/




























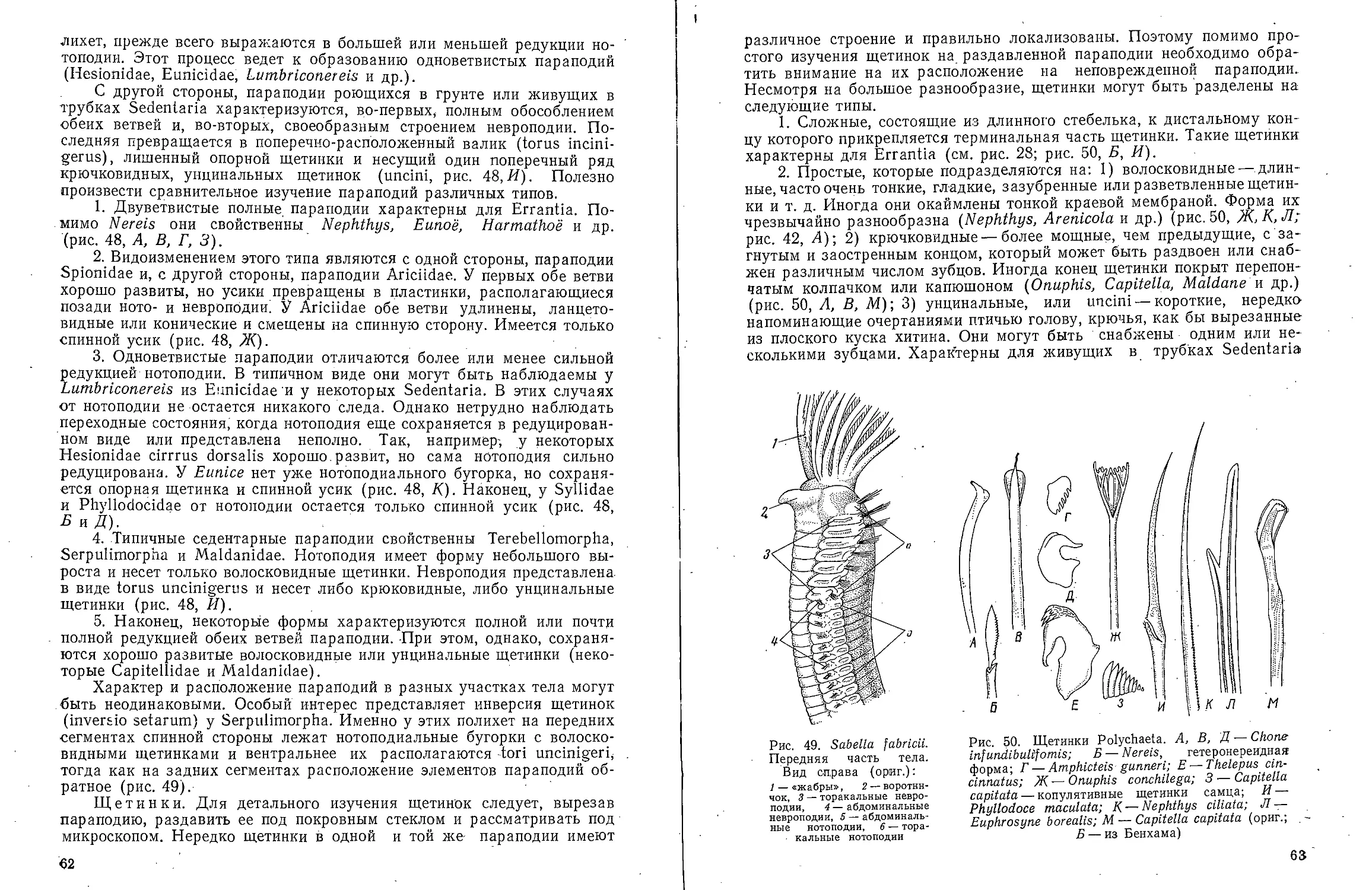

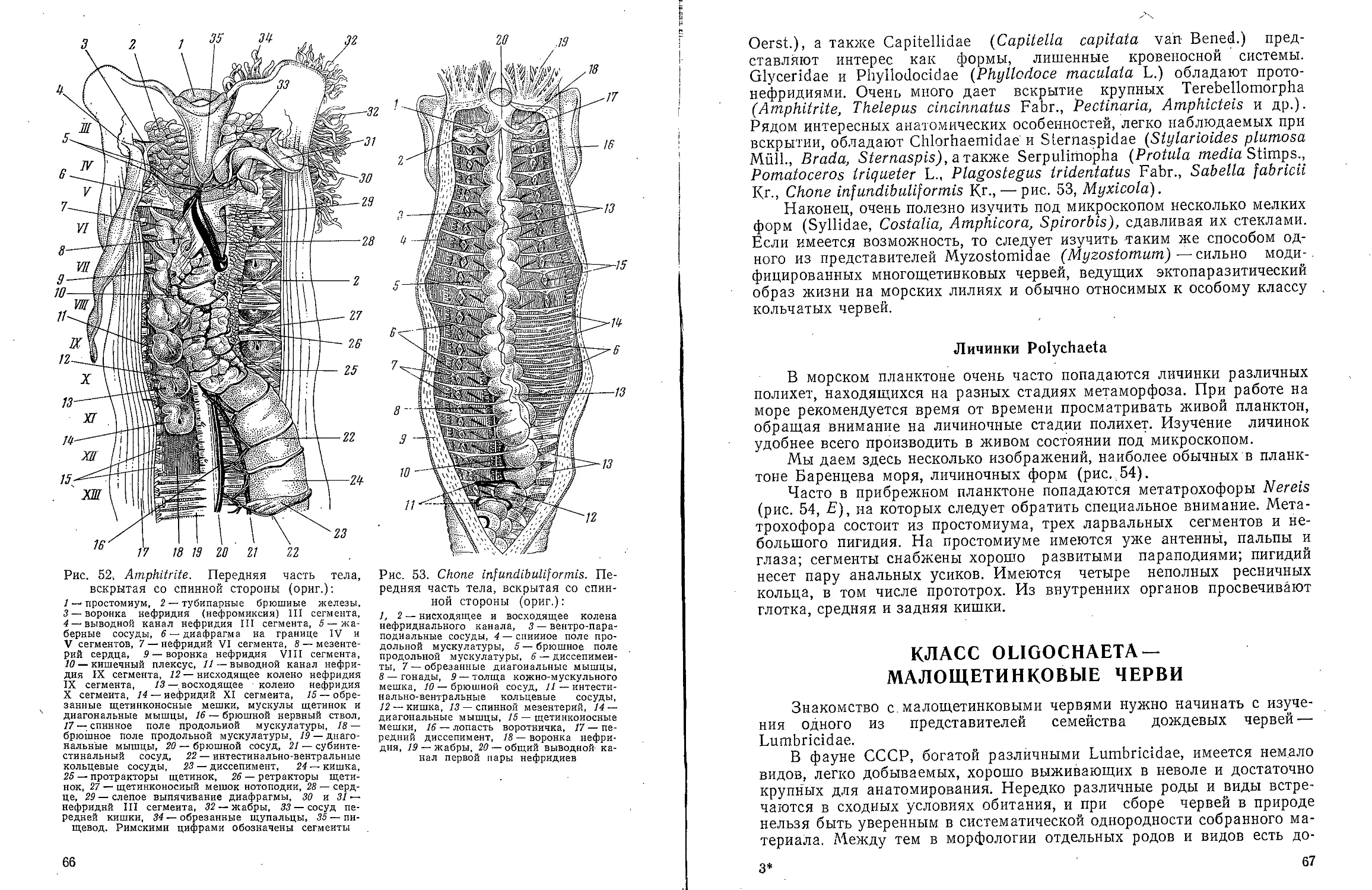
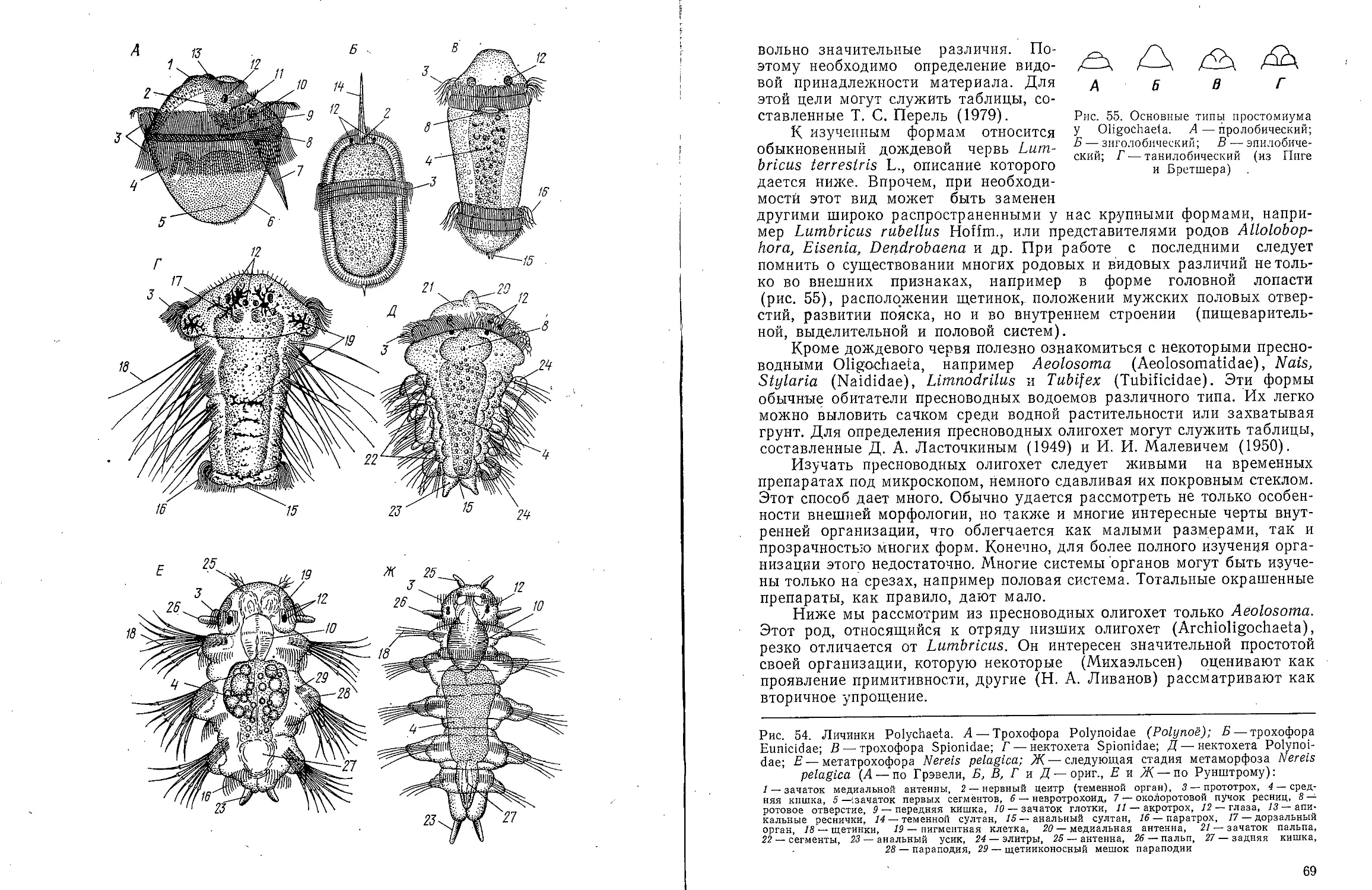


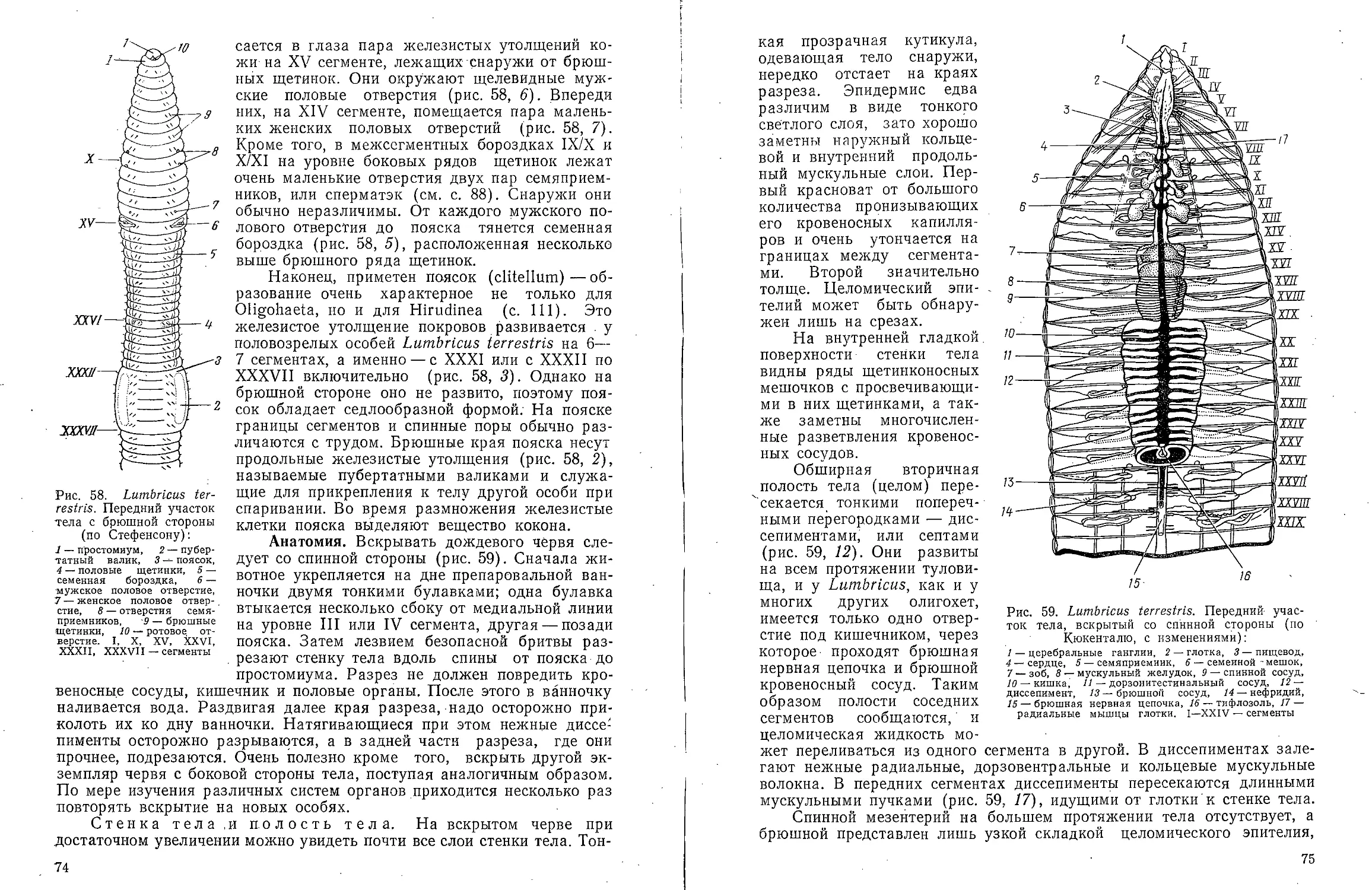





































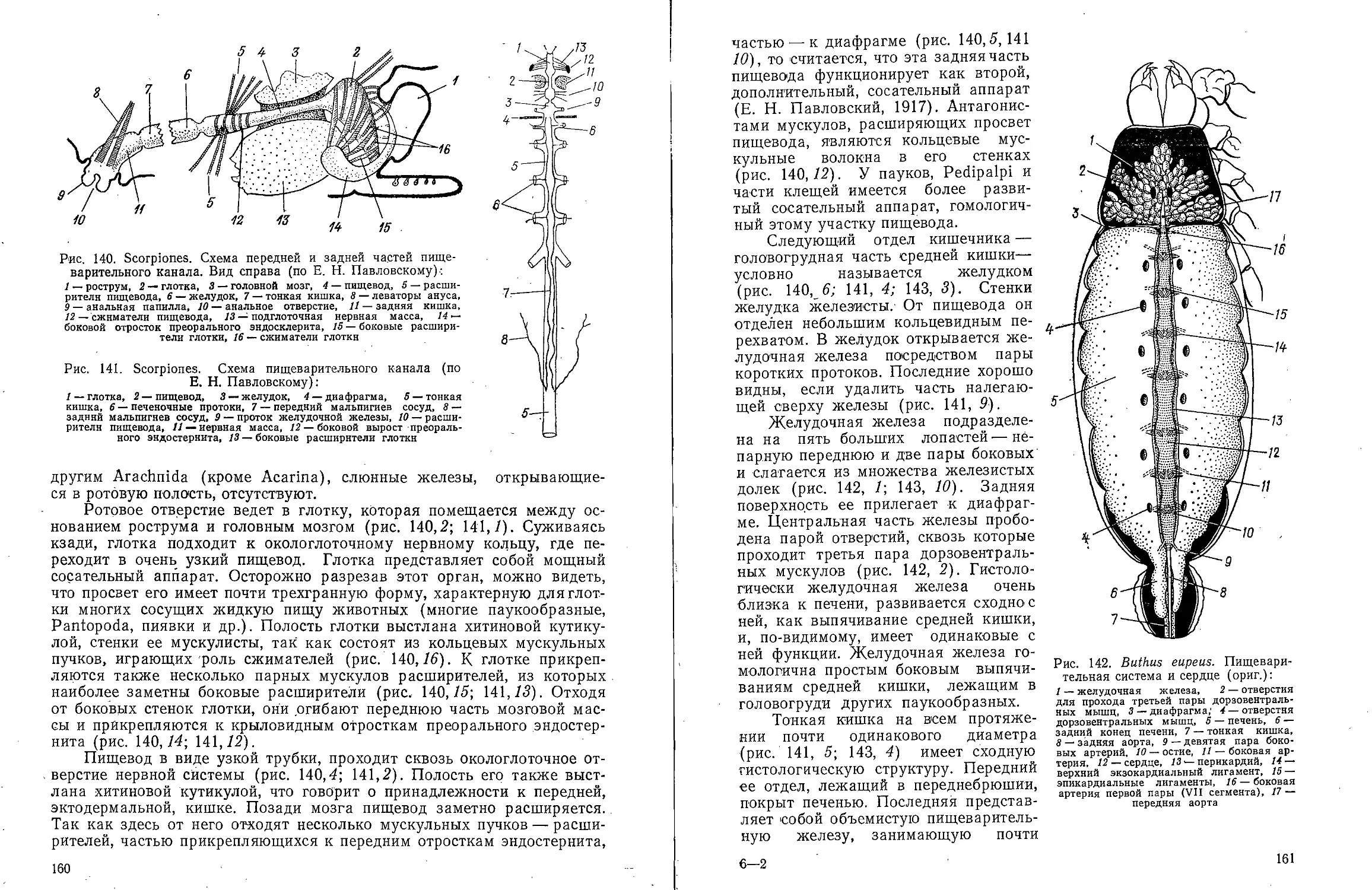
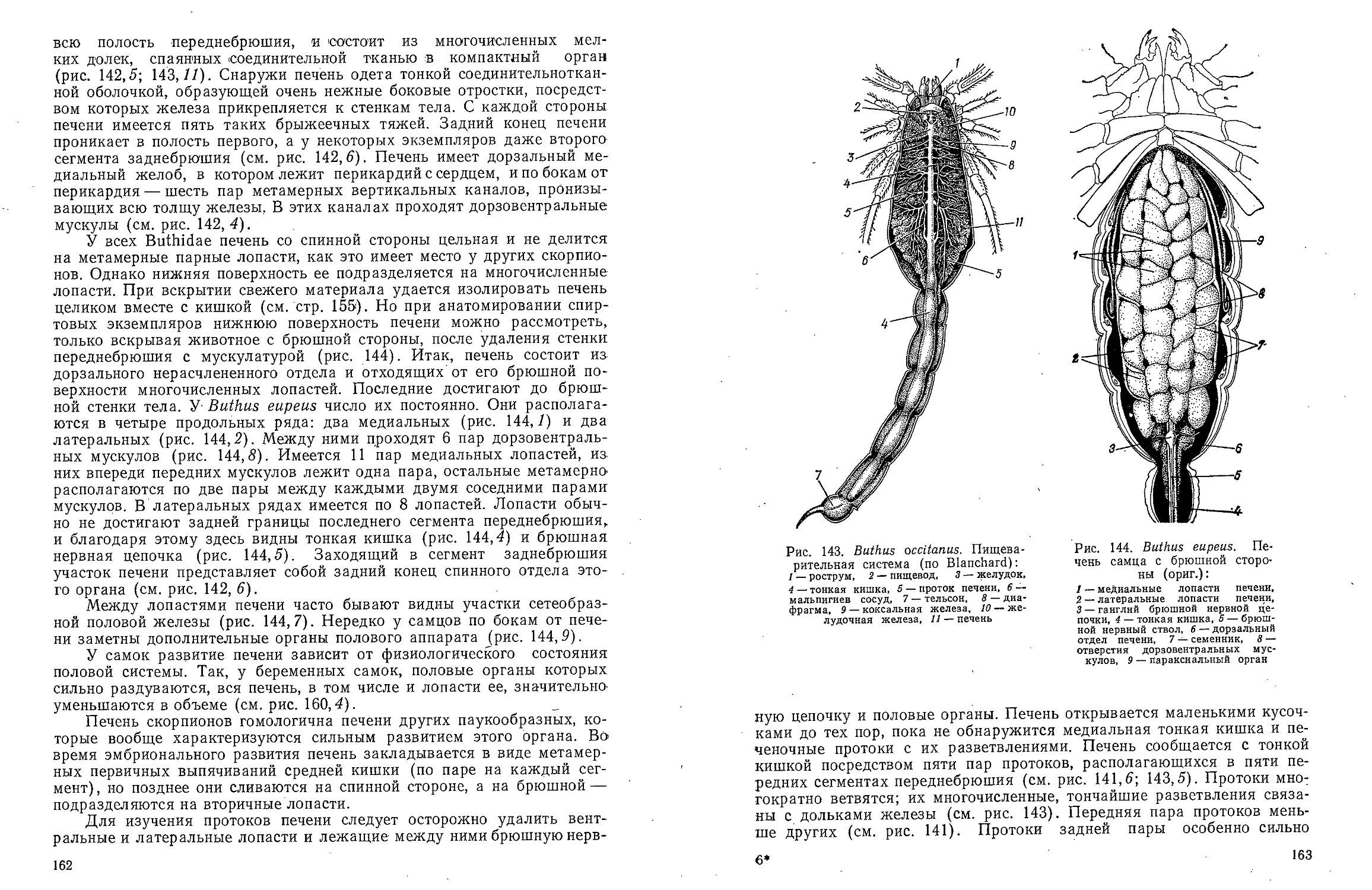













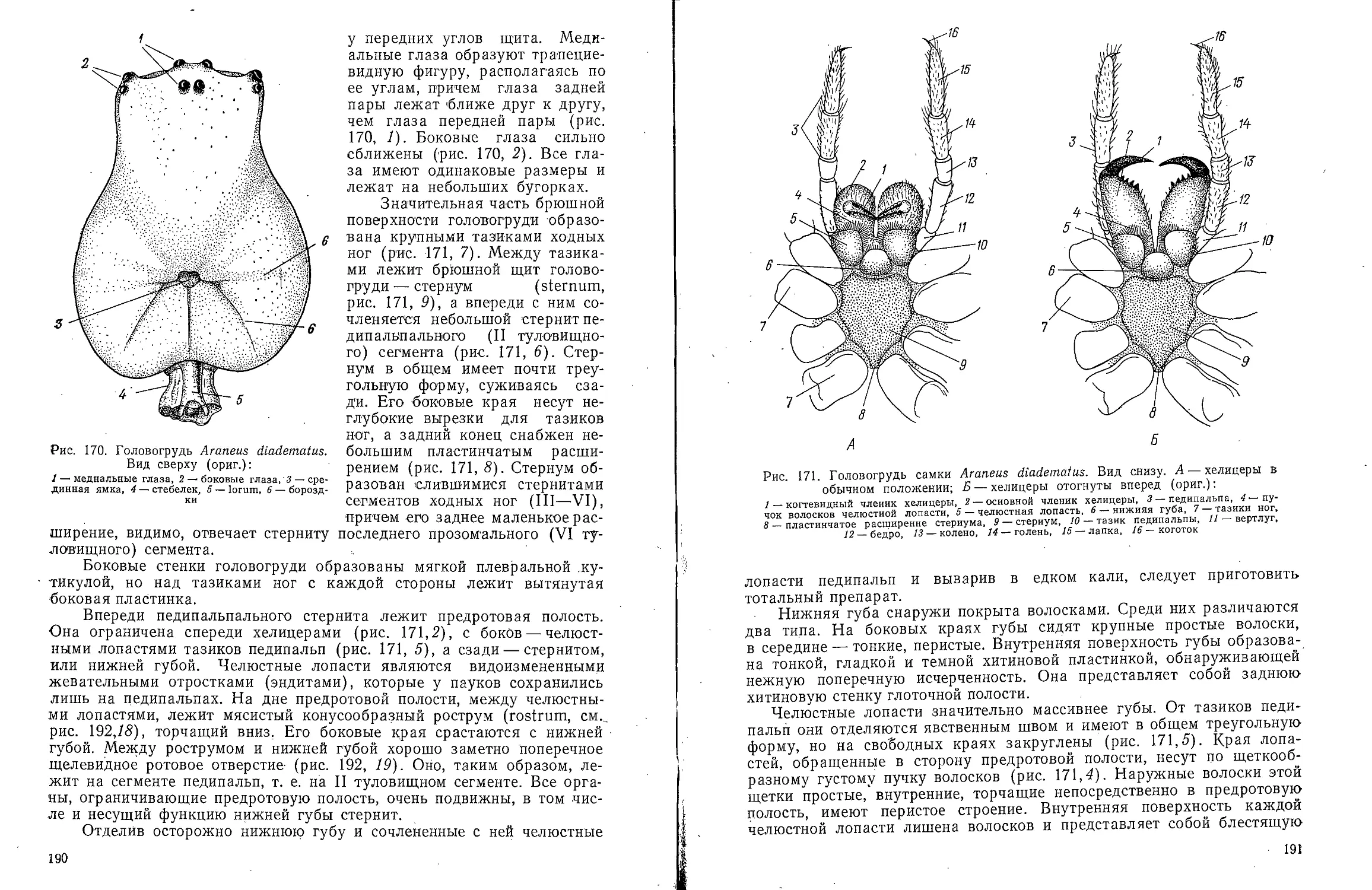















































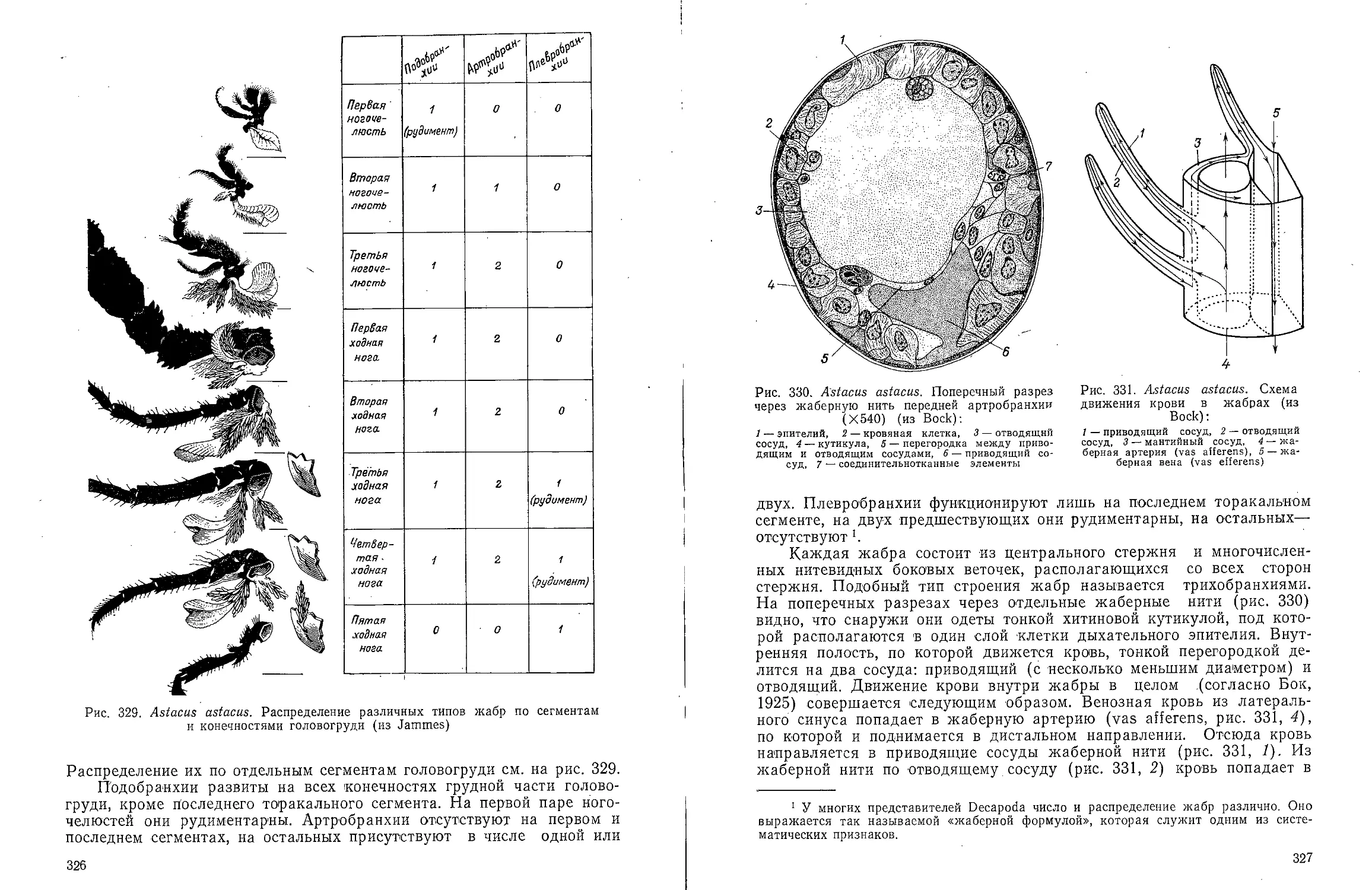


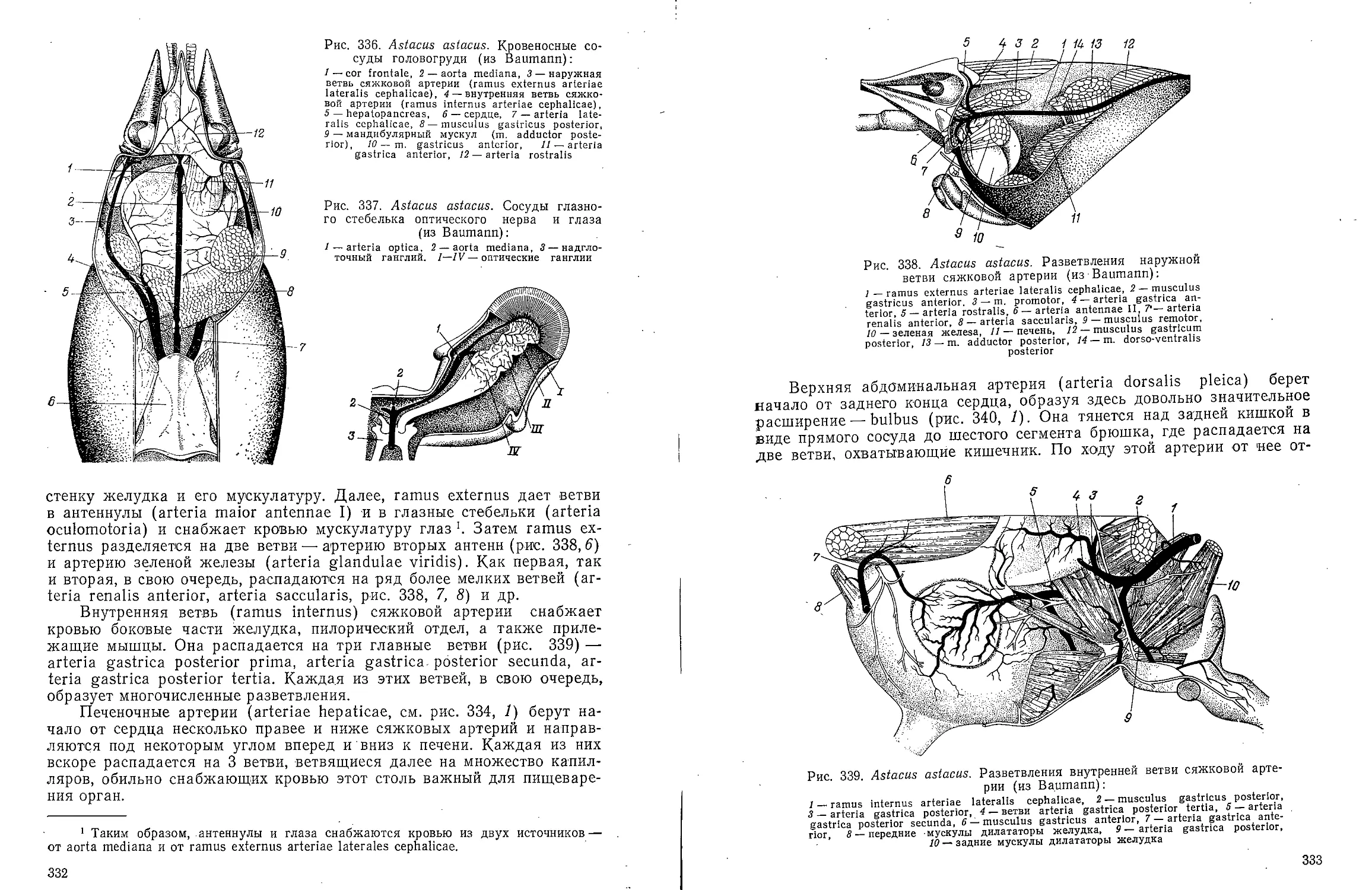
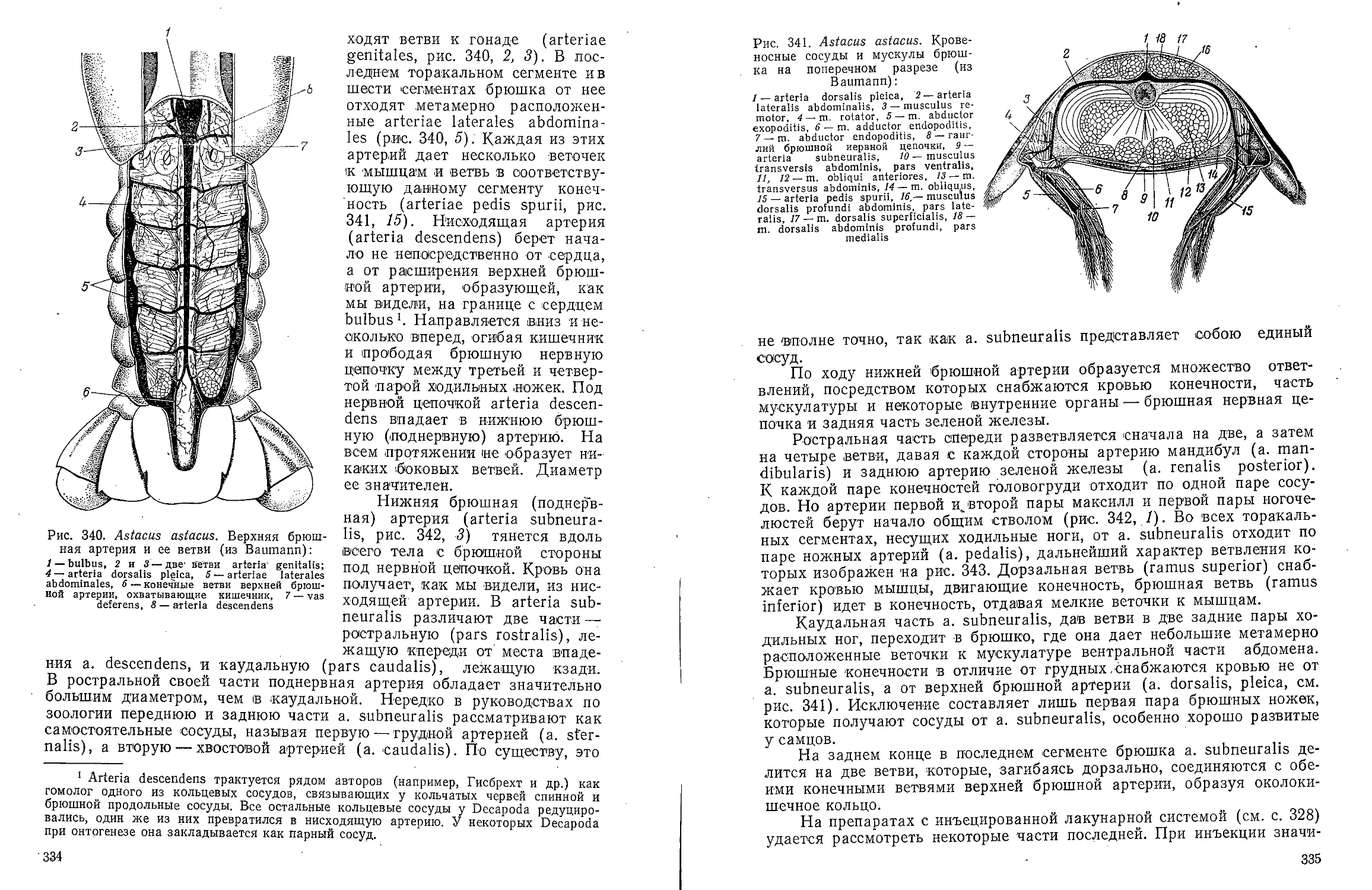



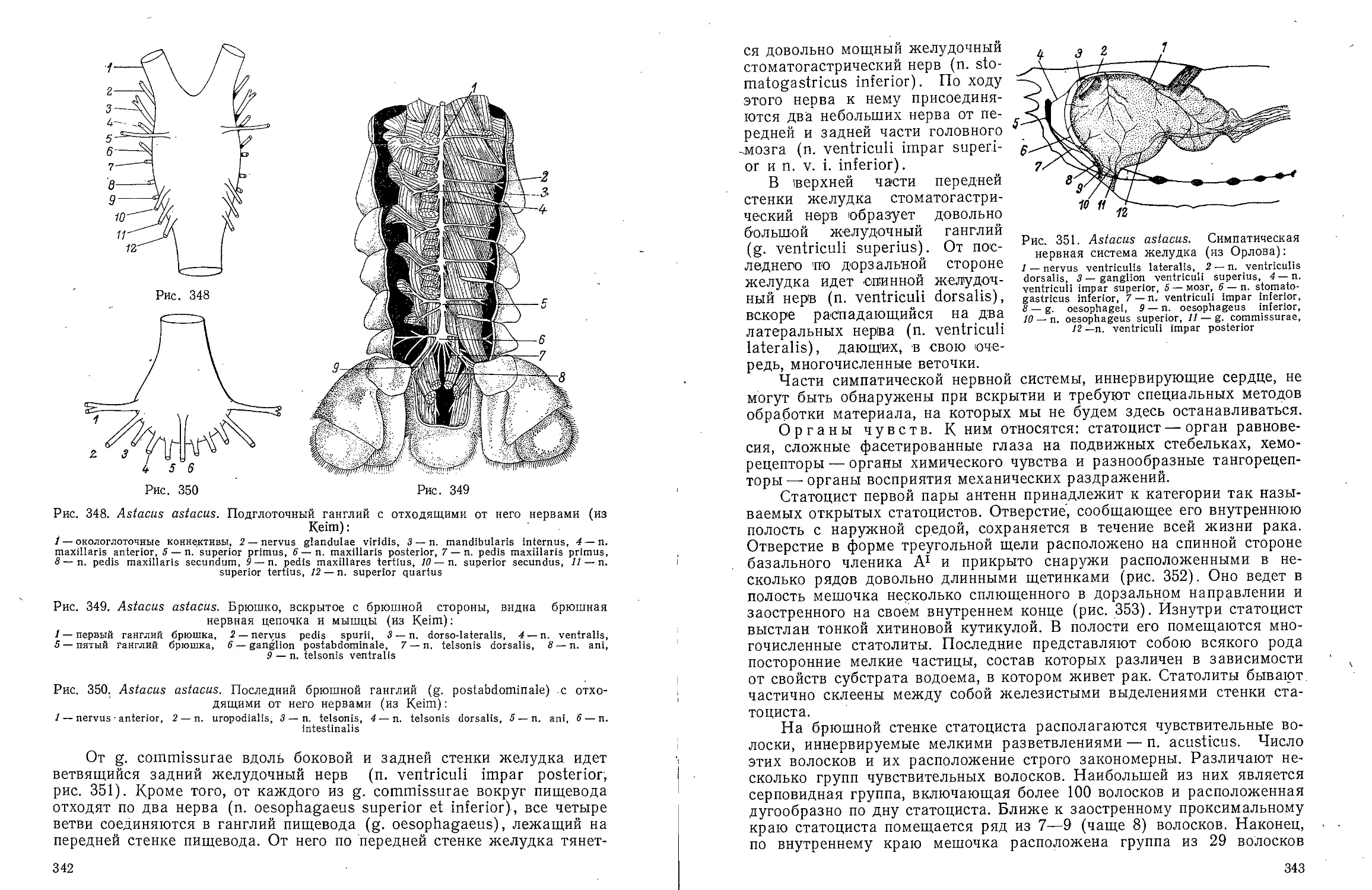























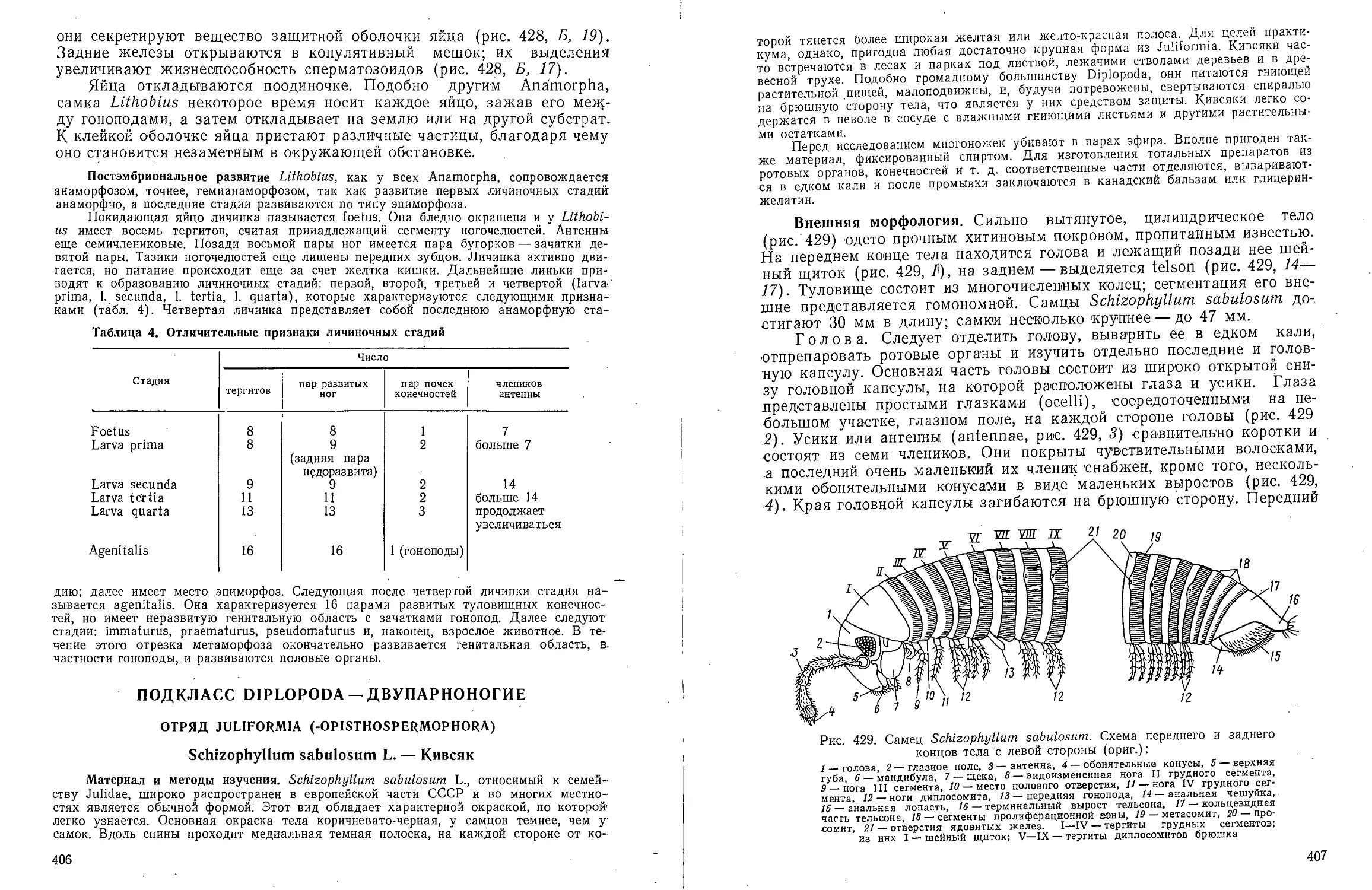

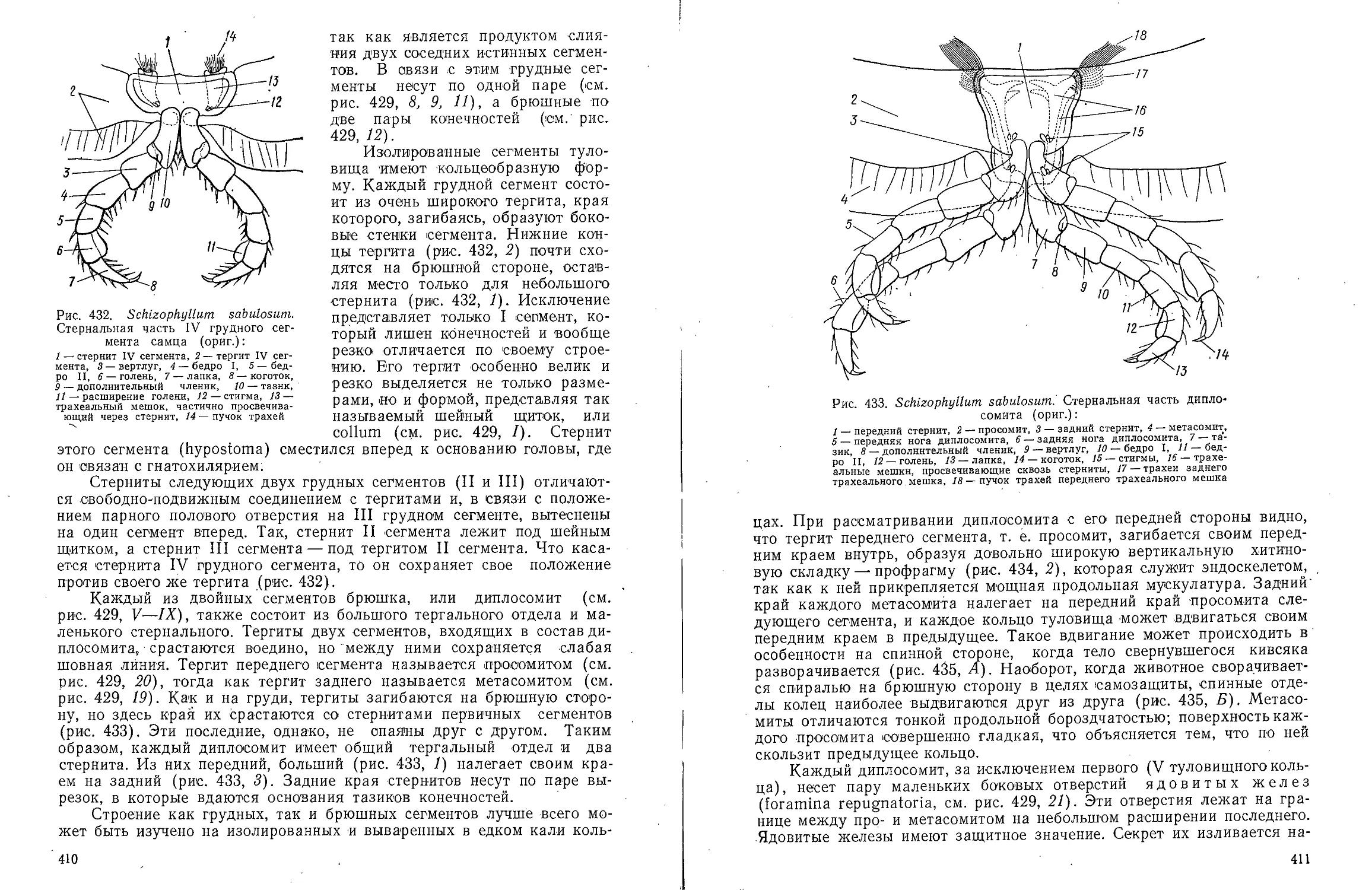





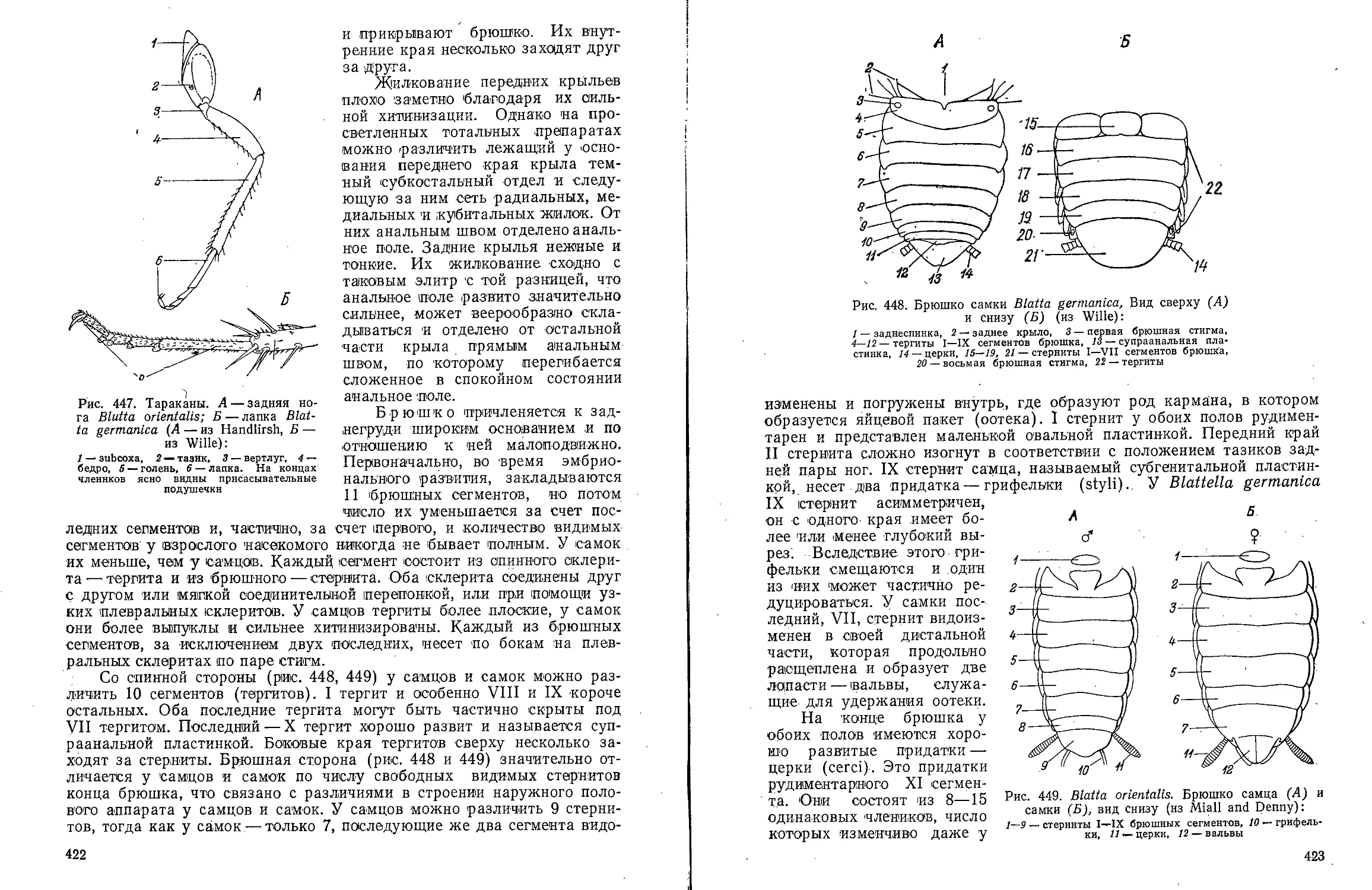
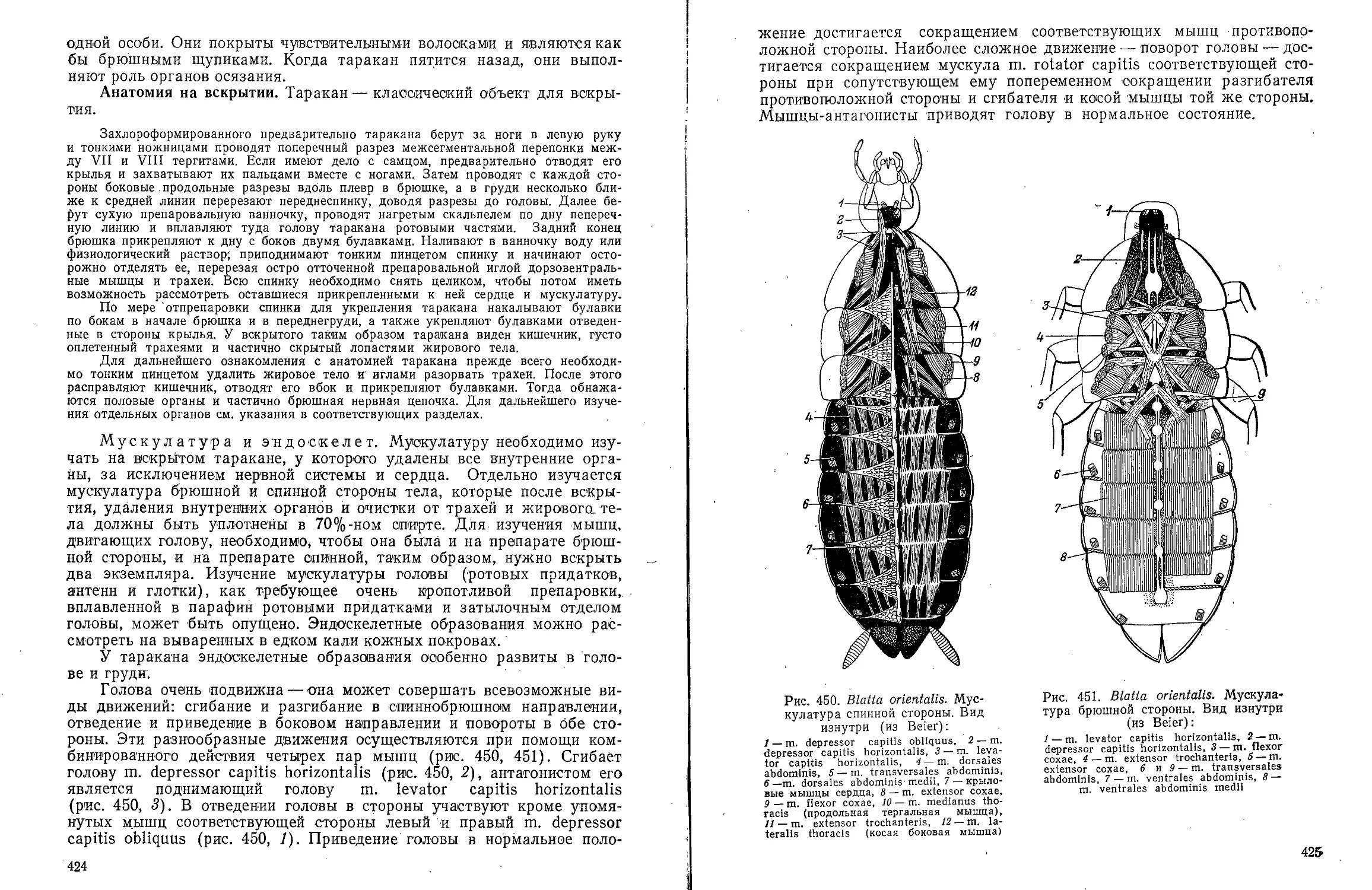

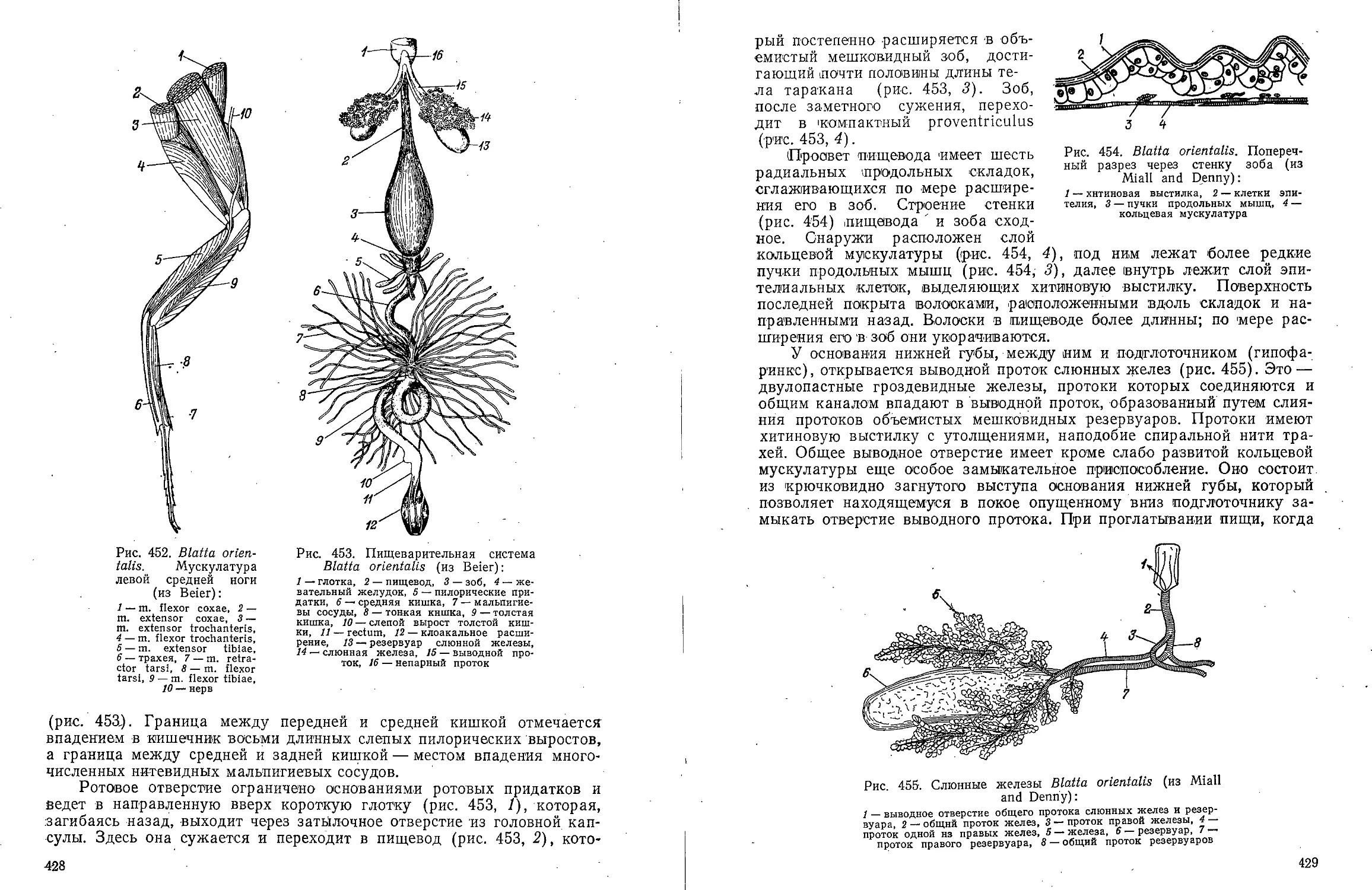

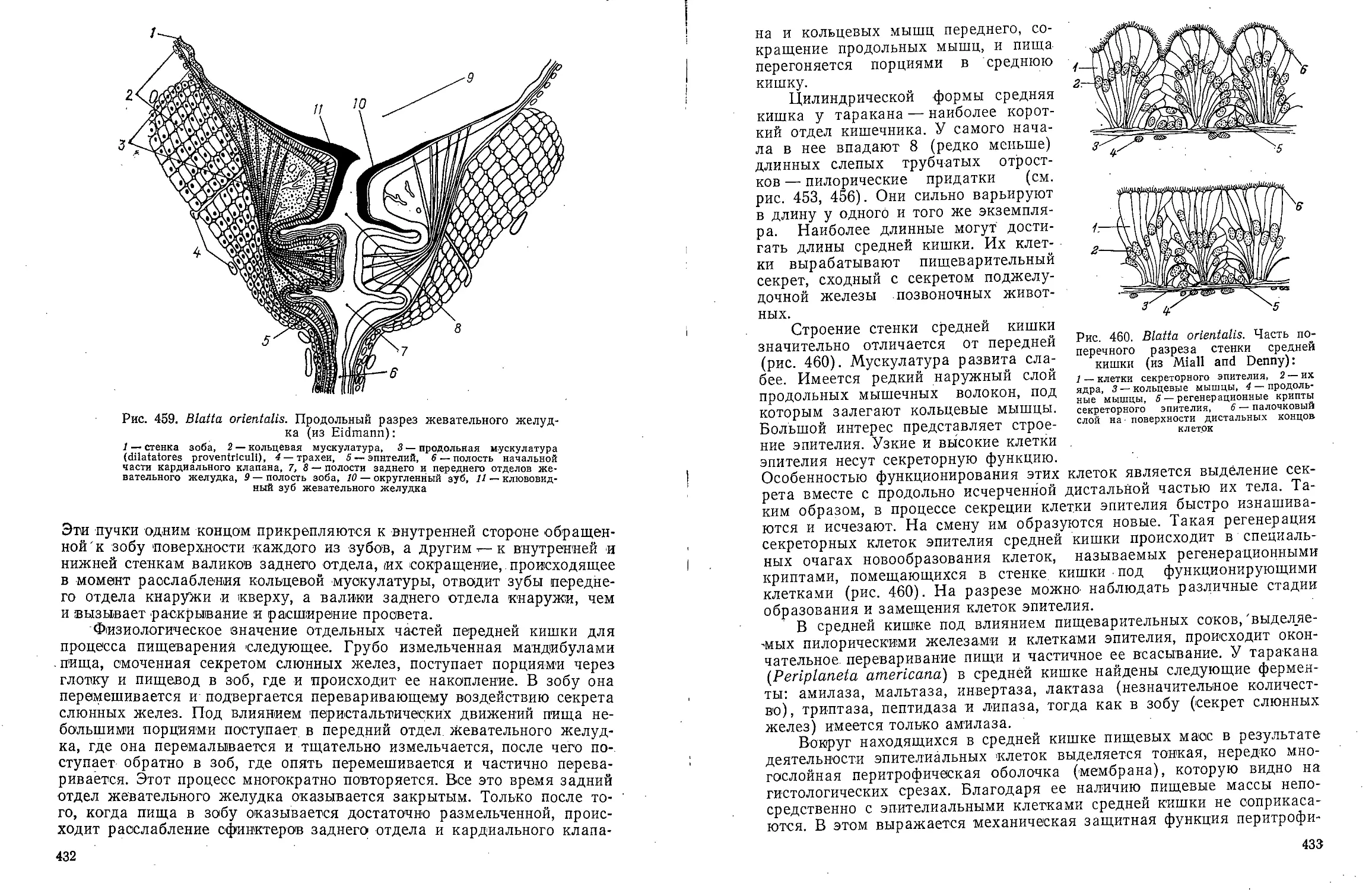
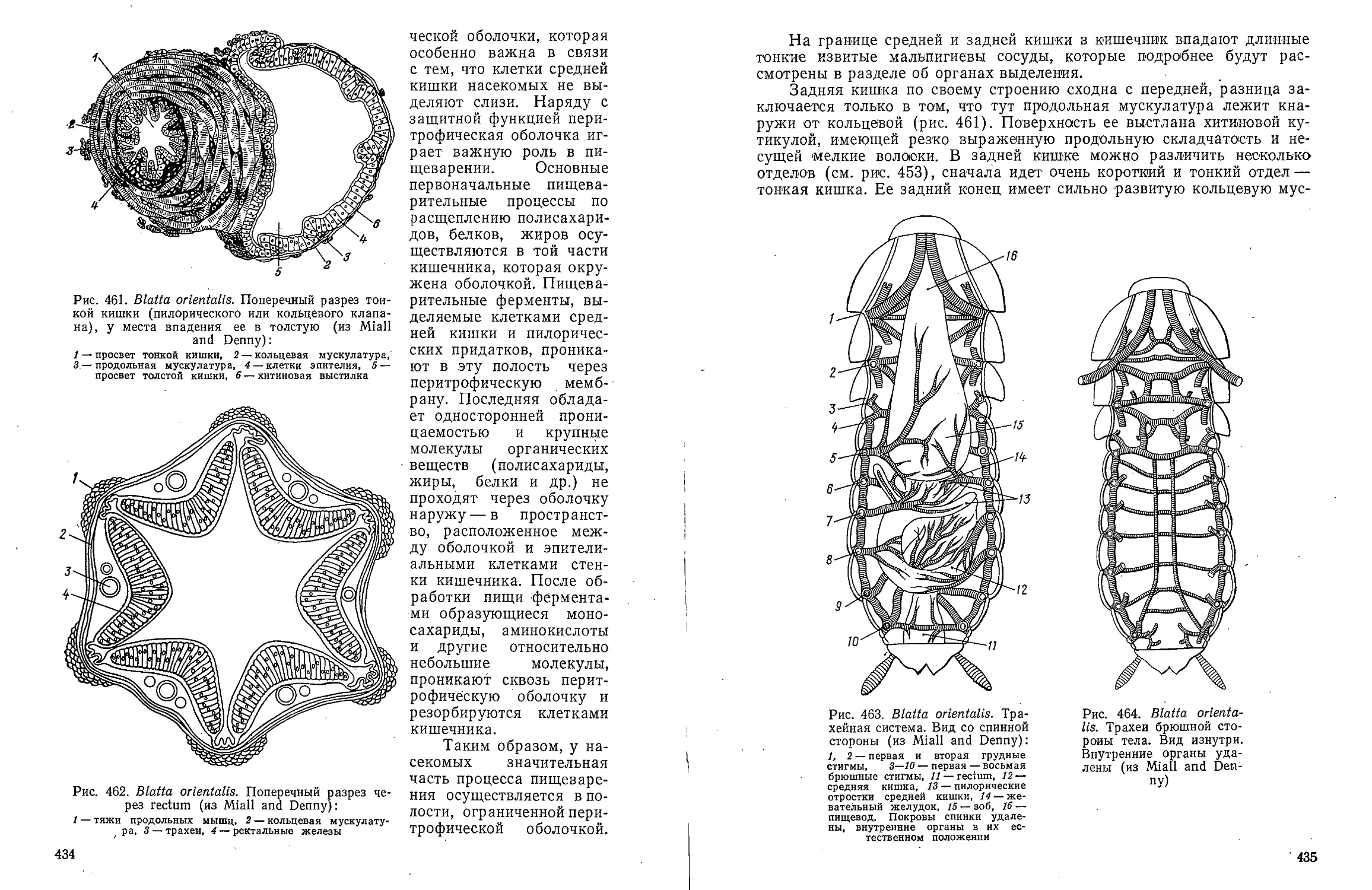





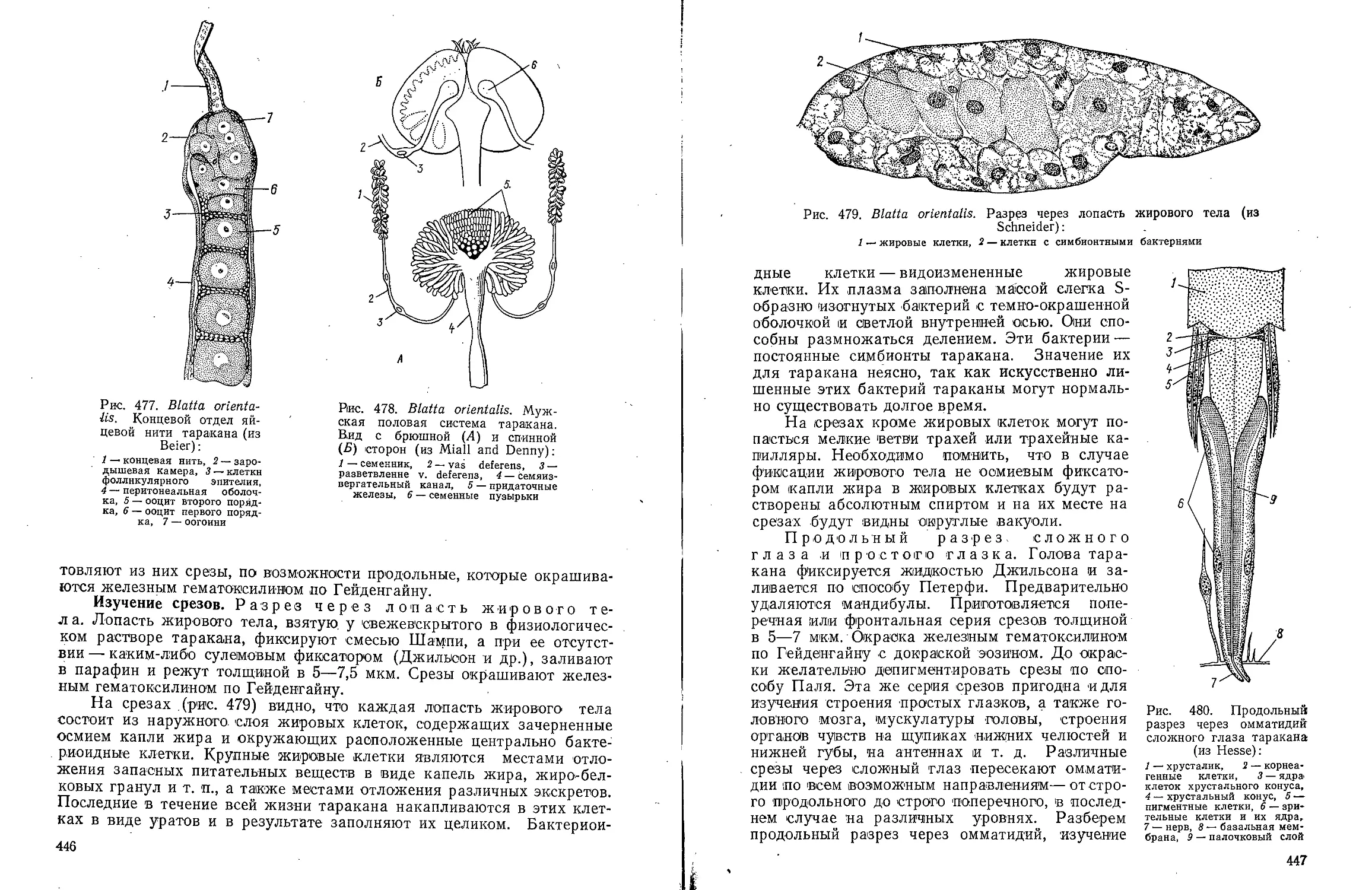























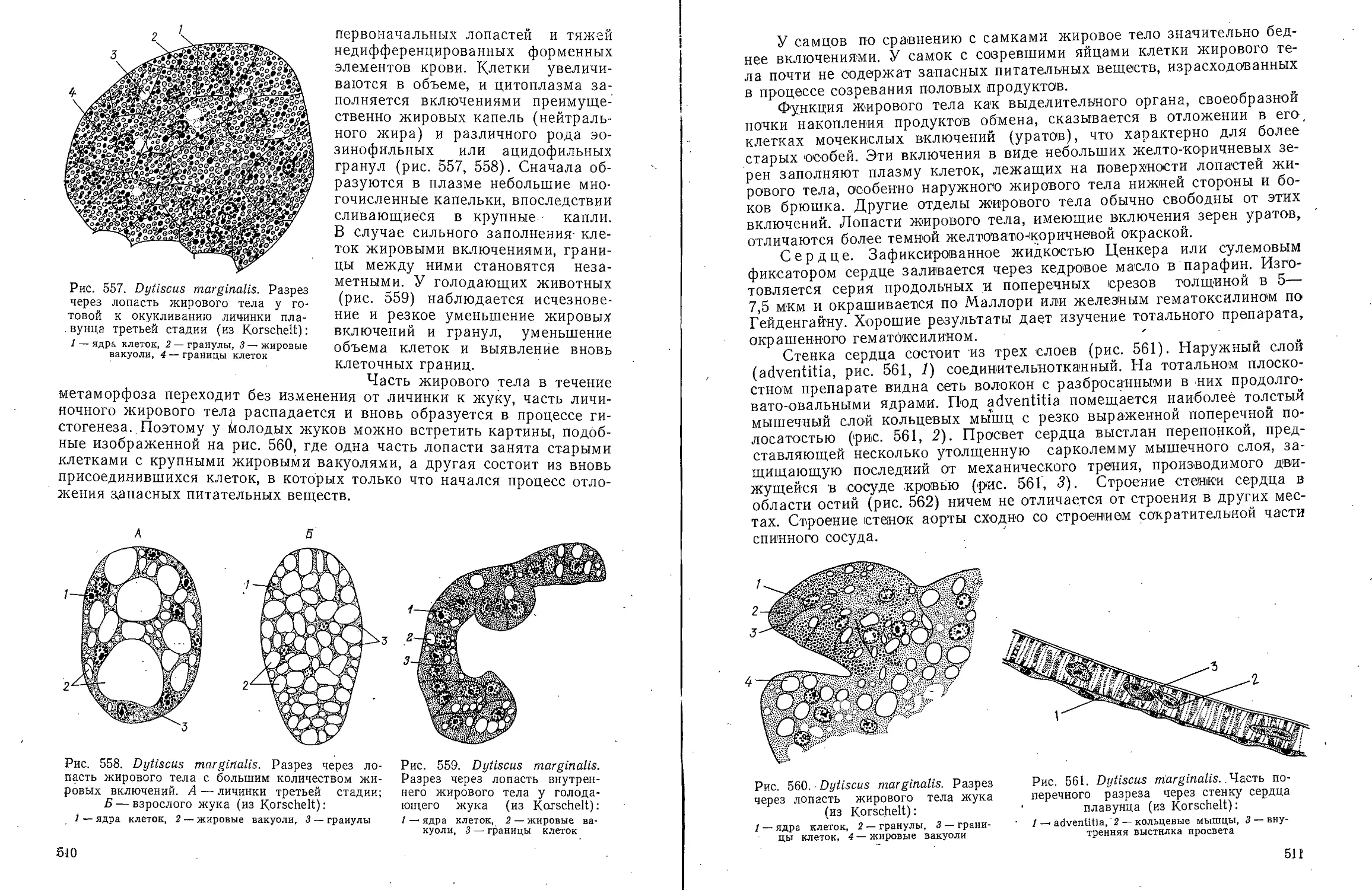

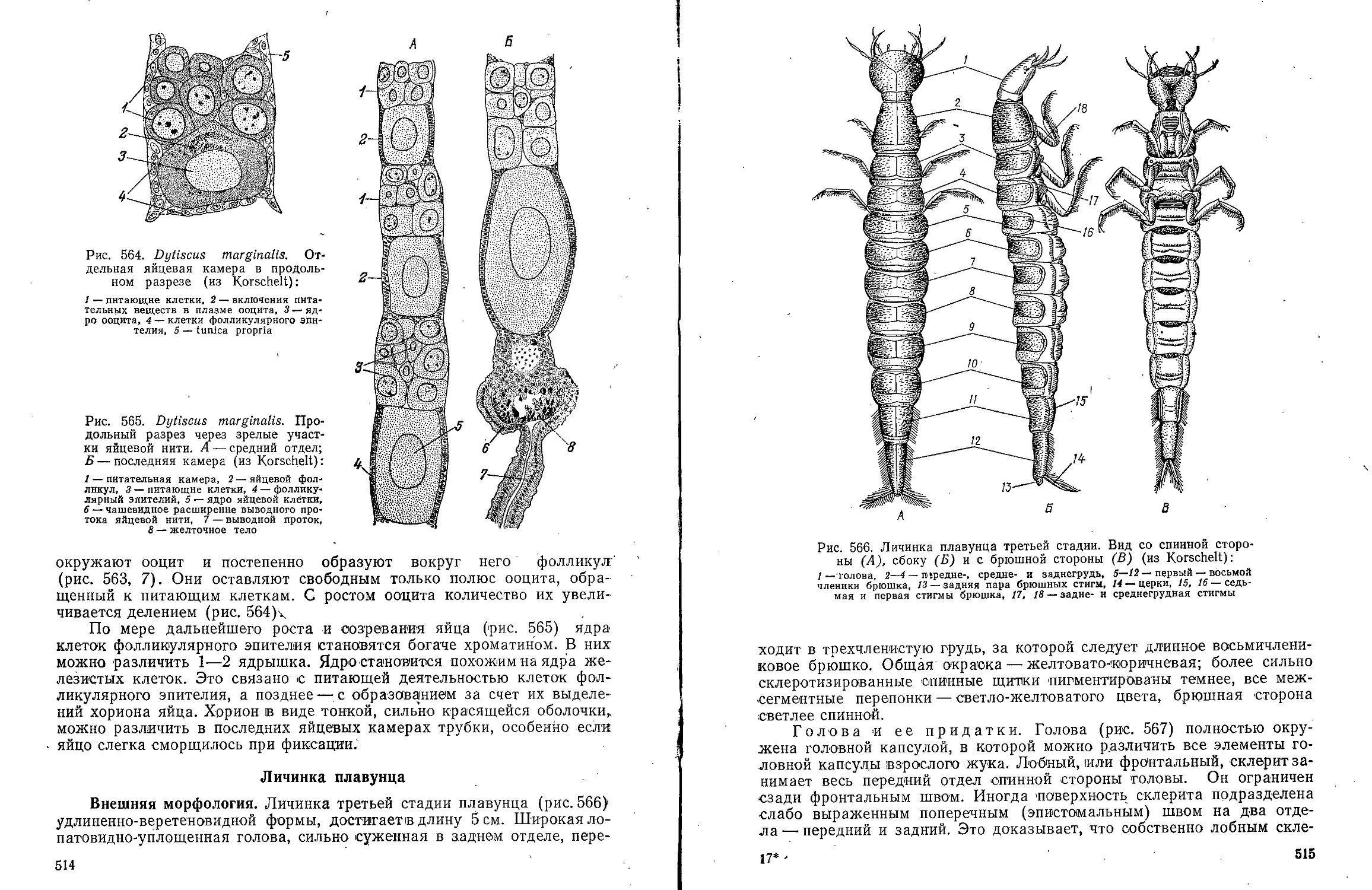



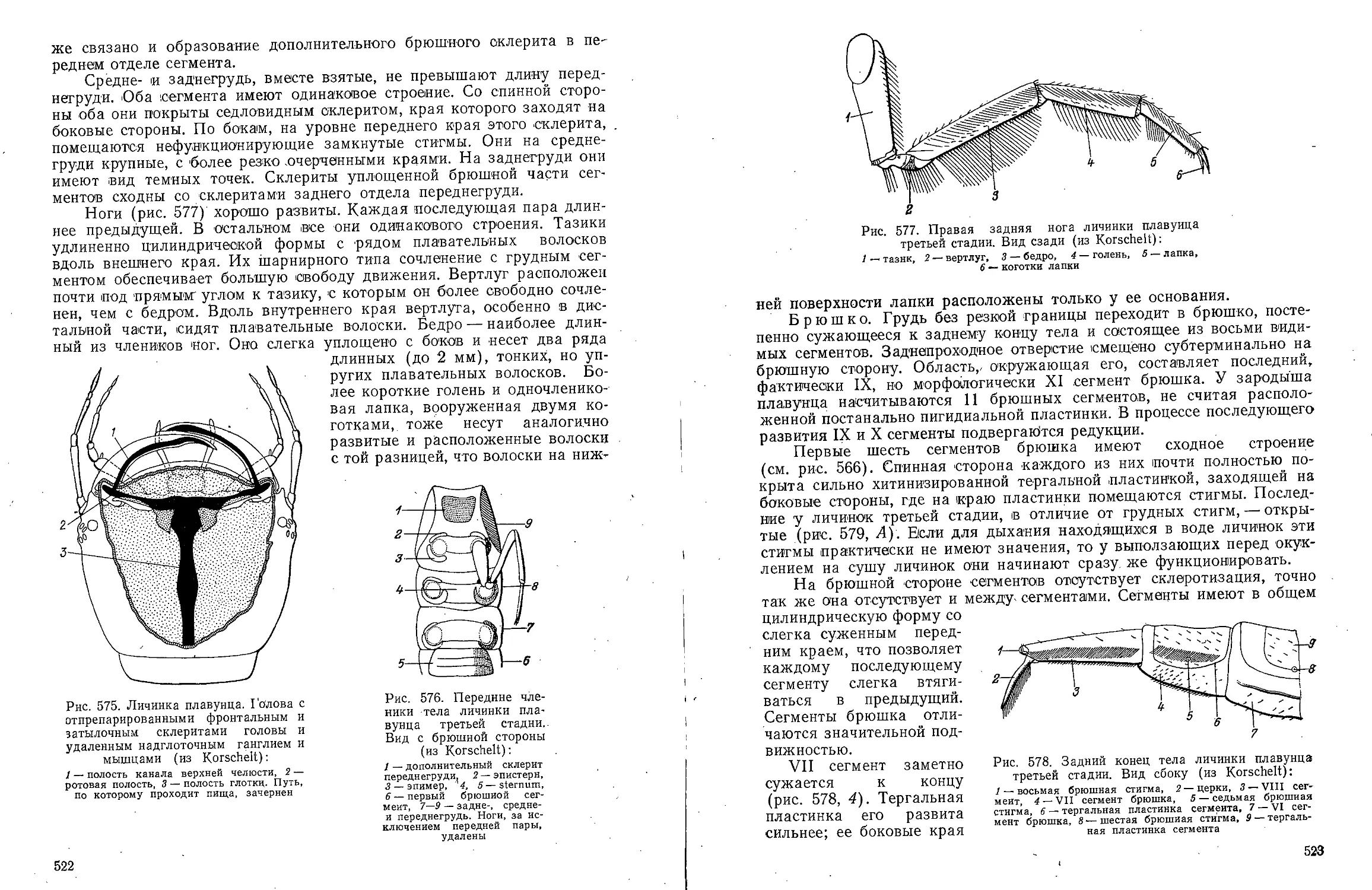
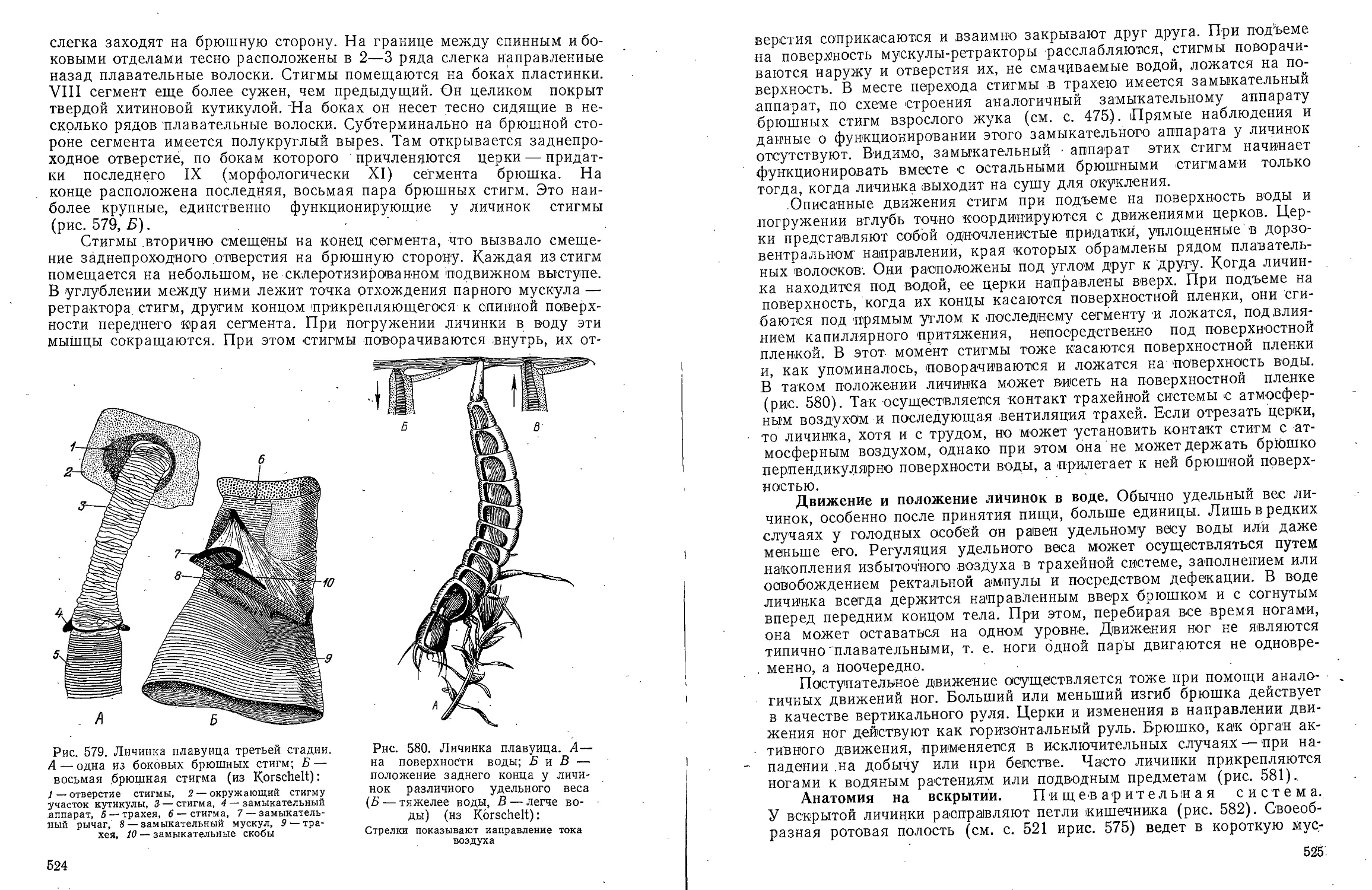












