/


































































































































































































































































































































































































































































































































































































































































































































































































































































































































Author: Ткаченко Б.И.
Tags: биология физиология анатомия




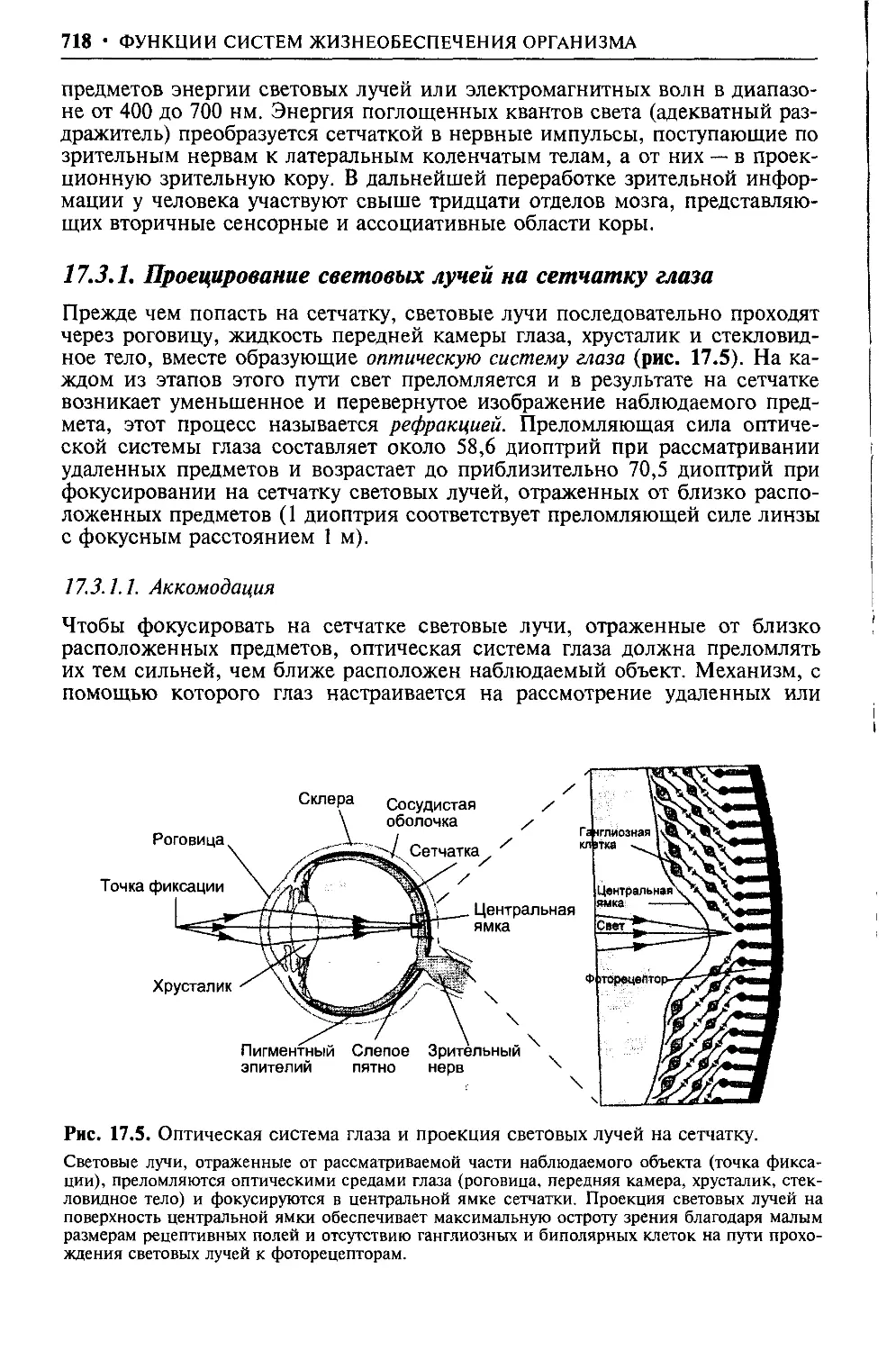
Text
ГЛАВА 1
Жидкие среды организма
Жизнедеятельность многоклеточного организма полностью зависит от ок-
ружающей среды, ее газового, водного, солевого состава, питательных ве-
ществ, температуры среды, в которой он эволюционировал и обитает, и т.
д. Именно внешняя среда в ходе эволюции сформировала видовые особен-
ности обмена веществ между организмом человека, животных и внешней
средой: алиментарного (обмен питательными веществами и продуктами их
метаболизма), газового, водно-солевого и др. Этот обмен между организ-
мом и внешней средой прямого влияния на клетки тканей организма не
оказывает, так как жидкость в межклеточных пространствах является той
промежуточной средой, через которую из внешней среды в клетки посту-
пают кислород, энергетические и пластические ресурсы, и, напротив, в
нее из клеток поступают продукты белкового, жирового, углеводного, со-
левого обмена и др. Из жидкости межклеточных пространств последние, с
кровью и лимфой в ходе кровообращения и лимфообращения, перемеща-
ются к органам, обеспечивающим выведение этих веществ из организма
(желудочно-кишечный тракт, почки, легкие, кожные покровы и др.). Та-
ким образом, для клеток организма человека и животных «внешней сре-
дой» обитания является внеклеточная жидкость, которую Клод Бернар на-
звал «внутренней средой организма» и рассматривал ее существование как
необходимое условие жизни клеток организма, не зависящей от изменений
внешней среды.
