pf) гг
(1П1.4)
Сферические частицы различного размера и(или) плотности будут про-
ходить одно и то же расстояние в центрифужной пробирке за разное время;
этот факт лежит в основе дифференциального центрифугирования. Посколь-
ку относительно большие частицы, например ядра и интактные клетки, осаж-
даются довольно быстро, их можно собрать и отделить в виде осадка после
центрифугирования суспензии в течение ограниченного времени при сравни-
тельно невысоких скоростях. Затем супернатантную суспензию подвергают
повторному кратковременному центрифугированию при более высоких ско-
ростях вращения ротора, после чего отделяют второй осадок, содержащий
митохондрии. Продолжая эту операцию, получают ряд фракций компонентов
клетки. Весь процесс схематично изображен на рис. 1П1.2.
В более сложных методах центрифугирования применяют жидкие среды
с градиентом плотности по высоте центрифужной пробирки. Эти методы ис-
24
Глава 1
пользуются также для предварительного разделения и фракционирования
более мелких компонентов клетки, например некоторых типов макромолекул.
Другие полезные методы разделения таких сложных смесей, основанные на
различиях химических свойств разделяемых компонентов, подробнее будут
рассмотрены в гл. 11.
В применении и интерпретации результатов фракциониро-
вания компонентов клеток методом центрифугирования имеется
ряд ограничений, хорошо освещенных в книге Малера и Кордеса
(Mahler Н. R., Cordes Е. Н., Biological Chemistry, 2nd ed.,
Harper and Row, Publishers, Inc., New York, 1971). Одна из
трудностей характерна для любых работ в области изучения
и применения микроорганизмов. Для того чтобы получить до-
статочное для последующих работ количество клеток, органои-
дов, биологически важных молекул и других компонентов кле-
ток, мы вынуждены использовать популяцию, т. е. большое число
индивидуальных клеток. Обычно принимается, что эта популя-
ция гомогенна, или, иными словами, все входящие в ее состав
микроорганизмы идентичны. В таких случаях популяция нужна
только для увеличения числа этих микроорганизмов с тем, что-
бы облегчить дальнейшие экспериментальные исследования.
Обычно, однако, входящие в состав популяции микроорга-
низмы в той или иной степени различны; такую популяцию
называют гетерогенной. Например, в популяции растущих кле-
ток имеются старые и молодые, большие и малые клетки, часто
различающиеся по биохимическому составу и активности. Если
же рассматривать структурные элементы клетки, то следует
отметить, что органоиды одного типа, например митохондрии,
находящиеся в одной и той же клетке, также несколько отли-
чаются друг от друга. Поэтому содержащая митохондрии кле-
точная фракция также представляет собой гетерогенную попу-
ляцию. При дальнейшем изучении таких смесей определяют,
по сути дела, некоторые усредненные характеристики популя-
ции клеток, и, следовательно, найденные параметры будут за-
висеть от состава популяции.
1.3. Важнейшие типы клеток
В этом разделе будет вкратце рассмотрена классификация
царства протистов, к которому относятся все живые существа
с очень простой биологической организацией в сравнении с рас-
тениями и животными. К царству протистов принадлежат все
одноклеточные, а также многоклеточные организмы, построен-
ные из клеток только одного типа. Напротив, растения и жи-
вотные отличаются большим разнообразием типов клеток.
В конце настоящего раздела мы приведем примеры раститель-
ных и животных клеток, которые могут быть выращены на
твердой или жидкой питательной среде.
Введение в микробиологию
25
В табл. 1.1 показано деление царства протистов на удобные
для наших целей таксоны. Последние различаются рядом ха-
рактеристик: источниками и способом получения энергии и пи-
тательных веществ, скоростями роста и выделения продуктов
жизнедеятельности, способами самовоспроизведения, способно-
стью к передвижению и средствами для его осуществления. Все
эти факторы очень важны с точки зрения практического ис-
пользования протистов. Не последнюю роль в классификации
Таблица 1.1. Классификация организмов царства протистов
различных типов организмов играют морфологические разли-
чия, т. е. различия в их форме и структуре. Морфология мик-
роорганизма влияет на скорость массопередачи питательных
веществ и может также в существенной степени определять
динамические свойства суспензий, содержащих этот организм.
Все приведенные выше данные свидетельствуют о том, что каж-
дый из перечисленных в табл. 1.1 таксонов микроорганизмов
следует рассматривать отдельно.
Искусство биологической классификации называется таксо-
номией. В этой классификации основной единицей является вид;
организмы одного вида характеризуются высокой степенью бли-
зости морфологических и биохимических свойств и в то же вре-
мя существенно отличаются по этим свойствам от организмов
близких видов. Биологический вид обозначается двумя латин-
скими словами, первое из которых начинается с прописной
буквы и обозначает род организма, а второе (часто носящее
26
Глава 1
описательный характер)—собственно вид. Например, кишечная
палочка — очень подробно изученная бактерия, обнаруженная
в кишечнике человека,— называется Escherichia (наименование
рода) coli (наименование вида). Латинское название выделяет-
ся курсивом; если из контекста ясно, о каком роде идет речь,
то наименование рода обычно сокращается до первой буквы,
например Е. coli.
В целях систематизации видов и родов организмов разрабо-
тана иерархическая система таксономии, в которой родственные
роды объединены в семейства, близкие семейства образуют от-
ряды, объединяющиеся в свою очередь в классы; далее классы
сгруппированы в отделы, или типы, и, наконец, близкие отделы
объединены в царства. Например, в табл. 1.1 указаны царство
протистов, отдел грибов и класс дрожжей. Часто, однако, раз-
личия в свойствах микроорганизмов выражены не слишком от-
четливо и подробная классификация становится в известной
степени искусственной и произвольной; особенно это относится
к бактериям и дрожжам.
1.3.1. Бактерии
Как мы уже упоминали в ходе предварительного изучения
прокариот, бактерии представляют собой относительно не-
большие организмы, обычно заключенные в жесткую оболочку.
У многих видов бактерий наружная сторона клеточной стенки
покрыта упругой, вязкой оболочкой, называемой капсулой или
слизистым слоем. Бактерии представляют собой одноклеточные
организмы; морфологически они могут быть разделены на три
основные группы (рис. 1.6). Большинство бактерий не способно
поглощать световую энергию, может самопроизвольно передви-
гаться и размножается путем деления на две дочерние клетки,
хотя из всех этих правил известно множество исключений.
Существует большое число подотделов бактерий; некоторые
основные типы бактерий и их характерные особенности пере-
числены в табл. 1.2. В столбце «реакция Грама» имеется в виду
реакция бактерий на относительно прямой и быстрый цветной
тест. В этом тесте клетки сначала окрашивают красителем
кристаллическим фиолетовым, затем обрабатывают раствором
иода и промывают спиртом. Клетки, сохраняющие после такой
обработки голубой цвет красителя, называют грамположитель-
ными\ потеря окраски свидетельствует о принадлежности бак-
терий к грамотрицательному типу. Многие характеристики бак-
терий хорошо коррелируют с этой цветной реакцией, отражаю-
щей существенные различия в структуре их оболочек.
При промышленном использовании микроорганизмов особен-
но важен вопрос, обязательна ли подача кислорода в питатель-
Введение в микробиологию
27
ную среду (гл. 8, 12 и 14). В аэробных процессах для питания
микроорганизмов подают кислород, обычно в виде воздуха.
К числу таких процессов относятся практически важные мик-
робиологические способы производства уксуса, некоторых ан-
тибиотиков и добавок к кормам для животных. Одна из основ-
ных трудностей в разработке таких процессов связана с ограни-
ченной растворимостью кислорода в типичных для этих систем
РИС. 1.6. Три формы бактерий.
водных средах (гл. 8). В анаэробных процессах, например
в производстве некоторых спиртов или при переработке орга-
нических отходов, микроорганизмы функционируют в отсутствие
кислорода.
В промышленном применении и в контроле бактериального
заражения не менее важна способность бактерий образовывать
в неблагоприятных условиях так называемые эндоспоры. По-
следние представляют собой «спящую» форму клетки, в которой
они без вредных для себя последствий переносят воздействие
повышенной температуры, радиации и ядохимикатов. Когда
споры оказываются в пригодной для их жизнедеятельности сре-
де, они превращаются в нормально функционирующие клетки.
В отличие от споровой формы это нормальное, биологически
активное состояние клеток часто называют вегетативной фор-
мой. Как свидетельствуют приведенные в табл. 1.2 данные, су-
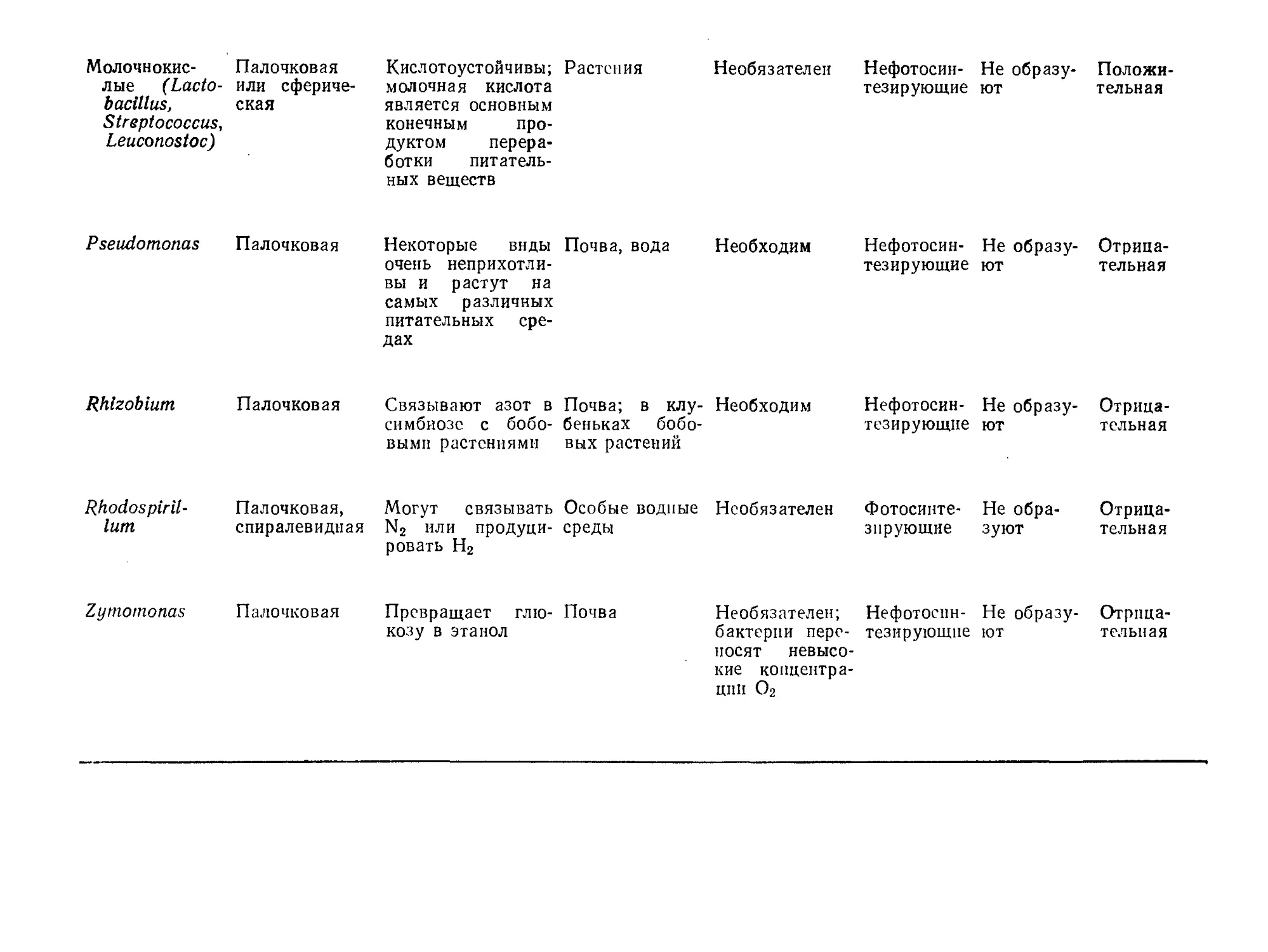
Таблица 1.2. Некоторые основные типы бактерий и их отличительные особенности
Тип бактерий Доминирующая морфологическая структура Некоторые особен- ности питания Обычная сфера обитания Потребность в кислороде для большинства видов Способность к фотосинтезу Способность к образова- нию спор Реакция Грама
Уксуснокис- Палочковая; Часто усваивают Разлагающие- Необходим Нефотосин- Не образу- Отрица-
лые бакте- рии (Aceto- bacter, Glu- conobacter) некоторые ви- ды Acetobacter образуют про- тяженные сли- зистые слои спирт; кислото- устойчивы ся растения тезирующие ют тельная
Bacillus Палочковая Универсальны; су- ществуют в самых различных пита- тельных средах Почва Необходим Нефотосин- тезирующие Образуют Положи- тельная
Clostridium Палочковая Для различных видов характер- ны различные тре- бования к пита- тельной среде Почва Большинство видов не пере- носят О2 Нефотосин- тезирующие Образуют Положи- тельная
Corynebacte- Неправильная Нетребовательны Почва, Необязателен, Нефотосин- Не образу- Положи-
rium форма; не раз- множаются де- лением; часто не способны к движению организм че- ловека но может ис- пользоваться при его нали- чии тезирующие ют тельная
Энтеробакте- Палочковая Несложные орга- Естественная Необязателен, Нефотосин- Не образу- Отрица-
рии или ко- либактерии (например, Е. coli) нические соедине- ния среда обита- ния некото- рых видов — кишечник выс- ших живот- ных но может ис- пользоваться при его нали- чии тезирующие ют тельная
Молочнокис- Палочковая Кислотоустойчивы; Растения
лые (Lacto- или сфериче- молочная кислота
bacillus, ская является основным
Streptococcus, конечным про-
Leuconostoc) дуктом перера- ботки питатель- ных веществ
Необязателен Нефотосин- Не образу- Положи-
тезирующие ют тельная
Pseudomonas Палочковая Некоторые виды очень неприхотли- вы и растут на самых различных питательных сре- дах Почва, вода Необходим Нефотосин- тезирующие Не ют образу- Отрица- тельная
Rhlzoblum Палочковая Связывают азот в симбиозе с бобо- выми растениями Почва; в клу- Необходим беньках бобо- вых растений Нефотосин- тезирующие Не ют образу- Отрица- тельная
Rhodospir il- ium Палочковая, спиралевидная Могут связывать N2 или продуци- ровать Н2 Особые водные Необязателен среды Фотосинте- зирующие Не обра- зуют Отрица- тельная
Zymomonas
Палочковая Превращает глю- Почва
козу в этанол
Необязателен; Нефотоспн- Не образу- Отрпца-
бактерпи пере- тезирующие ют тельная
носят невысо-
кие концентра-
ции О2
30
Глава 1
ществуют две основные группы спорообразующих бактерий.
Аэробные бактерии рода Bacillus чрезвычайно широко распро-
странены в природе и легко адаптируются в любых условиях.
Для нормально развивающихся в анаэробных условиях веге-
тативных форм некоторых видов Clostridium кислород детален,
однако споры этих бактерий устойчивы к действию кислорода.
Другие бактерии, вегетативные формы которых быстро погиба-
ют при 45°C, образуют споры, выдерживающие кипячение в во-
де в течение нескольких часов. Отсюда следует, что если мы
хотим убить микроорганизмы нагреванием {тепловой стерили-
зацией), то для уничтожения спорообразующих бактерий необ-
ходимы более высокие температуры — обычно кипячение под
давлением в автоклаве при температурах выше 120°C.
Мы не будем рассматривать здесь сине-зеленые водоросли
(цианобактерии), не имеющие большого промышленного зна-
чения. Следует отметить, однако, что активно участвующие в
кругообороте азота цианобактерии важны в общем кругообо-
роте веществ в водных экосистемах (гл. 14).
1.3.2. Дрожжи
Дрожжи составляют один из важных классов отдела гри-
бов. Грибы, как и бактерии, широко распространены в природе,
хотя обычно они живут в почве в относительно менее влажных
по сравнению с бактериями регионах. Грибы не способны
усваивать энергию солнечного света и, как правило, существу-
ют изолированно в виде отдельных одноклеточных организмов.
Хотя для большинства грибов характерна довольно сложная
морфология, дрожжи легко отличить по внешнему виду — обыч-
но они представляют собой отдельные небольшие клетки дли-
ной от 5 до 30 мкм и шириной от 1 до 5 мкм.
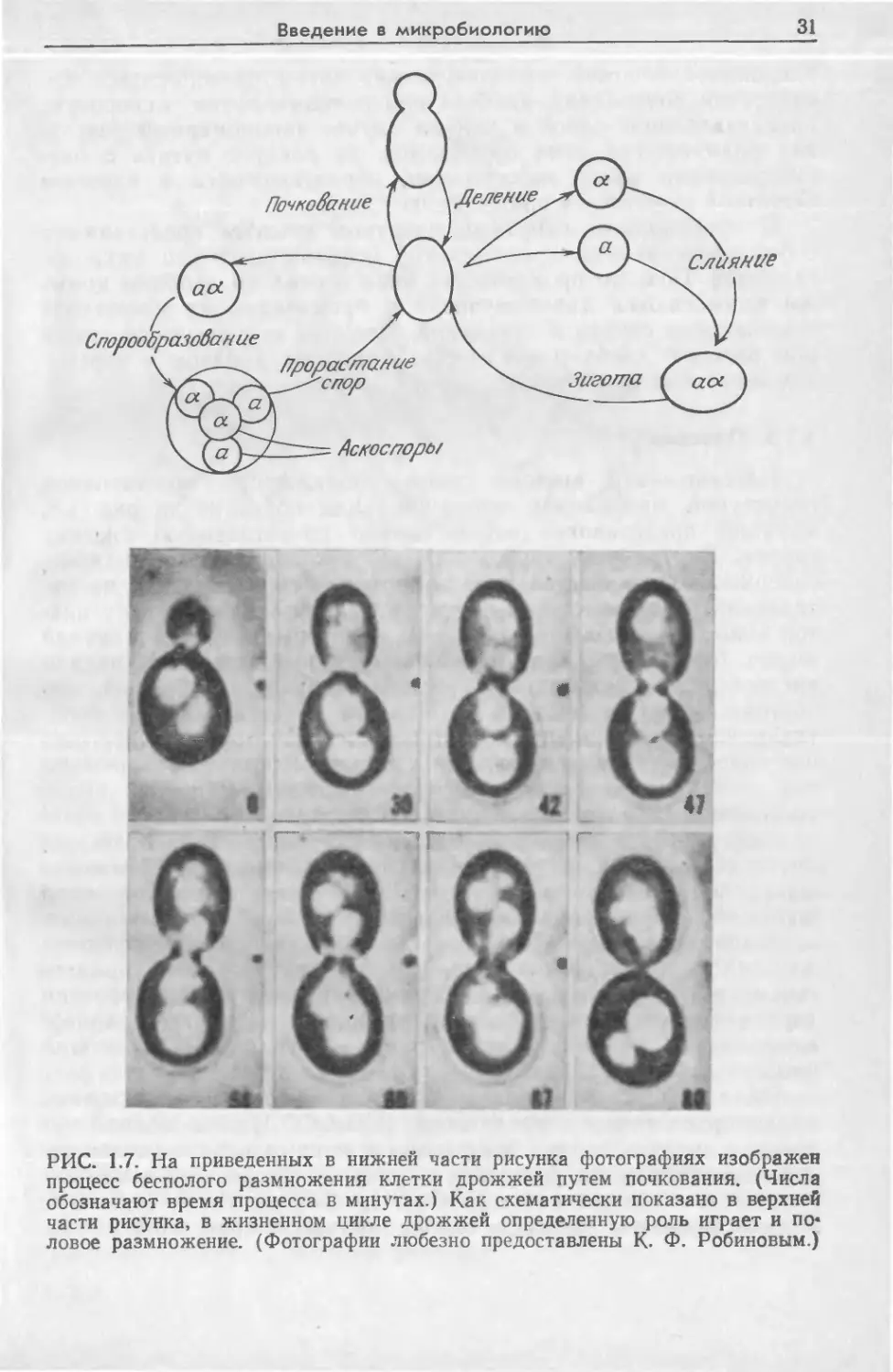
Дрожжи могут размножаться бесполым и половым путями;
схема бесполого размножения (посредством почкования и де-
ления) изображена на рис. 1.7. При почковании на родитель-
ской клетке сначала начинает расти небольшой отросток; от-
деление дочерней клетки от родительской происходит не сразу,
благодаря чему становится возможным образование колоний
дрожжевых клеток, состоящих из нескольких поколений. В ре-
зультате деления из одной клетки образуются две новые. По-
ловое размножение осуществляется путем слияния двух гапло-
идных, (имеющих одинарный набор хромосом) клеток, которое
сопровождается разрушением пограничной стенки и образова-
нием диплоидной (имеющей два набора хромосом) зиготы. Яд-
ро в диплоидной клетке может претерпевать одно или несколько
делений, в результате которых образуются аскоспоры\ каждая
из аскоспор в конце концов становится индивидуальной новой
Введение в микробиологию
31
РИС. 1.7. На приведенных в нижней части рисунка фотографиях изображен
процесс бесполого размножения клетки дрожжей путем почкования. (Числа
обозначают время процесса в минутах.) Как схематически показано в верхней
части рисунка, в жизненном цикле дрожжей определенную роль играет и по*
ловое размножение. (Фотографии любезно предоставлены К. Ф. Робиновым.)
32
Глава 1
гаплоидной клеткой, которая может затем размножаться по-
средством почкования, деления или половым путем. Аскоспоры,
представляющие собой в данном случае закономерный резуль-
тат размножения этих организмов, не следует путать с рас-
смотренными выше эндоспорами, образующимися в качестве
защитной реакции на враждебную среду.
В производстве спиртных напитков дрожжи представляют
собой единственный промышленно используемый тип микроор-
ганизмов. Помимо производства пива и вина анаэробные дрож-
жи применяются для получения в промышленных масштабах
технического спирта и глицерина. Дрожжи используются также
при выпечке хлеба и в качестве белковых добавок к кормам
для животных (см. гл. 12).
1.3.3. Плесени
Плесени — это высшие грибы, обладающие вегетативной
структурой, называемой мицелием. Как показано на рис. 1.8,
мицелий представляет собой сильно разветвленную систему
трубок. Внутри этих трубок находится подвижная цитоплазма,
содержащая множество ядер. Мицелий может состоять из не-
скольких типов родственных клеток. Длинные, тонкие нити кле-
ток мицелия называют гифами. В некоторых случаях мицелий
может быть очень плотным. Учитывая необходимость подачи
кислорода для нормальной жизнедеятельности плесеней, это
обстоятельство может вызвать большие затруднения в их куль-
тивировании, поскольку мицелий может оказывать существен-
ное сопротивление массопередаче. Эта проблема, как и необыч-
ные гидродинамические свойства суспензий мицелия, будут
подробнее рассмотрены в гл. 4 и 8.
Как и дрожжи, плесени не содержат хлорофилла и обычно
не способны передвигаться. Как правило, плесени размножа-
ются спорами половым или бесполым путем. Свойства спор
играют большую роль в классификации грибов.
С промышленной точки зрения наиболее важны плесени
Aspergillus и Penicillium (рис. 1.9). К числу основных продук-
тов метаболизма этих микроорганизмов относятся антибиотики
(продукты жизнедеятельности, убивающие некоторые микроор-
ганизмы или подавляющие их рост), органические кислоты и
биологические катализаторы.
Один из штаммов Aspergillus niger в нормальных условиях
продуцирует щавелевую кислоту (НО2ССО2Н), но если пита-
тельная среда обеднена фосфатами и ионами некоторых метал-
лов, например меди, железа и магния, то преимущественно об-
разуется лимонная кислота НООССН2С(ОН) (СООН)СН2СООН.
Эта особенность лежит в основе промышленного биохимиче-
Введение в микробиологию
33
РИС. 1.8. Структура мицелия плесеней. Условия в центре плотного мицелия
и в его периферийных участках могут существенно различаться.
ского способа производства лимонной кислоты. Таким образом,
плесень A. niger может служить интересным примером различия
в подходах к разработке и оптимизации биохимических и не-
биологических процессов. В биологических системах путем
сравнительно небольшого изменения состава питательной среды
иногда может быть достигнута значительно большая селектив-
ность.
Этот пример, как и приведенный ниже пример пенициллина,
показывает, насколько для специалиста в области биохимиче-
ской технологии важно знать строение клеток, их метаболизм
и функции; другие многочисленные подтверждения этого поло-
жения мы найдем в последующем изложении. Без учета основ-
ных свойств клеток и происходящих в них процессов все умение
инженера-технолога, проявляемое при разработке, проектиро-
вании и анализе различных промышленных биохимических про-
цессов, может оказаться совершенно бесполезным, поскольку
при этом не будут приниматься во внимание ключевые биоло-
гические свойства изучаемой системы.
3-537
34
Глава 1
Второе фундаментальное различие между микробиологиче-
скими и небиологическими процессами можно проиллюстриро-
вать на примере производства пенициллина. Основные успехи
в производстве пенициллина были достигнуты благодаря полу-
чению высокопродуктивных мутантов исходного штамма Penh
cilliutn путем ультрафиолетового облучения его спор (рис. 1.10).
Мутации могут вызываться различными агентами и часто при-
РИС. 1.9. Гифы Aspergillus и Penicillium двух промышленно важных пле-
сеней.
водят к увеличению выхода нужного продукта метаболизма на
несколько порядков. Рассматриваемая в гл. 6 технология ре-
комбинантных ДНК позволяет получать генетически видоизме-
ненные варианты некоторых организмов иным, целенаправлен-
ным и строго контролируемым путем. Таким образом стало
возможным, в частности, наладить производство некоторых про-
дуктов жизнедеятельности животных организмов (белков) с по-
мощью простых бактерий. Эти примеры говорят о той большой
роли, которую играет в биохимической технологии генетика.
В настоящее время генетике уделяется большое внимание как
в высших учебных заведениях и научных учреждениях, так и
в промышленных исследовательских центрах биологического
направления. Практическая важность разработки генетически
Введение в микробиологию
35
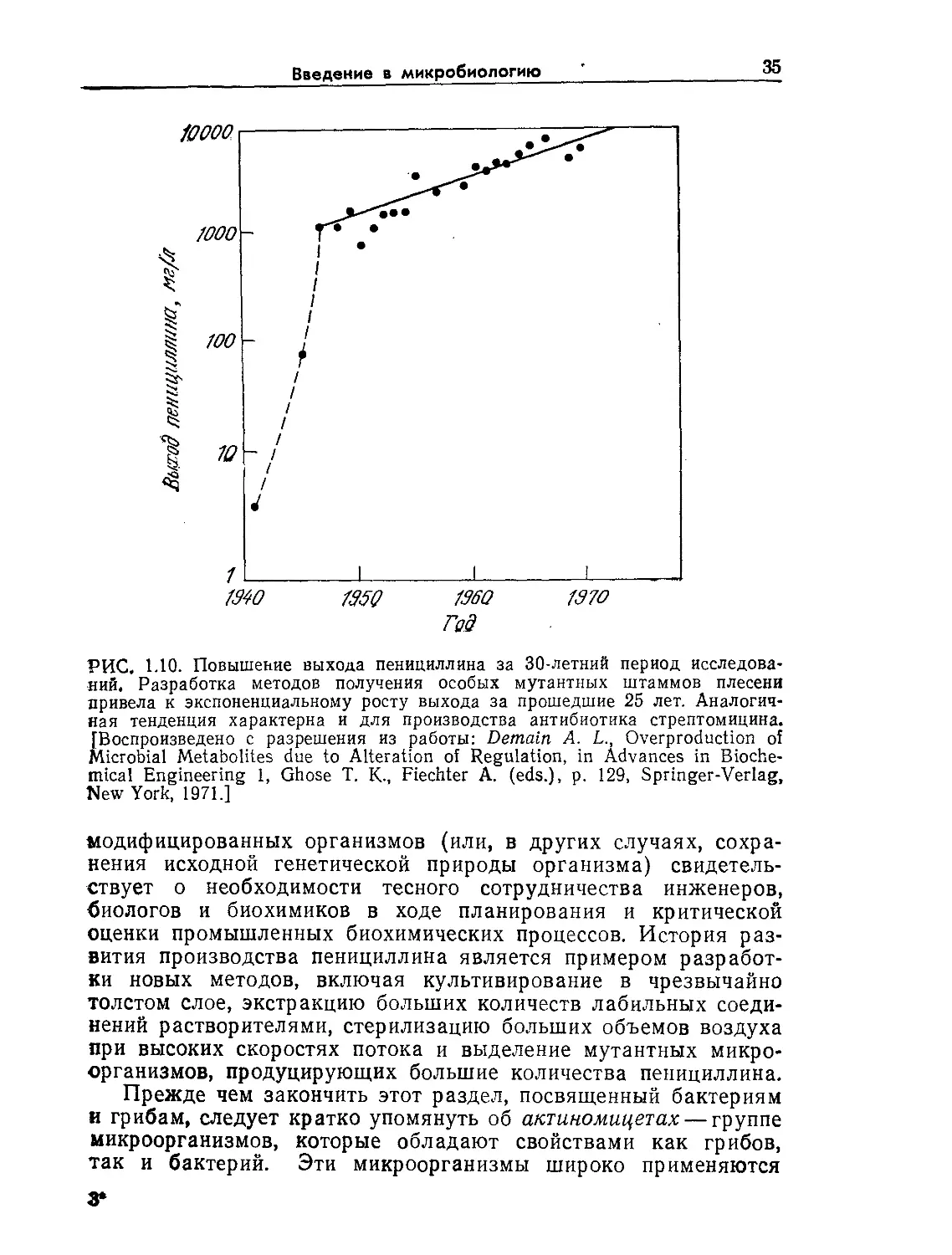
РИС. 1.10. Повышение выхода пенициллина за 30-летний период исследова-
ний, Разработка методов получения особых мутантных штаммов плесени
привела к экспоненциальному росту выхода за прошедшие 25 лет. Аналогич-
ная тенденция характерна и для производства антибиотика стрептомицина.
[Воспроизведено с разрешения из работы: Demain A. L., Overproduction of
Microbial Metabolites due to Alteration of Regulation, in Advances in Bioche-
mical Engineering 1, Ghose T. K-, Fiechter A. (eds.), p. 129, Springer-Verlag,
New York, 1971.]
модифицированных организмов (или, в других случаях, сохра-
нения исходной генетической природы организма) свидетель-
ствует о необходимости тесного сотрудничества инженеров,
биологов и биохимиков в ходе планирования и критической
оценки промышленных биохимических процессов. История раз-
вития производства пенициллина является примером разработ-
ки новых методов, включая культивирование в чрезвычайно
толстом слое, экстракцию больших количеств лабильных соеди-
нений растворителями, стерилизацию больших объемов воздуха
при высоких скоростях потока и выделение мутантных микро-
организмов, продуцирующих большие количества пенициллина.
Прежде чем закончить этот раздел, посвященный бактериям
и грибам, следует кратко упомянуть об актиномицетах — группе
микроорганизмов, которые обладают свойствами как грибов,
так и бактерий. Эти микроорганизмы широко применяются
3*
36
Глава 1
в производстве очень важных антибиотиков. Хотя формально
актиномицеты относят к бактериям, по способности образовы-
вать длинные, чрезвычайно разветвленные гифы они напоми-
нают грибы. Процессы производства антибиотиков с использо-
ванием актиномицетов и плесеней также имеют много общего.
Актиномицеты сближает с бактериями их восприимчивость к
заражению одними и теми же вирусами и к вирусным заболе-
ваниям. Вирусы мы вкратце рассмотрим ниже, в гл. 6.
1.3.4. Водоросли и простейшие
Эти относительно большие эукариоты обладают сложным
и высокоупорядоченным строением. Эвгленовые водоросли, на-
пример, передвигаются с помощью жгутиков, у них нет жесткой
оболочки, но имеется чувствительное к свету пятно, называе-
мое глазком. Последнее реагирует на свет и заставляет клетки
двигаться к более освещенному месту, что немаловажно для
жизнедеятельности этой водоросли, усваивающей, как и боль-
шинство других водорослей, световую энергию. Многие диато-
мовые (другой вид водорослей) имеют наружные двустворча-
тые оболочки (панцири) разнообразной формы, состоящие в ос-
новном из кремнезема. Эти панцири широко используются
в промышленности в качестве фильтрующего материала.
Повышенный интерес к водорослям обусловлен их потенци-
альной ценностью в качестве продукта питания или как добавки
к пищевым продуктам. В Японии, например, в настоящее время
работает несколько промышленных установок, на которых во-
доросли культивируют именно для этих целей. Кроме того,
в Азии в довольно широких масштабах в пищу употребляют
морские водоросли. Последние не являются микроорганизмами
и построены из множества однотипных клеток. Как и более
простые сине-зеленые водоросли, водоросли-эукариоты выпол-
няют важную функцию в круговороте веществ на земле (гл. 14).
В известном смысле водоросли можно рассматривать как
примитивные растения; точно так же простейших, не способных
усваивать солнечную энергию, можно считать примитивными
животными. Естественная среда обитания, морфология и актив-
ность простейших изменяются в довольно широких пределах.
Некоторые трипаносомы, например, являются переносчиками
серьезных заболеваний, включая африканскую сонную болезнь,
или трипаносомиаз. С другой стороны, простейшие Trichonympha
населяют кишечник термитов и помогают им переваривать дре-
весину. Амебы не обладают какой-либо определенной формой
и постоянно меняют свои внешние очертания, в то время как
для солнечников (Heliozoa) характерно наличие внутреннего
скелета и определенной формы.
Введение в микробиологию
37
Хотя простейшие в настоящее время не используются в про-
мышленном масштабе ни для производства клеточной массы,
ни для синтеза продуктов их жизнедеятельности, они наряду
с микроорганизмами играют большую роль в биологической
очистке сточных вод (гл. 14). С точки зрения микробиолога
эти процессы, широко применяющиеся во всем мире в городах
и на больших промышленных предприятиях, поразительно слож-
ны. Бытовые и промышленные сточные воды представляют со-
бой сложную смесь, в состав которой входят различные пита-
тельные вещества и самые разнообразные микроорганизмы; по-
этому для обработки стоков необходимо также большое число
различных протестов. Эти организмы конкурируют в потребле-
нии питательных веществ, уничтожают друг друга и взаимо-
действуют многими другими путями, характерными для неболь-
шой экологической системы. Подробнее вопросы взаимодействия
между различными видами будут рассмотрены и проанализи-
рованы в гл. 13.
1.3.5. Растительные и животные клетки
Многие вакцины и другие биохимикаты продуцируются в хо-
де роста животных клеток в реакторах, т. е. при выращивании
клеток вне организма животного. Совершенствование методов
культивирования тканевых клеток и разрабатываемые в послед-
ние годы методы генетической трансформации животных и рас-
тительных клеток открывают новые многообещающие пути для
их значительно более широкого промышленного использования.
Тканевые клетки можно выращивать почти в таких же реакто-
рах, какие используются для культивирования микроорганиз-
мов; по этой причине кинетика роста как тканевых, так и мик-
робных клеток и применяющиеся для этой цели биохимические
реакторы будут рассмотрены в одних главах (гл. 7 и 9 соответ-
ственно). Ниже мы вкратце обсудим важнейшие типы клеток
высших организмов, культуры которых могут быть выращены
в технологической аппаратуре независимо от организма живот-
ного или растения, от которого эти клетки были отобраны.
Если часть ткани животного (обычно полученную путем раз-
рушения межклеточных связей) поместить в соответствующую
питательную среду, то большинство типов клеток, в частности
клетки крови, погибнут в течение нескольких дней, недель или
месяцев. Другие клетки в этих условиях размножаются и дают
так называемую первичную линию клеток. Часто эти клетки
удается пассировать, т. е. перенести в свежую питательную
среду, где снова происходит размножение клеток, приводящее
ко вторичной линии клеток. Некоторые вторичные клетки, вы-
держивающие, по всей вероятности, неограниченное число пас-
38
Глава 1
сажей, называют стабильной, перманентной или установившейся
линией клеток.
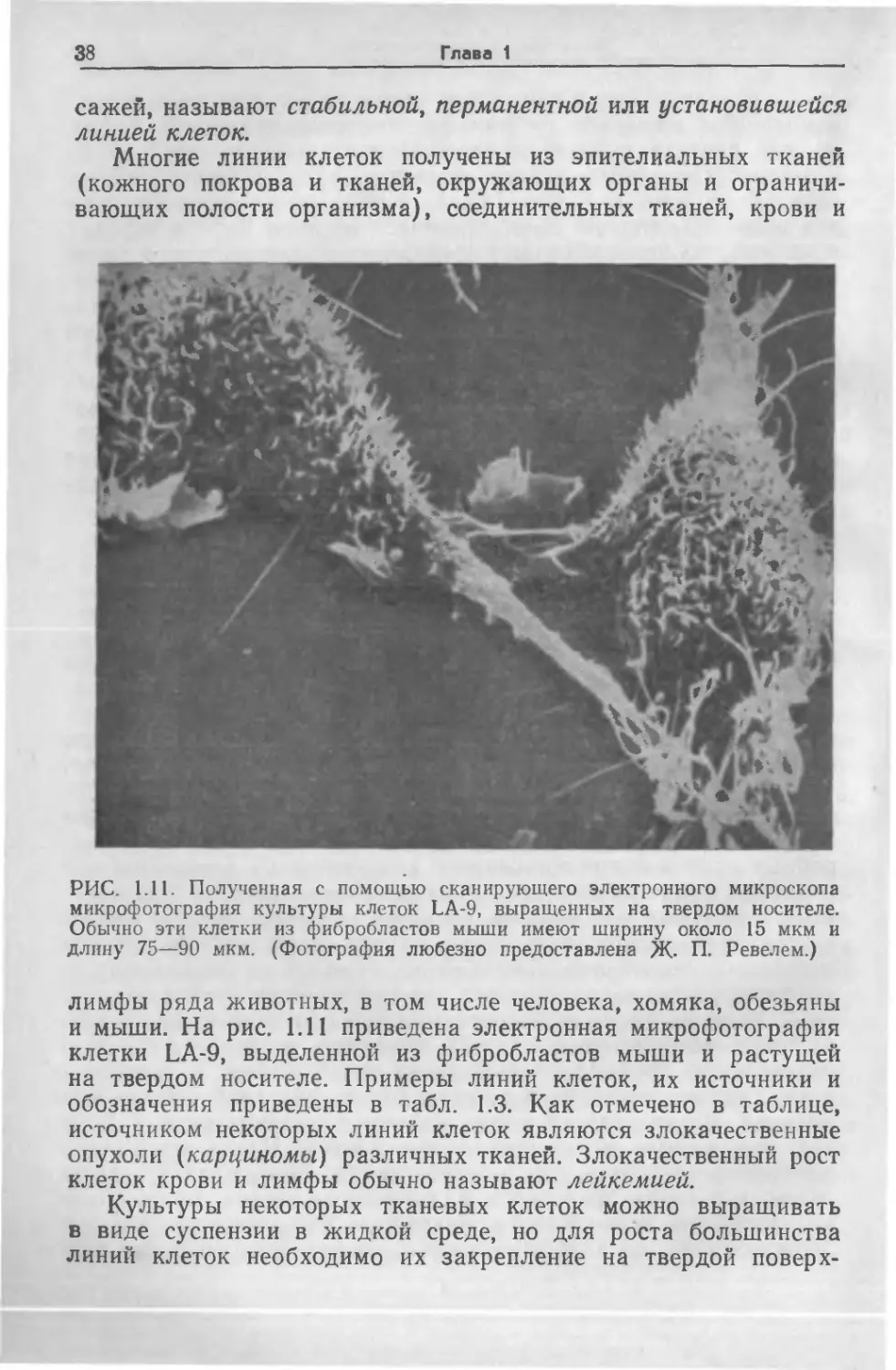
Многие линии клеток получены из эпителиальных тканей
(кожного покрова и тканей, окружающих органы и ограничи-
вающих полости организма), соединительных тканей, крови и
РИС. 1.11. Полученная с помощью сканирующего электронного микроскопа
микрофотография культуры клеток LA-9, выращенных на твердом носителе.
Обычно эти клетки из фибробластов мыши имеют ширину около 15 мкм и
длину 75—90 мкм. (Фотография любезно предоставлена Ж. П. Ревелем.)
лимфы ряда животных, в том числе человека, хомяка, обезьяны
и мыши. На рис. 1.11 приведена электронная микрофотография
клетки LA-9, выделенной из фибробластов мыши и растущей
на твердом носителе. Примеры линий клеток, их источники и
обозначения приведены в табл. 1.3. Как отмечено в таблице,
источником некоторых линий клеток являются злокачественные
опухоли (карциномы) различных тканей. Злокачественный рост
клеток крови и лимфы обычно называют лейкемией.
Культуры некоторых тканевых клеток можно выращивать
в виде суспензии в жидкой среде, но для роста большинства
линий клеток необходимо их закрепление на твердой поверх-
Введение в микробиологию
39
ности, что налагает серьезные ограничения на масштабы про-
мышленного производства вакцин и других биологических про-
дуктов на основе культур животных клеток. Метод культиви-
рования на микроносителях, который мы подробнее рассмотрим
в гл. 9, позволил значительно повысить производительность ре-
акторов (в расчете на единицу объема), предназначенных для
выращивания культур клеток, растущих только на носителях.
Культуры некоторых клеток растительного происхождения
можно выращивать также в виде каллюса (нароста недиффе-
Таблица 1.3. Примеры стандартных линий клеток животных и их
происхождение
Обозначение линии клеток Животное
Ткань
HeLa (CCL2)a
HLM
FS-4
MK2
CHO (CCL 61)
L-M (CCL 2)
Человек
Человек
Человек
Обезьяна
Китайский хомяк
Мышь
Карцинома шейки матки
Печень эмбриона
Фибробласт препуциума
Почки
Яичник
Соединительная ткань
а Обозначение CCL применяется Банком культур клеток Американской коллекции
типов культур (Cell Culture Repository, American Type Culture Collection, 12301 Park-
lawn Drive, Rockville, Maryland 20852 USA); эта организация является хранителем и
поставщиком линий клеток.
ренцированной ткани растения на твердой питательной среде)
или в виде суспензии агрегированных клеток. Поскольку рас-
тения продуцируют множество практически важных соединений,
в том числе душистые вещества, красители, лекарственные
средства и опиаты, использование культур растительных клеток
в будущем может оказаться весьма перспективным. Культуры
растительных клеток могут также служить весьма специфичны-
ми катализаторами ряда ценных реакций. Культуры раститель-
ных клеток могут оказаться полезными и в сельском хозяйстве,
например для регенерации целого растения. В то же время
будущему широкомасштабному применению культур раститель-
ных клеток должно предшествовать более тщательное изучение
общей биологии растений, а также особенностей и ограничений
процессов культивирования растительных клеток. Следует от-
метить, что можно культивировать также тканевые клетки на-
секомых и других беспозвоночных, однако в последующих гла-
вах при обсуждении культур тканевых эукариот мы ограни-
чимся клетками животного и растительного происхождения.
40
Глава 1
1.4. Перспективы дальнейшего изучения
Приведенные в этой главе краткие сведения о строении и
классификации клеток позволяют убедиться в обоснованности
и полезности клеточной теории в качестве основы биологической
науки. Здесь мы также неоднократно подчеркивали важность
фундаментальных основ биохимии для понимания процессов
биохимической технологии. И в последующих главах мы по-
стоянно будем уделять внимание основным положениям биоло-
гии клетки.
В гл. 2 будут рассмотрены основные типы химических соеди-
нений, которые клетка должна синтезировать для обеспечения
нормальной жизнедеятельности и размножения; гл. 3 и 4 посвя-
щены биологическим катализаторам, используемым клеткой для
осуществления химических реакций. Затем в гл. 5 мы изучим
последовательности реакций, необходимые для нормального
функционирования клеток, а гл. 6 будет посвящена главным
образом способам контроля и регулирования этих реакций и
проблемам генетики. В гл. 7 мы рассмотрим вопросы кинетики
роста микробных систем, а последующие главы будут посвя-
щены проблемам анализа, проектирования, контроля и оптими-
зации процессов биохимической технологии.
Упражнения
1.1. Выдающиеся микробиологи. Ознакомьтесь с биографией одного из
известных в прошлом микробиологов, например Роберта Гука, Антони ван
Левенгука, Ладзаро Спалланцани, Луи Пастера, Уолтера Рида, Д. И. Ива-
новского, П. Роуза, Теодора Шванна или М. Я. Шлейдена. Подготовьте крат^
кий обзор, указав в нем, какие технические и социальные трудности при-
шлось преодолеть этим ученым, какие методические достижения были (или
не были) достигнуты в ходе его работ, укажите, какое место занимали ме-
тоды индукции и дедукции в его исследованиях.
1.2. Экспериментальная микробиология. Микробиология располагает мно-
жеством простых и хорошо отработанных методик, которые, однако, могут
оказаться незнакомыми специалистам в области общей, органической и фи-
зической химии. Поскольку любое экспериментальное изучение базируется на
наблюдениях и измерениях, необходимо ознакомиться с методами практиче-
ского исследования, в том числе с обеспечиваемой ими точностью. Если вы
ранее не прошли курс лабораторных работ по микробиологии, то желатель-
но сделать это одновременно с изучением данного теоретического курса или
после него. При отсутствии такой возможности внимательно изучите разделы
краткого руководства по лабораторным работам (например, [7]), соответст-
вующие главам этой книги. При этом не забывайте высказывание Клода Бер-
нара: «Постановка эксперимента без четко поставленной цели равносильна
бесцельному блужданию». При изучении экспериментальных методов в лабо-
ратории или по руководству старайтесь сформулировать цель (или цели)
каждого эксперимента. Подумайте, какую дополнительную информацию мож-
но получить в ходе каждого эксперимента.
1.3. Методы изучения. Ознакомьтесь с кратким описанием методов мик-
41
В в е д ен ие в микробиологию
ппскопии в том числе с методами темного поля, фазового контраста, флуо-
ресцентной и электронной микроскопии. Сформулируйте относительные р
HMy^cTBj"^^nole^nocTapa^Jec^
минам, а при наличии нескольких терминов в одной строке постараит
дать сравнительную их оценку:
а)
б)
"S™XOaTVeS“°^3MaT„,ecKaa мембрана, эндоплазматический
ретикулум;
в) цитоплазма;
г) ядро, нуклеоид;
д) рибосома, митохондрия, хлоропласт;
е) морфология;
ж) спиральные, сферические и палочковые формы бактерий;
з) почкование, половое слияние, деление, спорообразование;
и) простейшие, водоросли, мицелий, амебы.
1.5. Идентификация и классификация, а) Начертите схему царства про-
тистов (по памяти).
б) Таксономическое деление микроорганизмов на виды в основном бази-
руется на результатах визуальных наблюдений с помощью оптического мик-
роскопа. Найдите изображения микроорганизмов Escherichia coli, Staphylo-
coccus aureus, Bacillus cereus и Spirillum serpens в справочных пособиях
[например, Краткий определитель бактерий Берги (М.: Мир, 1980) или «А Guide
to the Identification of the Genera of Bacteria» Скермана]. Перечислите отличи-
тельные черты любых двух видов из этих бактерий, которые позволили бы
идентифицировать их достаточно надежно. Начните с самых общих черт и по-
степенно переходите от семейства к более детализированным подразделениям
(подсемейству, роду) и, наконец, к виду.
1.6. Поэтапное изучение проблемы. Выберите интересную микробиологи-
ческую тему (процессы брожения в пивоварении, производство антибиотиков,
рост дрожжей, обработка сточных вод, микробиология почвы, вакцины, ква-
шение, производство сыра, экология озер, микробиология морской воды
и т. д.). Сначала ознакомьтесь с темой в том объеме, в каком она изложе-
на в общих руководствах, например, в книге «Encyclopedia of Chemical
Technology» Кирка и Отмера. Начертите схему технологического процесса
или явлений, происходящих в природной системе, укажите важнейшие участ-
вующие в этом процессе микроорганизмы, источники их питания, потоки
сточных вод или отходящие потоки. В ходе изучения курса каждую неделю
добавляйте к этой схеме одну-две страницы текста, в которых указывайте,
в частности, важность (или ее отсутствие) темы пройденной главы для дан-
ного процесса. По окончании курса подготовьте краткий доклад по вашей
теме и представьте его студентам вашей группы.
1.7. Промышленная микробиология. Краткое описание истории промыш-
ленной микробиологии и ряд предложений по ее развитию можно найти
в статье «Industrial Microbiology» \Demain A. L., Solomon N. A., Scientific
American, 245, 67—75 (1981)]. Прочтите эту статью и подготовьте краткий
(объемом около одной страницы) обзор об историческом развитии промыш-
ленной микробиологии, датах основных событий, продуктах и (или) процессах
этой отрасли промышленности и о видах применяющихся микроорганизмов.
1.8. Подвижность простейших. Предположим, что с помощью микроскопа
вы наблюдаете движение сферических простейших и можете определить их
размеры и скорость движения, выраженную в количестве длин организма,
преодолеваемых им в единицу времени.
а) Вычислите числа Рейнольдса для потока жидкости вокруг организма
для приведенных ниже размеров организмов и скоростей их движения при
условии, что жидкостью является вода при 20 °C и что поток жидкости
42
Глава 1
практически не подвергается возмущению жгутиками, слизистыми оболочка-
ми или вращением организмов.
Размеры: 10 мкм, 50 мкм, 100 мкм.
Скорости: 10 длин организма в секунду;
1 длина организма в секунду;
0,1 длины организма в секунду.
Какие режимы потоков характерны для этих движений клеток?
б) Для замедления движения микроорганизмов и, следовательно, для
облегчения наблюдений за ними в микроскопии часто применяют раствор
метилцеллюлозы. Как скажется на скорости движения клеток увеличение
вязкости окружающей их жидкости в 104 раз, если принять, что расходы
энергии на движение организма не меняются?
1.9. Пластинчатая разводка Е. coli. При выращивании Е. coli для
дальнейших экспериментов желательно иметь генетически гомогенную попу-
ляцию. Обычно для этой цели исходный бульон разбавляют до такой кон-
центрации, при которой на агаровых пластинках образуются отдельные ко-
лонии, образовавшиеся из одной клетки. Далее популяцию выращивают на
основе одной из таких колоний.
а) Предположим, что нам даны круглая агаровая пластинка радиусом
4 см, 1 мл бульона с концентрацией 1010 клеток на литр и 1 л стерильного
бульона для разбавления. Насколько следует разбавить бактериальный
бульон, чтобы на агаровой пластинке образовалось около 100 отдельных
колоний? (Примите, что пластинку радиусом 4 см можно покрыть 5 мл
раствора.)
б) Как вы смогли бы получить колонии из индивидуальных клеток, если
концентрация клеток в бульоне неизвестна?
1.10. Разделение с помощью центрифугирования. Рассмотрим разбавлен-
ную суспензию частиц типа А и типа В. Первоначально однородная суспен-
зия вращалась в центрифуге с угловой скоростью со в течение времени t.
Найдите уравнение, позволяющее определить, сколько частиц типа А оста-
лось в супернатантной суспензии после центрифугирования (по отношению
к исходной относительной концентрации f0 частиц А). Время t следует при-
нять достаточно малым, чтобы в суспензии остались частицы как типа А,
так и типа В.
1.11. Центрифугирование с угловым ротором. Рассмотрим схематично
изображенную на рис. 1У11.1 центрифугу, в которой пробирки расположены
под углом к оси вращения. Найдите время, за которое сферическая частица
РИС. 1У11.1.
Введение в микробиологию
43
радиусом R и плотностью р₽ пройдет расстояние от г=п до г=г2 при угло-
вой скорости вращения ротора <в, плотности жидкости р/ и ее вязкости у./,
если угол вращения составляет 0 от вертикали. Учтите, что стеклянная стен-
ка пробирки прилагает к частице вертикально направленную силу.
Литература
1. Sistrom IF. R., Microbial Life, 2d ed., Holt, Rinehart and Winston, Inc., N.Y.,
1969. В первых трех главах этого введения в микробиологию рассмотрено
большинство тем первой главы настоящего учебного пособия. В эту книгу
включены также сведения о метаболизме, росте и генетике микроорганиз-
мов, которые здесь будут рассмотрены в последующих главах.
2. Frobisher М., Fundamentals of Microbiology, 8th ed., W. B. Saunders Com-
pany, Phila., 1968. В этой книге описаны различные формы микробной жиз-
ни, а также классификация клеток, вирусы, проблемы стерилизации, имму-
нологии и применения микроорганизмов. Особое внимание уделено много-
численным связям между микроорганизмами и человеком.
3. Pelczar М. J., Jr., Reid R. D., Chan E. C. S., Microbiology, 4th ed., McGraw-
Hill, New York, 1977. Пособие для углубленного изучения микробиологии.
Детально рассмотрены грибы, водоросли, простейшие и вирусы, а также
промышленная микробиология и микробиология окружающей среды, физи-
ческие и химические методы регулирования размножения микроорганизмов.
4. Stanier R. Y„ Doudoroff М., Adelberg Е. A., The Microbial World, 4th ed.,
Prentice-Hall, Inc., Englewood Cliffs., N.J., 1975. Книга также довольно
глубокого содержания, написанная очень хорошим языком. В ней рассмот-
рены история микробиологии, классы микроорганизмов, вопросы симбиоза,
природы заболеваний, а также микробного метаболизма. Детально освеще-
на генетика на молекулярном уровне, включая проблемы мутаций и регу-
ляции.
5. The Living Cell, readings from Scientific American, W. H. Freeman and Com-
pany, San Francisco, 1965. Сборник статей из журнала «Scientific American»,
посвященных проблемам строения, энергетики, синтеза, деления и диффе-
ренциации клеток, а также ряду специальных вопросов, например коммуни-
кации, стимулированию и сокращению мышц. Несмотря на то что некото-
рые статьи отчасти устарели, этот тематически разнообразный и хорошо
иллюстрированный сборник все еще заслуживает большого внимания.
6. Paul J., Cell and Tissue Culture, 5th ed., Churchill Livingstone, Edinburgh,
1975. Великолепная исчерпывающая вводная монография, посвященная ли-
ниям животных клеток, методам и принципам культивирования клеток
и практическому использованию результатов биологических исследований.
Литература к разделу «Упражнения»
7. Crabtree К. Т., Hinsdill R. D., Fundamental Experiments in Microbiology,
W. B. Saunders Co., Phila., 1974.
Глава 2
Химические основы жизни
Любой организм должен синтезировать все химические сое-
динения, необходимые для жизнедеятельности и размножения
клеток. В последующих главах мы рассмотрим вопросы кине-
тики, энергетики и регулирования основных биохимических пу-
тей таких синтезов, однако, прежде чем приступить к этим
вопросам, необходимо ознакомиться с реагентами, продуктами
реакций, катализаторами и химическими регуляторами, которые
принимают участие в сложной сети происходящих в клетке
химических превращений.
Настоящая глава посвящена главным образом изучению
преобладающих в клетке высокомолекулярных соединений,
а также соответствующих небольших мономерных молекул, из
которых построены эти полимеры. Полимерные соединения
клетки делятся на четыре основных класса: жиры и липиды,
полисахариды (целлюлоза, крахмал и т. д.), носители инфор-
мации полидезоксирибонуклеиновые и полирибонуклеиновые
кислоты (ДНК, РНК), а также белки. Физико-химические свой-
ства этих соединений важны как для понимания функций клет-
ки, так и для рационального проектирования технологических
процессов с участием живых клеток.
В зависимости от строения различные биологические поли-
меры целесообразно подразделять на гомополимеры и сополи-
меры. Биологические гомополимеры построены из мономерных
единиц одного типа; в таком случае полимеры, содержащие
мономерные звенья одного типа, отличаются один от другого
прежде всего молекулярной массой и степенью разветвленности
полимерных цепей. Основная функция гомополимеров в клетке
заключается в создании структурных элементов, обладающих
необходимыми механической прочностью, химической инертно-
стью и достаточной проницаемостью. Кроме того, в виде гомо-
полимеров в клетках часто хранятся запасы питательных ве-
ществ-, 1 М раствор глюкозы, например, может храниться в ви-
де полимера гликогена, своеобразного резервного полисахарида
клетки, обеспечивающего снижение молярной концентрации
раствора в 10000 раз или даже в еще большей степени. По-
лимер является особенно удобной формой хранения питатель-
Химические основы жизни
45
ных веществ в тех случаях, когда клетке необходимо создать
их избыток без существенного изменения внутриклеточного ос-
мотического давления.
Сополимеры построены из нескольких различных мономер-
ных звеньев, число которых может достигать 20. Каждый из
таких полимеров имеет определенную молекулярную массу и
характерный мономерный состав; более того, остатки мономеров
соединены в строго определенной, генетически запрограммиро-
ванной последовательности.
Таблица 2.1. Элементный состав Е. со11л
Элемент Процентное содер- жание в расчете на массу сухого веще- ства Элемент Процентное содер- жание в расчете иа массу сухого веще- ства
Углерод 50 Натрий 1
Кислород 20 Кальций 0,5
Азот 14 Магний 0,5
Водород 8 Хлор 0,5
Фосфор 3 Железо 0,2
Сера 1 Все другие эле- 0,3
Калий 1 менты
а Данные из работы: Luria S. Е„ in The Bacteria, Gtmsalus I. C., Stanier R. Y.
(eds.), vol. 1, chap. 1, Academic Press, Inc., New York, 1960,
На приведенном в табл. 2.1 элементном составе Е. coli мож-
но видеть пример того набора химических элементов, из кото-
рых построены биополимеры. Преобладающие элементы (водо-
род, кислород, азот и углерод) образуют химические связи
путем заполнения своих внешних электронных оболочек одним,
двумя, тремя и четырьмя электронами соответственно. Это са-
мые легкие элементы периодической системы; за исключением
водорода, все они способны также образовывать кратные хи-
мические связи. Из этих элементов, а также в меньшей степени
из фосфора и серы построено огромное множество соединений,
в том числе и все четыре основных класса биополимеров.
Биохимические соединения, построенные из этих элементов,
отличаются не только большим разнообразием, но и высокой
устойчивостью; как правило, они очень медленно взаимодейству-
ют друг с другом, с водой или другими компонентами клетки.
Химические реакции с участием таких соединений ускоряются
биологическими катализаторами — особыми белками, называе-
мыми ферментами (напомним, что катализатором называется
вещество, повышающее скорость реакции, но само при этом
46
Глава 2
не подвергающееся необратимым изменениям). Таким образом,
клетка может регулировать как природу, так и скорость про-
исходящих в ней химических реакций путем изменения концент-
раций ферментов. Детали этих регуляторных механизмов будут
рассмотрены в гл. 6.
Фосфор и сера входят в состав органического вещества всех
живых существ, хотя и в относительно небольших количествах.
В клетках всегда присутствуют также ионы натрия, калия, маг-
ния, кальция и хлора, а следовые количества марганца, железа,,
кобальта, меди и цинка необходимы для активации определен-
ных ферментов. Для нормальной жизнедеятельности некоторых
организмов требуются также ничтожные количества бора, алю-
миния, ванадия, молибдена, иода, кремния, фтора и олова..
В общем случае для жизни необходимы по меньшей мере-
24 различных химических элемента.
Клетки живут в водной среде. Вода обладает целым рядом
весьма необычных свойств (высокой теплотой испарения, высо-
кой диэлектрической проницаемостью, способностью образовы-
вать при диссоциации кислоты и основания, склонностью к об-
разованию водородных связей), благодаря которым она явля-
ется чрезвычайно важным реагентом, участвующим во многих
катализируемых ферментами реакциях. Кроме того, проявляе-
мые биополимерами свойства в большой степени зависят от
свойств того растворителя, в среде которого они находятся; на
этом принципе основаны, в частности, многие процессы разде-
ления. взаимосвязь воды с другими обычными компонентами
клеток обсуждена Блюмом [12].
2.1. Липиды
Липидами называют соединения биологической природы,
растворимые в неполярных растворителях (бензоле, хлорофор-
ме, эфире и т. п.) и практически нерастворимые в воде. Из
такого определения следует, что липиды могут иметь различное
химическое строение и выполнять различные биологические
функции. Их относительно низкая растворимость в водных сре-
дах является причиной того, что липиды встречаются в основ-
ном в неводных биологических фазах, в особенности в клеточ-
ных мембранах и мембранах органоидов. К липидам относятся
жиры, представляющие собой резервы полимерного биологиче-
ского топлива, а также некоторые важные медиаторы биологи-
ческих процессов. Липиды также входят в состав более слож-
ных соединений, например липопротеинов и липосахаридов,
которые опять-таки в основном располагаются в биологических
мембранах клеток и во внешних оболочках некоторых вирусов.
Химические основы жизни
47
2.1.1. Жирные кислоты и родственные липиды
Насыщенные жирные кислоты представляют собой отно-
сительно простые липиды общей формулы СНз(СНг)яСООН.
В процессе биосинтеза углеводородная цепь жирных кислот
строится из идентичных мономерных звеньев с двумя атомами
углерода, поэтому жирные кислоты можно рассматривать как
не несущие информации биополимеры с концевой карбоксиль-
ной группой. В биологических системах п обычно принимает
четные значения от 12 до 20.
Полярная карбоксиль-
ная группа 1
Липидный монослой на границе
раздела раз воздух-вода
деполярная
'углеводородная
цепь
Липидная мицелла
в воде
РИС. 2.1. Некоторые устойчивые агрегированные состояния жирных кислот
в воде.
Ненасыщенные жирные кислоты образуются при замене на-
сыщенной (—С—С—) связи на двойную связь (—С=С—).
Например, олеиновая кислота является ненасыщенным анало-
гом стеариновой кислоты (п=16):
СНз (СН2) 16СООН СНз (СН2) 7СН=СН (СН2) 7СООН
стеариновая кислота олеиновая кислота
Углеводородная цепь придает этим соединениям гидрофоб-
ные свойства, но карбоксильная группа в высшей степени гид-
рофильна. Поэтому, когда жирная кислота находится на гра-
нице раздела фаз воздух — вода, небольшое ее количество
образует ориентированный мономолекулярный слой (монослой),
в котором полярные карбоксильные группы связаны водой,
а углеводородные цепи направлены в сторону воздушной фазы
(рис. 2.1). Это явление лежит в основе механизма действия
моющих средств, представляющих собой соли жирных кислот.
Образование мыльного монослоя значительно снижает поверх-
48
Глава 2
костное натяжение на границе воздух—вода, что резко повы-
шает способность раствора смачивать и очищать загрязненные
места.
О
II
Na+~O—С—(СН2)7НС=СН(СН2)7СНз
мыло — олеат натрия
Такого рода гидрофобно-гидрофильные молекулы липидов
обладают очень невысокой растворимостью; повышение кон-
центраций раствора выше того предела, который необходим
для создания монослоя, приводит к агрегированию избытка
растворенного вещества в виде сравнительно больших упоря-
доченных структур, называемых мицеллами (рис. 2.1). Движу-
щей силой этого процесса является уменьшение общей свобод-
ной энергии системы в процессе формирования мицелл из
раствора. Та или иная структура мицеллы диктуется увеличе-
нием числа энергетически выгодных контактов между группи-
ровками одинаковой степени гидрофобности (гидрофильности)
и соответствующим уменьшением числа взаимодействий между
гидрофобными и гидрофильными группировками. Известно, что
аналогичные взаимодействия между гидрофобными и гидро-
фильными участками одного биополимера являются причиной
существования полимерной цепи в одной предпочтительной кон-
формации. Такое поведение ДНК и белков мы вкратце рассмот-
рим позднее.
Жиры, выполняющие важную функцию внутриклеточного
топлива, представляют собой сложные эфиры, образующиеся
при конденсации жирных кислот с глицерином:
СН2ОН НО—ОС(СН2)„ — сн3
+
СНОН + НО—ОС(СН2)„:—сн3
+
СН2ОН НО—ОС(СН2)Лз—сн3
глицерин жирная кислота
СН2О—С-(СН2)„-сн,
о
II
СНО—С—(СН2)„,—СН3
о
II
СН2О-С-(СН2)„-СН3:
жир
В щелочной среде при нагревании жиры и другие липиды,
рассматриваемые в настоящем разделе, гидролизуются до гли-
церина и солей жирных кислот (мыла) — именно таким путем
мыла были впервые получены из животных жиров. Подобная
Химические основы жизни
49
реакция, обратная приведенной выше схеме синтеза жиров,
в пищеварительном тракте животных осуществляется при тем-
пературе тела и катализируется особыми ферментами, способ-
ными расщеплять жиры; микроорганизмы также продуцируют
такие ферменты, роль которых заключается в гидролизе неко-
торых жиров на более мелкие фрагменты, способные затем про-
никать в клетку через клеточные мембраны.
По строению (но не по выполняемой ими функции) жирам
близки фосфоглицериды. В молекулах последних остаток фос-
форной кислоты замещает один из концевых остатков жирной
кислоты. В результате получается соединение, в молекуле ко-
торого опять-таки имеются гидрофильные и гидрофобные ос-
татки, поэтому фосфоглицериды в достаточно высоких концент-
рациях также способны образовывать мицеллы. Если в раствор
фосфолипида поместить пластинку с небольшим отверстием, то
в последнем может сформироваться плоский бимолекулярный
слой (бислой) толщиной около 70 А (7-Ю-7 см) (рис. 2.2, а).
Биологические плазматические мембраны обычно содержат зна-
чительные количества фосфолипидов и других полярных липи-
дов. Кроме того, для плазматических мембран характерен хо-
рошо различимый на электронных микрофотографиях бимоле-
кулярный слой (рис. 2.2,6) примерно такой же толщины, как
и изображенный на рис. 2.2,а самопроизвольно образующийся
двойной фосфоглицеридный слой. По этой причине синтетиче-
ские бислойные липидные мембраны могут служить удобной
моделью для изучения основных особенностей тонких биологи-
ческих мембран.
По ряду физических свойств липидные бислойные мембраны
напоминают клеточные мембраны. Оба типа мембран имеют
высокое электрическое сопротивление и значительную электри-
ческую емкость, что в большой степени обусловливает непро-
ницаемость природных мембран для веществ, несущих электри-
ческий заряд, например фосфорилированных соединений. Это
свойство мембран в свою очередь позволяет клеткам хранить
запас заряженных питательных веществ и промежуточных про-
дуктов метаболизма, а также поддерживать высокую разность
концентраций небольших катионов (Н+, К+, Na+ и т. п.) внутри
клетки и вне ее.
Другие компоненты мембран и их роль в обмене веществ
между клеткой и средой будут рассмотрены в разд. 2.5 и 5.7.
Барьеры и каналы транспорта биохимически важных соедине-
ний определяют, какие вещества из числа участвующих в слож-
ной сети катализируемых ферментами процессов проникнут
в клетку, какие останутся в ней, а какие будут выведены из
клетки. Функции регулирования транспорта веществ обязатель-
ны для нормальной жизнедеятельности любой клетки; они так-
1—537
50
Глава 2
РИС. 2.2. а — самопроизвольное образова-
ние устойчивого фосфоглицеридного бислоя
отверстии перегородки, помещенной в со-
суд с раствором липида; б — по внешнему
виду самопроизвольно образующийся двой-
ной слой напоминает аналогичные структу-
ры клеточных мембран, хорошо различимые
на электронных микрофотографиях. [Рис.
2.2,6 воспроизведен с разрешения из рабо-
ты: Robertson J. В., Membrane Models:
Theoretical and Real, in The Nervous Sys-
tem, vol. 1: The Basic Neurosciences, To-
wer B. D. (ed.), p. 43, Raven Press, New
York.]
Химические основы жизни
51
же играют важную роль в технологических процессах, в кото-
рых клетки являются катализаторами.
Путем добавления небольшого количества различных ве-
ществ можно селективно изменять проницаемость как модель-
ных бимолекулярных, так и плазматических мембран относи-
тельно определенных ионов. В частности, было установлено,
что некоторые антибиотики и другие соединения, связывающие
катионы в комплексы, способны значительно усиливать пассив-
ный ионный транспорт в обеих типах мембран. Более сложные
процессы обусловливают повышение проницаемости клеточных
стенок живых организмов после их мягкой химической или
тепловой обработки. Это свойство клеток с успехом использу-
ется в процессах микробиологического производства веществ,
являющихся промежуточными соединениями в клеточном ме-
таболизме, а также в целях снижения содержания нуклеино-
вых кислот в клеточной массе, применяемой в качестве корма
для животных.
2.1.2. Жирорастворимые витамины, стероиды и другие липиды
Витаминами называют органические вещества, следовые ко-
личества которых необходимы для нормальной жизнедеятель-
ности клеток. Незаменимыми называют те витамины, которые
сам организм не может синтезировать; при их отсутствии в пи-
тании клетка погибает. (Этот факт послужил основой теста
на наличие или отсутствие какого-либо витамина путем выра-
щивания микроорганизмов в исследуемой среде.) Понятно, что.
водорастворимые витамины, например витамин С (аскорбино-
вая кислота), не являются липидами. В то же время витамины
А, Е, К и D не растворяются в воде, но растворимы в органи-
ческих растворителях, поэтому их относят к липидам. Биоло-
гическая роль жирорастворимых витаминов до конца не выяс-
нена; исключение составляет витамин А, необходимый для
предотвращения дневной слепоты (никталопии) у человека.
Интерес к микроорганизмам и пищевым продуктам как ис-
точникам витаминов обусловлен тем обстоятельством, что к чис-
лу незаменимых (или считающихся незаменимыми) для детей
и (или) взрослых относятся водорастворимые витамины тиамин,
рибофлавин, ниацин (никотиновая кислота), пантотеновая кис-
лота, биотин, фолиевая кислота, холин, а также липидные ви-
тамины A, D, Е и К. Многие из этих соединений могут синте-
зироваться различными микроорганизмами. Дрожжи, например,
продуцируют эргостерол — предшественник витамина D2 (каль-
циферола), превращающийся в него под действием солнечного
света. Жирорастворимый витамин К синтезируется микроорга-
низмами в пищеварительном тракте животных и человека; этот
4*
снз.
с=о
холестерин
в<
РИС. 2.3. Некоторые примеры структур стероидов.
он
пример является прекрасной иллюстрацией взаимной помощи,
оказываемой одним видом организмов другому (так называе-
мого комменсализма, рассматриваемого далее в гл. 13). Из-
вестно, что некоторые водорастворимые витамины необходимы
для перевода ряда ферментов в активную форму.
Стероиды представляют собой класс биологически важных
липидов, обладающих общим углеродным скелетом (рис. 2.3,а).
К стероидам относятся, в частности, гормоны, являющиеся
чрезвычайно мощными регуляторами скоростей биологических
Химические основы жизни
53
реакций; в тканях человека гормоны эффективны в концентра-
циях порядка 10-8 М. В настоящее время микроорганизмы
используются только для осуществления относительно неслож-
ных превращений, позволяющих из одних стероидов получать
другие, более ценные. Например, прогестерон можно превра-
тить в кортизон в две стадии: первая осуществляется микро-
биологическим путем, а вторая — химическим (рис. 2.3,6). Дру-
гие подобные примеры приведены в табл. 12.15. Структура
исходного прогестерона настолько сложна, что только опреде-
ленный фермент, продуцируемый одним или несколькими типа-
ми микроорганизмов, позволяет провести необходимую реакцию
с достаточной степенью селективности (т. е. с минимальным
образованием побочных продуктов). Широко известный стероид
холестерин (рис. 2.3, в) обнаружен практически только в мемб-
ранах животных тканей. Показано, что родственные стероиды
обладают способностью изменять проницаемость плазматиче-
ских мембран клетки.
Важным полимерным резервом питательных веществ для
ряда бактерий, в том числе Alcaligenes eutrophus, служит
поли-р-гидроксимасляная кислота (ПГМ), мономерное звено
которой имеет строение —ОСН(СН3)СН2СО—. Этот полимер
находится в клетках в виде гранул. В отсутствие достаточного
количества питательных веществ клетка деполимеризует ПГМ
с образованием растворимой, легко включающейся в метабо-
лический цикл p-гидроксимасляной кислоты. Физические и ме-
ханические свойства ПГМ позволяют изготовлять из нее упа-
ковочные материалы; если учесть, что ПГМ легко подвергается
биодеградации, то можно предположить, что этот биополимер
в будущем будет производиться в промышленных масштабах.
2.2. Сахара и полисахариды
Углеводами называют органические соединения общей фор-
мулы (СН2О)П, где п>3. Эти соединения, содержащиеся во всех
животных, растительных и микробных клетках, выполняют
функции как конструкционных элементов, так и резервных пи-
тательных веществ клетки. Формула (СН2О)П достаточно точно
отражает их состав и может использоваться в расчетах стехио-
метрии происходящих в клетке реакций и высвобождающейся
при этом энергии.
В биосфере вещества углеводной природы (включая крахмал
и целлюлозу) по массе превосходят все другие органические
соединения, вместе взятые. В процессе фотосинтеза диоксид
углерода под действием солнечного света превращается в про-
стые сахара, содержащие от 3 до 9 атомов углерода (подробнее
эти вопросы и другие проблемы биоэнергетики рассмотрены
54
Глава 2
Таблица 2.2. Моносахариды, обычно встречающиеся в биологических
системах
Альдозы (производные альде- Кетозы (производные ке-
гидов; префикс альдо-) тонов; префикс кето-)
Триозы (3 угле- СНО СН2ОН
родных атома) 1
неон с=о
сн2он СН2ОН
D-глицерииовый альдегид днгидроксиацетов
Пентозы (5 угле- СНО СН2ОН
родных атомов) 1 1 о
неон е=о
неон нс'он I
неон неон
сн2он сн2он
D-рибоза D-рибулоза
Гексозы (6 угле- СНО СНО СН2ОН
родных атомов) неон НОСН С=О
носн НОСН НОСН
неон неон неон
неон неон неон
СН2ОН СН2ОН СН2ОН
D-глюкоза D-манноза D-фруктоза
СНО
1 неон
1 носн
1 носн
неон
1 СН2ОН
D-галактоза
в гл. 5). Образующиеся моносахариды затем полимеризуются,
превращаясь в вещества, удобные для построения конструкци-
онных элементов клетки (целлюлоза) или для хранения резерва
сахаров (крахмал). Описанные процессы обеспечивают усвоение
солнечной энергии и ее накопление в виде определенных хими-
Химические основы жизни
55
ческих соединений для последующего использования. Подсчита-
но, что таким путем ежегодно трансформируется около 1018ккал
энергии, что составляет около 0,1% от всей падающей на землю
солнечной энергии. Само собой разумеется, что большая часть
этих ежегодно поступающих 1018 ккал в конце концов вновь
рассеивается в результате последующего окисления (в основ-
ном путем клеточного дыхания) до диоксида углерода.
2.2.1 D-Глюкоза и другие моносахариды
Моносахариды, или простые сахара, представляют собой
простейшие углеводы. Моносахариды содержат от трех до де-
вяти атомов углерода и выполняют роль мономерных звеньев
при построении не несущих генетической информации биополи-
меров с молекулярной массой до нескольких миллионов.
Глюкоза, как и другие простые сахара, представляет собой
производное полигидроксиуглеводорода (табл. 2.2) и в природ-
ных источниках всегда встречается в виде d-( + )-глюкозы, т. е.
в виде оптического изомера, вращающего плоскость поляриза-
ции света вправо.
Хотя D-глюкоза по распространенности намного превосходит
все другие моносахариды, в живых организмах встречаются и
иные простые сахара (табл. 2.2). Все они имеют альдегидную
или кетонную группу. В названии сахаров сначала обычно ука-
зывается природа этой группы, а затем уже число углеродных
атомов в цепи. Так, например, глюкоза называется альдогексо-
зой; символ d обозначает определенный оптический изомер,
в виде которого этот моносахарид практически всегда присут-
ствует в живых системах (относительно оптической активности
см. разд. 2.4).
В растворах D-глюкоза большей частью существует в виде
циклического соединения, так называемой пиранозы, образую-
щейся в результате взаимодействия альдегидной группы С-1
с гидроксилом при С-5 (обратите внимание на принятую систе-
му нумерации шести атомов углерода, а также на символы а
и р, обозначающие пространственное положение гидроксильной
группы при С-1).
56
Глава 2
Пятичленные сахара D-рибоза и дезоксирибоза являются основ-
ными компонентами мономерных звеньев нуклеиновых кислот
(ДНК и РНК) и других биологически важных соединений, ко-
торые мы вкратце рассмотрим позднее.
2.2.2. Дисахариды и полисахариды
Поскольку в растворах многие простые сахара существуют
преимущественно в циклической форме, они не дают реакций,
характерных для альдегидов или кетонов. В то же время
в D-гликопиранозном кольце гидроксильная группа при С-1
отличается от других гидроксилов большей реакционной спо-
собностью. Как показано ниже, на схеме реакции, эта гидрок-
сильная группа, занимающая a-положение, может вступать
в реакцию с гидроксильной группой при С-4 другого сахара,
в результате чего элиминируется молекула воды и образуется
оАД-гликозидная связь-.
он
<х-£-глюкоза
ос -D-глюкоза
конденсация
гидролиз
ос-7,4-гликозидная связь
-а-паль/поза
В результате конденсации двух молекул моносахаридов обра-
зуется дисахарид. Помимо мальтозы, образующейся из двух
молекул d-глюкозы, относительно широко распространены еле-
Химические основы жизни
57
дующие дисахариды:
(Х-В-глюкоза /-В-фруктоза
сахароза
/-Д-галактоза Ji-D-глюкйза
лактоза
Важнейший пищевой продукт, обыкновенный сахар, представ-
ляет собой сахарозу, которая обнаружена во всех фотосинте-
зирующих растениях. Сахароза легче других дисахаридов под-
вергается гидролизу; образующаяся при этом смесь моноса-
харидов глюкозы и фруктозы называется инвертным сахаром.
Относительно редкий, но важный дисахарид лактоза находится
только в молоке. Поскольку некоторые люди не переносят лак-
тозу и поэтому не могут употреблять в пищу молоко, в настоя-
щее время разрабатываются ферментативные процессы гидро-
лиза молочного сахара.
При дальнейшей полимеризации глюкозы могут образовы-
ваться новые 1,4-гликозидные связи. Амилоза представляет со-
бой неразветвленный полимер, построенный из остатков глюко-
зы с молекулярной массой от нескольких тысяч до полумил-
лиона:
а-/,4-гликозидные сдязи
Участок цепи амилозы f—ОН-группы
не показано!)
58
Глава 2
Резервный источник питания растений крахмал обычно содер-
жит около 20% амилозы, хотя эта величина может меняться
в довольно широких пределах. Гранулы крахмала достаточна
велики, и с помощью микроскопа их легко увидеть во многих
растительных клетках.
Амилозная фракция крахмала, как уже упоминалось, со-
стоит из нерастворимых в воде линейных полимеров; основная
же часть крахмала представляет собой амилопектин, также
полимер d-глюкозы, отличающийся от амилозы разветвленной
структурой*. В среднем на 25 остатков глюкозы приходится
один центр разветвления; разветвление цепи осуществляется
за счет образования гликозидной связи между гидроксилом
при С-1 одной цепи и гидроксилом при С-6 другой цепи:
Как правило, молекулы амилопектина больше молекул амило-
зы; их молекулярная масса составляет от 1 до 2 миллионов.
Амилопектин растворим в воде и, адсорбируя воду, может об-
разовывать гели. Частичный кислотный или ферментативный
гидролиз крахмала приводит к образованию различных раз-
ветвленных фрагментов амилопектина, называемых декстрина-
ми. Декстрины применяются в качестве загустителей и при
изготовлении паст. Конечно при частичном гидролизе образу-
ются также глюкоза, мальтоза и другие относительно неболь-
шие сахара. Таким путем из кукурузного крахмала получают
кукурузную патоку.
Резервным источником глюкозы в животных клетках, осо-
бенно в клетках печени и мышц, служат полимерные гранулы
гликогена, по степени разветвленности напоминающего амило-
пектин. В гликогене длина линейных участков между центрами
разветвления обычно составляет около 12 звеньев; таким об-
разом, молекулы гликогена разветвлены даже в большей сте-
пени, чем молекулы амилопектина. Молекулярная масса глико-
* См. статью: Green М. М., Blankenhorn G., Hart Н., «Textbook Errors. 123;
Which Starch Fraction is Water-Soluble, Amylose or Amylopectin?> J. Chem.
Educ., B52, 729 (1975).
Химические основы жизни
59
гена может составлять 5 миллионов и более. Гликоген выпол-
няет функции резервного источника энергии и в некоторых
микроорганизмах, в том числе в энтеробактериях.
2.2.3. Целлюлоза
Целлюлоза является главным структурным элементом всех
растительных клеток, от водорослей до деревьев, и самым рас-
пространенным органическим соединением на земле. Типичны-
ми примерами материалов, состоящих в основном из целлюлозы,
могут служить хлопок и древесина. Считается, что в биосфере
ежегодно образуется 1011 т целлюлозы. Каждая молекула цел-
люлозы представляет собой длинную неразветвленную цепь, по-
строенную из остатков d-глюкозы и имеющую молекулярную
массу от 50 000 до 1 миллиона и более.
Хотя в целлюлозе остатки глюкозы связаны 1,4-гликозидны-
ми связями, эти связи отличаются от тех, которые участвуют
в построении амилозы (сравните приведенную ниже структур-
ную формулу с формулой амилозы). Это на первый взгляд
небольшое различие имеет важные последствия. Гликозидные
связи типа а-1,4, характерные для крахмала и гликогена, легко
расщепляются (гидролизуются) ферментами множества микро-
организмов, растений и животных; напротив, лишь очень не-
многие живые существа способны гидролизовать £- 1,4-связи
целлюлозы. Одним из обычных продуктов ферментативного
гидролиза целлюлозы является дисахарид целлобиоза, молеку-
ла которой построена из двух остатков глюкозы, соединенных
{1-1,4-гликозидной связью.
Устойчивость целлюлозы к расщеплению как в природных,
так и в лабораторных условиях обусловлена не столько особен-
ностями ^-1,4-гликозидной связи, сколько кристаллической
с
Полиглюкозидная цепь целлю-
лозы
структурой целлюлозы и особой «упаковкой» ее молекул в био-
логических структурах. Как показано на рис. 2.4, для целлю-
лозы характерны внутрицепочечные водородные связи между
гидроксильной группой при С-3 и атомом кислорода пираноз-
ного кольца, а изредка также и водородные связи между от-
60
Глава 2
дельными цепями. Эти связи обусловливают формирование из
отдельных цепей целлюлозы больших надмолекулярных струк-
тур, называемых кристаллитами и легко различимых на элект-
ронных микрофотографиях.
Предложено несколько различных моделей кристаллической
структуры целлюлозы. Для наших целей достаточно привести
основные черты этих моделей, схематически изображенные на
рис. 2.5. Большая часть целлюлозы концентрируется в высоко-
РИС. 2.4. Схематическое изображение системы водородных связей между
остатками глюкозы в целлюлозе. Символом R обозначен центр возможной
химической модификации целлюлозы. Так, в метилцеллюлозе, ацетилцеллюло-
зе и карбоксиметилцеллюлозе R = CH3, СОСН3 и CHsCOONa соответственно.
упорядоченных кристаллических областях, в которых цепи или
фибриллы целлюлозы упакованы настолько плотно, что в них
с большим трудом проникают даже молекулы воды. По этой
причине целлюлоза не может растворяться в воде. Менее упо-
рядоченные участки, называемые аморфными, обычно составля-
ют около 15% микроструктуры целлюлозы. Аморфные участки
сравнительно легко гидролизуются, например, кислотами; кри-
сталлические области поддаются деструкции гораздо труднее.
Цепи молекул целлюлозы сгруппированы в микрофибриллы,
которые часто изображают в разрезе так, как это показано
на рис. 2.5, а. Здесь сплошные черточки обозначают плоскости
мономерных звеньев глюкозы, а прерывистыми черточками ука-
заны ориентации другого важного полисахарида, называемого
гемицеллюлозой. В древесине и других материалах целлюлоз-
ной природы молекулы гемицеллюлозы окружают кластеры
микрофибрилл; эти сложные элементы структуры в свою оче-
Химические основы жизни
61
РИС. 2.5. Строение микрофибрилл целлюлозы. [Воспроизведено из работы
Mulethaner К., Ann. Rev. Plant Physiol., 18, 1 (1967).]
редь окружены оболочкой из лигнина, упрочненной многочис-
ленными поперечными связями.
Материалы описанной структуры часто называют лигноцел-
люлозой. Приведенные в табл. 2.3 данные показывают, что
растительная биомасса, продукция целлюлозно-бумажной про-
мышленности и ее отходы содержат значительные количества
лигноцеллюлозы, в которой относительное содержание целлю-
лозы, гемицеллюлозы и лигнина может меняться в широких
пределах. Эти данные свидетельствуют также о том, что рас-
тительная биомасса является самовозобновляемым источником
Таблица 2.3. Относительное содержание (масс. %) целлюлозы,
гемицеллюлозы и лигиина в растительной биомассе, продукции
целлюлозно-бумажного производства и ее отходах
Материал
Целлюлоза
Гемицел-
люлоза
Лигнин
Древесина твердых пород
Древесина мягких (хвойных) пород
Трава
Листва
Волоски хлопкового семени
Газетная бумага
Отходы бумажного производства
40—55 24—40 18—25
45—50 25—35 25—35
25—40 25—50 10—30
15—20 80—85 ~0
80—95 5—20 ~0
40—55 25—40 18—30
60—70 10—20 5—10
62
Глава 2
не только материалов на основе целлюлозы, но и огромных
количеств другого потенциально ценного сырья. В связи с этим
ниже мы рассмотрим свойства гемицеллюлозы и лигнина.
Состав гемицеллюлоз изменяется в довольно широких пре-
делах в зависимости от природы растения. В общем случае
гемицеллюлозы представляют собой короткие разветвленные
полимеры, построенные из остатков пентоз (ксилозы и арабино-
зы) и некоторых гексоз (глюкозы, галактозы и маннозы). Эти
мономерные звенья, как правило содержащие большое коли-
чество ацетильных групп, связаны 1,3-, 1,6- и 1,4-гликозидными
связями. Основные черты химического строения гемицеллюлозы
твердой древесины показаны на следующей схеме:
3
А
— -X—X—х—X—X—X—X—X—X—X- -
3 2
А А
3 3 2 3
А А А ^.80—276
2
а
1
GA
4
М
X — ксилоза
GA — глюкуроновая кислота
А—ацетильная группа М — метоксильная группа
На этой схеме черточками обозначены £-1,4-гликозидные связи,
а цифры указывают порядковые номера атомов углерода, участ-
вующих в образовании различных связей.
Глюкуроновую кислоту мы ранее не упоминали. Она явля-
ется типичным представителем класса уроновых кислот, обра-
зующихся в результате селективного окисления одной из пер-
вичных гидроксильных групп моносахарида до карбоксильной
группы; в частности, глюкуроновая кислота представляет собой
производное глюкозы с карбоксильной группой в положении 6.
Гемицеллюлоза довольно легко поддается гидролизу с обра-
зованием растворимых соединений, например при обработке
разбавленной (0,05—3%-ной) серной кислотой или даже горя-
чей водой. Значительно более устойчива к гидролизу лигнино-
вая оболочка полисахаридных компонентов биомассы.
Лигнин представляет собой полифенол переменного состава.
Известное представление о степени сложности и гетерогенности
этого вещества дает модель структуры лигнина ели, приведен-
ная на рис. 2.6. Такая неупорядоченная комбинация различных
деталей структуры в высшей степени устойчива к действию хи-
мических и ферментативных агентов. Некоторые из применяю-
щихся в настоящее время методов переработки лигнина будут
Химические основы жизни
63
М2СОН
НС
ИСОН
Н2СОН
СО
сн2
МеО
Н2СОН
—СН (I
НС —
н2сон
со
н2сон сн2
° ' __[2У
^-^Ъме
он
(ОМе)05
Н2СОН^
МеОФоме н2сон нс —4^ОМе
О--------СН Нс----°
нс—
О Me
НС---О
Н2СОН ОМе
НС — о
НСО(С6Н10О5)пН
МеО
1
2
нсо н2сон
НС НС
II I
НС со
н2сон т
НС---1
неон
Н2С' 'СН
нс-сн
нс сн.
, Н2СОН
ОМе I
-СН
нс—о
ос^°-СН2
нс---СН
НС неон
оме^ОМе
он о
Н2СОН
О---СН Н2СОН Ц^оме
НСОН НС---о
МеоЦ^>
ОН
МеО
-о
МеО
ОМе
ОН
Н2СОН
НС—
неон
С^ОМе
ОН
ОМе
МЗН
сн
н2сон сн
—сн
6
ОМе
—О'
1
2
РИС. 2.6. Схематическое изображение на плоскости химического строений
лигнина ели. [Воспроизведено с разрешения из работы: Freudenberg К., Lig-
nin: Its Constitution and Formation from p-Hydroxycinnamyl Alcohols; Science,
148, 595 (1965). © 1965 by the American Association for the Advancement of
Science.]
вкратце рассмотрены в гл. 4. В заключение этого краткого
обзора химии и строения биомассы следует отметить, что про-
дукты частичного расщепления лигнина сами по себе являются
потенциально ценными химикатами и сырьем для химической
промышленности.
2.3. От нуклеотидов к РНК и ДНК
Информационный биополимер ДНК (дезоксирибонуклеино-
вая кислота) содержит всю генетическую информацию клетки.
При делении каждая дочерняя клетка наследует по меньшей
мере одну полную копию родительской ДНК, что позволяет
потомству сохранять все морфологические и функциональные
. ДезоксириОонуклеотады
(оВщая структура)
он
Основание
ОН Н
Фоарорная
кислота
W л
Сахар Основание
" V
РиВонуклеотиды
он он
Азотист&/е основания
Лирилшдаял/ I Лурины
а
б
Химические основы жизни
65
особенности, присущие данному виду. Генетическая информа-
ция в ДНК хранится в виде последовательности составляющих
ее полимерную цепь остатков нуклеотидов. Механизм передачи
информации и место процесса репликации ДНК в жизненном
цикле клетки, а также другие аспекты регуляции жизнедеятель-
ности клетки мы обсудим в гл. 6. Здесь же будут рассмотрены
химия мономеров нуклеиновых кислот, структура полимерной
ДНК и информация, закодированная в ее последовательности,
’йг* ’
2.3.1. Структурные элементы нуклеиновых кислот;
переносчик энергии и коферменты
Помимо их роли как структурных элементов нуклеиновых
кислот нуклеотиды и их производные представляют и самостоя-
тельный биологический интерес. Все нуклеотиды построены из
трех компонентов: фосфорной кислоты, альдопентоз рибозы или
дезоксирибозы и азотистого основания, обычно производного
пурина или пиримидина.
пурин
I II
НС. СН
пиримидин
Эти три компонента образуют два типа нуклеотидов, отли-
чающихся природой остатка альдопентозы (рис. 2.7). Рибонук-
леиновые кислоты (РНК) представляют собой полинуклеотиды,
содержащие остаток рибозы, а полимерные цепи дезоксирибо-
нуклеиновых кислот (ДНК) построены с участием моносахарида
дезоксирибозы. На рис. 2.7 изображены также химические фор-
мулы пяти азотистых оснований, входящих в состав нуклеотидов
ДНК и РНК. Три из этих оснований, аденин (А), гуанин (G)
и цитозин (С), типичны как для ДНК, так и для РНК. Напро-
тив, тимин (Т) входит в состав только ДНК, а родственное
пиримидиновое основание урацил (U) специфично для РНК.
Оба типа нуклеотидов представляют собой сильные кислоты,
что обусловлено наличием остатка фосфорной кислоты.
Отщепление фосфатной группы от 5'-углеродного атома
нуклеотида приводит к соответствующему нуклеозиду. Как по-
казано в табл. 2.4, названия нуклеозидов и нуклеотидов явля-
РИС. 2.7. а — общая структура рибонуклеотидов и дезоксирибонуклеотидов;
б — пять азотистых оснований, входящих в состав ДНК и РНК.
5—537
66
Глава 2
ются производными от названий соответствующих азотистых
оснований. Следует отметить, однако, что для нуклеотидов при-
меняется и другая номенклатура. Например, аденилат можно
назвать также аденозин-5'-монофосфатом. Номенклатура по-
следнего типа применяется обычно для производных нуклеози-
дов, у которых гидроксильная группа при С-5' этерифицирована
дифосфатной или трифосфатной группировкой, например адено-
зин-5'-трифосфат (АТР).
С биологической точки зрения особенно важен нуклеозид
аденозин, построенный из остатков рибозы и аденина. На
Таблица 2.4. Номенклатура нуклеозидов и нуклеотидов. Как показано
в последней строке, префикс «дезокси-» применяется для обозначения
соединений, содержащих остаток дезоксирибозы
Основание Нуклеозид Нуклеотид
Аденин (А) Цитозин (С) Гуанин (G) Урацил (U) Тимин (Т) Аденозин Цитидин Гуанозин Уридин Дезокситимидин Аденилат (АМР) Цитидил ат (СМР) Гуанилат (GMP) Уридилат (UMP) Дезокситимидилат (dTMP)
рис. 2.8 изображено строение аденозин-5'-монофосфата (АМР)
и ряда его важных производных. К АМР могут быть присоеди-
нены еще один или два остатка фосфорной кислоты, в резуль-
тате чего образуются ADP (аденозин-5'-дифосфат) и АТР со-
ответственно. При гидролизе соединяющих фосфатные группы
фосфодиэфирных связей высвобождается большое количество
энергии. Например, превращение АТР в ADP и фосфат при 3°С
и pH 7 (напомним, что рН=—lgaH, где ан — концентрация
ионов Н+ в растворе в моль/л) сопровождается изменением
стандартной энергии Гиббса, равным —7,3 ккал/моль.
Мы привыкли оценивать энергетический эффект реакций
прежде всего в единицах тепловой энергии, т. е. теплоты, одна-
ко клетка представляет собой, в сущности, изотермическую си-
стему, в которой, как правило, реализуются химические пути
трансформации энергии. Позднее, в гл. 6, мы значительно под-
робнее рассмотрим АТР в качестве основного переносчика хи-
мической энергии во всех клетках без исключения. По сути
дела, АТР является аккумулятором энергии, получаемой из пи-
тательных веществ или солнечного света, которая затем рас-
ходуется в биосинтезе полимеров, транспорте веществ через
мембраны и движении клеток. Дифосфаты и трифосфаты дру-
гих нуклеотидов также могут выполнять аналогичные функции
67
Химические основы жизни
ОН
НО —Р—О — СНг^ф^^Аденин
° \н Н/
л Н\1 IZH
Аденозинмонофосфат уМВГ
(АМР? он он
ОН он
Аденозиндифосфат
(ADP)
ОН ОН
ОН
НО —Р—О—Р—О—Р—О—СН2 ^Ох Аденин
Аденозинтрифосфат
(АТР)
Н
ОН . ОН
О
О
О
РИС. 2.8. Фосфаты аденозина. AMP, ADP и АТР участвуют в процессах пе-
реноса энергии в клетке, а циклический АМР выполняет регуляторные функ-
ции.
в химии клетки, но основными переносчиками энергии служат
все же аденозинфосфаты.
Циклическая форма АМР, содержащая внутримолекулярный
цикл с участием фосфатной группы (рис. 2.8), выполняет функ-
ции регулятора множества клеточных реакций, включая реакции
образования полисахаридов и резервных полимеров (жиров).
5*
68
Глава 2
Флавинадениндинуклеотид (FAD),
окисленная форма
NH2
Никотинамидадениндинуклеотид.
(NAD)
NH2
Аденин
Кофермент А
NH2
Аденин I
о он
но-p—он
Аденин |
ОН ОН
но-^=о
о
НО—Р=О
।
О
I II CH
но-т>=о
о
сн2
носн
Рибофлавин
о—
он он
♦ о
НО—Р=О 0Р03-в NADP+
HO—Р=0
О
I
сн,
I
сн3—с—сн3
но—in
о
I
но-р^о
реакционноспо-
собный центр
Н /
нс с—conh2
нс, сн
N+
никотинамид
НОСН
носн -
^н
""С—сн3
СН3
н
с=о
HN
СН,
СН2
с=о-
остаток
пантотеновой
кислоты
HN
ли g-меркапто-
,п2 этиленамин
СН2
ОН OH SH
РИС. 2.9. Три важных кофермента, являющиеся производными нуклеотидов.
Недостаток циклического АМР в тканях связывают с одним из
видов рака, т. е. состояния относительно неконтролируемого ро-
ста клеток.
Аденозинмонофосфат не только является структурным эле-
ментом нуклеиновых кислот, но и служит основой для построе-
ния ряда коферментов, химические формулы которых изобра-
жены на рис. 2.9. Кинетика ферментативных реакций рассмат-
ривается в следующей главе; здесь же достаточно отметить,
что коферментами мы называем органические соединения, не-
обходимые для активации некоторых ферментов, т. е. для их
перевода в ту форму, в которой они способны выполнять ката-
Химические основы жизни
69
литические функции. Поскольку практически все реакции
в клетке катализируются ферментами, изменение концентраций
коферментов представляет собой удобный способ регуляции
активности соответствующих ферментов в клетке и таким путем
изменения скорости некоторых внутриклеточных процессов.
2.3.2. Хранение биологической информации, ДНК и РНК
Как и полисахариды, полинуклеотиды образуются путем
конденсации мономеров. И в РНК, и в ДНК нуклеотиды соеди-
нены фосфатными связями, образующимися с участием гидрок-
сильных групп при С-3' и С-5' рибозы или дезоксирибозы.
В качестве примера на рис. 2.10 приведена структура одного
из тринуклеотидов. Клеточные молекулы ДНК невероятно ве-
лики: вся наследственная информация прокариот хранится в од-
ной молекуле ДНК с молекулярной массой порядка 2-Ю9.
В эукариотах ядро может содержать несколько больших мо-
лекул ДНК. Отрицательные заряды ДНК нейтрализуются дву-
зарядными катионами (в прокариотах) или основными амино-
кислотами (в эукариотах).
Следует подчеркнуть, что последовательность нуклеотидов,
как это показано на рис. 2.10, а, имеет направление или поляр-
ность, обусловленную тем обстоятельством, что на одном конце
цепи находится свободная гидроксильная группа при С-5', а на
другом — гидроксил при С-3'. На рис. 2.10,6 приведен удобный
способ изображения нуклеотидной последовательности в на-
правлении от 5'- к З'-концу. Часто применяют и еще более
короткие обозначения. Предположим, например, что основания
1, 2 и 3 на рис. 2.10,6 являются цитозином, аденином и тимином
соответственно; тогда та же самая нуклеотидная последова-
тельность может быть записана в виде рСрАрТ или даже еще
короче: CAT.
Как предположили в 1953 г. Джеймс Уотсон и Фрэнсис Крик,
молекула ДНК состоит из двух полинуклеотидных цепей, обра-
зующих двойную спираль (рис. 2.11). Имеющий форму пра-
вильной спирали каркас молекулы построен из остатков сахаров
и фосфатных групп. Внутри двойной спирали расположены пу-
риновые и пиримидиновые основания. Именно последователь-
ность четырех различных азотистых оснований в полинуклео-
тидной цепи содержит в себе всю генетическую информацию,
в частности данные о структуре синтезируемых рибонуклеино-
вых кислот и белков. Механизмы, регулирующие и использую-
щие генетическую информацию ДНК, мы рассмотрим в гл. 6.
Азотистое основание одной цепи двойной спирали взаимо-
действует с пространственно наиболее близко расположенным
СНгоН (]СНОвание /
$-//онце&ая группа
а.
он
группа
Схематическое изобра-
жение
Основа- Основа- Основа-
ние 1 нце2 ние 3
РИС. 2.10. а — конденсация нескольких
нуклеотидов с образованием цепи, свя-
/'занной фосфодиэфириыми связями; б —
упрощенное схематическое изображение
нуклеотидной цепи.
ОН
С вдхюфо-
► диз/рирных
сЗязях
*С и N
яоснойа-
к ниях
>р
РИС. 2.11. Изображенная несколькими способами структура двойной спирала
ДНК. На этом рисунке показана классическая структура Уотсона — Кри-
ка [7], называемая также В-ДНК; она представляет собой правую спираль,
в которой плоскости пар оснований перпендикулярны оси спирали. В нижней
части рисунка приведена схема, иллюстрирующая некоторые геометрические
параметры этой структуры. [Верхний и нижний рисунки воспроизведены с раз-
решения из работы: Yudkin М., Offord R., A Guidebook to Biochemistry, р. 52,
Cambridge University Press, London, 1971.]
72
Глава 2
РИС. 2.12. Комплементарные пары оснований аденин — тимин и гуанин —
цитозин, образующиеся за счет водородных связей и обладающие очень
близкими пространственными характеристиками, что способствует формиро-
ванию пар оснований внутри двойной спирали молекулы ДНК- (Из работы:
Лёви А., Сикевиц Ф., Структура и функция клетки. — М.: Мир, 1971, с. 164.)
основанием другой цепи строго определенным образом, так что
аденин специфично связывается только с тимином, а гуанин —
с цитозином. На рис. 2.12 показано замечательное геометриче-
ское сходство этих пар оснований, а также наличие двух и трех
водородных связей в парах оснований А—Т и С—G соответ-
ственно.
Две цепи ДНК имеют противоположную направленность
(5'—>-3' и 3'—>-5'). Поэтому, располагая информацией о нуклео-
тидной последовательности одной дезоксирибонуклеотидной це-
пи, например 5,CGAATCGTA3/, можно сделать вывод о строе-
нии соответствующего фрагмента двуспиральной молекулы
Химические основы жизни
73
ДНК; в нашем случае этот фрагмент будет иметь структуру
5'CGAATCGTA3'
3'GCTTAGCAT5'
К той же самой двуспиральной структуре мы пришли бы, если
бы нам была задана последовательность 5'TACGATTCG3'. Та-
ким образом, в информационном смысле последовательность
одной цепи определяет последовательность комплементарной
цепи, т. е. каждая цепь служит матрицей для другой.
Это характерное свойство ДНК является основой для син-
теза дочерних ДНК из исходной родительской молекулы ДНК —
процесса, называемого репликацией ДНК. Если две компле-
ментарные цепи ДНК разделяются и затем на каждой из цепей
в соответствии с правилами образования пар оснований стро-
ятся двойные спирали, то конечным продуктом такого процесса
будут две новые молекулы, каждая из которых идентична ис-
ходной двухцепочечной ДНК и содержит одну новую и одну
старую цепь (рис. 2.13). Таким образом, образование пар осно-
ваний составляет химическую основу считывания биологической
информации, закодированной в нуклеотидной последовательно-
сти ДНК. Справедливость общих принципов такого механизма,
схематично изображенного на рис. 2.13, была убедительно до-
казана экспериментами по выращиванию клеток Е. coli сначала
в среде, содержащей 15N, а затем в среде только с нормальным
изотопом азота HN. Содержащие различные изотопы ДНК мо-
гут быть разделены на ультрацентрифуге. Ультрацентрифуги-
рование и применение изотопных меток, в том числе радиоак-
тивных, являются важнейшими инструментами эксперименталь-
ных исследований в современной биохимии.
Другая важная деталь структуры ДНК—водородные связи,
с помощью которых формируются пары оснований (рис. 2.12).
Водородные связи способствуют стабилизации двухцепочечной
структуры ДНК, однако эта стабилизация не является ни по-
стоянной, ни необратимой. Как это показано на схеме репли-
кации (рис. 2.13), биологическая функция ДНК основана на
раскручивании спирали и разделении двух цепей. В ходе даль-
нейшего изучения молекулярной генетики (гл. 6) мы ознако-
мимся и с другими важными функциями одноцепочечных сег-
ментов ДНК.
Разделение двух цепей ДНК при нагревании является ос-
новой важного метода идентификации различных ДНК. По-
скольку пары оснований АТ связаны двумя водородными свя-
зями, а пары GC — тремя, то содержащие большее число пар АТ
участки ДНК плавятся (так условно называется процесс раз-
деления двух цепей) раньше, чем участки, обогащенные пара-
ми GC. Процесс плавления легко контролируется путем изме-
74
Глава 2
РИС. 2,13. Упрощенная схема репликации ДНК. По мере разделения цепей
родительской ДНК на них синтезируются комплементарные цепи, что приво-
дит к образованию двух дочерних молекул, идентичных родительской ДНК.
Обратите внимание на то, что каждая дочерняя молекула содержит одну
цепь родительской ДНК. (Из работы: Лёви А., Сикевиц Ф., Структура и
функция клетки.— М.: Мир, 1971, с. 169.)
рения поглощения раствора ДНК при 260 нм; одноцепочечные
днк поглощают сильнее, и процесс плавления ДНК регистри-
руется по увеличению общей абсорбции. Температурой плав-
ления, Тил, называют температуру, при которой изменение аб-
сорбции составляет 50% от разницы в абсорбции между
полностью двухцепочечной и полностью расплавленной (одно-
цепочечной) ДНК. Как и следовало ожидать, величина Тпл
коррелирует с содержанием GC. Например, ДНК из Е. coli,
содержащая 50% GC, имеет ТПЛ = 69 °C, а ДНК из Pseudomonas
aeruginosa с 68% GC характеризуется Тпл—76 °C. Содержание
GC в различных организмах меняется в широких пределах
(от 23 до 75%). Особенности нуклеотидного состава иногда
используются для идентификации организмов.
Химические основы жизни
75
Если раствор расплавленной ДНК охладить, то разделенные
комплементарные цепи вновь образуют структуру типа двойной
спирали (эту операцию называют отжигом). Аналогично два
различных одноцепочечных сегмента ДНК с частично компле-
ментарными последовательностями могут подвергаться гибри-
дизации с образованием сегмента двойной спирали. Гибриди-
зация является важнейшим экспериментальным приемом в био-
химии и технологии рекомбинантных ДНК (гл. 6). В завершение
этого краткого введения в физическую химию ДНК следует
отметить, что двухцепочечную ДНК можно также расплавить
или денатурировать путем добавления щелочи или кислоты, что
приводит к ионизации оснований. Можно упомянуть также, что
с точки зрения гидродинамики двухцепочечная ДНК в растворе
ведет себя подобно жесткому стержню, в то время как одно-
цепочечная ДНК по своему поведению напоминает неупорядо-
ченный полимерный клубок.
Как уже упоминалось выше, вся информация, необходимая
для выполнения клеткой ее важнейших функций, в том числе
роста и деления, в бактериях типа Е. coli содержится в одной
молекуле ДНК, которая имеет строение гигантского кольца.
В нативном состоянии ДНК из Е. coli существует в сверхспи-
рализованной форме, что отчетливо видно на электронных мик-
рофотографиях бактерии в виде несколько размытого нуклео-
тида (см. рис. 1.2). Эта большая основная ДНК клетки назы-
вается хромосомой бактерии; кольцевая хромосома Е. coli по-
строена из 4,7 миллиона пар оснований*.
В заключенном в мембрану ядре эукариот ДНК обычно
поделена между несколькими хромосомами; последние могут
быть намного больше хромосом прокариот. В состав хромосом
эукариот входят также небольшие основные белки, называемые
гистонами, которые составляют около половины массы хромо-
сомы. Этот комплекс нуклеиновых кислот и белков хромосом
эукариот называется хроматином. В хлоропластах и митохонд-
риях эукариотических клеток также имеются отдельные, более
мелкие молекулы ДНК-
Очень существен тот факт, что клетки могут содержать и
другие молекулы ДНК- Относительно небольшие ДНК, суще-
ствующие во многих бактериях и эукариотах, называют плаз-
мидами. Бактериальные плазмиды, например, представляют
собой кольцевые молекулы ДНК, содержащие от 4 до 50 тысяч
* Выраженную в тысячах оснований длину ДНК можно перевести в еди-
ницы длины, приняв, что 1 тысяче оснований соответствует длина 0,34 мкм.
Кроме того, молекулярная масса двухцепочечной ДНК с 1 тысячью оснований,
равна приблизительно 660 000 атомных единиц массы.
76
Глава .2
пар оснований. Биологические функции плазмид состоят в обе-
спечении клетки-хозяина полезными, но не самыми необходи-
мыми качествами, например устойчивостью к антибиотикам.
В биохимических исследованиях и в технологических процессах
плазмиды занимают одно из самых центральных мест, являясь
важным инструментом методов рекомбинантных ДНК. Как мы
увидим в гл. 6, модификация плазмид в лабораторных условиях
и последующее введение рекомбинантных плазмид в живые
клетки позволяют изменить их генетическую программу, с тем
чтобы заставить продуцировать новые соединения или более
эффективно расти.
Другие небольшие молекулы ДНК могут быть введены в жи-
вые клетки вирусами. Небольшие вирусы, способные заражать
бактерии, называют бактериофагами или, короче, фагами. По-
следние могут вызывать ряд трудностей в крупномасштабных
технологических процессах с использованием чистых бактери-
альных культур. В то же время изучение фагов позволило вне-
сти большой вклад в развитие молекулярной генетики. Кроме
того, фаги используются для создания стабильных рабочих кол-
лекций или библиотек сегментов ДНК. Фаг % Е. coli содержит
одну циклическую двухцепочечную молекулу ДНК, построенную
из 48,6 тысяч пар оснований. ДНК фага Т2 Е. coli состоит из
166 тысяч пар оснований.
Основная функция ДНК заключается в хранении инструк-
ций для синтеза молекул РНК определенной нуклеотидной
последовательности и длины; некоторые из этих РНК затем
направляют синтез различных специфических белков. Кодирую-
щий последовательность молекулы РНК. сегмент ДНК называ-
ется геном. Ниже мы вкратце расмотрим различные типы РНК,
а затем перейдем к обсуждению белков и тех структурных эле-
ментов, из которых они построены.
Во всех клетках имеются рибонуклеиновые кислоты трех
типов, четвертый тип характерен для некоторых вирусов. Моле-
кулы РНК построены из остатков рибонуклеотидов, углеводным
компонентом которых является рибоза (а не дезоксирибоза,
как в ДНК). Подобно ДНК, в РНК нуклеотиды расположены
в определенной последовательности. В сущности, нуклеотидная
последовательность РНК строится на основе той информации,
которая содержится в соответствующих сегментах ДНК- В пе-
редаче информации от ДНК к РНК основную роль опять-таки
играет образование пар оснований. Правила образования пар
оснований в этом случае аналогичны тем, которым подчиняется
процесс образования пар оснований двухцепочечных ДНК, за
исключением пары аденин-тимидин; при синтезе РНК вместо
нее формируется пара аденин-урацил:
Химические основы жизни
77
ДНК (матрица) РНК (синтезируе- мая на матрице) ДНК (матрица) РНК (синтезируе- мая на матрице)
Т А С G
А и G С
В нормальном цикле жизнедеятельности клетки участвуют раз-
личные РНК, выполняющие важную функцию прочтения и реа-
лизации генетической информации, заложенной в ДНК. Мат-
ричная, или информационная, РНК (мРНК или иРНК) компле-
ментарна последовательности оснований гена в ДНК- Каждая
молекула мРНК переносит определенную информацию от ДНК
к другому механизму биохимического аппарата клетки. По-
скольку объем этой информации меняется в довольно широких
пределах, то и величина молекул мРНК тоже может быть
различной; обычно цепь мРНК содержит 103—104 нуклеотидов.
РНК почти всегда имеют одноцепочечную структуру, тогда как
образование меж- и внутримолекулярных пар оснований часто
играет важную роль в структуре или функции РНК-
Содержащаяся в мРНК информация считывается в рибосоме
(рис. 1.2 и 1.3). До 65% рибосомы составляет рибосомальная
РНК (рРНК), которая в свою очередь может быть разделена
(например, центрифугированием) на РНК нескольких типов.
В частности, рибосома Е. coll содержит три различные РНК,
обозначаемые 23S, 16S и 5S, которые содержат около 3-103,
1,5-103 и 102 нуклеотидных остатков соответственно. В рибосо-
мах из цитоплазмы клеток-эукариот, с другой стороны, имеются
28S, 18S, 7S и 5S рРНК, а в рибосомах митохондрий и хлоро-
пластов обнаружены РНК, специфичные только для этих ор-
ганоидов.
Транспортные РНК (тРНК) отличаются наименьшей моле-
кулярной массой и содержат всего лишь от 70 до 95 нуклеотид-
ных остатков; они находятся в цитоплазме клетки и участвуют
в процессе реализации генетического кода в рибосоме.
В табл. 2.5 приведены основные свойства различных РНК бак-
терии Е. colt. Нуклеотидные последовательности и, следователь-
но, структуры и функции различных клеточных рРНК и тРНК,
как и мРНК, определяются последовательностями нуклеотид-
ных остатков соответствующих генов клеточной ДНК.
Конечный результат этого сложного процесса передачи и
трансляции информации состоит в синтезе молекулы белка.
Белки представляют собой материальное биохимическое выра-
жение генетической информации и инструкций, носителем ко-
торых является ДНК. Используя терминологию, принятую в тео-
рии управления процессами, можно сказать, что белки — это
78
Глава 2
Таблица 2.5. Свойства РНК Е colia
Тип Коэффи- циент Число нуклеотид- Содержание, %
РНК седи- Молекулярная масса иых остатков 'от всей РНК
мента- клетки)
ции
мРНК 6 S—25 S 25 000—1 000 000 75—3 000 ~2
TPHK-4S 23 000—30 000 75—90 16
рРНК 5 S —35 000 ~100
16 S —550000 ~1 5000 | 82
23 S 100 000 ~3 100
а Воспроизведено из работы: Lehninger A. L. Biochemistry, 2d ed., Table 12-3, Worth
Publishers, Inc., New York, 1975; есть перевод более раииего издания: Ленинджер А.,
Биохимия.— М.: Мир, 1976.
конечные элементы управления, которые передают командные
сигналы ДНК клеточным процессам. Ниже и в некоторых по-
следующих главах мы рассмотрим динамику и свойства этих
элементов управления.
2,4. Аминокислоты и белки
Белки представляют собой наиболее распространенные орга-
нические соединения клетки; обычно они составляют от 30 до
70% массы сухих веществ клеток. Все белки построены из че-
тырех самых распространенных биологических элементов — уг-
лерода, водорода, азота и кислорода. В среднем белки содер-
жат 50% С, 7% Н, 23% О и 16% N. Кроме того, в белках
имеется до 3% серы, которая играет важную роль в стабилиза-
ции трехмерной структуры почти всех белков за счет образо-
вания дисульфидных (S—S) связей между атомами серы, рас-
положенными в различных участках полимерной цепи. Моле-
кулярные массы этих неповторяющихся полимеров изменяются
в широких пределах от 6000 до миллиона и более. На рис. 2.14
изображены два основных типа пространственной структуры
белков — фибриллярный и глобулярный.
Преобладание в клетке веществ белковой природы неуди-
вительно, если принять во внимание разнообразие их биологи-
ческих функций (табл. 2.6). Главнейшая функция белков за-
ключается в катализе. Белковые катализаторы, называемые
ферментами, определяют скорость происходящих в клетке хи-
мических реакций. Ферменты локализуются в самых различных
участках клетки, а некоторые из них находятся в растворенном
или суспендированном виде в цитоплазме и таким образом рав-
номерно распределяются по всему объему клетки. Другие фер-
менты связаны с мембранами или существуют в виде ассоциа-
тов с другими веществами, с которыми они образуют надмоле-
Химические основы жизни
79
Одно цепочечный
йеной (миоглойин)
ЦК-
Тан-
тиларал-
лельный
складча-
тый лист)
РИС. 2.14. Два основных типа структур белков и их варианты — фибрилляр-
ные (а) и глобулярные (б).
Олигомерный белок,
построенный из четырех
субъединиц (гемоглобин)
S) Гло5улярнь/е оелки
кулярные агрегаты. Некоторые связанные с мембранами белки,
называемые пермеазами, помогают транспортировать специфи-
ческие питательные вещества в клетку.
Другие белки являются структурными элементами клеточных
мембран, а ряд белков выполняет двигательные функции. У мно-
гих одноклеточных организмов имеются небольшие образования,
по форме напоминающие волоски, которые называются жгути-
ками. Эти жгутики движутся под действием способных
сокращаться белков, обеспечивая тем самым перемещение всей
80
Глава 2
Таблица 2.6. Различные биологические функции белков
Белок Функция или источник выделения
Ферменты (биологические катали- заторы) глюкозоизомераза трипсин алкогольдегидрогеназа РНК-поли мер аза Регуляторные белки /ас-репрессор белок — активатор катаболизма ай Изомеризация глюкозы во фруктозу Гидролиз некоторых пептидов Окисление спиртов до альдегидов Катализ синтеза РНК Регуляция синтеза РНК Подавление синтеза РНК за счет катаболизма
интерфероны Индуцирование устойчивости к ви- русам
инсулин гормон роста быка Транспортные белки лактозопермеаза Регуляция метаболизма глюкозы Стимулирование роста и лактации Транспорт лактозы через клеточные мембраны
миоглобин гемоглобин Защитные белки крови позвоночных антитела Транспорт О2 в мышцах Транспорт О2 в крови Образование комплексов с чужерод- ными молекулами
тромбин Участие в механизмах свертывания крови
Токсины токсин Bacillus thuringiensis токсин ST Е. coll токсин Clostridium, botulinum Токсичен для насекомых Вызывает заболевания свиней Вызывает заражение пищевых про- дуктов
Резервные белки овальбумин казеин зеин Сократительные белки динеин Структурные белки коллаген гликопротеины эластин Яичный белок Белок молока Белок кукурузных зерен Реснички и жгутики Хрящи, сухожилия Стенки и оболочки клеток Связки
клетки. Другие нитевидные и трубчатые придатки, называемые
фимбриями, участвуют в инициировании связывания патоген-
ных бактерий с чувствительными к ним тканями.
Белки выделяют, очищают и характеризуют различными фи-
зическими и химическими методами. Методы разделения белков
(гл. 11) основаны на различиях в их молекулярных свойствах,
которые в свою очередь частично определяются природой со-
ставляющих их аминокислот; последним мы и посвятим следую-
щий раздел.
Химические основы жизни
8Г
2.4.1. Белковые аминокислоты и полипептиды
Мономерными звеньями полипептидов являются сс-аминокис-
лоты общей формулы
Н
h2n—С—соон
I
Таким образом, аминокислоты различаются природой группы R,.
связанной с а-углеродным атомом (соседним с карбоксильной
группой). Поскольку в общем случае все заместители у этого
атома углерода различны (за исключением глицина, у которого
R = H), этот углеродный атом асимметричен.
Многие биологически важные органические соединения,
в том числе сахара и аминокислоты, оптически активны, т. е.
обладают по меньшей мере одним асимметрическим атомом
углерода и поэтому могут существовать в двух формах, как
это показано ниже на примере аминокислот:
н
Г
H2N -с .-соон
R
L-форма
н
!
ноос- с- nh2
R
D-форма
Раствор одного изомера вращает плоскость поляризации
света вправо (правая или d-форма) или влево (левая или
Z-форма). Оптическая изомерия представляет собой чрезвычайно
важное явление, поскольку при отсутствии ферментов, превра-
щающих один изомер в другой, живые организмы могут усваи-
вать только один из изомеров. Ферменты, как правило, также
катализируют превращения только одного из оптических изо-
меров. Это свойство ферментов используется в промышленности
для разделения смесей рацемических ациламинокислот, когда
гидролизу подвергается только один из изомеров, так что в про-
дуктах реакции содержатся два существенно различающихся
и, следовательно, легче разделяемых вещества. Подробнее этот
процесс мы рассмотрим в гл. 4. Поскольку метод прямого фи-
зического разделения оптических изомеров дорог и неэффек-
тивен, подобные микробиологические и ферментативные (а так-
же химические) способы разделения могут оказаться гораздо
более выгодными.
Интересно, что в подавляющем большинстве белков найдены
только ь-изомеры аминокислот, о -Аминокислоты в природных
6—537
Циклические
Число атомов углерода в группе д
Jo 1 2 з 4
н— сн,— сн3 СН — сн3 сн3 \н—сн2— СНз
глицин (Gly; й) аланин (Ala; А) Валин (Vai; V) лейцин (Leu; L)
триптофан
(Ггр, или Try, W)
0 1 2
НО-СН2- I
СН--СН-
серин треонин
(Ser; Sj (Thr; Т)
Mb
\ '
с-сн,
о4
аспарагин
(Азп; Л9
~0
\
с-сн,-
о7/ '
аспарагиновая
кислота
(Asp; D)
Й5-СН2-
цистеир
(Cys‘}C)
СИ,—S—сн2—сн2—
метионин
(Met; М)
Ml,
\ '
с—сн2—сн2
o/z
глутамин
(Gin; 0.)
Фенилаланин
(Phe; Г)
сн,—сн,—сн —
I
СН3
изолейцин
(Не; I)
~о
С-СИ,-СН,- +H,N-(CH,).-
// i
О
глутаминовая лизин
кислота (Lys; К)
(Glu; Е)
h2N-C-NH-(CH2)3~
М12‘
аргинин
(Ary ; R)
НС=С-СН2-
4 HN NH
¥
н
гистидин
(His; Н)
н2
о
//
Другие
§
5s
I с-
H2C^N/ Xj| О'
Н
пролин
(Pro; Р)
(изображена веб
молекула)
Химические основы жизни
83
источниках встречаются редко; они были обнаружены в клеточ-
ных стенках некоторых микроорганизмов и в ряде антибиотиков.
Кислотная (—СООН) и основная (—NH2) группировки ами-
нокислот в водных растворах могут подвергаться ионизации.
Таблица 2.7. Величины рК (—1g К) концевых аминных и карбоксильных
групп, а также группировок в боковых цепях Ra
Аминокислота рК1 а-СООН рА'а a-NH2 pKR группа R
Глицин 2,34 9,6
Аланин 2,34 9,69
Лейцин 2,36 9,60
Серин 2,21 9,15
Треонин 2,63 10,43
Глутамин 2,17 9,13
Аспарагиновая кислота 2,09 9,82 3,86
Глутаминовая кислота 2,19 9,67 4,25
Гистидин 1,82 9,17 6,0
Цистеин 1,71 10,78 8,33
Тирозин 2,20 9,11 10,07
Лизин 2,18 8,95 10,53
Аргинин 2,17 9,04 12,48
а Воспроизведено из работы: Lehninger A. L., Biochemistry, 2d ed., Table 4-2, Worth
Publishers, Inc., New York, 1975; есть перевод более раннего издания: Ленинджер А.,
Биохимия.— М.: Мир, 1976.
Аминокислота несет положительный заряд (катион) при низ-
ких значениях pH и отрицательный заряд (анион) при высо-
ких pH. При некотором промежуточном значении pH амино-
кислота представляет собой биполярный ион (цвиттер-ион)
с нулевым результирующим зарядом. Эта величина pH назы-
вается изоэлектрической точкой и определяется природой за-
местителя R (табл. 2.7). В изоэлектрической точке аминокис-
лота под влиянием электрического поля не способна мигриро-
вать ни к аноду, ни к катоду, и, кроме того, ее растворимость
минимальна. На этих свойствах аминокислот основаны такие
методы разделения смесей, как ионный обмен, электродиализ
и электрофорез (гл. 11).
На рис. 2.15 приведены формулы 20 аминокислот, обычно
встречающихся в белках. Помимо характерных для всех ами-
РИС. 2.15. 20 белковых аминокислот.
6* '
84
Глава 2
нокислот карбоксильной и аминной (кроме пролина) групп не-
которые из них содержат ионизирующиеся группы и в заме-
стителе R. Для одних аминокислот типичны неполярные гидро-
фобные группы R, в других заместители R обладают гидро-
фильными свойствами. Как мы увидим в следующем разделе,
природа этих боковых цепей важна с точки зрения как функции
белка, так и его структуры.
Простые белки представляют собой полимеры, образующие-
ся путем конденсации одних только аминокислот. Реакция кон-
денсации, лежащая в основе синтеза белков, осуществляется
между аминогруппой одной аминокислоты и карбоксильной
группой другой и приводит к образованию пептидной связи-.
но но
I II г..- | ||
Н—С-т ОН + Н—N С—ОН
ZC\ ... XCZ
н R, Н R2
Пептидная связь частично имеет характер двойной связи, по-
этому шесть атомов (на схеме они лежат внутри прямоугольни-
ка, ограниченного штриховыми линиями) расположены в одной
плоскости. Запомните, что каждая аминокислота соединяется
с последующей пептидной связью, поэтому вся белковая цепь
может строиться с помощью одного фермента; в то же время
порядок расположения аминокислот определяется другими ме-
ханизмами (гл. 6).
Название образовавшегося в результате возникновения пеп-
тидной связи фрагмента (аминокислотного остатка) производят
от названия соответствующей аминокислоты путем добавления
окончания ил; например, остатками глицина и аланина явля-
ются глицил и аланил соответственно. Перечисление остатков
в олигопептиде начинают с конца, несущего свободную амино-
группу.
Полипептиды представляют собой сравнительно короткие
цепи, построенные из аминокислотных остатков (рис. 2.16). По-
нятно, что по мере увеличения длины цепи физико-химические
свойства полимера все в большей степени будут определяться
природой групп R аминокислотных остатков, а роль концевых
аминной и карбоксильной групп будет все менее и менее важ-
ной. Полипептидами принято называть относительно небольшие
Химические основы жизни
85
полиаминокислотные цепи. Многие полипептиды имеют большое
биологическое значение; в частности, к числу полипептидов
относится ряд гормонов, например инсулин, гормон роста и со-
матостатин.
Большие полиаминокислотные цепи называют белками-, гра-
ница между полипептидами и белками строго не определена;
обычно считают, что она лежит в пределах 50—100 аминокис-
лотных остатков. Поскольку средняя молекулярная масса ами-
соо"
р— f .
+
HN==\
СОО"
I
7-8
H3N\
н
к
сх
I
сн
I
сн2
6 сн2
II I
сн2
СН2
I
сн2
сн2
сн2
I
СН3
I Н з-
N I .СОО"
С^
I
Н
н о
сн
I
NH
NH
X 12-13
4*
nh2
Gly-Asp-Lys-Glu-Arg-His-Ala
РИС. 2.16. Гипотетический полипептид с различными ионизированными груп-
пами. Запомните, что перечисление аминокислотных остатков начинают с
N-конца последовательности. Числа возле каждой ионизированной группы
указывают соответствующие величины рК.
нокислотного остатка составляет около 120, то молекулярная
масса белков должна превышать 10 000; известны отдельные
белки с молекулярной массой более миллиона.
Аминокислотный состав какого-либо белка или смеси белков
может быть определен с помощью автоматического аминокис-
лотного анализатора. Полный гидролиз белка осуществляют
нагреванием в 6 н. НС1 в течение 10—24 ч при 100—120°C. При
этом с количественным выходом в виде гидрохлоридов обра-
зуются все аминокислоты (за исключением триптофана, аспара-
гина и глутамина); их разделяют и определяют количественно.
Для определения содержания триптофана можно применить ще-
лочной гидролиз. Результаты такого изучения белков из ки-
шечной палочки Е. coli приведены в табл. 2.8. Эти и ряд других
данных показывают, что не все 20 аминокислот (перечислен-
ных на рис. 2.15) входят в состав любого белка. Ни в одном
известном белке аминокислоты не содержатся в эквимолярных
количествах. Для любого конкретного белка, однако, относи-
86
Глава 2
тельные количества различных аминокислот представляют со-
бой строго определенные величины.
Аминокислоты не являются единственными компонентами
белков. В состав многих сложных белков входят другие орга-
нические или даже неорганические группировки, называемые
простетическими группами. Если белок построен только из ос-
татков аминокислот, его иногда, как уже упоминалось выше,
называют простым белком. Широко известный пример сложных
Таблица 2.8. Относительные количества различных аминокислот в белках
Е. coli3-
Аминокислота Относительное со- держание (содержа- ние аланина принято равным 100) Аминокислота Относительное со- держание (содержа- ние аланина принято равным 100)
Аланин 100 Треонин 35
Глутаминовая Пролин 35
кислота и глу- Изолейцин 34
тамин 83 Метионин 29
Аспарагиновая Фенилаланин 25
кислота и аспа- Тирозин 17
рагин 76 Цистеин 14
Лейцин 60 Триптофан 8
Глицин 60 Гистидин 5
Лизин 54
Серин 46
Валин 46
Аргинин 41
а Воспроизведено из работы: Lehninger A. L., Biochemistry, 2d ed.. Table 5-3, Worth
Publishers, Inc., New York, 1975; есть перевод более раннего издания: Ленинджер А.,
Биохимия. — М.: Мир, 1976.
белков представляет гемоглобин— переносчик кислорода в крас-
ных кровяных тельцах,— в состав молекулы которого входят
четыре группировки гема, представляющие собой железосодер-
жащие металлоорганические комплексы. В близком по струк-
туре, но меньшем по молекулярной массе миоглобине имеется
один гем. Каждая молекула фермента оксидазы с-аминокислот,
катализирующей дезаминирование ряда ь-аминокислот, содер-
жит два остатка флавинадениндинуклеотида (FAD) (см. рис. 2.9).
Как указывалось выше, простетической группой рибосом можно
считать РНК.
2.4.2. Структура белков
Способность белков выполнять множество специфических
функций (табл. 2.6) большей частью обусловлена тем обстоя-
тельством, что белки могут существовать в самых различных
Химические основы жизни
87
конформациях. В целях облегчения анализа строения белков принято подразделять их структуру на три уровня, а при нали- чии нескольких полипептидных цепей рассматривают еще и чет- вертый уровень. Приведенные в табл. 2.9 данные показывают,
Таблица 2.9. Структура белков
1. 2. 3. 4. Первичная Вторичная Третичная Четвертичная Последовательность аминокислотных остатков, соединенных пептидными связями Способ расположения полимерной цепи в прост- ранстве, обусловленный главным образом водо- родными связями между сравнительно близко расположенными в последовательности аминокис- лотными остатками Складывание и сгибание полипептидной цепи, обусловленное водородными, ионными и ковалент- ными (дисульфидными) связями, а также гидро- фобным и гидрофильным взаимодействиями Характер упаковки полипептидных цепей (субъ- единиц); структура стабилизируется теми же си- лами, что и третичная
что каждый структурный уровень определяется различными
факторами, которые в сумме обеспечивают все разнообразие
структур и функций белков.
2.4.3. Первичная структура
Под первичной структурой белка подразумевают свойствен-
ную ему последовательность аминокислотных остатков. Первым
белком, у которого удалось выяснить полную первичную струк-
туру, был инсулин (Сэнджер и сотрудники, 1955 г.). Теперь
можно считать доказанным, что для любого белка характерен
не только определенный аминокислотный состав, но и специфи-
ческая аминокислотная последовательность. Как мы увидим
ниже (разд. 6.1), эта последовательность определяется нуклео-
тидной последовательностью ДНК.
Детальное изучение методов определения аминокислотной
последовательности не входит в цели настоящей книги, поэтому
мы ограничимся только самыми общими сведениями. Существу-
ет много методов расщепления белка на различные полипептид-
ные фрагменты. Известны, например, ферменты, способные рас-
щеплять связи между строго определенными аминокислотными
остатками. Определить аминокислотную последовательность об-
разующихся относительно коротких полипептидных цепей мож-
но далее с помощью методов, разработанных Сэнджером и дру-
гими исследователями. Известные частично перекрывающиеся
последовательности фрагментов белка затем комбинируют та-
ким образом, чтобы в конце концов получить полную картину,
88
Глава 2
характеризующую аминокислотную последовательность всего
белка. На автоматических приборах для определения амино-
кислотной последовательности в белках (секвенаторах) с вра-
щающимся реакционным сосудом можно последовательно опре-
делить до 100 аминокислотных остатков в цепи; для опреде-
ления одного остатка требуется около 2 ч. Предложенный
недавно газофазный метод позволил значительно снизить коли-
чество белка, необходимое для определения его аминокислотной
последовательности. Например, для выяснения полной первич-
ной структуры полипептида, состоящего из 50 аминокислотных
остатков, на газофазном секвенаторе требуется 50—100 пико-
молей вещества, а на анализаторе с вращающимся сосудом
1—5 наномолей вещества. Как мы увидим позднее в гл. 6, ами-
нокислотная последовательность белка может быть выяснена
и косвенным путем на основе результатов экспериментального
определения нуклеотидной последовательности соответствующе-
го гена.
В настоящее время известна аминокислотная последователь-
ность многих полипептидов и белков. Пока что в белках не
было обнаружено повторяющихся участков с идентичными по-
следовательностями, хотя некоторые данные позволяют пред-
положить, что подобная картина возможна для ряда фибрил-
лярных белков. В качестве примера на рис. 2.17 приведена
аминокислотная последовательность фермента из яичного бел-
ка, называемого лизоцимом. Как мы уже неоднократно отме-
чали, нумерация аминокислотных остатков начинается с N-koh-
цевого, в данном случае лизина, и завершается С-концевым
остатком, которым в этом ферменте является лейцин.
Боковые цепи аминокислотных остатков взаимодействуют
друг с другом и с непосредственным окружением белка, тем
самым определяя геометрическую конфигурацию белковой мо-
лекулы. Складывание полипептидной цепи в строго определен-
ную трехмерную структуру приводит к белковой молекуле
сложной, запутанной формы типа изображенной на рис. 2.17, б
для случая лизоцима. Такое сложное пространственное строение
белка детерминируется в первую очередь его первичной струк-
турой.
Значимость первичной структуры белков становится особен-
но очевидной, если учесть, что она в свою очередь определяется
клеточной системой кодирования (разд. 6.1). Таким образом,
аминокислотная последовательность белков является как бы
тем звеном, которое соединяет главный командный центр клет-
ки, ДНК, со сложными, высокоспецифичными молекулами бел-
ков, осуществляющими и регулирующими разнообразные био-
химические процессы, необходимые для роста и выживания
клетки.
РИС. 2.17. а — аминокислотная последовательность или первичная структура
фермента яичного белка лизоцима; б — трехмерная (третичная) структура мо-
лекулы кристаллического лизоцима. [Воспроизведено с разрешения автора и
издательства из работы: Blake С. С. F., Structure of Hen Egg-White Lysozyme;
Nature, 206, 757 (1965).]
90
Глава 2
2.4.4. Пространственная структура белков;
вторичная и третичная структуры
Под вторичной структурой белков понимают относительное
расположение в пространстве аминокислотных остатков, зани-
мающих соседние положения в аминокислотной последователь-
ности. Напомним, что частичная двоесвязность амидной связи
обусловливает ее планарность. По этой причине вращение воз-
можно только вокруг двух из каждых трех связей пептидной
цепи (рис. 2.18), что ограничивает число конформаций, которые
может принимать короткий участок цепи.
РИС. 2.18. Планарная природа пептидной связи ограничивает вращение во-
круг связей пептидной цепи. (Воспроизведено из работы: Lehninger A. L.,
Biochemistry, 2d ed., р. 128, Worth Publishers, New York, 1975; есть перевод
более раннего издания: Ленинджер А., Биохимия. — М.: Мир, 1976.)
Как мы уже отмечали (рис. 2.14), существуют две основные
формы вторичной структуры белков: а-спираль и 0-структура
(0-слой). Считается, что конформация фибриллярных белков
волос, шерсти и ряда других объектов стабилизируется водо-
родными связями между атомами соседних аминокислотных
остатков. При свертывании белковой цепи в спираль возможно
образование таких же связей между группой —С=О одного
остатка и группой —NH другого, отделенного от первого тремя
аминокислотными остатками. Такое связывание без существен-
ной деформации связей и углов полипептидной цепи возможно
только при одной конформации, называемой а-спиралъю. Пред-
полагается, что в молекуле коллагена, самого распространен-
ного белка высших животных, имеются три а-спиральных участ-
ка, объединенных в одну суперспираль. Жесткие и относительно
малоэластичные молекулы коллагена выполняют механическую
(опорную) функцию. Коллаген обнаружен в коже, сухожилиях,
роговице глаза и многих других органах; он важен в иммоби-
Химические основы жизни
91
лизании ферментов и клеток и как составляющая часть био-
совместимых материалов.
Водородные связи не отличаются высокой прочностью, по-
этому а-спиральные структуры легко разрушаются. Белки мо-
гут терять спиральную структуру в водных растворах в резуль-
тате конкурентного образования водородных связей с молеку-
лами воды. Если волокна шерсти обработать паром и растянуть,
то белки шерсти принимают иную конформацию, называемую
^-структурой (fi-слоем, складчатым слоем). Последняя харак-
терна для нативных белков волокна шелка. ^-Структура также
стабилизирована водородными связями, однако в этом случае
они возникают между соседними параллельными цепями. Для
этой структуры характерны гибкость и в то же время высокая
устойчивость к растяжению.
Свойства вторичной структуры фибриллярных белков ис-
пользуются в пищевой промышленности. Некоторые пищевые
продукты растительного происхождения, например соевые бобы,
являются ценными источниками незаменимых аминокислот; в то
же время они не обладают консистенцией и текстурой мясных
продуктов. Для придания им этих качеств растворенные гло-
булярные белки перерабатывают в так называемый «тексту-
рированный белок» путем их трансформации в структуры, более
близкие к линейным.
В результате чрезвычайно трудоемких и сложных экспери-
ментов удалось определить полное пространственное строение
(третичную структуру) нескольких белков. Для этой цели вы-
ращивали кристаллы чистого белка, которые затем изучали
методом рентгеноструктурного анализа. В результате рассеяния
рентгеновских лучей на атомах образуется сложная дифракци-
онная картина, которая содержит информацию о деталях строе-
ния белка при разрешении до 2 А. Дешифровка этой информа-
ции в случае миоглобина (рис. 2.19) потребовала расчета
10 000 рядов Фурье, поэтому выяснение строения белков этим
методом в общем случае целесообразно только при условии
использования высокоэффективных компьютеров.
Как показано на рис. 2.17,6 и 2.19, третичная структура
белков, особенно глобулярных, чрезвычайно сложна. На ука-
занных рисунках без труда можно найти области со спиральной
вторичной структурой, а в молекуле лизоцима, кроме того, име-
ются и участки с ^-структурой. Каким путем стабилизируются
эти структуры и какой цели служит класс соединений, молекулы
которых обладают таким разнообразным строением?
Взаимодействия между боковыми группами R, удаленными
друг от друга в аминокислотной последовательности белков,
определяют характер складывания или изгиба белковой цепи
с образованием компактных конфигураций, типичных для гло-
92
Глава 2
булярных белков. Трехмерная структура белка формируется
в первую очередь за счет некоторых слабых взаимодействий,
в том числе ионных и водородных связей, а также гидрофобного
взаимодействия между неполярными группами R (рис. 2.20).
Подобно рассмотренным ранее мицеллам, образующимся из
липидов, у многих глобулярных белков гидрофобные остатки
сконцентрированы во внутренней части молекулы, а более гид-
рофильные группировки располагаются на ее поверхности. Та-
РИС. 2.19. Трехмерная структура миоглобина. Точками указаны положения
а-углеродных атомов 121 аминокислотного остатка в этом белке, выполняю-
щем функцию переносчика кислорода. В середине верхней части молекулы
более крупным кружком выделен атом железа, расположенный в центре един-
ственного гема белка. (Воспроизведено из работы: Edwards N. A., Hassall К. А.,
Cellular Biochemistry and Physiology, p. 57, McGraw-Hill Publishing Co., Ltd.,
London, 1971.)
кая конформация, иногда называемая моделью масляной капли,
вероятно, наиболее устойчива в естественной для нативных бел-
ков водной среде. (Чем, по вашему мнению, отличается от опи-
санной структура белка, входящего в состав клеточной мемб-
раны?)
В биохимической технологии со многих точек зрения чрез-
вычайно важно понять, что эти слабые взаимодействия легко
нарушаются при различных изменениях окружающей среды.
В биологических системах энергия водородной связи составляет
от 3 до 7 ккал/моль, а энергия ионной связи обычно равна
5 ккал/моль, поэтому уже умеренное нагревание может нару-
шить некоторые из этих связей. Изменение pH, ионной силы,
Химические основы жизни
93
РИС. 2.20. Различные типы связей и взаимодействий, стабилизирующих мо-
лекулярную структуру белка. (Из работы: Лёви А., Сикевиц Ф., Структура
и функция клетки.—М.: Мир, 1971, с. 245.)
физические воздействия и добавки органических веществ, оче-
видно, также могут нарушать третичную структуру белков, свой-
ственную им в нативной форме.
В создании устойчивых конформаций белков важную роль
играют также ковалентные связи, особенно часто образующие-
ся между двумя остатками цистеина. Дисульфидная связь обра-
зуется между двумя группами —SH путем элиминирования двух
атомов водорода:
—NH—СН—СО— —NH—СН—СО—
сн2 сн2
I I
SH —2Н S
+ ---> . | <- Дисульфидная связь
SH S
сн2 сн2
—NH—СН—СО— —NH—СН—СО —
Дисульфидные мостики играют роль поперечных связей в по-
липептидной цепи, а иногда они связывают две различные це-
пи; молекула инсулина, например, представляет собой две це-
пи, содержащие 21 и 30 аминокислотных остатков, соединенные-
94
Глава 2
двумя дисульфидными связями; третий дисульфидный мостик
образует внутреннюю поперечную связь в короткой цепи
(рис. 2.21). По сравнению с указанными выше слабыми связя-
ми дисульфидные ковалентные мостики более устойчивы к тер-
мическому воздействию. В то же время дисульфидные связи
легко восстанавливаются избытком сульфгидрильных соедине-
ний, например ^-меркаптоэтанолом.
в
Ser—Leu—'
t—-яCys-Vai
I NH» внутри- I
_____________) 1*2 целочеч- J
I ная связи Д I
Gly—Leu—Vai —Glu—Glu —Cvs Ser
•Туг
Glu—NH2
Leu
Glu
------------'I I Cys
Phe NH2 NH^_________
Vai Asp—Glu—His—L eu—Cys
----Ala
почечная
>язь
Asp—NH2
Туг
Gly
Se/
His'
Leu
xnh2
Cys—As/ J
Д Нежцедочв' чная
Туг—Leu—Vai —Cys—Gly—Glu
Vai. ।
Glu—Ala
[Ala—Lys—Pro
j Arg
^Gly
Phe'
РИС. 2.21. Дисульфидные мостики связывают две пептидные цепи, называе-
мые цепью А (21 аминокислотный остаток) и цепью В (30 остатков), в изобра-
женной здесь молекуле бычьего инсулина; инсулины других видов животных
построены аналогичным образом. Обратите внимание на третью внутрицепо-
чечную дисульфидную связь между двумя остатками короткой цепи А. (Вос-
произведено из работы: Reithel F. J., Concepts in Biochemistry, p. 237, McGraw-
Hill Book Co., New York, 1967.)
Конформация белка в существенной степени определяет его
биологическую активность. Многочисленные эксперименталь-
ные факты свидетельствуют о том, что во многих случаях бе-
лок способен выполнять свойственную ему функцию, только
находясь в определенной трехмерной структуре. Этот принцип,
в частности, положен в основу так называемой модели «замка
и ключа», достаточно наглядно объясняющей высокую специ-
фичность белковых ферментов, катализирующих биохимические
превращения. Известно, что данный фермент способен превра-
щать только определенные соединения, называемые субстрата-
ми. Согласно модели замка и ключа (рис. 2.22), фермент обла-
дает специфическим участком («замком»), который по своей
структуре комплементарен молекуле субстрата («ключу»), по-
этому с ферментом могут связываться (и, следовательно, под-
Химические основы жизни
95
вергаться дальнейшим каталитическим превращениям) только
субстраты, обладающие необходимой пространственной струк-
турой. Полученные в ходе изучения третичной структуры бел-
ков данные не только подтвердили эту гипотезу, но и помогли
глубже понять механизм каталитического эффекта ферментов;
на этой теме мы остановимся подробнее в следующей главе.
Неоднократно показано, что непосредственная связь между
пространственным строением белков и их высокоспецифически-
Активный
центр фермента
Комплекс (pep-
мент-субстрат
Продукты
Сбододный
рермент
РИС. 2.22. Упрощенная схема модели ферментативного катализа типа «замок
и ключ». Здесь форма каталитически активного центра фермента (замка)
комплементарна форме субстрата (ключа). (Воспроизведено из работы: Pelc-
zar М. J., Jr., Reid R. D., Microbiology, 3d ed., p. 158, McGraw-Hill Book Co.,
New York, 1972.)
ми взаимодействиями с другими веществами характерна также
для пермеаз, гормонов, антител и других белков.
Если белок находится в условиях, отличающихся от его
обычного биологического окружения, он может подвергаться
структурным изменениям, которые обычно сопровождаются по-
терей его функциональных свойств (рис. 2.23); этот процесс
называют денатурацией. Денатурация может быть вызвана,,
например, относительно небольшими изменениями pH и темпе-
ратуры раствора; в этом случае, как правило, она не сопро-
вождается разрушением ковалентных связей. При медленном
охлаждении разбавленного раствора денатурированного белка
до свойственной ему в природных условиях температуры часто
происходит обратный процесс, называемый ренатурацией, со-
провождающийся восстановлением функции белка. Как и в
случае многих других превращений, повышение температуры
способствует росту энтропии (разупорядоченности) и, следова-
тельно, разрушению глобулярных структур белков; охлаждение
благоприятствует осуществлению взаимодействий, обеспечива-
ющих компактную структуру белка и его ренатурацию.
96
Глава 2
Принимаемая белком и определяющая его свойства конфор-
мация характеризуется минимумом свободной энергии молеку-
лы. Пока что эту концепцию нельзя использовать для предска-
зания структуры и функции белка, если известна только его
аминокислотная последовательность. Она может, однако, ока-
заться полезной в тех случаях, когда необходимо высказать
обоснованные предположения об изменении свойств белка в
условиях конкретного технологического процесса (которые
часто существенно отличаются от условий существования на-
ЛатиВная КлуВок (неупорядо-
форма ценная структура)
<рорма
РИС. 2.23. Денатурированный под
влиянием тех или иных факторов бе-
лок часто может вновь принять на-
тивную биологически активную кон-
формацию, как только действие этих
факторов прекращается. Такого рода
эксперименты показывают, что пер-
вичная структура белка определяет
его вторичную и третичную структу-
ры. (Воспроизведено из работы: Leh-
ninger A. L., Biochemistry, 2d ed„
р. 62, Worth Publishers, New York,
1975; есть перевод более раннего из-
дания: Ленинджер А., Биохимия.—
М.: Мир, 1976.)
дивного белка) или в результате частичного изменения его
аминокислотной последовательности. В частности, эта концеп-
ция помогает понять, почему связанный с твердой поверхностью
фермент менее активен, чем тот же фермент в растворе.
В настоящей главе много внимания было уделено связи
между строением белков и их функцией. В то же время не сле-
дует забывать, что в силу слабости большинства взаимодейст-
вий и связей, стабилизирующих молекулы белков, реальные
конформации последних могут в существенной степени откло-
няться от некоторой характерной усредненной конформации.
Более того, эта способность белков в ряде случаев тесно свя-
зана с их биологической функцией подобно тому, как биологи-
ческая функция ДНК определяется ее способностью разделять-
ся в изотермических условиях внутри клетки на две цепи.
С точки же зрения биохимика-технолога относительная лег-
кость термической или химической денатурации многих белков
говорит в первую очередь о том, что ферментативные и клеточ-
ные процессы можно регулировать путем изменения, например
pH, температуры и ионной силы, только в довольно узких пре-
Химические основы жизни
97
делах. Денатурацию полезно учитывать и в некоторых других
случаях, например при выделении белков (гл. 11) ив конст-
рукциях стерилизаторов (гл. 7 и 9).
2.4.5. Четвертичная структура
и регулирование биологических процессов
Белки могут быть построены из нескольких полипептидных
цепей (субъединиц); из таких белков наиболее широко извес-
тен, по-видимому, гемоглобин. Под четвертичной структурой
белка понимают способ упаковки субъединиц. Как видно из
приведенных в табл. 2.10 данных, множество белков, в особен-
ности ферментов (обычно их названия оканчиваются суффиксом
-аза), представляют собой олигомеры и поэтому должны обла-
дать характерной четвертичной структурой. Считается, что чет-
вертичная структура стабилизируется теми же силами и связя-
ми, что и третичная. Иногда, как в случае инсулина, в обра-
зовании четвертичных структур участвуют дисульфидные свя-
зи, однако в большинстве приведенных в табл. 2.10 примеров
субъединицы удерживаются в олигомерной молекуле белка за
счет более слабых взаимодействий. Известно, что многие оли-
гомерные белки способны к самоконденсации\ так, например,
разделенные а- и [1-цепи гемоглобина в растворе быстро ассо-
циируют с образованием молекул интактного гемоглобина. Это
свойство белков чрезвычайно показательно в том смысле, что
оно свидетельствует (по меньшей мере в некоторых случаях)
об определяющей роли биохимического одномерного кода
ДНК, который характеризует не только первичную структуру
белков, но и посредством ее все высшие структурные уровни
белков, а следовательно, и их специфические биологические
функции.
Имеющиеся в настоящее время данные показывают, что
построение молекул некоторых белков из нескольких субъеди-
ниц обеспечивает выполнение по меньшей мере двух важных
биологических функций: во-первых, регулирует каталитическую
активность ферментов и, во-вторых, создает широкие возмож-
ности для построения родственных, но не идентичных молекул
из одного и того же набора субъединиц. Иллюстрацией послед-
ней функции могут служить белки, называемые изофермента-
ми или изозимами. Изоферменты представляют собой различ-
ные молекулярные формы фермента, катализирующие одну и
ту же реакцию в организмах одного вида. Существование изо-
ферментов само по себе может показаться ненужным, однако
на самом деле доступность параллельных, но различных ката-
литических процессов является важным элементом ряда систем
биохимической регуляции (гл. 3 и 5). Известно, что некоторые
изоферменты представляют собой олигомерные белки; описан
7—537
98
Глава 2
Таблица 2.10. Характеристики некоторых олигомерных белкова
Белок Молекулярная масса Число поли- пептидных цепей Число ди- сульфидных связей
Инсулин 5798 1+1 3
Рибонуклеаза 13 683 1 4
Лизоцим 14 400 1 5
Миоглобин 17 000 1 0
Папаин 20 900 1 3
Трипсин 23 800 1 6
Химотрипсин 24 500 3 5
Карбоксипептидаза 34 300 1 0
Гексокиназа 45 000 2 а
Такаамилаза 52 000 1 4
Альбумин сыворотки быка 66 500 1 17
Енолаза дрожжей 67 000 2 0
Гемоглобин 68 000 2+2 а
Алкогольдегидрогеназа из печени 78 000 2 а
Щелочная фосфатаза 80 000 2 4
Гемэритрин 107 000 8 а
Глицеральдегидфосфатдегидроге-
наза 140 000 4 а
Лактатдегидрогеназа 140 000 4 0
у-Глобулин 140 000 2-f-2 25
Алкогольдегидрогеназа из дрожжей 150 000 4 О'
Триптофансинтетаза 159 000 2+2
Альдолаза 160 000 4(?) 0
Фосфорилаза b 185 000 2
Треониндезаминаза из сальмонелл 194 000 4
Фумараза 200 000 4 0
Триптофаназа 220 000 8 4
Формилтетрагидрофолатсинтетаза 230 000 4
Аспартаттранскарбамилаза 310 000 4+4 0
Глутаматдегидрогеназа 316 000 6 0
Фибриноген 330 000 2+2+2
Фосфорилаза а 370 000 4
Миозин 500 000 2+3 0
[ГГалактозидаза 540 000 4
Рибулозодифосфаткарбоксилаза 557 000 24
а Воспроизведено с разрешения из работы: Лёви А., Сикевиц Ф., Структура и функ-
ция клетки. — М.: Мир, 1971.
случай изоферментов, построенных из пяти субъединиц только1
двух типов. Подобную конструкцию можно считать выгодной
и целесообразной, поскольку она позволяет синтезировать пять
различных белков всего лишь из двух полипептидных цепей.
2.5. Биохимические соединения смешанного строения
Существует много веществ биологического происхождения,
представляющих интерес с научной или практической точки
зрения, которые трудно отнести к одному из рассмотренных в
Химические основы жизни
99
предыдущих разделах классов соединений. Эти вещества име-
ют смешанное строение и сформированы из остатков липидов,
сахаров и аминокислот в различных сочетаниях. Как указано
в табл. 2.11, соединения смешанного строения выполняют ряд
важных биологических функций. В следующем разделе мы бо-
Таблица 2.11. Некоторые примеры биохимически важных соединений
смешанного строения, их основных элементов структуры и биологических
функций
Название Элементы структуры Локализация в клетке, функция
Пептидогликаны Связанные поперечными связями дисахариды и пептиды Стенки клеток бактерий
Протеогликаны Углеводы (95%) и белки (5%) Соединительные ткани
Гликопротеины Углеводы (1—30%), остальное — белки Находятся в различных объектах, в том числе в антителах; являются компонентами наружных оболочек эукариот
Гликолипиды Липиды, содержащие от 1 до 7 остатков сахаров Компоненты мембран
Липопротеины В липопротеинах плазмы от 1 до 50% белков, остальное — липиды Мембраны, компоненты плазмы крови
Липополисахариды Липид и в высшей сте- пени вариабельная оли- госахаридная цепь Наружные оболочки грамотрицательных бактерий
лее детально обсудим химию и строение наружных оболочек
различных клеток; здесь встречаются смешанные соединения,
а также специфические полисахариды и белки.
2.5.1. Клеточные стенки; пептидогликаны и липополисахариды
В разделе, посвященном липидам, мы уже говорили о том,
что клеточные мембраны играют жизненно важную роль в ре-
гуляции транспорта веществ в клетку и из клетки. В этом от-
ношении не менее важны и другие структурные элементы на-
ружных поверхностей клеток микроорганизмов и тканей. С точ-
ки зрения условий существования микроорганизмы, например
бактерии, живут в гораздо более изменчивом и менее контро-
лируемом окружении, чем клетки тканей животных, например
клетки печени, поэтому микроорганизмы должны обладать го-
раздо большей жесткостью, устойчивостью к физическим на-
грузкам и резким изменениям осмотического давления. Отсюда
следует, что по структуре наружные оболочки бактерий долж-
ны сильно отличаться от клеток животных.
7*
100
Глава 2
В процессах биохимической технологии оболочки клеток
представляют интерес в нескольких аспектах. Во-первых, свой-
ства наружных поверхностей клеток определяют их способность
к адгезии друг с другом, а также со стенками реакторов, тру-
бопроводов и сепараторов. Как мы увидим в последующих гла-
вах, такие явления необходимо учитывать при получении им-
мобилизованных клеточных катализаторов. Они влияют также
на работу микробных реакторов непрерывного действия и на
процессы отделения клеток от жидкости. Химические и меха-
нические характеристики оболочек определяют также устойчи-
Жгутик (спираль
диаметром (2-78 нм)
Ресничка (диа-
метром 4-35 нм)
•UM (толщиной-8 нм)
Грамположи-'
тельная
РИС. 2.24. Схематическое изображение структуры стенок грамположительных
и грамотрицательных бактерий. Обозначения: ЦМ — цитоплазматическая мем-
брана; ПГ — пептидогликан; ПП — периплазматическое пространство; ВМ —
внешняя мембрана. (Воспроизведено из работы: Stanier R. ¥., Adelberg Е. А.,
Ingraham J. L., Wheelis М. L., Introduction to the Microbial World, p. 128,
Prentice-Hall, Englewood Cliffs, N.Y. 1979.)
Капсула (различ-
ной толщина/)
ПГ (толщиной
J5-80 нм)
Грамотрица-
тельная
Капсула
(различной толщины)
ВМ (толщиной-8 нм)
КГ (толщиной -2 нм)
ПП (толщиной -7 нм)
ЦМ (толщиной-8 нм)
вость клетки к воздействию физических, ферментативных и хи-
мических факторов, что существенно в процессах выделения
компонентов клетки.
Как уже упоминалось выше при обсуждении реакции Грама,
структуры оболочек грамположительных и грамотрицательных
клеток сильно отличаются друг от друга (рис. 2.24). У тех и у
других оболочки состоят из нескольких слоев, но их положение,
толщина и состав далеко не идентичны. На этом рисунке не
показаны различные белки, находящиеся внутри клеточной
мембраны или закрепленные на ее поверхности; их мы рассмот-
рим ниже, в гл. 5, в ходе изучения транспорта веществ через
мембраны.
Грамположительные бактерии имеют одну мембрану, а
грамотрицательные клетки — две аналогичные мембраны. Меж-
ду наружной и цитоплазматической мембранами в грамотрица-
Химические основы жизни
101
тельных бактериях находится область, называемая периплазма-
тическим пространством или периплазмой. В периплазматичес-
ком пространстве, включающем от 20 до 40 процентов общей
массы клетки, находится ряд ферментов, а также белков, свя-
зывающих сахара и аминокислоты.
Как в грамположительных, так и в грамотрицательных бак-
териях непосредственно к внешней поверхности цитоплазмати-
ческой мембраны примыкает пептидогликановый слой. Пепти-
догликаны построены из остатков дисахарида [состоящего из
N-ацетилмурамовой кислоты (NAM) и N-ацетилглюкозамина
(NAG), связанных {3-1,4-гликозидной связью], мостикового пен-
тапептида, состоящего только из остатков глицина, и тетрапепти-
да. Строение последнего зависит от вида микроорганизма; в
Staphylococcus aureus, например, этот тетрапептид содержит ос-
татки ь-аланина, D-глутамина, ь-лизина и D-аланина (рис. 2.25,
а; обратите внимание на наличие редких D-аминокислот). Все
перечисленные элементы структуры пептидогликанов связаны
множеством поперечных связей, образуя как бы одну гигантс-
кую макромолекулу, окружающую всю клетку (рис. 2.25,6).
Фермент лизоцим, структура которого приведена на
рис. 2.17, является эффективным антибактериальным агентом.
В основе его бактерицидного действия лежит гидролиз глико-
зидных {3-1,4-связей между остатками NAM и NAG пептидогли-
кана, приводящий к разрушению и удалению пептидогликано-
вой оболочки бактерий и таким путем к разрыву (лизису) клет-
ки в нативных гипотонических растворах. В лабораторных ус-
ловиях в изотонической среде удается получить живые клетки,
не содержащие пептидогликановых оболочек. После обработки
лизоцимом в таких условиях образуются клетки, которые те-
ряют свойственные им очертания и принимают сферическую
форму; их называют сферопластами. Если известно, что кле-
точная стенка удалена полностью, то такие клетки называют
протопластами.
Важными компонентами внешней части наружных мембран
грамотрицательных бактерий являются липополисахариды. Не-
которые липополисахариды называют эндотоксинами, посколь-
ку они в высшей степени токсичны для животных. Токсичность
липополисахаридов — одна из причин того, почему заражение
крови Е. coll может быть чрезвычайно опасным. По этой же
причине при очистке белков, синтезируемых генетически транс-
формированной Е. coll, необходимо тщательное удаление эн-
дотоксинов.
Молекулы липополисахаридов внешней мембраны имеют
три участка: а), липидный компонент А, состоящий из шести
ненасыщенных жирных кислот, углеводородные цепи которых
проходят в мембрану; они связаны с остатком диглюкозами-
N-А цетил-
D- глюкозагшн
^Ацетилмурагювая
кислота
Пептидогликановые
цепи
РИС. 2.25. а — строение основных структурных элементов пептидогликанов;
б — схематическое изображение пептидогликана клеточной стенки грамположи-
тельной бактерии Staphylococcus aureus, содержащего большое количество по-
перечных связей. (Из работы: Ленинджер А., Основы биохимии, т. 1—3. — М.:
Мир, 1985, т. 1, с. 317.)
Химические основы жизни
103
на; б) центральную! олигосахаридную область, построенную из
остатков десяти моносахаридов; некоторые из них относятся к
числу редких сахаров; в) боковую О-цепь, состоящую из мно-
жества повторяющихся тетрасахаридных остатков. Централь-
ная область и О-цепь направлены от клетки в среду. Таким об-
разом, именно наружные боковые О-цепи взаимодействуют с
иммунной системой зараженного животного. Путем мутаций
бактерии могут достаточно быстро менять структуру О-цепей,
что является частью системы их антииммунной защиты.
На этом, однако, рассказ о структуре клеточных стенок
бактерий не кончается. У многих видов микроорганизмов внеш-
няя мембрана окружена капсулой или слизистым слоем, по хи-
мической природе представляющим собой полисахарид. Капсу-
ла одного из штаммов пневмококков (бактерий, вызывающих
пневмонию) построена из чередующихся остатков глюкозы и
глюкуроновой кислоты. Не имеющие такой полисахаридной
капсулы мутанты не обладают патогенными свойствами. Поли-
сахаридная капсула некоторых бактерий может растворяться
в среде. Производство внеклеточных полисахаридов является
важным промышленным процессом, который, однако, сущест-
венно осложняется неньютоновским характером течения жид-
кой среды (гл. 8). Слизистые слои, кроме того, принимают
участие при флокуляции бактерий, являющейся важным этапом
процессов обработки сточных вод методом активного ила.
Цитоплазматическая мембрана дрожжевых клеток состоит
из липидов, белков и полисахаридов, содержащих остатки
маннозы. Если двигаться дальше к внешней оболочке клетки,
то за цитоплазматической мембраной будет расположено пери-
плазматическое пространство, окруженное в свою очередь кле-
точной стенкой. В пекарских дрожжах, Saccharomyces cerevi-
siae, клеточная стенка содержит от 6 до 8% белков (в том чис-
ле несколько ферментов), а также приблизительно по 30% (по
массе) глюкана (полисахарида, построенного из остатков d-
глюкозы, соединенных {3-1,6-связями, а также поперечными
{3-1,3-связями) и маннана (полиманнозы с а-1,6-связями и сс-
1,2-боковыми цепями). При обработке S. cerevisiae ферментом,
гидролизующим глюкан, например 1,3-глюканазой, можно по-
лучать соответствующие протопласты. Такую обработку обыч-
но проводят при введении рекомбинантных молекул ДНК в
дрожжевые клетки.
Клеточные стенки дрожжей и многих плесеней содержат
хитин, макромолекула которого построена из остатков N-аце-
тилглюкозамина, соединенных гликозидными р-1,4-связями
(строение хитина показано на следующей странице).
Животные клетки, существующие обычно в строго контро-
лируемом изотоническом окружении, не имеют клеточных сте-
104
Глава 2
нок. Помимо фосфолипидов и белков в их плазматических
мембранах содержится от 2 до 10% углеводов. Последние об-
наружены на внешней поверхности всех клеток млекопитаю-
щих, изученных до настоящего времени; они связаны с липида-
ми и белками в виде гликолипидов и гликопротеинов соответ-
ственно.
scu/пин
2.5.2. Антитела и другие гликопротеины
Белки, содержащие ковалентно связанные остатки моноса-
харидов или коротких олигосахаридных цепей, называют гли-
копротеинами. Гликопротеины самых различных типов обнару-
жены в эукариотах и окружающей их среде. К числу глико-
протеинов относится ряд ферментов, например глюкозооксида-
за, продуцируемая Aspergillus niger. Упоминавшийся выше кол-
лаген— биологический опорный элемент—также представляет
собой гликозилированный белок. Гликопротеинами являются и
некоторые интерфероны, мощные противовирусные агенты. По
сути дела, большая часть белков эукариот, контактирующих с
окружением клетки или выделяемых ею в среду, представляет
собой гликопротеины. Некоторые гликопротеины уже стали
или, по всей вероятности, скоро станут ценными промышленны-
ми продуктами. Биосинтез гликопротеинов и их участие в
транспорте веществ мы рассмотрим в гл. 5 и 6.
К гликопротеинам относятся и антитела — важнейшее ору-
жие иммунологической защиты позвоночных (рис. 2.26). С по-
мощью обсуждаемых в гл. 6 методов слияния клеток можно
получать в значительных количествах гомогенные антитела в
организмах животных или в биологических реакторах, что по-
зволит в будущем широко применять антитела для диагности-
ческих целей, введения лекарственных препаратов и разделе-
ния биологически важных веществ. По этой причине нам ка-
жется целесообразным вкратце обсудить здесь происхожде-
ние, строение и функции антител.
В состав иммунной системы позвоночных входят В-клетки,
принадлежащие к одному из двух типов обнаруженных в ор-
РИС. 2.26. а — схематичное изображение структуры иммуноглобулина. Сим-
волами С и V обозначены постоянные и вариабельные части цепей соответст-
венно, а индексами Н и L — тяжелые и легкие цепи соответственно. Показаны
дисульфидные связи, а также место присоединения углеводного остатка (СНО)
к тяжелым цепям. Область ствола (Fc) и центр, связывающий антиген, рас-
положены на карбоксильном и аминном концах тяжелых цепей соответствен-
но. Специфичность связывания антигена и сродство к нему определяются
вариабельными частями как тяжелых, так и легких цепей; б — молекулярная
структура антитела IgG, иммуноглобулина G. [Воспроизведено с разрешения
из работы: Silverton Е. W„ Navia М. A., Davies D. R., Proc. Natl. Acad. Sci.,
74, 5142 (1977).]
106
Глава 2
ганизме лимфоцитов; в присутствии чужеродного вещества, ви-
руса или клетки (антигена) В-клетки дифференцируются и об-
разуют плазматические клетки, которые выделяют антитела.
Последние специфично связывают антигены (и близкие по
строению вещества); образующийся комплекс антиген-антитело
осаждается и выводится из организма.
Антитела представляют собой особый класс белков, назы-
ваемых иммуноглобулинами', свойственные иммуноглобулинам
общие детали структуры изображены на рис. 2.26. Молекула
иммуноглобулина состоит из двух идентичных, более длинных
«тяжелых» цепей, связанных друг с другом дисульфидными
связями, а нековалентными связями с двумя другими, также
идентичными, но более короткими «легкими» цепями. В наибо-
лее широко распространенном классе иммуноглобулинов, обо-
значаемом IgG, легкие и тяжелые цепи имеют молекулярные
массы 23 000 и 53 000 соответственно. C-Концевые участки как
легких, так и тяжелых цепей, практически не отличающиеся
по структуре в иммуноглобулинах одного типа, называют по-
стоянными участками. Область Fc, представляющая собой ствол
молекулы антитела, построена из С-концевых последовательно-
стей постоянных участков тяжелых цепей.
Дифференциация антител в пределах одного типа осущест-
вляется прежде всего в N-концевых участках цепей, называе-
мых вариабельными. Связывающий антиген активный центр
антитела (паратоп) формируется из вариабельных участкоз
легких и тяжелых цепей. Как и в случае каталитической актив-
ности ферментов, связывающие антиген центры в различных
антителах могут существенно различаться по своей специфич-
ности и сродству к антигенам.
2.6. Иерархия клеточной структуры
В предыдущих разделах мы рассмотрели основные типы
биологически важных соединений небольшой молекулярной
массы и построенных из них биополимеров. Хотя мы неодно-
кратно подчеркивали связь между химическим строением этих
веществ и их функциями в клетке, полезно еще раз обсудить
динамическую природу и компартментализацию этих функций.
По мере роста живой клетки в ней должны постоянно синтези-
роваться все рассмотренные в настоящей главе биополимеры.
Обычно питательная среда клетки состоит из сахаров, диокси-
да углерода, некоторых аминокислот, воды и ряда неорганичес-
ких ионов, а биополимеры и ряд необходимых для их построе-
ния мономеров, как правило, в сколько-нибудь значительных
количествах отсутствуют. Таким образом, клетка должна синте-
зировать все другие необходимые ей аминокислоты, нуклеино-
Химические основы жизни 107
'Клетка -
h
Органоиды
Ядро
Митохондрии
/поропласты
h
Надмолекулярные
комплексы (мол. масса
частиц (О6-/О9)
Ферментные комплексы
Рибосомы
Сократительные системы
Макромолекулы
(мол. масса
103-109)
Основные
Нуклеиновые белки
кислоты
д
темы гл. 2
Строительные
дроки (мол. масса
100-350)
Мононуклео- Амино-
тиды кислоты
Простые Жирные
сахара кислоты,
11 глицерин
I
Промежуточные Рибоза,
соединения (мол. карбамоил-
масса 50-250) (росрат
А-Не то- Фосфо- Ацетат,
кислоте/ пирубат, малонат
малат
Предшественники,
поступающие из среды
\(Мол. масса 18 44)
РИС. 2.27. Распределение описанных в настоящей главе биологически важ-
ных веществ в порядке усложнения их структуры и повышения степени орга-
низации. (Из работы: Ленинджер А., Биохимия. — М.: Мир, 1976, с. 22.)
108
Глава 2
вые кислоты, липиды, белки и другие вещества из имеющихся
простейших предшественников. Протекающие с поглощением
энергии процессы синтеза предшественников и образования
биополимеров рассматриваются в гл. 5.
В клетке существует множество надмолекулярных, структур.
Как мы уже видели, клеточная мембрана, например, представ-
ляет собой сложное сочетание молекул многих типов. Другим
примером могут служить рибосомы, являющиеся специфичес-
Связанные с кле-
точной мем-
браной фер-
менты
Полирибосома
Свободная рибосома
(белки и мРНН)
Связанная с клеточ-
ной мембраной
хромосома СДНН)
I ТПРНп
Свобод-
ные
фермен-
те/ (бел-
ка)
клеточная стен-
ка (полисахариды
и пептиды)
молеку-
влеточная мембра-
на (липиды и белки)
Небольшие
лы (аминокислоты,
нуклеотиды, прос-
тые сахара, гли-
церин, жирные кис-
лоты, ацетат, па-
ри ват, кетокислоты
и т. л.)
РИС. 2.28. Схема компартментализации описанных в настоящей главе био-
логически важных молекул в клетке кишечной палочки Е. coli.
ким комплексом нескольких различных белков и нуклеиновых
кислот. Во многих случаях ферменты, катализирующие не-
сколько последовательных химических реакций, объединены в
одном ферментном комплексе, в котором, по-видимому, обес-
печивается максимальная эффективность использования проме-
жуточных веществ. Последний по сложности уровень организа-
ции, непосредственно предшествующий самой клетке, занимают
органоиды типа митохондрий и хлоропластов. Различные уров-
ни сложности от простейших веществ до клетки схематически
изображены па рис. 2.27.
Схема локализации различных биологически важных ве-
ществ, рассмотренных в этой главе, в клетке микроорганизма-
прокариоты приведена на рис. 2.28, а общий химический со-
став Е. coli суммирован в табл. 2.12. Небольшие молекулы
Химические основы жизни
109
Таблица 2.12. Состав быстрорастущей клетки Е. со№
Содержание, % от общей Средняя мо- Приблизитель- ное число МО- Число различ-
Компонент лекулярная лекул в клет- ных типов
массы клетки масса ке
Н2О 70 18 4-Ю10 1
Неорганические ионы 1 40 2,5-Ю8 20
(Na+, К+, Mg2+, Са2+, Fe2+, С1-, РО43-, SO42-
и т. п.) Углеводы и их предше- 3 150 2-Ю8 200
ственники 3-107
Аминокислоты и их пред- 0,4 120 100
шественники 1,2-107
Нуклеотиды и их пред- 0,4 300 200
шественники 2,5-Ю7
Липиды и их предшест- 2 750 50
венники 1,5-Ю7
Другие небольшие моле- кулы (гемы, хиноны, 0,2 150 200
продукты распада пита- тельных веществ и т. п.)
Белки Нуклеиновые кислоты: 15 40 000 2,5-109 106 2000—3000
ДНК 1 4 1
РНК 16s рРнк 6 500 000 3-104 1
23S рРНК 1 000 000 3-104 1
тРНК 25 000 4-105 40
мРНК 1 000 000 103 1000
а Уотсон Дж., Молекулярная биология гена, —М.: Мир, 1978, с. 70.
типа аминокислот и простых сахаров, а также некоторые зна-
чительно большие молекулы, например ряд ферментов и тРНК,
равномерно распределены по всему объему цитоплазмы. Дру-
гие биополимеры локализованы в определенных участках внут-
ри клетки или на ее поверхности, например на клеточной мем-
бране.
Связь между различными ионами, питательными вещества-
ми, промежуточными соединениями и другими составными час-
тями клетки осуществляется с помощью разветвленной сети
химических реакций (гл. 5 и 6). Отдельные реакции этой сети
катализируются ферментами, которые обеспечивают их осу-
ществление в мягких условиях, способствующих, как это было
показано выше, сохранению нативной формы белков. Ряд ре-
акций, протекающих с поглощением энергии (гл. 5), а также
тенденция некоторых молекул к самопроизвольному образова-
нию локализованных в объеме клетки структурных элементов
по
Глава 2
обеспечивают образование мембран, ферментных комплексов и
органоидов.
Гл. 3 и 4 посвящены кинетике катализируемых ферментами
реакций и практическому использованию последних. Далее в
гл. 5—7 мы рассмотрим вопросы энергетики и стехиометрии в
клеточных процессах, проблемы генетики и регуляции сети про-
исходящих в клетке реакций, а также математические выраже-
ния, характеризующие кинетику роста популяций клеток. Пос-
ле этих вводных глав мы перейдем к основной теме книги —
изучению биохимических реакторов.
Упражнения
2.1. Химическое строение, а) Напишите полные или обобщенные химиче-
ские формулы перечисленных ниже веществ. Объясните (возможную) функ-
циональную роль различных группировок в молекулах этих веществ.
1. Жнр, фосфолипид, мицелла, витамин А.