Text
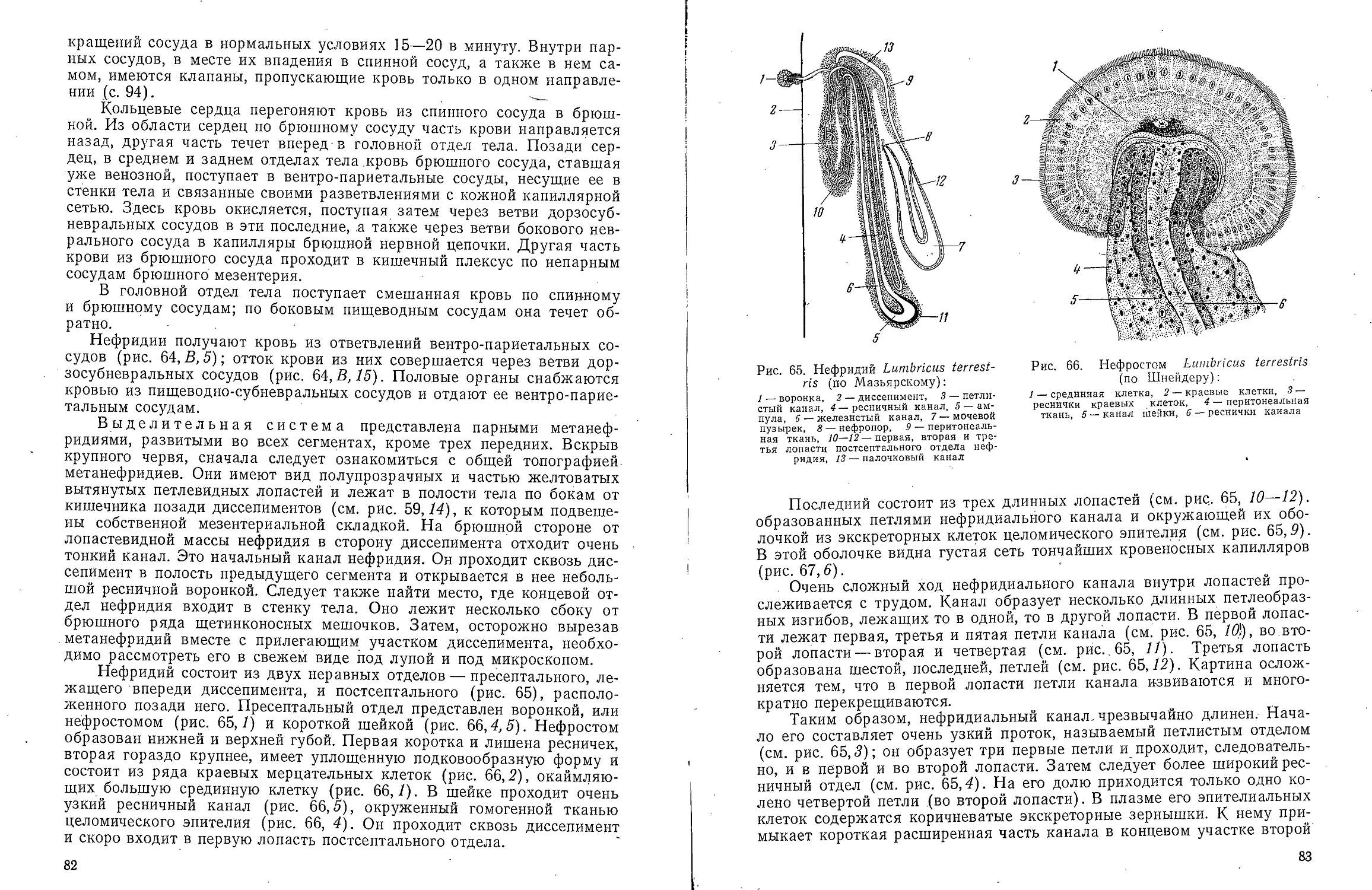
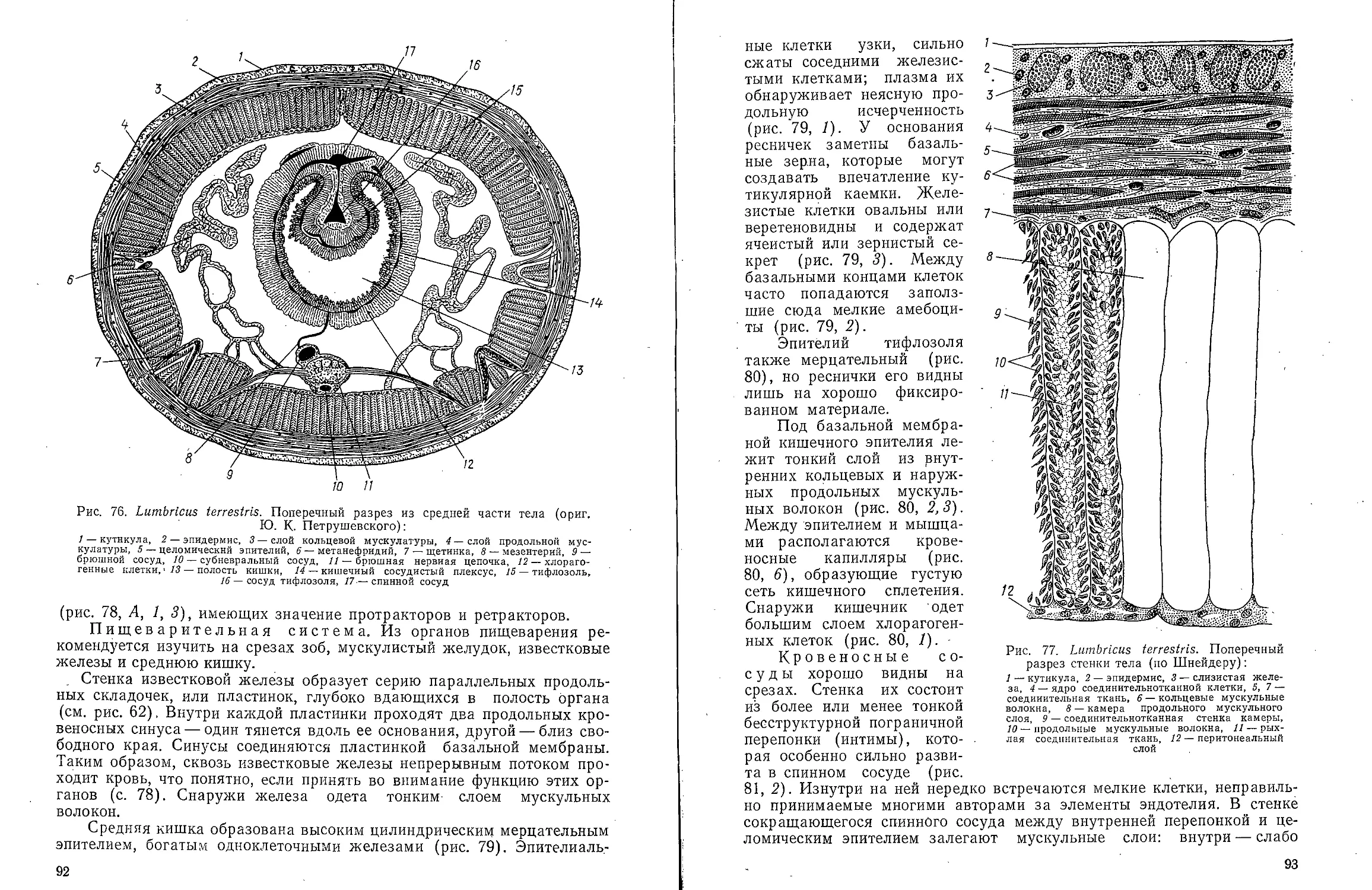
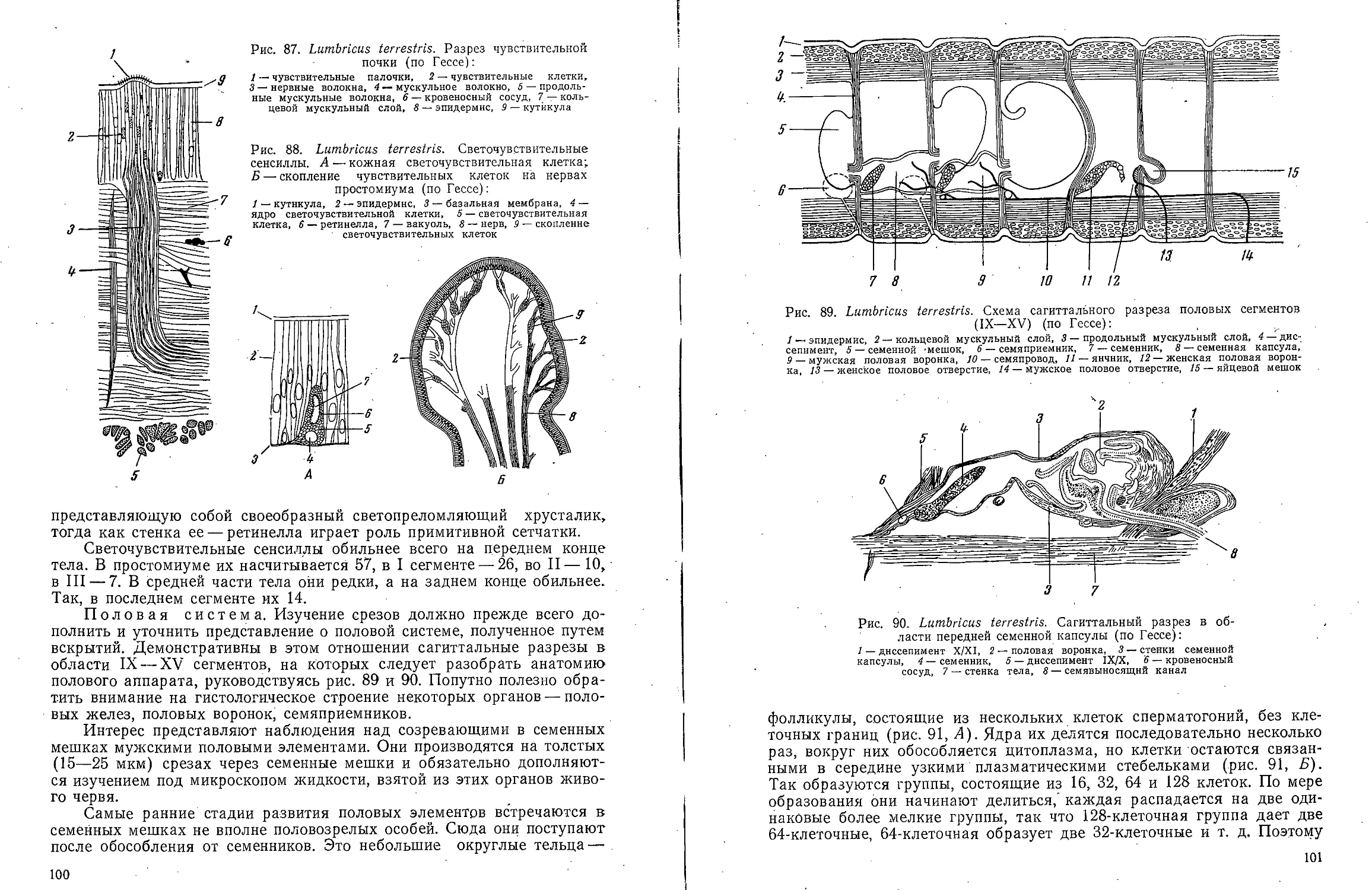
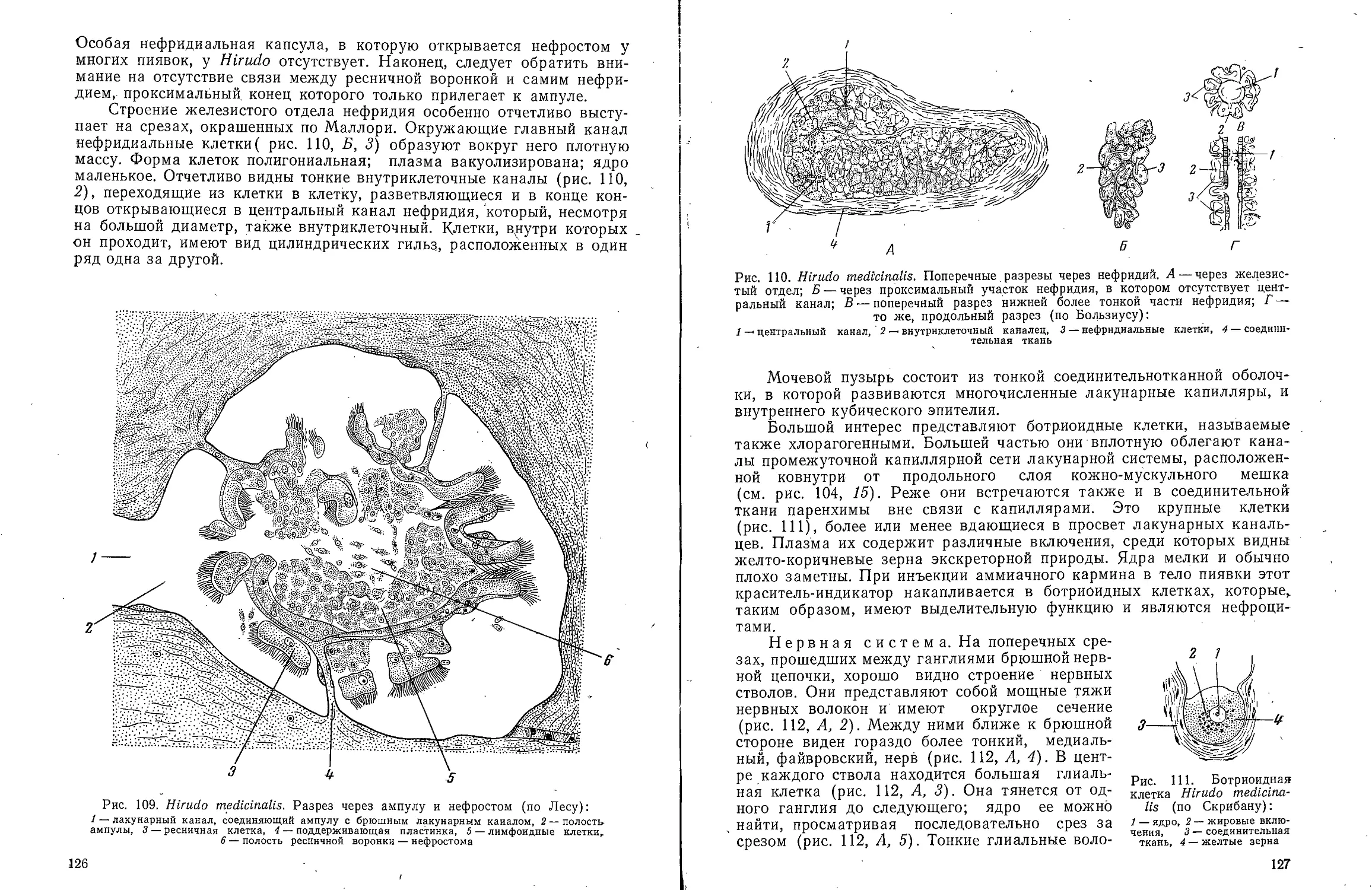
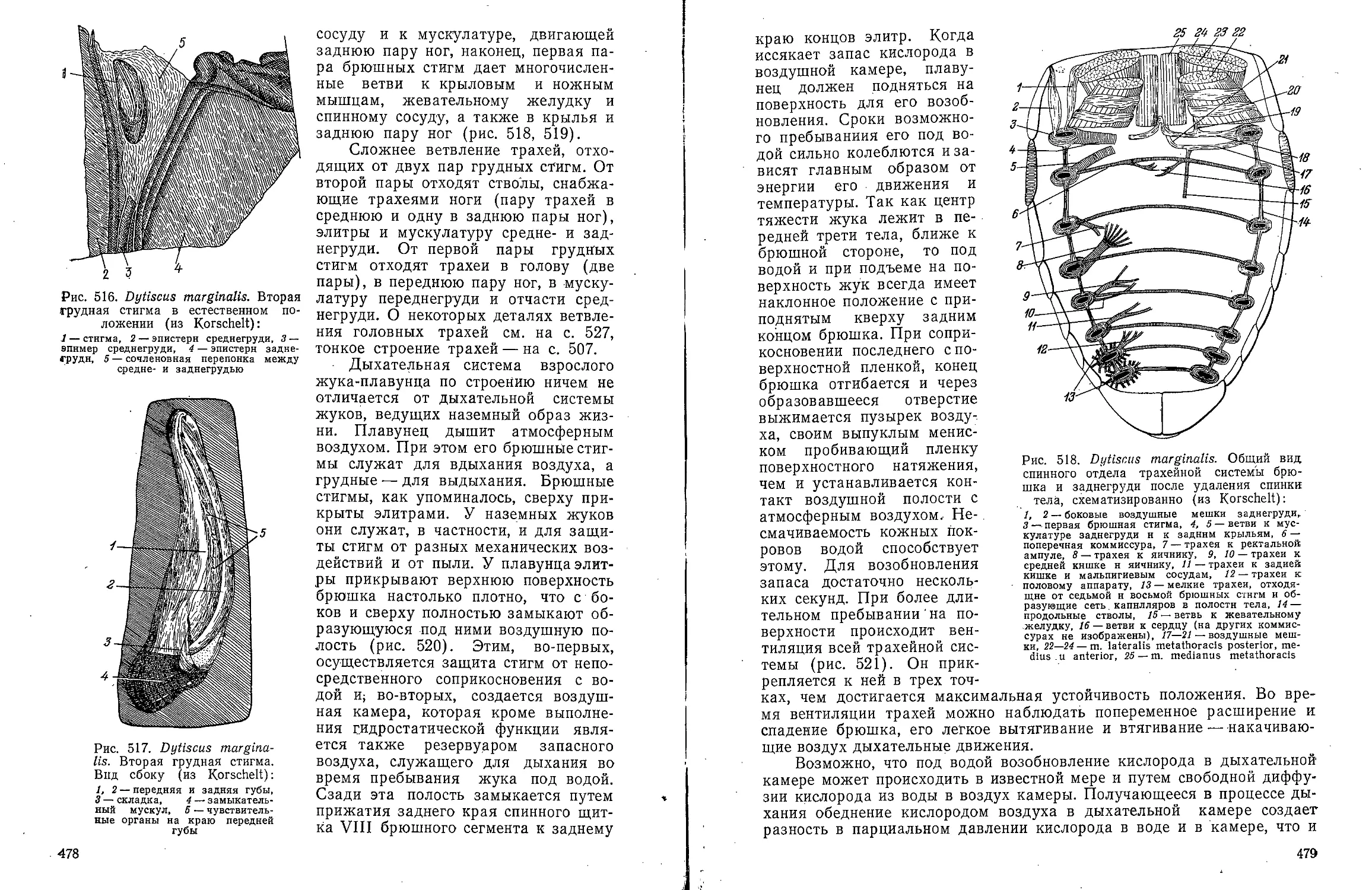
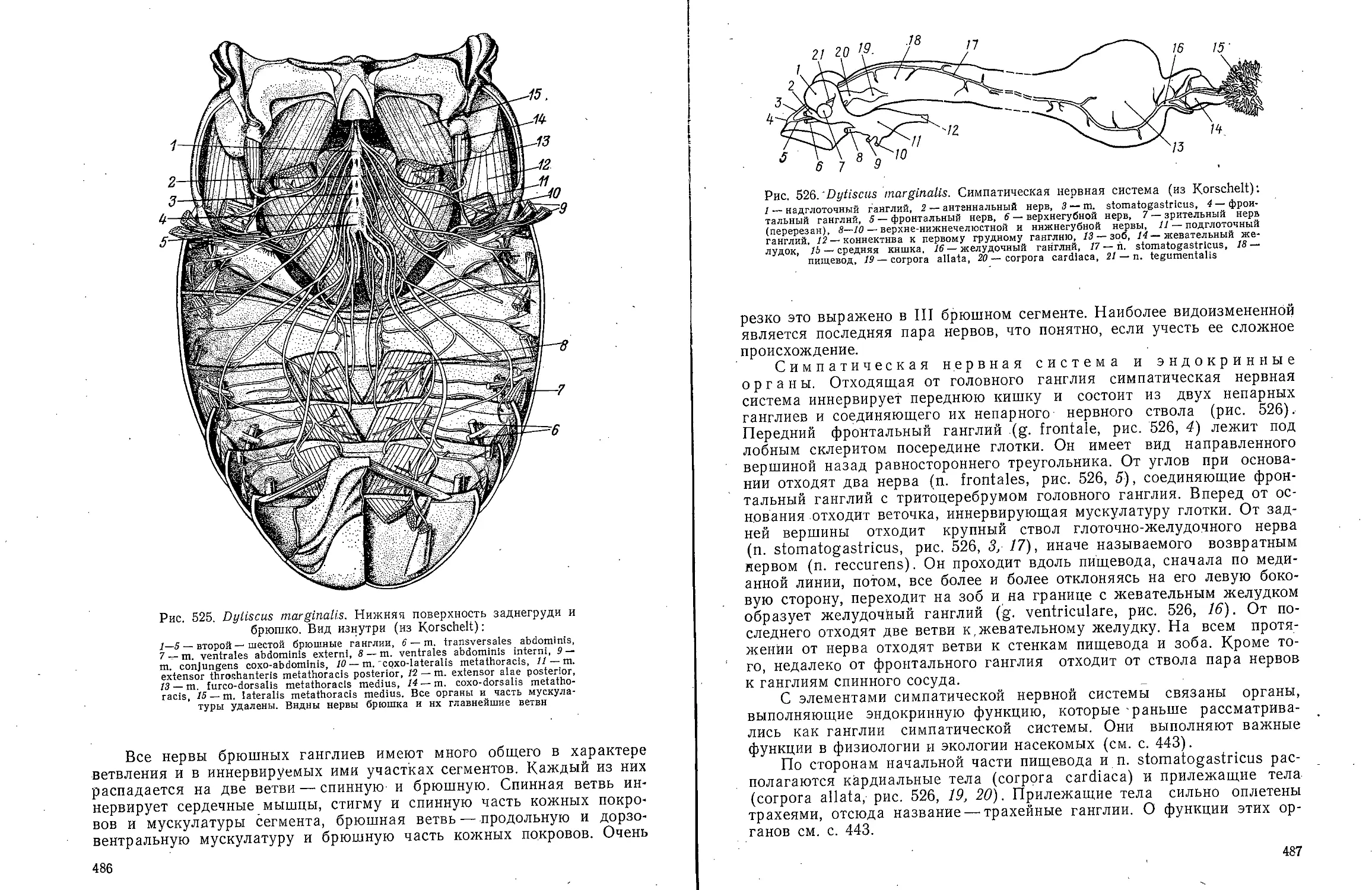
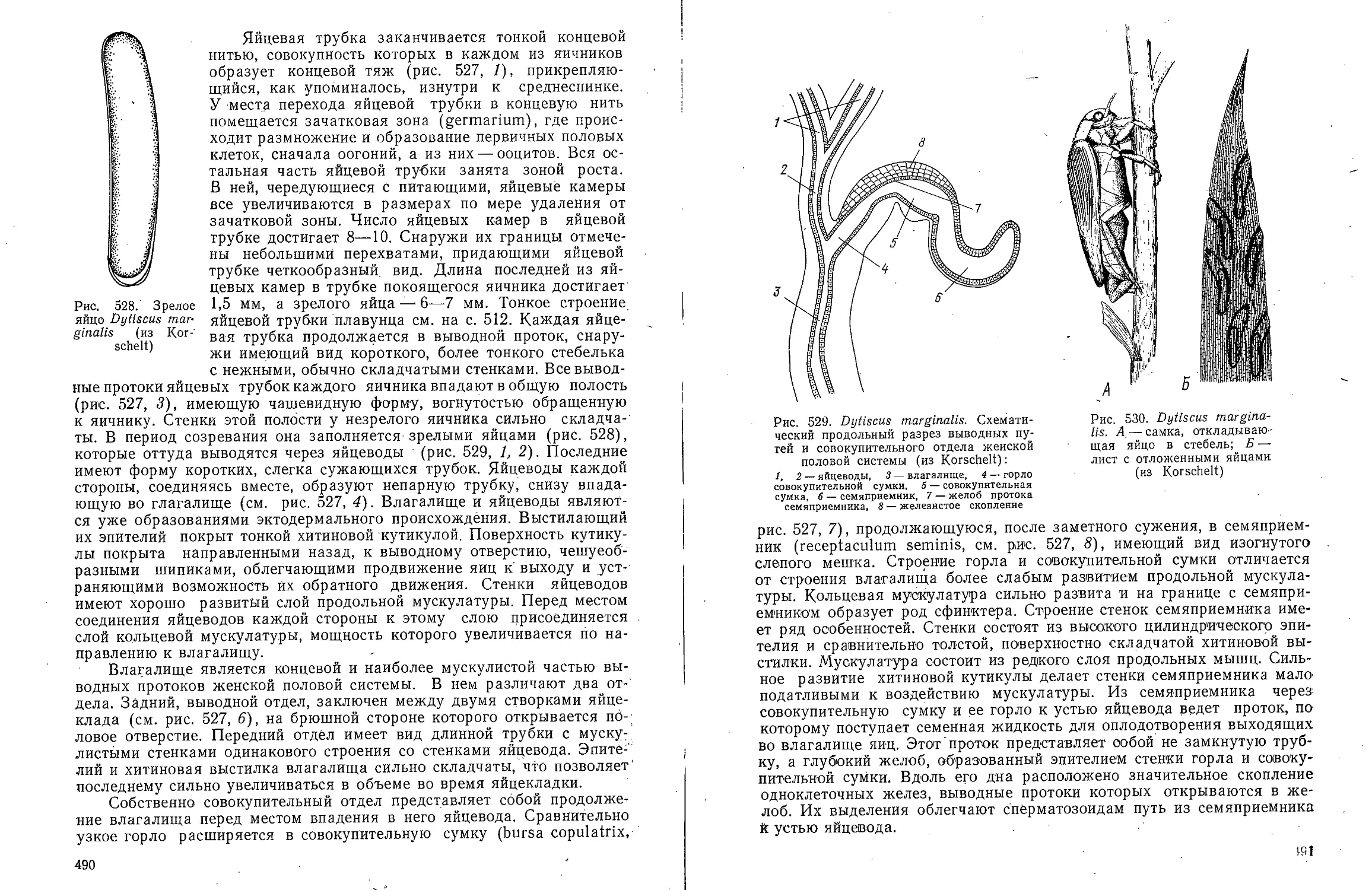
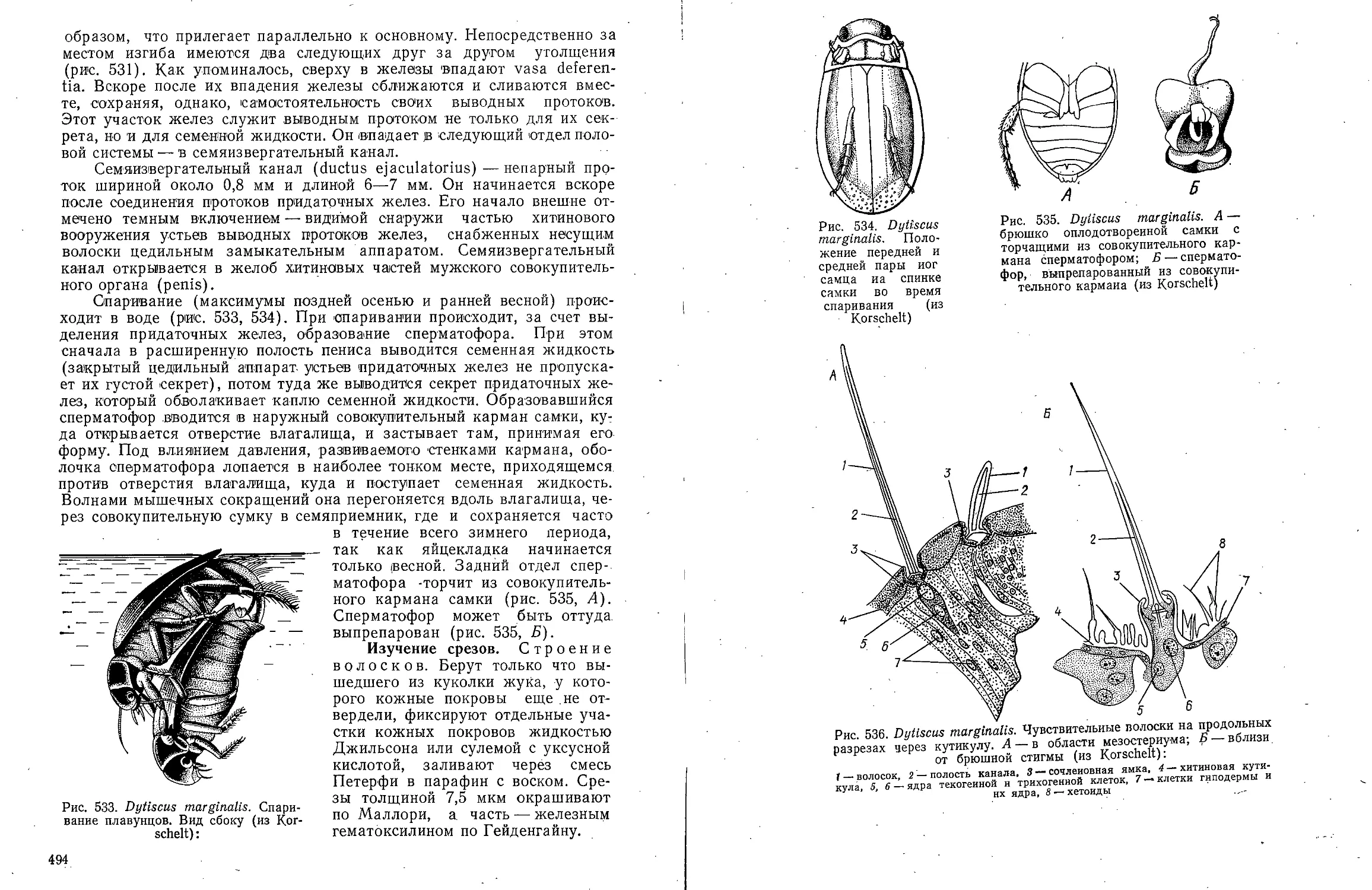
В ов А С Моичадскии
ЮИ не А. Л Стрелков
Боль ой
актикум
о зоочогаи
беспозвоночных
ебн*е * "Ив
А.В.Иванов, А.С. Мончадский,
Ю.И. Полянский, А.А.Стрелков
Большой
практикум
по зоологии
беспозвоночных
ТИПЫ: КОЛЬЧАТЫЕ ЧЕРВИ, ЧЛЕНИСТОНОГИЕ
ЧАСТЬ ВТОРАЯ
Издание третье, переработанное
и дополненное
Допущено
Министерством высшего и среднего специального
образования СССР
в качестве учебного пособия
для студентов биологических специальностей
университетов
МОСКВА <ВЫСШАЯ ШКОЛА> 1983
ББК 28.691
Б 79
УДК 592
Рецензент:
кафедра зоологии Львовского государственного университета
(зав. кафедрой д-р биол. наук, проф. В. И. Здун)
Большой практикум по зоологии беспозвоночных. Типы:
Б 79 Кольчатые черви, Членистоногие: Учеб. пособие для
студентов биолог, спед. ун-тов. Ч. 2/Иванов А. В., Мончадский А. С,
Полянский Ю. И., Стрелков А. А. — 3-е изд., перераб. и доп.—
М.: Высш. шк., 1983. — 543 с, ил.
В пер.: 1 р. 90 к.
Книга представляет собой третье издание учебного пособия к
специализированному студенческому лабораторному практикуму по зоологии беспозвоночных
животных, выпускаемому в трех частях. I часть — «Простейшие, Губки,
Кишечнополостные, Гребневики, Плоские черви, Немертины, Круглые черви» вышла в 1981 г.
В III части будут описаны Сипункулиды, Моллюски, Щупальцевые и Иглокожие.
Второе издание Практикума вышло в двух частях в 1958 г.
Рассмотрены внешняя и внутренняя морфология типичных представителей двух
типов — Кольчатые черви и Членистоногие. Описание объектов составлено jio
единому плану. Объекты подобраны таким образом, чтобы оин были по возможности
доступны.
2005000000—273 ББК 28.691
И 81—83
001(01)—83 592
© Издательство «Высшая школа», 1983
ПРЕДИСЛОВИЕ
Книга представляет собой третье издание
учебного пособия к специализированному студенческому
лабораторному практикуму по зоологии
беспозвоночных животных, выпускаемому в трех частях.
I часть «Простейшие, Губки, Кишечнополостные,
Гребневики, Плоские черви, Немертины, Круглые
черви» вышла в 1981 г. В III части будут описаны
Сипункулиды, Моллюски, Щупальцевые и
Иглокожие. Второе издание Практикума вышло в двух
частях в 1958 г.
Вторая часть «Большого практикума по зоологии
беспозвоночных» посвящена членистым животным
(Articulata в понимании Кювье): Кольчатым
червям (Annelida) и Членистоногим (Arthropoda).
Третье издание Практикума переработано и
дополнено в соответствии с новыми достижениями
науки. Особенно значительному изменению
подвергся раздел, посвященный Многощетинковым
кольчецам. Систематика приведена в соответствие с
новыми исследованиями, выполненными в основном
советскими зоологами. Библиографические ссылки
снабжены краткими аннотациями. Опыт
показывает, что они полезны для подбора литературы.
В настоящую книгу включены также два
представителя насекомых — таракан и жук-плавунец.
Это отнюдь не дублирует курса энтомологии.
Упомянутые насекомые рассматриваются в
зоологическом плане сравнительно с другими классами
членистоногих. В существующих энтомологических
практикумах таких подробных очерков морфологии
насекомых нет.
А. В. Ивановым написаны: • Кольчатые черви,
Паукообразные (кроме Клещей) и Многоножки;
А. С. Мончадским — Насекомые; Ю. И.
Полянским — Ракообразные; А. А. Стрелковым — Клещи.
Академик А. В. Иванов,
член-корреспондент АН СССР Ю. И. Полянский
Тип Annelida — Кольчатые черви
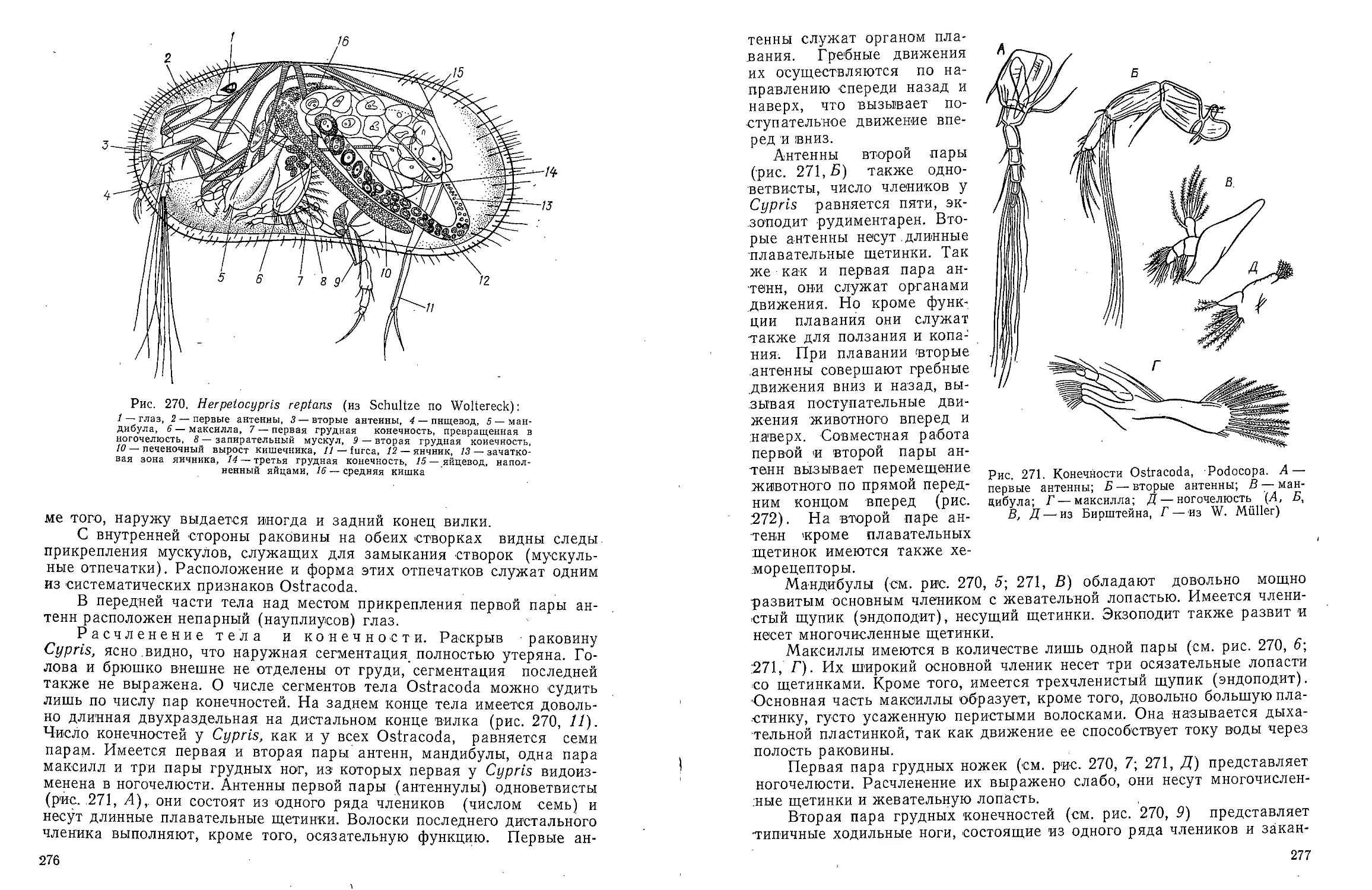
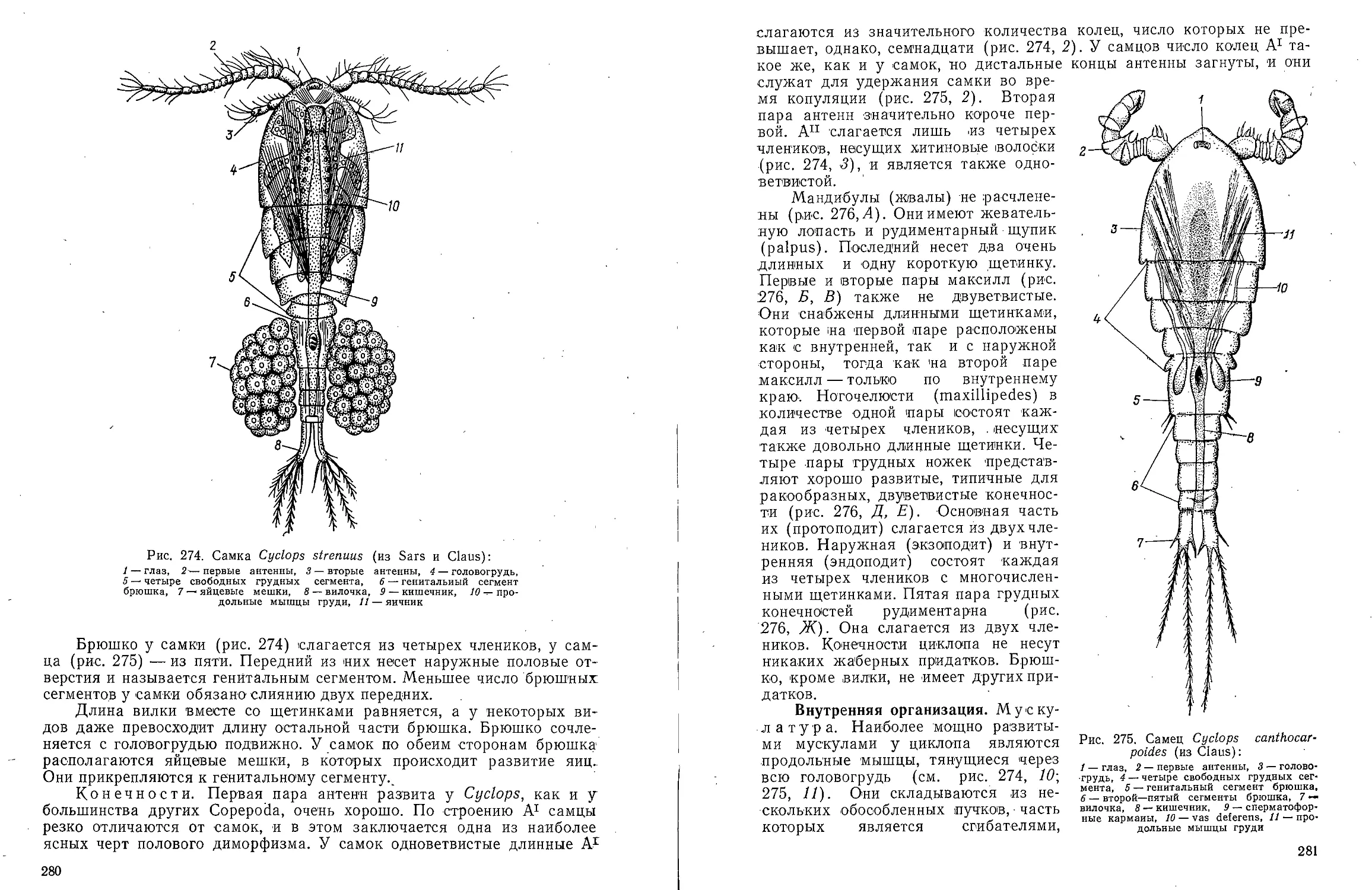
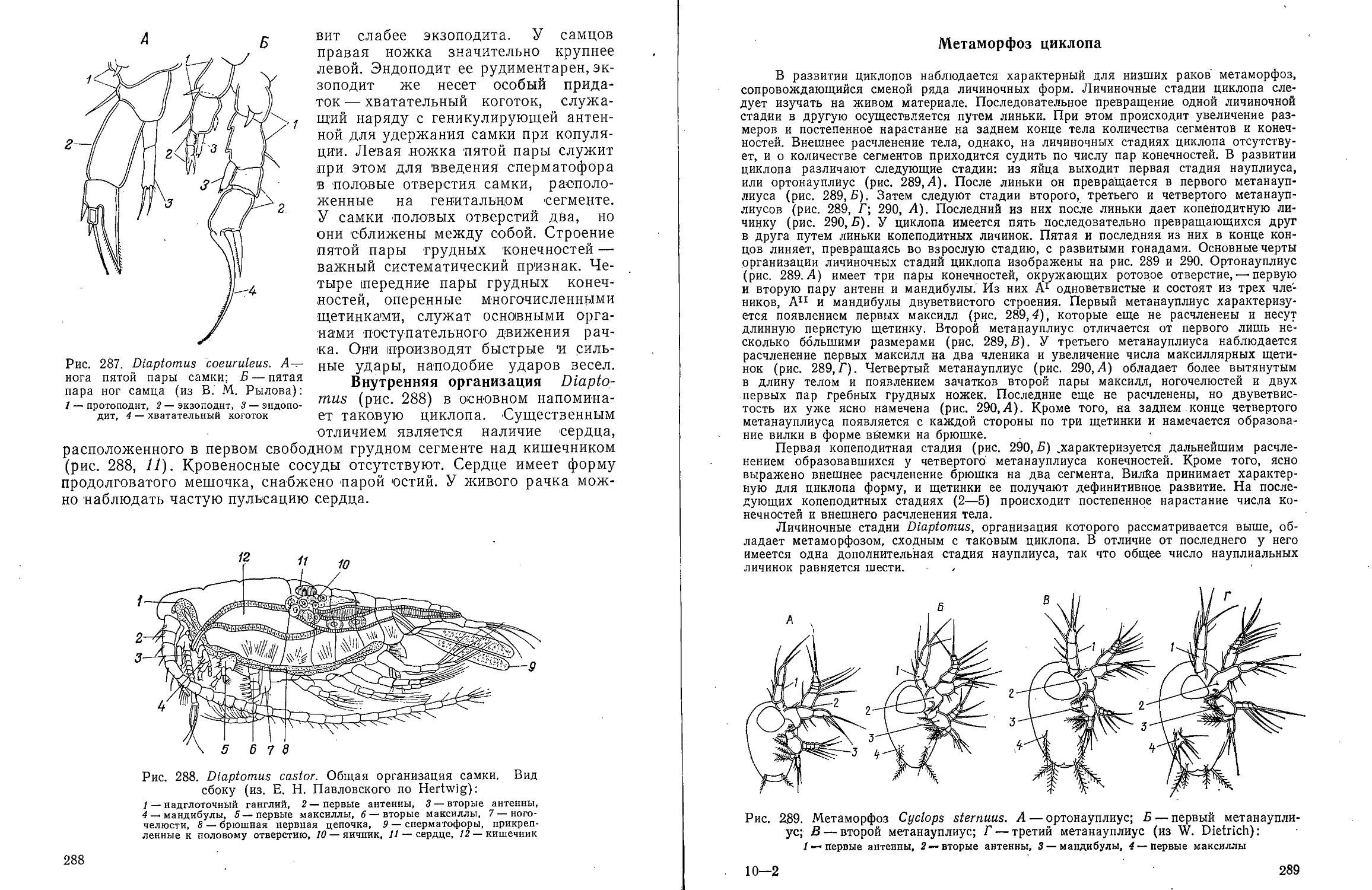
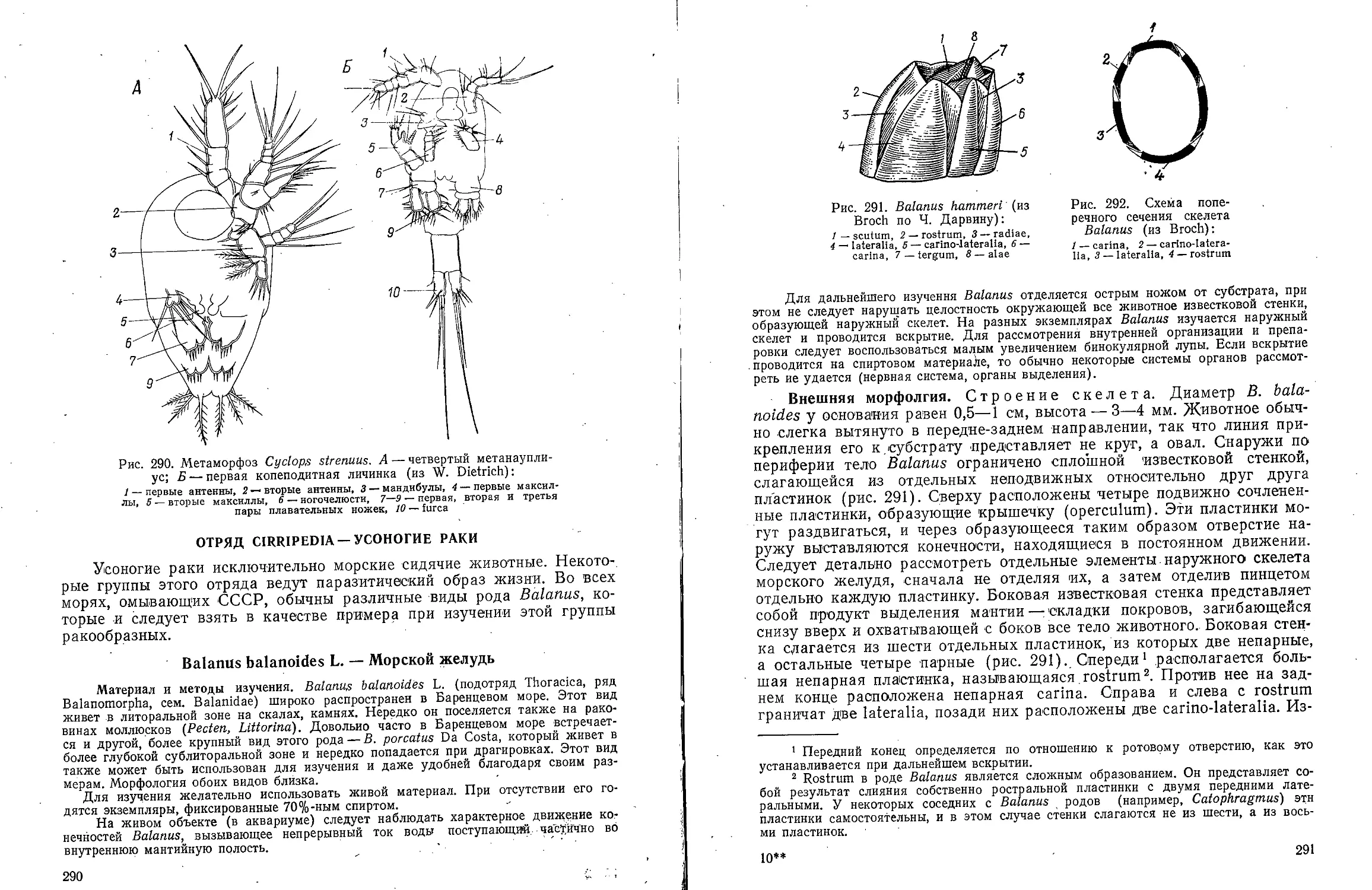
КЛАСС POLYCHAETA-
МНОГОЩЕТИНКОВЫЕ ЧЕРВИ
Для ознакомления с классом многощетинковых червей предлагаем
изучить представителей различных групп: из отряда Archiannelida —
Dinophilus vorticoides О. Schmidt и Polygordius ponticus Salensky, из
отряда Nereimorpha — Nereis pelagica L. и из отряда Drilomorpha —
Arenicola marina L.
Систематическое положение архианнелид до сих пор остается
спорным. Многие зоологи рассматривают эту группу как самостоятельный
примитивный класс аннелид. В настоящее время после их детального
изучения, представляется более вероятным, что это примитивные поли-
хеты, приспособившиеся к жизни в толще песка и приобретшие
некоторые упрощения организации (Ливанов, 1955; Германе, 1969; Орхаге,
1974). Среди них Dinophilidae олигомерны, тело их слагается только из
ларвальных сегментов, а целом лишен метамерного расчленения.
Общий характер метамерии, охватывающей ресничные пояски, кожные
железы, протонефридии и брюшные ганглии, отсутствие параподий,
щетинок и кровеносной системы и развитие без личинки трохофоры
резко отличают динофилид от остальных архианнелид и придают им
большой теоретический интерес.
Polygordiidae и другие родственные им полимерные архианнелиды
во многих отношениях занимают такое же место среди аннелид, как
и ланцетник среди хордовых, который, несмотря на наличие вторичных
приспособлений к роющему образу жизни, рассматривается как
прототип позвоночных.
Представители рода Nereis широко распространены почти во всех
морях СССР, в которых представлены несколькими легко
добываемыми видами. В частности, N. pelagica принадлежит к наиболее
распространенным и обычным формам. При отсутствии этого вида он может
быть легко заменен другим, так как организация разных видов
чрезвычайно близка. Труднее подобрать главный объект среди полихет,
ведущих седентарный образ жизни. На побережьях Баренцева и Белого
морей обычна Arenicola marina, она обитает в легко доступных местах
на литорали, где может быть добыта в больших количествах.
Разнообразие организации Polychaeta и сравнительно большое
морфологическое значение класса делают необходимым ознакомление со
строением еще и других представителей этой обширной группы.
л
ОТРЯД ARCHIANNELIDA- ПЕРВИЧНЫЕ КОЛЬЧЕЦЫ
Dinophilus vorticoides О. Schmidt
Материал и методы изучения. Dinophilus vorticoides — (сем. Dinophilidae) один
из обычных обитателей прибрежной зоны Белого и Баренцева- морей (Мурманский
берег). Собирается Dinophilus следующим образом. С небольших глубин с помощью
сачка или частой драги добываются мелкие водоросли и оставляются в покое в
сосуде с морской водой. Через некоторое время из водорослей выползают различные
мелкие беспозвоночные, в том числе Dinophilus, которые заметны простым глазом в
виде оранжевых червячков, напоминающих на первый взгляд мелких ресничных
червей, рабдоцелид. Хорошие результаты дает также просмотр водорослей в чашке
Петри под лупой или бинокуляром. Иногда, для того чтобы найти несколько экземпляров,
приходится просматривать большие количества проб,
нередко Dinophilus обнаруживаются сразу же. Особенно
хорошие сборы может дать просмотр водорослей Clado-
phora и Ectocarpus. По данным И. Г. Малиновой (1971),
на Мурманском побережье размножение Dinophilus
vorticoides происходит в апреле и мае1. Вышедшие из яиц
молодые червячки к середине июня или июля достигают
0,5—0,8 мм в длину и инцистируются, переходя в покою-
щееся состояние. Они выделяют вокруг себя защитную
цисту; тело их округляется, реснички исчезают, рот и
анус замыкаются, происходит некоторая дегенерация
тканей. Из цисты животные выходят только в феврале —
марте следующего года. После этого они испытывают
второй период роста и, достигнув к апрелю — маю
максимальной величины (1,2—1,5 мм), становятся
половозрелыми. Самки, отложив несколько кладок, погибают. На
западном побережье Швеции все этапы жизненного цикла
Dinophilus vorticoides наступают на 1—2 месяца раньше
(Иегерстен, 1944). На Белом море жизненный цикл ди-
нофилюса не изучался. В Японском море (побережье
залива Петра Великого) встречается Dinophilus gyrociliatus
О. Schmidt (рис. 1). Этот вид также' может быть
использован для целей большого практикума. Дальневосточный
Dinophilus, однако, принадлежит к группе видов с
особенно резким половым диморфизмом. Самцы у этой формы
рудиментарные, а в кладках самок есть яйца большие,
дающие самок, и мелкие, развивающиеся в самцов.
Изучение организации Dinophilus производится на
живых объектах методом витальных окрасок и на сериях
срезов. Хорошие результаты дает прижизненная окраска
метиленовым синим. Для этого живые черви помещаются
в слабый раствор красителя в морской воде. Через
несколько часов краска проникает в клетки покровов и
окрашивает секрет кожных желез и пигментные гранулы.
Специфически окрашиваются ресничные клетки, особенно
клетки мерцательных поясков и оральной области,
которые заполняются густыми зернышками
вишнево-фиолетового цвета. Позднее окрашиваются нефридии и эпителий
кишечника. При продолжительном выдерживании в мети-
леновом синем красится кожная мускулатура и
одновременно отдельные участки периферической нервной системы.
1 И. Г. Маликова определила мурманских особей как
Dinophilus taeniatus Harmer, но, судя по ее рисункам, это
скорее D. vorticoides. Мы считаем, что в Белом,
Баренцевом, а также в Северном морях (у берегов
Скандинавии) обитает один вид Dinophilus vorticoides О. Schmidt.
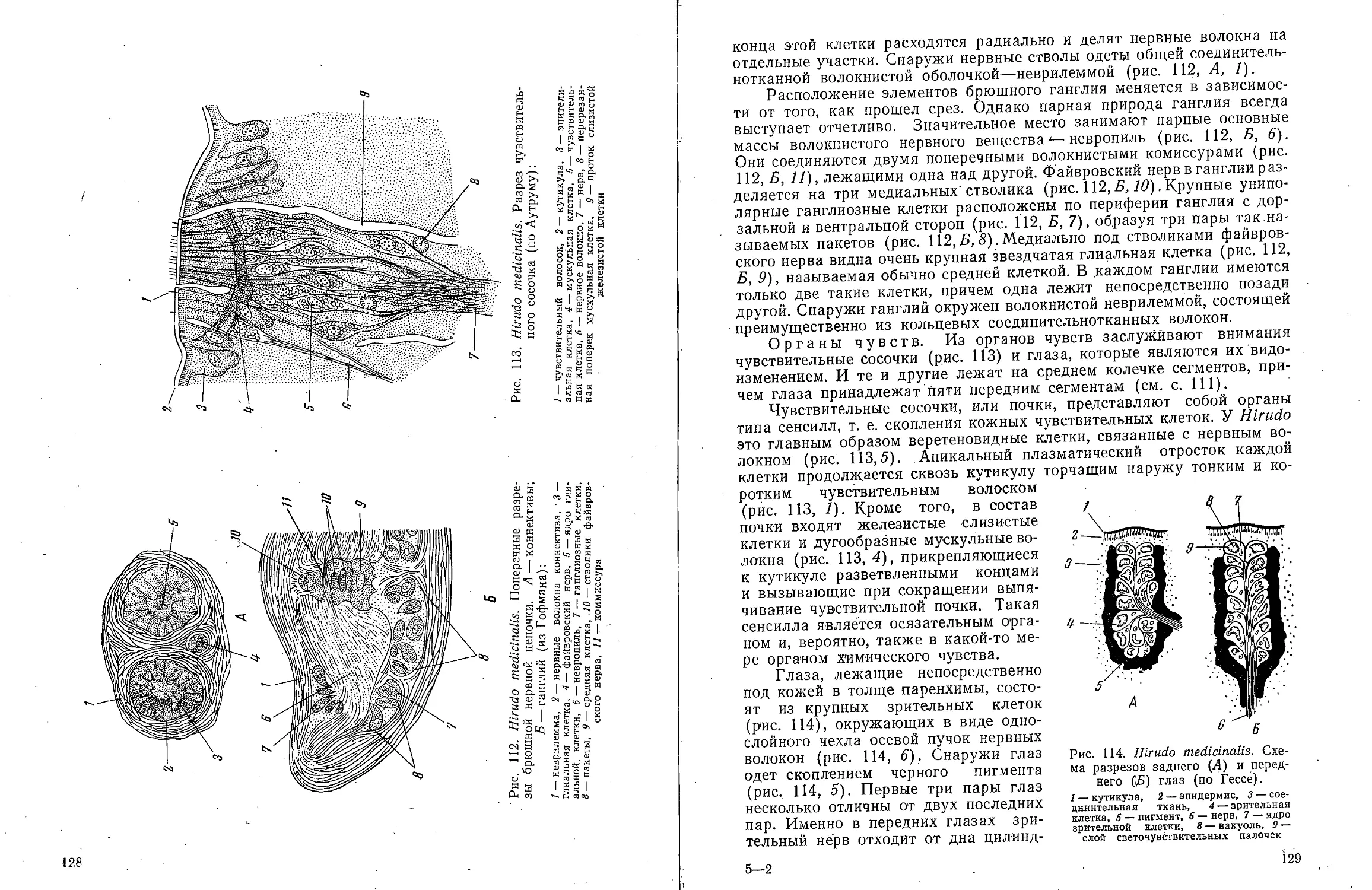
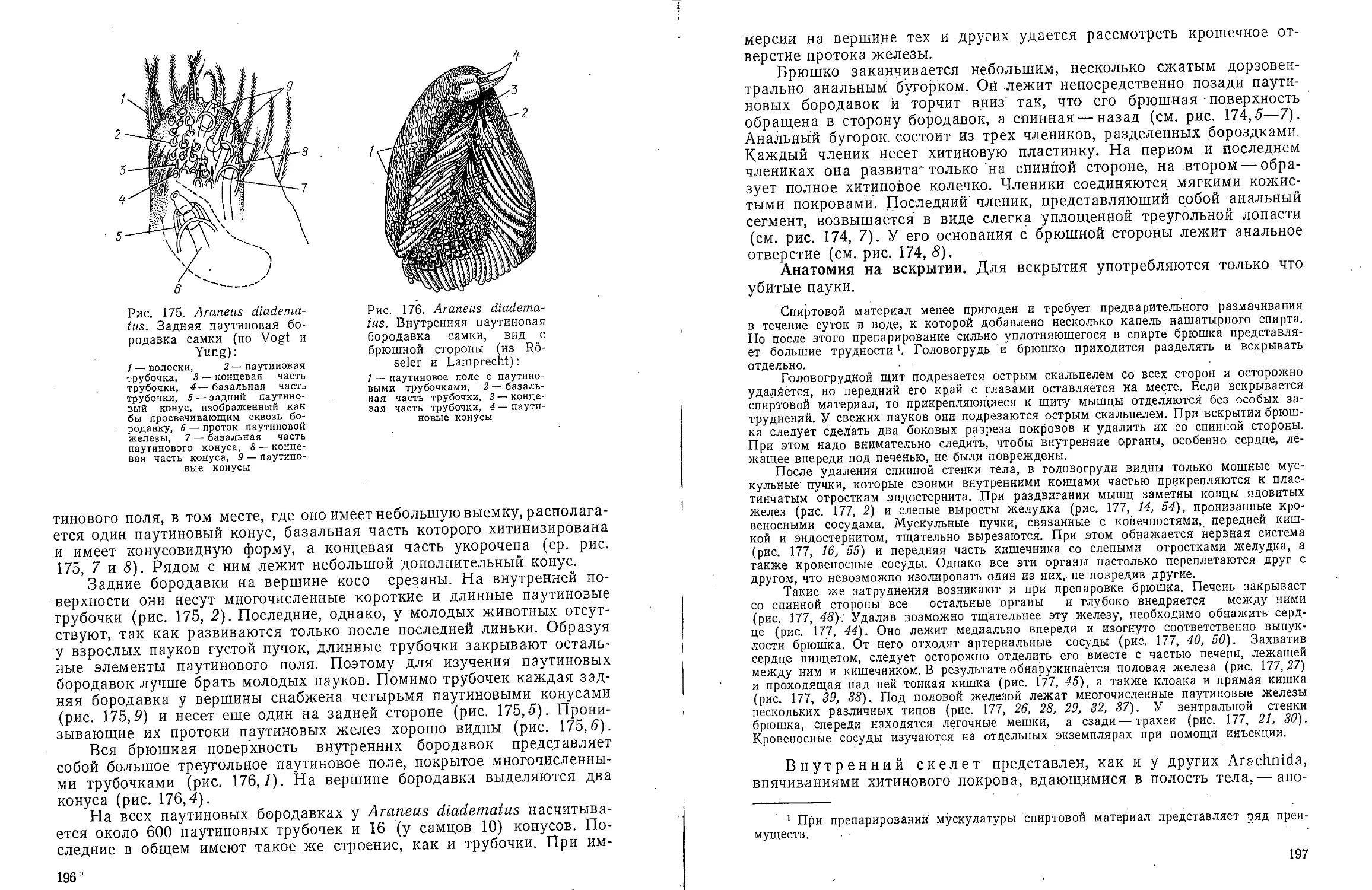
Рис. 1. Dinophilus
gyrociliatus. Самка. Вид со
спинной стороны (ориг.):
1 — слюнная железа, 2 —
отверстие глоточного
кармана, 3 — ротовое отверстие,
4 — глаз, 5 — осязательные
волоски, 5 — простомиум,
7 — преоральный ресничный
поясок, 8 — глоточный
карман, 9 — средняя кишка,
10 — яичник, И — задняя
кишка, 12 — анус, /У— пи-
гидий, 14 — ресничный
поясок V сегмента
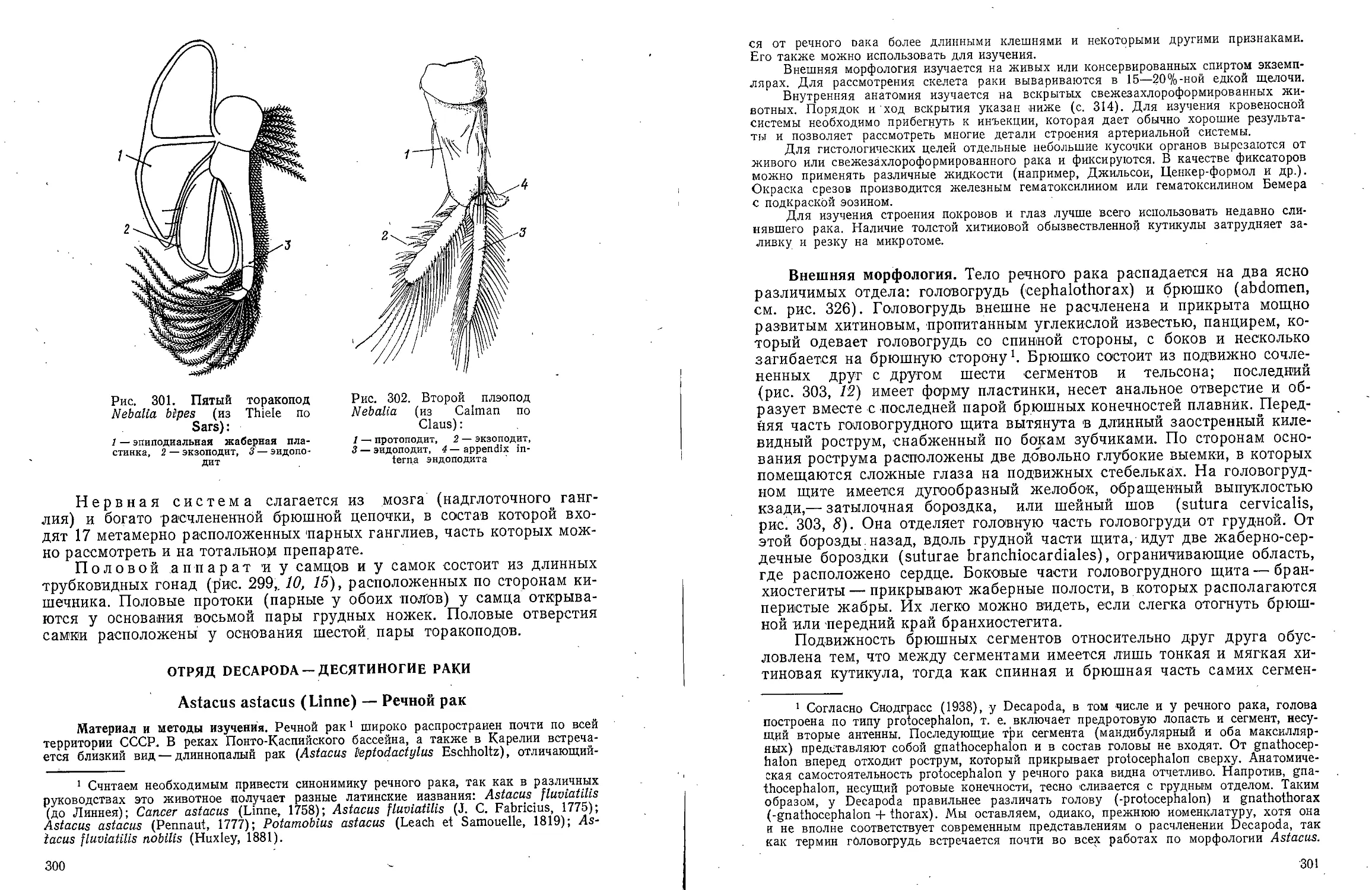
5
Животные, предназначенные для изготовления срезов, фиксируются холодной
или горячей смесью сулемы с уксусной кислотой, или раствором сулемы в морской
воде, или же подогретым абсолютным спиртом. Заливку можно производить обычным
способом через кедровое масло и парафин. Однако для лучшей ориентировки
рекомендуется заливка по Петерфи на целлоидиновых пластинках. Срезы окрашиваются
гематоксилином (например, гематоксилином Эрлиха) или кислым гемалауном Майера.
Хорошие результаты дает окраска in toto борным кармином.
Внешняя морфология. Тело червя продолговатое, цилиндрической
формы, в четыре раза длиннее своей ширины (рис.2). Длина 1,2—1,5 мм;
самцы несколько меньше самок. Метамерия в расположении нефридиев
и в строении нервной системы (см. ниже), так же как метамерное
расположение мерцательных поясков, слизистых желез и кольцевой
мускулатуры, указывает на сегментированность Dinophilus. Помимо просто-
миума (головного отдела), тело слагается из шести сегментов.
Простомиум (рис. 2, 4) закруглен спереди и отделен от туловища
ясной кольцевой бороздой. На заднем конце тело заметно сужено,
образуя небольшой хвостовой
отдел, или пигидий (рис. 2,
12). На простомиуме и туло-
вТище располагаются
поперечные ресничные пояски (рис.
2, 20). Хорошо видны пояски
при витальной окраске мети-
леновым синим. Одновременно
окрашивается довольно
широкая брюшная мерцательная
полоска, которая впереди
переходит в мерцательный
эпителий околоротовой области
(рис. 2,18). С ней же
соединяются и ресничные пояски
головы. Сзади брюшная
мерцательная полоска переходит в
ресничный покров хвостового
отдела. Каждый сегмент
снабжен двумя мерцательными
поясками, состоящими из одного
ряда ресничных клеток, и
только второй, головной, поясок
образован двумя рядами
клеток (рис. 2, 1). По характеру
расположения этих клеток он
чрезвычайно напоминает пре-
оральный мерцательный
поясок трохофоры. Передний
головной поясок прерван на
брюшной стороне. На спинной
стороне головного отдела ле-
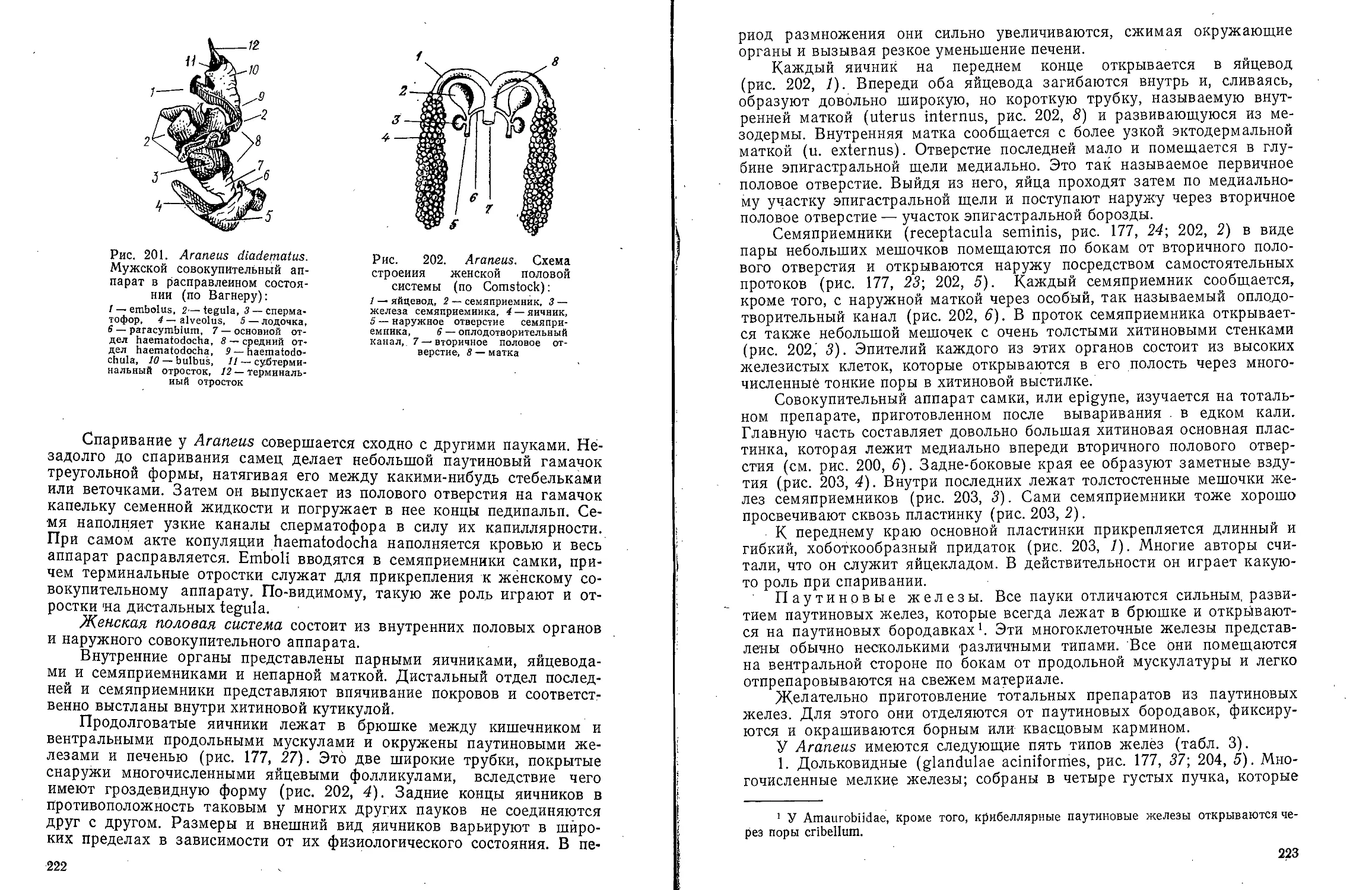
Рис. 2. Dinophilus vorticoides. A — самка со
спинной стороны; Б — самец с брюшной
стороны (ориг.):
/ — преоральный ресничный поясок, 2 — глаз, 3 —
апикальные осязательные волоски, 4 — простомиум,
5 — глоточный карман, 6 — слюнные железы, 7 —
средняя кишка, 8 — передний яичник, 9 — задний
янчник, 10 — задняя кишка, // — анальное отверстие,
12 — пигидий, 13 — хромофильные железы, 14 — ну-
хальный орган, 15 — язычок, 16 — отверстие глоточ-
ного кармана, Я —рот, 18 — брюшная ресничная по- ЖИТ Пара ГЛЗЗ (рИС. 2, 2). При
лоска, 19 — семенник, 20 — пояски III сегмента, 21— <„„, „„. .,„„„„„„ „ „. „„„ ;„
семенной пузырек, 22 - половое отверстие ООЛЬШОМ увеличении МИКрОСКО-
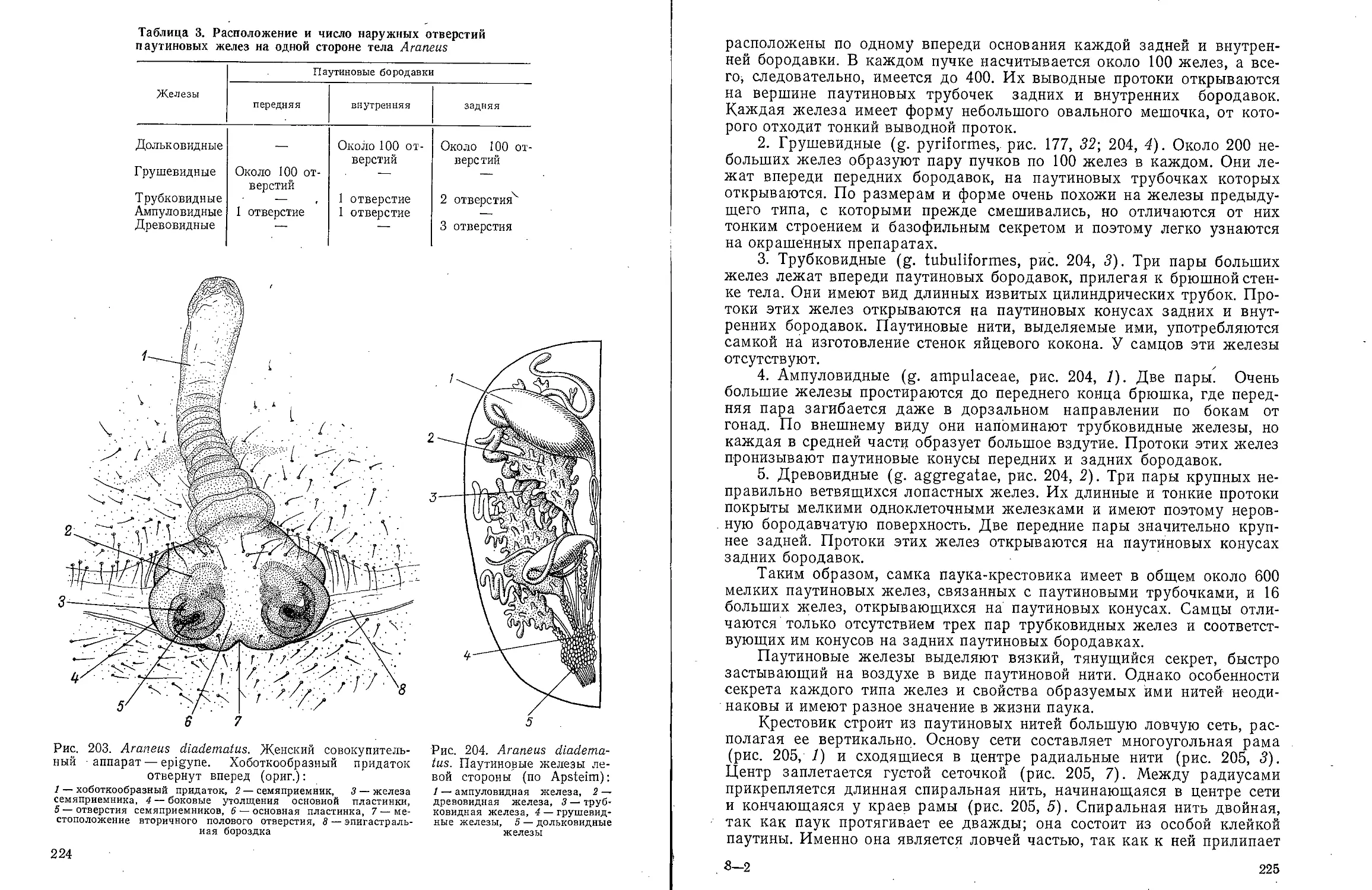
Рис. 3. Dinophilus vorticoides. Самцы, окрашенные метиленовым
синим. А — окрасились мерцательные пояски и кишечник; Б —
протонефридии и кишечник (по Шимкевичу):
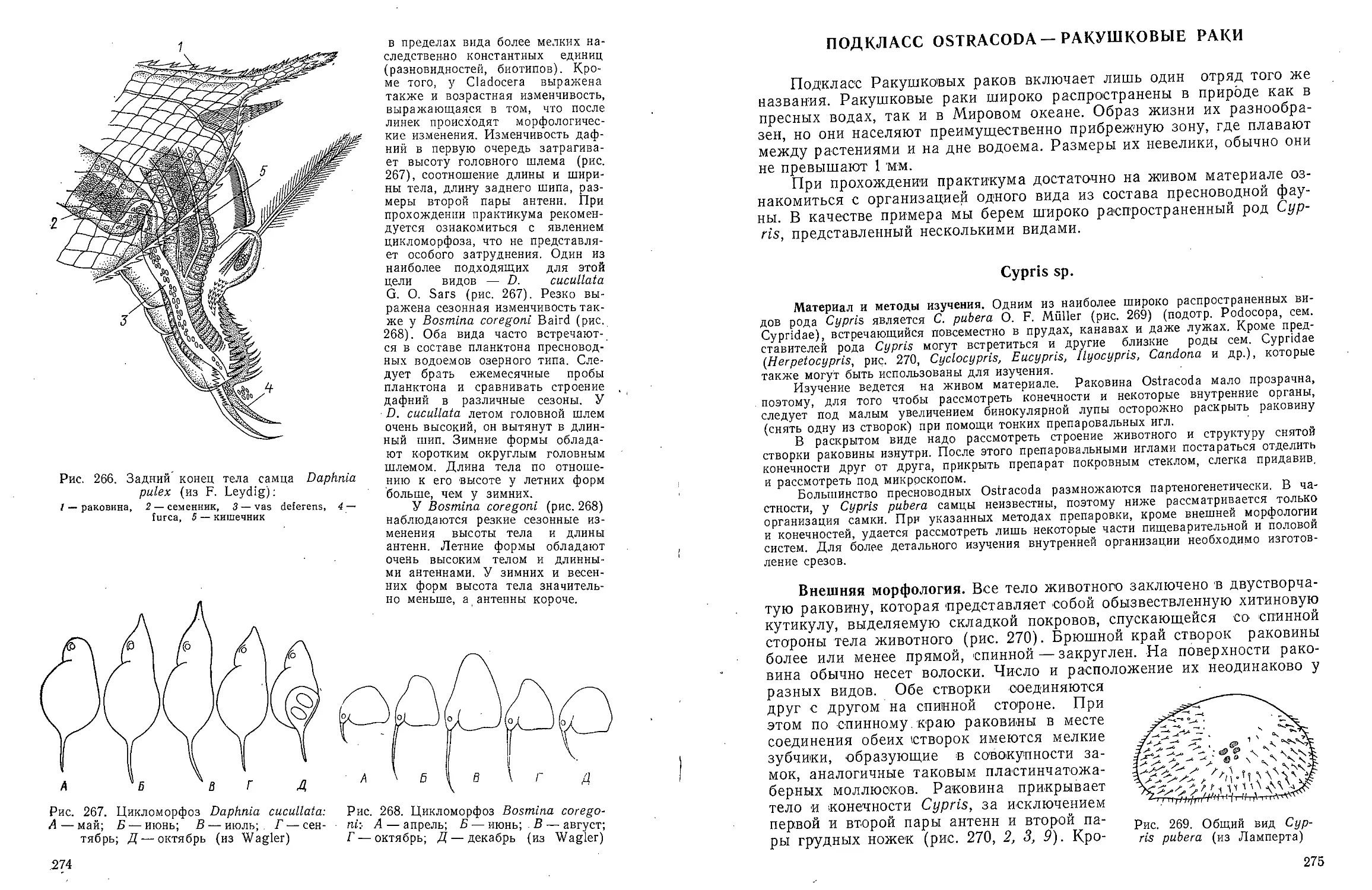
/ — бороздка, отграничивающая головной отдел, 2 — совокупительный
орган, 3 — анальное отверстие, 4 — семенные пузырьки, 5 — туловищный
отдел, 6 — ротовое отверстие, 7 — глаз, 8 — чувствительные волоски
головного отдела, 9 — протонефридии, 10 — задняя кишка, Л — передняя киш- .
ка, 12 — средняя кишка, 13 — семенник, 14 — мужское половое отверстие,
л1—л11 — мерцательные пояски, I—VI — сегменты тела
па видно, что каждый из них состоит из темного пигментированного
бокала и двух светопреломляющих телец.
На переднем крае головного отдела хорошо заметны осязательные
волоски (рис. 2,5), такие же волоски рассеяны на туловище и хвостовом
отделе. Небольшое ротовое отверстие Т-образной формы, оно
располагается на брюшной стороне на границе между головным отделом
и туловищем (рис. 2,17). Анальное отверстие лежит на спинной
стороне у основания хвостового отдела (рис. 2,11).
Хорошо красятся метиленовым синим, но и без витального
окрашивания заметны одноклеточные слизистые железы эпидермиса, а также
некоторые внутренние органы: передняя средняя и задняя кишки и
некоторые части полового аппарата (рис. 3). Живые черви имеют желто-
7
красный цвет, что зависит от
наличия пигмента в клетках
эпидермиса и эпителия
кишки.
Анатомия. При
ознакомлении с внутренней
организацией рекомендуется изучение
серий срезов дополнять
наблюдениями над животными,
окрашенными in vivo.
Необходимо приготовить серии
поперечных, фронтальных и
сагиттальных срезов.
Кожно-мускульн ы й
мешок. Все клетки
однослойного эпидермиса, за
исключением клеток
мерцательных поясков и медио-вентраль-
нои мерцательной полоски, лишены ресничек (рис. 4). На
брюшной стороне и головном отделе эпидермис гораздо выше, чем в других
местах; всюду он подстилается базальной мембраной. Между
обычными покровными клетками залегают бокаловидные одноклеточные хро-
мофильные железы (рис. 4, /). Содержимое их хорошо окрашивается
витальными красками, в частности метиленовым синим. Слизистые
железы развиты на брюшной стороне тела и особенно многочисленны в
хвостовом отделе. На брюшной стороне тела в толще эпидермиса лежат
крупные базальные клетки, не имеющие протоков (рис. 4,6). Их функция
неизвестна. Кроме этих желез имеются головные, или теменные, железы,
которые хорошо видны на фронтальных срезах. Это группы
одноклеточных железистых клеток, лежащие глубоко под эпидермисом в
передней части головного отдела (рис. 5,/). Их длинные выводные протоки
соединяются пучками, которые открываются наружу субвентрально на
уровне первого головного мерцательного пояска. Такими же
погруженными под эпидермис железами являются передние и задние губные
железы, заметные на поперечных срезах в области рта (рис. 5,14). Они
представляют собой комплексы одноклеточных желез.
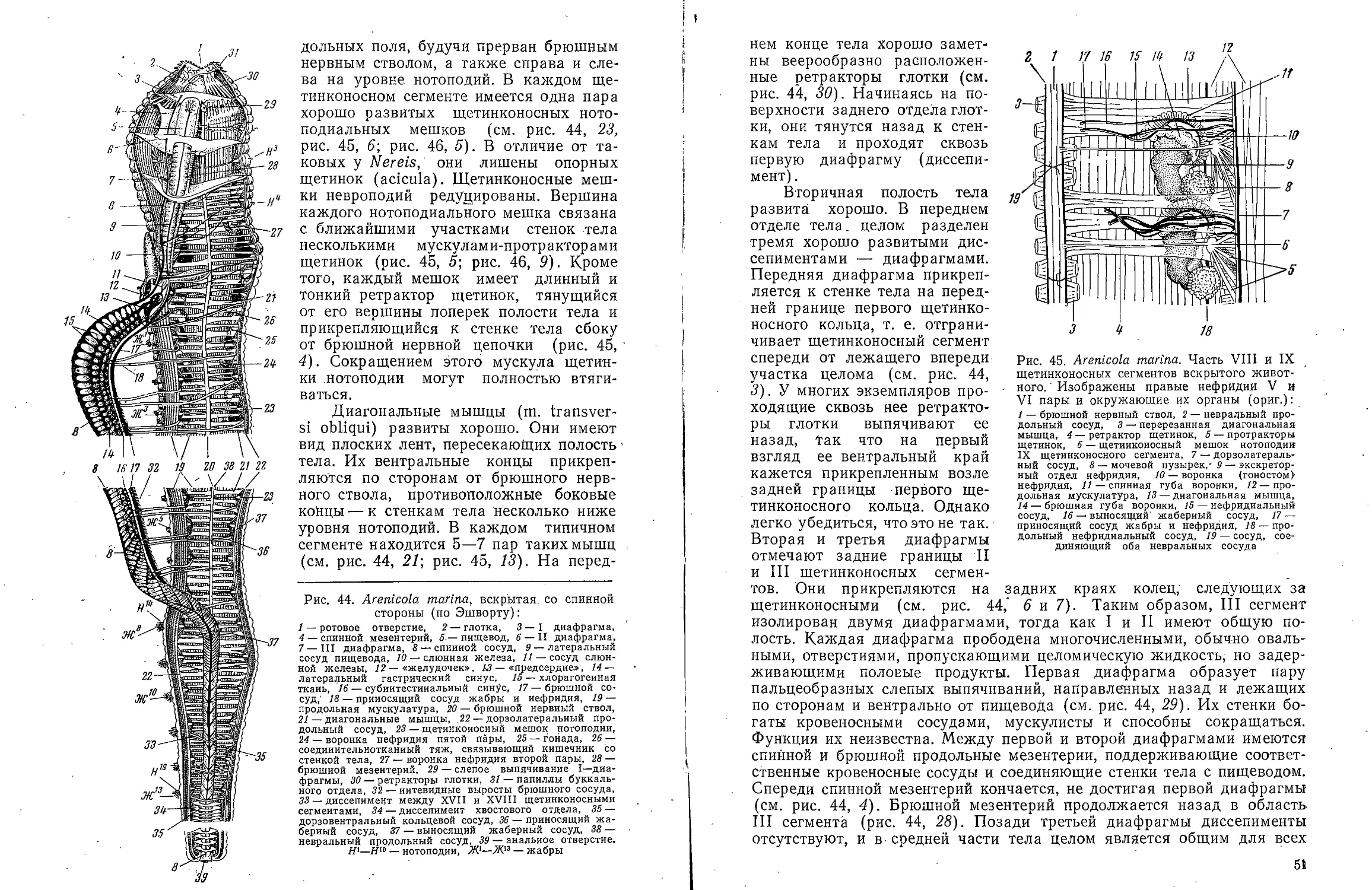
Мускулатура развита слабо. Под кожей залегают редкие
мышечные пучки различных направлений, которые можно наблюдать на
животных, окрашенных in vivo метиленовым синим. Каждый мускульный
пучок состоит из 3—4 тонких гладких мускульных волокон. Кольцевая
мускулатура лежит непосредственно под эпидермисом; ее мускульные
пучки немногочисленны. Расположение поперечных и продольных
мускульных волокон (а также продольных нервных стволов) показано на
рис. 6.
К мускулатуре кожно-мускульного мешка принадлежат также два
мощных продольных брюшных мускула, которые тянутся над брюшными
нервными стволами и отвечают вентральным продольным мышечным
полям других аннелид (рис. 6,10).. Гомологи спинной пары мышечных
полей у Dinophilus отсутствуют. В головном отделе волокна каждого
8
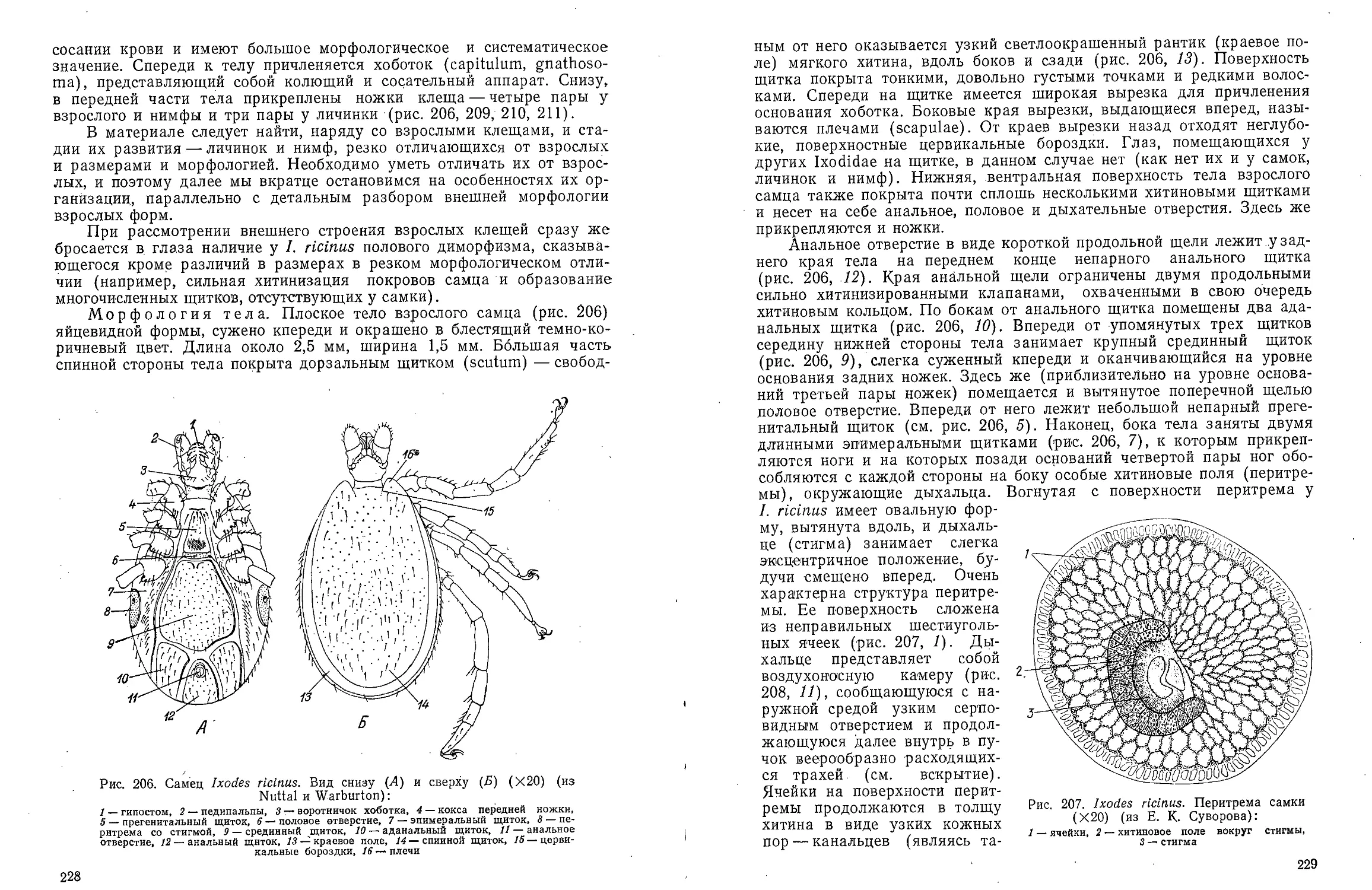
Рис. 4. Dinophilus vorticoides. Поперечный
разрез эпидермиса в латеро-вентралыюй
области туловища (по Иегерстену):
/ — хромофильные железы, 2 — прокутикула, 3 —
слизистые железы, 4 — брюшной нервный ствол,
5 — ядра, 6 — базальные клетки, 7 — базальная
мембрана, 8 — мерцательная клетка
брюшного мускула загибаются
внутрь, проходят вдоль заднего
края мозга, где взаимно
перекрещиваются и прикрепляются к
стенке тела позади глаз (см. рис. 5,9).
Кроме того, имеется еще пара
поперечных мускулов, пересекающих
головной отдел и соединяющих
стенки тела противоположных
сторон (рис. 5, З)1.
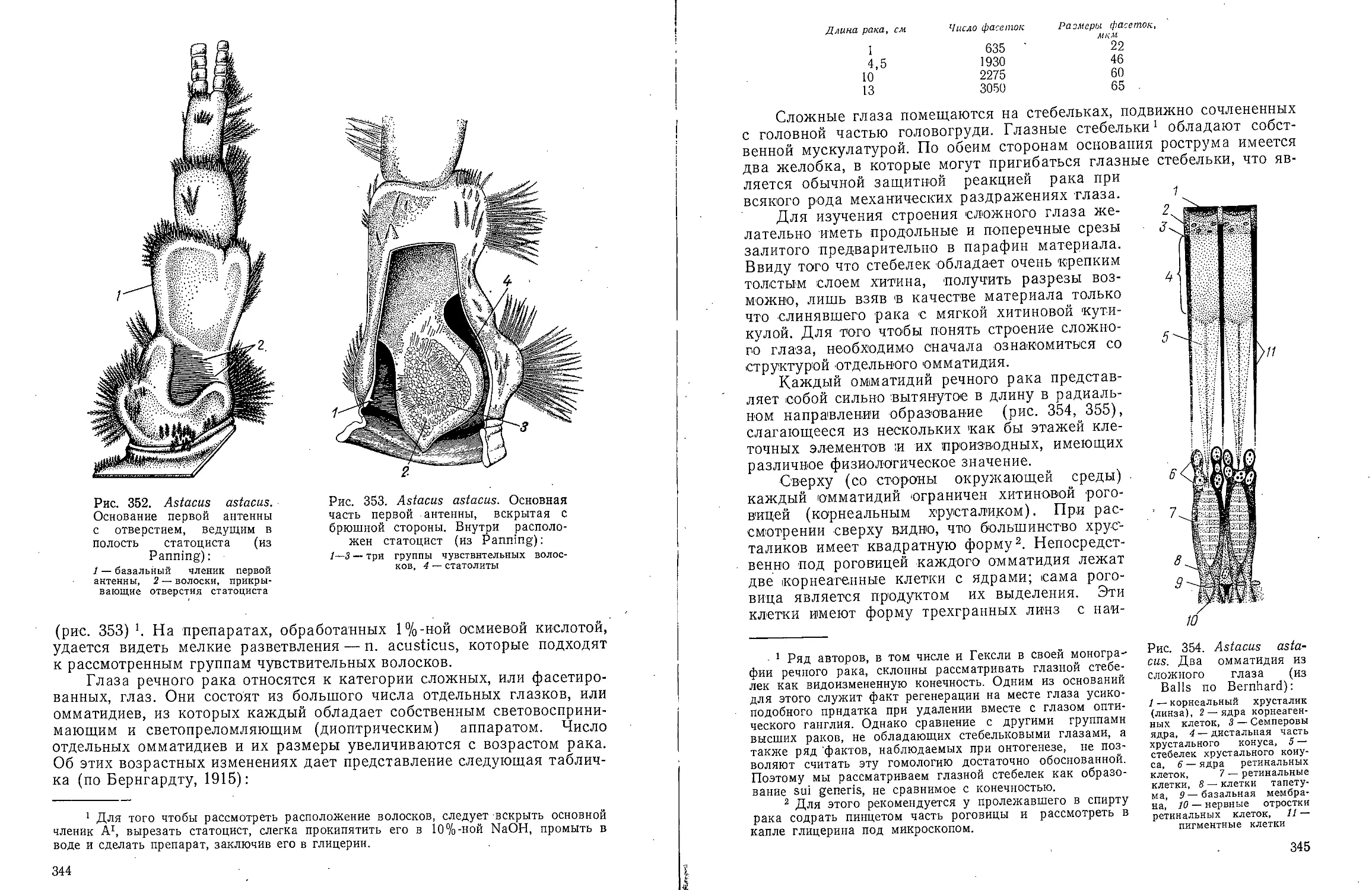
Полость тела. Динофили-
ды обладают хорошо развитой
вторичной полостью тела (целомом),
которая, однако, лишена
каких-либо признаков метамерии. Септы
(диссепименты) и мезентерии
отсутствуют. Целом состоит из пары
боковых (латеральных) мешков,
лежащих по бокам от средней
кишки и сообщающихся в средней
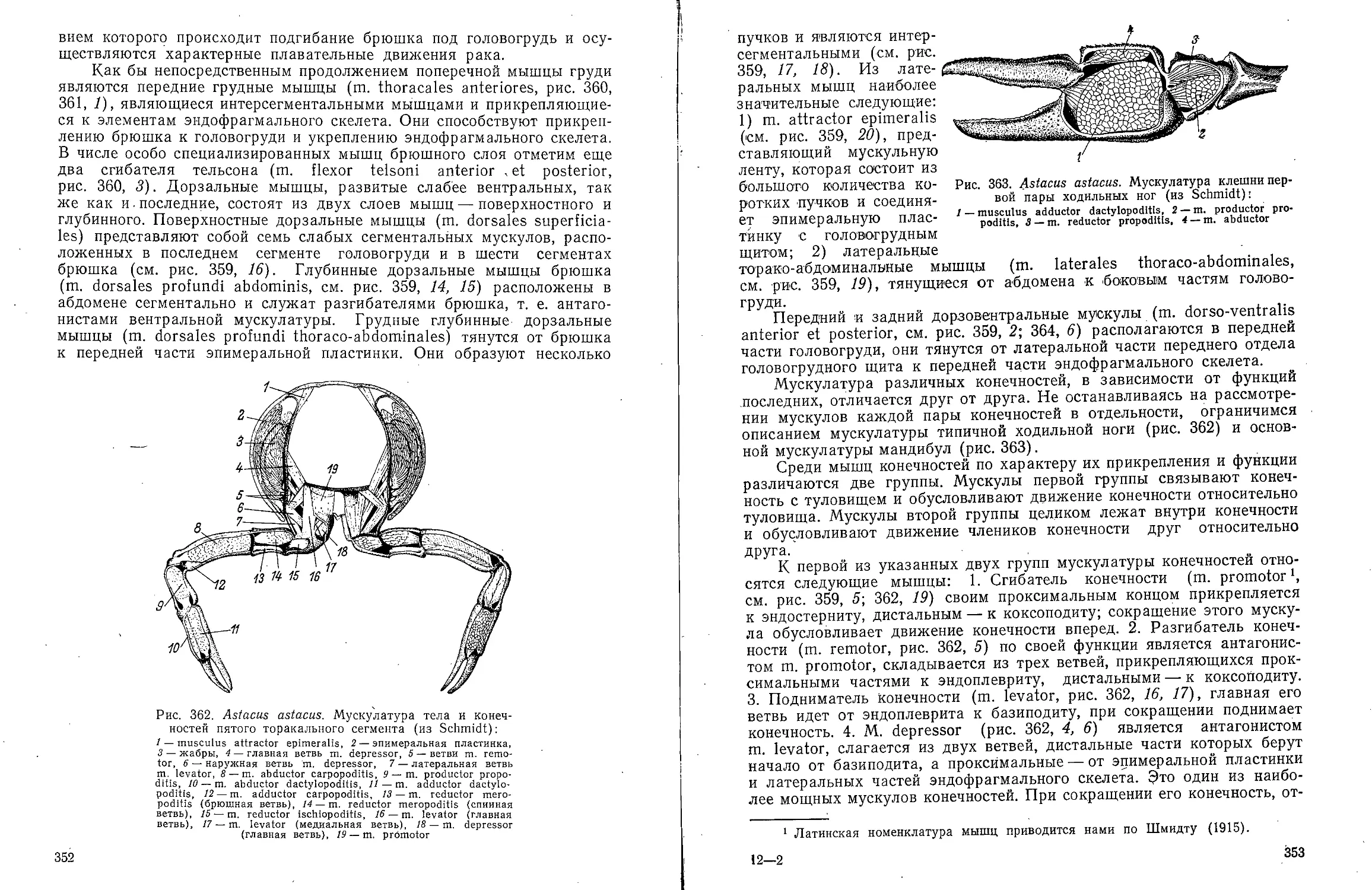
части тела с непарным брюшным
(вентральным) мешком; сзади и дор-
зально боковые мешки соединяются
друг с другом (рис. 7). Такая
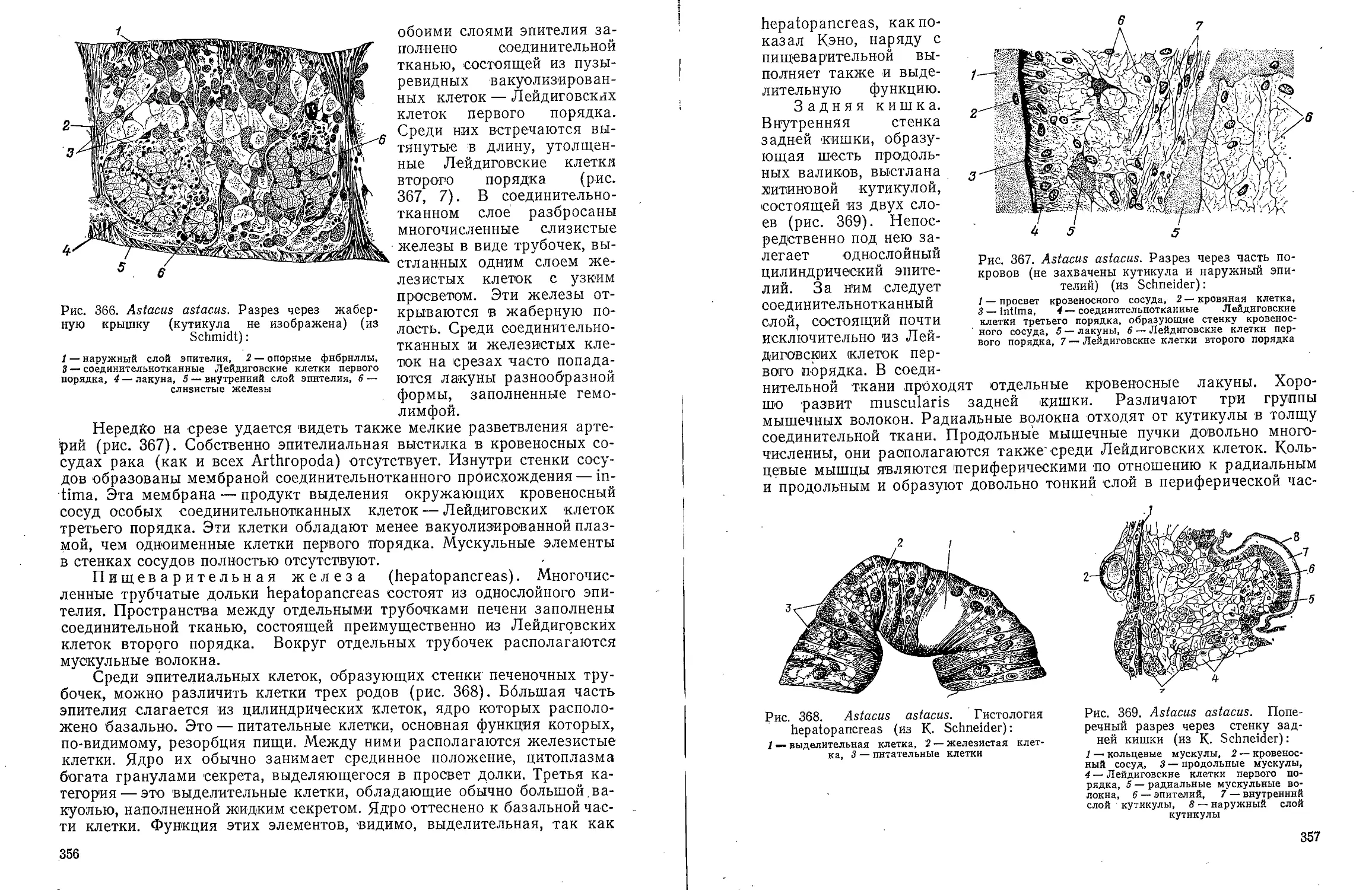
конфигурация целомической полости
объясняется, по-видимому тем,
что она развивается в промежутках
между внутренними органами,
вытесняя соединительную ткань
паренхимы. Стенка целома тонка и
имеет вид мембраны, которая на
первый взгляд представляется
неклеточной. На срезах, однако, на
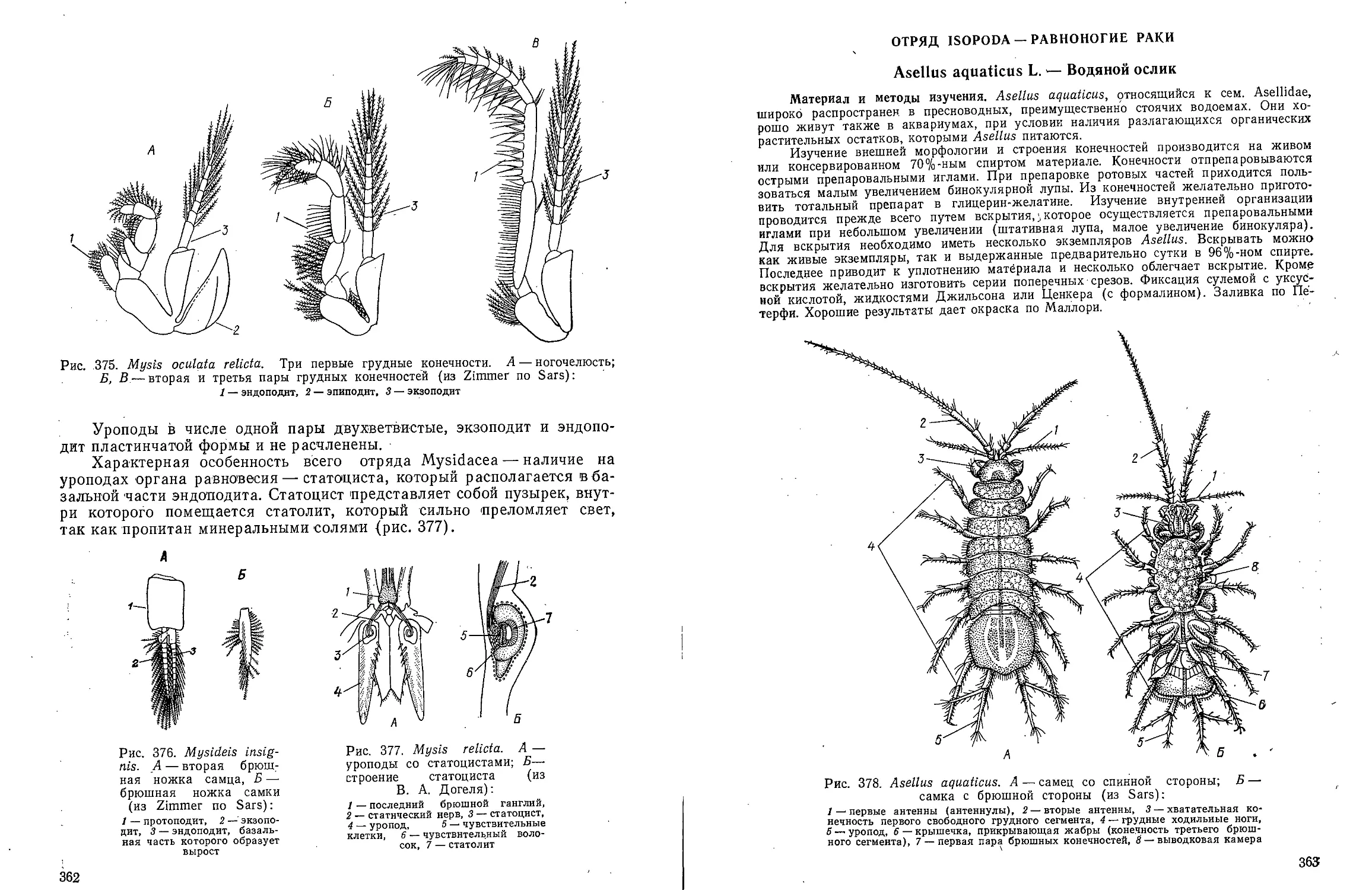
ней видны редкие линзообразно
уплощенные клетки с ядрами (Иегер-
стен, 1944).
Пищеварительная
система. Т-образное ротовое
отверстие, расположенное вентрально
на переднем крае первого
сегмента, окружено подвижными
кожными складками (см. рис. 2, 17),
причем обычно ясно выделяется
подковообразный валик,
ограничивающий рот с боков и сзади. Позади
1 В тканях различных органов
Dinophilus могут встретиться молодые и
взрослые стадии грегарины Clepsidrina dinophili.
12 В
Рис. 5. Dinophilus vorticoides. A —
фронтальный разрез передней . части
тела самки на уровне поперечных
мускулов головного отдела; Б~ то
же на уровне прикрепления
брюшной продольной мускулатуры в
области глаз; В — сагиттальный разрез
передней части тела самки (по Шим-
кевичу):
/ — теменные железы, 2 — бороздка,
отграничивающая головной отдел, 3 —
поперечные мускулы головного отдела,
4—пищевод, 5 — волокнистая часть головного
мозга, 6 — слой ганглиозных клеток головного
мозга, 7 — место прикрепления брюшных
продольных мышц, 8 — глаз, 9 —
перекрещивающиеся брюшные продольные
мышцы, 10 — брюшные продольные мышцы,
// — глотка, 12 — ротовое отверстие, 13 —
головной мозг, 14 — передняя губная
железа
рта лежит серповидное
поперечное отверстие
посторального органа — глоточного
кармана. Это довольно обширное
впячивание покровов, в
котором находится своеобразный
плотный язычок (см. рис. 5,
//). Стенки ротовой полости
способны сильно
выпячиваться, вследствие чего рот
оказывается в центре
подушкообразного возвышения покровов.
У фиксированных животных,
напротив, вся околоротовая
область втянута внутрь,
поэтому создается впечатление, что
глоточный карман
открывается непосредственно в ротовую
полость (см. рис. 5, В). Как
видно на поперечных срезах,
ротовая полость окружена
мускульными волокнами,
образующими ротовой сфинктер.
От стенок тела к пищеводу
проходят мускульные пучки.
Глоточный карман снабжен
ретракторами и парой
протракторов. Подвижный язычок
состоит из плотно
упакованных уплощенных мускульных
клеток, каждая из которых
содержит две системы
пересекающихся миофибрилл.
Пищевод (передняя
кишка) изогнут коленообразно
(см. рис. 7); его ресничный
эпителий богат
бокаловидными железистыми клетками.
С пищеводом связаны хорошо
заметные на срезах,
небольшие мешковидные придатки
неизвестного назначения,
находящиеся на боковых стенках
в задней его части, и группы
слюнных железистых клеток,
изливающие секрет через длинные протоки в просвет пищевода; они
расположены по бокам вдоль пищевода (см. рис. 2, А, 6).
Средняя кишка занимает значительную часть тела (рис. 6,2; 12, Л,
3). Ее эпителий, богатый желтыми и оранжевыми включениями, несет
Рис. 6. Dinophilus vorticoides. Схема
поперечного разреза в средней части тела (по Иегер-
стену из Бошана):
/ — семенник, 2 — кишка, 3 — продольные мускульные
волокна, 4 — боковой мешок целома, 5 — протонеф-
ридий, 6 — брюшной мешок, 7 — брюшная
мерцательная полоска, 8— добавочные нервные стволы, 9 —
брюшной нервный ствол, 10 — продольные брюшные
мускулы
Рис.,7. Dinophilus vorticoides. Схема
расположения целомических мешков самки. А —
медианный разрез; Б — парамедианный разрез (по
Иегерстену):
/ — брюшной мешок целома, 2— боковой мешок, 3—
соединение брюшного мешка с боковыми, 4 —
спинное соединение боковых мешков
10
Рис. 8. Dinophilus vorticoides. Расположение ресничных колец,
органов чувств и протонефридиев самки (по Иегерстену):
/ — апикальные волоски, 2—глаз, 3 — теменные волоски, 4 — нухальный орган,
5 — протонефридин, 6 — анус, 7 — отверстия нефридиев, 8 — рот
редкие реснички и содержит железистые клетки. Задняя кишка гораздо
меньшего размера (рис. 2, А, 10) образована ресничным эпителием.
Анальное отверстие лежит на спинной стороне у основания «хвостика»
(рис. 2,4,//).
Выделительная система. Органы выделения представлены
протонефридиями, причем у самца Dinophilus vorticoides их 4 пары
(см. рис. 3,5, 9), ау самки —6 пар (рис. 8, 5). Каждый нефридий
состоит из трех частей: тонкостенного лопастного терминального
пузырька, вдающегося в целом, промежуточного отдела и выводного канала
(рис. 9). В пузырьке находится пучок жгутов, в узком просвете
канала видны реснички. Полость терминального пузырька, по-видимому,
сообщается с выводным каналом лишь через неправильные вакуоли
промежуточного отдела (Иегерстен, 1944). Данные о наличии у
Dinophilus соленоцитов не подтвердились. Наружные
поры протонефридиев расположены латеро-вент-
рально (рис. 8, 7).
Нервная система. Центральная
нервная система изучается на срезах (желательно
изготовление графической реконструкции, для
чего наиболее удобны фронтальные срезы —
см. рис. 5, А). Демонстративны также
тотальные препараты, изготовленные по методу Же-
ребцова в модификации Котиковой (
гистохимическая реакция на холинэстеразы)1. На них
видны многие детали строения нервной системы,
ускользающие при изучении одних срезов.
Головной мозг лежит в простомиуме. От него отходят
окологлоточные коннективы, огибающие
пищевод и соединяющиеся на брюшной стороне тела
с вентральными нервными стволами, имеющими
вид волнообразно изогнутых тяжей (рис. 10). Рис g Протонефридии
Задние концы вентральных стволов соединены Dinophilus vorticoides
(по Иегерстену):
1 — терминальный отдел,
2 — промежуточный отдел,
1 См. СПИСОК литературы. 3— канал, 4 — жгуты
11
/4 / 75
Рис. 10. Dinophilus vorti-
coides. Нервная система
на препарате после
обработки на холинэстера-
зы. Вид с брюшной
стороны (А) и спереди (Б)
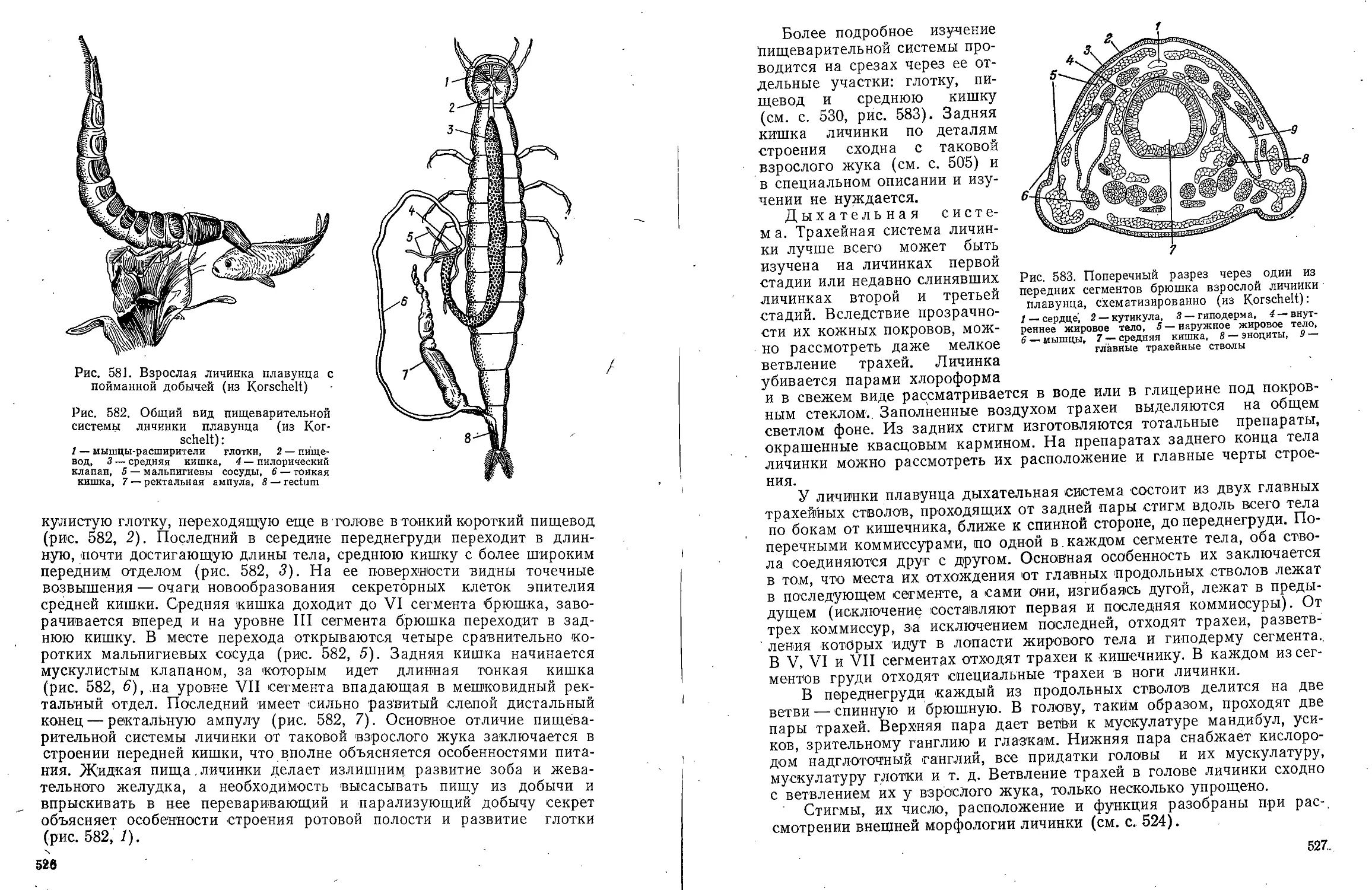
(по Е. А. Котиковой):
/ — глоточный ганглий, 2 —
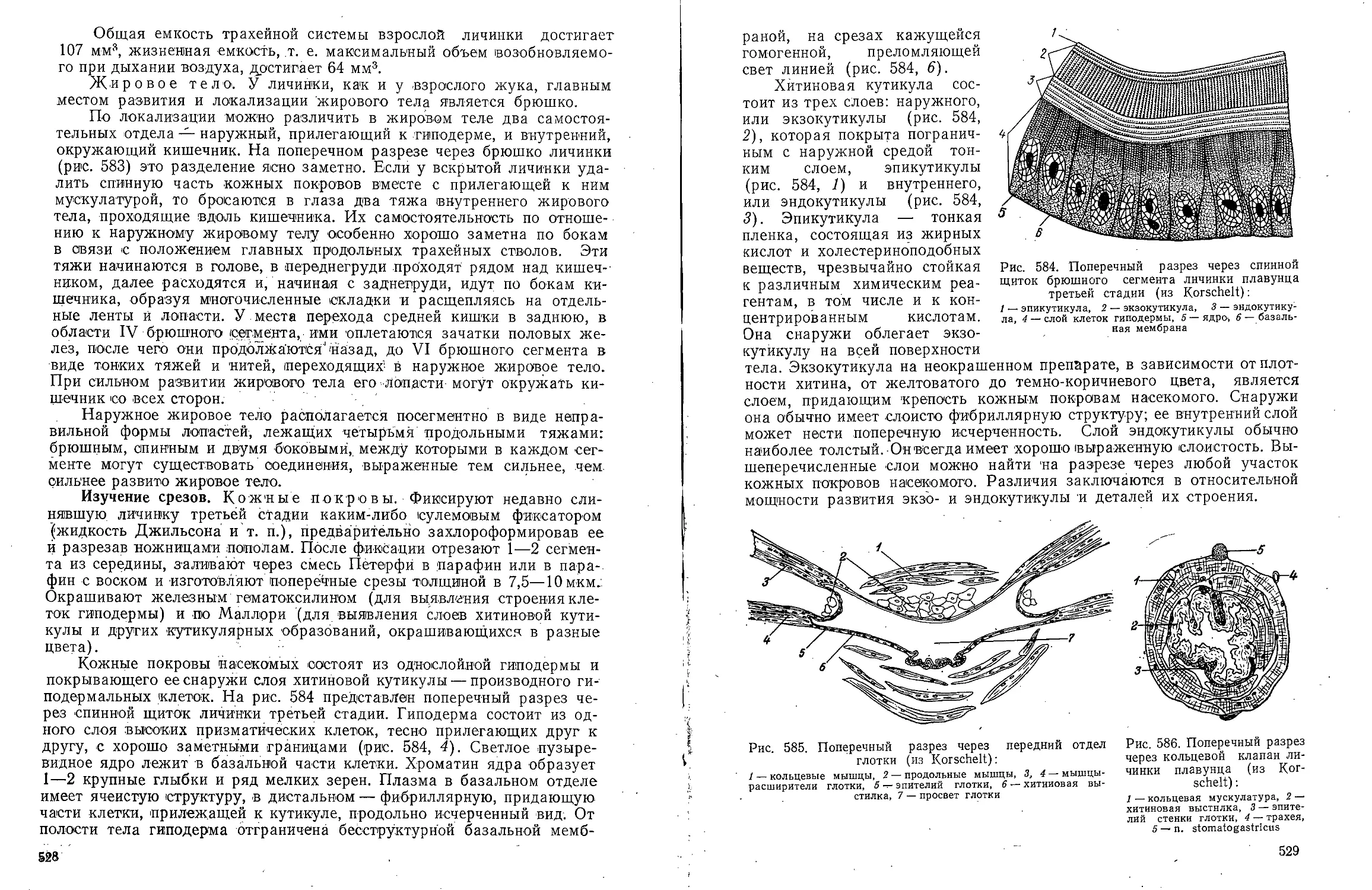
окологлоточный коннектнв,
3 — боковой нерв, 4 — глаз,
5 — мозг, 6 — глоточный
коннектив, 7 — брюшной
нервный ствол, 8 —
дополнительные нервные стволы,
9 — медиальный брюшной
ствол, 10 — спинной нерв,
// — ганглии, 12 — коммис-
сура, 13 — нервное кольцо,
14 — чувствительная нервная
сеть, 15 — глоточная ком-
миссура
нервным кольцом. Между стволами
расположены три очень тонких дополнительных
продольных ствола (рис. 10, 8). Имеется 6 пар
брюшных ганглиев, из них 4 передние пары с
поперечной коммиссурой. Между ганглиями
расположены парные ганглиозные расширения, от
которых отходит по нерву, загибающемуся на
спинную сторону.
Следует обратить внимание на пару
небольших ганглиев, лежащих в начальных
отделах окологлоточных коннектив. Это глоточные
ганглии (рис. 10, /), связанные коммиссурой; от
них начинаются довольно мощные глоточные
коннективы, идущие по стенке глоточного
кармана на спинную сторону. Вероятно это
элементы стоматогастрической нервной системы.
Мозг — цельный, нерасчлененный орган с
боковыми лопастями. На его передней и
боковых сторонах расположены нервные клетки,
центральная часть образована
преимущественно нервными волокнами (см. рис. 5, А, 6). От
мозга отходят боковые нервы, с ним
контактируют глаза (рис. 10, 4).
Органы чувств представлены глазами,
осязательными волосками и обонятельными
ямками — нухальными органами. Глаза лежат на
спинной стороне простомиума (см. рис. 2, А, 2);
по строению они напоминают глаза турбелля-
рий. Чувствительные волоски — длинные пучки
ресничек, правильно расположены главным
образом на головном отделе, спинной стороне,
боковых поверхностях тела и «хвостике». Самые
крупные осязательные волоски (апикальные)
находятся на переднем конце тела, дорзально
на задней границе головного отдела
(затылочные) и спинной поверхности хвостового отдела
(см. рис. 8, 1, 3). Нухальные органы хорошо
видны у особей обоих полов, после
выдерживания в растворе метиленового синего. Они в виде
небольших овальных мерцательных ямок
расположены дорзо-латерально непосредственно
впереди третьего ресничного пояска (см. рис. 2, А,
14; 8,4).
Половая система. У самцов есть пара
колбасовидных семенников, лежащих под
средней кишкой по бокам от брюшного целомическо-
го мешка. У зрелых особей они соединяются
друг с другом широкими перемычками спереди
и сзади (рис. 11, 2—3). Сзади семенники обра-
12
зуют боковые соединения (протоки) с
овальными семенными пузырьками,
лежащими в VI сегменте (рис. 11,5). В
семенниках и перемычках между ними у зрелых
самцов in vivo видны густые пучки
спермы, а в семенных пузырьках — «кишащий
клубок» сперматозоидов. Стенка семенных
пузырьков состоит из плоского эпителия.
От пузырьков отходят ресничные каналы,
открывающиеся в мужское половое
преддверие, в котором лежит конусовидный
совокупительный орган с внутренним
каналом, напоминающий выворачивающийся
пенис ресничных червей (рис. 11,8).
Преддверие открывается наружу брюшным
половым отверстием (рис. 11, 9). В
проксимальную часть пениса открываются
протоки зернистых железистых клеток, а на его
дистальной вершине — протоки
одноклеточных желез, выделяющих оформленный
игловидный секрет (рис. 12,11). При
спаривании пенис может значительно
выпячиваться наружу, благодаря сокращениям
специальных мускульных волокон.
Женская половая система
слагается из двух пар овальных яичников,
лежащих вентрально по бокам брюшного
целомического мешка (рис. 12, 5).
У зрелой самки в целомическом
мешке находится сперма самца, а позднее
происходит созревание ооцитов. Выпячивания
брюшного целомического мешка доходят
между яичниками почти до боковых стенок
тела (рис. 12, /). Яичники имеют хорошо
Рис. 11. Dinophilus vorticoi-
des. Мужская половая
система (ориг. рис. Ю. В. Мам-
каева):
/ — семенник, 2, 3 — переднее н
заднее соединения семенников,
4 — проток, ведущий в
семейной пузырек, 5 — семейной
пузырек, 5 — ресничный канал,
7 — мужское преддверие, 8 —
пеиис, 9 — половое отверстие,
10 — канал пениса, // — железы
с игловидным секретом, 12 —
зернистые железы
заметную клеточную стенку. Они
полностью обособлены от целома, в который ооциты попадают через
разрыв стенки яичников (Иегерстен, 1944). Таким образом, у Dinophili-
dae, как и у других аннелид, одной из важных функций целома
является половая функция. ,
Polygordius ponticus Salensky
Материалы и методы изучения. В окрестностях Севастополя и в других местах
по побережью Черного моря в изобилии встречается Polygordius ponticus Salensky
(сем. Polygordiidae), морфологически почти не отличимый от средиземноморского
P. neapolitanus Fraipont. Наиболее существенное отличие между этими видами
сводится к неодинаковому строению личинки трохофоры и различию в метаморфозе. Для
P. neapolitanus характерна трохофора типа exolarva, тогда как P. ponticus обладает
трохофорой типа endolarva. P. ponticus ведет роющий образ жизни в песчаных
грунтах, встречаясь на глубине от 10 до 20 м, откуда добывается драгой. Обычно
Polygordius встречается вместе с ланцетником (Amphioxus). Помимо Polygordius ponticus
13
Рис. 12. Dlnophilus vorticoides. Поперечные разрезы самки в средней части
туловища (А) и самца в области совокупительного органа (Б) (по Р. В.
Селивановой) :
/ — боковой мешок целома, 2 —эпидермис, 3 —средняя кишка, 4 — мезенхима, 5 —
яичник, 6 — брюшные продольные мускулы, 7 — брюшной нервный ствол, 8 — брюшной
продольный мускул, 9 — брюшной мешок целома, 10 — задняя кишка, 11 — семяпровод,
12 — железы с игловидным секретом, 13 — пенис, 14 — половое отверстие, 15 — мужское
преддверие, 16 — брюшиая ресничная полоска, 17 — канал пениса, 18 зернистые Железы
в морях СССР имеются представители и других семейств Archiannelida. В Черном
море обычен Protodrilus flavocapitatus Uljanin (сем. Protodrilidae) и Saccocirrus papillo-
cercus Bobrezky (сем. Saccocirridae). В Японском море (залив Петра Великого)
встречаются Polygordlus pacificus Uschida, Saccocirrus gabriellae Marcus и Protodrilus sp.
Если невозможно достать материал по Polygordlus, для ознакомления с
Archiannelida можно воспользоваться Protodrilus или Saccocirrus. В таком случае надо иметь
в виду, что эти формы во многом отличаются от Polygordlus.
Внешнюю морфологию Polygordlus предпочтительнее изучать на живых червях.
Для этой же цели полезно изготовить также несколько тотальных препаратов,
различно ориентировав на них объекты. Перед заключением в канадский бальзам
животные окрашиваются квасцовым или борным кармином. Фиксация живого материала
для изготовления .срезов производится' подогретой сулемой с уксусной кислотой,
жидкостью Джильсона, Ценкера и другими фиксаторами. Наилучшие результаты, однако,
дает способ фиксации, предложенный Вольтереком (концентрированный раствор
сулемы—1 ч., спирт 80%-ный—1 ч., ледяная уксусная кислота — 0,2 ч.). Для того чтобы
животные зафиксировались в вытянутом состоянии, предварительно в морскую воду
с живыми червями капля за каплей добавляется 50%-ный спирт. Постепенно
концентрация спирта в воде достигает такой степени, что черви остаются совершенно
неподвижными и вытянутыми. В таком состоянии их перекладывают в фиксатор. Заливку
следует производить в парафин. Срезы окрашиваются гематоксилином. Хорошие
результаты дает окраска слабым раствором кислого гемалауна Майера с последующей
окраской слабым водным раствором эозина.
Внешняя морфология. Сильно вытянутое, цилиндрическое тело
состоит, из многочисленных одинаковых сегментов (гомономная
сегментация), лишенных каких бы то ни было придатков (рис. 13). Все сегменты
U
Polygordius постларвальные; лар-
вальные сегменты отсутствуют1.
Брюшная сторона тела несколько
уплощена. Сегменты тела почти
одинаковой длины и ширины;
границы между ними отчетливы и
имеют вид кольцевых бороздок,
особенно хорошо заметных на
консервированных животных. Передний
конец червя уже, чем остальное
тело, и представляет собой
небольшую головную лопасть, или просто-
миум (prostomium, рис. 13, 1),
ясно отграниченный с брюшной
стороны поперечной бороздкой от
туловища. Простомиум снабжен
парой пальп и парой обонятельных
ямок. Глаза отсутствуют.
Цилиндрические пальпы направлены
вперед и прикрепляются к спинной
стороне переднего края головной
лопасти. Длина их едва превышает
длину простомиума (рис. 13, 5). На
поверхности пальпов хорошо
заметны редкие осязательные волоски.
Обонятельные ямки имеют
овальные очертания и расположены по
боками простомиума, у его задней
границы (рис. 13, 3).
Следующий за простомиумом
I сегмент, также входящий в
состав головного отдела, называется
ротовым сегментом, или метасто-
миумом (metastomium). Спереди
он ясно отграничен бороздкой, но
сзади лишен ясной границы (рис.
13, Б). На брюшной его стороне
впереди располагается широкое
поперечное ротовое отверстие.
Анальная лопасть, или пигидий (pygidi-
um), отличается вздутой формой и обычно резко отделена от впереди
лежащих сегментов кольцевой бороздой (рис. 13, 2). В средней
вздутой своей части она несет кольцевой ряд маленьких сосочков, на
которых открываются хвостовые же_лезы (рис. 13, 6). Анальное отверстие
расположено терминально (рис. 13, 7).
Рис. 13. Polygordlus neapoletanus.
А — внешний вид животного; Б —
передний конец тела с правой стороны;
В — задний конец тела с левой
стороны (по Фрэпону):
1 — простомиум, 2 — пигидий, 3 —
обонятельная ямка, 4 — место расположения
ротового отверстия, 5 т- пальп, 5 — анальные
папиллы с хвостовыми железами, 7 —
анальное отверстие, 8 — задние сегменты
туловища
1 О ларвальных и постларвальных сегментах см.: Иванов П. П. Общая и
сравнительная эмбриология. М., 1937, с. 199—202.
15
Общее число сегментов нередко превышает сотню; количество их
варьирует в широких пределах. Длина червя измеряется несколькими
сантиметрами(2,5—5 см). Самки крупнее самцов. Основная желтовато-
розоватая окраска живых червей несколько изменяется в период
половой зрелости, становясь у самок более яркой, а у самцов более бледной,
от цвета просвечивающих сквозь стенки тела половых продуктов.
Анатомия. Детальное ознакомление с внутренней организацией
Polygordius возможно только на сериях срезов. В дополнение к этому
. основному методу, следует внимательно изучить тотальные препараты,
а также исследовать несколько живых экземпляров под микроскопом
или бинокуляром. При этом живых червей следует сдавить между
двумя предметными стеклами.
Необходимо приготовить следующие серии срезов: поперечную
головного отдела, включая 2— 3 первых сегмента тела; поперечную
2 —3 сегментов из средней области тела; поперечную нескольких
задних сегментов и пигидия; фронтальные — головного, среднего и
заднего отделов тела и сагиттальную — головного отдела.
Изучение анатомии и гистологии производится путем
детального просмотра серий срез за срезом, сопоставления их и сравнения
поперечных серий с продольными, а также путем сравнения тоталь^
ных препаратов со срезами. При разборе серий срезов полезно делать
глазомерные графические реконструкции.
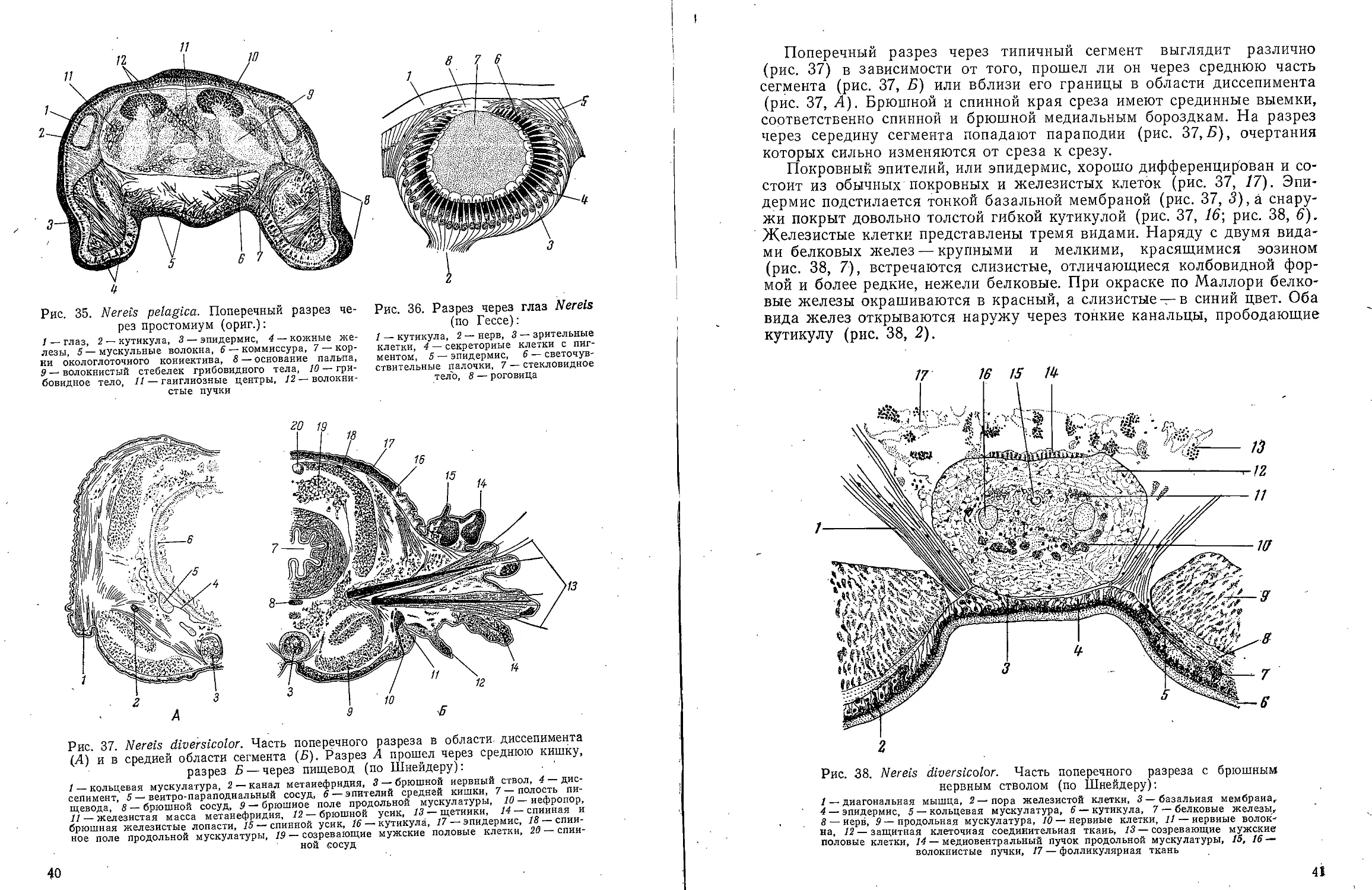
К о ж н о-м ускульный мешок. Строение кожно-мускульного
мешка хорошо видно на поперечных разрезах через среднюю область
тела (рис. 14). Он слагается из кожного эпителия и слоя продольной
мускулатуры. Брюшной нервный ствол тесно связан с кожей.
Однослойный кожный эпителий (эпидермис) снаружи покрыт тонкой
кутикулой, лишен ресничного покрова и подстилается отчетливой базаль-
ной мембраной. Следует обратить внимание на отсутствие невротро-
хоида, т. е. медиальной брюшной ресничной бороздки, характерной
для других представителей Archiannelida и личинок аннелид.
Эпидермис слагается из различных клетбчных элементов, отличающихся
строением ядер. В основном, однако, это более или менее уплощенные
цилиндрические клетки без ясных границ (рис. 14, /).
Немногочисленные кожные железы представлены двумя сортами клеток. Одни
из них представляют слизистые железы, разбросанные по всей
поверхности тела, другие, хвостовые, железы сосредоточены на
вздутой части пигидия. Именно каждый сосочек кольцевого вздутия имеет
свою группу хвостовых желез, выводные канальцы которых
открываются на его вершине (рис. 15, 3). Хвостовые железы выделяют
клейкий секрет, служащий для временного прикрепления червя к
субстрату.
Продольная мускулатура кожно-мускульного мешка образует
толстый слой, непрерывно продолжающийся из сегмента в сегмент
(см. рис. 14, 2; рис. 15, 8). Этот слой распадается на два спинных
и два брюшных пучка. Он разделен сверху и снизу мезентериями, а
по бокам — в местах прикрепления диагональных мышц. В головной
лопасти четыре мускульных пучка широко расставлены друг от друга.
Они кончаются у основания пальпов, не продолжаясь в них.
16
12 П
Рис. 14. Polygordius ponticus. A — поперечный
разрез в средней области туловища; Б — часть
поперечного разреза с брюшным нервным
стволом (по В. В. Заленскому):
/ — эпидермис, 2 — продольная мускулатура, 3 —
канал метанефридия, 4 — брюшной кровеносный сосуд,
5 — диагональные мышцы, 6 — брюшной нервный
ствол, 7 — гонада, 8 — ядро мышечной клетки, 9 —
перитониум, 10 — спинной кровеносный сосуд, 11 —
спннной мезентерий, 12 — средняя кишка, 13 —
медиальный пучок нервных волокон, 14 — опорные
клетки, 15 — латеральный пучок нервных волокон, 16 —
кишечная камера целома, П — нервные клетки, 18 —
нефридиальная камера целома
Рис. 15. Polygordius ponticus.
Фронтальный разрез через
задний конец тела (по В. В.
Заленскому) :
/ — средняя кишка, 2 — диссепи-
меиты, 3 — хвостовые железы, 4 —
пигидий, 5 — анальное отверстие,
6 — кольцевые мышцы, 7 — задняя
кишка, 8 — продольные мышцы,
9 — диагональные мышцы, 10 —
гонада
Кольцевая мускулатура кожно-мускульного мешка у Polygordius
отсутствует. Лишь в пигидий внутрь от узкого продольного слоя
имеется слабый слой кольцевых мускульных волокон (рис. 15, 6).
Положение этого слоя не позволяет гомологизировать его с
кольцевым слоем других аннелид. Мускульные клетки (миоЦиты)
продольного слоя своеобразного строения. Они представляют собой
заостренные на концах лентовидные элементы, сильно вытянутые в
продольном направлении. Плоские стороны соседних миоцитов прилегают
друг к другу. На поперечных срезах видно, что их наружные части
слегка ряейшреш»ц-^__вщЩ1енние куполообразно закруглены и вдаются
в поло£й^т^лжл<В«р^Ер£нд^| отделы состоят из тонкозернистой плаз-
к J-
БИБЯ-.ОТЕК
им. Ф. м. Достсйюк,г0
UU-Г АП. > м .-. _ .
agp/pps
17
мы, содержащей одно ядро (см. рис. 14, 8). Наружные — содержат
миофибриллы в форме пластинок, которые располагаются
перпендикулярно к поверхности животного и параллельно друг другу, подобно
листам книги. В каждом миоците залегает 10—12 таких пластинок
(см. рис. 14, 2). Строение миоцитов видно не на каждом срезе.
Сплошь и рядом внутренние отделы мускульных клеток склеиваются
вместе, сливаясь как бы в сплошной плазматический слой с ядрами
или образуя плазматическую сеть, сливающуюся с клетками перито-
неального эпителия. На поперечных срезах хорошо заметны косые
мускульные пучки, пересекающие полость тела. Это диагональные
мышцы (musculi transversi obliqui), не относящиеся к кожно-мускуль-
нбму мешку. Их брюшные концы прикрепляются в области брюшного
нервного ствола, а противоположные — к боковым стенкам тела (см.
рис. 14, 5; рис. 15, 9). Сравнивая ряд поперечных срезов одной
серии, нетрудно обнаружить их метамерное расположение. Наличие
диагональных мышц характерно для аннелид. В головном отделе
диагональные мышцы дифференцированы в пучки специальною значения.
Вся мускулатура Polygordius слагается из гладких мышечных волокон.
Полость тела. Хорошо развитая между стенками тела,
кишечником и другими органами полость представляет собой целом (см.
рис. 14). Соответственно наружной сегментации целом делится
многочисленными поперечными перегородками, диссепиментами (рис.
Рис. 16. Polygordius ponticus. Передний конец тела,
нарисованный с тотального препарата. Вид справа (А); с брюшной
(Б) и со спинной (В) сторон (по В. В. Заленскому):
/ — пальпы, 2 — нерв пальпа, 3 — пальпарный ганглий, 4 —
обонятельная ямка, 5 — пищевод, 6 — глоточная коммиссура, 7 — ротовое
отверстие, 8 — окологлоточный кониектив, 9 — верхняя губа, 10 — головной
карман, Я — церебральные ганглии, 12 — борозда на задней границе
головной лопасти, 13 — глотка, 14 — задний глоточный карман, 15 —
передний глоточный карман, 16 — обонятельный ганглий
18
15, 2) на ряд камер, или сомитов. Все сомиты разделены также на
одинаковые правые и левые половины продольной перегородкой,
проходящей в медианной плоскости — спинным и брюшным
мезентериями (см. рис. 14, //). Пересекающие целом диагональные мышцы
делят каждый сомит на центральную кишечную камеру, в которой
лежит кишка (см. рис. 14, 16), и две боковых нефридиальных камеры
(см. рис. 14, 18). В простомиуме целом отсутствует. У основания
пальпов и впереди церебральных ганглиев имеются узкие щелевидные
пространства первичной полости тела.
Стенки полости тела выстланы перитонеальным (целомическим)
эпителием, который одевает также все органы, лежащие в целоме
(кишечник, метанефридии, кровеносные сосуды). В клетках
париетального листка перитонеума нередко встречаются жировые капли
и скопления экскреторных зерен. Диссепименты и мезентерии состоят
из двух слоев перитонеального эпителия, между которыми встречаются
мускульные волокна, особенно сильно развитые в диссепиментах в
виде мускульного кольца, охватывающего кишечник.
Зона образования новых
сегментов расположена непосредственно
впереди пигидия. Здесь, как хорошо
видно на фронтальных срезах, всегда
имеется несколько молодых сегментов
и сегментов, находящихся в процессе
образования (рис. 15).
Пищеварительная систе-
м а. Кишечный канал представляет
собой прямую трубку, проходящую
через все тело; он слагается из
передней, средней и задней кишок.
Наиболее сложно устроена передняя кишка,
которая состоит из ротовой полости,
глотки и пищевода. На тотальных
хорошо просветленных препаратах
отчетливо видны главнейшие части
передней кишки. Ротовое отверстие
ограничено спереди пузыревидным
утолщением кожи — верхней губой
(рис. 16, 9). Передняя граница
последней образует глубокое впячивание
покровов — головной карман, которое
начинается впереди губы и имеет вид
длинного сильно уплощенного
кармана, простирающегося назад и в дор-
зальном направлении и кончающегося
слепо вблизи спинной стенки
головного отдела (рис. 16, 10; см. рис. 17, //).
Посредством впячивания покровов
верхняя губа отделена от головной
лопасти. Задняя граница рта образо-
Рис. 17. Polygordius ponticus.
Сагиттальный разрез передней части тела
(по В. В. Заленскому):
/ — нерв пальпа, 2 — пальпарный ганглий,
3 — церебральный ганглий, 4 —
обонятельный орган, 5 — глоточная мускулатура,
6 — эпидермис, 7 — продольная
мускулатура, 8 — нижняя губа, 9 — передний
глоточный карман, 10 — верхняя губа, 11 •—
• отверстие головного кармана
19
7 вана поперечным, изогнутым назад,
эктодермальным валиком — нижней
губой (см. рис. 17, 8), под кожей
которой даже на тотальных препаратах
заметен пучок поперечных
мускульных волокон (сжиматель нижней
губы) . Через широкое ротовое отверстие
видны стенки ротовой полости,
которая образует пару направленных
вперед выпячиваний, обычно хорошо
заметных вследствие того, что их
клеточные элементы интенсивно
окрашиваются. Это передние глоточные
карманы (рис. 16, 15). Стенки их в
передней части сильно утончаются, а
полости имеют характер каналов.
Позади нижней губы заметен
лежащий на брюшной стороне широкий
полуовальный толстостенный мешок —
глотка, просвечивающая сквозь
покровы (рис. 16, £, 13). При известных
положениях объекта видны боковые
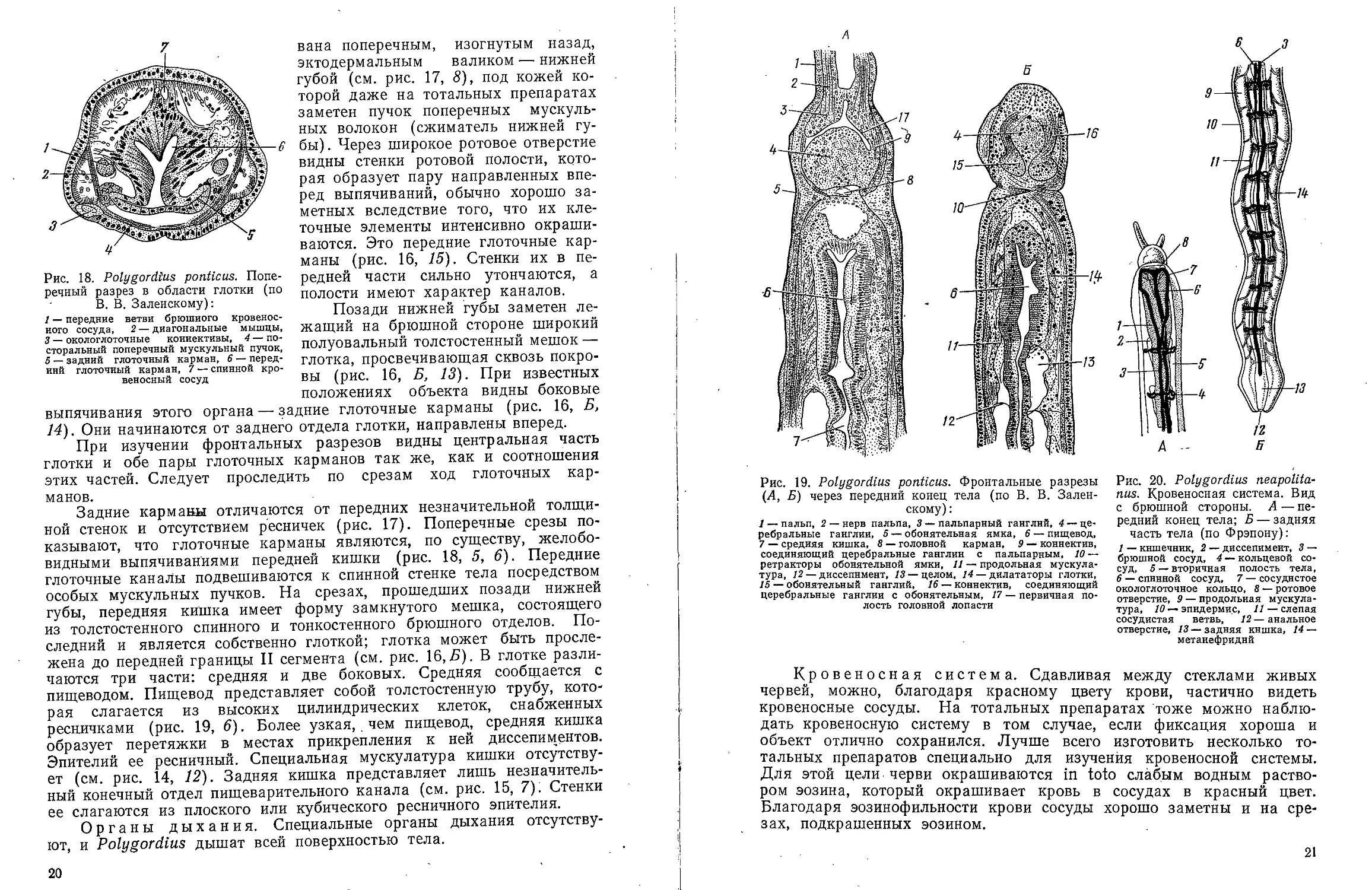
выпячивания этого органа — задние глоточные карманы (рис. 16, Б,
14). Они начинаются от заднего отдела глотки, направлены вперед.
При изучении фронтальных разрезов видны центральная часть
глотки и обе пары глоточных карманов так же, как и соотношения
этих частей. Следует проследить по срезам ход глоточных
карманов.
Задние карманы отличаются от передних незначительной
толщиной стенок и отсутствием ресничек (рис. 17). Поперечные срезы
показывают, что глоточные карманы являются, по существу, желобо-
видными выпячиваниями передней кишки (рис. 18, 5, 6). Передние
глоточные каналы подвешиваются к спинной стенке тела посредством
особых мускульных пучков. На срезах, прошедших позади нижней
губы, передняя кишка имеет форму замкнутого мешка, состоящего
из толстостенного спинного и тонкостенного брюшного отделов.
Последний и является собственно глоткой; глотка может быть
прослежена до передней границы II сегмента (см. рис. 16,5). В глотке
различаются три части: средняя и две боковых. Средняя сообщается с
пищеводом. Пищевод представляет собой толстостенную трубу,
которая слагается из высоких цилиндрических клеток, снабженных
ресничками (рис. 19, 6). Более узкая,. чем пищевод, средняя кишка
образует перетяжки в местах прикрепления к ней диссепиментов.
Эпителий ее ресничный. Специальная мускулатура кишки
отсутствует (см. рис. 14, 12). Задняя кишка представляет лишь
незначительный конечный отдел пищеварительного канала (см. рис. 15, 7). Стенки
ее слагаются из плоского или кубического ресничного эпителия.
Органы дыхания. Специальные органы дыхания
отсутствуют, и Polygordius дышат всей поверхностью тела.
20 •
Рис. 18. Polygordius ponticus.
Поперечный разрез в области глотки (по
В. В. Заленскому):
/ — передние ветви брюшного
кровеносного сосуда, 2 — диагональные мышцы,
3 — окологлоточные коннективы, 4 —
посторальный поперечный мускульный пучок,
5 — задний глоточный карман, 6 —
передний глоточный карман, 7 — спинной
кровеносный сосуд
Рис. 19. Polygordius ponticus. Фронтальные разрезы
(А, Б) через передний конец тела (по В. В.
Заленскому) :
/ — пальп, 2 — нерв пальпа, 3 — пальпарный ганглнй, 4 —
церебральные ганглии, 5 — обонятельная ямка, 6 — пищевод,
7 — средняя кишка, 8 — головной карман, 9 — коннектив,
соединяющий церебральные ганглин с пальпарным, 10 —
ретракторы обонятельной ямки, // — продольная
мускулатура, 12 — диссепнмент, 13 — целом, 14 — дилататоры глотки,
15 — обонятельный ганглий, 16 — коннектив, соединяющий
церебральные ганглии с обонятельным, 17 — первичная
полость головной лопасти
Рис. 20. Polygordius neapolita-
nus. Кровеносная система. Вид
с брюшной стороны. А —
передний конец тела; Б — задняя
часть тела (по Фрэпону):
/ — кншечннк, 2 — диссепимент, 3 —
брюшной сосуд, 4 — кольцевой
сосуд, 5 — вторичная полость тела,
6 — спннной сосуд, 7 — сосудистое
окологлоточное кольцо, 8 — ротовое
отверстие, 9 — продольная
мускулатура, 10 — эпидермис, // — слепая
сосудистая ветвь, 12 — анальное
отверстие, 13—задняя кншка, 14 —
метанефриднй
Кровеносная система. Сдавливая между стеклами живых
червей, можно, благодаря красному цвету крови, частично видеть
кровеносные сосуды. На тотальных препаратах тоже можно
наблюдать кровеносную систему в том случае, если фиксация хороша и
объект отлично сохранился. Лучше всего изготовить несколько
тотальных препаратов специально для изучения кровеносной системы.
Для этой цели черви окрашиваются in toto слабым водным
раствором эозина, который окрашивает кровь в сосудах в красный цвет.
Благодаря эозинофильности крови сосуды хорошо заметны и на
срезах, подкрашенных эозином.
21
Главными кровеносными сосудами являются продольные
спинной (vas dorsale) и брюшной (vas ventrale) сосуды, которые
проходят вдоль всего тела над и под кишкой (рис. 20, 6 и 3). Оба
продольных сосуда связаны друг с другом интерсегментальными
кольцевыми сосудами (vasa commissuralia, рис. 20, 4). От последних назад
отходят короткие слепые сосудистые ветви, направляющиеся к
половым железам (рис. 20, 11). На переднем своем конце брюшной сосуд
раздваивается, ветви его окружают глотку с боков и в головном
отделе соединяются со спинным сосудом (рис. 20, 7). Продольные
сосуды залегают в мезентериях, причем более объемистый брюшной
занимает почти весь брюшной мезентерий (см. рис. 14, 4). Эти
сосуды лишены собственных стенок, представляя собой лишь пространства
между обоими листками мезентериев, т. е. являются, по существу,
кровеносными синусами. Только в головном отделе они отделяются
от перитонеума и принимают характер настоящих сосудов. На
фронтальных разрезах через брюшной сосуд обнаруживаются редкие
кольцевые мускульные волокна, залегающие под клетками перитоне-
ального эпителия, прилегающего к сосудам. Спинной сосуд помимо
кольцевых имеет еще наружный продольный слой мускульных
волокон. По спинному сосуду кровь движется от заднего конца к
переднему, по брюшному — в обратном направлении. В переднем отделе
туловища кровь поступает по кольцевым сосудам из спинного сосуда
в брюшной, а в заднем — из брюшного в спинной сосуд.
Выделительная система. Органы выделения имеют экто-
дермальное происхождение и гомологичны протонефридиям, от
которых, однако, отличаются отсутствием клеток с мерцательным
пламенем (соленоцитов) и наличием ресничной воронки. Таким образом,
это истинные метанефридии, отличные от мезодермальных целомо-
дуктов. Они могут быть найдены только на очень хороших срезах.
Гораздо больше дает изучение живых червей под микроскопом.
Именно in vivo метанефридии обнаруживаются довольно легко по
мерцанию ресничек внутри нефридиального канала. Каждый сегмент
тела имеет свою пару метанефридиев, которые занимают передние
2/з сомита, располагаясь в его боковых частях и прилегая к
внутренней поверхности стенок тела (рис. 20, 14). Каждый метанефридии
состоит из воронки-, или нефростома (рис. 21, 1), открывающегося в
полость предыдущего сегмента, и прямого нефридиального канала
(рис. 21, 2). Нефростом лежит на передней стороне диссепимента.
У живых червей воронки различимы благодаря мерцанию ресничек.
Наружные отверстия, или нефропоры метанефридиев, не различимы.
Они лежат попарно на боковых поверхностях каждого сегмента. На
поперечных срезах иногда видно строение нефридиального канала.
Стенки его слагаются из многих клеток без ясных границ.
Нервная-систем а. Центральная нервная система Polygordius
состоит из головного мозга, окологлоточных коннективов и брюшного
нервного ствола.
Несмотря на слабое развитие органов чувств, головной мозг
имеет сравнительно сложное строение. Замечательной чертой
является ясное разделение головного мозга на три пары ганглиев,
22
Рис. 21.
Метанефридии Polygordius
neapolitanus (no
Фрэпону):
/ — нефростом, 2 —
нефрнднальный
канал
которые обнаруживаются на тотальных препаратах.
Все они лежат в простомиуме. Передняя часть
головной лопасти занята парой крупных пальпарных
ганглиев (см. рис. 16, 3). Позади них лежит большая
масса церебральных ганглиев (см. рис. 16, И), а
сзади и дорзальнее церебральных располагаются
небольшие обонятельные ганглии (см. рис. 16, А, 16).
Пальпарные ганглии представляют собой
компактные грушевидные массы, разделенные щелевидным
участком первичной полости тела (см. рис. 17, 2;
рис. 19, А, 2). Передние их концы соединяются под
эпидермисом. От переднего края каждого ганглия
берет начало мощный нерв, пронизывающий пальп
(см. рис. 17, 1; рис. 19, А, 2). Сзади от пальпарных
ганглиев по направлению к церебральным отходят
короткие коннективы, лучше всего заметные на
фронтальных срезах (см. рис. 19, А, 9).
Церебральные ганглии, занимающие большую
часть простомиума, представляют ганглиозную
массу, нередко разделенную на две симметричные
половины (см. рис. 19, 4). Небольшие участки боковых
поверхностей, а также передней брюшной поверхности церебральных
ганглиев тесно связаны с эпидермисом, образуя места фиксации мозга.
От этих ганглиев отходит пара нервов, иннервирующая переднюю
часть кишечника, что хорошо видно на срезах. Обонятельные ганглии
берут начало от боковых частей переднего отдела церебральной массы.
Далее они загибаются дугообразно назад и примыкают своими
задними концами ко дну обонятельных ямок, которые они иннервируют
(см. рис. 19, Б, 15). От церебральных ганглиев они отделены
значительными участками полости тела.
Гистологическая дифференциация мозга невысока. Во всех
ганглиях униполярные и биполярные нервные клетки располагаются по
периферии, тогда как Центральная часть занята волокнистым
веществом. В пальпарных. ганглиях нервные клетки сосредоточены
преимущественно на задних концах, а в церебральных — на передних (см.
рис. 19, А, 3 и 4).
Окологлоточные коннективы тесно связаны с эпидермисом, от
которого, однако, их немногочисленные клетки резко отграничены.
Начинаясь от боковых поверхностей церебральных ганглиев, коннективы
направляются назад. Далее, загибаясь на брюшную сторону, они
охватывают с обеих сторон переднюю кишку (см. рис. 16, А, 8) и
позади глотки соединяются с брюшным нервным стволом. Последний
представляет собой продукт слияния двух брюшных нервных стволов.
Он погружен в стенку тела, будучи тесно связан с эпидермисом (см.
рис. 14, А, 6). Ганглиозные клетки располагаются на всем
протяжении ствола, так что брюшные сегментарные ганглии отсутствуют.
На поперечном срезе брюшной нервный ствол выглядит как медио-
вентральное утолщение эпидермиса (см. рис. 14, Б). Он слагается из
трех волокнистых пучков, из которых медиальный намного меньше бо-
23
ковых (см. рис. 14, Б, 13> 15). Клетки эпидермиса, прилегающие
к нервному стволу, дифференцированы в качестве опорных и
защитных элементов (см. рс. 14, Б, 14). Они снабжены длинными
отростками, которые внедряются между волокнистыми пучками и окружают
нервный ствол снаружи. Собственно нервные клетки прилегают к
волокнистым пучкам с брюшной стороны, образуя довольно резко
очерченные клеточные группы. В поперечном разрезе они имеют
полулунную форму (см. рис. 14, Б, 17).
Органы чувств представлены пальпами, несущими
осязательную функцию, и обонятельными ямками. Глаза й статоцисты
отсутствуют. Пальпы Polygordius не имеют внутреннего целомического
канала (см. рис. 13, Б, 5). Обонятельные ямки, называемые также ну-
хальными органами, окаймлены светлым кожным валиком (см. рис. 13,
Б, 3). Более темное дно ямок выстлано чувствительными ресничными
клетками, связанными с обонятельными ганглиями. Это лучше всего
видно на фронтальных срезах. Дно обонятельных ямок способно
несколько погружаться внутрь, благодаря действию особых мускулов-
ретракторов. (см. рис. 19, Б, 10).
Половая система. Все виды Polygordius раздельнополы.
Гонады различимы только в период половой зрелости. Половые клетки
развиваются в перитонеуме, . образуя на поверхности диагональных
мышц более или менее заметные компактные клеточные группы —
гонады (см, рис. 14, Л, 7 и рис. 15, 10). Последние развиваются во всех
сегментах, за исключением нескольких передних и задних молодых
сегментов. Созревающие и вполне созревшие половые элементы
плавают в полостной жидкости сомитов и к моменту полного созревания
совершенно его переполняют. Выход половых продуктов
осуществляется через разрыв стенки тела.
Трохофора Polygordius ponticus Salensky
Онтогенетическое развитие Polygordius сопровождается
метаморфозом, причем личинкой является типичная трохофора.
Личинка Polygordius ponticus принадлежит к довольно обычным
планктонным формам Черного моря. Так, в окрестностях Севастополя
трохофоры встречаются в планктоне массами в течение лета (июнь —
сентябрь), реже — зимой. Собирание личинок Polygordius производится
планктонными сетками с судна или шлюпки. Изучение организации
рекомендуется, если возможно, производить на живом материале. Для
этого трохофору помещают в капле воды под покровное стекло на
восковых ножках и рассматривают в микроскоп.
Для изготовления тотальных препаратов достаточна фиксация
4%-ным раствором формалина. Затем трохофоры отмываются от
формалина в воде и окрашиваются квасцовым кармином. Просветление
производится в гвоздичном масле или, лучше, во избежание
сморщивания объектов, посредством постепенного замещения абсолютного
спирта кедровым маслом. Покровное стекло должно быть с восковыми
ножками, иначе личинки будут раздавлены. Восковые ножки помогают
также правильно ориентировать объект. Если покровное стекло едва
24
касается объекта, то, сдвигая его
в ту или иную сторону и
перекатывая таким образом личинку,
нетрудно добиться желаемой
ориентировки.
На тотальных препаратах
организация трохофоры может
быть изучена только в общих
чертах. Для изучения нервной
системы, мускулатуры,
выделительных органов (протонефриди-
ев), а также образования
мезодермы и органогенеза
необходимо применять специальную
методику и проводить исследование
на срезах. Здесь мы ограничимся
описанием того, что обычно
видно на тотальных препаратах
(рис. 22).
Известно два типа трохо-
фор, принадлежащие к
различным видам Polygordius и
называемые exolarva и endolarva.
Различие сводится главным
образом к тому, что у exolarva
тело будущего червя развивается
в форме торчащего свободно,
постепенно вырастающего
придатка на нижней половине
трохофоры, тогда как у endolarva оно
развивается внутри тела
личинки. Личинкой типа exolarva
обладает, например,
средиземноморский P. neapolitanus.
Endolarva свойственна P. lacteus
Schneider из Северного моря.
Трохофора P. ponticus также
относится к последнему типу.
Трохофора Polygordius ponticus почти шаровидной формы.
Экваториальный ресничный поясок — прототрох, делит тело личинки на два
полушария — верхнее, или эписферу, и нижнее, или гипосферу
(рис. 22, А, 13 и 14). Прототрох проходит над ротовым отверстием,
являясь преоральным мерцательным пояском (рис. 22, А, 3). В центре эпи-
сферы на верхнем полюсе личинки расположено хорошо заметное
утолщение — теменная пластинка, центральный орган личиночной нервной
системы (рис. 22, А, 1). На ней обычно различима пара пигментных
пятен — глазков. Довольно широкое ротовое отверстие расположено
непосредственно под прототрохом на брюшной стороне тела (рис. 22, А,
9). Анальное отверстие лежит на нижнем полюсе личинки в центре ги-
Рис. 22. Трохофора Polygordius ponticus,
нарисованная с "тотального препарата. А и
Б — последовательные стадии (ориг.):
/ — теменная пластинка, 2 — нервные клетки,
3 — прототрох, 4 — впячнвание вокруг зачатка
туловища, 5 — зачаток туловища, 6 — мезобласт,
7 — задняя кишка, 8 — средняя кишка, 9 —
ротовое отверстие, 10 — передняя кишка, Я —
впячнвание, в котором лежит зачаток туловища, 12 —
мембрана, замыкающая сзади впячивание, 13 —
эпнсфера, 14 — гипосфера, 15 — полость тела
25
посферы. Эктодерма личинки слагается из плоского эпителия, но клетки
прототроха довольно высоки. В области порошицы эктодерма образует
циркуманальное утолщение, которое несколько выдается над
поверхностью тела. Это возвышение представляет собой первый заметный
зачаток будущего туловища червя (рис. 22, А, 5). Сквозь стенки тела
просвечивают внутренние органы. Ротовое отверстие ведет в короткую
переднюю кишку, стенки которой состоят из довольно высокого
мерцательного эпителия (рис. 22, 10). Затем следует сравнительно объемистая
и тонкостенная средняя кишка (рис. 22, А, 8). Кишечник заканчивается
задней кишкой (рис. 22, А, 7). Передняя и задняя кишки имеют
эктодермальное происхождение.
Кишечник лежит в объемистой первичной полости тела (рис. 22, Б,
15). Далее, под клетками прототроха заметен ряд крупных клеток,
прилегающих к стенке тела изнутри. Это нервные клетки
экваториального нервного кольца (рис. 22, А, 2). По сторонам от задней кишки в
зачатке туловища червя легко обнаружить пару крупных светлых
клеток, представляющих собой мезобласты (рис. 22, 6). Во время
метаморфоза они образуют мезодермальные полоски.
На более поздней стадии видны следующие этапы метаморфоза.
Туловищный зачаток сильно разрастается, тогда как тело трохофоры
увеличивается очень незначительно. Зачаток туловища червя (рис. 22,
Б, 5) целиком помещается на нижней стороне личинки, будучи
погружен в особое впячивание стенок тела—амниотическую полость
(рис. 22, Б, И) и закрыт снизу специальной тонкостенной складкой
экдотермы, или мембраной (рис. 22, Б,12). Благодаря быстрому росту
туловищного зачатка и недостатку места внутри впячивания, стенки
зачатка образуют многочисленные складки (рис. 22, Б, 5).
Помимо трохофор, едва начавших метаморфоз, и средних стадий в
планктоне нередко попадаются поздние стадии метаморфоза. На них
кроме еще большего увеличения туловищного зачатка можно
наблюдать плотные зачатки щупалец, которые развиваются на поверхности
теменной пластинки. Голова взрослого червя образуется из эписферы,
причем теменная пластинка дает начало церебральным ганглиям, а все
туловище червя развивается из туловищного зачатка. Сближение этих
двух участков личинки при образовании тела взрослого животного
сопровождается элиминацией промежуточной зоны стенок тела.
ОТРЯД NEREIMORPHA
Nereis pelagica L.
Материал и методы изучения. Род Nereis (сем. Nereidae) в морях СССР
представлен несколькими видами. Из них наиболее доступен N. pelagica L., в изобилии
встречающийся в Баренцевом море на Мурмане, в Белом море и во всех
дальневосточных морях. На Мурмане N. pelagica обычен в биоценозе литотамния и среди
зарослей асцидий Phallusia.
При невозможности получить именно этот вид в качестве материала могут быть
использованы другие виды Nereis. Для вскрытий предпочтительнее более крупные
формы, например N. virens (Sars.) из Белого и Японского морей, тогда как для
фиксации с гистологическими целями следует выбирать мелких червей. В Баренцевом
море кроме N. pelagica встречается N. zonata Malmgren. В Черном и Азовском мо-
26
рях имеется несколько видов. Из ннх обычны N. cultifera (Gr.) и более крупный
N. diversicolor Mull., а также N. succinea (Leuckart) н N. zonata. В 1939—1941 гг.
N. diversicolor был пересажен из Азовского моря в Каспнй с целью увеличения
запасов живого корма для промысловых рыб. Это мероприятие удалось, и теперь этот
вид является обычным обитателем северного Каспия '.
Nereis cultifera жнвет в тонких трубках, которые легко покидает, N. succinea
обитает в U-образных норках.
Добываются Nereis сачком среди водорослей илн путем драгировок.
Внешняя морфология изучается на живых или консервированных в спирту
червях, а также на тотальных препаратах (параподии, щетинки). Изучение внутренней
организации производится путем вскрытий и.на срезах. Для фиксации с
гистологическими целями следует выбирать не слишком крупные .экземпляры. Рекомендуется
предварительно разрезать тело червя на несколько кусков. Хорошие результаты дает
фиксация подогретой сулемой с уксусной кислотой илн фиксация жидкостью Джильсона,
а также смесью Ценкера или Флеммннга. Срезы окрашиваются обычными
красителями: гематоксилином с докраской эозином по Маллори или борным кармином in toto.
Окраска срезов по' Маллори дает возможность изучить пограничные образования.
Для изготовления больших продольных срезов необходимо производить залнвку в
целлоидин.
Наблюдения над живыми животными. Следует обратить внимание
на характер движения Nereis. Передвигающийся по дну аквариума
червь совершает синусоидальные движения. Передний конец тела
остается вытянутым вперед, тогда как остальное туловище
волнообразно изгибается в горизонтальной плоскости, причем
волны пробегают спереди назад. Параподии
участвуют в движении как плавники. При этом обе
параподии одного сегмента движутся одновременно в
противоположных направлениях, в строгом соответствии с
волнообразными движениями тела. На гребне волны
они ударяют с силой назад, тогда как параподии на
другой стороне тех же сегментов движутся вперед
(рис. 23). При ударе в заднем направлении щетинки
параподии высовываются, что увеличивает их
рабочую поверхность, при обратном движении
втягиваются. Спасаясь от преследования, Nereis начинает
быстро плавать, при этом характер движения
сохраняется. Таким образом передвигаются многие Erran-
tia. Помимо того легко наблюдать дыхательные
движения Nereis. He меняя своего положения на дне
аквариума, червь многократно совершает
волнообразные колебания туловища в вертикальной плоскости,
что приводит к обновлению омывающей -его воды.
Иногда удается видеть, как животное выпячивает
ротовую (буккальную) область, и тогда на переднем
ее конце заметны две крючкообразные хитиновые
челюсти, служащие для схватывания добычи.
Процесс выворачивания и вворачивания буккального
отдела происходит очень быстро. Надавливая пальца-
)
)
$
1 См.: Сборник работ по акклиматизации N. succinea в
Каспийском море. М., 1952.
Рис. 23. Схема
синусоидального
движения Nereis (по
Будденброку).
Стрелки показывают
направление
движения параподнй
27
ми на передний конец живого
червя, можно заставить его
выпятить буккальный отдел.
Внешняя морфология. Для
изучения внешнего строения
пригоден как живой, так и
фиксированный материал.
Тело Nereis pelagica сильно
вытянуто и достигает в длину 6—
12 см. На Мурмане были
случаи нахождения экземпляров
более 20 см длиной.
Наибольшей ширины тело достигает в
передней и средней своей
части; задняя треть тела
постепенно суживается к заднему
концу (рис. 24).
Спинная,-более темноокрашенная, сторона
выпукла, брюшная
поверхность плоская с глубокой
медиальной продольной
бороздкой. Живые Nereis pelagica
окрашены в буро-зеленый
цвет, нередко с бронзовым
отливом и металлическим
блеском. На головной лопасти
между глазами иногда
имеется светлое пятно неправильной
формы. Все тело состоит из
80—100 сегментов.
Голова слагается из- двух
отделов, небольшого
переднего — простомиума, или
головной лопасти, и более крупного
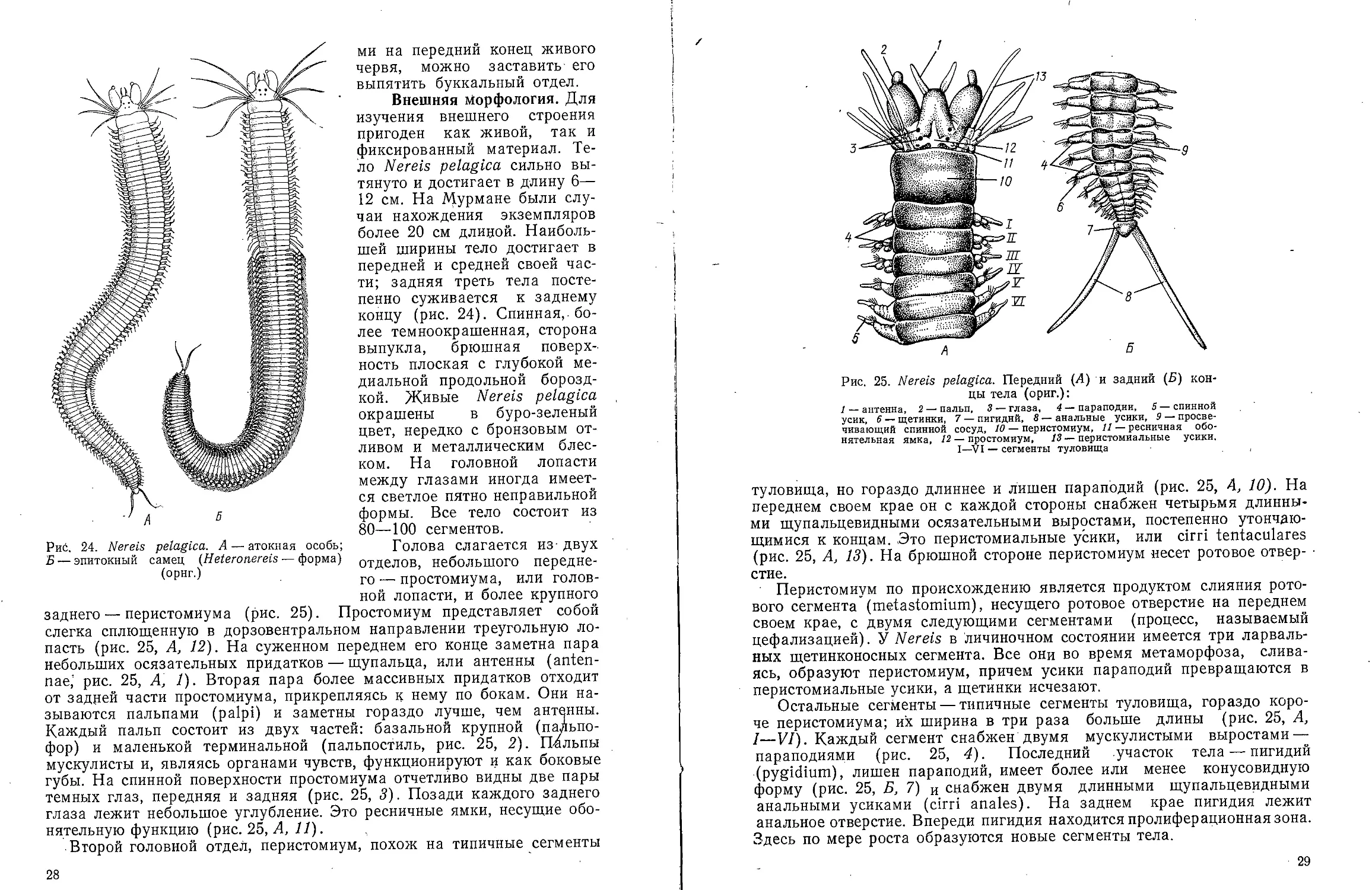
заднего — перистомиума (рис. 25). Простомиум представляет собой
слегка сплющенную в дорзовентральном направлении треугольную
лопасть (рис. 25, А, 12). На суженном переднем его конце заметна пара
небольших осязательных придатков — щупальца, или антенны
(antennae,' рис. 25, А, 1). Вторая пара более массивных придатков отходит
от задней части простомиума, прикрепляясь к нему по бокам. Они
называются пальпами (palpi) и заметны гораздо лучше, чем антенны.
Каждый пальп состоит из двух частей: базальной крупной (пальпо-
фор) и маленькой терминальной (пальпостиль, рис. 25, 2). Пальпы
мускулисты и, являясь органами чувств, функционируют и как боковые
губы. На спинной поверхности простомиума отчетливо видны две пары
темных глаз, передняя и задняя (рис. 25, 3). Позади каждого заднего
глаза лежит небольшое углубление. Это ресничные ямки, несущие
обонятельную функцию (рис. 25, А, 11).
Второй головной отдел, перистомиум, похож на типичные сегменты
Рис. 24. Nereis pelagica. A — атокная особь;
Б — эпитокный самец (Heteronereis — форма)
(орнг.)
28
Рис. 25. Nereis pelagica. Передний (А) и задний (Б)
концы тела (ориг.):
/ — антенна, 2 — пальп, 3— глаза, 4 — параподий, 5—спинной
усик, 6 — щетинки, 7 — пигиднй, 8 — анальные усики, 9 —
просвечивающий спинной сосуд, 10 — перистомиум, Я — ресничная
обонятельная ямка, 12 — простомиум, 13 — перистомиальные усики.
I—VI — сегменты туловища
туловища, но гораздо длиннее и лишен параподий (рис. 25, А, 10). На
переднем своем крае он с каждой стороны снабжен четырьмя
длинными щупальцевидными осязательными выростами, постепенно
утончающимися к концам. Это перистомиальные усики, или cirri tentaculares
(рис. 25, А, 13). На брюшной стороне перистомиум несет ротовое
отверстие.
Перистомиум по происхождению является продуктом слияния
ротового сегмента (metastomium), несущего ротовое отверстие на переднем
своем крае, с двумя следующими сегментами (процесс, называемый
цефализацией). У Nereis в личиночном состоянии имеется три ларваль-
ных щетинконосных сегмента. Все они во время метаморфоза,
сливаясь, образуют перистомиум, причем усики параподий превращаются в
перистомиальные усики, а щетинки исчезают.
Остальные сегменты — типичные сегменты туловища, гораздо
короче перистомиума; их ширина в три раза больше длины (рис. 25, А,
I—VI). Каждый сегмент снабжен двумя мускулистыми выростами —
параподиями (рис. 25, 4). Последний участок тела — пигидий
(pygidium), лишен параподий, имеет более или менее конусовидную
форму (рис. 25, Б, 7) и снабжен двумя длинными щупальцевидными
анальными усиками (cirri anales). На заднем крае пигидия лежит
анальное отверстие. Впереди пигидия находится пролиферационная зона.
Здесь по мере роста образуются новые сегменты тела.
29
Рис. 26. Nereis pelagica. Перед- Рис. 27. Nereis pelagica. Параподия атокной
ннй конец тела с выпяченным формы (орнг.):
буккальным отделом (орнг.): / — спинной усик, 2 —лопасти нотоподни, 3 — тол-
/—челюсть, 2 —буккальный отдел, стые гомогомфные щетннки, 4 — тонкие гомогомф-
3 — антенна 4 — простомиум 5 — пери- ные щетинки, 5 — тонкие гетерогомфные щетинки,
стомиум, 5 —обонятельная 'ямка, 7 — 5 —толстые гетерогомфные щетинки, 7 — лопасть
неглаза, 8 — перистомиальные усики,- вроподии, S —брюшиой усик, 9 — невроподия, 10 —
9 — пальп, 10 — парагнаты, И — линии, опорные щетинки, И — нотоподия
по которым происходит складывание
буккального отдела
Следует рассмотреть буккальный отдел, способный выворачиваться.
Надавливая пальцами на передний конец живого червя и вызвав его
выворачивание, следует перевязать ниткой животное поперек в
передней части тела. Этим предотвращается обратное вворачивание
буккального отдела. Имея под рукой лишь фиксированных червей, необходимо
выбрать особи, буккальный отдел которых выпятился при фиксации.
Вполне выпяченный буккальный отдел (рис. 26) имеет вид
широкого мешка, разделенного поперечной бороздкой на переднюю и заднюю
части. На его переднем крае лежит отверстие, ведущее
непосредственно в полость глотки (см. рис. 30, Б, 7). По бокам этого" отверстия торчит
пара мощных челюстей черного цвета. Челюсти крючкообразно
изогнуты внутрь остриями и на внутреннем крае снабжены несколькими
зубцами (рис. 26, 1). Они служат своего рад граблями,
загребающими пищу — детрит с различными мелкими организмами, а также
соскребывающими водоросли с камней1.
Кутикула, покрывающая буккальный отдел, образует местные
небольшие утолщения, зубчики или бляшки черноватого цвета,
называемые парагнатами (paragnathae, рис. 26, 10).
1 См.: Беклемишев К. В. Питание N. pelagica и функциональное значение его
челюстного аппарата. — Докл. АН СССР, 1950, т. 73, j4° б.
30
Далее следует детально изучить параподии. Для
этого острой бритвой или скальпелем вырезают один
из сегментов тела и рассматривают его под бинокуля-
ром или микроскопом. Можно изготовить тотальный
препарат, предварительно подкрасив вырезанный
кусок борным или квасцовым кармином. Параподии
Nereis имеют типичное строение. Каждая параподия
(рис. 27) представляет собой мускулистый боковой
вырост стенки тела и состоит из базальной нерасчле-
ненной части и двух ветвей: спинной (notopodium) и
брюшной ветви (neuropodium, рис. 27, 11 и 9).
Нотоподия на дорзальной стороне снабжена щупальце-
видным придатком — спинным усиком (cirrus dorsa-
lis, рис. 27, 1), который у многих полихет"
дифференцирован в качестве жабры. Сходный брюшной усик
(cirrus ventralis) имеется и на брюшной стороне нев-
роподии (рис. 27, 8). Каждая ветвь параподии
подразделяется на две вторичных лопасти и снабжена
щетинками (chaetae), концы которых торчат наружу
(рис. 27, 3—6). В качестве внутреннего скелета
параподия имеет две мощных хитиновых опорных
щетинки (acicula, рис. 27, 10).
Следует рассмотреть торчащие наружу щетинки
параподии при сильном увеличении микроскопа.
Щетинки Nereis pelagica (рис. 28) относятся к типу
сложных щетинок. Каждая состоит из длинной
стеблевой части, кончающейся бокаловидным углублением, со дна которого
поднимается терминальная часть, более или менее вытянутая. У одних
щетинок края воронковидного углубления одинаковой высоты; такие
щетинки называются гомогомфными (рис. 28, А). У других — гетеро-
гомфных щетинок, края воронки скошены, т. е. с одной стороны они
выше, чем с другой (рис. 28, Б). Кроме того, сами щетинки различаются
по толщине. Наряду с толстыми, грубыми, имеются более тонкие.
Терминальные части толстых щетинок коротки и массивны, тогда как у
тонких они вытянуты в длинную, слегка изогнутую, утончающуюся к
концу саблевидную пластинку, один край которой несет ряд тонких и
коротких волосков (рис. 28, А). Следует обратить внимание на
расположение этих форм щетинок на параподии (см. рис. 27, 3—6). Нотоподия
снабжена несколькими толстыми гомогомфными щетинками. Щетинки
невроподии более многочисленны и распадаются на два пучка. Из них
пучок, расположенный дорзальнее aciculum, слагается из тонких гомо-
гомфных и толстых гетерогомфных щетинок, тогда как пучок, лежащий
вентральнее aciculum, состоит из тонких и толстых гетерогомфных
щетинок.
Анатомия. С анатомией Nereis удобнее всего познакомиться путем
вскрытий. Однако очень много дает также наблюдение над живыми
мелкими экземплярами, которых следует рассматривать под бинокуля-
ром или микроскопом, сдавливая предметными стеклами. При таком
способе изучения помимо ряда анатомических особенностей можно наб-
Рис. 28. Щетинки
Nereis pelagica.
А — тонкая гомо-
гомфная; Б —
толстая гетерогомф-
ная (ориг.)
31
Рис. 29. Nereis pelagica. Передняя часть
тела, вскрытая со спинной стороны (ориг.):
1 — концевая ветвь спинного сосуда, проходящая
по поверхности буккального отдела, 2 — буккаль-
ный отдел, 3— протрактор буккального отдела,
4 — спинной сосуд, 5 — просвечивающие челюсти,
6 — глотка, 7 — передние ретракторы глотки, 8 —
слюнная железа (частично удалена), 9— задние
ретракторы глотки, 10 — пищевод, 11 — дорзоин-
тестииальные сосуды, 12 — параподио-дорзальиые
сосуды, 13 — спинной мезентерий, 14 — кишка,
15 — брюшной нервный ствол, 16 — брюшной
ганглий, 17 — вентро-параподиальные сосуды, 18 —
брюшное поле продольной мускулатуры, 19 —
железистый отдел метанефридия, 20— спинное поле
продольной мускулатуры, 21 — диагональные
мышцы, 22 — интерсегментарная бороздка,
просвечивающая сквозь стенку тела, 23 — ннтести-
нально-вентральные сосуды, 24 — щетинкоиосный
мешок, 25 — брюшной сосуд, 26 — протракторы
щетинок, 27, 28 — мембраны с сетью кровеносных
сосудов, 29 — линии, по которым происходит
складывание стенок буккального отдела, 30—-диссе-
пимент
людать выпячивание и впячивание
буккального отдела, пульсацию
спинного кровеносного сосуда,
движение параподийи пр.
Вскрывать следует наиболее крупные
экземпляры живыми или, лучше,
убив их слабым спиртом.
Вскрытие ведется в препаровальной
ванночке в морской воде.
Приколов Nereis ко дну ванночки
спинной стороной кверху ( одну
булавку следует втыкать в просто-
миум, другую — в заднюю часть
тела), разрезают стенку тела
вдоль, несколько справа от
спинной медиальной линии. Края раз-'
реза отворачиваются и
прикалываются ко дну ванночки. При
этом следует перерезать с
правой стороны мощные мышечные
пучки, прикрепляющиеся к
задней части глотки. Обратить
внимание на общее расположение
органов. Многие из них
располагаются метамерно
соответственно наружной сегментации (рис.
29).
Стенка тела,
мускулатура и полость тела. Кож-
но-мускульный мешок состоит из
наружного покровного эпителия,
одетого снаружи кутикулой, и из
мышечных слоев: наружного —
кольцевого и внутреннего —•
продольного. Последний
распадается на четыре мускульных ленты:
две брюшных и две спинных (см.
рис. 31, 9, 25).
На вскрытом животном
видны только продольные спинные и
брюшные мышечные ленты (рис.
29, 18, 20). Первые разделены
спинным мезентерием, и между
ними проходит спинной
кровеносный сосуд; между вторыми
проходит нервная цепочка. Спинные
мышечные ленты пересекаются
поперечными линиями, которые
представляют собой просвечива-
32
ющие сквозь стенку тела границы между сегментами (рис. 29, 22).
По бокам тела между брюшными и спинными лентами продольной
мускулатуры хорошо заметны метамерно расположенные щетинконос-
ные мешки параподий (рис. 29, 24), сквозь прозрачную стенку которых
видны лежащие в них acicula. Щетинконосные мешки связаны
специальными мускульными пучками — протракторами щетинок —со стенками
тела (рис. 29, 26). Далее хорошо заметны метамерные диагональные
мышцы (musculi transversi obliqui), характерные для полихет. У
Nereis эти лентовидные мышцы прикрепляются своими более широкими
брюшными концами по сторонам от брюшной нервной цепочки, а более
узкими латеральными концами к стенкам тела в области параподий
(рис. 29, 21). В каждом сегменте имеется одна пара таких мышц. Они
пересекают полость тела в косом направлении.
Полость тела представляет собой целом и пересекается
вертикальными нежными и прозрачными мембранами — мезентериями и диссепи-
ментами. Эти мембраны отсутствуют лишь в передних сегментах тела,
где подвижность кишечной трубки, связанная со способностью
буккального отдела выворачиваться наружу, исключает их развитие.
Спинной мезентерий начинается от спинной медиальной линии кожно-
мускульного мешка и прикрепляется к дорзальной поверхности
кишечника (рис. 29, 13). Брюшной мезентерий соединяет таким же образом
брюшную стенку тела с вентральной поверхностью кишечника.
Поперечно расположенные диссепименты являются неполными и связывают
только боковые поверхности кожно-мускульного мешка с кишкой. Они
проходят на границах между сегментами и связаны с поперечными
кровеносными сосудами. В местах прикрепления к стенкам тела
диссепименты уже, чем в местах прикрепления к кишке. Таким образом,
диссепименты делят полость тела на ряд камер, из которых каждая
отвечает полости одного сегмента, т. е. сомиту. Ниже и выше свободных
краев диссепиментов соседние камеры сообщаются друг с другом. Кроме
того, и сами диссепименты могут быть прободены небольшими
отверстиями. При вскрытии большая часть диссепиментов разрывается
(рис. 29, 30).-
Пищеварительная система. Пищеварительный канал,
начинаясь ротовым отверстием на перистомиуме, проходит через все тело
в виде прямой трубки, которая дифференцирована на буккальный
отдел, глотку, пищевод, среднюю и заднюю кишки.
Толстостенный и мускулистый буккальный отдел цилиндрической
формы (рис. 29, 2); на его поверхности заметны продольные и поперечные
линии, по которым происходит складывание стенок этого органа (рис.
29). Следующий отдел — глотка представляется как бы продолжением
буккального отдела (рис. 29, 6). Стенки ее очень толсты и мускулисты,
а полость узка. На границе между глоткой и буккальный отделом
просвечивают лежащие внутри челюсти (рис. 29, 5). В спокойном состоянии
глотка у Nereis pelagica достигает V сегмента. При выворачивании
буккального отдела сама глотка не выворачивается, но выдвигается далеко
вперед. При этом она оказывается внутри выпятившегося наружу
буккального отдела, а челюсти, лежащие по сторонам отверстия,
соединяющего буккальную полость с полостью глотки, оказываются на перед-
2—2
33
нем конце выпятившегося буккального
отдела (рис. 30). Выворачивание
последнего происходит в результате
сокращения мускулов протракторов
(рис. 30, А, 3) и давления полостной
жидкости, возникающего благодаря
сокращению мышц в стенках
передней части тела. Выворачивание
останавливается диафрагмой —
мускулистой пластинкой, охватывающей кругом
буккальный отдел и
прикрепляющейся к стенкам I сегмента. Впячивание
происходит благодаря сокращению-
диафрагмы, а также вследствие
сокращения мощных ретракторов глотки.
Эти последние соединяют задний
отдел глотки со стенками тела (см. рис.
29, 7, 9; рис. 30, 5). Движение
челюстей зависит от мощных мускулов,
лежащих в стенках глотки.
Отходящий от заднего конца
глотки пищевод представляет собой узкую-
цилиндрическую трубку (см. рис.
29, 10), продолжающуюся сзади в
среднюю кишку. В передний отдел
пищевода открывается пара
продолговатых слюнных желез, наружная
поверхность которых сильно складчата
(см. рис. 29, 8). Более широкая, чем
пищевод, средняя кишка представляет собой тонкостенную,
расширяющуюся в каждом сегменте трубку (см. рис. 29, 14). Наконец, сзади
кишечник оканчивается короткой задней кишкой.
Органы дыхания. Специальные органы дыхания у Nereis
pelagica отсутствуют. Дыхание происходит через кожу, причем
особенно большое значение для окисления крови имеют сосудистые сплетения
в параподиях (рис. 31, 6). Рассматривая живых мелких Nereis, сдавлен-
. ных между стеклами, можно видеть в параподиях эту респираторную'
сеть кровеносных сосудов.
Кровеносная система. Кровеносные сосуды хорошо заметны
благодаря красному цвету наполняющей их крови. Кровеносная система
замкнутая, состоит из двух главных продольных сосудов —
пульсирующего спинного (vas. dorsale, см. рис. 29, 4), несокращающегося
брюшного (vas ventrale, см. рис. 29, 25)— и периферических сосудов. В
каждом типичном сегменте кровеносная система имеет следующее строение..
Со спинным сосудом связаны две пары поперечных сосудов. Одна
несет кровь к спинной поверхности кишки (дорзоинтестинальные сосуды,.
рис. 31, 2; рис. 29, 11), где разветвляясь, образует капиллярную сеть в
стенках кишечника. Вторая" пара сосудов принимает кровь от спинной
стенки тела и главным образом от параподий (параподио-дорзальные
Рис. 30. Схема выпячивания
буккального отдела Nereis. A — буккальный
отдел н глотка втянуты; Б — бук-,
кальнын отдел вывернут, глотка
выдвинута вперед (по Лангу):
1 — мозг, 2 — буккальный отдел, 3 —
протракторы, 4 — глотка, 5 — ретракторы, 6 —
парагнаты, 7 — отверстие, ведущее в
полость глотки, 8 — челюсти
34
Рис. 31. Схема поперечного разреза Nereis через сегмент туловища (по
Бенхаму):
/—'спинной сосуд, 2 — дорзоинтестинальный сосуд, 3 — параподио-дорзальный
сосуд, 4 — полость тела, 5 — лопасти параподин, 6 — респираторная сеть кровеносных
сосудов, 7 — интестинально-вентральиый. сосуд, 8 —вентро-параподиальные сосуды,
9 — брюшное поле продольной мускулатуры, 10 — брюшная нервная цепочка, 11 —
брюшной сосуд, 12 — диагональная мышца, /3—параподнальный нерв, 14 — нефро-
пор, 15 — брюшной усик, 16 — метанефридий, 17 — невроподия, 18 —' опорная
щетинка, 19 — нотоподия, 20 — протрактор щетинок, 21 — спинной усик, 22 — нерв спинного
усика, 23 — цилиарный орган, 24 — кольцевая мускулатура, 25 — спинное поле
продольной мускулатуры, 26 — полость кишки. Слева зачернена кровеносная система,
справа изображена мускулатура параподий и параподиальный иерв
сосуды, см. рис. 29, 12; рис. 31, 3). С брюшным сосудом связаны три
пары поперечных сосудов. Одна из них принимает кровь от
сосудистого сплетения кишки (интестинально-вентральные сосуды, см. рис. 29,
23; рис. 31, 7), а две других несут кровь к брюшной стенке тела и пара-
подиям, именно к нотоподиям и невроподиям (вентро-параподиальные
сосуды, см. рис. 29, 17; рис. 31, 8). По спинному сосуду кровь течет от
заднего конца к переднему, по брюшному — в обратном направлении.
Поперечные сосуды частично связаны с диссепиментами. Сосудистый
комплекс одного типичного сегмента составляет ангиосомит — понятие,
введенное С. И. Тимофеевым1. Главные элементы ангиосомита
гомологичны у различных полихет.
В передних сегментах тела у Nereis расположение сегментарных
кровеносных сосудов отличается от описанной схемы. Здесь эти сосуды
особенно длинны. От обоих продольных сосудов они направляются назад к
передней части кишки, где на небольшом участке кишечной
поверхности сближается большое количество поперечных сосудов (см. рис, 29,
23). Такое отклонение от расположения сосудов в типичном сегменте
связано с подвижностью передней части пищеварительного тракта.
1 Тимофеев С. И. К морфологии Polychaeta. Сборник трудов Иркутского
университета, 1923, т. V.
2*
35
Спинной сосуд проходит вдоль всего тела, достигая простомиума; он
лежит на внутренней поверхности спинной стенки тела в промежутке
между продольными мышечными лентами кожно-мускульного мешка
(рис. 31, 1), Брюшной сосуд проникает в переднюю часть тела не столь
далеко, кончаясь в области глотки, к которой он прилегает с брюшной
стороны (см. рис. 29, 25). При ввороченном буккальном отделе и заднем
положении глотки передняя часть брюшного сосуда образует
многочисленные, изгибы, выравнивающиеся, когда буккальный отдел
выпячивается и глотка выдвигается вперед.
Оба продольных сосуда впереди разделяются на две конечные ветви,
которые связаны друг с другом на каждой стороне. Конечные
разветвления спинного сосуда длинны и сначала тянутся по спинной
поверхности буккального отдела назад (см. рис. 29,1). Оба продольных
сосуда, кроме того, отделяют две боковых веточки, которые
соединяются друг с другом на каждой стороне, как и конечные
разветвления.
Таким образом, между продольными сосудами имеется двойная связь,
которая, однако, не является непосредственной. А именно боковые
сосуды многократно ветвятся, образуя густые сосудистые сплетения,
расположенные в двух парах тонких мембран (см. рис. 29, 27, 28). Последние
лежат по бокам глотки в полости тела и соответствуют диссепиментам
типичных сегментов.
Выделительная система. В каждом сегменте, исключая
пять передних и несколько задних, лежит пара метанефридиев.
Благодаря малой величине эти органы обнаруживаются с трудом и лучше
заметны у крупных экземпляров. Они лежат в передней части сегмента
под диагональными мышцами перед входом в полости параподии
(см. рис. 29, 19).
Выделив осторожно метанефридий, следует^ рассмотреть его в
свежем состоянии под микроскопом или бинокуляром. Метанефридий
Nereis своеобразного строения, напоминают таковые некоторых
Oligochaeta. Каждый состоит из компактной железистой массы (рис.
32, 1), содержащей сильно извитой канал, мерцательный на большем
своем протяжении, но лишенный ресничек в своей последней петле
(рис. 32, 2). Эта последняя часть представляет собой выводной проток
органа, выходит из
железистой массы нефридия,
прободает стенку тела и
открывается нефридиаль-
ной порой (рис. 32, 3) у
основания невроподйи-
Ресничный канал также
обособляется от
железистой массы, вытягивается
вперед и, прободая дис-
сепимент, открывается в
полость предыдущего
сегмента воронкой или неф-
ростомом (рис. 32, 4).
Рис. 32. Метанефридий Nereis (по Бенхаму):
/ — железистая масса, 2 — нефридиальный канал, 3 — не-
фридиальная пора, 4 — нефростом
36
Края, или губы нефростома, снабжены ресничками. У Nereis cultifera
губы широкого нефростома покрыты бугорковидными отростками с
длинными пучками ресничек.
Нервная система состоит из церебральных, или надглоточных,
ганглиев, слитых в одну массу, из окологлоточных коннективов,
брюшной нервной цепочки и отходящих из этих отделов нервов. На вскрытом
черве хорошо видна брюшная нервная цепочка, являющаяся продуктом
слияния сблизившихся в поперечном направлении двух брюшных
нервных стволов и их ганглиев. Она проходит вдоль всего тела под кишкой,
прилегая к кожно-мускульному мешку в промежутке между
вентральными продольными мышечными лентами (см. рис. 29, 15). "В каждом
сегменте нервная цепочка образует легкое утолщение — брюшной
ганглий данного сегмента (см. рис. 29, 16). От каждого ганглия берут
начало нервы, из которых наибольшего развития достигает одна пара, ин-
нервирующая параподии. Строение и топографию переднего отдела
центральной нервной системы удобнее изучить на срезах (см. с. 39,43).
Органы чувств. Осязательными органами являются антенны,
а также усикп перистомиума, параподии и анальные. Органы
химического чувства (обонятельные) представлены парой пальпов и парой
ресничных ямок, лежащих дорзально в задней части простомиума (см.
рис. 25, 11). Кроме того, имеются две пары хорошо развитых глаз
(см. рис. 25, 3), строение которых разбирается ниже. Статоцисты у
Nereis отсутствуют.
Половая система. Встречающиеся большую часть года
экземпляры Nereis представляют собой неполовозрелые, лишенные половых
органов особи. С наступлением половой зрелости половые продукты
развиваются только в сегментах задней половины тела, которые
называются эпитокными сегментами в отличие от передних, атокных. В
течение короткого промежутка времени (несколько недель) эпитокные
сегменты сильно изменяются, причем особенно это изменение касается
параподии. Тело животного приобретает характерное резкое
разделение на два отдела, чем существенно отличается от неполовозрелых форм
(см. рис. 24, Б). Благодаря этому половозрелые формы прежде
относились даже к особому роду Heteronereis. Nereis раздельнополы, как
огромное большинство полихет, и гетеронереидные формы представлены
как самцами, так и самками. Неполовозрелые Nereis ведут придонный
образ жизни, тогда как гетеронереидные особи оживленно плавают на
поверхности моря.
Следует рассмотреть внешнюю морфологию гетеронереидной формы
и вскрыть ее заднюю эпитокную часть тела. На параподиях эпиток-
ных сегментов развиваются широкие вертикальные плавникообразные
лопасти (рис. 33, 1, 4, 6), щетинки заменяются своебразными
плавательными щетинками с веслообразно уплощенным терминальным
отделом. Изменения касаются также и передней атокной части тела:
глаза увеличиваются, антенны удлиняются, увеличивается несущая
чувствительные клетки поверхность усиков на передних сегментах у
самцов. Окраска становится ярче. Задний эпитокный отдел тела
приобретает беловатый оттенок у самцов и желтоватый или зеленоватый у
самок, так как мускулатура кожно-мускульного мешка претерпевает
37
Рис. 33. Nereis pelagica. Параподия гетероне-
реидной формы (по Элерсу):
/ — мембрана нотоподии, 2 — спинной усик, 3 —
щетинки, 4 — мембрана невроподии, 5 — брюшной усик,
£ —мембрана, связанная с усиком, 7 —опорные
щетинки
дегенеративные изменения и
сквозь стенки тела
просвечивает масса спермы или
яиц. Другие внутренние изг
менения в эпитокных
сегментах выражаются в
дегенерации кишечника, стенки
которого спадаются и
склеиваются вместе в тонкую
пластинку. Мускулатура па-
раподий, напротив, не
только не ослабляется, но даже
усиливается. Явление эпи-
токии обычно у Nereidae. и
хорошо известно у Nereis
pelagica и других видов.
Гонады у Nereis
дифференцируются к моменту
наступления половой
зрелости. Половые клетки
развиваются в целомическом
эпителии. Гонады прикрепляются к перитонеуму брюшной стенки тела
вблизи медианной линии посредине сегмента. Созревшие половые
клетки наполняют всю полость сегмента и выводятся наружу через разрыв
стенки тела. N. diversicolor — живородящая форма.
Изучение срезов. Следует приготовить серию поперечных и
фронтальных срезов через передний конец червя, а также через несколько
типичных сегментов тела.
Для ознакомления со строением и топографией переднего отдела
центральной нервной системы рекомендуется произвести реконструкцию
этой системы по срезам. Удобнее всего сделать фронтальную
реконструкцию по фронтальным срезам, хотя могут быть применены и другие
способы реконструкции. В качестве примера реконструкции мы даем
здесь изображение нервной системы Nereis virens, близкое к таковой
N. pelagica и других видов (рис. 34). От боковых сторон компактного
мозга (церебральных ганглиев, рис. 34, 5), лежащего в простомиуме,
отходят окологлоточные коннективы, начинаясь каждый тремя корнями
(рис. 34, 12). У основания перистомиальных усиков коннективы
раздваиваются и затем загибаются под передним концом пищеварительной
трубки на брюшную сторону, где соединяются с подглоточным
ганглием (рис. 34, 7). Брюшные нервные стволы и ганглии сближены в попе-,
речном направлении до полного слияния (рис. 34,8). От церебральных
ганглиев берут начало нервы, направляющиеся к буккальному отделу,
антеннам, пальпам, мускулатуре головы, ресничным обонятельным
ямкам и глазам. Перистомиальные усики'иннервируются от
окологлоточных коннективов (рис. 34, /3)..Перистомиум получает нервы от подгло-
точного ганглия. Каждый ганглий сегмента дает несколько пар нервов,
иннервирующих данный сегмент. Из них особенного развития достигает
одна пара, направляющаяся в пара подии (рис. 34, 11): У основания па-
38
раподий эти нервы образуют небольшой
параподиальный ганглий (рис. 34, 10).
Нужно разобрать гистологическое
строение церебральных ганглиев, что при
указанных обработках может быть сделано
лишь в общих чертах. Ганглиозные клетки
занимают периферические отделы мозга,
располагаясь вокруг волокнистой массы —
центрального тела. По Гольмгрену, мозг
Nereis имеет 26 ганглиозных центров,
между которыми правильно располагаются
участки волокнистого вещества (рис. 35).
Следует обратить внимание на пару крупных
скоплений ганглиозных клеток в передней
части мозга, называемых грибовидными
телами (corpora pedunculata или globuli, рис.
35,10). У Nereis pelagica каждое
грибовидное тело слагается из двух шапковидных
скоплений ганглиозных клеток,
волокнистые отростки которых, сходясь вместе,
образуют стебелек грибовидного тела.
Грибовидные тела представляют собой высшие
центры мозга; наличием их объясняют
способность Nereis к некоторым условным
рефлексам. Далее следует обратить внимание
на сгущения волокнистого вещества у
основания каждого нерва, иннервирующего
пальп, так называемые glomeruli, а также
на поперечные" волокнистые тяжи — ком-
миссуры (оптическая, стоматогастрическая
и др.).
На срезах удается изучить строение глаз
ставляет уже вполне замкнутый глазной пузырь, но еще не
отделившийся полностью от эпидермиса, впячиванием которого он является (рис. 36).
Место схождения краев глазного впячивания лишено пигмента и
представляет собой прозрачную роговицу (cornea), не разделенную еще на
настоящие внутреннюю к наружную роговицы (рис. 36, 8). В остальных
местах стенки глазного пузыря представляют собой сильно
пигментированную сетчатку (retina), которая слагается из двух сортов клеток —
зрительных, или ретинальных, и секреторных (рис. 36, 3 и 4). Более
многочисленные зрительные клетки не особенно густо окружены
пигментом; на своих внутренних концах каждая из них снабжена
светочувствительной «палочкой» (рис. 36, 6). Последняя представляет собой
трубочку с проходящим внутри тончайшим нервным волоконцем.
Секреторные клетки имеют густую пигментную обкладку. Их внутренние
концы выделяют секрет, различимый в виде нитей. За его счет
образуется стекловидное тело, занимающее полость глаза (рис. 36, 7). Ко дну
глаза подходит зрительный нерв (рис. 36, 2). У молодых Nereis глаза
представляют собой еще незамкнутые глазные бокалы без роговицы.
Рис. 34. Nereis pelagica. Схема
центральной нервной системы
переднего отдела тела (по Ха-
макеру):
1 — буккальный отдел, 2 — нерв бук-
кальиого отдела, 3 — грибовидное
тело, 4 — глаз, 5 — мозг, 6 —
обонятельная ямка, 7 — подглоточный
ганглнй, 8 — брюшные ганглин
сегментов, 9 — медиальный нерв, 10 —
параподиальный ганглий, 11 —
параподиальный нерв, 12 —
окологлоточный коннектив, 13 — перистоми-
альный уснк, 14 — нерв буккально-
го отдела, 15, П — нервы
мускулатуры стенок тела, 16 — нерв пальпа
. Каждый глаз Nereis пред-
39
Рис. 35. Nereis pelagica. Поперечный разрез
через простомиум (ориг.):
/ — глаз, 2 — кутикула, 3 — эпидермис, 4 — кожные
железы, 5 — мускульные волокна, 6 — коммиссура, 7 —
корни окологлоточного кониектива, 8 — основание пальца,
9 — волокнистый стебелек грибовидного тела, 10 —
грибовидное тело, // — гаиглиозные центры, 12 —
волокнистые пучки
Рис. 36. Разрез через глаз Nereis
(по Гессе):
1 — кутикула, 2 — нерв, 3 — зрительные
клетки, 4 — секреторные клетки с
пигментом, 5 — эпидермис, 6 —
светочувствительные палочки, 7 — стекловидное
телЪ, 8 — роговица
го w
Рис. 37. Nereis diversicolor. Часть поперечного разреза в области, диссепимента
(А) и в средней области сегмента (5). Разрез А прошел через среднюю кишку,
разрез 5 — через пищевод (по Шнейдеру):
I — кольцевая мускулатура, 2 — канал метаиефридия, 3 — брюшной нервный ствол, 4 — дис-
сепимент, 5 — веитро-параподиальный сосуд, б —эпителий среднгй кишки, 7 — полость
пищевода, 8 — брюшной сосуд, 9 — брюшиое поле продольной мускулатуры, 10 — иефропор,
II — железистая масса метаиефридия, 12 — брюшной усик, 13 — щетиики, 14 — спииная и
брюшная железистые лопасти, 15 — спинной усик, 16 — кутикула, 17 — эпидермис, 18 —
спинное поле продольной мускулатуры, 19 — созревающие мужские половые клетки, 20 —
спинной еосуд
40
Поперечный разрез через типичный сегмент выглядит различно
(рис. 37) в зависимости от того, прошел ли он через среднюю часть
сегмента (рис. 37, Б) или вблизи его границы в области диссепимента
(рис. 37, А). Брюшной и спинной края среза имеют срединные выемки,
соответственно спинной и брюшной медиальным бороздкам. На разрез
через середину сегмента попадают параподии (рис. 37,5), очертания
которых сильно изменяются от среза к срезу.
Покровный эпителий, или эпидермис, хорошо дифференцирован и
состоит из обычных покровных и железистых клеток (рис. 37, 17).
Эпидермис подстилается тонкой базальной мембраной (рис. 37, 3), а
снаружи покрыт довольно толстой гибкой кутикулой (рис. 37, 16; рис. 38, 6).
Железистые клетки представлены тремя видами. Наряду с двумя
видами белковых желез — крупными и мелкими, красящимися эозином
(рис. 38, 7), встречаются слизистые, отличающиеся колбовидной
формой и более редкие, нежели белковые. При окраске по Маллори
белковые железы окрашиваются в красный, а слизистые — в синий цвет. Оба
вида желез открываются наружу через тонкие канальцы, прободающие
кутикулу (рис. 38, 2).
17 16 15 1U-
Рис. 38. Nereis diversicolor. Часть поперечного разреза с брюшным
нервным стволом (по Шнейдеру):
/ — диагональная мышца, 2— пора железистой клетки, 3 — базальиая мембрана,
4 — эпидермис, 5 — кольцевая мускулатура, 6 — кутикула, 7 — белковые железы,
8 — иерв, 9 — продольная мускулатура, 10 — нервные клетки, 11 — нервные
волокна, 12 — защитная клеточная соединительная ткань, 13 — созревающие мужские
половые клетки, 14 — медиовентральный пучок продольной мускулатуры, 15, 16 —
волокнистые пучки, 17 — фолликулярная ткань
41
В средней области сегмента слизистые и крупные белковые железы
располагаются группами, или пакетами, которые достигают особенного
развития на параподиях. Здесь богатые ими спинная лопасть нотоподии
и брюшная невроподии получают название железистых лопастей (см.
рис. 37, 14).
Эпителий параподии образует глубокие тонкостенные впячивания,
щетинконосные мешки, в которых лежат щетинки. ч Стенки каждого
мешка прилегают к щетинкам и различимы как нежная мембрана,
местами утолщенная благодаря лежащим в ней большим ядрам. На
слепом конце мешка находятся клетки — образовательницы щетинок; ядра
их не отличаются от ядер других клеток. Щетинконосные мешки
глубоко вдаются в полость параподии и полость тела и окружены светлой
пузырчатой тканью, модифицированным перитонеумом.
Мускулатура кожно-мускульного мешка состоит из наружного
кольцевого и внутреннего продольного мышечных слоев. Кольцевой слой
лежит непосредственно под -эпидермисом (рис. 38, 5). Он прерывается
медиовентрально в области брюшной нервной цепочки, в тех местах,
где она примыкает непосредственно к эпидермису, а также с боков, в
местах прикрепления диагональных мышц к базальной мембране
покровов. В области параподии кольцевой слой распадается на ряд
мускульных пучков. Одни из них оканчиваются на дистальном конце параподии
или прикрепляются на поверхности щетинконосных мешков (ретракто-
ры щетинок), а другие соединяют стенки тела с внутренними концами
щетинконосных мешков (протракторы щетинок) (см. рис. 37, Б).
Продольная мускулатура кожно-мускульного мешка разделяется
на четыре мощные ленты, или поля, на пару вентральных и пару дор-
зальных (см. рис. 37, Б, 9, 18). Кроме того, медиовентрально на
брюшной нервной цепочке располагается пятая, слабо развитая продольная
мышечная лента (рис. 38, 14). Четыре большие мышечные ленты
прилегают к кольцевой мускулатуре только дорзально и вентрально;
боковые их края вдаются в полость тела. Каждая вентральная лента в
поперечнике имеет S-образную форму.
Диагональные мышцы прикрепляются к базальной мембране
эпидермиса по бокам от брюшной нервной цепочки (рис. 38, 1), пересекают
полость тела в косом направлении, проходят к основаниям параподии и
здесь прикрепляются к базальной мембране боковой стенки тела.
Мускулатура Nereis состоит из гладких мышечных волокон.
Конфигурация полости тела усложнена вдающимися в нее
продольными мышечными, лентами и пересекающими ее диагональными
мускулами. Последние делят полость тела на широкое пространство,
окружающее кишечник, — кишечную, или интестинальную, камеру и на две
вентральных — подиальных, или нефридиальных, камеры, к которым
принадлежат и полости параподии. Между боковыми стенками дорзаль-
ных мышечных лент и боковыми стенками тела остаются щелевидные
пространства целом а, пересекаемые цучками кольцевой мускулатуры,
которые внедряются в параподии и тянутся к щетинконосным мешкам —
побочным камерам. Местами стенки побочных камер впячиваются в
кожно-мускульный мешок в виде кожных каналов, сопровождающих
ответвления кровеносных сосудов. Вся полость тела, не исключая кож-
АО
ных каналов, выстлана плоским перитонеальным эпителием. Этот же
эпителий выстилает все органы, лежащие в полости тела. Вокруг
щетинконосных мешков перитонеум принимает характер пузырчатой
клеточной ткани. В полости тела, реже в мускулатуре и других тканях
встречаются амебовидные лимфатические клетки. Они имеют различную
форму и величину. Плазма их наполнена эозинофильными зернами.
В задней части каждого сегмента имеется пара утолщений перито-
■неального эпителия, покрытых ресничками и лежащих на дорзальных
мускульных лентах на границе "между кишечной и побочными камерами.
Это ресничные, или. цилиарные, органы, представляющие широко
раскрытую и сильно складчатую воронку (см. рис. 31,23).Эти органы
рассматриваются Гудричем как видоизмененные половые воронки (гоносто-
мы), свойственные ряду других полихет. Они не связаны с метанефри-
диями и несут функцию фагоцитарных органов.
Метанефридии лежат в нефридиальных камерах целома. На срезах
легко найти их компактную железистую массу (см. рис. 37, Б, 11).
Внутри последней отчетливо видны многократно перерезанные петли
нефридиального канала. Большая часть последнего железиста и
снабжена ресничками, но петля канала, продолжающаяся в конечный выводной
канал метанефридия, лишена ресничек. Просвет канала является
частью межклеточным и частью внутриклеточным. При инъекции в
полость тела живого червя карминокислого аммония вся железистая
масса метанефридия поглощает это вещество, приобретая красный цвет
(А. Ковалевский). Нетрудно проследить на срезах ход короткого
выводного канала. Он открывается наружу нефридиальной порой- вентрально
у самого основания параподии (см. рис. 37, Б, 10). Найти-воронку (не-
фростом) гораздо труднее. С метанефридиями связаны вентро-парапо-
диальные кровеносные сосуды.
Кровеносные сосуды на срезах хорошо заметны. Стенки
сокращающегося спинного продольного сосуда имеют кольцевые мускульные
волокна (см. рис. 37, Б, 20). Все сосуды в тех,местах, где они свободно
лежат в полости тела, снаружи одеты перитонеумом. По крайней мере,
часть из них имеет собственные эндотелиальные стенки; сосуды
содержат округлые кровяные тельца. Красный цвет крови зависит от
растворенного в ней гемоглобина.
Брюшная нервная цепочка расчленяется на ганглии сегментов и
связывающие их коннективы, лежащие интерсегментально. Ганглии
непосредственно прилегают к базальной мембране эпидермиса, а
коннективы отделены от нее кольцевой мускулатурой. Брюшная нервная
цепочка Nereis представляет собой продукт слияния двух продольных
нервных стволов с их сегментарными ганглиями. На срезах парная
природа нервной цепочки легко обнаруживается. Коннектив в
поперечном разрезе имеет округлые очертания. Он состоит из двух мощных
продольных пучков нервных волокон (рис. 38, 16), окруженных толстой
оболочкой, имеющей клеточную структуру (рис. 38, 12). Между
боковыми пучками волокон лежит еще маленький медианный пучок
(медиальный нерв, рис. 38, 15). Все волокнистые пучки одеты глиальной тканью,,
клетки которой непосредственно к ним прилегают. При окраске
железным гематоксилином глиальные волокна хорошо .заметны.
43
Коннективы переходят без резких границ в брюшной ганглий
сегмента. От ганглия отходят парные нервы. Легко обнаруживаются
наиболее мощные параподиальные нервы. В месте их отхождения ганглий
уплощен. Гистологически ганглий отличается от коннектива наличием
грушевидных униполярных ганглиозных клеток, погруженных в ткань
оболочки и расположенных группами латерально и снизу от
волокнистых пучков (рис. 38, 10). Отростки ганглиозных клеток входят в
волокнистые пучки, где перемешиваются с другими нервными волокнами.
У половозрелых животных в полости тела встречаются
многочисленные половые клетки (рис. 37, Б, 19; рис. 38, 13). Они лежат среди
пузырчатой ткани, соединяющейся местами с перитонеальным
эпителием, разращением которого эта ткань является (рис. 38, 17). Оогонии
сначала соединены группами, позднее самостоятельны. Семенные клетки
всегда связаны в грушевидные массы и имеют различный вид в
зависимости от стадии созревания.
ОТРЯД DRILOMORPHA
Arenicola marina L. — Пескожил
Материал и методы изучения. Arenicoh. marina L. (сем. Arenicolidae)
принадлежит к числу обычных литоральных форм Баренцева (Мурман) и Белого морей. В
Черном море этот род представлен A. branchialis Audouin et M. Edwards, а в Японском —
A. cristata Stimpson. Морфологически все эти виды близки, отличаясь друг от друга
лишь деталями. ,т, и * с ! + • \
Пескожил — типичный представитель седентарных полихет (Polycnaeta bettentanaj.
Этим названием, не имеющим систематического значения, принято обозначать много-
щетинковых червей, ведущих сидячий или роющий образ жизни, в отличие от форм
«бродячих», или эррантных (Polychaeta Errantia).
Arenicola marina в изобилии заселяет песчаные отмели, обнажающиеся во время
отлива. Животное ведет скрытный образ жизни в U-образиой норке (рис. 39). Конец
норки отмечен конусовидной кучкой грунта — экскрементами. Переднее колено целиком
заполнено рыхлым песком, который образует здесь песочный столбик. Животное
занимает горизонтальную часть норки, упираясь головой в нижний конец столбика,
тогда как его хвостовой отдел вдается в заднее вертикальное колено. В результате
перистальтического движения животного, вода промывает норку, входя через заднее
отверстие и фильтруясь сквозь песочный столбик. По мере того, как червь
заглатывает песок столбика, передний вертикальный канал норки заполняется сверху новыми
порциями поверхностного грунта. Червь никогда не покидает своей норки.
Добывание Arenicola производится во время отлива на песчанистых отмелях.
Черви выкапываются лопатой или «червеницей» — специальным орудием,
употребляемым1 рыбаками, которые собирают их для наживки.
A. marina заселяет не только обнажающуюся во время отлива литораль, но
встречается и глубже, в верхних-горизонтах зоны ламинарий. Здесь она представлена
особой разновидностью, отличающейся некоторыми морфологическими особенностями.
Изучение наружной морфологии Arenicola производится на живых или
консервированных в спирте животных. Перед вскрытием черви анестезируются 8%-ным
раствором хлористого магния в морской воде, в которой они должны быть выдержаны
некоторое время.
Наблюдения над живыми животными. Передвижение Arenicola
происходит иначе, чем у Nereis, и называется перистальтическим
движением, характерным для роющихся в грунте и живущих в трубках Sedenta-
ria. Вынутая из норки и положенная на песок дна аквариума Arenicola
позволяет наблюдать характер перистальтического движения. Прежде
ЛА
Рис. 39. Arenicola marina в норке (по Крюгеру).
Черные стрелки показывают ток воды через норку и песочный столбик,
светлые — движение песка столбика
всего на заднем конце туловища возникает энергичная волна
сокращения— несколько сегментов сильно вздуваются. Эта волна быстро
движется вперед, захватывая все новые и новые участки туловища
вплоть до IV или V сегмента. При этом полостная жидкость гонится
сзади вперед, благодаря ее давлению вбуравливание переднего конца
в грунт усиливается. После, волны сокращения продольной мускулатуры
кожно-мускульного мешка очень быстро возникает слабая волна
сокращения кольцевой мускулатуры, в особенности в передней трети тела.
В результате эта часть тела сильно вытягивается, дугообразно
изгибаясь над грунтом, а передний конец втыкается в песок. Так как при
этом выпячиваются и раздуваются буккальный отдел и глотка, то
частицы грунта раздвигаются. Быстро чередующиеся сокращения
продольной и кольцевой мускулатуры в комбинации с действием
выпячивающегося буккального отдела приводят к закапыванию червя. Хвостовой
отдел и три первых сегмента туловища в перистальтическом движении
не участвуют. При выпячивании буккального отдела и глотки сначала
выворачивается первый, затем на его конце выворачивается и глотка.
Впячивание происходит в обратном порядке.
Внешняя морфология. С внешним строением животного лучше
ознакомиться на свежеубитом экземпляре, хотя для этой цели годятся и
консервированные экземпляры. Вытянутое, червеобразное тело, круглое
в поперечном сечении, постепенно суживается к переднему концу, а сза:
45
ди вытянуто в более узкий хвостовой отдел (рис. 40, Л). Взрослые черви
имеют длину 150—200 мм, а иногда достигают и 300 мм. Наибольшая
ширина' 8—10 мм. Живые черви окрашены в желтовато-серый или
бурый цвет, но сильно пигментированные особи могут быть
темно-зеленого, почти черного цвета. В средней области тела бросаются в глаза
ярко-красные кустики жабр (рис. 40, Л, 7). Поверхность кожи
шероховатая, покрыта многочисленными мелкими бородавочками, или папил-
лами. Наружная сегментация выражена отчетливо, хотя и несколько
маскируется вторичной кольчатостью. Каждый типичный сегмент
подразделяется внешне на пять вторичных колец (anrnili, рис. 40, IV—XIX;
рис. 43, X). По сравнению с Nereis, бросается в глаза слабое развитие
простомиума и параподии.
Простомиум можно видеть, рассматривая передний конец
животного со спинной стороны (рис. 41). Это небольшое трехлопастное
образование,,, достигающее у взрослых животных 1 мм в длину и 2 мм в
ширину (рис. 40, 3; рис. 41, 3). Простомиум способен впячиваться вместе с
полукруглой обонятельной ямкой, которая лежит у его основания
(рис. 41, 9). Он лишен чувствительных придатков и снабжен глазами,
которые могут быть обнаружены лишь на срезах. Два передних
колечка, прилегающие сзади к простомиуму, представляют собой перистоми-
. ум, который также лишен каких бы то ни было придатков
(см. ркс. 40, 4; рис. 41, 4). С брюшной стороны на его переднем конце
расположено ротовое отверстие, хорошо.заметное, когда буккальный
отдел и глотка втянуты внутрь. Ротовое отверстие имеет вид полулунной
поперечной щели, спереди оно ограничено маленькой верхней губой и
со всех сторон окаймлено рядом папилл (см. рис. 40, Б, 13). Следующие
два кольца, по-видимому, являются сегментами, утратившими
параподии (см. рис; 40, 5; рис. 41, 5).
Все остальное тело червя может быть разделено на три области
(см. рис. 40, Л): 1) шесть передних сегментов с параподиямй, но без
жабр (сегменты I—VI); 2) 13 средних сегментов с параподиямй и с
жабрами (сегменты VII—XIX); 3) хвостовой отдел, лишенный и пара-
подий и жабр. Как уже упоминалось, каждый типичный сегмент состоит
из пяти колец. Одно из них — предпоследнее, или четвертое, несет
параподии (см. рис. 40, А; рис. 43).
Таким образом, граница между сегментами отвечает задней
границе кольца, следующего за' щетинконосным. Такое строение имеют все
16 щетинконосных сегментов, кроме трех передних. I сегмент слагается
из двух колец, причем параподии помещаются на переднем кольце
(см. рис. 40, I; рис. 4Г, /). II сегмент состоит из трех колец; среднее из
них —с параподиямй (см. рис. 40, Л и Б, II). Такое же строение имеет
следующий, III сегмент у разновидности из зоны ламинарий (см.
рис. 40, Л, III), но у литоральной разновидности в этом сегменте
впереди щетинконосного кольца имеется не одно, а два простых кольца, т. е^.
весь сегмент слагается из четырех колец (см. рис. 40, Б, III). Хвостовой
отдел, лишенный параподии и жабр, значительно варьирует у разных
экземпляров. Обычно, когда он вытянут, он подразделяется серией
перетяжек, которые отвечают границам сегментов (см. рис. 40, Л, 8).
На конце последнего сегмента лежит округлое анальное отверстие (см.
46
Рис. 40. Arenicola marina. А —
разновидность из зоны ламинарий, вид с
правой стороны, буккальный отдел и
глотка полностью выпячены; Б —
передний конец литоральной
разновидности, вид с брюшной стороны,
буккальный отдел и глотка втянуты (по
Эшворту):
/ — глотка, 2 — буккальный отдел, 3 — про-
стомнум, 4 — перистомиум, 5 — передние
сегменты, утратившие параподии, 6 — но-
топодия, 7 — жабра, 8 — хвостовой отдел,
9 — анальное отверстие, 10 — невроподин,
// — метастомиальная бороздка, 12—
брюшная бороздка, 13 — ротовое
отверстие. 1—XIX — щетинкоиосные сегменты
Рис. 41. Arenicola marina. Передний
конец, вид со спинной стороны, рук-
кальный отдел выпячен лишь
отчасти. Простомиум выпячен вполне
(по Эшворту): х
/ — первый щетиикоиосный сегмент, 2 —
буккальный отдел, 3 — простомиум, 4 —
перистомнум, 5 — передние сегменты,
утратившие параподии, 6 — нотоподия, 7 —
наружное отверстие статоциста, 8 —
метастомиальная бороздка, 9 — обонятельная
ямка
2
рис. 40, 9), из которого иногда может выпячиваться небольшой участок
задней кишки.
В онтогенезе из задней зоны роста образуется около 80 хвостовых
сегментов. Сегменты, которые лежат спереди, остаются неразвитыми,
короткими, они многочисленны. При утрате хвоста они быстро растут в
длину, восстанавливая нормальную длину хвостового отдела. Такое
приспособление к быстрой регенерации объясняется тем, что хвостовой
отдел пескожила часто отрывается хищниками, преимущественно
рыбами и птицами.
Параподии хорошо развиты на средних жаброносных сегментах, но
по сравнению с параподиямй Nereis значительно редуцированы. Каждая
47
параподия состоит из изолированных и различных
по строению нотоподий (спинная ветвь) и невропо-
дии (брюшная ветвь). Нотоподий — это конические
возвышения, расположенные дорзолатерально (см.
рис. 40, А, 6; рис. 43, 5). На закругленном конце
каждой помещается отверстие щетинконосного
мешка (см. рис. 43, 6); из него торчит пучок волосковид-
ных щетинок. Каждая невроподия (torus uncinige-
rus) слагается из двух, плотно прижатых друг к
другу, поперечных мясистых валиков, лежащих вентро-
латерально (см. рис. 40, А, 10; рис. 43, 3). Узкая
щель между валиками является отверстием
щетинконосного мешка; из нее выдаются кончики одного
ряда крючковидных щетинок. Параподиальные усики и
опорные щетинки (acicula) отсутствуют. Первые пять
пар невроподий малы, но в задней части
жаброносного отдела они сильно развиты, их брюшные концы
медиально почти соприкасаются (см. рис. 40, А, 10).
Отделив щетинки нотоподий и невроподий, следует
изучить их под микроскопом (рис. 42).
13 пар жабр располагаются на VII — XIX
сегментах (см. рис. 40, А, 7). Это нежные, сильно
разветвленные придатки, красный цвет которых зависит от
пронизывающих их кровеносных сосудов. Первая
пара жабр гораздо меньше остальных. У середины
жаброносного отдела эти органы достигают
наибольшего развития. Жабры прикрепляются
непосредственно позади нотоподий (рис. 43,1). Форма жабр
несколько различается у двух основных разновидно-
. стей A. marina. У литоральной формы они в виде
кустиков. Каждая жабра состоит из 8—12 радиально
расположенных^ жаберных стволиков.. (2—3 мм
длиной), древовидно ветвящихся. Основания стволиков связаны тонкой
мембраной. У A. marina из зоны ламинарий жабры развиты лучше.
От каждого из 11—14 стволиков (5—6 мм длиной) отходит до 12 пар
боковых чередующихся веточек. Последние разделяются
дихотомически, образуя 20—25 жаберных нитей (рис. 43, 1).
Вдоль всего тела по брюшной медиальной линии проходит
неглубокая бороздка. Она отмечает положение брюшного нервного ствола
(см. рис. 40, Б, 12). Несколько впереди от первого щетинконосного
кольца эта брюшная бороздка раздваивается. Ее ветви следуют по боковым
поверхностям перистомиума в переднеспинном направлении и
называются метастомиальными бороздками (см. рис. 40, Б, 11; рис. 41, 8).
Под ними залегают нервные окологлоточные коннективы.
Наружные отверстия шести пар нефридиев малы и могут быть
обнаружены с трудом. Они щелевидной формы и лежат несколько выше-
и сзади верхних концов невроподий на IV—IX сегментах (рис. 43, 4).
Живых червей можно заставить вывернуть буккальный отдел и
глотку, если надавливать пальцами на передний конец тела. Следует
48
Рис, 42. Щетинки
Arenicola marina.
А — волосковид-
ная щетинка
нотоподий; Б — крюч-
ковидная щетинка
невроподий (ориг.)
рассмотреть эти отделы
пищеварительного тракта в выпяченном
состоянии. У молодых особей они
обычно розового цвета, но у
взрослых сильно
пигментированы. Буккальный отдел
представляет собой мешковидный,
расширенный впереди, довольно
объемистый орган (см. рис. 40, Л,
2). В отличие от Nereis, у
Arenicola выворачивается также
часть глотки, которая
оказывается при.этом впереди
буквального отдела (см. рис. 40, 1).
Поверхность буккального отдела
несет несколько рядов изогнутых
папилл. Более или менее
округлая глотка покрыта мелкими
папиллами.
Анатомия. Arenicola
представляет собой прекрасный
объект для анатомического
вскрытия. Вскрывать следует только
что пойманных свежеубитых
хлороформом животных. Для этого
их помещают в банку с морской водой, в которой предварительно
разбалтывается небольшое количество хлороформа. При этом животные
постепенно наркотизируются, оставаясь в более или менее расслабленном
состоянии, что очень важно, так как иначе при вскрытии тело сильно
сокращается, а нередко и разрывается. Вскрывать следует в морской
воде со спинной стороны, приколов животное булавками ко дну
ванночки. Две булавки втыкаются в бока I сегмента, две другие — в бока
последнего жаброносного сегмента. Хвостовой отдел укрепляется одной
или двумя булавками. Разрез стенки тела рекомендуется производить
несколько сбоку (например, справа) от спинной медиальной линии.
Нередко через разрез выпячивается средняя часть кишечника, что
зависит от давления полостной жидкости и сокращения кожно-мускульно-
го мешка.
После ознакомления с общей топографией органов (рис. 44)
следует перейти к их систематическому изучению.
Стенка тела, мускулатура и полость тела. Кожно-
мускульный мешок состоит, как и у других полихет, из наружного
эпидермиса, покрытого кутикулой, лежащего под ним слоя кольцевых
мышечных волокон и внутреннего продольного мускульного слоя.
Последний хоройю виден на внутренней,поверхности стенок тела (рис. 44, 19;
рис. 45, 12). Он состоит из многочисленных продольных мускульных
пучков, вдающихся в полость тела в виде невысоких валиков (рис.
46, 6). Со стороны полости тела продольная мускулатура одета пери-
тонеальным эпителием. Этот мышечный слой разделяется на три про-
Рис. 43. Часть девятого сегмента и десятый
щетинконосный сегмент Arenicola marina
из зоны ламинарий! Вид справа (по Эш-
ворту):
/ — жабра, 2 — мембрана между основаниями
жаберных стволиков, 3 — невроподия, 4 — нефро- ■
пор, 5 — иотоподиальный бугорок, 6 — отверстие
щетинконосного мешка, 7 — основание обрезанной
' жабры
49
дольных поля, будучи прерван брюшным
нервным стволом, а также справа и
слева на уровне нотоподий. В каждом ще-
тинконосном сегменте имеется одна пара
хорошо развитых щетинконосных ното-
подиальных мешков (см. рис. 44, 23,
рис. 45, 6; рис. 46, 5). В отличие от
таковых у Nereis, они лишены опорных
щетинок (acicula). Щетинконосные
мешки невроподий редуцированы. Вершина
каждого нотоподиального мешка связана
27 с ближайшими участками стенок тела
несколькими мускулами-протракторами
щетинок (рис. 45, 5; рис. 46, 9). Кроме
того, каждый мешок имеет длинный и
тонкий ретрактор щетинок, тянущийся
от его вершины поперек полости тела и
прикрепляющийся к стенке тела сбоку
от брюшной нервной цепочки (рис. 45,
4). Сокращением этого мускула
щетинки нотоподий могут полностью
втягиваться.
Диагональные мышцы (m. transver-
si obiiqui) развиты хорошо. Они имеют
вид плоских лент, пересекающих полость
тела. Их вентральные концы
прикрепляются по сторонам от брюшного
нервного ствола, противоположные боковые
концы — к стенкам тела несколько ниже
уровня нотоподий. В каждом типичном
сегменте находится 5—7 пар таких мышц
(см. рис. 44, 21; рис. 45, 13). На перед-
Рис. 44. Arenicola marina, вскрытая, со спинной
стороны (по Эшворту):
/ — ротовое отверстие, 2 — глотка, 3 — I диафрагма,
4— спинной мезентерий, 5 —пищевод, 6— II диафрагма,
7—III диафрагма, 8 — спиииой сосуд, 9 — латеральный
сосуд пищевода, 10 — слюнная железа, // — сосуд
слюнной железы, 12 — «желудочек», i3 — «предсердие», 14 —
латеральный гастрический синус, 15 — хлорагогеиная
ткаиь, 16 — субиитестииальиый синус, П — брюшной
сосуд,' 18 — приносящий сосуд жабры и нефридия, 19 —
продольная мускулатура, 20 — брюшной нервный ствол,
21 — диагональные мышцы, 22 — дорзолатеральиый
продольный сосуд, 23 — щетинкоиосный мешок нотоподий,
24 — воронка нефридия пятой пары, 25 — гонада, 26 —
соединительнотканный тяж, связывающий кишечник со
стенкой тела, 27 — воронка нефридия второй пары, 28 —
брюшиой мезентерий, 29 — слепое выпячивание
I—диафрагмы, 30 — ретракторы глотки, 31 — папиллы буккаль-
ного отдела, 32 — нитевидные выросты брюшного сосуда,
33 — диссепимент между XVII и XVIII щетинконосными
сегментами, 34 — диссепимеит хвостового отдела, 35 —
дорзовентральиый кольцевой сосуд, 36 — приносящий
жаберный сосуд, 37 — выносящий жаберный сосуд, 38 —
невральный продольный сосуд, 39 — анальное отверстие.
Н1—На — нотоподий, Ж1—Ж13 — жабры
нем конце тела хорошо
заметны веерообразно
расположенные ретракторы глотки (см.
рис. 44, 30). Начинаясь на
поверхности заднего отдела
глотки, они тянутся назад к
стенкам тела и проходят сквозь
первую диафрагму
(диссепимент) .
Вторичная полость тела
развита хорошо. В переднем
отделе тела, целом разделен
тремя хорошо развитыми дис-
сепиментами — диафрагмами.
Передняя диафрагма
прикрепляется к стенке тела на
передней границе первого щетинко-
носного кольца, т. е.
отграничивает щетинкоиосный сегмент
спереди от лежащего впереди
участка целома (см. рис. 44,
3). У многих экземпляров
проходящие сквозь нее
ретракторы глотки выпячивают ее
назад, Так что на первый
взгляд ее вентральный край
кажется прикрепленным возле
задней границы первого ще-
тинконосного кольца. Однако
легко убедиться, что это не так.
Вторая и третья диафрагмы
отмечают задние границы II
и III щетинконосных
сегментов. Они прикрепляются на
щетинконосными (см. рис. 44, 6 я 7). Таким образом, III сегмент
изолирован двумя диафрагмами, тогда как I и II имеют общую
полость. Каждая диафрагма прободена многочисленными, обычно
овальными, отверстиями, пропускающими целомическую жидкость, но
задерживающими половые продукты. Первая диафрагма образует пару
пальцеобразных слепых выпячиваний, направленных назад и лежащих
по сторонам и вентрально от пищевода (см. рис. 44, 29). Их стенки
богаты кровеносными сосудами, мускулисты и способны сокращаться.
Функция их неизвестна. Между первой и второй диафрагмами имеются
спинной и брюшной продольные мезентерии, поддерживающие
соответственные кровеносные сосуды и соединяющие стенки тела с пищеводом.
Спереди спинной мезентерий кончается, не достигая первой диафрагмы
(см. рис. 44, 4). Брюшной мезентерий продолжается назад в область
III сегмента (рис. 44, 28). Позади третьей диафрагмы диссепименты
отсутствуют, и в средней части тела целом является общим для всех
2 1 17 Iff 15 Ш 13
Рис. 45. Arenicola marina. Часть VIII и IX
щетинконосных сегментов вскрытого
животного. Изображены правые нефридии V и
VI пары и окружающие их органы (ориг.):
/ — брюшной нервный ствол, 2 — невральный
продольный сосуд, 3 — перерезанная диагональная
мышца, 4 — ретрактор щетинок, 5 — протракторы
щетинок, 6 — щетинкоиосный мешок нотоподий
IX щетинконосного сегмента, 7 —
дорзолатеральиый сосуд, 8 — мочевой пузырек,- 9 —
экскреторный отдел нефридия, 10 — воронка (гоностом>
нефридия, Л — спинная губа воронки,
12—продольная мускулатура, 13 — диагональная мышца,
14 — брюшиая губа воронки, 15 — нефридиальный
сосуд, 16 — выносящий жаберный сосуд, 17 —
приносящий сосуд жабры и нефридия, 18 —
продольный нефридиальный сосуд, 19 — сосуд,
соединяющий оба невральных сосуда
задних краях колец; следующих за
51
Рис. 46. Arenicola marina. Передний конец животного, вскрытый со
спинной стороны. Глотка и первая диафрагма удалены (по Эшворту):
1 — простомиум, 2 — передняя лопасть церебрального гаиглия, 3 — статоцист,
4 — место прикрепления I диафрагмы, S — щетинкоиосиый мешок иотоподии I ще-
тиикоиосного сегмента, 6 — продольная мускулатура, 7 — брюшной нервный ствол,
8 — ретрактор щетинок, 9 — протракторы щетинок, 10 — окологлоточный коннек-
тив, // — нерв статоциста, 12 — задняя лопасть церебрального ганглия, 13 —
церебральная коммиссура ч
сегментов. Однако на некоторых поперечных кровеносных сосудах
(приносящие и выносящие жаберные сосуды) имеются рудименты дис-
сепиментов. Особенно хорошо они заметны в задних щетинконосных
сегментах, из которых два последних имеют почти полные диссепименты
(см. рис. 44, 33). В хвостовом отделе полость тела сравнительно узка
и пересекается спинным и брюшным мезентерием, а также
многочисленными диссепиментами (см. рис. 44, 34).
Так же как у Nereis, целом образует многочисленные каналообраз-
ные выпячивания, выстланные перитонеальным эпителием, которые
проникают в толщу кожно-мускульного мешка и нередко
сопровождаются кровеносными сосудами. При вскрытии эти кожные каналы
хорошо видны на переднем конце тела.
Пищеварительная система. Пищеварительный канал
состоит из буккального отдела, глотки, пищевода, средней кишки,
передняя часть которой несколько отличается строением и называется
«желудком», и задней кишки. Глотка хорошо заметна, она имеет округлые
очертания (см. рис. 44, 2). К заднему отделу глотки прикрепляются
многочисленные ретракторы. Пищевод представляет собой
цилиндрическую тонкостенную трубку, способную значительно растягиваться.
В спокойном состоянии задний его конец достигает VI щетинконосного
сегмента (см. рис. 44, 5). В заднюю часть пищевода открывается пара
продолговатых «слюнных» желез, обычно зеленоватого цвета. Каждая
52
из них имеет короткий проток, на котором сидит, как на стебельке
(см. рис. 44, 10). Следующий отдел — желудок — самая широкая и
объемистая часть пищеварительного тракта. Он покрыт желтыми хлораго-
генными клетками. На поверхности его хорошо заметна сложная сеть
кишечных кровеносных синусов и сосудов, которые делят поверхность
■органа на отдельные овальные площадки (см. рис. 44, 15). В области
XII или XIII сегмента желудок суживается и без резких границ
переходит в собственно кишку. Задний отдел последней представляет неясно
дифференцированную заднюю кишку, кончающуюся на конце тела
анальным отверстием (см. рис. 44, 39)..
У Arenicola, находящихся в естественных условиях, в норке,
буквальный отдел и глотка непрерывно то выпячиваются, то втягиваются.
При этом захватывается песок, который сначала поступает в пищевод.
'Проходя по пищеводу, песок смешивается со слизью, выделяемой
клетками, выстилающими эту часть кишечного тракта, а в задней части
пищевода подвергается действию секрета слюнных желез.
Образовавшаяся таким образом смесь,поступает далее в желудок, где к ней
примешиваются еще выделения слизистых и пищеварительных клеток
желудка. Вследствие сокращения мускулатуры стенок тела, а также
выпячивания и втягивания переднего отдела кишечного тракта желудок
совершает ритмические движения вперед и назад. Это облегчает
перемешивание песка и пищеварительных соков. Всасывание переваренных
питательных веществ происходит преимущественно в желудке. Таким
образом, Arenicola, подобно Lumbricus и Sipunculus, пропускает сквозь
кишечник большие количества грунта, изменяя его химические и
физические свойства. Пройдя сквозь кишечник, песок (экскременты)
выбрасывается из норки, образуя у заднего конца последней небольшую
кучку.
Органы дыхания. Каждая из 13 пар жабр представляет собой
вырост стенки тела, в который продолжается целом. Стенки жабры
очень тонки; они слагаются из наружного эпителия, под которым
залегает слой мускульных волокон. Полость жабры выстлана
перитонеальным эпителием. Между последним и мускульным слоем проходят
кровеносные сосуды. Целомическая полость жабры пересекается
мускульными тяжами; жабры способны к сильным сокращениям. Еще Мильн-
Эдвардс считал, что жабры Arenicola регулярно сокращаются,
выталкивая окисленную кровь в выносящие жаберные сосуды. Когда
Arenicola лежит в норке, не меняя места, по ее телу проходят волны
вздутий от заднего конца к переднему^ В результате этих дыхательных
движений вода в норке все время обновляется. При расширении жаб-
роносных сегментов жабры сжимаются, благодаря чему достигается
обновление в них крови. -
Кровеносная система. Наполненные красной кровью,
содержащей гемоглобин, кровеносные сосуды отчетливо видны на вскрытом
животном. Как и у Nereis, главными кровеносными сосудами
являются спинной и брюшной продольные сосуды.
Спинной сосуд (vas dorsale) начинается вблизи анального
отверстия к тянется над пищеварительным каналом до переднего конца, где
распадается на мелкие сосуды, и капилляры (см. рис. 44, 8). Ритмиче-
53
ски сокращаясь, он прогоняет кровь от заднего конца тела к
переднему. В хвостовом отделе спинной сосуд связан с брюшным посредством;
кольцевых сосудов, идущих по поверхности кишки. В каждом сегменте-
имеется одна пара таких дорзовентральных сосудов (см. рис."-44, 35).
В задней части жаброносного отдела в спинной сосуд открываются,
семь пар (от семи пар жабр) выносящих жаберных сосудов,
располагающихся метамерно и отвечающих параподио-дорзальным сосудам.
Nereis (см. рис. 44, 37). Между ближайшими парами выносящих
сосудов имеется две или три пары дорзовентральных сосудов. Впереди 7-й:
пары жабр спинной сосуд не принимает в себя выносящих жаберных,
сосудов. Здесь он тесно связан многочисленными соединениями с
кишечным кровеносным плексусом (кишечным синусом, см. рис. 44, 8)..
Впереди желудка в области пищевода спинной сосуд принимает
следующие парные сосуды от: 1) первых трех пар нефридиев и
прилегающих к ним участков кожно-мускульного мешка, 2) второй диафрагмы,
и стенок тела в области 2-й пары нотоподий, 3) стенок тела,
прилегающих ко второй диафрагме, и стенок тела в 1-й паре нотоподий.
Впереди первой диафрагмы спинной сосуд дает капиллярную сеть буккаль-
ного отдела, глотки с ее мускулатурой, стенок тела и т.д.
Из этих органов кровь собирается по мелким сосудам в переднюю-
часть брюшного сосуда (vas ventrale). Этот последний проходит под.
кишечником до заднего конца тела (см. рис. 44, 17). Он дает
небольшую ветвь — медиальный сосуд первой диафрагмы и ее мешков, а на-
уровне второй пары нотоподий-—одну пару сосудов к брюшному
нервному стволу и другую пару — к стенкам тела (вентро-париетальные
сосуды). Несколько сзади от него ответвляется медиальный сосуд
второй диафрагмы, а также сосуды к брюшному нервному стволу и
мелкие сосуды третьей диафрагмы и первой пары нефридиев. От этого
места брюшной сосуд тянется назад, снабжая кровью посредством
сегментарных боковых парных сосудов (вентро-париетальных) щетинконос-
ные мешки, стенку тела, нефридии и жабры (см. рис. 44, 18, 36; рис..
45, 17). В области кишки брюшной сосуд связан со спинным
посредством дорзовентральных сосудов. Вблизи анального отверстия он
распадается на капилляры.
На стенках кишки и особенно желудка развиты многочисленные-
кровеносные синусы, образующие сложный кишечный плексус. У
молодых экземпляров он представляет собой сеть мелких сосудов, но у~
взрослых эти сосуды превращаются в систему синусов. У старых
особей можно говорить о наличии почти сплошного кишечного синуса,,
расположенного между кишечным эпителием и перитонеумом,
покрывающим кишечник. Спинной сосуд всегда хорошо различим, хотя в
области желудка тесно связан с кишечным плексусом. На кишечном плек-
сусе дифференцированы, во-первых, два продольных синуса, лежащие-,
вентрально, и, во-вторых, пара продольных латеральных гастрических'
синусов. Первые называются субинтестинальными синусами. Они
начинаются на переднем конце желудка (см. рис. 44, 16) и могут
простираться до XII щетинконосного сегмента, позади которого постепенно-
исчезают. Субинтестинальные синусы принимают выносящие сосудьЕ
первых пяти пар жабр (см. рис. 44, 37; рис. 45, 16).
54
Таким образом, в области желудка все поперечные сегментарные
сосуды связаны либо с брюшным сосудом, либо с брюшной стороной
кишечного тракта. Кроме того, они здесь длиннее, чем в других
участках тела. Это связано с сильной подвижностью желудка, который
совершает постоянные ритмические движения в полости тела. В качестве
-приспособления, ограничивающего эти движения желудка и таким
образом предохраняющего сегментарные сосуды от возможного разрыва,
имеется пара прочных тонких соединительнотканных тяжей,
связывающих передний конец желудка с боковыми стенками гела. Они проходят
рядом с сегментарными сосудами, идущими от брюшного сосуда к
четвертой паре, метанефридиев и соседним с ними участкам кожно-мус-
жульного мешка (см. рис. 44, 26). Латеральные гастрические синусы
(см. рис. 44, 14) теряются позади середины желудка. Каждый
принимает кровь из спинного сосуда и субинтестинального синуса своей
стороны через плексус и образует впереди тонкостенное вздутие, называемое
«предсердием» (см. рис. 44, 13). С ним связан «желудочек»,
представляющий мешковидное расширение (см. рис. 44, 12). Итак, на границе
пищевода и желудка с каждой стороны имеется по «сердцу»,
состоящему из предсердия и желудочка. Желудочки обладают мускулистыми
стенками и, пульсируя, выталкивают кровь в брюшной сосуд. В
желудочках старых экземпляров встречается так называемое сердечное т-е--
ло. Латеральные синусы продолжаются в виде сосудов, вдоль
пищевода и, не достигая первой диафрагмы, распадаются на капилляры.
По сторонам брюшного нервного ствола проходят
сопровождающие его тонкие продольные невральные сосуды (см. рис. 44, 38;
рис. 45, 2). Они начинаются впереди в области окологлоточных кон-
нективов, где связаны с капиллярной сетью. В передних сегментах (от
II до IV включительно) с ними соединяются ветви сегментарных
сосудов, отходящих от брюшного сосуда. Сосуды стенок тела хорошо
развиты. Среди них выделяются дорзолатеральные продольные сосуды,
проходящие выше уровня нотоподий (см. рис. 44, 22; рис. 45, 7).
Кроме того, имеется еще одна пара продольных более тонких нефридиаль-
ных сосудов, которые тянутся тоже на внутренней поверхности стенок
тела, но ниже нотоподий и нефридиев. Они хорошо заметны впереди и
позади каждого нефридия (см. рис. 45, 18). Дорзолатеральные сосуды
становятся различными позади первой пары нотоподий и
простираются до заднего конца тела.
В каждом сегменте между продольными сосудами
кожно-мускульного мешка развиты следующие сегментарные поперечные сосуды:
1) короткие поперечные сосуды, соединяющие на каждой стороне дор-
золатеральный сосуд с нефридиальным; 2) кольцевые сосуды,
связывающие оба дорзолатеральных сосуда; они проходят по спинной
поверхности кожно-мускульного мешка; 3) сосуды, соединяющие на
каждой стороне нефридиальный сосуд с невральным; 4) короткие сосуды,
соединяющие невральные сосуды поверх брюшного нервного ствола
(см. рис. 45, 19).
Дорзолатеральные и нефридиальные сосуды снабжают кровью
кожно-мускульный мешок в спинной и боковых областях, тогда как
невральные сосуды снабжают нервный ствол.и брюшную область кож-
55
но-мускульного мешка. Все эти сосуды хорошо заметны без вскрытия
на маленьких молодых экземплярах, так как просвечивают сквозь
покровы.
Брюшной кровеносный сосуд на значительном протяжении покрыт
многочисленными темно-коричневыми нитями, развитие которых
зависит от величины и возраста животного (см. рис. 44, 32). Они
многочисленнее и лучше выражены у наиболее старых животных, у которых
развиваются и на других сосудах в жаброносном отделе. Каждая
такая нить представляет собой слепую веточку сосуда, покрытую хлораго-
генными клетками.
Выделительная система. Шесть пар крупных нефридиев,.
лежащих под диагональными мышцами, т. е. в нефридиальных камерах,
целома, принадлежат IV—IX сегментам (см! рис. 44). Их наружные
отверстия — нефропоры помещаются на поверхности щетинконосных
колец несколько дорзальнее и позади невроподий (см. рис. 43, 4).
Нефридий Arenicola представляет собой нефромиксий, т. е. каждый из них:
есть продукт слияния метанефридия с половой воронкой (гоностомом),,
причем настоящий нефростом не сохраняется. Это характерно для
многих полихет. Таким образом, воронка нефридия Arenicola
гомологична це нефростому Nereis, а цилиарному органу.
Следует вырезать один из нефридиев и рассмотреть его
внимательно в морской воде под бинокуляром. Каждый нефридий состоит из трех
частей: воронки (гоностом), среднего экскреторного отдела и задней
части — мочевого пузырька (см. рис. 45). Воронка велика. Она
открывается- в целом вытянутой щелью, которая обращена главным образом:
вперед и внутрь и ограничена спинной и брюшной губами (см. рис. 45„
11 и 14). Спинная губа воронки несколько крупнее брюшной, бахром-
чата, так как покрыта рядом лопастных придатков, содержащих
кровеносные полости, от чего зависит их красный цвет. Внутренняя
поверхность воронки одета ресничным эпителием. Экскреторный отдел
представляет собой тонкостенный вытянутый мешок, в который спереди
открывается воронка, а сзади он сообщается с полостью мочевого
пузырька. Благодаря наличию в клетках "эпителия многочисленных
коричневых гранул, он обычно имеет темно-коричневый цвет (см. рис.
45, 9). Мочевой пузырек окрашен в зеленоватые или коричневатые
тона. В наполненном состоянии форма его почти шаровидна, но при
сокращении, спадаясь, он становится розетковидным (см. рис. 45, 8)..
Воронка и передняя часть экскреторного отдела прикрепляются к
стенке тела тонким мезентерием; воронка, кроме того, удерживается в
своем положении одной из диагональных мышц. Воронки первой пары;
нефридиев располагаются на передней стороне третьей диафрагмы.
Каждый нефридий снабжается специальными кровеносными сосудами-
(см. с. 55). Нефридии Arenicola помимо экскреторной функции
выделительных органов несут функцию выводных половых путей.
Нервная система. Центральная нервная система состоит из-
надглоточных церебральных ганглиев, или мозга, окологлоточных кон-
нективов и брюшного нервного ствола. Чтобы обнаружить передний
отдел нервной системы, нужно удалить глотку и буккальный отдел и
осторожно препарировать сначала окологлоточные коннективы, начаа
56
с места соединения их с брюшным нервным стволом, а затем и мозг.
Обнаружить коннективы нетрудно: их положение отмечено снаружи
•метастомиальными бороздками.
Церебральные ганглии расположены в простомиуме и у крупных
экземпляров достигают 1 мм в длину. Каждый подразделяется на
переднюю и заднюю лопасти (рис. 46, 2, 12). Ганглии соединяются
широкой коммиссурой (рис. 46, 13). Передние лопасти посылают серию
дорзальных нервов к эпителию простомиум.а и несколько вентральных
нервов к верхней губе и околоротовому пространству. Окологлоточные
коннективы отходят от боковых поверхностей ганглиев (рис. 46, 10).
Несколько впереди третьего щетинконосного кольца они соединяются
с брюшным нервным стволом (рис. 46, 7). От них берут начало нервы
статоцистов (рис. 46, 11) буккального отдела и многочисленные нервы,
иннервирующие перистомиум и первые, лишенные параподий,
сегменты.
Брюшной нервный ствол (рис. 46, 7) представляет собой продукт
полного слияния двух нервных стволов. Обычно он отделен от
эпидермиса слоем кольцевой мускулатуры, но у некоторых особей в хвостовом
отделе и последних щетинконосных сегментах прилегает
непосредственно к эпидермису. Брюшные сегментарные ганглии отсутствуют; гангли-
озные клетки покрывают брюшной ствол по всему его протяжению.
В каждом сегменте от брюшного ствола отходит несколько пар
боковых нервов, из которых наиболее развиты параподиальные нервы. .
Органы чувств. Помимо рассеянных чувствительных клеток в
эпидермисе тела в папиллах буккального отдела и в других местах
имеются локализированные органы чувств: статоцисты, обонятельный
орган и'глаза.
Статоцисты видны при вскрытии. Их надо искать у наружного края
дорзолатеральных частей окологлоточных коннективов (рис. 46, 3).
Каждый статоцист является небольшим пузырьком, связанным с
наружной средой посредством узкого канала. Наружное отверстие
последнего ,лежит на перистомиуме вблизи пересечения метастомиальной
борозды с первой интераннулярной бороздой (см. рис. 41, 7). Под
микроскопом внутри статоциста видно несколько конкреций — слуховых
камешков, или отокониев, неправильной формы. Статоцисты удобно
наблюдать на молодых прозрачных экземплярах. При наблюдении in
vivo видно, что отоконии совершают постоянные колебательные
движения.
Обонятельный орган имеет вид U-образной ресничной ямки,
расположенной на переднем крае простомиума (см. рис. 41, 9). Постлар-
вальиая стадия обладает 2—5 парами глаз. Снаружи они заметны
только у очень молодых особей, а на срезах могут быть обнаружены
.лишь у экземпляров до 70 мм длиной. Они погружены в ганглиозные
клетки на спинной стороне мозга. У'взрослых особей найти глаза даже
на срезах очень трудно.
Половые органы. У половозрелых индивидуумов нетрудно
-обнаружить половые железы. Гонады расположены сегментарно по оя-
ной на поверхности каждого нефридия, исключая нефридии первой
пары. Они лежат позади задней части воронки и имеют вид маленьких
57
розоватых овальных телец размером от 0,4 до 1 мм. (см. рис. 44, 25).
Зрелые половые продукты свободно плавают в полости тела, нередка
скопляясь в нефридиальных камерах целома. Для выведения половых
продуктов наружу служат последние пять пар нефридиев.
Внешняя морфология различных Polychaeta
Благодаря высокой пластичности организации Polychaeta
приспособились к разнообразному образу жизни. В связи с этим и внешняя ш'
внутренняя морфология в различных отрядах, семействах и даже родах
варьирует в широких пределах.
В целях ознакомления с внешними морфологическими
особенностями мы рекомендуем изучить возможно большее число
представителей из разных отрядов и семейств. Это удобнее всего сделать, работая
на берегу моря, где можно получить различных полихет. Однако можно-
использовать и фиксированный материал. Следует обращать внимание
на характер сегментации, строение отдельных сегментов и параподий с
их придатками, характер и особенности прсстомиума и перистомиума
и их придатков, устройство буккального отдела и его вооружение и т.д.
Для определения полихет могут служить определители Фовелз
(1923 и 1927), Фридриха (1938) и П. В. Ушакова (1955).
Сегментация. За редкими исключениями сегментация
выражена отчетливо. Обычно тело слагается из многочисленных сегментов,
подобных друг другу (гомономная сегментация у Nephthydae, Glyceri-
dae, Syllidae, Eunicidae и др.). Но для некоторых форм, напротив,
характерно небольшое число сегментов (Spirorbis, Hesionidae). У Seden--
taria тело разделяется на грудной (thorax) и брюшной (abdomen)
отделы (гетерономная сегментация, рис. 47, Г). Иногда налицо особый
хвостовой отдел (Arenicola, Pectinaria). У видов Spirorbis на
нескольких сегментах между грудью и брюшком параподий отсутствуют.
В некоторых случаях гетерономность сегментации туловища
связана с различием в строении ларвальных и постларвальных сегментов.
Даже по чисто внешним признакам у некоторых взрослых полихет
можно отличить передние дарвальные сегменты от последующих
постларвальных (Chaetopterus, Spionidae, Serpulidae и др). Число ларваль-'
ных сегментов у полихет разных семейств варьирует от 3 до 13, но в
пределах одного семейства оно постоянно.
Простомиум. Головная лопасть хорошо развита у Errantia, но
у Sedentaria низведена до степени незначительного придатка. У Serpu-
limorpha она совершенно закрыта перистомиумом, что дало повод Бен-
хаму разделить Polychaeta на Phanerocephala и Cryptocephala. У
Errantia простомиум имеет более или менее округленную или коническую
форму (см. рис. 25, Л, 12). У Nephthys он квадратный, а у Glycera
конусовидный и подразделен поперечными бороздками на мелкие колечки
(рис. 47, Б, 2). Простомиум обычно снабжен характерными чувстви-
Рис. 47. Различные виды Polychaeta. A—Harmathoe imbricata; 5 —Передний конец.
тела. .Glycera capitata; В — Cirratulus; Г — Amphitrite; Д — Amphicora fabricia; E —
Spirorbis borealis (ориг):
1 — медиальная антенна, 2 — простомиум, 3 — пальп, 4 — спиииые усики, 5 — щетиики иевроподии„
6 — щетиики нотоподии, 7 —элитры, 8— аиальные уснки, 9 — боковые антенны, 10 — аитеины, 11—
5
тельными выростами и другими органами чувств, которые наиболее
полно выражены у Errantia (рис. 47, А и Б). Именно здесь могут быть
щупальца (antennae), пальпы (palpi), глаза и ресничные обонятельные
ямки или бороздки'. Наиболее характерны пальпы, встречающиеся
обычно в количестве одной пары. Они хорошо развиты /у Nereidae, Ро-
lynoidae, Syllidae и др. (см. рис. 25, 2; рис. 47, А, 3), но отсутствуют
у некоторых Errantia (например, Nephthydidae). Особое внимание еле-
дует обратить на видоизменения пальпов. Так, у Amphictenidae и Ат-
pharetidae пальпы представлены так называемыми ротовыми
щупальцами. Далее полые щупальца Chlorhaemidae являются гомологами
пальпов так же, как венец перистых «жабр» на переднем конце тела
Serpulimorpha (рис. 47, Д, 17; Е, 26).
Простомиальные глаза представлены чаще всего одной (Phyllodo-
cidae, Nephthys) или двумя парами (Polynoidae, Hesionidae, Syllidae,
Nereidae и др. (см. рис. 25, 3). У Sedentaria простомиальные^ глаза
встречаются реже (Spionidae. Myxicola, Amphicora и Др.). У Sabellidae
и Serpulidae нередко на жаберных нитях имеются жаберные глаза
(Dasychone, Potamilla и др.). Глаза полихет представляют все степени
усложнения, от простых светочувствительных пигментных пятен до
высокоразвитых органов зрения.
Рекомендуется произвести сравнительное изучение простомиума у
представителей различных отрядов и семейств:
1. Большинство Errantia (Nereidae, Polynoidae, Syllidae, Eunici-
dae, Phyllodocidae, Amphinomidae и др.) имеют крупный, хорошо
развитый простомиум округлой, четырехугольной, двулопастной
трапециевидной или конусовидной формы. Обычно имеется одна пара пальпов
и антенны в различном числе (см. рис. 25, 2; рис. 47, А, 1, 3, 9; Б, 10).
Phyllodocidae и Nephthydidae лишены пальпов.
2. У Spionidae простомиум удлинен, прямоугольной формы или
заострен спереди. Имеется пара очень длинных пальпов с целомичес-
ким каналом внутри; они служат для захватывания пищи и при
сильном раздражении легко отваливаются.
3. Иное строение имеет простомиум у Ariciidae, Maldanidae и
части Capitellidae. Он сильно упрощен, обычно имеет вид заостренного
спереди конуса, лишенного каких-либо придатков.
4. У Terebeliidae простомиум имеет характер широкой поперечной,,
гофрированной лопасти, несущей множество (до 100) тонких,
нитевидных очень длинных придатков. В совокупности они отвечают пальпам
других полихет. Высовываясь из трубки, они стелются во все стороны
по поверхности грунта. Брюшная сторона каждой нити имеет
мерцательную бороздку, по которой пищевые частицы подгоняются ко рту.
5. Наконец, у ряда Sedentaria, например, у Sabellidae и Serpulidae,
простомиум более или менее скрыт внутри перистомиума. Пальпы
представлены в виде сильно развитых перистых придатков — «жабр»,
выставляемых из трубки. Они состоят из более или менее многочисленных
жаберных лучей (radioli), несущих боковые веточки — пиннулы (pin-
1 Глаза и обонятельные органы у различных полихет могут быть развиты не
только на простомиуме, но и на других частях тела. Так, у Amphicora пара.глаз
расположена на заднем конце брюшка (см. рис. 47, Д).
60
nulae). Соответственно происхождению этих жабр от пары пальп,
жаберные лучи поднимаются от двух боковых дисковидных оснований
(рис. 47, Д, 17). У Serpulidae за счет головных жабр развиваются две
(Filigrana) или одна (Protula, Splrorbis) крышечка (operculum),
служащая для замыкания входа в трубку (рис. 47, Е, 28). У некоторых
видов Spirorbis крышечка снабжена особой выводковой камерой, в
которой происходит инкубация эмбрионов.
Головные жабры Serpulimorpha в первую очередь служат для
собирания пищи. Они покрыты ресничками, мерцание которых гонит
воду вместе со взвешенными в ней мелкими организмами к ротовому
отверстию червя.
Перистомиум. Следующий за головной лопастью отдел — пе-
ристомиум всегда выражен хорошо; на его брюшной стороне лежит
ротовое отверстие. Обычно перистомиум представляет собой несколько'
слившихся передних сегментов (цефализация). На перистомиуме могут
быть видоизмененные параподии (Phyllodocidae и др.) и в различном;
числе перистомиальные усики, или щупальца (cirri tentaculares, см..
рис. 25, 13).
Параподии. Для изучения параподии вырезаются из сегмента
(можно вырезать целиком и весь сегмент) и рассматриваются в
свежем состоянии под бинокуляром или микроскопом, или же
предварительно из них изготовляется тотальный препарат.
Основной формой являются двуветвистые и так называемые
«полные»'параподии, хорошо выраженные у Nereis (см. рис. 27) и Nephihys
(рис. 48, А). Кроме наличия ясных нотоподии (спинной ветви) и невро-
подии (брюшной ветви), сидящих на общем оснований, такие
параподии характеризуются присутствием опорных щетинок (acicula), одного-
спинного усика (cirrus dorsalis) и одного брюшного усика (cirrus vent-
ralis). Видоизменения, которым подвергаются параподии у многих по-
Рис. 48. Различные формы параподии Polychaeta. А — Nephthys; Б — Phyl-
lodoce; В — Polynoidae, параподия с элитрой; Г — то же с простым cirrus
dorsalis; Д — Syllis; E — Amphicteis; Ж — Aricia; 3—Glycera; И — Sabel-
la; К — Eunice (по Фовелю)
61
лихет, прежде всего выражаются в большей или меньшей редукции
нотоподии. Этот процесс ведет к образованию одноветвистых параподий
(Hesionidae, Eunicidae, Lumbriconereis и др.).
С другой стороны, параподий роющихся в грунте или живущих в
трубках Sedenlaria характеризуются, во-первых, полным обособлением
обеих ветвей и, во-вторых, своеобразным строением невроподии.
Последняя превращается в поперечно-расположенный валик (torus incini-
gerus), лишенный опорной щетинки и несущий один поперечный ряд
крючковидных, унцинальных щетинок (uncini, рис. 48,Я). Полезно
произвести сравнительное изучение параподий различных типов.
1. Двуветвистые полные, параподий характерны для Errantia.
Помимо Nereis они свойственны Nephthys, Еипо'ё, Harmathoe и др.
(рис. 48, А, В, Г, 3).
2. Видоизменением этого типа являются с одной стороны, параподий
Spionidae и, с другой стороны, параподий Ariciidae. У первых обе ветви
хорошо развиты, но усики превращены в пластинки, располагающиеся
позади ното- и невроподии! У Ariciidae обе ветви удлинены,
ланцетовидные или конические и смещены на спинную сторону. Имеется только
спинной усик (рис. 48, Ж).
3. Одноветвистые параподий отличаются более или менее сильной
редукцией нотоподии. В типичном виде они могут быть наблюдаемы у
Lumbriconereis из Eunicidae и у некоторых Sedentaria. В этих случаях
от нотоподии не остается никакого следа. Однако нетрудно наблюдать
переходные состояния, когда нотоподия еще сохраняется в
редуцированном виде или представлена неполно. Так, например-, у некоторых
Hesionidae cirrrus dorsalis хорошо.развит, но сама нотоподия сильно
редуцирована. У Eunice нет уже нотоподиального бугорка, но
сохраняется опорная щетинка и спинной усик (рис. 48, К). Наконец, у Syllidae
и Phyllodocidae от нотоподии остается только спинной усик (рис. 48,
БЯД). '
4. Типичные седентарные параподий свойственны Terebellomorpha,
Serpulimorpha и Maldanidae. Нотоподия имеет форму небольшого
выроста и несет только волосковидные щетинки. Невроподия представлена.
в виде torus uncinigerus и несет либо крюковидные, либо унцинальные
щетинки (рис. 48, И).
5. Наконец, некоторые формы характеризуются полной или почти
полной редукцией обеих ветвей параподий. При этом, однако,
сохраняются хорошо развитые волосковидные или унцинальные щетинки
(некоторые Capitellidae и Maldanidae).
Характер и расположение параподий в разных участках тела могут
быть неодинаковыми. Особый интерес представляет инверсия щетинок
(inversio setarum) у Serpulimorpha. Именно у этих полихет на передних
сегментах спинной стороны лежат нотоподиальные бугорки с волоско-
видными щетинками и вентральнее их располагаются -tori uncinigeri,
тогда как на задних сегментах расположение элементов параподий
обратное (рис. 49).
Щетинки. Для детального изучения щетинок следует, вырезав
параподию, раздавить ее под покровным стеклом и рассматривать под
микроскопом. Нередко щетинки в одной и той же- параподий имеют
62
различное строение и правильно локализованы. Поэтому помимо
простого изучения щетинок на раздавленной параподий необходимо
обратить внимание на их расположение на неповрежденной параподий.
Несмотря на большое разнообразие, щетинки могут быть разделены на
следующие типы.
1. Сложные, состоящие из длинного стебелька, к дистальному
концу которого прикрепляется терминальная часть щетинки. Такие щетинки
характерны для Errantia (см. рис. 28; рис. 50, Б, И).
2. Простые, которые подразделяются на: 1) волосковидные
—длинные, часто очень тонкие, гладкие, зазубренные или разветвленные
щетинки и т. д. Иногда они окаймлены тонкой краевой мембраной. Форма их
чрезвычайно разнообразна (Nephthys, Arenicola и др.) (рис.50, Ж, К, Л;
рис. 42, А); 2) крючковидные —более мощные, чем предыдущие, с
загнутым и заостренным концом, который может быть раздвоен или
снабжен различным числом зубцов. Иногда конец щетинки покрыт
перепончатым колпачком или капюшоном (Onuphis, Capitella, Maldane и др.)
(рис. 50, А, В, М)\ 3) унцинальные, или uncini —короткие, нередка
напоминающие очертаниями птичью голову, крючья, как бы вырезанные
из плоского куска хитина. Они могут быть снабжены одним или
несколькими зубцами. Характерны для живущих в трубках Sedentaria
Рис. 49. Sabella fabricii.
Передняя часть тела.
Вид справа (ориг.):
; — «жабры», 2 —
воротничок, 3 — торакальные
невроподии, 4 — абдоминальные
невроподии, 5 —
абдоминальные нотоподии, 6 —
торакальные нотоподии
Рис 50. Щетинки Polychaeta. А, В, Д — Chone
infundibultfomis; Б — Nereis, гетеронереидная
форма; Г — Amphicteis gunneri; E — Thelepus ctn-
cinnatus; Ж—Onuphis conchilega; 3 — Capitella
capitata — копулятивные щетинки самца; Я —
Phyllodoce maculata; K —Nephthys cihata; Л —
Euphrosyne borealis; M — Capitella capitata (ориг.;
Б — из Бенхама)
63
(Terebellomorpha, Serpulimorpha и др.), у которых они приурочены к
невроподиям — tori uncinigeri (рис. 50, Г, Д, Е).
Спинные усики (cirri dorsales). Следует обратить внимание на
различные модификации спинных усиков параподий. Строение и форма,
а также и функции этих органов подвержены большим колебаниям.
Наибольший интерес представляют их видоизменения у Phyllodocidae,
Polynoidae и Serpulidae. У Phyllodocidae (например, Phyllodoce) они
представляют собой сердцевидные более или менее округлые пластинки,
богатые железами и расположенные вертикально. Здесь они играют
роль плавников (см. рис. 48, Б).
У Polynolidae (Еипоё, Harmathoe, Lepidonotus и др.) эти органы
превращены в элитры — плоские широкие щитки, связанные со стенкой
тела коротким стебельком (см. рис. 48, В). Элитры располагаются че-
репицеобразно, закрывая всю или часть спинной поверхности, для
защиты которой они главным образом служат (см. рис. 47, А, 7). Некоторые
формы под ними вынашивают развивающиеся яйца. Элитры
расположены не на всех сегментах, так как сегменты, несущие их, правильно
чередуются с сегментами, снабженными обыкновенными спинными усиками
(см. рис. 48, Г). Характер такого чередования варьирует у разных форм,
являясь характерным и постоянным признаком. У Serpulidae спинные
и брюшные усики сегментов сравнительно короткой груди (thorax)
сливаются вместе, образуя с каждой стороны продольные, выдающиеся на
спинную сторону, торакальные мембраны.
Первичной функцией спинных усиков является функция
осязательная и химического чувства, но у ряда форм они играют роль жабр и в
связи с этим бывают-более или менее разветвлены.
Жабры. Большинство мелких форм, а также некоторые крупные
(Capitellidae, Maldanidae) лишены специальных дыхательных органов;
дыхание у них происходит всей поверхностью кожи. У ряда других форм
дыхание сосредоточено на определенных участках тела (Lumbriconereis,
параподий Nereis). Большинство же полихет обладает специальными
органами дыхания — жабрами разнообразной морфологической
природы. Нередко это модифицированные усики параподий (Eunice, Scolele-
pis, Arenicola, Cirratulus, Amphitrite и др.) (см. рис. 48, К; рис. 43, 1;
рис. 47, В, 4; рис. 47, Г, 17). Наиболее распространены истинные, или
кровяные, жабры, пронизанные кровеносными сосудами (приносящий и
выносящий жаберные сосуды), но у .Glyceridae и Capitellidae, у
которых кровеносная система отсутствует, имеются целомические, или
лимфатические, жабры. Внутрь таких жабр заходит участок полости тела.
Целомические жабры можно видеть у Glycera, где они представлены
небольшими выростами, расположенными над нотоподиями и способными
втягиваться в особое впячивание стенок тела (см. рис. 47, Б, 12;
рис.48, 3).
Вскрытие различных Polychaeta
Полихеты — прекрасные объекты для вскрытия. Для этой цели
годится любая достаточно крупная форма. Вскрытие производится на
живом или свежеубитом материале. При отсутствии живых полихет
€4
вскрытие фиксированных
червей полезно но дает гораздо
меньшие результаты. Перед
вскрытием черви убиваются
слабым спиртом или
хлороформом как это было указано
для Nereis и Arenicola (см.
с. 32, 49). Иногда перед
умерщвлением полезно
анестезировать животных сернокислым
магнием. Не имея
возможности описывать анатомию
различных полихет, возможных
объектов для вскрытия,
укажем лишь на наиболее
подходящие для этой цели и
интересные формы из числа
встречающихся на Мурмане.
Некоторые из них во вскрытом
состоянии изображены на рис.
51, 52, 53. Таковы, например,
крупные Polynoidae
Harmathoe imbricata L. (см. рис. 47,
А) и Еипоё nodosa Sars.,
интересные главным образом
строением пищеварительной
системы. Amphinomilae (Euph-
rosyne) замечательны
наличием тетраневральной нервной
системы и другими
особенностями. Eunicidae (Eunice nor-
vegica L. — рис. 51 и Onuphis
conchilega Sars.) интересны
наличием сложно устроенного
челюстного аппарата,
лежащего в особом, способном
выворачиваться мешке, и наличием
парного спинного сосуда.
У Nephthydidae (Nephthys сое-
са Fabr. или N. ciliata Mull.)
можно наблюдать протонефри-
дии, снабженные
многочисленными соленоцитами, и
связанные с ними цилиарные органы
(видоизмененные половые
воронки). Интерес представляет
также мускулатура и
кровеносная система Nephthys.
Glyceridae (Glycera capitata
Рис. 51. Eunice norvegica. Передняя часть
тела, вскрыта'я со спинной стороны (ориг.):
/ — простомиум, 2 — перистомиум, 3 — спинной
кровеносный сосуд, 4 — диагональная мышца, 5 —
вентральный сосуд пищевода, 6 — ретракторы глоточного
мешка, 7 — пищевод, 8 — брюшное поле продольной
мускулатуры, 9 — диссепимеиты, 10 — спинное поле
продольной мускулатуры, 11 —- парный спнниой
сосуд, 12 — спинной мезентерий, 13 — поперечный сосуд,
связывающий спинные сосуды, 14 — пульсирующая
ампула, 15 — субиитестинальный сосуд, 16 — экскре-
тофор, 17 — параподиальные железы, 18 — кишка,
19 — дорзо-субинтестинальный сосуд, 20 — выносящий
жаберный сосуд, 21 — приносящий жаберный сосуд,
22 — фагоцитарный орган, 23 — вентро-параподиаль-
ный сосуд, 24 — брюшиой сосуд, 25 — глоточный
мешок, 26 — челюстной аппарат, лежащий в глоточном
мешке, 27 — глаз, 28 — антенна
3-2
65
Рис. 52, Amphitrite. Передняя часть тела,
вскрытая со спинной стороны (ориг.):
/ — простомиум, 2— тубипарные брюшиые железы,
3— воронка нефридия (нефромиксия) III сегмента,
4— выводной канал нефридия III сегмента, 5—
жаберные сосуды, 6— диафрагма на границе IV и
V сегментов, 7 —нефридий VI сегмента, 8 —
мезентерий сердца, 9 — воронка нефридия VIII сегмента,
10 — кишечный плексус, // — выводной канал
нефридия IX сегмента, 12 — нисходящее колено нефридия
IX сегмента, 13 — восходящее колено нефридия
X сегмента, 14 — нефридий XI сегмента, 15 —
обрезанные щетинконосные мешки, мускулы щетинок и
диагональные мышцы, 16 — брюшной нервный ствол,
17 — спннное поле продольной мускулатуры, 18 —
брюшное поле продольной мускулатуры, 19 —
диагональные мышцы, 20 — брюшной сосуд, 21 — субинте-
стинальный сосуд, 22 — интестинально-вентральные
кольцевые сосуды, 23 — диссепимент, 24 — кишка,
25 — протракторы щетинок, 26 — ретракторы
щетинок, 27 — щетинконосиый мешок нотоподии, 28 —
сердце, 29 — слепое выпячивание диафрагмы, 30 и 31 —
нефридий III сегмента, 32 — жабры, 33 — сосуд
передней кишки, 34 — обрезанные щупальцы, 35 —
пищевод. Римскими цифрами обозначены сегменты
Рис. 53. Chone infundibuliformis.
Передняя часть тела, вскрытая со
спинной стороны (ориг.):
1, 2 — нисходящее и восходящее колена
нефриднального канала, 3 — вентро-пара-
подиальные сосуды, 4 — спиииое поле
продольной мускулатуры, 5 — брюшное поле
продольной мускулатуры, 6 — диссепимеи-
ты, 7 — обрезанные диагональные мышцы,
8 — гонады, 9 —- толща кожно-мускульного
мешка, 10 — брюшной сосуд, 11 —
интестинально-вентральные кольцевые сосуды,
12 — кишка, 13 — спинной мезентерий, 14 —
диагональные мышцы, 15 — щетинконосные
мешки, 16 — лопасть воротничка, 17 —
передний диссепимент, 18— воронка
нефридия, 19 — жабры, 20 — общий выводной
канал первой пары нефридиев
66
Oerst.), а также Capitellidae (Capitella capitata van Bened.)
представляют интерес как формы, лишенные кровеносной системы.
Glyceridae и Phyllodocidae (Phyllodoce maculaia L.) обладают прото-
нефридиями. Очень много дает вскрытие крупных Terebellomorpha
(Amphitrite, Thelepus cincinnatus Fabr., Pectinaria, Amphicteis и др.).
Рядом интересных анатомических особенностей, легко наблюдаемых при
вскрытии, обладают Chlorhaemidae и Slernaspidae (Siylarioides plumosa
Mull., Brada, Sternaspis), а также Serpulirhopha (Protula media Stimps.,
Pomatoceros triqueter L., Plagostegus iridentatus Fabr., Sabella fabricii
Kr., Chone infundibuliformis Kr.,— рис. 53, Myxicola).
Наконец, очень полезно изучить под микроскопом несколько мелких
форм (Syllidae, Costalia, Amphicora, Spirorbis), сдавливая их стеклами.
Если имеется возможность, то следует изучить таким же способом
одного из представителей Myzostomidae (Myzostomum)—сильно моди-.
фицированных многощетинковых червей, ведущих эктопаразитический
образ жизни на морских лилиях и обычно относимых к особому классу
кольчатых червей.
Личинки Polychaeta
В морском планктоне очень часто попадаются личинки различных
полихет, находящихся на разных стадиях метаморфоза. При работе на
море рекомендуется время от времени просматривать живой планктон,
обращая внимание на личиночные стадии полихет. Изучение личинок
удобнее всего производить в живом состоянии под микроскопом.
Мы даем здесь несколько изображений, наиболее обычных в
планктоне Баренцева моря, личиночных форм (рис.,54).
Часто в прибрежном планктоне попадаются метатрохофоры Nereis
(рис. 54, £), на которых следует обратить специальное внимание. Мета-
трохофора состоит из простомиума, трех ларвальных сегментов и
небольшого пигидия. На простомиуме имеются уже антенны, пальпы и
глаза; сегменты снабжены хорошо развитыми параподиями; пигидий
несет пару анальных усиков. Имеются четыре неполных ресничных
кольца, в том числе прототрох. Из внутренних органов просвечивают
глотка, средняя и задняя кишки.
КЛАСС OLIGOCHAETA —
МАЛОЩЕТИНКОВЫЕ ЧЕРВИ
Знакомство с.малощетинковыми червями нужно начинать с
изучения одного из представителей семейства дождевых червей —
Lumbricidae.
В фауне СССР, богатой различными Lumbricidae, имеется немало
видов, легко добываемых, хорошо выживающих в неволе и достаточно
крупных для анатомирования. Нередко различные роды и виды
встречаются в сходных условиях обитания, и при сборе червей в природе
нельзя быть уверенным в систематической однородности собранного
материала. Между тем в морфологии отдельных родов и видов есть до-
3*
67
вольно значительные различия. По- ^-^ /Л ^г-\ лл
этому необходимо определение видо- /~-\ А—А А—А А~^-\
вой принадлежности материала. Для А Б в Г
этой цели могут служить таблицы,
составленные Т. С. Перель (1979). Рис. 55. Основные типы простомиума
К изученным формам относится У Oligochaeta. Л — пролобический;
обыкновенный дождевой червь Lum- Я-зиголобический; В-эпилобиче-
«„,-,, . „, • т скии; Г — танилобическии (из Пиге
bncus terrestris L., описание которого и Бретшера)
дается ниже. Впрочем, при
необходимости этот вид может быть заменен
другими широко распространенными у нас крупными формами,
например Lumbricus rubellus Hoffm., или представителями родов Allolobop-
hora, Eisenia, Dendrobaena и др. При работе с последними следует
помнить о существовании многих родовых и видовых различий не
только во внешних признаках, например в форме головной лопасти
(рис. 55), расположении щетинок, положении мужских половых
отверстий, развитии пояска, но и во внутреннем строении
(пищеварительной, выделительной и половой систем).
Кроме дождевого червя полезно ознакомиться с некоторыми
пресноводными Oligochaeta, например Aeolosoma (Aeolosomatidae), Nais,
Stylaria (Naididae), Limnodrilus и Tubifex (Tubificidae). Эти формы
обычные обитатели пресноводных водоемов различного типа. Их легко
можно выловить сачком среди водной растительности или захватывая
грунт. Для определения пресноводных олигохет могут служить таблицы,
составленные Д. А. Ласточкиным (1949) и И. И. Малевичем (1950).
Изучать пресноводных олигохет следует живыми на временных
препаратах под микроскопом, немного сдавливая их покровным стеклом.
Этот способ дает много. Обычно удается рассмотреть не только
особенности внешней морфологии, но также и многие интересные черты
внутренней организации, что облегчается как малыми размерами, так и
прозрачностью многих форм. Конечно, для более полного изучения
организации этого недостаточно. Многие системы органов могут быть
изучены только на срезах, например половая система. Тотальные окрашенные
препараты, как правило, дают мало.
Ниже мы рассмотрим из пресноводных олигохет только Aeolosoma.
Этот род, относящийся к отряду низших олигохет (Archioligochaeta),
резко отличается от Lumbricus. Он интересен значительной простотой
своей организации, которую некоторые (Михаэльсен) оценивают как
проявление примитивности, другие (Н. А. Ливанов) рассматривают как
вторичное упрощение.
Рис. 54. Личинки Polychaeta. А — Трохофора Polynoidae (Palynoe); £ —трохофора
Eunicidae; В — трохофора Spionidae; Г — нектохета Spionidae; Д — нектохета
Polynoidae; Е — метатрохофора Nereis pelagica; Ж—следующая стадия метаморфоза Nereis
pelagica (Л —по Грэвели, Б, В, Г и Д— ориг., Е и Ж —по Рунштрому):
i — зачаток медиальной антенны, 2 —нервный центр (теменной орган), 3 — прототрох, 4 —
средняя кишка, 5—'.зачаток первых сегментов, 6 — невротрохоид, 7 — околоротовой пучок ресниц, 8 —
ротовое отверстие, 9 — передняя кишка, 10 — зачаток глотки, Л — акротрох, 12 — глаза, IS —
апикальные реснички, 14 — теменной султан, 15—анальный султан, 16 — паратрох, 17— дорзальный
орган, 18 — щетинки, 19 — пигментная клетка, 20 — медиальная антенна, 21 — зачаток пальпа,
22 — сегменты, 23 — анальный усик, 24 — элитры, 25 — антенна, 26 — пальп, 27 — задняя кишка,
28 — параподия, 29 — щетииконосный мешок параподии
69
ОТРЯД OPISTHOPORA
Lumbricus terrestris L. — Обыкновенный дождевой червь
Материал и методы изучения. Lumbricus terrestris, принадлежащий к сем.
Luinbricidae, распространен в европейской части СССР (за исключением нижнего
течения Дона и Волги). Этот вид приурочен главным образом к культурным почвам и
эбитает в вертикальных норках, глубина которых у крупных особей может
достигать 2 м.
Черви добываются выкапыванием из почвы или собираются под лежащими на
земле камнями, кусками гнилого дерева, старыми досками, кусками дерна в парках,
садах, огородах или на краю пашни. Увидев дождевого червя под перевернутым
камнем или доской, надо как можно быстрее прижать рукой к земле заднюю часть его
тела; уходящую в норку. Затем следует подкопать лопатой или совком оставшуюся в
норке часть тела. Этот способ ловли удается лишь при достаточной быстроте
действий, так как дождевые черви очень чувствительны к свету, сотрясению почвы и
другим раздражениям и почти мгновенно уходят в норку. В тех местах, где червей
много, их можно ловить также ночью, когда они выходят на поверхность почвы на
поиски пищи. Фонарь, которым приходится пользоваться, надо прикрывать. Хорошие
результаты дает химический способ ловли, основанный на применении слабого
(0,15%-ного) водного раствора 'марганцевокислого калия. Этим раствором поливается
почва там, где имеются дождевые черви. Последние появляются на поверхности земли
почти сразу же и нередко в большом, количестве. Немедленно они должны быть
обмыты водой для удаления марганцевокислого калия. Наконец, большое количество
червей и к тому же в короткое в,ремя можно собрать, шерекапывая кучи старого навоза,
компостные кучи или огородные грядки. Однако при этом попадаются представители
различных родов и видов; в навозе живет 'преимущественно Eisenia foetida
(Sav.).
В неволе дождевые черви содержатся в деревянном ящике с землей. Ящик
наполняется на три четверти чередующимися слоями огородной земли и прошлогодних
древесных листьев или компостной смесью, которая составляется из равных частей
садовой земли и измельченных опавших листьев, огородной ботвы и т. п. с
небольшой примесью конского или кроличьего навоза. На каждую сотню червей следует
брать 3—5 ведер земли. Затем почву слегка поливают водой и кладут в ящик
червей. Чтобы обеспечить лучшее сохранение влажности, землю полезно прикрыть
мешковиной, сложенной несколько раз. Закрытый крышкой ящик хранят в полутемном
месте при температуре 12—15°С. Земля по мере подсыхания слегка поливается. Для
подкормки червей время от времени в ящик подкладывают размельченные
подгнивающие листья, картофельные очистки или раздавленный вареный картофель.
Появляющуюся плесень, загнивающие остатки пищи и трупы червей следует немедленно
удалять.
Перед изучением внешней морфологии или перед вскрытием черви убиваются в
парах хлороформа или погружением в 15—20%-ный спирт. Перед фиксацией кишечник
животных необходимо освободить от земли, так как иначе резать на микротоме его
невозможно. Для этого небольших червей помещают на пару дней на влажную
фильтровальную бумагу в чашке Петри, которую следует держать в темном месте.
После того как значительная часть содержимого кишечника выйдет наружу,
животных пересаживают в сосуд с крахмалом смоченным водой. Через 2—3 дня черви
заполнят кишечник крахмалом, а остатки земли выйдут из кишки.
Для фиксации рекомендуется сулема с уксусной кислотой, жидкости Штиве или
Ценкера. Сначала червь погружается на 1—2 мин в фиксатор целиком, затем
извлекается и разрезается на дощечке лезвием безопасной бритвы на куски не более 1 см.
Предназначенные для заливки куски снова погружаются в фиксатор на 3—4 ч.
Заливка производится через ксилол или касторовое масло в парафин. Срезы
окрашиваются гематоксилином Бёмера, Делафильда или железным гематоксилином и
подкрашиваются эозином, смесью Маллори.
Необходимы: поперечная серия срезов переднего конца половой области (IX—
XV сегменты), области средней кишки и пояска, а также серия сагиттальных срезов
половой области и, наконец, несколько толстых (15—20 мкм) срезов через семенные
мешки не вполне зрелой и вполне зрелой особей.
70
Наблюдения над живыми, животными. Дождевой червь, ползущий
по горизонтальной поверхности, передвигается перистальтическими
движениями тела, то вытягивая его, то сокращая. Этот способ
движения характерен для кольчатых червей, ведущих роющий образ жизни
(см. также с. 44). Сокращения продольного и кольцевого мышечных
слоев кожно-мускульного мешка происходят попеременно. Щетинки,
которыми червь опирается при ползании по бумаге, производят
характерный легкий шорох.
Рытье норки и передвижение в ней, происходящее тоже посредством
перистальтических сокращений, можно наблюдать в ящике или банке с
землей или в специальном плоском садке со стеклянной стенкой.
Задняя стенка садка делается из доски или куска фанеры размером
примерно 40X50 см, передняя — из такого же по размеру стекла.
Между фанерой и стеклом вдоль нижней и боковых сторон садка
помещаются узкие деревянные прокладки с таким расчетом, чтобы расстояние
между фанерой и стеклом было около 5—10 мм. Все части садка
скрепляются деревянными скобками, имеющими форму вытянутой буквы П.
Садок умеренно наполняется влажной почвой, которая должна быть
уплотнена; на ее поверхность кладут дождевого червя.
Зарываясь в почву, червь сначала вытягивает передний конец тела
и изгибает его дугообразно так, что упирается им в грунт. Затем
передний конец немного внедряется в землю и, расширяясь, раздвигает ее
частицы. Это позволяет ему внедриться еще глубже и т. д. При таком
способе зарывания помимо перистальтических сокращений мышц
важная роль принадлежит и жидкости, находящейся в полости тела,
которая вследствие своей несжимаемости придает телу животного
достаточную упругость (опорная функция целома).
Расширение норки и уплотнение ее стенок производится вздутием
различных участков тела. В очень плотном грунте червь прокладывает
ход, поедая постепенно почву и пропуская ее через кишечник. Та часть
экскрементов, которая не может быть впрессована в стенку норки,
выбрасывается на поверхность грунта. Характерные засохшие землистые
кучки экскрементов дождевых червей часто можно видеть на
поверхности почвы в садах и парках; особенно заметны они на гладких
дорожках. Наблюдая червей в ящиках, банках или садках с землей,
легко заметить, что свою пищу — различные растительные остатки — они
затаскивают в норку, где и поедают. Черви ведут ночной образ жизни,
норку покидают только в темное время суток. Однако, разыскивая
пищу по соседству с норкой, они все же держат в норке задний конец тела
и при опасности мгновенно в ней исчезают.
Нетрудно заметить, что Черви не выносят сухости. Будучи оставлены
на открытом воздухе, они быстро становятся вялыми, сморщиваются,
темнеют и, наконец, погибают. При подсыхании они прежде всего
испытывают недостаток в кислороде, так как обладают только кожным
дыханием. В проточной или сменяемой дважды в сутки воде, достаточно
богатой кислородом, они могут прожить несколько дней, но быстро
гибнут в кипяченой (лишенной растворенного кислорода) воде.
Среди червей, достаточно долго содержавшихся в неволе, иногда
попадаются особи, находящиеся в процессе регенерации. У таких червей
71
на месте утраченного переднего или заднего конца тела имеется более
или менее развитый и обычно легко отличимый по толщине и цвету
регенерирующий участок (регенерат). Опыты по регенерации удаются
легко, но регенерация протекает медленно в течение нескольких месяцев.
Подопытные животные содержатся в стаканах или в банках с влажной
фильтровальной бумагой.
Внешняя морфология. Длинное цилиндрическое тело Lumbricus
terrestris суживается к переднему концу. Задний отдел тела заметно
уплощен и при жизни способен, лопатообразно расширяться; им червь
держится за края норки, когда выползает из нее на поверхность почвы.
На переднем конце тела впереди вентрального рта лежит небольшая
закругленная головная лопасть—простомиум (см. рис. 58, 1). В лупу
видно, что на спинной стороне медиальная часть простомиума глубоко
вдается в I сегмент и достигает передней границы II сегмента (танило-
бическая головная лопасть) (см. рис. 55, Г; рис. 56, 1).
Туловище состоит из многочисленных (110—180) сегментов, которые
в передней части тела значительно крупнее (длиннее), чем в задней.
Первый сегмент, лишенный щетинок, является ротовым сегментом (ме-
тастомиум), ротовое отверстие лежит на его переднем крае. В шести
или семи передних сегментах олигохет имеется как ларвальная, так и
постларвальная мезодерма. Однако есть все основания считать их лар-
вальными. В них помимо формирования ларвальных мезодермальных
полосок развиваются провизорные протонефридии и кишечник
образуется несколько иначе, чем в других сегментах. У некоторых олигохет эти
ларвальные сегменты и во взрослом состоянии отличаются особыми
чертами строения (подробнее см. П. П. Иванов, 1944).
Анальная лопасть, или пигидиум, которым заканчивается тело, очень
мал, лишен щетинок и несет большое, сжатое с боков, анальное
отверстие. Длина тела от 9 до 30 см.
Окраска живого червя темная — красно-бурая с фиолетовым
оттенком. При ярком освещении кутикула сильно иризирует, особенно на
переднем конце тела. Средняя и задняя области
туловища светлее, с темной спинной полоской.
Брюшная сторона заметно бледнее спинной.
У половозрелых червей выделяется более темная
область пояска (см. с. 74).
Некоторые особенности внешней
морфологии требуют внимательного изучения под бнно-
куляром.
Все сегменты, кроме ротового, снабжены
щетинками, представляющими все, что осталось
у олигохет от параподий. Каждый сегмент