1.1. Внутренняя среда организма
К внутренней среде организма относят все жидкости внеклеточного про-
странства: интерстициальную, или тканевую, жидкость, кровь, лимфу,
трансцеллюлярные жидкости, заполняющие полости организма и некото-
рых органов, — спинномозговую, внутриглазную, внутрисуставную (сино-
виальную) жидкость, жидкости серозных пространств (плевральную, пери-
кардиальную, перитонеальную). Жидкости внутренней среды организма
характеризуются постоянством физико-химических показателей — осмо-
лярности, pH, концентрации различных ионов и органических соедине-
ний, температуры Постоянны объемы, занимаемые различными жидкостя-
ми внеклеточного пространства. Для каждого показателя внеклеточной
жидкости эмпирически установлены средняя величина и границы ее нор-
мальных отклонений (табл. 1.1), а также крайние сдвиги границ величины
показателей внеклеточной жидкости, при которых может нарушаться жиз-
недеятельность клеток или организма в целом.
Показатели внутренней среды организма, диапазон границ отклонения
которых от средней величины мал (концентрация Na+, К+, pH), относят к
жестким константам. Показатели, границы отклонений которых от сред-
них величин имеют более значительный диапазон (концентрация глюкозы,
белки плазмы крови), относят к пластическим константам. Отклонения
20 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Таблица 1.1. Средние величины некоторых показателей внеклеточной жидкости и гра-
ницы их нормальных отклонений
Показатель внеклеточной жидкости Нормальная сред- няя величина по- казателя Нормальные гра- ницы отклонения показателя от сред- ней величины Крайние, опасные для жизни границы сдвига показателя
Na+, ммоль/л К+, ммоль/л 142 138-146 115-175
4,2 3,8-5,0 1,5-9,0
Бикарбонат натрия, ммоль/л 28 24-32 8-45
рн 7,4 7,35-7,45 6,9-8,0
Са, ммоль/ л 1,2 1,0-1,4 0,5-2,0
О2, мм рт. ст. СО2, мм рт. ст. 40 35-45 10-1000
40 35-45 5-80
средних величин показателей внеклеточной жидкости к их верхним или
нижним нормальным границам (см. табл. 1.1) могут быть обусловлены воз-
растом организма, социальными и профессиональными условиями, в ко-
торых находится человек, а также временем года и суток, географическими
и природными условиями, половыми и индивидуальными особенностями
организма (см. главу 23).
Выход за нормальные границы физико-химических и биологических
жестких констант внутриклеточной жидкости является или причиной,
или следствием заболевания организма. Поэтому для врача исследование
показателей внеклеточной жидкости (крови, спинномозговой жидкости
и др.) и их отклонений от нормальных величин имеет большую диагно-
стическую ценность, поскольку используется врачом при постановке ди-
агноза заболевания, позволяет по изменению этих показателей анализи-
ровать течение заболевания, оценивать эффективность применяемого ле-
чения.
Отклонения от нормальных границ физико-химических показателей,
объемов или давления жидкостей внутренней среды организма восприни-
маются рецепторами (осмо-, хемо-, волюмо- и барорецепторами), улавли-
вающими изменение ионного, газового, антигенного состава жидкостей,
их объема, оказываемого ими механического давления на стенки крове-
носных сосудов, что приводит к включению нервной, гормональной и им-
мунной систем в организацию биохимических, биофизических и физиоло-
гических регуляторных реакций, реализуемых различными системами ор-
ганизма (дыхания, кровообращения, крови, выделения, иммунной систе-
мой и др.), которые и устраняют сдвиги, возникшие во внутренней среде
организма.
При этом показатели внутренней среды контролируются в организме
механизмами, реализуемыми на уровне генома клеток, клеточном, ткане-
вом, органном, системном, и организменном. Например, сниженное со-
держание кислорода в крови уменьшает поступление его в клетки почек,
являющихся высокочувствительными к недостатку кислорода, что приво-
дит к активации в них участка генома, ответственного за синтез эритропо-
этиновой и-РНК. Биосинтез эритропоэтина клетками почек резко усили-
вается, костный мозг, стимулированный эритропоэтином, воспроизводит
больше эритроцитов, и с увеличением массы гемоглобина в крови возрас-
тает количество переносимого кровью кислорода. На данном примере
можно видеть, что изменение физиологической константы — содержания
кислорода в крови — включило механизмы, контролирующие этот пара-
1. Жидкие среды организма • 21
метр внутренней среды организма на уровне генома клеток почек, на тка-
невом уровне — в костном мозге и в системе крови — в целом.
У. Кэннон предложил обозначать постоянство внутренней среды орга-
низма, обеспечиваемое совокупностью физиологических реакций систем
организма, возникающих при действии на него внешних и внутренних
возмущающих влияний, термином «гомеостазис» («гомеостаз» — от греч.
homoios — подобный, stasis — неподвижность).
Гомеостазис есть состояние относительного и динамического постоян-
ства показателей внутренней среды организма, отклонение от которого
устраняется регуляторными механизмами.
1.2. Биологические свойства жидкостей, составляющих
внутреннюю среду организма
1.2.1. Вода как составная часть жидкостей организма
Основу жидкостей внутренней среды организма составляет вода. Общее
содержание воды в организме взрослого человека составляет около 60—
65 % от массы его тела. При этом на долю внутриклеточной (интрацеллю-
лярной) воды приходится 73, т. е. 40—45 % от массы тела, а остальное ко-
личество воды, составляющее 20—23 % от массы тела, распределено во
внеклеточном пространстве. Из них около 16 % содержится в составе меж-
клеточной жидкости (интерстициальная жидкость), 5 % — внутри сосудов,
т. е. интравазально, в составе плазмы крови, 2 % — в лимфатических сосу-
дах, в составе лимфы, 1—3 % воды от массы тела — в составе трансцеллю-
лярных жидкостей.
Между всеми водными секторами организма осуществляется постоян-
ный водный обмен (рис. 1.1)
Вода является растворителем всех органических и неорганических ве-
ществ, поступающих из внешней среды в организм и всасывающихся в же-
лудочно-кишечном тракте. Вода, входящая в состав крови, интерстициаль-
ной жидкости и лимфы, переносит растворенные в ней питательные вещест-
ва к клеткам тканей и продукты обмена — к органам выделения. В воде, вхо-
дящей в состав клеток организма, осуществляются все процессы обмена. Во-
да, благодаря высокой теплоемкости
регуляции, обеспечивая выведе-
ние тепла из организма (теплоот-
дачу) с помощью потоотделения,
испарения пота, тепловой одышки.
В крови, лимфе, трансцеллю-
лярных жидкостях, пищевари-
тельных соках вода находится в
свободном, т. е. не связанном с
органическими соединениями со-
стоянии. В интерстициальном
пространстве и клетках организма
большая ее часть находится в свя-
занном состоянии, т. е. связана с
белками (например, с молекулами
коллагена), другими органически-
ми соединениями. Поэтому вода
не вытекает при рассечении тка-
ней органов.
и теплопроводности, участвует в термо-
Рис. 1.1. Распределение и обмен воды ме-
жду разными секторами тела взрослого
человека (в процентах от массы тела).
А — вода плазмы крови; Б — интерстициальная
вода; В — внутриклеточная вода; Г — вода лим-
фы; Д — трансцеллюлярная вода. В процентах
указан объем воды, находящейся в секторе,
рассчитанный по отношению к массе тела.