2. D(+)-Глюкоза, пираноза, лактоза, крахмал, амилопектин, целлюлоза.
3. Рибонуклеотид, аденозинмонофосфат (АМР), аденозинтрифосфат
(АТР), никотинамидадениндинуклеотид (NAD), дезоксирибонуклеино-
вая кислота (ДНК).
4. Аминокислота, лизин, трипептид, белок.
5. Глобулярные и фибриллярные белки.
б) Начертите схему уровней строения белков, укажите важнейшие осо-
бенности каждого из уровнен (от первичной до четвертичной структуры).
2.2. Спектрофотометрия. Интенсивность поглощения света раствором при
определенных длинах волн измеряется с помощью спектрофотометра — при-
бора, имеющего очень большое значение в аналитической биохимии. Методом
спектрофотометрии определяют концентрации растворенных веществ, а по
характеру спектров поглощения их можно идентифицировать. Спектрофото-
метрическое определение концентрации основано на законе Бера — Ламберта:
А = 1g = атЬс
где А — поглощение или оптическая плотность; 10 — интенсивность падающего
света; I — интенсивность пропускаемого света; ат — молярный коэффициент
экстинкции; Ь — длина пути светового пучка; с — молярная концентрация
растворенного вещества.
Обычно длину волны падающего света выбирают равной ктах, при ко-
торой изучаемое растворенное вещество поглощает в наибольшей степени.
В табл. 2У2.1 приведены величины ^тах ряда биологически важных соедине-
Таблица 2У2.1
Соединение ^тах* им ат-10~3, л/(сммоль)
Триптофан 218 33,5
АТР 259 15,4
NAD+ 259 18
Рибофлавин 260 37
NADH 339 6,22
FAD 450 11,3
Химические основы жизни
111
ний и соответствующие молярные коэффициенты экстинкции (при длине све-
тового пути Ь = 1 см и pH 7). Другие необходимые данные можно найти
в справочниках; обратите внимание на то, что значения Ктах часто лежат
в ультрафиолетовой области.
а) Раствор АТР в кювете толщиной 1 см пропускает 70% падающего
света при 259 нм. Вычислите концентрацию АТР. Какова оптическая плот-
ность 5-10—5 М раствора АТР?
б) Предположим, что соединения А и В имеют одинаковые молярные
коэффициенты экстинкции, приведенные в табл. 2У2.2. Вычислите
Таблица 2У2.2
Длина волны, нм ат, л/(см-моль)
А в
340 14 000 7100
410 2900 6600
концентрации А и В в растворе, содержащем как А, так и В и обладающем
оптической плотностью (в кювете толщиной 1 см) 0,35 при 340 нм и 0,220
при 410 нм.
2.3. Молекулярные массы. Для оценки молекулярной массы методами
седиментации и диффузии часто пользуются уравнением Сведберга:
М= —
<0(1 — vp)
где R — газовая постоянная; Т — абсолютная температура; s — коэффициент
седиментации, с; 2D —коэффициент диффузии, см2/с; v — парциальный удель-
ный объем вещества, см3/г; р — плотность раствора, г/см3.
Проверьте молекулярные массы перечисленных в табл. 2У3.1 веществ
(7’=20°С). Примите р = 1,0.
Таблица 2У3.1
Вещество s -1013 D-107 А’ ,м
Лизоцим 1,91 • П,2 0,703 14 400
Фибриноген 7,9 2,02 0,706 330 000
Вирус карликовости 132 1,15 0,74 10 700 000
кустарниковых
2.4. Буферная емкость и pH растворов полимеров, а) Вычислите pH
0,2 М растворов серина, гидрохлорида серина, калиевой соли серина.
б) Начертите кривую титрования раствора гидрохлорида серина сильным
основанием (КОН); шкалы градуируйте в величинах pH и числе молей КОН,
добавленных к 1 молю серина соответственно.
в) Вычислите pH растворов линейных полипептидов, содержащих 5, 20
и 100 остатков серина, соответствующих гидрохлоридов и калиевых солей
при их процентной концентрации, равной концентрации 0,2 М. раствора гид-
рохлорида серина. Примите, что константа диссоциации не зависит от степе-
ни полимеризации.
г) Вычислите pH раствора моногидрохлорида лизина при его процентной
концентрации, равной концентрации 0,2 М раствора гидрохлорида серина.
112
Глава 2
Считайте константы равновесия реакций протонирования а-аминогруппы и
аминогруппы боковой цепи равными 8,91 -Ю-6 и 2,95-10-11 л/моль соответ-
ственно.
д) Вычислите pH 1%-ных (по массе) растворов целлюлозы и крахмала
(примите, что молекулярная масса крахмала очень велика, >106). Как
в общем случае влияют резервные полимеры на pH клетки?
2.5. Разделение аминокислот, а) Оцените, правильны ли следующие
утверждения, обоснуйте ваши ответы:
1. рК простой аминокислоты выражается формулой
р/= V2 (рКсоон+рКянг)
2. В диапазоне между рКсоон и рКин2 аминокислоты нейтральны.
б) При разделении аминокислот хроматографическими методами успеш-
но используются хроматографические носители, являющиеся сульфирован-
ными производными полистирола. Основные группы аминокислот взаимодей-
ствуют с отрицательно заряженными сульфогруппами, а гидрофобные участ-
ки аминокислот взаимодействуют с ароматическими группировками полимера.
Оцените, в каком (приблизительно) порядке будут элюироваться аминокисло-
ты при pH 3, 7 и 10 из колонки, содержащей:
1. Только сульфогруппы на инертном носителе.
2. Только полистирол.
3. Сульфированный полистирол с доступными в одинаковой степени суль-
фогруппами и фенильными кольцами.
в) Какие взаимодействия, гидрофильные или гидрофобные, будут играть
определяющую роль в процессе разделения смеси полипептидов и смеси бел-
ков на хроматографических носителях, перечисленных в предыдущем уп-
ражнении?
2.6. Физическая химия макромолекул, а) Используя свои знания, спе-
циальную литературу по химии и другие источники, опишите общие химиче-
ские процессы и (или) физические конформационные изменения, сопровож-
дающие поджаривание яичницы, приготовление теста, свертывание молока,
застывание желе, выдувание мыльного пузыря, кристаллизацию белка, дуб-
ление кожи, взбивание яичных белков.
б) Важными параметрами заменителей пищевых продуктов из микроб-
ных и других источников, например белков одноклеточных организмов, за-
менителей мяса растительного происхождения, являются степень их тексту-
рирования и реологические свойства (текучесть при приложении усилия).
Опишите в общих чертах, как бы вы организовали лабораторию испытаний
проб синтетической пищи. (Какую подготовку вы бы предусмотрели для
персонала лаборатории? Какие методы проверки вы бы организовали?)
2.7. Уравнение материального баланса; стехиометрия. Из химических
элементов для роста клеток необходимы в первую очередь углерод, кисло-
род, водород и азот. Предположим, что источниками углерода и азота явля-
ются СН2О и NH4+ соответственно:
aCH2O + jJNH4+ + £O2 —> CaHz,OcNd+nCO2+yH2O
новое клеточное
вещество
а) Опишите ограничения, налагаемые на коэффициенты a, f и т. д. в си-
лу сохранения элементного состава. Какая дополнительная информация
необходима для определения всех коэффициентов при заданном «составе»
клетки?
б) Выполните такое же задание для фотосинтезирующей системы:
aCO2+jJNH4+ + YH2O —> CaHftOcNd+eO2
в) Подробно опишите ряд экспериментов, с помощью которых вы могла
бы определить элементный состав клетки (коэффициенты a, b, с, d).
Химические основы жизни
113
2.8. Стехиометрия; аэробные и анаэробные микроорганизмы. Для облег-
чения расчетов потребности в питательных веществах, скоростей аэрации и
теплопередачи (гл. 5, 8) можно приписать клетке кажущуюся молекулярную
формулу и на этой основе записать общее стехиометрическое уравнение кле-
точного роста.
а) Вычислите формулу клетки дрожжей, которые растут на питательной
среде, содержащей сахар, если экспериментально для этого процесса опреде-
лен следующий баланс веществ (примите, что образуется 1 «моль» клеток):
100 г СбН12О6+5,1 г NHs+46,63 г О2 —>
—>- 43,23 г клеточной массы+41,08 г Н2О +67,42 г СО2
б) Какую формулу следует приписать клетке, если в приведенной выше
реакции образуется 0,30017 «моля» клеток?
в) При анаэробном брожении наряду с клеточной массой обычно обра-
зуется множество частично окисленных соединений. Исходя из «молекуляр-
ной формулы» клетки, найденной в предыдущем упражнении, вычислите не-
известные коэффициенты для следующих типичных уравнений реакций (здесь
коэффициенты указывают количество молей, а не массу):
0,55556 глюкоза+<х аммиак —>- р клеточная масса-)- 0,05697 глицерин
+ у бутанол (типичный
продукт метаболизма
спиртовой природы)
+ 8 янтарная кислота
(типичный продукт
метаболизма кислотной
природы)
+ 0,01164 Н2О+1,03076 СО»
1,00380 этанол
Скорости процессов, описанных в упражнениях 2.8а и 2.8в, легко регулиру-
ются путем изменения скорости аэрации, что позволяет получать дрожжи
и этанол в любом промежуточном соотношении (венский процесс) [Harrison
J. В., Adv. Ind. Microbiol., 10, 129 (1971)].
2.9. Клеточные и искусственные липидные двуслойные мембраны.
В табл. 2У9.1 приведены сравнительные характеристики природных и ис-
кусственных двуслойных липидных мембран.
Таблица 2У9.1а
Природные мембраны Искусственные липидные мем- браны
Толщина (А) по данным:
электронной микроскопии 40—130 60—90
реитгеноструктурного анализа 40—85
оптических методов 40—80
определения электрической емкости 30—150 40—130
Сопротивление, Ом/см2 102—105 103—109
Напряжение пробоя, В 100 100—550
Электрическая емкость, мкФ/см2 0,5—1,3 0,4—1,3
а Tien И. Т„ Bilayer Lipid Membranes: An Experimental Model for Biological Mem-
branes, in Chemistry of Biosurfaces, Hair M. L. (ed.), vol. 1, p. 239, Marcel Dekker,
New York, 1971.
8—-537
114
Глава 2
а) Почему два типа мембран имеют примерно одинаковую толщину?
(Постарайтесь дать подробный и обоснованный ответ.)
б) Выполнению каких функций клетки (вероятно) способствуют высокое
сопротивление и высокая электрическая емкость мембран?
в) Сравните напряжение пробоя порядка 1 В/A с отношением потенциа-
ла ионизации любого углеводорода, содержащего 10 или более атомов угле-
рода, к длине его молекулы. Обсудите полученные результаты.
2.10. Модель согласованных переходных состояний для связывания кис-
лорода гемоглобином. Согласно модели согласованных переходных состоя-
ний. неактивная форма белка То находится в равновесии с активной фор-
.мой Ro:
L
Ro < То L — константа равновесия
В свою очередь активная форма, в случае гемоглобина построенная из четы-
рех субъединиц, может связывать одну молекулу субстрата S каждой субъ-
единицей :
+S +S -]-S +S
Ro < Ri < •*- Ro < у Rs < R<
—S —S —S —S
Обозначим константу диссоциации комплекса гемоглобина с субстратом, об-
разующимся в каждой из приведенных выше стадий, символом Kos'-
„ _____ [S] [несвязанные центры]
Ds [связанные центры]
и вычислим для различных s/Kds и L отношения
№=•
концентрация связанных с S субъединиц
общая концентрация субъединиц
Постройте график зависимости у от sIKds при /.=9000. Найдите в биохими-
ческой литературе график зависимости количества связанного гемоглобином
О2 от парциального давления О2 и сравните с вашим графиком. В найденной
литературе прочтите о других кооперативных явлениях в биохимии.
2.11. Ферменты, построенные из нескольких субъединиц; вероятность
ошибки. На первый взгляд может показаться, что ферменты, состоящие из
нескольких субъединиц, например аллостерические ферменты, имеют более
сложное строение, чем одноцепочечные белки. Тем не менее относительная
вероятность ошибки при биосинтезе таких белков может оказаться меньшей.
а) Рассмотрим синтез двух белков, каждый из которых построен из
-850 аминокислотных остатков. Пусть первый белок состоит из одной поли-
пептидной цепи, а второй — из трех субъединиц с 200, 300 и 350 остатками.
Оцените относительное количество первого и второго белка с одной или бо-
лее ошибками, если вероятность ошибки одинакова для любого остатка и со-
ставляет 5-Ю-9 на остаток и если наличие двух ошибок в любой цепи
предотвращает ее дальнейший синтез или включение в активную структуру.
б) Как ни убедительны расчеты, приведенные в упражнении 2.11а, из-
вестно, что у ряда ферментов отщепление небольшой части аминокислотной
последовательности не сопровождается потерей активности н что одну и ту
же функцию может выполнять довольно большое количество белков с раз-
личными первичными структурами (см., например, различные структуры ци-
тохромов с в монографии: Ленинджер А., Основы биохимии, т. 1. — М.: Мир,
1985). Какие другие функции могут выполнять в клетке эти на первый взгляд
несущественные аминокислотные последовательности и остатки? (Вспомните
внутреннюю структуру клетки.)
Химические основы жизни
115
2.12. Обмен веществ. Для изучения энергетического и массового балан-
сов клеточных процессов необходимо знать, в какой последовательности
осуществляются химические превращения и в каких участках клетки проис-
ходит каждое из них. (Детальнее различные пути метаболизма мы изучим
в последующих главах.) На основе данных, приведенных на рис. 2.27 и 2.28,
начертите схему образования основных биополимеров и высших структурных
элементов клетки; стрелками укажите последовательность процессов и из-
менение мест их осуществления; считайте, что все структурные элементы
клетки образуются из простых предшественников, например О2, Н2О, NH4+
и глюкозы. В необходимых случаях укажите наличие (или отсутствие) про-
странственных ограничений для данных промежуточных продуктов мета-
болизма.
2.13. Соответствие биологически важных соединений. В этой главе были
рассмотрены мономерные соединения и биополимеры, играющие важнейшую1
роль в жизнедеятельности микроорганизмов. Прочтите книгу [12], в которой
с точки зрения истории развития планеты обсуждается вопрос, почему
именно эти соединения возникли в процессе эволюции. Перечислите основные
доводы Блюма в пользу пригодности воды, глюкозы и АТР для выполнения
их биологической роли.
2.14. Реакция Грама. Из всех цветных реакций в бактериологии чаще
всего используется реакция Грама. Перепишите из какого-либо руководства
по микробиологии стандартную методику реакции Грама. Обсудите основ-
ные пункты методики и механизм реакции Грама.
2.15. Внутриклеточные концентрации. Рассчитайте молярные концентра-
ции ДНК, различных РНК, аминокислот и их предшественников, липидов-
и их предшественников в клетке Е. coli. Определите общую молярную кон-
центрацию веществ в Е. coli и обсудите возможность применения термоди-
намических законов для разбавленных (идеальных) растворов к цитоплаз-
ме Е. coli.
2.16. Строение ферментов. В состав активного центра фермента карбок-
сипептидазы, содержащего 307 аминокислотных остатков, входят аргинин
(145), тирозин (248) и глутаминовая кислота (270) (числа обозначают по-
рядковый номер аминокислотного остатка, начиная с N-конца цепи). При-
нимая, что молекула фермента имеет структуру а-спирали, рассчитайте рас-
стояния между тремя указанными остатками. Какие необходимые для осу-
ществления каталитического эффекта структурные изменения происходят
в нативной молекуле карбоксипептидазы, если они приводят к сближению
этих трех остатков до расстояний порядка десятых долей нанометра?
2.17. Термодинамика складывания полипептидных цепей белков. Рас-
смотрим гипотетический белок, находящийся в водной среде. Предположим,
что часть молекулы этого белка имеет определенную вторичную структуру,,
а другая часть содержит Ser, Thr, Asn и два неполярных остатка. Определи-
те AG (ккал/моль белка) н оцените, произойдет ли складывание второй части
белка в следующих условиях:
а) Все полярные группы в последовательности образуют водородные
связи.
б) Все полярные группы, за исключением расположенных в боковых
цепях, образуют внутримолекулярные водородные связи.
в) Ни одна из полярных групп в последовательности не участвует в об-
разовании внутримолекулярных водородных связей.
Примите, что перемещение гидрофобного остатка из водной среды с ней-
тральным pH в неполярную внутреннюю область белка сопровождается
&G«—4 ккал/моль (в расчете на остаток) и что для образования водородной
связи между двумя любыми несвязанными полярными группами AG«
«—5 ккал/моль (в расчете на одну полярную группу). Учтите, что не уча-
ствующие во внутримолекулярных водородных связях остатки могут обра-
зовывать водородные связи с водой.
8*
116
Глава 2
Таблица 2У18.1
Агент Необходимый Ri Необходимый R?
Трипсин Lys, Arg Любой
Химотрипсин Phe, Тгр, Туг Любой
Пепсин Любой Phe, Тгр, Туг, Leu, Asp, Glu
Бромциан Met Любой
2.18. Определение аминокислотной последовательности белков. При ча-
стичном гидролизе полипептидных цепей белков образуются фрагменты, ами-
нокислотная последовательность которых может дать необходимую инфор-
мацию о первичной структуре всего белка. В табл. 2У18.1 перечислен ряд
.агентов, специфично гидролизующих определенные пептидные связи:
Другим важным инструментом определения аминокислотной последователь-
ности является реакция Сэнджера, в которой 2,4-динитрофторбензол реаги-
рует с N-концевой аминокислотой с образованием соответствующего динитро-
фенильного (DNP) производного.
Таблица 2У18.2
Обработка
Результат
Частичный кислотный гидролиз
Пепсин
Реакция Сэнджера
Определение С-концевой аминокисло-
ты
Пептидные фрагменты с последова-
тельностями Gly-Ile-Val-Glu-Glu,
Glu-Glu-Cys, Glu-Asp-Tyr, Ser-Val-
Cys, Ser-Leu-Tyr, Glu-Cys-Cys, Glu-
Leu-Glu, Cys-Asp, Leu-Tyr-Glu, Cys-
Cys-Ala, Tyr-Cys
При гидролизе одного из пептических
пептидов образовывались фрагмен-
ты Ser-Val-Cys, Ser-Leu
DNP-глицин
Аспарагиновая кислота
В табл. 2У18.2 приведен ряд экспериментальных результатов, получен-
ных в ходе изучения цепи А бычьего инсулина. Определите ее первичную
структуру. При полном кислотном гидролизе белковой цепи образуются Gly,
Ala, 2 Vai, 2 Leu, He, 4 Cys, 2 Asp, 4 Glu, 2 Ser и 2 Туг.
Литература
1. Страйер Л., Биохимия, т. 1— 3. — М.: Мир, 1984. Углубленный, написанный
очень понятным языком учебник, освещающий основные биохимические
концепции в свете последних достижений молекулярной биологии.
Химические основы жизни
117
2. Yudkin М., Offord R., A Guidebook to Biochemistry, Cambridge University
Press, London, 1971. Хороший вводный курс, в котором даются основы
биохимии с привлечением минимального количества деталей.
3. Лёви А., Сикевиц Ф., Структура и функция клетки. — М.: Мир, 1971.
Третья часть этой книги представляет собой великолепное короткое введе-
ние в биохимию; его особенно сильной стороной является большое внима-
ние, уделяемое экспериментальной проверке биологических теорий и мо-
делей.
4. Lehninger A. L., Biochemistry, 2d ed., Worth Publishers, New York, 1975;
есть перевод первого издания: Ленинджер А., Биохимия. — М.: Мир, 1976.
Великолепное пособие для углубленного изучения биохимии. Постоянное
обращение к примерам из области физиологии клетки и человека помогает
читателю лучше понять, запомнить и усвоить материал.
5. Ленинджер А., Основы биохимии, т. 1—3. — М.: Мир; 1985. Это не так
давно вышедшее издание предназначено для студентов; написанное в том
же стиле, что и предыдущая книга, оно в то же время отличается более
подробным изложением современных проблем и перспектив развития мо-
лекулярной биологии.
6. Mahler Н. R., Cordes Е. Н., Biological Chemistry, Harper and Row, Pub-
lishers, Inc., New York, 1971. Здесь более детально, чем в книгах Ленинд-
жера, рассмотрены химические и математические аспекты биохимии,
а в разделах, посвященных экспериментальным методам биохимии и мик-
робиологии, вопросы моделирования и транспорта веществ изложены
глубже и подробнее, чем в большинстве других книг.
7. Уотсон Дж. Молекулярная биология гена. — М.: Мир, 1978. Хотя в основ-
ном эта хорошо написанная и уже ставшая классической книга посвящена
молекулярной биологии, в ее начальных главах кратко изложены и основы
биохимии. Здесь убедительно показана важность слабых взаимодействий
и молекулярной динамики для структуры и функции биохимически важ-
ных соединений.
3. Wood W. В., Wilson J. Н., Benbow R. М., Hood L. Е., Biochemistry: A Prob-
lems Approach, 2d ed., Benjamin/Cummings Publishing Co., Menlo Park,
California, 1981. Краткое изложение основ биохимии, дополненное упраж-
нениями (и ответами на них).
9. Dowben R. М., General Physiology: A Molecular Approach, Harper and Row,
Publishers, Inc., New York, 1969. Эта интересно и увлекательно написанная
книга посвящена физиологии клетки, рассматриваемой с точки зрения ос-
нов физической химии.
10. Dickerson R. W., Geis I., The Structure and Action of Proteins, Harper and
Row, Publishers, Inc., New York, 1969. Написанная совместно биологом
и популяризатором науки, эта монография своеобразно рассматривает
различные функции белков, особенно связь между строением и функцией.
Ее полезно прочесть наряду с одной из перечисленных выше книг.
11. Уотсон Дж. Д., Двойная спираль. — М.: Мир, 1969. История открытия
структуры ДНК, поворотного пункта в развитии биологии. Здесь доста-
точно откровенно показан также сам процесс рождения открытий в совре-
менной науке.
12. Blum Н. F., Time’s Arrow and Evolution, 3d ed., Princeton University Press,
Princeton, N.J., 1968. Изучение биологического соответствия элементов и
соединений с точки зрения эволюции.
13. Fukui, Ishida, Microbial Production of Amino Acids, Kodansha Ltd., Tokyo,
and John Wiley and Sons, Inc., New York, 1972. Интересный обзор различ-
ных способов выделения промежуточных продуктов метаболизма, в дан-
ном случае аминокислот.
Глава 3
Кинетика катализируемых
ферментами реакций
В предыдущей главе мы узнали, что в клетке имеется мно-
жество химических соединений. Как они синтезируются и вза-
имодействуют друг с другом с достаточно высокими скоростями
при относительно низких температурах и давлениях? Как клет-
ка делает свой выбор, какие именно вещества следует в данный
момент ввести в реакцию, а какие молекулы подвергнуть де-
градации? Ответ на оба этих вопроса один — все реакции в
клетке осуществляются и регулируются путем ферментативно-
го катализа; ферменты же, как мы уже знаем, представляют
собой глобулярные белки.
Поворотным пунктом в развитии энзимологии был 1897 г.,
когда Бухнер впервые выделил активные ферменты из живых
клеток. Эта работа имела большое значение с двух точек зре-
ния. Во-первых, было показано, что работающие в живом ор-
ганизме катализаторы могут функционировать совершенно не-
зависимо от любого другого клеточного процесса; как мы уви-
дим в следующей главе, изолированные ферменты в настоящее
время находят самое широкое применение. Во-вторых, откры-
тие Бухнера стимулировало работы по выделению и очистке
индивидуальных ферментов. В чистом виде один из ферментов
впервые был выделен Самнером в 1926 г. Его изучение показа-
ло, что он является белком; сейчас известно, что все ферменты
представляют собой белки.
После основополагающих работ Самнера число известных
ферментов постоянно возрастало и в настоящее время уже
намного превысило 1500. Судя по количеству генетической ин-
формации, заложенной даже в таких простых организмах, как
Е. coli, можно уверенно предсказать, что в будущем будет
идентифицировано и охарактеризовано множество новых фер-
ментов. Действительно, единственная молекула ДНК, состав-
ляющая хромосому Е. coli, несет в себе информацию, достаточ-
ную для кодирования структур от 3000 до 4500 различных бел-
ков.
Поскольку в этой главе и позднее мы будем упоминать ряд
конкретных ферментов, здесь уместно сказать несколько слов
об их номенклатуре. К сожалению, не существует общих пра-
Кинетика катализируемых ферментами реакций
119
вил номенклатуры, применимых ко всем ферментам. В подав-
ляющем большинстве случаев в названии фермента отражается
,ег0 функция, а не строение', обычно к названию субстрата до-
бавляют суффикс -аза (например, уреазой называют фермент,
катализирующий разложение мочевины). Иногда тот же суф-
фикс добавляется к названию реакции, которую катализирует
данный фермент (алкогольдегидрогеназа, например, катализи-
рует окислительное дегидрирование спирта). Исключения из
этого правила представляют исторически сложившиеся назва-
ния ферментов (как правило, давно известных); сюда относят-
ся пепсин и трипсин из пищеварительного тракта человека,
используемый в сыроделии реннин и «старый желтый» фермент,
вызывающий появление коричневой окраски на срезах яблок.
Ферменты катализируют реакции шести основных типов, ко-
торые лежат в основе системы, рекомендованной Комиссией по
ферментам (КФ), классификации и условного цифрового обоз-
начения всех ферментов (табл. 3.1). Эта «официальная» систе-
ма позволяет выразить в табличной форме и классифицировать
выполняемые ферментами разнообразные функции, хотя наря-
ду с ней до сих пор широко используется и прежняя более тра-
диционная номенклатура ферментов.
Во избежание недоразумений давайте вспомним, что такое
катализатор вообще. Катализатором называют вещество, ко-
торое повышает скорость химической реакции, не претерпевая
при этом необратимых химических изменений. Повышая ско-
рость химической реакции, катализатор в то же время не на-
рушает равновесия реакции (рис. 3.1). Равновесные концент-
рации можно вычислить на основе одних лишь термодинами-
ческих свойств субстратов (напомним, что субстратом в био-
химии называют вещество, вступающее в катализируемую
ферментом реакцию) и продуктов реакции. Кинетика реакции,
однако, зависит от молекулярной динамики, и в настоящее вре-
мя ее невозможно достаточно точно предсказать, не распола-
гая экспериментальными данными.
Для изучения какой-либо реакции и разработки соответст-
вующего технологического процесса необходимо располагать
математическим выражением, определяющим скорость реакции
(т. е. число молей вещества, реагирующего в единицу времени
в единице объема) в зависимости от состава, температуры, дав-
ления и других параметров реакционной смеси. Если вы ранее
изучали каталитические реакции, то вам должны быть извест-
ны общие принципы вывода выражений, характеризующих
скорость реакций. Обычно прежде всего высказываются доста-
точно обоснованные соображения относительно элементарных
реакций, происходящих на молекулярном уровне. Затем с при-
влечением некоторых приближений, затрагивающих динамику
Таблица 3.1. Классификация ферментов в соответствии с рекомендациями
Международной комиссии по ферментам (указаны название класса
ферментов, кодовые номера Комиссии по ферментам, тип катализируемых
реакций)3
1. Оксидоредуктазы
(окислительно-восстановительные реакции)
1.1. Действуют на группу —СН—ОН
1.2. Действуют на группу —С=О
1.3. Действуют на группу —СН=СН—
1.4. Действуют на группу —СН—NH2
1.5. Действуют на группу — СН—NH—
1.6. Действуют на NADH, NADPH
2. Трансферазы
(перенос функциональных групп)
2.1. Переносят одноуглеродные остатки
2.2. Переносят альдегидные и кетонные группы
2.3. Переносят ацильные группы
2.4. Переносят гликозильные группы
2.7. Переносят фосфатные группы
2.8. Переносят группы, содержащие серу
3. Гидролазы
(реакции гидролиза)
3.1. Действуют на сложноэфирные связи
3.2. Действуют на гликозидные связи
3.4. Действуют иа пептидные связи
3.5. Действуют иа С—N-связи, отличающиеся от пептидных связей
3.6. Действуют на кислотноангидридные связи
4. Лиазы
(присоединение по двойным связям)
4.1. — С=С—
I
4.2. — С = О
I
4.3. — C = N—
5. Изомеразы
(реакции изомеризации)
5.1. Рацемазы
6. Лигазы
(образование связей, сопровождающееся расщеплением АТР)
6.1. Образуют С—О-связи
6.2. Образуют С—S-связи
6.3. Образуют С—N-связи
6.4. Образуют С—С-связи
а Воспроизведено из работы: Lehninger A. L„ Biochemistry, 2d ed., table 8-1, Worth
Publishers, Inc., New York, 1975; есть перевод первого издания: Ленинджер А., Биохи-
мия. — М.; Мир, 1976.
120
Кинетика катализируемых ферментами реакций
121
Некаталитическая реакция
Глюкоза-Г
фоарат
Глюноэо-6-
- фосфат
Высокая энергия
активации
Изменение стандар-
тной свободной
энергии реакции
AG°=-1,7S ккал
_’L_________________
Каталитическая реакция
AG°=-17S ккал
Глюкоза-f-
фосфагп
Гл/окозо-6-
фосфат
Низкая энергия
активаищи
Необходимый
для реакции
минимум энергии
Число молекул
Высокая
энергия
активации
Энергия,
Энергия, Только эти моле -
приходящаяся кулы обладают
” достаточной для
начала реакции
внутренней энер-
гией'. скорость
реакции низка
Средняя
внутренняя
энергия
Необходимый
для реанции
минимум
энергии
Низкая
энергия
активации
Средняя
внутренняя
энергия
Все эти молекулы
обладают достаточ-
ной для начала реак-
ции внутренней
энергией; скорость
реакции высока
Энергия,
приходящаяся
на одну
молекулу
на одну
молекулу
РИС. 3.1. Катализатор снижает энергию активации реакции и тем самым
обеспечивает возможность реакции фермента с молекулами субстрата, обла-
дающими малой внутренней энергией. [Воспроизведено с разрешения из ра-
боты: Lehninger A. L., Bioenergetics, 2d ed., р. 35, W. A. Benjamin, Inc., Menlo
Park, С A, 1971.]
одного или нескольких реакционноспособных промежуточных
соединений, и с использованием ряда простых математических
преобразований находят выражение для скорости суммарной
реакции. В настоящей главе мы воспользуемся точно таким же
подходом для оценки скоростей катализируемых ферментами
реакций.
Аналогия между синтетическими катализаторами и фер-
ментами не заканчивается на принципах моделирования кине-
тики реакций. Математические выражения, определяющие ско-
рости реакций, катализируемых этими двумя типами катализа-
торов, очень близки, а иногда даже идентичны. Это объясня-
ется тем, что в обоих случаях, как известно, в качестве проме-
122
Глава 3
жуточного соединения образуется тот или иной комплекс реаги-
рующего вещества (субстрата) с катализатором; общий
механизм каталитических процессов, естественно, приводит к
одинаковым выражениям для их скоростей. Позднее мы еще-
раз вкратце остановимся на этом вопросе.
В то же время важно не забывать и о существенных разли-
чиях между синтетическими катализаторами и ферментами; о
некоторых из таких различий мы уже упоминали. Подавляю-
щее большинство синтетических катализаторов неспецифична
в том смысле, что они могут катализировать аналогичные ре-
акции с участием самых разнообразных реагентов. Некоторые
ферменты также не отличаются высокой специфичностью, но
многие катализируют только одно превращение крайне ограни-
ченного числа субстратов. Обычно степень специфичности фер-
мента соответствует его биологической функции. Высокая спе-
цифичность нежелательна, например, для фермента, основная
задача которого заключается в гидролизе белков до небольших
пептидов и аминокислот. Напротив, фермент, катализирующий
изомеризацию одного конкретного соединения, должен быть в
высшей степени специфичным. Как уже упоминалось выше в
разд. 2.2.4, считается, что специфичность фермента обусловлена
его сложной объемной структурой, позволяющей сформировать
ответственный за каталитические свойства фермента активный
центр.
Другой отличительной чертой многих ферментов является
наличие кофакторов, необходимых для проявления'фермента-
тивной активности. Кофактором называют соединение небелко-
вой природы, которое связывается с неактивным белком (апо-
ферментом) с образованием каталитически активного комплек-
са. Последний биохимики часто называют голоферментом, мы
же чаще будем называть его просто ферментом. Существует
два различных типа кофакторов. Простейшими кофакторами
являются ионы металлов (табл. 3.2). Роль кофакторов могут
выполнять и сложные органические соединения, называемые
коферментами. В предыдущей главе мы уже упоминали кофер-
менты NAD, FAD и кофермент А(СоА); иногда кофактором
может быть и АТР. Часто кофакторы связываются с фермента-
ми довольно слабыми связями; в таких случаях существует
равновесие между ферментом, апоферментом и кофактором.
Вообще говоря, и прочно связанные с ферментом небелковые
элементы структуры также являются коферментами, как, на-
пример, гем в цитохроме с. Однако, как мы уже упоминали, ча-
ще такие необратимо связанные группы называют простетичес-
кими.
Приведенные в табл. 3.2 данные интересны и с той точки
зрения, что здесь содержатся сведения, к которым мы будем
Кинетика катализируемых ферментами реакций
123
неоднократно возвращаться при изучении процессов выделения
и применения ферментов. Обратите внимание на то, что рядом
с названиями некоторых ферментов указаны источники их вы-
деления; например, глюкозоизомераза из бактерии Bacillus
coagulans проявляет максимальную активность в присутствии
кобальтового кофактора. Для того чтобы точно определить,
о каком ферменте идет речь, во многих случаях необходи-
Таблица 3.2. Некоторые ферменты, содержащие в качестве кофакторов ионы
металлов или проявляющие активность только в присутствии ионов
металлов
Са2+ а-Амилаза (из поджелудочной железы свиньи) Коллагеназа Липаза
Со2+ Нуклеаза микрококков Глюкозоизомераза (из Bacillus coagulans; для активации необходим также Mg2+)
Cu2+ (Си+) Г алактозооксидаза Триозиназа
Fe2+ или Fe3+ Каталаза Цитохромы Пероксидаза
Mg2+ Дезоксирибонуклеаза (из поджелудочной железы быка)
Mn2+ Na+ Аргиназа Аденозинтрифосфатаза из плазматических мемб- ран (для активации необходимы также К+ и
Zn2+ Mg2+) Алкогольдегидрогеназа Щелочная фосфатаза Карбоксипептидаза
мо указывать и его происхождение. Дело в том, что другие
организмы также синтезируют ферменты, катализирующие
изомеризацию глюкозы во фруктозу, и поэтому их тоже назы-
вают глюкозоизомер азами. В то же время ферменты с одина-
ковыми названиями, но выделенные из разных организмов, час-
то имеют различные аминокислотные последовательности и по
этой причине различаются по свойствам и каталитической ак-
тивности. Например, для глюкозоизомеразы из В. coagulans
необходим ион Со2+, а глюкозоизомераза из мутантного штам-
ма того же организма при рН>8 активна и в отсутствие ко-
бальта. Надо иметь всегда в виду, что одно лишь название
фермента ничего не говорит о природе какого-либо конкретно-
го белка с определенными свойствами и соответствующими тех-
нологическими требованиями. Чтобы устранить возможность
Двусмысленного толкования, нужно указать полное название
продуцирующего этот фермент организма.
124
Глава 3
Как синтетические, так и биологические катализаторы в
ходе выполнения своей каталитической функции постепенно
утрачивают активность, однако ферменты в общем случае го-
раздо более недолговечны. Сложная и запутанная пространст-
венная структура ферментов, обусловливающая их необычно
Таблица 3.3. Сравнительная активность ферментов и синтетических твердых
катализаторов (N — число молекул субстрата, превращаемых одним
активным центром катализатора в 1 с)а
Фермент Субстрат или ката- лизируемая реакция Диапазон найденных зна- чений N при 0—37 °C
Рибонуклеаза Перенос фосфатной группы полинуклеоти- да 2—2-Ю3
Трипсин Гидролиз пептидов з-ю-3—1 -102
Папаин Гидролиз пептидов 8-Ю-2—1-Ю1
Бромелаин Гидролиз пептидов 4.10-3—5-10-*
Карбонилангидраза Обратимая гидратация карбонильных соеди- нений 8-Ю-1—6-Ю5
Фумаратгидратаза Обратимое превраще- ние малата в фумарат и воду 1-Ю3 (прямая реакция) З-Ю3 (обратная реак- ция)
Твердый катализатор Катализируемая реакция Температура, N °C
SiO2—А120з (импрегни- рованный) Крекинг кумола 3-10-8 25 2-Ю4 420
Декатионизированный цеолит Крекинг кумола ~103 25 ~Ю8 325
V2O5 Дегидрирование цикло- гексена 7-Ю"11 25 102 350
Обработанный Cu3Au Дегидрирование НСО2Н 2-Ю7 25 З-Ю10 327
AICI3- А120з Изомеризация н-гексана (в жидкой фазе) 1-10~2 25 1,5-Ю“2 60
а Воспроизведено из работы: Maatman R. W., Enzyme and Solid Catalyst Efficien-
cies and Solid Catalyst Site Densities, Catal. Rev., 8, 1 (1973).
высокие специфичность и активность, легко нарушается, что
приводит к потере ферментом его каталитических свойств. Раз-
личные пути инактивации ферментов и кинетика этих процес-
сов будут рассмотрены в разд. 3.7.
Часто утверждают, что ферменты более активны в том
смысле, что они повышают скорость реакций в большей степе-
ни, чем небиологические катализаторы. Степень активности ка-
тализатора обычно выражают числом оборотов, которое пред-
Кинетика катализируемых ферментами реакций 12S
ставляет собой число молекул субстрата, реагирующих с актив-
ным центром катализатора в единицу времени. Для сравнения
в табл. 3.3 приведены параметры ряда реакций, катализируе-
мых ферментами и синтетическими катализаторами. Оказыва-
ется, что в диапазоне температур, в котором ферменты наибо-
лее активны, они действительно повышают скорость реакций &
большей степени, чем большинство синтетических катализато-
ров. При более высоких температурах, однако, активность ис-
кусственных катализаторов обычно превосходит активность
ферментов. К сожалению, ферментативная активность не мо-
жет бесконечно возрастать при повышении температуры; на-
против, ферменты обычно теряют свою активность уже при
сравнительно низких температурах, часто лишь на несколько
градусов превышающих температуру живой клетки.
Характерной особенностью ферментативного катализа явля-
ется возможность его регуляции с помощью соединений не-
большой молекулярной массы. Некоторые ферменты «выключа-
ются» в присутствии определенных веществ; последними часто
оказываются конечные продукты последовательности реакций,,
в которой участвует регулируемый фермент. Эта особенность
ферментов играет большую роль в нормальном жизненном цик-
ле клетки. Некоторые аспекты кинетики ферментативных реак-
ций такого типа мы изучим в разд. 3.5, а в гл. 6 мы узнаем,,
как посредством изменения нормальных каналов регуляции
внутриклеточных реакций можно резко повысить эффектив-
ность промышленных биологических процессов.
Прежде чем приступить к моделированию кинетики фермен-
тативного катализа, мы рассмотрим имеющиеся эксперимен-
тальные данные о характере молекулярных превращений, ко-
торые действительно происходят в процессе катализируемых
ферментами реакций. На этой основе мы сможем высказать
обоснованные гипотезы о последовательности элементарных,
реакций и затем применить эти гипотезы для вывода матема-
тических выражений, характеризующих скорость реакций.
3.1. Фермент-субстратные комплексы
и механизм действия ферментов
В настоящее время не существует единой теории, объясня-
ющей необычно высокую специфичность и активность фермент-
ных катализаторов. В то же время в отношении небольшого
числа конкретных ферментов был выдвинут целый ряд вполне-
вероятных гипотез, подтвержденных экспериментальными дан-
ными. По-видимому, положенные в основу этих гипотез явле-
ния в совокупности и обусловливают специфические свойства,
ферментов. В настоящем разделе мы вкратце рассмотрим не-
126
Глава 3
которые из этих гипотез; более подробные сведения читатель
сможет найти в литературе, приведенной в конце главы. По-
скольку все рассматриваемые здесь гипотезы только частично
объясняют механизм действия ферментов, мы не будем пытать-
ся обобщить эти данные с целью создания единой теории фер-
ментативной активности.
Существование фермент-субстратных комплексов доказано
с помощью различных экспериментальных методов, в том чис-
ле рентгеноструктурного анализа, спектроскопических методов
и электронного парамагнитного резонанса. Субстрат связыва-
ется с ферментом в определенной области молекулы фермента,
называемой активным центром, где осуществляется катализи-
руемая ферментом реакция и образуются ее продукты. В свя-
зывании субстрата с ферментом и образовании комплекса иног-
да принимают участие слабые взаимодействия, рассмотренные
в разд. 2.4.3, а в некоторых случаях при этом образуются и
ковалентные связи. Как схематически показано на рис. 2.24 и
3.2, комплекс образуется тогда, когда субстратный «ключ» вхо-
дит в ферментный «замок». На рис. 3.2 особенно отчетливо
видно, что фермент-субстратный комплекс образуется с по-
мощью водородных связей между субстратом и группами, рас-
положенными в самых разных участках аминокислотной после-
довательности фермента.
Этот пример также наглядно иллюстрирует понятие об ак-
тивном центре. Молекула белка сложена таким образом, что
реакционноспособные группы в боковых цепях нескольких ами-
нокислотных остатков фермента образуют в высшей степени
специфичную, пространственно организованную конструкцию,
точно отвечающую конфигурации субстрата. В состав актив-
ных центров ферментов входят боковые цепи остатков Asp,
Cys, Glu, His, Lys, Met, Ser, Thr, а также концевые аминные и
карбоксильные группы. Поскольку в среднем возле субстрата
расположено около 20 таких групп (значительно меньше, чем
общее число аминокислотных остатков в молекуле фермента),
принято считать, что непосредственное участие в функциониро-
вании активного центра фермента принимает только его не-
большая часть. У больших ферментов может быть несколько
активных центров. Большинство аминокислотных остатков, не
входящих в активный центр, определяют характер складывания
полипептидной цепи (вторичную структуру) и пространственное
расположение одной части цепи относительно другой (третич-
ную структуру), в результате чего и создается активный центр
фермента (рис. 3.4).
Хотя ряд изложенных ниже более детальных гипотез вес
еще не лишен некоторых противоречий, следует подчеркнуть,
что понятия об активном центре и фермент-субстратиом комп-
РИС. 3.2. Схематическое изображение активного центра лизоцима. Жирными?
линиями выделена молекула гексасахаридного субстрата, большими и малень-
кими кружками обозначены атомы кислорода и азота соответственно, а штри-
ховыми линиями—водородные связи. [Воспроизведено с разрешения из ра-
боты: Yudkin М„ Offord R., A Guidebook to Biochemistry, р. 48, Cambridge-
University Press, London, 1971.]
128
Глава 3
лексе в настоящее время общеприняты и лежат в основе боль-
шинства теорий, объясняющих механизм действия ферментов.
В дальнейшем мы также будем пользоваться этими понятиями
при математическом анализе кинетики ферментативного ката-
лиза.
Два различных подхода к объяснению ферментативной ак-
тивности схематически отображены на рис. 3.3. Фермент может
связывать молекулы, двух субстратов таким образом, что реак-
РИС. 3.3. Фермент ускоряет реакцию за счет пространственного сближения
двух субстратов (эффект сближения) и их расположения под выгодным уг-
лом (эффект ориентации). (Воспроизведено с разрешения Д. Э. Кошланда.)
ционноспособные группы последних будут располагаться побли-
зости друг от друга и от каталитических групп фермента. Это,
естественно, будет способствовать ускорению химической реак-
ции; такой эффект известен под названием эффекта сближе-
ния. Предположим далее, что молекулы двух субстратов не
обладают сферической симметрией. В этом случае реакция про-
изойдет только в том случае, когда при сближении молекулы
располагаются в ориентации, обеспечивающей тесное взаимо-
действие реакционноспособных атомов или групп. Считается,
что ферменты связывают молекулы субстратов, так что по-
следние занимают особенно благоприятное положение, созда-
вая таким образом эффект ориентации, который и обеспе-
чивает ускорение реакции. Это явление, иногда называемое
также орбитальным управлением, вносит свой вклад в процесс
ферментативного катализа, однако количественную величину
соответствующего эффекта в общем случае пока еще опреде-
лить трудно.
Прежде чем приступить к краткому обзору химии фермента-
тивного катализа, следует упомянуть еще одну гипотезу, связан-
Кинетика катализируемых ферментами реакций
129
ную с геометрией фермента. Известно, что связывание субст-
рата сопровождается небольшим изменением пространственной
структуры некоторых ферментов. В частности, путем изучения
трехмерной структуры ферментов лизоцима и карбоксипепти-
дазы А в виде комплексов с субстратами и без субстратов бы-
ло показано, что конформации ферментов при связывании суб-
страта несколько изменяются. Такое индуцированное соответст-
вие фермента и субстрата может вносить свой вклад в процесс
ферментативного катализа. Предлагались также усложненные
варианты модели индуцированного соответствия, в которых в
ходе процесса последовательно образуется несколько промежу-
точных фермент-субстратных комплексов. Некоторая эластич-
ность и гибкость молекулы фермента может способствовать
тщательной пространственной подгонке его каталитических
групп и тем самым ускорению превращений каждого из проме-
жуточных соединений. Возможно, что индуцирование субстра-
том изменений в конформации активного центра характерно
для ферментативного катализа вообще, хотя эксперименталь-
ное обнаружение этого эффекта связано с рядом трудностей и
поэтому было осуществлено только в случае нескольких фер-
ментов.
Некоторые ферменты осуществляют реакции, хорошо извест-
ные химику-органику. Одной из таких реакций является общий
кислотно-основной катализ, в котором катализатор на одной
из стадий процесса захватывает или отдает протон. Этот тип
катализа, в частности, лежит в основе механизма действия од-
ного из немногих ферментов, для которых предложена доста-
точно убедительная полная последовательность элементарных
стадий каталитического процесса (рис. 3.4). Выделяемый из
поджелудочной железы химотрипсин представляет собой про-
теолитический (т. е. гидролизующий белки) фермент, специ-
фично расщепляющий пептидные связи, образованные карбок-
сильными группами остатков тирозина, триптофана и фенила-
ланина. Считается, что в реакции химотрипсинового катализа
как при отщеплении, так и при присоединении протонов роль
переносчика последних играет вода. В сущности, к общему
кислотно-основному катализу можно свести целый ряд очень
важных в химии клетки реакций, в том числе процессы присо-
единения к карбонильным соединениям и гидролиз сложных
эфиров.
В ферментативном катализе важную роль могут играть и
Другие факторы, например ковалентный катализ, деформация
связей, электростатический катализ, многофункциональный ка-
тализ и эффекты растворителя (вспомните структуру масляной
капли, приведенную в предыдущей главе). Детальное описание
этих эффектов можно найти в приведенной в конце главы ли-
9-537
Липептидныи
суде трат
нонк,
I II I I
R—С—C—N - CH-COO
I
+ NH3
Продукт
реакции 7
Продукт
реакции 2
I
CHj,
б—H-
H О Н R,
। LI ।
R—С—С—N—CH— COO
O
R, R
I I +
R NH, — СН —ООО- Н —С —NHj
I I
н—с—ЫН3 СООН
с=о
О
N^'NH
Ser 195 -------- His 57
Белок
CH;,
СН2
б—
I Г
СН -----
Белок
Белок
Ser 195 -----
Белок
His 57
Свободный
химотрипсин
A
Рцилгсиыотрик -
сгт
Свободный
химотрипсин
РИС. 3.4. а — предполагаемая последовательность реакций при расщеп-
лении пептидной связи химотрипсином. (Воспроизведено из работы:
Lehninger A. L., Biochemistry, 2d ed., рр. 229—231, Worth Publishers,
New York, 1975; есть перевод первого издания: Ленинджер А., Биохи-
мия.— М.: Мир, 1976, с. 203); б — структура химотрипсина. [Воспроиз-
ведено из статьи: Matthews В. W. и др., Three-Dimensional Structure of
Tosyl-a-Chymotrypsin; Nature, 214, 652 (1976)].
A. — Гидроксильная.группа Serl95 связана водородной связью с имид-
азольным атомом азота His 57.
Б. Протон очень быстро мигрирует от Serl95 к имидазольному атому
азота His57. Образуется временная водородная связь между имид-
азольным кольцом и субстратом, ориентирующая дипептид таким обра-
зом, что становится возможной нуклеофильная атака атома кислорода
серина на карбонильный атом углерода субстрата.
В. Аминоацильная группа отрывается от дипептида и переносится к
ферменту; одновременно образуется свободная С-концевая аминокис-
лота субстрата.
Г. Под действием Н2О аминоацильная группа отделяется от Serl95,
в результате чего регенерируется свободный фермент.
Кинетика катализируемых ферментами реакций
131
тературе; вероятно, они, как и рассмотренные в настоящем раз-
деле факторы, могут оказывать влияние на некоторые катали-
зируемые ферментами реакции. Поскольку в общем случае ре-
акции ферментативного катализа включают в себя множество
эффектов, неудивительно, что пока еще не удалось разработать
простую общую схему, которая позволила бы оценить их сум-
марное влияние и относительную значимость каждого из них.
К счастью, математические выражения для скоростей катали-
зируемых ферментами реакций можно вывести, опираясь лишь
на основную концепцию о фермент-субстратном комплексе как
основном промежуточном соединении.
3.2. Кинетика простых ферментативных реакций
с одним и двумя субстратами
В настоящем разделе мы попытаемся вывести математиче-
ские выражения, характеризующие скорости катализируемых
ферментами реакций. Естественно, важнейшим критерием пра-
вильности такого выражения будет соответствие вычислен-
ных и экспериментально найденных скоростей. Чтобы исклю-
чить возможные недоразумения, сначала мы в общих чертах
опишем некоторые экспериментальные методы определения ско-
ростей реакций.
Прежде всего определим, что мы имеем в виду под ско-
ростью реакции. Рассмотрим реакцию
S ---> Р
Скорость этой реакции в приближении квазиравновесного со-
стояния (раздел 3.2.1) определяется как
ds dp
dt dt
(3-1)
где символами s и р обозначены молярные концентрации ис-
ходного вещества S и продукта реакции Р соответственно. От-
сюда следует, что v выражается в числе молей в единице объе-
ма в единицу времени. Скорость реакции является интенсивной
величиной, имеющей определенное значение в каждой точке
реакционной смеси. Поэтому, если концентрации или другие
интенсивные переменные изменяются от точки к точке, то и
скорости реакции в этих точках будут различными. При экспе-
риментальном изучении кинетики часто используют реакторы
с эффективным перемешиванием, в которых обеспечивается од-
на и та же скорость реакции во всем объеме реакционной
смеси.
Как и при моделировании любых других технологических
процессов, слово «точка» в определении скорости реакции ис-
9*
132
Глава 3
пользуется не в строго геометрическом смысле. Напротив, здесь
под точкой подразумевается некоторый объем, в который вхо-
дит множество молекул, но который тем не менее очень мал
по сравнению с объемом всей реакционной смеси. Это на пер-
вый взгляд не столь существенное уточнение очень важно не
забывать при переходе от гипотетических моделей к конкрет-
ным биологическим системам. Например, при моделировании
молекулярных процессов в отдельной изолированной клетке
понятия о концентрации данного вещества Ai и скорости его
превращения в определенной точке могут оказаться вообще не-
применимыми, если в клетке содержится только небольшое чис-
ло молекул А]. К этому вопросу мы вернемся позднее, при
изучении кинетики роста клетки (гл. 7).
Типичный эксперимент по изучению кинетики ферментатив-
ных. реакций (т. е. реакций, катализируемых ферментами)
обычно проводят следующим образом. В нулевой момент вре-
мени растворы субстрата и соответствующего очищенного фер-
мента смешивают при эффективном перемешивании в закрытом
сосуде, в котором поддерживается постоянная температура и
который содержит буферный раствор, необходимый для под-
держания определенного значения pH. Далее через заданные
промежутки времени определяют концентрации субстрата
и (или) продукта реакции. Для этой цели применяют различ-
ные методы, в том числе спектрофотометрические, манометри-
ческие, электродные, поляриметрические, а также методы, свя-
занные с отбором проб. Обычно используют только данные о
начальных скоростях реакций. Поскольку условия реакции, в
том числе концентрации фермента и субстрата, точнее всего
известны при нулевом времени реакции, начальный наклон
кривой, отражающей изменение концентрации субстрата или
продукта реакции в зависимости от времени, определяется по
следующим данным:
= п|/=о, где е = е0, s=s0 и р = 0 (3.2)
/=о
На рис. 3.5 приведены данные, полученные в одном из экспе-
риментов описанного типа. Обратите внимание на то, что при
( = 0 в смеси уже имеется некоторое количество продукта реак-
ции; отсюда следует, что нулевое время на самом деле не яв-
ляется временем начала реакции. В этом заключается одна из
трудностей, присущих методу определения начальных скорос-
тей реакций; другие недостатки метода описаны в специальной
литературе по химической кинетике. Тем не менее метод на-
чальных скоростей позволяет с достаточной воспроизводи-
мостью определять ферментативную активность и концентрации
dp _______ ds
dt dt
Кинетика катализируемых ферментами реакций
133
веществ в первый момент реакции и поэтому с успехом может
применяться в практической работе.
Проблема воспроизводимости результатов определения фер-
ментативной активности важна со многих точек зрения, в том
числе и с точки зрения конструкций реакторов, в которых осу-
ществляются процессы с изолированными ферментами. В пре-
дыдущих разделах мы уже упоминали, что белки в условиях,
О 20 40 60 80 100 120 140
Ародолжителбность реакции, мин
РИС. 3.5. Экспериментальное определение скорости гидролиза 30%-го рас-
твора крахмала глюкоамилазой в реакторе периодического действия (60 °C,
е0=И 600 единиц, объем реактора I л).
отличающихся от свойственного им биологического окружения,
легко подвергаются денатурации; в этой связи неудивительно,
что изолированный фермент в «необычной» для него водной
среде может постепенно терять свою каталитическую актив-
ность (рис. 3.6). Известно, что такая инактивация характерна
для многих ферментов; в то же время во многих руководствах
по кинетике ферментативного катализа это явление в лучшем
случае только упоминается. Проблема постепенной потери ка-
талитической активности ферментами не столь существенна in
vivo (в интактном живом организме), когда снижение концент-
раций ферментов за счет инактивации компенсируется их био-
синтезом в необходимых количествах. Напротив, инактивацию
фермента in vitro (вне живой клетки) необходимо учитывать
как при изучении кинетики реакций, так и при проектировании
134
Глава 3
ферментативных реакторов. К этой теме мы обратимся еще
раз в разд. 3.7.
В подписи1 к рис. 3.5 (в скобках) нам впервые встречается
другая типичная для кинетики ферментативного катализа осо-
бенность. Обратите внимание на то, что количество фермента,
использовавшегося в этом эксперименте, выражено в неких
РИС. 3.6. Инактивация фермента в растворе. В течение 1 ч при 37 °C раствор
аспартазы теряет более 5% исходной активности. [Воспроизведено из статьи:
Tosa Т. et al., Continuous Production of L-Aspartic Acid by Immobilized Aspar-
tase, Biotech. Bioeng., 15, 68 (1973)].
«единицах». Что это за таинственные единицы и почему нельзя
количество фермента выразить более понятным и более опре-
деленным способом, например в молях или в единицах массы?
Сначала ответим на вторую часть вопроса. Количество фер-
мента обычно не выражают, например, в единицах массы, по-
скольку ферментные препараты, как правило, представляют
собой смесь различных белков, в которой данный фермент яв-
ляется всего лишь одним из нескольких компонентов. Содержа-
ние интересующего нас фермента в этой смеси часто неизвест-
но; более того, оно может быть различным в разных партиях
препарата. Чтобы охарактеризовать препарат, содержание фер-
мента обычно выражают в единицах каталитической активно-
сти, содержащейся в единице массы ферментного препарата.
Кинетика катализируемых ферментами реакций 135
Единицей или единицей активности называют количество
фермента, которое обладает определенной каталитической ак-
тивностью в заданных для этого фермента стандартных усло-
виях. Например, в эксперименте по гидролизу крахмала, ре-
зультаты которого приведены на рис. 3.5, за единицу активно-
сти глюкоамилазы принимали количество фермента, которое в
4%-ном растворе крахмала Линтнера при pH 4,5 и температу-
ре 60 °C образует 1 мкмоль глюкозы в 1 мин. Отсюда следу-
ет, что единица активности каждого конкретного фермента бу-
дет иметь свое специфическое определение и, более того, один
фермент может иметь несколько определений активности в за-
висимости от конкретных условий катализируемых им реакций.
Поэтому во избежание ошибок при интерпретации или исполь-
зовании результатов изучения кинетики реакций ферментатив-
ного катализа всегда следует тщательно проверять применяемое
в каждом случае определение единицы ферментативной актив-
ности. Конечно, вероятность ошибки резко снижается, если из-
вестна активность высокоочищенного фермента.
3.2.1. Уравнение Михаэлиса — Ментен
Предположим теперь, что в нашем распоряжении имеется
целый ряд экспериментальных данных по определению скорос-
ти ферментативной реакции (рис. 3.7) и перед нами стоит зада-
ча выразить эти данные в математической форме. Информация
типа показанной на рис. 3.7 часто позволяет сделать следую-
щие выводы качественного характера:
1. При относительно низких концентрациях субстрата реакция
имеет первый порядок по субстрату. (Напомним, что если
v=k-sn, где k — константа, то п называют порядком реак-
ции.)
2. По мере повышения концентрации субстрата порядок реак-
ции по субстрату постепенно снижается от единицы до нуля.
3. Скорость реакции пропорциональна общему количеству фер-
мента в реакционной смеси.
На базе экспериментальных данных такого рода в 1902 г. Ан-
ри предложил следующее выражение для скорости фермента-
тивной реакции:
g:=7m^ ’ где = (3.3)
В уравнении (3.3) учтены все три указанные выше закономер-
ности. Обратите внимание, что при s, равном Km, и = итах/2. Во
избежание недоразумений, связанных с употреблением в лите-
ратуре различных символов и сокращений, подчеркнем, что s
136
Главе 3
РИС. 3.7. Экспериментальное изуче-
ние кинетики катализируемых фер-
ментами реакций,
а — зависимость скорости реакции от
концентрации субстрата при посто-
янной концентрации фермента. [Вос-
произведено из работы: Laidler К. J.,
The Chemical Kinetics of Enzyme Ac-
tion, p. 64, The Clarendon Press,
Oxford, 1958, по данным, опублико-
ванным ранее в статье: Quellet L„
Laidler К. J., Morales M. F., Molecular
Kinetics of Muscle Adenosine Tripho-
sphate, Arch. Biochem. Biophys., 39,
37 (1942).]
6 — зависимость скорости реакции
(изменение оптической плотности при
200 нм за 1 мин) от концентрации
фермента при постоянной концентра-
ции субстрата. Из работы: Диксон М.,
Уэбб Э., Ферменты, т. 1—3. — М.:
Мир, 1982.)
обозначает концентрацию свободного субстрата в реакционной
смеси, а е0 — общую концентрацию фермента как в свободном,
так и в связанном виде (см. ниже).
Хотя Анри дал теоретическое толкование уравнению (3.3)
на базе гипотетического механизма реакции, его доказательст-
во, равно как и близкое обоснование, предложенное в 1913 г.
Михаэлисом и Ментен, в настоящее время считается в общем
случае не строгим. В то же время в этой главе мы неодно-
кратно и не без успеха будем применять общий ход рассужде-
ний Михаэлиса и Ментен (так и не нашедший до сих пор стро-
гого подтверждения) для разработки более сложных моделей
Кинетика катализируемых ферментами реакций 137
кинетики ферментативного катализа. Поэтому сначала мы рас-
смотрим основные положения гипотезы Михаэлиса — Ментен.
В качестве одного из исходных положений принимают, что
фермент Е и субстрат S, вступая в реакцию, образуют комп-
лекс ES, который затем диссоциирует на продукт реакции Р
и свободный (несвязанный) фермент Е:
S+E ES (3.4а)
Л-1
ES Р + Е (3.46)
В этом механизме учтены как рассмотренное выше образование
фермент-субстратного комплекса, так и регенерация катализа-
тора в исходной форме по завершении последовательности ре-
акций. Уравнения (3.4) в принципе верны, хотя и чрезмерно
упрощены.
Анри, а также Михаэлис и Ментен предположили, что реак-
ция (3.4а) обратима, и, следовательно, в соответствии с зако-
ном действующих масс мы можем записать:
= -4^- = Кт=-константа диссоциации (3.5)
(es) j «1 771
Здесь s, е и (es) обозначают концентрации S,E и ES соответ-
ственно. Кроме того, принимается, что процесс разложения
комплекса на продукт реакции и свободный фермент необра-
тим:
(з.б)
Поскольку весь фермент находится или в свободной, или в
связанной форме, то
e+(es)=eo (3.7)
где е0 — общая концентрация фермента в системе, известная
из исходных данных и равная количеству фермента, загружен-
ного в реактор. Теперь путем исключения (es) и е из трех по-
следних уравнений можно получить уравнение (3.3), в котором
vma*=k2e0. Обычно, говорят, что реакция характеризуется урав-
нением Михаэлиса — Ментен, если ее скорость может быть
описана уравнением (3.3), хотя в разработку и подтверждение
правильности этого выражения внесли не меньший вклад и
другие исследователи. Параметр итах называют максимальной
или предельной скоростью, а Кт известна как константа Миха-
элиса. Уравнение Михаэлиса — Ментен удачно описывает ки-
нетику многих катализируемых ферментами реакций, но тем не
менее оно не универсально. В последующих разделах и в уп-
ражнениях мы рассмотрим ряд модифицированных вариантов
138
Глава 3
РИС. 3.8. Изменение концентраций веществ в реакции
£+i k+2
Е + S <—ES -------> Р + Е при k+1 « « k+2
k-i
этого уравнения, которые точнее описывают конкретные фер-
ментативные реакции, протекающие в специфических условиях.
Бриггс и Холдейн пришли к уравнению (3.3) другим путем,
который, как было показано позднее путем экспериментального
изучения кинетики и математического анализа, носит наиболее
общий характер. Если реакция осуществляется в закрытом со-
суде при энергичном перемешивании, то уравнения материаль-
ного баланса для субстрата и фермент-субстратного комплекса
можно записать следующим образом:
и = —= k^e—k_l (es)
и
= kiSe—(£_t + k2) (es)
С учетом выражения (3.7) эти два уравнения представляют
собой систему двух обычных дифференциальных уравнений с
двумя неизвестными, s и (es). Понятно, что соответствующие
начальные условия при / = 0 выражаются следующим образом:
s(O) = so (es)(0)=0
Эти уравнения нельзя решить аналитически, но путем ин-
тегрирования с помощью компьютеров можно пайти зависимо-
сти концентраций S, Е, ES и Р от времени. Полученные та-
ким путем результаты приведены на рис. 3.8; очевидно, что
Кинетика катализируемых ферментами реакций 139
после короткого стартового периода можно с весьма большой
точностью принять, что
d(es)
dt
Ключевым пунктом анализа Бриггса и Холдейна явилось до-
пущение о правильности условия (3.8). Можно показать, что
для нашего случая катализируемой ферментом реакции в за-
крытой системе это допущение, называемое обычно приближе-
нием квазиравновесного состояния, справедливо при условии
достаточно малой величины отношения e0/s0. Если же началь-
ная концентрация субстрата сравнима с общей концентрацией
фермента, то допущение (3.8) может оказаться неверным.
В большинстве случаев, однако, количество ферментного ката-
лизатора намного меньше количества реагентов; в такой ситуа-
ции после стартового периода реакции выражение (3.8) мож-
но считать применимым с достаточно высокой точностью.
Если далее, следуя рассуждениям Бриггса и Холдейна, из
приведенных выше уравнений с помощью выражения (3.7)
исключить е и (es), то мы придем к уравнению
__________________k2e0s______ /о q\
dt [(fe-x + ^/M + s__________{ '
Таким образом мы получили выражение типа уравнения Миха-
элиса — Ментен, в котором
ишах=^2^о (3.10)
И
(ЗЛ1)
Обратите внимание на то, что здесь Кт по физическому смыслу
уже не является константой диссоциации.
Располагая математическим выражением для скорости ре-
акции типа уравнения Михаэлиса — Ментен, мы можем опре-
делить изменения концентраций реагентов во времени путем
интегрирования
ПРИ условии, что s(O) = so (3.12)
Таким путем получаем, что
^max^~so (3.13)
Разумеется, на практике это уравнение проще использовать для
140
Глава 3
расчета времени /, необходимого для достижения определен-
ных концентраций субстрата, а не наоборот.
Полезно сравнить ход реакции, предсказываемый уравне-
нием (3.13), с результатами, полученными без приближения
квазиравновесного состояния. Как показано на рис. 3.9, откло-
нения могут быть значительными, если общая концентрация
фермента приближается к s0, поэтому в таких случаях не сле-
РИС. 3.9. Вычисленные зависимости степени гидролиза эфира ацетил-Ь-фенил-
аланина химотрипсином от продолжительности реакции в реакторе периоди-
ческого действия. При больших величинах eo/so=a наблюдается значитель-
ное несоответствие между точным решением и решением в квазиравновесном
приближении. [Воспроизведено с разрешения из статьи: Lim Н. С., On Kinetic
Behavior at High Enzyme Concentrations, Amer. Inst. Chem. Eng. J., 19,
659 (1973).]
1—решение в квазиравновесном приближении; 2 — точное решение.
дует применять уравнение Михаэлиса — Ментен. На рис. 3.10
приведен другой пример, а именно зависимость эстеразной ак-
тивности от концентрации фермента. Здесь предсказываемая
моделью Михаэлиса — Ментен линейная зависимость справед-
лива при малых концентрациях фермента, но также не соблю-
дается при более высоких концентрациях. Запомните, что для
линейного участка кривой тангенс угла наклона равен
^2s/ (Кт +5) •
В некоторых случаях в зависимости от скоростей отдельных
стадий реакции приближение квазиравновесного состояния мо-
жет оказаться справедливым и при больших значениях eo/so-
В частности, такая ситуация могла бы возникнуть, если бы
комплекс ES диссоциировал гораздо быстрее, чем образовывал-
ся, т. е. если бы Кт была значительно большего. Одйако обыч-
Кинетика катализируемых ферментами реакций 141
но константа Михаэлиса очень мала и составляет от 10-2 до
10_5, поэтому такие случаи не типичны.
Отсюда следует, что если концентрация фермента сравнима
с So, то, как правило, у нас пет достаточных оснований для уп-
рощения модели кинетики соответствующего процесса с исполь-
зованием приближения квазистационарного состояния. Относи-
тельная концентрация фермента может достигать больших зна-
Содержание фермента, ед.
РИС. 3.10. При высоких начальных концентрациях фермента зависимость
эстеразной активности от концентрации эстеразы не подчиняется уравнению
Михаэлиса — Ментен. (Воспроизведено с разрешения из работы: Frobisher М„
Fundamentals of Microbiology, 8th ed., p. 61, W. B. Saunders Co., Philadelphia,
1968.)
чений, например, если работа ферментативного реактора про-
должается и. тогда, когда большая часть субстрата уже под-
верглась превращению. В таком случае s уменьшается до зна-
чений, сравнимых с во, и на последнем этапе процесса уравне-
ние Михаэлиса — Ментен может уже не отражать его реальной
кинетики. К счастью, в этот период реакция резко замедляет-
ся и поэтому, как правило, уже не представляет практического
интереса. Концентрация субстрата вблизи фермента может
быть незначительной и при проведении ферментативных реак-
ций на поверхности раздела фаз; реакции такого типа мы
рассмотрим в разд. 4.4. В таких случаях следует учитывать и
скорость переноса субстрата к границе раздела фаз. Ограни-
чения, свойственные модели Михаэлиса — Ментен, выявились
лишь в самое последнее время, и поэтому соответствующее
уравнение все еще широко используется при изучении и проек-
142
Глава 3
тировании ферментативных реакторов. Хотя в некоторых ситу-
ациях равновесное приближение и уравнение Михаэлиса —
Ментен нельзя считать достаточно точными, в целом они оказа-
лись очень полезными инструментами исследования, и в этом
качестве мы часто будем их применять и в дальнейшем изло-
жении.
Говоря о простом уравнении Михаэлиса — Ментен, нельзя
не упомянуть, что точно таким же математическим выражени-
ем широко пользуются для оценки скоростей многих реакций,
ускоряемых твердыми катализаторами. Уравнение (3.3) в хи-
мической технологии называется уравнением Ленгмюра — Хин-
шелвуда или Хоугена — Уотсона. Поскольку в химической и
нефтяной промышленности широко применяются реакции на
синтетических твердых катализаторах, анализу и проектирова-
нию таких каталитических реакторов было посвящено огромное
число работ. В большей части этих работ использовали урав-
нение Ленгмюра — Хиншелвуда; следовательно, их результаты
с успехом могут быть перенесены и на катализируемые фер-
ментами реакции, описываемые уравнением Михаэлиса —
Ментен. В последующих главах мы еще не раз встретимся с
аналогиями и сходством между классической химической и био-
химической технологиями, но в то же время узнаем и многие
особенности, характерные только для биологических процес-
сов.
3.2.2. Определение параметров
в уравнении Михаэлиса — Ментен
В своей первоначальной форме уравнение Михаэлиса —
Ментен (3.3) неудобно для определения кинетических пара-
метров утах и Кт. Как показано на рис. 3.7, по графику зави-
симости v от s довольно сложно определить точное значение
Утах. Путем ряда несложных преобразований уравнения (3.3)
нетрудно, однако, найти приведенные ниже уравнения, более
пригодные для изображения результатов в графической форме
и для графического определения параметров:
— -- ----(3.14)
и ушах ^тпах 5
— = ---— .S (3.15)
и ymax ушах
У = ашах—(зл6)
Каждое из приведенных уравнений отражает линейную за-
висимость одной величины от другой. В то же время при опре-
Кинетика катализируемых ферментами реакций
143
делении кинетических параметров по таким зависимостям сле-
дует принимать во внимание ряд свойственных им ограничений
и недостатков. В графическом выражении уравнения (3.14) в
виде зависимости 1/и от 1/s (известном под названием графика
Лайнуивера—Бэрка) четко разделены зависимая и независи-
мая переменные (рис. 3.11, а). Определяемые с наибольшей
РИС. 3.11. а — результаты экспериментального изучения ферментативной ак-
тивности пепсина в координатах Лайнуивера — Бэрка;
б — экспериментальные данные по изучению катализируемого химотрипсином
гидролиза метилового эфира дигидрокоричной кислоты в координатах Эди —
Хофсти. [Воспроизведено с разрешения из работы: Laidler К. J-, The Chemical
Kinetics of Enzyme Action, pp. 65—66, The Clarendon Press, Oxford, 1958.]
точностью значения скоростей реакций, близкие umax, концент-
рируются возле исходной точки, а менее точно определяемые
значения v удалены от нее и таким образом в наибольшей
степени влияют на наклон прямой Кт/атгх; поэтому в таких
случаях нельзя применять метод наименьших квадратов. Вто-
рое уравнение (3.15) обеспечивает более равномерное распре-
деление больших значений v и, таким образом, более точное
определение наклона кривой 1/итах, однако отрезок, отсекае-
мый прямой на координатной оси, часто слишком мал, что не
позволяет достаточно точно определить этим методом Кт-
В третьем методе применяется так называемый график Эди —
Хофсти, представляющий собой график зависимости и от v/s
(рис. 3.11,6); здесь наименее точно определяемая переменная
v является составной частью как функции, так и аргумента.
Приведенные выше соображения диктуют следующую стра-
тегию определения утах и Кт: сначала определяют ’ итах по
графику уравнения (3.14) (находят точную величину отрезка,
144
Глава 3
отсекаемого прямой на оси 1/и) или уравнения (3.15) (точно
определяют наклон прямой). Затем возвращаются к графику
зависимости v от s и находят Si/2, т. е. такую концентрацию
субстрата, при которой v равно vmsx/2. Как мы уже упоминали
при обсуждении уравнения (3.3), Кт по величине и размернос-
ти равно Si/2.
Важно представлять себе, в каких диапазонах могут изме-
няться эти кинетические параметры. В табл. 3.4 приведен ряд
параметров, характерных для различных ферментов. Обратите
внимание на то, что величина Л2, может изменяться в очень
широких пределах, а Кт, напротив, обычно имеет значения
2-Ю-3—10-10—3 М. Почти все приведенные в табл. 3.4 данные
были получены при умеренной температуре и почти нейтраль-
ном pH. Исключение составляет пепсин, основная биологичес-
кая функция которого сводится к гидролизу белков в кислой
среде желудка и который поэтому наиболее активен при низ-
ких значениях pH; эти значения, естественно, использовались и
при экспериментальном определении его кинетических па-
раметров. Модели, отражающие влияние pH и температу-
ры на кинетику ферментативных реакций, мы рассмотрим в
разд. 3.6.
3.2.3. Кинетика обратимых реакций, реакций с двумя
субстратами и с активацией фермента кофактором
Равновесие многих катализируемых ферментами реакций,
например реакций гидролиза биополимеров, сильно смещено в
сторону продуктов реакции, поэтому, как правило, такие пре-
вращения можно считать необратимыми. В других случаях,
например при изомеризации глюкозы во фруктозу под дейст-
вием фермента глюкозоизомеразы, может установиться равно-
весное состояние, требующее учета вклада обратной реакции.
В качестве простейшей модели обратимой ферментативной ре-
акции рассмотрим кинетику последовательных превращений
типа
S+E ES (3.17а)
k-i
ES Р + Е (3.176)
*-2
Эта последовательность элементарных реакций отличается
от последовательности реакций, предложенных Анри — Михаэ-
лисом— Ментен и выражаемых уравнениями (3.4), только
тем, что уравнение (3.176) теперь предусматривает образова-
ние комплекса ES из продукта реакции и свободного фермента.
Таблица 3.4. Кинетические параметры некоторых ферментативных реакций®
zss-ei
Фермент
Субстрат
Температу- рН с_, м_,
Пепсин
Трипсин
Химотрипсин
Карбоксипептидаза
Аденозинтрифосфатаза
Уреаза
Этиловый эфир карбобензокси-Е-глутамил-Е-ти- 31,6 4,0 0,00108 530
розина Карбобензокси-Ь-глутамил-Ь-тирозин 31,6 4,0 0,00141 560
Бензоил-Ь-аргининамид 25,5 7,8 27,0 480
Химотрипсиноген 19,6 7,5 2900 <770
Стурнн 24,5 7,5 13100 400
Эфир бензоил-Е-аргинина 25,0 8,0 26,7 12 500
Метилгидроциннамат 25,0 7,8 0,026 256
Метиловый эфир «Я-а-хлор-р-фенилпропионовой 25,0 7,8 0,135 83,3
кислоты
Метиловый эфир d-fJ-фенилмолочной кислоты 25,0 7,8 0,139 28,6
Метиловый эфир /-fj-фенилмолочной кислоты 25,0 7,8 1,38 100
Метиловый эфир бепзоил-Е-фенилаланипа 25,0 7,8 51,0 217
Этиловый эфир ацетил-Е-триптофана 25,0 7,8 30,7 588
Этиловый эфир ацетил-Б-тирозина 25,0 7,8 193,0 31,2
Этиловый эфир бензоил-Б-о-нитротирозина 25,0 7,8 3,27 90,9
Этиловый эфир бензоил-Б-тирозина 25,0 7,8 78,0 250
Этиловый эфир бензоил-Е-фенилаланина 25,0 7,8 37,4 167
Этиловый эфир бензоил-Е-метионина 25,0 7,8 0,77 1250
Бензоил-Е-тирозинамид 25,0 7,8 0,625 23,8
Ацетил-Б-тирозинамид 25,0 7,8 0,279 12,3
Карбобепзоксиглицил-Б-триптофан 25,0 25,0 7,5 8,2 89 94 196 164
Карбобензоксиглицил-Б-фенилаланин 25,0 7,5 181 154
Карбобензоксиглицил-Б-лейцин 25,0 7,5 10,6 37
АТР 25,0 7,0 104 79 000
Мочевина | 20,8 20,8 7,1 8,0 20 000 30 800 250 256
а Воспроизведено из работы: Laidler К. J., The Chemical Kinetics of Enzyme Action, p. 67, Oxford University Press, London, 1958.
146
Глава 3
Как и ранее, будем считать, что реакционная смесь нахо-
дится в закрытом сосуде и эффективно перемешивается. Тогда
на основе материального баланса различных форм фермента
[уравнение (3.7)] с учетом приближения квазистационарного
состояния [уравнение (3.8)] легко находим, что
7, = _ ds = dp _ (vs/Ks) S - (Vp/Kp) p /о 1 m
dt dt ~ l + slKs+plKp ' '
Здесь vs и Ks аналогичны wmax и Km в уравнениях (3.10) и
(3.11) соответственно, и
к»=-¥г <ЗЛ9>
Подавляющее большинство катализируемых ферментами
реакций протекает с участием по меньшей мере двух субстра-
тов. В то же время чаще всего, одним из субстратов является
вода, концентрация которой практически постоянна и обычно в
1000 или более раз превышает концентрации других субстра-
тов. Как мы покажем позднее в этом же разделе, в таких
случаях можно считать, что реакция протекает с одним субст-
ратом S, и, следовательно, ее кинетику можно изучать так, как
это было описано в предыдущем разделе. Кроме того,, описан-
ные ниже кинетические модели иногда могут объяснять и
влияние кофакторов на скорости ферментативных реакций.
По всей вероятности, во многих процессах с участием двух
субстратов могут образовываться тройные комплексы, в кото-
рых фермент одновременно связан с двумя субстратами. В та-
ком случае возможна, например, такая последовательность ре-
акций:
Константа
равновесия
диссоциации
Е +Sj ESr Kj
Е +S2 =рь ES2 К.2
(3.20)
ES1 + S2 ES1S2 К\2
ES2+S1 ESiSs К21
k
ESjSj --> P + E
так что
v=k{es\Si) (3.21)
Как и раньше, строчными буквами мы будем обозначать кон-
центрации, а такими же буквами в скобках — концентрацию
одной формы, например фермепт-субстратпого комплекса. Ес-
Кинетика катализируемых ферментами реакций
147
ли допустить, что первые четыре реакции (3.20) равновесны, то
получим
V —
ke0
, . к21 . К12 , х/2 (К2к21 + КтКтг)
1+~+ —+
(3.22)
Вывод этого выражения не представляет затруднений, если
учесть, что общая концентрация фермента е0 должна быть рав-
на сумме концентраций свободного фермента е и трех комп-
лексов ESj, ES2 и ESiS2. Как и в случае реакций с одним
субстратом, в ферментативных реакциях с двумя субстратами
можно применить приближение квазиравновесного состояния.
В общем случае, однако, в результате получается довольно
громоздкое уравнение с таким числом параметров, что оно ста-
новится непригодным для практического использования. Удов-
летворительное по ряду параметров приближение описывается
уравнением (3.22).
Уравнение (3.22) можно далее несколько упростить, если
принять во внимание, что условия равновесия требуют, чтобы
K±K12 = K2K2i (3.23)
Соответствующим преобразованием уравнения (3.22) можно
прийти к уже знакомой форме уравнений:
у ^*maxsi
Kl* H-Si
где
_______ ^g0S2
u max _i_ /г-
й2 I л 12
И
' _ ^21S2 + ММ2
S2 + М2
(3.24
(3.25)
(3.26)
Из последних трех уравнений следует, что если s2 постоянна, а
Si изменяется, то реакция будет подчиняться уравнению Миха-
элиса— Ментен. В то же время уравнения (3.25) и (3.26) по-
казывают, что кажущиеся максимальная скорость и константа
Михаэлиса зависят от концентрации S2.
Если считать приведенные выше последовательность реак-
ций и выражение для скорости реакции с двумя субстратами
правильными, то мы можем проверить справедливость уравне-
ния Михаэлиса — Ментен (3.3), допустив, что один субстрат
находится в большом избытке. Тогда y*max становится равной
keQ, a М* приближается к постоянному значению Следова-
0
148
Глава 3
тельно, реакцию с двумя субстратами при s2^>Ki2 можно рас-
сматривать как реакцию с одним субстратом.
На рис. 3.12 изображена схема механизма ферментативной
реакции с одним субстратом (или с двумя субстратами при
S2^>/Ci2) при участии кофактора, которым может быть ион ме-
талла или кофермент. Поскольку эта ситуация аналогична
только что рассмотренному нами двухсубстратному механизму,
здесь нет необходимости снова обращаться к допущению о рав-
новесных последовательных реакциях. Если принять, что суб-
Субстрат
+
Апофермент
(белок)
Фермент
кофермент
или ион
металла
Продукты
реакции
Свободный
фермент
Фермент -
субстратный
комплекс
РИС. 3.12. Схематическое изображе-
ние предполагаемого механизма фер-
ментативного катализа с участием
кофактора.
страт связывается только апофермент-коферментным комплек-
сом, то в конце концов мы получим
v =------*£o£s---- (3.27)
cs + Ks(c + Ke)
где с — концентрация кофактора. Если считать концентрацию
субстрата s постоянной, то это выражение преобразуется в
уравнение Михаэлиса — Ментен, отражающее зависимость ско-
рости реакции от концентрации кофактора с. Так, при невысо-
кой концентрации кофактора (с<бйКс) реакция будет иметь пер-
вый порядок по концентрации кофактора с. С другой стороны,
при с^Кс мы получим уравнение скорости реакции с одним
субстратом, которая не зависит от концентрации кофактора.
Если Si и S2 или С и Si связываются с ферментом в строго
определенном порядке, то соответствующее уравнение скорости
процесса можно получить, приняв, что Kij запрещенной реак-
ции приближается к бесконечно большой величине.
Кинетика катализируемых ферментами реакций 149
3.3. Определение констант скоростей элементарных
стадий ферментативной реакции
В разд. 3.2.2 мы рассмотрели различные графические способы определе-
ния параметров vmax и Кт, входящих в уравнение Михаэлиса — Ментеи. Как
показывает анализ Бриггса — Холдейна, эти параметры определяются кон-
стантами скоростей элементарных стадий реакции k\, k-i и k2. В то же вре-
мя из уравнений (3.10) и (3.11) следует, что знания параметров Кт и vmaX
еще недостаточно для определения скорости отдельных стадий реакции. Для
оценки последних, очевидно, необходимо найти какие-то иные, независимые
взаимосвязи между константами скоростей элементарных стадий и экспери-
ментальными данными.
Такие более фундаментальные кинетические параметры представляют ин-
терес по меньшей мере по двум причинам. Во-первых, они дают углублен-
ное представление о реальных процессах, происходящих в ходе катализируе-
мых ферментами реакций. Мы можем узнать, например, насколько быстро
субстрат взаимодействует с ферментом, и сравнить эти величины со скоростью
обратного процесса диссоциации комплекса ES. Во-вторых, как мы уже ви-
дели, упрощения, лежащие в основе уравнения Михаэлиса — Ментен, с до-
статочной точностью оправдываются только при относительно невысоких кон-
центрациях фермента. Если это условие не соблюдается, то изучение кинети-
ки ферментативной реакции возможно только при учете данных материаль-
ного баланса для s, р и (es), для чего в свою очередь необходимо знать
константы отдельных стадий реакции &_] и k2. По указанным причинам
нам представляется целесообразным рассмотреть здесь экспериментальные
методы определения этих параметров и полученные с помощью таких мето-
дов результаты.
В методе кинетики предстащионарного состояния основное внимание уде-
ляется короткому периоду (около 1 с или меньше) сразу после начала реак-
ции, когда приближение квазистационарного состояния еще неприменимо.
В течение этого периода концентрация субстрата изменяется незначительно,
что позволяет приближенно решить уравнения материального баланса для
(es) и р. Сравнение найденной таким образом зависимости р от t с экспери-
ментальными данными, а также найденные значения vmax и Кт дают всю
информацию, необходимую для расчета k-\ и k2. Экспериментальной ос-
новой этого метода является методика остановленного потока, разработан-
ная и развитая Б. Чансом с сотрудниками. Согласно этой методике, растворы
фермента и субстрата быстро смешивают в кювете спектрофотометра; спект-
рофотометрический метод позволяет контролировать изменения концентраций
исходных веществ и продуктов реакции, осуществляющиеся в течение не-
скольких миллисекунд.
Разработанные и широко применявшиеся Эйгеном и сотрудниками ре-
лаксационные методы дают возможность изучать реакции, завершающиеся
в течение нескольких наносекунд. Существует несколько вариантов этого
метода, но все они основаны на принципе возмущения равновесного (или
стационарного) состояния реакционной смеси путем искусственного скачко-
образного изменения некоторых условий реакции, например температуры,
давления или напряженности электрического поля. Последующий отклик ре-
акционной системы на это изменение регистрируют непрерывно. Как схема-
тически показано на рис. 3.13, отклик на скачкообразное изменение условий
реакции представляет собой не что иное, как переход в новое, почти рав-
новесное или стационарное состояние.
3.3.1. Релаксационные методы изучения кинетики
В этом разделе мы несколько подробнее остановимся на релаксационных
методах, основанных на скачкообразном изменении условий реакции. Теория
и практика релаксационных методов была разработана также в применении
150
Глава 3
Температура
Наложение
электри-
ческого
ноля или
микроволнового
импульса
при в=О
Реакционная
смесь (вравно-
весии при t<0)
Время, ~ 70 6 с
Концентрация
РИС. 3.13. В релаксационных методах небольшое скачкообразное изменение
условий реакции, например температуры, вызывает быстрый переход в новое
равновесное состояние.
к осциллирующим возмущениям условий реакций. Сначала рассмотрим рав-
новесие только между субстратом, ферментом и фермент-субстратным ком-
плексом:
E-J-S <—> ES
/Мы знаем, что для этой реакции
s + (es) =s0
и
е + (es) = е0
Поэтому для эффективно перемешиваемой реакционной смеси, находящейся
в реакторе периодического действия, уравнение материального баланса для$
-^- = ^_1(es)—k^e (3.28)
принимает форму
-^- = ^1(s0—$)—^s(e0 + s—s0)= f(s)
(3.29)
Если мы символом s* обозначим концентрацию s в равновесном состоя-
нии после скачкообразного изменения условий реакции, то $* можно опре-
Кинетика катализируемых ферментами реакций
151
делить, решив уравнение
f(s*)=O (3.30)
где f(s)—правая часть уравнения (3.29), равная dsfdt. В любом релаксацион-
ном эксперименте $ будет близко s*, и приближенно можно считать, что f(s)
равно первым членам в разложении Тейлора типа
df (s*)
f(s) («*) +-----— (s—s*)-1-члены порядка (s—s*)2 и высших порядков.
(3.31)
Здесь f(s*), согласно уравнению (3.30), равно нулю. Обозначив символом %
отклонение от равновесной концентрации
%=s —s* (3.32)
и принимая во внимание, что s* не зависит от времени, путем преобразова-
ния уравнений (3.29) — (3.32) можно получить линейное уравнение следую-
щего вида [если пренебречь членами второго и высшего порядков в уравне-
нии (3.31)]:
dt —R-i + ^т(ео — so_r2s*)lx (3.33)
Если система сначала находится в состоянии исходного равновесия, ко-
торое отвечает условиям, предшествующим скачкообразному возмуще-
нию, то
Х(О)=Дхо (3.34)
а затем
Х(/)=ДХов-^ (3.35)
где
=s k_± -|- k1(e0 — sQ -f- 2s*) =4 k_^ 1\(е* -{- s*) (3.36)
V
В уравнении (3.36) e* выражает концентрацию не связанного в комплекс
фермента в состоянии конечного равновесия. Уравнение (3.35) свидетельст-
вует о том, что т можно определить по наклону прямой, отражающей полу-
логарифмическую зависимость X(f) от ДХо. Другими словами, т представляет
собой время, в течение которого X(f) уменьшается до 37% от своего перво-
начального значения. Определив т, %* и е* (или е0 и з0), далее по уравне-
нию (3.36) можно найти взаимосвязь между kt и k-i, не зависящую от
уравнения равновесия. Таким путем можно вычислить как kt, так и k-i.
Этот метод нашел широкое применение в химической кинетике, в том
числе и в кинетике рассматриваемого здесь ферментативного катализа.
3.3.2. Некоторые результаты изучения кинетики
переходных состояний
В табл. 3.5 приведены значения kx и fe-i для ряда ферментов и субстра-
тов. Эти данные были получены с помощью описанных выше методов, а так-
же ряда других специальных методик, описанных в приведенной в конце
настоящей главы литературе. Нетрудно видеть, что константы скорости пря-
мой реакции субстрата с ферментом kt почти всегда близки к теоретическо-
Таблица 3.5. Константы скорости некоторых элементарных стадий
фермент-субстратных реакций3
Белок или фермент
Субстрат
fei, (М-с)-1
с-'
Фумараза Фумарат L-Малат >109 >Ю9 >4,5-104 >4-104
Ацетилхолинэстераза Ацетилхолин >10э
Уреаза Мочевина >5-10е
Миозин (аденозин- АТР >8-10е
трифосфатаза)
Гексокиназа Глюкоза 3,7-10® 1,5-10®
MgATP >4-10® >6-102
Р-Амилаза Амилоза >5,8-107
Алкогольдегидроге- NAD 5,3-10® 74
наза из печени NADH 1,1•107 3,1
NAD-Алкогольдегид- С2Н5ОН >1,2-104 >74
рогеназа из печени СН3СНО >3,7-103 >3,!
Малатдегидрогеназа N.ADH 6,8-10® 2.4-108
Желтый старый фер- Флавинмононуклео- 1,5-10® ~10’4
мент Каталаза тид Н2О2 5-10®
Н2О2-Каталаза Н2О2 1.5-107
Пероксидаза Н2О2 9-10® <1.4
Н2О2-Пероксида- Гидрохинон 2,5-10®
за(П)
Пероксидаза Цитохром с 1,2-10®
Глутамат-аспартат- Глутамат 3,3-107 2,8-10®
трансаминаза, Аспартат >107 >5-10®
альдегидная форма NH2OH 3,7-10® 38
Кетоглутарат, >5-108 >5-104
Глутамат-аспартат- оксалоацетат Оксалоацетат 7-Ю7 1,4-10®
трансаминаза, Кетоглутарат 2,1-Ю7 70
аминная форма
Альбумин из сыво- I-Нафтол-4- (4'-азо- 2-10® 35
ротки быка бензолазофенил)- арсоновая кисло- та (I) [Нафтол-З-сульфо-4- 3,5-105 2,5
у-Глобулин кролика (4/-азобензолазо)]- фениларсоновая кислота (II) 1-Нафтол-4- (4'-азо- 2,2-Ю7 50
(антитела к I и II) бензолазофенил)ар -
у-Глобулин кролика соновая кислота 2,4-Динитрофенил- 8,1-10’ 1,1
(антитела к III) Гемоглобин НЬ(О2)з лизин (III) о2 2-Ю7 36
Глобин Карбоксигем 7-107
а Воспроизведено из работы: Mahler Н. R., Cordes Е. Н„ Biological Chemistry,
2d ed., р. 322, Harper and Row, Publishers, Inc., New York, 1971.
152
Кинетика катализируемых ферментами реакций
153
му максимальному значению, составляющему для абсолютной скорости бимо-
лекулярных соударений в растворе от 109 до 1011 [(моль-с)/л]-1. Напротив,
обратный процесс протекает сравнительно медленно, и его скорость, как и
скорость диссоциации промежуточного фермент-субстратного комплекса на
продукт реакции и свободный фермент (см. значения k2 в табл. 3.4), в зна-
чительно большей степени зависит от природы реагирующих веществ.
Описанными выше методами была выявлена другая интересная особен-
ность ферментативного катализа, заключающаяся в образовании нескольких
промежуточных фермент-субстратных комплексов ES. В ряде случаев ком-
плекс претерпевает ряд последовательных конформационных превращений,
которые можно рассматривать как реакции изомеризации. Методами изуче-
ния стационарных состояний невозможно установить, образуется ли одно
или несколько промежуточных соединений, если скорость диссоциации по-
следнего промежуточного комплекса (до конечных продуктов реакции) мала
по сравнению со скоростью превращения комплексов, образующихся на пре-
дыдущих стадиях. В таких случаях ценная информация может быть полу-
чена релаксационными методами.
3.4. Другие типы зависимостей скорости
ферментативной реакции от концентрации
субстрата
Скорости не всех катализируемых ферментами реакций под-
чиняются описанному в разд. 3.2 уравнению Михаэлиса — Мен-
тен. В настоящем разделе мы рассмотрим некоторые наиболее
типичные отклонения. Отклонение первого типа, характерное
для регуляторных ферментов, можно объяснить связыванием
с ферментом нескольких субстратов. Далее мы опишем второй
тип отклонений от уравнения Михаэлиса — Ментен, когда суб-
страт представляет собой смесь различных реагирующих с фер-
ментом веществ, обладающих разными кинетическими свойст-
вами. Анализ отклонений второго типа поможет выявить ряд
проблем, с которыми приходится сталкиваться при работе с не-
достаточно очищенными или плохо охарактеризованными сме-
сями субстратов.
3.4.1. Активация и ингибирование ферментов субстратами
Характерная для некоторых ферментов S-образная форма
кривой зависимости скорости реакции от концентрации субст-
рата свидетельствует об эффекте активации (рис. 3.14). При
низких концентрациях субстрата связывание ферментом одной
молекулы субстрата повышает скорость связывания следующей
молекулы (с математической точки зрения это означает, что
на этом этапе реакции dvlds возрастает). Моделью такого про-
цесса может служить согласованное связывание субстрата
субъединицами белка. Предположим, что фермент, например,
олигомерной структуры имеет несколько связывающих субст-
рат центров и что связывание первой молекулы субстрата с
154
Глава 3
РИС. 3.14. Активация (а) и ингибирование (б) ферментативной реакции суб-
стратом.
ферментом изменяет структуру последнего таким образом, что
сродство оставшихся свободными центров по отношению к суб-
страту увеличивается. Математический анализ такой модели
аналогичен рассмотренному в упражнении 2.10 анализу модели
гемоглобина, и поэтому мы не будем повторять его в основ-
ном тексте.
Иногда в присутствии большого количества субстрата ско-
рость катализируемой ферментом реакции снижается при по-
-3 -2 -1
bgs
а
РИС. 3.15. Экспериментальное обнаружение субстратного ингибирования.
[Воспроизведено из работы: Laidler К. L, The Chemical Kinetics of Enzyme
Action, p. 71, The Clarendon Press, Oxford, 1958.
a — данные из статьи: Augustinsson К. D., Substrate Concentration and Speci-
ficity of Choline Ester Splitting Enzymes, Arch. Biochem., 23, 111 (1949);
6 — данные из статьи: Lumry R., Smith E. L., Glantz R. R., Kinetics of Carbo-
xypeptidase Action. I. Effect of Various Extrinsic Factors on Kinetic Parame-
ters, J. Amer. Chem. Soc., 73, 4330 (1951).]
Кинетика катализируемых ферментами реакций 155
вышении концентрации субстрата (рис. 3.15). Это явление на-
зывается субстратным ингибированием. Как показано на
рис. 3.15, в таких случаях скорость реакции v по мере повыше-
ния концентрации субстрата сначала возрастает, затем дости-
гает максимума и, наконец, снижается. Если s больше, чем s,
соответствующая итаХ) то снижение концентрации субстрата
вызывает повышение скорости реакции. Автокаталитический
характер реакций такого типа может оказывать существенное
влияние на работу биохимических реакторов.
Взаимосвязь между концентрацией субстрата и скоростью
ингибируемой субстратом реакции можно оценить количествен-
но с достаточно высокой точностью, используя подход Миха-
элиса и Ментен- и допустив, что фермент может связывать и
вторую молекулу субстрата, причем в результате присоедине-
ния S к ES образуется нереакционноспособное промежуточное
соединение. В этом случае в равновесном состоянии будут осу-
ществляться следующие последовательные реакции:
Константа
диссоциации
Е + S ES
k-i
ES + S ч—* ES2
k
ES ----> Е + Р
Ki например,
(медленная стадия)
(3.37)
(3.38)
На основе двух указанных равновесных реакций диссоциации
с учетом уравнения материального баланса различных состоя-
ний фермента, аналогичного уравнению (3.7), после неслож-
ных алгебраических преобразований можно получить следую-
щее выражение:
(3.39)
Экспериментальное определение параметров этого уравне-
ния не вызывает затруднений. Так, сначала путем изменения
е0 можно определить k, а затем, построив график зависимости
1/и от s, можно оценить тангенс угла наклона прямого участка
кривой, при высоких концентрациях субстрата равный 1//C2^f?o
(рис. 3.16). Наконец, Ki можно найти по уравнению (3.40);
здесь smax удовлетворяет уравнению (3.39) при dv/ds = Q.
$тах К К1К2
(3.40)
156
Глава 3
3.4.2. Взаимодействие одного фермента
с несколькими различными субстратами
Многие ферменты могут взаимодействовать с несколькими
субстратами. В таких случаях различные субстраты конкури-
руют за связывание с активным центром (здесь мы принимаем,
что роль активного центра для различных субстратов выполня-
ет один и тот же участок молекулы фермента). Наиболее из-
бонцентрация субстрата Ю~3
РИС. 3.16. Способ определения константы К.2 по наклону прямой зависимости
1/у от s. Приведены данные для гидролиза этилбутирата карбоксипептидазой
из печени овцы. (Из работы: Диксон М., Уэбб Э. Ферменты, т. 1—3. — М.:
Мир, 1982, т. 1, с. 99.)
вестными примерами ферментов такого типа являются гидро-
лизующие деполимеразы, которые расщепляют многие иден-
тичные связи, часто независимо от длины гидролизуемого по-
лимерного сегмента. В то же время кинетические параметры
реакций такого фермента с димером, тримером или олигоме-
ром могут заметно отличаться от параметров аналогичной ре-
акции с достаточно высокомолекулярными полимерными цепя-
ми. Примером таких ферментов может служить лизоцим, содер-
жащийся в яичном белке, слизистых выделениях и слезах. Ли-
зоцим гидролизует сложные мукополисахариды, в том числе и
муреины клеточных стенок, вызывая гибель грамположитель-
ных бактерий. Приведенные выше общие положения подтверж-
даются экспериментальными данными (табл. 3.6), где указа-
ны найденные скорости и константы диссоциации для каждого
Кинетика катализируемых ферментами реакций[57
субстрата лизоцима в отсутствие других субстратов. Нетрудно
видеть, что наибольшие отличия характерны для мономерного
и олигомерных субстратов.
В приведенных выше и в последующих примерах каждую
подвергающуюся гидролизу связь олигомера или полимера
можно рассматривать как индивидуальный субстрат. Более то-
го, полимерный субстрат типа крахмала на самом деле также
Таблица 3.6. Кинетические параметры реакции лизоцима с компонентами
клеточной стенки бактерий (сополимеры GlcNAc—MurNAc;
GlcNAc— N-ацетилглюкозамин, MurNAc—N-ацетилмурамовая кислота)3
Реакция
Тип реакции
Kij
1. E+Sz < fe-Gry; /—0 t+1 Ассоциация
S;= (GlcNAc—MurNAc), — олигомер
из i звеньев
kc
2. Сц —>• A/+Si-/; /=1, ..., i—1 Гидролиз комплекса
hv
3. Си —>- А/+Н2О Диссоциация промежу-
точного комплекса
4. Ai(+H2O) —>- E+S; Гидролиз промежуточ-
ного комплекса
5. Ai+S/ —>- E + S/+/ Трансгликозилирование
с образованием олиго-
мера из i+j звеньев
Кп=0,5 М-1; К21=6,3-10-4 М-1; К22=2000 М~'; К32=3,5-104 М"1; йс =
= 1,75 с-1; 6, = 100 с-1; йг=2,77-105 (М-с)-1.
а По данным работы: Chipman D. М., A Kinetic Analysis of the Reaction of Lyso-
zyme with Oligosaccharides from Bacterial Cell Walls; Biochemistry, 10, 1714 (1971).
представляет собой не индивидуальный субстрат в строгом
смысле слова, а смесь полимеров различной молекулярной мас-
сы, которые к тому же могут содержать мономер-мономерные
связи разных типов (разд. 2.2.2). На рис. 3.17 отражены раз-
личия в кинетике гидролиза разных крахмалов. Пример кине-
тики ферментативного гидролиза более четко охарактеризо-
ванной системы приведен на рис. 3.18. Такого типа реакции
используются в промышленности для разжижения растворов
крахмала амилазами. Аналогичные эффекты следует ожидать
и в других подобных реакциях, например, в случае гидролиза
белков протеолитическими ферментами. Прежде чем перейти
к количественной оценке конкуренции между различными суб-
стратами, следует подчеркнуть, что в приведенных выше при-
РИС. 3.17. Кривые зависимости скорости гидролиза различных фракций ами-
лозы под действием амилоглюкозидазы от времени (25 °C; pH 4,6; So=
=0,1 г/100 мл, ео=6,8-1О~8 М); молекулярные массы фракций амилозы около
1 650 000 (a), 1 100 000 (б) и 360 000 (в). [Воспроизведено с разрешения из
статьи: Hiromi К. et al., A Kinetic Method for the Determination of Number-
Average Molecular Weight of Linear High-Polymer by Using an Exo-Enzyme,
J. Biochem. (Tokyo), 60, 439 (1966).]
РИС. 3.18. Кривые зависимости скорости ферментативного гидролиза олиго-
меров и полимеров глюкозы под действием амилоглюкозидазы от времени
(15 °C; pH 5,15; s0 = 0,04 М; ео=2,82-10-7 М). [Воспроизведено из работы:
Ono S., et al., Kinetic Studies of Gluc-amylase; J. Biochem. (Tokyo), 55, 315
(1964),]
Кинетика катализируемых ферментами реакций
159
мерах определенную роль могут играть и другие факторы, в
частности различия в физических размерах и состояниях инди-
видуальных субстратов, а также в числе поддающихся гидро-
лизу связей в каждом из находящихся в реакционной смеси
субстратов.
Для некоторых гидролитических ферментов характерна се-
лективность в отношении различных участков цепи биополиме-
ра. Так, экзоферменты расщепляют концевые (или близкие к
концевым) связи цепи, а эндогидролазы катализируют гидро-
лиз внутренних связей цепей. При описании кинетики таких
систем важно учитывать оба типа ферментативных реакций, а
также «концентрации» концевых и внутренних связей в смеси
биополимеров. С таким подходом мы встретимся в следующей
главе при изучении ферментативного гидролиза целлюлозы.
Ниже мы рассмотрим один простой, но показательный при-
мер, иллюстрирующий некоторые типичные затруднения, кото-
рые встречаются при описании кинетики ферментативных ре-
акций со смесями субстратов. Пусть последовательности реак-
ций (3.41) и (3.42) отражают взаимодействие двух различных
субстратов с одним и тем же ферментом:
E + St ч=ь ESi; константа диссоциации
(3.41)
E + S2 ES2; константа диссоциации Л2
Медленные стадии:
ESx Е + Рг; ES2 ЕР2 (3.42)
Здесь подразумевается, что каждый субстрат связывает какую-
то часть фермента. Если мы далее, как и в наших предыдущих
анализах кинетики ферментативных реакций, используем урав-
нение общего материального баланса фермента и условия рав-
новесия, то получим следующие выражения для скоростей ре-
акций:
— =-----MA//Q------- (3 43)
d/ 1 + -(- s2//C2
— = =--------fe2gQS2/Kg-- ,344)
1 + Si/M 4- s2/^2
Из уравнений (3.43) и (3.44) следует, что если две реакции
катализируются одним и тем же ферментом, то скорости инди-
видуальных реакций в присутствии двух субстратов ниже, чем
в присутствии только одного субстрата. Этот факт успешно ис-
пользовался для идентификации ферментных препаратов; дей-
ствительно, таким путем можно выяснить, действует ли на оба
субстрата один и тот же фермент или же реакция каждого
субстрата катализируется своим ферментом.
160
Глава 3
В экспериментах и в технологических процессах обычно
трудно или даже невозможно определять и контролировать
концентрации всех компонентов реакционной смеси. Например,
изображенные на рис. 3.18 кривые отражают ход многих одно-
временно протекающих реакций, в том числе гидролиза маль-
тозы, мальтотриозы и т. д. Другими словами, в действительно-
сти изучают не индивидуальную реакцию, а некоторую гипоте-
тическую общую реакцию превращения полимеров глюкозы в
мономерную глюкозу под действием фермента амилоглюкозида-
зы:
амилоглюкозидаза
Полимеры глюкозы--------------► глюкоза
В этой реакции все субстраты, построенные из остатков глюко-
зы, объединены в некий воображаемый единый субстрат, кон-
центрация которого может быть определена сравнительно лег-
ко. Такого типа усреднение субстратов широко применяется во
всех областях химической кинетики. Рассмотрим, например,
каталитический крекинг газойля. В проектных разработках или
при анализе работы реактора этот процесс для удобства часто
изображают в виде следующей двустадийной реакции:
Газойль ---> бензин ---> газы
хотя в данном случае каждый из реагентов* очевидно, пред-
ставляет собой смесь соединений. В научно-технической литера-
туре по биологии и биохимической технологии при изучении и
моделировании биологических реакций, а также при проектиро-
вании соответствующих технологических процессов подобные
усреднения, хотя и далеко не всегда столь очевидные, встреча-
ются сплошь и рядом. В нашей книге мы постараемся тщатель-
но анализировать обоснованность таких и подобных упроще-
ний, применяющихся (обычно косвенным образом) при описа-
нии биологических систем. Неоправданные упрощения могут
привести к серьезным ошибкам как в теоретических исследо-
ваниях, так и в проектных разработках.
Приведем пример одной из таких ошибок. Предположим,
что 5т представляет собой общую концентрацию всех субст-
ратов в смеси:
~ st §2 (3.45)
Тогда суммарная скорость превращения Vr будет определяться
выражением
у __ ds-p _______f dsl . dsz \ _ g0 (kis1IK1 4- zg
dt \ dt dt J 1 -f- Si/Ki 4- Sn /Kt
Очевидно, что скорость vT зависит от отношения величин Si и
Кинетика катализируемых ферментами реакций
161
$2. Например, если суммарная концентрация субстрата имеет
определенную величину syo, то суммарная скорость Vt в об-
щем случае не имеет определенного значения, а в зависимости
от отношения si/s2 может изменяться в следующих пределах:
ит
sl=sTo
S2=0
gpfelsro
Ki + sTo
уменьшение si
------------>-
увеличение s2
VT
Si=0
S2=STo
^0^2$ 7* 0
+ ST0
(3-47)
Отсюда следует, что определенные экспериментальным пу-
тем кинетические параметры итах и Кт будут зависеть как от
состава смеси (отношения Si/s2), так и от общей концентрации
субстрата Sr. Точно так же параметры работы ферментативно-
го реактора будут изменяться при изменении относительных
количеств субстратов Si и S2, даже если общая концентрация
вводимых в реактор реагентов будет поддерживаться па по-
стоянном уровне.
3.5. Регуляция ферментативной активности
Изменять, или модулировать, каталитическую активность
ферментов могут не только субстраты, но и другие взаимодей-
ствующие с ферментами вещества; их называют модулятора-
ми, или эффекторами. Эффекторы могут быть обычными ком-
понентами клетки, но могут и проникать в клетку из среды
или действовать на изолированные ферменты. Основное внима-
ние в этом разделе будет уделено ингибиторам — веществам,
понижающим ферментативную активность. Следует, однако,
иметь в виду, что известны и случаи активации ферментов суб-
стратами.
Взаимодействие фермента с эффектором представляет собой
химическую реакцию и поэтому может быть полностью обрати-
мым, частично обратимым или практически необратимым.
К числу известных необратимых ингибиторов принадлежат
яды, например цианидный ион, инактивирующий ксантинокси-
дазу, а также группа соединений, называемых нервнопарали-
тическими ядами-, последние необратимо инактивируют холин-
эстеразы (ферменты, участвующие в системах передачи нерв-
ных импульсов и, таким образом, в моторной активности).
Если процесс ингибирования необратим, то кинетика ингиби-
торзависимых ферментативных реакций не подчиняется урав-
нению Михаэлиса — Ментен, основой которого является нали-
чие равновесия между свободной и связанной формами фер-
мента. Часто по мере полной инактивации все большего и
большего числа молекул фермента процесс необратимого ин-
гибирования прогрессирует во времени. В других случаях,
труднее поддающихся выявлению, фермент инактивируется
11—537
162
Глава 3
лишь частично и сохраняет каталитическую активность, хотя
и в меньшей степени по сравнению с чистым ферментом.
В разд. 2.6 мы уже говорили о замечательной способности
большинства микроорганизмов продуцировать множество раз-
нообразных сложных соединений из относительно небольшого
числа простых предшественников. Для достижения этой цели
необходима некая система очень эффективного распределения
поступающих предшественников по многим биосинтетическим
путям, ведущим к различным конечным продуктам метаболиз-
Z 'Треонин L-Изолейцин
СН3 сн3
I £, Е3 Еа Ёь ।
неон ---------- > Рт ——> р2----> р3----> Р4--СН,
I Г
hcnh2 | НССНз
соон I hcnh2
I I
I соон.
РИС. 3.19. Последовательность реакций биосинтеза L-изолейцина, ингибируе-
мая по принципу обратной связи; здесь активность фермента Ei (L-треонин-
дезаминазы) снижается в присутствии конечного продукта последовательно-
сти— L-изолейцина. (Р1=а-кетобутират; Р2=а-ацетогидроксибутират; Рз=
=аф-дигидроксиф-метилвалерат; Р4=а-кетоф-метилвалерат.)
ма. В большинстве случаев принцип оптимального усвоения до-
ступного исходного продукта требует синтеза любого из конеч-
ных продуктов только в строго необходимом количестве. Если,,
например, в клетке имеется достаточное количество данного
мономера, то его дальнейшее накопление должно быть прерва-
но, с тем чтобы ресурсы клетки могли быть направлены на син-
тез других соединений, количество которых в этот момент от-
носительно мало.
Одним из регуляторных механизмов, используемых клеткой
для достижения максимально эффективного усвоения питатель-
ных веществ, является обратимая регуляция ферментативной
активности. (Другой важный механизм управления будет рас-
смотрен в гл. 6, разд. 1.) Наиболее интересные примеры регу-
ляции ферментативной активности включают в себя сложную
сеть реакций с несколькими регуляторными петлями; эти при-
меры мы рассмотрим позднее, после того как изучим основы
химии клетки (гл. 5). Сейчас же нам будет достаточно приме
ра регуляции простой последовательности реакций. На рис. 3.19
изображена пятистадийная последовательность биосинтеза
Кинет ина катализируемых ферментами реакций
163
аминокислоты ь-нзолейцнна. Регуляция этой последователь-
ности реакций осуществляется путем ингибирования по прин-
ципу обратной связи, когда конечный продукт, ь-изолейцин, ин-
гибирует активность фермента, катализирующего первую ста-
дию последовательности. Таким образом, биосинтез конечного
продукта последовательности реакций будет тормозиться по ме-
ре его накопления.
Таблица 3.7. Некоторые типы обратимого ингибирования1
Тип ингибирования Механизм ингибирования
Ингибирующий
эффект
1а. Полное кон- курентное Ингибитор сорбируется активным Увеличение ка- центром, связывающим субстрат жущегося зна- чения Кт
б. Частичное конкурентное Ингибитор и субстрат связываются Увеличение кажу- различными центрами; связывание щегося значения ингибитора влияет на связывание Кт субстрата
Па. Неконкурент- ное Связывание ингибитора не влияет на Снижение vmax сродство субстрата к ферменту, но без изменения фермент-субстрат-ингибиторный Кт комплекс устойчив и далее не дис-
б. Неконкурент- ное социирует Тот же, что в ингибировании типа Снижение vmax Па, но фермент-субстрат-ингиби- без изменения торный комплекс распадается с об- Кт разованием нормального продукта реакции со скоростью, меньшей скорости диссоциации ES
III. Смешанное Изменение как Кт, так и vmax
а Из работы: Диксон М., Уэбб Э., Ферменты. — М.: Мир, 196S.
Поскольку катализируемые ферментами Е2—Е5 реакции на-
ходятся в состоянии равновесия, а первая реакция в клеточной
среде «необратима», то быстродействие такой регуляторной
системы очень высоко. По сути дела, большинство регулятор-
ных ферментов катализирует именно такие «необратимые» ре-
акции. С точки зрения биологии клетки очевидно, что такая
«природная» регуляция ферментативной активности должна
быть обратимой. Если, например, весь ь-изолейцин был израс-
ходован в процессе белкового синтеза, то ингибирование фер-
мента Е[ должно прекратиться и система вновь должна син-
тезировать необходимое количество этой аминокислоты.
В случае обратимого ингибирования изложенные в разд. 3.2
подходы к анализу кинетики оказались очень плодотворными.
Поскольку многие изолированные фермент-субстратные систе-
11*
164
Глава 3
мы подчиняются кинетике Михаэлиса — Ментен, принято клас-
сифицировать ингибиторы по их влиянию на параметры урав-
нения Михаэлиса — Ментен ymax и Кт.
Обратимые ингибиторы называют конкурентными, если в их
присутствии повышается значение Кт, a ymax не изменяется.
Вызываемый ингибиторами такого типа эффект можно частич-
но или полностью подавить путем повышения концентрации
субстрата. Неконкурентные ингибиторы, напротив, инактиви-
руют фермент или фермент-субстратный комплекс путем умень-
шения Утах фермента, но не влияют на величину Кт. В этом
случае повышение концентрации субстрата до сколь угодно вы-
сокого уровня уже не позволит повысить скорость реакции до
величины, характерной для неингибированного фермента. Ти-
пичными примерами неконкурентных ингибиторов являются
ионы тяжелых металлов, обратимо реагирующие с сульфгид-
рильными (—SH) группами цистеиновых остатков.
Известны несколько сочетаний и вариантов этих двух ос-
новных типов обратимых ингибиторов; некоторые из них при-
ведены в табл. 3.7. Чуть позже мы рассмотрим эксперименталь-
ные методы определения типа ингибирования, но прежде
вкратце обсудим некоторые теории, объясняющие механизмы
действия регуляторов ферментативной активности,, и соответст-
вующие экспериментальные данные.
3.5.1. Механизмы обратимой регуляции
ферментативной активности
Многие из известных конкурентных ингибиторов по своей
химической природе близки обычным субстратам; такие инги-
биторы называют субстратными аналогами. Полагают, что по-
добные ингибиторы имеют пространственную структуру типа
ключа, который может входить в «замочную скважину» актив-
ного центра фермента, однако «ключ» не вполне подходит к
данному «замку» и реакция не идет. Рассмотрим, например,
ингибирование процесса дегидрирования янтарной кислоты ма-
лоновой кислотой:
СООН
СН2
СН2
СООН
процесс катализируется
сукцинилдегидр огеназой
я конкурентно ингибируется
малоновой кислотой
янтарная
кислота
соон
I
сн
II
сн
соон
фумаровая
кислота
соон\
сн2 I
соон /
малоновая
кислота
В данном случае малоновая кислота может образовывать
комплекс с сукцинилдегидрогеназой, но на этом процесс оста-
навливается.
Кинетика катализируемых ферментами реакций
165
Конкурентное ингибирование лежит в основе механизма
действия одного из лекарственных сульфамидных препаратов —
сульфаниламида (стрептоцида). Структура последнего очень
близка структуре n-аминобензойной кислоты — важного вита-
мина многих бактерий. Сульфаниламид ингибирует фермент,
участвующий в превращении n-аминобензойной кислоты в фо-
лиевую кислоту, тем самым блокируя биохимический аппарат
бактерии, что приводит к ее гибели.
л-амииобеизойная сульфаниламид
кислота
При другом механизме, называемом аллостерической регу-
ляцией, поведение фермент-субстратной системы типично для
неконкурентного ингибирования. Считается, что механизм алло-
стерической регуляции вообще играет доминирующую роль в
неконкурентном ингибировании и активации. Название аллосте-
рический (имеющий другую форму) было дано этому механиз-
му сначала на том основании, что по своему строению многие
эффекторы ферментативной активности существенно отличают-
ся от субстратов. Отсюда был сделан вывод, что регуляторное
действие таких эффекторов основано на их связывании со спе-
цифическим регуляторным центром фермента, отличным от ак-
тивного центра, в котором осуществляется катализ превраще-
ний субстрата. Соответственно ферменты, обладающие цент-
рами регуляции и катализа, были названы аллостерическими
ферментами.
Аллостерическая регуляция может как ингибировать (сни-
жать), так и активировать (повышать) каталитическую способ-
ность фермента. Процесс аллостерической регуляции схемати-
чески изображен на рис. 3.20. Обратите внимание на то, что
фермент здесь изображен в виде двух субъединиц; известно,
что многие аллостерические ферменты и на самом деле явля-
ются олигомерными белками.
Наиболее убедительным доказательством справедливости
аллостерической теории ферментативной активности явилось
экспериментальное изучение аспартилтранскарбамоилазы
(АТС-азы). После разделения фермента на две субъединицы
выяснилось, что на обладающую каталитической активностью
166
Глава 3
большую субъединицу ингибитор интактного фермента СТР
(цитидинтрифосфат) не влияет. Напротив, меньшая субъеди-
ница каталитически не активна, но связывает СТР.
В отсутствие экспериментальных доказательств типа при-
веденных выше для АТС-азы нельзя быть уверенным в том,
что эффектор не связывается активным центром фермента.
С кинетической точки зрения, однако, представляет интерес
Каталитически
активное состоя-
ние (К форма)
Связь/вание субстрата
S и эффектора А стаби-
лизирует каталити-
чески активное состояние
Аллостеричес-
ка ингибирован-
ное состояние
Связывание алло-
стерического инги-
битора I стабили-
зирует ингибиро-
ванное состояние
„Принцип симметрии’ исключает воз-
можность смешанного T-R-состоя-
ния
РИС. 3.20. В симметричной модели аллостерической регуляции ферментатив-
ной активности связывание активатора А и субстрата S приводит к каталити-
чески активной R-форме фермента, а связывание ингибитора изменяет кон-
формацию всех субъединиц в олигомерном белке таким образом, что молеку-
ла белка принимает неактивную Т-форму. (Из работы: Лёви А., Сикевиц Ф.,
Структура и функция клетки. — М.: Мир, 1971, с. 306.]
только влияние эффектора на ряд параметров, в том числе на
сродство субстрата к ферменту, концентрацию несвязанного
фермента и (или) скорость распада комплекса. В следующем
разделе мы попытаемся найти математические выражения, ха-
рактеризующие эти эффекты количественно.
3.5.2. Анализ влияния обратимой регуляции
на кинетику ферментативных реакций
Основная ценность подхода Михаэлиса и Ментен к анализу
кинетики ферментативных реакций заключается в том, что он
позволяет количественно описать влияние эффекторов. Преж-
де всего допустим, что приведенная ниже последовательность
реакций достаточно точно отражает механизм взаимодействия
Кинетика катализируемых ферментами реакций 167
ингибитора и субстрата с ферментом при (полном) конкурент-
ном ингибировании. Здесь, как обычно, Е и S обозначают фер-
мент и субстрат соответственно, а символами I и EI обозна-
чены ингибитор и фермепт-ингибиторный комплекс соответст-
венно.
Равновесные стадии реакции:
Константа
диссоциации
E + S ES Ks
(3.48)
E+I EI Ki
Медленная стадия:
k
ES ----> E+P (3.49)
В этом случае фермент может связывать или только субстрат,
или только ингибитор. Поскольку часть фермента связана в
комплексе EI, не весь фермент участвует в каталитическом
превращении субстрата; следовательно, скорость реакции в
присутствии ингибитора замедляется.
На базе равновесных реакций (3.48) и уравнения общего
материального баланса фермента
е+ (es) 4- (ei) = (3.50)
скорость реакции можно выразить через общую концентрацию
фермента во и концентрации свободного субстрата и ингибито-
ра (s и i соответственно):
р == s .уд.) (полное) конкурентное ингибирование (3.51)
Сравнение с исходной формой уравнения Михаэлиса — Ментен
показывает, что параметр утах остался без изменений, а кажу-
щаяся константа Михаэлиса Ктарр в присутствии конкурентно-
го ингибитора возрастает:
<3-52)
\ Ki /
Другими словами, обусловленное конкурентным ингибитором
снижение скорости реакции может быть полностью компенси-
ровано путем соответствующего повышения концентрации суб-
страта; на максимально возможную скорость реакции конку-
рентный ингибитор не влияет.
Если последовательные реакции (3.48) и (3.49) дополнить
равновесными реакциями (3.53а) и (3.536), то можно получить
168
Глава 3
полезную модель (полного) неконкурентного ингибирования:
Константа
диссоциации
EI+S EIS к, (3.53а)
ES + I =₽ь EIS Ki (3.536)
Здесь ингибитор и субстрат могут одновременно связываться с
ферментом, образуя тройной комплекс EIS (фермент-ингиби-
тор-субстратный). В этой простейшей модели неконкурентного,
ингибирования мы принимаем, что связывание ингибитора не
влияет на сродство фермента к субстрату, а связывание субст-
рата не сказывается на сродстве фермента к ингибитору. От-
сюда следует, что Ks и Ki в уравнениях (3.53) идентичны со-
ответствующим константам диссоциации в уравнении (3.48).
Примем также, что комплекс EIS не диссоциирует с образова-
нием продукта реакции Р.
Используя уже известный нам прием выражения концентра-
ций комплексов через е0, s и I, можно получить следующее вы-
ражение для скорости реакции:
t> = s£e0/(^-H//G) (полное) неконкурентное ингибирование (3.54)
Нетрудно заметить, что в этом выражении константа Михаэли-
са осталась без изменений, а максимальная скорость реакции
снизилась до
уЯрр _ ^go_______
max 1 + i/Ki
(3.55)
Отсюда следует, что в присутствии неконкурентного ингибито-
ра добавление к реакционной смеси сколь угодно большого
количества субстрата не позволит повысить максимальную ско-
рость реакции до той величины, которая может быть достигну-
та в отсутствие ингибитора.
Конкурентное и неконкурентное ингибирование легко раз-
личить по графикам зависимостей кинетических параметров в
координатах Лайнуивера — Бэрка. В случае конкурентного ин-
гибирования добавление ингибитора приводит к увеличению от-
резка, отсекаемого прямой на оси 1/s, и не влияет на отрезок,
отсекаемый на оси 1/щ Напротив, при неконкурентном ингиби-
ровании возрастает только отрезок, отсекаемый на оси 1/v.
Рассмотрим в качестве примера приведенные на рис. 3.21 дан-
ные по гидролизу крахмала. Сравнивая графики зависимостей
1/у от 1/s в присутствии различных ингибиторов, а также без
ингибитора, нетрудно видеть, что а-декстрин является конку-
рентным ингибитором, а мальтоза и предельный декстрин
представляют собой неконкурентные ингибиторы.
Кинетика катализируемых ферментами реакций
169
РИС. 3.21. Конкурентное (а-декстрин) и неконкурентное (мальтоза, предель-
ный декстрин) ингибирование а-амилазы. (Воспроизведено с разрешения из
работы: Aiba S., Humphrey А. Е., Millis N., Biochemical Engineering, 2d ed.,
p. 101, University of Tokyo Press, Tokyo, 1974; есть перевод 1-го издания:
Аиба Ш., Хемфри А., Миллис Н„ Биохимическая технология и аппаратура. —
М.: Пищевая промышленность, 1975, с. 81.)
Как мы указывали в табл. 3.7, существуют и другие типы
ингибирования ферментативных реакций. Ниже мы в несколь-
ких словах обсудим, как можно их описать путем видоизмене-
ния рассмотренных выше моделей ингибирования. При частич-
ном конкурентном ингибировании (тип 16 в табл. 3.7) ингиби-
тор и субстрат соединяются с различными участками молекулы
фермента и ингибитор влияет на сродство фермента к субстра-
ту. Чтобы описать этот случай, добавим к последовательности
170
Глава 3
реакций (3.48) и (3.49) следующие стадии,. которые мы будем
считать равновесными:
Константа
диссоциации
EI + S EIS К*
(3.56)
ES + I EIS Ks{
и медленную стадию:
k
EIS ----> EI + Р (3.57)
(Обратите внимание: можно показать, что Kis = KsKsi/Ki.)
В другом случае неконкурентного ингибирования (типа Пб)
описанная выше модель видоизменяется с учетом возможности
образования продукта реакции из комплекса EIS; диссоциация
последнего по сравнению с комплексом ES протекает с мень-
шей скоростью:
EIS EI+ р (3.58)
Одну из моделей смешанного ингибирования, включающего в
себя полный конкурентный и частично конкурентный типы,
можно получить, опустив реакцию (3.57) в приведенной выше
модели частичного конкурентного ингибирования. Результаты
анализа всех этих типов ингибирования можно записать в фор-
ме уравнения Михаэлиса — Ментен:
vapp S
V __----—
s + C₽₽
(3.59)
В этом уравнении кажущиеся параметры t'maxa₽₽ и Ктарр зави-
сят от концентрации ингибитора и различных кинетических па-
раметров (табл. 3.8). На рис. 3.22 схематически показано вли-
яние разных типов ингибирования на различные формы графи-
ческого выражения данных по изучению кинетики ферментатив-
ных реакций.
Один из типов активации фермента эффектором может быть
описан моделью типа Пб при условии, что kl больше k. С точ-
ки зрения механизма процесса это означает, что скорость обра-
зования продукта реакции из комплекса EIS выше, чем из
комплекса ES. Таким образом, здесь вещество I выполняет
функцию активатора фермента.
Кинетика катализируемых ферментами реакций
171
3.6. Другие факторы, влияющие на ферментативную
активность
Прежде чем приступить к изложению темы этого раздела,
полезно напомнить, что основная цель настоящей главы заклю-
чается в описании скорости катализируемых ферментами реак-
ций с помощью математических выражений. Без последних мы
не сможем разработать проект реактора или спланировать
эксперимент с участием изолированных ферментов. Более того,
когда мы приступим к изучению кинетики клеточного роста, мы
узнаем, что и здесь приложимы различные аспекты кинетики
ферментативного катализа. В этой связи становится очевидной
Таблица 3.8. Параметры кинетики ферментативных реакций в присутствии
ингибиторов различных типов; отрезки, отсекаемые прямыми на осях
1/ц и 1/s
Тип ингибирования Отрезок на оси ординат Отрезок на оси абсцисс
1а. Полное конкурентное I 1
vmax Ks(l + */Ki)
16. Частичное конкурент- 1 1 + iKslKiKt»
ное vmax Ks(l + UKi)
Па. Полное неконку- 1 + i/Kj 1
рентное vmax
Пб. Частичное некой- 1 + '4Ki 1
курентное vmax + Ks
16 и Па. Смешанное 1 + iKslKiKis 1 + iKsIKiKis
vmax Ks(l + i/Kt)
важность тщательного изучения различных факторов, влияю-
щих на ферментативный катализ, в том числе количественной
оценки степени их влияния.
Мы уже знаем, что различные химические соединения, свя-
зываясь с ферментами, могут изменять скорость катализируе-
мых ферментами реакций, и в общих чертах представляем, как
это происходит. На каталитическую активность ферментов вли-
яют и многие другие факторы, которые могут изменять строе-
ние или химическую природу фермента. К числу таких факто-
ров относятся:
1. pH
2. Температура
3. Силы, действующие в текучих средах (гидродинамические
силы, гидростатическое давление и поверхностное натяжение)
4. Химические агенты (например, спирт, мочевина или перок-
сид водорода)
5. Облучение (свет, звук, ионизирующая радиация)
Неконкурентное
Конкурентное
Смешанное
РИС. 3.22. Влияние типа ингибирования на различные формы графического
выражения кинетики ферментативных реакций. Индексом i обозначены пара-
метры реакции в присутствии ингибитора, (Из работы: Диксон М., Уэбб Э.,
Ферменты, т. 2, с. 498, — М.: Мир, 1982.]
Кинетика катализируемых ферментами реакций 173
Иногда снижение каталитической активности, вызванное, на-
пример изменением pH, обратимо; в таких случаях возврат к
первоначальным условиям сопровождается восстановлением
активности фермента. В известном смысле такая ситуация ана-
логична рассмотренному случаю обратимого ингибирования;
небольшие изменения одного из перечисленных выше факторов,
по сути дела, только слегка сдвигают равновесие (или квази-
стационарное состояние), характерное для данной фермента-
тивной реакции. В общем случае отклонение от условий, ти-
пичных для биологического окружения нативного фермента,
должно быть относительно небольшим (или кратковременным);
в противном случае возрастает вероятность инактивации фер-
мента. Многие из перечисленных выше факторов мы обсудим в
следующем разделе, посвященном инактивации ферментов,
здесь же основное внимание будет уделено «обратимому» вли-
янию pH и температуры на каталитическую активность фер-
ментов.
Следует подчеркнуть, что граница между «обратимой» и
«необратимой» инактивацией белков не всегда четко определе-
на. Например, подвергнутый кратковременному нагреванию
фермент при охлаждении до свойственной ему «рабочей» тем-
пературы может полностью восстановить свою активность.
С другой стороны, более продолжительное нагревание при той
же температуре или столь же кратковременная термическая об-
работка при более высокой температуре могут привести к то-
му, что при последующем охлаждении активность фермента
восстановится лишь частично. Такое поведение белков вообще
и ферментов в частности становится понятным, если учесть
связь между их строением и функцией, влияние молекулярной
динамики на функцию белков и возможность разрыва некото-
рых слабых связей при изменении условий среды.
3.6.1. Влияние pH на кинетику ферментативных реакций
в растворах
На рис. 2.15 приведены структурные формулы различных
аминокислот, из которых построены все белки. Аминокислоты
обладают основными, нейтральными и кислотными группиров-
ками, поэтому интактный фермент при любом заданном pH
может содержать как положительно, так и отрицательно заря-
женные группы. Заряженные группировки часто входят в со-
став активных центров ферментов, так как в основе целого
ряда механизмов ферментативного катализа лежит катализ
кислотного или основного типа. Необходимым условием для
осуществления кислотного или основного катализа может быть
наличие определенного заряда на ионизируемых группах ак-
174
Глава 3
тивного центра. Отсюда следует, что каталитически активная
форма фермента существует только в одном строго определен-
ном состоянии ионизации и в зависимости от pH в нее может
превращаться большая или меньшая часть всего имеющегося в
смеси фермента. На рис. 3.23 показано влияние pH на актив-
ность некоторых ферментов; нетрудно видеть, что по мере по-
вышения pH каталитическая активность фермента достигает
максимума (при оптимальном pH) и затем снижается.
РИС. 3.23. Зависимости актив-
ности ферментов от pH и диа-
пазона максимальной активно-
сти фермента от его природы.
(Воспроизведено с разрешения,
из работы: Fruton J. S., Sim-
monds S., General Biochemistry,
p. 260, John Wiley and Sons,
Inc., New York, 1953.)
Полезное математическое выражение, связывающее актив-
ность фермента с pH, можно получить с помощью следующей
простой модели ионизации активного центра:
—н+ —н+
Е +=> Е- Е2- (3.60)
+н+ +н+
Ki
В этих кислотно-основных реакциях Е~ обозначает активную
форму фермента, а Е и Е2~ — неактивные формы, образующие-
ся соответственно при протонировании и депротонировании ак-
тивного центра Е~. Константы равновесия указанных реакций
обозначены символами К\ и Kz- Мы не будем здесь рассмат-
ривать другие состояния ионизации фермента, поскольку пред-
полагается, что депротопирование Е- уже полностью инактиви-
рует фермент.
Записав условия равновесия для двух ступеней ионизации в
виде
(3.6i>
Кинетика катализируемых ферментами реакций 175
Где /г+=[Н+], мы можем определить относительное количество
фермента, находящегося в активной форме. Если обозначить
общую концентрацию фермента через е0, так что
е0=е+е~+е2~ (3.62)
то активная фракция фермента у~ равна е~/е0 и определяется
уравнением
у~ =--------!------- (3.63)
J 1 + h+/Kt + K2/h+
Уравнение (3.63) представляет собой одну из pH-функций Ми-
хаэлиса. Две другие, у и у2", определяют относительные коли-
чества фермента, находящегося в кислой и основной формах со-
ответственно.
Зависимость функции у~ от pH выражается кривой такого
же типа, какие приведены на рис. 3.23 для экспериментально
найденных зависимостей ферментативной активности от pH.
Функция у~ имеет один максимум при следующем pH:
h+opt = V^^2 или (pH)opZ = 1/2(pK1 + pK2) (3.64)
где pKi = —IgKi. Функция у~ убывает равномерно и симметрич-
но по мере удаления pH от оптимального значения.
Реакции протонирования и депротонирования представляют
собой очень быстрые процессы по сравнению со скоростями
большинства других реакций в растворах. Поэтому можно при-
нять, что фракция фермента, находящегося в активном состоя-
нии, равна Г и в том случае, когда этот фермент выполняет
функцию катализатора. Отсюда следует, что выражение для
максимальной скорости реакции можно получить, взяв вместо
общей концентрации фермента е0 общую концентрацию его ак-
тивной формы еоу~:
»тах = КУ~ =------------------ (3.65)
° l + h+/K1 + K2/h+ v 1
Располагая данными по ферментативной активности при
различных pH, с помощью последнего уравнения можно опре-
делить параметры К) и К2. Действительно, pH максимальной
активности фермента связан с Ki и К2 уравнением (3.64). Оп-
ределяемая экспериментальным путем зависимость фермента-
тивной активности от pH устанавливает независимую взаимо-
связь между Ki и К2, позволяя таким образом определить оба
эти параметра.
Согласно расширенному толкованию уравнения Михаэли-
са—Ментен для случая простейшей катализируемой фермен-
том последовательности реакций [уравнение (3.4)], pH может
влиять и на константу Михаэлиса, Кот. В то же время если
176
Глава 3
субстрат не может существовать в нескольких ионизированных
состояниях, характеризующихся различным сродством к фер-
менту и если образование фермент-субстратных комплексов с
каждой из форм фермента не влияет на Ki и Кг, то в таком
случае можно показать, что Кт не зависит от pH. Эксперимен-
тальные данные показывают, что, как правило, Кт очень слабо
зависит от pH, поэтому на практике для выражения зависимо-
сти скоростей катализируемых ферментами реакций от pH
чаще всего пользуются только уравнением (3.65).
Никогда не следует забывать, что все приведенные выше
рассуждения могут оказаться несостоятельными, если речь идет
о значениях pH, значительно отличающихся от оптимальных.
В такой ситуации нарушение сил, стабилизирующих конформа-
цию нативного белка, может привести к его денатурации,
и тогда даже после восстановления оптимального (или близко-
го к оптимальному) pH быстрая реактивация фермента стано-
вится маловероятной или вообще невозможной.
3.6.2. Скорость ферментативных реакций и температура
При изучении любых проблем химической кинетики одним
из основных теоретических положений является уравнение Ар-
рениуса, выражающее зависимость константы скорости реакции
от температуры:
k=Ae~E«/RT (3.66)
где Еа — энергия активации реакции; R — газовая постоянная;
А — предэкспоненциальный множитель; Т — абсолютная темпе-
ратура.
При графическом изображении зависимость 1п& от 1/Т вы-
ражается прямой с тангенсом угла наклона — EaIR [конечно,
если соблюдается условие уравнения (3.66)]. Аррениусовской
зависимости константы скорости реакции от температуры под-
чиняются и многие реакции, катализируемые ферментами; при-
мером могут служить экспериментальные данные, представлен-
ные на рис. 3.24.
Следует подчеркнуть, однако, что температурный диапазон,
в котором были получены приведенные на рис. 3.24 данные,
очень узок. В этом эксперименте не рассматривались темпера-
туры, значительно превышающие обычные в биологических ус-
ловиях пределы. Что же произойдет, если мы попытаемся за-
ставить фермент катализировать процесс еще быстрее, подняв
температуру еще выше? Как показано на рис. 3.25, в большин-
стве случаев результат окажется очень печальным.
Денатурация большинства белков начинается в диапазоне
температур от 45 до 50 °C и завершается очень быстро при
Кинетика катализируемых ферментами реакций
177-
РИС. 3.24. Аррениусовская зависимость скорости ферментативной реакции
(катализируемый миозином гидролиз АТР) от температуры. [Воспроизведено
из работы: Laidler К. L, The Chemical Kinetics of Enzyme Action, p. 197, The
Clarendon Press, Oxford, 1958; данные из статьи Quellet L., Laidler K. J.,
Morales M. F., Molecular Kinetics of Muscle Adenosine Triphosphate, Arch. Bio-
chem. Biophys., 39, 37 (1952).]
55°C. Один из механизмов термической денатурации белков
очевиден: по мере повышения температуры атомы в молекуле
белка приобретают все более высокую энергию, в том числе ки-
нетическую, и в конце концов становится возможным разруше-
ние слабых связей, стабилизирующих глобулярную структуру
белка, что и приводит к его инактивации.
Термическая инактивация ферментов может быть обрати-
мой, необратимой или смешанной. Зависимость скорости реак-
ции ферментативного катализа от температуры в достаточно
широком диапазоне температур может быть описана с помо-
щью простой модели обратимой термической инактивации. Со-
гласно этой модели, мы принимаем, что неактивная (i) и ак-
тивная (а) формы фермента находятся в равновесии:
Еа Ef (3.67)
Константу равновесия этой реакции можно выразить следую-
12-587
178
Глава 3
РИС. 3.25. При высоких температурах, когда начинает доминировать процесс
термической инактивации фермента, аррениусовская зависимость скорости
реакции от температуры нарушается (приведен пример разложения H2O2 ка-
талазой). Сплошная кривая вычислена по уравнению (3.73) при следующих
значениях параметров: £=3,5 ккал/моль, A#d=55,5 ккал/моль, ASd=
= 168 ккал/(моль-К), ^ = 258 мм3/мин. [Воспроизведено из статьи: Sizer I. W.,
Temperature Activation and Inactivation of the Crystalline Catalase-Hydrogen
Peroxide System, J. Biol. Chem., 154, 461 (1944).]
щим уравнением:
— Kd = exp f- —=x exp f--~ exp ((3.68)
ea \ RT ) 4 RT J \ R J
В этом уравнении символами AGd, A//d и ASd обозначены сво-
бодная энергия, энтальпия и энтропия инактивации соответст-
венно.
Хотя изолированные водородные связи сравнительно слабы
(их энергия обычно изменяется в пределах от 3 до 7 ккал/
/моль), энтальпия инактивации ферментов A//d достаточно вы-
сока, составляя, например, для трипсина и лизоцима белка
куриных яиц соответственно 68 и 73,5 ккал/моль. Инактивация
этих ферментов сопровождается изменением энтропии, равным
+ 213 кал/(моль-К). Благодаря высокой энтальпии денатура-
ции уже небольшие изменения температуры существенно изме-
няют относительное количество активной формы фермента.
______Кинетика катализируемых ферментами реакций
179-
При таких высоких значениях AZZd фермент инактивируется
практически полностью в диапазоне тридцати градусов.
Поскольку весь фермент существует либо в активной, либо
в неактивной форме, т. е.
ea+ei = eo (3.69)
из уравнения (3.68) следует, что
е =____£о_
° 1 + Kd
(3.70)
Согласно теории переходного состояния, скорость реакций
(3.4) при высоких концентрациях субстрата может быть выра-
жена следующим образом:
Vmax = ва ' kiT} (3.71)
где
4(Г)»а(^е45,/«ГВДГ (3.72)
Здесь kB и h — константы Больцмана и Планка соответственно,
а а — коэффициент. Из уравнений (3.68), (3.70) — (3.72) следу-
ет, что
Утах ~ д$ /я -bHjRT (3.73)
1 -f- е и е и
В уравнении (3.73) коэффициент р включает се, kB, h, е0 и
exp(AS*/Z?).
Это уравнение и представляет собой математическое выра-
жение, описывающее схематично изображенное на рис. 3.25 по-
ведение систем фермент—субстрат. Показанная на этом рисун-
ке сплошная линия, проходящая через экспериментально най-
денные точки, фактически была вычислена по уравнению (3.73)
после определения параметров р, Е, &Sd и AZZd на основе тех
же экспериментальных данных. При больших значениях 1/7
тангенс угла наклона кривой приближенно равен —E/Z? (по-
грешность равна Т (К) и обычно не вносит большого вклада).
Тангенс угла наклона другого прямого участка кривой при
более высоких температурах равен приблизительно (&Hd—Е)/
/Z?. Величину ASd можно определить, используя тот факт, что
при температуре Ттах, когда lnumax принимает наивысшее зна-
чение,
х) = -.„£+/Гт”------ (3 74)
&па с. К/ max
[Это выражение легко получить, приняв <Z(ln vmzy^ jdT рав-
ным нулю.] Поскольку Тщах известна из измерений, а величины Е
12*
180
Г лава 3
и мы уже определили, то не представляет труда вычис-
лить правую часть уравнения (3.74). Теперь, зная величину
Kd{Tmzx), возвратимся к уравнению (3.68) и найдем ASd. На-
конец, подберем такое значение коэффициента 0, чтобы итах
[Ттах в уравнении (3.73)] стало равным измеренной величи-
не. При необходимости более точные значения параметров мож-
но получить методом последовательного приближения. В част-
ности, очень большое влияние на результаты может оказывать
значение Ттах. Конечно, от температуры зависят и другие па-
раметры уравнений скорости реакции, например константа Ми-
хаэлиса или ингибиторная константа. Если эти параметры счи-
тать константами равновесия, как это часто бывает в действи-
тельности, то мы должны прийти к температурной зависимости
типа той, которая выражается уравнением (3.68). Тогда гра-
фик зависимости 1п/( (или рК) от 1/Т будет представлять со-
бой прямую линию, по наклону которой можно определить
стандартную свободную энергию.
В некоторых случаях графическая зависимость в этих ко-
ординатах изображается не прямой линией, иными словами,
наклон линии меняется при изменении диапазона температур.
Причиной подобных осложнений могут быть неоправданные
упрощения в интерпретации Кт или Ki или даже в последова-
тельности катализируемых ферментом реакций. Напомним, на-
пример, что для случая простейшей последовательности реак-
ций (3.4) Кт представляет собой не что иное, как отношение
констант скоростей элементарных стадий реакции, выражаемое
уравнением (3.5). Если каждую из этих констант элементарных
стадий в свою очередь выразить в форме уравнения Аррениуса
[уравнение (3.66)] или в форме уравнения теории переходного
состояния [уравнение (3.72)], то получится весьма сложная
форма зависимости от температуры. Другим возможным источ-
ником затруднений в определении зависимости некоторых па-
раметров скорости реакции от температуры может служить
существование нескольких промежуточных фермент-субстрат-
ных комплексов (или комплексов фермента с продуктом реак-
ции).
Подобные соображения были положены в основу полезного
и довольно широко применяемого способа проверки корректно-
сти кинетических моделей и соответствующих гипотетических
последовательностей реакций. Если зависимость кинетических
параметров в аррениусовских координатах выражается не пря-
мой, то исходная модель или чрезмерно упрощена, или вообще
неверна.
Кинетика катализируемых ферментами реакций
181
3.7. Инактивация ферментов
Подавляющая часть опубликованных в литературе данных
по кинетике ферментативного катализа посвящена изучению
начальных скоростей реакций и обратимых изменений фермен-
тативной активности. В то же время скорость снижения катали-
тической активности является одной из основных характеристик
ферментов при их практическом использовании. В первую оче-
редь это относится к процессам, в которых фермент использу-
ется в течение длительного времени, например , в проточных ре-
акторах непрерывного действия (следующая глава будет по-
священа в основном методам иммобилизации ферментов, пред-
назначенным для их удерживания именно в таких реакторах).
В подобных ситуациях полезное время жизни ферментного био-
катализатора может определять экономическую целесообраз-
ность всего процесса.
Основное внимание в этом разделе будет уделено механиз-
мам и кинетике процессов, при которых фермент теряет свою
каталитическую активность; эти процессы, связанные с измене-
нием структуры и функции белков, важны и с практической
точки зрения. Так, например, белки часто осаждают из культу-
рального бульона путем изменения pH или ионной силы, что
сопровождается изменением конформации или химического со-
стояния белка. На последующих стадиях очистки белков в ка-
честве высокоспецифичных сорбентов иногда применяют анти-
тела; инактивация последних сопровождается потерей способ-
ности узнавать и адсорбировать определенный белок. К про-
блемам разделения мы вернемся в гл. 11, а сейчас рассмотрим
причины снижения ферментативной активности, кинетику этих
процессов и сопровождающие их эффекты.
3.7.1. Механизмы денатурации белков
и сопутствующие эффекты
Как мы уже знаем, структура белков стабилизуется за
счет слабых взаимодействий; таким путем обеспечивается
функционально важная молекулярная подвижность. С другой
стороны, слабость стабилизирующих сил предполагает, что
с энергетической точки зрения белки могут сравнительно легко
принимать несколько альтернативных и биологически менее
активных конформаций. Иллюстрацией этого свойства белков
могут служить приведенные на рис. 3.26 термодинамические
характеристики различных состояний а-лактальбумина. Не-
трудно видеть, что разность свободных энергий нативного и
полностью денатурированного состояний этого белка составля-
ет всего лишь 9,0 ккал/моль (хотя переход из одной конформа-
182
Глава 3
35 А
50 А
РИС. 3.26. Схематичное изображение ступенчатого разворачивания белковой
молекулы а-лактальбумина и соответствующие термодинамические данные.
[Воспроизведено с разрешения из статьи: Kuwajima К., A Folding Model of
a-Lactalbumin Deduced from the Three-state Denaturation Mechanism; J. Molec.
Biol, 114, 241 (1977).]
Обозначения: N — нативная конформация; A* — критически активированное
состояние; ID — частично неупорядоченная конформация; RC — неупорядо-
ченный клубок (полностью денатурированное состояние),
дни в другую может потребовать преодоления более значи-
тельных энергетических барьеров переходных состояний). По-
этому приходится признать, что стабильность нативной
структуры белков крайне низка.
В этой связи неудивительно, что сопровождающиеся сниже-
нием активности нарушения природной геометрической и хими-
ческой структуры белков могут быть вызваны множеством фи-
зических и химических факторов (табл. 3.9). Установив инди-
видуальные факторы, обусловливающие денатурацию белков,
следует в то же время учитывать, что скорость денатурации
белков определяется не каждым из этих факторов в отдельнос-
ти, а их сочетанием. Например, склонность белка к денатура-
ции при повышенных температурах может изменяться в очень
широких пределах в зависимости от pH раствора, а совместное
влияние определенных pH и температуры на белок в очень
большой степени зависит от его природы.
Таблица 3.9. Факторы, вызывающие денатурацию белков, и сопутствующие денатурации эффекты*
Фактор, иниции- Типы взаимодействий, связей и группи-
рующий денату- ровок, подвергающихся действию данного Движущая сила процесса денатурации Результат денатурации
рацию фактора
Физические
факторы
Нагревание Водородные связи Повышение содержания денатурированных Высоконеупорядочен-
конформаций благодаря усилению теп- ные конформации, аг-
лового движения и разрушению структу- регаты
ры растворителя
Необратимые изменения ковалентных свя-
зей (например, дисульфидный обмен)
Охлаждение Гидрофобные связи, сольватирован- Нарушение структуры растворителя, дегид- Агрегаты, неактивные
ные группы ратация мономеры
Механические Сольватированные группы, пустоты Изменение сольватации, разрушение пус- Высоконеупорядочен-
силы тот, разрыв связей ные конформации, не-
' активные мономеры
Радиация Функциональные группы (например, Уменьшение числа структурообразующих Высоконеупорядочен-
пептидные связи, группы SH остат- взаимодействий после фотоокисления или ные конформации, аг-
ков Cys) атаки свободными радикалами регаты
Химические
факторы
Кислоты Расположенные в глубине молекулы Снижение числа структурообразующих ион- Неупорядоченный клу-
иезаряженпыс группы (например, ных взаимодействий бок
His, пептидные связи)
Основания Расположенные в глубине молекулы Снижение числа структурообразующих ион- Неупорядоченный клу-
незаряженные группы [например, ных взаимодействий бок
Туг, Cys, (Cys)2]
органические Водородные связи Уменьшение числа структурообразующих Неупорядоченный клубок
соединения, водородных связей между водой и натив-
образующие ной конформацией
2 водородные
w связи
Продолжение табл. 3.9
Фактор, иниции- Типы взаимодействий, связей и группи-
рующий денатура- ровок, подвергающихся действию данного Движущая сила процесса денатурации
цию фактора
Результат денатурации
Соли Полярные и неполярные группы
Растворители Неполярные группы
«Высаливание» полярных групп в раствори-
тель с большей диэлектрической прони-
цаемостью и обратный эффект в отноше-
нии неполярных групп
Сольватация неполярных групп
Высокопеупорядоченные
конформации
Поверхностно- Гидрофобные участки (для всех ти- Образование частично нарушенных субст-
антивные вс- пов ПАВ) и заряженные группы руктур, в том числе мицеллоподобных
щества (для ионных ПАВ) участков
Оксиданты Функциональные группы (например, Снижение числа структурообразующих
в остатках Cys, Met, Try и др.) и (или) функциональных взаимодейст-
вий
Ионы тяжелых Функциональные группы (например, Маскирование групп, существенных для
металлов в остатках Cys, His и т. д.) структуры или функции белка
Хелатообразую- Катионы, необходимые для структу- Замещение лиганда или потеря катиона
щие агенты ры или функции белка
Высокоупорядоченная
конформация пептид-
ных цепей с преобла-
данием спиральных
участков
Частично неупорядочен-
ная конформация,
значительные спираль-
ные участки
Инактивированный фер-
мент, иногда неупоря-
доченная структура
Инактивированный фер-
мент
Инактивированный фер-
мент
Биологические
факторы
Протеазы Пептидные связи
Гидролиз концевых или внутренних пен- Олигопептиды,
тидных связей кислоты
амино-
в Воспроизведено с разрешения из работы: Schmid R. D., Stabilized Soluble Enzymes, Adv. Biochem. Eng., 12, 41 (1979).
Кинетика катализируемых ферментами реакций
185
Можно указать целый ряд различных, но часто взаимосвя-
занных свойств белков, которые изменяются под воздействием
перечисленных в табл. 3.9 физико-химических факторов.
К внешним проявлениям денатурации белков относится изме-
нение таких их свойств, как растворимость, тенденция к обра-
зованию гелей или к кристаллизации, а также нарушение би-
ологической активности (каталитической активности, способ-
ности связывать антитела) растворов белков. Точно так же, как
при денатурации важно прежде всего совместное действие
различных факторов, так и в сопровождающих денатурацию
эффектах главнейшую роль играет не каждый эффект в отдель-
ности, а их комбинация. Например, если фермент выделяют
осаждением, то существенно, чтобы условия осаждения не вы-
зывали необратимой потери активности фермента.
3.7.2. Моделирование и кинетика процессов инактивации
В простейшей модели инактивации молекулы активного
фермента (Ея) претерпевают необратимые структурные или
химические изменения, приводящие к неактивной форме (Ег):
Еа ----> Ef (3.75)
Скорость этой реакции Га пропорциональна концентрации ак-
тивной формы фермента:
rd = kdea (3.76)
Следовательно, в закрытой системе при эффективном переме-
шивании (примем, что в реакционной смеси нет ни субстрата,
ни продукта реакции, ни ингибитора, ни иного эффектора) из-
менение концентрации активной формы фермента во времени
будет описываться уравнением
' (3-77)
так что
1п[м^г]=-^ (3-78)
С этим выражением согласуются, в частности, приведенные на
рис. 3.27 экспериментальные данные по определению активно-
сти аденозинтрифосфатазы в различных условиях. Зависимость
константы инактивации kd от температуры может быть описа-
на теорией переходного состояния [например, уравнением
(3.72) при а=1] в форме, несколько отличающейся от урав-
нения Аррениуса [уравнение (3.66)] для узкого температурно-
го диапазона, представляющего интерес для биологических си-
186
Глава 3
стем. В табл. 3.10 приведены величины энергии активации Е
и энтропии AS* для денатурации ряда распространенных фер-
ментов. Как видно из приведенных в таблице данных, для про-
цессов денатурации белков характерны очень большие энергии
активации.
В отсутствие ферментативной активности жизнедеятель-
ность клетки невозможна. В некоторых случаях клетка поги-
бает уже после разрушения очень небольшой части внутрикле-
РИС. 3.27. Зависимость инактивации аденозинтрифосфатазы от времени при
различных pH и температуре. [Воспроизведено с разрешения из статьи: Pelle-
tier G. Е., Quellet L., Influence of Temperature and pH on Myosin Inactivation;
Can. J. Chem„ 39, 265 (1961).]
точных ферментов. В этой связи становится понятным, почему
для стерилизации (т. е. для уничтожения всех форм микроорга-
низмов) газов, жидкостей или твердых тел можно использо-
вать тепловую обработку. В гл. 7 мы узнаем детальнее, почему
лишь немногие ферменты (и соответственно немногие микро-
организмы) способны без вреда для себя переносить длитель-
ное нагревание.
Данные об инактивации ферментов обычно получают, вы-
держивая фермент некоторое время в вызывающих денатура-
цию условиях без субстрата, после чего действие денатурирую-
щих факторов снимают, создают некоторые стандартные усло-
вия, добавляют субстрат и определяют активность по
начальной скорости ферментативной реакции. В присутствии
субстрата скорость инактивации фермента может существенно
отличаться от скорости, определенной описанным выше путем,,
если, например, свободный фермент и фермент-субстратный
Кинетика катализируемых ферментами реакций
187
комплекс (или комплекс фермента с продуктом реакции)
инактивируются с различными скоростями или если субстрат
и (или) продукт реакции сами инициируют инактивацию фер-
мента. Одну из возможностей анализа таких систем мы пока-
жем на простом примере. Допустим, что связывание субстрата
с ферментом стабилизирует последний (подобный эффект в из-
вестной степени действительно наблюдался в ряде систем). Та-
ким образом, в нашем примере инактивации будет подвергать-
ся только свободный фермент.
Таблица 3.10. Энергия активации и энтропия денатурации ферментов®
Фермент pH Энергия актива- ции, ккал/моль Энтропия акти- вации AS*, э. ед./моль
Панкреатическая липаза 6,0 46,0 68,2
Трипсин 6,5 40,8 44,7
Пепсин 4,83 56—147 Нет данных
Аденозицтрифосфатаза 7,0 70 150,0
а Воспроизведено из работы: Laidler К. Bunting Р. S., The Chemical Kinetics
of Enzyme Action, 2d ed., p. 430, Oxford University Press, London, 1973.
Комбинируя такую модельную систему инактивации с про-
стой последовательностью элементарных реакций Михаэлиса и
Ментен [уравнение (3.4)], получим
Ea-f-S =Д±: ЕдБ Еа+Р (3.79а)
kd
Еа -Л Ег (3.796)
Если мы далее достаточно обоснованно допустим, что процесс
инактивации происходит значительно медленнее, чем реакции
(3.79а), то, привлекая приближение квазистационарного состоя-
ния для комплекса (EaS), получим следующее выражение для
скорости реакции:
k?ptot,as
V== Km + S
(3.80)
где etot.a — общая концентрация активного фермента как в сво-
бодной форме, так и в виде комплекса. Скорость изменения
^tot.a выражается уравнением
(3.81)
си>
188
Глава 3
Возвращаясь к расчетам кинетики квазистационарного состоя-
ния, применявшимся при выводе уравнения (3.80), мы можем
выразить еа через etot,a и параметры каталитической реакции;
тогда уравнение (3.81) приобретет такую форму:
de tot,a kdetot,a
~dT~ = 1 + s/Km
(3.82)
Из уравнений (3.80) и (3.82) следует, что скорости превраще-
ния субстрата и инактивации фермента взаимосвязаны. В част-
ности, скорость инактивации фермента зависит от концентра-
ции субстрата. С другой стороны, если Еа и (EaS) инактивиру-
ются с одинаковыми скоростями, то в условиях ферментативной
реакции активность фермента будет снижаться точно так же,
как и в отсутствие субстрата.
Обобщая эти рассуждения, нетрудно прийти к выводу, что
вследствие различия в скоростях инактивации разных форм
фермента (свободного фермента, разнообразных комплексов,
ионизированных состояний и т. д.) скорость общей реакции
зависит от любого параметра (концентраций субстрата и ин-
гибитора, pH и т. д.), влияющего на относительные количества
разных форм фермента. В частности, для описания сложной
зависимости скорости инактивации от pH была разработана
следующая модель:
-н+ —н+ -н+ —н+
р р р р
^01 -< — -Ьа2 ч Ьаз *-'а4
+н+ +Н+ г +н+ +н+
kd3,,
kdi
Kt
Ef Ej
На рис. 3.28 в качестве примера такого сложного процесса
изображена зависимость скорости инактивации рицина от pH
(рицин не является ферментом; содержание активной формы
белка определяли путем измерения растворимой фракции бел-
ка). Для некоторых других белков также наблюдался минимум
скорости инактивации при определенном значении pH.
Снижение ферментативной активности во времени не всегда
соответствует реакции первого порядка, т. е. уравнению (3.78).
Для ряда белков найдены нелинейные зависимости логарифма
активности от времени, иногда на соответствующих кривых
имеется по два различных прямых участка (рис. 3.29, а). Для
объяснения и анализа таких систем предложен ряд моделей.
Одна из них, включающая параллельные обратимую и необра-
Кинетика катализируемых ферментами реакций
189
pH
РИС. 3.28. Зависимость константы скорости реакции первого порядка инакти-
вации рицина от температуры и pH. [Воспроизведено с разрешения из статьи:
Levy М., Benaglio. А. Е., The Influence of Temperature and pH upon the Rate'
of Denaturation of Ricin, J. Biol. Chem., 186, 829 (1950).]
тимую инактивации, нашла применение при объяснении и ана-
лизе снижения активности некоторых ферментов:
В частности, эта модель, предполагающая кинетику первого'-
порядка для каждой из указанных элементарных реакций, ус-
пешно использовалась для интерпретации результатов изуче-
ния инактивации люциферазы, приведенных на рис. 3.29, а.
Кривая процесса инактивации при 45 °C, например, была по-
лучена расчетным методом с помощью параметров kdi=kd? =
= 1,02 и &г = 0,02 ч-1.
Потеря активности в растворах протеаз представляет собой
более сложный процесс, поскольку протеазы катализируют ре-
акцию собственного гидролиза. Эта особенность (автолиз)*
190
Глава 3
РИС. 3.29. а — инактивация препарата люциферазы при pH 6,8 и указанной
температуре. [Воспроизведено с разрешения из статьи: Chase А. М., Studies
on Ceil Enzyme Systems. IV. The Kinetics of Heat Inactivation of Cypridina
Luciferase, J. Gen. Physiol., 33, 535 (1950).] 6—инактивация а-химотрипсина
в растворе при pH 7,8, температуре 40 °C, концентрации Са2+ 10-3 М и на-
чальной концентрации фермента 7,3-10—7 М (Л), 3,65-10—6 М (Б), l,46-10-sM
(В) и 2,92-10-5 М. (Г). [Воспроизведено с разрешения из статьи: Kawamu-
га Y., Nakanishi К., Matsuno R., Kamikubo Т., Stability of Immobilized «-Chymo-
trypsin, Biotech. Bioeng., 23, 1219 (1981).]
должна учитываться и в соответствующей модели инактива-
ции. Приведенная ниже модель инактивации протеазы а-хи-
мотрипсина является вариантом модели (3.83), дополненной
стадией автолиза:
Еа Ег1 _^EZ2
к
Еа 4- Ел ;—> (ЕдЕл) ---> Еа 4-неактивные продукты гидролиза (пептиды)
(3.84)
Некоторые особенности этой последовательности реакций были
подтверждены химическими методами. Обратите внимание, в
частности, на то, что только обратимо инактивированная фор-
ма Е/l этого очень хорошо изученного фермента способна под-
вергаться атаке и гидролизу активной формой протеазы Ея.
Более глубокий анализ этой модели и ее применение для рас-
Кинетика катализируемых ферментами реакций
191
чета кривых, изображенных на рис. 3.29,6, предлагается в уп-
ражнении 3.14.
Завершая обзор кинетики инактивации ферментов, следует
упомянуть о подходах к анализу необратимых реакций инакти-
вации ферментов ядами. В простейшем случае имеем
Ев+яд ----> Е</ rd = kd-ea-(nR) (3.85)
Поскольку, однако, яды часто действуют на активный центр
фермента, доступ к которому может быть блокирован связан-
ным с ферментом субстратом, то в присутствии последнего ход
анализа следует опять-таки видоизменить. Очевидно, нам нуж-
но рассматривать процессы катализа, например реакцию
(3.79а), и отравления фермента (3.85) совместно, причем в
последнем уравнении Еа будет обозначать свободный, не свя-
занный в комплекс фермент. Подобные видоизмененные моде-
ли, которые мы не будем здесь рассматривать, могут быть соз-
даны и для процессов инактивации, описываемых уравнениями
(3.83) и (3.84).
3.7.3. Денатурация ферментов
под действием механических факторов
Механические воздействия могут нарушить сложную гео-
метрию молекул ферментов вплоть до их полной денатурации.
К числу таких воздействий относятся и гидравлические силы,
возникающие при движении потока жидкости. Эксперименты,,
позволяющие оценить влияние сдвига на ферментативную ак-
тивность, проводились путем истечения через капилляр и с по-
мощью вискозиметра с коаксиальными цилиндрами.
Обозначим символами 0 и у время воздействия сдвига и его
скорость соответственно (в случае истечения через капилляр под
Ч подразумевается усредненная по поперечному сечению капил-
ляра скорость). Изучение индуцированной сдвигом денатурации
каталазы и уреазы показало, что потеря активности определя-
ется произведением параметров 0у (рис. 3.30, а). Результаты
этих экспериментов еще раз свидетельствуют о том, что степень
инактивации фермента обусловливается сочетанием интенсивно-
сти денатурирующего фактора и времени его действия. В то же
время изучение лактатдегидрогеназы показало, что инактивация
этого фермента в потоке определяется напряжением сдвига, а не
его скоростью.
Приведенные на рис. 3.30, б данные убедительно свидетель-
ствуют о частичной обратимости процесса индуцированной сдви-
гом инактивации уреазы. Кривая А отражает скорость превра-
щения субстрата (мочевины) в заторможенном растворе уреа-
зы, а кривая Б получена путем определения скорости той же
192
Глава 3
РИС. 3.30. а — инактивация каталазы и уреазы под воздействием сдвига
(здесь различными символами обозначены разные скорости сдвига), б — кон-
версия мочевины раствором уреазы в состоянии покоя (Л), под воздействием
сдвига (скорость сдвига 1717 см-1) (5) и через разные периоды времени
после снятия сдвига (0,4 единицы уреазы в 1 мл раствора, pH 6,75, 23 °C).
[Результаты изучения каталазы воспроизведены с разрешения из статьи:
Charm S. Е„ Wong В. L., Enzyme Inactivation with Shearing, Biotech. Bioeng.,
12, 1103 (1970). Данные по изучению уреазы воспроизведены из работы:
Tirrell М., Middleman S., Shear Modification of Enzyme Kinetics, Biotech. Bio-
eng., 17, 299 (1975).]
реакции в вискозиметре с коаксиальными цилиндрами при ско-
рости сдвига 1717 с-1. Очевидно, что движение жидкости сопро-
вождается снижением активности фермента. Другие точки на
рис. 3.30, б были определены после снятия гидравлического воз-
действия различной продолжительности. В этих случаях фермен-
тативная активность после снятия сдвигового усилия сразу же
начинает возрастать, но прежнего уровня не достигает. Остаточ-
ное различие в активности было объяснено частичной необрати-
мой инактивацией.
Кинетика катализируемых ферментами реакций 193
Характерная для ферментов чувствительность к механиче-
ским воздействиям может налагать определенные ограничения
на допустимую величину гидравлического усилия в ферментных
реакторах, в которых для повышения скорости массопередачи
субстрата используется перемешивание, или в системах ультра-
фильтрации растворов ферментов, когда повышение пропускной
способности мембраны сопровождается увеличением напряже-
ния сдвига и растяжения непосредственно перед мембраной и
сразу после нее.
Денатурацию белков и, следовательно, инактивацию фер-
ментов часто вызывает и другое механическое воздействие — по-
верхностное натяжение. Поскольку поверхностное натяжение на
границе раздела между воздухом и чистой водой составляет
80 дин/см, пенообразование обычно вызывает денатурацию бел-
ков, адсорбированных на границе раздела фаз. Граница раздела
между двумя жидкими фазами характеризуется значительно
более низким поверхностным натяжением; считается, например,
что плазматическая мембрана клетки имеет поверхностное на-
тяжение порядка 1 дин/см или даже меньше. Очевидно, низкие
поверхностные натяжения не инактивируют ферменты, посколь-
ку, как хорошо известно, в таких плазматических мембранах
существуют многие активные белки. Пенное фракционирование
представляет собой метод разделения, основанный на концен-
трировании выделяемого вещества на границе раздела фаз по-
верхностно-активное вещество — воздух; в этих условиях белки
не денатурируют, так как поверхностно-активное вещество сни-
жает поверхностное натяжение в системе вода — воздух до ве-
личин порядка 1 дин/см.
В технологических процессах и иногда в лабораторных экспе-
риментах на скорость инактивации ферментов одновременно
влияют различные механические и химические факторы, напри-
мер окисление. Сложная природа проявления этих взаимозави-
симых факторов хорошо освещена в работе Томаса и Даннил-
ла*, в которой, в частности, тщательное изучение влияния сдви-
га на те же ферменты (см. рис. 3.30) привело к иным результа-
там и показало, что один только сдвиг вызывает незначительную
инактивацию ферментов. Кроме упоминавшихся факторов на
инактивацию ферментов могут оказывать влияние расширяю-
щиеся потоки, кавитация, локальный адиабатический нагрев,
следы металлов и поверхностная денатурация в полостях. Более
подробные сведения о проблеме денатурации читатель может
найти в приведенной в конце главы литературе. Нам же доста-
точно запомнить одно общее правило: если фермент in vitro на-
* Thomas С. R., Dunnill Р., Action of Shear on Enzymes: Studies with Cata-
lase and Urease, Biotechn. Bioeng., 21, 2279 (1979).
13-537
194
Глава 3
ходится по существу в такой же среде и в таких же условиях,
как и in vivo, он будет активен. При изменении любого из пара-
метров, характерных для естественных условий существования
фермента, появляется вероятность его инактивации.
Это правило находит многочисленные применения в биохи-
мической технологии. Так, если нам нужен фермент, активный
при экстремальных значениях pH или температуры, то целесо-
образно прежде всего попытаться найти организм, для которого
эти экстремальные условия являются нормальными. Часто та-
кой организм содержит ферменты, способные эффективно функ-
ционировать именно в таких необычных условиях. Показатель-
ным примером может служить обнаружение продуцируемых
микроорганизмами ферментов, обладающих устойчивостью в
щелочных средах; эти ферменты успешно применяются в качест-
ве добавок к стиральным порошкам, растворы которых обычно
имеют pH 9,0—9,5.
3.7.4. Методы стабилизации ферментов
Помимо поиска более устойчивых природных форм существу-
ет ряд методов повышения стабильности уже известных фермен-
тов. Эти методы можно разделить на три основные группы:
а) добавление стабилизирующих веществ к среде, в которой
хранится фермент или проводится ферментативная реакция;
б) химическая модификация растворимого белка;
в) иммобилизация белка на поверхности либо во всем объ-
еме нерастворимого твердого носителя или матрицы. В нашем
кратком обзоре основное внимание будет уделено первым двум
методам стабилизации, а иммобилизацию и ее влияние на ус-
тойчивость белков мы обсудим в следующей главе.
К веществам, повышающим устойчивость белков, относятся
субстраты, органические растворители и соли. Поскольку актив-
ный центр фермента часто представляет собой одновременно и
наиболее лабильный участок его молекулы, то присутствие суб-
страта может стабилизировать фермент путем закрепления ча-
сти молекулы белка в виде фермент-субстратного комплекса.
С другой стороны, известны случаи дестабилизации ферментов
соответствующими субстратами. Растворители типа многоатом-
ных спиртов, стабилизирующие некоторые ферменты, возмож-
но, повышают устойчивость внутримолекулярных водородных
связей белка.
При низких концентрациях солей (<0,1 М.) ряд катионов,
например Са2+, Zn2+, Mn2+, Fe2+, Мо2+ и Си2+, специфично взаи-
модействуют с особыми ферментами, называемыми металлофер-
ментами. Некоторые из перечисленных катионов являются ко-
факторами, и их присутствие стабилизирует фермент. Катион
Кинетика катализируемых ферментами реакций
195
Таблица 3.11. Примеры методов стабилизации ферментов
Фермент
Метод стабилизации
Результат стабилизации
Глюкоамилаза
Лактатдегидро
ген аз а
а-Амилаза
Химотрипсин
^-Галактозидаза
Добавление аналогов суб-
страта— глюкозы, глю-
конолактона
Добавление субстрата (лак-
тата) или эффектора
(фруктозодифосфата)
Добавление 50—70% сорби-
та
Добавление 50—90% гли-
церина
Добавление 5—10% этанола
или изопропанола
а-Амилаза (из
Bacillus caldoly-
ticus )
Трипсин
Добавление Са2+
Аспарагиназа
Глякогенфосфо-
рилаза
Папаин
Конденсация полиаланила
(около 10 аминокислот-
ных остатков) с амино-
группами белка
Введение сукцинильных
групп обработкой янтар-
ным ангидридом
Введение бутильных или
пропильных заместителей
обработкой альдегидом и
NaBH4
Образование поперечных
связей с помощью глу-
тарового альдегида
Повышение термической
устойчивости
Повышение термической
устойчивости; дестабили-
зация в присутствии дру-
гого субстрата (пирува-
та)
Повышение термической
устойчивости и устойчиво-
сти при хранении
Повышение устойчивости к
протеолизу
Повышение термической
устойчивости; метанол
или «-пропанол в тех же
концентрациях дестаби-
лизируют фермент
Значительное повышение
термической устойчиво-
сти
Повышение устойчивости к
протеолизу и тепловой
инактивации
Повышение устойчивости к
протеазам
Повышение термической
устойчивости
Повышение термической
устойчивости
Са2+ участвует в стабилизации третичной структуры ряда бел-
ков. Образуя ионные связи с двумя различными аминокислот-
ными остатками, ионы Са2+ могут выполнять функцию стабили-
зирующего мостика аналогично дисульфидным связям.
Примеры влияния различных веществ на стабильность фер-
ментов приведены в табл. 3.11, а более подробные сведения чи-
татель сможет найти в обзоре [4]. Следует подчеркнуть, что
влияние данного вещества или фактора на стабильность какого-
либо фермента не всегда означает, что это вещество или фактор
будут таким же образом действовать и на другие ферменты.
С другой стороны, стабилизирующий эффект часто проявляется
13*
196
Глава 3
только при определенном сочетании состава раствора и темпе-
ратуры; например, добавление небольших количеств ряда солей
стабилизирует ферменты, а более высокие концентрации тех же
солей обычно вызывают их денатурацию. Это правило относит-
ся и к органическим растворителям.
Для повышения стабильности ферментов сравнительно ус-
пешно использовался также такой важный инструмент биохими-
ческих исследований, как химическая модификация белков.
В одном из вариантов этого метода модификации подвергают
боковые цепи остатков некоторых аминокислот, например путем
ацилирования, восстановительного алкилирования или конден-
сации аминогрупп нативного белка с боковыми цепями поли-
аминокислот.
Другой вариант химических методов стабилизации белков
основан на применении бифункциональных реагентов, напри-
мер глутарового альдегида. Такие реагенты образуют попереч-
ные связи между аминогруппами белка и тем самым, во-первых,
затрудняют доступ протеаз к белку и, во-вторых, могут закреп-
лять активную конформацию белка. Для химической стабилиза-
ции могут применяться и диимиды, связывающие поперечными
амидными связями аминные и карбоксильные группы. Эти реа-
генты и соответствующие реакции мы рассмотрим подробнее в
ходе изучения их применения для иммобилизации ферментов.
3.8. Ферментативные реакции в гетерогенных
системах
До сих пор основное внимание мы уделяли поведению фер-
ментов в растворе, где и осуществляется их взаимодействие с
растворенными субстратами. Однако, как мы уже указывали,
такая ситуация не является единственно возможной. Так, на
рис. 2.27, где схематически изображена прокариотическая клет-
ка, было показано, что некоторые клеточные ферменты связаны
с мембранами клетки. Аналогичные структуры имеются и в эу-
кариотах; в митохондриях, например, со сложной системой внут-
ренних мембран связаны ферменты, отвечающие за очень слож-
ную последовательность реакций.
Как показано на рис. 3.31, в природе и в биохимической тех-
нологии возможны многие другие сочетания различных физиче-
ских состояний фермента и субстрата. Здесь мы рассмотрим ки-
нетику ферментативных реакций между растворами ферментов
и нерастворимыми субстратами, а в следующей главе изучим
реакции растворимых субстратов, катализируемые связанными
с носителями ферментами.
Если субстрат одновременно может существовать в несколь-
ких фазах, то иногда ферментативной реакции подвергается
К и н ет и ка катализируемых ферментами реакций
197
Состояние
фермента
Раствор
Генсониназа
Поверхность
Живность
Твердое
вещество
да
Твердое
вещество
Пденозинтрифо'
„ „ сфатаза
Поверх- ----------
ность Протеиназа на а£д
сорёщ
Сухое ве-
щество
с низ ной
влажностью
Состояние
субстрата
Раствор
Пепсин-
альбуминовь/е лленни
Гель
Заморо-
женное
состояние
' Газ
РИС. 3.31. Находящиеся в различных физических состояниях ферменты ката-
лизируют превращения различных форм субстратов. Классический случай
ферментативной реакции в растворе является только одним из множества
возможных вариантов фермент-субстратных взаимодействий. [Воспроизведе-
но из статьи: McLaren A. D„ Packer L., Some Aspects of Enzyme Reactions in
Heterogeneous Systems, Adv. Enzymol. Rel. Sub. Biochem., 33, 245 (1970).]