имеет 8 щетинок, сближенных попарно, — две
боковые и две брюшные пары. Таким образом, вдоль
Рис. 56. Lumbricus ter- тела тянутся 4 двойных ряда щетинок (рис. 58,
restris. Вид со спинной 9). Вытащив осторожно маленьким пинцетом
стороны (по Михаэль- щетинки из нескольких различных сегментов в
сенУ): передней области тела, следует рассмотреть их
1 — головная лопасть, 2 ■— тл **
спннные поры под микроскопом. Размеры щетинок болыиин-
72
ства сегментов около 1 мм (от 0,75 до 1,25 мм)
(рис. 57). Они желтоваты, слегка изогнуты
S-образно; дистальный конец их заострен.
В средней части щетинки, несколько ближе к ди-
сталыюму концу, имеется легкое утолщение,
называемое узелком (рис. 57, А, Г). Вещество
щетинок близко к хитину, разбухает в едком
кали и растворяется в крепкой серной кислоте.
На 2/3 своей длины щетинки погружены в
толщу кожно-мускульного мешка, но благодаря
сокращению мышечных пучков, связанных с
мешочком, в котором они сидят (с. 91), могут
несколько выдвигаться наружу или втягиваться.
Они дают опору телу, когда червь
передвигается в норке или ползет по поверхности почвы.
Среди брюшных щетинок, имеются, однако,
модифицированные, называемые половыми
щетинками, так как играют определенную роль при
спаривании. Таковы бороздчатые щетинки X,
XV, XXVI сегментов и пояска. Они в 2 раза
длиннее нормальных щетинок и снабжены
четырьмя бороздками в дистальном отделе,
вытянутом в тонкое острие (рис. 57, Б, В). Во время
спаривания они -втыкаются в тело партнера, и
по их желобкам в ранку проникает секрет
железистых клеток щетинконосного мешочка. Другую
группу половых щетинок составляют брюшные
щетинки VI — X сегментов. Они на 7з длиннее и
в три раза толще нормальных щетинок (рис. 57,
Г). При спаривании ими зажимается тело партнера.
На спинной стороне тела расположены очень маленькие непарные
медиальные спинные поры, посредством которых полость тела
сообщается с наружной средой (см. рис. 56, 2). Они отсутствуют только на
передних сегментах. Искать их следует в межсегментных бороздках; на
заспиртованных экземплярах они видны яснее; на только что убитых
червях их можно обнаружить по выступающей из них полостной
жидкости, слегка сдавливая тело животного. Как правило, спинные- поры
лежат у передней границы сегмента, но иногда, особенно в заднем
отделе тела, и на средней его части. Предполагают, что выходящая через
поры жидкость облегчает скольжение поверхности тела при
передвижении животного в норке. Считается также, что во время сбрасывания
кокона через передний конец тела (с. 89) истечение полостной жидкости
облегчает этот процесс, так как уменьшается давление в передних
сегментах. В задней части тела спинные поры несомненно служат для
выделения эскреторных телец (с. 76) из полости тела. Наконец, благодаря
спинным порам, подсыхающая поверхность тела может быстро
увлажняться, что важно для нормального кожного дыхания. Все эти функции,
разумеется, не исключают друг друга.
Парные половые отверстия расположены на брюшной стороне. Бро-
Рис. 57. Щетинки"
Lumbricus terrestris. A —
двигательная щетинка;
Б — половая щетинка из
области пояска; В — ее
поперечное сечение; Г—■
половая щетинка из
области VI—X сегментов
(по Фельдкампу)
73
сается в глаза пара железистых утолщений
кожи на XV сегменте, лежащих снаружи от
брюшных щетинок. Они окружают щелевидные
мужские половые отверстия (рис. 58, 6). Впереди
них, на XIV сегменте, помещается пара
маленьких женских половых отверстий (рис. 58, 7).
Кроме того, в межсегментных бороздках IX/X и
X/XI на уровне боковых рядов щетинок лежат
очень маленькие отверстия двух пар
семяприемников, или сперматэк (см. с. 88). Снаружи они
обычно неразличимы. От каждого мужского
полового отверстия до пояска тянется семенная
бороздка (рис. 58, 5), расположенная несколько
выше брюшного ряда щетинок.
Наконец, приметен поясок (clitellum) —
образование очень характерное не только для
Oligohaeta, но и для Hirudinea (с. 111). Это
железистое утолщение покровов развивается - у
половозрелых особей Lumbricus lerrestris на 6—
7 сегментах, а именно —с XXXI или с XXXII по
XXXVII включительно (рис. 58, 3). Однако на
брюшной стороне оно не развито, поэтому
поясок обладает седлообразной формой. На пояске
границы сегментов и спинные поры обычно
различаются с трудом. Брюшные края пояска несут
продольные железистые утолщения (рис. 58, 2),
называемые пубертатными валиками и
служащие для прикрепления к телу другой особи при
спаривании. Во время размножения железистые
клетки пояска выделяют вещество кокона.
Анатомия. Вскрывать дождевого червя
следует со спинной стороны (рис. 59). Сначала
животное укрепляется на дне препаровальной
ванночки двумя тонкими булавками; одна булавка
втыкается несколько сбоку от медиальной линии
на уровне III или IV сегмента, другая — позади
пояска. Затем лезвием безопасной бритвы
разрезают стенку тела вдоль спины от пояска до
простомиума. Разрез не должен повредить
кровеносные сосуды, кишечник и половые органы. После этого в ванночку
наливается вода. Раздвигая далее края разреза, надо осторожно
приколоть их ко дну ванночки. Натягивающиеся при этом нежные диссе-
тшменты осторожно разрываются, а в задней части разреза, где они
прочнее, подрезаются. Очень полезно кроме того, вскрыть другой
экземпляр червя с боковой стороны тела, поступая аналогичным образом.
По мере изучения различных систем органов приходится несколько раз
повторять вскрытие на новых особях.
Стенка тела,и полость тела. На вскрытом черве при
достаточном увеличении можно увидеть почти все слои стенки тела. Тон-
XXV/
ХШ
XXXVI—:
Рис. 58. Lumbricus ter-
restris. Передний участок
тела с брюшной стороны
(по Стефенсону):
1 — простомиум, 2 —
пубертатный валик, 3 -^ поясок,
4 — половые щетинки, 5 —
семенная бороздка, 6 —
мужское половое отверстие,
7 — женское половое
отверстие, 8 — отверстия
семяприемников, 9 — брюшные
щетинки, 10 — ротовое
отверстие. I, X, XV, XXVI,
XXXII, XXXVII —сегменты
74
кая прозрачная кутикула,
одевающая тело снаружи,
нередко отстает на краях
разреза. Эпидермис едва
различим в виде тонкого
светлого слоя, зато хорошо
заметны наружный
кольцевой и внутренний
продольный мускульные слои.
Первый красноват от большого
количества пронизывающих
его кровеносных
капилляров и очень утончается на
границах между
сегментами. Второй значительно
толще. Целомический
эпителий может быть
обнаружен лишь на срезах.
На внутренней гладкой
поверхности стенки тела
видны ряды щетинконосных
мешочков с
просвечивающими в них щетинками, а
также заметны
многочисленные разветвления
кровеносных сосудов.
Обширная вторичная
полость тела (целом)
пересекается тонкими
поперечными перегородками — дис-
сепиментами, или септами
(рис. 59, 12). Они развиты
на всем протяжении
туловища, и у Lumbricus, как и у
многих других олигохет,
имеется только одно
отверстие под кишечником, через
которое проходят брюшная
нервная цепочка и брюшной
кровеносный сосуд. Таким
образом полости соседних
сегментов сообщаются, и
целомическая жидкость
может переливаться из одного сегмента в другой. В диссепиментах
залегают нежные радиальные, дорзовентральные и кольцевые мускульные
волокна. В передних сегментах диссепименты пересекаются длинными
мускульными пучками (рис. 59, 17), идущими от глотки к стенке тела.
Спинной мезентерий на большем протяжении тела отсутствует, а
брюшной представлен лишь узкой складкой целомического эпителия,
Рис. 59. Lumbricus terrestris. Передний
участок тела, вскрытый со спннной стороны (по
Кюкенталю, с изменениями):
/ — церебральные ганглии, 2 — глотка, 3 — пищевод,
4—'Сердце, 5 — семяприемник, 6 — семейной-мешок,
7 — зоб, 8 — мускульный желудок, 9 — спинной сосуд,
10— кишка, 11 — дорзоиитестинальный сосуд, 12 —
диссепимент, 13—брюшной сосуд, 14 — нефридий,
15 — брюшная нервная цепочка, 16 — тифлозоль, 17 —
радиальные мышцы глотки. I—XXIV — сегменты
75
Рис. 60. Lumbricus terrestris. Клетки целомической жидкости. А — амебоцит с
многочисленными мелкими псевдоподиями; Б — амебоциты с листовидными псевдоподиями;
В — амебоциты с короткими псевдоподиями; Г—округлившиеся амебоциты;
Д—бактерии (ориг.)
соединяющей кишечник с брюшным кровеносным сосудом и нервной
цепочкой и не достигает брюшной стенки.
Средняя кишка и значительная часть кровеносных сосудов на
спинной стороне покрыты слоем, желтых хлорагогенных клеток, которые
представляют собой модификацию клеток целомического эпителия
(см. с. 95).
Целомическая жидкость водяниста и обычно мутновата (иногда
молочно-белая). Взяв каплю этой жидкости из переднего и заднего
участков тела, следует, исследовать ее под микроскопом. Содержащиеся
в ней свободные клетки представлены несколькими формами.
Особенно обильны крупные амебоциты с пальцевидными псевдоподиями,
которых так много, что клетка представляется как бы лохматой (рис.60,Л).
Нередко эти элементы собираются в большие комки. Встречаются также
амебоидные клетки' с меньшим количеством прозрачных
листовидных псевдоподий (рис. 60, Б), округлые клетки с мелкими
заостренными псевдоподиями (рис. 60,5) и, наконец, небольшие пузырчатые
шаровидные элементы с небольшим отчетливым ядром (рис. 60,Г).
Возможно, что все эти формы, за исключением последних, представляют
различные возрастные или физиологические состояния одних и тех же
амебоцитов. Между свободными клетками нередко можно видеть
большое количество бактерий (рис. 60, Д).
Главная функция амебоцитов — фагоцитоз твердых экскреторных
частиц и различных инородных тел. Убедиться в этом можно,
произведя инъекцию в полость тела мелкорастертой туши с помощью
обыкновенного шприца. Через несколько дней частицы туши окажутся
сосредоточенными в амебоцитах. Кроме того, в полостной жидкости
довольно часто встречаются экскреторные тельца. Это сравнительно
крупные (до 1—2 мм) комки, глыбки неправильной формы, состоящие из
множества амебоцитов и бурой зернистой массы и содержащие
выпавшие в полость тела щетинки, продукты распада хлорагогенных клеток
и даже паразитов — мелких нематод и цисты грегарин (рис. 61).
Таким образом, помимо упомянутого опорного значения (см. с. 71)
целомическая жидкость имеет и экскреторную функцию.
76
Рис. 61. Lumbricus terrestris. Экскреторные тельца из полости тела.
А—тельце с цистами грегарин; Б — тельце с щетинками (по Штоль-
те)
Пищеварительная система. Ротовое отверстие ведет в
небольшую складчатую ротовую полость, выстланную кутикулой. Со
стенками тела ее соединяют небольшие мускульные пучки. Далее следует
объемистая глотка (см. рис. 59,2), простирающаяся до передней
границы VII сегмента. Поверхность ее со стороны полости тела кажется
лохматой от' многочисленных прилегающих слюнных желез. От глотки
отходят длинные мускульные пучки (см. рис. 59,17), расходящиеся в
разные стороны, пересекающие диссепименты и прикрепляющиеся к
стенке тела. Те из них, которые тянутся вперед, играют роль
протракторов глотки, идущие назад являются ее ретракторами. Эти мышцы,
кроме того, служат расширителями глотки. Толстостенная глотка состоит
из нижнего довольно складчатого отдела, переходящего сзади в
пищевод, и из верхнего слепого глоточного кармана. Упомянутые слюнные
железы открываются в последний посредством тонких протоков. Это
крупные, колбовидные железистые клетки, секрет которых увлажняет
пищу и содержит ферменты, действующие на белки.
По наблюдениям Ч. Дарвина, пища сначала смачивается
выделениями передней части пищеварительного канала (слюнных желез).
Лишь после этого она заглатывается, причем ротовая полость и
отчасти глотка выворачиваются наружу, что достигается сокращением
протракторов и облегчается давлением полостной жидкости.
Пищевод снабжен перехватами в местах прикрепления диссепимен-
тов; он тянется до XV сегмента (см. рис. 59,3).
С пищеводом связаны три пары небольших известковых, или мор-
реновых, желез (рис. 62). Эти железистые выпячивания боковых
стенок пищевода лежат у Lumbricus terrestris в X, XI и XII сегментах.
Чтобы их рассмотреть, необходимо осторожно обрезать заслоняющие их
семенные мешки. Передняя пара желез имеет форму полусферических
мешков, две следующие пары сидят на боковых поверхностях пищевода
в виде коротких поперечных колбасовидных валиков. Железы покрыты
легко стирающимся слоем хлорагогенных клеток. Сквозь их стенки
просвечивают многочисленные продольные параллельные очень тонкие
кровеносные сосуды (синусы — см. с. 92). Осторожно разрезав
пищевод вдоль, следует убедиться, что передняя пара желез (рис. 62,5)
сообщается с его полостью на границе X и XI сегментов, тогда как две
77
s —
задние пары (рис. 62, 7, 8) лишены
сообщения с пищеводом. Они открываются в
полости передних желез (рис. 62, 4),
которые вероятно выполняют роль общих
резервуаров секрета. В железах находится
белая масса кристаллов углекислой
извести.
Функция известковых желез состоит в
удалении карбонатов, накопляющихся в
крови. Однако при известных условиях
известь желез может растворяться в крови,
образуя бикарбонаты и связывая избыточ-
. ную углекислоту. Излишки извести
поступают в пищевод. Здесь они, как показал
еще Ч. Дарвин, примешиваясь к пище,
нейтрализуют содержащиеся в ней гуминовые
кислоты.
Пищевод открывается в широкий
тонкостенный зоб сероватого цвета (см. рис.
59, 7), лежащий в XV и XVI сегментах.
В стенках зоба разветвляются
многочисленные кровеносные сосуды; полость его
выстлана кутикулярным эпителием.
Сзади к зобу примыкает широкий, бо-
чонковидный мускульный желудок (см. рис.
59, 8), представляющий самый задний
отдел эктодермальной кишки. Он помещается
в XVII, XVIII и XIX сегментах.
Поверхность его темно-красная от множества
очень тонких кольцевых кровеносных сосудов, проходящих
параллельно друг другу в поверхностном слое органа. Стенки его толсты и
мускулисты, полость выстлана кутикулой. В мускульном желудке
перетирается пища, чему способствуют довольно крупные кварцевые
песчинки,-обычно находящиеся в его полости.
Средняя кишка начинается вслед за желудком и тянется почти
до заднего конца тела. Это широкая, прямая трубка (см. рис. 59,10),
покрытая снаружи слоем хлорагогенных клеток. В местах
прикрепления диссепиментов она имеет небольшие перехваты, а между ними
образует метамерные расширения, покрытые поперечными складками.
На спинной стороне стенка кишки глубоко вдается в кишечную полость
в виде продольной складки — тифлозоля " (typhlosolis, см. рис. 59,16),
за счет которого значительно увеличивается пищеварительная и
всасывающая поверхность. Следует разрезать кишку вдоль, удалить и
отмыть ее содержимое и рассмотреть тифлозоль. Он заканчивается
значительно не доходя до заднего конца тела. В кишке пищевая масса
подвергается воздействию пищеварительных ферментов — протеолитиче-
ских, амилазы, переводящей крахмал в сахар — мальтозу, а также
липазы, расщепляющей жиры, и др. Здесь же происходит всасывание.
Короткая задняя кишка заканчивается анальным отверстием.
XI
хи
хш
XIV
Рис. 62. Lumbricus terrest-
ris. Продольный разрез
через пищевод и известковые
железы( по Харрингтону):
/ — просвет пищевода, 2 —
кровеносный сосуд, 3 — хлорагоген-
ные клетки, 4 — полость
передней известковой железы, 5 —
передняя известковая железа,
6 — кровеносные синусы
железы, 7 — средняя железа, 8 —
задняя железа, 9 — кристаллы
извести. X—XIV — сегменты
78
Рис. 63. Lumbricus terrestris. Схема кровеносной системы передней части тела. Вид
с левой стороны (ориг.):
1 — дорзоинтестинальные сосуды, 2 — заднее соединение бокового пищеводного сосуда со спинным
сосудом, 3 — сосуды задней известковой железы, 4 — спинной сосуд, 5 —переднее соединение
бокового пищеводного сосуда со спинным сосудом, 6 — вентро-парнетальные сосуды, 7 — боковой
пищеводный сосуд, 8 — сердце, 9 — брюшной сосуд, 10 — пищеводно-поднервные сосуды, It — боковой
невральный сосуд, 12 —"субневральный сосуд, 13 — сосуды мускульного желудка, 14 — вентро-ин-
тестинальные сосуды, 15 — дорзосубневральные сосуды. I—XXI — сегменты
Органы дыхания отсутствуют. Газообмен происходит через
кожу посредством густой сети капиллярных сосудов, залегающих в
поверхностных слоях стенки тела (с. 94).
Кровеносная система замкнута, как и у других кольчатых
червей. Сосуды залегают под целомическим эпителием кишечника,
диссепиментов, стенок тела и т. д. Капиллярные сосуды проходят в толще
органов и тканей. Кровь окрашена в красный цвет, что облегчает
наблюдение сосудов.
Для изучения кровеносной системы следует брать только что
убитых червей. Ванночку лучше наполнять не водой, а 20—30%-ным
спиртом, который несколько уплотняет ткани и стенки сосудов. Однако для
наблюдений над сокращением сосудов необходимо вскрывать живых
червей, лишь потерявших подвижность от недолгого пребывания в
слабом спирте. При препаровке следует остерегаться излишнего
повреждения сосудов. Хлорагогенные клетки, покрывающие многие сосуды и
затрудняющие их изучение, легко счищаются кисточкой, после чего
следует сменить спирт в ванночке.
Кровеносная система (рис. 63, 64) состоит из продольных и мета-
мерных сосудов. Среди последних различаются кольцевые, или коммис-
суральные, сосуды, соединяющие продольные, и не представляющие
полных колец поперечные сосуды. В стенках средней кишки между
кишечным эпителием и мускульными слоями залегает сосудистая
капиллярная сеть кишечного плексуса.
1. Продольные сосуды.
Спинной сосуд (vas dorsale, см. рис. 63,4) тянется над
пищеварительным каналом от одного конца тела до другого. Он вплотную
прилегает к кишечнику, за исключением области кольцевых сердец (см.
ниже), где связан с пищеводом узким мезентерием (рис. 64, Б, 12). Наи-
79
-18
Рис. 64. Lumbricus terrestris. Схемы ангиосомитов А—X сегмента; Б — VIII сегмента;
В — сегмента из средней части тела (ориг.):
1 — спинной сосуд, 2 — сердце, 3 — соединение бокового пищеводного сосуда со спинным сосудом,
4 — боковой пищеводный сосуд, 5 — вентро-париетальный сосуд, 6 — пищеводно-поднервный сосуд,
7 — боковой иевральный сосуд, 8 — субневральный сосуд, 5 — ветвь, несущая кровь из стенки тела
в боковой иевральный сосуд, 10 — брюшной сосуд, 11 — брюшной- мезентерий, 12 — спинной
мезентерий, IS — дорзоинтестинальный сосуд, 14 — нефридий, 15 — дорзосубневральиый сосуд, 16 — вен-
тро-интестинальный сосуд, 17 — кишечный плексус, 18 — сосуд тифлозоля, 19 — сосуд, соединяющий
тифлозолярный и спинной сосуды
большую толщину спинной сосуд имеет позади желудка и суживается
постепенно как впереди, так и сзади. Он пульсирует на всем
протяжении, прогоняя кровь от заднего конца к переднему.
Брюшной сосуд (vas ventrale, рис. 63,5) проходит под-кишечником,
будучи подвешен к нему мезентерием (рис. 64, Б, 11). Он тоньше
спинного и неспособен сокращаться. По нему кровь течет спереди назад.
В каждом сегменте брюшной сосуд связан с кишечным плексусом
тремя непарными вентро-интестинальными сосудами (рис. 63,14),
проходящими в брюшном мезентерии.
Боковые пищеводные сосуды (vasa extraoesophagealia, рис. 63,7)
развиты лишь в передних 12 сегментах. Эти парные сосуды тянутся
вдоль пищевода под целомическим эпителием. Хлорагогенных клеток
на них нет. Они начинаются в области глотки, образуясь слиянием
многочисленных разветвляющихся в ее стенках сосудов, и тянутся
назад до XII сегмента, где впадают в спинной сосуд (рис. 63,2), с
которым связаны также поперечной перемычкой в X сегменте (рис. 63, 5;
рис. 64, Л, 3). На своем пути они принимают метамерные сосуды,
выносящие кровь из стенки тела, пищевода и нефридиев, от них отходят
короткие сосуды к известковым железам (рис. 63,3). .
Субневральный сосуд (vas subneurale, рис. 63,12; рис. 64,8)
прилегает медиально к брюшной стороне нервной цепочки. Он тоньше
брюшного сосуда и в каждом сегменте соединен с парой кольцевых
сосудов ( рис. 63,15, рис. 64,15), несущих кровь в спинной сосуд.
80
Боковые невральные, или околоневральные, сосуды (vasa extraneu-
ralia, рис. 63,7/, рис. 64,7) очень тонкие парные сосуды,
.сопровождающие по бокам брюшную нервную цепочку. В середине каждого
сегмента они принимают тонкую веточку из стенки тела (рис. 64,5). Ко-
. роткие латеральные сосудики соединяют их с субневральный сосудом.
Сосуд тифлозоля, или надкишечный сосуд (vas supraintestinale,
рис. 64, В, 18), проходит внутри тифлозоля средней кишки, где залегает
в хлорагогенной ткани. Он собирает кровь из кишечного плексуса.
В каждом сегменте от него отходят 2—3 вертикальных сосуда
(рис. 64, В, 19), впадающие в спинной сосуд.
2. Кольцевые сосуды.
Кольцевые сердца (vasa dorso-ventro-commissuralia, рис. 63,8) в
количестве 5 пар 1 лежат в VII—XI сегментах, где соединяют спинной
и брюшной сосуды. Средняя часть их сильно расширена. Пульсируя,
они перекачивают кровь из спинного сосуда в брюшной.
Дорзосубневральные, или дорзопариетальные, сосуды (vasa dorso-
subneuro-commissuralia, s. vasa dorso-parietalia, — рис. 63,15;
рис. 64, В, 15) соединяют субневральный и спинной сосуды. Проходя по
задней стороне диссепиментов, они принимают несколько ветвей,
выносящих кровь из стенки тела, диссепиментов и из нефридиев. В 12
передних сегментах эти сосуды отсутствуют; здесь их заменяют пищеводно-
поднервные сосуды. По дорзосубневральным сосудам кровь течет в
спинной сосуд.
Пищеводно-поднервные сосуды (vasa extraoesophageo-subneuro-
coramissuralia, рис. 63,10; рис. 64, А, Б, 6) соединяют в передних
сегментах боковые пищеводные сосуды с субневральный и, следовательно,
не образуют полных колец.
3. Поперечные сосуды.
Дорзоинтестинальные сосуды (vasa dorso-intestinalia,—-рис. 63,/;
рис. 64, В, 13). В каждом сегменте, кроме 12 передних, в спинной сосуд
впадают две пары коротких дорзоинтестинальных сосудов, отходящих
от стенок кишечника. Они лежат свободно поверх кишки и одеты слоем
хлорагогенных клеток. По ним кровь течет из кишечного плексуса
в спинной сосуд.
Вентро-париетальные сосуды (vasa ventro-parietalia, — рис. 63,6;
рис. 64, В, 5) имеются во всех сегментах. Начинаясь от брюшного
сосуда, они проходят по передней стенке диссепиментов и снабжают
кровью стенки тела.
Кровообращение совершается следующим образом. В спинной
сосуд из кишечного плексуса через парные дорзоинтестинальные сосуды
и непарные сосуды тифлозоля поступает венозная кровь, обогащенная
питательными веществами. Сюда же попадает и кровь, богатая
кислородом, из дорзосубневральных сосудов, несущих ее из субневрального
хюсуда и из стенок тела. Таким обр-азом, в спинном сосуде венозная
кровь смешивается с артериальной. Вся эта масса смешанной крови
течет вперед по спинному сосуду благодаря его пульсации. Частота со-
1 Другие Lumbricidae могут обладать большим количеством сердец. Так, у АНо-
lobophora caliginosa (Sav.) их 6—7 пар.
81
крашений сосуда в нормальных условиях 15—20 в минуту. Внутри
парных сосудов, в месте их впадения в спинной сосуд, а также в нем
самом, имеются клапаны, пропускающие кровь только в одном
направлении (с. 94). -^-
Кольцевые сердца перегоняют кровь из спинного сосуда в
брюшной. Из области сердец по брюшному сосуду часть крови направляется
назад, другая часть течет вперед-в головной отдел тела. Позади
сердец, в среднем и заднем отделах тела кровь брюшного сосуда, ставшая
уже венозной, поступает в вентро-париетальные сосуды, несущие ее в
стенки тела и связанные своими разветвлениями с кожной капиллярной
сетью. Здесь кровь окисляется, поступая затем через ветви дорзосуб-
невральных сосудов в эти последние, а также через ветви бокового нев-
рального сосуда в капилляры брюшной нервной цепочки. Другая часть
крови из брюшного сосуда проходит в кишечный плексус по непарным
сосудам брюшного мезентерия.
В головной отдел тела поступает смешанная кровь по спинному
и брюшному сосудам; по боковым пищеводным сосудам она течет
обратно.
Нефридии получают кровь из ответвлений вентро-париетальных
сосудов (рис. 64, В, 5); отток крови из них совершается через ветви дор-
зосубневральных сосудов (рис. 64, В, 15). Половые органы снабжаются
кровью из пищеводно-субневральных сосудов и отдают ее вентро-парие-
тальным сосудам.
Выделительная система представлена парными метанеф-
ридиями, развитыми во всех сегментах, кроме трех передних. Вскрыв
крупного червя, сначала следует ознакомиться с общей топографией
метанефридиев. Они имеют вид полупрозрачных и частью желтоватых
вытянутых петлевидных лопастей и лежат в полости тела по бокам от
кишечника позади диссепиментов (см. рис. 59,14), к которым
подвешены собственной мезентериальной складкой. На брюшной стороне от
лопастевидной массы нефридия в сторону диссепимента отходит очень
тонкий канал. Это начальный канал нефридия. Он проходит сквозь дис-
сепимент в полость предыдущего сегмента и открывается в нее
небольшой ресничной воронкой. Следует также найти место, где концевой
отдел нефридия входит в стенку тела. Оно лежит несколько сбоку от
брюшного ряда щетинконосных мешочков. Затем, осторожно вырезав
. метанефридий вместе с прилегающим участком диссепимента,
необходимо рассмотреть его в свежем виде под лупой и под микроскопом.
Нефридий состоит из двух неравных отделов — пресептального,
лежащего впереди диссепимента, и постсептального (рис. 65),
расположенного позади него. Пресептальный отдел представлен воронкой, или
нефростомом (рис. 65,/) и короткой шейкой (рис. 66,4,5). Нефростом
образован нижней и верхней губой. Первая коротка и лишена ресничек,
вторая гораздо крупнее, имеет уплощенную подковообразную форму и
состоит из ряда краевых мерцательных клеток (рис. 66,2),
окаймляющих большую срединную клетку (рис. 66, 1). В шейке проходит очень
узкий ресничный канал (рис. 66,5), окруженный гомогенной тканью
целомического эпителия (рис. 66, 4). Он проходит сквозь диссепимент
и скоро входит в первую лопасть постсептального отдела.
82
Рис. 65. Нефридий Lumbricus terrest-
ris (по Мазьярскому):
1 — воронка, 2 — диссепимент, 3 —
петлистый канал, 4 — ресничный канал, 5 —
ампула, 6 — железистый канал, 7 — мочевой
пузырек, 8 — нефропор, 9 — перитонеаль-
ная ткань, 10—12—первая, вторая и
третья лопасти постсептального отдела
нефридия, 13 — палочковый канал
Рис. 66. Нефростом Lumbricus terrestris
(по Шнейдеру):
1 — срединная клетка, 2 — краевые клетки, 3 —
реснички краевых клеток, 4— перитонеальная
ткань, 5 — канал шейки, 6 — реснички канала
Последний состоит из трех длинных лопастей (см. рис.. 65, 10—12).
образованных петлями нефридиального канала и окружающей их
оболочкой из экскреторных клеток целомического эпителия (см. рис. 65,5).
В этой оболочке видна густая сеть тончайших кровеносных капилляров
(рис. 67,6).
Очень сложный ход нефридиального канала внутри лопастей
прослеживается с трудом. Канал образует несколько длинных
петлеобразных изгибов, лежащих то в одной, то в другой лопасти. В первой
лопасти лежат первая, третья и пятая петли канала (см. рис. 65, 10}),
во.второй лопасти — вторая и четвертая (см. рис. .65, 11). Третья лопасть
образована шестой, последней, петлей (см. рис. 65,12). Картина
осложняется тем, что в первой лопасти петли канала извиваются и
многократно перекрещиваются.
Таким образом, нефридиальный канал, чрезвычайно длинен.
Начало его составляет очень узкий проток, называемый петлистым отделом
(см. рис. 65,5); он образует три первые петли и проходит,
следовательно, и в первой и во второй лопасти. Затем следует более широкий
ресничный отдел (см. рис. 65,4). На его долю приходится только одно
колено четвертой петли (во второй лопасти). В плазме его эпителиальных
клеток содержатся коричневатые экскреторные зернышки. К нему
примыкает короткая расширенная часть канала в концевом участке второй
83
Рис. 67. Кровеносные сосуды нефридия Lumbricus terrestris
(по Бенхаму):
/ — воронка, 2 — брюшной сосуд, 3 —диссепнмент, 4 — брюшная
нервная цепочка, 5 — субневральиый сосуд, 6 — расширения капилляров
нефридия, 7 — вторая лопасть нефридия, 8 — первая лопасть нефридия,
9 — третья лопасть нефридия, 10 — выносящий сосуд нефридия, // —
дорзосубневральный сосуд, 12 — приносящий сосуд нефридия
лопасти, называемая ампулой (см. рис. 65,5). От ампулы начинается
железистый отдел (см. рис. 65,6). Перейдя из второй лопасти в первую,
он суживается и делает здесь полную петлю (пятую). Продолжение его
за пределами первой лопасти составляет палочковый отдел (см. рис.65,
13), получивший название от скоплений палочковидных телец в
клетках эпителия. Наконец, канал заканчивается U-образно изогнутым
мочевым пузырьком (см. рис. 65,7), открывающимся наружу нефропором
(см. рис. 65,8).
Спустя несколько дней после инъекции в полость тела
аммиачного кармина или мелкорастертой туши можно наблюдать
концентрацию этих красок в ресничном отделе нефридиального канала, что
доказывает его фагоцитарную роль.
Через воронку и начальную часть нефридия лишь выводится
жидкость, уносящая с собой экскреты. Выделение происходит
преимущественно через стенки нефридиального канала, извлекающего экскреторные
вещества из крови капиллярной сети органа. В выделении
участвуют также хлорагогенные клетки (с. 95). Накопив в плазме в
большом количестве буро-желтые или зеленоватые зерна, они отрываются
от эпителия и дегенерируют в целоме. Освобождающиеся при этом
экскреты выводятся затем нефридиями. Кроме того, выделительная роль
принадлежит амебоцитам, целомической жидкости (с. 76).
Продуктами выделения у дождевых червей являются мочевина,
аммиак и креатин.
Нервная система. Изучение нервной системы путем вскрытия
не представляет затруднений.
Церебральные ганглии, или мозг, помещаются в III сегменте над
самым началом глотки, т. е. из области простомиума вторично сдвину-
84
Рис.
ты назад. Это две
грушевидные тесно
спаянные ганглиозные массы
(рис. 68, 4), от которых
по бокам отходят
довольно толстые
окологлоточные коннективы (рис. 68,
7), соединяющиеся с под-
глоточным ганглием (рис.
68, 15) брюшной нервной
цепочки.
Простомиум
снабжается парой нервов,
отходящих от боковых
областей мозга (рис. 68, 1).
Вблизи своего основания
они разделяются надвое,
а затем раздваиваются
еще раз. Таким образом
на каждой стороне
получается четыре ветви. Из
них три иннервируют
простомиум (рис. 68, 3),
где разветвляются, а
одна направляется в I
сегмент (рис. 68, 2).
Помимо того, I сегмент
снабжается парой нервов,
берущих начало от
середины окологлоточных
коннективов (рис. 68, 5).
II сегмент получает две
пары нервов (большие и
маленькие-—рис. 68,5), отходящих тоже от окологлоточных
коннективов, но в том месте, где они соединяются с подглоточным ганглием.
От окологлоточных коннективов отходят, кроме того, стоматогастриче-
ские нервы (рис. 68, 16), иннервирующие кишечник.
Брюшная нервная цепочка гомономна, но передняя ее часть
характеризуется значительными вторичными изменениями, на которые
указывают места отхождения упомянутых нервов двух первых сегментов.
Именно ганглии I сегмента смещены на боковые стороны
окологлоточных коннективов; ганглии, принадлежащие II сегменту, переместились
на вентральную область коннективов, а ганглии III сегмента (слитые
вместе) сдвинуты в переднюю часть IV сегмента. Они-то и
представляют собой первый ганглий брюшной нервной цепочки, или подглоточный
ганглий (рис. 68,15). Таким образом, в IV сегменте имеются два
ганглия (рис. 68,14 и 15), принадлежащие двум соседним сегментам.
В остальном нервная цепочка Lumbricidae характеризуется
сильным сближением и слиянием брюшных стволов и ганглиев (рис. Q8,10),
68. Lumbricus terrestris. Нервная система.
Вид со спинной стороны (по Гессу):
/ — нерв простомиума и I сегмента, 2 — ветвь, иннерви-
рующая I сегмент, 3 — ветви, иннервирующие
простомиум, 4 — церебральный ганглий, 5 — нервы I сегмента,
6 — большой нерв II сегмента, 7 — окологлоточный кон-
нектив, 8 — боковые щетинки, 9 — брюшные щетинкн,
10 — брюшная нервная цепочка, // — задний нерв VI
сегмента, 12 — средний нерв VI сегмента, 13 — передний
нерв VI сегмента, 14 — ганглий IV сегмента, 15 —
ганглий III сегмента, 16 — стоматогастрический нерв. I—
VI — сегменты
85
так что ее парная
природа обнаруживается
отчетливо лишь на
поперечных срезах.
Другая особенность
выражается в очень слабом
анатомическом
обособлении ганглиев; они
имеют вид
незначительных и нерезко
отграниченных
расширений, лежащих в
задней половине своего
сегмента.
Каждый сегмент,
начиная с III, иннерви-
руется тремя парами
нервов. Передняя пара
(рис. 68, 13) отходит
впереди ганглиозного
расширения в передней
части сегмента. Две
другие пары (рис. 68,
//, 12) начинаются
рядом друг с другом от
ганглия. От заднего
нерва ответвляется
тонкая веточка, иннер-
вирующая диссепимент
(см. рис. 85,. 7).
Половая
система. Характерными
особенностями Oligo-
chaeta являются
гермафродитизм и малое
число сегментов,
содержащих гонады. У Lum-
bricus ierrestris,
подобно большинству' малощетинковых червей, имеются лишь две пары
семенников и одна пара яичников.
Мужской половой аппарат состоит из двух пар семенников,
лежащих в семенных капсулах, трех пар семенных мешков и двух пар
половых воронок с их протоками. На вскрытом дождевом черве из половых
органов прежде всего бросаются в глаза семенные мешки. Это три
пары больших мягких беловатых лопастей (рис. 69, 1, 3,4) по бокам от
пищевода. Удалив осторожно пищевод и раздвинув семенные мешки,
можно видеть, что на брюшной стороне они соединяются узкими
основаниями с двумя непарными мешками, называемыми семенными
капсулами (рис. 69,2). Последние служат особыми отделами целома X и
Рис. 69. Lumbricus terrestris. Схема половой
системы. На правой стороне семенные мешки
вырезаны, а семенные капсулы вскрыты (ориг.):
/ — передний семенной мешок, 2 — семенные капсулы,
3— средний семенной мешок, 4—задний семенной
мешок, 5 — диссепимент ХП/ХШ, 6 — днссепимент
XIII/XIV, 7 — часть диссепимента XIII/XIV, 8 — женская
половая вороика, 9 — женский половой проток, 10—
брюшная нервная цепочка, // — брюшиой ряд щетинок,
12 — боковой ряд щетинок, 13 — мужское половое
отверстие, 14 — семяпровод, 15 — яйдевой мешок, 16 — яичник,
П — семявыносящий проток, 18 — мужские половые
воронки, 19 — семенник, 20 — семяприемники. IX—XVI —
сегменты
86
XI сегментов и образуются разрастанием целомического эпителия.
Каждая капсула представляет собой тонкостенную широкую трубку,
простирающуюся на брюшной стороне своего сегмента от переднего
диссепимента до заднего. Поверх семенных капсул проходит брюшной
кровеносный сосуд, нервная цепочка тянется внутри капсул (рис. 69,10).
Семенные мешки развиваются как выпячивания диссепиментов.
Передние мешки (рис. 69,/), уступающие по размерам остальным и
отличающиеся яйцевидной формой, лежат в IX сегменте; они
представляют собой выпячивание диссепимента IX/X и открываются в переднюю
часть семенной капсулы X сегмента. Вторая пара мешков (рис. 69,5)
расположена в XI сегменте и образовалась за счет диссепимента X/XI.
Она тоже открывается в ту же капсулу, но не спереди, а сзади.
Наконец, самые крупные задние мешки (рис. 69,4), представляющие
разрастание диссепимента XI/XII, соединяются с задней частью капсулы
XI сегмента. Эти мешки заполняют не только полость XII сегмента, но,
оттесняя назад диссепимент ХП/ХШ, внедряются в XIII сегмент.
Далее следует удалить семенные мешки, осторожно вскрыть
стенку семенных капсул и тщательно промыть их полости с помощью
пипетки. Внутри капсулы помещаются семенники и половые воронки1.
Семенники в количестве двух пар (по паре в каждой капсуле)
представляют собой очень маленькие (не более 1 мм) гроздевидные
органы (рис. 69, 19), прикрепляющиеся к передней стенке капсулы
справа и слева от брюшной нервной цепочки. У зрелых червей
обнаружить их очень трудно, так как, отделяя большое количество половых
клеток, заполняющих семенные мешки, они сильно уменьшаются в
размерах. Зато у задней стенки капсул хорошо видны крупные, сильно
складчатые половые воронки (в каждой капсуле имеется одна пара)
(рис. 69, 18). Они иризируют от мерцания ресничек на их внутренней
поверхности и от множества двигающихся спермиев. От каждой
воронки начинается узкий семявыносящий канал (рис. 69, 17),
направляющийся вкось — назад и наружу и прободающий диссепимент. В XII
сегменте на каждой стороне оба семявыносящих канала соединяются в
общий семяпровод (рис. 69, 14). При вскрытии семяпроводы не видны,
так как залегают в соединительной ткани под целомическим эпителием.
Они тянутся назад до XV сегмента и на его брюшной стороне
открываются наружными мужскими половыми отверстиями.^.
Половые воронки..вместе с выводными протоками представляют
собой настоящие целомодукты, т. е. образования мезодермальной
природы, генетически не связанные с метанефридиями.
В молочно-белой жидкости из семенных мешков содержится масса
половых клеток на различных стадиях созревания, объединенных в
характерные группы. Изучение их, однако, удобнее производить на
срезах (с. 100). В семенные мешки половые клетки попадают из семенных
капсул, после того как они отделились от семенников. Зрелые спермин
1 Половые воронки легче наблюдать у Allolobophora, у которой семенные
капсулы отсутствуют, а семенники и воронки лежат в полости X и XI сегментов.
87
Рис. 70. Яичник Lumbricus ter-
restris (no Ю. К. Петрушевскому)
из семенных мешков поступают обратно
в семенные капсулы, откуда при
спаривании выводятся через воронки и
семяпроводы '.
Женский половой аппарат состоит
из нары яичников, пары половых
воронок с яйцеводами, являющихся целомо-
дуктами, и двух пар семяприемников.
Яичники помещаются в XIII сегменте,
где прикрепляются к переднему диссе-
пименту по бокам от брюшной нервной
цепочки (рис. 69, 16). Это сравнительно
маленькие грушевидные заостренные на
свободном конце органы. При
рассматривании отпрепарированного яичника
под микроскопом в нем видны довольно
крупные ооциты, содержащие большое
ядро с ядрышком и желточные зерна
(рис. 70).
Напротив яичников на заднем диссе-
пименте того же сегмента
располагаются женские половые воронки (см. рис.
69, 8), представляющие собой целомоду-
кты. От каждой воронки начинается короткий канал яйцевода,
прободающий диссепимент XIII/XIV и заканчивающийся женским половым
отверстием уже в XIV сегменте. Половые отверстия лежат кнаружи
от брюшных щетинок.
Над воронками расположена пара яйцевых мешков (см. рис. 69,
15). В отличие от семенных мешков это совсем незначительные
выпячивания диссепимента XIII/XIV. Они вдаются в полость XIV сегмента и
легко узнаются по обилию разветвляющихся в них кровеносных
капилляров. В них задерживаются развивающиеся ооциты, отделившиеся'
от яичников.
Семяприемники, или сперматеки, принимающие сперму партнера во
время копуляции, лежат в IX и X сегментах, по паре в каждом. Эти
небольшие шаровидные мешочки (см. рис. 69, 20) открываются наружу
коротким и узким протоком в межсегментных бороздках IX/X и X/XI.
Нередко в семяприемниках обнаруживается сперма с подвижными
сперматозоидами.
При копуляции черви прикладываются друг к другу брюшными
сторонами, так что передние концы их обращены в разные стороны
(рис. 71). Поясок каждой особи при этом расположен против области
IX—XI сегментов другого червя, т. е. против отверстий его
семяприемников. Прочное соединение обоих копулянтов в таком положении
обеспечивается выделением кожных желез в области пояска и в~-обла-
сти X, XI и XXVI сегментов, а также благодаря половым,^ щетинкам
1 J3 семенных мешках Lumbricidae часто встречаются грегарины из рода Мопо-
cystis на различных стадиях жизненного цикла (см.: Большой практикум по
зоологии беспозвоночных. Т. 1, М., 1981, с. 150).
Рис. 71. Lumbricus lerrestris. Две
копулирующие особи (по Грове и Коули):
/ — общая слизистая муфта, 2 — XV — сегмент
(с. 73). Кроме того, в начале
спаривания каждое животное
.выделяет на протяжении от
IX сегмента до конца пояска
быстро уплотняющийся
слизистый футляр. Будучи
независимыми друг от друга на
большем своём протяжении, эти
футляры сливаются в области
поясков, образуя здесь общие
муфты, охватывающие обоих
червей.
Каждая особь выделяет
из мужских половых отверстий
по капельке семенной
жидкости, которые сокращением мускулатуры кожно-мускульного мешка
гонятся по ее семенным бороздкам к области ее же пояска. Как мужские
половые отверстия,, так и семенные бороздки находятся на участке
тела, закрытом снаружи слизистым футляром, но в области пояска
внутри общей муфты семенная жидкость попадает на поверхность тела
партнера и- засасывается его семяприемниками. Поэтому
семяприемники разошедшихся после копуляции животных содержат чужую сперму.
Через некоторое время червь приступает к откладке кокона. Вокруг
передней части тела образуется такой же слизистый футляр, как при
копуляции. Железы пояска выделяют застывающий секрет,
формирующий стенку кокона, который охватывает поясок в виде несколько
расширенной посередине муфты. Затем в полость кокона выделяется
прозрачная белковая жидкость, которой в дальнейшем питаются зародыши.
После этого животное сбрасывает слизистый футляр и кокон через
головной конец. Когда кокон проходит мимо.женских половых отверстий,
в него откладывается от 1 до 3 яиц; почти сразу же в него поступает и
сперма при прохождении мимо отверстий семяприемников. Таким
образом, оплодотворение происходит в коконе.
Коконы откладываются в поверхностных слоях почвы. Края
отложенного кокона сжимаются, замыкая его спереди к сзади^ и благодаря
этому он приобретает характерную лимонообразную форму (рис. 72).
На переднем его конце некоторое время сохраняется остаток
слизистого футляра. Размеры кокона у Lumbricus terrestris 6—8 мм. Стенки его
сначала беловатые, через некоторое время желтеют.
Изучение срезов. Кожно-мускульный мешок. Кутикула,
покрывающая кожный эпителий снаружи, на срезах не обнаруживает
отчетливой структуры. Однако она легко отделяется после мацерации
червя в 30%-ном спирте в течение нескольких дней. Следует
рассмотреть небольшой ее кусочек под микроскопом. Кутикула имеет тонкую
волокнистую структуру, причем волоконца двух направлений
перекрещиваются под прямым углом (рис. 73, 1)\ по отношению к главной оси
тела они располагаются диагонально. Видны также крошечные
отверстия кожных желез (рис. 73, 2), расположенные в центре характерных
крестообразных утолщений.
89
ч^йшшг
Рис. 72. Lumbricus Рис. 73. Lumbricus terrest-
rubellus. Кокон с ris. Участок кутикулы из
средней части тела .(по
Серфонтэну):
1 — перекрещивающиеся
волоконца, 2 — отверстия кожных
желез в центре крестообразных
фигур
остатком
слизистой трубки на
одном конце (по
Вейдовскому)
Эпидермис представлен
цилиндрическим эпителием
(рис. 74), подостланным
тонкой базальной
мембраной (рис. 74, 5). Среди его
клеток рассеяны
многочисленные одноклеточные
железы — слизистые (рис. 74,
4), содержащие
грубозернистый ацидофильный секрет
и более редкие — белковые
(рис. 74, 2) с гомогенным
базофильным содержимым.
Выделения слизистых желез
увлажняют кожу, что
является необходимым условием
кожного дыхания, и
образуют слизистый футляр в
период размножения. Между
основаниями эпителиальных
клеток часто встречаются
мелкие клетки с темными
ядрами; это амебоциты,
проникшие сквозь базаль-
ную мембрану (рис. 74, 6).
В области пояска (cli-
tellum) покровы сильно
утолщены . за счет мощно
развитых клителлярных
желез (рис. 75). Из-за их
обилия эпидермис здесь плохо
заметен (рис. 75, 9), но по
толщине не особенно
отличается от эпидермиса в дру- '
гих местах тела, он только
оттеснен к периферии
толстым слоем желез. Этот суб-
эпителиальиый слой (рис.
75, 8) состоит из массы
длинных, трубковидных
клеток, глубоко погруженных под базальную мембрану эпидермиса
и достигающих кольцевой мускулатуры. Среди клителлярных желез
различаются: 1) слизистые клетки, сходные с таковыми обычного
эпидермиса (рис. 75, 2), они не погружены под эпителий; 2)
грубозернистые клетки (рис. 75, 3) с секретом окрашивающимся по Маллори в
оранжевый цвет; они продуцируют вещество стенок кокона; 3)
тонкозернистые клетки (рис. 75, 4), голубые при окраске по Маллори,
выделяющие белок кокона. Эти железы составляют главную массу; их
особенно длинные тела лежат субэпителиально и образуют пачки, ог-
Рис. 74. Эпидермис Lumbricus terrestris
(по Серфонтэну):
/ — кутикула, 2 —белковая клетка, 3 —эпидер-
мальная клетка, 4 — слизистая клетка, 5 — базаль-
ная мембрана, 6— амебоцит
90
раниченные врастающими в их толщу
тонкими соединительнотканными
прослойками (рис. 75, 6).
Под эпидермисом лежит довольно
тонкий слой кольцевой мускулатуры (рис.
76, 3). Он состоит из сильно вытянутых
веретеновидных мускульных волокон,
'между которыми развита соединительная ткань.
Клеточное тело мускульного волокна с
овальным ядром расположено в средней
его части, прилегая к нему сбоку.
Соединительнотканные клетки снабжены
отростками, ядра их невелики и округлы, плазма
содержит скопления белковых кристаллов
в виде сильно преломляющих свет телец,
известных под неправильным названием
бактероидов.
Следующий слой продольных мышц
гораздо толще (рис. 76, 4). Он разделен
на несколько продольных полей,
ограниченных четырьмя рядами щетинконосных
мешочков и спинной линией. Мускульные
волокна находятся в особых узких камерах
(рис. 77, 8), образованных тонкими
соединительнотканными перепонками и
расположенных параллельно друг другу и
перпендикулярно к поверхности тела. На
соединительнотканных перепонках этих камер
(рис. 77, 9) и располагаются довольно
правильными рядами мускульные волокна
(рис. 77, 10). Между волокнами в полости камер находится рыхлая
соединительная ткань с многочисленными ядрами (рис. 77, 11). Ядра
мускульных волокон редки и не всегда обнаруживаются.
Под продольной мускулатурой имеется тонкий слой соединительной
ткани. Со стороны целома стенка тела ограничена уплощенным целоми-
ческим эпителием, или перитонеумом, с довольно редкими ядрами
(рис. 77, 12).
Щетинконосные мешочки (рис. 78), в которых сидят щетинки,
представляют собой впячивания кожного эпителия. Наружная часть
мешочка образована более или менее высоким эпителием (рис. 78, Л, 7),
несущим кутикулу, но в глубине он становится очень низким, а
кутикула исчезает. Щетинка имеет нежную продольную волокнистую
структуру (рис. 78, А, о). Она представляет собой кутикулярное образование,
выделяемое единственной гиподермальной клеткой, которая изредка
сохраняется на внутреннем конце щетинки. Иногда рядом с щетиико-
носным мешочком лежит молодой замещающий мешочек, внутри
которого формируется новая замещающая щетинка (рис. 78, Б, 8) с
образующей клеткой у основания (рис. 78, Б, 10). К внутреннему концу
щетинконосного мешочка прикрепляется несколько мускульных пучков
Рис. 75. Эпидермис пояска Ei-
senia sp. (по Нейманну):
1 — кутикула, 2 — слизистые
клетки, 3 — грубозернистые клетки, 4 —
тонкозернистые клетки, 5 —
кольцевые мускульные волокна, 6 —
соединительнотканные перегородки,
7 — базальная мембрана эпителия,
8 — субэпителиальный железистый
слой, 9 — эпидермис
91
10 11
Рис. 76. Lumbricus terrestris. Поперечный разрез из средней части тела (ориг.
Ю. К. Петрушевского):
/ — кутнкула, 2 — эпидермис, 3 —слой кольцевой мускулатуры, 4 — слой продольной
мускулатуры, 5 — целомическнй эпителий, 6 — метанефридий, 7 — щетинка, 8 — мезентерий 9 —
брюшной сосуд, 10 — субневральный сосуд, Л — брюшная нервная цепочка, 12 — хло'раго-
генные клетки, ■ 13 — полость кишки, 14 — кишечный сосудистый плексус, 15 — тифлозоль,
16— сосуд тифлозоля, 17 — спинной сосуд
(рис. 78, А, 1, 3), имеющих значение протракторов и ретракторов.
Пищеварительная система. Из органов пищеварения
рекомендуется изучить на срезах зоб, мускулистый желудок, известковые
железы и среднюю кишку.
. Стенка известковой железы образует серию параллельных
продольных складочек, или пластинок, глубоко вдающихся в полость органа
(см. рис. 62), Внутри каждой пластинки проходят два продольных
кровеносных синуса — один тянется вдоль ее основания, другой — близ
свободного края. Синусы соединяются пластинкой базальной мембраны.
Таким образом, сквозь известковые железы непрерывным потоком
проходит кровь, что понятно, если принять во внимание функцию этих
органов (с. 78). Снаружи железа одета тонким- слоем мускульных
волокон.
Средняя кишка образована высоким цилиндрическим мерцательным
эпителием, богатым одноклеточными железами (рис. 79). Эпителиаль-
92
ные клетки узки, сильно
сжаты соседними
железистыми клетками; плазма их
обнаруживает неясную
продольную исчерченность
(рис.'79, /). У основания
ресничек заметны базаль-
ные зерна, которые могут
создавать впечатление ку-
тикулярной каемки.
Железистые клетки овальны или
веретеновидны и содержат
ячеистый или зернистый
секрет (рис. 79, 3). Между
базальными концами клеток
часто попадаются
заползшие сюда мелкие
амебоциты (рис. 79, 2).
Эпителий тифлозоля
также мерцательный (рис.
80), но реснички его видны
лишь на хорошо
фиксированном материале.
Под базальной
мембраной кишечного эпителия
лежит тонкий слой из
внутренних кольцевых и
наружных продольных
мускульных волокон (рис. 80, 2, 3).
Между эпителием и
мышцами располагаются
кровеносные капилляры (рис.
80, 6), образующие густую
сеть кишечного сплетения.
Снаружи кишечник одет
большим слоем хлорагоген-
ных клеток (рис. 80, 1). -
Кровеносные
сосуды хорошо видны на
срезах. Стенка их состоит
из более или менее тонкой
бесструктурной пограничной
перепонки (интимы),
которая особенно сильно
развита в спинном сосуде (рис.
81, 2). Изнутри на ней нередко встречаются мелкие клетки,
неправильно принимаемые многими авторами за элементы эндотелия. В стенке
сокращающегося спинного сосуда между внутренней перепонкой и це-
ломическим эпителием залегают мускульные слои: внутри — слабо
Рис. 77. Lumbricus terrestris. Поперечный
разрез стенки тела (по Шнейдеру):
1 — кутикула, 2 —эпидермис, 3 — слизистая
железа, 4 — ядро соединительнотканной клетки, 5, 7 —
соединительная ткань, 6 — кольцевые мускульные
волокна, 8 — камера продольного мускульного
слоя, 9 — соединительнотканная стенка камеры,
10 — продольные мускульные волокна, 11 —
рыхлая соединительная ткань, 12 — перитонеальный
слой
93
развитый продольный (рис.
81, 4), снаружи — довольно
мощный кольцевой (рис. 81,5).
Сосуды, лежащие в полости
тела, одеты снаружи, целоми-
ческим эпителием или хлора-
гогенными клетками.
Заслуживает. внимания
кожная сеть кровеносных
капилляров в стенке тела,
развитие которой связано с
кожным дыханием. Эти очень
тонкие сосуды особенно развиты
в кольцевом мускульном слое;
тончайшие петлеобразные
веточки их проникают даже в
толщу эпидермиса (рис. 82).
Очень интересны
горизонтальные разрезы через
спинной сосуд в области средней
кишки. На удачно прошедших
срезах хорошо видны
клапаны в месте впадения дорзоин-.
тестинальных и дорзосубнев-,
ральных сосудов (рис. 83, 4,
3) и клапаны в самом
спинном сосуде (рис. 83, 6), Они
образуются за счет клеток це-
ломического эпителия,
врастающих в сосуды.
Нефридии могут быть
подробно изучены на срезах.
В зависимости .от того, как
прошел разрез, видны то одни,
то другие части органа.
Следует найти на соседних срезах
различные отделы нефриди-
ального канала. Петлистый
канал состоит из вытянутых
клеток с редко расположенными
ядрами (рис. 84, 4). В его
просвете кое-где заметны
реснички. Ресничный канал легко
узнается по мелким
экскреторным зернам, наполняющим плазму его клеток, и по густому покрову
ресничек (рис. 84, 3). На внутренней поверхности стенки ампулы
(рис. 84, 2) хорошо заметен слой мелких телец — «бактероидов»,
которые прежде неправильно принимались за бактерий. В плазме
эпителиальных клеток палочкового отдела канала содержатся палочковид-
Рис. 78. Щетинконосный мешочек Lumbricus
sp. А— продольный разрез щетинконосного
мешочка; Б — разрез через основание
замещающего щетинконосного мешочка (А — из
, Шнейдера, Б — по Сайовичу):
/ — протракторы, 2 — основание мешочка с
разрезанной поперек мускулатурой, 3 —ретрактор, 4 — цело-,
мический эпителий, 5 — щетинка, 6 — стенка
мешочка, 7 — эпителий, 8 — основание замещающей
щетинки, 9 — боковая клетка, 10 — клетка — образователь-
ница щетинки
94
ные включения. Стенки мочевого
пузырька имеют собственную мускулатуру в
виде перекрещивающихся мускульных
волокон. Каналы окружены более или
менее толстым слоем перитонеальной
ткани (рис. 84, 5), пронизанной
многочисленными кровеносными капиллярами.
Хлорагогенные клетки (рис.
81, 1), являющиеся видоизменением
клеток целомического эпителия, образуют
неровный, часто очень высокий
эпителий на поверхности кишки и многих
сосудов.
Они неравной высоты, часто вздуты
на дистальном конце. Ядра
располагаются на разных уровнях; плазма
варьирует по виду. Обычно она переполнена
множеством зеленоватых экскреторных
зерен различного размера, но иногда
лишена их и вакуолизирована. Как
отмечалось (с. 84), хлорагогенные клетки
несут выделительную функцию. Однако,
кроме того, они служат -местом
накопления запасных питательных веществ
(гликогена) .
Нервная система. Прежде чем
ознакомиться с главными чертами-
гистологического строения брюшной нервной
цепочки, следует проследить области
иннервации сегментарных нервов.
В каждом сегменте, кроме трех передних, парные нервы,
начинающиеся от ганглиев, образуют нервные кольца, залегающие между
кольцевым и продольным мускульными слоями. Каждый нерв (рис. 85)
делится на две главные ветви; одна — короткая, идет на брюшную
сторону (рис. 85, 6), другая —длинная, проходит в спинном
направлении (рис. 85, 2). От них отходят веточки в толщу кольцевой
мускулатуры и к коже (рис. 85, /). Нервные кольца Lumbricidae на спинной
стороне незамкнуты.
- На поперечном разрезе через ганглий брюшной нервной цепочки
бросаются в глаза две большие массы волокнистого вещества —
невропиля (рис. 86, Б, 3), соединенные поперечной волокнистой коммиссурой
(рис. 86, Б, 5). Со спинной стороны и с боков к ним прилегают и частью
вдвигаются между внешними пучками нервных волокон мелкие плоские
или булавовидные клетки опорной ткани — невроглии (рис. 86, Б, 8).
Волоконца этих клеток, трудно различимые без специальной обработки,
примешиваются, к волокнам невропиля. В дорзальнои части ганглия
лежат три очень толстых продольных волокна, изолированные друг от
друга. Это гигантские волокна, или неврохорды (рис. 86, Б, 9). Гангли-
озные клетки (рис. 86, Б, 4) прилегают к массам невропиля с боков и
Рис. 79. Эпителий средней кишки
Lumbricus sp. (по Жозесру):
/ — ресничная клетка, 2 — амебоциты,
3 — железистая клетка
95
3
в
Рис. 80. Lumbricus terrestris. Поперечный разрез кишки в области тифлозоля
(по Шнейдеру):
/ — хлорагогенные клетки, 2 — продольные мускульные волокна, 3 — кольцевые
мускульные волокна, 4-—сосуд тифлозоля, 5 — хлорагогенная ткань внутри тифлозоля,
6—кровеносный плексус, 7 — кишечный эпителий, 8 — спинной сосуд, 9 — дорзоинтестинальный
сосуд
с брюшной стороны. Они крупные, колбовидной формы и снабжены
невритом и многочисленными плохо различимыми отростками. Плазма
их сильно окрашивается гематоксилином. Невриты проникают в невро-
пиль, где вскоре теряются среди других волокон. Ганглий окружен
рыхлой, волокнистой, облегающей тканью (рис. 86, Б, 2), волоконца
которой образуют вокруг неврохордов слоистые оболочки (рис. 86, Б,
11). Облегающая ткань содержит очень тонкие кровеносные сосуды и
покрыта снаружи невральной пластинкой. К невральной пластинке
прилегают продольные мускульные волокна (рис. 86, Б, 12). Наконец, от
полости тела ганглий отграничен одевающим его слоем целомического
эпителия (рис. 86, 5, 1).
96
Рис. 81. Lumbricus sp. Поперечный разрез спинного сосуда
(по Шнейдеру):
/-хлорагогенная клетка, 2 - пограничная перепонка, 3-кольцевой
мускульный слой, 4 — продольные мускульные волокна
2—
3—
Рис. 82. Lumbricus sp. Кровеносные капилляры
. кожи (по Леноссеку): Ч
1 — эпидермис, 2 — кольцевой слой мускулатуры, 3 —
продольный слой мускулатуры
Рис. 83. Lumbricus sp. Горизонтальный разрез
через спинной сосуд. Задняя часть сосуда находит-
, ся в состоянии сокращения (по Джонстону):
; _ диссепименты, 2 — спииной сосуд, 3 — дорзосубнев-
ральные сосуды, 4 — дорзоинтестинальные сосуды, 5 —
■ отверстия сосудов, несущих кровь из сосуда тифлозоля,
6 — клапаны
?
Рис. 85. Lumbricus terrestris. Схема нервного
кольца (по Гессу):
1 — кожный нерв, 2 — спинная ветвь, 3 — продольный
мускульный слой, 4 — кольцевой мускульный слой,
5 — эпидермис, 6 — брюшная ветвь, 7 — септальный
нерв, 8 — брюшная нервная цепочка, 9 — кншка, 10 —
тифлозоль
Картина значительно меняется, если разрез прошел через коннек-
тивы брюшной нервной цепочки. Здесь продольные тяжи нервных
волокон отделены друг от друга (рис. 86, А, 10), Над ними проходят три
неврохорда (рис. 86, А, 9). Ганглиозные клетки малочисленны (рис. 86,.
А, 4). С брюшной стороны нервной цепочки под целомическим
эпителием тянется субневральный кровеносный сосуд (рис. 86, 6), по бокам
проходят узкие боковые невральные сосуды.
Органы чувств у дождевых червей представлены группами
чувствительных клетак — сенсиллами, диффузно рассеянными главным
образом в коже. Таковы чувствительные почки и светочувствительные
клетки и их группы.
Чувствительные почки залегают в кожном эпителии на всем теле.
Особенно много их на простомиуме и, в частности, на переднем крае
рта, где их насчитывается около 1800. Развиты они также и в ротовой
полости. На передних 15 сегментах (не считая I сегмента) имеется
399 почек. К заднему концу тела количество их уменьшается. На
каждом сегменте они располагаются тремя поясками, соответственно ходу
нервных колец, причем больше всего их на брюшной стороне сегмента..
Каждая почка представляет собой группу вытянутых чувствительных
клеток (рис. 87, 2), лежащих в толще эпидермиса. Базальный конец
клетки связан с нервным волокном (рис. 87, 5), дистальный
продолжается в короткую чувствительную палочку (штифтик), проходящую
98
Рис. 84. Lumbricus sp.
Разрез, через лопасть постсеп-
тального отдела нефридия
(по Бенхаму):
/ — капилляр, 2 — ампула, 3 —
ресничный канал, 4 — петлистый
канал, 5 — леритонеальная ткань
Рис. 86. Eisenia rosea. Поперечные разрезы брюшной нервной цепочки. А —
разрез через коннективы; Б — разрез ганглия (по Шнейдеру):
/ — целомическнй эпителий, 2 — облегающая ткань, 3 — невропиль, 4 — ганглиозные
клетки, 5 — коммиссура, 6 — субневральный сосуд, 7 — волокна нерва, 8 — клетки невроглии,
9 — неврохорды, 10 — волокнистый пучок кониектива, // — волокнистая оболочка неврохорда,
12 — продольные мускульные волокна
сквозь кутикулу и слегка возвышающуюся над его поверхностью
(рис. 87, /). Среди веретёиовидных чувствительных клеток и вокруг них
лежат опорные клетки, окрашивающиеся обычно светлее.
Чувствительные почки рассматриваются как органы осязания и химического
чувства.
Светочувствительные клетки рассеяны поодиночке в толще
эпидермиса (рис, 88, Л, 5) или образуют небольшие групяы — сенсиллы — на
тонких разветвлениях кожных нервов. Эти группы легче всего
обнаружить на продольных разрезах через простомиум, где они особенно
многочисленны (рис. 88, Б, 9). Каждая клетка сенсиллы связана с
чувствительным нервным волокном и помимо ядра содержит округлую или
вытянутую и более или менее искривленную вакуоль (рис. 88, Л, 7),
4*
99
—«Й*-
Рис. 87. Lumbricus terrestris. Разрез чувствительной
почки (по Гессе):
1 —• чувствительные палочки, 2 — чувствительные клетки,
3 — нервные волокна, 4 — мускульное волокно, 5 —
продольные мускульные волокна, 6 — кровеносный сосуд, 7 —
кольцевой мускульный слой, 8 — эпидермис, 9 — кутикула
Рис. 88. Lumbricus terrestris. Светочувствительные
сенсиллы. А — кожная светочувствительная клетка;
Б — скопление чувствительных клеток на нервах
простомиума (по Гессе):
/ — кутикула, 2 — эпидермис, 3 — базальная мембрана, 4 —
ядро светочувствительной клетки, 5 — светочувствительная
клетка, 6 — ретинелла, 7 — вакуоль, 8 — нерв, 9 — скопление
светочувствительных клеток
представляющую собой своеобразный светопреломляющий хрусталик,
тогда как стенка ее — ретинелла играет роль примитивной сетчатки.
Светочувствительные сенсиллы обильнее всего на переднем конце
тела. В простомиуме их насчитывается 57, в I сегменте — 26, во II— 10,
в III — 7. В средней части тела они редки, а на заднем конце обильнее.
Так, в последнем сегменте их 14.
Половая система. Изучение срезов должно прежде всего
дополнить и уточнить представление о половой системе, полученное путем
вскрытий. Демонстративны в этом отношении сагиттальные разрезы в
области IX — XV сегментов, на которых следует разобрать анатомию
полового аппарата, руководствуясь рис. 89 и 90. Попутно полезно
обратить внимание на гистологическое строение некоторых органов —
половых желез, половых воронок, семяприемников.
Интерес представляют наблюдения над созревающими в семенных
мешках мужскими половыми элементами. Они производятся на толстых
(15—25 мкм) срезах через семенные мешки и обязательно
дополняются изучением под микроскопом жидкости, взятой из этих органов
живого червя.
Самые ранние стадии развития половых элементов встречаются в
семенных мешках не вполне половозрелых особей. Сюда они поступают
после обособления от семенников. Это небольшие округлые тельца —
100
// 12
Рис. 89. Lumbricus terrestris. Схема сагиттального разреза половых сегментов
(IX—XV) (по Гессе):
/ — эпидермис, 2 — кольцевой мускульный слой, 3 — продольный мускульный слой, 4 —' дис-
сепимент, 5 — семенной 'мешок, 6 — семяприемник, 7 — семенник, 8 — семенная капсула,
9 — мужская половая воронка, 10 — семяпровод, 11 — янчник, 12 — женская половая
воронка, 13 — женское половое отверстие, 14 — мужское половое отверстие, 15 — яйцевой мешок
Рис. 90. Lumbricus terrestris. Сагиттальный разрез в
области передней семенной капсулы (по Гессе):
1 — днссепимент X/XI, 2 — половая воронка, 3—стенки семенной
капсулы, 4 — семенник, 5 — днссепимент 1ХД, 6 — кровеносный
сосуд, 7 — стенка тела, 8 — семявыносящнй канал
фолликулы, состоящие из нескольких клеток сперматогоний, без
клеточных границ (рис. 91, А). Ядра их делятся последовательно несколько
раз, вокруг них обособляется цитоплазма, но клетки остаются
связанными в середине узкими плазматическими стебельками (рис. 91, Б).
Так образуются группы, состоящие из 16, 32, 64 и 128 клеток. По мере
образования они начинают делиться," каждая распадается на две
одинаковые более мелкие группы, так что 128-клеточная группа дает две
64-клеточные, 64-клеточная образует две 32-клеточные и т. д. Поэтому
101
Лк 4* W W J?ZJ
Рис 91 Lumbricus sp. Стадии созревания мужских половых клеток. А-фолликул-
Б — деление сперматогоний в фолликуле; В-двойная группа спеоматогонийГ
сперматосфера из 64 сперматоцитов ^порядка; Д-сперма^осфераТз 128 спГрмати^
£-сперматосфера из 64 сперматид, превращающихся в спермин (А и Б по Колкие
СУ, В—Е по Бюньон и Попову):
/ — цитофор, 2 — половые клетки, 3 — стебельки
102
очень часто встречаются группы двойные, находящиеся в процессе
разделения (рис. 91, В). Все эти группы встречаются вместе, вперемешку;
половые клетки в них остаются на стадии сперматогоний.
В семенных мешках вполне зрелых червей встречается множество
более поздних, различных стадий развития. После описанных фаз
размножения и разделения, наступает новая фаза деления ядер,
приводящая к образованию сперматосфер, или «морул» (рис. 91, Г).
Протоплазма клеток сливается в середине в одну общую безъядерную
плазматическую массу — цитофор (рис. 91, Д, 1); клетки (рис. 91, Д, 2)
располагаются вокруг цитофора, с которым связаны плазматическими
стебельками. В результате получаются сперматосферы, состоящие
сначала из 32 сперматоцитов I порядка, из 64 сперматоцитов II порядка и
даже из 128 сперматид (рис. 91, Д).
Наконец сперматиды дифференцируются в спермин,
освобождающиеся от цитофора, который погибает. Однако очень часто наблюдаются
отклонения от описанного развития, которые выражаются в сильной
изменчивости числа клеточных делений в фазе размножения клеток.
Поэтому фактически одни и те же стадии сперматогенеза встречаются
в сперматосферах с различным числом клеток.
Спермин имеют^ очень длинную тонкую головку и сравнительно
короткий хвостик и бывают короткими (длиной 40 мкм) и длинными
(80—86 мкм). Первые образуются в сперматосферах из 256 или
128клеток, вторые--в сперматосферах из 64 и даже из 32 клеток.
ОТРЯД PLESIOPORA
Aeolosoma hemprichii Ehr.
Материал и методы изучения. К наиболее обычным видам Aeolosoma относятся
A. hemprichii Ehr. A. variegation Vejd. A. quaternarium Ehr. Они встречаются в
разнообразных водоемах, причем A. variegatum обычно попадается среди водной
растительности и в иле, a A. quaternarium — на камнях и в колониях мшанок Plumatella. Это
мелкие, не более 5 мм, прозрачные формы с характерными желтыми, оранжевыми,
красноватыми или зеленоватыми кожными железками.
Aeolosoma hemprichii легко разводится и в лабораторных условиях живет
подолгу. Исходный материал для культуры можно взять непосредственно из природы
или же получить следующим образом. Верхний слой ила, взятого вместе с
растительными остатками в прибрежной части водоема, распределяют слоем 1—1,5 см в
большом кристаллизаторе, доливают водой и оставляют стоять при комнатной
температуре. Скоро на поверхности воды от разложения -растительных остатков
образуется бактериальная пленка, в которой через 10—15 дней появляются Aeolosoma и
другие пресноводные олигохеты.
Тем временем нужно подготовить среду для культуры. Для этого 0,15—0,2 г
хорошо очищенных зерен риса заливают водой, подогретой До 40°С в чашке Петри.
Зерна должны быть стекловидно-прозрачными и предварительно подсушиться. Вода
должна полностью их покрывать. Через 2—3 дня в чашку Пет.ри вносят пипеткой
мелких жгутиконосцев Chilomonas, Gonium или инфузорий Colpidium. Когда они
размножатся (через несколько дней), подсаживают и Aeolosoma. Далее время от
времени в культуру следует подкладывать небольшое количество^рисовых зерен. При
недостатке пищи Aeolosoma инцистируются.
Изучаются Aeolosoma в живом виде на временных препаратах под покровным
стеклом.
Внешняя морфология. Наряду с одиночными особями часто
встречаются находящиеся в процессе бесполого размножения цепочки из не-
103
скольких индивидов. Одиночные особи состоят не более чем из 7
сегментов. Установить количество сегментов проще всего по числу парных
пучков щетинок, так как межсегментные бороздки отсутствуют
(щетинок нет только на I сегменте).
Простомиум (рис. 92, /) довольно большой, расширен в виде
закругленной уплощенной лопасти, очень подвижен и может значительно
изменять форму, совершая ощупывательные движения. Он шире I
сегмента и легко от него отличается, несмотря на отсутствие резкой
границы. На его плоской брюшной стороне заметно мерцание низкого
ресничного покрова (рис. 92,22), при помощи которого животное скользит
по субстрату. Этот способ передвижения связан с малыми размерами
Aeolosoma и у других олигохет не встречается. У основания по бокам
простомиум несет пару маленьких обонятельных ресничных ямок
(рис. 92,2). Их довольно длинные реснички малоподвижны. Наконец,
простомиум усажен редкими очень тонкими неподвижными
осязательными волосками (рис. 92,2/).
Первый сегмент, или метастомиум (рис. 92,5), отделенный от
следующего ясной поперечной бороздкой, лишен щетинок, как у всех
олигохет. Все остальные сегменты снабжены четырьмя пучками щетинок
(рис. 92,6) — парой брюшных и парой спинных, причем каждый пучок
состоит из 3—5 длинных волосовидных щетинок, между которыми
имеются укороченные. Хорошо, видны щетинкон'осные мешочки (рис. 92,5),
вдающиеся в целом, и мускульные пучки (рис. 92,13), связывающие их
со стенками тела. Анальная лопасть (пигидиум) мала и не обособлена
от последнего сегмента.
Кожные железки (рис. 92,//), особенно обильные на простомиуме,
представляют собой округлые клетки эпидермиса, содержащие по
большой окрашенной капле секрета, который прежде ошибочно принимался
за жировое включение.
Анатомия. Отчетливо видна полость тела (целом), в которой при
сокращении кожно-мускульного мешка и кишки переливается
прозрачная целомическая жидкость. Следует отметить почти полное отсутствие
диссепиментов. Вместо них на границах сегментов развиты лишь
рыхлые пучки мускульных волокон (рис. 92,14). Настоящий диссепимент
имеется только между I и II сегментами (рис. 92,17). Спинной и
брюшной мезентерии отсутствуют.
Ротовое отверстие, окаймленное ресничками, лежит на переднем
крае метастомиума'и ведет в мускулистую глотку, (рис. 92,/5), полость
которой выстлана мерцательным эпителием. За глоткой следует узкий
и несколько извитой пищевод (рис. 92,15), за ним — довольно широкая
кишка в виде простой трубки (рис. 92, 7). В просвете кишки видно
энергичное мерцание ресничек-(рис. 92, 29), направленное сзади
наперед. Небольшое анальное отверстие лежит терминально на заднем
конце тела (рис. 92,30). Aeolosoma питается детритом, бактериями и
мелкими жгутиконосцами, которые загоняются в рот благодаря мерцанию
ресничек простомиума.
Кровеносная система видна прекрасно, несмотря на то что кровь
бесцветна. В стенках кишки имеются многочисленные лакуны,
образующие сеть кишечного плексуса (рис. 92,5), который спереди соединя-
104
Рис. 92. Aeolosoma hemprichii. Цепочка из четырех особей. Спинные
пучки щетинок не изображены (ориг.):
1 — простомиум, 2 — обонятельная ямка, 3 — метастомиум, 4 — бороздка между мета-
стомиумом и первым щетинконосным сегментом, 5 — щетинконосный мешочек, 6 —
брюшной пучок щетинок, 7 — кишка, 8 — кишечный кровеносный плексус, 9 — зона
почкования, 10 — третья дочерняя особь, Л — кожные железы, 12 — нефридий, 13 —
мускул щетииконосного мешочка, 14 — мускульные волокна, 15 — пищевод, 16 —
спинной сосуд П — диссепимент, 18 — глотка, 19 — полость глотки, 20 — мозг, 21 —
чувствительные волоски, 22 — реснички нижней поверхности простомиума, 23 — зона
почкования дочерней особи, 24 — первая дочерняя особь, 25 — ее глотка, 26 — ее
простомиум, ' 27 — анальное отверстие второй дочерней особи, 28 — вторая дочерняя особь,
29 — полость кишки, 30 — анальное отверстие первой дочерней особн
ется со спинным сосудом (рис. 92,16). Последний развит только в
передней части тела над пищеводом и глоткой. В простомиуме его ветви
соединяются с брюшным сосудом. Благодаря частым и очень
энергичным антиперистальтическим сокращениям кишки кровь из кишечного
сплетения проталкивается в спинной сосуд, который, наполняясь
кровью, постепенно расширяется до известных пределов, потом вдруг
сокращается. Таким образом, его пульсация происходит гораздо
медленнее, чем сокращения кишки. Кровь по нему движется вперед.
Брюшной сосуд можно хорошо рассмотреть только с брюшной
стороны, для чего надо найти на препарате соответственно
ориентированное животное. Он тянется вдоль всего тела, тесно прилегая к
кишечнику. Кольцевые сосуды отсутствуют.
Метанефридии развиты во всех сегментах, кроме метастомиума и
первого щетинконосного. Так как диссепимеНты отсутствуют, то они
прикрепляются к боковым стенкам кишечника. Нефридий начинается
очень маленькой воронкой и снабжен большим компактным
железистым отделом (рис. 92,12), прободеняым тонким петлистым каналом,
в котором заметно энергичное мерцание ресничек.
Нервная система замечательна своей примитивностью. Мозг лежит,
на спинной стороне в задней части простомиума, будучи тесно связан
с кожным эпителием (рис. 92,20). Окологлоточные коннективы и
широко расставленные брюшные нервные стволы, связанные поперечными
коммиссурами, целиком залегают в толще эпидермиса и на живых
червях не видны, поэтому прежде ошибочно считалось, что они вообще
отсутствуют.
Aelosoma размножается главным образом бесполым путем, и
половозрелые особи встречаются очень редко. Данные о половой системе
недостаточны и противоречивы. Наблюдения над ее строением in vivo
представляют трудности. Известно, что мужские и женские половые
клетки развиваются в области V—VII сегментов в полости тела.
Непарное женское половое отверстие лежит вентрально на VII сегменте.
Поясок (clitellum) развивается на брюшной стенке V—VII сегментов.
Бесполое размножение. Очень часто встречаются цепочки из
нескольких особей, возникающие в результате паратомии, т. е. бесполого
размножения посредством поперечного деления. Перед делением
животного позади последнего (VI или VII) щетинконосного сегмента
образуется короткая зона почкования (рис. 92,9), из которой одна за
другой формируются дочерние особи. Этот процесс' происходит
последовательно, так что в возникающей цепочке первая по времени
образования дочерняя особь оказывается сзади (рис. 92,24), а последняя —
передней ( рис. 92,10). Дочерние особи отделены друг от друга ясными
перетяжками и характеризуются неодинаковыми размерами тела и
щетинок, а также разным числом сегментов, в зависимости от возраста.
Когда формирование задней, старшей дочерней, особи заканчивается,
она отрывается. Однако еще до этого она сама образует собственную
зону почкования (рис. 92,23).
В каждой дочерней особи сначала формируется один сегмент,
который в дальнейшем становится III сегментом, затем образуется IV
сегмент и т. д. до VI (или VII). Возникшая таким образом, но еще непол-
106
ная особь, лишенная еще двух передних сегментов и простомиума,
может сама перейти к бесполому размножению. II, I сегменты и головная
лопасть образуются последними (рис. 92,25). Кишка, которая
незадолго до этого тянулась через все дочерние особи непрерывно, на этой
стадии прервана. На особи, лежащей перед закончившей свое
формирование, образуется анальное отверстие (рис. 92,27), а часть кишки,
лежащая между этими особями, дегенерирует. Сформированная дочерняя
особь оказывается связанной с предыдущей лишь небольшим участком
эпидермиса, который легко разрывается, и тогда задняя дочерняя особь
отделяется. От первой закладки новой особи до ее отрывания проходит
около 4 дней (при комнатной температуре). Хорошо питающиеся
животные образуют цепочки из 3—4 особей. Число сегментов в такой
цепочке может быть представлено формулой: 7 + 0+2+4+(5+1), где
7 — число щетинконосных сегментов материнского организма; 0 —
отвечает самой молодой дочерней особи, не имеющей еще щетинок;
следующие особи имеют соответственно 2 и 4 сегмента; задняя особь (5 +
+ 1) не имеет еще двух передних сегментов, но образовала уже свою
односегментную почку. На рис. 92 изображена цепочка, у которой
старшая дочерняя особь (рис. 92,24) сформировала уже передние сегменты
и простомиум (рис. 92,26) и готова оторваться.
КЛАСС HIRUDINEA-ПИЯВКИ
Удобным объектом изучения является медицинская пиявка — Hiru-
do medicinatis L., принадлежащая к семейству Hirudinidae отряда
челюстных пиявок — Gnathobdellea. Она широко распространена, достаточно
крупна для анатомирования и относится к числу хорошо
исследованных беспозвоночных. В крайнем случае она может быть заменена
обычной в пресноводных водоемах большой ложноконской пиявкой — Нае-
mopis sanguisuga L., относящейся к тому же семейству Hirudinidae.
Кроме того, желательно ознакомиться с организацией одного" из
представителей отряда хоботных пиявок —■ Rhynchobdellea, например
улитковой пиявкой—Glossiphonia complanata (L.) (сем. Glossiphoniidae).
ОТРЯД GNATHOBDELLEA—ЧЕЛЮСТНЫЕ ПИЯВКИ
Hirudo medicinalis L. — Медицинская пиявка
Материалы и методы изучения. Медицинские пиявки издавна используются с
лечебными целями, в связи с чем они продаются в аптеках. Нетрудно собрать их и
в природе. Этот вид широко распространен в южных областях европейской части
СССР (Воронежская, Курская, Саратовская, Ростовская области, Украина, Кавказ),
в Среднеазиатских советских республиках. Излюбленные места обитания — бедные
известью стоячие водоемы с илистым дном,, заросшие растительностью. Здесь в теплое
время года пиявок легко наловить простым сачком. Медицинские пиявки долго живут
в неволе, если их содержать в прохладном месте в непрозрачной (лучше фаянсовой)1
посуде, плотно завязанной полотном. Полезно положить в воду немного конского
волоса, о который животные могут счищать слизь. Для содержания 10 червей требуется
не менее литра воды. Через каждые 2—4 дня воду (следует менять, а посуду
тщательно мыть. Пиявки способны голодать долгое время. Однако при длительном
содержании их необходимо подкармливать сгустками свежей крови (полученной с бойии) или
подсаживать к ним на время живую лягушку.
новится неподвижным После этогосл^Т™361 Кр°ВЬ' 3аТеМ Успокаивается и ста-
слизь, покрывающую теао^зу^ениР^Г-ЩТ1ЛЬНО "ереТЬ полотенВДм обильную
тых животных. Из>чение внешней морфологии производится на свежеуби-
ж„дкЙЛТиГсГаИ1иРсуГемо™ЯсВгс&^^
удина. Перед фиксацией ■рекомендуется" Гегка S'n^Z"4"0™0 Ша~
ковому дну маленькой г-тркчяттй ™„„ растянув пиявку, приколоть ее к вое-
дольных срезов заливают^^целлоидин Л°' ДЛЯ ИЗготовле™я больших про-
M^^Xo^^S^^'^^'^^P" С Д°КраСК0Й эозшом "- по
кармином и осмиевой. Слотой Сле^т и и"еЛЫХ КуСКОВ пеРед заливкой борным
реднего конца, ^"ых ^^^^^^ГпЯГХ ^
тальные и сагиттальные срезы переднего и заднего концов тела ' ** Же Фр°Н"
Наблюдения над живыми животными. Представляет интеоес хяпяк
тер движения медицинской пиявки. Она может ползать плавав
конец, производить так называемые ДыхательныГдвижения *' **'
*шаган;я>>НПппа!пРШаеТСЯ СХ°ДН° С° ВСеми ДРУ™ пиявками путем
«шагания». Прочно- присосавшись к субстрату задней присоской чепвь
вытягивается, нащупывает передней присоской подходящее место пои
nSnTleS ппи°СГб0ДИВ 3*аДНЮЮ ПРИС0СКУ' прДик°рТлеяетМ:ТпоГаРди
передней. Тело при этом изгибается петлеобразно кверху Затем
освобождается передняя присоска, тело вновь вытягиваетсяи т. д (рис 93)
Плавает пиявка быстро, волнообразно
изгибая уплощенное тело в вертикальной
плоскости. Иногда она начинает совершать
волнообразные движения всем телом,
оставаясь при этом прикрепленной задней
присоской. Эти движения производятся для
обновления воды вокруг тела и являются
признаком того, что животное испытывает
недостаток в кислороде. Часто можно
видеть, как пиявки вылезают из воды и
присасываются к стенке сосуда. Это
вызывается резким недостатком кислорода в воде
и связано с наличием у пиявок чисто
кожного дыхания.
Голодные пиявки быстро
присасываются к коже живой лягушки и долго сосут
кровь. В нормальных условиях пиявка
всасывает за один прием от 3 до 15 г крови, в
Рис 93 Схема движения Him- результате чего сильно раздувается vrp-
do medicinalis (по Гертеру) личиваясь в массе в несколько раз.''После
108
Рис. 94. Hirudo medicinalis.
Вид со спинной стороны (по
Скрибану)
Рис. 95. Внешняя морфология
Hirudo medicinalis. Вид со
спинной (А) и, брюшной (Б)
сторон (по Скрибану, с
изменениями) : '
/ — простомиум, 2 — глаза, 3 —
чувствительное колечко, 4 —
анальное отверстие, 5 — задняя
присоска, 6 — женское половое
отверстие, 7 — мужское половое
отверстие, 8 — наружное
отверстие нефридия, 9 — передняя
присоска, 10 — ротовое
отверстие. I—XXVI — сегменты
однократного приема пищи пиявка может голодать до 15 месяцев,
за это время высосанная кровь полностью переваривается.
Пиявки время от-времени линяют, т. е. сбрасывают старую
кутикулу,- под которой оказывается новая. Особенно часто это происходит,
когда в аквариуме вода начинает портиться.
Размножение в неволе наблюдать трудно. В природе откладка
коконов происходит в июле— августе. Животное делает ход в сырой
земле над уровнем воды и откладывает в нем кокон яйцевидной формы,
размером с лесной орех. Коричневая оболочка кокона имеет губчатую
структуру; внутри него находится от 5 до 30 яиц.
Внешняя морфология. В спокойном состоянии тело медицинской
пиявки сильно вытянуто в длину (рис. 94). Спереди оно несколько уже.
Характерна сильная дорзовентральная сплющенность: спинная сторона
109
выпуклее брюшной. На переднем конце с брюшной стороны видна
передняя, или ротовая, присоска (рис. 95, Б, 9) с ротовым отверстием на
дне. Сзади тело заканчивается дисковидной задней присоской (рис. 95,
Б, 5), у основания которой дорзально лежит анальное отверстие
(рис. 95, А, 4). Длина взрослого животного в покое достигает 8—12 см,
ширина 12—20 мм. Спинная поверхность темная, зеленоватая, с тремя
парами коричневатых, ржаво-красных или красновато-желтоватых
полосок, на фоне которых заметны неправильные, но в общем метамер-
ные черные пятна. Окраска подвержена сильным индивидуальным
колебаниям. Брюшная сторона бледная — желтовато-зеленоватая, с
неправильными черными пятнами.
Подобно всем пиявкам Hirudo обладает вторичной кольчатостью
сегментов. Все тело животного разделено тонкими поперечными
бороздками на многочисленные кольца. Нормальный сегмент (в средней
области тела) состоит, как у большинства Gnathobdellea, из 5 колец.
Следует внимательно изучить внешнюю морфологию пиявки,
пользуясь лупой или бинокуляром. Необходимо иметь в виду, что^установ-
ление границ сегментов на поверхности тела требует большой
внимательности. Среднее (третье) кольцо каждого сегмента (рис. 95, А, 3)
узнается по наличию на его спинной стороне поперечного ряда
маленьких, округлых, окрашенных светлее окружающей кожи, возвышений —
чувствительных папилл, или сосочков. Поэтому третье кольцо сегмента
обычно называют чувствительным. Тщательно прослеживая
расположение чувствительных колец на теле, можно выяснить общее число
туловищных сегментов. Различению границ отдельных сегментов помогают
также повторяющиеся на определенных кольцах детали рисунка —
черные пятна. Наконец, на брюшной стороне по бокам второго кольца
каждого сегмента в средней области тела в хороший бинокуляр
заметны парные щелевидные отверстия нефридиев (рис. 95,5).
На головном отделе видны кольца передних сегментов, за счет
которых формируется передняя, или ротовая, присоска. Впереди лежит
неясно обособленная головная лопасть (рис. 95, А,1), затем следуют
7 узких колечек. Из них первые 2 кольца являются чувствительными
кольцами I и II сегментов (рис. 95,/,//), третье и четвертое колечки
принадлежат III сегменту (рис. 95,///), а пятое и шестое—IV
сегменту (рис. 95, IV). Каждый из этих четырех головных сегментов несет
пару глаз в виде небольших черных пятен (рис. 95, А, 2), представляющих
собой видоизменение чувствительных сосочков.
Ротовая присоска лежит на нижней стороне головного отдела. Ее
почти треугольное углубление обращено основанием назад, а
вершиной вперед (рис. 95, Б, 9). Края ее несколько уплощены и несут
радиальные бороздки — границы колечек передних сегментов. На дне
присоски в задней ее части хорошо заметно ротовое отверстие трехлучевой
формы (рис. 95, Б, 10). Сзади присоска ограничена IV сегментом.
Таким образом, этот орган слагается из головной лопасти и четырех
передних сегментов, которые, по-видимому, являются ларвальными
(П. П. Иванов, 1944).
Туловище состоит из 22 сегментов (с V по XXVI), которые сначала
постепенно расширяются, а сзади довольно быстро суживаются. Перед-
110
ние два туловищных сегмента (V и VI) трехкольчатые (рис. 95, V, VI).
Первый сегмент туловища несет пару глаз. Hirudo, следовательно,
обладает 5 парами глаз — 4 парами головных и 1 парой туловищных.
VII сегмент разделяется на 4 колечка (рис. 95, VII). Сегменты всей
средней области тела (с VII по XXII) состоят каждый из 5 колец.
Наконец, XXIII сегмент четырехкольчатый (рис. 95,XXIII), XXIV
—состоит из трех колечек (рис. 95, XXIV), а последние два — XXV и XXVI —
двухкольчатые (рис. 95,ХАУ, XXVI). На последнем сегменте лежит
анальное отверстие (рис. 95, А, 4), поэтому его иногда называют
анальным сегментом.
Задняя присоска куполовидна и обращена своей вогнутой стороной
на брюшную сторону (рис. 95, £,5). Диаметр ее в два раза меньше
наибольшей ширины тела. Она лишена каких-либо следов
сегментации, но сформирована семью задними сегментами.
Таким образом, в состав всего тела пиявки входят 33 сегмента
(4 передней присоски, 22 — туловища й 7 —задней присоски). Это
число постоянно для всех пиявок, если не считать отряда' щетинковых
пиявок (Acanthobdellea), обладающих 30 сегментами.
Следует также рассмотреть расположение выделительных и
половых отверстий. Маленькие щелевидные поры нефридиев в количестве
17 пар помещаются по бокам на брюшной стороне VI—XXII
сегментов (рис. 95, Б, 8). Непарное мужское половое отверстие (рис. 95,5, 7)
лежит медиально на брюшной поверхности X сегмента, между его
четвертым и пятым кольцами. Иногда из него выдается совокупительный
орган — циррус — значительной длины. Такое же положение, но на
следующем (XI) сегменте занимает непарное женское половое отверстие
(рис. 95, Б, 6).
В период половой зрелости оба половых сегмента, а также
предыдущий (IX) сегмент заметно "'отличаются по внешнему виду от
соседних, так как. в них развиваются особые клителлярные железы,
выделяющие вещество кокона. Эти три сегмента (IX, X и XI)
образуют область пояска и носят название клйтеллярных.
Наконец, необходимо обратить внимание на полное отсутствие
щетинок— характерную особенность для всех пиявок, кроме
примитивных— Acanthobdellea, у которых на пяти передних сегментах еще
сохраняются щетинки.
Анатомия. Умерщвленная пиявка укрепляется на дне
препаровальной ванночки спиной кверху. Одна булавка втыкается в заднюю
присоску, другая — в передний конец тела. Животное следует возможно
больше растянуть.
Вскрытие представляет ряд трудностей, так как рространства
между внутренними органами заполнены довольно плотной и прочной
соединительной тканью. Сначала острым скальпелем делается
неглубокий продольный разрез стенки тела несколько в стороне от медио-
дорзальной линии. Затем следует осторожно отпрепарировать
кишечник. Приподнимая пинцетом края разреза и отводя их осторожно в
стороны, одновременно подрезают кончиком скальпеля соединительную
ткань. Если при этом повреждается кишечник, то из него осторожно
выдавливается выступающая кровь и весь препарат промывается
111
Рйс. 96. Hirudo medicinalis. Общий вид
животного, вскрытого со спинной стороны. А—
пищеварительная и лакунарная системы; Б — нервная,
лакунариая и половая системы (по Гатчеку и
Кори):
1 — глотка, 2 — мускулатура глотки, 3 — нефридий, 4 —
желудок, 5 — боковой лакунарный канал, 6 — спннной
лакунарный канал, 7 — задний вырост желудка, 8 —
кишка, 9 — задняя кишка, 10 — задняя присоска, И —
анальное отверстие, 12 — железистый придаток кишки, 13 —
церебральные ганглии, 14 — предстательная железа, 15 —
придаток семенника, 16 — железистый отдел нефридия,
17 — мочевой пузырек, 18 — брюшной лакунарный канал
с брюшной нервной цепочкой, 19 — семяпровод, 20 —
семенные мешки, 21 — влагалище, 22 — яйцевой мешок с
яичником, 23 — мужской совокупительный орган
112
свежей водой. После
изучения кишечника таким
же образом отпрепаровы-
вают и остальные
системы органов (рис. 96, А,
Б).
Пищеварите л ь-
ная система.
Передний отдел
пищеварительного аппарата удобнее
изучить на отдельном
экземпляре пиявки.
Сначала необходимо внима-
. тельно рассмотреть под
бинокуляром дно
передней присоски с ротовым
отверстием, потом
рассечь острым скальпелем
передний конец тела по
медио-вентральной линии
(рис. 97, Б).
Передняя кишка
представлена ротовой
полостью и глоткой,
выстланными кутикулой, что
указывает на их эктодер-
мальную природу.
Трехлучевая форма ротового
отверстия (рис. 97, А, 2)
зависит от наличия трех
кожных складок— губ.
Одна из них
ограничивает рот с заднебрюшной
стороны, пара других —
спереди и с боков. Под
губами находится
небольшая ротовая полость, в
которую вдаются три
челюсти (характерный при- •
знак отряда Gnathobdel-
lea) в виде полукруглых
линзообразных беловатых
валиков (рис. 97, Б, 3,
4). Передняя челюсть
лежит в медианной
плоскости, две другие, вентро-
латеральные,
расположены к ней под углом в
120°. Челюсти мускулис-
Рис. 97. Hirudo medicinalis. Передняя часть пищеварительной системы. А — передний
конец тела с брюшной стороны; Б — он же, вскрытый вдоль; В — изолированная
челюсть; Г — зубчик челюсти с широкой стороны (А—В — из Гертера, Г — по Цину):
/ — передняя присоска, 2 —ротовое отверстие, 3 — латеро-вентральная челюсть, 4 — дорзальная
челюсть, 5 — брюшной лакунарный канал с брюшиой нервной цепочкой, 6 — продольная
мускулатура, 7 — полость глотки, 8 — радиальные мускулы глотки, 9 — ряд зубчиков, 10 — двигательные
мускулы челюсти
ты и несут по свободному килевидному краю ряд мелких выступающих
зубчиков (рис. 97,В, 9). Чтобы их рассмотреть, нужно вырезав одну из
челюстей, приготовить временный или постоянный тотальный препарат и
воспользоваться слабым увеличением микроскопа. Зубчики (рис. 97, Г)
состоят из органического вещества, пропитанного известью, и имеют
характерную форму сердцевидных пластиночек (размер около 80 мкм).
Когда животное присасывается передней присоской для сосания крови,
челюсти, выдаваясь между губами, прижимаются к коже жертвы. Ки-
левидный край челюсти представляет как бы тонкую пилочку и при
сокращении мускулатуры производит на коже жертвы разрез, действуя
наподобие круговой пилы.
Между зубчиками челюстей открываются протоки, слюнных желез.
Так как в слюне содержится гирудин, препятствующий свертыванию
крови, то ранка, прорезанная пиявкой, долго кровоточит и червь имеет
возможность достаточно насытиться. В том месте, где сосала пиявка,
на коже жертвы остается ранка характерной трехлучевой формы.
Роль нагнетательного аппарата при сосании играет глотка. Это
мускулистый орган веретеновидной формы (см. рис. 9Q,A,1). Особенно
сильно развита радиальная глоточная мускулатура, связывающая
глотку со стенками тела (см. рис. 96, А, 2; 97, Б, 8).
Высосанная кровь поступает в следующий отдел кишечника —
желудок. Этот орган, относящийся к средней кишке, объемист и тянется
почти до заднего конца тела (см. рис. 96,А,4). Он имеет 11 парных
боковых выростов (см. рис. 96, Л, 7). Впрочем, первая пара часто
бывает рудиментарной. Особенно велика задняя пара выростов,
проходящая по бокам вдоль следующего отдела средней кишки. Желудок
служит главным образом резервуаром, наполняемым пищей — кровью.
113
Переваривание и всасывание происходит преимущественно в
заднем отделе средней кишки, которая имеет вид тонкой трубочки (см.
рис. 96, А, 8). В своей начальной части она снабжена парой небольших
железистых придатков (см. рис. 96, А, 12), а сзади открывается в
несколько расширенную эктодерм ал ьную .заднюю кишку (см. рис. 96, Л,
9). Последняя открывается наружу анальным отверстием (см. рис. 96,
А, 11).
Лакунарная система. Hlrudo medicinalis, подобно всем Gna-
thobdellea, лишена истинной кровеносной системы, которая
функционально замещена остатками целома в виде узких каналов, образующих
лакунарную систему. Часть последней — спинной канал ■—
обнаруживается уже при препаровке кишечника. Для изучения всей остальной
части системы необходимо удалить желудок, который перерезается позади
глотки, захватывается пинцетом и осторожно отделяется. Довольно
прочную соединительную ткань4, связывающую кишечник с брюшными
стенками тела, следует подрезать острыми ножницами. Лакунарные
каналы легко узнаются, так как содержат красную кровь, цвет которой
зависит от растворенного в ней гемоглобина. Однако изучение лаку-
нарной системы путем анатомирования затрудняется тем, что часть
каналов при этом неизбежно повреждается, кровь вытекает из них и
они становятся незаметными. Приходится поэтому вскрывать несколько
пиявок, варьируя каждый раз способ вскрытия и отпрепаровывая те
части системы, которые не удалось рассмотреть на предыдущем
экземпляре животного. Вскрытие и изучение препарата производится под би-
нокуляром.
Лакунарная система состоит из четырех продольных каналов —
спинного, брюшного и двух боковых, соединенных множеством сильно
ветвящихся промежуточных лакун, сетью кожных лакун и, наконец,
кольцевыми коммиссурами.
Спинной канал (см. рис. 96, А, 6; рис. 98, 13), проходящий
медиально над кишечником, особенно часто .повреждается при -вскрытии.
От него отходят парные веточки (рис. 98,14), которые, сильно
разветвляясь, образуют густую кожную сеть лакун спинной стороны тела.
Брюшной канал (см. рис. 96, Б, 18; рис. 98,/) заключает в себе
брюшную нервную цепочку и образует небольшие расширения вокруг
ее ганглиев. Спереди вокруг окологлоточного нервного кольца и сзади
вокруг большого последнего ганглия он расширяется особенно сильно.
От каждого расширения ответвляется пара вентродорзальных каналов
(рис. 98, 7); они поднимаются на спинную сторону мимо боковых лаку-
нарных каналов, не соединяясь с ними, проходят между слепыми
выростами желудка к коже, где связаны с кожным сплетением лакун
(рис. 98,16).
Позади расширений с ганглиями отходят также тонкие парные
каналы (рис. 98,5), которые загибаются слегка на спинную сторону и
скоро расширяются на конце, образуя маленький пузырек, называемый
ампулой (рис. 98,5). Внутри ампулы находится ресничная воронка
нефридия (нефростом), и она, следовательно, представляет собой пе-
ринефростомальный участок целома. Имеется всего 11 пар ампул,
лежащих в средних сегментах тела (с XII по XXII), причем они прилегают
114
/ 17 16 15
9 10 11 12 13
Рис. 98. Hirudo medicinalis. Часть лакунарной системы щэавой
половины тела (по Гратиоле):
1 — брюшной лакунарный канал, 2 — его расширение, содержащее ганглий
брюшной нервной цепочки, 3 — латеро-вентральный канал, 4 — канал,
проходящий от ампулы в дорзальном направлении, 5—ампула, 6 — каналы,,
соединяющие ампулу с брюшным лакунарным каналом, 7 — вентро-дор-
зальный канал, 8 — свободный конец нефрндня, 9 — семяпровод, 10 —
боковой лакунарный канал, 11 — латеро-латеральный канал, 12 — латеро-
дорзальный сосуд, 13 — спинной лакунарный канал, 14 — канал, идущий
от спинного канала к коже, 15 — железистый отдел нефридия, 16 —
кожная сеть капилляров, 17 — мочевой пузырек
сверху к округлым семенным мешкам, кроме двух последних пар,
которые помещаются уже в сегментах, лишенных семенных мешков. В
шести передних нефридиальных сегментах ампулы отсутствуют. От каждой
ампулы отходит тонкий боковой сосудик, направляющийся к стенке
тела (рис. 98,4), где он соединяется с кожной сетью лакун.
Боковые лакунарные каналы (рис. 96,5; 98,10) проходят
симметрично с боков от кишечника. Это самые заметные и толстые каналы,,
отличающиеся к тому же беловатым цветом, что зависит от сильно
развитого в их стенках слоя кольцевых мускульных волокон. Они слегка
изгибаются волнообразно; передние и задние концы их сильно
разветвляются, образуя анастомозирующие капилляры.
От боковых каналов ответвляются следующие ветви:
1. Тонкие латеро-вентральные сосуды (рис. 98,5), представленные
18 парами. Каждый канал скоро раздваивается, а обе ветви его, широ-.
ко расходясь, направляются на брюшную сторону, где соединяются под.
брюшным лакунарным каналом с такими же ветвями с
противоположной стороны тела. Таким образом, посредством латеро-вентральных
сосудов боковые каналы связаны друг с другом. Латеро-вентральные со-
115.
суды дают мелкие веточки, оплетающие нефридии густой сетью
капилляров.
2. Очень толстые, но короткие латеро-латеральные каналы
(рис. 98,11), распадающиеся на густое капиллярное сплетение.
3. Латеро-дорзальные ветви (рис. 98,12), очень толстые длинные
стволы, чередующиеся с предыдущими, в количестве 17 пар. Они
направляются к средней области спинной стенки тела, ветвятся и
соединяются через систему капилляров с такими же ветвями
противоположной стороны.
Промежуточная сеть лакун чрезвычайно сложна. Она развита
между кожно-мускульным мешком й кишкой и состоит из многочисленных
капиллярных разветвлений латеро-вентральных и латеро-дорзальных
каналов. Капилляры имеют неправильно узловатый характер и
отличаются темно-коричневым цветом, так как покрыты ботриоидной тканью
(см. с. 127).
Кожная сеть капилляров (рис. 98,16) также очень сильно развита.
Ее разветвления лежат в коже между основаниями клеток эпидермиса
и связаны с ветвями латеро-латеральных, латеро-вентральных и
латеро-дорзальных каналов.
Кровообращение в лакунарной системе исследовано недостаточно.
В частности, неизвестно направление тока крови. Снабженные
кольцевой мускулатурой боковые каналы пульсируют попеременно, когда один
находится в состояний сокращения, другой наполнен кровью и
наоборот. Способны сокращаться-и ампулы, называемые иногда сердцами; в
их стенках также есть мускульные волокна. Все ампулы,
принадлежащие к одной стороне тела, сокращаются одновременно, причем
сокращения правых и левых ампул чередуются. Наконец, к сокращениям
способен и брюшной лакунарный канал.
Выделительная система. Выделительный аппарат
представлен нефридиями и ботриоидной тканью (с. 127). Нефридии в
количестве 17 пар (см. рис. 96, А, 3) расположены на вентральной стороне VI—
XXII сегментов; размеры их постепенно возрастают к заднему концу
тела. Один из нефридиев следует осторожно вырезать и внимательно
изучить на временном или постоянном тотальном препарате.
Ознакомление с тонкой структурой органа возможно лишь на срезах
(с. 125).
Каждый нефридий имеет вид длинного канала, образующего
характерную подковообразную петлю (рис. 99), заметно утолщенную в
середине. Она лежит между соседними слепыми выростами желудка.
На вентральной стороне ее выдается тонкий проксимальный конец
нефридия (рис. 99,8), связанный с ресничной воронкой (рис. 99,10).
Рядом от петли отходит тонкий проток, называемый экскреторным
каналом (рис. 99,4). Он направляется к большому сферическому мочевому
пузырьку (рис. 99,5) и открывается в него. Мочевые пузырьки (см.
рис. 96, Б, 17) лежат на брюшной стороне тела ковнутри от боковых
лакунарных каналов. Короткий выводной проток сообщает мочевой
пузырек с наружной средой. Наружные выделительные отверстия (нефро-
поры) в виде маленьких поперечных щелей (см. рис. 95, Б, 8) лежат
на брюшной стороне второго колечка сегмента.
116
На свободном проксимальном конце каждый нефридий прилегает
ж ресничной воронке. Последняя, называемая также ресничным
органом, лежит в ампуле — мешковидном расширении поперечного лаку-
ларного канала (с. 125). Шесть передних пар нефридиев имеют,
однако, свободные воронки, так как ампулы в их сегментах отсутствуют.
Проксимальный участок нефридия и следующая за ним петля
вместе обозначаются как железистый отдел органа (рис. 99,5,6, 8); у Hiru-
■dinidae железистый отдел отличается своеобразным строением. Он
слагается из массы нефридиальных клеток (см. .рис. ПО, Б,3), среди
которых проходит центральный канал нефридия. Очень сложный ход
этого канала представлен на рис. 99. В центральный канал открываются
многочисленные внутриклеточные канальцы (см. рис. 110,2),
пронизывающие плазму в толще железистого отдела и образующие густую сеть
(см. также с. 126). Свободный проксимальный конец нефридия
(железистого отдела) не имеет центрального канала, а его ветвящиеся
канальцы не сообщаются с ресничным органом. Таким образом воронка
•отделена от нефридия. Эта особенность характерна для пиявок.
Нефридии вместе с их
воронками имеют мезодер-
мальное происхождение. Из
эктодермы формируются
только мочевой пузырек и
выводной канал.
Нервная система.
Центральная нервная
система, состоящая из
церебральных ганглиев и брюшной
нервной цепочки, тщательно
отпрепаровывается,
окрашивается борным кармином, и
из нее приготавливается
тотальный препарат.
Церебральные ганглии
пиявок по сравнению с
таковыми у других аннелид
значительно сдвинуты
назад. У Hirudinidae они
помещаются в - VI сегменте
над глоткой (см. рис. 96, Б,
13). Связывающая их ком-
:миссура коротка. Коротки
и окологлоточные коннекти-
вы (рис. 100, 5),
соединяющие церебральные ганглии
vC большой передней гангли-
юзной массой (рис. 100, 6)
•брюшной нервной цепочки.
Затем следуют 21 простой
;ганглий цепочки (рис. 100,
Рис. 99. Нефридий Hirudo medicinalis (по
Берну)":
/ — эпидермис, 2'— нефропор, 3 — мочевой пузырек,
4 — экскреторный канал, 5, 6 — части железистого
отдела нефридия, 7 — центральный канал, 8—
свободный проксимальный отдел нефридия, 9 — полость
ресничного органа, 10 — ресничный орган
117
7) и, наконец, большая задняя ганглиозная
масса (рис. 96, Б).
В типичном ганглии брюшной нервной:
цепочки нервные клетки собраны в виде
характерных пакетов (рис. 100, 8), разделенных
неврилеммой. Каждый ганглий содержит 6
таких пакетов —2 пары боковых и два медио-
вентральных. Соседние ганглии соединены,
друг с другом тесно сближенными коннекти-
вами (см. рис. 112, А, 2), между которыми,
проходит очень тонкий, медиальный, так
называемый файвровский нерв (см. рис. 112, А,
4). От каждого ганглия отходят 2 пары
боковых нервов.
Анализ расположения и числа пакетов;
внутри сложных передней и задней ганглиоз-
ных масс брюшной нервной цепочки, а также
изучение областей иннервации отходящих or
них нервов, позволили установить их состав.
У Hirudinidae передняя ганглиозная масса-
состоит из 4 слившихся простых ганглиев, а
задняя — из 8, причем самый задний ганглий;
рудиментарен.
От боковой поверхности церебральных:
ганглиев отходит пара коротких стоматогаст-
рических нервов, заканчивающихся
придаточными ганглиями (рис. 100, /, 3). Последние
входят в состав рыхлого нервного колечка
симпатической нервной системы, лежащего
вокруг глотки впереди церебральных ганглиев
и окологлоточных коннективов. Оно иннерви-
рует челюсти; от него отходят многочисленные нервы,' образующие
сплетение в стенках глотки и продолжающиеся назад в виде рыхлога
нервного сплетения кишечника.
Половая система. Hirudo medicinatis гермафродит, как все
пиявки. Изучение доловой системы путем вскрытия не представляет-
трудностей. У пиявок, так же как у малощетинковых червей (с. 86),
половые железы лежат в половых мешках. Однако в связи с редукцией
целома, последние приняли характер совершенно обособленных
образований. Семенные мешки у Hirudinidae и многих других пиявок
представлены несколькими метамерными парами, но их множественность
есть результат вторичного расчленения одной пары первоначально
цельных мешков, которые в свою очередь возникли обособлением от целома
X сегмента.
У медицинской пиявки 9 пар (в виде исключения— 10 пар)
семенных мешков, лежащих на брюшной стороне XII—XX сегментов, ковнут-
ри от мочевых пузырьков (см. рис. 96, Б, 20). От каждого мешка с его
наружной стороны отходит поперечный тонкий семявыносящий канал
(рис. 101, Л, 7), открывающийся в продольный семяпровод своей сторо-
118
Рис. 100. Hirudo medici-
nalis. Нервная система,
церебральные ганглии и
подглоточная
ганглиозная масса (из Н. А.
Ливанова) : "
1 — челюстной ганглий сто-
матогастрической системы,
2 — нерв церебрального
ганглия, 3 — ганглий стомато-
гастрнческой системы, 4 —
церебральный ганглий, 5 —
окологлоточный коннектив,
6 — подглоточная
ганглиозная масса, 7 — ганглий
брюшной нервной цепочки, 8 —.
пакеты
Рис. 101. Половая система Hirudo medicinalis. A — передняя часть
мужской половой системы; Б — дистальная часть мужского полового
аппарата в оптическом разрезе (по Брандесу):
/ — базальная часть мужского совокупительного органа, 2 — семяизвергатель-
ный канал, 3 — придаток семенника, 4 — семяпровод, 5 — железистая часть
семяпровода, 6 — семенной мешок, 7 — семявыносящий канал, 8 — ганглий
брюшной нервной цепочки, 9 — влагалище, 10 -т яйцевой мешок, //— мужской
совокупительный орган, 12 — предстательная железа, 13 — семяпроводящий
канал, 14 — cornua, 15 — атрий
иы (рис. 101, Л, 4,5). Пара семяпроводов тянется вперед до X сегмента,
где они закручиваются в плотные клубочки, называемые придатками
семенников (epididymis, рис. 101, Л, 5). Они играют роль семенных
пузырьков, т. е. служат для накопления спермы; у неполовозрелых
животных они содержат секрет железистых стенок семяпроводов, а у
половозрелых— массы спермиев, соединенных пучками.
От каждого из этих органов отходит короткий, но довольно
широкий семяизвергательный канал (ductus ejaculatorius, — рис. 101, 2) с
толстыми мускулистыми стенками. Семяизвергательные каналы
открываются в базальную часть мужского совокупительного органа, которая
образована непарным железистым мешком, называемым атрием
(atrium, рис. 101, Б, 15). Атрий снабжен правым и левым отростками
(cornua, рис. 101, Б, 14). Толстая железистая масса, одевающая снаружи
лтрий, ^называется лредстательной железой (prostata, рис. 101, Б, 12).
От атрия отходит непарный совокупительный орган; он
направляется сначала назад, потом вперед, образуя характерную петлю
(рис. 101,11). Он мускулист, содержит внутри семяпроводящий канал
и прикреплен передним концом к. стенке тела. Вся его дистальная
половина способна выворачиваться наподобие пальца перчатки, и тогда
он выпячивается через половое отверстие далеко наружу.
Рекомендуется изучить под микроскопом содержимое семенного
мешка, взяв его от живой или свежеубитой пиявки.
119
У зрелых особей в жидкости семенных мешков плавают мужские
половые клетки различных стадий развития. Самые молодые стадии:
представлены одиночными клетками сперматогониями, которые очень-
рано отрываются от зачаткового эпителия стенок семенного мешка.
Наряду с ними встречаются 4, 6 и 8-клеточные стадии размножения,,
напоминающие морулу (рис. 102,Л). При дальнейшем размножении
сперматогонии обособляют большую центральную плазматическую
массу— цитофор (рис. 102, Б, 2), на поверхности которого сидят сами
клетки (рис. 102, Б, 1). Цитофор лишен ядер, в нем постепенно
накапливаются экскреты; после отделения от него половых клеток он погибает.
Образование сперматозоидов из сперматид сопровождается сильным
вытягиванием половых клеток и склеиванием их в характерные пучки,
в которых они расположены параллельно друг другу.
Женская половая система состоит из пары небольших яйцевых
(овариальных) мешков, лежащих в XI сегменте, и выводных путей.
Яйцевые мешки (рис. 103,5) представляют собой обособившиеся отделы
целома XII сегмента; в каждом свободно лежат два шнуровидных
компактных тяжа, образующие несколько петель. Это и есть яичники
(рис. 103, 1). Тонкостенные яйцевые мешки продолжаются в виде
коротких маток (рис. 103,2), которые, соединяются далее в непарный
яйцевод (рис. 103,5), образующий несколько петель и покрытый массой
одноклеточных слизистых, так называемых яйцеводных желез
(рис. 103,4). Яйцевод сообщается с широким петлеобразным
влагалищем (рис. 103,5), которое открывается наружу женским половым
отверстием. Половые отверстия лежат медиально на брюшной стороне —
мужское на X, а женское — на XI сегменте. ~~~
Рис. 102. Hirudo medicinalis. Co- Рис. 103. Hirudo medicirialis. Женский по-
зревающие мужские половые клет- ловой аппарат в оптическом разрезе (по
ки. А — сперматогонии в стадии Брандесу):
размножения; Б — образование ;-яичннк, г-матка, з- яйцевод, 4-яйцевод-
ЦИТОфора (по Иогансону): ные железы, 5 — влагалище, 5 —яйцевой мешок
1 — половые клетки, 2 — цитофор
120
Рис. 104. Hirudo medicinalis. Поперечный разрез (ориг. рис. Ю. "К. Петрушевского):
/ — кожный эпителий, 2 —кольцевая мускулатура, 3 — диагональная мускулатура, 4 — продольная
мускулатура, 5 — дорзовёитральный пучок мускульных волокон, 6 — боковой лакуиарный канал,
7 —боковой карман желудка, 8 — нефридий, 9 — семенной мешок, 10 — семяпровод, // —брюшной
лакуиарный канал с брюшной нервной цепочкой, 12 — желудок, 13 — лакунарный канал, 14 —
мочевой пузырек, 15 — ботрноидная ткань, 16 — спннной лакунарный канал
При спаривании совокупительный орган одной особи вводится во
влагалище другой, в связи с чем у Hirudinidae не образуются сперма-
тофоры, характерные для других пиявок (см. с. 140). Спаривание
происходит в воде и продолжается от 10—20 мин до 2—3 ч. Оно может
быть перекрестным, когда каждая особь одновременно играет роль
самца и самки, или односторонним. Между спариванием и откладкой
яиц проходит от нескольких дней до 3 месяцев. Откладка коконов
происходит в июле — августе. Стенки коконов формируются из белковых
выделений клителлярных желез, причем образуется муфта,
сбрасываемая затем через головной конец тела так же, как у дождевых червей
(с. 89).
Изучение срезов. Кожно-мускульный мешок. Поверхность
тела покрыта тонкой .слоистой, окрашивающейся гематоксилином,
кутикулой. Эпидермис отличается некоторыми, свойственными только
пиявкам, особенностями. Он состоит из слоя, высоких колбовидных или
цилиндрических клеток (см. рис. 105, 4), соприкасающихся друг с
другом только своими апикальными (периферическими) концами, тогда как
тела клеток, содержащие ядра, разъединены внедряющей между ними
соединительной тканью. Так как границы между апикальными
отделами клеток незаметны, то создается впечатление, что под кутикулой
лежит сплошная плазматическая безъядерная пластинка. Базальная
мембрана, в отличие от таковой других аннелид, не тянется прямоли-
1 2
■w
Э—
нейно под эпителием, но
образует между клетками
глубокие выпячивания, в
которые и заходят
элементы соединительной
ткани.
Кожные железы
представлены большими
погруженными в
соединительную ткань клетками.
Одни из них—белковые—
называются трубчатыми
железами. Они в виде
длинных, узких
извилистых трубок (см. рис. 105,.
1) проникают вплоть до
кожно-мускульного
мешка, и содержат базофиль-
ный зернистый или ваку-
олизированный секрет.
Слизистые железы (см.
рис' 105, 7) с тонкими
протоками имеют
ацидофильное грубозернистое
содержимое. В области
пояска в коже развиты
многочисленные клител-
лярные одноклеточные
железы. Каждая железа
имеет широкое клеточное
тело с крупным ядром и
узкий выводной канал.
В полость передней
присоски открываются
многочисленные специальные одноклеточные железы (см. рис. 108, 17),
являющиеся разновидностью трубчатых желез. Наконец, особенно
мощно развиты железы задней присоски, за их счет образуется
большая компактная железистая масса внутри присоски, тогда как их
многочисленные поры расположены на внутренней поверхности присоски.
Эти одноклеточные железы также относятся к группе трубчатых желез.
Важной особенностью кожи Hirudo является ее богатство лакунар-
ными каналами, что связано с дыхательной функцией кожи. Между ба-
зальными концами клеток эпидермиса находится густая капиллярная
сеть лакун, содержащая кровь (см. рис. 105, 3 и с. 116).
Под эпидермисом находится довольно толстый слой волокнистой
соединительной ткани — кутис кожи. В петлях волокон здесь находятся
многочисленные жировые1 и пигментные клетки. Последние легко
узнаются по желто-коричневым зернам пигмента в плазме (см. рис. 105,5).
Мускулатура стенок тела состоит из наружного кольцевого слоя
122
Рис. 105. Hirudo medicinalis. Поперечный разрез
стенки тела (по Шнейдеру):
J — белковая ■ железистая клетка, 2-— ядро эпидермаль-
ной клетки, 3 — лакунарные капилляры, 4 — клетка
кожного эпителия, 5 — пигментная клетка, 6 — дорзовент-
ральное мускульное волокно, 7 — слизистая железистая
клетка, 8 — соединительная ткань, 9 — волокно
кольцевого мускульного слоя, Л) —лакун арный сосуд, 11 —
пучок продольных мускульных волокон, 12 — дорзовент-
ральные мышцы
(рис. 104,2), следующего за ним диагонального (рис. 104,3) и
внутреннего очень толстого продольного слоя мускульных волокон
(рис. 104,4). Кроме того, сильно развиты пучки дорзовентральных
мышц (рис. 104,5), пересекающие паренхиму и продольный слой
кожно-мускульного мешка.
Мышечные волокна толстые. Структура их хорошо видна на
поперечном сечении и в особенности на волокнах продольного слоя. Они
напоминают как бы длинные трубки (рис. 105,11), образованные пе-.
риферическим слоем параллельных друг другу вытянутых вдоль
волокна миофибрилл. Центральная часть волокна заполнена саркоплазмой,
в которой лежит ядро. Между мускульными волокнами развита
соединительная ткань с капиллярными лакунами и пигментными клетками.
По форме различаются простые, ветвящиеся и сетеобразные
мускульные волокна. Передняя и задняя присоски имеют особую сильно
развитую систему мышц. Собственной мускулатурой обладают также
различные внутренние органы — челюсти, глотка, лакунарные сосуды, органы
полового аппарата и др.
Паренхима, заполняющая все пространство между внутренними
органами, представлена волокнистой соединительной тканью (рис. 105,8).
В прозрачном основном веществе ткани лежат сильно вытянутые
соединительнотканные клетки, снабженные одним или двумя длинными не-
ветвящимися отростками,, которые и придают паренхиме грубо
волокнистое строение. Плазма этих опорных клеток сильно вакуолизирована,
что зависит от растворения жировых капель при изготовлении
препарата; ядра небольшие, темноокрашивающиеся. Имеются, кроме того,
округлые жировые клетки и подвижные трудно различимые амебоциты
•с более короткими отростками.
Пищеварительная система. Гистологическое строение
челюстей удобнее изучать на поперечных разрезах через эти органы
(рис. 106). Челюсть, представляющая мускулистую складку переднего
края глотки, одета кутикулярным эпителием. Кутикула очень тонка
(рис. 106,8). Между зубцами на килевидном крае челюсти
открываются тонкие протоки слюнных желез (рис. 106,7), выделяющих гирудин.
Мускулатура состоит из поперечных, радиальных и продольных
периферических волокон (рис. 106,3—5). Середина челюсти занята
многочисленными выводными каналами слюнных желез (рис. 106,6). Кроме
перечисленных мышц к основанию челюсти прикрепляются еще две
труппы антагонистических мускульных пучков; сокращаясь они
двигают челюсть наподобие круговой пилы, в результате чего челюсть
прорезает кожу жертвы, когда пиявка питается.
Слюнные железы лежат в переднем отделе тела, где рассеяны в
соединительной ткани между кожно-мускульным мешком и покровами
(рис. 107, А, 4). Это многочисленные мелкие колбовидные железистые
клетки с очень длинными и тонкими протоками (рис. 107,5). Тела
клеток округлы; плазма их содержит большое количество мелких капель
секрета. Протоки объединяются в мощные пучки, направляющиеся
внутрь челюстей.
Строение глотки выясняется из сопоставления срезов различных
направлений. Полость ее выстлана однослойным эктодермальным эпи-
123
Рис. 106. Hirudo medicinalis.
Поперечный разрез челюсти (по Скрибану):
1 — наружное отверстие протоков слюнных
желез, 2 — утолщение кутикулы, 3 — продольные
мускульные клетки, 4 — радиальные
мускульные волокна, 5 — поперечные мускульные
волокна, 6 — протоки слюнных желез, 7 —
эпидермис, 8 — кутикула, 9 — концевые участки
протоков слюнных желез
Рис. 107. Слюнные железы
Hirudo medicinalis. A —
топография слюнных желез; Б —
отдельная железистая слюннаж
клетка (по Списсу):
/ — глотка, 2 — продольная
мускулатура, 3 —желудок, 4— скопление
слюнных желез, 5 — протоки желез,
входящие в челюсть, 6 — клеточный
проток, 7 — зерна секрета, 8 — ядро
железистой клетки
телием, снабженным кутикулой. Под эпителием лежит продольный
мускульный слой (рис. 108,5), состоящий из маленьких пучков волокон,
отделенных друг от друга радиальной мускулатурой. К' нему прилегает
кольцевой слой (рис. 108,6), между пучками которого тоже проходят
радиальные волокна (рис. 108,27). Последние не образуют
самостоятельного слоя; выходя из стенок глотки, они тянутся во все стороны и
внешними концами прикрепляются к коже переднего конца тела.
Просвет глотки в передней ее части имеет треугольную форму, а стенка
глотки соответственно приобретает трехлучевую симметрию.
Стенки желудка складчаты (см. рис. 104,12). Однослойный
эпителий, из которого они состоят, покрыт тонким палочковым слоем. В
плазме цилиндрических или кубических клеток находятся жировые
включения, растворяющиеся при изготовлении препаратов; на их месте
остаются небольшие вакуоли. На границе с кишкой расположен
мускульный сфинктер. Эпителий кишки толще, чем в желудке. Клетки здесь
значительно выше, но тоже снабжены палочковым слоем. Прямая
кишка состоит из цилиндрического ресничного эпителия.
Лакунарная система. Следует обратить внимание на
гистологическое строение продольных каналов лакунарной системы.
Спинной и брюшной каналы (см. рис. 104,16,11) тонкостенны; стенка их
состоит из тонкой, но прочной соединительнотканной перепонки, одетой
изнутри слоем эндотелиальных клеток. Стенки боковых каналов
(см. рис. 104,5) толстые и слагаются из слоев наружного мощного слоя
кольцевых мускульных волокон, соединительнотканного, внутреннего-
продольного мускульного и, наконец, эндотелиального.
124
/ — передний край ротовой присоски с отверстиями железистых клеток, 2 — полость
ротовой присоски, 3 — заднебоковая губа, 4—подглоточиая ганглиозная масса, 5 —
продольная мускулатура глотки, 6 — кольцевая мускулатура глотки, 7 — ганглий
брюшной нервной цепочки, 8 — пищевод, 9 — сфинктер, 10 — слюнные железы, // —
продольный мускульный слой, 12 — пучок слюнных протоков, 13 — церебральный
ганглнй, 14 — дорзальиая челюсть, 15 — переднебоковая губа, 16 — кольцевой
мускульный слой, П — железы передней присоски, 18 — глаз, 19 — вентро-латеральная
челюсть, 20 — нефридий, 21 — радиальные мускульные волокна глотки, 22 —
мускулатура челюсти
Выделительная система. На многих поперечных срезах
видны разрезы через различные отделы нефридиев. Следует найти
разрез через ампулу и изучить лежащий в ней нефростом, или ресничный
орган. Последний состоит из многих ресничных клеток (рис. 109,3),
образующих воронку вокруг мерцательного канала. Каждая клетка
несет снабженную ресничной бороздкой лопасть. В пространстве между
клетками ресничной воронки лежат многочисленные лимфоидные'
клетки (рис. 109,5). Со стенками ампулы воронка соединяется несколькими
соединительнотканными поддерживающими пластинками (рис. 109,4).
125
Особая нефридиальная капсула, в которую открывается нефростом у
многих пиявок, у Hirudo отсутствует. Наконец, следует обратить
внимание на отсутствие связи между ресничной воронкой и самим
нефридием, проксимальный конец которого только прилегает к ампуле.
Строение железистого отдела нефридия особенно отчетливо
выступает на срезах, окрашенных по Маллори. Окружающие главный канал
нефридиальные клетки( рис. ПО, Б, 3) образуют вокруг него плотную
массу. Форма клеток полигониальная; плазма вакуолизирована; ядро
маленькое. Отчетливо видны тонкие внутриклеточные каналы (рис. ПО,
2), переходящие из клетки в клетку, разветвляющиеся и в конце
концов открывающиеся в центральный канал нефридия, 'который, несмотря
на большой диаметр, также внутриклеточный. Клетки, внутри которых
он проходит, имеют вид цилиндрических гильз., расположенных в один
ряд одна за другой.
Рис. 109. Hirudo medicinalis. Разрез через ампулу и нефростом (по Лесу):
/ — лакунарный канал, соединяющий ампулу с брюшным лакунарным каналом, 2 —полость
ампулы, 3 —ресничная клетка, 4 — поддерживающая пластинка, 5 — лимфоидные клетки,.
6 — полость ресничной воронки — нефростома
126
Б Г
Рис. 110. Hirudo medicinalis. Поперечные разрезы через нефридий. Л —через
железистый отдел; Б — через проксимальный участок нефридия, в котором отсутствует
центральный канал; В —поперечный разрез нижней более тонкой части нефридия; Г —
то же, продольный разрез (по Бользиусу):
/ — центральный канал, 2 — внутриклеточный каналец, 3 — нефридиальные клетки, 4 —
соединительная ткань
Мочевой пузырь состоит из тонкой соединительнотканной
оболочки, в которой развиваются многочисленные лакунарные капилляры, и
внутреннего кубического эпителия.
Большой интерес представляют ботриоидные клетки, называемые
также хлорагогенными. Большей частью они вплотную облегают
каналы промежуточной капиллярной сети лакунарной системы,
расположенной ковнутри от продольного слоя кожно-мускульного мешка
(см. рис. 104, 15). Реже они встречаются также и в соединительной
ткани паренхимы вне связи с капиллярами. Это крупные клетки
(рис. 111), более или менее вдающиеся в просвет лакунарных
канальцев. Плазма их содержит различные включения, среди которых видны
желто-коричневые зерна экскреторной природы. Ядра мелки и обычно
плохо заметны. При инъекции аммиачного кармина в тело пиявки этот
краситель-индикатор накапливается в ботриоидных клетках, которые,,
таким образом, имеют выделительную функцию и являются нефроци-
тами.
Нервная система. На поперечных
срезах, прошедших между ганглиями брюшной
нервной цепочки, хорошо видно строение нервных
стволов. Они представляют собой мощные тяжи
нервных волокон и имеют округлое сечение
(рис. 112, А, 2). Между ними ближе к брюшной
стороне виден гораздо более тонкий,
медиальный, файвровский, нерв (рис. 112, А, 4). В
центре каждого ствола находится большая глиаль-
ная клетка (рис. 112, А, 3). Она тянется от
одного ганглия до следующего; ядро ее можно
найти, просматривая последовательно срез за
срезом (рис. 112, А, 5). Тонкие глиальные воло-
Рис. 111. Ботриоидная
клетка Hirudo
medicinalis (по Скрибану):
1 — ядро, 2 — жировые
включения, 3 — соединительная
ткань, 4 — желтые зерна
127
конца этой клетки расходятся радиально и делят нервные волокна на
отдельные участки. Снаружи нервные стволы одеты общей
соединительнотканной волокнистой оболочкой—неврилеммой (рис. 112, А, 1).
Расположение элементов брюшного ганглия меняется в
зависимости от того, как прошел срез. Однако парная природа ганглия всегда
выступает отчетливо. Значительное место занимают парные основные
массы волокнистого нервного вещества — невропиль (рис. 112, Б, 6).
Они соединяются двумя поперечными волокнистыми комиссурами (рис.
112, Б, 11), лежащими одна над другой. Файвровский нерв в ганглии
разделяется на три медиальных'стволика (рис. 112, Б, 10). Крупные
униполярные ганглиозные клетки расположены по периферии ганглия с дор-
зальной и вентральной сторон (рис. 112, Б, 7), образуя три пары так
называемых пакетов (рис. 112, Б, 8). Медиально под стволиками файвров-
ского нерва видна очень крупная звездчатая глиальная клетка (рис. 112,
Б, 9), называемая обычно средней клеткой. В .каждом ганглии имеются
только две такие клетки, причем одна лежит непосредственно позади
другой. Снаружи ганглий окружен волокнистой неврилеммой, состоящей
преимущественно из кольцевых соединительнотканных волокон.
Органы чувств. Из органов чувств заслуживают внимания
чувствительные сосочки (рис. 113) и глаза, которые являются их
видоизменением. И те и другие лежат на среднем колечке сегментов,
причем глаза принадлежат пяти передним сегментам (см. с. 111).
Чувствительные сосочки, или почки, представляют собой органы
типа сенсилл, т. е. скопления кожных чувствительных клеток. У Hirudo
это главным образом веретеновидные клетки, связанные с нервным
волокном (рис. 113,5). Апикальный плазматический отросток каждой
клетки продолжается сквозь кутикулу торчащим наружу тонким и
коротким чувствительным волоском
(рис. 113, 1). Кроме того, в состав
почки входят железистые слизистые
клетки и дугообразные мускульные
волокна (рис. 113, 4), прикрепляющиеся
к кутикуле разветвленными концами
и вызывающие при сокращении
выпячивание чувствительной почки. Такая
сенсилла является осязательным
органом и, вероятно, также в какой-то
мере органом химического чувства.
Глаза, лежащие непосредственно
под кожей в толще паренхимы,
состоят из крупных зрительных клеток
(рис. 114), окружающих в виде
однослойного чехла осевой пучок нервных
волокон (рис. 114, 6). Снаружи глаз
одет скоплением черного пигмента
(рис. 114, 5). Первые три пары глаз
несколько отличны от двух последних
пар. Именно в передних глазах
зрительный нерв отходит от дна цилинд-
Рис. 114. Hirudo medicinalis.
Схема разрезов заднего (Л) и
переднего (£) глаз (по Гессе).
/ — кутикула, 2 — эпидермис, 3 —
соединительная ткань, 4 — зрительная
клетка, 5 — пигмент, 6 — нерв, 7 — ядро
зрительной клетки, в — вакуоль, 9 —
слой светочувствительных палочек
5-2
J29
рического глазного бокала, составляя продолжение его оси (рис. 114,
Б, 6), тогда как в задних глазах он выходит сбоку (рис. 114, А, 6).
Строение зрительных клеток своеобразно и характерно для пиявок
и олигохет. Каждая клетка, кроме ядра, содержит большую вакуоль
(рис. 114, Б, 8) с белковой жидкостью, не окрашивающейся
гематоксилином. Стенка вакуоли покрыта слоем маленьких светочувствительных
палочек (рис. \\А,Б,9). Удаленный от источника света полюс
зрительной клетки связан с нервным волокном (рис. 114,Б). Следовательно,
глаз принадлежит к неинвертированному типу. Однако в задних глазах
(рис. 114,Л), те зрительные клетки, которые лежат проксимальнее
места выхода нерва, получают нервные волокна со стороны источника света
и являются инвертированными. Первичными у пиявок являются
глаза инвертированные, а отсутствие инверсии развилось вторично. В
задних глазах Hirudo процесс превращения инвертированных органов
зрения во вторично неинвертированные (или реинвертированные) не
дошел до конца.
ОТРЯД RHYNCHOBDELLEA —ХОБОТКОВЫЕ ПИЯВКИ
Glossiphonia complanata L. — Улитковая пиявка
Материал и методы изучения. Glossiphonia complanata, принадлежащая к сем.
Glossiphoniidae, широко распространена по всему СССР. Это обычный обитатель
пресноводных водоемов, заросших водной растительностью. Ее пищей являются
преимущественно мелкие пластинчатожаберные Sphaerium и Pisidium, а также
прудовики и другие уЛитки, в меньшей степени черви и личинки водных насекомых.
Внешняя морфология изучается на животных, убитых слабым спиртом или
10%-ным раствором азотной кислоты. Для изучения внутреннего строения
производятся вскрытия и приготавливаются тотальные препараты и срезы, поперечные и
продольные. Очень много дает также .изучение молодых живых особей, сдавленных
между стеклами. Разрезая на куски, фиксировать пиявок.следует жидкостью Буэна
или Флемминга или сулемой с уксусной кислотой. Срезы окрашиваются гемотокси-
лином Бёмера с эозином или по Маллори.
Внешняя морфология Glossiphoniidae,
к которым принадлежит улитковая пиявка,
отличаются широким, сильно уплощенным
телом (рис. 115). Длина Glossiphonia
complanata достигает 3 см, ширина до 1 см.
Передняя присоска имеет вид
треугольного углубления на брюшной стороне
переднего конца тела (см. рис. 117, 1).
Ротовое отверстие лежит на ее переднем крае
(см. рис. 117, 2). Задняя присоска
сравнительно невелика, дисконидна (см. рис. 117,
12); у ее основания на спинной стороне
заметно маленькое анальное отверстие
(рис. 116, 8).
В состав тела, как у других пиявок,
кроме Acanthobdellea, входят 33 сегмента.
. , . , За счет первых четырех образуется перед-
^.^ид^ТиГойстоЙы няя присоска,, за^ счет семи последних-
(по Аутруму) задняя. Типичный сегмент состоит из 3 ко-
130
лечек — признак, свойственный всем Glossiphoniidae, причем среднее
является чувствительным, так как несет три пары спинных
чувствительных сосочков (рис. 116, 11). Последние, таким образом, располагаются
тремя парными продольными рядами (рис. 116, 5, 9, 10). Сосочки
внутренних рядов крупнее (рис. 116, 5); пространства между ними
отмечены черными продольными линиями (рис. 116, 6). Иногда правильное
расположение сосочков 'нарушается, так как некоторые из них могут
отсутствовать или, напротив, развиты дополнительные на третьих
колечках сегментов. На трех передних сегментах за счет сосочков
образуется три пары сближенных медиальных глаз, которые заметны в
виде небольших пятен (рис. 116, 2).
Вторичная кольчатость Glossiphonia отличается от таковой Hirudo.
Впереди глаз различимы простомиум (рис. 116, У) и I сегмент в виде
простого колечка (рис. 116,7). За ними следует II сегмент с двумя
парами глаз, состоящий из двух неясно разграниченных колечек
(рис. 116,Я). III сегмент состоит из одного колечка, которое несет
третью пару глаз (рис. 116, III). Сегменты с IV по XXIII являются
полноценными, трехкольчатымл. XXIV сегмент редуцирован до двух колец,
а XXV до одного кольца. Наконец, XXVI сегмент состоит из двух
колечек; анус лежит на задней границе последнего колечка (рис. 116,8).
Мужское половое отверстие (рис. 117,5)' помещается на брюшной
стороне между X и XI сегментами, а женское (рис. 117,5) — на XI
сегменте между его вторым и третьим кольцами. Поясок (clitellum) у
Glossiphonia внешне не различим.
Анатомия. Пищеварительная система. Общая морфология
и топография пищеварительного тракта хорошо видны при простом
сдавливании живой пиявки между двумя стеклами, если кишечник ее
заполнен пищей. Многое видно и на тотальных препаратах. Вскрытие
затрудняется наличием прочной мускулатуры и соединительной ткани,
развитых между внутренними органами. Поэтому можно ограничиться
следующим простым приемом. После укрепления на дне ванночки
вскрывается спинная лакуна. Для этого острым скальпелем
разрезается стенка тела несколько сбоку от медиальной спинной линии. Под
довольно широкой спинной лакуной обнажается кишечник. Гистология
органов пищеварения изучается на срезах.
Ротовое отверстие ведет в длинную трубку — влагалище хоботка
(рис. 118,2), которое в задней своей половине расширяется, окружая
длинный хоботок (рис. 118, 3). Стенка влагалища представляет
продолжение кожного эпителия и изнутри одета тонкой кутикулой. Она
снабжена продольными мускульными волокнами, при сокращении
которых влагалище хоботка укорачивается, образуя многочисленные
кольцевые складки.
Хоботок, представляющий собой глотку, имеет вытянутую
цилиндрическую форму и трехгранный просвет, ограниченный кутикулярным
эпителием. Стенка его чрезвычайно мускулиста — в ней развиты
кольцевые, радиальные и продольные мышечные волокна. Кроме того,
имеются протракторы, расположенные вне хоботкового влагалища.
Благодаря их сокращению, хоботок способен выдвигаться наружу через
ротовое отверстие, в чем легко убедиться, если сильно сдавить пальцами
5*
131
Рис. 116. Сегментация Glos-
slphonia complanata (no
Аутруму):
/ г- простомиум, 2 — глаза, 3 —
уровень мужского полового
отверстия, 4 — уровень женского
полового отверстия, 5 —
внутренние чувствительные сосочкн,
6 — пигментные полоски, 7 —
задняя присоска, 8 — анальное
отверстие, 9 — латеральный ряд
чувствительных сосочков, 10 —
средний ряд чувствительных
сосочков, // — чувствительное
колечко. Римские цифры —
сегменты
Рис. 117. Glossiphonia complanata.
Расположение брюшной нервной
цепочки, нефридиальных капсул, иеф-
ридиальных и половых отверстий
(по Брумпту):
/ — передняя присоска, 2 — ротовое
отверстие, 3 — подглоточная ганглиозиая масса,
4 — первое иефридиальное отверстие, 5 —
мужское половое отверстие, 6 — женское
половое отверстие, 7 — иефридиальные
капсулы, в — иефридиальные отверстия, 9 —
гаиглии брюшной нервной цепочки, 10 —
14-е иефридиальное отверстие, 11 — задняя
ганглиозная масса, 12 — задняя прнс'оска
переднюю часть тела живой пиявки. Будучи очень упругим, хоботок
может проникать сквозь нежные покровы животных. Он же играет роль
сосательного органа, перекачивающего кровь жертвы в желудок.
Втягивание хоботка осуществляется сокращ&нием пары ретракторов,
соединяющих его основание со спинной стенкой тела.
132
На вершине хоботка
открываются слюнные железы (рис. 118, 4),
лежащие в соединительной ткани
вокруг него и пищевода. Это
многочисленные округлые железистые
клетки, снабженные очень
длинными и тонкими клеточными
протоками, которые образуют пучки,
пронизывающие хоботок по всей его
длине. Ядра слюнных клеток велики и
богаты хроматином, плазма
содержит многочисленные зерна секрета.
Пищевод — гибкая цилиндрическая
трубка (рис. 118, 5), соединяющая
хоботок с желудком. По бокам от
него проходят два ретрактора
.хоботка, внутри которых видны
протоки слюнных желез.
Желудок (рис. 118, 6),
принадлежащий уже к средней энтодер-
мальной кишке, объемист и
снабжен шестью парами боковых
слепых выростов (рис. 118, 7), из
которых задняя пара достигает
наибольшего развития.' От следующего,
заднего, отдела средней кишки он
отделен мускулистым сфинктером.
Собственно кишка (рис. 118, 9), в
которой происходит переваривание
и всасывание пищи, также
снабжена боковыми карманами в
количестве четырех пар (рис. 118, 8).
Задняя кишка состоит из переднего
ректального пузырька (рис. 118,
10), образованного ресничным
эпителием, и из прямой кишки
(рис. 118, 12), выстланной кутику-
лярным эпителием.
Лак унарна я система.
Для изучения лакунарных каналов применяется инъекция. Можно
получить демонстративные препараты простой инъекцией воздуха в
лакунарную систему. Еще лучшие результаты дает инъекция растертой
туши или окрашенной желатины в спинную или брюшную лакуну.
Инъецированные животные затем обезвоживаются и просветляются в
кедровом масле. На таких тотальных препаратах лакунарную систему
можно изучить гораздо быстрее и проще, чем на сериях срезов.
Лакунарная система хоботных пиявок является системой каналов
настоящего целома и полностью отделена от кровеносных сосудов,
которые еще сохраняются.
Рис. 118. Пищеварительная система
Glossiphonia complanata (по Брумпту):
/ — ротовое отверстие, 2 — влагалище хоботка,
3 — хоботок, 4 — слюнные железы и их
протоки, б — пищевод, 6 — желудок, 7 — слепой
вырост желудка, 8 — слепой вырост кишки,
9 — кишка, 10 — ректальный пузырек, 11 —
анальное отверстие, 12 — прямая кишка, 13 —
семенной мешок, 14 — семяпровод
133.
В широком спинном
продольном канале (р'ис. 119, 1),
проходящем над кишечником, лежит
спинной кровеносный сосуд (рис. 119,
12). В более обширном брюшном
канале (рис. 119, 11) помещаются
брюшной кровеносный сосуд и
лежащая под ним брюшная нервная
цепочка (рис. 119, 10, 9). В
передних и задних сегментах" оба канала
сливаются, образуя переднюю и
заднюю медиальные лакуны (рис.
119, 4). Боковые лакунарные
каналы (рис. 119, 6) тянутся вдоль
узких боковых краев тела и
отличаются небольшим диаметром.
Все четыре продольных канала
соединяются сложной системой
поперечных лакун, среди которых
различаются: метамерные вентральные
и дорзальные соединительные
лакуны (рис. 119, 8, 2), промежуточные
лакуны (рис. 119, 4), боковые
соединительные лакуны (рис. 119, 7),
кожные лакунарные капилляры (рис. 119, 3, 120, 1—12 и др.) Сильное
развитие лакун в коже показывает, что у хоботных пиявок, несмотря
на наличие настоящей кровеносной системы, лимфа лакун берет на
себя дыхательную функцию.
Кровеносная система. Кровеносные сосуды могут быть
изучены на живых мелких молодых, сжатых между двумя стеклами,
пиявках. Кроме того, они хорошо видны на срезах.
Замкнутая кровеносная система состоит из спинного и брюшного
сосудов (см. рис. 119,12,10; 121, 2,3) и из серии петлеобразных
капиллярных сосудов, соединяющих их спереди и сзади (рис. 121,5—7,12).
Настоящие кольцевые метамерные сосуды отсутствуют.
В области XII—XVIII сегментов спинной сосуд подразделен рядом
перетяжек на 15 метамерных камер. Между камерами имеются
клапаны, пропускающие кровь только вперед. На уровне заднего отдела
средней кишки спинной сосуд сильно расширяется и охватывает его це-.
ликом вместе с боковыми карманами, образуя соответственно 4 пары
слепых боковых расширений (рис. \2\,10). Благодаря этому вся кишка
омывается кровью.
Направление тока крови такое же, как у других аннелид. По
спинному сосуду кровь движется от заднего конца тела к переднему. Его
толстые стенки обладают собственной мускулатурой, и он играет роль
сердца. По брюшному сосуду кровь течет в обратном направлении.
Выделительная система G. complanata обладает 14
парами нефридиев. Передняя пара, принадлежащая VII сегменту,
рудиментарна и мала. Следующие две пары нормальны, но соединены друг с
Рис. 119. Glossiphonia complanata.
Схема лакунарной системы в средней
части тела (по Ока):
/—спннной лакунарный канал, 2 —
спинная соединительная лакуна, 3 — кожные
капиллярные лакуны, 4 — промежуточные
лакуны, 5 — кожная соединительная
лакуна, 6 — боковой лакунарный канал, 7 —
боковая соединительная лакуна, 8 —
брюшная соединительная лакуна, 9 — брюшная
нервная цепочка, 10 — брюшной
кровеносный сосуд, // — брюшной лакунарный
канал, 12 — спинной кровеносный сосуд.
Стрелка показывает направление
движения крови
134
Рис. 120. Glossiphonia complanata. Схема
лакунарной системы передней и задней
частей тела (по Ока):
/— передняя часть спинного кровеносного
сосуда, 2 — передние петлеобразные кровеносные
сосуды, 3 — брюшная соединительная лакуна, 4 —
медиальная лакуна, 5 — боковая соединительная
лакуна. 6 — промежуточные лакуны, 7 — спинная
соединительная лакуна, 8 — спинной кровеносный
сосуд, 9 — брюшной кровеносный сосуд, 10 —
брюшная нервная цепочка, 11 — боковой
лакунарный канал, 12 — задние петлеобразные
кровеносные сосуды. Черным цветом изображены
кровеносные сосуды, лежащие вне лакунарной
системы
Рис. 121. Hemiclepsis marginata. Схема
кровеносной системы (по Ока):
1 — раздвоенный передний конец спинного
сосуда, 2 —тонкая часть спинного кровеносного
сосуда, 3 —брюшной сосуд, 4 — капилляры хоботка,
5 — вторая пара передних петлеобразных
сосудов, б —третья пара петлеобразных сосудов, 7 —
четвертая пара петлеобразных сосудов, 8 —
боковой карман желудка, 9 — часть спинного сосуда,
разделенная на камеры, 10 — слепые мешки
спинного сосуда, //— анальное кольцо спинного
сосуда, 12 — задние петлеобразные сосуды
135
другом и с первой парой. В
половых сегментах (X и XI)
нефридии отсутствуют. Однако в
соединительной ткани здесь имеются
их рудименты в виде
изолированных нефр'идиальных клеток.
Следующие 11 сегментов (с XII по
XXII) имеют нормальные
нефридии.
Каждый нефридий состоит
из ресничной воронки — нефро-
стома (рис. 122, 6), связанной
узким каналом с нефридиальной
капсулой (рис. 122,5), и
длинного петлистого железистого
отдела (рис. 122,4). Внутри
последнего проходит иефридиаль-
ный канал, заканчивающийся
небольшим мочевым пузырьком
(рис. 122, 8), который
открывается наружу на брюшной
стороне второго колечка своего
сегмента. Железистый отдел не
соединяется с нефридиальной капсулой, а только тесно к ней прилегает
проксимальным концом. Двулопастная ресничная воронка (рис. 123,
А, Г, 1)~ вдается в брюшную лакуну (рис. 123, Г, 11), широко
открываясь в ее полость. Нефридиальная капсула лежит уже вне лакуны
(рис. 123, Г, 4).
Ресничная воронка состоит только из трех больших клеток,
причем две образуют ресничные лопасти (рис. 123, А, 2), а
одна—мерцательный канал стебелька воронки (рис. 123, А, 3). Капсула представляет
собой тонкостенный эпителиальный пузырек (рис. 123, Г, 4).
Воронка служит для собирания твердых и жидких экскретов и
перегруженных ими амебоидных фагоцитов, которые накапливаются в капсуле.
Нередко поэтому воронку и капсулу называют цилиофагоцитарным
органом.
При инъекции в спинной или брюшной лакунарный канал
растертой в воде туши или кармина частицы этих красок через сутки
накапливаются в капсулах (рис. 123, .Г, 12). Жидкие экскреты отсюда могут
просачиваться в каналы железистого отдела нефридия, который также
извлекает конечные продукты обмена из окружающих его тканей и ла-
кунарных каналов.
Железистый отдел нефридия состоит из одного ряда крупных
клеток (рис. 123,5,7), пронизанных центральным каналом (рис. 123,5),
от которого в тело каждой клетки отходят тончайшие разветвленные
канальцы (рис. \22>,В,10).
Ботриоидные клетки, несущие у пиявок выделительную функцию,
у G. complanata встречаются только в спинной, брюшной и
промежуточных лакунах, где они обычно видны вместе с амебоцитами. Они го-
Рис. 122. Glossiphoniidae. Схема
естественного расположения нефридия (по Ока):
/, 2 —первое и второе колечки сегмента, 3 —
третье колечко предыдущего сегмента, 4 —
железистый отдел нефридия, 5 — нефридиальная
капсула, 6 — воронка, 7 —проксимальный конец
железистого отдела, прилегающий к нефридиальной
капсуле, 8 — мочевой пузырек
136
5 В
■■ , '• Г- , f —f
Рис. 123. Glossiphonia complanata. Гистологическое строение нефридия. А —
воронка; Б — разрез проксимальной части нефридия; В — часть предыдущего
разреза при более сильном увеличении; Г—разрез через брюшной
лакунарный канал, в который были инъецированы кармин и взвесь бактерий (А, Б,
В — по Ока, Г — по А. Ковалевскому):
/ — вход в воронку, 2 — клетка левой лопасти воронки, 3 — ресничный канал воронки,
образованный одной клеткой, 4—стенка капсулы, 5— ядро клетки канала, б
—нефридиальная капсула с фагоцитами, 7 — проксимальная часть железистого отдела
нефридия, 8 — канал нефридия, 9 — ядро иефроцита, 10 — внутриклеточный каналец, 11 —
брюшиой лакунарный канал, 12 — скопление кармина и бактерий в капсуле, 13—
брюшной кровеносный сосуд, 14 — брюшная нервная цепочка, /5 — соединительная ткань
раздо крупнее последних и содержат в плазме так называемые кислые
зерна, поэтому обычно именуются кислыми клетками (рис. 124).
Нервная система отпрепаровывается сравнительно легко.
Желательно приготовить ее тотальный
препарат, окрасив борным или
квасцовым кармином. Строение ее
сходно с нервной системой
медицинской пиявки (с. 117). Нервное
кольцо (церебральные ганглии,
окологлоточные коннективы и подгло-
точный ганглий — см. рис. 117,3) —
лежат впереди хоботка, окружая
его влагалище.
Органы чувств. Из
органов чувств следует обратить
внимание на байеровские органы и глаза.
И те и другие легко обнаружить на
срезах. Первые встречаются в
большом количестве в коже на спинной
стороне тела (рис. 125, 1). Эти
Рис.
124. Кислые клетки Glossiphonia
complanata (по Скрибану):
/ — лакуна, 2 — соединительнотканная стенка
лакуны, 3 — соединительная ткань, 4— кислая
клетка, 5—плазма с кислыми зернами, 6 —
амебоцит
137
Рис. 125. Glossiphonia complanata.
Поперечный разрез стенки тела (по Байе-
РУ):
/ — байеровскне органы, 2 —эпидермис, 3 —
кутикула, 4 — грушевидная железистая
клетка, 5 — кольцевые мускульные волокна, 6 —
продольные мускульные волокна, 7 —
трубчатая железистая клетка, в'— лакуна, 9 —
желтая клетка, 10 — дорзовентральное мускульное
волокно, 11 — соединительная ткань, 12 —
подкожная соединительная ткань
кий и короткий семявыносящии
длинный семяпровод-'(рис. 128, 8)
своеобразные органы, принимаемые
за органы осязания, просты и
состоят всего из двух клеток.
Чувствительная клетка (рис. 126, 2) слегка
выдается над эпидермисом,
основание ее охвачено мускульной
клеткой (рис. 126, 4). Бокаловидные
глаза замечательны своей
простотой. Глаз состоит из нескольких
зрительных клеток (рис. 127, 1),
окруженных чашевидным скоплени-
ем пигмента (рис. 127, 3). Каждая
клетка на конце, прилегающем к
пигменту, содержит светлую
вакуоль со слоем светочувствительных
палочек (рис. 127, 4, ср. также
с. 130). Нервные волокна (рис.
127, 5) отходят от обращенных к
свету концов клеток, т. е. глаз
является инвертированным.
Половая система.
Органы полового аппарата
наблюдаются при вскрытии, за исключением
части мужских протоков, слишком
тонких и нежных; для их изучения
необходимы срезы.
Десять пар семенных мешков
(рис. 128, 4) лежат на брюшной
стороне тела между слепыми
выростами кишечника (начиная с XII
сегмента). От спинной поверхности
каждого мешка отходит очень тон-
проток (рис. 128, 7), впадающий в
своей стороны. Достигнув X сегмен-
Рис. 126. Glossiphonia
complanata. Байеровский орган на разрезе
(по Байеру):
/ — кутикула, 2 — чувствительная
клетка, з — кожный эпителий, 4 —
мускульная клетка
Рис. 127. Схема
строения глаза Glossiphonia
complanata (по Гессе):
/ — зрительная клетка, 2 —
ее ядро, 3 — пигмент, 4 —
вакуоль с палочковым
слоем, 5 — нерв
138
та, семяпроводы переходят в каналы
несколько большего диаметра, называемые
по их функции семенными пузырьками и
отвечающие придаткам семенников
челюстных пиявок (с. 119). Эти каналы
образуют большие петли (рис. 128, 9,10),
направленные назад, и легко
наблюдаются на вскрытии-, так как свободно
лежат в брюшной лакуне. В области X
сегмента семенные пузырьки переходят в
короткие мускулистые семяизвергатель-
ные каналы (рис. 128, 1), которые в
свою очередь продолжаются в виде
небольших железистых расширений — спер-
матофорных мешков. Последние
гомологичны отросткам атрия (cornua)
медицинской пиявки (с. 119). Внутри них
образуются хитиноидные стенки спермато-
форов. Наконец, сперматофорные мешки
соединяются под брюшной нервной
цепочкой в короткий непарный канал,
заканчивающийся мужским половым
отверстием на брюшной стороне между X
и XI сегментами. Совокупительный орган
отсутствует, что связано с развитием у
хоботных пиявок сперматофорного
оплодотворения. Женский половой аппарат
состоит из пары вытянутых овариальных
мешков (рис. 128, 3), лежащих в
брюшной лакуне, и выводных путей. В
полости каждого овариального мешка
свободно лежит нитевидный, извитой яичник,
утолщенный на заднем конце (рис.
128, 2). Овариальные мешки спереди
соединяются друг с другом над брюшной
нервной цепочкой и под ней, окружая ее
в виде кольца. На брюшной стороне это
кольцо сообщается с коротким непарным
влагалищем, которое открывается
наружу на XI сегменте.
В средней полосе СССР спаривание
у G. complanata происходит во второй
половине мая, чаще всего ночью.
Копулирующие пиявки прикрепляют друг к
другу где-либо наспинной стороне по
нескольку (3—4, реже до 6) сперматофо-
ров. Через 5—6 дней после этого пиявка
откладывает 3—8 крупных коконов,
содержащих в общем до 200 яиц, и при-
Рис. 128. Половая система
Glossiphonia complanata (по Брумп-
ту):
1 — семянзвергательный канал, 2 —
яичник, 3 — яйцевой мешок, 4 —
семенные мешки, 5 — часть брюшной
нервной цепочки, 6 — задняя присоска, 7 —
семявыносящие каналы, 8 —
семяпровод, 9 — нисходящее колено семенного
пузырька, 10 — его восходящее колено
Рис. 129. Сперма тофор
Glossiphonia complanata (по Бренту)
139
крывает их своим широким телом. Вылупляющаяся вскоре молодь
прикрепляется присосками к брюшной поверхности матери, которая
некоторое время «охраняет» свое потомство.
Сперматофор (рис. 129) состоит из двух веретеновидных, спаянных
вдоль, хитиноидных мешочков, содержащих сперму. Двураздельность
его объясняется тем, что один мешочек формируется в правом,
другой — в левом сперматофорном мешке. Оба мешочка открываются
раздельно на более широком конце сперматофора. Здесь же находится
скопление зернистой цементной массы, служащей для приклеивания
сперматофора к коже пиявки. Из сперматофора спермин проходят
через кожу пиявки в лакунарную систему и по ней достигают области
женских половых органов, в которые активно проникают по
окружающей их особой рыхлой проводящей ткани. Такой способ введения
спермы в тело партнера при спаривании называется гиподермальной
импрегнацией.
Тип Arthropoda — Членистоногие
Тип членистоногих содержит большое количество разнообразных
по организации групп. Однако при прохождении зоологического
практикума приходится несколько ограничить выбор материала. Это
выражается, с одной стороны, в том, что некоторые относительно мелкие
группы, не представляющие большого сравнительно-анатомического и
филогенетического интереса, в практикум не включаются (Tardigrada,
Pentastomida, Pantopoda). С другой стороны, классы Onychophora и
Xiphosura хотя и представляют большой интерес, нами также не
рассматриваются, так как добыть материал практически недоступно.
В пределах изучаемых групп некоторые объекты берутся как
основные и рассматриваются подробно (например, Artemla, Daphnia,
речной рак, скорпион, жук-плавунец). В отношении же ряда других
можно ограничиться изучением внешней морфологии и тотальных
препаратов.
Техника исследования членистоногих отличается своеобразием в
основном благодаря наличию у них наружного хитинового скелета.
Большое значение приобретает здесь метод вскрытия.
Техника изучения внешней морфологии
и анатомии членистоногих
Наличие у членистоногих твердого хитинового наружного скелета вызывает при
их изучении, во-первых, изменение относительного значения отдельных применяемых
методов исследования в их общем комплексе, во-вторых, модификацию некоторых из
них. Слабая проницаемость хитиновой кутикулы по отношению к ряду реактивов,
твердость и в то же время хрупкость хитина создают ряд специфических трудностей
и вызывают необходимость в добавочных операциях при фиксации, изготовлении
тотальных препаратов и при заливке и изготовлении срезов из большинства
членистоногих. С другой стороны, твердый наружный скелет дает возможность применения
в большей степени, чем при изучении других групп, методов вскрытия и препаровки
в целях изучения объекта или отдельных его частей in toto. Именно эти методы
должны применяться в первую очередь при изучении анатомии членистоногих. Изучение
срезов лишь дополняет в части знакомства с тонким строением данные, полученные
при вскрытии и на тотальных препаратах.
Техника изучения внешней морфологии (хитинового скелета) и изготовления
тотальных препаратов. Для изучения внешней морфологии крупных членистоногих не
требуется специальных методов обработки; оно может производиться как на свежем,
так и иа спиртовом или сухом материале. Если необходимо произвести расчленение
наружного хитинового скелета на отдельные склериты, то объект, с предварительно
разрезанными в нескольких местах межсегментными перепонками, вываривается в
едком кали' и после промывки в воде разнимается на отдельные склериты,
высушивается и монтируется путем наклейки иа картой или иа проволочном каркасе. Если у
животного имеются втянутые внутрь сегменты или их части (последние сегменты
брюшка у большинства насекомых или их придатки), то для их обнаружения и изу-
141
чения впрыскивается внутрь живого объекта шприцем спирт, формалин или другой
фиксатор, после чего, вследствие резкого увеличения внутреннего давления, все
втянутые и скрытые части выпячиваются наружу и становятся легко обозримыми
(способ, предложенный акад. Е. Н. Павловским).
Из мелких объектов или из отдельных отпрепарованных частей, конечностей
и т. п. приготовляются тотальные препараты. Препаровку конечностей или других
деталей удобнее проводить не на живых или свежеубитых объектах, а на спиртовом
материале или даже на просветленном в гвоздичном масле. Тогда мышцы становятся
более хрупкими, что облегчает выделение мелких деталей и уменьшает риск потерять
их при переводе через спирты возрастающей крепости.
Темнопигментированный и малопрозрачный объект необходимо выварить в
едком кали; иежные и мелкие объекты подвергаются действию 1—5%-ного раствора
без нагревания на срок до 1 сут, крупные и грубые — при нагревании в 5—20%-ном
растворе. Иногда необходимо полностью депигментировать объект. Для этого
служит диафанол (диоксидуксусная кислота). Приводим рецепт его изготовления: 150 г
порошковидной щавелевой кислоты смешивают с 40 г бертолетовой соли (не
растирать, может взорваться), помещают в колбе с 20 см3 дистиллированной воды в
водяную баню при 60°С. Образующиеся красно-коричневые пары С102', пропустив для
очистки через воду, отводят в сосуд с 500 см3 50%-ной уксусной кислоты, стоящий
во льду. Все соединения и газопроводные трубки должны "быть герметичны.
Полученная в сосуде жидкость желтого цвета сохраняется в склянке коричневого стекла,
герметически закупоренной пропарафинированной притертой пробкой, в темном
прохладном месте. Объекты помещают в темноте в диафанол из 65%-ного спирта. При
обесцвечивании диафанола его сменяют. После окончательной депигментации
необходима промывка в 65%-ном спирте. Для депигментации пригоден также указанный
ниже метод Паля (см. с. 145) при условии значительного удлинения сроков обработки.
Прозрачный объект с непигментированным хитином при заключении в бальзам
может просветлиться настолько, что его мелкие детали сделаются неуловимыми для
глаза. Этого избежать можно двумя путями. Во-первых, путем заключения в среду
менее преломляющую свет: глицерин, глицерин-желатин (см. т. 1, с. 17) или в гум-
миарабиковую смесь (дистиллированной воды 50,0 г, глицерина 20 см3, гуммиарабика
40,0 г, хлорал-гидрат 50 см3, кокаина 0,5 г). В последнюю смесь на предметном
стекле можно непосредственно помещать живые объекты и покрывать их покровным.
Полученный тотальный препарат может храниться долгое время. Если не работать
с живым материалом, то можно в гуммиарабиковой смеси хлорал-гидрат и кокаин
заменить 5 г кристаллической карболовой кислоты или несколькими кристалликами
тимола (против загнивания). Гуммиарабиковая смесь очень удобна для
приготовления тотальных препаратов из мелких Crustacea — Ectomostraca, их конечностей,
мелких клещей, личинок насекомых и т. п. Во-вторых, путем окраски. Наиболее простой
способ — окраска пикриновой кислотой, растворенной в ксилоле. Обезвоженный в
абсолютном спирте объект переносится в ксилол с пикриновой кислотой, где он
одновременно и просветляется и окрашивается. Значительно совершеннее метод
серебрения. В наиболее простом виде он применяется следующим образом. Объект из воды
погружается в 5%-ный раствор азотнокислого серебра на 5—10 мин, затем
ополаскивается водой для удаления избытка раствора серебра и помещается в 5%-ный
раствор едкого натра или кали на 5—15 мин или на более долгий срок, до появления
желто-коричневого окрашивания; затем следует промывка в воде, необходимы спирты
возрастающей крепости и гвоздичное масло, в котором отпрепаровываются
необходимые части.
Нередко приходится иметь дело со сморщенными сброшенными шкурками, с
высохшими сморщившимися нежными объектами и т. п. материалом. Для
расправления таких объектов их помещают в смесь равных объемов молочной кислоты и
96%-ного спирта на 5—10 мин, переводят в чистую молочную кислоту, где объект
быстро размягчается. Затем его переносят на предметное стекло и под бинокуляром
или лупой расправляют при помощи препаровальных игл. Придав нужное положение,
удаляют фильтровальной бумагой избыток молочной кислоты и прибавляют по
каплям абсолютный спирт. Когда объект уплотнится (1—2 мин), его на шпателе
осторожно переносят в абсолютный спирт, а после обезвоживания — в гвоздичное масло
1 Необходимо иметь в виду, что С102 ядовито, и требуется соблюдение правил
техники безопасности.
142
для просветления. Таким путем можно получать хорошие препараты из материала,
который при употреблении обычных методов считается негодным.
При изготовлении препаратов из цельных объектов надо всегда их надкалывать,
надрезать или даже разрезать пополам при переносе их во время обезвоживания из
70%-ного в 96%-ный спирт.
При заключении препаратов в бальзам часто бывает необходимо расположить
отдельные отпрепарованные части в определенном порядке (например, ротовые
органы, конечности и т. п.), который обычно нарушается при покрывании покровным
стеклом, вследствие возникающих в бальзаме токов. Для избежания этого можно
пользоваться указанным ниже методом заключения отпрепарованных органов в
целлоидиновую пленку (см. 144) или, что проще, на предметном стекле в- тонком слое
жидкого бальзама разложить отпрепарованные объекты в нужном порядке и,
прикрыв стекло от пыли, оставить на 15—30 мин, пока бальзам слегка не подсохнет;
после этого прибавить бальзама и прикрыть покровным стеклом. Последнее всегда
должно иметь восковые ножки, высота которых должна соответствовать толщине
объекта.
Вскрытие и препаровка членистоногих. Перед вскрытием животное должно быть
анестезировано хлороформом или эфиром. При отсутствии живого материала можно
пользоваться спиртовым. Для препаровки мускулатуры и нервной системы спиртовой
материал даже более удобен, но прочие органы, вследствие уплотнения тканей,
утрачивают гибкость, делаются ломкими и ие.поддаются расправлению.
Вскрытие и препаровка, особенно мелких объектов, производится под
бинокулярным препаровальным микроскопом или под препаровальной лупой. При выборе
увеличения начинающие обычно берут наибольшее. Это ошибка; необходимо
пользоваться наименьшим, при котором видны нужные^ детали. Пользование крыльями,
имеющимися при каждом бинокуляре или лупе,— обязательно; иначе руки будут на
весу и скоро начнут дрожать. Вскрытие и препаровка производятся при падающем
свете, предпочтительно — при естественном. Для усиления освещения объекта, если
нет специальных осветительных ламп, 'между источником света (окно, электрическая
лампочка) и объектом помещается обычная колба или стеклянный шар, наполненные
водой; при работе с искусственным светом вода слегка подкрашивается медным
купоросом в слабо голубоватый цвет. Необходимо помнить, что плохое освещение
объекта может явиться причиной неудачи работы. Если объект можно держать
пальцами, то все необходимые для вскрытия первоначальные разрезы (например, продольные
боковые разрезы на спинной стороне) необходимо сделать, держа его в руке.
Вскрытие производится в препаровальной ванне с восковым дном (см. т. 1,
с. 42). Мелкие объекты надо вскрывать в небольших плоскодонных чашечках, или
солонках с восковым дном, или в стеклах с углублением. Крупные и средней
величины объекты прикрепляются ко дну ванночки булавками. При вскрытии мелких и
средней величины объектов, имеющих достаточно твердый хитиновый покров иа
брюшной стороне, удобнее вплавлять их брюшной стороной в восковую массу дна
ванночки. Для укрепления мелких объектов удобно в солонку налить немного
расплавленного парафина, поместить на его поверхность живой объект спинной
стороной кверху и дать застыть. Мелкие объекты с мягкой кутикулой вскрывают, не
прикрепляя их к субстрату, а придерживая препаровальной иглой.
Для вскрытия и последующей препаровки необходимы кроме тонких (глазных)
ножниц и остроконечного пинцета препаровальные иглы. При препаровке это
самые нужные инструменты. Они приготовляются работающим самостоятельно путем
заточки на точильном бруске обыкновенных швейных игл. Наиболее пригодны иглы
двух родов: заточенные в форме очень острого конуса и в форме плоского с двух
сторон кинжала. Кроме того, очень удобны копьевидная игла и приготовляемая из
нее, путем стачивания полукругом концевого острия, лопатовидная препаровальная
игла. Иногда удобны иглы с согнутым под углом концом. Для очень тонких
вскрытий и препаровки употребляют «минуции» — наиболее тонкие из энтомологических
булавок. Иглы должны быть острыми (их необходимо часто подтачивать) и
чистыми (обезжиривать, протирая ваткой с хлороформом или 96%-ным спиртом); к
грязным иглам прилипают части препарируемых органов, что создает большие неудобства.
Вскрытие для знакомства с общей анатомией производится под водой, сменяе- *
мой по мере загрязнения. Если объект вскрывается для последующей препаровки и
\ фиксации отдельных органов, вместо воды надо пользоваться физиологическим
раствором или другими изотоническими жидкостями (Рингера и т. п., см. т. 1, с. 8).
143
Так как поверхность хитина, особенно у наземных форм, не смачивается водой,
мелкие объекты необходимо перед вскрытием на мгновение опустить в 96%-ный спирт;
после этого они без труда могут быть погружены в воду для вскрытия. При
препаровке надо держать препаровальные иглы в пальцах свободно, не сжимая их. В
противном случае уже через 1—2 мин утомленные пальцы начинают дрожать и движения
становятся неточными.
Давать общую схему вскрытия в порядок препаровки отдельных органов
членистоногого—■ бесполезно: схема и порядок будут слишком общи. Поэтому,
ограничиваясь здесь сделанными замечаниями, отсылаем к систематическому обзору
членистоногих, где для каждого объекта указаны необходимые приемы и порядок
вскрытия и препаровки.
Приготовление тотальных препаратов из отпрепарованных органов (способ
Е. Н. Павловского). 1. Отпрепарованные органы переносят на чистое предметное
стекло с помощью шпателя (крупные объекты), лопатовидной или копьевидной иглы
(мелкие объекты). Часто для этого удобно пользоваться «ушком». Во избежание
прилипания переносимого объекта к шпателю или иглам, последние должны быть
тщательно обезжирены.
2. На стекло прибавляют физиологический раствор и под бинокуляром или
препаровальной лупой, с помощью препаровальных игл, расправляют орган, после чего
тонкой пипеткой или полоской фильтровальной бумаги осторожно удаляют
физиологический раствор, стараясь не сдвинуть с места расправленный орган.
3. Приводят стекло в строго горизонтальное положение и осторожно, по каплям,
чтобы ие сдвинуть с места объект, прибавляют жидкость Карнуа (см. т. I, с. 15) до
тех пор, пока объект не будет полностью покрыт фиксатором. Фиксация длится 1—
2 мин для мелких' объектов и 5—10 мин для крупных (длиной свыше 1—1,5 см); в
последнем случае фиксатор необходимо сменить. При осторожной работе объект при
фиксации приклеивается к стеклу.
Далее осторожно сливают фиксатор и для промывки наливают 96°/о-ный спирт,
—который сменяют 2—3 раза.
4. На предметное стекло наливают затем несколько капель смеси равных
объемов абсолютного спирта и серного эфира, сливают и быстро наливают 0,25%-ный
раствор целлоидина (коллодия или фотоксилина) в смеси равных объемов абсолютного
спирта и серного эфира. Легким покачиванием стекла равномерно распределяют
раствор вокруг объекта и оставляют на воздухе до начала загустения пленки (1—
3 мин), когда игла после прикосновения начинает оставлять на ней след. После этого
стекло переносят в 85—90%-ный спирт до следующего утра, а затем в 70—75%-иый
спирт для длительного хранения.
5. Для приготовления постоянного препарата стекло переносят в воду,
окрашивают объект квасцовым кармином или гематоксилином (Бёмера, Деляфильда, Эрли-
ха). При последующем проведении через спирты возрастающей крепости надо иметь
■ в виду, что абсолютный спирт растворяет целлоидиновую пленку. Поэтому лучше
выдержать стекло в 96%-ном спирте и сменить его лишний раз, а в .абсолютном'спирте
держать не более 1—2 ч.
После просветления в ксилоле препарат заключают в бальзам под стекло с
восковыми ножками. Если при просветлении в ксилоле на пленке остаются
молочно-белые непрозрачные пятна — это признак, что препарат недостаточно обезвожен и его
снова надо перевести иа 30—45 мин в абсолютный спирт. Чтобы избежать вредного
действия абсолютного спирта на целлоидиновую пленку, можно для просветления
употреблять карбол-ксилол, куда стекло можно переносить непосредственно из
96%-ного спирта.
Так как длительное хранение в спирте на предметных стеклах иногда
(например, в экспедиционных условиях) бывает громоздко, можно наклеивать объекты на
покровные стекла или в спирте снимать со стекла целлоидиновую пленку вместе с
объектом и хранить только ее. В последнем случае надо брать более крепкий
раствор целлоидина (1—2%-ный), наливать его на стекло в большем количестве, чтобы
пленка была более толстая, и, после 1—2 мин пребывания залитого целлоидином
препарата на открытом воздухе, поместить его для отвердения целлоидина в пары
хлороформа. После отвердения стекло переносят в спирт, где с помощью лезвия от
безопасной бритвы снимают пленку вместе с объектом.
Фиксация. При фиксации членистоногих необходимо помнить, что их хитиновая
кутикула плохо проницаема для большинства реактивов и красок. Поэтому в целые
144
объекты фиксатор проникает внутрь очень медленно, и притом разные его
компоненты — с неодинаковой скоростью. Это влечет неравномерную и плохую фиксацию
отдельных тканей и органов, а у объектов с мягкой кутикулой — сморщивание и
деформацию. Во избежание этого при фиксации необходимо накладывать (мелкие) и
надрезать или разрезать (крупные) объекты.
Перед фиксацией объекты желательно анестезировать хлороформом или
эфиром, чтобы они не съеживались под влиянием судорожных сокращений мышц.
Особенно это необходимо для объектов с мягкой хитиновой кутикулой (личинки
насекомых, животные непосредственно после линьки и т. п.), у которых во время фиксации
сокращения мускулатуры вйзывают выпячивание наружу частей кишечника и других
органов из проколов и разрезов в кожных покровах. Хорошие анатомические и
гистологические картины дают различные формалиновые, сулемовые, пикриновые и
осмиевые фиксаторы, а также и жидкость Карнуа (см. т. I, с. 10—15). После всех
этих фиксаторов для получения хороших обзорных анатомических препаратов с
окрашенными в различные цвета отдельными тканями пригодна окраска срезов по Мал-
лори (см. т. I, с. 34—35). После осмиевых фиксаторов для получения хорошей
окраски по Маллори необходимо срезы (из воды) обработать по методу Паля (1)
0,25%-ный марганцевокислый калий — 2—3 мин, 2) короткое ополаскивание в воде,
3) смесь равных объемов 1%-ной щавелевой кислоты и 1%-ного серноватистокисло-
го калия — 4—5 мин, 4) промывка в воде — 20—30 мин).
Заливка и приготовление срезов. Приготовление срезов из членистоногих,
особенно с твердой хитиновой кутикулой, сопряжено с большими трудностями; хитин
обычно крошится, рвет срезы, в результате чего не удается получить не только
полную серию срезов, но и один хороший срез. Кроме того, вследствие плохой
проницаемости кутикулы, отдельные участки объекта, особенно удаленные от места разреза,
могут недостаточно обезводиться и не пропитаться парафином, следствием чего яв--
ляется выкрашивание этих участков при резке на микротоме. Во избежание этих
трудностей необходимо подготовляемые к заливке мелкие объекты при переносе из
70%-ного в 96%-ный спирт разрезать пополам, а крупные надрезать. Если объект
удлиненный, то места, удаленные от разреза, надо надколоть.
Заливать надо всегда в целлоидин-парафии по Петерфн (см. т. I, с. 23). При
этом желательно употреблять более крепкий (3—6%-ный) раствор целлоидина.
Объекты в растворе целлоидина следует выдерживать не менее 4—6 дней. Заливать
следует не в чистом парафине, а в смеси с пчелиным воском (9 ч. парафина, 1 ч. воска;
нагреть до появления паров при тщательном помешивании и охладить). Держать
объекты в термостате в парафине с воском надо не более 30—40 мин. При такой за;
ливке для объектов с умеренно твердым хитином возможность неудач исключается.
Заливка объектов в смесь парафина с воском имеет еще одно важное
преимущество. Она дает возможность с одного блока получать срезы различной толщины —
от 2—5 мкм до 50—70 мкм. Это важно для получения с одного объекта толстых
срезов. В целях изучения общей топография органов и тканей (особенно мускулатуры)
и промежуточных между толстыми — тонких срезов для параллельного изучения
гистологического строения отдельных деталей. Так как точка плавления смеси
парафина с воском несколько ниже точки плавления чистого парафина, то при получении
тонких срезов (5—6 мкм и тоньше) надо парафиновый блок время от времени
охлаждать, смачивая его 'бока эфиром или 96%-ным спиртом, и дуть на него, чтобы
ускорить испарение. Наоборот, для получения толстых срезов приходится блок слегка
подогревать, придвигая к нему сверху электрическую лампочку. При работе со
срезами неодинаковой толщины надо наклеивать их соответственно на разные
предметные стекла. В противном случае при окраске тонкие срезы могут быть уже
перекрашены, в то время как 'толстые еще не успевают полностью прокраситься.
Рекомендуемые обычно методы размягчения хитина путем химического растворения
сообщающих ему твердость включений (инкрустов) органического и неорганического
характера (диафанол или соответствующие ему сернокислый' и азотнокислый препараты,
щавелевая вода и т. п.) здесь не рассматриваются, поскольку они действуют
неблагоприятно на структуру тканей и не имеют преимущества по сравнению с описанным
способом заливки.
Объекты с хитином, пропитанным углекислой известью (многие ракообразные,
некоторые паукообразные и как исключение насекомые), необходимо до заливки
выдерживать в подкисленном спирте (0,5—2%-ных азотной или соляной кислот) до
размягчения на ощупь их кожных покровов.
145
Подтип Chelicerata — Хелицеровые
КЛАСС ARACHNIDA—ПАУКООБРАЗНЫЕ
В качестве представителей паукообразных рассматриваются
скорпион (Buthus eupeus), паук (AraneuS diadematus) и клещ (Ixodes ri-
cinus). Подробное изучение организации скорпиона представляет
интерес в том отношении, что этот отряд является наиболее примитивной
и древней группой среди современных арахнид. Паук-крестовик
издавна считается классическим объектом изучения паукообразных. Кроме
того, широкое распространение и многообразие форм пауков
заставляет остановиться на организации этой группы достаточно подробно.
Организация клеща (Ixodes ricinus) представляет интерес как пример
глубоко специализированной группы арахнид, являющихся вместе с
тем паразитическими организмами.
ОТРЯД SCORPIONES —СКОРПИОНЫ
Buthus eupeus (С, Koch) — Скорпион пестрый
Материалы и методы изучения. Buthus eupeus (С. Koch), или скорпион пестрый
(см. Buthidae), принадлежит к наиболее обычным видам скорпионов, встречающихся
в СССР. Он распространен в Закавказье, на юго-востоке европейской части СССР
и в Средней Азии. Скорпионы ведут ночной образ жизни, выползая из своих убежищ
лишь с наступлением темноты на охоту за насекомыми и другими мелкими
животными. В местностях, где они обычны, их можно собрать, разбирая кучи камней, под
отдельными камнями, в трещинах глинобитных строений, в развалинах и т. п. Обычно
скорпионы живут одиночно, на некотором расстоянии друг от друга, никогда не
образуя скоплений из нескольких особей. Ввиду наличия у них ядовитого жала, для
ловли следует употреблять длинный пинцет. Живых скорпионов собирают порознь в
отдельные баночки или в ящик с отделениями, так как они могут поедать друг друга.
Buthus eupeus, как -и другие представители семейства Buthidae, живет в
неволе по нескольку месяцев. Для кормления скорпионов рекомендуются мухи,
личинки мучного жука Tenebrio и другие насекомые, а также пауки и мелкие дождевые
черви. От тараканов скорпионы обычно отказываются. Влаголюбивые скорпионы
Euscorpius выносят неволю Гораздо хуже.
Анатомически Buthus eupeus довольно близок к другим видам этого рода.
Поэтому он может быть заменен В. caucasicus (Nord.), обитающим в Закавказье,
Азербайджане и Средней Азии, и В. crassicauda (Oliv.) из юго-восточной части
Закавказья или даже представителями других родов и семейств. Из числа последних
наиболее обыкновенны Euscorpius italicus (Her.) (сем. Chactidae) из западного
Закавказья и крымский Е. taurtcus (С. Koch). Несколько видов Buthidae обитают
также у нас в Средней Азии (Psammobuthus, Liobuthus, Ammalobuthus и др.)1.
1 Для определения скорпионов ом. работу. А. А. Бялыницкого-Бирули «Фауна
России».
146 '
Изучение внешней морфологии производится на животных, убитых хлороформом
или эфиром, или на спиртовом материале. Вскрытие предпочтительно делать на
свежем материале, по пригодны и фиксированные скорпионы.
Для изучения кровеносной системы применяется инъекция. Для изготовления
срезов через печень, легкие и т. д. эти органы вырезаются и фиксируются подогретой
сулемой с уксусной кислотой. Мозг и глаза рекомендуется фиксировать жидкостью
Тройана ' или Карнуа. Для изготовления разрезов через все тело следует выбирать
самые маленькие, молодые экземпляры и, обрезав предварительно конечности,
фиксировать сулемой с уксусной кислотой. Перед заливкой в парафин, хитин
размягчается диафанолом, а затем объект проводится через жидкость Петерфи.
Срезы окрашиваются обычными красками — борным кармином in toto перед
заливкой, железным гематоксилином. Для нервной системы рекомендуется гематоксилин
Делафильда.
Внешняя морфология. Вытянутое тело скорпиона разделяется на
небольшую, покрытую общим хитиновым щитом головогрудь (cepha-
lothorax, рис. 130, А, 1) и длинное, сегментированное брюшко
(abdomen). Последнее в свою очередь подразделяется на передний, более
широкий отдел — переднебрюшие (praeabdomen, рис. 130, 21) и
суженный отдел — заднебрюшие (postabdomen, рис. 130, 20). Тело
заканчивается несущей искривленное острие анальной лопастью, или тельсо-
ном (telson, рис. 130, А, 19).
Длина тела скорпиона пестрого достигает 5 см; самцы несколько
меньше самок. Основной цвет хитинового покрова желтый, но
головогрудь и переднебрюшие со спинной стороны нередко с черноватыми
полосками и пятнами. Последний сегмент заднебрюшия обычно
темный, особенно на брюшной стороне.
Как у всех Scorpionoidea и у большинства Chelicerata, в состав
головогруди входят акрон (асгоп), отвечающий головной лопасти
(prostomium) аннелид, и шесть сегментов, соответственно шести парам
головогрудных конечностей: хелицер, педипальп и четырех пар ходных
ног. Из них четыре передних сегмента ларвальные; их мезодерма
развивается иначе, чем в остальных постларвальных сегментах. Эти
ларвальные сегменты гомологичны у всех Chelicerata и отвечают четырем
сегментам протасписовои личинки трилобитов, образующим голову
взрослых трилобитов. Таким образом, две последние пары ходных ног
принадлежат уже передним постларвальным сегментам. Следующий,
относящийся уже к брюшку сегмент, — рудиментарный, лишенный
конечностей VII сегмент, появляющийся в эмбриональном развитии. Но
его присутствие у взрослых животных выражается только в наличии
принадлежащей ему пары боковых артерий и лигаментов сердца,
тогда как в других системах органов и в наружном скелете от него
не остается никаких следов. В состав переднебрюшия входят семь
сегментов (с VIII по XIV). Из них шесть передних несут сильно
видоизмененные конечности (половые крышечки, гребневидные придатки,
легкие). XIV сегмент, так же как все следующие пять сегментов
заднебрюшия (с XV по XIX), лишен конечностей или их дериватов. Между
последним сегментом заднебрюшия и тельсоном лежит анальное
отверстие.
1 В состав жидкости Тройана входят: 5%-ный водный раствор двухромовокисло-
го калия — 7 частей, формалин — 1 часть, уксусная кислота — 1 часть.
147
Рис. 130. Buthus eupeus. Вид со спинной (А) и брюшной сторон (Б) (по-
А. А. Бялыницкому-Бируле):
1 — головогрудь, 2 — хелицера, 3 —вертлуг педипальпы, 4 — бедро педипальпы, 5 —
подвижный палец, 6 — ладонь, 7 — голень педипальпы, 8 — нога второй пары, 9 — нога
третьей пары, 10 — тазик, // —вертлуг, 12 — бедро, 13 — голень, 14 — основной
членик лапки, 15 — первый членик лапки, 16 — второй членик лапки, 17 — коготки,
18 — ядовитая игла, 19 — тельсои, 20 — заднебрюшне, 21 — переднебрюшие, 22 —
анальное отверстие, 23 — шпоры, 24 — стигма, 25 — стерннт XII сегмента, 26 ~ зубчики,
27 — средние пластинки, 28 — краевые пластинки, 29 — тазик ноги четвертой пары,
30 — половая крышечка, 31 — стернум, 32 -- педипальпа, 33 — неподвижный палец,
34 —тазик педипальпы, 35 — челюстная лопасть ноги первой пары, 36 — челюстная
лопасть ноги второй пары, 37 — тазик ноги второй пары, 38 — тазик ноги третьей
пары, 39 — гребневидный орган, 40 — четковидные пластинки, 41 — метастернум.
VIII—XIX — сегменты туловища
148
Итак, если считать VII рудиментарный сегмент, в состав тела
помимо акрона и тельсона входят 19 сегментов.
Наряду с подразделением тела скорпионов на головогрудь,
переднебрюшие и заднебрюшне, существует другое, не совпадающее с
ним, но применимое ко всем Chelicerata разделение на pro-, meso- и
metasoma. Первое деление наиболее распространено среди
систематиков, второе основывается на большом сравнительно морфологическом
(главным образом эмбриологическом) материале. К переднему отделу
тела — prosoma кроме акрона относятся шесть упомянутых сегментов
головогруди (рис. 130, А, 1), VII рудиментарный сегмент представляет
собой первый сегмент mesosoma. Последний сегмент переднебрюшия.
(XIV сегмент), который у Chelicerata никогда не имеет конечностей
даже в зародышевом состоянии и в этом отношении сходен с
сегментами заднебрюшия, вместе с этими последними составляет задний отдел
metasoma. Следовательно, граница между meso- и metasoma проходит
между VI и VII сегментами переднебрюшия (рис. 130, А, граница
между XIII и XIV сегментами).
Головогрудь невелика; она занимает только 7в—!/э часть
тела. На ней располагаются глаза, ротовое отверстие и шесть пар
конечностей. Впереди лежат хелицеры, затем следуют педипальпы и далее
четыре пары ходных ног.
Спинной щит головогруди (сагарах) представляет собой продукт
полного слияния акрона и тергитов (дорзальных щитков) шести
следующих сегментов. Он трапециевидной формы, заметно расширяется
кзади; углы его равномерно закруглены (рис. 131). Поверхность щита
неровна, сильно скульптурирована, покрыта бороздками, ямками и
гребешками с характерными рядами бугорков, а также редкими
волосками. Последние особенно хорошо заметны на переднем крае, где они
крупнее и, по некоторым данным, являются чувствительными.
Примерно посредине спинного щита расположена пара круглых выпуклых и
довольно крупных медиальных глаз (рис. 131,2). Близ каждого
переднего угла щита лежит группа из пяти боковых глаз (рис. 131,/). Из
них три передних и внешних глаза крупнее. Два маленьких задних
глаза мало заметны, так как легко смешиваются с окружающими
хитиновыми бугорками. Пара медиальных глаз свой- _„_„^^
ственна всем Chelicerata; число боковых у 1^-£"'/'"\"\~\к
различных скорпионов варьирует от двух до ^Ж ( J% } т.
пяти пар. 2 Кк у|||Т,|</ (Шк
Бока головогруди, или ее плевральные |7ЖА11« ||1
области, покрыты мягкой, кожистой кутику- I/ VTWlllw I \f\
к/IP Я «Г1' *? £ ' А\ ill
Брюшная поверхность головогруди почти 1^^Ът0''- "№ЧШ?'^Ж.
целиком образована тазиками, или коксами Wf^\ ^'к\т^^^^Ж
(coxae), педипальп (рис. 132,/) и ходных ног i|^^^=Ai=2?^^3
(рис. 132, 5, 6, 8, 9). Только сзади, между та- ^继
зиками третьей и четвертой пары ног сохра- Рис. 131. Сагарах But-
няется небольшая хитиновая пластинка почти hus sp. (по А. А. Бялы-
треугольной формы — метастом, или стернум иицкому-Бируле):
(sternum, рис. 132, 10), ~ результат слияния '~бок°1™™13?к*1~меда"
149
стернитов (брюшных щитков) V и
VI сегментов. В состав стернума
входит также и задняя часть стер-
нита IV сегмента (Павловский,
1917). Из передних стернитов
сохраняются принадлежащие III и
II сегментам, но снаружи они не