22 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Суточная потребность человека с массой тела 70 кг в воде составляет
2,5 л, из них 1,2 л поступает в организм человека в виде питьевой воды,
1 л с пищей, 0,3 л образуется при окислении жиров, белков и углеводов.
Такое же количество воды (2,5 л) ежесуточно выводится из организма: с
мочой до 1,5 л, с потом 0,5 л, с выдыхаемым воздухом 0,4 л, с калом 0,1 л.
Вода в организме человека обновляется за 1 мес, а внеклеточная — за
1 нед. Недостаточное поступление воды в организм человека (например,
если поступление воды в организм меньше ее объема, интенсивно выде-
ляемого с потом на жаре) приводит к уменьшению воды в крови, интер-
стициальном пространстве. Это состояние называется дегидратацией. Де-
гидратация вызывает сгущение крови, повышение ее вязкости и, как след-
ствие, нарушение кровообращения. Дегидратация, достигающая 20 % от
массы тела, может привести к летальным последствиям.
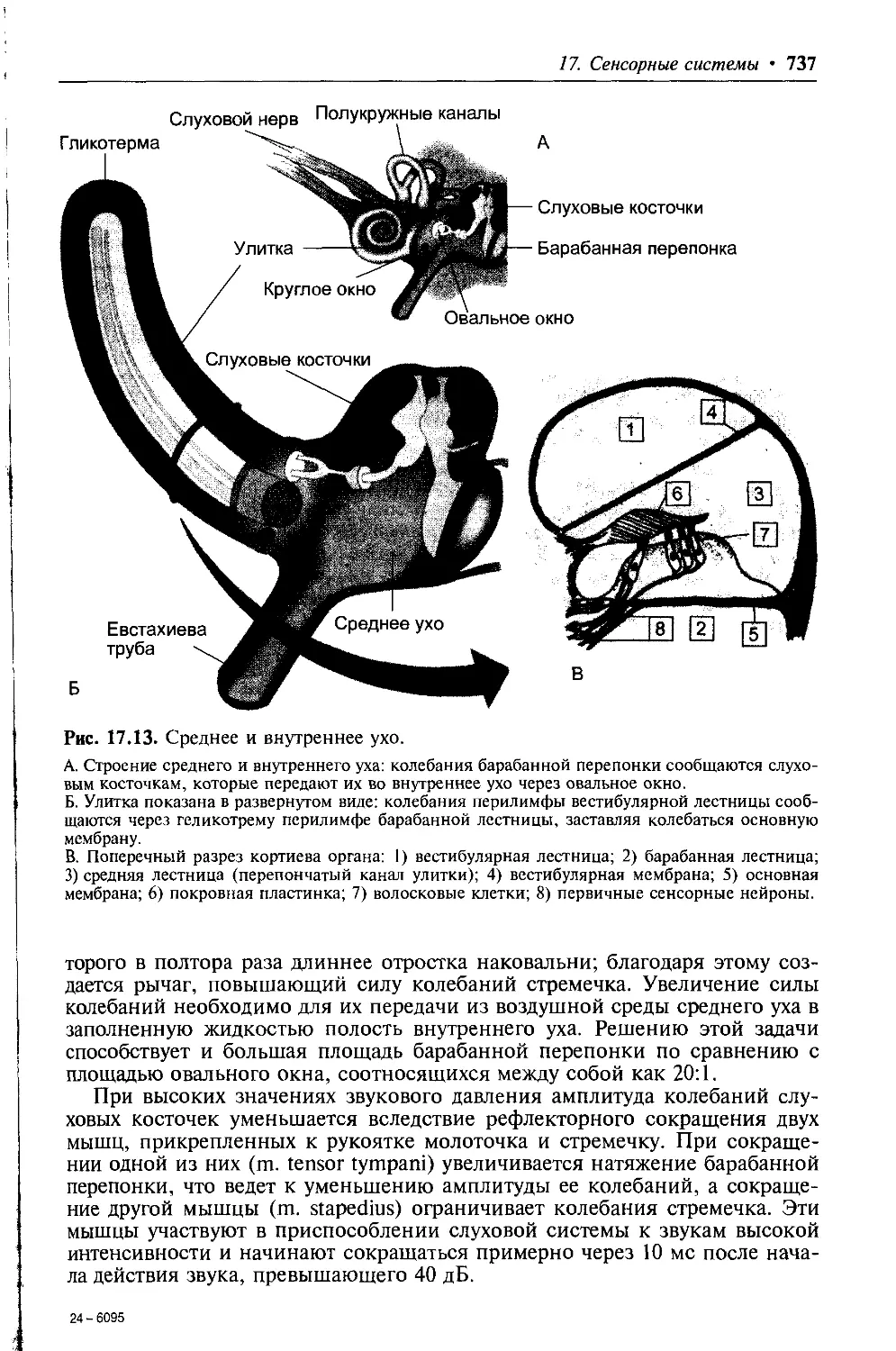
1.2.2. Гистогематические барьеры
Между кровью и внеклеточным пространством находятся образования, по-
лучившие название гистогематических барьеров, отделяющие плазму кро-
ви от внеклеточной жидкости разных тканей организма. Последняя отде-
ляется от внутриклеточной жидкости мембранами клеток. Гистогематиче-
ские барьеры и мембраны клеток обладают избирательной проницаемо-
стью для ионов и органических соединений. Поэтому электролитный и ор-
ганический составы плазмы крови, внеклеточной и внутриклеточной жид-
кости различаются между собой.
По особенностям проницаемости для белков на уровне кровь — ткань
все гистогематические барьеры делят на три группы: изолирующие, час-
тично изолирующие и неизолирующие. К изолирующим барьерам относят
гематоликворный (между ликвором и кровью), гематонейрональный, гема-
тотестикулярный (между кровью и тестикулами), гематоэнцефалический
(между кровью и мозговой тканью) и гематоофтальмический (между кро-
вью и внутриглазной жидкостью), барьер хрусталика глаза. К частично изо-
лирующим относятся барьеры на уровне желчных капилляров печени, коры
надпочечников, пигментного эпителия глаза между сосудистой и сетчатой
оболочками, щитовидной железы, концевых долек поджелудочной железы
и гематоофтальмический барьер на уровне цилиарных отростков глаза. Не-
изолирующие барьеры хотя и позволяют белку проникать из крови в интер-
стициальную жидкость, однако ограничивают его транспорт в микроокру-
жение и цитоплазму паренхиматозных клеток. Такие барьеры существуют
в миокарде, скелетных мышцах, мозговом слое надпочечников, околощи-
товидных железах.
Структурным элементом гистогематических барьеров является стенка
кровеносных капилляров. Морфологические и функциональные особенно-
сти клеток эндотелия капилляров — размер пор в их мембране, наличие
фенестр, наличие межклеточного основного вещества, цементирующего
щели между эндотелиоцитами капилляров и толщина базальной мембраны
определяют проницаемость барьера для воды и растворенных в ней моле-
кул веществ различных размеров и строения. Содержащиеся в крови веще-
ства (вода, кислород, СО2, глюкоза, аминокислоты, мочевина и др.) могут
проникать через барьер двумя путями (рис. 1.2): трансцеллюлярно (через
клетки эндотелия) и парацеллюлярно (через межклеточное основное веще-
ство).