только та его часть, которая находится в растворе. Примером
могут служить данные, приведенные на рис. 3.32. Поскольку все
вещества в той или иной степени растворимы в воде, то неболь-
шое количество субстрата всегда будет находиться в растворе
и, следовательно, подвергаться действию фермента. В то же вре-
мя скорость этого процесса может быть настолько мала, что с
практической точки зрения он не будет представлять никакого
интереса. Теперь рассмотрим некоторые примеры другого рода.
Одним из таких примеров может являться реакция гидро-
лиза метилбутирата под действием панкреатической липазы—
фермента, образующегося в пищеварительном тракте человека
и способного расщеплять жиры. В отличие от предыдущего при-
мера здесь реакция не идет до тех пор, пока в реакционной
смеси не сформируется нерастворимая форма субстрата в виде
небольших капель (рис. 3.33). Очевидно, этот фермент способен.
198
Глава 3
концентрация субстрата
РИС. 3.32. Ферментативная реакция осуществляется только с растворенной
формой субстрата. (Из работы: Диксон М., Уэбб Э., Ферменты, т. 1—3. — М.:
Мир, 1982, т. 1, с. 122.]
Ллощадь тежразной поверхности, спг/пп
О ОД 7,0 7,5 2,0
Общая концентрация тетилбцтирата (отно-
тетто нась/щсния)
РИС. 3.33. Реакция на поверхности раздела двух жидких фаз. [Воспроизве-
дено с разрешения из статьи: Sarda L., Desnuelle Р., Action de la Lipase Pan-
creatique sur les Esters en Emulsion, Biochim. Biophys. Acta, 30, 513 (1958).]
проявлять свою активность только на границе раздела двух
жидких фаз. Учитывая возможность денатурации фермента за
счет поверхностного натяжения, интересно отметить, что, как и
в случае упоминавшегося выше метода пенного фракционирова-
ния в присутствии детергентов, поверхностное натяжение на
жировых каплях может существенно снизиться вследствие ад-
сорбции желчных кислот — природных поверхностно-активных
веществ, выделяемых в пищеварительный тракт.
Кинетика катализируемых ферментами реакций 199
Другие ферменты активны в отношении как растворимой, так
и нерастворимой форм субстрата. Так, например, было показа-
но, что протеолитический фермент трипсин способен расщеплять
как свободный лизоцим, так и лизоцим, адсорбированный на
поверхности каолинита. Сам лизоцим также может взаимодей-
ствовать с растворимыми и «нерастворимыми» субстратами. Как
мы уже отмечали, лизоцим активно разрушает стенки бактери-
альных клеток; в то же время он способен катализировать и
расщепление растворимых олигомеров, образующихся из поли-
меров клеточной стенки (разд. 3.4.2).
РИС. 3.34. Зависимость скорости разруше-
ния твердого субстрата (тиогеля) от кон-
центрации во фермента в растворе. [Дан-
ные из статьи: Tsuk A. G., Oster G., Deter-
mination of Enzyme Activity by a Linear
Measurement, Nature (London), 190, 721
(1961).]
Взаимодействие растворенного фермента с нерастворимым
субстратом путем адсорбции на поверхности последнего может
быть описано с помощью интересного варианта рассмотренных
ранее кинетических уравнений. В отличие от предыдущих слу-
чаев, когда скорость реакции возрастала пропорционально об-
щей концентрации фермента, здесь при повышении концентра-
ции фермента скорость реакции сначала возрастает, а затем
приближается к некоторой предельной величине. Такое поведе-
ние фермент-субстратной системы наглядно иллюстрируется
приведенными на рис. 3.34 данными, отражающими кинетику
гидролиза твердого блока белка (в данном случае тиогеля —
сшитого поперечными связями желатина) под действием трип-
сина.
Разработку достаточно обоснованной модели для кинетики
гетерогенной реакции мы начнем с допущения, обратного тому,
которое было положено в основу анализа скоростей фермента-
тивных реакций в растворе — теперь мы будем считать, что
фермент адсорбируется на субстрате. Обозначив символом А
200
Глава 3
вакантные центры на поверхности субстрата, мы можем напи-
сать следующее уравнение:
Е4-А ЕА (3.86)
&des
Если общее число молей центров адсорбции на поверхности суб-
страта в расчете на единицу объема реакционной смеси принять
равным а0, то
aQ=a+(ea) (3.87)
Из уравнения (3.87) и уравнения равновесной адсорбции фер-
мента (3.86) следует, что
(еа) = —-----, где Ка = -^- (3.88)
Ад + е kads
Теперь остается только допустить, что реакция завершается не-
обратимым расщеплением комплекса ЕА. Следовательно,
v = ks(ea) = -~~- (3.89)
Лд+ е
По условиям модели е обозначает концентрацию свободного
фермента, которая в начале эксперимента связана с общей кон-
центрацией фермента е0 соотношением
ео = е+(еа) (3.90)
Если начальная концентрация фермента значительно превыша-
ет начальную концентрацию субстрата (со^Яо), то с хорошей
степенью приближения можно считать, что
е0~е (3.91)
Поэтому
v __ k3a0e0
К а + ео
Для реакций с твердыми субстратами ситуации, когда ео^>ао,
ни в коей мере не являются исключением. Например, результа-
ты, приведенные на рис. 3.34, были получены в эксперименте,
в котором отношение е0/а0 составляло около 4000. Этим гетеро-
генные реакции резко отличаются от реакций в растворах, ког-
да s0 обычно значительно больше, чем е0.
Из уравнения (3.92) следует, что график зависимости 1/v от
1/е0 (в двойных обратных координатах Лайнуивера — Бэрка)
должен представлять собой прямую линию. Как показано на
рис. 3.35, так оно и есть на самом деле. Здесь прямая на
рис. 3.35, а выражает в другой форме те же данные, которые
Кинетика катализируемых ферментами реакций
201
РИС, 3.35. а — зависимость ско-
рости деструкции твердого субст-
рата (тиогеля) (1/и) от концент-
рации трипсина (1/е0) в растворе
в координатах Лайнуивера—Бэр-
ка. б — зависимость скорости де-
струкции нерастворимого субстра-
та (частиц поли-р-гидроксибути-
рата) раствором фермента (де-
полимеразы из Р. lemoignei). [Вос-
произведено из работы: McLa-
ren A. D., Packer L., Some Aspects
of Enzyme Reactions in Heteroge- .. ,
neous Systems; Adv. Enzymol. Rel.
Sub. Biochem, 33, 245 (1970).]
были приведены на рис. 3.34, а на рис. 3.35,6 отражены анало-
гичные результаты, полученные для другой системы раствори-
мый фермент — нерастворимый субстрат.
Многие источники питательных веществ для микроорганиз-
мов представляют собой твердые частицы (в потоках сточных
вод, озерах, хранилищах компоста и т. д.). Очевидно, что ад-
сорбции и транспорту растворенных питательных веществ через
клеточные мембраны должен предшествовать гидролиз этих
частиц внеклеточными ферментами. Точно так же гидролиз цел-
люлозы ферментами типа целлюлаз требует предварительного
расщепления нерастворимых частиц, поэтому надо полагать, что
рассмотренная в этом разделе кинетика гетерогенных фермента-
тивных реакций окажется пригодной для расчетов реакторов с
такого типа субстратами.
В заключение следует подчеркнуть, что область применения
рассмотренной кинетической модели не ограничивается тверды-
202
Глава 3
ми субстратами; она использовалась и для описания нераство-
римых жидких субстратов, диспергированных в растворе фер-
мента (например, системы, кинетика которой изображена на
рис. 3.33). Кроме того, здесь мы не принимали во внимание раз-
личий в концентрациях реагентов в основной массе жидкой
фазы и на границе раздела фаз. Последнюю проблему мы рас-
смотрим детально в следующей главе.
Упражнения
3.1. Определение Кт и vmax. В табл.-ЗУ 1.1 приведены начальные скорости
катализируемой ферментом реакции при различных концентрациях субстрата.
а) Определите vmax и Кт методом Лайнуивера — Бэрка.
б) Определите те же величины графически в координатах Эди—Хофсти.
в) Вычислите стандартные отклонения наклонов прямых и отсекаемых
на координатных осях отрезков для каждого метода.
Таблица ЗУ1.1
s, моль/л v, моль/(л-мин)-103 S, моль/л V, моль/(л-мин) Ю3
4,1-10-s 177 4,9-10~5 80
9,5-10“4 173 1,06-IO’3 67
5,2-10“4 125 5,1-10-® 43
1,03-10’4 106
3.2. Ферментативная реакция в реакторе периодического действия. Фер-
мент с Km=l-10_3 М. инкубировали с субстратом при начальной концентра-
ции последнего 3-10~3 М. Через 2 мин прореагировало 5% субстрата. Какое
количество субстрата будет трансформировано в течение 10, 30 и 60 мин?
3.3. Кинетика ферментативных реакций при участии ферментов с не-
сколькими активными центрами. Предположим, что фермент обладает двумя
активными центрами, так что субстрат трансформируется в продукт реакции
в соответствии со следующей последовательностью элементарных реакций:
kt kz
E4-S (ES) (ES)+S <=> (ESS)
Й-1 Д-2
ks ki
(ESS) ----> (ES)P (ES) ----------> E + P
Приняв условия квазистационарного состояния для (ES) и (ESS), найдите
выражение, определяющее скорость образования Р.
3.4. Образование нескольких фермент-субстратных комплексов. В ходе
катализируемых ферментами реакций иногда промежуточно образуется не-
сколько комплексов. Найдите соответствующие выражения для скорости
реакции, применив а) равновесный подход Михаэлиса и б) приближение
квазистационарного состояния для комплексов, если имеет место следующая
последовательность реакций:
&3 kb
S+E (ES)X (ES)2---------------------> Р + Е
kz kt
____________Кинетика катализируемых ферментами реакций_____________203
3.5. Релаксационная кинетика с синусоидальными возмущениями. Как
показано на рис. ЗУ5.1, если параметры реагирующей системы, находящейся
в равновесии (или в стационарном состоянии), подвергаются небольшому
возмущению по синусоидальному закону, то и концентрации реагирующих
веществ также изменяются по такому же закону. Найдите уравнения, связы-
РИС. ЗУ5.1.
вающие наблюдаемые флуктуации концентраций с кинетическими парамет-
рами, для реакции
А <— В г ^k^ — k^b
kz
если средняя температура изменяется в соответствии с уравнением
7’=T0(l+asin cot), где а<1
Покажите, как вы будете определять ki и k2> если известны отклики а и Ь
иа периодические возмущения.
3.6. Обратимые реакции. Предположим, что мы изучаем обратимую ре-
акцию:
k\ kz
E + S (ES) E-f-P
k-1 k.z
а) Покажите, что равновесие будет значительно смещено вправо только
При УСЛОВИИ kik2^k-ik-2-
б) Покажите, что параметры vs, vp, Ks и Кр не являются независимыми.
в) При каких условиях графическое решение уравнения задачи (б) в ко-
ординатах Лайнуивера — Бэрка даст полезные результаты?
г) Путем интегрирования dpjdt для приведенной последовательности
реакций определите зависимость р от t и значения р при равновесии
( Р
At s= Bp + С In [ 1 — -т—
\ "еЧ
д) Покажите, что В=0, если Ks=Kp-
3.7. Инактивация ферментов. Пусть фермент необратимо инактивируется
в соответствии с уравнением (3.77).
204
Глава 3
а) Покажите, что в таком процессе изменяется только vmax, но не Кт.
б) Покажите, что для ферментов, действующих на нерастворимые суб-
страты [уравнение (3.92)], можно ошибочно найти решение, противополож-
ное решению задачи (а), если не принять во внимание изменение концентра-
ции активной формы фермента.
3.8. Зависимость от pH. В результате катализируемой ферментом необра-
тимой реакции генерируются протоны в соответствии со следующим урав-
нением:
H2O + E+S+ —> E+SOH+H+
Допустим, что е~ — концентрация активной формы фермента и что е~/е=1,0
при рН=6,0=р/<|, е2-/е~ = 1,0 при рН= 10,0=pA'2.
а) Покажите, что скорость реакции при pH 7,0 приближенно выражается
уравнением
sKj.
" = (Ks + ^+V ™еА=1Н+1
б) Путем интегрирования предыдущего уравнения покажите, что измене-
ние s во времени осуществляется по закону
s
a In — + P(s0 — s) + 6(s20 — s2) t= vmt
a0
где а и {J — параметры, зависящие от начальных pH и концентрации
субстрата.
3.9. Кинетика ингибирования. Пестицид ингибирует активность данного
фермента А, и, следовательно, последний может применяться для определе-
ния пестицида в пробах неизвестного состава.
а) В лабораторных экспериментах измерены начальные скорости реакции;
результаты измерений приведены в табл. ЗУ9.1. Каким ингибитором является
Таблица ЗУ9.1
s, моль/л V, моль/(л-мин) • 103
В отсутствие ингибитора В 10~5 М растворе ингибитора
3,3-10~4 56 37
5,0-10~4 71 47
6,7-10~4 88 61
1,65-10-3 129 103
2,21-10-3 149 125
этот пестицид, конкурентным или неконкурентным? Определите величины Кь
Vmax К Кт.
б) Раствор фермента (50 мл), описанного в задаче (а), смешали с 50мл
8-Ю-4 М раствора субстрата и 25 мл пробы; начальная скорость реакции
составила 18 мкмоль/(мин-л). Какова концентрация ингибитора в пробе, если
последняя не содержит других ингибиторов (кроме пестицида) и субстратов?
3.10. Кинетика ферментативных реакций, протекающих с участием иони-
зирующегося кофактора. Для проявления каталитической активности фермен-
ту необходим связанный кофактор. Связь кофактора с ферментом очень
прочная. Необходимая для проявления активности группа кофактора имеет
рК, равное р/С; кофактор активирует фермент только в депротонированной
форме. Первые значения рК активного центра фермента по обе стороны от
Кинетика катализируемых ферментами реакций 205
pH максимальной активности фермента равны pKi и рК9. Найдите (или
просто напишите) соответствующие выражения для скорости превращения
индивидуального субстрата в продукт реакции при следующих условиях:
а) рКс| < IpAi < I рЛз 1
б) p/CiI < I$Кс < (рКг!
в) p/Cc=p/Ci
3.11. Субстратная активация. Найдите уравнения, описывающие скорость
ферментативной реакции при активации субстратом:
Ks
Е + S , ES
KSA S< K's к.
E + S ,-----ESa ESaS -----esa + p
s
Как обычно, примите, что а) промежуточные соединения находятся в квази-
стационарном состоянии и б) концентрация субстрата значительно превы-
шает концентрацию фермента.
3.12. Выделение тепла в процессе ферментативных реакций. Максималь-
ное повышение температуры в реакторе полного вытеснения можно опреде-
лить с помощью уравнения теплового баланса превращения субстрата при
его однократном прохождении через реактор.
а) Учитывая, что количество выделившейся теплоты больше количества
теплоты, переданной жидкой среде (или равно ему), выразите максимальное
повышение температуры в катализируемой одним ферментом реакции через
теплоту реакции на моль реагента &НГ, относительное количество прореаги-
ровавшего реагента б, концентрацию реагента на входе в реактор s0, тепло-
емкость ЖИДКОСТИ Ср И &Т=Т выход—Г вход.
б) Покажите, что для гидролиза 20%-ного раствора лактозы на 80%
[Д//г=—7100 кал/(г-моль)] максимальное повышение температуры состав-
ляет всего лишь несколько градусов.
в) Если тонкостенный ферментный реактор снабжен охлаждающей ру-
башкой, то температуру в средней линии выхода приближенно можно опре-
делять по уравнению
У* =- ^BblS0A ^стенка г
^вход — Т’стенка 2 У <xt
где z— длина колонны, t — время пребывания реакционной массы в реакторе
и a.=ktlpCp, где kt — общая теплопроводность, р — плотность слоя насадки
и erf — функция погрешности (таблицы значений этой функции для различ-
ных аргументов z!2}at можно найти в любом справочнике). Покажите, что
для условий, описанных в части (б) настоящего упражнения, Т* быстро при-
ближается к единице (вычислите Т* при z=l, 2, 3, 4 дюйма). (Pitcher W. Н.,
Immobilized Enzymes for Industrial Reactors, p. 151, Academic Press, New
York, 1975.)
3.13. Уравнение Хилла для кооперативного связывания. Для описания
кооперативного (п>1) связывания субстратов (S) с олигомерными белками
(Еп) часто применяется следующая простая модель:
nS-)-Ert =?=> ErtSrt
Найдите простой графический метод определения п, если известен ряд экспе-
риментально определенных значений Еп8п.
3.14. Инактивация а-химотрипсина. Рассмотрим процессы тепловой дена-
турации и автолиза а-химотрипсина, одновременно протекающие в соответ-
ствии с уравнением (3.84), где К — константа равновесия реакции между Еа
206
Глава 3
и Ен, а Кт — константа диссоциации комплекса ЕаЕц. Принимая, что
(вавн) <С£а+ец, определите зависимость концентрации потенциально активно-
го фермента е (е=еа+ец) от времени в реакторе периодического действия.
Литература
Во всех перечисленных в конце гл. 1 руководствах по биохимии дается
и введение в кинетику ферментативного катализа. Особенно подробно и иа
современном уровне эта проблема освещена в книге Малера и Кордеса (ссыл-
ка [4] в гл. 2). Более подробную информацию читатель может найти в сле-
дующих книгах и статьях.
1. Bernhard A., Structure and Function of Enzymes, W. A. Benjamin, New York,
1968. Короткое, написанное очень доступным языком введение в энзимо-
логию.
2. Диксон М„ Уэбб Э., Ферменты, т. 1—3. — М.: Мир, 1982. Самое полное
однотомное руководство по химии и биологии ферментов, являющееся
настольной книгой для любого энзимолога. В нем имеется обширная глава
(200 страниц), посвященная кинетике ферментативного катализа. Кроме
того, в монографии рассмотрены многие другие проблемы энзимологии, при-
ведена большая таблица ферментов и катализируемых ими реакций, дана
обширная библиография.
3. Laidler К. J„ Bunting Р. S., The Chemical Kinetics of Enzyme Action, 2d ed.,
Oxford University Press, London, 1973. В этой монографии более подробно
рассмотрена кинетика ферментативного катализа и представлен обширный
экспериментальный материал, подтверждающий различные математические
выражения, предложенные для определения скоростей реакций.
4. Schmid R. D., Stabilized Soluble Enzymes, in Advances in Biochemical Engi-
neering, Vol. 12: Immobilized Enzymes II, p. 41, Ghose T. K-, Fiechter A.,
Blakebrough N. (eds.), Springer-Verlag, Berlin, 1979. Превосходная обзор-
ная статья, в которой дан глубокий анализ причин и факторов, вызываю-
щих денатурацию белков, а также великолепный обзор методов повышения
стабильности ферментов.
5. Johnson F. Н., Eyring Н., Polissar М. J., The Kinetic Basis of Molecular
Biology, John Wiley and Sons, Inc., New York, 1954. Великолепная моно-
графия, в которую включено множество данных о влиянии температуры,
pH, давления и ингибиторов на ферментативную и другие виды биологиче-
ской активности, а также интерпретации этих данных. Особое внимание
уделено применению термодинамики и теории абсолютных скоростей
реакций.
6. Жоли М., Физическая химия денатурации белков. — М.: Мир, 1968. Хотя
в эту монографию, естественно, не включены многие последние данные,
в ней рассмотрены все основные явления, связанные с причинами, прояв-
лениями, обнаружением и изучением денатурации белков. Все эти явления
рассматриваются с точки зрения химика.
7. McLaren A. D., Packer L., Some Aspects of Enzyme Reactions in Heteroge-
neous Systems, Advan. Enzymol., 33, 245 (1970). Превосходная обзорная
статья, в которой рассмотрено множество гетерогенных ферментативных
реакций, протекающих в биологических системах в природных условиях.
Эта работа посвящена важной, но относительно мало изученной теме пове-
дения нерастворимых субстратов, а также связанных ферментов в клетке
и почве.
Глава 4
Применение реакций,
катализируемых ферментами
В этой главе мы рассмотрим некоторые примеры практиче-
ского применения ферментов и изучим различные типы и мето-
ды получения иммобилизованных ферментных катализаторов,
позволяющих использовать ферментативные реакции в непре-
рывных процессах. Поскольку кинетические параметры таких
биокатализаторов зависят как от массопередачи, так и от при-
роды химических реакций, важно знать совместное влияние
этих двух факторов на свойства катализаторов.
Все применяющиеся в практической работе ферменты полу-
чают из природных источников (табл. 4.1). Хотя ферменты син-
тезируются в любой живой клетке, тот или иной определенный
фермент обычно целесообразно выделять только из одного ис-
точника— какого-либо растения, животного или микроорганиз-
ма. Некоторые ферменты, например, продуцируются только в
организмах животных. Выделенные из животных ферменты, од-
нако, могут иметь сравнительно высокую стоимость, как, напри-
мер, реннин из желудка теленка, а их доступность может опре-
деляться, например, спросом на говядину или баранину. Выде-
ление растительных ферментов (например, папаина из папайи)
обычно проще, но доступность соответствующего сырья также
зависит от потребности в данных пищевых продуктах. Методы
получения микробных ферментов позволяют легко расширить
масштабы производства. Как будет показано далее в гл. 6, тех-
нология рекомбинантных ДНК представляет собой новый метод
получения самых разнообразных ферментов (в том числе и та-
ких, которые в нормальных условиях не синтезируются микро-
организмами или перманентными линиями клеток) с помощью
бактерий, дрожжей и культур клеток. Более того, благодаря
быстрому по сравнению с растениями и животными самовоспро-
изведению микроорганизмов микробиологические процессы зна-
чительно проще приспособить к колебаниям в спросе на фер-
менты. С другой стороны, ферменты, применяющиеся в произ-
водстве пищевых продуктов или лекарственных препаратов,
могут производиться только с помощью заведомо безопасных
микроорганизмов.
Таблица 4.1. Примеры ферментов, применяемых в промышленности8
Фермент Источник фермента Применение Примечания Степень промы ш- ленной важности
Диастаза Солод Амилазы (разжижающие крахмал) Способствующее пищеварению сред- а-Амилазная и Р-амилаз- +++
Такадиастаза Aspergillus oryzae ство; добавка к хлебопродуктам; производство патоки Способствующее пищеварению сред- ная активности Содержат много других +++
Амилаза Bacillus subtilis ство; добавка к хлебопродуктам; производство патоки Расшлихтовка тканей; производство ферментов, в том числе . протеазы, рибонуклеазу Неочищенные препараты со- 4—1—h
Кислотоустойчивая Aspergillus niger патоки, глюкозы, этанола (броже- ние) Способствующее пищеварению сред- держат протеазы Максимальная активность +
амилаза Амилоглюкозидаза Rhizopus niveus, A. ство Амилазы (осахаривающие крахмал) ni- Производство глюкозы при pH 4—5 +++
Трипсин ger, Endomycopsls fi- bull ger Протеазы из организмов животных и растений Поджелудочная железа Применяется в медицине, а также +++
Пепсин животных Желудок животных для мягчения мяса, осветления пива Способствующее пищеварению сред- +++
а-Химотрипсин Желудок животных ство; мягчение мяса Применяется в медицине +++
Реннин Желудок теленка Производство сыра 4-4-4-
4—537
Фермент Источник фермента Применение Примечания Степень промыш - ленной важно- сти
Панкреатическая протеаза Папаин Бромелаин, фицин Протеаза Протеаза Поджелудочная железа животных Папайя Ананас, инжир A. oryzae A. tiiger Способствующее пищеварению сред- ство; производство моющих средств; мягчение кожи; обесвола- шиваиие; улучшение качества кор- мов Способствующее пищеварению сред- ство; применяется для осветления пива, мягчения мяса Способствующее пищеварению сред- ство; применяется для мягчения мяса, осветления пива Протеазы из микроорганизмов Осветление и вкусовая обработка сакэ Производство кормов; способствую- Кислотоустойчивая протеа- ++ +++ ++ + ++
Протеаза В. subtilis щее пищеварению средство Производство детергентов; разруше- за, оптимум активности при pH 2—3 Максимальная активность ++
Протеаза Streptomyces griseus пне желатиновых пленок (при реге- нерации серебра); мягчение мяса; производство рыбных гидролизатов Производство детергентов; разруше- при pH 7,0 Максимальная активность ++
Варидаза Streptococcus sp. ние желатиновых пленок (при реге- нерации серебра); мягчение мяса; производство рыбных гидролизатов Применяется в медицине при pH 8,0 Производится компанией ++
Стрептокиназа Streptococcus sp. Lederle Профнбрииолизцн
КЗ
о
Продолжение табл. 4.1
Фермент
Источник фермента
Применение
Примечания
Степень
промыш-
ленной
важнос-
ти
Некоторые другие промышленно важные ферменты
Глюкозоизомераза Lactobacillus brevis, Ba- cillus coagulans, Arth- robacter simplex, Acti- noplanes missourensis Превращение глюкозы во фруктозу Производится компаниями Novo, ICI, Gist Brocades +++
Пенициллиназа B. subtilis, Bacillus ce- reus Разрушение пенициллина Производится компаниями Takamine, Schenley +
Глюкозооксидаза Aspergillus niger (Dee 0, Dee G) Связывание кислорода или деструк- ция глюкозы в различных пищевых продуктах; производство яичного порошка Производится Takamine компанией +
Penicillium chrysogenum Определение глюкозы Производится Nagase Со. компанией +
Гиалуронидаза Липаза Животные, бактерии Поджелудочная железа, плесень (Rhizopus) Применяется в медицине Способствующее пищеварению сред- ство; улучшает вкусовые качест- ва молочных продуктов + + +
Цитохром с Дрожжи (Candida) Применяется в медицине Производится Sankyo Со. компанией +
Каталаза Кератиназа Streptomyces fradiae Стерилизация молока Обесволашивание шкур животных Производится Merck Со. компанией +
Примечания
Фермент
Применение
Источник фермента
Степень
промыш-
ленной
важнос-
ти
5'-Фосфодиэстер аза Penicillium citrinum, S. griseus, В. subtilis Производство 5'-нуклеотидов (ино- зиновой и гуаниловой кислот) Производится компаниями —Ы- Yamasa Со., Takeda Со.
Аденилатдеаминаза A. oryzae Превращение АМР в IMP Содержится в такадиастазе -f-
Микробный реннин Mucor sp. Производство сыра Производится компанией -j—f- Meito Sangyo Со.
Нарингиназа Лакказа Целлюлаза Инвертаза Aspergillus niger Coriolus versicolor Trichoderma koningi Trichoderma viride Saccharomyces cerevisiae Устранение горького привкуса соков цитрусовых Высушивание лаков Способствующее пищеварению сред- ство Гидролиз целлюлозы Предотвращение кристаллизации са- хара в производстве кондитерских изделий; производство шоколада, высококачественной мелассы Производится компанией Rohm and Haas Максимальная активность при pH 4,6 Смесь ферментов
Пектиназа Sclerotina liber Una Coniothyrium, diplodiella, Aspergillus oryzae, A. niger, A. flavus Осветление и повышение выхода со- ков Разрушение пектина, концентриро- вание кофе Производится компаниями —h+ Sankyo (скраза), Rohm and Haas (пектипол), Ta- kamine, Haas Delete Ta- kamine Производится компаниями Takamine (пектиназа-кла- раза), I. G. Farbcn (фильтрагол)
а Из работы: Arima К., Microbial Enzyme Production, in Global Impacts of Applied Microbiology, Starr M. P. (cd.), pp. 278—279, John
Wiley and Sons, Inc., New York, 1964.
212
Глава 4
Хотя все применяющиеся в настоящее время ферменты по-
лучают из природных источников, в настоящей главе мы рас-
смотрим использование ферментов только вне живого организ-
ма. Все биологические катализаторы подразделяют на две ка-
тегории— внеклеточные и внутриклеточные ферменты. К первой
категории относятся ферменты, выделяемые клеткой в среду, где
они расщепляют питательные полимерные вещества до низко-
молекулярных соединений, которые могут проникать в клетку
через клеточную стенку. Внутриклеточные ферменты в нормаль-
ных условиях сконцентрированы в объеме клетки и в среду не
транспортируются; для их выделения необходимо дезинтегриро-
вать клетки путем размола, растирания, лизиса или каким-либо
другим способом.
Для некоторых областей применения ферментов необходимы
относительно чистые препараты. Например, глюкозооксидаза,
применяющаяся для обессахаривания яиц (в производстве яич-
ного порошка) не должна содержать протеолитических фермен-
тов, а протеазы, вводимые внутримышечно домашнему скоту
перед забоем для мягчения мяса не должны содержать никаких
соединений, которые могли бы вызвать какую-либо сильную фи-
зиологическую реакцию. Относительно чистые ферменты приме-
няются в клинической диагностике и в процессах, связанных с
производством и обработкой пищевых продуктов.
Кинетика ферментативных реакций, как правило, изучалась
на наиболее чистых препаратах ферментов. Как мы уже указы-
вали в гл. 3, в ходе таких исследований, кроме того, применя-
лось минимальное число субстратов (лучше всего один, если
только это возможно), а также строго контролировались пара-
метры среды в отношении содержания активаторов (Са2+, M.g2+
и т. п.), кофакторов и ингибиторов. Результаты таких исследо-
ваний наиболее достоверно отражают кинетику ферментативных
реакций.
В то же время многие из применяемых в промышленности
препаратов ферментов очищены в гораздо меньшей степени.
Как правило, они содержат ряд ферментов с различными ката-
литическими свойствами. Кроме того, в большинстве случаев
эти препараты используются для обработки таких материалов,
которые даже отдаленно не напоминают чистый субстрат или
синтетическую среду строго определенного состава типа рас-
смотренных в гл. 3. Следует отметить, что одновременное дей-
ствие нескольких ферментов может быть более эффективным,
чем последовательная обработка рядом индивидуальных фер-
ментов. Несмотря на все эти сложности, такие препараты фер-
ментов с кинетической точки зрения все же намного проще ин-
тактного живого организма, из которого они были выделены.
В этой связи представляется целесообразным сначала рассмот-
Применение реакций, катализируемых ферментами 213
реть применяемые в промышленности ферментные препараты,
а затем перейти к анализу путей клеточного метаболизма и к
изучению промышленного применения биохимической техно-
логии.
4.1. Применение гидролитических ферментов
Катализируемые гидролитическими ферментами реакции ле-
жат в основе не только таких очевидных макроскопических про-
цессов, как порча пищевых продуктов, ожижение крахмала или
обработка сточных вод, но и в химии процессов дозревания не-
спелых плодов, лизисе (автолизе) отмерших клеток, созревания
мяса, производства сыра, предотвращении образования осадка
в пиве, текстурировании кондитерских изделий, обработке ран
или расшлихтовке текстиля. Настоящий раздел будет посвящен
изучению ферментов, применяемых в указанных и других про-
изводствах, а также источников их выделения.
В табл. 4.2 приведена общая классификация важнейших гид-
ролитических ферментов. Три основные группы гидролаз рас-
щепляют сложноэфирные, гликозидные и различные связи с
участием атома азота соответственно. Широко применяется и бо-
лее детальная классификация ферментов внутри каждой из этих
трех групп. Для наших целей достаточно указать, что, напри-
мер, некоторые ферменты катализируют гидролиз самых разно-
образных гликозидных связей, а другие могут расщеплять толь-
ко один-единственный олигомер глюкозы. Таким образом, на-
звание фермента само по себе еще не определяет его субстрат-
ную специфичность.
Здесь следует еще раз подчеркнуть, что названия большинст-
ва ферментов связаны с катализируемыми ими реакциями, а не
с их химической структурой*. Поскольку в этом случае не су-
ществует правила «один фермент — одна реакция», то катали-
зирующие одну и ту же реакцию ферменты, выделенные из раз-
личных растительных или животных источников, не всегда будут
иметь одну и ту же структуру, и поэтому кинетика соответст-
вующих реакций также может быть различной. Отсюда следу-
ет, что максимальная скорость реакции, константа Михаэлиса,
pH оптимума стабильности и активности и другие свойства фер-
мента будут определяться его природой и, следовательно, ис-
точником его выделения.
Многие гидролазы компартментализованы в тех или иных
* Классификация ферментов в соответствии с рекомендациями Комиссии
по ферментам приведена в табл. 3.1. Более подробно вопросы классификации
ферментов изложены в книге: Диксон М., Уэбб Э., Ферменты, т. 1—3, гл. 5,
М.: Мир, 1982.
214
Глава 4
Таблица 4.2. Гидролитические ферменты3
Фермент
Субстрат
Продукты гидролиза
Эстеразы:
Липазы Глицериды (жиры) Глицерин и жирные кис- лоты
Фосфатазы:
Лецитиназа Лецитин Холин, Н3РО4 и жирные кислоты
Пектинэстераза Метиловый эфир пекти- Метанол и полигалакту-
на роновая кислота
Карбогидразы:
Фруктозидазы Сахароза Фруктоза и глюкоза
а-Глюкозидазы Мальтоза Глюкоза
(мальтаза)
^-Глюкозидазы Целлобиоза Глюкоза
(целлобиаза)
^-Г алактозидазы Лактоза Галактоза и глюкоза
(лактаза)
Амилазы Крахмал Мальтоза или глюкоза и мальтоолигосахари- ДЫ
Целлюлаза Целлюлоза Целлобиоза
Цитаза Простые сахара
Полигалактуроиаза Полигалактуроновая кислота Галактуроновая кислота
Ферменты, гидролизую- щие азотсодержащие со- единения:
Протеиназы Белки Полипептиды
Полипептидазы Белки Аминокислоты
Дезаминазы:
Уреаза Мочевина СО2 и NH3
Аспарагиназа Аспарагин Аспарагиновая кислота в NH3
Деаминазы Аминокислоты Карбоновые кислоты и
NH3
а Weiser Н. Н., Practical Food Microbiology and Technology, p. 37, Avi Publishing
Co., Westport, Conn., 1962.
структурных элементах клетки, отделенных от цитоплазмы мем-
бранами. Очевидно, что такая локализация гидролаз защищает
важные биополимеры цитоплазмы от деструкции. Грамположи-
тельные бактерии выделяют в среду множество гидролаз.
У грамотрицательных бактерий надежным хранилищем для раз-
личных гидролаз служит ограниченное двумя мембранами пери-
плазматическое пространство наружной оболочки. В клетках
Применение реакций, катализируемых ферментами
215
эукариот гидролазы могут локализоваться в ограниченных мем-
браной особых органоидах — лизосомах, в периплазме (в мик-
роорганизмах типа дрожжей) или выделяться в среду. Большин-
ство используемых в промышленности гидролитических фермен-
тов представляет собой внеклеточные продукты жизнедеятель-
ности микроорганизмов.
В то же время некоторые гидролазы обнаружены и в цито-
плазме. Здесь они участвуют в метаболических циклах и тем
самым выполняют важную функцию, помогая клетке полностью
использовать ее внутренние ресурсы. Внутриклеточные гидрола-
зы играют большую роль в методах генетической инженерии
(гл. 6).
Поскольку вода является универсальным субстратом, кон-
центрация которого составляет около 55 М, и поскольку гидро-
литические ферменты обычно катализируют реакции деграда-
ции, например превращение крахмала в сахара, белков в поли-
пептиды, липидов в глицерин, жирные кислоты и фосфат, то ос-
новой для последующего изложения нам послужат различные
типы полимерных субстратов. Следует отметить, однако, что в
некоторых разработанных недавно методах синтеза полипепти-
дов в средах с низким содержанием воды природные протеазы
использовались в качестве конденсирующих агентов. Очевидно,
в специфических условиях протеазы могут иногда катализиро-
вать и обратную реакцию синтеза.
4.1.1. Гидролиз крахмала и целлюлозы
Находящие широкое применение амилазы представляют со-
бой ферменты, которые гидролизуют гликозидные связи в крах-
мале и родственных полимерах глюкозы. Чтобы понять различие
между двумя основными типами амилаз, мы должны вспом-
нить, что крахмал состоит из линейных полимеров глюкозы, на-
зываемых амилозой, и разветвленного полимера (амилопекти-
на). Последний значительно более растворим по сравнению с
амилозой и способен резко повышать вязкость растворов крах-
мала. При обработке а-амилазой вязкость растворов снижается
за счет неупорядоченного разрыва любых сс-1,4-гликозидных
связей; по этой причине ос-амилазу часто называют разжижаю-
щим крахмал ферментом*. Напротив, ^-амилаза может атако-
вать а-1,4-связи только на невосстанавливающих концевых
участках полимерной цепи, поэтому при гидролизе линейных по-
* По-видимому, а-амилаза неспецифично разрывает гликозидные связи
только в субстратах с достаточно длинной полимерной цепью; в случае более
коротких олигомеров наблюдается известная специфичность.
216
Глава 4
лимеров этим ферментом всегда образуется мальтоза. По этой
причине {3-амилазу называют также осахаривающим ферментом.
При обработке крахмала {3-амилазой в конечном итоге образу-
ется смесь мальтозы и декстринов, представляющих собой фраг-
менты молекул крахмала с концевыми 1,6-связями. Эти связи
{3-амилаза не гидролизует.
Другой осахаривающий фермент, амилоглюкозидаза (имею-
щая много других названий, в том числе глюкоамилаза), ата-
кует прежде всего невосстанавливающие концевые а-1,4-связи
крахмала, гликогена, декстринов и мальтозы; амилоглюкозида-
за гидролизует также а-1,6-связи, но со значительно меньшей
скоростью. Если целью процесса является получение глюкозы,
а не мальтозы, то применяют смесь ферментов или последова-
тельную обработку а-амилазой и глюкоамилазой; такие процес-
сы применяются на спиртоводочных заводах (в отличие от пи-
воваренных предприятий), в производстве концентрированных
растворов глюкозы (кукурузной патоки) и кристаллической
глюкозы. Подсчитано, что в 1971 г. в США таким методом было
получено 0,61 млрд, кг глюкозы. Использование амилаз в про-
изводстве глюкозы и других областях (табл. 4.3) обусловливает
очень большую практическую значимость этих ферментов. От-
носительные количества а- и {3-амилаз, применяемые в каждом
конкретном случае, определяются конечной целью процесса.
Амилазы получают из самых различных источников. Это и
неудивительно, поскольку крахмал представляет собой обычную
форму резервного углевода для самых различных форм жизни.
Амилазы продуцируются рядом бактерий и плесеней; в качест-
ве примера практически важного фермента можно привести
амилазу из Clostridium acetobutylicum — микроорганизма,
участвующего в микробиологическом превращении полисахари-
дов в бутанол и ацетон. Препараты амилазы, используемые че-
ловеком для пищевых целей, обычно получают из зерновых
культур, в первую очередь из ячменя, пшеницы, ржи, овса, ку-
курузы, сорго и риса. В амилазах зерна отношение осахариваю-
щей и разжижающей ферментативных активностей зависит не
только от типа растения, но и от состояния зерен, в первую
очередь от того, проросли ли зерна или нет. В производстве со-
лода (размягченных проросших зерен ячменя), применяемого в
пивоварении, непроросшие зерна ячменя хранят в условиях вы-
сокой влажности и соответствующей температуры, способствую-
щих их быстрому прорастанию, причем одновременно в них воз-
растает содержание а-амилазы. Затем проросшие зерна мед-
ленно высушивают в печи; в сухом препарате амилазная актив-
ность не проявляется, но ферменты не подвергаются необрати-
мой инактивации. Обладающий огромной гидролитической ак-
тивностью высушенный и измельченный солод используют затем
Применение реакций, катализируемых ферментами
217
Таблица 4.3. Области наиболее широкого применения амилазных
ферментов®
Производство
Применение
Производство глюкозы и
патоки
Пивоварение
Хлебопечение
Производство фруктовых
соков
Бумажная промышлен-
ность
Текстильная промышлен-
ность
Кондитерская промыш-
ленность
Крупномасштабное производство сахаров путем
полного или частичного гидролиза крахмала
амилоглюкозидазой или а-амилазой
Превращение размолотых зерен крахмала в маль-
тозу (субстрат дисахаридной природы, пригод-
ный для дальнейшего дрожжевого брожения)
Заквашивание теста: превращение части крахма-
ла в дисахариды, образующие при последующем
ферментативном распаде Диоксид углерода
Осветление путем гидролиза нерастворимых
фракций крахмала
Снижение вязкости раствора крахмала под дей-
ствием а-амилазы, предшествующее нанесению
раствора на целлюлозную основу (бумага с ре-
гулируемой массой)
Шлихтование: а-амилаза снижает вязкость рас-
творов крахмала, применяющихся затем для
усиления волокон основы
Расшлихтовка: обработка а- и ^-амилазами уда-
ляет шлихт из тканого материала; это обеспечи-
вает равномерность последующего окрашива-
ния и необходимую текстуру ткани
Производство кондитерских изделий необходимой
консистенции
а Из работы: Weiser Н. Н., Practical Food Microbiology and Technology, p. 37,
Avi Publishing Co., Westport, Conn., 1962.
для превращения крахмала в сахара, которые впоследствии мо-
гут быть переработаны путем дрожжевого брожения.
Другие перечисленные в табл. 4.1 карбогидразы также рас-
щепляют гликозидные связи. Сообщалось, например, что комби-
нированное применение пуллуланазы, которая селективно рас-
щепляет 1,6-гликозидные связи боковых цепей, и амилазы повы-
днает выход глюкозы из крахмала. В производстве мороженого
обычно используют лактазу, расщепляющую лактозу на более
сладкие моносахариды глюкозу и галактозу. Родственный фер-
мент инвертаза гидролизует сахарозу и полисахариды, содержа-
щие p-D-фруктофуранозильные связи. Свое название этот фер-
мент получил после того, как было установлено, что гидролиз
сахарозы в растворах и образование глюкозо-фруктозного рас-
твора сопровождается изменением направления вращения плос-
-Кости поляризации света. Частично или полностью гидролизо-
218
Глава 4
ванный раствор сахарозы обладает двумя ценными для конди-
терской промышленности качествами — он несколько более сла-
док, чем исходный раствор, и не кристаллизуется при упарива-
нии до значительно более высоких концентраций.
Изучению ферментативного гидролиза целлюлозы посвящено
очень много работ, проводившихся в различных странах. Преж-
де чем приступить к изложению этой темы, следует подчеркнуть,
что ферментные препараты, используемые для деполимеризации
целлюлозы и обычно называемые целлюлазой, представляют
собой сложную смесь многих ферментов. Более того, природа
входящих в состав целлюлазы ферментов и их относительные
количества зависят от типа микроорганизма, из которого была
выделена целлюлаза, а в некоторых случаях и от процесса по-
лучения ферментного препарата. Как мы уже упоминали в гл. 2
при обсуждении структуры лигноцеллюлозы, вся биомасса и
промышленные отходы различных производств различаются по
ряду свойств, в том числе по степени кристалличности и удель-
ной поверхности, а также по химическому составу. Некоторые
методы предварительной обработки позволяют модифицировать
эти субстраты и тем самым снизить их устойчивость к гидро-
лизу. Таким образом, скорость гидролиза целлюлозных мате-
риалов и выходы продуктов в каждом конкретном процессе за-
висят от совокупности нескольких факторов: во-первых, от при-
роды и свойств субстрата, во-вторых, от результатов предвари-
тельной обработки и, в-третьих, от степени активности и специ-
фичности индивидуальных ферментов, входящих в состав цел-
люлазы.
В настоящее время наиболее тщательно изучены и охаракте-
ризованы целлюлазные системы, продуцируемые грибами
Trichoderma. Как показано на рис. 4.1, в состав этих систем
входят три типа ферментов, действующих на различные суб-
страты и образующих различные продукты реакции. На этом
же рисунке указаны пути ингибирования индивидуальных фер-
ментов по принципу обратной связи. Кинетика каждой стадии
этой сложной системы реакций подчиняется уравнению Михаэ-
лиса— Ментен со строго конкурентным или неконкурентным ин-
гибированием. Целлюлазы с различными свойствами и активно-
стями продуцируют и многие другие микроорганизмы, в том чис-
ле плесени Fusarium solani, Aspergillus niger, Penicillium funi-
colsum, Sporotrichum pulverulentum, Cellulomonas sp., Clostri-
dium thermocellum и Clostridium thermosaccharolyticum.
В табл. 4.4 приведены дополнительные сведения о целлюлаз-
ной системе из Т. viride. Обратите внимание на то, что для опре-
деления специфических активностей применяются различные
«стандартные» субстраты. Немаловажны и размеры молекул
ферментов, сравнимые с размерами микрофибрилл природной
Применение реакций, катализируемых ферментами
219
целлюлозы. Скорость гидролиза зависит главным образом от
степени кристалличности целлюлозы. Соответствующие выраже-
ния, описывающие кинетику гидролиза с учетом степени кри-
сталличности, мы опишем ниже, в примере 4.1; сейчас же вкрат-
це обсудим химические и физические методы предварительной
обработки лигноцеллюлозных материалов.
РИС. 4.1. Схема взаимодействия субстратов, целлюлаз и продуктов реакции
в ходе биологической деградации целлюлозы (-> основная реакция;---------> по-
бочная реакция;------> эффекты ингибирования). [Воспроизведено из статьи:
Lee У. Н., Fan. L. Т., Properties and Mode of Action of Ce lulase, in Advances in
Biochemical Engineering, Fiechter A. (ed.), 17, 101 (1980).]
Изучение методов химической обработки древесины дает
представление о влиянии лигнина на устойчивость целлюлозных
материалов к ферментативному гидролизу. В бумажной про-
мышленности для растворения лигнинов и пектинов измельчен-
ную древесину подвергают сульфитной варке (обработке вод-
ным раствором гидросульфита, содержащим свободный SO2)
или сульфатной варке (обработке водным раствором NaOH и
Na2S) при повышенных температуре и давлении (крафт-про-
цесс). Путем такой обработки четырех различных пород древе-
сины в различных условиях были получены целлюлозные мас-
сы с различным содержанием лигнина, которые затем были об-
работаны серной кислотой для гидролиза веществ целлюлозной
природы. Результаты этого эксперимента (рис. 4.2) наглядно
220
Глава 4
показывают, что способность целлюлозы к гидролизу в первую
очередь зависит от количества оставшегося лигнина. Для раз-
рушения лигнина применяют и другие методы, в том числе обра-
ботку газообразным SO2, минеральными кислотами и специфи-
ческими ферментами, которые продуцируются рядом грибов, на-
пример Sporotrichum pulverulentum и Pleurotus ostreatus.
Структуру целлюлозы, в том числе такие ее характеристики,
как степень кристалличности, удельная поверхность, степень по-
лимеризации, можно изменить с помощью предварительной об-
Таблица 4.4. Свойства различных типов ферментов в целлюлазной
системе Т. viride
Свойства ферментов Типы ферментов
Эндо-р-1,4- глюкаиаза Экзо-|3-1,4-цел- лобиогидролаза Р-Глюкозндаза (целлобиаза)
Стандартный субстрат Карбоксиме- тилцеллюло- за (КМЦ) Авицельа Целлобиоза
Ингибиторы Целлобиоза Глюкоза (целлобиоза) ?
Ориентировочная молеку- лярная масса 12 500—52 000 46 000 76 000
Расчетный диаметр сфери- ческой молекулы, А 34—64 62 76
а Выпускаемая промышленностью кристаллическая целлюлоза, прошедшая стадию
кислотной обработки.
работки, например путем размола на шаровой или роликовой
мельнице, 'у-облучения, пиролиза, обработки кислыми или ще-
лочными реагентами. В качестве примера рассмотрим кинетику
изменения степени кристалличности целлюлозы при кислотной
обработке древесины; сведения об изменении других свойств
целлюлозы читатель может найти в приведенной в конце главы
литературе.
Как показано на рис. 4.3, а, влияние предварительной обра-
ботки на кристалличность лучше всего контролировать методом
рентгеноструктурного анализа. Последний позволяет определить
параметр, называемый индексом кристалличности (CrI) и рас-
считываемый по эмпирическим формулам; в нашем случае
(рис. 4.3, а) эта формула имеет следующий вид:
Сг1(%) =
I(am)
1(002) .
.100%
(4.1>
1 —
где I(am) (am — аморфный) и 1(002)—интенсивности дифрак-
ции при 20=18,5 и 22,5° соответственно. Значения CrI не явля-
Применение реакций, катализируемых ферментами
221
ются точной мерой содержания кристаллической фракции; тем
не менее индекс CrI удобен для оценки изменения степени кри-
сталличности и средних размеров кристаллитов в различных
целлюлозных материалах. Приведенные на рис. 4.3, а экспери-
ментальные данные наглядно показывают, что при многократной
РИС. 4.2. Результаты экспериментального изучения влияния степени делигни-
фикации на гидролизуемость целлюлозы из четырех различных пород древе-
сины. [Воспроизведено с разрешения из статьи: Baker A. J., Effect of Lignin
on the in vitro Digestibility of Wood Pulp, J. Anim. Sci., 37, 768 (1973).]
обработке на роликовой мельнице степень кристалличности цел-
люлозы снижается.
Ранее, при обсуждении структуры целлюлозы и активности
целлюлазных ферментных систем, мы уже упоминали, что ско-
рость гидролиза аморфных и паракристаллических участков
целлюлозы должна быть выше скорости гидролиза кристалли-
ческой целлюлозы. Отсюда следует, что снижение степени кри-
сталличности целлюлозы должно сопровождаться повышением
скорости ее ферментативного гидролиза. Действительно, такая
зависимость была обнаружена, в частности, при изучении гидро-
лиза тех же целлюлозных материалов, которые исследовались
РИС. 4.3. Изменение характера дифракции рентгеновского излучения целлю-
лозой типа авицель после многократных пропусканий через двухроликовую
мельницу (на кривых указано число пропусканий) (а); после обработки цел-
люлазой из Т. reesei (на кривых указано время инкубирования в часах) (б).
Для расчета индекса кристалличности CrI использовались интенсивности при
углах дифракции 20 = 18,5 и 22,5°. [Воспроизведено с разрешения: а) из
статьи Ryu D. D. Y„ Lee S. В., Tassinari Т„ Масу С., Effect of Compression
Milling on Cellulose Structure and on Enzymatic Hydrolysis Kinetics; Biotech.
Bioeng, 24, 1047 (1982); б) из статьи: Lee S. B„ K.im I. H„ Ryu D. D. Y„
Taguchi H., Structural Properties of Cellulose and Cellulase Reaction Mecha-
nism; Biotech. Bioeng., 25, 33 (1983).]
Применение реакций, катализируемых ферментами
223
в эксперименте, изображенном на рис. 4.3, а, под действием
суммарного препарата целлюлаз из Trichoderma reesei MCG-77
(табл. 4.5). В табл. 4.5 приведены кажущиеся средние значения
параметров в обычном уравнении Михаэлиса — Ментен; их оп-
ределяли, допустив, что при измерении начальной скорости ре-
акции целлобиоза в смеси отсутствовала.
Расчеты изменения скорости ферментативного гидролиза
целлюлозных материалов во времени осложняются тем обстоя-
Таблица 4.5. Влияние числа пропусканий через роликовую мельницу
на кристаллическую структуру целлюлозы типа авицель и средние
кинетические параметры ее гидролиза под действием целлюлазы
из Т. reeseia
Число пропусканий CrI % Размер крн-° uappmax, Kapp/n. oappmax/Каррпг> ’ сталлитов, А мкг/(мл-мнн) мг/мл ч—1
0 81 38
4 71 31 19,6 31,3 0,038
10 61 21 22,2 20,0 0,067
20 37 10 23,1 14,8 0,110
35 17 7 19,3 7,7 0,150
а Данные из статьи: Ryu. D. D. Y„ Lee S. B., Tassinari T., Macy C., Effect of
Compression Milling on Biotech. Bioeng., 24, 1047 Cellulose (1982). Structure and on Enzymatic Hydrolysis Kinetics,
тельством, что в ходе таких процессов меняется также относи-
тельное содержание кристаллической и аморфной фракций суб-
страта (рис. 4.3,6). В приведенном ниже примере мы рассмот-
рим модифицирование принципов моделирования, изложенных в
гл. 3, в применении к системам такого рода.
Пример 4.1. Влияние степени кристалличности целлюлозы на скорость ее
ферментативного гидролиза*. Для изучения гидролиза целлюлозы фермент-
ными системами типа целлюлазы разработано несколько математических мо-
делей кинетики этих процессов. В одной из них наибольшее внимание уде-
ляется влиянию кристалличности; в этом подходе принимается, что фермен-
ты целлюлазы (их сумма обозначается здесь символом Е) адсорбируются
на целлюлозе в состоянии £*:
Е
kad^
<kd
Е*
(4П1.1)
Допуская, что адсорбция фермента носит равновесный характер и что кон-
центрация фермента в растворе достаточно низка (на практике £q<0,5 мг
белка в 1 мл реакционной смеси), получим
e*=tKDe (4П1.2)
* Ryu D. D. Y., Lee S. B., Tassinari T.t Macy C., Effect of Compression
Milling on Cellulose Structure and on Enzymatic Hydrolysis Kinetics, Biotech.
Bioeng., 24, 1047 (1982).
224
Глава 4
Описание модели завершается еще двумя допущениями, согласно которым
адсорбированный фермент катализирует гидролиз аморфной (Sa) и кристал-
лической (Sc) целлюлозы в соответствии со следующими уравнениями парал-
лельно осуществляющихся реакций:
kia kza
E*4-Se E*Se ------------> Е* + Р (4П1.3)
й-1а
E* + Sc <=> E*SC--------> Е*+Р (4П1.4)
k-ic
причем адсорбированная и свободная целлюлазы связываются с инертным
материалом целлюлозного субстрата, а также с продуктом реакции соответ-
ственно:
kix
E* + Sx ;<=> E*SX
kli
E+P EP
k-ii
(4П1.5)
(4П1.6)
Применив ко всем фермент-субстратным комплексам приближение квази-
стационарного состояния и суммируя концентрации всех форм фермента, по-
лучим следующее выражение для начальной скорости гидролиза чистой цел-
люлозы (sx=0) при низкой концентрации фермента и в отсутствие продукта
реакции (р=0):
__ dS _ ^max so
v dt s=so,P=O,sx=O- ^PP + s0 (4nL7)
Здесь s (s=so4-Sc) —общая концентрация целлюлозы. Зависимость кажущих-
ся максимальной скорости и константы Михаэлиса от начального индекса
кристалличности Crlo [CrIo=sco/(Sao+sCo)] и констант скоростей элементарных
реакций можно выразить следующим образом:
___ ^шах,a (1 Сг10) + Ощах,сСг10 (KalKo)
Umax= 1 - Сг10 + (Ка/Кс) СП0 (4П1.8)
%т= 1 СП IK <СН~ (4Ш>9)
1 Сг10 -]- (ixalixc)
тде
^max,a = ^2a^0 И Vmax.c = ^2с<?0
Ka=kl + KD)^k-ia + k2a')/KDkia (4П1.10)
v iv ____ (k-ia + ^аа)
Лв/Лр_ kla(k_lc+k2c)
Выражения (4П1.8) и (4П1.9) свидетельствуют о зависимости начальной
•скорости реакции от начальной кристалличности субстрата Сг10, причем кон-
станта скорости реакции псевдопервого порядка omaxapp/^Cmapp и величина,
обратная кажущейся константе Михаэлиса, 1/Ктарр> линейно зависят от
Сг10; это дает возможность проверить соответствие описанной модели экспе-
риментальным данным. Построенные на основе результатов эксперимента
Применение реакций, катализируемых ферментами
225
О 50 ЮО
Степень кристалличности CrIQ7 %
РИС. 4П1.1. Экспериментальное определение зависимости кажущихся кинети-
ческих параметров цтахарр и Ктарр от начальной степени кристалличности суб-
страта Сг10 для авицеля (А, •), хлопка (X, О) и целлюлозы типа солка-
флок (С, □). Цифры указывают число предварительных пропусканий субстра-
та через роликовую мельницу. Характер дифракции рентгеновского излучения
субстратами представлен на рис. 4.3,а. [Воспроизведено из статьи: Ryu D. D. Y.,
Lee S. В., Tassinari Т., Масу С., Effect of Compression Milling on Cellulose
Structure and on Enzyme Hydrolysis Kinetics, Biotech. Bioeng., 24, 1047
15-537
226
Глава 4
графики зависимостей указанных кинетических параметров от начальной
степени кристалличности субстрата для ряда образцов целлюлозы приведены
на рис. 4П1.1. Большинство данных хорошо согласуется с описанной мо-
делью; наибольшее отклонение обнаружено в случае константы скорости ре-
акции псевдопервого порядка для целлюлозы типа солка-флок.
Эти результаты весьма поучительны в том отношении, что они могут
служить примером вывода полезных математических выражений для интер-
претации кинетических данных на базе обоснованной гипотезы о природе
происходящих в системе превращений. Приведенное выше выражение для
начальной скорости несколько громоздко, однако уже из механизма про-
цесса очевидно, что начальная скорость гидролиза должна зависеть от всех
параметров, входящих в состав этого уравнения. Более того, соответствую-
щее выражение для зависимости скорости реакции гидролиза целлюлозы
в реакторе периодического действия от времени будет, очевидно, еще слож-
нее, поскольку в нем необходимо учитывать и ингибирование процесса про-
дуктом реакции [см. уравнение (4.П1.6)]. Наконец, при расчете изменения
концентрации субстрата во времени следует учитывать и тот факт, что суб-
страт представляет собой смесь кристаллической и аморфной форм, гидро-
лизующихся с различными скоростями. Следовательно, описывающее кинети-
ку гидролиза выражение должно отражать два параллельных процесса с дву-
мя различными субстратами, причем эти процессы взаимосвязаны, поскольку
относительные количества аморфной и кристаллической целлюлозы также не
постоянны во времени, о чем свидетельствует изменение индекса кристал-
личности.
Этот пример иллюстрирует только некоторые из тех многочисленных
затруднений, которые встречаются при моделировании биологических субстра-
тов и катализаторов. Как мы еще не раз увидим в последующих главах и
разделах, общий принцип создания моделей подобных сложных систем сво-
дится к выбору такой биохимической или механистической схемы и соответст-
вующих математических выражений, которые отражали бы изменение только
ключевых переменных, представляющих основной интерес в конкретном ана-
лизе. Так, в нашем примере мы рассматривали некоторый идеализированный
субстрат целлюлозной природы. Такое допущение позволяет оценить влияние
кристалличности на ход гидролиза, определить соответствующие кинетические
параметры и рассчитать закономерность изменения кристалличности субстра-
та в реакторе периодического действия. Все приведенные экспериментальные
данные были получены с использованием одного и того же целлюлазного
ферментного комплекса, поэтому описанная модель вообще не учитывает
влияние индивидуальных ферментов, входящих в состав комплекса, и их от-
носительное содержание в комплексе. Очевидно, при сравнении с данными,
полученными в ходе изучения целлюлаз из других источников, возникнет
необходимость в учете относительного содержания различных ферментов
в комплексе и их активностей. В идеальном варианте в результате ряда тща-
тельных экспериментальных исследований и разработки соответствующих ма-
тематических моделей типа рассмотренной выше мы в конце концов смогли
бы получить разностороннюю информацию о происходящих в системе цел-
люлоза— целлюлаза превращениях и на этой основе разработать некоторую
общую модель, которая более полно отражала бы свойства субстратов и
ферментов этой системы.
В завершение этого обзора, посвященного проблеме исполь-
зования целлюлозы, следует упомянуть, что существуют и дру-
гие физические и химические методы деградации целлюлозы.
Так, в последние годы были разработаны усовершенствованные
процессы кислотного гидролиза целлюлозы, отличающиеся при-
менением более высоких температур и сокращенным временем
Применение реакций, катализируемых ферментами
227
обработки и позволяющие получать необходимые продукты де-
градации при минимальном количестве побочных веществ. Дру-
гой многообещающий метод переработки целлюлозы базируется
на ее пиролитической деградации. Изучаются, разрабатываются
или уже применяются также методы переработки лигноцеллю-
лозы, основанные на шнековой экструзии, обработке гетероген-
ными катализаторами, газификации в псевдоожиженном слое и
на самом старом способе переработки путем сжигания.
4.1.2. Протеолитические ферменты
Существует множество ферментов, селективно атакующих
азотсодержащие соединения, особенно белки. Эти ферменты на-
ходят и разнообразные практические применения. Как и амила-
зы, протеазы разделяются на ферменты, гидролизующие конце-
вые группы (экзопептидазы), и ферменты, расщепляющие внут-
ренние пептидные связи (эндопептидазы).
Поскольку ферменты, эти важнейшие катализаторы всех жи-
вых организмов, сами являются белками, неудивительно, что
протеолитические ферменты часто синтезируются сначала в не-
активной форме. Неактивная форма фермента или хранится в
клетке, или транспортируется от места ее синтеза к тому цент-
ру, где этот фермент необходим в активной форме; в частности,
в клетке транспортируются неактивные формы пепсина, трипси-
на, химотрипсина и карбоксипептидазы. Активация этих протео-
литических ферментов осуществляется по одному из двух путей
(рис. 4.4). Интересно отметить, что активация пепсина и трип-
сина происходит автокаталитически: неактивный предшествен-
ник фермента является субстратом для активной формы, поэто-
му в результате реакции образуется большее количество актив-
ной формы фермента. Интересен и тот факт, что после превра-
щения неактивного трипсина или пепсина в активную форму
дальнейшее автокаталитическое расщепление фермента не наб-
людается. Для активации второй группы ферментов, обычно
экзопептидаз, необходимы ионы одного или нескольких опреде-
ленных металлов. Стандартный метод определения потребности
ферментов и других компонентов клетки в ионах металлов за-
ключается в изучении изменения активности раствора фермента
в процессе диализа.
В промышленных масштабах протеазы выделяют из организ-
мов животных (из поджелудочной железы), высших растений
(из соков и латексов), а также из дрожжей, плесеней и бакте-
рий. Некоторые из протеаз и области их применения перечисле-
ны в табл. 4.1.
Свободные протеазы применяются в основном в производст-
ве моющих средств, в химической чистке, при мягчении мяса,
15*
228
Глава 4
в сыроделии (только реннин), дублении, извлечении серебра из
фотографических пленок и бумаг, производстве препаратов,
способствующих пищеварению, а также в медицине при лечении
воспалительных процессов и вирулентных ран. В качестве доба-
вок к моющим средствам ферменты применялись начиная уже
с 1913 г., однако особенно резкий рост потребления протеаз в
производстве детергентов наблюдался в конце 1960-х гг. Фер-
а) Из предшественников:
Н+, пепсин
Пепсиноген --------> Пепсин + X
мол. масса мол. масса мол. масса
42 000 38 000 -4000
Трипсиноген
мол. масса
3*000
трипсин
------------>
энтерокиназа
Трипсин
мол. масса
-34 000
трипсин
Химотрипсиноген ---------->• Химотрипсин
трипсин
Прокарбоксипептидаза —--------> Карбоксипептидаза
б) В присутствии ионов металлов:
Глицилглициндипептидаза (неактивная) + Со2+ + Mg2+ ---->
----> Глицилглициндипептидаза (активная)
Лейцинаминопептидаза (неактивная) + Мп2+ (или Mg2+) ----->
----> Лейцинаминопептидаза (активная)
РИС. 4.4. Активация протеолитических ферментов, а—образование активных
ферментов из предшественников; б — образование активных ферментов в при-
сутствии ионов металлов.
менты, способствующие удалению загрязнений белковой приро-
ды, представляют собой смесь нейтральной и щелочной бакте-
риальных протеаз, активных в интервале pH от 6,5 до 10,0 при
температурах от 30 до 60 °C. Наибольшего уровня производство
ферментов для этих целей достигло в 1969 г., когда с фермент-
ными добавками выпускалось 30—75% всех моющих средств в
Западной Европе и 40% в США. Затем, однако, Федеральная
комиссия по торговле США высказала опасение, что такие де-
тергенты оказывают вредное воздействие на здоровье человека,
и в 1970—1971 гг. производство бактериальных протеолитиче-
ских ферментов резко сократилось. Последовавшее позднее сня-
тие предупреждения Федеральной комиссии и внедрение усовер-
шенствованных методов производства, при которых образование
ферментных аэрозолей сводится к минимуму (путем «парафини-
рования»), способствовали частичному восстановлению спроса
Применение реакций, катализируемых ферментами 229
на бактериальные протеазы для детергентов. В 1980 г. в США
таких протеаз было произведено на сумму около 6 млн. долл.
Различные методы мягчения разделанных мясных туш с по-
мощью специальных агентов основаны на протеолитической ак-
тивности относительно недорогих и устойчивых к действию теп-
ла растительных протеаз папаина и бромелаина. Процесс «со-
зревания» целых туш обычно проводят до их разделки и
расфасовки путем контролируемого автолиза при температуре
около 15 °C с одновременным облучением ультрафиолетом, ко-
торый выполняет роль гермицидного препарата и предотвраща-
ет рост нежелательных .микроорганизмов на поверхности туш.
Сырые ферментные препараты поджелудочной железы раз-
личных животных содержат все пищеварительные протеазы, в
том числе трипсин, а также липазы и амилазы. Эти смеси, обла-
дающие мощной гидролитической активностью, используются
для обезволашивания (удаления волос) шкур животных и одно-
временного гидролиза других неколлагеновых белков. Посколь-
ку пепсин гидролизует и коллаген (фибриллярный белок, яв-
ляющийся основой кожного покрова животных), его нельзя
применять при выделке кож.
В молочной промышленности широко применяется только
один фермент — реннин. Реннин отщепляет гликопептид от рас-
творимого казеината кальция и таким образом превращает его
в относительно малорастворимый параказеинат кальция, осаж-
дающийся в виде творожного сгустка. Превращать казеинат
кальция в параказеинат могут и другие протеазы, однако ката-
лизируемый ими протеолиз обычно на этом не останавливается,
а поскольку продукты более глубокой деградации казеина обла-
дают большей растворимостью, то творожный сгусток в этих
условиях не образуется. Нехватка животного реннина стимули-
ровала разработку методов получения аналогичных бактериаль-
ных ферментов, и последние уже применяются в сыроделии. Для
получения реннина теленка с помощью микроорганизмов при-
меняются также методы генетической инженерии.
Протеолитические ферменты применяются в медицине и кли-
нической практике в качестве способствующих пищеварению
средств, а также при лечении тяжелых ран. Поскольку фермен-
ты представляют собой белки, то способствующие пищеварению
ферменты применяют только в капсулах, предохраняющих фер-
менты от кислой среды желудка, которая неминуемо вызвала
бы их денатурацию. Из перечисленных в табл. 4.1 ферментов
нет ни одного, который был бы выделен из организма человека.
Введение протеаз животных человеку (а трипсин свиньи, на-
пример, отличается от трипсина человека) применяется также
для подавления воспалительных процессов в тканях; реакция
иммунной системы человека минимальна в случае высокоочи-
230
Глава 4
щенных кристаллических препаратов ферментов. В отличие от
живых мертвые клетки обычно не способны защищать себя от
действия протеаз. На этом различии основано применение рас-
творов протеаз для лечения вирулентных или сочащихся ран;
протеазы способствуют размягчению омертвевшей ткани и кле-
ток, облегчая тем самым дренаж ран и ускоряя их заживление.
Протеолитические ферменты, особенно трипсин, подавляют
воспалительные процессы и опухоли, связанные с внутренними
повреждениями и инфекциями. Их действие, вероятно, основано
на растворении тромбов и осадков внеклеточных белков или на
локальной активации других защитных систем организма, вы-
полняющих те же функции, или на обоих этих факторах одно-
временно. Протеолитические ферменты тормозят развитие или
полностью излечивают ряд тяжелых инфекционных заболева-
ний, приводящих к накоплению в легких слизи.
4.1.3. Применение эстераз
Ферменты этой группы расщепляют сложноэфирные связи с
образованием спирта и кислоты или катализируют обратную ре-
акцию:
R'COOR2+H2O R'COOH+R2OH
Наиболее важной подгруппой эстераз являются липазы, гидро-
лизующие жиры до глицерина и жирных кислот:
Н2С—OCOR1 Н2СОН RXCOOH
| липаза I +
НС—OCOR2 -------> НСОН + R2COOH
I 1 +
Н2С—OCOR3 Н2СОН R3COOH
липид
глицерин жирные
кислоты
Панкреатическая липаза, выделяемая в пищеварительный тракт
после нейтрализации желудочной кислотности, расщепляет не-
переваренные жиры до жирных кислот; при этом образуются и
промежуточные продукты гидролиза — моно- и диглицериды.
Как мы уже упоминали в гл. 3, нерастворимость высокомолеку-
лярных жиров служит причиной того, что липазы проявляют за-
метный каталитический эффект только на границе раздела фаз
вода — липид (см. рис. 3.33).
Специфичность липаз, так же как и специфичность второй
группы эстераз, называемых алиэстеразами, невысока. Липазы
активны при гидролизе высокомолекулярных жиров и не дейст-
вуют на жиры, построенные из карбоновых кислот с короткой
углеводородной цепью. Последние, напротив, легко гидролизу-
ются алиэстеразами, не активными по отношению к высокомо-
Применение реакций, катализируемых ферментами
231
пекулярным жирам. Активность липаз повышается в присут-
ствии поверхностно-активных веществ (например, желчных кис-
лот, которые стабилизируют высокую удельную поверхность
жиров в эмульсиях) и ионов кальция, которые, очевидно, свя-
зывают образующиеся жирные кислоты путем осаждения или
образования комплексов. Связывание жирных кислот выводит их
из сферы реакции; в противном случае они действуют как ин-
гибиторы.
В аэробных процессах очистки сточных вод, происходящих в
природных условиях и применяющихся в методе активного ила
(гл. 14), для поддержания необходимых форм жизни нужен по-
стоянный приток достаточного количества кислорода. Поскольку
транспорт кислорода через жировые пленки затруднен, то на
соответствующих установках приходится постоянно освобождать
поверхность аэрируемых вод от таких пленок. Содержащую
большое количество жиров собранную жидкость затем инкуби-
руют с микроорганизмами, способными продуцировать внекле-
точные липазы. Образование нефтяных или жировых пленок
на больших площадях природных водоемов приводит к резкому
снижению концентрации растворенного кислорода и таким обра-
зом к гибели всех микроорганизмов и более сложных форм жиз-
ни, не способных существовать в анаэробных условиях, если
только эти организмы не могут пробиться через липидный слой.
В мясной промышленности путем обработки отрубов липаз-
ными препаратами, вызывающими частичный гидролиз жиров,
можно производить относительно нежирные мясные продукты.
В процессе переработки пищевых продуктов или бытовых и
промышленных отходов водостоки иногда забиваются смесями
нерастворимых веществ, содержащими белки, жиры и углеводы.
Для предотвращения или ликвидации таких пробок успешно
применяются смеси протеаз, различных карбогидраз и липаз.
Ко второй группе эстераз относят ферменты, называемые
фосфатазами. Фосфатазы являются неотъемлемыми элемента-
ми внутриклеточных биосинтетических путей метаболизма, в ко-
торых используется химическая свободная энергия, высвобож-
дающаяся при окислении клеточного топлива. Данные о прак-
тическом применении фосфатаз вне живых клеток отсутствуют.
4.1.4. Ферментные смеси, пектиновые ферменты
и другие пути использования ферментов
Вместо индивидуальных ферментов часто удобнее или целе-
сообразнее использовать их смеси. Компоненты смеси могут
принадлежать к одному и тому же типу (а- и ^-амилазы или
трипсин и химотрипсин) или к разным классам ферментов
(экстракты поджелудочной железы, представляющие собой
232
Глава 4
смесь трипсина, липазы и амилазы). Так, использование специ-
альных смесей различных амилаз позволяет повысить выход
осахаренного крахмала, являющегося сырьем для производства
спирта путем спиртового брожения, а смесь амилаз с меньшим
содержанием р-амилазы, позволяющую понизить вязкость рас-
творов крахмала без существенного осахаривания, целесообраз-
нее применять, например, при расшлихтовке текстиля. Экстрак-
ты поджелудочной железы животных содержат смесь пищевари-
тельных ферментов, которая способна выполнять многие функ-
ции ферментного комплекса панкреатической жидкости челове-
ка. По меньшей мере один препарат, выпускающийся в США
в промышленном масштабе в качестве добавки к моющим сред-
ствам, содержит смесь бактериальных протеаз и амилаз; сооб-
щалось, что этот препарат эффективнее протеаз при удалении
некоторых типов загрязнений.
Окисление галактозы до галактуроновой кислоты с после-
дующей поликонденсацией приводит к образованию полигалак-
туроновых кислот. Последние являются основным структурным
элементом ряда природных материалов растительного проис-
хождения, называемых пектинами. В пектинах карбоксильные
группы часто этерифицированы до карбометоксильных. Фермент
пектинметилэстераза (пектинэстераза, гидролизующая пектаза)
гидролизует карбометоксильные группы, а полигалактуроназы
(пектиназы, пектолазы) расщепляют гликозидные связи в пек-
тинах. Основными источниками ферментов первого типа служат
фрукты, а ферменты второго типа получают главным образом
из грибов.
Пектиновые ферменты наиболее широко применяются в двух
отраслях пищевой промышленности — в производстве соков и
виноделии. После первичной переработки фруктов и овощей по-
лучаются очень вязкие соки, что вполне приемлемо для соков
из томатов и цитрусовых, но часто нежелательно для яблочно-
го и других соков. Путем контролируемого частичного гидроли-
за пектинов удается получить продукт с необходимой теку-
честью, но в то же время достаточно вязкий, чтобы не происхо-
дило осаждения нерастворимых веществ. В производстве яблоч-
ного сока применяют более глубокий гидролиз пектинов, благо-
даря чему облегчается последующее фильтрование, обеспечи-
вающее необходимую прозрачность сока. Если в дальнейшем
предполагается использовать соки в производстве желеобраз-
ных продуктов, то их обрабатывают только пектинэстеразой.
Образующиеся полигалактуроновые кислоты затем желатини-
зируют ионами кальция.
Другой важной областью применения пектиновых ферментов
является виноделие. Добавление смеси пектиновых ферментов
к раздавленному винограду приводит к повышению выхода сока,
Применение реакций, катализируемых ферментами 233
более эффективному экстрагированию красящих веществ из
кожуры, а также ускоряет процессы фильтрования и отжимки.
Обработка пектиновыми ферментами уже перебродившего вино-
градного сока также дает положительный эффект, связанный
с ускорением процесса отделения вина от дрожжей и осадка,
повышением его прозрачности и стабильности (в основном за
счет снижения концентрации суспендированных белков в гото-
вом вине). Таким образом, как в первой, так и во второй обла-
сти применения пектиновые ферменты способствуют главным
образом достижению необходимой вязкости и фильтруемости
Таблица 4.6. Перспективные или находящиеся в стадии разработки
области применения гидролаз
Фермент Процесс
Пенициллинацилаза Получение исходного сырья для производства полусинтетических пенициллинов из пеницилли- на G
Лактаза Гидролиз лактозы, содержащейся в молоке, сы- воротке
Рибонуклеаза Получение 5'-нуклеотидов из РНК
Декстраназа Удаление зубных бляшек
Изоамилаза Производство мальтозы из крахмала
Кератиназа Обработка шерсти, щетины, кожи
Танназа Деструкция дубильных кислот в пищевых про- дуктах
полупродуктов, что положительно сказывается на экономично-
сти процессов и на внешнем виде готовой продукции.
В будущем, возможно, пектиновые ферменты будут приме-
няться и для обработки древесины хвойных пород. Свежесруб-
ленная древесина таких пород, например норвежской ели, не
поддается пропитке химическими консервантами. Как оказалось,
предварительная обработка пектиновыми ферментами улучшает
проницаемость древесины и повышает таким образом эффектив-
ность химической пропитки.
В ограниченных масштабах применяются и другие гидроли-
тические ферменты; кроме того, разрабатываются новые много-
обещающие процессы с использованием гидролаз (табл. 4.6).
4.2. Другие области применения
ферментативных реакций в растворах
Рассмотренные в предыдущих разделах сферы применения
гидролаз доминировали в прошлом и преобладают сейчас в био-
химической технологии с использованием ферментов. В то же
234
Глава 4
Таблица 4.7. Ферменты, применяющиеся в медицине
Фермент Характерная область применения
Трипсин Противовоспалительное средство; применяется при обработке ран
Глюкозооксидаза Лизоцим (белок, не гид- ролизующийся трип- сином, химотрипсином и папаином) Гиалуронидаза (из се- менников быка) Определение глюкозы в крови и моче Рекомендуется при лечении некоторых язв, кори, рассеянного склероза, ряда кожных заболева- ний и послеоперационных инфекций (антибак- териальное средство) Гидролизует полигиалуроновую кислоту (содер- жащийся в организме человека полимер, пре- пятствующий транспорту различных веществ); вводится совместно с другими лекарственными препаратами для облегчения их диффузии (в частности, с антибиотиками, адреналином, ге- парином, местными анестетиками в хирургии и стоматологии)
Пищеварительные фер- менты (смеси амилаз, липаз, протеаз и цел- люлаз) Стрептокиназа Стрептодорназа Способствуют процессу пищеварения Противовоспалительное средство Противовоспалительное средство; кроме того, гидролизует ДНК и тем самым снижает вяз- кость экссудатов (воспалительных выпотов) из ран
Пенициллиназа Разрушает аллергенные формы пенициллинов у чувствительных к ним больных
Урокиназа Предотвращение и рассасывание тромбов в кро- веносных сосудах
Активатор тканевого плазминогена Аспарагиназа Растворение тромбов в кровеносных сосудах Противоопухолевое средство
время в пищевой, фармацевтической и биохимической отраслях
промышленности применяются и другие ферментативные про-
цессы. Более того, как мы увидим в последующих разделах этой
главы, в последние годы открылись новые возможности исполь-
зования ферментов.
4.2.1. Применение ферментов в медицине
Свободные или внеклеточные ферменты находят все более и
более широкое применение в медицине. В табл. 4.7 приведен
краткий перечень самых распространенных и наиболее перспек-
тивных в медицине ферментов.
В защитных жидкостях организма, например в слизи носовой
полости и слезах, содержится фермент лизоцим, гидролизующий
мукополисахариды клеточных стенок ряда (грамположитель-
Применение реакций^ катализируемых ферментами 235
ных) бактерий. Этот фермент применяется в качестве антибак-
териального средства; по-видимому, он выполняет и другие
каталитические функции.
Начальные стадии ряда заболеваний, а также внутренние
травмы сопровождаются понижением или повышением концен-
траций некоторых ферментов в таких легко поддающихся ана-
лизу жидкостях организма, как лимфа, кровь или моча. Наличие
данного фермента легко установить с помощью соответствующе-
го субстратного теста. Таким образом, определение фермента-
тивной активности в жидкостях организма можно использовать
в качестве средства диагностики. Данные об изменении концен-
трации тех или иных ферментов в сыворотке крови дают полез-
ную информацию при диагностировании целого ряда заболева-
ний, связанных с нарушениями функций сердца, поджелудочной
железы, патологическими изменениями в мышечных и костных
тканях, а также с ростом злокачественных опухолей.
Фермент ь -аспарагиназа катализирует гидролиз ь-аспара-
гина:
Ь-Аспарагин + Н2О —> L-аспартат+NHs
Для некоторых типов опухолевых клеток ь-аспарагин является
незаменимым питательным веществом; ь-аспарагиназа разру-
шает аспарагин и тем самым может тормозить рост таких кле-
ток. Известно, что Е. coli продуцирует две различные аспараги-
назы, из которых только одна проявляет антилимфомную ак-
тивность.
Как мы уже отмечали выше, проницаемость клеточных мем-
бран и составляющих их структурных элементов для многих ве-
ществ сравнительно низка; в случае клеток высших животных
это свойство мембран обусловлено наличием очень вязкой поли-
гиалуроновой кислоты. Эффективность некоторых лекарствен-
ных препаратов и местных анестетиков, например в стоматоло-
гии, повышается при одновременном введении гиалуронидазы,
частично разрушающей этот мембранный барьер.
Некоторые одноклеточные организмы продуцируют пеницил-
линазу (или ^-лактамазу) и таким образом защищают себя от
летальных доз пенициллиновых антибиотиков. Организм чело-
века не вырабатывает пенициллиназу, поэтому аллергическую
реакцию на введенный больному пенициллин можно снять путем
инъекции раствора пенициллиназы, которая в идеальном вари-
анте превращает этот антибиотик в вещество, не вызывающее
аллергии.
Глюкозооксидаза катализирует окисление глюкозы до глю-
коновой кислоты:
Глюкоза+ О2+Н2О —> глюконовая кислота + Н2О2
236
Глава 4
Поскольку образующийся при этом пероксид водорода легко
обнаружить с помощью какого-либо индикатора, глюкозоокси-
дазу можно использовать в качестве чувствительного и специ-
фичного реагента на глюкозу, определяемую, например, в крови
или моче (в частности, в случае сахарного диабета). Другие
области применения биологических катализаторов в медицине
мы рассмотрим в ходе изучения иммобилизованных ферментов.
4.2.2. Применение негидролитических ферментов
в промышленности и перспективы
расширения сферы их использования
Только что упоминавшийся фермент глюкозооксидаза при-
меняется и в тех случаях, когда возникает необходимость в де-
струкции глюкозы или в связывании кислорода. Так, сухой яич-
ный порошок при хранении темнеет в результате взаимодейст-
вия глюкозы с белками. Добавление глюкозооксидазы предот-
вращает эту реакцию, обычно называемую браунинг-реакцией
или реакцией Майяра. Обескислороживание с помощью глюко-
зооксидазы применяется также в производстве и при хранении
безалкогольных напитков на основе апельсинового сока, консер-
вированных напитков, порошкообразных сухих пищевых про-
дуктов, майонеза, соусов для салатов и расфасованного сыра.
В последнем случае в присутствии кислорода образуются про-
дукты окисления, имеющие неприятный запах; для связывания
кислорода в упаковку с сыром добавляют не только глюкозо-
оксидазу, но и глюкозу. Такие ферментные добавки в течение
длительного времени связывают кислород, проникающий через
упаковочные материалы, и тем самым способствуют увеличению
срока годности расфасованных пищевых продуктов, так как при
температуре их хранения ферментативная активность сохраня-
ется достаточно долго.
Образующийся в процессе катализируемой глюкозооксида-
зой реакции пероксид водорода обладает бактерицидными свой-
ствами. Если же его наличие по каким-либо причинам нежела-
тельно, то в систему добавляют еще один фермент — каталазу,
которая катализирует реакцию:
2Н2О2 —> 2Н2О + О2
В присутствии каталазы эта реакция протекает очень быстро
[фтах/ео= 1,2-107 (с-М)~1, ^п=10-7 М]. Способность каталазы
быстро разлагать пероксид водорода используется в некоторых
средствах для крашения волос.
В табл. 4.8 перечислены некоторые другие области приме-
нения негидролитических ферментов; соответствующие процес-
сы находятся в стадии разработки или промышленного внедре-
Применение реакций, катализируемых ферментами 237
Таблица 4.8. Некоторые разработанные в последние годы области
применения нсгпдролитических ферментов и наиболее перспективные
сферы их использования3
Производство L-яблочной кислоты из фумаровой с применением фумаразы
Производство L-аспарагиновой кислоты из фумаровой кислоты с помощью
аспартазы
Производство АТР из аденина в присутствии ферментов из микроорганизмов
Производство NAD из аденина и никотинамида в присутствии ферментов из
микроорганизмов
Производство фруктозы из глюкозы с помощью глюкозоизомеразы
Производство фруктозы из сорбита с помощью сорбитдегидрогеназы
Разделение DL-аминокислот с помощью иммобилизованной аминоацилазы
Производство L-тирозина и ь-дигидроксифенилаланина с помощью тирозин —
фенол-лиазы
Производство L-дигидроксифенилаланина из соответствующей а-кетокислоты
с помощью трансаминазы
Ферментативный синтез L-триптофаиа из индола, пировиноградной кислоты
и аммиака
Ферментативный синтез L-лизина из а-амино-8-капролактама
Трансформация насыщенных жирных кислот в полиненасыщенные жирные
кислоты с помощью десатуразы жирных кислот
Производство галактонолактозы из сыворотки с помощью глюкозооксидазы
и галактозооксидазы
а Воспроизведено нз работы: Pye Е. К., Wingard L. В. (eds.), Enzyme Engineering,
р. 365, Plenum Press, New York, 1973.
ния. Одним из перспективных ферментов, в частности, являет-
ся глюкозоизомераза, превращающая глюкозу во фруктозу. По-
скольку этот и ряд других приведенных в таблице процессов
часто осуществляются с помощью иммобилизованных фермен-
тов, мы рассмотрим их подробнее в следующих разделах.
4.3. Технологические процессы
с участием иммобилизованных ферментов
Под иммобилизацией понимают способы трансформации
ферментов, обеспечивающие возможность их использования в
непрерывных процессах. Необходимость в иммобилизации мо-
жет возникнуть по целому ряду причин. Во-первых, иммобили-
зованные индивидуальные ферменты удерживаются в реакторе,
в то время как растворимые ферменты обычно переходят в
смесь, содержащую продукт реакции. В результате ценные фер-
менты необратимо теряются, и в реактор приходится вводить
дополнительные количества биологического катализатора; к тому
же фермент может оказаться нежелательной примесью и его
придется отделять от конечного продукта реакции. Во-вторых,
как мы уже упоминали в гл. 3, по сравнению с растворимыми
биологическими катализаторами иммобилизованные ферменты
238
Глава 4
часто сохраняют активность в течение более длительного перио-
да (рис. 3.6). В-третьих, иммобилизованные ферменты можно
локализовать в соответствии с последовательностью осуществ-
ляемых ими каталитических превращений, что будет способст-
вовать повышению эффективности многостадийного процесса.
Указанные характеристики иммобилизованных ферментов
свидетельствуют о том, что их применение целесообразно в слу-
чае переработки больших количеств субстрата и (или) в случае
высокой стоимости фермента. Кроме того, возможность скон-
центрировать фермент в определенном объеме и придать ему
любую форму позволяет найти новые уникальные сферы приме-
нения иммобилизованных биологических катализаторов. Сле-
дующий раздел будет посвящен изучению методов иммобилиза-
ции ферментов, а затем мы рассмотрим пути применения иммо-
билизованных ферментов и перспективы их дальнейшего
промышленного использования.
4.3.1. Иммобилизация ферментов
Существует много методов иммобилизации ферментов. Как
мы увидим ниже, от метода иммобилизации во многом зависят
и свойства полученного биокатализатора. Таким образом, вы-
бор конкретного способа иммобилизации зависит от целого ряда
факторов и параметров каталитического процесса, в том числе
от общей каталитической активности, эффективности использо-
вания катализатора, его способности подвергаться инактивации
и регенерации и, конечно, от его цены. В связи с проблемой за-
хоронения отходов производства, а также в зависимости от
предполагаемой области использования получаемых продуктов
следует принимать во внимание и токсичность реагентов, при-
меняемых в реакциях синтеза иммобилизованных ферментов.
Все методы иммобилизации ферментов можно разделить на
две группы: химические методы, основанные на создании кова-
лентных связей между ферментом и носителем, и физические
методы, основанные на более слабых взаимодействиях или на
включении ферментов в какую-либо оболочку или матрицу
(рис. 4.5). Ферменты можно превратить в иммобилизованную
форму посредством адсорбции на различных носителях
(табл. 4.9); целесообразность применения этого метода иммо-
билизации обусловлена тем обстоятельством, что в ряде случа-
ев катализатор удается сравнительно просто регенерировать
путем десорбции отработанного фермента и адсорбции новой
порции активного биокатализатора. Свойства иммобилизованно-
го фермента определяются как свойствами фермента, так и ха-
рактеристиками носителя или матрицы и методикой иммобили-
зации.
Применение реакций, катализируемых ферментами
239
а) Свимические методы
М = Водонерастворимая матрица
Е = Фермент
Связывание фермента
с патрицей ковалент-
ными связями
Ферментная
матрица с
по пере чными
связями
8) физические методе/
Образование поперечных
связей неэ/сду молекулами
рерпента с помощью
поли/рункционаленого
реагента
ZZZ1 ZZZZZZZZZ2 VZZZZZ2ZZ3 VZZZZ
Ое 0е Ge
ZZZZZZZZ2 VZZZZZZZ2 ZZZZZZZZ2 ZZ
Ферменты, включенные 6
пористые пустотелые
волокна
Фермент, включенный
б нерастворимую геле-
вую матрица
Ферменты, включенные в скрученные
волокна
S
Фермент, включен-
ный в микрокал-
сулу
РИС. 4.5. Некоторые методы иммобилизации ферментов: химические (а); фи-
зические (б).
Выбор носителя для химической или адсорбционной иммо-
билизации фермента зависит прежде всего от свойств его по-
верхности. Способен ли фермент адсорбироваться на этой по-
верхности? Есть ли у носителя функциональные группы, связы-
вающие фермент? Если поверхность носителя не удовлетвори-
тельна в этом отношении, то можно ли химически модифициро-
вать или иным образом трансформировать поверхность носите-
ля, чтобы облегчить связывание фермента? В табл. 4.10 перечне-
240
Глава 4
Таблица 4.9. Носители, применяемые для иммобилизации ферментов
методом адсорбции, и типы взаимодействий между ферментом и носителем
Взаимодействие
Адсорбенты
Физическая адсорбция Активированный уголь, силикагель, оксид алю- миния, крахмал, глина, стекло Модифицированные носители: таннинаминогек-
Ионное связывание силцеллюлоза, конканавалин А — сефароза Катионообменники: карбоксиметилцеллюлоза, ам- берлит CG-50, дауэкс 50 Анионообменники: диэтиламиноэтилцеллюлоза, ди- этиламиноэтилсефадекс, поли аминополистирол, амберлит IR-45
лены некоторые носители, используемые . для иммобилизации
ферментов путем создания ковалентных связей, и приведены
функциональные группы, участвующие в этих связях. В качест-
ве носителей иммобилизованных ферментов применяются также
керамические материалы, стекло и оксиды ряда металлов.
Описание методики ковалентной иммобилизации фермента
часто начинается со стадии модификации или активации по-
верхности носителя. В случае неорганических носителей для
этой цели широко применяется силанизация, т. е. обработка по-
Таблица 4.10. Нерастворимые носители, применяемые для ковалентного
связывания ферментов (указаны участвующие в связывании
функциональные группы, расположенные на поверхности носителей)
Носители природного происхождения
Синтетические носители
Целлюлоза (—ОН)
Карбоксиметилцеллюлоза
(—СООН)
Агароза (сефароза) (—ОН)
Декстран (сефадекс) (—ОН)
Полиакриламиды и их производные
(биогель, энзакрил и др.) (арома-
тические аминогруппы)
Поли аминополистирол (—NH2)
Сополимеры малеинового ангидрида
верхности кремнийорганическими реагентами (например,
(CH3CH2O)3Si (CH2)3R, где R чаще всего NH2). Обработанный
таким способом носитель содержит на поверхности аминогруп-
пы, которые (как и аминогруппы других носителей) можно пере-
вести в альдегидные группы конденсацией с глутаровым альде-
гидом или в ариламинные группы с помощью п-нитробензоил-
хлорида, или в карбоксильные группы конденсацией с янтарным
ангидридом. Другой способ модификации поверхности носителей
основан на введении гибких, сравнительно длинных цепей (на-
пример, остатка w-пропиламина). Ввиду отсутствия пространст-
венных препятствий последующее присоединение фермента к
Применение реакций, катализируемых ферментами 24Г
этим гибким группировкам позволяет в основном сохранить на-
тивную структуру фермента, что в свою очередь способствует
сохранению и стабилизации его каталитической активности.
С помощью подобных модификаций можно также изменять гид-
рофобность или гидрофильность поверхности носителя.
На рис. 4.6 суммированы типичные реакции иммобилизации
ферментов с участием функциональных групп на поверхности
носителей, в том числе перечисленных в табл. 4.10. Следует от-
метить, что каждая из указанных реакций протекает только-
в строго определенных условиях, в частности при заданных ве-
личинах pH, ионной силы и концентраций реагентов. Подробные
сведения об условиях конкретных реакций, в том числе обшир-
ные табличные данные о различных методах иммобилизации
ферментов на различных носителях, можно найти в литературе,,
приведенной в конце главы. Химическая природа функциональ-
ных групп на поверхности носителя определяет и функциональ-
ные группировки белка, с участием которых он образует кова-
лентные связи. Благодаря наличию достаточно большого числа
методов поверхностной модификации носителей иммобилизацию
какого-либо определенного фермента на конкретном носителе
обычно можно осуществить несколькими способами. Конденса-
ция фермента с носителем, очевидно, не должна затрагивать
аминокислотные остатки, входящие в активный центр фермен-
та или расположенные вблизи него.
Решая вопрос о выборе носителя и способе модификации его
свойств, нельзя забывать и о взаимодействиях между поверх-
ностью носителя и реакционной смесью, благодаря которым не-
посредственно примыкающие к носителю, а следовательно, и к
ферменту слои реакционной смеси могут существенно отличаться
по своим свойствам от основного объема реакционной смеси.
Например, вокруг частиц носителя с ионизированными группа-
ми образуется зона с повышенной концентрацией противопо-
ложно заряженных ионов, что в свою очередь приводит к су-
щественному изменению зависимости наблюдаемой каталитиче-
ской активности от pH жидкой фазы в сравнении с аналогичной
зависимостью для ферментативной реакции в растворе (рис. 4.7).
Аналогично гидрофобные или гидрофильные свойства носителя
будут влиять на локальные концентрации растворенных ве-
ществ и растворителей в соответствии с их гидрофобностью (или.
гидрофильностью).
Другая важная функция поверхности носителя связана с ее
прямым или косвенным влиянием на молекулярное окружение
фермента. В какой-то степени фермент всегда взаимодействует
с поверхностью носителя на молекулярном уровне и в то же
время контактирует с реакционной средой, на которую так же„
как только что было показано, влияет носитель. Молекула фер-
16-537
Диазо-метод
Активация вромцианом
\~он . \ о
:а + cnbf--------> Й )с = nh
/-он /-О
НО
O)-N{ I "° (о) I! Л -(o') N = N-^O^E
\
Также группы NH2
и имидазольное
кольцо г ист и на
о
k-O-C-NHE
О zc = NH + ENH;--> Я
/-О /он
Концевая амино-
группа или е-амино-
группа лизина
Карводи имидный метод
Метод алкилирования
* *
NH*
I
R
он
^иБсЯГ^ОСОСНгВг
диоксане
^-ОСОСН2Вг +□-£-» ^-ОСОСН2Е
Аминная, /ренальная
или сульфгидрильная
группа
О
II °
C-NH-E + II
С
/ \
R'NH HNR
R
Можно также осуществлять
конденсацию группы NHZ на
носителе с группой СООН на ферменте
РИС. 4.6. Некоторые методы ковалентного связывания ферментов с носителями (Е — фермент).
Применение реакций, катализируемых ферментами
243-
РИС. 4.7. Различные зависимости каталитической активности от pH свобод-
ного фермента и фермента, иммобилизованного на ионизированном носителе.
Субстратом является этиловый эфир ацетил-L-тирозина. [Воспроизведено с
разрешения из статьи: Goldstein L. et al., A Water-Insoluble Polyanionic Deri-
vative of Trypsin. II Effect of the Polyelectrolyte Carrier on the Kinetic Behavior
of Bound Trypsin, Biochemistry, 3, 1913 (1964). (C) American Chemical Society.]
1 — химотрипсин (свободный); 2 — ЕМА-химотрипсин (иммобилизованный на
отрицательно заряженном носителе — сополимере этилена с метакриловой кис-
лотой) ; 3 — полиорнитилхимотрипсин (иммобилизованный на положительно
заряженном носителе).
мента построена таким образом, что она принимает определен-
ную конформацию и проявляет соответствующую активность
только в свойственной ей природной среде. Очевидно, что путем
подбора носителя для иммобилизации фермента можно попы-
таться как сохранить свойственное данному ферменту окруже-
ние, так и направленно изменить его с тем, чтобы повлиять на
активность, а также на селективность и стабильность фермента.
В настоящее время большое внимание уделяется изучению взаи-
16*
244
Глава 4
модействий ферментов с их окружением на молекулярном уров-
не и выяснению возможности использования таких взаимодейст-
вий для углубления наших знаний о поведении ферментов в
природных условиях и для оптимизации соответствующих био-
каталитических процессов.
На активность и другие параметры иммобилизованного фер-
мента оказывают влияние также физические и механические
свойства носителя. Пористость носителя и распределение пор по
размерам определяют количество фермента, которое может
быть иммобилизовано на этом носителе, и доступность субстра-
та для молекул фермента, связанных с внутренними поверхно-
стями. Механическая прочность носителя в существенной степе-
ни влияет на возможность его использования в реакторах опре-
деленных конструкций. Например, в реакторе с перемешиваемой
взвесью твердого катализатора нецелесообразно использовать
носители, склонные к механическому истиранию, а легко сжи-
маемые материалы нельзя применять в насадочных колоннах
крупномасштабных производств. Важна и способность носителя
к набуханию. Другие сведения о физических свойствах различ-
ных носителей, а также о способах их изучения можно найти
в приведенной в конце главы литературе.
Для создания ковалентных связей между молекулами фер-
ментов можно использовать некоторые реагенты, содержащие
две (или более) функциональные группы. Из таких реагентов
чаще всего применяют глутаровый альдегид, образующий меж-
молекулярные (и, возможно, внутримолекулярные) связи с уча-
стием аминогрупп фермента:
Е—NH2 + ОНС(СН2)3СНО + H2N—Е'
—2Н2О
Е—N=HC(CH2)3CH=N—Е'
В качестве бифункциональных реагентов используют также бис-
диазобензидин, циаиурхлорид и гексаметилендиизоцианат. Сши-
тые поперечными связями ферменты обычно представляют со-
бой желатинообразную массу, механические свойства которой
не отвечают требованиям большинства процессов. Как показано
на рис. 4.5, адсорбция фермента на носителе и последующая его
сшивка бифункциональными реагентами способствуют улучше-
нию механических свойств катализатора и облегчают доступ
субстрата к ферменту.
Метод иммобилизации фермента путем его включения вы-
годно отличается относительной независимостью от природы
фермента (и, следовательно, возможностью применения одной
методики для иммобилизации различных ферментов), а также
Применение реакций, катализируемых ферментами
245
Е Фермент + сн, = сн 1 CONH2 Акриламид (мономер) N, л/'-Метилен5исак- риламид СН, = СН 1 с=о + NH сн2
NH
С=О
I
сн=сн2
полимеризация
K,S2O8
(инициатор)
DMAPN
Ф (ускоритель)
NH
СО CONHj
I I
—сн2— сн—сн2—сн—сн2— сн—сн2—сн—сн2—
СО со
NH NH
СНг Фермент NH । СН, 1 NH
СО СО
—сн2— сн—сн2 — сн—сн2—сн—сн2—сн—сн2—
CONH2 CONH —
РИС. 4.8. Метод иммобилизации фермента путем его включения в полимер.
В данном случае полимерная матрица представляет собой полиакриламид,
сшитый поперечными связями.
сравнительно небольшими нарушениями нативной структуры
фермента. Методы включения можно разделить на две группы:
к первой группе относят методы, основанные на включении фер-
мента в полимерную матрицу, а ко второй — методы, в которых
раствор фермента отделяют мембраной, проницаемой для суб-
стратов и (или) продуктов реакции, но непроницаемой для фер-
мента.
Чаще всего фермент иммобилизуют в трехмерной полимер-
ной решетке полиакриламидного геля путем полимеризации ак-
риламида в присутствии сшивающих мономеров и фермента (Е)
(рис. 4.8). Как указано на рисунке, инициатором полимеризации
может быть персульфат калия (КгЗгОз), а ее ускорителем —
₽-диметиламинопропионитрил (DMAPN). Сообщалось, что с по-
246
Глава 4
мощью этих реагентов можно получать гели иммобилизован-
ных ферментов с диаметром пор от 100 до 400 нм, которые на-
дежно удерживают многие ферменты, имеющие обычно диаметр-
молекул от 300 до 2000 нм.
В альтернативном способе иммобилизации раствор фермента
отделяют мембраной таким образом, чтобы фермент постоянно-
находился в объеме, ограниченном этой полупроницаемой мем-
браной. Биологическим прототипом этого метода иммобилиза-
ции являются лизосомы, в которых сконцентрированы гидроли-
тические ферменты, вызывающие при попадании в цитоплазму
мгновенную гибель клетки. В одном из вариантов этого способа,
называемом микрокапсулированием, ферменты заключают в-
крохотные капсулы диаметром до 300 мкм. Капсулы заключены
в сферические мембраны с порами, через которые в любом на-
правлении могут транспортироваться небольшие молекулы суб-
стратов и продуктов реакции и в то же время не могут прони-
кать ферменты и другие высокомолекулярные соединения.
Известны два метода изготовления полупроницаемых микро-
капсул. В первом методе формируют постоянную полимерную
мембрану. Для этой цели на границе раздела двух фаз — орга-
нической и водной, содержащей фермент, проводят реакцию
сополимеризации. Если в качестве одного из реагентов взять
водонерастворимый мономер, а в качестве другого—мономер,
обладающий заметной растворимостью в той и другой фазах,,
то их сополимеризация будет осуществляться только вблизи
границы раздела фаз. Устойчивые полимерные микрокапсулы,
можно получить также путем коацервации, т. е. посредством,
выделения из раствора полимера в виде микрокапель новой,
жидкой фазы, обогащенной полимером. Неустойчивые микро-
капсулы получают путем эмульгирования водного раствора фер-
мента в присутствии поверхностно-активного вещества; при
этом образуются капсулы, заключенные в мембрану из поверх-
ностно-активного вещества, которые можно затем добавлять к.
водному раствору субстрата.
Оба метода микрокапсулирования выгодно отличаются высо-
кой удельной поверхностью катализатора (характерная величи-
на составляет 2500 см2 на 1 мл суспензии фермента) и возмож-
ностью повышения специфичности — иногда удается изготовить
мембраны, селективно пропускающие одни субстраты и задер-
живающие другие. В принципе эти методы применимы к любым
ферментам. В то же время мембрана представляет собой опре-
деленное препятствие для массопередачи, поэтому «коэффици-
ент эффективности» заключенных в микрокапсулу ферментов,
может быть очень низким. Кроме того, эти методы вообще не-
применимы, если размеры молекулы субстрата сравнимы с раз-
мерами молекулы фермента.
Применение реакций, катализируемых ферментами
247
Для иммобилизации ферментов можно использовать и уст-
ройства, основанные на ультрафильтрации через полупроницае-
мые мембраны, которые не мешают обмену молекулами неболь-
шого диаметра между раствором фермента и соседним раство-
ром. На рис. 4.9 изображена принципиальная схема непрерыв-
ной ультрафильтрации, где, как и в случае микрокапсулирова-
ния, фермент удерживается в определенном объеме. В то же
Фермент и другие
высокомолекулярные
вещества
Субстрат
Реактор
Мембрана
Насос
Фермент,
.субстрат а
продукт
реакции
л
Блок ультра-
фильтрации
Фильтрат,
содержащий
продукт
реакции
РИС. 4.9. Локализация фермента в масштабе всего реактора. Полупроницае-
мая мембрана удерживает фермент и другие высокомолекулярные соединения
в реакторе.
время в отличие от метода микрокапсулирования в этом случае
площадь поверхности, разделяющей два раствора, невелика, и к
тому же здесь возникает опасность индуцированной потоком
денатурации фермента. Скорость массообмена через мембрану
довольно низка, что может лимитировать скорость всего про-
цесса. К преимуществам метода иммобилизации ферментов с
помощью полупроницаемых мембран можно отнести возмож-
ность применения практически любого фермента и смесей любых
ферментов, а также высокомолекулярных или нерастворимых
субстратов (по отношению к которым связанные с полимерными
носителями ферменты обычно малоэффективны). Кроме того,
в реакциях гидролиза полимеров через полупроницаемые мем-
браны проникают только низкомолекулярные соединения, а все
вещества с относительно большой молекулярной массой остают-
ся в растворе фермента. На этом принципе основан интересный
248
Глава 4
метод регулирования молекулярно-массового распределения про-
дуктов реакции на выходе из реактора.
Изучен целый ряд вариантов принципиальной схемы, изо-
браженной на рис. 4.9. Так, вместо реактора с перемешиванием
можно использовать трубчатый реактор. Блок с одной полупро-
ницаемой мембраной можно заменить на устройство с множест-
вом параллельных пустотелых нитей, в котором один поток,
направляется во внутреннюю полость нитей, а другой — в омы-
Таблица 4.11. Сравнение основных характеристик различных методов
иммобилизации ферментов3
Метод связывания с носителем
Характеристики
Сшнвка би-
Физическая Ионное свя- Ковалентное функциональ- Методы
адсорбция зыванне связывание ными реагеи- включения
тами
Методика иммо- Простая Простая
билизации
Ферментативная активность Низкая Высокая
Субстратная Не изме- Не изме-
специфичность няется няется
Связывающие силы Слабые Умеренные
Регенерация Возмож- на Возможна
Область приме- Узкая Умеренная
нения
Стоимость процес- Низкая Низкая
са иммобилиза-
ции
Сложная
Высокая
Может из-
меняться
Сильные
Невозмож-
на
Умеренная
Высокая
Сложная
Умеренная
Может из-
меняться
Сильные
Невозмож-
на
Узкая
Умеренная
Слож-
ная
Высокая
Не из-
меня-
ется
Сильные
Невоз-
мож-
на
Широ-
кая
Низкая
а Воспроизведено с разрешения из работы: Immobilized Enzymes, Research апс»
Development, Chibata I. (ed.), p. 72, Kodansha Ltd., Tokyo, 1978.
вающий нити объем. Процесс ультрафильтрации и его коли-
чественную трактовку мы рассмотрим подробнее в гл. 11.
Важнейшие характеристики различных методов иммобили-
зации ферментов суммированы в табл. 4.11. Здесь под «фер-
ментативной активностью» понимается собственная каталитиче-
ская активность иммобилизованных ферментов по отношению к
активности тех же ферментов в растворе. В общем случае мето-
ды химической иммобилизации снижают активность ферментов,,
поскольку создающиеся в процессе иммобилизации ковалентные
связи могут в какой-то мере нарушать „ третичную структуру,
свойственную нативным белкам. С другой стороны, ковалентные
связи обеспечивают надежное, прочное связывание фермента.
Применение реакций, катализируемых ферментами
249
а иногда снижают скорость его инактивации и изменяют спе-
цифичность в нужном направлении. Иммобилизация ферментов
путем их включения или адсорбции обычно в меньшей степени
сопровождается нарушением их структуры, и поэтому свойства
иммобилизованных такими методами ферментов мало отлича-
ются от их свойств в растворах. В то же время не надо забы-
вать, что все сказанное выше относится только к собственным
свойствам ферментов, а на практике в силу эффектов массопе-
редачи соответствующие параметры иммобилизованного фермен-
та могут иметь совершенно другие значения. Этот важный во-
прос мы изучим подробнее в разд. 4.4. Диффузионные эффекты
обычно сильнее проявляются в случае сшитых поперечными свя-
зями и включенных ферментных катализаторов и в меньшей сте-
пени— у ферментов, иммобилизованных на носителях.
4.3.2. Промышленные процессы
Катализаторы на основе иммобилизованных ферментов уже
используются в той или иной мере в ряде крупномасштабных
промышленных производств. К числу наиболее важных областей
применения иммобилизованных ферментов относятся производ-
ство сиропа с высоким содержанием фруктозы из кукурузного
крахмала и производство ь-аминокислот путем разделения
рацемических смесей (состоящих из ь- и D-изомеров). Кроме
того, в производстве полусинтетических пенициллинов применя-
ют иммобилизованную пенициллинацилазу. С экономической
точки зрения наиболее важна первая из упомянутых областей.
Сахар (сахарозу) нельзя заменить d-глюкозой, поскольку
глюкоза менее сладка. К тому же кристаллизация концентриро-
ванных растворов глюкозы может затруднить их последующую
переработку и хранение. Эти осложнения в значительной мере
могут быть устранены, если глюкозу частично изомеризовать во
фруктозу с помощью фермента глюкозоизомеразы:
Н ОН
С
н—с—он
...I....
но—с—н
I
н—с—он
I
н—с—он
I
сн2он
D-глюкоза
глюкозоизо-
мер аз а
------------>.
Ч------------
i СН2ОН
i I
= с=о
.....
но—с—н
н-с-он
н-с—он
Ан2он
D-фруктоза
250
Глава 4
При 50 °C константа равновесия этой реакции близка к еди-
нице, и изменение температуры практически не влияет на нее,,
поскольку теплота реакции изомеризации составляет всего лишь,
около 1 ккал/моль. Поэтому продуктом реакции является смесь-
глюкозы и фруктозы в отношении приблизительно 1 : 1. Такая
смесь значительно более сладка, чем чистая глюкоза, и может
с успехом заменять сахар в самых различных областях, в том
Осахаривание
Сырой
крахмал
Ожижение
~| йвй—______> Очистка с
| j помощью
Фильтрование че- ванного ?цгля
рез слои фильтру-
ющего материала
концентри-
рование
Ионный
обмен
Изомеризация
Подготовка
загрузочной
смеси
Очистка с
помощью
активи-
рованного
угля
Ионный
обмен
Упаривание
Охлаж-
] де кие
Продукт
РИС. 4.10. Схема процесса производства кукурузного сиропа с высоким со-
держанием фруктозы с помощью глюкозоизомеразы. [Воспроизведено из
статьи: Harden J. D. On-Line Control Optimizes Processing; Food Eng., 44, 59
(1972).]
числе в производстве безалкогольных напитков, приготовлении
различных пищевых продуктов и в хлебопекарной промышлен-
ности. Еще более сладкие смеси можно получать путем хрома-
тографического разделения (гл. 11) продуктов изомеризации,
приводящего к обогащению смеси фруктозой.
Глюкозоизомераза представляет собой внутриклеточный
фермент, продуцируемый рядом микроорганизмов, из которых
используются главным образом некоторые штаммы Arthrobac-
ter и Streptomyces. Необходимость дезинтеграции клеток доста-
точно мягкими методами, не вызывающими необратимую инак-
тивацию фермента, приводит к существенному повышению стои-
мости глюкозоизомеразы по сравнению, например, с внеклеточ-
ными гидролазами. К тому же глюкозоизомераза очень чувстви-
тельна к ряду ингибиторов. Оба этих фактора предполагают
Применение реакций, катализируемых ферментами 251
целесообразность иммобилизации глюкозоизомеразы и ведения
процесса в строю контролируемых условиях. Изучены различ-
ные методы иммобилизации глюкозоизомеразы, в том числе и
метод, основанный на применении содержащих фермент целых
клеток, закрепленных в свою очередь в коллагене или с помо-
щью какого-либо другого флокулирующего и связывающего
агента. Такой прием может служить примером еще одного под-
хода к иммобилизации внутриклеточных ферментов, заключаю-
щегося в иммобилизации продуцирующих этот фермент клеток
(обычно обработанных с целью снижения сопротивления клеточ-
ных мембран массообмену) или лизатов таких клеток. К пробле-
ме иммобилизации клеток мы еще вернемся в гл. 9.
На рис. 4.10 приведена схема процесса производства куку-
рузного сиропа с высоким содержанием фруктозы, основанного
на использовании иммобилизованной глюкозоизомеразы. Здесь
необходимость множества операций разделения и обработки
промежуточных продуктов между стадиями осахаривания и изо-
меризации диктуется свойствами ферментных систем. Так, для
повышения термической устойчивости сх-амилазы, применяемой
для гидролиза крахмала (обычно этот процесс проводят при
температуре около 105°C), добавляют ионы кальция. Послед-
ние, однако, ингибируют глюкозоизомеразу, и поэтому их свя-
зывают ионообменными смолами прежде, чем декстрозный рас-
твор поступит в реактор изомеризации.
Пример 4.2. Параметры процесса изомеризации глюкозы в реакторе
в присутствии иммобилизованной глюкозоизомеразы*. Для выбора типа ка-
тализатора, конструкции реактора и условий каталитического процесса в ре-
акторе потребовалось тщательное изучение множества взаимосвязанных па-
раметров. В табл. 4П2.1 перечислены некоторые из параметров и критериев,
которые учитывались ранее при проектировании реакторов для получения
кукурузного сиропа с высоким содержанием фруктозы с помощью иммоби-
лизованной глюкозоизомеразы. В табл. 4П2.2 указаны основные свойства
катализатора, выбранного в результате анализа перечисленных параметров
в ходе разработки проекта компанией Н. J. Heinz Company. Выбранный раз-
мер частиц катализатора удовлетворяет двум взаимоисключающим требова-
ниям: с одной стороны, частицы катализатора достаточно малы, чтобы ско-
рость диффузии не лимитировала скорость всего процесса, а с другой — их
размеры достаточно велики, чтобы удалось свести к минимуму перепад дав-
лений на реакторе колонного типа со слоем иммобилизованного фермента.
Найденные оптимальные условия реакции суммированы в табл. 4П2.3.
Обратите внимание на необходимость высокого содержания глюкозы в по-
ступающей в реактор смеси; присутствие в последней более 10% олигосаха-
ридов существенно снижает активность фермента. Большой диапазон време-
ни контакта катализатора с субстратом объясняется постепенной инактива-
цией катализатора. С течением времени активность фермента падает и при
постоянной скорости потока реагентов степень превращения субстрата по-
* Venkatasubramanian К., Harrow Н. S., Design and Operation of a Com-
mercial Immobilized Glucose Isomerase Reactor System, Ann. N.Y. Acad. Sci.,
326, 141 (1979).
252
Глава 4
Таблица 4П2.1. Параметры, изучавшиеся в ходе проектирования реактора
с иммобилизованной глюкозоизомеразойа
Биохимические параметры
1. Активность
2. Стабильность фермента в процессе работы реактора (время жизни ката-
лизатора) и зависимость инактивации от времени
3. Производительность в используемом диапазоне времени жизни
4. Оптимальная концентрация субстрата
5. Влияние концентрации олигосахаридов
6. Влияние растворенного кислорода
7. Минимальное и максимальное время контакта с субстратом
8. Образование побочных продуктов реакции
9. Чувствительность к изменению pH и температуры
10. Устойчивость при хранении
11. Вымывание фермента
12. Рост микроорганизмов (если таковой вообще наблюдается)
13. Характеристики потока на выходе из реактора (состав, цвет, запах, со-
держание белков, pH и т. п.).
Механические параметры
1. Размер, форма частиц и их распределение по размерам
2. Насыпная масса в сухом и влажном виде
3. Набухание
4. Сжимаемость
5. Когезия
6. Истирание частиц
Гидромеханические параметры
1. Перепад давлений
2. Тип потока (восходящий или нисходящий)
3. Уплотнение слоя
4. Осевая дисперсия и каналообразование
5. Распределение времени пребывания
6. Расслаивание
7. Отношение длины к диаметру
8. Минимальная скорость начала псевдоожижения
а Воспроизведено с разрешения из статьи: Venkatasubramanian К.. Harrow Н. S.,
Design and Operation of a Commercial Immobilized Glucose Isomerase Reactor System,
Ann. N.Y. Acad. Sci., 326, 141 (1979).
степенно снижается. Поэтому для обеспечения необходимого качества про-
дукции по мере инактивации фермента время его контакта с субстратом уве-
личивают путем уменьшения скорости потока. Соответствующие математиче-
ские выражения, позволяющие количественно оценить взаимосвязь между
активностью, временем контакта и степенью превращения для различных
типов ферментных катализаторов, мы рассмотрим в гл. 9.
Размеры реактора колонного типа определяются гидромеханическими
свойствами слоя катализатора. При нисходящем потоке реакционной смеси
слой ферментного иммобилизованного катализатора способен сжиматься под
давлением; в результате сопротивление потоку возрастает. На рис. 4П2.1
изображена зависимость перепада давлений на колонне от высоты слоя
катализатора при максимальной скорости потока, допустимой в случае све-
жего катализатора. Если перепад давления на колонне превышает
0,2 кг/см2, то сопротивление колонны резко возрастает. Отсюда следует, что
максимальная высота слоя катализатора должна составлять около 4,6 м.
Учитывая известное время контакта катализатора с субстратом и заданную
производительность всей установки, нетрудно определить размеры необходи-
Применение реакций, катализируемых ферментами
253:
Таблица 4П2.2. Физические и каталитические свойства иммобилазы —
катализатора с глюкозоизомеразной активностью (продукт фирмы 1С1)а
Форма катализатора
Внешний вид
Размер частиц, меш
Объемная плотность в сухом состоя-
нии, г/см3
Объемная плотность во влажном
состоянии, г/см3
Характерный размер пор, мкм
Активность6, ед./г
Производительность
Объем пустот в слое, %
Сухие гранулы
Окрашен в желто-коричневый цвет
Номинальный размер 12X20 (ци-
линдрические гранулы диаметром5
около 1 мм, длиной около 2,5 мм)
0,64—0,72
0,2 (±10%)
0,2
Не менее 0,04
В среднем до 907 кг 42%-ного обо-
гащенного фруктозой сиропа Ha-
О.45 кг фермента в течение 1000 ч
Около 45%
а Воспроизведено из статьи: Venkatasubramanian К., Harrow Н. S., Design and:
Operation of a Commercial Immobilized Glucose Isomerase Reactor System, Ann. N. Y.
Acad. Sci., 326, 141 (1979).
б Единица активности определяется как количество катализатора, способное пре-
вратить 10-9 молей субстрата в 1 мин при 60 °C и pH 8,0.
Таблица 4П2.3. Условия реакции изомеризации глюкозного сиропа
в реакторе колонного типа в слое иммобилизованного фермента3
Содержание сухого вещества, %
Содержание глюкозы в исходной
смеси, %
Требования к исходной смеси
pH
Падение pH
Температура, °C
Активатор
Время контакта фермента с субст-
ратом, ч
40—45
93—96
Смесь необходимо очистить фильт-
рованием, обработкой активирован-
ным углем и методом ионного об-
мена
8,2—8,5
На 0,2—0,4
60
0,0004 М Mg2+
0,5—4
а Воспроизведено с разрешения из статьи: Venkatasubramanian К., Harrow Н. S.,
Design and Operation of a Commersial Immobilized Glucose Isomerase Reactor Svstem,
Ann. N.Y. Acad. Sci., 326, 141 (1979).
мых колонных реакторов и их число. В силу упоминавшихся выше процес-
сов инактивации катализатора целесообразно устанавливать несколько ко-
лонн, так чтобы они инактивировались (и заменялись) последовательно; это
позволит максимальным образом использовать катализатор и в то же время
обеспечить постоянную высокую производительность установки в целом.
Потребность в ь-аминокислотах для производства пищевых,
продуктов и в медицинских целях постоянно возрастает. В свя-
зи с этим большое внимание уделяется разработке как микро-
биологических, так и химических методов получения ь-амино-
254
Глава 4
РИС. 4П2.1. Зависимость перепада давления в колонне от высоты слоя ката-
лизатора; колонна с нисходящим потоком раствора заполнена гранулами им-
мобилизованной глюкозоизомеразы. [Воспроизведено с разрешения из статьи:
Venkatasubramanian К., Harrow Н. S., Design and Operation of a Commercial
Immobilized Glucose Isomerase Reactor System; Ann. N.Y. Acad. Set, 326,
1141 (1979).]
кислот. Недостатком химических методов является рацемиче-
ская природа синтетических аминокислот. В общем случае
о-изомеры не имеют питательной ценности, поэтому желатель-
но получать только физиологически активные ь-аминокислоты.
Указанная цель была достигнута в процессе, разработанном
фирмой Tanabe Seiyaku Со., Ltd. (Осака, Япония), в котором
впервые в промышленном масштабе применялись иммобилизо-
ванные ферменты и результаты которого были опубликованы в
печати. Основой процесса является метод разделения оптических
изомеров в соответствии со следующей реакцией, катализируе-
мой ферментом аминоацилазой:
NHCOR1 NH2 NHCOR1
| аминоацилаза | I
DL-R—СН— СООН + Н2О------------> L-R—СН—СООН + D-R—СН—СООН
DL-ациламиио- Ь-амииокислота D-ациламино-
кислота кислота
Применение реакций, катализируемых ферментами
255
РИС. 4.11. Реакторы колонного типа с иммобилизованным ферментом, при-
меняющиеся фирмой Tanabe Seiyaku Со., Ltd. в производстве L-аминокислот.
(Фотография любезно предоставлена доктором И. Шибатой из фирмы Tanabe
Seiyaku Со., Ltd.)
Эту реакцию осуществляют в реакторе колонного типа с иммо-
билизованной аминоацилазой (рис. 4.11), после чего основной
продукт реакции, ь-аминокислоту, отделяют от негидролизован-
ной D-ациламинокислоты, используя их различные растворимо-
сти. Затем D-ациламинокислоту рацемизуют до оь-ациламино-
кислоты, которая снова поступает в колонну с аминоацилазой.
Общая схема процесса приведена на рис. 4.12.
Чтобы найти оптимальную форму иммобилизованного фер-
мента для этого процесса, Шибата, Тоса и их сотрудники из
фирмы Tanabe Seiyaku провели обширные исследования, ре-
зультаты которых частично изложены в опубликованной ими
статье [16]. В табл. 4.12 суммированы их выводы о свойствах
256
Глава 4
РИС. 4.12. Технологическая схема процесса с применением иммобилизованной
аминоацилазы, реализованного фирмой Tanabe Seiyaku. [Воспроизведено с
разрешения из работы: Chibata I. et al., Preparation and Industrial Application
of Immobilized Aminoacylases, in Fermentation Technology Today, Terui G. (ed.),
p. 387, Society of Fermentation Technology, Japan, Osaka, Japan, 1972.]
различных форм иммобилизованных ферментов. Эти данные еще
раз говорят о том, что при подборе катализатора для промыш-
ленного использования помимо начальной активности необходи-
мо учитывать и многие другие факторы. В рассматриваемом слу-
чае в конечном итоге исследователи остановились на амино-
ацилазе, иммобилизованной ионными связями на DEAE-сефа-
дексе, в силу высокой активности, простоты получения, возмож-
ности регенерации и устойчивости такого катализатора. В 1978г.
представители компании Tanabe Seiyaku сообщили, что этот ка-
тализатор используется уже более пяти лет без какого-либо
механического разрушения и снижения (связывающей) актив-
ности.
В 1983 г. было опубликовано сообщение о первом промыш-
ленном процессе с применением ферментов, иммобилизованных
полупроницаемыми мембранами. В данном случае речь шла о
процессе превращения Ы-ацетил-оь-метионина в u-метионин с
помощью ацилазного фермента. Изучены возможности приме-
нения иммобилизованных ферментов во многих других химико-
Применение реакций, катализируемых ферментами
257
Таблица 4.12. Свойства различных форм аминоацилазы (субстрат — ацетил-
ВЬ-метионин)а
Иммобилизованные аминоацилазы
Свойства Нативная ами- Ионное свя- Ковалентное связывание (с иодацетил- целлюлозой) Фермент, включенный в полиакрил- амидный гель
ноацилаза зывание (на DEAE-сефа- дексе)
Оптимальный pH 7,5—8,0 7,0 7,5—8,5 7,0
Оптимальная темпера- 60 72 55 65
тура, °C
Энергия активации, 6,9 7,0 3,9 5,3
ккал/моль
Оптимальная концент- 0,5 0,5 0,5 0,5
рация Со2+, мМ.
Кт, мМ 5,7 8,7 6,7 5,0
Отах, МКМОЛЬ/Ч 1,52 3,33 4,65 2,33
Методика получения Связывающие силы Возможность регенера- ции а Воспроизведено из статьи: Mort Т., Простая Слабые Возможна Sato Т., Tosa Сложная Сильные Невозмож- на Т., Chibata Сложная Сильные Невозмож- на Studies on Im'
mobilized Enzymes X: Preparation and Properties of Aminoacylase Entrapped into Acryl-
amide Gel-Lattice, Enzymologia, 43, 213 (1972).
технологических процессах (табл. 4.13). Некоторые из этих воз-
можностей уже реализованы в промышленности, другие нахо-
дятся на стадиях разработки и внедрения.
4.3.3. Применение иммобилизованных ферментов
в медицине и химическом анализе
В настоящее время известно уже более 120 врожденных ме-
таболических заболеваний человека; многие из этих дефектов
связаны с отсутствием активности у какого-либо одного кон-
кретного фермента, имеющегося в организме здоровых людей.
Например, фенилкетонурия (болезнь, приводящая к задержке
умственного развития), как полагают, вызывается недостатком
фермента, превращающего фенилаланин в тирозин. В настоящее
время терапевтический способ лечения фенилкетонурии сводит-
ся к специальной диете, не содержащей фенилаланина. Альтер-
нативным способом лечения могло бы быть введение недостаю-
щего фермента, однако фермент с такой же активностью, выде-
ленный из животных, вызывает резкую иммунологическую реак-
цию организма человека. Эту проблему, возможно, удастся ре-
шить, заключив фермент в микрокапсулы, волокна или гель.
Вполне вероятно, что иммобилизованный таким образом фер-
мент уже не вызовет защитной реакции иммунной системы, и в
17—537
258
Глава 4
Таблица 4.13. Применение иммобилизованных ферментов в химических
процессах и потенциальные сферы их использования®
Тип катализируе- мой реакции Иммобилизованный фермент Область применения
Окисление-восста- новление Оксидаза L-аминокис- лот 3-Тирозиназа Д'-Гидрогеназа Флавопротеиноксидаза Производство D-амииокислот Производство L-дигидрокси- фенилалаиина Производство L-тирозина Производство преднизолона N-Окисление лекарственных препаратов, содержащих аминные или гидразиновые
Миграция групп Гидролиз Лиазные реакции (асимметриче- ский синтез) Изомеризация Декстрансукраза Фосфорилаза Полинуклеотидфосфо- рилаза Карбаматкииаза Рибонуклеаза а-Амилаза Глюкоамилаза Целлюлаза Инвертаза Лейцинаминопептидаза Карбоксипептидаза Папаин Пенициллинамидаза Амииоацилаза Аденозинмонофосфат- деаминаза Аспартаза Триптофаназа D-Оксииитрилаза Г люкозоизомераза Производство декстрана Полимеризация глюкозы Получение полинуклеотидов Регенерация АТР Синтез тринуклеотидов Производство глюкозы Производство глюкозы Производство глюкозы Производство инвертного са- хара Разделение dl-аминокислот иа оптические изомеры Разделение DL-аминокислот на оптические изомеры Гидролиз казеина Производство 6-аминопеницил- лановой кислоты Синтез пенициллинов и цефа- лоспоринов Разделение DL-аминокислот на оптические антиподы Получение б'-инозиновой кис- лоты Производство L-аспарагиновой кислоты Производство L-триптофана Производство нитрила D-мин- дальной кислоты Производство фруктозы
а Воспроизведено Development, Chibata с разрешения из работы: Immobilized Enzymes, Research and I. (ed.), p. 164, Kodansha, Ltd., Tokyo, 1978.
то же время небольшие молекулы субстрата смогут контактиро-
вать с ферментом, проникая через стенки микрокапсулы, волок-
на или гель. Защищенные мембранами ферменты не подверга-
ются действию антител, но в то же время концентрация биока-
тализаторов на поверхности природных мембран снижает
Применение реакций, катализируемых ферментами 259
эффективность массообмена, а следовательно, и скорость ути-
лизации субстрата.
Вариант изложенного выше подхода был положен в основу
одного из предполагаемых проектов компактной искусственной
почки. Согласно этому проекту, уреазу и ионообменную смолу
или активированный уголь помещают в одну микрокапсулу; об-
разующийся в процессе разложения мочевины аммиак адсорби-
руется внутри микрокапсулы:
диффузия в
Мочевина -------------> мочевина
микрокап-
сулу
уреаза
----> НСОз~+^Н4+ —*- адсорбция на смо-
ле или на активи-
рованном угле
Из попыток применения иммобилизованных ферментов в
мелкомасштабном производстве следует отметить их использо-
вание для трансформаций стероидов (разд. 2.1.2). Так, напри-
мер, применяемый при лечении артрита кортизол можно полу-
чать из дешевого предшественника 11-дезоксикортизола в ко-
лонне с иммобилизованной 1 ip-гидроксилазой, затем кортизол
можно перевести в еще более ценный лекарственный препарат
преднизолон в реакторе со слоем иммобилизованной Д^дегид-
рогеназы. Обратите внимание на чрезвычайно высокую специ-
фичность этих ферментов. В настоящее время подавляющее
иммодилизо-
ванная Д -де-
, гидрогеназа
преднизолон
17*
260
Глава 4
из\
могут
РИС. 4.13. Электрод для опре-
деления мочевины; в этом
электроде уреаза иммобилизо-
вана в геле, нанесенном на по-
верхность стеклянного элект-
рода.
большинство трансформаций стероидов в промышленном мас-
штабе осуществляют микробиологическим путем.
Иммобилизованные ферменты уже сейчас широко применя-
ются в аналитической биохимии, и сфера их использования в
ближайшие годы, безусловно, еще больше расширится. Одним
ь электроды с иммобилизованными
ферментами, позволяющие осущест-
влять непрерывный контроль (мо-
ниторинг) за низкими концентра-
циями биохимически важных ве-
ществ. Так, в электроде для опреде-
ления мочевины (рис. 4.13) иммоби-
лизованная уреаза разлагает моче-
вину на ионы, которые могут быть
обнаружены обычными электрохи-
мическими методами. Электроды с
иммобилизованными ферментами
позволяют, как это показано схема-
тически на рис. 4.14, автоматизиро-
вать стандартные биохимические
анализы. Такая автоматизированная
система может применяться, напри-
мер, для определения концентраций
глюкозы или лактата с помощью
иммобилизованной глюкозооксидазы
или лактатдегидрогеназы.
На таком же принципе основаны
конструкции ферментных электро-
дов, предназначенных для опреде-
ления многих других биологически
важных соединений (табл. 4.14).
Для изучения ферментативных ре-
акций в мембранах недавно был
разработан также метод флуоримет-
рии на поверхностях, позволяющий непосредственно определять
концентрации различных ферментов, субстратов и кофакторов.
В приведенной в конце главы литературе читатель найдет об-
ширный дополнительный материал, детальнее освещающий эту
проблему.
Использование иммобилизованных биохимических соедине-
ний в аффинной хроматографии мы рассмотрим в гл. 11. Этот
метод, основанный на чрезвычайно высоком сродстве определен-
ных веществ, находящихся в растворе, к иммобилизованному
соединению, позволяет выделять, очищать и анализировать ин-
гибиторы ферментов, кофакторы, антигены, антитела и другие
вещества.
буферный
или конт-
рольный
раствор
0,2 мл/мин
Коллектор
ярой
Ч г 0,8 мл,/пин
1-мл шприц
с иммобили-
зованным в
геле фер-
ментом
Нейлоно-
вая сетка
/в мл/мин
Цветной
реагент
Ре г истрируюшее
устройство
Избыток
злюата"*
За
Стандартные
рас творог
мл/мин
Фотометричес-
кие ячейки
РИС. 4.14. Схема автоматической системы для определения глюкозы (с по-
мощью иммобилизованной глюкозооксидазы) или лактата (с помощью иммо-
билизованной лактатдегидрогеназы). [Воспроизведено с разрешения из статьи:
Hicks G. Т., Updike S. J., The Preparation and Characterization of Lyophilized
Polyacrylamide Enzyme Gels for Chemical Analysis, Anal. Chem., 38, 726 (1966).
© American Chemical Society.]
Таблица 4.14. Некоторые соединения, которые можно определять
с помощью электродов с иммобилизованными ферментами.
Ацетальдегид
Ацетилхолин
D-Аланин
L -Аланин
Алифатические иитросоединения
Щелочная фосфатаза
L -Аргинин
D -Аспартат
Бензальдегид
Холинэстераза
Креатин
Креатинфосфокиназа
L-Цистеин
Дегидрогеназы
Диамины
2-Дезокси-D-глюкоза
Этанол
Формальдегид
L-Галактонолактон
D-Галактоза
D-Глюкоза
D-Глутамат
L-Гулоно-А-лактон
Гипоксантин
D-Лактоза
Лактатдегидрогеназа
L-Лактоза
NADH
Пенициллин
Некоторые пестициды
L-Фенилаланин
Фосфат
Сульфат
L-Триптофан
L-Тирозин
Мочевина
Мочевая кислота
262
Глава 4
4.3.4. Утилизация и регенерация кофакторов
Наряду с применением в аффинной хроматографии кофакто-
ры представляют интерес и с точки зрения их способности обес-
печивать активность ферментов. В отсутствие кофакторов толь-
ко два класса ферментов из шести способны проявлять катали-
тическую активность. Очевидно, что для широкомасштабного
промышленного использования ферментов четырех других клас-
сов необходимо располагать эффективными способами получе-
ния, разделения и выделения достаточных количеств органиче-
ских кофакторов. Более того, для эффективной утилизации и
регенерации коферментов необходимы специальные реакторы и
каталитические установки. К настоящему времени многие из
перечисленных задач еще не решены, хотя в этой области и про-
водятся интенсивные исследования, направленные на реализа-
цию больших потенциальных практических и научных возмож-
ностей тщательно отработанной технологии кофакторов. Здесь
мы вкратце рассмотрим только некоторые из проблем, возни-
кающих при проектировании фермент-коферментных реакторов;
более подробные сведения по вопросам применения кофакторов
можно найти в соответствующей литературе, приведенной в кон-
це главы.
Если фермент и кофермент помещены в полые волокна с по-
лупроницаемыми стенками, через которые могут проникать и
субстрат, и продукты реакции, то ферментативную реакцию
можно провести без потери фермента и кофермента. На
рис. 4.15 приведена схема лабораторного реактора, основанно-
го на таком принципе. Очевидно, что при достаточной активно-
сти и стабильности каталитической системы подобная схема
может быть взята за основу и при проектировании более круп-
номасштабного процесса. В таком реакторе, в частности, изу-
чали протекающую в присутствии NAD двустадийную реакцию
окисления этанола:
алкогольдегидрогеиаза
Этанол + ЫАО+------------------* ацетальдегид+NAD Н + Н+
альдегиддегидрогеназа
Ацетальдегид4-;ЫАО+------------------- ацетат+ЫАОН-|-Н+
Очевидно, что этот процесс может протекать непрерывно только
при условии постоянной регенерации NAD+ путем окисления
NADH; для этой цели в реакционную смесь, находящуюся внут-
ри полых волокон, добавляли фермент диафоразу (из сердца
свиньи). Диафораза катализирует окисление NADH кислоро-
дом, причем в процессе регенерации кофактора NAD+ образу-
ется также Н2О2. Поскольку пероксид водорода инактивирует
многие ферменты, ферментную систему дополняют еще фермен-
том— каталазой. Понятно, что такой подход может быть рас-
Применение реакций, катализируемых ферментами
263
пространен и на другие ферментативные реакции. В то же вре-
мя если молекулярная масса субстрата близка молекулярной
массе кофермента (коферментов) и ферментов, то потребуется
некоторая модификация схемы. (Объясните, почему.) Если же
и молекулы кофермента относительно невелики, то устранить
их потерю за счет ультрафильтрации можно, например, путем
Проточная''
кювета
РИС. 4.15. Схема лабораторного реактора с полыми полупроницаемыми во-
локнами, в котором осуществляется непрерывный процесс, катализируемый
фермент-коферментной системой.
ковалентного связывания с растворимым полимером типа поли-
этиленгликоля, декстрана или полилизина.
Теперь рассмотрим такие катализируемые фермент-кофер-
ментной системой процессы, в которых ферменты и (или) соот-
ветствующие коферменты связаны с нерастворимыми носителя-
ми. Любой катализатор такого типа должен обеспечивать, во-
первых, физический контакт фермента с коферментом и, во-вто-
рых, возможность регенерации кофактора. Можно наметить по
меньшей мере три различных подхода к решению такого типа
задач.
1. Можно иммобилизовать кофермент, а фермент и все необхо-
димые для регенерации вещества вводить в растворенном
виде. Тогда в реакционной смеси на выходе из реактора бу-
дут содержаться все реагенты, за исключением кофермента.
264
Глава 4
2. Можно иммобилизовать фермент, а кофермент оставить в
растворе; следовательно, он перейдет и в продукты реакции.
Другие ферменты и субстраты, необходимые для регенерации
кофермента, можно вводить в реактор в виде раствора.
В альтернативном варианте процесс регенерации может осу-
ществляться в отдельном реакторе, в котором также могут
использоваться иммобилизованные ферменты.
3. Можно, наконец, соединить молекулы фермента и кофермен-
та длинной гибкой цепью и затем иммобилизовать этот комп-
лекс фермент—кофермент («кофермент на привязи»).
Первый и второй подходы были реализованы в лабораторных
экспериментах, а возможности третьего подхода пока еще толь-
ко изучаются. При переходе к промышленному производству
выбор между первым и вторым подходами будет определяться
целым рядом технологических и экономических факторов, из ко-
торых важнейшими являются легкость осуществления, эффек-
тивность и стоимость процессов выделения и повторного исполь-
зования фермента, кофермента и (или.) системы регенерации
последнего. Если, например, повторное использование ни одно-
го из этих компонентов реакционной смеси не представляется
возможным, то первый подход может оказаться более целесо-
образным, поскольку большинство коферментов намного дороже
любого другого компонента смеси.
Высокая эффективность процессов утилизации и регенерации
кофакторов в живых клетках подсказывает принцип альтерна-
тивного подхода к разработке процессов, катализируемых фер-
мент-коферментными системами. Этот подход заключается в
иммобилизации живых клеток и последующем использовании
природных систем синтеза и регенерации кофакторов. Практи-
ческая реализация этого внешне заманчивого подхода, однако,
пока что была не слишком успешной. В частности, до сих пор
не решены проблемы обеспечения стабильности иммобилизован-
ных клеток и введения необходимых фермент-коферментных си-
стем в легкоиммобилизуемые, активные клетки. Последняя
проблема в принципе может быть решена методами генетиче-
ской инженерии, позволяющими внедрить в геном подходящей
клетки ген, синтезирующий необходимый фермент; тогда клет-
ка будет выполнять в основном функции хранилища кофактора
и соответствующей системы его регенерации. Другое принци-
пиальное затруднение, возникающее при работе с иммобилизо-
ванными клетками, связано с регуляцией транспорта через обо-
лочку клетки. Точно так же, как и в случае фермент-кофермент -
ной каталитической системы, иммобилизованной полупроницае-
мой мембраной, живая клетка должна пропускать внутрь клет-
ки субстраты, выделять во внеклеточную среду продукты реак-
Применение реакций, катализируемых ферментами
265
ции и в то же время не должна терять ферменты, кофакторы и
другие вещества, необходимые для нормального функциониро-
вания внутриклеточных каталитических систем.
4.4. Кинетика реакций,
катализируемых иммобилизованными
ферментами
Для проектирования реакторов с иммобилизованными фер-
ментными катализаторами необходимо достаточно хорошо знать
кинетические свойства иммобилизованных ферментов. Наблю-
даемые каталитические свойства отдельной частицы катализа-
тора на основе иммобилизованного фермента или реактора,
в котором ферменты удерживаются полупроницаемой мембра-
ной, определяются взаимосвязанным сочетанием двух процес-
сов— транспорта субстрата и каталитической ферментативной
реакции. В этом разделе основное внимание будет уделено изу-
чению взаимосвязи между массопередачей и каталитической ре-
акцией и ее влияния на каталитическую активность отдельной
частицы иммобилизованного фермента.
Наше изучение путей взаимодействия массопередачи и хими-
ческих процессов, определяющего общую активность, инактива-
цию, и другие параметры иммобилизованного ферментного ката-
лизатора, мы начнем со схемы, приведенной на рис. 4.16. На
этой схеме изображено поперечное сечение тонкого слоя иммо-
билизованного фермента, контактирующего с обеих сторон с
раствором субстрата. Вдали от катализатора концентрация суб-
страта и другие параметры, например pH, имеют те же значе-
ния, что и во всей реакционной среде. Эти параметры легко
определить обычными методами аналитической химии.
Поскольку субстрат трансформируется внутри иммобилизо-
ванного фермента и здесь же образуется продукт реакции, меж-
ду раствором вдали от катализатора и активными центрами им-
мобилизованного фермента возникают градиенты концентраций.
В частности, субстрат должен транспортироваться из раствора
к наружной поверхности катализатора. В отсутствие переме-
шивания этот процесс осуществляется за счет молекулярной
диффузии. Обычно скорость транспорта субстрата к поверхно-
сти катализатора повышают путем перемешивания или создания
потока его раствора. Если активный фермент находится только
на поверхности иммобилизованного фермента или если субстрат
не может проникнуть внутрь частицы катализатора, то нам
следует учитывать только эту внешнюю массопередачу. Часто,
однако, фермент равномерно распределен по всему объему про-
ницаемой для субстрата матрицы. В таком случае каталитиче-
266 Глава 4
ОК и дная----
.(раза
Граничнош
слой или
стоячая пленка
Слой иммоби---
лизоданного.
фе/змента
[Транспорт субстрата через
^наружный граничный слой
Диффузия субстрата 0 слой
катализатора
Диффузия продукта реакции
из катализатора
Транспорт продуктов реакции
0 жидкую среду
яд (субстрат)
ПАРАМЕТРЫ
козакрициенту
•Ц.ру концентрация
'субстрата ц продукта
реакции 0 жидкой среде
Толщина
слоя ката-~
лизатора 2L
Козффициен'ть/
- масса передачи
(*з’ кр)
Собственные
кинетические.
//параметры
РИС. 4.16. Схема массопередачи и химических превращений в двустороннем
слое иммобилизованного фермента. Указанные в верхней части рисунка явле-
ния обусловливают специфические профили изменения концентраций субстра-
та и продукта реакции, изображенные в средней части рисунка. Эти профили
могут быть описаны количественно с помощью параметров, перечисленных
в нижней части схемы.
'атализи- С
руемая
ферментом-
реакция
ПРОФИЛИ
ИЗМЕРЕНИЕ
КОНЦЕНТРАЦИИ
Средняя линия слоя
р0 (продукт
еакции)
ская активность большей частью сосредоточена и распределена
внутри гранулы катализатора, и субстрат, очевидно, должен
диффундировать в гранулу до некоторого активного центра, где
и осуществляется реакция. Тот же путь, но в обратном направ-
лении, должны проходить и продукты реакции. В таких случаях
необходимо учитывать как внешнюю массопередачу, так и про-
цессы диффузии внутри частицы катализатора.
Как показано на рис. 4.16, реакция в слое иммобилизованно-
го фермента протекает со скоростью, определяемой концентра-
циями внутри этого слоя. В силу упоминавшихся выше градиен-
Применение реакций, катализируемых ферментами
267
тов концентрации локальные скорости реакции изменяются в
зависимости от положения центра реакции, а общая скорость
превращения субстрата представляет собой сумму всех локаль-
ных скоростей трансформации субстрата внутри проницаемого
катализатора. В стационарном состоянии эта общая скорость
равна скорости транспорта субстрата к катализатору. Очевидно,
что в общем случае скорость всего процесса зависит как от
скорости транспорта, так и от собственной скорости каталити-
ческой реакции.
Вывод математических выражений, позволяющих описать эти-
взаимосвязанные процессы, и критериев, дающих возможность
оценить их относительную важность, явился большим достиже-
нием химической технологии. Эти взаимосвязанные процессы,
играют очень большую роль в катализируемых иммобилизован-
ными ферментами и иммобилизованными клетками реакциях;
в то же время их роль часто не учитывалась как в теоретиче-
ских исследованиях, так и в разработке путей практического при-
менения иммобилизованных биокатализаторов. Поэтому мы
рассмотрим здесь достаточно подробно основные принципы и
методы математического моделирования процессов на иммоби-
лизованных ферментах, а также его результаты. Еще более де-
тально эта тема освещена в учебных пособиях по химической
технологии и в ряде монографий.
4.4.1. Влияние внешнего сопротивления массопередаче
Чтобы читателю было легче усвоить некоторые основные по-
нятия и специфическую терминологию, мы начнем с самого про-
стого варианта. Предположим, что фермент иммобилизован
только на наружных поверхностях носителя, имеющего форму
пластины. В этом случае мы должны рассматривать только
массопередачу из раствора к поверхности носителя и реакцию
на этом носителе.
Одна из традиционных моделей, которую биохимики назы-
вают диффузионным слоем Нернста, а технологи — стоячей
пленкой или граничным слоем, приводит к следующему выра-
жению для потока (выраженному в молях в единицу времени
на единицу поверхности) субстрата из жидкой фазы (биохи-
мики иногда называют ее пулом) к границе раздела фаз:
Ns = fes(s0 — s) (4.2)
Здесь s и So — концентрации субстрата на границе раздела фаз
и в жидкой фазе соответственно, a ks — коэффициент массопере-
дачи. Последний зависит от физических свойств среды, а также
от гидродинамических условий вблизи границы раздела фаз и
может быть определен по известным корреляциям, рассматрц-.
268
Глава 4
ваемым в гл. 8. В частности, ks возрастает при повышении ско-
рости потока через реактор колонного типа с насадкой иммоби-
лизованного фермента.
В стационарном состоянии субстрат не может аккумулиро-
ваться на поверхности катализатора, поэтому скорость поступ-
ления субстрата за счет массопередачи должна быть равна ско-
рости его расхода в реакции на поверхности катализатора. Если
допустить, что скорость v ферментативной реакции на поверх-
ности (выраженная в молях в единицу времени на единице по-
верхности) подчиняется уравнению Михаэлиса — Ментен, то по-
лучим
ks(sl>-s')=v = -^- (4.3)
Л/п "Г s
Число необходимых для описания системы параметров (ks, s0,
Umax и Кт) можно уменьшить до двух (Da и к) путем введения
следующих безразмерных переменных:
х = Da=-^- х = (4.4)
Sfl k^Sg S о
С учетом этих параметров уравнение материального баланса
по субстрату примет вид
1 ~х. =___(4.5)
Da х 4- х
где 0^х^1,0.
Здесь необходимо подчеркнуть физический смысл парамет-
ра Da, числа Дамкелера:
р __ Ртах __ максимальная скорость реакции (4 6)
&ss0 максимальная скорость массопередачи ' ' 1
Так, если Da значительно меньше единицы, то максимальная
скорость массопередачи намного превышает максимальную ско-
рость реакции (низкое сопротивление массопередаче). Если же
сопротивление массопередаче велико, то этот фактор ограничи-
вает скорость всего процесса, a Da принимает значения, намного
превышающие единицу. Эти случаи известны как лимитируемый
реакцией режим и лимитируемый диффузией режим соответ-
ственно.
Алгебраическим преобразованием уравнения (4.5) легко по-
лучить квадратное уравнение, позволяющее определить х ана-
литическим путем:
где
P=Da + x-1 (4-8)
Применение реакций, катализируемых ферментами
269
В уравнении (4.7) знаки + и — применяются при [}>0 и р<0
соответственно. Если [3 = 0, то х = Используя это выражение
для s/s0, мы можем с помощью правой или левой части уравне-
ния (4.7) оценить безразмерную наблюдаемую' скорость реак-
ции y/fmax- Следует помнить, что в общем случае зависимость v
от So не выражается уравнением Михаэлиса — Ментен. Кроме
того, Кт уже нельзя считать равным Sy2, когда v ровно в два
раза меньше наблюдаемой максимальной скорости реакции
»пшарр, поскольку величина Si/2 зависит от Da.
Тем не менее параметр sy2 часто называют кажущейся кон-
стантой Михаэлиса (Ктарр) и используют для оценки влияния
массопередачи. Если перед исследователем стоит только задача
определения Si/2, такое упрощение может оказаться удобным, но
в общем случае оно может привести к серьезным ошибкам. На-
пример, можно прийти к выводу, что наблюдаемой скорости ре-
акции соответствует следующее некорректное уравнение:
,,ЭРР S
гтах5о
СРР+*о
(4.9)
В каком-то конкретном случае это уравнение может даже удов-
летворительно описывать наблюдаемую кинетику процесса, но
в целом оно неверно, поскольку в нем не учтена зависимость
Ктарр от свойств жидкой среды и гидродинамических свойств
слоя вблизи границы раздела фаз. Как показывают эксперимен-
тальные данные, такая зависимость может резко изменять ки-
нетику процесса (рис. 4.17).
В химической технологии влияние массопередачи на общую
скорость реакции традиционно выражают через коэффициент
эффективности rj, физический смысл которого определяется сле-
дующим образом:
_________наблюдаемая скорость реакции_________ ,, , qx
скорость, которая наблюдалась бы в отсутствие_' ' ’
сопротивления массопередаче, т. е. при условии
равенства концентраций субстрата на поверхности
катализатора (s) и в растворе ($о)
Следовательно, в нашем случае
х/(х + х)
1/(х+1)
(4.Н)
Поэтому т]^1, и, таким образом, в общем случае повышение
сопротивления массопередаче будет сопровождаться снижением
наблюдаемой активности катализатора. На рис. 4.18 приведены
результаты эксперимента, подтверждающие такую зависимость.
270
Глава 4
РИС. 4.17. Экспериментальное обнаружение взаимосвязи между скоростью
трансформации субстрата 6 [6= (s; — se)/sf, где Sf и se — концентрации суб-
страта в исходной смеси и в продуктах реакции соответственно] и скоростью
потока [указана цифрами на кривых (мл/ч)] в реакторе со слоем иммобили-
зованного фермента (изучалась реакция гидролиза этилового эфира бензоил-
L-аргинина фицином на карбоксиметилцеллюлозе). Если бы кинетика процес-
са подчинялась уравнению Михаэлиса — Ментен, то при потоке идеального
вытеснения наклон всех кривых был бы одинаков и равен Кт- Изменение
наклона с изменением скорости потока свидетельствует о значительном влия-
нии массопередачи на кинетику процесса в целом. [Воспроизведено с разре-
шения из статьи: Lilly М. D., Hornby W. Е„ Crook Е. М., The Kinetics of Car-
boxymethylcellulose— Ficin in Packed Beds, Biochem. J., 100, 718 (1966).]
Если параметр Da близок нулю (очень медленная по сравне-
нию с максимальной скоростью массопередачи реакция), то, со-
гласно уравнению (4.5), х приближается к единице, и, следова-
тельно, для лимитируемого реакцией режима (Da->0)
Т1=1 ^maxSo., (4.12)
В этом случае кинетика процесса не отличается от истинной,
Применение реакций, катализируемых ферментами
271
собственной кинетики реакции на границе раздела фаз жид-
кость— твердое тело. Если приготовлен новый катализатор, со-
держащий иммобилизованный фермент на поверхности, то всегда
следует экспериментально оценить, как методика иммобилиза-
ции отразилась на каталитических свойствах фермента. Необхо-
димое для этой цели определение параметров Umax и Кт должно
РИС. 4.18. Экспериментальный метод обнаружения эффектов внешней диф-
фузии; здесь v* — скорость реакции при высоких скоростях потока через
насадочную колонну (изучалась реакция гидролиза 4-Ю-4 М раствора га-нит-
роанилида М-бензоил-оь -аргинина иммобилизованным трипсином при pH 8
и 25 °C). [Воспроизведено с разрешения из работы: Ford J. R. et al., Recircu-
lation Reactor System for Kinetic Studies of Immobilized Enzymes, in Enzyme
Engineering, Wingard L. B., Jr. (ed.), Wiley-Interscience, New York, 1972.]
проводиться в условиях, когда Da<Cl; только тогда будут уст-
ранены маскирующие диффузионные эффекты.
Сведения о собственной кинетике ферментативной реакции
необходимы также при разработке и проектировании реакторов
с иммобилизованными ферментами, поскольку только таким пу-
тем можно достаточно надежно учесть влияние свойств жидкой
среды, формы носителя фермента и характеристик смешения.
Разработано много типов экспериментальных реакторов, пред-
назначенных для изучения реакций обычного гетерогенного ка-
тализа в лимитируемом реакцией режиме. Некоторые из таких
реакторов применяли и для исследования кинетики реакций, ка-
тализируемых иммобилизованными ферментами (рис. 4.19).
В целях минимизации сопротивления массопередаче во всех этих
реакторах вблизи катализатора создавались высокие скорости
потока (большие ks, малые Da). Что касается иммобилизован-
272
Глава 4
ных ферментов, то такой подход имеет несколько недостатков.
Во-первых,^как мы уже упоминали в разд. 3.7.3, гидродинамиче-
ское воздействие может вызвать частичную или даже полную
денатурацию ферментов, связанных с носителем. Во-вторых, ме-
ханическое перемещение одних частиц катализатора относи-
тельно других может приводить к потере фермента за счет ис-
тирания.
Проточная
K/ofema
РИС. 4.19. Реактор с рециркуляцией для изучения влияния внешней массопе-
редачи на кинетику реакций, катализируемых иммобилизованными фермен-
тами.
В системах, в которых скорость процесса зависит как от хи-
мической реакции, так и от массопередачи, лимитируемый диф-
фузией режим возникает тогда, когда утах значительно больше
fess0, т. е. при Da^>l. Для лимитируемого диффузией режима
(Da->oo, % ограничен) путем ряда преобразований уравнения
(4.7), включая разложение в ряд квадратного корня, можно по-
лучить следующие выражения:
14-х
Da
(4.13)
Таким образом, при очень больших значениях Da реакция имеет
первый порядок по общей концентрации субстрата и ее ско-
рость v совершенно не зависит от собственных параметров ката-
лизатора Утах И В такой ситуации собственные кинетические
параметры иммобилизованных ферментов вообще не проявляют-
ся. В лимитируемом диффузией режиме, например, при задан-
Применение реакций, катализируемых ферментами
273
ном So наблюдаемая активность постоянна, даже если ферменты
на границе раздела фаз на самом деле теряют активность,
в частности за счет изменения в неблагоприятную сторону тем-
пературы, pH или других условий процесса. Поэтому изучение
скорости денатурации иммобилизованного фермента должно
проводиться только в условиях, максимально приближенных к
лимитируемому реакцией режиму.
4.4.2. Моделирование диффузии
и реакций внутри частицы катализатора
Как мы уже не раз упоминали выше, обычно ферменты им-
мобилизуют путем связывания с внутренними поверхностями по-
ристых носителей или путем включения в матрицы, через кото-
рые может диффундировать субстрат. В таких системах для
расчета наблюдаемой скорости трансформации субстрата необ-
ходимо знать профиль изменения его концентрации в частице
катализатора. Для этого в свою очередь прежде всего надо най-
ти уравнение материального баланса в стационарном состоянии
для тонкого слоя проницаемого катализатора. Предположим,
что фермент иммобилизован в грануле, форму которой можно
считать сферической. Тогда необходимый тонкий слой будет
заключен между двумя концентрическими сферами радиусов г
и r+dr соответственно (рис. 4.20). Толщина этого слоя настоль-
ко мала, что все условия внутри него можно считать постоян-
ными независимо от положения.
Символами 2)es и v обозначают эффективный коэффициент
диффузии субстрата и локальную скорость его трансформации
соответственно. Оба этих параметра принципиально отличаются
от аналогичных параметров для процессов в растворе; сущест-
венна и количественная разница между ними. Рассмотрим сна-
чала эффективный коэффициент диффузии, учитывая, что ско-
рость диффузии любых веществ через носитель зависит от сле-
дующих факторов:
1. Часть поперечного сечения частицы катализатора занята мо-
лекулами твердого носителя и поэтому не может использо-
ваться для диффузионного транспорта субстратов (соответ-
ствующий параметр — пористость частицы ер).
2. Сеть пор переплетена очень сложным образом, поэтому диф-
фузия может происходить только в разрешенных, часто ме-
няющихся направлениях (соответствующий параметр — ко-
эффициент извилистости т).
3. Поры могут иметь очень небольшой диаметр, сравнимый с
размерами молекул субстрата (ограниченная диффузия; соот-
ветствующий параметр КР!Кг)-
18-537
274
Глава 4
Проницаемая
гранула ката-
лизатора
Уравнение материального баланса для стационарного состояния в тонком
слое, ограниченном радиусами г и r+dr, можно записать в следующем виде:
(“ dr •4яг2)
I es dr /
Г \ /
= и-4лгМг
r+dr
Если принять, что эффективный коэффициент диффузии субстрата Sbes по-
стоянен, то, разделив на 4ndr, получим
ds I ds
^~L~r
dr
В пределе при dr-^Q
r2v
или
РИС. 4.20. Вывод уравнения стационарного состояния сферической проницае-
мой гранулы иммобилизованного фермента с помощью концепции тонкого
слоя.
Теперь мы можем выразить эффективный коэффициент диффу-
зии через эти параметры:
(4.14)
Здесь 3)s0 — коэффициент диффузии субстрата в жидкой реак-
ционной среде. Параметр пористости ер необходимо определять
экспериментально для каждого конкретного носителя. Для коэф-
фициента извилистости обычно принимают значения, располо-
женные в диапазоне от 1,4 до 7. Что касается ограниченной
диффузии, то соответствующий параметр KplKr в первом при-
ближении можно оценить, воспользовавшись следующим урав-
нением:
Кр 7 । У
V к Гр)
(415)
Применение реакций, катализируемых ферментами 275
где rs и гр— (эквивалентный) радиус молекул субстрата и ха-
рактеристический радиус поры соответственно. Неопределен-
ность всех параметров в правой части уравнения (4.14) являет-
ся причиной того, что найти достоверное значение 3)es расчет-
ным путем очень трудно. Поэтому предпочтительнее оценивать
Фе5 на основе экспериментального изучения общей кинетики
процесса. Этот метод мы рассмотрим позднее, после анализа
взаимного влияния диффузионных и химических процессов
внутри частицы иммобилизованного фермента.
Следует отметить также, что массопередача внутри частицы
может зависеть от химии ее внутренних поверхностей и от на-
личия ионизированных групп. Более того, если в реакции участ-
вуют или образуются ионизированные соединения, то внутри
частицы катализатора могут создаваться градиенты электриче-
ского потенциала, которые в свою очередь будут изменять ско-
рости транспорта заряженных веществ. Методы анализа и уче-
та этих факторов читатель может найти в литературе, приведен-
ной в конце главы.
Если допустить, что собственная кинетика локальной фер-
ментативной реакции с участием иммобилизованного фермента
подчиняется уравнению Михаэлиса — Ментен:
V = . (4.16)
Лт + s
то параметр максимальной скорости в данном случае можно оп-
ределить как произведение нагрузки носителя ферментом eimm
(количество микромолей фермента в 1 г носителя) на удельную
активность иммобилизованного фермента <7E,imm (число микро-
молей субстрата, превращаемых в 1 с 1 мкмолем фермента) и
на плотность частиц рр (количество граммов носителя в единице
его объема):
^тах — бтт ' Рр' ^E.imm (4.17)
Напомним, что иммобилизация может изменять структуру фер-
мента и (или) его молекулярное окружение, и, следовательно,
величины <7E,imm и Кт для иммобилизованного фермента могут
отличаться от соответствующих величин для фермента в раство-
ре. Как мы уже отмечали в разд. 4.4.1, собственную кинетику
фермента желательно изучать в условиях минимального вклада
в общую кинетику системы эффектов, обусловленных массопе-
редачей. Критерий для таких лимитируемых реакцией условий
мы найдем в ходе приведенного ниже анализа.
Уравнение материального баланса по субстрату в стационар-
ном состоянии, вывод которого приведен на рис. 4.20, представ-
18*
276
Глава 4
ляет собой обычное дифференциальное уравнение второго по-
рядка:
d2s
dr2
2 ds \ ,
-----т— = V (s)
г dr 4 '
(4-18)
Изображенную на рисунке модель необходимо дополнить гра-
ничными условиями. Профиль изменения концентрации в части-
це катализатора почти всегда симметричен относительно цент-
ра сферы, т. е.
4- =0 (4.19)
иг г=0
В этом анализе мы допускаем, что концентрация субстрата на
наружной поверхности гранулы равна концентрации субстрата
So в жидкой среде, омывающей гранулу. (Случаи одновременно-
го сопротивления массопередаче в граничном слое и внутри
частицы будут рассмотрены в следующем разделе.) Следова-
тельно,
s]r=« = s0 (4.20)
Наблюдаемая общая скорость v0 утилизации субстрата час-
тицей катализатора равна поступлению субстрата в гранулу за
счет диффузии. Величина и0, выраженная в молях на единицу
объема гранулы в единицу времени, составляет
(4'21)
Здесь символами Vp и Ар обозначены объем частицы и площадь
ее наружной поверхности соответственно. Как и в предыдущем
разделе, такие скорости мы будем выражать в виде безразмер-
ного параметра, определяющего влияние диффузионных эффек-
тов. Коэффициент эффективности ц определяется как
__ v0 ____________наблюдаемая скорость (Л
v (so) скорость, которая наблюдалась бы при_' '
отсутствии градиентов концентраций в
грануле
К сожалению, если в соответствии с уравнением (4.16) и из-
меняется нелинейно, то определить коэффициент эффективно-
сти аналитическим путем непросто. Для этого необходимо
решить в цифровой форме задачу о граничном значении [уравне-
ния (4.18) — (4.20)] и затем по уравнению (4.21) найти и0. Та-
кие вычисления достаточно сложны и трудоемки, поэтому мы
постараемся выразить результаты в наиболее простой и в то же
время достаточно общей форме. Для этого преобразуем приве-
Применение реакций, катализируемых ферментами
277
денные выше уравнения в эквивалентные безразмерные выра-
жения.
Введем параметры s = s/s0 и r = rlR‘, тогда уравнение (4.18)
можно записать в следующей форме:
dr2 г dr S)esso 1 + ₽s
Здесь безразмерные параметры Ф и £ определяются как
Л —— 1Л8 = 4- (4.24)
Соответствующие уравнению (4.23) безразмерные граничные
условия равны
(4-25)
Физический смысл параметра Ф, называемого модулем Тила,
заключается в том, что квадрат модуля Тила представляет со-
бой частное от деления скорости реакции первого порядка
R3(ymax/Km)s0 на скорость диффузии R0essQ. Параметр насы-
щения является мерой отклонения локальных скоростей от ки-
нетики первого порядка, причем большие значения £ свидетель-
ствуют о том, что порядок реакции приближается к нулевому.
Выразим через эти безразмерные переменные коэффициент
эффективности
т) = (426)
зф2[1/(1 + ₽)1
Согласно уравнениям (4.23) и (4.25), s является функцией толь-
ко г, Ф и р, и поэтому (dsldfyr=\ зависит только от £ и Ф. Учи-
тывая уравнение (4.26), отсюда следует, что т] зависит только
от модуля Тила и параметра насыщения:
Т)=ЛМ) (4.27)
При использовании выражения (4.27) для оценки коэффи-
циента эффективности возникает проблема, связанная с тем, что
собственные параметры кинетики каталитической реакции итах
и Кт часто неизвестны. С помощью простых преобразований
можно показать, однако, что при определении т] можно обойтись
и без первого параметра итах. Действительно, из уравнений
(4.26) и (4.21) следует, что
0 =
9т].0е58о
(1+Р)Т'2
Подстановка этого выражения в правую часть уравнения (4.27)
278
Глава 4
приводит к уравнению (4.28), отражающему неявную зависи-
мость между т], £ и новым безразмерным наблюдаемым моду-
лем ф, который определяется следующим образом:
ф = —f-Хг-У (4.28)
<®esso \ Ар /
(Обратите внимание на то, что Ф зависит только от наблюдае-
мой общей скорости и0 и не зависит от собственных кинетиче-
ских параметров каталитической реакции.) Решение этого не-
РИС. 4.21. Коэффициенты эффективности иммобилизованных ферментных
катализаторов с собственной кинетикой, описываемой уравнением Михаэли-
са— Ментен (j3=s0/Km)- Определение безразмерного наблюдаемого модуля
Ф дано в уравнении (4.28).
явного уравнения позволяет определить т) как функцию от
и наблюдаемого модуля Ф:
П=£(Ф, (4.29)
На рис. 4.21 приведены графики зависимости т] от Ф, выра-
жаемые уравнением (4.29) при (3 ->0 и (1—>оо. Поскольку коэф-
фициенты эффективности для промежуточных значений Кт/$ъ
располагаются между кривыми для этих двух предельных слу-
чаев, нетрудно видеть, что коэффициент rj сравнительно не чув-
ствителен к последнему собственному параметру Kmlso.
Прежде чем перейти к изучению аналитических решений, ос-
нованных на приближенной оценке кинетики процессов, нам
нужно найти выражение для т), применимое при больших значе-
ниях Ф (или Ф) и кинетике, выражаемой уравнением Михаэли-
Применение реакций, катализируемых ферментами
279
са — Ментен. Если модуль Ф достаточно велик (Ф^З; см.
табл. 4.15), то субстрат быстрее утилизируется ферментом, чем
диффундирует внутрь гранулы катализатора. В такой ситуации,
когда скорость всего процесса определяется скоростью диффу-
зии, можно допустить, что весь субстрат трансформируется в
граничащем с наружной поверхностью тонком слое гранулы ка-
тализатора, поэтому эффектом кривизны (Zlr-dsfdr) в уравне-
Таблица 4.15. Критерий для оценки влияния эффектов массопередачи
на кинетику всего процесса
Критерий Значение f] Стадия, определяющая скорость процесса Влияние эффектов массопередачи
Ф<0,3 Химическая реакция Незначительное
Ф>3 ос ф-1 Диффузия Большое
нии (4.18) можно пренебречь. Отсюда следует, что
d2s
I -------= и
es dr2
(4.30)
Если в уравнение (4.30) вместо d2sjdr2 подставить правую часть
уравнения (4.31)
d2s _ 1 d f ds \2
dr2 ~ 2 ds [ dr )
и затем интегрировать no s, то мы получим следующее выра-
жение:
ds I
dr |г=д
so
9 Г
— v(s)ds
->es J
sc
1/2
(4-32)
Здесь sc и s0 — концентрации субстрата в центре частицы (г = 0)
и на ее поверхности (r=R) соответственно.
В случае лимитируемой диффузией реакции st~0, что по-
зволяет вычислить интеграл в уравнении (4.32). Совместное ре-
шение уравнений (4.21), (4.22) и (4.32) приводит к следующему
выражению для коэффициента эффективности, справедливому
при достаточно больших значениях Ф или Ф\
2
S)es
__
большие Ф, ф R
(общая скорость
процесса опре-
деляется диф-
фузией)
so
j V (s) ds
0
У (So)
1/2
(4.33)
280
Глава 4
Преобразование этого уравнения для частного случая кинетики,
выражаемой уравнением Михаэлиса — Ментен (4.16), дает
’>b»i=v + (4.34)
Продолжая изучение кинетики, выражаемой уравнением Ми-
хаэлиса— Ментен, отметим, что если допустить первый порядок
реакции (это справедливо при то, согласно данным, при-
веденным на рис. 4.21,
v = ks (4.35)
где k=vma-x.lKm, и т] принимает консервативное (низкое) значе-
ние, достаточно близко приближающееся к истинному значению.
Воспользовавшись линейной зависимостью (4.35), мы можем
решить аналитическим путем уравнения модели лимитируемого
диффузией процесса; таким образом получим
4=v(vh—Л-) (4-36>
В этом выражении модуль Тила Ф равен
= (4.37)
Нр
С помощью уравнения (4.37) можно получить изображенную
на рис. 4.21 кривую, отвечающую реакции первого порядка, если
использовать справедливую в такой ситуации зависимость:
ф = П02 (4.38)
Другой предельный случай кинетики Михаэлиса — Ментен
характерен для реакций, порядок которых приближается к нуле-
вому (s^>/(m); такую ситуацию можно определить следующим
образом (&0 = Umax):
u = |fe0=c°nst при s>0 39
( 0 при $=0
Совместное решение задачи о граничном значении [уравнения
(4.18)—(4.20)] и функции утилизации фермента показывает, что
концентрация субстрата s зависит от радиуса г следующим об-
разом:
s=s0--$-(R2-r!) (4-40)
Это выражение справедливо при всех неотрицательных значе-
ниях s, т. е. в интервале от r=R до некоторого критического ра-
диуса r=Rc, определяемого путем решения уравнения (4.18)
Применение реакций^ катализируемых ферментами
281
совместно с уравнением (4.20) при следующих граничных ус-
ловиях:
s=0 и -^- = 0 при r=Rc
Это решение приводит к кубическому уравнению
но Rc:
/ Re \3 3 / Rc\2 . 1 3£>ess0
\ R ) 2 \ R ) Ф 2 ~ k0R2
(4.41)
относитель-
(4.42)
Таким образом, если уравнение (4.42) имеет положитель-
ный корень (Rc/R), величина которого меньше единицы, то внут-
ри гранулы катализатора существует некоторая область (от
г=0 до r = Rc), в которой s = 0 и у = 0. В таких случаях реакция
протекает только во внешнем слое частицы (Rc<Zr^R), так что
~n(R3-R3c)k0
--------4-------
--------nR3k0
3
(4-43)
Если не существует имеющего физический смысл решения урав-
нения (4.42), то т] = 1 и скорость трансформации субстрата во
всем объеме гранулы катализатора одинакова.
Другим важным следствием изображенных на рис. 4.21 за-
висимостей является малое влияние геометрии частицы катали-
затора на взаимозависимость т] и Ф. Например, коэффициент
эффективности реакции первого порядка в пластине определя-
ется выражением
Здесь Ф определяется по уравнению (4.37) при VPMP, равном
половине толщины пластины. Эта функция во всем диапазоне
значений Ф отличается от функции в уравнении (4.36) не более
чем на 10%. Наибольшие отличия наблюдаются при Ф, близком
единице; с возрастанием или убыванием Ф эта разница быстро
уменьшается. Незначительное влияние геометрии частиц ката-
лизатора на функцию т] = f(0) часто позволяет использовать сле-
дующие эмпирические корреляции для коэффициента эффектив-
ности пластины иммобилизованного фермента с собственной ки-
нетикой, описываемой уравнением Михаэлиса — Ментен:
j tg 0 / Tld _j \
л = , Ф ) приц^! (4 45)
T]d--f -----------1 ) ПРИ 1
w Ф tg T|rf J
282
Глава 4
Здесь r]d—асимптотическое значение коэффициента эффектив-
ности, определяемое по уравнению (4.34). Эти корреляции, пер-
воначально разработанные для катализаторов с частицами,
имеющими форму пластин, можно применять и к другим сим-
метричным геометриям частиц, используя в формулировке мо-
дуля Тила в качестве параметра характеристической длины
частное от деления объема частицы на ее наружную поверх-
ность.
Малая чувствительность функции г] к изменениям порядка
реакции и геометрии частиц катализатора явилась основой
удобного общего критерия, позволяющего различать лимитируе-
мые реакцией и лимитируемые диффузией процессы. Этот кри-
терий (табл. 4.15) сводится к единственному параметру Ф, ко-
торый, как мы знаем, зависит только от наблюдаемых перемен-
ных. Подробнее этот критерий рассмотрен в работе [21].
В небиологическом катализе параметр г] обычно рассматри-
вают как меру эффективности использования катализатора.
Если т] существенно отличается от единицы, то общая скорость
процесса может быть повышена путем измельчения катализато-
ра; при этом Ф уменьшается и, следовательно, возрастает т).
Такой подход может оказаться полезным и в биологическом ка-
тализе; во всяком случае, он указывает на наличие связи между
размером частиц иммобилизованного фермента и максимальной
эффективностью процесса.
Непосредственное изучение собственной кинетики также воз-
можно только в тех случаях, когда фактор эффективности бли-
зок единице. На катализаторе с достаточно малыми размерами
частиц (см. упражнение 4.5) можно экспериментально опреде-
лить характер изменения скорости катализируемой иммобилизо-
ванным ферментохм реакции и найти значения соответствующих
кинетических параметров. Эта информация необходима для рас-
чета наблюдаемой общей кинетики реакции на более крупных
частицах катализатора. Поскольку в водных растворах эффек-
тивные коэффициенты диффузии субстрата и (или) общие кон-
центрации субстрата обычно невелики, то лимитируемый диф-
фузией режим, как правило, обусловливается сравнительно
большими размерами частиц катализатора, что может быть не-
обходимым или для снижения перепада давлений и сопротивле-
ния потоку в реакторах с неподвижным слоем катализатора,
или для предотвращения разрушения частиц катализатора в
реакторах с псевдоожиженным слоем и со взвесями катализато-
ра. Иногда лимитируемый реакцией режим трудно воспроизве-
сти даже .в лабораторных условиях. В таких случаях данные о
собственных кинетических параметрах иммобилизованного фер-
мента приходится получать описанным выше аналитическим
путем на основе экспериментальных данных, усложненныхдиф-
Применение реакций2 катализируемых ферментами 283
фузионными эффектами. Как это показано ниже на конкретном
примере, экспериментальное изучение кинетики реакций, ката-
лизируемых иммобилизованными ферментами, позволяет оце-
нить и эффективный коэффициент диффузии субстрата.