видны. Позади стернума лежат
парные половые крышечки (рис.
130, Б, 30), относящиеся уже к
VIII туловищному сегменту, т. е.
ко второму сегменту переднебрю-
шия. Тазики педипальп первой и
второй пар ног на внутренней
стороне несут челюстные отростки, или
лопасти, тазики двух задних пар
ног лишены их. Челюстные лопасти
велики и образуют значительную
часть брюшной поверхности
головогруди (рис. 132, 3, 4). Характерно,
что челюстные лопасти второй
пары ног вытянуты вперед по
направлению к ротовому отверстию и
внедряются между" отростками
первой пары ног (рис. 132, 3).
Челюстные лопасти представляют собой
не что иное, как жевательные
отростки (эндиты), свойственные
всем низшим Chelicerata и Trilobita,
у которых они располагались на
всех ходных ногах. У скорпионов,
однако, они не имеют жевательной функции, хотя и ограничивают
предротовую полость.
Необходимо отрезать ножницами челюстные лопасти первой и
второй пары ног в том месте, где они отходят от тазиков. Лучше это'
сделать только на одной стороне (рис. 132). При этом открывается
предротовая полость, в глубине которой лежит ротовое отверстие.
Предротовая полость представляет собой узкое пространство,
ограниченное дорзально хелицерами, направленными параллельно друг
другу вперед (рис. 132,17,19), с боков — коксами педипальп (рис. 132, 7),
а вентрально — налегающими друг на друга челюстными лопастя-
мя ног (рис. 132,3,4). От задней стенки предротовой полости отходит
довольно большой, килеобразный, направленный вперед мускулистый
вырост — ротовой конус, или рострум (rostrum, рис. 132,16). У самого
его основания, между подходящими сюда челюстными лопастями
первой пары ног и педипальп, помещается очень маленькое ротовое
отверстие (рис. 132,75). Таким образом, оно лежит между сегментами
педипальп и первой пары ходных ног. Внутренние поверхности тазиков
педипальп, образующие боковые стенки предротовой полости; представ-
Рис. 132. Buthus eupeus, вид голо-
груди снизу. Левые первая и вторая
ноги удалены (ориг.):
/ — тазик педипальпы, 2 — вертлуг педи-
пальпы, 3 — челюстная лопасть второй
ноги, 4 — челюстная лопасть первой ноги,
5 — тазик первой ноги, б — тазик второй
ноги, 7 — вертлуг второй ноги, 8 — тазик
третьей ноги, 9 — тазик четвертой ноги,
10 — стернум, 11 — предротовая полость,
12 — основание второй ноги, 13 — ротовое
отверстие, 14 — боковой край карапакса,
15 — основание первой ноги, 16 — рострум,
17 — первый членик хелицеры, 18 —
боковая стенка предротовой полости, 19 —
второй членик хелицеры, 20 — конечный
членик хелицеры — подвижный палец, 21 —
фурка, 22 — неподвижный палец хелицеры
150
ляют выпуклые, мясистые подушечки, покрытые тонкой и гибкой
кутикулой (рис. 132,18) и в этом отношении сходные с рострумом.
Своими мягкими покровами они резко отличаются от соседних сильно хи-
тинизированных частей тазиков педипальп и ходных ног. Подушечки
несут густые и довольно длинные волоски. Такие же волоски
покрывают рострум и нижнюю поверхность хелицер.
Таким образом, стенки предротовой полости образованы
подвижными, но более или менее плотно примыкающими друг к другу
поверхностями соседних органов. В предротовую полость поступает сильно
размельченная и обильно смоченная выделениями пищеварительных
желез (вероятно, главным образом печени) добыча, которая здесь
высасывается. (Скорпионы питаются только соками своей добычи, и
твердые пищевые частицы отбрасываются.) Снабженный сложной
специальной мускулатурой рострум при этом играет роль поршня;
сокращаясь, он создает в полости отрицательное давление! Волоски, в изобилии
покрывающие рострум и соседние органы, смачиваются и как бы
«законопачивают» щели между отдельными частями стенок предротовой
полости, препятствуя проникновению воздуха извне, что важно для
более полного высасывания добычи.
Антенны I у Chelicerata отсутствуют, и передняя пара конечностей
представлена хелицерами (chelicerae, см. рис. 130, Д 2), которые
гомологичны антеннам II трилобитов и ракообразных, т. е. первой паре
посторальных конечностей. Хелицеры лежат впереди и над ротовым
отверстием и прикрепляются к передней мягкой, кожистой стенке
головогруди под краем спинного щита так, что последний нависает над ними.
Хелицеры состоят из трех члеников. Первый короткий членик скрыт
под передним краем головогрудного щита (рис. ,132,77), второй —
толстый, яйцевидной формы (рис. 132,75) на своем передневнутреннем
крае вытянут в крючковидный отросток, называемый неподвижным
пальцем ( рис. 132,22). Третий членик, или подвижный палец,
прикреплен на передне-наружном крае второго и вместе с неподвижным
пальцем составляет клешню хелицеры (рис. 132,20). Подвижный палец на
конце раздвоен, образуя два серповидных острия вилочки, или фурки
(furca, — рис. 132,27), в которую вкладывается конец неподвижного
пальца, когда клешня сжимается. Внутренние края обоих пальцев
вооружены крепкими дополнительными хитиновыми зубцами. Нижняя
поверхность хелицер покрыта густыми и длинными щетинками. В покое
из-под головогрудного щита торчат только передние части хелицер, но
они способны значительно выдвигаться вперед, в чем легко убедиться,
осторожно вытягивая их пинцетом. Поочередно действуя хелицерами,
скорпион разрывает свою добычу и медленно перемалывает ее в
кашицеобразную массу, которая затем высасывается в предротовой полости.
Педипальпы (pedipalpi, см. рис. 130, А, 3—7) значительно толще и
длиннее всех прочих конечностей. Они служат для осязания,
схватывания и удерживания добычи, поднесения ее к хелицерам и для чистки
хелицер. Педипальпы состоят из шести члеников: тазика (соха —
рис. 132,7), вертлуга (trochanter, см. рис. 130, А, 3), бедра (femur,
ем. рис. 130, А, 4), голени (tibia, см. рис. 130, Л, 7), основного (basitar-
sus, см. рис. 130, А, 6) и конечного (telotarsus, рис. 130, Л, 5) члеников
151
Рис. 133. Четвертая левая нога
Buthus eupeus (по А. А. Бялыницкому-
Бируле):
/ — вертлуг, 2 — бедро, 3 — голень, 4 —
основной членик лапки, 5 — шпоры, 6 —
первый членик лапки, 7 — второй, членик
лапки, 8 — коготковый членик, 9 — шип,
10 — коготки
лапки. Последний, по-видимому,
представляет, собой результат слияния
нескольких члеников. Тазики подвижны.
Два последних членика образуют
мощную клешню, причем основная
часть ее — ладонь представлена
предпоследним члеником, вытянутым на
передневнутреннем углу в длинный
неподвижный палец клешни (см.
рис. 130, Б, 33). Последний членик
играет роль подвижного пальца и
прикреплен на внешнем углу ладони (см.
рис. 130, А, 5). Пальцы у Buthus
eupeus слегка изогнуты; на внутренней
стороне они заострены в лезвия, вдоль
которых расположены мелкие
хитиновые бугорки, группирующиеся в
косые, параллельные друг другу ряды.
Членики педипальп несут ряды
бугорков и слабо развитые гребешки. На
них рассеяны волоски, а на ладонях и особенно на пальцах, кроме
того, многочисленные чувствительные волоски, или трихоботрии.
Ходные ноги постепенно увеличиваются в каудальном направлении.
Самая маленькая — первая пара, самая мощная — четвертая. Все они
состоят из восьми члеников, из которых первые пять те же, что и на
педипальпах, шестой и седьмой называются первым и вторым
члениками лапки (tarsus I и II, рис. 133, 6,7), а восьмой представлен коготко-
вым члеником (praetarsus, рис. 133, <§). Последний очень мал и почти
скрыт в углублении на вершине лапки. На нижней стороне он
вооружен крепким, но коротким шипом (рис. 133,5), к его верхнему краю
подвижно прикреплены два острых коготка (рис. 133,/0). При ходьбе
скорпион опирается на землю лишь двумя последними члениками.
Тазики ходных ног отличаются характерной особенностью,
свойственной многим паукообразным, — они неподвижны, так как входят в
состав брюшной стенки головогруди. Ограниченную подвижность
сохраняют лишь коксы первой пары ног. Первые четыре членика несут
только редкие волоски, остальные снабжены многочисленными щетинками.
Кроме того, конец второго членика лапки вооружен одной, а конец
первого членика — двумя подвижными и заостренными шпорами.
Переднебрюшие состоит из семи широких и несколько
уплощенных сегментов. Каждый сегмент сверху прикрыт твердым
хитиновым спинным щитком, или тергитом (см. рис. 130, Л, VIII—XIV). С
каждой стороны от тергита отчленяется по одному маленькому отогнутому
вниз боковому щитку. Бока переднебрюшия покрыты тонкой и мягкой,
морщинистой хитиновой кутикулой, соединяющей тергиты со стерни-
тами. Передние тергиты самые короткие и имеют прямоугольную фор-~
му, последний, седьмой, самый длинный; форма его трапециевидная
(см. рис. 130, А, XIV). На поверхности тергитов имеются продольные
бугорчатые ребрышки.
152
Все сегменты переднебрюшия, кроме первого, обладают также
хорошо выраженными стернитами. Стернит второго сегмента (IX
туловищный) представлен небольшой, вытянутой в поперечном направлении
пластинкой — метастернумом (metasternum, см. рис. 130, Б, 41),
расположенной между гребневидными органами. Стерниты всех следующих
сегментов (X—XIV) велики и занимают всю их брюшную сторону. Они
имеют форму прямоугольников, кроме заднего — трапециевидного
(см. рис. 130, Б).
Сегменты переднебрюшия, за исключением последнего, снабжены
сильно видоизмененными конечностями. Лишенный стернита передний
сегмент (VIII туловищный) несет пару небольших половых крышечек,
прикрывающих половое отверстие (см. рис. 130, Б, 30). Замечательно,
что у всех Chelicerata половое отверстие всегда лежит на этом
сегменте. Половые крышечки находятся непосредственно впереди метастерну-
ма и гомологичны конечностям VIII сегмента. У самцов Buthus
eupeus, как и у многих других скорпионов, под половыми крышечками по
бокам от полового отверстия лежат небольшие хитинизированные
сосочки, половые папиллы, которые, по-видимому, играют роль мужских
совокупительных органов.
Конечности следующего, второго сегмента представлены
гребневидными придатками (см. рис. 130, Б, 39), своеобразными органами,
которые свойственны только скорпионам. Каждый состоит из
суживающегося к дистальному концу стержня, который прикрепляется к боковому
краю метастернальной пластинки, и сидящих на его заднем краю,
подобно зубьям гребня, подвижных зубчиков, или пластинок. Стержень
слагается из трех рядов неподвижно спаянных друг с другом пластинок.
Передний край образован большими, удлиненными краевыми
пластинками (см. рис. 130, Б, 28); во втором ряду лежат округлые средние
(см. рис. 130, Б, 27); третий ряд составляют многочисленные мелкие
четковидные пластинки (см. рис. 130, Б, 40). К последним
прикрепляются подвижные, образующие гребень зубчики, которые слегка колено-
образно изогнуты, по внешнему краю несколько расширены и отчасти
налегают друг на друга (см. рис. 130, Б, 26). Гребневидные органы
имеются у обоих полов, но у самцов они несколько длиннее и шире и
несут большее количество зубчиков.
На четырех следующих сегментах (с X по XIII) располагается по
паре легких, мешки которых часто просвечивают сквозь стерниты на
их боках. У заднего края каждого легкого видна поперечная щель,
окруженная невысоким хитиновым валиком, — дыхальце, или стигма
(см. рис. 130,24). Легкие скорпионов гомологичны жаброносным
конечностям их предков (Eurypterida), они располагаются на этих же
сегментах.
Принадлежность заднего, седьмого, сегмента переднебрюшия к ме-
тазоме видна не только в отсутствии конечностей, но и в общей форме
и характере скульптуры, особенно на его стерните. Расположенные
здесь продольные гребешки напоминают таковые на заднебрюшных
сегментах (см. рис. 130,5).
Заднебрюшие — это самая узкая и длинная часть тела
(см. рис. 130, А, 20). Все пять сегментов его отличаются полным слия-
153
ш-
UK-
ШЕ-
-— тель-
-XIX —
нием своих тергитов и стернитов так, что
каждый имеет вид сплошного хитинового
кольца. Передний край каждого кольца
имеет выемку (хорошо видно на брюшной
стороне), в которую входит округлый
выступ предыдущего кольца. Подобное
сочленение 'придает 'всему заднебрюшию особую
подвижность и позволяет животному
поднимать эту часть тела вверх, загибая ее
дугой над спиной.
Анальное отверстие лежит на брюшной
стороне между пятым сегментом заднебрю-
шия и тельсоном, под самым основанием
последнего (см. рис. 130, Б, 22; 134,/). Оно
окружено четырьмя мясистыми лопастями
несколько вывороченной наружу задней
кишки и прикрывается полукруглой,
морщинистой хитиновой лопастью заднего края
тельсона.
Т е л ь с о н сильно вздут у своего
основания и загибается на спинную сторону
(рис. 134, 2), заканчиваясь острой,
искривленной назад иглой, или жалом (рис. 130,
А, 18; 134,5). В тельсоне лежит пара
ядовитых желез, протоки которых
открываются на вершине иглы двумя маленькими отверстиями.
Анатомия на вскрытии. Вскрытие скорпиона производится в
препаровальной ванночке в воде или, если анатомируется свежеубитое
животное, в физиологическом растворе.'
Вначале разрезаются острыми ножницами плевральные стенки переднебрюшия
и головогруди.,Разрезы проводятся по сторонам тела, вдоль под боковыми щитками и
под краями спинного щита головогруди. Оба разреза соединяются поперечными — под
передним краем головогрудного щита и позади последнего тергита переднебрюшия.
Затем пинцетом слегка приподнимается передний край головогрудного щита. При
этом обнаруживается, что к щиту изнутри прикрепляются многочисленные мышечные
пучки, которые следует осторожно и тщательно отделить острым скальпелем.
Действуя так же и далее, следует отделить до.рзальную стенку переднебрюшия. Эту в
общем трудную операцию удобнее производить вне воды, до прикалывания животного
к дну ванночки, и если, проделать ее аккуратно и тщательно, то внутренние органы
и мускулатура не повреждаются. Наконец, разрезаются с боков кольца заднебрюшия
и снимается спинная стенка с этой части тела. В результате обнажается дорзальная
мускулатура, которая закрывает все внутренние органы. Только в головогруди видна
часть так называемой желудочной железы, а в заднебрюшии спинная аорта и лежа-,
щая под ней кишка '.
Далее необходимо вырезать мускульные пучки в головогруди и удалить
мускульную стенку переднебрюшия и дорзальные мышцы заднебрюшия, после чего
обнажается часть пищеварительной системы и сердца. При анатомировании свежего ма:
териала печень, разрываясь, мутит воду. Прибавление к воде небольшого количества
спирта препятствует помутнению.
Рис. 134. Конец заднебрюшия
Buthus eupeus. A — сбоку, Б —
с брюшной стороны (по А. А.
Бялыницкому-Бируле):
/ — анальное отверстие, 2
сон, 3 — ядовитая игла. XVI-
сегменты туловища
1 При недостаточно аккуратном вскрытии большая часть мускулатуры брюшка
остается на снимаемых спинных покровах.
154
Полость тела разделена поперечной мускулистой перегородкой, диафрагмой,
проходящей между головогрудью и переднебрюшием (см. рис. 138, 6; 142, 3).
Впереди от диафрагмы сразу бросается в глаза большая пятилопастная
желудочная железа, занимающая всю заднюю половину головогруди (рис. 142, 1). Пе-
реднебрюшие заполнено объемистой коричневатой или сероватой печенью (рис. 142,5).
Медиально над печенью помещается вытянутое, трубковидное сердце (рис. 142, 12),
лежащее в тонкостенном иерикардиальном мешке (рис. 142, 13). Многие детали
сердца хорошо видны. Спереди оно продолжается в переднюю аорту (рис. 142, 17), а
сзади от него отходит задняя аорта (рис. 142, 8). Передняя аорта проходит сквозь
диафрагму и направляется вперед и вниз под желудочную железу так, что видно
только основание этого сосуда. Спинная аорта тянется над кишечником до заднего
конца тела. По бокам от перикардия печень прободается шестью парами дорзовент-
ральных мускулов (рис. 142, 4). Пара таких же мускулов проходит сквозь
желудочную железу (рис. 142, 2).
Дальнейшее вскрытие сводится к постепенному и осторожному удалению
органов, лежащих сверху. Сначала удаляется сердце, желудочная железа и печень.
Последняя отрывается маленькими кусочками, и препарат время от времени промывается
водой. При вскрытии только что умерщвленного скорпиона печень легко уд'аляется
целиком. Для этого надо перерезать кишку сзади диафрагмы, захватить пинцетом,
спереди всю массу печени и, осторожно подергивая, отделить ее от нижележащих
органов. Таким же образом можно изолировать передний участок кишечника с
желудочной железой.
В головогруди обнажаются передняя часть кишечника, эндостернит, коксальные
железы, и головной мозг. У основания брюшных лопастей печени проходит связанная
с ней средняя кишка. Глубже лежит половая система, а еще вентральнее — брюшная
нервная цепочка. Наконец, после удаления всех внутренностей становятся видными
мускулы, связанные с брюшной стенкой тела, и легкие.
В некоторых случаях удобнее производить вскрытие с брюшной стороны.
Особенно это рекомендуется прн изучении на спиртовом материале печени, ее протоков
и средней кишки, а также мальпигиевых сосудов.
Наибольшие трудности представляет препарирование глотки, надглоточной и
подглоточной нервных масс и особенно тонкостенных и нежных кровеносных сосудов.
Изучение нервной системы должно предшествовать изучению кровеносных сосудов.
Внутренний скелет скорпионов, как и у других
паукообразных и Xiphosura, состоит из образований двоякого происхождения.
С одной стороны, имеются аподемы, или эндосклериты, т. е. глубокие
впячивания наружного хитинового покрова, с другой стороны — эндо-
хондриты, или скелетные образования мезодермальной природы (Шим-
кевич, 1893; Переяславцева, 1907), не связанные с наружным хитином.
К числу последних относится сложный по форме лежащий в
головогруди эндостернит.
Для изучения аподем, головогрудь без педипальп и ног и без
спинного щита вываривается в 10%-ном едком кали. После промывания
хорошо заметны: 1. Коксальные аподемы — пластинчатые выросты,
отходящие от первого членика хелицер и тазиков первой и второй пары ног
(рис. 135,70). Они глубоко вдаются в полость тела и служат Местом
прикрепления мощной мускулатуры. 2. Медиальные, непарные,
аподемы. Впереди лежит преоральный эндосклерит с двумя большими
боковыми выростами (рис. 135,13). Позади него виден посторальный
эндосклерит в виде короткой хитиновой пластинки (рис. 135,75). В области
стернума лежит карино-стернальная аподема.
Интерес представляет посторальный эндосклерит, который
представляет собой видоизмененный стернит III сегмента (сегмента первой
пары ходных ног). Он образует задний край ротового отверстия; с бо-
155
Рис. 135. Buthus eupeus. Аподемы
головогруди (ориг.):
/ — хелицера, 2 — педипальпа, 3 — тазик
первой ноги, 4 — тазик второй ноги, 5 —
тазик третьей ноги, 6—тазик четвертой
ноги, 7 — гребенчатый орган, 8 — легкое,
9 — стернит, 10 — коксальные аподемы,
// — половая крышечка, 12 — стернум,
13 — боковой вырост преорального эндо-
склерита, 14— аподема хелицеры, 15 —
посторальный эндосклерит
Рис. 136. Эндостернит головогруди
Buthus eupeus. Вид сзади (А) и
сверху (Б) (ориг.):
/ — субневральная дужка, 2 —
пластинчатые выросты
ков к нему прикрепляются базаль-
ные углы тазиков педипальп
первой и второй ходных ног.
На этом же препарате следует
ознакомиться с особенностями
соединения головогруди с переднебрю-
шием, которое на цельных
скорпионах рассмотреть не удается.
Мягкая соединительная кожа между
этими двумя отделами глубоко
вдается в тело между тазиками
последней лары ходных ног в виде
большой складки.
Эндохондриты состоят из
волокнистой, хрящеподобной ткани и
при кипячении в едком кали легко
разрушаются. Поэтому приходится
их отпрепаровывать особо.
Эндостернит лежит в головогруди под
передним отделом кишечника и
выделяется обычно с обрывками
прикрепляющихся к нему
диафрагмы и мышц. Это бесцветное,
довольно широкое кольцо с четырьмя
парами пластинчатых выростов
(рис. 136, А а Б). Оно окружает
подглоточную нервную массу (рис.
137). Представляет интерес
нижняя часть кольца, называемая суб-
невральной дужкой (рис. 136, А,1).
Она поддерживает нервную систему
и отвечает подобной же дужке
эндостернита у Limulus. На верхней
дуге эндостернита имеется медиг
альный ' желобок для пищевода и
сосудов. Сзади эндостернит связан
с диафрагмой.
В переднебрюшии лежит над-
гребешковый эндохондрит. Он
принадлежит IX сегменту, являясь его
эндостернитом, и имеет вид
маленькой поперечной пластинки,
расположенной- вблизи брюшной
стенки тела над метастернумом. К
нему прикрепляются дорзовентраль-
ные мускулы и мускулатура
гребенчатых органов.
Мускулатура. В теле
скорпиона имеется не менее 150 (боль-
156
шей частью парных) мышц, не считая
мускулатуры конечностей.
Сравнительно с .сегментированным
брюшком, лишенным двигательных конечностей,
головогрудь представляет сложный отдел тела.
Соответственно и мускулатура головогруди по
дифференциации и расположению резко
отличается от таковой брюшка, где она целиком Рис т. Scorpiones. Поло-
носит метамерныи характер. жение эндостернита относи-
Мезозома и метазома различаются по тельно нервной системы (из
характеру своей мускулатуры. Versluys и Demoll):
Вся туловищная мускулатура может быть т0~нТяТеНр0вйная0массаГзп-АсУб-
раЗДеЛена На Следующие ГРУППЫ: невральная дужка, 4 — эндо-
I. Продольная мускулатура. II. Дорзо- стернит
вентральные мускулы. III. Мускулы,
прикрепляющиеся к конечностям. IV. Мускулы, прикрепляющиеся к непарным
элементам внутреннего скелета (энд остер ниту, надгребешковому эндо-
хондриту, преоральному эндооклериту и др.). V. Мускулатура глотки,
перикардия и других внутренних органов 1.
Следует обратить внимание на наиболее заметные мышцы первых
трех групп.
I. Продольная мускулатура наиболее примитивна в мезозоме. Здесь
каждый типичный сегмент имеет три пары спинных и четыре пары
брюшных мускулов. Это лентовидные продольные и косые мышцы.
На спинной стороне лежат:
1. Передне-задние мускулы (m. antero-posteriores, рис. 138,5). Они
прилегают к нижней поверхности тергита, прикрепляясь у переднего
края следующего тергита. При сокращении сближают соседние терги-
ты, т. е. укорачивают переднебрюшие.
2. Косые тергальные мускулы (m. tergales obliqui — рис. 138,8).
Прикрепляются спереди к тергиту, а сзади к переднему краю
следующего тергита. Ретракторы сегментов.
3. Боковые спинные мускулы (m. laterodorsales, рис. 138,10).
Прикрепляются сходно с предыдущими. Контракторы и дилататоры
брюшка.
На брюшной стороне в первых, двух сегментах мезозомы (VIII и
IX сегменты) продольная мускулатура сильно видоизменена. В
остальных же четырех сегментах имеются:
1. Средние передне-задние мускулы (m. antero-posteriores. mediani,
рис. 139,4). Проходят от переднего края стернита к переднему краю
следующего. Действие их аналогично таковому передне-задних
мускулов.
2. Боковые передне-задние мускулы (m. antero-posteriores laterales,
рис. 139,5). Соединяют стерниты двух соседних сегментов.
3. Косые стернальные мускулы (m. sternales obliqui, рис. 139,6).
1 Некоторые мышцы этой группы рассматриваются ниже при описании
соответствующих органов. Особый интерес представляют перикардиовентральные мускулы
(см. с. 167—168).
157
Рис. 138. Buthus eupeus. Спинная
мускулатура туловища (ориг.):
I — первый дорзовеитральный мускул, 2 —
медиальный глаз, 3 — второй дорзовент-
ральиый мускул, 4 — третий
дорзовеитральный мускул, 5 — четвертый
дорзовеитральный мускул, 6 — диафрагма, 7 —
пятый дорзовеитральный мускул, 8 — косой
тёргальный мускул, 9 — передне-задний
мускул, 10 — боковой спинной мускул,
II — дорзовеитральный мускул, 12 —
дорзовеитральный мускул метазомы, 13 —
передне-задний мускул метазомы, 14 —
косой мускул метазомы, 15 — поверхностный
дорзовеитральный мускул, 16 —
желудочная железа, 17 — дорзококсальные
мускулы ног, 18 — дорзодейтомеритные мускулы
педипальпы, 19 — дорзококсальные
мускулы педипальпы, 20 — дорзоэндосклеритиые
мускулы хелицеры, 21 — дорзококсальные
мускулы хелицеры
158
Проходят от стернита косо назад к
межсегментной сочленовной коже.
4. Задне^тигмальные мускулы
(m. poststigmatici, рис. 139, 7).
Проходят косо позади и кнутри от
легочных мешков и стигм. Прикрепляются
к стерниту и сочленовной коже. Из
этих мышц в прозоме имеются только
спинные, именно пара передне-задних
(см. рис. 138, 9) и пара косых тергаль-
ных (см. рис. 138, 10); брюшные
мышцы здесь отсутствуют.
В каждом сегменте метазомы
сохраняются по четыре пары мускулов:
сверху — передне-задние (см. рис. 138,
13) и косые (см. рис. 138, 14), а
снизу— средние передне-задние (рис. 139,
4) и косые стернальные (рис. 139, 6).
Однако они здесь имеют характер
мощных толстых пучков и иначе
называются. В первом метазоматичес-
ком сегменте (XIV) лежит еще, кроме
того, пара мощных поверхностных
дорзовентральных мышц (m. superfi-
ciales dorso-ventrales, см. рис. 138,15).
Среди мышц метазомы имеются
элеваторы, ретракторы и экстензоры,
обеспечивающие сильную подвижность
этого отдела тела.
II. Дорзовентральные мускулы
(m. dorso-ventrales) представлены
одиннадцатью парами. При
сокращении они сплющивают тело в спинно-
брюшном направлении.
В прозоме имеется четыре пары.
Все они начинаются от головогрудно-
го щита и направляются вниз.
Передняя пара (см. рис. 138, /) вентрально
прикрепляется к мягкой коже между
хелицерами, вторая пара (см.
рис. 138, 3)—к преоральному эндо-
склериту, третья (см. рис. 138, 4)
проходит сквозь желудочную железу к
эндостерниту. К нему же
прикрепляется и четвертая пара (см. рис. 138,
5), проходящая впереди диафрагмы.
Остальные дорзовентральные
мускулы принадлежат брюшку. Пара,
отходящая от первого тергита мезо-
зомы (см. рис. 138, 7), тянется по
задней стороне диафрагмы, прободает ее и
прикрепляется к задней части эндостер-
нита. Следующая пара проходит через
печень от второго тергита к надгребеш-
ковому эндохондриту. Далее следует пять
пар мускулов (см. рис. 138, // и 12),
принадлежащих четырем задним
сегментам мезозомы и первому сегменту
метазомы. Они соединяют тергит со стерни-
том своего сегмента и, как предыдущая -
пара, прободают печень.
III. Мускулы, двигающие
конечностями и прикрепляющиеся к их
основаниям, лежат в прозоме. Наиболее заметны
из них следующие группы:
Мускулы хелицер.
1. Дорзо-коксальные (m. dorso-coxa-
les chelicerae, см рис. 138, 21). Две
пары. Прикрепляются к первому членику
хелицер.
2. Дорзо-эндосклеритные (m. dorso-
endosclerites chelicerae, см. рис. 138, 20).
Две пары. Идут к эндосклеритам (апо-
демам) хелицер.
Мускулы педипалвп.
1. Дорзо-коксальные (m. dorso-coxa-
les pedipalpi, см. рис. 138, 19).
Соединяются с тазиками.
2. Дорзо-дейтомеритные (m. dorso-
deutomerites pedipalpi, см. рис. 138, 18).
Тянутся в основание педипальп, где
прикрепляются к вертлугу.
Мускулы ходных ног.
1. Дорзо-коксальные мышцы первой,
второй, третьей и четвертой пар ног (т.
dorso-coxales pedis, см. рис. 138, 17).
Прикрепляются к тазикам
соответствующих конечностей.
Пищеварительная система.
Кишечный канал представляет собой
прямую трубку небольшого диаметра и
дифференцирован на глотку, пищевод,
желудок, тонкую и заднюю кишки.
Желудочная железа открывается в желудок, который является первым
отделом средней, энтодермальной, кишки. Печень открывается в
переднюю половину тонкой кишки. Первые три отдела кишечника
помещаются в головогруди, остальные—в брюшке, причем задняя кишка
ограничена последним сегментом заднебрюшия. У скорпионов, подобно
Рис. 139. Buthus eupeus.
Брюшная мускулатура
туловища (ориг.):
/ — мускул половой крышечки, 2 —
надгребешковый эндохондрит, 3 —
брюшные концы дорзовентральных
мускулов, 4 — средний передне-
задний мускул, 5 — боковые
передне-задние мускулы, 6 — косые
стериальные мускулы, 7 — задне-
стигмальный мускул, 8 —
латеральный мускул, 9 — стигма, 10 —
легкое
159
Рис. 140. Scorpiones. Схема передней и задней частей
пищеварительного канала. Вид справа (по Е. Н. Павловскому)-.
/ — рострум, 2 — глотка, 3 — головной мозг, 4 — пищевод, 5 —
расширители пищевода, 6 — желудок, 7 — тонкая кишка, 8 — леваторы ануса,
9 — анальная папилла, 10 — анальное отверстие, Я — задняя кишка,
12 — сжнматели пищевода, 13 — подглоточная нервная масса, 14 —
боковой отросток преорального эндосклерита, 15 — боковые
расширители глотки, 16 — сжиматели глотки
Рис. 141. Scorpiones. Схема пищеварительного канала (по
Е. Н. Павловскому):
/ — глотка, 2 — пищевод, 3 — желудок, 4 — диафрагма, 5 — тонкая
кишка, 6 — печеночные протоки, 7 — передний мальпигиев сосуд, 8 —
задний мальпигнев сосуд, 9 — проток желудочной железы, 10 —
расширители пищевода, Л —нервная масса, 12 — боковой вырост
преорального эндостернита, 13 — боковые расширители глотки
другим Arachnida (кроме Acarina), слюнные железы,
открывающиеся в ротовую полость, отсутствуют.
Ротовое отверстие ведет в глотку, которая помещается между
основанием рострума и головным мозгом (рис. 140,2; 141,/). Суживаясь
кзади, глотка подходит к окологлоточному нервному кольцу, где
переходит в очень узкий пищевод. Глотка представляет собой мощный
сосательный аппарат. Осторожно разрезав этот орган, можно видеть,
что просвет его имеет почти трехгранную форму, характерную для
глотки многих сосущих жидкую пищу животных (многие паукообразные,
Pantopoda, пиявки и др.). Полость глотки выстлана хитиновой
кутикулой, стенки ее мускулисты, так как состоят из кольцевых мускульных
пучков, играющих роль сжимателей (рис. 140,16). К глотке
прикрепляются также несколько парных мускулов расширителей, из которых
наиболее заметны боковые расширители (рис. 140,75; 141,73). Отходя
от боковых стенок глотки, они огибают переднюю часть мозговой
массы и прикрепляются к крыловидным отросткам преорального
эндостернита (рис. Ш,14; 141,72).
Пищевод в виде узкой трубки, проходит сквозь окологлоточное
отверстие нервной системы (рис. 140,4; 141,2). Полость его также
выстлана хитиновой кутикулой, что говорит о принадлежности к передней,
эктодермальной, кишке. Позади мозга пищевод заметно расширяется.
Так как здесь от него отходят несколько мускульных пучков —
расширителей, частью прикрепляющихся к передним отросткам эндостернита,
160
частью — к диафрагме (рис. 140,5,141
10), то считается, что эта задняя часть
пищевода функционирует как второй,
дополнительный, сосательный аппарат
(Е. Н. Павловский, 1917).
Антагонистами мускулов, расширяющих просвет
пищевода, являются кольцевые
мускульные волокна в его стенках
(рис. 140,72). У пауков, Pedipalpi и
части клещей имеется более
развитый сосательный аппарат,
гомологичный этому участку пищевода.
Следующий отдел кишечника —
головогрудная часть средней кишки—
условно называется желудком
(рис. 140, 6; 141, 4; 143, 3). Стенки
желудка железисты.- От пищевода он
отделен небольшим кольцевидным
перехватом. В желудок открывается
желудочная железа посредством пары
коротких протоков. Последние хорошо
видны, если удалить часть
налегающей сверху железы (рис. 141, 9).
Желудочная железа
подразделена на пять больших
лопастей—непарную переднюю и две пары боковых'
и слагается из множества железистых
долек (рис. 142, 7; 143, 10). Задняя
поверхность ее прилегает к
диафрагме. Центральная часть железы пробо-
дена парой отверстий, сквозь которые
проходит третья пара дорзовентраль-
ных мускулов (рис. 142, 2).
Гистологически желудочная железа очень
близка к печени, развивается сходное
ней, как выпячивание средней кишки,
и, по-видимому, имеет одинаковые с
ней функции. Желудочная железа
гомологична простым боковым
выпячиваниям средней кишки, лежащим в
головогруди других паукообразных.
Тонкая кишка на всем
протяжении почти одинакового диаметра
(рис. 141, 5; 143, 4) имеет сходную
гистологическую структуру. Передний
ее отдел, лежащий в переднебрюшии,
покрыт печенью. Последняя
представляет собой объемистую
пищеварительную железу, занимающую почти
Рис. 142. Buthus eupeus.
Пищеварительная система и сердце (ориг.):
/ — желудочная железа, 2 — отверстия
для прохода третьей пары дорзовентраль-
ных мышц, 3 —диафрагма; 4 — отверстия
дорзовентральных мышц, 5 — печень, 6—
задний конец печени, 7 — тонкая кишка,
8 — задняя аорта, 9 — девятая пара
боковых артерий, 10 — остие, 11 — боковая
артерия, J2 —сердце, 13^ перикардий, 14 —
верхний экзокардиальный лигамент, 15 —
эпикардиальные лигаменты, 16 — боковая
артерия первой пары (VII сегмента), 17 —
передняя аорта
6—2
161
всю полость переднебрюшия, и состоит из многочисленных
мелких долек, спаянных соединительной тканью в компактный орган
(рис. 142,5; 143,//). Снаружи печень одета тонкой
соединительнотканной оболочкой, образующей очень нежные боковые отростки,
посредством которых железа прикрепляется к стенкам тела. С каждой стороны
печени имеется пять таких брыжеечных тяжей. Задний конец печени
проникает в полость первого, а у некоторых экземпляров даже второго
сегмента заднебрюшия (см. рис. 142,6). Печень имеет дорзальный
медиальный желоб, в котором лежит перикардий с сердцем, и по бокам от
перикардия — шесть пар метамерных вертикальных каналов,
пронизывающих всю толщу железы, В этих каналах проходят дорзовентральные
мускулы (см. рис. 142,4).
У всех Buthidae печень со спинной стороны цельная и не делится
на метамерные парные лопасти, как это имеет место у других
скорпионов. Однако нижняя поверхность ее подразделяется на многочисленные
лопасти. При вскрытии свежего материала удается изолировать печень
целиком вместе с кишкой (см. стр. 155). Но при анатомировании
спиртовых экземпляров нижнюю поверхность печени можно рассмотреть,
только вскрывая животное с брюшной стороны, после удаления стенки
переднебрюшия с мускулатурой (рис. 144). Итак, печень состоит из
дорзального нерасчлененного отдела и отходящих'от его брюшной
поверхности многочисленных лопастей. Последние достигают до
брюшной стенки тела. У- Buthus eupeus число их постоянно. Они
располагаются в четыре продольных ряда: два медиальных (рис. 144,/) и два
латеральных (рис. 144,2). Между ними проходят 6 пар дорзовентраль-
ных мускулов (рис. 144,5). Имеется 11 пар медиальных лопастей, из.
них впереди передних мускулов лежит одна пара, остальные метамерно
располагаются по две пары между каждыми двумя соседними парами
мускулов. В латеральных рядах имеется по 8 лопастей. Лопасти
обычно не достигают задней границы последнего сегмента переднебрюшия,
и благодаря этому здесь видны тонкая кишка (рис. 144,4) и брюшная
нервная цепочка (рис. 144,5). Заходящий в сегмент заднебрюшия
участок печени представляет собой задний конец спинного отдела
этого органа (см. рис. 142, 6).
Между лопастями печени часто бывают видны участки сетеобраз-
ной половой железы (рис. 144,7). Нередко у самцов по бокам от
печени заметны дополнительные органы полового аппарата (рис. 144,5).
У самок развитие печени зависит от физиологического состояния
половой системы. Так, у беременных самок, половые органы которых
сильно раздуваются, вся печень, в том числе и лопасти ее, значительно,
уменьшаются в объеме (см. рис. 160,4).
Печень скорпионов гомологична печени других паукообразных,
которые вообще характеризуются сильным развитием этого органа. Во
время эмбрионального развития печень закладывается в виде
метамерных первичных выпячиваний средней кишки (по паре на каждый
сегмент), но позднее они сливаются на спинной стороне, а на брюшной —
подразделяются на вторичные лопасти.
Для изучения протоков печени следует осторожно удалить
вентральные и латеральные лопасти и лежащие между ними брюшную нерв-
162
ж
Рис. 143. Buthus occitanus.
Пищеварительная система (по Blanchard):
/ — рострум, 2 — пищевод, 3 — желудок,
4 — тонкая кишка, 5 — проток печени, 6 —
мальпигиев сосуд, 7 — тельсон, 8 —
диафрагма, 9 — коксальная железа, 10 —
желудочная железа, И — печень
Рис. 144. Buthus eupeus.
Печень самца с брюшной
стороны (ориг.):
/ — медиальные лопасти печени,
2 — латеральные лопасти печени,
3 — ганглнй брюшной нервной
цепочки, 4 — тонкая кишка, 5 —
брюшной нервный ствол, 6 — дорзальный
отдел печени, 7 — семенник, 8 —
отверстия дорзовентральных
мускулов, 9 — параксиальный орган
ную цепочку и половые органы. Печень открывается маленькими
кусочками до тех пор, пока не обнаружится медиальная тонкая кишка и
печеночные протоки с их разветвлениями. Печень сообщается с тонкой
кишкой посредством пяти пар протоков, располагающихся в пяти
передних сегментах переднебрюшия (см. рис. 141,6; 143,5). Протоки мно:
гократно ветвятся; их многочисленные, тончайшие разветвления
связаны с дольками железы (см. рис. 143). Передняя пара протоков
меньше других (см. рис. 141). Протоки задней пары особенно сильно
6*
163
развиты. Они направляются назад и
принадлежат всей задней половине печени (см. рис. 143,
5).
Физиологическая роль печени велика, так
как в ее полостях происходит переваривание
пищи, а в клетках печеночного эпителия —
внутриклеточное пищеварение. Особые клетки печени
секретируют пищеварительные ферменты. Кроме
того, в ней происходит резорбция и отложение
Рис 145. Euscorpius car- запасных питательных веществ. Наконец, печень
pathicus. Легкое после несет также отчасти и экскреторную функцию,
вываривания в едком ка- В последнем сегменте переднебрюшия в тон-
ли. Вид со спинной сто- кую кишку открываются две пары тонких маль-
роны (из Kastner): пигиевых сосудов (см. рис. 141, 7, 8; 143, 6).
1 — легочные пластинки, 2 — у, ^ J ч г ' '
преддверие,- з-стернит В заднебрюшии тонкая кишка несколько
расширяется (см. рис. 143, 4). В последнем сегменте
-заднебрюшия она переходит в заднюю кишку, от
которой отделяется слабой кольцевой бороздкой. Задняя, или прямая,
кишка коротка (см. рис. 140, //). Полость ее выстлана хитиновой
кутикулой. К концу ее прикрепляется пара косых мускулов — леваторы
ануса (см. рис. 140, 8), при сокращении которых вся область
заднепроходного отверстия с анальными лопастями втягивается внутрь.
Органы дыхания. Четыре пары легких располагаются на
брюшной стороне X—XIII сегментов. Их щелевидные наружные
отверстия, стигмы, или дыхальца (stigmata, см. рис. 130, Б, 24; 139,5), лежат
по бокам на соответствующих стернитах. Каждая стигма открывается
в заднюю часть довольно объемистого легочного мешка. Таким образом
легочные мешки лежат целиком впереди стигм (см. рис. 139,10).
От передней стенки легочного мешка отходит большое .число
(около 7ТГ—80) широких, очень тонких и нежных легочных пластинок
(рис. 145,/). Они представляют собой полые внутри, сильно
уплощенные складки легочной стенки.-В их полость заходит кровь из венозных
синусов, тогда как воздух проникает сзади между ними. Наличие их
увеличивает общую дыхательную поверхность легкого. Нежность и
тонкость их стенок обеспечивает быстрое окисление крови.
Легочные пластинки разделяются узкими щелевидными
пространствами (рис. 146, А, 4), располагаясь параллельно друг другу,
наподобие листов книги, почему английские авторы нередко называют легкие
паукообразных «booklungs». Легочные пластинки заполняют почти все
легкое, оставляя свободной лишь самую заднюю его часть, называемую
преддверием легкого (см. рис. 145,2; 146,6). Расположение их не
вполне горизонтально; они пересекают полость легкого в косом
направлении, так как их наружные края лежат несколько ниже краев,
обращенных внутрь. Пластинки связаны со стенками легкого не только спереди»
но и по бокам, и только задние их края остаются свободными.
Поэтому если смотреть на вскрытый легочный мешок сзади, то пластинки,
как таковые, не видны, но бросаются в глаза многочисленные,
параллельные друг другу, узкие, щелевидные отверстия, ведущие в
пространства между пластинками (рис. 146, А, 4). Узкие перегородки между со-
164
седними щелевидными
отверстиями представляют
собой задние свободные края
легочных пластинок
(рис. 146, Л, 5).
Стенки легкого, которое
является глубоким впячива-
нием кожных покровов, а
также поверхности легочных
пластинок, выстланы тонкой
хитиновой кутикулой.
Изучить детали строения
легкого можно, предварительно
выварив его в 10%-ном
растворе едкого кали. При
этом ткани, находящиеся
внутри каждой легочной
пластинки и связывающие
оба ее хитиновых листка,
разрушаются. Поэтому
последние легко расходятся при
вдвигании между ними кончика препаровальной иглы со стороны
передней стенки легкого. Каждое щелевидное, легко расширяемое иглой,
пространство между двумя соседними листками представляет собой,
таким образом, внутреннюю полость одной легочной пластинки
(рис. 146, 8).
Легочные пластинки скорпионов снабжены особыми утолщениями
хитина (хитиновыми столбиками, гребешками и т. д.), которые
препятствуют слипанию соседних пластинок, обеспечивая наличие
постоянного свободного пространства между ними. У Buthus eupeus пластинки
легких покрыты тончайшими хитиновыми гребешками, образующими
на их поверхности очень нежную сеточку. Такую же роль играют и
мельчайшие хитиновые шипики на свободных краях пластинок. Судя по
тому, что на них могут оседать мелкие посторонние частицы,
попадающие в легкие с воздухом, эти шипики служат также фильтром,
предохраняющим легочные пластинки от загрязнения. Все эти хитиновые
образования легко наблюдать под микроскопом на изолированной
пластинке.
Легочные пластинки скорпионов очень похожи на жаберные
лепестки Limulus, а сами легкие гомологичны жаброносным конечностям
водных Chelicerata, видоизменение которых они и представляют
(MacLeod, 1884, Kingsley, 1885, 1893, и др.).
Обновление воздуха внутри легкого осуществляется посредством
вентиляции и диффузии. К задней стенке преддверия легкого
прикрепляется небольшой мускул (расширитель преддверия — гомолог
внутреннего бр.анхиального мускула Limulus). При его сокращении стигма
открывается, а полость преддверия расширяется. При расслаблении
мускула стенка преддверия возвращается в прежнее положение, а
стигма закрывается. Таким образом, полость преддверия хорошо' вентили-
Рис. 146. Opisthophthalmus. Схема строения
легкого. А — средняя часть; Б — боковой
участок легкого (по Kastner):
1 — наружные покровы, 2 — внутренняя поверхность
стенки легочной пластинки, 3 —
соединительнотканный столбик, 4 — щелевидный вход в пространство
между двумя легочными пластинками, 5 — задний
свободный край легочной пластинки, 6 ~ преддве- /
рие, 7 — стигма, 8 — внутренняя полость легочной
пластинки
165
руется. В узких пространствах между легочными пластинками воздух
обновляется посредством диффузии. Благодаря пульсации крови в
синусе, омывающем легкое, ритмически сужаются и расширяются лаку-
нарные пространства легочных пластинок, и сами пластинки при этом
совершают слабые колебательные движения, что, конечно, способствует
обновлению воздуха между ними.
Кровеносная система скорпионов, сравнительно с таковой
у других паукообразных, развита очень сильно, что связано с наличием
локализированных органов дыхания.
Сердце может быть изучено путем вскрытий живых или даже
консервированных скорпионов, но для препарирования кровеносных
сосудов требуется инъекция.
Животное убивается в парах хлороформа и предварительно
вскрывается в физиологическом растворе. Осторожно удаляются тергиты пе-
реднебрюшия начиная с шестого по второй включительно. При этом
дорзовентральные мышцы и лигаменты сердца подрезаются острым
скальпелем. Инъекция производится обыкновенным шприцем с очень
тонкой иглой. В качестве инъекционной краски можно взять
разведенную берлинскую лазурь, растертую тушь или карминовую глицерин-
желатиновую массу.
Артериальную систему следует инъецировать через сердце, втыкая
иглу шприца в его стенку наискось, острием вперед. При этом, однако,
сосуды заднебрюшия наливаются плохо, и для их выявления следует
сделать инъекцию на другом экземпляре в задний конец сердца,
направляя острие иглы назад.
Венозная система инъецируется через брюшные венозные синусы,
что гораздо труднее, так как приходится, не вскрывая скорпиона,
вводить иглу наугад между стернитами переднебрюшия несколько кнутри
от легочных отверстий. Эта операция удается не сразу, и для нее
обычно требуется несколькб скорпионов.
Инъецированное животное фиксируется затем в течение
нескольких часов 95%-ным спиртом и перекладывается на сутки в 75%-ный
спирт, после чего его молено анатомировать. Однако прослеживание
сосудов облегчается, если после фиксации, обезвоженное в абсолютном
спирте животное просветляется в гвоздичном масле. Препарирование
производится под бинокуляром. Все органы и ткани, мешающие
изучению сосудов, осторожно удаляются с помощью игл и остроконечного
пинцета.
Сердце сильно вытянутое, трубковидное тянется на спинной
стороне почти через все переднебрюшие — от переднего края первого терги-
та до середины седьмого (рис. 142, 12; 147, 5; 148, 24). Оно заключено в
несколько более широкий мешковидный, тонкостенный перикардий
(см. рис. 142,13), вместе с которым лежит в медиальном желобке
печени. На спинной стороне сердца заметны семь пар метамерно
расположенных остий в биде косых, щелевидных, маленьких отверстий,
сообщающих полости перикардия и сердца (см. рис. \42,10; 151,75).
Первые пять пар остий лежат на уровне задних краев соответствующих
тергитов, шестая пара — несколько впереди заднего края, а седьмая —
ближе к переднему краю своего тергита. Смещение задних пар остий
166
вперед — явление вторичное (они
закладываются интерсегментарно)
и связано с укорочением сердца на
заднем конце. Наличие остий
придает сердцу вид семикамерного
органа, но в действительности оно не
разделено ни перетяжками, ни
внутренними перегородками и полость
его сплошная '.
Через остий кровь может
проходить только в одном
направлении, а именно из перикардия в
сердце. Это обусловлено наличием
клапанов. Края каждого отверстия
заворачиваются внутрь, образуя
уплощенную вороночку, которая и
играет роль клапана. Рассмотреть
клапан можно не только на срезах,
но и в бинокуляр, вырезав
соответствующий участок сердечной
стенки.
Сердце подвешивается
посредством восьми метамерно
расположенных групп парных лигаментов,
т. е. соединительнотканных
эластичных тяжей. Каждая группа, за
исключением передней, состоит из
четырех пар лигаментов,
расположенных на уровне остий. Самые
короткие из них — эпикардиальные
лигаменты (ligamenta epicardialia, см.
рис. 142, 15), которые отходят на
спинной стороне сердца. Вторая,
более длинная пара начинается
кнаружи от первой на краях остий и
тянется в дорзолатеральном
направлении. Это верхние наружные
лигаменты (1. exocardialia superio-
res, см. рис. 142, 14). Третья пара
представлена горизонтальными
нижними наружными лигаментами (1.
exocardialia inferiores), которые часто неправильно принимаются за
крыловидные мышцы сердца.
Четвертая пара называется гипокардиальными лигаментами (1. hy-
pocardialia). Они начинаются от брюшной стенки сердца, тянутся,
несколько расходясь в брюшном направлении, и сопровождают перикар-
Рис. 147. Buthus occitanus. Сердце и
артериальные сосуды (по Blanchard):
/ — артерии хелицеры, 2 — артерия педи-
пальпы, 3 — артерия ноги, 4 — передняя
аорта, 5 — сердце, 6 — печень, 7 — экзо-
кардиальный лигамент, 8 — боковая
артерия 9-й , пары, 10 — тонкая кишка, 11 —
боковая артерия, 12 — остие, 13 — толстая
артерия
1 В ряде учебников и руководств ошибочно указывается, что сердце скорпионов
состоит' из. восьми камер, из которых каждая несет пару остий.
167
Рис. 148. Схема организации скорпиона (по Newport из Kastner):
1 — хелицеры, 2 — артерия педипальпы, 3 — нерв педипальпы, 4 — артерия ноги, 5 — под-
глоточная нервная масса, 6 — проток желудочной железы, 7— основание ноги, 8— стигма,
9 — ганглий XII сегмента, 10 — проток печени, И — спинальная артерия, 12 — брюшной
нервный ствол, 13 — легкое, 14 — тонкая кишка, 15 — боковая артерия XV сегмента, 16 —
передняя узелковая ветвь, П— подузелковый сосуд, 18—задняя узелковая ветвь, 19
—-'межузелковый сосуд, 20 — анальное отверстие, 21 — ядовитая железа, 22 — задняя аорта, 23—
боковая артерия, 24 — сердце, 25 — остие, 26 — передний клапан, 27 — передняя аорта, 28 — диа-
фрагмальная артерия, 29 — медиальный глаз, 30 — головной мозг, 31 — глазная артерия, 32 —
зрительный нерв, 33 — боковые глаза, 34 — хелицеральная артерия, 35 — хелицеральный нерв
дио-вентральные мускулы (рис. 149,2). Дистальные концы всех лига-
ментов связаны с соединительной тканью тела или прикрепляются к
базальной мембране гиподермы. Первая группа лигаментов
прикрепляется к переднему концу сердца впереди первой пары остий и состоит
только из эпи- и гипокардиальных лигаментов. Она принадлежит
рудиментарному прегенитальному (VII) сегменту.
Лигаменты пересекают перикардий и делят его полость на области,
которые называются перикардиальными синусами, хотя и сообщаются
широко друг с другом в тех местах, где лигаменты отсутствуют.
Функциональное значение лигаментов отнюдь не ограничивается их опорной
ролью, они имеют также важное значение в работе сердца. Систола
сердца осуществляется сокращением кольцевых мускульных волокон,
залегающих в его стенках. Сокращаясь, сердце преодолевает
сопротивление лигаментов и растягивает их. Когда мышцы сердца-
расслабляются, то эластичные лигаменты возвращают стенки сердца в прежнее
положение, т. е. наступает диастола. Осторожно удалив у живого
скорпиона несколько тергитов, нетрудно наблюдать сокращения сердца. Оно
сокращается целиком, как одно целое. Число сокращений у Buthus
около 60 в минуту.
Метамерность сердца подчеркивается еще сегментарным
расположением боковых сосудов, связанных с ним и с перикардием. Под каж-
168
дой парой остий от брюшной поверхности
сердца берет начало пара боковых артерий,
которые, как и у других паукообразных
(см. 216), проходят в вентральном
направлении между лопастями печени. Но позади
диафрагмы имеется еще одна пара боковых
артерий (см. рис. 142, 16; 151, 17), а от заднего
конца сердца отходят 'парные сосуды,
впадающие на брюшной стороне в спинальную
артерию (см. рис. 142, 9; 148, 15; 151, 12). Они
представляют собой самую заднюю
видоизмененную пару боковых артерий. Итак, всего
имеется девять пар боковых артерий, причем
передняя пара, очевидно, принадлежит
прегенитальному (VII) сегменту, а задняя,
девятая,— первому сегменту заднебрюшня (XV
туловищному), хотя и лежит в предыдущем
XIV сегменте. Кроме того, непосредственно
против остий в перикардий открываются семь
пар легочных вен (табл. 1).
Наличие лишних, по сравнению, с остия-
ми, передних лигаментов и артерий, а также
задних артерий указывает на то, что у
предков скорпионов сердце было гораздо длиннее
и распространялось далее как вперед, так и
назад, захватывая большее'число сегментов.
Лигаменты и артерии VII, редуцированного
сегмента, по-видимому, представляют собой
все, что от него осталось у скорпионов.
Артериальная система. Спереди сердце
продолжается в переднюю аорту, сзади от
него берет начало задняя аорта. Все сосуды,
отходящие от сердца, в том числе метамерные боковые артерии, имеют
у своего основания клапаны, пропускающие кровь только из сердца;
Если осторожно вскрыть основание аорт, то можно рассмотреть в бино-
куляр их клапаны. Клапаны боковых артерий видны только на срезах.
Передний клапан лежит в самом начале передней аорты,, в том
месте, где она проходит через, диафрагму. Он представляет собой косой
складкообразный вырост, направленный вперед и вниз. Спинной и
боковые края его прикреплены к стенке сосуда, а брюшной край свободен
(см. рис. 148, .26). При расширении сердца свободный край клапана
прижимается к вентральной стенке аорты, закрывая вход в сердце.
Клапан задней аорты лежит в ее основании и устроен иначе. Он имеет вид
воронки, обращенной узким концом назад и приросший своим
широким концом к стенке сосуда.
Передняя аорта (aorta anterior) проходит сквозь диафрагму и
тянется вперед и вниз под пищеводом до головного мозга (см. рис. 148,
27). Близ своего начала она ответвляет пару маленьких артерий,
снабжающих кровью желудочную железу. Возле задней поверхности голов-
Рис. 149. Androctonus.
Сердце с брюшной стороны
(по R. Lankester):
1 — передняя аорта, 2 — гипо-
кардиальный лигамент, 3 —
боковая артерия, 4 — дорзовеит-
ральный мускул, 5 — дорзовент-
ральный мускул XIV сегмента,
6 — боковая артерия последней
пары, 7 — основание задней
аорты, 8 — перикардий с
сердцем
169
Таблица 1. Сегментарная принадлежность всех образований, связанных с сердцем
Головогрудь
V VI
Переднебрюшие
VII 1 VIII 1 IX
LA
OLAV OLAV
X 1 XI 1 XII XIII
OLAV
OLAV
OLAV
OLAV
XIV
OLAV
Заднебрю-
шие
XV
A
XVI
О—остии, L — лигаменты, А—боковые артерии, V — легочные вены.
ного мозга аорта дает последовательно несколько сосудов. Сначала
ответвляется непарная верхняя мозговая артерия (a. cerebralis superior),
которая несет кровь в головной мозг. Несколько впереди начинаются
мощные парные головные артерии (a. cephalicae), огибающие мозг с
боков, и непарная нижняя мозговая артерия (a. cerebralis
inferior).Последняя проходит над пищеводом под головной мозг и входит в него
снизу. Под корнем нижней мозговой артерии аорта делится на две
мощные, но короткие толстые артерии (arteriae crassae, см. рис. 147,73),
охватывающие пищевод с боков и лежащие на верхней стороне подгло-
точной нервной массы. Каждая головная артерия (a. cephalica) впереди
мозга образует две ветви. Одна из них проходит в хелицеры рядом с
хелицеральными нервами и дает ответвления к мускулатуре
головогруди (см. рис. 147,/; 148,34). Другая — несет кровь к глазам и назад к
диафрагме (рис. 149,3/ и 28). Впереди мозга обе задние ветви
головной артерии соединяются сосудистой коммиссурой (ramus supercerebra-
lis).
Толстые артерии дают начало пяти парам крупных артерий
конечностей (a. pedipalpales, рис. 147,2; 148,2; 150,1 и a. pedales, 147,3;
148,4; 150,2), которые тянутся рядом с соответствующими нервами и
снабжают кровью мускулатуру. Кроме
того, толстые артерии связаны друг с
другом короткими поперечными
сосудистыми мостиками (rami transversales,
рис. 150, 6). Последние снабжают
кровью подглоточную нервную массу, к
которой прилегают сверху. Распределение
их ветвей внутри нервной массы может
быть изучено при препарировании
инъецированного и просветленного
материала и представляет большой интерес с
точки зрения метамерии нервной и
кровеносной систем. Однако оно становится
понятным после ознакомления с более
примитивными отношениями между
артериями и ганглиями брюшной нервной
цепочки.
От заднего, лежащего на
подглоточной нервной массе, поперечного
сосудистого мостика начинается большой меди- -
Рис. 150. Scorpiones. Артерии,
отходящие от arteriae crassae, вид
со спинной стороны (из Kastner):
1 — педипальпальная артерия, 2 —
педальные артерии, 3 — толстая артерия,
4 — спинальная артерия, 5 —
межузелковые артерии, отходящие от спиналь-
ной артерии, 6 — поперечные артерии,
7 — межузелковые артерии, отходящие
от поперечных артерий
170
альный сосуд — спинальная артерия (a. spinalis, рис. 150, 4; 148, 11),
которая проходит над брюшной нервной цепочкой до последнего
брюшного ганглия, где разделяется на две ветви, сопровождающие нервы
этого ганглия. Позади диафрагмы она ответвляет многочисленные
парные сосуды к различным органам брюшка.'Но особенно интересны
короткие непарные сосуды, которые отходят от спинальной артерии к
свободным брюшным ганглиям. У каждого ганглия эта артерия дает
две непарные ветви:
1. Переднюю узелковую ветвь (ramus ganglionaris anterior, см.
рис. 148,76), которая идет вертикально вниз по передней стороне
нервного узла между коннективами.
2. Заднюю узелковую ветвь (г. g.. posterior, см. рис. 148,78),
проходящую вниз по задней поверхности ганглия. На нижней стороне
ганглия обе узелковые ветви соединяются медиальным продольным подузел-
ковым сосудом (ramus subganglionaris, см. рис. 148, 17).
Таким же образом снабжаются сосудами все свободные ганглий,
кроме седьмого, заднего, состоящего из слившихся нервных узлов XVIII
и XIX сегментов. Этот ганглий имеет не только передний и задний
узелковые сосуды, но, кроме того, прободен коротким вертикальным
межузелковым сосудом (г. interganglionaris, см. рис. 148,19), проходящим
в соединительнотканной прослойке между обоими составляющими его
невромерами. Межузелковый сосуд представляет собой продукт
слияния задней узелковой ветви, принадлежащей ганглию XVIII сегмента
с передней узелковой ветвью ганглия XIX сегмента.
Совершенно сходно, вследствие сближения и слияния четырех
передних ганглиев брюшка, образовались вертикальные межузелковые
сосуды в задней части подглоточной нервной массы (рис. 150,5)..
И здесь межузелковые сосуды отходят от нижней поверхности
спинальной артерии и разделяют невромеры VIII—XI сегментов.
Каждый межузелковый сосуд на нижней стороне подглоточной
массы дает подузелковую ветвь, как. и на свободных брюшных
ганглиях. Подузелковые ветви отдельных невромеров соединяются друг с
другом в продольный медиальный сосуд, проходящий по
нижней.стороне нервной массы.
Подобным же образом снабжается сосудами и передний отдел
подглоточной нервной массы, состоящий из слившихся ганглиев педипалыт
и четырех пар ходных ног. Но здесь межузелковые сосуды
начинаются не от спинальной артерии, а от поперечных сосудистых мостиков
(рис. 151,7).
Все задние узелковые и межузелковые сосуды дают по небольшой
непарной стернальной ветви (г. sternalis), которая направляется вниз
к стенке тела.
Сосудистая система подглоточной нервной массы усложняется еще
наличием коксальных ветвей (г. coxales), расположенных на ее
нижней стороне и соединяющих стернальные ветви с артериями
конечностей.
Задняя аорта (a. posterior, см. рис. 142,8; 147,9; 148,22)
представляет собой большой сосуд, проходящий над кишечником от заднего
конца сердца через все заднебрюшие. По пути она дает парные ветви к
171
Рис. 151. Scorpiones. Главнейшие вены переднебрю-
шия. Вид с левой стороны (по Kastner):
/ — передняя аорта, 2 — тонкая кишка, 3 — диафрагма, 4 —
проток печени, 5 — легочные вены первого легкого, 6 —
легкое, 7 — стигма, 8 — циркумпульмональный синус, 9 —
вентральный синус, 10— легочные вены четвертого легкого, // —
брюшная нервная цепочка, 12 — девятая боковая артерия,
13 — задняя аорта, 14 — боковая артерия, 15 — остие, 16 —
перикардий с сердцем, П — первая боковая артерия
стенкам кишки и мускулатуре (см. рис. 147). Ее конец проходит в тель-
сон, где она разветвляется на поверхности ядовитых желез.
Из девяти пар боковых артерий (a. laterales, см. рис. 142, //, 16;
148,23) передние восемь погружаются в печень, где дают
многочисленные разветвления, пронизывающие весь орган. Артерии девятой пары
(рис. 148,75; 151,12), несомненно, также являются видоизмененными
боковыми артериями, ибо имеют у основания собственные клапаны.
Они загибаются по бокам кишечника вниз и открываются в спинальную
артерию1.
В таком длинном, проходящем почти через все тело сосуде, как
спинальная артерия, нагнетающее действие сердца должно в конце
концов затухать. По-видимому, именно в связи с этим у скорпионов
установилось вторичное соединение этого сосуда с сердцем посредством
девятой пары боковых артерий, благодаря чему в спинальной артерии
создается дополнительное давление, достаточное для проталкивания
крови до конца.
Несколько расширенные основания боковых артерий, снабженные
клапанами, гомологичны кардио-целомическим отверстиям,
свойственным некоторым другим паукообразным (Pedipalpi).
Венозная система. У скорпионов, как и у других Arthropoda,
кровеносная система незамкнутая.
Артерии/ сильно ветвятся, образуя густую сеть тонких сосудов
(см. рис. 147). Эти последние открываются в лакуны и синусы, часть
которых обладает собственными соединительнотканными стенками и
заслуживает название вен. В головогруди и переднебрюшии из лакун
венозная кровь собирается в пару продольных вентральных синусов (si-
1 Некоторые авторы неправильно считают последнюю пару боковых артерий ла^
теральными ветвями задней аорты.
172
nus ventrales, рис. 151,9). В сегментах переднебрюшия, имеющих
легкие, синусы расширяются, образуя легочные, или циркумпульмональ-
ные, синусы (s. circumpulmonales, рис. 151,8), которые омывают
легочные мешки. Заходя из них в лакунарные пространства легочных
пластинок, кровь окисляется. Из легочных синусов окисленная кровь
поступает в перикардий по семи парам легочных вен (v. pulmonales, см. рис.
151, 5, 10). Эти вены проходят поверхностно в каждом сегменте
переднебрюшия между продольной мускулатурой и покровами, соединяя
синусы легких с полостью перикардия. Так как количество легочных вен
и легких не совпадает, то с передней парой легких связаны три пары
легочных вен (в VIII, IX и X сегментах — рис. 151, 5), а с задней — две
лары (вены XIII и XIV сегментов — рис. 151, 10). Вторая и третья пары
легких имеют по одной паре вен (рис. 151).
При каждом сокращении сердца давление в перикардиальной
полости падает, что вызывает приток крови через легочные вены.
Существенную роль в кровообращении играют также перикардио-вентраль-
ные мускулы (m. pericardio-ventrales), которые в количестве семи пар
прикрепляются своими дорзальными концами к брюшной стенке
перикардия. Первый сегмент переднебрюшия (VIII туловищный) лишен
этих мышц, последний (XIV) — содержит две пары, причем задняя
принадлежит первому сегменту заднебрюшия (XV туловищному). Они
проходят в брюшном направлении несколько впереди дорзовентральных
мускулов, их брюшные концы связаны с соединительной тканью стенки
вентральных венозных синусов.
В сегментах, в которых имеются легкие, они прикрепляются к
стенкам легочных синусов. Строение перикардио-вентральных мускулов
своеобразно, так как в верхних и нижних своих отделах они полые, но
средняя их часть лишена полости, поэтому они не могут проводить кровь
из брюшных синусов в перикардий, как может показаться на первый
взгляд. По-видимому, они сокращаются согласованно с сердцем и в том
же ритме. При этом они поднимают верхнюю стенку вентральных и
легочных синусов и насасывают в них кровь.
Вероятно, и движение стенок тела (тергитов и стернитов) также
способствует циркуляции крови.
Выделительная система. Органы выделения представлены
коксальными железами, мальпигиевыми сосудами и лимфатическими
железами.
Коксальные железы в числе одной пары лежат в головогруди под
боковыми лопастями желудочной железы, у основания третьей и
четвертой пары ног (см. рис. 153,9). Их задняя поверхность прилегает к
диафрагме. Каждая железа состоит из концевого целомического мешочка
(sacculus, рис. 152,/) и очень длинного мезодермального
экскреторного канала, многочисленные петли которого закрывают мешочек почти
со всех сторон. Только в передней части железы, где К'ней подходит
коксальная артерия, ответвляющаяся от артерии третьей пары ног
(рис. 152, 3), мешочек не закрыт петлями канала. Самый канал состоит
из нескольких частей различного строения. От мешочка отходит
короткий и неясно выраженный собирательный канал (рис. 152,7); он
переходит в наиболее длинный участок — так называемый лабиринт
17.3
(рис. 152, 5). Затем следует небольшое
расширение—■ мочевой пузырек (рис. 152, 2).
Наконец весь орган заканчивается коротким
выводным каналом, который выстлан
хитиновой кутикулой и является
производным эктодермы (рис. 152, 4). Наружные
отверстия коксальных желез лежат в
глубине бороздок, между тазиками третьей
пары ходных ног и стернумом. Они очень
малы, и у Buthus eupeus без приготовления
^аГой жеГеГТпо Buxton срезов обнаружить их не удается. По 'мне-
из Kastner): нию многих авторов, они имеются только у
/ — мешочек, 2- мочевой пузырек, ЭмбрИОНОВ И МОЛОДЫХ ЖИВОТНЫХ.
но! к0акнаалл,ь5-Жй.*в-3-" Коксалыше железы у Chelicerata - (
рагма, 7 — собирательный канал. ЭТО ВИДОИЗМеНеННЫе ЦеЛОМОДуКТЫ, И у СКОр-
v-vi сегменты пионов принадлежат V сегменту. Во время
развития зародыша целомодукты
закладываются в III, IV, V и VI, а также VIII сегментах, но в коксальные
железы превращается только одна пара, лежащая в V сегменте.
Остальные редуцируются, за исключением целомодуктов VIII (генитального)
сегмента, из которых формируются половые протоки.
Мальпигиевы сосуды хорошо видны при вскрытии, но
отпрепарировать их целиком, особенно на свежем материале, очень трудно. Их
четыре, и открываются они в кишку в VII сегменте переднебрюшия. Обе
пары представляют собой очень тонкие трубочки, которые благодаря
белому цвету хорошо выделяются на фоне темной печени. Передняя
пара тянется над кишечником вперед к диафрагме и образует боковые
веточки, которые теряются в массе печени (см..рис. 141,7; 143,6).
Задняя пара гораздо короче; от места впадения в кишку они расходятся
в стороны, разветвляются и, по-видимому, анастомозируют с
концевыми вбтвями передней пары (см. рис. 141,5).
Мальпигиевы сосуды скорпионов тесно связаны с печенью, залегая
в соединительнотканной оболочке лопастей этого органа. В печени
происходят интенсивные процессы пищеварения, накопление запасных
питательных веществ и вообще происходит энергичный обмен веществ.
Здесь же происходит усиленная экскреция отчасти посредством
печеночных клеток. Поэтому предполагают, что мальпигиевы сосуды
являются выделительными органами, обслуживающими специально только
печень, тогда как экскреция в других частях тела осуществляется кок-'
сальными и лимфатическими железами. Мальпигиевы сосуды
скорпионов, так же, как и у других Arachnida, имеют энтодермальное
происхождение и не гомологичны эктодермальным мальпигиевым сосудам
насекомых. В связи с этим следует обратить внимание на то, что у
скорпионов они открываются в среднюю, энтодермальную, кишку очень
далеко от начала задней, эктодермальной, кишки.
Лимфатические железы развиты неодинаково у различных
скорпионов. У Buthus eupeus они представлены несколькими лимфатическими
узелками неправильной формы, расположенными над брюшной
нервной цепочкой, и всегда легко обнаруживаются при препарировании
174
нервной системы (рис. 153, 4). Эти органы
представляют собой разросшиеся дорзальные. выпячивания
стенок артерии,' проходящей над нервной цепочкой,
с которой они связаны, и несут кроветворную и
фагоцитарную функции. Свойственные некоторым родам
скорпионов особые лимфоидные железы, связанные с
диафрагмой, у Buthidae отсутствуют.
В экскреторной функции коксальных и
лимфатических желез можно убедиться, произведя опыт с
инъекцией в полость тела соответствующих веществ.
Для изучения роли коксальных желез следует
применить растворы красок индикаторов — индиго-
кармина и отдельно на других экземплярах
скорпионов — аммиачного кармина. Для наблюдения над
лимфатическими железами впрыскивают взвесь
твердых частиц — тушь или порошок кармина, растертых
в воде. Во всех случаях инъекция делается
стерильной пастеровской пипеткой. Живой скорпион
предварительно укрепляется на куске пробки путем
обкалывания его булавками и заостренными дужками
проволоки. Особенно тщательно укрепляется одна из
ходных ног, в которую затем и производится впрыски-
Рис. 153. Buthus
eupeus.
Лимфатические железы (по
Е. Н.
Павловскому) :
/ — диафрагма, 2--—
/-, а ганглии, 3 — нервный
ВЭНИе. ОТЛОМИВ КОНЧИК ПИпеТКИ, Следует Набрать В ствол,
^-лимфатические железы
нее краску и вращательным движением ввести ее под
мягкую сочленовную перепонку между двумя
члениками ноги. Из пипетки краска выдувается ртом. Если она не выходит,
то пипетку следует несколько подать назад и повернуть вокруг оси.
Через несколько минут после извлечения пипетки из ноги скорпион
освобождается и пересаживается в садок. Если инъекция произведена
аккуратно, то кровь из ранки не выступает. Инъецированных скорпионов
следует вскрывать через сутки.
Результаты опыта видны невооруженным глазом. При введении
индигокармина окрашиваются лабиринты коксальных желез, которые,
таким образом, выделяют кислые вещества. Аммиачный кармин и,
следовательно, вещества с щелочной реакцией выделяются только
мешочками коксальных желез'1.
Твердые частицы туши и кармина накапливаются в лимфатических
железах, которые через сутки, после введения этих веществ, становятся'
черными или красными и, следовательно, играют роль фагоцитарных
органов, или почек накопления.
Нервная система. Из всех Arachnida скорпионы
отличаются наименее концентрированной нервной системой.
Центральная нервная система состоит из головного мозга и
длинной брюшной нервной цепочки, доходящей до четвертого сегмента зад-
небрюшия. Передние ганглии нервной цепочки слиты в одну объемис-
1 По некоторым данным, аммиачный кармин выделяется также мальпигиевыми
сосудами и поглощается нефроцитами — крупными клетками, рассеянными
преимущественно на брюшной стенке тела у основания ног и на органах нервной системы.
175
тую подглоточную массу, которая вместе с головным мозгом лежит
приблизительно в середине головогруди.
Головной мозг, лежащий над пищеводом, смещен несколько назад,
так как помещается не впереди, а непосредственно над передним
отделом подглоточной массы (рис. 154, Л и Б,1). На его верхней стороне
имеется неглубокая медиальная бороздка. Окологлоточные коннективы
очень коротки и массивны.
От передней поверхности головного мозга отходят следующие
нервы:
1. Зрительные нервы медиальных глаз (п. optici mediales, рис. 154,
10). Самые толстые нервы мозга; берут начало от его переднего
верхнего края.
2. Нервы боковых глаз (п. optici laterales, рис. 154,5).
Начинаются несколько ниже предыдущих.
3. Непарный нерв рострума (п. rostralis, рис. 154,8).
4. Нервы хелицер (п.. chelicerales,
рис. 154, 7). Пара больших нервов,
отходящих от нижнего отдела мозга, почти
от окологлоточных коннективов. У их
основания начинаются дополнительные
хелицеральные нервы.
С головным мозгом связана также
симпатическая нервная система,
представленная двумя парами тонких нервов,
начинающихся от задней части мозга.
Латерально отходит пара боковых
нервов (п. laterales, рис. 154, Б, 14), они ин-
нервируют желудочную железу и печень,
а также легкие. Вторая пара — интести-
нальные нервы (п. intestinales, рис. 154,
Б, 13) — сопровождает кишечник вплоть
до заднебрюшия.
Подглоточная нервная масса лежит
под пищеводом и над стернумом,
будучи окружена кольцом эндостернита. Она
значительно крупнее и шире головного
мозга и разделяется на широкий,
большой передний и узкий, маленький
задний отделы (рис. 1.54, 2).
От переднего отдела спереди берут
начало мощные нервы педипальп (п. ре-
dipalpales, рис. 154, 6; 155, 20). Затем
следуют четыре пары нервов ходных ног
(п. pedales, рис. 154, 5; 155, 23).
Следовательно, эта часть подглоточной массы
состоит из пяти пар слившихся друг с
другом ганглиев, принадлежащих пяти
сегментам головогруди (со II по VI).
У основания нервов педипальп отходит
Рис. 154. Мозг Euscorpius сагра-
thicus. Вид с правой (А) и
спинной сторон (Б) (по Gottlieb):
/ — головной мозг; 2 — подглоточная
нервная масса, 3 — брюшные
коннективы, 4 — иерв VIII сегмента, 5
—педальные иервы, 6 — педипальпальный
нерв, 7 — хелицеральный иерв, 8 —
ростральный нерв, 9 — нерв боковых
глаз, 10 — нервы медиальных глаз,
//— иерв первого и второго легких,
12—нерв гребенчатого органа, 13—
интестинальный нерв, 14 —
латеральный нерв
176
пара маленьких дополнительных
нервов. Кроме того, следующие
четыре невромера дают начало,
по меньшей мере, четырем
парным дорзальным нервам.
Последние иннервируют мускулатуру
головогруди и переднюю аорту,
а также связаны нервными
анастомозами с непарным-эпикарди-
альным нервом (п. epicardicus), ~
который проходит медиально над
сердцем, содержит ганглиозные
клетки и иннервирует этот орган
(Kastner, 1940)1.
От небольшого заднего
отдела подглоточной массы
начинаются нервы генитального, VIII
сегмента (нервы мускулатуры
диафрагмы, см. рис. 154, Б, 4),
гребенчатых органов (см. рис.
154, 12) и двух передних пар
легких (см. рис. 154, 11), т. е. в
состав этой части нервной массы
входят ганглии четырех передних
сегментов переднебрюшия (с VIII
по XI). Невромер
рудиментарного прегенитального (VII)
сегмента у взрослых скорпионов
отсутствует. Таким образом,
подглоточная нервная масса есть
результат слияния девяти невроме-
ров, ее метамерный состав см. в
табл. 2.
На протяжении -брюшной
нервной цепочки, лежащей в
брюшке, имеются семь ганглиев.
Из них три передние лежат в пе-
реднебрюшии, остальные
помещаются в заднебрюшии (см. рис.
148 и 155). Все элементы
нервной цепочки непарны. Но
первично-парная природа ганглиев и
коннективов, их связывающих, у
скорпионов еще- очень хорошо
видна на протяжении всей цепочки,
Чем ближе к заднему концу, тем
1 По данным других авторов (Police,
от задней поверхности головного мозга.
Рис. 155. Buthus occitanus. Нервная
система in situ (no Blanchard):
/ — хелицера, 2— педипальпа, 3 — ходиые
ноги, 4 — циркумпульмональный синус, 5 — дор-
зовентральный мускул, 6 — тельсон, 7 —
кишка, 8 — ганглий XVIII и XIX сегментов, 9 —
ганглий XVII сегмента, 10 — нервные кокиек-
тивы, // —ганглий XVI сегмента, 12 —
ганглий XV сегмента, 13 — ганглий XIV сегмента,
14 — ганглий XIII сегмента, 15 — гаиглий
XII сегмента, 16 — подглоточная нервная
масса, 17 — головной мозг, 18 — нерв ноги, 19 —
хелицеральный нерв, 20 — педипальпальный
нерв, 21 — медиальные глаза, 22 — рострум,
23 — нерв ногн, 24 — боковой нерв, 25 —
легочный нерв, 26 — иерв мускулатуры
за исключением ее передней части,
яснее эта парность, зависящая от
1902), эпикардиальный нерв берет начало
177
Таблица 2. Метамерный состав подглоточной нервной массы
Невромеры
1
2
3
4
5
6
7
8
9
Сегменты
туловища
II
III
IV
V
VI
VIII
IX
X
XI
Иннервируемые органы
Педипальпы
Ходные ноги I
» » II
» » III
» » IV
Мускулатура диафрагмы
Гребенчатые органы
Легкие 1
» II
просвечивающих волокнистых масс ганглиев и волокнистых пучков кон-
нективов.
Первый свободный ганглий принадлежит V сегменту брюшка и
соответственно иннервирует третью пару легких, но у Buthus eupeus
лежит на границе III и IV брюшных сегментов (рис. 155,15). Следующий
ганглий помещается в передней части VI сегмента брюшка и снабжает
нервами четвертую пару легких (рис. 155,14).. Третий- ганглий лежит в
VII сегменте и обслуживает органы этого сегмента (рис. 155,13)1.
Каждый из четырех следующих ганглиев лежит в передней части
своего сегмента (с I по IV сегмент заднебрюшия — рис. 155,8,9,11).
Последний ганглий образован слиянием ганглиев IV и V сегментов
заднебрюшия и дает пару больших нервов, которые соответственно иннер-
вируют эти два сегмента и тельсон. Этот последний своего ганглия не
имеет.
От каждого ганглия отходят две пары нервов,, но на каждой сторо-
не два нерва сливаются в один большой нерв (рис. 155,24). Каждый
такой нерв двух передних ганглиев дает заднюю веточку к легкому
(рис. 155,25) и более крупную боковую ветвь к мускулатуре и стенке
тела своего сегмента (рис. 155, .26). Сходно разветвляются и нервы двух
задних невромеров подглоточной нервной массы.
Органы чувств у Buthus eupeus представлены осязательными
щетинками, рассеянными на разных частях тела, и в том числе трихо-
ботриями, чувствительными органами гребенчатых придатков,
медиальными и боковыми глазами. Однако тонкое гистологическое строение
всех этих органов может быть изучено только на срезах. (Строение
глаз. см. на с. 184—186.)
Трихоботрии расположены на ладонях и пальцах педипальп и
представляют собой волоски, сидящие в окаймленных, круглых ямках.
Строение их недостаточно известно. Волосок поднимается со дна
чашевидного углубления хитина; у его основания лежат чувствительные
клетки, происходящие за счет гиподермы. По мнению одних авторов,
1 Положение передних трех ганглиев по отношению к сегментам брюшка и
легким варьируе\ у различных скорпионов и, по данным Laurie (1896), неодинаково у
близкородственных родов и даже видов и сходно у форм неродственных. В заднебрю-
шии локализация четырех нервных узлов постоянна у всех скорпионов.
178
трихоботрии имеют
слуховую функцию, другие
считают их осязательными
органами.
Гребенчатые придатки —
характерные органы
скорпионов, отсутствующие у
других паукообразных.
Внешнее строение их
описано на с. 153. Будучи
сильно измененными
конечностями первого постгениталь-
ного сегмента, они
обладают сложной мускулатурой.
Наиболее развиты мускулы,
прикрепляющиеся к их
основному членику и
головогруди и двигающие всем
органом в разных
направлениях. Внутри самого
гребневидного придатка
имеются серии мышц, двигающие
отдельными его частями. На
гребневидных придатках
расположены
чувствительные органы и чувствительные палочки. Последние во множестве
собраны на особых чувствительных полях на зубчиках гребневидных
придатков. Каждая «алочка связана со сложным многоклеточным
чувствительным аппаратом.
Функциональное значение гребешков до сих пор неизвестно. Им
приписывалась функция органов осязания и органов обоняния. Это
мнение, основанное на анализе структуры чувствительных аппаратов
гребешков, наиболее вероятно.
Половая система. Для скорпионов характерно отсутствие
резко выраженного полового диморфизма. В частности, у Buthus eupeus
самцы почти не отличаются от самок. Небольшие различия
выражаются лишь в несколько меньших размерах самцов и касаются
незначительных отличий в форме клешни педипальп и величине гребневидных
придатков.
Мужской половой аппарат состоит из пары семенников и парных
дополнительных органов. Семенники помещаются в переднебрюшии и
достигают задними концами передней границы XIV сегмента. Каждый
семенник состоит из двух продольных семенных трубок, соединенных
четырьмя более короткими поперечными (рис. 156, //). Нередко
семенники более или менее анастомозируют друг с другом (рис. 157).
Короткие семяпроводы (рис. 156,10) передними концами связаны
с параксиальными органами, которые имеют вид вытянутых пластинок
(рис. 156,8). Каждый параксиальный орган загибается на конце
наружу и вперед, образуя тонкий придаток (flagellum, рис. 156,5). Сквозь
179
Рис. 156. Buthus eupeus. Мужская половая
система. Левый семенник удален (по Е. Н.
Павловскому):
/ — половое отверстие, 2 — передняя придаточная
железа, 3 — овальная железа, 4 — расширение
семяпровода, 5 — цилиндрическая железа, 6 — семенной
пузырек, 7—стержень параксиального органа, 8 —
параксиальный орган, 9—flagellum, 10 — семяпровод,
11 — семенник
Рис. 157. Buthus eupeus.
Различные вариации в степени
слияния семенников (по Е. Н.
Павловскому)
стенки параксиальных органов
просвечивают лежащие в них твердые и темные
стержни (рис. 156, 7). Передние концы
параксиальных органов соединяются друг с
другом и открываются в половое отверстие.
Сюда же открывается пара передних
придаточных желез (glandulae accessoriae,
рис. 156, 2). Перед впадением в полость
параксиального органа каждый семяпровод
образует небольшое расширение (рис. 156,
4). В последний открывается
мешковидный семенной пузырек (vesicula seminalis,
рис. 156, 6), величина которого зависит от
наполнения его семенем. Впереди от
расширения семяпровода лежит овальная
железа (glandula ovalis, рис. 156, <3). К
параксиальному органу с внутренней стороны
прилегает довольно большая, вытянутая
цилиндрическая железа (g. cylindracea,
рис. 156, 5).
Функциональное значение всех этих
органов выяснено недостаточно. По одним
данным, стержни параксиальных органов
при спаривании, по-видимому,
высовываются наружу и вводятся в половое отверстие
самки. Каждый стержень имеет -желобо-
видную бороздку, по которой, вероятно,
стекает семенная жидкость. Однако
возможно, что роль совокупительных органов
играют и наружные половые папиллы,
лежащие под половыми крышечками по
бокам от мужского полового отверстия. ■
Женский половой аппарат. Скорпионы
живородящи, и яйца развиваются в
половых органах самки. Женская половая
система имеет различный вид в зависимости
. от ее физиологического состояния. Яичник
у всех скорпионов непарный и занимает то
же 'положение, что и семенник у самцов. У небеременной самки Buthus
eupeus (и других видов этого рода) яичник состоит из трех продольных
трубок, соединяющихся пятью парами поперечных (рис. 158, 5). На
поверхности трубок сидят многочисленные фолликулы различных
размеров (рис. 158, 6). В каждом из них находится один ооцит или яйцо.
Боковые продольные трубки продолжаются вперед в виде коротких
яйцеводов (рис. 158, 4), которые, не доходя до полового отверстия,
образуют по расширению, называемому семяприемником (receptaculum
seminis, рис. 158, 3). Соединяясь друг с другом впереди,
семяприемники образуют короткое влагалище (рис. 158, 2), открывающееся наружу
женским половым отверстием (рис. 158, 1).
Рис. 158. Buthus eupeus.
Женская половая система (по
Е. Н. Павловскому):
/ — половое отверстие, 2 —
влагалище, 3 — семяприемник, 4 — яйцевод,
5 — яичник, 6 — фолликулы
180
Иногда в половом отверстии и
влагалище обнаруживается твердое
органическое вещество темного цвета. Это так
называемый сперматоклейтрум (spermatocleith-
rum), играющий роль пробочки,
замыкающей половое отверстие после спаривания.
По-видимому, он является затвердевшим
выделением параксиальных органов самца.
Развитие эмбрионов происходит в
трубках яичника, куда попадают зрелые яйца,
выскальзывая из яйцевых фолликулов К
Поэтому у беременных самок трубки
яичника приобретают четковидную форму.
Эмбрионы, сильно увеличиваясь в
размерах, растягивают стенки половых трубок
и создают ряд вздутий, разделенных
узкими перехватами. На вздутиях обычно
видны еще мелкие фолликулы с молодыми
ооцитами, которые дадут потомство в
следующий сезон размножения. К концу
развития эмбрионов шаровидные вначале
утолщения яичника становятся овальными,
а трубки яичника значительно удлиняются.
Несмотря на наличие эмбриональных
оболочек, которые окружают зародышей,
сквозь стенки вздутий просвечивают их
темные глаза и сегментированное тело
<ряс. 159, 3).
У самок в конце беременности лопасти-печени, между которыми
лежат половые трубки, сильно уменьшаются в объеме (рис. 160,4).
Самка Buthus eupeus рождает от 14 до 16 детенышей
молочно-белого цвета и некоторое время носит их на спине. Новорожденные
скорпионы имеют в общем такое же строение, как и взрослые, и
постэмбриональное развитие сводится в основном к росту, который
сопровождается линьками, и к окончательному формированию половой системы.
Ядовитый аппарат. Для ознакомления со строением
ядовитого аппарата необходимо вскрыть тельсон, вдоль, а также приготовить
грубые поперечные разрезы бритвой через этот участок тела.
Пара мешковидных ядовитых желез (g. veneniferae, рис. 161,7)
занимает почти всю полость тельсона. Каждая железа имеет овальную
форму и сзади постепенно сужается в длинный выводной канал,
который проходит внутри жала (рис. 161,А,2). Каналы открываются
наружу у самого острия жала отдельными вытянутыми маленькими
отверстиями (рис. 161, А, 4). Выводные каналы выстланы тонкой хитиновой
кутикулой. Как видно на поперечных разрезах, стенки ядовитых желез
складчаты (рис. 161, Б, 1), и каждая железа окружена изнутри и свер-
Рис. 159. Buthus occitanus.
Половая система беременной
самки (по Blanchard):
/ — половое отверстие, 2 — яйцевод,
3 —' эмбрион
1 Исключением являются представители семейства Scorpionidae, у которых
бедные желтком яйца развиваются в фолликулах.
181
ху толстым слоем поперечных мускульных
волокон (рис.. 161, Б, в). При сокращении этих мышц,
секрет желез с силой выбрасывается наружу.
Изучение срезов. Серия поперечных срезов-
должна помочь разобраться в некоторых
анатомических деталях. Особенно ценные дополнения
к вскрытию дают разрезы через головогрудь в
области глотки и эндостернита и разрезы через
переднебрюшие, захватывающие легкие и
легочные вены. Кроме того, следует обратить
внимание на гистологическое строение некоторых
органов, в первую очередь головного мозга, глаз,,
легких и пищеварительных желез.
Головной мозг удобнее изучать на
горизонтальных, фронтальных разрезах, а легкие—■ на
сагиттальных, пересекающих легочные
пластинки.
Головной мозг Chelicerata, в отличие ог
Crustacea и Insecta, состоит только из proto- и
tritocerebrum. Второй отдел мозга — deutocereb-
rum, дающий у других членистоногих нервы к
антеннам I, отсутствует.
Proto- и tritocerebrum внешне не разделены,
и границы между ними можно наметить только'
на срезах. Весь головной мозг, а также
короткие окологлоточные коннективы состоят из
центрального волокнистого вещества и периферической коры
ганглиозных клеток. Однако в волокнистом веществе заметны отдельные
сгущения нервных волокон, которые 'вместе с прилегающими к ним
группами и скоплениями нервных клеток периферической ганглиозной ко-.
ры являются чувствительными и ассоциативными центрами.
В верхней большей части мозга, представляющей собой протоцереб-
рум, имеется несколько таких центров. Расположение их волокнистых
Рис. 160. Buthus occita-
nus. Половая система
беременной самки in situ
(по Blanchard):
/ — половое отверстие, 2 —
яйцевод, 3 —эмбрион, 4 —
лопасти печени
Рис. 161. Ядовитый аппарат Buthus sp. A — тельсон,
вскрытый вдоль; Б — поперечный разрез через
тельсон (А — из Kastner, Б — по Е. Н. Павловскому):
/ — ядовитые железы, 2 — проток ядовитой железы, 3 —
жало, 4 — отверстие протока, 5 — стенка тельсона, 6 —
мускульная мантия
182
/ а
Рис. 162. Схема расположения
чувствительных и ассоциативных
центров в головном мозге
скорпионов. Вид сверху (из Hanst-
rom):
/ — зрительный нерв срединного глаза.
2 — его нервная масса, 3 — зрительный
нерв боковых глазков, 4,5, 6 — его
яервные массы, 7 — globulus,
грибовидное тело, 8 — зрительная коммиссура,
■9—glomerulus, 10— центральное тело,
//.—мостик, 12 — волокнистый пучок
нерва срединного глаза
Рис. 163. Euscorpius carpathicus. Разрез
через боковой глаз (по Scheuring):
/ — хрусталик, 2 — переходные клетки ириса, 3 —
кутикула, 4 — гиподерма, 5 — соединительная
ткаиь, 6 — зрительные клетки, 7 — рабдом, 8 —
базальная мембрана, 9 — преретинальная
мембрана
участков показано на рис. 162, который является схемой нескольких
горизонтальных разрезов через мозг. Разобраться в строении этой
части мозга можно, только тщательно изучая серии срезов и сравнивая
один с другим.
Дорзально и по бокам, впереди лежат зрительные центры. Нервы
медиальных глаз (рис. 162,7) связаны с овальными зрительными
массами, которые слагаются из маленькой шапочки мелких ганглиозных
клеток, сильно красящихся гематоксилином, и густого волокнистого
узелка (рис. 162,2). От последнего тянется пучок волокон (рис. 162, ./2)
назад к третьей паре зрительных центров боковых глаз (рис. 162,6).
Эти центры соединяются длинной дугообразной зрительной коммиссу-
рой (рис. 162,8). Боковые глаза имеют три пары зрительных центров,
нз которых только передняя (рис. 162,4) обладает богатыми
хроматином ганглиозными клетками, тогда как другие (рис. 162,5 и 6) имеют
обкладку из клеток, не отличимых от обыкновенных ганглиозных
клеточных элементов. У дорзального заднего края мозга лежит большая
волокнистая масса в форме полумесяца. Это так называемое
центральное тело (corpus centrale, рис. 162,70). К нему принадлежит мантия
•сильно красящихся нервных клеток. Центральное тело — важнейший
ассоциативный орган мозга, так как связан волокнистыми пучками,
хорошо заметными без специальной обработки препарата, со всеми
центрами мозга, в том числе с тритоцеребрумом и с подглоточной нервной
массой. В средней части мозга лежат волокнистые пучки,
принадлежащие грибовидным телам (corpora pedunculata). Последние состоят из
183
пары шапочек сильно красящихся мелких нервных клеток, которые
образуют середину дорзальной части ганглиозной коры мозга (globulin
рис. 162,7), и упомянутых волокнистых пучков. Грибовидные тела
связаны с третьей парой зрительных центров боковых глаз, центральным,
телом, окологлоточными коннективами и соединяются друг с другом
поперечной волокнистой коммиссурой, называемой мостиком и лежащей,
впереди центрального тела (рис. 162, J7).
Тритоцеребральный отдел мозга образует его самую нижнюю часть
и содержит хелицеральные ганглии и центры симпатической нервной:
системы. Соответственно от тритоцеребрума берут начало спереди
нервы хелицер и ростральный нерв, а сзади —■ латеральные и кишечные-
нервы. Хелицеральные ганглии лежат на границе мозга и ганглиозных.
окологлоточных коннективов и соединяются поперечной волокнистой
коммиссурой, которая проходит под пищеводом, т. е. посторально.
Кроме того, хелицеральные ганглии связаны еще одной преоральной стома-
тогастрической коммиссурой.
Глаза скорпионов очень интересны, так как принадлежат к
переходному типу, соединяя в себе черты строения простых и сложных
глаз Arthropoda. Видимо, они образовались в результате слияния
(интеграции) отдельных омматидиев фасеточных глаз предков (Eurypteroi-
dea). Медиальные глаза стоят ближе к фасеточным, нежели боковые-
Боковые глаза устроены проще. Это бокаловидные глаза, т. е.
развивающиеся как простое впячивание наружного эпителия, почему и
состоят из одного слоя клеток.
Сильно утолщенная над глазом хитиновая кутикула покровов
вдается внутрь глазного бокала, образуя выпуклую линзу, или хрусталик
(рис. 163, 1). Дно глазного бокала представляет собой сетчатку, или.
ретину. Между последней и хрусталиком лежит тонкая преретинальная
мембрана (рис. 163,9). Сетчатка состоит только из вытянутых
зрительных клеток с ядром у основания (рис. 163,6). Но они располагаются
группами по нескольку клеток, образуя многочисленные просто
устроенные ретинулы, не отграниченные друг от друга пигментом. Каждая,
зрительная клетка на стороне, обращенной внутрь ретинулы, несет раб-
домер, или светочувствительную палочку1, а рабдомеры всех клеток
ретинулы сливаются в один общий рабдом (рис. 163,7). Пигмент
находится в самих зрительных клетках. На краях сетчатка постепенно
переходит в гиподерму, клетки которой здесь сильно пигментированы и
образуют вокруг глаза темное кольцо (iris, рис. 163,2). Сетчатка
подостлана базальной мембраной (рис. 163,8). Волокна зрительного нерва
подходят к базальным концам клеток, т. е. глаз является неинвертиро-
ванным. Боковые глаза скорпионов, по-видимому, служат для
рассматривания предметов вблизи, тогда как удаленные предметы
рассматриваются медиальными глазами.
Строение медиальных глаз легче понять, если сначала обратиться
к их эмбриональному развитию. Каждый глаз образуется из плоского*
1 Рабдомеры фасеточных глаз членистоногих представляют собой результат
слияния каймы светочувствительных штифтиков, которыми заканчиваются отдельные невро-
фибриллы зрительных клеток.
184
Рис 164 Scorpiones. Схема развития медиального глаза. А, Б,
ледовательные стадии развития (из П. П. Иванова):
■ зрительный нерв, 3 — сетчатка, 4 — постретннальный слой,
иый слой
/ — хрусталик, 2 -
Г — ПОС-
- корнеоген-
карманообразного впячивания гиподермы (рис. 164, А, Б), которое
затем ложится под кожей, образуя трехслойную складку (рис. 164, £,У ).
Наружный слой складки дает выделяющий хрусталик корнеогенныи
слой, получивший у Arachnoidea название «стекловидного тела»
(рис 164, Л 5). Второй слой дифференцируется в сетчатку (рис. 1Ь4,
Г,3), а третий образует так называемый постретннальный слои
(рис! 164, Г, 4).
Сетчатка состоит из многочисленных ретинул, а каждая ретинула
■слагается из пяти зрительных клеток с их рабдомерами. Пять рабдоме-
ров сливаются вместе, образуя
пятигранный рабдом (рис. 165, 8).
Рассмотреть пятилучевое строение
отдельной ретинулы и рабдома можно,
конечно, только на фронтальных
разрезах (рис. 165, Б). Между
зрительными 'клетками кое-где видны особые
пигментные клетки.
Каждая зрительная клетка имеет
V-образную форму, перегибаясь
пополам, причем перегиб обращен в
сторону стекловидного тела, а концы
клетки — к постретинальному слою.
Обращенное внутрь ретинулы
суженное колено клетки образует рабдомер,
тогда как наружное колено
грушевидно расширено и содержит ядро.
Нервное волокно подходит к основанию
этой расширенной части клетки^
Таким образом, светочувствительный КО- рис 155. Медиальный глаз Euscorpi-
нец клетки С рабдомерОМ повернут ОТ us carpathicus. A — разрез глаза;
источника света, как в инвертирован- с- —' -■ *"""тям™ пя*пе"
ных глазах, но волокно зрительного
нерва связано с удаленным от света
концом клетки, как в глазах неинвер-
тированных. Это обстоятельство дало
Б — ретинула на фронтальном
разрезе сетчатки (по Scheuring):
\ — хрусталик, 2 — стекловидное тело, 3 —
кутикула, 4 — гиподерма, 5 — сетчатка, 6 —
постретннальный слой, 7 — зрительная
клетка, 8 — рабдом, 9 — зрительный иерв,
10 — зериа- пигмента
185
повод некоторым авторам (Versluys und
Demoll, 1922) рассматривать медиальные
глаза скорпионов как вторично
видоизмененные инвентированные глаза '.
Форму зрительных клеток и детали их
строения без специальных методов
исследования изучить не удается.
Легкие. На сагиттальном разрезе
через легкое следует обратить внимание
на общее строение этого органа, а
также на тонкую структуру отдельных
легочных листков (рис. 166). Стенки
последних образованы очень тонкой
хитиновой кутикулой {рис. 166, 5), под которой
лежит сильно уплощенный слой
гиподермы с редкими ядрами (рис. 166, 6).
Лакунарные пространства легочных
листков кое-где пересекаются
соединительными столбиками, которые связывают
обе стенки листка и, видимо, имеют со-
(рис. 167, 1). Кроме того, внутри
легочных листков встречаются отдельные клеточные элементы крови. Задние
свободные края листков несут хитиновые шипики (рис. 166, 8), а на
наружной поверхности их хитиновых стенок видны перерезанные
гребешки (рис. 167, 7), образующие характерную для Buthus сетчатую
микроскульптуру (см. с. 165).
Печень и желудочная железа. Железистый эпителий
печени состоит из клеток двух типов: питательных, или резорбирующих
(рис. 168, Л, 5), и ферментных (рис. 168, А, 4). Первые несколько
крупнее, ядра их занимают центральное, или базальное, положение (рис. 168,
А, 7), плазма переполнена крупными гомогенными шарообразными
включениями разного размера (рис. 168, А, 2). Включения имеют
белковую природу и окрашиваются кислыми красками. Часто, кроме того,
плазма содержит небольшие скопления пигментных зерен (рис. 168,
А, 6).
Ферментные клетки не столь обильны и более или менее сдавлены
окружающими их питательными клетками. Плазма их также содержит
крупные ацидофильные включения, но зернистой структуры (рис. 168,
А, 3). Между этими включениями видно множество более мелких—ба-
зофильного характера в форме зернышек или нитей. Эти клетки секре-
тируют пищеварительные ферменты и отвечают ферментным клеткам
печени других Arachnida и Crustacea.
Снаружи печень одета рыхлой соединительнотканной оболочкой,
состоящей из звездчатых разветвленных клеток, анастомозирующих
своими отростками (рис. 168, А, 1). Нередко в них встречаются крупные
1 Боковые глаза Pedipalpi и глаза Pseudoscorpiones инвертированного типа.
186
Рис. 166. Buthus caucasicus.
Разрез через легочные пластинки (по
Е. Н. Павловскому):
/ — скульптура на поверхности
пластинки, 2 — лакунарная полость
пластинки, 3 — легочная пластинка, 4 —
пространство между легочными
пластинками, 5—кутикула, 6 — утолщение
гиподермы, 7 — утолщение кутикулы,
8 — шипы на задних краях пластинок
единительнотканную природу
ацидофильные шаровидные включения, сходные
■с таковыми питательных клеток.
Под действием секрета ферментных клеток
пища переваривается в полости печени, но затем
фагоцитируется питательными клетками или,
будучи резорбирована ими, уплотняется в
характерные белковые шары, в которых, видимо,
переваривается окончательно. Образовавшиеся
при этом экскреторные зерна выталкиваются
обратно в полость печени.
Очень сходное гистологическое строение
имеет и желудочная железа и даже стенка средней
кишки.
В желудочной железе Buthus eupeus
обнаружены липаза, пепсин, химозин и трипсин, а
в печени, кроме того, каталаза и амилаза.
Маль!пигиевы сосуды. . В
соединительной ткани, окружающей печень, между ее
лопастями попадаются разрезы через мальпиги-
евы сосуды. Мелкие разветвления этих органов,
которые нигде за пределы соединительной ткани
не выходят, образованы сильно уплощенным
эпителием с дистально расположенными ядрами
(рис. 168, Б, 9). Плазма имеет, характерную,
перпендикулярную к базальной мембране исчер-
ченность. Клетки лишены включений, которые
обычны в эпителии мальпигиевых сосудов
насекомых.
Рис. 167. Teuthraustes.
Горизонтальный разрез
через легкое (по Е. Н.
Павловскому):
I — соединительные
столбики, 2 — внутренние полости
легочных пластинок,
сообщающиеся с окружающей
орган полостью циркум-
пульмонального синуса, 3 —
часть преддверия, 4 —
пространства между соседними
легочными пластинками
Рис. 168. Buthus eupeus. Разрезы через печень
(А) и мальпигиев сосуд (Б) (по Е. Н.
Павловскому) :
/ — соединительнотканная оболочка печени, 2 —
шарообразные белковые включения, 3 — ацидофильные
включения, 4 — ферментная клетка, 5 — резорбирующая
клетка, 6 — пигментные зерна, 7 — ядро резорбирующей
клетки, 8 — ядро ферментной клетки, 9 — эпителий маль-
пигиева сосуда
187
ОТРЯД ARANEI —ПАУКИ
Araneus (-Epeira)diadematus L. — Паук-крестовик
Материал и методы изучения. Araneus diadematus L., или паук-крестовик (сем,
Araneidae подотряда Labidognatha), обычен в европейской части СССР. Крупные
половозрелые самки встречаются в августе и сентябре в лесах, парках, во дворах и
старых строениях, на заборах, где плетут большие ловчие сети из паутины, на которых
они большей частью сидят. Более мелкие самцы находятся обычно вблизи гнезд
самок под корой деревьев, в трещинах строений и т. д. Яйца откладываются самкой
осенью в паутиновых яйцевых коконах, которые подвешиваются в укромном уголке.
Молодь выходит из яиц весной, и в течение лета до августа встречаются только
молодые, неполовозрелые особи.
Помимо A. diadematus в нашей фауне обычен еще целый ряд видов этого рода.
Все они очень близки по своей организации и отличаются главным образом только
деталями строения совокупительных органов и поэтому с успехом могут быть
использованы.
Сбор пауков, предназначенных для изучения внешней морфологии и
мускулатуры, производится сразу в спирт. Для изучения внутренней организации путем
вскрытий, особенно для инъекции кровеносной системы, нужны живые пауки. Анатомия
может быть изучена полно путем вскрытий, но желательно и дополнительное изучение
серий срезов. Для заливки следует выбирать не слишком крупные (меньше 10 мм)
экземпляры, которые фиксируются подогретой сулемой с уксусной кислотой или
жидкостью Буэна. Перед фиксацией паукам обрезаются ноги. Заливку следует
производить в целлоидин или парафин через жидкость Петерфи. Толстые срезы не менее
15 мкм окрашиваются обычными красками.
Внешняя морфология. Тело состоит из сравнительно
небольшой головогруди (cephalothorax, рис. 169,7), несущей конечности, и
крупного несегментированного брюшка (abdomen, рис. 169,5).
Последнее соединяется с головогрудью посредством узкого и короткого сте- ~
белька (petiolus, см. рис. 177,20).
Размеры тела подвержены значительным индивидуальным
колебаниям и варьируют у самцов от 10 до 11 мм, у самок от 20 до 25 мм в
длину. Обильные сероватые волоски покрывают все тело, в
особенности брюшную поверхность, головогрудь и конечности. На спинной
стороне брюшка волоски развиты гораздо слабее. Хитин покровов бурого
цвета. Характерна окраска спинной стороны тела. На темно-буром
сетчатом фоне брюшка выделяются большие белые пятна и волнистые
полоски, окаймленные бурой каемкой. Белые пятна образуют более или
менее ясную крестообразную фигуру. Низ брюшка и головогруди тем- '
но- бурый, брюшко — с двумя продольными белыми полосками.
Середина члеников ног со светлыми перевязками. У самцов эта окраска
выражена не столь ясно.
В состав головогруди входят акрон и шесть сегментов прозомы
(четыре ларвальных и два постларвальных), подобно большинству
других Chelicerata. Соответственно головогрудь несет шесть пар
конечностей— хелицеры, педипальпы и четыре пары ходных ног.
Стебелек образован первым абдоминальным (VII туловищным)
сегментом. Пауки происходят от предков с расчлененным, 12-членис-
тым брюшком, но у всех высших пауков Labidognatha, к которым
принадлежит Araneus, брюшко состоит лишь из пяти передних сегментов
мезозомы (с VII по XI), сливающихся полностью, и-последнего
анального (XVIII). Остальные брюшные сегменты развиты лишь в зароды- •
188
Рис. 169. Самка Araneus diadematus. Вид со спинной стороны
(ориг.):
1 - головогрудь, 2 — хелицеры, 3 — педипальпы, 4 — ходные ноги, 5 —
брюшко, 6 — хнтинизированные площадки
шевом состоянии и у взрослых животных редуцируются, и только XII
и XIII представлены частично невромерами, входящими в состав под-
глоточной нервной массы.
Спинной щит головогруди (сагарах, рис. 170) — результат слияния
акрона и всех тергитов прозомы. Его передний и боковые края
загибаются вниз. Несколько позади середины щит имеет небольшое
медиальное вдавление в виде плоской ямки (рис. 170, 3); от нее начинаются
несколько слабо намеченных, радиальных желобков (рис. 170, 6),
которые отвечают местам прикрепления мускулов сосательного желудка
и конечностей.
Впереди головогрудной щит несет восемь глаз, расположенных в
два поперечных ряда. Четыре глаза лежат медиально, а две пары —
189
у передних углов щита.
Медиальные глаза образуют
трапециевидную фигуру, располагаясь по
ее углам, причем глаза задней
пары лежат 'ближе друг к другу,
чем глаза передней пары (рис.
170, 1). Боковые глаза сильно
сближены (рис. 170, 2). Все
глаза имеют одинаковые размеры и
лежат на небольших бугорках.
Значительная часть брюшной
поверхности головогруди
образована крупными тазиками ходных
ног (рис. 171, 7). Между
тазиками лежит брюшной щит
головогруди — стернум (sternum,
рис. 171, 9), а впереди с ним
сочленяется небольшой стернит
педипальп а льно го (II
туловищного) сегмента (рис. 171, 6).
Стернум в общем имеет почти
треугольную форму, суживаясь
сзади. Его боковые края несут
неглубокие вырезки для тазиков
ног, а задний конец снабжен
небольшим пластинчатым
расширением (рис. 171, 8). Стернум
образован 'слившимися стернитами
сегментов ходных ног (III—VI),
причем его заднее маленькое
расширение, видимо, отвечает стерниту последнего прозомального (VI
туловищного) сегмента.
Боковые стенки головогруди образованы мягкой плевральной
кутикулой, но над тазиками ног с каждой стороны лежит вытянутая
боковая пластинка.
Впереди педипальпального стернита лежит предротовая полость.
Она ограничена спереди хелицерами (рис. 171,2), с боков —
челюстными лопастями тазиков педипальп (рис. 171, 5), а сзади — стернитом,
или нижней губой. Челюстные лопасти являются видоизмененными
жевательными отростками (эндитами), которые у пауков сохранились
лишь на педипальпах. На дне предротовой полости, между
челюстными лопастями, лежит мясистый конусообразный рострум (rostrum, см...
рис. 192,75), торчащий вниз. Его боковые края срастаются с нижней
губой. Между рострумом и нижней губой хорошо заметно поперечное
щелевидное ротовое отверстие- (рис. 192, 19). Оно, таким образом,
лежит на сегменте педипальп, т. е. на II туловищном сегменте. Все
органы, ограничивающие предротовую полость, очень подвижны, в том
числе и несущий функцию нижней губы стернит.
Отделив осторожно нижнюю губу и сочлененные с ней челюстные
Рис. 170. Головогрудь Araneus diadematus.
Вид сверху (ориг.):
/ — медиальные глаза, 2 — боковые глаза, 3 —
срединная ямка, 4 — стебелек, 5 — lorum, 6 — борозд-
190
Рис. 171. Головогрудь самки Araneus diadematus. Вид снизу. А — хелицеры в
обычном положении; Б — хелицеры отогнуты вперед (ориг.):
1 - когтевидный члеиик хелицеры, 2-основной членик хелицеры, 3- педипальпа,
4—пучок волосков челюстной лопасти, 5 - челюстная лопасть, 6 — нижняя губа, / —тазики ног,
8 - пластинчатое расширение стериума, 9 — стериум, 10 - тазик педипальпы, 11 - вертлуг,
12— бедро 13 — колено, « — голень, 15 — лапка, 16 — коготок
лопасти педипальп и выварив в едком кали, следует приготовить
тотальный препарат.
Нижняя губа снаружи покрыта волосками. Среди них различаются
два типа. На боковых краях губы сидят крупные простые волоски,
в середине — тонкие, перистые. Внутренняя поверхность губы
образованна тонкой, гладкой и темной хитиновой пластинкой, обнаруживающей
нежную поперечную исчерченность. Она представляет собой заднюю
хитиновую стенку глоточной полости.
Челюстные лопасти значительно массивнее губы. От тазиков
педипальп они отделяются явственным швом и имеют в общем треугольную
форму, но на свободных краях закруглены (рис. 171,5). Края
лопастей обращенные в сторону предротовой полости, несут по щеткооб-
разному густому пучку волосков (рис. 171,4). Наружные волоски этой
щетки простые," внутренние, торчащие непосредственно в предротовую
полость, имеют перистое строение. Внутренняя поверхность каждой
челюстной лопасти лишена волосков и представляет собой блестящую
191
хитинизированную поверхность. Передний, косо срезанный, край этой
поверхности снабжен рядом чрезвычайно мелких хитиновых зубчиков.
Рострум хорошо виден при раздвигании челюстных лопастей в
■стороны. Он прикрывает ротовое отверстие спереди и нередко называется
верхней губой. Задняя его поверхность, ограничивающая вертикальную
глотку, образована голой, плотной хитиновой пластинкой, на других
местах покровы этого органа состоят из мягкой, кожистой кутикулы и
несут волоски. Рострум очень подвижен, так как обладает собственной
сложной мускулатурой.
Итак, предротовая полость, сходная с таковой скорпионов,
ограничена подвижными частями окружающих органов. Волоски, торчащие
в ее просвет и обильно развитые у входа, играют важную роль в
процессе принятия пищи. Пауки, подобно другим Arachnida, питаются
исключительно жидкой пищей, высасывая свою добычу (насекомых).
Убитая хелицерами добыча сначала разминается этими органами и
челюстными лопастями и затем подвергается действию секрета
желудочных отростков и желез, залегающих в челюстных лопастях и рост-
руме. Под влиянием сильно действующих ферментов этих выделений
все мягкие ткани добычи разжижаются. Таким образом, пищеварение
начинается уже вне тела паука. Жидкая пища затем всасывается в
предротовую полость, куда она поступает уже в силу капиллярности
волосков. Существенную роль, .однако, играют также сосательные
движения рострума и челюстных лопастей. В кишечник пища
насасывается действием сосательного, желудка. В результате от добычи остаются
только хитиновые покровы.
Передняя пара конечностей — хелицеры (chelicerae, рис. 171,1,2)
прикрепляется впереди рта на брюшной стороне головогруди. Эти
мощные, но короткие двучленистые придатки у самцов несколько уже ' и
длиннее. В покоящемся положении хелицеры располагаются
вертикально, будучи направлены вниз, параллельно друг другу (рис. 171, А, 1, 2).
Сверху при этом видны только их основания. Но они довольно
подвижны и* могут отгибаться вперед и вверх (рис. 171, Б, 1,2). Толстый
основной членик хелицеры имеет почти цилиндрическую форму, хотя
у основания заметно вздут. На вершине у внешнего края он сочленен
с острым, когтевидно изогнутым, черным конечным члеником
(рис.171,/). Этот последний двигается только в одной плоскости и
может, прогибаясь внутрь, вкладываться, подобно лезвию перочинного
ножа, в бороздку на основном членике. Края бороздки вооружены
хитиновыми зубцами. На концах когтевидных члеников открываются
протоки пары ядовитых желез, лежащих в передней части головогруди.
Хелицеры служат для умерщвления добычи и для защиты.
Вторая пара конечностей — педипальпы (pedipalpi, рис. 171,5)
значительно длиннее хелицер, но в 3—4 раза короче ходных ног, на
которые очень похожи. У самцов педипальпы на концах сильно
вздуты и своеобразно изменены, так как функционируют в качестве
совокупительных органов. (Строение их рассматривается ниже — с. 220).
Педипальпы самки состоят из шести члеников: тазика (соха, рис. 171,
10) с его челюстной лопастью, вертлуга (trochanter, рис. 171,17),
бедра (femur, рис. 171, 12), колена (patella, рис. 171, 13), голени (tibia,
192
рис. 171, 14) и лапки (tarsus рис. 171, 15).
Последний членик постепенно суживается к
концу, обильно покрыт различной длины
чувствительными волосками и заканчивается
гребневидным коготком (рис. 171, 16), очень похожим
на такие же коготки ходных ног. Самка держит
педипальпы горизонтально, вытягивая их
вперед. Главная функция педипальп
чувствительная.
Четыре пары ходных ног имеют одинаковое
строение, различаясь только своей длиной.
Третья пара несколько короче всех других (см.
рис. 169, 4). Каждая нога состоит из семи
члеников, причем первые пять носят те же
названия, что и на педипальпах, шестой называется
пяткой (metatarsus, см. рис. 197, А, 6), а
седьмой лапкой (tarsus, см. рис. 197, А, 7).
Для изучения коготков, расположенных на
концах ног, необходимо приготовить несколько
тотальных препаратов тарзального членика,
различно его ориентировав. Каждая нога
заканчивается парой больших гребенчатых коготков
(рис. 172, 1). Между ними прикрепляется'
изогнутый крючкообразно непарный коготок с
одним зубцом у основания (рис. 172, 2).
Несколько проксимальней от этих трех коготков имеются
еще несколько зазубренных шипов, среди
которых выделяются два особенно мощных, слегка изогнутых й-ооразно
(рис. 172, 3). Ноги задней пары, кроме того, на нижней стороне лапкй
у ее вершины несут по одному прямому шипу. Шипы и коготки служат
для передвижения по тенетам. Коготки задних ног и прямые шипы
также употребляются для натягивания паутиновых нитей при
постройке тенет. При лазании по вертикальным паутинавым нитям
функционирует главным образом передняя пара ног, нить при этом пропускается
между крючковидным коготком и S-образными щетинками. Когда паук
бежит по поверхности, то опирается на концы гребневидных коготков.
Стебелек, прикрепляющийся к головогруди не на заднем ее конце,
а несколько дорзально, снабжен маленькими спинной и брюшной
пластинками. Продолговатая спинная пластинка (lorum, см. рис. 170, 5)
имеет характерную седловидную форму, будучи изогнута задним
концом кверху, соответственно изгибу самого стебелька, который
соединяется с нижней поверхностью брюшка. Боковые края пластинки
загибаются вниз, а передний и задний концы глубоко вырезаны. Лорум
представляет собой не что иное, как тергит VII, прегенитального
сегмента, за счет которого, образуется стебелек. V-образная • брюшная
пластинка (plagula) гораздо меньше. Она направлена двумя концами
назад и, видимо, отвечает стерниту VII сегмента.
Брюшко у самок гораздо крупнее головогруди и имеет округло-
яйцевидную форму. У самцов оно более вытянуто и гораздо меньше.
Рис. 172. Araneus dlade-
matus. A — конец
тарзального членика ноги
снизу; Б — то же сбоку
после удаления волосков
(А — из Roseler и Lamp-
recht, Б — по Nielsen):
/ — гребенчатые коготки,
2 — крючковидиый коготок,
3 — S-образный шип, 4 — за- '
зубренные шипы, 5 —
волоски
7-2
193
Рис. 173. Брюшко самки Araneus dladema-
tus. Вид снизу (по Kastner):
/ — место прикрепления стебелька, 2 — легочная
крышечка, 3 — стигма правого легкого, 4 — эпи-
гастральиая бороздка, 5—стигма трахей, 6 —
паутиновые бородавки, 7 — аиальиый бугорок,
8 — epigyne
Рис. 174. Araneus diadematus. Паутиновые
бородавки и анальный бугорок (из Ger-
hardt и Kastner):
/ — colulus, 2—передняя паутиновая бородавка,
3 — задняя паутиновая бородавка, 4 —
внутренняя паутиновая бородавка, 5 — первый членик
анального бугорка, 6 — второй членик анального
бугорка, 7 — третий членик анального бугорка,
8—местоположение анального отверстия, 9 —
паутнновое поле передней бородавки
194
Место соединения со стебельком
лежит на нижней стороне
брюшка, отступя от переднего края не
менее чем на 7з его длины
(рис. 173, 1). Вследствие этого,
во-первых, вентральная
поверхность брюшка значительно
меньше дорзальной и, во-вторых,
широким передним краем оно
налегает на головогрудь сверху,
закрывая заднюю половину голово-
грудного щита (см. рис. 169, 5).
Все брюшко покрыто мягким
хитиновым покровом, за
исключением небольших хитинизиро-
ванных площадок в местах
прикрепления мышц (см. рис. 169,
6) и так называемых легочных
крышечек (рис. 173, 2).
На вентральной стороне
брюшка, позади стебелька,
лежит непарное половое отверстие,
принадлежащее второму
абдоминальному (VIII туловищному)
сегменту. У взрослых самцов оно
имеет вид небольшой поперечной
щели, ограниченной спереди и
сзади сильно хитинизированны-
ми складками. У самок в области
полового отверстия
дифференцируется совокупительный аппарат
сложного строения — эпигине
(epigyne, рис. 173, 8). Он
состоит из основной хитиновой
пластинки с парными утолщениями и
хитинизированного
червеобразного придатка (описание см. на
с. 223).
На уровне полового
отверстия покровы брюшка образуют
поперечную борозду, называемую
эпигастральной (рис. 173, 4),
которая заканчивается парой
боковых щелевидных легочных
отверстий, или стигм (stigmata,
рис. 173, <3). Впереди последних
лежат щитковидные хитиновые
пластинки — легочные крышечки,
резко отличающиеся от окружа-
9
ющих покровов темной окраской, наличием поперечных морщинок и
отсутствием волосков ('рис. 173, 2). Непосредственно под ними лежат
легкие, которые, таким образом, помещаются несколько впереди
полового отверстия, почти по бокам от места прикрепления стебелька.
Легкие принадлежат VIII половому сегменту, гомологичны первой паре
легких низших четырехлегочных пауков и крышечке (operculum) Li-
mulus и не находят себе гомологов у скорпионов, у которых первая
пара легочных мешков лежит на X сегменте '.
На заднем конце брюшка с вентральной стороны лежат заметные
простым глазом паутиновые бородавки (рис. 173,5). Впереди них
помещается непарная стигма трахей, сзади — анальное отверстие. Для
изучения этих органов отрезают заднюю часть брюшка, вываривают в
едком кали и рассматривают при слабом увеличении. Кроме того,
необходимо приготовить тотальные препараты изолированных
паутиновых бородавок. Перед паутиновыми бородавками видно медиальное
дыхальце, или стигма (stigma, рис. 173,5), в форме небольшой
поперечной щели. Это отверстие ведет в трахеальное,преддверие —
непарный мешочек, в который открываются две пары трахей. Как
показывает эмбриональное развитие этих органов, они образуются на IX
туловищном сегменте. На этом же сегменте у низших пауков (Mesothelae
и Orthognathia) лежит вторая пара легких.
У самого основания передних паутиновых бородавок хорошо
заметна небольшая непарная хитинизированная и волосистая папилла
(colulus, рис. 174, 1) —остаток cribellum. Этот хорошо развитый у
некоторых пауков (Amaurobiidae) орган у Araneus находится в
рудиментарном состоянии2.
Паутиновые бородавки представлены тремя парами — большими
передними и задними и маленькими внутренними (рис. 174,2,3,4).
Последние лежат между задними бородавками. Передние и задние
бородавки состоят из двух члеников, соединенных мягкой сочленовой
кожей, и, как показывает их история развития, представляют собой
сильно видоизмененные ножки X и XI туловищных сегментов. Внутренние
бородавки во время эмбрионального развития отделяются от зачатков
задней пары и самостоятельными конечностями не являются.
Бородавки снабжены паутиновыми трубочками и паутиновыми
конусами, которыми заканчиваются протоки многочисленных, лежащих
в брюшке паутиновых желез. На передних бородавках трубочки
сосредоточены на вершине небольшого конечного членика. Пространство,
занятое ими, имеет почковидную форму и называется паутиновым
полем (рис. 174,9). Здесь на каждой бородавке располагается около
сотни паутиновых трубочек. Каждая из них состоит из двух частей — более
толстой, базальной, которая как бы срезана на конце, и более тонкой,
концевой части (ср. рис. 175,3 и 4). Последняя является
продолжением протока железы, хорошо заметного на препарате, не слишком долго
вываривавшемся в едком кали. Кроме того, с внутренней стороны пау-
1 У некоторых пауков (Caponiidae) на месте легких развиваются трахеи.
2 У Amaurobiidae cribellum представляет ситовидную пластинку, через поры
которой выделяется паутина. У этих же пауков на предпоследнем членике задних ног
имеется продольный гребневидный ряд волосков — calamistrum.
7*
195
Рис. 175. Araneus diadema-
tus. Задняя паутиновая
бородавка самки (по Vogt и
Yung):
/ — волоски, 2 — паутииовая
трубочка, 3 — концевая часть
трубочки, 4—базальная часть
трубочки, 5 — задний паутино-
вый конус, изображенный как
бы просвечивающим сквозь
бородавку, 6 — проток паутиновой
железы, 7 — базальная часть
паутинового конуса, 8 —
концевая часть конуса, 9 — паутино-
вые конусы
Рис. 176. Araneus diadema-
tus. Внутренняя паутиновая
бородавка самки, вид с
брюшной стороны (из R6-
seler и Lamprecht):
1 — паутиновое поле с паутино-
выми трубочками, 2 —
базальная часть трубочки, 3 —
концевая часть трубочки, 4 — паути-
новые конусы
тинового поля, в том месте, где оно имеет небольшую выемку,
располагается один паутиновый конус, базальная часть которого хитинизирована
и имеет конусовидную форму, а концевая часть укорочена (ср. рис.
175, 7 и 8). Рядом с ним лежит небольшой дополнительный конус.
Задние бородавки на вершине косо срезаны. На внутренней
поверхности они несут многочисленные короткие и длинные паутиновые
трубочки (рис. 175, 2). Последние, однако, у молодых животных
отсутствуют, так как развиваются только после последней линьки. Образуя
у взрослых пауков густой пучок, длинные трубочки закрывают
остальные элементы паутинового поля. Поэтому для изучения паутиновых
бородавок лучше брать молодых пауков. Помимо трубочек каждая
задняя бородавка у вершины снабжена четырьмя паутиновыми конусами
(рис. 175,9) и несет еще один на задней стороне (рис. 175,5).
Пронизывающие их протоки паутиновых желез хорошо видны (рис. 175,6).
Вся брюшная поверхность внутренних бородавок представляет
собой большое треугольное паутиновое поле, покрытое
многочисленными трубочками (рис. 176,/). На вершине бородавки выделяются два
конуса (рис. 176,4).
На всех паутиновых бородавках у Araneus diadematus
насчитывается около 600 паутиновых трубочек и 16 (у самцов 10) конусов.
Последние в общем имеют такое же строение, как и трубочки. При им-
196
мерсии на вершине тех и других удается рассмотреть крошечное
отверстие протока железы.
Брюшко заканчивается небольшим, несколько сжатым дорзовен-
трально анальным бугорком. Он лежит непосредственно позади
паутиновых бородавок и торчит вниз так, что его брюшная ■ поверхность
обращена в сторону бородавок, а спинная — назад (см. рис. 174,5—7).
Анальный бугорок, состоит из трех члеников, разделенных бороздками.
Каждый членик несет хитиновую пластинку. На первом и последнем
члениках она развита"только на спинной стороне, на втором —
образует полное хитиновое колечко. Членики соединяются мягкими
кожистыми покровами. Последний'членик, представляющий собой анальный
сегмент, возвышается в виде слегка уплощенной треугольной лопасти
(см. рис. 174, 7). У его основания с брюшной стороны лежит анальное
отверстие (см. рис. 174, 8).
Анатомия на вскрытии. Для вскрытия употребляются только что
убитые пауки.
Спиртовой материал менее пригоден и требует предварительного размачивания
в течение суток в воде, к которой добавлено несколько капель нашатырного спирта.
Но после этого препарирование сильно уплотняющегося в спирте брюшка
представляет большие трудности1. Головогрудь и брюшко приходится разделять и вскрывать
отдельно.
Головогрудной щит подрезается острым скальпелем со всех сторон и осторожно
удаляется, но передний его край с глазами оставляется на месте. Если вскрывается
спиртовой материал, то прикрепляющиеся к щиту мышцы отделяются без особых
затруднений. У свежих пауков они подрезаются острым скальпелем. При вскрытии
брюшка следует сделать два боковых разреза покровов и удалить их со спинной стороны.
При этом надо внимательно следить, чтобы внутренние органы, особенно сердце,
лежащее впереди под печенью, не были повреждены.
После удаления спинной стенки тела, в головогруди видны только мощные
мускульные' пучки, которые своими внутренними концами частью прикрепляются к
пластинчатым отросткам эндостернита. При раздвигании мышц заметны концы ядовитых
желез (рис. 177, 2) и слепые выросты желудка (рис. 177, 14, 54), пронизанные
кровеносными сосудами. Мускульные пучки, связанные с конечностями, передней
кишкой и эндостернито.м, тщательно вырезаются. При этом обнажается нервная система
(рис. 177, 16, 55) и передняя часть кишечника со слепыми отростками желудка, а
также кровеносные сосуды. Однако все эти органы настолько переплетаются друг с
другом, что невозможно изолировать один из них, не повредив другие.
Такие же затруднения возникают и при препаровке брюшка. Печень закрывает
со спинной стороны все остальные органы и глубоко внедряется между ними
(рис. 177, 48): Удалив возможно тщательнее эту железу, необходимо обнажить
сердце (рис. 177, 44). Оно лежит медиально впереди и изогнуто соответственно
выпуклости брюшка. От него отходят артериальные сосуды (рис. 177, 40, 50). Захватив
сердце пинцетом, следует осторожно отделить его вместе с частью печени, лежащей
между ним и кишечником. В результате обнаруживается половая железа (рис. 177,27)
и проходящая над ней тонкая кишка (рис. 177, 45), а также клоака и прямая кишка
(рис. 177, 39, 38), Под половой железой лежат многочисленные паутиновые железы
нескольких различных типов (рис. 177, 26, 28, 29, 32, 37). У вентральной стенки
брюшка, спереди находятся легочные мешки, а сзади — трахеи (рис. 177, 21, 30).
Кровеносные сосуды изучаются на отдельных экземплярах при помощи инъекции.
Внутренний скелет представлен, как и у других Arach.nida,
впячиваниями хитинового покрова, вдающимися в полость тела,— апо-
1 При препарирований мускулатуры спиртовой материал представляет ряд
преимуществ. ' ■ -
197
Рис. 177. Схема организации Araneus (no Comstock):
/ — глаза, 2 — ядовитая железа, 3— хелицеральный нерв, 4 — артерия педипальпы, 5 — педипальпа,
6 — нерв педипальпы, 7 — проток ядовитой железы, 8 — основной членик хелнцеры, 9 — артерия
хелицеры, 10— коготок, Л — когтевидиый членик хелицеры, 12— верхняя и нижняя губы, 13 —
нерв ноги, 14 —• боковые ветви желудочных выростов, 15 — артерия иогн, 16 — подглоточная
нервная масса, 17 — truncus peristomacalis, IS — желудок, 19 — нервный ствол, 20 — стебелек, 21 —
легкое, 22—стигма легкого, 23 — отверстие семяприемника, 24 — семяприемник, 25 — матка и половое
отверстие, 26 — древовидная железа, 27 — янчник, 28 — ампуловидиая железа, 29 — трубковидная
железа, 30 — трахеальное преддверие, 31 — стигма трахей, 32 — грушевидные железы, 33 — передняя
паутиновая бородавка, 34 — внутренняя паутиновая бородавка, 35—задняя паутииовая бородавка,
36 — анальное отверстие, 37 — дольковидные железы, 38 — задняя кишка, 39 — клоака, 40 — задняя
аорта. 41 — эпикардиальные лигаменты, 42 — мальпигиевы сосуды, 43 — проток печени, 44—■
сердце, 45 — расширение тонкой кишкн, 46—остие сердца, 47 — перикардий, 48 — разветвления печени,
49 — легочная вена, 50 — передняя аорта, 51 — сосательный желудок, 52 — мускулатура
сосательного желудка, 53 — пищевод, 54 — вырост желудка, 55 — головной мозг, 56 — зрительные нервы,
S7 — глазная артерия
демами и мезодермальными эндохондритами. Аподемы
обнаруживаются очень легко — вывариванием вскрытых головогруди и брюшка в
едком кали. Нежные эндохондриты отпрепаровываются без
предварительного вываривания, если изучается свежий материал, или, если
материал сильно уплотнен спиртом, после непродолжительного
кипячения в слабом растворе едкого кали.
В головогруди аподемы развиты очень слабо. Спинной щит в
области медиальной ямки имеет небольшую непарную аподему, к которой
прикрепляются мускулы сосательного желудка. На брюшной стороне
головогруди небольшие треугольные пластинчатые аподемы развиты
лишь на тазиках педипальп.
В стебельке задние углы тергальной пластинки (lorum) снабжены.
маленькими аподемальными отростками (рис. 178, /; 180, 42). Ниже
их на границе стебелька и брюшка лежат две пары очень маленьких
аподем (рис. 178, 21), к которым прикрепляются медиальные
продольные мускулы брюшка. Но особенно интересны вентральные аподёмы,
так как они расположены на задних границах брюшных сегментов.
Это. довольно значительные, языкообразные или пластинчатые впячи-
вания интерсегментарной кожи VIII—IX сегментов. Передняя из них
непарная широкая аподема помещается на уровне легочных стигм,
(рис. \78,20). Эпигастральная борозда представляет собой наружное
щелевидное отверстие этой аподемы, которая принадлежит VIII сег-
198
менту. Следующая, тоже
непарная аподема IX
сегмента лежит позади трахей
(рис. 178, 18), и, наконец,
две пары пластинчатых
аподем, принадлежащих X и
XI сегментам, помещаются
позади передних и задних
паутиновых бородавок (рис.
178, 15, 16).
Эндохондриты
представлены главным образом
брюшными элементами —
эндостернитами (endoster-
па). Из спинных эндохонд-
ритов (endoterga)
сохраняется лишь одна пара в
основании брЧошка.
Самый крупный,
непарный эндостернит,
отвечающий таковому Limulus и
других Arachnida, лежит в
головогруди между
сосательным желудком и под-
глоточной нервной массой.
Это довольно широкая,
большая бесцветная
пластинка, несколько вогнутая
сверху, >с глубокой
вырезкой, обращенной в сторону
головного мозга (рис. 179,
А я Б). С 'каждой стороны
край эндостернита образует
два вертикальных
пластинчатых киля— спинной и
брюшной (рис. 179, 2, 3).
Края этих килей вытянуты
в пластинчатые выросты.
На брюшном киле видны четыре слабо развитых, неправильно
зазубренных отростка (рис. 179, 4). Сп'инной киль щесет столько же верхних
отростков, направленных кверху, параллельно друг другу, и, кроме
того, один внутренний (рис. 179, 5, 6). К верхним отросткам
прикрепляются мощные мускулы, связывающие эндостернит со спинным щитом.
От брюшной поверхности эндостернита и от нижних его отростков
отходят мышцы к стернальной пластинке (Sternum). Спинная поверхность
эндостернита связана мощными мускульными пучками с сосательным
желудком.
В брюшке единственная пара эндотергитов лежит непосредственно
над задними концами тергальной пластинки стебелька (рис. 178, 3;
Рис. 178. Araneus sp. Аподемы брюшка и
органы дыхания (ориг.):
/ — аподемальиые отростки lorum, 2 — утолщения
кутикулы в местах прикрепления мышц, 3 — эндотергит
стебелькового сегмента, 4 — lorum, 5 — место прикрепления
стебелька, 6 — легкое, 7 — преддверие легкого, 8 —
внутренняя трахея, 9 — наружная трахея, 10 — трахеальиое
преддверие, // — передняя паутиновая бородавка, 12 —
задняя паутиновая бородавка, 13 — внутренняя
паутиновая бородавка, 14 — анальное отверстие, 15 — аподемы
XI сегмента, 16 — аподемы X сегмента, 17 — colulus, 18 —
аподемы IX сегмента, 19 — место прикрепления
вентральных продольных мускулов, 20 — аподема VIII
сегмента, 21 — аподемы VII стебелькового сегмента
19Э
180, 2). К ним прикрепляются
несколько парных мускулов. Небольшие
эндостерниты располагаются метамерно на
вентральной стороне. Имеются четыре
пары медиальных и столько же
латеральных эндостернитов, связанных друг с
другом многочисленными мышцами. Они
принадлежат VIII—XI сегментам,
причем каждый сегмент имеет по одной
паре медиальных и по паре латеральных
элементов (рис. 180,8,24,26,35,38).
Медиальные эндостерниты лежат над
аподемами соответственных сегментов,
т. е. интерсегментарно. Передние
медиальные, эндостерниты связаны друг с
другом поперечным сухожильным тяжем
(рис. 180, 38; 182, 12).
Мускулатура. Препарирование
мускулов удобнее производить на
спиртовом, предварительно размягченном в
воде материале. Если от
продолжительного пребывания в спирте материал
сильно уплотнен, то хорошие результаты
дает вываривание в 5%-ном растворе
едкого кали. Перед вывариванием следует
удалить спинной щит головогруди и
покровы со спинной стороны брюшка.
Остатки размягченных тканей и органов
удаляются тщательной промывкой;
остается только мускулатура, которая дольше противостоит разрушающему
действию щелочи.
В головогруди ряд мышц, прикрепленных к спинному щиту,
соединяют его с эндостернитом. В середине располагаются мускульные
пучки сосательного желудка (см. рис. 177,52; 191, 2; 192,4), а впереди,
медиально, подниматели глотки (см. рис. 191,5; 192,9). Другие
мышцы своими нижними концами прикрепляются к конечностям.
Несколько мускульных пучков соединяют головогрудь с брюшком.
В стебельке рядом с кишечником проходит пара продольных мускулов,
которые спереди начинаются от эндостернита а головогруди, а сзади
подходят к спинным.мускулам брюшка. Более тонкие парные
мускульные тяжи тянутся на брюшной стороне стебелька и сзади
прикрепляются к аподемам VII сегмента.
Гораздо интереснее мускулатура брюшка, так как она сохраняет
метамерный характер и дает ясное представление о сегментарном
составе этого отдела тела.
Все мускулы брюшка могут быть разделены на следующие группы.
I. Дорзальные мускулы. Две пары мощных мышц, которые
соединяют эндотергиты со спинными покровами. Из -них передняя пара
коротка и направлена вперед (рис. 180,/; 181,18). Задняя пара проходит
Рис. 179. Araneus reau-muri. Эн-
достернит головогруди. Вид
сверху (А) и сбоку (Б) (из Gerhardt
и Kastner):
/ — передняя вырезка, 2 — брюшной
киль, 3 — спинной киль, 4 — отростки
брюшного киля, 5 — отростки спинного
киля, 6 — внутренний вырост
200
в дорзальном
направлении, сходно с дорзо-
вентральными
мускулами (см. ниже), с
которыми может быть
легко смешана. Места
прикрепления этих
мускулов на спине
хорошо видны после
удаления стенок тела (см.
рис. 183, А, Б, 8). Они
тянутся рядом с
сердцем по бокам от' него,
почему нередко
называются латеро-карди-
альными мышцами
(latero-cardiales, рис.
180, 4; 181, 20). Обе
нары спинных
мускулов, видимо,
представляют собой остатки ме-
тамерной дорзальной
мускулатуры предков. . •
II. Вентральные
мускулы.
Многочисленные мышечные пучки,
разделяющиеся в свою
очередь на несколько
групп:
1. Продольные
медиальные мускулы (т.
medio - longitudinales,
рис. 180, 16, 21, 25;
181, 6, 9, 32). Парные
мышцы, соединяющие
медиальные
эндостерниты VIII—IX
сегментов. Передняя пара,
относящаяся к VIII
сегменту, спереди
прикрепляется к боковым
аподемам VII
сегмента (рис 180, 39; 181,
13; 182, 5) и закрыта
сверху особыми
мускульными пучками,
которые передними
концами фиксируются на
верхних атюдемах тер-
/ . -43
Рис. 180. Araneus sp. Мускулатура брюшка. Вид со
спинной стороны (ориг.):
;_ передний дорзальный мускул, 2 — эндотергит VII сегмента,
3 — легочная вена, 4 — латеро-карднальный мускул, 5 — цир-
кумпульмональный синус, 6 — передний продольный
латеральный мускул, 7 —легкое, 8 — латеральный эндостернит VIII
сегмента, 9 — открыватель стигмы, 10 — поперечный мускул VIII
сегмента, 11 — мускул, соединяющий медиальный эндостернит
VIII с тергитом VII, 12 — дорзовентральный мускул VIII, 13 —
латеральный продольный мускул IX, 14 — косой мускул IX,
15 — задний конец продольного медиального мускула,
связанный с покровами, 16 — продольный медиальный мускул IX,
П — латеральный продольный мускул X, 18 — мускул передней
паутнновой бородавки, 19 — мускулы задней паутнновой
бородавки, 20 — латеральный продольный мускул XI, 21 —
медиальный продольный мускул X, 22 — анальный мускул, 23 —
■ анальное отверстие, 24 — медиальные эндостерниты XI, 25 —
медиальный продольный мускул XI, 26 — латеральный
эндостернит XI, 27 — дорзовентральный мускул X, 28 — задняя паутнно-
вая бородавка, 29 — передняя паутиновая бородавка, 30 —
латеральный эндостернит X, 31 — косой мускул X, 32 — мускул
передней паутнновой бородавки, 33 — поперечный мускул IX,
34 — дорзовентральный мускул IX, 35 — латеральный
эндостернит IX, 36 — косой мускул IX, 37 — продольный мускул,
соединяющий медиальные эндостерниты VIII и X, 38 — медиальные
эндостерниты VIII, 39— продольный медиальный мускул VIII,
40 — дорзовентральный мускул VII, 41 — поверхностные
мускулы, 42 — задние аподемы brum, 43 — поперечный мускул,
соединяющий задние аподемы lorum
201
1 8 9 wll ПЯ 14 15
гита стебелька (рис. 180, 11;
181, 15).
2. Кроме того, ближе к
брюшной стенке тела лежат
продольные мускулы,
связанные своими задними концами
частью с эндостернитами и
частью с аподемами и
брюшными покровами. Из них одна
пара проходит под эндостерни-
там« IX, соединяя эндостерни-
ты VLII и X сегментов;
(рис. 181, 8; 182, <§).
Непосредственно к стенке тела
прикрепляются задние концы
нескольких парных мускулов
(рис. 180, 15; 181, 5, 10),
места прикрепления которых
хорошо видны на брюшных
покровах (см. рис. 178, 19).
3. Продольные
латеральные мускулы (m. latero-longi-
tudinales, см. рис. 180, 17, 20).
Четыре пары проходят между
латеральными
эндостернитами. Из них две передние пары
отличаются некоторыми
особенностями. Первая пара,
принадлежащая VIII сегменту,
проходит над легочными
мешками и сильно скошена (см.
рис. 180, 6), что связано с
сужением брюшка впереди у
стебелька. Вторая пара (IX сегмента) в передней своей половине
редуцирована и не доходит до латеральных зндостернитов VIII сегмента (см..
рис. 180, 13).
4. Поперечные мускулы (m. transversales) соединяют медиальные
и латеральные эндостерниты своего сегмента. К ним относятся длинные
поперечные мышцы VIII сегмента, проходящие параллельно задним
краям легочных мешков (см. рис. 180,10), и короткие, скошенные
мускулы IX сегмента (см. рис. 180,33). В X и XI и частично в IX
сегментах эти мышцы своими внутренними концами прикрепляются не к
эндостернитам, а у основания паутиновых бородавок, для движения
которых они служат (см. рис. 180,18,32). Возможно, что к этой же
группе принадлежат анальные мускульные пучки, проходящие позади
задней кишки (см. рис. 180,22).
5. Косые мускулы (m. obliqui, см. рис. 180, 14, 31, 36). Три пары в
IX и X сегментах. Они соединяют медиальные эндостерниты с
латеральными следующего сегмента.
Рис. 181. Araneus. Схема расположения
сердца, мускулатуры и зндостернитов в брюшке.
Вид с правой стороны (ориг.):
/ — анальное отверстие, 2 — задняя паутииовая
бородавка, 3 — передняя паутиновая бородавка, 4 — эндо-
стернит X сегмента, 5 — мускул X, прикрепляющийся
к покровам, 6 — медиальный продольный мускул X,
7 — медиальный эндостернит IX, 8 — мускул,
соединяющий эндостерниты VIII и X, 9 — медиальный
продольный мускул IX, 10 — мускул XI,
прикрепляющийся к покровам, // — эпигастральная щель, /2 —
медиальный эндостернит VIII, 13 — медиальный
продольный мускул VIII, 14 — дорзовентральныи
мускул VII, 15 — мускул, соединяющий медиальный
эндостернит VIII с тергнтом VII, 16 — аподема терги-
та VII (lorum), П — эндостергнт VII, 18— передний
дорзальный мускул, 19 — остие сердца VIII, 20 —
латеро-кардиальный мускул, 21 — остие сердца IX,
22 — эпикардиальный лигамент, 23 — гипокардиаль-
ный лигамент IX, 24 — сердце, 25 —
дорзовентральныи мускул VIII, 26 — остие сердца X, 27 — гипокар-
диальный лигамент X, 28 — гипокарднальныи
лигамент VIII, 29 — дорзовентральныи мускул IX, 30 —
дорзовентральныи мускул X, 31 — гипокардиальный
лигамент XI, 32—медиальный продольный мускул XI,
33 — медиальный эндостернит XI
202
III. Дорзовентральные мускулы (m. dorso-
ventrales). Четыре пары. Первая пара очень
мала, располагается на задней границе
стебелька и принадлежит стебельковому сегменту.
Начинается она на брюшной стенке тела и
проходит вверх к верхним аподемам lorum (см. рис.
181, 14). Три следующие пары принадлежат
VIII, IX и X сегментам и снизу прикрепляются
к соответственным медиальным эндостернитам,
а верхними концами к спинной стенке тела. Они
развиты неодинаково: передняя пара очень
мощная (см. рис. 180, 12; 181, 25), следующая
несколько слабее (см. рис. 180, 34; 181, 29), зад-
ня-я очень тонка (см. рис. 180, 27; 181, 30).
В XI сегменте дорзовентральные мускулы
полностью редуцированы.
У низших пауков с сегментированным
брюшком (Liphistiidae) дорзовентральные мускулы
занимают интерсегментарное положение и
верхние их 'концы лежат в одной плоскости с остия-
ми сердца того же сегмента. Но у Labidognatha
спинные концы мускулов вторично смещаются,
а сердце укорачивается, и вследствие этого
первичные отношения мышц и остий нарушаются.
У Araneus наиболее мощные дорзовентральные
мускулы VIII сегмента прикрепляются на епине
по бокам от остий X сегмента (см. рис. 181, 25;
189, 6; 190, 11).
IV. Открыватели легочных стигм. Пара
широких косых мышечных лент, которые
связывают задние стенки легочных мешков с брюшной
стенкой тела (см. рис. 180, 9).
V. Поверхностные мускулы.
Многочисленные, очень своеобразные мускульные пучки,
начинающиеся от брюшной стенки тела (см.
рис. 180," 41) и проходящие между печенью и
гиподермой на спинную сторону. Здесь они
постепенно сливаются друг с другом, образуя
сравнительно немного радиальных тяжей. Эти
последние лежат в неглубоких бороздках печени и
в конце концов подходят к выходящим сквозь
печень на спинную сторону дорзальным
концам латерокардиальных и дорзовентральных
мускулов, а также лигаментов сердца, с
которыми они, видимо, прочно связаны (рис. 183, А,
Б, 7; 189, А, Б, 10).
Пищеварительная система. Передняя кишка
дифференцирована на глотку, пищевод и сосательный желудок. Средняя кишка
разделяется на желудок и тонкую кишку, причем первый лежит еще в
Рис. 182. Araneus sp.
Часть брюшной
мускулатуры (ориг.):
/ — аподемальные выросты
lorum, 2 — lorum, 3 —
задняя граница стебелька, 4 —
боковые аподемы стебелька,
5 — продольный медиальный
мускул VIII сегмента, 6 —
поперечный мускул VIII, 7 —
косой мускул IX, 8 —
продольный мускул,
соединяющий эндостерниты VIII и
X, 9 — медиальный
эндостернит X, 10 — продольный
медиальный мускул XI,
// — эндостернит XI, 12 —
сухожильный тяж, 13 —
медиальный эндостернит VIII.
Мускулы, соединяющие
аподемы lorum с
эндостернитами VIII сегмента, а также
часть медиальных
мускулов IX и X сегментов,
удалены
203
головогруди, а вторая помещается в брюшке, где заканчивается
расширением — клоакой, которая сообщается с очень короткой задней
кишкой. Желудок снабжен парными слепо замкнутыми на концах
железистыми выростами. Тонкая кишка принимает протоки объемистой
печени, целиком лежащей в брюшке.
Все отделы кишечника, находящиеся в головогруди, могут быть
легко изолированы целиком. Для этого, срезав спинной щит бритвой,
надо выварить головогрудь в течение часа в слабом растворе едкого
кали. После тщательной промывки, которая удаляет растворенную и
размягченную мускулатуру, передняя часть кишечника оказывается
совершенно открытой.
Ротовое отверстие имеет вид поперечной щели и ограничено
спереди рострумом, а сзади стернитом педипальпального сегмента,
играющими роль верхней и нижней губ. Ротовое отверстие ведет в
объемистую глотку, которая располагается вертикально, сплющена в передне-
заднем направлении и, вследствие этого, имеет щелевидныи просвет
(см. рис. 191, 18; 192, 21). Ее передняя и задняя стенки выстланы
широкими хитиновыми пластинками, связанными друг с другом по бокам
глотки очень тонкой и нежной, прозрачной кутикулой. На спинной
стороне глотка открывается в тонкий, расположенный горизонтально
пищевод. Обе глоточные пластинки имеют одинаковые размеры, но
отличаются по структуре. Передняя, граничащая с рострумом, снабжена
медиальным желобком, постепенно углубляющимся по мере
приближения к пищеводу, внутрь которого он продолжается (рис. 184, /). Темный
цвет пластинки зависит от просвечивающей железистой гиподермы.
Наружный край снабжен перистыми волосками (рис. 184,2). Задняя
пластинка глотки, прилегающая к нижней губе, очень тонка,
прозрачна, слегка выпукла и обнаруживает тонкую поперечную исчерченность.
Под ней лежит темная железистая гиподерма.
Глотка снабжена мощной мускулатурой. От ее верхнего конца
отходит пара больших (треугольных поднимателей глотки, которые дор-
зально прикрепляются к щиту головогруди (см. рис. 191, 8; 192, 9).
Антагонистами их являются расширители глотки — несколько
мускульных пучков, из которых наиболее заметна одна пара,
начинающаяся тоже от верхнего отдела глотки и проходящая параллельно ее
стенкам вниз к области нижней губы.
Тонкий трубчатый пищевод отходит от глотки назад под прямым
углом (см. рис. 177, 53; 192, 22). Пройдя сквозь окологлоточное
нервное кольцо, под головным мозгом, он соединяется с сосательным
желудком. Полость пищевода выстлана хитиновой кутикулой, очень
тонкой и нежной на брюшной стороне и образующей толстый продольный
желобок на спинной стороне.
Сосательный желудок лежит позади мозга над подглоточной
нервной массой на пластинке эндостернита. В общем он имеет форму
трехгранной пирамиды, обращенной основанием вперед (см. рис. 177, 51;
192, 3). Боковые стенки его сильно вогнуты внутрь (рис. 185). К
стенкам сосательного желудка прикрепляются мощные мускульные пучки,
расходящиеся в разные стороны. Следует осторожно отделить этот
орган и "приготовить тотальный препарат в глицерин-желатине. Полость
205
Рис. 184. Araneus diadematus. Передняя
глоточная пластинка (по Vogt и Yung):
/ — медиальный желобок, г —перистые
волоски, 3 — спинной желобок пищевода
Рис. 185. Пауки Labidognatha. Схема
строения сосательного желудка (из Ger-
chardt и Kastner):
/~ спинная пластинка, 2—пищевод, 3 —
брюшная пластинка, 4 — боковая пластинка,
5 — задний конец, переходящий в - среднюю
кишку
сосательного желудка имеет трехгранную, почти Т-образную форму и
ограничена хитиновой кутикулой, которая утолщается в четыре
продольные пластинки: широкую спинную (рис. 185, /), две боковые
(рис. 185, 4) и желобовидную брюшную (рис. 185,3). Пластинки
соединяются друг с другом нежной гибкой кутикулой. От спинной
пластинки отходят мощные мускулы, которые верхними концами
прикрепляются к спинному щиту головогруди (см. рис. 177, 52; 191, 2; 192, 4).
Боковые и брюшная пластинки связаны мышцами с эндостернитом.
Помимо этих мускулов-расширителей сосательный желудок снабжен
мантией кольцевых сжимателей. Он представляет собой очень мощный
сосательный аппарат.
Следующий отдел кишечника — желудок имеет энтодермальное
происхождение и состоит из железистого эпителия, лишенного
внутренней кутикулы. Сам желудок невелик (см. рис. 177,18), но его боковые
стенки образуют пару больших, разветвленных выпячиваний, которые
гомологичны желудочной железе скорпионов и головогрудным
железистым придаткам, других паукообразных. Эти железистые выросты
загибаются на спинную сторону и вперед, помещаясь сверху и по
бокам от сосательного желудка (см. рис. 177, 54). Здесь между ними
проходят мощные пучки спинных мышц сосательного желудка, а
боковая стенка каждого из них дает четыре широкие боковые ветви. Далее
выросты желудка проходят над головным мозгом и, суживаясь,
заканчиваются под задними концами ядовитых желез (см. рис. 177,2).
Боковые ветви выростов загибаются вниз, внедряются в тазики ходных
206
ног между мышцами, здесь резко загибаются назад и на брюшную
сторону и заканчиваются слепо между подглоточной нервной массой
и стернумом (см. рис. 177, 14).
Желудок продолжается в цилиндрическую тонкую кишку, которая
проходит через стебелек в брюшко, где тянется в виде-медиальной
беловатой трубки, изгибаясь характерной дугой кверху более или
менее параллельно выпуклой спинной поверхности брюшка и несколько
расширяясь (см. рис. 177, 45; 192, 29). Она лежит в массе печени
между сердцем и половой железой и на своем пути принимает протоки
печени. Задний конец тонкой кишки открывается в широкий мешок-—
клоаку—грушевидный мешок, занимающий около 7б части брюшка,
с толстым закругленным передним концом, суживающимся кзади
(см. рис. 177, 39). Кишка открывается в клоакальный мешок с его
брюшной стороны, задняя часть кишки проходит под нижней стенкой
мешка, будучи настолько тесно с ним связана, что ее трудно отделить.
В клоаке почти всегда лежат крупные, черные веретенообразные
экскременты. У места впадения тонкой кишки в клоаку открываются две
пары мальпигиевых сосудов белого цвета (см. рис. 177,42).
Сзади клоака сообщается с очень короткой задней кишкой
(см. рис. 177,38), выстланной хитиновой кутикулой и
заканчивающейся анальным отверстием (см. рис. 177, 36).
Печень представляет собой сплошную массу, не разделяющуюся на
правую и левую доли (см. рис. 177, .9), но по происхождению является
парным органом. Наличие нескольких пар печеночных протоков
указывает, кроме того, на 'метамерную природу этой железы (ом. рис. 177,43).
У многих пауков имеется непарный брюшной проток, впадающий в
кишку впереди, и четыре парных протока. Но у Araneus кроме этих
первичных протоков развивается еще несколько вторичных: изменчивой
формы. Коричневатая ткань печени слагается из бесчисленных мелких
железистых долек, что видно хорошо уже при небольшом увеличении
лупы или бинокуляра. Печень занимает большую часть брюшка,
окружая все органы, за исключением легочных мешков. Полости долек
сообщаются друг с другом и связаны в одну массу рыхлой
соединительной тканью.
Наружная поверхность печени покрыта неглубокими бороздками,
более обильными на брюшной стороне; в них лежат тонкие подкожные
поверхностные мускульные волокна, соединяющиеся на спинной
стороне с дорзовентральными мускулами и лигаментами сердца, которые
прободают печень (см. рис. 183, 7).
Жидкая пища из сосательного желудка поступает в слепые
мешки желудка и в полости печени. В этих органах она переваривается и
всасывается, причем главную роль играет печень.
Эпителий последней состоит, как и у других паукообразных, из
ферментных и резорбирующих, или питательных, клеток. Первые
выделяют ферменты, поступающие в просвет печени. Питательные клетки
воспринимают частицы пищевой кашицы с ферментами и переполнены
вакуолями, в которых продолжается переработка пищевых веществ.
Непереваренные остатки в виде темных экскреторных шаров и
конкреций выталкиваются в просвет печени, откуда затем выносятся в тонкую
207
5
Рис. 186. Aranei. Схема строения легкого. А— наружная; Б — средняя и
В — внутренняя часть легкого (по Kastner):
/ — легочная крышечка, 2 — наружный угол легочного мешка, 3 — щелевидное
пространство между соседними легочными пластинками, 4 — полость преддверия, 5 —
стигма, 6 — внутренний угол легкого, 7 — внутреннее лакунарное пространство
легочной пластинки, 8 — задний, свободный край легочной пластинки, 9 —
соединительнотканные столбики между стенками легочной пластинки, 10 — внутренняя поверхность
легочной пластинки
кишку и далее в клоаку. В печени пауков обнаружены сильный протео-
литический фермент, близкий к трипсину, липаза и_амилаза.
Органы дыхания. Подобно большинству Labidognatha, паук-
крестовик имеет пару легких и две пары трахей. Легкие лежат на
вентральной стороне брюшка впереди эпигастральной борозды и
принадлежат VIII туловищному сегменту, на котором, кроме того,
помещается половое отверстие. Трахеи открываются наружу общей непарной
стигмой впереди паутиновых бородавок и относятся к следующему,
IX сегменту.
Стигмы (stigmata), или дыхальца, легких имеют вид поперечных,
щелевидных отверстий; лежащих на краях эпигастральной борозды
позади легочных крышечек (см. рис. 173, 3). С внутренней стороны они
ничем не отделены от борозды, непосредственное продолжение которой
представляют. Соответственно и преддверие легкого, в которое
открывается каждая стигма, на внутреннем своем крае непосредственно
соединяется с узкой полостью эпигастральной складки и через нее
сообщается с полостью преддверия противоположной стороны.
Преддверия легких, таким образом, являются как бы местными
расширениями полости эпигастральной складки (см. рис. 178, 7; 186,
4), а стигмы — боковыми участками эпигастральной щели. В то же
208
Рис. 187. Araneus diadematus. Разрез легкого.
Легочные пластинки перерезаны поперек (по
Kastner):
/ — печень, 2 — циркумпульмональный синус, 3 —
мускульные пучки, 4 — легочные пластинки, 5 —
складкн хитина легочной крышечки, в — легочная
крышечка,. 7 — стигма, 8 — преддверие, 9 —
пространства между соседними легочными пластинками,
10 —■ хитиновые столбики на задней стенке
преддверия
время преддверия легких
являются задними отделами
легочных мешков. Последние
уплощены дорзовентрально,
особенно спереди, имеют
треугольную форму и одной из
вершин обращены вперед (см.
рис. 180,7). Они окружены
тонкостенными циркумпульмо-
нальными кровеносными
синусами (см. рис. 180,5), передние
концы которых, суживаясь,
образуют легочные вены (см.
рис. 180,3). По общему плану
строения легкие пауков и
скорпионов схожи. Большая часть
легкого заполнена
многочисленными (у взрослых самок до
100) легочными пластинками,
свободной от них остается
только задняя часть легочного мешка, т. е. преддверие (рис. 178, 7;
186, 4). Пластинки представляют собой тонкие уплощенные складки
стенок легкого (см. рис. 178, 6). У Araneus они имеют форму
вытянутых продольно равнобедренных треугольников, обращенных короткой
стороной назад, и располагаются параллельно друг другу и почти
параллельно брюшной поверхности тела. Хитиновая кутикула,
покрывающая обе стороны пластинок, непосредственно переходит в хитиновую
стенку легочного мешка. Остаются свободными только задние края
пластинок, обращенные в сторону преддверия (рис. 186, 8). Между
ними, на вскрытом легком, видны многочисленные параллельные друг
другу щели, ведущие в промежутки между легочными пластинками
(рис. 186,3).
Для более полного ознакомления с легкими желательно сделать
дополнительные наблюдения на продольных срезах, пересекающих
легочные пластинки поперек (рис. 187). При большом увеличении видна
структура пластинок. Каждая из них ограничена очень тонкой
хитиновой кутикулой (рис. 188, 4), которая, однако, на заднем свободном краю
пластинки значительно утолщается (рис. 188, //). На верхней стороне
каждая пластинка несет маленькие хитиновые столбики, слегка
расширенные на концах (рис. 188, 7). Они не позволяют соседним
пластинкам слипаться, благодаря чему в промежутках между пластинками
всегда находится воздух. На задних краях пластинок концы столбиков
соединяются друг с другом дужками, образуя характерное арочное
окаймление (рис. 188, 10). Хитиновые столбики покрывают и заднюю
стенку преддверия легкого (см. рис. 187, 10). Под кутикулой
пластинок лежит очень тонкий слой гиподермы с редкими ядрами,
утолщающийся в области заднего края (рис. 188, 2, 8). Узкие лакунарные
пространства легочных пластинок (рис. 186, 7; 188, 3) кое-где
пересекаются соединительнотканными мостиками, соединяющими обе стенки
209
Рис. 188. Araneus diadematus. Разрез через легочные пластинки
(по Kastner):
/ — амебоциг, 2 — гиподерма, 3—'лакунарное пространство внутри
легочной пластинки, 4 — хитиновая кутикула, 5 — легочные пластинки, 6 —
промежутки между легочными пластинками, 7 — хитиновые столбики, 8 —
утолщение гиподермы, 9 — вход из преддверия в пространство между
пластинками, 10 — задний край пластинки, // —утолщение хитина, 12—.
арочное окаймление края пластинки
(см. рис. 186, 9). Эти пространства наполнены кровью, которая
заходит сюда из окололегочного синуса, и газообмен совершается через
тонкие стенки пластинок. Легкое прочно фиксируется на брюшной
стенке тела посредством мелких мускульных пучков (см. рис. 187, 3),
прикрепляющихся главным образом к глубоким складкам хитина,
легочной крышечки (см. рис. 187, 5).
Обновление воздуха внутри легких происходит благодаря
вентиляции, которая осуществляется работой особых мускульных пучков —
открывателей стигм. Эти последние спереди прикрепляются к задней
стенке преддверия легкого над краем стигмы и тянутся назад и внутрь
к брюшной стенке тела (см. рис. 180, 9). Сокращаясь, они тянут назад
заднюю стенку преддверия и открывают стигму. При этом полость
преддверия резко расширяется и в него заходит воздух извне. При
расслаблении мускула задняя стенка преддверия, вследствие своей
эластичности, занимает прежнее положение, при этом объем преддверия
уменьшается, а стигма закрывается. Обновление воздуха между
легочными пластинками совершается главным образом через диффузию.
Трахеи лежат на брюшной стороне тела (см. рис. 178, 8, 9). Эти
нежные тонкостенные трубочки выстланы тонким прозрачным слоем
хитина. Трахеи внутренней пары в передней трети слегка расширяются
(см. рис. 178, 8); более длинные боковые трахеи на всем протяжении
210
имеют одинаковый диаметр (см. рис. 178, 9), Обе пары открываются
в небольшое общее преддверие (см. рис, 178, 10), которое сообщается
с внешней средой через непарную стигму (см. рис. 173, 5). При
рассматривании трахей под микроскопом видно, что они лишены
спиральной опорной хитиновой нити, столь характерной для трахей насекомых
и некоторых паукообразных (Solifugae).
Происхождение обеих пар трахей различно. Наружная пара есть
видоизменение второй пары легких низших четырехлегочных пауков, и
отвечает первой паре легких скорпионов. Внутренняя пара
образовалась из парных аподемальных выростов на задней границе IX сегмента.
Кровеносная система. Сосудистая система изучается
посредством инъекции через сердце. Для этого у только что
умерщвленных пауков обнажается сердце. Вскрытие следует производить в
физиологическом растворе. Инъекция делается шприцем с очень тонкой
иглой, а в качестве краски употребляется берлинская лазурь или
карминовая желатиновая масса. Игла шприца вводится в сердце
наискось острием вперед. После инъекции объект уплотняется в спирте.
Хорошие результаты дает просветление в гвоздичном масле1.
Ознакомившись с морфологией кровеносной системы, полезно
произвести наблюдение под бинокуляром или микроскопом на живых
только что вылупившихся из яиц паучках Araneus и других форм. На них
сквозь прозрачные стенки тела обычно хорошо видны сокращения
сердца и все кровообращение.
Сердце представляет собой мускулистую трубку, которая
располагается в передней части брюшка, над кишечником, будучи покрыта
довольно толстым слоем печени (рис. 177, 44; 189, 2). Оно сильно
изогнуто соответственно передней выпуклости брюшка, а при
рассматривании сверху, со спинной стороны, видна только задняя его половина
(рис. 189, А, 2). Сердце лежит в тонкостенном перикардии (рис. 177,
47; 189, 1; 190, 2), который, в свою очередь, окружен узким лакунар-
ным пространством, граничащим с печенью. На его спинной стороне
легко обнаруживаются три пары остий — маленькие, щелевидные,
косые отверстия. Первая пара лежит на переднем конце (рис. 189, Б, 20;
190, 17), вторая —позади середины (рис. 189, А, 5; 190, 16) и третья —
несколько впереди его заднего конца (см. рис. 189, А, 7; 190,15). В
области остий от брюшной стороны сердца отходят тонкие
соединительнотканные тяжи — гйпокардиальные лигаменты (ligamenta hypocardia-
lia, см. рис. 181, 28, 27, 23). Они проходят сквозь печень, между
внутренними органами на брюшную сторону и здесь прикрепляются к
медиальным эндостернитам VIII, IX и X сегментов. Кроме того, имеется
еще четвертая пара гипокардиальных лигаментов, берущая начало от
заднего конца сердца (см. рис. 181, 31). Она прикрепляется к эндо-
стернитам XI сегмента.
Очевидно, сердце имеет сегментарное строение и слагается из
участков, принадлежащих трем брюшным сегментам (VIII, IX и X),
1 Ввиду того что у всех Labidognatha расположение и развитие кровеносных
сосудов сходно, можно Araneus заменить другими более крупными формами, например
тарантулом — Trochosa.
211
О. у К
О ~ о л
I Я м 2
.. I S "> з
— ч> >> Й
П.
о
О)
t- * 1> I
5 S Л I
о- ч 2, к oj^<
О) >.nuS
с w ё; р я „-
ы s и ч §
- ° Й5
§S&[|£
О- Ч J.^g'M
о ^ л я '
•я ait»*
2 S 1 S о>^
2 « 5 s в .
C£S ft.д,
« § i 5ls
00 So «
-3 W „ я О c
Si «''■о»
ел 3 «sk
o. ft? 3 _ «
и « « 5 <u
3 e.«!«
§ «Sl||
.*4I'
F4 (U
0) ««S3 I
2, o,H S 5
o T s .a
~- « ЗЙ I i-
■ £ oi.s
О «« я В «
s ftg x ч
^ я S ft
^ «;»«
s s ш а
£ft§5
" l S
I « I S
a,
и рудиментарной, недоразвитой части,
лежащей позади третьей пары остий и
относящейся к XI сегменту. Сравнение с сердцем
низших пауков, обладающих сегментированным
брюшком (Liphistiidae), показывает, что
остии и гипокардиальные лигаменты, так же
как и дорзовентральные мускулы,
первоначально помещаются на задней границе своего
сегмента, т. е. интерсегментарно. У Araneus,
как у всех высших пауков, сердце сильно
укорачивается, теряя свои задние кардиомеры,
что, конечно, связано с редукцией задних
брюшных сегментов. Параллельно с этим
места прикрепления дорзовентральных мышц на
спине сдвигаются назад, а первая пара остий
вперед ближе к легким. Первичное
сегментарное расположение этих органов на спинной
стороне нарушается (см. рис. 181).
В области остий сердце образует!
расширения, между которыми лежат еще вторичные'
расширения. В результате оно приобретает
вид органа, состоящего как бы из пяти мета-,
мер (рис. 190). Некоторые авторы именно так
трактовали метамерию сердца высших пауков,
причем отсутствие остий на четвертом и
пятом метамерах сердца неправильно
рассматривалось как результат редукции.
Помимо гипокардиальных лигаментов в
прикреплении сердца участвуют и другие
тяжи соединительнотканной природы. Сверху к
сердцу прикрепляются в количестве девяти,
пар короткие эпикардиальные лигаменты
(1. epicardialia, рис. 181,22; 189,15; 190, 6).
Они подвешивают сердце к спинным
покровам, причем места-их прикрепления к
кутикуле располагаются медиально (рис. 183, 1).
На каждый полный кардиомер их приходится
четыре пары. От краев остий отходят верхние
экзокардиальные лигаменты (1. exocardialia
superiores). Соответственно числу остий они
представлены тремя парами — в VIII, IX и
X сегментах (189, 13; 190, 4). Но, кроме того,
по паре сходных с ними лигаментов
начинается от обоих вторичных расширений сердца
(рис. 189, 14; 190, 5). От боковых стенок
сердца отходят нижние экзокардиальные
лигаменты (1. exocardialia inferiores),
принимавшиеся прежде за сердечные мышцы. Они
представлены семью длинными, прочными.тя-
- ,1 * .
Рис. 190. Сердце Araneus
sp. Вид сверху (ориг.):
/ — передняя аорта, 2 —
перикардий, 3 — полость
перикардия, 4 — верхние
экзокардиальные лигаменты, лежащие на
уровне остий, 5 —
дополнительные верхние экзокардиальные
лигаменты, 6 —
эпикардиальные лигаменты, 7 — сердце,
8 — латеро-кардиальный
мускул, 9 — нижние
экзокардиальные лигаменты, 10— боковые
артерии, 11 — дорзовентральный
мускул VIII сегмента, 12 —
дорзовентральный мускул IX, 13 —
задняя аорта, -14 — непарный
задний нижний экзокардиаль-
иый лигамент, 15 — остие X,
16 — остие IX, П — остие VIII,
18 — легочная вена. VIII—XI —
1 кардиомеры
213
15 ie 11
Рнс. 191. Agelenidae. Артерии
головогруди. Вид слева (по Causard из
Gerhard и Kastner):
/ — передняя аорта, 2 — мускулатура
сосательного желудка, 3 —- сосательный
желудок, 4 — truncus peristomacalis, 5 —-
толстая артерия, 6 — головная артерия, 7—
головной мозг, 8—подниматель глотки,
9 — глазная артерия, 10 — зрительный нерв,
И,— глаза, 12 — хелицеральная артерия,
13 — хелицеральный нерв, 14 — хелицера,
15 — губная артерия, 16 — верхняя губа
(рострум), 17 — ротовое отверстие, 18 —
глотка, 19 — артерия педипальпы, 20 —
педальные артерии, 21 — подглоточная
нервная масса, 22 — спннальная артерия, 23 —
брюшной нервный ствол, 24 — стебелек
fS.fs 20
Рис. 192. Tegenaria. Медиальный
разрез головогруди. Изображены
артерии правой стороны (из Gerhard
и Kastner):
/ — передняя аорта, 2 — желудок, 3 —
сосательный желудок, 4 — мускулатура
сосательного желудка, 5 —truncus
peristomacalis, 6 — толстая артерия, 7 —
головная артерия, 8 — головной мозг, 9 —
подниматель глотки, 10 — глазная артерия,
// — зрительный нерв, 12 — глаз, 13 —
ядовитая железа, 14 — хелицеральная
артерия, 15 — проток ядовитой железы, 16 —
хелицера, 17 — хелицеральный нерв, 18 —
рострум, 19 — ротовое отверстие, 20 —
нижняя губа, 21 — глотка, 22 — пищевод,
23 — межузелковые артерии, 24 — невроме-
ры подглоточной нервной массы, 25 — эн-
достернит, 26 — спинальная артерия, 27 —
стебелек, 28 — нервный ствол, 29 — тонкая
кншка
жами, которые выходят на спинную сторону тела на некотором
расстоянии от сердца (рис. 183, А, 3; 189, А, 11; 190, 9). Здесь они
прикрепляются к кутикуле. В VIII сегменте они отсутствуют, зато на заднем
конце сердца имеется пара, принадлежащая XI сегменту (рис. 190, 9),
и еще один такой же непарный лигамент (рис. 190, 14).
Функция лигаментов Araneus сходна с таковой у скорпионов (см.
с. 168). Они служат для фиксации сердца, а также, вследствие своей
растяжимости и эластичности, обеспечивают расширение сердечной
трубки после каждой систолы.
Остии сердца снабжены клапанами, которые пропускают кровь
только из перикардия в сердце. Это небольшие складочки, свисающие
от краев остий в полость сердца. Во время систолы они автоматически
замыкаются. Осторожно вскрыв стенку сердца, их можно рассмотреть
в бинокуляр.
Артериальные сосуды. От сердца берут начало: впереди — передняя
аорта, сзади — задняя аорта и три пары боковых артерий. Последние
принадлежат IX, X и XI сегментам; из них две передние пары
начинаются ниже остий, а третья отходит от заднего конца сердца (рис. 189,
А, 12; 190, 10). В начале этих сосудов имеется клапан, пропускающий
кровь только из сердца. Из этих клапанов сравнительно легко можно
наблюдать лишь клапан передней аорты, для чего надо вскрыть осно-
214
вание сосуда. Он
представляет собой 'косую направленную
вперед складку спинной
стенки аорты.
Передняя аорта (aorta
anterior, рис. 177, 50; 190, 1)
проходит сквозь стебелек в
головогрудь, где тянется вперед
над кишкой. Затем она
разветвляется на два одинаковых
ствола (trunci peristomaca-
les, рис. 177, 17; 191, 4;
192, 5), которые проходят по
бокам сосательного желудка
до задней стороны головного
мозга. Здесь каждый ствол
делится на две ветви —
головную и толстую артерии.
Головная артерия (а. сер-
haiica, рис. 191, 6; 192, 7)
тянется по бокам мозга вперед.
Она дает несколько ветвей к
мозгу, которые проходят
частью по его поверхности,
частью в соединительнотканной
пластинке, разделяющей
отделы мозга (протоцеребрум и
тритоцеребрум). Далее каждая головная артерия разделяется на
глазную артерию (a. oculorum, рис. 191, 9; 192, J0), несущую ировь к
переднему краю головогруди и к глазам, и артерию хелицер (a. cheliceralis,
см. рис. 191, 12; 192, 14).
Толстые артерии (a. crassae) загибаются вниз. Они очень коротки
и кончаются на верхней стороне подглоточной нервной массы (рис. 191,
5; 192, 6). Здесь они разделяются на артерии конечностей — педипальп
и ходных ног (a. pedipalpales, рис. 191^/2; 193, 2; 177, 4 и a. pedales,
рис. 191, 20; 177, 15; 193, 4). Обе a. crassae соединяются друг с другом
поверх подглоточного мозга посредством шести поперечных ветвей (га-
mitransversales, рис. 193, И). Из них пять первых ответвляют по
одной медиальной межузелковой артерии (a. interganglionaris, рис. 192,
23), которая проходит на брюшную сторону нервной массы, залегая в
соединительнотканной прослойке между двумя соседними невромерами.
Передний из этих межузелковых сосудов проходит под пищеводом
сквозь окологлоточное нервное кольцо и разделяет, таким образом,
хелицеральный и педипальпальный ганглии. Четыре других — проходят в
пограничных соединительнотканных прослойках между ганглиями пе-
дипальпального сегмента и четырех сегментов ходных ног.
Последняя поперечная ветвь дает начало мощному продольному
сосуду — спйнальной артерии (a. spinalis, см. рис. 191, 22; 192, 26),
которая тянется назад над задней частью подглоточной нервной массы.
Рис. 193. Agelena. Подглоточная нервная
масса с ее сосудами. Вид сверху (по
Causard из Gerhard и Kastner):
/ — рострум, 2 — педипальпальная артерия, 3 —
толстая артерия, 4 — педальные артерии, 5 —
спинальная артерия, 6 — брюшной нервный ствол,
7 — стебелек, 8 — подглоточная нервная масса,
9 — педальные нервы, 10 — педипальпальный нерв,
// — поперечные артерии
215
Следующие семь вертикальных межузелковых сосудов, проходящих
между невромерами сегментов брюшка, берут начало на ее нижней
стороне (см.. рис. 192, 23). Спинальная артерия проходит затем по нижней
стороне нервных стволов, направляющихся в брюшко от заднего конца
подглоточной нервной массы.
Органы брюшка снабжаются кровью через слабо развитую заднюю
аорту (a. posterior, см. рис. 177, 40; 190, 13), называемую также
хвостовой артерией (a. caudalis), и три парные боковые артерии (a. latera-
les, см. рис. 190, 10). Все эти сосуды глубоко внедряются в массу
печени, где многократно ветвятся.
Венозная сосудистая система. Из мелких разветвлений
артериальных сосудов кровь изливается в многочисленные мелкие лакуны и
щели. Затем она омывает внутренние органы, становится венозной и
собирается наконец в. более крупные венозные синусы.
В головогруди венозная кровь собирается на брюшной стороне,,
причем главную массу составляет кровь, вытекающая из конечностей
и нервной системы. Отсюда кровь течет по парным синусам стебелька
в брюшко, где поступает в брюшной венозный синус (sinus ventralis),
расположенный между лентами продольных мышц. В брюшке венозная
кровь в конце концов собирается в этот же синус. Из последнего часть
крови затем поднимается по наружной стороне печеночных долек к
перикардию, другая, большая часть поступает в окололегочные синусы
(s. circumpulmonales, см. рис. 180, 5). Из легких артериальная кровь
проходит по паре легочных вен (v. pulmonales, см. рис. 177, 49; 180, 3;
190, 18) в переднюю часть перикардия, откуда уже попадает в сердце
через, остии.
Кровь пауков бесцветна.
Выделительная система. Экскреторную функцию несут
коксальные железы, мальпигиевы сосуды, клоака, нефроциты и
гуанинов ые клетки.
Коксальные железы — видоизмененные целомодукты, как и у
других Arachnida. У всех Labidognatha имеется лишь одна пара,
принадлежащая III сегменту туловища и открывающаяся у основания первой
пары ходных ног. У Araneus, однако, коксальные железы рудиментарны
и малы, поэтому при вскрытии их трудно обнаружить. Каждая железа
состоит из двух частей — одна отвечает мешочку (sacculus, рис. 194,/),
другая является сильно редуцированным лабиринтом (рис. 194, 2).
Нахождение коксальных желез
облегчается, если за сутки до вскрытия
живому пауку ввести в полость тела
раствор аммиачного кармина, который
выделяется мешочками. Инъекция
производится в одну из ходных ног
стерильной пастеровской пипеткой или
шприцем с очень тонкой иглой.
Рудиментарное состояние коксальных
желез у паука-крестовика многие
авторы объясняют тем, что сильно
развитые паутиновые железы извлекают из
у~\_
Ж
Рис. 194. Aranei. Схема строения
' коксальной железы (по Buxton):
/ — мешочек, 2 — лабиринт, 3 — выводной
проток,. 4 — отверстие коксальной железы.
III—VI — сегменты головогруди
216
Рис. 195. Мальпигиевы сосуды
Araneus diadematus. Вид со
едины (из В. А. Догеля):
/ — мальпигиев сосуд, 2 — тонкая
кишка, 3 — клоака
тела экскреты и утилизируют их при
образовании вещества паутины.
Две пары мальпигиевых сосудов энто-
дермального 'происхождения открываются в
клоаку с брюш-ной стороны. Это тонкие,
белые, сильно разветвляющиеся трубочки,
которые за пределы брюшка не выходят
(рис. 177, 42; 195, /). Конечные
раз-ветвления их заканчиваются слепо. Оба сосуда
на каждой стороне сливаются в очень
короткий общий проток.
Продуктом выделения является гуанин.
При инъекции в полость тела неитральрота
мальпигиевы сосуды окрашиваются.
Выделительную функцию выполняет
также клоака — большое расширение
средней кишки (рис. 177, 39; 195, 3).
Гистологическое строение стенок клоаки
напоминает структуру мальпигиевых сосудов, и физиологически эти органы
чрезвычайно близки. Стенки клоаки, как и "мальпигиевы сосуды, извлекают
из окружающей их крови гуанин и выводят его наружу.
Нефроциты — специальные выделительные клетки отличаются
своей необычайно крупной величиной. Это самые крупные, не считая яиц,
клеточные элементы в теле паука. Они располагаются в головогруди,
преимущественно на брюшной стороне тела, в частности на нижней
поверхности нервной системы, и легко обнаруживаются при простом
вскрытии после инъекции живому пауку аммиачного кармина.
Нефроциты содержат сферические, концентрически слоистые конкреции
гуанина, которые затем освобождаются в растворенном виде и
выделяются мальпигиевыми сосудами и коксальными железами.
Наконец, выделительными являются также ярко-белые гуаниновые
клетки, скопления которых легко обнаруживаются на поверхности
печени на спинной стороне брюшка. Гуаниновые клетки развиваются из
питательных клеток печени и, следовательно, имеют энтодермаяьную
природу. Плазма их густо набита белыми кристаллическими
конкрециями гуанина, которые выделяются затем в полость печени, откуда
гуанин в растворенном виде выносится в клоаку и через заднюю
кишку — наружу. -
Нервная система. Все г'англии брюшной нервной цепочки
слиты в одну большую подглоточную нервную массу, которая
помещается на брюшной стороне в головогруди между слепыми выростами
желудка и эндостернитом (рис. 177, 16; 196, 4). На ее передней части
располагается надглоточный, или головной, мозг. Он невелик и на
верхней стороне несет ясную медиальную бороздку (рис. 177, 55; 196,
/). С подглоточной массой мозг соединяется короткими, но толстыми,
ганглиозными коннективами. Между ними проходит пищевод.
Надглоточный мозг дает несколько парных нервов. От верхнего
переднего края берут начало толстые зрительные нервы (п. optici,
см. рис. 177, 56; 195, 21), основания которых тесно сближены в меди-
217
альной плоскости. Нервы хелицер (п.
chelicerae, см. рис. 177, 3) начинаются
под ними от нижнего отдела мозга и
проходят над пищеводом.
Подглоточ'ная нервная масса
состоит из двух неравных частей: передней —
очень широкой и звездообразной,
слагающейся из ганглиев головогруди
(рис. 196, 4), >и задней —очень узкой и
вытянутой, которая образована
ганглиями брюшка (рис. 196, 5). Передняя
часть подглоточной массы образует пять
парных баковых лопастей (рис. 193, 8;
196, 4). От них начинаются мощные
нервы конечностей. Таким образом, эта
часть подглоточной массы имеет
отчетливую наружную сегментацию, и ее ме-
тамериый состав совершенно ясен. В
состав ее входят ганглии педипальпально-
го и четырех следующих сегментов.
Задний суженный участок
подглоточной массы состоит из семи слившихся
брюшных невромеров, которые
обнаруживаются только на срезах. Нервные
волокна, отходящие от них, . соединяются
сзади в мощный нервный ствол,
тянущийся в брюшко (рис. 196, 7). Здесь он
делится на два ствола, которые проходят
с внутренней стороны продольных
брюшных мускулов (рис. 196, 8). Сначала
каждый из этих стволов ответвляет
боковой нерв VII сегмента,'т. е. стебелька,
и затем в основании брюшка
последовательно дает три ветви. Первые две ветви
принадлежат VIII сегменту. Из них
одна тянется вдоль медиальной стороны
легкого (рис. 196, 19), а другая — в
косом направлении над легким к
латеральному эндостерниту VIII сегмента
(рис. 196, 18). Третья ветвь относится к
IX сегменту и проходит вдоль брюшной
стенки тела назад (рис. 196, 17).
Главные нервные стволы достигают медиальных эндостернитов
IX сегмента, где разделяются каждый на три ветви: нерв передних пау-
тиновых бородавок( рис. 196, 16), нерв задних и внутренних бородавок
(рис. 196, 15) и нерв XII сегмента (рис. 196, 14), направляющийся к
нижней стенке клоаки и к области анального отверстия.
Судя по числу невромеров, входящих в состав подглоточной нервной
массы, должна быть еще одна пара нервов для следующего (XIII) сег-
Рис. .196. Нервная система Tege-
naria (по Kastner):
/ — головной мозг, 2 — педнпальпаль-
ный нерв, 3 — нерв ноги первой пары,
4 — головогрудная часть подглоточной
нервной массы, 5 — абдоминальная
часть подглоточной нервной массы, 6—
нерв ногн четвертой пары, 7 —
брюшной нервный ствол, 8 — ветвь
брюшного нервного ствола, 9 — легкое, 10 —
основание передней паутиновой
бородавки, 11 — основание задней
паутиновой бородавки, 12 — задняя паутнновая
бородавка, 13 — анальный бугорок,
/4 — нерв XII сегмента, IS — нерв
задней и внутренней бородавки, 16 — нерв
передней бородавки, 17 — нерв IX
сегмента, 18 и 19 — нервы VIII сегмента,
20 — пищевод, 21 — зрительный нерв
218
мента, который, однако, не имеет ни наружных хитиновых, ни
мускульных элементов.
Органы чувств представлены осязательными волосками,
лировидными органами, тарзальными органами и глазами. Их тонкое
строение может быть изучено только на срезах.
Осязание имеет большое значение в жизни пауков — при бегании
по паутине, устройстве тенет, ловле добычи, спаривании и т. д.
Соответственно с этим осязательные волоски многочисленны и рассеянны
на всех частях тела, особенно на конечностях. Среди них трихоботрии
имеют довольно сложное строение. Они подвижно прикреплены на дне
ямки хитиновых покровов и снабжены у своего основания группой
чувствительных клеток, посылающих тонкие отростки в самый волосок.
При помощи лировидных органов, по-видимому, определяется
растяжение и изгибание покровов, давление на покровы, постановка ног
и т. д., т. е. они служат органами механического чувства. Эти
своеобразные органы представлены одиночными «чувствительными щелями»
и более сложными, собственно лировидными, органами. И те и другие
очень малы (от 50 до 160 мкм длиной) и рассеяны по всему телу,
причем лировидные органы точно локализированы. На рис. 197, А
показано распределение этих органов на одной из ног у самки Araneus undatus
Oliver. У самок их несколько больше, чем у самцов, а вообще их очень
много (самка A. undatus имеет около 2500 одиночных щелей и 132лиро-
в'идных органа).
Чувствительные щели имеют вид маленьких, вытянутых,
ромбовидной формы ямок на хитине (рис. 197, А, 10). Дно их образовано тонкой
хитиновой перепонкой, к которой прикрепляется отросток
чувствительной клетки. Лировидные органы представляют собой совокупность
параллельных друг другу чувствительных щелей (рис. 197, Б) и в
принципе имеют сходное с ними строение. .
Тарзальные органы располагаются у обоих полов на педипаль-
пах и ногах, по одному на верхней стороне тарзального членика ближе
к его основанию (рис. 198, 2). Они хорошо заметны в сильную лупу
или бинокуляр на поверхности членика в виде крошечных ямок, окайм-
Рис. 197. Самка Araneus undatus. А — расположение лировидных
органов на задней стороне ноги второй пары; Б — лировидный
орган (по Vogel):
/ — тазик, 2 — вертлуг, 3 — бедро, 4 — колено, 5 — голень, 6 — пятка, 7 —
лапка, 8 — коготки, 9 — лировидные органы, 10 — чувствительные щели,
//— место отростка чувствительной клетки
г219
* f I—J
w—*—*-y
5
Рис. 198. Araneus diadematus.
Тарзальный орган (по Blumen-
thal):
/ — пятка, 2 — тарзальный орган, 3 —
лапка, 4 — гребневидные коготки, 5 —
крючковидный коготок
Рис. 199. Araneus diadematus.
Схема строения мужской
половой системы (по Jogt и Yung):
J — семенной мешок, 2— семяпровод,
3 — половое ■ отверстие, 4 — семенник
ленных очень тонким кольцевидным хитиновым валиком. На дне ямки
лежит несколько чувствительных папилл, в которых имеются
воспринимающие раздражение клетки. По-видимому, это органы обоняния.
Наконец, подобно большинству пауков, Araneus обладает
четырьмя парами развитых глаз, расположенных в два поперечных ряда на
переднем краю головогруди. Они имеют различное строение. Передние
медиальные глаза относятся к инвертированному- типу, тогда как все
остальные неинвертированы. Передние медиальные глаза, кроме того,
замечательны тем, что к каждому прикрепляется особый глазной
мускул, который, сокращаясь, сдвигает сетчатку и, видимо, служит для
аккомодации.
Половая система. Мужская половая система носит парный
характер. Семенники имеют вид продолговатых мешочков и лежат в
передней трети брюшка на вентральной стороне под печенью. Их
задание концы несколько расширены (рис. 199, 4). Спереди они постепенно
суживаются, переходя без резкой границы в тонкие семяпроводы
(рис. 199, 2), которые достигают уровня эпигастральной щели. Здесь
оба протока загибаются внутрь и назад и открываются в переднюю
часть небольшого мускулистого непарного семенного мешка (рис. 199,
1). Последний открывается мужским половым отверстием на дне
эпигастральной щели.
В качестве совокупительных органов функционируют своеобразно
измененные педипальпы. У молодых самцов, не проделавших еще
последней линьки, педипальпы, так же как и у самок, состоят из шести
члеников, но последний членик сильно вздут и лишен коготка.
Окончательного развития совокупительный аппарат достигает лишь после
последней линьки, превращаясь в чрезвычайно сложное образование
(рис. 201).
Совокупительный аппарат изучается на тотальном препарате. В
покоящемся состоянии все его многочисленные части тесно прилегают
друг к другу, будучи собраны на вогнутой стороне видоизмененного
тарзального членика. В таком состоянии изучить его строение невоз-
220
Рис. 200. Aranei. Схема строения
мужского совокупительного
аппарата (по Вагнеру):
1 — alveolus, 2 — лодочка, 3 —
haematodocha, 4 — bulbus, 5 — каналы в
стенке сперматофора, 6 — сперматофор,
7 — embolus, 8 — отверстие
сперматофора
можно. Поэтому сначала следует
основательно выварить его в едком кали и
затем осторожно расправить и заключить в
глицерин-желатин или в канадский
бальзам.
Понять его строение у Araneus легче,
зная более простое, строение
совокупительного аппарата некоторых других
пауков (например, Segestria, Attus или
Thomisus). Схема такого примитивного
копулятивного органа представлена на
рис. 200. Он прикрепляется на дне
особого углубления — alveolus (рис. 200, 1)
на шестом, тарзальном членике, который,
называется лодочкой, или cymbium
(рис. 200, 2). Собственно копулятивный