Трансцеллюлярный транспорт веществ может быть пассивным (т. е. по
концентрационному или электрохимическому градиенту без затрат энер-
1. Жидкие среды организма • 23
Трансцеллюлярный обмен
Парацеллюлярный обмен
Л
Эр
Вода
Газы: О2, СО2
Мочевина
Глюкоза
Электролиты
Белки плазмы
Клетки крови
Бактерии
Вода
Газы: О2, СО2
Мочевина
”•.> Глюкоза
Электролиты
* Белки плазмы
(пиноцитоз)
Рис. 1.2. Транспорт веществ через стенку капилляров.
Эр — эритроциты, ЭК — эндотелиальные клетки, Л — лейкоциты.
гии) и активным (против градиента с затратами энергии). Трансцеллюляр-
ный перенос веществ осуществляется и с помощью пиноцитоза, т. е. про-
цесса активного поглощения клетками пузырьков жидкости или коллоид-
ных растворов. Парацеллюлярный транспорт, или перенос веществ через
межклеточные щели, заполненные основным веществом, окутывающим
волокнистые структуры фибриллярного белка, возможен для молекул раз-
ных размеров (от 2 до 30 нм), поскольку в капиллярах размеры межклеточ-
ных щелей неодинаковы. Базальная мембрана капилляров разных органов
имеет неодинаковую толщину, а в некоторых тканях прерывиста. Эта
структура барьера играет роль молекулярного фильтра, пропускающего мо-
лекулы определенного размера. В состав базальной мембраны входят гли-
козаминогликаны, способные уменьшать степень полимеризации и адсор-
бировать ферменты, повышающие проницаемость барьера. Снаружи в ба-
зальной мембране располагаются отростчатые клетки — перициты. Точных
сведений о функции этих клеток нет, предполагается, что они выполняют
опорную роль и продуцируют основное вещество базальной мембраны.
Основные функции гистогематических барьеров — защитная и регуля-
торная. Защитная функция заключается в задержке барьерами перехода
вредных веществ эндогенной природы, а также чужеродных молекул из
крови в интерстициальную среду и микроокружение клеток. При этом не
только сама сосудистая стенка с ее избирательной проницаемостью, но и
ячеисто-коллоидные структуры интерстиция, адсорбируя такие вещества,
24 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
препятствуют их поступлению в микросреду клеток. Если же произошло
проникновение крупномолекулярных чужеродных веществ в интерстици-
альное пространство и они не подверглись здесь адсорбции, фагоцитозу и
распаду, то такие вещества поступают в лимфу, а не в клеточное микроок-
ружение. Лимфа в этом плане представляет собой как бы «вторую линию
обороны», поскольку содержащиеся в ней антитела, лимфоциты и моно-
циты обеспечивают обезвреживание чужеродных веществ.
Благодаря регуляторной функции гистогематические барьеры контроли-
руют состав и концентрацию молекул различных соединений в интерсти-
циальной жидкости, изменяя проницаемость барьеров для ионов, пита-
тельных веществ, медиаторов, цитокинов, гормонов, продуктов метаблиз-
ма клеток. Таким образом гистогематические барьеры регулируют поступ-
ление различных веществ из крови в интерстициальную жидкость и свое-
временный отток продуктов клеточного обмена из межклеточного про-
странства в кровь.
Проницаемость гистогематических барьеров изменяется под влиянием
вегетативной нервной системы (например, симпатические влияния
уменьшают их проницаемость). Изменяют проницаемость гистогематиче-
ских барьеров в сторону как увеличения, так и уменьшения циркулирую-
щие в крови гормоны (например, кортикостероиды уменьшают прони-
цаемость гематоэнцефалического барьера), тканевые биологически актив-
ные вещества (биогенные амины — серотонин, гистамин, гепарин и др.),
ферменты (гиалуронидаза и др.), образуемые как самими эндотелиальны-
ми клетками, так и клеточными элементами интерстициального про-
странства. Например, гиалуронидаза — фермент, вызывающий деполиме-
ризацию гиалуроновой кислоты основного вещества межклеточных про-
странств. Поэтому при его активации резко повышается проницаемость
барьеров; серотонин — снижает их проницаемость, гистамин повышает
ее; гепарин — ингибирует гиалуронидазу и в результате уменьшения ее
активности снижает проницаемость барьеров; цитокиназы — активизиру-
ют плазминоген, и усиливая растворение фибриновых волокон, повыша-
ют проницаемость барьера. Повышают проницаемость барьеров метабо-
литы, вызывающие сдвиг pH в кислую сторону (например, молочная ки-
слота).
Проницаемость гистогематических барьеров зависит и от химического
строения молекул переносимых веществ, их физико-химических свойств.
Так, для растворимых в липидах веществ гистогематические барьеры более
проницаемы, поскольку такие молекулы легче проходят через липидные
слои мембран клеток.
1.2.3. Внутриклеточная жидкость
Во внутриклеточной жидкости преобладающими катионами являются ка-
лий (150 мэкв/л) и магний (40 мэкв/л), содержится большое количество
ионов НРО; (100 мэкв/л) и белков анионпротеинатов (т. е. молекул белка,
имеющих отрицательный заряд) (55 мэкв/л). Столь высокие концентрации
ионов калия во внутриклеточной жидкости связаны с их участием в био-
синтезе белков и углеводов, магния — участием более чем в 300 энзимных
внутриклеточных реакциях. Фосфатные ионы и анионпротеинаты входят в
состав основных буферных систем, поддерживающих pH внутриклеточной
жидкости. Осмотическое давление во внутриклеточной и внеклеточной
жидкости примерно равно, что поддерживает постоянство объемов воды в
этих секторах. Поэтому важнейшим следствием поддержания постоянства
1. Жидкие среды организма • 25
осмотического давления внеклеточной жидкости являетсй стабильность
объема воды, содержащейся в клетках организма.
В отличие от внеклеточной жидкости, физико-химические показатели
которой строго поддерживаются на постоянном уровне регуляторными ме-
ханизмами, что и обеспечивает оптимальные условия для деятельности
клеток организма, физико-химические показатели внутриклеточной жид-
кости характеризуются весьма широким диапазоном колебаний, которые
обусловлены уровнем функциональной активности клеток. Так, при пере-
ходе клетки из ее нормального состояния в состояние возбуждения или
торможения ее активности концентрация ионов — К, Mg, Са — в жидкой
среде цитоплазмы резко изменяется. Например, концентрация ионов ка-
лия, являющаяся «жесткой» константой внеклеточной жидкости во внут-
риклеточной жидкости во время активации клеточной функции может из-
меняться от 115 до 150 мэкв/л. Концентрация ионов кальция в цитоплазме
клетки, находящейся в состоянии физиологического покоя, составляет
10-7—10~8 моль/л, а при действии возбуждающего активность клетки сиг-
нала (медиатора, гормона) концентрация Са++ в цитоплазме возрастает до
10~5—10~6 моль/л, т. е. в 20 раз. В то же время даже незначительное увели-
чение концентрации ионов Са во внеклеточной жидкости — с 1,2 до
1,4 ммоль/л, включает регуляторные механизмы, восстанавливающие нор-
мальную концентрацию Са++ во внеклеточной жидкости.
1.2.4. Интерстициальная, или тканевая, жидкость
Около '/6 объема тела человека составляет пространство, расположенное
между клетками организма, стенками кровеносных и лимфатических сосу-
дов. Оно называется интерстициальным, или интерстициумом, а жидкость,
заключенная в границах, образованных с одной стороны мембранами кле-
ток, с другой — стенками кровеносных и лимфатических капилляров, на-
зывается интерстициальной, или тканевой, жидкостью. Как заметил К.