Пример 4.3. Определение эффективного коэффициента диффузии субстра-
та и собственных кинетических параметров катализатора на основе иммоби-
лизованного фермента. В качестве исходного положения примем, что собст-
венная кинетика каталитической реакции подчиняется уравнению Михаэли-
са— Ментен. Характерные для данного катализатора собственные кинетиче-
ские параметры Umax и Кт, а также эффективный коэффициент диффузии
субстрата 2Des можно определить по результатам двух серий кинетических
экспериментов, в одной из которых используется катализатор со сравнитель-
но большими частицами (обеспечивающими большие значения Ф и, следова-
тельно, лимитируемые диффузией условия), а во второй — катализатор с го-
раздо более мелкими частицами (достаточно малыми, чтобы скорость транс-
формации субстрата определялась скоростью реакции). В каждой серии
экспериментов необходимо определить значения v0, отвечающие различным
s0, и затем выразить эти зависимости в графической форме в координатах
Эди — Хофсти (зависимость vo/so от и0; см- разд. 3.2.2). При достаточно боль-
ших значениях s0 реакция имеет нулевой порядок во всем объеме частиц
катализатора и, следовательно, коэффициент эффективности равен единице,
а кривые для больших и малых частиц катализатора совпадают [это говорит
о том, что в случае катализатора с большими частицами условие (4.42)
не выполняется и что ограниченная растворимость субстрата не лимитирует
эксперименты в диапазоне s03>/<m]. Таким образом, независимо от размера
частиц кривые будут отсекать на оси абсцисс отрезок, равный итах-
С другой стороны, если внешняя концентрация субстрата и общая ско-
рость реакции невелики, то собственный порядок каталитических реакций
будет приближаться к единице. Тогда для частиц малого размера (лимити-
руемый реакцией режим) v0/s0 равно vmaX/Km, а для больших частиц иммо-
билизованного фермента, т. е. для лимитируемого диффузией режима:
v0 __п ^тах _ 3 1/ Umax^ss МПЗ 1)
so S°Z<Km 1 ^рвый Krn R ? Кт { ’
<Ь порядок
Следовательно, кривая зависимости u0/s0 от и0 будет отсекать на оси ординат
в случае мелких частиц отрезок, равный итлх1Кт, а в случае больших ча-
стиц— отрезок, определяемый уравнением (4П3.1). Таким образом, зная
размер больших частиц катализатора и величины трех отрезков, отсекаемых
на осях координат кривыми зависимости uq/sq от и0, можно определить итах>
Кт И SDes-
На рис. 4П3.1 представлены результаты применения описанной методики
к а-химотрипсину, иммобилизованному на активированной бромцианом се-
фарозе 4В. Определенные таким образом удельная активность иммобилизо-
ванного фермента (<?е — частное отделения Umax на число молей иммо-
билизованного активного фермента в единице объема катализатора) и Кт
составляют 213 мкмоль субстрата (АТЕЕ, этилового эфира N-ацетил- L-тиро-
зина) на 1 мкмоль активного фермента в секунду и 2,6 мМ соответственно.
Эти значения существенно отличаются от соответствующих параметров того
же фермента в растворе, составляющих 311 мкмоль АТЕЕ на 1 мкмоль ак-
тивного фермента в секунду (удельная активность) и 0,73 мМ (Кт)', отсюда
следует, что иммобилизация заметно изменяет каталитические свойства фер-
мента. Оценка эффективного коэффициента диффузии АТЕЕ в этом катали-
заторе приводит к величине 3,8-10-6 см2/с. Столь малое значение 3)es и до-
вольно высокая активность иммобилизованного фермента предполагают
284
Глава 4
РИС. 4П3.1. Зависимость So/^o от v0 для а-химотрипсина, иммобилизованного
на больших (У? = 60 мкм; ф) и маленьких (R = 10 мкм; О) гранулах активи-
рованной BrCN сефарозы 4В. [Экспериментальные данные заимствованы и»
работы: Clark D. S., Bailey J. Е., Structure-Function Relationships in Immobi-
lized Chymotrypsin Catalysis, Biotech. Bioeng., 25, 1027 (1983).]
существенные диффузионные ограничения даже в случае катализатора с ча-
стицами радиусом 60 мкм. Определение модуля Ф этого катализатора дает
величину 2,6.
При создании катализаторов на основе чрезвычайно доро-
гих и высокоактивных металлов на носителях обычно стремят-
ся локализовать собственно катализатор в тонком наружном
слое частиц носителя. Очевидно, такой же подход применим и
при разработке катализаторов на основе иммобилизованных
ферментов: используя максимально допустимые для данного-
процесса размеры частиц носителя, можно обходиться мини-
мальным количеством фермента. Существует несколько мето-
дов, принципиально пригодных для получения таких катализа-
торов. Недавно было показано, что в катализаторах, полученных
путем импрегнирования пористого носителя раствором фермен-
та, распределение биокатализатора внутри частицы носителя
может быть в высшей степени неравномерным. Как мы увидим
Применение реакций^ катализируемых ферментами 285
в упражнении 4.10, неравномерное распределение фермента
влияет на кажущуюся активность и характер инактивации им-
мобилизованного ферментного катализатора.
Прежде чем перейти к проблеме одновременного сопротивле-
ния массопередаче в растворе и внутри частиц катализатора,
следует подчеркнуть, что, как и в случае ферментативных реак-
ций в растворе, кинетика катализируемой иммобилизованным
ферментом реакции может отличаться от кинетики необратимо-
го превращения одного субстрата, описываемой уравнением Ми-
хаэлиса— Ментен. Достаточно полно вопросы анализа кинетики
других типов реакций с участием иммобилизованных ферментов!
освещены в приведенной в конце главы литературе; здесь же
мы ограничимся несколькими замечаниями общего характера.
Прежде всего следует отметить, что в случае обратимой реак-
ции минимальная концентрация субстрата внутри частицы ката-
лизатора равна его равновесной концентрации. Следовательно,
при вычислении асимптотического коэффициента эффективности,
например по уравнению (4.33), нижний предел интегрирования
должен быть равен равновесному значению. Интересно, что в
силу различных коэффициентов диффузии разных веществ сте-
хиометрические соотношения внутри частицы катализатора мо-
гут отличаться от стехиометрии такого же процесса в растворе.
Это явление подробнее рассмотрено в упражнении 4.8.
Изучение систем, в которых скорость процесса определяется
как диффузией, так и самой реакцией, мы завершим кратким
анализом любопытной ситуации, когда субстрат является инги-
битором собственной реакции иммобилизованного фермента.
Если внешняя концентрация субстрата s0 превышает концентра-
цию субстрата, соответствующую максимальной скорости (см.
разд. 3.4.1), то внутри частицы катализатора снижение концен-
трации субстрата вызовет увеличение локальной скорости реак-
ции до значений, превышающих скорость реакции на внешней
поверхности частицы. В этой ситуации при определенных усло-
виях можно наблюдать коэффициенты эффективности, превы-
шающие единицу. Такое на первый взгляд парадоксальное, на
в сущности легко объяснимое поведение возможно для любой
автокаталитической реакции, т. е. такой системы, в которой ско-
рость реакции возрастает во времени.
4.4.3. Одновременное сопротивление массопередаче
в граничном слое
и внутри частицы катализатора
В двух предыдущих разделах мы рассматривали особые слу-
чаи взаимного влияния массопередачи и химической реакции,,
схематически изображенные на рис. 4.16. В общем случае, одна-
286
Глава 4
ко, субстрат должен сначала пройти через граничный слой или
стоячую пленку, а затем диффундировать в частицу катализато-
ра, где и происходит его превращение. Одна из целей нашего
анализа этой наиболее общей ситуации будет заключаться в
изыскании путей использования рассмотренных выше упрощен-
ных моделей. Другими словами, нам нужно знать, когда доми-
нирует внутреннее сопротивление массопередаче, в каких слу-
чаях определяющую роль играет внешнее сопротивление и ког-
да нужно учитывать оба типа сопротивления.
Решение поставленных задач целесообразно начать с изуче-
ния простой модели иммобилизованного ферментного катализа-
тора с геометрией пластины и кинетикой собственной реакции
первого порядка. В таком случае уравнение материального ба-
ланса для субстрата внутри частицы в стационарном состоянии
можно записать следующим образом:
<4-46»
•с условием симметрии для средней линии
ds
~dx
= 0
х=0
(4-47)
На внешней поверхности пластины (x = L) субстрат не аккуму-
лируется. Транспорт через эту поверхность внутрь частицы ка-
тализатора путем диффузии равен транспорту в обратном на-
правлении через граничный слой; следовательно, граничное ус-
ловие выражается следующим уравнением:
(4«)
ал x=L
Совместное решение уравнений (4.46) — (4.48) дает возмож-
ность определить коэффициент эффективности ф, который в
.нашем случае может быть записан в следующей форме:
п =____________________ (4.49)
1,8 ф[1 +(ф tg ф)/ВЦ 1 1
В этом выражении модуль Тила Ф имеет тот же физический
смысл, что и ранее [см. уравнение (4.37); для геометрии пласти-
ны VPMp = L]. Здесь мы ввели новый важный параметр Bi, на-
зываемый числом Био и определяемый как
удельная скорость транспорта
g.__ ksL __через граничный слой_______ м ggj
&es удельная скорость диффузии
в частице катализатора
Применение реакций, катализируемых ферментами
287
Таким;
глася-
сопро-
Уравнение (4.49) удобнее преобразовать в другую форму:
_L = _L_|__|L (4.51)
Здесь т] (i] = tg0/0)—коэффициент эффективности катализато-
ра в отсутствие сопротивления транспорту в граничном слое.
Величину, обратную коэффициенту эффективности, можно рас-
сматривать как меру сопротивления превращению субстрата,,
обусловленного требованиями к транспорту последнего. ”
образом, уравнение (4.51) отражает известное правило,
щее, что общее сопротивление равно сумме отдельных
тивлений.
Уравнение (4.51) позволяет ответить на вопрос, при
условиях можно пренебречь одним из сопротивлений. Если
т=^-=тг«1 <4’52>
каких;
то можно пренебречь сопротивлением внешнего граничного слоя.
Если же kL)ks намного больше единицы, то можно не принимать
во внимание внутреннее сопротивление частицы катализатора.
Во всех промежуточных случаях необходимо учитывать оба со-
противления. Обратите внимание, что при больших значениях
Ф наблюдается обратная пропорциональная зависимость т]$
от Ф2. Альтернативный критерий, не требующий знания собст-
венной константы скорости, основан на построении графиков за-
висимости r]s от Ф для различных Bi; при Bi^ 100 влияние
внешнего сопротивления незначительно.
Рассмотренные принципы анализа применимы и к иммобили-
зованным ферментам с другой геометрией частиц и с другой
собственной кинетикой; эти случаи подробно рассмотрены в
приведенной в конце главы литературе. Мы же ограничимся
замечанием, что оценка типов массопередачи, оказывающих
большое влияние на скорость всего процесса, всегда базируется’
на сравнительном изучении собственной скорости реакции, диф-
фузионных эффектов в частице катализатора и внешней массо-
передачи.
4.4.4. Влияние ингибиторов, температуры и pH
на каталитическую активность
и инактивацию иммобилизованных ферментов
В предыдущей главе были рассмотрены математические вы-
ражения, описывающие процессы инактивации ферментов и
влияние различных параметров (концентраций ингибиторов и-
активаторов, температуры, pH, ионной силы и т. д.) на фермен-
288 Глава 4
тативную активность в растворах. В общем случае можно ожи-
дать, что эти выражения будут пригодны и для описания зави-
симости собственной активности иммобилизованного фермента
от различных параметров. В то же время, как мы уже не раз
отмечали, иммобилизация может сопровождаться изменением
собственных свойств фермента. Так, иммобилизованные фер-
менты могут иметь другие константы скорости реакций; более
того, иммобилизация может даже потребовать изменения фор-
мы уравнений, описывающих влияние параметров реакции на
собственную активность фермента. Для определения отклика
собственной каталитической активности иммобилизованного
фермента на изменения параметров реакции необходимо тща-
тельное изучение кинетики реакции с применением описанных
выше методов, позволяющих изучать только собственное кине-
тическое поведение системы, а не сочетание скоростей массопе-
редачи и каталитических реакций.
В настоящем разделе мы прежде всего постараемся под-
черкнуть то влияние, которое процессы массопередачи могут
оказывать на взаимосвязь между кажущейся общей каталити-
ческой активностью иммобилизованного фермента и условиями
среды. Все необходимые данные могут быть получены путем
изучения кинетики реакций первого порядка. Прежде всего сле-
дует запомнить, что наблюдаемая общая кинетика реакций с
участием иммобилизованного фермента, функционирующего в
лимитируемом реакцией режиме, не отличается от собственной
локальной кинетики этой ферментативной реакции. Следова-
тельно, изучаемая в этом режиме реакция будет иметь первый
порядок, а константа скорости процесса будет равна собствен-
ной константе скорости ферментативной реакции. В случае же
иммобилизованного ферментного катализатора, функционирую-
щего в лимитируемом диффузией режиме, кажущийся порядок
реакции также будет первым, но кажущаяся константа скорости
будет равна квадратному корню из собственной константы ско-
рости ферментативной реакции. [В общем случае если реакция
протекает по /7-му порядку, то кажущийся порядок этой реакции
в лимитируемых диффузией условиях будет равен (п+1)/2.]
Таким образом, кажущаяся энергия активации составит лишь
половину истинной энергии активации. Точно так же и влияние
изменения любых параметров реакции на наблюдаемую общую
кинетику процесса будет меньшим, чем действительный эффект
.изменения этих же параметров на локальную, собственную ки-
нетику ферментативной реакции. Например, если процессы
инактивации, введение ингибитора или изменение pH уменьша-
ют константу скорости собственно ферментативной реакции в
4 раза, то в лимитируемых диффузией условиях наблюдаемая
-общая скорость процесса уменьшится только в 2 раза.
Применение реакций, катализируемых ферментами
289
Из всего сказанного следует, что при изучении влияния па-
раметров реакции на кинетику катализируемых иммобилизо-
ванными ферментами реакций следует быть предельно внима-
тельным. Если не соблюдены лимитируемые реакцией условия
или если экспериментальные данные, полученные в условиях,
когда диффузия вносит определенный вклад в течение процесса,
не переработаны с целью получения сведений о собственной ки-
нетике ферментативной реакции, то любые выводы о зависимо-
сти активности иммобилизованного фермента от условий про-
цесса будут иметь ценность только для данной партии катали-
затора, только в данных условиях реакции и никоим образом
не будут описывать собственное поведение иммобилизованного
фермента. В таком случае любые изменения скорости потока
раствора субстрата через слой катализатора, размера его час-
тиц, структуры пор, содержания или распределения фермента в
частице катализатора будут сопровождаться изменениями най-
денных ранее зависимостей активности катализатора от пара-
метров процесса. Этот факт еще раз подчеркивает важность и
необходимость четкого разграничения собственных свойств им-
мобилизованного фермента от тех его характеристик, которые
обусловлены эффектами массопередачи.
К сожалению, это обстоятельство не учитывалось во многих
опубликованных в литературе работах, посвященных изучению
зависимостей характеристик иммобилизованных ферментных
систем от условий реакции. Поскольку в большинстве случаев
иммобилизованные ферменты функционируют в условиях, ког-
да диффузионные эффекты в заметной степени влияют на ско-
рость процесса или даже определяют ее, то многие имеющиеся
в литературе экспериментальные данные по определению скоро-
стей инактивации и влияния pH и температуры на кинетику
реакций, очевидно, не описывают собственные свойства иммоби-
лизованных ферментов.
В то же время имеется множество фактов, свидетельствую-
щих об изменении собственной кинетики инактивации фермен-
тов после их иммобилизации. Предлагались и постулировались
различные механизмы и интерпретации этого явления; некото-
рые из гипотез, которые мы вкратце здесь рассмотрим, выдер-
жали экспериментальную проверку. Во-первых, вследствие им-
мобилизации ферменты удерживаются в относительно фиксиро-
ванном в пространстве положении, что уменьшает вероятность
таких взаимодействий между молекулами фермента, которые
способствуют их инактивации за счет агрегации или автолиза
(в случае протеолитических ферментов). Во-вторых, наличие
нескольких связей молекулы фермента с носителем затрудняет
нарушение третичной структуры белка. Сообщалось, что попыт-
ки создать комплементарную поверхности фермента микрострук-
19—537
290
Глава 4
туру носителя путем включения фермента в гель сопровожда-
лись резким повышением его стабильности. Аналогично иммо-
билизация стабилизирует активную форму ферментов, состоя-
щих из нескольких субъединиц, и тем самым может замед-
лять инактивацию, обусловленную диссоциацией олигомерных
белков.
Повышенная устойчивость иммобилизованных ферментов мо-
жет быть также обусловлена благоприятным местным окруже-
нием, в меньшей степени инактивирующим белок, чем окруже-
ние в растворе. В качестве примеров можно привести носители,
поверхности которых обладают буферными свойствами, или сни-
жают местную концентрацию кислорода, или адсорбируют фер-
ментные яды, или катализируют превращение денатурирующих
веществ (например, Н2О2) в безвредные соединения. Вероятно,
на стабилизацию иммобилизованных ферментов могут оказы-
вать влияние и другие факторы, которые будут выяснены в ходе
последующих работ по изучению фундаментальных свойств им-
мобилизованных ферментов и взаимосвязи между их структурой
и функцией.
4.5. Заключение
Мы познакомились с основными направлениями промышлен-
ного применения ферментных катализаторов, узнали, как фер-
менты можно иммобилизовать для использования в непрерыв-
ных процессах, и изучили особенности кинетики реакций, ката-
лизируемых иммобилизованными ферментами. Теперь обратимся
к рассмотрению природных многоферментных систем, характер-
ных для живых клеток, в которых осуществляются сложнейшие
разветвленные последовательности катализируемых фермента-
ми реакций, часто обладающие элементами обратной связи.
Следующая глава посвящена стехиометрии и термодинамике
взаимосвязанных клеточных реакций. Затем в гл. 6 будут рас-
смотрены системы регуляции и контроля этих последовательно-
стей реакций, а в гл. 7 — кинетика роста популяций клеток.
Прежде чем перейти к изучению биологических реакторов
(гл. 9), в гл. 8 рассмотрим основные проблемы массопередачи
и энергообмена. Если же читателя интересуют прежде всего
реакторы для осуществления ферментативных процессов, то он
может сразу ознакомиться с разд. гл. 9, в которых описывают-
ся основные проблемы анализа и проектирования биологических
реакторов. В частности, разд. 9.1.2 и 9.1.4 посвящены исключи-
тельно анализу ферментативных реакторов, а несколько тем из
разд. 9.6 по многофазным реакторам имеют непосредственное
отношение к биологическим реакторам с иммобилизованными
ферментами.
Применение реакций, катализируемых ферментами
291
Упражнения
4.1. Нерастворимые субстраты. В результате смешения водной суспензии
однородных по размерам частиц желатина (полиглицина) и порошкообразно-
го протеолитического фермента получена смесь, содержащая а объемных ча-
стей желатина и е молей фермента в литре суспензии.
а) Приняв, что первоначальный размер частиц равен d0 и что единствен-
ным продуктом реакции является глицин, найдите выражения, описывающие
зависимость скорости образования глицина и изменения размера частиц от
времени. Примите также, что скорость реакции при s0<^e0 выражается урав-
нением Михаэлиса — Ментен, т. е.
скорость ___ keosa
объем е0 + КЕ
где sa — площадь поверхности субстрата на единицу объема реакционной
смеси.
б) Из проявленной фотопленки необходимо выделить серебро путем фер-
ментативного гидролиза связывающего серебро желатина. Определите мини-
мальное время контакта фотопленки с раствором фермента, за которое гид-
ролизуется весь желатин.
4.2. Автокатализ. Можно принять, что в периодическом процессе кинетика
автокаталитической активации пепсина (рис. 4.4) описывается уравнением
Михаэлиса — Ментен.
а) Покажите, что максимальная скорость активации достигается при
„ — %s (so + ео — s)
As 4" S
б) Покажите, что ход реакции во времени описывается уравнением
в) Для указанной автокаталитической системы начертите график зави-
симости 1/и от 1/s в координатах Лайнуивера — Бэрка. График какой другой
зависимости был бы более удобным?
4.3. Ферментативные реакции с несколькими субстратами. В промышлен-
ных ферментных реакторах в реакционной смеси часто присутствует несколь-
ко субстратов одновременно.
а) Покажите, что в системе с двумя субстратами скорость превращения
Si и S2 выражается уравнением
и что таким образом каждый субстрат является конкурентным ингибитором
другого субстрата.
б) Найдите общее выражение, описывающее общую скорость реакции
одного фермента с т субстратами одновременно.
4.4. Кинетика гидролиза целлюлозы. Гидролиз целлюлозы протекает
с участием солюбилизирующего фермента, фермента, образующего дисаха-
рид целлобиозу, и гидролизующей этот дисахарид ^-глюкозидазы (рис. 4.1).
Первые два фермента ингибируются целлобиозой (будем считать, что целло-
биоза представляет все олигомерные ингибиторы), поэтому упрощенная схе-
19*
292
Глава 4
ма реакций будет выглядеть следующим образом:
ki
{GaJ + Ej ----> EHGd
El + G2 ----> E1G2
Ei [G2] ----> Ej + G2
Здесь [G2] и G2— нерастворимая целлюлоза и растворимая целлобиоза со-
ответственно, a Ei — фермент, катализирующий наиболее медленную стадию
образования целлобиозы.
а) Можно допустить, что в отсутствие ^-глюкозидазы глюкоза не обра-
зуется. Найдите соответствующие выражения, исчерпывающе описывающие
зависимость g2 от времени для случаев конкурентного и неконкурентного
ингибирования.
б) Покажите, что по графику зависимости g2/t от (1 Д) In (s0/g2) можно
различить системы без ингибирования, с конкурентным ингибированием и не-
конкурентным ингибированием. Почему в этом случае лучше применять g2,
а не концентрацию непрореагировавшего субстрата? [Экспериментально пока-
зано, что описанная система ингибируется неконкурентно; см. статью: Ho-
well 1. A., Stuck 1. D., Kinetics of Solka floc Hydrolysis by Trichoderma viride
Cellulase: Biotech. Bioeng., 17, 873 (1975).]
4.5. Подбор биокатализатора для лимитируемого реакцией режима. Эти-
ловый эфир ацетил-L-тирозина (АТЕЕ) гидролизуется иммобилизованным
а-химотрипсином с удельной объемной скоростью 18,4 мкмоль АТЕЕ в 1 см3
за 1 с. Найдено, что эффективный коэффициент диффузии АТЕЕ равен при-
мерно 3,8-10~6 см2/с. Биокатализатор с каким радиусом сфер следует выб-
рать для изучения кинетических свойств иммобилизованного фермента в ли-
митируемом реакцией режиме?
4.6. Маскирование инактивации иммобилизованного фермента массопере-
дачей. Рассмотрим иммобилизованный фермент, необратимо теряющий свою
активность в соответствии с уравнением (3.78). Примем, что превращение
субстрата является реакцией приблизительно первого порядка.
а) Предположим, что фермент иммобилизован на наружной поверхности
непроницаемого для субстрата твердого носителя и что при t=0 число Дам-
келера велико. Начертите графики зависимости коэффициента эффективности
и скорости реакции от времени. Какие ошибочные выводы относительно влия-
ния иммобилизации на стабильность фермента можно сделать, если не ввести
соответствующие поправки на эффекты массопередачи?
б) Ответьте на тот же вопрос в другой ситуации, когда фермент иммо-
билизован внутри проницаемой для субстрата пластины. Примите, что внеш-
нее сопротивление массопередаче относительно невелико.
4.7. Гистерезис; влияние pH на иммобилизованный папаин. Изучение
pH-электрода, иа поверхность которого нанесен папаин, показывает, что за-
висимость генерируемого иа поверхности электрода pH (внутреннего pH)
от pH в растворе при различных направлениях изменения pH выражается
разными кривыми. [В данном случае изучалась реакция гидролиза этилового
эфира бензоиларгинина (ВАЕЕ) до соответствующей кислоты; см. рис.
в статье: Naparstek A., Romette 1. L., Kernevez J. Р., Thomas D., Nature, 249,
490 (1974).]
а) Приняв, что so^Km, слой фермента очень тонок и активность папаи-
на максимальна при pH 6,0, покажите, что это явление может быть обуслов-
лено внешним сопротивлением массопередаче.
б) Кинетические константы этой реакции, катализируемой растворимым
ферментом, при pH 6,0 и 20 °C равны: /г2=19 международных единиц (1 меж-
дународная единица равна количеству микромолей ВАЕЕ, гидролизуемых
1 мг фермента в 1 мин), /С1 = 5-10—3 М. Найдите величину коэффициента
Применение реакций, катализируемых ферментами
293
массопередачи иона водорода и соответствующее указанным константам
значение рК2 фермента, если на поверхность электрода нанесено а мг/см2
фермента и активность иммобилизованного фермента составляет только 6%
активности фермента в растворе. (Во всех случаях s>Km.)
в) По найденной величине рК2 и приведенным в предыдущей части за-
дачи параметрам вычислите коэффициенты эффективности при различных
внешних pH; постройте график зависимости коэффициента эффективности от
внешнего pH. Иногда коэффициент должен превышать единицу, а при неко-
которых pH задача должна иметь несколько решений. [См. также работу:
Bailey J. Е., Chow М. Т. С., Biotech. Bioeng., 16, 1345 (1974).]
4.8. Стехиометрия реакций внутри частицы катализатора. Внутри пори-
стой частицы иммобилизованного фермента осуществляется катализируемая
ферментом реакция
ccSi + ^Sj -—>- 7P1 + SP2
где а, [3, 7 и б — стехиометрические коэффициенты.
а) Используя уравнения материального баланса для субстратов и про-
дуктов реакции, найдите выражения, определяющие концентрации всех ве-
ществ внутри частицы катализатора через концентрацию S[ (в той же точке
частицы), концентрации на наружной поверхности частицы, а также эффек-
тивные коэффициенты диффузии субстратов и продуктов реакции.
б) Покажите, что реагент, ограниченно растворимый в реакционной сре-
де, будет лимитировать скорости реакций внутри частицы иммобилизованного
фермента.
4.9. Обратимая изомеризация глюкозы в насадочной колонне. Промыш-
ленно важный процесс изомеризации глюкозы во фруктозу осуществляют
в реакторе колонного типа с неподвижным слоем иммобилизованной глюко-
зоизомеразы; кинетика катализируемой этим ферментом реакции описывается
уравнением Михаэлиса — Ментен для обратимых реакций [уравнение (3.18)].
а) Покажите, что подстановка в указанное уравнение s=s—se (se — рав-
новесная концентрация) приводит к простому уравнению Михаэлиса — Мен-
тен (относительно s). Как в этом уравнении будут выражены кажущиеся
константа Михаэлиса (КаРР) и максимальная скорость (царр)?
б) Изменение 7 в зависимости от положения в одномерном неподвижном
слое катализатора с пористостью е и при средней скорости потока иг может
быть описано уравнением
I ds \ ,
e«z —— t= — (1 — е)ц
\ dz )
с одним краевым условием: s=so—se при z=0. Путем интегрирования этого
выражения найдите другое уравнение, определяющее параметры работы реак-
тора через объемную производительность (т), определяемую как т=
=L(1—e)/e«2 [частное от деления объема реактора (катализатора) на объ-
емную скорость потока].
в) Рассмотрим медленно инактивирующийся иммобилизованный фермент,
находящийся в реакторе с неподвижным слоем катализатора, описанным в
части «б» настоящей задачи. Если процесс инактивации подчиняется зако-
ну г(активн.) =£(t=O)exp(—М), найдите по измененным соответствую-
щим образом данным предыдущей задачи функцию f[s0, ym,app, s(Z-)], гра-
фик зависимости которой от времени t позволил бы определить как т,
так и kd.
4.10. Иммобилизованный ферментный катализатор с неравномерным рас-
пределением фермента. Приняв, что катализируемая реакция имеет пер-
вый порядок, сравните общие скорости и коэффициенты эффективности,
294
Глава 4
когда иммобилизованный в пористой пластине фермент, имеющий одно и
то же число единиц ферментативной активности,
а) распределен равномерно по всему объему пластины;
б) сконцентрирован и равномерно распределен во внешнем слое пла-
стины вблизи наружной поверхности катализатора, причем во внутренней
области, толщина которой равна половине толщины пластины, фермент
совершенно отсутствует.
4.11. Одновременная оценка параметров кинетики реакции и массопе-
редачи. Фермент иммобилизован на поверхности непористого твердого тела.
Приняв, что внешним сопротивлением массопередаче нельзя пренебречь и
что собственная кинетика ферментативной реакции описывается уравнением
Михаэлиса — Ментен:
а) найдите выражение, четко определяющее коэффициенты в коорди-
натах Лайнуивера — Бэрка; с помощью этого уравнения выразите кажу-
щиеся максимальную скорость (wmaxapp) и константу Михаэлиса (/Старр)
через реальные переменные Wmax, Кт и ks (коэффициент массопередачи);
б) покажите, как можно графическим путем определить параметры
Стах, Кт и ks, если имеются данные для достаточно большого диапазона
концентраций субстратов.
4.12. Кинетика реакций иммобилизованных ферментов с разделением
зарядов. Концентрацию заряженного вещества (субстрата, ингибитора) на
заряженной матрице, например на ферментной мембране, можно выразить
следующим уравнением:
[С + ] мембрана = [С + ] растпорСге'‘>?,^Г
п0 д2а _
‘ Ki<x ‘ h0
где z— ионный заряд субстрата или ингибитора; е — заряд электрона;
ф— электростатический потенциал мембраны (ф (раствор) =0). Покажите,
что кажущиеся величины Ks и Ki определяются выражением
— К, г. .e~ze^/RT
' (ист)
если рассматривать концентрации в объеме всего раствора. Допуская, что
в мембране имеются е, е~, е2~ (е~— активная форма фермента), покажите,
что максимальная начальная скорость реакции (если пренебречь влиянием
массопередачи) достигается в случае матрицы-носителя фермента с ф, оп-
ределяемым уравнением
Sq \ / К%
№
где а=еге^/ЯТ и ho— [Н+] в основной массе раствора.
4.13. Микрокапсулирование; ^-галактозидаза. Гидролиз лактозы [5-галак-
тозидазой ингибируется продуктом реакции, и поэтому уравнение, выражаю-
щее скорость этой реакции, имеет форму
V =---------------------
Кщ (1 + РIKi) + s
С целью облегчения процесса выделения ^-галактозидазы из молока рас-
твор фермента заключен в микрокапсулы из нитроцеллюлозы диаметром
около 30 мкм.
а) Постройте график в координатах Лайнуивера — Бэрка для р=0;
0,5; 1,5; 5 и 10 мМ (е0=75 мг в 100 мл) при условии, что пт=^рво,
Ар = 0,57 мкмоль/(мг фермента в минуту), /(т=0,54 мМ, Ki= 1,5 мМ.
Применение реакций, катализируемых ферментами
295
б) На основе графика части «а» задачи, не привлекая приведенное
в условии уравнение, постройте графики зависимости 1/о от 1/з для перио-
дического процесса при следующих начальных условиях:
До, мМ 0,5 5,0 10,0 5,0
ро, мМ 0 0 0 5,0
в) Покажите, что даже при максимально возможной скорости реакции
в микрокапсуле в системе, описанной в части «а», скорость процесса лимити-
руется реакцией, т. е. диффузия субстрата и продукта реакции осуществля-
ются быстрее, чем сама реакция [см. статью: Wadiak D. Т., Carbonell R. G.,
Kinetic Behavior of Microencapsulated ^-Galactosidase, Biotech. Bioeng., 17,
1157 (1975)1.
4.14. Мембранный биореактор. Фермент удерживается в проточном
биореакторе полупроницаемой мембраной. Активный фермент инактивирует-
ся в ходе реакции первого порядка с константой скорости kd. Постоянство
состава продуктов реакции обеспечивают путем постепенного введения ак-
тивного фермента в реактор, чтобы концентрация активного фермента
в нем была постоянной, равной еа. На мембране концентрируется гелеоб-
разный слой фермента, увеличивающий сопротивление потоку раствора,
поэтому с целью обеспечения постоянной скорости потока / реакционной
смеси через мембрану приходится непрерывно регулировать падение дав-
ления на мембране. В то же время необходимая скорость потока / может
быть достигнута только в том случае, если общая концентрация фермен-
та £tot (сумма активной и неактивной форм) не превышает некоторое мак-
симально допустимое значение, определяемое уравнение:*!
emax = es exp (— J/k^
где es — концентрация насыщения фермента и ke — коэффициент массопере-
дачи фермента.
а) Определите зависимость общей концентрации фермента в реакторе
от времени.
б) Если скорость потока падает ниже /, то процесс следует остано-
вить, а реактор подвергнуть чистке. Определите максимальное время не-
прерывной работы реактора tmax как функцию Л
в) Найдите значение J, позволяющее получить максимальное количе-
ство продукта реакции за период непрерывной работы реактора (0<t<
c^max) при е0=5 кг/м3, £s = 100 кг/м3, &е=0,017 мм/с, kd~0,018 сут~!.
г) Решите задачу «в» при ео=50 кг/м3 (все другие параметры не из-
меняются) .
д) Как влияет константа скорости инактивации kd на величину J, оп-
ределяемую по условиям задачи «в»? На какие другие важные с эконо-
мической точки зрения параметры влияет kd?
Литература
В большинстве работ, приведенных в конце гл. 3, содержится материал,
относящийся и к темам настоящей главы. Кроме того, многие проблемы тех-
нологии ферментов рассмотрены и в ряде других литературных источников,
к числу которых относятся:
296
Глава 4
1. Tauber Н., The Chemistry and Technology of Enzymes, John Wiley and Sons,
Inc., New York, 1949.
2. LJnderkofler L. A., Manufacture and Use of Industrial Microbial Enzymes,
Bioeng. Food Process. CEP Symp. Ser. no. 69, 62, 11 (1966).
3. Faith IF. T„ Neubeck С. E., Reese E. T., Production and Application of Enzy-
mes, Advan. Biochem. Eng., 1, 77 (1971).
4. Arima K., Microbial Enzyme Production, in Global Impacts of Applied Micro-
biology, Starr M. P. (ed.), John Wiley and Sons, Inc., New York, 1964.
5. Wolnak B. et al., The Present and Future Technological Status of Enzymes,
U.S. Dept. Commerce Doc., PB-219636, Natl. Tech. Inf. Serv., December
1972.
6. Aunstrup K-, Enzymes of Industrial Interest: Traditional Products, in Annual
Reports on Fermentation Processes, Perlman D. (ed.), vol. 2, Academic Press,
New York, 1978.
7. Godfrey T., Reichelt J., Industrial Enzymology, Macmillan Publishers Ltd.,
London, 1983.
В перечисленных ниже монографиях и сборниках содержится ценная ин-
формация о получении, идентификации и применении иммобилизованных фер-
ментов:
8. Guilbault G. G.. Enzvmatic /Methods of Analysis, Pergamon Press, New York,
1970.
9. Zaborsky 0. R., Immobilized Enzymes, CRC Press, Cleveland, Ohio, 1973.
10. Insolubilized Enzymes, Salmona M„ Saronio C., Garatlin S. (eds.), Raven
Press, New York, 1974.
11. Olson A. C., Cooney C. L., Immobilized Enzymes in Food and Microbial
Processes, Plenum Press, New York, 1974.
12. Immobilized Enzymes for Industrial Reactors, Messing R. A. (ed.), Academic
Press, Inc., New York, 1975.
13. Immobilized Enzymes, Mosbach K- (ed.). Methods in Enzymology, vol. XLIV
(Colwick S. P., Kaplan N. O., eds.-in-chief), Academic Press, New York,
1976.
14. Immobilized Enzymes, Research and Development, Chibata I. (ed.), Halsted
Press, New York, 1978.
15. Characterization of Immobilized Biocatalysts, Buchholz K-, (ed.), DECHEMA
Monographs No. 1724—1731, vol. 31, Verlag-Chemie, Weinheim, 1979.
16. Chibata I., Tosa T., Sato T., Mori T., Matsuo Y., Preparation and Industrial
Application of Immobilized Aminoacylases, in Fermetation Technology Today,
Terui G. (ed.), p. 383, Society of Fermentation Technology, Japan, 1972.
17. Wang S. S., King C.-K-, The Use of Coenzymes in Biochemical Reactors in
Advances in Biochemical Engineering, vol. 12, Ghose T. K-, Feichter A.,
Blakebrough N. (eds.), p. 119, Springer Verlag, New York, 1979.
18. Klibanov A. M., Stabilization of Enzymes by Immobilization, Analyt. Bio-
chem., 93, 1 (1979).
Опубликованы две ценные обзорные статьи, посвященные целлюлозе и
«целлюлазе»:
19. Lee Y. И., Fan L. Т., Properties and Mode of Action of Cellulase; Kinetics of
Hydrolyses of Insoluble Cellulose by Cellulase, in Advances in Biochemical
Engineering, vol. 17, p. 101, 131, Fietcher A., (ed.), Springer Verlag, New
York, 1980.
Применение реакций, катализируемых ферментами 297
В приведенных ниже работах подробно рассмотрена взаимосвязь между
химическими реакциями и массопередачей. В некоторых из них основное вни-
мание уделено биологическим системам, а в других — синтетическим катали-
заторам. Приведенный в последних работах материал, как мы уже указыва-
ли, можно после небольших изменений в терминологии и обозначениях отнести
и к биологическим системам.
20. Atkinson В., Biochemical Reactors, Pion Ltd., London, 1974.
21. Weisz P. U7, Diffusion and Chemical Transformation: An Interdisciplinary
Excursion, Science, 179, 433 (1973).
22. Satterfield C. N., Mass Transfer in Heterogeneous Catalysis, MIT Press,
Cambridge, Mass., 1970.
23. Petersen E. E., Chemical Reaction Analysis, Prentice-Hall, Inc., Englewood
Cliffs, N.J., 1965.
24. Aris R., The Mathematical Theory of Diffusion and Reaction in Permeable
Catalysts, vol. 1, The Theory of the Steady State, vol. 2, Questions of Uni-
queness, Stability and Transient Behavior, Clarendon Press, Oxford, 1975.
В многотомном издании под общим названием Enzyme Engineering пуб-
ликуются доклады, представленные на проводящихся раз в два года конфе-
ренциях по технологии ферментов, организуемых инженерным фондом. Пер-
вый том был опубликован издательством Wiley-Interscience (New York, 1972);
последующие тома выпускались издательством Plenum (New York), а также
публиковались в журнале Annals of the New York Academy of Sciences.
Глава 5
Стехиометрия и энергетика
метаболических превращений
Живая клетка представляет собой сложный химический ре-
актор, в котором осуществляется более 1000 независимых ка-
тализируемых ферментами реакций. Тем не менее биологичес-
кие системы подчиняются тем же основным законам сохране-
ния вещества и энергии и тем же принципам термодинамики,
которые хорошо знакомы нам по процессам химической техно-
логии. В настоящей главе мы будем рассматривать клетку
именно как химический реактор, концентрируя внимание на
стехиометрических соотношениях, процессах выделения или по-
глощения энергии, характерных для живых организмов.
Совокупность всех осуществляющихся в клетке химических
превращений называется метаболизмом. На рис. 5.1 приведена
упрощенная схема некоторых наиболее важных процессов ме-
таболизма в бактерии Escherichia coli. Нетрудно видеть, что
метаболические реакции большей частью объединяются в по-
следовательности, называемые путями метаболизма, причем
связь между ними осуществляется, во-первых, через цикличес-
кие, закрытые пути, в которых продукт последней стадии явля-
ется исходным веществом для первой стадии, и, во-вторых, по-
средством ответвлений от основных путей, соединяющих одну
последовательность реакций с другой. Все стрелки на рис. 5.1
обозначают одну или несколько катализируемых ферментами
реакций, в результате которых одни компоненты клетки (мета-
болиты) превращаются в другие.
Отдельные клетки и их популяции представляют собой бо-
лее упорядоченную по сравнению с окружающей средой систе-
му, а упорядочение любой системы требует затрат энергии.
Сейчас мы уже более или менее детально знаем, каким обра-
зом необходимая для этого энергия извлекается из окружаю-
щей клетку среды. С другой стороны, — это, вероятно, наибо-
лее важно с точки зрения биохимической технологии, — изуче-
ние энергетического обмена помогает понять и объяснить ос-
новные различия между функционированием клеток в присут-
ствии и в отсутствие кислорода. Как мы уже отмечали, такие
условия называются аэробными и анаэробными соответственно.
Одни клетки (облигатные, или строгие, анаэробы) вообще не
Фенилаланин
Шикимат-S- ®Г-— >► Триптофан
j Тирозин
Шикимовая кислота
Компоненты
клеточной одолочки
-------Пути синтеза аминокислот
--------Пути си теза нуклеиновых кислот
-------Рута синтеза ЛТР
.......Гута синтеза липидов
Галактоза
Зритрозо-в-®
[ f Гликоген
Уридин-® ®-глк>юэа \
Путь Эмвдена-Мейергофа-Парнаса
Рибулоза-5-®-^- 6-Фосфоглюкуроновая —^Глюкозо
I кислота
Глюкоза
Глюкоза-в-®
Клеточная
мембрана
4
Нуклеиновые
нислоты
I Ривозо-5-®
! 1
I । !----х----1 ,
\®-Ри5озо-5-®) Пуриновые нуклеотиды^
| 'рэ-Пири миди новые —и' ^Глицин ч
| ! нуклеотиды к----------------^-Серин—<------
। Оротовая кислота Цистеин
Лизин \ваТин
' н | "
\Лолуаль^егид аспа- I Ацетолак- -*
,^аеиновой кислоты I тат
| fАспарагин I Лейцин /
Изоле йцин-^
Фруктоза-Г®-в-®
Аланин
в- дрсроглиц^оино--*— Дигидроксиацетон-®
--------3- Фосфоглицериновая
кислота
I
—------Фосфоенолпировиноградная
------------кислота-
Пируват ар
Аспартат
I
„ *
Гомосе^ин
*• Метионин
---*- Треонин-----— оС-КетобутарЪгп
Аргинин
Глутамин -,-Длутамат
Пролин '
S- Амииолевулинатт
Гем
I
_1
Цитрат
цис-Акош.
Цикл ТНК
(никл ь
Кребса )
Оксало-
рГ ацетат
Малат
Фумарат
укцинат
А
Сукцинил -СоА
цинат
сГКетоглу--
тарам
Фосфолипид.ы
Ф осратидиловая
кислота
i
I
Глицерин-®-
Триглицериды
(Ясиры) '
Бутирил- СоА • • •
Лактат у
Ацетил-^оК- *Малой ил- СоА
-^Дыхатемна:. ^ед-т*-н о
л t
Дыхательные ферменты
РИС. 5.1. Схематическое изображение некоторых основных путей метаболизма бактерии Е. coli. (Уотсон
Дж., Молекулярная биология гена. — М.: Мир, 1978.)
300
Глава 5
используют свободный кислород, другим клеткам (облигатным
аэробам) кислород необходим, а третьи клетки могут расти
как в присутствии кислорода, так и без него. Метаболически
универсальную третью группу клеток называют факультативны-
ми анаэробами; к ним относятся, в частности, хорошо извест-
ные дрожжи.
Микроорганизмы усваивают два вида энергии — световую и
химическую. Фототрофами называют организмы, для которых
основным источником энергии является свет, а хемотрофами —
организмы, получающие энергию в результате деградации оп-
ределенных питательных веществ. В соответствии с природой
последних хемотрофы подразделяют далее на ряд подгрупп.
Так, литотрофы окисляют неорганические вещества, а органо-
трофы для извлечения энергии используют органические пита-
тельные вещества. Несколько конкретных примеров мы приве-
дем после того, как детальнее ознакомимся с основами пита-
ния клеток.
Полученная тем или иным способом энергия обычно хранит-
ся и транспортируется (внутри клетки) в виде удобных высоко-
энергетических промежуточных соединений типа АТР (разд.
2.3.1). Клетка использует эту энергию для выполнения следую-
щих трех функций: химического синтеза больших или сложных
молекул, (т. е. для роста); транспорта ионных и нейтральных
веществ в клетку, из клетки, а также в клеточные органоиды
и из них; механической работы, связанной с делением и пере-
движением клеток. Ни один из этих процессов не происходит
спонтанно, а их результатом является повышение свободной
энергии клетки. По этой причине они осуществляются только
совместно с другими реакциями, сопровождающимися боль-
шим по абсолютной величине понижением свободной энергии.
К этой теме мы вернемся в разд. 5.1.
Эффективность использования свободной энергии в биосин-
тезе довольно высока и обычно превышает 20%. Транспорт
веществ также происходит с участием АТР; этот процесс уни-
кален в том отношении, что он обеспечивает перенос неболь-
ших молекул и ионов через мембраны против градиента кон-
центрации даже в тех случаях, когда отношение концентраций
транспортируемого вещества по обе стороны мембраны со-
ставляет 105. Механическая работа, очевидно, совершается при
делении клеток, а также при движении бактерий и простей-
ших. В мышечной активности животных и движении спермато-
зоидов также участвует АТР; прямое превращение химической
свободной энергии в механическую работу без участия таких
посредников, как электричество или горячие газы, представля-
ет собой еще одно уникальное свойство живых систем. Частич-
но теряющаяся в ходе таких процессов химическая энергия
Стехиометрия и энергетика метаболических превращений 301
превращается в тепловую, что необходимо учитывать в техно-
логических процессах с участием растущих популяций клеток.
Чтобы расти и самовоспроизводиться, клетка должна по-
треблять вещества, необходимые для построения мембран,
белков, хромосом, стенок и других структурных элементов. Из
основных химических элементов для жизни необходимы преж-
де всего углерод, азот, сера и фосфор; водород и кислород мо-
Таблица 5.1. Важнейшие источники химических элементов
Химический элемент Источник
Углерод Азот Сера Фосфор СО2, сахара, белки, жиры Белки, NH3, NO3~ Белки, SO42- РО43-
гут быть получены из компонентов среды или в некоторых
случаях из воды. Типичные источники этих элементов приведе-
ны в табл. 5.1, а на рис. 5.2 изображена схема их утилизации
некоторой воображаемой бактерией. Этот организм можно
было бы назвать гетеротрофом, поскольку он получает необхо-
димый углерод из органического вещества. Другие микроорга-
низмы могут использовать более простые питательные вещест-
ва; так, аутотрофы всю свою потребность в углероде удовлет-
Синтез низко-
молекулярных
соединений
биосинтез
макромолекул/
Расщепление
питательный:
teiyecmB
। 1
мн;
। мн;
‘.Углерод-
ные сне-\
। летпь/ ,
isoH ;
Глюкоза р'о’- S02-
Гексоз-
амины
Амино-
кислоты
уНуклео-
I тиды
\Мем5рана\
Мукопептиды
Нлеточная\
ооолочка ।
\белки
~60 %
_] PH К ^\Раег£орины^Ридосомб11
\Ю-20 %—
(ДНК ,-------------,
j 2- % I Хромосома |
РЧС 5.2. Схематическое изображение биосинтеза макромолекулярных соеди-
нений бактерией из несложных питательных веществ. [Воспроизведено с раз-
решения из работы: Biochemistry of Bacterial Growth, Mandelstam J„
McQuillen K., (eds.), 2d ed., p. 4, Blackwell Scientific Publications, Oxford,
302
Глава 5
воряют за счет СОг- Объединив эту градацию с указанной вы-
ше классификацией организмов в соответствии с источниками
энергии, мы можем определить совокупность признаков, описы-
вающих метаболизм клетки. Соответствующая номенклатура
и примеры организмов каждого типа приведены в табл. 5.2.
Использование разными клетками различных источников энер-
гии п углерода говорит о том, что и химизм протекающих в них
процессов неодинаков. В настоящей главе основное внимание
уделено изучению наиболее общих путей метаболизма, харак-
Таблица 5.2. Классификация и номенклатура от источников углерода и энергии организмов в зависимости
Источник Источник энергии
углерода Химический Световой
Органические соединения СО2 Хемогетеротрофы (высшие животные, простейшие, грибы и большинство бактерий) Хемоаутотрофы (некото- рые бактерии) Фотогетеротрофы (некоторые бактерии, ряд водорослей- эукариот) Фотоаутотрофы (высшие рас- тения, водоросли-эукариоты, сине-зеленые водоросли и не- которые бактерии)
терных для самых разных типов живых клеток; важные про-
блемы дифференциации путей метаболизма обсудим позднее.
На рис. 5.2 происходящие в клетке реакции разделены на
три группы: деградация питательных веществ, биосинтез ве-
ществ небольшой молекулярной массы и биосинтез полимер-
ных макромолекул. Как мы уже отмечали, каждая реакция
катализируется специфическим ферментом; последние, таким
образом, выполняют важнейшую функцию, определяя, какие
реакции и с какой скоростью будут протекать в клетке.
Чтобы оценить важность регуляции относительных скорос-
тей реакций, полезно взглянуть на химию клетки с другой сто-
роны. Многие находящиеся в клетке вещества (метаболиты)
могут одновременно подвергаться действию нескольких фер-
ментов, которые, например, могут окислять, восстанавливать
или конденсировать их с другими веществами. В результате
происходящие в клетке последовательности реакций пересека-
ются и перекрываются самым замысловатым образом, как это
показано, в частности, на рис. 5.1. Обратите внимание, напри-
мер, на то, что конечный продукт пути Эмбдена — Мейергофа
(называемого также путем Эмбдена — Мейергофа — Парнаса
или ЭМП) пируват может далее использоваться в пяти раз-
личных путях биосинтеза. Хотя приведенная на рис. 5.1 схема
может на первый взгляд показаться очень сложной, нельзя
Стехиометрия и эн»ргетика метаболических превращений 303
забывать, что па самом деле здесь изображена лишь ничтож-
ная доля из приблизительно 1000 происходящих в клетке реак-
ций. К тому же на рис. 5.1 не указано, что многие стадии этих
путей метаболизма обратимы; обратимость реакций обеспечи-
вает возможность как синтеза, так и деградации биооргапичес-
ких соединений. Несколько позже в этой же главе мы более
подробно рассмотрим некоторые из этих путей метаболизма,
а также контролирующие и регулирующие их ферментативные
механизмы.
На рис. 5.1 и 5.2 отсутствует и другая важная деталь. Мы
не показали здесь пути выведения, конечных продуктов метабо-
лизма из клетки. Многие из этих'веществ не нужны для жиз-
недеятельности клетки, а другие, например антибиотики или
внеклеточные ферменты, выполняют определенную функцию.
Некоторые из этих продуктов метаболизма (спирты, органи-
ческие кислоты, аминокислоты, антибиотики и многие другие)
представляют большую ценность для человека, что экономичес-
ки оправдывает их промышленное производство с помощью вы-
ращивания культур клеток. Для таких целей обычно стремятся
найти неэффективный в биологическом смысле штамм клеток,
который продуцирует интересующее нас вещество в количест-
вах, намного превышающих потребности организма. По этой
причине в настоящей главе мы попытаемся также проследить
пути взаимопревращений различных химических соедине-
ний внутри клетки, с тем чтобы понять, что можно произво-
дить с помощью живых организмов. Хотя здесь и в
последующих главах мы лишь коснемся этой темы, даже этого
поверхностного взгляда будет достаточно, чтобы оценить неве-
роятную сложность и почти неисчерпаемое разнообразие про-
исходящих в биологическом мире химических превращений.
Химический состав микроорганизмов или животных клеток
одного типа изменяется в сравнительно узких пределах, и ме-
таболическая активность клетки подчиняется обычным прин-
ципам стехиометрии и термодинамики. Так, для синтеза задан-
ного количества клеточного вещества требуется определенное
количество углерода, азота и кислорода из непосредственного
окружения клетки. Далее, если нам известна химическая фор-
ма, в которой эти элементы потребляются клетками из среды,
мы можем установить дополнительные ограничения, связываю-
щие количество утилизованных клетками питательных веществ
с массой образовавшихся клеток вообще и определенных про-
дуктов биосинтеза в частности. Кроме того, поскольку синтез
всех необходимых для построения новых клеток веществ тре-
бует затрат энергии, то необходимо учитывать и источники
этой энергии. Это в свою очередь налагает соответствующие
ограничения на количество используемой клеткой химической
304
Глава 5
или световой энергии, так как клетка усваивает энергию в виде
АТР в результате определенной последовательности реакций.
По сути дела, уже одни ограничения материального балан-
са и принципы термодинамики могут служить достаточной ос-
новой для выработки математических выражений, в весьма уп-
рощенном виде описывающих клеточную активность; в этом
подходе, рассматриваемом подробнее в разд. 5.10, совершенно
не учитываются данные о внутреннем химизме происходящих
в клетке процессов, а изучаются только параметры, характе-
ризующие рост клеточной массы, потребление клетками субст-
ратов и продуцирование определенных продуктов метаболизма.
Такой подход дает возможность получить уравнения, позволяю-
щие по ряду легко измеряемых величин сделать определенные
выводы о некоторых необходимых условиях работы биологи-
ческого реактора и проверить соответствие экспериментальным
данным.
Изучение материала мы начнем с фундаментальных меха-
низмов, лежащих в основе жизнедеятельности клетки, и затем
перейдем к конечным результатам процессов, осуществляющих-
ся в соответствии с этими механизмами. Прежде всего более
или менее подробно проанализируем роль АТР, универсального
переносчика энергии в клетке, а также типы реакций, в кото-
рых АТР поглощает и отдает энергию. В этих же разделах мы
рассмотрим важные окислительно-восстановительные процессы
и способствующие их осуществлению в клетке переносчики
электронов (NADH и NADPH), затем перейдем к изучению
некоторых путей катаболизма, связанных с разложением пита-
тельных веществ (из которых основное внимание будет уделе-
но глюкозе), определИхМ энергетику и стехиометрию этих про-
цессов и вкратце рассмотрим также механизмы фотосинтеза.
Далее с той же точки зрения мы изучим некоторые пути био-
синтеза различных веществ в клетке, после чего перейдем к
рассмотрению взаимосвязи между путями метаболизма, в кото-
рых генерируются энергия и электроны, и использующими их
последовательностями реакций. Наконец, мы проанализируем
общие стехиометрические соотношения, характерные для кле-
точного роста, и в завершение главы рассмотрим стехиометрию
образования продуктов метаболизма и ее связь со стехиометри-
ей клеточного роста.
5.1. Принципы термодинамики
Чтобы определить, пойдет ли данная химическая реакция
в клетке в прямом или обратном направлении, нам придется
принять ряд упрощений, поскольку полный анализ всех путей
метаболизма практически нереален. Прежде всего отметим, что
Стехиометрия и энергетика метаболических превращений
305
в случае элементарной химической реакции
аА+₽В ------------------------> VC + 6D (5.1)
изменение свободной энергии AG' может быть записано в виде
AG' = AG°'4-flTln(^^ (5.2)
Как и в предыдущих главах, строчными буквами а, b ц т. д.
мы будем обозначать молярные концентрации соединений А,
В и т. д. Поскольку концентрации веществ в биологических
растворах обычно невысоки, то в уравнении (5.2) вместо ак-
тивностей можно подставить концентрации реагентов. Надст-
рочный индекс (') над символами AG означает, что все расче-
ты относятся к водным растворам при pH 7; это позволяет не
включать концентрации воды и Н+ в последний член уравне-
ния (5.2), даже если Н2О и Н+ участвуют в реакции (5.1). Та-
ким образом, стандартное изменение свободной энергии
AGO/ обозначает изменение свободной энергии в реакции (5.1)
в нейтральном водном растворе, когда концентрации всех дру-
гих реагентов и продуктов реакции равны 1 М (см. [1]).
В закрытой системе реакция будет протекать слева напра-
во только в том случае, если AG' отрицательно. Отсюда сле-
дует, что в состоянии равновесия AG' равно нулю; таким обра-
зом мы получим хорошо известное выражение для равновесной
реакции:
AG0, = —7?? 1П (5.3)
где
C^egd^eq
a^eq^eq
K'eq
(5.4)
Запомните, что если в реакции участвуют вода или Н+, то их
концентрации не входят в правую часть уравнения (5.4); эти
параметры уже учтены в Л'еч (при pH 7).
В последующем обсуждении время от времени мы будем
считать отрицательные значения AGO/ свидетельством в пользу
того, что реакция идет слева направо. Такое допущение, без-
условно, справедливо только в самом первом приближении,
поскольку, во-первых, клетка не представляет собой закрытую
систему и, во-вторых, концентрации реагентов обычно не дости-
гают 1 М. Полезность этого допущения для понимания основ-
ных принципов биоэнергетики хорошо освещена в книге Ле-
нинджера [1], рекомендуемой для дополнительного чтения.
В то же время следует отчетливо представлять себе и те
ограничения, которые накладывает такой приближенный под-
ход. В сложной метаболической схеме типа изображенной на
20—537
306
Глаза 5
рис. 5.1 направление основного пути реакции в метаболизме
клетки не всегда можно установить путем изучения изолиро-
ванной реакции. Рассмотрим, например, реакцию между двумя
изомерами, являющимися промежуточными соединениями при
расщеплении глюкозы по пути Эмбдена — Мейергофа:
СНО сн2он
СНОН -------> СО AG°'=s — 1830 кал/моль (5.5)
СН2О—Р СН2О—Р
глицеральде- дигидроксиаце-
гид-3-фосфат тонфосфат
Здесь Р — фосфатная группа. Отрицательная величина измене-
ния свободной энергии обусловливает смещение равновесия в
сторону дигидроксиацетона до отношения 22:1. Однако, как
показано на рис. 5.1, в пути Эмбдена — Мейергофа глице-
ральдегидфосфат постоянно вовлекается в другую последова-
тельность реакций, приводящую в конце концов к пирувату, в
результате чего равновесие смещается и реакция (5.5) проте-
кает справа налево.
Многие биологические реакции и процессы, связанные с
энергообменом, включают стадии окисления-восстановления
типа
Аох 4~ Вгес1 < - Ared Вох (5.6)
Для определения направления таких реакций часто использу-
ют параметр, называемый изменением стандартного потенциа-
ла ДЕ®' и определяемый следующим образом:
Д£°” “£Длга/лге<1>—£°'<ви/вге<1> (5-7)
Здесь ^о,(яох/а red) — стандартный электродный потенциал по-
луреакции
АОх “Ь 2с" -> Ared (5.8)
В качестве точки отсчета для таких электродных потенциалов
применяют водородный электрод, потенциал которого при дав-
лении водорода 1,01 -105 Па и активности ионов гидроксония в
растворе, равной 1, принимают равным нулю:
2Н+ + 2е- Н2 £о=0,00В (рН = 0) (5.9)
Изменения свободной энергии и соответствующие изменения
электродного потенциала связаны соотношением
ДЕ' = — n!FEE' (5.10)
Здесь п — число участвующих в реакции электронов, а —
константа Фарадея [££' = 23,062 ккал/(В • моль) ]. Согласно
уравнению (5.10), слева направо протекают только реакции с
Стехиометрия и энергетика метаболических превращений
307
положительными значениями А£'. Если необходимо определить
значение ДЕ' для нестандартных условий, можно воспользо-
ваться уравнениями (5.2) и (5,10).
5.2. Взаимосвязанность метаболических реакций;
АТР и NAD
В гл. 3 и 4 мы рассматривали кинетику реакций в системах
с изолированными ферментами, а также практическое примене-
ние свободных ферментов и ферментных препаратов. Большин-
ство упоминавшихся в гл. 3 работ по изучению кинетики фер-
ментативного катализа и большая часть приведенных в гл. 4
примеров практического применения ферментов относятся к
гидролазам, которые по определению расщепляют или разла-
гают большие молекулы на более мелкие с участием воды в
качестве второго субстрата. Такие процессы деградации со-
провождаются снижением свободной энергии системы и поэто-
му в закрытой системе происходят спонтанно. В настоящем
разделе мы рассмотрим механизмы, которые позволяют откры-
той системе, а именно живой клетке, сочетать реакции, проте-
кающие с выделением энергии (экзэргонические), с реакциями
и другими функциями, осуществляющимися только с затратой
определенного количества энергии (эндэргонические процессы).
5.2.1. АТР и другие фосфаты
Ранее мы уже упоминали о строении и некоторых свойствах
аденозинтрифосфата (АТР; см. разд. 2.3.1 и рис. 2.8), где, в
частности, отмечали, что ферментативный гидролиз АТР до
ADP и неорганического фосфата сопровождается большим от-
рицательным изменением свободной энергии:
АТР + Н2О —>- ADP + Pz AG°'=—7,3 ккал/моль (5.11)
Здесь Pi — неорганический фосфат. Таким образом, в ходе гид-
ролиза АТР выделяется большое количество энергии, а в об-
ратной реакции присоединения фосфата к ADP энергия может
запасаться и храниться в виде АТР для последующего исполь-
зования. Давайте рассмотрим, как реализуется последний про-
цесс в условиях сопряженных химических реакций.
Понятие об общем промежуточном соединении можно про-
иллюстрировать на примере другой последовательности реак-
ций, являющейся частью пути метаболизма Эмбдена — Мейер-
гофа — Парнаса. Превращение альдегида в карбоновую кис-
лоту (в водном растворе) сопровождается понижением свобод-
ной энергии примерно на 7000 кал/моль. Как показано на
рис. 5.3, в изолированной системе эта свободная химическая
энергия полностью рассеялась бы в растворе. В живой клетке,
20*
308
Глаза 5
однако, этого не происходит. При биохимическом окислении
глюкозы превращение глицеральдегид-3-фосфата в соответст-
вующую карбоновую кислоту (3-фосфоглицерат) сопровожда-
ется регенерацией АТР из ADP (см. реакцию 2 на рис. 5.3).
Таким образом, обусловленное окислением альдегида уменьше-
ние свободной энергии благодаря участию ферментативной си-
стемы клетки сочетается с одновременной регенерацией АТР.
1) Окисление альдегида до карбоновой кислоты в изолированной системе
(в водном растворе)
RCHO + H2O —>- 2H+RCOO_+H+ AGi0'»*—7 ккал/моль
2) Те же реакции, сопряженные с регенерацией АТР (окисление глюкозы)
RCHO + HPO42-+ADP3- —2H + RCOO-+ATP4- AG2o/~0 ккал/моль
3) Вычитая из реакции 2 реакцию 1, получим
ADP3- + HPO?~ + H+ АТР4~ + Н2О AG3o,«+7 ккал/моль
4) Элементарные стадии реакции 2:
о~ н н О
-I I I II 2-
О-Р-О-С-С-С-Н + НТО. —
II I I
о н он
/ О- НН о- \
: _ I II I
--*- 2Н + Н2О + : О-Р-О-С-С-С-О-Р-О
II I I П II
\ о н он о о
; О—Р—О —С—С—С—О—Р —О- ; + ADP3
II I I II II
\ О Н ОНО О
--*- RCOO" + АТР4-
РИС. 5.3. Примеры реакций с общим промежуточным соединением.
Поскольку в результате реакции 2 свободная энергия практи-
чески не изменяется, то высвобождающаяся при окислении гли-
церальдегид-3-фосфата свободная энергия, очевидно, трансфор-
мируется в так называемую высокоэнергетическую (макроэрги-
ческую) фосфатную связь аденозинтрифосфата.
Последовательность элементарных реакций, в действитель-
ности происходящих в клетке, изображена в нижней части
рис. 5.3. К наиболее существенным характеристикам этих двух
последних реакций относятся, во-первых, наличие общего про-
межуточного соединения, являющегося продуктом первой реак-
ции и реагентом во второй, и, во-вторых, большая свободная
энергия реакции гидролиза (отщепления фосфатной группы)
Стехиометрия и энергетика метаболических превращений
309
промежуточного фосфорилированного соединения по сравнению
с гидролизом АТР. Равновесие последней стадии смещено
вправо, т. е. в сторону образования продукта реакции; таким
образом, этот этап метаболизма глюкозы одновременно пред-
ставляет собой и один из нескольких центров регенерации
АТР, необходимого для эндергонических реакций. Регенерация
АТР осуществляется путем превращения частично метаболизи-
рованного питательного вещества в высокоэнергетическое фос-
форилированное промежуточное соединение, которое затем в
результате катализируемой ферментом реакции передает фос-
фатную группу ADP.
Фосфорилирование различных соединений, в том числе и
ADP, используется клеткой для нескольких целей. Во-первых,
фосфорилированные соединения представляют собой удобное
хранилище значительной части свободной энергии, выделяю-
щейся в процессе окисления клеточного топлива. Свободная
энергия гидролиза некоторых соединений, называемых фосфат-
ными донорами, превышает AGO/ гидролиза АТР (например,
величины AGO/ для гидролиза фосфоенолпирувата и 1,3-дифос-
фоглицерата равны —14,8 и —11,8 ккал/моль соответственно).
Следовательно, гидролиз этих соединений может использовать-
ся для фосфорилирования ADP. Аналогично гидролиз АТР
может быть связан с фосфорилированием некоторых соедине-
ний, в результате которого образуются «низкоэнергетические»
фосфаты; величины AGO/ расщепления последних ниже соот-
ветствующей величины для реакции гидролиза АТР (например,
AGO/ гидролиза глюкозо-6-фосфата и глицеро-1-фосфата состав-
ляют —3,3 и —2,20 ккал/моль соответственно).
Вторая важная функция фосфорилирования в жизни клеток
связана с неспособностью силыюионизированных органических
веществ проникать через клеточные мембраны. Благодаря это-
му обстоятельству заряженные фосфорилированные соедине-
ния, являющиеся промежуточными соединениями во многих
путях метаболизма, остаются внутри клетки, что в свою оче-
редь позволяет получить максимум энергии и химического
сырья из питательных веществ. Обычно дефосфорилирование
является последней стадией метаболического пути; неионизи-
рованные отходы жизнедеятельности клетки могут выводиться
в среду.
5.2.2. Окисление и восстановление;
сочетание с превращением NAD
Мы только что рассмотрели роль АТР как переносчика фос-
фатных групп, связывание и отщепление которых сопровожда-
ется поглощением или выделением довольно значительного ко-
310
Глава 5
личества энергии. В этом разделе мы вкратце проанализируем,
как в живой клетке происходят процессы окисления и восста-
новления и как они связаны с метаболизмом АТР.
Прежде всего вспомним, что окисление какого-либо соеди-
нения означает, что его молекулы теряют электроны, а присо-
единение электронов есть не что иное, как восстановление. При
биохимическом окислении органическое вещество обычно теря-
ет электроны в виде атомов водорода; следовательно, термины
окисление и дегидрирование здесь являются синонимами. Точ-
но так же восстановление или присоединение электронов обыч-
но осуществляется путем гидрирования. Примером могут слу-
жить реакции восстановления пировиноградной кислоты и окис-
ления молочной кислоты:
+2Н (восстановление
ртт ПИрОВИНОГрадНОЙ z-тт
3 КИСЛОТЫ) 3
СО -... ................... снон
I —2Н (окисление молоч- <
соон ““°™’ соон
пировино- молочная
градная кислота
кислота
(Пировиноградная кислота представляет собой по сути дела
то же, что пируват на рис. 5.1. Пируватом называют ионизиро-
ванную форму кислоты СНзСОСОО-, преобладающую при
биологических pH.)
Роль переносчиков пар атомов водорода, высвобождающих-
ся в процессах окисления и расходующихся в реакциях восста-
новления, выполняют производные нуклеотидов, в первую оче-
редь никотинамидадениндинуклеотид (NAD) (см. рис. 2.9) и
его фосфорилированное производное NADP. Ранее эти соедине-
ния относили к коферментам, поскольку их присутствие обычно
обязательно в процессах ферментативного окисления или вос-
становления. Если, например, необходимы атомы водорода, то
никотинамидная группировка восстановленной формы NAD
может отдать их в процессе окисления:
NADH
\/н О
11
нс с—с—nh2
II II
НС\ сн
NAD*
— 2 Н (окисл}
+ 2 Н (восст.)
R
восстановленная
форма
форма
Стехиометрия м энергетика метаболических превращений 311
Процесс окисления легкообратим, поэтому NAD может и при-
соединять электроны (атомы Н), если последние высвобожда-
ются в процессах окисления других соединений. Окисленную и
восстановленную формы NAD мы будем обозначать далее так,
как и на приведенной выше схеме, символами NAD+ и
и NADH соответственно.
Как переносчик электронов NAD выполняет две основные
функции, первая из которых аналогична функции АТР и за-
ключается в переносе восстанавливающих эквивалентов (элект-
ронов или атомов IT), высвобождающихся в ходе распада пи-
тательных веществ, к биосинтетическим реакциям. Необходи-
мость в восстановлении часто обусловливается тем обстоя-
тельством, что степень окисления непосредственных продуктов
биосинтеза не всегда соответствует степени окисления веществ,
необходимых для построения структурных элементов клетки.
Как мы уже отмечали, степень окисления углерода в клетке
приблизительно соответствует углеводам (СН2О)П, поэтому
аутотрофным организмам, использующим в качестве источника
углерода СО2, для успешной ассимиляции углерода необходимо
обладать большой восстанавливающей способностью:
СО2+4Н —> (СН2О)+Н2О
Хотя восстанавливающие эквиваленты в наибольшей степе-
ни используются в процессах ассимиляции углерода, усвоение
серы и азота также часто требует участия восстановительных
реакций. Чтобы оценить эти потребности, можно принять, что
клетка содержит азот в основном в степени окисления аммиака
(NH3), а серу — главным образом в форме, аналогичной суль-
фидному иону (S2-). Тогда для утилизации, например, сульфа-
та, в качестве источника серы клетке потребуется значительная
восстанавливающая способность, о чем свидетельствует соот-
ветствующее уравнение реакции:
SO42- + 8H —> S2~+4H2O
NAD и родственные пиридиновые нуклеотиды, также выпол-
няющие функцию переносчика атомов водорода, участвуют и в
образовании АТР в аэробном метаболизме. В разд. 5.3 мы под-
робнее изучим процесс соединения атомов водорода из NADH
с кислородом в последовательности реакций, известной под на-
званием дыхательной цепи. Выделяющейся в этой последова-
тельности реакций окисления энергии достаточно для превра-
щения трех молекул ADP в АТР.
Интересно, что во всех известных биологических системах,
включая анаэробные, аэробные и фотосинтезирующие орга-
низмы, АТР является основным инструментом аккумулирова-
ния и хранения энергии окисления или света, необходимой для
312
Глава 5
осуществления эндэргонических процессов в клетке. В после-
дующих разделах этой главы мы проследим за ходом процео
сов утилизации АТР и переноса электронов во все более и бо-
лее сложных путях метаболизма в анаэробных, аэробных и
фотосинтезирующих системах. Анализ уравнений окислительно-
восстановительных реакций позволит нам найти стехиометриче-
ские ограничения, налагаемые составом питательной среды и
клеток, путями метаболизма и природой конечных метаболи-
тов. Поскольку свободная энергия окисления клеточного топли-
ва в конечном счете хранится в АТР, то эффективность утили-
зации АТР в различных клеточных процессах может служить
удобной мерой относительного количества энергии, потребляе-
мой клеткой в этих процессах. Более того, изучение АТР по-
зволяет логически увязать процессы построения углеродных
скелетов и связанные с ними изменения свободной энергии. На-
конец, сравнение энергетики различных биологических систем
дает возможность понять основу высокой термодинамической
эффективности их биосинтетического аппарата.
5.3, Катаболизм углерода
Процессы расщепления питательных веществ с целью извле-
чения энергии называются катаболизмом. В качестве источника
углеродсодержащих питательных веществ в подавляющем боль-
шинстве случаев используются углеводы, хотя некоторые мик-
роорганизмы могут также утилизировать аминокислоты, углево-
дороды и другие соединения. Иллюстрацией разнообразия ми-
ра микробов может служить тот факт, что практически любой
углевод или родственное соединение может усваиваться тем
или иным микроорганизмом. Большинство микроорганизмов,
способных утилизировать углеводы, может перерабатывать и
наиболее распространенный моносахарид глюкозу. В следую-
щем разделе мы изучим основные пути деградации глюкозы.
Существует по меньшей мере семь различных путей катаболиз-
ма глюкозы. Природа микроорганизма определяет, какой из
этих механизмов реализуется и какие конечные продукты об-
разуются. При этом основное внимание мы будем уделять
участию рассмотренных выше процессов переноса энергии и
электронов, общей стехиометрии метаболического пути, а так-
же идентификации важнейших на данном пути промежуточных
соединений.
5.3.1. Метаболический путь Эмбдена — Мейергофа — Парнаса
Изображенный на рис. 5.1 метаболический путь Эмбдена —
хМейергофа — Парнаса (ЭМП) включает десять катализируе-
мых ферментами стадий, исходным веществом для которых яв-
Стехиометрия и энергетика метаболических превращений
313
н,он
5 .
но
ОН
глюкоза
Н
гексокиназа
ОН
ОН
<АТР
CHpPOj’
нЬон
К ОН Н и глюкоз о-о- фосфат
но\—<он
Н ОН
фоссрогексо-
изопераза , ,
I
3-фосфоглицериновыи
альдегид_ +
глицвральдв - р~1ЧЛи
гид-3- фосфат-
дегидрогеназа ip^NADH
снррог
НСОН 1,3-дифос<ро-
6-ОРоГ глицерат
II 11
СН,ОРО32‘
I ,О\СН7ОН
(<н но>
н у—г он
Он н
фруктоза -6-(рос-
<рат
фОСфОфРУ -
чтокиназа <_ дрр
СНрРОз
I
кн но;.
Н>—{'ОН
он н
, 1/-ADP
3-фоссрогли- V
цераткиназа д-рр
СНгОРО
з неон
5 000
Ц фОсфогли-\
~ церотутазаХ
СНгОН
н^оро*'
СОО’
2-
3
3-сроссрогЛи-
церат
З-фосрогли-
церат
альдолаза
СНрРОз'
<рруктозо -1,6-
дифосфат
1- Н
снррОз с=о
0 = 0 ИСОН
J с^враза J“*u
снгон СНгОРО3
дигадроксиацеток- рикобый^^ег'ид
(poctpam I
СН2
II ' г-
с-о- ро3
Соо“ фосфоенол-
камаза L
Л^АТР
Сг1о
I
с= о лирудат
ООО-
н
I
РИС. 5.4. Путь Эмбдена — Мейергофа — Парнаса (ЭМП). Обратите внимание
на то, что из каждой шестиуглеродной молекулы субстрата (глюкозы) обра-
зуются два промежуточных соединения с тремя атомами углерода, каждое из
которых претерпевает последовательные превращения, изображенные на пра-
вой стороне рисунка.
ляется глюкоза, а конечным продуктом последовательности ре-
акций— пируват (рис. 5.4). В настоящем разделе мы рассмот-
рим этот наиболее детально изученный путь катаболизма угле-
рода, обращая особое внимание на энергетику и процессы вос-
становления.
Анализ приведенной на рис. 5.4 последовательности реакций
пути ЭМП показывает, что каждая стадия представляет собой
314
Глава 5
простую реакцию изомеризации, расщепления кольца или миг-
рации небольших групп, например атомов водорода или фос-
фатной группировки. В конечном счете из одного моля глюко-
зы образуются два моля пирувата. Первая и третья реакции
осуществляются совместно с гидролизом АТР; спонтанно в не-
обходимом направлении эти реакции не протекают. Две другие
стадии связаны с высвобождением свободной энергии, доста-
точной для фосфорилирования ADP; эти две реакции осуществ-
ляются дважды в ходе превращения каждой молекулы исход-
ной глюкозы, поэтому конечным итогом пути ЭМП является
фосфорилирование ADP. Как показано на рис. 5.4, дегидриро-
вание глицеральдегид-3-фосфата сочетается с восстановлением
NAD+, причем эта реакция происходит дважды в ходе превра-
щения каждой молекулы глюкозы. Таким образом, общую сте-
хиометрию пути метаболизма ЭМП можно выразить следую-
щим уравнением:
CsH12O6+2Pz+2ADP + 2NAD+ —»
—2C3H4O3+2ATP + 2(NADH+H+) (5.12)
В результате превращения глюкозы в пировиноградную кисло-
ту клетки обеспечиваются резервами химической энергии и
восстанавливающих эквивалентов. Накапливание энергии за
счет превращения этого или какого-либо другого субстрата на-
зывается фосфорилированием на уровне субстрата.
Метаболический путь ЭМП выполняет и другую важную
функцию, связанную со снабжением исходными веществами ря-
да биосинтетических последовательностей реакций. В качестве
биосинтетических субстратов (предшественников) использу-
ется не только пируват, но и ряд промежуточных веществ
этого пути метаболизма (см. рис. 5.1). По этой причине мета-
болический путь ЭМП называют (как и некоторые другие пути
метаболизма) амфиболическим, подразумевая при этом, что
он обеспечивает клетку как источниками энергии, так и исход-
ными веществами.
В клетках мышечных тканей и молочнокислых бактериях,
а также в ряде других клеток путь ЭМП не завершается на
этапе пирувата, а удлиняется еще на одну стадию:
C3H4O3+NADH+H+ —> C3H6O3 + NAD+
Вся последовательность реакций, начинающаяся с глюкозы и
завершающаяся молочной кислотой, называется гликолизом.
Интересно сравнить изменение свободной энергии в гликолизе:
Глюкоза+2P/+2ADP —> 2лактат + 2АТР + 2Н2О
AG°' = —32 400 кал/моль (5.13)
Стехиометрия и энергетика метаболических превращений
315
с изменением свободной энергии расщепления
Глюкоза —2 молочная кислота
ОДНОЙ ГЛЮКОЗЫ
AG°'=—47 000 кал/моль (5.14)
Из уравнений (5.13) и (5.14) следует, что в виде высоко-
энергетических фосфатов сохраняется энергия, равная
14,6 ккал/моль глюкозы (или 7,3 ккал/моль АТР). Кажущая-
ся эффективность переноса свободной энергии равна 14/47Х
Таблица 5.3. Концентрации промежуточных веществ, участвующих
в процессе гликолиза в эритроцитах человека, и соответствующие
изменения стандартной свободной энергии. Обратите внимание на то, что
концентрации всех вешеств существенно отличаются от значения 1М,
используемого при определении стандартной свободной энергии. Значения
AG0/ соответствуют стадиям, в которых участвует указанное в данной
строке промежуточное соединениеа
Концентра-
ция, мкМ
5000
83
14
31
138
18,5
118
29,5
23
51
2900
1850
138
1000
Промежуточное соединение
Изменение стан-
дартной свобод-
ной энергии Дб°',
ккал/моль
Глюкоза
Глюкозо-6-фосфат (G6P)
Фруктозо-6-фосфат (F6P)
Фруктозо-1,6-дифосфат (FDP)
Дигидроксиацетонфосфат (DHP)
Глицеральдегнд-З-фосфат (GAP)
З-Фосфоглицерат (3PG)
2-Фосфоглицерат (2PG)
Фосфоенолпируват (РЕР)
Пируват (Руг)
Лактат (Lact)
АТР
ADP
Фосфат
—4,0
-4-0,4
—3,40
-4-5,73
4-1,83
—3,0
-Н ,06
4-0,44
—7,5
—6,0
аИз книги: Lehninger A. L., Biochemistry, 2d ed., table 16.1, Worth Publishers, Inc.,
New York, 1975; есть перевод первого издания: Леиинджео А., Биохимия,—М.: Мир,
1976.
X100 — 31 %. Введение поправок на реальные концентрации и
преобладающие in vivo pH (см. табл. 5.3) показывает, что эта
оценка существенно занижена и истинная эффективность со-
ставляет около 53%. Причина столь существенного разли-
чия между кажущейся и действительной эффективностями
становится понятной после коррекции приведенных в табл. 5.3
величин AG0/ в соответствии с реальными концентрациями (см.
упражнение 5.4); результаты коррекции показывают, что Д6'
примерно равно нулю для всех стадий, за исключением трех
(превращение глюкозы в G6P, F6P в FDP и РЕР в Руг;
расшифровка сокращений дана в табл. 5.3). Таким обра-
316
Глава 5
зом, большинство элементарных реакций в последовательности
ЭМП являются практически равновесными и, следовательно,
обратимыми. Значение неравновесных стадий для всего процес-
са мы поймем после изучения ферментативной регуляции пути
ЭМП (разд. 5.7.2).
5.3.2. Другие пути катаболизма углеводов
Прежде всего рассмотрим две другие последовательности
реакций, в которых осуществляется катаболизм глюкозы. Пен-
тозофосфатный цикл или пентозофосфатный путь (называемый
также гексозомонофосфатным путем или шунтом) начинается
с окисления глюкозофосфата:
Глюкозо-6-фосфат-гКАЭР+ —>• 6-фосфоглюконат4^АРРН + Н+
Основной задачей пентозофосфатного пути является обеспече-
ние клетки NADPH, выполняющим роль переносчика электро-
нов в биосинтетических реакциях. Схема всех происходящих на
этом пути реакций довольно сложна (см. стр. 456 в работе
[1]), но в целом стехиометрию пентозофосфатного пути можно
выразить следующим довольно простым уравнением:
Глюкоза-Ь 12NADP+ + 7H2O+ATP —>
—> 6CO2+P,-+12(NADPH + H+)+ADP (5.15>
Таким образом, конечным результатом пентозофосфатного пу-
ти является полное окисление глюкозо-6-фосфата до СОг с пе-
редачей всех электронов (Н) NADP.
В указанном пути потребляется АТР. Чтобы обеспечить
клетку АТР, пентозофосфатный цикл может быть прерван на
стадии, когда общая стехиометрия процесса выражается урав-
нением
3 Глюкоза+ 6NADP++ АТР —>•
—> 2 фруктозо-6-фосфат+глицеральдегид-3-фосфат+ЗС02+
+ 6(NADPH + H+) +ADP + Pi (5.16)
Если дальнейшие превращения фруктозо-6-фосфата и глице-
ральдегид-3-фосфата до пирувата будут осуществляться по пу-
ти ЭМП, то на этом пути произойдет и фосфорилирование
6 молекул ADP (в расчете на 3 молекулы глюкозы). В резуль-
тате из моля глюкозы образуется 5/3 моля АТР, но общий вы-
игрыш в энергии будет все же меньше, чем в метаболическом
пути ЭМП.
В то же время пентозофосфатный путь имеет и свои пре-
имущества, обусловленные прежде всего тем обстоятельством,
что в нем в качестве промежуточных веществ образуются ри-
Стехиометрия и энергетика метаболических превращений
317
бозо-5-фосфат и эритрозо-4-фосфат, являющиеся важными
предшественниками в синтезе пуринов и пиримидинов в клетке.
В метаболическом пути ЭМП такие соединения не образуются.
По этой причине микроорганизмы типа молочнокислых бакте-
рий, располагающие для катаболизма углеводов только путем
ЭМП, в анаэробных условиях могут расти только на сложных
питательных средах, в состав которых входят пентозы, пурины
и пиримидины. Напротив, бактерия Е. coli даже в анаэробных
условиях способна расти и на более простых средах, поскольку
в ней реализуются одновременно и путь ЭМП, и пентозофос-
фатный путь (в некоторых условиях, например, 75% углеводов
катаболизируется по пути ЭМП, а 25%—по пентозофосфатно-
му пути). Этот факт является иллюстрацией общего принципа,
согласно которому клетки могут катаболизировать данное пи-
тательное вещество несколькими путями одновременно, что, по
всей вероятности, обеспечивает их оптимальный рост за счет
удовлетворения потребностей как в энергии, так и в веществах-
предшественниках.
Последний из рассматриваемых здесь путей катаболизма
глюкозы называется путем Энтнера — Дудорова (ЭД). Общая
стехиометрия этой последовательности реакций выглядит сле-
дующим образом:
Глюкоза + АТР+NADP+ —>
—> глицеральдегид-3-фосфат+пировиноградная кислота +
+ADP+NADPH+H+ (5.17)
Если учесть, что последующее превращение глицеральдегид-3-
фосфатд в пируват по той же схеме, что и в пути ЭМП, со-
провождается фосфорилированием двух молекул ADP, то ста-
новится очевидным, что с энергетической точки зрения путь
ЭД, в котором образуется моль АТР из моля глюкозы, не
очень выгоден. Прежде чем перейти к изучению катаболизма
других питательных веществ, следует подчеркнуть, что описан-
ные выше основные пути катаболизма глюкозы далеко не ис-
черпывают возможности бактерий.
Ранее мы упоминали, что в качестве источников углерода
микроорганизмы могут утилизировать различные соединения.
С детальным анализом этой проблемы читатель может ознако-
миться в работе [3]; мы же ограничимся несколькими приме-
рами, иллюстрирующими только основные пути катаболизма
отличных от глюкозы веществ.
Утилизация других сахаров часто начинается с их превра-
щения с помощью одной или нескольких реакций в глюкозу
или в одно из промежуточных соединений в путях ее катабо-
лизма. В качестве одного из примеров на рис. 5.5 приведена
последовательность реакций, посредством которых Е. coli на-
318
Глава 5
Путь
ЗИП
РИС. 5.5. Катаболизм лактозы Е. coli (UDPG — уридиндифосфатглюкоза;
UDPGal — уридиндифосфатгалактоза).
чинает ассимиляцию дисахарида лактозы. Особенность этого
пути катаболизма заключается в специфическом механизме,
обеспечивающем утилизацию галактозы. Обратите внимание на
циклический характер утилизации уридиндифосфата (UDP),
который выполняет таким образом функции катализатора.
В качестве второго примера можно привести утилизацию фрук-
тозы некоторыми бактериями; в этом случае в результате двух
катализируемых ферментами реакций фруктоза превращается
в соответствующий 1,6-дифосфат, который затем может быть
включен в путь ЭМП. Утилизация бактериями пентоз обычно
начинается с последовательности реакций, в результате кото-
рых образуется ксилулозо-5-фосфат; последний может быть
включен в пентозофосфатный путь, а может претерпевать рас-
щепление по фосфокетолазному пути (рис. 5.6).
Стехиометрия и энергетика метаболических превращений
319
Чрезвычайно интересны и разнообразны процессы утилиза-
ции микроорганизмами и других органических соединений, в
частности углеводородов; любопытны и происходящие при
этом химические превращения. Здесь мы ограничимся упоми-
нанием о существовании в мире микроорганизмов ферментных
систем, способных окислять углеводороды с изменением или без
изменения их углеродного скелета, превращать углеводородные
субстраты в аминокислоты, липиды, витамины, другие компо-
АТР ADP
Рибоза
к у
рибозо~Р-росрат
ксилулозо - J- cpocpcmt
Ацрталросрагп
NADH
NAD
2ADP
2 АТР
и
пируват
Рибоза + NAD + + ADP + 2 Р, —пируват ^ацетилростат +
+ATP+NADH + H*
РИС. 5.6. Упрощенная схема катаболизма рнбозы по фосфокетолазному пути.
ненты микробных клеток и иные соединения. Более того, неко-
торые микроорганизмы способны расщеплять токсичные веще-
ства, например фенолы и полихлорбифенилы. Очевидно, такие
микроорганизмы и синтезируемые ими ферменты потенциально
очень важны как катализаторы специфических химических пре-
вращений, которые могут оказаться полезными в решении ряда
проблем охраны окружающей среды. Введение в биохимию пу-
тей метаболизма углеводородов и соответствующих ферментов
дано в превосходной книге Доуэлла [3].
5.4. Дыхание
Дыханием называется сопровождающийся выделением энер-
гии процесс, в котором органические или восстановленные не-
органические соединения окисляются в результате взаимодей-
320
Глава 5
ствия с неорганическими веществами. Как показано в табл. 5.4,
в различных бактериях процесс дыхания осуществляется с уча-
стием различных восстановителей и окислителей. Если окисли-
телем является не кислород, то процесс называют анаэробным
дыханием, а термин аэробное дыхание применяют в тех типич-
ных для эукариот и многих бактерий случаях, когда роль
окислителя выполняет Ог. Напомним, что литотрофы исполь-
зуют неорганические восстановители. Примеры литотрофов
Таблица 5.4. Восстановители и окислители в процессах дыхания бактерий3
Восстановитель Окислитель Продукты реакции Организмы
Н2 О2 H20 Водородные бактерии
Н2 so?- H2O + S2- Desulfovibrio
Органические вещества 02 C.O2+ H2U Многие бактерии, все растения и животные
NH3 02 N O2~ 4" H2O Нитрифицирующие бак-
NO.- 02 no3-+h2o Нитрифицирующие бак- терии
Органические вещества N03- n2+(JO2 Денитрифицирующие бактерии
Fe2+ 02 Fe3+ Ferrobacillus (железобактерии)
S2- 02 SO?- + H2O Thiobacillus (серные бактерии)
а Из книги: and Winston, Inc. Sistrom W. R., Microbial Life, 2d ed., , New York, 1969. table 4-2, р. 53, Holt, Rinehart
также приведены в табл. 5.4. По имеющимся в настоящее вре-
мя данным все литотрофы являются аутотрофами, так как ис-
точником углерода для них служит СОг-
В большинстве форм дыхания органические вещества окис-
ляются под действием кислорода. Далее в настоящем разделе
мы будем рассматривать только такую форму дыхания и, упот-
ребляя термин «дыхание», будем подразумевать только аэроб-
ный процесс. Удобно подразделять весь процесс дыхания на
две фазы, в первой из которых органические вещества окисля-
ются до СОг, а атомы водорода (электроны) захватываются
NAD. В следующей фазе атомы водорода участвуют в ряде по-
следовательных химических превращений, в результате которых
из ADP регенерируется АТР, а сами атомы водорода соединя-
ются с кислородом, образуя воду. Ниже мы рассмотрим эти
две фазы более детально.
Стехиометрия и энергетика метаболических превращений
32!
5.4.1. Цикл трикарбоновых кислот
Сначала в дополнение к разд. 5.3, посвященному метабо-
лизму углеводов, отметим, что все описанные там пути метабо-
лизма, ведущие к пирувату, могут реализоваться и в процессе
дыхания, а свойственные собственно дыханию реакции начи-
наются со стадии образования пирувата. В то же время в ме-
таболизме дыхания пируват не восстанавливается до какого-
либо конечного продукта с помощью образовавшихся в процес-
се расщепления глюкозы атомов водорода; последние исполь-
зуются для других целей, которые мы вкратце здесь также
рассмотрим. Более того, в процессе превращения пирувата в
производное уксусной кислоты (ацетил-СоА) генерируется до-
полнительное количество универсального восстановителя
NADH:
пируватдегидрогеназный комплекс
CH3COCOOH + NAD+4-COA—SH-------------------------->
—у СНзСО—S—CoA+CO2+NADH+H+ (5.18)
Ацетил-СоА (см. рис. 2.9) является также ключевым промежу-
точным веществом в катаболизме аминокислот и жирных кис-
лот; следовательно, три класса органических соединений могут
окисляться до ацетил-СоА.
Первая фаза этого окисления осуществляется в замкнутой
последовательности реакций, называемой циклом трикарбоно-
вых кислот (ТКК), циклом Кребса или циклом лимонной кисло-
ты (рис. 5.7). Обратите внимание на то, что два оставшихся ато-
ма углерода пирувата (один теряется ранее в виде СО2 при об-
разовании ацетил-СоА) включаются в цикл путем образования
карбоновой кислоты с 6 углеродными атомами из ацетил-СоА и
С4-кислоты. В дальнейших стадиях цикла ТКК, однако, два ато-
ма углерода выделяются в виде СО2, и таким образом в этой
первой фазе дыхания потребляются все углеродные атомы ис-
ходного пирувата.
Общая стехиометрия цикла ТКК выглядит следующим об-
разом:
С3Н4Оз+АОР + Р; + 2Н2О + ЕАО+4МАО+ —>
—>3CO2+ATP + FADH2+4(NADH + H+) (5.19)
Может показаться, что цикл ТКК (в таком виде, как он
изображен на рис. 5.7) выполняет только каталитическую
функцию, поскольку здесь не указано никакого иного источника
углерода, кроме субстрата. На самом же деле, как показано
на рис. 5.1, цикл ТКК очень важен и в качестве источника
предшественников для последующих биосинтетических реак-
ций. Следовательно, некоторые промежуточные соединения
21—537
•хоЕЭия xrasoHogdBMHdi ггииП ’Z'S DHd
'd + JQV
yog -i/TiHnhx/fy
Q>
II
VODS-D
огн
гНЭ
-HD
HOOD
(qvn *)
-QD HSV°3
< \ V s
\( 0) DUlOlfO/lM
uogodaw/ftssoway-io •
HOOD
HSVQD<->
ашо^эпя uDHtfzMWtf
HOOD
"HD
"HD
I
HOOD
<avj .*)
HZ 4-
(*□) Ю/УОУЭПУ
кпдос/пь/Яф
HOOD
Oz
0=9
HOOD гНО-ОгН
'^•гоо
z->(aVN Л-)
Нс
nii/CMVnX ifDHHOJjlWVlM
HOOD
tfOMMWnj/- I
i/wnft ‘njgatfy u-ynh) HOHD
Х/9ООУ00 . I
-dw/ncfaj /swift HOOD"HD — DH
HOOD
t
( 9g) ПШОУЭПЛ XDHMOAMW
HOOD
"HD
. HOOD"HD —D—OH
(7g) ашо/гэпу |
уоуз/1м^одэ//9до‘т HOOD
HOOD
г ' ОгН
n^j \
> I----\
HODH , V ч O=D
I (avN i
HOOD j_{£ HOOD У is/vaA/aaa>oy-2mwany
VOD-DCHD
II
О
A (qvn )
HD
HD
I
HOOD
Ь V
HOOD
"HD
V°DSH
г.
HSVO0->
ои/о&эпя #омро&гонпдо1/иу
HOODDfHD
II
О
$ eaeuj
Стехиометрия и энергетика метаболических превращений
323
этого цикла постоянно отбираются для целей биосинтеза и их
количества должны возобновляться. Эта цель достигается пу-
тем синтеза щавелевоуксусной кислоты из пирувата или другой
кислоты с тремя углеродными атомами. В некоторых микро-
организмах недостаток промежуточных веществ в цикле ТКК
возмещается за счет глиоксилатного цикла, в котором в ре-
зультате конденсации двух молекул ацетата образуется моле-
кула сукцината.
5.4.2. Дыхательная цепь
Оставив на время метаболизм соединений углерода, давай-
те проследим за реакциями, в которых атомы водорода окисля-
ются до воды, — именно этот процесс в основном обеспечивает
аэробные клетки энергией. В каждом единичном цикле ТКК
высвобождаются четыре пары атомов водорода, три из которых
переносятся к NAD, а одна, образующаяся при дегидрировании
янтарной кислоты, как показано на рис. 5.7, переносится к фла-
винадениндинуклеотиду (FAD) (см. рис. 2.9). Высвобождаю-
щиеся в цикле ТКК восстанавливающие эквиваленты частично
используются в биосинтетических реакциях, а частично — в ре-
генерации АТР.
В последующем обсуждении регенерации АТР в процессе
дыхания мы будем уделять особое внимание такой ситуации,
когда в реакциях дыхательной цепи участвуют все атомы во-
дорода, образующиеся при расщеплении глюкозы. Именно та-
ким путем создается большая часть энергетических запасов
клетки в виде АТР. На рис. 5.8 в сокращенном виде приведена
последовательность реакций в дыхательной цепи, а также их
связь с процессом полного расщепления пирувата через ацетил-
СоА в цикле ТКК (Кребса). На этой схеме символами FPi и
FP2 обозначены два различных флавопротеина, которые пред-
ставляют собой ферменты, содержащие FAD и участвующие в
транспорте электронов. Электроны от NADH (NADH = FPJ
передаются коферменту Q (обозначенному на рисунке симво-
лом Q); в этом процессе передача каждой пары электронов со-
провождается фосфорилированием одной молекулы ADP до
АТР. Электроны, высвобождающиеся в цикле ТКК в ходе де-
гидрирования сукцината, передаются при посредстве FAD (в
FP2) непосредственно коферменту Q.
Начиная с этого момента судьба всех электронов одинако-
ва: все они взаимодействуют с рядом цитохромов — гем-содер-
жащих белков, обозначенных на рис. 5.8 символами Ь, с, а и
аз. В ходе этого взаимодействия каждая пара электронов гене-
рирует две молекулы АТР. Процесс регенерации АТР в дыха-
тельной цепи называется окислительным фосфорилированием.
21*
'Пируват
Аиртил-Тл\
Жирные
кислоты
Амино- -
кислоты
'—Цикл тринар&онобых—'
'—Транспорт электронов 8 дыхательной цепи--'
кислот
РИС. 5.8. Фосфорилирование ADP в дыхательной цепи с участием высокоэнергетических электронов высво-
бождающихся при окислении углеводов, жирных кислот и аминокислот (FPi —NADH; FP2 — сукцинатде-
гидрогеназа; Q — кофермент Q; Ь, с, а и а3 — соответствующие цитохромы). [Воспроизведено из работы- Left-
mnger A., Bioenergetics, 2d ed., р. 74, W. A. Benjamin, Inc., Palo Alto, CA, 1974.] H
Стехиометрия и энергетика метаболических превращений 325
В конце концов атомы водорода вступают в реакцию с раство-
ренным кислородом, в результате чего в качестве второго ко-
нечного продукта окисления образуется вода. Если проследить
за изменениями свободной энергии на отдельных стадиях ды-
хательной цепи, нетрудно заметить, что АТР регенерируется во
всех случаях, когда1, уменьшение свободной энергии превышает
величину 7,3 ккал/моль, необходимую для фосфорилирования
ADP.
Таким образом, все происходящие в дыхательной цепи реак-
ции в конечном счете описываются следующими уравнениями:
NADH+H+-P/2O2+3ADP+3P, —* NAD++4H2O+3ATP (5.20)
И
FADH2+V2O2+2ADP + 2P,- —> FAD + 3H2O+2ATP (5.21)
Потенциальные возможности дыхания в отношении обеспе-
чения клетки энергией намного превосходят возможности гли-
колиза, поскольку изменение стандартной свободной энергии
AG0' в реакции
Глюкоза + 6О2 —>• 6СО2+6Н2О (5.22)
составляет —686 ккал/моль. Можно проверить, насколько эф-
фективно используется этот богатый источник энергии в живых
аэробных организмах. Для сравнения с реакцией (5.22) допус-
тим, что глюкоза полностью окисляется до СО2 и Н2О по ме-
таболическим путям ЭМП, ТКК и в дыхательной цепи. При
этом не следует забывать, однако, что путь ЭМП и цикл ТКК
являются амфиболическими и что, следовательно, часть проме-
жуточных соединений постоянно отвлекается, включаясь в био-
синтетические пути метаболизма. Поэтому на самом деле глю-
коза не всегда претерпевает именно эти превращения, и при-
веденные ниже расчеты позволяют оценить только высший пре-
дел выхода АТР из глюкозы в аэробной клетке.
Путем сложения уравнения (5.12): (путь ЭМП) с умножен-
ным на два уравнением (5.19) (цикл ТКК), умноженным на
десять уравнением (5.20) [окисление всех NADH + H+, образу-
ющихся по пути ЭМП (дважды), и реакций цикла ТКК (четыре
раза)] и умноженным на два уравнением (5.21) [окисление
FADH2] получим
СбН12Об+38АОР+38Р;+6О2 —* 6СО2+З8АТР+44Н2О (5.23)
Поскольку изменение стандартной свободной энергии при гид-
ролизе АТР равно —7,3 ккал/моль, то изменение свободной
энергии в реакции (5.23) составит приблизительно
AGO,«(38 моль ATP/моль глюкозы) (7,3 ккал/моль АТР) =
=—277 ккал/моль глюкозы (5.24)
326
Глава 5
Эта величина в 19 раз превышает энергию, усваиваемую клет-
кой в ходе гликолиза. Как и в гликолизе, эффективность про-
цесса дыхания очень высока:
277
Эффективность утилизации энергии =>-40 % (5.25)
686
Если ввести поправки на нестандартные концентрации в клет-
ке, то мы получим поразительно высокую степень эффективно-
сти, превышающую 70%. Большая часть не использованной в
процессе регенерации АТР энергии рассеивается в виде тепло-
ты, которая должна тем или иным способом отводиться, чтобы
температура не вышла за рамки допустимых для клетки физио-
логических пределов.
Поскольку в практической термодинамике химические ма-
шины обычно не изучаются, здесь может оказаться полезной
аналогия с классическим примером сжатия газа в цилиндое
с поршнем. Если газ сжимают быстро, то большое количество
энергии теряется в виде теплоты и поэтому не может быть ис-
пользовано при последующем расширении. Аналогичным про-
цессом является совершенно неэффективное сжигание глюкозы
на воздухе. К идеальному обратимому процессу приближается
сжатие газа путем очень медленного движения поршня, когда
выделяется минимальное количество тепла. Точно так же и
процессы в живой клетке приближаются к обратимым и мак-
симально эффективным с точки зрения извлечения энергии бла-
годаря тому, что окисление глюкозы осуществляется в несколь-
ко стадий, каждая из которых характеризуется относительно
небольшим изменением свободной энергии.
Описанные выше реакции в дыхательной цепи типичны для
обычной эукариотической клетки. В случае прокариот транс-
порт электрона к атому кислорода может сопровождаться фос-
форилированием менее трех молекул ADP. Кроме того, цито-
хромный путь может несколько видоизменяться в зависимости
от вида прокариоты, а на эффективность фосфорилирования
могут влиять условия роста клеток. Для оценки эффективности
усвоения энергии в дыхательной цепи используется особый па-
раметр, отношение P/О, равное числу актов фосфорилирования
в расчете на один усвоенный атом кислорода.
Позднее в этой главе мы узнаем, каким образом степень
окисления субстрата и биосинтетические потребности клетки
совместно налагают определенные ограничения на стехиомет-
рию амфиболических путей. Прежде, однако, мы завершим наш
обзор путей метаболизма как источников энергии, вкратце рас-
смотрев фотосинтез — процесс преобразования энергии, без
которого была бы невозможна жизнь на земле.
Стехиометрия и энергетика метаболических превращений
327
5.5. Фотосинтез и его связь
с первичным источником энергии
В большинстве типов дыхания атомы водорода постоянно
переносятся от топлива к кислороду; этот процесс сопровожда-
ется выделением энергии. Фотосинтез представляет собой, по
сути дела, обратный процесс, в котором осуществляется улав-
ливание световой энергии, используемой для превращения ди-
оксида углерода в глюкозу и ее полимеры. Фотосинтез являет-
ся первичным поставщиком энергии для всей биосферы. В фо-
тосинтезе усваивается энергия единственного значимого
источника энергии в Солнечной системе — самого солнца. Дру-
гая жизненно важная функция фотосинтеза заключается в его
участии в круговороте углерода и кислорода; именно благодаря
фотосинтезу осуществляется восстановление углерода, окислен-
ного в ходе дыхания:
Фотосинтез: 6СО2+6Н2О+свет —>• С6Н12Об + 6О2 (5.26)
5.5.1. Аккумулирование световой энергии
В прокариотах (цианобактериях, называемых также сине-
зелеными водорослями; зеленых серобактериях; пурпурных се-
робактериях) фотосинтез осуществляется в многослойных мем-
бранах, а в эукариотах (водорослях, растениях) центром фото-
синтеза являются особые органоиды, называемые хлоропласта-
ми (рис. 1.5). И те и другие системы содержат хлорофилл —
сложное органическое соединение, сильно поглощающее свет
в видимой области спектра (рис. 5.9). Энергия одного фотона
Ер зависит от его частоты (или длины волны) следующим об-
разом:
Е = hv = h-^- (5.27)
где h — константа Планка; v — частота фотона; X — длина вол-
ны фотона; с — скорость света в среде. Два основных пика в
спектре поглощения хлорофилла (рис. 5.9) соответствуют энер-
гиям фотонов 43,5 ккал/моль (650 нм) и приблизительно
67 ккал/моль (430 нм). Обе полосы поглощения, очевидно, со-
ответствуют свободной энергии, значительно превосходящей
необходимый для фосфорилирования ADP минимум.
Известны две различные системы аккумулирования световой
энергии и сопровождающих этот процесс химических реакций,
называемые фотосистемами I и II. Фотосистемы I и II активи-
руются светом с длиной волны более и менее 680 нм соответст-
венно. Обе системы обнаружены во всех фотосинтезирующих
328
Глава 5
РИС. 5.9. Максимумы в спектре поглощения хлорофилла а {нижняя кривая)
связаны с максимумами поглощения в спектре зеленых листьев {верхняя кри-
вая) и со спектром фотосинтетической активности листьев {средняя кривая).
Соответствующие пигменты листьев способствуют повышению эффективности
процесса утилизации света в диапазоне 500—600 нм по сравнению с чистым
хлорофиллом. {Ленинджер А., Основы биохимии, т. 1—3. — М.: Мир, 1985.)
1—спектр поглощения; 2 — спектр фотосинтетической активности; 3 — хло-
рофилл а.
организмах, выделяющих кислород. Напротив, фотосинтезирую-
щие бактерии, не выделяющие кислород, содержат только
фотосистему I.
5.5.2. Транспорт электронов и фотофосфорилирование
В фотосистемах I и II поглощение двух световых квантов
(фотонов) хлорофиллом и другими пигментами приводит к
возбуждению электрона в реакционном центре (например,
Р700 в случае фотосистемы I; см. рис. 5.10). Обладающий до-
статочно высокой энергией электрон включается в цепь элект-
ронного транспорта. Затем электрон «падает» на низший
энергетический уровень (этот процесс изображен на рис. 5.10
как повышение значения стандартного электродного потенциа-
ла Е°'). В фотосистеме I электрон в конечном счете исполь-
зуется для восстановления NADP+, а возбужденные электроны
фотосистемы II передаются фотосистеме I, причем в процессе
их транспорта каждая пара возбужденных электронов участ-
вует в фосфорилировании одной молекулы ADP. Таким обра-
зом, общая стехиометрия реакций, индуцируемых фотонами в
Стехиометрия и энергетика метаболических превращений
329
РИС. 5.10. Инициированное поглощением света возбуждение электронов (вер-
тикальные стрелки) и их транспорт в фотосинтезирующих системах. (Ленинд-
жер А., Основы биохимии, т. 1—3. — М.: Мир, 1985.)
фотосинтезирующих эукариотах, выглядит следующим образом:
H2O+4/iv+NADP++ADP+P/ —>
—> NADPH+H+ + >/2O2+(ATP+H2O) (5.28)
В используемом диапазоне энергий светового возбуждения
только при совместной работе двух фотосистем вода может
служить донором (Ео/ = + 0,82 В), a NADP+—акцептором (Е°' =
= —0,32 В) электронов. Уравнение (5.10) объясняет, почему,
например, в бактериях, использующих в качестве донора элек-
тронов H2S, фотосинтез может осуществляться только фотосис-
темой I: это связано с величиной стандартного электродного
потенциала восстановления серы Е°', равного —0,23 В.
Поглощенная световая энергия может участвовать в регене-
рации дополнительных количеств АТР по пути циклического
фотофосфорилирования, изображенному на рис. 5.10 штриховой
линией. В этом случае возбужденные электроны транспортиру-
330
Глава 5
ются от Р700 к Р430, затем к цитохрому &5вз и снова к Р700;
на последнем этапе этого пути электроны принимают участие
в фосфорилировании ADP по следующему простому уравне-
нию:
З/iv+ADP+P,- АТР+Н2О (5.29)
Использование накопленных рассмотренными выше путями
энергии и восстанавливающих эквивалентов для синтеза глюко-
зы из СО2 и Н2О мы опишем в следующем разделе. Здесь же
отметим, что изменение свободной энергии в реакции фотосин-
теза
6СО2+6Н2О —> С6Н12О6+6О2 (5.30)
равно +686 ккал/моль. Поскольку в любой из фотосистем для
возбуждения одного электрона необходимы два фотона и по-
скольку генерирование одной молекулы О2 сопровождается пе-
реносом четырех электронов, то образование 6О2 требует по-
глощения 48 квантов света. Согласно уравнению (5.27), энер-
гия фотонов зависит от длины их волны. Если принять % = 700 нм,
то минимальное количество световой энергии, необходимой
для инициирования реакции (5.30) с помощью фотосинтезирую-
щей системы растений, составит 48-41 = 1968 ккал/моль глюко-
зы. Следовательно, в первом приближении эффективность ути-
лизации поглощенного света будет равна 686: 1968 = 0,35 = 35%.
Действительную эффективность фотосинтеза в растениях и мик-
роорганизмах мы рассмотрим позднее, в ходе обсуждения об-
щей стехиометрии метаболизма.
5.6. Биосинтез
Во вводных разделах мы отмечали, что большинство процес-
сов в мире микроорганизмов характеризуются 1) утилизацией
субстратов или питательных веществ, 2) ростом клеток и
3) выделением в среду продуктов их метаболизма. Биосинтез
связан со всеми этими тремя сторонами жизнедеятельности
клеток. Необходимость в питательных веществах определяется
потребностями клетки в веществах-предшественниках, запасах
химической энергии и восстанавливающих эквивалентах. Неко-
торые продукты биосинтеза клетка выделяет в среду. Наконец,
скорость биосинтеза определяет скорость образования новых
компонентов клетки и, таким образом, темпы клеточного роста.
Скорости роста клеток изменяются в очень широких пределах;
так, бактерии Е. coll могут делиться каждые 20 мин, жизнен-
ный цикл клеток печени крысы составляет от двух до трех ме-
сяцев, а нервные клетки взрослого человека вообще не само-
воспроизводятся. Даже в последнем случае, однако, необходи-
Стехиометрия и энергетика метаболических превращений
331
мость в биосинтезе не отпадает; здесь он нужен для поддержа-
ния жизнедеятельности клеток и восстановления повреждений.
Для биосинтетических целей клетка использует химические
ресурсы, образующиеся в процессах катаболизма. Как прави-
ло, реакции синтеза термодинамически невыгодны и протека-
ют только одновременно с гидролизом АТР до ADP или АМР.
При гидролизе образующегося в последнем случае пирофосфа-
та (Р-Р) выделяется дополнительная свободная энергия
(AG0/~7 ккал/моль), также использующаяся для «запуска»
реакций синтеза. Поскольку питательные вещества обычно
окислены в большей степени, чем необходимые клетке соеди-
нения, то для биосинтеза последних нужны и восстанавливаю-
щие эквиваленты.
5.6.1. Биосинтез низкомолекулярных соединений
Биологически важные низкомолекулярные соединения в ос-
новном являются мономерами и используются клеткой для по-
строения биополимеров. Для этой цели необходимо в общей
сложности примерно 70 различных соединений: 4 рибонуклео-
тида, 4 дезоксирибонуклеотида, 20 аминокислот, около 15 мо-
носахаридов и около 20 жирных кислот и других предшествен-
ников липидов. Кроме того, в реакциях второго типа должны
синтезироваться АТР, NAD, другие соединения-переносчики и
коферменты. Все эти вещества называют центральными про-
межуточными метаболитами. В настоящем разделе мы рас-
смотрим некоторые основные пути биосинтеза промежуточных
метаболитов и организацию этих путей.
Аминокислоты можно разделить на четыре группы, разли-
чающиеся химическим строением и природой биосинтетических
предшественников. Как мы увидим позднее в ходе обсуждения
регуляции метаболизма, синтез аминокислот каждой группы
осуществляется с помощью разветвленной последовательности
регулируемых реакций. Биосинтез всех аминокислот начинает-
ся с промежуточных соединений в метаболизме углерода (см.
схему на рис. 5.11).
Живые клетки усваивают азот путем его включения в ами-
нокислоты глутамин и глутаминовую кислоту. Сначала при
взаимодействии аммиака с а-кетоглутаровой кислотой (одним
из промежуточных веществ в цикле трикарбоновых кислот) об-
разуе^я глутаминовая кислота:
глутаматдегидрогеназа
НООС (СН2) 2СОСООН+NH4+ + NADH ---------------*
—> HOOC(CH2)2CHNH2COOH + NAD+ + H2O (5.31)
Глутаминовая кислота может связать еще один аммониевый
332
Глава 5
Семейство аромати-
ческих аминокислот
и гистидина
Семейство аспарагиновой Семейство глутаминовой
кислоты кислоты
РИС. 5.11. Семейства аминокислот и их предшественники — углеводы.
(Стент Г., Кэлиндар Р., Молекулярная генетика. — М.: Мир, 1981.)
Стехиометрия и энергетика метаболических превращений
333
ион и превратиться в глутамин:
глутаминсин таза
НООС (СН2) 2CHNH2COOH +NH4++АТР-----------►
—> H2NOC(CH2)2CHNH2COOH+ADP+P<+H+ (5.32)
AG°'=—3,9 ккал/моль
Вторая реакция, происходящая с поглощением метаболической
энергии, осуществляется при недостатке аммиака в среде.
В некоторых бактериях с участием NADH происходит непо-
средственное аминирование пирувата до аланина, а другие
бактерии способны превращать фумарат в аспартат. Ряд микро-
организмов усваивает азот в виде нитрата NO3- и свободного
азота N2; сейчас известно, однако, что эти питательные вещест-
ва сначала трансформируются в аммиак, который затем и ас-
симилируется клетками по описанным выше путям.
Все другие аминокислоты образуются из глутамата или на
основе его углеродного скелета, или путем переноса аминогруп-
пы к другим молекулам. Например, глутамат превращается в
пролин в результате двух последовательных катализируемых
ферментами реакций и одной неферментативной гидролитичес-
кой реакции по общему уравнению:
C5NH8O4--I-ATP + 2(NADPH-|-H+) —>
—> C5NH8O2-+ADP-|-P<-|-2NADP+ + H2O (5.33)
В большинстве организмов аланин и аспартат образуются пу-
тем переаминирования глутамата:
Глутамат+оксалоацетат —> а-кетоглутарат+аспартат
Глутамат+пируват —> а-кетоглутарат+аланин
Все реакции переаминирования происходят только в присутст-
вии кофермента пиридоксальфосфата, производного витамина
В6 (пиридоксаля).
В завершение этого краткого обзора биосинтеза аминокис-
лот на рис. 5.12 приведены некоторые отдельные стадии и об-
щая схема биосинтеза аминокислот семейства аспарагиновой
кислоты; здесь обращает на себя внимание тот факт, что боль-
шинство стадий протекает с участием АТР или NADPH. На
этом рисунке штриховыми линиями обозначены регуляторные
механизмы, которые мы обсудим позднее.
Сколько-нибудь детальное изучение путей биосинтеза нук-
леотидов выходит за рамки тематики этой книги. Источники
предшественников этих соединений указаны на рис. 5.13. Более
подробные сведения читатель может найти в литературе.
В биосинтезе жирных кислот и других предшественников
липидов роль исходных веществ выполняют ацетил-СоА и гли-
церин. Чтобы еще раз подчеркнуть участие АТР и NADPH в
334
Глава 5
сосг с
I
сн2
I
I
I
I
I
I
I
I
I
I
I
I
I
I
i
I
СОО-
Аспартап?
о
И
с—о ©
HCNHJ р-Асларлш/г-Р
I
СОО’
ll^-NADPH + Н+
^~»NADP+ + Р,
нс=о
сн2
I J3 -/Толуальдегид
HCNHJ аспарагиновой
| кислоппн ' *
СОО’ t
NADPH + Н-
сн2он
сн2
HCNHJ
СОО’
АТР
Гокосеран-Н>
Г ^ADP
I * -
| СН2О ®
Г ।
СИ2
HCNHJ
СОО’ Гопосерил-
-Р '
“S
I
I
I
I
I
I
I
I
I
I
I
I
I
I
I
I
Met
J
I сн3
I I
неон
I
j HCNHJ
1 соо’ Треонин —>—>це
>___________________I 4у_________,1
в биосинтезе, мы несколько
подробнее рассмотрим путь
биосинтеза самой обычной из
жирных кислот — пальмитино-
вой СН3(СН2) мСООН. Первая
стадия этого пути заключается
в карбоксилировании ацетил-
СоА, в результате чего обра-
зуется малонил-СоА:
Ацетил-СоА 4~ АТР + СО2 -*•
ацетил-СоА —
карбоксилаза
малонил-СоА
(HOOCCH2COSCOA)
+ ADP + P; (5.34)
Далее из семи молекул мало-
нил-СоА и одной молекулы
ацетил-СоА образуется паль-
митат. Этот процесс осуществ-
ляется ступенчато, а его конеч-
ный результат можно выра-
зить следующим уравнением:
Ацетил-СоА+7 малонил-СоА+
+ 14(NADPH+H+) —>
—>- пальмитат+ 7CO2+8HSCoA-I-
+ 14NADP++6H2O (5.35)
Обратите внимание на то, что
в этом процессе в соответст-
вии с уравнением (5.34) на
каждую молекулу образующе-
гося малонил-СоА расходуется
одна молекула макроэргичес-
кого фосфата.
В завершение этого крат-
кого обзора биосинтеза рас-
смотрим пути образования
глюкозы и родственных соеди-
нений. Мы уже знаем, что
продукты катаболизма глюко-
зы являются важнейшими
Рис. 5.12. Схема реакций, в ходе которых аспартат превращается в лизин,
гомосерин, метионин, треоиин и изолейцин. (Воспроизведено с разрешения из
работы: Wood W. В., Willson J. Н., Benbow В. М., Hood L. Е., Biochemistry,
A Problems Approach, 2d ed., р. 294, Benjamin/Cummings Publishing Co.,
Menlo Park, CA, 1981.)
Стехиометрия и энергетика метаболических превращений 335
предшественниками в биосинтезе аминокислот, нуклеотидов
и жирных кислот. Следовательно, организмы, растущие на дру-
гих источниках углерода, например СО2, должны превращать
их в глюкозу или в один из ближайших продуктов ее метабо-
лизма. Кроме того, при избытке соединений углерода (по срав-
нению с другими питательными веществами) многие клетки
превращают глюкозу в резервные углеводы и хранят их для
последующего использования.
Глюкоза
РИС. 5.13. Схема превращения веществ-предшественников в нуклеотиды и
дезоксирибонуклеотиды. ГВоспроизведено с разрешения из работы: Bioche-
mistry of Bacterial Growth, Mandelstam J., McQuillan K- (eds.), 2d ed., p. 33,
Blackwell Scientific Publications, Oxford, 1973.]
Глюконеогенезом называют синтез глюкозы в хемотрофах.
Интересно, что в синтезе глюкозы из пирувата участвуют все
промежуточные соединения, типичные для метаболического пу-
ти ЭМП. Еще более важен тот факт, что все ферменты, катали-
зирующие практически равновесные превращения в пути ЭМП,
катализируют обратимые реакции и в биосинтезе глюкозы. В то
же время в биосинтезе глюкозы реакции фосфорилирования и
дефосфорилирования несколько отличаются от аналогичных ре-
акций в гликолизе и катализируются другими ферментами. Эти
биосинтетические реакции, изображенные на рис. 5.14 (снизу
вверх), происходят спонтанно и сопровождаются уменьшением
свободной энергии. Общая стехиометрия глюконеогенеза пока-
зывает, что в целом процесс сопровождается поглощением энер-
Глюкоза
ГенсекиназаР \
дтр ( Хй-Глюкозо-б-росфатаза
Глюкоза-6- фосфат
Фосфофру ктокина-
за (активируется
АМР или AD5^ инги-
бируется АТР)
Фруктозо-6-фосфат
Дифосрофруктозо -
фосфатаза (инги-
бируется ADP или АМР)
Фруктозо-1,6. - дифосфат
, 3-Фосфоглицериново/й альдегид
1,З-Дисросфогли церат
3- Фосфоглицерат
2 - Фоссроглицерат
Ряд реакции, —
активируемых
ацетил- СоА
РИС. 5.14. Расщепление глюкозы по пути гликолиза (сверху вниз) и биосин-
тез глюкозы (снизу вверх). Для этих двух путей метаболизма характерна
идентичность ряда реакций, особенно близких к равновесным. В то же время
неравновесные реакции в зависимости от их направления катализируются
различными ферментами (большей частью аллостерическими), что позволяет
независимо регулировать процессы синтеза и расщепления глюкозы. (Воспро-
изведено из работы Lehninger A., Bioenergetics, 2d ed., р. 129, W. A. Benjamin,
Inc., Palo Alto, CA, 1974.)
Стехиометрия и энергетика метаболических превращений
337'
гии и не является повторением пути ЭМП в обратном направле-
нии (GTP — гуанозинтрифосфат):
2 nHpyBaT + 4ATP + 2GTP + 2NADH + 2H++4H2O —
—> ivii0K03a + 2NAD+ + 2GDP-|-4ADP + 6Pz (5.36)
Наиболее важная биосинтетическая реакция — это синтез
глюкозы из СО2 в растениях. Без этого процесса невозможно су-
ществование биосферы в том виде, в каком мы привыкли себе ее-
представлять. Кроме того, масштабы и эффективность этих ре-
акций определяют объемы возобновляемых ресурсов углеродсо-
держащего сырья, служащего топливом и исходным материа-
лом в биохимических и химических процессах.
Выше были описаны световые реакции фотосинтеза: погло-
щение света хлорофиллом и другими пигментами и формирова-
ние потока возбужденных электронов, генерирующего АТР и
NADPH. Далее мы рассмотрим темновые реакции фотосинтеза,
названные так потому, что они протекают и в отсутствие света
до тех пор, пока в клетке имеется достаточный запас АТР и
NAD PH.
Первоначальное включение в цикл СО2 осуществляется в ре-
акции с рибулозо-1,5-дифосфатом:
рибулозодифосфаткарбокси л аза
Рибулозо-1,5-дифосфат + СО2---------------------”
—> 2 (глицеральдегид-3-фосфат) (5.37)
Образующийся на этой стадии глицеральдегид-3-фосфат может
далее претерпевать несколько превращений, в частности вклю-
чаться в аналогичную глюконеогенезу последовательность реак-
ций, в результате которой образуется глюкоза. В реакциях цик-
ла Калвина (рис. 5.15) регенерируется рибулозо-1,5-дифосфат,
который может снова ассимилировать СО2 в соответствии с ре-
акцией (5.37). В отличие от цикла ТКК, представляющего со-
бой, по сути дела, источник АТР и NADH, в цикле Калвина рас-
ходуются как АТР, так и NADPH. Поскольку в одном цикле
Калвина утилизируется одна молекула СО2, то для синтеза глю-
козы необходимо повторить этот цикл шесть раз; тогда общее
уравнение синтеза глюкозы будет выглядеть следующим обра-
зом:
6CO2+12NADPH + 12H+ + 18ATP+12H2O —>
—> C6H12O6+12NADP+ + 18ADP + 18PZ (5.38)
5.6.2. Синтез макромолекулярных соединений
Как показано на общей карте путей метаболизма, приведен-
ной во введении к настоящей главе, из мономерных веществ-
предшественников далее должны строиться полимерные компо-
22-537
338
Глава 5
ненты клетки. Для этой цели опять-таки требуется большое ко-
личество метаболической энергии, поскольку любая реакция
конденсации сопровождается повышением свободной энергии.
Более того, чрезвычайно большие запасы свободной энергии яв-
ляются той движущей силой, которая резко смещает общее рав-
новесие в сторону образования необходимых полимеров. Это осо-
бенно важно в случае биосинтеза полимеров строго определен-
Ридулозо -5- (pocipam 3- Фосфуглицерино -
новг/й алоаегид
Фосфатаза,
зиисгераза, трата >
кетолаза, альдолаза
а другие рер/зенть/
фосфат
РИС. 5.15. Упрощенная схема синтеза глюкозы из СО2 в цикле Калвина.
ного строения, например нуклеиновых кислот и белков с задан-
ными последовательностями.
На рис. 5.16 показано, что хранящаяся в фосфатных связях
АТР энергия при посредстве трифосфатов других нуклеотидов
утилизируется в биосинтезе четырех классов биополимеров. Для
биосинтеза макромолекулярных соединений типично сочетание
реакции конденсации с гидролизом двух высокоэнергетических
фосфатных связей, причем сначала нуклеозидтрифосфат пре-
вращается в нуклеозидмонофосфат и пирофосфат, который так-
же подвергается гидролизу. Таким путем высвобождается при-
мерно в 2 раза больше свободной энергии (около 14 ккал/моль),
чем при превращении нуклеозидтрифосфата в соответствующий
дифосфат. Та или иная модификация этого общего механизма
реализуется в биосинтезе липидов, РНК, ДНК и гликогена.
Стехиометрия и энергетика метаболических превращений 339
Здесь опять-таки используется принцип общих промежуточных
соединений, хотя и в несколько усложненной форме. Так, про-
цесс присоединения остатка глюкозы к растущей молекуле гли-
когена осуществляется в шесть стадий, четыре из которых яв-
ляются последовательными и, следовательно, связаны общими
промежуточными соединениями (табл. 5.5).
АТР
Полисахариды
Порфирины
Целлюлоза
Бел ни
РИС. 5.16. Участие высокоэнергетических трифосфатов нуклеозидов в различ-
ных путях биосинтеза. (Воспроизведено с разрешения из работы: Lehninger А.,
Bioenergetics, 2d ed., р. 136, W. A. Benjamin, Inc., Palo Alto, 1975.)
Естественно, синтез информационных полимеров (РНК, ДНК
и белков) представляет собой гораздо более сложный процесс.
И в том и в другом случае, однако, наращиванию полимерной
цепи предшествует активация мономера. В схему синтеза РНК
и ДНК нуклеотиды входят в виде нуклеозидтрифосфатов, а при-
соединяется к соответствующей полимерной цепи монофосфат;
реакция сопровождается образованием пирофосфата. Таким
образом, для присоединения одного мономерного остатка необ-
ходимо затратить 14 ккал/моль энергии. Активация аминокислот
в ходе синтеза белков осуществляется в соответствии со следу-
ющей схемой:
Аминокислота + АТР —>- аминоацил-АМР + Р — Р (5.39)
(активированная
аминокислота)
22*
340
Глава 5
Кроме того, процесс присоединения аденилата аминокислоты к
пептидной цепи сопровождается гидролизом GTP; следователь-
но, для присоединения одного аминокислотного остатка к белко-
вой цепи необходимо гидролизовать в общей сложности три вы-
сокоэнергетические фосфатные связи. В гл. 6 мы рассмотрим ме-
ханизмы биосинтеза заданных последовательностей мономерных
остатков в ДНК, РНК и белках.
Таблица 5.5. Последовательные стадии биосинтеза гликогена в клетках
млекопитающих (в бактериях и растениях вместо UDP-глюкозы в качестве
промежуточного соединения образуется АТР-глюкоза)а
гексокиназа
1. Глюкозан-АТР -----------> глюкозо-6-фосфат + ADP
фосфоглюкомутаза
2. Глюкозо-6-фосфат --------------глюкозо-1 -фосфат
_ _ , , т UDP-глюкозопиро -
3. Глюкозо-1-фосфат4-UTP фОСфорндаза-------UDP-глюкоза-}- пирофосфат
гликогенсинтетаза
-4. UDP-глюкоза+гликогенп---------------гликогенп+1 + UDP
иуклеотиддифосфокнназа
5. ATP-UDP------------------------> ADP + UTP
пирофосфатаза
6. Пирофосфат+ Н2О------------>- 2 фосфат
Суммарная реакция:
Глюкоза + 2АТР + гликоген,, 4- Н2О----> 2ADP,+2Pi + гликогенп +1
а Воспроизведено из работы: Lehninger A. L„ Bioenergetics, 2d ed., р. 140, W. A. Ben-
jamin, Inc., Palo Alto, Ca., 1971.
5.7. Транспорт через клеточные мембраны
Нормальная жизнедеятельность клетки возможна только при
условии регулируемого транспорта ионов и молекул между
клеткой и средой. Клеточные мембраны обладают селективной
проницаемостью по отношению к различным компонентам и со-
ставным частям клетки и тем самым регулируют транспортные
процессы, выполняющие ряд важных функций. Во-первых, бла-
годаря процессам транспорта pH и состав внутриклеточной сре-
ды поддерживаются на одном уровне в достаточно узком диапа-
зоне, обеспечивающем необходимую активность ферментов.
Кроме того, мембранный транспорт регулирует объем клетки,
обеспечивает доступ и нужную концентрацию питательных ве-
ществ, удаление токсичных соединений.
Известны по меньшей мере три способа транспорта через
клеточные мембраны: пассивная диффузия, сопряженный транс-
порт и активный транспорт. Независимо от механизма способ-
Стехиометрия и энергетика метаболических превращений
34!
ность данной мембраны транспортировать данный субстрат ча-
сто выражают как проницаемость мембраны К, которую рас-
считывают по уравнению
/< = ^_1п-Се~.сА.
At се — ct (t)
(5.40)
где V — объем клетки; А— наружная поверхность клетки; се —
наружная концентрация субстрата; сг0 — начальная внутрикле-
точная концентрация субстрата; сг(7) —внутриклеточная кон-
центрация субстрата через время t. Из уравнения (5.40) следует,
что проницаемость имеет ту же размерность, что и скорость
(сантиметров в секунду). Как мы увидим далее в ходе реше-
ния упражнений, лежащие в основе этого уравнения допущения
ограничивают его применимость.
5.7.1. Пассивная диффузия и сопряженный транспорт
Пассивная диффузия представляет собой транспорт вещест-
ва через мембрану из области с высокой концентрацией в об-
ласть с низкой концентрацией (рис. 5.17). Скорость диффузии
пропорциональна общей движущей силе, которой в данном
случае является разность концентраций по обе стороны мемб-
раны. Термодинамический анализ показывает, что пассивная
диффузия спонтанна, поскольку транспорт вещества из обла-
сти с концентрацией с2 в область с концентрацией щ сопровож-
дается изменением свободной энергии ДО0, равным
Д(?0 = ^7'1п — (5.41)
Так как Ci меньше с2, то Д(?° в случае пассивной диффузии всег-
да отрицательно. Если транспортируемое вещество заряжено, то
уравнение (5.41) следует видоизменить следующим образом:
ДС° = ^Т1п-^-4-71^'Дф (5.42)
где Zi — заряд транспортируемых молекул; — число Фарадея
[23,062 ккал/(В-моль)]; Дф— разность потенциалов на мемб-
ране, В.
В силу специфичности структуры плазматических мембран,
окружающих любые клетки и органоиды эукариот, не все соеди-
нения проникают через мембрану одинаково легко. Существует
корреляция между скоростью диффузии большинства сравни-
тельно объемных молекул и их растворимостью в липофильных
средах (рис. 5.18). Это и неудивительно, поскольку основой
плазматических мембран, как полагают, является главным об-
342
Глава 5
Внеклеточ- Клеточная Внутрек-
ная среда мембрана ний объем
а) Диффузия (пассивный
транспорт)
Внекле- 'Внутрен-
точная Клеточная ний объем
среда мембрана клетки
б) Сопряженный транспорт
Внеклеточ - Кле то чна я Внутренний
ная среда мембрана объем клетки
8) Активный транспорт
РИС. 5.17. Различные механизмы транспорта через мембрану: а — диффузия;
б — сопряженный транспорт; в — активный транспорт (S — транспортируемое
вещество; С — белок-переносчик; CS—комплекс).
разом липидный бислой (вспомните рис. 2.2). Аномально высо-
кую скорость диффузии воды и других очень малых молекул
объясняет другая модель, предусматривающая наличие в мемб-
ранах пор небольшого диаметра.
Для ионизированных и полярных веществ характерна очень
низкая растворимость в липофильных растворителях, поэтому
такие вещества практически не транспортируются через мемб-
рану за счет пассивной диффузии. Важность этого обстоятель-
ства мы уже не раз подчеркивали: промежуточные метаболиты
большей частью представляют собой ионизированные соедине-
ния и поэтому удерживаются внутри клетки. Впрочем, некою-
Стехиометрия и энергетика метаболических превращений
343
рые полярные вещества очень легко преодолевают мембранный
барьер.
Одним из механизмов, объясняющих такое аномальное пове-
дение полярных веществ, является механизм сопряженного
транспорта. Как схематично показано на рис. 5.17, согласно это-
<5
I
I
fO's
Метанол
t
Вода
Триметил-
цитрат
Цианамид г- Валерамид
• Вцтирамид
Пропионамид • нДДмм •Диацетин
^^ЭтилоВыи эфир глицерина
* Метилодб/й эфир глицерина
^»Диметилмочедина
• Моноацетин
Этилмо чедина
Тиомочедина
• Диэтилмалонамид
Лантанид
Мочебина
fO'6
Форманид*
Ацетамид •
Этиленгликоль
Метилмочебин
ф Глицерин
КГ* Ю~3 Ю'г Ю'7
Коэффициент распределения 6 системе масло/Вода
РИС. 5.18. Способность многих веществ проникать через мембрану водорослей
Chara коррелирует с их растворимостью (коэффициентом распределения)
в липофильных растворителях. Размеры точек примерно пропорциональны
размерам соответствующих молекул. (По данным Коллайдера, 1947.)
му механизму субстрат на наружной поверхности мембраны со-
единяется с молекулой-переносчиком-, образующийся комплекс
далее «диффундирует» к другой поверхности мембраны и там
расщепляется, выделяя перенесенную молекулу во внутренний
объем клетки. Такой способ биологического транспорта имеет
ряд характерных особенностей. Так, скорость транспорта веще-
ства в этом случае изменяется в зависимости от разности кон-
центраций не линейно, а достигает некоторого максимума (уров-
ня насыщения), после которого дальнейшее повышение разности
концентраций вещества не изменяет скорость его транспорта.
344
Глава 5
Зная уравнение Михаэлиса—Ментен и другие математические
модели кинетики ферментативного катализа, нетрудно понять,
почему изображенный на рис. 5.17, б процесс назван сопряжен-
ным транспортом.
Другие важные свойства сопряженного транспорта также на-
поминают особенности ферментативного катализа; так, через
мембрану транспортируются только определенные вещества,
а специфические ингибиторы замедляют процесс. Специфичность
и кинетические характеристики свидетельствуют о том, что носи-
тели представляют собой белки. Многие из носителей удалось
выделить и охарактеризовать; их называют пермеазами. Веро-
ятно, самым известным примером сопряженной диффузии явля-
ется транспорт глюкозы в эритроцитах (красных кровяных тель-
цах) человека. Подробные сведения об этой системе и дополни-
тельную информацию о сопряженном транспорте читатель мо-
жет найти в литературе.
5.7.2. Активный транспорт
Как мы уже показывали на рис. 5.17, для активного транс-
порта характерно, во-первых, движение вещества против его хи-
мического (или электрохимического) градиента, т. е. из области
с низкой концентрацией в область с высокой концентрацией, и,
во-вторых, как показывает уравнение (5.41), необходимость за-
трат метаболической энергии. Например, если соединение, не
несущее заряда, транспортируется из среды, где его концентра-
ция равна 0,001М, в клетку, где его концентрация составляет
0,1 М, то
AG°=1,98 кал/(моль-град) (298 град) (In 100) =
= 2,72 ккал/моль
Следовательно, для реализации этого процесса необходимо за-
тратить по меньшей мере указанное количество энергии. Актив-
ный транспорт также осуществляется при участии специфиче-
ских пермеаз.
Активный транспорт лежит в основе передачи нервных им-
пульсов, но клетки используют этот механизм в гораздо более
широких масштабах. Так, почти все клетки обладают системами
активного транспорта (ионными насосами), позволяющими под-
держивать необходимые концентрации ионов КЛ, Na+ и воды
внутри клетки. Эти системы перемещают ионы Na+ из клетки,
а ионы К+ в клетку, что позволяет компенсировать пассивную
диффузию этих ионов. Указанные системы транспорта взаимо-
связаны и сопряжены с гидролизом АТР (рис. 5.19). В красных
кровяных тельцах роль натрий-калиевого насоса выполняет оли-
Стехиометрия и энергетика метаболических превращений 345
гомерный белок, называемый Na+—К+-АТРазой. Как показано
на рис. 5.19, этот белок располагается в толще плазматической
мембраны и пронизывает ее насквозь; такие белки называют
трансмембранными.
Вторая обычная группа систем активного транспорта обес-
печивает поступление в клетку молекул питательных веществ,
например глюкозы и аминокислот, со скоростями, значительно
превышающими скорость их транспорта за счет пассивной диф-
фузии. В клетках высших животных активный транспорт глюко-
Внутракле- Клеточная Внеклеточная
точное про- те/чорана среда
странство
РИС. 5.19. Na+—К+-насос (Na+—К+-АТРаза) транспортирует Na+ из клетки
и К+ в клетку; движущей силой этого процесса является гидролиз АТР.
зы совмещен с транспортом Na+ в клетку, а удаление Na+ из
клетки осуществляется параллельно с гидролизом АТР под дей-
ствием Na+—К+-АТРазы. Весь процесс схематично изображен
на рис. 5.20. В бактериях для активного транспорта глюкозы ис-
пользуется другой процесс, называемый транслокацией групп.
В этом процессе глюкоза высвобождается во внутриклеточное
пространство в виде высокоэнергетического глюкозо-6-фосфата,
с трудом проникающего через мембраны (рис. 5.21). Считается,
что этот процесс является той стадией, скорость которой опре-
деляет скорость роста ряда клеток.
Движущей силой многих систем транспорта в бактериях слу-
жит поток протонов через плазматическую мембрану. Основная
роль потока протонов в энергетике бактерий описывается хеми-
осмотической теорией, разработанной Митчеллом в 1961 г. На
рис. 5.22 изображена схема совместного транспорта протонов
и лактозы в процессе дыхания бактерии Е. coli. Протоны и
лактоза совместно транспортируются в клетку лактозопермеа-
зой (продуктом транскрипции гена у оперона lac). Одновре-
346
Глава 5
Мем&рана
РИС. 5.20. Ионы Na+ и глюкоза
транспортируются из среды в клетку
одним и тем же пассивным перенос-
чиком. Низкая внутриклеточная кон-
центрация Na+ поддерживается за
счет активного транспорта. (Воспро-
изведено с разрешения из работы:
Lehninger A., Bioenergetics, 2d ed.,
р. 205, W. A. Benjamin, Inc., Palo
Alto, CA, 1974.)
внеклеточ- Клеточная Внутриклеточ-
ная среда мемора на ное пространство
РИС. 5.21. Фосфорилирование глюкозы в клеточной мембране создает посто-
янную движущую силу для транспорта глюкозы через мембрану и удержи-
вает глюкозу (в виде производного) внутри клетки. Здесь G — глюкоза; Ен —
мембранный фермент; Е! — цитоплазматический фермент; НРг — цитоплазма-
тический белок; РЕР — фосфоенолпируват. (Воспроизведено с разрешения из
работы: Lehninger A. L., Biochemistry, 2d ed., р. 799, Worth Publishers, New
York, 1975; есть перевод первого издания: Ленинджер А., Биохимия. — М.:
Мир, 1976, с. 715.)
менно в дыхательной цепи протоны вместе с потоком электро-
нов выводятся из клетки. В целом pH внутри клетки выше,
чем в непосредственном ее окружении.
Эукариотические клетки должны обладать целым комплек-
сом систем транспорта, поскольку концентрации многих веществ
Стехиометрия и энергетика метаболических превращений
347
в органоидах поддерживаются на уровне, отличающемся от кон-
центрации этих веществ в цитоплазме. Этот факт лишний раз
говорит о том, что любая клетка, и особенно эукариотическая,
представляет собой не однородную смесь множества компонен-
тов, а высокоорганизованную, вплоть до молекулярного уровня,
систему.
Разработано много математических моделей активного тран-
спорта, но сейчас мы не будем их рассматривать. Более подроб-
РИС. 5.22. Сочетание активного транспорта лактозы с транспортом протонов
по градиенту электронного потока дыхательной цепи.
ные сведения читатель может найти в приведенной в конце гла-
вы литературе и упражнениях.
Прежде чем перейти к изучению потоков и регуляторных
систем в последовательностях метаболических реакций, полез-
но несколько полнее и детальнее рассмотреть структуру и ор-
ганизацию клеточной мембраны. На рис. 5.23 схематично изо-
бражено строение клеточной оболочки бактерии; нетрудно ви-
деть, что ее структура значительно сложнее описанной в гл. 2
упрощенной модели, включающей липидный бислой и белковую
оболочку. На рисунке показано, что в толще мембраны нахо-
дится множество белковых молекул; одни из них располага-
ются у наружной или внутренней поверхности мембраны, другие
молекулы пронизывают всю толщу мембраны. Эти белки
(пермеазы и белки-рецепторы) узнают специфические соеди-
нения в среде и контактируют с регуляторными системами
клетки, обеспечивая ее ответную реакцию на изменения в сре-
де. В случае бактерий белковые компоненты мембран позволя-
348
Глава 5
ют клеткам быстро приспособиться к изменившимся условиям
окружения. Поверхностные рецепторы высших эукариотиче-
ских клеток участвуют в процессах клеточного узнавания, иг-
рающих большую роль как в организации тканей, так и в им-
мунной реакции клеток.
Л4ембраны в общем случае асимметричны; их наружные и
внутренние поверхности содержат различные компоненты и
выполняют различные функции. Липиды и разнообразные
РИС. 5.23. Детали строения клеточной оболочки Е. coli. Здесь белые глобу-
лярные объекты на поверхности и в толще клеточных мембран обозначают
отдельные белки (НМ — наружная мембрана; ПГ — пептидогликан; ПП —
периплазматическое пространство; ЦМ — цитоплазматическая мембрана).
(Воспроизведено с разрешения из работы: Inouye М., Bacterial Outer Mem-
branes, John Wiley, New York, 1979.)
мембранные белки диффундируют в плоскости мембраны очень
быстро; по некоторым оценкам скорость диффузии липидов в
ряде мембран составляет 10-8 см2/с.
5.8. Организация и регуляция метаболизма
Подобно любому химическому предприятию, клетка долж-
на обладать системами регулирования сложного переплетения
осуществляющихся в ней химических реакций. Эти системы
должны обеспечивать соответствующее потребностям клетки
поступление материалов, энергии и электронов, а также высо-
кую эффективность использования этих ресурсов. Кроме того,
последовательности и циклы реакций должны быть организо-
ваны таким образом, чтобы они поддавались эффективному
контролю и регулированию. В этом разделе мы изучим спосо-
бы регуляции потоков веществ в многочисленных разветвлен-
ных и замкнутых путях метаболизма клетки путем модуляции
Стехиометрия и энергетика метаболических превращений
349-
активности ряда ключевых ферментов и, кроме того, рассмот-
рим основные особенности структуры метаболической сети.
Чтобы оценить важность и эффективность этих структур и си-
стем регуляции, достаточно ознакомиться с метаболической ак-
тивностью бактерий Е. coli. В богатой питательными вещества-
ми среде эти клетки делятся каждые 20 мин; при этом осуще-
ствляется огромное количество химических реакций с порази-
Таблица 5.6. Биосинтетическая активность в процессе 20-минутного цикла
деления клеток Е. со11'л
Химический компонент Содержание (в рас- чете на массу сухо- го вещества), % Ориентировочная мо- лекулярная масса Число молекул в 1 клетке
днк 5 2 000 000 000 1
РНК 10 1 000 000 15 000
Белки 70 60 000 1 700 000
Липиды 10 1000 15 000 000
Полисахариды 5 200 000 39 000
Число молекул АТР, Относительное коли-
Химический Число молекул, син необходимых для чество необходимой
компонент тезируемых в 1 с синтеза соответст- биосинтетической
вующего компонен- энергии, %
та в 1 с
днк 0,00083 60 000 2,5
РНК 12,5 75 000 3,1
Белки 1400 2 120 000 88,0
Липиды 12 500 87 500 3,7
Полисахариды 32,5 65 000 2,7
а Lehninger А. L., Bioenergetics, 2d ed., р. 123, W. A. Benj amin, Inc., Palo Alto,
Са., 1965.
тельными точностью, производительностью и сбалансированно-
стью (табл. 5.6).
В качестве примера достаточно рассмотреть белковый син-
тез. Приведенные в таблице данные показывают, что в среднем
в клетке синтезируется 1400 молекул белков в секунду. По-
скольку белки представляют собой сравнительно большие био-
полимеры, при синтезе каждого из которых создается в среднем-
около 300 ковалентных связей, то в среднем в одной клетке за
секунду образуется около 420 000 пептидных связей. Более то-
го, так как белки являются информационными полимерами, со-
ставляющие их мономерные звенья должны быть соединены
только в строго определенной последовательности.
За такую огромную скорость синтеза белков живой клетке
приходится расплачиваться существенной частью образующейся
в процессе метаболизма химической энергии. Почти всю свою
350
Глава 5
энергию бактерия вкладывает в биосинтез, причем около 88%
ее расходуется на синтез белков. Из данных табл. 5.6 следует
также, что на цели биосинтеза расходуется около 2,5 миллиона
молекул АТР в секунду. Поскольку всего в клетке Е. coli со-
держится приблизительно пять миллионов молекул АТР, то
этого запаса хватит клетке всего лишь на две секунды жизне-
деятельности. Этот факт дает возможность оценить те огром-
ные скорости регенерации АТР, которые необходимы для нор-
мального функционирования .клетки, а также важность регуля-
ции скорости образования АТР в соответствии с потребностями
клетки. Аналогичные выводы могут быть сделаны и в отношении
других важных участников процессов первичного метаболизма
NADH и NADPH.
5.8.1. Ключевые точки пересечения
и разветвления метаболических путей
Анализ путей метаболизма как сети направленных реакций
позволяет выявить ключевые элементы их организации. Нераз-
ветвленная последовательность метаболических реакций харак-
терна для ряда взаимосвязанных превращений, осуществляю-
щихся на данном пути. Исходные и конечные вещества нераз-
ветвленного пути представляют собой или исходные питатель-
ные вещества, или конечные продукты метаболизма (которые в
свою очередь могут задерживаться в клетке или выделяться в
среду), или, наконец, исходные вещества для других путей ме-
таболизма, т. е. точки разветвления. Обычно первая после точ-
ки разветвления метаболического пути необратимая (т. е. име-
ющая существенное по абсолютной величине отрицательное
AG' при внутриклеточных условиях) реакция является аллосте-
рически регулируемой стадией. В этом случае посредством ре-
гуляции поступления исходных веществ клетка распределяет
метаболиты, используемые одновременно для нескольких це-
лей. Со структурной точки зрения эти метаболиты представля-
ют собой исходные точки или узловые пункты метаболической
ветви. Отсюда следует, что точки разветвления и модулируе-
мые необратимые реакции являются теми критическими эле-
ментами, которые определяют все стороны химической актив-
ности клетки. К этому вопросу мы еще не раз вернемся в по-
следующих разделах и главах, а изложенные выше принципы
будем использовать в упрощенной технологической интерпрета-
ции клетки как химического реактора.
Прежде всего обратим внимание на важность схем разветв-
ленных реакций с самой общей точки зрения. Оказывается, что
все типы метаболической активности клетки зависят от рас-
пределения потоков трех основных реагентов: глюкозо-6-фосфа-
Стехиометрия и энергетика метаболических превращений
351
Глюкоза
//
Путь ЭМП
(низкая конц.
АТР и С-со-
держащих
предшест-
венников) 11
Оксалоацетагп
Пируват
Глюнозо-б-гросрат
а <6S *
dC3 Л
rj7u,<o8eH
Рибозо-в-фоссрат.
Лактат
К циклу ТКК
(синтез АТР и
С - содержащих
предшествен-
ников)
Кетоны,
холесте- ~
рин
(Низкая конц.
АТР и С2-пред-
шественников)
Алании
Синтез жирных
кислот _
---------=— Жирные
----------кислоты
^-Окисление
Ацетил- Со А
и
Цикл ТКК
(низкая
конц. АТР;
f
со2
РИС. 5.24. Схема пересечения основных путей метаболизма углерода. В клет-
ках млекопитающих превращение ацетил-СоА в пируват не происходит.
та, пирувата и ацетил-СоА. На рис. 5.24 схематично изображе-
ны пересечения различных путей метаболизма, в которых эти
реагенты являются исходными или конечными веществами.
Объем потоков по каждому из путей регулируется клеткой в за-
висимости от метаболических потребностей и ее состава в дан-
ный момент.
В качестве более частного примера можно проанализировать
разветвленные пути биосинтеза аминокислот, изображенные на
рис. 5.12. Здесь также обращает на себя внимание наличие уз-
ловых точек, являющихся исходными пунктами для прямых по-
следовательностей реакций, которые далее в свою очередь так-
532
Глава 5
же разветвляются. В следующем разделе мы рассмотрим лока-
лизацию регуляционных центров в этой части метаболической
сети.
5.8.2. Регуляция метаболизма на ферментативном уровне
Аллостерические ферменты регулируют распределение пото-
ков веществ по различным путям метаболизма. Роль регуля-
торных сигналов, которые управляют активностями этих фер-
ментов, выполняют концентрации АТР, NADH, NADPH, а так-
же некоторых ключевых соединений-предшественников и про-
межуточных веществ. Влияние некоторых из этих сигналов на
разделение потоков в точках разветвления в общих чертах по-
казано на рис. 5.24. Здесь мы рассмотрим некоторые наиболее
показательные примеры регуляции метаболизма, а также осу-
ществляющийся иногда координированный контроль нескольких
путей метаболизма.
Очевидно, что концентрация АТР в клетке должна поддер-
живаться на необходимом уровне. Мерой энергетического со-
стояния клетки может служить аденилатный энергетический
заряд [4], определяемый как отношение концентрации АТР к
сумме концентраций AMP, ADP и АТР:
. [АТР] + 0,5 [ADP]
Энергетически» заряд- [Амр) * |Д№) * [Дтр] (5.43)
В большинстве клеток энергетический заряд изменяется в пре-
делах от 0,87 до 0,94.
Чтобы понять, как достигается столь высокая степень посто-
янства концентрации АТР, рассмотрим сначала путь ЭМП
(рис. 5.25). Первичным регуляторным ферментом здесь служит
фосфофруктокиназа, которая активируется ADP и ингибирует-
ся АТР. Таким образом, этот фермент замедляет поток мета-
болитов по пути ЭМП, если энергетический заряд высок, и на-
оборот. Аналогично действует пируваткиназа, активируемая
АМР. Вспомним, что фосфорилирование глюкозы, а также ре-
акции фруктозо-6-фосфата и фосфоенолпирувата представляют
собой единственные три необратимые реакции пути ЭМП. Те-
перь мы можем понять причину этого явления: на первой ста-
дии образуется ионизированное производное, прочно удержи-
ваемое внутри клеточной оболочки, а две другие необрати-
мые стадии представляют собой регуляторные пункты.
Энергетический заряд влияет и на активность цикла ТКК-
Изоцитратдегидрогеназа активируется ADP и ингибируется АТР.
Поскольку одна из основных функций метаболических путей ЭПМ
и ТКК заключается в фосфорилировании ADP, что приводит к
повышению энергетического заряда клетки, нетрудно видеть, что
Стехиометрия и энергетика метаболических превращений
353
Глюкоза
Путь
ЭМП
г₽р ^,-лтре
ФосфОфруктокиназа
(Фоссрориларо-
ванае на уров- ।
не субстрата) Пируваткиназа-^- —АМР ©
РЕР
ADP©
Пируват
Пируват-дегидрогеназнглй _ ЛТ„ _
комплекс А I г у
Ацетил- Сок
фитратсинтазо — АМР©
Пи ню иная кислота
(Давелевоуксус-
ная кислота
_ Изолитонная кислота
ADP © /
Изоцитратдегидро -
С ген аза
Цикл ТКК
ИТР 0
<х- Кетоглутаровая
кислота
Л
NADH
I
Окислительное фосфорилирование
I
АТР
РИС. 5.25. Регуляция метаболического пути ЭМП и цикла ТКК аденозинфос-
фатами. Штриховые стрелки указывают аллостерический эффект в отношении
данного фермента; знаком © обозначена активация фермента, знаком © —
его ингибирование.
23—537
35 4
Глава 5
основным типом регуляции в схеме на рис. 5.25 являются согла-
сованные и координированные формы ингибирования по прин-
ципу обратной связи.
Для управления поступлением веществ-предшественников и
процессами биосинтеза, очевидно, необходимы другие системы
регуляции. Цитрат, например, служит аллостерическим модуля-
тором нескольких ферментов. Он повышает ингибиторный эф-
фект АТР в отношении фосфофруктокиназы. Этот факт отражает
амфиболическую природу путей ЭМП и ТКК, благодаря которой
в любом из них при высоком энергетическом заряде и поступле-
нии достаточного количества питательных веществ могут исполь-
зоваться соединения с шестью атомами углерода. Цитрат явля-
ется также мощным активатором первой реакции в биосинтезе
жирных кислот [уравнение (5.34)].
Центром регуляции путей биосинтеза обычно являются ста-
дии, следующие непосредственно за точкой разветвления. При-
мером такой регуляции могут служить реакции биосинтеза
аминокислот семейства аспарагиновой кислоты, изображенные
на рис. 5.12, где штриховые линии обозначают пути ингибиро-
вания по принципу обратной связи. Выгоды разветвленной по-
следовательности реакций с несколькими регулируемыми ста-
диями очевидны: таким путем в результате ограниченного чис-
ла реакций образуется несколько конечных продуктов, причем
путь синтеза каждой аминокислоты регулируется независимо.
На первый взгляд может показаться, что в этой системе есть
один дефект: высокие концентрации лизина и треонина долж-
ны, вероятно, замедлять синтез гомосерина и метионина. На
самом же деле этого не происходит, поскольку превращение
аспартата в аспартил-р-фосфат катализируют три различных
фермента. Активность одного из этих ферментов не регулирует-
ся, второй ингибируется лизином, а третий ингибируется трео-
нином. Различные ферменты, катализирующие одну и ту же
реакцию, называются изозимами или изоферментами. Точно
так же превращение полуальдегида аспарагиновой кислоты в
гомосерин катализируют два изофермента, один из которых не
регулируется, а второй ингибируется треонином. Эти факты по-
могают понять, как осуществляется действенный и эффектив-
ный контроль сети реакций при любом сочетании избыточных
концентраций продуктов метаболизма.
Целью того или иного процесса биохимической технологии
может быть производство больших количеств определенных ме-
таболитов. В таком случае стремление добиться максимальной
производительности в отношении этих соединений, очевидно,
вступает в противоречие с системами жизнеобеспечения и регу-
ляции нативного организма. Чтобы заставить клетку продуци-
ровать необходимые метаболиты достаточно эффективно (с точ-
Стехиометрия и энергетика метаболических превращений 355
ки зрения человека, но не «с точки зрения» клетки), часто пы-
таются модифицировать регулирующие метаболизм ферменты
таким образом, чтобы свойственные нативной клетке системы
регуляции не срабатывали и происходило перепроизводство
нужных метаболитов. На этой проблеме мы еще остановимся
в следующей главе в ходе изучения прикладной генетики.
Здесь же нам осталось рассмотреть только стехиометрические
и энергетические аспекты сочетания различных метаболических
путей, а также суммарную стехиометрию утилизации субстра-
тов, клеточного роста, образования метаболитов и выделения
теплоты. В качестве введения к этой теме мы познакомимся с
природой конечных продуктов метаболизма, выделяемых клет-
кой в среду.
5.9. Конечные продукты метаболизма
Клетки выделяют в среду множество различных соединений.
Во многих случаях эти соединения образуются в результате про-
цессов, сопровождающихся выделением энергии. Типичными
примерами конечных продуктов метаболизма являются спирты
и органические кислоты. Другие продукты метаболизма, напри-
мер антибиотики, токсины, алкалоиды и факторы роста, пред-
ставляют собой более сложные соединения, выполняющие опре-
деленные функции в жизнедеятельности клетки. Такие соеди-
нения, наличие которых не является необходимым для роста
культуры клеток в чистой питательной среде, называют вторич-
ными метаболитами. В настоящем разделе мы вкратце рассмот-
рим некоторые пути метаболизма, ведущие от промежуточных
метаболитов к конечным продуктам метаболизма, а также ти-
пы последних в зависимости от природы продуцирующих их
клеток и условий культивирования.
5.9.1. Продукты анаэробного метаболизма (брожения)
Прежде всего рассмотрим различные конечные продукты
анаэробного метаболизма глюкозы. Многие организмы перера-
батывают глюкозу в пируват по путям Эмбдена — Мейерго-
фа— Парнаса, Энтнера — Дудорова или по пентозофосфатно-
му шунту. Дальнейшие пути от пирувата к конечным продук-
там метаболизма могут существенно различаться. Экономиче-
ски наиболее важен процесс превращения пирувата в этанол
(спиртовое брожение), осуществляющийся по следующей схеме:
пируватдекарбоксилаза
Пируват -* ацетальдегид 4-СОг
алкогольдегидрогеназа
Ацетальдегид-}-NADH 4-Н+------------------ этанол + NAD-1- (5.44)
23*
356
Глава 5
Таблица 5.7. Конечные продукты метаболизма углеводов в бактериях по пути Эмбдена — Мейергофа — Парнаса®
Тип брожения Основные продукты мета- болизма пировиноградной кислоты Бактерии, осуществляющие данный тип брожения
1. Молочнокислое Молочная кислота Молочнокислые бактерии ро- дов Streptococcus, Pediococ- cus, Lactobacillus (некоторые виды)
2. Смешанное Молочная, уксусная, ян- Различные энтеробактерии, на-
кислотное тарная, муравьиная (или СО2 и Н2) кисло- ты, этанол пример Escherichia, Salmo- nella, Shigella, Proteus, Yer- sinia
а. Бутандиоль- Как при брожении ти- Aerobacter, Serratia, Aeromonas,
ное па 2, а также бутан- диол-2,3 Bacillus polymyxa
3. Маслянокислое Масляная кислота, ук- сусная кислота, СО2 и н2 Многие анаэробные спорообра- зующие бактерии (Clostri- dium')', некоторые неспорооб- разующие анаэробы (Butyri- bacterium)
а. Ацетоно-бу- Как при брожении типа Некоторые спорообразующие
тиловое 3, а также бутанол, этанол, ацетон и изо- пропанол анаэробы (Clostridium)
4. Пропионово- кислое Пропионовая кислота, уксусная кислота, ян- тарная кислота, СО2 Propionibacterium, Veillonella
а Stonier ft. У., Doudoroff М., Adelberg Е. А., Prentice-Hall, Inc., Englewood Cliffs, N. J., 1970. The Microbial World, 3d ed., p. 183,
В этих реакциях, как и в рассматриваемых ниже других
процессах брожения углеводов, образующийся в ходе биосин-
теза пирувата NADH вновь окисляется. Поскольку в отличие от
дыхания при анаэробном метаболизме электроны не переносят-
ся к внешнему акцептору, то процессы утилизации субстратов
и образования продуктов брожения могут быть описаны обыч-
ными уравнениями окислительно-восстановительных реакций.
Другим примером участия NADH в образовании продуктов ана-
эробного метаболизма является молочнокислое брожение, в хо-
де которого пируват в одну стадию превращается в лактат:
лактатдегидрогеназа /е
Пируват+ЫАЭН4-Н+-------------------- лактат+NAD-1- 1ОЛО7
В табл. 5.7 перечислены другие типы анаэробного метабо-
лизма углеводов, начинающиеся с превращения в пируват по
пути ЭМП, а на рис. 5.26 приведена схема соответствующих
последовательностей реакций. Следует подчеркнуть, что при-
рода и относительные количества конечных продуктов броже-
Стехиометрия и энергетика метаболических превращений
357
РИС. 5.26. Конечные продукты (обведены кружком) анаэробного метаболиз-
ма пирувата в микроорганизмах. Буквами обозначены микроорганизмы, осу-
ществляющие указанные реакции: А — молочнокислые бактерии (Streptococ-
cus, Lactobacillus); В — Clostridium propionicunv, С — дрожжи, уксуснокислые
бактерии, Zymomonas, Sarcina ventriculi, Erwinia amylovora-, D — энтеробак-
терии (аэрогенные колибактерии); E — клостридии; F—аэробактерии; G —
дрожжи; H — клостридии (виды butyricum, acetobutylicum); I — пропионово-
кислые бактерии. [Воспроизведено с разрешения из работы: Biochemistry of
Bacterial Growth, Mandelstam J., McQuillen K. (eds.), 2d ed., p. 166, Black-
well Scientific Publications, 1973.]
358
Глава 5
ния зависят как от самого организма, так и от условий его
культивирования. Например, при спиртовом брожении 5%-ного
раствора глюкозы в присутствии дрожжей Saccharomyces се-
revisiae изменение pH от 3,0 до 7,0 приводит к снижению вы-
хода этанола от 171,5 до 149,5 ммоль (в расчете на 100 ммоль
исходной глюкозы) и к повышению выхода глицерина с 6,16
до 22,2 ммоль (также из 100 ммоль глюкозы). При добавлении к
питательной среде сульфита выход глицерина повышается еще
больше; основанный на этом типе брожения способ получения
глицерина широко применялся в годы первой мировой войны.
В заключение этого краткого обзора процессов брожения
следует отметить, что некоторые организмы способны превра-
щать находящиеся в питательной среде аминокислоты в другие
органические соединения. Подобные процессы метаболизма иг-
рают важную роль в производстве спиртных напитков, посколь-
ку образующиеся из азотсодержащих соединений высшие али-
фатические спирты придают напиткам соответствующие вкус и
аромат.
5.9.2. Частичное окисление и его конечные продукты
В аэробных микроорганизмах в большинстве случаев ко-
нечными продуктами дыхания являются вода и диоксид угле-
рода. В то же время некоторые аэробные микроорганизмы,
а также любые другие аэробы в аномальных условиях окисля-
ют органические вещества неполностью; в таких случаях в
клетках или в среде накапливаются продукты частичного окис-
ления. Поскольку ряд процессов частичного микробиологиче-
ского окисления используется в промышленном масштабе, мы
вкратце рассмотрим здесь некоторые из них.
Иногда направление частичного окисления определяется со-
ставом питательной среды микроорганизма. Так, в разд. 1.3.3
мы уже упоминали о способе производства лимонной кислоты с
помощью плесени A. niger. Выход лимонной кислоты может
быть значительно увеличен путем повышения концентрации
сахара и снижения концентрации ионов железа (кофактора
фермента, использующего в качестве субстрата лимонную кис-
лоту) (табл. 5.8). Выращивая другие грибы и аэробные бакте-
рии в аномальных условиях, можно получать другие промежу-
точные соединения цикла ТКК, например а-кетоглутаровую и
фумаровую кислоты.
Хорошо известна склонность уксуснокислых бактерий к не-
полному окислению. Бактерии родов Acetobacter и Glucono-
bacter окисляют этанол до уксусной кислоты, а в отсутствие
этанола бактерии Acetobacter могут окислять уксусную кислоту
Стехиометрия и энергетика метаболических превращений
359
Таблица 5.8. Зависимость выхода лимонной кислоты3, образующейся
из сахара в погруженной культуре Aspergillus niger, от концентрации
солей железа6
Содержание железа (в виде FeCl3), мг/л Выход лимонной кислоты, г на 100 г гексоз Содержание железа (в виде FeCl3), мг/л Выход лимонной кислоты, г иа 100 г гексоз
0,0 67,00 1,00 76,00
0,05 73,0 2,00 71,00
0,50 88,0 5,00 57,0
0,75 79,0 10,00 39,0
а Состав среды; 40%-ный раствор сахарозы, очищенный ионным обменом до со-
противления 3,6 МОм и разбавленный до 14,2%; КН2РО4 (0,014%); MgSO<-7H2O (0,1%);
(NH4)2CO3 (0,2%); НС1 до pH 2,6 (из патента США 2970084).
б Microbial Technology, table 8-1, Peppier H. J. (ed.), Reinhold Publishing Corpo-
ration, New York, 1967.
далее до CO2. Напротив, бактерии Gluconobacter не способны
метаболизировать уксусную кислоту; известно, что у некоторых
видов этих бактерий отсутствуют ферменты, участвующие в
цикле ТКК- Образующийся из этанола под действием Acetobac-
ter конечный продукт (раствор уксусной кислоты) называется
винным уксусом. В табл. 5.9 перечислены другие конечные про-
дукты метаболизма в уксуснокислых бактериях.
5.9.3. Синтез вторичных метаболитов
В соответствующих условиях микроорганизмы и другие
клетки синтезируют вторичные метаболиты. Обычно во время
быстрого роста клеток эти вещества в сколько-нибудь значи-
тельных количествах не образуются. Известно множество вто-
ричных метаболитов, имеющих самое разнообразное строение и
различные биологические активности. В настоящее время иден-
тифицировано, например, около 6000 различных антибиотиков.
Как Bacillus subtilis, так и Streptomyces griseus продуцируют
более 50 антибиотиков!
На рис. 5.27 изображены различные вторичные метаболиты
и наиболее общие пути их биосинтеза. Нетрудно видеть, что ис-
ходными веществами здесь являются первичные метаболиты.
Кро-ме того, первичные метаболиты и энергетический заряд
клетки служат важнейшими регуляторами путей биосинтеза
вторичных метаболитов. Мы еще вернемся к этой теме в ходе
изучения катаболитной репрессии в гл. 6 и 7. Другим важным
параметром регуляции биосинтеза вторичных метаболитов яв-
ляется средний уровень концентрации фосфатов.
360
Глава 5
СНз
бутандиол-2,3
н н онон
Таблица 5.9. Находящие практическое применение частично окисленные
продукты метаболизма уксуснокислых бактерий®
СН3СН2ОН+О2 —> СНзСООН+НзО
этанол уксусная кислота
СН3СН2СН2ОН+О8 —> СНзСН8СООН+Н8О
(пропанол пропионовая кислота
(НзС)2СНОН+‘/2О2—> (НзС)2С=О+Н2О
изопропанол ацетон
снаон снаон
CHOH+V2O2—>• С=О-Н2О
<!:наон с!:н2он
глицерин дигидроксиацетон
СНз СНз
(СНОН)2+1/2О2—> снон+н2о
с=о
I
СНз
ацетоин
н он он
НОСН2—С—С—С—С—CH2OH+V2O8—>НОСН2—С—с—с—с—СН8ОН+Н2О
Till и I I I
ононн н о онн н
маннит фруктоза
н н онн н н онн
НОСНг—С—С—С—С—СНО + ’/2О2—►НОСНг—С—С—С—С—СООН+Н2О
ОНОНН он ононн он
глюкоза глюконовая кислота
н н онн н онн
НОСН2—С—С—С—С—СООН+*/2О2 —> носн2—с—с—с—с—соон+н2о
Illi II I I I
ононн он о онн он
глюконовая кислота 5-кетоглюконовая кислота
а Startler R. У., Doudoroff М., Adelberg Е. A., The Microbial World, 3d ed., p. 211,
Prentice-Hall, Inc., Englewood Cliffs, N. J., 1970.
5.10. Стехиометрия клеточного роста
и образования продуктов метаболизма
Хотя рост клеток представляет собой чрезвычайно сложный
и изящный процесс, он, естественно, подчиняется законам со-
хранения материи. В ходе метаболических процессов в клетке
Нойедая кислота
Неомицин
Гентамицин
Панамицин
Тобрамицин
риоостамицин
Паромомицин
Стрептомицин
Спектиномицин
Фосфаты еенсоз
Нуклеозид» ЩыаРМ
ДНК, РНК А
I
----^Фоараты пентоз------>• Фосфаты тетроз
Шикимовая кислота
Пенициллин
Церамицин
Цефалоспорин
Аспартат —
Серин
Г ”
Цистеин
Валин -*
Лизергиновая кислота
Эрголин
Кандицидин
Коринецины
Хлорамфеникол
Пиоцианик
Наводи о цин
Антрамицин
Сибиромицин
-Пинномицин
Рубрадирин
Ацетил-Со К
Оксалоацетат
а-Амино -
адипат
Сукцинат
Пируват
Пирролнитрин
Индолмицин
Актиномицин
Глиотоксин
Цитрат
Аконитат-
Жирные кислоте/
брсфелбдин
а - Кетоглута- Итак окат
pan? '
Изопреноиды
Гиббереллины
Стероиды
Наротиноиды
Трислоровые кислоты
Поликетиды
I
Орселлиновая кислота
в-Метилсалицилат
Патулин
Гризеофулсвин
Афлатоксин
Эритромицин
тетрациклин
ри/рамицин
Стрептоварицин
Стрептолидигин
РИС. 5.27. Схема путей метаболизма, ведущих к вторичным метаболитам. Жирными стрелками выделены катализируемые
ферментами последовательности реакций, в результате которых образуются вторичные метаболиты. [Воспроизведено с раз-
решения из работы: Malik V. S., Microbial Secondary Metabolism, Trends Biochem. Sci., 5, 68 (1980).]
362
Глава 5
перегруппировываются атомы углерода, азота, водорода, кисло-
рода и других жизненно важных химических элементов, рас-
смотренных в гл. 2, но общее количество каждого из этих эле-
ментов, включенное в структуры клетки, в точности равно коли-
честву, взятому клеткой из питательной среды. Кроме того,
количество образующегося метаболита или выделившейся в про-
цессе роста клеток теплоты часто пропорционально количеству
поглощенного клеткой субстрата или количеству другого про-
дукта метаболизма, например СО2. В этом разделе мы рассмот-
рим некоторые достаточно жесткие ограничения, налагаемые на
баланс веществ, а также ряд полезных приближенных эмпири-
ческих стехиометрических соотношений. Как мы увидим, эти
стехиометрические аспекты имеют самое непосредственное отно-
шение к проблемам биохимической технологии, где они должны
учитываться при решении самых различных вопросов от выбора
состава питательной среды до выбора систем автоматизирован-
ного контроля и способа охлаждения биологических реакторов.
Кроме того, как и в анализе любых реакторов, стехиометриче-
ские соотношения определяют материальный баланс веществ в
биореакторе и позволяют наиболее эффективным и целенаправ-
ленным путем использовать специфику кинетики реакций (гл. 7).
5.10.1. Общая стехиометрия клеточного роста;
состав среды и коэффициенты выхода
В процессе роста клетки потребляют субстраты, которые
обеспечивают их энергией и исходными веществами для синте-
за новой клеточной массы. С самой общей точки зрения для
реализации этого процесса необходимо, чтобы в питательной
среде находились все химические элементы, требующиеся для
формирования новой клеточной массы, и чтобы свободная энер-
гия усвоенных субстратов превышала свободную энергию обра-
зующихся клеток и продуктов метаболизма (рис. 5.28)*. Дру-
гими словами, свободная энергия образующихся веществ долж-
на быть меньше свободной энергии утилизированных субстратов.
Кроме того, питательные вещества должны иметь соответст-
вующее молекулярное строение, определяемое специфичностью
клеточных ферментов, участвующих в процессах катаболизма и
биосинтеза. В то же время при изучении роста клеток как еди-
ного процесса (а именно с этой точки зрения мы будем изучать
его в большей части настоящего раздела) привлечение деталей
* В настоящем разделе продуктами метаболизма мы будем называть ор-
ганические вещества, отличающиеся от продуктов жизнедеятельности клеток,
выделяемых ими в среду. Под питательными веществами или субстратами мы
будем подразумевать вещества, потребляемые клеткой из среды в процессе
роста или образования продуктов метаболизма.
Стехиометрия и энергетика метаболических превращений 363
механизмов метаболических превращений может оказаться не
только ненужным, но и вредным. Основной смысл изображенно-
го на рис. 5.28 процесса роста клеток как единого явления за-
ключается в том, что ему свойственны определенные стехиомет-
рические ограничения независимо от того, какой конкретный
механизм или какие метаболические пути использует «система»
клеток для достижения цели «реакции» — роста клеток. Поэтому
для решения вопроса о пригодности тех или иных питательных
РИС. 5.28. Клеточный рост как единый процесс. Под «системой» здесь подра-
зумевается определенное количество клеток. В процессе роста «система» вы-
полняет функции катализатора, превращая субстраты в новые клетки и про-
дукты метаболизма.
веществ мы просто будем рассматривать только те питательные
вещества, которые заведомо являются подходящими субстрата-
ми для роста изучаемого штамма клеток.
Простейшая форма стехиометрических ограничений, накла-
дываемых на состав питательной среды, в краткой форме мо-
жет быть выражена следующим образом: если мы хотим выра-
стить общую массу клеток (X) (прописными буквами в скобках
мы будем обозначать общую массу компонента, указанного в
скобках) и если весовая доля элемента i в клетке равна Wi, то
минимально необходимое общее содержание элемента i во всех
субстратах должно составлять wq(X). В общем случае подобные
вычисления лучше выполнять в расчете на массу сухого клеточ-
ного вещества. При выполнении таких расчетов для различных
элементов необходимо знать их относительное содержание в
клетке. Примером могут служить довольно подробные данные
об элементном составе бактерии Е. coli, приведенные в табл.
2.1, а в табл. 5.10 указано содержание основных элементов (С,
Таблица 5.10. Элементный состав некоторых микроорганизмов. Символом ц, обозначена удельная скорость
роста, равная массе клеток, образующихся из данной массы клеток в единицу времени (см. гл. 7)а
Микроорганизм Лимитирующее питательное вещество Состав, масс. % Эмпирическая химическая формула «Молекул яр- пая> масса, отвечающая эмпирической формуле
-1 Н N О Р S Зола
Ц. ч С
Бактерия 53,0 7,3 12,0 19,0 '"*г4 8 СН1,б6бМо,2оОо,27 20,7
Бактерия 47,1 7,8 13,7 31,3 СНгМо.гбОо.б 25,5
Aerobacter aero- 48,7 7,3 13,9 21,1 8,9 CHi,78No,240o,33 22,5
genes
Klebsiella aero- Глицерин 0,1 50,6 7,3 13,0 29,0 CH1.74N0.22O0,43 23,7
genes K. aerogenes Глицерин 0,85 50,1 7,3 14,0 28,7 CHi .73N0.24O0.43 24,0
Дрожжи 47,0 6,5 7,5 31,0 8 СН^ббМолзОо^о 23,5
Дрожжи 50,3 7,4 8,8 33,5 CHijsNqjsOo.b 23,9
Дрожжи 44,7 6,2 8,5 311,2 1,08 0,6 CHi,64No,ieOo,52Po,OlSo,005 26,9
Candida utilis Глюкоза 0,08 50,0 7,6 11,1 31,3 CH1.82N0.i9O0.47 24,0
C. utilis Глюкоза 0,45 46,9 7,2 10,9 35,0 CHi,84No,2O0.se 25,6
C. utilis Этанол 0,06 50,3 7,7 11,0 30,8 CHi,82|No,190o,46 23,9
C. utilis Этанол 0,43 47,2 7,3 11,0 34,6 CH i,84No,2O0.se 25,5
® Воспроизведено с разрешения из работы: Atkinson. В., Mavituna F., Biochemical Engineering and Biotechnology Handbook, p. 120,
Macmillan Publishers Ltd., Surrey, England, 1983.