аппарат состоит из двух основных
отделов: проксимального — haematodocha
(рис. 200, 3) и дистального — bulbus
(рис. 200, 4). Haematodocha
представляет складчатый мешок из нежного хитина, прикрепляющийся на дне
ямки — alveolus. Bulbus имеет хитинизированные стенки и заканчивается
тонким носиком — отростком—embolus (рис. 200, 7). На его вершине
лежит маленькое отверстие, ведущее в длинную хитиновую трубочку,
слепо замкнутую на конце и называемую семенным каналом, или спер-
матофором (рис. 200, 6). В нее набирается сперма перед спариванием.
Внутри haematodocha находится большая наполненная кровью лакуна,
соединяющаяся с лакунами внутри лодочки.
Дри спаривании самец мускульными сокращениями брюшка вгоняет
кровь в эти лакуны, и haematodocha сильно раздувается, расправляя
совокупительный аппарат. Лакунарная полость haematodocha
сообщается также с полостью сперматофора через тонкие поры в его стенках
(рис. 200, 5). Находящаяся под давлением кровь проникает сквозь эти
поры в канал сперматофора и выталкивает семя через отверстие на
конце embolus.
Все эти основные отделы имеются и в копулятивном аппарате
Araneus. Однако вследствие их сложной дифференциации, развития
дополнительных хитиновых частей и отростков весь аппарат приобретает
необычайно сложное строение. Главнейшие усложнения сводятся к
следующему: haematodocha очень велика и распадается на три отдела —
базальный, прикрепленный на дне alveolus (рис. 201, 7), средний
(рис. 201, 8) и дистальный, на котором лежит bulbus с embolus и
который обозначается как haematodochula (рис. 201, 9). В среднем отделе
развиваются дополнительные хитиновые пластинки — tegula (рис. 201,2).
Сперматофор длинен и глубоко внедряется в haematodocha (рис. 201,3).
Рядом с bulbus на haematodochula расположены хитиновые отростки —
терминальный и субтерминальный апофизы (рис. 201, 11, 12). Cymbium
ладьевидно изогнут (рис. 201, 5) и у основания несет загнутый
крючковидный отросток (paracymbium, рис. 201, 6).
22Ь
Рис. 201. Araneus diadetnatus.
Мужской совокупительный
аппарат в расправленном
состоянии (по Вагнеру):
/ — embolus, 2—-tegula, 3— сперма-
тофор, 4 — alveolus, 5 — лодочка,
S — paracymbium, 7 — основной
отдел haematodocha, 8 — средний
отдел haematodocha, 9 — haematodo-
chula, 10— bulbus, //—- субтерми-
нальный отросток, 12 —
терминальный отросток
Рис. 202. Araneus. Схема
строения женской половой
системы (по Comstock):
/ — яйцевод, 2 — семяприемник, 3 —
железа семяприемника, 4 — яичник,
5— наружное отверстие
семяприемника, 6 — оплодотворительный
канал,. 7— вторичное половое
отверстие, 8 — матка
Спаривание у Araneus совершается сходно с другими пауками.
Незадолго до спаривания самец делает небольшой паутиновый гамачок
треугольной формы, натягивая его между какими-нибудь стебельками
или веточками. Затем он выпускает из полового отверстия на гамачок
капельку семенной жидкости и погружает в нее концы педипальп.
Семя наполняет узкие каналы сперматофора в силу их капиллярности.
При самом акте копуляции haematodocha наполняется кровью и весь
аппарат расправляется. Emboli вводятся в семяприемники самки,
причем терминальные отростки служат для прикрепления к женскому
совокупительному аппарату. По-видимому, такую же роль играют и
отростки на дистальных tegula.
Женская половая система состоит из внутренних половых органов
и наружного совокупительного аппарата.
Внутренние органы представлены парными яичниками,
яйцеводами и семяприемниками и непарной маткой. Дистальный отдел
последней и семяприемники представляют впячивание покровов и
соответственно выстланы внутри хитиновой кутикулой.
Продолговатые яичники лежат в брюшке между кишечником и
вентральными продольными мускулами и окружены паутиновыми
железами и печенью (рис. 177, 27). Это две широкие трубки, покрытые
снаружи многочисленными яйцевыми фолликулами, вследствие чего
имеют гроздевидную форму (рис. 202, 4). Задние концы яичников в
противоположность таковым у многих других пауков не соединяются
друг с другом. Размеры и внешний вид яичников варьируют в
широких пределах в зависимости от их физиологического состояния. В пе-
222
риод размножения они сильно увеличиваются, сжимая окружающие
органы и вызывая резкое уменьшение печени.
Каждый яичник на переднем конце открывается в яйцевод
(рис. 202, 1). Впереди оба яйцевода загибаются внутрь и, сливаясь,
образуют довольно широкую, но короткую трубку, называемую
внутренней маткой (uterus internus, рис. 202, 8) и развивающуюся из
мезодермы. Внутренняя матка сообщается с более узкой эктодермальной
маткой (u. externus). Отверстие последней мало и помещается в
глубине эпигастральной щели медиально. Это так называемое первичное
половое отверстие. Выйдя из него, яйца проходят затем по
медиальному участку эпигастральной щели и поступают наружу через вторичное
половое отверстие — участок эпигастральной борозды.
Семяприемники (receptacula seminis, рис. 177, 24; 202, 2) в виде
пары небольших мешочков помещаются по бокам от вторичного
полового отверстия и открываются наружу посредством самостоятельных
протоков (рис. 177, 23; 202, 5). Каждый семяприемник сообщается,
кроме того, с наружной маткой через особый, так называемый
оплодотворительный канал (рис. 202, 6). В проток семяприемника
открывается также небольшой мешочек с очень толстыми хитиновыми стенками
(рис. 202,' 3). Эпителий каждого из этих органов состоит из высоких
железистых клеток, которые открываются в его полость через
многочисленные тонкие поры в хитиновой выстилке.
Совокупительный аппарат самки, или epigyne, изучается на
тотальном препарате, приготовленном после вываривания . в едком кали.
Главную часть составляет довольно большая хитиновая основная
пластинка, которая лежит медиально впереди вторичного полового
отверстия (см. рис. 200, 6). Задне-боковые края ее образуют заметные
вздутия (рис. 203, 4). Внутри последних лежат толстостенные мешочки
желез семяприемников (рис. 203, 3). Сами семяприемники тоже хорошо
просвечивают сквозь пластинку (рис. 203, 2).
К переднему краю основной пластинки прикрепляется длинный и
гибкий, хоботкообразный придаток (рис. 203, 1). Многие авторы
считали, что он служит яйцекладом. В действительности он играет какую-
то роль при спаривании.
Паутиновые железы. Все пауки отличаются сильным,
развитием паутиновых желез, которые всегда лежат в брюшке и
открываются на паутиновых бородавках1. Эти многоклеточные железы
представлены обычно несколькими различными типами. Все они помещаются
на вентральной стороне по бокам от продольной мускулатуры и легко
отпрепаровываются на свежем материале.
Желательно приготовление тотальных препаратов из паутиновых
желез. Для этого они отделяются от паутиновых бородавок,
фиксируются и окрашиваются борным или квасцовым кармином.
У Araneus имеются следующие пять типов желез (табл. 3).
1. Дольковидные (glandulae aciniformes, рис. 177, 37; 204, 5).
Многочисленные мелкие железы; собраны в четыре густых пучка, которые
1 У Amaurobiidae, кроме того, крибеллярные паутиновые железы открываются
через поры cribellum.
223
Таблица 3. Расположение и число наружных отверстий
паутиновых желез на одной стороне тела Araneus
Железы
Дольковидные
Грушевидные
Трубковидные
Ампуловидные
Древовидные
Паутиновые бородавки
передняя
Около 100
отверстий
1 отверстие
внутренняя
Около 100
отверстий
1 отверстие
1 отверстие
задняя
Около 100
отверстий
2 отверстия
3 отверстия
Рис. 203. Araneus diadematus. Женский
совокупительный аппарат — epigyne. Хоботкообразный придаток
отвернут вперед (ориг.):
/ — хоботкообразный придаток, 2 — семяприемник, 3 — железа
семяприемника, 4— боковые утолщения основной пластинки,
5 — отверстия семяприемников, 6 — основная пластинка, 7 —
местоположение вторичного полового отверстия, 8 — эпигастраль-
иая бороздка
Рис. 204. Araneus
diadematus. Паутиновые железы
левой стороны (по Apsteim):
/ — ампуловидная железа, 2 —
древовидная железа, 3— труб-
ковидная железа, 4 —
грушевидные железы, 5 — дольковидные
железы
224
расположены по одному впереди основания каждой задней и
внутренней бородавки. В каждом пучке насчитывается около 100 желез, а
всего, следовательно, имеется до 400. Их выводные протоки открываются
на вершине паутиновых трубочек задних и внутренних бородавок.
Каждая железа имеет форму небольшого овального мешочка, от
которого отходит тонкий выводной проток.
2. Грушевидные (g. pyriformes, рис. 177, 32; 204, 4). Около 200
небольших желез образуют пару пучков по 100 желез в каждом. Они
лежат впереди передних бородавок, на паутиновых трубочках которых
открываются. По размерам и форме очень похожи на железы
предыдущего типа, с которыми прежде смешивались, но отличаются от них
тонким строением и базофильным секретом и поэтому легко узнаются
на окрашенных препаратах.
3. Трубковидные (g. tubuliformes, рис. 204, 3). Три пары больших
желез лежат впереди паутиновых бородавок, прилегая к брюшной
стенке тела. Они имеют вид длинных извитых цилиндрических трубок.
Протоки этих желез открываются на паутиновых конусах задних и
внутренних бородавок. Паутиновые нити, выделяемые ими, употребляются
самкой на изготовление стенок яйцевого кокона. У самцов эти железы
отсутствуют.
4. Ампуловидные (g. ampulaceae, рис. 204, 1). Две пары. Очень
большие железы простираются до переднего конца брюшка, где
передняя пара загибается даже в дорзальном направлении по бокам от
гонад. По внешнему виду они напоминают трубковидные железы, но
каждая в средней части образует большое вздутие. Протоки этих желез
пронизывают паутиновые конусы передних и задних бородавок.
5. Древовидные (g. aggregatae, рис. 204, 2). Три пары крупных
неправильно ветвящихся лопастных желез. Их длинные и тонкие протоки
покрыты мелкими одноклеточными железками и имеют поэтому неров-
. ную бородавчатую поверхность. Две передние пары значительно
крупнее задней. Протоки этих желез открываются на паутиновых конусах
задних бородавок.
Таким образом, самка паука-крестовика имеет в общем около 600
мелких паутиновых желез, связанных с паутиновыми трубочками, и 16
больших желез, открывающихся на паутиновых конусах. Самцы
отличаются только отсутствием трех пар трубковидных желез и
соответствующих им конусов на задних паутиновых бородавках.
Паутиновые железы выделяют вязкий, тянущийся секрет, быстро
застывающий на воздухе в виде паутиновой нити. Однако особенности
секрета каждого типа желез и свойства образуемых ими нитей
неодинаковы и имеют разное значение в жизни паука.
Крестовик строит из паутиновых нитей большую ловчую сеть,
располагая ее вертикально. Основу сети составляет многоугольная рама
(рис. 205, 1) и сходящиеся в центре радиальные нити (рис. 205, 3).
Центр заплетается густой сеточкой (рис. 205, 7). Между радиусами
прикрепляется длинная спиральная нить, начинающаяся в центре сети
и кончающаяся у краев рамы (рис. 205, 5). Спиральная нить двойная,
так как паук протягивает ее дважды; она состоит из особой клейкой
паутины. Именно она является ловчей частью, так как к ней прилипает
8-2
225
попавшее в сеть насекомое.
Рядом с сетью где-нибудь в
укромном уголке паук делает себе
примитивное логовище, или гнездо»
в котором сидит, поджидая
добычу. Между 'центром сети и
убежищем натягивается особая
сигнальная нить. По дрожанию этой
нити паук чувствует, когда в сеть
попадает добыча.
Для построения сети
употребляется секрет всех паутино-
вых желез, кроме трубковидных,
которые выделяют паутину для
яйцевого кокона. Рама и
радиусы делаются из сравнительно
толстых и прочных сухих паути-
новых нитей, выделяемых ампу-
ловидными железами. При этом
несколько желез действуют
одновременно, выделяя каждая по
одной нити, но они соединяются
вместе капельками жидкого
секрета и образуют более толстый
«кабель». В зависимости от
числа действующих одновременно
желез (двух или четырех)
толщина и прочность последнего
изменяется. Грушевидные железы
выделяют пучки тонких и
прочных паутинок, служащих для
прикрепления концов радиальных
.нитей к окружающим
предметам—стволам деревьев, ветвям и т. п. Основу клейких спиральных
нитей образуют, по-видимому, паутиновые нити, выделяемые дольковид-
ными железами. На эту основу накладывается в виде капелек липкий
секрет древовидных желез.
Ядовитые железы помещаются в передней части головогруди
на спинной стороне, располагаясь параллельно друг другу. Их задние
концы прилегают к спинной стенке тела (см. рис. 177, 2). -Это большие
цилиндрические мешки с характерной косой исчерченностью на
поверхности, которая зависит от наличия наружной мускульной мантии из
косых спиральных волокон. Неровный железистый эпителий желез не
образует глубоко вдающихся в полость складок, что хорошо видно на
органе, разрезанном вдоль. От передних концов желез отходит по
тонкому выводному протоку (см. рис. 177, 7), который проходит в хелице-
ру и открывается у вершины когтевидного членика маленьким
округлым отверстием.
Рис. 205. Ловчая сеть Araneus (из Qerhadt
и Kastner):
/ — верхняя нить рамы, 2 — нить рамы второго
порядка, в — радиальная ннть, 4 — рама, 5 —
спиральная клейкая нить, 6 — ловчая зона, 7 —
сетчатый центр, 8 — свободная зона
226
ОТРЯД ACARI — КЛЕЩИ
Ixodes ricinus L. — Собачий клещ
Материал и методы изучения. Ixodes ricinus L. относится к семейству Ixodidae
подотряда Parasitiformes и широко распространен в лесной зоне Евразии, к- Северу
почти до Белого моря; на юге же распространение ограничивается зоной лесостепи.
Паразитирует на различных домашних животных (лошади, крупном и мелком
рогатом скоте, собаке, кошке и других, вплоть до птицы), присасывается также и к
диким животным, часто к ежам и особенно к грызунам (встречается и на ящерицах).
Хозяином собачьего клеща может быть и человек. Относясь к треххозяинным клещам,
/. ricinus в стадии личинки нападает на скот и особенно на мелких диких
млекопитающих (например, на ежей). В нимфу сытая личинка линяет на земле, и нимфа
должна снова присосаться к хозяину и питаться перед окончательной линькой,
которая также происходит на земле. Взрослые клещи (самцы и самки) перед копуляцией
в третий раз нападают на хозяина, питаются, копулируют до или во время сосания
и отпадают для откладки яиц на земле. Поэтому кроме нахождения клещей иа
животных их случайно можно собрать и во время энтомологических экскурсий, когда
голодные личинки, нимфы и взрослые ползают по земле, по траве и кустарнику в
поисках хозяина. Личинки, нимфы и взрослые попадаются преимущественно в первую
половину лета, хотя сроки развития чрезвычайно варьируют, и от полугода,
требующегося для завершения всего жизненного цикла, этот срок может удлиниться до
нескольких лет. Клещи присасываются к телу животного хозяина под шерстью в
местах с наиболее тонкой кожей (в пахе, у корня хвоста, на мошонке самцов, на ушах)
и внедряются глубоко и прочно своим хоботком, вооруженным шипами. Последние
направлены назад, назад же отгибаются и подвижные членики хелицер и при
насильственном снимании клеща настолько прочно удерживаются в коже, что хоботок
нередко отрывается от тела клеща и остается в коже; поэтому требуется
осторожность при снимании клещей, и лучше предварительно ослабить клеща, смазав его
керосином, бензином, маслом с целью закупорки дыхалец. Тогда расслабляются
мышцы, управляющие внедрением хелицер, и' клеща легче сиять неповрежденным.
Захватывать клеща при этом можно и пинцетом и просто пальцами.
Личинок для изучения можно получать в большом количестве, заставив
насосавшуюся самку отложить яйца в пробирке с увлажненным воздухом (внутри
следует держать полоску фильтровальной бумаги, постоянно смачиваемую). Развитие
должно идти также во влажной атмосфере, и при комнатной температуре вылупле-
ние личинок начинается иа 40-й день после откладки яиц.
Внешняя морфология может изучаться на материале, консервированном в
70%-ном спирте. Если стремиться сохранить расправленные ножки, то клещей
следует убивать горячей водой (около 80°С) и потом перекладывать в спирт. Ряд
деталей внешней морфологии удобнее разобрать на готовых тотальных препаратах,
которые изготовляются из отдельных частей тела после их выварки в едком кали.
Можно изготовить препараты и из целого клеща, но для лучшего рассмотрения обеих
сторон их следует отделить друг от друга, подрезая край тела лучше всего острой
копьевидной иглой. При этом хоботок следует оставить со спииной половинкой, а ножки
и дыхальца — с брюшиой. После разъединения каждую половинку необходимо очис-
. тить от внутренностей и мышц и заключить обычным способом в канадский бальзам.
Внутреннее строение предлагается изучать путем вскрытия живого объекта,
которого замаривают предварительно эфиром или хлороформом. Из отпрепарованных
\ органов изготовляют также тотальные препараты, приклеивая их к предметному
стеклу по способу Павловского (см. с. 144).
Внешняя морфология. Тело изучаемого клеща овальной, или яйце-
( видной, формы, в отличие от тела других паукообразных, слито в одно
сплошное целое, на котором не удается найти и следов сегментации,
подразделения на типичные для других паукообразных прозому, мезо-
зому и метазому. Поверхность тела покрыта хитиновой кутикулой, об-
I разующей местами резкие утолщения (щитки), которые в противо-
I положность остальной поверхности тела не способны растягиваться при
227
сосании крови и имеют большое морфологическое и систематическое
значение. Спереди к телу причленяется хоботок (capitulum, gnathoso-
ma), представляющий собой колющий и сосательный аппарат. Снизу,
в передней части тела прикреплены ножки клеща — четыре пары у
взрослого и нимфы и три пары у личинки (рис. 206, 209, 210, 211).
В материале следует найти, наряду со взрослыми клещами, и
стадии их развития — личинок и нимф, резко отличающихся от взрослых
и размерами и морфологией. Необходимо уметь отличать их от
взрослых, и поэтому далее мы вкратце остановимся на особенностях их
организации, параллельно с детальным разбором внешней морфологии
взрослых форм.
При рассмотрении внешнего строения взрослых клещей сразу же
бросается в глаза наличие у /. ricinus полового диморфизма,
сказывающегося кроме различий в размерах в резком морфологическом
отличии (например, сильная хитинизация покровов самца и образование
многочисленных щитков, отсутствующих у самки).
Морфология тела. Плоское тело взрослого самца (рис. 206)
яйцевидной формы, сужено кпереди и окрашено в блестящий
темно-коричневый цвет. Длина около 2,5 мм, ширина 1,5 мм. Большая часть
спинной стороны тела покрыта дорзальным щитком (scutum) — свобод-
Рис. 206. Самец Ixodes ricinus. Вид снизу (А) и сверху (Б) (Х20) (из
Nuttal и Warburton):
/ — гипостом, 2 — педипальпы, 3 — воротничок хоботка, 4 — кокса передней ножки,
5 — прегенитальный щиток, 6 — половое отверстие, 7 — эпимеральный щиток, 8 — пе-
рнтрема со стигмой, 9 — срединный щиток, 10 — аданальный щиток, Я — анальное
отверстие, 12 — анальный щнток, 13 — краевое поле, 14—спииной щиток, 15— церви-
кальные бороздки, 16 — плечи
228
ным от него оказывается узкий светлоокрашенный рантик (краевое
поле) мягкого хитина, вдоль боков и сзади (рис. 206, 13). Поверхность
щитка покрыта тонкими, довольно густыми точками и редкими
волосками. Спереди на щитке имеется широкая вырезка для причленения
основания хоботка. Боковые края вырезки, выдающиеся вперед,
называются плечами (scapulae). От краев вырезки назад отходят
неглубокие, поверхностные цервикальные бороздки. Глаз, помещающихся у
других Ixodidae на щитке, в данном случае нет (как нет их и у самок,
личинок и нимф). Нижняя, вентральная поверхность тела взрослого
самца также покрыта почти сплошь несколькими хитиновыми щитками
и несет на себе анальное, половое и дыхательные отверстия. Здесь же
прикрепляются и ножки.
Анальное отверстие в виде короткой продольной щели лежит у
заднего края тела на переднем конце непарного анального щитка
(рис. 206, 12). Края анальной щели ограничены двумя продольными
сильно хитинизированными клапанами, охваченными в свою очередь
хитиновым кольцом. По бокам от анального щитка помещены два ада-
нальных щитка (рис. 206, 10). Впереди от упомянутых трех щитков
середину нижней стороны тела занимает крупный срединный щиток
(рис. 206, 9), слегка суженный кпереди и оканчивающийся на уровне
основания задних ножек. Здесь же (приблизительно на уровне
оснований третьей пары ножек) помещается и вытянутое поперечной щелью
половое отверстие. Впереди от него лежит небольшой непарный
прегенитальный щиток (см. рис. 206, 5). Наконец, бока тела заняты двумя
длинными энимеральными щитками (рис. 206, 7), к которым
прикрепляются ноги и на которых позади оснований четвертой пары ног
обособляются с каждой стороны на боку особые хитиновые поля (перитре-
мы), окружающие дыхальца. Вогнутая с поверхности перитрема у
/. ricinus имеет овальную
форму, вытянута вдоль, и
дыхальце (стигма) занимает слегка
эксцентричное положение,
будучи смещено вперед. Очень
характерна структура перитре-
мы. Ее поверхность сложена
из неправильных
шестиугольных ячеек (рис. 207, 1).
Дыхальце представляет собой
воздухоносную камеру (рис.
208, 11), сообщающуюся с
наружной средой узким
серповидным отверстием и
продолжающуюся далее внутрь в
пучок веерообразно
расходящихся трахей (см. вскрытие).
Ячейки на поверхности перит-
ремы Продолжаются В толщу рис. 207. Ixodes ricinus. Перитрема самки
хитина в виде узких кожных <х20) (из Е- к> сУв°Р°ва):
/ / — ячейки, 2 — хитиновое поле вокруг стигмы,
пор — канальцев (являясь та- з-стигма
229
ким образом расширением последних
на поверхности тела). Система пор на
перитреме считается органом чувств
(обоняния?), ибо в их области в
гиподерме залегают чувствительные
клетки (рис. 208, 6).
Тело взрослой самки резко
отличается от тела самца прежде всего
размерами. Правда, голодная самка
немного больше самца (длина ее
тела 4 мм, рис. 209, А, Б). Но при
сосании крови, благодаря тому, что на
теле сам'юи из твердых щитков есть
только небольшой спинной щиток,
остальная же поверхность покрыта
мягким растяжимым покровом, тело ее
сильно раздувается (рис. 209, В) и
может достигнуть в длину до 11 мм
при ширине 7 мм и более (у самца
сосание крови почти не отражается на
изменении формы тела).
Дорзальный щиток у голодной
самки покрывает спереди всего одну
треть длины овального тела (рис.209,
А). Форма щитка почти пятиугольная
с округленными углами. Цвет темно-
коричневый. Так же как и у самца,
ясно выражены плечи и слабо
намечены цервикальные бороздки.
Поверхность щитка тонко пунктирована и
покрыта редкими короткими
волосками.
На остальном, теле, за
исключением расположенных на боках перитрем, . хитинизированных
щитков нет, и тело одето тонким, гибким растяжимым
хитином с нежной поперечной штриховкой, обусловленной
наличием ряда утолщенных хитиновых полосок. Хитиновый покров
пронизан многочисленными порами, снабженными волосками и
выполняющими роль органов чувств. Цвет этот покров имеет чаще всего серый,
иногда красноватый, в зависимости от степени упитанности. На верхней
поверхности мягких покровов (так называемом alloscutum), вдоль тела
проходят бороздки: боковые, передние и задние (маргинальные) — по
краю тела, одна срединная (медианная) и по бокам от нее две задние
(парамедианные) —все три в задней части тела (рис. 209, В).
Спереди же от краев щитка назад тянутся передние парамедианные
(рис. 209, В, 13), Длинная половая бороздка огибает спереди половое
отверстие, расположенное на уровне оснований задних ножек, и ее
концы, расходясь в стороны, достигают заднего края тела. Анальное
отверстие также огибается спереди анальной бороздкой (рис. 209, Б, 8).
Рис. 208. Ixodes ricmus.
Вертикальный разрез через перитрему в
области дыхальца (Х240) (из Falke,
1931):
/ — воздушная полость в ячейке, 2 —
поры, 3 — замыкательная хитиновая
пластинка в области стигмы, 4 — стигма, 5 —
мышечный пучок, 6 — органы чувств, 7 —
центральный замыкательный мускул, 8 —
трахеи, 9 — главный изогнутый трахеаль-
ный ствол 10 — дорзальный
мускул-замыкатель, // — воздухоносная камера
230
В
Рис. 209. Самка Ixodes ricinus. A — голодная, слабо насосавшаяся, вид сверху; Б —
то же, вид снизу; В — сильно насосавшаяся, вид сверху (Х8) (ориг.):
/ — хоботок, 2 — щиток, 3 — аллоскутум, 4 — половое отверстие, 5 — перитрема, 6 — половая
бороздка, 7 — анальное отверстие, 8 — анальная бороздка, 9 — медианная бороздка, 10 — задняя
парамедиаиная бороздка, // -■- задняя боковая бороздка, 12—передняя боковая бороздка, 13 —
передняя парамедиаиная бороздка
Нимфа по внешнему виду напоминает самку, но значительно
мельче по размерам (длина около 1,5 мм), и у нее дорзальный щиток
относительно крупней, занимает почти половину длины тела (рис. 210).
На нижней стороне обращает внимание отсутствие полового отверстия,
но половая бороздка уже есть (рис. 210, 3), так же как и анальная.
Последняя, как и у взрослой самки, огибает анальное отверстие
спереди. У нимфы четыре пары ножек и дыхальца на боках тела.
Личинка (рис. 211) также в общем походит на самку, но обладает
только тремя парами ножек и в длину достигает всего 1 мм.
Дорзальный щиток, подобно нимфе, относительно велик, личинка лишена трахе-
альной системы, следовательно, у нее нет и дыхалец; нет также и
полового отверстия. Снизу видна только анальная бороздка
(рис. 211, А,1).
При сосании тело у нимфы и личинки способно раздуваться.
Конечности. В общих чертах устройство ножек одинаково во
всех стадиях, и для примера рекомендуется разобрать их строение у
самца. Расположены ножки двумя расходящимися кзади рядами в
пределах передней половины тела (рис. 206). Каждая из них
неподвижно прирастает к соответствующему эпимеральному щитку при помощи
базального членика — коксы. Последние у /. ricinus несут отростки и
бугорки. Коксы первой пары оттянуты с внутренней стороны в длинный
шип, налегающий на коксу П. На остальных коксах, на их задне-внеш-
них углах есть небольшие шипы (вернее, бугорки). Такие же бугорки
231
гг^я'
Рис. 210. Нимфа Ixodes ricinus. Вид сверху (А) и
снизу (Б) (Х25) (из Nuttal и Warburton):
/ — щиток, 2 — перитрема со стигмой, 3 — половая бороздка,
4 —• анальное отверстие, 5 — анальная бороздка
Рис. 211. Личинка Ixodes ricinus. Вид снизу (А) и сверху (Б)
(Х40) (из Nuttal и Warburton):
/ — анальная бороздка, 2 — щиток
сидят и на задне-внутрен-
них углах кокс II и III
(рис. 212, 2). Следующие
членики ножки подвижно
сочленены друг с другом и
называются: вертлуг
(trochanter), бедро (femur),
голень (tibia), переднелапка
(praetarsus) и лапка
(tarsus) (рис. 212). Все бедра
и три задние лапки
подразделяются на две части —
образуется неподвижное
сочленение
(псевдоартикуляция). Лапка кончается
тонким расчлененным
придатком с двумя крепкими,
загнутыми острыми
коготками, у основания которых
вентрально прикреплена
подушечка с
присасывательной вогнутой поверхностью
(рис. 213, 4). Благодаря
коготкам и подушечкам, клещ
свободно передвигается по
поверхностям любого
наклона, в том числе и по совершенно гладким (стеклу). Передняя лапка
с дорзальной стороны несет орган Галлера (рис. 213, 1; 214) в виде
двух ямок, лежащих рядом и затянутых бесцветной кожицей. Внутри
ямок видны несколько волосков. Предполагаемая функция органа
Галлера — обоняние, но высказывались предположения и о его функции в
качестве органа равновесия.
Хоботок клеща содержит части, гомологичные ротовым частям
других паукообразных: около ротового отверстия находятся и хелицеры
и педипальпы, видоизмененные и объединенные в колюще-сосущий
аппарат. Своим основанием хоботок вклинивается в переднюю вырезку
дорзального щитка и, составляя передний конец тела, называется
capitulum.
Разберем во всех деталях устройство хоботка у самца (у самки,
нимфы и личинки ниже отметим отличия в некоторых деталях строения
одинаковых, в общих чертах, частей).
Основание хоботка представляет собой воротничок — хитиновую
пластинку в виде кольца. Снизу воротничок в средней своей части
продолжается вперед в виде длинного, суженного, разделенного бороздой
на две половины колющего выроста — гипостома (рис. 215, 2): У самца
гипостом усажен с брюшной стороны шестью-семью поперечными
рядами шипов, из которых наиболее крупные боковые шипы средней части.
Продольной бороздой шипы распределяются на две боковые группы.
Концы шипов направлены назад, благодаря чему при внедрении хобот-
Рис. 212. Ixodes ricinus. Ножки самца с левой
стороны. Вид снизу (Х20) (ориг.):
/ — край тела, 2 — коксы с шипами и бугорками, 3 —
вертлуги, 4 — бедра, 5 — голени, 6 — переднелапки, 7 —
лапкн, 8 — придатки лапок с присасывательными
подушками и коготками, 9 — орган Галлера
233
ка в кожу хозяина клещ крепко
держится на ней. Опереди по бокам воротничка
прикреплены два щупика — пальпы
(рис. 215, 3), широкие членистые
придатки, едва выдающиеся за передний
конец гипостома и имеющие на
внутренней поверхности вогнутость, что позволя-
Рис. 213. Ixodes. Л —конец первой
ножки, вид сверху; 5 —конец одной из
задних ножек, вид сбоку (Х100) ; (из
Lahille, 1905):
/ — орган Галлера, 2 — лапка, 3—придаток
лапки, 4 — присасывательная подушечка, 5 —
коготки
Рис. 214. Ixodes ricinus. Орган
Галлера на передней ножке, про-.
дольный разрез (Х200) (из
Samson) :
/ — чувствительный бугорок впереди
чувствительных ямок, 2 — передняя и
3 —задняя чувствительные ямкн, 4 —
чувствительные волоски, 5 — нерв
Рис. 215. Хоботок самца Ixodes ricinus.
Вид сверху, со спинной стороны (А) и
снизу (Б) (Х70) (из Nuttal и Warburton):
/ — конечный членик хелицер, 2 — гипостом, 3 —
пальпы, 4 — воротничок хоботка, 5 — плечи
щитка, 6 — футляр для хелицер, I—IV— 1-й — 4-й
члеиики пальп
234
Рис. 216. Ixodes ricinus.
A—конечный членик хелицер самца;
Б — то же самки (Х150); В —
гипостом самки с внутренней
стороны с ротовым желобком (А и
Б из Nuttal, Warburton; В — из
Schulze):
/ — продольные гребни с зубцами, 2 —
базис членика, 3 — основной членик
хелицеры
ет им при складывании плотно
обхватывать 'С боков колющий аппарат. Пальпы
состоят из четырех члеников. В основании
каждой лежит узкий короткий членик,
подвижно сочлененный со следующим отделом
пальп. Два следующих членика, самые
длинные и широкие, неподвижно
сочленяются друг с другом. Четвертый членик
ничтожных размеров, прикреплен вентрально
(рис. 215, /V) в -специальной ямке третьего
членика и сверху незаметен. В спокойном
состоянии клещ держит пальпы сомкнуты-'
ми, прижатыми к хоботку; при сосании же
они отгибаются в стороны под прямым
углом, предоставляя сосущему аппарату как
можно глубже внедряться в кожу хозяина.
Пальпы усажены немногочисленными
крепкими волосками. Воротничок с гипостомом
и пальпами соответствует (гомологичен)
педипальпам других паукообразных.
Дорзальный край воротничка оттянут
вперед в парные, короткие (в половину
длины гипостома) придатки, загибающиеся
своими боками вниз и образующие два
трубчатых футляра (рис. 215, 6), внутри
которых находится пара свободно
движущихся хелицер (рис. 215, 1; 217, 2).
Основной членик хелицеры вдается далеко в
глубь тела и действием специальных мышц
может втягиваться внутрь и, наоборот,
высовываться наружу. Второй, конечный
членик (палец, крючок, рис. 215, 1) короток,
заострен, изогнут' вбок и по наружному
краю несет несколько сильных острых
зубцов, направленных назад и расположенных
в два неравных ряда на особых продольных
гребнях (рис. 216, 1). При высовывании
хелицер этот членик действием
специальной мускулатуры, лежащей внутри полого
основного членика хелицер (рис. 217, 2),
может отгибаться в сторону, что и
осуществляется в момент присасывания клеща к
хозяину.
Ротовая полость заключена между футлярами хелицер и
гипостомом. Последний с внутренней стороны несет продольный желобок (рис.
216, В). Уже у основания футляров хелицер гипостом срастается с ними,
и ротовая полость полностью замыкается и превращается в плоскую
трубку (рис. 217, 6), куда открывается ротовое отверстие. Тут же, немного
кзади, впадают и протоки двух слюнных желез (рис. 217, 7).
Рис. 217. Ixodes ricinus.
Поперечные разрезы через хоботок
самки. А — иа уровне
футляров хелицер; Б — на уровне
первого членика педипальп;
В — через основание воротника
хоботка (ХЗОО) (А и Б— из
Samson, В — из Ruser):
/ — футляр хелицеры, 2 — хелице-
ра, 3 — педипальпа, 4 — ротовая
бороздка гипостома, 5 — субхели-
церная пластинка, 6 — ротовая
полость, 7 — проток слюнной железы,
8 — мускулы-дилататоры глотки
(верхние), 9 — мускулы
констрикторы глотки, 10 — дилататоры
глотки (нижние), 11 — боковые
мускулы, 12 — просвет глотки, 13 —
хитин воротника
235
Присасываясь к коже хозяина, клещ боковыми-движениями
пальцев хелицер, -выдвинутых вперед, раздирает кожу и в образовавшуюся
ранку вводит гипостом и футляр хелицер. Крючья гипостома и загнутые
вбок пальцы хелицер позволяют клещу прочно держаться в коже. Весь
хоботок обладает подвижностью и может загибаться книзу.
У самки гипостом раза в полтора длиннее, чем и самца, уже и
усажен многочисленными мелкими зубцами, расположенными с каждой
стороны от срединной борозды в три продольных ряда (рис. 218, Л). С
верхней стороны на основании хоботка видны так называемые поровые
поля, грушевидного очертания (рис. 218, А, 6), отсутствующие у самца.
Хитин в их области пронизан массой пор и канальцев, представляющих,
подобно порам на перитреме, органы чувств. Кроме того, у самки у
основания хоботка с дорзальной стороны следует отыскать щелевидное
отверстие органа Женэ, способного выпячиваться наружу и
выделениями своих стенок обволакивающего откладываемые яйца, которые
благодаря этому склеиваются в комки.
Ротовые части нимфы и личинки по своей конфигурации
напоминают таковые у самки (рис. 218, Б, В). Отличие: 1) меньшее количество
рядов зубцов (по два с каждой стороны) и 2) отсутствуют поровые
поля.
Анатомия на вскрытии. Изучение внутреннего строения выгоднее
начать с самки — более крупной и, следовательно, удобной для
вскрытия.
Вскрывать можно также и других клещей Ixodidae, и в южных областях
Советского Союза могут быть с успехом использованы крупные самки, например Нуа-
lomma, Dermacentor, Boophilus, длина тела которых в насосавшемся состоянии мо-
236
жет достигнуть 2 см и более1 У них в общем
анатомия будет сходна с таковой /. ricinus. Различия
наблюдаются в мелких, несущественных деталях.
Для вскрытия желательно брать самок /.
ricinus, не слишком насосавшихся или даже голодных
(последние, впрочем, менее выгодны, ибо у иих
половая система может быть недоразвита).
Заморенного клеща перед вскрытием
рекомендуется опустить на несколько мгновений в 96%-ный
спирт и сразу перенести в ванночку с
физиологическим раствором для вскрытия (покровы клеща
плохо смачиваются водой, и он всплывает иа
поверхности жидкости; погружение в спирт облегчает
смачивание и предохраняет от всплывания). Ванночка
для вскрытия — это небольшая чашечка Коха с
восковым дном. Дальнейший ход вскрытия изложен
по Е. Н. Павловскому (1928).
Вскрытие начинается с того, что, придерживая
клеща иглой или пинцетом, удаляют ноги и остро
отточенной копьевидной или лопатовидной иглой
обрезают край тела. Сильно напившиеся самки могут
быть вскрыты по всей окружности тонкими
ножницами. Дыхальца должны быть обрезаны сверху
таким образом, чтобы оставить их в нижней части
клеща. Далее клеща следует приколоть к воску
спинкой кверху тончайшими энтомологическими
булавками, втыкая их в передней и задней части
хитинового покрова брюшной половинки. Захватив
пинцетом край спинного хитина, следует осторожно
поднимать его, обрывая пучки дорзовентральных
мышц. Эти мышцы составляют две симметричные
боковые передние и задние группы, парамедианиые
передние и задние и одну срединную (медианную)
группу (рис. 219). Их наличие при насасывании крови приводит к формированию у
самки на дорзальной стороне вышеупомянутых борозд (с. 230). Кроме того,
развиваются пучки мышц и в передней части тела клеща — мощные ретракторы хелицер,
тянущиеся от заднего края щитка к хелицерам и основанию хоботка. Разрыву мышц
следует помогать иглами. Снимая верхний хитин с клеща, одновременно удаляем
пленочку жирового тела, покрывающую внутренние органы, и также удаляем и сердце.
Последнее очень мало, плохо заметно, и во избежание его потери следует сразу же
внимательно рассмотреть внутреннюю сторону снятого спиниого хитинового покрова.
Затем начинается разбор и препаровка внутренних органов (рис. 220), что
лучше всего вести под бинокуляром. Удаляются по возможности дольки жирового
тела, разрываются трахейные трубочки, связывающие органы друг с другом, и
начинается распутывание ветвей кишечника, которые у сосавшей самки достигают большой
длины, загибаются концами вентрально, и требуется большая осторожность, чтобы
сохранить их в целости. Однако стенки кишечника неизбежно прорываются, из него
выходит пищевое содержимое — кровь хозяина. Струей из тонкой пипетки эту кровь
удаляют, и время от времени приходится сменять физиологический раствор в
ванночке. Одиовремеиио в передней части тела выпутываются из жирового тела
слюнные железы и распутываются длинные трубки двух мальпигиевых сосудов. Тут же
следует отпрепаровать и железистые дольки субскутальиой железы (органа Женэ) в
виде двух пар трубок, сходящихся к общему резервуару у основания хоботка
(рис. 220, 4). Освобождение трахейных пучков производится таким образом: игла
подводится под каждый из них снизу и поднимается осторожно кверху — тонкие
веточки обрываются и весь трахиальный пучок освобождается и отводится в сторону.
Далее следует изготовить тотальные препараты из расправленных внутренних
органов. Для этого ветви кишечника приподнимают и осторожно отводят вперед и в
сторону и копьецем вырезают вокруг анального отверстия квадратный кусочек
хитина, который вместе с задней кишкой и ректальным пузырем с мальпигиевыми
сосудами отводится вперед. Передняя кишка перерезается около нервного ганглия, и
Рис. 219. Ixodes ricinus.
Расположение дорзовентральных
мышц самки — мест
прикрепления их аподем к спинному
хитину (ориг.):
/ — щиток, 2 — передние
парамедианиые мышцы, 3 — передние
боковые мышцы, 4 — задние боковые
мышцы, 5 — задние
парамедианиые мышцы, 6 — срединные
(медианные) мышцы
237
Рис. 220. Вскрытая упитанная самка Ixodes ricinus (X15) (ориг.):
/ — хоботок, 2 — поровые поля, 3 — щиток, 4 — орган Женэ, 5 — слюнная железа, 6 — боковые
отростки средней кишки, 7 — средняя кишка, 8 — ректальный пузырь, 9 — левый мальпигиев сосуд,
10— яичник, 11— правый мальпнгиев сосуд, 12 — яйцевод, 13—трахеи, 14 — нервная масса, 15 —
пищевод
весь кишечник выводится из тела клеща. Нижняя хитиновая стенка тела
перерезается сразу за хоботком впереди от полового отверстия, и таким образом
освобождаются слюнная и субскутальная железы. Наконец, отпрепаровываются и
расправляются половые железы и перерезается влагалище около полового отверстия. При
отделении полового аппарата отпрепаровывают по пути и нервную систему,
сконцентрированную впереди, рядом с влагалищем.
Все отпрепарованные органы осторожно переносятся шпателем на предметное
стекло, очищаются от жирового тела, расправляются и обрабатываются методом
Е. Н. Павловского.
Пищеварительная система. От ротового отверстия (в
области воротничка хоботка у основания гипостома) начинается глотка с
плотными, выстланными внутри хитином стенками. Просвет глотки
имеет крестообразную форму, и к ее стенкам со стороны полости тела
прикрепляется серия мышц (см. рис. 217), действием которых
осуществляется сосательная работа глотки. Тянется глотка до нервной массы, где
238
она резко утончается, и переходит в узкий, сплющенный, тонкостенный
пищевод. Последний S-образно изгибается в вертикальном
направлении, пронизывает нервную массу насквозь (рис. 220, 14) и по выходе
из нее сразу же впадает в центральный отдел средней кишки, который
тянется вдоль тела в виде вытянутого мешка. От этого центрального
отдела в разные стороны отходят кишечные отростки, слепые рукава
(рис. 220,6). Развиты они различно, в зависимости от степени насасыва-
ния клещом крови, и у крупных самок, переполненных кровью, концы их
загибаются вентрально, перепутываются друг с другом и с прочими
внутренними органами. Расположение боковых отростков следующее.
От переднего конца центрального мешка в обе стороны расходятся
по одному рукаву, раздвоенному на конце. От задней части
центрального отдела берут начало две пары главных рукавов. Указанные
разветвления могут отдавать дополнительные боковые отростки при
переполнении кишечника кровью, и правильность их расположения может
нарушаться. Стенки кишечника обладают способностью сокращаться,
причиной чего является наличие в стенке кишечника густой сети продольных
и поперечных мышечных волокон. Общее количество крови, которое
высасывает самка /. ricinus, приблизительно равно 400 мг.
Центральный мешок резким пережимом отделен от ректального
пузыря, который снизу прилегает к заднему концу средней кишки (рис.
220, 8). Содержимое ректального пузыря беловатого цвета от
переполняющих его выделений из мальпигиевых сосудов, впадающих с обеих
сторон в ректальный пузырь с вентральной стороны. Тонкая задняя
кишка с хитиновой выстилкой внутри сообщает ректальный пузырь с
наружной средой и открывается' наружу анальным отверстием. К
хитиновому кольцу, окружающему последнее, подходят со спинной стороны
два пучка мышц, способные своими сокращениями втягивать слегка
это кольцо внутрь, что приводит к сближению и замыканию анальных
клапанов.
Как было указано выше, в ротовую полость сразу позади
ротового отверстия открываются два протока слюнных желез. В
отпрепарированном состояний эти железы лежат в виде гроздьев железистых
долек с обеих сторон от внутренних органов (рис. 220, 5). Концы
слюнных желез достигают уровня перитрем. Внутри каждой железы
проходит дихотомически ветвящийся проток и обычно, кроме главного
скопления долек, с каждой стороны обособляется небольшая добавочная
лопасть (рис. 221, 4). Ягодообразные альвеолы (концевые мешочки
железы) чередуются с одноклеточными дольками. Протоки железы
выстланы внутри хитиновой спиральной ленточкой, что делает их
похожими на трахеи (рис. 221, 3). Степень развития слюнных желез
зависит от упитанности клеща. У самки, начавшей питаться, они развиты
максимально — их альвеолы и дольки находятся в
напряженном*состоянии (переполнены секретом). Насосавшиеся же самки имеют
железы уже опорожненные. Секрет слюнных желез, впрыснутый при
сосании в ранку, производит анестезирующее действие (укусы клещей
нечувствительны), сопровождаемое местным воспалением кожи хозяина.
Примешиваясь к всосанной крови, этот секрет оказывает антикоагули-
рующее действие.
239-
Рис. 221. Ixodes ricinus. Отпрепарированные
органы Женэ и левая слюнная железа. Вид с
брюшной стороны (Х20) (из Е. Н. Павловского):
/ — хоботок, 2 — железистые дольки органа Женэ, 3 —
слюнной проток, 4 — дольки слюнной железы
Рис. 222. Ixodes ricinus.
Сердце и сосуды самки.
Вид с брюшной стороны
(ХЗО) (из Samson):
1 — головная аорта, 2 —
перерезанные сосуды ног,
3 — нервный синус, 4 —
аорта, 5 — сердце, 6 — остии
Органы дыхания. Трахеальная система начинается от
дыхалец (стигм), лежащих по бокам тела на перитремах. Щель стигмы
серповидной формы (см. рис. 207, 3), пронизывает толщу сплошного
хитинового поля в центре перитремы и продолжается в сферическую
дыхательную полость (см. рис. 208, /У), от которой и берут свое начало
трахеи. Последние пучком многочисленных, - расходящихся- в разные
стороны трубочек начинаются от короткого, изогнутого ствола,
непосредственно отходящего от дыхательной полости. Этот ствол снабжен
системой мышц (вентральных и дорзальных, см. рис. 208, 7, 10) и
функционирует в качестве замыкательного аппарата. Давлением крови
производится сжатие стенки ствола и уменьшается ток воздуха внутрь
трахеальной системы. При сокращении же упомянутых мышц
увеличивается просвет ствола и воздух беспрепятственно проникает внутрь.
Трахеи своими мельчайшими разветвлениями оплетают все внутренние
органы. Стенки их выстланы спиральной нитью хитина (это следует
рассмотреть под микроскопом, выделив несколько трубочек на
предметное стекло), чего, впрочем, лишены тончайшие их окончания уже
непосредственно в пределах органов. Движение воздуха в трахеях,
вероятно, вызывается работой дорзовентральных мышц тела. Подобное
мощное развитие трахей у Ixodidae представляет исключение у
других паукообразных, кроме сольпуг.
Кровеносная система. Органы кровообращения у I. ricinus
развиты очень слабо, и при вскрытии с большим трудом удается рас-
240
смотреть их элементы. Сердце прикреплено к спинному хитину двумя
большими пучками мышц около линии, проведенной между перитре-
мами. Оно имеет овальную форму, вытянуто поперек (рис. 222, 5) и в
стадии сокращения имеет около У4 мм в длину, при наполнении же
доходит до 0,5 мм. Сердце сплющено сверху вниз. Кровь из полости тела
поступает в него через две пары вентрально расположенных остий
(рис. 222, 6). От них в полость сердца вдаются трубочки, выполняющие
роль клапанов. От сердца вперед над центральным резервуаром
кишечника идет аорта — тонкий прямой сосуд, который вокруг нервной
системы расширяется в объемистый синус (рис. 222, 3). Из синуса кровь
изливается в полость тела через переднее отверстие — остаток головной
аорты. Через четыре пары боковых сосудов, внутри которых проходят
нервы к ногам, кровь проходит в полость ног.
Выделительная система. .Как уже было упомянуто, в
ректальный пузырь впадают два мальпигиевых сосуда (рис. 220, 9, 11).
Каждый из них, сильно извиваясь, уходит вперед и у упитанных самок
в расправленном состоянии может достигнуть 2 см в длину. Окраска
сосудов— белая. По ходу сосудов наблюдаются варикозные утолщения-
места скопления зерен секрета гуанина. При прорыве стенок сосудов
зернышки могут выпадать из просвета, и, рассматривая их под
микроскопом, можно обнаружить концентрическую слоистость.
Нервная система. Центральная нервная система в виде
компактной бесцветной массы лежит вентрально впереди от полового
отверстия (рис. 220, 14). Передний край нервной массы закруглен, задний
прямой. Проходящим сквозь нее пищеводом нервная масса (мозг)
подразделяется на верхний отдел —
надглоточный ганглий и на нижний
отдел — слившиеся вместе остальные
нервные ганглии тела.
Окологлоточные коннективы, благодаря тому что
весь мозг сплюснут дорзовентралыно,
очень коротки и широки. От мозга
отходят 11 пар нервов (рис. 223).
Спереди прежде всего следует найти три
пары тонких нервов, идущих к хели-
церам, педипальпам и к глотке со"
слюнными железами. С боков мозг
отдает четыре пары самых мощных
нервов, иннервирующих ножки. Наконец,
от заднего края мозга отходят четыре
пары нервов, иннервирующих органы
пищеварения и половую систему.
Поверхность нервной массы
покрыта тонкой соединительнотканной
неврилеммой, сквозь которую при
рассматривании мозга под микроскопом
просвечивают сферические,
поверхностно залегающие, нервные клетки.
Половая система. Половые
Рис. 223. Центральная нервная
система Ixodes ricinus (X50) (из
Pagenstecher):
/ — глоточный нерв, 2 — нерв к хели-
церам, 3 — нерв к педипальпам, 4—7 —
нервы к ножкам, 8 — нервы к половой
системе и кишечнику, 9 — пищевод,
10 — подглоточная масса, 11 —
надглоточная масса
241
f4
.X»
4
i
S/ч
■у
S
железы варьируют в
степени своего
развития, в зависимости от
упитанности самки.
Поэтому следует
проследить степень их
развития у голодной и
насосавшейся крови
самки.
У голодной самки
непарный яичник в
виде тонкого, длинного
полого внутри рукава
переплетается с
прочими внутренними
органами и вместе с
отходящими от его концов
яйцеводами образует
в расправленном виде
кольцо" (рис."224, Б).
Однако следует иметь
в виду, что непарность
яичника —явление
вторичное. У нимфы
половые железы
закладываются в виде парных
зачатков, которые
опаиваются своими кау-
дальными концами и
образуют
вышеописанного вида яичник. На
его поверхности в
данной стадии видны
многочисленные еще
мелкие оогонии. Оба
яйцевода тянутся вперед,
загибаются потом
назад и, сблизившись
около середины тела,
сливаются вместе,
образуя вздутый
двурогий 'непарный отдел —
матку (рис. 224, 3). От матки вперед отходит узкая
соединительная трубка, впадающая в объемистое влагалище, вытянутое вдоль
тела. Влагалище пережимом отграничивается от преддверия (atrium,
рис. 224, 7), открывающегося наружу половым отверстием. В области
пережима во влагалище впадают две придаточные трубчатые железы
(рис. 224, 5). Их зернистый секрет вероятно служит своеобразной
пробкой, закупоривающей вход во влагалище после оплодотворения.
#*V/
Рис. 224. Ixodes ricinus. Женский половой
аппарат у самок различной упитанности. А —
выводная часть половой системы при большом
увеличении (ХЗО); Б — слабо насосавшаяся самка
после оплодотворения (Х10); В — самка сильно
упитанная (Х10) (из Е. Н'. Павловского):
/ — яичник, 2 — яйцевод, 3 — матка, 4 — семяприемник,
5 — добавочные железы, 6 — влагалище, 7 — преддверие
242
Рис. 225. Копуляция у Ixodes ricinus (X12) (из
Brumpt)
С другой стороны, этому
секрету приписывается
значение смазки,
облегчающей выход яиц из
влагалища. Здесь же
около пережима
прикреплены пучки мышц,
направленные вентрально.
Влагалище и соединительный
каналец выстланы тонкой
хитиновой пленкой.
Наконец, в области
пережима 'выпячивается
довольно крупный,
пузыревидный, непарный,
направленный вершиной вперед
мешок — семяприемник
(рис. 224, 4).
У самки,
насосавшейся крови (а
следовательно, уже и оплодотворенной, что происходит у /. ricinus после линьки
нимфы, но чаще еще до питания), яичник достигает окончательного
развития и сильно увеличивается как в длину, так и в толщину
(рис. 224, В). На его поверхности — масса уже созревших яиц, часть
из них уже уходит в просвет яичника и выносится в яйцеводы, которые
приобретают четковидный характер. Матка у описываемой самки
непосредственно переходит во влагалище (исчезает соединительный
каналец), и отличие между этими двумя отделами можно обнаружить
только потому, что в матке нет хитиновой выстилки.
При копуляции (рис. 225) самец длительное время (несколько
часов) сидит на брюшке самки, удерживаясь за нее своими ножками и
прицепляясь также с помощью хоботка, вводимого периодически в по-,
ловое отверстие самки. Последняя манипуляция приводит также к
расширению полового отверстия и к выпрямлению обычно перегнутого
влагалища (Е. Н. Павловский, 1940), что должно привести к
облегчению введения сперматофора. Далее самец, видимо, прикладывает свое
половое отверстие к половому отверстию самки, и туда вводится
шейка сперматофора. Последний имеет колбовидную форму, и в
описанных случаях его резервуар свисает наружу. Его шейка, вероятно,
разрывается повторным введением хелицер самца в половое отверстие
самки. Содержимое сперматофора при этом перекачивается в
семяприемник, где вокруг массы сперматозоидов возникает новая
оболочка— образуется внутренний сперматофор, или сперматодоза. За время
питания самок (до двух недель) сперматозоиды выхоДятиз сперматодо-
зы и по яйцеводам попадают в яичник, где и совершается
оплодотворение.
Откладка яиц идет своеобразным способом, причем в этом
процессе принимают участие и хоботок самки и орган Женэ. При
откладке яиц хоботок подгибается вентрально и гипостом прячется"~за выпя-
243
Рис. 226. Ixodes ricinus. Mo- Рис. 227. Ixodes ricinus. Половая система самца.
мент откладки яйца — выпя- Вид сбоку (по Е. К. Суворову):
чеИНЫЙ орган Женэ, охватив- / — семенники, 2 — семенной мешок, 3 — семяпровод, 4 — се-
ШИЙ выделенное ИЗ ПОЛОВОГО мянзвергательный канал, 5 — половое отверстие, 6 — непар-
отверстия ЯЙЦО (из Nuttal И пая добавочная железа. I—IV—парные добавочные железы
Warburton):
1 — орган Женэ, 2 — ножки, 3 —
яйцо, 4 — пальпы, 5 — гипостом,
6 — половое отверстие
ченное наружу преддверие, из которого и показывается яйцо.
Одновременно выпячивается резервуар органа Женэ в виде двойного мешка
(рис. 226, 1). Его лопасти охватывают яйцо и обволакивают клейким
секретом, благодаря чему отложенные яйца склеиваются друг с
другом в характерный для Ixodidae комок. Движением хоботка кверху
яйцо удаляется из полового отверстия, и их скопление часто
покрывает воротничок хоботка и дорзальный щиток. Вероятно также
выделения органа Женэ предохраняют яйца от высыхания. Кладка /.
ricinus может содержать несколько тысяч яиц.
Вскрытие самца. В общих чертах вскрытие самца ведется так же,
как и самки. Из внутренних органов особое внимание, конечно,
следует обратить на половую систему, так как остальные органы в
общем устроены сходно с таковыми самки, но большинство из них
развиты слабее. При вскрытии самца бросается в глаза отсутствие органа
Женэ.
Наконец, следует отпрепарировать и половую систему (рис. 227).
Два длинных семенника тянутся снизу под кишечником вдоль тела, и
их слегка вздутые вершины направлены назад. Семяпроводы,
представляющие непосредственное продолжение семенников вперед, в области
нервного ганглия загибаются назад, и их короткие колена вливаются
в семяизвергательный канал (рис. 227, 4). Тут же в семяизвергательный
канал впадает серия (пакет) придаточных желез, среди которых
выделяется своими крупными размерами дорзально расположенный
семенной мешок (рис. 227, 2). Недалеко от полового отверстия в
семяизвергательный канал вливается небольшой непарный придаток (рис. 227,5).
Остальные четыре категории придаточных желез являются парными, и
выделяемый ими секрет вероятно участвует в образовании сперма-
тофора.
У взрослого-самца семенники наполнены уже готовой спермой.
Созревание же мужских половых клеток идет у нимф.
244
Подтип Branchiata — Жабродышащие
КЛАСС CRUSTACEA —РАКООБРАЗНЫЕ
Класс Ракообразные характерен чрезвычайным многообразием как
внешней морфологии, так и внутренней организации относящихся к
нему форм. Поэтому при прохождении практикума необходимо
ознакомиться с довольно большим количеством представителей этого класса.
Еще недавно всех ракообразных делили на два покласса: низших
(Entomostraca) и высших раков (Malacostraca). Однако организация
ракообразных, относящихся к низшим ракам, настолько многообразна
и филогенетические пути их настолько различны, что объединение их
в один подкласс низших раков .оказалось совершенно искусственным.
Бывший подкласс Entomostraca в настоящее время разбит на
несколько подклассов, тогда как высшие раки (Malacostraca) представляют
естественную группу, характеризующуюся единством метамерно-
го состава и рядом других признаков. Класс Ракообразные делится на
5 подклассов: Branchiopoda — Жаброногие раки, Cephalocarida — Це-
фалокариды, Maxillopoda — Максиллоподы, Ostracoda — Ракушковые
раки, Malacostraca — Высшие раки. Большинство названных
подклассов распадаются на многочисленные отряды.
ПОДКЛАСС BRANCHIOPODA —ЖАБРОНОГИЕ РАКИ
ОТРЯД ANOSTRACA —ЖАБРОНОГИЕ
Среди представителей этого отряда в качестве основных объектов
для изучения лучше всего взять Artetnia или Branchipus, так как
благодаря их прозрачности и небольшим размерам на живых экземплярах
или тотальных препаратах, удается рассмотреть довольно подробно их
•строение.
Материал и методы изучения. По своей организации Artetnia salina и Branchipus
stagnalis весьма близки между собой и могут быть рассмотрены вместе. A. salina —
■космополитический галобионтный вид, живущий в солоновато-водных водоемах.
В СССР встречаются в Крыму, в районе Каспийского моря, в Западной Сибири. Arte-
mia хорошо разводятся в аквариумах. Яйца их вместе с илом легко выдерживают
высыхание. Для того чтобы вывести из яиц Artemia, их заливают водой из солонова-
товодного водоема, которая может быть заменена следующим искусственно
приготовленным составом: воды—1 л; NaCl —50 г; 5% Na2HP04—10 см3; 5% KN03 —
10 см3; 2,5% MgS04—10 см3; 5% NH4N03—10 см3; 5% СаС12—10 см3; 1% KI —
10 см3 К
1 Приведенный состав воды для Artemia предложен М. М. Исаковой-Кео.
245
Для длительного существования Artemia в аквариуме их следует регулярно
подкармливать. Пищей служат протококковые водоросли, простейшие. При отсутствии
указанной пищи Artemia можно подкармливать дрожжами. При этом раки
вылавливаются из аквариума широкой пипеткой и пересаживаются в небольшие сосуды
(солонки, чашки Петри), где предварительно делается взвесь из дрожжевых клеток.
Через полчаса — час Artemia вновь переносятся в аквариум.
В подобной среде Artemia в Ленинградском университете живут и размножаются,
годами. Чтобы вывести Artemia из яиц, отложенных в аквариумах, яйца следует в
течение 1—2 сут продержать в пресной воде, после чего перенести в солоноватую. Эти.
организмы выносят без вреда для себя значительное колебание солености. Branchipus-
slagnalis встречается в средней и южной полосе европейской части СССР, а также в
Сибири. Живет преимущественно в мелких пересыхающих водоемах, появляется в
весенние месяцы. Для изучения лучше всего использовать живой материал, что вполне
доступно в отношении A. salina в любое время года, если ее разводить в аквариумах..
При отсутствии такового можно использовать тотальные препараты, слегка подкрасив-,
объект квасцовым кармином. Конечности отпрепаровать иглами под бинокуляром и.
сделать препарат, заключив их в глицерин-желатин. Дополнительно к изучению
объекта in toto следует изготовить серию поперечных срезов. В качестве фиксатора
можно взять жидкость Флемминга. Сулемовые фиксаторы дают худшие результаты.
Окраска срезов — железным гематоксилином с эозином.
Примечание. Phyllopoda Anostraca относятся к числу довольно редких животных.
Поэтому возможно, что при прохождении практикума не удастся добыть ни Artemia,.
ни Branchipus. В таком случае можно использовать другие виды и роды этой группы,,
так как их организация в общем довольно однотипна. Широко распространенным в
СССР является Prislicephalus josephinae (Grub), встречающийся в средней полосе
европейской и азиатской части СССР, а также распространенный далеко на Север
вплоть до Архангельска и Карелии. Нередки также Chirocephalopsis birostratus (Dyb),.
Steptocephalus corvicornis (Wagi) и некоторые другие.
Ниже, при рассмотрении организации Artemia и Branchipus, мы укажем
некоторые морфологические отличия указанных форм, затрагивающие в первую очередь
строение головных придатков.
Изучение живых объектов или тотальных препаратов. Artemia
salina и Branchipus stagnalis, так же как и другие виды Branchiopoda
Anostraca, обладают вытянутым, ясно сегментированным телом
(рис. 228, 229). В. stagnalis имеет в длину 10—12 мм, A. salina — 8 —
10 мм. Тело распадается на четыре отдела. Передний отдел — голова
(рис. 231, Г) образует монолитную капсулу, несущую верхнюю губу,,
глаза, первую и вторую пару антенн. Морфологически голова
Anostraca соответствует предротовой лопасти — акрону (гомолог простомиума
Annelides), несущей первую пару антенн (антеннулы) и слившейся с
первым постантеннулярным сегментом, несущим вторую пару антенн.
Такой тип строения головы называется protocephalon. Второй отдел —
gnathocephalon — состоит из свободного сегмента мандибул (жвал) и-
двух слившихся между собой сегментов первой и второй пары мак-
силл. Таким образом в состав gnathocephalon входят три сегмента1..
Третьим отделом является грудь (называемая также thorax или ра-
1 Описание расчленения Phyllopoda Anostraca показывает, что широко
распространенное представление о том, что все ракообразные имеют голову, состоящую из
пяти слившихся сегментов, неточно. На самом деле protocephalon и gnathocephalon
представляют собой обособленные морфологически отделы. Наличие обособленного-
protocephalon имеет место не только у Phyllopoda Anostraca, но и у большинства
высших раков, в том числе и у Decapoda. У некоторых отрядов, как низших, так и
высших раков (Copepoda, Leptostraca, Isopoda, Amphipoda), protocephalon сливается;
с gnathocephalon, образуя составную голову — syncephalon. Все эти отношения
рассматриваются ниже при изучении представителей отрядов.
246
Рис 228 Самка Chirocephalus grubei (А); самцы Branchipus stagnalis (Б) и Branchi-
necta sp (В) (А - из Wagler, Б и В - из Gestaecker и Ortmann):
/-нопяоный глаз 2-печеночный вырост кишечника, 3 - первая пара антенн, 4 -вторая пара
' придатки, 15 — семенник
reion) состоящая из 11 сегментов и несущая плавательные конечности.
Четвертый отдел — брюшко, лишено конечностей, оно слагается из
девяти сегментов. Два первых (передних) сегмента брюшка сливаются
в половой сегмент. Задний брюшной сегмент несет вилочку (turcaj,
Аорма которой характерна для отдельных видов. Она состоит из двух
удлиненных придатков, на которых расположены многочисленные
волоски1 (рис. 228, 229, 230).
Конечности. Первая пара антенн (антеннулы) располагается
со спинной стороны у переднего края головы. Эти антенны нитевидны,
1 Для Artemia salina характерна широкая изменчивость формы iurca в замш-
мости от концентрации солей в окружающей среде. Как показали ™™?™™™^
работы Шманкевича (1875), Гаевской (1916), увеличение концентрации солеи
приводит к постепенной редукции вилочки вплоть до ее полного исчезновения.
247
одноветвисты и слагаются из двух члеников (см. рис. 228, 229). На них
располагаются чувствительные волоски-эстетаски, являющиеся хемо-
рецепторами (увидеть их можно лишь под микроскопом).
Вторая пара (Ап) антенн, которая развита значительно сильнее
первой пары, у самцов и самок имеет совершенно различную
структуру (рис. 231, 232). Вместе с тем различие в строении А11 у самцов
представляет собой один из существенных систематических признаков.
И у самцов и у самок Ап одноветвисты. У самок (рис. 231, Б, Г) они
представляют собой относительно небольшие крючковидные
образования, несущие немногочисленные волоски.
. У самцов А11 видоизменены в хватательный орган, служащий для
удержания самки во время копуляции. У Artemia salina (рис. 231, А)
они состоят из двух члеников, причем дистальныи из них сплющен
дорзовентрально, заострен на конце и направлен внутрь. У Branchipus
stagnalis строение их еще более сложно. Оба основных членика
сливаются между собой и с головой. Дистальные членики направлены
вперед. Кроме того, к основным членикам
прикрепляются еще особые подвижные
нитевидно-заостренные придатки,
выдающиеся вперед дальше дистальных концов
самих антенн. Их роль при копуляции еще
недостаточно выяснена. У Pristicephatus
josephinae и Chirocephalopsis birostratus
антенны имеют отличную от описанной для
Branchipus и Artemia форму, как это
показано на рис. 232.
Мандибулы, расположенные по
сторонам ротового отверстия, лишены щупикта.
Они представляют довольно массивные
хитиновые пластинки, зазубренные по своему
внутреннему краю (рис. 233).
Первая (рис. 233) и особенно вторая
пара максилл развиты значительно слабее
Рис. 230. Artemia salina. Различные
степени развития furca в зависимости от
концентрации солей. В
концентрированных растворах furca почти не
выражена (на рисунке слева). По мере
ослабления концентрации обе ветви furca
развиты все сильнее (из Wagler)
^Pиc. 229. Artemia salina. Вид
с брюшной стороны (из Я. А.
Бирштейна):
1 — непарный глаз, 2 — первые
антенны, 3 — парный сложный глаз,
4 — грудные конечности, 5 —
яйцевой мешок, 6 — брюшко, 7 — furca
248
КУ (fiagna-liГд-EutalMpus vSis(аЛ из Daday, Д-из Snodgrass):
Stagnant,, м ^ „„'.,* глаз 3-вторая пара антенн, 4-нитевидные придатки,
1 - первая ™?^1%%£Г£2^*Т/Т- protoce°phaSlon. P//_W - сегменты тела
мандибул Они представлены небольшими хитиновыми пластинками.
ТрТдные ножки Artemia и Branchipus, так же как и всех. Anostra-
са поедставляют собой типичные листовидные конечности. Все 11 пар
их имеют сходную структуру и отличаются друг от друга только
размерами^ НожкиУпереднихУдУвух грудных сегментов несколько меньше
Рис 232 Chirocephalopsis birostratus (А) и Pristicephalus jostphinae (Б),
вид'головы самца сверху (А - из Daday; Б - ориг. по рис. С. С.
Смирнова):
1- первая пара антенн, 2-сложный глаз, -3- вторая пара антенн
249
Рис. 233. Конечности Branchipus. A -—
мандибула; Б — первые максиллы; В—
пятая грудная конечность. (из Gestaec-
ker и Ortmann):
/ — экзиты (эпиподиальные жабры), 2 — эндн-
ты с плавательными волосками
остальных. Каждая ножка
сплющена в передне-заднем направлении.
■Ясное расчленение на отдельные
членики отсутствует. Общая
подвижность конечности
обусловливается тем, что хитиновая кутикула
относительно тонка и эластична.
Конечность .полая, ее эластичность
обусловлена тем, что гемолимфа,
заполняющая полость, находится
под довольно значительным
давлением. С внутреннего и наружного
края конечность образует несколько
выростов, имеющих форму
лопастей (рис. 233, В). По внутреннему
краю располагаются пять так
называемых эндитов, несущих
многочисленные длинные волоски,
способствующие плаванию. По
наружному краю располагаются три эк-
'зита, называющиеся также эпипо-
дитами (рис. 233, В, 1). Эти
придатки лишены волосков, покровы их очень тонки, они богато
снабжаются кровью и в функциональном отношении являются жабрами. Из
них два проксимальных имеют форму пластинок, дистальный же
является мешковидным.
Мускулатура. При рассмотрении Artemia или Branchipus in
toto хорошо видны некоторые основные системы мышц. Более полное
представление об их структуре и расположении получается при
изучении поперечных срезов (с. 256). Имеются парные спинная и брюшная
продольные мышцы, тянущиеся от максиллярного сегмента до заднего
конца тела. В каждом сегменте груди, кроме того, видны метамерно
расположенные дорзовентральные мышцы, идущие от спинной части
сегмента к его брюшной стенке. В грудных конечностях отчетливо
видны мышечные пучки в количестве трех пар в каждой ножке. Одни из
этих мышц берут начало в спинном отделе, другие — в брюшной части
каждого сегмента. В голове особенно мощно развиты мускулы
мандибул и второй пары антенн.
Пищеварительная система. Органы пищеварения удается
без труда рассмотреть как на живых объектах, так и на тотальных
препаратах. Ротовое отверстие, прикрытое спереди верхней губой,
помещается на нижней стороне головы. Оно ведет в короткий пищевод
эктодермального происхождения, выстланный изнутри хитиновой
кутикулой. На границе между ним и средней кишкой, следующей за
пищеводом, имеется клапан, состоящий из волосков. Энтодермальная
средняя кишка в виде прямой трубки тянется через все тело животного и
переходит в заднем конце брюшка в короткую эктодермальную заднюю
кишку, в свою очередь открывающуюся анальным отверстием на
последнем абдоминальном сегменте. Передний конец средней кишки об-
250
разует два небольших розетковидных
слепых выроста (ом. рис. 228, 2; 235,
3), лежащих в голове. Эти выросты
.называются печеночными.
Кровеносная система.
Длинное трубиовидное сердце
расположено над кишечником, оно тянется
от максиллярного сегмента до заднего
конца брюшка. На живом объекте при
небольшом увеличении микроскопа
удается наблюдать пульсацию сердца,
гонящего бесцветную гемолимфу
сзади наперед. В каждом сегменте
имеется пара остий. Стенки сердца очень
тонки. Они обладают сократимостью
благодаря наличию мускульных
элементов. При рассматривании сердца
со спинной стороны, особенно в
области брюшка, удается увидеть, что
оно слагается из отдельных камер,
расположенных метамерно по
продольной оси животного (рис. 234).
В каждом сегменте имеются тонкие
тяжи соединительнотканного
характера, при помощи которых сердце
подвешено к стенке тела. На заднем
конце последняя камера несет небольшое
непарное отверстие, представляющее
собой остию. В головном отделе
гемолимфа изливается из сердечной
трубки- в окружающие ткани, откуда
передвигается по брюшному синусу к
жабрам (рис. 235). Кроме сердечной
трубки никаких других морфологически оформленных кровеносных
сосудов нет.
Выделительная система представлена парой максиллярных
желез, хорошо видимых на живых экземплярах при небольшом
увеличении микроскопа (рис. 228, 13). Они лежат по бокам тела во втором
максиллярном сегменте. Каждая железа слагается из небольшого цело-
мического мешочка и нефридиального канальца, образующего три
петли. Дистальный конец его открывается в небольшой тонкостенный
пузырь, в свою очередь коротким каналом открывающийся наружу у
основания второй пары максилл.
Нервная система. У Artemia и Branchipus, как и у всех
Branchiopoda, наиболее примитивный среди членистоногих тип строения
нервной системы, так называемая брюшная нервная лестница. Ее
удается рассмотреть in vivo под микроскопом, лучше всего на молодых
неполовозрелых экземплярах, довольно сильно придавив объект
покровным стеклом.
Рис. 234. Branchipus. Четыре задние
камеры сердца молодого
неполовозрелого экземпляра со спинной
стороны (из С. Claus):
1 — продольный мускул тела, 2
—соединительнотканные тяжи, на которых
подвешено сердце, 3 — остии, 4 — мускульные
клетки стенки сердца, 5 — задняя
непарная остия
251
Рис. 235. Artemia salina. Передний конец тела. Вид со
спинной стороны (из С. Claus):
1 — непарный глаз, 2— сложный парный глаз, 3—печеночные
выросты кишечника, 4— максиллярная железа, 5 — передняя
часть сердца, 6 — продольные мускулы тела, 7 — дорзальные
мускулы мандибул. Стрелками показано направление движения
кровн
Головной мозг (надглоточный ганглий) располагается в передней
части головы, он вытянут в поперечном направлении (рис. 236). От
него отходят мощные оптические нервы в стебельки сложных глаз и
нервы к первой паре антенн. Непосредственно к мозгу прилежит непарный
фронтальный глаз. От мозга берут начало окологлоточные коннективы,
огибающие пищевод. На них располагаются ганглии второй пары
антенн.
Подглоточного нервного узла, иннервирующего ротовые части, как
это имеет место у огромного большинства членистоногих, здесь не
образуется. Ганглии мандибул и обеих пар максилл обособлены и входят
в состав брюшной нервной лестницы (рис. 237). Одноименные ганглии,
относящиеся к одному сегменту, расставлены на довольно значительное
расстояние и связаны коммиссурами. Следующие друг за другом
ганглии соединяются коннективами. Каждый из одиннадцати грудных,
несущих конечности, сегментов имеет одну пару ганглиев, от которых
отходят периферические нервы. В первых двух, слившихся в половой
сегмент, метамерах брюшка имеется две пары ганглиев. В
последующих абдоминальных сегментах небольшие ганглии имеются лишь в
первых двух, следующих за половым, сегментах. Остальные сегменты
252
Рис. 236. Branchipus stagnalis. Фронтальный срез через надглоточный ганглий (из
С. Claus):
1 — оптический нерв, 2 — антеннальный нерв, (А1)
лишены ганглиев, и по их брюшной стороне тянутся два нервных
тяжа — продолжение брюшной нервной лестницы.
Органы чувств. Хорошо развиты сложные глаза,
расположенные на конце длинных подвижных глазных стебельков. Основные
элементы строения глаз удается рассмотреть in vivo или на тотальных
Рис. 237. Максиллярная область Artemia salina (из С. Claus):
/ — ганглий мандибул, 2 — мускулы мандибул, 3 — продольный мускул, 4 —
ганглий первых максилл, 5—первая максилла, 6 — вторая макснлла, 7 —
ганглий первой пары грудных ножек, 8 — ганглий вторых максилл
253
Рис. 239. Brinchipus.
Участок среза через
сложный глаз при
большом увеличении (из
С. Claus):
1 —■ хитиновая - прозрачная
«утикула, 2 — хрустальный
конус, 3 — днстальные части
ретинальных клеток, 4 — ре-
тинальные клетки, между
отдельными омматиднями
прослойки пигмента, 5 — ба-
зальная мембрана,
6^нервные волокна, отходящие от
базальной части
ретинальных клеток
Рис. 238. Branchipus. Фронтальный
срез через сложный глаз (из С.
Claus):
/ — хрустальные конусы, 2 — слой
ретинальных и пигментных клеток, 3—
нервные волокна, отходящие от ретинальных
клеток, 4 — мускул глазного стебелька,
5 — мозг, 6 — оптический нерв, 7 —
оптический ганглий, 8 —ретинальный ганглий
препаратах, которые дополняются изучением
срезов. Сложный глаз слагается из большого
количества отдельных глазков, или омматидиев
-(рис. 239). Снаружи он покрыт прозрачной
хитиновой кутикулой, которая, однако, не
распадается на отдельные фасетки, а является
сплошной. Вслед за кутикулой следует слой
прозрачных клеток (рис. 238, 239), образующих так
называемые хрустальные конусы отдельных
омматидиев. Хрустальные конусы представляют
собой диоптрическую, светопреломляющую часть
омматидиев. Далее вглубь следует
пигментированная часть глаза, состоящая из
светочувствительных (ретинальных) и пигментных клеток.
При рассматривании глаза in toto, благодаря
наличию большого количества пигмента,
увидеть ретинулы отдельных омматидиев не
удается. Они сливаются в сплошную темную
массу.
В центре ретинальных клеток каждого ом-
матидия (по продольной их оси) располагается
светочувствительная палочка — рабдом.
От базальной части ретинальных клеток
берет начало оптический нерв, хорошо видимый
in vivo. Он направляется к мозгу. По ходу пег-
vus opticus располагается оптический ганглий.
Непарный глаз располагается на передней
части головы впереди мозга. Он лишен
светопреломляющего аппарата (рис. 228, 1). В состав
его входят два тесно прилежащих друг к другу
пигментных бокала, окруженных светочувстви-
254
Рис. 240. Branchipus. Непарный
глаз молодого экземпляра (из
С. Claus):
/ — пигментный бокал, 2 —
светочувствительные клетки, 3 — боковые нервы,
4 — центральный нерв
тельными клетками. От непарного
глаза берут начало 3 нерва (один
срединный и два боковых), которые
соединяют его с мозгом (рис. 240,
3,4).
Кроме глаз, у живого объекта
следует рассмотреть органы химического
чувства — чувствительные волоски-эс-
тетаски, расположенные, как указано
(с. 248), на первой паре антенн (на
дистальной их части).
Половая система. Муоюская
половая система. Парные семенники
представляют собой прямые трубки,
располагающиеся по сторонам
кишечника, начиная от анального сегмента
и далее назад. У Artemia salina они тянутся лишь до абдоминального
сегмента (рис. 241, А). У видов рода Branchipus семенники длиннее и
могут простираться назад вплоть до предпоследнего сегмента брюшка
(рис. 241, £).
В передней части семенники переходят в семяпроводы (vas
deferens). По ходу последних иногда образуется небольшое расширение —
семенной пузырек (рис. 241, 4). Конечные отделы мужских половых
протоков открываются на вершине хитиновых парных копулятивных
органов (рис. 241, 3), форма которых у видов Anostraca различна.
Женская половая система. Парные трубчатые яичники занимают
в теле самки такое же положение, как семенники у самца. У Artemia
они более короткие, чем у Branchipus. В передней части каждого
яичника в половом сегменте от него отходит в медиальном направлении
яйцевод, образующий петлю (рис. 242).
Оба яйцевода открываются в непарный довольно объемистый
отдел женской половой системы — матку. В области матки брюшная
стенка генитального сегмента образует выпячивание, представляющее
собой яйцевой мешок (рис. 242). На дистальном конце при помощи
небольшого протока (vagina) он открывается непарным отверстием
наружу.
В конечные отделы яйцеводов открываются довольно объемистые
железы (рис. 242, 4), фолликулы которых также располагаются в ге-
нитальном сегменте. За счет секрета этих желез выделяется оболочка
яиц, заполняющих обычно в значительном количестве матку.
У Artemia salina, в отличие от Branchipus и большинства других
Anostraca, наблюдается иногда партеногенетическоё развитие яиц.
Изучение срезов. На серии поперечных срезов следует,
исходя из изучения Artemia или Branchipus in toto, уточнить некоторые
анатомические отношения отдельных систем органов, а также
ознакомиться с гистологическим строением их.
Покровы слагаются из тонкой хитиновой кутикулы (рис. 243, 244)
и подлежащих эпителиальных гиподермальных клеток. В цитоплазме
последних имеются опорные фибриллы, проходящие от базальной к
255
Рис. 241. Мужской по- Рис. 242. Женский половой алпа-
ловой аппарат Artemia рат Branchipus grubei (из Gestae-
salina (А) и Branchipus ker и Ortmann):
grubei (Б) (из Dybonsky ; —яйцевод, 2 — яичиик, 3 — яйцевой
И Leydig): мешок, 4 — железа
1 — семенник, 2 — vas
deferens, 3—penis, 4 — vesicula
seminalis
дистальной части клетки. Своеобразное строение имеют покровы в
области жаберных пластинок (экзиты грудных конечностей). Кутикула
здесь чрезвычайно тонка (рис. 245), эпителиальные клетки сильно
уплощены. В центральной части жаберной пластинки имеется просвет,
наполненный гемолимфой с отдельными амебоидными клетками
(рис. 245). Между обоими слоями эпителия расположены опорные
фибриллы, благодаря наличию которых внутренний просвет жаберной
пластинки не спадается.
На срезах через торакальный отдел видно взаимное расположение
отдельных систем органов. На брюшной стороне имеется довольно
глубокая бороздка, по сторонам ее широким основанием берут начало
конечности, полость которых не отграничена от полости тела.
Значительную часть поперечного разреза занимают хорошо
развитые поперечнополосатые мышцы. Со спинной и с брюшной стороны
проходят парные продольные мышцы, причем каждая спинная мышца
распадается на деэ параллельно идущих пучка (сн, рис. 243, 9). Вид-
256
Рис. 243. Branchipus stagnalis.. Поперечный срез в области груди (по К. Schneider):
1 — гиподерма, 2 — дорзовентральный (сагиттальный) мускул, 3 — брюшной продольный мускул,
4 — лимфатические клетки, 5 — дыхательная пластинка, 6 — мускулы конечностей, 7 — брюшная
нервная лестница, 8 — трансверсальный мускул, 9 — спинные (дорзальные) мускулы, 10 — средняя
кишка, 11 — сердце
ны также дорзовентральные (боковые) мышцы, образующие широкий
пучок, и трансверсальные мышцы. Последние одним концом
прикрепляются ко дну брюшной бороздки, другим — к боковым стенкам тела.
Наконец, мышечные пучки направляются в конечности.
Средняя кишка (см. рис. 243, 10) выстлана изнутри однослойным
эпителием.
Сердце, лежащее над кишкой (см. рис. 243, 11), обладает очень
тонкими стенками, состоящими из одного слоя клеток мускульного
характера, эндотелий отсутствует. В полости тела разбросаны отдельные
довольно многочисленные лимфатические клетки. По обеим сторонам
9—2
257
'^5
Рис. 244. Покровы Bran-
chipus stagnalis (из
К. Schneider):
1 — хитиновая кутикула, 2 —
ядро гиподермальной
клетки, 3 — опорные фибриллы,
4— примыкающий к
гиподерме мускул, 5 — ядро
мускульной клетки
Рис. 245. Branchipus
stagnalis. Поперечный
срез через дыхательную
пластинку (экзит
грудной конечности) (из
К- Schneider):
1 — кутикула, 2 — ядро
гиподермальной клетки, 3—
лимфоцит, 4 — опорные
фибриллы
брюшной бороздки располагаются элементы брюшной нервной
лестницы. Ганглии находятся под некоторым углом друг к другу,
соединяющая их коммиссура огибает брюшную бороздку (см. рис.
243,7).Нервные клетки расположены по периферии ганглия.
При изучении срезов через головной отдел Branchipus или Artemia
обратить внимание на строение мозга (см. рис. 236).
Нервные клетки, расположенные группами, локализуются по
периферии. Центральные части мозга слагаются преимущественно из
нервных волокон. В центре находится особо дифференцированный отдел,
так называемое центральное тело. Следует проследить отхождение от
мозга глазного нерва, нервов первой пары антенн и окологлоточных
коннективов.
На срезах, захвативших сложный глаз, можно рассмотреть
гистологические элементы последнего. Хорошо видны клетки хрустального
конуса (см. рис. 239). Подними лежат ретинальные. клетки,
охватывающие суживающиеся базальные части хрустальных конусов. В
состав ретинулы каждого омматидия входит семь ретинальных клеток, из
которых две недоразвиты. Между ретинулами отдельных омматидиев
располагаются пигментные клетки. У основания ретинальной части
глаза проходит тонкая базальная мембрана. Нервные отростки,
отходящие от ретинальных клеток, входят в состав оптического нерва.
На срезах, прошедших через сегмент второй пары максилл,
следует отыскать выделительную железу (см. с. 251), стенки канала
которой слагаются из железистых клеток.
258
ОТРЯД PHYLLOPODA —ЛИСТОНОГИЕ РАКИ
Отряд Phyllcpoda объединяет три подотряда: Notostraca — Щитни,
Conchostraca — Раковинные листоногие раки и Cladocera — Ветвисто-
усые раки. В отношении представителей первых двух подотрядов мы
ограничимся изучением их внешней морфологии, тогда как ветвисто-
усых раков, составляющих значительную часть пресноводного
зоопланктона, рекомендуем изучить подробно.
Подотряд Notostraca — Щитни
В качестве примера Phyllopoda Notostraca можно взять щитня
(Apus cancriformis Schaeffer) или же виды близкого к нему рода
Lepidurus Leach. Щитень один из наиболее крупных представителей листо-
ногих раков. Его длина 6 см. Встречается спорадически,
преимущественно в средней и южной полосе СССР, в мелких пересыхающих
водоемах и просто лужах, образовавшихся после дождя. Появление его
носит более или менее внезапный характер, и столь же внезапно он
исчезает. Объясняется это тем, что яйца щитня могут без вреда
выкосить длительное высыхание и развиваются лишь при наступлении
благоприятных условий. Самцы встречаются значительно реже самок.
Lepidurus arcticus встречаются в северной полосе вплоть до Арктики.
Тело Apus (и Lepidurus)
слагается из головы и большого а 5
числа сегментов, без резкого
разграничения на грудной и
брюшной отделы (рис. 246). Число
сегментов в отделах Notostraca
варьирует. У некоторых видов
Apus оно достигает 42 (не считая
головы). Со спинной стороны
голова и большая часть туловища
прикрыты двускатным широким
щитом К В передней части его
расположены два глаза. Задний
конец тела выдается за границу
щита, так что vпоследние 10—
15 сегментов щитом не
прикрыты. Задний сегмент несет вилку
(furca), состоящую из двух
длинных придатков, длина которых
превышает длину тела. При рас-
1 У Notostraca protocephalon и
gnathocepha'lon довольно тесно
сливаются. Со спинной стороны граница
между ними, однако, заметна (рис. 246, Л, 5).
Щит в большей части образуется за счет
максиллярных сегментов, но передняя
часть его принадлежит protocephalon.
Рис. 246. Apus cancriformis — (А) и
Lepidurus macrurus (Б), вид со спинной
стороны (из С.-С. Смирнова):
1 — глаза, 2 — нитевидные придатки первой пары
грудных ножек, 3 — брюшко, 4 — вилка, 5 —
бороздка, отделяющая область protocephalon от
манднбулярного сегмента
9*
259
смотрении щитня с брюшной стороны видны
многочисленные конечности. Первая пара антенн представлена короткими
нитевидными придатками (рис. 246, 2, 3). Вторая пара рудиментарна и у
некоторых видов отсутствует. Из ротовых частей хорошо развиты
мандибулы (рис. 247, 4), обладающие зазубренным внутренним краем.
Первая и вторая пара максилл (рис. 247, 5) представляют небольшие
пластинчатые образования. Челюсти Notostraca лишены щупиков.
Остальные конечности являются типичными листовидными ножками.
Их размеры спереди назад постепенно уменьшаются, задние сегменты,
выходящие за границу спинного щита, лишены конечностей. Общее
число конечностей превосходит 40 пар, т. е. больше числа сегментов,
несущих конечности. Это объясняется тем, что сегменты,
расположенные позади одиннадцатого (на котором открываются наружу половые
отверстия как у самцов, так и у самок), имеют более чем по одной
паре (до пяти) конечностей.
Каждая листовидная ножка. Apus (рис. 248) по своему
медиальному краю несет пять эндитов. По латеральному краю расположены два
экзита. Из них проксимальный — эпиподит представляет мешковидную
жабру. Дистальный в форме пластинки, он играет существенную роль
при плавании щитня. Первая пара листовидных ножек Apus и Lepidu-
rus обладает тремя нитевидными придатками (см. рис. 246, 247),
далеко выдающимися за границы щита и при рассматривании животного
сверху производящими впечатление антенн. У самок на 11-й паре
конечностей за счет видоизменения экзитов образуется яйцевая камера, куда
поступают откладываемые самкой яйца.
Рис. 247. Lepidurus glacialis. Передний
конец брюшной стороны (из Caiman):
1 — верхняя губа, 2 — первые антенны, 3 —
вторые антенны, 4 — мандибула, 5 — первая
и вторая максиллы, 6— первая пара грудных
иожек с нитевидными придатками
260
Рис. 248. Листовидная
ножка Apus cancriformis .(из
Caiman):
/ — дистальный экзит
(плавательная пластинка), 2—
проксимальный экзит (эпиподиальная
жабра), 3 — эндиты
Подотряд Conchostraca — Раковинные ластоногие раки
В качестве представителя Conchostraca можно взять разные виды
родов Esther ia Ruppel', а также Lynceus br achy иг us О. F. M.,
встречающихся в пресноводных водоемах Европы, в том числе и в водоемах
СССР. Характерная особенность этой группы — наличие двустворчатой
раковины, напоминающей таковую пластинчатожаберных моллюсков.
Размеры большинства видов Conchostraca:— около 4—8 мм (рис. 249,
250). Род Esther ia отличается от других родов этой группы наличием
на спинной стороне раковины ясно выраженной верхушки. Раковина
пропитана известью и закрывает почти все тело животного вместе с
головой и конечностями. Наружу выдаются лишь антенны и задний
конец брюшка, несущий вилку. Головной отдел несет ростральный
выступ и непарный глаз (рис. 249). В передней части тела расположен
мощный мускул аддуктор, служащий для замыкания створок
раковины. Створки раковины малопрозрачны, поэтому для ознакомления с
морфологией животного одну из створок нужно осторожно удалить.
Тело животного слагается из значительного числа сегментов
(рис. 250), нерезко отграниченных от головы и снабженных
листовидными конечностями. Вилка состоит из двух шиловидных придатков. Ан-
теннулы короткие, одноветвистые, зазубренные по краю. Вторая пара
антенн мощно развита, двуветвиста, несет многочисленные волоски и
служит органом движения (см. рис. 249). Мандибулы и обе пары
максилл лишены щупиков.
Число листовидных конечностей у многих видов различно, в
зависимости от числа сегментов. В пределах Conchostraca оно варьирует от
10 до 32. Подобно тому как это имеет место и у других Phyllopoda, у
Conchostraca конечность несет эпиподиальную мешковидную жабру,
зкзит и обычно пять эндитов (рис. 251). В отношении строения неко-
Рис. 249. Estheria obliqua. Л—раковина; Б — самец с удаленной створкой
(из Caiman):
/ — глаз, 2 — ростральный вырост головы, 3 — вторая антенна, 4 — передние грудные
ножки с хватательными придатками, 5 — листовидные ножки, 6 —- f urea
1 В современной систематике Phyllopoda род Estheria разбит на несколько
родов. Мы сохраняем, однако, прежнюю номенклатуру, принятую в большинстве
учебных руководств.
261
Рис. 250. Самец Lynceis brachyurus (из
С. С. Смирнова по Sars):
1 — глаз, 2 — кишечник, 3 — furca, 4 — грудные
ножкн, б — передние грудные ножки с
хватательными придатками, 6 — первые антенны, 7 —
вторые антенны
Рис. 251. Листовидная
конечность Estheria obli-
qua (из Caiman):
/ — дистальный экзит, 2 —
проксимальный экзит (эпи-
поднальная жабра), 3 — эн-
диты
торых конечностей у Conchostraca наблюдается половой диморфизм.
У самцов одна или две (см. рис. 249) передних пары конечностей
снабжены специальными хватательными придатками (видоизмененные эн-
диты), используемыми как хватательный орган при копуляции. У
самок 9, 10 и 11-я пары несут длинные нитевидные придатки. Последние
служат для направления яиц в выводковую камеру, расположенную на
спинной стороне раковины. Эти придатки представляют собой сильно
вытянутые в длину экзиты.
Подотряд Cladocera — Ветвигтоусые раки
Многочисленные представители этого отряда входят в состав
планктона пресных вод. Их удается находить в природе начиная с
момента вскрытия водоемов до ледостава. Зимой огромное большинство
Cladocera находится в состоянии зимних яиц. В водоемах озерного
типа некоторые виды сохраняются в течение всей зимы. В условиях
лаборатории не представляет труда содержать их всю зиму в
аквариумах. Чаще других встречаются представители родов Daphnia, Simo-
cephalus, Bosmina.
Для изучения морфологии наиболее удобным объектом служит
обычная пресноводная форма — Daphnia putex De Geer.
Кроме изучения морфологии, желательно произвести, некоторые
наблюдения за размножением и цикломорфозом Cladocera.
262
Daphnia pulex De Geer — Обыкновенная дафния
Материал и методы изучения. Daphnia pulex относится к сем. Daphnidae и
является одной из обычнейших форм в небольших пресноводных водоемах. Встречается
по всей территории СССР. Зиму хорошо выживает в аквариумах при условии
равномерной комнатной температуры и наличии пищи. Для питания D. pulex рекомендуем
периодически прибавлять в аквариум небольшие количества бульона, наличие
которого способствует развитию бактерий.
Для получения живого материала зимой можно использовать следующий прием.
Из водоема, в котором летом встречались дафнии и имеются зимние яйца, небольшое
количество льда и ила перенести в аквариум в теплое помещение. При этом вскоре из
зимних яиц развиваются дафнии, которые приступают к партеногенетическому
размножению. В большинстве случаев приходится иметь дело с партеногенетическими
самками, так как самцы появляются относительно редко. В условиях культуры изменением
внешних факторов удается вызвать появление самцов и образование зимних яиц и
эфиппий (см. с. 272). D. pulex относится к полициклическим видам — появление
самцов и оплодотворение имеет место у нее несколько раз в течение года. Поэтому
иногда самцов, а также самок с зимними яйцами удается находить в природе не только
поздней осенью, но и весной и летом.
Изучение строения дафнии проводится на живом объекте. Применяя различные
увеличения микроскопа благодаря небольшой величине и прозрачности объекта,
удается рассмотреть ее строение. К изготовлению тотальных препаратов и срезов
прибегать нет необходимости. Вместо D. pulex можно использовать и других
представителей этого рода, в частности наиболее крупную из всех дафний — D. magna Straus/
D. longispina Sars, D. cucullata Sars и др. D. cuculiata встречается преимущественно
в водоемах озерного типа и. представляет хороший объект для знакомства с явлением
цикломорфозов.
Наблюдения за движением и размножением дафний. Поместив
несколько экземпляров дафний в высокую энтомологическую пробирку,
можно, воспользовавшись лупой, наблюдать характер и механизм их
движения. Оно осуществляется характерными скачками, причем
органом поступательного движения является вторая пара антенн, мощно
развитая у Cladocera. Периодически совершающиеся движения антенн
носят характер коротких сильных ударов, осуществляемых в
направлении спереди назад. При каждом ударе антенн дафния перемещается
вверх и вперед (рис. 252). При этом продольная ось животного,
которая в начале скачка занимала вертикальное положение, становится
косо, так как голова перемещается несколько книзу в то время, как
задний конец приподнимается наверх. Дафния как бы стремится
перевернуться через голову. После
удара антенн наступает период покоя.
Животное начинает медленно
опускаться вниз. Продольная ось
дафнии вновь занимает вертикальное
положение. Антенны при этом
отводятся назад в исходное положение,
причем их основной членик
становится почти перпендикулярно
поверхности тела, а обе ветви с
волосками широко расставлены.
Антенны при этом, увеличивая общую п осо гл ' „ „
r ' J . J Рис. 252. Изменение положения про-
удельную поверхность животного, ДОЛьной оси дафнии при движении
замедляют скорость его погруже- (из Wagler no Woltereck)
263
ния. Через некоторое время
удар антенн повторяется
вновь. Указанная форма
движения дафнии
определяет ее зигзагообразный путь
в воде. При этом в
зависимости от
продолжительности указанных периодов и
частоты ударов антенн
животное либо поднимается,
либо постепенно опускается
вниз 1.
Дафнии питаются
мельчайшими органическими
частицами, взвешенными в
воде (бактерии, детрит,
мелкие водоросли и простейшие
и т. п.), которые попадают
в кишечник благодаря
деятельности ротовых частей и
грудных конечностей,
обусловливающих своими
движениями постоянный ток
воды и создающих фильтр,
удерживающий взвешенные
в воде частицы. Если в
воду, в которой живут даф-
•нии, прибавить мелко
растертую тушь или кармин, то
под лупой или бинокуляром
можно наблюдать довольно
быстрое заполнение
кишечника.
Вполне доступны также
наблюдения над темпом
размножения и плодовитостью
дафний 'при партеногенезе.
Партеногенетические самки
высаживаются по одиночке
в стаканчики или.
кристаллизаторы и содержатся при
комнатной температуре и достаточном количестве пищи. Каждая самка
D. pulex при благоприятных условиях среды дает до 40 потомков
(D. magna — до 80).
Добиться появления самцов удается путем понижения
температуры (ниже 10°С) и ухудшения условий питания.
1 Подробный анализ движения дафнии дан в работе: WoUereck R. Uber Funktion,
Herkunft und Entstehungsursache der sog. «Schwebefortsatze» pelagischer Cladoce-
ren. — Zoologica, Bd. 67, 1913.
Рис. 253. Daphnia pulex (из О. Storch):
J — первые антенны, 2 — мозг, 3 — науплиальный
глаз, 4 —сложный глаз, 5 —вторые антенны, 6 —
второй печеночный вырост, 7 — фронтальный орган,
8 — мандибула, 9 — максиллярная железа, 10 —
сердце, // — выводковая камера, 12— латеральная
складка, 13 — экзит пятой пары грудных конечностей,
14 — яичник, IS — спинной вырост, 16 — кишечник,
17 — экзоподит четвертой пары конечностей, 18 —
щетинки четвертой пары конечностей, 19 — эпиподит
четвертой пары конечностей, 20 — экзоподит третьей
пары конечностей, 21 — щетники третьей пары
конечностей, 22 — вторая пара конечностей, 23 — первая
пара конечностей
264
Организации самки и самца значительно отличаются друг от
друга, поэтому их удобнее рассмотреть отдельно.
Строение самки Daphnia pulex. Внешняя морфология.-Тело
дафнии несколько сплющено с боков. Благодаря этому на препарате
животное оказывается обычно лежащим на боку. Голова, имеющая
шлемовидную форму, клювообразно вытянута на брюшную сторону
(рис. 253), она несет обе пары антенн, непарный сложный глаз и
простой (науплиусов) глаз. Все остальное тело дафнии заключено в
прозрачную двустворчатую раковину, охватывающую с боков грудной,
несущий конечности, и брюшной, лишенный конечностей, отделы '.
Конечности также располагаются в полости раковины. На заднем конце
со спинной стороны раковина продолжается в форме острого шипа.
Ясно выраженной наружной сегментации у дафнии не наблюдается, о
числе сегментов можно судить только по числу конечностей. Брюшной
отдел тела загибается на брюшную сторону и заканчивается
характерной для огромного большинства Branchiopoda вилочкой (furca). Кроме
того, на спинной стороне брюшка имеется две довольно длинные
щетинки, выдающиеся назад из полости раковины (рис. 253). На спинной
стороне тела дафнии расположена выводковая камера, обычно
заполненная развивающимися эмбрионами. На заднем конце она
замыкается двумя особыми выростами, которые отгибаются при выходе
развившихся эмбрионов из выводковой камеры наружу.
Конечности. Первая пара антенн развита очень слабо (см.
рис. 253, /) (А1 сильнее развиты у самцов — см. с. 273). Они
представляют собой короткие нерасчлененные придатки, расположенные у
основания клюва головы. На дистальном конце А1 несут пучок
чувствительных волосков, являющихся, вероятно, хеморецепторами. Вторая
пара антенн, напротив, мощно развита (рис. 253, 5) и является
основным органом движения дафнии. Каждая антенна слагается из
массивной основной части (протоподита) и двух ветвей — четырехчленистой
наружной (экзоподита) и трехчленистой внутренней (эндоподита). Обе
ветви несут длинные перистые плавательные щетинки.
Мандибулы (рис. 253, 8) представляют собой довольно массивные
нерасчлененные вытянутые в длину пластинки, расположенные по
сторонам ротового отверстия. На своем дистальном конце мандибулы
несут многочисленные зубчики (рис. 254). Обе пары
мансилл развиты чрезвычайно слабо, носят
рудиментарный характер, разглядеть их, в силу малых
размеров и прозрачности, затруднительно.
Пять пар грудных конечностей (рис. 255) имеют
в общем листовидный характер, так же как и.-у
Phyllopoda. Но все они отличаются друг от друга и
специализированы для выполнения определенных
функций. Их следует отпрепаровать тонкими
препаровальными иглами под бинокуляром.
1 Раковина Cladocera в большей своей части развивается за
счет максиллярной области. Морфологически граница между
protocephalon и gnathocephalon не выражена.
Рис. 254.
Мандибула Daphnia
pulex (из Lilljeborg)
265
Pc. 255. Daphnia pulex. Грудные конечности I—V (из Schultze no Beh-
ning):
/—эпиподиты, 2 — экзиты, 3 — эндиты, 4 — максиллярный вырост второй пары
грудных ножек
Каждая из пяти пар грудных конечностей обладает внутренними и
наружными выростами — эндитами и экзитами. Один из' экзитов
каждой из пяти пар представляет собой небольшую эпиподиальную жабру.
Первая пара имеет наименьшие размеры и несет довольно длинные
перистые волоски (рис. 255, /). Вторая пара характеризуется наличием
особого максиллярного выроста, расположенного у основания
конечности и снабженного одной длинной и рядом более коротких щетинок
(рис. 255, II). Максиллярный вырост со щетинками играет важную
роль в захвате пищи (см. ниже). Кроме того, вторая пара конечностей
несет и на своем дистальном конце длинные перистые щетинки. Третья
и четвертая пара грудных конечностей являются наиболее крупными
из всех (рис. 255, III—IV). Характерную особенность их строения
представляет наличие с внутренней стороны множества длинных тонких
щетинок, расположенных почти вплотную друг к другу и образующих
фильтр, служащий для удержания мельчайших
взвешенных в воде частиц. Характерной чертой
пятой (рис. 255, V) пары конечностей является
наличие длинной загибающейся назад крючко-
видной щетинки. В совокупности грудные
конечности образуют довольно сложный насасыва-
тельный и фильтрующий аппарат. Они находятся
в непрерывном движении в течение всей жизни
животного. Механизм действия этого аппарата
обстоятельно изучен Шторхом. Вода из
наружной среды проникает в полость, ограниченную
со спинной стороны брюшной стенкой тела и с
боков—конечностями и створками раковины,
через брюшную щель. Последняя образуется
благодаря раздвиганию конечностей. При
замыкании брюшной щели вода направляется назад
Рис. 256. Схема, показы- и выходит наружу с брюшной стороны в задней
еающая направление то- треТи раковины (рис. 256, 257). Попадая в по-
L^1eBч0„AoЫcтerliPИдaфнBииЖeH(из Л0СТЬ' Расположенную между конечностями, ,во-
О. Storch) да при сближении последних фильтруется через
266
Рис. 257. Схема действия фильтрационного аппарата дафнии. А—Г —
последовательные стадии поступления и фильтрации воды (из О. Storch):
/ — четвертая пара грудных ножек, 2 — третья пара грудных ножек, 3 — волоски
третьей пары грудных ножек, 4 — волоскн четвертой пары грудных ножек. Стрелки
показывают направление тока воды
тонкий фильтр, образованный волосками третьей и четвертой пары
ножек (рис. 255, III—IV). Мельчайшие взвешенные в воде частицы,
служащие дафнии нищей, при этом задерживаются. Они собираются в
желобок, образуемый брюшной стенкой тела. Здесь формируется.пищевой
комочек. Его дальнейший путь к ротовому отверстию осуществляется
благодаря действию максиллярного выроста 'второй пары конечностей,
который проталкивает его вперед по направлению к голове. Проходя
мимо мандибул, пищевой комочек размельчается, после чего, благодаря
глотательному движению, попадает в пищевод.
Мускулатура. На живом' объекте хорошо видны мускулатура
второй пары антенн и мандибул. Движения Ап осуществляются
действием четырех пар мышц (рис. 258). Три из них прикрепляются к
спинной части головного шлема, одна — проходит под кишечником. Их
проксимальные концы расширены. Они тянутся в полость антенн и
продолжаются в обе ветви их. Мандибулы обладают двумя парами сильных
мышц, из которых задняя
значительно крупнее передней.
Все мышцы дафнии являются
поперечнополосатыми.
Пищеварительная
система. Ротовое отверстие
расположено между
мандибулами и прикрыто спереди
верхней губой,
представляющей довольно большую
хитиновую пластинку. Оно ведет в
узкий эктодермальиый
пищевод (переднюю кишку, см.
рис. 253), обладающий муску- Рис. 258. Daphnia magna. Мышцы второй
листыми стенками. Пищевод- паРы антенн (из Klotzsche):
пткпьтяется R энтпттрпмяпктгтп '-сердце, 2 - мышцы мандибул, 3-б-пер-
и1к.рьшае1сн и сшшдермальную вый — четвертый парные мускулы вторых антенн
267
Рис. 259.
молимфы
среднюю кишку, тянущуюся в виде
прямой трубки почти до заднего конца
животного, где она загибается на брюшную
сторону. На границе между передней и
средней кишкой располагается клапан,
препятствующий переходу пищи из
средней кишки в переднюю.
Средняя кишка в голове, недалеко
от места своего соединения с пищеводом,
образует два небольших слепых
выпячивания, называемых печеночными
отростками (см. рис. 253, 6). В задней
части брюшка средняя кишка переходит в
заднюю (эктодермальную). Граница
между этими отделами анатомически
почти не различима. Анальное отверстие
расположено на заднем конце брюшка
между ветвями вилочки. Форма
кишечного канала дафнии выступает особенно
отчетливо, если животное подкормить
тушью.
Кровеносная система.
Мешковидное сердце, имеющее лишь одну
пару остий по бокам и выходное
отверстие на переднем конце, расположено на
спинной стороне в максиллярном сегмент
.те (см. рис. 253, 10). Хорошо видны
отдельные мускульные клетки, слагающие
сердце и расположенные наподобие
долек апельсина. На живой дафнии очень
хорошо, даже при малом увеличении микроскопа, видны частые
сокращения сердца (до 200—250 раз в минуту). Кровь (гемолимфа)
—бесцветна, в ней встречаются лишь отдельные амебоидные клетки.
Благодаря почти полной прозрачности тела дафнии, возможно, особенно на
молодых экземплярах, проследить основные пути движения крови в
полости тела, которое осуществляется вполне закономерно (рис. 259).
Направление основных токов определяется наличием в полости тела
двух мембран. Одна из них (дорзальная) охватывает спереди сердце и
тянется назад над кишечником. Вторая— (брюшная) проходит под
кишечником. Ее средняя часть прикрепляется к брюшной стенке тела,
благодаря чему образуется правый и левый брюшные синусы. В
области сердца брюшная мембрана поднимается кверху до уровня дорзаль-
ной бтенки кишечника. Общая схема циркуляции крови показана на
рис. 259. Кровь из сердца направляется в голову, омывает головной
мозг, откуда главная ее масса поступает в правый и левый брюшной
синус. Отсюда она направляется в конечности, где обогащается
кислородом. Из конечностей кровь по брюшному синусу поступает в задний
конец тела, далее переходит на спинную сторону и движется вперед к
сердцу.
Пути движения ге-
в теле дафнии.
Основные токи показаны
стрелками (по Horonard из Schult-
ze):
1 — дорзальная мембрана,
2—вентральная мембрана, 3— загнутая
на спннную сторону часть
вентральной мембраны
268
4
г
з-
и-
6-
Рис. 260. Схема нервной
системы Cladocera (из
Giesbrecht):
/ — оптическая лопасть, 2 —
иерв первой пары антенн,
3 — нервы второй пары
антенн, 4 — ганглий мандибул,
5 — ганглий первой пары
грудных ножек, 6 — ганглий
пятой пары грудных ножек'
Выделительная система. У D. ри-
lex имеется хорошо развитая парная максилляр-
ная железа, лежащая в сегменте второй пары
максилл (см. рис. 253, 9). Аналогично тому, как
это имеет место у Artemia (с. 251), она состоит
из целомичеокого мешочка, открывающегося при
посредстве нефростома в выделительный канал.
Последний образует тройную петлю. Конечный
суженный отдел его (выводной канал):
открывается наружу у основания второй пары максилл.
Все указанные отделы выделительной железы
хорошо видны in vivo на небольших прозрачных
экземплярах дафний.
Нервная система. На живом объекте
хорошо виден надглоточный ганглий, или мозг,
располагающийся в голове несколько кпереди от
основания второй лары антенн (см. рис. 253,'£)■
От него отходят нервы к глазу и фронтальным
органам. Окологлоточные коннективы огибают
с обеих сторон пищевод. В их задней части
расположены ганглии, иннервирующие вторую пару
антенн. Таким образом, у Cladocera, так же как и у Phyllopoda,
наблюдаются примитивные отношения в том смысле, что ганглий второй
пары антенн еще не входит в состав мозга в противоположность тому,
что имеет место у огромного большинства других ракообразных.
Брюшная нервная лестница слагается из парного ганглия
мандибул и пяти пар ганглиев, соответственно числу грудных конечностей
{рис. 260). Одноименные ганглии соединяются парными коммиссурами.
Специальные ганглии, соответствующие максиллам, отсутствуют.
В брюшко, лишенное конечностей, нервная лестница не продолжается,
туда тянутся лишь от последней пары ганглиев парные нервы. Части
брюшной нервной лестницы, лежащие под кишечником, in vivo видны
не очень отчетливо, благодаря конечностям и максиллярной железе. Тем
не менее на молодых, прозрачных экземплярах рассмотреть ее вполне
возможно.
Органы чувств. У дафнии имеется непарный сложный глаз,
простой, 'или науплиусов, глаз, фронтальные органы и хеморецепторы,
или эстетаски. Последние располагаются на первой паре антенн. Все
эти органы могут быть рассмотрены на живом объекте (см. рис. 253).
Непарный сложный глаз дафний, расположенный в передней части
головы, обладает значительными размерами и подвижен '. Последнее
обусловлено, с одной стороны, тем, что к глазу прикрепляются
специальные глазные мышцы, а с другой — особым положением глазного
яблока. Вокруг глаза имеется полость, к стенкам которой он как бы
подвешен при помощи небольших лигаментов (рис. 261). Расположенная
1 Непарный сложный глаз в онтогенезе образуется путем сближения и
последующего слияния двух зачатков. У некоторых видов Cladocera следы этой
первоначальной парности сохраняются.
269
над глазом наружная
хитиновая кутикула не образует
фасеток. В состав
сложного глаза дафний входит
обычно 22 омматидия.
Общая схема строения омма-
тидиев та же, что у Anost-
гаса (см. с. 254). In vivo
хорошо видны хрустальные
конусы отдельных оммати-
диев, обладающие выпуклой
формой. По их числу можно
судить о количестве омма-
тидивв, входящих в состав
сложного глаза.
Рассмотреть отдельные элементы
ретинальной части (рети-
нальные и пигментные
клетки), благодаря наличию
большого количества
пигмента, на живом объекте
не удается. От глаза к
мозгу отходит хорошо развитый
оптический нерв. Простой
(науплиусов) глаз
небольших размеров. В виде
небольшого пигментного
пятнышка он располагается на
нижней стороне головы и
почти прилежит к мозгу
(см. рис. 253, 3). При большом увеличении, придавив животное,
удается рассмотреть, что глаз складывается из пигментного бокала и
небольшого числа прилежащих к нему чувствительных клеток, собранных
в четыре группы.
Фронтальные органы, функция которых остается неясной,
расположены также в голове. У D. pulex имеются парные фронтальные органы
на спинной и брюшной сторонах головы (см. рис. 253, 7). Нервы от них
идут к передней части мозга.
Эстетаски (хеморецепторы), как уже отмечалось (с. 265),
расположены на дистальном конце первой пары антенн. Обычно каждая
антенна несет девять чувствительных волосков. От их основания в
проксимальном направлении отходят нервные волоконца. Последние, в свою
очередь, соединены с группой биполярных нервных клеток,
расположенных у основания антенн. От них идет нерв к мозгу. Все это удается
рассмотреть при больших увеличениях на мелких прозрачных
экземплярах. Иногда полезно бывает слегка подкрасить дафнию слабым
раствором метиленового синего.
Половая система. Парные яичники лежат по сторонам
средней кишки в грудном отделе (см. рис. 253,14). На заднем конце от каж-
Рис. 261. Строение сложного глаза Daphnia
magna (из Klotzsche):
1 — хитиновая кутикула, 2 — камера, окружающая глаз,
3— хрустальные конусы омматидиев, 4 — лигамент, на
котором подвешен глаз, 5 — мышцы глаза
207
Рис. 262. Строение яичиика Daphnia. A — яичник, продуцирующий летние
яйца; Б — яичник, продуцирующий летние яйца и одно зимнее яйцо; В —
яичиик с зимними яйцами (из Schultze):
/_ четверная группа, 2 — зона размножения, 3 — зимиее яйцо
дого яичника отходит короткий яйцевод, который открывается в
выводковую камеру, расположенную на спинной стороне тела. В яичнике на
заднем конце его расположена зона размножения, состоящая из мелких
оогониальных клеток, размножающихся путем деления. В остальной
части яичника располагаются яйцевые клетки на разных стадиях
роста. В яичнике происходит образование летних партеногенетических или
зимних яиц. Последние развиваются только после оплодотворения.
Иногда образование того и другого типа яиц может протекать
одновременно (рис. 262). Летние яйца дафний относительно мельче
зимних, они более прозрачны, так как содержат многочисленные капельки
жира, тогда как зимние яйца более темные, благодаря наличию
значительного количества желтка. В яичнике, где происходит формирование
летних яиц, клетки располагаются характерными четверными группами
(рис. 262, А). В пределах каждой такой группы лишь одна клетка
заканчивает свой рост и превращается в яйцо, способное к дальнейшему
£7.1,
развитию. Остальные три по мере роста собственно яйцевой клетки
постепенно уменьшаются в размерах, так как их цитоплазма
поглощается растущим яйцом. При образовании зимних яиц также образуются
четверные группы. Но кроме трех клеток растущим ооцитом
поглощаются клетки и соседних с нею четверных групп, что связано с большими
размерами зимних яиц по сравнению с летними. Оплодотворение зимнего
яйца происходит уже после выхода его из яичника. Размеры зрелых яиц
у D. pulex по Вейсманну следующие: зимнее яйцо — 380-210 мкм,
летнее яйцо 250-190 мкм.
У самок, продуцирующих зимние яйца, на спинной стороне вокруг
выводковой камеры образуется темно-пигментированное седлышко, или
эфиппий (см. рис. 286), внутри которого и помещаются яйца.
Одновременно в эфиппий у D. pulex имеются два яйца. Седлышко
сбрасывается дафнией во время линьки. Оно не опускается на дно, а плавает в
воде благодаря наличию в стенке эфиппия особых наполненных газом
хитиновых камер. Строение эфиппия довольно сложно. Он
представляет собой видоизмененную часть обеих створок раковины, прилежащих
к выводковой камере. Наружная стенка раковинки при образовании
эфиппия сильно утолщается благодаря деятельности подлежащих
эпителиальных покровных клеток. В стенке эфиппия имеется множество
мелких камер, причем каждая камера образуется одной покровной
клеткой. Внутренняя полость камер заполняется газом. Благодаря
наличию указанной структуры, при рассматривании эфиппия сбоку,
стенки его кажутся мелкоячеистыми (рис. 263). Сбрасывание эфиппия
Рис. 264. Самец Daphnia
pulex (из F. Leydig):
1 — первые антенны, 2 — вторые
антенны, 3 — кишечник, 4 —
сердце, 5—семенник, 6 — furca,
7 — жгутовидиый придаток
первой пары грудных конечностей
Рис. 263. Эфиппий. А — Daphnia pulex; Б — Daphnia
magna (из Schultze) •
необязательно сопровождается
гибелью самки, которая может
последовательно образовать
несколько .седлышек.
У различных видов дафний
и других Cladocera эфиппий
имеют различную структуру, что
является одним из
систематических признаков.
Строение самца D. pulex.
Самцы D. pulex меньших
размеров. Длина самца меньше
таковой самки приблизительно на
'/з1- Голова имеет несколько
Рис. 265. Самец Daphnia pulex. A — первые
антенны; Б — первая пара грудных ног
(А — из F. Leydig, Б — из P. Schultze):
1 — чувствительные волоски, 2 — крючковндно
загнутая терминальная щетинка, 3— ганглий, 4 —
нерв, 5 — жгутовидиый придаток
иную форму, чем у самки, так
как не образуется заостренного
на конце «клюва» (рис. 264).
Бросающимся в глаза отличительным
признаком служит отсутствие выводковой камеры, благодаря чему
спина у самца не выпуклая. Имеются также существенные различия в
строении конечностей. Первая пара антенн у самца развита гораздо
сильнее и является двучленистой (рис. 265). При этом кроме
чувствительных волосков А1 снабжены особым крючковидно загнутым на
конце придатком, который'способствует удержанию самки при копуляции.
Первая пфа грудных ног самца (см. рис. 264, 7) в отличие от самки
имеет два особых придатка — крючковидный и жгутовидиый, которые
выполняют роль хватательных аппаратов при копуляции. Строение
других конечностей, так же как и внутренняя организация, у обоих полов
сходно за исключением органов размножения. Семенники (парные)
занимают в теле самца то же положение, что и яичники у самки. Их
задний конец продолжается в длинный vas deferens, тянущийся вдоль
всего брюшка рядом с кишечником. Мужские половые отверстия
(каждый vas deferens открывается наружу самостоятельно)
располагаются на заднем конце брюшка, вблизи анального отверстия, по
отношению к которому они несколько сдвинуты на спинную сторону
(рис. 266).
При копуляции задний конец брюшка самца вводится в
выводковую камеру самки, где и происходит оплодотворение поступивших туда
же зимних яиц. Сперматозоиды дафнии, так же как и всех Cladocera,
не имеют типичной жгутиковой формы. Они снабжены плазматическими
выростами, напоминающими псевдоподии.
Наблюдения над цикломорфозом. Некоторые, особенно пелагические, формы
Cladocera обладают резко выраженной изменчивостью. С одной стороны, изменчивость
г эта выражается в закономерной смене форм по временам года и может быть названа
цикломорфозом. С другой стороны, представители одного и того же.вида из разных
водоемов нередко морфологически отличаются между собой, что указывает на наличие
1 У некоторых других видов дафний самцы бывают еще мельче. Например, у
D. magna сайец примерно в 2,5 раза короче самки.
273
Рис. 266. Задний конец тела самца Daphnla
pulex (из F. Leydig):
/ — раковина, 2 — семенник, 3 — vas deferens, 4 —
furca, 5 — кишечник
в пределах вида более мелких
наследственно константных единиц
(разновидностей, биотипов).
Кроме того, у Cladocera выражена
также и возрастная изменчивость,
выражающаяся в том, что после
линек происходят
морфологические изменения. Изменчивость
дафний в первую очередь
затрагивает высоту головного шлема (рис.
267), соотношение длины и
ширины тела, длину заднего шипа,
размеры второй пары антенн. При
прохождении практикума
рекомендуется ознакомиться с явлением
цикломорфоза, что не
представляет особого затруднения. Один из
наиболее подходящих для этой
цели видов — D. cucullata
G. О. Sars (рис. 267). Резко
выражена сезонная изменчивость
также у Bosmina coregoni Baird (рис.
268). Оба вида часто
встречаются в составе планктона
пресноводных водоемов озерного типа.
Следует брать ежемесячные пробы
планктона и сравнивать строение
дафний в различные сезоны. У
D. cucullata летом головной шлем
очень высокий, он вытянут в
длинный шип. Зимние формы
обладают коротким округлым головным
шлемом. Длина тела по
отношению к его высоте у летних форм
больше, чем у зимних.
У Bosmina coregoni (рис. 268)
наблюдаются резкие сезонные
изменения высоты тела и длины
антенн. Летние формы обладают
очень высоким телом и
длинными антеннами. У зимних и
весенних форм высота тела
значительно меньше, а антенны короче.
Рис. 267. Цикломорфоз Daphnia cucullata: Рис. 268. Цикломорфоз Bosmina corego-
А — май; Б — июнь; В — июль; . Г — сен- fit:- А — апрель; Б — июнь; . В — август;
тябрь; Д — октябрь (из Wagler) Г — октябрь; Д — декабрь (из Wagler)
274
ПОДКЛАСС OSTRACODA —РАКУШКОВЫЕ РАКИ
Подкласс Ракушковых раков включает лишь один отряд того же
названия. Ракушковые раки широко распространены в природе как в
пресных водах, так и в Мировом океане. Образ жизни их
разнообразен, но они населяют преимущественно прибрежную зону, где плавают
между растениями и на дне водоема. Размеры их невелики, обычно они
не превышают 1 мм.
При прохождении практикума достаточно на живом материале
ознакомиться с организацией одного вида из состава пресноводной
фауны. В качестве примера мы берем широко распространенный род
Cypris, представленный несколькими видами.
Cypris sp.
Материал и методы изучения. Одним из наиболее широко распространенных
видов рода Cypris является С. pubera О. F. Milller (рис. 269) (подотр. Podocopa, сем.
Cypridae), встречающийся повсеместно в прудах, канавах и даже лужах. Кроме
представителей рода Cypris могут встретиться и другие близкие роды сем. Cypridae
(Herpetocypris, рис. 270, Cyclocypris, Eucypris, Ilyocypris, Candona и др.), которые
также могут быть использованы для изучения.
Изучение ведется на живом материале. Раковина Ostracoda мало прозрачна,
поэтому, для того чтобы рассмотреть конечности и некоторые внутренние органы,
следует под малым увеличением бинокулярной лупы осторожно раскрыть раковину
(снять одну из створок) при помощи тонких препаровальных игл.
В раскрытом виде надо рассмотреть строение животного и структуру снятой
створки раковины изнутри. После этого препаровальными иглами постараться отделить
конечности друг от друга, прикрыть препарат покровным стеклом, слегка придавив,
и рассмотреть под микроскопом.
Большинство пресноводных Ostracoda размножаются партеногенетически. В
частности, у Cypris pubera самцы неизвестны, поэтому ниже рассматривается только
организация самки. При указанных методах препаровки, кроме внешней морфологии
и конечностей, удается рассмотреть лишь некоторые части пищеварительной и половой
систем. Для более детального изучения внутренней организации необходимо
изготовление срезов.
Внешняя морфология. Все тело животного заключено 'В
двустворчатую раковину, которая представляет собой обызвествленную хитиновую
кутикулу, выделяемую складкой покровов, спускающейся со спинной
стороны тела животного (рис. 270). Брюшной край створок раковины
более или менее прямой, спинной — закруглен. На поверхности
раковина обычно несет волоски. Число и расположение их неодинаково у
разных видов. Обе створки соединяются
друг с другом на спинной стороне. При
этом по спинному, краю раковины в месте
соединения обеих створок имеются мелкие
зубчики, образующие в совокупности
замок, аналогичные таковым пластинчатожа- З^^- -* "//a f "\\N^N$
берных моллюсков. Раковина прикрывает ^3^<C^f Л^'^л4^:^
тело и конечности Cypris, за исключением ^T-nrUfrrrVitrHr^h^rr^^^
первой и второй пары антенн и второй па- Рис 269 0б й с
ры грудных ножек (рис. 270, 2, 3, 9). Кро- ris pubera (из Ламперта)
275
Рис. 270. Herpetocypris reptans (из Schultze no Woltereck):
/ — глаз, 2— первые антенны, 3— вторые антенны, 4 — пищевод, 5
—мандибула, 6 — максилла, 7 — первая грудная конечность, превращенная в
ногочелюсть, 8— запирательный мускул, 9 — вторая грудная конечность,
10 — печеночный вырост кишечника, /} — furea, 12 — янчник, 13 —
зачатковая зона яичника, 14 — третья грудная конечность, 15— яйцевод,
наполненный яйцами, 16 — средняя кишка
ме того, наружу выдается иногда и задний конец вилки.
С внутренней стороны раковины на обеих створках видны следы
прикрепления мускулов, служащих для замыкания створок
(мускульные отпечатки). Расположение и форма этих отпечатков служат одним
из систематических признаков Ostracoda.
В передней части тела над местом прикрепления первой пары
антенн расположен непарный (науплиусов) глаз.
Расчленение тела и конечности. Раскрыв раковину
Cypris, ясно.видно, что наружная сегментация, полностью утеряна.
Голова и брюшко внешне не отделены от груди, сегментация последней
также не выражена. О числе сегментов тела Ostracoda можно судить
лишь по числу пар конечностей. На заднем конце тела имеется
довольно длинная двухраздельная на дистальном конце вилка (рис. 270, 11).
Число конечностей у Cypris, как и у всех Ostracoda, равняется семи
парам. Имеется первая и вторая пары антенн, мандибулы, одна пара
максилл и три пары грудных ног, из которых первая у Cypris
видоизменена в ногочелюсти. Антенны первой пары (антеннулы) одноветвисты
(рис. .271, А),, они состоят из одного ряда члеников (числом семь) и
несут длинные плавательные щетинки. Волоски последнего дистального
членика выполняют, кроме того, осязательную функцию. Первые ан-
276
тенны служат органом
плавания. Гребные движения
их осуществляются по
направлению спереди назад и
наверх, что вызывает
поступательное движение
вперед и вниз.
Антенны второй пары
(рис. 271, Б) также одно-
ветвисты, число члеников у
Cypris равняется пяти, эк-
зоподит рудиментарен.
Вторые антенны несут .длинные
плавательные щетинки. Так
же как и первая пара
антенн, они служат органами
движения. Но кроме
функции плавания они служат
также для ползания и копа:
ния. При плавании 'вторые
антенны совершают гребные
.движения вниз и назад,
вызывая поступательные
движения животного вперед и
наверх. Совместная работа
первой и второй пары ан-
тенн вызывает перемещение рщ; ш Конечйости Ostracoda, Podocopa. A-
животного ПО прямой перед- первые антенны; Б — вторые антенны; В — ман-
НИМ КОНЦОМ вперед (рис. дибула; Г —максилла; Д — ногочелюсть (А, Б,
272). На второй паре ан- В, Д —из Бирштейна, Г —из W. Muller)
тенн «кроме плавательных
щетинок имеются также хе-
морецепторы.
Мандибулы (см. рис. 270, 5; 271, В) обладают довольно мощно
развитым основным члеником с жевательной лопастью. Имеется
членистый щупик (эндоподит), несущий щетинки. Экзоподит также развит и
несет многочисленные щетинки.
Максиллы имеются в количестве лишь одной пары (см. рис. 270, 6;
271, Г). Их широкий основной членик несет три осязательные лопасти
со щетинками. Кроме того, имеется трехчленистый щупик (эндоподит).
Основная часть максиллы образует, кроме того, довольно большую
пластинку, густо усаженную перистыми волосками. Она называется
дыхательной пластинкой, так как движение ее способствует току воды через
полость раковины.
Первая пара грудных ножек (см. рис. 270, 7; 271, Д) представляет
ногочелюсти. Расчленение их выражено слабо, они несут
многочисленные щетинки и жевательную лопасть.
Вторая пара грудных конечностей (см. рис. 270, 9) представляет
типичные ходильные ноги, состоящие из одного ряда члеников и закан-
277
чивающиеся когтевидным
придатком. Вместе со вторыми
антеннами они служат для
передвижения по субстрату (рис. 273).
Третья шара грудных ножек
у Cypris и других Cypridae
называется чистильными ногами
(см. рис. 270, 14). Они загнуты
внутрь раковины и служат для
очищения внутренней
поверхности створок ют ^посторонних
частиц.
Внутренняя организация. Из
.внутренних органов на Cypris с
удаленной створкой раковины
удается 'рассмотреть только
некоторые части пищеварительной
и половой системы.
Ротовое отверстие
помещается между ротовыми
конечностями, спереди прикрыто верхней,
сзади — нижней губой. Оно
ведет ш узкий эктодермальный
пищевод (см. рис. 270, 4),
открывающийся в расширенную среднюю
кишку (см. рис. 270, 16). В
последнюю открываются длинные
трубчатые железистые,
придатки — печеночные отростки (см,
рис. 270, 10). Они направлены назад и вниз и залегают между
двумя листками боковых складок кожи. Средняя кишка переходит в
эктодермальную заднюю кишку, открывающуюся анальным отверстием
на заднем конце, за местом отхождения вилки. Женский половой
аппарат располагается в задней половине тела Cypris (см. рис. 270, 12, 13) г
Парные яичники имеют форму трубок. Их начальный отдел обычно
загнут на спинную сторону. В этом отделе яичника находится зона
размножения, в которой 'происходят деления ооганий. Границы между
клетками здесь отсутствуют и образуется синцитий. Остальная часть
яичника заполнена растущими ооцитами. Значительная часть яичника
расположена в боковых складках покровов непосредственно под
раковиной. Яичники переходят в широкие яйцеводы, наполненные обычно
многочисленными яйцевыми клетками. Яйцеводы посредством коротких
vagina открываются наружу. Половое отверстие помещается на
брюшной стороне между последней парой конечностей и вилкой.
Кровеносная и дыхательная система у Cypris отсутствуют,
элементы нервной системы не видны. ,
Из мышц более или менее отчетливо виден мускул — запиратель
створок (см. рис. 270, 8), а также иногда мышцы, двигающие
мандибулы.
Рис. 272. Схема плавательных движений
Ostracoda. Стрелками показано
направление движений первой и второй пары
антенн и направление, по которому
перемещается животное (из P. Schultze)
Рис. 273. Схема передвижения Cypris по
субстрату (из P. Schultze)
278
ПОДКЛАСС MAXILLOPODA — МАКСИЛЛОПОДЫ
ОТРЯД COPEPODA —ВЕСЛОНОГИЕ РАКИ
Веслоногие большей частью планктонные рачки, как и ветвисто-
усые раки. В отличие от последних Copepoda входят в большом
количестве и в состав морского планктона. Добывание живого материала
не представляет затруднений даже в зимнее время. В качестве
основного объекта для изучения берутся представители рода Cyclops Muller,
относящегося к подотряду Podoplea. В качестве дополнительного
объекта следует изучить организацию рода Diaptomus Westwood, как
представителя подотряда Gymnoplea.
Copepoda, в частности Cyclops, удобный объект для изучения
метаморфоза. На живом материале следует изучить все личиночные
стадии циклопа или Diaptomus, начиная со стадии науплиуса и кончая
половозрелым животным.
Cyclops sp.
Материал и методы изучения. В пресных водах встречаются разные виды рода
Cyclops, общее число которых в пределах СССР более тридцати. Для изучения могут
быть использованы многие виды, мелкие различия между которыми, затрагивающие
строение вилки пятой пары грудных' конечностей, число колец первой пары антенн и
некоторые другие, для морфологических целей не столь существенны.
Все виды Cyclops небольших размеров, длина их варьирует в среднем от 1 до
5 мм. Наиболее часто в мелких водоемах встречаются Cyclops fuscus Jur., С. atbidus
Jur., С. strenuus Fischer, С. leuckarti Claus, C. viridis Iur.
Материал добывается при помощи планктонной сетки или простым
зачерпыванием воды из пруда. Циклопы хорошо живут и размножаются в аквариумах, особенно
•если их подкармливать инфузориями. Изучение строения циклопа проводится на
живом объекте. Под микроскопом хорошо видны не только внешняя морфология, но и
■внутренняя организация. Конечности отпрепаровываются тонкими иглами под средним
■увеличением бинокулярного микроскопа. При этом лучше взять экземпляры,
предварительно консервированные спиртом или формалином. Из отпрепарованных конечностей
рекомендуется изготовить тотальный препарат в глицерин-желатине.
В качестве дополнения к изучению живого объекта желательно изготовить серию
поперечных срезов (заливка по Петерфи). Материал фиксируется какой-либо
сулемовой смесью. Окрашивается по Маллори или гематоксилином с эозином.
Внешняя морфология. Тело циклопа ясно сегментировано,
образования типа раковины отсутствуют. Тело распадается на овальную (при
рассматривании сверху) головогрудь и узкое удлиненное брюшко,
заканчивающееся вилочкой, снабженной длинными перистыми
щетинками (рис. 274,275).
Головогрудь слагается из пяти члеников. Из. них первый занимает
по длине примерно половину головогруди и, как можно судить по
конечностям, представляет собой голову1, слившуюся с первым грудным
■сегментом. Далее назад следуют четыре ясно разграниченных грудных
сегмента. Из них три передних широки, задний же по ширине своей
лишь немногим превосходит брюшко.
1 Голова Copepoda представляет собой результат слияния protocephalon и gnatho-
■cephalon, т. е. является syncephalon (см. с. 246).
279
Рис. 274. Самка Cyclops strenuus (из Sars и Claus):
1 — глаз, 2— первые антенны, 3 — вторые антенны, 4 — головогрудь,
5 — четыре свободных грудных сегмента, 6 — генитальиый сегмент
брюшка, 7 — яйцевые мешки, 8 — вилочка, 9 — кишечник, 10 -*-
продольные мышцы груди, 11 — яичник
Брюшко у самки (рис. 274) слагается из четырех члеников, у
самца (рис. 275) — из пяти. Передний из них несет наружные половые
отверстия и называется генйтальным сегментом. Меньшее число брюшных
сегментов у самки обязано слиянию двух передних.
Длина вилки вместе со щетинками равняется, а у некоторых
видов даже превосходит длину остальной части брюшка. Брюшко
сочленяется с головогрудью подвижно. У самок по обеим сторонам брюшка
располагаются яйцевые мешки, в которых происходит развитие яиц.
Они прикрепляются к генитальному сегменту..
Конечности. Первая пара антенн развита у Cyclops, как и у
большинства других Copepoda, очень хорошо. По строению А1 самцы
резко отличаются от самок, и в этом заключается одна из наиболее
ясных черт полового диморфизма. У самок одноветвистые длинные Аг
280
слагаются из значительного количества
вышает, однако, семнадцати (рис. 274, I
кое же, как и у самок, но дистальные
служат для удержания самки во
время копуляции (рис. 275, 2). Вторая
пара антенн значительно короче
первой. Ап слагается лишь из четырех
члеников, несущих хитиновые волоски
(рис. 274, 3), и является также одно-
ветвистой.
Мандибулы (жвалы) не
расчленены (рис. 276, Л). Они имеют
жевательную лопасть и рудиментарный щупик
(palpus). Последний несет два очень
длинных и одну короткую щетинку.
Первые и вторые пары максилл (рис.
276, Б, В) также не двуветвистые.
Они снабжены длинными щетинками,
которые на первой паре расположены
как с внутренней, так и с наружной
стороны, тогда как 'на второй паре
максилл — только по внутреннему
краю. Ноточелюсти (maxillipedes) в
количестве одной пары состоят
каждая из четырех члеников, . несущих
также довольно длинные щетинки.
Четыре пары грудных ножек
представляют хорошо развитые, типичные для
ракообразных, двуветвистые
конечности (рис. 276, Д, Е). Основная часть
их (протоподит) слагается из двух
члеников. Наружная (экзоподит) и
внутренняя (эндоподит) состоят каждая
из четырех члеников с
многочисленными щетинками. Пятая пара грудных
конечностей рудиментарна (рис.
276, Ж). Она слагается из двух
члеников. Конечности циклопа не несут
никаких жаберных придатков.
Брюшко, кроме вилки, не имеет других
придатков.
Внутренняя организация.
Мускулатура. Наиболее мощно
развитыми мускулами у циклопа являются
продольные мышцы, тянущиеся через
всю головогрудь (см. рис. 274, 10;
275, 11). Они складываются из
нескольких обособленных пучков, • часть
которых является сгибателями,
колец, число которых не пре-
). У самцов число колец А1 та-
концы антенны загнуты, и они
Рис. 275. Самец Cyclops canthocar-
poides (из Claus):
/ — глаз, 2 — первые антенны, 3 —
головогрудь, 4 — четыре свободных грудных
сегмента, 5 — генитальиый сегмент брюшка,
6 — второй—пятый сегменты брюшка, 7 —
вилочка, 8 — кишечник, 9 — сперматофор-
ные карманы, 10 — vas deferens, И —
продольные мышцы груди
281
Рис. 276. Cyclops strenuus. A — мандибула; Б — первая максилла;
.В —вторая максилла; Г — ногочелюсть; Д— первая грудная
плавательная ножка; Е—- четвертая грудная плавательная ножка; Ж
—питая грудная конечность (из Sars) :■'
1 — протоподит, 2 — экзоподит, 3 — эндоподит
часть — разгибателями тела. Из других мускулов на живом объекте
видны еще продольные мускулы в брюшке, а также метамерно
расположенные мелкие мышцы в головогруди, направляющиеся в конечности.
Для более детального ознакомления с мускулатурой циклопа
необходимо изучение срезов.
Пищеварительная система. Расположенное на нижней
стороне головы между ротовыми конечностями ротовое отверстие
прикрыто двумя хитиновыми складочками — более крупной верхней и
нижней губой. Рот ведет в довольно узкий эктодермального происхождения
пищевод, идущий вверх и назад. Пищевод открывается в энтодермаль-
ную среднюю кишку, тянущуюся через всю головогрудь и часть брюшка
(см. рис. 274, 9; 275, 8). Та часть кишечника, которая залегает в
головогруди, довольно сильно расширена. Ее нередко называют желудком.
Средняя кишка у циклопа лишена каких бы то ни было выростов и
придатков. В брюшке она переходит в короткую эктодермальную
заднюю кишку, которая анальным отверстием, расположенным на заднем
конце брюшка, открывается наружу. У живых объектов кишечник
обычно бывает наполнен пищей, благодаря чему хорошо виден. От
переднего отдела кишечника к спинной стенке тела тянутся особые мускулы.
Благодаря их сокращению кишечник может подниматься и опускаться.
Эти сокращения совершаются более или менее ритмически. Последнее
282
способствует движению в полости тела полостной
жидкости. Специальные органы кровообращения у
циклопа отсутствуют.
Выделительная система представлена
парой максиллярных желез, расположенных в виде
двух клубочков в сегменте второй пары максилл.
Разглядеть их удается с большим трудом.
Нервная система (рис. 277) на живом объекте
видна плохо, благодаря тому что со спинной стороны
она прикрыта кишечником и гонадой, с брюшной же
рассмотрению ее мешают 'конечности. Тем не менее
можно видеть расположенный в голове впереди
пищевода мозг (надглоточный ганглий), от которого
отходят окологлоточные коннективы к брюшной
нервной цепочке. В последней отдельные ганглии
морфологически 'плохо отграничены друг от друга. Нервная
цепочка в брюшко не заходит, она лежит только в
головогруди. Из органов чувств хорошо виден
наживой объекте непарный (науплиусов) глаз (см. рис.
274, 275), расположенный со спинной стороны
передней части головы. При большом увеличении можно
рассмотреть некоторые детали его строения. Он
слагается из трех пигментных бокалов, с которыми
связаны светочувствительные ретинальные клетки
(рис. 278).
На первой паре антенн, кроме того,
располагаются чувствительные волоски, являющиеся, вероятно,
хеморецепторами.
Половая система. Половые органы как у самца, так и у
самки хорошо видны на живом объекте.
Мужская половая железа — непарная. Семенник лежит на
спинной стороне в передней трети головогруди ( рис. 279, /). От него берут
начало парные семяпроводы (vas deferens), в начальный отдел
которых открываются протоки двух больших дополнительных желез
(рис. 279, 3). В геяитальном сегменте каждый из семяпроводов
образует расширение — сперматофорный карман (рис. 279, 4). Каждое из
этих расширений одним отверстием открывается наружу. У циклопа,
так же как и у других Copepoda, сперматозоиды склеиваются в
довольно крупные сперматофоры, которые хранятся в сперматофорных
карманах, откуда выводятся во время копуляции.
Женский половой аппарат, так же как и мужской,
характеризуется непарной половой железой. Яичник расположен на спинной
стороне в передней части головогруди. Справа и слева от него отходят
яйцеводы, начальные отделы которых сильно расширены и образуют
слепые выпячивания (см. рис. 274, 11). Яйцеводы продолжаются назад
вплоть до генитального сегмента, где двумя отверстиями (справа и
слева) открываются наружу. Конечные отделы яйцеводов тонкие. Кроме
того, в генитальном сегменте помещается семяприемник, в который
попадают сперматофоры при копуляции. Семяприемник имеет самостоя-
Рис. 277. Схема
нервной системы
Copepoda Gymnop-
lea (из Gies-
brecht):
/ — непарный глаз,
2 — нервы антеннул,
3 — нервы антенн, 4 —
нервы мандибул, 5 и
6 — нервы первой и
второй пары максилл,
7 — нервы первой
пары торакопод, 8 и 9 —
нервы пятой и шестой
пары торакопод
283
Рис. 278. Науплиусов
глаз Eucalanus (из Gies-
brecht по Grenadier):
/ — ретииальные клетки, 2 —
нерв к фронтальному
органу, з — nervus opticus, 4—
медиальный глазной бокал,
5 — латеральный глазной
бокал
нослоиного эпителия,
ка. Над кишечником
тельное непарное отверстие на брюшной стороне
генитального сегмента. Двумя короткими
протоками он соединяется с конечными отделами
яйцеводов (рис. 280). Оплодотворенные яйца у
циклопов склеиваются выделениями яйцеводов.
в два мешка, которые выдаются наружу из
отверстий яйцеводов; самка носит их вплоть до-
выхода из яиц личинок — науплиусов. В этом
можно видеть одну из форм заботы о потомстве.
Изучение срезов. В дополнение к изучению-
организации циклопа in vivo желательно
изготовить и рассмотреть поперечные срезы. На срезе
через грудную часть тела циклопа (рис. 281)
следует рассмотреть тонкую хитиновую кутикулу
(рис. 281, /) и лежащую -под ней гиподерму.
Полость тела обширна и заполнена ■ гемолимфой.
В полости.тела видны перерезанные в
различных направлениях поперечнополосатые мышцы
(рис. 281, 3, 5). Часть их идет в конечности,
часть является продольными мышцами тела.
В центральной части среза хорошо виден
кишечник (рис. 281, 7), стенки которого состоят из од-
Под кишечником лежит брюшная нервная цепоч-
залегает гонада, от которой идут протоки, лежа-
Рис. 279. Мужской половой аппарат Cyclops.
Вид сбоку (А) и со спинной стороны (Б)
(из С. Claus):
/ — семенник, 2 — vas deferens, 3 — дополнительная
железа, 4 — сперматофорный карман
Рис. 280. Женский
половой аппарат
Cyclops (из Gies-
brecht):
/ — янчник, 2 —
яйцевод, 3 — наружное
отверстие яйцевода,
4 — семяприемник,
5 — сперматофоры
284
Рис. 281. Cyclops sp. Поперечный
разрез в области головогруди
(ориг.):
/ — хитиновая кутикула, 2 — гиподерма,
3—продольные мышцы, 4—полость тела,
5 — мышцы конечностей, 6 — нервная
цепочка, 7 — кишечник, 8 — яйцевые клетки
в яйцеводе, 9 — яичник
щие по сторонам кишечника.
У самки (рис. 281, 8, 9) в
яичнике отчетливо видны
ооциты на разных стадиях
роста. Яйцевые клетки в
яйцеводах богаты желтком (рис.
281, 8).
Diaptomus sp.
Материал и методы изучения. В пресных водах СССР встречается около 40
видов рода Diaptomus (отр. Gymnoplea, сем. Diaptornidae), относящегося к числу
наиболее распространенных веслоногих рачков пресноводного планктона. Наиболее
обычные виды — D. coeruleus Fischer, D. castor Jur. и др.
Следует рассмотреть живые или фиксированные спиртом экземпляры при
помощи бинокулярной лупы, ограничиваясь главным образом изучением внешней
морфологии. Конечности надо отпрепаровать тонкими иглами и рассмотреть под микроскопом.
На живом экземпляре можно наблюдать своеобразный характер движения Diaptomus.
Внешняя морфология.
Расчленение тела. Тело
Diaptomus, так же как и
циклопа, распадается на
головогрудь и брюшко. Головогрудь
относительно длиннее, чем у
циклопа (рис. 282, 283). На
переднем ее конце со спинной
стороны расположен непарный
глаз. Головогрудь слагается
из пяти или шести члеников.
Передний представляет
голову, слитую с первым грудньГм
сегментом. Задний (пятый)
сегмент головогруди нередко
несет след слияния из двух
сегментов (рис. 282) или же
пятый и шестой сегменты
свободны (рис. 283).
Брюшко Diaptomus
короче, чем у циклопа. У самок
большинства видов оно
состоит из трех сегментов. Первый
из них — генитальный сегмент.
Рис. 282. Diaptomus ЬасйЩег. А — самка с
яйцевым мешком, вид со спинной стороны; В —
первая антенна самца; В— вторая антенна
самки Diaptomus castor (из В. М. Рылова по
G. Sars):
/ — двучленнстый протоподит, 2 — эндоподит, 3 —
экзоподит
285
несущий половое отверстие, значительно
длиннее остальных. Он обычно снабжен двумя
боковыми шипами. У самцов брюшко из яяти
сегментов. Уменьшение числа сегментов брюшка у
самки происходит за счет их слияния. Брюшко
заканчивается относительно короткой вилочкой,
несущей по пять перистых щетинок с каждой
стороны.
У самки к генитальному сегменту
прикрепляется один (а не два, как у циклопа) яйцевой
мешок (рис. 282).
Конечности. Первая пара антенн
отличается у Piaptomus большой длиной. У
некоторых видов они превосходят по длине все тело
вместе с вилкой. Первые антенны служат
органом плавания. Они слагаются из одного ряда
члеников (колец), несущих,волоски, число колец
в .каждой антенне равно 24—25.
У самцов правая антенна первой пары
является геникулирующей — она видоизменена и
приспособлена для удержания самки во время
копуляции. В связи с этим на ней развиваются
особые крючковидные шипики, а некоторые
членики расширены. Детали строения
геникулирующей антенны у Diaptomus — один из важных
Рис. 283. Diaptomus sp. систематических признаков при определении ви-
Вид сбоку (из Е. Н. дов (рис. 282, Б). Вторая пара антенн двувет-
Павловского по Storch): виста. Основная часть их состоит из двух чле-
/ — место прнчленения пер
вых антенн, 2 — верхняя
губа, 3 — вторые антенны, 4—
мандибулы, 5 — первые мак-
сил'лы, 6 — вторые максил-
лы, 7 — ногочелюсть, 8 —
грудные плавательные ножки
се-
ников, эндоподит двучленистыи, экзоподит■
мичленистый (рис. 282, В).
Ротовые части Diaptomus состоят из пары
мандибул, двух пар максилл и пары ногочелюг
стей. Их строение изображено на рис. 284, а
расположение на брюшной стороне головы
рачка — на рис. 285. Из особенностей строения ротовых частей
следует отметить, что мандибулы сохраняют двуветвистый тип строения, их
основной членик превращается в жевательную пластинку. Вблизи
ротового отверстия кроме конечностей располагаются верхняя
и нижняя губы (рис. 285, 3, 4). Первая пластинка непарная,
вторая— парная.
Вторая пара антенн, щупики мандибул и дистальные части
максилл находятся в постоянном колебательном движении (согласно Штор-
ху, они производят до 300 колебательных движений в минуту), что
обусловливает постоянный ток-воды к ротовому отверстию. При этом
питательные частицы задерживаются щетинками околоротовых
конечностей и направляются в рот. Описанное вибрирующее движение
ротовых частей следует рассмотреть на живом экземпляре. Наряду с током
воды оно обусловливает медленное скользящее поступательное
движение животного.
286
Рис. 284. Ротовые конечности Diaptomus castor (из В. М. Рылова по различным
авторам):
/ — мандибула, 2 —первая максилла, 3 —вторая максилла, 4 — ногочелюсть
Грудные конечности присутствуют в числе пяти пар (см. рис. 283).
Из них четыре передние пары обладают типичным двуветвистый
строением (рис. 286). Пятая пара устроена различно у самок и самцов
(рис. 287). У самок обе ножки пятой пары одинаковы, эндоподит раз-
Рис. 285. Diaptomus sp.
Передняя часть тела. Вид снизу (из
Е. Н. Павловского по Storch):
1 — первая антенна, 2—вторая
антенна, 3 — верхняя губа, 4 — нижняя
губа, -5 — ногочелюсть, 6 — вторая
максилла, 7—первая максилла,
8 — мандибула
Рис. 286. Diaptomus castor.
Плавательные ножки первой (А) и
четвертой (Б) пары (из В. М. Рылова по
Sars):
/ — протоподнт, 2 — экзоподит, 3 —
эндоподит
287
г—,
вит слабее экзоподита. У самцов
правая ножка значительно крупнее
левой. Эндоподит ее рудиментарен, эк-
зоподит же несет особый
придаток— хватательный коготок,
служащий наряду с геникулирующей
антенной для удержания самки при
копуляции. Левая ножка пятой пары служит
при этом для введения сперматофора
в половые отверстия самки,
расположенные на генитальном сегменте.
У самки половых отверстий два, но
они сближены между собой. Строение
пятой пары грудных конечностей —
важный систематический признак.
Четыре шередние пары грудных
конечностей, оперенные многочисленными
щетинками, служат основными
органами поступательного движения
рачка. Они производят быстрые и
сильные удары, наподобие ударов весел.
Внутренняя организация
Diaptomus (рис. 288) в основном
напоминает таковую циклопа. 'Существенным
отличием является наличие сердца,
расположенного в первом свободном грудном сегменте над кишечником
(рис. 288, 11). Кровеносные сосуды отсутствуют. Сердце имеет форму
продолговатого мешочка, снабжено парой остий. У живого рачка
можно наблюдать частую пульсацию сердца.
Рис. 287. Diaptomus coeuruleus. А—
нога пятой пары самки; 5 — пятая
пара ног самца (из В. М. Рылова):
/ — протоподнт, 2 — экзоподнт, 3 —
эндоподит, 4 — хватательный коготок
7 8
Рис. 288. Diaptomus castor. Общая организация самки. Вид
сбоку (из. Е. Н. Павловского по Hertwig):
/ — надглоточный ганглий, 2—первые антенны, 3 —вторые антенны,
4 —. мандибулы, 5—первые максиллы, 6 — вторые максиллы, 7 — ного-
челюсти, 8 — брюшная нервная цепочка, 9 — сперматофоры,
прикрепленные к половому отверстию, 10 — яичник, 11 — сердце, 12 — кишечник
288
Метаморфоз циклопа
В развитии циклопов наблюдается характерный для низших раков метаморфоз,
сопровождающийся сменой ряда личиночных форм. Личиночные стадии циклопа
следует изучать на живом материале. Последовательное превращение одной личиночной
стадии в другую осуществляется путем линьки. При этом происходит увеличение
размеров и постепенное нарастание на заднем конце тела количества сегментов и
конечностей. Внешнее расчленение тела, однако, на личиночных стадиях циклопа
отсутствует, и о количестве сегментов приходится судить по числу пар конечностей. В развитии
циклопа различают следующие стадии: из яйца выходит первая стадия науплиуса,
или ортонауплиус (рис. 289,Л). После линьки он превращается в первого
метанауплиуса (рис. 289,Б). Затем следуют стадии второго, третьего и четвертого метанауп-
лиусов (рис. 289, Г; 290, Л). Последний из них после линьки дает копеподитную
личинку (рис. 290,5). У циклопа имеется пять последовательно превращающихся друг
в друга путем линьки копеподитных личинок. Пятая и последняя из них в конце
концов линяет, превращаясь во взрослую стадию, с развитыми гонадами. Основные черты
организации личиночных стадий циклопа изображены на рис. 289 и 290. Ортонауплиус
(рис. 289. Л) имеет три пары конечностей, окружающих ротовое отверстие, — первую
и вторую пару антенн и мандибулы/ Из них А1 одноветвистые и состоят из трех чле'-
ников, А11 и мандибулы двуветвистого строения. Первый метанауплиус
характеризуется появлением первых максилл (рис. 289,4), которые еще не расчленены и несут
длинную перистую щетинку. Второй метанауплиус отличается от первого лишь
несколько большими размерами (рис. 289,5). У третьего метанауплиуса наблюдается
расчленение первых максилл на два членика и увеличение числа максиллярных
щетинок (рис. 289,Г). Четвертый метанауплиус (рис. 290,Л) обладает более вытянутым
в длину телом и появлением зачатков второй пары максилл, ногочелюстей и двух
первых пар гребных грудных ножек. Последние еще не расчленены, но двуветвис-
тость их уже ясно намечена (рис. 290,Л). Кроме того, на заднем конце четвертого
метанауплиуса появляется с каждой стороны по три щетинки и намечается
образование вилки в форме выемки на брюшке.
Первая копеподитная стадия (рис. 290,5) ^характеризуется дальнейшим
расчленением образовавшихся у четвертого метанауплиуса конечностей. Кроме того, ясно
выражено внешнее расчленение брюшка на два сегмента. Вилка принимает
характерную для циклопа форму, и щетинки ее получают дефинитивное развитие. На
последующих копеподитных стадиях (2—5) происходит постепенное нарастание числа
конечностей и внешнего расчленения тела.
Личиночные стадии Diaptomus, организация которого рассматривается выше,
обладает метаморфозом, сходным с таковым циклопа. В отличие от последнего у него
имеется одна дополнительная стадия науплиуса, так что общее число науплиальных
личинок равняется шести.
Рис. Я89. Метаморфоз Cyclops sternuus. A — ортонауплиус; 5 — первый
метанауплиус; В — второй метанауплиус; Г — третий метанауплиус (из W. Dietrich):
/ — Первые антенны, 2 — вторые антенны, 3 —мандибулы, 4 — первые максиллы
10—2
289
Рис. 290. Метаморфоз Cyclops strenuus. A — четвертый метанаупли-
ус; Б — первая копеподитная личинка (из W. Dietrich):
1 — первые антенны, 2 —вторые антенны, 3 — мандибулы, 4 — первые максил-
лы, 5 — вторые максиллы, 6 — ногочелюсти, 7—9 — первая, вторая и третья
пары плавательных ножек, 10 — furca
ОТРЯД CIRRIPEDIA —УСОНОГИЕ РАКИ
Усоногие раки исключительно морские сидячие животные.
Некоторые группы этого отряда ведут паразитический образ жизни. Во всех
морях, омывающих СССР, обычны различные виды рода Balanus,
которые -и следует взять в качестве примера при изучении этой группы
ракообразных.
Balanus balanoides L. — Морской желудь
Материал и методы изучения. Balanus balanoides L. (подотряд Thoracica, ряд
Balanomorpha, сем. Balanidae) широко распространен в Баренцевом море. Этот вид
живет в литоральной зоне на скалах, камнях. Нередко он поселяется также на
раковинах моллюсков (Pecten, Littorina). Довольно часто в Баренцевом море
встречается и другой, более крупный вид этого рода — В. porcatus Da Costa, который живет в
более глубокой сублиторальной зоне и нередко попадается при драгировках. Этот вид
также может быть использован для изучения и даже удобней благодаря своим
размерам. Морфология обоих видов близка.
Для изучения желательно использовать живой материал. При отсутствии его
годятся экземпляры, фиксированные 70%-ным спиртом.
На живом объекте (в аквариуме) следует наблюдать характерное движение
конечностей Balanus, вызывающее непрерывный ток воды поступающий ..частично во
внутреннюю мантийную полость, - '
290
Рис. 291. Balanus hammed (из Рис. 292. Схема попе-
Broch по Ч. Дарвину): речного сечения скелета
/ - scutum, 2 — rostrum, 3 — radiae, Balanus (из Broch):
4 - lateralia, 5 — carino-lateralia, 6 - ,_ carina, 2 - carlno-latera-
canna, 7 —tergum, 8 — alae lia, 3 —lateralia, 4 —rostrum
Для дальнейшего изучения Balanus отделяется острым ножом от субстрата, при
этом не следует нарушать целостность окружающей все животное известковой стенки,
образующей наружный скелет. На разных экземплярах Balanus изучается наружный
скелет и проводится вскрытие. Для рассмотрения внутренней организации и
препаровки следует воспользоваться малым увеличением бинокулярной лупы. Если вскрытие
проводится на спиртовом материале, то обычно некоторые системы органов
рассмотреть ие удается (нервная система, органы выделения).
Внешняя морфолгия. Строение скелета. Диаметр В.
balanoides у основания равен 0,5—1 см, высота — 3—4 мм. Животное
обычно слегка вытянуто в передне-заднем направлении, так что линия
прикрепления его к.субстрату представляет не круг, а овал. Снаружи по
периферии тело Balanus ограничено сплошной 'известковой стенкой,
слагающейся из отдельных неподвижных относительно друг друга
пластинок (рис. 291). Сверху расположены четыре подвижно
сочлененные пластинки, образующие крышечку (operculum). Эти пластинки
могут раздвигаться, и через образующееся таким образом отверстие
наружу выставляются конечности, находящиеся в постоянном движении.
Следует детально рассмотреть отдельные элементы наружного скелета
морского желудя, сначала не отделяя их, а затем отделив пинцетом
отдельно каждую пластинку. Боковая известковая стенка представляет
собой продукт выделения мантии—:складки покровов, загибающейся
снизу вверх и охватывающей с боков все тело животного. Боковая
стенка слагается из шести отдельных пластинок, из которых две непарные,
а остальные четыре парные (рис. 291). Спереди1 располагается
большая непарная пластинка, называющаяся rostrum2. Против нее на
заднем конце расположена непарная carina. Справа и слева с rostrum
граничат две lateralia, позади них расположены две carino-lateralia. Из-
1 Передний конец определяется по отношению к ротовому отверстию, как это
устанавливается при дальнейшем вскрытии.
2 Rostrum в роде Balanus является сложным образованием. Он представляет
собой результат слияния собственно ростральной пластинки с двумя передними
латеральными. У некоторых соседних с Balanus родов (например, Catophragmus) этн
пластинки самостоятельны, и в этом случае стенки слагаются не из шести, а из
восьми пластинок.
10**
291
Рис. 293. Схема метаморфоза
личинки Lepas. Л—В
—последовательные стадии (из А. В. Ива.нова
по Korschelt-Heider):
1 — полость мантии, 2 — кишечник, 3 —
abdomen, 4— грудные конечности, 5 —
ротовое отверстие, 6 — непарный глаз,
7 — парный глав, 8 — первые антенны
8
вестковые пластинки, слагающие стенку, особенно в своей базальной
части, обладают бороздками, идущими сверху вниз. В местах
соединения отдельных пластинок они налегают друг на друга на некотором
довольно значительном протяжении (рис. 292). Части пластинок, которые
при этом направлены в сторону мантийной складки, называются alae,
те же, которые занимают периферическое положение,— radiae (рис.292).
Крышечка слагается из четырех известковых пластинок, образующих
две пары. Передние называются scutum, задние — tergum1 (рис. 291,
1.7):
Конечности. Прежде чем приступить к вскрытию Balanus,
следует, воспользовавшись лупой, рассмотреть его с нижней стороны,
которой животное прикрепляется к субстрату. Если отделение от
субстрата произведено достаточно аккуратно, то в центре подошвы можно
видеть два небольших нерасчлененных придатка, представляющих сильно
редуцированную первую пару антенн. Их присутствие на подошве
объясняется тем, что свободноплавающая личинка Cirripedia (циприсовид-
ная личинка) прикрепляется к субстрату головным концом.
Для того чтобы понять ориентировку основных частей тела
Balanus, так же как других Cirripedia, необходимо представить себе, как
происходит метаморфоз личинки. Циприсовидная личинка усоногих
получила свое название за внешнее сходство, с Ostracoda (род Cypris).
Она снабжена двустворчатым головным щитом (рис. 293). При
метаморфозе личинка прикрепляется к субстрату передним концом, который
и превращается в подошву. Ее продольная ось первоначально проходит
параллельно брюшному краю, задний же конец тела направлен кверху.
После прикрепления личинки происходит процесс изгибания
продольной оси тела в передней его части таким образом, что брюшная
поверхность поворачивается кверху, т. е. в сторону, противоположную субстра-
1 На внутренней, обращенной к телу поверхности scutum и tergum имеются
характерные складки и гребни, которые служат для прикрепления мышц.
292
Рис. 294. Balanus tintinnabulum. iU 13
Боковая часть стенки удалена
(из Ч. Дарвина):
1 — scutum, 2 — rostrum, 3 — m.
adductor scutorum, 4 — m. depressor scutorum
rostrale, 5 — яйцевод, 6 — яичник, 7 —
prosoma, 8 — полость мантнн, 9 —
грудной отдел, 10 — m. depressor tergorum,
И — carina, 12 — tergum, 13 — грудные
конечности, 14 — ротовое отверстие
4-
5
ту. Соответственно спинная сторона становится почти параллельной
подошве. В результате конечности Balanus (так же как и других
Cirripedia) 'направлены 'наверх, и все животное, за исключением головного
конца, превращенного в подошву, как бы лежит на спине (рис. 293,
294).
Часть тела, ближе к которой расположено ротовое отверстие и
где помещается rostrum, названа передней. Мантийная скл.адка,
окружающая все тело, образуется в виде выроста'подошвы. Вторая4 пара
антенн отсутствует. Сквозь мантийную складку, прилежащую к
подошве, просвечивает парный яичник, залегающий в толще самой складки
(рис. 294, 6). У Balanus он очень сильно развит. Отделив часть его,
следует рассмотреть под микроскопом яйцевые клетки, которые имеют
овальную форму и богаты желтком. Парные яйцеводы, открывающиеся
у основания первой пары грудных конечностей и образующие особые
расширения, обычно наполненные яйцами, могут быть видны лишь при
вскрытии. Вскрытие следует начать, осторожно удалив с одной
стороны известковые пластинки стенки. Непосредственно под ними залегает
мантия, прикрывающая собственно тело животного, лежащее во
внутренней (мантийной) полости. Отрезав небольшой кусочек мантии,
следует рассмотреть ее внутреннюю стенку под бинокулярной лупой. Она
покрыта многочисленными складками, которые увеличивают
внутреннюю поверхность мантии и представляют собой жабры.
После удаления мантии с одной стороны^ видно общее
расположение основных частей тела и некоторые мышцы (рис. 294). Брюшная
сторона животного повернута кверху и дугообразно изогнута, причем
выпуклая (спинная) сторона дуги направлена к субстрату.
Происхождение подобной ориентировки тела находит свое объяснение в
метаморфозе личинки, описанном в общих чертах выше. К переднему концу
тесно прилежат передние пластинки крышечки — scutum.
293
Ротовое отверстие,
окруженное ротовыми
частями, помещается на вершине
небольшого хоботка (рис.
294, 14). Та часть тела,
которая расположена кпереди
от ротового хоботка,
называется prosoma. Сзади от
нее «а брюшной стороне
помещаются шесть 'пар усо-
видных конечностей (рис.
294, 13). Несущий их отдел
тела называется грудным.
На заднем конце
помещается длинный жгутовидный
penis (рис. 295, 22). Длина
его превосходит длину
конечностей. На животном с
удаленной стенкой мантии
видны некоторые мышцы,
относящиеся к мантийному
комплексу мускулатуры и
служащие для закрывания
крышечки. В 'переднем
конце проходит парный muscu-
lus depressor scutum rostrale (рис. 294, 4), который одним концом
прикрепляется к scutum, а другим — к основанию стенки в области
ростральной пластинки.
На заднем конце расположен также парный- musculus depressor
tergorum (рис. 294, 10), который обусловливает замыкание tergum.
Одним концом он прикрепляется к tergum, а другим — к основанию хтен-
ки в области задней непарной пластинки — carina. Кроме этих мышц
имеется еще парный depressor scutorum lateralis, проходящий в
боковых частях мантии и тянущийся от scutum -к основанию стенки. Он
удаляется при вскрытии вместе с боковыми частями мантии. Если
удалить, осторожно подрезав ножницами, одну из двух scutum, то
становится виден мощный мускул-аддуктор (рис. 294, 3; 295, 3),
соединяющий обе пластинки scutum и служащий для замыкания щели между
ними.
После общего ознакомления с расположением основных частей
тела Balanus следует на отдельном экземпляре отпрепаровать и
рассмотреть конечности. После того как отделен ротовой хоботок
препаровальными иглами под бинокулярной лупой, изолируются отдельные ротовые
части и рассматриваются под микроскопом (рис.296). Спереди ротовое
отверстие прикрыто пластинкой — верхней губой (labrum), снабженной
парой нерасчлененных щупиков (рис. 296, 1). Гомология их с
ротовыми частями других раков неясна.
Мандибулы (рис. 296, 2) представляют собой нерасчлененные
хитиновые пластинки с зубчатым краем. Нерасчлененная пара максилл
-Рис. 295. Balanus sp. Расположение
внутренних органов (из Caiman no Gravel):
/ — отверстие, ведущее в мантийную полость, 2 —
scutum, 3 — т. adductor scutorum, 4— ростральный
синус, 5 — яйцевод, 6 — rostrum,' 7— m. depressor
scutorum rostrale, 8 — яйцевые клетки мантийной
полости, 9 — яичник, 10 — цементные железы, 11 —
семенники, 12 — желудок, 13 — надглоточный ганглий,
14—-vesicula seminalis, 15 — первые антенны, 16 —
задняя кишка, 17 — брюшная нервная масса, 18 —
мантийная полость, 19 — m. depressor tergorum, 20 —
carina, 21 — анальное отверстие, 22— penis, 23 —
грудные конечности, 24 — ротовое отверстие
294
1 — labrum с пальцами, 2 — мандибула, 3—первая максилла, 4 — вторая максилла
снабжена длинными волосками (рис. 296). Более сложную
конфигурацию имеют вторые максиллы. Они также не расчленены, но в каждой
из них можно различить две лопасти, покрытые волосками (рис. 296,4).
Шесть пар грудных конечностей, называющихся cirri, или усоножками,
имеют типичное двуветвистое строение (рис. 297). Основная часть (про-
топодит) слагается из двух члеников. Экзоподит и эндоподит
многочленистые и, за исключением первой пары, почти равной длины. Они
покрыты многочисленными длинными волосками.
Внутренняя организация. На отдельном экземпляре Balanus
производится вскрытие тела рачка. Тонкими ножницами или скальпелем
проводится разрез по средней линии спинной стороны, отделяются
покровы. Отдельные органы отпрепаровываются иглами. Мышцы тела
Balanus хорошо развиты и довольно многочисленны. Они распадаются
на две основные группы. Ближе к спинной стороне расположены
длинные мускулы — разгибатели (m. extensores) тела, ближе к брюшной
стороне — мускулы-сгибатели (m. flexores).
Пищеварительный канал. Ротовое отверстие ведет в
короткий тонкий эктодермальный пищевод, проходящий косо вперед и на
спинную сторону (рис. 295). Он открывается в объемистую энтодер-
мальную среднюю кишку, называющуюся нередко желудком
(см. рис. 295, 12). В переднюю часть этого отдела кишечника
открываются довольно многочисленные трубчатые пищеварительные железы.
Средняя кишка переходит в эктодермальную заднюю кишку, которая
тянется по спинной стороне животного, загибается на брюшную сторо-
Рис. 297. Грудные конечности Balanus balanoid.es (из Broch)
295
ну и открывается анальным отверстием на
заднем конце тела (см. рис. 295, 21).
Специальные органы
кровообращения у Balanus отсутствуют. Кровь циркулирует
по лакунам, установить расположение которых
при вскрытии, однако, не удается. Обычно не
видны органы выделения,
представленные парой макоиллярных желез.
Нервная система, которую можно
обнаружить при ъскрытии живого Balanus,
состоит из следующих частей. Впереди пищевода
помещается надглоточный ганглий, или мозг
(рис. 298, 4, 7). Пер'едний отдел мозга — proto-
cerebrum вынесен 'вперед и соединяется с deuto-
cerebrum длинной коннективой. Окологлоточное
нервное кольцо соединяет мозг с подглоточным
■ганглием. В тесной связи с последним стоят
слившиеся в общую массу ганглии брюшной
цепочки (рис. 298, 10). Таким образом, нервная
система Balanus обнаруживает сильную
концентрацию. Из органов чувств следует отметить
многочисленные чувствительные (осязательные)
волоски, располагающиеся «а конечностях.
Кроме того, у Balanus имеются сильно
редуцированные глаза, которые в виде двух маленьких
пигментных пятнышек располагаются в передней
части тела( рис. 298, /).
Balanus, как /и огромное большинство Cirri-
pedia, — гермафродиты.
Мужской половой аппарат развит
у Balanus очень сильно. Семенники,
образующие многочисленные фолликулы, располагаются
в передней половине тела (см. рис. 295, 11),
где они заполняют все промежутки между
органами. От семенников отходят два семяпровода,
кишечнику назад. Они образуют расширение,
наполненное сперматозоидами, — vesiculae seminalis (см. рис. 295, 14).
У основания penis оба семяпровода соединяются в непарный семяиз-
вергательный канал (ductus ejaculatoris), проходящий -через длинный
penis и открывающийся половым отверстием на его дистальном конце.
Женская половая система частично уже рассмотрена
(см. рис. 294). Желательно отыскать яйцеводы, проходящие в передней
части тела и открывающиеся у основания первой пары конечностей.
Несмотря на гермафродитизм, у Balanus обычно осуществляется
перекрестное оплодотворение. Морские желуди обычно живут
группами. Во время копуляции penis одной особи вводится в мантийную
полость другой, расположенной по соседству. В некоторых случаях
спермин просто выпускаются в воду и попадают в мантийную полость
соседа вместе с током воды.
Рис. 298. Нервная
система Balanus tintlnnabu-
lum (из Broch no Gru-
bel):
/ — глаз, 2 — нерв желудка,
3 — оптический нерв, 4 —
protocerebrum, 5 — нервы
подошвы, 6 — нерв,
соединяющий proto- и deutocerebrum,
7 — deutocerebrum, 8 —
пищевод, 9 — нерв мускула
аддуктора, 10 — первый
ганглий брюшной нервной
массы, 11 — нерв первой пары
грудных конечностей, 12 —
нервы второй — шестой пар
грудных конечностей, 13 —
нерв пениса, 14 и 15—нервы
мантни
идущие параллельно
296
ПОДКЛАСС MALACOSTRACA— ВЫСШИЕ РАКИ
ОТРЯД LEPTOSTRACA —ТОНКОПАНЦИРНЫЕ РАКИ
Организация Leptostraca представляет значительный интерес. Эта
небольшая группа ракообразных относится к высшим ракам и несет
некоторые черты низших ракообразных, что дает основание некоторым
зоологам выделять Leptostraca в отдельный подкласс. В качестве
объекта для изучения можно взять Nebalia bipes, встречающуюся в наших
северных морях. При прохождении практикума достаточно
ограничиться рассмотрением внешней морфологии и некоторых черт внутренней
организации, видимых на тотальных препаратах.
Nebalia bipes (О. Fabricius)
Материал и методы изучения. Nebalia bipes (сем. Nebaliidae — единственное в
отряде Leptostraca) встречается изредка в Баренцевом море на разнообразных
глубинах (от 1 до 200 м). Живет на илистом и песчаном грунте, чаще всего ее можно
найти в ризоидах ламинарий.
Для изучения используется живой материал или консервированный 70%-ным
спиртом. Под бинокулярной лупой рассматривается внешняя морфология рачка,
после чего отпрепаровываются конечности. Желательно также изготовить тотальный
препарат, слегка подкрасив его квасцовым кармином.
Внешняя морфология. Nebalia bipes — небольшие рачки, длина их
не, превышает 1,5 см. Самки встречаются чаще самцов. Половой
диморфизм выражается в том, что у самцов значительно длиннее антенны,
менее высокий панцирь, несколько больше длина ветвей вилки
(рис. 299).
Характерной особенностью строения Nebalia является наличие
двустворчатого хитинового панциря, прикрывающего часть головы', грудь
и первые четыре сегмента брюшка. Панцирь очень тонкий, прозрачный.
Обе створки соединяются мощным мускулом-замыкателем,
расположенным в передней трети тела (рис. 299, 13). Спереди к панцирю причле-
няется длинный рострум. По сторонам его на голове (расположены
подвижные стебельчатые сложные глаза. Сквозь панцирь можно различить
восемь грудных сегментов, несущих каждый по паре листовидных
конечностей. Брюшко слагается из семи разграниченных и подвижно
сочлененных друг с другом сегментов и тельсона. В этом заключается
одно из существенных отличий отряда Leptostraca от всех Malacostraca,
у которых брюшко всегда слагается не более чем из шести члеников и
тельсона. Тельсон у Nebalia несет хорошо, развитую длинную вилку,
состоящую из двух ветвей (рис. 299, 8).
Конечности. Первая и вторая пара антенн Nebalia являются
длинными членистыми придатками. Антеннулы одноветвисты. Основная
часть их состоит из четырех довольно крупных члеников, несущих
многочисленные волоски, часть которых является хеморецепторами.
Четвертый членик антеннул снабжен особым листовидным придатком,
который обильно покрыт волосками. Дистальная часть антеннул состоит
1 Голова Leptostraca морфологически представляет собой syncephalon (protocepha-
lon + gnathocephalon, см. с. 246). Панцирь развивается за счет максиллярной
области головы.
297
Рис. 299. Nebalia bipes. A — самец; Б — самка (из Thiele по Claus):
1 — rostrum, 2 — первые антенны, 3 — глаза, 4 — щупик мандибулы, 5 — вторые
антенны, 6 — грудные ножки, 7 — брюшные ножки, 8 — furca, 9 — кишечник,
10 — семенник, 11 — сердце, 12 — жгутовидный придаток максилл, 13 — замы-
кательный мускул панциря, 14 — выделительная антеннальная железа, 15 —
яичник
из многочисленных мелких члеников, постепенно утончающихся
(рис. 299,2). , , ~
Вторая пара антенн также одноветвиста. Основная часть их
включает 3 членика, за которыми следует длинный жгутовидный отдел,
особенно длинный у самцов. Вокруг ротового отверстия, помещающегося
на нижней части головы, располагаются ротовые части (рис. 300).
Спереди рот прикрыт довольно крупной складкой — верхней губой,
сзади— двулопастной нижней губой (рис. 300, 4, 6). Мандибулы обладают
длинным трехчленистым щупиком (рис. 300, 2). Основная часть их
снабжена жевательными лопастями (рис. 300, Б). Характерным
является строение первой пары максилл (рис. 300, 5). Их основная часть
двучленистая. К ней причленяется длинный, покрытый волосками
жгутовидный придаток, очищающий внутреннюю поверхность панциря от
посторонних частиц, проникающих с током воды. Вторая пара максилл
двуветвистая. Их основная двучленистая часть обильно покрыта
волосками (рис. 300, 8). Восемь пар грудных конечностей. (торакоподов),
полностью прикрытых панцирем, имеют сходное строение (рис. 301).
298
К основному членику их
причленяется листовидный эпиподит,
выполняющий функцию жабры.
Экзоподит также листовидный и
нерасчлененный. Эндоподит,
несущий многочисленные перистые
волоски, слагается из четырех
члеников1.
Первые четыре пары брюшных
конечностей (плэолодов)
представляют собою типичные
плавательные ножки (рис. 302). Они двувет-
вистые, выдаются наружу из
створок гаанциря и несут волоски. Эти
конечности снабжены хорошо
развитой мускулатурой. Их активные
движения обусловливают плавание
рачка. Конечности пятого и
шестого абдоминальных сегментов
развиты слабо, они 'представляют собой
маленькие одноветвистые
придатки, покрытые короткими
волосками. Седьмой брюшной сегмент
лишен конечностей. Тельсон, как уже
отмечалось, несет вилку.
Внутренняя организация. На
тотальных препаратах, а тем более
на живом материале, удается обычно рассмотреть некоторые черты
внутренней организации Nebalia (см. рис. 299).
Передний отдел кишечного канала образует жевательный
желудок, открывающийся наружу при помощи короткого пищевода.
Жевательный желудок переходит в кишку, которая в виде прямой трубки
тянется до тельсона, где анальным отверстием открывается наружу.
Кишечник образует несколько слепых железистых выростов. От начала
средней кишки вперед отходят две пары коротких слепых отростков,
назад через всю грудь и переднюю половину брюшка тянутся три пары
железистых выростов, имеющих форму трубок.
Сердце представляет собой длинную трубку, лежащую над
кишечником и тянущуюся от максиллярного сегмента до четвертого
членика брюшка. Другие кровеносные сосуды отсутствуют. В сердце
(см. рис. 299, 11) видна одна пара крупных остий, расположенных в
шестом торакальном сегменте. Впереди от них метамерно расположено
еще шесть пар очень мелких остий. Кзади от шестого грудного
сегмента остий отсутствуют.
В области антенн лежит в форме клубочка парная
антеннальная железа — орган выделения. Рудиментарные максиллярные
железы обычно не видны.
1 Наряду с эпиподиальными жабрами у Nebalia функцию дыхания выполняет
также внутренняя поверхность панциря.
Рис. 300. Nebalia bipes. A — передняя
часть с брюшной стороны; Б —
изолированные мандибулы (из Thiele no Sars):
/— место причленения первых антенн
(удалены), 2 — щупик мандибул, 3 — место
прикрепления вторых антеин (удалены), 4 — верхняя
губа, 5 — первые максиллы со жгутовидный
придатком, 6~ нижняя губа, 7—замыкатель-
ный мускул панциря, 8 — вторая максилла, 9 —
жевательная пластинка мандибул, 10 —
мускул мандибул
299
Рис. 301. Пятый торакопод
Nebalia blpes (из Thiele no
Sars):
/ — эпиподиальная жаберная
пластинка, 2 — экзоподит, 3'— эидопо-
дит
Рис. 302. Второй плэопод
Nebalia (из Caiman по
Claus):
/ — протоподит, 2 — экзоподит,
3 — эидоподит, 4 — appendix
interna эндоподита
Нервная система слагается из мозга (надглоточного
ганглия) и богато расчлененной брюшной цепочки, в состав которой
входят 17 метамерно расположенных 'парных ганглиев, часть которых
можно рассмотреть и на тотальном препарате.
Половой аппарат иу самцов и у самок состоит из длинных
трубковидных гонад (р'ис. 299,. 10, 15), расположенных по сторонам
кишечника. Половые протоки (парные у обоих подов) у самца
открываются у основания восьмой пары грудных ножек. Половые отверстия
самки расположены у основания шестой пары торакоподов.
ОТРЯД DECAPODA —ДЕСЯТИНОГИЕ РАКИ
Astacus astacus (Linne) — Речной рак
Материал и методы изучения. Речной рак1 широко распространен почти по всей
территории СССР. В реках Понто-Каспийского бассейна, а также в Карелии
встречается близкий вид — длиннопалын рак (Astacus I'eptodactylus Eschholtz), отличающий-
1 Считаем необходимым привести синонимику речного рака, так как в различных
руководствах это животное получает разные латинские названия: Astacus fluviatilis
(до Линнея); Cancer astacus (Linne, 1758); Astacus fluviatilis (J. С Fabricius, 1775);
Astacus astacus (Pennaut, 1777); Potamobius astacus (Leach et Samouelle, 1819); As-
lacus fluviatilis nobilis (Huxley, 1881).
300
ся от речного оака более длинными клешнями и некоторыми другими признаками.
Его также можно использовать для изучения.
Внешняя морфология изучается на живых или консервированных спиртом
экземплярах. Для рассмотрения скелета раки вывариваются в 15—20%-ной едкой щелочи.
Внутренняя анатомия изучается на вскрытых свежезахлороформированных
животных. Порядок и ход вскрытия указан ниже (с. 314). Для изучения кровеносной
системы необходимо прибегнуть к инъекции, которая дает обычно хорошие
результаты и позволяет рассмотреть многие детали строения артериальной системы.
Для гистологических целей отдельные небольшие кусочки органов вырезаются от
живого или свежезахлороформированного рака и фиксируются. В качестве фиксаторов
можно применять различные жидкости (например, Джильсои, Ценкер-формол и др.).
Окраска срезов производится железным гематоксилином или гематоксилином Бемера
с подкраской эозином.
Для изучения строения покровов и глаз лучше всего использовать недавно
слинявшего рака. Наличие толстой хитиновой обызвествленной кутикулы затрудняет
заливку и резку на микротоме.
Внешняя морфология. Тело речного рака распадается на два ясно
различимых отдела: головогрудь (cephalothorax) и брюшко (abdomen,
см. рис. 326). Головогрудь внешне не расчленена и прикрыта мощно
развитым хитиновым, пропитанным углекислой известью, панцирем,
который одевает головогрудь со спинной стороны, с боков и несколько
загибается на брюшную сторону1. Брюшко состоит из подвижно
сочлененных друг с другом шести сегментов и тельсона; последний
(рис. 303, 12) имеет форму пластинки, несет анальное отверстие и
образует вместе с последней парой брюшных конечностей плавник.
Передняя часть головогрудного щита вытянута в длинный заостренный киле-
видный рострум, снабженный по бокам зубчиками. По сторонам
основания рострума расположены две довольно глубокие выемки, в которых
помещаются сложные глаза на подвижных стебельках. На головогруд-
ном щите имеется дугообразный желобок, обращенный выпуклостью
кзади,— затылочная бороздка, или шейный шов (sutura cervicalis,
рис. 303, 5). Она отделяет головную часть головогруди от грудной. От
этой борозды.назад, вдоль грудной части щита, идут две жаберно-сер-
дечные бороздки (suturae branchiocardiales), ограничивающие область,
где расположено сердце. Боковые части головогрудного щита—бран-
хиостегиты — прикрывают жаберные полости, в которых располагаются
перистые жабры. Их легко можно видеть, если слегка отогнуть
брюшной или передний край бранхиостегита.
Подвижность брюшных сегментов относительно друг друга
обусловлена тем, что между сегментами имеется лишь тонкая и мягкая
хитиновая кутикула, тогда как спинная и брюшная часть самих сегмен-
1 Согласно Снодграсс (1938), у Decapoda, в том числе и у речного рака, голова
построена по типу protocephalon, т. е. включает предротовую лопасть и сегмент,
несущий вторые антенны. Последующие три сегмента (мандибулярный и оба максилляр-
ных) представляют собой gnathocephalon и в состав головы не входят. От gnathocep-
halon вперед отходит рострум, который прикрывает protocephalon сверху.
Анатомическая самостоятельность protocephalon у речного рака видна отчетливо. Напротив,
gnathocephalon, несущий ротовые конечности, тесно сливается с грудным отделом. Таким
образом, у Decapoda правильнее различать голову (-protocephalon) и gnathothorax
(-gnathocephalon + thorax). Мы оставляем, однако, прежнюю номенклатуру, хотя она
и не вполне соответствует современным представлениям о расчленении Decapoda, так
как термин головогрудь встречается почти во всех работах по морфологии Astacus.
301
12
Рис. 303. Самка Astacus astacus (из Thiele no Sars):
/"•— рострум, 2 —первые антенны, 3—-вторые антенны, 4 — экзоподит
(squama) вторых аитени, 5 — стебельчатый глаз, 6 — первая пара
ходильных йог, 7 — вторые — пятые ходильные ноги, 8 — sutura cervicalis,
9 — sutura brauchlocardialls, 10 — брюшко, // — последняя (шестая)
пара брюшных ножек, 12 — тельсон
тов одета прочными обызвествленными хитиновыми пластинками,
образующими наружный скелет (строение его подробно рассмотрено на
с. 311). Брюшко лишь в малой степени может сгибаться на спинную
сторону, тогда как на брюшную сторону оно способно подгибаться под
головогрудь, что происходит, например, при плавании рака. Это
зависит отчасти от того, что спинные пластинки (тергиты) наружного
скелета абдоминальных сегментов широки (считая по продольной оси
животного) и крышеобразно налегают друг на друга, тогда как брюшные
пластинки (стерниты) узки, и между соседними стернитами имеются
широкие промежутки, ограниченные снаружи тонкой эластичной
хитиновой кутикулой. Относительная ширина брюшка по отношению к
головогруди раз'лична у самцов и у самок (рис. 304). У самцов более
302
Рис. 304. Самец (А) и самка (Б) Astacus astacus. Вид с
брюшной стороны (из Huxley):
/ _ первые антенны, 2 — вторые антенны, 3 — третья пара иогочелюстей,
4—8 — ходильные ноги (торакоподы), 9 —половое отверстие, 10 —
вторая пара брюшных ножек, И — 3—5-я пары брюшных ножек, 12 —
шестая пара брюшных ножек, 13 — апиз, 14 — первая пара брюшных
ножек, 15 — стернит пятого грудного сегмента, 16 — стернит четвертого
грудного сегмента, 17 — развивающиеся яйца, прикрепленные к
брюшным ножкам
узкое брюшко, у самок оно шире. У самцов ширина его несколько
меньше ширины головогруди, у самок превышает последнюю.
Перевернув речного рака брюшной стороной кверху, следует
рассмотреть общее расположение конечностей. Более детально строение
отдельных конечностей изучается при изоляции и препаровке их.
Спереди по сторонам рострума расположена первая пара сяжков, или
антенн (антеннулы, рис. 304, 7), в дистальной своей части каждая антен-
нула несет два жгутовидных придатка. Общая длина их невелика. По
303
сторонам антеннул и несколько кзади от них причленяется вторая пара
сяжков, характеризующаяся длинным жгутовидным придатком. Далее
назад, на брюшной стороне головогруди расположено ротовое
отверстие, окруженное ротовыми конечностями, служащими для удержания,
ощупывания и размельчения пищи. Число конечностей равно шести
парам (одна пара верхних челюстей, или мандибул, две пары нижних
челюстей, или максилл, и три пары ногочелюстей). Рассмотреть
строение ротовых частей без их препаровки не удается.
К грудной части головогруди причленяется пять пар ходильных
ног (торакоподы), служащих для передвижения рака по субстрату.
Из них три первые пары несут клешни, две задние лишены их.