Бернар, это «внутреннее море», в котором живут клетки.
Структура интерстиция представлена сетью коллагеновых и эластиче-
ских волокон, филаментов протеогликанов. Коллагеновые волокна пред-
ставляют собой белок, образуемый фиброцитами соединительной ткани.
Масса коллагеновых волокон составляет 6 % массы тела, а общая поверх-
ность этих волокон превышает 1 000 000 м2. Сеть этой своеобразной кол-
лагеновой «губки» накапливает в интерстиции воду и электролиты, осо-
бенно натрий. Пучки волокон коллагена простираются вдоль всего интер-
стиция и обеспечивают механическую прочность (сопротивление) тканей.
К плотным структурам интерстиция относятся также филаменты протеог-
ликанов, очень тонкие и едва различимые в световом микроскопе. Их
свернутые спиралью молекулы на 98 % состоят из гликозаминогликанов —
гиалуроновой кислоты, хондроитинсульфатов А, В и С, а также белка. Мо-
лекулы протеогликанов и гликозаминогликанов имеют отрицательный за-
ряд (анионы), благодаря чему поддерживается ионное равновесие с катио-
нами интерстициальной жидкости.
Интерстициальная жидкость заключена в основном в мельчайших про-
странствах между филаментами протеогликанов и имеет характер геля. По-
этому ее еще называют тканевым гелем. Таким образом, филамент проте-
огликанов интерстиция формирует первую, коллоидную, или гелеподобную,
фазу интерстиция, которая благодаря высокой гидрофильности связывает
или освобождает воду под влиянием ферментов и биологически активных
веществ (гиалуронидаза, гепарин, гистамин и др.). Быстрый транспорт мо-
26 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
лекул воды, О2, СО2, электролитов, питательных веществ, экскретов клеток
между кровеносными капиллярами и клетками тканей обеспечивается про-
стой диффузией через гель этих соединений. Скорость диффузии указан-
ных веществ от стенок капилляров до клеток на расстоянии до 50 мкм осу-
ществляется за несколько секунд. Вторая фаза интерстиция — водная, в ви-
де свободной жидкости, «текущей» по тонким «каналам» вдоль коллагено-
вых волокон, составляет не более 1 % интерстициальной жидкости. При
развитии отека (т. е. скопления воды и электролитов в межклеточном про-
странстве) содержание свободной жидкости в интерстициальном простран-
стве резко увеличивается, а «каналы» оказываются резко расширенными.
В обеих фазах интерстициального пространства у взрослого человека со-
держится в среднем 11 — 12 л жидкости, т. е. около 16 % массы тела (см.
рис. 1.1).
В ионном составе интерстициальной жидкости преобладают ионы нат-
рия (142—144 мэкв/л) и ионы хлора (120 мэкв/л). Высокая суммарная кон-
центрация данных ионов определяет величину осмотического давления
интерстициальной жидкости. Поэтому при уменьшении концентрации Na
в плазме крови и интерстициальной жидкости (например, при недостаточ-
ности коры надпочечников уменьшается секреция гормона альдостерона,
усиливающего реабсорбцию Na+ в канальцах почек и Na+ в больших коли-
чествах с мочей выводится из организма) в интерстициальной жидкости
появляется «осмотически свободная вода», которая выводится из организ-
ма через почки, а также по осмотическому градиенту диффундирует в
клетки и вызывает их набухание. При увеличении же концентрации Na+ в
интерстициальной жидкости (например, вследствие избыточного поступле-
ния NaCl в организм с соленой пищей) ее осмотическое давление повыша-
ется, вода задерживается в интерстициальном пространстве, что приводит
к развитию отеков. Концентрация К+ в интерстиции (3,8—5 ммоль/л) в
30 раз меньше, чем во внутриклеточной жидкости. Это «жесткая» констан-
та интерстициальной жидкости, и ее сдвиги вызывают нарушение функ-
ций клеток. Так, например, увеличение концентрации К+ в интерстици-
альной жидкости миокарда (следствие гиперкалиемии — увеличения кон-
центрации К+ в плазме крови) уменьшает соотношение концентраций —
К+ внутриклеточный / К внеклеточной жидкости, что приводит к деполя-
ризации мембраны, нарушает восстановление мембранного потенциала
клеток миокарда. В результате замедляется проведение возбуждения в сер-
дечной мышце, что может вызвать остановку сердца. Жесткими константа-
ми являются и содержание Mg++ (0,75—1,2 ммоль/л) и Са++ (0,8—1,2
ммоль/л) во внеклеточной жидкости. Оба иона участвуют в поддержании
нервно-мышечной возбудимости. Например, ионы Mg++ влияют на транс-
порт К+ через мембрану клетки и увеличение их концентрации во внекле-
точной жидкости (следствие гипермагниемии) угнетает возбудимость нерв-
ной системы, скелетных мышц. Напротив, уменьшение концентрации
Mg++ или Са++ в крови вызывает повышение нервно-мышечной возбуди-
мости.
В интерстициальной жидкости содержится кислород, большое количест-
во питательных веществ для клеток — глюкозы, аминокислот, жирных ки-
слот, в ней содержится и СО2, поступающий из клеток и диффундирующий
из интерстиция в кровь для удаления из организма, продукты белкового ме-
таболизма клеток (мочевина, креатин, креатинин и др.). Из интерстициаль-
ной жидкости продукты обмена поступают в кровь и транспортируются ею
к органам выделения — желудочно-кишечному тракту, почкам, которыми и
выводятся из организма. Через поры капилляров соматического типа — их
1. Жидкие среды организма • 27
стенка представлена непрерывной базальной мембраной и эндотелиальным
слоем, в которых имеются поры, шириной от 6 до 7 нм (в легких, коже) —
в интерстиций способно выходить небольшое количество белков плазмы
крови. Много большее количество их поступает в интерстициальное про-
странство через стенку капилляров синусоидного типа, представленных, на-
пример, в печени и имеющих эндотелиальный слой с фенестрами, базаль-
ную мембрану — с перерывами и в результате такой структурной организа-
ции — прерывистую стенку с большими просветами. Поэтому содержание
белка неодинаково в интерстициальной жидкости разных тканей: оно низ-
кое в подкожной ткани, в легких — 0,5—2 г/л. Однако в лимфе, оттекающей
от интерстиция печени, в который из капилляров синусоидного типа посту-
пает большое количество белка, содержание последнего достигает 55—60 г/л.
Общее количество белка во всем объеме интерстициальной жидкости
организма (в 11—12 л) достигает 330—360 г. Отсюда, концентрация белка в
1 л интерстициальной жидкости составляет около 30 г/л и создает колло-
идно-осмотическое (онкотическое) давление, равное 8 мм рт. ст., являю-
щееся силой, удерживающей жидкость в интерстиции.
Все белки из интерстициальной жидкости возвращаются обратно в
кровь только через лимфатическую систему. По пути кровь—лимфа—кровь
за сутки рециркулирует от 50 до 100 % белка.
Давление интерстициальной жидкости оказывается на 3 мм рт. ст.
меньше атмосферного. Причиной отрицательного давления интерстици-
альной жидкости по отношению к атмосферному давлению является по-
стоянный отток жидкости из интерстиция по лимфатическим сосудам.