Стехиометрия и энергетика метаболических превращений 365
N, О и Н) в клетках целого ряда микроорганизмов. Хотя эле-
ментный состав клеток несколько изменяется в зависимости от
вида микроорганизма и даже от состава питательной среды в
случае одного и того же штамма, в целом эти данные вполне
надежно определяют элементный состав клеточной массы.
Смысл приведенных в этой таблице эмпирических формул кле-
ток и их применение мы рассмотрим в следующем разделе.
На самом деле определить необходимый для эффективного
роста клеток состав среды значительно сложнее, поскольку из-
ложенный выше основной стехиометрический принцип не учиты-
вает, во-первых, то, что часть химических элементов субстра-
тов входит в выделяемые в среду продукты метаболизма, во-
вторых, ограничений, накладываемых скоростями реакций,
в-третьих, особых метаболических свойств данного штамма кле-
ток, обусловленных ингибирующими эффектами каких-либо пи-
тательных веществ или продуктов метаболизма. Первую при-
чину осложнений мы рассмотрим позднее в этом же разделе,
а вторую обсудим сейчас. Затем мы перейдем к изучению эко-
номических коэффициентов — простейших параметров, с помо-
щью которых можно количественно оценить стехиометрию роста
клеток.
Хотя в этом разделе основное внимание мы постараемся
уделять именно стехиометрии, при разработке подходов к упро-
щению стехиометрических выражений клеточного роста и обра-
зования продуктов метаболизма нам придется принимать во вни-
мание и кинетические факторы. Число питательных веществ в сре-
де обычно очень велико, и учесть все эти вещества в анализе
стехиометрии или кинетики процесса практически невозможно.
Поэтому обычно стараются описать систему упрощенно, на базе
ограниченного числа лимитирующих процесс ключевых соедине-
ний или элементов. Найти стехиометрически лимитирующее со-
единение можно на основе стехиометрии клеточного роста. Для
этого необходимо определить, какой субстрат первым полно-
стью израсходуется в «реакции» клеточного роста, изображен-
ной на рис. 5.28. Лимитирующие соединения других типов мож-
но идентифицировать путем изучения влияния состава среды на
рост клеток. Для данного штамма в данных условиях (темпера-
тура, pH, присутствие других питательных веществ) лимитиру-
ющий скорость роста компонент питательной среды можно най-
ти экспериментально следующим образом. Предположим, что
клетки растут в определенной среде в заданных условиях. В этой
системе резко повышают концентрацию одного из компонентов
среды и затем определяют изменение скорости роста клеток.
Часто состав среды подбирают так, чтобы скорость клеточного
роста лимитировал только один компонент, а относительно не-
большие (по сравнению с базовыми значениями) изменения кон-
366
Глава 5
центраций всех других компонентов среды практически не влия-
ли бы на скорость роста клеток. В принципе, однако, возможны
и такие варианты, когда скорость клеточного роста определяют
несколько субстратов одновременно.
Компонент, лимитирующий скорость клеточного роста, и сте-
хиометрически лимитирующий компонент не обязательно дол-
жны быть одним и тем же веществом. Другими словами, в оп-
ределенных условиях скорость роста клеток может ограничи-
вать одно соединение, но рост клеток в периодическом процес-
се может прекратиться в силу истощения запасов другого со-
единения. В то же время в различных математических выраже-
ниях, описывающих рост микроорганизмов количественно, та-
кая возможность часто игнорируется, что может приводить к из-
вестным трудностям при изучении режима биореакторов и не-
правильным выводам при интерпретации их характеристик. Не
забывая о возможных осложнениях такого рода, мы тем не ме-
нее будем считать, что клеточный рост лимитирует только одно
питательное вещество. Если речь идет о конкретной системе и
определенной среде, то всегда можно экспериментально прове-
рить справедливость допущения о том, что лимитирующий ско-
рость роста и стехиометрически лимитирующий компоненты яв-
ляются одним и тем же веществом.
Часто оказывается, что общее количество клеточной массы,
образующейся в процессе клеточного роста, пропорционально
массе утилизированного клетками субстрата (обычно углерод-
содержащего предшественника, источника энергии или кислоро-
да) (рис. 5.29). Экономическим коэффициентом называют соот-
ветствующее соотношение:
(5-46)
Следует отметить, что размерность экономического коэффици-
ента определяется способом выражения количества клеточной
массы и количества субстрата (например, в единицах массы или
в числе молей); этот факт всегда следует учитывать при исполь-
зовании опубликованных в литературе значений У. При одновре-
менной утилизации из среды нескольких различных субстратов
Sj можно определить несколько соответствующих экономических
коэффициентов клеточного роста Yx/si В табл. 5.11 приведены
экономические коэффициенты для роста ряда микроорганизмов
в средах, отличающихся природой источника углерода; здесь
цифры в одной строке отвечают разным способам измерения ко-
личества утилизированного субстрата (в граммах, молях или в
граммах углерода в 1 моль субстрата).
Если для данной культуры клеток в данной питательной сре-
де экономический коэффициент приблизительно постоянен, то
Стехиометрия и энергетика метаболических превращений
367
уравнение (5.46) позволяет определить изменение концентрации
клеточной массы по изменению концентрации субстрата, и на-
оборот. Иными словами, одну из этих переменных можно выра-
зить через другую и при расчетах в ходе проектирования реак-
торов одну переменную можно не учитывать. Это стехиометри-
ческое соотношение может пригодиться и при контроле работы
биологических реакторов; в данном случае концентрация суб-
страта может служить одновременно и мерой концентрации
РИС. 5.29. Зависимость общего количества образующейся биомассы от коли-
чества утилизированного субстрата в случае роста бактерий Aerobacter aero-
genes в аэробных условиях (а) и бактерий Propionibacterium pentosaceum
в анаэробных условиях (б) на различных источниках углерода. [Воспроиз-
ведено с разрешения из работ: a) Hadjipetrou L. Р., Gerrits J. Р., Ten-
lings F. A. G., Stouthamer А. Н., Relation between Energy Production and
Growth of Aerobacter aerogenes, J. Gen. Microbiol., 36, 139 (1964); 6) Bau-
•chop T., Elsden S. R„ Growth of Microorganisms in Relation to their Energy
Supply, J. Gen. Microbiol., 23, 457 (I960).]
клеточной массы, если только известны обе соответствующие
исходные величины. Это чрезвычайно удобно, поскольку обычно
концентрации субстратов определять значительно легче, чем
концентрацию биомассы. На практике удобнее всего определять
концентрации веществ, покидающих зону аэробного биореакто-
ра, например СО2. В этом случае в качестве основы для уста-
новления зависимостей между различными переменными био-
технологических процессов также используются стехиометриче-
ские соотношения. Однако, прежде чем обратиться к изучению
таких зависимостей, необходимо упомянуть о вариабельности
экономического коэффициента и о понятии метаболизма под-
держания.
Нет никакой гарантии, что найденный эмпирическим путем
экономический коэффициент, представляющий собой кажуще-
368
Глава 5
Таблица 5.11. Экономические коэффициенты для аэробного роста ряда
микроорганизмов на различных источниках углерода. Здесь символом
Ух/о2 обозначен коэффициент, выраженный в граммах клеточной массы
в расчете на 1 г усвоенного О2а
yx/s
Микроорганизм Субстрат г/г г/моль г/г (угле- рода) 7Х/О г/г
Aerobacter aerogenes Мальтоза 0,46 149,2 1,03 1,50
Маннит 0,52 95,5 1,32 1,18
Фруктоза 0,42 76,1 1,05 1,46
Глюкоза 0,40 72,7 1,01 1,П
Candida utilis Глюкоза 0,51 91,8 1,28 1,32
Penicillium chrysogenum Глюкоза 0,43 77,4 1,08 1,35
Pseudomonas fluorescens Глюкоза 0,38 68,4 0,95 0,85
Rhodopseudomonas sphe-
roides Глюкоза 0,45 81,0 1,12 1,46
Saccharomyces cerevisiae Глюкоза 0,50 90,0 1,25 0,97
Aerobacter aero genes Рибоза 0,35 53,2 0,88 0,98
Сукцинат 0,25 29,7 0,62 0,62
Глицерин 0,45 41,8 1,16 0,97
Лактат 0,18 16,6 0,46 0,37
Пируват 0,20 17,9 0,49 0,48
Ацетат 0,18 10,5 0,43 0,31
Candida utilis Ацетат 0,36 21,0 0,90 0,70
Pseudomonas fluorescens Ацетат 0,28 16,8 0,70 0,46
Candida utilis Этанол 0,68 31,2 1,30 0,61
Pseudomonas fluorescens Этанол 0,49 22,5 0,93 0,42
Klebsiella sp. Метанол 0,38 12,2 1,01 0,56
Methylomonas sp. Метанол 0,48 15,4 1,28 0,53
Pseudomonas sp. Метанол 0,41 13,1 1,09 0,44
Methylococcus sp. Метан 1,01 16,2 1,34 0,29
Pseudomonas sp. Метан 0,80 12,8 1,06 0,20
Pseudomonas sp. Метан 0,60 9,6 0,80 0,19
Pseudomonas methanica Метан 0,56 9,0 0,75 0,17
а Воспроизведено с разрешения из работы: Nagai S., Mass and Energy Balances
for Microbial Growth Kinetics, in Advances in Biochemical Engineering, vol. 11, Gho-
se T. K., Fiechter A., Blakebrough N. (eds.), Springer-Verlag, New York, p. 53, 1979.
еся стехиометрическое соотношение, всегда постоянен для дан-
ного организма в данной среде. Действительно, эксперимен-
тально установлено, что часто экономический коэффициент из-
меняется при изменении скорости роста клеток. Чтобы понять
причину этих изменений, иногда полезно разбить процесс ути-
лизации субстрата на три части: превращение субстрата в кле-
точную массу (ассимиляция), обеспечение энергии для клеточ-
ного синтеза и обеспечение энергии для поддержания культу-
ры. Под энергией поддержания здесь подразумеваются энерге-
тические затраты клетки, необходимые для ее выживания или
сохранения в определенном (потенциально) жизнеспособном
состоянии, но не обеспечивающие клеточный рост и образова-
Стехиометрия и энергетика метаболических превращений
369
ние новых клеток. Эта энергия расходуется на активный
транспорт ионов и других веществ через клеточные мембраны
и на возобновление деградирующих составных частей клетки.
В случае хемогетеротрофов один и тот же субстрат выпол-
няет функции источника; энергии и углерода, поэтому суммар-
ную утилизацию субстрата можно выразить следующим урав-
нением:
(AS) — (А^)ассимиляция “Ь (А^)энергия роста “Ь (А^)энеРгия поддержания (5.47)
Разделив это уравнение на (АХ), получим
1 (AS) ассимиляция , (AS) энергия роста (А3)энергня поддержания
Ух/s ” (ДХЦ
(АХ)
;(ДГ)
(5.48)
Истинный экономический коэффициент клеточного роста АХ/
/(AS)ассимиляция относительно постоянен и представляет собой
стехиометрически четко определенную величину. Этого, однако,,
нельзя сказать об общем коэффициенте Ух/s, что связано с за-
висимостью различных (AS) в правой части уравнения (5.48)
от условий роста. Так, в быстро растущей популяции клеток
большая часть субстрата расходуется на ассимиляцию и рост.,
в то время как в популяции клеток, находящейся в стационар-
ном состоянии или покое, субстрат часто расходуется исключи-
тельно на поддержание (в этом случае Ух/з = 0).
В следующем разделе мы рассмотрим другие, более строгие
стехиометрические выражения для клеточного роста с учетом
уравнений материального баланса по углероду, водороду, азо-
ту и кислороду. Кроме того, мы познакомимся с более деталь-
ным подходом к моделированию клеточного роста, при котором
рост клеток все еще рассматривается как единый процесс, но-
вместе с этим учитывается ведущая роль NADH и АТР в ме-
таболизме роста.
5.10.2. Материальный баланс по элементам и клеточный рост
Рост клеток и связанные с ним метаболические процессы
можно описать такими же уравнениями, какие используются
для изображения обычных химических реакций. Для этого
прежде всего необходимо найти «химическую формулу» сухого
клеточного вещества.; Если известен элементарный состав дан-
ного штамма, растущего в определенных условиях (см., напри-
мер, данные табл. 5.10), то не представляет труда найти и от-
носительные значения коэффициентов в эмпирической формуле
клетки СеНаСШ. Для того чтобы клетке отвечали одна строго
определенная формула и соответствующая «молекулярная мас-
24—537
370
Глава 5
са», удобно принять 9=1 и затем в соответствии с элементным
составом найти значения а, и б. Приведенные в табл. 5.10
«формулы» клеток получены именно таким путем. Один С-моль
клеток определяют как количество клеток, в котором содержит-
ся один грамм-атом (12,011 г) углерода и которое отвечает
эмпирической формуле клетки при 0=1.
В качестве простейшего примера анализа стехиометрии
клеточного роста на основе материального баланса по элемен-
там рассмотрим прежде всего рост клеток в аэробных условиях,
когда помимо новых клеток, СО2 и Н2О не образуется никаких
иных продуктов метаболизма. Если источники углерода и азота
мы обозначим формулами СНхО^ и H/OmNn соответственно, то
уравнение клеточного роста будет выглядеть следующим обра-
зом:
a'CHxO, + &,O2+c'H/OmN„ —>- CHaOpNe+dT^O-f-e'CC^ (5.49)
Здесь коэффициент при формуле клетки принят равным еди-
нице, что не отражается на общности уравнения, поскольку
стехиометрические коэффициенты можно умножать или делить
на любое одно и то же число.
Из уравнения (5.49) следует, что пять неизвестных стехио-
метрических коэффициентов a', b', с1, d' и е1 связаны четырьмя
уравнениями:
С: а'=1+е'
Н: a'x + c'l = a + 2d'
О: a'y-f-2b' + c'm=$+d'+2e'
N: с'п=8
Еще одно уравнение можно получить, если экспериментальным
путем определить дыхательный коэффициент (RQ) реакции
клеточного роста. Дыхательным коэффициентом называют мо-
лярное (или объемное) отношение выделяемого при дыхании
СО2 к поглощенному О2:
число молей выделившегося СО2
Дыхательный коэффициент = RQ = число молей поглощенного о2
Для реакции роста клеток, выраженной уравнением (5.49),
RQ = e'/&' (5.52)
Таким образом, пять уравнений (5.52) и (5.50) содержат пять
неизвестных стехиометрических коэффициентов, и если извес-
тен RQ, то можно определить все эти коэффициенты. Возмож-
ные затруднения, связанные с очень большими колебаниями
значений стехиометрических коэффициентов при относительно
небольших погрешностях в измерении RQ, мы рассмотрим в
упражнении 5.16.
Стехиометрия и энергетика метаболических превращений 371
В общем случае химическая формула элементного состава
клетки и стехиометрические коэффициенты в уравнении (5.49)
зависят от состава питательной среды и условий, в которых
находится клетка. Не следует забывать, что эта зависимость
имеет описательный характер и в используемом здесь смысле
отражает суммарный эффект множества независимых химичес-
ких реакций. Чтобы получить какое-то представление о влия-
нии среды или условий клеточного роста на стехиометрию это-
го процесса и связать это влияние с обсужденными выше пу-
тями метаболизма, можно, например, разделить общую реак-
цию роста на несколько реакций, отвечающих различным
метаболическим функциям (но не обязательно специфическим
путям метаболизма).
Теперь мы несколько более детально рассмотрим стехио-
метрию роста хемогетеротрофпого организма в аэробных усло-
виях. С каждой реакцией мы будем связывать реакции генери-
рования и утилизации АТР. Ниже последние записаны отдель-
но и не включены в уравнения соответствующих реакций с уча-
стием соединений углерода, водорода, кислорода и азота из-за
ряда практических затруднений, связанных со стехиометрией
АТР; вкратце мы рассмотрим и эти затруднения.
Диссимиляция источника энергии:
аСНхОу 4- (коэффициент) • Н2О 4- -у- aNAD+ >
Vc
---> aCO24~-y- a (NADH 4-Н+)
esa(ADP + Pi) —* ssa(ATP+H2O)
Окислительное фосфорилирование:
2b(NADH+Н+)+Ю2 —»- 2ftNAD++2ftH2O
2ft (P/О) (ADP-f-Pi) —> 2ft (P/О) (ATP+HaO)
Биосинтез:
(5.53а)
(5.536)
(5.54а)
(5.546)
б 1 ( б \
(I + о)СНхО^ + — H/OmNn 4- -у I ув - (1 + о) - — yN\ (NADH4-H+)—
(5.55а)
CHaOpNfi + суСО2 4- (коэффициент)-Н2О
MWB
^г(арр + р<)
1 АТР
(5.556)
(5.56)
—I (АТР -г Н2О)
1 АТР
Поддержание и рассеяние энергии:
с(АТР + Н2О) —* c(ADP + Pz)
Взаимосвязь между этими реакциями можно установить, если
допустить, что в рассматриваемых здесь условиях не накапли-
ваются ни АТР, ни NADH, и, следовательно, синтез этих соеди-
нений должен уравновешиваться их утилизацией.
24*
372
Глава 5
Приведенная выше усложненная, но более близкая к дей-
ствительности система химических уравнений, отражающих
клеточный рост, получена ценой резкого увеличения числа сте-
хиометрических параметров. Заметьте, что в уравнениях (5.53) —
(5.56), во-первых, стехиометрические коэффициенты отражают
баланс по элементам в каждой отдельной реакции и, во-вто-
рых, стехиометрический коэффициент биомассы CHaO₽N6 при-
нят равным единице. Поэтому коэффициенты, а, & и с в других
реакциях отражают, по сути дела, степень протекания этих ре-
акций относительно реакции роста клеток (биосинтеза). Сте-
хиометрические коэффициенты для воды в двух случаях обо-
значены словом «коэффициент», а не каким-либо специальным
символом, поскольку вовлечение воды в реакцию или ее выде-
ление в результате реакции обычно практически не изменяет
ее общего количества в клетке. Символом е$ обозначено число
актов фосфорилирования в расчете на атом углерода, участву-
ющий в процессах катаболизма.
Параметрами ув, 7s и уд обозначены степени восстало елей-
ности биомассы, источника углерода и источника азота соот-
ветственно. Степень восстановленности у соединения с брутто-
формулой CHrOsNv определяется как у = 4 +г—2s—3v. Нетруд-
но видеть, что степень восстановленности СОг, НгО и NH3
равна нулю.
Коэффициент Удтртах обозначает массу клеток, образующих-
ся в результате усвоения 1 моль АТР, при условии, что АТР
расходуется только на цели биосинтеза. Предпринимались по-
пытки оценить величину Удтртах по известным элементному со-
ставу клеток и путям биосинтеза [13]. Эти оценки показали,
что, например, при росте Е. coll на глюкозе и неорганические
солях Удтртах составляет 28,8 г клеток на 1 грамм-моль АТР.
Для перехода от единиц массы к мольным единицам в уравне-
ние (5.556) включена молекулярная масса клеток, обозначен-
ная символом MWb-
Как мы уже отмечали выше, P/О обозначает число актов
фосфорилирования ADP в расчете на один атом усвоенного
кислорода; этот параметр характеризует эффективность окисли-
тельного фосфорилирования и в общем случае зависит от ус-
ловий культивирования клеток. Расчетами на основе среднего
элементного состава микроорганизмов показано, что теорети-
ческие максимальные значения P/О составляют 2,25, 2,50 и 3,00
при использовании в качестве основного питательного вещества
ацетата, малата и глюкозы соответственно.
Прямое экспериментальное определение параметра P/О за-
трудняется тем обстоятельством, что изучать опытным путем
отдельно пути синтеза или утилизации АТР крайне сложно;
к тому же относительные количества образующегося и исполь-
Стехиометрия и энергетика метаболических превращений 373
зующегося в каждом пути АТР зависят от условий культивиро-
вания клеток. Основная трудность здесь обусловлена частич-
ной утилизацией АТР в процессах транспорта веществ через
мембраны, в биосинтезе соединений, далее разлагаемых внутри-
клеточными гидролазами, и в других недостаточно изученных
процессах (их иногда называют бесполезными циклами). Все
перечисленные источники потребления АТР, сгруппированные
здесь в одну «реакцию» (5.56) под общим названием «поддер-
жание и рассеяние энергии», естественно, оказывают влияние
на другие процессы генерирования и утилизации АТР.
Поскольку количество вновь синтезированного АТР должно
быть равно количеству утилизированного АТР, то любые изме-
нения в относительном количестве АТР, участвующего в «ре-
акции» (5.56) (это количество отражает коэффициент с), по-
влекут за собой и изменения в количествах АТР, принимаю-
щего участие в других реакциях. Экспериментальные данные
показывают, что потребность в АТР для поддержания клетки
определяется не стехиометрическими факторами, а скоростями
метаболических процессов, поэтому количественную оценку
этих затрат мы отложим до изучения кинетики клеточ-
ного роста в гл. 7.
Здесь целесообразно упомянуть, что экспериментально най-
денные величины выхода биомассы в расчете на количество
утилизированного АТР в анаэробных условиях обычно весьма
мало зависят от типа субстрата и вида микроорганизма и в
среднем величина Татр составляет 10,7 г клеточной массы на
1 моль АТР. У хемогетеротрофных организмов, как правило,
значительно больше субстрата используется для выработки
энергии, чем для роста биомассы. Так, в одной из работ было
показано, что при росте пекарских дрожжей Saccharomyces
cerevisiae в анаэробных условиях в богатой глюкозой среде
(глюкоза являлась источником углерода) 98% усвоенного уг-
лерода расходовалось на выработку энергии и только 2% —на
рост клеточной массы. Определить с достаточной точностью
количество синтезируемого АТР в условиях аэробного роста
оказалось крайне сложно. Имеющиеся данные говорят о том,
что величина отношения P/О составляет от 0,5 до 1,8, т. е. зна-
чительно ниже теоретического значения. Изучение бактерии
Aerobacter cloacae показало, что при ее росте в аэробной мини-
мальной среде 55% глюкозы ассимилируется во вновь синте-
зированной биомассе, а 45% расходуется для выработки энер-
гии. Эти данные свидетельствуют о том, что в аэробных усло-
виях выход клеточной массы в расчете на количество
утилизированного субстрата обычно выше, чем при анаэроб-
ном росте. Например, коэффициент Ух/s (масса клеток в грам-
мах, образовавшихся из 1 моль глюкозы) Streptococcus faeca-
374
Глава 5
lis в глюкозной среде составляет 21,5 в анаэробных условиях и
58,2 при аэробном росте В следующем разделе мы изучим
влияние такого распределения субстрата на стехиометрию об-
разования процессов метаболизма.
5.10.3. Стехиометрия процессов образования
продуктов метаболизма
Множество конечных продуктов метаболизма выделяется
клеткой в питательную среду или аккумулируется в клетке.
В соответствии с особенностями стехиометрии процессы обра-
зования продуктов метаболизма удобно подразделить на четы-
ре перечисленных ниже типа; первые три типа отвечают клас-
сификации путей анаэробного метаболизма, впервые предло-
женной в 1955 г. Гэйденом-младшим.
1. Основной продукт метаболизма является результатом пер-
вичного энергетического обмена. Примером может служить
образование этанола в ходе анаэробного роста дрожжей.
2. Образование основного продукта метаболизма не связано
непосредственно с энергетическим обменом, .как, например,
при биосинтезе лимонной кислоты при аэробном культиви-
ровании плесеней.
3. Конечный продукт является вторичным метаболитом. В ка-
честве примера можно привести биосинтез пенициллина, об-
разующегося в аэрируемой культуре плесени.
4. Биотрансформация. Здесь конечный продукт метаболизма
образуется из субстрата в результате одной или несколь-
ких реакций, катализируемых клеточными ферментами. При-
мером может служить гидроксилирование стероидов.
Процессы первого типа характеризуются относительно про-
стой стехиометрией, что связано со сравнительно постоянной
величиной отношения количества образующегося метаболита
к количеству накопленной клеточной массы и утилизированно-
го субстрата. В этом случае процессы утилизации субстрата,
синтеза биомассы и образования конечного продукта метабо-
лизма формально можно выразить одним уравнением химичес-
кой реакции, если, например, к правой части уравнения (5.49)
добавить продукт метаболизма СН^О®:
o/CHzOi; + b'O% + c'HzOmNn —>
—> CHaOfjNa 4" е^СОгД/'CHyOw (5.57)
(Аналогичным путем можно выразить стехиометрию синтеза
азотсодержащих продуктов метаболизма.)
Нетрудно получить и выражение для соответствующего эко-
номического коэффициента, если учесть, что в этом параметре
Стехиометрия и энергетика метаболических превращений
375
количества веществ должны быть выражены в молях. Так, если
субстратом является глюкоза, молекула которой содержит
шесть атомов углерода, то в реакции (5.57) будет участвовать
а'/§ молекул глюкозы. Аналогично если в молекуле продукта
метаболизма содержится пр атомов углерода, то в результате
реакции (5.57) образуется Y/пр его молекул. Следовательно, мо-
лярный коэффициент Ур/s (число молей продуктов метаболиз-
ма, образующегося из 1 моль утилизированного субстрата) со-
ставляет
• _ V
P/s “ а'
ns
Пр
(5.58)
Здесь ns — число атомов углерода в молекуле субстрата.
Простая стехиометрия уравнения (5.57) неприменима к ме-
таболитам второго типа, количество которых не обязательно
должно быть пропорциональным количеству утилизированного
субстрата или приросту клеточной массы. В этом случае сте-
хиометрию процесса можно выразить только с помощью от-
дельного уравнения реакции образования продукта метаболиз-
ма. Интересно проследить за стехиометрией реакций, уравне-
ния которых получены путем введения стадии образования
продукта метаболизма в приведенные выше уравнения реакций
роста клеток (5.53) — (5.56):
Образование продукта метаболизма:
dCHxOy + (коэффициент) -Н2О + (ys — zyp) NAD+ ->
---> dCH^O^H-d(l — z) СО2 +d-j-(ys —гур) (NADH + H+) (5.59a)
epd(ADP+P,) —> epd(ATP+H2O) (5.596)
Здесь коэффициентом z обозначена часть углерода субстрата,
используемая для образования продукта метаболизма и най-
денная экспериментально в последнем. Генерируемое в процес-
се биосинтеза продукта метаболизма число молекул АТР
обозначено коэффициентом ер; этот коэффициент может прини-
мать и отрицательные значения, если в процессе образования
продукта метаболизма АТР не синтезируется, а, напротив, рас-
ходуется. Аналогично если коэффициент (у5—гур) имеет отри-
цательное значение, то это означает, что в процессе образова-
ния продукта метаболизма используются восстанавливающие
эквиваленты NADH (или NADPH; на принятом здесь довольно
невысоком уровне детализации механизмов путей метаболизма
нет смысла различать эти два агента).
376
Глава 5
Способ описания стехиометрии образования метаболитов 3
и 4 зависит от природы субстратов и продуктов метаболизма.
В этих случаях образование последних обычно никак не свя-
зано с клеточным ростом. Накопление вторичных метаболитов
определяется кинетическими факторами и активностью клеток.
Изучение стехиометрии образования продуктов метаболизма
позволяет сделать ряд важных с точки зрения инженера-техно-
лога выводов. Например, с помощью стехиометрических расче-
тов можно оценить максимально возможный выход продукта
метаболизма, а эти данные в свою очередь могут оказаться
очень полезными в предварительном экономическом анализе
технологического процесса (см. гл. 12). В качестве первого
примера рассмотрим анаэробный процесс превращения глюко-
зы в этанол. (Приведенные выше реакции аэробного метабо-
лизма могут быть применены и для изучения анаэробных про-
цессов, если во всех реакциях опустить О2 и стадии окисли-
тельного фосфорилирования.) Очевидно, что оптимальным ва-
риантом является использование всего субстрата только для
образования продукта метаболизма. Если принять, что рост
клеток пренебрежимо мал, то уравнение (5.57) в нашем случае
принимает очень простую форму:
СНгО —>• УзСОг + г/зСНзОо.Б
(5.60)
Отсюда следует, что в идеальном варианте продукт метаболиз-
ма будет содержать две трети углерода утилизированного
субстрата. В соответствии с уравнением (5.58) высший предел
молярного коэффициента составит: (Ур/5)тах = (2/з)(6/2) = 2.
Такой же подход может быть использован и в случае обра-
зования метаболитов всех других типов, в том числе и при од-
новременном биосинтезе нескольких продуктов метаболизма.
Предельное значение выхода данного продукта метаболизма
можно определить, приняв, что одновременно с ним не образу-
ются новые клетки или другие метаболиты. Например, по урав-
нению реакции биосинтеза пенициллина из соединений-пред-
шественников:
1,5 глюкоза + Н25О4+2МНз+фенилацетат —►
—► пенициллин G+CO2+8H2O
(5.61)
легко определить максимально возможный выход пенициллина,
составляющий 1,2 г на 1г глюкозы. В реальных промышлен-
ных процессах выходы намного ниже (около 0,1 г/г), что сви-
детельствует о возможности дальнейшего совершенствования и
технологического процесса, и микроорганизма-продуцента. Ма-
лая величина реального выхода продукта метаболизма по срав-
Стехиометрия и энергетика метаболических превращений 377
нению с максимальной расчетной величиной указывает на то,
что значительное количество субстрата расходуется на рост
клеток, на их поддержание или на синтез других веществ.
Изучение зависимости между энергетическим обменом и
биосинтезом продуктов метаболизма в том виде, в каком эти
процессы выражаются уравнениями (5.53) — (5.56) и (5.59),
может дать полезные сведения о влиянии затрат на поддержа-
ние клеток на выходы продуктов метаболизма. Предположим,
что в анаэробных условиях в среде, содержащей в качестве
источника азота аммиак, из глюкозы образуется этанол. Един-
ственным источником АТР здесь является субстратное фосфо-
рилирование [уравнение (5.536)]. В такой ситуации относи-
тельно высокие затраты субстрата на поддержание клеток
приведут к тому, что сравнительно большие количества глюко-
зы будут расходоваться в реакциях диссимиляции. Это в свою
очередь приведет к регенерации большего количества NADH,
не использующегося в сколько-нибудь значительных количест-
вах в реакции биосинтеза [уравнение (5.55а)], поскольку сте-
пени восстановленности субстрата и биомассы примерно оди-
наковы. Восстанавливающие эквиваленты будут использоваться
в образовании продукта метаболизма по уравнению реакции
(5.59а), что приведет к его накоплению в среде, поскольку
степень восстановленности этанола выше степени восстановлен-
ности субстрата (глюкозы). Напротив, в аэробных условиях,
когда целью процесса часто является синтез метаболитов треть-
его или четвертого типов, повышение расхода субстрата на
поддержание клеток приводит к снижению выхода продукта
метаболизма за счет интенсификации утилизации субстрата при
выработке энергии.
Очевидно, метаболические превращения можно описывать
не только уравнениями (5.53) — (5.56) и (5.59). Эти уравнения
должны быть модифицированы, например, в тех случаях, когда
одновременно утилизируется несколько источников углерода
и (или) азота или когда продукт метаболизма содержит азот.
Стехиометрические уравнения могут описывать и превращения
по какому-либо определенному пути метаболизма. Действи-
тельно, сочетание стехиометрического анализа и эксперимен-
тального определения количеств использованного субстрата и
образовавшегося метаболита успешно применялось для оценки
потоков веществ по параллельным и разветвляющимся путям
метаболизма. Более детально альтернативные стехиометричес-
кие уравнения рассмотрены в упражнениях, поскольку принци-
пы и методы их построения очень близки принципам, изло-
женным в этом разделе. Обсуждение стехиометрии мы продол-
жим в следующем разделе, посвященном проблеме выделения
тепловой энергии в метаболических процессах.
378
Глава 5
5.10.4. Стехиометрия энергетического обмена;
оценка количества выделяющейся теплоты
и соответствующих экономических коэффициентов
В клетках химическая энергия используется очень эффек-
тивно, но, как и в любом другом реальном процессе, часть
энергии субстрата выделяется в виде теплоты, что, в
частности, обусловливает необходимость охлаждения биологи-
ческих реакторов с культурами клеток. Клетки генерируют теп-
лоту в первую очередь за счет процессов связанного с клеточ-
ным ростом метаболизма и энергообмена. Следовательно, мож-
О2 + , СО, , Н2о
Дп^, ккал/г окисленного
субстрата
ДНС, ккал/г
полностью
окисленных
клеток
Ш Сгорание
клеточной
/чассь/
И Синтез кле-
точного беще-
стба б процессе
дыхания
,Н2О + СО2 + клетки сшкроорганизноб-
РИС. 5.30. Ориентировочный тепловой баланс утилизации субстрата.
но предполагать наличие примерно пропорциональной зависи-
мости между количеством выделившейся теплоты и количест-
вом утилизированного высокоэнергетического субстрата.
В этой связи был введен особый параметр — коэффициент
Уд (отношение количества образовавшейся клеточной массы в
граммах к количеству выделившейся теплоты в килокалориях),
аналогичный рассмотренным выше другим экономическим ко-
эффициентам. Если количество клеточной массы (в граммах),
продуцируемой из одного грамма утилизированного субстрата,
мы обозначим символом У5, а теплоты сгорания субстрата и
клеточной массы (в килокалориях на грамм) — символами
\HS и ЛНС соответственно, то взаимосвязь между этими вели-
чинами будет выражаться следующим уравнением:
- YS6HO
Это уравнение вытекает из приближенного энергетического ба-
ланса между двумя путями полного окисления субстрата в
случае аэробного роста, схематично изображенного па рис. 5.30.
Стехиометрия и энергетика метаболических превращений
379
Если окислителем в основном является кислород, то разность
между количеством теплоты, выделяющейся при полном окис-
лении одного грамма субстрата (A/7S), и теплотой сгорания
клеток (и сухого остатка внеклеточной жидкости), выросших
на таком же количестве субстрата (У5АЯС), будет приблизи-
тельно равна количеству теплоты, выделяющейся в процессе
роста клеток (а также выделения Н2О и СО2) на одном грамме
утилизированного субстрата.
Оценить точность допущений, положенных в основу уравне-
ния (5.62) и схемы на рис. 5.30, можно на примере данных, от-
носящихся к росту дрожжей на неразветвленных парафиновых
углеводородах. Определенные экспериментальным путем вели-
чины А/Д и АЯс составляют 11,4 и 4,7 ккал/г соответственно;
отсюда по уравнению (5.62) находим, что
— = В * * 11-400- —4700
Ул ys
Непосредственное измерение У5 и Уд дало следующие резуль-
таты f 16]:
Ys, найдено Ч Вычислено по уравнению Найдено (5.62)
1,25 1,03 1,09 4400 4640 6400 6060 5800 5830
В отсутствие экспериментальных данных теплоту сгорания
клеток (как и любых других материалов) можно определить на
основе следующего эмпирического факта. При переносе элект-
ронов от соединения со степенью восстановленности к соеди-
нениям е нулевой степенью восстановленности (например, СО2
или Н2О) выделяется количество энергии, равное К-^s, где К —
коэффициент, величина которого изменяется в пределах от 26
до 31 ккал на один химический эквивалент окисляемого веще-
ства. Поскольку молекулярный кислород О2 в процессе дыха-
ния принимает четыре электрона, то поглощение моля кислоро-
да должно сопровождаться выделением 104—124 ккал теплоты.
На рис. 5.31 приведены экспериментальные данные для двух
бактерий, дрожжей и плесени, выросших на различных средах.
Разброс результатов довольно велик, но зависимость скорости
тепловыделения от скорости поглощения кислорода просмат-
ривается достаточно отчетливо.
В качестве примера оценим теплоту сгорания клеток Pseu-
domonas fluorescens, растущих на глюкозной среде. Для этого
380
Глава 5
РИС. 5.31. Экспериментально найденные скорости тепловыделения и погло-
щения кислорода в процессе роста ряда микроорганизмов в различных сре-
дах (Glu — глюкозная среда, Mol — среда на основе мелассы, Sbm — среда
на основе соевых бобов). Наклон прямой регрессионной зависимости равен
124±3 ккал/моль О2. [Воспроизведено с разрешения из работы: Cooney С. L.,
Wang D. I. С., Mateles R. I., Measurement of Heat Evolution and Correlation
with Oxygen Consumption during Microbial Growth, Biotech. Bioeng., 11,
269 (1968)].
прежде всего запишем уравнение реакции сгорания клеток в
расчете на экспериментально найденную «брутто-формулу»,
приняв, что продуктами сгорания являются СО2, Н2О и N2:
СН^деЫо.гоОо,27+1,280г —* СОг+ОДОМг+О^ЗНгО (5.63)
клетки
Далее определим теплоту, выделяющуюся при сгорании бакте-
рий по уравнению (5.63), приняв теплоту сгорания в расчете
на 1 моль О2 равной 104 ккал:
(1,28 моль О2)(Ю4 ккал/моль О2) _ fi 41 ..ял/г
[12+(1,66)(1) + (0,20)(14)+(0,27)(16)] г ’ 7 '
При этом1 мы не учли, однако, что сухое вещество клеток со-
держит и золу. Если в сухом клеточном веществе содержится
10% золы, то теплота сгорания клеток будет равна
М »(0,90) (6,41 ккал/г) = 5,8 г сух()г0 клеХго „ешееты <5'6S>
Значение Уд зависит от природы как микроорганизма (влия-
Таблица 5.12. Экономические коэффициенты для бактерий, растущих на средах с различными источниками
Субстрат Ys> г клеток/г субстрата Уо2. г клеток/г утилизированного О; Уд, г клеток/ккал
Малат 0,34 1,02 0,30
Ацетат 0,36 0,70 0,21
Эквиваленты глюкозы крахмал, целлюлоза) (меласса, 0,51 1,47 0,42
Метанол 0,40 0,44 0,12
Этанол 0,68 0,61 0,18
Изопропанол 0,43 0,23 0,074
н-Парафины 1,03 0,50 0,16
Метан 0,62 0,20 0,061
а Из работы. Abbott В. J., Clamen A., The Relationship of Substrate, Growth Rate, and Maintenance Coefficient to Single Cell Protein
Production, Biotech. Bioeng., 15, 117 (1973).
382
Глава 5
ющего на АЯС), так и утилизируемого субстрата (влияющего
на А7Л). В табл. 5.12 приведен ряд вычисленных значений ко-
эффициентов Ys в зависимости от природы субстрата. Обрати-
те внимание на то, что в общем случае углеводороды обеспе-
чивают большее тепловыделение, чем частично окисленные со-
единения [Уд(СН4)<Уд(СНзОН); УА(«-алканы)<УА(глюкоза)].
Следовательно, более восстановленные субстраты в большей сте-
пени требуют отвода тепла от биореактора. Влияние энергетиче-
ской стехиометрии на экономику процессов рассмотрено в гл. 12.
В процессах брожения или очистки сточных вод субстрата-
ми часто являются смеси множества веществ с различными
теплотами сгорания. Сервици и Боуген [18] отметили, что сте-
хиометрическое отношение т0 (число молей О2, необходимого
для полного окисления 1 моль субстрата) для ряда соедине-
ний пропорционально стандартной свободной энергии сгорания
AG0. Например, для углеводов, промежуточных соединений
цикла ТКК и некоторых продуктов гликолиза
AG° =— 116m0 ккал/моль субстрата (5.66)
а для ароматических соединений, спиртов и алифатических
кислот
AG°= — 104mo ккал/моль субстрата (5.67)
[Хаттори (ссылка [4] в гл. 13) отмечал, что если известна ве-
личина химической потребности в кислороде (ХПК), выражен-
ная в граммах кислорода на моль, то вместо то в этих урав-
нениях можно применять ХПК/32.] Усредненную свободную
энергию полного окисления смеси субстратов можно определить
суммированием свободных энергий окисления всех индивиду-
альных субстратов:
AG° = ккал/моль (5.68)
где i—субстраты; Мг-— число молей i-ro субстрата; AG/ —
свободная энергия полного окисления i-ro субстрата.
Оказалось также, что экономический коэффициент У про-
цесса, в котором участвуют несколько видов одноклеточных
организмов, можно определить так же, как и в случае описан-
ных выше процессов с участием микроорганизмов одного вида
(бактерий или дрожжей). Сервици и ^Боуген [18] определили
средние экономические коэффициенты Ys (выраженные в грам-
мах клеток на моль субстрата) для активного ила (см.
гл. 14), содержащего множество микроорганизмов и большое
число субстратов, и нашли (рис. 5.32), что
ys = 0,108AG° г клеток/моль субстрата (5.69)
Стехиометрия и энергетика метаболических превращений 385
При данном У5 все другие, величины, входящие в уравнение
(5.62), легко определить экспериментально; следовательно, ив
этом случае можно вычислить Уд.
Следует ожидать, что величина коэффициента в уравнении
(5.69) зависит от вида микроорганизма, поэтому опять-таки
желательно экспериментальное определение этого коэффициен-
та по отношению к участвующему в процессе организму (или
организмам). В то же время, предполагая существование про-
РИС. 5.32. Зависимость среднего экономического коэффициента роста актив-
ного ила от средней свободной энергии окисления субстратов. [Воспроизве-
дено из работы: Servizi J. A., Bogan R. Н., Thermodynamic Aspects of Biologi-
cal Oxydation and Synthesis, J. Water Pollut. Control Fed., 36, 607 (1961).]
порциональной зависимости между Ys и AG°, Сервици и Боуген
исходили из линейной^зависимости между средним экономичес-
ким коэффициентом У5 и числом Л%Тр молекул АТР, синтези-
руемых в ходе утилизации 1 моль субстрата. Поскольку У5
есть не что иное, как количество клеток (в граммах), синтези-
руемых из 1 г субстрата, то коэффициент пропорциональности
между Ys и Адтр представляет собой массу клеток (в граммах),
синтезируемых в процессе утилизации 1 моль АТР. Известно,
что последняя величина приблизительно постоянна для боль-
шого числа анаэробов, но изменяется в гораздо более широких
пределах в случае аэробных организмов (разд. 5.9.2). В от-
сутствие более надежных данных уравнение (5.69) в общем
случае может служить основой для оценки систем, состоящих
из нескольких субстратов и микроорганизмов. Точность такой
оценки, по-видимому, не превышает ±20—30%, о чем наглядно-
384
Глава 5
свидетельствуют корреляционные данные, приведенные на
рис. 5.32.
Упоминавшаяся выше взаимосвязь между числом участвую-
щих в процессе электронов и внутренней энергией вещества
может быть использована для оценки максимально возможного
выхода органического вещества из органического субстрата.
Долю электронов, переходящих в процессе реакции от субст-
рата к продуктам метаболизма, t,p можно определить по урав-
нению
(число участвующих в процессе электронов в 1 г продукта метаболизма)
(число участвующих в процессе электронов в 1 г субстрата)
X
'число граммов продукта метаболизма
число граммов субстрата
(5.70)
Последний член правой части уравнения (5.70) представляет
собой экономический коэффициент YP/S. Число участвующих в
процессе электронов в 1 г органического вещества можно выра-
зить как оу/12, где о—массовая доля углерода в соединении
и 7 — степень восстановленности этого углерода. Подставив
это выражение в уравнение (5.70), получим
(5-71)
Поскольку энергии субстрата и продукта метаболизма при-
близительно пропорциональны содержанию в них участвую-
щих в реакции электронов, то параметр можно интерпрети-
ровать и как энергетический коэффициент выхода продукта
метаболизма из субстрата. В этом случае, очевидно, коэффи-
циент Zp не должен превышать единицы. Следовательно, верх-
ний предел выхода продукта метаболизма Yp/smax можно оп-
ределить по уравнению (5.71) при £₽=1:
<5-72)
Значения Yp/smax для нескольких субстратов и продуктов мета-
болизма приведены в табл. 5.13.
5.10.5. Стехиометрия фотосинтеза
Оценка выходов продуктов реакции фотосинтетического
восстановления СОг до углеводов важна как в процессах произ-
водства продуктов питания, так и для определения скоростей
воспроизводства исходного сырья при получении биомассы.
Известные сведения о последовательности реакций фотосинтеза
и сопутствующих потоках электронов позволяют сделать вы-
Стехиометрия и энергетика метаболических превращений
385
вод, что теоретически возможная эффективность фотосинтеза
(выраженное в процентах отношение свободной энергии глюко-
зы к свободной энергии поглощенного излучения при 700 нм)
составляет 35%. Конечно, эта величина представляет собой
предельное значение, только теоретически достижимое в иде-
альных условиях. Здесь не учтены такие факторы, как утили-
зация в фотосинтезе только части солнечной радиации (соот-
ветствующий понижающий коэффициент равен приблизительно
0,43), отражение света и затенение листьев (понижающий ко-
эффициент 0,80 или меньше), темновое дыхание (соответству-
Таблица 5.13. Максимальные значения выходов продуктов метаболизма
по массе ymaxp/s (выраженные в граммах продукта метаболизма на 1 г
субстрата), вычисленные по уравнению (5.72)а
Продукт метаболизма Субстрат
н-Алканы Этанол Глюкоза
Лимонная кислота 4,55 2,77 1,42
Полисахариды 3,21 1,96 1,00
Уксусная кислота 3,21 1,96 1,00
Лизин 2,23 1,36 0,70
Стрептомицин — — 0,66
Этанол 1,64 — 0,51
Триглицериды 1,18 0,72 0,37
а Воспроизведено с разрешения из работы: Eroshin V. К.. Minkevich !. G., On the
Upper Limit of Mass Yields of an Organic Product from an Organic Substrate, Biotech.
Bioeng., 24, 2263 (1982).
ющий коэффициент может изменяться в широких пределах;
более или менее типичная величина составляет 0,67). Умножив
все эти коэффициенты на теоретическую эффективность фото-
синтеза, можно определить реальную (высшую) эффективность,
составляющую около 8%.
Соответствующее количество биомассы (в виде крахмала),
продуцируемое на единице площади (м2) в единицу времени
(например, за 1 сут), можно оценить следующим образом:
(продуцируемая биомасса (г крахмала) А
---------------м2".Сут------------[(поток солнечной радиации,
ккал/(м2-сут)] (эффективность фотосинтеза Х0,01)Х
/1 моль глюкозыА (180—18) г крахмала , г -„ч
Х| 686 ккал Г 1 моль глюкозы
Интенсивность солнечной радиации зависит от времени года и
географической зоны. Например, средняя интенсивность солнеч-
ной радиации в США составляет 3930 ккал/(м2-сут), а на
юго-западе США в летний период достигает 6775 ккал/(м2Х
25—537
386
Глава 5
Хсут). Последний сомножитель уравнения (5.73) учитывает
потерю одной молекулы воды каждой молекулой глюкозы,
включаемой в полимерную цепь крахмала.
Если имеются экспериментальные данные, характеризую-
щие продуктивность процесса образования биомассы и интен-
сивность солнечной радиации, то по уравнению (5.73) можно
рассчитать действительную эффективность фотосинтеза в рас-
тениях и водорослях (табл. 5.14). Найденные таким путем
Таблица 5.14. Экспериментально найденные среднегодовые и максимальные
значения эффективности и продуктивности фотосинтеза в некоторых
растениях3
Среднегодовые Найденные максимальные
Растение Выход, г/(м2-сут) Эффектив- ность, % Выход, г/(м2-сут) Эффектив- ность, %
Сз-Растения 23
Люцерна 8 0,7 1,4
Сахарная свекла 9 0,8 31 1,9
Хлорелла СгРастения 28 1,7
Сахарный тростник 31 2,8 38 2,4
Сорго 10 0,9 51 3,2
Кукуруза 52 3,2
а Воспроизведено с разрешения из работы: Bassham J. A., Increasing Crop Pro-
duction Through More-Controlled Photosynthesis; Science, 197, 630 (1977).
величины лежат в пределах от 0,8 до 3,2%, т. е. намного ниже
теоретической оценки. Обратите внимание также на то, что
эффективность в группе фотосинтезирующих так называемых
С4-растений значительно выше, чем в группе Сз-растений. Ни-
же мы вкратце рассмотрим биохимическую основу этого раз-
личия.
Явление фотодыхания (светового дыхания), т. е. поглоще-
ние кислорода и одновременное выделение СОг на свету, пред-
ставляет собой важный процесс, влияющий на общую эффек-
тивность роста фотосинтезирующих организмов. В этом про-
цессе происходящие в растении реакции фиксации углерода
приводят к промежуточному соединению — рибулозо-1,5-дифос-
фату (см. разд. 5.6.1), который далее распадается следующим
образом:
рибулозодифосфат-карбоксилаза
Рибулозо-1,5-дифосфат+Ог —------------------------
—>- фосфогликолят+3-фосфоглицерат+2Н+ (5.74)
Затем гидролизуется фосфогликолят, образуя гликолят, кото-
Стехиометрия и энергетика метаболических превращений 387
рый в свою очередь, в органоидах растительной клетки превра-
щается в глицин и другие вещества,
В этом окислительном процессе не образуется АТР и не
высвобождаются электроны; следовательно, здесь расходуется
энергия фотосинтеза. В то же время реакция окисления (5.74)
ингибируется высокими концентрациями СОг. В клетках С4-
растений, в которых цикл Калвина (рис. 5.15) любопытным
образом видоизменен, поддерживается относительно высокая
концентрация СОг. В непосредственно контактирующих с воз-
духом клетках мезофилла С4-растений осуществляется ряд
химических превращений, которые суммарно можно выразить
следующим уравнением:
Клетки мезофилла:
Пируват + СО2+АТР + 2Н2О —> малат +АТР + 2Р; (5.75)
Образующийся таким образом малат диффундирует в соседние
клетки обкладки проводящего пучка, в которых реализуются
реакции цикла Калвина. Здесь включенный в малат СО2 вы-
деляется в соответствии с реакцией
Малат —> пируват + СОг (5.76)
В конечном счете в этом метаболическом цикле для под-
держания обеспечивающей минимум фотодыхания высокой кон-
центрации СОг вблизи от рибулозодифосфат-карбоксилазы рас-
щепляются две высокоэнергетические фосфатные связи. Отсюда
следует, что фотосинтез 1 моль глюкозы дополнительно требу-
ет 12 моль АТР. Таким образом в С4-растениях энергия, оче-
видно, расходуется на два конкурирующих процесса: цикл ре-
акций транспорта СОг и усиленное фотодыхание (в отсутствие
цикла транспорта СОг). Интересно отметить, что энергия акти-
вации реакции фотодыхания (5.74) превышает энергию актива-
ции реакции карбоксилирования (5.37), которая также ката-
лизируется рибулозодифосфат-карбоксилазой. Возможно, имен-
но этот факт может объяснить, почему С4-растения шире рас-
пространены в тропиках, а Сз-растения преобладают в умерен-
ной зоне.
5.11 Заключение
Теперь мы знаем, что в живой клетке происходит множест-
во чрезвычайно сложных химических превращений. Несмотря
на всю сложность химии клеточных процессов, в них очень час-
то можно обнаружить некоторые ключевые соединения и реак-
ции, общие для всех путей клеточного метаболизма. Вероятно,
самыми важными процессами такого типа являются транспорт
энергии при посредстве АТР (рис. 5.33) и транспорт восста-
25*
388
Глава 5
навливающих эквивалентов в виде NAD. Мы узнали также,
что стехиометрические соотношения на разных уровнях могут
использоваться для выяснения взаимосвязей между различны-
ми химическими превращениями, являющимися частью кле-
точного метаболизма.
В следующей главе мы закончим наше краткое изучение
биологии и биохимии живой клетки, а затем приступим к труд-
. Микроорганизм»/ ------------
гетеротрофы х >//r-
Ъ atbUlJrxOfe лиС'
_______________2____лото/, ими-
'\нохислоты
\CSem, Н20,
^Растения артотроры qq
Животные 2 3
гетеротрсиры\ |
I Кища
С
Полисахариды,
жиро/, белки
Ферменты
Клетка
Сахара,
жирные
аминокислоты
РИС. 5.33. Схема потоков энергии в клетке и ее окружении. {Леви А., Сикв‘
виц Ф., Структура и функция клетки. —М.: Мир, 1971.)
ной проблеме анализа, изучения и проектирования технологиче-
ских процессов, в которых используются биологические реак-
ции.
Упражнения
5.1. Равновесие. Стандартное изменение свободной энергии в реакции
фосфоенолпируват ADP—>-пируват + АТР равно —7,50 ккал. Найдите
концентрации всех веществ, если исходная смесь содержала 6,0 мМ ADP,
6,0 мМ фосфоенолпирувата, 6,0 мМ АТР и не содержала пирувата.
5.2. Функции химических элементов. Назовите важнейшую биологическую
функцию (или функции) в клетке и доминирующую при выполнении каждой
из функций химическую форму для каждого из перечисленных в табл. 5У2.1
элементов и соединений. Действительно ли каждый из содержащихся в золе
элементов имеет определенную биохимическую функцию? [См. Frey С. N-*
Ind. Eng. Chem., 22, 1154 (1930).]
Стехиометрия и энергетика метаболических превращений 389
Таблица 5У2.1. Ориентировочный состав золы, образующейся при полном
окислении дрожжей
Компонент Ориентировочное содержание, % Компонент Ориентировочное содержание, %
Пентоксид фосфора 50 Оксид кремния 1
Оксид калия 35 Оксид натрия 1
Оксид магния 5 Триоксид серы 0,5
Оксид кальция 1 Хлор, железо Следы
5.3. Стандартное изменение свободной энергии. Изменение свободной
энергии реакции ДО связано с электродными потенциалами Ei и Е2 инди-
видуальных полуреакций следующим уравнением:
AG = — nF&E=—п$~{Е{ —Е2)
где п — число электронов, мигрирующих при превращении одной молекулы,
5Г— константа Фарадея [t7"«23 ккал/(В-моль)], a Ei (i=I, 2) определяет-
ся по уравнению Нернста:
RT . si, red
-~г In-7-------
Sj>ox
Ei = Eio
где — концентрация реагента в окисленной или восстановленной форме
и Ею — электродный потенциал при Si, Геа/$г, ох = 1,0 по отношению к водо-
родному полуэлементу:
Si,red Si,ох + пе~ E0^=Q
Н2 —► 2Н++2е~ Ео=О по определению
Электродные потенциалы некоторых необходимых реакций приведены
в табл. 5У3.1.
Таблица 5У3.1а
Реакция иа электроде
Ацетат + СО2+2Н+ —> пируват+2Н2О
Ацетоацетат+2Н+ —>- р-гидроксибутират
Пируват+2Н+ —>- лактат
S + 2H+-^-H2S
V2O2+2H+—>Н2О
NAD(P)4- + 2H4-—NAD(P)H+H+
FAD + 2H+—>-FADH2 (свободный кофермент)
Хлорофилл4- —>- хлорофилл* + е~
(фотосинтез в эукариотах, нециклическая часть)
Хлорофилл4- —>- хлорофилл
(фотосинтез в эукариотах, нециклическая часть)
МОз_ + 2Н+ —>-NO2_ + H2O
SO42~+2H+ —> SO32-+H2O
Ацетат+2Н+ —> ацетальдегид + Н2О
Ацетальдегид+2Н+ —>- этанол
Е0, В
2 —0,70
2 —0,27
2 —0,19
2 —0,23
2 0,82
2 —0,32
2 —0,18
1 —0,2
1 0,9
2 0,42
2 0,48
2 —0,60
2 —0,20
а Данные нз работы: Wood W. В., Wilson J. Н„ Benbow R. М., Hood L. Е„ Bio-
chemistry: A Problems Approach, рр. 190—191» W. A. Benjamin, Inc., Palo Alto, CA, 1974.
390
Глава 5
Воспользовавшись приведенными в таблице стандартными электродны-
ми потенциалами, выясните, возможны ли следующие реакции в стандарт-
ных условиях:
а) перенос электрона от возбужденного хлорофилла,
б) окисление ^-гидроксибутирата кислородом,
в) окисление ацетальдегида ацетальдегидом (дисмутация или радикаль-
ное диспропорционирование).
5.4. Изменение свободной энергии в клетке, а) На базе приведенных
в табл. 5.3 данных вычислите изменение свободной энергии для каждой
стадии гликолиза в эритроцитах человека.
б) По полученным вами данным и величине свободной энергии гидро-
лиза АТР до ADP в тех же условиях докажите необходимость АТР для
инициирования отдельных стадий, протекающих с его участием.
в) По данным предыдущих расчетов, определите общее изменение сво-
бодной энергии AG' в процессе гликолиза в клетке.
5.5. Кинетика взаимосвязанных реакций. В клетке одновременно осуще-
ствляется множество реакций, однако часто кажущаяся кинетика последо-
вательности реакций определяется очень небольшим числом стадий.
а) Покажите, что в стационарном состоянии математическое выраже-
ние, определяющее скорость образования Р в последовательности реакций,
формально отвечающей схеме Михаэлиса — Ментен
E + S =Ft ES —>- Е+Р
не отличается от соответствующего выражения для последовательности
равновесных реакций:
S + E] S2+E1
S2+E2 ч* S3+E2
S3 + E3 E3S3 —> Ез+Р
б) Приведенные на рис. 5.4 десять последовательных реакций можно
выразить только тремя кинетическими уравнениями, описывающими
М &2 &з
А -----> В -----> С -----> D (лактат)
Назовите вещества А, В и С.
Таблица 5У5.1
Субстрат
ДО°', ккал/моль Субстрат
ДО0', ккал/моль
Фосфоенолпируват —12,8 Глюкозо-1-фосфат —5,0
1,3-Дифосфоглицерат —11,8 Фруктозо-6-фосфат —3,8
Фосфокреатинин —10,5 Глюкозо-6-фосфат —3,3
Ацетилфосфат —10,1 З-Фосфоглицерат —3,1
АТР (концевая —7,3 Г лицеро-1 -фосфат —2,3
связь) —
в) В табл. 5У5.1 приведены стандартные свободные энергии гидроли-
за ряда фосфатов. Как изменится вычисленная вами (в упражнении 5.4)
разность свободных энергий для трех указанных в пункте б стадий, если
в клетке отношение ATP/ADP уменьшится в 10 раз (при постоянной сум-
марной концентрации ATP-j-ADP)? Какой полезный (или вредный) для клет-
ки эффект будет результатом этого изменения?
________Стехиометрия и энергетика метаболических превращений 391
5.6. Регенерация АТР. Благодаря сопряжению с гидролизом АТР до
ADP или АМР в клетке осуществляется множество реакций с неблагоприят-
ными константами равновесия. Применение для аналогичного синтеза сво-
бодных ферментов потребовало бы в конечном счете переноса фосфатной
группы (для регенерации АТР) от системы с более высокой свободной
энергией гидролиза.
а) Какова стоимость 1 фунта каждого из соединений, перечисленных
в табл. 5У5.1, свободная энергия гидролиза которых больше AG' АТР?
(Данные можно взять из любого каталога биохимических реактивов.)
б) Какой должна быть концентрация фосфорилированного соединения
(глюкозо-1-фосфата, фруктозо-6-фосфата, глицеро-1-фосфата), чтобы из
ADP образовывался АТР в концентрациях, указанных в табл. 5.3?
5.7. Регенерация NAD в ферментативных трансформациях стероидов. Ме-
тод трансформации стероидов в присутствии 20{}-гидроксистероид-дегидроге-
иазы (20JJ-HSDH) [Cremonesi Р. et al., Enzymatic preparation of 20p-Hydro-
xysteroids in a two phase system, Biotech. Bioeng., 17, 1101 (1975)] состоит из
трех стадий:
Гидрирование стероида:
206-HSDH
KopTH3OH4-,NADH + H+ < NAD+ +дигидрокортизон (1)
Окисление этанола:
алкогольдегидрогеназа
c2h5oh+nad+ ; ............ CH3CH0+NADH+H+ (2)
Связывание ацетальдегида:
CH3CHO+NH2NHCONH2 —> CH3CHNNHCONH2 (3)
Константа равновесия протекающих совместно реакций (1) и (2) близка
единице.
а) Оцените термодинамическую вероятность превращения кортизона,
если принять, что константа равновесия реакций (1) и (2) равна единице
и что константа равновесия реакции (3) (которая в состоянии равновесия
также должна быть обратимой) составляет 10, 103, 105, 1010 (в единицах
обратных концентраций). Выразите результаты в виде графической зависи-
мости степени превращения кортизона от 1g К.
б) Поскольку стероиды относительно малорастворимы в воде (в 1 л
насыщенного водного раствора содержится от 10~4 до 10-5 моль стерои-
да), для увеличения общего количества кортизона в системе добавляют
органические растворители, образующие двухфазную систему. Таким обра-
зом, органическая фаза, содержащая относительно высокие концентрации
стероида (в 100 мл бутилацетата растворяется 0,160 г кортизона), выпол-
няет функцию резервного источника, постоянно подпитывающего водную
фазу. Оцените максимально возможную степень превращения кортизона,
приняв, что объемная доля бутилацетата в водно-бутанольной эмульсии
равна е, растворимость кортизона в воде равна 10-4 моль/л, дигидрокорти-
зон в воде растворим в той же мере, что и кортизон, а в бутилацетате
нерастворим и что А реакции (3) составляет 10, 103 или 105. Выполните те
же расчеты, приняв, что растворимость дигидрокортизона в 100 мл бутил-
ацетата равна 0,160 г.
в) Перечислите термодинамические механизмы, использующиеся в этом
процессе для осуществления реакции (1).
5.8. Проницаемость. Выведите уравнение (5.40). Обоснуйте все до-
пущения.
5.9. Материальный баланс веществ, участвующих в сети взаимосвязан-
ных реакций. Предположим, что у вас есть рассеянный друг, который.
392
Глава 5
к несчастью, не слишком хороший экспериментатор. На лабораторных за-
нятиях он выполнял задачу по брожению, однако забыл взвесить добавлен-
ную глюкозу и определить содержание спирта в образовавшейся смеси.
Он установил, что в результате брожения образовались перечисленные
в табл. 5У9.1 вещества. Ему известно, что вся глюкоза (СбЕЦгОе) окис-
ляется по пути ЭМП до пировиноградной кислоты (СНзСОСООН) и что
никакие другие соединения, кроме этанола, ие должны образовываться. Он
просит вас помочь ему как-нибудь разобраться в этой путанице. Что вы
должны ответить ему на вопрос о том, сколько молей этанола образова-
лось? (Обоснуйте ваш ответ,)
Таблица 5У9.1
Вещество Количество, моль Вещество Количество, моль
Молочная кислота СНзСНОНСООН 10 Диоксид углерода СО2 15
Уксусная кислота 5 Водород Н2 10
СНзСООН
5.10. Определение понятия «жизнь». Выскажите свои соображения о не-
обходимости (дли ее отсутствии) каждого слова в следующем определении
жизни [Perrett J., New Biol., 12, 68 (1952)]: «Жизнь —это потенциально са-
мовозмущающаяся открытая система связанных органических реакций, ката-
лизируемых постадийно и практически изотермично сложными и специфиче-
скими органическими катализаторами, которые продуцируются этой же систе-
мой». Более поздние определения понятия «жизнь» можно найти, например,
в книге: Bernal J. D., Theoretical and Mathematical Biology, p. 96, Blaisdell
Publ. Co., N.Y., 1965.
5.11. Свободная энергия и перенос электронов. Внутриклеточные соедине-
ния-предшественники обычно представляют собой ионы; с другой стороны,
перенос свободной энергии в гликолизе, цикле ТКК, митохондриальных и
других метаболических процессах тесно связан с миграцией электронов. От-
сюда следует, что энергетика внутриклеточных процессов может оцениваться
в каких-либо единицах, отражающих явления переноса электронов. Мак-Кар-
ти в статье Energetics of Organic Matter Degradation [Water Pollution Micro-
biology, Mitchell R. (ed.), p. 91, Wiley-Interscience, New York, 1972] предложил
использовать особый параметр — коэффициент У« (количество клеток в грам-
мах, отвечающее 1 химическому эквиваленту образующихся клеток), опреде-
ляемый как
где С — количество клеток в граммах, отвечающее 1 химическому эквива-
ленту в полуреакции их окисления; h — число электронов, действительно
переносимых от молекулы-донора, деленное на число химических эквивален-
тов в моле (в соответствующей электродной полуреакции), обычно Л=1,0;
А — отношение количества химических эквивалентов донора электронов
(источника энергии) к числу химических эквивалентов образовавшихся
клеток.
Стехиометрия и энергетика метаболических превращений
393
Если клетке отвечает формула CsHzO2N и полуреакция окисления вы-
ражается уравнением
V 20C5H7O2N +8/20Н2О = V 5СО2+'/20НСО3- +
+1/20NH4++H+4-e-
то вычисленное значение С составляет 5,65.
а) Проверьте правильность вычисленного значения С.
б) Проверьте правильность приведенных в табл. 5У11.1 значений А
и Уе по другим данным той же таблицы.
Таблица 5У11.1. Определение коэффициентов Уе по энергетическим
параметрам окисления субстрата (/<=0,6; источник азота — аммиак)3
Донор электронов Акцептор электронов XGr6, ккал AGp, ккал А (вычислено) Ye (вычисле- но), г/экв
Ацетат О2 —25,28 1,94 0,71 7,96
NO3- (восстановле- ние до N2) —23,74 1,94 0,76 7,43
SO42- —1,52 1,94 11,8 0,48
СО2 —0,85 1,94 21,1 0,27
Глюкоза о2 —28,70 —1,48 0,38 14,90
СО2 —4,26 —1,48 2,58 2,19
Этанол о2 —26,27 0,95 0,58 9,76
СО2 —1,83 0,95 8,3 0,67
а McCarty Р. L., in Water Pollution Microbiology, Mitchell R. (ed.), p. 107, Wiley-
Interscience, New York, 1972.
6 Продукты реакции и реагенты в стандартных условиях, за исключением pH
(рН-7).
Если К всегда (как в табл. 5У14.1) принимать равным 0,60, то расчет-
ные значения А отличаются от экспериментальных значений (для 25 сис-
тем, приблизительно половина из которых анаэробные, а другие — аэроб-
ные) обычно не более чем на 50%.
Необходимое в определении Уе (см. выше) значение А находят по
уравнению
А = -AGP/K+AGc + AGn/fr
tfAG,
где AGp — свободная энергия, необходимая для превращения веществ, яв-
ляющихся источником углерода в клеточном синтезе, в промежу-
точные соединения;
AGn — свободная энергия, необходимая для превращения неорганических
соединений — источников азота в аммиак (степень окисления ко-
торого соответствует степени окисления азота в веществах
клетки);
AGp — свободная энергия, необходимая для превращения промежуточ-
ных соединений (источников углерода) и аммиака в компоненты
клетки (с учетом эффективности процессов AGc?s7,5 ккал на
1 химический эквивалент клеток);
394
Глава 5
К— средняя эффективность переноса свободной энергии (в гетеротро-
фах или аутотрофных бактериях /<=0,4—0,8);
ДО, — изменение свободной энергии при окислении 1 химического экви-
валента субстрата.
При указанной выше стехиометрии клетки в зависимости от источника
азота значение AG,, составляет 0 (аммиак), 3,25 (нитрит), 4,17 (нитрат),
3,78 (Na).
5.12. Сопряженный транспорт. Предположим, что компонент А нераство-
рим в мембране толщиной L и что его концентрация на наружной и внут-
ренней поверхностях мембраны равна а\ и а^. Существующий только в мемб-
ране носитель В образует на поверхности мембраны комплекс АВ. Прини-
мая, что реакция комплексообразования равновесна на любой поверхности
мембраны (но не внутри ее) и что константы равновесия равны Ki и К2,
определите поток А от поверхности 1 к поверхности 2, если коэффициент
диффузии АВ равен S). При каких условиях (если такие условия вообще
возможны) будет происходить активный транспорт А?
5.13. Стехиометрия клеточного роста и экономические коэффициенты,
а) Приняв, что клетки могут превращать в биомассу две трети (по массе)
углеродсодержащего субстрата (углеводорода или глюкозы), вычислите
«стехиометрические» коэффициенты утилизации гексадекана или глюкозы:
Гексадекан:
С16Нз4+а1О2+ааНН3 —>-
--Pl (С4,4Н7,з|Ыо,8б02,2) + Р2СО2+ Р3Н2О
Глюкоза:
СеН^Об + а^Ог+а'гХ’Нз —>- [},1(С4,4Н7,з|Мо,8б02,2) + [К2СО2+(/зН2О
б) Вычислите три коэффициента Ys, Уо, Уд. Обратите внимание на то,
что Уо и Уд для углеводорода меньше соответствующих коэффициентов для
глюкозы даже при условии равных эффективностей превращения углерода
в биомассу.
5.14. Метаболические реакции in v it г о. Экстракт дрожжей инкубируют с
200 ммоль D-глюкозы, 20 ммоль АТР, 2 ммоль NAD+ и 20 ммоль фосфата
(экстракт содержит все ферменты, необходимые для превращения глюкозы
в этанол). Какова будет концентрация глюкозы и этанола в состоянии равно-
весия? Как можно довести процесс брожения до конца?
5.15. Энергетическая эффективность накопления глюкозы в виде гликоге-
на. Сравните количество энергии, необходимой для включения одной моле-
кулы глюкозы в гликоген, с энергией, выделяющейся при последующей дегра-
дации одного остатка глюкозы в процессе а) брожения и б) дыхания. Какие
энергетические условия способствуют накоплению гликогена?
5.16. Особые случаи стехиометрии метаболизма. Для оценки стехиометри-
ческих коэффициентов в процессах метаболизма часто предлагалось опреде-
лять дыхательный коэффициент [RQ; уравнение (5.51)]. а) Найдите систему
двух линейных алгебраических уравнений с двумя неизвестными а и Ь, вхо-
дящими в уравнения (5.53) и (5.54) прис? [NADH+H+]/d/=0; используйте при
этом определение RQ. б) Найдите условия, при которых матрица коэффициен-
тов в уравнениях для а и Ь становится сингулярной (т. е. имеет нулевой оп-
ределитель); объясните физический смысл таких условий, в) Сообщалось, что
при росте дрожжей (у=4,14) на глюкозе (у$=4,0) и аммиаке (1№=0) свя‘
занное с биосинтезом выделение СО2 невелико (о=0,095). Вычислите а и b
для значений RQ, равных 1,05 и 1,06. Объясните практическую ценность
применения RQ для определения стехиометрии метаболизма в этой си-
стеме [12].
Стехиометрия и энергетика метаболических превращений
395
Литература
Биоэнергетика
В качестве дополнительной литературы рекомендуются ссылки [4—6]
в гл. 2, а также следующие источники:
1. Lehninger A. L., Bioenergetics, 2d ed., W. A. Benjamin, Inc., New York,
1971.
2. Peusner L., Concepts in Bioenergetics, Prentice-Hall, Inc., Englewood Cliffs,
N.J., 1974. В этой монографии при изучении биоэнергетики значительно
больше внимания уделяется термодинамике.
Метаболизм
Рекомендуется вся литература, приведенная в гл. 2, а также следующие
источники:
3. Doelle Н. W., Bacterial Metabolism, 2d ed., Academic Press, New York.
Великолепный обзор важнейших метаболических процессов в различных
бактериях.
4. Atkinson D. Е., Cellular Energy Metabolism and Its Regulation, Academic
Press, New York, 1977. Основное внимание в этой очень глубокой моно-
графии уделяется роли энергетических факторов в регуляции метаболизма.
Стехиометрия клеточного роста и образования продуктов метаболизма
5. Atkinson В., Mavituna F., Biochemical Engineering and Biotechnology Hand-
book, Macmillan Publishers Ltd., Surrey, England, 1983. В этом справочнике
собраны превосходно подобранные и удачно расположенные многочислен-
ные данные и сведения по теме настоящей главы и по другим интересным
вопросам.
6. Pirt S. L, Principles of Microbe and Cell Cultivation, Blackwell Scientific
Publications, Oxford, 1975. Здесь хорошо объясняются понятие об эконо-
мических коэффициентах и многие другие аспекты клеточного роста.
7. Herbert D., Stoichiometric Aspects of Microbial Growth, p. 1 in Continuous
Culture 6: Applications and New Fields, Dean A. C. R„ Ellwood D. C.,
Evans C. G. T., Melling J. (eds.), Ellis Horwood Ltd., Chichester, England,
1976. Здесь дано четкое объяснение понятия о С-моле и обсуждается зна-
чение этого понятия.
8. Poels J. A., Simple Model for the Energetics of Growth on Substrates with
Different Degrees of Reduction, Biotech. Bioeng., 22, 33 (1980).
9. Poels J. A., Bioengineering Report: Application of Macroscopic Principles to
Microbial Metabolism Biotech. Bioeng., 22, 1437 (1980).
10. Erickson L. E., Minkevich I. G., Eroshin V. K. Application of Mass and Ener-
gy Balance Regularities in Fermentation, Biotech. Bioeng., 20, 1595 (1978).
11. Nagai S., Mass and Energy Balances for Microbial Growth Kinetics, p. 49,
in Advances in Biochemical Engineering, v. 11, Ghose T. K., Fiechter A.,
Blakebrough N. (eds.), Springer-Verlag, Berlin, 1979.
12. Stephanopoulos G., San К. У., Grosz P., Studies on On-Line Bioreactor
Identification I—IV; Biotech Bioeng., 26, 1176, 1189, 1198, 1209 (1984).
13. Stouthamer A. H., A Theoretical Study on the Amount of ATP Required for
the Synthesis of Microbial Cell Material; Anton, van Leeuwenhoek, 39, 545
(1973).
14. Minkevich I. G., Mass-Energy Balance for Microbial Product Synthesis —
Biochemical and Cultural Aspects, Biotech. Bioeng., 25, 1267 (1983).
15. Cooney C. L., Acevedo F., Theoretical Conversion Yields for Penicillin Syn-
thesis, Biotech. Bioeng., 19, 1449 (1977).
396
Глава 5
Выделение теплоты в процессах метаболизма
См. приведенные выше ссылки [5, 6, 8, 9, 11, 14] и [4] в гл. 14, а также
следующую литературу:
16. Abbott В. J., Clamen A., The Relationship of Substrate, Growth Rate, and
Maintenance Coefficient to Single Cell Protein Production, Biotech. Bioeng.,
15, 117 (1973).
17. Kanazawa M., The Production of Yeast from n-Paraffins, p. 438 in Single
Cell Proteins II, Tannenbaum S., Wang D. I. C. (eds.), MIT Press, Camb-
ridge, MA, 1975.
18. Servizi J. A., Bogan R. H., Thermodynamic Aspects of Biological Oxidation
and Synthesis, J. Water Poll. Control Fed., 36, 607 (1961).
Глава 6
Молекулярная генетика
и системы регуляции
В предыдущих главах мы рассмотрели кинетику фермента-
тивного катализа и общие черты некоторых наиболее распрост-
раненных путей метаболизма. На любом химическом предпри-
ятии синтезируют ограниченное число веществ, а стационар-
ный режим работы реакторов (к которому обычно и стремят-
ся) требует минимального регулирования. Напротив, такой
сложнейший химический комбинат, каким является живая
клетка, постоянно находится в переходном состоянии. В клетке
по мере прохождения различных периодов и этапов жизненно-
го цикла непрерывно меняются концентрации и относительные
количества субстратов, ферментных катализаторов, РНК, ко-
факторов и других компонентов. Настоящая глава посвящена
изучению синтеза и регуляции систем, управляющих клеточ-
ной динамикой. Эта тема тесно связана с проблемами вирус-
ного заражения культур клеток, естественных и вызванных ис-
кусственным путем мутаций, генетических трансформаций и
универсальности некоторых культур в отношении используемых
питательных веществ.
6.1. Молекулярная генетика
Прежде чем начать изучение генетики клетки и экспрессии
генетической информации на молекулярном уровне, мы долж-
ны вспомнить рассмотренные в гл. 2 состав и строение ДНК,
трех разновидностей РНК и белков. Чтобы оценить важность
рассматриваемых ниже механизмов, полезно напомнить, что
белки, особенно ферменты, непосредственно определяют харак-
тер функционирования клетки путем регуляции осуществляю-
щихся в ней химических реакций. Следовательно, в известной
степени набор клеточных ферментов (типы ферментов и кон-
центрации каждого из них) определяют метаболические про-
цессы, т. е. природу и особенности осуществляющихся в клетке
последовательных и параллельных реакций. Другие белки
также выполняют важные для клетки функции (вспомните
табл. 2.6). Очевидно, что любая дочерняя клетка должна по-
лучать от родительской клетки всю наследственную информа-
398
Глава 6
цию, необходимую для синтеза тех же самых белков, которые
присутствуют в исходной клетке. Механизмы передачи этой
информации изучает генетика. В первом разделе настоящей
главы мы узнаем, каким образом нуклеотидная последователь-
ность неповторяющегося и, следовательно, информационного
биополимера, дезоксирибонуклеиновой кислоты (ДНК), опре-
деляет ход белкового синтеза в клетке.
6.1.1. Процессы экспрессии генов
Мы уже знаем, что молекула ДНК представляет собой
очень длинную двойную спираль, состоящую из двух цепей.
Обе цепи представляют собой полимеры, построенные из четы-
рех нуклеотидов, отличающихся друг от друга природой азо-
тистого основания [аденина (А), гуанина (G), цитозина (С) и
тимина (Т)]. Две цепи в двойной спирали ДНК имеют комп-
лементарные последовательности так, что А в одной цепи всег-
да связывается с Т другой; аналогично связываются G и С.
Геном называют участок ДНК, кодирующий определенный
белок. Экспрессия гена, в результате которой формируется
аминокислотная последовательность соответствующего белка,
представляет собой двустадийный процесс; промежуточным
продуктом этого процесса является матричная (или информа-
ционная) рибонуклеиновая кислота (мРНК или иРНК). На
первом этапе экспрессии гена, называемом транскрипцией,
сложный олигомерный фермент РНК-полимераза катализирует
синтез мРНК на матрице, роль которой выполняет ген. На
втором этапе экспрессии гена на основе содержащейся в
нуклеотидной последовательности мРНК информации синтези-
руется пептидная цепь с соответствующей аминокислотной
последовательностью. В этом процессе, называемом трансля-
цией, участвуют различные компоненты клетки, в том числе
центры и регуляторы пептидного синтеза — рибосомы, а также
несколько различных транспортных РНК (тРНК), которые пе-
реносят химически активированные аминокислоты к месту
синтеза пептидной цепи в соответствии с диктуемой мРНК по-
следовательностью. Ниже мы рассмотрим эти процессы, а так-
же генетический код, являющийся связующим звеном между
нуклеотидной и аминокислотной последовательностями. В на-
шем обсуждении мы будем рассматривать только основные мо-
менты и по необходимости опустим немаловажные в ряде си-
туаций детали; поэтому читателю настоятельно рекомендуется
подробнее ознакомиться с проблемой экспрессии генов в при-
веденной в конце главы литературе.
Транскрипция начинается со связывания фермента РНК-по-
лимеразы с сигнальной нуклеотидной последовательностью,так
Молекулярная генетика и системы регуляции 399
называемым промотором, располагающейся на одной из комп-
лементарных цепей ДНК. Связанная с РНК-полимеразой ну-
клеотидная цепь становится таким образом матрицей ДНК
для синтеза молекулы мРНК- После частичного расплетания
двойной спирали ДНК вблизи от комплекса РНК-полимеразы
с промотором полимераза начинает двигаться вдоль матричной
цепи ДНК в направлении 3Z->5Z. При построении комплемен-
тарной ДНК последовательности одноцепочечной молекулы
мРНК соблюдается правило образования пар нуклеотидов
ДНК 5' Г —Я— GCCTTTATGGAAATTATACGTTAAAGC —3'
3' 1 —!!~ CGGAAATACCTTTAATATGCAATTTCG —11—5'
Транскрипция PH К- полимераза
мРНК 5' — GCC UUU AUG GAA AUU AUA CGU UAA AGC—3'
Правило образования пар
нркпео/пидоб'.К Т С G ДНК
U A G С мРНК
РИС. 6.1. Схематическое изображение транскрипции матричной цепи ДНК
(здесь транскрибируется нижняя цепь) при участии фермента РНК-полимера-
зы, в результате чего синтезируется молекула соответствующей мРНК с ком-
плементарной нуклеотидной последовательностью.
ДНК—РНК (вспомните разд. 2.3.2 или см. рис. 6.1). Синтези-
руемая мРНК располагается антипараллельно матричной цепи
ДНК; следовательно, мРНК синтезируется от 57-конца к З'-кон-
цу. В приведенном на рис. 6.1 примере матрицей является ниж-
няя цепь ДНК- Транскрипция прекращается, т. е. синтез
мРНК останавливается в тот момент, когда РНК-полимераза
достигает специфической нуклеотидной последовательности
матричной цепи {терминатора), сигнализирующей об оконча-
нии синтеза. Прежде чем перейти к рассмотрению следующих
стадий, следует отметить, что РНК-полимераза связывается с
ДНК и неспецифично в центрах, не являющихся промоторами.
Неспецифично связанная полимераза не инициирует и не осу-
ществляет транскрипцию.
Длина молекулы мРНК изменяется от приблизительно 300
до 3000 нуклеотидных остатков и более. Обычно одна мРНК
соответствует одному гену в цепи ДНК, но в некоторых случа-
ях молекула мРНК несет генетическую информацию о группе
родственных и тесно связанных генов — так называемом опе-
роне (см. разд. 6.1.4). Как мы уже отмечали при обсуждении
400
Глава 6
энергетики биосинтеза в гл. 5, построение цепи РНК осуществ-
ляется путем отщепления пирофосфатной группировки от ну-
клеозидтрифосфатов. Следовательно, образование каждой
фосфодиэфирной связи РНК сопровождается расщеплением
двух высокоэнергетических фосфатных связей. Это способст-
вует тому, что реакция конденсации протекает «до конца» и
свидетельствует о важности синтеза этих биополимеров для
нормального функционирования клетки.
Прежде чем перейти к описанию процесса трансляции, нам
нужно несколько более детально ознакомиться с транспортны-
ми РНК. Известно, что каждая разновидность тРНК может
транспортировать только одну соответствующую ей аминокис-
лоту. На рис. 6.2 изображена вторичная структура аланил-
тРНК из дрожжей. Присоединение аланильного остатка к со-
ответствующей тРНК требует предварительной активации ала-
нина путем его превращения в аланил-АМР, как это отраже-
но в уравнении реакции (5.39). Активированный аланиновый
остаток затем связывается с соответствующей тРНК при учас-
тии фермента, специфичного как в отношении аминокислоты,
так и в отношении тРНК. Таким образом обеспечивается свя-
зывание аланина именно с аланиновой тРНК-
Теперь мы можем перейти к обсуждению природы специ-
фичности тРНК. В основании нижней петли молекулы тРНК
имеется последовательность из трех нуклеотидов, называемая
антикодоном. Этот элемент структуры присутствует во всех
изученных тРНК- Последовательность оснований антикодона
комплементарна трехнуклеотидному сегменту мРНК, называе-
мому кодоном. Изучение биохимии нуклеиновых кислот пока-
зало, что каждый кодон представляет собой «слово» в генети-
ческом послании, т. е. каждый кодон соответствует одной ами-
нокислоте. Поскольку химический алфавит РНК состоит всего
лишь из четырех букв (четырех оснований А, С, G и U), а
всего должно быть не менее 20 «слов» (по числу аминокислот),
то для передачи генетической информации должен существо-
вать специальный «язык» или код. Как это ни удивительно, но
генетический код был полностью расшифрован; более того, ока-
залось, что он практически универсален для всех живых орга-
низмов— бактерий, растений и животных (рис. 6.3).
Процесс трансляции начинается со связывания меньшей из
двух субъединиц рибосомы с молекулой мРНК с участием спе-
цифической последовательности, называемой связывающим ри-
босому сайтом. Этот сайт обычно расположен на определенном
расстоянии, различном у прокариот и эукариот, «выше» кодо-
на AUG, с которого почти всегда начинается считывание ин-
формации о последовательности полипептидной цепи. Здесь
термин «выше» означает, что этот сайт расположен в направ-
Молекулярная генетика и системы регуляции
401
Неодычнь/е основания
у Пседдоуридин
I Инозин
UHZ Дигиарруридин
Г РиЗогпикидин
GMe Метилгуанозин
£Мег Динётилгуано-
1Ме ' Метилинозин
РИС. 6.2. Антикодон аланиновой тРНК узнает комплементарный кодон из
трех оснований в мРНК. (Уотсон Дж., Молекулярная биология гена. — М.:
Мир, 1978.)
лении 5'-конца от кодона AUG. Последовательность кодонов,
определяющая аминокислотную последовательность белковой
цепи, начинается с кодона AUG и идет в направлении 5'—>3'.
Далее мРНК образует комплекс со всей рибосомой, и к это-
му комплексу присоединяются тРНК, несущие аминокислоты,
которые отвечают двум первым кодонам мРНК. В соответст-
вий с заданной аминокислотной последовательностью между
карбоксильной группой первой аминокислоты и аминогруппой
второй образуется пептидная связь. Затем рибосома смещается
26-537
402
Глава 6
в направлении 3' на один кодон; при этом высвобождается
тРНК первой аминокислоты, после чего с комплексом связыва-
ется третья аминокислота в виде соответствующей аминоацил-
тРНК и присоединяется к пептидной цепи, в результате чего
последняя удлиняется еще на один остаток. Этот процесс удли-
нения полипептидной цепи (рис. 6.4) продолжается вплоть до
терминирующего кодона (см. рис. 6.3); в этот момент пептид-
Второе основание
Bepffoe U C A C Третье основание
основание Phe Ser Tyr Cys и
Phe Ser Tyr Cys С
U ) Leu Ser Stop Stop А
Leu Ser Stop Trp 9
' Leu Pro His Arg и
С Leu Pro His Arg с
Leu Pro Gin Arg А
Leu Pro Gin Arg G
r lie Thr Asn Ser и
A -s He Thr Asn Ser с
He Thr Lys Arg А
Met Thr Lys Arg G
Г Vai Ala Asp Gly и
G < Vai Ala Asp Gly С
Vai Ala Glu Gly А
tVal Ala Glu Gly G
РИС. 6.3. Генетический код. Три кодона (Stop) сигнализируют о прекращении
синтеза пептидной цепи. Нетрудно видеть, что замена одного основания кодо-
на (особенно третьего) часто приводит к близкой аминокислоте (принцип
надежности).
ная цепь отделяется, а комплекс рибосомы и мРНК разруша-
ется. Таким образом, построение полипептидной цепи начина-
ется с N-концевого остатка и завершается С-концевым амино-
кислотным остатком.
На рис. 6.5 процесс трансляции изображен с другой точки
зрения; здесь основное внимание уделено передаче информации
от нуклеотидной последовательности мРНК к соответствующей
аминокислотной последовательности полипептида. Образовав-
шаяся в результате транскрипции (рис. 6.1) гипотетическая
мРНК транслируется в соответствующий полипептид. Обратите
внимание на то, что здесь тринуклеотидная последовательность
AUG выполняет две функции, указывая, во-первых, на начало
ДМР
Покидающий ло-
ве рхносто матрицы
переносчик
Образование пептидной
связи при участии
пептидсинтазб/
’-О NH2
\ I
\ C-CH—R.
\ гл PH К, взаимодействуя
с соответствующим антико-
доном, доставляет на место
сборки следующую аминокислоту
'Рядом находятся
гие тРНК с соответ-
ствующими аминокислот-
ными остатками
, ~ — -• Аниноацил-
Транспорт- ЧАМР гдАмишхщил-№Р синтаза
ная РНК ' X. О | 2
S- — СН R.
АТР Н—О
Аминокислота
бел-
ковая цель, лона еще
связанная с движу-
щейся рибосомой
РИС. 6.4. Трансляция — реализация генетической информации мРНК в первичной структуре белка — осуществляется пу-
тем ряда специфических взаимодействий между мРНК и тРНК иа рибосоме. (Воспроизведено из работы: Edwards N. А.,
Hassall К. A., Cellular Biochemistry and Physiology, p. 342, McGraw-Hill Publishing Company Ltd., London, 197J.)
404
Глава 6
кодирующей последовательности.для белка и, во-вторых, коди-
руя метионин (Met) как N-концевой аминокислотный остаток
полипептида. Заметьте также, что не вся молекула мРНК не-
сет в себе информацию о структуре синтезируемого белка.
Выше мы уже упоминали о предшествующем участку копиро-
вания связывающем рибосому сайте; другие нуклеотидные по-
следовательности, в том числе кодирующие последовательности
со своими инициирующими и терминирующими кодонами, мо-
гут быть расположены ниже (в направлении 5'->3') кодирую-
щей области.
[*рнк]
5'—GCCUUUAUGGAA-AUUAUACGUUAAAGC—Зг
Иниции- Термини-
рующим руюёций кодон
кодон
Трансляция (Рибосома)
\6еяок |
Met—G1u—Не — Не —Arg
N- конце'вой с -концевой оста-
оста ток спок
РИС. 6.5. Схема передачи (трансляции) информации, содержащейся в нук-
леотидной последовательности мРНК, аминокислотной последовательности
соответствующего полипептида на рибосоме. Здесь изображена та же мРНК,
что и на рис. 6.1,
Здесь полезно рассмотреть процесс экспрессии гена также
с точки зрения регулирующих этот процесс элементов, посколь-
ку контролирование последних немаловажно для успешного ре-
шения задач генетической инженерии. На рис. 6.6 в самом
общем виде приведены важнейшие функции регуляторных по-
следовательностей в процессе передачи генетической информа-
ции. Слева и справа от гена расположены последовательности
промотора и терминатора, контролирующие начало (инициа-
цию) и окончание (терминацию) транскрипции. ДНК должна
содержать также регуляторные сигналы трансляции (связыва-
ющий рибосому сайт, кодон-инициатор и терминирующий ко-
дон), которые должны транскрибироваться в мРНК. Наконец,
часть мРНК, комплементарная структурному гену ДНК, кото-
рый кодирует определенную аминокислотную последователь-
ность, должна использоваться для направления белкового син-
теза в ходе трансляции.
Часто одна мРНК связывается с несколькими рибосомами.
Полирибосомы (полисомы), представляющие собой сложные
агрегаты из нескольких индивидуальных рибосом, часто обла-
дают высокоупорядоченной структурой. В полирибосомах одно-
Молекулярная генетика и системы регуляции
405
временно осуществляется считывание генетической информа-
ции с различных участков мРНК, что существенно ускоряет
белковый синтез и обеспечивает высокую эффективность этого
процесса.
Как мРНК, так и тРНК, и рибосомальные рибонуклеиновые
кислоты (рРНК) синтезируются на матрице — определенном
участке цепи ДНК. Таким образом, ДНК содержит в закоди-
Структурнб/й еен (ко-
дирующая структуру бел-
ка последовательность)
Связывающее———*---------------
' рибосому пос-
Пплмптпп'/,-^ателмос'Л6 Иници- _ Терпи -
' ///с// и шир ирующии Ниру/О-—\
кодон щий
кодон
Термина-
г- тор
Р RB
Транскрипция
V
РНК
Трансляция
V
Белок
РИС. 6.6. Регулирующие и информационные последовательности ДНК направ-
ляют процессы транскрипции и трансляции (экспрессию гена).
рованном виде информацию не только о первичной структуре
белков, но и о строении различных веществ, с помощью которых
осуществляется белковый синтез.
Здесь полезно напомнить, что взаимодействия между ами-
нокислотными остатками полипептидной цепи определяют вто-
ричную, третичную и даже четвертичную структуры белков
(разд. 2.4). Следовательно, слабые взаимодействия между ато-
мами и группами играют большую роль на всех этапах хране-
ния генетической информации, ее передачи и реализации.
Прежде чем перейти к вопросу о механизмах передачи ге-
нетической информации от поколения к поколению и о факто-
рах, нарушающих этот процесс, следует упомянуть о различи-
ях в механизмах синтеза белков в эукариотах и прокариотах.
В клетках бактерий синтез мРНК и трансляция мРНК в белки
на рибосомах осуществляются почти в одном и том же центре
406
Глава 6
нуклеоида. В эукариотах транскрипция происходит в ядре.
Затем молекулы мРНК претерпевают модификацию (описан-
ную в следующем разделе), диффундируют через поры ядер-
ной мембраны в цитоплазму и образуют комплексы с рибосо-
мами, где и происходит процесс трансляции. В эукариотах и в
меньшей степени в прокариотах продукт трансляции часто пре-
терпевает модификацию, результатом которой является окон-
чательная, функционально-активная молекула белка. Пост-
трансляционные модификации белков рассматриваются в
разд. 6.1.3. Наконец, следует отметить, что в эукариотах и про-
кариотах процесс синтеза белка различно протекает и во вре-
мени; подробнее этот вопрос мы рассмотрим в разд. 6.5.
6.1.2. Расщепленные гены и модификация мРНК в эукариотах
Матричная РНК эукариот модифицируется путем присоеди-
нения к 5'-концу особого структурного элемента, называемого
кэпом или кэп-структурой (остаток У'-метилгуанозина); кэп-
структура повышает устойчивость мРНК к действию фосфатаз
и нуклеаз. Кроме того, к З'-концу большинства мРНК эукариот
присоединяется последовательность поли-А, содержащая от
150 до 200 остатков дезоксиаденилата. Этот факт можно ис-
пользовать при выделении эукариотических мРНК, сорбируя их
на носителях, содержащих ковалентно-связанные олигонуклео-
тидные цепи поли-Т.
В настоящее время можно считать твердо установленным,
что многие эукариотические гепы представляют собой чередо-
вание последовательностей, кодирующих структуру белка и не
несущих информацию о белковой структуре. Последователь-
ности первого типа иногда называют экзонами, а некодирую-
щие последовательности — интронами или промежуточными
последовательностями. Последние, как следует из их назва-
ния, располагаются между экзонами и разделяют кодирующие
последовательности участками, которые на первый взгляд со-
держат ненужную избыточную информацию. Очевидно, что в
эукариотических клетках должны , существовать механизмы,
распознающие экзоны и интроны и обеспечивающие в процессе
экспрессии гена непосредственную трансляцию одних только
экзонов.
Основные структурные элементы эукариотического гена и
главные этапы его экспрессии изображены на рис. 6.7. В верх-
ней части рисунка расположен гипотетический сегмент струк-
турного эукариотического гена, содержащий экзоны и интро-
ны. Во время транскрипции матричная цепь ДНК направляет
синтез мРНК (первичного транскрипта), комплементарной всей
Молекулярная генетика и системы регуляции
407
Экспресси-
руемая пос- Декодирую"
ледоЗатель- щая после -
нос то ' оователь -
(экзон) кость (интрон) ДНК
CAG ATGTATGGG GTACTAAAGTAG CACTTTTAG GTCATCCTGA
АТС ТАСАТАССС CATGATTTCATC GTGAAAATC CAGTAGGACT
Транскрипция
Терзанный
транскрипт
(me^tyrXglyXjwsXph^ Стоп Пептид
РИС. 6.7. В процессе экспрессии генов эукариот первичный транскрипт мРНК
подвергается сплайсингу; в результате образуется зрелая (активная) мРНК,
участвующая в трансляции. Здесь изображены гипотетические укороченные
последовательности.
последовательности матрицы, т. е. содержащей последователь-
ности как экзонов, так и интронов.
Далее следует характерная, по всей вероятности только для
эукариот, стадия сплайсинга. В процессе сплайсинга участки
мРНК, отвечающие интронам, вырезаются, а оставшиеся по-
следовательности, комплементарные последовательностям эк-
зонов, соединяются в том же порядке, в каком они располага-
лись в первичном транскрипте. Образующаяся зрелая мРНК
затем транслируется описанным выше путем, в результате чего
синтезируется соответствующий данному гену белок.
Имеющиеся в настоящее время экспериментальные данные
говорят о том, что почти все эукариотические гены содержат
интроны. Впрочем, известно и несколько исключений из этого
правила, в том числе гены сс-интерферона человека. Интроны
могут быть очень большими; возможно, что в состав интро-
нов входит более половины всех нуклеотидных остатков ДНК.
408
Глава 6
Ко времени написания настоящей главы биологическая
функция интронов оставалась невыясненной. Возможно, что
интроны участвуют в регуляции экспрессии эукариотических
генов. С точки зрения эволюции организмов интроны могут
выполнять роль инструмента для осуществления относительно
крупных изменений генов посредством включения или исклю-
чения последовательности целого интрона или экзона. Предпо-
лагалось также, что экзоны кодируют довольно крупные уча-
стки (домены) первичной структуры белков и что перестановка
экзонов дает клетке возможность синтезировать новые белки
и таким путем оценивать их функциональную пригодность.
С технологической точки зрения с интронами связаны извест-
ные затруднения при экспрессии эукариотические генов в про-
кариотических клетках-хозяевах, поскольку последние не об-
ладают аппаратом сплайсинга РНК- В разд. 6.3, посвященном
технологии рекомбинантных ДНК, мы увидим, как можно пре-
одолеть это затруднение.
6.1.3. Посттрансляционная модификация белков
Образующийся в результате трансляции мРНК полипептид
часто еще не является зрелой, биологически активной формой
белка. Иногда после трансляции от белка отщепляется N-
концевой остаток метионина. В других случаях происходит
окисление двух остатков цистеина, приводящее к образованию
дисульфидной связи — важного фактора в закреплении третич-
ной структуры белка (гл. 2). Присоединение к боковым цепям
аспарагина, серина и треонина углеводных остатков приводит
к гликопротеинам. В химической модификации (процессинге)
полипептидов используются также реакции гидроксилирова-
ния, фосфорилирования и ацетилирования.
У некоторых полипептидов на N-конце имеется короткая
(15—30 остатков) последовательность гидрофобных аминокис-
лот. Подобные сигнальные последовательности играют большую
роль в транспорте белков через клеточные мембраны. Секре-
тируемые белки, как правило, содержат такие последователь-
ности. К названию белков с сигнальными последовательностя-
ми обычно добавляют префикс «пре» (например, прелизоцим).
Сигнальная последовательность отщепляется в процессе
транспорта пребелка через мембрану, в результате чего образу-
ется функционально активный белок (в нашем примере — ли-
зоцим).
Белки могут содержать и другие аминокислотные последова-
тельности, отщепляющиеся при их превращении в функциональ-
но активную форму. Наряду с упомянутыми выше механизма-
ми секреции белков такие элементы структуры позволяют клет-
Молекулярная генетика и системы регуляции 409
ке осуществлять белковый синтез и активацию специфических
белков в различных точках внутри клетки или вне ее. Молеку-
ла препроинсулина, например, построена из N-концевой сиг-
нальной последовательности (24 аминокислотных остатка), за
которой следует последовательность из 21 остатка (цепь В),
затем еще из 21 остатка (цепь С) и, наконец, С-концевой учас-
ток из 30 остатков (цепь А). Сигнальная последовательность
отщепляется в ходе секреции; образующийся проинсулин (це-
пи А—С—В) далее расщепляется под действием ферментов до
инсулина, состоящего из соединенных двумя межцепочечными
дисульфидными связями цепей А и В. В инсулине между цепя-
ми А и В пептидных связей нет (рис. 6.8).
6.1.4. Индукция и репрессия; регуляция белкового синтеза
Мы уже знаем, что живые клетки обладают сложными си-
стемами управления, обеспечивающими высокую эффективность
использования материальных и энергетических ресурсов. Из
предыдущих глав мы узнали также, что активация и ингиби-
рование ферментативной активности метаболитами позволяют
направлять эти промежуточные вещества по необходимым
путям в сложной сети клеточных реакций. В этом разделе мы
рассмотрим другой уровень регуляции, принципиально отли-
чающийся от указанных выше. Активация и ингибирование
влияют на каталитическую активность уже имеющихся в клет-
ке ферментов, но не изменяют количества последних. Таким
образом, эти способы регуляции действуют на уровне фермен-
тов. Напротив, рассматриваемые здесь механизмы регуляции —
индукция и репрессия — изменяют скорости синтеза белков (и,
следовательно, количества содержащихся в клетке ферментов)
и осуществляются на уровне гена. По-видимому, сходство меж-
ду этими двумя типами регуляции сводится лишь к их чувст-
вительности по отношению к низкомолекулярным соединениям.
, Здесь основное внимание мы будем уделять регуляции бел:
нового синтеза на генетическом уровне в бактериях, поскольку
именно в этих организмах лучше всего изучены молекулярные
механизмы этих процессов. Аналогичные механизмы опериру-
ют и в клетках высших растений и животных, однако последние
располагают также гораздо менее изученной системой регуля-
ции, направляющей дифференцировку клеток в процессе раз-
вития организма.
Прежде чем углубиться в детали механизмов индукции и
репрессии, полезно попытаться оценить роль регуляции белко-
вого синтеза в жизни микроорганизмов. В гл. 1 мы уже упоми-
нали, что бактерии представляют собой изолированные, само-
стоятельно существующие клетки, и поэтому они практически
410
Глава 6
в
Сигнальная
последовательность
н,н-|Т~ ~
Транспорт через
мембрану знооплазмати чес-
каео ретикулума , f
У комплексу Голь-
джи, резервным
гранулам
Инсулин
s—-----------------s Цепь А
Gly-Ile-Val-Glu-Gln-Cys-Cys-Ala-Ser-Val-Cys-Ser-Leu-Tyr-GIn-Leu-Glu-Asn-Tyr-Cys-Asn
5 | 10 15 | ?1
S /S
s s цепь В
I f „
Phe-Val-Asn-Gln-His-Leu-Cys-Gly-Ser-His-Leu-Val-Glu-Ala-Leu-Tyr-Leu-Val-Cys-Gly-Glu-Arg-GIy-Phe-PhO
5 10 15 20 24 |
Thr- Lys- Pro-Thr-Tyr
30 . ,
РИС. 6.8. Активная форма инсулина образуется в результате нескольких пост-
трансляционных превращений, в том числе в результате отщепления сигналь-
ной последовательности, образования дисульфидных связей и отщепления
цепи С. На рисунке приведена аминокислотная последовательность цепей А
и В инсулина человека.
Молекулярная генетика и системы регуляции
411
не влияют па внешнее окружение. Отсюда следует, что бакте-
рии должны обладать очень высокой способностью к адапта-
ции; возможность эффективно функционировать в самых раз-
личных условиях может оказаться решающей для их выжива-
ния.
Многие бактерии и в самом деле обладают высокой при-
способляемостью. Они могут синтезировать ферментные систе-
мы, способные эффективно утилизировать самые различные пи-
тательные вещества. Примером особенно универсального в этом
отношении микроорганизма может служить бактерия Pseudo-
monas multivorans, которая в качестве единственного источни-
ка углерода может использовать более 90 различных соедине-
ний, в том числе углеводы и их производные, жирные кислоты,
дикарбоновые и другие органические кислоты, первичные спир-
ты, аминокислоты и другие азотсодержащие соединения и даже
циклические соединения, например фенол. Обычно для усвоения
каждого питательного вещества необходим свой фермент. Сле-
довательно, бактерия должна нести генетическую информацию
о структуре всех необходимых ферментов. Вся содержащаяся
в хромосомах клетки генетическая информация называется
генотипом клетки.
Однако такой бактерии не нужны одновременно все фермен-
ты, способные усваивать любые питательные вещества, а син-
тез лишних белков связан с непроизводительными затратами
энергии и промежуточных метаболитов. Поэтому в данной сре-
де в целях максимальной эффективности в бактериях претер-
певает экспрессию (т. е. выражается в синтезе конкретных бел-
ков) только часть всей генетической информации. Под феноти-
пом понимают внешний облик организма. Учитывая изложен-
ные выше факторы, можно сказать, что данный фенотип обу-
словливается сочетанием генотипа организма и условий среды
(его окружения). Конститутивные ферменты синтезируются не-
зависимо от состава и условий среды.
Напротив, скорость биосинтеза индуцибельных ферментов
зависит от окружения клетки. Типичным примером такой зави-
симости может служить синтез fl-галактозидазы. Этот фермент
катализирует гидролиз дисахарида лактозы на составляющие
его моносахариды глюкозу и галактозу. Указанная реакция не-
обходима, если клетка вынуждена использовать лактозу в ка-
честве питательного вещества, поскольку в последующих реак-
циях могут использоваться только моносахариды. Клетка бакте-
рии Е. coli регулирует синтез fl-галактозидазы в соответствии с
потребностью в этом ферменте. С появлением в среде лактозы
клетка начинает синтез фермента; этот процесс ускоряется по
мере повышения концентрации лактозы вплоть до максимально
возможного, полностью индуцированного уровня (рис. 6.9). Та-
412
Глава 6
ким образом, в конечном счете именно субстрат индуцирует об-
разование фермента.
Аналогичная ситуация складывается в случае репрессибель-
ных ферментов. Например, Е. coll может синтезировать любые
ферменты, необходимые для синтеза всех 20 аминокислот из бо-
лее простых предшественников; в то же время если в питатель-
ной среде содержатся нужные аминокислоты, то соответствую-
РИС. 6.9. Концентрация ^-галактозидазы в клетке возрастает по мере повы-
шения концентрации индуктора (лактозы) в питательной среде. Повышение в
среде концентрации репрессора (гистидина) уменьшает содержание в клетке
ферментов, катализирующих биосинтез репрессора, {Уотсон Дж., Молекуляр-
ная биология гена. — М.: Мир, 1978.)
щие ферменты не образуются в клетке в сколько-нибудь зна-
чительных количествах. В этом случае конечный продукт пути
биосинтеза подавляет синтез ферментов, катализирующих от-
дельные стадии этого же пути. Это явление также изображено
на рис. 6.9, где в качестве примера приведен синтез гистидина.
На рис. 6.10 приведены модели процессов индукции и реп-
рессии, разработанные Моно с сотрудниками во Франции и от-
ражающие внутреннее сходство этих двух процессов. В обеих
случаях экспрессия гена регулируется на уровне транскрипции
и ген-регулятор продуцирует белок, взаимодействующий со спе-
цифической последовательностью ДНК, называемой операто-
ром. Если белок-модулятор связан с последовательностью опе-
ратора, то РНК-полимераза не взаимодействует с промотором;
в результате блокируется процесс транскрипции и структурный
ген не экспрессируется.
Молекулярная генетика и системы регуляции
413
В модели индукции (рис. 6.10, а) регуляторный ген проду-
цирует молекулу репрессора, который может предотвращать
синтез фермента. Если в среде присутствует индуктор, то он
связывается с репрессором с образованием неактивного комп-
лекса, не влияющего на последующую транскрипцию ДНК-
В присутствии корепрессора
Апорепрессор \ РНК-тти-
мераза
В отсутствие индуктора {
Последователь - Последовательность
кость промотора оператора
В присутствии индуктора
I « Г
В отсутствие корепрессора
I R I р ’I. о I 8 ЩНК
Репрессору
Индук-
тор
рн к - поли - Транскрип-
мераза ,, ция
Неактивный
репрессор
мРНК
Трансляция
Щ РНК - поли-
'Апорепрессор ^еРа?а
'1рансн.-
рипция
мРНК
Фермент
Трансляция
ф^ферненпт
РИС. 6.10. Слева-, индуктор (например, галактоза) инактивирует репрессор-
таким образом, что последний не связывается с соответствующей последова-
тельностью оператора и не блокирует транскрипцию структурного гена (на-
пример, гена ^-галактозидазы). Справа: корепрессор (например, гистидин)
взаимодействует с апорепрессором, образуя активный репрессор, который,
связываясь с оператором, может блокировать экспрессию структурного гена;
таким путем регулируется синтез репрессибельных ферментов. [Воспроизве-
дено из работы: Demain A. L., Theoretical and Applied Aspects of Enzyme
Regulation and Biosynthesis in Microbial Cells, in Enzyme Engineering, Win-
gard L. B., Jr. (ed.), Interscience, New York, 1972.]
В модели репрессии (рис. 6.10, б) регуляторный ген должен об-
разовывать комплекс с другой молекулой, в результате чего и
возникает репрессор. В отсутствие корепрессора (в приведен-
ном здесь примере гистидина) белковый синтез не останавлива-
ется.
Подобные регулируемые промоторы могут одновременно
контролировать синтез нескольких ферментов. При индуцирова-
нии синтеза ^-галактозидазы, например, одновременно стимули-
руется и синтез двух других белков, одним из которых является
галактозидпермеаза, участвующая в активном транспорте ^-га-
лактозидов. Здесь налицо логическая связь между функциями1
414
Глава 6
этих двух белков, поэтому координация их биосинтеза представ-
ляется вполне оправданной. Группа регулируемых совместно ге-
нов называется опероном; в частности, только что упомянутую
систему называют Zac-опероном. Количественный математиче-
ский анализ регуляторных свойств Zac-промотор а-оператора мы
рассмотрим позднее, в разд. 7.5.3.
Хотя в нашем обсуждении основное внимание уделяется ре-
гуляции синтеза ферментов в бактериях, аналогичные механиз-
мы используются для контроля синтеза других белков в бакте-
риях и в клетках высших растений и животных. В общем слу-
чае следует всегда иметь в виду, что синтезируемый клеткой на-
бор белков и связанные с этим биологические и каталитические
активности могут изменяться и часто действительно изменяются
в ответ на изменения состава и условий среды. Эта способность
к адаптации обусловливает целый ряд проблем при анализе ки-
нетики клеточных реакций и при расчете биореакторов; такие
проблемы совершенно нетипичны для технологических процес-
сов с использованием синтетических катализаторов.
Существует несколько вариантов контроля на генетическом
уровне. Рассмотреть здесь все эти варианты не представляется
возможным, однако следует упомянуть о катаболитной репрес-
сии. Если бактерии Е. coli культивируют в среде, содержащей
глюкозу, и другой источник углерода, труднее включающийся в
схему метаболизма, например лактозу, то преимущественно
усваивается глюкоза. В этих условиях лактоза не индуцирует
синтез ^-галактозидазы. В быстро растущей на глюкозе клетке
ингибируется образование циклического АМР. Концентрация
цикло-АМР (рис. 2.8) оказывает влияние на активность промо-
тора Zac-оперона таким образом, что при низких внутриклеточ-
ных концентрациях цикло-АМР индукция блокируется или
ослабляется. Поскольку продукты катаболизма глюкозы снижа-
ют концентрацию цикло-АМР, то такой способ регуляции назы-
вают катаболитной репрессией.
Следует подчеркнуть, что катаболитная репрессия может осу-
ществляться и в отсутствие глюкозы. Обычно бактерия, расту-
щая на смеси источников углерода, селективно утилизирует наи-
более выгодный (т. е. обеспечивающий наивысшую скорость
роста) из них и катаболически подавляет утилизацию менее
выгодных питательных веществ. Что же произойдет, когда исто-
щится первый источник углерода? (См. гл. 7, разделы, посвя-
щенные кинетике процессов брожения.)
Катаболитная репрессия играет важную роль и в синтезе
продуктов метаболизма. При быстром росте клеток часто не
синтезируются ферменты, необходимые для образования вторич-
ных метаболитов. Отсюда следует, что перед технологом-биохи-
миком может возникнуть очень интересная и важная задача
Молекулярная генетика и системы регуляции
415
РИС. 6.11. Упрощенная схема репликации ДНК. По мере разделения роди-
тельских цепей на каждой из них формируется комплементарная цепь, в ре-
зультате чего образуются две идентичные родительской дочерние молекулы.
Заметьте, что каждая дочерняя молекула содержит одну цепь родительской-
ДНК. (Лёви А., Сикевиц Ф., Структура и функция клетки. — М.: Мир, 1971.)
обеспечения минимальной катаболитной репрессии и макси-
мального накопления вторичных метаболитов. Пути решения
этой задачи мы обсудим в гл. 7, 9 и 10.
6.1.5. Репликация ДНК и мутации
Поскольку ДНК содержит всю информацию, необходимую
для развития и жизнедеятельности клетки, то последняя долж-
на обладать практически безошибо.чно функционирующим ме-
ханизмом копирования ДНК. При самовоспроизведении клеток
каждая из дочерних клеток должна получить полную генетиче-
скую информацию в виде ДНК. Мы уже отмечали, что двуспи-
ральная модель структуры ДНК обеспечивает точное воспроиз-
ных метаболитов. Отсюда следует, что перед технологом-биохи-
миком может возникнуть очень интересная и важная задача
416
Глава 6
лементарная цепь, и в результате образуются две идентичные
молекулы ДНК, каждая из которых содержит по одной цепи
исходной ДНК- По-видимому, такая схема позволяет свести к
минимуму ошибки в процессе репликации.
На самом деле процесс репликации ДНК осуществляется бо-
лее сложным путем, чем это показано на рис. 6.11. Синтез ДНК
представляет собой сложный ступенчатый процесс; особый фер-
мент, называемый ДНК-no лимеразой, легко строит дочерние
цепи в направлении б'->3'. Этот процесс без затруднений про-
исходит на одной из родительских цепей в направлении 3'-*
Таблица 6.1. Число хромосом в клетках различных
организмов в нормальном состоянии
Организм Число хромосом
Прокариоты (гаплоидные) Бактерии Эукариоты (диплоидные) Клевер красный Пчела медоносная Пекарские дрожжи (Saccharomyces cerevisiae) Лягушка Гидра Кошка Крыса Человек Курица 1 14 16 17 26 30 38" “ 42 46 78
На другой цепи ДНК-полимераза подбирает фрагменты до-
черней цепи в направлении 5'->3' родительской цепи, а кова-
лентное связывание фрагментов осуществляет другой фер-
мент— ДНК-лигаза. Таким образом происходит и синтез вто-
рой дочерней цепи в направлении З'-’-б'.
Здесь мы вкратце рассмотрим также различия в хранении
информации и репликации между прокариотическими и эука-
риотическими ДНК. В нечетко отграниченном нуклеотиде про-
кариот имеется только одна хромосома (носитель генетической
информации), содержащая кольцевую двойную спираль ДНК.
Длина этой огромной молекулы составляет 1,2 мм, толщина
около 20 ангстрем, а ее молекулярная масса равна примерно
2,8-109. Этого достаточно для кодирования приблизительно
2000 различных белков. Кольцевая природа ДНК Е. coli впер-
вые была выяснена в ходе генетических исследований, направ-
ленных на определение относительного расположения отдельных
генов. Хромосома эукариот построена из молекулы ДНК, свя-
занной с белками и, возможно, с небольшим количеством РНК.
Молекулярная генетика и системы регуляции
417
Как показано в табл. 6.1, эукариотические клетки обычно со-
держат несколько хромосом. Некоторые эукариоты, например
дрожжи, могут быть или гаплоидными (т. е. содержащими толь-
ко по одной хромосоме каждого типа), или диплоидными,
в каждой клетке которых содержится по две хромосомы каж-
дого типа (за исключением, быть может, половых хромосом).
Ген дикого типа
— А Т—Т—С—G—А—С—1 —G—Т—А—С—G— '
— Т—A—A—G—С—T--G- -Л —С — А—Т — G- -С —
Мнсерция ।
— А—Т—Г—С—G — A—G— С— Т—G—Т—А—С—G —
— Т — А—А—G—С — Т—С — G— А—С—А—Т — G—С-
Деяеиия ? А
— А — Т—Т—С—Ci—С—Т— G—Т—А—С—G—
—т — А—А—G — С—G—А—С—А—Г—G—С--
ч
Грани ария г
— А— Т — Т—С—g4-G-\-C—Т— G—Т—А—С— G —
: ! : : । : 1 • j :
г—Т—А—А—G—C-^C-Z-G — А—С—Л — Т G С
\ /
ч >
Трансверсия
—А—Т—Т—С —G-At-A-C—Т—G—Т—А—С—G —
; : i ; । I = : : i j :
—T'^A—A—G—С-т-A-t-G—A—С—A—T—G—C--
4/
РИС. 6.12. Различные типы мутаций в последовательности пар оснований мо-
лекулы ДНК.
Мутацией называют изменение структуры ДНК, передаю-
щееся последующим поколениям. На молекулярном уровне му-
тация есть не что иное, как изменение нуклеотидной последова-
тельности ДНК. Некоторые возможные варианты мутаций изо-
бражены на рис. 6.12. В какой-то степени мутация представля-
ет собой спонтанный процесс, которому ДНК подвергается по-
стоянно. Скорость спонтанных мутаций, однако, невелика и со-
ставляет в среднем 1 ошибку на миллион дупликаций гена.
Серьезность ошибки при копировании гена зависит от ее
природы. При так называемой миссенс (изменяющей смысл)
мутации кодон одной аминокислоты изменяется таким образом,
что соответствующее место в аминокислотной последовательно-
сти белка занимает другая аминокислота. Этот тип мутации
приводит, например, к аномальному гемоглобину, характерно-
27—537
418
Глава 6
му для человека, страдающего серповидно-клеточной анемией;
в этом конкретном случае в аномальном белке вместо Glu по-
является остаток Vai. Согласно генетическому коду (рис. 6.3),
остаток Glu кодируют кодоны GAA и GAG, а валину соот-
ветствуют кодоны GUA и GUG. Отсюда следует, что замена
всего лишь одного основания может приводить к тяжелому на-
следственному заболеванию. Другие изменения в кодонах мо-
гут приводить, например, к терминирующему кодону и тем са-
мым вызывать преждевременное прекращение пептидного син-
теза; мутации такого типа называются бессмысленными (нон-
сенс-мутациями).
Предлагалось несколько механизмов спонтанных мутаций.
Во-первых, нуклеотидные основания ДНК могут существовать в
нескольких структурно различных формах, называемых тауто-
мерными. Предполагается, что изображенные на рис. 2.7 струк-
туры доминируют в нативной ДНК; вместе с тем не исключает-
ся возможность и других таутомеров, образование пар основа-
ний с которыми может приводить к ошибкам. Другая возмож-
ная причина спонтанных мутаций связана с функционированием
ферментов, необходимых для синтеза и репарации (восстанов-
ления повреждений в структуре) ДНК. Наконец, некоторые
промежуточные соединения в нормальном метаболизме клетки,
например пероксиды, азотистая кислота, формальдегид, явля-
ются мутагенами, т. е. веществами, способными индуцировать
химические мутации ДНК.
Действие химических мутагенов на ДНК широко изучалось
путем выращивания клеток в среде, обогащенной такими аген-
тами. К числу мутагенов относятся, например, соединения,
структура которых близка структуре обычных оснований ДНК.
В силу близости их строения и химических свойств в процессе
биосинтеза ДНК в нуклеотидную цепь может быть включен не
обычный нуклеотид, а его аналог. Другие типы мутагенов и спо-
собы проявления ими мутагенной активности приведены в
табл. 6.2.
Второй общей причиной мутации является радиация. В ча-
стности, ДНК интенсивно поглощает ультрафиолетовое излуче-
ние, и при достаточной дозе это излучение довольно быстро
убивает большинство клеток, а выжившие клетки подвергаются
очень глубокой мутации. Любая клетка имеет в своем распоря-
жении ферментативные механизмы для репарации ДНК, по-
врежденной ультрафиолетовым излучением. Эти ферменты до-
вольно сложным путем заменяют поврежденный сегмент, со-
держащий ковалентно-связанные пиримидиновые остатки.
В биохимической технологии явление мутации важно с не-
скольких точек зрения. Вспомните, например, рис. 1.10, на ко-
тором показано, как с помощью мутаций можно изменить при-
Молекулярная генетика и системы регуляции
419
роду микроорганизма, чтобы он стал более полезным для ка-
кой-либо определенной цели. Мутагены и ультрафиолетовое из-
лучение часто используются для создания мутантов протестов,
потенциально обладающих более высокой продуктивностью.
В любой работе по изысканию новых штаммов микроорганиз-
мов важно иметь в своем распоряжении эффективные способы
идентификации и выделения мутантов, обладающих специфиче-
скими свойствами. В табл. 6.3 перечислены некоторые основ-
ные методы, применяющиеся для этой цели. Другие подходы к
Таблица 6.2. Химические мутагены и соответствующие способы проявления
мутагенной активности3
Химический агент Мутагенный эффект
Аналоги оснований Включение в ДНК вместо обычных
оснований
Азотистая кислота Дезаминирование пуриновых и пири-
мидиновых оснований ДНК
Профлавин, акридиновый оранжевый Интеркалярное включение в стэкинг-
комплексы оснований ДНК
Алкилирующие агенты Модификация пуриновых оснований
ДНК
а Из монографии: Startler R. У., Doudoroff М., Adelberg Е. A., The Microbial World,
р. '418, Prentice-Hall, Inc., Englewood Cliffs, N. J., 1970.
генетической трансформации продуцирующих организмов мы
рассмотрим в разд. 6.2 и 6.3.
С другой стороны, мутации могут создавать в биотехнологии
и ряд затруднений. Для успеха микробиологического промыш-
ленного процесса часто необходимы чистые штаммы микроор-
ганизмов, обладающие хорошо известными характеристиками.
Вместе с тем никогда нельзя исключать возможность мутаций
в таких культурах, поэтому необходима регулярная проверка
их генетической гомогенности. Другие практические проблемы,
связанные с мутациями и иными типами генетической неста-
бильности, мы рассмотрим в последующих главах.
6.1.6. Общая схема потока информации в клетке
Известная пословица гласит, что лучше один раз увидеть,
чем сто раз услышать. Рис. 6.13, на котором в компактном ви-
де изображена схема каналов регуляции и передачи информа-
ции в клетке, еще раз подтверждает эту истину. На схеме чет-
ко видна взаимосвязь между регуляцией и передачей информа-
ции. Обратите внимание на то, что система контроля построена
27*
Таблица 6.3. Методы селекции и обнаружения различных мутантов
бактерий*
Тип мутанта
Методы селекции
Обнаружение
Способный использовать
в качестве источника
углерода соединение,
не утилизирующееся
диким штаммом
Посев па агаровой пла-
стинке, содержащей
указанное соединение
как единственный ис-
точник углерода
Метод посева на пла-
стинках абсолютно се-
лективен; только иско-
мый мутант образует
колонии
Устойчивый к химиче-
ским ингибиторам, на-
пример к пенициллину,
стрептомицину, сульф-
амидным препаратам,
красителям
Посев на агаровой пла-
стинке, содержащей
ингибитор
Метод посева на пла-
стинках абсолютно се-
лективен; только иско-
мый мутант образует
колонии
Устойчивый к бактерио-
фагу
Посев на агаровой пла-
стинке, предварительно
обработанной суспензи-
ей фага
Метод посева на пла-
стинках абсолютно се-
лективен; только ис-
комый мутант образу-
ет колонии
Ауксотрофные (требую-
щие один или несколь-
ко факторов роста, не
являющихся необхо-
димыми ДЛЯ дикого
штамма)
Инкубирование в отсут-
ствие фактора роста,
но в присутствии пени-
циллина; клетки дико-
го типа размножаются
и большей частью по-
гибают; ауксотрофные
мутанты в отсутствие
фактора роста не раз-
множаются и выжива-
ют (пенициллин уби-
вает только активно
растущие клетки)
Посев на агаровой пла-
стинке в отсутствие
фактора роста; после
образования колоний
дикого типа их отмеча-
ют, а затем добавля-
ют слой агара, содер-
жащего фактор роста;
ауксотрофы образуют
колонии только после
добавления фактора
роста
Неспособный утилизиро-
вать данный углевод
Применим описанный вы-
ше пенициллиновый
метод с использовани-
ем в качестве единст-
венного источника уг-
лерода указанного уг-
левода
Посев на агаровой пла-
стинке, содержащей
указанный углевод и
химический индика-
тор, который изменяет
окраску в присутствии
осуществляющих бро-
жение клеток (напри-
мер, кислотно-основ-
ной индикатор); му-
тантные колонии вы-
деляются специфиче-
ской окраской
Молекулярная генетика и системы регуляции
421
Тип мутанта Методы селекции Обнаружение
Неспособный синтезиро- Инкубируют суспензию Испытание колонии на
вать ДНК при повы- клеток при 42 °C в те- способность к росту
шейной температуре чение 15 мин, добавля- ют 5-бромурацил (5-BU) и продолжают инкубирование еще 60 мин, облучают све- том с Х = 310 нм; при этом клетки, ДНК ко- торых содержит 5-BU, погибают, а клетки, ДНК которых не под- вергалась репликации при 42 °C, выживают при 42 °C, например, методом перепечатки
Чувствительные к ульт- Инфицируют культуру Приготовление суспеи-
рафиолетовому облу- слегка инактивирован- зий из изолированных
чению (неспособные к ным ультрафиолетом колоний; определение
репарации поврежде- бактериофагом; нор- выживания при раз-
ний, индуцированных мальные клетки вое- личных дозах ультра-
ультрафиолетом) станавливают индуци- рованные ультрафиоле- том повреждения фага и впоследствии погиба- ют от того же фага; неспособные репари- ровать повреждения клетки выживают фиолетового излучения.
а Stonier R. У., Doudoroff М., Adelberg Е. A., The Microbial World, 3nd ed., p. 450, Prentice-Hall, Inc., Englewood Cliffs, N. J., 1970.
по каскадному типу: активация и ингибирование (на уровне
ферментов) позволяют клетке быстро приспособиться к кратко-
временным изменениям в химическом балансе клетки, в то вре-
мя как индукция и репрессия (на уровне генов) дают возмож-
ность изменить всю картину метаболизма в тех случаях, когда
однажды изменившиеся условия и состав среды далее стабили-
зируются на достаточно длительный период.
6.2. Модификация структуры клеточной ДНК
Не только мутации, но и многие другие процессы могут из-
менять генетический материал клетки. С точки зрения промыш-
ленной биотехнологии эти процессы могут иметь как положи-
тельный, так и отрицательный эффекты. С одной стороны, лю-
бой метод изменения структуры клеточной ДНК может быть по-
лезен для получения более продуктивных штаммов. С другой
422
Глава 6
РИС. 6.13. На этой схеме, изображающей информационные потоки в клетке,
можно выделить различные уровни систем регуляции, передачи и переработки
информации. (Лёви А., Сикевиц Ф., Структура и функция клетки. — М..: Мир,
1971).
стороны, неожиданная модификация определенной ДНК может
вызвать крайне нежелательные последствия для промышленно-
го биотехнологического процесса.
Большая часть приведенного ниже материала относится толь-
ко к бактериям, причем в большинстве случаев соответствую-
щие исследования проводились на Е. coli. Прежде всего рас-
смотрим методы слияния клеток, позволяющие осуществлять ге-
нетические модификации, а разд. 6.4 посвящен методам с при-
менением рекомбинантных ДНК и их использованию. Мы не
будем рассматривать сложные специфические процессы половой
рекомбинации, которые оперируют во многих эукариотах. Крат-
кое введение в эту тему в контексте промышленно важных мик-
роорганизмов можно найти в книге Эландера [4]-
Молекулярная генетика и системы регуляции
423
6.2.1. Вирусы и фаги; лизогения и трансдукция
Мы знаем, что вирусы вызывают целый ряд заболеваний че-
ловека, в том числе обычные респираторные заболевания, грипп,
оспу, полиомиелит, желтую лихорадку, свинку, корь и многие
другие. Они способны поражать и бактерии; вирусное зараже-
ние может крайне неблагоприятно влиять на различные про-
мышленные биологические процессы с участием бактерий, в ча-
стности в сыроделии и производстве антибиотиков. Вирусы, за-
ражающие бактерии, называют бактериофагами или просто
фагами.
Вирусные частицы построены из белков и нуклеиновой кис-
лоты и могут включать, кроме того, липопротеины. Основная
функция вирусных белков заключается в создании оболочки
(капсида), защищающей вирусную нуклеиновую кислоту,
а иногда в адсорбции вируса на поверхностных рецепторах жи-
вых клеток. Вирусное заражение и его последствия обусловли-
ваются именно нуклеиновой кислотой ДНК или РНК. В табл.
6.4 перечислены свойства ряда вирусов; вирусная нуклеиновая
кислота может быть одно- или двухцепочечной, линейной или
кольцевой.
Вне живой клетки определенного типа вирус представляет
собой инертную частицу, не способную к самовоспроизведению.
Вирус размножается только после проникновения в клетку-хо-
зяина. Таким образом, вирусы являются типичными паразита-
ми. Короткая сводка основных различий между вирусами и жи-
выми клетками приведена в табл. 6.5.
Теперь рассмотрим типичный «жизненный цикл» вирусов на
примере инфицирующего бактерию Е. coli вируса, называемого
фагом Л. Фаг прикрепляется к клеточной стенке и затем вводит
свою ДНК в клетку. Начиная с этого момента возможны два
альтернативных пути. В одном из этих путей, называемом лизо-
генией, ДНК фага (ее в этом случае называют профагом) внед-
ряется в хромосому бактерии. В этом случае клетка нормаль-
но живет и самовоспроизводится, одновременно копируя про-
фаг и создавая таким образом новые лизогенные клетки. По-
добные фаги, находящиеся в лизогенном взаимодействии с
клеткой-хозяином, называют умеренными.
Другой возможный вариант возникновения заражения клетки
фагом называется литическим циклом; он неизбежно приводит
к гибели клетки-хозяина. Инфекция литическими фагами, на-
пример фагом Т2, всегда приводит к литическому циклу. Во вре-
мя литического цикла фаг находится в так называемом веге-
тативном состоянии, когда ДНК фага, по сути дела, контроли-
рует все клеточные процессы. Прежде всего фаговая ДНК за-
ставляет рибосомы синтезировать ферменты, разрушающие
Таблица 6.4. Характеристики некоторых вирусов"
Тип вируса
Размеры, нм Форма, состав и другие сведения о вирусе
Вирусы животных:
Кубические:
вирус полиомиелита
30 Построен из одной спиральной молекулы РНК с мол. массой около
2-10°, которая окружена молекулами белка, образующими поверх-
ность икосаэдра; наружной мембраны не имеет; молекулярная мас-
са частиц 10-10s
Спиральные:
вирус гриппа
100 Построен из одной молекулы РНК (мол. масса около 2-Ю6); спираль-
ная молекула нуклеопротеина окружена липопротеиновой оболочкой;
молекулярная масса частиц 100-106
Вирусы растений:
Палочкообразные:
вирус табачной мозаики 300X15 Вирусная частица, имеющая палочкообразную форму, построена из
1 молекулы РНК (мол. масса около 2-Ю6), связанной с белковыми
молекулами; нуклеопротеидный комплекс имеет спиральную струк-
туру; мол. масса частиц 39-Ю6
Сферические:
вирус карликовости томата
30 Икосаэдр; содержит 16% РНК (мол. масса 1,6-106) и белок; мол. мас-
са частиц 9•1О6
Продолжение
Тип вируса Размеры, нм Форма, состав и другие сведения о вирусе
Вирусы насекомых: вирус тутового шелкопряда 280X40 Собственно вирус имеет палочкообразную форму и содержит около 8% ДНК (в расчете на массу сухих веществ), но in vivo вирусные палочки включены в полиэдрические кристаллические агрегаты диа- метром 0,5—15 мкм
Бактериофаги: С двухцепочечпой ДНК: Т-четный Е. coll головка отросток (хвост) нити отростка 90X60 100X25 130X2,5 Внешне напоминает головастика; содержит ДНК (с мол. массой 130- •106) в головке; некоторые из белков фага способны сокращаться, а длинные нити участвуют в присоединении к клетке-хозяину (мол. масса частиц 250-106)
С одноцепочечной кислотой:- С ДНК: 0X174 Е. coli нуклеиновой 22 Додекаэдр, построенный из 12 субъединиц; содержание ДНК (мол. масса 1,6-106) 25% (в расчете па массу сухих веществ); мол. масса частиц 6,2 - 106
с РНК: f2 Е. coli 20 Многогранник; состоит из РНК (3-10~12 мкг/вирус) и белка; содержа- ние нуклеиновой кислоты, вероятно, такое же, как в фаге 0X174
а
Aiba S„ Humphrey A. E„ .Millis N, F., Biochemical Engineering, 2nd cd., p. 19, Academic Picss, Inc., New York, 1973.
426
Глава 6
ДНК клетки-хозяина (эта особенность, впрочем, присуща не
всем вирусным инфекциям) и содействующие многократному
копированию фаговой ДНК. Затем синтезируются белки, необ-
ходимые для формирования интактных частиц фага. Эти белки
спонтанно соединяются с ДНК фага, образуя зрелые высокоор-
ганизованные частицы бактериофага. По завершении процесса
самосборки множества новых фагов начинается синтез фермен-
та лизоцима. Как мы уже знаем, этот фермент разрушает му-
реин клеточных стенок бактерий. Таким путем осуществляется
лизис, в результате которого клетки разрушаются, высвобож-
дая множество фагов.
Таблица 6.5. Сравнение свойств вирусов и клеток
Вирус Клетка
Содержит нуклеиновую кислоту только одного тина Содержит очень ограниченное число ферментов Воспроизводится путем самосборки компонентов (нуклеиновых кислот и белков), синтезируемых клеткой- хозяином Содержит ДНК и РНК Содержит тысячи ферментов Самовоспроизводится строго опреде- ленным, регулируемым путем
Уже этот краткий обзор природы фагов достаточно ясно по-
казывает, почему фаговая инфекция промышленных культур
бактерий может представлять собой серьезную проблему. Час-
тицы фагов очень малы, в соответствующей среде они могут
размножаться очень быстро, и в то же время в состоянии отно-
сительного покоя — лизогении их обнаружение может быть не-
простой задачей.
Иногда в процессе репродукции фага в клетке-хозяине обра-
зуются фаговые частицы, содержащие небольшой фрагмент
(примерно от 1 до 2%) хромосомы клетки-хозяина. Когда такой
трансдуцирующий фаг вводит свою ДНК в другую бактерию,
перенесенная из первой клетки ДНК может подвергнуться крос-
синговеру с фрагментом хромосомы нового хозяина. Результа-
том этого процесса, называемого трансдукцией, может быть
изменение генетических характеристик клетки — нового хозяи-
на фага. Обычно кроссинговер осуществляется только между
фрагментами ДНК, содержащими гомологичные нуклеотидные
последовательности, которые контролируют сходные характери-
стики клеток. Например, в результате трансдукции фаг может
перенести небольшой фрагмент ДНК, обеспечивающий усвоение
лактозы, от утилизирующего лактозу штамма Е. coli к другому
Молекулярная генетика и системы регуляции
427
а) Фаг заражает
клетку
5) Некоторые из образо-
вавшихся после зара-
жения <рагов несут
в севе фрагмент клеточ-
ной хромосоме/ ; фаги
высвобождаются в ре-
зультате лизиса клетки
в) Фаг, содержащий фраг-
мент бактериалоной
- хромосомы, заражает
другую клетку
г) Рекомбинация между сег- .
ментом бактериальной ДНК
которую несет фаг, и. гомоло-
гичным сегментом ДНК второй
клетки
д) Генетически
модифицирован^
кая Га к терпя
РИС. 6.14. Схематическое изображение модификации бактериальной хромосо-
мы путем трансдукции — процесса, протекающего при участии бактериофагов.
штамму Е. coli, неспособному выполнять эту операцию. На рис.
6.14 приведена упрощенная схема трансдукции и кроссинговера
сегментов ДНК-
Известно, что трансдукцию легко претерпевают многие акти-
номицеты. В ходе изучения трансдукции в Streptomyces griseus
с помощью фагов удалось передать способность синтезировать
антибиотик стрептомицин от продуцирующего к непродуцирую-
щему штамму. Некоторые из подвергшихся трансдукции куль-
тур обладали большей продуктивностью в отношении антибиоти-
ка, чем даже исходные синтезирующие штаммы.
428
Глава 6
6.2.2. Трансформация и конъюгация в бактериях
Под трансформацией понимают процесс переноса генетиче-
ской информации из одной клетки в другую в виде свободной
ДНК. В этом процессе фрагмент двухцепочечной ДНК прони-
кает в клетку-реципиент, находящуюся в специфическом состоя-
нии, когда она способна включать чужеродную ДНК (такие
клетки называют компетентными). Если перенесенный фрагмент
аналогичен фрагменту ДНК реципиента, то хромосома послед-
него быстро трансформируется путем кроссинговера.
Механизм трансформации позволяет включать в живые бак-
терии не только фрагменты хромосом, но и плазмиды. В разд.
2.3 мы узнали, что плазмидой называется молекула ДНК, суще-
ствующая и реплицирующаяся независимо от бактериальной
хромосомы. Плазмиды обычно представляют собой относитель-
но небольшие кольцевые молекулы с молекулярной массой по-
рядка 106—108 (рис. 6.15). Трансформация с помощью плазмид
является одним из основных приемов в методе направленного
изменения структуры ДНК, рассматриваемом в разд. 6.3.
Плазмиды обычно не выполняют каких-либо существенных
функций в нормальных условиях жизнедеятельности клетки, но
в то же время могут обеспечивать клетку рядом полезных
(в специфических условиях) качеств. Установлено, например,
что плазмиды, называемые R-факторами (факторами устойчи-
вости), обусловливают устойчивость бактерий к антибиотикам.
Плазмидам близки так называемые эписомы — молекулы
ДНК, которые могут или быть составной частью клеточной хро-
мосомы, или существовать независимо от нее. Один из наиболее
известных примеров эписом — фактор F (фактор фертильности),
характеризующий клетки Е. coli при их конъюгации. В процес-
се конъюгации Е. coli содержащие F-фажтор клетки (Е+-клет-
ки) передают его клеткам, не имеющим фактора F ^“-клет-
кам); иногда и часть хромосомы Е+-клетки также передается
F^-клетке. Аналогичным образом могут передаваться и R-фак-
торы. Впрочем, некоторые Е+-мутанты, обозначаемые Hfr, ведут
себя по-иному; они не инфицируют Е~-клетку F-фактором, а пе-
редают ей большую часть своей хромосомы, которая в резуль-
тате кроссинговера с хромосомой Е~-клетки образует генетиче-
ски модифицированные штаммы.
Многочисленными экспериментами показано, что хромосома
Е+-клетки проникает в Е--клетку в линейном порядке в виде
нити; если процесс прерывать через различные промежутки вре-
мени, то в Е~-клет.ках будут находиться участки хромосомы
различной длины. Последующее сравнение характеристик попу-
ляций клеток-реципиентов, содержащих различные количества
хромосомного материала из Е+-клеток, позволяет выяснить рас-
Молекулярная генетика и системы регуляции
429
РИС. 6.15. Электронная микрофотография плазмиды pSClOl (Х230 000). Эта
кольцевая ДНК, существующая и реплицирующаяся независимо от бактери-
альной хромосомы, была важным инструментом в первых экспериментах с ре-
комбинантными ДНК. (Микрофотография предоставлена д-ром С. Н. Коуэном.)
положение генетического материала в хромосоме Е+-клетки.
Именно таким путем были получены детальные генетические
карты типа изображенной на рис. 6.16. На этой карте обозначе-
ны относительные положения характерных для различных му-
тантов генов. Например, символ thr указывает ген, характер-
ный для мутанта, 'который растет только в присутствии треони-
на. Расшифровка многих подобных обозначений приведена в
монографии Уотсона (ссылка [7] в гл. 2).
6.2.3. Слияние клеток
Глубокие генетические изменения, включающие кроссинго-
вер генетического материала различных видов, могут быть до-
стигнуты путем слияния клеток разных типов. Как показано на
схеме слияния клеток микроорганизмов (рис. 6.17), первой ста-
430
Глава 6
РИС. 6.16. Генетическая карта Е. coll К12, на которой показаны относитель-
ные положения различных генов в кольцевой хромосоме. [Воспроизведено из
статьи: Taylor A. L., Trotter С. D., Revised Linkage Map of E. coli, Bacteriol.
Rev., 31, 332 (1967).]
дней этого сложного процесса является получение протоплас-
тов— клеток, лишенных наружных стенок и окруженных только
плазматическими мембранами. Для этой цели применяют раз-
личные ферменты (гидролазы клеточных стенок), а процесс гид-
ролиза проводят в строго гипотонической среде, чтобы свести к
минимуму разрушение протопластов под влиянием осмотиче-
ского давления. Инкубирование протопластов приводит к реге-
нерации клеточных стенок и восстановлению нормальной кле-
точной морфологии. Это обстоятельство играет решающую роль
во всем процессе, поскольку в результате слияния протоплас-
тов различных штаммов нам, очевидно, необходимо в конце
Молекулярная генетика и системы регуляции
431
Плетки ,
(штамм 1 а~ Ь~ с~)
н
Обработка
литическим рерментом\
Плетки
(штамм 2 cP е~
Обработка
литическим ферментом -
Протопласты
Протопласты
Лосев на пластинках с ми-
нимальной средой для про-
дерни на обратную мутацию
и с полной средой для опреде-
ления жизнеспособности
Лосев на пластинках с ми-
нимальной средой для про-
верки на обратную мутации
и с полной средой для опре-
деления жизнеспособности
Гибридные протопласты
(не менее /• (0 s клеток
каждого штамма)
Обработка
полиэтиленгликолем
Промывка и (или) разводе -'
ние гипертоническим раст-
вором
Реверсия протопласта#
в минимальной среде
Реверсия протопластов
в полной среде
РИС. 6.17. Основные этапы работы по получению и выделению новых штам-
мов путем слияния протопластов двух различных исходных штаммов. [Воспро-
изведено из статьи: Peberdy J. F., Protoplast Fusion —A Tool for Genetic
Manipulation and Breeding in Industrial Microorganisms Enzyme Microb
Technol., 2, 25 (1980).]
концов получить организм, который можно было бы далее
культивировать в обычных условиях, в том числе и в крупно-
масштабном промышленном производстве. Слияние протоплас-
тов обычно осуществляют в растворе полиэтиленгликоля (ПЭГ),
который индуцирует этот процесс, а для выделения продуктов
слияния протопластов применяют методы селекции, аналогич-
ные использующимся в традиционной генетике микроорганиз-
мов (табл. 6.3).
432
Глава 6
Описанным путем успешно получали гибридные клетки раз-
личных штаммов Penicillium, Aspergillus, Streptomyces, Cepha-
losporium, дрожжей и бактерий. Межвидовые гибриды получе-
ны, в частности, между различными видами Penicillium, Asper-
gillus, между Saccharomyces cerevisiae и Saccharomyces diasta-
ticus, между Candida tropicalis и Saccharomyces fibuligera. Гиб-
риды существенно различающихся видов недостаточно стабиль-
ны и через несколько поколений часто превращаются в один из
родительских видов, однако в целом метод слияния протоплас-
тов представляет собой очень интересный способ существенной
перестройки генетического материала клетки в одну стадию.
В настоящее время проводится изучение возможности слияния
протопластов быстро растущих штаммов Streptomyces с мутант-
ными высокопродуктивными, но медленно растущими штам-
мами.
Метод слияния клеток может применяться и для получения
гибридов клеток растительных или животных организмов. При-
мером могут служить имеющие уже сейчас большое практиче-
ское значение и очень перспективные гибридомы, образующие-
ся при слиянии продуцирующих антитела клеток лейкоцитов с
клетками миеломы (рака кожи) мыши или другого животного.
Каждая гибридомная клетка синтезирует только один тип анти-
тел, поэтому полученная из одной гибридомы культура клеток
продуцирует моноклональные антитела. Напротив, если антите-
ла получают путем введения в организм, например кролика,
белка или другого антигена, то этот организм одновременно
синтезирует множество антител, специфичных в отношении раз-
личных участков молекулы антигена. Моноклональные антите-
ла уже в настоящее время представляют собой важный инстру-
мент исследования и служат основой для применяемых сейчас
или разрабатываемых многочисленных методов диагностирова-
ния заболеваний. В иммобилизованной форме моноклональные
антитела являются чрезвычайно специфичными аффинными ад-
сорбентами, пригодными для выделения и очистки антигенов в
лабораторных и промышленных масштабах. В перспективе мо-
ноклональные антитела могут оказаться очень ценными агента-
ми для интроскопии опухолей и транспорта лекарственных пре-
паратов к определенным типам клеток организма.
Небольшие количества моноклональных антител удобно по-
лучать путем инокуляции гибридомных клеток в брюшную по-
лость иммунологически совместимой мыши. Развивающаяся
опухоль выделяет моноклональные антитела в окружающие
опухоль жидкости организма; концентрация антител в послед-
них достигает 1—20 мг/мл. Гибридомы можно выращивать и в
культуре. В настоящее время большое внимание уделяется раз-
работке и внедрению в производство новых конструкций реак-
Молекулярная генетика и системы регуляции
433
торов, позволяющих повысить плотность клеток и их продуктив-
ность в отношении антител, а также поиску путей снижения
стоимости моноклональных антител при их крупномасштабном
производстве.
6.3. Применение результатов исследования генетики
микроорганизмов и популяций мутантов
в промышленности
Достижения молекулярной биологии и результаты изучения
закономерностей систем регуляции клеточных процессов име-
ют важные практические приложения. Некоторые из этих при-
ложений, связанные с возможностью определения положения
нужного или вредного гена и с последующими попытками раз-
личными методами модифицировать этот ген, были рассмотре-
ны в предыдущем разделе. Другой подход основан на индуци-
ровании мутации в более продуктивный штамм; о результатах
такого подхода мы уже упоминали в гл. 1 в связи с повышени-
ем выхода пенициллина за счет использования особых штам-
мов, полученных путем ряда последовательных мутаций. В за-
ключительной части настоящего раздела мы рассмотрим ряд
других микробиологических процессов, основанных главным об-
разом на применении специально выведенных мутантных
штаммов.
Часто повышенная продуктивность мутантного микроорга-
низма легко объясняется изменениями в основных системах ре-
гуляции клетки. Понимание механизмов, контролирующих био-
синтез необходимого продукта метаболизма, важно еще и по-
тому, что только на этой основе можно принять обоснованное
решение об обеспечивающем максимальную продуктивность
составе питательной среды и о способах ее контроля. Влияние
среды на продуктивность клеток мы и рассмотрим в первую
очередь.
6.3.1. Системы клеточной регуляции;
влияние состава среды
Существуют два подхода к решению проблемы повышения
продуктивности клеток путем изменения состава среды: во-пер-
вых, можно ввести в среду индуцирующие вещества и, во-вто-
рых, попытаться снизить количество репрессора. На первый
взгляд оба этих подхода довольно очевидны, однако их практи-
ческое использование может быть связано с рядом трудностей.
Чрезвычайно эффективными индукторами биосинтеза фер-
ментов помимо самого субстрата могут быть не подвергающие-
8—537
434
Глава 6
ся метабо. ическим превращениям аналоги субстратов. Напри-
мер, галактозиды, в частности изопропил-^-Э-тиогалактозид,
могут повышать удельную ^-галактозидазную активность Е. co-
li более чем в 1000 раз. Аналоги других субстратов эффективны
в отношении соответствующих ферментов; так, N-ацетилацет-
амид повышает амидазную активность, метициллин ускоряет
биосинтез пенициллин-р-лактамазы, а малоновая кислота спо-
собствует повышению малеатизомеразной активности.
Известно, что во многих практически важных процессах био-
синтез продукта метаболизма может тормозиться за счет ката-
болитной репрессии. Доступность быстро усваиваемого субст-
рата, например сравнительно концентрированных растворов
глюкозы, стимулирует рост клеток, но в то же время тормозит
синтез антибиотиков пенициллина, митомицина, бацитрацина и
стрептомицина. Выход пенициллина значительно повышается
в условиях искусственной диауксии (рис. 7.14), т. е. при добав-
лении к глюкозной питательной среде медленно метаболизирую-
щегося сахара типа лактозы. В таких условиях биомасса растет
на глюкозе, а синтез пенициллина осуществляется по мере ус-
воения лактозы. Тот же самый эффект может быть достигнут и
другим путем, если глюкозу медленно добавлять к питательной
среде.
В качестве других примеров репрессоров, концентрации ко-
торых легко регулируются путем изменения состава среды, мож-
но привести неорганический фосфат, подавляющий синтез фос-
фатазы в Е. coli и нуклеазы в Aspergillus quernicus, а также
аммиак, подавляющий биосинтез уреазы в Proteus retgeri. На-
пример, путем снижения концентрации фосфатного иона в пи-
тательной среде можно повысить содержание щелочной фосфа-
тазы в Е. coli от практически нулевого уровня до примерно 5%
от массы всех белков клетки. Аналогично снижение концентра-
ции аминокислот или сульфатного иона существенно ускоряет
синтез протеаз бактериями или водорослями Aspergillus niger
соответственно.
Труднее поддерживать низкую концентрацию репрессора,
если последний синтезируется микроорганизмом. Один из воз-
можных подходов к решению этой задачи сводится к модифи-
цированию клеточной мембраны таким образом, чтобы репрес-
сор легко диффундировал из клетки в среду. Такой подход ис-
пользуется, в частности, в очень эффективном производстве
глутаминовой кислоты с участием Corynebacterium glutamicum,
когда выделение глутаминовой кислоты в среду индуцируется
снижением концентрации биотина. В присутствии избытка био-
тина концентрации других аминокислот внутри клетки достига-
ют максимума. Хотя механизм этого эффекта не известен, не-
которые экспериментальные факты свидетельствуют о том, что
Молекулярная генетика и системы регуляции
435
биотин влияет на фосфолипидные компоненты клеточной мемб-
раны.
В следующем разделе мы рассмотрим другие методы борьбы
с отрицательными эффектами репрессоров.
6.3.2. Применение ауксотрофных мутантов
Ауксотрофные мутанты (методы их селекции и идентифика-
ции приведены в табл. 6.3) отличаются отсутствием ферментатив-
ной активности, необходимой для осуществления одной или не-
скольких стадий в каком-либо из биосинтетических путей. В ре-
зультате ауксотрофы не способны синтезировать конечный про-
I---------Г
Фермент а ] Фермент
II-------------1
— Фермент с Фермент d ।
С о ооооо D--------—*-Е
РИС. 6.18. Для повышения выхода промежуточных продуктов какого-либо
пути метаболизма можно использовать ауксотрофные мутанты. В приведен-
ном здесь гипотетическом примере у мутанта отсутствует фермент с и, следо-
вательно, не синтезируется репрессор Е. (Воспроизведено из работы: De-
main A. L., Overproduction of Microbial Metabolites and Enzymes due to Alte-
ration of Regulation, in Advances in Biochemical Engineering 1, Ghose T. K.,
Fiechter A. (eds.), p. 120, Springer-Verlag, New York, 1971.)
дукт этого пути метаболизма. Такой мутант может выжить
только при наличии в питательной среде несинтезируемого ме-
таболита. Например, триптофановый ауксотроф будет расти
только в присутствии триптофана. Если штамм сам синтезирует
триптофан, то его называют прототрофным по триптофану.
Поскольку отсутствующие метаболиты вводятся в питатель-
ную среду, а не синтезируются ауксотрофом, то концентрации
этих метаболитов регулирует, по сути дела, не микроорганизм,
а инженер-технолог. С практической точки зрения такая ситуа-
ция может быть очень удобной, если несинтезируемый продукт
метаболизма является репрессором. Как показано на рис. 6.18,
неспособные к синтезу репрессора ауксотрофные мутанты мож-
но заставить перепроизводить* тот или иной промежуточный
* Под «перепроизводством» здесь понимается повышенная скорость био-
синтеза вещества по сравнению с оптимальной скоростью, необходимой для
роста клетки. Оптимальная для клетки скорость биосинтеза, однако, не обес-
печивает выход, достаточно высокий для промышленного производства. Имен-
но поэтому в промышленности используют мутантные штаммы с нарушенными
системами регуляции.
28*
436
Глава 6
продукт метаболизма. Поддерживая в среде низкую концентра-
цию репрессора Е, можно свести к минимуму процессы ингиби-
рования по принципу обратной связи и подавления активности
ферментов (участвующих в данном пути метаболизма). При этом
промежуточное соединение С (обычно являющееся субстратом
фермента с, отсутствующего в данном мутанте) образуется в го-
раздо более высокой концентрации, чем в нативном организме.
Аспартат
।---------------Аспартокиназа
। Аспартиаироссрат
flam/албдегид аспарагиновой кислоты
I i/ icfiUiju
О
о
РИС. 6.19. Мутант С. glutamicum, у которого отсутствует фермент гомосерин-
дегидрогеназа, продуцирует повышенные количества L-лизина. (Воспроизве-
дено из работы: Demaiti A. L., Overproduction of Microbial Metabolites and
Enzymes due to Alteration of Regulation, in Advances in Biochemical Engi-
neering 1, Ghose T. K., Fiechter A., (eds.), p. 122, Springer-Verlag, New York,
1971.)
Как мы уже указывали в конце предыдущего раздела, в подоб-
ной ситуации может возникнуть потребность в дополнительных
операциях, обеспечивающих диффузию промежуточного веще-
ства С через клеточную мембрану в среду.
На рис. 6.19 изображен механизм, лежащий в основе про-
мышленного способа производства ь-лизина с участием ауксо-
трофного мутанта С. glutamicum. В высокопродуктивном мутан-
те отсутствует фермент гомосериндегидрогеназа, поэтому инги-
бирование синтеза лизина треонином (через аспартокиназу)
у этого мутанта не наблюдается. Поскольку ауксотрофный му-
тант не синтезирует ни треонин, ни метионин, то обе эти амино-
кислоты должны добавляться к питательной среде.
Молекулярная генетика и системы регуляции 437
Сравнение регуляции путей биосинтеза аминокислот семей-
ства аспарагиновой кислоты в С. glutamicum (рис. 6.19) и в
Е. coli (рис. 5.12) приводит к важному выводу: организмы, об-
ладающие сходными путями биосинтеза, не обязательно имеют
идентичные системы регуляции. Обратите внимание, в частно-
сти, на то, что на схеме рис. 5.12 лизин автоингибирует свой
собственный биосинтез с помощью реакции, ведущей к дигид-
ропиколиновой кислоте. В диком (немутантном) штамме С. glu-
tamicum соответствующая стадия ингибирования отсутствует.
Обнаруженная в Corynebacterium аспартокиназная система, по-
видимому, отличается от системы Е. coli тем, что первая подав-
ляется только при совместном присутствии лизина и треонина
(согласованное или поливалентное ингибирование по принципу
обратной связи), а изоферменты аспартокиназы в Е. coli инги-
бируются одним лизином. Именно по этим причинам с промыш-
ленной точки зрения выгоднее использовать Corynebacterium.
Аналогичный подход успешно применялся при разработке
высокоэффективных микробиологических процессов производст-
ва вкусовых веществ— пуриновых нуклеотидов гуанозинмоно-
фосфата (GMP), инозинмонофосфата (IMP) и ксантинмонофос-
фата (ХМР). Как показано на рис. 6.20, минимизация концент-
раций АМР и GMP с помощью ауксотрофного мутанта Brevi-
bacterium ammoniagenes позволяет интенсифицировать биосин-
тез инозина и гипоксантина. Концентрация последних в среде
возрастает при добавлении небольших количеств (около 10 мкг/л)
ионов марганца (Мп2+). Имеющиеся в настоящее время дан-
ные показывают, что марганец изменяет проницаемость клеточ-
ных мембран.
6.3.3. Мутанты с модифицированными системами регуляции
Более выгодные с точки зрения их промышленного примене-
ния штаммы микроорганизмов получали также, модифицируя
системы регуляции иа уровне ферментов и (или) генов с по-
мощью иных генетических трансформаций и селекции. Мутанты
такого типа можно использовать для повышения выходов про-
дуктов метаболизма и ферментов.
Чтобы добиться перепроизводства метаболита, обладающего
ингибиторными и (или) репрессорными свойствами в своем пу-
ти биосинтеза, необходимо искать мутантные организмы, алло-
стерические ферменты и опероны которых нечувствительны к
присутствию метаболита. Часто такие мутанты удается, выде-
лить с помощью антиметаболитов — токсичных аналогов изу-
. чаемых метаболитов. В содержащей антиметаболит среде нор-
мальные клетки не растут, поскольку антиметаболит подавляет
или ингибирует биосинтез необходимого метаболита, не будучи
438
Глава 6
Фосууоибозилппророс<рат I
Фосрорибозилакин
^Гипоксантин 1
(9реакций)
Инозин
Инозинмоносросфат
' (IMP) \
Аденила -
сукцинат
' Аденозинмонофоссрат
(АМР)
Ксантинионофосфат
(ХМР)
Гуанозин/нзно/россрат.
(GMP)
Аденозинтриросрат
(АТР)
а) Регуляция биосинтеза нуклеотидов
Фосрорибозилпироросрат
Гипоксантин
Фосрорибозиламин
Инозин |
Инозинноноростат
блокирование z±z i блокирование
Аденило-
сукцинат
ХМР
AMP GMP
5) Накопление инозина и гипоксантина'
РИС. 6.20. В нормальной клетке АМР и GMP ингибируют последовательность
реакций биосинтеза нуклеотидов по принципу обратной связи на нескольких
стадиях (я). В мутанте, не способном синтезировать АМР и GMP, концен-
трацию этих веществ при соответствующем составе среды можно поддержи-
вать на достаточно низком уровне, обеспечивая таким путем повышение выхо-
да инозина и гипоксантина (б). (Аябя Ш., Хемфри А., Миллис Н., Биохими-
ческая технология и аппаратура. — М.: Пищевая промышленность, 1975.)
Молекулярная генетика и системы регуляции
439
способным выполнять функции несинтезирующегося метаболита
или заменять его в последующих путях метаболизма. Напротив,
картина биосинтеза и скорости реакции у штаммов с нарушен-
ной системой регуляции по принципу обратной связи в присут-
ствии антиметаболита не изменяются, поэтому такие штаммы
нормально развиваются и в среде, содержащей антиметаболит.
В табл. 6.6 перечислены некоторые продукты жизнедеятельно-
Таблица 6.6. Продукты метаболизма и соответствующие антиметаболиты,
применение которых в селекции микроорганизмов позволяет выделить более
продуктивные мутантыа
Продукт метаболизма Аитиметаболит Продукт метаболизма Антиметаболит
Аргинин Канаванин Фенилаланин п-Фторфенилала- нин Тиенилаланин Тирозин п-Фторфенилала- нин Тиенилаланин D-Тирозин Триптофан 5-Метилтрипто- фан 6-Метилтрипто- фан Валин а-Аминобутират Изолейцин Валин Лейцин Трифторлейцин 4-Азалейцин л Detnain A. L., Overproduction of Mier lation, in Advances in Biochemical Engineei p. 113, Springer-Verlag, New York, 1971. Треонин а-Амино-8-гидр- оксивалерат Метионин Этионин Норлейцин а-Метил метионин ь-Метионин-оь- сульфоксимин Гистидин 2-Тиазолаланин 1,2,3-Триазол-3- аланин Пролин 3,4-Дегидропролин Аденин 2,6-Диаминопурин Урацил 5-Фторурацил i ohiai Metabolites due to Alteration in Regu- ing, vol. 1. Ghose T. K., Fiechter A. (eds.).
сти микроорганизмов, выходы которых можно повысить таким
путем.
Устойчивость к антиметаболитам часто обусловлена различ-
ными изменениями в системах регуляции, например частичным
или полным восстановлением активности ферментов, каталити-
ческие свойства которых в нормальном пути биосинтеза подав-
ляются. В частности, с помощью антиметаболитов можно так-
же вести поиск мутантов с необычно высокими концентрациями
биосинтезирующих ферментов. Подобный подход, например, мо-
жет оказаться целесообразным при разработке процессов про-
изводства ферментов, используемых затем в других биосинтети-
ческих процессах in vitro.
Для конститутивных мутантов характерно то, что биосинтез
индуцибельных или репрессибельных ферментов не зависит от
наличия или отсутствия индуктора или репрессора. Путем не-
440
Глава 6
пользования конститутивных мутантов можно повысить выходы
следующих ферментов: ^-галактозидазы, каталазы, фосфатаз,
протеаз, гомосериндегидрогеназы, инвертазы, гистидазы, пени-
циллиназы и амидазы.
В приведенном выше обсуждении мы уделяли основное вни-
мание различным подходам к повышению выхода интересующих
нас веществ. Те же самые молекулярно-биологические подходы
могут служить основой для поиска путей биосинтеза производ-
ных нормальных метаболитов, а также совершенно новых ве-
ществ, которые, возможно, найдут применение в фармакологии
и в других областях. Эта тема вкратце рассмотрена в обзорах
[4—6]. Примером практически важного процесса может слу-
жить биосинтез тетрациклинового антибиотика 6-деметилтетра-
циклина мутантньш штаммом Streptomyces aureofaciens. 6-Де-
метилтетрациклин более устойчив в кислых средах, чем обыч-
ный тетрациклин, и поэтому является одним из основных анти-
биотиков тетрациклиновой группы, производящихся в промыш-
ленном масштабе.
6.4. Технология рекомбинантных ДНК
В конце 70-х годов в биохимии был разработан ряд новых
методологических подходов, ознаменовавших начало эры гене-
тической инженерии. Методы рекомбинантных ДНК, перевер-
нувшие наши представления о возможностях биотехнологии в
сфере удовлетворения потребностей человека, позволяют на-
правленно конструировать генетический материал, вводить его
в живые клетки и с помощью последних реализовать эту гене-
тическую информацию. Кроме того, благодаря технологии ре-
комбинантных ДНК значительно ускорились темпы расширения
и развития наших знаний о функциях ДНК, организации ге-
нов, регуляции их экспрессии, первичной структуре белков, что
в свою очередь создает основу для понимания природы различ-
ных заболеваний и борьбы с ними. Возможность введения лю-
бых сегментов ДНК в клетки позволяет создавать промышлен-
ные микроорганизмы, способные синтезировать ценнейшие
белки. В этом разделе мы уделим основное внимание примене-
нию генетической инженерии в промышленности и использова-
нию в качестве организма-хозяина бактерии Escherichia coli.
Прежде чем перейти к изучению основных приемов и мето-
дов генетической инженерии, целесообразно рассмотреть общие
принципы процесса и познакомиться со специальной терминоло-
гией. На рис. 6.21 изображена упрощенная схема клонирования
фрагмента ДНК. Под клонированием здесь мы понимаем при-
готовление колонии генетически идентичных клеток, которые
содержат интересующий нас сегмент ДНК- Этот сегмент, на-
Молекулярная генетика и системы регуляции
441
званный на рис. 6.21 «чужеродной ДНК», получают путем раз-
резания большого участка ДНК на фрагменты с помощью спе-
цифичных эндонуклеазных ферментов или путем синтеза —
химического или ферментативного. Чужеродную ДНК соединя-
ют (in vitro) с вектором — специальным инструментом для вве-
дения чужеродной ДНК в бактериальную клетку. В нашем при-
мере вектор представляет собой бактериальную плазмиду, не-
Z/7 vitro
[7Получение чужеродной ДНК
2 Соединение чужеродной ДНК с плазмидным вевтором
Иден,77ири-
кация кло-
нированной
ДНК
РИС. 6.21. Основные этапы клонирования фрагментов чужеродной ДНК.
сколько модифицированную с целью облегчения процесса кло-
нирования. Затем молекулу рекомбинантной ДНК (комплекса
чужеродной ДНК с плазмидой) вводят в ДНК клетки бактерии,
используя механизм трансформации. После этого с помощью
описанных ниже методов идентифицируют клон, содержащий
плазмиду с чужеродной ДНК.
Клонирование фрагмента ДНК дает возможность получить
его в количествах, достаточных для детального изучения и при-
менения в качестве реагента в последующих биохимических и
генетических работах. В качестве примера использования кло-
нированных ДНК для аналитических целей можно указать на
ход работ по изучению интерферона из фибробластов (IFN-p)
в конце 70-х годов. Этот белок был чрезвычайно труднодоступен,
поэтому даже с помощью самых чувствительных приборов для
442
Глава 6
определения аминокислотной последовательности удавалось
выяснить природу только нескольких N-концевых аминокислот-
ных остатков белка. Однако вскоре после клонирования гена
интерферона из фибробластов была определена его нуклеотид-
ная последовательность, из которой с учетом генетического ко-
да непосредственно вытекала и аминокислотная последова-
тельность соответствующего белка! В разд. 6.4.4 мы рассмотрим
примеры использования клонированных ДНК для получения
штаммов микроорганизмов, продуцирующих большие количе-
ства чужеродных белков, а следующий раздел будет посвящен
изучению ряда ферментов, без которых невозможна технология
рекомбинантных ДНК-
6.4.1. Ферменты для расщепления и сшивки ДНК
Успехи генетической инженерии во многом связаны с выделе-
нием и последующим промышленным производством ряда фер-
ментов, с помощью которых можно разрезать, видоизменять и
соединять молекулы ДНК в лабораторных условиях. Особое ме-
сто среди этих ферментов занимают рестрикционные эндонук-
леазы, способные узнавать и расщеплять специфические нуклео-
тидные последовательности в молекулах ДНК- Например, рест-
риктаза EcoRI специфична по отношению к последовательности
шести нуклеотидных остатков в двойной спирали следующего
типа:
!
---G—А—А—Т—Т—С-----
--С—Т—Т—A—A—G------
t
Эта рестриктаза расщепляет каждую цепь между остатками А
и G по связям, отмеченным стрелками. В результате образуют-
ся два фрагмента:
---G А—А—Т—Т—С-----
---С—Т—Т—A—A G---
Обратите внимание на то, что каждый фрагмент содержит ко-
роткую одноцепочечную последовательность, называемую «лип-
ким концом», поскольку эти две последовательности компле-
ментарны и, следовательно, имеют тенденцию к связыванию
друг с другом за счет водородных связей между парными осно-
ваниями. Реализация этой тенденции зависит от эксперимента-
тора, который посредством изменения pH или температуры мо-
жет добиться образования водородных связей или их разру-
шения.
К настоящему времени идентифицированы более 100 раз-
Молекулярная генетика и системы регуляции
443
Таблица 6.7. Некоторые рестрикционные
эндонуклеазы, соответствующие сайты узнавания
и расщепляемые связи (указаны стрелками)
Фермент. Сайт узнавания
BamHl — G-- G—А—Т—С—С—
—С—С—Т—A—G—G-—
t
В gill —A—G—А—Т—С—Т—
—Т—С—Т—A—G—А—
t
EcoRI — G—А—А—Т—Т—С—
—С—Т—Т—A—A—G—
t
I
Haelll — G—G—С—С—
—С—С—G—G—
t
I
Hindlll — A—A—G—С—T—T—
—T—T—C—G— A—A—
t
I
tfpal — G—T—T—A—A—C—
—С—A—A—T—T—G—
t
I
Pstl — С—T—G—C—A—G—
—G—A—C—G—T—C—
t
личных ферментов, обладающих рестриктазной активностью.
В табл. 6.7 перечислены наиболее часто используемые рестрик-
тазы, соответствующие сайты узнавания и гидролизуемые ими
фосфодиэфирные связи. Нетрудно заметить, что не все рестрик-
тазы образуют липкие концы. Рестриктаза Hpal, например,
расщепляет специфическую последовательность шести нуклео-
тидных остатков таким образом, что образуются только двухце-
почечные фрагменты с «тупыми концами».
Номенклатура рестрикционных ферментов базируется на на-
звании организма, из которого данная рестриктаза была впер-
вые выделена. Первая прописная буква в обозначении рестрик-
тазы соответствует первой букве названия рода организма,
а последующие две или три строчные буквы берутся из первых
444
Глава 6
двух или трех букв названия вида этого организма. Наконец,
римская цифра обозначает порядковый номер фермента, отра-
жающий хронологию его выделения из данного организма. Так,
рестриктаза Bglll представляет собой вторую рестриктазу, об-
наруженную в бактерии Bacillus globigii. Для уточнения при-
роды штамма, продуцирующего данный фермент, в название
рестриктазы иногда включают дополнительные буквы; так, ре-
стриктаза EcoRI была выделена из Е. coli RY13.
Ценность рестрикционных ферментов в технологии рекомби-
нантных ДНК обусловливается их высокой специфичностью,
т. е. способностью расщеплять только строго определенные нук-
леотидные последовательности. Поскольку вероятность наличия
в молекуле ДНК специфических сайтов узнавания, содержащих
от 4 до 6 нуклеотидных остатков, не очень велика, то образую-
щиеся в результате гидролиза рестриктазами фрагменты ДНК
обычно имеют довольно большую молекулярную массу (как
правило, несколько сотен нуклеотидных остатков). Это доста-
точно много для того, чтобы образовавшиеся фрагменты несли
ценную генетическую информацию, и одновременно довольно
мало для того, чтобы их можно было изучать и модифицировать
in virto.
Объем настоящего учебного пособия позволяет лишь вкрат-
це упомянуть другие важные ферменты, применяющиеся в тех-
нологии рекомбинантных ДНК. В первую очередь следует отме-
тить ДНК-лигазу. Предположим, что мы создали такие усло-
вия, когда липкие концы, образовавшиеся в результате расщеп-
ления различных молекул ДНК под действием EcoRI, «отжига-
ются», т. е. сближаются и связываются за счет образования
пар оснований. Образовавшиеся ДНК не будут, однако, кова-
лентно замкнутыми, поскольку между остатками А и G в обеих
цепях отсутствуют фосфодиэфирные связи. Последние могут
быть вновь созданы с помощью ДНК-лигазы, катализирующей
конденсацию З'-гидроксильной группы с б'-фосфатной группи-
ровкой.
Таким путем, например, отожженные фрагменты
---G А—А—Т—Т—С-----
---с—Т—Т—A—A G----
ковалентно сшиваются ДНК-лигазой:
---G—А—А—Т—Т—С-----
---С—Т—Т—A—A—G-----
В заключение отметим другие ферменты, катализирующие
синтез олигонуклеотидов (рис. 6.22). На матричной цепи ДНК
фермент ДНК-полимераза1 катализирует построение компле-
ментарной цепи в 5'—> З'-направлении. Для осуществления этого
Молекулярная генетика и системы регуляции
445
Матрица
G Т A G. С
III I II х
3' Синтез помпне-
тентарной
цепи
ОВратная транскриптаза
мРНК $ +
- CAUGGCUAAA
Дезохсирийо-
• нунпеотид-
Нб/й прайтер
„ ~ со сВоооВнои
Синтез котипе пентарной 3 - О Н - гпиптн
цени ДНК в направлении 3'^3'
\НонцеВая трансфераза |
^Свободная ОН-группа •
+ dGTP ----*-
О _ — - г
- G(G)nG 3'
З'
РИС. 6.22. Характерные реакции некоторых синтезирующих ферментов, ис-
пользуемых при получении рекомбинантных ДНК.
процесса необходимо присутствие всех четырех дезоксирибо-
нуклеотидов, а также праймера, обладающего свободной
З'-гидроксильной группой. Обратная транскриптаза (ревертаза;
РНК-зависимая ДНК-полимераза) выполняет задачу, потреб-
ность в решении которой возникает сравнительно редко; этот
фермент как бы обращает обычный порядок передачи генетиче-
ской информации, синтезируя цепь ДНК, комплементарную
матрице мРНК. Соединение различных фрагментов ДНК часто
облегчается после их удлинения на комплементарные гомопояи-
мерные цепи (хвосты); в присутствии только одного нуклеотид-
трифосфата фермент концевая трансфераза способен много-
446
Глава 6
кратно присоединять этот нуклеотид к концевой З'-ОН-группе
молекулы ДНК.
Некоторые другие ферменты, играющие большую роль в ме-
тодологии генетической инженерии, мы упомянем ниже, а сле-
дующий раздел будет посвящен требованиям, предъявляемым к
векторам рекомбинантных ДНК.
6.4.2. Векторы для Escherichia coli
Векторами называют молекулы ДНК, обеспечивающие ам-
плификацию фрагмента ДНК в растущей популяции клеток.
Чтобы вектор отвечал всем предъявляемым к нему в процессе
клонирования требованиям, он должен обладать следующими
свойствами:
1. Способностью реплицироваться в клетке-хозяине.
2. Способностью включать чужеродные ДНК различной моле-
кулярной массы без нарушения способности к репликации.
3. Легкостью введения в клетку-хозяина после включения чуже-
родной ДНК.
4. Наличием генетического маркера селекции, обеспечивающего
быструю положительную селекцию клеток, содержащих век-
тор.
5. Наличием только одного сайта, подвергающегося расщепле-
нию одной или несколькими рестрикционными эндонуклеа-
зами.
Для клонирования ДНК в Е. coli применяли два класса векто-
ров— плазмиды и бактериофаги. Здесь основное внимание бу-
дет уделено плазмидным векторам.
Как мы уже упоминали выше, при введении плазмид в клет-
ки Е. coli используется механизм трансформации. Поскольку
трансформированная клетка Е. coli может включить только од-
ну плазмиду, а для последующего выделения, идентификации и
применения интересующих нас последовательностей ДНК необ-
ходим клон, состоящий из большого числа идентичных клеток,
содержащих плазмиды, то плазмида обязательно должна обла-
дать способностью к репликации в растущей бактериальной
клетке. Для этого в свою очередь нужно, чтобы в состав плаз-
миды входил участок начала репликации — нуклеотидная по-
следовательность (около 600 пар оснований в случае плазмид
Е. coli типа Со1Е-1), направляющая и регулирующая процесс
репликации таким образом, чтобы каждая клетка содержала
достаточное число копий плазмид (обычно около 30).