Особенно мощно развиты клешни первой пары ходильных ног. Они служат
для охватывания, удержания и расщипывания пищи, а также являются
органами защиты рака. На основном членике третьей пары ходильных
ног помещаются наружные половые отверстия самки. У самцов они
расположены на первом (проксимальном) членике пятой пары
ходильных ног (рис. 304, 9). Брюшко несет типичные для ракообразных дву-
ветвистые небольшие конечности (плэоподы), числом пять, дистальная
часть которых состоит из двух ветвей. Шестая пара брюшных ножек
(уроподы, рис. 303, 11; 304, 12) сильно сплющена в дорзовентральном
направлении и образует вместе с тельсоном (см. рис. 303, 12),
лишенным конечностей, плавник.
В отношении строения двух передних пар брюшных конечностей
самцы и самки резко отличаются друг от друга. У самцов они
видоизменены в копулятивный аппарат, хорошо развиты и подогнуты
вперед под задний отдел головогруди (рис. 304, 10, 14). У самок первая
пара брюшных ножек недоразвита и присутствует в виде небольшого
придатка. Вторая же пара имеет обычное для плэоподов двуветвистое
строение.
Суммируя внешние морфологические отличия между самцами и
самками речного рака, можно свести их в следующую табличку (по
Павловскому).
Название частей Самец Самка
Брюшко Не шире головогруди Шире головогруди
Половые отверстия У основания пятой пары У основания третьей пары
ходильных ног ходильных ног
Ножки'брюшка (плэоподы) Две передние пары разви- Передняя пара рудиментар-
ты сильнее и образуют на, остальные развиты
копулятивный аппарат одинаково
Весной на брюшных ножках самок нередко можно обнаружить
развивающиеся яйца или молодых, недавно вышедших из яиц рачков.
В вынашивании ,на брюшке развивающихся яиц и молоди и
выражается забота о потомстве (рис. 304, 17). Хитиновый наружный скелет
рака обладает темно-грязно-зеленой окраской, обязанной наличию
особого пигмента — цианокристаллина. При опускании раков в горячую
воду изменение их окраски в красную происходит от того, что цианокрис-
таллин переходит в красный зооэритрин.
Рост рака сопровождается периодическими линьками. Только что
слинявший рак, сбросивший старый хитиновый панцирь, обладает мяг-
304
кими, не обызвествленными покровами1.
В этот период своей жизни раки совершенно
беззащитны, в связи с чем они особенно
тщательно -прячутся под камнями и в норах.
Мелкие молодые раки линяют несколько раз в
году, крупные половозрелые раки — самки —■
один раз в году (обычно в начале лета),
самцы— два раза в год.
Строение конечностей. Конечности отпре-
паровываются пинцетом, начиная от задних
брюшных ножек и кончая антеннулами. При
этом следует соблюдать большую
аккуратность, чтобы не повредить некоторые,
особенно нежные конечности, прилежащие к
ротовому отверстию. Рассмотрение строения
конечностей лучше вести спереди назад.
В основе строения конечностей
десятиногих раков, как и вообще большинства рако-
- образных, лежит двуветвистый тип, с
которым мы уже отчасти встречались при
изучении других подклассов.
Конечности речного рака, несущие разнообразные
функции, представляют собой (за
исключением антеннул) различные модификации двувет- подите /// — эк'зоподит (из
вистой конечности. Общая схема строения ко- Schmidt):
нечности Astacus дана на рис. 305. Основная £~ipo™4 - ьсВДаи"" 2-
ЧЭСТЬ КОНечНОСТИ — protOpodit СОСТОИТ ИЗ Двух meropodit 6 -carpopodit,7 —
члеников — coxopodit и basipodit. Coxopodit ргоро '' 8 ~~ ас у оро '
нередко несет вырост — epipodit. К basipodit'y
причленяются обе ветви конечности — exopodit и endopodit. Экзоподит
(у Decapoda почти всегда более короткая ветвь) состоит из одного
проксимального членика и кольчатого жгутовидного придатка. Эндо-
подит в типичном случае слагается из пяти члеников (ischiopodit,
meropodit, carpopodit, propodit, dactylopodit).
Первая пара сяжков, или антенн (antennulae)
(рис. 306, /). Основная часть состоит из трех члеников, среди которых
проксимальный значительно длиннее двух остальных. Со спинной
стороны он несет прикрытое щитинками отверстие, ведущее в полость ста-
тоциста (см. с. 343). Дистальная часть антеннул состоит из двух
кольчатых нитей, из которых наружная более длинная2. На ней располага-
Рис. 305. Схема строения
конечности речного рака.
/ — протоподит, // —■ эндо-
1 Состав (по Бальеку) хитинового панциря рака следующий: 48%' СаС03; 6,8%
Саз(Р04)2; 40,6% органических веществ, остальное-—небольшие количества солей Mg,
Al, Fe.
2 Несмотря на то что антеннулы дзуветзистые, сравнивать эти зетви с экзо-
подитом и эндоподитом других конечностей нельзя по ряду причин. Одной из них
является то обстоятельство, что А1 — конечности преоральные и филогенетически не
могут быть выведены на параподий Polychaeta, тогда как двуветвистость остальных
конечностей ракообразных сравнивают с двуветвистостью параподий многощетинковых
кольчецов. А1 принадлежат предротовой лопасти — асгоп (см. с. 246).
305
ются чувствительные волооки, служащие, вероятно, органами
химического чувства.
Вторая пара сяжков, или антенн (рис. 306, Б). Прото-
подит двучленистый. У основания на нижней поверхности коксоподита
открывается отверстие зеленой железы — органа выделения рака.
Экзоподит представлен треугольной нерасчлененной пластинкой — чешуей
(squama). Эндоподит чрезвычайно длинный, в виде многочленистого
жгутовидного придатка. Два первых членика больше остальных и
подвижно сочленены друг с другом. Вторая пара антенн служит органом
осязания.
Верхние челюсти, или жвалы (рис. 306, В). По
сравнению с общей схемой строения двуветвистой конечности они
видоизменены значительно в направлении редукции ряда частей. Жвалы —
главный орган откусывания и размельчения пищи. Сострят они из широкой
обызвествленной прочной хитиновой пластинки, зазубренной по
внутреннему краю, и короткого трехчленистого щупика (palpus).
Мандибулы обладают мощно развитыми мышцами (с. 354). Жевательная
пластинка соответствует протоподиту. Щупик некоторыми авторами гомоло-
гизируется с эндоподитом. Однако, согласно Гисбрехту, эта гомология
является сомнительной 1.
Первая пара нижних челюстей (maxillae I, рис. 306, Г).
В отличие от массивных жвал, максиллы очень тонкие, относительно
слабо хитинизированные образования. Каждая нижняя челюсть
слагается из двух пластинок, представляющих собой коксоподит и базиподит
основной части конечности. По наружному краю базиподита,
направленному к ротовому отверстию, расположены многочисленные
хитиновые зубчики. Базиподит несет нерасчлененный листовидный щупик,
соответствующий, вероятно, эндоподиту. Экзоподит отсутствует.
Пластинки первых максилл, в особенности базиподит, принимают участие в
откусывании и размельчении пищи.
Вторая пара нижних челюстей (maxillae II, рис. 306,Д).
Коксоподит и базиподит тонкие листовидные, каждый из них делится
продольной щелью на две лопасти. По краю несут многочисленные
волоски. Эндоподит нерасчлененный в виде длинного щупальцевидного
придатка. Экзоподит срастается с придаткам коксоподита (эпиподи-
том) и образует удлиненную лодочку (scaphognatit), усаженную по
краям волосками. Лодочка играет существенную роль в процессе
дыхания2. У живого рака она находится в непрерывном движении,
благодаря которому вода вычерпывается из жаберной полости.
Таким-образом, скафогнатит способствует постоянному току воды через
жаберную полость. Движения его легко наблюдать, если у живого рака
срезать ножницами переднюю часть жаберной крышки.
Первая пара ногочелюстей (maxillipedes I, рис. 306, Е).
Оба членика протоподита (коксоподит и базиподит) листовидны. Эпи-
1 При онтогенезе по-Гисбрехту мандибулы закладываются как типичная двувет-
вистая конечность. Но з дальнейшем экзо- и эндоподит редуцируются, и щупик
возникает вновь как новообразование.
2 Некоторые авторы склонны рассматривать весь скафогнатит как
видоизмененный экзоподит, считая, что эпиподит у вторых максилл отсутствует.
306
Рис. 306. Конечности Astacus astacus. А— первая антенна (антеннула); Б —
вторая антенна; В — мандибула; Г — первая максилла; Д — вторая максилла;
Е — первая ногочелюсть; Ж —вторая ногочелюсть; 3— третья ногочелюсть;
И — ходильная нога второй пары (из Huxley):
/ — эндоподит вторых антенн, 2 —экзоподит (squama) вторых аитеии, 3 — щупик мандибул,
4 — жевательная пластинка мандибул, 5 — эндоподит первых максилл, 6 — дистальиая
жевательная лопасть (basipodit) первых максилл, 7 — проксимальная жевательная лопасть
(coxopodit) первых максилл, 8 — эндоподит вторых максилл, 9 — дистальная лопасть
(basipodit) вторых максилл, 10 — проксимальная лопасть (coxopodit) вторых максилл, 11 —
скафогнатит, 12 — экзоподит первых ногочелюстей, 13 — эндоподит первых ногочелюстей,
14, 15 — жевательные лопасти первых ногочелюстей, 16 — эпнподит, П — экзоподит вторых
ногочелюстей, 18 — эндоподит вторых ногочелюстей, 19 — basipodit, 20— coxopodit, 21 —
эпиподит вторых ногочелюстей, 22 — жабры вторых ногочелюстей, 23 — экзоподит третьих
ногочелюстей, 24 — эндоподит третьих ногочелюстей, 25 — эпнподнт третьих ногочелюстей, 26 —
жабра, 27 —нити коксоподита, 28 — эпиподит второй пары ходильных ног, 29 — ischiopodit,
30 — meropodit, 31 — carpopodit, 32 — propodit, 33 — dactylopodit
подит хорошо выражен. Эндоподит в форме небольшого двучленистого
придатка. Экзоподит развит значительно сильнее эндоподита. Состоит
из довольно длинного проксимального членика и членистого
жгутовидного придатка. Первая пара ногочелюстей принимает участие в удер-
307
жании и ощупывании пищи. Эпиподйт вместе со скафогнатитом второй
пары максилл участвует в вычерпывании воды из жаберной полости.
Двуветвистый тип строения выражен здесь более полно, чем у обеих
пар максилл.
Вторая пара ногочелюстей (maxillipedes II, рис. 306, Ж)
имеет типичное для двуветвистой конечности строение. По размерам
несколько крупнее первой пары ногочелюстей. Протоподит двучлепис-
тый. От коксоподита отходит нежная, слабо хитинизированная
пластинка — эпиподйт. К нему прикрепляется вполне развитая перистая
жабра, лежащая в жаберной полости. Эндоподит выражен хорошо,
слагается из пяти характерных для этого отдела члеников. Самым
крупным из них в данном случае является meropodit. По внутреннему краю
членики эндоподита усажены многочисленными волосками. Экзоподит
довольно длинный, дистальная часть его образует многочленистый
жгут, усаженный волосками.
Третья пара ногочелюстей (maxillipedes III, рис. 306, 3).
Так-же как и вторая пара, имеет типичное двуветвистое строение. По
размерам — самая большая среди ногочелюстей, приближается к
размерам ходильных ног. Подобно тому как и у второй ногочелюсти, кок-
соподит имеет листовидный эпиподйт, к которому прикрепляется жаб)ра.
Кроме того, эпиподйт несет пучок длинных волосков. Эндоподит
хорошо развит, пятичленист и по размерам значительно превосходит
экзоподит. В отличие от второй пары ногочелюстей, здесь самым большим
члеником является ischiopodit, который своей базальной частью
срастается с базиподитом. Экзоподит обладает довольно длинным основным
члеником и членистым жгутом. Как протоподит, так и эндоподит несут
по внутреннему краю многочисленные волоски.
Вторая и третья пары ногочелюстей служат главным образом для
удержания пищи и подачи ее ко рту при питании рака. Кроме того,
эндоподит третьей ногочелюсти является органом, служащим для'
очистки глазных стебельков и антеннул от оседающих на них посторонних
частиц.
Первая пара ходильных ног (pereiopodae I, рис. 307).
Первая пара ходильных ног, так же как и остальные четыре пары,
совершенно лищена экзоподита. Благодаря этому они не двуветвистые,
а одноветвистые. Первая пара ходильных ног самая мощная из всех.
Она состоит из семи члеников, в числе которых два членика протопо-
дита и пять члеников эндоподита. Но проксимальный членик
эндоподита (ischiopodit) срастается с базиподитом, так что сочленение между
ними отсутствует. Коксоподит несет эпиподйт и жабру. Проподит
образует в дистальном направлении вырост, у основания которого причле-
няется дактилоподит. В совокупности образуется мощно развитая
клешня — орган защиты и захвата пищи. Вместе с тем первая пара
ходильных ног выполняет и локомоторную функцию при ползании
животного.
Вторая — пятая пары ходильных ног (pereiopodae II—
V, рис. 306, И). Вторая и третья пары ходильных ног, подобно первой,
несут клешни, образованные проподитом и дактилоподитом. Размеры
клешней, однако, значительно меньше, чем у первой пары. Четвертая и
308
-Рис. 307.
astacus.
ходильная
нечность
Astacus
Первая
ко-
(из
пятая пары лишены клешней, у них дактилоподит
является непосредственным продолжением проподита,
с которым он подвижно сочленен. Все ходильные яо-
ги состоят из семи члеников. В отличие от первой
пары базиподит и ischiopodit не срастаются. Вторая,
третья и четвертая нары несут эпиподиты (придатки
ноксоподитов), волосковидные щетинки и жабры.
Пятая пара ходильных ног лишена эпиподита,
жабр и щетинок. Ходильные ноги —■ локомоторные
органы рака, они служат для ползания по
субстрату.
Представляет интерес характер сочленения
отдельных члеников ходильных ножек между собой.
В пределах каждого сочленения движение
осуществляется лишь в одной плоскости. Но плоскости
движения различных члеников не совпадают между собой.
Каждый последующий членик двигается под
известным определенным углом тго отношению к
предыдущему. В.результате вся конечность в целом
приобретает подвижность в различных направлениях.
Брюшные ножки — и л э о и о д ы первой —
пятой пары (см. рис. 304). В отношении
строения двух первых пар плэопод наблюдается, как
отмечалось выше, диморфизм между самцами и
самками. Поэтому (мы рассмотрим сначала строение
третьей пары 'брюшных ножек, которые одинаковы у обоих
полов. По размерам плэоподы значительно меньше
ходильных ножек. Они имеют ясно выраженное
двуветвистое «строение, но обладают неполным числом
члеников (рис. 308, Г). Протоподит слагается из
короткого коксоподита, причленяющегося ю брюшку, и более длинного ба-
зиподита. Эндоподит и экзоподит хорошо выражены, причем первый
несколько длиннее второго. Эндоподит состоит из одного длинного
членика и членистого жгутовидного придатка. Экзоподит слагается из более
короткого 'Основного членика и также членистого жгута. Как
протоподит, так -и обе ветви конечности покрыты многочисленными
хитиновыми волосками. У самки третья пара илэоподов несколько крупнее, чем
у самцов.
Четвертая и пятая пары брюшных ножек устроены так же, как и
третья пара, и не требуют специального описания.
Первая и вторая пары плэопод у самца превращены в копулятив-
ный орган и по строению резко отличаются от всех остальных
брюшных ножек. Обе пары у живого рака подогнуты под брюшко,
направлены вперед и тянутся до брюшной пластинки (sternum) последнего
торакального сегмента. Первая пара плэопод представляет собой нерас-
члененные придатки (рис. 308, А), образовавшиеся, вероятно, в
результате слияния элементов протоподита и эндоподита. Края их загнуты,
в результате чего внутри конечности образуется замкнутый желобок,
служащий для продвижения сперматозоидов самца при копуляции.
Netz):
1 — coxopodit, 2 — ba-
sipodit, 3 —
ischiopodit, 4 — meropodit,
5 — carpopodit, 6 —
propodit, 7 — dactylo-
podit
309
Рис. 308. Брюшные конечности Astacus astacus. Плэоподы—первой
пары самца '(А); самки (Б); второй пары самца (В); третьей пары
самки (Г) и уроподы шестой пары брюшных конечностей (Д) (из
Huxley):
/ — экзоподит, 2 — эидоподит
Вторая пара плэоподов самца, в большей степени, чем первая,
сохраняет тип строения двуветвистой конечности (рис. 308, В). Протоподит
двучленистый. Эндоподит развит хорошо. Его основной членик в дис-
тальной части образует выемку, в которой причленяется небольшой
жгутовидный членистый придаток. Дистальная часть основного
членика эндоподита имеет с наружной стороны выемку в форме желобка и
заканчивается воронкообразным расширением. Экзоподит развит
значительно слабее эндоподита, состоит из небольшого основного членика
и расчлененной жгутовидной части. Первая и вторая пары плэопод у
самки устроены иначе, чем у самца. Первая пара рудиментарна, в 3—
4 раза меньше остальных плэопод (рис. 308, Б), экзоподит у нее не
развит. Вторая пара имеет вполне типичное двуветвистое строение
подобно третьей — пятой парам.
Закончив рассмотрение конечностей головогруди и плэоподов, сле'-
дует обратить внимание на характер сочленения между отдельными
члениками конечностей. Можно различить два основных типа
сочленений — простой сустав и шарнирный сустав. Первый свойствен плэопо-
дам, сочленению жгутов антеннул с основным члеником, а также
отдельных колец жгутов антеннул/и антенн. Второй находим главным
образом в сочленении отдельных члеников ходильных ног. При простом
сочленении между обоими соединяющимися члениками имеется тонкая,
слабо хитинизирйванная и не пропитанная известковыми слоями
эластичная кожица. Ширина ее по всей поверхности сочленения более или
менее одинакова. Простое сочленение позволяет производить движения
в различных направлениях.
В шарнирном суставе (рис. 309) ширина суставной кожицы не
везде одинакова. С двух сторон один из члеников образует зубовидный
вырост, который подвижно сочленяется с небольшой выемкой на
соответствующем выросте соседнего членика. Этим создается возможность
движения лишь в одной плоскости, как это и отмечалось в отношении
отдельных члеников ходильных ног. Но в отличие от простого сочлене-
310
Л
Рис. 309. Шарнирный тип со- Рис. 310. Astacus astacus. Третий сег-
членения члеников конечное- мент брюшка самки. Вид сзади, (из
тей. А — схема; Б — действи- Huxley):
тельный ВИД сочленения / — tergum, 2 — pleuron, 3 — eplmeron, 4 —
третья брюшиая иожка, 5 — sternum
ния, за счет уменьшения направлений, движения, создается большая
прочность сочленения и достигается большая сила при движении.
Шестая пара брюшных конечностей — уроподы
(см. рис. 308, Д). Все элементы ее сплющены в одной плоскости.
Вместе с тельсоном уроподы, как указывалось, образуют плавник.
Протоподит состоит лишь из одного сильно укороченного членика. Эндоподит —
широкая нерасчлененная пластинка. Экзоподит поперечной бороздкой
разделен на две части, по размерам несколько больше эндоподита.
Уроподы по краю несут многочисленные щетинки.
Таким образом, общее число пар конечностей речного рака
равняется девятнадцати.
Скелет. Речной рак, так же как и все членистоногие животные,
обладает наружным хитиновым скелетом. В результате довольно
сложных процессов, имеющих место при онтогенезе, некоторые части
наружного скелета смещаются довольно глубоко внутрь тела, что у речного
рака имеет место в отношении некоторых вентральных элементов
скелета головогруди. Выше, при общем обзоре внешней морфологии
речного рака, уже были отмечены основные части наружного скелета; голо-4
вогрудный щит (сагарах) и наружный скелет сегментов брюшка.
Для более детального изучения скелета рака следует использовать
материал, вываренный в 10—20%-ной едкой щелочи и промытый в
воде, причем должны быть удалены все мягкие части.
Скелетные элементы брюшка имеют более простое строение, чем
головогруди. Поэтому изучение строения скелета удобнее начать с
рассмотрения отдельного членика брюшка.
Со спинной стороны каждый сегмент прикрыт выпуклой широкой
спинной пластинкой — tergum (рис. 310). С брюшной стороны между
точками прикрепления конечностей расположена более узкая (считая
по продольной оси животного) брюшная пластинка — sternum.
Кнаружи от конечностей лежат надбедрия — epimera. Размеры этой части
311
скелета невелики. По бокам каждого
брюшного сегмента (справа и слева)
располагаются два крыловидных
выроста боковой стенки — боковины, или
pleuron. Они представляют собой
продолжение с одной стропы tergum, а с
другой — epimeron.
В головогрудном отделе спинные
пластинки — terga утрачивают внешне
видимое метамерное расположение и
входят в состав головогрудного щита
(сагарах), боковые части которого
(бранхиостегиты) представляют собой
сильно развитые и слившиеся pleura.
Брюшная же часть скелета и в этом
отделе сохраняет метамерное
расположение, особенно в трудной части
головогруди.
Здесь, однако, к основным
элементам брюшного скелета — стер-
нальным пластинкам, присоединяется
сложная система перекладин,
соединяющих между собою следующие друг
за другом метамеры и служащих
точками прикрепления мышц. Указанные элементы, образующие в
совокупности сложный эндофрагмальный скелет, представляют собой не
что иное, как результат впячивания в'нутрь складок кутикулы, т. е.
систему аподэм. Во время линьки сбрасывается также и кутикула эндо-
фрагмы, хотя она и является но своему топографическому положению
в теле внутренним скелетом. Изучение этой части скелета следует
вести на поперечных разрезах (макроскопически) через середину
головогруди (рис. 311) и на изолированном путем кипячения в растворе
NaOH цельном эндофрагмальном скелете (рис. 312, 313).
На поперечном {разрезе через грудь (лучше в области второй или
третьей пары грудных ножек) видно расположение скелетных
элементов грудного отдела cephalothorax (рис. 311). По средней брюшной
линии между местами причленения конечностей располагается вытянутая
в продольном направлении узкая грудина — sternum. В области
головогруди sternum значительно уже, чем в брюшке. По обеим сторонам
sternum отходят выросты — эндостерниты. Они поднимаются от
грудины вверх и почти смыкаются по средней линии брюшка, образуя канал
(canalis sternalis), в котором лежит брюшная нервная цепочка.
Кнаружи от эндостернитов располагаются примыкающие непосредственно к
ним эндоплевриты (рис. 313, 4, 5). От последних вверх к спинной
части отходят тонкие хитиновые пластинки, гомологичные надбедриям
брюшных сегментов — эпимеры. Они отграничивают жаберную полость,
расположенную между ними и бранхиостегитами, от полости тела..
Эпимеры на спинной стороне непосредственно переходят в стенку
сагарах.
Рис. 311. Astacus astacus.
Поперечный разрез груди за основанием
первой пары ходильных ног (из
Huxley):
I — спинка (tergum), 2 — жаберная полость,
3— иадбедрие (epimeron), ограничивающее
жаберную полость со стороны тела, 4 —
эидоплеврит, 5 — эндостериит, 6 — сапаНз
sternalis, 7 —грудина (sternum), S—бран-
хиостегит
312
На изолированном
эндофрагмальном
скелете (рис. 312) видно, что
эндостерниты и
эндоплевриты образуют решетку,
на месте отверстий
которой причленяются
конечности. В совокупности
перекладины этой решетки,
тянущиеся от sternum к
epimeron, называются
arthrophragmae.
Sternum в разных
сегментах головогруди
выражена неодинаково.
Наиболее широкими
являются стерниты трех
последних сегментов
головогруди (рис. 312, 12—
14).
Коксоподиты
конечностей (правой и левой),
причленяющиеся к
туловищу, в этой области
довольно далеко
расставлены друг от друга, если
сравнивать с двумя
первыми парами ходильных
ног.
Стернит самого
заднего (XIII) сегмента
головогруди отличается
еще следующими особенностями. Между ним и предыдущим
сегментом (XII) образуется довольно глубокое впячивание (складка-). Сам
стернит' благодаря этому приобретает некоторую подвижность, что
несвойственно остальным стернитам головогруди. Стерниты IX и X
сегментов головогруди, несущие две первые пары ходильных ног,
представляют собою тонкие, вытянутые в длину пластинки (рис. 312, *
10, 11). В сегментах, несущих три пары ногочелюстей (шестую,
седьмую и восьмую), стерниты являются также узкими короткими
пластинками (рис. 312, 7—9). Стерниты двух максиллярных сегментов (IV и
V) сливаются в общую известковую пластинку, вытянутую в
поперечном направлении (рис. 312, 6).
В области ротового отверстия сзади имеется кутикулярный вырост,
образующий две лопасти (рис. 312,5). Это нижняя губа, или metastoma.
В предротовой области скелетные элементы загибаются кверху
(рис. 314). Из отдельных частей скелета здесь следует отметить
следующие. Спереди в ротовое отверстие вдается верхняя губа (labrum,
рис. 314, 6), несущая несколько хитиновых обызвествленных утолще-
313
Рис. 312. Astacus astacus. Эндофрагмальный скелет
и стернальные элементы. Вид с брюшной (Л) и со
спинной (Б) сторон (из Huxley):
/ — место причленения глазного стебелька, II—XIV — места
причлеиения конечностей головогруди (от первых антеин до
пятой пары ходильных ног); / н 2 — эпимер и стериит
первых антени, 3 — стернит сегмента вторых аитени, 4 —
верхняя губа (labrum), 5 — нижняя губа (metastoma), 6 —
стернит сегмента первой и второй пары максилл, 7 — стернит
сегмента первой пары ногочелюстей, 8 — стерннт сегмента
второй пары ногочелюстей, 9 — стериит сегмента третьей
пары ногочелюстей, 10—14 — стерииты сегментов первой —
пятой пары ходильных ног, 15 — эндоплевриты, 16 —
эндостериит
Рис. 313. Astacus astacus. Строение скелета на сагиттальном
разрезе (из Netz):
/ — эпимеральиая пластинка, 2 — линия прикрепления эпимеральной пластинки
к сагарах, 3 — рострум, 4 — эндоплеврит, 5 — эидостериит, 6 — сочленовая
ямка, 7 — тергнты сегментов брюшка, 8 — сочленовный бугорок
ний. Перед нею располагается довольно широкая прочная пластинка
(рис. 314, 5) (вероятно, стёрнит сегмента антенн), называемая episto-
ma. Впереди нее имеется узенькая пластинка — стернит антеннулярно-
го сегмента.
Наконец, в области антеннул и глазных стебельков довольно
хорошо развиты боковые обызвеетвленные части хитинового скелета,
которые представляют собой эпимеры двух передних сегментов
(см. рис. 312, 2). Они непосредственно прилежат к области-причлене-
ния глазных стебельков.
Изучение скелета следует
закончить рассмотрением скелета
конечностей. На перерезанных
поперек ходильных ножках ясдо
видно, что прочный хитиновый
скелет образует цилиндр, внутри
которого проходят отдельные
пучки мышц. Местами хитиновая
кутикула заворачивает внутрь,
образуя сухожилия, называемые у
членистоногих аподэмами,
служащие для прикрепления отдельных
мышц. Особенно прочным
является скелет клешней первой пары
ходильных ног.
Анатомия на вскрытии.
Умерщвленные в парах хлороформа
раки вскрываются со спинной
стороны. Тонкая кожица,
соединяющая брюшко и. головогрудь,
на спинной стороне, разрушается
скальпелем (брюшко при этом
отгибается книзу). После этого
Рис. 314. Astacus astacus. Строение
скелета
на
сагиттальном разрезе (из
Huxley):
/ — мышцы, соединяющие эпимеры с голово-
грудиым щитком, 2 —рострум, 3 — место
применения антеннул, 4 — место причленения
вторых антени, 5 — стернит сегмента вторых
антенн, 6 — верхняя губа, 7 — мандибула, 8 —
нижняя губа, 9 — стернит сегментов первых н
вторых максилл, 10— стернит сегментов
первой и второй пары ногочелюстей, // — стернит
сегмента третьей пары ногочелюстей, 12 — эн-
достерннт, 13 — эндоплеврит, 14 —
эпимеральиая пластинка
314
/£
\\: i/S
шт^
Рис. 315. Astacus astacus.
Схема, показывающая способ
вскрытия рака. Стрелками
указаны направления разреза (из
Jammes):
под головогрудный щит вводится ветвь
анатомических ножниц и делаются два
продольных разреза, как указано на рис. 315.
Поперечный разрез делается тотчас позади глаз.
Затем осторожно снимается спинная часть
сагарах. Тупой стороной скальпеля следует
при этом отделять хитин от подлежащих
тканей. Проводятся два продольных
разреза вдоль брюшка со спинной стороны,
поперечный разрез делается на VI
абдоминальном сегменте.
Тергиты снимаются пинцетом вместе с
дорзальной поверхностной мускулатурой
абдомена, и таким путем обнажается
задняя часть кишечника. Отдельные системы
органов для более детального изучения
необходимо изолировать. Изучение вскрытого
рака ведется в ванночке под водой.-
Расположение внутренних
органов вскрытого рака. После
удаления спинной части головогрудного щита под
ним остается слой гиподермы,
представляющий собой пленку красно-фиолетового
цвета. Ее нужно осторожно удалить пинцетом
и рассмотреть общую картину
расположения внутренних органов (рис. 316).
На спинной стороне в околосердечной сумке лежит сердце,
имеющее вид пятиугольного мешочка. На спинной стороне его бывает видна
пара остий и отходящая вперед передняя аорта, или глазная артерия
(aorta mediana). В переднем же 'направлении под некоторым углом к
продольной оси животного отходят сяжковые артерии (arteria
lateralis cephalicae, рис. 316, 14, 15). От заднего конца сердца отходит в
брюшко брюшная, или задняя, артерия (arteria dorsalis pleica,
рис. 316, 13).
Под сердцем залегает половая железа (яичник, или семенник) и
видны половые протоки.
В передней части головогруди расположен объемистый желудок
(рис. 316, 2), состоящий из передней кардиальной и задней пилориче-
ской частей. От него назад проходит в виде прямой трубки кишка,
лежащая ниже сердца и тянущаяся вплоть до тельсона, где она
кончается анальным отверстием. Между сердцем и желудком большая часть
полости тела занята сильно развитой объемистой желтовато-'бурой
печенью (hepatopancreas). По сторонам передней части кардиального
отдела желудка у основания второй пары антенн лежат зеленые, или
антеннальные, железы, являющиеся органами выделения (рис. 316, 3).
Наконец, при общем осмотре вскрытого рака следует отметить
некоторые наиболее мощно развитые мышцы. По сторонам пилоричеекой
части желудка лежат два мощных аддуктора мандибул (musculus
adductor posterior mandibulae). Впереди желудка — парные передние, поза-
315
ди — парные задние мышцы желудка.
Особенно мощно развиты мышцы
брюшка, образующие здесь сложную систему
мускулов сгибателей (в нижней части
брюшка) и разгибателей (в спинной
части).
Нервная система, лежащая на
брюшной стороне, при вскрытии не
видна. Для рассмотрения ее необходимо
удалить большинство других органов, о
чем будет сказано ниже. Ознакомившись
с общей картиной расположения
внутренних органов речного рака, следует
приступить к более детальному изучению
отдельных систем органов.
Пищеварительная система
речного рака, как и других Decapoda,
устроена сложно. Она слагается из
передней, средней и задней кишок.
Передняя и задняя кишки — эктодермального
происхождения и выстланы изнутри
хитиновой кутикулой. Средняя кишка — эн-
тодермальная. Передняя кишка
распадается на пищевод (oesophagus) и
объемистый желудок (рис. 317), Средняя
кишка коротка. Она образует короткую
дорзальную слепую кишку (рис. 317, 7),
и в нее открываются протоки мощно
развитой пищеварительной железы,
называемой печенью (hepatopancreas, см.
рис. 316, 6). Задняя кишка — прямая
трубка.
Для изучения пищеварительного
аппарата рекомендуется изолировать весь
кишечник. Для этого перерезают
пищевод, подводя ножницы под желудок и
заднюю кишку близ анального
отверстия, и осторожно отлрепаровывают
кишечник. Изучение мускулатуры
желудка проводится на вскрытом раке, у которого удален толовогрудный щит
и с одной стороны срезаны жабры и epimeron. Разобраться в сложной
структуре желудка можно, отделив его от кишечника и сняв пинцетом
поверхностную мускулатуру. Кроме того, необходимо вскрыть полость
желудка в его передней кардиальной части с брюшной стороны. Пйло-
рическую часть следует разрезать как в продольном, так и в
поперечном направлениях. Чтобы детальнее рассмотреть обызвествленные
скелетные элементы желудка, можно рекомендовать кратковременное
вываривание в едкой щелочи с последующей промывкой в воде.
Ротовое отверстие помещается с брюшной стороны в передней час-
Рис. 316. Самец Astacus
astacus. Общая картина вскрытия
(из Jammes):
/ — передние мускулы желудка,
2 — желудок, 3 — выделительная
железа, 4 —• мускул мандибул, 5 ~
задние мускулы желудка, 6 —
hepatopancreas, 7 — жабры, 8 — сердце,
9 — семенник, 10 — уаз deferens,
11 — задняя кишка, 12 — мускулы
брюшка, 13 — arteria dorsalis pleica,
14 — arteriae laterales cephalicae,
15 — aorta mediana
316
5 *
Рис. 317. Astacus astacus. Сагиттальный разрез, показывающий
расположение основных частей органов пищеварения (из Huxley):
/ — пищевод, 2 — кардиальная часть желудка, 3 — передние мышцы желудка,
4 — срединный зуб, 5 —.пилорическая часть желудка, 6 — задние мышцы желуд-
ка_ 7 — слепой дорзальный вырост средней кишки, 8 — задняя кишка, 9 —
отверстие, ведущее в проток hepatopancreas
ти головогруди. Оно окружено ротовыми конечностями и ограничено
спереди верхней губой, сзади — нижней губой, или метастомой
(рис. 318), скелетные элементы которых рассмотрены (см. с. 313) при
изучении скелета.
Пищевод представляет собой короткую трубку, идущую вверх к
спинной стороне. Стенки его мускулисты. Внутренний просвет имеет
форму продольно вытянутой щели, расширяющейся у передней и
задней стенки (рис. 319). Пищевод выстлан хитиновой кутикулой.
Имеется пять пар мышц дилататоров пищевода (musculus dilatatores
oesophagi), служащих для расширения его внутреннего просвета при приеме
пищи. Эти мышцы с одной стороны прикрепляются к стенке пищевода,
а с другой — к прилежащим частям хитинового скелета. Спинной конец
пищевода открывается в Желудок.
2 1
Рис. 318. Astacus astacus. Ротовое отверстие
снизу (Х7) (из Ringel):
/ — верхняя губа (labrum), 2 — чувствительные волоски,
расположенные на верхней губе, 3 — мандибула, 4 —
пластинки нижней губы (paragnathae), несущие волоски,
5 — боковые складки начала пищевода, 6 — брюшиые
складкн, 7 — спинная складка начала пищевода
Рис. .319. Astacus astacus.
Нижняя часть пищевода в
поперечном разрезе (Х4), видны
парные мускулы — дилататоры
(из Ringel):
/ — musculi dilatatores oesophagi
anteriores, 2 —m. d. oe. latero-infe-
riores, 3 — m. d. oe. posteriore3, 4 —
просвет пищевода
317
Рис. 320. Astacus astacus.
Мышцы желудка. Вид со спинной
стороны (хЗ) (из Ringel):
/ — musculus gastricus anterior, 2 —
m. gastricus posterior, 3 — m. cardio-
pyloricus dorsalis
Желудок распадается на
объемистый передний кардиалнный отдел (pars
cardiaca) и меньших размеров задний
пилорический отдел (pars pylorica,
см. рис. 317).
Прежде чем рассмотреть внутреннее
строение этих отделов, ознакомимся с
расположением главнейших мышц
желудка, обусловливающих его
значительную 'подвижность, которая имеет
большое физиологическое значение, так как
в желудке речного рака происходит
дополнительное измельчение пищи при
помощи имеющихся здесь сложных
хитиновых образований.
Мускулатуру желудка можно
разделить на две группы мышц. Одни из
них, называемые наружными мышцами,
прикрепляются одним концом к стенкам
желудка, а другим — к стенкам тела.
Другие — внутренние мышцы, целиком
входят в состав стенок желудка. Среди
наружных мышц наиболее мощные
передние и задние мышцы желудка (musculi gastrici anteriores et poste-
riores, рис. 320). Передние прикрепляются внутренним концом к
спинной стенке кардиальной части желудка, наружным — у основания рост-
рума. Задние мышцы желудка берут начало со спинной стороны
кардиальной и пилорической части и тянутся юазад, прикрепляясь к
спинному щиту в области затылочной борозды. Указанные мышцы играют
основную роль в работе желудка как жевательного аппарата. Их
сокращение приводит в действие «желудочную мельницу», состоящую из
хитиновых зубов, (строение которых будет рассмотрено ниже.
Кроме.,этих наиболее мощных мышц желудка имеется еще
значительное количество более слабо развитых мышечных пучков,
прикрепляющихся преимущественно к нижней части желудка. Четыре пары
мускулов дилататоров (расширителей) служат для расширения
кардиальной части полости желудка. Из них верхняя передняя пара (m. dila-
tatores gastrici antero-superiores, рис. 321, 2) идет от передней стенки
желудка 'постепенно сужающимся пучком к переднему концу, где
прикрепляется недалеко от основания глазных стебельков. Передняя
нижняя пара (m. dilatatores gastrici antero-inferiores) — от дна передней
кардиальной части желудка к epistoma (рис. 321, 4). Передняя
латеральная пара дилататоров (m. dilatatores gastrici latero-anteriores,
рис. 321, 7) отходит сбоку передней нижней части желудка и
прикрепляется к epistoma. Задняя латеральная пара дилататоров (m.
dilatatores gastrici latero-posteriores, рис. 321, 9) отходит от дна кардиальной
части желудка и также идет к epistoma. Пилорическая часть желудка
обладает тремя парами- дилататоров. Верхние дилататоры (m.
dilatatores pylorici superiores) идут от середины спинной части пилоруса к
318
головогрудному щиту, где прикрепляются близ затылочной борозды
(рис. 321, 22). Две пары нижних дилататоров (m. dilatatores pylorici
inferiores) являются длинными, но тонкими мышечными пучками,
тянущимися от нижней стенки пилоруса к области максилл, где
прикрепляются при помощи длинных аподэм (рис. 321, И, 13).
Внутренние мышцы желудка довольно многочисленны. Они
выполняют различные функции, из которых основными являются следующие:
1. Часть их представляет собой антагонистов описанных мускулов-ди-
лататоров, они сжимают желудок, способствуя жевательной функции
«желудочной мельницы». 2. Другая группа мышц стенок желудка
способствует своими сокращениями переходу пищи из кардиальной в пи-
лорическую часть. 3. Ряд мышц стенок пилоруса образует пресс,
сжатием которого отжимаются жидкие составные части пищи от более
крупных частиц.
К первой из указанных трех категорий относятся передние карди-
альные мышцы (m. cardiaci anteriores, рис. 321, 6), спинные кардио-4
пилорические мышцы (т. cardio-pylorici dorsales, см. рис. 320, 3),
латеральные желудочные мускулы (m. laminae lateralis, рис. 321, 10).
Из второй группы удается рассмотреть мускул-иодниматель
кардио-пилорической перегородки (m. levator valvulae cardio-pyloricae
posterior, рис. 321, 23). v
К третьей категории относится ряд мускульных пучков,
образующих сфинктер пилоруса (m. sphincter pylorici anterior, medianus,
posterior, рис. 321, 19, 20 я 21), а также несколько продольных мелких
мускулов пилоруса (m. pylorici longitudinales, т. pylorici obliquus, m.
pylorici latero-anteriores, рис. 321, 16, 17, 18). Кроме указанных мышц в
7 8 9 10 11 12 13 Щ 15
Рис. 321. Astacus astacus. Мышцы желудка (ХЗ) (из Ringel):
1 — т. gastrici anteriores, 2 — т. dilatatores gastrici antero-superiores, 3 — жерновок, 4 — т.
dilatatores gastrici antero-inferiores, 5 — т. dilatator oesophagi antero-superior, 6 — m. cardiacii anterior,
7 — m. dilatatores gastrici latero-anteriores, 8 — m. dilatatores oesophagi latero-superiores, 9 — m.
dilatatores gastrici latero-posteriores, 10 — m. laminae lateralis, //— m. dilatator pylorici infero-interior,
12 — m. cardio-inferomedianum, 13 — m. dilatator pylorici infero-exterior, 14 — m. cardio-pyloricu3
lateralis exterior, 15 — m. pylorici anteroventrales, 16 — m. pyloricus obliquus, 17 — m. pylorici latero-
anteriores, 18 —m. pylorici longitudinales, 19 —m. sphincter pylorici posterior, 20 — m. sphincter
pylorici mediana, 21 — ra. dilatator pylorici anterior, 22 — m. dilatator pylorici superior, 23 — m.
levator valvulae cardio-pyloricae posterior, 24 —m. gastricus posterior, 25 — m. intercalarium posterior,
26 — m. intercalarium anterior
319
/
Рис. 322. Astacus astacus. Хитиновый
скелет желудка. Вид снаружи (Х5) (из
Ringel):
/ — cardio-prosuperomedianum, 2 — cardio-pro-
superolateralia, 3 — жерновок, 4 — cardio-la-
minalateralis, 5 — cardio-inferolateralla superio-
res, 6 — cardio-inferolateralia inferiores, 7 —
железистый фильтр пилорической части
желудка, 8 — pylorico-metasuperolateralia, 9 —
pylorico-mesosuperomedianum, 10 — pylorico-
intercalare superius, // — pylorico-mesosupero-
laterale, 12— cardio-intercalare superius, 13 —
pylorico-prosuperomedianum, 14 — dens
lateralis, 15 — cardio-mesosuperolaterale, 16 — cardio-
intercalare medianum, 17 — cardio-intercalare
anterius
стенке желудка есть еще несколько мелких мышечных пучков, на
которых мы не будем останавливаться (см. рис. 321) х.
Кардиальная часть желудка представляет объемистый мешок.
Пищевод открывается в нее снизу (рис. 317, 322). Изнутри эта часть
желудка имеет плотную эластическую хитиновую выстилку, которая
сбрасывается во время линьки. В передней части кардиального отдела
желудка не образуется сколько-нибудь значительных обызвествлений
хитина. Но в период, предшествующий линьке, здесь откладываются
особые известковые образования — так называемые жерновки, имеющие
форму белых круглых лепешек (рис. 322, 3). Содержащаяся в них
известь растворяется в период линьки и употребляется на пропитывание
известью хитинового панциря. Спинная и задне-боковые части
кардиального отдела обладают сложной системой хитиновых пластинок,
пропитанных известью, и зубцов, направленных внутрь желудка.
Вся эта сложная система хитинового вооружения образует в
совокупности аппарат, служащий для размельчения пищи и получивший
название «желудочной мельницы». Было 'бы, однако, неправильно
думать, что размельчение пищи — единственная функция кардиального
отдела желудка речного рака. Из печени в него поступают пищевари-
1 Для изучения указанных мелких мышц рекомендуется пользоваться
бинокулярной лупой.
Рис. 323. Astacus astacus. Жевательный
желудок, вскрытый с брюшной стороны
(Х5) (из Ringel):
/ —cardio-prosuperomedianum, 2—cardio-meso-
superomedianum, 3 — cardio-prosuperolateralia,
4 — dens lateralis, 5 — cardio-lamina lateralis,
6 — dens mediana, 7 —воронка, 8
—железистый фильтр, 9 — утолщение стенки пресса
пилорической части желудка, 10 — дорзальная
складка, отделяющая кардиальную часть от
пилорической
320
тельные соки (благодаря этому содержимое
передней части желудка обычно окрашено в
коричневый цвет) и здесь, наряду с чисто
механическим размельчением, начинается также и
переваривание пищи.
Для изучения отдельных элементов
«желудочной мельницы» кардиальный отдел следует
вскрыть с брюшной стороны. Укажем
главнейшие составные части хитинового вооружения
переднего отдела желудка (рис. 322, 323, 324). На
спинной стороне лежит довольно большая
широкая срединная известковая пластинка (cardio-
prosuperomedianum1, рис. 322, 1; 323, 1; 324, 1).
От нее назад и вниз отходит вторая пластинка,
имеющая форму рукоятки (eardio-mesosuperome-
dianum, рис. 323, 2; 324, 3). В этом месте
образуется складка спинной стенки желудка. К этой
пластинке, под углом, подвижно нричленяётся
еще небольшая известковая пластиночка,
идущая назад 'И наверх к пилорическому отделу.
Эта пластинка (cardio-metasuperomedianum, рис.
324, 5) на -своем обращенном вперед конце
несет двухлопастный прочный хитиновый зуб (dens
mediana, рйс. 323, 6; 324, 6), вдающийся в
полость желудка. Задним концом она сочленяется
с широкой довольно .большой пластинкой,
прикрывающей сверху пилорический отдел желудка
(pylorico-prosuperomedianum, рис. 322, 13; 324, 8).
От этой же пластинки вниз и вперед справа и слева отходят два
массивных известковых образования (рис. 322, 14), имеющих форму не
совсем правильных треугольников. На своей расширенной стороне они
несут мощно развитые латеральные зубы (dens lateralis, рис. 323, 4),
вдающиеся в полость кардиальной части желудка и направленные
острыми концами к срединному зубу. Каждый из латеральных зубов
является довольно сложным образованием. Он несет три острых зубчика,
за которыми следует плоская жевательная поверхность с
многочисленными поперечными бороздками. Эта часть латерального зуба по
структуре несколько напоминает коренной зуб слона. Кроме трех хорошо
развитых зубов имеются еще два боковых слабо развитых зубчика,
расположенных несколько ниже латеральных зубов. Они сидят на
небольших известковых пластинках (cardio-intercalare medianum,
рис. 322, 16). Рядом с ними имеются многочисленные волоски.
Действием прикрепленных к желудку мышц (главным образом musculi
gastrici anteriores et posteriores, см. с. 318) происходит сближение всех
трех зубов в одной точке и осуществляется измельчение пищи. При
1 Латинские названия отдельных элементов желудочной мельницы введены М. Рин-
гель (1924), при этом само название указывает на положение данного элемента в
желудке.
Рис. 324. Astacus
astacus. Изолированные
наиболее крупные скелетные
элементы желудка (из
Huxley, терминология по
Ringel):
/— дорзальная пластинка
(cardio-prosuperomedianum),
2 — cardio-prosuperalatera-
lia, 3 — cardio-mesosupero-
medianum, 4 — сочленовная
поверхность, 5 —
cardio-metasuperomedianum, б — dens
mediana, 7— латеральные
зубы (den3 lateralia), 8 —
pylorico-prosuperomedianum
11—2
321
расслаблении мышц зубы отодвигаются друг от друга. Последнее
достигается главным образом благодаря эластичности сочленений
рассмотренных скелетных элементов и общей эластичности хитиновой
выстилки желудка.
Кроме отмеченных отдельных известковых элементов хитинового
вооружения кардиальной части желудка имеется еще несколько
мелких и крупных пластинок, из которых отметим только некоторые, ясно
видимые при вскрытии. От большой спинной кардиальной пластинки
справа и слева располагаются небольшие треугольные боковые карди-
альные пластинки (cardio-prosuperolateralia, рис. 322, 2; 323, 3). По
обеим сторонам сбоку лежат довольно большие латеральные пластинки
(cardio-laminae laterales, рис. 322, 4; 323, 5), несущие на своей
внутренней поверхности многочисленные шипики и волоски. Кзади от
них располагаются две длинные саблевидные пластинки, 'вытянутые в
дорзовентральном направлении (eardio-inferolateralia superius et infe-
rius, рис. 322, 5, 6). Еще некоторые мелкие скелетные элементы
кардиальной части желудка указаны на рис. 322.
Пилорическая часть желудка, будучи по размерам значительно
меньше кардиальной, обладает еще более сложной, чем последняя,
структурой. Ее физиологическая функция сводится в значительной
мере к отделению, жидких и мелко измельченных компонентов пищи,
подвергшейся уже обработке в кардиальной части желудка, от более
крупных частиц. Первые попадают из пилорической части в среднюю
кишку и печень. Вторые непосредственно направляются в заднюю
кишку и в дальнейшем выводятся наружу Ч
Отверстие из кардиальной части желудка в пилорическую
представляет собою узкую щель дельтообразной формы. Это обусловлено
тем, что на границе между двумя указанными отделами в полость
желудка с брюшной стороны вдается в виде треугольного языка
мускулистая кардио-пилорическая перегородка (valvula cardio-pylorica, см.
рис. 323, 10; 325, 1; 327, 2), а со спинной —две боковые дорзальные
складки. Благодаря этому в пилорус не могут попадать очень крупные
частицы пищи и поступление пищи осуществляется относительно
небольшими порциями. В пилорической части различают следующие
отделы: 1) преддверие, 2) отстойную камеру и пресс, 3) фильтр средней
кишки, 4) фильтр железистой средней кишки, 5) воронку.
Конфигурация всех этих отделов сложна (рис. 325). Для изучения их следует
вскрыть пилорус в различных направлениях и изготовить поперечные
срезы.
Через узкую щель пища из кардиальной части желудка попадает
в небольшой передний отдел — преддверие (рис. 325, 2). Оттуда она
поступает в полость пресса (рис. 325, 3), которая сильно вытянута в-
дорзовентральном направлении. Стенки этой части пилоруса толстые
и наряду с соединительной тканью имеют сильно развитые мускулы.
Здесь пищевые массы подвергаются довольно сильному давлению со
1 Строение и физиология желудка речного рака служат объектом значительного
количества исследований. Наиболее полными и обстоятельными работами являются
исследования М. Рингель (1924), приведенные в списке литературы.
322
Рис. 325. Astacus astacus. Сагиттальный разрез пилориче1
ского отдела желудка (из Jordan):
/ — кардио-пилорическая перегородка, 2 — преддверие, 3 —
отстойная камера и пресс, 4 — фильтр железистой средней кишки, 5 —
отверстие, ведущее в hepatopancreas, б — задняя кишка, 7 — воронка,
8 — хитиновый клапан, препятствующий обратному поступлению
пищи в желудок, 9 — слепой дорзальный вырост средней кишки
стороны стенок пресса и от них отжимаются
жидкие и полужидкие фракции. Полость пресса
далее назад продолжается в хитиновую трубку,
имеющую форму воронки (см. рис. 325, 7),
которая своим заостренным концом
непосредственно ведет в полость задней кишки (см. рис. 325,6),
минуя среднюю кишку и пищеварительную
железу hepatopancreas. Через воронку крупные
частицы пищи, которая не подверглась измельчению
ротовыми частями и «желудочной мельницей»,
непосредственно попадают в заднюю кишку и
вместе с каловыми массами выводятся наружу.
Из задней кишки назад в желудок вновь пища
попадать не может, так как сама воронка
образует клапан, препятствующий движению пищи в
обратном направлении.
Жидкие фракции пищи из пресса
двигаются в двух направлениях. Go опинной стороны
полость пресса узкой щелью сообщается с
фильтром средней кишки. На границе между этими
отделами имеется клапан, слагающийся из
многочисленных длинных хитиновых волосков,
препятствующих прохождению твердых частиц.
Полость фильтра средней кишки — седловидной
формы, она охватывает со спинной стороны
полость пресса (рис. 327, 2). В фильтр средней
Рис. 326. Astacus
astacus. Пилорическая часть
желудка с брюшной
стороны (Х15) (из Ringer):
/ —■ известковая пластинка
cardio-inferomediana, 2 —
складка, отделяющая cardi-
um от pylorus (valvula
cardio-pylorica), 3— фильтр
железистой части кишечника,
4 — клапаны воронки, 5 —
известковая пластинка pylo-
rico-proinferomediana
кишки пища может проникнуть
не только из пресса, но и
непосредственно из преддверия (ом.
рис. 325). На этом пути также
располагаются хитиновые
волоски. Из фильтра средней кишки
пища далее направляется в
короткую среднюю кишку и в
небольшую дорзальную слепую
кишку, где имеет место
резорбция. Однако большая часть пищи
проделывает иной путь и
попадает в пищеварительную железу
hepatopancreas. С брюшной
стороны июлость пресса имеет щеле-
видное отверстие, ведущее в
фильтр железистой части
средней кишки (рис. 327, 5). Он
является более обширным, чем
спинной фильтр, и полость его
охватывает полость пресса
справа и слева с брюшной стороны.
При рассматривании пилору-
са с брюшной стороны (рис. 326)
правый и левый фильтры
железистой части кишечника выступают в форме двух довольно объемистых
ореховидных тел. В этой части пилоруса располагается сложный
цедильный фильтровальный аппарат. Со шинной стороны в полость его
вдаются многочисленные хитиновые волоски, брюшная стенка образует
продольно идущие хитиновые ребрышки, которые продолжаются далее
в полость в виде волосков. В задней части правой и левой половины
полости железистого фильтра открываются отверстия, ведущие в полость
пищеварительной железы (см. рис. 325, 5).
Наличие описанных сложных фильтрующих аппаратов
обеспечивает поступление в hepatopancreas лишь жидких частей пищи. Последнее
физиологически является существенным, так как стенки печени
чрезвычайно нежны и повреждение их неразмельченными элементами
пищи легко могло бы привести к прободению стенок. В стенках пилори-
ческой части желудка располагается несколько известковых пластинок,
имеющих опорное значение. Их расположение и название см. на
рис. 322.
Средняя кишка речного рака очень коротка. Ее длина равняется
приблизительно Vi3 длины задней кишки. На ее спинной стороне, как
уже отмечалось, открывается короткая слепая кишка (см. рис. 325, 9).
Пищеварительная железа, называемая также hepatopancreas, или
печенью, имеет большое значение в пищеварении речного рака. Она
состоит из двух лопастей (правой и левой), из которых каждая
самостоятельным отверстием сообщается с задней частью железистого
фильтра.
Рис. 327. Astacus astacus. Поперечный
разрез pylorus (из Jordan):
1 — слепой дорзальный вырост средней кишки,
2 — боковая часть фильтра средней кишки,
3 — отстойная камера, 4 — мускулистые
стенки пилорического отдела, 5 — фильтр
железистой средней кишки
324
Рис. 328. Astacus astacus. A—■ жабры в естественном положении после удаления бран-
хиостергита; Б — жабры после удаления подобранхий (из Huxley):
1 — подобранхий четвертой пары ходильных ног, 2 — эпиподиты, образующие пластинки,
расположенные между подобранхиями, 3 — подобранхий второй пары ногочелюстей, 4— скафогнатит, 5 —
первая ногочелюсть, 6 — плевробранхий последнего торакального сегмента, 7 — верхний
артробранхий четвертой пары ходильных ног, 8 — верхний артробранхий третьей пары ногочелюстей, 9 —
нижний артробранхий третьей пары ногочелюстей, 10 — нижний артробранхий четвертой пары
ходильных ног
Печень слагается из большого количества отдельных трубочек;
общая внутренняя поверхность ее велика. Непосредственно в полость
этих трубочек поступает жидкая пища. Как показал ряд
физиологических исследований, железистые клетки печени выделяют ферменты,
расщепляющие жиры, белки и углеводы. Кроме того, непосредственно
здесь же происходит резорбция пищи. Фагоцитарной деятельности
клеток печени и внутриклеточного пищеварения не наблюдается.
Изложенные данные о физиологии печени речного рака ясно
показывают, что по своей функции она не сравнима с одноименным
органом позвоночных животных. Более правильным для функции этого
органа является название hepatopancreas.
Задняя кишка — длинная, прямая трубка, тянущаяся вплоть до
анального отверстия. Стенки задней кишки обладают хорошо развитой
мускулатурой. Ее внутренняя стенка образует шесть продольных
складок, довольно глубоко вдающихся в полость кишки. Эти складки ясно
видны как на вскрытом кишечнике (см. рис. 317, 8) так и на
поперечных разрезах. Поскольку задняя кишка эктодермального
происхождения, она имеет изнутри тонкую хитиновую выстилку.
Органы дыхания. Органами дыхания речного рака служат
жабры, расположенные в головогруди в жаберных камерах,
отграниченных от наружной среды головогрудным щитом (бранхиостегитом);
а от внутренних органов и полости тела — эпимером (см. с. 312 и
рис. 311).
Жабры речного рака в большинстве связаны с конечностями. Они
располагаются в восьми грудных сегментах — от первой пары
ногочелюстей и до пятой пары ходильных ног включительно. У речного рака,
так же как и у других Decapoda, по отношению к конечностям и стенке
тела различают три типа жабер. Подобранхий прикрепляются
непосредственно к основному членику конечностей (коксоподиту),
артробранхий — к месту сочленения конечностей с туловищем, наконец,
плевробранхий прикрепляются непосредственно к туловищу (рис. 328).
325
5 2 2=
Iff
ш
III
"go
Третья
лодная
нога
Четвертая ■
ладная
нога
Пятая
ходная
нога
1
(рудимент)
1
1
1
1
1
1
0
0
1
2
2
2
г
2
0
0
0
0
0
0
1
(рудимент)
1
(рудимент)
1
Рис. 329. Astacus astacus. Распределение различных типов жабр по сегментам
и конечностями головогруди (из Jammes)
Распределение их по отдельным сегментам головогруди см. на рис. 329.
П'одобранхии развиты на всех конечностях грудной части
головогруди, кроме последнего торакального сегмента. На первой паре ного-
челюстей они рудиментарны. Артробранхии отсутствуют на первом и
последнем сегментах, на остальных присутствуют в числе одной или
326
Рис. 330. Astacus astacus. Поперечный разрез Рис. 331. Astacus astacus. Схема
через жаберную нить передней артробранхии движения крови в жабрах (из
(Х540) (из Воск): Воск):
1 — эпителий, 2 — кровяная клетка, 3 — отводящий 1 — приводящий сосуд, 2 — отводящий
сосуд, 4 — кутикула, 5 — перегородка между приво- сосуд, 3 — мантийный сосуд, 4 —
жалящим и отводящим сосудами, 6 — приводящий со- верная артерия (vas afferens), 5 — жа-
суд, 7 — соединительнотканные элементы верная вена (vas efferens)
двух. Плевробранхии функционируют лишь на последнем торакальном
сегменте, на двух предшествующих они рудиментарны, на остальных—
отсутствуют'.
Каждая жабра состоит из центрального стержня и
многочисленных нитевидных боковых веточек, располагающихся со всех сторон
стержня. Подобный тип строения жабр называется трихобранхиями.
На поперечных разрезах через отдельные жаберные нити (рис. 330)
видно, что снаружи они одеты тонкой хитиновой кутикулой, под
которой располагаются ъ один слой клетки дыхательного эпителия.
Внутренняя полость, по которой движется кровь, тонкой перегородкой
делится на два сосуда: приводящий (с несколько меньшим диаметром) и
отводящий. Движение крови внутри жабры в целом (согласно Бок,
1925) совершается следующим образом. Венозная кровь из
латерального синуса попадает в жаберную артерию (vas afferens, рис. 331, 4),
по которой и поднимается в дистальном направлении. Отсюда кровь
направляется в приводящие сосуды жаберной нити (рис. 331, 1). Из
жаберной нити по отводящему сосуду (рис. 331, 2) кровь попадает в
1 У многих представителей Decapoda число и распределение жабр различно. Оно
выражается так называемой «жаберной формулой», которая служит одним из
систематических признаков.
327
периферический, окружающий весь стержень жабры, так называемый
мантийный сосуд (рис. 331, 3), отделенный перегородкой от
приводящей жаберной артерии, занимающей более центральное положение. Из
этого сосуда кровывновь проникает еще в одну жаберную нить, откуда
уже поступает' в жаберную вену (vas efferens, рис. 331, 5), связанную
с рассмотренными бранхио-кардиальными сосудами. Таким образом,
кровь, обогащаясь кислородом в жабрах, проходит обычно не через
одну, а через две жаберные нити.
К подобранхиям прилежат непосредственно эпиподиальные
пластинки (см. рис. 328, А, 2), которые располагаются в жаберной полости
друг за другом, как листы книги, отделяя один жаберный пучок от
следующего за ним.
Постоянный ток воды через жаберную полость осуществляется, как
указывалось, благодаря деятельности скафогнатита (с. 306).
Внутрь жаберной полости вода проникает у речного рака через
семь отверстий (с каждой стороны), образующихся у основания
грудных конечностей, начиная со второй пары ногочелюстей, где головогруд-
ный щит не плотно прилегает к стенке тела и основаниям ножек,
оставляя просветы.
Кровеносная система. При вскрытии видны лишь
немногие части кровеносной системы — перикардий, сердце и главнейшие
отходящие от него сосуды. Для более детального изучения кровеносной
системы необходимо прибегнуть к инъекции (см. т. I, с. 43).
Инъекция артериальной системы производится путем введения
иглы шприца в сердце, через которое - и наполняются артерии. Чтобы
установить, в какой момент следует прекратить давление поршня,
полезно прибегнуть к следующему приему. Перед инъекцией надо
отрезать часть одной из ходильных ножек. Наполнение сосудов вести до
того момента, когда окрашенная массаначнет выступать из поперечного
разреза. Это будет указывать на наполнение артерий конечностей, а
вместе с ними и других мелких разветвлений кровеносной системы. Для
инъекции взять свеженаркотизированных или живых раков, острым
скальпелем или ножницами вырезать в хитине окошечко над сердцем,
снять перикардий. После этого поместить рака в теплую воду (чтобы
в сосудах не сразу застывала желатина). Инъекцию начинать еще при
слабом сокращении сердца. После окончания инъекции до начала
препаровки рака помещают на несколько часов или на сутки в 4%-ный
раствор формалина. После формалина объект полезно перенести на 2—
3 ч в 90°-ный спирт для уплотнения органов. При препаровке
инъецированный рак переносится в воду. Осторожно снимается головогрудный
щит. Последовательно отыскиваются артерии. При этом приходится
удалить некоторые органы, чтобы рассмотреть нижележащие.
Инъекция лакунарной системы может быть произведена только
частично, так как лакуны, не имеющие собственных стенок, легко
пропускают инъекционную массу в просветы между органами. Для инъекции
поступают следующим образом. С одной стороны только что
усыпленного рака снимают жаберную крышку (не повредить жабры!). В
крышке головогрудного щита просверливают над сердцем небольшое
отверстие. Вводят иглу шприца в перикардий (не в сердце!) и осторожным
328
Рис. 332. Схема кровеносных сосудов и главнейших
лакун у Decapoda macrura (из Blass по Gegenbanr):
/ — сердце, 2 — перикардий, 3 — передняя аорта или глазная
артерия (aorta mediana-arteria opthalmica), 4 — сяжковая
артерия (arteria lateralis cephalica-arteria antennularis), 5 —нижняя
. брюшиая (поднервная) артерия (a. subneuralis), 5 —брюшной
венозный синус, 7 — приносящие жаберные сосуды, 8 —
нисходящая артерия, 9 — верхняя брюшная артерия (a. dorsalis plei-
са), 10 — жаберно-'сердечные каналы. Печеночные артерии на
схеме не изображены
давлением шприца наполняют полость перикардия и впадающие в него
лакуны. При этом наполняются в случае удачной инъекции также
выносящие сосуды жабр. Препарат уплотняется в формалине. В
дальнейшем жабры отсекаются, и тогда ста>новятся видны некоторые
элементы лакунарной системы (рис. 332, 10).
Прежде чем остановиться на подробном рассмотрении отдельных
частей кровеносной системы речного рака, напомним кратко общий ход
кровообращения.
Кровеносная система не замкнута. Кровь в сердце проникает из
перикардиальной полости через три пары остий. При систоле остии
замыкаются и кровь поступает в артерии. От сердца отходят следующие
основные артерии: 1) непарная глазная артерия (aorta mediana l = а.
ophthalmica) идет прямо от сердца вперед к голове; 2) парные сяжко-
вые артерии (arteriae laterales cephalicae = аа. antennariae) проходят
от сердца вперед в направлении второй пары сяжков; 3) парные
печеночные артерии (arteriae hepaticae) идут от сердца под углом вниз к
печени; 4) непарная верхняя брюшная артерия (arteria dorsalis plei-
са) — от сердца назад по спинной стороне брюшка; 5) непарная
нисходящая артерия (arteria descendens) —от заднего конца сердца вниз на
брюшную сторону, где она впадает в брюшную артерию (arteria
subneuralis).
Из конечных разветвлений артерий кровь непосредственно
изливается в просветы между органами и движется по лакунам. Венозная
кровь собирается в большом вентральном венозном синусе, лежащем на
вентральной стороне головогруди. Из него кровь проходит по коротким
1 Терминология сосудов речного рака запутана. Во многих работах находим
различные названия для одних и тех же сосудов. Нами применяется терминология
Баумана (1921), данная им в монографии «Кровеносная система Potamobius». Иногда
приводим также наиболее распространенные синонимы.
329
•^
/,
5
6-—
7—
8-—
сл
JBy
■p' Iff I
^5;' ;'Г№"'''
W-F-Jtt' I
- vE§ai':rV:'v':-'; / g
/МииЖ^
Рис. 333. Astacus astacus. Сердце,
вскрытое со спинной стороны. Стенка
перикардия частично снята (из Ваи-
mann):
1 — aorta mediana, 2 — arteria lateralis
cephalicae, 3 — ligamentum dorsale laterale,
4—1. dorsale medianum, 5 — 1. ostii dorsa-
Нз anterior, 6 — 1. ostii dorsalis posterior,
7 — 1. dorsale medianum, 8 — 1. latero poste-
rius, 9 — 1. ventralia, 10 — дорэальные остин
каналам в два латеральных синуса,
расположенных вдоль головогруди
непосредственно у основания жабр. Из
латеральных синусов кровь попадает
в жабры, откуда через жаберно-сер-
дечные каналы поступает в перикар-
диальную полость и далее через ос-
тии в сердце.
Исходя из общей схемы
кровеносной системы речного рака, следует
детально проследить разветвления
отдельных артерий и частично
рассмотреть некоторые элементы лакунар-
ной системы.
Сердце, расположенное на
спинной стороне в полости
околосердечной сумки, является мускулистым,
мешочком, имеющим не совсем
правильную пятиугольную форму.
Осторожно приподняв сердце
пинцетом, можно увидеть несколько
лигаментов, при помощи которых оно
прикрепляется к стенкам перикардия.
Бауман описывает 2 непарных и
6 парных лигаментов. Их
расположение изображено на рис. 333, 334.
При вскрытии дорзальные лига-
менты разрушаются, и удается
видеть лишь латеральные и брюшные
лигаменты. Три пары остий
расположены следующим образом:
дорзальные остии (рис. 333, 10) — на спинной
стороне несколько кпереди от
середины сердца, латеральные остии (см.
7
Рис. 334. Astacus astacus (из Baumann):
/ — arteria hepatica, 2 — ligamentum latero-anterius, 3
—вентральные остии, 4 — 1. ventralia, 5 — луковица (bulbus) arteriae
dorsalis pleicae, б — 1. posterius, 7 — 1. dorsale medianum, 8 —
латеральные остни, 9 — 1. dorsale posterius
330
рис. 334, 8) — по бокам сердца, вентральные
остии (рис. 334, 3) — с брюшной стороны в
передней трети сердца.
Сняв полости сердца (рис. 335), его
спинную стенку, можно увидеть внутреннее
строение сердца и. расположение мускульных
пучков. В сердце имеется лишь одна камера, но
косо идущим вдоль брюшной стенки
мускульным тяжем она подразделяется (не
полностью) на две части, на которых левая
несколько больших размеров, чем правая.
Глазная артерия (aorta mediana)
начинается от переднего конца сердца и идет, не
ветвясь, до переднего края жевательного
желудка (рис.336). Здесь она образует мускулистое
расширение — cor frontale (рис. 336, 1),
являющееся дополнительным пропульсаторным
органом. Вскоре после этого на брюшную
сторону отходит непарная мозговая артерия
(arteria cerebri). Дойдя до надглоточного ганглия,
она распадается сначала на две ветви,
которые, в свою очередь, ветвятся, оплетая
мелкими капиллярами надглоточный ганглий.
Несколько далее кпереди от aorta
mediana отходят две крупные артерии в глазные
стебельки (arteriae opticae, рис. 337, /),
снабжающие кровью главным образом
оптический нерв ,и ганглии. После отхождения этих артерий aorta mediana,
ставшая значительно тоньше, распадается на две парные ветви, из
которых одни идут к мышцам, а другие представляют собою малую
артерию первых антенн (arteria minor antennalis prima). Все это удается
рассмотреть на инъецированном объекте, если осторожно снять
панцирь в области рострума и глазных стебельков, а также удалить
хитиновую стенку последних.
, Сяжковые артерии (arteriae laterales cephalicae) снабжают кровью
антеннулы, антенны, органы выделения и желудок, образуя множество
разветвлений. От сердца они берут начало по сторонам от aorta
mediana двумя мощными стволами (рис. 336, 7). На уровне приблизительно
переднего конца гонады каждая из них делится на две большие
ветви — наружную и внутреннюю (ramus externus et ramus internus, см.
рис. 336, 3, 4). Первая из них направляется к голове, вторая — к
желудку.
На хорошо инъецированных препаратах можно проследить
дальнейшие разветвления обеих ветвей. Ramus externus дает несколько
мелких веточек к мускулам мандибул, но до.уровня переднего отдела
желудка от нее не берут начало крупные артерии. Здесь же на
спинную сторону отходят общим стволом артерия рострума (arteria rostra-
lis) и передняя артерия желудка (arteria gastrica anterior, рис. 338, 4,
5). Первая снабжает кровью полость рострума, вторая — переднюю
Рис. 335. Astacus
astacus. Сердце, вскрытое со
спинной стороны. Видны
косо идущие пучки
мышечных волокон (из
Baumann)
331
Рис. 336. Astacus astacus. Кровеносные
сосуды головогруди (из Baumann):
1 — cor frontale, 2 — aorta mediana, 3 — наружная
ветвь сяжковой артерии (ramus externus arteriae
lateralis cephalicae), 4 — внутренняя ветвь
сяжковой артерии (ramus internus arteriae cephalicae),
5 — hepatopancreas, 6 — сердце, 7 — arteria
lateralis cephalicae, 8 — musculus gastricus posterior,
9 — мандибулярный мускул (m. adductor
posterior), 10 — m. gastricus anterior, 11 — arteria
gastrica anterior, 12 — arteria rostralis
Рис. 337. Astacus astacus. Сосуды
глазного стебелька оптического нерва и глаза
(из Baumann):
/ — arteria optica. 2 — aorta mediana, 3—
надглоточный ганглий. I—IV—оптические ганглии
стенку желудка и его мускулатуру. Далее, ramus externus дает ветви
в антеннулы (arteria maior antennae I) и в глазные стебельки (arteria
oculomotoria) и снабжает кровью мускулатуру глаз '. Затем ramus
externus разделяется на две ветви—-артерию вторых антенн (рис. 338,5)
и артерию зеленой железы (arteria glandulae viridis). Как первая, так
и вторая, в свою очередь, распадаются на ряд более мелких ветвей
(arteria renalis anterior, arteria saccularis, рис. 338, 7, 8) и др.
Внутренняя ветвь (ramus internus) сяжковой артерии снабжает
кровью боковые части желудка, пилорический отдел, а также
прилежащие мышцы. Она распадается на три главные ветви (рис. 339) —
arteria gastrica posterior prima, arteria gastrica- posterior secunda,
arteria gastrica posterior tertia. Каждая из этих ветвей, в свою очередь,
образует многочисленные разветвления.
Печеночные артерии (arteriae hepaticae, см. рис. 334, /) берут
начало от сердца несколько правее и ниже сяжковых артерий и
направляются под некоторым углом вперед и вниз к печени. Каждая из них
вскоре распадается на 3 ветви, ветвящиеся далее на множество
капилляров, обильно снабжающих кровью этот столь важный для
пищеварения орган.
1 Таким образом, антеннулы и глаза снабжаются кровью из двух источников —
от aorta mediana и от ramus externus arteriae laterales cephalicae.
332
Рис. 338. Astacus astacus. Разветвления наружной
ветви сяжковой артерии (из Baumann):
i —ramus externus arteriae lateralis cephalicae, 2 —musculus
gastricus anterior. 3 — m. promotor, 4 — arteria gastrica
anterior, 5 — arteria rostralis, 6 — arteria antennae II, T— arteria
renalis anterior, 8 — arteria saccularis, 9 — musculus remotor,
10 — зеленая железа, 11 — печень, 12 — musculus gastricum
posterior, 13 —m. adductor posterior, 14 — m. dorso-ventralis
posterior
Верхняя абдоминальная артерия (arteria dorsalis pleica) берет
начало от заднего конца сердца, образуя здесь довольно значительное
расширение — bulbus (рис. 340, 1). Она тянется над задней кишкой в
виде прямого сосуда до шестого сегмента брюшка, где распадается на
две ветви, охватывающие кишечник. По ходу этой артерии от нее от-
Рис. 339. Astacus astacus. Разветвления внутренней ветви сяжковой
артерии (из Baumann):
1 — ramus internus arteriae lateralis cephalicae, 2— musculus gastricus posterior,
3 — arteria gastrica posterior, 4 — ветви arteria gastrica posterior tertia, 5 — arteria
gastrica posterior secunda, 6 — musculus gastricus anterior, 7 — arteria gastrica
anterior, 3 — передние мускулы дилататоры желудка, 9—arteria gastrica posterior,
10 — задние мускулы дилататоры желудка
333
ходят ветви к гонаде (arteriae
genitales, рис. 340, 2, 3). В
последнем торакальном сегменте ив
шести сегментах брюшка от нее
отходят метамерно
расположенные arteriae laterales abdomina-
les (р.ие. 340, 5). Каждая из этих
артерий дает несколько веточек
ж мышцам и ветвь в
соответствующую данному сегменту
конечность (arteriae pedis spurii, рис.
341, 15). Нисходящая артерия
(arteria descendens) берет
начало не непосредственно от сердца,
а от расширения верхней
брюшной артерии, образующей, как
мы видели, на границе с сердцем
bulbus1. Направляется вниз и
несколько вперед, огибая кишечник
и прободая брюшную нервную
цепочку между третьей и
четвертой парой ходильных .ножек. Под
нервной цепочкой arteria
descendens впадает в нижнюю
брюшную (лоднервную) артерию. На
всем протяжении we образует
никаких боковых ветвей. Диаметр
ее значителен.
Нижняя брюшная
(поднервная) артерия (arteria
subneuralis, рис. 342, 3) тянется вдоль
всего тела с брюшной стороны
под нервной цепочкой. Кровь она
получает, как мы видели, из
нисходящей артерии. В arteria
subneuralis различают две части —
ростральную (pars rostralis),
лежащую кпереди от' места
впадения a. descendens, и каудальную (pars caudalis), лежащую кзади.
В ростральной своей части поднервная артерия обладает значительно
большим диаметром, чем в каудальной. Нередко в руководствах по
зоологии переднюю и заднюю части a. subneuralis рассматривают как
самостоятельные сосуды, называя первую — грудной артерией (a. ster-
nalis), а вторую — хвостовой артерией (a. caudalis). По существу, это
1 Arteria descendens трактуется рядом авторов (например, Гисбрехт и др.) как
гомолог одного из кольцевых сосудов, связывающих у кольчатых червей спинной и
брюшной продольные сосуды. Все остальные кольцевые сосуды у Decapoda
редуцировались, один же из них превратился в нисходящую артерию. У некоторых Decapoda
при онтогенезе она закладывается как парный сосуд.
Рис. 340. Astacus astacus. Верхняя
брюшная артерия и ее ветви (из Baumann):
У —bulbus, 2 и 3— две- ветви arteria' genitalis;
4—arteria dorsalis pleica, 5 — arteriae laterales
abdominales, 6 — конечные ветви верхней
брюшной артерии, охватывающие кишечник, 7 — vas
deferens, 8— arteria descendens
334
Рис. 341. Astacus astacus.
Кровеносные сосуды и мускулы
брюшка на поперечном разрезе (из
Baumann):
/ — arteria dorsalis pleica, 2 — arteria
lateralis abdominalis, 3 — musculus re-
motor, 4— m. rotator, 5 — m. abductor
exopoditis, 6 — m. adductor endopoditis,
7— m. abductor endopoditis, 8—
ганглий брюшной нервной цепочки, 9 —
arteria subneuralis, 10 — musculus
iransversis abdominis, pars ventralis,
11, 12 — m. obliqui anteriores, 13 — m.
transversus abdominis, 14 —- m. obliquus,
15 — arteria pedis spurii, 16,— musculus
dorsalis profundi abdominis, pars
lateralis, 17 — m. dorsalis superficialis, 18 —•
m. dorsalis abdominis profundi, pars
medialis
не вполне точно, так как a. subneuralis представляет собою единый
сосуд.
По ходу нижней брюшной артерии образуется множество
ответвлений, посредством которых снабжаются кровью конечности, часть
мускулатуры и некоторые внутренние органы — брюшная нервная
цепочка и задняя часть зеленой железы.
Ростральная часть спереди разветвляется сначала на две, а затем
на четыре ветви, давая с каждой стороны артерию мандибул (a. man-
dibularis) и заднюю артерию зеленой железы (a. renalis posterior).
К каждой паре конечностей головогруди отходит по одной паре
сосудов. Но артерии первой и^второй пары максилл и первой пары ногоче-
люстей берут начало общим стволом (рис. 342, .7)- Во всех
торакальных сегментах, несущих ходильные ноги, от a. subneuralis отходит по
паре ножных артерий (a. pedalis), дальнейший характер ветвления
которых изображен на рис. 343. Дорзальная ветвь (ramus superior)
снабжает кровью мышцы, двигающие конечность, брюшная ветвь (ramus
inferior) идет в конечность, отдавая мелкие веточки к мышцам.
Каудальная часть a. subneuralis, дав ветви в две задние пары
ходильных ног, переходит в брюшко, где она дает небольшие метамерно
расположенные веточки к мускулатуре вентральной части абдомена.
Брюшные конечности в отличие от грудных,снабжаются кровью не от
a. subneuralis, а от верхней брюшной артерии (a. dorsalis, pleica, см.
рис. 341). Исключение составляет лишь первая пара брюшных ножек,
которые получают сосуды от a. subneuralis, особенно хорошо развитые
у самцов.
На заднем конце в последнем сегменте брюшка a. subneuralis
делится на две ветви, которые, загибаясь дорзально, соединяются с
обеими конечными ветвями верхней брюшной артерии, образуя
околокишечное кольцо.
На препаратах с инъецированной лакунарной системой (см. с. 328)
удается рассмотреть некоторые части последней. При инъекции значи-
335
Рис. 342. Astacus astacus.
Брюшная (субневральная)
артерия и ее разветвления
(из Baumann):
1 — общая ветвь arteria subneu-
ralis к первым и вторым мак-
силлам и первой паре ногоче-
люстей, 2 — артерия третьей
пары ногочелюстей, 3 — arteria
subneuralis, 4 — артерия пятой
пары ходильных йог, 5 —
абдоминальная часть a. subneuralis
тельное количество инъекционной массы
скопляется с брюшной стороны головогруди.
Это наполняются 'брюшной и латеральный
венозные синусы, из которых кровь
поступает в жабры. В случае удачной инъекции
в осевой части жабр становится виден
отводящий сосуд, ino которому кровь
направляется от жабр к перикардиальной сумке.
Отчетливо видны кардио-'бра'нхиальные
сосуды. У речного рака с каждой стороны от
жаберных пучков к околосердечной сумке
поднимается шесть бранхио-кардиальных
сосудов (рис. 344). Они лежат
непосредственно под epimeron и просвечивают через
его стенку. Сосуды, соответствующие
жабрам ногочелюстей и первой пары
ходильных ног, соединяются вместе и
открываются в перикардий общим стволом. Сосуды,
идущие от жабр вторых, третьих и
четвертых пар ходильных ног, открываются в
перикардий порознь. Перед впадением в
перикардий они соединяются анастомозом с
сосудом, лежащим кзади. В задний из
сосудов 'Попадает также кровь от жабры
пятой пары ходильных ног. Таким образом, с
каждой стороны бранхио-кардиальные
сосуды открываются в перикардий четырьмя
стволами.
Выделительная система.
Органы выделения речного рака представлены
одной иарой антеннальных желез, часто
называемых также зелеными железами,
благодаря присущему им зеленоватому
оттенку. Они соответствуют ло своему развитию
и положению в теле сегменту вторых
антенн.
Филогенетически антеннальные
железы следует рассматривать как
видоизмененную пару метанефридиев. Они хорошо видны при вскрытии по
сторонам передней части желудка, особенно после удаления последнего.
Следует изолировать зеленую железу (лучше на уплотненном спиртом
материале), а также изготовить срезы для ознакомления с гистологией.
Антеннальная железа слагается из небольшого целомического
мешочка и выводного канала (рис. 345), дистальный конец которого
расширяется в мочевой пузырь. Последний открывается при помощи
короткого канала отверстием, расположенным на основном членике
второй пары антенн, наружу. . • -
Целомический мешочек по сравнению с другими отделами железы
относительно небольших размеров. Отверстие, ведущее из него в канал,
336
Рис 343 Astacus astacus. Сосуды и нервы пятого торакального сегмента
(из Keim):
/ _ arteria subneuralis, 2- ramus superior■ 3-^amus; inferior 4 - пР°Д°™е_га-
mus inferior. /-™™%*™Т- „.VedTu^W-^krnus
Рис 344. Astacus astacus. Основные сосуды лакунарной
системы (из Huxley):
1 - перикардий, 2 - кардио-броихиальные сосуды
337
называется нефростомом. Канал,
соединяющий целомический мешочек с мочевым
пузырем, распадается 'на ряд отделов
(рис. 345). Неф ростом ведет
непосредственно в расширенную пузыревидную часть —
зеленый канал. Его внутренняя стенка
образует многочисленные складки и выросты
в просвет канала, значительно
увеличивающие выделительную поверхность.
Зеленый канал при посредстве
короткого и узкого прозрачного канала ведет в
белый канал (рис. 345, 3). Последний
довольна длинен, образует несколько петель.
Его внутренняя стенка, так же как и
зеленого канала, покрыта многочисленными
складками. Белый канал открывается в
гладшетенный мочевой пузырь — самый
большой отдел выделительного органа.
В естественном положении в теле рака
мочевой пузырь 'прикрывает сверху все
остальные отделы антеннальной железы,
которые лежат под ним, будучи свернуты в
клубочек.. При этом целомический мешочек
прилежит с внутренней стороны к
мочевому .пузырю (рис. 345, 4). Выделительными
частями зеленой железы являются зеленый,
прозрачный и белый каналы, которые в
совокупности называются лабиринтами. В
мочевом пузыре собираются жидкие экскреты
перед выведением их наружу.
Нервная система. Центральная
нервная система речного рака хорошо
видна при вскрытии после удаления всех
внутренних органов. В области головогруди она лежит в canalis sternalis,
образованном отростками эндостернитов (см. с. 312), которые нужно
осторожно удалить.
Для изучения нервной системы лучше употреблять раков,
выдержанных перед вскрытием в течение суток в 80%-ном спирте.
Симпатическая нервная система, значительная часть которой
располагается на желудке, рассматривается до удаления последнего. Для
рассмотрения отдельных нервов, отходящих от ганглиев, следует вое
пользоваться малым увеличением бинокулярной лупы. Брюшная
нервная цепочка может быть изолирована целиком, из нее следует
изготовить тотальный препарат в канадском бальзаме. Надглоточный и под-
глоточный ганглии необходимо фиксировать отдельно и изготовить
серии фронтальных срезов для изучения их гистологии.
Центральная нервная система речного рака состоит из следующих
основных частей: надглоточного ганглия, или мозга, подглоточного
ганглия, связанного с мозгом окологлоточными коннективами, пяти то-
Рис. 345. Astacus astacus. Ан-
геннальная железа. А —
схематизированный продольный
разрез в естественном положении;
Б — в расправленном виде (из
Balss):
1 — мочевой пузырь, 2 —
выделительная пора, 3— белый канал, 4 —
прозрачный канал, 5 — зеленый
канал, 6 — целомический мешочек
338
Рис. 346. Astacus astacus. Общая
топография нервной системы (из Keim):
1 — надглоточный ганглий (мозг), i—
окологлоточные коннективы, 3—5 —
внутренняя, средняя и наружная ветви нерва
первой пары ходильных ног, 6 — подглоточ-
ный ганглий, 7 — первый грудной ганглий
брюшной нервной цепочки, 8 —
продольная щель брюшной нервной цепочки, через
которую проходит artcria descendens, 9 —
пятый грудной ганглнй, 10 — первый
брюшной ганглий, 11—13 — нервы, отходящие от
брюшных ганглиев, 11 — nervus pedis spu-
rii, 12 — n. dorso-lateralis, 13 — n. ventra-
lis, 14 — пятый брюшной ганглий, 15 —
последний (шестой) брюшиой ганглий
(ganglion postabdominale), 16 — нерв,
идущий к антеннальной железе (п. glandulae
viridis), 17 — ganglion oesophagei
(симпатической нервной системы)
ракальных ганглиев и шести
абдоминальных ганглиев (рис.
346). Все ганглии образуют в
совокупности брюшную
нервную цепочку и соединены друг
с другом коннективами.
Каждый ганглий брюшной цепочки
является, по существу,
парным, образовавшимся в
результате слияния двух
ганглиев— правого и левого. Эта
па/рность видна отчетливо на
тотальных препаратах. Точно
так же парными по
происхождению являются и коннективы между ганглиями. На просветленных
тотальных препаратах эту парность легко обнаружить, так как нервные
волокна, составляющие коннектив, ясно слагаются в два продольных
пучка. Нервные клетки сосредоточены в ганглиях, коннективы состоят
исключительно из нервных волокон. Коннектив между третьим и
четвертым торакальными ганглиями образует довольно значительную
продольную щель, через которую проходит arteria descendens (см. с. 334).
После ознакомления с общей топографией центральной нервной
системы следует перейти к более детальному рассмотрению отдельных
ганглиев и отходящих от них нервов.
Надглоточный ганглий, или мозг, располагается впереди пищевода
в области причленения антенн (рис. 347). Внешне на нем не заметно
расчленения, лишь передние углы несколько вытянуты в глазные ло-
пдети (lobi optici)1.
От головного мозга, отпрепаровав прилежащие части наружного
скелета и мышцы, можно видеть отхождение пяти пар нервов. От пе-
1 Мозг Astacus, как и мозг большинства других Arthropoda, слагается из трех
отделов — proto-, deuto- и tritocerebrum. При изучении срезов эти отделы могут быть
обнаружены, но анатомически расчленение мозга речного рака не выражено.
339
редней части мозга отходят
мощные оптические нервы 1
(nervus opticus—pedun-
culus lobi optici по Ханстро-
му), .идущие в глазные
стебельки и иннервирующие
глаза (рис. 347, /). Рядом
с п. opticus отходит
гораздо 'более тонкий
глазодвигательный нерв (п. oculomo-
toris), иннервирующий
мускулатуру, глазного
стебелька (-рис. 347, 2). Следующей
парой нервов, отходящих с
нижней стороны мозга,
являются нервы первой пары
антенн (п. antennalis pri-
Рис. 347. Astacus astacus. Надглоточный ганг- mus, р1ИС. 347, 3). В ПОЛОС-
лий с отходящими от него нервами (из Keim): ти последних они дают спе-
/ — nervus opticus, 2 — п. oculomotorius, 3 — п. an- ЦИаЛЬНуЮ ВвТВЬ К СТаТОЦИС-
tennalis primus, 4 —п. tegumentarius, 5 — п. antenna- /r-ni n" n г-м
lis secundus, 6 — окологлоточные коннектнвы Ту (СЛуХОВОИ Нер.В П. aCU-
sticus) и ряд мелких
веточек, иннервирующих антен-
нулы. Четвертой парой нервов головного мозга являются покровные
нервы (п. tegumentarius), иннервирующие гиподерму в области рострума,
антенн и боковых частей головного отдела головогруди (рис. 347,4).
Наконец, пятая пара —■ нервы второй пары антенн (п. antennalis secundus)
берут начало от задней боковой части мозга. Они иннервируют
антенны и вскоре после своего отхождения от мозга распадаются на две
ветви, из которых одна направляется в экзоподит, а вторая — в эндо-
подит2.
Окологлоточные коннективы, отходящие от заднего конца мозга,
огибают с обеих сторон пищевод и тянутся довольно далеко назад. По
ходу их на уровне пищевода расположены два небольших ганглия —
ganglion commissurae, от которых берут начало элементы
симпатической нервной системы, которую мы рассмотрим ниже.
Подглоточный ганглий (ganglion infraoesophageum) лежит на
уровне первой пары ходильных ног (см. рис. 346). Он представляет
собою результат слияния ганглиев брюшной цепочки, соответствующие
сегментам мандибул, обеих максилл и трех пар ногочелюстей, являясь
таким образом довольно сложным образованием. Указанное строение
подглоточного ганглия становится совершенно ясным при изучении
отходящих от него нервов, число которых равняется 11 парам. Из них
семь пар отходят от вентральной поверхности ганглия и четыре — от
1 Латинские названия нервов нами даются по . работе В. Кейм (1915). Все
отклонения от этой терминологии специально оговариваются.
2 Сравнительно-анатомически от protocerebrum отходят п. opticus я п. oculomo-
toris, от deutocerebrum — п. antennalis primus, от tritocerebrum — п. tegumentaris я
a. antennalis secundus.
340
дорзальной (рис. 348). Первой парой нервов, отходящих с брюшной
стороны ганглия, являются нервы зеленой железы (п. glandulae viri-
dis), иннервирующие железистую часть выделительного органа. За
ними следует шесть пар нервов ротовых конечностей: п. mandibularis in-
ternus, п. maxillaris anterior, n. maxillaris posterior, п. pedis maxillaris
primus, n. pedis maxillaris secundus, n. pedis maxillaris tertius. Эти
нервы проходят в конечности, а также дают ветви к мышцам,
обусловливающим движение данной пары конечностей.
Со спинной стороны подглоточного ганглия отходит четыре пары
нервов (рис. 348) — п. superins primus, secundus, tertius et quartus, йн-
нервирующих в основном дорзальную и дорзовентральную
мускулатуру головогруди. Последующие пять ганглиев брюшной нервной цепочки
(ganglion thoracale primum — quintum) имеют сходную структуру, и
из них каждый принадлежит лишь одному сегменту. Ганглий
четвертый и пятый несколько сближены между собой. С каждой стороны от
грудных ганглиев отходит по два нерва, один из них (п. thoracalis) ин-
нервирует мускулатуру соответствующего сегмента груди, тогда как
другой (п. pedalis) иннервирует конечность. Последний, в свою очередь,
делится на три ветви, из которых две направляются к мышцам
конечности, а одна непосредственно в конечность (см. рис. 343). N. thoracalis
у самцов в последнем грудном сегменте, а у самок в сегменте третьей
пары ходильных ног дает специальную веточку (п. genitalis) к
половому отверстию.
Из шести брюшных ганглиев (g. abdominales) пять имеют
одинаковое строение и соответствуют каждый одному брюшному сегменту,
шестой же (последний) имеет более сложную структуру в связи с тем,
что иннервирует кроме шестого сегмента абдомена еще и тельсон
(рис. 349). С каждым g. abdominales связаны три пары нервов. Одна
из них (п. pedis spurii) направляется к конечностям (плэоподам).
Вторая (п. dorso-lateralis) иннервирует глубокие слои мускулатуры
брюшка. Третья (п. ventralis) берет начало не от самого ганглия, а от ком-
миссуры и иннервирует брюшные части мускулатуры.
Шестой брюшной ганглий (ganglion postabdominale, рис. 350)
несколько больших размеров, чем предыдущие пять. От него отходят
пять парных и один непарный нерв. Парные нервы — следующие: п.
anterior — к мышцам, п. uropodialis — в уроподы; п. telsoni ventralis
иннервирует частью уроподы, частью тельсон; п. telsoni dorsalis —
чувствительные волоски тельсона; п. ani иннервирует область анального
отверстия. Непарным нервом, отходящим от g. postabdominale является
кишечный нерв (п. intestinalis), загибающийся на спинную сторону и
тянущийся вдоль значительной части кишечника. Видеть обычно
удается лишь его начало. Этот нерв относится к симпатической нервной
системе, к рассмотрению которой мы и переходим.
Основные элементы симпатической нервной системы берут начало
от двух g. commissurae, расположенных по ходу окологлоточных кон-
нективов на уровне пищевода. Рассмотреть все части симпатической
системы без специальной методики удается далеко не всегда. Тем не
менее мы отметим ее основные компоненты, так как многие из них все же
могут быть обнаружены при вскрытии.
341
Рис. 348. Astacus astacus. Подглоточный ганглий с отходящими от него нервами (из
Keim):
/ — окологлоточные коннективы, 2 — nervus glandulae viridis, 3 — п. mandibularis internus, 4 — п.
maxillaris anterior, 5 — n. superior primus, 6—n. maxillaris posterior, 7 — n. pedis maxillaris primus,
8 — n. pedis maxillaris secundum, 9 — n. pedis maxillares tertius, 10 — n. superior secundus, 11 — n.
superior tertius, 12 — n. superior quartus
Рис. 349. Astacus astacus. Брюшко, вскрытое с брюшной стороны, видна брюшная
нервная цепочка и мышцы (из Keim):
/ — первый ганглий брюшка, 2 — nervus pedis spurii, 3 — п. dorso-lateralis, 4— п. ventralis,
5 — пятый ганглий брюшка, 6— ganglion postabdomihale, 7 — п. telsonis dorsalis, 8— п. ani,
9 — п. telsonis ventralis
Рис. 350. Astacus astacus. Последний брюшной ганглий (g. postabdominale) .с
отходящими от него нервами (из Keim):
1 — nervus-anterior, 2 — п. uropodialis, 3 — п. telsonis, 4— п. telsonis dorsalis, 5 — п. ani, 6 — п.
intestinalis
От g. commissurae вдоль боковой и задней стенки желудка идет
ветвящийся задний желудочный нерв (п. ventriculi impar posterior,
рис. 351). Кроме того, от каждого из g. commissurae вокруг пищевода
отходят по два нерва (п. oesophagaeus superior et inferior), все четыре
ветви соединяются в ганглий пищевода (g. oesophagaeus), лежащий на
передней стенке пищевода. От него по передней стенке желудка тянет-
342
ся довольно мощный желудочный
стоматогастрический нерв (п. sto-
matogastricus inferior). По ходу
этого нерва к нему
присоединяются два небольших нерва от
передней и задней части головного
-мозга (п. ventriculi impar
superior и п. v. i. inferior).
В верхней части передней
стенки желудка
стоматогастрический нерв образует довольно
большой желудочный ганглий
(g. ventriculi superius). От
последнего 'по дорзальной стороне
желудка идет сйинной
желудочный нерв (п. ventriculi dorsalis),
вскоре распадающийся на два
латеральных нерва (п. ventriculi
lateralis), дающих, в свою
очередь, многочисленные веточки.
Части симпатической нервно;
могут быть обнаружены при вскрытии и требуют специальных методов
обработки материала, на которых мы не будем здесь останавливаться.
Органы чувств. К ним относятся: статоцист — орган
равновесия, сложные фасетированные глаза на подвижных стебельках, хемо-
рецепторы — органы химического чувства и разнообразные тангорецеп-
торы—-органы восприятия механических раздражений.
Статоцист первой пары антенн принадлежит к категории так
называемых открытых статоцистов. Отверстие, сообщающее его внутреннюю
полость с наружной средой, сохраняется в течение всей жизни рака.
Отверстие в форме треугольной щели расположено на спинной стороне
базального членика А1 и прикрыто снаружи расположенными в
несколько рядов довольно длинными щетинками (рис. 352). Оно ведет в
полость мешочка несколько сплющенного в дорзальном направлении и
заостренного на своем внутреннем конце (рис. 353). Изнутри статоцист
выстлан тонкой хитиновой кутикулой. В полости его помещаются
многочисленные статолиты. Последние представляют собою всякого рода
посторонние мелкие частицы, состав которых различен в зависимости
от свойств субстрата водоема, в котором живет рак. Статолиты бывают,
частично склеены между собой железистыми выделениями стенки ста-
то цист а.
На брюшной стенке статоциста располагаются чувствительные
волоски, иннервируемые мелкими разветвлениями — п. acusticus. Число
этих волосков и их расположение строго закономерны. Различают
несколько групп чувствительных волосков. Наибольшей из них является
серповидная группа, включающая более 100 волосков и расположенная
дугообразно по дну статоциста. Ближе к заостренному проксимальному
краю статоциста помещается ряд из 7—9 (чаще 8) волосков. Наконец,
по внутреннему краю мешочка расположена группа из 29 волосков
Рис. 351. Astacus astacus. Симпатическая
нервная система желудка (из Орлова):
1 — nervus ventriculis lateralis, 2 —п. ventriculis
dorsalis, 3—ganglion ventriculi superius, 4 — n.
ventriculi impar superior, 5 — мозг, 6 — п. stomato-
gastricus inferior, 7 — n. ventriculi impar inferior,
8 — g. oesophagei, 9 — n. oesophageus inferior,
10 — n. oesophageus superior, 11 — g. commissurae,
12 —n. ventriculi impar posterior
системы, иннервирующие сердце, не
343
Рис. 352. Astacus astacus.
Основание первой антенны
с отверстием, ведущим в
полость статоциста (из
Panning):
1 — базальный членик первой
антенны, 2 — волоски,
прикрывающие отверстия статоциста
Рис. 353. Astacus astacus. Основная
часть первой антенны, вскрытая с
брюшной стороны. Внутри
расположен статоцист (из Panning):
1—3 — три группы чувствительных
волосков, 4 — статолиты
(рис. 353) К На препаратах, обработанных 1%-ной осмиевой кислотой,
удается видеть мелкие разветвления — п. acusticus, которые подходят
к рассмотренным группам чувствительных волосков.
Глаза речного рака относятся к категории сложных, или фасетиро-
ванных, глаз. Они состоят из большого числа отдельных глазков, или
омматидиев, из которых каждый обладает собственным световосприни-
мающим и светопреломляющим (диоптрическим) аппаратом. Число
отдельных омматидиев и их размеры увеличиваются с возрастом рака.
Об этих возрастных изменениях дает представление следующая
табличка (по Бернгардту, 1915):
Для того чтобы рассмотреть расположение волосков, следует вскрыть основной
членик А1, вырезать статоцист, слегка прокипятить его в 10%-ной NaOH, промыть в
воде и сделать препарат, заключив его в глицерин.
344
рака, см
1
4,5
10
13
Число фасеток
635 "
1930
2275
3050
Ра
омсры
фасеток
мкм
22
46
60
65 .
Сложные глаза помещаются на стебельках, подвижно сочлененных
с головной частью головогруди. Глазные стебельки' обладают
собственной мускулатурой. По обеим сторонам основания рострума имеется
два желобка, в которые могут пригибаться глазные стебельки, что
является обычной защитной реакцией рака при
всякого рода механических раздражениях глаза.
Для изучения строения сложного глаза
желательно иметь продольные и поперечные срезы
залитого предварительно в парафин материала.
Ввиду того что стебелек обладает очень крепким
толстым слоем хитина, получить разрезы
возможно, лишь взяв в качестве материала только
что -слинявшего рака с мягкой хитиновой
кутикулой. Для того чтобы понять строение
сложного глаза, необходимо сначала ознакомиться со
структурой отдельного омматидия.
Каждый омматидий речного рака
представляет собой сильно вытянутое в длину в
радиальном направлении образование (рис. 354, 355),
слагающееся из нескольких как бы этажей
клеточных элементов :и их производных, имеющих
различное физиологическое значение.
Сверху (со стороны окружающей среды) ■
каждый омматидий ограничен хитиновой
роговицей (корнеальным хрусталиком). При
рассмотрении сверху видно, что большинство
хрусталиков имеет квадратную форму2.
Непосредственно под роговицей каждого омматидия лежат
две корнеагенные клетки с ядрами; сама
роговица является продуктом их выделения. Эти
клетки имеют форму трехгранных линз с наи-
- ' Ряд авторов, в том числе и Гексли в своей
монографии речного рака, склонны рассматривать глазной
стебелек как видоизмененную конечность. Одним из оснований
для этого служит факт регенерации на месте глаза усико-
подобного придатка при удалении вместе с глазом
оптического ганглия. Однако сравнение с другими группами
высших раков, не обладающих стебельковыми глазами, а
также ряд 'фактов, наблюдаемых при онтогенезе, не
позволяют считать эту гомологию достаточно обоснованной.
Поэтому мы рассматриваем глазной стебелек как
образование sui generis, не сравнимое с конечностью.
2 Для этого рекомендуется у пролежавшего в спирту
рака содрать пинцетом часть роговицы и рассмотреть в
капле глицерина под микроскопом.
Рис. 354. Astacus
astacus. Два омматидия из
сложного глаза (из
Balls no Bernhard):
/ — корнеальный хрусталик
(линза), 2 — ядра корнеаген-
ных клеток, 3 — Семперовы
ядра, 4 — дистальная часть
хрустального конуса, 5 —
стебелек хрустального
конуса, 6 — ядра ретинальных
клеток, 7 — ретинальные
клетки, 8 — клетки тапету-
ма, 9—базальная
мембрана, 10 — нервные отростки
ретинальных клеток, // —
пигментные клетки
345
меньшей толщиной в центре
омматидия. Далее в глубине
омматидия лежит длинное
прозрачное, сильно
преломляющее свет образование —
хрустальный конус. Последний
складывается из четырех
расположенных рядом по
продольной оси клеток, ядра
которых (так называемые Сем-
пероъы ядра) лежат в дис-
тальной части конуса,
непосредственно под корнеагенны-
ми клетками (рис. 354, 3). Ди-
стальная часть конуса
образует выпуклый купол,
проксимальный же отдел постепенно
суживается и заостряется. Эта
часть хрустального конуса
называется стебельком. Рогови-
да, корнеагенные клетки и
хрустальный конус образуют в
совокупности диоптрическую
часть омматидия.
Основание стебелька
хрустального конуса
непосредственно окружается в своей
проксимальной части ретинальны-
ми клетками,
представляющими световоспринимающий,
перципирующий отдел
омматидия. В составе каждого омматидия восемь вытянутых по продольной оси
ретинальных клеток, из которых одна развита слабее остальных. Их
совокупность образует ретинулу омматидия. Ядра этих клеток
располагаются в их верхней дистальной части (рис. 354, 6), окружающей стебелек
хрустального конуса. На внутренней стороне каждой ретинальной
клетки, обращенной к центру ретинулы по ее продольной оси, имеется
особо дифференцированная часть в форме гребешка, представляющая собой
рабдомер. Рабдомеры всех ретинальных клеток данного омматидия,
сходясь в центре, образуют рабдом. Рабдомеры представляют собой,,
как показал Гессе, палочковидно утолщенные окончания неврофибрил-
лей. На продольном разрезе через омматидий рабдом поперечно
исчерчен. Это обусловлено тем, что направления фибриллей,
входящих в состав рабдомеров, на разных уровнях не одинаковы. На
срезе они оказываются перерезанными то в продольном, то в
поперечном направлении. Нижние, проксимальные части светочувствительных
ретинальных клеток вытянуты в форме нервных отростков, которые
прободают базальную мембрану и входят в состав оптического нерва.
Внутри ретинальных клеток заключается темный пигмент.
Рис. 355. Astacus astacus. Продольный разрез
сложного глаза и глазного стебелька (нз
Balls, no Bernhard):
1 — базальвая мембрана, 2 — мелкие ганглиозные
клетки, окружающие оптические ганглии, 3 — ветви
глазной артерии, 4 — musculus oculi adductor, 5 —
nervus opticus, 6 — Лейдиговские
соединительнотканные клетки, 7 — ретина. /—IV — четыре оптических
ганглия
346
Кроме ретинулы пигмент находится и в некоторых других клетках.
По бокам каждого омматидия лежат две сильно вытянутые в длину
главные пигментные клетки, тянущиеся от корнеагенных клеток вплоть
до ретинулы. Благодаря наличию в них темного пигмента, отдельные
омматидий оказываются изолированными друг от друга.
У основания ретинальных клеток между отдельными ретинулами
вклиниваются пигментные клетки особого типа — клетки тапетума,
заполненные не темным, а, желтоватым пигментом. В совокупности эти
клетки образуют отражательную поверхность — тапетум. Если
рассматривать целиком срез через сложный глаз речного рака, то все
указанные элементы, располагающиеся на одинаковых уровнях в соседних
омматидиях, образуют несколько концентрических слоев полусферы
(рис. 355). Нервные отростки ретинальных клеток входят в состав
мощного оптического нерва, проходящего по центральной части глазного
стебелька. В той части нерва, которая прилежит непосредственно к
глазу, располагаются следом друг за другом четыре оптических ганглия.
Из них два дистальных в виде медулярных пластинок, между которыми
располагается прослойка из нейроглиальных элементов, тогда как два
проксимальных яйцевидной формы. Кроме того, вокруг указанных
ганглиев располагается обкладка из нервных клеток.
На продольном разрезе через глаз и стебелек кроме
рассмотренных уже элементов видны мышцы глазного стебелька, а также
перерезанные веточки сосудов. Снаружи стебелька—мощно развитая
кутикула, под ней —гиподерма. Между гиподермой, сосудами, мышцами и
нервами располагается соединительная ткань, состоящая главным
образом из довольно крупных Лейдиговских клеток и промежуточного
вещества.
Органы химического чувства сосредоточены почти исключительно ■
на антеннулах, на их дорзальной (наружной) ветви. При
рассматривании А1 при малом увеличении микроскопа1 ясно видны
многочисленные чувствительные волоски, которые и являются хеморецепторами.
Эти волоски, называемые также эстетами, полые внутри. У их
основания под гиподермой располагаются биполярные нервные клетки, один
отросток которых направлен к волоску, а другой — к нервному
окончанию.
Ряд физиологических данных указывает на то, что хеморецепторы
у речного рака локализуются не только на антеннулах, но также и в
других частях тела. В частности, внутренняя поверхность бранхиосте-
гита богата нервными окончаниями и, согласно ряду физиологических
экспериментов, воспринимает химические раздражения. Однако
морфологически оформленные органы химического чувства известны пока
лишь на антеннулах в форме описанных эстетов.
В качестве тангорецепторов—органов восприятия механических
раздражений у речного рака, как и у других Crustacea, следует
рассматривать многочисленные волоски, разбросанные по всему телу и
особенно многочисленные на конечностях.
1 Лучше предварительно антенну выварить в 10—15%-ном растворе едкой
щелочи.
347
Рис. 356. Astacus astacus.
Мужской половой аппарат (из
Huxley):
1 — парная часть семенника, 2 —
непарная часть семенника, 3 — vas
deferens, 4— ductus ejaculatoris
Половая система. Мужской
половой аппарат расположен в головогруди.
Семенник, по происхождению парный, у
взрослого самца ^представляют собой
непарную железу, образовавшуюся в
результате слияния двух зачатков.
Первоначальная парность семенника сохраняется в его
переднем, направленном к голове отделе,
который состоит из двух лопастей. Задний
отдел семенника — непарный (рис. 356), он
образует лишь одну вытянутую в
продольном направлении лопасть. Семенник лежит
на спинной стороне под сердцем над
кишечником. Если кусочек семенника
расщепить иглами в воде, то видно, что он
состоит из многочисленных трубочек,
оканчивающихся мешковидными расширениями.
В трубочках семенника происходят
процессы созревания половых клеток.
Семяпроводы (vasa deferentia) — парные.
Они берут начало в месте соединения
парных и непарной лопастей семенника и
образуют многочисленные петли и извивы
(рис. 356, 3). Их проксимальная, прилежащая к семеннику часть
гораздо тоньше дистальной. Первая является железистой частью
семяпроводов, вторая обладает мускулистыми стенками и представляют
собой семяизвергательный канал (ductus ejaculatorius). Мужские
протоки открываются наружу на ко'ксоподитах пятой пары ходильных ног.
Перенеся содержимое семяпроводов в каплю воды и изучая под
микроскопом, следует рассмотреть строение своеобразных аберантных
спермиев речного рака (рис. 357). Они дисковидной формы и
снабжены длинными лучеобразно расходящимися от центра отростками.
Центральный диск на одной стороне несет хитиновую капсулу, другая же
его поверхность соответствует головке.
Строение женского полового аппарата сходно с мужским. Яичник,
занимающий в теле рака то же положение, что и семенник, в передней
части является двухлопастным, в задней — однолопастным (рис. 358).
В нем видны многочисленные яйцевые клетки, находящиеся на разных
стадиях роста. Справа и слева от средней части яичника отходят
короткие яйцеводы, не образующие петель и открывающиеся наружу на
коксоподитах третьей пары ходильных ног.
Оплодотворение у речного рака протекает своеобразно.. При
копуляции самец и самка обращены друг к другу брюшными сторонами.
Поступающая из мужских половых отверстий сперма по желобку
первой пары брюшных ножек, превращенных, как мы видели, в копулятив-
ный аппарат, переносится на брюшко самки, где прикрепляется в
области sternum восьмого сегмента груди. Здесь она застывает в виде
молочно-белой массы и в женские половые протоки непосредственно не
проникает. Оплодотворение яиц совершается позже, в момент яйцеклад-
348
ки самки. При этом"самка подгибает брюшко под головогрудь. В
образованное таким путем замкнутое пространство выделяется особая
жидкость, которая растворяет вещество, склеивающее сперматозоиды.
После этого выводятся наружу созревшие яйцевые клетки, которые и
оплодотворяются сперматозоидами'внетела самки в образованной путем
подгибания брюшка камере. После этого оплодотворенные яйцеклетки
выделением липкого секрета женских половых путей приклеиваются к
брюшным ножкам самки, где и протекает их развитие.
У речного рака копуляция обычно происходит-поздней осенью и в
начале зимы. Перед копуляцией самка всегда линяет. Развитие яиц
падает на зимние месяцы. Некоторое время самка носит на брюшке
вылупляющуюся молодь, в этом выражается забота о потомстве.
Мускулатура речного рака сравнительно-анатомически
относится к категории локализованной мускулатуры; мышцы
дифференцированы на отдельные пучки, имеющие строго определенное положение и
точки прикрепления в теле животного. В отличие от низших раков
мускулатура Astacus более сложна. Мы рассмотрим только некоторые
наиболее важные в физиологическом отношении группы мышц: 1) Мышцы
тел"а, обусловливающие движение его частей; 2) мышцы конечностей,
при помощи которых конечность перемещается в отношении туловища;
3) мышцы члеников конечностей; 4) мышцы внутренних органов.
Последняя категория была нами рассмотрена при изложении строения
отдельных систем органов.
Изучение мускулатуры осуществляется при вскрытии, причем для
рассмотрения мышц последовательно удаляются внутренние органы.
Кроме того, необходимо произвести большими острыми ножницами
сагиттальный разрез рака начиная от тельсона и кончая рострумом.
После этого удалить внутренние органы, тогда расположение большинства
349
Рис. 359. Astacus 'astacus. Мускулатура, видимая на сагиттальном разрезе (из
Schmidt):
1— musculus adductor posterior, 2— m. dorso-ventralis anterior, 3— m. promotor antennae II,
4 — m. remotor antennae II, 5 — m. promotor ходильных ног, 6 — m. thoracales anteriores, 7—
m. ventrales superficiales thoracis, 8— m. transversus thoracis, 9 — m. ventralis superficialis
thoraco-abdominalis, 10 — m. obliqui anteriores, Л — m. ventralis profundi, 12 — m. transversi
abdominis, 13 — m. obliqui anteriores fasciae dorso-medianus, 14 — m. dorsales profundi, 15 —
m. dorsales profundi abdominis, pars mediana, 16 — m. dorsales superficiales, /7 — m. dorsales
profundi, 18 — m. dorsales profundi thoraco-abdominales, pars lateralis, 19 —m. Iaterales,
20 — m. attractor epimeralis
/
основных мышц выступает отчетливо (рис. 359). Для изучения
мускулатуры конечностей следует вырезать вместе с прилежащими частями
эндофрагмального скелета одну из ходильных ножек. Тоненькими
острыми ножницами снять с одной стороны хитиновый скелет, отпрепаро-
вав подлежащие мышцы.
Кроме приведенной группировки мышц в пределах туловищной
мускулатуры, с которой и нужно начинать изучение, различают
следующие группы мускулов: 1) сегментальные, которые принадлежат лишь
одной метамере или соединяют два сегмента между собой, 2)
интерсегментальные, тянущиеся через несколько метамер и 3) дорзовентраль-
ные, идущие в спинно-брюшном направлении. По положению в теле
среди мышц туловища различают мышцы спинные, брюшные и
латеральные.
Туловищная мускулатура сильнее развита в брюшке, чем в
головогруди, что связано с большой подвижностью брюшка и подвижностью
сочленения его отдельных сегментов. Головогрудные же сегменты, как
известно, неподвижны в отношении друг друга. У речного рака
наиболее развитой частью туловищной мускулатуры являются вентральные
мышцы. И в головогруди и в брюшке они образуют два слоя —
поверхностный (слабо развитый) и внутренний (мощный). Большинство этих
мышц относится к категории сегментальной мускулатуры.
Поверхностных брюшных шесть пар в головогруди (musculi ventrales superficiales
thoracis, рис. 359, 7) и пять пар в брюшке (m. v. s. abdominis).
Внутренние мышцы брюшного слоя образуют несколько систем
мускулов, проходящих в различных направлениях. Из них на
сагиттальном разрезе при вскрытии видны поперечные мышцы груди (один мус-
350
Рис. 360. Astacus astacus.
Мускулатура задней части головогруди и
брюшка (поверхностные мышцы
удалены (из Schmidt):
/—musculus thoracales anteriores, 2 —
m. obliqui anteriores et posteriores
(расположены сегментально), З — m. flexor tel-
soni anterior et posterior, 4 — m. transversi
abdominis (расположены сегментально)
Рис. 361. Astacus astacus.
Положение передних грудных и поперечных
брюшных мышц. Вид с брюшной
стороны (из Schmidt):
/ — musculus thoracales anteriores, 2 —гл.
transversus thoracis, 3 — дорзальиая часть
m. transversi abdominis, 4 — вентральная
часть m. transversi abdominis
кул) и брюшка (шесть мускулов) (m. transversus thoracis и m.
transversi abdominis, рис. 359, 8, 12; 360, 4; 361, 2—4). Передние (семь
мускулов) и .задние (пять мускулов) косые мышцы брюшка (m. obliqui
anteriores et т. о. posteriores, рис. 359,10; 360,2), наиболее мощные
мускулы абдомена. Они проходят в переднем и заднем направлениях косо
по отношению как к сагиттальной, так и к латеральной плоскостям
животного. Рассмотренные мускулы брюшка (поверхностные, поперечные
и косые) образуют в совокупности мощный сгибатель абдомена, дейст-
351
вием которого происходит подгибание брюшка под головогрудь и
осуществляются характерные плавательные движения рака.
Как бы непосредственным продолжением поперечной мышцы груди
являются передние грудные мышцы (m. thoracales anteriores, рис. 360,
361, 1), являющиеся интерсегментальными мышцами и
прикрепляющиеся к элементам эндофрагмального скелета. Они способствуют
прикреплению брюшка к головогруди и укреплению эндофрагмального скелета.
В числе особо специализированных мышц брюшного слоя отметим еще
два сгибателя тельсона (m. flexor telsoni anterior , et posterior,
рис. 360, 3). Дорзальные мышцы, развитые слабее вентральных, так
же как и. последние, состоят из двух слоев мышц — поверхностного и
глубинного. Поверхностные дорзальные мышцы (m. dorsales superficia-
les) представляют собой семь слабых сегментальных мускулов,
расположенных в последнем сегменте головогруди и в шести сегментах
брюшка (см. рис. 359, 16). Глубинные дорзальные мышцы брюшка
(m. dorsales profundi abdominis, см. рис. 359, 14, 15) расположены в
абдомене сегментально и служат разгибателями брюшка, т. е.
антагонистами вентральной мускулатуры. Грудные глубинные дорзальные
мышцы (m. dorsales profundi thoraco-abdominales) тянутся от брюшка
к передней части эпимеральной пластинки. Они образуют несколько
Рис. 362. Astacus astacus. Мускулатура тела и
конечностей пятого торакального сегмента (из Schmidt):
/ — musculus attractor epimeralis, 2— эпимеральная пластинка,
3 — жабры, 4 — главная ветвь m. depressor, 5 — ветви m.
remoter, 6 — наружная ветвь m. depressor, 7 — латеральная ветвь
m. levator, 8 — т. abductor carpopoditis, 9 —- т. productor propo-
ditis, 10— т. abductor dactylopoditis, Jl — т. adductor dactylo-
poditis, 12— m. adductor carpopoditis, 13 —m. reductor mero-
poditis (брюшная ветвь), 14 — m. reductor meropoditis (спииная
ветвь), 15 — m. reductor ischiopoditis, 16 —m. levator (главная
ветвь), 17 — m. levator (медиальная ветвь), 18 — ra. depressor
(главная ветвь), 19 — m. promoter
352
Рис. 363. Astacus astacus. Мускулатура клешни
первой пары ходильных ног (из Schmidt):
/ — musculus adductor dactylopoditis, 2 — m. productor pro-
poditis, S — m. reductor propoditls, 4 — m. abductor
пучков и являются
интерсегментальными (см. рис.
359, 17, 18). Из
латеральных мышц наиболее
значительные следующие:
1) m. attractor epimeralis
(см. рис. 359, 20),
представляющий мускульную
ленту, которая состоит из
большого количества
коротких лучков и
соединяет эпимеральную
пластинку с головогрудным
щитом; 2) латеральные
торано-абдоминалвные мышцы (m. laterales thoraco-abdominales,
см. рис. 359, 19), тянущиеся от абдомена к боковым частям
головогруди.
Передний и задний дорзовентральные мускулы (m. dorso-ventralis
anterior et posterior, см. рис. 359, 2; 364, 6) располагаются в передней
части головогруди, они тянутся от латеральной части переднего отдела
головогрудного щита к передней части эндофрагмального скелета.
Мускулатура различных конечностей, в зависимости от функций
.последних, отличается друг от друга. Не останавливаясь на
рассмотрении мускулов каждой пары конечностей в отдельности, ограничимся
описанием мускулатуры типичной ходильной ноги (рис. 362) и
основной мускулатуры мандибул (рис. 363).
Среди мышц конечностей по характеру их прикрепления и функции
различаются две группы. Мускулы первой группы связывают
конечность с туловищем и обусловливают движение конечности относительно
туловища. Мускулы второй группы целиком лежат внутри конечности
и обусловливают движение члеников конечности друг относительно
друга.
К первой из указанных двух групп мускулатуры конечностей
относятся следующие мышцы: 1. Сгибатель конечности (m. promotorl,
см. рис. 359, 5; 362, 19) своим проксимальным концом прикрепляется
к эндостерниту, дистальным — к коксоподиту; сокращение этого
мускула обусловливает движение конечности вперед. 2. Разгибатель
конечности (m. remotor, рис. 362, 5) по своей функции является
антагонистом m. promotor, складывается из трех ветвей, прикрепляющихся
проксимальными частями к эндоплевриту, дистальными — к коксоподиту.
3. Подниматель конечности (m. levator, рис. 362, 16, 17), главная его
ветвь идет от эндоплеврита к базиподиту, при сокращении поднимает
конечность. 4. М. depressor (рис. 362, 4, 6) является антагонистом
m. levator, слагается из двух ветвей, дистальные части которых берут
начало от базиподита, а проксимальные — от эпимеральной пластинки
и латеральных частей эндофрагмального скелета. Это один из
наиболее мощных мускулов конечностей. При сокращении его конечность, от-
Латинская номенклатура мышц приводится нами по Шмидту (1915).
12—2
353
Рис. 364. Astacus astacus (из Schmidt):
/ — musculus adductor anterior, 2—m. adductor
lateralis,' 3 — m. adductor posterior, 4 — m. compressor
endophragmatis, 5 —m. dorso-ventralis anterior, 6 —
m. dorso-ventralis posterior, 7 — m. abductor minor,
8 — m. abductor major
гибается книзу, а
рак,.стоящий на конечностях,
приподнимается кверху.
Остальные мышцы
конечностей относятся ко
второй .группе, т. е. соединяют
отдельные членики
конечностей друг с другом. Сюда
относятся следующие
отдельные мускулы,
изображенные на рис. 362. М. ге-
ductor ischiopoditis (рис.
362, 15) идет от базяподита
к ишжшодиту, 'вызывает
перемещение' ишиоподита
назад. М. reductor meropoditis-
(рис. 362, 13, 14) состоит
из дорзальной и
вентральной ветвей, тянется от
ишиоподита к мероподиту,.
обусловливает движение ме-
роподита назад. М. abductor
carpopoditis (р:ис. 362, 8) обусловливает движение картюподита кверху;,
его антагонистом является га. adductor carpopoditis (рис. 362, 12)..
Движение п'роподита вперед вызывает т. productor propoditis
(рис. 362, 9), антагонистом которого является m. reductor
propoditis.
Наконец, два сильно развитых мускула вызывают движения
клешни. М. abductor dactylopoditis — мышца, раскрывающая клешню. М.
adductor dactylopoditis — сжиматель клешни. Он особенно мощно развит
в первой паре ходильных ног (рис. 362, 10, 11; 363).
Мускулатура мандибул менее многообразная, чем мускулатура
ходильных ног, что связано с малой расчлененностью этой пары ротовых
конечностей. Удалив желудок и рассматривая мандибулы со стороны
полости тела, можно обнаружить основные мускулы жвал. Все они
парные, так же как и сами мандибулы (рис. 364).
Раздвигание мандибул достигается сокращением двух пар мышц,
(m. abductor mandibulae major и т. а. т. гтпог.рис. 364, 7, 8),
прикрепляющихся дистальной частью к внутренней поверхности жвал,
проксимальной — к боковым частям скелета головы. Сжимание
мандибул, играющее важную роль в измельчении пищи, достигается
действием трех пар мускулов, из которых самая мощная задняя пара (m.
adductor posterior mandibulae, см. рис. 359, 1; 364, 3). Узким концом эти
мышцы прикрепляются к жвалам, тянутся от последних кверху по
сторонам желудка и широкой частью прикрепляются к спинной части са-
гарах впереди затылочной борозды. Менее мощно развита передняя пара
аддукторов (m. adductor anterior mandibulae, рис. 364, 1), состоящая
из двух листовидных мускулов, тянущихся от мандибул почти прямо-
вперед. Наконец, третьей парой мышц является состоящий также из
354
Рис. 365. Astacus
astacus. Разрез через
кутикулу панциря
головогруди (из Schneider):
/ — наружный слой
кутикулы, 2 — гомогенный слой,
3 — пигментный слой, 4 —
основной слой
двух пучков m. adductor lateralis mandibulae,
тянущийся от жвал .к 'боковым частям
головного отдела панциря (рис. 364, 2).
Кроме рассмотренных мышц в толще
самих мандибул залегают два небольших
мускула, обусловливающих движение щупика.
Изучение гистологических препаратов.
Для того чтобы полнее представить себе
■строение и функцию отдельных органов
речного рака, желательно рассмотреть еще
некоторые гистологические препараты.
Кутикула, покровы и
соединительная ткань. Для изучения покровов
речного рака следует воспользоваться
боковыми складками сагарах — жаберными
крышками (бранхиостегитом). Жаберная крышка
образуется в процессе онтогенеза как складка
покровов. Поэтому она несет два слоя
внешнего эпителия ■— один наружный и другой внутренний, смотрящий в
полость жаберной камеры. Между обоими слоями эпителия находится
■слой соединительной ткани.
Небольшой кусочек жаберной крышки фиксировать жидкостью
Ценкера (с формалином) или жидкостью Перени. Лучше взять
недавно слинявшего рака. Декальцинировать в 1%-ной хромовой кислоте и
перед заливкой в парафин длительно (около месяца) выдержать в
растворе целлоидина по Петерфи.
Хитиновая кутикула мощно развита на обращенной наружу
стороне жаберной крышки. На срезах ясно видна продольная исчерченность
кутикулы, слагающейся из нескольких слоев (рис. 365). Самый
наружный слой незначительной толщины (около 1 мкм), за ним следует
более темно-окрашенный гомогенный слой, также незначительной
толщины (около 7 мкм). Далее внутрь следует мощный пигментный слой, в
котором локализован характерный для.речного рака пигмент,
окрашивающий его панцирь в темный цвет.
Наконец, самым мощным, занимающим около половины толщины
всей кутикулы, является внутренний —г основной слой. Пигментный и
основной слой в свою очередь обладают продольной исчерченностью,
так как состоят из большого количества вторичных слоев. Кроме
продольной в кутикуле наблюдается и поперечная исчерченность;
последняя обязана наличию особых опорных фибрилл, являющихся
непосредственным продолжением опорных фибрилл, подстилающих кутикулу
клеток гиподермы. В толще кутикулы нередко на срезах попадаются
канальцы, идущие от внутренних слоев к наружным. Эти канальцы
сообщаются с лимфатическими пространствами соединительнотканного
слоя. Кутикулу подстилает однослойный эпителий (гиподерма, рис. 366).
Клетки его, особенно с наружной стороны жаберной крышки, сильно
вытянуты в длину, почти веретенообразно. Внутри гиподермальных
клеток имеются опорного характера фибриллы, которые в дистальном
направлении продолжаются в толщу кутикулы. Пространство между
12*
355
Рис. 366. Astacus astacus. Разрез через
жаберную крышку (кутикула не изображена) (из
Schmidt):
/ — наружный слой эпителия, 2 — опорные фнбрнллы,
S — соединительнотканные Лейдиговские клетки первого
порядка, 4 — лакуна, 5 — внутренний слой эпителия, 6 —
слизистые железы
обоими слоями эпителия
заполнено соединительной
тканью, состоящей из
пузыревидных вакуолизирован-
ных клеток — Лейдиговских
клеток первого порядка.
Среди них встречаются
вытянутые в длину,
утолщенные Лейдиговские клетки
второго порядка (рис.
367, 7). В
соединительнотканном слое разбросаны
многочисленные слизистые
железы в виде трубочек,
выстланных одним слоем
железистых клеток с узким
просветом. Эти железы
открываются в жаберную
полость. Среди
соединительнотканных и железистых
клеток на срезах часто
попадаются лакуны разнообразной
формы, заполненные
гемолимфой.
Нередко на срезе удается 'видеть также мелкие разветвления
артерий (рис. 367). Собственно эпителиальная выстилка в кровеносных
сосудах рака (как и всех Arthropoda) отсутствует. Изнутри стенки
сосудов образованы мембраной соединительнотканного происхождения — in-
tima. Эта мембрана — продукт выделения окружающих кровеносный
сосуд особых соединительнотканных клеток — Лейдиговских клеток
третьего порядка. Эти клетки обладают менее вакуолизированной
плазмой, чем одноименные клетки первого порядка. Мускульные элементы
в стенках сосудов полностью отсутствуют.
Пищеварительная железа (hepatopancreas).
Многочисленные трубчатые дольки hepatopancreas состоят из однослойного
эпителия. Пространства между отдельными трубочками печени заполнены
соединительной тканью, состоящей преимущественно из Лейдиговских
клеток второго порядка. Вокруг отдельных трубочек располагаются
мускульные волокна.
Среди эпителиальных клеток, образующих стенки печеночных
трубочек, можно различить клетки трех родов (рис. 368). Большая часть
эпителия слагается из цилиндрических клеток, ядро которых
расположено базально. Это — питательные клетки, основная функция которых,
по-видимому, резорбция пищи. Между ними располагаются железистые
клетки. Ядро их обычно занимает срединное положение, цитоплазма
богата гранулами секрета, выделяющегося в просвет долки. Третья
категория— это выделительные клетки, обладающие обычно
большой.вакуолью, наполненной жидким секретом. Ядро оттеснено к базальной
части клетки. Функция этих элементов, видимо, выделительная, так как
356
hepatopancreas, как
показал Кэно, наряду с
пищеварительной
выполняет также и
выделительную функцию.
Задняя кишка.
Внутренняя стенка
задней кишки,
образующая шесть
продольных валиков, выстлана
хитиновой кутикулой,
состоящей из двух
слоев (рис. 369).
Непосредственно под нею
залегает однослойный
цилиндрический
эпителий. За ним следует
соединительнотканный
слой, состоящий почти
1? состоящий
исключительно из Лей
Рис. 367. Astacus astacus. Разрез через часть
покровов (не захвачены кутикула и наружный
эпителий) (из Schneider):
/ — просвет кровеносного сосуда, 2 — кровяная клетка,
3 —intlma, 4 — соединительнотканные Лейдиговские
клетки третьего порядка, образующие стенку
кровеносного сосуда, 5 — лакуны, 6 — Лейдиговские клетки
первого порядка, 7 — Лейдиговские клетки второго порядка
диговсиих клеток
первого порядка. В
соединительной ткани проходят отдельные кровеносные лакуны.
Хорошо развит muscularis задней кишки. Различают три группы
мышечных волокон. Радиальные волокна отходят от кутикулы в толщу
соединительной ткани. Продольные мышечные пучки довольно
многочисленны, они располагаются также'среди Лейдиговских клеток.
Кольцевые мышцы являются периферическими по отношению к радиальным
и продольным и образуют довольно тонкий слой в периферической час-
Рис. 368. Astacus astacus. Гистология
hepatopancreas (из К. Schneider):
/ — выделительная клетка, 2 — железистая
клетка, 3 — питательные клетки
Рис. 369. Astacus astacus.
Поперечный разрез через стенку
задней кишки (из К. Schneider):
1 — кольцевые мускулы, 2 —
кровеносный сосуд, 3 — продольные мускулы,
4 — Лейдиговские клетки первого
порядка, 5 — радиальные мускульные
волокна, 6 — эпителий, 7 — внутренний
слой кутикулы, 8 — наружный слой
кутикулы
357
Рис. 370. Astacus astacus. Поперечный разрез через брюшной ганглий (из
К- Schneider):
i — дорзальная коммиссура, 2 — вентральная коммиссура, 3 — гигантские продольные
волокна, 4 — лакуны, 5 — латеральный нерв, 6 — нервные клетки, 7 — перииеурнум
ти стенки кишки. Кнаружи от кольцевых мышц располагается тонкий
периферический слой соединительной ткани (tunica externa),
непосредственно граничащий с полостью тела. В этом слое проходят мелкие
разветвления сосудов, а также нервные волокна.
Ганглии брюшной цепочки. В ганглиях брюшной нервной
цепочки можно видеть следующие гистологические элементы (рис.370):
нервные клетки, нервные волокна, соединительную ткань (perineurium)
и мелкие кровеносные лакуны.
На поперечном разрезе через ганглий в его дорзаЛьной половине
видны продольные нервные волокна, расположенные в три этажа (дор-
зальный, средний и брюшной нейропиль). В состав спинного пучка
входят два волокна, расположенных справа и слева, обладающих, по
сравнению с другими волокнами, чрезвычайно большим диаметром. Это так
называемые «гигантские» волокна: Между пучками продольных
волокон располагаются две поперечные коммиосуры — дорзальная и
вентральная. С боков от ганглия отходят латеральные нервы к периферии,
состоящие в основной своей массе из нервных волокон.
Нервные клетки располагаются в брюшной половине ганглия. Они
образуют правую и левую группы, в чем сказывается парное
происхождение ганглиев. Нервные клетки различной величины, на некоторых из
них удается видеть отхождение нервного волокна. Последние частью
входят в состав продольных нервных пучков, частью — в состав
латеральных нервов.
358
Соединительнотканные клетки типа Лейдиговских клеток особенно
многочисленны в брюшной части ганглия. Они окружают нервные
клетки и располагаются между нервными волокнами. Кровеносные лакуны
многочисленны. Они располагаются между соединительнотканными
элементами, преимущественно в латеральных частях ганглия.
ОТРЯД MYSIDACEA —МИЗИДЫ
Большинство видов отряда Mysidacea — морские пелагические
формы. Часть их ведет придонный образ жизни. Некоторые виды перешли
к пресноводному образу жизни,.встречаясь в нижнем и среднем
течении Волги, Днепра, Дона ив северных реках и озерах. В пределах СССР
встречается около 60 видов мизид, наиболее богато ими Каспийское
море. Ряд видов живет также в Черном, Азовском и Баренцевом морях.
При прохождении практикума достаточно ограничиться изучением
внешней морфологии представителей этого отряда, отличающегося от
рассмотренных Decapoda рядом характерных особенностей. В реках
добывают мизид тралом _или драгой на песчаных местах с быстрым
течением.
Mysis Latreille
Материал и методы изучения. Одна из наиболее дбступных форм для
изучения— Mysis ocutata (Fabr.) (сем. Mysidae, подсем. Mysinae). Этот вид встречается
в Баренцевом море. Близка к нему пресноводная М. oculata relicta Loven, живущая в
Онежском и Ладожском озерах и в других пресноводных водоемах севера
европейской части СССР. Вместо указанного вида с успехом можно использовать формы,
живущие в Понто-Каспийском бассейне, как, например, Mesomysis kowalewskyi Czern.,
Caspiomysis knipowitschi Sars, Metamysis strauchi Sars, разные виды рода Paramysls
и др. Все они относятся к сем. Mysidae и отличаются друг от друга только
некоторыми деталями строения.
Изучение ведется на живом или консервированном спиртом материале.
Конечности отпрепаровываются препаровальными иглами и рассматриваются под бинокуляром
или при малом увеличении микроскопа.
Внешняя морфология. Длина половозрелых М. oculata 1,5—2 см.
Тело распадается на головогрудь и брюшко (рис. 371). С головой, в
отличие от Decapoda, прочно срастаются лишь два передних (у некоторых
видов Mysidae — три передних) сегмента. Остальные шесть грудных
сегментов ясно отграничены друг от друга. Имеется хорошо
выраженный головогрудный панцирь (сагарах). Хитин его очень мягок. На
спинной стороне он срастается лишь с первыми двумя грудными
сегментами, слившимися с головой. Остальные сегменты груди, хотя и
прикрыты им, но не прирастают к сагарах.. На заднем конце головогрудного
щита со спинной стороны имеется довольно глубокая вырезка, тогда
как боковые части его выдаются назад. В передней трети проходит
поперечная бороздка. Голова несет два больших стебельчатых
сложных глаза, расположенных по бокам от рострума. Кроме того, на
голове расположены длинные первая и вторая пары антенн.
Брюшко длиннее головогруди. Оно слагается из ясно
отграниченных друг от друга шести сегментов и тельсона, последний имеет
форму пластинки и вместе с последней парой брюшных конечностей
359
Рис. 371. My sis oculata relicta. Вид сбоку (А) и со спинной
стороны (Б) (из Я. А. Бирштейна):
1 — первая антенна, 2 — вторая антенна, 3 — грудная часть cephalotho-
rax, несущая двуветвистые конечности, 4 — брюшко, б — уроподы
(уроподами) образует плавник.
Сегменты брюшка обладают
подвижностью относительно друг друга, и все
брюшко в целом может совершать
плавательные движения, быстро
подгибаясь под головогрудь.
Конечности. Первая пара
антенн хорошо развита. Каждая из этих
антенн слагается из основной части,
состоящей из трех члеников и двух
многочленистых жгутовидных ветвей
(рис. 372, А). У самцов, кроме того,
на третьем членике основной части ан-
теннул есть особый придаток
(processus masculinus), несущий многочис-
Рис. 372. Mesomysis inserta. A — первые
антенны; Б —вторые антенны (из Sars):
1 — эндоподнт, 2 — экзоподит
ленные чувствительные волоски (рис. 373). Вторая
пара антенн (рис. 372, Б) обладает трехчлениетым
протоподитом. Эндоиодит — в форме длинного жгута,
слагающегося на- множества колец. Экзоподит имеет
форму вытянутой в длину пластинки с
многочисленными щетинками на конце.
Мандибулы и обе пары максилл (рис. 374)
развиты хорошо. Мандибулы несут трехчленистый
щупик (palpus, рис. 374, Б). Первая пара максилл
.(рис. 374, В) лишена щупика, трехчлениста и имеет
два эндйта с волосками. Вторые максиллы (рис.
374, Г) также трехчленисты и несут двучленистый
щупик (palpus). Все грудные конечности имеют
типичное двуветвистое строение (рис. 375). Из них
первая пара укорочена и представляет собой ногоче-
люсть (рис. 375, А). Остальные семь пар служат
типичными шлавательными ножками (рис. 375, Б, В).
Основные части двух задних шар грудных конечное- вые "антенны сам-
тей несут у самок особые пластинки — оостегиты, в да (из Zimmer):
совокупности образующие выводковую камеру, в ко- /1^5,Г0СнесущийПащё"
торой вынашиваются развивающиеся яйца1. Брюш- тиики, г —три чле-
/ \ ника основной части
ные конечности (тлэоподы) присутствуют в
количестве шести пар. У самцов они сохраняют типичное
двуветвистое строение (рис. 376, А). У самок же в сем. Mysidae
наблюдается частичная их редукция—они превращаются в одноветвистые
нерасчлененные придатки (рис. 376, Б).
Рис. 373. Leptomy-
sis gracilis. Пер-
Рис. 374. Caspiomysis knipowitschi. A — жевательная часть мандибул, Б — щупик
мандибул, В —первая максилла, Г — вторая максилла (из Sars)
1 У представителей сем. Mysidae выводковая камера складывается из двух пар
оостегитов, у других семейств Mysidacea число оостегитов может быть больше,
достигая иногда семи пар.
361
Рис. .375. Mysis ocutata relicta. Три первые грудные конечности. А — ногочелюсть;
Б, В — вторая и третья пары грудных конечностей (из Zimmer no Sars):
1 — эндоподнт, 2 — эпиподнт, 3 — экзоподит
Уроподы в числе одной пары двухветвистые, экзоподит и эндопо-
дит пластинчатой формы и не расчленены.
Характерная особенность всего отряда Mysidacea — наличие на
уроподах органа равновесия — статоциста, который располагается вба-
зальной части эндотюдита. Статоцист представляет собой пузырек,
внутри которого помещается статолит, который сильно 'Преломляет свет,
так как пропитан минеральными солями {рис. 377).
Рис. 376. Mysideis insig-
nis. A — вторая брюш:
ная ножка самца, Б —
брюшная ножка самки
(из Zimmer no Sars):
1 — протоподит, 2 —'
экзоподит, 3 — эндоподит, базаль-
ная часть которого образует
вырост
Рис. 377. Mysis relicta. A —
уроподы со статоцистами; Б—
строение статоциста (из
В. А. Догеля):
/ — последний брюшной ганглий,
2 — статический нерв, 3 — статоцист,
4 — уропод, 5 — чувствительные
клетки, 6 — чувствительный
волосок, 7 — статолит
362
ОТРЯД ISOPODA —РАВНОНОГИЕ РАКИ
Asellus aquaticus L. — Водяной ослик
Материал и методы изучения. Asellus aquaticus, относящийся к сем. Asellidae,
широко распространен, в пресноводных, преимущественно стоячих водоемах. Они
хорошо живут также в аквариумах, при условии наличия разлагающихся органических
растительных остатков, которыми Asellus питаются.
Изучение внешней морфологии и строения конечностей производится на живом
или консервированном 70%-ным спиртом материале. Конечности отпрепаровываются
острыми препаровальными иглами. При препаровке ротовых частей приходится
пользоваться малым увеличением бинокулярной лупы. Из конечностей желательно
приготовить тотальный препарат в глицерин-желатине. Изучение внутренней организации
проводится прежде всего путем вскрытия,;, которое осуществляется препаровальными
иглами при небольшом увеличении (штативная лупа, малое увеличение бинокуляра).
Для вскрытия необходимо иметь несколько экземпляров Asellus. Вскрывать можно
как живые экземпляры, так и выдержанные предварительно сутки в 96%-ном спирте.
Последнее приводит к уплотнению материала и несколько облегчает вскрытие. Кроме
вскрытия желательно изготовить серии поперечных срезов. Фиксация сулемой с
уксусной кислотой, жидкостями Джильсона или Ценкера (с формалином). Заливка по Пё-
терфи. Хорошие результаты дает окраска по Маллори.
Рис. 378. Asellus aquaticus. A — самец со спинной стороны; Б —
самка с брюшной стороны (из Sars):
/ — первые антенны (антеннулы), 2 — вторые антенны, 3 — хватательная
конечность первого свободного грудного сегмента, 4 — грудные ходильные ноги,
5 — уропод, 6 — крышечка, прикрывающая жабры (конечность третьего
брюшного сегмента), 7 — первая пара брюшных конечностей, 8 — выводковая камера
363
Рнс. 379. Asellus aquaticus. Схе- Рис. 380. Asetlus aquattcus. Передняя часть
ма поперечного разреза в области тела. Вид сбоку (из Sars):
свободных грудных сегментов (из / — первая антенна, 2 — вторая антенна, 3 — верхняя
С С Четверикова): губа, 4 — щупик мандибул, 5 —мандибула, 5
—язычок, 7 — первая максилла, 8 — вторая максилла, 9 —
1—-тергнт, Z —место причленения ко- ногочелюсть, 10 — первая грудная конечность, 11 —
нечности, 3—стернит глаз
Внешняя морфология. Тело водяного ослика сплющено 1в дорзовент-
ральном направлении. Семь грудных сегментов, начиная со второго,
свободные, первый же слит с головой (рис. 378). Брюшные сегменты,
за исключением двух передних, сливаются в общий нерасчлененный
отдел. На основании эмбриологических данных принимается, что в
состав брюшка Asellus входит шесть сегментов, как и у других Malacost-
гаса. Между двумя передними брюшными сегментами граница видна
отчетливо, остальные же четыре сегмента полностью слиты и прикрыты
сверху общей хитиновой пластинкой. Голова и первый сегмент груди
имеют прочный наружный хитиновый скелет без каких бы то ни было
следов расчленений (типичный syncephalon). По обе стороны головы
в передней ее части располагается по четыре небольших простых
глазка. Длина каждого из грудных сегментов примерно в 3 раза меньше
ширины. Скелет грудных сегментов слагается из спинной (тергит) и
брюшной (стернит) пластинок (рис. 379).
Конечности. Обе пары антенн (сяжков) одноветвисты. Первая
пара антенн (антеннулы, рис. 378, 380) относительно коротка,
состоит из трех основных более крупных члеников и 11—14 мелких колец.
Вторая пара антенн отличается значительной длиной, почти
равной длине тела, каждая из них слагается из пяти основных
члеников и большого количества мелких колец.
Ротовые конечности располагаются вокруг ротового
отверстия, помещающегося с нижней стороны головы. Спереди оно прикрыто
верхней губой, представляющей собою хитинизированную складку
покровов (рис. 380, 3).
Верхние челюсти, или мандибулы, развиты довольно мощно и
являются наиболее массивной частью среди ротовых частей водяного
ослика. Каждая из мандибул состоит из основной части, двух
жевательных лопастей и четырехчленистого щупика (рис. 380, 5; 381, А). Одна
364
Рис. 381. Asellus aquaticus. Конечности головы. А — верхняя губа и мандибулы; Б—
первая максилла; В— вторая максилла; Г — ногочелюсти (А, Б, В — из Sars, Г — из
С. С. Четверикова):
1 — верхняя губа, 2 — щупик мандибул, S — жевательные лопасти, 4 — мускул — аддуктор мандибул
из жевательных лопастей сильно хитинизирована. Она служит главным
образом для размельчения пищи. В строении мандибул наблюдается
некоторая асимметрия. На левой челюсти имеется дополнительный вырост
верхней лопасти, который отсутствует на правой.
Первая и вторая пары максйлл (рис. 380, 7, 8; 381, Б, В) нерас-
членены. Первая пара снабжена двумя лопастями, вторая — тремя,
несущими многочисленные волоски.
Ногочелюсти (maxillipedes) присутствуют у Asellus в
количестве одной пары (рис. 380, 9; 381,/), в соответствии с числом
грудных сегментов, слившихся с головой (см. рис. 378). Каждая
ногочелюсть складывается из двух основных члеников (coxopodit и basipod.it)
и пятичленикового щупика, который соответствует, по-видимому, эндо-
подиту. Наиболее крупным элементом ногочелюсти является базиподит,
несущий пластинчатую лопасть и многочисленные волоски. Правая и
левая ногочелюсти у живого Asellus соединены между собой особыми
волосками, расположенными по внутреннему краю базиподита. Кроме
указанных частей каждая ногочелюсть несет еще по два эпиподиаль-
ных придатка, из которых наружный развит довольно сильно и
снабжен волосками. Он называется обычно боковой пластинкой.
Грудные ножки (торакоподы). В соответствии с числом
свободных сегментов, у водяного ослика имеется семь пар таракопод. Из
них первая пара отличается от остальных размерами, структурой и
функцией. Она представляет собой хватательную конечность, тогда как
остальные шесть пар являются ходильными ножками. Все
торакоподы — одноветвисты, экзоподит у них полностью редуцирован. Каждая
грудная конечность складывается из семи члеников (рис. 382), так же
как у большинства других групп высших раков,1 но у первой пары,
несущей хватательную функцию, коксоподит сливается с туловищем
(рис. 382, А), так что остается лишь шесть свободных члеников.
Размеры грудных конечностей возрастают спереди назад, так что
седьмая пара торакоподов почти в 2 раза длиннее первой. Три передние
1 Номенклатура члеников та же, что и у речного рака (см. с. 305).
36&
Рис. 382. Asellus aquaticus. Грудные
конечности первой (А) и второй (Б) пары (из.
С. С. Четверикова):
/ — coxopodit, 2 — basipodit, 3 — ischiopodit, 4 —
meropodit, 5 — carpopodit, 6 — propodit, 7 — dacty-
lopodit
Рис. 383. Asellus aquaticus. Схема
поперечного разреза самки в области передних
свободных грудных сегментов (из
С. С. Четверикова):
/ — оостегиты, образующие выводковую камеру
пары грудных ножек в естественном состоянии направлены под
некоторым углом к продольной оси тела животного вперед, четвертая обычно*
ориентирована прямо вниз, три задние пары направлены назад.
Характерно положение отдельных члеников конечностей в отношении
медиальной плоскости животного в естественном состоянии, когда водяной
ослик передвигается на грудных ножках по субстрату, как это
показано на рис. 379. Базиподиты—самые крупные членики конечностей —
направлены внутрь, ишиоподиты занимают почти вертикальное положение,,
меро- и карпоподиты направлены наружу почти параллельно
субстрату, наконец, про- и дактилотюдит расположены вертикально. Благодаря
указанному расположению частей конечностей достигается большая
механическая прочность при движении.
У самок в период размножения у основного членика четырех
передних пар грудных конечностей развиваются довольно большие тонкие
хитиновые пластинки (рис. 383) которые, тесно соприкасаясь друг с
другом, образуют в совокупности выводковую камеру (см. рис. 378, Б).
В ней происходит эмбриональное развитие отложенных самкой ,яиц.
Брюшко у самцов несет шесть пар конечностей, у самок — лишь
пять пар.
Абдоминальные придатки сильно специализированы и
связаны с функциями размножения и дыхания. Самцы отличаются от
самок структурой первой и второй пары абдоминальных ножек, тогда
как остальные пары конечностей устроены у обоих полов одинаково.
Первая пара брюшных конечностей у самцов (рис. 384, А) состоит и»
двух сплющенных в одной плоскости пластинок. Базальная пластинка
несет хитиновые крючочки, при помощи которых обе пластинки соеди-
366
Рис. 384. Asellus aquaticus. Брюшные конечности. А — первая и вторая пары брюшных
ножек самца; Б— первая пара брюшных ножек самки; В — третья пара брюшных
ножек, образующая operculum; Г, Д — четвертая и пятая брюшные ножки, превращен-
. ные в жаберные лепестки (из Sars):
1 — экзоподит, 2 — эндоподит
няются друг с другом. Дистальный членик снабжен многочисленными
перистыми волосками. Вторая пара брюшных конечностей самца
своеобразной формы (рис. 384, А); каждая из них состоит из основного
сплющенного в одной плоскости членика, богато снабженного
мускулатурой, и двух дистальных члеников, независимо друг от друга причле-
няющихся к основному. Внутренний из них несет крючковидный
вырост. Эта пара конечностей играет, по-видимому, существенную роль
во время копуляции при направлении спермы к половому отверстию
самки. Более точно функция их, однако, до сих пор не установлена.
У самок брюшные ножки первой пары состоят только из одного
членика, и каждая из них (рис. 384, Б; 385, 4) представляет собой овальную
пластинку с многочисленными перистыми волосками. Вторая пара у
самок отсутствует.
Третья, четвертая и пятая пары брюшных ножек выполняют
функцию дыхания. Каждая из этих конечностей состоит из короткого
основного членика и двух пластинок, соответствующих, видимо, экзо- и эндо-
подиту (см. рис. 384, В, Г, Д). Жаберные ножки причленяются к
брюшку на небольшом расстоянии друг от друга, благодаря чему отдельные
пластинки прикрывают одна другую. Первая пара жаберных ножек
обладает относительно крупным экзоподитом, довольно сильно хитинизи-
рованным, несущим перистые волоски и поделенным поперечным швом
на два отдела. Он прикрывает все остальные жаберные ножки,
образуя operculum (рис. 385). Вторая и третья пары брюшных ножек
меньше operculum. Стенки их очень тонки, они богато снабжаются кровью
и выполняют функции жабр. Последняя пара брюшных придатков
представляет собой уроподы. Каждая из этих конечностей слагается
из основного довольно длинного членика (рис. 385, 6) и двух нерас-
члененных ветвей. На уроподах в значительном количестве
расположены волоски. На дистальном конце конечностей они образуют целый
пучок. Среди волосков, расположенных на уроподах, некоторые являются
чувствительными и выполняют осязательную функцию.
367
Рис. 385. Asellus aquaticus.
Задний отдел тела самки. Вид
с брюшной стороны (из Sars):
1 — кишечник, . 2 — выделительные
(Ценкеровские) тельца, 3 —
основная часть шестой пары ходильных
йог, 4 — первая пара брюшных
конечностей, 5 — крышечка, образуе-
"" третьего брюш-
6 — уроподы
Мая конечностью
ного сегмента.
Рис. 386. Asellus
aquaticus. Изолированная
пищеварительная система
(из Sars):
/ — жевательный желудок,
2 — средняя кишка, 3 —
печеночные придатки, 4—
задняя кишка
Анатомия на вскрытии. Для вскрытия желательно использовать не
менее четырех экземпляров водяных осликов. На одном экземпляре от-
препаровывается пищеварительная система, на втором и третьем —
половой аппарат (следует взять самца и самку), на четвертом — нервная
система. Органы выделения и кровообращения изучаются на
поперечных срезах. Тергиты осторожно отпрепаровьтваются иглами. Из
изолированных органов желательно изготовить тотальные препараты,
приклеив, их к предметным или покровным стеклам (указания по технике
вскрытия см. на с. 144).
Органы пищеварения. При хорошо проведенном вскрытии
удается выделить весь комплекс органов пищеварения (рис. 386), хотя
очень часто печеночные отростки благодаря нежности тканей
обрываются. Довольно короткий пищевод открывается в желудок, нерезко
поделенный на передний, кардиальный, и задний, пилорический, отделы.
Пищевод и желудок целиком помещаются в головном отделе
животного. Оба эти отдела эктодермального происхождения и имеют
внутреннюю хитиновую выстилку. Средняя кишка представляет прямую, не-
368
сколько суживающуюся к заднему концу трубку. В брюшке она
переходит в короткую эктодермальную заднюю кишку, открывающуюся
наружу анальным отверстием на заднем конце тела.
Две пары длинных печеночных отростков тянутся вдоль всего тела
животного по сторонам средней кишки. Они открываются в желудок с
нижней стороны на границе его со средней кишкой (рис. 386, 3).
После ознакомления с общим строением и топографией отдельных
частей 'пищеварительного аппарата Asellus следует изолировать
желудок и рассмотреть его под бинокулярной лупой или при малом
увеличении микроскопа. Желудок обладает, как и у большинства Malacostra-
са, сложной структурой, так как за счет его стенок развиваются
жевательный и цедильный аппараты. Стенки желудка достаточно прозрачны,
что позволяет рассматривать его не вскрывая. Следует дополнить
рассмотрение желудка in toto путем изучения срезов (см. с. 372).
В кардиальной части желудка располагается два боковых
мускулистых утолщения стенки, покрытых многочисленными хитиновыми
зубчиками (рис. 387, 2). В кардиальной же части желудка с брюшной
стороны имеется продольное утолщение брюшной стенки, снабженное
хитиновыми волосками, образующими первый фильтр (рис. 387, 3).
Далее кзади в пилорической части желудка располагаются два боковых
утолщения, между которыми лежит мощно развитый медиальный
валик (рис. 387, 6). Образующиеся между ними две продольные
бороздки богато снабжены волосками, образующими второй фильтр.
Медиальный валик второго фильтра на заднем своем конце вдается в
полость желудка в виде язычка. Под ним открываются оба отверстия
печеночных отростков. Пища в них может попасть, лишь пройдя через
оба описанные фильтра, которые задерживают все твердые пищевые
частицы. Кроме указанных частей имеются два кольцевидных
утолщения, также снабженных волосками. Пища, не прошедшая сквозь
фильтры, непосредственно поступает в среднюю кишку.
Нервная система. Для того чтобы рассмотреть строение
нервной системы, следует удалить на отдельном экземпляре Asellus все
части пищеварительной и половой систем. Препаровальными иглами
можно изолировать все ганглии брюшной нервной цепочки вместе с
надглоточным ганглием и, приклеив к стеклу, изготовить из них
тотальный препарат. Надглоточный ганглий, или мозг (рис. 388),
водяного ослика значительных размеров. Он несколько вытянут в поперечном
направлении. Хорошо выражены оптические лопасти, от которых
отходят глазные нервы.
Окологлоточные коннективы, огибающие с обеих сторон пищевод,
соединяют мозг с тгодглоточным ганглием, расположенным в задней
части головы. Он представляет собой результат слияния парных
ганглиев сегментов головы, несущих ротовые части, а также сегмента,
несущего ногочелюсти. Ротовые конечности иннервируются от подглоточ-
ного ганглия.
В каждом из свободных грудных сегментов имеется по одному
ганглию (общее число 7), ясно обнаруживающему свою парную
природу (рис. 388). От каждого торакального ганглия отходит пара коннек-
тивов к ганглию следующего сегмента, а также периферические нервы.
369
Рис. 387. Asellus aquaticus.
Жевательный желудок in toto
(из Zimmer no Sochoenichen):
1 — утолщение стенок пищевода в
месте перехода в кардиальный
отдел желудка, 2 — боковые
мускульные валики кардиума с
хитиновыми зубчиками (жевательный
аппарат), 3 — первый фильтр, 4 —
боковые утолщения пнлорической
части, 5 — кольцевые складки
пнлорической части (laminae annulares),
6 — медиальный валик второго (пи-
лорического) фильтра, 7 —
отверстие, ведущее в печеночные
придатки, 8 — пластинчатые выросты
желудка, вдающиеся в среднюю
кншку
Рис. 388. Нервная система
Asellus aquaticus. (из Sars):
1 — мозг, 2 — подглоточный
ганглий, 3 — первый свободный
грудной ганглнй, 4 — последний
свободный грудной ганглий, 5 — ганглий
брюшка, 6 — нервы уроподов
К заднему торакальному ганглию непосредственно примыкает нервная
масса, представляющая результат слияния абдоминальных узлов
нервной цепочки. В ней обычно удается рассмотреть две перетяжки,
подразделяющие ее на три расположенные друг; за другом части. От
брюшного ганглия отходят многочисленные нервы к периферии, иннервиру-
ющие брюшные конечности. Один длинный непарный нерв отходит
непосредственно назад к анальному отверстию, где он распадается
сначала на две, а затем на большее количество мелких веточек. Два
мощных нерва отходят от заднего конца брюшного ганглия к уроподам.
Половая система. Парные семенники (рис. 389)
располагаются в пяти задних торакальных сегментах. Каждый семенник состоит
из трех лопастей, из которых каждая самостоятельно открывается в
семяпровод (vas deferens). Парные семяпроводы образуют в задней тре-
370
Рис. 389. Мужская
половая система Asellus
aquaticus (из Sars):
1 — семенник, 2 —
семяпровод, 3 — половое отверстие
Рис. 390. Женская
половая система Asellus
aquaticus (из Sars):
/ — ovarium, 2 — яйцевод,
3 — половое отверстие
ти небольшие расширения. Они открываются двумя самостоятельными
отверстиями в задней части последнего торакального сегмента, на
границе с брюшком. Дистальная часть семяпроводов снабжена хитиновым
вооружением в форме трубочки, которое следует рассматривать как
penis.
Парные яичники (рис. 390) представляют две почти прямые
трубки одинакового диаметра на всем своем протяжении. Они
располагаются со спинной стороны по сторонам кишечника вдоль всего
торакального отдела, задние их части заходят в abdomen. Примерно на
расстоянии 2/3 от передних концов яичников берут начало короткие яйцеводы,,
которые открываются наружу на брюшной стороне пятого грудного
сегмента. Женские наружные половые отверстия имеют форму щелей.
Ценкеровские органы. Кроме рассмотренных систем
органов при вскрытии обычно удается отчетливо видеть в гаолости тела
особые характерные образования, связанные, как предполагают, с
выделительной функцией- и получившие название Ценкеровских органов. Это
более или менее округлые тела, обычно окрашенные в
желтовато-зеленый тон, располагающиеся по сторонам кишечника в задней половине
груди и в брюшке (см. рис. 385, 2). Высказано (предположение,
физиологически еще недостаточно обоснованное, что Ценкеровские органы
представляют собой органы выделения, функционирующие по типу так
называемых почек накопления.
Изучение срезов. В дополнение к вскрытию следует разобрать
серию поперечных срезов через Asellus, причем изучение нужно начинать
371
с переднего конца, постепенно
переходя к грудному отделу и
брюшку.
ПокрО'вы Asellus
состоят из хитиновой слоистой
кутикулы и подлежащего слоя
гиподермы, 'состоящего из
эпителиальных клеток. В
гиподерме, преимущественно на
спинной стороне, имеются
включения зерен пигмента.
Мускулатура —
типичная поперечнополосатая.
Расположение основных групп
мышц в общих чертах
напоминает таковые речного рака.
Поэтому на описании
мускулатуры водяного ослика мы
специально останавливаться
не будем.
На срезах через голову
следует рассмотреть строение
различных частей желудка,
которые отмечены при
изучении вскрытого Asellus. На
срезах, прошедших через кар-
диальную часть, отчетливо
видны боковые утолщения с хитиновыми зубчиками и передние
медиальный и боковые брюшные валики, щетинки которых образуют первый
фильтр (рис. 391, 1, 2). Далее кзади, в пилоричеокой части, на
поперечном разрезе видны латеральный валик и две бороздки между ними
(рис. 391,4). Эти образования, снабженные хитиновыми волосками,
формирующими второй фильтр, ограничивают две продольно идущие
бороздки, которые в задней части желудка соединяются вместе,
образуя общую полость. От нее берут начало печеночные придатки
(рис. 391, 7). Со спинной стороны задней части пилорического отдела
внутрь желудка вдается складка в виде пластинки, залегающей над
медиальным и латеральным валиками. На срезах через торакальную
часть хорошо видны средняя кишка и печеночные придатки (рис. 392,
В, 15, 21). Стенки последних слагаются из крупных железистых
клеток.
Из элементов кровеносной системы на срезах следует
отыскать главные сосуды и сердце, исходя из общей схемы строения
кровеносной системы Isopoda, изображённой на рис. 393.
На срезах через голову и переднюю часть груди видны передняя
аорта (aorta cephalica, см. рис. 392, 12) и брюшная стернальная
артерия (aorta sternalis), лежащая под нервной цепочкой, а также
латеральные артерии (arteriae laterales). Сердце, лежащее в двух
последних сегментах груди, в передней половине брюшка имеет довольно зна-
Рис. 391. Asellus aquaticus. I—VII—
последовательные срезы через желудок (сравнить с
рис. 387) (из Zimmer no Schoenichen):
/ — утолщение брюшной части карднального отдела,
образующее передний фильтр, 2 — боковые валнкн
карднума, 3— боковые утолщения (валнкн)
пилорического отдела, 4— медиальный валнк второго
фильтра, образующий в задней части язычок
(срезы VI, VII), 5 — хитиновая пластинка, вдающаяся
в начальный отдел средней кишки, 6 — энтодермаль-
ный эпителий средней кишки, 7 — начальная часть
печеночных отростков
372
Рис. 392. Поперечные разрезы самки Asellus aquaticus. A -^ в области надглоточного
ганглия (мозга); Б —- максиллярного сегмента; В— седьмого свободного
торакального сегмента; Г —средней части брюшка (ориг.):
/ — хитиновая кутикула, 2 — эпителий, 3 — начальный отдел пищевода, 4 — мышцы головы 5 —
хитиновая, аподэма, 6 — надглоточный ганглий, 7 ~ брюшная нервная цепочка, 8 — эндоскеле'т
передней части головогруди, 9 — нефрндиальный канал максиллярной железы, 10 — целомическнй мешок
максиллярной железы, // — жевательный желудок, 12 — aorta cephalica, 13 — продольные спинные
мышцы, 14 — янчник, 15 ~ печеночные отростки, 16 — ганглий брюшной нервной цепочки. 17 —
■брюшные продольные мышцы, 18 - пернкарднальная полость, 19 — arteria lateralis, отходящая-от
•сердца, 20 — сердце,21 — средняя кишка, 22 — кровеносные лакуны брюшка, 23 — крышечкГ 24 —
жаберные лепестки (брюшные иожкн), 25 — Ценкеровский орган
373
жы:
г*
11 'T\v
3 4
Рис. 393. Схема кровеносной системы Isopoda (из Zimmer):
■сердце, 2 — aorta cephalica, 3 — arteria lateralis anterior, 4 — arteria sternalis, 5 — arteria
lateralis posterior
чительный диаметр (см. рис. 392, 20) и помещается непосредственно-
над кишечником.
При последовательном просмотре серии срезов удается обнаружить-
также отходящие от него латеральные артерии (см. рис. 392, 19),
несущие кровь в конечности и анастомозирующие со етернальной
артерией. На заднем конце сердце замкнуто слепо. Из лакунарной системы:
обычно бывают видны латеральные лакуны брюшка, богато
снабженные кровью (см. рис. 392, Г, 22).
На срезах через abdomen следует рассмотреть гистологическое-
строение жаберных ножек (см. рис. 392, 24), образующих ряд,
лежащих друг под другом пластинчатых жабр. Хитиновая кутикула,
здесь тонка и нежна; в толще жабр, слагающихся из клеток
эпителиального характера, имеются многочисленные лакуны, наполненные
гемолимфой.
Органы выделения,
представленные маисиллярными железами
(рис. 394), располагаются в области
головы, где их следует отыскать на
срезах >. Каждая железа состоит из целоми-
чесгого мешочка, нефридиального
канала !и дистального отдела — мочевого
канальца, открывающегося наружу у
основания мансилл (см. рис. 392, 9, 10)..
Нефридиальные канальцы образуют
петли, которые (и видны на срезах
перерезанными в нескольких местах (см.
рис. 392, 9). . . .
Из отделав нервной системы
на срезах через переднюю половину го-
лавы следует отыскать надглоточный
ганглий, или мозг (см. рис. 392, 6)„
Рис. 394. Oniscus sp.
Схематизированный разрез через
максиллярную железу (из
Zimmer no Ter-Poghossians):
/ — целомический мешочек, 2 —
замыкающие клетки, 3 — нефридиаль-
ный канал, 4—мочевой канал, 5 —
наружное отверстие
1 При вскрытии максиллярные железы
обычно рассмотреть не удается. Поэтому изучение
органов выделения Asellus осуществляется на
срезах.
374
В нем видны крупные нервные клетки, нервные волокна. При
рассмотрении ганглиев и коннективов брюшной нервной цепочки отметить
парный характер того и другого, а также наличие нервных клеток только
в ганглиях. Коннективы состоят исключительно из нервных волокон.
Половые железы хорошо видны на срезах (см. рис. 392, 14),
они лежат со спинной стороны по сторонам кишечника. В семенниках
отыскать разные стадии сперматогенеза. В яичниках отметить разные
стадии роста ооцитов. Крупные ооциты Asellus обладают большим
количеством желтка.
ОТРЯД AMPHIPODA—БОК.ОПЛАВЫ
Gammarus pulex (L.)— Бокоплав-блоха
Материал и методы изучения. Gammarus pulex, относящийся к подотряду Gam-
maridea, сем. Gammaridae, часто встречается в пресноводных — текучих и стоячих
водоемах, Gammarus не живет в загрязненных прудах с заиленным дном. Это
типичное придонное животное. Для изучения кроме G. pulex с успехом может быть
использован морской вид—G. locusta, встречающийся в больших количествах в
прибрежной зоне почти во всех морях, омывающих СССР. Строение указанных видов сходно.
Внешняя морфология изучается на живом или консервированном спиртом
материале. Конечности отпрепаровываются тонкими иглами под бинокулярной лупой.
Изготовляется тотальный препарат небольшого экземпляра бокоплава. Из конечностей
также желательно изготовить тотальный препарат в глицерин-желатине.
Внутренняя анатомия изучается путем вскрытия свежезахлороформированных
животных.
На вскрытом животном не удается рассмотреть выделительную и кровеносную
■систему. Поэтому в качестве дополнения желательно изготовить поперечные разрезы.
Техника фиксации и окраски та же, что для Asellus (с. 363). Ввиду того что боко-
плавы довольно сильно дугообразно изогнуты, их спина выпукла, а брюшная
поверхность вогнута, продольная ось их тела представляет дугу. Это затрудняет
ориентировку при изготовлении поперечной серии срезов. Можно рекомендовать перед
заливкой разрезать бокоплава поперек на три части, и каждую из них резать
самостоятельно. Таким путем можно получить более или менее правильно ориентированную
поперечную серию.
Внешняя морфология. Тело Gammarus довольно сильно сплющено
с боков и изогнуто дугообразно на брюшную сторону. Поэтому
название бокоплав вполне соответствует действительному положению
животного, когда оно передвигается но субстрату. Длина половозрелых
экземпляров 1—1,5 см, причем самцы крупнее самок.
Голова (syncephalon), подобно таковой Asellus, сливается с
первым сегментом груди. Остальные семь грудных сегментов свободно
сочленяются между собой (рис. 395).
Брюшко слагается из шести свободно сочлененных между собой
сегментов, снабженных конечностями, и не резко отграничено от груди.
К шестому абдоминальному сегменту причленяется небольшой двураз-
дельный тельсон. По сторонам передней части головы, у основания
первой пары антенн расположены глаза (рис. 395, 8). По своему
строению это сложные глаза, но наружная кутикула их не фасетирована
(см. рис. 406, 7). Четыре передних торакальных сегмента несут сильно
развитые боковые пластинки — этшмеры. На остальных грудных и трех
первых брюшных сегментах эпимеры развиты слабее. В пределах брюш-
375
7
Рис. 395. Самка Gammarus pulex (из Sars):
1 — первые антенны, 2 — вторые антенны, 3'— эпнмеры первых грудных
сегментов, 4 — развивающиеся яйца, 5 — задний отдел брюшка (uroso-
ma), 6 — передний отдел брюшка (metasoma), 7 —семь свободных
торакальных сегментов, 8 — глаз
ка различают два отдела. Передние три сегмента напоминают сегменты
груди, несут плавательные двуветвистые конечности. Этот отдел
брюшка называется метазома. Задние, три абдоминальные сегмента
значительно меньше, чем в метазоме, не несут эпимеров, конечности их иной
структуры. Этот отдел брюшка называется урозома.
Конечности. Первая пара антенн (рис. 395, 1) длиннее, чем
вторая пара. Основная часть ее состоит из трех члеников, к которым
причленяется длинная, состоящая из многих колец дистальная часть.
Кроме нее имеется очень небольшая трехчленистая вторая ветвь.
Вторая пара антенн (рис. 395, 2) несколько короче первой, одноветвиста.
На члениках ее диетальной части помещаются особые булавовидно
утолщенные образования—calceoli. Функция их неясна.
Среди ротовых конечностей наиболее массивные верхние челюсти,,
или мандибулы (рис. 396, А). Они состоят из основной части и двух
лопастей. Одна из лопастей сильно хитинизирована. Она служит
жевательной лопастью, приспособленной для измельчения пищи. Кроме
того, каждая мандибула несет трехчленистый щупик (palpus). Первая
пара нижних челюстей, или максилл (рис. 396, Б), имеет три лопасти,
несущие на своем дистально'м конце хитиновые зубчики и волоски.- И*
них наружная лопасть, видимо, соответствует щупику. Вторая пара
нижних челюстей (рис. 396, В) слагается из основной части и двух
лопастей, несуших многочисленные щетинки. Ногочелюстей (maxillipedes)
у Gammarus лишь одна пара. Она соответствует тому грудному
сегменту, который сливается с головой. Каждая ногочелюсть состоит из
шести члеников. Первый и второй проксимальные членики несут жева-
376
Рис. 396. Ротовые конечности Gammarus pulex. A — мандибулы; Б — первая
максилла; В — вторая максилла; Г — ногочелюсти (из Sars):
1 — щупик мандибул, 2 — жевательные лопастн
тельные лопасти. Первые членики правой и левой ногочелюстей
срастаются между собой (рис. 396, Г)1
Семь пар конечностей, причленяющиеся к свободным торакальным
сегментам, одноветвистые (экзоподит полностью редуцирован) и
слагаются из семи члеников, как и у большинства других Malacostraca.
Коксоподит тесно срастается с боковыми пластинками-эпимерами и с
туловищем. Две первые пары грудных конечностей самые маленькие
{рис. 395, 397, А). В области ишиоподита и карпоподита они изогнуты
под острым углом вперед. Конечности эти служат главным образом не
для движения, а для удержания пищи, поэтому могут быть названы
гнатоподами. Их предпоследний членик (проподит) сильно расширен,
последний же членик (дактилоподит) когтевиден и действием
мускулатуры может пригибаться к проподиту. Грудные конечности (торако-
поды) с третьего по седьмой построены более или менее сходно. Они
постепенно увеличиваются в размерах по направлению спереди назад
(см. рис. 395) и являются двигательными конечностями. Задние три
пары их несколько отогнуты назад.
Основной членик всех торакоподов, кроме первой пары (гнатопо-
дов), несет по одной жаберной пластинке (лепестку), направленной
внутрь и прикрытой с боков эпимерами (рис. 397). Общее число
жаберных лепестков равняется таким образом шести парам. Каждый из них
представляет собой тонкостенную овальную пластиночку, внутренние
лакуны которой богато снабжаются кровью. У самок, кроме того, кок-
соподиты второй, третьей, четвертой и пятой пары торакоподов несут
особые тонкие хитиновые пластинки, прикрепляющиеся над
жаберными лепестками и заворачивающиеся внутрь. В совокупности эти
пластинки, называемые оостегитами, образуют выводковую камеру, в
которой самка вынашивает развивающиеся яйца (рис. 397, Б; 404, 9).
Конечности переднего отдела брюшка (metasoma) в количестве
трех пар представляют собой типичные двуветвистые ножки (рис.398).
377
Рис. 397. Грудные конечности Gammarus pulex. A — первой пары; Б — второй пары;
В—третьей пары; Г — седьмой пары (из Sars):
1 — эпнмер, 2— оостегнт, 3—жаберная пластинка
Каждая из них состоит из основной (нерасчлененной) части —■ прототю-
дита и двух многочленистых ветвей — экзоподита и эндоподита,
несущих многочисленные перистые волоски. Конечности эти являются
плавательными. Кроме того, их постоянное движение -способствует
циркуляции воды, омывающей жаберные лепестки. Четвертая и пятая пары
брюшных ножек (уроподы), причленяющиеся к двум передним
сегментам заднего отдела брюшка (urosoma), также двуветвистые (рис. 395,
398). В отличие от плавательных ножек их экзо- и эндоподит не
расчленены и вместо волосков несут лишь короткие хитиновые шип-ики.
Уроподы являются прыгательны-
ми конечностями. Их резкие
движения вызывают перемещения
животного, напоминающее
скачки. Они направлены назад, почти
параллельно продольной оси
брюшка. Наконец, последняя
пара брюшных конечностей
занимает терминальное положение,
тогда как тельсон смещен на
спинную сторону. Эти конечности
также двуветвистые, причем их не-
расчлененные экзоподит и
эндоподит снабжены
многочисленными щетинками.
Тельсон (рис. 398) глубокой
вырезкой разделен на две
лопасти, которые несут
немногочисленные волоски.
Рис. 398. Брюшные конечности Gamma- Анатомия на вскрытии. Для
rus pulex А-плавательная брюшная вскрытия следует взять нескОль-
ножка; Б — задний отдел брюшка с v ,- тг
уроподами и тельсоном (из Sars): и0 экземпляров бокоплавов. Ко-
1 — уроподы, 2-тельсон нечности предварительно удаля-
378
Рис. 399. Схема организации Gammarus. I — голова, // — семь свободных
торакальных сегментов, /// — metasoma, IV—urosoma (из Cussans, с изменениями):
1 — глаз, 2— первая антенна, 3— вторая антенна, 4— мозг, 5 — антеннальная железа, 6 — ногоче-
люсть, 7 — гнатоподы, 8 — брюшная нервная цепочка, 9 — торакоподы, 10 — брюшные конечности,
// — уроподы, 12 — тельсон, 13 — задняя кншка, 14 — задние слепые выросты кишечника, 15 —
средняя кишка, 16 — aorta posterior, 17 — передние печеночные придатки, 18 — teste3, 19 — сердце, 20 —
передний слепой непарный вырост кишечника, 21 — aorta anterior (cephalica)
тотся. Общий ход вскрытия примерно такой же, как это указано для
водяного ослика (с. 368). Отдельные системы органов при этом
изолируются, из них желательно изготовить тотальные препараты.
Осторожно действуя препаровальными иглами, удаляя
последовательно кольца наружного скелета, следует попытаться прежде всего
выделить, не нарушая цельности, пищеварительную систему Gammarus.
•Общее расположение органов представлено на рис. 399.
Пищеварительная система (рис. 400). Ротовое отверстие
ведет в короткий пищевод, открывающийся в желудок (эктодермаль-
ный), снабженный, как и у других Malacostraca, жевательным и
цедильным аппаратом. За желудком следует средняя кишка (энтодер-
мальная), представляющая собой прямую трубку, тянущуюся до
третьего брюшного сегмента. Средняя кишка переходит в заднюю,
выстланную хитиновой кутикулой и открывающуюся наружу на заднем конце
-брюшка. В задний конец желудка открываются две пары длинных
цилиндрических печеночных отростков, тянущихся через весь грудной
отдел вплоть до середины брюшка. На спинной стороне передней части
средней кишки имеется короткий слепой вырост, проходящий над
задней частью желудка. На границе между средней и задней кишкой в
нее открываются с дорзальной стороны еще две слепые кишки,
продолжающиеся вперед примерно до шестого грудного сегмента. .
Желудок следует рассмотреть, изолировав его отдельно/при
малом увеличении микроскопа. Строение напоминает таковое у Asellus
379
Рис. 400. Gammarus pulex.
Изолированная пищеварительная
система (из Sars):
1 — непарный дорзальиый вырост
передней части средней кншки, 2 —
передние печеночные придатки, 3 —
средняя кишка, 4 — задние печеночные
придатки, 5 — rectum, 6 — яичник
380
(с. 369). В вытянутом в длину желудке
можно различить не (резко
отграниченную переднюю кардиальную и заднюю
пилорическую части (рис. 401). На
границе между пищеводом и желудком
справа и слева располагаются мускулистые
утолщения стенок, несущие
многочисленные щетинки и волоски. Этот аппарат
препятствует обратному попаданию
пищи из желудка в пищевод. Далее в
желудке располагаются еще две пары
хитиновых пластинок, несущих по краю
волоски. Передняя из них лежит в карди-
альиой части желудка, задняя заходит в
пилоричеокий отдел.
На брюшной стенке пилорической
части помещается углубление, на
дне которого располагается валик
(рис. 401, 2). По сторонам его
образуются две довольно глубокие бороздки. Со
стороны боковых стенок они ограничены
мускулистыми утолщениями. В
совокупности эти образования, снабженные
многочисленными волосками (рис. 401, 3),
образуют фильтр. В своей задней части
медиальный валик продолжается в
язычок, снабженный многочисленными
волосками и свободно вдающийся в
полость заднего отдела желудка. В глубине
задней части 'боковых бороздок берут
начало печеночные трубки. Справа и
слева они начинаются отверстием,
ведущим в одну трубку, вскоре, однако,
распадающуюся на две ветви. Таким
образом, (пища, так же как и у водяного
ослика, может попасть в печеночные
трубки, функционально и
морфологически соответствующие hep atop ancreas
речного рака, только пройдя через
фильтр. Последний пропускает жидкую
и очень мелко размельченную пищу.
Нервная система. Удалив при
вскрытии 'на отдельном экземпляре
Gammarus пищеварительную и половую
системы, следует выделить брюшную
нервную цепочку и иадглоточный ганглий и,
приклеив к покровному или
предметному стеклу (с. 144), изготовить
тотальный препарат.
Рис. 401. Вскрытый
желудок Gammarus pulex
(из Sars):
/,— утолщение стенки
переднего отдела кардиальной
части желудка, 2 —
мускулистые валики пилорической
части, 3 — язычок
Центральная нервная система бокоплава
состоит из большего числа ганглиев, чем у
водяного ослика, что связано с отсутствием слияния
сегментов брюшка. Общее число ганглиев
(включая моэг) равняется 13 (рис. 402).
Надглоточный ганглий, или мозг, лежащий
в передней части головы, слагается из двух
ясно различимых парных лопастей. С нижней
стороны от него отходят нервы к первой и второй
паре антенн, а с боков — короткие оптические
нервы к глазам.
Короткие окологлоточные коннективы,
огибающие пищевод, соединяют мозг с подглоточ-
ньгм ганглием. Последний состоит из двух
частей. Передний отдел его иннервирует
-мандибулы и. обе пары максилл, от заднего отходят
нервы к ногочелюстям. Таким образом, задний
отдел подглоточного ганглия принадлежит, строго
говоря, первому трудному сегменту,
слившемуся с головой. Каждый из свободных торакальных
сегментов несет парный ганглий брюшной
цепочки, соединяющийся 'С последующим парными
коннективами.
В переднем отделе брюшка (metasoma) каждый сегмент имеет
парный ганглий. Все ганглии заднего отдела брюшка (urosoma)
сливаются вместе в общую нервную массу. Как от самих ганглиев
нервной "цепочки, так и от связывающих их коннективов, отходят нервы к
конечностям, мускулатуре и внутренним органам. ■ Нервы, отходящие
от коннективов, связаны с нервными клетками, входящими в состав
ганглиев, так как коннективы нервных клеток не несут. От заднего
ганглия брюшной цепочки отходят нервы, иннервирующие конечности,
мускулатуру урозомы и заднюю кишку. Из элементов симпатической
нервной системы удается обнаружить парные нервы, отходящие от
заднего конца окологлоточных коннективов и иннервирующие желудок:
Половая система. Мужские половые органы (семенники)
парные, лежат в грудных сегментах по сторонам и несколько выше
средней кишки (рис 399, 18; 403, А). От заднего конца каждого
семенника отходит короткий семяпровод (vas deferens), открывающийся
у основания последней пары грудных ножек. Передняя часть каждого
семенника состоит из клеток, находящихся на разных стадиях
сперматогенеза. В задней его части помещаются преимущественно зрелые
спермин.
Женские половые органы (яичники) также парные, располагаются
в грудных сегментах по сторонам средней кишки (рис. 403, В; 400, 6)
и несколько дорзально от нее. У живого животного яичники
выделяются своей темной окраской. Каждый яичник состоит из немногих рядов
довольно крупных яйцевых клеток-ооцитов. От каждого яичника,
отступя на Уз от его заднего конца, отходит короткий яйцевод,
открывающийся наружу у основания пятой пары грудных конечностей.
381
Рис. 403. Половые органы Gam-
marus pulex. A — мужские; Б ■—
женские (из Sars):
/ — testes, 2— vas deferens, 3
—ovarium, 4 — яйцевод
Рис. 402. Нервная система Gam-
marus pulex (из Sars):
/ — мозг, 2 — подглоточный гаиглий,
3 — ганглии свободных грудных
сегментов, 4—гаиглий сегментов метазо-
мы, 5 —ганглий, образовавшийся в
результате слияния ганглиев урозомы
Изучение срезов. На
поперечной серии срезов,
которую следует
'Просмотреть начиная с головного
отдела до брюшка,
необходимо разобраться в
топографии органов,
руководствуясь общей схемой
организации Gammarus,
изображенной на рис.
399, 404. Главное
внимание надо обратить на те
системы органов,
которые не удается
обнаружить при вскрытии.
Покровы
Gammarus 'СОСТОЯТ ИЗ ХИТИНОВОЙ
кутикулы и подлежащих
шподёр'мальных
эпителиальных клеток.
На срезах через
головной отдел следует
рассмотреть строение
желудка, основные части
хитинового вооружения
которого отмечены при
изучении его in toto.
Отыскать место отхожде-
ния от желудка
печеночных трубок и дорзальной
передней слепой кишки.
На срезах через
головной отдел у основания
второй пары антенн
рассмотреть расположенную
здесь парную
выделительную антенналь-
ную железу (рис.
405, 406, 5). Она состоит
из целомическопо
мешочка, длинного извитого не-
фридиального канальца и
выводного канала. Цело-
мический мешочек и
выводной канал лежат у
самого основания антенн,
нефридиальный же
каналец помещается в
полости головы. На границе
между целомическим ]мешочком и нефриди-
альным канальцем расположены особые
замыкающие крупные клетки и кольцевые
мышцы, образующие сфинктер. Нефридиальный
каналец образует многочисленные извивы,
которые на срезах оказываются перерезанными
в различных направлениях. Стенки целомиче-
ского мешочка состоят из одного слоя
плоских эпителиальных клеток, стенки же нефри-
диального канальца — из более высоких
эпителиальных 'клеток железистого характера,
расположенных также в один слой.
На срезах видны .некоторые части
кровеносной системы. У Gammarus сердце
(см. рис. 399, 19) представляет длинный
сосуд, тянущийся ото опинной стороне с
середины второго до середины шестого
торакального юегмента и несущий три пары остий. От
сердца к голове кровь направляется по
передней аорте. Кроме того, от переднего конца
сердца отходит пара головных артерий. От
сердца назад отходит задняя аорта (см. рис.
399, 16), тянущаяся по спинной стороне
груди и брюшка над кишечником. Из сосудов
кровь собирается в обширный брюшной синус,
откуда направляется к жабрам. Из жабр она
по каналам попадает в перикардиальный
синус, окружающий сердце, откуда через остии проникает в сердце. На
срезах видны сердце и отходящие от него сосуды (см. рис. 404, 1).
Сердце оказывается лежащим в полости, представляющей собой
перикардиальный синус, обычно наполненный свернувшейся гемолимфой. На
срезах хорошо виден мозг и элементы брюшной нервной цепочки.
Следует рассмотреть гистологическое строение глаза. Последний
состоит из большого количества омматидиев, из которых каждый
слагается из типичных элементов — хрустального конуса, ретинальных и
пигментных клеток (рис. 406, 7). Снаружи глаз прикрыт прозрачной
хитиновой кутикулой, не поделенной на фасетки. Таким образом,
хитиновая cornea у каждого отдельного омматидия отсутствует.
Кроме указанных органов, на срезах хорошо видны гонады и.
мускулатура.
Метаморфоз и личинки высших раков (Malacostraca)
Онтогенез во многих отрядах высших раков протекает без превращений, и
личиночные стадии отсутствуют. Но у большинства представителей отрядов Decapoda и у
всех Euphausiacea и Stomatopoda имеется ясно выраженный метаморфоз,
сопровождающийся сменой ряда личиночных форм.
Наибольшей сложности достигает метаморфоз у морских десятиногих раков,,
большинство личинок которых ведет планктонный образ жизни. Для изучения
личиночных форм Decapoda следует воспользоваться пробами планктона из Черного или
385
Рис. 404. Gammarus pulex.
Схема поперечного разреза
самки в области третьего-
грудного сегмента (и»
Sars):
/ — сердце, 2 — яичник, 3 —
средняя кишка, 4 — передние
печеночные придатки, 5 — брюшная;
нервная цепочка, 6 — эпимеры,
7 — грудная конечность, 8 —
жаберная пластинка, 9 — оостегиг
Рис. 405. Gamrriarus pulex. A — продольный разрез ан-
теднальной железы; Б — конечный отдел антеннальной
железы (из Reibisch no Vejdovsky):
/ — целомический мешочек, 2 — сфинктер, замыкающий вход в
нефрндиальиый канал, 3 — замыкательные клетки, 4 — иефриди-
альиый каиал, 5 —кутикула, 6 — гиподермальный эпителий,
7 — мезодермальные клетки иефридиальиого канала
Баренцева морей, взятыми в различное время года, преимущественно летом. Stomato-
poda в наших морях, за исключением Японского моря, почти не встречаются, так что
практически приходится иметь дело почти исключительно с личинками Decapoda.
Материал и методы изучения. Фиксированные 4%-ным формалином или 70%-ным
спиртом пробы морского планктона просматриваются под бинокулярной лупой.
Попадающиеся, наряду с другими организмами, личинки высших раков, отбираются
пипеткой. Изучение их ведется в спирту, воде или же предварительно изготовляются
тотальные препараты в глицерин-желатине.. При работе на морских биологических
станциях желательно использовать свежие пробы планктона с живыми объектами.
Пробы берутся обычным планктонным сачком с мелкими или средними