Интерстициальное пространство содержит клетки соединительной тка-
ни — фибробласты и фиброциты, тучные клетки, макрофаги и лимфоциты,
которые секретируют в микросреду клеток биологически активные соеди-
нения (ферменты, гепарин, биогенные амины, простагландины, лейко-
триены, цитокины и др.), поддерживающие нормальное функциональное
состояние интерстиция. Клетки интерстициального пространства осущест-
вляют фагоцитоз, иммунную защиту интерстиция.
Микросреда клеток — это часть интерстициального пространства, непо-
средственно прилегающего к поверхности клеток, толщиной порядка 10—
20 нм, играет основную роль в обмене веществ через ее мембрану.
Микросреда клеток отличается от среды общего интерстициального
пространства более высокой концентрацией поступающих из крови в ин-
терстиций аминокислот и жирных кислот, используемых в пластических и
энергетических процессах в клетке; медиаторов, гормонов и антигенов, ре-
гулирующих клеточные функции (пролиферацию, дифференциацию, мета-
болизм, синтез и секрецию антител и др.). Обмен воды и молекул между
микросредой клеток и общим интерстициальным пространством происхо-
дит под влиянием градиентов сил гидростатического, онкотического и ос-
мотического давления, электрокинетических и электростатических потен-
циалов. В создании последнего участвуют гликозаминогликаны, форми-
рующие отрицательный заряд на поверхности мембран клеток.
Находящиеся в микросреде гуморальные факторы — нейромедиаторы,
гормоны, метаболиты, цитокины, связываясь с их мембранными клеточ-
ными рецепторами, осуществляют физиологическую регуляцию различных
функций клеток: процессов пролиферации и дифференцировки клеток, их
метаболизма, например синтеза и продукции ими белков, гликопротеидов,
липидов и других продуктов, что поддерживает постоянство структуры ор-
ганов и тканей организма, обеспечивает приспособительную реакцию кле-
ток к изменениям внешней среды.
28 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
1.3. Плазма крови как внутренняя среда организма
Кровь состоит из плазмы и клеток (форменных элементов) — эритроцитов,
лейкоцитов и тромбоцитов, находящихся во взвешенном состоянии в плаз-
ме крови (см. главу 7). Плазма крови состоит из воды (около 90 % массы),
солей или электролитов (0,9 %), углеводов, липидов (1,1 %), органических
кислот и оснований, промежуточных продуктов обмена, витаминов и бел-
ков (8 % массы плазмы). Плазма является для клеток крови жидкой сре-
дой, обеспечивающей их нормальную жизнедеятельность.
Функции плазмы крови. Транспортная функция (дыхательная, питатель-
ная, интегративно-регуляторная и экскреторная) — обеспечивает перенос
газов О2 СО2, а также веществ, необходимых для жизнедеятельности как
клеток крови, так и других тканей организма (глюкозы, аминокислот, ви-
таминов и др.) или регуляции их функций (гормоны, цитокины) обеспечи-
вает перенос веществ, подлежащих удалению из организма (например,
продуктов белкового обмена — креатина, креатинина, мочевины и др.). За-
щитная функция — связывает и нейтрализует токсичные вещества, попа-
дающие в организм или образующиеся в нем, связывает и разрушает ино-
родные белковые молекулы и чужеродные клетки, в том числе и инфекци-
онного происхождения. Клетки крови (лейкоциты, тромбоциты) секрети-
руют в плазму пептидные молекулы — цитокины, активирующие регенера-
цию различных тканей организма (кроветворной, костной, кровеносных
сосудов и др.).
Объем циркулирующей крови, включающий форменные элементы и плаз-
му крови, у взрослого человека (нормоволемия) составляет 6—8 % от массы
тела (4—6 л) или 62,4 ± 7,8 мл/кг массы тела. Из них на долю циркули-
рующей плазмы крови приходится 34,2 ± 4,5 мл/кг массы тела, формен-
ных элементов крови — 28,2 ± 4,0 мл/кг. После избыточного приема воды
объем плазмы крови может повышаться (гиперволемия), а при тяжелой фи-
зической работе (например, в жарких цехах металлургического производст-
ва) и связанном с ней избыточным потоотделением — снижаться (гиповоле-
мия). Процентная часть объема, занимаемая клетками крови, от общего
объема крови называется гематокритом. Гематокрит в норме (при нормо-
цитемии, т. е. при нормальном содержании форменных элементов в кро-
ви) у мужчин равен 44—48 %, у женщин — 41—45 %.
Электролиты и белки плазмы крови формируют ее осмотическое и он-
котическое давление, коллоидную стабильность, влияют на процесс свер-
тывания крови и фибринолиза, на агрегатное (т. е. жидкое) состояние кро-
ви, ее суспензионную устойчивость (т. е. поддержание клеток крови во
взвешенном состоянии), удельный вес и вязкость.
1.3.1. Электролитный состав плазмы крови
Электролитный состав плазмы крови (он соответствует таковому и в сыво-
ротке крови — под последней понимают плазму крови, лишенную фибри-
ногена) представлен катионами Na+, К+, Са++, Mg++, Fe+++, Zn++ и Cu++,
анионами СГ, НСО7, НРО7, HSO7 органических кислот и анионпротеина-
тами (1 г белков плазмы соответствует 0,208 мэкв/л) (табл. 1.2).
Суммарная величина анионов и катионов в плазме крови, выраженная
в мэкв/л, примерно одинакова. Нарушение этого равновесия может возни-
кать при расстройствах кислотно-основного баланса.
Содержание натрия, калия и кальция в плазме крови относится к жест-
ким гомеостатическим константам. Содержание электролитов в плазме
1. Жидкие среды организма • 29
Таблица 1.2. Содержание основных электролитов и некоторых микроэлементов (Fe, Zn
и Си) в плазме крови (средняя величина)
Катионы мг/л мэкв/л Анионы мг/л мэкв/л
Na+ 3260 142 СГ 3600 101,5
К+ 190 4,8 HCOJ 1500 24,6
Са++ 100 5 ро; 100 1,9
Mg++ 21 1,8 Протеины 72 000 15
Fe"+ 1 0,05 Органические кислоты 270 5
Zn++ 3 0,09 Другие анионы — 5
Cu++ 1 0,03
Общее количество 154 Общее количество 154
крови зависит от величины их поступления в организм с пищей и водой и
выведения из него с мочой, калом и потом, а также перераспределения ио-
нов между клетками и внеклеточной средой. Гомеостазис катионов натрия
и калия поддерживается изменением пищевого поведения человека (боль-
шее или меньшее потребление соли), ренин-ангиотензин-альдостероновой
системой (усиливает реабсорбцию натрия в почках и выведение К+ с мо-
чой) и натрийуретическим гормоном предсердий (усиливает выведение
Na+ с мочой). На уровень ионизированного кальция в плазме крови влия-
ют гормоны: околощитовидных желез (паратирин — задерживает выведе-
ние Са++ с мочой, повышает уровень Са++ в крови), щитовидной железы
(кальцитонин и его аналоги — усиливают выведение с мочой Са++), почек
(кальцитриол — усиливает всасывание Са++ в кровь из кишечника).
Минимум 15 микроэлементов содержатся в плазме крови (железо, медь,
кобальт, марганец, цинк, хром, стронций и др.). Они необходимы для
обеспечения нормального метаболизма клеток и их функций, поскольку
входят в состав ферментов, катализируют их действие. Ионы меди, цинка
участвуют в процессах образования клеток крови (гемопоэзе), а ионы ко-
бальта, железа — в синтезе гемоглобина.