Немаловажны и генетические маркеры селекции. Если, на-
пример, в плазмиду включить ген устойчивости к антибиотику,
то быстрая положительная селекция содержащих плазмиды кле-
Молекулярная генетика и системы регуляции 447
ток нс вызовет затруднений. Для этого достаточно вырастить
клетки на среде, содержащей антибиотик в такой концентрации,
которая приведет к гибели всех не имеющих плазмиды клеток,
но в то же время обеспечит рост клеток, содержащих плазми-
ды, а следовательно, и ген устойчивости к антибиотику. Другой
общий подход к селекции заключается в использовании мутант-
ной клетки-хозяина, у которой отсутствует фермент, необходи-
мый для роста в определенной среде, и в одновременном введе-
нии соответствующего этому ферменту нормального гена в ре-
Hind Ш
BeoRl
Ват HI
у ивоета к
астициллану
Участок кача-
ларепликации
Рои I
Psi I
WI
Рои Д
\ Ген устой-
чивости к
тетрациклину
p8R322
РИС. 6.23. Генетическая карта плазмиды pBR322. Показаны локусы^ генов
и ряд сайтов рестрикции. Расшифрована последовательность всех 4363 пар
оснований плазмиды.
комбинантную плазмиду. В этом случае экспрессия гена плаз-
миды комплементирует генетический дефект клетки-хозяина.
В технологии рекомбинантных ДНК маркеры селекции в
плазмидах могут выполнять две различные функции. Во-первых,
методы положительной селекции, позволяющие быстро иденти-
фицировать содержащие вектор колонии, резко интенсифициру-
ют лабораторные исследования. Во-вторых, при последующем
выращивании содержащих плазмиду клеток давление отбора
(обусловленное, например, введением в питательную среду ан-
тибиотика) сводит к минимуму ^конкуренцию со стороны любых
не содержащих плазмиду клеток, которые могут возникать вхо-
де роста популяции.
Плазмидные векторы для клонирования в Е. coli особенно
широко изучались в последние годы. К числу наиболее попу-
лярных относится плазмида pBR322 (рис. 6.23), содержащая
ряд единичных сайтов рестрикции, гены устойчивости к тетра-
циклину и ампициллину, а также участок начала репликации,
обеспечивающий амплификацию плазмиды в клетке. Здесь под
амплификацией (экстракопированием) понимается существен-
448
Глава 6
ное повышение количества плазмид по отношению к хромосом-
ной ДНК. В случае pBR322 и родственных плазмид амплифи-
кация становится возможной благодаря тому обстоятельству,
что в отличие от хромосомы репликация плазмиды в клетке-хо-
зяине может осуществляться и тогда, когда белковый синтез
остановлен. Поэтому добавление к культуральной жидкости ин-
гибитора белкового синтеза, например хлорамфеникола, приво-
дит к синтезу от 30 до 1000 и более копий плазмиды, а следо-
вательно, и клонируемой ДНК. Это позволяет получить послед-
нюю в количествах, достаточных для ее идентификации и после-
дующего использования в качестве реагента при конструирова-
нии других ДНК. В плазмиде pBR322 и родственных векторах
могут клонироваться сегменты чужеродных ДНК, содержащие
до 15 тысяч пар оснований.
В качестве клонирующего вектора широко изучался также
бактериофаг К, который сохраняет способность к репликации и
литические функции после делеции 25% генома дикого типа;
это позволяет клонировать сегменты чужеродной ДНК, содер-
жащие до 12 тысяч пар оснований. Значительно большие сег-
менты ДНК (до 50 тысяч пар оснований) удается клонировать с
помощью гибридных плазмидно-фаговых X векторов, называе-
мых космидами; последние могут быть введены в головку и
хвост фага X. По этой причине векторы-бактериофаги предпоч-
тительнее в тех случаях, когда возникает необходимость в кло-
нировании библиотек, состоящих из целого генома эукариоты
или из ее очень больших фрагментов. Полезные векторы полу-
чали также из фагов с одноцепочечным геномом, например из
фага КПЗ. Получаемые с их помощью одноцепочечные клоны
особенно удобны для определения нуклеотидных последователь-
ностей ДНК по методу Сэнджера. Следующий раздел будет по-
священ методам выяснения последовательностей нуклеотидных
остатков и другим методам идентификации клонированных ДНК.
6.4.3. Идентификация клонированных ДНК
Предположим, что рекомбинантные плазмиды удалось вклю-
чить в популяцию клеток Е. coll и что содержащие плазмиды
клетки удалось обнаружить на агаровых пластинках в селектив-
ной среде. В этом разделе мы вкратце рассмотрим методы
скрининга полученных таким путем клонов и детального изуче-
ния инсерций (вставок) ДНК из интересующих нас клонов.
Прежде всего нам нужно отличать клоны, содержащие
плазмиды с инсерциями чужеродной ДНК, от клонов, в состав
которых входят только исходные плазмиды; это может быть до-
стигнуто с помощью инсерционной инактивации в процессе кло-
нирования. Предположим, например, что фрагменты ДНК вклю-
Молекулярная генетика и системы регуляции 449
чены в сайт плазмиды pBR322, расщепляемый рестриктазой
Pstl. Поскольку инсерция в этом центре разрывает ген устойчи-
вости к ампициллину, то содержащие рекомбинантные плазми-
ды клетки будут чувствительны к ампициллину и устойчивы к
тетрациклину. Напротив, клетки с плазмидами без инсерции бу-
дут устойчивы как к ампициллину, так и к тетрациклину.
В этом случае клоны, содержащие рекомбинантные плазми-
ды, можно обнаружить следующим образом. Сначала клетки
выращивают на пластинках в присутствии тетрациклина; таким
путем избавляются от существенного фона не содержащих
плазмиды клеток. Затем применяют метод реплик (отпечат-
ков) : войлочную подушечку или нитроцеллюлозную фильтро-
вальную бумагу прижимают сначала к исходной матричной
пластинке, а затем ко второй пластинке (реплике), получая на
ней точную копию распределения колоний на матричной пла-
стинке. Если реплика обработана ампициллином, то колонии
клеток, содержащих плазмиды с инсерциями в гене устойчиво-
сти к ампициллину, растут на матричной пластинке, но не рас-
тут после перепечатки.
Поиск в клоне специфических нуклеотидных последователь-
ностей может быть осуществлен с помощью ряда методик, ос-
нованных на гибридизации денатурированной (т. е. одноцепо-
чечной) ДНК из клона с особым нуклеотидным зондом, обычно
меченным радиоактивным изотопом 32Р. Нуклеотидная последо-
вательность зонда (которым может быть как ДНК, так и мРНК)
комплементарна части последовательности искомого сегмента
ДНК- Зонды мРНК получают путем выделения мРНК из кле-
ток, обогащенных интересующей исследователя генетической
информацией, а зонды ДНК выделяют на одной из первых ста-
дий клонирования или получают прямым химическим син-
тезом.
В методе гибридизации колонии in situ на нитроцеллюлоз-
ной Фильтровальной бумаге, лежащей на чашке Петри с пита-
тельной средой, выращивают реплику клонов. Затем клетки
разрушают лизоцимом, а ДНК денатурируют обработкой NaOH,
после чего нитроцеллюлозную бумагу отделяют, обрабатывают
зондом, промывают и экспонируют с рентгеновской пленкой.
На пленке появляются темные пятна в тех местах, где произош-
ла гибридизация зонда с комплементарными последовательно-
стями ДНК; эти клоны и представляют интерес для дальнейше-
го изучения.
Плазмидную ДНК, которую можно выделить ультрацентри-
фугированием в градиенте плотности хлорида цезия, далее об-
рабатывают рестриктазой. Полученные фрагменты ДНК образу-
ют своеобразную картину («отпечатки пальцев») для данной
плазмидной ДНК. Фрагменты могут быть разделены в соответ-
29—537
450
Глава 6
ствии с их молекулярными массами методом электрофореза в
агарозном геле (0,5—1,5%); для обнаружения фрагментов ДНК
часто используют флуоресцентный краситель бромистый эти-
дий, специфичный по отношению к двухцепочечным нуклеино-
вым кислотам. Путем сравнительного анализа длины фрагмен-
тов, образующихся при действии других рестриктаз и при со-
вместном расщеплении несколькими рестрикционными эндонук-
леазами, получают рестрикционную карту, характеризующую
относительные положения различных сайтов рестрикции. По-
добные карты чрезвычайно полезны для проверки правильности
включения ДНК в заданные участки плазмиды и в качестве
основы для дальнейшей работы с рекомбинантными плазми-
дами.
Упоминавшиеся выше меченные радиоактивными изотопами
зонды можно использовать и для идентификации определенных
фрагментов, образующихся при обработке рестриктазами. В ме-
тоде блоттинга по Саузерну фрагменты ДНК в геле денатури-
руют щелочью и на гель накладывают нитроцеллюлозную плен-
ку. Сверху помещают фильтровальную бумагу, в которую пере-
ходит буферный раствор; одновременно фрагменты ДНК пере-
ходят на нитроцеллюлозную пленку. ДНК ковалентно связыва-
ется с нитроцеллюлозой при 80°C. Затем связанные денатури-
рованные фрагменты ДНК обрабатывают, как было описано
выше, зондом, экспонируют с рентгеновской пленкой и таким
образом идентифицируют полосы на электрофореграмме, соот-
ветствующие комплементарным зонду нуклеотидным последо-
вательностям.
Современные экспериментальные методы биохимии ДНК на-
столько чувствительны, что нескольких нанограммов ДНК, выде-
ленных из полосы на гелевой электрофореграмме, достаточно
для клонирования или для дальнейшего изучения. Очевидно,
что исчерпывающая идентификация фрагмента ДНК должна
включать в себя определение его нуклеотидной последователь-
ности. В 70-е годы были разработаны два метода быстрого, на-
дежного и эффективного определения последовательностей нук-
леотидных оснований в ДНК — Максама — Гилберта и Сэнд-
жера. Эти методы оказали громадное воздействие на биологи-
ческие науки вообще и на биотехнологию в частности.
В методе Максама — Гилберта двойную спираль ДНК сна-
чала метят, присоединяя радиоактивную метку к одному концу
каждой цепи. Затем ДНК денатурируют и препарат одной из
двух цепей разделяют на четыре аликвотные части, каждую из
которых обрабатывают специфичным реагентом. В результате
каждой из таких обработок селективно разрушается одно осно-
вание (или два), а цепи ДНК в этих центрах расщепляются.
Для успеха метода существенно, чтобы это разрушение было
Молекулярная генетика и системы регуляции
451
Радиоактивная
тетка
Олигонуклеотид
g|a|t|c |а|т |g
Параллельная обработка различными
.реагентами х
Разрушает Разрушает Разрушает Разрушает
Изображение злектрофореераммь/
на рентгеновской пленке
РИС. 6.24. Определение нуклеотидной последовательности методом Макси-
ма — Гилберта. При обработке изучаемой последовательности четырьмя спе-
цифическими реагентами, разрушающими различные основания, образуется
смесь фрагментов, молекулярные массы которых зависят от положения рас-
щепленного основания. Расположение полос на четырех дорожках гелевой
электрофореграммы позволяет считывать нуклеотидную последовательность
непосредственно с геля; считывание сверху вниз соответствует направлению
последовательности к меченому концу исходного олигонуклеотида.
Полоса/ на электро-
фореграмме отде-
ча/от
частичным; в идеальном варианте желательно, чтобы каждая
цепь расщеплялась бы только в одном центре. Затем образо-
вавшиеся в результате таких четырех параллельных расщепле-
ний фрагменты изучают на достаточно протяженном полиакрил-
амидном геле. Полосы в геле обнаруживают с помощью чув-
ствительной к рентгеновскому излучению фотопленки. Опреде-
ляемую нуклеотидную последовательность считывают непосред-
ственно с четырех параллельных дорожек гелевой электрофоре-
граммы (рис. 6.24). Таким методом в одном эксперименте без
труда можно выяснить последовательность около 200 нуклео-
29*
452
Глава 6
тидных остатков. Метод определения нуклеотидных последова-
тельностей по Сэнджеру рассмотрен в работе [21]; обычно его
применяют при изучении более крупных фрагментов ДНК.
Часто возникает необходимость в скрининге из множества
различных клонов одного клона (или нескольких клонов), обла-
дающих способностью к экспрессии определенного белка.
В большинстве случаев для решения такой задачи прибегают к
помощи антитела, специфичного по отношению к данному бел-
ку. В антитела вводят радиоактивную метку (например, 1251}
и раствором антител обрабатывают изучаемые колонии на пла-
стинке; затем их промывают и с помощью рентгеновской плен-
ки обнаруживают искомые колонии, содержащие данный белок..
В другом варианте антитела связывают с ферментом и антиге-
ны обнаруживают по свойствам этого фермента. Идентифициро-
вать колонии, содержащие искомый белок, можно, например,,
с помощью хромогенного субстрата (т. е. субстрата, изменяю-
щего свою окраску или окрашивающегося после обработки
ферментом). Более чувствительные иммунохимические методы,
с помощью которых можно обнаружить белки в концентрациях
1—5 молекул в клетке Е. coli, описаны в приведенной в конце:
главы литературе.
Объем и тематика этой книги не позволяют обсудить де-
тально все методы идентификации и изучения клонов; все же-
надо надеяться, что даже приведенные здесь данные дают
представление о том, насколько трудоемкой, длительной и уто-
мительной может быть работа по скринингу клонов и по конт-
ролю каждого шага в построении новых рекомбинантных мо-
лекул. Хотя для специалиста в области биохимической техно-
логии больший интерес представит тема следующего раздела,,
посвященного экспрессии клонированных генов, следует отда-
вать себе отчет в том, что основным препятствием в программе^
синтеза чужеродного белка в Е. coli или другой клетке-хозяине-
скорее всего будет именно клонирование гена. В то же время
успехи методических разработок и развитие их автоматизации:
будут содействовать внедрению «типовых методик» молекуляр-
ной генетики, что в свою очередь создаст основы для существен-
ного расширения набора доступных и интересных генов и регу-
ляторных последовательностей.
6.4.4. Экспрессия эукариотических белков в Е. coli
Первыми примерами успешного применения генетической
инженерии в биотехнологии были синтезы ряда ценных белков-
эукариот в клетках Е. coli. В этом разделе мы прежде всего
рассмотрим методы получения генов, пригодных для экспрессии
эукариотических белков, затем обсудим требования, предъяв-
Молекулярная генетика и системы регуляции
453
ляемые к вектору экспрессии, и завершим раздел рядом заме-
чаний, касающихся проблем скрининга продуктов экспрессии
генов, а также устойчивости генов и соответствующих белков.
Если вернуться к рис. 6.7 и соответствующему разделу тек-
ста, нетрудно понять, что прямая экспрессия генов эукариот
Е. coli связана с одним принципиальным препятствием. Прока-
риоты вообще и Е. coli в частности не обладают биохимиче-
ским механизмом, способным осуществлять сплайсинг РНК-
Поэтому эукариотический ген, включенный непосредственно в
Е. coli (при условии рассматриваемой ниже соответствующей
бактериальной регуляции экспрессии), не будет нормально экс-
прессироваться. В Е. coli будут транслироваться последова-
тельности интронов эукариотического гена мРНК и синтезиру-
ющийся полипептид в общем случае будет резко отличаться от
нормального эукариотического белка.
Очевидно, нам нужно каким-то образом приготовить бакте-
риальный ген, отвечающий эукариотическому белку, т. е. такой
ген, который содержал бы только нуклеотидную последова-
тельность, кодирующую первичную структуру белка, и не имел
бы ни одного интрона. Эта цель может быть достигнута двумя
принципиально различными путями. Один из них сводится к хи-
мическому синтезу заданной нуклеотидной последовательно-
сти. Хотя здесь мы не можем углубляться в детали химическо-
го синтеза генов (интересующийся этой проблемой читатель мо-
жет найти все необходимые сведения в литературе), все же
необходимо подчеркнуть, что синтетическая химия ДНК пока
еще не позволяет осуществлять прямой синтез всего гена.
Обычно синтезируют олигонуклеотиды с частично перекрыва-
ющимися последовательностями, которые обладают способно-
стью к самосборке в полную двухцепочечную нуклеотидную по-
следовательность за счет образования водородных связей в па-
рах оснований. В качестве примера на рис. 6.25 приведен ген,
кодирующий последовательность 21 аминокислотного остатка
цепи А инсулина человека; этот ген был собран из 12 различ-
ных олигодезоксирибонуклеотидов, построенных из 10—15 нук-
леотидных остатков. При планировании синтеза гена следует
принимать во внимание и ряд других требований, связанных с
клонированием и экспрессией, в том числе свойства кодонов,
отсутствие сигналов терминации, наличие сайтов рестрикции.
Возможность химического синтеза генов или их фрагментов
создает базу для чрезвычайно перспективной области белковой
инженерии. В принципе уже сейчас мы можем заменить любую
аминокислоту в белке, с тем чтобы попытаться повысить биоло-
гическую активность или технологическую устойчивость данно-
го белка. Более того, мы можем заставить клетку синтезировать
новые (или, во всяком случае, неизвестные нам) аминокислот--
454
Глава 6
ные последовательности и таким путем создавать новые ката-
лизаторы, лекарственные препараты и компоненты пищевых
продуктов. К сожалению, в настоящее время мы знаем так ма-
ло о связи между первичной структурой и функцией белков, что
возможности улучшения свойств белков путем их модификации
пока что остаются практически нереализованными.
В другом пути получения бактериального гена, соответству-
ющего эукариотическому белку, исходят из зрелой эукариоти-
ческой матричной РНК, уже прошедшей стадию сплайсинга.
Ген цепи А
1234567 S 9 10 11 12 13 14 15 16 17 18 19 20 21
Met Glv Ik Vai Glu Gin Счя Суя Thr Scr lie Суя Ser Leu Туг Gin Leu Glu Asn Туг Cys Asn
£c'>R1A, a2 A3 A4 As A6 Стоп-кодоны
AATrCATGGGCATCGTTGAACAGTGTTGCACTTCTATCTGCTCTCTTTACCAGCTTGAGAACTACTGTAACTAATAG
gtacccgtagcaacttgtcacaacgtgaagatagacgagagaaatggtcgaactcttgatgaCattgattatcci
РИС. 6.25. Ген, отвечающий цепи А инсулина человека (аминокислотная по-
следовательность приведена в верхней части рисунка). Этот ген был синте-
зирован нз 12 олигодезоксирибонуклеотидов At—А12. [Воспроизведено из
статьи: Crea R„ Kraszewski A., Hirose Т., Itakura К., Chemical Synthesis of
Genes for Human Insulin, Proc. Natl. Acad. Sci. (USA), 75, 5765 (1978).]
Как схематично показано на рис. 6.26, соответствующая необхо-
димому белку мРНК сама может выполнять роль матрицы, на
которой особый фермент, обратная транскриптаза, может синте-
зировать комплементарную цепь ДНК. Цепи мРНК и ДНК за-
тем разделяют, на цепи ДНК с помощью ДНК.-полимеразы
строят комплементарную цепь, после чего специфическим фер-
ментом нуклеазой S1 отщепляют небольшой петлевой одноце-
почечный участок. В результате образуется молекула двухцепо-
чечной комплементарной ДНК (кДНК). К последней, как пока-
зано на рис. 6.26, с помощью концевой трансферазы присоединя-
ют гомополимерные последовательности дезоксицитидина (dC).
Таким путем создаются предпосылки для последующего клони-
рования, предпочтительно с плазмидным вектором pBR322, рас-
щепленным рестриктазой PstI и имеющим гомополимерные по-
следовательности (dG).
Для эффективной экспрессии структурному гену должны
предшествовать нуклеотидные последовательности, обеспечива-
ющие инициацию транскрипции (промотирующая последова-
тельность) и начало трансляции (связывающий рибосому сайт
Молекулярная генетика и системы регуляции
455
и стартовый кодон); за структурным геном должен следовать
терминирующий кодон, означающий прекращение трансляции, и
терминатор транскрипции (вспомните рис. 6.6). В векторы экс-
прессии входят все эти регуляторные последовательности, уча-
сток начала репликации, а также по меньшей мере один мар-
кер селекции. С помощью клонирования и синтетическими ме-
тодами получено большое число регулирующих экспрессию по-
мРНК, coomffem-
Обратная транскриптаза, ходимоми бета/
нрклеозидтри/рос/раты,
Ф оииго-а1Т
А А А А А
_________ Т Т Т Т Т
| ДНК-тмитераза
ААААА
^Нуклеаза Sf
^Дбухцемчечная %
нат/тетентарная
ДНК (кДНк)
ААААА
ТТТТТ
\концебая транарераза, dCTP
сссссс
АААААССССССС
РИС. 6.26. Схема синтеза двухцепочечной ДНК с нуклеотидной последова-
тельностью, комплементарной нуклеотидной последовательности молекулы
мРНК (волнистая линия). В результате синтеза получают пригодную для
клонирования молекулу комплементарной ДНК (кДНК).
следовательностей, пригодных для включения в векторы экс-
прессии. Чаще других используются промоторы lac, trp и XpL.
Как правило, желательно, чтобы применяющийся промотор
был регулируемым, т. е. включался только при каком-либо из-
менении параметров среды, например при добавлении индукто-
ра, инактивации репрессора или при изменении температуры.
В хорошо отработанной системе экспрессии содержание белка,
отвечающего клонированному гену, может достигать 70% от
всех клеточных белков, хотя обычно эта величина составляет
от 10 до 25%. Поскольку продукт экспрессии не выполняет ни-
какой полезной для клетки-хозяина функции, столь серьезное
отвлечение биосинтезирующего аппарата клетки на синтез не-
456
Глава 6
РИС. 6.27. а — электронная сканирующая микрофотография рекомбинантных
клеток Е. coli, продуцирующих химерный белок политриптофан-проинсулин.
Рекомбинантные клетки морфологически отличаются от нормальных клеток
Е. coli (на вставке, Х5300) удлиненной формой и наличием утолщений; б —
электронная просвечивающая микрофотография рекомбинантных клеток Е. co-
li, продуцирующих химерный белок Р-галактозидаза—цепь А инсулина
(X17500); в — рекомбинантные клетки Е. coli, продуцирующие политрипто-
фан-проинсулин (Х30 000); г—в клетках Е. coli, не содержащих плазмид,
белковые включения отсутствуют (Х30 000). [Воспроизведено с разрешения
из статьи: Williams D. С., Van Frank R. М„ Muth W. L., Burnett I. P., Cyto-
plasmic Inclusion Bodies in Escherichia coli Producing Biosynthetic Human Insu-
lin Protein; Science, 215, 687 (1982).]
нужных клетке белков может сказаться на скорости роста кле-
ток и даже на их жизнеспособности. Поэтому в практической
работе, как правило, условия культивирования в реакторе ме-
няют таким образом, чтобы сначала экспрессия клонированного
гена была практически подавлена и обеспечивался в основном
быстрый рост клеточной массы вплоть до достижения необхо-
димой плотности клеток, а затем с максимальной скоростью
осуществлялась бы экспрессия.
Молекулярная генетика и системы регуляции 457
Когда в Е. coli концентрация чужеродных белков достигает
высокого уровня, они аккумулируются в клетке в виде плотных
образований (рис. 6.27). Последние представляют собой глав-
ным образом чужеродный белок, но содержат и небольшие ко-
личества бактериальных белков, причем чужеродный белок
обычно находится в денатурированном состоянии и клетки, со-
держащие плотные образования, часто имеют искаженную фор-
му. Поэтому, хотя высокая концентрация белка в той или иной
степени облегчает процессы выделения, его дальнейшая стаби-
лизация и превращение в активную форму обусловливают не-
обходимость дополнительных технологических стадий.
Конечная продуктивность рекомбинантной культуры может
лимитироваться обусловленной тем или иным фактором неста-
бильностью белка. Бактерия Е. coli содержит внутриклеточные
протеазы, способные каким-то образом узнавать небольшие
аномальные белки и разрушать их. В обычных клетках эти про-
теазы выполняют полезную функцию, разрушая ошибочно син-
тезированные белки до аминокислот, которые затем могут вновь
использоваться для синтеза белков; в рекомбинантных систе-
мах подобные протеазы могут быстро расщепить весь чужерод-
ный для клетки белок. Для повышения устойчивости последне-
го часто методами генной инженерии синтезируют гибридный
белок, состоящий из белка Е. coli, к которому (как правило,
к С-концу) присоединен чужеродный белок. Такой подход при-
менялся для стабилизации ряда относительно небольших поли-
пептидов, в том числе соматостатина (14 аминокислот), цепей
А (21 аминокислота) и В (30 аминокислот) инсулина человека.
В этих случаях роль соединительного звена между белковым
фрагментом Е. coli и пептидом человека выполнял остаток ме-
тионина; при последующей обработке бромцианом, расщепля-
ющим полипептидные цепи по остаткам метионина, образовы-
вался активный пептид человека. (К счастью, ни один из пере-
численных полипептидов не содержал остатков метионина!)
На рекомбинантных популяциях могут сказываться две раз-
личные формы генетической нестабильности. Во-первых, опре-
деленная часть вновь образующихся клеток может не содер-
жать плазмид. В этом случае благодаря так называемой сегре-
гационной неустойчивости не несущие плазмид клетки могут
подавлять рост содержащих плазмиды продуктивных клеток,
если только система не находится под давлением отбора. Одна-
ко даже под давлением отбора сегрегационная нестабильность
снижает общую скорость роста и продуктивность рекомбинант-
ной популяции. Другой тип нестабильности — структурная не-
устойчивость — приводит к неспособности клеток синтезировать
активный белок. Причина этой нестабильности может заклю-
чаться в мутации структурного гена или регуляторных нуклео-
458
Глава 6
тидных последовательностей. К сожалению, эта форма неста-
бильности не поддается регуляции путем давления отбора.
6.4.5. Генетическая инженерия с участием других клеток-хозяев
£. coli-—излюбленный объект подавляющего большинства
работ по клонированию и генетической инженерии, поскольку
этот организм наиболее детально изучен на молекулярном уров-
не. В то же время Е. coli практически не известна в микробио-
логической промышленности; будучи грамотрицательной бакте-
рией, Е. coli содержит в своей наружной оболочке токсичные
липополисахариды (заражение крови Е. coli приводит к ле-
тальному исходу в 50% случаев) и не секретирует белки в сре-
ду. К тому же как прокариота Е. coli не способна осуществлять
характерные для эукариотических клеток сплайсинг и посттранс-
ляционную модификацию белков. В силу этих и ряда других
причин большой интерес представляет разработка систем кло-
нирования и экспрессии в других, отличных от Е. coli организ-
мах. При этом во всех случаях важно иметь в виду следующие
факторы, которые могут препятствовать экспрессии чужеродно-
го гена:
1. Разрушение чужеродных ДНК или РНК нуклеазами клетки-
хозяина.
2. Несрабатывание механизма репликации вектора.
3. Низкая активность промотора или терминатора транскрип-
ции.
4. Неполный сплайсинг мРНК.
5. Неэффективная трансляция.
6. Гидролиз протеазами.
Кроме того, необходима эффективная методика, обеспечиваю-
щая введение вектора в клетку-хозяина с высоким выходом.
Известны методы клонирования и экспрессии генов в не-
скольких бактериях. Из последних наиболее хорошо изучена,
по-видимому, Bacillus subtilis — грамположительный, непато-
генный, непаразитический микроорганизм, использовавшийся
до начала эры генетической инженерии в микробиологической
промышленности для производства ряда ферментов и полипеп-
тидных антибиотиков. В. subtilis секретирует некоторые свои
белки в среду. В генетической инженерии последнее свойство
особенно удобно, поскольку секретируемые белки обычно не за-
грязнены большим количеством очень близких внутриклеточных
белков. К тому же секреция белка в среду в принципе позволя-
ет достичь более высокой концентрации синтезируемого белка
по сравнению с внутриклеточными белками. (Каков верхний
Молекулярная генетика и системы регуляции 459
предел количества внутриклеточного белка на единицу объема
культуральной жидкости?)
В В. subtilis можно клонировать ряд плазмид и бактерио-
фагов. Для введения в В. subtilis чужеродной ДНК могут при-
меняться методы трансформации, трансдукции и слияния про-
топластов. В В. subtilis удалось успешно осуществить экспрес-
сию нескольких белков млекопитающих, в том числе инсулина
и интерферонов, однако вплоть до момента написания этой гла-
вы извлечение чужеродных белков из В. subtilis оставалось
проблематичным.
Технология клонирования применима и к ряду штаммов
Pseudomonas и Streptomyces. Путем сочетания генетической ин-
женерии с методами мутации и селекции удалось получить
штаммы Pseudomonas с совершенно новым механизмом метабо-
лизма; эти штаммы, например, могут расти на обычно токсич-
ных хлорированных углеводородах как на единственном источ-
нике углерода. Большое промышленное значение различных ви-
дов Streptomyces обусловило интенсивное изучение возможно-
стей применения методов генетической инженерии к этим мик-
роорганизмам. Целью этих работ является поиск штаммов, ко-
торые были бы более продуктивными в отношении имеющих
практическое значение ферментов или синтезировали бы новые
полусинтетические или гибридные антибиотики.
Последние примеры заслуживают несколько более детально-
го обсуждения. Здесь мы впервые сталкиваемся с метаболиче-
ской инженерией. В целом генетическая инженерия позволяет
не только синтезировать белки, представляющие интерес как
таковые, но и вводить в живую клетку особые ферменты, регу-
ляторные белки, пермеазы и вообще любые другие белки. Та-
ким путем мы можем придать клетке совершенно новую фер-
ментативную, регуляторную или транспортную активность, най-
ти которую среди нативных организмов или получить путем
случайного мутагенеза было бы крайне маловероятно. Отсюда
следует, что теперь мы в состоянии строго определенным и на-
правленным путем перестраивать и реконструировать отдель-
ные участки сети метаболических превращений клетки. В перс-
пективе именно в этом направлении в основном и будет разви-
ваться генетическая инженерия. Пока что достижению опреде-
ленных результатов мешает недостаток данных о деталях кон-
кретных важных путей метаболизма и о стадиях, лимитирую-
щих скорость превращений по этим путям. К тому же наши зна-
ния о регуляции всей метаболической сети явно недостаточны
для того, чтобы можно было попытаться предсказать влияние
изменений в одном пути на ход других. Наконец, экспрессия
большого числа новых генов в рекомбинантных клетках требу-
ет разработки новых приемов генетической инженерии.
460
Глава 6
В последние годы удалось добиться больших успехов в ге-
нетической инженерии эукариотических клеток. Особенно ин-
тенсивно изучались дрожжи Saccharomyces cerevisiae-, этот мик-
роорганизм выгодно отличается мощной генетической системой,
способной к прямой экспрессии как некоторых эукариотических,
так и ряда прокариотических генов, а также способностью
трансформироваться с помощью чистой ДНК. Дрожжи также
осуществляют по меньшей мере некоторые из типичных для
эукариот посттрансляционных модификаций и секретируют не-
которые белки в среду. Например, рекомбинантные S. cerevi-
siae продуцируют антиген австралийского гепатита В, который
по степени гликозилирования и агрегации, по-видимому, не от-
личается от антигенов, обнаруженных в организме страдающих
этим заболеванием людей. Генетически модифицированные
дрожжи синтезируют и секретируют в среду иммунный интер-
ферон человека (INF-7).
Клонирование и экспрессия генов в клетках млекопитающих
пока что осуществлялись только с помощью векторов на основе
вируса SV40. Геном этого вируса (ковалентно замкнутая коль-
цевая ДНК) может реплицироваться в клетках млекопитающих
как самостоятельно, так и после включения в хромосомы клет-
ки-хозяина. Процесс экспрессии клонированного гена обеспечи-
вается наличием в вирусе SV40 как промоторов, так и участка
начала репликации, функционирующими в клетках млекопита-
ющих.
Наиболее удобным для клонирования и идентификации
ДНК остается все же быстрорастущий организм Е. coli, к кото-
рому применимо большинство методик генетической инженерии.
Поэтому, если конечной целью являются многократная репли-
кация вектора и экспрессия его генов в другом организме, час-
то оказывается удобным прибегнуть к помощи так называемо-
го челночного вектора, который может реплицироваться как в
Е. coli, так и в другом организме. Очевидно, что челночный век-
тор должен обладать двумя участками начала репликации —
по одному на каждый организм-хозяин. На рис. 6.28 приведена
схема челночного вектора, предназначенного для продуцирова-
ния иммунного интерферона (INF-7) в клетках обезьян. В этом
векторе фрагмент из 342 пар оснований, содержащий участок
начала репликации и «поздний» промотор из SV40. связан с
фрагментом кДНК, в состав которой входят кодирующая по-
следовательность пре-INF-^, а также с фрагментом pBR322,
включающим ген устойчивости к ампициллину и участок нача-
ла репликации Е. coli. Трансфекция (инфекция клеток чистой
вирусной ДНК) трансформированных клеток обезьян линии
COS-7 через 3—4 дня привела к INF-7-активности порядка
50—100 единиц в 1 мл культуральной жидкости. Этот пример
Молекулярная генетика и системы регуляции
461
может служить иллюстрацией тех преимуществ, которыми об-
ладают генетически модифицированные клетки-хозяева млеко-
питающих в синтезе белков млекопитающих, когда все стадии
биосинтеза белков и их секреции должны с максимально дости-
жимой точностью воспроизводить соответствующие стадии,
осуществляющиеся при синтезе и транспорте этих белков in
vivo.
Успешно развиваются также работы по введению чужерод-
ных генов в целые организмы растений и животных и их после-
Промотор
SV^O (Вероятная) сиг-
нальная последе-
/ дательность
Последова/пель-
ность, кодирую:
щая IFN-y
Участок нача-
ла репликации
SV40
Фрагмен.
кДНК
Ген устойчивос-
ти к ампи-
циллину
Участок начала
репликации pBR322
РИС. 6.28. Схема челночного вектора для Е. coli и клеток обезьяны, осуще-
ствляющего экспрессию иммунного интерферона в клетках обезьяны и его
секрецию [23].
дующей экспрессии. Хотя анализ методологии этих работ не
входит в тематику настоящей книги, их потенциальная значи-
мость для человека очевидна. Открывающие новые перспекти-
вы научные и технические достижения в то же время стимули-
ровали широкие дискуссии (в том числе и в литературе) о воз-
можности неблагоприятного воздействия биотехнологии на об-
щество [24].
6.4.6. Заключение
Приведенный в табл. 6.8 перечень основных этапов синтеза
белка с помощью рекомбинантных ДНК может служить крат-
ким обзором темы настоящего раздела. Запомните, что клони-
рование необходимого гена и его экспрессия эффективно вы-
полняются как отдельные последовательные операции. Следует
еще раз подчеркнуть необходимость постоянного контроля лю-
бых превращений ДНК путем определения нуклеотидной после-
462
Глава 6
довательности и изучения фрагментов, образующихся при рас-
щеплении рестриктазами.
Объем нашей книги позволяет изложить тему этого раздела
только в очень сжатой форме. Принимая во внимание ту боль-
шую роль, которую будет играть технология рекомбинантных
ДНК в будущем, технологу-биохимику необходимо значительно
глубже изучить основы молекулярной биологии и соответствую-
щей методологии. Знание этих отраслей науки необходимо при
знакомстве с современной биологической литературой, а также
Таблица 6.8. Последовательность этапов работы по клонированию гена
и экспрессии соответствующего белка методами технологии
рекомбинантных ДНК
1. Получение ДНК
2. Введение ДНК в плазмидный вектор
3. Трансформация
4. Обнаружение необходимого клона
5. Выращивание культуры и выделение плазмид
6. Определение нуклеотидной последовательности клонированного фраг-
мента ДНК
7. Конструирование и построение плазмиды для экспрессии
8. Трансформация
9. Обнаружение необходимого клона
10. Выращивание культуры и выделение плазмид
И. Проверка нуклеотидной последовательности ДНК
12. Трансформация
13. Выращивание культуры для производства белка
при разработке в сотрудничестве с молекулярными биологами
оптимальных организмов и процессов. Факторы, определяющие
продуктивность рекомбинантных популяций (уровни экспрессии
гена, активности внутриклеточных протеаз, генетическая ста-
бильность и т. п.), зависят от нуклеотидной последовательности
вектора, генетических свойств клетки-хозяина, состава среды,
конструкции биореактора и условий его работы. Для понимания
этих зависимостей, а также различных взаимосвязей в генетиче-
ски видоизмененном организме необходимы совместные усилия
специалистов в области биохимической технологии и молеку-
лярной биологии.
6.5. Рост и самовоспроизведение изолированной
клетки
Ростом организма можно назвать упорядоченное увеличение
массы всех составляющих этот организм веществ. Растущий
одноклеточный организм в конце концов претерпевает деление.
Следовательно, рост популяции микроорганизмов сопровожда-
ется как увеличением числа клеток, так и возрастанием массы
Молекулярная генетика и системы регуляции
463
всего клеточного вещества; для количественного математиче-
ского анализа клеточного роста можно использовать любой из
этих параметров. На практике применяют несколько единиц
измерения клеточной массы. Так, масса сухих высокомолеку-
лярных веществ учитывает только высокомолекулярные компо-
ненты клетки. Под общей массой сухих веществ понимают не
только высокомолекулярные, но и низкомолекулярные вещества
клетки (за исключением воды). Наконец, клеточным объемом
РИС. 6.29. События, происходящие с момента рождения новой клетки до ее
деления, могут быть представлены в виде циклической (а) или линейной (б)
карт клеточного цикла. Буквами D и А обозначены деление клетки и начало
синтеза ДНК соответственно.
или общей клеточной массой называют массу (или объем) всех
веществ клетки, в том числе и воды.
В следующей главе мы попытаемся понять закономерности
роста множества клеток в питательной среде. В этом же разде-
ле перед нами стоит более простая задача. Здесь мы рассмот-
рим основные события клеточного цикла, под которым пони-
мают период времени с момента образования данной клетки в
результате деления материнской клетки до последующего деле-
ния дочерней клетки. Изучение клеточного цикла должно пред-
шествовать теме гл. 7 по двум причинам: во-первых, таким об-
разом мы сможем оценить трудность задачи анализа событий
при одновременном росте и делении множества клеток, и, во-
вторых, знание основ клеточного цикла поможет в разработке
необходимых моделей, которые потребуются нам в гл. 7.
Жизненный цикл изолированной клетки можно схематично
изобразить в виде циклической или линейной карты (рис. 6.29).
В любом случае точкой отсчета служит момент деления клетки.
464
Глава 6
Эту точку обычно помещают на правом конце линейной схемы
клеточного цикла, а левый конец, как правило, отвечает самой
молодой дочерней клетке. Для описания относительной хроно-
логии других важных событий клеточного цикла, например на-
чала и прекращения синтеза ДНК, удобен особый параметр а,
называемый возрастом клетки или возрастом клеточного цикла.
В клеточном цикле величина этого параметра обычно изменя-
ется от нуля (момента рождения клетки) до tD, времени деления
дочернего организма (для делящихся клеток); период от нуля
до tD называют периодом удвоения. В этом вводном обсуждении
мы принимаем, что деление клетки приводит к образованию
двух дочерних идентичных клеток. Это допущение справедливо'
для простого бинарного деления, но при почковании, как это
можно показать на примере пекарских дрожжей S. cerevisiae,
могут возникать и другие ситуации.
6.5.1. Методы эксперимента;
цитометрия в потоке и синхронные культуры
Читатель, интересующийся методами экспериментального
изучения клеточного цикла во всех деталях, может найти соот-
ветствующие сведения в монографии Митчисона [25]; мы же
вкратце опишем только принципы основных методик. Наиболее
очевидный подход, заключающийся в наблюдении жизненного
цикла изолированной клетки, имеет весьма лимитированные
возможности. Этот подход требует много времени, а получаемая
таким путем информация ограничена, поскольку проводить из-
мерения на одной клетке крайне затруднительно. Поэтому та-
кой подход имеет существенное значение только в особых слу-
чаях.
Очевидно, любые биохимические анализы проще выполнять,
когда в распоряжении экспериментатора имеется множество кле-
ток. К сожалению, популяция клеток в суспензии с очень боль-
шой степенью вероятности будет гетерогенной в том смысле, что
различные клетки будут находиться в разных периодах клеточ-
ного цикла; одни клетки только что выйдут из стадии деления,,
другие, зрелые клетки вплотную подойдут к процессу деления,
а третьи будут занимать какое-либо промежуточное положение.
Обычными методами в такой популяции можно определить-
лишь некоторые усредненные характеристики, но не свойства
отдельных клеток.
Для быстрой оценки некоторых свойств индивидуальных
клеток в гетерогенной популяции можно использовать методы-
цитометрии в потоке. В этих методах пробу клеток разбавляют
и иногда обрабатывают тем или иным способом, чтобы ввести-
метку в определенные биохимические компоненты клетки. Затем!
Молекулярная генетика и системы регуляции 465
пробу (разбавленную суспензию клеток) пропускают через
ячейку, в которой измеряют характеристики индивидуальных
клеток по мере их прохождения. К числу таких характеристик
может относиться полный объем индивидуальной клетки, опре-
деляемый по рассеянию света или по изменению удельного со-
противления потока. Рассеяние света дает также ценную ин-
формацию о морфологии изолированой клетки и ее внутренней
структуре. Для проточной цитометрии разработано большое
число различных флуоресцентных маркеров, позволяющих оп-
ределять общее содержание клеточной ДНК, двухцепочечной
ДНК, белков, определенных ферментов, компонентов клеточной
РИС. 6.30. Зависимость отно-
сительного содержания ДНК
от относительного числа кле-
ток в популяции пекарских
дрожжей (Saccharomyces сеге-
visiae). Относительную интен-
сивность флуоресценции ДНК
индивидуальных клеток, окра-
шенных иодистым пропидием,
измеряли методом проточной
цитометрии в популяции 105
клеток.
оболочки. Применяющиеся для этих целей приборы и устройст-
ва мы рассмотрим в разд. 10.3.2.
Для измерения в потоке одной характеристики индивиду-
альной клетки требуется всего лишь около 1 мкс, поэтому ана-
лиз, например, 105 клеток длится только несколько минут. По-
лученные таким способом данные обычно изображают в виде
частотного распределения измеренного параметра индивидуаль-
ных клеток, т. е. в виде зависимости относительного числа кле-
ток (частоты в пробе) от интенсивности рассеяния света или
флуоресценции одной клетки. В качестве примера на рис. 6.30
приведены экспериментальные результаты оценки содержания
ДНК в одной клетке среди популяции Saccharomyces cerevisiae,
выросшей в проточном реакторе (см. гл. 7). Ценность таких дан-
ных заключается в том, что они непосредственно характеризуют
гетерогенность популяции клеток. К тому же часто удается свя-
зать форму кривой частотного распределения с важными пара-
метрами или характеристиками роста и деления индивидуаль-
ной клетки.
Остроумные методики синхронизации культур позволяют
применять и обычные методы биохимического анализа для изу-
чения клеточного цикла. Современные методики селекции син~
30—437
466
Глава 6
хронных культур позволяют получать популяции клеток, нахо-
дящихся на одной стадии клеточного цикла. Для этого клетки
иммобилизуют на твердом носителе, а только что возникшие
дочерние клетки смывают по мере их образования. Таким обра-
зом, в любой данный момент с колонки смываются клетки толь-
ко одного возраста, т. е. синхронные. В другом методе суспен-
зию клеток центрифугируют в градиенте плотности; находящие-
ся на различных стадиях жизненного цикла клетки различают-
ся размерами и плотностью и поэтому разделяются. Затем зо-
ну определенной плотности, представляющую собой гомоген-
ную популяцию, помещают в питательный раствор и выращи-
вают.
Зреня, ч
РИС. 6.31. Рост синхронной культуры
дрожжей Schizosaccharomyces pombe
сопровождается ступенчатым ростом
числа клеток. (Воспроизведено из ра-
боты: Mitchison J. М., Vincent W. S.,
A Method of Making Synchronous
Cell Cultures by Density Gradient
Centrifugation, in Cell Synchrony,
Cameron I. L„ Padilla G. M. (eds.),
p. 328, Academic Press, New York,
1966.)
Если мы затем проследим за изменением концентрации кле-
ток во времени, то мы вправе ожидать ступенчатого увеличения
числа клеток, поскольку число клеток должно удваиваться в
каждом клеточном цикле. Приведенные на рис. 6.31 экспери-
ментальные результаты могут служить иллюстрацией такого
поведения синхронной популяции. Оказывается, что по мень-
шей мере в первых двух циклах популяция клеток остается го-
могенной и синхронизированной; в любой момент цикла все
клетки находятся примерно на одной и той же стадии развития.
Как мы уже отмечали в гл. 1, гомогенные популяции являются
очень удобным объектом для экспериментального исследова-
ния, поскольку они обеспечивают резкое (в миллионы раз) по-
вышение интенсивности сигнала, отражающего характеристики
индивидуальных клеток. Таким путем можно детально изучить
многие особенности клеточного цикла.
Другой часто применяемый подход к получению синхрон-
ных культур называют индуцированной синхронизацией. В этом
подходе популяцию вынуждают расти синхронно путем перио-
дического изменения условий окружения. Однако поскольку та-
кие изменения могут сопровождаться нарушением нормального
метаболизма клеток, то для изучения физиологии клеток пред-
почтительнее методы селекции синхронных культур.
Молекулярная генетика и системы регуляции 467
Обратите особое внимание на то, что, согласно приведен-
ным на рис. 6.31 данным, по мере роста популяции синхронность
постепенно исчезает; первоначально ступенчатая зависимость
числа клеток от времени затем сменяется более плавной кривой.
Причина этого кроется в потере популяцией гомогенности; тен-
денция к гетерогенности обусловлена стохастической природой
роста и размножения клеток. Дело в том, что продолжитель-
ность клеточного цикла не является строго фиксированной, оп-
ределенной величиной для всех клеток одного типа. Так, две
первоначально идентичные дочерние клетки могут делиться не
одновременно. В то же время продолжительность среднего
жизненного цикла для популяций, состоящих из миллиардов
клеток, представляет собой строго определенную величину, ко-
торая имеет большое значение при проектировании и конст-
руировании реакторов (гл. 7).
В следующих двух разделах мы изучим картины клеточного
роста и синтеза высокомолекулярных соединений в хорошо ис-
следованной прокариоте Е. coli и эукариотическом организме
Saccharomyces cerevisiae. Основное внимание в каждом из этих
разделов мы будем уделять связи между репликацией ДНК и
делением клетки. Очевидно, что делению родительской клетки
на две дочерних должен предшествовать синтез геномов, содер-
жащих в себе всю генетическую информацию. Следовательно,
мы вправе ожидать тесной связи между процессами реплика-
ции ДНК и деления клетки; в противном случае данный вих
был бы нежизнеспособен.
6.5.2. Клеточный цикл Е. coll
Простая прокариота Е. coll размножается бинарным деле-
нием; в богатой питательными веществами среде число клеток
этого организма может удваиваться менее чем за 20 мин. Бак-
терии Е. coli имеют палочкообразную форму, длина которой
может служить надежной мерой объема и массы клетки. Не-
посредственным изучением изменения длины отдельных клеток
Е. coli под микроскопом показано, что в процессе клеточного
цикла длина бактерий возрастает постоянно; зависимость дли-
ны клетки от времени удовлетворительно описывается возра-
стающей экспоненциальной функцией. В течение клеточного
цикла аналогично изменяется и суммарное содержание всех
белков.
В то же время содержание ДНК в течение клеточного цик-
ла Е. coli изменяется довольно сложным образом; по характе-
ру этой зависимости Е. coli резко отличается от эукариотиче-
ских клеток. Координация процессов синтеза ДНК и деления
30*
468
Глава 6
клеток в случае Е. coli зависит от ряда регуляторных и кине-
тических параметров, которые мы и рассмотрим ниже.
В самых различных условиях роста, когда деление осуще-
ствляется через различные промежутки времени, период реп-
ликации хромосомы Е. coli остается приблизительно постоян-
ным (около 40 мин). В процессе синтеза ДНК* скорость дви-
жения репликационной вилки (участка молекулы ДНК, в кото-
ром осуществляются разделение цепей и синтез ДНК) также
остается постоянной. Следовательно, скорость синтеза ДНК
прямо пропорциональна числу активных репликационных вилок.
Репликация ДНК всегда начинается в определенном центре
хромосомы, называемом участком начала репликации. Две вил-
ки движутся в противоположных направлениях от участка на-
чала репликации вокруг циклической хромосомы Е. coli и схо-
дятся в момент окончания процесса репликации.
Регуляция процесса деления клеток связана с синтезом
ДНК; бактериальная клетка делится примерно через 20 мин
после того, как репликационные вилки достигают конца хромо-
сомы. Как показано на рис. 6.32, ситуация в целом остается
сравнительно простой, если Е. coli растет в среде, позволяющей
клеткам делиться в среднем раз в час. В этом случае синтез
ДНК осуществляется с постоянной скоростью в течение первых
сорока минут клеточного цикла, и в оставшиеся двадцать ми-
нут, предшествующие делению, клетка не синтезирует ДНК.
Росту клетки Е. coli с пятидесятиминутным клеточным циклом
(например, при слегка повышенной температуре) отвечает бо-
лее сложная картина синтеза ДНК; в этом случае новый набор
репликационных вилок начинает синтез ДНК за 10 мин до де-
ления клетки. Следовательно, в течение последних 10 мин этого
клеточного цикла ДНК синтезируется вдвое быстрее, чем в лю-
бой момент медленного (часового) цикла. Нетрудно видеть, что
кинетика синтеза ДНК в Е. coli и, вероятно, в некоторых дру-
гих бактериях в общем случае довольно сложна и характери-
зуется чередованием периодов спада и повышения синтетиче-
ской активности; характер чередования зависит от возраста
клетки и продолжительности клеточного цикла.
Относительно зависимости синтеза РНК от времени клеточ-
ного цикла имеется немного данных. Предполагается, что син-
тез РНК осуществляется непрерывно. В то же время показано,
что для фракции РНК, представляющей собой, по-видимому,
мРНК, характерны флуктуации. Следовательно, и скорость син-
теза индивидуальных белков может изменяться циклически.
Такое поведение и в самом деле было обнаружено в ряде
* В настоящем разделе под синтезом ДНК мы подразумеваем только
синтез ДНК хромосомы; репликацию плазмид мы рассмотрим позднее.
Молекулярная генетика и системы регуляции 469
экспериментальных работ, посвященных главным образом изу-
чению ферментов, активность которых может быть индуциро-
вана пли подавлена. Синтез большого числа индуцибельных
ферментов в бактерии можно стимулировать в любой момент
клеточного цикла. В синхронной культуре скорость синтеза ин-
дуцибельных ферментов изменяется ступенчато, что, вероятно,
обусловлено удвоением числа соответствующих этим ферментам
О
О 10 20 30 40 60
Возраст клетки, тин
ВродбУжителбность /меточного цикла 60пин
40
го
40-60
О
Возраст к ле тли
20 30
60
/Зродолтсителоность клеточного •
цикла 60 /чин
РИС. 6.32. Зависимость скорости синтеза ДНК и структуры хромосомы
в Е. coli от возраста клетки и продолжительности периода ее удвоения.
В правой части рисунка схематично изображена половина хромосомы Е. coli
от участка начала репликации до участка завершения репликации. Черными
точками обозначены репликационные вилки.
генов в момент репликации ДНК. В случае репрессии фермен-
тов имеется несколько возможностей. Если бактерии растут в
условиях, когда конечный продукт участка метаболического пу-
ти подавляет синтез фермента в максимальной или в мини-
мальной степени, то скорость синтеза фермента остается посто-
янной в течение промежутка времени, равного продолжитель-
ности клеточного цикла, а затем возрастает вдвое. Для проме-
жуточных уровней подавления характерно регулярное периоди-
ческое изменение скорости синтеза фермента. Некоторые экс-
периментальные данные позволяют предположить, что эти пе-
риодические изменения обусловлены устойчивыми колебания-
ми в системе регуляции, функционирующей по принципу обрат-
ной связи.
470
Глава 6
Зазраст штш, ч
а) К/гелгки пь/ши L -929
РИС. 6.33. а — Увеличение массы клетки ткани мыши в течение клеточного
цикла. [Воспроизведено из статьи: Killander D.t Zetterberg A., Quantitative
Cytochemical Studies on Interphase Growth. I. Determination of DNA, RNA
and Mass Content of Age Determined Mouse Fibroblasts in vitro and of Inter-
cellular Variation in Generation Time, Exp. Cell Res., 38, 272 (1965).]
6 — рост массы Amoeba proteus в течение клеточного цикла; каждая кривая
отвечает одной из шести различных индивидуальных клеток. [Воспроизведено
из статьи: Prescott М., Relations Between Cell Growth and Cell Division. I. Re-
duced Weight, Cell Volume, Protein Content, and Nuclear Volume of Amoeba
proteus from Division to Division, Exp. Cell Res., 9, 328 (1955).]
Молекулярная генетика и системы регуляции 471
6.5.3. Клеточный цикл эукариот
Прежде всего нам следует вспомнить, что в природе не су-
ществует типичной эукариотической клетки. В то же время мо-
дель такой воображаемой клетки оказывается очень полезной
при обсуждении черт, свойственных большинству эукариот;
именно в этом смысле мы будем использовать понятие о типич-
ной эукариоте в настоящем разделе.
Возможные различия между разными типами эукариотиче-
ских клеток легко оценить, если, например, проанализировать
увеличение клеточной массы в процессе жизненного цикла кле-
ток мыши и амебы (рис. 6.33). В то время как масса сухих ве-
ществ клеток мыши возрастает во времени линейно или экспо-
ненциально, скорость роста клеток амебы постепенно уменьша-
ется и в момент деления становится практически равной нулю.
Второй вариант более типичен для эукариот вообще. Обратите
внимание также на шкалу времени — клеточный цикл этих
эукариот длится около суток. Впрочем, некоторые эукариоти-
ческие организмы, например дрожжи, в оптимальных условиях
могут делиться примерно раз в час.
Для клеточного цикла эукариот характерна (по сравнению с
прокариотами) высокая степень дифференциации между от-
дельными периодами цикла. Обычный цикл эукариотической
клетки изображен на рис. 6.34. Он подразделяется на четы-
ре периода: Gi, S, G2 и М; относительные продолжительности
этих периодов указаны на рисунке. В периоде Gi происходит
активный синтез белков и РНК, а ДНК не синтезируется. В сле-
дующем периоде S осуществляется репликация хромосом. Функ-
ция периода G2 пока еще достаточно достоверно не выяснена.
Вслед за этим периодом начинается деление клеток (период
7И). Сложный, хорошо координированный процесс деления эу-
кариот называют митозом. В митозе два набора хромосом раз-
деляются и распределяются по дочерним клеткам. В медленно
растущих или нерастущих культурах клетки находятся в со-
стоянии покоя, обозначаемом символом Go.
В отличие от характерной для прокариот линейной времен-
ной зависимости синтез ферментов в эукариотах почти всегда
осуществляется периодически. В определенный момент клеточ-
ного цикла наблюдается резкое повышение концентрации того
или иного фермента. Имеющиеся экспериментальные данные
позволяют предположить, что в основе этого явления лежит по-
следовательный характер транскрипции ДНК.
В качестве примера, иллюстрирующего структуру клеточно-
го цикла эукариот и асимметричное деление клеток, вкратце
рассмотрим жизненный цикл размножающихся почкованием
472
Глава 6
РИС. 6.34. Последовательность событий в жизненном цикле типичной эука-
риотической клетки.
Цикл деления клети
Цикл почкования
Старт
Появление
Отпочковав-
шаяся дочер-
няя клетка
Период Gf
Деление
клеток
esp—Материнс-
--------| кая клетка
Синтез Период &г и деле-
ДНК (S) ^\^ние ядра (М)
Я верны и цикл
РИС. 6.35, Клеточный цикл почкующихся дрожжей 5. cerevisiae.
Молекулярная генетика и системы регуляции 473
дрожжей S. cerevisiae. Этот микроорганизм широко применяется
в пивоварении, производстве пищевых продуктов и генетической
инженерии. Как показано на рис. 6.35, дочерняя клетка растет
на материнской в виде почки. Затем, после деления, материн-
ская клетка быстро возвращается в начальную фазу цикла поч-
кования, в то время как обычно сравнительно небольшая до-
черняя клетка должна еще в течение какого-то времени расти;
только после этого она достигает типичного для начала цикла
почкования состояния («старт»).
Начало периода S примерно совпадает с появлением поч-
ки. Все непочкующиеся клетки находятся в периоде Gp Если
обратиться к изображенному на рис. 6.30 распределению ДНК
в индивидуальных клетках S. cerevisiae, можно достаточно
обоснованно предположить, что первый пик отвечает клеткам с
одним геномом, т. е. клеткам, находящимся в периоде Gb Вто-
рой пик отвечает клетками с двумя геномами, которые могут на-
ходиться в одном из двух периодов — G2 или М. Математиче-
ский анализ распределений подобного типа позволяет быстро
определить относительные времена пребывания клеток S. cerevi-
siae в периодах Gb S и G2-f-M и таким образом изучать влия-
ние генетических изменений и среды на регуляцию клеточного
цикла.
В этом разделе мы ознакомились с некоторыми особенностя-
ми клеточных циклов. В сущности, в настоящее время о них
имеется чрезвычайно мало данных. Как показало изучение кле-
точного метаболизма (гл. 5), для эффективной утилизации пи-
тательных веществ клетке необходима тщательнейшая коорди-
нация чрезвычайно сложной сети химических реакций. Биохи-
мия этих реакций известна достаточно детально, однако мы
почти ничего не знаем о том, как они осуществляются in vivo.
На рис. 6.36 приведены интересные данные, свидетельствующие
о наличии сложных закономерностей в распределении отдель-
ных взаимосвязанных реакций во времени. Следовательно, хи-
мические реакции в клетке локализованы не только в простран-
стве (например, за счет иммобилизации фермента на поверхно-
сти мембраны), но и во времени. Все эти сложные эффекты вы-
нуждают нас проявлять крайнюю осторожность, если перед на-
ми стоит задача математического анализа кинетики роста живых
клеток.
Этим мы заканчиваем наше введение в биологические науки.
Вооруженные полученными в этой и предыдущих главах зна-
ниями, мы должны быть готовы к изучению технологии процес-
сов с участием живых клеток. Основное внимание в последую-
щих главах мы уделим анализу, моделированию и проектиро-
ванию биологических процессов. В гл. 7 рассмотрена кинетика
клеточного роста.
474
Глава 6
РИС. 6.36. Сложный характер изменения относительного содержания свобод-
ных аминокислот в течение клеточного цикла в Chlorella pyrenoidsa. [Воспро-
изведено с разрешения из статьи: Hare Т. A., Schmidt R. R., Nitrogen Metabo-
lism During Synchronous Growth of Chlorella: II. Free-, Peptide-, and Protein-
Amino Acid Distribution, J. Cell Physiol., 75, 73 (1970).]
Упражнения
6.1. Частота мутаций. Мутации приводят к новым видам, мутантам. По-
скольку ДНК представляет собой всего лишь полимерную молекулу, в ходе
мутаций она, вероятно, претерпевает какие-либо химические и физические
изменения точно так же, как и молекулы любого другого соединения.
а) Исходя из приведенных на рис. 6У1.1,а данных, вычислите энергик>
активации мутации Ет и предэкспоненциальный множитель k°m в уравнении
km—k°mexp( — Em/RT). Сравните вычисленные величины с аналогичными па-
раметрами для более простых соединений, претерпевающих химические прев-
ращения. (Эти данные вы можете взять из любого учебного пособия по ор-
ганической химии.)
б) Поглощение излучения подчиняется закону Бера, согласно которому
каждый компонент мишени поглощает одну и ту же часть излучения, падаю-
щего на этот компонент; следовательно, 7(х) =/об~ах. На рис. 6У1.1,б изо-
Молекулярная генетика и системы регуляции
475
Тем/гера/пура, °с
РИС. 6У1.1. Скорость мутаций Е. coli возрастает при повышении температу-
ры и дозы ионизирующего излучения, а — влияние температуры на частоту
мутаций his~ (неспособных синтезировать гистидин бактерий) в his+ (синте-
зирующие гистидин организмы); б — влияние дозы рентгеновского излуче-
ния на превращение организмов met-2- (не синтезирующих метионин) в му-
танты met-2+ (синтезирующие метионин). (Воспроизведено из работы: Sa-
ger R„ Ryon F. J., Cell Heredity: An Analysis of the Mechanisms of Hereditary
at the Cellular Level, John Wiley and Sons, Inc., New York, 1961.)
а I
бражена линейная зависимость частоты мутаций от интенсивности излуче-
ния Д. При каких условиях все клетки могли получить одинаковую дозу
радиации? Можно ли ожидать линейную зависимость частоты мутаций от
дозы, если не все клетки получат одинаковую дозу радиации?
в) Приведенные на рис. 6У1.1 данные относятся к популяциям Е. coli,
мутирующим от his~ (неспособных синтезировать гистидин) к his+
(рис. 6У1.1,а) и от met-2~ (неспособных синтезировать метионин)
к met-2+. Если любые изменения ДНК осуществляются с такой же вероят-
ностью, как и в указанных случаях, какова общая скорость мутации ДНК
Е. coli? Обоснуйте ваш ответ. Характерно ли для этих мутаций фенотипиче-
ское проявление?
6.2. Репарация и термодинамика мутаций. Если в процессе участвует
большое число организмов, то кинетику мутаций можно описать теми же
параметрами, какие используются в кинетике обычных химических реакций.
а) В популяции бактерий с концентрацией клеток З-Ю7 мл-1 (объем
культуры 1000 л) мутация гена gi в ген g2 происходит с частотой 10~s на
.деление клетки, а обратная мутация g2 в gi — с частотой 10-6 на деление
жлетки. Вычислите «равновесные» концентрации каждого мутанта.
476
Глава 6
б) Поскольку в клетках, способных ферментативным путем репариро-
вать повреждения ДНК, имеется соответствующий специфический фермент,
можно предположить, что в таких случаях имеет место и равновесие между
нормальной и поврежденной ДНК:
Нормальная ДНК < > поврежденная ДНК
k-i
ферменты
репарации
Предположим, что ДНК повреждается еще и ультрафиолетовым излучением;
константа скорости этой реакции равна kl. Покажите, что в этом случае рав-
новесная популяция поврежденной ДНК повышается от ki/iki+k-i) до
(^i+^1)/(^i+^1+A—i).
в) Покажите, что после отключения источника ультрафиолетового излуче-
ния (при условии, что Кт репарационного фермента значительно превышает
общую концентрацию ДНК) средняя концентрация поврежденной ДНК воз-
вратится к исходному уровню со скоростью, пропорциональной разности кон-
центраций поврежденной ДНК в отсутствие и при наличии ультрафиоле-
тового излучения.
6.3. Упрощенная модель кинетики действия репрессора. Дж. М. Смит
предложил упрощенную схему сети кинетической регуляции, изображенную
на рис. 6У3.1.
РИС. 6У3.1. Упрощенная модель механизма действия репрессора. (Воспроиз-
ведено из работы: Smith J. М., Mathematical Ideas in Biology, fig. 30, Cam-
bridge University Press, London, 1971.)
а) Приняв, что концентрации P и гена стационарны, разработайте кине-
тическую модель, описывающую (после решения соответствующих уравнений)
зависимость концентраций РНК, М и Z от времени. Считайте, что скорость
потери РНК пропорциональна ее концентрации. Постарайтесь, насколько это
только возможно, приблизиться к полному решению для переходного со-
стояния.
б) Основываясь на материале этой и предыдущих глав, укажите, долж-
на ли в данном случае приниматься во внимание диффузия, и отметьте не-
достатки модели.
6.4. Мутация и генетический код. В мутантных белках обнаружены ука-
занные ниже замены аминокислот. Объясните, какие кодоны подверглись.
Молекулярная генетика и системы регуляции
477
мутации в каждом случае, и укажите тип мутации.
6.5. Организация внутриклеточных превращений; многоферментные систе-
мы. Многие реакции в клетке протекают путем превращения Sj в S3 через
промежуточное соединение 5г, причем по сравнению с Si и 5з равновесная
концентрация S2 мала:
ki k%
Si v..... S2 S3
fe_l Й_2
Если Зз участвует в следующей катализируемой ферментом реакции, то об-
ратимостью второй реакции можно пренебречь.
а) Приняв, что для каждого фермента и что Si и S2, Ei и Е2
распределены в объеме равномерно, покажите, что скорость реакции в ста-
ционарном состоянии определяется следующим выражением:
Скорость = -т—пттЬтть- —
б) Покажите, что если Ei закреплен на одной стороне проницаемой пла-
стины, а Е2 — на другой, то скорость реакции в стационарном состоянии бу-
дет подчиняться уравнению
Скоростью
Покажите также, что эта величина меньше, чем s2,equiiS)/L, где L — расстоя-
ние между поверхностями пластины, a s2,equii — концентрация S2, находяще-
гося в равновесии с Si при концентрации последнего si.
в) Обычно скорость поглощения кислорода равна 10~8 моль/(с-см3)..
Приняв, что скорости каждой из стадий внутреннего метаболизма примерно-
одинаковы, вычислите максимальное расстояние между Ei и Е2, при котором
была бы возможной указанная скорость поглощения кислорода, если величи-
на $2.equii составляет от 10~4 до 10“12 моль/см3. [Weisz Р. В., Enzymatic Reac-
tion Sequences and Cytological Dimensions, Nature, 195, 772 (1962).]
478
Глава 6
6.6. Организация внутриклеточных превращений; синтез и утилизация
АТР. В интактной клетке имеются локализованные центры (органоиды),
призванные снабжать всю клетку определенными соединениями. В этой связи
представляется интересным рассмотреть сперматозоиды быка, в которых
АТР синтезируется только в промежуточном отделе сперматозоида, а жгутик
расходует значительную часть АТР для движения.
а) Приняв, что реакция утилизации АТР в любой точке жгутика имеет
нулевой порядок и что АТР распределен в промежуточном отделе равно-
мерно в концентрации с0, покажите, что изменение концентрации АТР в жгу-
тике может быть описано уравнением с=1—z(l+02/2) + 02z2/2, где 0—мо-
дуль Тила жгутика, z — безразмерный параметр, характеризующий расстояние
от жгутика, и., с — безразмерный параметр, характеризующий концентрацию.
б) Каждый движущийся сперматозоид потребляет кислород для движе-
ния со скоростью 3,7—5,0-10-18 моль/с. С помощью указанных ниже допол-
нительных данных вычислите минимальное содержание АТР в сперматозои-
де, необходимое для поддерживания его транспорта в жгутик за счет
диффузии.
Коэффициент диффузии АТР (определенный с учетом содержания воды
в жгутике и извилистости) 3,6-10-6 см2/с
Объем промежуточного отдела 1,3-10—12 см3
Длина жгутика 5-Ю-3 см
Площадь поперечного сечения жгутика (без учета оболочки н фибрилл)
Выход АТР в расчете на поглощенный О2 6%
в) Найдено, что в среднем в одном сперматозоиде содержится
200-10~18 моль АТР. Утверждалось, что от одной трети до половины всего
АТР расходуется митохондриями клетки. Оцените результаты ваших расче-
тов в части «б» этой задачи с точки зрения необходимости (или необязатель-
ности) учета других, отличных от пассивной диффузии механизмов транспор-
та для обеспечения соответствующего распределения АТР из митохондрий,
локализованных в промежуточном отделе. [Neva А. С„ Rikmenspoel R., Diffu-
sion of АТР in Sperm Flagella, J. Theoret. Biol., 26, 11 (1970).]
6.7. Теория мишени. Чтобы объяснить с молекулярно-биологической точ-
ки зрения потерю активности спорами, вирусами или клетками, была предло-
жена гипотеза, согласно которой «в каждой клетке и в каждом организме
существует ряд «мишеней» и изменение активности является результатом
неупорядоченного поражения этих мишеней». (Smith. J. М., Mathematical Ideas
in Biology, p. 87, Cambridge University Press, London, 1971.) Рассмотрим сле-
дующую ситуацию.
Предположим, что содержащая N мишеней клетка облучена дозой К
частиц. Пусть вероятность поражения данной мишени данной частицей рав-
на р; очевидно, что величина р очень мала.
Вероятность того, что данная мишень не будет поражена данной части-
цей, равна 1— р.
Отсюда следует, что вероятность того, что данная мишень не будет по-
ражена ни одной из К частиц, составляет
(1—
а) Учитывая, что при большом К и малом р Кр~ (К—1)р~ (К—2)р, до-
кажите справедливость указанного приближения для этого типичного слу-
чая (большое К, малое р).
б) Покажите, что если относительное количество поврежденных клеток
(под повреждением здесь подразумевается, что клетка поражена один раз
или более) невелико, то «относительное количество поврежденных клеток
равно NKp, т. е. пропорционально дозе». Если же, напротив, осталось мало
неповрежденных клеток, то доля s последних определяется выражением
In s= — NKp-, подтвердите справедливость этого уравнения.
Молекулярная генетика и системы регуляции 479
в) Если клетка повреждается только после нескольких поражений, то для
анализа такой ситуации можно использовать аналогичный подход. «Бактерия
поражается идентичными бактериофагами; число поражающих одну бактерию
бактериофагов равно г, каждый бактериофаг содержит N существенных
генов. Жизнеспособный бактериофаг может образовываться путем рекомби-
нации при условии, что в каждом из N локусов один ген не поврежден»
(см. приведенную выше ссылку). Покажите, что если «вероятность непораже-
ния каждой мишени мала», то Ins — N In г — NpK (здесь под мишенями подра-
зумевается число копий бактериофага г в клетке в момент облучения).
г) Покажите, что результаты предыдущего упражнения 6в согласуются
с данными, изображенными графически на рис. 6У7.1, а; оцените все возмож-
РИС. 6У7.1. Экспериментальная зависимость заражаемости S бактерий бак-
териофагом Т7 (а) и бактериофагом Т2 (б) от дозы облучения при различной
продолжительности периода между заражением и облучением. [По материа-
лам статьи: Benzer S., Resistance to Ultraviolet Light as an Index to the
Reproduction of Bacteriophage, J. Bacteriol., 63, 59 (1952).]
ные параметры. Какие явления могут обусловливать поведение системы, гра-
фически представленное на рис. 6У7.1.6?
6.8. Судьба мутантного гена в растущей популяции. В популяциях мута-
ции происходят часто. Жизнеспособность мутантных организмов поразитель-
но высока, а устойчивость мутантных генов низка. Для размножающейся
половым путем популяции AI диплоидных организмов в момент появления
мутантного гена (аллеля) частота мутации равна 1/2Лг. Общее число организ-
мов в популяции N превышает число организмов, участвующих в размно-
жении Ne.
а) Для селективно нейтральной мутации (когда мутант не имеет ни
преимуществ, ни недостатков по сравнению с исходным штаммом) среднее
число поколений, предшествующих исчезновению мутантного гена, опреде-
ляется выражением ti = 2(Ne/N)in2N, а среднее число поколений, предшест-
вующих закреплению гена (т. е. состоянию, когда частота гена равна 1,0)
составляет tf—4Ne. Здесь величины ti и tf усреднены только по параметрам
исчезновения и закрепления гена соответственно. Приняв Ne/IV=0,5 и время
480
Глава 6
удвоения равным 1 ч, вычислите ti и tf для популяции с концентрацией 107
клеток в 1 мл при объеме культуры 1 л.
б) Разброс значений tf мал, а значений ti велик по отношению к их сред-
ним величинам; этот факт вы можете подтвердить расчетом, взяв за основу
следующие выражения:
var tt « 4,58№е var tf * ----(2 i In 2N \
' N \ N J
в) Вероятность закрепления мутации в популяции можно оценить с по-
мощью уравнения
1 -ехр[ — 2CVg/7V)gx ]
1 — exp ( — iNgSj
где $i — селективное преимущество мутантного гена по сравнению с исходным
геном. Покажите, что для типичных условий популяции микроорганизмов
u=2s\Ne/N. Следовательно, даже если селективное преимущество велико
(S( = 1 % = 10-2), вероятность того, что мутант в конце концов станет домини-
рующим в данной популяции, невелика (Kimura М., Ohta Т., Theoretical
Aspects of Population Genetics, Princeton University Press, Princeton, N.J.,
1971).
6.9. Естественный отбор. Относительную приспособленность w данного
генотипа х можно определить как отношение выживаемости генотипа х
к выживаемости х среднего генотипа популяции. (Под выживаемостью пони-
мают отношение числа организмов, участвующих в размножении, к числу
родившихся в каждом поколении.) Предположим, что приспособленность
генотипа х линейно зависит от среднего выражения фенотипа _так, что
w(x) = l—а(х—х), где а — мера интенсивности влияния разности (х—х) на
w(x) (а>0). По определению g(x)—нормализованное распределение ис-
ходного генотипа [ f g(x)dx—1,0]. Через одно поколение распределение ста-
новится иным: g'(x)=w(x)g(x).
а) Выразите х' через среднее значение х и генетическую дисперсию о2.
[Напомним, что среднее х' = f xg(х)dx, а дисперсия а2^ { (х— x)2g(x)dx.]
б) Покажите, что в одном поколении изменение х пропорционально о2,
т. е. «скорость эволюции пропорциональна генетической дисперсии популяции»
(основная теорема естественного отбора; см. работу: Wilson Е. О., Bos-
sert W. И., Primer of Population Biology, pp. 79—83, Sinauer Associates, Stam-
ford, Conn., 1971).
в) Уилсон и Боссерт утверждают, что величина (х'— х) «некоторым об-
разом пропорциональна дисперсии генотипа», даже если w нелинейно за-
висит от х. Выберите ряд обоснованных зависимостей w=f(х) и покажите
справедливость (или некорректность) этого утверждения.
6.10. Кратные сайты оператора; регуляция промотора ЛРл. Белок-репрес-
сор промотора фага ХРд может связываться с тремя различными сайтами
оператора, обозначенными символами OR1, OR2 и OR3. Как показано на
рис. 6У10.1, связывание репрессора с любым сайтом (за исключением О«3)
блокирует промотор PR. Допустим, что реакция связывания репрессора
с оператором равновесна и что константы равновесия реакций связывания
репрессора с тремя сайтами составляют Ki, К2 и Кз соответственно.
а) Найдите общее уравнение, выражающее доступную для связывания
с РНК-полимеразой долю промоторов Pr через константы равновесия, общее
количество белка-репрессора и последовательности PROR в клетке.
б) Оцените долю доступных промоторов в клетке, содержащей 25 после-
довательностей RrOr и 70 молекул репрессора. Примите Ki, К2 н Кз равны-
ми 125-108, 1,25 10s и 1' 109 М~‘ соответственно. Каков физический смысл
полученного вами результата?
Молекулярная генетика и системы регуляции
481
Pr
г---------------
I
L____________________________________________________
РИС. 6У10.1. Относительные положения промотора PR и трех операторов О»/.
Or 2 и Or3 в фаге X. н
в) Решение предыдущих упражнений основывалось на принципах клас-
сической термодинамики. Статистическая термодинамика позволяет точнее
оценить взаимодействия небольшого числа различных молекул в популяции
клеток. Найдите решения задач 6.10а и 6.106 в свете теории, изложенной
в статье: Berg О. G., Blomberg С., Mass Action Relations in vivo with Applica-
tion to the Lac Operon; J. Theoret. Biol., 67, 523 (1977)..
6.11. Скорости репликации ДНК в прокариотах и эукариотах, а) Для
осуществления репликации необходимо расплетание двойной спирали ДНК.
Предположим, что в ДНК имеются два центра расплетания (две реплика-
ционные вилки). Определите скорость вращения в центрах расплетания
(в числе оборотов в минуту) ДНК Е. coli, если вся хромосома синтезируется
за 41 мин. б) Период S линии клеток млекопитающих с геномом, содержа-
щим 1,1 м двухцепочечной ДНК, длится 4,8 ч. Если по скорости синтеза
ДНК эти клетки не отличаются от Е. coli, сколько репликационных вилок
должно функционировать одновременно?
6.12. Рекомбинация фрагментов ДНК. Две двухцепочечные молекулы
ДНК, 1 и 2, расщеплены рестрикционной эндонуклеазой EcoRI; одна цепь
каждой из этих ДНК изображена ниже. Образовавшиеся фрагменты смеша-
ны, рекомбинированы и обработаны ДНК-лигазой, катализирующей образова-
ние ковалентных связен. Перечислите все возможные продукты реакций.
ДНК 1: (5/)CGATAGAATTCAGTCAA(3/)
ДНК 2: (5/)GAATTCGGTTCGAATTCG(3/)
6.13. Карты рестрикции. Относительные положения сайтов рестрикции во
фрагментах ДНК определяют путем их частичного или полного расщепления
рестриктазами и последующей оценки молекулярных масс образующихся
фрагментов методом электрофореза в геле. Фрагмент ДНК Hind III, содер-
жащий 3000 пар оснований, обработан EcoRI, В gill и смесью этих фермен-
тов. Электрофорез в геле показал, что при этом образуются два фрагмента,
содержащие 1400 и 1600 пар оснований, три фрагмента с 400, 900 и 1700
парами оснований и четыре фрагмента с 400, 500, 900 и 1200 парами основа-
ний соответственно. Определите карту рестрикции Hind III.
6.14. Праймер для кДНК интерферона. Для обеспечения максимальной
вероятности клонирования кДНК, кодирующей аминокислотную последова-
тельность интерферона нз фибробластов (IFN-^), полезно воспользоваться
олиго дезоксирибонуклеотидным праймером, комплементарным 5'-концевой
кодирующей последовательности мРНК. N-Концевая последовательность бел-
ка выражается формулой Met—Ser—Туг—Asn. Перечислите все возможные
соответствующие нуклеотидные последовательности мРНК и набор отвечаю-
щих им праймеров. Будьте внимательны при идентификации 3'- и 5'-концов
в каждом из праймеров.
31-537
482
Глава 6
6.15. Продуцирование внутриклеточных белков рекомбинантными клетками.
Оценка емкости биореактора, необходимой для продуцирования определен-
ного количества аккумулирующегося в клетках продукта экспрессии клониро-
ванного гена, не представляет затруднений. Предположим, что рекомбинант-
ный штамм Е. coli аккумулирует чужеродный белок, продукт экспрессии
плазмидного гена, в концентрации до 30% от общего белка клетки. Если
максимально допустимая плотность клеток равна 109 клеток/мл, сколько
белка можно выделить из 1 л объема биореактора?
Литература
Каждая из работ, указанная в гл. 1 и 2, содержит ценные сведения, имею-
щие непосредственное отношение к той или иной теме настоящей главы. Кро-
ме того, рекомендуется следующая литература:
1. Стент Г. С., Кэлиндар Р., Молекулярная генетика. — М.: Мир, 1981'.
Прекрасно описаны основные понятия молекулярной генетики в историче-
ском аспекте; особое внимание уделено подтверждению принятых в настоя-
щее время гипотез.
2. Lewin В., Genes, 2d ed., John Wiley and Sons, New York, 1985. Хорошо из-
ложенный, очень подробный и глубокий обзор, посвященный природе ге-
нов, их репликации и функциям.
3. Албертс Б., Брэй Д„ Льюис Дж., Рафф М., Робертс К., Уотсон Дж. Д.,
Молекулярная биология клетки, т. 1—5. — М.: Мир, 1986—1987. Биология
клетки, изложенная с точки зрения молекулярной биологии, представлена
как одна из самых интересных отраслей биологических наук.
Введение в проблемы мутагенеза и прикладную генетику (с точки зрения
микробиологической промышленности) дается в следующей литературе:
4. Elander R. Р., Applications of Microbial Genetics to Industrial Fermentation,
in Fermentation Advances, Perlman D. (ed.), Academic Press, Inc., New
York, 1969.
5. Demain A. L., Overproduction of Microbial Metabolites and Enzymes Due to
Alteration of Regulation, Adv. Biochem. Eng., 1, 113 (1971).
6. Dobrazanski W. T., Microbial Genetics in Pharmacy, Chem. Br., 10, 386
(1974).
7. Biology of Industrial Microorganisms, Demain A. L., Solomon N. A. (eds.).
Benjamin Cummings and Co., Menlo Park, CA, 1984. В этом недавно опуб-
ликованном сборнике суммированы данные о метаболизме, системах регу-
ляции и генетике микроорганизмов, используемых в настоящее время в
промышленности. Одна из очень немногих книг, в которых детально рас-
смотрена микробиология плесеней и Streptomyces.
В приведенных ниже статьях рассмотрены гибридомы и методы слияния
клеток, применяемые при синтезе моноклональных антител:
8. Peberdy J. F., Protoplast Fusion — A Tool for Genetic Manipulation and
Breeding in Industrial Microorganisms, Enzyme and Microbial Tech., 2,
25 (1980).
9. Milstein C.y Monoclonal Antibodies, Scientific American, 243, 66 (1980).
10. Boyd J. E., James Д., McClelland D. B. L., Human Monoclonal Antibodies —
Production and Potential, Trends in Biotech., 2, 70 (1984).
Отдельные примеры практического применения генетики микроорганизмов рас-
смотрены в следующих статьях из сборника Fermentation Advances, Perl-
man D. (ed.), Academic Press, Inc., New York, 1969:
11. Pardee A. B., Enzyme Production by Bacteria, p. &
Молекулярная генетика и системы регуляции 483
12. Veda К., Some Fundamental Problems of Continuous L-Glutamic Acid Fer-
mentations, p. 43.
13. Furuya A., Misawa M., Nara T., Abe S., Kinoshita S., Metabolic Controls of
Accumulations of Amino Acids and Nucleotides, p. 177.
В связи с ростом научных и технических достижений в последние годы публи-
куется все больше работ, посвященных технологии рекомбинантных ДНК.
В приведенном ниже перечне они расположены по возрастающей степени
сложности — от вводных, общих обзоров до подробного описания методов
и результатов исследований.
14. Cohen S. N., The Manipulation of Genes, Scientific American, 233, 24
(1975).
15. Gilbert IF., Villa-Komaroff L., Useful Proteins from Recombinant Bacteria,
Scientific American, 242, 74 (1980).
16. Wetzel R., Applications of Recombinant DNA Technology, American Scientist,
68, 664 (1980).
17. Watson J. D., Tooze J., Kurz D. T., Recombinant DNA: A Short Course,
W. H. Freeman and Co., New York, 1983.
18. Glover D. M., Genetic Engineering: Cloning DNA, Chapman and Hall, New-
York, 1980.
19. Old R. W„ Primrose S. B., Principles of Gene Manipulation, 2d ed., Univer-
sity of California Press, Berkeley, 1981.
20. Rodriguez R. L„ Tait R. C., Recombinant DNA Techniques: An Introduction,
Addison-Wesley Publishing Co., Reading Massachusetts, 1983.
21. Recombinant DNA, Methods in Enzymology, vols. 68 (1979), 100, 101
(1983), Academic Press, New York.
22. Sanger F., Nicklen S., Coulson A. R., DNA Sequencing with Chain-Termina-
ting Inhibitors, Proc. Natl. Acad. Sci. USA, 74, 5463 (1977).
23. Gray P. IF. et al., Expression of Human Immune Interferon cDNA in E. coli
and Monkey Cells, Nature, 295, 503 (1982).
Научные и общественные дискуссии, посвященные технологии рекомби-
нантных ДНК, отражены в книге:
24. Watson J. D., Tooze J., The DNA Story, W. M. Freeman and Co., San Fran-
cisco, 1981.
Работы, посвященные изучению клеточных циклов вообще и циклов Е. coll
и S. cerevisiae в частности, обобщены в следующей литературе:
25. Mitchison J. М., The Biology of the Cell Cycle, Cambridge University Press,
London, 1971.
26. Ingraham J. L., Maalee O., Neidhardt F. C., Growth of the Bacterial Cell,
Sinauer Associates, Inc., Sunderland, Massachusetts, 1983.
27. Hartwell L. H., Saccharomyces cerevisiae Cell Cycle, Bact. Rev., 38, 164
(1974).
28. Lievense J. C., Lim H. C., The Growth and Dynamics of Saccharomyces
cerevisiae, in Annual Reports on Fermentation Processes, vol. 5, p. 211,
Tsao G. T. (ed.)., Academic Press, New York, 1982.
31
Глава 7
Кинетика процессов
утилизации субстрата,
образования продуктов метаболизма
и биомассы в культурах
клеток
Если небольшое количество живых клеток поместить в рас-
твор, содержащий все необходимые питательные вещества, то
при определенных температуре и pH клетки будут расти. В за-
висимости от морфологии клетки процесс ее роста может проис-
ходить двумя путями. Одноклеточные организмы претерпевают
в процессе роста деление, и поэтому увеличение их биомассы
(массы живых организмов) сопровождается увеличением числа
клеток. Основное внимание в настоящей главе будет уделено
именно этому типу роста; здесь мы встретимся с проблемой ро-
ста популяции. Совершенно по-иному происходит рост плесне-
вых грибов — в этом случае по мере роста организма возраста-
ют в первую очередь длина и число гиф мицелия. Таким обра-
зом, в культуре растущей плесени увеличиваются размеры ор-
ганизмов и их концентрация, но не обязательно число орга-
низмов.
С клеточным ростом тесно связаны два других процесса: по-
глощение клеткой некоторых веществ из окружения и выделе-
ние конечных продуктов метаболизма клетки в среду. Как мы
увидим ниже, в процессе роста скорости этих процессов изме-
няются в очень широких пределах. В общем случае предска-
зать эти скорости не представляется возможным; вместе с тем
весь многолетний опыт говорит о том, что среди множества из-
вестных процессов с участием культур клеток достаточно часто
встречается только ограниченное число типичных путей утили-
зации субстрата и образования продуктов жизнедеятельности
клеток. Изучив сначала эти типичные пути, мы будем лучше
подготовлены к решению новых задач, связанных с примене-
нием культур растущих клеток.
Необходимая и достаточная точность (а следовательно, и
сложность) математического описания кинетических свойств
системы в свою очередь зависит от ее сложности и предпола-
гаемой области применения математических выражений.
Прежде чем перейти к анализу конкретных процессов и соот-
ветствующих математических выражений, описывающих их ки-
нетику, мы должны иметь определенное представление о раз-
Образование биомассы в культурах клеток 485
нообразии реакций и процессов в популяции клеток. В нашем
обсуждении основное внимание будет уделено клеточному рос-
ту, но аналогичные принципы, концепции и ход рассуждений
применимы и к другим аспектам изучения химических процес-
сов в клетке.
Начнем с самого сложного случая — с анализа растущей
популяции клеток в самом общем виде. Затем рассмотрим, ка-
кие упрощения можно допустить в этом анализе и в каких си-
туациях эти упрощения оправданны. На рис. 7.1 перечислены не-
Среда (окружение) Китателбнь/е вещества Копуляция клеток
• Многокомпонентностб • Реакции драотборе •Кислотно-основное рад коде сие • Изменение pH, температуры и других параметров г •Изменение реологических сооисто (Вязкости •Многофазноето (Г-Ж, Ж~Ж, Г-Ж-Ж) • Объемная неоднородность • Многокомпонент- ное те • Гетерогенность популяции клеток • Множественность реакций . • Системы онутрен ней регуляции . •Способность к адап /нации • Стохастические факторы •Генетические изменения
Субстраты Ироду к те/
метаболизма Ге плота
Механические^
Воздействия '
РИС. 7.1. Некоторые из основных параметров, явлений и взаимодействий,
определяющих кинетику роста популяции клеток.
которые параметры, явления и взаимодействия, влияющие на
кинетику роста популяции клеток. При этом прежде всего сле-
дует усвоить, что во всех случаях нужно учитывать две взаимо-
действующие системы — биологическую фазу, состоящую из по-
пуляции клеток, и фазу, окружающую эту популяцию, или сре-
ду. Клетки поглощают питательные вещества и превращают
субстраты (взятые из среды) в продукты метаболизма. Клетки
генерируют теплоту, а температура среды в свою очередь опре-
деляет температуру клеток. Среда оказывает механическое воз-
действие на клетки посредством гидростатического давления
или гидродинамических эффектов, а также путем изменения
вязкости среды вследствие накопления клеточной массы и про-
дуктов жизнедеятельности клеток.
Что касается среды, то из ряда ее важнейших характери-
стик прежде всего следует отметить многокомпонентность: во-
первых, она должна содержать все необходимые для роста кле-
ток питательные вещества, и, во-вторых, в ней накапливаются
(по мере роста клеток) различные конечные продукты клеточ-
ного метаболизма. Химические реакции могут происходить и в
среде, что приводит к модификации продуктов метаболизма,
486
Глава 7
как, например, при гидролизе пенициллина. Часто клетки по-
требляют или продуцируют вещества, изменяющие кислотность
среды; взаимосвязь клеточных процессов с кислотно-основным
равновесием определяет pH среды, который в свою очередь
влияет на клеточную активность и на процессы транспорта.
В ходе клеточных реакций могут происходить изменения темпе-
ратуры, ионной силы, pH и реологических свойств бульона во
времени.
Часто среда представляет собой многофазную систему, со-
стоящую из жидкой фазы и диспергированных в ней пузырьков
газа или из двух несмешивающихся жидких фаз, а иногда из
одной газовой и двух жидких фаз. К числу последних достиже-
ний в разработке биореакторов относятся методы одновремен-
ного проведения клеточных реакций и процессов разделения пу-
тем введения в среду особых веществ, способствующих обра-
зованию дополнительной жидкой или твердой фазы. Наконец,
в силу большого объема биологического реактора, высокой вяз-
кости и неньютоновской природы бульона в отдельных его точ-
ках могут создаться разные условия. Все эти параметры и пере-
менные среды существенно влияют на кинетику клеточного роста.
Что же касается наиболее важных особенностей популяции
клеток, то, как мы уже знаем из предыдущих глав, каждая
индивидуальная клетка представляет собой сложную много-
компонентную систему, которая даже на клеточном уровне да-
лека от гомогенности. В каждой клетке одновременно осуществ-
ляется множество химических реакций, управляемых сложным
комплексом систем регуляции. Благодаря этим системам клет-
ка может изменять (и изменяет на самом деле) скорость и да-
же тип происходящих в ней химических реакций в зависимости
от условий и состава среды. Длительное культивирование кле-
точной популяции может сопровождаться накоплением спон-
танных мутаций; кроме того, особенности технологического
процесса могут налагать давление отбора, приводящее к мед-
ленным изменениям генетической природы штамма. С другой
стороны, в культуре растущих клеток всегда наблюдается суще-
ственная гетерогенность популяции, т. е. в любой момент, в лю-
бом, даже самом малом объеме культуры индивидуальные
клетки различаются по возрасту (одни только что образовались
в результате деления, другие уже достигли состояния зрело-
сти, а третьи находятся в стадии деления) и, следовательно, по
биохимической активности. Из предыдущей главы, посвящен-
ной особенностям клеточного цикла, мы знаем, что различаю-
щиеся по возрасту клетки часто обладают различными метабо-
лическими функциями и активностями.
Очевидно, практически невозможно создать кинетическую
модель, которая учитывала бы все параметры и факторы, пе-
Образование биомассы в культурах клеток
487
речисленные на рис. 7.1. Поэтому теперь рассмотрим ряд при-
ближений, которые позволяют упростить эту картину и найти
математические выражения, описывающие кинетику роста по-
пуляции клеток. Во-первых, что касается среды, то обычно до-
пускают, что, за исключением одного компонента, все ее со-
ставляющие присутствуют в таких высоких концентрациях, что
их изменения практически не отражаются на общих скоростях
процессов. Таким образом, один компонент среды становится
питательным веществом, лимитирующим скорость клеточного
роста, и при анализе влияния состава среды на кинетику роста
клеток мы должны учитывать концентрацию только этого ком-
понента. Иногда возникает необходимость в учете некоторых
других составляющих среды, например накапливающегося в
среде ингибитора, в противном случае наше описание кинетики
было бы слишком далеким от истинной картины. В отношении
других параметров среды часто можно допустить, что их изме-
нения не влияют в сколько-нибудь существенной степени на ки-
нетику роста микроорганизмов за тот период, в течение которо-
го проводится типичный эксперимент или осуществляется ти-
пичный процесс, если эти изменения не превышают обычных
для этого эксперимента или процесса пределов. К тому же си-
стемы контроля и регулирования биореакторов могут поддер-
живать на постоянном уровне целый ряд параметров среды,
в том числе pH, температуру и концентрацию растворенного
кислорода. Впрочем, в некоторых случаях для адекватного опи-
сания требуемого диапазона кинетического поведения может
возникнуть необходимость учета в модели многокомпонентно-
сти и нескольких параметров среды.
На рис. 7.2 суммированы основные принципы различных
приближений и способов описания, которые могут быть полез-
ны при математическом анализе клеточной фазы системы. Со-
гласно этой схеме, разработанной Фредриксоном и Цучия, под-
ходы к анализу микробиологических систем классифицируются
в соответствии с числом компонентов, используемых при описа-
нии клеток, а также в зависимости от того, рассматриваются ли
клетки как гетерогенная популяция различных объектов (как
оно и есть на самом деле) или как популяция некоторых усред-
ненных клеток (в этом случае клетки принципиально не отли-
чаются от какого-либо компонента в растворе). Многокомпо-
нентные модели клеток называют структурированными, а одно-
компонентные— неструктурированными. Описание популяции
клеток с учетом их гетерогенности называют сегрегированным
подходом, а в несегрегированном описании рассматривают
только усредненные свойства клеток. Как показано на рис. 7.2,
реальная ситуация отвечает структурированной сегрегирован-
ной системе. Если гетерогенность клеток не влияет в сущест-
488
Глава 7
венной степени на кинетику исследуемых процессов, то можно
принять «приближение усредненной клетки» и тем самым упро-
стить способ описания от сегрегированного до несегрегирован-
ного. В так называемом состоянии «сбалансированного роста»
вся синтетическая активность клеток скоординирована таким
образом, что на средний состав клеток пролиферация популя-
ции не влияет. В этом случае допустимо использовать модели,
не учитывающие многокомпонентную природу клеток. Как мы
Реструктурированный
Структурированный
Максимально упрощенный с косой
Приближение
Популяция клеток сбалансированного Многокомпонент-
рассматрибаетс я ----------------- ное описание усред-
как однокомпонентное роста пенной клетки
растворенное бещестбо
Приближений'
-—усредненной -----
“клетки”
Приближение
к усредненной—
клетки”
Однокомпонентное
описание; индиди-
дуально/е клетки
гетерогенны
Приближение Многокомпо г. .
сбалансированного нентное дпи-
роста сание; йндиПи-
дуальные клет-
ки гетерогенны
Реальная ситуация
РИС. 7.2. Различные способы описания кинетики роста популяции клеток.
увидим ниже, при анализе и описании роста популяций клеток
обычно принимают наиболее идеализированную ситуацию, т. е.
несегрегированную, неструктурированную модель.
С другой стороны, мы познакомимся с рядом таких ситуа-
ций, когда выгоднее рассматривать биологическую фазу как
более сложную систему. Так, в нестационарном состоянии (ти-
пичном, в частности, для периодических микробиологических
процессов) условия сбалансированного роста даже в первом
приближении соблюдаются только в течение сравнительно ко-
роткого промежутка времени. Хорошо известно, что в ходе пе-
риодического процесса в очень широких пределах могут изме-
няться как клеточный состав популяции, так и скорости и типы
реакций, осуществляющихся в этой популяции, и тогда могут
пригодиться более детальные модели. Кроме того, структуриро-
ванные модели позволяют отражать непосредственно в матема-
Образование биомассы в культурах клеток
489
тических выражениях известные особенности сети клеточных
биохимических реакций. Аналогично не представляющее осо-
бых затруднений включение в модель кинетики клеточного ро-
ста особенностей клеточного цикла (сегрегированный подход)
способствует повышению ценности и расширению диапазона
применимости модели. Важнейшими элементами сегрегирован-
ных моделей являются кинетика и особенности регуляции роста
индивидуальной клетки.
Живые клетки представляют собой чрезвычайно малые си-
стемы, поэтому количество любого химического компонента в
них крайне ограничено. Действительно, приведенные в табл. 5.6
величины очень малы по сравнению с обычным для химика
числом молекул порядка 1023. Предельным случаем является
ДНК, которая в медленно растущей бактерии содержится толь-
ко в количестве одной молекулы; очевидно, что в этом случае
само понятие «концентрация ДНК в клетке» становится весьма
неопределенным. То же самое можно сказать о содержащихся
в следовых количествах ионах, органоидах и многих других
компонентах клетки. Хотя в некоторых из рассматриваемых ни-
же моделей мы будем описывать внутриклеточные события как
непрерывные, следует отдавать себе отчет в том, что такое опи-
сание является всего лишь удобным приближением типичной
усредненной клетки в популяции клеток (см. ниже). Реакции и
процессы массопередачи, в которых участвует ограниченное
число молекул, должны рассматриваться как случайные собы-
тия. Для описания таких событий создавались стохастические
модели популяций клеток, не имеющих существенных преиму-
ществ по сравнению с более простыми детерминистическими
моделями. Предсказуемость поведения популяции клеток можно
проиллюстрировать, например, оценив точность детерминисти-
ческого описания очень малой популяции, в частности ти-
пичного инокулята (посевного материала; обычно около 105 кле-
ток). Пусть совокупность продолжительностей клеточных цик-
лов индивидуальных клеток описывается нормальным распреде-
лением:
P(()=__Uexp ..-('-О»
2о2
Примем i=l, а стандартное отклонение о равным 0,5. Тогда мы
можем сказать, что с коэффициентом доверительности 0,95 про-
должительность клеточного цикла отдельной клетки составляет
Л)дна клетка — t±l,96o= 1,0±0,98
Далее рассмотрим суспензию, содержащую достаточно боль-
шое число клеток, например m клеток. Пусть каждая из пг кле-
ток растет независимо от других клеток, т. е. клеточная суспен-
490 Глава 7
зия содержит т независимых образцов. Тогда 95%-ный довери-
тельный предел времени удвоения численности этой популяции
будет равен 2опОпУляция, где
- 0,98л
^популяция-- п /—
у т
Следовательно, с возрастанием т неопределенность времени
удвоения численности популяции быстро уменьшается. Приняв,
например, как и раньше, о = 0,5 и t —1,0, легко найти, что
95%-ные доверительные пределы для t в случае популяции из т
клеток составляют
т 95%-ный доверительный предел для t
1 Z= 1+0,98
ю4 /= 1 ±0,0098
10s /=1 ±0.000098
В сущности, этот несложный расчет еще раз подтверждает
известное положение, что, как и в любых стохастических про-
цессах при относительно большом числе событий (например,
в химических реакциях или в явлениях, связанных с транспор-
том жидкостей), изменения характеристик популяции могут
быть предсказаны с достаточно большой точностью, даже если
стандартные отклонения характеристик индивидуальных клеток
велики. Так, рост инокулята, приводящий к повышению кон-
центрации с 104 до 108 клеток в 1 мл, очевидно, обеспечивает
достаточное усреднение по всем стадиям роста, что позволяет с
высокой точностью определить время удвоения численности по-
пуляции. Точно так же можно достаточно обоснованно гово-
рить о скорости синтеза ДНК в типичной клетке популяции,
хотя в каждой индивидульной клетке синтезируется только од-
на или две молекулы ДНК со скоростью, которая в данный мо-
мент может резко отличаться от средней величины.
Невозможно обсуждать кинетику в отсутствие данных о кон-
струкции реактора для измерения скоростей процессов и оцен-
ки их кинетики. В следующем разделе мы вкратце рассмотрим
соответствующие материальные балансы для двух типов иде-
альных биореакторов. Затем мы приступим к анализу кинетики
клеточного роста, начиная с ситуации, схематично изображен-
ной в верхнем левом углу рис. 7.2, т. е. с простейших моделей
кинетики клеточного роста, утилизации субстрата и образова-
ния продуктов метаболизма. В последующих разделах главы
мы изучим другие ситуации, также изображенные на рис. 7.2,
и попытаемся оценить важность различных концептуальных и
математических подходов к анализу кинетики роста популяции
Образование биомассы в культурах клеток
491
клеток. В общем случае разработка кинетической модели попу-
ляции клеток является своего рода искусством, требующим, во-
первых, учета окончательной цели, для которой эта модель
предназначается, во-вторых, тщательно продуманного и обосно-
ванного выбора основных переменных и параметров, влияющих
на наиболее важные процессы, и, в-третьих, известной концеп-
туальной и математической гибкости, что необходимо для пере-
вода качественных характеристик системы на язык примени-
мых в практической деятельности математических выражений.
7.1. Идеальные реакторы
для изучения кинетики клеточного роста
Получить полезную информацию о кинетике роста популя-
ции микроорганизмов практически невозможно, если условия в
разных участках объема реактора неидентичны. Поэтому жела-
тельно изучать кинетику только в реакторах с полным переме-
шиванием. Здесь мы изучим реакторы с полным перемешиванием
периодического действия и проточные (непрерывного действия)
реакторы. Другие типы реакторов, в том числе реакторы пе-
риодического действия с добавлением субстрата и реакторы с
неполным перемешиванием, будут рассмотрены в гл. 9.
7.1.1. Идеальный реактор периодического действия
Основой многих биохимических процессов является перио-
дический рост популяций клеток, при котором после посева
инокулята живых клеток в жидкую среду к последней в тече-
ние всего периода роста ничего не добавляется и из нее ничего
не выделяется (за исключением, быть может, некоторых га-
зов). Обычно в таком реакторе по мере роста популяции из-
меняются концентрации питательных веществ, клеток и про-
дуктов метаболизма.
Материальный баланс по компоненту i показывает, что ско-
рость его накопления, выраженная в виде производной общего
количества компонента i по времени, должна быть равна ско-
рости его образования, обусловленного химическими реакция-
ми в реакторе. Таким образом,
d
dt
объем \ /молярная концентрация
культуры J \ компонента i
/ объем
(.культуры
число образовавшихся в реакции молей i
единица объема культуры-единица времени
или
^-(VRc<)=VR-r„
(7.1)
(7.2)
492
Глава 7
где VR — объем культуры; с, — количество молей i в единице
объема культуры; гд. — количество образовавшихся в реакции
молей i в единице объема культуры за единицу времени.
Если в реактор не добавляют и из него не отбирают ника-
ких жидких веществ и если потерями жидкости с потоком газа
можно пренебречь, то Vr постоянно и уравнение (7.2) упроща-
ется:
Следует отметить, что аналогичными уравнениями может быть
выражено не только изменение количества молей, но и массы
или концентрации (плотности) компонента. Если же компонент
i входит в состав газового потока, поступающего в реактор (или
выходящего из реактора), то указанные выше уравнения долж-
ны быть дополнены слагаемыми, отражающими результирую-
щую скорость поступления компонента i с газовой фазой.
Из уравнения (7.3) следует, что определение скорости из-
менения концентрации компонента i позволяет непосредственно
определить общую скорость образования i в реакциях (в том
числе внутриклеточных), которые происходят в реакторе пе-
риодического действия. В общем случае скорость образования
Гц зависит от состояния клеточной популяции (состава, морфо-
логии, распределения по возрасту и т. д.) и всех параметров
среды, оказывающих влияние на скорости реакций в клетках и
среде. В то же время, как мы уже упоминали во введении к
этой главе, обычно кинетику клеточного роста описывают
упрощенно, принимая во внимание только наиболее важные
параметры. В последующих разделах мы увидим, каким обра-
зом стехиометрия реакций и их скорости связаны со скоростью
образования продуктов жизнедеятельности клеток.
7.1.2. Идеальный проточный реактор
с полным перемешиванием (ПРПП)
На рис. 7.3, а изображены основные узлы лабораторного
проточного реактора с полным перемешиванием (ПРПП), а на
рис. 7.3, б указаны основные обозначения, используемые при
моделировании и анализе такого рода реакторов. Если эти ре-
акторы предназначены для изучения роста культур клеток, то
их часто называют хемостатами. Как показано на этих рисун-
ках, перемешивание осуществляется с помощью мешалки, по-
тока восходящих пузырьков газа или того и другого. Примем,
что в ПРПП перемешивание культуральной жидкости осущест-
вляется настолько эффективно, что каждая фаза содержимого
реактора вполне однородна по составу, т. е. концентрации лю-
Образование биомассы в культурах клеток
493
S
РИС. 7.3. Проточный реактор с полным перемешиванием (ПРПП), предназна-
ченный для непрерывного культивирования популяций клеток, а — основные
элементы лабораторного ПРПП, предназначенного для изучения растущей
популяции клеток; б—обозначения, обычно применяемые при моделировании
и анализе таких реакторов.
бых компонентов в любой из фаз одинаковы во всем объеме ре-
актора. Как показано на схеме, отсюда следует важный вывод
о том, что состав вытекающего потока не отличается от соста-
ва содержимого реактора.
Полное перемешивание должно обеспечивать и одинаковую
концентрацию растворенного кислорода во всем объеме жидкой
фазы. Этот факт особенно важен при анализе аэрируемых
ПРПП, поскольку отсюда следует, что в большинстве случаев
мы можем изучать происходящие в реакторе процессы незави-
симо от конструкций аэратора или мешалки. Если система
аэрации обеспечивает такую концентрацию растворенного кис-
494
Глава 7
лорода, которая не лимитирует клеточный рост в ПРПП, то
анализ кинетики клеточного роста мы можем рассматривать
как независимую проблему. Аналогичные соображения приме-
нимы и к проблемам теплопередачи, которые могут возникать,
в ходе роста микроорганизмов. Систему можно считать изотер-
мической, если реактор снабжен устройствами для полного пе-
ремешивания, эффективного отвода тепла и регулирования тем-
пературы на заданном уровне; тогда можно изучать процессы
микробиологических превращений в изотермических условиях.
В большей части последующего материала мы будем принимать-
все указанные допущения.
В стационарном состоянии, когда концентрации всех компо-
нентов в реакторе не изменяются во времени, к любому компо-
ненту системы применимо следующее уравнение:
"Скорость бавления до-" в ре- — "Скорость вывода из -J- "Скорость зоваиия обра-' в ре- = 0 (7-4)
.актор .реактора .акторе
Обозначив, как и раньше, символом Уя общий объем культуры
в реакторе, указанное уравнение стационарного состояния
можно записать в следующем виде:
Ftcif — сд+ VRrfi = Q (7.57
где F — объемная скорость потока раствора питательных ве-
ществ и вытекающего потока; cif — молярная концентрация
компонента i в потоке питательных веществ; а— концентрация
компонента i в реакционной смеси и вытекающем потоке.
Преобразовав уравнение (7.5),
= cif) = D(Ci—cif) (7.6)
можно без труда определить скорость образования компонента
I, измерив его (стационарные) концентрации на входе в реак-
тор и на выходе из него. Введенный в уравнение (7.6) пара-
метр D называют скоростью разведения и определяют в виде
D= -Т- (7.7)
'R
Этот параметр определяет время пребывания или скорость
переработки в реакторе и равен числу объемов жидкой фазы
реактора, проходящей через него в единицу времени. Параметр
D представляет собой величину, обратную более обычным в
химической технологии параметрам среднего времени пребыва-
ния или среднего времени удерживания. Здесь, однако, мы бу-
дем пользоваться общепринятым в литературе по биохимической?
технологии понятием о скорости разведения.
Из сравнения уравнений (7.3) и (7.6) нетрудно заметить»
Образование биомассы в культурах клеток 495
что кинетика процессов в ПРПП проще, чем в реакторе пери-
одического действия; действительно, здесь нет необходимости
в определении зависимости концентрации от времени и в по-
следующем дифференцировании полученных данных. Изучение
кинетики роста популяции клеток в этих условиях имеет еще
одно преимущество, заключающееся в том, что в ПРПП клетки
могут приспособиться к стационарным условиям и таким путем
перейти в состояние сбалансированного или почти сбалансиро-
ванного роста. Этим самым создается реальная возможность
для обеспечения относительно определенного, воспроизводимо-
го состояния клеточной популяции; в случае периодических
процессов с участием микроорганизмов решить эту задачу зна-
чительно труднее. С другой стороны, эксперименты по изуче-
нию роста клеток в периодических процессах могут быть вы-
полнены в небольших сосудах, размещенных на термостатиро-
ванной качалке; оборудование для ПРПП значительно слож-
нее и дороже. Состояние стационарности в биологических
ПРПП может быть достигнуто только через несколько часов
или даже дней, что существенно повышает опасность внесения
загрязнений, обесценивающих результаты эксперимента. Нако-
нец, в крупномасштабном производстве в ряде случаев, веро-
ятно, более целесообразными окажутся периодические процес-
сы, отличающиеся непостоянным, несбалансированным ростом,
различными метаболическими процессами и активностями в
разные периоды процесса; для таких производств кинетические
модели, основанные на стационарных условиях в ПРПП, могут
оказаться вообще непригодными. Отсюда следует, что экспе-
риментальному изучению кинетики роста популяций клеток
микроорганизмов и высших организмов и разработке соответ-
ствующих математических выражений должно предшествовать
точное определение предполагаемой сферы применения этих
выражений. Это требование является основой для разработки
программ экспериментального и математического моделирова-
ния.
В следующем разделе мы рассмотрим простейшие модели
роста популяций клеток. При этом основное внимание мы бу-
дем уделять кинетике роста клеток в ПРПП, поскольку основ-
ные принципы построения кинетических уравнений были раз-
работаны и наиболее полно развиты именно в экспериментах
с хемостатами.
7.2. Кинетика сбалансированного роста
Моделирование и анализ кинетики роста популяций клеток
мы начнем с неструктурированных моделей, в которых биофаза
характеризуется только величиной клеточной массы или чис-
496
Глава 7
ленной концентрацией. Суммарную скорость роста клеточной
массы гх часто выражают как цх, где х — клеточная масса в
единице объема культуры, а ц— удельная скорость роста кле-
ток, выраженная в единицах обратного времени. В этих пара-
метрах уравнение материального баланса клеточной массы для
ПРПП в стационарном состоянии будет выглядеть следующим
образом:
Dxf^kD—ц)х (7.8)
Часто поступающий в проточный реактор поток раствора
питательных веществ состоит только из стерильного питатель-
ного вещества, так что х^ = 0. В таком случае из уравнения
(7.8) следует, что отличная1 от нуля популяция клеток может
существовать в стационарном состоянии только при
(7.9)
т. е. тогда, когда удельная скорость клеточного роста равна
скорости разведения. Если выполняется условие уравнения
РИС. 7.4. Как следует из уравнения (7.9), при постоянном ц концентрация
клеточной массы в проточной культуре неопределенна (В. lineus при 26 °C и
£> = 0,417 ч-1). [Воспроизведено из статьи: Finn R. К., Wilson R. Е., Population
Dynamics of a Continuous Propagator for Microorganisms, J. Agri. Food Chem.,
2, 66 (1954). © American Chemical Society.]
(7.9), то при стерильном питательном веществе уравнение
(7.8) уже не определяет х. Неопределенность уровня популя-
ции была подтверждена в экспериментах с культурой Bacil-
lus linens в проточном реакторе. В этих экспериментах стацио-
нарное состояние было достигнуто через 6 ч (рис. 7.4), после
чего культуру подвергали двум последовательным разведениям,
при которых часть содержимого реактора, состоящего из попу-
ляции клеток и среды, удаляли и замещали чистой средой.
Образование биомассы в культурах клеток
497
После каждого такого разведения через некоторое время в сис-
теме устанавливалось новое стационарное состояние, характе-
ризующееся иной концентрацией клеточной массы.
7.2.1. Уравнение Моно для кинетики клеточного роста
Приведенная на рис. 7.4 зависимость характерна только для
тех случаев, когда удельная скорость клеточного роста не за-
висит от х и s. [Здесь под s мы понимаем массовую концентра-
цию субстрата, лимитирующего клеточный рост.] Система
становится определенной, если скорость роста клеток лимити-
руется каким-либо питательным веществом.
Прежде чем перейти к обсуждению деталей зависимости
скорости роста от подачи питательных веществ, рассмотрим об-
Таблица 7.1. Примеры синтетических и сложных среда
Дополнительные ингредиенты
Основные компоненты,
общие для всех сред
(минеральная основа)
Среда 1® Среда 2® Среда 3® Среда 4В
Вода (1 л), NH4C1 Глюкоза1, Глюкоза (5 г), К2НРО4 (1 г), (1 г) (5 г), NH4CI (1 г), MgSO4-7H2O (200 мг), NH4CI (1 г) никотиновая FeSO4-7H2O (10 мг), кислота СаС12 (10 мг), (0,1 мг) следовые компоненты Глюкоза (5 г), дрожже- вой экст- ракт (5 г)
(Мп, Mo, Си, Со, Zn)
в виде солей неорга-
нических кислот (по
0,02—0,5 мг каждой)
а Startler R. У., Doudoroff М„ Adelberg Е. A., The Microbial World, 3d ed., p. 79,
Prentice-Hall, Inc., Englewood Cliffs, N. J., 1970.
® Синтетическая,
в Сложная.
г Если среда стерилизуется в автоклаве, то глюкозу следует стерилизовать отдельно
и вводить асептически. Сахара при нагревании в присутствии других веществ, особенно
фосфатов, подвергаются частичному разложению, в результате которого образуются
соединения, очень токсичные для ряда микроорганизмов.
щие теоретические и практические пути создания сред для
культивирования клеток. В зависимости от состава различают
два типа сред. Синтетическими называют среды с четко опре-
деленным химическим составом. Как показано в табл. 7.1, та-
кие среды создаются на основе раствора неорганических солей,
к которому добавляют необходимые источники углерода, азота
и энергии, а также все незаменимые витамины. Неорганические
соли служат не только источником ионов, необходимых для
нормального функционирования клеток, но и выполняют бу-
32—53Т
498
Глава 7
ферную функцию, сглаживая большие флуктуации pH в про-
цессе роста популяций. Сложные среды содержат материалы
неопределенного состава. Например, среда 4 в табл. 7.1 отно-
сится к сложным, поскольку точный химический состав дрож-
жевого экстракта неизвестен. В состав других сложных сред
входят мясной бульон, кровяной бульон, кукурузный экстракт
и сточные воды.
Основное требование, предъявляемое к среде, заключается
в обеспечении высокой скорости роста клеток и(или) синтеза
РИС. 7.5. Зависимость удельной скорости роста Е. coli от концентрации лими-
тирующего рост питательного вещества в глюкозной среде (а) и триптофано-
вой среде (для мутанта, нуждающегося в триптофане) (б). (Воспроизведено
с разрешения издательства из работы: Stanier R. Y., Doudoroff М., Adel-
berg Е. A., The Microbial World, 3d ed., p. 315, Prentice-Hall, Inc., Englewood
Cliffs, N.J., 1970.)
продуктов их жизнедеятельности. Отсюда, однако, отнюдь не
следует (как можно было бы предположить чисто интуитивно),
что все питательные вещества нужно подавать в большом из-
бытке. Во-первых, избыточная концентрация питательного ве-
щества может ингибировать рост клеток или даже привести к
их гибели. Во-вторых, если клетки растут слишком интенсив-
но, то накапливающиеся конечные продукты метаболизма мо-
гут нарушить нормальные биохимические процессы в клетках.
Поэтому обычно общий рост клеток лимитируют, ограничивая
количество одного питательного вещества в среде.
Если концентрация одного незаменимого компонента среды
изменяется, а концентрации всех других компонентов остаются
без изменения, то, как это показано на рис. 7.5, зависимость
скорости клеточного роста от концентрации незаменимого пи-
тательного вещества обычно выражается кривой типа гипербо-
лы. Математическое выражение, отражающее функциональную
Образование биомассы в культурах клеток 49£>'
зависимость удельной скорости клеточного роста у, от концент-
рации незаменимого питательного вещества, было предложено
Моно в 1942 г. Уравнение Моно имеет ту же форму, что и
уравнение изотермы адсорбции Ленгмюра (1918 г.) и обычное
уравнение скорости катализируемой ферментом реакции с од-
ним субстратом, предложенное Анри (1902 г.) и Михаэлисом
и Ментен (1913 г.), и устанавливает, что
у = -НтахД. (7.Ю)
As + s
В этом уравнении у,тах — максимальная скорость роста, дости-
гаемая при s^>Ks и постоянных концентрациях всех других не-
заменимых питательных веществ. Ks — это такая концентрация
лимитирующего клеточный рост питательного вещества, при
которой удельная скорость роста вдвое ниже максимального
значения. В первом приближении можно считать, что Аз пред-
ставляет собой промежуточную зону между диапазоном низ-
ких концентраций, при которых у, линейно зависит от s, и диа-
пазоном высоких концентраций, когда у, становится независи-
мым от s. Как показано на рис. 7.5, значения Ks для штаммов
Е. coli, растущих в лимитируемых глюкозой и триптофаном
средах, составляют 0,22-10—4 М и 1,1 нг/мл соответственно.
Поскольку мы уже знакомы с биохимией клетки, для нас
должно быть очевидно, что уравнение Моно, скорее всего, чрез-
мерно упрощено. Как, однако, это часто бывает и в других об-
ластях технологии, относительно простое уравнение иногда
удовлетворительно описывает определенные зависимости, хотя
физический смысл параметров модели неизвестен или, возмож-
но, даже, вообще не существует. Впрочем, в некоторых специ-
фических ситуациях уравнению Моно можно приписать опре-
деленный физический смысл. Одна из наиболее показательных
ситуаций такого типа складывается тогда, когда скорость кле-
точного роста лимитируется скоростью транспорта через мем-
брану при участии пермеаз (вспомните разд. 5.7).
В уравнении Моно (7.10) привлекает его простота, однако
применение этого простого выражения требует от исследова-
теля большой осторожности. Во-первых, величина Ks часто
довольно мала, поэтому s обычно намного больше Ks и выра-
жение s/(Ks + s) можно рассматривать как адекватное выраже-
нию для расчета отклонений у, от ymax по мере снижения кон-
центрации s. Это соотношение предполагает также, что удель-
ная скорость роста у, не равна нулю при любых отличных от
нуля концентрациях лимитирующего скорость клеточного роста
компонента. В общем случае данное условие нельзя считать,
доказанным для s<^Ks.
32*
500
Глава 7
Если скорость роста популяции клеток связана с концентра-
цией лимитирующего клеточный рост питательного вещества
математическим выражением типа уравнения Моно, то отсюда
следует, что между условиями работы реактора и кинетически-
ми и стехиометрическими параметрами популяции клеток
должны существовать определенные зависимости. Для выявле-
ния этих зависимостей можно начать с уравнения материаль-
ного баланса по лимитирующему клеточный рост субстрату,
связанного с балансом по, клеточной массе, поскольку ц зави-
сит от s. В балансе по субстрату мы будем использовать эко-
номический коэффициент Ух/s (см. разд. 5.10.1):
Г масса образующихся клеток
масса утилизированного субстрата
(7.11)
Тогда уравнение материального баланса по субстрату для ста-
ционарного состояния можно записать в следующей форме:
Если в это выражение подставить ц из уравнения (7.10), то
получим, что
D^~ <7Л2>
В этом случае уравнение материального баланса по клеточной
массе будет выглядеть следующим образом:
----D)x+£>*,= 0 (7.13)
Ks 4-s J
Уравнения (7.12) и (7.13) часто называют моделью Моно для
хемостата.
Если, как это обычно бывает, питательные вещества сте-
рильны (xf = 0), то преобразованием этих уравнений легко най-
ти соответствующие выражения для х и s:
•^стерильные = Г x/s
питательные
вещества
^стерильные = _.. х ,
питательные ах
вещества
Уравнения (7.14) и (7.15) отражают зависимость х и s от
скорости потока (D = F/V) в стационарном состоянии. Для
очень медленных скоростей потока при заданном объеме D-*-0
и, следовательно, s также приближается к нулю. Поскольку
почти весь вводимый в реактор субстрат поглощается клетка-
(7.14)
t Hmax — D
Образование биомассы в культурах клеток
501
ми, то концентрация клеточной массы на выходе из реакторах
будет равна $fУх/s.
По мере постепенного повышения D возрастает и s: сначала
пропорционально D, а затем, по мере приближения D к ртах,
еще быстрее. Точно таким же образом снижается и концентра-
ция клеточной массы — сначала в линейной зависимости от D,
а затем, при D->-pimax, с большей скоростью. При приближении
РИС. 7.6. Зависимости концентрации субстрата s в продуктах, концентрации
клеточной массы х и скорости образования клеток Dx от скорости разведе-
ния непрерывной культуры D, вычисленные на основе модели Моно для хемо-
стата (|Лтах=1 Ч-1, Ks = o,2 г/Л, Ух/5 = 0,5, S,' = 10 Г/л).
D к ртах в какой-то момент х становится равной нулю; это оз-
начает, что скорость разведения D только что превысила мак-
симально возможную скорость роста и решение уравнения
(7.14) в условиях стационарного состояния возможно только
при х = 0. Потеря всех клеток в стационарном состоянии, назы-
ваемая вымыванием, происходит, когда D больше Лтах; послед-
ний параметр можно определить по уравнению (7.14) при х =
= 0:
Л =_HmaxSf (7
max Ks + sf V-10'
На рис. 7.6 приведены отражаемые уравнениями (7.14) и
(7.15) графики зависимостей концентраций субстрата и клеточ-
ной массы от D для следующего ряда значений параметров:
Ртах=1,0ч~1 Ух/3=0,5 Ks = 0,2 г/л 5/=10г/л
502
Глава 7
Заметьте, что вблизи точки вымывания реакционная смесь ста-
новится очень чувствительной к колебаниям D; даже неболь-
шое изменение D сопровождается относительно большим изме-
нением х и (или) s.
Эту зависимость необходимо иметь в виду в тех случаях,
когда целью непрерывного микробиологического процесса явля-
ется производство биомассы. Скорость продуцирования клеток
в единице объема реактора равна Dx; на кривой зависимости
Dx от D (рис. 7.6) имеется резкий максимум. Максимальную
скорость образования клеток можно вычислить, решив диффе-
ренциальное уравнение
-d(£.x) =0 (7.17)
aD ' '
в котором для описания х как функции от D используется
уравнение (7.14). Решение уравнения (7.17) дает
•^макс.произв. = Птах j/" (7-18)
Если, как это часто бывает на практике, Sf^>Ks, то
D
макс.произв. приближается к цтах и, следовательно, находится
вблизи точки вымывания. В такой ситуации, изображенной, в
частности, на рис. 7.6, как правило, чтобы избежать диапазона
наибольшей чувствительности, целесообразно не добиваться
максимальной скорости образования биомассы. Вообще при
решении любых проблем оптимизации в поисках сравнительно
легко регулируемых параметров важно не забывать практиче-
ские аспекты сочетания чувствительности, регулируемости и на-
дежности.
При изучении закономерностей образования конечных про-
дуктов непрерывных микробиологических процессов введем еще
один коэффициент Ур/х, представляющий собой отношение
у ____масса образующегося продукта метаболизма jgv
р^х прирост биомассы ' ’ '
Изучив в гл. 5 стехиометрию образования продуктов метабо-
лизма, мы уже знаем, что коэффициент Ур/х постоянен при
микробиологических процессах типа I, но в других процессах
изменяется. Применение этого коэффициента позволяет запи-
сать уравнение материального баланса для продукта жизне-
деятельности клеток в стационарном состоянии в форме
D(pf-p)+yP/xpx=0 (7.20}
Отсюда следует, что
, Ур/хн*
p=pi+——
(7.21)
Образование биомассы в культурах клеток
503
Вместе с приведенными выше уравнениями, выражающими за-
висимость ц и х от параметров процесса, уравнение (7.21)
позволяет вычислить концентрацию продукта в выходящем из
реактора потоке. Производительность реактора по отношению
к продукту равна pD и при постоянном Ур/х максимальна,
если D имеет значение, определяемое уравнением (7.18). Сле-
довательно, при попытках добиться максимальной производи-
РИС. 7.7. Экспериментально найденные параметры роста непрерывной куль-
туры (Л. aerogenes) качественно согласуются с моделью Моно. (Воспроизве-
дено из работы: Herbert D., A Theoretical Analysis of Continuous Culture Sys-
tems, in Society of Chemical Industries Monograph 12, p. 247, London, 1961.)
тельности процесса также необходимо учитывать фактор чув-
ствительности. [При каком значении D величина pD макси-
мальна, если YP/x=f (D) ?]
В качестве примера на рис. 7.7 приведены результаты экспе-
риментального изучения роста культуры бактерий Aerobacter
aerogenes в проточном реакторе. Экспериментальные результа-
ты и описанная выше простая математическая модель качест-
венно вполне согласуются. В то же время найденная скорость
образования клеточной массы и найденная концентрация суб-
страта остаются приблизительно постоянными в значительно
более широком диапазоне условий, чем следует из рассчитан-
ных по уравнению Моно данных (рис. 7.6). Кроме того, модель
Моно не согласуется с экспериментальными данными при очень
504
Глава 7
низких и очень высоких скоростях разведения. В следующем
разделе мы изучим эти две экстремальные ситуации и попыта-
емся понять, почему в таких случаях модель Моно неприме-
нима.
7.2.2. Влияние эндогенного метаболизма
и метаболизма поддержания на кинетику клеточного роста
Приведенные на рис. 7.8 данные для культуры A. aerogenes
показывают, что при малых скоростях разведения концентра-
ция клеток заметно снижается. Аналогичное поведение наблю-
далось и в случае кормовых дрожжей Torula utilis. Эту особен-
Ofya/тшая мор ос теразведения D~’ у
РИС. 7.8. Снижение концентрации клеточной маесы (культура A. aerogenes)
в глицериновой среде в проточном реакторе при уменьшении скорости раз-
ведения противоречит модели Моно. (Воспроизведено из работы: Herbert D.,
Continuous Culture of Microorganisms: Some Theoretical Aspects, in Continuous
Culture of Microorganisms: A Symposium, p. 48, Publishing House of the
Czechoslovakia Academy of Sciences, Prague, 1958.)
ность, противоречащую модели Моно для хемостата, можно
объяснить, если в модели учесть возможность эндогенного ме-
таболизма. Под эндогенным метаболизмом подразумеваются
происходящие в клетке реакции, в которых расходуются клеточ-
ные вещества. Чтобы учесть этот эффект, дополним уравнение
Моно слагаемым —kex:
г __ Мтиах5*
s+tfs
(7.22>
Образование биомассы в культурах клеток
505
РИС. 7.9. Согласующаяся с уравнением (7.24) линейная зависимость удель-
ной скорости дыхания бактерии A. aerogenes от скорости разведения в про-
точном реакторе. (Воспроизведено из работы: Herbert D., Continuous Culture
of Microorganisms: Some Theoretical Aspects, in Continuous Culture of Micro-
organisms: A Symposium, p. 49, Publishing House of the Czechoslovakia Aca-
demy of Sciences, Prague, 1958.)
Обратите внимание на то, что слагаемое —kex в уравнении
(7.22) можно интерпретировать и как скорость гибели клеток.
Модифицированное уравнение Моно согласуется и с други-
ми экспериментальными данными. Например, если скорость
дыхания аэробной культуры пропорциональна скорости утили-
зации субстрата, т. е.
г __ PM-maxsx /7
rresp-"s+^-
то из уравнений (7.8), (7.22) и (7.23) следует, что удельная
скорость дыхания равна
-^=₽(D+W (7.24)
Приведенные на рис. 7.9 экспериментальные данные хорошо
согласуются с уравнением (7.24).
Наблюдаемая зависимость экономического коэффициента
Y от D также подтверждает справедливость уравнения (7.22)
для скорости клеточного роста. Если скорость поглощения суб-
506
Глава 7
страта равна
- __ 1____Ртах^
s У'х/s s+Ks
(7.25>
где У'х/s — «истинный» коэффициент, то из уравнений (7.22)
и (7.25) и определения Ух/s (напомним, что Ух/s — это стехио-
метрический коэффициент, равный частному от деления об-
щей массы образовавшихся клеток на общую массу поглощен-
ного субстрата, поэтому при условии стерильности питатель-
ных веществ —rs = Dx!Yx/s) следует, что
. ДУ'х/s
X/S' D -}- kg
(7.26)
Экспериментально показано, что для ряда микроорганизмов за-
висимость Ух/s от D выражается именно таким уравнением.
Другая возможность, рассмотренная вкратце при обсужде-
нии стехиометрии клеточного роста в разд. 5.10.1, заключается
в параллельной утилизации субстрата как для клеточного
роста, так и для других энергетических потребностей клетки
(метаболизм поддержания). В этом случае скорость утилиза-
ции субстрата определяется выражением
— rs = y?x/s + тх (7.27)
где m—удельная скорость потребления субстрата, расходуе-
мого в метаболизме поддержания. Если принять, что гх равно
цх, то
у _ У x/s^
D mY x/s
(7.28)
Таким образом мы получили точно такую же функциональную
зависимость Ух/s от D, как и в случае модели эндогенного ме-
таболизма [уравнение (7.26)].
Как показано на рис. 7.10, при высоких скоростях разведе-
ния поведение культуры клеток в проточном реакторе может
существенно отклоняться от поведения, предсказываемого мо-
делью Моно для хемостата. Здесь не только слишком велика
ошибка в определении концентрации клеток вблизи точки вы-
мывания, но и вся популяция клеток может существовать при
скоростях разведения, значительно превышающих предсказыва-
емую теорией критическую величину. Более того, по мере
приближения D к критическому максимальному значению эко-
номический коэффициент снижается. Одна из возможных при-
чин этого несоответствия может заключаться в относительно
высокой концентрации субстрата, характерной для высоких
скоростей разведения. В таких условиях субстрат часто не ог-
Образование биомассы в культурах клеток
507
РИС. 7.10. Экспериментальные
данные по росту Aerobacter cloa-
cae в проточном реакторе, свиде-
тельствующие об отклонении кон-
центрации клеточной массы от ну-
левого значения при скоростях
разведения, превышающих вычис-
ленную критическую величину.
[Воспроизведено из статьи: Her-
bert D. et al., The Continuous Cul-
ture of Bacteria: A Theoretical and
Experimental Study, J. Gen. Micro-
biol., 14, 601 (1956).]
0,0
0,2
0,6
Обратная критическая
скорость разведения, сог-
L/ласно результатам изучения
'периодической культуры
раоуая Г,Гд v ।____________,
0 1 Z 3 в 5
Обратная скорость разведения D~, у/
раничивает рост клеток, и составляющие популяцию клетки
могут изменить характер метаболизма, выбрав какой-либо
другой лимитирующий фактор среды. Еще одной причиной
может быть недостаточно эффективное перемешивание; этот
вопрос мы изучим в гл. 9.
7.2.3. Другие уравнения кинетики клеточного роста
Предлагались и другие математические выражения, опре-
деляющие удельную скорость клеточного роста; в ряде случаев
эти выражения лучше соответствуют экспериментальным дан-
ным. Например, Теиссье, Мозер и Контуа предложили следую-
щие уравнения: Уравнение Теиссье: H = Hmax(l—е S'K&) (7.29)
Уравнение Мозера: Ц = Цтах( 1 4" KsS *") (7.30)
Уравнение Контуа: S И—Птах (7.31)
По сравнению с уравнением Моно алгебраическое решение
двух первых уравнений клеточного роста затруднительно.
В уравнение Контуа входит кажущаяся константа Михаэлиса,
508
Глава 7
пропорциональная концентрации биомассы х, поэтому по мере
увеличения плотности популяции максимальная скорость роста
должна уменьшаться и в конце концов ц станет пропорцио-
нальной х~1.
Те или иные компоненты среды, например субстрат или про-
дукт метаболизма, могут снижать удельную скорость клеточно-
го роста. Эндрюс предложил уравнение, учитывающее инги-
бирование роста клеток субстратом [11]:
s
И = Hmax ks+s+s2/k. (7.32)
Примером процесса, ингибируемого продуктом жизнедея-
тельности клеток, может служить спиртовое брожение; Аиба„
Шода и Нагатани для описания анаэробного брожения глюко-
зы в присутствии дрожжей предложили функцию роста сле-
дующего типа [9, 10]:
П-33)
Ар Р
Возможны случаи, когда рост клеток лимитируют два (или
более) субстрата одновременно. Хотя имеется очень мало дан-
ных, характеризующих такие процессы, можно предположить^
что каждое из лимитирующих питательных веществ подчиняет-
ся уравнению Моно:
и = Umax-77-4---^4---••• (7.34)
В отсутствие более убедительных данных, подтверждающих
это уравнение, его можно рассматривать как своего рода кри-
терий, позволяющий установить, что рост клеток действительно
определяется несколькими питательными веществами.
7.2.4. Влияние других параметров среды
на кинетику клеточного роста
При сбалансированном росте для характеристики кинетики
роста популяции клеток достаточно только одного параметра
ц (или времени удвоения численности популяции id = ln2/p).
По этой причине параметр удельной скорости роста ц широко
используется для описания влияния среды на поведение попу-
ляции. Рассмотрим прежде всего влияние температуры; извест-
но, что живые организмы могут существовать в диапазоне тем-
ператур примерно от —5 до 95 °C. Принято классифицировать
прокариоты в соответствии с диапазоном температур их роста.
Как показывают приведенные в табл. 7.2 данные, для каждого
класса микроорганизмов характерна некоторая оптимальная
Образование биомассы в культурах клеток 509
температура, при которой скорость роста достигает максимума,
а также верхняя и нижняя температурные границы, вне кото-
рых популяция вообще не способна к росту.
Приведенные на рис. 7.11 кривые свидетельствуют об очень
сильном влиянии температуры на рост Е. coli. Обратите вни-
мание на то, что в координатах Аррениуса классическое арре-
ниусовское поведение наблюдается только при низких темпе-
ратурах, а по мере приближения температуры к той границе,
при которой бактерии начинают погибать, скорость их роста
резко падает. Бросается в глаза сходство зависимостей кле-
Таблица 7.2. Классификация микроорганизмов в соответствии с зависимостью
скорости их роста от температуры3
Температура, °C
Группа микроорганизмов
Минимальная Оптимальная Максимальная
Термофильные от 40 до 45 от 55 до 75 от 60 до 80
Мезофильные от 10 до 15 от 30 до 45 от 35 до 47
Психрофильные
облигатные от —5 до 5 от 15 до 18 от 19 до 22
факультативные от —5 до 5 от 25 до 30 от 30 до 35
аStonier R. У., Doudoroff М„ Adelberg Е. A., The Microbial World, 3d ed., p. 3161.
Prentice-Hall, Inc., Englewood Cliffs, N. J., 1970.
точного роста (рис. 7.11) и каталитической активности фер-
ментов (рис. 3.25) от температуры. В этой связи неудивитель-
но, что зависимость удельной скорости клеточного роста от
температуры часто можно выразить в виде уравнения типа
(3.73). Очевидно, в диапазоне относительно низких температур
возрастание метаболической активности клетки при повыше-
нии температуры связано с повышением каталитической актив-
ности входящих в ее состав ферментов. Когда же температура
достигает того предела, при котором начинается денатурация
термически наиболее лабильных важных для жизнедеятельнос-
ти клетки белков, то клетка погибает. В ряде случаев эта ги-
потеза была подтверждена генетическими методами; показано,
в частности, что мутация одного гена может существенно из-
менить максимальную температуру, которую способен выдер-
жать микроорганизм.
Поскольку конформация белка и его активность зависят от
pH, мы вправе ожидать, что величина pH будет оказывать су-
щественное влияние на процессы клеточного транспорта, ско-
рости реакций, а следовательно, и скорость роста клеток.
Обычно скорость роста бактерий максимальна в диапазоне pH
от 6,5 до 7,5. В качестве примера на рис. 7.12 приведена зави-
время удвоения популяции,
РИС. 7.11. а — при повышении температуры скорость роста клеток Е. coll до
определенного момента возрастает, но при слишком высокой температуре
клетки погибают; б — та же зависимость в аррениусовских координатах. (Вос-
произведено с разрешения издательства из работы: Stanier R. Y., Doudo-
roff М., Adelberg Е. A., The Microbial World, 3d ed., pp. 316, 317, Prentice-Hall,
Inc., Englewood Cliffs, N.J., 1970.)
РИС. 7.12. Влияние температуры и pH на время удвоения Е. coll. [Воспроиз-
ведено с разрешения из работы: Methods in Microbiology, Norris J. R., Rib-
bons D. W. (eds.), vol. 2, Academic Press, New York, 1970.]
Образование биомассы в культурах клеток
511
симость скорости роста Е. coli от pH. Впрочем, из этого пра-
вила есть и исключения. Так, ацидофильные бактерии растут
при pH 2,0. Обычно отклонение pH на 1,5—2 единицы в любую
сторону от оптимальной величины приводит к практически пол-
ному прекращению роста (рис. 7.12). Дрожжи лучше всего рас-
тут в диапазоне pH от 4 до 5, а оптимальный pH для роста
плесневых грибов обычно равен 5—7. Для дрожжей и плесеней
характерен довольно широкий диапазон роста — приблизитель-
но от pH 3,0 до pH 8,5. Как показывают приведенные на
рис. 7.12 данные, при повышении температуры pH максималь-
ной скорости клеточного роста обычно возрастает.
На удельную скорость роста микроорганизмов влияют так-
же термодинамическая активность среды (зависящая в свою
очередь от растворенных в ней веществ) и гидростатическое
давление. Скорость роста аэробных организмов, очевидно, в
существенной степени зависит от концентрации растворенного
кислорода. Влияние потребляемого клетками кислорода на ки-
нетику их роста мы рассмотрим при изучении аэрации реакто-
ров в гл. 8.
7.3. Кинетика клеточного роста
в переходном состоянии
В определенные периоды роста культур клеток в реакторах
периодического действия или после возмущений в проточных
реакторах непрерывного действия популяции клеток находятся
в переходном состоянии, кинетика которого может оказаться
весьма сложной. Для описания различных вариантов роста
клеток в переходном состоянии могут потребоваться разные
типы математических моделей кинетики роста. Здесь мы рас-
смотрим несколько примеров кинетики клеточного роста в пе-
реходном состоянии, причем основное внимание уделим наибо-
лее общему случаю роста популяции в реакторе периодического
действия. Другие аспекты кинетики роста клеток в переходном
состоянии мы обсудим в ходе изучения динамики биореакторов
в гл. 9.2.
7.3.1. Основные фазы роста клеток
в реакторах периодического действия
Как показано на рис. 7.13, в типичном периодическом про-
цессе число живых клеток изменяется во времени. После лаг-
фазы (фазы задержки, латентной фазы), в течение которой
численность популяции практически не изменяется, наступает
период быстрого роста, когда число клеток возрастает во вре-
мени экспоненциально. Часто эту стадию роста культур клеток
512
Глава 7
в реакторе периодического действия называют логарифмичес-
кой фазой (лаг-фазой); мы предпочитаем более точный тер-
мин— экспоненциальный рост, который и будем использовать
в последующем изложении. Понятно, что в закрытом сосуде
клетки не могут размножаться бесконечно, и вслед за фазой
экспоненциального роста наступает стационарная фаза. В этот
момент число клеток достигает максимума. В конце концов
•стационарная фаза сменяется фазой отмирания, когда число
.клеток начинает снижаться. Здесь также часто наблюдается
РИС. 7.13. Кривая роста культуры в реакторе периодического действия,
экспоненциальное снижение, численности популяции живых ор-
ганизмов.
В микробиологических процессах важную роль может иг-
рать любая из фаз. Так, при разработке процесса одной из его
задач может быть сокращение до минимума продолжительнос-
ти лаг-фазы и достижение максимальной длительности фазы
экспоненциального роста (и скорости роста клеток в этой фа-
зе); вторая цель в свою очередь достигается путем искусствен-
ного торможения перехода к стационарной фазе. Часто очень
важно к концу процесса добиться максимальной плотности по-
пуляции клеток, что возможно только при условии учета всех
переменных, влияющих на фазы роста культуры, В последую-
щем изложении мы будем рассматривать каждую фазу от-
дельно и в соответствующие моменты обсудим математические
модели наиболее важных явлений. Это обсуждение мы продол-
жим в разд. 7.3.2, где рассмотрим математические модели
всего процесса клеточного роста в реакторе периодического
действия.
Образование биомассы в культурах клеток 513
Продолжительность лаг-фазы, наступающей после инокуля-
ции свежей среды, зависит как от изменений в составе пита-
тельных веществ (если такие изменения имеют место), так и
от возраста и массы инокулята. Сопровождающее перенос кле-
ток из одной среды в другую резкое изменение условий может
оказывать на живые клетки разностороннее действие. Во-пер-
вых, как мы уже знаем, системы контроля и регуляции фер-
ментативной активности включают и адаптационные механиз-
мы; в частности, сталкиваясь с новым питательным веществом,
клетка начинает его усваивать только после синтеза новых фер-
ментов. Поэтому, например, перенос находящейся в экспонен-
циальной фазе культуры из глюкозной среды в лактозную всег-
да сопровождается периодом с незначительной скоростью деле-
ния клеток; в этот период в клетках синтезируются ферменты
и кофакторы, участвующие в пути метаболизма лактозы. (Что
произойдет, если выращенную на лактозе культуру перенести
в глюкозо-лактозную среду?) Аналогично лаг-фаза может на-
ступить и после изменения концентраций питательных веществ.
Если новая питательная среда богаче лимитирующим клеточ-
ный рост питательным веществом, то в течение некоторого вре-
мени это вещество будет расходоваться не на рост клеток, а на
повышение концентраций метаболизирующих ферментов. Сни-
жение концентрации питательного вещества может и не сопро-
вождаться лаг-фазой; в этом случае экспоненциальный рост
возобновляется сразу же после снижения концентрации, но
скорость клеточного роста снижается.
Для активации многих внутриклеточных ферментов необхо-
димы те или иные низкомолекулярные соединения (витамины,
кофакторы) или ионы металлов (активаторы), которые могут
проникать через клеточные мембраны. Перенос небольшого
объема культуры клеток или инокулята в значительно боль-
ший объем среды вызовет обратную диффузию этих необходи-
мых для ферментативного катализа веществ в среду, если в
последней они отсутствуют или если среда резко отличается от
инокулята по своей ионной силе. В таком случае опять-таки
скорость роста упадет соответственно понижению концентрации
активаторов в клетке, что в свою очередь, повлечет за собой
лаг-фазу на период «включения» новых механизмов синтеза
этих активаторов. Если в описанных процессах снижаются кон-
центрации незаменимых активаторов (ионов и витаминов, ко-
торые не могут синтезироваться в клетке), то общий уровень
клеточной активности должен снизиться необратимо.
На длительность лаг-фазы очень большое влияние оказыва-
ет фаза роста, в которой находится инокулят, полученный в
свою очередь путем культивирования клеток в небольшом реак-
торе периодического действия. Объем инокулята также являет-
33-537
514
Глава 7
ся важной переменной; в частности, как мы уже упоминали,
она влияет на количество диффундирующих в среду веществ,
например витаминов и активаторов. Так, перенос популяции
молодых клеток в среду, богатую промежуточными метаболи-
тами (например, аминокислотами), не сопровождается лаг-фа-
зой; напротив, при переносе того же самого инокулята в среду
с сульфатом аммония клетки будут обедняться жизненно важ-
ными промежуточными веществами за счет их транспорта в
раствор. Если в момент переноса культура находится в фазе
экспоненциального роста, то в среде инокулята уже должны
содержаться в значительных концентрациях продукты проме-
Время, ч
! Утилизация । Утилизация^.
I глюкозы Г ксилозы ' I
РИС. 7.14. Если среда содержит оди-
наковые начальные концентрации
глюкозы и ксилозы, то в реакторе
периодического действия наблюдает-
ся диауксия — двухфазный рост куль-
туры Е. coli. (Воспроизведено с раз-
решения издательства из работы:
Startler R. У., Doudoroff М„ Adel-
berg Е. A, The Microbial World, 3d
ed., p. 308, Prentice-Hall, Inc., Engle-
wood Cliffs. N.J., 1970.)
жуточного метаболизма, поэтому сопровождающее перенос
разведение окажет меньший эффект. Перенос в аммониево-
сульфатную среду старой культуры (находящейся в стационар-
ной фазе или близкой к ней) приведет к более длительной лаг-
фазе.
Если среда содержит несколько источников углерода, то
иногда можно наблюдать ряд последовательных лаг-фаз
(рис. 7.14). Это явление, называемое диауксией {двухфазным
ростом), обусловливается изменением метаболического меха-
низма в процессе роста. При диауксии клетки сначала усваи-
вают один источник углерода, а затем, когда это питательное
вещество истощается, клеткам приходится переключать свою
активность с роста на «перестройку», необходимую для усвое-
ния нового источника углерода. В основе такой последователь-
ной утилизации питательных веществ, возможно, лежит явле-
ние катаболитной репрессии, рассмотренное в разд. 6.1.4.
Образование биомассы в культурах клеток
515
Попытки свести к минимуму время роста культуры и всего
процесса обычно включали сокращение продолжительности
лаг-фазы, типичной для любого периодического процесса с
участием микроорганизмов. Из уже рассмотренного нами ма-
териала и ряда других данных (описанных в приведенной в
конце главы литературе) вытекают следующие общие прин-
ципы:
1. Культура инокулята должна обладать максимальной актив-
ностью, а в момент введения в среду инокулят должен на-
ходиться в фазе экспоненциального роста.
2. Среда, в которой выращивается инокулят, по своему соста-
ву должна быть близкой к составу среды крупномасштаб-
ного микробиологического процесса.
3. Для предотвращения обусловленных диффузией потерь не-
заменимых промежуточных веществ и активаторов целесооб-
разно применять инокулят большего объема (до 5—10% от
объема новой среды).
К концу лаг-фазы популяция микроорганизмов приспосаб-
ливается к новым условиям. Теперь клетки могут быстро раз-
множаться, и клеточная масса (или число живых клеток) уд-
ваивается через определенные промежутки времени. Увеличе-
ние числа клеток в этот период может быть описано уравне-
нием
dx 1 dx irt or- \
= или —(7.35a)
причем
X = X0 при Z = Zlag (7.356)
Таким образом, скорость увеличения х пропорциональна х. Ин-
тегрируя уравнения (7.35), получим
In—— =р(£—Zlag) или х = f>^iag (7.36)
Отсюда следует, что время, необходимое для удвоения числен-
ности популяции клеток, составляет
= (7-37)
г
Как и в случае роста клеток в стационарном состоянии в
ПРПП, для описания популяции в ходе экспоненциального
роста в периодическом процессе необходим только один пара-
метр ц (или to). В первом приближении можно считать, что
в этой фазе рост периодической культуры сбалансирован. Сле-
довательно, путем изучения роста культуры клеток в реакторе
33*
516
Глава 7
периодического действия можно получить полезные данные о
кинетике сбалансированного роста, если только это изучение
ограничено фазой экспоненциального роста.
Экспоненциальный рост заканчивается, когда одна из важ-
ных переменных процесса (например, концентрация питатель-
ного вещества или токсина) достигает значения, не обеспечива-
ющего максимальную скорость роста клеток. Истощение запа-
сов лимитирующего рост питательного вещества может при-
вести к очень резкому изменению скорости клеточного роста,
поскольку в экспоненциальной фазе общая скорость утилиза-
ции питательного вещества возрастает очень быстро. Чтобы
найти описывающее это явление приближенное математическое
выражение, предположим, что до установления стационарной
фазы скорость потребления питательного вещества А пропор-
циональна массовой концентрации живых клеток:
= -V (7.38)
Допустим далее, что экспоненциальный рост продолжается до
тех пор, пока не установится стационарная фаза, и что экспо-
ненциальный рост начинается в нулевой момент времени. Тогда
x = xoeuf (7.39)
где хо — массовая концентрация живых клеток в момент нача-
ла экспоненциального роста.
Если в нулевой момент времени концентрация А равна flo,
то из уравнений (7.38) и (7.39) следует, что А будет полностью
утилизировано, когда
= х0) (7.40)
Г
Здесь Xs—массовая концентрация популяции в тот момент,
когда вещество А истощено и популяция переходит в стацио-
нарную фазу. Следовательно, xs представляет собой максималь-
ную концентрацию биомассы в периодическом процессе (ср.
рис. 7.13). Преобразованием уравнения (7.40) можно получить
уравнение, выражающее максимальную концентрацию биомас-
сы в момент истощения питательного вещества:
^==хо + -г~ао - (7-41)
Линейная зависимость xs от начальной концентрации пита-
тельного вещества ао во многих случаях наблюдалась экспери-
ментально (хо часто настолько мало, что им можно прене-
бречь). Пример такого поведения популяции клеток приведен
на рис. 7.15. В то же время график зависимости ns (численной
Образование биомассы в культурах клеток
517
О 2 4 6
Концентрация фруктозы, нг/мл
лактозы, г/л тартрата аммония, г/л
Влияние концентрации питательного вещества
на суммарную популяцию A. aerogenes
5 6
РИС. 7.15. Зависимость максимальной концентрации биомассы в периодиче-
ском процессе от исходной концентрации лимитирующего рост питательного
вещества: а — Pseudomonas sp. во фруктозной среде; б — A. aerogenes в лак-
тозной среде; в — A. aerogenes в среде на основе тартрата аммония. (Воспро-
изведено с разрешения издательства из работы: a) Stanier R. Y„ Doudoroff М„
Adelberg Е. A., The Microbial World, За ed., р. 313, Prentice-Hall, Inc., Engle-
wood Cliffs, N.J., 1970; б, в) Dean A. C. R„ Hinshelwood C., Growth, Function
and Regulation in Bacterial Cells, p. 72, Oxford University Press, London,
1956.)
плотности клеток в стационарной фазе) бактерии A. aeroge-
nes от концентрации лактозы заметно отличается от зависимос-
ти, предсказываемой уравнением (7.41). Очевидно, на начало
стационарной фазы и максимальную концентрацию биомассы
могут влиять и другие факторы.
518
Глава 7
Если в культуре накапливается токсин, тормозящий ско-
рость роста клеток (по сравнению с ростом в экспоненциаль-
ной фазе), то для описания поведения такой системы может
оказаться полезным уравнение следующего типа (с/ — концент-
рация токсина):
-^ = b[l-f(c,)l
В частном случае линейной зависимости скорости роста от
Ct это уравнение приобретает такой вид:
=S(1_te,) (7.42)
Здесь b — константа. Логично предположить, что скорость об-
разования токсина зависит только от х и пропорциональна х:
= (7.43)
Поэтому Ct^q^xdt (если с; = 0 при / = 0), а уравнение клеточ-
ного роста преобразуется следующим образом:
t \
-^- = kx[\—bq^xdt\ (7.44)
' 6 /
Величина мгновенной эффективной удельной скорости роста
geff равна
1- (7.45)
о /
Эта величина уменьшается во времени с возрастающей ско-
ростью, так что
-*2- = -АЬ?х<0 (7.46)
Рост клеток прекращается, когда
t
= f (7Л7)
О
Согласно уравнению (7.42), рост клеток прекращается толь-
ко тогда, когда Ct достигает значения, равного 1/&. Разведение
данной среды, содержащей токсин, или введение непитатель-
ного вещества, связывающего этот токсин, должно сопровож-
даться возобновлением клеточного роста и, следовательно, уве-
Образование биомассы в культурах клеток
519
личением максимальной концентрации биомассы xs в стацио-
нарной фазе. Если рост сдерживается истощением питательного
вещества, то разведение раствором, пе содержащим питатель-
ных веществ, не влияет на xs. Этими зависимостями можно
пользоваться в качестве критериев для предварительной оцен-
ки причин снижения скорости и прекращения клеточного роста.
Более точные критерии сформулировать труднее, поскольку,
как мы увидим позднее, рост популяции клеток в условиях ис-
тощения основного питательного вещества замедляется не-
сколько раньше, чем это вещество полностью утилизируется, а
скорость роста клеток в присутствии токсина может стать неиз-
Нититиру/ощии рантор-.
накопление токсичных
веществ д___________
Лимитирующии фактор:
истощение с ростра та
Начальная концентрация питательного
вещества
РИС. 7.16. Взаимосвязь между начальной концентрацией питательного веще-
ства и максимальной концентрацией биомассы в периодической культуре. При
высоких концентрациях питательных веществ, когда они не определяют мак-
симальную концентрацию биомассы, фактором, лимитирующим концентрацию,
может стать накопление токсичных веществ.
меримо малой задолго до того момента, когда dx/dt станет
равным нулю.
На рис. 7.16 изображена предсказываемая теорией зависи-
мость максимальной концентрации биомассы от исходной
концентрации данного питательного вещества. Постепенное
снижение концентрации питательного вещества в конце концов
приводит к линейной зависимости максимальной концентрации
биомассы от начальной концентрации питательного вещества.
В этом случае отсутствие фазы экспоненциального роста, оче-
видно, вызвано недостатком питательного вещества. Напротив,
повышение начальной концентрации питательного вещества
может привести к тому, что величина xs в конце концов пере-
станет быть зависимой от а0; возможно, это объясняется нако-
плением токсичных веществ или влиянием другого лимитирую-
щего клеточный рост питательного вещества.
Изучая всю популяцию, мы не должны забывать о судьбе
индивидуальных клеток. В общем случае популяции всегда
520
Глава 7
негомогенны, и кривая роста популяции в периодическом про-
цессе отражает лишь некоторый усредненный параметр чрезвы-
чайно сложной системы. Во время фазы экспоненциального
роста, например, некоторые клетки делятся и порождают но-
вые, молодые клетки, а другие в тот же самый момент растут
и созревают. Поскольку клетки разного возраста обычно име-
ют различные размеры и химический состав, мы можем рас-
сматривать клетки одного возраста как некоторое «вещество».
С этой точки зрения рост одного вида микроорганизма приво-
дит к популяции, содержащей множество различных «веществ».
Гетерогенность популяции становится особенно заметной
в стационарной фазе и фазе отмирания. Так, в стационарной
фазе некоторые клетки делятся, а некоторые погибают. Мерт-
вые клетки часто подвергаются лизису (разрушению и раст-
ворению); в результате углеводы, аминокислоты и другие ком-
поненты клеток попадают в среду и становятся питательными
веществами для живых членов популяции. Такой каннибализм
способствует сохранению массы популяции в стационарной фа-
зе. В конце концов, однако, истощение питательных веществ и
накопление токсинов приводит к тому, что популяция не мо-
жет поддерживать далее свое существование и наступает фа-
за отмирания.
Фазе отмирания было посвящено сравнительно небольшое
число исследований, возможно, вследствие того, что многие
промышленные микробиологические периодические процессы
заканчиваются задолго до наступления этой фазы. Обычно счи-
тается, что гибель популяции подчиняется экспоненциальному
закону:
х=х,е~^ (7.48)
В этом уравнении t означает время, прошедшее с начала фазы
отмирания. В основу уравнения (7.48) положено допущение о
том, что в любое время погибает одна и та же доля живых
клеток.
Одно из объяснений экспоненциального уменьшения числен-
ности популяции в фазе отмирания заключается в том,, что в
культуре клеток летальный исход является случайным событи-
ем и гибель данной клетки определяется только вероятностью
этого события. Недостатком такого объяснения является пре-
небрежение историей популяции. Дин и Хиншелвуд предполо-
жили, что в стабильной и отмирающей популяциях не только
живые клетки существуют за счет отмерших, но и конкурирую-
щие участки метаболического механизма клетки в борьбе за
дефицитные промежуточные соединения также существуют
один за счет другого [3]. Если принять это предположение,
можно показать, что скорость гибели клеток должна подчи-
Образование биомассы в культурах клеток
521
няться экспоненциальному закону. Другие модели снижения
численности популяции мы обсудим в разд. 7.7.
Чтобы лучше понять закономерности роста популяции кле-
ток в периодическом процессе и создать основу для вывода
соответствующих математических выражений, целесообразно,
рассмотреть ряд результатов экспериментального изучения из-
менений массы и химического состава популяции в периодиче-
ских процессах. Следует опять-таки подчеркнуть, что эти ре-
зультаты относятся не к индивидуальной клетке, а к совокуп-
ности множества клеток, составляющих популяцию в данный
момент.
Дин, Хиншелвуд и другие исследователи посвятили много
работ изучению изменений в составе культуры бактерий
A. aerogenes. В частности, ими было показано, что в среде,,
содержащей глюкозу, лаг-фаза сокращается до минимума,,
если инокулят взят из популяции в стационарной фазе, насту-
пившей после истощения запасов глюкозы. Полученные данные
изображены на, рис. 7.17. Обратите внимание на то, что на
ранних этапах цикла роста на кривых изменения средней
массы клетки и отношения РНК/ДНК наблюдаются резкие
максимумы. Хотя количество ДНК в расчете на единицу кле-
точной массы и отношение массы белков к массе РНК оста-
ются относительно постоянными, по этим данным в совокуп-
ности с некоторой другой информацией нетрудно воспроизвести
картину активизации метаболической активности клетки, на-
правленной на использование повышенных количеств пита-
тельных веществ в новой среде и на синтез больших количеств
диффундирующих промежуточных метаболитов.
На рис. 7.18 представлены данные, иллюстрирующие другое
интересное явление, к которому мы обратимся еще раз при
изучении скоростей образования конечных продуктов жизне-
деятельности клеток. Средняя концентрация РНК в клетке
возрастает пропорционально скорости роста популяции. Следу-
ет подчеркнуть, что такая зависимость наблюдается только тог-
да, когда изменение скорости роста обусловлено вариациями
в составе питательной среды. Если же скорость клеточного рос-
та изменяется, например, за счет понижения или повышения
температуры, то в фазе экспоненциального роста концентрация
РНК, по-видимому, остается практически на одном уровне.
В завершение этого раздела мы несколько подробнее рас-
смотрим свойства популяций клеток и характер изменения
активностей ферментов в течение клеточного цикла. В фазе
экспоненциального роста A. aerogenes концентрации двух фер-
ментов с гидрогеназной активностью изменяются очень мало,
в то время как удельная активность аспарагиндеаминазы
варьирует в довольно широких пределах. Первоначальное па-.
522
Глава 7
РИС. 7.17. Изменения характеристик клеток в ходе роста A. aerogenes в пе-
риодическом процессе. (Воспроизведено с разрешения из работы: Dean А. С. R.,
Hinshelwood С., Growth, Function and Regulation in Bacterial Cells, p. 87—89,
Oxford University Press, London, 1966.)
депие активности этого фермента приписывается разбавлению
инокулята, а повышение активности в конце клеточного цикла
обусловлено, по-видимому, снижением pH среды на этой ста-
дии процесса.
Этот факт лишний раз свидетельствует о том, что периоди-
ческую культуру клеток следует рассматривать как сложную
единую систему, состоящую из популяции клеток и жидкой сре-
ды переменного состава. В известном смысле рост популяции
в периодическом процессе можно считать функцией начальных
характеристик не только клеток, но и среды, поэтому любые
Образование биомассы в культурах клеток
523
Среднее обратное бремя удбоения
клетки, 1/(мин-АО3)
РИС. 7.18. Зависимость содержания РНК от скорости роста A. aerogenes для
различных источников углерода. (Воспроизведено из работы: Dean А. С. R.,
Hinshelwood С., Growth, Function and Regulation in Bacterial Cells, p. 92,
Oxford University Press, London, 1966.)
параметры популяции после начала процесса зависят от каж-
дой из этих двух фаз. Таким образом, в общем случае до из-
вестной степени необоснованно рассматривать свойства одной
лишь популяции клеток в периодическом процессе роста, по-
скольку эти свойства определяются взаимодействием между
средой и популяцией.
7.3.2. Неструктурированные модели клеточного роста
в периодических процессах
В простейшем подходе к моделированию роста культуры
клеток в периодическом процессе мы допускаем, что скорость
роста клеточной массы во времени является функцией одной
лишь клеточной массы, т. е.
(7.49)
Как мы скоро увидим, из этого уравнения отнюдь не следует,
что мы должны пренебрегать любыми изменениями, происходя-
щими в ходе клеточного роста в среде.
Одной из простейших моделей типа (7.49) является закон
Мальтуса, который математически можно выразить следующим
образом:
/(х) = цх
(7.50)
524
Глава 7
где ц — константа. Нетрудно видеть, что таким же образом
выражается уже известная нам закономерность накопления
клеточной массы в фазе экспоненциального роста. Предсказа-
ние Мальтуса о неизбежности рокового конца человечества в
результате неконтролируемого роста народонаселения не оправ-
далось (или, быть может, пока еще не оправдалось?); так же
и в случае популяций микроорганизмов вслед за фазой экспо-
ненциального роста наблюдается переход к стационарной по-
пуляции.
РИС. 7.19. Логистическая кривая (&>0; Р>0).
Эта теория получила дальнейшее развитие в работах Ферл-
хурста (1844 г.), а также Перла и Рида (1920 г.), которые при
анализе роста популяции стали учитывать ингибирующий фак-
тор. Допустив, что ингибирование пропорционально х2, эти ис-
следователи предложили уравнение
-^ = ^(1-₽х) х(О) = хо (7.51)
Это выражение представляет собой уравнение Риккати; его ин-
тегрирование не представляет затруднений и приводит к урав-
нению логистической кривой:
Как показано на рис. 7.19, логистическая кривая имеет S-об-
разную форму и приводит к стационарной популяции с макси-
мальной концентрацией биомассы xs=l/$.
Одно из возможных объяснений логистической кривой сво-
дится к допущению, что скорость продуцирования токсина про-
Образование биомассы в культурах клеток
525
порциональна скорости роста популяции:
det dx ~dt~a~dT (7.53)
И если (7.54)
то Сг = а(х —Xo) (7.55)
Обычно Хо пренебрежимо мало по сравнению с х; в таких слу-
чаях подстановка уравнения (7.55) в (7.42) приводит к урав-
нению типа (7.51).
В основу неструктурированных моделей другого типа, опи-
сывающих приближающиеся к стационарному состоянию попу-
ляции, положен принцип истощения лимитирующего рост пита-
тельного вещества. Если, как и при выводе уравнения Моно,
допустить, что p = p(s) и что общий экономический коэффици-
ент Ух/s постоянен, то уравнения материального баланса для
питательного вещества и клеточных веществ можно объединить
в одном уравнении типа (7.49) (см. упражнение 7.14).
Недостатком логистического уравнения является пренебре-
жение фазой снижения массы (численности) популяции после
того, как стационарная популяция исчерпала все свои ресурсы.
Этот фактор учтен в модели, разработанной в начале этого
столетия Вольтеррой. В модели Вольтерры уравнение (7.50)
дополняется интегральным членом типа
t
§ K(t,f)x(r)dr (7.56)
о
Физический смысл выражения (7.56) в первом приближении
можно интерпретировать как влияние истории популяции
/((/, г) на скорость ее роста. Выражение (7.56) отражает зави-
симость скорости роста во время t от всех предыдущих значе-
ний плотности популяции. Если К не зависит от t, то выраже-
ние (7.56) можно рассматривать как отражение влияния ком-
понента культуры, изменение концентрации которого подчиня-
ется закону
^-=|Х(/)х(01 с(0) = 0 (7.57)
Предположим, что постоянно и равно Ко, и дополним
уравнение (7.51) выражением (7.57), отражающим предыду-
щую историю популяции. Тогда изменение массы или плотности
526
Глава 7
РИС. 7.20. Решение уравнения Вольтерры при различных значениях парамет-
ра X, отражающего историю популяции клеток,
популяции во времени можно описать следующим уравнениемз
t
dx
dT
= kx(\—Px) + 7<0 \x(r)dr
x(0)=xo (7.58)
о
В соответствии с уравнением (7.58) Ко для ингибитора имеет
отрицательное значение, а для соединения, промотирующего
клеточный рост, — положительное. Это уравнение можно ре-
шить численно; пример такого решения приведен на рис. 7.20.
При отрицательных Ко масса популяции достигает максимума
и затем уменьшается.
Описанные выше неструктурированные модели клеточного
роста имеют несколько недостатков. В частности, в них не на-
ходит отражения лаг-фаза, не учтены влияющие на рост клеток
различные переменные, а также данные о метаболизме клетки
и его регуляции. В последующих разделах мы попытаемся най-
ти связи между сложной биохимией клетки и явлениями, на-
блюдаемыми при росте популяции в периодическом процессе.
Образование биомассы в культурах клеток
527
7.3.3. Рост филаментозных организмов
В предыдущих разделах настоящей главы мы рассматрива-
ли популяции микроорганизмов, в которых увеличение биомас-
сы сопровождается повышением числа клеток. По-другому
складывается ситуация при росте плесневых грибов и других
филаментозных организмов; здесь по мере роста культуры из-
меняется как масса, так и морфология гранул или суспензии
плесени. Экспериментальное изучение роста погруженных куль-
тур в периодических процессах показало, что количество био-
массы возрастает во времени не экспоненциально, а с меньшей
скоростью, так что масса популяции приблизительно пропорци-
ональна третьей степени времени. Чтобы объяснить эту зави-
симость, нужно начать изучение с одно- и двумерных культур
плесневых грибов. В случае одномерной культуры скорость
увеличения длины колонии остается постоянной во времени, а
для культур плесеней на поверхности (двумерных) характерна
постоянная скорость увеличения радиуса культуры.
Экстраполируя эти результаты на сферическую гранулу,
растущую в погруженной культуре, предположим, что
-^- = kg = const (7.59)
где R — радиус гранулы. Поскольку биомасса М равна
М = (7.60)
то из уравнений (7.59) и (7.60) следует, что
= = <7-61)
Подставляя в уравнение (7.61) значение R из уравнения
(7.60), получим
= 7М2/3 (7.62)
где
7 = /гя[36лр]1''3 (7.63)
Интегрирование уравнения (7.62) при начальной биомассе
Мо дает
М = [МО1/3 + -4-Y (7.64)
\ I
Поскольку Мо по сравнению с М обычно очень мало, то урав-
528
Глава 7
Штамм
Соетав среде/
Р«
Пооерхностно-
антивные
вещества
Штамм
Состав среде/
Перемешивание
pH
во/щ.. инокулята
Перемешивание
pH
Поверхностно -актив-
ные вещества
Твердые частице/
РИС. 7.21. Факторы, определяющие образование гранулы (а) и ее структу-
ру (б) в процессе роста мицеллярных организмов. [Воспроизведено с разре-
шения из статьи: M.etz В., Kossen N. W. F., Biotechnology Review: The Growth
of Molds in the Form of Pellets. — A Literature Review, Biotech. Bioeng., 19,
781 (1977).]
нение (7.64) свидетельствует об упоминавшейся выше пропор-
циональной зависимости М от t3.
Полный анализ роста филаментозного организма должен
учитывать также кинетику образования гранул. Последние
возникают в результате агломерации спор и последующего
роста или в результате разрастания отдельной споры. Имею-
Образование биомассы в культурах клеток
529
щиеся данные показывают, что на образование гранул влияют
самые различные свойства организма и среды (рис. 7.21). Об-
щая модель кинетики образования гранулы пока еще не раз-
работана, поскольку сложные механизмы этого явления недос-
таточно изучены и не вполне понятны.
При анализе роста уже образовавшихся гранул описанная
модель может рассматриваться только в качестве очень грубо-
го приближенного описания. На кинетику роста гранул долж-
ны оказывать влияние их размер, морфология и внутренняя
структура, а эти факторы в свою очередь определяются сово-
купностью таких параметров, как интенсивность перемешива-
ния, концентрация гранул, свойства организма и состав среды
[12]. Кинетика роста гранул часто зависит и от взаимосвязей
типа диффузия — реакция, рассмотренных нами в разд. 4.4.
7.4. Структурированные модели кинетики
клеточного роста
Неструктурированные модели описывают только количест-
во биологической фазы. Такие модели не учитывают и никоим
образом не отражают состав биофазы, т. е. то, что можно
назвать ее качественными характеристиками. Если в процессе
роста существенно изменяется состав клеточной популяции и
если это изменение влияет на кинетику клеточного роста, то
анализ таких систем возможен только с помощью структури-
рованных моделей. Поскольку ни в одной модели практически
невозможно учесть материальные балансы всех компонентов
клетки, то при создании структурированной модели (по необхо-
димости приближенной) мы, очевидно, должны тщательно ото-
брать ключевые переменные и процессы, играющие важнейшую
роль в планируемом применении этой модели. Как мы пока-
жем на ряде примеров в следующих разделах, структурирован-
ные модели могут быть построены на основе различных кон-
цепций.
В структурированных моделях в качестве переменных био-
фазы обычно применяются массовая (х/) или молярная (с;)
концентрации в единице объема биофазы [15]. При условии
полного перемешивания уравнение материального баланса по
компоненту / можно записать в следующем виде:
-4-Г—— VdXCj1=-^— VRxrf.-]——CDxCj (7.
di L P« K J Pc * J Pc
где pc — плотность клеток, равная массе клеток в единице
объема клеток; —молярная скорость образования компо-
нента / [(число молей /) • (время)-1 • (единица объема кле-
34—537
530
Глава 7
ток)-1]; ФЛ — масса клеток, вводимых в реактор в единицу
времени; — объем культуры; х — концентрация клеточной
массы [(масса клеток) • (единица объема культуры)-1]; с/—
(число молей /) • (единица объема клеток)-1. В последнем сла-
гаемом уравнения (7.65) принимается, что вводимые в реактор
(например, в рецикле) или выводимые из реактора клетки на-
ходятся в том же состоянии, что и популяция клеток в реакто-
ре. Если это условие не выполняется (например, во втором или
одном из последующих реакторов в каскаде ПРПП), то урав-
нение баланса должно быть соответствующим образом моди-
фицировано.
Если допустить, что плотность клеток рс и объем культуры
VR не изменяются во времени, то после дифференцирования
уравнения (7.65) получим
<7-66’
Для реактора периодического действия Фх равно нулю, а выра-
жение в скобках в правой части уравнения (7.66) есть не что
иное, как удельная скорость роста ц; следовательно, для этого
случая уравнение (7.66) преобразуется следующим образом:
= (7.67)
Физический смысл rf. ясен; выражение—цс, отражает сниже-
ние концентрации, обусловленное разведением в процессе роста
популяции клеток. Интересно, что оценка правой части уравне-
ния (7.66) для ПРПП при стерильности питательных веществ
опять-таки приводит к уравнению (7.67).
7.4.1. Компартментальные модели
В простейших структурированных моделях предусматривает-
ся компартментация биомассы в небольшом числе компонентов.
Иногда эти компоненты имеют приближенное биохимическое
толкование, например синтетический компонент (РНК и пред-
шественники) и структурный компонент (ДНК и белки).
Компартменты могут быть также определены как ассимилиру-
ющий компонент и синтетический компонент. В другой интер-
претации при создании простых компартментальных моделей
используется понятие об узких местах метаболизма.
Использование структурированных моделей с небольшим
числом переменных оправданно также с точки зрения некото-
рых принципов динамики систем. Каждый тип реакций и транс-
портных явлений, осуществляющихся в популяции клеток (мо-
лекулярные соударения, химические реакции, диффузия, кру-
Образование биомассы в культурах клеток
531
гооборот РНК, синтез белков,, увеличение числа клеток, завер-
шение периодического процесса, спонтанные мутации), харак-
теризуется своим временем релаксации, т. е. временем, необ-
ходимым для перехода в стационарное состояние после возму-
щения (вспомните разд. 3.1). Время релаксации может изме-
няться от долей секунды до нескольких часов. С точки зрения
динамики систем в моделировании наиболее важна взаимо-
связь между диапазоном времени, в течение которого соверша-
ются изменения в среде (те), и спектром времен релаксации
внутриклеточных процессов. Можно допустить, что клеточные
процессы, характеризующиеся быстрой реакцией на изменения
в среде (т. е. небольшой величиной отношения времени релак-
сации к те), находятся в квазистационарном состоянии
(разд. 3.2.1). В другом крайнем случае, когда время релаксации
некоторых клеточных процессов велико по сравнению с те, эти
клеточные процессы можно считать «замороженными» в на-
чальном состоянии. Примером могут служить глубокие генети-
ческие изменения, которые практически никогда не осущест-
вляются в ходе одного периодического процесса.
Установлено, что в случае многих сложных систем только
два или три времени релаксации находятся в том же времен-
ном диапазоне, что и изменения в среде; этот факт еще раз:
подтверждает обоснованность приближенного описания дина-
мики сложных систем с помощью упрощенной модели с двумя
или тремя переменными. В то же время часто трудно или да-
же невозможно связать некоторые из этих переменных с изме-
ряемыми физическими количественными характеристиками
системы. Ниже мы рассмотрим некоторые примеры простых
компартментальных кинетических моделей роста клеточной по-
пуляции.
Уильямс предложил двукомпартментальную модель, которая
удивительно точно описывает некоторые стороны динамики
клеточного роста в периодическом процессе [16]. Основные по-
стулаты этой модели формулируются следующим образом:
1. Синтетический компонент (1) продуцируется в результате
поглощения внешнего питательного вещества S с экономиче-
ским коэффициентом У (отношение массы компонента 1 к
массе субстрата). Реакция образования синтетического кле-
точного компонента имеет первый порядок по общей плот-
ности клеток х (отношению клеточной массы к объему куль-
туры) и по массовой концентрации питательного вещества
(отношению массы субстрата к объему культуры).
2. Структурно-генетический компонент клетки (2) продуциру-
ется из компонента 1 со скоростью, пропорциональной
pip2 (р£— отношение массы i к единице объема клеток).
34*
532
Глава 7
3. Для деления клетки необходимо и достаточно удвоение
компонента 2. Следовательно, численная плотность клеток
пропорциональна концентрации компонента 2 в культуре.
4. Биомасса построена только из компонентов 1 и 2.
Для реактора периодического действия при условии полного
перемешивания эти же постулаты можно выразить в математи-
ческой форме:
-|-=—rkisx (7.68)
- #- = ^ (7-69)
- ^- = ^1s(p1 + p2)—РР1 (7.70)
^- = ^PA-PP2 (7-71)
Принимая во внимание, что
y, = &is (7.72)
и
Pi + p2 = Pc = const (7.73)
уравнение (7.71) можно записать в следующей форме:
- ^-=(7.74)
где f2 = P2/pc представляет собой долю клеточной массы, состо-
ящей из компонента 2. С помощью двух уравнений этой моде-
ли s можно выразить через х. Если таким путем преобразовать
уравнение (7.69), то мы получим выражение, по форме совпа-
дающее с уравнением (7.51). Следовательно, изменение х бу-
дет подчиняться логистическому уравнению типа (7.52).