номерами мельничного газа.
Личинки Decapoda, относящихся к различным семействам, отличаются друг от
друга некоторыми деталями строения. Однако определение видов Decapoda по
личинкам представляет большие затруднения и почти еще не разработано. Поэтому при
изучении метаморфоза Decapoda вполне достаточно ограничиться рассмотрением
наиболее характерных личинок, не определяя ближе их видовую принадлежность. В
соответствии с этим описание личинок дается нами ниже: по фазам метаморфоза.
Нужно иметь в виду,' что у отдельного вида никогда не бывает всех возможных для
Decapoda личиночных фаз.
Науплиус. Типичный науплиус, соответствующий таковому Copepoda и
некоторых других раков, у Decapoda встречается редко. У большинства десятиногих
раков личинка выходит из яйца на более поздних стадиях метаморфоза. Тем не
менее некоторые креветки, относящиеся к сем. Peneidae, начинают свой метаморфоз со
стадии науплиуса. Последний (рис. 407, А) имеет типичное для этой стадии
строение — три пары конечностей (две пары антенн и мандибулы) и нерасчлененное тело.
Конечности двуветвисты и несут длинные плавательные щетинки. Анальное отверстие
отсутствует. Науплиус в результате линьки превращается в метанауплиуса, который
384
Рис. 406. Gammarus pulex. Поперечный разрез в области головы (ориг.):
/ — кутикула, 2 — гиподерма, 3 — полость тела, 4 — мозг, 5 — перерезанные петли нефри-
диального канала антеннальной железы, 6 — хрустальные конусы глаза, 7 — ретинальиые
клетки глаза, 8 — мышцы
отличается от только что рассмотренной стадии зачатками еще трех или четырех пар
конечностей, следующих за мандибулами (обе максиллы и 1—2 ногочелюсти).
Протозоея (Protozoea). Тело этой личиночной стадии распадается на
головогрудь и брюшко (рис. 407,5). Нерасчлененная головогрудь покрыта тонким
панцирем и имеет два сложных глаза. Имеются обе пары антенн, мандибулы, максиллы и
две.пары ногочелюстей. Все эти конечности, кроме А1 и мандибул, двуветвистые.
Длинное брюшко на конце раздвоено, и обе его ветви несут плавательные щетинки.
Брюшко или совсем нерасчленено, или сегментация его уже намечена, но плэоподы
отсутствуют. У некоторых Decapoda имеются две стадии Protozoea, отличающиеся
друг от друга размерами и степенью расчлененности брюшка.
Зоеа (Zoea) представляет собой наиболее типичную для Decapoda личинку.
Очень многие десятиногие раки выходят из яйца именно на этой стадии. Зоеа
отличается от протозоеа большей степенью расчлененности тела и большим числом
конечностей (рис. 408, 409).
Головогрудь несет сложные глаза на стебельках. Сегментация грудного отдела
намечена, грудные конечности заложены, но еще недоразвиты. Брюшко
сегментировано, но несет лишь одну (заднюю) пару хорошо развитых конечностей. Зоеа крабов
13—2
385
Рис. 409. Zoea краба Maja. Рис. 410. Мизидная личинка Pandalus (из'
Обозначения см. на рис. 408 Giesbrecht no Sars):
(ИЗ Giesbrecht ПО Claus) / — первые антенны, 2 —вторые антенны, 3 —
грудные конечности, 4 — брюшные конечности, 5 — уро-
поды, 6 — тельсои
Рис. 411. Phyllosoma Palinurus (из Gies- Рис. 412. Megalopa краба Portunus
brecht по Claus): (из Giesbrecht no Claus):
/ — первые аитеины, 2 — вторые антенны, 3 — груд- / — вторые антенны, 2 — грудные коиечно-
иые конечности, 4 — abdomen сти, 3 — сегменты брюшка с конечностями
(Brachyura) характеризуется наличием на головогруди длинного щипа (рйс. 409).
Мизидная личинка (рис. 410) обладает хорошо развитым головогрудным щитом
с рострумом. Грудные конечности двуветвистые. Брюшные ножки — плэоподы, также
имеются. Сегментация брюшка выражена вполне ясно, отдельные сегменты подвижно
сочленены друг с другом. На мизидной стадии личинки Decapoda претерпевают
обычно несколько линек. При этом происходит увеличение размеров. У большинства
Brachyura и Anomura личинка, соответствующая мизидной, обладает не двуветвистыми
торакоподами, а состоящими из одного ряда члеников. Подобная личиночная стадия
называется метазоеа (Metazoea).
Phyllosoma — своеобразное видоизменение мизидной стадии, свойственное сем.
Scyllaridea (рис. 411). Она характеризуется прозрачным листовидным телом, длинными
двуветвистыми торакоподами и чрезвычайно коротким, сегментированным брюшком.
- Megalopa — еще одна особая личиночная форма для крабов (рис. 412). Эта
личинка ведет придонный образ жизни. Ее головогрудиой отдел и конечности в общем
соответствуют тому, что имеется и у взрослого животного. Но в отличие от
последнего брюшко вытянуто в длину, явственно сегментировано и несет конечности.
13* *
Подтип Tracheata — Трахейные
КЛАСС MYRIAPODA-МНОГОНОЖКИ
Основным объектом может служить камнелаз Lithobius из
подкласса Chilopoda. Виды этого рода, в частности L. forficatus L.,
широко распространены в пределах СССР. Однако, ввиду того, что Diplopo-
da отличаются рядом существенных особенностей, совершенно
необходимо знакомство с организацией и этой группы. Наиболее удобны для
этой цели кивсяки из семейства Julidae, например Schizophyllum sabu-
losum L. Ниже приводится описание внешней морфологии этой формы.
Разбор внутренней организации опущен, так как методы вскрытия не
разработаны, а изготовление срезов представляет большие трудности.
Желательно также ознакомиться с организацией представителей
подклассов Symphyla и Pauropoda,
ПОДКЛАСС CHILOPODA—ГУБОНОГИЕ
ОТРЯД LITHOBIOMORPHA
Lithobius forficatus L. — Камнелаз, или Костянка
Материал и методы исследования. Lithobius forficatus L. (ceM.Lithobiidae)
принадлежит к самым обычным представителям наземной фауны беспозвоночных
европейской части СССР. Встречается с ранней весны до поздней осени под камнями,,
лежачими стволами деревьев, опавшими листьями, под корой старых пней и т. п.
Эта очень подвижная, быстро бегающая многоножка питается разными мелкими
насекомыми, паукообразными и червями, за которыми охотится ночью. Днем камнела»
прячется в темные укромные уголки. Зиму проводит в почве. В неволе Lithobius
выживают хорошо, если содержатся в сосуде с влажным мхом или гнилыми
древесными листьями. В качестве корма рекомендуются мухи или мелкие кусочки свежего или
даже вареного мяса.
Вместо L. forficatus для целей практикума можно взять любой другой
достаточно крупный вид этого рода.
Перед изучением организации многоножек убивают в парах эфира. Для
изучения хитиновых частей животные вывариваются в растворе едкого кали, который
растворяет внутренние органы. Если нужно при этом сохранить трахеи, стенки которых
разрушаются в концентрированном растворе, то Животное оставляют на некоторое
время в слабом растворе едкого кали. В этом случае, для лучшего проникновения
реактива внутрь тела, полезно, сделав один-два надреза на боках тела, лёгким,
давлением выдавить через них жировое тело и содержимое кишечника, которое не всегда
просветляется едким кали. Изготовленные таким образом тотальные препараты
отчетливы и пригодны для изучения при больших увеличениях.
Благодаря сравнительно большим размерам тела камнелаз вполне пригоден для
анатомического вскрытия.
388
Внешняя морфология. Тело
Lithobius состоит из крупной, ясно
обособленной головы и
сегментированного туловища, в состав
которого входят 19 сегментов (рис. 413).
Как у всех Chilopoda и сходно с
Insecta, в образовании головы
принимают участие: асгоп,
отвечающий prostomium аннелид, и четыре
сегмента: интеркалярный, мандибу-
лярный и два максиллярных1.
Сегментация туловища отличается
сравнительно большой гомоном-
ностью, хотя три последних
сегмента меньше других и имеют особое
строение. Все головные и три задние
сегменты Chilopoda по развитию
мезодермы рассматриваются как
ларвальные. Туловищные сегменты
(I—XVI) являются постларвальны-
ми2.
Каждый туловищный сегмент,
кроме двух последних, снабжен
парой конечностей. Длина тела
достигает 2,5—3 см. Хитиновые покровы
блестящие, коричневато-бурого
цвета.
Голова. Для изучения
внешней морфологии головы и ее
придатков следует, отделив голову от
туловища, выварить ее в растворе
едкого кали и заключить в
канадский бальзам. Необходимо, кроме
того, выварив голову от другого
экземпляра, отпрепарировать иглами
ротовые части и приготовить
тотальные препараты челюстей и
головной капсулы в
глицерин-желатине.
1 Некоторые авторы считают в составе
головы еще антеннальный и преантенналь-
ный сегменты. По-видимому, целомические
мешки, появляющиеся во время развития
впереди интеркалярного сегмента,
представляют собой продукт вторичного
разделения целома головы зародыша и не
отвечают особым головным сегментам.
2 Относительно ларвальных и постлар-
вальных сегментов у губоногих см.
Иванов П. П. (1940).
Рис. 413. Lithobius forficatus. Вид со
спинной стороны (ориг.):
/ — голова, 2 — глазное поле с глазками, 3 —
полоска интерсегмеитарной кожи, 4 — йога
XVI сегмента, 5—анальное отверстие, 6 —
стигмы, 7 — мягкая кутикула плевры, 8— нога
VIII сегмента, 9 — нога II сегмента, 10 —ко-
гочелюсть, 11 — аитениа, I—XIX — тергиты
сегментов туловища
389
Рис. 414. Lithobius forficatus. Головная
капсула после удаления челюстей. Вид
снизу (из Attems):
/ — наличник, 2 — первый членик антенны, 3 —
орган Темешвари, 4 — глазки, 5—
дополнительная пластинка, 6 — опорная пластинка,
7 — верхняя губа, 8 — срединная вырезка
верхней губы с пятью зубцами, 9 — передний
бугорок наличника
Рис. 415. Lithobius forficatus. Голова и
I туловищный сегмент с брюшной стороны
(из Roseler и Lamprecht):
/ — срединная вырезка верхней губы, 2 —
передний бугорок наличника, 3 — наличинк, 4 — когте-
вой членик иогочелюсти, 5 — голень, 6 — бедро,
7 — максилла II, 8 — вертлуг II, 9— вертлуг I,
10 — тазик, 11 — зубцы иа переднем крае тазика,
12 — максилла I, 13 — мандибула, 14 —канал
ядовитой железы, 15 — верхняя губа, 16 — антенна
Голова имеет округлые очертания и заметно сплющена в дорзо-
вентральном направлении. Хитиновые покровы головы образуют
головную капсулу (рис. 413, /, 414). Спинная поверхность капсулы
уплощена, без ясных швов и других следов слияния отдельных частей. На
боках лобной поверхности головной капсулы прикрепляются длинные
усики, или антенны (antennae), состоящие из многочисленных
цилиндрических члеников (см. рис. 413, //). Последние покрыты
осязательными волосками и чувствительными шипиками. Усики постепенно
утончаются к концам. Базальные более мощные членики усиков
сочленяются с головой на дне особых ямок. Непосредственно позади
основания каждой антенны находится глазное поле, на котором заметно
около 20 простых глаз (ocelli), расположенных в три-четыре
продольных ряда (рис. 413, 2; 414, 4). Возле передне-брюшного края каждого
глазного поля заметно небольшое окаймленное отверстие особого
органа чувств ■— органа Темешвари (рис. 414, 3).
При рассматривании головы с брюшной стороны, сразу же позади
основания антенн нетрудно заметить подковообразную хитиновую
пластинку (рис. 414, /; 415, <?). Она неподвижно спаяна с головной
капсулой, часть которой она представляет. Это — наличник (clypeus). На
своем лобном краю наличник несколько выдается вперед в виде
небольшого бугорка, снабженного неподвижными щетинками (рис. 414, 9;
415,2).
390
Рис. 416. Lithobius forficatus. Верхняя губа, эпифаринке и склериты мандибулярного
сегмента левой стороны. Вид с внутренней стороны верхней губы (ориг.):
/ — медиальный зубец верхней губы, 2 — вырезка верхней губы, 3 —боковой зубец верхней губы,
4 — внутренняя пластинка верхней губы, 5 — чувствительные перистые щетиики, 6 — перепончатая
стенка ротовой полости, 7 — пластинка с чувствительными шипами, 8 — отверстия желез, 9 —
передняя стенка начала пищевода, 10 — дополнительная пластинка, // — место сочленовного бугорка
мандибулы, 12 — опорная пластинка, 13 — край головной капсулы, 14— поля чувствительных
щетинок верхней губы и эпифариикса
На брюшной стороне головы располагается ротовое отверстие,
окруженное ротовыми органами: верхней губой (рис. 415, 15), верхними
челюстями (см. рис. 415, 13), первой и второй парами нижних челюстей
(рис, 415, 12 и 7). Эти органы представляют собой более или менее
измененные конечности соответственных головных сегментов, за
исключением верхней губы, которая является хитинизированнои складкой
_ брюшной -стенки головы.
Верхняя губа (labrum) лежит непосредственно позади наличника.
Она имеет дугообразную форму, выгнута вперед и спаяна с
наличником (см. рис. 414, 7). Верхняя губа состоит из двух пластинок:
наружной, которая, собственно, и видна снаружи при рассматривании
головы с брюшной стороны, и внутренней, лежащей дорзальнее первой,
т. е., при рассматривании головы снизу, под наружной пластинкой. Обе
пластинки сходятся друг с другом под острым углом на заднем крае
верхней губы, образуя как бы лезвие, нависающее спереди над
ротовым отверстием. Средняя часть этого края снабжена перистыми
щетинками, которые расположены несколькими тесными рядами
(рис. 416, 14). По-видимому, они несут осязательную функцию; такие
же щетинки встречаются на других частях ротового аппарата. Верхняя
губа имеет ясную срединную вырезку (рис. 414, 8; 416, 2) с пятью
мощными зубцами черного цвета; один из них занимает медиальное
391
положение (рис. 416, 1), другие парны и лежат по сторонам от
среднего зубца (рис. 416, <?). Верхняя губа граничит с передней стенкой
ротовой полости, представляющей собой эпифаринкс (epipharynx).
Чтобы рассмотреть эпифаринкс необходимо выделить его вместе с
верхней губой и заключить в глицерин-желатин. Изучая препарат со
стороны внутренней пластинки верхней губы, можно видеть, что
передняя стенка ротовой полости (рис. 416, 6) соединяется с labrum
(рис. 416, 4) и образована тонкой прозрачной и бесцветной хитиновой
перепонкой, на которой заметны волосистые поля (рис. 416, 14). В
задней части эпифаринкса видны две нежные хитиновые пластинки,
усаженные рядом направленных назад шипов чувствительного характера
(рис. 416, 7).
Позади верхней губы, по бокам от ротового отверстия лежат
верхние челюсти, или жвалы (mandibulae), служащие для размельчения
пищи. Каждая челюсть представляет собой вытянутую хитиновую
пластинку, расширенную дистально (рис. 417). Проксимальный отдел
челюсти сильно вытянут и искривлен. Жевательная поверхность верхней
челюсти снабжена мощными хитиновыми зубцами черного цвета
(рис. 417, 7). Когда жвалы смыкаются над ротовой областью, зубцы
одной челюсти входят в промежутки между зубцами другой. Позади
зубцов заметна густая щетка из мелких зубчиков и жестких волосков
(рис. 417, 6), которая представляет как бы продолжение ряда
жевательных зубцов. На другой стороне от зубцов находятся густые ряды
жестких щетинок. В верхне-наружном краю этого пучка имеется около
20 длинных, изогнутых желтоватых, перистых или зазубренных
гребневидно волосков (рис. 417, /).
Мандибулы у Lithobius сохраняют следы первичной двучленисто-
сти. Именно каждая челюсть неполно подразделяется косой щелью
(рис. 417, 3) на проксимальный и диетальный членики. На нижней
поверхности дистального отдела заметен небольшой сочленовный бугорок
с закругленной вершиной. Посредством него верхняя челюсть
сочленяется с особыми хитиновыми трехветвистыми пластинками —
склеритами мандибулярного сегмента. Последних имеется две пары. Одна пара
лежит по бокам от верхней губы—опорные пластинки (см. рис. 414, 6;
416, 12), другая пара позади верхней губы — дополнительные
пластинки (см. рис. 414, 5; 416, 10). На каждой стороне обе пластинки
подвижно соединены друг с другом, а опорная связана мягкой кутикулой и
связками с наличником и верхней губой и сочленена с боковым краем
головной капсулы. Внутренние ветви опорных пластино'к
поддерживают также гипофаринкс (см. ниже). Сочленовные бугорки верхних
челюстей входят в промежутки между двумя внутренними отростками
опорных пластинок (см. рис. 416, 11). Дополнительные пластинки
вдаются в полость головы и представляют собой, таким образом, аподемы,
т. е. внутренний скелет, и, по-видимому, отвечают части такового
насекомых (см. с. 455).
К мандибулярному сегменту у Chilopoda относится гипофаринкс
(hypopharynx) — непарная посторальная лопасть вентральной стенки
головы, ограничивающая ротовую полость сзади. Гипофаринкс
представляет собой, по-видимому, видоизменный стернит мандибулярного
392
Рис. 417. Lithobius forficattis.
Верхняя челюсть (из Attems):
/ — гребневидные волоски, 2 —
дистальный отдел, 3 — щель между
отделами челюсти, 4 —
проксимальный отдел, 5 — сочленовный
бугорок, 6 — щетка зубчиков и жестких
волосков, 7 — хитиновые зубцы
Рис. 418. Lithobius forficatus.
Гипофаринкс. Вид с передне-
брюшной стороны (ориг.):
/ — лопасть гипофаринкса, 2 — ряд
крупных шипов, 3 — язычок,
покрытый перистыми волосками, 4'—
спинная стенка начальной части
пищевода
сегмента. Этот орган обнаруживается при препаровке под первой парой
нижних челюстей, между ними и ротовым отверстием. Выделив
гипофаринкс, следует заключить его в глицерин-желатин и рассмотреть с
передней стороны (рис. 418). Это довольно крупная лопасть с вырезкой
на вершине, густо покрытая мелкими шипиками и зубцами. На
боковых краях лопасти выделяются ряды более крупных шипов (рис. 418,
2). У основания гипофаринкса заметен небольшой язычок, покрытый
длинными перистыми волосками (рис. 418, 3).
Следующие две пары ротовых конечностей очень напоминают ход-
ные ноги. Нижние челюсти I, или максиллы I (maxillae I),
представляют собой трехчленистые конечности. Их основные^членики
(кокситы) листовидно расширены в поперечном направлении и спаяны
медиально друг с другом, но шов, разделяющий их, сохраняется
(рис. 419, 3). Передние внутренние углы этих члеников вытянуты в
большие отростки (эндиты,, рис. 419, 4), которые сходясь друг с другом,
образуют подобие нижней губы, лежащей сразу же позади
гипофаринкса, и ограничивающей рот сзади. У основания челюстей лежит узкая
поперечная хитиновая пластинка — стернит первого максиллярного сег-
393
Рис. 419. Максиллы I Lithobius forficatus Рис. 420. Максиллы II Lithobius for-
(из Attems): ficatus (ориг.):
/ и 2 — члеиики челюсти, 3 — коксит, 4 — эидит, / — чувствительные перистые щетинки, 2 —
5 — чувствительные перистые волоски коготок, 3 — коицевой членик, 4 — членики
челюсти, 5 — coxosternum
мента. Концевые членики, а также передние отростки основных
члеников максилл I снабжены чувствительными волосками и перистыми
щетинками (рис. 419, 5). При помощи максилл I животное поддерживает
добычу, убитую ногочелюстями (см. ниже).
Вторая пара нижних челюстей, или максиллы II (maxillae II),
еще более напоминают настоящие ножки. Основные членики этих
конечностей сливаются со стернитом второго максиллярного сегмента в
вытянутую в поперечном направлении пластинку (coxosternum,
рис. 420, 5). К боковым краям этой пластинки прикрепляются трехчле-
нистые челюсти (рис. 420, 3, 4). Все членики покрыты чувствительными
волосками, а концевые, кроме того, имеют по Овальному полю перистых
щетинок (рис. 420, /) и снабжены каждый одним коготком (рис. 420,
2). Роль максилл II сводится к ощупыванию добычи и поднесению ее
к ротовому отверстию и жвалам.
Туловище. Из 19 сегментов, составляющих туловище, первый и
три задних резко отличаются от остальных (см. рис. 413). Первый
сегмент снабжен парой мощных ногочелюстей, прикрывающих отчасти
голову с брюшной стороны. Три задних сегмента очень малы. Из них
только один XVII сегмент снабжен короткими ножками, остальные два
безногие, XVIII сегмент несет половое отверстие; вместе с
предыдущим он составляет генитальную область. Последний, XIX сегмент —
анальный, он несет Т-образую порошицу. ,
Остальные туловищные сегменты в основном имеют сходное
строение. Каждый снабжен парой ног и имеет хитиновый спинной щиток —
тергит (см. рис. 413, /—XIX), а также брюшной щиток, называемый
стернитом. Оба щитка на боках сегмента связаны плеврами,
образованными мягкой складчатой кутикулой (см. рис. 413, 7). На плеврах
заметны хитинизированные пластинки, расположенные вокруг основа-
394
ния ножек и в верхне-задних углах возле дыхалец. Складчатость
плевральных участков зависит от степени наполнения кишечника; когда
кишечник набит пищей, плевры расправляются.
Однако между сегментами средней, области туловища полного
сходства нет, что бросается в глаза с первого взгляда (см. рис. 413).
Различие заключается главным образом в величине тергитов. Часть сег-
.ментов отличается более крупными тергитами. Таковы II, IV, VI, VIII,
IX, XI, XIII, XV сегменты. Остальные имеют укороченные тергиты.
Дыхальца (стигмы) расположены также не на всех сегментах, а
только на шести крупных (на IV, VI, IX, XI, XIII и XV, рис. 413, 6).
Первый сегмент, снабженный ногочелюстями, имеет короткий
тергит и слабо развитые плевры. Стернит этого сегмента отсутствует, так
как вся его брюшная сторона занята спаянными друг с другом
основными члениками ногочелюстей.
Значительные отличия в развитии склеритов свойственны также
трем задним сегментам. Оба сегмента генитальной области у обоих
полов имеют общий тергит. Впереди него лежит очень укороченный
тергит XVI сегмента. Сзади на конце туловища заметен тергит
анального сегмента, соединенный с генитальной областью довольно длинной
полоской интерсегментарной кожи (см. рис. 413, <?). При
рассматривании заднего конца тела с брюшной стороны- позади 16-го стернита
виден довольно большой стернит
прегенитального (XVII)
сегмента (рис. 421, 10), который
снабжен парой направленных
назад половых ножек (рис. .
421, 9) (у самцов они
рудиментарны). Стернит гениталь-
ного (XVIII) сегмента
редуцирован до степени
незначительной хитиновой пластинки и
трудно различим. Половое
отверстие лежит под
основаниями (половых ножек на
брюшной стороне полового сегмента.
Конечности тулови-
щ а. Ходные; или бегательные,
ноги (II—XVI сегментов)
состоят из семи члеников (рис.
422). Основной членик — тазик
(соха), следующие вертлуг I
(trochanter), вертлуг II (ргае-
femur), бедро (femur), голень
(tibia), лапка (tarsus) и когот-
ковый членик (praetarsus).
На вершине последнего
имеется изогнутый коготок, у
основания которого заметен
маленький дополнительный кого-
39S
Рис. 421. Lithobius forficatus. Задний конец
тела самки, вид с брюшной стороны (из R6-
seler и Lamprecht):
I — стернит XVI сегмента, 2 — тазик, 3 — вертлуг I,.
4 — вертлуг II, 5— бедро, 6— аиальиый сегмент, 7 —
подвижный зубец, 8 — аиальиое отверстие, 9 —
половая ножка, 10 — стернит прегенитального сегмента,
II — коксальиые поры. Стрелка указывает положение
полового отверстия
ток (рис. 422, 10). Все членики конечностей, так же как склериты
сегментов, покрыты редкими волосками; вершина каждого членика
ножки, кроме того, вооружена несколькими острыми подвижными шипами..
Задние туловищные ножки отличаются большей длиной, а когда
животное находится в покое, обращены назад (см. рис. 413). Ноги XVI
сегмента особенно длинны и служат главным образом для • осязания
(см. рис. 413, 4).
На брюшной стороне каждого тазика четырех задних пар бега-
тельных ножек заметен ряд из 5—-8 овальных отверстий, лежащих на
дне продольного желобка. Это так называемые коксальные поры
(рис. 421, //; 422, 1). Каждая пора окружена утолщенным хитиновым
кольцом; дно ее составляет очень тонкая бесструктурная хитиновая
перепонка. По некоторым данным коксальные поры представляют собой
отверстия особых желез, выделяющих бесцветный клейкий секрет.
Передняя пара туловищных конечностей, принадлежащая I
сегменту, известна под названием ногочелюстей (maxillipedes, см.
рис. 415, 4—6, 8—10). Каждая ногочелюсть состоит из шести члеников
(coxa, trochanter, praefemur, femur, tibia и tarsungulum). Основные
членики ногочелюстей, или тазики, содержащие мощную мускулатуру,
очень велики; они срастаются друг с другом медиально, хотя шов
между ними остается, и занимают всю брюшную сторону I сегмента (см.
рис. 415, 10). Каждый из них образует прямоугольный вырост,
направленный вперед и усаженный по краю черными коническими зубцами
(см. рис. 415, 11), число которых увеличивается с возрастом животного
и достигает семи. Из следующих члеников ногочелюсти вертлуг I
(trochanter, см. рис. 415, 9) очень мал, тогда как вертлуг II
(praefemur, см. рис. 415, 8) достигает значительных размеров. Конечный
членик (tarsungulum, представляющий собой результат слияния tarsus и
praetarsus), заострен и когтевидно изогнут внутрь (см. рис. 415, 4).
На поверхности tarsungulum заметны слабые продольные бороздки со
слегка возвышенными краями. После вываривания в едком кали
видно, что внутри когтевого членика проходит тонкий канал ядовитой
железы (см. рис. 415, 14), которая лежит в этом и отчасти в предыдущем
членике. Канал открывается на конце когтевого членика
маленьким.отверстием.
Рис. 422. Нога XIV сегмента Lithobius forftcatus (из At-
tems),:
/ — коксальные поры, 2 — стернит XIV сегмента, 3 — тазик, 4 —
вертлуг I, 5 —вертлуг II, 6 — бедро, 7 —голень, 8 — лапка, 9 —
коготковый членнк, 10 — дополнительный коготок, // — коготок
396
Ногочелюсти прикрывают снизу значительную часть головы, а их
крючковидные концы сходятся друг с другом впереди ротового
отверстия. Ядовитые ногочелюсти являются органами нападения и защиты;
ими схватывается и умерщвляется добыча.
Половые ножки, или гоноподы, расположенные на прегенитальном
(XVII) сегменте, у самок развиты довольно хорошо, тогда как "у
самцов редуцированы до степени небольших двучленистых бугорков.
Гоноподы самки представляют собой короткие трехчленистые конечности,
торчащие назад параллельно друг другу (см. рис. 421, 9). Их основные
членики на своих внутренних краях несут небольшие бугорки,
вооруженные маленькими подвижными зубцами (см.. рис. 421, 7), которые
служат для поддержания яиц при их откладке. Конечными члениками
гонопод самка обхватывает пенис самца при копуляции.
Анатомия на вскрытии. Вскрывается убитый эфиром Lithobius.
Вскрытие производится со спиниой стороны под водой, причем животное
прикрепляется двумя булавками к дну ванночки. При помощи остро отточенного тонкого
скальпеля, копьевидной иглы или острых маленьких ножниц делаются два боковых
разреза стенки тела вдоль всего туловища. Для этого инструмент втыкается
где-нибудь сбоку под одним из тергитов, а затем рассекаются покровы вперед и назад.
При этом острие инструмента нельзя погружать глубоко в тело, но надо вести по
внутренней стороне тергитов, что нетрудно контролировать глазом, так как хитиновые
покровы прозрачны. Часто уже при первом надрезе из него выпячиваются внутренние
органы, поэтому необходимо соблюдать осторожность, чтобы их не повредить. Оба
продольных разреза соединяются спереди и сзади короткими поперечными. Затем
спиниые покровы осторожно снимаются и анатомирование продолжается при помощи
препаровальных игл. Вскрытие удобно производить при слабом увеличении. После
ознакомления с общим расположением внутренних органов следует перейти к
изучению отдельных систем органов.
Пищеварительная система. Кишечник представляет
собой прямую трубку, которая делится на три отдела: переднюю кишку,
состоящую из ротовой полости и пищевода, среднюю и заднюю кишки.
На границе между двумя последними отделами в кишечник
открывается пара длинных трубочек — мальпигиевы сосуды. Имеются слюнные
железы.
Пищевод тоньше, чем средняя кишка; кзади он несколько
расширяется (рис. 423, /). На его поверхности заметна продольная исчерчен-
ность, зависящая от внутренней складчатости. Изнутри пищевод,
который имеет эктодермальное происхождение, выстлан хитиновой
кутикулой — продолжением хитиновых покровов тела.
Средняя кишка в виде сравнительно широкой прямой трубки
(рис. 423, 2). Это самый длинный отдел кишечника. Иногда в
некоторых местах кишки наблюдаются вздутия или неровности, но они
зависят от частиц пищи, находящихся в просвете кишки.
Задняя, или прямая, кишка тянется от места впадения мальпигие-
вых сосудов до конца туловища, постепенно суживаясь кзади
(рис. 423, 3). Поверхность ее покрыта поперечными кольцевыми
складочками. Подобно пищеводу, задняя кишка имеет эктодермальное
происхождение и обладает внутренней хитиновой выстилкой.
Что касается слюнных желез (называемых также максиллярными
железами), то это довольно крупные органы (до 8 мм длиной),
лежащие по бокам от пищевода (рис. 423, 6) и обычно окрашенные в яркий
397
Рис. 423 Пищеварительная система
Lithobius forficatus (по Plateau с изме
нениями):
™,£ИЩТ°А- 2-сРеДняя кишка, 3- задняя
ев сосуд f3 ™°е отвеРстие- 5-мальпигн
ев сосуд, 6 -слюнная железа, 7 - проток
слюнной железы
Рис. 424. Lithobius forficatus. Стигма
после вываривания в едком кали (из
Vogt и Yung):
/-отверстие стигмы, 2-перитрема,
3-защитная щетинка, 4 - поверхностные трахеи
5-стигмальный склерит, 6-задние трахеи
7-трахеальный мешочек, а - поперечное
трахеи
*И^еТ0ВЫЙ ^ВеТ- К°Р°ткие и тонкие протоки желез открываются в об-
жел™ lTShohf,rCm- ФУНКЦИЯ ЭТИХ Ж6Ле3 Неясна" Помимо слюнных
лТз?лежащ^в1головГСЯ "** ** МрЫ Неб0ЛЬШИХ мак™Рных же-
кухояосиьти\^аЯИЯ У многоножек представлены трахеями,
возили Тигм (^Ля?яТаМИ'/0Т°РЬ1е откРываютс* посредством дыхалец,
или стигм (stigmata),.на боковых поверхностях сегментов
1рахеальная система Lithobius изучается на тотальных гтпрпяпя
тах, монтированных в глицерин-желатине. Предварительно живот&е
выдерживается в течение нескольких суток в" слабом растворяемого
учас?кТхаГу1Тх°ШХ^3^тетТНЬ1у5НИ Р3™3™^* на плевральных
учаиках iv, VI, IX, XI, XIII и XV сегментов (см. рис 413 6) Кяжттяа
гТщиаткЛаеЖКосо «^Г™ б°^™°^™ во'звышении^и^иГво-
го щитка. Косо расположенное щелевидное отверстие стигмы
398
(рис. 424, /) окружено хитиновым утолщением — перитремой
(рис. 424, 2). Несущий стигму склерит (рис. 424, 5) снабжен
неподвижными щетинками защитного значения (рис. 424, 3). При
рассматривании в профиль видно, что стигма ведет в небольшой, выстланный
хитином мешочек (рис. 424, 7), стенки которого покрыты большим
количеством мелких зубчиков, торчащих в полость мешочка. Эти зубчики
препятствуют проникновению в трахеальную систему мелких
посторонних тел. От мешочка отходит пучок из нескольких трахеальных
стволов, которые очень скоро начинают ветвиться, расходясь в разных
направлениях (рис. 424, 6, 8). От боковых, стенок мешочка, независимо
от главных трахей, берут начало очень тонкие поверхностные трахеи,
которые разветвляются вблизи стигмы (рис. 424, 4).
Трахеальные стволы, принадлежащие разным стигмам", не
соединяются друг с другом, отсутствуют как продольные, так и поперечные
интерстигмальные анастомозы. Расположение трахей задних пяти пар
дыхалец довольно просто. От каждой стигмы всегда начинаются две
группы стволов: поверхностные трахеи, дающие ветви в поперечном
направлении к противоположной стороне сегмента (рис. 424, 8, 425, 7), и.
более глубокие трахеи, которые загибаются назад и несут воздух в
заднюю часть сегмента и в следующий сегмент (рис. 424, 6).
Отдельные тонкие продольные ответвления можно проследить на большом
протяжении; они захватывают несколько сегментов.
Особенной мощностью отличаются трахеи первой пары стигм
(рис. 425). Из них выделяются две пары толстых стволов, посылающие
свои ветви в передние сегменты, ногочелюсти и голову. Пара таких
стволов занимает дорзальное положение (рис. 425, 4), другая пара
расположена ближе к брюшной стенке тела (рис. 425, 5). Оба спинных
ствола сближаются друг с другом на уровне III сегмента. В этом
месте они отделяют боковые ветви в третью пару туловищных
конечностей (рис. 425, //). Далее спинные стволы расходятся, а на уровне
II сегмента дают ветви в ногочелюсти (рис. 425, 14). В голове каждый
ствол разделяется на пучок более мелких трахей, идущих к глазам,
головному мозгу и другим органам (рис. 425, 2). От дорзальных же
стволов берут начало тонкие трахеи антенн (рис. 425, 16). Брюшные
продольные стволы первой пары стигм также достигают головы. По
пути они дают ветви во вторую пару туловищных конечностей
(рис. 425, 12), ногочелюсти (рис. 425, 13) и, наконец, разветвляются
на пучки более мелких трахей (рис. 425, 15). Иногда брюшные стволы
перекрещиваются в голове так, что от правого снабжаются левые
органы головы (нервная система, ротовые части) и наоборот
(рис. 425, 3).
Все трахеальные стволы многократно ветвятся, образуя в общем
густое сплетение мелких трахей, оплетающее внутренние органы.
Мелкие трахеи, так же как и главные, нигде не образуют анастомозов.
Некоторые органы имеют особенно густую трахеальную сеть. Таковы
пищевод, задняя кишка, брюшная нервная цепочка. Тонкие
поверхностные трахеи сопровождают сердце вдоль всей его длины.
Трахеи имеют сходное строение с. таковыми насекомых. Они
представляют собой глубокие впячивания покровов, изнутри выстланы хи-
399
тином и имеют поддерживающую
спиральную нить, завитки которой лежат
очень тесно. Хитиновая выстилка
трахеи выделяется тонким эпителиальным
слоем — matrix, богатым ядрами. Этот
слой можно видеть, рассмотрев в
микроскоп кусочек трахеи в свежем
состоянии под покровным стеклом в
воде или глицерине.
Мускулатура. Перед
вскрытием мускулатура должна быть
уплотнена. Для этого животному,
убитому эфиром, впрыскивают в полость
тела 96%-ный спирт. Игла шприца
вводится наискось между двумя
сегментами. Полезно предварительно
обрезать усики у основания. Затем
животное оставляется на час в 96%-ном
спирте.
Следует обратить внимание наме-
тамерный характер распределения
мускулов в сегментах туловища.
Мускулатура головы 'и заднего отдела
тела резко отличается от таковой
туловищных сепментов. Ротовые
конечности имеют собственную сложную
мускулатуру.
Мышечные волокна, как у всех
Arthropoda, имеют
поперечнополосатую исчерченность, что легко
наблюдать, рассматривая кусочек свежей
мускульной ткани под микроскопом.
Кровеносная система. Не»
замкнутая кровеносная система
состоит из дорзального сердца и
вентрального продольного сосуда. Тонкое
продолжение сердца, называемое
головной аортой (aorta cephalica), несет
кровь в голову. Здесь аорта соединяется с околопищеводным
сосудистым кольцом (мандибулярное кольцо), от которого отходят парные
короткие сосуды к мандибулам и гипофаринксу. Передний конец сердца,
кроме того, связан вторым сосудистым кольцом (максиллипедное
кольцо) с брюшным сосудом. От второго кольца берут начало артерии ного-
челюстей. Мэксиллы I и II получают кровь через ветви брюшного
сосуда. От последнего также ответвляются сосуды ног.
У молодых экземпляров с прозрачным хитином можно непосредст- .
венно наблюдать сквозь спинные покровы сердце в виде медиальной
светлой, местами расширенной полоски. Если животное лишь усыплено
эфиром, то можно видеть пульсацию сердца (около 80 сокращений в
Рис. 425. Lithobius forficatus.
Трахейная система передней пары стигм
.с препарата, обработанного едким
кали (из Vogt и Yung):
7 — антеннальная ветвь брюшного трахе-
ального ствола, 2 — головные ветви
спинного ствола, 3 — перекрест брюшных
стволов, 4 — спинной ствол, 5 — брюшной
ствол, 6 — короткая поверхностная трахея,
7 —. поперечная трахея, 8 — задняя трахея,
9 — стигма, 10 — трахея ноги четвертой
пары, 11 — трахея ногн третьей пары, 12 —
трахея ноги второй пары, 13 — ногочелюст-
ная ветвь брюшного ствола, 14 — ногоче-
люстная ветвь спинного ствола, 15 — ветви
левого брюшного ствола, 16 —
антеннальная ветвь спинного ствола
минуту). Если осторожно вырезать всю спинную стенку тела, то, после
удаления жирового тела и части дорзальных мышц, обнаруживается
.сердце. Оно тянется вдоль всего тела от I до прегенитального сегмента,
будучи прикреплено к внутренней поверхности спинной стенки тела.
Сердце состоит из 15 камер, которые заметны как небольшие
расширения, по одному в каждом сегменте. Все камеры сердца одинаковы,
кроме передней и задней, и разделены интерсегментарными клапанами.
В середине сегмента между двумя клапанами лежит пара отверстий —
остий. Передний конец сердца через головную аорту открывается в
головной синус, задний сообщается с каудальным синусом.
Непосредственно над сердцем тянется узкий спинной сердечный синус (sinus cordis
dorsalis), а по бокам расположены продольные «перикардиальные»
синусы. Все эти полости представляют собой участки первичной
полости тела. Брюшной сосуд проходит между брюшной нервной цепочкой
и кишкой. Спереди он сообщается с сердцем и с головным синусом.
Сзади брюшной сосуд соединяется с сердцем третьим (ректальным)
сосудистым кольцом и связан с каудальным синусом. При впрыскивании
в полость тела растертой в воде туши иногда брюшной сосуд
наполняется этой краской. В таком случае, после вскрытия и удаления
жирового тела, кишечника и половых органов, его можно видеть над
брюшной нервной цепочкой.
Кровообращение у Lithobius совершается следующим образом.
Вследствие давления тергитов кровь из спинного сердечного синуса
поступает через остии в сердце. По сердцу кровь течет от заднего
конца к переднему из камеры в камеру, благодаря волнам сокращений
сердечных камер, которые начинаются сзади и распространяются
вперед. Из большого головного синуса кровь течет по брюшному сосуду
назад, но значительная ее часть, минуя этот сосуд, проходит в лаку-
нарные пространства туловища. Сзади кровь собирается в каудальный
синус, из него затем поступает в заднюю камеру сердца и дорзальный
сердечный синус. Кровь бесцветна и содержит небольшое количество
лейкоцитов.
Выделительная система. Органы выделения представлены
парой мальпигиевых сосудов. Выделительную функцию выполняют
также жировое тело и лимфатические тяжи и видоизмененные целомодук-
ты — максиллярные железы. Мальпигиевы сосуды — длинные, тонкие
трубки белого цвета, тянущиеся по бокам кишечника до головы (см.
рис. 423, 5). Они открываются в кишечник на границе средней и
задней кишок и происходят от эктодермальной задней кишки.
Эпителиальные клетки мальпигиевых сосудов содержат конкреции мочевой
кислоты.
Жировое тело, как у насекомых, заполняет полость тела между
внутренними органами. Жировое тело имеет вид неправильной формы
лопастей или тяжей буроватого цвета, перекрещивающихся друг с
другом в различных направлениях и пронизанных тонкими разветвлениями
трахей. Гистологически жировое тело представляет синцитий с
многочисленными ядрами и содержит включения: жировые капли, хромо-
фильные пигментные зерна, экскреторные зерна и зерна запасных
питательных веществ. Последние три категории неотличимы по виду друг
401
от друга. Функция жирового тела заключается не только в экскреции,
но и в накоплении запасных питательных веществ.
Вместе с жировым телом в полости тела лежит ретикулярная
соединительная ткань, богатая фиолетовыми волокнами, она легко
узнается по цвету. Встречается она всегда в непосредственной близости от
брюшной нервной цепочки, под боковыми краями тергитов, вдоль
боковых стенок тела, а также в основаниях ног.
Лимфатические тяжи, или ацидофильные нити Ковалевского,
развиты вокруг, слюнных желез и образуют неправильные, часто
перешнурованные или лопастные ленты. В цитоплазме клеток лимфатических-
тяжей находятся вакуоли жидкости и пигментные зерна.
У Chilopoda доказано наличие пары видоизмененных целомодук-
тов — максиллярных почек, или желез. У Lithobius forficatus они
помещаются в голове. Каждая максиллярная железа открывается двумя
каналами, отверстия которых лежат медиально от оснований максилл I
и максилл II. '
Наличие в пузыревидном отделе каждой железы остатков двух
воронок и двух выводных каналов показывает, что у Lithobius эти
железы представляют собой продукт слияния двух пар целомодуктов.
Экскреторная функция этих ортанов несомненна. По положению анатомии
и гистологии они близки к максиллярным железам Crustacea и
лабиальным почкам Diplopoda и Insecta Apterygota.
Нервная система. Препарирование нервной системы очень
облегчается, если только что убитое, вскрытое со спины животное
оставить в воде на двое суток. За это время нервная, ткань не подвергается
разложению, тогда как прочие органы мацерируются. В результате
нервная система может быть выделена без труда со всеми ганглиями
и главнейшими нервами.
Нервная система состоит из парного головного мозга, лежащего
над глоткой (церебральные ганглии) и брюшной нервной цепочки.
Головной мозг Myriapoda, как у большинства членистоногих,
состоит из трех более или менее слившихся парных отделов: protocereb-
rum, deutocerebrum и tritocerebrum. Первый из них иннервирует глаза
и принадлежит acron. Deutocerebrum иннервирует антенны и относится
также к acron. Наконец, tritocerebrum принадлежит интеркалярному
сегменту.
У Lithobius передняя часть мозга, отвечающая protocerebrum,
сильно вытянута в поперечном направлении и состоит из пары больших
грушевидных фронтальных лопастей (lobi frontales, рис. 426, 3; 427,17),
спаянных медиально друг с другом. Их боковые участки, называемые
оптическими лопастями (1. optici, рис. 426, 4), дают начало мощным
зрительным нервам (п. optici, рис. 426, 6). Спереди от оптических
лопастей отходят нервы органа Темешвари (рис. 426, 5). От
фронтальных лопастей начинаются тонкие нервы (рис. 426, 17), связанные с
парой небольших округлых телец, которые лежат в полости головы и
называются мозговыми железами, или фронтальными органами
(рис. 426, 16). Природа и функции последних неизвестны.
Deutocerebrum представлен парой вытянутых в продольном направлении антен-
нальных лопастей, или ганглиев (1. antennales, рис. 426, /), которые
402
Рис. 426. Головной мозг Lithobius
forficatus. Вид с брюшной стороны (по
Fahlander с изменениями):
/ — антеннальная лопасть, 2 — чувствительный
антеннальный нерв, 3 — фронтальная лопасть, 4 —
оптическая лопасть, 5 — нерв органа Темешвари,
6 — зрительный нерв, 7 — трнтоцеребральная
область, 8— стоматогастрический нерв, 9—
свободная трнтоцеребральная коммнссура, 10 —
окологлоточный коннектив, 11 — нервы ногочелюсти,
12 — нервы максиллы II, 13— нервы максиллы I,
14 — нервы мандибулы, 15 — стоматогастрический
ганглий, 16 — фронтальный орган, П — нерв
фронтального органа, 18 — нерв верхней губы, 19 —
двигательный антеннальный нерв, 20 —
стоматогастрический мостик
Рис. 427. Нервная система Lithobius forfi,-
catus. Вид со спинной стороны (ориг.):
/ — стоматогастрический мостик, 2 —
чувствительный антеннальный нерв, 3 — двигательный
антеннальный нерв, 4 — глазные нервы, 5 — нерв
фронтального органа, 6—окологлоточный коннектив,
7 — подглоточный ганглнй, 8 — ганглий III
сегмента, 9 — коннектив, 10 — нерв конечности, 11 —
ганглий XVI сегмента, 12 — ганглиозная масса
XVII и XVIII сегментов, 13 — нервы ногочелюстей,
14 — нервы максиллы II, 15 — нерв максиллы I,
16 — нерв мандибулы, П — фронтальная лопасть,
18 — антеннальная лопасть, 19 —
стоматогастрический нерв
лежат под protocerebrum и срастаются с ним своими верхними
поверхностями. Сзади антеннальные лопасти без заметных границ сливаются
с парным tritocefebrum (рис. 426, 7). В состав последнего входит ганг-
лиозный стоматогастический мостик (pons cerebri, рис. 426, 20),
который имеет характер короткой и толстой коммиссуры и состоит из
слившегося с мозгом фронтального ганглия симпатической нервной
системы. Кроме того, задние участки этого отдела мозга связаны тонкой
тритоцеребральной коммиссурой (рис. 426, 9).
Сзади тритоцеребрум соединяется с подглоточным ганглием
брюшной нервной цепочки посредством коротких окологлоточных коннекти-
вов (рис. 426, 10).
От передних концов антеннальных лопастей отходят
чувствительные и двигательные нервы усиков (рис. 426, 2 к 19). От тритоцереб-
рального отдела иннервируется верхняя губа, принадлежащая интерка-
лярному сегменту (рис. 426, 18). Наконец, от стоматогастрическото
мостика берет начало медиальный стоматогастрический нерв (п. reccu-
rens, рис. 426, 8), идущий назад над кишкой и сходный с висцеральным
непарным нервом (п. ventriculi impar inferior) ракообразных (см.
с. 343). Основание этого нерва образует небольшой
стоматогастрический ганглий (рис. 426, 15).
Брюшная нервная цепочка содержит 17 парных ганглиев (рис. 427,
7—12). Все они связаны друг с другом продольными коннективами
(рис. 427, 9). Первый из них — подглоточный ганглий (рис. 427, 7)
лежит в I сегменте и представляет собой сложный ганглий, продукт
слияния ганглиев четырех сегментов: мандибулярного, двух максиллярных
и I туловищного. Соответственно он иннервирует жвалы, первую и
вторую пару нижних челюстей и ногочелюсти. Ганглии прегенитального
и генитального сегментов сливаются также, образуя общий
терминальный ганглий (рис. 427, 12). Анальный сегмент не имеет своего ганглия.
От каждого брюшного ганглия отходят боковые нервы к конечности и
к мускулатуре данного сегмента.
Органы чувств у Lithoblus представлены глазами, органами
Темешвари, фронтальными органами и многочисленными
осязательными щетинками на различных,участках тела. Кроме того, по-видимому,
имеются обонятельные щетинки на усиках. Строение всех этих органов
может быть изучено только на срезах.
Органы Темешвари лежат между основаниями усиков и глазами
(см. рис. 414, 3) и иннервируются от оптических лопастей. Внешне они
различимы в виде округлых двухконтурных хитиновых колечек,
окружающих участок нежной кутикулярной перепонки. В середине
последней имеется маленькое отверстие. Непосредственно под перепонкой
находятся круглые группы чувствительных клеток. Тонкие дистальные
выросты этих клеток прилегают к внутренней поверхности хитиновой
перепонки.
Функция органов Темешвари неизвестна; возможно, как считает
Ферхефф, они являются органами слуха.
Половая система. Все многоножки раздельнополы. Половой
диморфизм у Lithobius выражен слабо и заключается в различиях
строения генитальной области, в частности в неодинаковом развитии гоно-
404
под у самцов и' самок.
Гонады непарны, но
происходят из (парного зачатка.
Мужская половая
система непарна. Очень
вытянутый, образующий одну
петлю семенник (рис. 428,
А, 1) лежит над кишкой.
Его свободный передний
колец вытянут в нитевидный
вырост (рис. 428, А, 9).
Задний конец семенника
открывается в небольшое
пузыревидное расширение (рис.
-428, А, 4), стенки которого,
■сильно выпячиваясь вперед,
образуют два длинных
мешковидных семенных
пузырька (vesiculae seminales,
рис. 428, А, 2). В пузырьках
содержится молочно-белая
.зернистая масса со
сперматозоидами. От
пузыревидного расширения отходят
две ветви семяпровода (va-
sa deferentia, рис. 428, А, 5).
огибающие с боков заднюю
кишку и соединяющиеся
под ней в непарный семяиз-
вергательный канал (ductus
•ejaculatorius, рис. 428, А, 6).
.Этот последний открывается половым отверстием на брюшной стороне
генитального сегмента. В семяиэвергательный канал открываются
протоки двух пар придаточных желез (рис. 428, А, 3, 8). Они имеют
вытянутую форму и дольчатое строение. Семяизвергательный канал
заканчивается совокупительным органом (penis), который представляет собой
небольшой конусовидный, слабо хитинизированяый придаток (рис. 428,
А, 7).
Женская половая система имеет сходное строение. Мешковидный
яичник (рис. 428, Б, 10) сзади сообщается с непарным яйцеводом
{рис. 428, Б, 11), который вскоре раздваивается, огибая кишечник
(рис. 428, Б, .12). Под задней кишкой ветви яйцевода снова
соединяются (рис. 428, Б, 13). Конечный отдел яйцевода открывается в копу-
лятивный мешок (atrium genitale, рис. 428, Б, 14). Под кишечником
лежит пара продолговатых семяприемников (receptacula seminis,
рис. 428, Б, 18); их протоки раздельно впадают в заднюю непарную
часть яйцевода. Как и у самца, имеются две пары гроздевидных
вытянутых придаточных желез. Из них передние железы открываются рядом
,с семяприемниками в яйцевод и являются скорлуповыми железами;
Рис. 428. Половая система Lithobius forficatus.
А — мужская, Б — женская (по Schaufler):
/ — семенник, 2 — семенной пузырек, 3 — передняя
придаточная железа, 4 — пузыревидное расширение
полового протока, 5 — парная ветвь семяпровода, 6 —
семяизвергательный канал, 7 —пенис, 8 — задние придаточные
железы, 9 — нитевидный вырост семенника, 10 — яичник,
Л — непарный яйцевод, 12 — парная ветвь яйцевода,
13 — задний непарный отдел яйцевода, 14 — область ко-
пулятивиого мешка, 15 — половое отверстие, 16 —
половая ножка, 17 — задняя придаточная железа, 18 —
семяприемник, 19 — передняя придаточная железа
405
они секретируют вещество защитной оболочки яйца (рис. 428, Б, 19).
Задние железы открываются в копулятивный мешок; их выделения
увеличивают жизнеспособность сперматозоидов (рис. 428, Б, 17),
Яйца откладываются поодиночке. Подобно другим Anarnorpha,
самка Lithobius некоторое время носит каждое яйцо, зажав его
между гоноподами, а затем откладывает на землю или на другой субстрат.
К клейкой оболочке яйца пристают различные частицы, благодаря чему
оно становится незаметным в окружающей обстановке.
Постэмбриональное развитие Lithobius, как у всех Anarnorpha, сопровождается
анаморфозом, точнее, гемианаморфозом, так как развитие первых личиночных стадий"
анаморфно, а последние стадии развиваются по типу эпиморфоза.
Покидающая яйцо личинка называется foetus. Она бледно окрашена и у
Lithobius имеет восемь тергитов, считая принадлежащий сегменту ногочелюстей. Антенны
еще семичлениковые. Позади восьмой пары ног имеется пара бугорков — зачатки
девятой пары. Тазики ногочелюстей еще лишены передних зубцов. Личинка активно
двигается, но питание происходит еще за счет желтка кишки. Дальнейшие линьки
приводят к образованию личиночных стадий: первой, второй, третьей и четвертой (larva/
prima, I. secunda, 1. tertia, 1. quarta), которые характеризуются следующими
признаками (табл. 4). Четвертая личинка представляет собой последнюю анаморфную ста-
Таблица 4. Отличительные признаки личиночных стадий
Стадия
Foetus
Larva prima
Larva secunda
Larva tertia
Larva quarta
Agenifalis
Число
тергнтов
8
8
9
11
13
16
пар развитых
ног
8
9
(задняя пара
недоразвита)
9
11
13
16
пар почек
конечностей
1
2
2
2
3
1 (гоноподы)
члеников
антенны
7
больше 7
14
больше 14
продолжает
увеличиваться
дию; далее имеет место эпиморфоз. Следующая после четвертой личинки стадия
называется agenitalis. Она характеризуется 16 парами развитых туловищных
конечностей, но имеет неразвитую генитальную область с зачатками гонопод. Далее следуют
стадии: immaturus, praematurus, pseudomaturus и, наконец, взрослое животное. В
течение этого отрезка метаморфоза окончательно развивается генитальная область, а
частности гоноподы, и развиваются половые органы.
ПОДКЛАСС DIPLOPODA —ДВУПАРНОНОГИЕ
ОТРЯД JULIFORMIA (-OPISTHOSPERMOPHORA)
Schizophyllum sabulosum L. — Кивсяк
Материал и методы изучения. Schizophyllum sabulosum L., относимый к
семейству Julidae, широко распространен в европейской части СССР и во многих
местностях является обычной формой: Этот вид обладает характерной окраской, по которой
легко узнается. Основная окраска тела коричневато-черная, у самцов темнее, чем у
самок. Вдоль спины проходит медиальная темная полоска, на каждой стороне от ко-
406
торой тянется более широкая желтая или желто-красная полоса. Для целей
практикума, однако, пригодна любая достаточно крупная форма из Juliformia. Кивсяки
часто встречаются в лесах и парках под листвой, лежачими стволами деревьев и в
древесной трухе. Подобно громадному большинству Diplopoda, они питаются гниющей
растительной пищей, малоподвижны, и, будучи потревожены, свертываются спиралью
на брюшную сторону тела, что является у них средством защиты. Кивсяки легко
содержатся в неволе в сосуде с влажными гниющими листьями и другими
растительными остатками.
Перед исследованием многоножек убивают в парах эфира. Вполне пригоден
также материал, фиксированный спиртом. Для изготовления тотальных препаратов из
ротовых органов, конечностей и т. д. соответственные части отделяются,
вывариваются в едком кали и после промывки заключаются в канадский бальзам или глицерин-
желатин.
Внешняя морфология. Сильно вытянутое, цилиндрическое тело
(рис. 429) одето прочным хитиновым покровом, пропитанным известью.
На переднем конце тела находится голова и лежащий позади нее
шейный щиток (рис. 429, /)» на заднем — выделяется telson (рис. 429, 14—
17). Туловище состоит из многочисленных колец; сегментация его
внешне представляется гомономной. Самцы Schizophyllum sabulosum
достигают 30 мм в длину; самки несколько крупнее — до 47 мм.
Голова. Следует отделить голову, выварить ее в едком кали,
отпрепаровать ротовые органы и изучить отдельно последние и
головную капсулу. Основная часть головы состоит из широко открытой
снизу головной капсулы, на которой расположены глаза и усики. Глаза
представлены простыми глазками (ocelli), 'сосредоточенными на
небольшом участке, глазном поле, на каждой стороне головы (рис. 429
2). Усики или антенны (antennae, рис. 429, 3) сравнительно коротки и
■состоят из семи члеников. Они покрыты чувствительными волосками,
а последний очень маленький их членик снабжен, кроме того,
несколькими обонятельными конусами в виде маленьких выростов (рис. 429,
4). Края головной капсулы загибаются на брюшную сторону. Передний
Рис. 429. Самец Schizophyllum sabulosum. Схема переднего и заднего
концов тела с левой стороны (ориг.):
/ — голова, 2— глазное поле, 3—антенна, 4 — обонятельные конусы, 5 — верхняя
губа, 6 — мандибула, 7 —щека, 8 — видоизмененная нога II грудного сегмента,
9—нога III сегмента, 10— место полового отверстия, И— нога IV грудного
сегмента, 12 — ноги диплосомита, 13— передняя гонопода, 14 — анальная чешуйка, -
15—анальная лопасть, 16 — терминальный вырост тельсона, 17—кольцевидная
чаггь тельсона, 18 — сегменты пролиферационной еоны, 19 — метасомит, 20 — про-
сомит, 21 — отверстия ядовитых желез. I—IV — тергиты грудных сегментов;
из ннх I — шейный щиток; V—IX — тергиты диплосомитов брюшка
407
край образует поперечную, снабженную зубцами пластинку,
называемую верхней губой (labrum, рис. 429, 5). Задний край головной
капсулы образует выросты, уходящие в глубь головы, и служащие для
прикрепления мощной мускулатуры. Наконец, с боковыми краями капсулы
подвижно сочленены хитиновые пластинки, представляющие собой
внутренний скелет головы — тенториум.
На брюшной стороне головы лежит ротовое отверстие,
ограниченное с боков мощными верхними челюстями и прикрытое сзади второй
парой нижних челюстей, так называемым гнатохилярием.
Первая пара нижних челюстей у Diplopoda отсутствует '.
Верхние челюсти, жвалы, или мандибулы (mandibulae), сильно
развиты и имеют очень сложное строение. Каждая челюсть состоит из
щеки (gena, рис. 430, 2) и подвижно сочлененной с ней премандибуляр-
ной части (praemandibula, рис. 430, 1). В состав щеки, которая
представляет большую проксимальную часть челюсти, входят несколько
крупных хитиновых пластинок. Щека связана мягкой кожей с головной
капсулой и гнатохилярием.
Премандибулярный отдел состоит из большого основного членика
(рис. 430, 6) с теркой, т. е. пластинкой, усаженной мелкими
палочковидными шипами (рис. 430, 8), среднего членика (рис. 430, 5),
подвижного главного зубца (рис. 430, 4), внутреннего зубца (рис. 430, 3)
и пластинки, несущей ряды острых крючковидных зубцов (рис. 430, 9).
Гнатохилярий (gnathochilarium) представляет собой большой
непарный орган сложного строения, прикрывающий снизу значительную
часть головы и верхних челюстей. Основание гнатохилярия образовано
небольшой вытянутой в поперечном направлении пребазилярной
пластинкой (praebasilare, рис. 431, 7). К ней опереди прилегают парные
подбородочные пластинки (mentum secundarium, рис. 431, 6), латераль-
но от которых помещаются маленькие основные пластинки (cardines,
рис. 431, 5). Большая часть гнатохилярия образована двумя крупными
пластинками (mentostipites, рис. 431, 4), которые соприкасаются
медиально друг с другом в задней своей половине. Между передними
отделами mentostipites лежат соприкасающиеся друг с другом довольно
крупные язычковые пластинки (lamellae linguales, рис. 431, 3),
связанные впереди с непарным, так называемым центральным телом
(рис. 431, )). Между язычковыми пластинками лежит треугольный
promentum (рис. 431, 9). На диетальнЫх концах lamellae linguales и
mentostipites располагаются короткие щупики: пара внутренних
(рис. 431, 11) и две пары наружных (рис. 431, 10). На концах щупиков
видны небольшие обонятельные конусы (рис. 431, 2). Отдельные части
гнатохилярия снабжены, кроме того, чувствительными волосками.
Своим основанием гнатохилярий связан с широкой непарной пластинкой
(hypostoma, рис. 431, 8), которая является стернитом I туловищного
сегмента.
1 Некоторые авторы считают, что гнатохилярий гомологичен первой паре нижних
челюстей, тогда как вторая пара максилл отсутствует. При этом предполагается, что-
в состав головы двупарноногих кроме акрона входят не четыре, а три сегмента
(см.: Догель В. А. Зоология беспозвоночных. М., 1981, с. 332).
408
Гнатохилярий представляет собой результат слияния конечностей
второго максиллярного сегмента (максилл II) с элементами стернита
этого же сегмента. По мнению некоторых авторов, пока еще
непроверенному эмбриологически, praebasilare есть только отчленившаяся
задняя часть стернита. Остальная часть стернита разделилась и
образовала promentum, парный mentum secundarium и частью вошла в
состав mentostipites. Эти последние образовались в результате слияния
стернальных элементов со стволиками (stipites), которые, в свою
очередь, рассматриваются как гомологи тазиков. Lamellae linguales
отвечают самим конечностям. Щупики, несущие органы чувств, суть
отчленившиеся участки тех частей, на которых они сидят. Наконец, cardines,
по-видимому, отвечают плевритам второго максиллярного сегмента.
Туловище состоит из неопределенного и большого числа
сегментов (более 40). По строению главным образом брюшной стерналь-
ной части сегментов, туловище делится на два неравных отдела:
передний— грудь (thorax, см. рис. 429, /—IV), состоящий из четырех
истинных сегментов, и задний — брюшко (abdomen,— см. рис. 429, V, VI,
VII и т. д.), к которому относятся все остальные сегменты. Каждое
кольцо брюшка представляет собой двойной сегмент, или диплосомит,
Рис. 430. Мандибула Schizoph-
yllum sabulo.sum (ориг.):
1 — премандибулярный отдел, 2 —
щека, 3— внутренний зубец, 4—
главный зубец, S — средний членик
преманднбулярного отдела, 6 —
основной членик преманднбулярного
отдела, 7 — пластннкн щекн, 8 —
терка, 9 — ряды гребневидных
зубцов
Рис. 431. Гнатохилярий Schizoph-
yllum sabulosum (ориг.):
1 — центральное тело, 2 —
обонятельные конусы, 3 — язычковая пластинка,
4 — mentostipes, 5 — основная
пластинка, 6 — подбородочная пластинка, 7 —
пребазилярная пластинка, 8 — гипосто-
ма, 9 — promentum, 10 — наружные
щупики, // — внутренний щупик
409
Рис. 432. Schizophyllum sabulosum.
Стернальная часть IV грудного
сегмента самца (ориг.):
/ — стернит IV сегмента, 2 — тергит IV
сегмента, 3 — вертлуг, 4 — бедро I, 5 —
бедро II, 6 — голень, 7 — лапка, 8—коготок,
9 — дополнительный членик, 10 — тазнк,
// — расширение голени, 12— стигма, IS —
трахеальный мешок, частично
просвечивающий через стернит, 14 — пучок трахей
так как является продуктом
слияния двух соседних истинных
сегментов. В авязи с этим грудные
сегменты несут по одной паре (см.
рис. 429, 8, 9, И), а брюшные по
две пары конечностей (см/ рис.
429, 12).
Изолированные сегменты
туловища имеют кольцеобразную
форму. Каждый грудной сегмент
состоит из очень широкого тергита, края
которого, загибаясь, образуют
боковые стенки сегмента. Нижние
концы тергита (рис. 432, 2) почти
сходятся на брюшной стороне,
оставляя место только для небольшого
стернита (рис. 432, 1). Исключение
представляет только I сегмент,
который лишен конечностей и вообще
резко отличается по своему
строению. Его тергит особенно велик и
резко выделяется не только
размерами, «о и формой, представляя так
называемый шейный щиток, или
collum (см. рис. 429, /). Стернит
этого сегмента (hypostoma) сместился вперед к основанию головы, где
он связан с гнатохилярием.
Стерниты следующих двух грудных сегментов (II и III)
отличаются свободно-подвижным соединением с тергитами и, в связи с
положением парного полового отверстия на III грудном сегменте, вытеснены
на один сегмент вперед. Так, стернит II сегмента лежит под шейным
щитком, а стернит III сегмента — под тергитом II сегмента. Что
касается стернита IV грудного сегмента, то он сохраняет свое положение
против своего же тергита (рис. 432).
Каждый из двойных сегментов брюшка, или диплосомит (см.
рис. 429, V-—IX), также состоит из большого тергального отдела и
маленького стернального. Тергиты двух сегментов, входящих в состав
диплосомита, срастаются воедино, но между ними сохраняется слабая
шовная линия. Тергит переднего сегмента называется просомитом (см.
рис. 429, 20), тогда как тергит заднего называется метасомитом (см.
рис. 429, 19). Как и на груди, тергиты загибаются на брюшную
сторону, но здесь края их срастаются со стернитами первичных сегментов
(рис. 433). Эти последние, однако, не опаяны друг с другом. Таким
образом, каждый диплосомит имеет общий тергальный отдел и два
стернита. Из них передний, больший (рис. 433, 1) налегает своим
краем на задний (рис. 433, 3). Задние края стернитов несут по паре
вырезок, в которые вдаются основания тазиков конечностей.
Строение как грудных, так и брюшных сегментов лучше всего
может быть изучено на изолированных и вываренных в едком кали коль-
410
Рис. 433. Schizophyllum sabulosum. Стернальная часть дипло-
сомита (ориг.):
/ — передний стернит, 2 — просомит, 3 — задний стернит, 4 — метасомит,
5 — передняя нога диплосомита, 6 — задняя нога диплосомита, 7 — та'-
зик, 8 — дополнительный членик, 9— вертлуг, 10 — бедро I, // —
бедро II, 12 — голень, 13 — лапка, 14 — коготок, 15 — стигмы, 16 — трахе-
альные мешкн, просвечивающие сквозь стерниты, 17 — трахеи заднего
трахеального. мешка, 18— пучок трахей переднего трахеального мешка
цах. При рассматривании диплосомита с его передней стороны видно,
что тергит переднего сегмента, т. е. просомит, загибается своим
передним краем внутрь, образуя довольно широкую вертикальную
хитиновую складку — профрагму (рис. 434, 2), которая служит эндоскелетом,
так как к ней прикрепляется мощная продольная мускулатура. Задний'
край каждого метасомита налегает на передний край -просомита
следующего сегмента, и каждое кольцо туловища может вдвигаться своим
передним краем в предыдущее. Такое вдвигание может происходить в
особенности на спинной стороне, когда тело свернувшегося кивсяка
разворачивается (рис. 435, А). Наоборот, когда животное
сворачивается спиралью на брюшную сторону в целях самозащиты, спинные
отделы колец наиболее выдвигаются друг из друга (рис. 435, Б). Метасо-
миты отличаются тонкой продольной бороздчатостью; поверхность
каждого просомита совершенно гладкая, что объясняется тем, что по ней
скользит предыдущее кольцо.
Каждый диплосомит, за исключением первого (V туловищного
кольца), несет пару маленьких боковых отверстий ядовитых желез
(foramina repugnatoria, см. рис. 429, 21). Эти отверстия лежат на
границе между про- и метасомитом на небольшом расширении последнего.
Ядовитые железы имеют защитное значение. Секрет их изливается на-
411
Рис. 434. Schizophyllum sabulosum.
Изолированный диплосомит. Вид
спереди (ориг.):
/ — тергальная часть, 2— профрагма, 3—
трахеальные мешки, 4 — стернальная часть,
5 — передняя нога, в — задняя нога, 7 —
пучки трахей
Рис. 435. Четыре диплосомита Julidae прк
распрямленном (А) и- свернутом на
брюшную сторону туловище (Б) (из Attems):
/ — просомит, 2 — метасомит
ружу, когда животное сворачивается спиралью, и у кивсяков обладает
характерным запахом и содержит иод и синильную кислоту.
На стернитах лежат дыхальца, или стигмы. Они располагаются
впереди основания ножек несколько латерально (см. рис. 432, 12;
433, 15), Каждый стернит имеет одну пару стигм. Поэтому брюшные
кольца несут две пары стигм, тогда как грудные снабжены только од- .
ной парой. Как можно видеть на препаратах, изготовленных из
вываренных в едком кали стернальных отделов туловищных колец, каждая
стигма ведет в небольшой треугольный трахеальный мешок, выстланный
хитином (рис. 432, 13; 433, 16; 436, 7 и 8). От переднего края этого
мешка отходят пучки многочисленных тонких трахей (рис. 436, 9, 10).
Особое внимание следует обратить на положение парного
полового отверстия. Оно лежит на дне брюшного виячивания на III грудном
сегменте (см. рис. 429, 10). Последний сегмент туловища (telson)
несет заднепроходное отверстие и лишен стигм, ядовитых желез и
конечностей. Он состоит из кольцевидной части (см. рис. 429, 17), на
нижне-заднем краю которой сидит непарная анальная чешуйка (см.
рис. 429, 14), тогда как ее боковые края снабжены парными анальными
лопастями (см. рис. 429, 15). Между чешуйкой и лопастями находится
заднепроходное отверстие. Кольцевидная часть анального сегмента
образует спинной, торчащий назад, непарный вырост, которым у Schi- .
zophyllum заканчивается тело (см. рис. 429, 16). На конце этого
выроста, так же как на анальных лопастях, видны чувствительные волоски.
Некоторые авторы считают кольцевидное основание анального
сегмента за особый преанальный сегмент. Однако эта точка зрения не
412
подтверждается ни
анатомическими, ни эмбриологическими
данными.
Между последним дипло-
сомитом и тельсояом
расположена зона образования новых
сегментов (ом. рис. 429, 18).
Здесь находится несколько
колец, еще не вполне
сформированных и лишенных
конечностей. Постэмбриональное
развитие Diplopoda совершается
по типу анаморфоза, который у
Julidae не заканчивается даже
тогда, когда животное
достигает половой зрелости, т. е.
совершается 'в течение всей
жизни. Из яйца выходит
личинка с шестью сегментами —
четырьмя грудными, двумя
брюшными — и тельсоном
(рис. 437, А). Имеется три
пары ног (на II, III и IV
сегментах, рис. 437, А, 4). Брюшные
кольца несут по две пары
зачатков ног (рис. 437, А, 3).
В дальнейшем с каждой
линькой число брюшных колец и
конечностей увеличивается за
счет формирования новых
Рис. 436. Schizophyllum mediterraneum.
Трахеальные мешки и трахеи второго диплосомита
(по Krug):
1 — передний стернит, 2 — задний стернит, 3 — тазик
передней ноги, 4 — тазик задней ноги, б — задняя
стигма, 6 — передняя стигма, 7— задний
трахеальный мешок, 8 — передний трахеальный мешок, 9 —
толстые трахеи заднего трахеального мешка, 10 —
пучки тонких трахей переднего трахеального мешка,
// — сухожильный тяж
сегментов в пролиферационной зоне
(рис. 437, Б).
Конечности туловища представлены ходными ногами,
которые расположены на брюшной стороне почти всех сегментов.
Лишены ножек только I грудной сегмент (сегмент шейного щитка),
сегменты пролиферационной зоны и тельсон. В связи со смещением вперед
ш ш
Рис. 437. lulus sp. A — личинка по вылуплении из яйца; Б — вторая личиночная
стадия того же животного (из Verhoeff):
/ — голова, 2 — тельсон, 3 — зачатки брюшных ног, 4— грудные ноги, 5 — антенна, 6 —
кишка, 7 — ядовитая железа, 8 — брюшные ноги. I—IV — грудные сегменты, V—X — диплосо-
миты брюшка
413
стернитов двух передних грудных сегментов конечности этих сегментов
также переместились вперед. Соответственно с этим, под collum
прикрепляются ножки, по происхождению принадлежащие II сегменту
(см. рис. 429, 8), а на II сегменте — ножки, принадлежащие III
сегменту (см. рис. 429, 9). Таким образом, брюшная поверхность III
сегмента освобождена, и на ней располагается парное половое отверстие.
Грудные сегменты несут по одной паре ног, диплосомиты брюшка
соответственно своей двойной природе —две пары ног (см. рис. 429, 12).
Типичная ножка состоит из семи члеников: вытянутого тазика (соха,
см. рис. 432, 10), вертлуга (trochanter, см. рис. 432, 3), бедра I (femur,
рис. 432, 4), бедра II (postfemur, см. рис. 432, 5), голени (tibia, см.
рис. 432, 6), лапки (tarsus, см. рис. 432, 7) и подвижного коготка (см.
рис. 432, 8). Кроме того, между тазиком и вертлугом находится очень
маленький дополнительный членик, который .представляет собой
участок, отчленившийся от тазика (см. рис. 432, 9). Все членики подвижно
сочленены друг с другом и несут редкие волоски. На препаратах,
приготовленных без предварительного вываривания в едком кали, видна
внутренняя мускулатура конечности.
Ножки самца отличаются наличием на голени внутреннего
пластинчатого расширения (см. рис. 432, 11). Помимо того у самцов
особое строение имеют передняя грудная пара ног, а также обе пары
второго диплосомита, или VII туловищного кольца..
Передние ножки самца чрезвычайно малы и имеют вид загнутых
внутрь крючков (см. рис. 429, 8). Каждая ножка состоит из
сросшегося со стернитом, вытянутого в поперечном направлении тазика
(рис. 438, 2) и из трех подвижных члеников, снабженных волосками.
Ножки второго диплосомита, половые ножки, или гоноподы, погруже-
Рис. 438. Schizophyllum sabu- Рис. 439. Schizophyllum sabulosum. Гонопо-
losum. Видоизмененная ножка ды самца. А — передняя; Б — задняя гоко-
второго грудного сегмента сам- поды (А — ориг., Б — по Schubart)
ца (ориг.):
/ — конечный крючковидный
членик, 2 — тазик
414
ны в особое карманообразное впячивание покровов и чрезвычайно
сильно модифицированы. Чтобы их рассмотреть, необходимо, осторожно
отпрепарировав их иглами, приготовить тотальный препарат в глицерин-
желатине.
У Schizophyllum sabulosum передняя пара гонопод имеет вид не-
расчлененных широких, суженных на концах и несколько
искривленных пластинок (рис. 439, Л). Вторая пара имеет сложное строение. На
дистальном конце каждой видоизмененной конечности видны четыре
выроста характерной для этого 'вида формы (рис. 439, Б).
Строение гонопод самца у Diplopoda чрезвычайно разнообразно
и отличается даже у близких форм, этому признаку придается важное
значение в систематике.
Гоноподы способны выпячиваться наружу и служат для
перенесения спермы к половым отверстиям самки во время копуляции. Перед
копуляцией самец изгибает переднюю часть тела на брюшную сторону,
сближая III и VII кольца. При этом гоноподы принимают из половых
отверстий сперматофоры; сперма поступает в особые бутылкообразные
ямки задних гонопод. При копуляции оба животных прилегают друг к
другу брюшными поверхностями так, что VII кольцо самца находится
против III кольца самки. Самка, видимо, держится челюстями закрюч-
ковидные передние ножки самца. Гоноподы самца играют роль
'совокупительных органов.
КЛАСС INSECTA—НАСЕКОМЫЕ
Практическая важность и многообразие насекомых давно
привели к выделению энтомологии в качестве самостоятельной дисциплины.
Поэтому большой практикум по беспозвоночным не ставит в этом
разделе задачу замены энтомологического практикума, в частности
ознакомления с представителями отдельных отрядов насекомых. Это —
дело большого практикума по энтомологии. Наша задача — получение
общих представлений о насекомых как об одном из классов Arthropo-
da и овладение методами их морфологического и анатомического
исследования. Это может быть достигнуто подробным изучением одного-
двух объектов. Выполнив это и имея опыт прохождения большого
практикума, учащийся без труда может перейти к изучению любого
другого представителя класса насекомых как зоологического объекта.
Выбор объектов определяется доступностью материала, степенью
его изученности, наличием специальной литературы. Таракан —
классический объект изучения насекомых. Он рассматривается как пример
насекомого с неполным превращением. Вторым объектом служит жук-
плавунец, насекомое с полным превращением, уровень изученности
которого особенно высок.
Во избежание повторений некоторые отделы изложены более
подробно для одного объекта и менее подробно для другого. Например,
мускулатура тела и строение сложных глаз подробно описываются для
таракана и менее подробно для плавунца.
415
ПОДКЛАСС ECTOGNATHA —ОТКРЫТОЧЕЛЮСТНЫЕ,
ИЛИ НАСТОЯЩИЕ НАСЕКОМЫЕ
ОТРЯД BLATTOIDEA —ТАРАКАНЫ
Blatta orientalis L. — Черный таракан
Материалы и методы изучения. Тараканы (относятся к сем. Blattidae)
поселяются в жилище человека. Пойманных тараканов можно содержать (зимой — в теплом
месте) в садках с проволочными сетчатыми стенками или в ящиках с сетчатой,
плотно закрывающейся крышкой, на дно которых помещается в несколько слоев бумага
и стружки. Их кормят размоченным в воде хлебом с прибавлением небольшого
количества сырых овощей, а для питья ставят мелкий сосуд с водой.
В основном описывается черный таракан, но иногда привлекаются и другие
близкие виды: пруссак (Blattebla germanica L.) и американский таракан (Periplaneta arne-
ricana L.).
Изучение внешней морфологии (рис. 440) проводится на сухих или
заспиртованных, а также на вываренных в едком кали тараканах. На последних лучше всего
видны детали расчленения и отдельные склериты, входящие в состав грудных и
брюшных сегментов. Из придатков (усики, ротовые части, ноги и т. д.)
изготовляются тотальные бальзамные препараты. Так же лучше всего поступить и со склеритами
груди и брюшка.
Голова и' ее придатки. Голова таракана гипогнатического
типа, с направленными вниз и немного назад ротовыми придатками,
сильно уплощенная в передне-заднем направлении. Она очень
подвижно сочленена с переднегрудью и сверху частично прикрыта
выступающим передним краем переднеспинки, под который может быть втянута
почти полностью.
Голова окружена сильно хитинизированной головной капсулой
(epicranium), открытой сзади и снизу, к которой подвижно причленя-
Рис. 440. Blatta orientalis. A — самец; Б — самка (из Kukenthal)
416
ются усики (antennae) и видоизмененные конечности части вошедших
в ее состав первичных сегментов — ротовые придатки (trophi).
В головной капсуле таракана (рис. 441) различают несколько
отделов. Переднюю лицевую часть -головы занимает фронто-клипеальный
отдел. Он состоит из лобного, или фронтального, склерита (frons,
рис. 441, А, 6) и примыкающего к нему снизу наличника (elypeus,
рис. 441, А, 7), которому подвижно причленяется верхняя губа (lab-
rum). Лобный склерит отделяется от наличника не
резко выраженным эпистомальным швом (рис. 441, А, 10), концы
которого упираются во внутренние углы оснований -верхних челюстей.
Боковые границы лобного склерита идут к нижнему краю сложных
глаз, огибают с внутренней стороны ямки основания антенн и
переходят в лобный, или фронтальный, шов (рис. 441, А, 3), отделяющий
лицевой фронто-клипеальный отдел головной капсулы от теменного
(vertex, рис. 441, А, 2). Последний по средней линии разделяется ко-
рональным швом (рис. 441, А, 1) на два париетальных или теменных
склерита, 'баковые границы которых примыкают к сложным глазам.
Позади (рис. 442) эти склериты окружают узкий изогнутый затылочный
склерит (occiput, рис. 442, 3), отделенный от них затылочным швом.
Затылочный склерит сверху и с боков окружает затылочное отверстие
(foramen occipitale, рис. 442, 4). С нижней стороны затылочное
отверстие замыкается основанием нижней губы. Боковые отделы головы,
расположенные под сложными глазами и отчасти позади них,
называются щеками (genae, см. рис. 441, В, 11), в заднем отделе они
переходят в париетальные склериты.
Многочисленные попытки установления гомологии между отдель:
ными склеритами и швами головной капсулы и сегментами, вошедши-
S
Рис. 441. Голова Blatta orientalis. Вид спереди (Л), сверху
(Б) и сбоку (В) (из Miall.and Denny):
/ _ корональный шов, 2 — темя, 3 — фронтальный шов, 4 — глазок,
5 — усик, 6 — лобный склерит, 7 — наличник, 8 — верхняя челюсть,
9 — нижняя губа, 10 — эпистомальный шов, Л — щека, 12 — ямка
усика, 13 — сложный глаз, 14 — шейный склерит, 15 — затылок,
16 — темя, П — лоб, 18 — верхняя губа, 19 — нижнечелюстной
щупик, 20 — ннжнегубной щупик
14—2
417
ми (в состав головы и границами между ними,
пока с достоверностью установили, что фрон-
то-клилеальный отдел, ограниченный сверху
фронтальным швом, и область сложных глаз,
принадлежат к простомиальной области
первичной головы. Основания антенн, положение
которых на голове насекомых изменчиво,
относятся ж тому же сегменту. Для всех же
остальных отделов головной капсулы пока
гомологии установить не удалось.
Голова соединяется с грудью при помощи
танкой шейки, образованной за счет
разрастания межоегментальной перепонки,
соединяющей голову с переднегрудью. Ее мягкие
стенки укреплены восемью цервикальными
склеритами (см. рис. 441, Б, 14): двумя
спинными, двумя брюшными и четырьмя боковыми.
Ротовые органы таракана — грызуще-
жующего типа. Они состоят из хорошо
подвижной непарной верхней губы (labrum),
пары верхних челюстей (mandibulae), пары
нижних челюстей (maxillae) и нижней' губы
(labium, рис. 443).
Верхняя губа образует переднюю стенку
ротовой полости и прикрывает спереди
ротовые органы (см. рис. 441). Это не конечность.
По происхождению верхняя губа является
кожной складкой, отчленившейся от просто-
миального отдела первичной головы. У таракана она, посредством двух
боковых сочленений и соединительной перепонки, подвижно сочленена
с наличником. Верхняя губа может двигаться вверх и вниз, по
отношению к наличнику, и слегка отводиться вперед. Она приводится в
движение двумя парами мышц, отходящих от лобного склерита. Имеет
форму пластинки с выпуклым, закругленным свободным нижним
краем. Бе дистальный 'отдел несет многочисленные чувствительные волоски
и ямки. На внутренней стороне имеются направленные назад волооки.
С их помощью верхняя губа поддерживает откусываемые
мандибулами кусочки пищи, в чем легко убедиться, отрезав ее у живого
таракана и наблюдая после этого за действием его ротовых частей.
Верхние челюсти (рис. 443, А) являются мощными сильно хитини-
зированными темными пластинками, внутренний край которых несет
сильно развитые зубцы, число и величина последних варьируют, но
зубцы одной стороны всегда плотно входят в промежутки между зубцами
другой. Они служат для откусывания пищи, или ее соскребывания с
субстрата. Зубцы, лежащие ближе к основанию, уплощены и
расширены. Они служат для перетирания пищи. Мандибулы двумя суставами
сочленены с головной капсулой и могут двигаться щипцеобразно одна
по отношению к другой. Каждая из них приводится в движение двумя
мышцами — отводящей и приводящей. Наиболее мощная последняя
Рис. 442. Голова Blatta ori-
entalis. Вид сзади (из Miall
and Denny):
/ — корональный шов, 2 —
затылок, 3 — затылочный склерит,
4 — затылочное отверстие, 5 —
сложный глаз, 6 —
париетальный склерит, 7 — car do, 8 —
stipes, 9 — нижнечелюстной
щупик, 10 — наружная лопасть,
// — внутренняя лопасть, 12 —
щупнк нижней гуйы, 13 —
наружная лопасть, 14 —
внутренняя лопасть, 15 — подподборо-
док
418
прикрепляется широким
основанием в затылочном отделе головы.
Нижние челюсти (макоиллы,
рис. 443, Б) состоят из основного
членика и сидящих на нем двух
подвижных лопастей и щупика.
Основной членик разделяется на
проксимальный отдел — cardo^
подвижно (Причленяющийся к голове, и
дистальиый стволик — stipes. Оба
отдела подвижно сочленены друг с
другом. Они состоят из выпуклых
наружных склеритов, соединенных
перепонкой о головной капсулой.
Наружная лопасть (galea)
слабо хютинизи'роеана и на конце
несет многочисленные волооки.
Внутренняя лопасть (lacinia) заострена
на дистальном конце, образуя два
темно хитинизированных загнутых
внутрь зубца, у основания которых
имеется вырез. На ее внутренней
стороне расположены в два ряда
длинные слегка изогнутые щетинки.
Внутренняя лопасть активно
участвует в раощипьгвании пищи, тогда
как наружная играет роль органа
очистки. Пятичлениковый щупик
(palpus maxillaris) подвижно
сочленен с наружным краем stipes. Его
базальный членик очень короткий,
дистальный членик на внутренней стороне несет в продолговатом
углублении чувствительный орган сложного строения (рис. 475) —орган
вкуса.
Нижняя губа (labium, рис. 443, В) с нижней стороны здмыкает
ротовую полость. Она состоит из крупного базального склерита,
замыкающего с брюшной стороны головную капсулу,— подподбородка
(submentum). К нему примыкает не полностью твердо хитинизирован-
ный отдел — подбородок (mentum), несущий две пары терминальных
придатков — наружные и внутренние лопасти (paraglossa и glossa), и
пару нижнегубных трехчлениковых щупиков. Уже одно внешнее
строение нижней губы свидетельствует о ее происхождении путем слияния
по средней линии базальныхотделов вторыхмаксилл. Слившиеся кардо
образовали подподбородок, а стипес — подбородок. Терминальные
части соответственно гомологичны наружным и внутренним лопастям и
щупику максилл. Лопасти нижней губы слабо хитинизированы. Их
функция — не размельчение пищи, как у внутренних лопастей нижних
челюстей, а слизывание жидкой пищи. Нижнегубной щупик на конце
имеет хорошо развитое чувствительное поле и несет сходную с нижне-
Рис. 443. Ротовые органы Blatta orien-
talis. A — верхние челюсти (слева —
изнутри, справа — снаружи); Б — нижняя
челюсть; В — нижняя губа (из Miall
and Denny):
Б — 1 — cardo, 2 — stipes, 3 — щупик, 4 —
наружная лопасть, 5 —внутренняя лопасть; В —
1 — подподбородок, 2 — подбородок, 3 —
щупик, 4 — наружная лопасть, 5 — внутренняя
лопасть
14**
419
челюстным щупиком функцию. Подтлоточник (hypopharynx, язык)
представляет собой непарное образование, лежащее внутри у
основания нижней губы и являющееся результатом видоизменения нижней
стенки начала глотки. Он состоит из двух S-образно выгнутых боковых
хитиновых полосок, между которыми находится срединное языкообраз-
ное утолщение, в дистальном отделе покрытое многочисленными
мелкими волосками. Подтлоточник принимает участие в слизывании жидкой
пищи. Более существенна его роль в проглатывании пищи, которую он
проталкивает в глотку, двигаясь спереди и снизу назад и вверх.
Антенны. Кроме ротовых придатков, на голове таракана имеется
пара усиков, или антенн. Они щетинковидно-нитевидной формы, у
самцов немного длиннее, чем у самок, и состоят из 75—90 члеников, из
которых три основных крупнее остальных (рис. 444). Отдельные членики
при помощи сочленовных перепонок подвижно соединены друг с
другом. Их поверхность покрыта направленными вперед упругими
щетинками и чувствительными колбочками. Антенны—органы осязания и
обоняния таракана.
Грудь и ее придатки. Грудь состоит из трех сегментов:
передне-, средне- и заднегрудного (pro-, meso- и metathorax). У самок
сверху каждый сегмент покрыт спинным щитком (тергиты). У самцов
спинные щитки средне- и заднегруди развиты слабее, они тонкие,
полупрозрачные, так как сверху прикрыты кожистыми передними
крыльями (рис. 445). Спинной щиток переднегруди (переднеспинка) —
наибольший. Оверху он почти целиком прикрывает головку. Средне- и
заднеспинка самок по величине значительно уступают переднеспинке:
они уже и короче. Склериты брюшной стороны (стерниты) и боков,,
вследствие мощного развития коксальных члеников ног, очень малы
, 3 JJ 20 49 18 П 16 151>f
1 1 ?1 I 1 \ \ \ \ k iS
Рис. 444. Blatta
orientalis. Членики основания
усиков самца (А) и
самки (Б) (из .Miall and
Denny)
Рис. 445. Blatta orientalis: Тело самки (А) и
самца (Б) Вид сбоку (из Miall and Denny):
1, 2, 3 — передне-, средне-, заднеспинка, 4 — элитры, 5—
И — стерниты II—VIII сегментов брюшка, 12 — стерниг
IX сегмента брюшка (виден лишь у самца), 13 — тергиг
X сегмента, 14 — церки, 15—21 — тергиты сегментов
брюшка (у самца часть нх прикрыта элнтрами), 22 — грнфель-
кн (имеются только у самца).
420
(рис. 446). Брюшной щиток переднегруди —
простернум — является продольно вытянутой
простой пластинкой (рис. 446, 2), мезостернум—
в виде более широкой пластинки, у самцов
иногда, а у самок почти всегда продольно
разделенной на два склерита (рис. 446, 4). Метастернум
у обоих полов разделен на два склерита (рис.
446, 6). Позади стернитов, особенно на.средне-
и-заднегруди, находится уступ, к бокам которого
причленяются тазики .ног. В средней части этих
уступов помещаются (промежуточные склериты:
в переднегруди — в виде небольшой, вытянутой
поперечно пластинки, в средне- ш заднегруди—
в шде трехконечных вилок (furca, рис. 446, 5
и 7), с короткой передней и длинными
боковыми ветвями. К 'ним изнутри прикрепляются
мышцы. Боковые, или плевральные склериты (эпис-
терны), развиты очень слабо. Они в виде
небольших пластинок лежат между основаниями
тазиков и стерштами.
Ноги таракана—-резко выраженного бега-
тельного типа. Все три пары имеют сходное
строение. Передние — наиболее короткие,
задние— наиболее длинные. Тазики (коксы, соха,
рис. 447) сильно развиты, длинные, широкие и
уплощенные, почти одной длины с бедрами. Сих
величиной связано слабое развитие стернитов.
Вследствие мягкости окружающих их покровов
они обладают значительной подвижностью.
Между тазиками /и плевральными склеритами
помещается небольшой субкоксальный членик. За
тазиками следует маленький, короткий вертлуг
(trochanter), неподвижно сочлененный с последующим более тонким1
стройным бедром (femur). Голени (tibia) приблизительно одной
длины с бедрами и, как и последние, покрыты
многочисленными шипами. Лапка (tarsus) ■— пятичлениковая, причем первый членик
значительно длиннее остальных. Конечный членик лапки вооружен дву-'
мя коготками с присоской (pulvillum) между ними. На вершинах пер-'
вых четырех члеников могут находиться небольшие подушечки
(рис. 447). Бег — основной тип передвижения таракана. При этом пе>
редняя и задняя ноги одной стороны двигаются одновременно со
средней ногой другой стороны. Кроме бега разной скорости, тараканы
могут совершать и прыжки, главную роль при этом в качестве отталки-
вателей играют задние ноги.
Крылья. На средне- и заднегруди таракана помещаются крылья,
функционирующие, и то в слабой степени, только у самцов. У
последних передняя пара более сильно хитинизирована и превращена в род
надкрыльев, прикрывающих и защищающих более нежные и
перепончатые задние крылья. В спокойном состоянии крылья направлены назад
Рис. 446. Blatta orientalis.
Склериты брюшной
стороны сегментов груди (из
Miall and Denny): ■
1 — шейные склериты, 2 —
простернум, 3 —
промежуточный склерит
переднегруди, 4 — мезостернум, 5 —
вилка ■ среднегруди, 6 —
метастернум, 7 — вилка
заднегруди, 8> 9 — добавочные
склериты, места отхожде.-
ния аподем внутреннего
скелета
421
- задняя но-
■ лапка Blat-
и прикрывают брюшко. Их
внутренние края несколько заходят друг
за друга.
Жилкование передних крыльев
плохо заметно благодаря их
сильной хитинизации. Однако на
просветленных тотальных .препаратах
можно (различить лежащий у
основания переднего края крыла
темный субкостальный отдел и
следующую за ним сеть радиальных,
медиальных <и кубитальных 'жилок. От
них анальным швом отделено
анальное поле. Задние крылья нежные и
тонкие. Их жилкование сходно с
таковым элитр с той разницей, что
анальное иоле (развито значительно
сильнее, может веерообразно
складываться и отделено от остальной
части крыла прямым анальным
швом, по 'которому перегибается
сложенное в спокойном состоянии
анальное ноле.
Брюшко таричленяется к зад-
негруди широким основанием и по
отношению к ней малоподвижно.
Першоначально, во время
эмбрионального развития, закладываются
11 брюшных сегментов, но потом
число их уменьшается за счет
последних сегментов и, частично, за счет первого, и количество видимых
сегментов' у взрослого насекомого никогда не бывает полным. У самок ,
их меньше, чем у самцов. Каждый сегмент состоит из спинного
склерита— терпита и из брюшного — етернита. Оба склерита соединены друг
с другом или мялкой соединительной перепонкой, или при (помощи
узких плевральных склеритов. У самцов тер гиты более плоские, у самок
они более выпуклы и сильнее хитинизированы. Каждый из брюшных
сегментов, за исключением двух последних, несет по бокам на
плевральных склеритах по паре стигм.
Со спинной стороны (рис. 448, 449) у самцов и самок можно
различить 10 сегментов (тергитов). I тергит и особенно VIII и IX короче
остальных. Оба последние тергита могут быть частично скрыты под
VII тергитом. Последний — X тергит хорошо развит и называется суп-
раанальной пластинкой. Боковые края тергитов сверху несколько
заходят за стерниты. Брюшная сторона (рис. 448 и 449) значительно
отличается у самцов и самок по числу свободных видимых стернитов
конца брюшка, что связано с различиями в строении наружного
полового аппарата у самцов и самок. У самцов можно различить 9
стернитов, тогда как у самок — только 7, последующие же два сегмента видо-
Рис. 447. Тараканы. А ■
га Blutta orientalis; Б -
ta germanica (A — из Handlirsh, Б —
из Wille):
/ — subcoxa, 2 — тазик, 3 — вертлуг, 4 —
бедро, 5 — голень, 6 — лапка. На концах
члеников ясно видны присасывательные
подушечки
422
Рис. 448. Брюшко самки Blatta germanica, Вид сверху (А)
и снизу (Б) (из Wille):
; — заднеспинка, 2 — заднее крыло, 3 — первая брюшная стигма,
4—12 — тергиты I—IX сегментов брюшка, 13 — супраанальная
пластинка, 14 — церки, 15—19, 21 — стерниты I—VII сегментов брюшка,
20 — восьмая брюшная стигма, 22 — тергиты
изменены и погружены внутрь, где образуют род кармана, в котором
образуется яйцевой пакет (оотека). I стернит у обоих полов
рудиментарен и представлен маленькой овальной пластинкой. Передний край
II стернита сложно изогнут в соответствии с положением тазиков
задней пары ног. IX стернит самца, называемый субгенитальной
пластинкой, несет два придатка — грифельки (styli).. У Blattella germanica
IX стернит асимметричен,
он с одного- края имеет
более 'или менее глубокий
вырез. Вследствие этого
грифельки смещаются и один
из них может частично
редуцироваться. У самки
последний, VII, стернит
видоизменен в своей дистальной
части, которая продольно
расщеплена и образует две
лопасти — вальвы,
служащие для удержания оотеки.
На конце брюшка у
обоих полов имеются
хорошо развитые придатки —
церки (cerci). Это придатки
рудиментарного XI
сегмента. Они состоят из 8—15
одинаковых члеников, число
которых изменчиво даже у
о*
1—
о
6—
7—
£1Г>
\ Jr
Рис. 449. Blatta orientalis. Брюшко самца (А) и
самки (Б), вид снизу (из Miall and Denny):
1—9 — стерниты I—IX брюшных сегментов, 10 —
грифельки, 11 — церки, 12 — вальвы
423
одной особи. Они покрыты чувствительными волосками и являются как
бы брюшными щупиками. Когда таракан пятится назад, они
выполняют роль органов осязания.
Анатомия на вскрытии. Таракан — классический объект для
вскрытия.
Захлороформированного предварительно таракана берут за ноги в левую руку
и тонкими ножницами проводят поперечный разрез межсегментальной перепонки
между VII и VIII тергитами. Если имеют дело с самцом, предварительно отводят его
крылья и захватывают их пальцами вместе с ногами. Затем проводят с каждой
стороны боковые , продольные разрезы вдоль плевр в брюшке, а в груди несколько
ближе к средней линии перерезают переднеспинку, доводя разрезы до головы. Далее
берут сухую препаровальную ванночку, проводят нагретым скальпелем по дну пепереч-
ную линию и вплавляют туда голову таракана ротовыми частями. Задний конец
брюшка прикрепляют к дну с боков двумя булавками. Наливают в ванночку воду или
физиологический раствор; приподнимают тонким пинцетом спинку и начинают
осторожно отделять ее, перерезая остро отточенной препаровальной иглой дорзовентраль-
ные мышцы и трахеи. Всю спинку необходимо снять целиком, чтобы потом иметь
возможность рассмотреть оставшиеся прикрепленными к ней сердце и мускулатуру.
По мере отпрепаровки спинки для укрепления таракана накалывают булавки
по бокам в начале брюшка и в переднегруди, а также укрепляют булавками
отведенные в стороны крылья. У вскрытого таким образом таракана виден кишечник, густо
оплетенный трахеями и частично скрытый лопастями жирового тела.
Для дальнейшего ознакомления с анатомией таракана прежде всего
необходимо тонким пинцетом удалить жировое тело и иглами разорвать трахеи. После этого
расправляют кишечник, отводят его вбок и прикрепляют булавками. Тогда
обнажаются половые органы и частично брюшная нервная цепочка. Для дальнейшего
изучения отдельных органов см. указания в соответствующих разделах.
Мускулатура и эндоскелет. Мускулатуру необходимо
изучать на вскрытом таракане, у которого удалены все внутренние
органы, за исключением нервной системы и сердца. Отдельно изучается
мускулатура брюшной и спинной стороны тела, которые после
вскрытия, удаления внутренних органов и очистки от трахей и жирового,
тела должны быть уплотнены в 70%-ном спирте. Для изучения мышц,
двигающих голову, необходимо, чтобы она была и на препарате
брюшной стороны, и на препарате спинной, таким образом, нужно вскрыть
два экземпляра. Изучение мускулатуры головы (ротовых придатков,
антенн и глотки), как требующее очень кропотливой препаровки,
вплавленной в парафин ротовыми придатками и затылочным отделом
головы, может быть опущено. Эндоскелетные образования можно
рассмотреть на вываренных в едком кали кожных покровах.
У таракана эндоскелетные образования особенно развиты в
голове и груди:
Голова очень подвижна — она может совершать всевозможные
виды движений: сгибание и разгибание в сшшнобрюшном Направлении,
отведение и приведение в боковом направлении и повороты в обе
стороны. Эти разнообразные движения осуществляются при помощи
комбинированного действия четырех пар мышц (рис. 450, 451). Сгибает
голову m. depressor capitis horizontals (рис. 450, 2), антагонистом его
является поднимающий голову m. levator capitis horizontalis
(рис. 450, 3). В отведении головы в стороны участвуют кроме
упомянутых мышц соответствующей стороны левый и правый m. depressor
capitis obliquus (рис. 450, 1). Приведение головы в нормальное поло-
424
жение достигается сокращением соответствующих мышц
противоположной стороны. Наиболее сложное движение — поворот головы —
достигается сокращением мускула m. rotator capitis соответствующей
стороны при сопутствующем ему попеременном сокращении разгибателя
противоположной стороны и сгибателя и косой мышцы той же стороны.
Мышцы-антагонисты приводят голову в нормальное состояние.
Рис. 450. Blatta orientalis.
Мускулатура спинной стороны. Вид
изнутри (из Beier):
/ —m. depressor capitis obliquus, 2 —m.
depres3or capitis horizontalis, 3 — m.
levator capitis horizontalis, 4 — m. dorsales
abdominis, 5 —m. transversales abdominis,
5— m. dorsales abdominis'medii, 7 —
крыловые мышцы сердца, 8 — т. extensor coxae,
9 — m. flexor coxae, 10 —m. medianus
thoracis (продольная тергальная мышца),
11 — m. extensor trochanteris, 12 — m.
lateralis thoracis (косая боковая мышца)
Рис. 451. Blatta orientalis.
Мускулатура брюшной стороны. Вид изнутри
(из Beier):
1 — т. levator capitis horizontalis, 2 — m.
depressor capitis horizontalis, 3 —m. flexor
coxae, 4 — xa. extensor trochanteris, 5 — m.
extensor coxae, 6 и 9 — m. transversales
abdominis, 7 — m. ventrales abdominis, 8 —
m. ventrales abdominis medii
425
Наиболее проста мускулатура брюшка. На брюшной и спинной
стороне каждого сегмента проходят продольные мышцы (рис. 450, 4),
соединяющие передний край предыдущего сегмента с передним краем
последующего. На спинной стороне они помещаются на I—VII
сегментах, на брюшной стороне—на II—VI. На брюшной стороне эти
мышцы (m. ventrales abdominis interni, рис. 451, 7) лежат сплошным слоем
и развиты значительно сильнее, чем на опинной (m. dorsales abdominis,
рис. 450, 4). Кроме того, стерниты брюшка соединены между собой
короткими косыми мышцами (m. ventrales abdominis medii). Такие же
мышцы (т. dorsales abd. medii) соединяют тергиты между собой. Все
эти мышцы соединяют задний край предыдущего сегмента с передним
краем последующего. Система сегментальных продольных мышц
вызывает телескопическое втягивание последующих сегментов брюшка в
предыдущие, косые межсегментальные мышцы являются их
антагонистами. По бокам каждого сегмента дорзовентрально проходят
поперечные мышцы, соединяющие тергиты и стерниты каждого сегмента
между собой (m. transversales abdominis, рис. 451, 6 и 9). При их
сокращении брюшко сжимается в дорзовентральном направлении. Это
движение играет важную роль при вентиляции трахей, вызывая акт
выдыхания. ,
Мускулатура грудных сегментов значительно отклоняется от более
примитивного и упрощенного, вследствие отсутствия конечностей, типа
мускулатуры брюшка. Самки таракана совсем не летают, а самцы
обладают этой способностью в очень слабой степени. В связи с этим
специальное летательные мышцы у таракана отсутствуют, и движение
крыльев у самцов зависит от сокращения собственно мускулатуры
грудных сегментов. У представителей высших отрядов Ectognatha
(жуки, двукрылые, /перепончатокрылые и др.) отсутствие специальных
летательных мышц крыльев и зависимость движения последних от
сокращения продольных и дорзо'вентральных мышц груди с полным правом
рассматривается как прогрессивное явление, ведущее к большему
совершенству полета. Подобные отношения у таракана — представителя
одного из примитивных отрядов Ectognatha, надо рассматривать как
следствие редукции специальной летательной мускулатуры, связанное
с переходом от летающего к бегающему образу жизни, а движение
крыльев под косвенным влиянием мышц сегментов груди •— как
связанное с этой редукцией вторичное явление. Таким образом, у таракана
особенности развития мускулатуры грудных сегментов связаны только
с мускулатурой нот, за исключением переднегруди, продольная
мускулатура которой частично видоизменена в уже рассмотренную нами
мускулатуру, двигающую голову.
Спинная мускулатура грудных сегментов (см. рис. 450) очень
бедна сравнительно с таковой брюшных сегментов. В каждом сегменте
имеются продольные тергальные мышцы и косые боковые мышцы,
идущие от середины спинки каждого сегмента к бокам переднего края
спинки последующего.
Брюшная мускулатура груди сложнее и сильнее- видоизменена
сравнительно с мускулатурой брюшных сегментов. Брюшные мышцы
отходят от трубчатых аподем, являющихся внутренними отростками
426
промежуточных склеритов, лежащих между стернитами грудных
сегментов. От-передней аподемы, лежащей между передне- и среднегрудью
отходят три парные мышцы и одна непарная медиальная. Последняя
идет назад и прикрепляется к средней аподеме. Из парных мышц
передние расходятся по диагонали к основаниям тазиков передних ног,
средние направлены вбок к переднему краю оснований тазиков
средних ног, задние — отходят к концам боковых ветвей вилки среднего из
промежуточных склеритов. Этот последний образует среднюю аподему,
от которой отходят 4 пары мышц. Передняя пара проходит
диагонально вперед к заднему краю основания тазиков передних ног, вторая
пара идет вбок к переднему краю оснований тазиков задних ног, третья
пара проходит диагонально назад к концам боковых ветвей вилки
заднего из промежуточных склеритов, четвертая пара направлена назад
и прикрепляется к переднему краю второго стернита брюшка.
Задняя вилка не имеет аподемы. От ее основания вперед по
диагонали расходится пара мышц к боковым ветвям средней вилки, а
назад— пара диагональных мышц к Переднему краю второго стернита
брюшка. Кроме того, имеется пара мышц, соответствующая трансвер-
сальным тергостериальным мышцам брюшка, она отходит тоже от
основания задней вилки к бокам первого брюшного тергита. Наконец,
пара мышц соединяет боковые ветви задней вилки с серединой заднего
отдела второго стернита брюшка.
Мышцы, соединяющие аподемы передней и средней вилки с
тазиками ног, при своем сокращении поворачивают их. Это имеет место при
быстром беге таракана, позволяя ему значительно увеличивать его
«шаги». Мышцы, отходящие от боковых ветвей вилок, сокращаясь,
вызывают более тесное сближение стернальных склеритов с вилками, что
усиливает их значение как опорных образований для тазиков ног.
Мускулатура ноги изображена на рис. 452.
Пищеварительная система. Исследование
пищеварительной системы производится (прежде всего на тотально отпрепарированном
кишечнике. Отдельные участки кишечника фиксируют частями каким-
либо сулемовым фиксатором (Джильсон или др.) для последующей
заливки и изготовления поперечных срезов (окраска по Маллори). Из
слюнных желез и мальпигиевых сосудов необходимо приготовить
окрашенные квасцовым кармином или гематоксилином тотальные
препараты. Proventriculus (этот отдел передней кишки называют также
жевательным, или .мышечным, желудком) вываривается в едком кали, после
чего изготовляются тотальные препараты его зубов. Кроме микро-
томных срезов из фиксированного proventriculus можно сделать
несколько толстых поперечных срезов при помощи лезвия безопасной
бритвы (под лупой или малым увеличением бинокуляра) и приготовить
из них неокрашенные тотальные препараты1.
Все три главных отдела кишечника — передняя, средняя и задняя '
кишка — у таракана "внешне хорошо отграничены друг от друга
1 В дальнейшем при описании пищеварительной, выделительной и нервной систем
таракана излагаются параллельно анатомические и гистологические данные, что
позволяет более полно осветить некоторые физиологические вопросы.
427
■8
Рис. 452. Blatta orien-
talis. Мускулатура
левой средней ноги
(из Beier):
1 — m. flexor coxae, 2 —
m. extensor coxae, 3 —
m. extensor trochanteris,
4 — m. flexor trochanteris,
5 — m. extensor tibiae,
6 — трахея, 7 — m.
retractor tarsi, 8 — m. flexor
tarsi, 9 — m. flexor tibiae,
10 — нерв
Рис. 453. Пищеварительная система
Blatta orientalis (из Beier):
1 — глотка, 2 — пищевод, 3 — зоб, 4 —
жевательный желудок, 5 — пилорические
придатки, 6 — средняя кишка, 7—мальпигие-
вы сосуды, 8 — тонкая кншка, 9 — толстая
кишка, 10 — слепой вырост толстой
кишки, 11 — rectum, 12 — клоакальное
расширение, 13 — резервуар слюнной железы,
14 — слюнная железа, 15 — выводной
проток, 16 — непарный проток
(рис. 453,). Граница между передней и средней кишкой отмечается
впадением в кишечник восьми длинных слепых пилорических выростов,
а граница между средней и задней кишкой — местом впадения
многочисленных нитевидных мальпигиевых сосудов.
Ротовое отверстие ограничено основаниями ротовых придатков и
ведет в направленную вверх короткую глотку (рис. 453, 1), которая,
•загибаясь назад, выходит через затылочное отверстие из головной
капсулы. Здесь она сужается и переходит в пищевод (рис. 453, 2), кото-
428
Рис. 454. Blatta orientalis.
Поперечный разрез через стенку зоба (из
Miall and Denny):
1 — хитиновая выстилка, 2 — клетки
эпителия, 3 — пучки продольных мышц, 4 —
кольцевая мускулатура
рый постепенно расширяется в
объемистый мешковидный зоб,
достигающий почти половины длины
тела таракана (рис. 453, 3). Зоб,
после заметного сужения,
переходит в 'компактный proventriculus
(рис. 453, 4).
(Просвет пищевода имеет шесть
радиальных продольных складок,
сглаживающихся по мере
расширения его в зоб. Строение стенки
(рис. 454) пищевода ' и зоба
сходное. Снаружи расположен слой
кольцевой мускулатуры (рис. 454, 4), под ним лежат более редкие
пучки продольных мышц (рис. 454, 3), далее внутрь лежит слой
эпителиальных клеток, выделяющих хитиновую выстилку. Поверхность
последней покрыта волосками, расположенными вдоль складок и
направленными назад. Волоски в пищеводе более длинны; по мере
расширения его в зоб они укорачиваются.
У основания нижней губы, между ним и подглоточником (гипофа-
ринкс), открывается выводной проток слюнных желез (рис. 455). Это —
двулопастные гроздевидные железы, протоки которых соединяются и
общим каналом впадают в выводной проток, образованный путем
слияния протоков объемистых мешковидных резервуаров. Протоки имеют
хитиновую выстилку с утолщениями, наподобие спиральной нити
трахей. Общее выводное отверстие имеет кроме слабо развитой кольцевой
мускулатуры еще особое замыкательное приспособление. Оно состоит,
из крючковидно загнутого выступа основания нижней губы, который
позволяет находящемуся в покое опущенному вниз подглоточнику
замыкать отверстие выводного протока. При проглатывании пищи, когда
Рис. 455. Слюнные железы Blatta orientalis (из Miall
and Denny):
1 — выводное отверстие общего протока слюнных желез и
резервуара, 2 —общий проток желез, 3—проток правой железы, 4 —
проток одной нз правых желез, 5 —железа, 6 — резервуар, 7 —
проток правого резервуара, 8 — общий проток резервуаров
429
подглоточник приподнимается вверх,
отверстие протока открывается и из него
выходит капля слюны, омачивающая
пищу. Слюнные железы хорошо
оплетаются трахеями и богато иннервированы.
Иннервация идет из различных центров:
из 'симпатической нервной системы — от
'возвратного нерва и от бокового нерва,
отходящего от «оннективов между под-
глоточным и проторакальным ганглиями,.
Секрет содержит амилазу, способную
переводить крахмал в сахар.
Интересно строение жевательного
желудка (proventriculus). Он имеет
округло-коническую форму с обращенной
назад вершиной, конец которой вытянут
и вдается в просвет средней кишки в
виде цилиндрической трубки—кар ди а ль-
ного «лапана (valvula cardiaca).
Proventriculus (рис. 456, 3) разделен на два
отдела — передний и задний —
различные по строению и функции
(proventriculus anterior и posterior, рис. 456, 7 и
8). Передний отдел, вследствие мощного
развития кольцевой мускулатуры,
сильно вздут. Это вздутие снаружи резко
отграничивает его от зоба. Хитиновая
кутикула (переднего отдела сильно
утолщается и образует шесть больших, радиаль-
но расположенных зубов (рис. 457, 1),
концы которых направлены в просвет.
Зубы по форме бывают двух родов: три из
них имеют заостренный клювовидный
выступ (рис. 458), остальные
притуплены и имеют округленный край (рис.
459, 10). Эти различия в строении
обусловливают наиболее плотное и полное
соприкосновение поверхностей зубов при
перетирании пищи. У недавно слинявших
особей поверхность зубов покрыта
маленькими бородавчатыми возвышениями,
которые в процессе функционирования
постепенно стираются, и через некоторое
время поверхность становится гладкой.
Между основаниями зубов лежит
система продольных складок (рис. 457, 4), усаженных короткими волосками.
В каждом промежутке между зубами можно различить три главные
складки — более крупную срединную (рис. 457, 10), заходящую в
задний отдел жевательного желудка, и две боковые (рис. 457, 8), в проме-
Рис. 456. Blatta orientalis.
Схематический продольный разрез через
переднюю и начало средней кишок
(из Eidmann):
/ — пищевод, 2 — зоб, 3 — жевательный
Желудок, 4 — кардиальный клапан, 5 —
средняя кишка, 6 — пнлорические
придатки, 7 и 8- задний н передний
отделы жевательного желудка
430
5 6
Рис. 457. Blatta orientalis. Поперечный разрез через передний отдел жевательного
желудка. А —общий вид, Б — система складок между основаниями зубов (из Eidmann):
/ — зуб 2—продольная мускулатура, 3 —кольцевая мускулатура, 4 — система складок между
основаниями зубов, 5 —эпителий, б — хитиновая кутикула, 7 —основание зуба, 8 — боковая
складка, 9 — дополнительная складка, 10 — срединная складка
жутках между ними и между основаниями 'зубов лежат более мелкие
дополнительные складки (рис. 457, 9). Присутствие этих складок
обеспечивает большую подвижность зубов и лучшие растяжимость и
сжатие стенок жевательного желудка в целом.
К описанному переднему отделу непосредственно примыкает
задний. Если первый по функции является жевательным аппаратом, то
последний несет запирательную функцию. Соответственно зубам
переднего отдела тут имеется шесть радиальных валикообразных выступов.
Они.покрыты, хотя и толстым, но мягким хитином, поверхность
которого несет многочисленные направленные назад волоски. Далее кзади
начинаются еще шесть более мелких валиков, переходящих в складки
кардиального клапана. Между ними лежат складки, являющиеся
продолжениями главных срединных складок переднего отдела. При сжатии
кольцевой мускулатуры вся эта система валиков и складок сжимается
и, вследствие их эластичности, обеспечивает'
герметическое закрывание просвета
кишечника.
Мускулатура жевательного желудка
развита очень сильно (рис. 457, 459). Она состоит, как
и в других отделах передней кишки, из
наружного мощно развитого слоя кольцевых мышц
и внутреннего слоя продольных. Кольцевая
мускулатура особенно сильно развита в переднем
отделе, наиболее слабо в кардиальном клапане.
Продольная мускулатура дифференцирована
так, что часть пучков проходит по всей длине
жевательного желудка от зоба до конца карди- „,.„„,.. . , ,-
ального клапана, большая же часть пучков спе- ^458. В1аШ=Ш,
циализована в качестве особых мышц-расши- Ных зубов жевательного
рителей (dilatatores poventriculi, рис. 459, 3). желудка (из Eidmann)
431
Рис. 459. Blatta orientalis. Продольный разрез жевательного
желудка (из Eidmann):
1 — стенка зоба, 2 — кольцевая мускулатура, 3 — продольная мускулатура
(dilatatores proventricull), 4 — трахеи, 5 — эпителий, 6 —полость начальной
части кардиального клапана, 7, 8 — полости заднего и переднего отделов
жевательного желудка, 9 — полость зоба, 10 — округленный зуб, 11 —
клювовидный зуб жевательного желудка
Эти пучки одним концом прикрепляются к внутренней стороне
обращенной'к зобу поверхности каждого из зубов, а другим т- к внутренней и
нижней стенкам валиков заднего отдела, их сокращение, происходящее
в момент расслабления кольцевой мускулатуры, отвадит зубы
переднего отдела кнаружи и кверху, а валики заднего отдела кнаружи, чем
и вызывает раскрывание и расширение просвета.
Физиологическое значение отдельных частей передней кишки для
процесса пищеварения следующее. Грубо измельченная мандибулами
-пища, смоченная секретом слюнных желез, поступает порциями через
глотку и пищевод в зоб, где и происходит ее накопление. В зобу она
перемешивается и подвергается переваривающему воздействию секрета
слюнных желез. Под влиянием перистальтических движений пища
небольшими порциями поступает в передний отдел, жевательного
желудка, где она перемалывается и тщательно измельчается, после чего
поступает обратно в зоб, где опять перемешивается и частично
переваривается. Этот процесс многократно повторяется. Все это время задний
отдел жевательного желудка оказывается закрытым. Только после
того, когда пища в зобу оказывается достаточно размельченной,
происходит расслабление сфинктеров заднего отдела и кардиального клапа-
432
на и кольцевых мышц переднего,
сокращение продольных мышц, и пища
перегоняется порциями в среднюю
кишку.
Цилиндрической формы средняя
кишка у таракана — наиболее
короткий отдел кишечника. У самого
начала в нее впадают 8 (редко меньше)
длинных слепых трубчатых
отростков — пилорические придатки (см.
рис. 453, 456). Они сильно варьируют
в длину у одного и того же
экземпляра. Наиболее длинные могут
достигать длины средней кишки. Их
клетки вырабатывают пищеварительный
секрет, сходный с секретом
поджелудочной железы позвоночных
животных.
Строение стенки средней кишки
значительно отличается от передней
(рис. 460). Мускулатура развита
слабее. Имеется редкий наружный слой
продольных мышечных волокон, под
которым залегают кольцевые мышцы.
Большой интерес представляет
строение эпителия. Узкие и высокие клетки
эпителия несут секреторную функцию.
Особенностью функционирования этих клеток является выделение
секрета вместе с продольно исчерченной дистальной частью их тела.
Таким образом, в процессе секреции клетки эпителия быстро
изнашиваются и исчезают. На смену им образуются новые. Такая регенерация
секреторных клеток эпителия средней кишки происходит в
специальных очагах новообразования клеток, называемых регенерационными
криптами, помещающихся в стенке кишки под функционирующими
клетками (рис. 460). На разрезе можно наблюдать различные стадии
образования и замещения клеток эпителия.
В средней кишке под влиянием пищеварительных соков,
'выделяемых пилорическими железами и клетками эпителия, происходит
окончательное переваривание пищи и частичное ее всасывание. У таракана
(Periplaneta americana) в средней кишке найдены следующие
ферменты: амилаза, мальтаза, инвертаза, лактаза (незначительное
количество), триптаза, пептидаза и липаза, тогда как в зобу (секрет слюнных
желез) имеется только амилаза.
Вокруг находящихся в средней кишке пищевых масс в результате
деятельности эпителиальных клеток выделяется тонкая, нередко
многослойная перитрофическая оболочка (мембрана), которую видно на
гистологических срезах. Благодаря ее наличию пищевые массы
непосредственно с эпителиальными клетками средней кишки не
соприкасаются. В этом выражается механическая защитная функция перитрофи-
433
Рис. 460. Blatta orientalis. Часть
поперечного разреза стенки средней
кишки (из Miall and Denny):
/ — клетки секреторного эпителия, 2 — их
ядра, 3 — кольцевые мышцы, 4 —
продольные мышцы, 5 — регенерационные крипты
секреторного эпителия, 6 — палочковый
слой на поверхности дистальных концов-
клеток
Рис. 461. Blatta orientalis. Поперечный разрез
тонкой кишки (пилорического или кольцевого
клапана), у места впадения ее в толстую (из Miall
and Denny):
/ — просвет тонкой кишки, 2 — кольцевая мускулатура,
3 —продольная мускулатура, 4 — клетки эпителия, 5 —
просвет толстой кишки, 6 — хитиновая выстилка
Рис. 462. Blatta orientalis. Поперечный разрез
через rectum (из Miall and Denny):
/ — тяжи продольных мышц, 2 — кольцевая
мускулатура, 3 —трахеи, 4 — ректальные железы
ческой оболочки, которая
особенно важна в связи
с тем, что клетки средней
кишки насекомых не
выделяют слизи. Наряду с
защитной функцией пери-
трофическая оболочка
играет важную роль в
пищеварении. Основные
первоначальные
пищеварительные процессы по
расщеплению
полисахаридов, белков, жиров
осуществляются в той части
кишечника, которая
окружена оболочкой.
Пищеварительные ферменты,
выделяемые клетками
средней кишки и пилоричес-
ских придатков,
проникают в эту полость через
перитрофическую
мембрану. Последняя
обладает односторонней
проницаемостью и крупные
молекулы органических
веществ (полисахариды,
жиры, белки и др.) не
проходят через оболочку
наружу — в
пространство, расположенное
между оболочкой и
эпителиальными клетками
стенки кишечника. После
обработки пищи
ферментами образующиеся
моносахариды, аминокислоты
и другие относительно
небольшие молекулы,
проникают сквозь
перитрофическую оболочку и
резорбируются клетками
кишечника.
Таким образом, у
насекомых значительная
часть процесса
пищеварения осуществляется в
полости, ограниченной пери-
трофической оболочкой.
434
На границе средней и задней кишки в кишечник впадают длинные
тонкие извитые малыгагиевы сосуды, которые подробнее будут
рассмотрены в разделе об органах выделения.
Задняя кишка по своему строению сходна с передней, разница
заключается только в том, что тут продольная мускулатура лежит
кнаружи от кольцевой (рис. 461). Поверхность ее выстлана хитиновой
кутикулой, имеющей резко выраженную продольную складчатость и
несущей мелкие волооки. В задней кишке можно различить несколько
отделов (см. рис. 453), сначала идет очень короткий и тонкий отдел —
тонкая кишка. Ее задний конец имеет сильно развитую кольцевую мус-
Рис. 463. Blatta orientalis.
Трахейная система. Вид со спинной
стороны (из Miall and Denny):
1, 2 — первая и вторая грудные
стигмы, 3—10 — первая — восьмая
брюшные стигмы, 11 — rectum, 12 —
средняя кишка, 13 — пилорические
отростки средней кишки, 14 —
жевательный желудок, IS — зоб, 16 —
пищевод. Покровы спинки
удалены, внутренние органы в их
естественном положении
Рис. 464. Blatta
orientalis. Трахеи брюшной
стороны тела. Вид изнутри.
Внутренние органы
удалены (из Miall and
Denny)
435
кулатуру, образующую род сфинктера, или кольцевого клапана
(рис. 461). При ее сокращении 'Просвет тонкой кишки может
герметически закрываться. Далее идет широкая и длинная толстая кишка,
имеющая у дистального конца слепой вырост (соесшп), вслед за которым
она переходит в короткий ректальный отдел, имеющий перед анальным
отверстием пузыревидное клоакальное расширение с резко
выраженными шестью продольными складками — ректальными железами
(рис. 462). Непереваренные и неусвоенные остатки пищи скопляются
в клоакальном пузыре и выводятся в виде экскрементов наружу. Цвет
и консистенция их зависят от состава пищи и содержания воды в
организме.
Органы дыхания. Изучение трахейной системы надо
производить на специально вскрытом для этого насекомом, а не при общем
ознакомлении с анатомией таракана, так как в этом случае при
выделении отдельных органов приходится, в первую очередь, разрывать
оплетающие их трахеи. Порядок вскрытия остается тем же.
Трахейная система хорошо развита. При
помощи 10 пар стигм она сообщается с
атмосферным воздухом. В груди имеются две пары
стигм — одна на средне-, другая на заднегруди,
соответственно между основаниями первой,
второй и третьей пар ног. Восемь пар брюшных
стигм расположены на плеврах I—VIII
брюшных сегментов. Грудные стигмы несколько
отличаются по строению от брюшных. У стигм
имеются специальные запирательные
приспособления, приводимые в действие при помощи особых
мышц.
По бокам тела проходят две пары
продольных главных трахейных стволов — спинные и
брюшные (рис. 463—465). Оба ствола каждой
стороны соединены между собой посегментно
расположенными соединительными трахеями, от
которых отходят короткие поперечные стволы к
стигмам. В брюшке в каждом сегменте от
каждого из главных стволов отходит поперечная
ветвь. На брюшной стороне эти ветви
соединяются с ветвями противоположной стороны и при
помощи продольных ветвей друг с другом. Такое
расположение трахей обеспечивает, снабжение
кислородом брюшной нервной цепочки, ганглии
которой лежат в ячеях, образованных
поперечными и продольными ветвями трахей. На
спинной стороне поперечные ветви трахей каждой
Рис 465. Blatta orienta- стороны соединены друг с другом субмедиаль-
Ш. Трахеи спинной сто- ными продольными стволами, но не имеют со-
роны тела. Вид изнутри, единения с соответствующими ветвями другой
Внутренние органы уда- п я J о rj
лены (из Miall and Den- стороны. В свободном от трахеи пространстве
пу) вдоль'медиальной линии тела помещается спин-
436
Рис. 466. Blatta orientalis.
Ветвление трахей в голове
таракана. Вид сбоку (из
Miall and Denny)
ной сосуд. В груди поперечные соединения
имеются и на спинной стороне.
От этих главных трахей посегментно
отходят более мелкие ветви, разветвления
которых оплетают внутренние органы и
мышцы. Особенно хорошо оплетен
трахеями кишечник. Две пары главных
трахейных стволов продолжаются в голову
(рис. 466), где брюшные стволы дают
ветви в ротовые придатки и антенны, а
спинные — в головной мозг и глаза.
В груди от спинных главных
трахейных стволов берут начало трахеи,
заходящие в крылья и образующие их жилки. Это
лучше всего можно рассмотреть на
просветленных в глицерине зачатках крыльев
личинок таракана (рис. 467). От каждого
из спинных стволов в средне- и заднегруди
отходят по две ветви, которые соединяются
вместе, дают заходящий в зачаток крыла
общий ствол, дальнейшие разветвления
которого образуют сеть будущих жилок
крыла взрослого насекомого.
Кровеносная система, как и у
всех Arthropoda, незамкнутая, и кровь
только часть своего пути проходит в замкнутых
каналах или полостях пульсирующих органов. Сердце (рис. 468), или
спинной сосуд, в виде тонкой длинной трубки, лежит по средней линии
непосредственно под слоем спинных продольных мышц. Трубка поделена на
камеры, из которых две лежат в груди, а 10 или 11 — в брюшке. В
переднем направлении трубка в виде неразделенной на камеры аорты
продолжается в голову. Каждая камера имеет пару боковых
отверстий — остий, замыкание которых происходит при помощи клапанов.
Сердце прикрепляется к тергитам спинки при помощи строго
сегментально расположенных плоских крыловых мышц. Сокращение их
вызывает диастолу сердца и насасывание в него гемолимфы. Последующее
их расслабление и сокращение мышц сердца вызывает систолу.
Сокращение сердца идет волной от заднего конца к переднему и прогоняет
кровь через аорту вперед в голову. Там она попадает в сложную
систему синусов, в совокупности обеспечивающих снабжение кровью
ротовых придатков, головного мозга, глаз и т. д. Для обеспечения
кровообращения в длинных и тонких антеннах у таракана имеются добавочные
пульсирующие ампулы, лежащие у основания антенн и прогоняющие
туда кровь. Из головы кровь двумя потоками — брюшным, проходящим
над нервной системой и. спинным, огибающим головной мозг и
попадающим в околосердечный синус, — направляется в полость тела груди
и брюшка, откуда она насасывается в сердце при следующей диастоле.
Изучение сердца таракана надо производить на отпрепарованной
целиком спинке. Ее лучше всего зафиксировать в расправленном виде,
437
хотя бы в 70%-ном спирте.
После этого при помощи
препаровальных игл под бинокуляром
отпрепаровывают сердце вместе
с крыловыми мышцами. Из него-
приготовляется окрашенный
квасцовым кармином, или
гематоксилином тотальный препарат.
Выделительная систе-
м а. Выделительную функцию у
таракана несут мальпигиевы
сосуды, перикардиальные клетки,,
нефроцйты и жировое тело.
Мальпигиевы сосуды, как:
упоминалось, в виде тонких
извитых трубочек впадают в
кишечник на границе средней и задней
Рис. 467. Ветвление трахей в зачатках кишки. У взрослого таракана их
крыльев у личинкимследней стадии чисд0 дости/ает 80—100, у
ЛИЧИНОК их меньше. Они впадают в;
кишечник шестью радиально
расположенными пучками. Каждый сосуд является слепо замкнутой на
конце трубочкой с просветом, окруженным слоем крупных плоских
полигональных клеток. Снаружи клетки покрыты бесструктурной базальной:
мембраной, к которой примыкает сеть тонких поперечнополосатых
диагональных мышечных волокон, разветвляющихся и анастомозирующих.
друг с другом. Их сокращения вызывают червеобразные движения маль-
пигиевых сосудов в полости тела. Поверхность клеток, обращенная в.
просвет, несет палочковидную исчерченность, подобно клеткам
секреторного эпителия средней кишки. В плазме их можно наблюдать
внутриклеточные каналы и зерна экскретов. Содержимое просвета бывает
заполнено зернами кристаллов мочекислого натрия и аммония, щавелевокислого-
кальция и сферитами лейцина. В зависимости от физиологического
состояния мальпигиевы сосуды прозрачны или окрашены в
желтовато-белый цвет. Индигокармин, инъецированный в полость тела, может быть,
обнаружен в клетках малвпигаевых сосудов.
Мальпигиевы сосуды — главные органы .выделения, выводящие &
просвет кишечника продукты обмена и лишнюю воду.
Перикардиальные клетки, лежащие поодиночке по бокам
околосердечной полости, тоже несут выделительную функцию. Аналогичную
функцию выполняют нефроцйты. У Blatta orlentalis они расположены-
двумя небольшими группами у основания передней пары ног и двумя:
крупными скоплениями в голове около основания нижней губы.
Нефроцйты и перикардиальные клетки могут поглощать впрыснутый в
полость тела аммиачный кармин и откладывать его в своей плазме^
Их специфической ролью является удаление из крови различных
ядовитых для организма веществ. Фагоцитарные органы и фагоциты у
таракана, по крайней мере у взрослых особей, до сих пор не
обнаружены.
438
—1
Наконец, наряду с другими, несет
выделительную функцию и жировое
тело. Оно сильно развито в виде
многочисленных лопастей и тяжей в теле,
особенно в брюшке. Лопасти
жирового тела богато оплетены трахеями."
Их строение см. на с. 447. Жировое
тело таракана характерно тем, что
•оно состоит из клеток:
периферических, имеющих, наряду с
экскреторной, функцию накопления запасных
питательных веществ, и центральных,
играющих роль симбионтного органа.
Отложенные в клетках жирового тела
продукты обмена (ураты)
накапливаются в них в течение всей жизни
таракана и никуда из них не выводятся.'
В выделении продуктов обмена у
таракана, как и у других насекомых,
большое значение имеют многие
жизненные процессы, не имеющие
прямого отношения к выделительной
функции. К таким косвенным процессам
выделения можно отнести
сбрасывание хитиновой кутикулы во время
линьки. Состоящая из азотистых
веществ, пропитанная различными
органическими (пигменты) и
неорганическими включениями (инкрусты)
хитиновая кутикула с полным правом
может рассматриваться как экскрет.
К последним в известной мере
относятся и выделения различных желез.
Центральная нервная
система. Исследование нервной
системы производится методом препаровки.
У вскрытого таракана удаляются
пищеварительные и половые органы и
жировое тело. Затем таракана
помещают в 70%-ный спирт для
уплотнения и на следующий день препаруют под бинокуляром. Препаровка
головного мозга трудна. Отрезанную голову, после обработки 70%-ным
спиртом, вплавляют ротовыми частями в парафин так, чтобы она
была наклонена затылочным отверстием косо книзу. Отламывают по
кусочкам лобный и теменной отделы головы между глазами, после чего
обнажается головной мозг.
Изучение деталей его строения проводится на фронтальных
разрезах через голову, у которой перед заливкой надо удалить мандибулы.
Окраска железным гематоксилином по Гейденгайну с докраской эози-
Рис. 468. Схема сердца Blatta orien-
talis. Вид сбоку (А) и снизу (Б) (из
Miall and Denny):
1 — аорта, 2 — крыловые мышцы в груди,
3 — первая крыловая мышца брюшка, 4 —
тер гиты брюшных сегментов, 5— остия,
6 — концевая камера сердца, 7 — трахеи,
оплетающие околосердечную перегородку
439
ном дает удовлетворительные результаты.
Для препаровки симпатической нервной
системы и corpora allata выделяют сначала
передний отдел кишечника вместе с головой и,
после уплотнения в 70%-ном спирте,
вплавляют голову ротовыми частями в парафин так,,
чтобы она была наклонена несколько вперед,
находят под бинокуляром на спинной
поверхности зоба висцеральные ганглии, следуют по
ходу нервов вперед к голове, в которой
осторожно удаляют затылочный и теменной
отделы.
Центральная нервная система таракана
по степени расчлененности сравнительно
примитивна. Она состоит (рис. 469) из
церебрального (надглоточного) ганглия — головного
мозга, и брюшной нервной цепочки.
Последняя слагается из подглоточного ганглия, трех
грудных и шести ясно отделенных друг от
друга брюшных ганглиев в отличие от
плавунца, у которого они сближены. У зародыша
таракана закладываются 11 брюшных
ганглиев, но в процессе развития задние сливаются
вместе и образуют сложный шестой брюшнок
ганглий, лежащий в начале VII сегмента
брюшка.- Поперечные коммиссуры, соединяю^
- щие одноименные парные ганглии, сильно
укорочены и внешне не выражены. Парные
продольные коннективы выражены хорошо,
особенно в груди.
Наиболее сложное строение имеет
головной мозг таракана. Он лежит над глоткой в
передней части головы. В нем, как и у всех
других насекомых, можно различить три
отдела — прото-, дейто- и тритоцеребрум (рис.
470—472).
Протоцеребрум по величине занимает
наибольшую часть головного мозга. В нем
можно различить следующие отделы.
Центральную и верхнюю его части занимают
грибовидные тела (corpora pedunculata)
сложного строения. Грибовидные тела
рассматриваются как центр инстинктивной деятельности,
таракана. По бокам от них помещаются
зрительные доли (ganglion opticum), от которых;
отходит к сложным фасетчатым глазам толстый, короткий зрительный'
нерв (п. opticus). Между грибовидными телами, в срединной части,,
находится видимое только на срезах центральное тело, соединенное со
зрительными долями, грибовидными телами, обонятельными долями, к
Рис. 469. Blatta qrlentalis.
Общий вид нервной системы
(из Miall and Denny):
1 — надглоточный ганглий, 2 —
окологлоточная коннектива, 3 —
подглоточный ганглий, 4, 5 —
передне- н заднегрудной ганглии,
6—11 — первый — шестой
брюшные ганглии, 12 — нерв к цер-
кам, 13, 16 и 18 — нервы к
задним, средним и передним
ногам, 14 — нерв к задним
крыльям, 15 — нерв ко второй грудной
стигме, 17 — нерв к передним
крыльям, 19 — антеннальный
нерв, 20 — зрительный нерв
440
Рис. 470. Blatta orientalis. Надглоточный и
подглоточный ганглии. Вид спереди (из Breitschnei-
der):
/ — грибовидное тело, 2 — нерв к простым глазкам, 3 — if
зрительные доли, 4 — зрительный нерв, 5 — протоцере-
■бральные доли, 6 — дейтоцеребрум, 7 — антеннальный
яерв, 8 — двигательный иерв антенны, 9 —
тритоцеребрум, 10 — фронтальный нерв, //— тритоцеребральная
комиссура, 12 — подглоточный ганглий, 13 —
верхнечелюстной нерв, 14 — нижнегубной нерв, 15 —
нижнечелюстной нерв, 16 — верхнегубной нерв, 17 — фронтальный
ганглий
даже с окологлоточными коммиссурами.
Центральное тело важнейший
ассоциативный и зрительный центр. Над центральным телом находится pars
intercerebralis, от которой отходят нервы, иннервирующие простые
глазки. Пространство по бокам между грибовидными телами и
зрительными долями, занято протоцеребральными долями. Сзади и
спереди они охватывают центральные части протоцеребрума и соединяются
таким путем друг с другом. Их главная масса состоит из проводящих
волокон, образующих главнейшие пучки, которые соединяют отдельные
части мозга между собой. Здесь же происходит и перекрест
одноименных нейронов. Заслуживает упоминания наличие пучков,
непосредственно соединяющих зрительные доли с двигательными нервами антенн,
что допускает прямую двигательную реакцию антенн под влиянием
зрительных раздражений.
Дейтоцеребрум примыкает непосредственно снизу к протоцереб-
ральным долям протоцеребрума и помещается уже по бокам глотки.
Главную его массу кроме проводящих путей образуют, обонятельные
доли (lobus olfactorius) от которых отходят к антеннам мощные
обонятельные нервы — проводники воспринимаемых антеннами различных
(обонятельные, тактильные, температурные и т. д.) раздражений.
Отсюда же отходят нервы к мускулатуре антенн. Дейтоцеребрум каж-
2
Рис. 471. Blatta orientalis. Схема
внутреннего строения надглоточного
ганглия (из Breitschneider):
1 — центральное тело, 2 — грибовидное
тело, 3 — зрительные доли, 4 — зрительный
иерв, 5 — протоцеребральные доли, 6 —
обонятельные доли, 7 — антеннальный
нерв, 8 — фронтальный нерв, 9 —
верхнегубной нерв, 10 — тритоцеребральная ком-
миссура, Л — подглоточный ганглий, 12 —
глотка, 13 — п. stomatogastricus
441
дои стороны связан друг с другом
специальной коммиссурой (антеннальная коммиссура),
проходящей внутри через протоцеребральные
доли.
Непосредственным продолжением книзу
является тритоцеребрум (рис. 470). Этот
отдел мозга развит еще слабее предыдущего.
Отсюда отходят пара чувствительных
верхнегубных нервов (п. labralis), иннервирующих
верхнюю губу и лобный склерит, и пара
фронтальных нервов (п. frontalis), отходящих
вперед и соединяющихся над глоткой в
фронтальный ганглий (g. frontale), от которого
берет начало симпатическая нервная система
таракана. Вместе с фронтальным нервом
проходит двигательная ветвь верхнегубного
нерва. Обе части тритоцеребрума соединены
проходящей под глоткой тритоцеребральной
коммиссурой от которой отходят нервы к
мышцам-расширителям глотки.
Подглоточный ганглий соединен с трито-
церебрумом короткими окологлоточными кон-
нективами (рис. 472). Он состоит из трех пар
ганглиев — верхне-, нижнечелюстного и
нижнегубного, которые иннервируют
соответствующие ротовые придатки. Отходящие от них
нервы — смешанные, чувствительные и
двигательные. От подглоточного ганглия отходят также тонкие нервы к
слюнным железам.
Грудные ганглии (см. рис. 469) имеют сходное строение. От каж-'
дой пары отходят мощные нервы к ногам, разветвляющиеся по своему
ходу и иннервирующие отдельные мышцы. Грудные ганглии являются
центром, координирующим локомоторную деятельность ног; Средне- и
заднегрудные ганглии дают нервы, иннервирующие соответственно
переднюю (надкрылья) и заднюю пары крыльев. От этих же нервов
отходят ветви к грудным стигмам. Брюшные (1—6) ганглии значительно
меньше грудных. От каждой пары отходят нервы к мускулатуре и
стигмам 'соответствующего сегмента. Последняя пара заметно крупнее
остальных, так как она является результатом слияния шести (6—11)
последних пар первичных 'брюшных ганглиев. От шестой пары ганглиев
назад отходит пара толстых нервов, скоро распадающихся на ряд
ветвей, которые иннервируют мускулатуру заднего конца брюшка,
половой аппарат и церки. Сложный характер последнего ганглия брюшка
виден на фронтальных срезах через него. Но и таким путем нельзя
найти следы всех шести составляющих его первичных ганглиев. Это же
относится и к ветвям, на 'которые разделяются отходящие от него
нервы.
Симпатическая нервная система и эндокринные
органы. Симпатическая, или висцеральная, нервная система таракана
Рис. 472. Blatta orientalis.
Надглоточный и
подглоточный ганглии. Вид с левой
стороны (из Breitschneider):
1 — грибовидное тело, 2 —
зрительный нерв, 3 — нерв к
простым глазкам, 4 — антенналь-
ный нерв, б — двигательный
нерв антенны, 6 — фронтальный
ганглий, 7 — фронтальный нерв,
8 — верхнегубной нерв, 9 —
подглоточный ганглий, 10—12 —
верхне-нижнечелюстной и
нижнегубной нервы, 13 — нерв к
слюнным железам, 14 — коннек-
тивы к переднегрудному
ганглию, 15 — п. stomatogastricus
442
расположена на спинной стороне пищевода и
зоба (рис. 473). От фронтального ганглия отходит
назад непарный глоточно-желудочный нерв
(п. stomatogastricus. s. recurrens). Он идет
непосредственно над глоткой, проходит сквозь
окологлоточное кольцо и продолжается далее над
пищеводом, давая к нему нервные веточки,
регулирующие перистальтические движения
мускулатуры последнего. На уровне зоба нерв
оканчивается небольшим зобным ганглием, имеющим
форму треугольника, обращенного назад
основанием. От углов последнего отходят назад два
нерва, охватывающие в косом направлении зоб
и оканчивающиеся на нижней стороне
жевательного желудка. Видимо и эти нервы регулируют
движения мускулатуры зоба и жевательного
желудка.
На уровне глотки, по бокам п.
stomatogastricus, частично прикрытые задним отделом
головного мозга, лежат две пары железистых
органов — кардиальные тела — corpora cardiaca
(рис. 473, 4) и прилежащие тела — corpora alla-
ta (рис. 473, 5). Ранее эти структуры считали
ганглиями симпатической нервной системы.
В настоящее время установлена их сложная
комплексная природа. Эти тела образуют ретро-
церебральный комплекс нейрогемальных и
железистых органов. В этом комплексе
откладывается нейросекрет, продуцируемый нейросекретор-
ными клетками разных отделов мозга, а также
секретируются гормоны, в том числе ювениль-
ный гормон, регулирующий разные стороны
жизнедеятельности насекомых — обмен веществ, некоторые
поведенческие реакции, а также размножение и развитие.
Органы чувств. Органы зрения таракана в виде сложных,
или фасетчатых, глаз, имеющих почковидную форму (см. рис. 441),
расположены по бокам 'головы, позади и над основаниями антенн.
У самцов Bl. orientalis они заметно крупнее, чем у самок. Глаза
состоят из большого числа отдельных омматидиев эуконного типа.
Кроме сложных глаз у таракана имеется пара простых глазков
(ocellae), .помещающихся у основания антенн (см. рис. 441). И в
развитии глазков у В1. orientalis наблюдается половой диморфизм.
Тонкое строение сложных глаз и глазков должно быть изучено на
срезах.
Органами осязания, обоняния и температурного чувства являются
у таракана антенны, или усики, на которых расположены специальные
сенсиллы (рис. 474); к ним подходят мелкие ответвления антенналь-
ного нерва. Это, по-видимому, органы обоняния, тогда как для
осязания служат многочисленные волоски на теле члеников антенн. Различ-
Рис. 473. Blatta
orientalis. Передний отдел
симпатической нервной
системы (из Deegener):
/ — фронтальный ганглий,
2 — фронтальный нерв, 3 —
п. stomatogastricus, 4 —
corpora cardiaca, 5 — corpora
allata, 6 — нервы к слюнным
железам
443
ные волоски, покрывающие церки, тоже имеют
кроме осязательной и хорошо выраженную
обонятельную функцию.
Специальные органы слуха пока не
найдены. Но так как тараканы хорошо реагируют на
различные звуки, возможно, что различные
покрывающие поверхность их тела волоски,
вибрируя под влиянием звуковых волн, выполняют эту
функцию.
Наконец, органы вкуса, наряду с осязанием,
мы находим на ротовых придатках таракана,
особенно на щупиках нижних челюстей и нижней
Рис. 474. Periplanetaame- rvgbI (рис 475)
Hfa^H?eflVvonXCth)^ По лов ая" систем а. Женские половые
/ — чувствительный штнф- органы (рис. 476) состоят из парных сравнитель-
тик:, 2-<кутик™а'„ з-гип°- но крупных яичников, каждый из которых имеет
дерма, 4 — скопление чувст- rJ \ > ^
вительных клеток, 5 — нерв восемь или больше яйцевых трубок наиболее
примитивного у насекомых паноистического
типа. Концевые нити трубок соединяются вместе в общую концевую нить.
В каждой яйцевой трубке вслед за концевой нитью помещается
зачатковая зона (germarium, рис. 477). Образующиеся там оогонии
путем многократного деления дают яйцеклетки — ооциты. Последние,
продвигаясь, располагаются в линейном порядке и растут. Каждый ооцит
окружается цилиндрическими клетками фолликулярного эпителия,
происходящими из клеток эпителия зачатковой зоны. Этот отдел яйцевой
трубки называется желточной зоной (vitellarium). Ооцит, окруженный
фолликулярным эпителием, образует отдельную яйцевую камеру, или
фолликул, внутри которого и происходит рост ооцита. Приток
питательных веществ, за счет которых идет рост ооцита и накопление в
нем желтка, происходит только посредством эпителия фолликула, за
счет выделений которого образуется и наружная оболочка созревшего
яйца — хорион. По мере созревания и роста яйца фолликул
продвигается к основанию трубки. После созревания яйца клетки фолликула
дегенерируют и образуют так называемое остаточное, или желточное,
тело (corpus luteum), которое выталкивается последующим яйцом при
его продвижении.
В период вынашивания самкой яйцевого кокона рост ооцитов в.
яичнике приостанавливается. Эта задержка, как показали недавние
исследования, есть результат воздействия веществ, выделяемых в
гемолимфу проксимальным отделом яичника, т. е. теми его частями, где
находились фолликулы отложенных яиц, образующие желточное тело.
Яйцевые трубки открываются в короткие парные яйцеводы,
соединяющиеся в непарный отдел, называемый также маткой. Матка
открывается в наружные половые органы, представляющие собой род
кармана, на спинной стенке которого открывается отверстие семяприемника.
Этот карман произошел в результате впячивания внутрь тела VIII и
IX стернитов брюшка, которые и образовали спинную стенку
кармана, тогда как его слепой конец и брюшная стенка образованы
межсегментальной перепонкой VIII стернита и VII стернитом.
444
В матку, сразу позади места впадения в нее яйцеводов, впадают
придаточные железы. Это сильно разветвленные и извитые трубчатые
железы, секрет которых идет на образование яйцевого кокона (оотеки).
Мужские половые органы (рис. 478) состоят из парных семенников,
состоящих из многочисленных мелких округлых фолликулов,
расположенных вдоль проксимального конца длинных парных семяпроводов
(vasa deferentia). Семяпроводы по своему ходу дважды образуют
небольшие разветвления, сразу же вновь соединяющиеся. Сливаясь,
семяпроводы образуют непарный еемяизвергательный канал (ductus
ejaculatorius), идущий к наружным совокупительным придаткам.
Семяпроводы перед слиянием образуют расширения — семенные
пузырьки. В них впадают многочисленные придаточные железы в форме
слепых трубочек, более крупных по бокам, тонких к середине. Все они у
половозрелых особей заполнены спермой. Семенники могут быть
обнаружены только у молодых, недавно слинявших самцов. Они быстро
дегенерируют, и их трудно найти в окружающих лопастях жирового
тела. .
Для изучения мужских и женских половых органов их отпрепаро-
вывают вместе с выводными путями, зарисовывают в расправленном
виде и приготовляют окрашенный квасцовым кармином или
гематоксилином тотальный бальзамный препарат. Фиксируют яичники и изго-
445
Рис. 477. Blatta
orientalis. Концевой отдел
яйцевой нити таракана (из
Beier):
1 — концевая нить, 2 —
зародышевая камера, 3 — клетки
фолликулярного эпителия,
4 — перитонеальная
оболочка, 5 — ооцит второго
порядка, 6 — ооцит первого
порядка, 7 — оогоини
Рис. 478. Blatta orientalis.
Мужская половая система таракана.
В,ид с брюшной (А) и спинной
(Б) сторон (из Miall and Denny):
1 — семенник, 2 — vas deferens, 3 —
разветвление v. deferens, 4 — семяиз-
вергательный канал, 5 — придаточные
железы, 6 — семенные пузырьки
товляют из них срезы, по возможности продольные, которые
окрашиваются железным гематоксилином по Гейденгайну.
Изучение срезов. Разрез через лопасть жирового те-
л а. Лопасть жирового тела, взятую у 'свежев'скрытого в
физиологическом растворе таракана, фиксируют смесью Шампи, а при ее
отсутствии— каким-либо сулемовым фиксатором (Джильсон и др.), заливают
в парафин и режут толщиной в 5—7,5 мкм. Срезы окрашивают
железным гематоксилином по Гейденгайну.
На срезах (рис. 479) видно, что каждая лопасть жирового тела
состоит из наружного слоя жировых клеток, содержащих зачерненные
осмием капли жира и окружающих расположенные центрально бакте-
риоидные клетки. Крупные жировые клетки являются местами
отложения запасных питательных веществ в виде капель жира,
жиро-белковых гранул и т. п., а также местами отложения различных экскретов.
Последние в течение всей жизни таракана накапливаются в этих
клетках в виде уратов и в результате заполняют их целиком. Бактериои-
446
Рис. 479. Blatta orientalis. Разрез через лопасть жирового тела (из
Schneider):
/ — жировые клетки, 2 — клетки с симбионтными бактериями
дные клетки — видоизмененные жировые
клетки. Их плазма заполнена массой слегка S-
образно изогнутых бактерий с темно-окрашенной
оболочкой и светлой внутренней осью. Они
способны размножаться делением. Эти бактерии —
постоянные симбионты таракана. Значение их
для таракана неясно, так как искусственно
лишенные этих бактерий тараканы могут
нормально существовать долгое время.
На срезах кроме жировых клеток могут
попасться мелкие ветви трахей или трахейные
капилляры. Необходимо помнить, что в случае
фиксации жирового тела не осмиевым
фиксатором капли жира в жировых клетках будут
растворены абсолютным спиртом и на их месте на
срезах будут видны округлые вакуоли.
Продольный разрез. сложного
глаза и простого глазка. Голова
таракана фиксируется жидкостью Джильсона и
заливается по способу Петерфи. Предварительно
удаляются мандибулы. Приготовляется
поперечная или фронтальная серия срезов толщиной
в 5—7 мкм. Окраска железным гематоксилином
по Гейденгайну с докраской эозином. До
окраски желательно депигментировать срезы по
способу Паля. Эта же серия срезов пригодна и для
изучения строения простых глазков, а также
головного мозга, мускулатуры головы, строения
органов чувств на щупиках нижних челюстей и
нижней губы, на антеннах и т. д. Различные
срезы через сложный глаз пересекают оммати-
дии по всем возможным направлениям—от
строго продольного до строго поперечного, в
последнем случае на различных уровнях. Разберем
продольный разрез через омматидий, изучение
Рис. 480. Продольный
разрез через омматидий
сложного глаза таракана
(из Hesse):
1 — хрусталик, 2 — корнеа-
генные клетки, 3 — ядра
клеток хрустального конуса,
4 — хрустальный конус, 5 —
пигментные клетки, 6 —
зрительные клетки и их ядра,
7 — нерв, 8 — базальная
мембрана, 9 — палочковый слой
447
которого надо дополнить изучением поперечных разрезов на разных
уровнях.
В сложном глазе таракана насчитывается до 1800 омматидиев.
Каждый омматидий (рис. 480) состоит из двух частей:
светопреломляющей и воспринимающей. Хитиновая кутикула тела на месте глаза
лишена пигмента, т. е. прозрачна. Над каждым омматидием она
образует двояковыпуклое линзоподобное утолщение — корнеальную линзу,
или хрусталик (рис. 480, /), под которой лежит слой видоизмененных
клеток гиподермы — линзообразующих, или корнеагенных, клеток
(рис. 480, 2). К линзе изнутри примыкает хрустальный конус
(рис. 480, 4), образованный четырьмя клетками (Семперовские
клетки), сливающимися вместе, ядра которых лежат на стороне конуса,
обращенной к хрусталику. По бокам хрустальный конус окружен
пигментными клетками, изолирующими отдельные омматидий друг от
друга (рис. 480, 5). Далее вглубь расположена ретинула —
воспринимающий отдел глаза (рис. 480, 6). Она состоит из семи зрительных клеток,
основания которыхпродолжаются в нервное волокно, образуя
зрительный нерв (рис. 480, 7). Дистальные концы клеток охватывают
суженный отдел хрустального конуса. Ядра зрительных клеток
располагаются двумя группами. Четыре ядра помещаются у конца хрустального
конуса, три — у середины ретинулы, т. е. дистального конца последней
достигают только четыре клетки. Обращенные внутрь ретинулы
участки клеток по всей длине образуют особо дифференцированный
поперечно исчерченный слой тесно прилегающих друг к другу палочек,
имеющий вид гребня — рабдомер (рис. 480, 9). Рабдомеры всех клеток
образуют светочувствительный стержень — рабдом. Цитоплазма зри-
Рис. 481. Фронтальный разрез через простой глазок таракана
(из Link):
/ — лннза хрусталика, 2 — хитиновая кутикула, 3 — гиподерма, 4 — корне-
агенные клетки, 5 — зрительные клетки, 6 — клетки тапетума, 7 — нерв
448
тельных клеток ретинулы богата мелкозернистым пигментом, могущим
перемещаться вдоль ретинулы.
Кроме сложных глаз у таракана имеются простые глазки, или
оцеллы, помещающиеся у основания антенн (см. рис. 441). На
продольном разрезе (рис. 481) видим слабо утолщенную прозрачную кутикулу,
образующую плоско выпуклый хрусталик (рис. 481, /). Ее подстилают
видоизмененные гиподермальные — корнеагенные клетки (рис. 481, 4).
Под ними лежит многослойное скопление зрительных клеток
неправильно полигональной формы, соединенных в группы по 2—4 клетки
(рис. 481, 5). Каждая из групп образует палочковидный рабдом. Раб-
домы не ориентированы по отношению к падающим лучам света, а
лежат в беспорядке. Вся масса зрительных клеток окружена, как
чашей, слоем тапетума (рис. 481, 6), который пронизан нервными
волокнами глазного нерва.
ОТРЯД COLEOPTERA —ЖЕСТКОКРЫЛЫЕ, ИЛИ ЖУКИ
Dytiscus marginalis L. — Плавунец окаймленный
Материал и методы изучения. Плавунец окаймленный относится к подотряду
Adephaga, сем. Dytiscidae. Это один из обычных обитателей пресноводных стоячих
водоемов, начиная от поросших высшей водной растительностью ям до прудов н озер.
Добыча материала не представляет затруднений: взрослых жуков и их личинок
ловят обычным водным сачком, быстро проводя им вдоль поверхности воды
преимущественно среди водной растительности. Содержать пойманных жуков, и особенно
личинок, следует поодиночке, иначе они будут нападать, друг на друга. Аквариумы со
взрослыми жуками следует держать закрытыми. Подкармливать жуков можно
кусочками мяса, головастиками и т. п., личинок — головастиками, мелкими водными
насекомыми. Весной можно найти в значительном числе личинок младших стадий.
Позднее встречаются только более взрослые личинки. Жуков можно ловить круглый год.
Зимой в пруду прорубается прорубь, к которой вскоре приплывают взрослые жуки.
Если вода подо льдом содержит в заметном количестве (ощутимом по запаху)
сероводород, то численность собирающихся к проруби жуков может быть значительной.
Взрослые личинки к концу лета выползают на берег для окукления в земле вдоль
берегов водоемов, где поздней осенью (куколки) н ранней весной (готовые к выходу
жуки) их можно найти в земляных коконах под камнями, упавшими деревьями, в
земле.
Изучение внешней морфологии проводится как на целых (сухих или
заспиртованных) жуках, так и на вываренных в едком кали. На последних следует провести
расчленение груди и брюшка на сегменты, для более детального изучения отдельных
склеритов, препаровку придатков головы, ног, элитр и крыльев, из которых
изготовляются тотальные препараты в канадском бальзаме.
Внешняя морфология. Тело плавунца (рис. 482, 483) состоит, как
и у всех насекомых, из трех отделов: головы (caput), груди (thorax) и
брюшка (abdomen). Эти три отдела резко отграничены друг от друга.
Однако, как следствие плавающего образа жизни, они настолько тесно
соединены друг с другом, что тело жука приобретает резко
выраженную обтекаемую форму, уплощенную в дорзовентральном направлении
и лишенную заметных выемок и углублений. Широкая голова глубоко
и плотно вдвинута в переднегрудь, членики груди соединены в
единый грудной комплекс и без резкой границы переходят в относительно
короткое, закругленное назади брюшко, слитое с грудью. Сочленение
между отделами тела таково, что совершенно исключает возможность
15—2
449
Рис. 482. Самка Dytiscus marginalis. Рнс. 483. Самка Dytiscus marginalis.
Вид со спинной стороны (из Ког- Вид с брюшной стороны (из Ког-
schelt): schelt):
/ — голова, 2 — глаз, 3 — усик, 4 — перед- / — глаз, 2 — переднегрудь, 3 — брюшной
няя нога, 5 — переднеспинка, 6 —щиток щиток заднегрудн (metasternum), 4 — та-
среднегрудн (scutellum), 7—средняя нога, зик задней ногн, 5—10 — стерниты II—VII
8 — надкрылье, 9 — задняя нога брюшных сегментов, Я — вертлуг задней
йоги, 12 — отросток тазнка задней ноги,
13 — край надкрылья (эпиплевра), 14 —
эпнстерн заднегруди, 15 — средняя нога,
16 — среднегрудь, 17 — передняя йога, 18 —
усик
их движения относительно продольной оси тела. Все это сообщает
общей форме тела полную стабильность и, наряду с ее обтекаемостью,
служит приспособлением к плаванию.
Голова и ее придатки. В состав головы, так же как это
имеет место и у многоножек (с. 389), входят предротовая лопасть
(асгоп) и четыре сегмента (интеркалярный,1 верхнечелюстной,
нижнечелюстной и нижнегубной). Глаза и верхняя губа относятся к предро-
товой лопасти, по существу не являющейся истинным сегментом, как
не содержащей при своей закладке мезодерм альных зачатков. Все
головные сегменты слились в сформированной голове настолько полно,
что все попытки гомологизации отдельных участков головы тем или
иным из них до сих пор оставались безуспешными. Голова покрыта
наружной головной капсулой (epicranium), по бокам которой лежат
1 Интеркалярный, или постантеннальный, сегмент соответствует сегменту вторых
антенн, которые у насекомых отсутствуют.
450
Рис. 484. Голова Dytiscus marginalis. Вид
сверху (из Korschelt):
1 — верхние челюсти, 2— нижние челюсти, 3 —
верхняя губа, 4 — наличник, 5 —лобный склерит, 6 —
темя, 7 — корональный шов, 8 — щека, 9 — сложный
глаз, 10 — уснк, // — нижнечелюстной щупнк
глаза; она несет придатки
головных сегментов —
антенны и ротовые органы.
Кроме того, в голове
имеются внутренние скелетные
образования — эндоскелет.
Головная капсула. На
спинной стороне головной
капсулы имеется более
темно 'Пигментированный
рисунок, подверженный
индивидуальным вариациям (рис.
484, 485). В центре
помещается треугольное пятно,
основанием обращенное
вперед. Линии, соединяющие
вершины углов при
основании этого треугольника с
передне-боковыми углами
головы, ограничивают
лобный, или фронтальный, склерит (frons, рис. 484, 5). Соответствующий
этим линиям и-абразный фронтальный шов выражен слабо. Впереди
резко выраженным эпиетомальным швом от фронтального склерита
отделяется наличник (clypeus, рис. 484, 4), к которому причленяется
верхняя губа (labrum). Последняя морфологически не является
конечностью головы, но, поскольку она функционально связана с ротовыми
конечностями, ее описание будет помещено ниже.
Кроме лба и наличника, в пределах которых у различных
насекомых возможно проведение гомологии, все остальные отделы головной
капсулы насекомых, по отмеченным причинам (с. 450), гомологизации
не поддаются. Поэтому терминология этих отделов имеет чисто
описательно-топографическое значение.
От вершины треугольного пятна, по средней линии, начинается
корональный шов (рис. 484, 7), делящий верхний и зданий отделы
головы на две симметричные половины. Теменной отдел (vertex,
рис. 484, 6) помещается по обе стороны коронального шва. Назади он
переходит в затылочный отдел (occiput, рис. 486, 6), а по бокам в
щеки (genae, рис. 486, 5). Последние, огибая сзади глазную область,
заходят на брюшную сторону, до расположенного по средней линии
горла (gula, рис. 486, 4). Горло является видоизмененным, вошедшим в
(Jv4)
Рис. 485. Dytiscus marginalis. Вариации рисунка верхней поверхности головы
(из Korschelt):
/ — верхняя губа, 2 — наличник, 3 — лобный склерит, 4 — глаз, 5 — корональный шов
15*
451
Рис. 486. Голова Dytiscus
marginalis. Внд с брюшной
стороны (нз Korschelt):
/ — верхние челюсти, 2 —
нижние челюсти, 3 — нижняя губа,
4 — gula, 5 — щеки, 6 —
затылочный склерит, 7 — затылочное
отверстие, 8 — глаз, 9 я 10 —
ннжнечелюстной и нйжнегубной
щупики
Рис. 487. Dytiscus marginalis. A—
усик; 5 — верхняя губа (внд
изнутри) (из Korschelt):
1 — основной членик жгутика, 2 — зса-
pus, 3 — pedicellus
452
состав головной капсулы, подподбород-
ком (submentum) нижней губы.
Затылочное отверстие (foramen occipitale)
оказывается у плавунца, таким образом,
замкнутым и с брюшной стороны, это
придает особую прочность головной
капсуле. Сильно выпуклые сложные глаза
(строение см. на с. 447), имеющие
почковидную форму, расположены по бокам
головы впереди от щек.
Усики (antennae) помещаются
впереди глаз. Они состоят из 11 члеников.
Базальный членик (scapus, рис. 487, 2}
подвижно сочленен с головной капсулой
при помощи шаровидного сустава. Его
расширенное шаровидное основание,
отграниченное от остальной части
суженной шейкой, лежит в специальной уси-
ковой ямке. Второй членик (pedicellus,
рис. 487, 3) относительно короткий и
имеет цилиндрическую форму.
Остальные 9 члеников, одинакового строения и
величины, образуют жгутик (funiculus).
Все они слегка булавовидно вздуты на
дистальном конце, где расположены
многочисленные чувствительные ямки,
особенно развитые на конечном членике.
На усиках расположены органы
тактильного чувства и обоняния. В покое усики
направлены вдоль головы назад, огибая
внутренний край глаз.
Движение усика в целом
достигается действием трех мышц, отходящих по
кругу от трех точек шаровидной головки-
базального членика, другими концами
они прикрепляются к внутреннему
скелету головы. Их попеременное
сокращение вызывает круговые движения, а
сокращение одной или двух из них
одновременно — сгибание в любую сторону.
Внутри базального членика
расположены два антагонистических мускула,
сокращения которых могут сгибать и
разгибать второй членик усика вместе со
жгутиком относительно первого. На
окрашенном квасцовым кармином тотальном
препарате продольно разрезанного
основания усика можно рассмотреть оба эти
мускула.
Рис. 488. Верхняя челюсть
Dytiscus marginalis. Вид
изнутри (А) и снаружи (Б) (из
Korschelt):
1 — суставная ямка, 2 — хитиновая
аподема мускула-сгибателя, 3 —
суставной отросток
Ротовые органы, плавунца.
Плавунец — хищник. Ротовые органы
грызущего типа сильно развиты и, по
расположению на голове, направлены вперед,
т. е. они принадлежат к прогнатическому
типу, в отличие от свисающих вниз ги-
погнатического типа ротовых органов
таракана и ряда других всеядных или
растительноядных насекомых. В состав
ротовых органов входят верхняя губа
(labrum), верхние челюсти (мандибулы,
mandibulae), нижние челюсти (максил-
лы, maxillae) и нижняя губа (labium).
Верхняя губа (рис. 487, Б) имеет
форму узкой, поперечно вытянутой
непарной пластинки, верхняя поверхность
которой слегка выдается из-под
наличника, с которым она соединяется сочленовной перепонкой. Она слабо
подвижна: может немного приподниматься при помощи двух отходящих
от фронтального склерита мускулов (m. levatores labri), ее опускание
происходит самопроизвольно, при расслаблении этих мышц'.
Нижняя поверхность верхней губы переходит в ротовую полость,
образуя на ее верхнем оводе надглоточник (epipharynx). Верхняя губа
прикрывает опереди ротовую полость и, возможно, играет роль при
ощупывании пищи. В срединном углублении ее переднего края
помещаются густо сидящие волоски, а с внутренней стороны, несколько
отступя от края, расположены в один ряд 20—30 направленных вперед и
внутрь чувствительных щетинок. На эпифаринмсе сидит пара крупных
штифтиков.
Верхние челюсти (рис. 488) развиты особенно сильно. Их верхняя
поверхность выпуклая, а .нижняя, прилегающая к нижним челюстям —
плоская. На последней проходит ряд коротких крепких шипов,
отграничивающих зубы челюсти от собственно ее тела. Мандибула на
конце несет острый двойной зуб с- режущим краем, у его основания
помещается небольшой непарный зуб.
Мандибулы соединены с головной капсулой двойным шарнирным
сочленением, допускающим их движение только в одной, фронтальной
плоскости. На нижнем крае мандибулы с внутренней (нижней)
стороны имеется округлый выступ, входящий в соответствующую ямку на
краю щек. С наружной стороны нижнего края имеется углубление,
куда входит выступ, отходящий с внутренней стороны головной капсулы
у границы лобного склерита и наличника, у места прикрепления
усика. Движение мандибулы происходит при действии двух
антагонистических мышц. Отведение мандибулы в сторону вызывается
сокращением отводящего мускула (m. extensor mandibulae). Он отходит от ее
1 Мускулатуру ротовых органов можно рассмотреть путем препаровки
вплавленной брюшной стороной в дно парафиновой ванночки, предварительно заспиртованной
головы. Иглами осторожно обламывают склериты спинной стороны головной капсулы.
453
Рис. 489. Нижняя
челюсть Dytiscus margina-
Us. Вид снаружи (из
Korschelt):
/ — наружная лопасть, 2 —
внутренняя лопасть, 3 — car-
do, 4 — stipes, 5 — щупик
основания позади от точек сочленения и
прикрепляется к нижней задней поверхности
боковых отделов головной капсулы. Мышца,
приводящая мандибулу, наиболее мощная из всех
мышц головы (m. flexor mandibulae). Она
отходит от плоской хитиновой аподемы (рис. 488,2),
которая обычно отпрепаровывается вместе с
мандибулой, а другим концом прикрепляется к
верхней и задней поверхности головы капсулы,
заполняя этот отдел ее целиком.
Нижние челюсти (рис. 489) вместе с
мандибулами ограничивают с боков ротовую полость.
Они уплощены в дорзовентральном направлении
и хорошо расчленены. Можно различить
следующие части: основной членик (cardo), стволик
(stipes), от последнего отходят нижнечелюстной
щупик (palpus maxillaris) и наружная и
внутренняя лопасти (lobus externus и I. internus или
соответственно — galea и lacinia). Cardo при-
членяется к головной капсуле у переднего края
щек, суженной частью вдаваясь внутрь полости
головной капсулы. К его расширенному дис-
тальному концу причленяется stipes. На его конце, ближе к
наружному краю находится специальное возвышение, к которому
причленяется нижнечелюстной щупик. Он состоит из четырех члеников, из
них наиболее короткий — базальный, а последний несет большое
количество чувствительных кутикулярных образований. На конце членика
расположены скопления осязательных штифтиков, среди которых
разбросаны более крупные одиночные вкусовые. Функцию вкусовых
органов несет скопление особых чашевидных образований на наружной
поверхности дистальной трети последнего членика. Кроме них на
поверхности члеников щупика расположены многочисленные чувствительные
ямки.
Внутри второго и третьего членика имеются мышцы, сгибающие
при своем сокращении членики внутрь, разгибание происходит за счет
эластических свойств хитиновой мембраны сочленений.
От внутреннего края stipes отходят сильно развитая внутренняя
и более слабая наружная лопасти. Это важнейшие части максиллы.
Внутренняя лопасть — в виде S-образно изогнутой пластинки,
оканчивающейся тонким заостренным зубцом. Вдоль ее внутреннего края
сидят длинные крепкие щетинки. Наружная лопасть состоит из двух
подвижных члеников и неподвижно соединенного с stipes базального. Она
имеет характер щупика. Последний членик густо усажен
чувствительными органами. Максиллы обладают способностью к сложным
движениям, в связи с чем их мускулатура сильно дифференцирована. Мак-
силлу в целом двигают четыре мышцы. Из них одна (m. extensor
maxillae) отводит ее в сторону, а остальные три могут сгибать ее внутрь
в разных направлениях (m. flexor maxillae posterior, anterior и
superior). Экстензор и первый из флексоров отходят от cardo, а другие два
454
Рис. 490. Нижняя губа Dytiscus mar-
ginalis. Вид снаружи (из Korschelt):
/ — язычок (ligula), 2 —наружные лопасти
(paraglossae), 3 — щупик, 4 — подбородок
(mentum), S —gula
флексора — от stipes. К головной
капсуле они прикрепляются в заднем
отделе около затылочного отверстия.
Когда зубцы внутренних лопастей
вонзаются в схваченный мандибулами
кусок пищи, наружные лопасти,
лежащие нормально в продолговатом
углублении внутренних, попадают в
образовавшуюся рану, служат они, по-
видимому, органами вкуса.
Нижняя губа (рис. 490) замыкает
ротовую полость снизу и тесно
прилегает к основаниям максилл. Ее ба-
зальная часть (submentum), как уже
упоминалось (с. 451), вошла в состав
головной капсулы в виде горла (gu-
1а). Последнее хорошо заметным швом
отделено от подбородка (mentum) —
вытянутой в поперечном направлении пластинки, имеющей на переднем
крае глубокий вырез, в котором помещается соединенная с mentum
сочленовной перепонкой пластинка praementum. Последняя несет пару
трехчленистых нижнегубных щупиков, пару небольших имеющих вид
густо покрытых волосками'щеточек — наружных лопастей (paraglossae)
и крупную подвижную срединную лопасть — язычок (ligula),
образовавшуюся в результате полного слияния обеих внутренних лопастей
(glossae).
Мускулатура нижней губы состоит из мышц, приводящих в
движение отдельные ее элементы, а не всю губу в целом. От середины
основания mentum отходит парный мускул (m. levator labii), при
сокращении которого приподнимается вся пластинка mentum. Далее
имеются три пары мышц, двигающих наружные лопасти и язычок, и,
наконец, пара мышц двигает нижнегубные щупики.
Как уже указывалось, плавунец питается всякого рода животной
пищей. Его нападению подвергаются головастики, мелкие лягушки,
насекомые, водные моллюски, олигохеты и т. д. Он питается любым
живым или мертвым животным, если только может его схватить,
удержать. Добыча схватывается передними и средними ногами и
подносится к голове. Мелкие животные, вроде молодых головастиков,
заглатываются целиком, от более крупных плавунец при помощи верхних
челюстей отрывает куски величиной около 1—2.мм. Затем они
передаются нижним челюстям, которые продвигают эти куски в глотку.
Все время добыча ощупывается нижнечелюстными и нижнегубными
щупиками.
Внутренний скелет головы. Твердый наружный скелет — хитиновая
кутикула — служит местом прикрепления мускулатуры тела и его
придатков. Под воздействием регулярно повторяющихся сокращений особо
мощных мышц, в местах их отхождения, на кутикуле образуются
специальные внутренние гребни и отростки (аподемы), в совокупности
составляющие внутренний скелет. Эти эндоскелетные образования име-
455
ют большое функциональное значение. Они не только укрепляют
склериты, к которым прикрепляются мышцы (разного рода внутренние
гребни), но и увеличивают площадь прикрепления мышц без
увеличения этой площади на поверхности склерита, т. е. позволяют к данной
точке склерита приложить большую силу в виде мышцы с большим
количеством мышечных волокон. Кроме того, эндоскелетные
образования создают систему рычагов, позволяющую изменять первоначальное
направление мышечных воздействий, что способствует изменению и
усложнению производимых движений.
Особенно сложен эндоскелет головы, где к перечисленным
функциям прибавляется и функция, поддерживающая глотку и передний
отдел пищевода. Эндоскелет можно обнаружить на вываренной в едком
кали голове. Она вплавляется в парафин ротовыми частями и брюшным
отделом, после чего у нее удаляются затылочный и теменной отделы и
фронтальный склерит. На деталях строения этого сложного
образования мы останавливаться не будем.
Грудь (thorax) состоит из трех сегментов — передне-, средне- и
заднегрудного, иначе называемых передне-, средне- и заднегрудью
(pro, meso- и metathorax). Сегменты груди несут органы движения
жука— ноги, расположенные на каждом из сегментов, и крылья — на
средне- и заднегруди. Передняя пара крыльев утратила свой
перепончатый характер и в виде жестких надкрылий, или элитр, прикрывает
сверху задние крылья и все тело жука. Переднегрудь подвижна и
выделяется своим сильно развитым спинным" щитком. Среднегрудь тесно
соединена с заднегрудью. Последняя развита наиболее сильно, что
связано с наибольшей активностью движения задних ног и крыльев и с
соответствующим развитием мускулатуры;
Переднегрудь (рис. 491) состоит из спинного щитка (переднеспин-
ка, pronotum), боковых частей (propleurae) и брюшного щитка
(presternum) и несет переднюю пару ног. Переднеопинка (рис. 491, /)
сверху представляет собой склерит трапециевидной формы, лежащий
между головой и надкрыльями. По краю она окаймлена желтоватой
полоской, что является видовым
признаком для Dytiscus mar-
ginalis. На ее поверхности
рассеяны чувствительные ямки,
слабее выраженные и
встречающиеся в меньшем числе у
самцов. Боковые края перед-
неспинки, загибаясь вниз под
острым углом, граничат с
остальными склеритами. Про-
плевры (рис. 491, 2), хорошо
видимые снизу, состоят из
одного склерита. Они сужаются
Рис. 491. Dytiscus marginalis. Переднегрудь. в медиальном направлении, И
Вид с брюшной стороны. Ноги удалены (из их внутренние края принима-
Korschelt): ют участие в образовании
; — переднеспинка, 2 — propleurae, 3 — presternum, _ , ;„ „ _ » _
4 -ямки сочленения тазиков передних ног ТаЗИКОВЫХ ЯМОК ПерВОЙ Пары
456
Рис. 492. Dytiscus marginalis. Склериты
спинной стороны среднегруди (из Korschelt):
/ — praescutum, 2 — промежуточные склериты
сочленения надкрыльев, 3 — надкрылья, 4 — scutum, 5 —
scutellum
Рис. 493. Dytiscus marginalis.
Склериты брюшиой стороны
среднегруди. Вид снаружи (из Ког-
Vhelt):
/ —эпистерн (mesepisternum), 2 — эпи-
мер (mesepimeron), 3 — стернум (meso-
sternum)
ног. Простернум (рис. 491, 3) тоже в виде одного склерита. Его
характерной особенностью является присутствие вентрального выступа,
разделяющего тазиковые ямки и продолжающегося назад в виде крюч-
ковидного отростка, конец которого входит в соответствующее
раздвоение метастернума. Это является приспособлением для более плотного
взаимного укрепления сегментов груди. Передний край простернума
несет вытянутые боковые крыловидные отростки, срастающиеся по всей
длине с проплеврами и достигающие краев переднеспинки. По бокам,
в складке мягкой перепонки, соединяющей передне- и среднегрудь,
помещается передняя пара стигм.
Среднегрудь (рис. 492) — наиболее слабо развитый сегмент
груди— тесно соединена с заднегрудью. Сверху она прикрыта задним
краем переднеспинки и основаниями отходящих от нее надкрыльев, так
что из всех скелетных элементов среднеапинки видимым остается
только ее задний отдел — лежащий между основаниями надкрыльев
треугольный щиток (scutellum). Если согнуть переднеспинку вперед,
становится видимой остальная часть среднеспинки (praescutum). Можно
увидеть, что основание щитка вдается в ее срединную часть и делит
ее на два симметричных склерита, к боковым частям которого
посредством нескольких небольших склеритов подвижно причленены
надкрылья. Боковые плевральные щитки (рис. 493) имеют неправильно
прямоугольную форму. Их узкие основания тесно прилегают к
брюшному склериту среднегруди (mesosternum, рис. 493, 3) и затем
поднимаются косо вверх, так что их плоскости располагаются под углом к
медиальной плоскости тела. Этим они образуют глубокие брюшно-боко-
вые вырезы в теле жука, как бы разделяющие его на передний и
задний отделы. В этих вырезах при плавании помещаются передние ноги.
Боковые щитки при помощи дугообразного шва, проходящего по
диагонали, резко разделены на два склерита: один, прилегающий к
среднеспинке,— эпимер (mesepimeron, рис. 493, 2) и другой,
прилегающий к брюшному щитку,— этшетерн (mesepisternum — рис. 493, /).
Расположенный впереди эпистерн имеет вид равнобедренного треуголь-
457.
Рис. 494. Dytiscus marginalis. Склериты спинной
стороны заднегруди. Вид снаружи (из Korschelt):
/ — praescutum, 2 — склериты сочленения основания крыльев,
3 — scutellum, 4— pseudonotum, 5 — scutum
ника, основание которого соединено с боковым отростком мезостернума,
а свободная (медиальная) сторона ограничивает обращенный вперед
край кольца среднегруди. Брюшной щиток среднегруди (mesosternum)
образован одним склеритом. Его центральная часть, имеющая форму
трапеции, по бокам несет два отростка. Задний край мезостернума
принимает участие в образовании тазиковых ямок второй пары ног, чем
и объясняется его форма.
Заднегрудь (рис. 494) —наиболее крупный из отделов груди. В ней
можно различить те же скелетные элементы, что и в предыдущих двух
сегментах с той разницей, что здесь они подвижно сочленены друг с
другом. Эта подвижность обусловлена прежде всего тем, что между
боковыми краями заднеспинки и боковыми хитиновыми склеритами
причленяются задние крылья, движение которых под влиянием
летательных мышц оказывается возможным только как прямое следствие
движения склеритов заднеспинки и боков. Нижняя часть заднегруди
значительно больше по площади, чем верхняя, и образует
непосредственный переход к стершиам брюшка.
Заднеспинка (metanotum) имеет вид вытянутого в поперечном
направлении 'прямоугольника. В ней можно различить следующие отделы.
Впереди лежит узкая тонкая полоса (praescutum, рис. 494, /),
вдающаяся посередине внутрь щитка заднеспинки в виде двух
расходящихся медиальных ветвей, пространство между которыми остается
перепончатым. Боковые отростки praescutum на концах резко расширены и
сочленены посредством промежуточных склеритов с основанием
крыльев. Большую часть заднеспинки занимает щиток (scutum, рис. 494, 5).
Каждая из его половин, лежащих кнаружи от медиальных ветвей
praescutum, подразделена поперечным швом на два склерита. Их боковые
края тоже сочленены с промежуточными склеритами основания
крыльев. Задний щиток (scutellum, рис. 494, 3) имеет вид выпуклой полосы,
замыкающей сзади заднеопинку. Посередине она несет языковидный
отросток, вдающийся в пространство между склеритами scutum.
Боковые ветви его соединены с основанием югальной области крыла.
Позади scutellum лежит соединенная с ним перепонкой хитиновая
полоса — ложношшнка (pseudonotum, рис. 494, 4). Морфологически
она не является частью заднеспинки. Это — склеротизированная склад-
458
ка межсегментальной перепонки, вторичного происхождения. Боковые
склериты заднегруди, как и в среднегруди, состоят из эпистерна и эпи-
мера. Эпистерн имеет форму прямоугольного треугольника,
вклиненного между эпимером среднегруди и metasternum заднегруди. . Его
задняя вершина доходит до тазиков задних ног. Эпимер представлен
маленьким склеритом, причленяющимся сзади к эпистерну. К верхнему
краю эпистерна причленяется продолговатый склерит, который
постепенно становится тоньше и непосредственно переходит в мембрану
крыла. Эта область называется околокрыловой (parapleurae).
Брюшные склериты заднегруди ясно видимы с брюшной стороны.
Metasternum вместе с ограничивающими его сзади тазиками,задней
пары ног и эииетернами образует брюшную сторону заднегруди, Спереди
он граничит с Мезостернумом. По его средней линии проходит шов,
спереди образующий вилообразный выступ, куда входит крючковидный
отросток простернума переднегруди. По бокам от него лежат
полушаровидные' выемки, образующие заднюю половину тазиковых ямок
средней пары ног.
Ноги. Типичное строение имеют только первая и вторая пары
ног у самки. Соответствующие пары у самцов несколько видоизменены
для схватывания и удержания самок ври копуляции. Третья пара ног
у обоих полов является органом плавания, в связи с чем и
претерпевает некоторые видоизменения.
Начнем описание с типичных передних ног самки (рис. 495, А).
Продолговатый тазик (соха) помещается в специальной тазиковой-
ямке. К его суженному дистальному концу причленяется вертлуг
(trochanter), имеющий форму равнобедренного треугольника, прилегающего
основанием к тазику, а одной из боковых сторон — к бедру (femur) —
наиболее сильно развитому из члеников ноги. На внутреннем крае его
дистальной трети имеется глубокая борозда, куда при складывании ноги
входит голень. Поверхность бедра покрыта короткими волосками.
Голень (tibia) сочленяется с бедром, слегка изогнутым внутрь более
тонким основанием. Как и бедро, она уплощена дорзовентрально. Ее
поверхность покрыта мелкими волосками. На наружном крае сидят
длинные, гибкие волоски, а на дистальном конце — шипы, из которых два
очень крупные. Лапка (tarsus) состоит из пяти члеников. Из них
первый членик немного шире остальных и имеет цилиндрическую форму,
а пятый удлинен и на конце вооружен двумя сильно развитыми
коготками.
Передние ноги самца (рис. 495, Б) сильно видоизменены по
сравнению с ногами самки. Бедро заметно длиннее и слегка изогнуто.
Голень, сразу после изгиба тонкого основания, достигает предельной
ширины. Расположение и развитие волосков и шипов иное, чем у самки.
Наибольшие изменения претерпевают первые три членика лапок. В
совокупности они образуют округлую пластинку (рис. 495, В), на
которой помещаются две крупные и около 40 более мелких сидящих на
стебельках присосок. Это — приспособление для схватывания и
удержания самки во время совокупления.
Вторая пара ног самки (рис. 496, А) в общих чертах сходна с
первой. Тазики имеют шаровидную форму, а голени не так расширены.
459
Рис. 495. Dytiscus marginalis. Передняя левая нога. А — самки; Б — сам- f
ца; В — лапка самца при большом увеличении (из Korschelt):
/ — тазик, 2 — вертлуг, 3 — бедро, 4 — голень, 5—9 — первый — пятый члеиики лапки,
10 — коготки
Края их, так же как и члеников лапок, покрыты многочисленными
длинными, тонкими волосками. У самца (рис. 496, Б) эти ноги
отличаются большей длиной за счет удлинения бедер и особенно лапок.
Первые три членика последних на нижней стороне несут большое
число (свыше 1000) гусго сидящих мелких стебельчатых присосок.
Третья пара ног (рис. 497) у обоих полов имеет одинаковое
строение и является главным органом движения в воде. Их очень сильно
развитые тазики неподвижны. Они срослись с мегастернумом и вошли в
состав наружного скелета заднегруди. Соответственно тазиковые
ямки отсутствуют (см. рис. 483). Косо расположенные тазики сходятся
своими суженными дистальными концами у средней линии тела и
образуют продольный шов, по бокам которого отходят направленные
назад отростки с раздвоенными и заостренными концами. Под ними, в
середине заднего края тазиков, лежит место сочленения ног,
осуществляемое посредством вертлугов. Их изогнутый базальный отдел
входит в имеющееся там отверстие в тазиках и расширяется там в
листовидную аподему, к которой прикрепляются мышцы, приводящие в
движение ноги при плавании. К середине бокового края вертлуга причле-
няется вытянутое и изогнутое в виде шейки основание бедра, в
остальном напоминающее этот членик второй пары ног. Голень веслообразно
расширена и несет многочисленные волоски и щетинки. Членики лапки
сильно видоизменены. Они уплощены и постепенно сужаются к дис-
тальному концу. По краям они оторочены рядом длинных упругих
волосков.
Благодаря особенностям сочленения, задние ноги плавунца могут
двигаться только в горизонтальной плоскости. При отведении ног впе-
460
Рис. 496. Dytiscus marginalis. Средня? Рис. 497- Задняя левая нога
левая нога самки (А) и самца (Б) (из Dytiscus marginalis (из Ког-
Korschelt). Обозначения см. на рис. 495. schelt):
/ — вертлуг, 2 — бедро, 3 —
голень, 4—8 — первый—пятый
членики, лапки, S —коготки
ред голень и лапка сгибаются под прямым углом, что уменьшает
сопротивление при движении. Затем следует толчок, при котором они
выпрямляются и действуют как настоящие весла. Волоски^
оторачивающие лапку и голень, при этом расправляются, увеличивая почти в
три раза гребную поверхность, в то время как при отведении они
плотно прилегают к членикам. Передние ноги в плавании участия не
принимают. В это время они сложены и помещаются в углублениях
между передне- и среднегрудью. Средняя пара ног играет роль
вертикальных рулей, движения которых изменяют глубину плавания. Роль
горизонтального руля играют задние ноги.
Такое глубокое приспособление задних ног к плаванию сказывается
на передвижении жука на суше. Хотя ритм передвижения ног при
ходьбе и беге сходен у плавунца с его ближайшими наземными родичами—
жужелицами (сем. Carabidae), но при этом он не подымает свое тело,
а в значительной мере волочит, его по .земле. Задние .ноги и тут
играют наиболее важную роль, являясь своего рода отталкивателями—
благодаря их действию жук продвигается вперед. Обе передние пары
ног имеют подчиненное значение. При передвижении на суше важную
роль имеют шипы на концах голеней. Они служат для упора и
отталкивания от субстрата. Бег у плавунца не равномерный, как у других
461
бегающих насекомых, а совершается толчками, иногда переходящими
в прыжки.
Крылья. Несмотря на приспособление к жизни и плаванию в
воде, крылья плавунца хорошо развиты, и он не утратил способности
совершать перелеты часто на значительные расстояния.
Передняя пара крыльев, как и у всех жуков, видоизменена в
твердые надкрылья, или элитры. Они имеюг неправильно треугольную
форму. Слабо, выпуклое узкое основание граничит с переднеспинкой,
огибает щиток среднеспинки и переходит в прямой внутренний край,
оканчивающийся у вершины. Вдоль наружного края тянется желтоватая