1.3.2. Осмотическое и онкотическое давление плазмы крови
Концентрация ионов, выраженная в мэкв/л (или ммоль/л), позволяет рас-
считать осмотическое давление раствора (в данном случае плазмы крови),
создаваемое растворенными в ней ионами. Один ион-грамм вещества соз-
дает 6,023 • 1023 частиц в растворе (число Авогадро) и формирует в 1 л воды
осмотическое давление в 1 осмоль. Следовательно, одна тысячная ион-
грамма (милли-ион-грамм) создает давление, равное 1 миллиосмолю
(мОсм). Отсюда:
Осмотическое давление в милиосмолях = Количество вещества в мэкв/Ва-
лентность.
Например, концентрация Na+ в плазме равна 142 мэкв/л, его валент-
ность — 1, создаваемое им осмотическое давление — 142:1 = 142 мОсм/л.
Концентрация СГ в литре плазмы составляет 101,5 мэкв. Следователь-
но, его осмомолярность равна около 102 мОсм/л плазмы. Отсюда следует,
что величина осмотического давления плазмы, равная 304 мОсм/л, созда-
ется в основном ионами Na и С1 и изменение их концентрации в плазме
крови наиболее выражение изменяет ее осмомолярность.
Неионизированные вещества в плазме крови также поддерживают ос-
30 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
мотическое давление. Так, каждый грамм глюкозы в литре плазмы создает
давление в 5,6 мОсм.
Хотя содержание белков в плазме крови достигает 70—80 г/л, их кон-
центрация в ней весьма невелика, около 0,8 ммоль/л, поэтому осмотиче-
ское давление, формируемое белками, равно 0,8 мОсм/л или 14 мм рт.ст.
В плазме содержится 42 г альбуминов, т. е. 0,0006 моля, и 28 г глобулинов,
что составляет 0,0002 моль/л. Однако плазма крови с указанным количест-
вом белков создает осмотическое давление, эквивалентное 25—28 мм рт.ст.
Объясняется это тем, что белки, будучи отрицательно заряженными иона-
ми, удерживают вблизи своих молекул большое число положительно заря-
женных ионов (катионов, в основном ионов натрия). Эти добавочные ка-
тионы увеличивают величину осмотически активных веществ в плазме
крови и делают ее осмотическое давление примерно на 50 % больше, чем
создаваемое только белками (эффект равновесия Даннона). Осмотическое
давление, создаваемое белками плазмы, получило название коллоидно-ос-
мотического или онкотического давления. В основном оно создается альбу-
минами плазмы. Онкотическое давление белков плазмы крови обеспечива-
ет постоянное поступление воды в кровеносные капилляры из тканей,
так как интерстициальная жидкость имеет более низкое онкотическое дав-
ление (8 мм рт.ст.), чем плазма крови (25—28 мм рт.ст.). При заболеваниях
почек, при которых имеет место потеря альбуминов с мочой (альбумину-
рия), онкотическое давление плазмы крови резко снижается, жидкость за-
держивается в тканях, у больного возникают отеки.
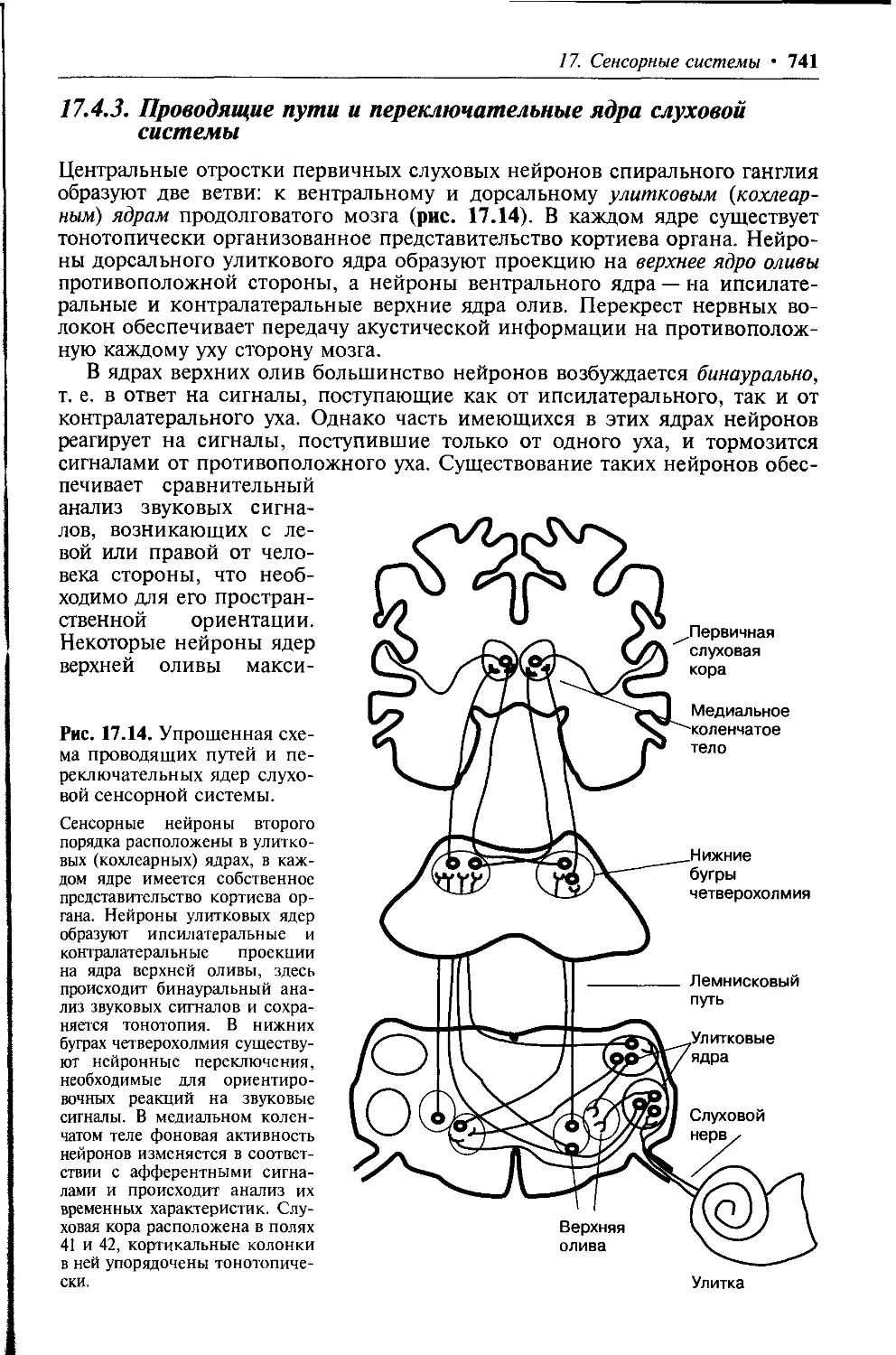
1.3.3. Обмен воды между плазмой крови и интерстициальной
жидкостью
В организме человека между плазмой крови и интерстициальной жидко-
стью имеет место постоянный обмен водой. Он возможен благодаря тому,
что в артериальном конце капилляра гидростатическое давление крови (си-
ла, выталкивающая воду из сосуда,— Ркапил) равно 30 мм рт.ст. и превосхо-
дит онкотическое давление плазмы (т. е. силу, удерживающую воду в сосу-
де, равную 25—28 мм рт.ст.). Выходу жидкости из капилляра в интерсти-
ций способствуют также отрицательное давление интерстициальной жид-
кости, равное —3 мм рт. ст., и притягивающее воду в интерстиций колло-
идно-осмотическое давление этой жидкости, равное 8 мм рт.ст. Таким об-
разом, общее давление сил, движущих жидкость из сосуда в интерстици-
альное пространство (фильтрационное давление), составляет 41 мм рт.ст. и
преодолевает силу, удерживающую жидкость в сосуде, равную 25—28 мм
рт.ст. Следовательно, фильтрационное давление, перемещающее жидкость
из капилляра в ткань, составляет:
^фильтрационное 41 ММ рТ. СТ. (Ркапил. Ринтерстиц. жидкости Рколл-осм. интерстиц. жидкости
Рколл.-осм. плазмы) 13 16 ММ рТ.СТ.,
где
Ркапил. = 30 ММ рт. СТ.,
Ринтерстиц. жидкости 3 ММ рТ. СТ.,
Рколл-осм. интерстиц. жидкости $ ММ рТ. СТ.,
Рколл.-осм. плазмы ^5 28 ММ рТ. СТ.