Предложенный Уильямсом подход применялся для модели-
рования роста популяции клеток в периодическом процессе с
использованием инокулята, находящегося в стационарной фазе
в среде с истощенным питательным веществом, что соответст-
вует pi = 0. Полученные таким путем результаты приведены на
рис. 7.22; они хорошо воспроизводят некоторые часто наблю-
даемые на практике особенности роста культур микроорганиз-
мов в периодическом процессе, включая:
1. Лаг-фазу, в течение которой возрастают размеры клеток.
2. Фазу экспоненциального роста, в которой размер клеток
достигает максимума.
Образование биомассы в культурах клеток
533
РИС. 7.22. Моделирование роста популяции клеток с использованием иноку-
лята, находящегося в стационарной фазе, по Уильямсу. Ординаты показывают
изменение во времени безразмерных параметров: концентрации субстрата
(s/so), концентрации биомассы (х/Узо), численности популяции (fix/Yso).
Здесь отражено также изменение во времени относительной массы клетки
(—l + 1/fa). (Параметры: &i=0,0125; А2Рс=0,025; У=0,5; хо=О,О5; s0=l;
f2(0) = l.)
3. Изменения в составе клеток в цикле роста. Поскольку такие
изменения, очевидно, происходят даже в экспоненциальной
фазе, отсюда следует, что рост клеток идеально не сбалан-
сирован ни в одной из фаз роста.
4. Стационарную фазу с относительно небольшими клетками.
Обратите особое внимание на то, что изменения во времени
плотности популяции по массе и по численности клеток непро-
порциональны; иными словами, масса индивидуальной клетки
зависит от скорости роста и даже от предыстории скорости
роста. Аналогичные результаты были получены в ходе деталь-
ного изучения роста популяций Е. coli и S. cerevisiae [30, 31].
Приведенные данные свидетельствуют о том, что динамику из-
менений клеточной массы часто можно описать с достаточно
хорошим приближением относительно простыми математичес-
кими моделями; напротив, расчеты динамики численности по-
пуляции более сложны.
534
Глава 7
На рис. 7.23 схематично изображены некоторые модифици-
рованные варианты описанной выше модели. В двухкомпарт-
ментальной модели Хардера и Роуэлса компонент G соответст-
вует ферментам, которые катализируют превращение субстрата
в промежуточные соединения, использующиеся при построении
биополимеров; составные части
РИС. 7.23. Схемы двух- и трехком-
партментальных моделей, описанных
Хардером и Роуэлсом [17].
клетки характеризуются компо-
нентом К, который образуется
из питательного вещества S со
скоростью, зависящей от ко-
личества G [17]. В этой прос-
той двухкомпонентной клетке
взаимопревращение G и К пу-
тем полимеризации и деполи-
меризации отвечает состоянию
метаболизма поддержания.
Составные части трехкомпо-
нентной клетки (рис. 7.23) ха-
рактеризуются более четко
определенной биохимической
природой; здесь К обозначает
РНК, G — белок, a R — дру-
гие компоненты биомассы
[17]. В последнем случае ме-
таболизм поддержания также
отражен как кругооборот ком-
понентов К и G.
По сравнению с некоторы-
ми из рассматриваемых ниже
более сложных структуриро-
ванных моделей эти неболь-
шие компартментальные моде-
ли относительно просты ма-
тематически и содержат не-
большое число кинетических
параметров. Их с умеренным успехом применяли для описания
различных состояний несбалансированного клеточного роста.
С другой стороны, отсутствие в ряде случаев четкого биохими-
ческого определения компонентов затрудняет оценку модели и
задачу нахождения параметров. При работе с такими моделями
мы не можем воспользоваться известными нам данными о
путях метаболизма клетки, регуляции молекулярных процессов
и закономерностях клеточного цикла. Эти данные можно при-
менять для моделирования кинетики клеточного роста, только
располагая глубоко структурированной моделью клетки, в ко-
торой учитывалось бы значительно большее число компонен-
тов, внутриклеточных реакций и взаимодействий.
Образование биомассы в культурах клеток
535
7.4.2. Метаболические модели
В этом разделе мы рассмотрим и изучим особенности двух
различных структурированных моделей кинетики клеточного
роста, включающих некоторые стороны клеточного метаболиз-
ма. На примере этих моделей мы познакомимся с некоторыми
преимуществами и недостатками, свойственными всем более
детальным моделям. Эти примеры покажут нам также, что по
мере включения в модель все большего числа биологических
деталей она становится более специфичной для определенного
организма или процесса. При создании метаболической кинети-
ческой модели определенной системы необходимо учитывать
ключевые детали метаболизма, иногда известные из научных
данных или биотехнологической литературы. В известном смыс-
ле мы может сказать, что, чем более детализированной стано-
вится наша модель, тем больше мы должны знать об изучае-
мом организме a priori. В противном случае задача выбора ки-
нетических уравнений и значений параметров становится слиш-
ком неопределенной, причем неизвестные параметры модели
нельзя определить на базе имеющихся (ограниченных) экспе-
риментальных данных. Хотя некоторые из описываемых здесь
и других высокоструктурированных моделей, которые мы обсу-
дим позднее, относятся к индивидуальной клетке, обычно при-
нимаемый детерминистический принцип предполагает описание
поведения усредненной клетки в большой популяции клеток.
Сначала мы изучим метаболическую модель, разработанную
Бийкерком и Холлом [18], а также Памментом, Холлом и Бар-
фордом [19] для аэробного роста почкующихся дрожжей
S. cerevisiae. В основу этой модели положены перечисленные
ниже допущения, большая часть которых соответствует извест-
ным данным о закономерностях клеточного цикла, метаболизма
и регуляции экспрессии ферментов организма.
Принятые в модели допущения
1. Индивидуальные изолированные клетки не рассматрива-
ются.
2. Лимитирующий рост клеток субстрат S является одновре-
менно источником углерода и энергии; символом Е обозна-
чен этанол.
3. Биомасса состоит из двух частей, А и В.
4. Масса А обеспечивает поглощение субстрата и снабжение
клетки энергией, масса В — синтез клеток и их деление.
5. Накопление энергии и продуктов метаболизма происходит в
ходе фазы Gi клеточного цикла, осуществляется массой А
и описывается превращением массы А в массу В [см. при-
веденные ниже уравнения (7.75) и (7.76)].
536
Глава 7
6. Репликация ДНК, митоз и деление клеток (фазы S, G2 и М)
происходят в течение определенного времени, осуществляют-
ся массой В и описываются превращением массы В в массу
А [см. приведенное ниже уравнение (7.77)].
7. Все участвующие в процессе брожения ферменты сгруппиро-
ваны и обозначены символом Ef. Аналогично сгруппированы
все ферменты дыхательного пути, обозначенные Ег.
8. В каждой ферментной системе биосинтез ферментов регули-
руется двумя различными путями. Во-первых, ферменты про-
дуцируются со скоростями, пропорциональными интенсивно-
сти потока метаболитов, проходящего через эту ферментную
систему. Во-вторых, ферменты каждой системы продуциру-
ются еще и в процессе адаптации клетки к изменяющимся
условиям; здесь скорость их биосинтеза пропорциональна
разности между имеющейся концентрацией фермента и
«контрольной» величиной (eF и eR для процессов брожения
и дыхания соответственно). «Контрольные» концентрации
ферментов пропорциональны интенсивности потока метабо-
литов, который был бы достигнут при нелимитирующих кон-
центрациях ферментов.
9. Скорости гликолиза и дыхания линейно зависят от е^е? и
от erleR соответственно.
10. Гликолитические ферменты продуцируются в результате
работы дыхательного пути (таким путем образуются фер-
менты для глюконеогенеза, биосинтеза и утилизации внут-
риклеточных резервных углеводов).
В таком виде клеточный рост можно описать с помощью
следующих стехиометрических уравнений [здесь и далее индек-
сами W обозначены переменные, выраженные в единицах мас-
сы; в других случаях подразумевается число молей или единиц
активности (для ферментов)]:
Брожение:
ГА
Л^ + a^Sw + Ef ---* 2Вц/ + a2Ew -|- Ef 4- СО2 (7.75)
Дыхание:
Aw + a^EwE О2+ Ef-E Ef ----> + Er + Ef + CO2 (7.76)
Деление:
BV Av (7.771
Скорости Га, Гв и Гс можно выразить следующими уравнениями:
_ кгаз ( ef \
А Ks-f-s eF J
(778)
Образование биомассы в культурах клеток
537
~ /^2^В / В г \
в Ке+в J
гс = КЪ
«Контрольные» концентрации ферментов равны
khk2,ae
R Ке+в
__ kfk^s k^ae
ер ~ Ks + s + \£+е
(7.79)
(7.80)
(7-81)
(7.82)
Скорости биосинтеза ферментов систем брожения и дыхания
принимаются равными соответственно
(7f) ферменты брожения — Г А Ч" С Ч" k^Ep — Ef) (7.83)
(7/Оферменты дыхания= гв+к6(ер — ег) (7.84)
В этих уравнениях символами ег и Ef обозначены активности
ферментов в единице объема культуры. Следовательно, как
мы уже упоминали, такая структурированная модель основана
не на «собственных» или внутриклеточных концентрациях.
Путем сравнения результатов моделирования и эксперимен-
тальных данных по росту культуры дрожжей в периодическом
процессе были определены параметры модели, значения которых
Таблица 7.3. Параметры математической
модели роста S. cerevisiae
в аэробных условиях [19]
Я1 5,95 г/г Ks 0,50 г/л
Й2 2,50 г/г Ке 0,02 г/л
d3 1,50 г/г ki 0,00 ч-1
kl 5,00 ч-1 k$ 0,75 ч-1
^2 0,26 ч-1 k6 0,225 ч-
К 0,50 ч-1 1,33 ч-1
kf 2,37 ча Sf 9,20 г/л
kh 0,244 ча
а Значения н kh вычислены по другим па-
раметрам модели с помощью уравнений, приве-
денных в работе [19].
приведены в табл. 7.3. Применение этой кинетической модели к
экспериментам по росту непрерывной культуры в стационарном
состоянии потребовало незначительного изменения (от 1,79 до
1,50 в периодическом и непрерывном процессах соответственно)
только одного параметра а3 — экономического коэффициента
роста за счет дыхания. Причиной его изменения отчасти послу-
жила известная физиология этого организма, в частности необ-
ходимость глюконеогенеза при росте периодической культуры
538
Глава 7
РИС. 7.24. а — рассчитанные путем
моделирования зависимости плотно-
сти клеточной массы х (сплошная
кривая) и удельных активностей фер-
ментов ef/x и ег/х (прерывистые кри-
вые) от скорости разведения D (ч-1)
для случая роста непрерывной культу-
ры S. cerevisiae в стационарном со-
стоянии в ПРПП. Зачерненными
квадратиками указаны результаты
экспериментального определения х\
б — результаты эксперименталь-
ного определения удельных активно-
стей ферментов брожения, NADP+-
глутаматдегидрогеназы (GluDH—
NADP+) и алкогольдегидрогеназы
(ADH); в — результаты эксперимен-
тального определения удельных ак-
тивностей ферментов малатдегидро-
геназы (MDH) и изоцитратлиазы
(ICL), участвующих в дыхании.
[Воспроизведено с разрешения из
статьи: Pamment N. В., Hall R. J.,
Barford J. R., Mathematical Modeling
of Lag Phases in Microbial Growth,
Biotech. Bioeng., 20, 345 (1978).]
на этаноле и отсутствие такой необходимости при росте проточ-
ной культуры на глюкозе (в ПРПП).
На рис. 7.24, а изображены графики зависимости плотности
клеточной массы х(х = а + Ь) и удельных активностей фермен-
тов брожения (ef/x) и дыхания (ег/х) от скорости разведения
для случая роста непрерывной культуры S. cerevisiae в стацио-
нарном состоянии; эти зависимости вычислены с помощью па-
раметров, перечисленных в табл. 7.3. Зачерненными квадрати-
ками на этом рисунке обозначены результаты эксперименталь-
ного определения х, хорошо согласующиеся с расчетными дан-
ными. Обратите внимание на то, что здесь зависимость х от
D как для экспериментальных, так и для расчетных данных ।
резко отличается от аналогичной упрощенной зависимости, по- ?
лученной на базе неструктурированной модели Моно (рис. 7.6)
На рис. 7.24, б приведены результаты экспериментального опре-
О б р а з о вание биомассы в культурах клеток 539
деления зависимости удельной активности двух ферментов, уча-
ствующих в метаболизме брожения (ЫАОР+-глутаматдегидро-
геназы и алкогольдегидрогеназы) от скорости разведения в
ПРПП. Эти зависимости качественно согласуются с расчетными
данными для е^/х модели. На кривой зависимости удельной ак-
тивности двух участвующих в дыхании ферментов (малатдегид-
рогеназы и изоцитратлиазы) от скорости разведения экспери-
ментально обнаружен максимум (рис. 7.24, в); модель предска-
зывает наличие максимума в зависимости ег/х от D.
Эти результаты, как и ряд других, приведенных в работе
[18], убедительно демонстрируют некоторые из преимуществ,
свойственных более структурированному описанию роста попу-
ляции клеток. Рассматриваемая модель успешно применялась
для описания периодического роста S. cerevisiae с добавлени-
ем субстрата (см. разд. 9.1.1); она также хорошо отражает
лаг-фазу в экспериментальных периодических процессах.
В качестве завершающего примера рассмотрим разработан-
ную Шулером и сотрудниками модель индивидуальной клетки
Е. coli [24]; из всех предложенных до настоящего времени мо-
делей роста микроорганизмов она наиболее удачна и высоко-
структирована. На рис. 7.25 схематично изображены учитывае-
мые в этой модели метаболиты, биополимеры и реакции, осу-
ществляющиеся, как предполагается, в клетке Е. coli В/г А.
Прерывистыми линиями здесь обозначены информационные по-
токи, регулирующие кинетику реакций. В модель включены ре-
акции образования клеточной оболочки, поэтому продолжи-
тельность клеточного цикла является величиной, предсказывае-
мой моделью, а не исходным параметром, как в большинстве
других структурированных моделей. Поскольку эта модель
разрабатывалась специально для описания роста клеток в ус-
ловиях ограниченного количества источника углерода или азо-
та, в ней учтены! многие структурные элементы клетки, участ-
вующие в транспорте и ассимиляции этих веществ. Приведен-
ные на рис. 7.25 реакции учитывают также синтез и утилиза-
цию АТР.
На рис. 7.25 не показаны важные детали модели, связан-
ные с регуляцией инициации синтеза ДНК. В течение очень ко-
роткого периода («взрывной синтез») синтезируется белок-реп-
рессор RP; он нейтрализуется белком-антирепрессором ARP,
который синтезируется со скоростью, пропорциональной скорос-
ти построения клеточной оболочки. При достаточно низкой кон-
центрации RP начинается транскрипция, которая приводит к
короткому участку РНК, необходимому для инициации репли-
кации.
К сожалению, объем нашей книги не позволяет описать эту
модель достаточно подробно; дополнительные данные читатель
540
Глава 7
РИС. 7.25. Схема высокоструктурированной модели индивидуальной клетки,
описывающей рост Е. coli В/гА на среде, содержащей глюкозу и соли аммо-
ния. [Воспроизведено с разрешения из работы: Shuler М. L., Domach М. М„
Mathematical Models of the Growth of Individual Cells, in Foundations of Bio-
chemical Engineering, Blanch H. W., Papoutsakis E. T., Stephanopoulos G.
(eds.), p. 101, American Chemical Society, Washington, 1983.]
Ai — ион аммония; A2 — глюкоза (и соединения, образующиеся из нее в клет-
ке) ; W — конечные продукты (СО2, Н2О и ацетат) энергетического обмена,
выделяемые в процессе аэробного роста; Pi — аминокислоты; Р2 — рибонук-
леотиды; Р3 — дезоксирибонуклеотиды; Р4—предшественники клеточной обо-
лочки; Mi — белки (как в цитоплазме, так и в клеточной оболочке);
M2rti — незрелая «стабильная» РНК; M2RTm — зрелая «стабильная» РНК
(рРНК и тРНК; во всех случаях принимается, что 85% составляет рРНК);
М2М — матричная - РНК; М3 — ДНК; М4 — небелковая часть клеточной обо-
лочки (принимается, что она содержит 16,7% пептидогликанов, 47,6% липи-
дов и 35,7% полисахаридов); М5 — гликоген; PG — ppGpp; Ei — ферменты,
участвующие в превращении Р2 в Р3; Е2, Е3— соединения, участвующие в об-
разовании клеточной оболочки и регуляции синтеза ее компонентов; GLN —
глутамин; Е4 — глутаминсинтетаза; * — вещества, находящиеся вне клетки.
может найти в работе [20]. Эта модель включает около 100
стехиометрических и кинетических параметров; почти все эти
параметры могут быть определены на основе опубликованных
результатов изучения биохимии Е. coll. Насколько велика ин-
формация, вложенная в эту модель, настолько же велики нее
возможности.
Модель достаточно точно описывает время инициации реп-
ликации хромосомы и другие важнейшие особенности клеточ-
ного цикла в широком диапазоне скоростей роста. На рис. 7.26
приведены графики зависимости в двойных обратных коорди-
натах удельной скорости роста отдельной клетки от концентра-
ции лимитирующего рост питательного вещества (в данном
случае иона аммония), полученные экспериментально (точки)
и расчетным путем на базе описанной модели (сплошная ли-
ния). Обратите внимание на то, что здесь и результаты модели-
Образование биомассы в культурах клеток 541
рования, и экспериментальные данные свидетельствуют о суще-
ствовании нескольких механизмов утилизации иона аммония;
это проявляется в изменении наклона при очень больших кон-
центрациях питательного вещества (малых величинах 1/s). Вы-
численные путем моделирования закономерности изменения
скорости клеточного роста, содержания в клетке гликогена и
размера клетки (рис. 7.27) хорошо согласуются с эксперимен-
f/s, (мг азота/а/)'г
РИС. 7.26. Зависимость удельной скорости лимитируемого источником азота
роста Е. coli В/г А от концентрации иона аммония (сплошная линия — мо-
дельные расчеты; жирные точки — экспериментальные данные). [Воспроизве-
дено с разрешения из работы: Shuler М. L„ Domach М. М., Mathematical Mo-
dels of the Growth of Individual Cells, in Foundations of Biochemical Engi-
neering, Blanch H. W., Papoutsakis E. T., Stephanopoulos G. (eds.), p. 101,
American Chemical Society, Washington, 1983.]
тальными результатами. Рассмотренная нами ранее модель
Уильямса предсказывает увеличение размеров клеток при вы-
соких скоростях роста; в отличие от нее описываемая высоко-
структурированная модель, учитывающая гораздо большее чис-
ло биохимических особенностей и деталей метаболизма клетки,
показывает, что размер клеток зависит не только от скорости
их роста, но и от того, какое питательное вещество лимитиру-
ет скорость роста.
7.4.3. Моделирование клеточного роста
как оптимального процесса
Изучение роста микроорганизмов на смесях различных суб-
стратов показывает, что обычно организмы быстрее усваивают
субстрат, обеспечивающий наиболее высокую скорость роста.
542
Глава 7
РИС. 7.27. Сравнение результатов, предсказываемых моделью изолированной
жлетки, с экспериментальными данными. Приведены расчетная кривая (У)
изменения объема клетки при лимитируемом глюкозой росте (2, 3 — экспери-
ментальные данные); то же (4) при лимитируемом ионом аммония росте (5 —
экспериментальные данные); расчетная кривая (6) изменения процентного
содержания гликогена (7 — экспериментальные данные). [Воспроизведено с
разрешения из работы: Shuler М. L„ Domach М. М., Mathematical Models of
the Growth of Individual Cells, in Foundations of Biochemical Engineering,
Blanch H. W„ Papoutsakis E. T., Stephanopoulos G„ (eds.), p. 101, American
^Chemical Society, Washington, 1983.]
На молекулярном уровне в основе такого поведения микроор-
ганизмов лежат процессы индукции, репрессии, ингибирования
и активации, рассмотренные в гл. .3 и 6. Их можно положить в
основу соответствующих моделей кинетики роста (разд. 7.5.3),
но для этого нужно учитывать множество параметров и деталь-
ных сведений о механизме процессов. В альтернативном под-
ходе, разработанном Рамкришной и сотрудниками, влияние
регуляторных клеточных процессов описывается как результат
оптимизации клеточного роста [21, 22]. В основе этого подхода
лежит постулат о том, что в результате естественного отбора и
эволюции биологические системы приобрели способность ис-
пользовать среду оптимальным образом. С точки зрения такой
.кибернетической модели действие регуляторных систем метабо-
Образование биомассы в культурах клеток
54Э-
лизма можно описать как решение задачи! оптимального расп-
ределения ресурсов, обеспечивающего максимальную скорость
роста.
Основанный на законе соответствия для распределения ре-
сурсов алгоритм оптимизации оказался вполне пригодным для
моделирования клеточного роста на смешанных субстратах.
Если отдача от утилизации ресурса h равна 0,(Z() и если
полная отдача (сумма всех 0г) максимизирована при условии,,
что сумма всех Z, равна постоянной величине, то должен удов-
летворяться следующий закон соответствия:
^®к==_^к=_^з_==... (7.85)
иХ 1 б/Х 2 d Z з
Предположим, что скорость роста популяции зависит от
внутриклеточных концентраций е2, • ряда ферментов, необ-
ходимых для ассимиляции субстратов Si, S2, - соответственно.
В таком случае, например, вклад субстрата 5г- в образование
биомассы может быть описан уравнением'
гх..=г^ (7.86)'
где
'<= ^7$^ (7.87)
Весовой коэффициент щ определяется условиями закона соот-
ветствия для энергетического выхода процессов потребления
различных субстратов, которые предполагают, что
f. = vd 1С1
1
(7.88)
где V/ — энергия, высвобождающаяся при утилизации единицы
массы субстрата /. Применение закона соответствия к распре-
делению клеточных ресурсов для синтеза ферментов приводит
к следующему выражению для скорости синтеза фермента Ег-:
= (7-89)‘
где
(7.90)‘
Важным преимуществом этой модели является возможность
экспериментального определения всех кинетических параметров
в реакциях с индивидуальными субстратами. Метод оптимиза-
ции учитывает все аспекты кинетики роста популяции микроор-
ганизмов в среде с несколькими субстратами. На рис. 7.28
544
Глава 7
приведено сравнение расчетных и экспериментальных данных
по росту культуры бактерии Klebsiella oxyloca в среде, содер-
жащей арабинозу и лактозу. Соответствующие параметры мо-
дели, определенные в отдельных экспериментах, можно найти
в работе [22]. В данном случае модель хорошо отражает про-
должительную лаг-фазу между периодом роста на лактозе и
периодом более медленного роста на арабинозе. Изучение
других моделей этого класса показало, что расчетные и экс-
РИС. 7.28. Сравнение экспериментальных (х) и полученных с помощью кибер-
нетической модели расчетных (-----) данных для двухфазного периодическо-
го роста Klebsiella oxyloca в арабинозно-лактозной среде. [Воспроизведено
с разрешения из статьи: Kampala D. S., Ramkrishna D., Tsao G. T., Cybernetic
Modeling of Microbial Growth on Multiple Substrates, Biotech. Bioeng., 26, 1272
(1984).]
периментальные данные не только качественно, но и количест-
венно хорошо согласуются и в случае трехфазного роста микро-
организмов на трех субстратах, как в периодическом процессе,
так и в стационарной и переходной фазах непрерывного про-
цесса в ПРПП. Большой интерес представило бы распростра-
нение кибернетического подхода на другие методы оптимиза-
ции и процессы образования продуктов жизнедеятельности
клеток. Кибернетические модели являются интересной альтер-
нативой моделей, в которых метаболизм рассматривается как
упрощенная сеть химических реакций.
7.5. Кинетика образования продуктов метаболизма
Кинетика образования популяциями клеток продуктов мета-
болизма может быть описана аналогично росту клеточных по-
пуляций. Здесь также возможны структурированный и неструк-
Образование биомассы в культурах клеток 545
турированный подходы. Как мы увидим в разд. 7.5.3, модели ки-
нетики белкового синтеза можно разработать и на молекуляр-
ном уровне, если воспользоваться известными в настоящее вре-
мя данными по регуляции молекулярных процессов.
7.5.1. Неструктурированные модели
Кинетика образования продуктов жизнедеятельности клеток
наиболее проста в тех случаях, когда образование метаболитов
и утилизация субстрата или клеточный рост связаны просты?л
стехиометрическим отношением. Тогда скорость образования
продуктов метаболизма можно выразить следующими уравне-
ниями:
% = -Ур/sQ, (7.91)
ИЛИ
% = (7.92)
соответственно. Такие случаи типичны для микробиологических
процессов типа I, рассмотренных в разд. 5.10.3. Примером мо-
жет служить спиртовое брожение, кинетика которого в перио-
дическом процессе отражена на рис. 7.29. Такого типа кинети-
ку образования продуктов жизнедеятельности иногда называют
сопряженной с ростом.
Во многих микробиологических процессах, особенно если
речь идет о вторичных метаболитах, продукт жизнедеятельнос-
ти клеток не образуется в сколько-нибудь значительных коли-
чествах в первых фазах периодического процесса вплоть до на-
чала стационарной фазы или даже несколько позже, как, на-
пример, процесс биосинтеза пенициллина (рис. 7.30). Иногда
в таких случаях кинетика образования продукта жизнедеятель-
ности клеток удовлетворительно описывается простой, не со-
пряженной с ростом моделью, в которой скорость образования
метаболита принимается пропорциональной не скорости роста
клеток, а их концентрации.
В ставшей классической работе Льюдикина и Пайрета*, по-
священной изучению молочнокислого брожения в присутствии
бактерии Lactobacillus delbruekii, было показано, что в кинети-
ку образования продукта метаболизма вносят вклад как сопря-
женные, так и не сопряженные с ростом факторы:
rfp=arfx+px (7.93)
* Leudeking R., Piret E. L., A Kinetic Study of the Lactic Acid Fermenta-
tion, J. Biochem. Microbiol. Technol. Eng., 1, 393 (1959).
35-537
546
Глава 7
/. Рост меток, г/(л- v)
8. Синтез спирта, г/(о- о)
С. Утилизация сахара, г/(л-ч)
Время процесса брожения, ч
РИС. 7.29. Для процесса спиртового брожения характерна простая сопряжен-
ная с клеточным ростом кинетика образования продукта метаболизма. На ри-
сунке указано изменение во времени биомассы, массы образовавшегося спир-
та и утилизированного сахара (а); объемных и удельных скоростей образова-
ния биомассы, утилизации субстрата и синтеза продукта метаболизма (спир-
та) (б, б). [Воспроизведено из работы: Leudeking R. Fermentation Process
Kinetics, in Biochemical and Biological Engineering, Blakebrough N. (ed.),
vol. 1, p. 203, Academic Press, Inc., (London) Ltd., London, 1967.]
Образован не биомассы в культурах клеток
547
0,5
0,4
Q3
0,2
0,1
О
-0,1
0,20
\ 0,15
о,ю-
0,05
25-
20-
Ъ 15-
х'
10-
5-
0,020-
0,015 -
0,010-
0,005 ~
О -
00012 -
0,0010 -
0,0008-
0,0006 -
0,0000 -
0,0002 -
О 20 00 60 80 100 120 740
8ремя ферментации, ч
РИС. 7.30. Усложненная кинетика биосинтеза вторичного метаболита пеницил-
лина. Указано изменение во времени биомассы, массы образовавшегося пени-
циллина и утилизированного сахара (а); объемных и удельных скоростей
образования биомассы, синтеза пенициллина, утилизации сахара и кислорода
(б, в). [Воспроизведено из работы: Leudeking R., Fermentation Process Kine-
tics, in Biochemical and Biological Engineering, Blakebrough N (ed.) vol 1
p. 205, Academic Press, Inc., (London) Ltd., London, 1967.]
35*
548
Глава 7
Такое уравнение скорости образования продукта метаболизма
с двумя параметрами, часто называемое уравнением Льюдики-
на — Пайрета, оказалось чрезвычайно полезным при интерпре-
тации экспериментальных результатов самых различных мик-
робиологических процессов. Именно такую форму зависимости
следует ожидать тогда, когда изучаемое вещество является ко-
нечным продуктом метаболического пути, связанного с выде-
лением энергии (например, в некоторых процессах анаэробного
брожения). В этих случаях первое и второе слагаемые правой
части уравнения (7.93) можно рассматривать как меру энер-
гии, расходуемой на клеточный рост и на поддержание клеток
соответственно (см., например, работу [5]).
Пример 7.1. Последовательное определение параметров простого периоди-
ческого процесса брожения. Рассмотрим модельный периодический процесс
брожения, в котором клеточный рост описывается логистическим уравнением
(7.51), а образование продукта метаболизма — уравнением Льюдикина —
Пайрета (7.93):
dp dx , о
-dF = a-dr + ?x (7Пк|>
Кинетику утилизации субстрата можно выразить следующим уравнением,
в котором учтены превращение субстрата в клеточную массу и продукт ме-
таболизма, а также его расход на поддержание клеток:
ds 1 dx 1 dp
dt Y'x/s dt Y'p/s &
(7П1.2)
Подстановка уравнения (7П1.1) в (7П1.2) позволяет записать уравнение ма-
териального баланса по субстрату:
4-=-v4—<7П1-з)
где
1 ( а
i"x/s +
Ч = (7П1.4)
Эта и другие аналогичные модели позволяют описать многие практически
важные микробиологические процессы, в том числе и такие, в которых обра-
зуется несколько конечных продуктов метаболизма (25]. К преимуществам
этих моделей можно отнести и тот факт, что их параметры могут быть оп-
ределены последовательно с помощью ряда специально разработанных удоб-
ных графических методов. Ниже мы вкратце рассмотрим эти методы.
Преобразованием уравнения (7.52) можно получить следующее выра-
жение:
In
x(t)jxs
l — x(f)/xs
= kt—In
%s_____
x0
1
(7П1.5)
Нетрудно видеть, что
фику зависимости
определив экспериментально xs, можно затем по гра-
1п
x(t)/xs 1
1 —х (0/xs J
Таблица 7П1.1. Значения параметров модели кинетики процессов биосинтеза внеклеточных
полисахаридов различными микроорганизмами3
Параметры образования продукта метаболизма
Продукт процесса или микроорганизм
Р а
Ксантан 0,155 г продукта/г биомассы в час 1,83 г продукта/г биомассы
Пуллулан (pH 4,5) 0 89% (по массе/(г в сутки в 100 мл)
Пуллулан (pH 5,5) 0 135% (по массе)/(г в сутки в 100 мл)
Пуллулан (pH 6,5) 0 110% (по массе)/(г в сутки в 100 мл)
Альгиновая кислота 0 1,60 г продукта/г биомассы
Pseudomonas sp. 10-3 г продукта/единиц биомассы в час 0
Продукт процесса или микроорганизм Параметры утилизации субстрата Параметры образо- вания биомассы
V п k, я-1
Ксантан 2,0 г субстрата/г биомассы 0,284 г субстрата/г биомассы в час 0,15
Пуллулан (pH 4,5) — — 1,12
Пуллулан (pH 5,5) — — 0,89
Пуллулан (pH 6,5) — — 1,12
Альгиновая кислота 6,6 г субстрата/г биомассы 0,015 г субстрата/г биомассы в час 0,12
Pseudomonas sp. 0,165% (масса/объем)/единиц биомассы 2,8-10-2 % биомассы (масса/объем)/единнц 0,31
в Воспроизведено с разрешения из статьи: Ollis D. F., A Simple Batch Fermentation Model: Theme and Variations. Annals N.Y. Acad.
Sci., 413, 144 (1983).
550
Глава 7
от t найти k (тангенс угла наклона прямой) и х0 (отрезок, отсекаемый пря-
мой на осп ординат). Что касается параметров кинетики образования продук-
та жизнедеятельности клеток, то из уравнения Льюдикина — Пайрета для
периодической культуры в стационарном состоянии следует
(.dp/^Юстационарная фаза
xs
(7П1.6)
Интегрированием уравнения (7П1.1) при х, определяемом уравнением (7.51),
получаем
Р(0—Ро~1—^-0— ekt) =а[х(0— Хо] (7П1.7)
Построив график зависимости левой части уравнения (7П1.7) от [х(0—х0],
можно определить а. Аналогичным путем можно найти параметры у и т|
в уравнении скорости утилизации субстрата.
В табл. 7П1.1 приведены найденные таким путем параметры кинетики
четырех различных микробиологических процессов, в результате которых
образуются внеклеточные полисахариды. Эти данные показывают, что про-
цессы образования внеклеточных биополимеров по своим кинетическим ха-
рактеристикам могут быть сопряжены с клеточным ростом (пуллулан и поли-
альгинат), не сопряжены с ним (биополимер из Pseudomonas sp.) или иметь
смешанный характер (ксантан).
Таблица 7.4. Классификация микробиологических процессов
по Дейндоерферуа
Тип процесса
Описание
Простой Превращение питательных веществ в продукты метаболизма осуществляется без накопления промежуточных веществ и характеризуется
Совместный строго определенной стехиометрией Превращение питательных веществ в продукты метаболизма не сопровождается накоплением промежуточных веществ, но стехиометрия про- цесса может изменяться
Последовательный Превращение питательных веществ в продукт ме- таболизма сопровождается накоплением про-
Ступенчатый межуточного соединения Превращению питательных веществ в продукт ме- таболизма предшествует полное превращение в промежуточное соединение, или питательные ве- щества селективно превращаются в продукт ме- таболизма в каком-либо определенном порядке
а Deindoerfer F. И., Adv. Appi. Microbiol., 2, 321 (1960).
Временная зависимость концентрации продукта метаболиз-
ма в ходе периодического процесса может быть очень слож-
ной, причем в среде могут накапливаться и подвергаться даль-
Образование биомассы в культурах клеток
551
Рис. 7.31. Образование различных веществ в ходе периодического роста
Cl. acetobutylicum при pH 5; приведены экспериментально найденные кон-
центрации глюкозы (7), клеточной массы (2), уксусной кислоты (3), масля-
ной кислоты (4}, ацетона (5), бутанола (6), этанола (7). [Воспроизведено
с разрешения из статьи: Costa J. М„ Moreira A. R., Growth Inhibition Kinetics
for the Acetone-Butanol Fermentation, ACS Symposium Series, 207, 501 (1983).]
нейшим превращениям несколько веществ. Различные возмож-
ные ситуации отражены в классификации микробиологических
процессов, предложенной Дейндоерфером (табл. 7.4). В неко-
торых случаях сложная кинетика образования продукта жиз-
недеятельности клеток может отражать изменения в механизме
метаболизма клетки. Показательным примером (важным каке
точки зрения промышленного использования, так и в истори-
ческом аспекте) такой кинетики может служить синтез ацетона
и бутанола бактерией Clostridium acetobutylicum (рис. 7.31).
В первой фазе периодического процесса глюкоза превращается
в уксусную и масляную кислоты, которые впоследствии перера-
батываются (наряду с глюкозой) в ацетон и бутанол. Образо-
вание продуктов метаболизма клетки может сопровождаться
также химическими превращениями метаболитов в среде, как,
например, при спонтанном гидролизе пенициллина. Описание
кинетики таких сложных процессов может потребовать вклю-
чения этих дополнительных реакций в схему математической
модели.
552
Глава 7
7.5.2. Химически структурированные модели кинетики
образования продуктов жизнедеятельности клеток
В сравнении с изучением кинетики роста индивидуальной
клетки и популяций клеток в проблеме кинетики процессов об-
разования продуктов метаболизма разработке структурирован-
ных моделей посвящено относительно небольшое число работ.
Можно ожидать, однако, что развитие структурированных мо-
делей клеточного роста в ближайшем будущем потребует раз-
работки ряда таких моделей. В этом разделе мы рассмотрим
интересный процесс образования одного из множества вторич-
ных метаболитов, на примере которого познакомимся с рядом
важных аспектов процессов микробиологического синтеза анти-
биотиков, а также с новыми подходами к моделированию ки-
нетики таких процессов.
Синтез ряда микробных антибиотиков и других продуктов
вторичного метаболизма ингибируется высокими концентрация-
ми фосфатного иона. Поскольку для роста клеток фосфат со-
вершенно необходим, очевидно, должен существовать некото-
рый оптимальный уровень его концентрации. Действительно,
такой оптимальный уровень начальной концентрации фосфата
наблюдался, например, при биосинтезе алкалоидов Claviseps
purpurea (рис. 7.32).
Приведенные на рис. 7.32 экспериментальные данные хоро-
шо моделируются следующими уравнениями кинетики клеточ-
ного роста периодической культуры и образования продукта
жизнедеятельности [26].
Рост клеток:
= М1 - exp (-K.Pi)} x-k^ (7.94)
CLi>
Концентрация фосфата в среде:
%— -Ь, V (7-95)
Концентрация фосфата в клетке:
-%- “ (Г₽/Х + Р‘) [ 1 - exp ((7-96)
Образование алкалоидов:
da _ » К3х
dt Кз + р?
(7.97)
В этой модели удельная скорость роста описывается уравнени
Образование биомассы в культурах клеток
553
РИС. 7.32. Изменение во времени плотности биомассы (ПБ) С. purpurea (•,
г сухого вещества в 1 л) и суммы продуцируемых алкалоидов (СА) (О,
мг/л) при различных начальных концентрациях фосфата. Кривые отражают
соответствующие расчетные данные, полученные с помощью описанных в тек-
сте математических моделей. [Воспроизведено с разрешения из статьи: Pazou-
tova S., Votruba J., Rehdcek Z., A Mathematical Model of Growth and Alkaloid
Production in the Submerged Culture of Claviceps purpurea-, Biotech. Bioeng.,
23, 2837 (1981).]
ем Теиссье, в котором одной из независимых переменных явля-
ется внутриклеточная концентрация фосфата pi. (Эта перемен-
ная равна числу граммов КН2РО4, содержащемуся в 1 г био-
массы, и отражает состав клетки. Все другие переменные этой
модели, описывающие компоненты системы, измеряются в
граммах на 1 л культуры.) Выражение k2x2 отражает скорость
лизиса клеток.
В результате лизиса фосфат выделяется в среду; как пока-
зывает второе слагаемое правой части уравнения (7.95), его
количество пропорционально содержанию фосфата в клеточной
массе Ур/х и внутриклеточной концентрации фосфата pi. Пер-
вое слагаемое той же части того же уравнения описывает ак-
554
Глава 7
тивнып транспорт фосфата в клетку; кинетика этого процесса
характеризуется наличием режима насыщения. Уравнение ма-
териального баланса по внутриклеточному фосфату [уравнение
(7.96)] является вариантом уравнения (7.67); здесь учтено, что
скорость его образования равна разности между скоростью
транспорта фосфата в клетку и скоростью включения фосфата
в компоненты клетки. В уравнении скорости образования про-
дуктов принято во внимание ингибирование процесса фосфатом
в том виде, в каком этот эффект учитывался ранее при моде-
лировании репрессии фермента фосфатазы.
Таблица 7.5. Значения параметров в структурированной модели кинетики
биосинтеза алкалоидов в С. purpurea^
Соответствующий дове-
рительный предел
Параметр
Размерность
Значение
kl сутки-1 0,5 0,058
k2 л/г в сутки 0,016 0,0024
kz сутки-1 0,0575 0,017
ki мг/г в сутки 6,028 1,69
Ур/IL — 0,0025 0,0024
Kt — 1,87-10"4 1,73-10~4
K2 г/л 4,29-Ю-4 6,67-10—4
Кз — 4,65-10-4 9,84-10-5
а Воспроизведено с разрешения из статьи: Pazoutova S., Vortruba J., Rehacek Z.,
A /Mathematical Model of Growth and Alkaloid Production in the Submerged Culture
of Claviceps purpurea, Biotech. Bioeng., 23, 2837 (1981).
Сплошные кривые на рис. 7.32 были вычислены с помощью
описанной модели, а значения соответствующих параметров пе-
речислены в табл. 7.5. Эта структурированная модель хорошо
отражает закономерности изменения биомассы и массы алкало-
идов во времени, в том числе наличие максимума на кривой
накопления суммы алкалоидов, но в то же время при высоких
концентрациях фосфата ее точность недостаточно высока. С по-
мощью такой модели, в частности, было показано, что макси-
мальное количество суммы алкалоидов продуцируется при на-
чальной концентрации фосфата в среде 0,17 г/л. Математичес-
кое моделирование различных методик введения фосфата (в
том числе добавления фосфата по частям в различные фазы
процесса) показало, что таким путем нельзя добиться сущест-
венного повышения выхода алкалоидов по сравнению с обыч-
ным периодическим процессом при оптимальной начальной кон-
центрации фосфата в среде. В других случаях, которые мы
рассмотрим в последующих главах, периодические процессы
с добавлением субстрата оказываются очень полезными.
Образование биомассы в культурах клеток
555
7.5.3. Моделирование кинетики образования продуктов
метаболизма на основе механизмов
молекулярных превращений; генетически
структурированные модели
Чем точнее описание кинетики в моделц отражает действи-
тельно осуществляющиеся в клетке химические события, тем
более универсальной становится наша модель. Здесь под «уни-
версальностью» мы понимаем возможность получения удовлет-
ворительных результатов в условиях, отличающихся от исполь-
зуемых в ходе разработки модели и определения ее парамет-
ров. Механизмы белкового синтеза, имеющего большое значе-
ние в производстве ферментов, гормонов и других практически
важных полипептидов, изучены настолько глубоко, что стано-
вится возможной разработка моделей на уровне молекулярных
событий и взаимодействий. Подобные модели должны быть в
высшей степени универсальными и полезными в оптимизации
параметров среды и генетической природы организма.
Материальный баланс по мРНК, образующейся при транс-
крипции данного гена G, можно выразить следующим уравне-
нием (здесь квадратные скобки служат для обозначения мо-
лярных концентраций данного компонента в клетке, выражен-
ных в молях в единице объема клетки) [27]:
d [М™* ]~ = МIG1 — kd 1мРНК] — И [мРНК] (7.98)
В этом уравнении kp — константа общей скорости транскрип-
ции, a kd—константа скорости инактивации и(или) деструкции
мРНК; допускается, что оба последних процесса имеют первый
порядок. В уравнении (7.98) учтен также коэффициент эффек-
тивности утилизации промотора ц, отражающий модуляцию
транскрипции оператором. Во многих случаях, представляющих
практический интерес, оценка т] в единицах концентраций бел-
ка-эффектора и индуктора или репрессора является наиболее
важным этапом всего моделирования.
Аналогично можно записать и уравнение материального
баланса по внутриклеточному продукту трансляции, т. е. по
исследуемому белку:
- k£ [мРНК]-ke [Р] - р [Р] (7.99)
Здесь скорость синтеза белка пропорциональна концентрации
кодирующей данный белок мРНК и эффективности ее утили-
зации на рибосомах (можно достаточно обоснованно предпо-
ложить, например, что изменение нуклеотидной последователь-
ности в связывающем рибосому центре приведет к изменению
556
Глава 7
величины £). В этом случае мы опять принимаем, что реакция
инактивации белка имеет первый порядок.
При сбалансированном росте производные от [мРНК] и
[Р] по времени равны нулю, и при этом условии из уравнений
(7.98) и (7.99) следует, что
<7ЛОО>
Это уравнение непосредственно выражает концентрацию внут-
риклеточного белка через параметры, характеризующие экс-
прессию гена и мРНК, а также стабильность белка в клетке.
Таблица 7.6. Значения3 параметров модели кинетики экспрессии генов
в Е. coli5
Размерность
Параметр
Значение
k р 2400/(233|л~2 + 78) мин-1
kq 3600а/(82,5ц,-’ + 145) а= 1 при jx>ln 2 а=ц/1п2 при р,<1п 2 мин-1
kd 0,46 мин-1
ke 0,07 мин-1
а В таблице приведены средние значения, которые могут потребовать уточнений
при расчете скорости экспрессии определенного гена и синтеза соответствующего белка.
Для удельной скорости роста ц здесь принята размерность ч—*.
“Данные из статьи: Lee S. В., Bailey J. Е., Analysis of Growth Rate Effects on
Productivity of Recombinant Escherichia coli Populations, Biotech. Bioeng., 26, 66 (1984).
В табл. 7.6 приведены характерные средние значения парамет-
ров описанной модели для синтеза белка в Е. coli, отражаю-
щие найденные зависимости параметров скоростей транскрип-
ции и трансляции от скорости роста.
Имеющиеся в настоящее время данные свидетельствуют о
том, что во многих случаях скорость экспрессии генов в Е. co-
li лимитируется стадией транскрипции. Следовательно, скорость
экспрессии регулируемого оператором гена зависит от эффек-
тивности транскрипции этого гена, которая в свою очередь оп-
ределяется совместным влиянием модулирующих агентов на
сайты оператора и связывания РНК-полимеразы. В качестве
примера рассмотрим систему промотор-оператор гена lac
Е. coli, причем основное внимание будем уделять совместному
влиянию /ас-репрессора, индуктора и ДНК на экспрессию гена
и, следовательно, на ^-галактозидазную активность [28].
Количество продуцируемого фермента пропорционально эф-
фективности утилизации промотора т), которая в свою очередь
пропорциональна вероятности того, что операторный сайт О не
Образование биомассы в культурах клеток
557
(7.101)
связан с белком-репрессором R. Таким образом,
1 J А [О]0
Здесь [О]о — общая концентрация операторных сайтов в клет-
ке Е. coli. Дробное выражение в правой части уравнения (7.101)
можно определить, если учесть взаимодействия между R, О,
другими неспецифическими сайтами ДНК (D), индуктором I;
предполагается, что все эти взаимодействия равновесны:
R+O RO KA (7.102a)
R+D RD Kb (7.1026)
R + I RI Kc (7.102b)
RI + O RIO KD (7.102r)
RI+D RID Ke (7.102д)
Оператор О может существовать в трех различных формах,
концентрации которых можно связать следующим уравнением
материального баланса:
[O]0=[O] + [RO] + [RIO] (7.103)
Аналогично уравнение материального баланса по R можно за-
писать в виде
[R]o= [R] + IRI] + [RO] + [RD] + [RIO] + [RID] (7.104)
Поскольку концентрация [D] неспецифично связывающих сай-
тов ДНК выше общей концентрации R, мы можем принять, что
[D]«[D]o (7.105)
Используя приближение (7.105), равновесную природу реакций
(7.102), уравнение материального баланса (7.104) и пренебре-
гая членами с множителем [0]о/[Д]о, нетрудно прийти к сле-
дующему уравнению:
ГР1 _ „___________1 + Кв [Р]о + Кс [Ц(1 + Ke [DJo)______
1 J A 1 + KB[D]O + KC[I] (1 + Ke[D]0) + (Ka4-KcKd[I])[R1o
(7.106)
Опубликованные в литературе экспериментальные данные поз-
воляют приближенно, с точностью не ниже одного порядка ве-
личин, оценить параметры этого уравнения [28]:
[D]o=4-IO-2 М
Кв=1-103 М-1
Kd = 2*109M-1
Ка=2-1012 М-1
Кс=Ы07М-!
Ке-1,5-104 М-1
(7.107)
Оценка параметров этой математической модели требует
обширных экспериментальных данных, но вместе с тем модели-
558
Глава 7
рование с учетом молекулярных взаимодействий имеет и свои
преимущества. Так, ее параметры имеют строгое физико-хими-
ческое толкование и часто могут быть определены в отдельных
экспериментах in vitro. Более того, модель такого уровня обла-
дает особой характеристикой, называемой генетической струк-
турой. Под генетической структурой в данном случае понима-
ется четкая зависимость определенных параметров модели от
генетической природы; так, изменение нуклеотидной последова-
тельности /яс-оператора должно сопровождаться изменением
параметров Ка и Ко, характеризующих взаимодействия 1ас-
оператора с белком-репрессором. По мере развития и углубле-
ния наших знаний о механизмах взаимодействий в системе
ДНК—белок, вероятно, мы сможем учитывать влияние опреде-
ленных изменений нуклеотидной последовательности на соот-
ветствующие константы связывания ДНК с белком расчетным'
путем. Тогда разработанная на молекулярном уровне кинети-
ческая модель позволит осуществлять картирование нуклеотид-
ной последовательности, отражающее продуктивность популя-
ции клеток. Потенциальные возможности такого подхода для
рациональной количественной оптимизации генетической моди-
фикации организма (или, по меньшей мере, клонированных
участков ДНК) очевидны.
В качестве примера рассмотрим применение уравнения
(7.106) для расчета влияния концентрации репрессора на ско-
рости индуцированной и неиндуцированной транскрипции 1ас-
промотора-оператора хромосомы Е. coli. Необходимые экспери-
ментальные данные, в том числе внутриклеточные концентрации
репрессора для каждого из мутантов, известны для му-
тантных штаммов i~, iq и isq Е. coli. Эти величины можно не-
посредственно использовать при моделировании; они не влия-
ют на другие параметры модели. Как показано в табл. 7.7, мо-
дель удовлетворительно отражает влияние внутриклеточной
концентрации репрессора как в присутствии индуктора, так и
без индуктора. В приведенной в конце главы литературе можно,
найти другие примеры моделирования с учетом регуляции на
молекулярном уровне.
7.5.4. Кинетика образования продуктов жизнедеятельности
филаментозными организмами
Кинетика образования продуктов метаболизма и утилизации
субстрата плесневыми грибами и другими филаментозными ор-
ганизмами обычно очень сложна. Типичным примером может
служить биосинтез пенициллина в периодическом процессе. Как
показано иа рис. 7.30, существуют три различных режима ус-
воения субстрата. В течение первых 20 ч происходит быстрая
Образование биомассы в культурах клеток
559
утилизация сахара, сопровождающаяся активным ростом пле-
сени. В стационарной фазе утилизация субстрата замедляется,
а скорость образования пенициллина достигает максимума. За-
тем скорость усвоения сахара опять возрастает и удерживается
на высоком уровне вплоть до его истощения.
Аналогичная картина наблюдается и в ходе биосинтеза
других антибиотиков, хотя каждый процесс имеет и свою спе-
цифику. В производстве стрептомицина, например, максималь-
Таблица 7.7. Вычисленные и найденные экспериментальным путем величины
^-галактозидазной активности, регулируемой системой /ас-промотор-оператор,
для дикого типа Е. coli и трех мутантов (i~, i4, isq) с различными
внутриклеточными концентрациями /ас-репрессора R ([R]o=2-lO~8 М)а
Генотип
Концентрация
репрессора
P-Галактозидазная активность®
Индуктор Литература
Вычислено Найдено
0 — 113 100—140 [331
1“ 0 + 113 100—130 [331
i+ [Ri 0 0,1 <0,1 [331
i+ [R 0 + 100 100 1331
i« 10 R 0 — 0,011 0,01—0,014 [34|
1ч 10 R 0 + 62 65 [351
50 lR 0 — 0,002 0,003—0,004 [34 J
Is Q 50 [R’ 0 + 23 25 [35]
аВоспроизведено с разрешения из статьи: Bailey J. Е., Hjortso М., Lee S. В.,
Srienc F., Kinetics of Product Formation and Plasmid Segregation in Recombinant Mic-
robial Populations, Annals N.Y. Acad. Sci., 413, 71 (1983).
® Относительно индуцированного дикого типа i+ (100%) при концентрации индук-
тора 10—3 М.
пая скорость синтеза антибиотика достигается (по сравнению
с пенициллином) позднее, а заключительная фаза ускоренной
утилизации сахара не наступает вообще. Процесс продуциро-
вания антибиотика окситетрациклина филаментозным микроор-
ганизмом Streptomyces rimosus интересен тем, что здесь резуль-
тат всего процесса зависит и от морфологии организма.
В табл. 7.8 суммированы основные стадии и особенности этого
процесса.
Ниже перечислены некоторые особенности образования
продуктов жизнедеятельности, характерные только для фила-
ментозных организмов и найденные экспериментальным путем:
1. Максимальный выход продукта метаболизма достигается в
определенной фазе периодического процесса при условии
оптимальной начальной концентрации субстрата. Повышение
концентрации субстрата приводит к ускорению роста, но
при этом субстрат расходуется в основном на образование
560
Глава 7
Таблица 7.8. Основные фазы периодического процесса биосинтеза
окситетрациклина в погруженной культуре Streptomyces rimosus*
Фаза Особенности
1. Лаг-фаза
2. Рост первичного ми-
целия
3. Фрагментация пер-
вичного мицелия
4. Рост вторичного ми-
целия
5. Стационарная фаза
Продолжительность около 90 мин, если объем
инокулята мал; метаболическая активность не
обнаруживается
Продолжительность от 10 до 25 ч в зависимости
от инокулята; интенсивны дыхание, синтез нук-
леиновых кислот и другие метаболические про-
цессы; концентрация пировиноградной кисло-
ты максимальна; антибиотик не образуется
Продолжительность около 10 ч; рост мицелия
прекращается, интенсивность дыхания и синте-
за нуклеиновых кислот снижается, концентра-
ция пировиноградной кислоты падает до очень
низкого уровня
Продолжительность около 25 ч; в этой фазе мас-
са вторичного мицелия в 2—4 раза превышает
биомассу в фазе 2; гифы мицелия значительно
утоньшаются; быстро возрастает скорость син-
теза антибиотика, возобновляется синтез нук-
леиновых кислот, а интенсивность дыхания про-
должает снижаться; сахара и азот (аммиак)-
быстро истощаются; концентрация пировино-
градной кислоты может немного повыситься.
В этой фазе рост клеток прекращается, а метабо-
лическая активность снижается; антибиотик еще
какое-то время синтезируется, но скорость син-
теза сравнительно низка
а Luedeking R., in Biochemical and Biological Engineering, Blakebrough N. (ed.)„
vol. 1, p. 208, Academic Press, Inc., New York, 1967; см. также Doskacil Sikyta B.r
Kasparova J., Poskocilova D., Zajicek J., J. Gen. Microbiol., 18, 302 (1958).
биомассы, а метаболит синтезируется в очень малых кон-
центрациях. Если же концентрация субстрата слишком низ-
ка, то образующееся незначительное количество биомассы не
в состоянии синтезировать антибиотик в достаточных кон-
центрациях даже в период максимальной скорости его био-
синтеза.
2. Образование продукта метаболизма максимально при мини-
мальном ветвлении активно растущих гиф инокулята. С дру-
гой стороны, уменьшение степени ветвления повышает про-
должительность лаг-фазы и, следовательно, длительность
всего процесса. Очевидно, должна существовать некоторая
оптимальная степень ветвления гиф инокулята.
3. Поскольку плесени и другие мицелиальные микроорганизмы
являются аэробами, казалось бы, что максимальная интен-
сивность перемешивания погруженной культуры должна спо-
Образование биомассы в культурах клеток
561
собствовать эффективному переносу кислорода к плесени и,
таким образом, росту последней. В случае пенициллина, од-
нако, было установлено, что максимальные выходы антибио-
тика достигаются при некоторой промежуточной интенсив-
ности перемешивания. Этому факту было дано несколько
объяснений. Известно, что на морфологию плесени влияют
механические воздействия; возможно, что более энергичное
)
Гиры
Ассимиляция м Истощение
метионина и N глюкозе/
глюкозы Ьз
$ ф= Индукция эндогенным
N метионином
N А----Натадолитная
pi репрессия глюкозой
Фермент (рерменты)^синтези-
} ЦОС
РИС. 7П2.1. Схема морфологически различных форм плесени С. acremonium,
их превращений и синтеза цефалоспорина С (ЦСС). [Воспроизведено с раз-
решения из статьи: Matsumura М., Imanaka Т., Yoshida Т., Taguchi Н., Mode-
ling of Cephalosporin С Production and Its Application to Fed-Batch Culture,
J. Ferment. Tech. (Japan), 59, 115 (1981).]
перемешивание способствует ветвлению гиф, которое, как
считается, снижает скорость биосинтеза антибиотика. Не
исключено также, что существует некоторая оптимальная
скорость подачи кислорода, но ввиду сложности системы
подтвердить это предположение экспериментальным путем
чрезвычайно трудно. (Влияние перемешивания мы рассмот-
рим подробнее в гл. 8.)
Пример 7.2. Морфологически структурированная модель кинетики био-
синтеза цефалоспорина С [14]. Экспериментальное изучение биосинтеза анти-
биотика цефалоспорина С (ЦСС) в глубинной культуре плесени Cephalospo-
rium acremonium показало, что в ней существуют три морфологически раз-
личные формы микроорганизмов: гифы (h), набухшие фрагменты гиф (s)
и артроспоры (а). В зависимости от состава среды эти формы могут взаимо-
превращаться, как это схематично показано на рис. 7П2.1. На этом же ри-
сунке отмечено, что ЦСС преимущественно продуцируется набухшими фраг*
36—537
562
Глава 7
ментами гиф. Скорость синтеза ЦСС непосредственно связана с активностью
соответствующих ферментов, которые индуцируются внутриклеточным метио-
нином и подавляются глюкозой.
Эти сведения послужили основой для приведенной ниже структурирован-
ной модели биосинтеза ЦСС в периодической культуре:
Рост и дифференциация плесени:
dt - K^g хн $\g,m)xh kDxh dts =$(g,m)xh у (g)xs —k^xs (7П2.1) (7П2.2)
dta -y(g)xs kDxa (7П2.3)
Здесь
№.'") = (*н + -к^)(~к7<7Г) (7П2.4)
У (g) = ^21 + ^22 ( <7П2-5>
Обратите внимание на то, что в этой модели общая биомасса разделена на
три морфологически различных типа, наблюдающиеся в эксперименте. Массо-
вые концентрации глюкозы и метионина в среде обозначены символами g
и m соответственно.
Модель утилизации субстрата подразумевает, что глюкоза и метионин
усваиваются только морфологическими формами h и s.
Утилизация глюкозы и метионина:
dg _________IhngXh_______VmgXs
dt ^h/g (g + Kg) g + Kg
dm j r j т
~dT = ~Uhxh — UsXs
(7П2.6)
(7П2.7)
где
• Umhm
h~ тфКщ
Umsfn
m + Km
(7П2.8)
С помощью уравнений (7П2.1) — (7П2.8) были рассчитаны характеристики
роста клеток и утилизации субстрата в периодическом процессе, причем для
лучшего соответствия расчетных данных результатам эксперимента парамет-
ры модели уточняли методом симплексной оптимизации.
Поскольку в регуляции экспрессии ферментов, синтезирующих ЦСС, осо-
бую роль играет внутриклеточный метионин, то кинетическая модель хими-
чески структурирована таким образом, чтобы можно было раздельно оце-
нить влияние концентраций метионина пць, tnts и m,a (во всех случаях в чис-
ле микромолей в 1 г клеток) в морфологических формах ft, s и а соответст-
венно. При этом использовались следующие уравнения материального балан-
са по метионину:
Образование биомассы в культурах клеток
563
Внутриклеточный метионин
, ушах „
= '4 + mih (7П2.9)
(7П2.10)
= — ksamia + V (f—* (tnis — mtа) (7П2.11)
up Ла
При выводе этих уравнений принималось, что скорость биосинтеза метионина
обратно пропорциональна его внутриклеточной концентрации и что присутст-
вие глюкозы в среде повышает скорость утилизации метионина в белковом
синтезе. Средняя внутриклеточная концентрация метионина равна
nii
xhmih + xsmis + xamia
(7П2.12)
x
где
X — Xh + Xs + Xa
(7П2.13)
Теперь модель описывает все основные стороны процесса: рост клеток, ути-
лизацию субстрата и концентрацию внутриклеточного метионина. Сравнением
найденных и расчетных значений mi можно определить новые параметры, по-
являющиеся в уравнениях (7П2.9) — (7П2.11). Эти параметры приведены во
втором столбце (группа 2) табл. 7П2.1.
В модели допускается, что скорость синтеза антибиотика зависит от ак-
тивности синтезирующих ЦСС ферментов е (выраженной в мг ЦСС в час
в расчете на 1 г клеток), которая описывается следующим уравнением:
Синтез ферментов
de 1 / VmpmicXc \ / xi, \
— = — ( mii + ^ (Л,-») е (7П2.14)
где
1 + (x/an) g
1 + (х/ап) (1 4- n) gn
(7П2.15)'
Параметр Q является мерой индуцируемой глюкозой катаболитной репрес-
сии. Подстрочный индекс (/—Ь) указывает на развитие процесса в период
t—tr, это отражает наличие лаг-фазы между индукцией и экспрессией гена.
Наконец, скорость образования ЦСС в этой модели описывается уравнением
Образование антибиотика
JP=exs~kPDp (7П2.16)
Параметры уравнений (7П2.15) и (7П2.16) приведены в третьем столбце
(группа 3) табл. 7П2.1.
На рис. 7П2.2 изображены экспериментальные (точки) и расчетные
(кривые) данные, отражающие клеточный рост, утилизацию субстратов и об-
разование антибиотика в периодическом процессе. Обратите внимание на тоь
36*
Таблица 7П2.1. Параметры математической модели, описывающей биосинтез цефалоспорина С (ЦСС)а
Группа 1 Группа 2 Группа 3
(параметры роста клеток и (параметры накопления эндогенного (параметры образования ферментов,
утилизации субстрата) метионина) синтезирующих ЦСС, и биосинтеза ЦСС)
pm = 0,069 ч-1 Итахй,5уп = 8,7 мкмоль Met/(г клеток-ч) VmE=l,2 мг ЦСС/(г клеток-ч)
Kg = 0,45 г клеток/г Glu £зл = 7,1 ч-1 Kg =12,0 мкмоль Met/r клеток
7<g=0,05 г GIu/л Vrmaxh,utii=48,6 ч-1 4г=13,0 ч
feu = 0,015 ч-1 Vmaxs,syn = 6,6 мкмоль Met/(г клеток-ч) к=0,29
£12=0,024 ч-1 £зз= 1,4 ч-1 а=0,15
£2i = 0,004 4-’ Vmax3,util = 2,2 ч-1- Т|= 1,6
£22=0,028 Ч-* £за=7,8 ч-1 л=5,1
Km—0,001 г Met/л £ed=0,38 ч~*
vm=0,023 г Glu/(г клеток-ч) £pd=0,012 ч—1
C/mh=0,0012 г Met/(r клеток-ч)
Um,= 0,0098 г Met/(г клеток-ч)
£о=0,014 ч“*
в Восппонзведено с разрешения из статьи: Matsumura М., Imanaka Т., Yoshida Т., Taguchi Н., Modeling of Cephalosporin С Production
and Its Application to Fed-Batch Culture, J. Ferment. Tech. (Japan), 59, 115 (1981).
Образование биомассы в культурах клеток
565
РИС. 7П2.2. Расчетные (сплошные кривые) и экспериментальные (точки) дан-
ные, отражающие развитие процесса биосинтеза цефалоспорина С (ЦСС)
в периодической культуре С. acremonium (символом Mi обозначены данные
для внутриклеточного метионина). [Воспроизведено с разрешения из статьи:
Matsumura М„ Imanaka Т., Yoshida Т„ Taguchi Н., Modeling of Cephalospo-
rin С Production and Its Application to Fed-Batch Culture, J. Ferment. Tech.
(Japan), 59, 115 (1981).]
что концентрация набухших фрагментов гиф сначала возрастает, а затем
снижается, а артроспоры образуются в основном на поздних стадиях процес-
са. Модель очень хорошо отражает своеобразный характер изменения во вре-
мени концентрации внутриклеточного метионина. В то же время разработка
и использование такой сложной модели с параметрами, определение которых
очень трудоемко, оправданны только в том случае, если эта модель приме-
нима для изучения культивирования в достаточно широком диапазоне усло-
вий. Описанная модель вполне отвечает этому требованию. Так, на рис.
7П2.3 приведены рассчитанные с помощью этой модели зависимости коли-
чества синтезируемого ЦСС (в единице объема) а) от начальной концент-
Началбная концентрация
глюкозы, г/л
а
Время введения метионина, ч
РИС. 7П2.3. Моделирование влияния начальной концентрации глюкозы (а)
и времени введения метионина (б) на синтез ЦСС. [Воспроизведено с разре-
шения из работы: Matsumura М., Imanaka Т., Yoshida Т„ Taguchi Н., Modeling
of Cephalosporin С Production and Its Application to Fed-Batch Culture, J Fer-
ment. Tech. (Japan), 59, 115 (1981).]
566
Глава 7
рации глюкозы и б) от времени введения метионина в культуру. В полном
соответствии с экспериментальными данными на первой кривой имеется чет-
ко выраженный максимум; вторая кривая свидетельствует о целесообразности
введения метионина на ранних стадиях процесса, что также подтверждается
экспериментом. Эта модель помогла также предсказать, что в периодическом
процессе одновременное введение метионина и глюкозы способствует значи-
тельному повышению выхода ЦСС, в то время как раздельное (во времени)
введение этих питательных веществ менее эффективно. Последующие экспе-
рименты подтвердили и эти расчетные данные.
7.6. Сегрегированные модели кинетики клеточного
роста и образования продуктов метаболизма
Обычно популяции клеток гетерогенны в том смысле, что
составляющие популяцию индивидуальные клетки различаются
по размерам, возрасту, скорости роста и другим свойствам.
Чтобы модель кинетики роста популяции клеток в большей
степени приближалась к реальной ситуации, она должна отра-
жать и распределение клеток по различным категориям, обла-
дающим разными биохимическими активностями. Другим пре-
имуществом такого подхода к моделированию является воз-
можность установления связи между особенностями клеточного
цикла и другими свойствами изолированной клетки, с одной
стороны, и общими характеристиками популяции — с другой.
Основной недостаток сегрегированных моделей обусловлен их
математической сложностью. В этой связи в настоящем разде-
ле основное внимание мы будем уделять распределению по
возрастам и коррелированным распределениям популяций кле-
ток, размножающихся бинарным делением. Вывод уравнений
материального баланса популяций, использующихся в сегреги-
рованных моделях, и решение уравнений модели для случая
некоторых микробных систем приведены в работах [29—31].
Частотная функция возраста клеток Ща) позволяет опреде-
лить относительное количество клеток в возрасте от а до a + da
как W(a)da. В этом разделе символом а мы будем обозначать
возраст клетки, считая, что нулевой возраст соответствует
только что возникшим в результате деления клеткам, а деля-
щиеся клетки имеют возраст tD, равный продолжительности
клеточного цикла или времени удвоения индивидуальной клет-
ки. Для сбалансированного экспоненциального роста популя-
ции основанное на возрасте клеток уравнение баланса популя-
ции будет выглядеть следующим образом:
-d-Wd{aa) = — pF (а) (7.108)
Здесь р — общая удельная скорость роста популяции. По-
скольку все клетки популяции имеют возраст от б до tD, то F
Образование биомассы в культурах клеток
567
должно также удовлетворять условию
f W(d)da=] (7.109)
о
В результате деления не всегда образуются две жизнеспо-
собные клетки. Причиной может быть старение клеток или их
гибель, а также рост содержащей неустойчивые плазмиды по-
пуляции в селективной среде. Если символом 0 обозначить долю
жизнеспособных дочерних клеток, образующихся в результате
бинарного деления жизнеспособной клетки, то уравнение ба-
ланса, связывающее количества делящихся и новых жизнеспо-
собных клеток, можно записать в следующей форме:
№(О) = (2 —0)TOz>) (7.110)
Решение уравнения (7.108) с учетом условий (7.109) и
(7.110) дает
(7Ш>
Из уравнений (7.110) и (7.111) следует приведенное ниже вы-
ражение для зависимости между скоростью роста ц и време-
нем генерации клетки:
и^.1п(Я.Г.е)_ (7.112)
Поскольку среднее время удвоения популяции (средняя продол-
жительность клеточного цикла) to равна
iz> = ln2/p. (7.113)
то
Время генерации отдельной клетки _ /д _ In (2 — 0) 114)
Время удвоения популяции Id In 2
Таким образом, если 0 не равно нулю, то to меньше to. Это
относительно простое соотношение иллюстрирует очень важный
общий принцип: общие кинетические свойства популяции не
обязательно должны соответствовать кинетическим парамет-
рам на уровне индивидуальной клетки.
Во многих случаях для расчета различных частотных функ-
ций и усредненных свойств популяции, имеющих конкретное
физическое содержание, можно использовать функцию плот-
ности возраста, которую нельзя непосредственно определить.
Предположим, например, что синтез некоторых компонентов
клетки или продуктов ее жизнедеятельности осуществляется с
постоянной скоростью в определенный период клеточного цикла
568
Глава 7
от ах до Яг- Установлено, в частности, что в эукариотах ДНК
синтезируется только в периоде S. Сообщалось также, что не-
которые ферменты синтезируются рядом организмов только в
течение очень узкого временного интервала клеточного цикла.
В таких случаях скорость образования продукта жизнедеятель-
ности клеток можно выразить уравнением
V(a)da
ai
Для рассмотренной выше ситуации получим
/ о________________________А \
Г1>2 = -—— [е-^1—е-^2]
(7.115)
(7:116)
Для любого количественного параметра клеточного роста х
(например, клеточной массы, содержания белка, объема, воз-
можно также содержания определенного продукта метаболиз-
ма), непосредственно связанного с возрастом клеточного цикла
а (иными словами, если а можно выразить или определить как
функцию от х), частотную функцию для а можно преобразо-
вать в частотную функцию для х в соответствии с уравнением
Wx(x)=W(a(x))
da (х)
(7.117)
Предположим, например, что в течение клеточного цикла масса
отдельной клетки возрастает во времени по линейному закону:
т{а) = ak
(7.118)
так, что
т — т0
(7.119)
Если в уравнение (7.117) подставить значения W из (7.111)
и а из (7.119), то получим
b(m_mo)
К (т) = (-ndr) 4г ехр
(7.120)
Здесь опять-таки следует подчеркнуть различие между кине-
тикой роста отдельной клетки и популяции клеток. Масса
отдельной клетки может возрастать в соответствии с уравне-
нием (7.118) (такая зависимость для некоторых организмов
была обнаружена и экспериментально), в то время как общая
масса популяции этих же клеток возрастает экспоненциально,
например в соответствии с уравнением (7.37), описывающим
Образование биомассы в культурах клеток 569
кинетику сбалансированного роста популяции в периодическом
процессе. Одно из преимуществ сегрегированного подхода к
моделированию кинетики клеточного роста заключается в
возможности выяснения непосредственной взаимосвязи между
параметрами кинетики и регуляции роста индивидуальной
клетки и параметрами кинетики роста популяции клеток.
Если допустить, что плотность клеточного вещества посто-
янна, то уравнение (7.120) можно также интерпретировать как
функцию распределения клеток по объему, которое можно оп-
ределить экспериментально с помощью счетчика Коултера или
методами проточной цитометрии по рассеянию света (см.
гл. 10). На практике, однако, чаще измеряют количественные
характеристики популяции в целом. Эти характеристики можно
вычислить с помощью соответствующих частотных функций и
уравнения
х = J xWx(x)dx (7.121)
о
В других подходах к моделированию гетерогенность попу-
ляции клеток можно рассматривать с иных точек зрения. Пред-
ложено несколько математических моделей образования про-
дуктов жизнедеятельности в популяциях микроорганизмов, учи-
тывающих длительные эффекты старения. В модели Пинг Шу
удельная скорость образования продукта жизнедеятельности
q
клеток выражается как SAe-*/9, где 0 — общее] время пре-
i=i
бывания клеток в реакторе [32]. Тогда концентрацию продукта
жизнедеятельности в момент времени t периодического процес-
са можно вычислить по уравнению
2 Ate~kiQ'dQ' Ы0 (7.122)
«=1 /
Здесь 7И(0, t)—частотная функция в момент времени t кон-
центрации клеточной массы, время пребывания которой в ре-
акторе равно 0. Поскольку клеточное вещество, имеющее в мо-
мент t возраст 0, образуется за время t—0, отсюда следует,
•что
M(Q,t)=rAt — 0) (7.123)
Эта модель отличается известной универсальностью и пригод-
на для описания различных механизмов накопления продуктов
метаболизма, наблюдаемых в периодических микробиологичес-
ких процессах.
570
Глава 7
7.7. Кинетика тепловой гибели
клеток и спор
Активность клеток, спор и вирусов в воздухе или жидких
средах может снижаться в результате их разрушения (под
воздействием тепла, радиации, химических агентов или меха-
нических сил), при механическом отделении (фильтрованием
или центрифугированием) или за счет ингибирования (при пе-
реохлаждении, высушивании, обезвоживании, воздействии хи-
мических реагентов). Жидкости стерилизуют в основном путем
нагревания (в промышленности) или хлорирования (для быто-
вых нужд), а для стерилизации воздуха применяют главным
образом фильтрование. В двух последних процессах большую
роль играют эффекты массопередачи, рассматриваемые в сле-
дующей главе; этот раздел посвящен кинетике инактивации
клеток и спор.
На рис. 7.33 изображены результаты экспериментального
изучения воздействия повышенной температуры на вегетатив-
ные клетки и споры. В общем случае по сравнению со спорами
клетки погибают при нагревании гораздо быстрее. Это и понят-
но, если учесть, что образование эндоспор, как мы уже отмеча-
ли ранее, представляет собой защитный механизм, позволяю-
щий некоторым клеткам переносить неблагоприятные условия.
Тепловая обработка приводит также к инактивации вирусов и
бактериофагов, поэтому использующаяся в микробиологической
промышленности тепловая стерилизация снижает как числен-
ность популяции жизнеспособных микроорганизмов, так и кон-
центрацию (титр) вирусов в подаваемом в реактор потоке пи-
тательных веществ.
Прежде чем перейти к изучению уравнений, описывающих
скорость снижения численности популяции в процессе стерили-
зации, необходимо сделать ряд замечаний. Гибель каждой кон-
кретной клетки, по-видимому, обусловливается термической де-
натурацией одного или нескольких жизненно важных для этой
клетки белков, например ферментов. Кинетика подобных ко-
оперативных превращений столь сложных молекул может изме-
няться во времени самым неожиданным образом. К тому же
скорость молекулярных процессов, приводящих в конце концов
к гибели клетки,, зависит от состава среды, в том числе и от
концентрации растворителя. Например, как показано в
табл. 7.9, температура, при которой наступает коагуляция
(денатурация с последующей интенсивной необратимой сшив-
кой молекул денатурированного белка) альбумина яич-
ного белка, повышается с уменьшением содержания в системе
воды.
Образование биомассы в культурах клеток
571
Доля вй/онгившиа: Кле/тмн
время нагревания, мин
РИС. 7.33. Результаты экспериментального определения скорости тепловой
гибели Е. coli и тепловой инактивации вегетативных спор Bacillus subti-
lis и вегетативных спор В. stearothermophilus. [Воспроизведено из ра-
бот: а) Аиба Ш., Хемфри А., Миллис Н., Биохимическая технология и аппа-
ратура.— М.: Пищевая промышленность, 1975; б, в) Burton Н„ Jayne-Wil-
liams D., Sterilized Milk, in Recent Advances in Food. Science, vol. 2: Proces-
sing, Hawthorn J., Leitch J. M. (eds.), p. 107, Butterworths & Co., Publishers
Ltd., London, 1962.]
a — скорость гибели E. coli в буферном растворе; б — скорость инактивации
спор Bacillus subtilis', в — скорость инактивации спор Bacillus stearothermo-
philus.
Эти данные и ряд других экспериментально обнаруженных
фактов свидетельствуют о том, что в случае сравнительно обез-
воженных организмов и структур (например, вирусов или
спор) их инактивацию правильнее рассматривать как двухэтап-
ный процесс, при котором сначала происходит гидратация, а
затем, возможно, и денатурация. Такому механизму отвечает
572
Глава 7
Таблица 7.9. Зависимость температуры коагуляции альбумина от содержания
водыа
Содержание воды, % Ориентировочная Содержание воды, Ориентировочная температура % температура коагуляции, °C 70 коагуляции, °C
50 56 5 149
25 76 0 165
15 96
а Данные из работы: Frobisher М., Fundamentals of Microbiology, р. 259, W. В. Sa-
unders Company, Phila., 1968.
зависимость скорости гибели от относительной влажности, об-
наруженная в случае бактериофагов [34]. Совместное влияние
нескольких инактивирующих факторов не обязательно носит
аддитивный характер; так, совместные обезвоживание и тепло-
вая обработка могут быть менее эффективными, чем можно
было бы предположить, судя по эффективности каждого из
этих факторов в отдельности.
Как мы упоминали выше, популяция состоит из множества
клеток различного возраста. Природа клеточной стенки и отно-
сительная важность любого из путей метаболизма зависят от
возраста, данной клетки и культуры в целом. Следовательно,
устойчивость клеток к тепловой или индуцированной иными
факторами инактивации будет зависеть от их истории, т. е. от
довольно неопределенного фактора, который далеко не всегда
удается охарактеризовать количественно. В частности, клетки
популяции, находящейся в фазе экспоненциального роста, име-
ют относительно проницаемые стенки, что способствует эффек-
тивному обмену растворенными веществами между внутрикле-
точным объемом и средой. Если эти растворенные вещества
влияют на устойчивость белков, то можно ожидать, что их
присутствие окажет влияние и на рост клеток в экспоненци-
альной фазе. Поэтому наиболее надежные данные о поведении
популяции могут быть получены только в результате экспери-
ментов в условиях, максимально приближенных к условиям
предполагаемого практического проведения процесса.
Анализ кинетики скорости гибели клеток мы начнем с заме-
чания о том, что линейная зависимость логарифма доли остав-
Образование биомассы в культурах клеток
573
шихся в живых клеток от времени (рис. 7.33) указывает на
первый порядок процесса снижения численности популяции
жизнеспособных клеток п:
= (7.124)
Отсюда следует, что при постоянном kd
n(t)—nae~kJ (7.125)
Здесь по — концентрация спор или вегетативных клеток при
t = 0. Наклон прямой полулогарифмической зависимости равен
— kd; график зависимости In^d от обратной температуры также
представляет собой прямую, поэтому зависимость kd от темпе-
ратуры может быть выражена уравнением аррениусовского
типа:
kt = kd„e-EJ^ (7.126)
Для многих спор и вегетативных клеток величина параметра
Ed лежит в диапазоне от 50 до 100 ккал/моль. Другой ранее
применявшийся параметр Dr, равный 2,303/^d и называемый
временем децимальной редукции (десятичным коэффициентом
снижения численности популяции), представляет собой не что
иное, как время, в течение которого численность популяции
жизнеспособных клеток снижается в 10 раз.
Приведенные выше кинетические уравнения справедливы
только при том условии, что количество спор или клеток в по-
пуляции достаточно велико с точки зрения статистики
(разд. 7.1). По мере снижения числа клеток или спор (и) ве-
роятность отклонений от величин, предсказываемых указанны-
ми уравнениями, возрастает, поскольку, например, при нор-
мальном распределении стандартное отклонение возрастает
пропорционально 1/п. Из предыдущего уравнения детермини-
стической модели следует, что оставшаяся жизнеспособной доля
популяции (если допустить, что в данных летальных условиях
роста клеток не происходит) определяется уравнением
Доля жизнеспособных =4 =* e~kdl (7.127)
клеток
Если судьба каждого организма не зависит от других организ-
мов, если организмы не самовоспроизводятся и если летальные
574
Глава 7
условия одинаковы для каждого организма, то стохастический
анализ процесса стерилизации показывает, что вероятность на-
личия в популяции N жизнеспособных организмов в любой мо-
мент времени t равна [33]
Рк(t> = W-V)iwi (e~V)A'(1 )<Л,°~Л,> (7Д28)
Здесь No — численность жизнеспособных организмов в стери-
лизуемой жидкости непосредственно, перед началом стерилиза-
ции.
В этОхМ уравнении параметр kd имеет тот же смысл, что и
константа скорости в уравнении (7.124); в стохастической мо-
дели kd можно интерпретировать как величину, обратную сред-
ней продолжительности жизни организма. По мере того как
число организмов снижается до некоторой сравнительно малой
величины, допущение о гомогенной популяции, достаточно пол-
но описываемой одним параметром kd, становится все менее и
менее обоснованным. Так, приведенные на рис. 7.34 данные,
характеризующие поведение Staphylococcus aureus в нейтраль-
ном фосфатном буфере, свидетельствуют об отклонении от по-
ведения, предсказываемого стохастическим распределением,
в положительную сторону; следовательно, большей устойчи-
востью к тепловой обработке отличается небольшая доля попу-
ляции.
Иногда допускается, чтобы после стерилизации оставалось
довольно значительное количество жизнеспособных организмов
(так, в пастеризованном молоке высшего качества концентра-
ция живых бактерий не должна превышать 30 000 клеток в
1 мл). В других ситуациях, например при выращивании чистых
культур, к стерилизации предъявляются значительно более
жесткие требования. В таких случаях важно оценить вероят-
ность гибели популяции, т. е. вероятность инактивации всех
организмов. Подставляя в уравнение (7.128) N = 0, полу-
чаем
Вероятность гибели = Ро(t)= 1—(1—e~kdi )N° (7.129)
Отсюда следует, что вероятность выживания по меньшей, мере
одного организма составляет
1— ро(0 = (1— е~^)Н0 (7.130)
Обычно A^o^l, поэтому
Образование биомассы в культурах клеток
575
О 2 4 6 8 f(HZ К Ю18 20 22 24
Время нагрела кия, мия
РИС. 7.34. Тепловая инактивация S. aureus. [Воспроизведено с разрешения
из статьи: Walker G. С., Harmon L. G., Thermal Resistance of Staphylococcus
aureus in Milk, Whey, and Phosphate Buffer, Appl. Microbiol., 14, 584 (1966).]
1—po(/)=l— e~Nt (7.131)
где
(7.132)
Величину (1—Po) можно интерпретировать как долю стерили-
заций, которые не должны приводить к полной гибели всех
организмов.
Изучив метаболизм клетки, мы узнали, что живые клетки
часто располагают несколькими альтернативными путями ус-
воения энергии и осуществления биосинтеза. Наличие таких
резервных систем повышает шансы организма на выживание
в неблагоприятных условиях. Статистический анализ показы-
вает, что с учетом существования резервных систем числен-
ность популяции должна составлять
576
Глава 7
ИЖ)]=Щ1 —{1 —exp[EdU)fl}')“ (7.133)
где со — число типов жизненно важных субклеточных структур;
г— число единиц каждой структуры в каждом организме;
Ad (0—средняя удельная скорость разрушения структуры каж-
дого типа.
Обратите внимание на то, что это уравнение можно
выразить через ряд убывающих экспоненциальных функций
и, следовательно, в полулогарифмических координатах боль-
шие участки соответствующей графической зависимости не
должны представлять собой прямую линию.
Самый простой подход к анализу популяции из т различ-
ных типов клеток (спор или вирусов) также сводится к допу-
щению о независимости видов организмов; тогда суммарная
популяция жизнеспособных организмов будет равна сумме ин-
дивидуальных популяций, выражаемых уравнениями типа
(7.125) с соответствующими индивидуальными коэффициентами
kdi'.
т т
= nt (0 = 2 V •“ (7-134)
1 = 1 Z=1
И снова график этой зависимости в полулогарифмических ко-
ординатах не должен представлять собой прямую. Далее, зна-
чения индивидуальных параметров kat не всегда известны; не-
которые методы оценки верхнего и нижнего пределов значений
n(t)/nQ по начальным данным обсуждены Хатчинсоном и Лас-
сом*. Достоверность таких оценок зависит от точности исход-
ных данных о гибели популяции и требует определения второй
производной d2n]dt2 при t = Q.
7.8. Заключение
Как мы упоминали во введении к настоящей главе, конечной
целью изучения кинетики клеточного роста является математи-
ческое количественное описание совместного влияния генетиче-
ской природы организмов и среды на скорости процессов, осу-
ществляющихся в популяции этих организмов. В настоящее
* Hutchinson Р., Luss D., Lumping of Mixtures with Many First-Order
Reactions, Chem. Eng. J., 1, 129 (1970).
Образование биомассы в культурах клеток 577
время мы располагаем целым рядом частных моделей, одни из
которых учитывают незначительные генетические изменения,
другие — небольшие отклонения в составе среды, а третьи опи-
сывают гетерогенность клеток культуры. Учитывая ограничен-
ную применимость каждой из таких моделей, специалист в
области биохимической технологии должен прежде всего четко
определить предполагаемую сферу использования данной кине-
тической модели и в соответствии с этим придать ей необхо-
димую форму и глубину.
Громадные успехи фундаментальных биологических наук и
вычислительной техники в будущем позволят создать более на-
дежные, более глубокие и более механистические модели кине-
тики клеточного роста.
В этой главе мы изучили зависимости между ростом био-
массы, утилизацией субстрата и образованием продуктов жиз-
недеятельности клеток. Экспериментальные данные помогли
нам выяснить основные характеристики роста популяций кле-
ток и филаментозных организмов. Мы рассмотрели также ма-
тематические модели роста популяций микроорганизмов, не-
обходимые для анализа и проектирования технологических
процессов с их участием.
Гл. 9 посвящена вопросам расчета, проектирования и анали-
за различных биологических реакторов периодического и непре-
рывного действия; при обсуждении этой темы мы будем опи-
раться в основном на рассмотренные в настоящей главе мате-
матические выражения, описывающие скорость процессов.
Прежде чем перейти к анализу биологических реакторов,
нужно изучить кинетику еще одного класса явлений, а именно
процессов транспорта в макроскопических системах. Здесь мы
имеем в виду транспорт кислорода, хлора и других газов в
жидкостях, использующийся, например, при аэрации и хлориро-
вании. Такие транспортные явления в основе своей тесно свя-
заны с характерными для биореакторов процессами естествен-
ной или вынужденной конвекции. Последние в свою очередь
невозможно рассматривать без учета потребляемой биореакто-
ром мощности. Транспортные явления необходимо учитывать к
при масштабировании биореакторов. В стерилизации и других
процессах биохимической технологии большую роль играет так-
же теплообмен. Все эти проблемы и ряд других вопросов мы
рассмотрим в следующей главе.
37—537
578
Глава 7
Упражнения
7.1. Кинетика периодического процесса брожения. Натуральный виноград-
ный сок„ содержит почти все питательные вещества, необходимые для роста
дрожжей. Выполните следующую лабораторную работу по брожению.
а) Приобретите или соберите сами простейшую установку для получения
пива или вина. Для получения вина вам потребуются виноградный сок
(в этом эксперименте необходим сок, не содержащий нераствориалых твердых
частиц), сахар, дрожжи, сосуд емкостью 3,8 л, небольшой химический стакан
и отводная трубка, соединенная с водяным затвором.
б) Проведение процесса брожения. Суспендируйте активные дрожжи
в 100 мл теплой кипяченой воды; смесь тщательно перемешайте, чтобы в сус-
пензии не оставалось комков дрожжевой массы. В ферментер (стерилизован-
ный кипящей водой или раствором сульфита натрия) поместите 0,95 л вино-
градного сока, 1,35 кг сахара и добавьте дистиллированной воды (с темпе-
ратурой чуть выше комнатной) до трех четвертей объема ферментера. Про-
следите за тем, чтобы над жидкостью осталось достаточно места для не-
большого ареометра, и предусмотрите, чтобы ареометр можно было вводить
через крышку ферментера. Добавьте дрожжевую суспензию, плотно закройте
сосуд и энергично встряхивайте в течение 30 с. В реакционную смесь опусти-
те ареометр. Закройте ферментер пробкой с одним отверстием, в которое
вставлена соединенная с водяным затвором отводная трубка: уровень воды
в затворе должен быть примерно на 1 см выше уровня отводной трубки.
в) Регистрируйте ход ферментации во времени (в течение недели), кон-
тролируя: 1) скорость выделения СО2 при постоянной разности высот отвод-
ной трубки и водяного затвора (придумайте способ определения объема од-
ного пузырька СО2); 2) показания ареометра (проследите за тем, чтобы не
нарушалась герметичность системы).
г) Выразите результаты в графической форме; обсудите графики в связи
с кажущимися периодами клеточного роста и зависимостью между скоростя-
ми образования этанола и СОг (четко сформулируйте все допущения). Зная
количество введенного в реакционную смесь сахара (содержащегося в вино-
градном соке п кристаллической сахарозе), массу сухого клеточного вещест-
ва в конце процесса и результаты предыдущего упражнения (пункт в), со-
ставьте уравнение материального баланса по углероду; сформулируйте приня-
тые в уравнении допущения.
д) Считается, что очень чувствительными измерительными устройствами
являются органы чувств человека. Попробуйте продукт вашего эксперимента
на запах и вкус!
7.2. Зависимость скорости клеточного роста от температуры. Джонсон,
Эйринг и Полиссар (The Kinetic Basis of xMolecular Biology, John W iley and
Sons, N.Y., 1954) предложили следующее уравнение для зависимости удель-
ной скорости роста ц Е. coli от температуры в диапазоне от 18 до 43 ЭС:
. Д/Ji \
аГехр — "
___________1 \ RT !
" ~ / AS АН, \
1 т ехр ПГ -
где а = 0?3612е24’04; Д/7) = 15 ккал/моль; Д/72=150 ккал/моль; Д5 = 476,46 кал/
/(г-моль-К).
а) Начертите график этой функции в виде зависимости In ц от 1/Т.
б) Покажите, что это уравнение можно представить в виде произведения
двух функций, форма которого предполагается графиком упражнения 7.2а.
Как можно объяснить физический смысл этих двух функций и значений при-
веденных выше параметров?
Образование биомассы в культурах клеток
579
в) Какие связанные с необратимой инактивацией допущения приняты
в этой интерпретации зависимости р, от 77
7.3. Кинетика микробиологических процессов с одним и несколькими
субстратами. Культура растет на простой среде, содержащей 0,3% (масса/
/объем) глюкозы. В момент времени /=0 культуру (как инокулят) разбавля-
ют большим объемом идентичной стерильной среды. Процесс роста культуры
контролируют по изменению оптической плотности (OD) при 420 нм во вре-
мени; полученные данные приведены в столбце 1 табл. 7У3.1. В столбце 2
Таблица 7У3.1а
Оптическая плотность Оптическая плотность
Время, ч (2) Время, ч (1) (2)
(1)
0 0,06 0,06 4,5 0,44б 0,43
0,5 0,08 0,06 5,0 0,526 0,48
1,0 0,11 0,06 5,5 0,526 0,50
1,5 0,14 0,07 6,0 0,52
2,0 0,20 0,10 6,5 0,30б
2,5 0,26 0,13 7,0 0,42б
3,0 0,37 0,18 7,5 0,506
3,5 4,0 0,49 0,356 0,26 0,32 8,0 0,506
а W. В. Из работы: Biochemical Reasonin Benjamin, Inc., Menlo Park, Calif., g. Kerridge D., 1972. Tipton К. (eds.), prob. 39,
б Определению оптической плотности предшествовало разведение пробы равным
объемом не содержащей клеток среды.
той же таблицы указано изменение OD этой же культуры, выросшей в слож-
ной среде и разбавленной питательным раствором, содержащим 0,15% глю-
козы и 0,15% лактозы (масса/объем). Допустим, что OD (при 420 нм) про-
порциональна концентрации клеточно/f массы, причем OD, равная 0,175, соот-
ветствует концентрации 0,1 мг клеток (в расчете на массу сухого вещества)
в 1 мл. Рассчитайте максимальную удельную скорость роста цтах, продол-
жительность лаг-фазы /lag и общие экономические коэффициенты Y (выра-
женные числом граммов клеток на 1 г субстрата) при условии, что субстрат
всегда утилизируется полностью. Объясните форму кривых роста для каждо-
го случая.
7.4. Кинетика инактивации и гибели клеток; хлорирование. Предложено
несколько уравнений, описывающих скорость инактивации или гибели клеток;
опубликованное одним из первых уравнение предусматривало экспоненциаль-
ное снижение во времени численности активных спор возбудителя сибирской
язвы в присутствии 5%-ного фенола [Chick Н., Investigation of the Laws of
Disinfection, J. Hyg., 8, 698 (1908)]. Соответствующее уравнение инактивации
dN!dt= —kN часто называют законом Чика. Применялись и три другие ма-
тематические модели:
Логистическая:
I2 :
Замедленная:
--^-^kN+k'N (No-N)
dN
-~dT-ktN
dN k
~ dt 1 + at n
37*
580
Глава 7
В трех последних уравнениях кажущаяся константа скорости процесса пер-
вого порядка изменяется во времени.
а) Проинтегрируйте каждое из приведенных уравнений; покажите, какую
форму будут иметь кривые соответствующих графических зависимостей и как
с помощью этих графиков можно определить параметры каждой из моделей.
б) В табл. 7У4.1 приведены данные, отражающие зависимость степени
инактивации популяции Е. coll от времени и концентрации хлора. Соответст-
вуют ли эти данные какому-либо из приведенных выше уравнений?
Таблица 7У4.1. Зависимость скорости инактивации клеток Е. coli от концентрации хлора и времени экспозиции (pH 8,5; 2—5°C); указана доля (в %) клеток, сохранивших жизнеспособность®
Концентрация С1, мг/л Длительность экспозиции, мин
0,5 2 5 10 20
0,14 52 II 0,7
0,07 80 56 30 0,5 0
0,05 95 85 65 21 0,31
а Из работы: Fair G. М., Geyer J. С., Okan D. A., Water and Wastewater Engi-
neering, рр. 31—39, John Wiley а nd Sons. Inc., New York, 1968.
в) Для данного дезинфицирующего агента время, в течение которого
гибнет или инактивируется определенная часть популяции, выражается урав-
нением (концентрация)0 (время) = const. При инактивации хлором (в виде
НОС1) величина а равна 0,86 для Е. coli, 3 для аденовирусов, 1 для вируса
полиомиелита и 2 для вируса А Коксаки. Объясните, почему можно было
(или нельзя) ожидать подобную изменчивость значений а.
7.5. Кинетика клеточного роста и образования продуктов жизнедеятель-
ности клеток. Конак предложил обобщенное логистическое уравнение [Ап
Equation for Batch Bacterial Growth, Biotech. Bioeng., 17, 271 (1975)]:
1 dN r I -V V I i N \b
ма+b dt \ у / \ N /
00 \ 4 00 / \ 00 /
где N<x> — масса популяции в стационарной фазе; а, b — константы; N — кле-
точная масса.
а) Покажите, что максимальная скорость роста достигается при
N/Nca = a/ и что ее величину можно определить по уравнению
dN kNa+ba^bb
dt max (a + 6)a+b
б) Покажите, как вы могли бы определить каждый из параметров мо-
дели, используя указанные уравнения и наброски соответствующих графиков.
в) Покажите, что путем подстановки Р вместо только в левую часть
уравнения и подбора соответствующих а и Ь можно получить выражение,
описывающее скорость образования продукта жизнедеятельности клеток
в соответствии с сопряженным с ростом, несопряженным с ростом или сме-
______________Образование биомассы в культурах клеток 58
шанным механизмами, отвечающими рассмотренной в этой главе модели
Льюдпкина и Пайрета.
7.6. Пути клеточного метаболизма и кинетика образования продуктов
жизнедеятельности клеток, а) Объясните, почему часто труднее описать ко-
личественно кинетику образования продуктов жизнедеятельности, чем кинети-
ку утилизации субстрата или образования биомассы.
б) Изучите литературу, описывающую биохимию, методы и технологию
микробиологического производства какого-либо антибиотика, витамина или
аминокислоты. Составьте краткий обзор, в котором опишите стадии микро-
биологического синтеза, укажите (если имеются соответствующие данные)
медленные и равновесные стадии, а также наличие процессов ингибирования
или активации, индукции или репрессии ферментов. В заключение на основа-
нии последовательности реакций биосинтеза предложите (или обсудите уже
описанную в литературе) кинетику образования этого метаболита.
7.7. Модуль Тила клетки. Показано, что модули Тила многих клеток и
клеточных органоидов (мембран, митохондрий, рибосом п т. п.) равны или
чуть меньше единицы [Weisz Р. В., Science, 179, 433 (1973)]. Объясните при-
чину этого явления с количественной точки зрения, допуская, что клетки мо-
гут продуцировать ферменты в очень широком диапазоне концентраций. Для
простоты примите, что в системе присутствует только один фермент. В ваших
рассуждениях учтите объемную скорость, общую скорость роста, энергетиче-
ские и другие затраты в процессе синтеза фермента. Обратите внимание на
то, что указанная величина модуля Тила предполагает, что определенная
в отсутствие внешнего сопротивления массопередаче кинетика клеточных
процессов (гл. 4) будет отражать истинную кинетику этих процессов, как и
предполагалось во всех разделах гл. 7.
7.8. Кинетика двойного ингибирования. Показано, что рост Candida
utilis на ацетате натрия ингибируется субстратом (ацетатом) и pH [Jack-
son J. V., Edwards V. Н., Biotech. Bioeng., 17, 943 (1975)].
а) Выразите скорость роста С. utilis через соответствующие константы,
а также концентрации ионов водорода и всего субстрата при следующих до-
пущениях:
1. Зависимость степени ингибирования субстратом от общей концентра-
ции субстрата аналогична неконкурентному ингибированию фермента
субстратом и может быть описана уравнением Эндрюса [уравне-
ние (7.32)].
2. Максимальная скорость роста цтах является аналогичной функцией
от концентрации ионов водорода.
3. Субстрат проявляет ингибиторную активность только в протонирован-
ной форме, т. е. существует равновесие HS (активный ингибитор)^
—S~ (неактивный ингибитор)+Н+.
б) Покажите, как кривая зависимости 1/р от 1/s (в координатах Лай-
нуивера — Бэрка) будет изменяться в зависимости от pH. Как можно опре-
делить параметры этой модели?
Аналогично pH влияет и на ряд процессов, использующихся при обра-
ботке сточных вод. Пример кинетики анаэробного микробиологического про-
цесса приведен в разд. 14.4.6.
7.9. Кинетика Моно в ПРПП. Рассмотрим организм, кинетика роста ко-
торого подчиняется уравнению Моно при ртах = 0,5 ч~1 и К$=2 г/л.
а) При какой скорости разведения D будет обеспечена максимальная
общая скорость образования клеточной массы, если принять, что культура
находится в стационарном состоянии, содержимое проточного реактора
полностью перемешивается, в культуре клетки не гибнут и So = 5O г/л, У=1
(г клеточной массы/г субстрата)?
б) Сколько последовательных реакторов одинаковой емкости потребует-
ся для снижения концентрации субстрата до 1 г/л при найденном в преды-
дущем упражнении значении D?
582
Глава 7
7.10. Особенности расчета ПРПП при ингибировании клеточного роста,
а) Постройте график зависимости х и s от D
________________
Гх^
где s0= 10 г/л, хо=О, Ks — 1 г/л, 7<, = 0,01 г/л, 1=0,05 г/л, ртах = 0,5 ч~‘,
Ух/з=0,1 г клеток/г субстрата. На том же чертеже постройте графики зави-
симостей х и s при 1 = 0.
б) Предположим, что ПРПП был предназначен для работы в отсутствие
ингибиторов. Найдите уравнения, определяющие отношения (s/«i) и xDI(xD)t,
т. е. отношения предсказываемых моделью концентрации субстрата и скорости
роста биомассы к соответствующим наблюдаемым величинам. Как присутст-
вие ингибитора влияет на поведение реактора в отношении (хО)тах и вы-
мывания?
в) Повторите решение упражнений 7.10, а и 7.10,6 при условии, что i
зависит от х (i = x'/10).
7.11. Экономический коэффициент. Если при моделировании кинетики
клеточного роста учитывается эндогенный метаболизм или энергия поддер-
жания клеток, то соответствующее уравнение (если допустить, что субстрат
имеется в избытке) можно записать в следующей форме:
1 1
— — Их=» — — р.Х — kex
где Y' — экономический коэффициент клеточного роста (количество граммов
клеток, образующихся из 1 г субстрата, расходуемого на рост клеток);
ke— количество граммов субстрата, расходуемого на энергию поддержания
1 г клеток; У — кажущийся экономический коэффициент (количество грам-
мов субстрата, расходуемого на образование 1 г клеточной массы).
Зависимость экономического коэффициента от скорости разведения (в двой-
ных обратных координатах) изображена на рис. 7У11.1.
РИС. 7У11.1. Зависимость экономического коэффициента от скорости разведе-
ния в двойных обратных координатах. [Воспроизведено из работы: Kirsh.
Sykes, Prog. Ind., Microbiol., 9, 155 (1971); данные заимствованы из работы:
Tempest D. W., Herbert D., Phipps P. J., in Microbial Physiology and Continuous
Culture, Powell E. O. et al. (eds.), HMSO, London, 1967.]
Образование биомассы в культурах клеток
583
РИС. 7У12.1. Схема и стехиометрия биосинтеза лимонной кислоты (АсСоА —
ацетил-СоА; CIT — цитрат; G6P — глюкозо-6-фосфат; GLU — глутамат; ICT —
изоцитрат; MAL— малат; ОАА — оксалоацетат; OGT — а-кетоглутарат;
PYR — пируват; SUC — сукцинат; GOX — глиоксилат). [Воспроизведено с
разрешения из работы: Aiba S., Matsuoka М., Identification of Metabolic Model:
Citrate Production from Glucose by Candida lipolytica, Biotech. Bioeng., 21,
1373 (1979).]
а) Покажите, что для ПРПП х и s связаны соотношением
DY' , .
Xt= keY' + D (S° S)
б) Каким уравнением в этом случае должны быть связаны ke, Y' и D?
в) Удовлетворяют ли этой модели данные по росту A. aerogenes иа гли-
цериновой среде, изображенные иа рис. 7У11.1? Обоснуйте ваш ответ коли-
чественно.
7.12. Стехиометрия и скорости метаболических превращений. На
рис. 7У12.1 приведена упрощенная схема биосинтеза лимонной кислоты
584
Глава 7
в Candida lipolytica. На схеме символами обозначены важнейшие стехиомет-
рические и кинетические параметры основных реакций в цикле ТКК и в гли-
оксилатном цикле. Здесь Vi — удельные скорости потоков углерода по раз-
личным метаболическим путям; фсаг, фрго и аг — стехиометрические коэффи-
циенты; р, — удельная скорость роста; v — удельная скорость утилизации глю-
козы; Qi — скорость образования определенных продуктов метаболизма.
fAiba S., Matsuoka М., Biotech. Bioeng., 21, 1373 (1979).]
а) Допустив, что все метаболиты находятся в квазистационарном состоя-
нии, найдите выражения, связывающие собственные скорости элементарных
реакций с наблюдаемыми общими скоростями процессов утилизации глюко-
зы, образования СОг, цитрата и изоцитрата.
б) Запишите уравнение общего материального баланса по углероду.
Зависит ли это уравнение от выражений, найденных вами при решении
упражнения 7.12, а?
в) Для оценки важности отдельных путей метаболизма в биосинтезе
цитрата были предложены две модели, в одной из которых отсутствует гли-
оксилатный цикл (о7 = 0), а во второй блокирован цикл ТКК (Уз = 0). С по-
мощью найденных вами в двух предыдущих упражнениях уравнений и экс-
периментальных данных, приведенных в таблице 1 статьи Аибы и Мацуоки,
вычислите относительные внутриклеточные метаболические потоки для ука-
занных двух моделей при £> = 0,122 и 0,0769 ч-1. Какая из двух моделей,
судя по результатам ваших расчетов, представляется более обоснованной?
7.13. Зависимость максимальной продуктивности и вымывания от скоро-
сти разведения, а) С помощью уравнений (7.16) и (7.18) покажите, что отно-
шение DmolDvio (т0— максимальная продуктивность; w0 — вымывание) за-
висит только от (Sf/Ks) и определяется выражением
Dm0/Dw0= [1 + (1 -Нг/Кг)-"2]-1
б) Покажите, что в предельных случаях предыдущее уравнение можно
преобразовать следующим образом:
1
2
4 Ks
при (Sf/Ks) « 1
D„!D„ ~ 1 - [1 - 4- (44] При (S'/Ks) » 1
в) Выскажите ваши соображения относительно стабильности работы
реактора и возможности его регулирования в предельных случаях, описан-
ных в пункте б.
г) Найдите выражения для Ур/х=/(^>) и соответствующей производной
df/dD, справедливые при максимальной продуктивности (pD)max.
7.14. Кинетика ингибирования. Если рост микроорганизма ингибируется
летучим продуктом его жизнедеятельности (например, этанолом), то скорость
роста клеток можно повысить за счет постоянного вакуумирования прост-
ранства над культуральной жидкостью.
а) Покажите, что для периодического процесса, описываемого уравне-
нием (7.33), временная зависимость концентрации субстрата для суммарнон
реакции S—>-0,3 Р-{-клеточная масса выражается уравнением
ds — [*o+rX/s(s0 —*)] s __________К/________
dt e KX/S p'max W ^4- 0,3 (s0 —s)
где x— концентрация биомассы (г/л). Экономический коэффициент >xs
считайте постоянным.
б) Путем интегрирования приведенного выше уравнения найдите выра-
жение, определяющее отношение x(t)lxi(t), где х и xi — концентрации био-
массы в отсутствие и в присутствии ингибитора соответственно.
Образование биомассы в культурах клеток
585
в) Определите отношение х (/)/*; (О (см. пункт б) при x0=10~6 г/мл,
Ух/з=0,1 г клеток/г субстрата, /<s=0,22 г/л, ртах=0,408 ч-1, /0 = 16 г/л для
двух концентраций субстрата: so = 5,O г/л и so=7O г/л (примите, что молеку-
лярная масса S втрое больше молекулярной массы Р).
7.15. Лаг-фаза в периодическом процессе, а) Показано, что при росте
Aerobacter aerogenes в периодическом процессе в аммониево-сульфатной
среде продолжительность лаг-фазы (/iag) приблизительно пропорциональна
объему инокулята и обратно пропорциональна концентрации клеток в ино-
куляте [3]. Предложенная Дином и Хиншелвудом [3] модель объясняет
этот факт тем, что лаг-фаза завершается тогда, когда концентрация опреде-
ленного вещества А в клетке достигает порогового значения с'. Эти исследо-
ватели предположили также, что в лаг-фазе концентрация с вещества А
изменяется во времени следующим образом:
с —a V+a'nat + a"t (1)
где V — объем инокулята; а — концентрация вещества А в единице объема
инокулята; по — число клеток в единице объема культуры (значение п0 мож-
но считать постоянным, поскольку скорость роста клеток в лаг-фазе ничтож-
но мала); а’ — среднее повышение концентрации вещества А (обусловленное
его синтезом другими клетками) в единицу времени в расчете на одну
клетку; а” — повышение концентрации вещества А за счет внутриклеточного
синтеза. С помощью уравнения (1) определите /iag и обсудите полученный
вами результат в связи с экспериментально наблюдаемой зависимостью.
Проанализируйте допущения, принятые при выводе уравнения (1).
б) Известно, что на длительность лаг-фазы влияет состояние популяции
инокулята. Предположим, что инокулят содержит п0 жизнеспособных клеток
и nd мертвых клеток и что рост жизнеспособных клеток начинается сразу
же после инокуляции и происходит с постоянной удельной скоростью р.
Определите зависимость рарр (наблюдаемой кажущейся удельной скорости
роста, оцениваемой по общей концентрации клеток) от времени. Опираясь на
полученные результаты, выскажите ваши соображения о влиянии жизнеспо-
собности инокулята и числа клеток в нем на кажущуюся длительность
лаг-фазы.
7.16. Рост популяции микроорганизмов. Найден новый микроорганизм,
при делении которого из каждой родительской клетки образуются три дочер-
ние. По приведенным ниже экспериментальным данным, отражающим рост
популяции, вычислите среднее время между двумя последовательными деле-
ниями клеток.
t, ч Масса сухого клеточного вещества, г/л । f’ 4 Масса сухого клеточного вещества, г/л
0 0,10 1,5 0,34
0,5 0,15 2,0 0,51
1,0 0,23
7.17. Кинетика биотрансформаций. Связанный с твердой поверхностью
монослой клеток катализирует превращение субстрата S в продукт Р, при-
чем рост клеток незначителен. Предположим, что скорость утилизации суб-
страта единицей поверхности описывается уравнением
где
и =4
^maxs'
/Cs + S
К,-=Л,о(1+р/ЛО
586
Глава 7
поскольку Р ингибирует реакцию. В указанных уравнениях s и р— концент-
рации S и Р соответственно на поверхности.
Концентрации S и Р в растворе вдали от поверхности равны s0 и р0
соответственно. Коэффициенты массопередачи S и Р равны hs и hp соответ-
ственно.
Выразите скорость образования Р на единице поверхности через наблю-
даемые концентрации в растворе s0 и р0, если из каждого моля S образует-
ся Ур/s молей Р.
7.18. Математические модели образования продуктов жизнедеятельности
клеток и метаболизма поддержания. Неоднократно высказывались пред-
положения, что модель Льюдикина—Пайрета, связывающая клеточный
рост с образованием продукта метаболизма [уравнение (7.93)]:
Г}Р=ар,х+$х
и разработанная Пертом [Proc. Royal Soc., Series В, 163, 224—231 (1965)]
модель, отражающая затраты энергии на поддержание:
1 1 m
Y ~ Y' + |л
где У— наблюдаемый экономический коэффициент в расчете на субстрат;
Y'— теоретический (без учета расхода на метаболизм поддержания) эконо-
мический коэффициент; m — коэффициент метаболизма поддержания (г суб-
страта/г клеток в час), эквивалентны. Согласны ли вы с этим предположе-
нием? Обоснуйте ваш ответ.
7.19. Структурированная модель клеточного роста Уильямса. Рассуждая
так, как рассуждал Уильямс, разделим биомассу на две части. В первую
часть мы включим промежуточные соединения, ферменты и другие компонен-
ты, участвующие в образовании структурного и генетического материала
клетки; это будет эквивалентом синтетической части биомассы в модели
Уильямса. Вторую часть составит структурный и генетический материал
биомассы. Предположим, что доли первой и второй частей биомассы состав-
ляют и f2 от общей биомассы; сумма ft и /2 равна единице.
Запишем уравнения модели в собственной форме:
dfi _______Wх/2 .. т
dt -a Kx-j-s K2 + /x
dfz p Wifa ..f
где
U—a_A^2__ i (R _ 1 \_WJ2________
a — количество синтетической части биомассы, образующейся при утили-
зации одной единицы массы лимитирующего скорость роста суб-
страта;
Р— количество структурной (генетической) биомассы, образующейся при
утилизации одной единицы массы синтетической биомассы.
а) Рассмотрим сбалансированный рост, при котором Л=/2=0; такое со-
стояние может быть достигнуто в том случае, если s будет постоянным до-
статочно длительное время. Каким образом ц зависит от s в состоянии сба-
лансированного роста?
б) Покажите, что модель Моно будет приблизительно соответствовать
состоянию сбалансированного роста, если К2^>1 и Чему (прибли-
женно) в этом случае будут равны ft и f2?
Образование биомассы в культурах клеток 587
в) Пусть и х2=с[2, где c=xt+x2. Напишите дифференциальные
уравнения для xj, х2 и s, описывающие систему хемостата. Напишите также
дифференциальное уравнение для плотности популяции л; примите, что 7
[количество структурного (генетического) материала в клетке) постоянно.
г) Постройте графики зависимостей 1g с от t, 1g п от t и т (средний раз-
мер клеток in=cln) от t для клеточного роста в периодическом процессе при
двух различных начальных значениях fi : fs — 0,40 и fi = 0,05. И в первом,
и во втором случае примите, что Со=2-10~3 г/л и so=5,O г/л. При построе-
нии используйте следующие константы: а = р = 0,50; &i = 6,0 ч-1; &2 = 3,0 ч-1;
/<i=0,2 г/л; /<2=0,25 г/л; масса клетки 1,1 -10—13 г.
д) С помощью параметров, перечисленных в пункте 7.19, г, постройте
график, отражающий изменение ц в зависимости от s для состояния сбалан-
сированного роста. На том же чертеже постройте график зависимости ц от
s в соответствии с уравнением Моно. Константы и Ks в уравнении Моно
подберите таким образом, чтобы ц асимптотически приближалось к постоян-
ной величине при больших значениях s и чтобы производная от ц по з при
s=0 имела бы одно и то же значение и в модели Моно, и в общей модели.
е) Постройте графики зависимости с, s и т от скорости разведения для
стационарного хемостата при s/ = 5,0 г/л.
ж) Как будут изменяться ci, s и т при «сдвиге» условий в хемостате от
0=0,1 ч-1 до 0=0,3 ч—1 и от 0=0,3 ч"1 до 0=0,1 ч-1? Примите, что
«сдвигу» предшествует стационарное состояние сбалансированного роста.
з) Обсудите полученные вами расчетные данные в сравнении с действи-
тельным поведением популяций бактерий.
Литература
В перечисленных ниже журналах публикуются результаты работ, посвя-
щенных изучению кинетики роста популяций клеток, а также многим другим
аспектам биохимической технологии и прикладной биологии: Biotechnology and
Bioengineering, Enzyme and Microbial Technology, Biotechnology Letters, Jour-
nal of Fermentation Technology, Journal of Chemical Technology and Biotechno-
logy, Journal of Applied Chemistry and Biotechnology, Process Biochemistry,
Trends in Biotechnology, Agricultural and Biological Chemistry, Applied Micro-
biology, Applied Biochemistry and Microbiology и другие. Многотомное издание
Advances in Biochemical Engineering and Biotechnology (ранее Advances in
Biochemical Engineering) публикует великолепные обзорные статьи, в которых
часто рассматриваются перспективы развития биохимической технологии. Об-
зорные и оригинальные статьи печатают также Annual Reports on Fermenta-
tion Processes, публикуемые в Annals of the New York Academy of Science
издания, посвященные биохимической технологии, и ежегодники Progress in
Industrial Microbiology, Society for General Microbiology Symposium.
1. Biochemical and Biological Engineering Science, Blakebrough N. (ed.),
vol. 1, Academic Press, Inc., New York, 1967. Здесь содержится полезный
материал справочного характера по конструкциям биологических реакто-
ров. Особенно рекомендуется гл. 6 (Luedeking R., Fermentation Process
Kinetics).
2. Аиба Ш„ Хемфри А., Миллис Н., Биохимическая технология и аппарату-
ра.— М.: Пищевая промышленность, 1978. В этом углубленном пособии
по биохимической технологии рассматривается кинетика периодических
процессов (гл. 4), подробно обсуждены непрерывные процессы (гл. 5),
а глава 9 посвящена проблемам стерилизации жидкостей.
3. Dean А. С. R„ Hinshelwood С. N., Growth, Function and Regulation in Bac-
terial Cells, Oxford University Press, London, 1966. Помимо методов мате-
матического анализа метаболических превращений в этой монографии рас-
смотрено множество экспериментальных данных по другим проблемам кле-
точного роста и его регуляции. Эта книга настоятельно рекомендуется
588
Глава 7
в качестве дополнительной литературы; ее единственным недостатком являет-
ся необоснованный скептицизм по отношению к молекулярной биологии.
4. Microbial Growth, Dawson Р. S. S. (ed.), Dowden, Hutchinson and Ross,
Inc., Stroudsburg, Pa., 1974. Превосходный сборник множества уже став-
ших классическими работ (в том числе оригинальной работы Моно), посвя-
щенных изучению роста микроорганизмов.
5. Roels J. A., Energetics and Kinetics in Biotechnology, Elsevier, Amsterdam,
1983. В этой монографии хорошо описаны как кинетические модели роста
микроорганизмов и образования продуктов их жизнедеятельности, так и
связанные проблемы химической термодинамики.
6. Foundation of Biochemical Engineering: Kinetics and Termodynamics in
Biological Systems, ACS Symposium Series 207, Blanch H. W., Papoutsa-
kis E. T., Stephanopoulos G. (eds.), American Chemical Society, Washington,
D.C., 1983. Довольно полная подборка обзорных статей по кинетике раз-
личных процессов от ферментативных реакций до роста популяций клеток.
Очень полезно ознакомиться со следующими обзорными статьями по ма-
тематическому моделированию клеточного роста:
7. Tsuchiya Н. М., Fredrickson A. G., Aris R., Dynamics of Microbial Cell Popu-
lations; Adv. Chem. Eng., 6, 125 (1966).
8. Fredrickson A. G., Megee R. D„ III, Tsuchiya H. M. Mathematical Models
for Fermentation Processes, Adv. Appl. Microbiol., 23, 419 (1970).
В приведенных ниже работах рассматриваются проблемы угнетения ро-
ста микроорганизмов продуктами метаболизма и субстратами:
9. Aiba S., Skoda М., Naga.ta.ni М., Kinetics of Product Inhibition in Alcohol
Fermentation, Biotech. Bioeng., 10, 845 (1968).
10. Aiba S., Shoda M., Reassessment of the Product Inhibition in Alcohol Fer-
mentation, J. Ferment. Technol. Japan, 47, 790 (1969).
11. Andrews J. F., A Mathematical Model for the Continuous Culture of Micro-
organisms Utilizing Inhibitory Substrates, Biotech. Bioeng., 10, 707 (1968).
Следующие статьи посвящены росту филаментозных организмов и обра-
зованию продуктов их жизнедеятельности.
12. Metz В.. Rossen N. IF. F., The Growth of Molds in the Form of Pellets —
A Literature Review, Biotech. Bioeng., 19, 781 (1977).
13. Megee R, D., Kinoshita S., Fredrickson A. G., Tsuchiya H. M., Differentiation
and Product Formation in Molds, Biotech. Bioeng., 12, 771 (1970).
14. Matsumura M„ I manuka T., Yoshida T., Taguchi H., Modeling of Cephalo-
sporin C Production and Its Application to Fed-batch Culture, J. Ferment.
Technol. Japan, 59, 115 (1981).
Co структурированными моделями клеточного роста можно ознакомиться
в работах [5—8], а также в следующих статьях:
15 Fredrickson A. G., Formulation of Structured Growth Models, Biotech.
Bioeng.. Г8, 1481 (1976).
16 Williams F. M., A Model of Cell Growth Dynamics, J. Theoret. Biol., 15, 190
(1967).
17. Harder A., Roels J. A., Application of Simple Structured Models in Bioen-
gineering, in Advances in Biochemical Engineering, vol. 21, Fiechter A. (ed.),
p. 55, Springer-Verlag, Berlin, 1982.
18. Bijkerk A. H. E., Hall R. J., A Mechanistic Model of the Aerobic Growth of
Saccharomyces cerevisiae, Biotech. Bioeng., 19, 267 (1977).
Образование биомассы в культурах клеток
589
19. Pamment N. В., Hall R. J., Barford J. Р., Mathematical Modeling of Lag
Phases in Microbial Growth, Biotech. Bioeng., 30, 349 (1978).
20. Shuler M. L., Domach M. M., Mathematical Models of the Growth of Indivi-
dual Cells, in Foundations of Biochemical Engineering, Blanch H. W., Papout-
sakis E. T., Stephanopoulos G. (eds.), p. 101, American Chemical Society,
Washington, D.C., 1983.
21. Ramkrishna D., A Cybernetic Perspective of Microbial Growth, in Founda-
tions of Biochemical Engineering, Blanch H. W., Papoutsakis E. T., Stepha-
nopoulos G. (eds.), p. 161, American Chemical Society, Washington, D.C.,
1983.
22. Kampala D. S., Ramkrishna D., Tsao G. T., Cybernetic Modeling of Microbial
Growth on Multiple Substrates, Biotech. Bioeng., 26, 1272 (1984).
Кинетика образования продуктов метаболизма рассмотрена в следующих
статьях:
23. Gaden Е. L., Chem. Ind. Rev., 154 (1955), J. Biochem. Microbiol. Technol.
Eng., 1, 413 (1959).
24. Deindoerfer F. H., Adv. Appl. Microbiol., 2, 321 (1960).
25. Ollis D. F., A Simple Batch Fermentation Model: Theme and Variations,
Ann. N.Y. Acad. Sci. USA, 413, 144 (1983).
26. Pazoutova S., Votruba J., Rehdcek Z., A Mathematical Model of Growth and
Alkaloid Production in the Submerged Culture of Claviseps purpurea, Bio-
tech. Bioeng., 23, 2837 (1981).
27. Lee S. B., Bailey J. E., Analysis of Growth Rate Effect of Productivity of
Recombinant Escherichia coli Populations, Biotech. Bioeng., 26, 66 (1984).
28. Lee S. B., Bailey J. E„ Genetically Structured Models for lac Promoter —
Operator Function in the Escherichia coli Chromosome and in Multicopy
Plasmids: lac Operator Function, Biotech. Bioeng., 26, 1372 (1984).
В перечисленных ниже статьях описаны сегрегированные кинетические мо-
дели.
29. Ramkrishna D., Statistical Models of Cell Populations, Adv. in Biochem. Eng.,
11, 1 (1979).
30. Nishimura Y„ Bailey J. E., On the Dynamics of Cooper — Helmstetter —
Donachie Procaryote Populations, Math. Biosci., 51, 505 (1980).
31. Hjortso M. A., Bailey J. E„ Steady-State Growth of Budding Yeast Popula-
tions in Well-Mixed Continuous Flow Microbial Reactors, Math. Biosci., 60,
235 (1982).
32. Shu P., Mathematical Models for the Product Accumulation in Microbial
Processes, J. Biochem. Microbiol. Technol. Eng., 3, 95 (1961).
Великолепное введение в проблему стерилизации дано в статье:
33. Blakebrough N., Preservation of Biological Materials Especially by Heat
Treatment, in Biochemical and Biological Engineering Science, vol. 2, Blake-
brough N. (ed.), Academic Press, Inc., New York, 1968.
Факторы, влияющие на устойчивость организмов к стерилизации, обобще-
ны в гл. 20 и 21 монографии Фробишера (ссылка [2] в гл. 1), а проблема
обезвреживания фагов рассмотрена в статье:
34. Hango et al., Phage Contamination and Control, in Microbial Production of
Amino Acids, Kodansha Ltd., Tokyo, and John Wiley and Sons, Inc., New
York, 1973.
Глава 8
Процессы переноса
в биотехнологических системах
В ходе изучения материала предыдущих глав мы постепен-
но переходили от сравнительно простых (молекулярных) сис-
тем ко все более и более сложным — сначала к отдельным
клеткам, затем к культурам, содержащим миллионы или мил-
лиарды клеток в миллилитре. Физическая разобщенность ком-
понентов системы (питательных веществ, клеток, продуктов
метаболизма) повышает вероятность того, что на общую ско-
рость превращений растворенных (или суспендированных) в
реакционной массе веществ будет влиять не столько скорость
химических реакций, сколько те или иные процессы физическо-
го переноса. Действительно, согласно данным Вейца, для кле-
ток и входящих в их состав каталитических комплексов харак-
терны модули Тила, приблизительно равные единице [1]; ины-
ми словами, клетки и субклеточные структуры функционируют
с максимальной скоростью практически без диффузионных ог-
раничений. Поэтому если мы, например, в каком-то микробио-
логическом процессе повысим количество углеродсодержащих
питательных веществ, то аэробные клетки смогут их полностью
усвоить только тогда, когда в непосредственной близости от
клеток будет поддерживаться достаточно высокая концентра-
ция кислорода. Если учесть, что растворимость кислорода в
водных средах очень мала, то представляется вполне вероят-
ным, что выполнить это условие можно будет только путем по-
вышения эффективности массообмена в системе газ — жид-
кость и таким путем обеспечить ускоренную подачу кислорода
к клеткам.
Очевидно, граница, разделяющая области с преобладанием
аэробных или анаэробных процессов, определяется локальной
концентрацией кислорода, коэффициентом диффузии Ог и ло-
кальными скоростями дыхания в аэробной области. В случае
облигатных аэробов, например плесеней в мицелиальных гра-
нулах или тканевых клеток в злокачественных опухолях, эта
граница одновременно разделяет жизнеспособные клетки от от-
мирающих; она определяет также глубину области с преобла-
данием аэробных процессов в поверхностных слоях озер и
разделяет сосуществующие колонии аэробов от анаэробных
Процессы переноса в биотехнологических системах 591
популяций бактерий в частицах почвы. Началом современных
работ по изучению роли массообмена кислорода в биологичес-
ких процессах можно считать сороковые годы нашего столетия,
когда в разгар второй мировой войны разворачивалось про-
мышленное производство пенициллина. В настоящее время
установлено, что массообмен кислорода играет большую роль
и в многочисленных природных процессах, в том числе и в об-
условленной окислительными процессами порче пищевых про-
дуктов и эвтрофикации озер, связанной или с неадекватной
аэрацией системы природными источниками кислорода, или с
повышенной концентрацией некоторых веществ, например фос-
фатов или нитратов.
В микробиологических процессах большую роль могут иг-
рать и другие ограниченно растворимые газы. Так, в производ-
стве белков одноклеточных организмов в качестве субстратов
применяют метан и другие углеводороды. В этом важном про-
цессе культуральная жидкость должна постоянно насыщаться
как кислородом, так и метаном со скоростью, достаточной для
удовлетворения потребностей микроорганизмов. Выведение ме-
тана из раствора является важным этапом в анаэробной обра-
ботке отходов; последней стадией этого процесса является де-
карбоксилирование карбоновых кислот (в первую очередь ук-
сусной кислоты) до соответствующих алканов.
Диоксид углерода генерируется почти в каждом микробио-
логическом процессе. Несмотря на высокую растворимость
СО2, связанные с массообменом взаимопревращения между га-
зообразным СО2 и различными формами растворенного диок-
сида углерода (СО2, Н2СО3, НСОз-, СОз2-) заметно влияют
на pH; этот факт немаловажен при регулировании pH в про-
цессах с участием неустойчивых к кислотам анаэробных орга-
низмов, в которых выделяются одновременно и СО2, и СН4.
Такие процессы мы рассмотрим в гл. 14.
Массообмен в системах жидкость — жидкость играет боль-
шую роль в производстве белков одноклеточных организмов из
жидких углеводородов, а также при выделении продуктов мик-
робиологических процессов, например при экстракции органи-
ческими растворителями культуральной жидкости или ее филь-
трата в производстве фармацевтических препаратов (гл. 11).
Биотрансформация возобновляемого природного сырья (на-
пример, отходов сельскохозяйственного и лесохимического про-
изводства типа целлюлозных, гемицеллюлозных и лигниновых
фракций), используемого в качестве сырья в различных микро-
биологических процессах, обычно включает стадии (солюбили-
зация биомассы, ожижение, гидролиз), скорость которых лими-
тируется доступной для реагентов площадью поверхности
твердого субстрата и скоростями диффузии растворенных ве-
592
Глава 8
ществ. Кроме того, массообмен между жидкой и твердой фаза-
ми играет определенную роль в различных сорбционных и хро-
матографических методах выделения и очистки веществ, а так-
же в транспорте растворенного кислорода к гранулам плесени
или к пленкам и гранулам иммобилизованных клеток.
При высокой плотности популяции клеток массообмен мо-
жет стать фактором, определяющим скорость всего процесса.
Такая ситуация характерна для самых разнообразных процес-
сов — от лабораторных сосудов, перемешиваемых встряхивани-
РИС. 8.1. Основные стадии переноса кислорода от газового пузырька в скоп-
ление клеток.
ем, до громадных промышленных реакторов, предназначенных
для производства пенициллина или внеклеточных биополимеров
(например, ксантана) и установок для обработки сточных вод
методом активного ила. Отсюда следует, что при разработке и
проектировании биологических реакторов инженер должен
четко знать, какой фактор определяет скорость процесса—яв-
ления переноса или кинетика биологических превращений.
Часто явления диффузии растворенного вещества и переноса
количества движения связаны самым тесным образом; в еще
более сложном варианте эти явления связаны и с кинетикой
химических реакций. Проблема взаимосвязи между диффузией
и реакцией была рассмотрена нами в гл. 4; с этой точки зрения
активность ферментов и клеток (в виде, например, коэффициен-
та эффективности) может быть полностью описана с помощью
двух параметров — модуля Тила и параметра насыщения
Ks/s0. К сожалению, в подавляющем большинстве случаев яв«
Процессы переноса в биотехнологических системах
593
ления массообмена самым тесным образом связаны с передачей
количества движения, т. е. с механикой жидкостей и газов; дей-
ствительно, этой теме посвящена большая часть литературы по
химической технологии. Здесь мы ограничимся изложением
основных концепций и таблицами формул, применяемых для
расчета или оценки коэффициентов массопередачи растворен-
ных веществ.
В заключительном кратком разделе этой главы будут рас-
смотрены примеры процессов, в которых на поведение биологи-
ческих систем существенное влияние оказывает (через неодно-
родное распределение температуры) другое важное явление
переноса — теплообмен. К числу таких примеров относятся
относительно экзотермические микробиологические процессы,
в том числе производство винного уксуса в реакторе со струй-
ным течением, обработка сточных вод, саморазогрев куч ком-
поста (а также бытовых и других отходов) и иные процессы
с участием твердой фазы.
8.1. Массообмен между газовой
и жидкой фазами в клеточных системах
Физическая основа рассматриваемого в этой главе явления
массообмена схематически представлена на рис. 8.1. Ограни-
ченно растворимый газ, чаще всего кислород, переносится от
его источника (например, поднимающихся воздушных пузырь-
ков) в содержащую клетки жидкую фазу. (Аналогичная карти-
на наблюдается и в случае любого другого ограниченно раство-
римого субстрата, например жидких углеводородов в микроби-
ологических способах переработки углеводородов.) На этом
пути кислород должен преодолеть ряд сопротивлений массооб-
мену, относительные величины которых зависят от гидродина-
мики пузырьков (капель), температуры, клеточной активности
и плотности культуры, состава раствора, ряда явлений на гра-
нице раздела фаз и от других факторов.
Общее сопротивление массообмену складывается из от-
дельных сопротивлений в следующих частных процессах:
1. Диффузии газа из ядра газовой фазы к границе раздела фаз
газ — жидкость.
2. Переносе через границу раздела фаз газ — жидкость.
3. Диффузии растворенного вещества через примыкающий к
пузырьку относительно неперемешиваемый слой жидкости в
перемешиваемую жидкую фазу.
4. Переносе растворенного вещества через жидкую фазу ко
второму относительно мало перемешиваемому жидкому
слою, окружающему клетки.
38—537
594
Глава 8
5. Переносе через второй относительно неперемешиваемый
слой, окружающий клетки.
6. Диффузии в скопления клеток, мицелий или в частицу
почвы.
7. Переносе через оболочку клетки во внутриклеточное прост-
ранство (реакционный центр).
Все эти типы сопротивлений изображены на рис. 8.1. Когда
организм существует в виде отдельных клеток, шестой тип со-
противления отпадает. Для клеток микроорганизмов характер-
на тенденция к адсорбции на границах раздела фаз. Так,
клетки могут концентрироваться преимущественно вблизи гра-
ницы, разделяющей газовый пузырек и жидкую фазу. Тогда
диффундирующий растворенный кислород преодолевает только
одну неперемешиваемую область; отпадает необходимость и в
переносе через жидкую фазу, предшествующем сопротивлению
пятого типа. В такой ситуации концентрация Ог, растворенного
в жидкой фазе, не отражает скорость его подачи для дыхания
микроорганизмов.
Точно так же в процессах микробиологических трансформа-
ций других ограниченно растворимых субстратов, например
капель углеводорода, часто наблюдалась адсорбция клеток на
границе раздела (или вблизи нее), отделяющей углеводород-
ную фазу от водной. Математическое моделирование работы
реакторов в таких ситуациях мы рассмотрим в гл. 9.
На рис. 8.2 изображены различные варианты физического
контакта между газовой и жидкой фазами. Все эти варианты
можно разделить на две группы, в первой из которых массо-
обмен совершается за счет свободного подъема пузырьков газа
или опускания жидких или твердых частиц, а во второй — за
счет движения жидкости под действием приложенных к ней
внешних сил, отличных от сил гравитации (принудительная
конвекция). Между этими двумя группами нет четкой грани-
цы, и, например, смешение газа с жидкостью в медленно пере-
мешиваемой полупроточной системе может в равной степени
обусловливаться как свободным подъемом пузырьков газа, так
и механическим перемешиванием. Важность гидродинамики в
процессах биохимической технологии требует изучения взаимо-
связи между течением жидкости и массообменом. Прежде чем
приступить к изучению этой проблемы, однако, необходимо сде-
лать несколько замечаний общего характера и познакомиться
с рядом терминов и определений, принятых в теории массооб-
мена.
Процессы переноса в биотехнологических системах 595
Таблица 8.1. Растворимость 02 при давлении 1 атм О2 в воде при различных
температурах и в растворах солей или кислот при 25 °Са
Температура, °C Растворимость О2 в воде, ммоль/л
0 2,18
10 1,70
15 1,54
20 1,38
25 1,26
30 1,16
35 1,09
40 1,03
Растворимость О2 в водных растворах при 25 °C
Растворимость О2, ммоль/л
Концентрация электролита, М
НС1
NaCl
0,0 1,26 1,26 1,26
0,5 1,21 1,21 1,07
1,0 1,16 1,12 0,89
2,0 1,12 1,02 0,71
I
а Из работы: International Critical Tables, vol. Ill, p. 271. McGraw-Hill Book Co.,
New York, 1928; Todt F„ Elektrochemische Sauerstoffmessungen, W. de Guy and Co..
Berlin, 1958.
8.1.1. Основные положения теории массообмена
Растворимость кислорода в водных средах при давлении
воздуха 1 атм и комнатной температуре равна приблизительно
10 миллионным долям (млн-1) (табл. 8.1). Популяция дрож-
жевых клеток в период активного дыхания может потреблять
кислород со скоростью около 0,3 г О2 в час (в расчете на 1 г
сухого клеточного вещества). Максимальную скорость утилиза-
ции кислорода прц плотности популяции 109 клеток/мл можно
оценить, допустив, что объем клетки составляет 10-10 мл, из
которых 80% составляет вода. Тогда абсолютная потребность
в кислороде составит
( -----(Ю9 клеток/мл) (Ю’10 мл)х
(г сухой биомассы)-ч J v '
,. о / чх /л п г сухой биомассы \
Х(1 г клеточной массы/см3) 0,2----•—----:----- =
v ’ у г клеточной массы /
= 6 • 10-3 г/(мл • ч) = 6 г О2/(л • ч)
38'
596
Глава 8
//' Отдельно подни-
мающиеся
пузырьки
Газ
3) Ступенчатый
противото чный
реактор
7)£стест8енная азрация в турбулентном
режиме
а
Процессы переноса в биотехнологических системах
597
Жидкость
Воздух
7)/7олупериодический процесс
(поток Воздуха д постоянном
объеме жидкости)
Воздух
2) Уепрерыонь/й процесс (пос-
тоянные потоки жидкой
разь/ и воздуха)
4) Реактор с мешалкой и отра -
жательными перегородками
(перегородки часто применяют-
ся ив реакторах типа 7 и 2)
Воздух
д)Ресколбко пропеллерных
мешапок на одном Верти-
кальном дану (полупериода-
чески й или непрерб/днб/й
процесс)
5
В) Ступенчатый
протидоточ- Воздух
нь/й процесс
РИС. 8.2. Различные способы контакта между газовой и жидкой фазами: а —
свободно поднимающиеся пузырьки газов, свободно опускающиеся твердые
частицы и капли жидкости; б — механическое перемешивание.
598
Глава 8
Следовательно, в период активного дыхания популяция клеток
потребляет кислород со скоростью, которая соответствует при-
близительно 750-кратному насыщению кислородом в течение
часа. Поскольку запасы растворенного кислорода довольно
скудны, то популяция жизнеспособных клеток может сущест-
вовать только при условии непрерывного введения кислорода в
жидкую среду. Эта задача далеко не проста, так как низкая
растворимость кислорода является причиной малой разности
его концентраций, которая представляет собой главную дви-
жущую силу переноса кислорода из одной фазы в другую.
В случае ограниченно растворимых веществ, например кис-
лорода и углеводородов в водных средах, зависимость между
двумя равновесными концентрациями на границе раздела фаз
Cgi (со стороны газовой фазы) и сц (со стороны жидкой фазы)
обычно можно выразить уравнением, отражающим закон ли-
нейного распределения, например закон Генри:
Mcu = cgi (8.1)
Указанное уравнение справедливо, если скорость обмена рас-
творенным веществом через границу раздела фаз значительно
выше общей скорости массообмена; обычно это условие удов-
летворяется, поскольку при давлении воздуха 1 атм и 25 °C
скорость столкновений молекул Ог с поверхностью равна при-
близительно 1024 на квадратный сантиметр в секунду, что на-
много превышает общую потребность в кислороде в приведен-
ном выше примере типичного микробиологического процесса.
В стационарном состоянии скорость переноса кислорода к
поверхности раздела фаз газ — жидкость равна скорости его
транспорта через пленку со стороны жидкой фазы (рис. 8.1).
Если и Ci — концентрации кислорода в газовой и жидкой фа-
зах соответственно, то мы можем записать равенство двух ско-
ростей переноса:
Поток кислорода = число молей Ог/(см2-с) =
= kg(cg—- cig) = kite и — ci) (8.2)
со стороны со стороны
газовой фазы жидкой фазы
Здесь ki и kg — коэффициенты массоотдачи в жидкой и газовой
фазах соответственно.
Поскольку при изучении массообмена непосредственно из-
мерить концентрации на границе раздела фаз не удается, то
количественно массообмен обычно выражают через общий ко-
эффициент массопередачи Ki и общую движущую силу (раз-
ность концентраций) а*—ci, где ci* — концентрация растворен-
Процессы переноса в биотехнологических системах
599
ного вещества в жидкой фазе, находящейся в равновесии с га-
зовой фазой:
Mct*=cg (8.3)
С помощью этих параметров можно выразить плотность потока
растворенного вещества:
Плотность потока = KAct*— ci) (8.4)
Из уравнений (8.1) — (8.4) следует хорошо известное выра-
жение, связывающее общий коэффициент массопередачи Ki с
физическими параметрами двухпленочной модели kg, ki и Л1:
= ~kT (8'^
В случае ограниченно растворимых веществ М намного боль-
ше единицы. Кроме того, kg обычно намного больше ki. Отсю-
да следует, что Ki приблизительно равно ki, и, таким образом,
практически все сопротивление массопередаче сосредоточено ня
стороне пленки, обращенной к жидкой фазе.
Скорость переноса кислорода в расчете на единицу объема
реактора Qoa определяется как
<20г = скорость поглощения кислорода =
_ (плотность потока) (площадь поверхности раздела фаз) _
объем жидкой фазы в реакторе
= ^(Q*—сг)^-=^а'(сг*—сг) (8.6)
Здесь a'=A/V— отношение площади поверхности раздела фаз
газ—жидкость к объему жидкой фазы; в уравнении (8.6) принято
только что рассмотренное приближение Ki~ki. Поскольку в
этой главе основное внимание уделяется процессам аэрации, то
в дальнейшем мы будем рассматривать главным образом пере-
нос кислорода и поэтому в качестве коэффициента массопере-
дачи вместо Ki будем применять ki. Входящим в некоторые
уравнения символом а обозначают площадь поверхности раз-
дела фаз газ — жидкость в расчете на единицу объема биоло-
гического реактора (т. е. на единицу объема дисперсной систе-
мы газ — жидкость). Находящийся над жидкой фазой газ не
учитывается.
Запомните, что Qo2 представляет собой параметр, отража-
ющий локальную объемную скорость поглощения Ог; средняя
объемная скорость утилизации кислорода (выраженная в мо-
лях за единицу времени в единице объема) <2о2 во всем объе-
600
Глава 8
ме жидкой фазы V определяется как
V
(8.7)
о
В общем случае (?о3 равна Qo2 тогда и только тогда, когда в
любой точке объема реактора одинаковы и гидродинамические
условия, и отношение площади поверхности раздела фаз к объ-
ему, и концентрация кислорода.
Наблюдаемая средняя скорость массообмена определяется
целым рядом явлений и факторов, в том числе потребляемой
реактором мощностью (в расчете на единицу объема), реоло-
гией жидкой фазы и дисперсной системы, характеристиками
барботирующего устройства, общей структурой течений в реак-
торе. На рис. 8.3 изображены многочисленные связи между
наблюдаемой средней скоростью переноса и обусловливающими
этот перенос явлениями. Поскольку мы обычно не располагаем
важнейшими фундаментальными данными о скоростях коалес-
ценции и повторного диспергирования, о распределении га-
зовых пузырьков по размерам и времени их пребывания в ре-
акторе, то, как правило, мы вынуждены разрабатывать необ-
ходимые уравнения на основе соответствующих усредненных
величин: среднем размере пузырьков, газосодержании (объем-
ной доли газа, диспергированного в жидкости), среднем вре-
мени пребывания пузырьков газа и жидкой фазы в реакторе и
т. Д.
Приведенные в табл. 8.1 данные говорят о том, что Ci* за-
висит от температуры и состава среды. Зависимость от состава
среды усложняется, если растворенный газ может реагировать
с жидкой фазой. Такая ситуация типична, например, для диок-
сида углерода, который может существовать в жидкой фазе
в любой из четырех форм: СОг, Н2СОз, НСОз- и СО32-. Соот-
ветствующие константы равновесия при 25°C:
д- = [Н+][НСО3-] = 10„6>3 м (8.8)
[СО21+ [Н8СОз1
К = = 10-10,25 М (8 9)
2 [НСО3-]
показывают, что общая концентрация Ст растворенного в виде
СО2 углерода в очень сильной степени зависит от pH:
ст=> 1СО21 + [Н2СО31 + [НСО3~] + [СОз2-] =
Ч1+[£г+Лй¥] (810)
Процессы перенос а в биотехнологических системах
601
РИС. 8.3. Зависимость между интенсивностью перемешивания и скоростью
переноса газа. [Воспроизведено с разрешения из работы; Resnick W„ Gal-
Ог В., Adv. Chem. Eng., 7, 295 (1968).]
Эта зависимость в графической форме представлена на рис. 8.4;
нетрудно видеть, что при pH<5 практически весь углерод на-
ходится в виде растворенного молекулярного СОг, бикарбонат
доминирует в диапазоне 7<рН<9, а карбонат — при рН>11.
Через границу раздела газ — жидкость переносится только рас-
творенный СОг; скорость его переноса может быть описана
уравнением (8.2).
602
Глава 8
РИС. 8.4. Равновесные концентрации СОг, НСО3_, СОз2- и Н2СО3 в растворе
(ст — общая концентрация СО2 во всех четырех формах; рсо2=10-3’5 атм
(что соответствует парциальному давлению СО2 в воздухе); pH доводили до
заданного значения сильной кислотой или сильным основанием). (Из работы:
Stamm W., Morgan J. J., Aquatic Chemistry, p. 127, John Wiley and Sons,
N.Y., 1970.)
В нейтральной или основной среде большую роль играют
как эффекты массообмена, так и химические процессы. Так,
скорость обратимой реакции (8.11) очень высока:
Н2СО3 НСО3-+Н+ (8.11)
ТС (Т = 28°С) = -1H+HHCQ3-] = 2 5.10-4 моль/л (8J2)
[Н2со3]
Напротив, реакция (8.13) протекает значительно медленнее
(^1 = 20 с-’; ^=0,03 с-1 при 25°C):
Н2СО3 СО2+Н2О .(8.13)
Следовательно, в зависимости от условий стадией, определяю-
щей скорость перехода СО2 в газовую фазу, может быть или
химическая реакция [уравнение (8.13)], или физический про-
цесс [СО2(раствор)->СО2(газ) ].
Процессы переноса в биотехнологических системах 603
8.1.2. Скорость утилизации кислорода
в процессах клеточного метаболизма
Проектируя аэробные биологические реакторы, мы с по-
мощью определенных математических выражений, с большим
или меньшим приближением описывающих интересующую нас
ситуацию, часто пытаемся выяснить, является ли самой мед-
ленной стадией процесса перенос кислорода (или другого лими-
тирующего клеточный рост вещества) или его утилизация клет-
ками. Максимально возможная скорость массообмена опреде-
ляется путем простой подстановки в указанные выше уравне-
ния с/ = 0; при этом условии весь поступающий в раствор кис-
лород быстро усваивается клетками. В предыдущей главе мы
установили, что максимальная скорость утилизации кислорода
составляет хцтах/Уо2, где х—плотность культуры клеток, а
Уо2—отношение числа грамм-атомов углерода в образующихся
клеточных углеродсодержащих соединениях к числу молей усво-
енного кислорода.
Если kia'ci* намного больше хцтах/Ео2, то, очевидно, основ-
ным препятствием дальнейшему повышению скорости утилиза-
ции кислорода является метаболизм микроорганизмов и ско-
рость процесса ограничивается биохимическими факторами.
В другом крайнем случае, когда kia'ci*<^x\imzx/Yo2, кажущееся
значение ci приближается к нулю и скорость процесса в реак-
торе должна определяться явлениями переноса.
В действительности ситуация несколько сложнее. В стацио-
нарном состоянии скорости утилизации и абсорбции кислорода
должны быть равны
Qo2 = абсорбция =а утилизация =4 л'<?о.2
(8.14)
Ог
Если известна зависимость ц от ci, то с помощью уравнения
(8.14) можно определить с/ и, следовательно, скорость утили-
зации кислорода.
В общем случае существует некоторая критическая концент-
рация кислорода в жидкой фазе, при достижении которой все
клетки насыщаются Оз и количество последнего превышает
минимум, необходимый для немедленного захвата всех элект-
ронных пар, участвующих в реакциях дыхательной цепи; в этой
ситуации, следовательно, скорость процесса будет лимитиро-
вать какой-то иной биохимический механизм (гл. 5). Напри-
мер, если зависимость удельной скорости роста клеток ц от
концентрации кислорода подчиняется уравнению Моно, то
-к£+с1 (8.15)
604
Глава 8
Общее решение уравнений такого типа было дано в разд. 4.4.1,
но здесь для наглядности примем, что величина с/ значи-
тельно меньше Ci*. Для биологических реакторов такая ситуа-
ция довольно обычна. Если С/СС/*, то С/ можно выразить сле-
дующим образом:
сг=сг*
max
_ 1 /Х|Хтах
(8.16)
Если найденная таким путем величина cz больше критического
значения ссг (около 3/Со2), то скорость утилизации кислорода
микроорганизмами лимитируется каким-либо иным фактором,
например низкой концентрацией другого субстрата, даже если
средняя концентрация растворенного кислорода в жидкой фазе
значительно ниже уровня насыщения. Величины критических
концентраций кислорода для различных организмов лежат в
диапазоне от 0,003 до 0,05 ммоль/л (табл. 8.2) или от 0,1 до
Таблица 8.2. Типичные значения со2,сг в присутствии субстрата3
Организм
Температура, °C
СО2> сг.
ммоль/л
Azotobacter vinelandii 30 0,018—0,049
Е. coli 37,8 0,0082
15 0,0031
Serratia marcescens 31 -0,015
Pseudomonas denitrificans 30 -0,009
Дрожжи 34,8 0,0046
20 0,0037
Penicillium chrysogenum 24 -0,022
30 -0,009
Aspergillus oryzae 30 -0,020
а Из работы: Finn. R. К.., р. 81 in Biochemical and Biological Engineering Science,
Blakebrough N. (ed.), vol. 1, Academic Press, Inc., New York, 1967.
10% максимальной растворимости Ог (табл. 8.1), или от 0,5
до 50% конц. насыщения кислородом воздуха. Очевидно, что
массообмен кислорода особенно важен в случае организмов,
для которых характерны большие значения критической кон-
центрации кислорода, например плесеней Penicillium.
Различные факторы могут влиять на общую потребность
микроорганизма в кислороде хц/Уо2, которая в свою очередь
определяет необходимые в проектировочных работах минималь-
ные значения kia' [уравнение (8.14)]. К числу наиболее важ-
ных факторов такого типа относятся вид клеток, фаза роста
культуры, источники углерода, pH среды, тип изучаемого мик-
Процессы переноса в биотехнологических системах
605
РИС. 8.5. Скорость утилизации кислорода периодической культурой Myrothe-
cium verrucaria. [Воспроизведено из статьи: Darby R. Т., Goddard D. R., Am. J.
Bot., 37, 379 (1950).]
робиологического процесса; последний фактор включает пара-
метры утилизации субстрата, образования биомассы или выхо-
да продукта жизнедеятельности микроорганизмов (гл. 7).
В приведенных на рис. 8.5 результатах определения скорос-
ти утилизации кислорода в периодическом процессе удельная
потребность в Ог достигает максимума в начале экспоненциаль-
ной фазы, а х возрастает и на более поздних этапах роста.
Произведение х\ь, а следовательно, и общая потребность в кис-
лороде максимальны в конце экспоненциальной и в самом на-
чале стационарной фазы; таким образом, скорость утилизации
кислорода достигает максимума позднее, чем удельная ско-
рость роста.
На скорость утилизации кислорода большое влияние оказы-
вает природа углерод содержащего питательного вещества. Глю-
коза, например, в общем случае метаболизируется быстрее, чем
другие углеводы. Найдено, что максимальная скорость потреб-
ления кислорода Penicillium составляет 4,9, 6,7 п 13,4 моль/(л -ч)
606
Глава 8
в случае роста плесени на лактозе, сахарозе и глюкозе соответ-
ственно [2].
Суммарная потребность клетки в кислороде складывается из
потребностей на поддержание жизнедеятельности клеток, окис-
лительные реакции в процессе дыхания (используемые для ро-
ста клеток и для целей биосинтеза) и окисление субстратов в
соответствующие конечные продукты метаболизма. При изуче-
нии стехиометрии метаболизма (гл. 5) мы видели, что скорость
утилизации кислорода, расходуемого на клеточный рост, обычно
непосредственно связана с количеством потребляемого субстра-
та, являющегося источником углерода. Кроме того, более вос-
становленные субстраты, например метан и высшие углеводоро-
ды, требуют усиленной утилизации кислорода клетками по
сравнению с субстратами типа глюкозы, степень восстановлен-
ности атомов С в которых приблизительно та же, что и в ком-
понентах клетки. Так, коэффициенты Уо2/с [т. е. отношение чис-
ла молей утилизированного кислорода к числу молей метаболи-
зированного субстрата (в расчете на одноуглеродный субстрат)]
равны 1,34, 1,0 и 0,4 для типичных микроорганизмов, растущих
на метане, парафиновых углеводородах и углеводах соответст-
венно.
Кислород может также выполнять роль реагента, участвую-
щего в биотрансформациях. В производстве 5-кетоглюконовой
кислоты из глюкозы, например, на первой стадии роста перио-
дической культуры Acetobacter происходит окисление части
содержащейся в среде глюкозы до глюконовой кислоты; при
этом С>2 используется как для клеточного роста, так и для син-
теза продукта метаболизма. После истощения глюкозы рост
клеток прекращается, а глюконовая кислота превращается в
5-кетоглюконовую кислоту по следующему уравнению реакции:
CsHisOyr’/гОг —>- CgHioOz+HaO (8.17)
В заключительной стадии процесса осуществляется только ука-
занная реакция, и потребление кислорода связано с образовани-
ем продукта метаболизма в соответствии со стехиометрией
уравнения (8.17).
8.2. Определение скорости переноса кислорода
В идеальном варианте скорость переноса кислорода должна
измеряться непосредственно в биологическом реакторе, содержа-
щем как питательную среду, так и изучаемую популяцию (или
популяции) клеток. Такой эксперимент, однако, возможен толь-
ко при соблюдении всех требований, предъявляемых к работе
со средой и посевным материалом, и всех предосторожностей,
необходимых для предотвращения случайного заражения и за-
Процессы переноса в биотехнологических системах
607
трязнения среды и культуры; поэтому для изучения эффектов
массообмена такой подход слишком сложен и трудоемок. В свя-
зи с этим скорость переноса кислорода обычно изучают на
синтетических системах, не содержащих живых организмов;
свойства таких систем должны примерно соответствовать свой-
ствам системы в биологическом реакторе. Основной целью этих
экспериментов является выяснение зависимости kta' от гидро-
динамики системы.
Для того чтобы синтетическая среда достаточно точно соот-
ветствовала изучаемой культуре клеток, необходимо, чтобы у
них были идентичными:
1. Вязкость и другие реологические характеристики (см. разд.
8.8).
2. Сопротивление массообмену на границе раздела газ — жид-
кость.
3. Коалесценция пузырьков.
4. Растворимость кислорода и его коэффициент диффузии.
В общем случае пригодность синтетической (бесклеточной)
системы для изучения и приближенного описания биологичес-
кого процесса зависит от степени выполнения указанных усло-
вий. Эксперименты по изучению поглощения кислорода чистой
водой, например, не удовлетворяют практически ни одному из
этих условий. Ниже мы вкратце рассмотрим влияние вязкости
жидкой среды, поверхностно-активных веществ и характеристик
перемешивания на скорость переноса кислорода.
8.2.1. Определение kta' с помощью химических реакций
газообразных веществ с жидкостями
Обращаясь к путям переноса кислорода, изображенным на
рис. 8.1, нетрудно заметить, что если кислород поглощается с
достаточно большой скоростью в химической реакции, проис-
ходящей в жидкой фазе, то с^О. В таком случае, определив
скорость химической реакции в жидкой фазе, равную k;a'c}*,
нетрудно найти величину kia'. Во многих ранних работах по
изучению массообмена в качестве поглотителя Ог использовали
сульфит натрия, окисляющийся в присутствии катализаторов
(ионов некоторых металлов, например Со2+) до сульфата:
катализатор
SO32- + 1/2О2 ----SO42- (8.18)
Кинетика реакции окисления сульфитного иона (в растворе)
до сульфатного довольно сложна. Порядок реакции по кисло-
роду и сульфиту зависит от природы катализатора и его кон-
центрации, что свидетельствует о своеобразной природе элемен-
608
Глава 8
тарных стадий, приводящих к отражаемому уравнением (8.18)
общему простому результату. Независимо от порядка реакции
в жидкой пленке, окружающей каждый пузырек газа, химиче-
ская реакция будет протекать в незначительной степени (и,
следовательно, ситуация будет близка к изображенной на рис.
8.1 схеме) только при условии, что общая скорость реакции в
пленке пренебрежимо мала по сравнению со скоростью массо-
передачи ki(c* — с). Если — толщина пленки, участвующей в
процессе массопередачи, то в математической форме этот кри-
терий можно выразить следующим образом:
С X скорость I пленка < kt (с* — с) (8.19)
Скорость реакции в пленке будет ниже скорости, отвечающей
концентрации сульфита и уровню насыщения кислородом в
жидкой фазе (с*, сульфитжидк); выраженное с помощью пара-
метра (с*, сульфитЖИдк), который можно определить экспери-
ментально или расчетным путем, условие низкой скорости реак-
ции в пленке можно сформулировать следующим образом:
^Хскорость (с*, сульфитжидк) <kl (с* — с) (8.20)
«Толщина» пленки массопередачи равна
» ®о2
Если допустить, что реакция имеет порядок ai по кислороду и
«2 по сульфиту, то получим неравенство
•®о2 IMc*)ai (сульфит)*2]
2 —С)
(8.21)
(8.22)
ki
Отсюда следует, что
kl
®о2 №г (с*)а1(сульфит)аз 1/2
с* — с
(8.23)
катализа-
этих условиях
В эксперименте Данквертса применялся кобальтовый
тор в концентрации 10~5 М (известно, что при
af = 2), а достаточно высокая концентрация сульфита (напри-
мер, 0,5 М) обеспечивала а2 = 0. Тогда при с<^с* неравенство
(8.23) преобразуется в выражение
k^lSD^krC^
Принимая ^о2= 1,6-Ю-5 см2/с, &r=0,85> 108 см3/(г-моль-с) (для
кобальтового катализатора) и с* = 1,35-10~7 г-моль/см3, получим
^/^>0,01 см/с
(8.24)
Процессы переноса в биотехнологических системах
609
В случае менее эффективного катализатора (меньшей величи-
ны kr) ki также уменьшится.
Позднее в этой же главе мы познакомимся с некоторыми
уравнениями, показывающими, что для больших пузырьков в
воде /г(^0,04 см/с, а для малых пузырьков &гл;0,01 см/с.
Следовательно, указанное неравенство устанавливает
минимальный размер пузырьков, при котором сох-
раняется справедливость приведенных выше выражений.
В случае мелких и медленно поднимающихся пузырьков газа
скорость реакции в пленке, окружающей эти пузырьки, будет
достигать заметных величин. Аналогично увеличение диаметра
пузырьков и повышение вязкости среды (по сравнению с во-
дой) приведут к снижению коэффициента массопередачи; учиты-
вающий этот пленочный эффект соответствующий коэффициент
можно вычислить, если известны константа скорости реакции и
ее порядок [3—5].
Для процесса окисления сульфитного иона характерны и
другие интересные особенности. Так, константа скорости kr
зависит, во-первых, от природы катализатора и его концентра-
ции, во-вторых, от ионной силы раствора, в-третьих, от наличия
загрязнений, влияющих на каталитическую активность, в-чет-
вертых, от pH раствора. Например, в присутствии солей кобаль-
та в концентрации 10~5 М при 20 °C повышение pH от 7,50 до
8,50 сопровождается увеличением kr в 10 раз. (В реакции гене-
рируются Н+, поэтому для поддержания постоянного pH к сме-
си нужно добавлять основание.)
Несмотря на все эти осложнения, в литературе описаны
примеры, когда определенная сульфитным методом для данных
барботера, скорости перемешивания и т. д. величина kia' доста-
точно точно соответствовала значениям kfi', найденным в кон-
кретном микробиологическом процессе. (Впрочем, известны и
обратные случаи.) Если допустить, что приведенная на рис. 8.1
схема отражает изучаемый биологический процесс, то на плен-
ке, окружающей пузырьки газа со стороны жидкости, практи-
чески не должно происходить поглощения О2. Следовательно,
любая химическая система, адсорбирующая О2 и предназначен-
ная для моделирования поведения культурального бульона,
должна помимо прочих предъявляемых к ней требований удов-
летворять и приведенному выше основному неравенству (8.19).
Если же растущие клетки концентрируются в тонком слое жид-
кости, окружающем пузырьки газа, то может возникнуть необ-
ходимость в применении иной химической модели.
Скорость переноса кислорода можно измерять и другими
методами. Если экспериментальная система представляет собой
строго периодическую культуру, когда в систему не вводят и
из нее не выводят ни жидкие, ни газообразные компоненты, то
39—537
610
Глава 8
<?о2 можно найти путем определения изменения объема или
давления газа во времени. Кроме того, как мы увидим в следую-
щем разделе, определению kta' помогает и оценка Q.
Если газ постоянно вводится в жидкую фазу и постоянно
выводится из нее, то для определения (>о2 можно использовать
следующее уравнение материального баланса по газообразному
кислороду:
^о2 ~ [^(вход) (Ро2)(вход) g(выход) (Ро2)(выХод)]/^Т (8.25)
Здесь Fg — объемная скорость потока газа, а /?о2 — парциаль-
ное давление О2. [При каких допущениях справедливо уравне-
ние (8.25)? Применимы ли в общем случае эти допущения к
биотехнологическим процессам?]
Как следует из уравнений (8.6) и (8.7) и соответствующего
обсуждения, найденные таким образом значения (?о2 можно
использовать для определения kia' только тогда, когда условия
в любой точке объема реактора одинаковы и, следовательно,
локальные скорости утилизации кислорода равны соответствую-
щей средней скорости. Поэтому при лабораторном изучении эф-
фектов массообмена с целью проектирования биологических
реакторов часто используются реакторы с перемешиванием
[рис. 8.2,6 (2)].
Если проблема обеспечения однородности условий в объеме
реактора решена и если известны Ci'* и Ci, то по уравнению
(8.6) можно определить kia'. Значение ct* можно найти в спра-
вочной литературе, например в табл. 8.1, а непосредственное
определение ct в настоящее время можно выполнить (даже в
чистых культурах микроорганизмов) с помощью полярографи-
ческого стерилизуемого кислородного электрода. Принцип дей-
ствия такого электрода, вырабатывающего ток, сила которого
пропорциональна локальному парциальному давлению раство-
ренного кислорода, описан в гл. 10. Там же рассматривается
другой метод определения kia', основанный на определении из-
менения концентрации О2 с помощью кислородного электрода.
В различных реакторах и в естественных процессах локаль-
ная скорость переноса кислорода может меняться в зависимо-
сти от пространственного положения данной точки. Если такие
изменения характерны для реактора, в котором измеряются
скорости массопередачи, то найденное усреднепнное (по объему
данного реактора) значение kia' уже нельзя непосредственно
использовать в операциях масштабного переноса, в которых
результаты лабораторных экспериментов переносятся на аппа-
раты значительно большего объема. Методы масштабного
переноса мы рассмотрим позднее в этой же главе. Изменения
концентрации растворенного О2 в «гомогенной» фазе (жидкой
Процессы переноса в биотехнологических системах 611
фазе, грануле плесневых грибов, пленке микроорганизмов
и т. д.) определяли с помощью миниатюрных кислородных зон-
дов, диаметр сенсорной головки которых составляет около
10 мкм. Это устройство использовалось, в частности, для изуче-
ния локальных профилей концентрации кислорода в гранулах
плесени и определения коэффициентов диффузии в скоплениях
микроорганизмов.
8.3. Массообмен с участием свободно
поднимающихся и свободно падающих тел
Скорость массообмена между различными зонами системы
определяется уравнениями, описывающими сохранение массы
всей системы, ее отдельных компонентов (например, кислорода)
и баланс количества движения. Если эти уравнения выражены
через относительные величины, отражающие изменение расстоя-
ний, скоростей и концентраций, и если основной движущей си-
лой переноса жидкого или газообразного компонента является
разность плотностей между двумя контактирующими фазами,
то в конечных уравнениях появляются три безразмерных пара-
метра (три критерия подобия), называемые числами Грасгофа,
Шервуда и Шмидта; при описании массообмена эти числа опре-
деляются следующим образом:
Число Грасгофа: Gr = Pz ^pz—(8.26a)
Й2/
Число Шервуда: Sh*=-^— (8.266)
Число Шмидта: Sc = —— (8.26в)
P/#o2 v '
Здесь D — характерная размерность и щ — вязкость непрерыв-
ной фазы. Можно ожидать, что в случае конвективного движе-
ния выражения для коэффициентов массопередачи будут вклю-
чать только эти три критерия подобия.
8.3.1. Коэффициенты массопередачи
для газовых пузырьков и потоков пузырьков
Коэффициент массопередачи для пузырька газа представля-
ет собой коэффициент пропорциональности между общей плот-
ностью потока пузырьков и общей движущей силой массопере-
дачи (с* — ct). Локальная плотность потока на границе раздела
фаз газ — жидкость равна —3)o2(dc/dz)z^=o (справедливо при
низких скоростях массопередачи), где z— координата, измерен*
39*
612
Глава 8
ная в жидкой фазе. Таким образом,
*< = 77^77 <8-27)
С1 С1 й 02 г-0
или в безразмерных параметрах:
kiD _ - 1
®о2 I — Cl
z=0
(8.28)
Относительная (безразмерная) концентрация с вблизи гра-
ницы раздела фаз может быть найдена путем решения уравне-
ний переноса:
c = f(z, Sh, Sc, Gr) (8.29)
Используя это выражение для оценки производной (dc/dz);=0
в уравнении (8.28), в принципе можно выразить искомый коэф-
фициент массопередачи ki через число Шервуда:
Sh = ^- = ?;(Sc,'Gr) (8.30)
Таким образом, безразмерный коэффициент массопередачи Sh
является функцией только двух параметров — Sc и Gr. Здесь
D — характерный диаметр пузырьков.
В литературе описаны и другие выражения для коэффициен-
тов массопередачи для случая опускающихся или поднимающих-
ся пузырьков, капелек или твердых частиц, в которых применя-
лись иные критерии подобия, например число Рейнольдса (Re =
= pzZ)zz/p,z) или число Пекле (Ре = нО/^о2). В этих безразмерных
параметрах и— скорость пузырьков газа относительно скоро-
сти жидкости. И в первом и во втором случае выражение ско-
рости и через разность плотностей Др= (рг—pg) приводит в
соответствии с уравнением (8.30) к зависимости коэффициента
массопередачи только от Gr и Sc.
Массопередачу из изолированной сферы с жесткой границей
раздела фаз (эта модель достаточно точно описывает систему,
состоящую из небольших пузырьков в культуральной жидкости
в присутствии поверхностно-активных веществ) можно опреде-
лить теоретически для случая Re=pzZ)w/pz<^l и Pe=wZ)/^o2^>l-
(Таким образом, uDl^^l^piDulpi, откуда следует, что
Цг/рг^о2 = 3с>1. Может ли быть справедливым обратное нера-
венство?) В водных средах, кинематическая вязкость которых
(v = pz/pz) составляет приблизительно 10~2см2/с, а ^о2 равен
примерно 10~5 см2/с, число Шмидта обычно имеет величину по-
Процессы переноса в биотехнологических системах
613
рядка 103. Следовательно, при Re=10-1—10-2 применимо полу-
ченное расчетным путем уравнение
Sh= 1,01Ре1/3 = 1,01 (иБ>/$О2)1/3 (8.31)
Для малых чисел Рейнольдса, при которых применимо урав-
нение (8.31), конечная скорость ut сферы определяется уравне-
нием
__ D2&pg /о оо\
Подстановка ut из уравнения (8.32) в уравнение (8.31) дает
sh= I.oif «^-У'3= 1,01 (
18р./£>о2 J 18р.2/ J Р/<0о2 У
= O.SOGr^Sc1^ (8.33)
[Выражение (£>3Apg)/p,/S>02 известно также как число Рэлея
Ra.] Обратите внимание на то, что здесь, как и следовало ожи-
дать, Sh = f(Gr, Sc).
В случае больших чисел Рейнольдса для одиночного нецир-
кулирующего сферического пузырька в ламинарном режиме
предложено другое уравнение:
Sh = 2,0 + 0,60Re,/2Sc'/a Re»l (8.34)
Интересно, что здесь Sh изменяется пропорционально квадрат-
ному (а не кубическому) корню из параметра относительной
скорости; это свидетельствует об изменении гидродинамического
режима. Опять-таки, подстановка вместо и выражения для соот-
ветствующей конечной скорости приведет к Sh=/(Gr, Sc).
Во многих промышленных реакторах с барботажем воздуха
(рис. 8.2) воздушные пузырьки группируются в потоки, в кото-
рых они соприкасаются настолько тесно, что течение в дисперс-
ной системе и массопередачу вблизи границы раздела фаз
газ — жидкость уже нельзя достаточно точно описать моделью,
учитывающей гидродинамические характеристики и массопере-
дачу для изолированного пузырька. Согласно данным Калдер-
бэнка и Му-Янга, адсорбцию ограниченно растворимых газов
в жидкостях, реагирующих с этими газами, можно описать с
помощью всего лишь двух уравнений [10]. Существуют два
принципиально различных режима массопередачи с участием
потоков пузырьков; граница между этими режимами определя-
ется критическим диаметром пузырьков Dc. В отсутствие поверх-
ностно-активных веществ Z)c«2,5 мм. Более крупные пузырьки
обычно возникают в перемешиваемых реакторах с чистой водой
и в колоннах с ситчатыми тарелками. Пузырьки меньшего диа-
метра характерны для колонн с тарелками из спеченного ма-
40—537
614
Глава 8
териала, а также для перемешиваемых сосудов, содержащих
водные растворы гидрофильных веществ.
При D<ZDc = 2,b мм
Sh = -^- = 0,31Gr1/3Sc1/3= 0,3Ra1/3 (8.35>
®o2
При D~>Dc = 2,b мм
Sh = = 0,42GrV3ScV2 (8.36>
Из уравнений (8.33) и (8.35) следует, что в потоке пузырь-
ков при одних и тех же значениях Sc и Gr коэффициент массо-
передачи приблизительно на 20% ниже по сравнению с изоли-
рованными пузырьками с неподвижной поверхностью. Показано,,
что уравнение (8.36) справедливо и для эрлифтных систем
(рис. 8.2, а), если вместо коэффициента 0,42 ввести коэффици-
ент 0,50.
Изменение показателя степени числа Шмидта в уравнении
(8.36) [по сравнению с уравнением (8.35)] свидетельствует об
изменении гидродинамического режима. В случае ньютоновских
жидкостей, вязкость которых постоянна и не зависит от скоро-
сти сдвига (обусловленной скоростью перемешивания, скоро-
стью движения пузырьков и другими факторами), переход от
области D<ZDC к области D^>DC сопровождается изменением
формы пузырьков от почти сферической (небольшие пузырьки)
до полусферической и грибообразной. Более подробно вопрос о
гидродинамике потоков пузырьков рассмотрен в работе [10].
На величину Dc влияют поверхностно-активные вещества; сооб-
щалось, что в некоторых средах Dc достигает величины 7,0 мм.
В некоторых неньютоновских жидкостях, которые мы рассмот-
рим позднее, переход от одного режима массопередачи к друго-
му совершается более плавно; в таких системах нет резкого
скачка, характерного для ньютоновских жидкостей.
Результаты изучения массопередачи в случае малых частиц
показывают, что по мере снижения разности плотностей Др
число Шервуда приближается к низшему пределу 2,0. В случае
суспензий отдельных клеток, их скоплений, хлопьевидных обра-
зований и т. д., а также в случае эмульсий газойля или других
углеводородов более точно значение числа Шервуда определя-
ется уравнением
Sh = = 2,0 + 0,3 lRa1/3 (8.37)
<=^о2
или
(8.38)
“^02 1) | Р/Ц)., v
Процессы переноса в биотехнологических системах
615
Отсюда следует, что относительный вклад чисто диффузионного
эффекта (Ар = 0, &г = 2,02>о2/£) уменьшается по мере увеличе-
ния размеров частиц. Для изолированной клетки 2&OJD равно
приблизительно 0,1 см/с, в то время как второе слагаемое, явля-
ющееся производным от числа Рэлея, равно 0,01 см/с; следова-
тельно, массопередача вблизи поверхности клетки близка к
массопередаче сферы в относительно неперемешиваемой среде.
Большие размеры хлопьев, пленок и т. п. обусловливают значи-
тельное повышение относительного вклада второго слагаемого.
8.3.2. Определение площади поверхности раздела фаз
и газосодержания дисперсной системы
Определив по той или иной из приведенных выше формул^,
мы далее должны найти площадь поверхности раздела фаз а',
приходящуюся на единицу объема. Значение а' можно оценить
расчетным путем по известному диаметру отверстий барботи-
рующего устройства и по другим данным о конструкции реак-
тора, а также путем фотографирования процесса или иными
методами. Если время пребывания пузырька в реакторе равно
tb, объемная скорость потока в расчете на одно отверстие бар-
ботера Fo, а общее число (одинаковых) отверстий п, то площадь
поверхности границы раздела фаз в единице объема а' (если
пренебречь коалесценцией и изменением D в результате изме-
нения гидростатического давления или абсорбции) определяется
уравнением
(8.39)
.объем 0 ° nD3/6 V D v
Ниже мы рассмотрим параметры правой части уравнения
(8.39), в частности, детально изучим физические факторы, оп-
ределяющие размер пузырьков, и на этой основе оценим воз-
можность определения размера пузырьков расчетным путем как
функции от параметров работы реактора, конструкции смеси-
теля и свойств жидкой среды. Многие из приведенных здесь,
концепций, относящихся к системам с поднимающимися отдель-
ными пузырьками или потоком пузырьков, применимы и для
изучения процессов переноса в реакторах с механическим пере-
мешиванием.
Размер пузырьков в биологических реакторах определяют
три важнейших взаимосвязанных фактора; аналогичные факто-
ры играют основную роль и в формировании дисперсии второй
ограниченно растворимой жидкой фазы. К этим факторам
относятся механизм образования пузырьков, их распад на мень-
шие пузырьки и коалесценция (слияние пузырьков). Образова-
ние пузырьков обусловлено неустойчивостью поступающего в
40*
516
Глава 8
жидкую фазу газового потока, что приводит к разрыву единой
струи на отдельные пузырьки. Распад пузырьков зависит от кон-
куренции между поверхностным натяжением, стабилизирую-
щим пузырек, и локальными гидромеханическими силами,
стремящимися его разрушить.
Вероятность коалесценции пузырьков определяется свойства-
ми поверхности раздела фаз газ — жидкость. В обычных для
процессов биохимической технологии водных системах на
коалесценцию влияют в первую очередь растворенные в жидкой
фазе вещества, например жирные кислоты, полиспирты, элект-
ролиты, кетоны. Добавление этих веществ подавляет коалесцен-
цию. В последующем обсуждении мы рассмотрим два крайних
случая: коалесцирующие дисперсные системы (например, воз-
дух— чистая вода) и некоалесцирующие дисперсные системы
(например, воздух — водный раствор электролита).
Наш анализ мы начнем с изучения процесса образования
пузырьков газа при его поступлении с объемной скоростью Fo
через единственное отверстие диаметром d. Символом Do мы
будем обозначать начальный диаметр образующегося пузырь-
ка. В этой ситуации можно различать два режима. При низких
скоростях потока газа в отверстии формируется последователь-
но один пузырек за другим. Простейший подход к анализу тако-
го режима основан на изучении равновесия сил, приложенных
к покидающему отверстие пузырьку. Это происходит в тот
момент, когда выталкивающая сила (nD03&pg)/6 уравновешива-
ется удерживающей силой nod:
=6 (8-«)
Подробнее теория этого вопроса изложена в работе [21].
При некоторой критической скорости потока газа Ео* в от-
верстии барботера наблюдается переход от отдельных пузырь-
ков газа к непрерывной струе. Методы точного расчета Fo* в
настоящее время не разработаны; известно, однако, что крити-
ческая скорость потока газа должна находиться в диапазоне
V-П5Г-с F* < 20’4 /т (см3/с) (8Л1)
Если скорость газового потока выше Fo*, то газовые пузырьки
образуются путем разрыва газовой струи вследствие неустой-
чивости поверхности раздела фаз. Разработанная Рэлеем теория
устойчивости предсказывает, что диаметр пузырьков, образую-
щихся в результате разрыва газовой струи при ламинарном
течении жидкой фазы, приближенно должен быть равен
Z>o=rf(12jt/0,485)1/3 = 4,27d (8.42)
Процессы переноса в биотехнологических системах
617
при условии, ЧТО [21]
op/d
тг
36
(8.43)
В процессах биохимической технологии это условие обычно
выполняется.