полоса, характерная для вида Dytiscus marginalis. Начиная от
основания, наружный край загибается вниз, образуя полоску (эпиплевру),
прикрывающую бока и исчезающую по направлению к вершине. У
самцов поверхность надкрыльев почти гладкая. Имеются лишь точечные
углубления, расположенные тремя продольными линиями. Надкрылья
самок несут на поверхности 9 продольных ребрышек, постепенно
сглаживающихся к вершине. Встречаются, однако, самки и с гладкими
надкрыльями.
Задние крылья (рис. 498) плавунца служат органами полета. Их
перепончатая поверхность несет сеть жилок, среди которых
преобладают продольные, частично анастомозирующие друг с другом,
соединенные небольшим числом поперечных жилок. Другой особенностью
жилкования является полная или частичная редукция концевых часте'й
жилок, особенно у вершины крыла. У основания крыла отходят две
жилки, идущие вдоль переднего края крыла, частично сливаясь друг
с другом,— это костальная и субкостальная жилки (vena costalis и
v. subcostalis, рис. 498, 11 и 12). Немного отступая от края, проходит
радиальная жилка (v. radialis, рис. 498, 9), доходящая почти до
конца второй трети длины крыла. Связанная с ней система ответвлений,
Рис. 498. Dytiscus marginalis. Жилкование крыла (из Korschelt, с
изменениями):
1 — югальная жилка, 2, 3 — анальные жилки, 4, S — кубитальные жилки, 6 —
ветви медиальной жилки, 7 — ветви радиальной жилки, 8 — радио-медиальная
ячейка, 9 — радиальная жилка, 10— медиальная жилка, // —субкостальная
жилка, 12 — костальная жилка
462
направленных к вершине крыла (первая, вторая и третья радиальные
жилки) в значительной степени редуцирована.
Такой же редукции подвергалась и следующая из основных жилок
крыла насекомых — медиальная жилка (v. medialis, рис. 498, 10), у
которой редуцирован прилегающий к основанию крыла отдел, а
сохранился дистальный с системой ответвлений. Поверхность крыла по
диагонали пересекается сильно развитой кубитальной жилкой (v. cubitalis,
рис. 498, 4, 5), которая, не доходя до заднего края, загибается по
направлению к переднему и сливается с ветвями медиальной жилки,
образуя совместно с ними замкнутую кубитальную ячейку. Впереди от
нее лежит незамкнутая радио-медиальная ячейка. Далее к
внутреннему краю крыла лежит система анальных жилок (v. analis, рис. 498,
2 и 3), соединяющихся друг с другом. Наконец, поперек основания
крыла, у его внутреннего края проходит югальная жилка (v. jugalis,
рис. 498, /). Жилкование крыльев у плавунца развито сравнительно
слабо, и ряд жилок претерпевает редукцию. Это в значительной мере
можно поставить в связь с тем, что в покое крылья находятся в
сложенном состояний. При этом они складываются не только вдоль, но и
поперек. В покое передний край крыла лежит вдоль края тела и
полностью прикрывается надкрыльями. В таком положении крылья
оказываются значительно длиннее надкрыльев, чем и обусловлена
необходимость их поперечного складывания. При складывании весь базальный
отдел крыла загибается вдоль средней из анальных жилок на нижнюю
сторону, а вершина крыла вдоль радио-медиальной и кубитальной
ячеек загибается наверх. При этом все эти три плоскости веерообразно
складываются продольно.
Непосредственно с поверхности воды плавунец взлететь не может.
Для взлета он должен или вылезти на сушу, на какое-нибудь
возвышение почвы или камень, или забраться на торчащее из воды растение.
Его удельный вес, нормально близкий к удельному весу воды, слишком
велик для полета. Поэтому полету предшествует ряд действий,
направленных на облегчение веса. Прежде всего жук через заднепроходное
отверстие освобождается от жидкого содержимого ректальной ампулы
(см. с. 468). После этого происходит заполнение воздухом трахейных
мешков дыхательной системы. Они помещаются преимущественно в
груди и в заполненном состоянии значительно уменьшают удельный вес
этого наиболее тяжелого из отделов тела. Это длится 1—2. мин, после
чего жук, сделав несколько шагов вперед, высоко приподнимается на
средних ногах, так что передние оказываются в воздухе, приподнимает
надкрылья, отводит их в стороны, расправляет крылья и взлетает.
Полет плавунца — медленный, способность маневрировать и быстро
изменять направление движения — очень слабая. Внезапный порыв ветра
или встречающееся на пути препятствие прекращают полет. Дальность
полета измеряется многими километрами. Спуск плавунца скорее
напоминает падение. После длительного полета плавунец не сразу может
погрузиться в воду. Прежде должно быть произведено выравнивание
его удельного веса путем заглатывания воды.
Лет плавунцов происходит весной и особенно осенью, после
захода солнца, преимущественно в лунные- ночи.
463
Рис. 499. Брюшко Dytiscus marginalis. Вид сбоку Рис. 500. Dytiscus marginalis.
(из Korschelt): Стигмы (из Korschelt):
/■-тергит I сегмента брюшка, 2—8— стерниты II—VIII / — плевральная перепонка, 2 —
сегментов брюшка, 9 — анальная или пнгидиальная пла- вторая брюшная стигма (первая не
стника, 10 — последняя (восьмая) брюшная стигма, //— изображена), 3—8 — стерниты И—
17 — тергиты VIII—II сегментов брюшка VII сегментов брюшка, 9 —
плевральная пластинка, 10 — тергит
VIII сегмента, // — межсегментная
сочленовная перепонка, 12—18 —
тергиты VII—I сегментов
Брюшко. Исследование эмбрионального развития плавунца
показывает, что первоначальное число закладывающихся сегментов
брюшка равно 11, -не считая пигидиальной пластинки. Однако быстро число
их у зародыша уменьшается до 10, у личинки можно различить 9
сегментов, а у взрослого жука число видимых сегментов уменьшается до
восьми.
В каждом нормальном сегменте брюшка (рис. 499, 500) можно
различить шинной щиток — тергит, брюшной щиток — стернит и два
боковых щитка — плевральные пластинки. Видимые после отведения в
стороны надкрыльев и крыльев 8 тергитов принадлежат I—VIII
сегментам брюшка, видимые с брюшной стороны 6 стернитов относятся
ко II—VII сегментам. У I сегмента брюшка отсутствуют плевральные
пластинки, а стернит сильно редуцирован и тесно связан с заднегрудью.
В VIII сегменте тоже отсутствуют плевральные пластинки, а
стернит видоизменен в половой клапан, прикрывающий половую
щель.
Тергиты образуют сравнительно слабо хитинизированную
подвижную спинную поверхность брюшка. Они соединены между собой и с
плевральными пластинками мягкой сочленовной перепонкой, от
которой нерезко отграничены. В боковой сочленовной перепонке
помещаются стигмы, из которых передние четыре пары лежат в ней свободно,
а каждая из последующих все теснее связана с тергитом
соответствующего сегмента, восьмая пара стигм лежит, включенная в передний
край восьмого тергита. Последний прикрывает невидимую сверху
анальную, или пигидиальную, пластинку.
464
Стерниты сильно хитинизированы. Второй, третий и четвертый
стерниты плотно соединены друг с другом; пятый, шестой и седьмой
соединены между собой сочленовными перепонками. Второй стернит состоит
из двух склеритов, посередине разъединенных пространством, которое
занято отростками граничащего с ним метаетернума. Третий стернит
посередине переднего края имеет выемку, обеспечивающую
подвижность вертлугов задних ног. Боковые края стернитов утолщены и
слегка загнуты кверху. Они сильнее всего развиты на втором стерните и
постепенно уменьшаются кзади. Развитие плевральных пластинок
находится в обратной зависимости по отношению к развитию боковых
краев стернитов. Они наиболее ужи у второго стернита и наиболее
широки у шестого, плевральные пластинки у седьмого стернита развиты
только в переднем отделе сегмента. В связи с наличием сочленений,
подвижным оказывается только задний конец брюшка — пятый —
седьмой стерниты, из которых каждый предыдущий налегает своим задним
краем на последующий. Склериты последующих сегментов участвуют
в образовании яйцеклада и копуляционного аппарата.
Анатомия на вскрытии. Dytiscus удобен для изучения анатомии на
вскрытии.
Для вскрытия берут свежеубитых в парах эфира, хлороформа или в слабом
(20—30%) спирте жуков. В крайнем случае можно вскрыть жуков, хранившихся
в 70%-ном спирте, но у таких жуков внутренние органы вследствие уплотнения
утрачивают гибкость и при расправлении легко ломаются. Для препаровки же нервной
системы и мускулатуры спиртовый материал, наоборот, более удобен. Вскрытие
производится в восковой ванночке, под водой или в физиологическом растворе, если отпре-
парованные органы будут фиксироваться для срезов или окрашенных тотальных
препаратов. Жука держат в левой руке, а правой приподнимают и отводят вбок и
вперед надкрылья и крылья, обнажая верхнюю поверхность брюшка. Конец тонких
ножниц вставляют в промежуток между предпоследним (VII) и последним (VIII) из
видимых сегментов брюшка и проводят два продольных боковых разреза вдоль
границы более темной части спинной поверхности брюшка так, чтобы более светлая краевая
часть вместе со стигмами лежала бы кнаружи от разрезов. Разрезы проводят до
заднегруди. Чтобы не повредить внутренние органы, необходимо держать ножницы
и вести разрезы под возможно более острым углом к горизонтальной плоскости жука.
Затем проводят поперечный разрез в области последнего из видимых брюшных
сегментов. Далее помещают жука в восковую ванночку и прикрепляют его двумя
булавками, косо вкалывая их по бокам между заднегрудью и I брюшным сегментом.
Отведенные в стороны крылья и надкрылья тоже прикалываются булавками. Вместо
прикалывания чрезвычайно удобно вплавлять жука брюшной стороной тела в дно
восковой или парафиновой ванночки.
При помощи тонкого пинцета и плоско заточенной острой препаровальной иглы
отпрепаровывают, приподнимают и отворачивают вперед спинную часть брюшка. Иглой
разрезают прикрепляющиеся к ней мышцы. При осторожной препаровке спинной
сосуд, отделенный от спинки, остается лежать на поверхности внутренних органов.
Если это не удалось, то спинной сосуд остается прикрепленным к внутренней
поверхности спинки при помощи крыловых мышц.
После отпрепаровки спинки брюшка укрепляют задний конец жука, косо втыкая
две булавки в бока заднего отдела' брюшка, и продолжают вскрытие. Проводят
вперед продольные боковые разрезы в груди по направлению к внутренним краям
сложных глаз. Ножницы встречают сопротивление со стороны более плотных хитиновых
склеритов спинки груди. К последним прикрепляются мощные мышцы, которые
необходимо осторожно перерезать поперек. Это удобнее сделать тонкими ножницами.
При разрезе и освобождении спинки подвижно сочлененного с грудью первого
сегмента надо соблюдать большую осторожность. С особой осторожностью производят
продольные разрезы спинной части головной капсулы, ведя их параллельно
внутреннему краю глаз.
465
Для рассмотрения внутренних органов и их взаимного расположения необходимо
удалить частично прикрывающие нх сверху и находящиеся между ними лопасти жи-
■ рового тела, а также разрушить густую сеть оплетающих органы трахей. Это
достигается при помощи тонкого пинцета и препаровальных игл, для удаления крупных
грудных мышц можно пользоваться ножницами, стараясь не повредить пищевод и
нервную систему. Спинной сосуд, вместе с доходящей до головы аортой, снимают
целиком, при удалении жирового тела, и рассматривают отдельно. После этого
обнажаются извивы пищеварительного канала и отчасти половые органы. При
распутывании петель кишечника и его придатков надо соблюдать осторожность, чтобы не
повредить темно-окрашенные длинные тонкие, сильно извитые мальпигиевы сосуды и
свернутые в клубок желтоватые анальные, или пигидиальные, железы.
Пищеварительный канал, по его расправлении, не отрезают у рта и заднепроходного отверстия, а
отводят вбок и закрепляют булавками. После этого полностью обнажаются половые
органы. При их препаровке следить, чтобы не оборвать концевые нити яичников у
самок и очень нежные семяпроводы у самцов. После расправления половых органов
приступают к препаровке нервной системы. Брюшная цепочка оказывается уже почти
полностью обнаженной благодаря всем предшествующим манипуляциям. Значительные
трудности представляет препаровка надглоточного ганглия. Он скрыт довольно
глубоко под мускулатурой головы, которую нужно аккуратно удалить. Препаровку
надглоточного ганглия надо вести, пользуясь средним увеличением бинокулярной
препаровальной лупы.
При препаровке мускулатуры и нервной системы, требующих длительной работы,
лучше всего не прикалывать жука, а вплавлять его в дно ванночки. После вскрытия
и вплавления его заливают 70%-ным спиртом и, прикрыв, оставляют на сутки, после
чего нервы и мышцы выступают значительно резче и препаровка их становится-более
удобной.
Жировое тело взрослого жука, сохраняя в целом имеющееся
у личинки (с. 528) разделение на наружной (париетальный) и
внутренний (висцеральный) слои, имеет, однако, ряд особенностей,
преимущественно связанных с развитием мускулатуры в грудных
сегментах. Так, париетальный слой развит почти исключительно в
брюшке.
При удалении спинной части кожных покровов жука (рис. 501)
могут быть обнаружены тяжи висцерального слоя. Как и у личинки,
они начинаются в голове, где образуют непарную лопасть,
прикрывающую сверху надглоточный ганглий. В грудных сегментах, особенно
в переднегруди, они полностью прикрывают сверху кишечник. В
середине среднегруди они обнаруживают парное строение. Далее кзади они
расходятся и идут по бокам зоба вплоть до средней кишки. В месте
расхождения от них отходят лопасти, у самок прикрывающие сверху
концевые отделы яичников, а от боковых тяжей идут лопасти,
подстилающие задние отделы яичников. При удалении кишечника
обнаруживаются лежащие под ним лопасти и тяжи внутреннего жирового тела
(висцерального слоя). Общее расположение их указывает, что
жировое тело помимо своего основного назначения — служить местом
отложения запасных питательных веществ и выполнять выделительную
функцию — имеет еще одно важное значение: его рыхлые, мягкие
лопасти защищают внутренние органы от механических воздействий при
сокращении мускулатуры и при резких движениях жука.
Наружное жировое тело сосредоточено почти исключительно в
брюшке. Только в середине брюшной стороны переднегруди имеется
лопасть постоянной лировидной формы, зависящей от расположения
мышц. В брюшке наружное жировое тело расположено посегментно.
466
На спинной стороне брюшка,
что хорошо заметно на отпре-
парованной целиком спинке,
его сплетение имеет вид
лестницы, снизу изолирующей
спинной сосуд и крыловые
мышцы от остальных органов
(рис. 502). Детали строения
лопастей и функции жирового
тела см. на с. 509.
Пищеварител ь н а я
система. У вскрытого (см.
с. 465) жука расправляются
путем перерезки оплетающих
их трахей петли кишечника,
удаляются излишние лопасти
жирового тела, после чего
можно рассмотреть внешнее
строение пищеварительной
системы (рис. 503).
Как и у всех насекомых,
кишечник плавунца состоит из
трех главных отделов —
передней, средней и задней кишки,
различающихся не только по
анатомии и функции, но и по
происхождению. Передняя и
задняя кишки — эктодермаль-
ного происхождения,
средняя — энтодермального.
Передняя кишка
начинается ротовой полостью. От нее
отходит мускулистая глотка
(рис. 503, /), ведущая в
пищевод (рис. 503, 2), характерный
заметной снаружи продольной
складчатостью. Пищевод,
расширяясь, переходит на уровне
границы груди и брюшка в
объемистый зоб (рис. 503,13),
переходящий после сужения
в мускулистый жевательный желудок (рис. 503, 12). Последний
полушаровидной формы и как бы вдвинут своим широким основанием
в суженный отдел зоба, вследствие чего и лежит асимметрично. Более
тонкий, короткий отдел, заключающий в себе кардиальный клапан,
соединяет жевательный желудок со средней кишкой.
Средняя кишка состоит из двух отделов (рис. 503, 5, 6). Передний,
более длинный и широкий, густо покрыт многочисленными слепыми
трубчатыми выростами — пищеварительными железами, придающими
Рис. 501. Dytiscus marginalis. Вскрытая самка
с удаленными кожными покровами спинной
стороны (из Korschelt):
/ — непарная лопасть внутреннего жирового тела, 2 —
парные тяжи жирового тела, прикрывающие сверху
зоб н яичник, 3 — задний отдел зоба, 4 —
жевательный желудок, 6 — лопасть жирового тела,
подстилающая яичннк, 6 — средняя кишка, 7 — тонкая кишка,
8 — янчннк, 9 — мальпигиевы сосуды
467
Рис. 502. Dytiscus marginalis. Рис. 503. Dytiscus marginalis. Пищевари-
Спинные покровы брюшка, вид тельная система в расправленном виде (из
изнутри (из Korschelt): Korschelt):
/ — сердце, 2— лопасти жирового тела, / — глотка, 2— пищевод, 3 — пилорический кла-
окружающне сердце, 3 — крыловые пан, 4 — мальпнгиевы сосуды, 5 — задний отдел
мышцы, 4 — спинные продольные мыш- средней кишкн, 6 — передний отдел средней кнш-
цы брюшка ки, 7 — тонкая кишка, 8 — ректальная ампула,
9 — rectum, 10 — пигидиальная железа, 11 —
резервуар пигидиальной железы, 12 — жевательный
желудок, 13 — зоб, 14 — надглоточный ганглий
ему ворсистый вид. Он переходит в почти вдвое более короткий тонкий
задний отдел, поверхность которого покрыта более короткими
бугорчатыми слепыми выростами.
В месте перехода средней кишки в заднюю в кишечник впадают
четыре мальпигиевых сосуда (рис. 503, 4). Задняя кишка тоже состоит
из двух резко различных отделов. Сначала идет длинная тонкая
кишка, которая почти в полтора раза длиннее всех остальных отделов
кишечника. Ода начинается коротким мускулистым участком усеченно
конической формы с направленной назад вершиной. Тут расположены
сфинктеры, запирающие вход в заднюю кишку. Тонкая кишка сбоку
впадает в мешковидно расширенный ректальный отдел задней кишки.
Она впадает не в концевую его часть, а ближе к заднепроходному
отверстию, поэтому его концевая часть образует объемистый слепой
отросток (рис. 503, 8), сильно растяжимый и достигающий в состоянии
наполнения свыше половины длины тела жука. О его функции см.
с. 463 и 506.
468
Общая длина кишечника
почти в 3 Ч2 раза превышает
длину тела жука. Подобное
соотношение скорее
характерно для растительноядного
животного, чем для хищника,
каковым является плавунец.
Такое сильное развитие
кишечника в длину, возможно,
объясняется тем, что жуки, будучи
хищниками, далеко не всегда,
особенно в зимнее время или
в случаях перенаселения ими
водоема, могут найти себе
пищу. Поэтому пойманная
добыча поглощается в избыточном
количестве, про запас.
Приспособлением к этому помимо
объемистого зоба и мощно
развитого секреторного
аппарата средней кишки и
является удлинение всасывающего
отдела кишечника, которым, в
частности, служит длинная
тонкая кишка. Все это
позволяет плавунцу поглощать сразу большое количество пищи и
переваривать и усваивать ее длительное время.
Чисто топографически с ректальным отделом кишечника связаны
пигидиальные железы. По бокам ректального отдела задней кишки, в
области трех последних брюшных сегментов лежат парные
пигидиальные железы (рис. 504). Каждая из желез состоит из длинного тонкого
слабо ветвящегося трубчатого секреторного отдела. Стенки трубки
заняты железистыми клетками, а в ее центре проходит осевой отводящий
канал, впадающий в мешковидно расширенный резервуар, где
сохраняется секрет железы. От резервуара отходит .слабо изогнутый выводной
проток, открывающийся наружу, сбоку от заднепроходного отверстия.
Секрет пигидиальных желез представляет ■ собой эмульсию — взвесь
мелких шаровидных жироподобных капелек в жидкости. Двойному
характеру секрета соответствует два типа железистых клеток. Жиропо-
добное вещество секрета выделяется клетками наружного слоя желез,
жидкое содержимое — клетками внутреннего слоя.
По своей, функции пигидиальные железы принадлежат к числу
смазочных желез. Их жироподобный маслянистый секрет выводится на
заднем конце тела наружу и попадает прежде всего на концы элитр
и верхнюю часть последнего сегмента брюшка, вследствие чего их
поверхность не смачивается водой. Воздушная полость, находящаяся под
элитрами, как раз в этом месте открывается наружу (см. с.-478), и
именно задний конец брюшка является местом установления контакта
воздушной полости с атмосферным воздухом. Поэтому секрет пигидй-
Рис. 504. Dytiscus marginalis.
Пигидиальная железа слева в естественном
положении и. справа расправленная (из Korschelt):
/ — место впадения тонкой кишкн в ректальную
ампулу, 2 — ректальная ампула, 3 —
пигидиальная железа, 4 — ее выводной проток, 5 — rectum,
£—секреторный отдел, 7 —резервуар, 8 — тонкая
кишка
469
альных желез имеет огромное значение в.приспособлении плавунца к:
дыханию атмосферным воздухом в условиях водной среды, препятствуя
попаданию воды в воздушную полость и облегчая установление ее
контакта с атмосферой.
После внешнего ознакомления с пищеварительной системой
плавунца необходимо более детальное изучение ее главнейших отделов.
Последнее необходимо вести на срезах через отдельные участки
кишечника. У взрослого жука необходимо изучить препараты срезов через::
пищевод (с. 499), зоб (с. 499), жевательный желудок (с. 500), карди-
альный клапан (с. 501), среднюю кишку (с. 501), привратник в месте
перехода средней в заднюю кишку, тонкую кишку (с. 504) и rectum
с ректальной ампулой (с. 506). Все отделы задней кишки жука
построению сходны с таковыми личинки, вследствие чего, если это
окажется почему-либо удобным, для их изучения можно пользоваться и:
личиночным материалом. Строение средней кишки жука, в частности:
крипт, представляет собой дальнейшее развитие крипт личинки.
Поэтому строение средней кишки надо сначала изучить у личинки (с. 531) „
а потом у взрослого жука (с. 501).
Органы выделения. Основные органы выделения плавунца—
мальпигиевы сосуды (см. с. 508), выделяющие из организма в просвет
задней кишки мочевую кислоту, ее соли и излишнюю воду (рис. 503).
Клетки жирового тела (см. с. 509) и перикардиальные клетки (с. 473))
играют роль «почек накопления», они улавливают и накопляют в
своей цитоплазме различные вредные вещества, находящиеся в крови в;
растворенном или во взвешенном состоянии.
Кровеносная система. Для изучения кровеносной системьк
у захлороформированного жука отрезают элитры и крылья и
вплавляют спинной стороной в дно парафиновой препаровальной ванночки-
Вскрытие ведут в физиологическом растворе поваренной соли.
Отделяют боковыми разрезами стерниты брюшка и груди, удаляют их и
закрывающие спинной сосуд внутренние органы (кишечник, половые
органы). Пользуясь бинокуляром, можно, при некоторой осторожности,,
без труда обнажить с помощью препаровальных игл всю кровеносную-
систему в ее естественном положении, вплоть до окончания под
надглоточным ганглием аорты (рис. 505). Наблюдают пульсацию сосуда,,
после чего фиксируют препарат жидкостью Ценкера и лишь затем
отделяют его от спинки. Таким путем можно получить окрашенный
квасцовым кармином или гематоксилином постоянный препарат. При
отсутствии живых жуков можно тем же путем вскрыть и заспиртованных..
Для рассмотрения только пульсирующей части кровеносной
системы у захлороформированного, жука отделяют спинку брюшка вдоль-
плевральных частей сегментов, перерезают ее поперек на границе с
заднеспинкой, снимают ее и прикалывают в ванночку с
физиологическим раствором.
Для срезов лучше всего заливать в парафин через кедровое
масло. Срезы, толщиной около 5 мкм, окрашиваются железным
гематоксилином по Гейденгайну. .
Сердце, или спинной сосуд плавунца, как и у всех насекомых,
состоит из помещающегося в брюшке собственно пульсирующего отдела,»
470
f
{ снабженного боковыми отверстиями, и из
•его продолжения — заходящей через грудь
в голову аорты. В брюшке спинной сосуд
проходит непосредственно под покровами
•спинки в виде прямой трубки, заключенной
.в особую околосердечную (перикардиаль-
ную) полость. В груди, при переходе в
аорту, он сужается и пролегает над кишечни-
жом, вместе с которым проникает в голову,
'.проходит под надглоточным ганглием и
-оканчивается в лобном отделе.
На сердце свежеотпрепарованном и
отделенном от покровов спинки, помещенном
:в физиологический раствор, можно
наблюдать сокращения в течение значительного
времени (свыше 2 ч). Последовательные
волны сокращений одна за другой
проходят вдоль сердца от заднего конца к
переднему короткими перехватами. Следующие
.-за ними расширения захватывают более
.крупные участки сердца.
Задний конец сердца слепо замкнут.
Кзади он сужается в форме воронки, конец
'■которой, в зависимости от состояния систо^
лы или диастолы, или нитевидно заострен,
или расширен и укорочен. Расположенная
в брюшке часть сердца имеет по бокам
восемь пар щелевидных отверстий — остий
(рис. 505, 4), через которые при диастоле
нагнетается кровь в полость сердца.
Сердечная трубка несколько расширена в
области остий и имеет небольшие перехваты
в промежутках между ними. Остий
представляют собой щёлевидные отверстия, их
загнутые внутрь края образуют
двухстворчатые клапаны, препятствующие крови
изливаться через них наружу во время систолы. Строение стенки сердца
и остий см. на с. 511.
На границе между первым брюшным сегментом и заднегрудью
•оканчивается пульсирующий отдел спинного сосуда и окружающая его
перикардиальная полость. Место перехода сердца в аорту напоминает
сужение бутылки у горлышка. В задне- и среднегруди от аорты вверх
-отходят каналы (рис. 506, 4, 5, 7, 8) слепо оканчивающиеся
расширениями, снабженными мускулатурой. Это — ампулы, дополнительные
пульсирующие органы спинного сосуда, помогающие сердцу прогонять
тсровь из брюшка в голову.
В области пульсирующего отдела спинного сосуда у плавунца
расположены восемь пар крыловых мышц (см. рис. 505, 3), Они
располагаются посегментно и образуют основу околосердечной перегородки,
Рис. 505. Dytiscus marginalis.
Кровеносная система. Вид
изнутри на спинную сторону
вскрытого с брюшной стороны
жука (из. Korschelf):
/ — аорта, 2 — начало аорты, 3 —
крыловые мышцы, 4 — остнн, 5 —
задняя камера сердца, 6 — восьмая
брюшная стигма; 7 — m. transver-
sales abdominis, 8— первая
брюшная стигма, 9 — m. lateralis meta-
thoracis posterior, 10 — т. medianus
metathoracis, 11 — спинная ветвь
головной трахеи, 12 — надглоточный
ганглий. Внутренние органы,
жировое тело н трахеи удалены
471
Рис. 506. Dytiscus marginalis. Схематический сагиттальный разрез
грудного отдела аорты (из Korschelt):
1—3 — спинка задне-, средне- и переднегруди, 4 — среднегрудная ампула, 5 — ее
канал, 6 — аорта, 7 — канал заднегрудной ампулы, 8 — заднегрудная ампула.
Стрелки показывают направление движения крови
разделяющей в брюшке полость тела жука на две неравные части:
нижнюю, в которой расположены внутренние органы, и верхнюю —
околосердечную полость. Сократительные элементы находятся только в
боковых выступах крыловых мышц. От места прикрепления по бокам
тергитов брюшка эти сократительные элементы расходятся
веерообразно в медиальном направлении (рис. 507), разделяясь на все более
мелкие волокна. Не доходя половины пути до средней линии, они
утрачивают поперечную исчерченность и превращаются в эластические
волокна, образующие в срединной части септы сложную сеть (рис. 508).
Как мышечные, так и сеть эластических волокон заключены в
волокнистую соединительную ткань, составляющую основу септы.
Рис. 507. Dytiscus marginalis. Участок тотального препарата
околосердечной перегородки сердца (из Korschelt):
'/ — перикардиальные клетки, 2 — крыловые мышцы, 3 — зона перехода
сократительных волокон крыловых мышц в эластические, 4 — остии, 5 — ядра соединительной
оболочки сердца (adventitial
472
Физиологическое значение
околосердечной перегородки многообразно.
Прежде всего она служит
механической защитой пульсирующего сердца
от нижележащих внутренних органов
(кишечник, гонады, ректальный
пузырь). Далее она служит для
укрепления положения сердца в
околосердечной полости (рис. 509). Последнее
при помощи эластических волокон
подвешено между спинной
поверхностью брюшка и околосердечной
перегородкой. Наконец, ее присутствие
определяет направление токов крови
в брюшке, не допуская застоя в
каком-либо из его отделов.
В околосердечной полости
располагаются клетки перикардиальной
ткани. Последняя окружает сердце и
располагается на поверхности околосердечной перегородки. Клетки
образуют многоядерные синцитии самой разнообразной формы. Если в
полость тела жука впрыснуть 0,2—0,4 см3 1%-ного раствора аммиачного
кармина, или индигокармина, или взвесь туши в физиологическом
растворе, то уже через 1,5—2 ч после инъекции при вскрытии можно
обнаружить красную окраску клеток перикардиальной ткани, а через
12—24 ч вся краска исчезает из крови и откладывается в этих клетках
в виде включений. Функция перикардиальных клеток напоминает, с
одной стороны, функцию клеток жирового тела, в которых
откладываются мочекислые соединения, т. е. функцию почек накопления, с другой
стороны, им можно приписать и фагоцитарную функцию. Если
правилен взгляд на единство происхождения форменных элементов крови
насекомых, клеток жирового тела и клеток перикардиальной ткани, то
эти функции последних являются вполне естественными, тем более что
их положение на пути токов крови, устремляющейся в сердце, тоже го-
Рис. 508. Dytiscus marginalis. Сеть
эластических волокон в срединной
части околосердечной перегородки
(из Korschelt):
/ — ядра, 2 — поперечные, 3 —
продольные эластические волокна
Рис. 509. Dytiscus marginalis. Часть поперечного разреза спинного отдела
брюшка. Соединение сердца с тергитами брюшка (из Korschelt):
1— эластические волокна, 2—сердце, 3 — трахея, 4 — внутренний гребень тергита, 5
—крыловые мышцы, 6 — стенка тергита сегмента брюшка
473
ворит за это. Они являются, таким образом, своеобразным
фильтрующим аппаратом, очищающим кровь от различных инородных тел перед,
поступлением ее в сердце.
Органы дыхания. Для исследования дыхательной системы
необходимо применить метод препаровки. Вплавленного брюшной
стороной в парафин или приколотого жука вскрывают, и под водой отпрепа-
ровывают главные трахеи. Для изучения хитиновых частей стигм
вырезают последние с участком кутикулы и с отходящими от них
главными трахеями, после обработки их в едком кали изготовляют
тотальные препараты. Для изучения мускулатуры замыкательного
аппарата, отпрепарованную вместе с кутикулой и участком трахей
стигму фиксируют спиртом, после чего очищают ее от лопастей жировога
тела, а кутикулу — от гиподермы, чтобы сильно красящаяся гиподерма
не маскировала более слабо красящиеся мышцы замыкательного
аппарата. После окраски квасцовым кармином изготовляют тотальные
препараты. Для срезов лучше фиксировать стигмы только что
вышедших из куколок жуков. Материал заливают через смесь Петерфи в
парафин, изготовляют сагиттальные срезы толщиной 7,5—10 мкм и
окрашивают их по Маллори.
У взрослого жука 10 пар стигм: две пары в груди и восемь пар в
брюшке. Если о принадлежности брюшных стигм I—VIII сегментам
брюшка в литературе нет никаких разногласий, то межсегментное
положение стигм в груди вызвало ряд разногласий в приурочении их к
тому или иному из грудных сегментов. Перемещение грудных стигм в
промежутки между сегментами — явление вторичное, связанное с
особенностями развития и расположения мускулатуры ног и крыльев.
Главнейшие мышцы расположены сегментально, и их сильное
развитие не допускает возможности отхождения крупных трахей от стигм,
если бы они оставались расположенными тоже сегментально, как у
зародыша. В процессе эмбрионального
развития плавунца грудные стигмы закладываются
на средне- и заднегруди, вследствие чего
грудные стигмы жука следует считать морфогене-
тически принадлежащими к средне- и задне-
грудному сегментам.
О расположении стигм сказано при
разборе внешней морфологии жука (с. 464). По»
строению различают три типа стигм: 1) все
брюшные, 2) передняя грудная, и 3) задняя
грудная.
Все брюшные стигмы имеют более или
менее сильно вытянутую эллиптическую
форму (рис. 510). Стигма окружена хитиновой
рамкой — перитремой, ограничивающей ее
отверстие. Внутренние края перитремы несут
Рис. 510. Dytiscus margina- древовидно ветвящиеся кустистые волоски,
lis. Брюшная стигма. Вид обычно на стороне, обращенной вперед, более
снаружи (из Korschelt): длинные, чем на задней. Их концы образуют
перн^еПмы?Д2Н-ЙволоскиНфиль?ра УЗКуЮ ЩвЛЬ. В СОВОКУПНОСТИ ВОЛОСКИ образу-
474
ют род фильтра,
защищающего полость стигмы и
трахейной системы от попадания
туда мелких взвешенных в
воздухе частиц пыли. Внутренний
'край перитремы
продолжается вглубь и образует стенки
•стигмы, окружающие ее
полость.
К стигме непосредственно
"примыкает трахея. В месте
перехода ее в трахею
расположен замыкательный аппарат
(рис. 511, А). Строение его
■следующее: прилегающий к
трахее нижний край стигмы
утолщен с одной стороны и
■образует хитиновую дугу,
всегда остающуюся неподвижной,
другая половина нижнего края
хитинизирована слабее и не
утрачивает подвижности и
эластичности (замыкательная скоба, рис. 511, 6). Выше ее боковая
поверхность стигмы несет хитиновую складку, продолжающуюся в виде
отростка в полость тела (рукоять замыкательного рычага, рис. 511,./),
другой отросток идет от основания, рукоятки рычага к замыкательной
скобе нижнего края стигмы (замыкательный рычаг, рис. 511, 2).
Замыкательный мускул (рис. 511, 3) соединяет рукоять рычага с одним из
неподвижных концов хитиновой дуги основания стигмы. При его
сокращении (рис. 511, Б) рукоять и весь рычаг приходят в движение,
последний давит на замыкательную скобу, которая прижимается к
неподвижной дуге основания стигмы и замыкает отверстие последней. При
расслаблении замыкательного мускула все части замыкательного
аппарата, вследствие их упругости, приходят в исходное положение.
На рис. 512 для иллюстрации описанных взаимоотношений
представлены два разреза через брюшную стигму: один — на уровне
рукояти замыкательного рычага, другой —• на уровне прикрепления
замыкательного мускула к стенке стигмы.
Первая грудная стигма (рис. 513) приподнята над поверхностью
перепонки, соединяющей передне- и среднегрудь, вследствие сильного
развития наружных стенок стигмы (рис. 514). Вдоль верхнего края
наружные стенки загибаются внутрь и образуют перитрему. Таким
образом, отверстие стигмы кажется погруженным внутрь. Древовидно
ветвящиеся хитиновые волоски, как это видно на разрезе, расположены
не только по ее краю, но и по всей внутренней поверхности полости.
Строение и действие замыкательного аппарата видны из рис. 515.
Вторая грудная стигма (рис. 516, 517) лежит в перепонке между
верхними боковыми краями эпистерн средне- и заднегруди, скрытая
под пластинкой эпимер среднегруди. Она имеет форму сильно вытяну-
Рис. 511. Dytiscus marginalis. Брюшная
стигма с замыкательный аппаратом. Вид сбоку
(А) и схема действия замыкательного
аппарата (Б) (из Korschelt):
1 — стенка стигмы, 2 — замыкательный рычаг, 3 —
замыкательный мускул, 4 — рукоять рычага, 5 — пери-
трема, 6 — замыкательная скоба, 7 — трахеи, 8 —
неподвижная точка, вокруг которой происходит
движение
475
L-5-
Рис. 512. Dytiscus marginal
lis. Разрез через брюшную
стигму на уровне рукояти
замыкательного рычага (А)
и прикрепления
замыкательного мускула (Б) (из Ког-
schelt):
/ — края пернтремы, 2 —
неподвижная хитиновая дуга
основания стигмы, 3 — полость
стигмы, 4 — стенка трахеи, 5 —
рукоять рычага, 6 — замыкатель-
ный мускул, 7 — гиподерма
того овала. Перитрема на обоих концах образует частично
прикрывающие их глубокие складки, из которых нижняя развита особенно
сильно. Слегка изогнутое щелевидное отверстие ограничено двумя
эластичными изогнутыми губами, имеющими пластинчатую форму.
Замыкание щели стигмы
осуществляется путем выпрямления
обеих губ. При этом
объемлющая створка одной губы своим
краем находит плотно на
другую и замыкает стигму.
Выпрямление обеих губ
происходит под действием
замыкательного мускула, одним
концом прикрепляющегося к
нежной подкововидной скобе,
соединяющей нижние края обеих
губ, и отходящего от внутрен-.
него основания хитиновой
складки. Здесь мы имеем
совершенно другой тип
замыкания стигм — губной затвор,
принципиально отличающийся
от сжимающего типа замыка-
Рис. 513. Dytiscus marginalis. Первая грудная тельного аппарата брюшных и
стигма. Вид сбоку (из Korschelt): ofi грудной стигм.
/ — пернтрема, 2 —волоски фильтра, 3 —наружная г\л •
стенка стигмы, 4 — трахеи Общая Схема ■ ТраХвИНОИ
476
системы жука-плавунца следующая.
Все стигмы каждой стороны
соединяются между собой продольными
трахеями, в совокупности образующими
продольные трахейные стволы.
Последние в груди и переднем отделе
брюшка образуют расширения —
воздушные мешки. От каждой из
брюшных стигм отходят спинные коммиссу-
ры, соединяющие продольные стволы
между собой (рис. 518, 6). От каждой
из брюшных стигм отходит, кроме
того, сильно ветвящаяся ветвь, к
различным внутренним органам
(висцеральная ветвь). Так, эти трахеи у
последних двух пар стигм подходят к
выводным путям половой системы, копу-
лятивному аппарату, задней кишке и
мальпигиевым сосудам, шестая пара
дает ветвь к тонкой кишке и тоже к
мальпигиевым сосудам, пятая пара —
трахею, ветви которой оплетают
среднюю кишку и яичник, четвертая
пара — ветвь, распадающуюся на массу
мелких трахей, оплетающих яйцевые
трубки; третья пара снабжает
трахеями слепой вырост ректальной ампулы;
вторая пара дает ветви к спинному
Рис. 514. Dytiscus marginalis.
Продольный разрез через первую груд'
ную стигму на уровне отхожде-
ния замыкательного мускула (из
Korschelt):
/ — перитрема, 2 — место отхождения
замыкательного мускула, 3 — наружная
стенка стигмы, 4 — замыкательная
скоба, 5, 8 — сочленовная перепонка
между передне- и среднегрудью, 6 —
полость трахеи, 7 — складка неподвижт
ной хитиновой дуги основания стигмы
^^^*- х
Рис. 515. Dytiscus marginalis. Замыкательный аппарат первой грудной
стигмы (А) и схема его действия (Б) (из Korschelt):
/ — чувствительные органы края перитремы, 2, 3 —хитиновые дужки, заменяющие
рукоять рычага, 4 — трахея, 5 — замыкательная скоба, 6 — замыкательный рычаг, 7 —
наружная стенка стигмы, 8 — замыкательный мускул
477
Рис. 516. Dytiscus marginalis. Вторая
грудная стигма в естественном
положении (из Korschelt):
J — стигма, 2 — эпистерн среднегруди, 3 —
эпнмер среднегруди, 4 — эпистерн
заднегруди, 5 — сочленовная перепонка между
средне- и заднегрудью
Рис. 517. Dytiscus
marginalis. Вторая грудная стигма.
Вид сбоку (из Korschelt):
1,2 — передняя и задняя губы,
3 — складка, 4 — замыкатель-
ный мускул, 5 —
чувствительные органы на краю передней
губы
сосуду и к мускулатуре, двигающей
заднюю пару ног, наконец, первая
пара брюшных стигм дает
многочисленные ветви к крыловым и ножным
мышцам, жевательному желудку и
спинному сосуду, а также в крылья и
заднюю пару ног (рис. 518, 519).
Сложнее ветвление трахей,
отходящих от двух пар грудных стигм. От
второй пары отходят стволы,
снабжающие трахеями ноги (пару трахей в
среднюю и одну в заднюю пары ног),
элитры и мускулатуру средне- и зад-
негруди. От первой пары грудных
стигм отходят трахеи в голову (две
пары), в переднюю пару ног, в
мускулатуру переднегруди и отчасти
среднегруди. О некоторых деталях
ветвления головных трахей см. на с. 527,
тонкое строение трахей — на с. 507.
- Дыхательная система взрослого
жука-плавунца по строению ничем не
отличается от дыхательной системы
жуков, ведущих наземный образ
жизни. Плавунец дышит атмосферным
воздухом. При этом его брюшные
стигмы служат для вдыхания воздуха, а
грудные — для выдыхания. Брюшные
стигмы, как упоминалось, сверху
прикрыты элитрами. У наземных жуков
они служат, в частности, и для
защиты стигм от разных механических
воздействий и от пыли. У плавунца
элитры прикрывают верхнюю поверхность
брюшка настолько плотно, что с
боков и сверху полностью замыкают
образующуюся под ними воздушную
полость (рис. 520). Этим, во-первых,
осуществляется защита стигм от
непосредственного соприкосновения с
водой и; во-вторых, создается
воздушная камера, которая кроме
выполнения гидростатической функции
является также резервуаром запасного
воздуха, служащего для дыхания во
время пребывания жука под водой.
Сзади эта полость замыкается путем
прижатия заднего края спинного
щитка VIII брюшного сегмента к заднему
478
краю концов элитр. Когда
иссякает запас кислорода в
воздушной камере,
плавунец должен подняться на
поверхность для его
возобновления. Сроки
возможного пребываниия его под
водой сильно колеблются и
зависят главным образом от
энергии его движения и
температуры. Так как центр
тяжести жука лежит в
передней трети тела, ближе к
брюшной стороне, то под
водой и при подъеме на
поверхность жук всегда имеет
наклонное положение с
приподнятым кверху задним
концом брюшка. При
соприкосновении последнего с
поверхностной пленкой, конец
брюшка отгибается и через
образовавшееся отверстие
выжимается пузырек
воздуха, своим выпуклым
мениском пробивающий пленку
поверхностного натяжения,
чем и устанавливается
контакт воздушной полости с
атмосферным воздухом.
Несмачиваемость кожных
покровов водой способствует
этому. Для возобновления
запаса достаточно
нескольких секунд. При более
длительном пребывании'на
поверхности происходит
вентиляция всей трахейной
системы (рис. 521). Он
прикрепляется к ней в трех
точках, чем достигается максимальная устойчивость положения. Во
время вентиляции трахей можно наблюдать попеременное расширение и
спадение брюшка, его легкое вытягивание и втягивание—
накачивающие воздух дыхательные движения.
Возможно, что под водой возобновление кислорода в дыхательной
камере может происходить в известной мере и путем свободной
диффузии кислорода из воды в воздух камеры. Получающееся в процессе
дыхания обеднение кислородом воздуха в дыхательной камере создает
разность в парциальном давлении кислорода в воде и в камере, что и
Рис. 518. Dytiscus marginalis. Общий вид
спинного отдела трахейной системы
брюшка и заднегруди после удаления спинки
тела, схематизированно (из Korschelt):
/, 2 — боковые воздушные мешки заднегруди,.
3 — первая брюшная стигма, 4, 5— ветви к
мускулатуре заднегруди н к задним крыльям, 6 —
поперечная коммиссура, 7 — трахея к ректальной
ампуле, 8 — трахея к яичнику, 9, 10 — трахеи к
средней кншке н яичнику, // — трахеи к задней
кишке и мальпигиевым сосудам, 12 — трахеи к
половому аппарату, 13 — мелкие трахеи,
отходящие от седьмой н восьмой брюшных стнгм и
образующие сеть. капилляров в полости тела, 14 —
продольные стволы, 15 — ветвь к жевательному
желудку, 16 — ветви к сердцу (на других коммис-
сурах не изображены), 17—21 — воздушные
мешки, 22—24 — m. lateralis metathoracis posterior, me-
dius u anterior, 25 — т. medianus metathoracis
479
Рис. 519. Dytiscus marginalis. Брюшко и заднегрудь после
удаления спинки. Жировое тело и поперечные трахейные коммис-
суры, кроме последних двух, удалены (из Korschelt):
/ — m. medianus metathoracis, 2—4 — m. laterales metathoracis posterior,
medius и anterior, 5 — m. cexodorsalis metathoracis, 6 — воздушный мешок,
7, д — трахеи к стенкам зоба, 8 — первая брюшная стигма, 10 —
продольный ствол, И — вторая брюшная стигма, 12 — m. extensor throchanteris
metathoracis posterior, 13 — so6, 14 — ректальная ампула, 15 — третья
брюшная стигма, 16 — ветви к ректальной ампуле, П — яичник, 18 —
трахейный ствол к яичнику, 19 — трахеи к средней кишке, 20 — трахея к
задней кншке н мальпигиевым сосудам, 21 — rectum, 22 — задняя кишка,
23 — мальпигиевы сосуды, 24 — средняя кишка, 25 — жевательный
желудок, 26 — ветвь к жировому телу, 27—31 — воздушные мешки
Рис. 520. Dytiscus marginalis. Схемати- Рис. 521. Dytiscus marginalis, вен-
ческий поперечный разрез через брюшко тилирующий трахеи на поверхио-
плавунца (из Korschelt): сти воды (из Korschelt).
/ — элитра, 2 — воздушная камера, 3 — брюшная
стигма
определяет направление процесса диффузии. Можно наблюдать, что
находящийся под водой жук выдавливает из-под заднего края элитр
воздух так, чтобы его поверхность соприкасалась с водой. Подобный
путь получения кислорода не может в течение сколько-нибудь
длительного времени покрывать потребность жука в кислороде, и воздушное
дыхание является основным.
Центральная нервная састема. Нервная система жука-
плавунца состоит из надглоточного ганглия (ganglion supraoesopha-
geum, рис. 522, /), называемого также головным или церебральным, и
из брюшной нервной цепочки. Последняя начинается лежащим в голове
сложным подглоточным ганглием (g. infraoesophageum, рис. 522, 2),
соединенным с надглоточным окологлоточными коннективами.4- За ним
идут три ганглия сегментов груди — передне-, средне- и заднегрудной
ганглии (g. prothoracis, g. mesothoracis и g. metathoracis, рис. 52.2,
3—5). Продольные коннективы, соединяющие эти ганглии между собой
и с подглоточным ганглием, становятся по направлению спереди назад
все короче, так что средне- и заднегрудной ганглии сильно сближены.
■ К' заднегрудному ганглию непосредственно примыкает первый брюшной
(рис. 522, 6). Коннективы, их соединяющие, настолько укорочены, что
один кажется продолжением другого. Наиболее длинные коннективы
соединяют первый и второй брюшные ганглии, за которым следуют на
близком расстоянии друг от друга третий — шестой брюшные ганглии.
Последний, шестой брюшной ганглий — сложный. Он произошел в
результате слияния 6—11 брюшных ганглиев, закладывающихся во время
эмбрионального развития у зародыша плавунца.
Головной ганглий (рис. 523) помещается в середине головы
непосредственно над глоткой, впереди мышц-сгибателей мандибул, в виде
поперечно вытянутой массы, разделенной посередине неглубокой бо-.
роздой на два полушария. По бокам, сужаясь, он переходит в мощный
я короткий глазной нерв, расширяющийся в прилегающий к сложным
глазам глазной ганглий. В головном ганглии различают три отдела,
однако внешне разграничить их трудно. Это видно, как следует, только
16—2
481
на срезах. Отделы головного
ганглия называются прото-,
дейто- и тритоцеребрумом. ■
Протоцеребрум занимает
весь верхний отдел головного
ганглия. Ему принадлежит
главная масса обоих верхних
полушарий. Их основная часть
составлена из больших
зрительных долей, от которых
отходят зрительные нервы
(п. opticus, рис. 524, 5). В про-
тоцеребруме заложены также
грибовидные тела, обычно
рассматриваемые как главные
центры двигательной и
психической деятельности жука.
Кроме того, от его заднего
края, прилегающего к
пищеводу, отходят нервы к ганглиям
симпатической нервной
системы, в частности к ганглию
спинного сосуда.
Дейтоцеребрум занимает
нижнюю часть полушарий.
Сверху видны только передние
края обонятельных долей, у
места отхождения идущих к.
антеннам обонятельных
нервов. При рассматривании
сбоку, а особенно снизу,
полностью видны утолщения
обеих обонятельных долей (1оЫ
olfactorii), соединенные друг с
другом внутренней коммиссу-
рой. Кроме обонятельного
нерва (п. olfactorius s. antennalis,
рис. 524, 4) от обонятельных
долей отходит пара
дополнительных нервов (п. antennalis
accessorius, рис. 523, 12),
иннервирующих мускулатуру
антенн. Наконец, по бокам отходят два нерва (п. tegumentalis, см. рис.
523, 5), иннервирующие главным образом верхний отдел (темя)
головной капсулы.
Тритоцеребрум примыкает снизу к предыдущему отделу и
занимает нижний отдел головного ганглия и большую часть окологлоточных
коммиссур. Оба его узла соединены коммиссурой (commissura transver-
salis s. annulus oesophageus), проходящей под пищеводом между око-
Рис. 522. Dytiscus marginalis. Общий вид
нервной системы жука (А) и личинки (Б) (из
• Blanchard):
/ — надглоточный ганглий, 2 — подглоточный ганглнй,
3—5 — передне-, средне- и заднегрудной ганглии, 6 —
13 — первый — восьмой брюшные ганглии
482
логлоточными
коллективами, недалеко от места
их соединения с
подглоточный ганглием. От
переднего отдела тритрце-
ребрума,
непосредственно под обонятельными
долями, отходит пара
нервов, каждый из которых
сразу же распадается на
два — верхнегубной нерв
(п. labralis, см. рис. 523,
16), иннервирующий
верхнюю губу и область
наличника, а также
частично мускулатуру
глотки, • и фронтальный нерв
(п. frontalis), дающий
ответвления к остальной
мускулатуре глотки, и
ветвь к расположенному
поверх пищевода
фронтальному ганглию
(g. frontale, см. рис.
523, /).
Подглоточный ганглий (рис. 524, 10) — сложный ганглий,
возникший в результате слияния трех пар нервных узлов верхнечелюстного,
нижнечелюстного и нижнегубного сегментов, значительно меньше
надглоточного, верхняя поверхность его сравнительно плоская, нижняя —
выпуклая. Он заметно сужается кзади. Окологлоточные коннективы
впадают в него по бокам верхней поверхности переднего отдела.
Помещается он непосредственно под пищеводом, над центральной частью
внутреннего скелета головы.
От нижнего переднего угла подглоточного ганглия, по бокам от
места впадения окологлоточных коннективов отходит пара
верхнечелюстных нервов (п. mandibularis, рис. 524, 6), иннервирующих
верхние челюсти и их мускулатуру. Сзади них, ближе к брюшной
стороне, отходит пара нижнечелюстных нервов (п. maxillaris, рис. 524,7),
иннервирующих нижние челюсти, их придатки и мускулатуру.
Непосредственно позади, но еще более смещенная на брюшную сторону,
отходит пара нижнегубных нервов (п. labialis, рис. 524, 8), которая ин-
нервирует нижнюю губу с придатками и мускулатуру, а также область
горла (gula), по происхождению тесно связанную с нижней губой
(см. с. 451). У места выхода нижнегубных нервов подглоточный
ганглий достигает наибольшей ширины. Далее кзади он сужается и
переходит в коннективы, соединяющие его с первым грудным ганглием.
В этом Отделе от него отходят две пары нервов, в совокупности
иннервирующие мышцы, двигающие голову в целом (п. jugularis primus и
п. jug. secundus, рис. 524, // и 49). Первый отходит позади нижнегуб-
16 15 1ii- а
Рис. 523. Dytiscus marginalis. Голова,
отпрепарированная со спинной стороны (из Korschelt):
/ — фронтальный ганглий, 2 — надглоточный ганглнй,
3 — оптический ганглий, 4 — глаз, 5 — п. tegumentalis,
6 — m. flexor mandibulae, 7 —глотка, 8 - п. stomatoga-
•stricus, 9—m. flexor antennae, 10 —m. entensor antennae,
11 — антеннальный нерв, 12 — п. antennalis accessorius,
13 — мышцы — расширители глотки, 14 — фронтальное
сплетение, 15 — кольцевая мускулатура глотки, 16 —
верхнегубной нерв
16*
483
Рис. 524. Dytiscus tnarginalisj Медианный сагиттальный разрез через голову и грудь.
Кишечник удален (из Korschelt):
/ — надглоточный ганглий, 2 — фронтальный ганглий, 3 — верхнегубной нерв, 4 — антеннальный
нерв, 5 —зрительный нерв (перерезан), 6 — верхнечелюстной нерв, 7 — ннжнечелюстной нерв,
8 — нижнегубной нерв, 9 — m. levator glossae, 10 — подглоточный ганглий, 11 — левый п. jugularis
primus, 12—14 — передне-, средне- н заднегрудной ганглнй, 15 — первый брюшной ганглий, 16, 39—
m. lateralis metathoracis anterior, 17 — коннектнва брюшной цепочки, 18, 22 — второй — шестой
брюшные ганглнн, 23—25 — m. extensor trochanteris metathoracis medius, inferior и posterior,
26—30 — нервы восьмого — четвертого брюшных сегментов, 31 — сердце, 32—36 — нервы третьего —
первого брюшных ганглиев, 37 —т. lateralis metathoracis medius, 38 — m. medianus metathoracis,
40— аорта, 41 — n. commissuralis, 42— m. retractor prothoracis, 43— m. extensor coxae mestothoracis,
44, 45 — m. mesonoti inferior и superior, 46 — m. flexor coxae mesothoracis, 47 — m. depressor capitis
obliquus, 48 — m. depressor prothoracis, 49 — правый п. jugularis primus, 50 — rotator capitis superior,
51 — глотка, 52 — corpus allatum, 53 — m. flexor maxilae, 54 — сосудистый ганглнй, 55 — n. tegu-
mentalis, 56 — мышцы-расширителн глотки
ного нерва от середины боковой стороны ганглия, второй, более
мощный, — непосредственно у места отхождения коннективов к грудным
ганглиям.
Все три грудных ганглия, вместе с подглоточным, образуют
направленную назад, проходящую в горизонтальном направлении цепочку.
Переднегрудной ганглий лежит у заднего края, среднегрудной — в
центре и заднегрудной — у переднего края соответствующего им сегм.ента.
Такое положение каждого из них определяется расположением
внутренних скелетных образований (вильчатые апофизы эпистернумов
каждого из сегментов), на которых лежат ганглии. Это обстоятельство, в
свою очередь, влияет на точки отхождения нервов от данного ганглия
к отдельным органам сегмента, так как нервы к данному органу
отходят от ближайшей к нему точки ганглия. Так, смещение заднегрудного
ганглия вперед вызвало перемещение точек отхождения нервов на
задний отдел ганглия. Сходство и различия в строении грудных га'нглиев
обусловлены сходством и различиями в строении грудных сегментов, в
первую очередь их придатков. Соответственно, иннервация ног во всех
трех сегментах одинакова.
Общими для каждого из грудных ганглиев являются шесть пар
нервов. Первый — грудинный нерв (п. sternalis) иннервирует стерналь-
484
ный склерит каждого из сегментов. Второй — передний тазиковый нерв
(п. coxalis anterior) — иннервирует в переднегруди главным образом
мышцы-разгибатели тазиков, а также мышцы, поднимающие голову, в
среднегруди — разгибатели тазиков и мышцы, поднимающие элитры, в
заднегруди, где тазики неподвижны — некоторые из крыловых мышц и
мышцу, двигающую эпистерны по отношению к sternum, так
называемые выдыхательные мышцы. Третий — задний тазиковый нерв (п.
coxalis posterior) — иннервирует в переднегруди сгибатели тазика и
вертлуга, в среднегруди — мышцы, опускающие элитры, разгибатели
тазика и сгибатели и разгибатели вертлуга, в заднегруди — крыловые
мышцы (как видоизмененные тазиковые) и мышцы, сгибающие вертлуг.
Четвертый — нижний тазиковый нерв (п. coxalis inferior) —иннервирует
в переднегруди мышцы-разгибатели тазика и сгибатели и разгибатели
вертлуга, в среднегруди — сгибатели тазика и вертлуга и в
заднегруди—сгибатели и разгибатели вертлуга. Пятый—нерв разгибателя
вертлуга (п. extensoris trochanteris) — во всех трех сегментах
иннервирует мышцы — разгибатели вертлуга. Шестой — ножной нерв (п. ischia-
dicus) во всех трех сегментах иннервирует членики ног и мышцы ниже
вертлуга.
Кроме этих нервов, общих всем трем грудным ганглиям, от
некоторых из них отходят только характерные для них нервы. От средне-
грудного ганглия отходит пара нервов к элитрам (п. elytri), иннерви-
рующих их поверхность, а от заднегрудного отходит пара крыловых
нервов (п. alae), иннервирующих, помимо некоторых мышц, связанных
с. движением крыльев, хордотональные органы последних. От
коннективов, соединяющих грудные ганглии между собой, отходит еще по
паре нервов.
Первый брюшной ганглий примыкает к заднегрудному настолько
тесно, что соединяющие их коннективы, внешне неразличимы. Он,
таким образом, лежит в середине заднегруди. Две пары отходящих от
него нервов иннервируют I и II сегменты брюшка. Это доказывает
сложность ганглия, возникшего в результате слияния ганглиев I и
II брюшных сегментов.
Длинные коннективы соединяют первый брюшной ганглий с
остальной частью брюшной нервной цепочки, лежащей целиком в облас-,
ти заднегруди (рис. 524, 525). Она состоит из пяти ганглиев. Первые
четыре, т. е. второй ■— пятый брюшные ганглии (морфологически это
будут третий — шестой), соединены друг с другом короткими коннек-
тивами. Последний (шестой) брюшной ганглий тесно прилегает к
предыдущему, почти сливаясь с ним. Он тоже является сложным
ганглием, образовавшимся в результате слияния ганглиев VII, VIII и
последующих сегментов брюшка. От каждого из ганглиев отходит пара
нервов. Только от последнего отходят две пары. Чем ближе лежит
ганглий к иннервируемой им области, тем скорее начинают ветвиться
отходящие от него нервы. Нервы, отходящие от второго — пятого
брюшных ганглиев, иннервируют III—VI брюшные сегменты. Первая из двух
пар нервов последнего (шестого) ганглия иннервирует VII сегмент
брюшка, вторая — половой аппарат и всю заднюю кишку (п. procto-
daeogenitalis).
485
Рис. 525. Dyliscus marginalis. Нижняя поверхность заднегруди и
брюшко. Вид изнутри (из Korsche.lt):
1—5— второй — шестой брюшные ганглии, 6 — m. transversales abdominis,
7 — т. ventrales abdominis extern!, 8— т. ventrales abdominis interni, 9 —
m. conjungens coxo-abdominis, 10— m. cqxo-lateralis metathoracis, // — m.
extensor throshanteris metathoracis posterior, 12 — m. extensor alae posterior,
13— m. furco-dorsalis metathoracis medius, 14 — m. coxo-dorsalis
metathoracis, 15 — m. lateralis metathoracis medius. Все органы и часть
мускулатуры удалены. Вндны нервы брюшка и нх главнейшие ветвн
Все нервы брюшных ганглиев имеют много общего в характере
ветвления и в иннервируемых ими участках сегментов. Каждый из них
распадается на две ветви — спинную и брюшную. Спинная ветвь ин-
нервирует сердечные мышцы, стигму и спинную часть кожных
покровов и мускулатуры сегмента, брюшная ветвь—■ продольную и дорзо-
вентральную мускулатуру и брюшную часть кожных покровов. Очень
486
Рис. 526.'Dytiscus marginalis. Симпатическая нервная система (из Korschelt)-.
/ — надглоточный ганглий, 2 — антеннальный нерв, 3 —m. stomatogastricus, 4 —
фронтальный ганглнй, 5 — фронтальный нерв, 6 — верхнегубной нерв, 7 — зрительный нерв
(перерезан), 8—10 — верхне-нижнечелюстной и ннжнегубной нервы, // — подглоточный
ганглий, 12 — коннектнва к первому грудному ганглню, 13 — зоб, 14—жевательный
желудок, 1Ь — средняя кншка. 16—желудочный ганглнй, 17 — h. stomatogastricus, 18 —
пищевод, 19 — corpora allata, 20—corpora cardiaca, 21 — n. tegumentalis
резко это выражено в III брюшном сегменте. Наиболее видоизмененной
является последняя пара нервов, что понятно, если учесть ее сложное
происхождение.
Симпатическая нервная система и эндокринные
органы. Отходящая от головного ганглия симпатическая нервная
система иннервирует переднюю кишку и состоит из двух непарных
ганглиев и соединяющего их непарного нервного ствола (рис. 526).
Передний фронтальный ганглий (g. frontale, рис. 526, 4) лежит под
лобным склеритом посередине глотки. Он имеет вид направленного
вершиной назад равностороннего треугольника. От углов при
основании отходят два нерва (п. frontales, рис. 526, 5), соединяющие
фронтальный ганглий с тритоцеребрумом головного ганглия. Вперед от
основания отходит веточка, иннервирующая мускулатуру глотки. От
задней вершины отходит крупный ствол глоточно-желудочного нерва
(п. stomatogastricus, рис. 526, 3, 17), иначе называемого возвратным
нервом (п. reccurens). Он проходит вдоль пищевода, сначала по
медианной линии, потом, все более и более отклоняясь на его левую
боковую сторону, переходит на зоб и на границе с жевательным желудком
образует желудочный ганглий (g. ventriculare, рис. 526, 16). От
последнего отходят две ветви к,жевательному желудку. На всем
протяжении от нерва отходят ветви к стенкам пищевода и зоба. Кроме
того, недалеко от фронтального ганглия отходит от ствола пара нервов
к ганглиям спинного сосуда.
С элементами симпатической нервной системы связаны органы,
выполняющие эндокринную функцию, которые -раньше
рассматривались как ганглии симпатической системы. Они выполняют важные
функции в физиологии и экологии насекомых (см. с. 443).
По сторонам начальной части пищевода и п. stomatogastricus
располагаются кардиальные тела (corpora cardiaca) и прилежащие тела
(corpora allata, рис. 526, 19, 20). Прилежащие тела сильно оплетены
трахеями, отсюда название — трахейные ганглии. О функции этих
органов см. с. 443.
487
Органы чувств. Оплошной твердый наружный хитиновый
покров жука сам по себе не является хорошим (проводником различных
внешних раздражений, поэтому разнообразные кожные чувствительные
органы распространены по всей поверхности тела. Среди них имеются
различно развитые чувствительные волоски, щетинки и кутикулярные
сенсиллы, являющиеся результатом специальных видоизменений
чувствительных волосков. Строение последних см. на с. 495. Они
воспринимают различные механические и химические раздражения.
Расположение сенсилл различного типа неодинаково на поверхности тела жука.
Особенно богато представлены кожные чувствительные органы на
голове. Сенсиллы, воспринимающие различные химические раздражения,
расположены на ротовых частях и усиках. Последние несут также
органы равновесия. На концах нижнечелюстных и нижнегубных щупиков
расположены и осязательные сенсиллы.
Органы химического чувства (обонятельные и вкусовые сенсиллы)
жука обладают значительной тонкостью восприятия. Капля крови,
попавшая в аквариум, быстро приводит жука в возбуждение и
привлекает его. При этом он усиленно вибрирует усиками и щупиками. При
схватывании добычи последние играют важную роль в ее ощупывании.
На поверхности груди и брюшка расположены преимущественно
сенсиллы, воспринимающие различные механические раздражения.
Органы химического чувства имеются в небольшом количестве и связаны
преимущественно с половой деятельностью жука. Они развиты в
точках соприкосновения жуков во время спаривания. В этих же местах
особенно многочисленны и осязательные сенсиллы. Соответственно
наблюдается половой диморфизм в распределении их. Так, на передне-
спивке и элитрах самок они развиты силь'нее. На поверхности вертлу-
гов располагаются особые сенсиллы, воспринимающие силу давления
воды или воздуха при плавании и полете. На крыльях тоже имеются
аналогичные. сенсиллы. Особенно богато представлены чувствительные
органы на элитрах.
На вторам -членике усиков помещается своеобразный орган
сложного строения — Джонстонов орган. Это усложненный хордотональный
орган, воспринимающий различные сотрясения внешней среды,
возможно вызываемые также и звуковыми волнами.
Органами зрения жука являются хорошо развитые сложные глаза,
насчитывающие небольшое число омматидиев — до 9000 (в глазе
стрекозы их насчитывается до 25 000). Принимая во внимание, что
резкость изображения в сложном глазе зависит от числа фасеток,
воспринимающих предмет, а число их изменяется обратно пропорционально
квадрату расстояния от предмета, дальность видения плавунца мала.
Поэтому зрение играет в жизни жука, в частности в поимке добычи,
подчиненную роль, сравнительно с другими чувствами, особенно с
химическими (обоняниеми вкусом) и механическими.
Так как исследование строения сложного^глаза у плавунца натал-
кивается на известные трудности, связанные с крепостью хитина
головы, то изучение этого органа лучше проводить у таракана (с. 447), тем
более что глаза обоих объектов принадлежат к одному типу эуконных
фасетчатых глаз.
488
Половая -система. Женские
половые органы. Нижняя часть
брюшка самки (рис. 527) занята женской
половой системой. Хитиновые части
яйцеклада занимают последние два
сегмента целиком, а частично вдаются
еще вперед до задней границы IV
сегмента брюшка. Вследствие этого
примыкающий к ним влагалищный
отдел загибается вправо и вниз, а этот
изгиб вызывает смещение яичников и
других отделов тоже вправо. Таким
образом, половая система занимает в
брюшке несколько асимметричное
положение. Передняя половина яичников
лежит между мощными мышцами
задней пары ног, а частично — и на
них, вследствие чего несколько
приподнята кверху. В таком положении
яичники закрепляются прежде всего
концевым тяжем, который направляется
вперед и вверх, проходит под спинным
сосудом через заднегрудь и
прикрепляется в среднегруди изнутри к сред-
неспийке. Многочисленные трахеи,
оплетающие яичник, тоже содействуют
прочному закреплению его положения
в теле жука. Они берут начало от
четвертой и пятой пары брюшных стигм
и оплетают яйцевидные трубки
яичников и их выводные протоки. Кроме того, яичники окружены лопастями
жирового тела, образующими вокруг них род перитонеальной оболочки,
Женская половая система состоит из четырех отделов: 1) половой
железы—яичников, 2) выводных путей, 3) совокупительного аппарата,
4) яйцеклада.
Яичники плавунца в период покоя (с июня — июля по февраль —
март) имеют форму укороченных с заднего конца и вытянутых с
переднего веретеновидных образований. Весной начинается интенсивное
созревание яиц, вследствие чего их объем увеличивается почти
вдвое.
Каждый яичник состоит из яйцевых трубок, число которых
варьирует от 38 до 49 штук, так что в тзбоих яичниках общая сумма их не
превышает 90. Яйцевые трубки плавунца принадлежат к мероистическо-
му полиграфному типу строения. В яйцевых трубках. этого типа из-
первичных половых клеток образуются кроме ооцитов первоначально
" ничем не отличающиеся от них питающие клетки, за счет содержимого
которых происходит дальнейший рост и созревание яйца. Тут, таким,
образом, за каждым яйцевым фолликулом следует питательная
камера, содержащая питающие или желточные клетки.
Рис. 527. Dytiscus marginalis.
Женская половая система в брюшке в
естественном положении (из Ког-
schelt):
/ — концевой тяж яичннка, 2 — яичник, 3 —
чашевидная полость, 4 — влагалище, 5 —*
мускулатура яйцевода, 6 — яйцеклад, 7 —
совокупительная сумка, 8 — семяприемник,
9 — перерезанные поперек мышцы задней
пары ног
489
Яйцевая трубка заканчивается тонкой концевой
нитью, совокупность которых в каждом из яичников
образует концевой тяж (рис. 527, /),
прикрепляющийся, как упоминалось, изнутри к среднеспинке.
У места перехода яйцевой трубки в концевую нить
помещается зачатковая зона (germarium), где
происходит размножение и образование первичных половых
клеток, сначала оогоний, а из них — ооцитов. Вся
остальная часть яйцевой трубки занята зоной роста.
В ней, чередующиеся с питающими, яйцевые камеры
все увеличиваются в размерах по мере удаления от
зачатковой зоны. Число яйцевых камер в яйцевой
трубке достигает 8—10. Снаружи их границы
отмечены небольшими перехватами, придающими яйцевой
трубке четкообразный. вид. Длина последней из
яйцевых камер в трубке покоящегося яичника достигает
Рис. 528. Зрелое 1,5 мм, а зрелого яйца — 6—7 мм. Тонкое строение
яйцо Dytlscus таг- яйцевой трубки плавунца см. на с. 512. Каждая яйце-
ginallsach^ Ког" вая трубка продолжается в выводной проток,
снаружи имеющий вид короткого, более тонкого стебелька
с нежными, обычно складчатыми стенками. Все
выводные протоки яйцевых трубок каждого яичника впадают в общую полость
(рис. 527, 3), имеющую чашевидную форму, вогнутостью обращенную
к яичнику. Стенки этой полости у незрелого яичника сильно складча-'
ты. В период созревания она заполняется зрелыми яйцами (рис. 528),
которые оттуда выводятся через яйцеводы (рис. 529, /, 2). Последние
имеют форму коротких, слегка сужающихся трубок. Яйцеводы каждой
стороны, соединяясь вместе, образуют непарную трубку, снизу
впадающую во глагалище (см. рис. 527, 4). Влагалище и яйцеводы
являются уже образованиями эктодермального происхождения. Выстилающий
их эпителий покрыт тонкой хитиновой кутикулой. Поверхность
кутикулы покрыта направленными назад, к выводному отверстию,
чешуеобразными шипиками, облегчающими продвижение яиц к' выходу и
устраняющими возможность их обратного движения. Стенки яйцеводов
имеют хорошо развитый слой продольной мускулатуры. Перед местом
соединения яйцеводов каждой стороны к этому слою присоединяется
слой кольцевой мускулатуры, мощность которого увеличивается по
направлению к влагалищу.
Влагалище является концевой и наиболее мускулистой частью
выводных протоков женской половой системы. В нем различают два от-'
дела. Задний, выводной отдел, заключен между двумя створками
яйцеклада (см. рис. 527, 6), на брюшной стороне которого открывается
половое отверстие. Передний отдел имеет вид длинной трубки с муску-.
листыми стенками одинакового строения со стенками яйцевода. Эпите:"
лий и хитиновая выстилка влагалища сильно складчаты, что позволяет'
последнему сильно увеличиваться в объеме во время яйцекладки.
Собственно совокупительный отдел представляет собой
продолжение влагалища перед местом впадения в него яйцевода. Сравнительно
узкое горло расширяется в совокупительную сумку (bursa copulatrix,
490
Рис. 529. Dytiscus marginalis.
Схематический продольный разрез выводных
путей и совокупительного отдела женской
половой системы (из Korschelt):
/, 2 — яйцеводы, 3 — влагалище, 4 — горло
совокупительной сумки, 5 — совокупительная
сумка, 6 — семяприемник, 7 — желоб протока
семяприемника, 8 — железистое скопление
Рис. 530. Dytiscus
marginalis. А—самка,
откладывающая яйцо в стебель; Б —
лист с отложенными яйцами
(из Korschelt)
рис. 527, 7), продолжающуюся, после заметного сужения, в
семяприемник (receptaculum seminis, см. рис. 527, 8), имеющий вид изогнутого
слепого мешка. Строение горла и совокупительной сумки отличается
от строения влагалища более слабым развитием продольной
мускулатуры. Кольцевая мускулатура сильно развита и на границе с
семяприемником образует род сфинктера. Строение стенок семяприемника
имеет ряд особенностей. Стенки состоят из высокого цилиндрического
эпителия и сравнительно толстой, поверхностно складчатой хитиновой
выстилки. Мускулатура состоит из редкого слоя продольных мышц.
Сильное развитие хитиновой кутикулы делает стенки семяприемника мало'
податливыми к воздействию мускулатуры. Из семяприемника через
совокупительную сумку и ее горло к устью яйцевода ведет проток, по
которому поступает семенная жидкость для оплодотворения выходящих
во влагалище яиц. Этот проток представляет собой не замкнутую
трубку, а глубокий желоб, образованный эпителием стенки горла и
совокупительной сумки. Вдоль его дна расположено значительное скопление
одноклеточных желез, выводные протоки которых открываются в
желоб. Их выделения облегчают сперматозоидам путь из семяприемника
й устью яйцевода.
т
Яйцекладка производится в ткани водных растений (Sagittaria,
Acorns, Potamogeton, Iris и др.). Положение тела самки при откладке
яиц видно на рис. 530. Острый конец яйцеклада прорезает кутикулу и
внедряется в ткани (в губчатую паренхиму листьев или в область
проводящих пучков стеблей). Процесс откладки яйца длится около 10 мин,
после чего жук поднимается на поверхность воды для дыхания. Яйца
откладываются всегда поодиночке. Исследования показали, что
главное биологическое значение подобного выбора места для яйцекладки
заключается в богатстве ассимилирующей ткани растения кислородом,
необходимым для развития яйца. Механическая защита и
предохранение яйца от высыхания являются факторами 'меньшего значения.
Одна самка может отложить от 500 до 1000 яиц (максимум 1500).
В период яйцекладки в течение суток одна самка откладывает в
среднем около 10 яиц (максимум 30).
Мужские половые органы. У самца задний отдел нижней стороны
брюшка занят копуляционным аппаратом, доходящим почти до
заднего края шестого брюшного сегмента. Весь передний отдел брюшка
занят семенниками и очень сильно развитыми придаточными железами.
В расправленном виде мужская половая система представлена на
рис. 531. Она состоит из пяти отделов: 1) из парных семенников
•(рис. 531, /); 2) половых
протоков, состоящих из отводящих
каналов, дистальная часть
которых служит в качестве
резервуара для хранения семени,
и выводящих протоков (рис.
531, 2, 5); 3) придаточных
желез (рис. 531, 7); 4) семяизвер-
гательного канала (рис. 531,3)
и 5) копулятивного аппарата
(рис. 531, 4). Первые два
отдела — мезодермального
происхождения, остальные •— эк-
тодермального.
Многочисленными трахеями и перитонеаль-
ной оболочкой в теле
плавунца удерживаются первые два
отдела в своем положении в
виде поперечно вытянутого
клубка на уровне VI—VII
сегментов брюшка. Осторожной
препаровкой можно без труда
расправить органы мужской
Рис. 531. Dytiscus marginalis. Мужская по- половой системы,
ловая система в расправленном виде (из Семенник в период интен-
Korschelt): сивного созревания половых
/.— семенник, 2 — vesicula seminalis, 3— семяиз- rmn/TVKTnF! ттртпм и пгрн^ю
верительный канал, 4- копулятнвный аппарат и продуктов леТОМ И ОСеНЬЮ
его мускулатура, 5 — vas efferens, 6 — мышечный
тяж, соединяющий концы придаточных желез,
7 — придаточные железы
492
представляет собой эллипсоидной формы тело
около 9 мм длиной и 6 мм в толщину и
ширину. Это сильно извитая в клубок, тонкая трубка,
изгибы которой хорошо заметны сквозь
покрывающую ее перитонеальную оболочку. Тонкие
разветвления трахей богато оплетают семенник.
В расположении извивов семенника наблюдается
известная закономерность . Его слепой концевой
отдел вместе с прилегающей к нему частью
трубки занимает всегда брюшную сторону
клубка, так что vas efferens отходит всегда со
спинной стороны. Общая длина расправленной
трубки семенника достигает 30—40 см. Его
расправление сопряжено с трудностями: необходимо
разрушить более плотные трахеи, оплетающие
извивы нежной трубки. Под лупой или при
малом увеличении бинокуляра содержимое
семенников кажется наполненным светлой зернистой
массой. В просвете трубки второй половины
семенника можно различить более плотную
желтоватую ось — скопления дегенерирующих
элементов, служащих для питания.
В период зимнего покоя семенник сильно
отличается от описанного функционирующего
семенника. Концевой отдел — зародышевая зона—
имеет нормально заполненный вид на
протяжении около 2 см. Вся остальная часть трубки
имеет спавшиеся, сморщенные стенки.
Половые протоки начинаются выходящей
дорзально из клубка семенника трубкой,
начальный отдел которой длиной в 4—6 мм еще заключен в
перитонеальную оболочку (vas efferens). Далее эта трубка образует клубок,
играющий роль семенного пузырька (vesicula seminalis), в котором
сохраняется семя. Поэтому этот отдел, в противоположность
семеннику, осенью и зимой сильно заполнен поступившим из семенника
семенем. Длина его достигает 15—17 см. Из этого клубка на брюшной
стороне выходит проток длиной в 3—4 мм — vas deferens. Каждый из
протоков впадает со спинной стороны (рис. 532) в лежащую под ним
придаточную железу соответствующей стороны. У места перехода они
сужаются вследствие сильного сокращения кольцевой мускулатуры,
замыкающей просвет и не допускающей выхода семени.
Придаточные железы имеют вид двух оканчивающихся слепо
трубок, достигающих в функционирующем состоянии 3—5 см длины и
1 мм ширины. Весной и в первую половину лета, когда копуляция
отсутствует, они не функционируют и бывают .значительно тоньше. Их
слепые концы соединены друг с другом тонким мышечным тяжем.
В «каждой железе различают два отдела: концевой, длиной 9—12 мм
молочно-белого цвета, слабо прозрачный и слегка более расширенный,
и основной, желтовато-белого цвета. Концевой отдел изогнут таким
Рис. 532. Dytiscus
marginalis. Выводные пути
мужской половой
системы (из Korschelt):
I — vas deferens, 2 —
придаточная железа, 3 — семяиз-
вергательный канал, 4—
penis, 5 — парамеры
493
образом, что прилегает параллельно к основному. Непосредственно за
местом изгиба имеются два следующих друг за другом утолщения
(рис. 531). Как упоминалось, сверху в железы впадают vasa deferen-
tia. Вскоре после их впадения железы сближаются и сливаются
вместе, сохраняя, однако, самостоятельность своих выводных протоков.
Этот участок желез служит выводным протоком не только для их
секрета, но и для семейной жидкости. Он впадает jb следующий отдел
половой системы — в семяизвергательный канал.
Семяизвергательный канал (ductus ejaculatorius) — непарный
проток шириной около 0,8 мм и длиной 6—7 мм. Он начинается вскоре
после соединения протоков придаточных желез. Его начало внешне
отмечено темным включением — видимой снаружи частью хитинового
вооружения устьев выводных протоков желез, снабженных несущим
волоски цедильным замыкательным аппаратом. Семяизвергательный
канал открывается в желоб хитиновых частей мужского
совокупительного органа (penis).
Спаривание (максимумы поздней осенью и ранней весной)
происходит в воде (рис. 533, 534). При спаривании происходит, за счет
выделения придаточных желез, образование сперматофора. При этом
сначала в расширенную полость пениса выводится семенная жидкость
(закрытый цедильный аппарат устьев придаточных желез не
пропускает их густой секрет), потом туда же выводится секрет придаточных
желез, который обволакивает каплю семенной жидкости. Образовавшийся
сперматофор вводится в наружный совокупительный карман самки,
куда открывается отверстие влагалища, и застывает там, принимая его
форму. Под влиянием давления, развиваемого стенками кармана,
оболочка сперматофора лапается в наиболее тонком месте, приходящемся,
против отверстия влагалища, куда и поступает семенная жидкость.
Волнами мышечных сокращений она перегоняется вдоль влагалища,
через совокупительную сумку в семяприемник, где и сохраняется часто
в течение всего зимнего периода,
так как яйцекладка начинается
только весной. Задний отдел спер-,
матофора -торчит из
совокупительного кармана самки (рис. 535, А).
Сперматофор может быть оттуда
выпрепарован (рис. 535, Б).
Изучение срезов. Строение
волосков. Берут только что
вышедшего из куколки жука, у
которого кожные покровы еще не
отвердели, фиксируют отдельные
участки кожных покровов жидкостью
Джильсона или сулемой с уксусной
кислотой, заливают через смесь
Петерфи в парафин с воском.
Срезы толщиной 7,5 мкм окрашивают
по Маллори, а часть — железным
гематоксилином по Гейденгайну.
Рис. 533. Dytiscus marginalis.
Спаривание плавунцов. Вид сбоку (из Ког-
schelt):
494
Рис. 534. Dytiscus
marginalis.
Положение передней и
средней пары йог
самца иа спинке
самки во время
спаривания (из
Korschelt)
Рис. 535. Dyiiscus marginalis. A —
брюшко оплодотворенной самки с
торчащими из совокупительного
кармана сперматофором; 5
—сперматофор, выпрепарованный из
совокупительного кармана (из Korschelt)
Рис. 536. Dytiscus marginalis. Чувствительные волоски на продольных
разрезах через кутикулу. Л-в области мезостериума; Б -вблизи.
е г от брюшной стигмы (из Korschelt).
кула, а, нар нх ядра_ 8„хетоиды
Чувствительные волоски развиты на поверхности склеритов груди.
На любом срезе можно найти картины, сходные с изображенными на
рис. 536. Видно, что у основания волоока хитиновая кутикула
прерывается, образуя вокруг него сочленовную ямку, или теку (theca,
рис. 536, 5), хитин которой отличается от окружающей его кутикулы.
Сверху такая сочленовная ямка выглядит 'в виде кольца. От теки к
основанию волоска отходит перепонка, укрепляющая его в ямке.
Волосок внутри имеет (полость (рис. 536, 2), которая через отверстие в теке
сообщается с нижележащими клетками. Каждый волосок образован
двумя клетками. Одна из них образует сочленовную ямку, другая —
волосок. Плазма последней может заходить в полость волоска. Эти обе
клетки — текогенная и трихогенная —отличаются от соседних клеток
гиподермы (рис. 536, 5, 6). Если волосок выполняет железистую функ-
В Г
Ркс. 537. Dytiscus marginalis. Различного типа сенсиллы на продольных разрезах через
кутикулу. А — куполовидные органы на наружной поверхности верхней губы; Б —
вкусовой орган на гипофаринксе; В — чувствительный штифтик на внутренней лопасти
нижней челюсти; Г — чувствительный штифтик на переднеспинке (из Korschelt):
/ — сенсилла, 2 —■ хитиновая кутикула, 3 — сочленовная ямка, 4 — клетки гиподермы, 5 — концевое
окончание, 6 — чувствительные клетки, 7 — ядра неврилеммы, 8 — нервное волокно, 9 — поровые
каналы кутикулы
496
цию, то трихогенная клетка бывает сильно
развита и ее тело может вдаваться в
полость тела насекомого. Если волосок
выполняет чувствительную функцию, то к
вытянутой трихогенной клетке может
подходить нервное окончание, обнаружить
которое можно часто лишь путем применения
специальных методов (окраска метилен- рис. 5З8. Dytiscus marginalis.
блау, серебрение). Расположение отпугивающих
У плавунца наблюдается чрезвычайно желез переднего края передне-
большое разнообразие типов кожных чув- спинки- ВвдмЖри (и3 К°Г"
СТВИТеЛЬНЫХ Образований. Все ОНИ ЯВЛЯЮТ- ,_железы, ,1 положение отвер-
СЯ ВИДОИЗМеНеНИЯМИ ОПИСаННОГО ТИПа ЧувСТ- стия выводного протока, 3-перед-
вительного' волоска. Видоизменения затра- неспинка
гивают как строение и форму самого
волоска, так и теки. На рис. 537 дано несколько типов видоизмененных
чувствительных волосков (сенсилл).
С этим типом волосков не надо смешивать хетоиды (см. рис. 536,8),
которые представляют собой шиловидные или волосковидные отростки
кутикулы — скульптурные образования ее поверхности.
Отпугивающие железы. Методика — см. предыдущий
препарат. Наряду с огромным количеством одноклеточных желез,
рассеянных по всей поверхности тела плавунца, в переднегруди, под передним
краем переднеопинки расположены две сложные отпугивающие
железы (рис. 538). Их секретом является молочно-белая жидкость,
появляющаяся по краям грудного отдела у чем-либо обеспокоенного
(например, взятого в руку) жука. Скопления железистых клеток окружают
объемистый резервуар. Протоки отдельных клеток каждого скопления
образуют общий проток, впадающий самостоятельно в резервуар.
Каждая из железистых клеток по строению напоминает обычные кожные
одноклеточные железы, поэтому специальное описание последних не
приводится.
На рис. 539 и 540 представлен сагиттальный разрез через передний
отдел переднеопинки жука и часть его при большом увеличении.
В каждой из клеток, кроме крупного пузыревидного ядра с
хорошо заметным ядрышком, виден сложный внутриклеточный секреторный
аппарат, имеющий резервуар и самостоятельный выводной проток.
Стенки резервуара радиалыно исчерчены.
Пи ще в ар я те л ьн а я система. Берут овежеотпрепарованный
в физиологическом растворе кишечник жука и фиксируют сулемовым
фиксатором (жидкость-Джильсона и т. п.). Затем разрезают кишечник
на куски, заливают через смесь Петерфи в парафин.или в смесь
парафина и 10%-ного пчелиного воска. Куски для заливки берут длиной
около 1—1,5 ом. Изготовляют поперечные срезы из всех
нижеперечисленных отделов толщиной 7,5—10 мкм и окрашивают их частью
гематоксилином по Гейденгайну с последующей докраокой эозином,
частью —по Маллори. Два-три кусочка средней кишки, дл,я получения
хороших гистологических картин, фиксируют осмиевым фиксатором
(Флемминг, Шампи).
497
Рис. 539. Dytiscus marginalis. Участок отпугивающей железы
при большом увеличении (из Korschelt):
/ — хитиновая кутикула, 2 — гиподерма, 3 — железистые клетки, 4 — их
ядра, 5 — секреторный внутриклеточный аппарат, 6 — хитиновая
трубочка выводного протока, 7 — мышечные волокна
Рис. 540. Dytiscus marginalis. Сагиттальный разрез через
передний отдел передиеспинки на уровне отверстия
выводного протока отпугивающих желез (из Korschelt):
1 — кутикула, 2 — гиподерма, 3 — железистые . клетки, 4 — полость
резервуара, 5 — выводной проток, 6 — его устье, 7 — мышечные
пучки, 8 — лопасть жирового тела, 9 — трахея, 10 —
одноклеточные кожные железы
498
Рис. 541. Dytiscus marginalis. Поперечный раз- Рис. 542. Dytiscus marginalis. Учас-
рез через пищевод (из Korschelt): ток поперечного разреза через пище-
/ —хитиновая выстилка, 2 — эпителий, 3 — продоль- ВОД При большом увеличении (из Ког-
ная мускулатура, 4 — наружный слой кольцевых schelt):
мышц ' Обозначения см. рис. 541
Поперечный разрез через пищевод (рис. 541, 542). Пищевод
представляет собой трубку с почти округлым, слегка дорзовентрально
уплощенным контуром, с широким просветом
стого слоя
продольно-складчатой хитиновой выстилки (inti-
ma, рис. 541, 1) и из
подстилающего ее очень низкого
однослойного эпителия (рис. 541,
2), окруженного тонкой ба-
зальной мембраной.
Мускулатура пищевода слабо развита.
Она состоит из внутреннего
слоя тесно расположенных
продольных мышечных пучков
и наружного слоя кольцевых
мышц (рис. 541, 3, 4).
Хитиновая выстилка несет тонкие
продольные складки,
увеличивающиеся по мере расширения
пищевода при переходе его в
зоб.
Поперечный разрез через
зоб (рис. 543, 544). Зоб
отличается от пищевода
значительно более широким просветом.
Выстилающая изнутри его
поверхность хитиновая кутикула
Его стенка состоит из тол-
Рис. 543. Dytiscus marginalis. Поперечный
разрез через зоб (из Korschelt):
1 — хитиновая выстилка, 2 — эпителий, 3, 4 —
продольная н кольцевая мускулатура, 5 — трахен
499
Рис. 544. Dytiscus marginalis.
менее толста, чем в пищеводе, но
располагается более глубокими и многочисленными
складками. Ее поверхность несет тонкую
продольную исчерченность. Подстилающий
ее в один слой эпителий состоит из
сравнительно плоских клеток. Он окружен базаль-
ной мембраной. Продольная мускулатура
расположена преимущественно в складках,
куда проникают мелкие ответвления трахей
(рис. 543, 5). Расположенная снаружи
кольцевая мускулатура не заходит в
складки. Можно наблюдать точки прикрепления
5-часток поперечного разреза отдельных пучков кольцевой мускулатуры
через зоб при большом увели- ■ J J J J r
чении (из Korschelt):
1,2 — продольная и кольцевая
мускулатура, 3 — эпителий, 4 —
хитиновая выстилка
к стенкам складки. Этим достигается более
тесная связь мышечного слоя с собственно
стенкой зоба, что важно, если принять во
внимание значительное изменение объема
при наполнении зоба пищей и
последующие изменения его формы во время измельчения ее в жевательном
желудке. Снаружи к кольцевой мускулатуре прилегают более крупные
ветви трахей.
Жевательный желудок (proventriculus, рис. 545—547). Вследствие
интенсивной хитинизации зубов жевательного желудка, изготовление
микротомных срезов через 'него сопряжено с большими трудностями и
обычно кончается неудачей. Поэтому изучение строения жевательного
желудка лучше вести и а разрезах, произведенных острой бритвой
(можно лезвием безопасной бритвы) от руки. Для этого выделенный
жевательный желудок слегка зажимают между двумя пластинками
бузины и разрезают в сагиттальной плоскости. Для общего знакомства
с его, строением достаточно провести два разреза: через пластинки
первого и второго порядка, для чего нужно иметь два жевательных
желудка. Изучение строения производится под бинокуляром. Только
после изучения сагиттальных разрезЬв^можно приступить к более
сложным —поперечным.
Жевательный желудок имеет форму воронки, широкое отверстие
которой направлено в сторону зоба, а узкое обращено к средней
кишке. Внутренние -стенки его образуют систему продольных и поперечных
■складок, края которых обращены в просвет и вооружены сложно
устроенными хитиновыми зубами. Мощно развитая наружная кольцевая и
более слабая продольная мускулатура (рис. 547, 2, 3; 548) своими
сокращениями вызывает измельчение и перетирание пищи при помощи
зубов.
От зоба жевательный желудок отделен родом крышки, вдающейся
в его полость. Крышка образована восемью продольными, радиально
расположенными складками, поверхность которых сильно хитинизиро-
вана в виде треугольных пластинок. Четыре из них сильнее развиты и
частично перекрывают расположенные между ними пластинки. Это,
-соответственно, пластинки первого и второго порядка (см. рис. 546, 1,2).
Такое восьмилучевое строение, обусловленное восемью продольными
500
складками, характерно для строения
жевательного желудка плавунца на всем его протяжении,
вплоть до места перехода в среднюю кишку.
Сходящиеся в центре концы пластинок образуют
небольшое крестообразное отверстие,
соединяющее зоб с'жевательным желудком и плотно
закрывающееся при сокращении мускулатуры. На
сагиттальном разрезе через жевательный
желудок видно, что этой системе из четырех пар
пластинок соответствует такая же система
жевательных зубцов, являющихся непосредственным
продолжением пластинок книзу. Весь этот
передний отдел жевательного желудка представляет
собой кольцевую складку, поделенную
продольными складками на восемь частей.
За этой первой кольцевой складкой следует
вторая, оставляющая свободным узкий
промежуток между ними, который может полностью
замыкаться при сокращении продольных и
кольцевых мышц и расширяться при их
расслаблении. Вторая кольцевая складка поделена на
восемь частей, образующих вторую систему из
четырех пар зубов. Вслед за ними продольные
складки уплощаются и образуют заднюю камеру
жевательного желудка. Последняя, сужаясь,
переходит в более тонкий отдел, оканчивающийся
кардиальным клапаном, ведущим в ср-еднюю
кишку.
Поперечный разрез средней кишки.
Строение средней кишки у взрослого жука очень
сходно со строением личинки. Оно представляет
собой дальнейшее развитие секреторного
аппарата личинок (см. ниже). Трубчатые крипты жука
(рис. 549) настолько увеличиваются, что их
свободные концы проходят сквозь редкую сеть
внутренней кольцевой и наружной продольной
мускулатуры и придают средней кишке
снаружи своеобразный ворсистый вид. Общая
длина трубки достигает 1 мм, но
собственно крипта занимает ее незначительную
самом дне лежит очаг размножения, в котором
Рис. 545. Dytiscus
marginalis. Схема сагиттально-
го разреза через
переднюю кишку,
показывающая отношения между
ее отделами (из
Korschelt) :
/ — пищевод, 2, 3 — зоб, 4 —
жевательный желудок, 5 —
его задняя камера, 6 —
отверстие кардиального
клапана, 7 — стенка средней
кишки
концевую часть. На
можно найти частые
картины митозов. Сравнительно мелкие клетки имеют неправильно
полигональную форму с неясно различимыми границами. Под влиянием
возрастающего давления размножающихся и растущих клеток дна
крипты последние становятся более вытянутыми и плоскими,
располагаются рядами и проталкиваются в дистальном направлении к
выводному отверстию. При этом они увеличиваются в размерах, начинают
секретировать в просвет крипты и приобретают вид, размерыи форму
настоящих клеток эпителия средней кишки (рис. 550). Так, отдельная
501
Рис. 546. Жевательный желудок Dytiscus marginalis. A — вид со стороны
зоба; Б — сагиттальный разрез (из Korschelt):
1, 2— пластинки I и II порядка, 3— стенка зоба, 4 — начальный отдел кардиального
клапана
крипта из очага размножения превращается в функционирующую
трубчатую пищеварительную железу.
В средней кишке пища не прикасается 'непосредственно к эпителию.
Она отделена от него бесструктурной, складчатой оболочкой — перит-
рофической мембраной. Последняя непрерывно образуется клетками
эпителия средней кишки в области кардиального клапана и, как футляр,
обволакивает содержимое кишки. Перитрофическая мембрана вместе
Рис. 547. Dyiiscus marginalis. Сагиттальный разрез через
жевательный желудок в области расположения пластинок I
порядка (из Korschelt):
; — стенка зоба, 2— пучки продольных мышц, 3 — кольцевая
мускулатура, 4 — эпителиальные клетки, 5 — пограничная складка между
первой н второй системами зубцов, 6 — полость задней камеры
жевательного желудка
502
Рис. 548. Dytiscus marginalis. Сагиттальный разрез
жевательного' желудка в области расположения пластинок
II порядка (из Korschelt):
/ — стенка зоба, 2 — хитиновая кутикула, 3 — кольцевая
мускулатура, 4 — клетки эпителия, 5 — режущий край зубца, 6,7 — полости
задней и передней камер
Рис. 549. Dytiscus marginalis.
Продольный разрез трубчатых
крипт плавунца (часть
поперечного разреза средней кишки,
мускулатура не изображена)
(из Korschelt):
7 — нормальные клетки
секреторного эпителия, 2 — вакуоли секрета,
3 — очаг размножения секреторных
клеток на дне крипты
Рис. 550. Dytiscus marginalis. Часть
поперечного разреза стенки средней
кишки между основаниями трубчатых крипт
(из Korschelt):
/ — вакуоли секрета вместе с участками
клеток в просвете кишечника, 2 — секреторные
клетки, 3 — их ядра, 4 — базальная перепонка,
5 — продольные мышечные волокна; 6 —
палочковый слой
503
с пищей попадает затем в заднюю кишку, где при формировании
экскрементов исчезает в ректальном ее отделе. Перитрофическая
мембрана служит приспособлением, защищающим нежный эпителий средней
кишки от возможных механических повреждений пищевыми частицами.
Она легко проницаема как для пищеварительного секрета, так и для
жидких продуктов переваренной пищи.
Поперечный разрез на границе перехода средней кишки в заднюю
и в области привратника (pylorus). Короткий начальный отдел задней
кишки характеризуется "рядом особенностей. Его передняя часть
отличается сильно развитыми складками, заходящими в просвет
кишечника, в совокупности образующими клапан привратника (valvula pylori-
са). В этой же области впадают в кишечник -и четыре мальпигиевых
сосуда. На рис. 551 видно, что эпителий этого отдела отличается от
эпителия средней кишки отсутствием всяких следов секреторной
функции в виде вакуолей, зернистости в плазме и палочкового слоя. Его
принадлежность к задней кишке сказывается в наличии тонкой хитино-
Рис. 551. Dytiscus marginalis. Поперечный разрез на границе
перехода средней кишки в заднюю (из Korschelt):
/ — эпителий, 2 — хитиновая выстилка, 3 — базальная мембрана, 4 —
продольные мышцы, 5 —трахея, 6 — кольцевые мышцы (сфинктеры),
окружающие устья мальпигиевых сосудов, 7 — мальпигиевы сосуды, 8 —
крипты средней кишки, 5 — складки клапана привратника
504
вой выстилки (intima) на его
поверхности, обращенной в
просвет кишечника. Эпителий
образует сложные складки.
Базальная мембрана отделяет
эпителий от сильно развитой
продольной мускулатуры,
связывающей этот отдел со
средней кишкой,-так как там
лежат точки отхождения этих
мышц. Сокращения этих мышц
действуют расширяюще на
клапан. Сеть кольцевых мышц
развита слабо. Тут же видны
места впадения мальпигиевых
сосудов (см. с. 508).
За клапаном следует
собственно область привратника
(рис. 55S), снабженная
мощной мускулатурой, особенно
сфинктерами, сокращения
которой образуют запирающий
аппарат клапана. Эпителий, с
более толстой, чем в
предыдущем отделе, хитиновой
выстилкой, образует шесть радиально расположенных глубоких складок,
вдающихся в просвет кишечника настолько далеко, что они почти
соприкасаются в центре. Мускулатура состоит из сильно развитого слоя
кольцевых мышц и лежащих кнаружи от них продольных,
расположенных шестью пучками, соответственно просветам между
складками.
. Поперечный разрез тонкой кишки. Привратник постепенно
переходит в тонкую кишку. Слой кольцевых мышц становится слабее, пучки
продольных мышц — тоньше, клетки эпителия крупнее и малочислен-
нее, а хитиновая выстилка толще (рис. 553, А). На протяжении первой
половины длины тонкой кишки сохраняются все шесть продольных
складок, около ее середины три из них уплощаются и исчезают. Вторая
половина тонкой кишки (рис. 553, Б) имеет только три складки.
Обращают на себя внимание слоистость хитиновой выстилки, поперечная
исчерченность плазмы клеток эпителия, крупные округлые или
овальные их ядра с грубозернистыми хроматиновыми включениями и
заметно развитая базальная мембрана.
Немного не доходя до места впадения в ректальный пузырь, в
промежутках между складками появляются снова еще три складки.
Плазма эпителия утрачивает исчерченность, ядра становятся меньше,
хитиновая выстилка — тоньше, кольцевая мускулатура—более мощной
(рис. 553, В). Это клапан, запирающий выход из тонкой кишки. В
месте перехода в ректальный пузырь она несколько вдается в полость
последнего.
Рис. 552. Dytiscus marginalis.
Поперечный разрез через заднюю кишку в
области привратника (из Korschelt):
1 — хитиновая выстилка, 2 — эпителий, 3 — тяжи
продольных мышц, 4 — кольцевая мускулатура, 5 —
трахеи
505
Рис. 553. Dytiscus marginalis. Поперечный разрез через тонкую кишку (из Korschelt):
в начальном отделе (А), середине (Б), в конце (В). Обозначения • см. на рис. 552
Строение эпителия тонкой кишки, развитие складок, сильно
увеличивающих поверхность, указывает на то, что в тонкой кишке
происходит интенсивное всасывание переваренной в средней кишке пищи.
Строение ректального пузыря, слепого выроста и прямой кишки.
Стенка ректального отдела кишечника характерна сильно развитой
складчатостью. В ректальном пузыре (рис. 554, Б) складки более
мелкие, но расположены теснее, что допускает значительное увеличение
его объема. Эпителий с мелкими немногочисленными ядрами развит
слабо и тонким слоем лежит под 'Продольно исчерченной хитиновой
выстилкой. Мускулатура состоит из разреженного внутреннего слоя
кольцевых мышц и из семи продольных радиально расположенных пучков,
начинающихся от вершины слепого выроста. Из них только шесть
доходят до прямой кишки. Ректальный пузырь, постепенно сужаясь,
переходит в прямую кишку
(рис. 554, А), складки ее
стенки не так
многочисленны, но крупнее. В
остальном она мало чем
отличается от
ректального пузыря.
Уже упоминалось об
одной из функций
ректального пузыря — аэро-
и гидростатической-
К этой функции
необходимо прибавить об его
роли как защитного
органа, откуда через задне-
Рис. 554. Dytiscus marginalis. Поперечный разрез че- проходное отверстие жук
рез прямую кишку (А) и стенку ректального пузы- с силой выпрыскивает
ря (Б) (из Korschelt): экскременты в момент
1 — хитиновая выстилка, 2 — эпителий, 3 — кольцевая мус- J
кулатура, 4 —продольные мышцы Приближения ОПаСНОСТИ.
506
У личинок ректальный пузырь развит еще значительнее. В
наполненном состоянии его конец может достигать головы. Такое заполнение
осуществляется путем нагнетания в него воды через ротовое отверстие
и всегда бывает приурочено к моменту линьки и короткому послели-
ночному периоду.
Процессу сбрасывания старой шкурки предшествует момент ее
разрыва (по швам в головной капсуле и по средней линии спинной
стороны в груди). Это может осуществиться при повышении внутреннего
•давления, что достигается насасыванием некоторого количества воды
в ректальный пузырь через рот. Сравнительно твердый хитиновый
покров личинки слабо растяжим и в недостаточной степени допускает
увеличение размеров личинки в течение стадии, когда интенсивное питание
личинки сопровождается значительным приростом живого вещества.
Поэтому для только что слинявшей личинки биологической
необходимостью является искусственное увеличение объема, вызывающее
полное расправление еще мягкой неинкрустированной после линьки
кутикулы, доведение кожных покровов до максимального объема, до того,
пока их хитин не успел отвердеть. Это и достигается заполнением
ректального пузыря водой до максимально возможного объема. После
отвердения хитина излишек воды выводится наружу постепенно, по мере
поступления пищи в организм личинки.
Аналогичную функцию — расправления хитина — выполняет
ректальный-пузырь и у жука, сразу после выхода его из куколки. Только
тут он заполняется не водой, а воздухом, видимо, путем его
заглатывания через рот, так как и в кишечнике в это чвремя встречаются
пузырьки воздуха.
Трахеи. Для изучения тонкого строения трахеи необходимо взять
участок одного, из крупных стволов. Рассмотрение такого участка
трахеи, еще наполненного воздухом, под покровным стеклом в воде или
в глицерине дает возможность при большом увеличении увидеть ход и
строение спиральных нитей. На тотальном препарате из такого участка,
окрашенном квасцовым кармином или гематоксилином, можно
разглядеть ядра эпителиального слоя трахеи. На любом срезе через
насекомое можно найти перерезанные в разных плоскостях трахеи и изучить
строение их стенки (наиболее удобны срезы, окрашенные по Маллори).
Строение трахей отражает их происхождение как парных,
сегментарных впячиваний эктодермы. Их строение поэтому сходно со
строением кожных покровов насекомых. Стенка трахеи состоит из
однослойного трахейного эпителия, соответствующего гиподерме-кожных покровов,
и выделяемой им хитиновой выстилки, являющейся
непосредственно продолжением хитиновой кутикулы покровов. Эпителий состоит из
крупных плоских клеток неправильно полигональной формы с
большими округлыми ядрами, имеющими хорошо заметное ядрышко. В
хитиновой выстилке можно различить при большом увеличении три слоя:
прилегающий к эпителию слой эндокутикулы, спиральные утолщения,
соответствующие экзокутикуле кожных покровов, и тонкий
бесструктурный слой эпикутикулы, выстилающий полость трахеи. Трахейные
капилляры спиральных экзокутикулярных утолщений не имеют.
Последние отсутствуют и в воздушных мешках.
507
A
Б
Рис. 555. Dytiscus marginalis. Поперечный разрез через мальпигиевы
сосуды. А — в проксимальном отделе; Б — в дистальной части (из Ког-
schelt):,
1 — tunica . peritonealis, 2 — tunica propria, 3 — трахея, 4 — секретирующие клетки,
5 — ядра клеток эпителия сосудов, 6 — зернышки экскретов, 7 — палочковый слой
Ма л ьп и г и ев ы сосуды открываются в кишечник в области
привратника самостоятельными, но сближенными попарно
отверстиями. Они достигают значительной длины. Сначала они поднимаются
вдоль средней кишки, затем загибаются и тесно оплетают петли тонкой
кишки. Сосуды, в свою очередь, богато оплетены тонкими
ответвлениями трахей.
Мальпигиевы сосуды окрашены в коричневый цвет вследствие
заполняющих их темно-коричневых зернистых включений; только у места
впадения в кишечник они бесцветны. Этот бесцветный начальный отдел
отличается и меньшей толщиной и гладкими стенками, тогда как
остальная часть сильно извита и на поверхности имеет различные
вздутия. Концевой отдел заканчивается тупо и по строению не отличается
от., остальных. У только что вскрытого животного можно наблюдать
пульсацию и сокращения сосудов.
На срезах (рис. 555, Б) строение пигментированного дистального
отдела мальпигиевых сосудов следующее. Снаружи сосуды окружены
перитонеальной оболочкой (tunica peritonealis, рис. 555, 1),
обладающей маленькими вытянутыми ядрами. В ней заключены сократительные
волокна, проходящие преимущественно кольцеобразно. К
перитонеальной оболочке прилегает базальная мембрана (tunica propria,
рис. 555, 2). Эпителий сосудов состоит из крупных клеток с
гигантскими ядрами, границы между клетками, хотя и существуют, но слабо
заметны.. Ядра (рис. 555, 5) обычно неправильно овальной формы с
1—2 ядрышками и с сетью хроматиновых зерен. Плазма клеток,
особенно около ядра, заполнена большим количеством сильно преломля-
508
ющих свет желтоватых зернышек. Они скопляются в части клетки,
обращенной к просвету, заполняют ее, отрываются вместе с окружающим
их участком плазмы и попадают в просвет. Свободная поверхность ■
клеток покрыта тонким палочковым слоем (рис. 555, 7).
Короткий проксимальный отдел отличается по строению от
остальной части мальпигиевых сосудов (рис. 555, А). Перитонеальная
оболочка развита значительно сильнее. Ее сократительные элементы
образуюттут, видимо, род запирательного клапана, регулирующего
освобождение сосудов от накапливающихся в их просвете продуктов
выделения— жидкого содержимого с плавающими в нем зернами. Клетки
эпителия в этом отделе значительно меньшей величины, но
функционально они ничем не отличаются от крупных клеток дистального
отдела. Просвет значительно шире, чем в дистальном отделе.
Жировое тело. У вскрытого животного пинцетом отрывают
несколько лопастей жирового тела и бросают в фиксатор. При обычной
фиксации (жидкости Джильоона, Ценкера и др.) жировые включения
в клетках растворяются при последующем проведении через спирты
. возрастающей крепости, и на месте капель жира остаются сохраняющие
их форму вакуоли. При фиксации осмиевыми смесями (крепкий Флем-
минг, еще лучше — Шампи) жировые включения в значительной
степени сохраняются. При этом они часто могут маскировать другие
включения. Поэтому лучше всего часть материала фиксировать осмием, а
часть — каким-либо сулемовым фиксатором. Из залитого в парафин
материала приготовляют срезы толщиной 4—7,5 мкм и окрашивают их
железным гематоксилином по Гейденгайну с докраской эозином.
Несколько стекол желательно окрасить по Маллори для выявления
различных включений. Необходимо иметь в виду,'что жировое тело — очень
своеобразная ткань как по происхождению, так и по функции.
Поэтому в зависимости от возраста особи, ее физиологического Состояния в
настоящем и особенностей ее прошедшего развития картины состояния
жирового тела могут быть очень различны. Это пока в достаточной
мере изучено лишь для немногих объектов.
Плавунец не, принадлежит к их числу. Поэтому далеко
не всегда есть возможность приурочить
определенные картины жирового тела к тому или
иному возрастному или физиологическому состоянию
насекомого.
У молодых личинок внутреннее жировое тело
(рис. 556) имеет вид клеточных тяжей,
состоящих из сравнительно небольшого числа
округлых клеток с крупными ядрами, хорошо
заметными границами, с цитоплазмой, почти
лишенной включений, с редкими вакуолями. Тяжи
располагаются около трахей, так как все процессы Рис- 556. Dytiscus margU
отложения, переработки и выделения запасных nalis- Внутреннее жиро-
питательных веществ требуют хорошего снабже- вунца^первоПтадии 7из
ния кислородом. Korschelt):
Постепенно С ВОЗраСТОМ КОЛИЧеСТВО КЛетОК /-клетки жирового тела,
увеличивается за счет оседания вокруг таких *~ "вакуоли?'4-7рГхея°вые
509
первоначальных лопастей и тяжей
недифференцированных форменных
элементов крови. Клетки
увеличиваются в объеме, и цитоплазма
заполняется включениями
преимущественно жировых капель
(нейтрального жира) и различного рода эо-
зинофильных или ацидофильных
гранул (рис. 557, 558). Сначала
образуются в плазме небольшие
многочисленные капельки, впоследствии
сливающиеся в крупные капли.
В случае сильного заполнения
клеток жировыми включениями,
границы между ними становятся
незаметными. У голодающих животных
(рис. 559) наблюдается
исчезновение и резкое уменьшение жировых
включений и гранул, уменьшение
объема клеток и выявление вновь
клеточных границ.
Часть жирового тела в течение
метаморфоза переходит без изменения от личинки к жуку, часть
личиночного жирового тела распадается и вновь образуется в процессе
гистогенеза. Поэтому у молодых жуков можно встретить картины,
подобные изображенной на рис. 560, где одна часть лопасти занята старыми
клетками с крупными жировыми вакуолями, а другая состоит из вновь
присоединившихся клеток, в которых только что начался процесс
отложения аапасных питательных веществ.
Рис. 557. Dytiscus marginalis. Разрез
через лопасть жирового тела у
готовой к окукливанию личинки пла-
. вунца третьей стадии (из Korschelt):
/ — ядра клеток, 2 — гранулы, 3 —жировые
вакуоли, 4 — границы клеток
Рис. 558. Dytiscus marginalis. Разрез через
лопасть жирового тела с большим количеством
жировых включений. А — личинки третьей стадии;
Б — взрослого жука (из Korschelt):
1 — ядра клеток, 2 — жировые вакуоли, 3 — гранулы
Рис. 559. Dytiscus marginalis.
Разрез через лопасть
внутреннего жирового тела у
голодающего жука (из Korschelt):
/ — ядра клеток, 2 — жировые
вакуоли, 3 — границы клеток
510
У самцов по сравнению с самками жировое тело значительно
беднее включениями. У самок с созревшими яйцами клетки жирового
тела почти не содержат запасных питательных веществ, израсходованных
в процессе созревания половых продуктов.
Функция жирового тела как выделительного органа, своеобразной
почки накопления продуктов обмена, сказывается в отложении в его
клетках мочекислых включений (уратов), что характерно для более
старых особей. Эти включения в виде небольших желто-коричневых
зерен заполняют плазму клеток, лежащих на поверхности лопастей
жирового тела, особенно наружного жирового тела нижней стороны и
боков брюшка. Другие отделы жирового тела обычно свободны от этих
включений. Лопасти жирового тела, имеющие включения зерен уратов,
отличаются более темной желтовато-коричневой окраской.
Сердце. Зафиксированное жидкостью Ценкера или сулемовым
фиксатором сердце заливается через кедровое масло в парафин.
Изготовляется серия продольных и поперечных срезов толщиной в 5—
7,5 мкм и окрашивается по Маллори или железным гематоксилином по
Гейденгайну. Хорошие результаты дает изучение тотального препарата,
окрашенного гематоксилином.
Стенка сердца состоит из трех слоев (рис. 561). Наружный слой
(adventitia, рис. 561, 1) соединительнотканный. На тотальном
плоскостном препарате видна сеть волокон с разбросанными в них
продолговато-овальными ядрами. Под adventitia помещается наиболее толстый
мышечный слой кольцевых мышц с резко выраженной поперечной по-
лосатостью (рис. 561, 2). Просвет сердца выстлан перепонкой,
представляющей несколько утолщенную сарколемму мышечного слоя,
защищающую последний от механического трения, производимого
движущейся в сосуде кровью (рис. 561, 3). Строение стенки сердца в
области остий (рис. 562) ничем не отличается от строения в других
местах. Строение стенок аорты сходно со строением сократительной части
спинного сосуда.
Рис. 560.- Dytiscus marginalis. Разрез Рис. 561. Dytiscus marginalis. Часть по-
через лопасть жирового тела жука перечного разреза через стенку сердца
(из Korschelt): ' плавунца (из Korschelt):
/ — ядра клеток, 2 — гранулы, 3 — грани- • / — adventitia, 2 — кольцевые мышцы, 3 — вну-
цы клеток, 4 — жировые вакуоли тренняя выстилка просвета
511
Рис. 562. Dytiscus marginalis. Поперечный разрез через сердце. А — в
момент систолы; Б — на уровне первой пары остий (из Korschelt):
/ — эластические волокна, 2 — стенка околосердечной полости, 3 — adventitia и ее
ядра, 4— кольцевые мышцы, 5 — перикардиальиые клетки, 6 — остии с клапанами
Яйцевая нить. У вскрытой самки плавунца быстро выделяют
яичник, перерезав ножницами трубку яйцевода и концевой тяж, и
помещают ,в фиксирующую жидкость (лучше всего в осмиевый
фиксатор— жидкость Флемминга или Шамтш). Отдельные яйцевые нити
заливают через смесь Петерфи в парафин. Продольные срезы толщиной
б 4—6 мкм окрашивают железным 'гематоксилином, по Гейденгайну.
Последующая докраска эозином необязательна. Изучение препарата
начинают с концевого отдела яйцевой нити (рис. 563).
, Концевая нить отделена от зародышевой камеры несколькими
слоями поперечно расположенных веретеновидных клеток (рис. 563, 1, 2).
Под ними начинается зародышевая зона. В ней можно различить дня
рода клеток; более мелкие клетки с плохо видимыми границами, с
маленькими овальными ядрами с обычно хорошо'заметным ядрышком, и
более крупные клетки с хорошо выраженными границами, с более
крупными богатыми хроматином ядрами. Клетки первого типа —
эпителиальные клетки, идентичны с клетками концевой нити, из которых впо-
512
следствии образуется эпителий яйцевых
фолликулов, в зародышевой камере они расположены
без порядка. Клетки второго типа — первичные
половые клетки — оогонии. По направлению от
концевого отдела они быстро увеличиваются в
размерах и митотически делятся (рис. 563, 3).
За этим первым отделом зародышевой
камеры, характеризующимся размножением и
ростом оогоний, следует второй, в котором
происходит образование питающих клеток. В этом
отделе, наряду с рассеянными повсюду
одиночными эпителиальными клетками, встречаются
клеточные скопления, отдельные клетки которых
лишь частично отделены границами друг от
друга. Среди них одна из клеток имеет более
крупное и богатое хроматином ядро. Это ооцит, а
более мелкие клетки, образовавшиеся в
результате неравного деления оогоний, —•
питающие клетки. Один ооцит обычно окружен
15 питающими клетками (рис. 563, 5). Связь
ооцита с питающими клетками .осуществляется
путем образования цитоплазматических
мостиков, образующихся через отверстия в их
клеточных оболочках. В этом клеточном .скоплении
ооцит расположен впереди, а питающие
клетки в виде розетковидной массы — позади
него.
В месте перехода зародышевой зоны в зону
роста клетки заметно увеличиваются- в
размерах. Сильно заполненные хроматином темные
ядра ооцитов становятся светлее и принимают
типичный вид зародышевого пузырька. Наоборот,
ядра питающих клеток становятся темнее
вследствие увеличения в них количества хро'матино-
вых зерен. Клетки тоже увеличиваются в разме- •
ре. Накопление хроматина в ядрах стоит в
связи с усилением их секреторной деятельности, как1
питающих ооцит клеток. По цитоплазматичес-
ким мостикам в плазму ооцита устремляются
токи из плазмы питающих клеток. В плазме
ооцита, особенно в ее части, обращенной к
питающим клеткам, происходит накопление зернистых
включений — результат переработки
притекающих питательных веществ (рис. 563, 6).
В результате роста диаметр ооцита
становится почти равным диаметру яйцевой трубки.
Вследствие этого ооциты вместе с розеткой пи--.
тающих клеток располагаются в линейном
порядке. К этому времени эпителиальные клетки
щ
\\
щд
ш
-%
ex.
Рис, 563. Dytiscus
marginalis. Продольный разрез
через концевой .'отдел
яйцевой нити; (из
Korschelt) :
/ — концевая нить, 2 —
граница между концевой иитью
и зародышевой камерой, 3 —
растущие и митотически
делящиеся оогонин, 4 —
розетки питающих клеток, 5 —
ядра растущих ооцитов, 6 —
включения питательных
веществ в плазме растущих
ооцитов, 7 — начало
образования яйцевого "фолликула,
8 — наружная оболочка
(tunica propria) яйцевой нити,
. 9 — эпителиальные клетки
17—2
513
Рис. 564. Dytiscus marginalis.
Отдельная яйцевая камера в
продольном разрезе (из Korschelt):
/ — питающие клетки, 2 — включения
питательных веществ в плазме ооцита,
3—ядро ооцита, 4 — клетки фолликулярного
эпителия, 5 — tunica propria
Рис. 565. Dytiscus marginalis.
Продольный разрез через зрелые
участки яйцевой нити. А ■— средний отдел;
Б — последняя камера (из Korschelt):
1 — питательная камера, 2— яйцевой
фолликул, 3 — питающие клетки, 4 —
фолликулярный эпителий, 5 — ядро яйцевой клетки,
б — чашевидное расширение выводного
протока яйцевой нити, 7 — выводной проток,
8 — желточное тело
окружают ооцит и постепенно образуют вокруг него фолликул
(рис. 563, 7). Они оставляют свободным только полюс ооцита,
обращенный к питающим клеткам. С ростом ооцита количество их
увеличивается делением (рис. 564)\
По мере дальнейшего роста и созревания яйца (рис. 565) ядра
клеток фолликулярного эпителия становятся богаче хроматином. В них
можно различить 1—2 ядрышка. Ядро становится -похожим на ядра
железистых клеток. Это связано с питающей деятельностью клеток
фолликулярного эпителия, а позднее — с образованием за счет их
выделений хориона яйца. Хорион ив виде тонкой, сильно красящейся оболочки,
можно различить в последних яйцевых камерах трубки, особенно если
яйцо слегка сморщилось при фиксации.
Личинка плавунца
Внешняя морфология. Личинка третьей стадии плавунца (рис.566)
удлиненно-веретеновидной формы, достигаете длину 5 см. Широкая ло-
патовидно-уплощенная голова, сильно суженная в заднем отделе, пере-
514
Рис. 566. Личинка плавунца третьей стадии. Вид со спииной
стороны (А), сбоку (Б) и с брюшной стороны (В) (из Korschelt):
1 — голова, 2—4 — передне-, средне- и заднегрудь, 5—12 — первый — восьмой
членики брюшка, 13 — задняя пара брюшных стигм, 14 — церки, 15, 16 —
седьмая и первая стигмы брюшка, 17, 18 — задне- и среднегрудная стигмы
ходит в трехчленистую грудь, за которой следует длинное восьмичлени-
ковое брюшко. Общая окраска — желтовато-коричневая; более сильно
склеротизированные спинные щитки 'пигментированы темнее, все
межсегментные перепонки —светло-желтоватого цвета, брюшная сторона
светлее спинной.
Голова « ее придатки. Голова (рис. 567) полностью
окружена головной капсулой, в которой можно р.азличить все элементы
головной капсулы взрослого жука. Лобный, или фронтальный, склерит
занимает весь передний отдел спинной стороны головы. Он ограничен
сзади фронтальным швом. Иногда 'поверхность склерита подразделена
слабо выраженным поперечным (эпистомальным) швом на два
отдела —■ передний и задний. Это доказывает, что собственно лобным скле-
17*-
515
ритом личинки является толь-
ко задний его отдел. Передний
же гомологичен наличнику.
Вдоль дугообразно
выпуклого переднего края лобного
склерита сидят расположенные
в 2—3 ряда уплощенные
волоски (рис. 568). Строение
сочленовной ямки, в которой
сидит своим основанием каждый
из них, таково, что ее верхний
край выступает в виде
козырька далеко за основание
волоска. Вследствие этого, при
внешнем давлении на них
спереди, они могут отгибаться
только вниз. Это надо
рассматривать как специальное
приспособление, помогающее
удержанию схваченной
мандибулами добычи, которая при
этом прижимается к
переднему краю лобного склерита'.
Отгибающиеся назад волоски
своими плоскозаостренными
концами мешают добыче
выскользнуть вперед. На
поверхности лобного склерита
рассеяны короткие чувствительные
волоски, располагающиеся в
два ряда. Одиночные
волоски сидят вдоль фронтального
шва.
От середины последнего отходит корональный шов, достигающий
заднего края головы. Он делит.всю постфронталвную часть головной
капсулы на две симметричные половины. Центральную часть занимает
теменной отдел, на боках переходящий в щеки. Их передне-боковые
отделы доходят до уровня боковых углов фронтального склерита и
прикрывают сверху основания мандибул. Кнаружи от последних лежат
точки отхождения усиков, а гоозади — глаза. Пооледние принадлежат
по своему строению к типу археомм (их строение см. с. 534). Около
Личиночных глаз у взрослых личинок могут просвечивать зачатки
сложных глаз взрослого жука в виде двух пятен. Кроме усиков и
мандибул из-под переднего края головы видны щупики нижних челюстей
и нижней губы..
'Начиная от глаз щеки сужаются, сначала постепенно, а затем
образуют резкий перехват, отделяющий шею от собственно головы. На
границе помещается поперечный затылочный шов, отделяющий сильно
развитый в виде воротничка шейный отдел — затылочный склерит —
Рис. 567. Голова личинки плавунца третьей,
стадии. Вид сверху (из Korschelt):
/ — нижнегубной щупик, . 2 — верхняя челюсть, 3 —
нижняя челюсть, 4 — усик, 5 — наличник, 6 — лоб,
7 — просвечивающий зачаток сложного глаза, 8 —
глазок, 9 — фронтальный „шов, 10— темя, 11 *=-
корональный шов, 12 — затылочный шов, 13 — затылочный
склерит, 14 — щеки, 15 — уплощенные волоскн
переднего края,головы
516
Рис. 568. Личинка плавунца.
Уплощенные волоски переднего края
головы. А — фронтальный вид;
Б — сагиттальный разрез (из
Korschelt) :
1 — клетки гиподермы, 2 — хитиновая
кутикула, 3 — волосок, 4 — выступ
сочленовной ямки
Рис. 569. Голова личинки плавунца третьей
стадии. Вид с брюшной стороны (из
Korschelt) :
/ — нижняя губа, 2 — нижняя челюсть, , 3 —
верхняя челюсть, 4 — усик, 5 — глазок, 6 — следы
прикрепления задинх ветвей внутреннего скелета, 7 —
щеки, 8 — затылочный шов, 9 — затылочное
отверстие, 10 — затылочный склерит, // —
продольный шов
от постфронтального-отдела. Этот склерит в голове взрослого жука не
выражен в качестве самостоятельного склерита, а входит в состав утол-'
щения, окружающего затылочное отверстие.
С брюшной стороны голова личинки плавунца (рис. 569)
отличается от головы взрослого жука помимо формы и своим строением.
Затылочный склерит образует на брюшной стороне глубокий вырез,
позволяющий личинке сгибать голову почти под прямым углом к груди.
Область горла (gula) сильно, развита, но швы, отделяющие ее от щек,
отсутствуют. Их местоположение отмечено точками 'прикрепления к
головной капсуле задних ветвей внутреннего скелета головы, ясно
заметными снаружи (рис. 569, 6).
Посередине проходит продольный шов, начинающийся от
затылочного отверстия и доходящий до основания нижней губы. Этот шов
вторичного'происхождения и имеет приспособительное значение,
связанное с особенностями линьки. При линьке головная капсула лопается
по фронтальному и корональному швам. Однако упругость склеритов
теменной части головы при узком, сравнительно с шириной.головы,
затылочном отверстии, не дает возможность личинке освободить голову
от шкурки без добавочных приспособлений. По этому шву. во .время
линьки шкурка головной капсулы или сгибается, что увеличивает
свободный промежуток между разошедшимися по швам дорзальными
склеритами, или иногда даже лопается, вследствие чего обе половинки
головной капсулы расходятся в стороны и затылочное отверстие
увеличивается в ширину, позволяя свободно вытащить сквозь него голову,
Основания.ротовых придатков сдвинуты вперед, к переднему краю
517
головы. Они сидят на узком уступе, сверху прикрытом передним краем
лобного склерита.
Усики (антенны, рис. 570) отходят от боков головы между
основаниями мандибул и глазами. У личинки первой стадии они состоят из
трех палочковидных члеников. У личинки третьей стадии каждый из
этих члеников у основания оказывается 'вторично перешнурованным.
Концевой членик несет на вершине маленький палочковидный
чувствительный орган. Подобное вторичное расчленение делает длинные и
тонкие усики значительно более гибкими и подвижными—существенное
преимущество при ощупывании схваченной добычи.
Ротовые придатки (рис. 571) личинки плавунца чрезвычайно
видоизменены по расположению, строению и функции сравнительно с
взрослым жуком. Как мы увидим ниже, это находится в прямой связи со
способом питания и схватывания добычи. Если у жука ротовые
придатки расположены по кругу, ограничивая ротовую полость, то у
личинки они расположены в два сильно сближенных параллельных ряда.
Таким образом, максиллы лежат не под, а кнутри от мандибул, и не
над, а кнаружи от нижней губы. Еще более значительны отклонения в
строении и функции. '
Верхняя губа как подвижное самостоятельное образование,
принимающее активное участие в работе ротовых придатков, отсутствует. За
ее гомолог принимается участок головной капсулы, ограниченный
уступом, от которого отходят ротовые придатки, и передним краем
лобного склерита. Она, таким образом, составляет нижнюю стенку
козырька, образуемого передним краем лобного склерита над
основаниями ротовых придатков. На ее поверхности по бокам, расположены
овальные железистые поля. Это участки, покрытые мягким хитином и
\)
А Б В
Рис. 571. Передний край головы личинки плавунца третьей
стадии. Вид с брюшной стороны (из Korschelt):
/ — верхняя челюсть, 2 — ее сочленение с головной капсулой, 3 —
основание усика, 4—■ cardo, 5 — железистое поле, 5 — stipes, 7 —
наружная лоиасть, 8 — нижнегубной щупик, Я — pfaementum
Рис. 570. Левый усйк
личинки плавунца
третьей (А), второй (5)
и первой (В) стадий.
Вид сверху (из
Korschelt)
несущие многочисленные па-
пиллы, крючки и зубчики. На
их поверхности наблюдается
присутствие клейкого секрета.
О возможном функциональном
значении железистых полей
будет сказано ниже.
Верхние челюсти —
мандибулы (рис. 572) — серповидно
изогнутой формы, с вытянутым
в виде острой иглы концом.
Они сильно хитинизированы,
очень упруги и крепки. На
внутренней поверхности,
немного отступя от конца
мандибулы, имеется продолговато
овальная щель, ведущая в
канал, проходящий по всей
длине мандибулы и
открывающийся у ее основания. Этот канал
морфологически не является
замкнутой с боков трубкой, а
представляет собой глубокую
складку на внутренней
поверхности мандибулы с сильно
сближенными краями (рис.
572, Б), что позволяет ей функционировать в качестве канала.
Мандибулы причленены к голове при помощи двуглавого
шарнирного сочленения и могут двигаться только в одной горизонтальной
плоскости. Они приводятся в движение двумя мощными
мышцами-антагонистами, прикрепляющимися к хитиновым аподемам у основания
мандибул (рис. 572, 4). Особенно сильно развит сгибатель (m. flexor
mandibulae), отходящий от спинной и брюшной стенок заднего отдела
головы. Его антагонист — разгибатель (m. extensor mandibulae)
отходит от бокового края заднего отдела головы.
Мандибулы личинки являются одновременно органами
схватывания и удержания добычи и органами, посредством которых
происходит питание личинки. Кожные покровы схваченной добычи
прокалываются концами мандибул, которые погружаются глубоко в тело жертвы.
В это же время через канал мандибул внутрь тела личинка
впрыскивает секрет, служащий не только для 'Парализования добычи, но и для
ее переваривания. Через некоторое время, не выпуская добычу,
личинка всасывает через те же каналы мандибул -*нереварившееся жидкое
содержимое тела добычи. У личинок плавунца мы встречаемся с
редким случаем внекишечного пищеварения. Строение мандибул является
одним из приспособлений к подобному типу питания.
В покое мандибулы сложены так, что их наружные края ложатся
параллельно изгабу переднего края головы, а концы соприкасаются с
поверхностью железистых полей верхней губы. Секрет этих полей сма-
Рис. 572. Личинка плавунца третьей стадии.
А — правая верхняя челюсть, Б — разрез
через верхнюю челюсть (из Korschelt):
1 — выводное отверстие канала, 2 — щель канала, 3 —
входное отверстие канала, 4 — аподемы сгибателя и
разгибателя верхней челюсти, 5 — щетинки краев
канала, 6 — полость канала, 7 — полость верхней
челюсти
519
f
/-*
зывает концы мандибул, а шипики и
волоски-полей счищают с их поверхности различные
посторонние частицы.
Нижние челюсти личинки — максиллы
помещаются между основаниями мандибул и нижней
губой. Внешне они представляют собой
типичные щупа льды (рис. 573). Однако, несмотря на
такое сильное отклонение от нормального типа
строения, в них можно найти все характерные
для максилл элементы. Основной членик (cardo,
рис. 573, 3) очень мал, в виде небольшой
пластинки он помещается с внешней стороны на'
основании стволика (stipes, рис. 573, 2) и
участвует в сочленении максиллы с головной капсулой,
т. е. сохраняет свое нормальное положение.
Стволик имеет удлиненную палочковидную
форму с слегка расширенными концами. Прямым
его продолжением является нижнечелюстной
щупик, состоявший первоначально из трех
члеников, у каждого из которых, подобно членикам
усика, вторично отшнурован базальный отдел.
Образовавшиеся, таким образом, шесть
члеников щупика становятся все тоньше в дистальном
направлении. Конечный членик на вершине
несет чувствительные органы, а на теле члеников
сидят в небольшом, но обычно постоянном
числе, чувствительные волоски. С внутренней
стороны вершины стволика, у основания щупика
сидит шипообразный рудимент наружной лопасти (lobus externus,
рис. 573, 5) нижних челюстей. Внутренняя лопасть полностью
редуцирована. Этот шип во время питания прижимается к схваченной добыче,
погружается в нее; он, возможно, является органом вкуса. Щупик
служит, видимо, органом осязания.
Нижняя губа (labium, рис: 574) занимает срединное положение
между нижними челюстями. Она тоже значительно видоизменена. Под-
подбородок (submentum) вошел в виде горла (gula) в состав головной
капсулы (см. с. 451). Подбородок (mentum) рудиментарен, в виде узг
кой хитиновой полосы у основания нижней тубы. Хорошо развит лишь
пластинчатый praementum, имеющий удлиненно прямоугольную.форму.
На внешних углах он несет палочковидные нижнегубные щупики,
состоящие у личинки третьей стадии из четырех члеников, а у- личинок
младших стадий — только из двух. Нижнегубные щупики, подобно ниж-
нечелюстным, имеют осязательную функцию.
У личинки плавунца, таким образом, из ротовых придатков
прямое участие :в процессе питания принимают только мандибулы. Осталь^
ные придатки в большей или меньшей степени видоизменены и
редуцированы и сохранили лишь чувствительную, преимущественно
осязательную функцию.
Описанные уплощение и расширение головы личинки, связанные с
Рис. 573. Правая нижняя
челюсть личинки
плавунца третьей стадии (нз
Korschelt):
1 — первый членик щупика,
2 — stipes, 3 — cardo, 4 —
аподемы сгибателя и
разгибателя нижней челюсти, 5 —
наружная лопасть
520
особенностями движения
мандибул, а также линейное
расположение ротовых частей
имеют следствием то, что ротовое
отверстие личинки принимает
форму узкой щели,
проходящей по всей ширине головы,
от одного основания
мандибулы до другого. При этом
внутренние части оснований
мандибул оказываются
погруженными в углы ротового
отверстия. Уже упоминалось, что в
процессе питания личинка
сначала впрыскивает через-
каналы мандибул в свою добычу
пищеварительный секрет, а
потом тем же путем всасывает
обратно переваренное
жидкое ее содержимое.
Наличие открытой ротовой
лость с внешней средой, делало бы эти процессы невозможными.
Поэтому, как глубокое приспособление к подобному типу питания у
личинок плавунца ротовая щель замкнута по всей своей длине, за
исключением углов- рта у оснований мандибул (рис. 574). Замыкание
настолько плотно, что сохраняется- на сброшенных личиночных шкурках.'
У только что слинявшей личинки оно отсутствует, но в это время она
не питается. Замыкание наступает при отвердении хитина головы.
Если отпрепаровать фронтальный склерит, и склериты затылочного
отдела головы, можно увидеть соотношения между ротовой полостью,
основаниями мандибул и глоткой (рис 575).
Грудь и ее придатки. Сегменты груди сходны между собой
(рис. 566, 576). Наиболее сильно развита переднегрудь (prothorax,
рис. 576, 9). Она имеет форму суженного кпереди усеченного конуса,
выпуклого на спинной стороне и уплощенного с 'брюшной. Вся спинная
сторона' и бока сегмента покрыты седловидным хитиновым щитком,
нижние края которого слегка загибаются на брюшную сторону. На
спинной стороне по средней линии тела проходит продольный шов, по
которому лопается кутикула во время линьки. На брюшной стороне в
переднем отделе помещается склерит, спереди примыкающий к горлу,,
а с боков — к краям спинного щитка (рис. 576, /). У последующих
сегментов груди он отсутствует. К нему прикрепляются мышцы,
сгибающие голову личинки. Места прикрепления их заметны снаружи.
Позади склерита, между основаниями передней пары ног, лежит поперечный
склерит (sternum, рис. 576, 4, 5). Основания тазиков ног окружены
двумя склеритами, соответствующими эпимеру и эпистерну (рис. 576,
2, 3). Сильное развитие переднегруди, сравнительно с-последующими
сегментами груди, необходимо поставить в связь с развитием хорошо'
дифференцированной мускулатуры, двигающей голову личинки, С этим
Рис. 574. Личинка плавунца. А —
сагиттальный разрез через передний отдел головы
личинки по средней линии; Б — замкнутые края
ротовой полости при большом увеличении (и»
Korschelt):
/ — лоб, 2 — наличник, 3 — плоские волоски переднего
края, 4 — нижняя губа, 5 — ее щупик, 6 — гипофа-
ринкс, 7 — полость глотки, 8 г- ротовая полость, 9 —
эпифаринкс
щели, сообщающей ротовую по-
521
же связано и образование дополнительного брюшного склерита в
переднем отделе сегмента.
Средне- и заднегрудь, вместе взятые, не превышают длину перед-
негруди. Оба сегмента имеют одинаковое строение. Со спинной
стороны оба они покрыты седловидным склеритом, края которого заходят на
боковые стороны. По бокам, на уровне переднего края этого склерита,
помещаются нефункционирующие замкнутые стигмы. Они на средне-
груди крупные, с 'более резко .очерченными краями. На заднегруди они
имеют вид темных точек. Склериты уплощенной брюшной части
сегментов сходны со склеритами заднего отдела переднегруди.
Ноги (рис. 577) хорошо развиты. Каждая последующая пара
длиннее предыдущей. В остальном ©се они одинакового строения. Тазики
удлиненно цилиндрической формы с рядом плавательных волосков
вдоль внешнего края. Их шарнирного типа сочленение с грудным
сегментом обеспечивает большую свободу движения. Вертлуг расположен
почти под прямым" углом к тазику, с которым он более свободно
сочленен, чем с бедром. Вдоль внутреннего края вертлуга, особенно в дис-
тальной части, сидят плавательные волоски. Бедро — наиболее
длинный из члеников ног. Оно слегка уплощено с баков и несет два ряда
длинных (до 2 мм), тонких, но
упругих плавательных волосков.
Более короткие голень и одночленико-
вая лапка, вооруженная двумя
коготками, тоже несут аналогично
развитые и расположенные волоски
с той разницей, что волоски на ниж~
Рнс. 575. Личинка плавунца. Голова с
отпрепарированными фронтальным и
затылочным склеритами головы и
удаленным надглоточным ганглием и
мышцами (из Korschelt):
/ — полость канала верхней челюсти, 2 —
ротовая полость, 3 — полость глотки. Путь,
по которому проходит пища, зачернен
Рис. 576. Передние
членики тела личинки
плавунца третьей стадии..
Вид с брюшной стороны
(из Korschelt):
/ — дополнительный склерит
переднегруди, 2 — эпистерн,
3 — эпимер, 4, 5 — sternum,
6 — первый брюшиой
сегмент, 7—9 — задне-, средне-
и переднегрудь. Ноги, за
исключением передней пары,
удалены
522
Рис. 577. Правая задняя нога личинки плавунца
третьей стадии. Вид сзади (из Korschelt):
1 — тазнк, 2 — вертлуг, 3 — бедро, 4 — голень, 5 — лапка,
6 — коготки лапки
ней поверхности лапки расположены только у ее основания.
Брюшко. Грудь без резкой границы переходит в брюшко,
постепенно сужающееся к заднему концу тела и состоящее из восьми
видимых сегментов. Заднепроходное отверстие смещено субтерминально на
брюшную сторону. Область,, окружающая его, составляет последний,
фактически IX, но морфологически XI сегмент брюшка. У зародыша
плавунца насчитываются 11 брюшных сегментов, не считая
расположенной постанально пигидиальной пластинки. В процессе последующего
развития IX и X сегменты подвергаются редукции.
Первые шесть сегментов брюшка имеют сходное строение
(см. рис. 566). Спинная сторона каждого из них почти полностью
покрыта сильно хитинизированнои тергальнои пластинкой, заходящей на
боковые стороны, где на краю пластинки помещаются стигмы.
Последние у личинок третьей стадии, в отличие от грудных стигм, —
открытые (рис. 579, А): Вели для дыхания находящихся в воде личинок эти
стигмы практически не имеют значения, то у выползающих перед
окуклением на сушу личинок они начинают сразу, же функционировать.
На брюшной стороне сегментов отсутствует склеротизация, точно
так же она отсутствует и между* сегментами. Сегменты имеют в общем
цилиндрическую форму со
слегка суженным
передним краем, что позволяет
каждому последующему
сегменту слегка
втягиваться в предыдущий.
Сегменты брюшка
отличаются значительной
подвижностью.
VII сегмент заметно
сужается к концу
(рис. 578, 4). Тергальная
пластинка его развита
сильнее; ее боковые края
Рис. 578. Задний конец тела личинки плавунца
третьей стадии. Вид сбоку (из Korschelt):
1 — восьмая брюшная стигма, 2 — церки, 3 — VIII
сегмент, 4 — VII сегмент брюшка, 5 — седьмая брюшиая
стигма, 6 — тергальная пластинка сегмента, 7 — VI
сегмент брюшка, 8 — шестая брюшиая стигма, 9 —
тергальная пластинка сегмента
523
слегка заходят на брюшную сторону. На границе между спинным и
боковыми отделами тесно расположены в 2—3 ряда слегка направленные
назад плавательные волоски. Стигмы помещаются на боках пластинки.
VIII сегмент еще более сужен, чем предыдущий. Он целиком покрыт
твердой хитиновой кутикулой. На боках он несет тесно сидящие в
несколько рядов плавательные волоски. Субтерминально на брюшной
стороне сегмента имеется полукруглый вырез. Там открывается
заднепроходное отверстие, по бокам которого причленяются церки —
придатки последнего IX (морфологически XI) сегмента брюшка. На
конце расположена последняя, восьмая пара брюшных стигм. Это
наиболее крупные, единственно функционирующие у личинок стигмы
(рис. 579, Б). ■ ' ■
Стигмы вторично смещены на конец сегмента, что вызвало
смещение заднепроходного отверстия на брюшную сторону. Каждая из стигм
помещается на небольшом, не еклеротизироваином подвижном выступе.
В углублении между ними лежит точка отхождения парного мускула —
ретрактора, стигм, другим концом прикрепляющегося к сливной
поверхности переднего края сегмента. При погружении личинки в воду эти
мышцы сокращаются. При этом стигмы поворачиваются .внутрь, их от-
Рнс. 580. Личинка плавунца. А—
на поверхности воды; Б и В —
положение заднего конца у
личинок различного удельного веса
(Б — тяжелее воды, В — легче
воды) (нз Korschelt):
Стрелки показывают направление тока
воздуха
524
Рис. 579. Личинка плавунца третьей стадии.
А— одна из боковых брюшных стигм; Б —
восьмая брюшная стигма (из Korschelt):
1 — отверстие стигмы, 2 — окружающий стигму
участок кутикулы, 3 —стигма, 4 — замыкательный
аппарат, 5 — трахея, 6 — стигма, 7 —
замыкательный рычаг, 8 — замыкательный мускул, 9 —
трахея, 10 — замыкательные скобы
верстия соприкасаются и взаимно закрывают друг друга. При подъеме
на поверхность мускулы-ретракторы расслабляются, стигмы
поворачиваются наружу и отверстия их, не смачиваемые водой, ложатся на
поверхность. В месте перехода стигмы ,в трахею имеется замыкательный
аппарат, по схеме строения аналогичный замыкательному аппарату
брюшных стигм взрослого жука (см. с. 475). Прямые наблюдения и
данные о функционировании этого замыкательного аппарата у личинок
отсутствуют. Видимо, замыкательный • аппарат этих стигм начинает
функционировать вместе с остальными брюшными стигмами только
тогда, когда личинка <выходит на сушу для окукления.
Описанные движения стигм при подъеме на поверхность воды и
погружении вглубь точно координируются с движениями церков.
Церки представляют собой одночленистые придатки, уплощенные в дорзо-
вентральном' направлении, края которых обрамлены рядом
плавательных 'волосков: Они расположены под углом друг к другу. Когда
личинка находится под водой, ее церки направлены вверх. При подъеме на
поверхность, когда их концы касаются поверхностной пленки, они
сгибаются под прямым углом к последнему сегменту и ложатся, под
влиянием капиллярного притяжения, непосредственно под поверхностной
пленкой. В этот момент стигмы тоже касаются поверхностной пленки
и, как упоминалось, поворачиваются и ложатся на поверхность воды.
В таком положении личинка может висеть на поверхностной пленке
(рис. 580). Так осуществляется контакт трахейной системы с
атмосферным воздухом и последующая вентиляция трахей. Если отрезать церки,
то личинка, хотя и с трудом, но может установить контакт стигм с
атмосферным воздухом, однако при этом она не может держать брюшко
перпендикулярно поверхности воды, а прилетает к ней брюшной
поверхностью.
Движение и положение личинок в воде. Обычно удельный вес
личинок, особенно после принятия пищи, больше единицы. Лишь в редких
случаях у голодных особей он равен удельному весу воды или даже
меньше его. Регуляция удельного веса может осуществляться путем
накопления избыточного воздуха в трахейной системе, заполнением или
освобождением ректальной ампулы и посредством дефекации. В воде
личинка всегда держится направленным вверх брюшком и с согнутым
вперед передним концом тела. При этом, перебирая все время ногами,
она может оставаться на одном уровне. Движения ног не являются
типично'плавательными, т. е. ноги одной пары двигаются не
одновременно, а поочередно.
Поступательное движение осуществляется тоже при помощи
аналогичных движений ног. Больший или меньший изгиб брюшка действует
в качестве вертикального руля. Церки и изменения в направлении
движения ног действуют как горизонтальный руль. Брюшко, как орган
активного движения, применяется в исключительных случаях — при
нападении .на добычу или при бегстве. Часто личинки прикрепляются
ногами к водяным растениям или подводным предметам (рис. 581).,
Анатомия на вскрытии. Пи щев арителын а я система.
У вскрытой личинки расправляют петли кишечника (рис. 582).
Своеобразная ротовая полость (см. с. 521 ирис. 575) ведет в короткую мус-
525.
Рис. 581. Взрослая личинка плавунца с
пойманной добычей (из Korschelt)
Рис. 582. Общий вид пищеварительной
системы личинки плавунца (из
Korschelt) :
/ — мышцы-расширители глотки, 2 —
пищевод, 3 — средняя кишка, 4 — пилорический
клапан, 5 — мальпигиевы сосуды, 6 — тонкая
кишка, 7 — ректальная ампула, 8— rectum
кулистую глотку, переходящую еще в голове в тонкий короткий пищевод
(рис. 582, 2). Последний в середине переднегруди переходит в
длинную, почти достигающую длины тела, среднюю кишку с более широким
передним отделом (рис. 582, 3). На ее поверхности видны точечные
возвышения — очаги новообразования секреторных клеток эпителия
средней кишки. Средняя кишка доходит до VI сегмента брюшка,
заворачивается вперед и на уровне III сегмента брюшка переходит в
заднюю кишку. В месте перехода открываются четыре сравнительно
коротких мальпигиевых сосуда (рис. 582, 5). Задняя кишка начинается
мускулистым клапаном, за которым идет длинная тонкая кишка
(рис. 582, 5),.на уровне VII сегмента впадающая в мешковидный рек-
талкный отдел. Последний имеет сильно развитый слепой дистальный
конец — ректальную ампулу (рис. 582, 7). Основное отличие
пищеварительной системы личинки от таковой взрослого жука заключается в
строении передней кишки, что вполне объясняется особенностями
питания. Жидкая пища,личинки делает излишним развитие зоба и
жевательного желудка, а необходимость высасывать пищу из добычи и
впрыскивать в нее переваривающий и парализующий добычу секрет
объясняет особенности строения ротовой полости и развитие глотки
(рис. 582,/).
526
Более подробное изучение
пищеварительной системы
проводится на срезах через ее
отдельные участки: глотку,
пищевод и среднюю кишку
(см. с. 530, рис. 583). Задняя
кишка личинки по деталям
строения сходна с таковой
взрослого жука (см. с. 505) и
в специальном описании и
изучении не нуждается.
Дыхательная систе-
м а. Трахейная система
личинки лучше всего может быть
изучена на личинках первой
стадии или недавно слинявших
личинках второй и третьей
стадий. Вследствие
прозрачности их кожных покровов,
можно рассмотреть даже мелкое
ветвление трахей. Личинка
убивается парами хлороформа
и в свежем виде рассматривается в воде или в глицерине под
покровным стеклом.. Заполненные воздухом трахеи выделяются на общем
светлом фоне. Из задних стигм изготовляются тотальные препараты,
окрашенные квасцовым кармином. На препаратах заднего конца тела
личинки можно рассмотреть их расположение и главные черты
строения.
У личинки плавунца дыхательная система состоит из двух главных
трахейных стволов, проходящих от задней пары стигм вдоль всего тела
по бокам от кишечника, ближе к спинной стороне, до переднегруди.
Поперечными коммиссурами, по одной в.каждом сегменте тела, оба
ствола соединяются друг с другом. Основная особенность их заключается
в том, что места их отхождения от главных продольных стволов лежат
в последующем сегменте, а сами они, изгибаясь дугой, лежат в
предыдущем (исключение составляют первая и последняя коммиосуры). От
трех коммиссур, за исключением последней, отходят трахеи,
разветвления котбрых идут в лопасти жирового тела и гиподерму сегмента.,
В V, VI и VII сегментах отходят трахеи к кишечнику. В каждом из
сегментов груди отходят специальные трахеи в ноги личинки.
В переднегруди каждый из продольных стволов делится на две
ветви — спинную и брюшную. В голову, таким образом, проходят две
пары трахей. Верхняя пара дает векви к мускулатуре мандибул,
усиков, зрительному ганглию и глазкам. Нижняя пара снабжает
кислородом надглоточный ганглий, все придатки головы и их мускулатуру,
мускулатуру глотки и т. д. Ветвление трахей в голове личинки сходно
с ветвлением их у взрослого жука, только несколько упрощено.
Стигмы, их число, расположение и функция разобраны при рас-,
смотрении внешней морфологии личинки (см. с. 524).
Рис. 583. Поперечный разрез через один из
передних сегментов брюшка взрослой личинки
плавунца, схематизированно (из Korschelt):
/ — сердце', 2 — кутикула, 3 — гиподерма, 4 —
внутреннее жировое тело, 5 — наружное жировое тело,
6 — мышцы, 7 — средняя кишка. 8 — эноциты, 9 —
главные трахейные стволы
527...
Общая емкость трахейной системы взрослой личинки достигает
107 mms, жизненная емкость, т. е. максимальный объем
'возобновляемого при дыхании воздуха, достигает 64 мм3.
Ж яровое тело. У личинки, как и у взрослого жука, главным
местом развития и локализации жирового тела является брюшко.
По локализации можно различить в жировом теле два
самостоятельных отдела — наружный, прилегающий к гиподерме, и внутренний,
окружающий кишечник. На поперечном разрезе через брюшко личинки
(рис. 583) это разделение ясно заметно. Если у вскрытой личинки
удалить спинную часть кожных покровов вместе с прилегающей к ним
мускулатурой, то бросаются в глаза два тяжа внутреннего жирового
тела, проходящие вдоль кишечника. Их самостоятельность по
отношению к наружному жировому телу особенно хорошо заметна по бокам
в связи с положением главных продольных трахейных стволов. Эти
тяжи начинаются в голове, в переднегруди проходят рядом над кишеч--
ником, далее расходятся и, начиная с заднепруди, идут по бокам
кишечника, образуя многочисленные складки и расщепляясь на
отдельные ленты И лопасти. У места перехода средней кишки в заднюю, в
области IV ■ брюшного сегмента, ими оплетаются зачатки половых
желез, после чего они продолжаются' назад, до VI брюшного сегмента в
виде тонких тяжей и нитей, переходящих в наружное жировое тело.
При сильном развитии жирового тела его ■■■лопасти- могут окружать
кишечник со всех сторон.
Наружное жировое тело располагается посегментно в виде
неправильной формы лопастей, лежащих четырьмя продольными тяжами:
брюшным, спинным и двумя боковыми, между которыми в каждом
сегменте могут существовать' соединения, выраженные тем сильнее, чем.
сильнее развито жировое тело.
Изучение срезов. Кожные покровы. Фиксируют недавно
слинявшую личинку третьей стадии каким-либо сулемовым фиксатором
(жидкость Джильсона и т. п.), предварительно захлороформировав ее
и разрезав ножницами пополам. После фиксации отрезают 1—2
сегмента из середины, заливают через смесь Петерфи в парафин или в
парафин с воском и изготовляют поперечные срезы толщиной в 7,5—Юмкм.:
Окрашивают железным гематоксилином (для выявления строения
клеток гиподермы) и по Маллори (для выявления слоев хитиновой
кутикулы и других кутикулярных образований, окрашивающихся в разные
цвета).
Кожные покровы насекомых состоят из однослойной гиподермы и
покрывающего ее снаружи слоя хитиновой кутикулы — производного ги-
подермальных клеток. На рис. 584 представлен поперечный разрез
через спинной щиток личинки третьей стадии. Гиподерма состоит из
одного слоя высоких призматических клеток, тесно прилегающих друг к
другу, с хорошо заметными границами (рис. 584, 4). Светлое
пузыревидное ядро лежит в базальной части клетки. Хроматин ядра образует
1—2 крупные глыбки и ряд мелких зерен. Плазма в базальном отделе
имеет ячеистую структуру, в дистальвом — фибриллярную, придающую
части клетки, прилежащей к кутикуле, продольно исчерченный вид. От
полости тела гиподерма отграничена бесструктурной базальной мемб-
§28
раной, на срезах кажущейся
гомогенной, преломляющей
свет линией (рис. 584, 6).
Хитиновая кутикула
состоит из трех слоев: наружвого,
или экзокутикулы (рис. 584,
2), которая покрыта
пограничным с наружной средой
тонким слоем, эпикутикулы
(рис. 584, 1) и внутреннего,
или эндокутикулы (рис. 584,
3). Эпикутикула — тонкая
пленка, состоящая из жирных
кислот и холестериноподобных
веществ, чрезвычайно стойкая
к различным химическим
реагентам, в том числе и к
концентрированным кислотам.
Она снаружи облегает экзо-
кутикулу на всей поверхности
тела. Экзокутикула на неокрашенном препарате, в зависимости от
плотности хитина, от желтоватого до темно-коричневого цвета, является
слоем, придающим крепость кожным покровам насекомого. Снаружи
она обычно имеет слоисто фибриллярную структуру; ее внутренний слой
может нести поперечную исчерченность. Слой эндокутикулы обычно
наиболее толстый. Он всегда имеет хорошо'выраженную слоистость.
Вышеперечисленные слои можно найти на разрезе через любой участок
кожных покровов насекомого. Различия заключаются в относительной
мощности развития экзо- и эндокутикулы и деталей их строения.
Рис. 584. Поперечный разрез через спинной
щиток брюшного сегмента личинки плавунца
третьей стадии (из Korschelt):
; — эпикутикула, 2 — экзокутикула, 3 — эндокутику-
ла, 4 — слой клеток гиподермы, 5 —ядро, 6— базаль-
ная мембрана
Рис 585 Поперечный разрез через передний отдел Рис. 586. Поперечный разрез
глотки (из Korschelt):
/ — кольцевые мышцы, 2 — продольные мышцы, 3, 4 — мышцы-
расширители глотки, б -г- эпителий глотки, 6 — хитиновая
выстилка, 7 — просвет глотки
через кольцевой клапан
личинки плавунца (из
Korschelt) :
; — кольцевая мускулатура, 2 —
хитиновая выстилка, 3 —
эпителий стенки глотки, 4 — трахея,
5 — п. stomatogastricus
529
Рис. 587. Поперечный разрез через задний
отдел глотки личинки плавунца, схематизиро-
ванйо (из Korschelt):
1 — кольцевая мускулатура, 2 — продольные мышцы,
3— брюшные расширители глотки, 4 — внутренний
скелет головы, 5 — подглоточиый ганглий, 6 — трахея,
7 — п. stomatogastricus
Рис. 588. Поперечный разрез через задний
отдел глотки около пищевода личинки
плавунца (из Korschelt):
/ — кольцевая мускулатура, 2 — расширитель глотки,
3 — эпителий, 4, 6 — продольные мышцы, 5 —
хитиновая выстилка, 7 — п. stomatogastricus
Пищеварительная система.
Фиксируют сулемовым
фиксатором отрезанную голову
личинки первой или
второй стадии или только
что слинявшей личинки
3-й стадии, заливают
через смесь Петерфи в
парафин, изготовляют серии
поперечных срезов
толщиной в 10 мкм и
окрашивают по Маллори.
У вскрытой личинки
выделяют в
физиологическом растворе кишечник,
фиксируют сулемовым
фиксатором, разрезают
на куски длиной до 1,5 см,
заливают через смесь
Петерфи в парафин и
изготовляют срезы толщиной
в 7,5 мкм по нескольку из
каждого отдела. Окраска
железным
гематоксилином и по Маллори. Для
получения более хороших
картин строения средней
кишки желательна
фиксация ее осмиевым
фиксатором (жидкость Флем-
минга, Шампи).
Поперечный разрез
глотки (рис. 585—588).
Короткая глотка целиком
помещается в голове
личинки. Она
подразделяется на передний и
задний отделы, граница
между которыми лежит на
уровне расположения
окологлоточных коннективов
и над- и подглоточного
ганглиев. Строение
глотки личинки и взрослого
жука в общем сходно, но
мускулатура глотки
личинки развита
значительно сильнее. Можно раз-
530
личить сильную кольцевую мускулатуру,
расположенную кнутри от нее продольную,
самостоятельную в переднем и заднем отделах, и
систему мышц-расширителей глотки — спинных в
переднем ее отделе и спинных и брюшных — в
заднем.
В переднем отделе глотки хитиновая
выстилка брюшной стороны значительно толще
(рис. 585, 6).,Подстилающий слой эпителия сое-
тоит из плоских клеток.^ Он окружен тонкой ^3 ^ ЙЗИЕ
бесструктурной базальнои мембраной. Спинной ЧИНки плавунца (из
продольный мускул лежит в глубокой срединной Korschelt):
складке, вследствие чего просвет глотки имеет i — кольцевые мышцы, г —
т т * • I г ос i\ продольные мышцы, 3 —хи-
U-обраЗНуЮ форму (рИС. 585, 7). тиновая выстилка, 4- эпи-
Снаружи расположены кольцевые мышцы телий
(рис. 585, 1). К боковым складкам глотки
подходят мышцы-расширители глотки, прикрепляющиеся к головной
капсуле (рис. 585, 3, 4). В результате их сокращения она принимает
вид, изображенный на рис. 585. Последующее сужение просвета
достигается сокращением кольцевой и продольной мускулатуры при
одновременном расслаблении мышц-расширителей. Таким образом, передний
отдел глотки представляет собой достаточно мощный насасывательный
аппарат.
В месте перехода переднего отдела в задний глотка заметно
сужается и округляется (рис. 586). Тут бросается в глаза отсутствие
продольных мышц, так как продольная мускулатура переднего отдела уже
окончилась, а заднего отдела — еще не началась. Очень сильно
развитая кольцевая мускулатура обеспечивает при сокращении плотное
замыкание просвета. На спинной стороне можно заметить перерезанный
поперек нервный тяж симпатической нервной системы (рис. 587, 7).
Задний отдел глотки (рис. 587, 588) 'несколько отличается от
переднего общей конфигурацией просвета, строением поверхности
хитиновой еыстилки, несущей направленные назад ширики (рис. 588, 5),
волооки и зубчики, а также наличием брюшных мышц-расширителей,
прикрепляющихся к внутреннему скелету головы. Под ним расположен
подглоточный ганглий (рис. 587, 5). Задний отдел глотки тоже
представляет собой действующий самостоятельно сильный
насасывательный аппарат, перекачивающий жидкое содержимое в пищевод. .
Поперечный разрез через пищевод (рис. 589). Пищевод имеет вид
короткой трубки, несколько расширяющейся кзади. Хитиновая
выстилка его несет многочисленные продольные складки. Мускулатура
(кольцевая— снаружи, продольная — 'внутри, между складками) развита
сравнительно слабо и не лежит сплошным слоем.
Поперечный разрез через среднюю кишку (рис. 590—593). Вся
поверхность средней кишки выстлана слоем тесно сидящих удлиненных
цилиндрических клеток с хорошо заметными, особенно на гематоксили-
новых препаратах, границами между ними. Только у неда'вно
слинявших личинок, вследствие растяжения у них стенок кишечника,
связанного с механизмом линьки, клетки эпителия становятся кубическими
531
Рис. 590.. Поперечный разрез через среднюю кишку личинки
плавунца (нз Korschelt):
/ — продольные мышцы, 2 — кольцевые мышцы, 3 — клетки эпнтелня, 4 ■—
трахеи, 5 — крипты
Рис. 591. Личинка плавунца первой стадии.
Поперечный разрез через стенку средней кишки. А —
начальный период накопления секрета; Б — начало
выделения секрета (из Korschelt):
/ — клетки эпителия, 2 —ядро, 3 — вакуоли секрета, 4 —
крипты,- 5 — продольные мышцы, 6 — кольцевые мышцы, 7 —
палочковый слой, 8 — выделяющиеся в просвет кишки капли
секрета
или слегка уплощенными.
Крупные ядра округлы
или
продолговато-овальной формы. Обращенная
в просвет кишки
поверхность клеток несет
оторочку из тонких палочек.
Такой вид имеют
покоящиеся, несекретирующие
клетки. У некоторых сек-
ретирующих клеток
наблюдается образование
одной или нескольких
вакуолей, лежащих обычно
впереди ядра, в части,
обращенной к просвету.
Вместе с ядром вакуоль
передвигается ближе к
дистальному концу, пока
532
А
В
Рис. 592. Разрезы через среднюю кишку на разных стадиях-
развития личинки плавунца. А — конец первой стадии (продольный
разрез); Б — начало второй стадии; В — конец второй стадии
(из Korschelt):
1 — палочковый слой, 2 — клетки эпителия, 3 — ядро, 4 — базальиая
мембрана, 5 — кольцевые мышцы, 6 — продольные мышцы, 7 — крипта, 8 —
очаг размножения клеток эпителия на дне крипты, 9 — вакуоли секрета
не достигает палочкового слоя, растворяет его
и выводится в просвет кишечника. При этом
типе секреции, по-видимому, не происходит
уничтожения секретирующей клетки.
Гораздо более распространен другой тип
секреции, ведущий в результате к распаду и
гибели секретирующей клетки. Под
палочковым слоем происходит накопление зернистого
■секрета, вызывающее набухание, вздутие и
выпячивание в просвет кишки всего дисталь-
ного отдела клетки. Последний постепенно от-
"шнуровывается, приобретает форму капли и
в результате выталкивается в просвет
кишечника. Часто при этом увлекается и ядро, и
■клетка погибает. Подобная гибель клеток
происходит все время на всей поверхности
средней кишки. Параллельно этому все время
происходит и процесс новообразования клеток
эпителия. Он происходит за счет особых, ле-
Рнс. 593. Поперечный
разрез через среднюю кншку
личинки плавунца третьей
стадии (из Korschelt):
J — клетки эпителия, 2 —
секреторный отдел крипты, 3 — очаг
размножения на дие крипты,
4 — продольные мышцы, 5 —
кольцевые мышцы
жащих под эпителием, на базаль-
ной перепонке, клеток. Эти клетки,
образуют под эпителием особые
скопления — очаги
новообразования, или крипты. Путем митотичес-
ких делений эти клетки
размножаются, растут и замещают
отработанные и погибающие клетки эпителия.
У личинок первой стадии крипты
(рис. 593, 2) состоят из небольшого-
числа клеток и почти не выступают
Рис. 594. Голова личинки плавунца сбо- из плоскости эпителия. Но скоро
ку. Видно расположение глазков (из интенсивное размножение клеток в-
Korschelt) v
' криптах начинает превышать их
расход на восстановление
клеток эпителия и к концу первой — началу второй стадии дно-
крипты выступает из плоскости эпителия и вдается в слой:
мускулатуры, а на конце, обращенном в просвет кишки,,
раздвигает клетки эпителия (рис. 592, Б). Наиболее дистально
расположенные клетки крипты сильно удлиняются, и крипта на разрезе
напоминает распускающуюся почку. Уже на этой стадии наиболее дис-
тальные клетки крипты начинают секретировать. К концу второй стадии,
(рис. 592, В) дно крипты, содержащее зону размножения клеток,
настолько увеличивается, что образует слепое выпячивание, а ее дисталь-
ный отдел уже содержит клетки, ничем не отличающиеся от клеток
эпителия и секретирующие наравне'с ними. У личинок третьей стадии это
увеличение крипт еще значительнее (рис. 593). Однако для личинок
является характерным, что дно такой выросшей крипты никогда не
выходит за границу мускульного слоя средней кишки. Последний состоит из
внутреннего редкого слоя кольцевых мышц и такого же наружного —
продольных (рис. 593, 4, 5).
Органы зрения. Приготовляют серию поперечных срезов
через фиксированную сулемовым фиксатором и залитую в парафин по-
способу Петерфи голову личинки плавунца.
Срезы окрашивают железным гематоксилином с последующей до-
краской эозином.
Чтобы лучше разобраться на срезах, необходимо предварительно,
когда голова находится в жидкости Петерфи, познакомиться с
расположением глаз на голове личинки. При рассмотрении головы сбоку
(рис. 594) видно, что глазки, в количестве шести с каждой стороны,
расположены на боковых сторонах головы непосредственно позади
основания усиков. Две верхние пары глазков имеют верхнее и боковое поле
зрения, нижняя пара ориентирована на брюшную сторону. Так как
все глазное поле и помещающиеся на нем глазки несколько
приподняты над окружающей поверхностью головы, передние три глазка
улавливают падающие спереди лучи, а задние — падающие сзади.
Нижние глазки имеют округлые линзы, верхние — овальные, средние
занимают промежуточное положение. В остальном их строение
одинаково.
534
Рис. 595. Продольный разрез через спинной глазок личинки плавунца. А —
разрез поперек глазной щели; Б — срединный разрез вдоль глазной щели (из
Korschelt):
J — хрусталик, 2 — стекловидное тело, 3 —^кутикула, 4 — гиподерма, 5 — пигментные клетки,
6— глазная щель, 7 — горизонтально расположенные палочки ретннальных клеток, 8 — нерв,
9 — крупные ретииальные клетки, 10 — их вертикально расположенные палочки
На продольных срезах (рис. 595) видна обращенная наружу
двояковыпуклая линза (хрусталик, рис. 595, /) — непосредственное
продолжение окружающей глаз хитиновой кутикулы (рис. 595, 3).
Подстилающие ее цилиндрические клетки гиподермы удлиняются и погружаются
внутрь, образуя бокал рецепторной части глазка, стенки которого
состоят из одного слоя клеток. Краевые клетки этого бокала, граничащие
•с хрусталиком, заполнены пигментом (рис. 595, .5). Их дистальныекон-
ды выделяют лежащее под хрусталиком стекловидное тело (corpus vit-
теит, рис. 595, 2). Более центрально лежащие клетки 'бокала образуют
щелевидное впячивание. Ретинальные клетки- (рис. 595, 9), лежащие на
дне этого впячивания, несут вертикально ' расположенные палочки
(рис. 595, 10). Именно к этим клеткам подходят окончания
зрительного нерва (рис. 595, 8). У клеток,, лежащих по бокам щели, палочки
расположены горизонтально (рис. 595, 7).
По строению глазки личинки плавунца принадлежат к
примитивному типу глазков — к археоммам. Это — однослойные неинвертиро-
'35
ванные бокальчатые глазки. С их помощью личинка может отличать
не только свет от темноты, но и направление световых лучей. Дальность
видения невелика. Личинка различает изменения в характере
освещения на расстоянии не более 5—10 см. "Движения добычи различаются
только на расстоянии 0,5—4 см от линзы. У личинок младших стадий
это расстояние еще меньше. Эти особенности находятся В1
непосредственной связи со способом ловли добычи. Личинка плавунца — хищник,,
подстерегающий добычу, сидя неподвижно среди водной растительности.
Она бросается на проплывающую мимо добычу, но бросок,
совершаемый при этом, обычно не превышает нескольких сантиметров. Этим и
определяется зрительная способность глазков.
;
ЛИТЕРАТУРА'
Кольчатые черви — Annelida
Многощетинковые кольчецы — Polychaeta, Archiannellda
Ливанов Н. А, Класс полихет (Polychaeta). Руководство' по зоологии. М., 1940,
т. 2, с. 10—136.
В руководстве дано полное изложение морфологии, экологии и систематики мно-
гощетинковых кольчецов.
Мамкаев Ю. В., Селиванова Р. В. Исследование организации Dinophilus vorticoi-
des с использованием электронной микроскопии.—Тр. Зоол. ин-та АН СССР, 1979,
т. 84, с. 73—83.
Мейер Э. Организация трубчатых червей сем. Serpulidae и Hermellidae как
результат их сидячего образа жизни. — Тр. об-ва естествоиспытателей, 1893, т. 26,
вып. 3, с. 1—131.
Классическое морфологическое исследование сндячнх полихет. Несколько
устарело, но и до снх пор является основой при изучении аннелид.
Свешников В. А. Морфология личинок полихет. М., 1978. Хорошо
иллюстрированная монография, необходимая при изучении живого материала на море.
Ушаков П. В. Многощетинковые черви Дальневосточных морей СССР. М., 1955,
сер. Определители по фауне СССР, издаваемые ЗИН АН СССР, № 56.
Полный определитель, включает общий очерк морфологии и биологии полихет.
Цетлин А. Б. Практический определитель многощетинковых червей Белого моря.
М., 1980. ■
Пособие необходимо при работе на Белом и Баренцевом морях.
Schimkewitsch W. M. Zur Kenntnis des Baues und der Entwicklung des Dinophilus
von Weifien Meere. Zeitschr. wiss. Zool. Bd. 59, 1895.
Основное исследование по морфологии Dinophilus, несколько устарело, но и
ныне служит основной работой при изучении этой группы полихет.
Fauvel P. Classe des Annelides, Polychetes. Traite de zoologie publ. P. Grasse,
Paris, 1959, p. 15—196.
Полное, богато иллюстрированное, изложение организации полихет и краткий
обзор систематики. Основное руководство в мировой литературе.
Kaestner A. Stamm Annelida. — In: Lehrbuch der speziellen Zool. Bd. I, T. I, Jena,
1965, s. 476—576.
Хорошо составленный, с многочисленными иллюстрациями очерк организации и
системы типа кольчатых червей. ' '
Krugen. F. Bau und Leben des Wattwurmer Arenicola marina Helgolander Wiss.
Meeresunters'. Bd. 22, 1971. S. 149—200.
Обстоятельная работа, посвященная морфологии и образу жизнн Arenicola
marina.
Малощетинковые черви— Oligochaeta
Малевич И. И. Собирание и изучение дождевых червей-почвообразователей. Сер.
В помощь работающим на полезащитных лесных полосах, № 6. 1950.
Полезное пособие, содержащее сведения о биологии, распространении дождевых
1 Общие сводки и руководства по зоологии беспозвоночных приведены в I томе
настоящего руководства: «Большой практикум по зоологии беспозвоночных
(простейшие, губки, кишечнополостные, низшие черви)». М., 1981, с. 493.
537
червей и указания по собиранию, вскрытню, содержанию в неволе. Приведены также
определительные таблицы.
Перель Т. С. Распространение и закономерности распределения дождевых червей,
фауны СССР. М., 1979, 270 с.
. Обширное, глубокое фаунистическое и систематическое исследование
распространения дождевых червей по территории СССР. Особое внимание обращено на роль-
дождевых червей в почвообразовании.
Чекановская О. В. Водные малощетинковые черви фауны СССР. Сер.
Определители по фауне СССР, издаваемые ЗИН АН СССР, 1962, № 78.
Работа, посвященная олнгохетам, содержит не только обзор системы малощетин-
ковых кольчецов, но также подробный очерк внешней н внутренней морфологии.
Avel M. Classe des Annelides Oligochetes. Traite de Zoologie publ. P. Grasse.
Paris, 1959, p. 225—470.
Подробный очерк организации, экологии, систематики олнгохет. Основное
руководство в мировой литературе. Богато иллюстрировано.
Stephenson J. The Oligochaeta. Oxford, 1930.
Одна из полных монографий с обширной морфологической частью и
многочисленными рисунками.
Veijdovsky F. System und Morphologie der Oligochaeten. Prag. 1884, S. 166.
Основное классическое монографическое исследование, положившее начало
современному изучению олнгохет. Точные описания, прекрасно иллюстрировано.
Пиявки — Hirudinea
Ливанов Н. A. Acanthobdella pelidina. Морфологическое исследование. — Зап.
Казанского ун-та, 1905, т. 22, с. 1—271.
Классическая монография по Acanthobdella — наиболее примитивной форме
пиявок.
Лукин Е. И. 1976. Пиявки, т. I. Пиявки пресных и солоноватоводных водоемов.
Нов. сер. Фауна СССР, изд. ЗИН АН СССР, № 109.
Обстоятельное монографическое исследование строения, экологии и систематики
пиявок. Одна из фундаментальных работ по изучению пиявок. х-
Заленский В. Д. Исследование по морфологии и систематике Hirudinea.
Организация Ichthyobdellidae. СПб., 1915.
Подробная монография, богато иллюстрированная.
Ливанов Н. А. Класс пиявок (Hirudinea).— В кн.: Руководство по зоологии-,,
т. II. М., 1940, с. 205—257.
Подробное описание морфологии, экологии и системы пиявок.
Haraut H., Grasse P.-P., Classe des. Annelides Achetes ou Hirudinees ou Sangsues,
Traite de Zoologie publ. P. Grasse. Paris, 1959. pp. 471—593.
Подробный, богато иллюстрированный очерк строения, экологии, систематик»
пиявок.
Членистоногие —Arthropoda
Паукообразные — Arachnoidea
Бялыницкий-Бируля А. А. Паукообразные. Скорпионы. Фауна России. Петроград.,
1917, т. I, вып. 1.
Подробное описание фауны скорпионов СССР. Определительные таблицы. В
общей части интересна гл.ава о географическом распространении и образе жизни
скорпионов.
Иванов А. В. Пауки, их строение и образ жизни и значение для человека. JL,.
1965.
Павловский Е. Н. Материалы к сравнительной анатомии и истории развития
скорпионов. Петроград, 1917.
Одна из основных работ по анатомии скорпионов. В гистологической части
несколько устарела. -
Павловский Е. Н. Ядовитые животные СССР. Л., 1931.
Подробные данные о скорпионах и ядовитых пауках.
Паразитология человека/Под ред. Г. С. Первомайского и В. Я. Подоляна. Л.,
1974.
538
В книге приведен обстоятельный очерк арахноэнтомологни, подробно
рассмотрены клещи — переносчики различных заболеваний.
Тыщенко В. П. Определитель пауков Европейской части СССР. Л., 1971.
Книга необходима при сборе пауков в природе. _ .
Andre M. Ordre des Acarienes Traite de Zoologie Publ. par. P. Grasse. Pans,
1949, vol. 6, p. 794—892. ,, „ „ , _, .
Millot J. Ordre des Araneides. Traite de Zoologie Publ. par. P. Grasse. Pans,
1949, Vol. 6, p. 589—744. , „ ,, _ _ „
Millot J. Vachon M., Ordre des Scorpions. Traite de Zoologie. Publ. par. P. P.
Grasse. Paris, 1949, vol 6, p. 386—437.
Указанные работы, опубликованные в многотомном руководстве по зоологии,
содержат наиболее полное изложение морфологии, экологии, систематики арахннд
Kaestner A., Unterstamm Chelicerata. — In: Lehrbuch der speziellen Zool. Bd. 1,
T. I, 1965, S. 628—784.
Один из лучших подробных учебников зоологии. Хорошо иллюстрирован. Может
служить основным руководством на практических занятиях.
Ракообразные — Crustacea
Гурьянова Е. Ф. Бокоплавы СССР и сопредельных вод. Сер. Определители по
фауне СССР. Л., 1951, № 41.
Обширная обобщающая работа по систематике бокоплавов. Содержит
подробные сведения по экологии, зоогеографии. Рассмотрен вопрос о происхождении морской
■фауны северных морей СССР.
Мкнуйлова Е. Ф. Ветвистоусые рачки фауны СССР. Сер. Определители по фауне
СССР. Л., 1964, № 88.
Пособие полезно при определении ветвнетоусых рачков. Содержит краткий очерк
морфологии, сведения по экологии, цикломорфозу.
Определитель пресноводных беспозвоночных европейской части СССР (планктон,
бентос)/Под ред. Л. А. Кутиковой и Я. И. Старобогатова. Л., 1977.
В книге на с. 213—259 помещены определительные таблицы и рисунки Cladocera
и Copepoda.
Рылов В. М. Пресноводные Calanoidea СССР. Сер. Определители пресных вод
СССР. Л., 1930, вып. 1.
Рылов В. М. Свободноживущие веслоногие ракообразные (Eucopepoda). Сер.
Пресноводная фауна Европейской России. М. 1922.
Darwin Ch. A. Monograph of the Subclass Cirripedia, 1—2. Roy. Society London,
1851—1853.
Классическая монография Ч. Дарвина и до сих пор является основной работой
по Cirripedia. Она включает систематику и биологию этой группы, а также данные
анатомического характера.
Hexley Th. L'ecrivisse, introduction a l'etude de la zoologie. 1880. Serie Bibliotheque
scientifique internationale.
Классическая монография речного рака, не утерявшая своего значения и до сих
лор.
Wesenberg — Lund С. Contributions to the biology and morphology of the genera
TDaphnia. Kobenhavn, 1926.
. Ценное исследование, посвященное в основном экологии Cladocera. Подробно
разбирается вопрос о жизненном цикле.
Серия специальных монографий по анатомии речного рака опубликована
сотрудниками Korschelt'a, работавшими в Зоологическом институте Марбургского
университета. К этой серии относятся следующие работы:
Baumann H. Das Gefafisystem von Astacus fluviatilis. Zeitschr. wiss Zool., Bd. 118,
1919. • ,
Bernhards H. Die Complexaugen von Astacus fluviatilis. Zeitschr. wiss. Zool., Bd.
116, 1916.
Bock. F. Das Respirationsapparat von Astacus fluviatilis. Zeitschr. wiss. Zool.,
Bd. 124, 1925.
Janisch E. Das Enddarm von Astacus fluviatilis. Zeitschr. wiss. Zool., Bd. 121,
1923.
Keim W. Das Nervensystem von Astacus fluviatilis (Potamobius astacus L.)
Zeitschr. wiss. Zool, Bd. 113, 1915.
539
Netz W. Das Skelett des Flufikrebses. Marburg, 1917.
Panning A. Die Statocyste von Astacus fluviatilis und ihre Beziehungen zu dem sie
umgebenden Gewebe. Zeitschr. wiss. Zool., Bd. 123, 1924.
Ringel M. Zur Morphologie des Vorderdarmes (Schlund und Magen) von Astacus
fluviatilis. Zeitschr. wiss. Zool., Bd. 123, 1924.
Schmidt W. Die Muskulatur von Astacus fluviatilis. Zeitschr. wiss. Zool. Bd. 113,.
1915.
В приведенных монографиях дается детальное анатомическое и отчасти
гистологическое описание. Работы хорошо иллюстрированы.
Многоножки — Myriapoda
Иванов П. П. Эмбриональное развитие сколопендры в связи с эмбриологией и
морфологией Tracheata. Изв. АН СССР. Сер. биол., 1940, № 6.
Эмбриологическое исследование Chilopoda, в котором, разбирается вопрос о лар-
вальных и постларвальных сегментах Tracheata, имеющий фундаментальное значение-
для понимания морфологии не только этой группы, но и всех Arthropoda.
Fahlander К. Beitrage zur Anatomie und systematischen Einleitung der Chilopoden..
Zoologiska Bidrag, Bd. 17, 1938.
В работе описываются покровы и эндоскелет, железы, максиллярные почки,
кровеносная система,_ половые органы, нервная система и органы. Темешвари. Один иа
объектов исследования — Lithobius forficatus. В последних главах разбирается стома-
тогастрическая- система Tracheata, вопрос о метамерии головы Tracheata,
рассматривается систематическое разделение Chilopoda. Работа снабжена многочисленными'
фотографиями срезов.
Seifert В. Anatomie und Biologie des Diplopoden Strongylosoma pallipes. Zeitschr.
f. Morph. u. Okol., Bd. 25, 1932.
Одна из основных работ по организации Diplopoda. В первой части подробно»
разбирается анатомия, во второй — биология.
- Voges E. Myriapodenstudien Zeitschr. wiss. Zool., Bd. 116, 1916.
Морфологическое исследование трахейной системы, в частности стигм и стиг-
мального замыкательного аппарата.у Lithobius, Scutigera^Julidae, Polydesmus, Glome-
ridae. Кроме того, разбирается морфология головы Diplopoda.
Насекомые — Insecta
Кузнецов Н. Я. Класс насекомых (Insecta, Hexapoda). Руководство по зоологии,
Т. III, ч. 2. М.,.1951, с. 168—458.
В некоторых частях несколько устаревшее подробное изложение морфологии и.
систематики насекомых. Много внимания уделено вопросам экологии и физиологии,
Кузнецов Н. Я- Основы физиологии насекомых. Л., т. I, 1948; т. II, 1953.-
Очень подробное, но несколько устаревшее изложение физиологии насекомых..
В вышедших томах рассмотрены процессы обмена веществ, пищеварения, выделения,,
газообмена, секреции. Не затронуты нервно-мышечная физиология и органы чувств.
Павловский Е. Н. Методы ручного анатомирования насекомых. Л., 1957.
Тыщенко В. П-. Основы физиологии насекомых. Л., ч. I, 1976; ч. II, 1977.
В работе дано современное представление об основах физиологии насекомых.
Рассмотрение функций ведется в тесной связи .с морфологией органов.
Холодковский Н. А. Курс энтомологии теоретической и прикладной. СПб, 1912.
Этот двухтомный курс энтомологии, во многих частях устаревший, продолжает
оставаться классическим произведением, содержащим обширный фактический
материал, мастерски изложенный .и богато иллюстрированный. ' г '
Шванвич Б. Н. Курс общей энтомологии. М.-Л., 1949.
Подробный курс энтомологии. Особое внимание уделено вопросам морфологии,
насекомых. Рассмотрены также вопросы экологии и поведения. Подробно освещены
вопросы окраски насекомых и ее значения. Изложена оригинальная система
насекомых.
Шванвич Б. Н. Введение в энтомологию. Л., 1959.
Кратко изложены основы энтомологии. В. книге много внимания уделено
экологии и практическому-значению насекомых. Дана систематика и характеристика
.отрядов. ' ■ ' ...
Kaestner A. Lehrbuch der speziellen Loologie, Bd. I. Wirbellosein, 3 —Insecta.
A. Allgemeiner Teil. Jena, 1972, S. 277—907; Spezieller Teil, 1973, S. 277—907.
Оба тома представляют подробный курс энтомологии, включающий общую
энтомологию и систематику. Прекрасно иллюстрированы.
Korschelt E. Bearbeitung einheimischer Tiere. Erste Monographie: Der Gelbrand
Dytiscus marginalis L. Bd. I u. II. Leipzig, 1924.
Классическая монография жука-плавунца, охватывающая вопросы строения,
развития, биологии, систематики и т. д. Библиография в некоторых частях несколько
ЛТ'РТ'Я ПР ТТо
Miall L., Denny A. The Structure of Life History of the Cockroach (Periplaneta
orientalis). An Introduction to the Study of Insects. London, 1886.
Классическая монография черного таракана. Во многом устарела, но и в наши
дни заключает важный материал для подробного изучения морфологии таракана.
Snodgrass R. E. Principles of Insect Morphology. N-Y —London., 1935.
Широко известная работа английского энтомолога, в которой особенно
подробно рассмотрен вопрос о сегментации членистоногих (в -том числе насекомых) и
обсуждается вопрос метамерии головного отдела членистоногих.
ОГЛАВЛЕНИЕ
Предисловие . . 3
Тип Annelida — Кольчатые черви 4
Класс Polychaeta — Многощетинковые черви 4
Отряд Archiannelida — Первичные кольчецы 5
Polygordius ponticus Salensky 13
Трохофора Polygordius ponticus Salensky 24
Отряд Nereimorpha 26
Nereis pelagica L 26
Отряд Drilomorpha 44
Arenicola marina L.—Пескожил 44
Внешняя морфология различных Polychaeta . 5.8
Вскрытие различных Polychaeta . 64
Личинки Polychaeta ' 67
Класс Oligochaeta — Малощетинковые черви 67
Отряд Opisthopora 70
Lumbricus terrestris L. — Обыкновенный дождевой червь 70
Отряд Plesiopora , 103
Aeolosoma hemprichii Ehr , J 03
Класс Hirudinea — Пиявки 107
Отряд Gnathobdellea — Челюстные пиявки '07
Hirudo medicinalis L.—Медицинская пиявка 107
Отряд Rhynchobdellea — Хоботковые пиявки 130
Glossiphonia complanata L.—-Улитковая пиявка 130
Тип Arthropoda — Членистоногие 141
Техника изучения внешней морфологии и анатомии членистоногих 141
Подтип Chelicerata — Хелицеровые 146
Класс Arachnida — Паукообразные 146
Отряд Scorpiones — Скорпионы 146
Buthus eupeus (С. Koch) — Скорпион пестрый 146
Отряд Aranei — Пауки . 188
Araneus(-Epeira)diadematus L.—Паук-крестовик 188
Отряд Acari — Клещи 227
Ixodes ricinus L. — Собачий клещ 227
Подтип Branchiata — Жабродышащие 245
Класс Crustacea — Ракообразные 245
Подкласс Branchiopoda — Жаброногие раки 245
Отряд Anostraca — Жаброиогие 245
Отряд Phyllopoda — Листоногие раки 259
Подотряд Notostraca — Щитни 259
Подотряд Conchostraca — Раковинные листоногие раки 261
Подотряд Cladocera — Ветвистоусые раки 262
Daphnia pulex de Geer — Обыкновенная дафния 263
Подкласс Ostracoda — Ракушковые раки 275.
Cypris sp 275
Подкласс Maxillopoda — Максиллоподы 279
Отряд Copepoda — Веслоногие раки 279
Cyclops sp ' 279
Diaptomus sp 285
Метаморфоз циклопа ., 289
~ Отряд Cirripedia — Усоногие раки 290
542
Balanus balanoides L. — Морской желудь 290
Подкласс Malacostraca — Высшие раки %*'
Отряд Leptostraca — Тонкопаицириые раки f^'
Nebalia bipes (О. Fabricius) **'
Отряд Decapoda — Десятиногие раки 300
Astacus astacus (Linne) — Речной рак 300
Отряд Mysidacea — Мизиды ЗЬУ
Mysis Latreille Зой
Отряд Isopoda — Равноногие раки • ■ 3bd
Asellus aquaticus L. — Водяной ослик- ЗоЗ
Отряд Amphipoda — Бокоплавы З/о
Gammarus pulex L. — Бокоплав-блоха . З/о
Метаморфоз и личинки высших раков (Malacostraca) Зад
Подтип Tracheata — Трахейные 3°»
Класс Myriapoda — Многоножки 38»
Подкласс Chilopoda — Губоногие , 3»»
Отряд Lithobiomorpha 3»»
Lithobius forficatus L. — Камнелаз, или Костянка . . Зой
Подкласс Diplopoda — Двупарноиогие -. . . 40Ь
Отряд Juliformia (-Opisthospermophora) 40»
Schizophyllum sabulosum L. — Кивсяк 40b
Класс Insecta — Насекомые '. 41b
Подкласс Ectognatha — Открыточелюстные, или Настоящие насекомые . . . 41b
Отряд Blattoidea — Тараканы jib
Blatta orientalis L. — Черный таракан . 41b
Отряд Coleoptera — Жесткокрылые, или Жуки 449
Dytiscus fnarginalis L. — Плавунец окаймленный 449
Личинка плавунца 514
Литература 5^
Артемий Васильевич Иванов, Александр Самойлович Мончадский,
Юрий Иванович Полянский, Александр Александрович Стрелков
Большой практикум по зоологии беспозвоночных
Типы: Кольчатые черви, Членистоногие, Часть вторая
Заведующий редакцией А. Г. Гаврилов. Редактор Т. А. Рыкова. Младшие редакторы Е. В. Бурова,
Е. Г. Самойло. Художник Н. Ю. Бабикова. Художественный редактор Т. А. Коленкова.
Технический редактор Т. Д. Гарина. Корректор Л. Э. Ганненко
ИБ № 4223
Изд. № Е-417. Сдано в набор 21.12.82. Подп. в печать 10.06.83. Формат 70X90'/t6. Бум. тип. № 2.
Гарнитура литературная. Печать высокая. Объем 39,78 усл. печ. л. Усл. кр.-отт. 39,78, 42,49 уч.-
изд. л. Тираж 10 000 экз. Зак. № 2. Цена 1 р. 90 к.
Издательство «Высшая школа», 101430, Москва, ГСП-4, Неглинная ул., д. 29/14.
Ярославский полиграфкомбинат Союзполиграфпрома при Государственном комитете СССР по делам
издательств, полиграфии и книжной торговли. 150014, Ярославль, ул. Свободы, 97.