При введении газового потока в вязкую культуральную
среду основное сопротивление образованию новых пузырьков
оказывает не столько поверхностное натяжение пузырьков,
сколько вязкость жидкой среды. Если пузырьки образуются, то
отношение диаметров пузырька и отверстия барботера (D/d)
определяется уравнением
(= 3,23Re0_0,1Fr00’21 (8.44)
Здесь относящиеся к отверстию барботера числа Рейнольдса и
Фруда задаются уравнениями (8.45) и (8.46) соответственно:
Re0 =
4pcF0*
зт d
Fr0 =
(Л)*)2
d5g
(8.45)
(8.46)
Иногда в дисперсной системе газ — жидкость пузырьки газа
имеют меньший диаметр, чем на выходе из распределяющего
газ устройства. Причиной этого явления может быть неустой-
чивость пузырьков под действием сил, прилагаемых к ним дви-
жущейся непрерывной фазой. Для диспергированной жидкой
или газовой фазы в непрерывной жидкой фазе максимальный
размер (диаметр) свободно поднимающихся (опускающихся)
или перемещаемых за счет перемешивания пузырьков или ка-
пель диспергированной фазы определяется равновесием проти-
водействующих сил:
1. Динамического давления т (суммы разностей напряжения
сдвига и нормального напряжения), стремящегося придать
каплям вытянутую форму и в конце концов разделить их на
меньшие капли; этому процессу препятствуют две указанные
ниже силы.
2. Сил поверхностного натяжения o/D частицы, стремящихся
вновь придать капле сферическую форму, отвечающую мини-
мальной поверхностной энергии.
3. Сопротивлением диспергированной фазы деформации, обус-
ловленным^ вязкостью; это сопротивление пропорциональ-
но Pd^|_|V'r/pd (здесь индекс d отражает принадлежность
параметра к диспергированной фазе).
618
Глава 8
В системах газ — жидкость сила 3 пренебрежимо мала по
сравнению с силой 2. В дисперсных системах типа жидкость —
жидкость сила 3 должна быть относительно большой, но, как мы
увидим позднее в примере 9.2, даже в этом случае силы поверх-
ностного натяжения играют главную роль.
Эффект двух последних факторов уменьшается пропорцио-
нально D~?, где р — положительное число. Отсюда следует, что
при некоторой критической величине диаметра Dc динамическое
давление преодолеет два противодействующих сопротивления и
пузырьки или капли распадутся на меньшие частицы. Критиче-
скому диаметру, очевидно, должно соответствовать следующее
равенство:
° । п-i т 1у2
ГП{ ~гГ -ГГПМЛ) 1 - = Т
1 D Pd
(8-47)
где mi и m2 — константы.
Если силы поверхностного натяжения намного превосходят
силы, обусловленные вязкостью диспергированной фазы, то при
критическом диаметре пузырьков
mt= т, или т^т-у-, или Dc — (8.48)
Согласно уравнениям (8.48) максимальный размер пузырька
равен частному от деления произведения безразмерной констан-
ты mi и поверхностного натяжения на динамическое давление.
Следовательно, повышение динамического давления приведет
к распаду пузырьков все меньшего диаметра.
В теоретических или эмпирических уравнениях, описываю-
щих взаимосвязь между максимальным диаметром устойчивого
пузырька (или капли) и свойствами жидкой среды и течения,
обычно применяется критерий подобия, основанный на уравне-
ниях (8.48). Так, число Вебера We определяется как
We = x-y (8.49)
Критическое число Вебера Wec есть не что иное, как значение
We при D = DC\ согласно уравнению (8.48), Wec является харак-
терной константой. Экспериментальным путем и теоретическими
расчетами показано, что для системы чистый воздух — BOAaWec
приблизительно равно единице (точнее, Wec=l,05).
Чтобы с помощью изложенных концепций и уравнений вы-
числить максимальный диаметр устойчивых пузырьков, нам
необходимо определить значения динамического давления т,
соответствующие различным режимам течения. Для свободно
Процессы переноса в биотехнологических системах
619
поднимающихся пузырьков т определяется по уравнению
А
т=_Р^Х (8.50)
Здесь ш — конечная скорость пузырьков, которую в случае сфе-
рических пузырьков можно определить по уравнению (8.32).
В сложном турбулентном течении определение динамического
.давления затруднено. Турбулентность следует ожидать, напри-
мер, в барботажных колоннах вблизи аэрирующего сопла.
Впервые столкнувшись здесь с этой проблемой, мы перейдем к
обсуждению некоторых связанных с турбулентностью понятий
общего характера, которые полезны и при рассмотрении ряда
последующих тем.
В разработанной А. Н. Колмогоровым и другими исследова-
телями статистической теории турбулентности поле турбулент-
ного течения рассматривается как результат суперпозиции вих-
рей или флуктуаций скоростей, характеризующихся масштабом
длины или частотой флуктуаций и величиной. Наиболее круп-
номасштабные вихревые элементы или первичные вихри имеют
масштаб основного течения. Эти первичные вихри нестабильны
и распадаются на меньшие, которые в силу своей неустойчиво-
сти распадаются на еще более мелкомасштабные вихри и так
далее. Этот каскад превращений крупномасштабных вихрей в
мелкомасштабные сопровождается потоком кинетической энер-
гии, пока в конце концов она не рассеется в виде теплоты. По-
ток кинетической энергии через каскад вихрей приводит к тому,
что направленный характер первичных вихрей (зависящий от
геометрии реактора, конструкции сопел, мешалок и т. п.) посте-
пенно затухает. Теория Колмогорова утверждает, что мелко-
масштабные вихри статистически не зависят от первичных
вихрей и локально изотропны (пространственно однородны).
Масштаб длины Хо наиболее мелкомасштабных вихрей, которые
рассеивают энергию турбулентности, определяется уравнением
а Н/3^4 ( Р \ /о с|\
где P/Vi — потребляемая мощность, приходящаяся на единицу
объема.
При изучении влияния турбулентности обычно используют
усредненные во времени параметры. Так, среднеквадратичная
скорость zzrms=<H2(/)>1/2 (скобки <> обозначают усреднение во
времени мгновенных флуктуаций скорости) отражает среднюю
величину изменений локальных скоростей. Для масштабов
длины I, намного меньших масштабов первичных вихрей и на-
много больших Хо, зависимость среднеквадратичной скорости
620
Глава 8
вихрей от характерного масштаба I определяется выражением
и I = ccf Р У/3
ггп|вихр и масштаба I \ Vl I I Pl
(8.52)
где а — константа.
Возвращаясь к проблеме нахождения выражения для дина-
мического давления, которое могло бы быть затем использова-
но для определения числа Вебера, отметим, что мы можем вос-
пользоваться уравнением
Р/ (Mrms n
|вихри масштаба Dc
(8.53)
Это выражение является мерой турбулентного напряжения
сдвига, действующего на пузырек диаметром Dc. Из уравнений
(8.49), (8.52) и (8.53) получаем
п ' а°’6
D. = a --------------
С (P/Vi)^pi°^
(8-54)
Здесь а' — константа. Таким образом, повышение потребляемой
мощности на единицу объема приводит к уменьшению макси-
мального размера устойчивых пузырьков.
Существенно, что, согласно теории изотропной турбулентно-
сти, приходящаяся на единицу объема потребляемая мощность
является важнейшим параметром, определяющим как масштаб
образующихся вихрей, так и интенсивность турбулентных флук-
туаций скорости, сравнимых по масштабу длины с размерами
пузырьков и капель. Локальная изотропная турбулентность
представляет собой теоретическое явление, далеко не всегда
наблюдаемое на практике; тем не менее нам важно не забы-
вать физическую интерпретацию механизма передачи энергии
(энергии сжатого газа или механического перемешивания) га-
зовым пузырькам, каплям, хлопьям или гранулам мицелия.
Кроме того, уравнения (8.52) и (8.54) предполагают, что вели-
чина P/Vi будет оказывать существенное влияние на коэффици-
енты массопередачи в барботажных колоннах и резервуарах с
перемешиванием. Особо следует подчеркнуть еще один важный
аспект этой теории: основную роль играет количество энергии,
приходящееся на единицу объема, а не способ ее передачи. Не
играет большой роли, например, передается ли энергия потоком
сжатого газа или перемешивающим устройством, не существен-
на и конструкция последнего и т. д. Это обобщение опять-таки
носит теоретический характер, однако в ряде случаев теорети-
ческие и соответствующие экспериментальные данные хорошо
согласуются.
Рассмотрев независимо процессы коалесценции, образования
и распада пузырьков, мы можем теперь перейти к изучению раз-
Процессы переноса в биотехнологических системах
621
личных возможных типов взаимодействия этих процессов в
барботажной колонне. Сначала предположим, что в рассматри-
ваемой двухфазной дисперсной системе медленно происходит
коалесценция пузырьков. Если начальный диаметр пузырьков
Do меньше максимального диаметра устойчивых пузырьков Dc,.
характерного для условий наибольшего динамического давления
(в барботажной колонне обычно в области формирования пу-
зырьков), то характерный размер пузырьков будет равен Do
(табл. 8.3). Если же Do больше Dc, то следует ожидать распада
пузырьков на более мелкие с характерным диаметром Dc.
Таблица 8.3. Зависимость характерного диаметра пузырьков D от их
тенденции к коалесценции и соотношения между диаметром пузырьков
в момент образования пузырьков (Dc) (Do) и максимальным диаметром устойчивых
Начальные условия Некоалесцирующая система Коалесцирующая система
D. <DC D«D0 Переход к равновесной?
Dq Dc дисперсной системе D « Dc D аг Dc (барботаж, перемешива- (локальное равновесие) ние)
С другой стороны, если коалесценция происходит быстро, то
образующиеся пузырьки будут сливаться и увеличиваться в-
размерах до тех пор, пока их диаметр не превысит Dc. Тогда
начнется обратный процесс распада пузырьков. В этом случае
начальный размер пузырьков Do может сохраняться только в
области барботера и практически не влияет на действительный
размер пузырьков в реакторе. Учитывая, что флуктуации скоро-
стей в турбулентном режиме, а следовательно, и Dc в общем
случае различны в разных точках реактора, для каждой точки
коалесцирующей системы характерна тенденция к равновесию
между коалесценцией и дроблением пузырьков так, что в конеч-
ном итоге образуются пузырьки с диаметром Dc.
Эти соображения необходимо учитывать при проектирова-
нии соответствующих аппаратов. В некоалесцирующей системе
рассеяние энергии, расходуемой на диспергирование газа, наи-
более эффективно в зонах образования и диспергирования
пузырьков. Если же процессы коалесценции играют большую
роль, то рассеяние энергии, расходуемой на диспергирование
газа, происходит более или менее равномерно во всем объеме-
жидкой фазы. Как мы увидим позднее (гл. 9), эти теоретиче-
ские выводы послужили основой для разработки, изучения иг
«22
Глава 8
использования перемешивающих и смесительных устройств
для биореакторов самых разнообразных конструкций; по этой
причине они и некоторые другие выводы качественного харак-
тера в настоящее время, возможно, являются наиболее ценны-
ми с практической точки зрения результатами рассмотренных
выше теоретических работ. Действительно, такие характеристи-
ки реакторов, как пространственная неоднородность структуры
течений, особенности турбулентных течений, объемные доли
газа и жидкой фазы, в большинстве случаев настолько сложны,
что их количественная оценка расчетным путем крайне затруд-
нена и получить необходимые численные данные обычно удается
только с помощью эмпирических уравнений. При выборе пос-
ледних, однако, мы не должны забывать только что рассмот-
ренную физическую основу явлений. Так, уравнение, описыва-
ющее коалесцирующую систему чистый воздух — вода, вероят-
нее всего, будет неприменимо к процессу, в котором участвует
относительно некоалесцирующая двухфазная система.
Теперь, возвращаясь к параметрам уравнения (8.39), опре-
деляющего площадь поверхности раздела фаз в единице объе-
ма а', рассмотрим время пребывания пузырьков tb в реакторе.
Этот параметр можно найти путем интегрирования скорости
подъема пузырька по высоте реактора hr:
hr
, _ f* dz ~ _hr_
b “ J «6 (2) ~ ~t
0
(8.55)
Здесь в правом выражении приближенно принято, что скорость
подъема пузырька равна его конечной скорости. В случае от-
дельных мелких пузырьков при небольших числах Рейнольдса
конечную скорость можно определить, воспользовавшись урав-
нением (8.32). Если же речь идет о больших сферических или
полусферических пузырьках диаметром D в ньютоновской жид-
кости, то в соответствующих расчетах следует использовать
значение конечной скорости, определяемое уравнением
щ = 0,711 (^D)V2 = 22,26 VD см/с (8.56)
Расчет характерной скорости движения пузырьков в потоке
более сложен, поскольку, во-первых, одни пузырьки влияют на
движение соседних и, во-вторых, в процессе движения пузырь-
ки могут коалесцировать или распадаться на более мелкие
пузырьки. Сравнивая уравнения (8.33) и (8.35), можно видеть,
-что для идентичных Sc и Sh в первом приближении
щ (поток пузырьков) ~]У3 0,31
ut (отдельный пузырек) 0,39
(8.57)
Процессы переноса в биотехнологических системах
623
«, следовательно,
И|(поток пузырьков) ~0,50 и/(отдельный пузырек) (8.58)
Для любой реальной дисперсной системы характерно то или
иное распределение пузырьков по размерам. В связи с этим
возникает проблема определения характерного или среднего
размера пузырьков. В уравнениях (8.35) — (8.38) под диаметром
пузырька D подразумевается усредненный по поверхности диа-
метр или средний диаметр по Заутеру Dsmi
Г)
8т SmjD2]
(8.59)
Здесь ntj — число пузырьков диаметром Dj.
Входящее в уравнение (8.39) выражение nF^tb отражает
общий объем пузырьков в реакторе. Отношение объема всех
пузырьков к объему реактора (объема газа к объему реактора)
называют газосодержанием Н. Если величина Н известна из
других лабораторных или заводских данных или из литератур-
ных источников (пример 8.1), то ее можно подставить в урав-
нение
а = Н~- (8.60)
В расчетах с привлечением уравнения материального балан-
са только жидкой фазы, например в уравнениях (8.14) и (8.15),
удобнее пользоваться параметром а', представляющим собой
отношение площади поверхности раздела фаз к объему жид-
кой фазы, а также параметром а — отношением площади по-
верхности раздела фаз к общему объему реакционной смеси
(жидкость4-газ). Эти два параметра, определяющие величину
межфазной поверхности, связаны через газосодержание Н:
а'(Л — Н)=а (8.61)
Будьте внимательны при использовании опубликованных в ли-
тературе уравнений массопередачи; проверьте, какой параметр
применялся в оригинальной работе — kta или kfi'.
Пример 8.1. Эмпирические уравнения для определения газосодержания.
Барботажная колонна*-.
Я/(1 — H)4=0,20(Bo)’/8(Ga)l/i2Fr (8П1.1)
где Во — число Бонда (Bo=gd<2Pc/o); Ga — число Галилея (Ga=gd?/«e2);
Fr — число Фруда (Fr=«G/Vgd<); ио — поверхностная скорость газа; dt —
диаметр колонны (реактора).
♦ Chakravarty М„ Begum S., Singh Н. D., Barrah J. N., Iyengar Al. S.,
Biotech. Bioeng. Symp., 4, 373 (1973).
624
Глава 8
Лабораторная газлифтная колонна*-.
Газосодержание в трубе подачи газа (с барботером):
(ц— №2о)2’75 + 1.61 — о
10"4и°’88
(8П1.2)
Газосодержание в кольцеобразном зазоре:
Н21= 1,23-10'2
Г 74,2 — 0 1
79,3 — о
(8П1.3)
Газосодержание над отражательной перегородкой;
Я3=7,5-1О-Зм0188 (8П1.4)
Общее газосодержание колонны:
< /7= О,ООЗи0,88
Здесь gz — вязкость жидкой фазы при температуре колонны, сП; Цн2о — вяз-
кость воды при температуре колонны, сП; о — поверхностное натяжение на
границе раздела фаз газ — жидкость, дин/см; и — поверхностная скорость га-
за, см/с; ylmt — площадь сечения трубы подачи газа, см2; Лапп — площадь се-
чения кольцеобразного зазора, см2.
Лабораторная газлифтная колонна**
В трубе подачи газа:
_____________
1,065ы^ + Uy
= доля свободного объема
(8П1.5)
где Ud—поверхностная скорость газа в трубе подачи;
__________объемная скорость потока газа___
объемная скорость потока смеси (газ+жидкость)
Г32 см/с у <0,43
“V= 1257 (у-0,43)+ 32 ?>0,43
Реактор с перемешиванием***
При условии, что диаметр реактора приблизительно равен высоте И
Re,0-7 (М,£),/и)°>3<2-104,
i/"~u~ р°'4р0-2 ( и V/a
|/ — Уя+О,О15ао и fl0=»l,44 J
При Re,0-7 (Л7г£+/и)°.3>2-104
Н =* + 0,015ог
Uq г
lg
=i 1,95-10-5Rei0.7
* Chakravarty M., Begum S., Singh H. D., Barrah J. N., Iyengar Л1. S.,
Biotech. Bioeng. Symp., 4, 373 (1973).
*♦ Hatch R. T., Ph. D. Thesis in Food Science and Nutrition, p. 150, Massa«
chusetts Institute of Technology, Cambridge, Mass., 1973.
Calderbank P. H., Trans. Inst. Chem. Eng., 36, 443 (1958).
Процессы переноса в биотехнологических системах
625
где do, 01 — площадь поверхности раздела фаз в единице объема культуры;
Re,— число Рейнольдса мешалки (Re( = pM.Di2/pic); и — поверхностная ско-
рость газа (для незаполненного реактора); — скорость подъема пузырьков.
Смысл символов Nlt Di, Р, р, а расшифрован в тексте (см. разд. 8.4).
Реакторы с перемешиванием*
Для системы вода — воздух данные могут быть выражены уравнением
/ р \0,4
Нт- и1/2=7,63Я + 2,37
где Р — мощность в л. с.; V — объем жидкой фазы без учета газа, м3; и —
поверхностная скорость, м/ч; Н — объемная доля газовой фазы (при 0,02 <
<Я<0,2).
8.4. Массообмен путем принудительной конвекции
Для достижения экономически оправданных скоростей про-
цессов роста биомассы, утилизации субстрата или образования
продуктов жизнедеятельности клеток часто возникает необхо-
димость в эффективном перемешивании дисперсной системы
вода — воздух. В этом разделе основное внимание снова будет
уделено зависимостям между соответствующими переменны-
ми, которые позволяют определить коэффициент массоотдачи
kt и (или) а или а', т. е. отношения площади поверхности разде-
ла фаз к соответствующему объему.
8.4.1. Общие принципы и основные критерии подобия
Основная цель механического перемешивания заключается
в повышении (иногда очень значительном) интенсивности сме-
шения фаз по сравнению с естественной конвекцией, обуслов-
ленной движением частиц свободно поднимающейся или опус-
кающейся диспергированной фазы. Эта цель может быть достиг-
нута за счет следующих эффектов:
1. Высокое динамическое давление вблизи конца лопасти ме-
шалки или другого перемешивающего устройства приводит к
образованию небольших пузырьков и тем самым к увеличе-
нию локальных величин а'. Если соответственно не возрастает
и скорость коалесценции во всем объеме жидкой фазы, то
в результате повышается и среднее объемное значение а'.
2. Культуральная среда может содержать суспензию твердой
или другой жидкой фазы, что приводит к расслаиванию
(осаждению) в реакторе. Механическое перемешивание обес-
печивает более однородное диспергирование этих фаз в не-
* Richards J. W„ Prog. Ind. Microbiol., 3, 143 (1961).
626
Глава 8
прерывной жидкой фазе. В случае эмульсий углеводородов в
уравнение для ki входит выражение, пропорциональное корню
кубическому из разности плотностей фаз (рн2о — Рнс)'Л
[см. уравнение (8.38)]; обусловленный этим обстоятельством
невысокий коэффициент массопередачи типичен для микро-
биологических процессов, лимитируемых углеводородным
субстратом, и может быть повышен за счет перемешивания.
3. Когда газовые пузырьки определенного диаметра находятся
в эффективно перемешиваемом реакторе, ki заметно не изме-
няется при повышении или снижении потребляемой реакто-
ром мощности, поскольку относительная скорость пузырьков
газа или капель жидкости определяется в основном разно-
стью плотностей. (Объясните, почему.) В то же время обус-
ловленная перемешиванием турбулентность уменьшает
диаметр пузырьков D и таким образом повышает а' при дан-
ном газосодержании; кроме того, изменение D отразится и
на величине kt.
4. Перемешивание может способствовать снижению максималь-
ного размера частиц не слишком плотного мицелия, слизи-
стых слоев микроорганизмов, скоплений плесневых грибов,
что приведет к уменьшению модуля Тила микроорганизмов
(разд. 4.4), а также к более однородному распределению
микроорганизмов в жидкой фазе. С другой стороны, известны
случаи, когда относительно высокие скорости перемешивания
приводили к снижению выхода необходимых продуктов кле-
точной жизнедеятельности вследствие нарушения структуры
клеточных или внеклеточных ферментов, влияния перемеши-
вания на морфологическое развитие и дифференциацию кле-
ток и т. д.
5. Суспензия клеток в жидкости может быть настолько вязкой,
что только механическое перемешивание позволит добиться
заметного диспергирования газа в жидкой среде (эту проб-
лему мы рассмотрим подробнее в разд. 8.8).
При принудительном перемешивании действие приложенных
механических сил обусловливает некоторую характерную ско-
рость, относительно которой можно оценивать масштабы дру-
гих движений в системе. В случае перемешивания лопастной
мешалкой существуют два масштаба — среднеквадратичная
флуктуация скорости жидкости wrms и окружная скорость ме-
шалки щ, пропорциональная NiDi, где JVf— скорость вращения
мешалки в оборотах за единицу времени, a Di — диаметр ло-
пастной мешалки.
Если в качестве характерной скорости использовать «rms, то
уравнения материального баланса для всей системы, отдельных
ее компонентов и баланс количества движения можно выразить
Процессы переноса в биотехнологических системах
627
через следующие безразмерные параметры (критерии подобия):
Число Шервуда:
Число Шмидта:
Число Рейнольдса:
Число Фруда:
Sh =
kjD
^о2
Sc=jiz/pzS>o2
Re—piDuxms/pt
Fr = u2rms/gD
(8.62a)
(8.626)
(8.62b)
(8.62r>
В альтернативном подходе в качестве характерного пара-
метра перемешиваемых систем можно взять диаметр лопастной
мешалки Д-; тогда характерной скоростью будет ее окружная
скорость NiDi. Индекс i говорит о том, что в данном случае ос-
новой масштабирования является скорость вращения мешалки,
а не параметры газообразных, жидких или твердых компонен-
тов дисперсной системы. В этом случае соответствующие числа
Рейнольдса и Фруда выражаются уравнениями
Ref = и Fn = (8.63)
И/ Р-/ ё '
Число Фруда имеет и другие определения. Так, для массо-
передачи в суспензии «нейтрально плавающих» частиц предла-
галось следующее выражение:
Fr _
gL
(8.64)
где L — высота реактора. Поскольку число Фруда отражает
зависимость динамики свободной поверхности от механического
перемешивания, то в нем необходимо учесть и расстояние от
поверхности дисперсной системы до дна реактора. Если в пере-
мешиваемой системе имеются две фазы (непрерывная и диспер-
гированная) с разными плотностями, например эмульсия
углеводорода в водной среде, то может пригодиться еще одно
модифицированное определение числа Фруда:
Fl"две фазы
pN2iD2i
SpgL
(8.65)
Очевидно, что во всех случаях при работе с литературными
данными необходимо внимательно следить как за формой са-
мих уравнений, так и за определениями применяемых в них
критериев подобия; нельзя использовать или цитировать урав-
нение, не определив все входящие в него критерии подобия.
628
Глава 8
8.4.2. Уравнения для определения коэффициентов
массопередачи и площади поверхности раздела фаз
Коэффициент массопередачи газов зависит главным образом
от гидродинамических свойств окружающей пузырек пленки
жидкости, которые на протяжении большей части времени пре-
бывания пузырька в свою очередь определяются в основном
выталкивающей силой (естественной конвекцией) и турбулент-
ностью, т. е. числом Рейнольдса. Поэтому в уравнения, описы-
вающие поведение свободно поднимающихся пузырьков, чаще
всего включают число Рейнольдса в форме уравнения (8.62в).
В достаточно больших реакторах, снабженных отражатель-
ными перегородками для обеспечения максимальных скоростей
перемешивания в непрерывной фазе (гл. 9), влияние эффектов
свободной поверхности (числа Фруда) становится несуществен-
ным. Газообмен на свободной поверхности может играть боль-
шую роль в небольших лабораторных биореакторах, но с уве-
личением объема реакторов этот эффект практически исчезает.
В отсутствие поверхностного влияния безразмерные решения
для полей скорости и концентрации дают
c = f(z, Re, Sc, Sh) (8.66)
Поэтому число Шервуда зависит только от Re и Sc:
Sh = = g (Re. Sc)
Калдербэнк предложил уравнение [11]
Sh (турбулентная аэрация) = 0,13Sc1/3Re3/4 (8.67)
С помощью уравнения (8.52) можно показать, что зависимость
&z(Sh) от мощности, потребляемой единицей объема реактора,
Р/V определяется выражением
Sh ос (А)1" (8.68)
Таким образом,
k, = 0,13 ( а3^ <P/V.LY/4 Sc"2'3 (8.69)
1 PiD J
В турбулентном режиме, когда становится применимым уравне-
ние (8.69), с повышением Р быстро снижается влияние этого
параметра на зависимость ki от Р:
Процессы переноса в биотехнологических системах
629
Познакомившись с процессами распада пузырьков, мы впра-
ве надеяться, что нам удастся найти математические выраже-
ния для зависимости максимального диаметра устойчивых пу-
зырьков Dc от о и переменных, определяющих динамическое
напряжение. Поскольку эта величина во многих случаях очень
близка характерному среднему истинному диаметру пузырьков
Dsm, неудивительно, что уравнения для Dsm имеют аналогичную
форму. Включение в эти уравнения дополнительных выраже-
ний, учитывающих газосодержание Н и вязкость диспергиро-
ванной фазы цс;, указывает на важные, но неопределяющие
вклады других факторов. Упоминавшаяся необходимость бли-
зости коалесцирующих свойств в экспериментальной системе,
послужившей основой для вывода эмпирических уравнений, и
в изучаемой системе сохраняет свою силу и в случае уравнений,
приведенных в примере 8.2.
Пример 8.2. Уравнения для определения максимального (D,) и средне-
го по Заутеру (Dsm) диаметров пузырьков или капель.
Экспериментально найденные значения Dc для свободно поднимающихся
пузырьков описываются уравнением*
/ а \1/2
Dc= Н,452-10-2-д^-1 см (8П2.1)
Что касается реакторов с перемешиванием, можно привести следующие
результаты экспериментов по изучению зависимости Dsm от мощности, по-
требляемой единицей объема реактора:
Система жидкость — жидкость**:
о0-8 п 5 ( щ- \°>25
ГО (8П22)
Система газ — жидкий электролит**'.
О0.6 а л
D-°2-25 p,»(W
Система газ — водный спирт***:
0,25
гтО.б / II \
Dim = 1,90——--------------Я0’65 | -1£.]о>25
Р/0-2 (P/V)^ щ J
Система газ — вязкая жидкость****:
о0-6 ( ц/ \о,1
п ____0 7---------------_Щ_
зт (р/V)0’^0’2 I Hg '
(8П2.3)
(8П2.4)
(8П2.5)
* Ни S„ Kinlner R. С., The Fall of Single Liquid Drops Through Water,
AlChE J., 1, 42 (1955).
** Calderbank P. H„ Trans. Inst. Chem. Eng., 36, 443 (1958).
*** McDonough J. A., Tomme IF. Holland C. D.. Formulation of Interfacial
Areas in Immiscible Liquids by Orifice Mixers, AIChE J., 6, 615 (1960).
**** Bhavaraju S. M., Russell T. IF. F., Blanch H. IF., Design of Gas Sparged
Devices for Viscous Liquid Systems, AIChE J., 24, 454 (1978).
630
Глава 8
Параметры лопастной
мешалки для реакторов с перемешиванием [26]:
Р
— « const
PiN3iD5i
D3i
при Rez > Ю4
£>c /о \о,б
—- — const -------------
Dt k N2iD3iPiJ
(8П2.6)
(8П2.7)
Для турбулентного течения в трубе [11]
(const) (We-XRe7X) -
~ (const) f тдЛ---------Г’6 ("Д---------У’1 (8П2.8)
\ Р/ \и / -^труба / \ сДруба^гтаР/ /
где
|______2______'l 1 г>Л __________ ^труба^гтвР/ /огтп п\
WeTpy6a - и ReTpy6a— (8П2.9)
\ Р/ <«*> Отруба / И/
Для течения через отверстие в трубе (определено в точке, отстоящей на
0,3 м от выхода из трубы)*
= 21 //VaiWe“p’v«.2Re^5 (“10’
Здесь WeiPy6a и ReTPy6a определяются уравнениями (8П2.9).
При данных Dc или DSm и газосодержании Н величину а можно вычис-
лить по уравнению (8.60). В сложных случаях, например в большинстве
реакторов с перемешиваемой двухфазной системой, изображенных на
рис. 8.2, Н следует измерять непосредственно в реакторе или определять по
эмпирическим уравнениям, разработанным специально для реактора данной
конструкции. Ряд таких уравнений был приведен в примере 8.1.
8,5. Определение kia' и мощность,
потребляемая реакторами с перемешиванием
и с барботажными устройствами
В этом разделе мы суммируем результаты различных экспе-
риментов, посвященных изучению и методам определения объ-
емного коэффициента массоотдачи kia'. Поскольку непосредст-
венно определить а' не удается, то экспериментальные данные
часто выражают именно в виде параметра kia'. Кроме того,
этот сложный параметр в некоторых случаях позволяет учесть
эффекты, обусловленные значительным временем пребывания
небольших пузырьков в очень вязких средах. В таких случаях
пузырьки уже практически не содержат кислорода и поэтому не
вносят никакого вклада в перенос О2. По этой причине, напри-
мер, определенные оптическими методами величины а' могут не
* McDonogh J. А., Тотте W. J., Holland С. D., Formulation of Interfacial
Areas in Immiscible Liquids by Orifice Mixers, AlChE J., 6, 615 (I960).
Процессы переноса в биотехнологических системах 631
отражать действительную удельную площадь поверхности разде-
ла фаз пузырьков, содержащих кислород.
Мы уже отмечали, что повышение потребляемой удельной
мощности может приводить к уменьшению размера газовых пу-
зырьков и таким образом к увеличению площади поверхности
раздела фаз. Здесь мы опишем методы расчета потребляемой
реактором мощности с помощью параметров, определяющих ха-
рактеристики барботажных и перемешивающих устройств. Кро-
ме того, мы рассмотрим довольно распространенный вариант
перемешивания барботажными устройствами и механическими
мешалками одновременно.
Многие методы изучения массообмена в системах газ — не-
вязкая жидкость в реакторах с перемешиванием рассмотрены
в обзоре Ван’т Риета [9]. Результаты большого числа экспери-
ментов в различных реакторах с применением мешалок разных
конструкций с точностью ±20—40% можно описать следующи-
ми уравнениями:
Реактор с перемешиванием, вода, коалесцирующая система:
^a=2,6.10-2^M(^s)0’5 с-1 (8.71)
(VC2600 л; 500<Р/П< 10 000 Вт/м3)
Реактор с перемешиванием, вода, некоалесцирующая система:
^а=2,0.10-3^О’70.у0’2 с’1 (8.72)
(2<7<4400 л; 500<Р/И<10 000 Вт/м3)
Здесь ugs — поверхностная скорость газа, равная частному от
деления объемной скорости подаваемого потока газа на произ-
ведение поперечного сечения реактора и газосодержания. Под
каждым уравнением указаны диапазоны объема реактора и объ-
емного потребления энергии, в которых справедливы эти вы-
ражения.
Необходимо подчеркнуть, что в соответствии с рассмотренны-
ми нами выше основными положениями влияния турбулентности
на массопередачу эти уравнения справедливы (с указанной
точностью ±20—40%) для самых различных перемешивающих
устройств (турбинных, лопастных, пропеллерных и стержневых
мешалок, самоиндуцирующихся устройств) и разного числа ме-
шалок. Положение мешалки в реакторе также не играет суще-
ственной роли, если только ее расстояние от дна реактора не
слишком мало (меньше ее диаметра), что может снижать сте-
пень рассеяния мощности, или если мешалка не расположена
слишком близко к поверхности, что сопровождается захватом
воздуха и снижением потребляемой мощности.
632
Глава 8
Аналогично изучение массообмена в барботажных колоннах
показало:
Барботажная колонна, вода, коалесцирующая система-.
£za = 0,32(«gS)0’7 (8.73)
Для некоалесцирующей системы в барботажной колонне общих
зависимостей не существует, поскольку конструкция барботера
влияет на величину kia'.
Поскольку определение потребляемой мощности или поверх-
ностной скорости газа может быть затруднено, в качестве при-
меров приведем несколько эмпирических уравнений других ти-
пов. Так, для переноса газа в барботажной колонне Акита и Йо-
шида предложили следующее уравнение [12]:
= 0,6 (Sc)1/2Bo°’62Ga0’31//1’1 (8.74)
Здесь в числах Бонда (Во^£>2рс/о) и Галилея (Ga=gd/3/zzz2)
учтен диаметр колонны dt. Показано, что это уравнение приме-
нимо при rf/^60 см (0,6 м) и может использоваться при dt>
>60 см, если в уравнении (8.74) dt принять равным 0,6 м.
Белло и другие для определения объемного коэффициента
массоотдачи kia в эрлифтной колонне предложили уравне-
ние [13]:
, __ 0,0005-(P/V)0-8
(1 + Лй/Л)
(8.75)
Здесь Аа/Аг — отношение площадей сечения зон опускания и
подъема дисперсной системы, P/V— отношение мощности, рас-
ходуемой на аэрацию, к объему. Этим уравнением можно поль-
зоваться, но в нем не учитывается тот факт, что массообмен,
газосодержание и другие параметры не одинаковы в различных
зонах эрлифтного аппарата.
Любые эмпирические уравнения имеют ограниченный диапа-
зон применения. Так, предсказываемое уравнением (8.75) боль-
шое влияние Р/V на kia' не выполняется при достаточно малых
P/V (P/V<1), поскольку в таких условиях динамика пузырь-
ков газа в жидкой фазе определяется главным образом естест-
венной конвекцией, т. е. их свободным подъемом.
Для деления и последующего смешения жидкой фазы, а так-
же для поддержания восходящего потока пузырьков и снижения
количества или полного устранения больших пузырьков и воз-
душных пробок можно применять неподвижные перемешиваю-
щие устройства. Для определения массопередачи в таких ап-
паратах Ванг и Фан предложили следующее уравнение [14]:
М = 0,12н,°-«( ,.99^47[, ) с- (8.76)
Процессы переноса в биотехнологических системах 633
Отрсинсаггкугбная Поверхность
перееороака жи&ностц
РИС. 8.6. Средние скорости течения (выраженные в долях от окружной ско-
рости лопасти мешалки, равной 5,2 фута в секунду) (1,6 м/с) и структура цир-
куляции воды в резервуаре высотой 12 дюймов (30,5 см) при скорости пере-
мешивания 200 об/мин. (McCabe W. L., Smith J. С., Unit Operations in Chemi-
cal Engineering, 3d ed„ p. 234. McGraw-Hill, New York, 1976.)
Здесь Ug и ui — поверхностные скорости газа и жидкости (в
см/с) соответственно.
Потенциальным недостатком любого эмпирического уравне-
ния является допущение о равномерном рассеянии энергии и
(или) постоянной величине kia' в зоне смешения, в которой из-
мерялись параметры, лежащие в основе данного уравнения,
и в изучаемом или проектируемом реакторе. В этой связи полез-
но рассмотреть изменения средней скорости течения, обнаружен-
ные экспериментально в стандартном перемешиваемом реакторе
с отражательными перегородками. Кривые на рис. 8.6 указыва-
ют структуру циркулирующих течений, а числа — локальные
средние скорости в долях от окружной скорости мешалки.
В ряде работ по изучению барботажных колонн было пока-
зано, что величина kia' существенно зависит от положения точ-
41-537
634
Глава 8
ки измерения этого параметра в объеме колонны. На рис. 8.7
приведены результаты экспериментального определения аксиаль-
ного распределения растворенного кислорода в барботажной
колонне и в трехфазной системе —псевдоожиженном слое, со-
держащем частицы диаметром 0,1 см. И в том и в другом слу-
чае вблизи барботера имеется входная зона, в которой скорости
переноса кислорода относительно велики; при увеличении рас-
Расс таяние от ffapffomepa, с/ч
a ff
РИС. 8.7. Экспериментально найденные (/) и рассчитанные с помощью мате-
матической модели (2) зависимости концентрации растворенного кислорода
от расстояния до барботера для барботажной колонны («/ = 7,5 см/с, ug=
= 28 см/с) (а) и трехфазного псевдоожиженного слоя, содержащего частицы
диаметром 0,1 см (щ = 7,5 см/с, ug=20 см/с) (б). Кривые отражают общий
характер зависимостей. [Воспроизведено с разрешения из работы: Alvarez-
Cuenca М., Nerenberg М. A., in Advances in Biotechnology, vol. 1, p. 477,
Moo-Young M. (ed.), Pergamon Press, 1980.]
стояния от барботера скорость переноса кислорода резко па-
дает. В основе этого явления, вероятно, лежит переход от пу-
зырьков одного размера, определяемого параметрами барботе-
ра, к пузырькам другого диаметра, зависящего от равновесия
между коалесценцией и дроблением пузырьков. Значения, обо-
значенные на рис. 8.7 символом □, вычислены с помощью ма-
тематической модели, в которой принимается, что поток газа
в режиме полного вытеснения проходит через две зоны с раз-
личными объемными коэффициентами массопередачи. Установле-
но, что граница между двумя зонами располагается приблизи-
тельно в 33 см над барботером. В другой модели для барботаж-
ной колонны иной конструкции принимается, что сначала (пер-
вые 27,6 см от барботера) kia' убывает, а затем принимает по-
стоянное значение.
Процессы переноса в биотехнологических системах
635
Определенные затруднения могут возникать при операциях
масштабирования. Так, упоминавшаяся выше входная зона вно-
сит очень большой вклад в небольших лабораторных системах,
а в высоких колоннах большого объема она составляет лишь
незначительную часть общего объема.
Теперь перейдем к рассмотрению методов расчета мощно-
сти, необходимой для достижения заданных скоростей ввода
газа в барботажную колонну и механического перемешивания.
Здесь мы обсудим только общие принципы этих методов и не-
которые уравнения; более детально вопросы учета потерь энер-
гии в элементах аппаратов рассмотрены в приведенной в конце
главы литературе. Для барботажной колонны мощность, необхо-
димая для сжатия газа до степени, обеспечивающей его барбо-
таж с объемной скоростью Fo при давлении р\, выражается
уравнением
Ря = PZo Г 1п — + а — °-] (8.77)
8 °L {MW) Р2 2 J v ’
Здесь p2—давление в верхней части резервуара, «о — скорость
газа в отверстии барботера, а а — доля кинетической энергии
газа, передаваемая жидкости; обычно а равен приблизитель-
но 0,06.
Расход мощности на перемешивание неаэрируемых жидко-
стей зависит от плотности и вязкости жидкой среды р/ и ц/, ско-
рости вращения мешалки Ni и ее диаметра Di, а также от ко-
эффициента торможения мешалки Cdc. Зависимость последнего
параметра от числа Рейнольдса мешалки должна быть разной
для каждого режима течения — ламинарного, переходного или
турбулентного. На рис. 8.8, а представлены результаты широко
известных работ Раштона, Костича и Эверетта по изучению
мешалок трех различных конструкций [26]. Эти результаты при-
ведены в виде зависимости безразмерного параметра — числа
мощности Рпо [определяемого уравнением (8.78)] от числа
Рейнольдса мешалки Re,:
?п0 = piN^D^t (8.78)
В турбулентном режиме потребляемая мощность не зависит от
Re,, т. е.
PocNpDj5 Рпо = const
Напротив, в ламинарном течении эти зависимости описываются
выражениями
Р ос N2tD\ или Рп0 ос —L-
41»
Число Рейнолбдса Re=D< и> p/j.i
5 5
РИС. 8.8. а — зависимость числа мощности от числа Рейнольдса для меша-
лок различных конструкций. [Из работы: Аиба Ш., Хемфри А., Маллис Н.,
Биохимическая технология и аппаратура. — М.: Пищевая промышленность,
1975; данные заимствованы из статьи Rushton J. Н., Costich Е. W., Eve-
rett Н. J., Power Characteristics of Mixing Impellers, part 2, Chem. Eng. Prog.,
46, 467 (1950).]
6 — зависимость коэффициента трения f потока в трубе от числа Рейнольдса
(z— относительная высота неровностей поверхности трубы). [Воспроизведе-
но с разрешения из работы: McCabe W. L„ Smith J. С., Unit Operations of
Chemical Engineering. McGraw-Hill, New York, 1954; данные заимствованы нз
статьи: Moody L. F., Trans. ASME, 66, 671 (1944).]
Процессы переноса в биотехнологических системах 637
В каждом конкретном случае константа пропорциональности оп-
ределяется конструкцией перемешивающего устройства. Обра-
щает на себя внимание сходство между кривыми на рис. 8.8, а
и графиком зависимости коэффициента трения f от числа Рей-
нольдса для течения в трубе. В последнем случае коэффициент
трения пропорционален 1/Re в ламинарном потоке (число мощ-
РИС. 8.9. Зависимость отношения мощности, расходуемой на перемешивание,
в аэрируемых (Ра) и неаэрируемых (Р) реакторах от Na (см. текст): а —
плосколопастная турбинная мешалка (8 лопастей); б — крыльчатый диск
(8 крыльев); в — крыльчатый диск (6 крыльев); г — крыльчатый диск
(16 крыльев); д — крыльчатый диск (4 крыла); е — лопастная мешалка. [Вос-
произведено с разрешения из статьи: Ohyama Y., Endoh К.., Power Characte-
ristics of Gas-Liquid Contacting Mixers, Chem. Eng. Japan, 19, 2 (1955).]
ности также пропорционально 1/Re/), а в турбулентном режи-
ме f стремится к почти постоянному значению, большему в слу-
чае труб с шероховатой внутренней поверхностью. Аналогично
по мере того, как геометрия мешалки становится менее «глад-
кой», Рпо для турбулентного режима достигает более высокого
постоянного значения (рис. 8.8, а); эта ситуация, однако, ослож-
няется влиянием стенок резервуара и отражательных перегоро-
док на определяемую величину потребляемой мощности Р.
Если реактор с перемешиванием одновременно аэрируется,
то потребление мощности на перемешивание снижается. На
рис. 8.9 приведены кривые зависимости Ра/Р (отношения по-
требляемой мощности в аэрируемом реакторе к мощности, по-
требляемой неаэрируемым реактором) от безразмерного пара-
638
Глава 8
метра Na (относительной скорости аэрации):
N _
а NiD\
(8.79)
Здесь Fg — объемная скорость газа.
За исключением наиболее быстро изменяющихся участков
соответствующих кривых, эти зависимости могут быть описаны
уравнением
Pq(Na) —Ра (ЛГа=>оо) _ -mNa ,g «дч
P-Pa{Na^) ' ' ’
где m — коэффициент. Мичел и Миллер предложили другое
уравнение, применимое и в случае турбулентной аэрации ненью-
тоновских жидкостей [28]:
п , / P^NlD^l W5 , ( pt (ДЛ£)3 )0,44 \0,4б
Ра = т------—- =т ——— --------
\ pg0’56 / k Na°’56 У
(8.81)
Здесь т' — коэффициент. В уравнениях (8.80) и (8.81) символом Р
обозначена мощность, потребляемая неаэрируемым реактором,
которая рассчитывается по формулам, приведенным ранее в этой
же главе.
Из указанных уравнений следует, что при постоянных М-
и Dt потребляемая мощность снижается при повышении Na
(т. е. при увеличении потока воздуха Fg). Этот эффект частично
обусловлен снижением средней плотности перемешиваемой дис-
персной системы. В то же время эффективность смешения си-
стемы при возрастании Na уменьшается.
В биологических реакторах некоторых конструкций переме-
шивание осуществляется с помощью струи жидкости. В этом
случае рассеиваемую мощность можно определить по следую-
щему уравнению [29]:
p = (8.82)
Здесь Dj — диаметр струи.
8.6. Массообмен через свободные поверхности
Перенос газов через свободные поверхности систем газ —
жидкость (рис. 8.10) играет большую роль в обеспечении кис-
лородом культур клеток животных и в удалении из них СОг-
Поверхностный массообмен важен также при росте культур во
встряхиваемых сосудах и в небольших перемешиваемых био-
реакторах, предназначенных для культивирования микроорганиз-
мов. Перенос газов через свободную поверхность жидкости
существенен для реаэрации естественных водных потоков и для
дыхания аэробных организмов в приповерхностных слоях морей
Процессы переноса в биотехнологических системах
639
без захвата
воздуха
С захватом
воздуха
5
ТРердь/й
носитель
Связанная с
поверхностью
плен на микроорганизмов
в
РИС. 8.10. Различные способы массообмена через свободные поверхности:
а — неподвижная поверхность (спокойное озеро); б — реакторы с перемеши-
ванием; в — стекающая пленка; г — турбулентный водный поток.
и озерных биоценозах. Массообмен через свободные поверхно-
сти жидкостей играет большую роль в различных микробиологи-
ческих процессах в слоевых реакторах со струйным течением
жидкости, применяющихся, в частности, в производстве винного
уксуса и обработке сточных вод. В перечисленных случаях глу-
бина проникновения кислорода зависит от масштаба вихрей
вблизи поверхности жидкости. Много работ посвящено изучению
массообмена с участием стекающей пленки жидкости, хотя да-
леко не все полученные результаты применимы в микробиологи-
ческих процессах. Массообмен в стекающих пленках жидкостей
мы и рассмотрим в первую очередь.
Для стекающей пленки жидкости с толщиной Л, длиной L
и шириной W при нулевой начальной концентрации растворен-
ного газа интегрированная по площади скорость абсорбции газа
выражается уравнением
Общая скорость абсорбции (число молей в единицу времени) =
= WLU ( 4а°;"т» Г (8.83)
* \ JlL )
640
Глава 8
где Иглах — скорость свободной поверхности. При выводе этого
уравнения допускалось, что концентрация растворенного газа
вблизи твердой поверхности всегда равна нулю; иными словами,
за время стекания пленки растворенное вещество не успевает
проникнуть в пленку на всю ее толщину [5].
В этом случае число Рейнольдса определяется как
цтах^/гР
Hz
где роль масштаба длины играет гидравлический радиус Rh:
h 1 rrrr
Rh =---------« — , если h « W
n 2W 4-2/i 2
Из определения коэффициента массоотдачи следует:
Общая скорость абсорбции = й/(с/*—• ci) WL (8.84)
Если Ci намного меньше cz*(cz«0), то уравнения (8.83) и (8.84)
можно записать в форме Sh = f (Sc, Re):
Sh = = 2b (ScRe)V2 (8.85)
^o2
Здесь h — масштаб длины для числа Шервуда и b—(Ljhy/2.
Таким образом, Sh пропорционально Re1/2.
Ливански и другие изучали абсорбцию СОг жидкими плен-
ками, стекающими по наклонной поверхности известной площа-
ди; в качестве абсорбирующих жидкостей использовали воду,
суспензии водорослей и питательные среды [18]. Для всех трех
систем значение ki при Re=(7—8) • 103 было одинаково; при
различных числах Рейнольдса изучали только суспензии водо-
рослей, для которых турбулентный режим течения наиболее ве-
роятен. Полученные результаты можно описать уравнением
£z = 4-10“5Re2/3 при 2000<Re^8000
В турбулентных водных потоках большое значение приобре-
тает масштаб циркулирующих вихрей, поскольку именно он
определяет толщину слоя, в который вода переносит свежие пор-
ции почти насыщенного раствора газа с поверхности в жидкую
фазу. Если мы представим себе изображенный на рис. 8.11 цир-
кулирующий вихрь длиной и глубиной А, то среднее число Шер-
вуда для массообмена в турбулентном режиме по аналогии
с уравнением (8.28) можно определить так:
dw
£>о, J \ дг Jz=o
1 о
Процессы переноса в биотехнологических системах
641
/7о fence.кость потопа
л-----
РИС. 8.11. Схема вихрей, циркулирующих вблизи свободной поверхности
жидкости.
где z и w_—безразмерные координаты, выраженные в масшта-
бе Л, a ki — усредненное по длине вихря значение коэффи-
циента.
Скорость массообмена в конечном счете локально зависит от
0о2, а также от скорости, с которой жидкость вблизи поверх-
ности обновляется за счет циркуляции. Согласно предложенно-
му одним из первых уравнению Хигби, коэффициент массоотда-
чи на поверхности системы газ — жидкость в случае одинаково-
го времени пребывания элементов жидкости равен*
Т/2 (8.86)
L ЛТ J
Это уравнение было модифицировано Данквертсом, чтобы его
можно было применять при том или ином распределении време-
ни пребывания элементов жидкости на поверхности; оказалось,
что и в этом случае ki пропорционально (®о2)1/2. Высказыва-
лось предположение, что время обновления т турбулентного по-
тока равно отношению его глубины h к средней скорости <uw>:
h
Тпоток-"<М_
Отсюда следует, что
Ь _ / {“w) \1/а
1 л h J
Если ширина потока равна W, то площадь поверхности раздела
фаз в единице объема потока составляет
Г(1) _ 1
1^(1)(й) ~ h
* Higbie R., Trans. AIChE, 35, 365 (1935).
642
Глава 8
Таким образом,
~ "4^”) (уравнение О’Коннора — Даббинса) (8.87)
Последнее уравнение довольно успешно применялось для описа-
ния скоростей реаэрации обедненных кислородом озер и рек.
Б другом уравнении учтено изменение ki в зависимости от поло-
жения w в вихре:
^=l,46(-^°2“rms )1/2 (8.88)
Здесь Urms — среднеквадратичная скорость циркуляции вихря.
В общем случае есть все основания считать Л и «rms пропор-
циональными средней глубине и средней скорости потока соот-
ветственно при одном и том же коэффициенте пропорциональ-
ности:
ТХглубина = Л уХсредняя скорость потока = «rms
Известно, что на перенос газа в очень большой степени влия-
ет волновое движение на границе раздела фаз вода —воздух.
В отличие от рассмотренных выше случаев здесь основными
факторами являются параметры течения газа, например сред-
няя и турбулентная компоненты скорости ветра, определяющего
характер океанских волн. Эта тема, однако, слишком сложна
для настоящего учебного пособия; интересующийся этой пробле-
мой читатель может ознакомиться с ней подробнее в приведен-
ной в конце главы литературе.
8.7. Другие факторы, влияющие на kta'
Из определений ki и а' и анализа факторов, влияющих на
толщину зоны сопротивления массообмену вблизи поверхностей
пузырьков и капель, следует, что величины ki и а' будут зави-
сеть от изменения коэффициента диффузии растворенного в
жидкой фазе газа ^о2> вязкости непрерывной фазы цс и сопро-
тивления массообмену на границе раздела фаз газ — жидкость.
«Вязкость» жидкости может изменяться в зависимости от ско-
рости сдвига; такое неньютоновское поведение имеет довольно
большое значение и поэтому будет рассмотрено отдельно
в разд. 8.8. Здесь же мы уделим внимание другим факторам.
8.7.1. Определение коэффициентов диффузии
Коэффициент диффузии небольших молекул в растворителях
невысокой молекулярной массы удобно оценивать (обычно
с точностью не менее 10—15%) с помощью уравнения Уилки —
Процессы переноса в биотехнологических системах
643
Чанга:
g) = 7,4.10-8-^-aM)1/2 см2/с (8.89)
Здесь М — молекулярная масса растворителя; Vm—молекуляр-
ный объем растворителя при нормальной температуре его кипе-
ния, см3/г-моль; щ— вязкость жидкости, сП.
Параметр ха представляет собой коэффициент ассоциации рас-
творителя; величина ха равна 2,6 для воды, 1,9 для метанола,
1,5 для этанола и 1,0 для бензола, эфира и гептана.
Коэффициент диффузии зависит от ионной силы (как и рас-
творимость; см. табл. 8.1) и концентраций растворенных ве-
ществ, влияющих на вязкость раствора. Если при изменении
Таблица 8.4. Коэффициенты диффузии О2 в пленках микроорганизмов
Биомасса Реактор 55. CM2/c-10b (найдено) 55, в % от 55 в н2о
Слизистые слои бактерий Вращающаяся труба 1,5 70
Погруженная пластина 0,04 2
Zoogloea ramige- ra Реактор с псевдоожи- женным слоем 0,21 8
вязкости раствора характер взаимодействия растворителя с рас-
творенным веществом не изменяется, то для внесения поправок
на изменение вязкости раствора (при постоянной температуре)
по сравнению, например, с чистой водой может оказаться полез-
ным уравнение
Как показано в табл. 8.4, коэффициенты диффузии кислорода
в скоплениях микроорганизмов обычно меньше, чем в чистой во-
де (2,25-10~5 см2/с). В ходе изучения влияния других перемен-
ных недавно было показано, что при увеличении времени пре-
бывания микроорганизмов в реакторе и отношения С/N утили-
зируемых субстратов коэффициент диффузии О2 в скоплениях
микроорганизмов, образующихся при переработке отходов,
уменьшается по сравнению с соответствующей величиной для
чистой воды в 5—20 раз.
8.7.2. Ионная сила
Достаточно точное разделение влияния ионной силы раствора
на все составляющие факторы оказалось затруднительным. Изу-
чение ньютоновских жидкостей привело к следующему уравне-
644
Глава 8
нию для массопередачи Оз в отсутствие химического взаимо-
действия:
Kfi — K
( Ра У [ Fg / Р/°’633^О22/3 \
k Vi М A J к О°.6р,;1/3 )
(8.90)
где Vi — объем жидкости; Ра — мощность, расходуемая на аэра-
цию; Л— площадь поперечного сечения реактора, перпендику-
лярного потоку, текущему со скоростью Fg. Параметры X, п и т
в зависимости от ионной силы / (/ = 1/2SZz2ci-, где i — тип иона,
Zi — его заряд) были найдены эмпирическим путем:
л ion 28,71' Т, I, если 0 < 1 < 0,40 г-ион/л
18,9— ’ , где Г =
0,276 4-/ (0,40, если /> 0,40 г-ион/л
Го лп I 0,862/' r, г п л
°Л0 + ~0.274 4-/- ПРИ 1 = 1 < °’4
0,90 при I > 0,4
т монотонно возрастает от 0,35 при / = 0 до 0,39 при /^0,4.
Здесь Kia выражено в с-'1, Ра — в фунт-сила-футах в минуту,
Vi—в кубических футах, Fg—в кубических футах в секунду,
А — в квадратных футах, другие параметры — в единицах СГС.
8.7.3. Поверхностно-активные вещества
Как мы упоминали в гл. 2, многие вещества биологического
происхождения по своей природе амфипатичны, т. е. содержат
как в высшей степени гидрофильные, так и гидрофобные участ-
ки, для которых характерна тенденция к концентрированию на
поверхностях раздела газ — жидкость и жидкость — жидкость.
На различных фазах роста клетки выделяют вещества, обла-
дающие поверхностно-активными свойствами (например, поли-
пептиды); иногда эти вещества вызывают вспенивание реакци-
онной массы аэрируемых реакторов. Добавление веществ, подав-
ляющих пенообразование, также влияет на сопротивление по-
верхности раздела массообмену; обычно это влияние противо-
положно действию поверхностно-активных веществ. _
Адсорбция поверхностно-активных веществ на поверхности
раздела фаз представляет собой спонтанный процесс; при этом
свободная энергия, а следовательно, и поверхностное натяже-
ние о на границе раздела понижаются. Из эмпирических урав-
нений, приведенных в примере 8.2, следует, что в присутствии
поверхностно-активных веществ величины Dsm и Dc должны сни-
жаться; это приведет к увеличению поверхности раздела фаз
в единице объема а'.
Повышение а' может компенсироваться влиянием пленок по-
верхностно-активных веществ на коэффициент массоотдачи ki.
Процессы переноса в биотехнологических системах 645
Адсорбция поверхностно-активных веществ повышает жесткость
и снижает подвижность поверхности раздела фаз. Рассматривае-
мое здесь снижение ki, как полагают, обусловлено одним из двух
механизмов (или обеими механизмами одновременно): 1) под-
вижность жидкой фазы вблизи границы раздела понижена в си-
лу относительно малой подвижности самой границы раздела;
поэтому различные теории массообмена, базирующиеся на оцен-
ке скоростей обмена малыми элементами жидкости между по-
верхностью и жидкой фазой, предсказывают уменьшение коэф-
фициента массопередачи (подробнее см. в работах [3, 4]);
2) подобно клеточной мембране, молекулярная пленка должна
обладать собственным сопротивлением, которое может вызвать
отклонения от предполагаемого равновесия газ — жидкость на
поверхности раздела фаз.
Добавление 10 млн-1 лаурилсульфата натрия (ЛСН) сни-
жает ki переноса кислорода на 56% (по отношению к чистой
воде). Дальнейшее повышение концентрации поверхностно-ак-
тивного вещества не изменяет kt. Площадь поверхности раздела
фаз в единице объема а' медленно возрастает при повышении
концентрации ЛСН от 0 до 75 млн-1, a kia' минимально при
концентрации ЛСН около 10 млн-1.
Найдено, что в реакторе с турбинным аэратором повышение
концентрации поверхностно-активного вещества сопровождается
монотонным возрастанием kta'. Анализ опубликованных в лите-
ратуре данных об изменении отношения а' в присутствии по-
верхностно-активного вещества к а' в его отсутствие и соответст-
вующего отношения величин kta' показал , что в присутствии
4 млн-1 ЛСН а' возрастает в 4 раза, a kia'—только на 15%;
отсюда следует, что ki уменьшается приблизительно на 71%.
Наблюдавшееся в двух указанных сериях экспериментов сни-
жение ki согласуется с результатами других исследований. В слу-
чае массообмена ряда ограниченно растворимых газов добавле-
ние поверхностно-активного вещества до постоянного ki приво-
дит к снижению этого параметра в среднем на 60%. Проведен-
ное Бенедеком и Хейдегером изучение влияния ЛСН показало,
что турбинная аэрация в присутствии этого агента соответствует
области, промежуточной между двумя областями, описываемыми
приведенными выше уравнениями (8.35) и (8.36) [33]. Анало-
гичный результат получен Данквертсом для случая аэрации
с помощью решетчатых тарелок. Эти результаты должны еще
раз предостеречь читателя от переоценки приведенных в данной
главе многочисленных эмпирических уравнений, и хотя послед-
ние, безусловно, полезны, при возможности следует использо-
вать экспериментальные данные, полученные в условиях и аппа-
ратах, максимально близких изучаемому процессу.
646
Глава 8
8.8. Неньютоновские жидкости
Для жидкостей отношение напряжения сдвига к градиен-
ту скорости du/dy определяется как вязкость (подстрочный
индекс v помогает отличить обозначение вязкости от символа
рассмотренного ранее коэффициента эффективности -q). Таким
образом,
„ du
Ts dy
(8.91)
Для ньютоновских жидкостей зависимость т от du/dy графи-
чески выражается прямой линией, проходящей через точку пере-
сечения осей координат. В условиях установившегося течения
растворов полимерных соединений и (или) дисперсных систем
с диспергированной твердой или жидкой фазой наблюдались
РИС. 8.12. Зависимость напряжения сдвига от скорости сдвига для ньютонов-
ских и наиболее типичных моделей неньютоновских жидкостей: 1 — пластич-
ная модель Бингама; 2— уравнение Кассона; 3 — дилатантная модель; 4 —
ньютоновская жидкость; 5—псевдопластичная. [Roels J. A., van den Berg J..
Voncken R. M., The Reology of Mycelial Broth, Biotech. Bioeng., 16, 181 (1974).]
Процессы переноса в биотехнологических системах 647
различные типы неньютоновского поведения; наиболее типичные
формы такого поведения изображены на рис. 8.12, где вместо
dujdy используется скорость сдвига 7.
8.8.1. Математические модели неньютоновских жидкостей
и соответствующие параметры
Частными случаями общего уравнения Оствальда — Вейля,
или степенного закона для жидкостей, являются дилатантное,
ньютоновское и псевдопластичное поведение:
ts = —i]o I 7 = — (кажущаяся вязкость) (7) =
= -тМ (8.92)
где п
>1 — дилатантная система
= 1 — ньютоновская жидкость
<1 — псевдопластичная жидкость
В некоторых случаях движение происходит только тогда,
когда напряжение сдвига превышает некоторую пороговую ве-
личину то. Для пластичных жидкостей Бингама применимо урав-
нение
Т5 = т0 —i]o I 7 ln-IY> где n=l, to=#O, Ts>t0 (8.93)
При ограниченном то и п<Л соответствующая уравнению
(8.93) кривая принимает тот же вид, что и кривая, отвечающая
уравнению Кассона [уравнение (8.94)]:
т? = то'/г — К(7,/г) (8.94)
При последующем изучении коэффициентов массопередачи,
потребляемой мощности, перемешивания и т. д. мы будем назы-
вать суспензии микроорганизмов или другие жидкие (газожид-
костные) системы (кажущимися) псевдопластичными, ньютонов-
скими и т. д. и приведем уравнения, связывающие параметры
соответствующей модели с параметрами исследуемой системы.
Описание типов течения жидкостей на рис. 8.12 относится к их
поведению при установившемся сдвиге. В нестационарных усло-
виях, возникающих, например, при ступенчатом изменении ско-
рости сдвига, часто наблюдается изменение кажущейся вязко-
сти во времени; переходные состояния в такой системе могут
быть описаны только более структурированной моделью (в из-
вестном смысле аналогичной рассмотренным в гл. 7 структури-
рованным моделям кинетики клеточного роста). По-видимому,
подобные переходные состояния точнее описывают перемешива-
ние с помощью турбинных мешалок и турбулентное смешение
648
Глава 8
в неньютоновских системах. Разработка более структурирован-
ных моделей, однако, представляет собой довольно трудную за-
дачу; вообще эта область механики довольно малоизученна.
Здесь мы ограничимся замечанием, что наше понимание факто-
ров, ответственных за неньютоновское поведение, и их матема-
тическое описание значительно уступают теоретическим разра-
боткам других проблем, рассмотренных в предыдущих разделах
этой главы.
Неньютоновское поведение может быть характерным по
меньшей мере для двух типов систем: 1) суспензий небольших
частиц и 2) растворов высокомолекулярных соединений. Оче-
видно, граница между этими двумя типами систем стирается,
если диаметр молекул полимеров превышает 50—100 А или
если размер частиц снижается до величин менее микрометра.
8.8.2. Суспензии
Различные теории предсказывают, что разбавленная суспен-
зия частиц сферической формы должна сохранять свойства
ньютоновской системы, причем эффективная вязкость т]и,ец сус-
пензии выражается уравнением
=1+-|-^ + Ь^+... (8.95)
Здесь Ф—объемная доля твердых частиц, а b имеет значение
от 6 до 8. При плотности культуры микроорганизмов около
109 клеток в миллилитре и диаметре клеток 3-10~4 см Ф равно
приблизительно 5-10~3; очевидно, влияние клеток в этом случае
ничтожно мало. Более высокие Ф характерны для операций
фильтрования, в частности, при сушке фильтрованием (в ходе
выделения продукта из культуральной жидкости), а также для
микробиологических процессов, в которых образуются рассмот-
ренные в предыдущих главах гранулы плесени или мицелий.
Для суспензий с высокой долей твердых частиц может на-
блюдаться то или иное напряжение пластического течения: так,
например, водные взвеси частиц ядерного топлива диаметром
порядка микрометра удовлетворительно описываются пластиче-
ской моделью Бингама. Уравнению Бингама подчиняются и си-
стемы с мицелием Streptomyces griseus, причем отклонения
наблюдаются только при очень низких скоростях сдвига; этот
факт может оказаться немаловажным в операциях аэрации. Из
уравнения (8.50) следует, что максимальное напряжение сдвига
для поднимающегося пузырька связано с конечной скоростью
его подъема. Таким образом, в бингамовских жидких средах до-
статочно мелкий пузырек газа не будет влиять на окружаю-
щую жидкость и останется закрепленным в одном элементе
Процессы переноса в биотехнологических системах
649'
жидкой фазы довольно долгое время, иными словами, в этих
условиях газовый пузырек будет циркулировать в реакторе вме-
сте с жидкой фазой, а не подниматься, как это свойственно
пузырькам большего диаметра.
Уравнение Кассона успешно применялось для описания тока
крови в кровеносных сосудах. Красные кровяные тельца обра-
зуют скопления, размер которых при повышении сил сдвига
уменьшается. Как показывает изображенная на рис. 8.12 кри-
вая, соответствующая уравнению Кассона, повышение скорости
сдвига сопровождается снижением кажущейся вязкости ц-.
Если в микробиологическом процессе образуются хлопья, то
мы вправе ожидать, что обусловленные перемешиванием силы
сдвига будут регулировать средний размер этих хлопьев, и, сле-
довательно, уравнение Кассона может оказаться полезным и
при описании процессов с участием хлопьевидных скоплений
микроорганизмов.
При изучении изменений во времени реологических свойств
микробиологических систем, использовавшихся в производстве
пенициллина, вязкость систем в широком диапазоне скоростей
сдвига определяли с помощью турбинной мешалки, а не обыч-
ным методом с вращающимся цилиндром. Указывалось, что та-
кой метод определения вязкости имеет два преимущества; во-
первых, перемешивание предотвращает разделение фаз, которое-
могло бы существенно повлиять на надежность полученных
результатов, поскольку в отсутствие перемешивания на стенке
цилиндра формируется тонкий «бесклеточный» слой жидкости;
во-вторых, скорость сдвига, индуцируемого мешалкой, непосред-
ственно связана со скоростью ее вращения и, как было показа-
но ранее, не зависит от реологии жидкой среды [38; стр. 188—
189]. В этой работе было показано, что уравнение Бингама и
степенная модель не отражают экспериментально полученных
результатов во всем изученном диапазоне скоростей сдвига.
В связи с этим было предложено модифицированное уравнение
Кассона
= (М^ ( 1 + °'69(У,1',‘~11 W'2) (8.96)
Здесь Мх — величина, пропорциональная напряжению сдвига,
a Л4т0=64/);3/2лЛ; Nt — скорость вращения мешалки (оборотов
в минуту); Л—отношение скорости сдвига к скорости вращения
мешалки (постоянная величина). Экспериментальные данные
хорошо согласуются с уравнением (8.96), хотя еще лучшее соот-
ветствие можно получить при замене второго слагаемого в пра-
вой части этого уравнения на
0,193 (Л4т0)°-75 + 0,1 w v/2
(Л4то)1/2 (
42—537
650
Глава 8
Приведенные результаты важны с трех точек зрения. Во-пер-
вых, в этих работах реологические свойства изучались в на-
столько широком диапазоне скоростей сдвига, что указанные
уравнения позволяют дискриминировать модели, дающие одина-
ковые результаты в более узких диапазонах скоростей сдвига.
Во-вторых, эти результаты показали, что степенная модель и
модель Бингама менее всего применимы в диапазоне низких
скоростей сдвига. В то же время именно этот диапазон с очень
большой степенью вероятности наиболее важен при определении
эффективности смешения в реакторе, и, следовательно, распро-
странение на него степенной модели и модели Бингама привело
бы к большим погрешностям (проблемы перемешивания в связи
с распределением времени циркуляции и времени пребывания
жидкости рассматриваются в гл. 9). В-третьих, как было пока-
зано, модифицированное уравнение Кассона предсказывает
существование коэффициента, зависящего от морфологии (фор-
мы) плесневых грибов, т. е. от отношения длины гиф к их диа-
метру. Этот вывод (и результаты других теоретических разра-
боток для несферических частиц) в принципе могут послужить
основой для учета морфологии микроорганизмов (опытных дан-
ных) в расчетах конструкций реакторов, тем более что в литера-
туре опубликовано множество данных, относящихся к изучению
морфологии различных видов организмов в различных условиях.
При описании влияния гиф плесеней степенная модель спра-
ведлива, по-видимому, в диапазоне высоких скоростей сдвига.
Она удовлетворительно описывает и поведение целлюлозной
пульпы; так, 4%-ная суспензия целлюлозной пульпы в воде хо-
рошо описывается степенной моделью с параметрами п = 0,575
и т|о = О,418 фунт-сила (секунда)"/квадратный фут. Изучение
переноса кислорода суспензиями пульпы применялось для моде-
лирования некоторых микробиологических процессов; такие мо-
дели в принципе могут оказаться полезными, если суммарное
поглощение кислорода лимитируется массообменом при высоких
скоростях сдвига. В то же время приведенные выше данные
заставляют усомниться в корректности этих моделей в тех слу-
чаях, когда перемешивание системы влияет на поглощение Ог
микроорганизмами.
В проточных системах с несколькими последовательно соеди-
ненными реакторами (гл. 9) порядковый номер реактора в из-
вестной мере выполняет функции шкалы времени и определяет
фазу роста популяции, в связи с чем содержимое реакторов
может различаться по реологическим параметрам. Например,
при культивировании Candida utilis в системе из двух реакторов
при больших скоростях течения условия во втором реакторе
становятся такими, что удельная скорость роста биомассы на-
много превышает удельную скорость деления клеток и, как след-
Процессы переноса в биотехнологических системах 651
ствие, наблюдается увеличение среднего размера клеток [41].
На возможную взаимосвязь между реологией и морфологией
указывает и наблюдавшееся изменение морфологии отдельных
клеток и их скоплений.
8.8.3. Растворы высокомолекулярных веществ
Некоторые микроорганизмы выделяют в среду полисахариды
и родственные биополимеры, а также их производные, например
ксантан, полиальгиновую кислоту и пуллулан. Ксантан широко
применяется для придания необходимой вязкости продукции
пищевой промышленности и стабилизации суспензий, а в буду-
щем, возможно, будет использоваться и при разработке нефтя-
ных месторождений. Выделяемые в среду биополимеры прида-
ют культуральному бульону свойства неньютоновской жидкости;
обычно поведение таких жидких сред описывается уравнением
(8.92) при (псевдопластичное поведение).
В периодическом процессе изменение концентрации биополи-
меров во времени приводит к тому, что свойства жидкой среды,
а следовательно, и параметры явлений переноса (массо- и теп-
лообмена) также изменяются во времени. Поскольку в уравне-
ниях, описывающих поведение неньютоновских жидкостей, ко-
эффициенты переноса обычно выражают через параметры сте-
пенной модели п и т|о, то естественно попытаться найти опреде-
ленную зависимость между концентрацией биополимера и этими
реологическими параметрами.
В случае ксантана эта зависимость выражается уравнением
Ло==Д.[р]в (8.97)
Здесь В«2,5, [Р]—концентрация полимера.
При росте некоторых внеклеточных биополимеров показатель
степени п и вязкость т|о изменяются во времени в соответствии
со следующим уравнением:
In-qo(/) = C+D-n(Z) (8.98/
Если зависимость [Р] от t известна из какой-либо биологиче-
ской кинетической модели, то уравнения (8.97) и (8.98) позво-
ляют без особых затруднений описать и зависимости n(t) и
По[Р(О]. Последние в свою очередь могут быть использованы
для расчета коэффициентов тепло- и массопередачи, а также па-
раметров смешения и потребляемой мощности [39, 40].
42*
652
Г лава 8
8.8.4. Потребляемая мощность и массообмен
в биотехнологических процессах
с неньютоновскими жидкостями
Мгновенное значение числа Рейнольдса Re/ мешалки, со-
гласно Калдербэнку [И], можно определить по уравнению
ре'. = / п \п
1 0,1/Q \ 6п + 2 J
(8.99)
Здесь Кс — индекс вязкости, равный напряжению сдвига т при
скорости сдвига 1 с-1.
РИС. 8.13. Зависимость числа мощности от числа Рейнольдса Re/ (см. текст)
в неаэрируемых системах для трех различных типов мешалок. [Из статьи:
Taguchi Н„ Miyamoto S., Power Requirements in Non-Newtonian Fermentation
Broth, Biotech. Bioeng., 8, 43 (1966).]
Как показано на рис. 8.13, для неаэрируемой, неньютоновской
культуральной жидкости Endomyces число мощности [уравне-
ние (8.78)] зависит от числа Рейнольдса Re/. Эту зависимость
можно выразить уравнением
р _ ь /р е' л* f У / \г
п0 - «(Re i) J
(8.100)
где Di — диаметр лопастной мешалки; DT — диаметр реактора;
Процессы переноса в биотехнологических системах
653
If — ширина лопасти мешалки; k, х, у и z зависят от диапазона
Re/ следующим образом:
Re/
<10 10-50 >50
k 32 11 9
X —0,9 —0,4 —0,05
у — 1,7 —1,7 —1,2
Z 0,4 0,5 0,9
Обращает на себя внимание сходство кривых, приведенных на
рис. 8.13, с данными Раштона и с кривыми зависимости коэф-
фициента трения в трубе от числа Рейнольдса (рис. 8.8, а, б).
Этими же исследователями показано, что в случае процессов
с аэрацией турбулентному режиму при Re/>50 удовлетворяет
уравнение Мичела и Миллера, применявшееся ранее для ньюто-
новских жидкостей:
F 0,56
г ё
Pg°4
(8.101)
Оказалось также, что ламинарный и второй режимы (Re/</50)
с удовлетворительной степенью приближения можно описать
тем же выражением с меньшим показателем степени:
р / РЧ^ \°,2~
& \ F °.56 /
\rg /
(8.102)
Две кривые пересекаются при P2M£>i3//7g°’56 = 2-10-2; коэффи-
циент пропорциональности в небольшой степени зависит от кон-
струкции перемешивающего устройства.
Разведение мицелиальных культур небольшим количеством
(10—25%) воды заметно изменяет ki и число мощности. Отсю-
да следует, что в непрерывных биологических процессах ско-
рость разведения может в существенной степени изменять вели-
чину потребляемой мощности.
Влияние популяции микроорганизмов зависит от ее физиче-
ского состояния (отдельные суспендированные клетки, скопле-
ния клеток, клетки, локализованные вблизи поверхности частиц
или пузырьков и т. д.) и характера воздействия микроорганиз-
мов на свойства системы. Показано, что при повышении кон-
центрации биомассы в мицелиальной культуре Aspergillus niger
от 0,02 до 2,5% kia снижается на 90%. С другой стороны, на-
блюдалось повышение скорости массообмена Ог в присутствии
суспензий Candida intermedia, Pseudomonas ovalis или частиц
оксида алюминия диаметром 0,3 мкм. С помощью ингибиторов
654
Глава 8
окислительного фосфорилирования (гл. 5) установлено, что
сходство эффектов клеток и оксида алюминия не ограничивает-
ся качественными характеристиками; оба фактора повышают
скорость массообмена примерно на 40% (по сравнению с водой),
причем вызываемый клетками эффект не зависит от их жизне-
способности. Недавно было показано, что влияние этих агентов
объясняется изменением гидродинамических условий вблизи
границы раздела газ — жидкость, сопровождающимся сниже-
нием сопротивления массообмену в жидкой пленке, окружаю-
щей пузырьки газа [44].
При высоких скоростях сдвига, необходимых для смешения многофаз-
ных систем и обеспечения необходимых скоростей массообмена, часто наблю-
дается снижение активности микроорганизмов. Так, серьезное снижение жиз-
неспособности популяции относительно большой клетки, простейшего Tetrahy-
тепа, при механическом повреждении клеток начинает проявляться при ско-
рости сдвига выше 1200 с-1. В этих экспериментах максимальная скорость
сдвига (характеризуемая окружной скоростью мешалки) оказалась более
важной переменной, чем число Рейнольдса или мощность, потребляемая еди-
ницей объема реактора.
Влияние механического перемешивания может проявляться и иным обра-
зом; выше мы уже упоминали, что при перемешивании может снижаться
размер существующих микробных скоплений. Тагучи и Йошида* рекомендо-
вали разделять экспериментально наблюдаемое снижение размеров гранулы
мицелия на два процесса: 1) отщепление значительно меньших частиц от
больших гранул; 2) непосредственное разрушение сферической формы гра-
нул. Оказалось, что обусловленное первым процессом уменьшение диамет-
ра D частиц во времени подчиняется закону
~= -k^D^D5’7 (8.103)
Второй процесс можно описать уравнением первого порядка:
-^-^^kiNp (8.104)
Здесь Np — число неразрушенных частиц. Зависимость коэффициента ско-
рости разрушения частиц kr от D, Ni и Di можно выразить таким урав-
нением:
kr= + (const) (D3. W'W'72) (8.105)
Тагучи и др. предположили, что последнее уравнение можно согласовать
с теоретическими данными, если предположить, что разрыв гранул опреде-
ляется не столько вязкостным, сколько турбулентным напряжением и что
устойчивость гранул непосредственно связана с их прочностью на разрыв,
определяемой экспериментально.
Что касается скорости переноса кислорода в суспензиях целлюлозной
пульпы, то здесь, как было показано, важны конструкции перемешивающе-
го устройства и реактора; эти данные согласуются с результатами изучения
аэрируемых и неаэрируемых суспензий Endomyces, опубликованными ранее
* Taguchi Н„ Yoshida Т., J. Ferment. Technol., 46, 814 (1968).
Процессы переноса в биотехнологических системах
655
D2Th
Тагучн и Миямото [42]. В 1,6 %-ной суспензии пульпы произведение kia
описывается уравнением
kia
-=0,113
Nt
1,437 1 пят ( Dt
(8J06)
где t — характерное время выравнивания концентраций в реакторе; h — высо-
та слоя жидкой фазы; W — ширина лопасти мешалки. Физический смысл
параметров Di, DT, L указан выше.
Подводя итог, можно отметить, что в общем случае коэффи-
циенты массопередачи и величина площади поверхности разде-
ла фаз в единице объема определяются целым рядом факторов,
в том числе размерами пузырьков и клеток, реологическими и
другими свойствами жидкой фазы (и газожидкостной системы),
конструкцией перемешивающего устройства и реактора, потреб-
ляемой мощностью. При проектировании и расчете биореакторов
необходимо принимать во внимание как все перечисленные фак-
торы, так и параметры кинетики клеточного роста (рассмотрен-
ные в предыдущей главе), а также параметры процесса смеше-
ния в реакторе и тип последнего (гл. 9).
Очевидно, желательно добиться такого уровня знаний, когда
можно было бы охватить все эти вопросы; в то же время надо
отдать себе отчет в том, что все изложенные в этом разделе
теории и соответствующие уравнения представляют собой всего
лишь введение в обширную область знаний, необходимую для
проектирования биореакторов на основе одних лишь теоретиче-
ских разработок. В заключение мы рассмотрим пример, иллю-
стрирующий некоторые сложные зависимости, природу которых
еще предстоит выяснить. В этом примере рассмотрено изменение
во времени соотношений между параметрами воздушных пу-
зырьков, диспергированного субстрата и клеток в процессе роста
Candida petrophilum на w-гексадекане (рис. 8.14). Заслуживает
внимания описание этой своеобразной системы газ—жидкость—
жидкость—клетка, данное авторами работы*:
«В первом периоде процесса маслянистые капли относительно велики,
и клетки концентрируются в основном вблизи этих капель, а не воздушных
пузырьков. Воздушные пузырьки неустойчивы и легко обновляются. Значе-
ние kia можно поддерживать на максимальном уровне, зависящем только
от конструкции реактора. В течение второго периода маслянистые капли
уменьшаются в размере, а клетки адсорбируются на их поверхности, обра-
зуя плотные хлопья. Для хлопьев характерна тенденция к связыванию с по-
верхностью воздушных пузырьков, но перемешивание легко разрушает эти
образования.
По мере развития процесса величина kta постоянно снижается. Третий
период процесса одновременно является вторым полупериодом фазы логариф-
мического роста, в котором дрожжи продолжают быстро расти, хотя под
* Mimura A., Takeda I., Wakasa R., Some Characteristic Phenomena of
Oxygen Transfer in Hydrocarbon Fermentation, Biotech. Bioeng. Symp. 4, pt. 1,
467 (1973).
656
Глава 8
микроскопом уже не видны маслянистые капельки, В этот момент kta дости-
гает минимального в течение всего процесса значения... [Воздушные] пу-
зырьки покрываются слоем дрожжевых клеток и сближаются, образуя скоп-
ления, в которых отдельные пузырьки разделены слоями биомассы. Подоб-
ные образования, плавающие на поверхности культуральной жидкости, очень
устойчивы. В четвертом периоде н-парафин полностью истощается. Клетки
распределяются равномерно по всему объему культуральной жидкости.
В этом периоде культуральная жидкость по своим свойствам, вероятно,
РИС. 8.14. Виды взаимодействия между воздушными пузырьками (В), ка-
пельками н-гексадекана (О) и дрожжевыми клетками (С) на различных ста-
диях периодического роста культуры дрожжей Candida petrophilum (1 — лаг-
фаза; 2 — первая половина экспоненциальной фазы; 3 — вторая половина
экспоненциальной фазы; 4 — после истощения запасов н-гексадекана). [Из
статьи: Mimura A., Takeda I., Wakasa R., Some Characteristic Phenomena of
Oxygen Transfer in Hydrocarbon Fermentation, in Adv. Microbial Eng., Siky-
ta B., Prokop A., Novak M. (eds.), part 7, p. 467, Wiley-Interscience, New York,
1973.]
близка культуральной жидкости, образующейся при брожении углеводов,
a kia возрастает до начального уровня».
Математическая модель, учитывающая ряд особенностей
процессов такого типа, будет рассмотрена в гл. 9 (пример 9.2).
8.9. Масштабный переход и массообмен
Как указывал Олдшу, при изменении объема реактора или.
скорости вращения мешалки различные параметры, которые мо-
гут влиять на произведение kia, изменяются по-разному*.
1. Турбулентное число Рейнольдса Rec определяет wrms и, таким
образом, коэффициент массоотдачи ki пузырьков:
Re — — Y'3 (8.107)
И k P T
* Oldshue J., Biotech. Bioeng., 8, 3 (1966).
Процессы переноса в биотехнологических системах
657
2. Окружная скорость лопасти мешалки TiNiDt определяет мак-
симальную скорость сдвига у, которая в свою очередь влияет
как на максимальный размер устойчивых пузырьков, так и
на размер хлопьев скоплений микроорганизмов (разд. 8.3),
а также может вызывать повреждения жизнеспособных кле-
ток (разд. 8.8.4).
3. Мощность, расходуемая на единицу объема, Р/V через Ref
определяет коэффициент массоотдачи и размеры частиц дис-
пергированной фазы. В ламинарном и переходном режимах
аэрации, согласно рис. 8.8, б,
PocNpD?
В турбулентном режиме число мощности постоянно, следова-
тельно,
РосМ?р.5
Тогда, считая объем реактора V пропорциональным Dp, по-
лучим
р с*, IN? аэрация в ламинарном или переходном режиме
V \NpDp аэрация в турбулентном режиме
4. Расход мощности в процессе аэрации равен
' Р2
Ра = т'
Таким образом,
-^2-» т'
V
^а°,58
Р2
у2,22
(8.108)
(8.109)
(NjD^)0^ чма
Последнее выражение позволяет определить мощность двига-
теля, необходимую для осуществления данного микробиоло-
гического процесса в данном биореакторе.
5. Если содержимое реактора эффективно перемешивается с по-
мощью внутреннего перемешивающего устройства, то система
в реакторе должна обладать характерным временем циркуля-
ции. Скорость рециркуляции потока жидкости Ft в области
перемешивающего устройства изменяется пропорционально
площади поперечного сечения n,Dp, а средняя скорость ме-
шалки в реакторе пропорционална NiDi. Таким образом,
д дг (время-1)
V D3t v ’
Величина представляет интерес с той точки зрения, что она
обратно пропорциональна времени, которое жидкость может
провести вне зоны гомогенизирующего действия перемешива-
ющего устройства.
658
Глава 8
Все перечисленные параметры влияют на ход процесса, и
все они по-разному зависят от режима перемешивания. В этой
связи возникает вопрос: какой (или какие) из этих параметров
должен быть взят за основу при масштабном переходе? Здесь
под основой для масштабного перехода мы понимаем простой
или сложный количественный параметр, который, будучи спра-
ведливым для реактора меньшего объема, может быть без из-
менения перенесен и на реакторы большего объема. Например,
если в качестве условия масштабного перехода мы выберем по-
стоянный расход мощности на единицу объема, то при механи-
ческом перемешивании в турбулентном режиме это будет озна-
чать, что
N^D^N^D2» (8.110)
Здесь индексы 1 и 2 обозначают параметры в небольшом и
большом реакторах соответственно. В ранних работах по изу-
чению промышленных способов получения пенициллина в каче-
стве условия масштабного перехода биореакторов принимался
постоянный расход мощности на единицу объема (около 1 л. с.
на 379 л) при сходной конструкции большого и малого реак-
торов. Как показано на рис. 8.15, а, такое условие масштабного
перехода обеспечивает достаточно постоянный выход пеницил-
лина в реакторах емкостью от 5 до 757 л. В то же время изме-
нение величины Р/V приводит к тому, что выход пенициллина
колеблется в зависимости от масштаба реактора.
Другим часто используемым условием масштабного перехода
является постоянство объемного коэффициента массоотдачи kia.
На рис. 8.15, б представлены кривые зависимости выхода про-
дукта жизнедеятельности бактерий витамина Вп от соответст-
вующих значений kiap при различных масштабах. Здесь в слож-
ный параметр kiap включено общее давление кислорода, что
позволяет учесть увеличение движущей силы переноса кислоро-
да при повышенном давлении, что типично для промышленных
биореакторов большого объема. В зависимости выхода витами-
на Bj2 от ktap достигнута удовлетворительная корреляция, хотя
выходы в небольших лабораторных реакторах обычно ниже, чем
в промышленных реакторах большого объема. Обратите внима-
ние также на то, что в отличие от рассмотренных ранее ситуа-
ций, когда клеточный рост лимитировался переносом кислорода
и возникала необходимость в более высоких величинах kia, на
кривой зависимости выхода витамина Bj2 от ktap имеется отчет-
ливый максимум. Это объясняется, по-видимому, тем, что в ре-
акторах различного объема при одном и том же kia другие па-
раметры перемешивания и течения жидкой фазы (например,
Процессы переноса в биотехнологических системах
659
о V = 7,6 л, металла типа Михсхо с ллосхими лопастями
1 - aV =420 л, меи/алха типа михсхо с плосхималопастями
о К=22,7 м3, мешалха типа Михсхо сплосхими лопастями
+V -45,5м3г мешалха :типа Михсхо с плосхими лопастями
---—I---1---J—— : т _________т—- — г
О 7Z34 667S
^а'р, г-моль П2/(мл-у)(х7О4)
6 Г
РИС. 8.15. а — зависимость выхода пенициллина от потребляемой мощности
в реакторах различного объема [по данным Gaden Е., Sci. Rep. 1st Super.
Samta, 1, 61 (1961)]; б — зависимость выхода витамина Bt2 (мкг/г) от слож-
ного параметра массопередачи ktap (см. текст) [по данным Bartholomew W. Н
Adv. Appl. Microbiology, 2, 289 (1960) ]
660
Глава 8
максимальная скорость сдвига или время циркуляции) в общем
случае могут различаться.
Из сказанного следует, что, осуществляя масштабный пере-
ход при одном постоянном условии, мы не должны забывать
о том, что при этом меняются другие характеристики переме-
шивания и структура течений. Этот факт особенно наглядно ил-
люстрируют данные Олдшу, который изучал изменения пара-
метров процесса при увеличении объема биореактора с переме-
шиванием от 80 до 10000 л*. При таком масштабном переходе
Таблица 8.5. Взаимосвязь между различными параметрами при масштабном
переходе3
Параметр Реактор малого объема (80 л) Реактор большого объема (104 л)
P 1,о 125 3125 25 0,2
P/V 1,0 1,0 25 0,2 0,0016
Ni 1,0 0,34 1,0 0,2 0,04
Di 1,0 5,0 5,0 5,0 5,0
Fi 1,0 42,5 125 25 5,0
F,V 1,0 0,34 1,0 0,2 0,04
NdDi 1,0 1,7 5,0 1,0 0,2
Re< 1,0 8,5 25 5,0 1,0
a Oldshue S. 3 (1966). У., Fermentation ^Mixing Scale-up Techniques, , Biotech. Bioeng., 8,
Di и V возрастают в 5 и 125 раз соответственно. Величина 1,0
в каждой колонке табл. 8.5 указывает, что данный сложный па-
раметр имеет одно и то же значение и в малом, и в большом
реакторе. Значения каждого параметра нормализованы по от-
ношению к 80-литровому реактору, поэтому в столбце «реактор
малого объема» все численные значения параметров равны 1,0.
Из приведенных в таблице данных следует, что масштабный
переход при постоянном Р/V сопровождается повышением мак-
симальной скорости сдвига на 70% (NiDi= 1,7), а времени цир-
куляции— примерно в три раза. С другой стороны, масштабный
переход при постоянном времени циркуляции Fi/V требует по-
вышения мощности в большом реакторе в 3125 раз!
Различные зависимости важнейших явлений переноса от
конструкции перемешивающего устройства приводят к тому, что
решение задачи масштабного перехода в случае реакторов с пе-
ремешиванием становится своего рода искусством. При этом
мы должны стараться выбрать в качестве условия переноса та-
* Oldshue S. Y., Fermentation Mixing Scale-up Techniques, Biotech. Bioeng..
8, 3 (1966).
Процессы переноса в биотехнологических системах
661’
кой параметр, который в данном биопроцессе является опреде-
ляющим. Эта проблема в свою очередь может быть связана
с рядом трудностей, обусловленных тем обстоятельством, что
явления переноса, зависящие от размеров мешалки и скорости
ее вращения, могут непредсказуемым образом влиять на чувст-
вительные к механическим воздействиям клетки.
Общую скорость поглощения кислорода в реакторе находят,
исходя из kia и соответствующего описания конструкции реак-
тора. Если жидкая фаза по своему составу гомогенна (однород-
на) и если пузырьки равномерно распределяются во всем объе-
ме раствора, то скорость массообмена определяется простым
выражением:
Vkta(c*— Ciiq)e = скорость поглощения Ог, моль/с
(8.111)
Здесь подстрочный индекс е указывает, что в данном случае
имеется в виду состав отходящих газов (если мы допустим, что
для реактора характерно полное перемешивание, то состав фазы
на выходе из реактора не отличается от состава этой фазы в ре-
акторе; гл. 9).
Если в реакторе пузырьки поднимаются в режиме полного
вытеснения, но перемешивающее устройство все еще обеспечи-
вает полное перемешивание жидкой фазы, то с* меняется в за-
висимости от положения пузырька в реакторе. Мгновенная по-
теря кислорода пузырьком при подъеме последнего на высоту
dz составляет
dpo2 1 объем газа (Дифференциальный
n/iaz & RT ~~ объем реактора \ объем реактора J
(скорость изменениях
концентрации I (8.112,у
Ог в пузырьке /
или
— kiakci* — Cb)Adz (8.113)
или скорости переноса кислорода из газовой фазы в жидкую.
Поскольку po2=Mci* (здесь М — константа в уравнении закона
Генри), то
Н dPp2 НМ dc? ^—к,а(с^—сь) (8.114):
RT dt RT dt
При постоянной скорости иъ подъема пузырьков dt = dzjub
и зависимость с* от z описывается уравнением
In (Q* ~ Сй) г = -kiaRT г (8.115>
(С/*-С6)вход иь
€62
Глава 8
ИЛИ
W - сЛ - (с,* - с„)км ехр (-k-^p-} (8.116)
\ ПГЛЩ) J
Следовательно, общая скорость массообмена в объеме Ah равна
h
С kfl (с,*-с„) (г) Adz = (с,« - сь) гх,адЛ Г1 — exp f Y
J 7?г L \ НМиь
(8.117)
Взаимное влияние перемешивания, кинетики микробиологиче-
ских процессов и массообмена мы рассмотрим в оставшихся гла-
вах этой книги.
8.10. Теплообмен
В биологических реакторах необходимость в отводе теплоты
или в обогреве культуральной жидкости может возникнуть по
следующим причинам.
1. Подаваемый в реактор питательный раствор желательно сте-
рилизовать нагреванием в проточном стерилизаторе или
в стерилизаторе периодического действия. Температура сте-
рилизации должна обеспечивать за общее время пребывания
гибель практически всех микроорганизмов (разд. 9.9.4).
2. Если количество выделяющейся в процессе переработки суб-
страта теплоты недостаточно для поддержания температуры
на заданном уровне, то реактор необходимо обогревать. Такая
ситуация типична, например, для процессов переработки от-
ходов в анаэробных условиях, оптимальная температура для
которых составляет 55—60 °C (разд. 14.4.7).
3. При превращении субстрата выделяется избыточное (по отно-
шению к оптимальным условиям, обеспечивающим, например,
жизнеспособность живых клеток) количество теплоты, кото-
рое необходимо отводить. Такая ситуация характерна для
большинства процессов с участием микроорганизмов.
4. Необходимость в дополнительном нагревании возникает так-
же в процессах сушки суспензий клеток.
Здесь мы рассмотрим первые три случая, связанные с жиз-
неспособностью организмов. Что касается сушки, то этот физи-
ческий процесс детально обсуждается в большинстве книг по
химической технологии.
Теплообмен между культуральной жидкостью и другой сре-
дой конструктивно осуществляется несколькими путями, в том
числе с помощью внешних рубашек, помещаемых в культураль-
ную жидкость трубчатых спиральных теплообменников (змееви-
Процессы переноса в биотехнологических системах
663
ков), циркуляции реакционной смеси через теплообменные ап-
параты, а также посредством упаривания или конденсации воды
и других летучих компонентов жидкой фазы, содержащей клет-
ки. Примеры способов теплообмена представлены на рис. 8.16.
Теплообмен, очевидно, играет определенную роль и в поддержа-
нии разницы температур между термически стратифицированны-
РИС. 8.16. Примеры теплообменных аппаратов и естественных процессов теп-
лообмена: а — реактор с рубашкой; б — реактор с внутренним змеевиком; в —
циркуляция реакционной массы через теплообменный аппарат; а — фазовые
переходы; д — естественные температурные колебания.
Гру/ш
д '
ми естественными водоемами и почвой; установившиеся зоны
с определенными диапазонами температур определяют соответ-
ствующие биологические ниши для совокупностей видов живых
организмов. В настоящем разделе основное внимание мы будем
уделять теплообмену в биологических реакторах.
Если принять, что скорости передачи и изменения других
форм энергии пренебрежимо малы, то основное уравнение теп-
лообмена в стационарном состоянии будет отражать связь меж-
ду скоростью отвода (с помощью какого-либо теплообменного
аппарата) и скоростью выделения теплоты. Следовательно,
Общая скорость выделения теплоты = скорость отвода теплоты = hA&T
(8.118)
664
Глава 8
Здесь АТ— характерная разность температур между содержи-
мым реактора и охлаждающей или нагревающей жидкостью;
А — площадь поверхности теплообмена; h — общий коэффициент
теплообмена.
Как и в случае массообмена, наибольшее сопротивление теп-
лопередаче оказывает относительно неподвижный тонкий слой
жидкости вблизи твердой стенки, разделяющей нагреваемую и
нагревающую жидкие среды, поскольку жидкая фаза обычно
эффективно перемешивается и, следовательно, ее можно считать
приближенно изотермичной. В этом разделе нас будут интере-
совать прежде всего вывод уравнений общего энергетического
Таблица 8.6. Примерные величины коэффициентов теплопередачи /га
h, ккал/(м2-ч-град)
Естественная конвекция
Г азы 3—20
Жидкости 100—600
Кипящая вода 1000—20 000
Принудительная конвекция
Газы 10—100
Вязкие жидкости 50—500
Вода 500—10 000
Конденсирующиеся пары 1000—100 000
а Из работы: Grober Н., Erk S., Grigull U., Warmedbertragung, 3d ed., p. 158,
Springer-Verlag, Berlin, 1955.
баланса и обзор математических выражений, определяющих h
для различных систем нагревания, охлаждения и стерилизации,
которые представляют интерес в микробиологических процес-
сах. Методы оценки тепловой нагрузки, сопровождающей рост
микроорганизмов, мы уже рассмотрели в гл. 5.
Высокие коэффициенты теплопередачи (табл. 8.6) кипящей
воды и конденсирующихся паров (обычно водяного пара) дела-
ют эти теплоносители особенно удобными в процессах стерили-
зации (разд. 9.9.4). Если необходимы более низкие температуры
(например, в обогреваемых аппаратах для анаэробной перера-
ботки), то в таких случаях удобнее применять воду с темпе-
ратурой ниже 100 °C. Для вязких жидкостей характерно более
высокое (по сравнению с водой) сопротивление теплопередаче;
как и в случае массообмена, это связано с пониженной скоро-
стью обмена между жидкой фазой и пленкой вблизи стенки,
а также с пониженной теплопроводностью вязких жидкостей
(аналогично малому коэффициенту диффузии Ог).
Изучение изображенных на рис. 8.16 принципов теплообмена
указывает на ряд проблем теплопередачи в биохимических ре-
Процессы переноса в биотехнологических системах
665
акторах. В системах с наружной охлаждающей или обогреваю-
щей рубашкой площадь теплопередающей поверхности Л про-
порциональна квадрату диаметра реактора (или диаметра ло-
пастной мешалки) D?. В то же время если мы хотим, чтобы
скорость микробиологических процессов и потребление мощно-
сти на единицу объема оставались постоянными, то степень
нагрева или охлаждения реактора должна быть пропорциональ-
ной Di3. Поэтому сосуды с рубашкой, обеспечивающие изотерми-
ческие условия в лабораторных экспериментах, при резком по-
вышении объемов реакторов часто приходится заменять на ре-
акторы с внутренними или внешними обогревающими (напри-
мер, при анаэробной переработке) или охлаждающими
(в частности, при переработке углеводородов в белок однокле-
точных организмов) змеевиками.
Понятно, что наличие системы внутренних трубопроводов из-
меняет картину перемешивания, структуру скоростей течения
жидкости и, возможно, скоростей коалесценции пузырьков газа.
Сложность такой ситуации еще раз напоминает нам о необхо-
димости экспериментального определения параметров только
в реакторах, конструкции которых в максимальной степени со-
ответствуют конструкции реактора, в котором будет осущест-
вляться крупномасштабный процесс; в процессах с участием
микроорганизмов априорное проектирование реакторов приве-
дет к еще более неопределенным результатам, чем в случае
более простых систем. (Проблемы перемешивания в реакторах
рассматриваются в тексте и в упражнениях гл. 9.) В частности,
приведенные в разд. 8.10.1 уравнения показывают, что коэффи-
циент теплопередачи h [выраженный в ккал/(м2-ч)] изменяется
при переходе от одной охлаждающей спирали, перпендикуляр-
ной течению жидкости, к устройству из нескольких располо-
женных в шахматном порядке рядов спиралей, также перпенди-
кулярному течению жидкости. Следовательно, первый ряд труб
изменяет характер потока, омывающего последующие ряды
труб.
В случае очень больших реакторов с большой тепловой на-
грузкой (например, при росте бактерий на метаноле в реакторе
объемом 1500 м3) внутренние змеевики уже не могут обеспечить
необходимый отвод теплоты. В таких случаях приходится при-
бегать к циркуляции через внешний теплообменник или через
теплообменник, являющийся составной частью реактора петле-
образной конструкции. Этот пример показывает, что иногда вы-
сокие тепловые нагрузки вкупе с другими факторами (напри-
мер, высоким расходом мощности на аэрацию и перемешивание)
обусловливают необходимость разработки биореакторов особых
конструкций, резко отличающихся от обычных резервуаров
с перемешиванием. Различные конструкции реакторов, способы
43—537
666
Глава 8
смешения и принципы перемешивания вкратце будут рассмот-
рены в гл. 9.
Известно, что скорости выделения и отвода теплоты позволя-
ют с достаточной точностью оценить тепловой баланс системы
при условии, что мы четко понимаем физическую основу соот-
ветствующих расчетов и учитываем обеспечиваемую этими рас-
четами степень достоверности результатов. В этом разделе мы
обсудим пути оценки потребности биотехнологических систем
в теплообмене (аналогичной потребности в кислороде); следую-
щий раздел будет посвящен проблеме определения коэффициен-
та теплопередачи h.
Определение потребности системы в нагревании или охлаж-
дении начинается с анализа общего энергетического баланса.
При постоянном давлении в системе с незначительными изме-
нениями потенциальной и кинетической энергии энергетический
баланс можно рассчитать через изменения энтальпии, в том
числе теплоты химических превращений, фазовых переходов
(испарения, конденсации), тепловых потоков, сопровождающих
процессы массообмена, теплопередачи от второй жидкости, ис-
пользуемой для охлаждения или нагревания. Введем следующие
обозначения:
Qmet — скорость выделения теплоты в процессах роста и
поддержания жизнедеятельности клеток;
Qag —скорость выделения теплоты за счет механического
перемешивания реакционной массы;
Qgas —скорость выделения теплоты в результате аэрации;
Qacc — скорость накопления теплоты;
Qexch—скорость теплопередачи в окружение реактора или
к теплоносителю теплообменного аппарата;
Qevap — скорость потери теплоты за счет испарения;
Qsen —скорость приращения энтальпии за счет разности
теплосодержания потоков на входе в реактор и на
выходе из него.
Тогда
Qmet + Qag 4“ Qgas = Qacc 4” Qexch 4“ Qevap4“ Qsen (8.1 19)
Куни, Ванг и Мателес [52] применяли это уравнение для расче-
та Qmet на основе экспериментального определения Qacc путем
контроля изменения температуры в практически изолированном
реакторе. В таких условиях величины Qevap и Qsen очень малы,
a Qexch ВНОСИТ большой вклад ПО сравнению С разностью Qacc —
Qag, хотя последние параметры сами по себе выше Qexch- Как
мы только что отмечали, Q9Ce контролировали калориметриче-
ски, a Qag вычисляли для каждого, значения скорости потока га-
Процессы переноса в биотехнологических системах 667
за и скорости вращения мешалки по уравнению Мичела и Мил-
лера [уравнение (8.81)].
При расчете стационарного режима работы реактора Qacc
принимается равным нулю, хотя при программированном изме-
нении температуры, необходимом для достижения оптимального
выхода в периодическом процессе (разд. 10.7), здесь могут воз-
никнуть дополнительные осложнения. Параметр Qag для аэри-
руемых или неаэрируемых систем определяется с помощью рас-
смотренных ранее уравнений для расчета потребляемой реакто-
ром мощности.
Если мы пренебрегаем Qevap (хотя этот параметр может быть
существенным в реакторах со струйным течением), а также по-
ка не учитываем Qsen, то единственным важным параметром
остается Qmet- Методы оценки и определения величины Qmet
были рассмотрены в гл. 5. При расчете реакторов большого
объема заданные рабочая температура и параметры течения оп-
ределяют Qevap и Qsen, а скорость вращения мешалки и ее диа-
метр определяют Qag (скорректированный с учетом выбранной
скорости аэрации). Конструкция барботера и скорость потока
газа определяют Qsas- Что касается оставшихся параметров
Qacc и Qexch, то независимо от того, находится ли реактор в изо-
термических условиях или нет, в каждый момент
Qexch = Qmet 4" Qag 4" Qgas Qacc (Qsen 4" Qevap) (8.120)
Нагрузка на- Выделение теплоты Накопление Отвод теплоты ины-
гревающего теплоты ми путями (не че-
или охлажда- рез твердые поверх-
ющего устрой- ности теплообмен-
ства ника)
Последнее уравнение устанавливает величину теплопередачи,
необходимую для поддержания заданной температуры или оп-
ределенной скорости накопления теплоты (если оно вообще не-
обходимо).
Мы можем использовать уравнение (7.35а), описывающее
мгновенную скорость образования клеточной массы в единице
объема реактора периодического действия:
(7.35а)
Соответствующая мгновенная скорость выделения теплоты в
микробиологическом процессе Qmet (теплота/время), очевидно,
будет определяться уравнением
Qmet = УреакторИ* (8.121)
Здесь Уд—коэффициент (г клеток/ккал), рассмотренный в
разд. 5.10.4. Там же были представлены методы определения Уд,
43*
668
Глава 8
а также иллюстративный материал, свидетельствующий об уве-
личении выделения метаболической теплоты при утилизации бо-
лее восстановленных субстратов (табл. 5.12).
Соответствующее уравнение для изотермического стационар-
ного процесса в проточном реакторе выглядит следующим об-
разом:
Удфте1 = VреакторЦ-^= (So $) Ух/sF — Xke (8.122)
(So-S) yxlsD —у^—\ IГд (8.123)
у реактор J I
Как мы упоминали выше, Ух/s может зависеть от возраста
культуры в периодическом процессе и скорости разведения Z>
в проточной системе. Зависимость Ух/s (а следовательно, и Уд)>
4met
V реактор
Таблица 8.7. Зависимость расходов на производство белка одноклеточных
организмов от природы субстрата3
Субстрат Расходы на производство 1 фунта (450 г) клеток, в центах
Субстрат Перенос О2 Охлаждение Всего
Малеат (отходы) 0 0,46 0,75 1,2
Эквиваленты глюкозы 3,9 0,23 0,54 4,7
(меласса)
Парафины 4,0 0,97 1,4 6,4
тётанбл 5,0 1,2 1,9 8,1
Метан 1,6 3,3 3,7 8,6
Этанол 8,8 0,75 1,3 11,0
Изопропанол Н,6 2,7 3,1 17,4
Ацетат 16,7 0,62 1,1 18,4
а Abbott В. J., Clamen
tenance Coefficient to Single
A., The Relationship of Substrate, Growth Rate, and Main-
Cell Protein Production, Biotech. Bioeng., 15, 117 (1973).
от затрат на поддержание клеточного метаболизма отражена
в уравнении (7.26).
Выполненные в 1973 г. Абботтом и Кламеном экономические
расчеты показывают, что при производстве биомассы бактерий
расходы как на теплообмен, так и на массообмен (перенос кис-
лорода) составляют значительную часть общей стоимости про-
дукции (табл. 8.7) [43].
8.10.1. Уравнения теплообмена
Из уравнения (8.118) и общего теплового баланса [уравне-
ние (8.119)] процессов нагревания, охлаждения и стерилизации
следует следующее общее уравнение для расчета теплопе-
редачи:
Qexch —ЛАДУ
(8.118а)
Процессы переноса в биотехнологических системах 669
Чтобы это уравнение можно было использовать в практических
расчетах, необходимо найти выражение для определения коэф-
фициента общей теплопередачи h, аналогичное рассмотренным
ранее выражениям для коэффициента общей массопередачи в
системе газ — жидкость. Условие непрерывности теплового пото-
ка требует, чтобы в стационарном состоянии для теплопередачи
через плоскую стенку толщиной Lw, разделяющую культураль-
ную жидкость с температурой Тжидк, i и нагревающую или ох-
лаждающую жидкость с температурой Тжидк. 2, удовлетворялось
уравнение
Ь {']' __т ч_______у ( ^стенкад Т’стенка.й \
"’wi х1 жидк,1 1 стенка. 1/ — Ks т ~—— —
\ J
~ (^стенка,2 жидк.г) ккал/(см2- ч) (8.124)
Здесь ks — теплопроводность стенки, выраженная в ккал/(см-
•с-град). Если в это уравнение ввести коэффициент общей теп-
лопередачи Л, определяемый как
тепловой П0Т0К=й(Тжидк,1 —Тжидк.2) (8.125)
то уравнение (8.124) можно преобразовать в
-J- = —-—|—-—|------— (плоская стенка) (8.126)
Л ks ^№2
По аналогии с уже изученной_нами массопередачей общее со-
противление теплопередаче \/h равно сумме трех сопротивле-
ний. При теплопередаче через цилиндрическую трубчатую по-
верхность нагревающих или охлаждающих змеевиков площадь
поверхности (в поперечном сечении) не остается постоянной,
а возрастает пропорционально расстоянию от внутренней по-
верхности трубы. В такой ситуации уравнение для h необходи-
мо модифицировать следующим образом:
-J— = —+ ln (<W)- 4---------— (стенка трубы) (8.127)
2kS hid(
Здесь di и d0 — наружный и внутренний диаметры трубы соот-
ветственно. Обратите внимание на индекс о при коэффициен-
те Ла, указывающий на то, что основой для расчета площади
теплопередачи является наружный диаметр трубы. Теплопро-
водность ks твердых тел зависит от их природы. Так, при 100 °C
ks равно 0,908 кал/(с-см-К) для меди и 0,107 кал/(с-см-К)
для стали; при понижении температуры ks медленно возрастает.
Величины ks для различных материалов, применяющихся в теп-
лообменниках, можно найти в любой технической справочной
литературе.
Изучение переноса количества движения и теплоты на лю-
бой из границ раздела жидкость — твердое тело позволяет опре-
670
Глава 8
делить соответствующие коэффициенты теплопередачи на гра-
ницах раздела фаз hwl, hw2 или h0, hi в уравнениях (8.126) и
(8.127) соответственно. Если такие индивидуальные коэффи-
циенты изменяются в зависимости от положения на поверхности
теплопередачи, то общий локальный коэффициент теплопереда-
чи определяется уравнениями типа (8.126) и (8.127), а для
расчета общего количества передаваемой теплоты необходимо
интегрирование по всей поверхности теплопередачи.
При изучении теплопередачи в системе жидкость — твердое
тело применяются следующие критерии подобия:
Число Нуссельта: к т hd NU=“7-- kf (8.128a)
Число Прандтля: Рг = kf (8.1286)
Число Бринкмана: Br= l^2 kf (Т’жидк — Т'стенка) (8.128b)
Число Фруда: т-. U2 Fr = —-г gd (8.128г)
Число Рейнольдса: Re = -^ (8.128д)
Здесь kf — теплопроводность жидкости, кал/(с*см *град); Ср —
теплоемкость, кал/(г-град); d—расстояние (или диаметр тру-
бы), см; ц— вязкость, П; и — скорость, см/с; g — постоянная
силы тяжести, г-см/с2.
Как указано в работе [5], число Бринкмана представляет со-
бой количество теплоты, участвующей в вязком рассеянии, отне-
сенное к теплопередаче за счет теплопроводности; в наших усло-
виях при теплопередаче через поверхность теплообменника этой
величиной можно пренебречь. (Число Бринкмана может быть
большим вблизи лопасти мешалки.) Аналогично в реакторах
с отражательными перегородками или с мешалкой, смещенной
относительно оси реактора, число Фруда обычно пренебрежимо
мало.
Коэффициент теплопередачи h, выраженный в виде безраз-
мерного критерия подобия Nu, является функцией от Рг и Re:
Nu=f(Pr, Re) (8.129а)
При ламинарном течении в прямых трубах теплопередача зави-
сит и от отношения длины трубы L к ее диаметру d (Lid), по-
этому предлагались и уравнения типа
Nu=f fPr.Re-^] (8.1296)
Процессы переноса в биотехнологических системах 671
Изменение температуры обусловливает изменение различных
свойств жидкости вблизи от поверхности теплообмена. Из этих
свойств важнейшим является вязкость, поэтому отношение
рь/ро, где ць и цо — вязкость жидкости при температуре жидкой
фазы и температуре стенки соответственно, также следует учи-
тывать в уравнении:
Nu=HPr,Re,4-,— 1 (8.129в)
\ “ Но /
Так как h (а следовательно, и Nu) можно определять и как
локальный коэффициент переноса, и как коэффициент, усреднен-
ный по поверхности теплопередачи одним из нескольких спосо-
бов, то при использовании опубликованных в литературе вели-
чин Nuioc или Nuav следует применять ДТюс или ДЛу соответ-
ственно.
Для теплопередачи при турбулентном течении жидкостей,
вязкость которых близка вязкости воды, предложено следующее
уравнение, полезное как в случае нагревания, так и при охлаж-
дении реакционной массы *:
Nu = ~ = 0,023Re0,8PrM (8.130)
rv
Это уравнение оказалось справедливым при
104^Re< 1,2-105 (турбулентный режим)
0,7^Рг<120 (для всех жидкостей, кроме расплавленных
металлов)
L/d>-№ (длинные трубы)
В модифицированном Сейдером и Тэйтом** виде последнее
уравнение применимо и при больших разностях температур; оно
оказалось полезным для определения теплопередачи вязкими
жидкостями, например маслами:
Nu== 0,027Re°’8Pr°’33 f (8.131)
« \ Но J
Если жидкая фаза неоднородна по плотности, то в теплообмен
вносит большой вклад и естественная конвекция; в этих случаях
в соответствующие уравнения вводят число Грасгофа:
Gr = —^Рар (8.132)
* McAdams W. Н., Heat Transmission, 3d ed., р. 152, McGraw-Hill Book
Company, New York, 1954.
*♦ Seider F. E. N., Tate G. E., Heat Transfer and Pressure Drop of Liquids
in Tubes, Ind. Eng. Chem., 28, 1429 (1936).
672
Глава 8
Тогда уравнение для Nu при течении жидкости в горизонталь-
ных трубах принимает вид*
Nu = 175Г—PrRe+0,04f4-GrPry,75l1/3f—Y’14 (8.133)
L L \ L /JX^0/ '
(В случае вертикальных труб вместо отношения вязкостей при-
меняют коэффициент 1,0, а константу 0,04 заменяют на 0,0722.)
Теплопередача неньютоновскими жидкостями описывается
другими уравнениями. Так, для псевдо пластических жидкостей
;(разд. 8.8) применяли два уравнения**:
Nu = —
k
V3r т]0 (жидкость)
3+ 1/n
(стенка) I 4
1 -10,14
т
(8.134а)
или
Nu = = 1,75 КеРгУ/3 ( Зга,+- У/3 (8.1346)
Здесь rja — кажущаяся вязкость [уравнение (8.92)], определен-
ная при температуре жидкой фазы или стенки. Обратите внима-
ние на то, что для ньютоновской жидкости (л=1) при неболь-
шой разности температур между жидкой фазой и стенкой урав-
нение (8.1346) сводится к уравнению (8.133). В уравнении
(8.134а) изменение вязкости в зависимости от температуры уч-
тено в той же мере, что и в уравнении (8.133).
В принципе теплообменные аппараты могут иметь самые раз-
личные поверхности и режимы потоков. Некоторые уравнения
для ряда конкретных ситуаций приведены в примере 8.3; другие
можно найти в обычных руководствах по теплообмену и в гл. 3
монографии Чарма (ссылка [30] в гл. 9). Ряд примеров расчета
коэффициентов теплопередачи и нагрузок теплообменных аппа-
ратов рассмотрен в упражнениях. Иногда культуральная жид-
кость образует осадок на поверхности нагревателя; это приво-
дит к загрязнению системы, изменению коэффициента теплопе-
редачи стенки во времени и увеличению h со стороны жидкости.
Пример 8.3. Уравнения теплопередачи
Теплопередача естественной конвекцией от вертикальной плоскости или
от поверхности цилиндра***:
Nu —7— =5 с (GrPr)a
rv
(8П3.1)
* Martinelli R. C., Boelter L. M., AIChE Mtg., 1942 (цитировано no
McAdams \W. H., см. выше).
•* Charni S. E., Merill E. W., Heat Transfer Coefficients in Straight Tubes for
Pseudoplastic Food Materials in Streamline Flow, Food Res., 24, 319 (1959).
*** Ring W. J., Free Convection, Meeh. Eng., 54, 347 (1932).
Процессы переноса в биотехнологических системах
673
Здесь L — длина стенки или диаметр цилиндра. Все параметры рассчитаны
При (7'жидк“}_7'стенка )/2, а также при
3,5- 107<GrPr< 1012 с=0,13, а = 1/2 (турбулентный режим)
104<GrPr<3,5-107 с=0,55, а=1/4 (ламинарный режим)
Теплопередача в пространстве между соосными цилиндрами*
Ламинарный режим:
hd ( d0 \о>8 ( d \°>«
Nus —(RePr)°.« GrM (8П3.2)
где do, d{ — наружный и внутренний диаметры.
Турбулентный режим:
, 0,23Рг-2/з (uft/u0)0>w
h ('Л(^-й)/иь]0’2' наружная стенка
CPG I (0,023)(0,87)(do/d/)0-63 (8ПЗ.З)
r"l(d0-d-)-G/^-------- внутренняя стенка
Течение под действием силы тяжести по горизонтальным трубным по-
верхностям
( w \1/з
Л 65 ~ j I британских тепловых единиц в час (8П3.4)
\ W о / на фуТ на градус Фаренгейта
w
если "2^7 < 525
Здесь w — скорость течения жидкости; L — длина трубы; d0 — наружный
диаметр.
Турбулентное течение в трубах**:
Nu = ~ = 0,023Reo,8Prd (8П3.5)
rv
f0,4 для нагревания
& =« •!
(0,3 для охлаждения
Течение, перпендикулярное изолированному цилиндру***:
Nu=—r— = (Рг) (0,35+ 0,56Re°’52) (8П3.6)
rv
Течение, перпендикулярное ряду труб, расположенных на расстоянии
двух диаметров одна от другой***:
Nu=™= o21Re7nO,6pri/3 (8П3.7)
rv
Здесь Rem —Re при Umax, а расстояние 2d определяется следующим обра-
зом:
* Monrad С. С., Pelton J. F., in McAdams W. H., Heat Transmission,
McGraw-Hill Book Company, New York, 1954.
** Dittus F. W., Boelter С. M. K., Univ. Calif. Publ. Eng., 2, 443 (1930);
(цитировано no McAdams W. H„ Heat Transmission, см. выше).
*♦* Charm S. E., Fundamentals of Food Engineering, 2d ed., Chap. 4, Avi
Publishing, Westport, Conn., 1971.
674
Глава 8
напрайление ш
/печения *“ “ |
2d
Ф
Расположенные в шахматном порядке последовательные ряды труб тако-
го же типа*:
Применяется уравнение (8П3.7), причем коэффициент 0,21 заменяется на
0,27 при 3 рядах, на 0,30 при 5 рядах и на 0,33 при 10 и более рядах труб.
Проблемам массо- и теплообмена посвящено огромное число
работ. В настоящей главе мы рассмотрели только основные
принципы определения параметров, представляющих наиболь-
ший интерес для биохимической технологии. Как указано в не-
которых из перечисленных в конце этой главы литературных
источников общего характера, к настоящему времени опублико-
вано очень большое число эмпирических и полуэмпирических
уравнений для определения тепло- и массопередачи в самых
разных условиях. В практической работе всегда следует исполь-
зовать уравнения, в наибольшей степени отвечающие изучаемо-
му процессу; кроме того, во всех случаях необходимо учитывать
области применимости уравнений.
8.11. Стерилизация газов и жидкостей фильтрованием
В предыдущих разделах мы обсуждали стерилизацию жид-
костей нагреванием. Однако высокие температуры могут способ-
ствовать разрушению некоторых компонентов среды, и, кроме
того, тепловая стерилизация газов нецелесообразна по экономи-
ческим соображениям. Альтернативным способом стерилизации,
пригодным как для газов, так и для жидкостей, является филь-
трование, позволяющее с помощью фильтров отделить все не-
желательные жизнеспособные клетки, а иногда и вирусы.
Фильтры изготавливаются из пористого фарфора, спрессо-
ванного асбестового волокна или из синтетических микропори-
стых полимерных мембран. Первые два материала в настоящее
время представляют интерес только с исторической точки зре-
ния; сейчас почти всегда для стерилизации фильтрованием при-
меняют полимерные микропористые мембраны. Такие фильтры
широко используются для стерилизации газов (в потоке) или
разбавленных жидких суспензий, поступающих в биореактор.
Мембранные фильтры имеют следующие преимущества:
* Charm S. Е., Fundamentals of Food Engineering, 2d ed., Chap. 4, Avi
Publishing Westport, Conn., 1971.
Процессы переноса в биотехнологических системах
675
1. Получаемые из устойчивых гелей пористые мембраны имеют
чрезвычайно малый разброс пор по диаметрам и благодаря
этому обстоятельству эффективно задерживают любые части-
цы, размер которых превышает определенную величину (кон-
тролируемую путем изменения размеров пор).
2. Высокая пористость (часто достигающая 70—80%) и малая
толщина (около 100 мкм) обеспечивают низкое сопротивле-
ние течению жидкости или газа и тем самым высокую про-
пускную способность мембранных фильтров по отношению к
растворителю (воде). Так, мембрана с фильтрующей по-
верхностью 10 см2 и порами диаметром 0,2 мкм способна
пропускать 1 л жидкости за 2—3 мин.
3. Все фильтрующие материалы мембран (в том числе нитро-
целлюлоза, ацетилцеллюлоза, винильные полимеры, полиами-
ды и фторуглеводороды) выдерживают стерилизацию паром,
устойчивы по отношению к большинству водных суспензий и
ко многим органическим веществам.
4. Качество мембраны заводского изготовления легко прове-
ряется с помощью суспензии жизнеспособных микроорганиз-
мов, обладающих практически одинаковыми размерами. На-
пример, пористые фильтры с диаметром пор 0,22 мкм про-
веряют с помощью суспензии бактерий Pseudomonas aerugi-
nosa, a Serratia marcescens может служить эффективным ин-
струментом для проверки мембран с порами диаметром
0,45 мкм. Штаммы вирусов можно применять для контроля
фильтров с очень мелкими порами; недостатком этого метода
является отсутствие отработанных методик культивирования
вирусов.
Рассмотренные в этом разделе фильтры применяют для от-
деления следовых количеств твердых частиц от газов или жид-
костей с целью стерилизации последних (в производстве фар-
мацевтических препаратов) или с целью отделения патогенных
организмов (в производстве напитков). Фильтрование как спо-
соб выделения твердых тел из концентрированных суспензий,
например культуральных жидкостей, мы рассмотрим в гл. 11.
Упражнения
8.1. Коэффициенты диффузии кислорода в растворах белков. Штреве
[53] указывал, что предложенное еще в 1881 г. Дж. К. Максвеллом
(Treatise on Electricity and Magnetism, vol. 1, 3d ed.) уравнение для диффузии
через жидкость, содержащую сферические частицы, упрощается до приведен-
ного ниже выражения, если эти частицы непроницаемы:
S) 2(1-/)
А 2 + /
где — кажущийся коэффициент диффузии в суспензии; S>o — кажущийся
676
Глава 8
коэффициент диффузии в чистой жидкости; f — объемная доля частиц. Он
нашел, что указанное уравнение хорошо согласуется с экспериментальными
данными, если /=fP+fь, где [р — объемная доля белка, a fb — объемная доля
воды, адсорбированной на поверхности молекул белка. Приняв, что молеку-
ла белка по своим размерам отвечает гемоглобину (сфероид 65X55X55 А),
вычислите и начертите график зависимости W-^о от fp (не от f) при
условии, что молекулы белка окружены 0, 1 или 2 монослоями иммобилизо-
ванной воды (диапазон /р от 0,1 до 0,5). Сравните полученные вами резуль-
таты с приведенными ниже экспериментальными данными для метгемогло-
бина; обсудите результаты сравнения.
<2>/^о 0,69 0,43 0,17
fp 0,1 0,2 0,4
8.2. Коэффициент массоотдачи. Определите ki для следующих условий:
объем жидкой фазы 10 л
диаметр турбинной мешалки 10 см
диаметр сосуда 50 см
скорость вращения мешалки 200 оборотов в мин
коэффициент бинарной диффузии воздух — среда 0,5-10—5 см2/с
скорость подачи воздуха 2 л/мин
плотность среды 1,2 г/см3
вязкость среды 0,1 г/(см-с)
8.3. Перенос кислорода в отсутствие перемешивания. Рассмотрим непере-
мешиваемый аэрируемый хемостат объемом 0,5 л, в дне которого имеется
10 отверстий. Какая удельная скорость роста клеток будет поддерживаться
в хемостате, если диаметр каждого отверстия равен 1 мм, скорость подачи
воздуха через одно отверстие составляет 5 мл/мин и если кислород лимити-
рует скорость роста клеток. Коалесценцией и дроблением пузырьков воздуха
можно пренебречь; считайте, что среда разбавлена в такой степени, что по
своему поведению не отличается от чистой воды. В расчетах примите следую-
щие значения параметров: цтах=0,5 ч-1; Ks = 0,l мМ; о=72 дин/см; g =
= 980 см/с2; Цгаз=2-10_4 г/(см-с); S>=0,5-10~5 см2/с; ЦжИдк = 10-2 г/(см-с);
ргаз = 1,4 г/л; //£=10 см; Уо/х = 1 г О2/г клеток; х=1,0 г клеток/л.
8.4. «Изменение» kia с температурой. Теория обновления поверхности
предусматривает, что коэффициент массоотдачи изменяется пропорциональ-
но ^1/2. Для случая диффузии в жидкостях, согласно уравнению Стокса —
Эйнштейна, 5>p,/T=const. Следовательно, ki должно быть пропорциональ-
ным (Т/ц)1/2.
а) Найдите в любом справочнике данные по зависимости вязкости воды
от температуры и вычислите изменение отношения ki(T)!ki (Т=15°С) в диа-
пазоне от 15 до 60° С; изобразите результаты в графической форме.
б) Равновесная концентрация растворенного кислорода зависит от тем-
пературы (табл. 8.1). Приняв, что отношение площади поверхности раздела
фаз к объему а не зависит от температуры, вычислите изменение 7s
s= [kiac*(T)/kiac*(T==\5°C)] от температуры в том же диапазоне, что и в
упражнении 8.4 а; изобразите результаты в графической форме.
Какое, по вашему мнению, практическое значение может иметь показан-
ное в этом упражнении постоянство величины 7? Постоянство 7 подтвержде-
но экспериментально [54].
8.5. Реактор периодического действия; зависимость клеточного роста от
массообмена. Периодическая культура микроорганизмов растет при 35 °C.
Методом окисления сульфитом натрия показано, что kia'Ci*=0,l моль/(л-ч).
Процессы переноса в биотехнологических системах
677
бремя удвоения численности популяции в периоде экспоненциального роста
составляет 30 мин, экономический коэффициент по кислороду равен 0,6 г
клеток/г Ог.
а) Вычислите удельную скорость клеточного роста р, в экспоненциаль-
ной фазе.
б) С помощью уравнения (8.14) рассчитайте концентрацию растворен-
ного кислорода ci при увеличении клеточной массы (начальное значение
Хо=Ю”6 г/мл). Постройте график зависимости ы от х. При какой концентра-
ции клеточной массы концентрация ci должна стать равной нулю?
в) В реальном процессе никогда не становится равной нулю. Вместо
этого при низких концентрациях растворенного кислорода р становится функ-
цией ci, на что указывает, например, уравнение (8.15). С помощью уравне-
ния (8.15) и приведенных выше значений параметров вычислите зависимость
ci от х. Примите К.ог равным 0,05 ммоль/л. (Удобнее по заданным величи-
«ам ci вычислять х.)
г) При с/>0,9 Ci* процесс лимитируется клеточным ростом, а при
С/<0,1 Ci* скорость процесса ограничивается массообменом. С помощью урав-
нения (8.15) и графика, полученного при решении упражнения 8.5в, опреде-
лите диапазоны значений х, которые соответствуют режимам, лимитируемым
ростом и массообменом.
Л) Правая часть уравнения (8.15) представляет собой dx/dt. На том же
чертеже, на котором вы решили упражнение 8.5в, постройте график зависи-
мости dxjdt от х. Каким выражением определяется х при (dx/d/)max?
8.6. Влияние давления. Перенос кислорода интенсифицируется при по-
вышении его парциального давления путем простой замены воздуха на чи-
стый кислород. Предлагалось также применять повышенное давление. К со-
жалению, рост микроорганизмов может подавляться кислородом за счет, на-
пример, накопления в клетке активных форм кислорода, которые могут по-
давлять внутриклеточные превращения, нуждающиеся в локальных
восстановительных условиях.
а) Допустим, что зависимость скорости клеточного роста от концентра-
ции кислорода можно описать уравнением
„ _ ______Hmaxg/_____
И /Co2+g/ + g2//^i
или
р___________с______
Птах 1 + с + ус2
где у = /Со2/К; и c^ci/Ko^. Покажите, что удельная скорость клеточного рос-
та будет максимальной при с=у~|/2.
б) На основе уравнения (8.15), модифицированного путем включения
приведенного выше выражения (отражающего ингибирующее действие кис-
лорода), выведите уравнение, которое будет предсказывать давление кисло-
рода рог, обеспечивающее максимальную скорость роста при любом значе-
нии х. Поскольку pmax=const, определите также соответствующую зависи-
мость ро2 от времени.
8.7. Секреция продукта клеточного метаболизма в среду, а) В микро-
биологическом процессе синтеза L-аспартата последний образуется в концент-
рации 1, 2, 3 и 4 г/л через 1, 2, 3 и 4 ч после начала процесса соответствен-
но. Если при /=0 добавить поверхностно-активное вещество (цетилпириди-
нийхлорид), то концентрация L-аспартата повысится до 12, 22, 30 и 35 г/л
соответственно. Можете ли вы объяснить, какой основной процесс (или про-
цессы) лежит в основе накопления L-аспартата в среде в отсутствие и в
678
Глава 8
присутствии поверхностно-активного вещества? (Данные взяты из ра-
боты [55].)
б) Накопление аминокислот в среде некоторыми штаммами Е. coll про-
исходит в результате двух одновременно протекающих конкурирующих яв-
лений—пассивного транспорта из клетки и активного транспорта в клетку.
Напишите уравнение скорости накопления в среде аминокислоты, если этот
процесс определяется двумя указанными явлениями. Опишите ряд методов,
экспериментального определения начальных скоростей накопления аминокис-
лот; с помощью меченных 14С аминокислот эти методы должны обеспечивать
определение всех параметров предложенного вами уравнения скорости
процесса.
Мутации, устраняющие катаболитную репрессию и подавляющие актив-
ный транспорт, могут привести к тому, что накопление продукта метаболиз-
ма будет лимитироваться только его транспортом (см. [56]).
8.8. Мицелий, иммобилизованный на микробусинах. Адсорбция спор
штамма Penicillium chrysogenum на пористых частицах диаметром 300—
500 мкм применялась для создания иммобилизованного мицелиального ката-
лизатора, характеристики которого ниже сравниваются с характеристиками
культуры суспензии мицелия. Обе культуры были выращены в одном и том
же биореакторе типа барботажной колонны.
Характеристика Суспензия клеток Иммобилизо- ванные клетки
Vmax, Г/Л 17,0 29,0
Ртах, г/л 2,0 5,5
&/аДс, ммолей ОгДл-ч-атм) 50—100 100—350
Базисный параметр — расход мощности, кВт/м3 2,3 2,3
Расход энергии на перенос кислорода, кг О2/кВтч 0,21 0,48
Удельный расход энергии, кВтч/г пенициллина G 0,12 0,07
а) Приняв (приближенно), что kt пропорционален ц~1/2, оцените отноше-
ние вязкостей суспендированной и иммобилизованной культур. Чем, по ваше-
му мнению, обусловлена пониженная вязкость культуральной жидкости
с большей концентрацией биомассы?
б) Оцените сравнительную стоимость культуры в суспензии и иммобили-
зованной культуры. В каком варианте процесс более выгоден с экономиче-
ской точки зрения [57]?
8.9. Методы масштабного перехода. В табл. 8.5 приведены различные
условия масштабного перехода (например, постоянная величина P/V).
а) Проанализируйте положительное (или отрицательное) влияние на
другие переменные процесса, если масштабный переход выполняется при по-
стоянстве P/V, Nt, Ft/V, NiDi или Re/.
б) Высказывалось мнение, что масштабный переход лучше всего выпол-
нять при постоянных kta и скорости сдвига жидкости (окружной скорости
мешалки), в частности, при D//T«0,25—0,4 и NiDi=0,5 м/с. С помощью
приведенных в этой главе уравнений покажите, как при этих условиях мас-
штабного перехода будут изменяться P/V, Nt, Fi/V и Re/ [58].
8.10. Параметры масштабного перехода в процессах аэрации, а) Соглас-
но многим уравнениям, при аэрации с перемешиванием Sh = aRe/miScm2. По-
кажите, что при одинаковом размере пузырьков одинаковые значения kt в двух
различных сосудах, например в маленьком (I) и большом (II), возможны
только в том случае, если изменение скорости вращения мешалки, выражен-
ной в числе оборотов в минуту Nt, при масштабном переходе описываете»
Процессыпереноса в биотехнологических системах
679
следующим выражением:
АМН) _Г £>,(!)
^•(1)
L Di (II) J
б) Покажите, что из этого уравнения следует приведенное ниже равен-
ство, справедливое при постоянном kr.
(Р/У)п г Ое(1) Т-3/От1
(P/Юг L Di (II) J
Какой частный вывод следует из последнего уравнения, если принять во вни-
мание рассмотренное в тексте уравнение для турбулентного режима?
в) Если размер пузырьков зависит от параметров перемешивания, какие
ограничения налагаются на отношения M(II)/M(I), (P/I/)n/(P/I/)i?
8.11. Работа барботажной колонны, а) Определите величины а, Н и kt
для барботажной колонны при следующих условиях:
скорость подачи газа 20 н. кубических футов (0,566 м3) в минуту
скорость подачи жидкой фазы 25 галлонов (94,633 л) в минуту (вода)
внутренний диаметр колонны 16 дюймов (40,64 см)
средний диаметр пузырьков D 0,25 дюйма (0,64 см)
б) Предлагалось иное уравнение для потока пузырьков (массопередача
в жидкость или в систему из двух жидкостей) [59]:
Sh «= 2,0 + 0,0187
Re0,484<;c0,339
Dg1/3
,2)2/3
\ 0,07241,61
Сравните это уравнение с приведенной в тексте формулой. Еще раз рассчи-
тайте kt и оцените разницу (в процентах) между двумя способами опреде-
ления Sh.
8.12. Реаэрация в естественном водном потоке (быстрины н спокойные
участки). Движущийся водный поток приближенно можно описать как чере-
дование глубоких и мелких участков одинаковой ширины. Пусть «глубокие»
и «мелкие» участки имеют глубину hD, hs и длину Id, Is соответственно.
а) Определите отношение коэффициентов массопередачи кислорода в глу-
боких и неглубоких участках.
б) Не подразделяя организмы на отдельные виды, опишите математиче-
ски процесс утилизации субстрата аэробными организмами в указанной си-
стеме из глубоких и мелких участков, считая, что скорость роста ограничи-
вает перенос кислорода. Четко укажите принятые допущения.
в) Найдите математические выражения (при необходимости приняв уп-
рощающие допущения), определяющие долю общего роста микроорганизмов
в спокойных участках и долю общего переноса кислорода, осуществляюще-
гося в быстринах.
8.13. Упрощенное описание реаэрации в водном потоке (уравнение Стри-
тера— Фелпса). В условиях, когда осаждение органических веществ, реак-
ции в осадке и потеря органических летучих веществ не играют большой
роли, реаэрацию в водном потоке можно описать как простое явление типа
потока идеального вытеснения. Уравнение материального баланса по орга-
ническому веществу S в потоке, движущемся со скоростью и, можно записать
в таком виде:
ds ds
~dT=a -“'^T-HmaxS
Для материального баланса по кислороду предложено уравнение
дс0 - Vi
=» ~ h ^О2 (с О2 — СО2) — Pmax5"o/S +
Н-скорость фотосинтеза — скорость дыхания водорослей — скорость осаждения
680
Глава 8
Здесь в правой части уравнения первое слагаемое отражает градиент кон-
центраций кислорода, второе — поступление Ог за счет массообмена, а
третье — расход Ог в процессах окисления с участием микроорганизмов.
а) Покажите, что
/ z \
s (z) 5z=o ехР I Ртах и I
если микроорганизмы, окисляющие питательные вещества, получают кисло-
род в избытке.
б) Покажите, что в стационарном состоянии в отсутствие фотосинтеза,
дыхания водорослей и осаждения профиль концентрации кислорода можно
описать уравнением Стритера — Фелпса:
^0/sPmaxsz=o Г / г \
' °’ - '°'<г> = io2-rO;SH»ax Н ~ "
/ ^z \1 I ^OZZ \
— ехР I------— I + (С*О2 - СОа.-=о) ехР I------— )
где Е'о2 = Ео2//; £о2— коэффициент массопередачи кислорода, усредненный
по вихрю; I — глубина потока.
в) Покажите, что в модели Стритера — Фелпса параметр с*о2 — co2(z),
называемый дефицитом кислорода, имеет единственный минимум при
Y0/SPmaxsz=o [ 2 \
с*Оа сОа (2) =* ехР I УЬ/SPmax I
к Оа \ /
Какое расстояние z соответствует этой точке максимального дефицита кис-
лорода? Выведите аналогичные выражения для случая, когда необходимо
учитывать фотосинтез, дыхание водорослей и осаждение, осуществляющиеся
с постоянными скоростями.
8.14. Теплообмен; размер пузырьков. Рассмотрим процесс, стехиометрия
которого дана в упражнении 5.13.
а) Предположим, что периодический аэробный микробиологический про-
цесс проводится в цилиндрическом резервуаре диаметром 1,83 м, снабжен-
ном охлаждающим змеевиком длиной 39,6 м и диаметром 2,54 см, причем
расстояние между витками змеевика составляет 3,08 см. Допустим далее,
что вся теплопередача осуществляется через змеевики и что барботер под-
держивает достаточно высокое значение Kia, при котором рост клеток не-
лимитируется кислородом. Какой должна быть минимальная, обеспечивающая
требуемую теплопередачу скорость вращения мешалки (в оборотах в мину-
ту), если средняя скорость жидкости в направлении, перпендикулярном тру-
бам змеевика, составляет 10% от скорости конца лопасти мешалки, средняя,
температура охлаждающей жидкости равна 18 °C и реактор должен работать
при температуре не выше 28 °C? Повторите расчеты для концентраций 106,.
107, 108 и I09 клеток в миллилитре [диаметр однолопастной мешалки
4,5 фута (1,37 м), толщина лопасти 1,2 дюйма (3,05 см), высота 6 дюймов-
(15,24 см)].
б) При плотности 109 клеток/мл скорость аэрации такова, что клетки
поглощают 10% поступающего кислорода. Из-за неудачной конструкции бар-
ботера размер пузырьков слишком велик. Какая скорость мешалки (в обо-
ротах в минуту) необходима для того, чтобы пузырьки имели адекватный
диаметр? Не даст ли лучшие результаты установленная непосредственно нал.
барботером вторая, значительно меньшая, но высокоскоростная юпастна?»
мешалка, например мешалка с параметрами £>2 = 0,2 Di, #2=10 jVi?
Процессы переноса в биотехнологических системах 681:
8.15. Критерии подобия. л-Теорема Бэкиигема. В процессах теплопередачи
при принудительном течении жидкости по поверхности трубы для определе-
ния коэффициента теплопередачи (й) со стороны жидкости важны следую-
щие параметры: характерный диаметр трубы D, скорость течения жидко-
сти и, вязкость жидкости ц (в пуазах), плотность р, удельная теплоемкость
при постоянном давлении Ср [в кал/(моль-град)] и теплопроводность жид-
кости kf [в кал/(с-см-град)]. л-Теорема Бэкингема устанавливает, что,
«функциональное соотношение между q величинами, элементы которых пред-
ставимы в виде комбинаций из р фундаментальных элементов, можно
записать, как функцию q—р безразмерных групп». В теплопередаче фунда-
ментальными элементами являются масса т, длина I, время t и темпе-
ратура Т.
а) Выразите ft, D, и, ц, р, Ср и k через указанные элементы, т. е. в виде
malbtcTd.
б) Четыре безразмерные группы обычно образуются из следующих кри-
териев подобия: Nu (Nu==ftD/&/), Re (Re=pDu/p), Рг (РггэСрр,/йД
и числа Стантона St (St = ft/zzCpp). Выскажите свои соображения. Что, по.
вашему мнению, отражает число Стантона?
8.16. Жидкости, подчиняющиеся степенному закону; гидролиз крахмала.
Нагревание 1%-ной (масса/объем) суспензии амилопектина в воде приводит
к псевдопластичному раствору, который должен подчиняться уравнению
т=т]у'1, где п<1,0. При гидролизе этого раствора ос-амилазой в реакторе пе-
риодического действия наблюдались следующие изменения параметров си-
стемы во времени [60]:
Т|, дин-сп/см2 0,32 0,26 0,20 0,14 0,10 0,03
п 0,73 0,75 0,78 0,83 0,85 0,98
а) Покажите, что эти данные можно описать уравнением То=тПол. где
То, Уо — константы.
б) С помощью соответствующих графиков покажите, что все кривые
зависимостей т от у при любой степени гидролиза пересекаются в од-
ной точке.
в) Как будет изменяться во времени потребляемая мощность, если ско-
рость вращения мешалки будет постоянной? Можно ли использовать этот
параметр для постоянного неавтономного контроля хода периодического
процесса?
8.17. Зависимость потребляемой мощности от скорости вращения переме-
шивающего устройства в неньютоновских жидкостях. Для неньютоновских
жидкостей с показателем степени п<1 в уравнении степенного закона ско-
рость сдвига у можно считать пропорциональной скорости вращения лопаст-
ной мешалки N, [61].
а) Покажите, что для жидкости, находящейся между двумя соосными
цилиндрическими поверхностями, скорость сдвига на внешнем цилиндре (при
изменении скорости вращения внутреннего цилиндра) изменяется в соответ-
ствии с выражением (dr/dNi) <xNin~i.
б) Покажите, что число Рейнольдса Re=£>;2M;p/T]0, где Т|г. — кажущаяся
вязкость (зависящая от скорости сдвига), пропорционально Ni2~n.
в) Для указанного выше числа Рейнольдса зависимость числа мощности
(PgdDfNfp) от числа Рейнольдса совпадает с аналогичной зависимостью
для ньютоновских жидкостей (или соответствующая кривая располагается
чуть ниже кривой для ньютоновской жидкости). Если Ро пропорционально.
44—537
682
Глава 8
Re“, покажите, что потребляемая мощность пропорциональна Nis+a^~n'>.
г) Как с помощью полученной в предыдущих упражнениях информации
вы могли бы определить коэффициент пропорциональности между f и Nt
для неньютоновской жидкости?
8.18. Фазы процессов микробиологической переработки углеводородов,
а) На рис. 8.14 изображены основные фазы процесса микробиологической
переработки углеводорода по данным Мимуры и др. Найдите математические
выражения, описывающие скорости клеточного роста и утилизации субстрата
в каждой фазе периодического процесса. Для каждой «фазы» приведите ко-
личественные характеристики, определяющие природу основного сопротивле-
ния клеточному росту, например растворимость углеводорода, транспорт кис-
лорода, клеточный метаболизм и т. д.
б) Назовите причины, почему система, состоящая из клеток, капелек
углеводорода, воздушных пузырьков и растворителя, принимает каждую из
упомянутых авторами структур. Какие эксперименты напрашиваются в связи
с результатами этих наблюдений? Как вы могли бы доказать предложенные
гипотезы или выбрать наиболее обоснованную из них?
8.19. Микробиологические процессы с двумя (газообразными) субстра-
тами. Хэймер и др. [SCP Production from Methane, р. 362 in Single Cell Pro-
tein II, Tannenbaum S. R., Wang D. I. C. (eds), MIT Press, Cambridge, Mass.,
1975] указывали, что рост клеток при утилизации метана в присутствии
кислорода в проточном, постоянно барботируемом реакторе можно описать
двойным уравнением Михаэлиса — Ментен:
С1_______С2
rx=^lhnaxX К1+С1 К2 + с2
здесь индексы 1 и 2 указывают параметры, относящиеся к кислороду и мета-
ну соответственно.
а) Допустив, что экономические коэффициенты У1 и Y2 (г клеток из г
субстрата i; i—1,2) постоянны, напишите уравнения материального баланса
по биомассе и субстратам 1 и 2 в стационарном состоянии при условии сте-
рильности питательных веществ. Коэффициент общей массопередачи примите
равным Киа-, считайте, что как жидкая, так и газовая фазы полностью пе-
ремешиваются.
б) Повышение скорости разведения в конце концов приводит к вымыва-
нию. Покажите графически или аналитически, что вымывание происходит
при D около 0,72 ч-1 при следующих значениях параметров: К\ = Кг^
= 5-10-4 г/л; У1 = 1,25 г клеток/г О2; У2=2,0 г клеток/г субстрата; Киа=
= Kz2<2=100 ч-1; Ci* = 0,015 г/л; с2*=0,007 г/л; р,Шах=0,8 ч-1.
Литература
Диффузия:
1. Weisz Р., Diffusion and Chemical Transformation: An Interdisciplinary
Excursion, Science, 179, 433 (1973).
Потребность культур клеток в кислороде и растворимость кислорода:
2. Finn R. К., Agitation and Aeration, in Biological Engineering Science,
Blakebrough N. (ed.), vol. 1, p. 69, Academic Press, Inc., New York, 1967.
Взаимосвязь между диффузией через жидкие пленки и химическими реак-
циями:
Процессы переноса в биотехнологических системах 68&
3. Данкверте П. В., Газо-жидкостные реакции. — М.: Химия, 1973.
4. Астарита Дж., Массопередача с химической реакцией. — Л.: Химия, 1971.
5. Берд Р., Стьюарт В., Лайтфут Е., Явления переноса. — М.: Химия, 1974.
Культуральные жидкости:
6. Taguchi Н., The Nature of Fermentation Fluids, Adv. Biochem. Eng., 1
(1971).
Проблемы масштабного перехода в микробиологических процессах:
7. Biotech. Bioeng., 8 (1966), Gaden Е. L., Jr. (ed.) (весь том).
Примеры массообмена в микробиологических реакторах:
8. Moser A., in Proc. Int. Symp. Adv. Microb. Eng., 1, 295—580 (1973).
9. Van’t Riet K-, Trends in Biotechnology, Mass Transfer in Fermentation, 1,
(4), 113 (1983).
Уравнения для определения переноса кислорода:
10. Calderbank Р. Н., Moo-Young М., The Continuous Phase Heat and Mass
Transfer Properties of Dispersion, Chem. Eng. Sci., 16, 39 (1961).
11. Calderbank P. H., Mass Transfer in Fermentation Equipment, in Biochemical
and Biological Engineering Science, Blakebrough N. (ed.), vol. 1, Academic
Press Inc., New York, 1967.
12. Akita K, Yoshida F., Bubble Size, Interfacial Area, and Liquid-Phase Mass
Transfer Coefficient in Bubble Columns, I & EC Process Des. Develop., 13,
84 (1974).
13. Bello R. A., Robinson C. W., Moo-Young M., Mass Transfer and Liquid
Mixing in External Circulating Loop Contactors, Adv. Biotech., 1, 547 (1981).
14. Wang К. B., Fan L. T., Mass Transfer in Bubble Columns Packed with
Motionless Mixers, Chem. Eng. Sci., 33, 945 (1978).
Потребность в кислороде в периодических процессах:
15. Darby R. Т., Goddard D. R., Studies of the Respiration of the Mycelium of
the Fungus Myrothecoum verracaria, A. J. Bot„ 37, 379 (1950).
Определение kia с помощью кислородного электрода:
16. Wernan W. C., Wilke C. R., New Method for Evaluation of Dissolved Oxygen
Response for Кьа Determination, Biotech. Bioeng., 15, 571 (1973).
Абсорбция водными пленками:
17. Briffaud J., Engasser M., Growth and Excretion Kinetics in a Trickle-Flow
Fermentor, Biotech. Bioeng., 21, 2093 (1979).
18. Livansky K, Prokes B„ Kihrt F., Benes V., Some Problems of CO2 Absorption
by Algae Suspensions, Biotech. Bioeng. Symp., 4, p. 513 (1973).
Стехиометрия утилизации метана:
19. Klass D. L„ landolo J. J., Knabel S. J., Key Process Factors in the Microbial
Conversion of Methane to Protein, CEP Symp. Ser., [93], 65, 72 (1969).
44*
684
Глава 8
Уравнения для определения необходимого диаметра отверстий:
20. van Krevelen D. W., Hoftijzer P. J., Studies of Gas-Bubble Formation: Calcu-
lation of Interfacial Area in Bubble Contactors, Chem. Eng. Prog., 46, 29
(1950).
21. Schugerl K.., Lucke J., Bubble Column Bioreactors, in Advances in Biochemb
cal Engineering, Ghose T. K., Fiechter A., Blakebrough N. (eds.), vol. 7,
p. 1, Springer-Verlag, Berlin, 1977.
Фотографическое определение a, H, D Заутера:
22. Calderbank P. H., Rennie J., Int. Symp. Distill. (Inst. Chem. Eng.), 1960.
Зависимость формы больших пузырьков от скорости [уравнение (8.36)]:
23. Davies R. М., Taylor G. I., Proc. Roy. Soc., A200, 375 (1956).
Массопередача через свободную поверхность к стекающей пленке жидкости
(см. работу [5]).
Массопередача через свободную поверхность к турбулентному водному по-
току:
24. Fortescue G. Е., Pearson J. R. A., On Gas Absorption into a Turbulent
Liquid, Chem. Eng. Sci., 22, 1163 (1967).
25. O’Connor D. J., Dobbins W., The Mechanism of Reaeration in Natural
Streams, J. Sanit. Eng. Div., Proc. ASCE, 82, SA6 (1966).
Зависимость мощности турбины от Re перемешивающего устройства:
26. Rushton J. Н., Costich Е. W„ Everett H. J., Power Characteristics of Mixing
Impellers, pt. 2, Chem. Eng. Prog., 46, 467 (1950).
Потребляемая мощность в аэрируемых и неаэрируемых процессах;
27. Ohyama Y., Endoh К., Power Characteristics of Gas-Liquid Contacting Mi-
xers, Chem. Eng. Japan, 19, 2 (1955).
28. Michel B. J., Miller S. A., Power Requirements of Gas-Liquid Agitated Sys-
tems, AIChE J., 8, 262 (1962).
Перемешивание с помощью струи жидкости:
29. Blenke H., Loop Reactors, in Advances in Biochemical Engineering, Gho-
se T. K., Fiechter A., Blakebrough N. (eds.), vol. 13, p. 121, Springer-Verlag,
Berlin, 1979.
Литературу по уравнениям с числом Вебера см. в работе [11].
Коэффициенты диффузии кислорода в пленках микроорганизмов:
30. Matson J. V., Characklis W. G., Oxygen Diffusion through Microbial Aggre-
gates, 77th AIChE Meet., Pittsburgh, June 1973.
Влияние ионной силы на kia:
31. Robinson C. W., Wilke C. R., Oxygen Absorption in Stirred Tanks: A Corre-
lation for Ionic Strength, Biotech. Bioeng., 15, 755 (1973).
Процессы переноса в биотехнологических системах 685
Поверхностно-активные вещества и массообмен:
32. Eckenfelder W. W., Jr., Barnhart Е. L., The Effect of Organic Substances on
the Transfer of Oxygen from Air Bubbles into Water, AIChE J., 7, 631
(1961).
33. Benedek A., Heideger W. J., Effect of Additives on Mass Transfer in Turbine
Aeration, Biotech. Bioeng., 13, 663 (1971).
34. Bull D. N., Kempe L. L., Influence of Surface Active Agents on Oxygen
Absorption to the Free Interface in a Stirred Fermentor, Biotech. Bioeng., 13,
529 (1971).
35. Aida S., Toda K., The Effect of Surface Active Agents on Oxygen Absorption
in Bubble Aeration I, J. Gen. Appl. Microbiol., 7, 100 (1963).
.36 . Maney К. H., Okun D. A., Effect of Surface Active Agents on the Rate of
Oxygen Transfer, Adv. Biol. Waste Treat., Ill (1963); см. также работы
МакКеоуна и Окуна, Тимсона и Данна, а также Карбера в этом же источ-
нике.
Реология микробных бульонов:
37. Leduy A., Marson A. A., Corpal В., A Study of the Rheological Properties of
a Non-Newtonian Fermentation Broth, Biotech. Bioeng., 16, 61 (1974). (Изу-
чение культуры A. pullulans.)
38. Reels J. A., van den Berg J., Voncken R. M., The Rheology of Mycelial
Broths, Biotech. Bioeng., 16, 181 (1974). (Изучение культуры Penicillium.)
39. Thompson N., Ollis D. F., Evolution of Power Law Parameters for Xanthan
and Pullulan Batch Fermentations, Biotech. Bioeng., 22, 875 (1980).
40. Chang H. T., Ollis D. F., Generalized Power Law for Polysaccharide Solu-
tions, Biotech. Bioeng., 24, 2309 (1982).
Изучение морфологии в связанных последовательно реакторах:
41. Vrana D., Some Morphological and Physiological Properties of Candida uti-
lis Growing «Hypertrophically» in Excess of Substrate in a Two-Stage Con-
tinuous Cultivation, Biotech. Bioeng. Symp., 4, 161 (1973).
Определение потребляемой мощности в неньютоновских биологических средах:
42. Taguchi Н., Miyamoto S., Power Requirement in Non-Newtonian Fermenta-
tion Broth, Biotech. Bioeng., 8, 43 (1966).
Концентрация клеток и массообмен:
43. Brierley М. R., Steel R., Agitation-Aeration in Submerged Fermentation,
pt. 2: Effect of Solid Dispersed Phase on Oxygen Absorption in a Fermentor,
Appl. Microbiol., 7, 57 (1959). (A. niger.)
44. Andrews G. F., Fonta J. P., Marrota E., Stroeve P., The Effects of Cells on
Oxygen Transfer Coefficients, Chem. Eng. J., 29, B39, B47 (1984).
Перемешивание и повреждение клеток. См. работу [7], а также:
45. Midler М., Finn R. К, A Model System for Evaluating Shear in the Design
of Stirred Fermentors, Biotech. Bioeng., 8, 71 (1966).
Суспензии целлюлозной пульпы:
46. Blakebrough N., Sambamurthy K, Mass Transfer and Mixing Rates in Fer-
mentation Vessels, Biotech. Bioeng., 8, 25 (1966).
686
Глава 8
Фильтрование через волокнистые фильтры:
47. Friedlander S. К-, Aerosol Filtration by Fibrous Filters, in Biochemical and
Biochemical Engineering Science, Blakebrough N. (ed.), vol. 1, p. 49,.
Academic Press, New York, 1967.
48. Air Filtration, Davies C. N. (ed.), Academic Press, New York, 1973.
Фильтрование через мембраны:
49. Dwyer J. L., Filtration in the Food, Beverage, and Pharmaceutical Industries,
p. 121, in Filtration: Principles and Practices, Part II, Marcel Dekker, New
York, 1979.
Теплопередача; анализ параметров; см. стр. 396 и далее в работе [5], а также*
50. Brown A. I., MacroS. М., Introduction to Heat Transfer, pp. 85—95, McGraw-
Hill Book Company, New York, 1958.
Экономические проблемы теплопередачи в микробиологических процессах:
51. Abbott В. J., Clamen A., The Relationships of Substrate. Growth Rate, and
Maintenance Coefficient to Single Cell Protein Production; Biotech. Bioeng.,
15, 117 (1973).
Баланс энтальпии в микробиологических процессах:
52. Cooney С. L., Wang D. I. C., Mateles R. I., Measurement of Heat Evolution
and Correlation with Oxygen Consumption during Microbial Growth, Bio-
tech. Bioeng., 11, 269 (1968).
Упражнения:
53. Stroeve P., On the Diffusion of Gases in Protein Solutions, Ind. Eng. Chem.
Fundam., 14, 140 (1975).
54. Aiba S., Koizumi J., Ru J. S„ Mukhopadhyay S. N., The Effect of Tempera-
ture on kia in Thermophilic Cultivation of Bacillus stearothermophilus, Bio-
tech. Bioeng., 26, 1136 (1984).
55. Fukui C., Ishida, Microbial Production of Amino Acids, Kodansha Ltd., To-
kyo and John Wiley, New York, 1972.
56. Rancourt D. E„ Stephenson J. T., Vickell G. A., Wood J. M., Proline Excre-
tion by Escherichia coli K12, Biotech. Bioeng., 26, 74 (1984).
57. Gbewonyo K-, Wang D. I. C., Enhancing Gas-Liquid Mass Transfer Rates in
Non-Newtonian Fermentations by Confining Mycelial Growth to Microbeads
in a Bubble Column, Biotech. Bioeng., 25, 2873 (1983).
58. Oldshue J., Fermentation. Mixing Scale-Up Techniques, Biotech. Bioeng., 8,
3 (1966).
59. Hughmark G. A., Holdup and Mass Transfer in Bubble Columns, Ind. Eng.
Chem. Process Des. Dev., 6, 218 (1967).
60. Cruz A., Kinetics and Shear Viscosity of Enzyme Hydrolysed Starch Pastes,
Ph. D. thesis, Princeton University, Princeton, N.J., 1976.
61. Metzner A. B„ Feehs R. H., Ramos H. L., Otto R. E„ Tuthill J. D., Agitation
of Viscous Newtonian and Non-Newtonian Fluids, AIChE J., 7, 3 (1961).
Оглавление
От переводчика................................................... 5
Предисловие авторов ........................................... 8
Глава 1. ВВЕДЕНИЕ В МИКРОБИОЛОГИЮ................................11
1.1. Биофизика и клеточная теория............................13
1.2. Строение клеток...........................................14
1.2.1. Прокариотические клетки.............................14
1.2.2. Эукариотические клетки..............................17
1.2.3. Фракционирование клеток.............................21
1.3. Важнейшие типы клеток.....................................24
1.3.1. Бактерии............................................26
1.3.2. Дрожжи..............................................30
1.3.3. Плесени.............................................32
1.3.4. Водоросли и простейшие..............................36
1.3.5. Растительные и животные клетки......................37
1.4. Перспективы дальнейшего изучения..........................40
Упражнения.....................................................40
Литература.................................................43
Глава 2. ХИМИЧЕСКИЕ ОСНОВЫ ЖИЗНИ.................................44
2.1. Липиды....................................................46
2.1.1. Жирные кислоты и родственные липиды.................47
2.1.2. Жирорастворимые витамины, стероиды и другие липиды 51
2.2. Сахара и полисахариды.....................................53
2.2.1. D-Глюкоза и другие моносахариды.....................55
2.2.2. Дисахариды и полисахариды...........................56
2.2.3. Целлюлоза...........................................59
2.3. От нуклеотидов к РНК и ДНК................................63
2.3.1. Структурные элементы нуклеиновых кислот; переносчик
энергии и коферменты.......................................65
2.3.2. Хранение биологической информации, ДНК и РНК . . 69
2.4. Аминокислоты и белки......................................78
2.4.1. Белковые аминокислоты и полипептиды................81
2.4.2. Структура белков....................................86
2.4.3. Первичная структура.................................87
2.4.4. Пространственная структура белков; вторичная и третичная
структуры..................................................90
2.4.5. Четвертичная структура и регулирование биологических
процессов...................................................97
2.5. Биохимические соединения смешанного строения..............98
2.5.1. Клеточные стенки; пептидогликаны и липополисахариды 99
2.5.2. Антитела и другие гликопротеины...................104
688
Оглавление
2.6. Иерархия клеточной структуры .............................. 106
Упражнения................................................НО
Литература.............................................ПО
Глава 3. КИНЕТИКА КАТАЛИЗИРУЕМЫХ ФЕРМЕНТАМИ РЕАК-
ЦИИ .......................................................118
3.1. Фермент-субстратные комплексы и механизм действия ферментов 125
3.2. Кинетика простых ферментативных реакций с одним и двумя суб-
стратами .....................................................131
3.2.1. Уравнение Михаэлиса — Ментен...................135
3.2.2. Определение параметров в уравнении Михаэлиса — Ментен 142
3.2.3. Кинетика обратимых реакций, реакций с двумя субстрата-
ми и с активацией фермента кофактором . . . . . 144
3.3. Определение констант скоростей элементарных стадий фермента-
тивной реакции................................................149
3.3.1. Релаксационные методы изучения кинетики .... 149
3.3.2. Некоторые результаты изучения кинетики переходных со-
стояний ...................................................151
3.4. Другие типы зависимостей скорости ферментативной реакции от
концентрации субстрата ...................................... 153
3.4.1. Активация и ингибирование ферментов субстратами . . 153
3.4.2. Взаимодействие одного фермента с несколькими различны-
ми субстратами.............................................156
3.5. Регуляция ферментативной активности.........................161
3.5.1. Механизмы обратимой регуляции ферментативной актив-
ности .....................................................164
3.5.2. Анализ влияния обратимой регуляции на кинетику фермен-
тативных реакций...........................................166
3.6. Другие факторы, влияющие на ферментативную активность 171
3.6.1. Влияние pH на кинетику ферментативных реакций в рас-
творах ....................................................173
3.6.2. Скорость ферментативных реакций и температура . . 176
3.7. Инактивация ферментов..................................... 181
3.7.1. Механизм денатурации белков и сопутствующие эффекты 181
3.7.2. Моделирование и кинетика процессов инактивации . . 185
3.7.3. Денатурация ферментов под действием механических фак-
торов .....................................................191
3.7.4. Методы стабилизации ферментов.................194
3.8. Ферментативные реакции в гетерогенных системах .... 196
Упражнения.......................................................202
Литература....................................................206
Глава 4. ПРИМЕНЕНИЕ РЕАКЦИЙ, КАТАЛИЗИРУЕМЫХ ФЕР-
МЕНТАМИ ...................................................207
4.1. Применение гидролитических ферментов........................213
4.1.1. Гидролиз крахмала и целлюлозы..........................215
4.1.2. Протеолитические ферменты..............................227
4.1.3. Применение эстераз.....................................230
4.1.4. Ферментные смеси, пектиновые ферменты и другие пути ис-
пользования ферментов......................................231
4.2. Другие области применения ферментативных реакций в растворах 233
4.2.1. Применение ферментов в медицине........................234
4.2.2. Применение негидролитических ферментов в промышленно-
сти и перспективы расширения сферы их использования 236
Оглавление
689
4.3. Технологические процессы с участием иммобилизованных фермен-
тов ........................................................237
4.3.1. Иммобилизация ферментов..................................238
4.3.2. Промышленные процессы....................................249
4.3.3. Применение иммобилизованных ферментов в медицине и хи-
мическом анализе......................................257
4.3.4. Утилизация и регенерация кофакторов......................262
4.4. Кинетика реакций, катализируемых иммобилизованными фермен-
тами ..........................................................265
4.4.1. Влияние внешнего сопротивления массопередаче . . . 267
4.4.2. Моделирование диффузии и реакций внутри частицы ката-
лизатора .............................................273
4.4.3. Одновременное сопротивление массопередаче в граничном
слое и внутри частицы катализатора...................285
4.4.4. Влияние ингибиторов, температуры и pH на каталитическую
активность и инактивацию иммобилизованных ферментов 287
4.5. Заключение....................................................290
Упражнения.......................................................291
Литература......................................................295
Глава 5. СТЕХИОМЕТРИЯ И ЭНЕРГЕТИКА МЕТАБОЛИЧЕСКИХ
ПРЕВРАЩЕНИЙ.................................................298
5.1. Принципы термодинамики......................................304
5.2. Взаимосвязанность метаболических реакций; АТР и NAD . . 307
5.2.1. АТР и другие фосфаты..................................307
5.2.2. Окисление и восстановление; сочетание с превращением
NAD....................................................309
5.3. Катаболизм углерода...................................312
5.3.1. Метаболический путь Эмбдена — Мейергофа — Парнаса 312
5.3.2. Другие пути катаболизма углеводов...............316
5.4. Дыхание...............................................319
5.4.1. Цикл трикарбоновых кислот.......................321
5.4.2. Дыхательная цепь................................323
5.5. Фотосинтез и его связь с первичным источником энергии . . 327
5.5.1. Аккумулирование световой энергии.................327
5.5.2. Транспорт электронов и фотофосфорилирование . . . 328
5.6. Биосинтез.............................................330
5.6.1. Биосинтез низкомолекулярных соединений .... 331
5.6.2. Синтез макромолекулярных соединений.............337
5.7. Транспорт через клеточные мембраны............................340
5.7.1. Пассивная диффузия и сопряженный транспорт . . . 341
5.7.2. Активный транспорт......................................344
5.8. Организация и регуляция метаболизма...........................348
5.8.1. Ключевые точки пересечения и разветвления метаболиче-
ских путей.....................................................350
5.8.2. Регуляция метаболизма на ферментативном уровне . . 352
5.9. Конечные продукты метаболизма.................................355
5.9.1. Продукты анаэробного метаболизма (брожения) . . . 355
5.9.2. Частичное окисление и его конечные продукты . . . 358
5.9.3. Синтез вторичных метаболитов............................359
5.10. Стехиометрия клеточного роста и образования продуктов мета-
болизма .......................................................360
5.10.1. Общая стехиометрия клеточного роста; состав среды и
коэффициенты выхода............................................362
5.10.2. Материальный баланс по элементам и клеточный рост 369
5.10.3. Стехиометрия процессов образования продуктов метабо-
лизма .........................................................374
690
Оглавление
5.10.4. Стехиометрия энергетического обмена; оценка количества
выделяющейся теплоты и соответствующих экономических
коэффициентов . . ,...............................378
5.10.5. Стехиометрия фотосинтеза.............................384
5.11. Заключение.................................................387
Упражнения.......................................................388
Литература...................................................395
Глава 6. МОЛЕКУЛЯРНАЯ ГЕНЕТИКА И СИСТЕМЫ РЕГУЛЯЦИИ 397
6.1. Молекулярная генетика.......................................397
6.1.1. Процессы экспрессии генов..............................398
6.1.2. Расщепленные гены и модификация мРНК в эукариотах 406
6.1.3. Посттрансляционная модификация белков..................408
6.1.4. Индукция и репрессия; регуляция белкового синтеза . . 409
6.1.5. Репликация ДНК и мутации...............................415
6.1.6. Общая схема потока информации в клетке.................419
6.2. Модификация структуры клеточной ДНК.........................421
6.2.1. Вирусы и фаги; лизогения и трансдукция.................423
6.2.2. Трансформация и конъюгация в бактериях.................428
6.2.3. Слияние клеток.........................................429
6.3. Применение результатов исследования генетики микроорганизмов
и популяций мутантов в промышленности...........................433
6.3.1. Системы клеточной регуляции; влияние состава среды 433
6.3.2. Применение ауксотрофных мутантов.......................435
6.3.3. Мутанты с модифицированными системами регуляции . . 437
6.4. Технология рекомбинантных ДНК...............................440
6.4.1. Ферменты для расщепления и сшивки ДНК .... 442
6.4.2. Векторы для Escherichia coli......................446
6.4.3. Идентификация клонированных ДНК...................448
6.4.4. Экспрессия эукариотических белков в Е. coli .... 452
6.4.5. Генетическая инженерия с участием других клеток-хозяев 458
6.4.6. Заключение........................................461
6.5. Рост и самовоспроизведение изолированной клетки .... 462
6.5.1. Методы эксперимента; цитометрия в потоке и синхронные
культуры....................................................464
6.5.2. Клеточный цикл Е. coli................................467
6.5.3. Клеточный цикл эукариот................................471
Упражнения.......................................................474
Литература....................................................482
Глава 7. КИНЕТИКА ПРОЦЕССОВ УТИЛИЗАЦИИ СУБСТРАТА, ОБ-
РАЗОВАНИЯ ПРОДУКТОВ МЕТАБОЛИЗМА И БИОМАС-
СЫ В КУЛЬТУРАХ КЛЕТОК.........................................484
7.1. Идеальные реакторы для изучения кинетики клеточного роста 491
7.1.1. Идеальный реактор периодического действия .... 491
7.1.2. Идеальный проточный реактор с полным перемешиванием
(ПРПП)......................................................492
7.2. Кинетика сбалансированного роста .......................... 495
7.2.1. Уравнение Моно для кинетики клеточного роста . . . 497
7.2.2. Влияние эндогенного метаболизма и метаболизма поддер-
жания на кинетику клеточного роста ........................ 504
7.2.3. Другие уравнения кинетики клеточного роста .... 507
7.2.4. Влияние других параметров среды на кинетику клеточного
роста.......................................................508
7.3. Кинетика клеточного роста в переходном состоянии . . . • 511
________________________ Оглавление _______________ 691
7.3.1. Основные фазы роста клеток в реакторах периодического
действия....................................................5ц
7.3.2. Неструктурированные модели клеточного роста в периоди-
ческих процессах...........................................523
7.3.3. Рост филаментозных организмов........................527
7.4. Структурированные модели кинетики клеточного роста . . 529
7.4.1. Компартментальные модели.............................530
7.4.2. Метаболические модели................................535
7.4.3. Моделирование клеточного роста как оптимального процесса 541
7.5. Кинетика образования продуктов метаболизма .......... 544
7.5.1. Неструктурированные модели...........................545
7.5.2. Химически структурированные модели кинетики образова-
ния продуктов жизнедеятельности клеток.....................552
7.5.3. Моделирование кинетики образования продуктов метаболиз-
ма на основе механизмов молекулярных превращений; ге-
нетически структурированные модели.........................555
7.5.4. Кинетика образования продуктов жизнедеятельности фила-
ментозными организмами.....................................558
7.6. Сегрегированные модели кинетики клеточного роста и образова-
ния продуктов метаболизма ..................................... 566
7.7. Кинетика тепловой гибели клеток и спор....................570
7.8. Заключение................................................576
Упражнения.....................................................578
Литература..................................................587
Глава 8. ПРОЦЕССЫ ПЕРЕНОСА В БИОТЕХНОЛОГИЧЕСКИХ СИ-
СТЕМАХ ......................................590
8.1. Массообмен между газовой и жидкой фазами в клеточных си-
стемах .........................................................593
8.1.1. Основные положения теории массообмена...............595
8.1.2. Скорость утилизации кислорода в процессах клеточного
метаболизма................................................603
8.2. Определение скорости переноса кислорода ................. 606
8.2.1. Определение kia' с помощью химических реакций газооб-
разных веществ с жидкостями................................607
8.3. Массообмен с участием свободно поднимающихся и свободно
падающих тел...............................................611
8.3.1. Коэффициенты массопередачи для газовых пузырьков и по-
токов пузырьков............................................611
8.3.2. Определение площади поверхности раздела фаз и газосо-
держания дисперсной системы................................615
8.4. Массообмен путем принудительной конвекции.................625
8.4.1. Общие принципы и основные критерии подобия ... 625
8.4.2. Уравнения- для определения коэффициентов массопередачи
и площади поверхности раздела фаз..........................628
8.5. Определение kia' и мощность, потребляемая реакторами с пере-
мешиванием и с барботажными устройствами........................630
8.6. Массообмен через свободные поверхности....................638
8.7. Другие факторы, влияющие на kia'..........................642
8.7.1. Определение коэффициентов диффузии..................642
8.7.2. Ионная сила.........................................643
8.7.3. Поверхностно-активные вещества......................644
8.8. Неньютоновские жидкости..........................• • •
8.8.1. Математические модели неньютоновских жидкостей и со-
ответствующие параметры....................................647
8.8.2. Суспензии . . . .............................648
692 Оглавление
8.8.3. Растворы высокомолекулярных веществ................651
8.8.4. Потребляемая мощность и массообмен в биотехнологиче-
ских процессах с неньютоновскими жидкостями 652
8.9. Масштабный переход и массообмен...........................656
8.10. Теплообмен...............................................662
8.10.1. Уравнения теплообмена.............................668
8.11. Стерилизация газов и жидкостей фильтрованием .... 674
Упражнения.....................................................675
Литература.............................................. 682
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформ-
лении, качестве перевода и другие просим присы-
лать по адресу:
129820 ГСП, Москва, И-110, 1-й Рижский пер.,
д. 2, издательство «Мир».
Учебное издание
Джеймс Э. Бейли, Дэвид Ф. Оллис
ОСНОВЫ биохимической инженерии
ч. I
Зав. редакцией академик О. А. Реутов
Зам. зав. редакцией 3. Ф. Ходецкая
Научный редактор Б. М. Комарова
Младший научный редактор И. И. Землячева
Художник А. Д. Смеляков
Художественный редактор М. Н. Кузьмина
Технический редактор Е. В. Алехина
Корректор В. И. Киселева
ИБ № 6715
Сдано в набор 08.09.88. Подписано к печати 28.02.89. Формат
60Х901/ц. Бумага книжно-журиальная. Печать высокая. Гарни-
тура Литературная. Объем 21,75 бум. л. Усл. печ. л. 43,5. Усл.
кр.-отт. 43,5. Уч.-изд. л. 43,89. Изд. № 3/5890. Тираж 7500 экз.
Зак. 537. Цена 9 р. 10 коп.
Издательство <МИР» В/О «Совэкспорткнига» Государственного
комитета СССР по делам издательств, полиграфии и книжной
торговли. 129820, ГСП, Москва, И-110, 1-й Рижский пер., 2.
Московская типография № 11 Союзполиграфпрома при Государ-
ственном комитете СССР по делам издательств, полиграфии и
книжной торговли. 113105, Москва, Нагатинская ул„ д. 1.
Фотографировал Семенюченко Владимир
chem vova@mail. univ. kiev. ua; vova2002@mail. ru
Дж.Бейли, Д.Оллис
основы
биохимической
инженерии