В результате в артериальной части капилляра имеет место выход воды,
составляющий 0,5 % плазмы крови, в интерстициальное пространство. На-
против, в венозной части капилляра, где гидростатическое давление крови
1. Жидкие среды организма • 31
Рис. 1.3. Обмен воды между плазмой крови в капилляре и интерстициальной жид-
костью.
— сила, направляющая движение воды из сосуда в интерстициальное пространство; j — си-
ла, удерживающая воду в сосуде; — направление движения воды. 1 — Ргшцюстаг. в артериаль-
ном КОНЦе КаПИЛЛЯра 30 ММ рт.ст., 2 Роик.плахмы крови 28 ММ рт.ст., 3 Рннтерстиц. жидкости
-3 мм рт.ст.; 4 — Рколл..осм. „„„ретин.жидкости = 8 мм рт. ст.; 5 — Рг1иростат. в венозной части капилляра
= 10 мм рт. ст.
снижается до 10 мм рт.ст и становится меньше онкотического, т. е. силы,
возвращающей жидкость в сосуд (25—28 мм рт. ст.), вода интерстициаль-
ной жидкости диффундирует в капилляр (см. рис. 1.3). Таким образом, си-
ла, возвращающая жидкость в сосуд из интерстиция, составляет:
Рколл.-осм. плазмы (25 28 ММ рТ.СТ.) (^капил. (Ю ММ рТ.СТ.) Т Ринтерстиц. жидкости
( 3 ММ рТ.СТ.) 4" Рколл.-осм. интерстиц. жидкости ММ рТ.СТ.) 4 7 ММ рТ. СТ.
Возврат воды из интерстиция в капилляр составляет 90 % от объема
жидкости, профильтровавшейся в артериальной части капилляра. Осталь-
ное количество жидкости возвращается из интерстиция в кровоток по
лимфатическим сосудам.
Осмотическое давление влияет на распределение воды не только между
кровью и интерстициальной жидкостью, но и между последней и клетками
организма. Это положение демонстрирует влияние гипертонического и ги-
потонического растворов поваренной соли на гидратацию эритроцитов.
Так, эритроциты, помещенные в гипертонический раствор, в результате
дегидратации сморщиваются, а в гипотоническом растворе они набухают и
лопаются, что используется для определения их осмотической резистент-
ности. Разрушение эритроцитов называется гемолизом. Концентрация Na-
Cl, при которой начинается гемолиз единичных эритроцитов, характеризу-
ет их минимальную осмотическую резистентность (обычно 0,5—0,4 %
NaCl), а концентрация NaCl (обычно 0,34—0,3 % NaCl), лизирующая все
эритроциты, — максимальную осмотическую резистентмостъ.
32 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
1.3.4. Продукты белкового обмена, углеводы и липиды плазмы
крови
Остаточный, или белковый, азот крови — это азотсодержащие продукты
белкового катаболизма (мочевина, мочевая кислота, креатин, свободные
аминокислоты, индикан). Величина остаточного азота (в норме 14,3—
28,6 ммоль/л) отражает эффективность выделения продуктов белкового
обмена через почки. Повышение остаточного азота и его компонентов в
крови является важным диагностическим показателем нарушения их экс-
креторной функции.
Концентрация мочевины в плазме крови в норме составляет 300 ±75 мг/л,
т. е. около 50 % небелкового азота плазмы. Мочевина — это конечный
продукт деградации белков, который подлежит выведению из организма.
Увеличение ее концентрации в плазме крови, так же как и креатинина
(в норме 9—11 мг/л плазмы), отмечается при почечной недостаточности.
Мочевая кислота — это конечный продукт расщепления основных бел-
ков. В норме ее концентрация составляет 50 мг/л плазмы. Ее увеличение в
плазме крови характерно для подагры.
Содержание свободных аминокислот достигает 500 мг/л плазмы. Умень-
шенное их содержание в сыворотке или плазме крови имеет место у боль-
ных с анемиями и объясняется усиленным использованием аминокислот в
костном мозге для синтеза гемоглобина.
Углеводы плазмы крови — более 90 % приходится на глюкозу — главный
источник энергии для клеток. Глюкоза растворима в воде, способна к мем-
бранному транспорту (она свободно проходит через мембрану эритроцита,
поэтому ее концентрация в единице объема воды в плазме и в эритроци-
те — одинакова). Глюкоза используется клетками для обеспечения их энер-
гией, образуемой при аэробном или анаэробном гликолизе. Глюкозы боль-
ше в плазме артериальной крови, чем в венозной, так как она непрерывно
используется клетками тканей. В целом, ее уровень зависит от всасывания
из желудочно-кишечного тракта, поступления из депо (гликоген печени),
новообразования из аминокислот и жирных кислот (глюконеогенез), ути-
лизации тканями и депонирования в виде гликогена.
У человека содержание глюкозы определяют натощак в цельной крови.
В норме ее количество составляет 0,8—1,2 г/л цельной крови. Гомеостазис
глюкозы регулируется вегетативной нервной системой и многочисленными
гормонами (инсулин, глюкагон, адреналин, глюкокортикоиды и др.). На-
пример, увеличение концентрации глюкозы в плазме крови активирует
секрецию инсулина, стимулирующего ее утилизацию клетками тканей, ис-
пользование глюкозы при синтезе молекул гликогена и жира. Увеличение
глюкозы в плазме крови (гипергликемия) характерно для сахарного диабе-
та, при заболевании которым в организме больного имеет место недоста-
точность инсулина.
Общее содержание липидов в плазме крови достигает 5—8 г/л. Разделен-
ные с помощью электрофореза (или ультрацентрифугированием), они об-
наруживают на липидограмме гетерогенный состав и включают холесте-
рин, фосфолипиды, триглицериды, жирные кислоты. Взаимодействие этих
липидов с белками — апопротеинами формирует липопротеины высокой
плотности (ЛПВП), состоящие на 50—60 % из белков; липопротеины низ-
кой плотности (ЛПНП), очень богатые жирами и лишь на 20—25 % со-
стоящие из белков и, наконец, липопротеины очень низкой плотности
(ЛПОНП), на 90 % состоящие из липидов. ЛПНП и ЛПОНП переносят от
печени к другим тканям липиды, а ЛПВП транспортируют избыточный
1. Жидкие среды организма • 33
холестерин из тканей в печень, где он и его эфиры превращаются в желч-
ные кислоты и выводятся с желчью в кишечник, а затем — из организма.
Среди органических кислот в плазме крови в наибольших количествах
представлены молочная (80—100 мг/л), пировиноградная (3—8 мг/л) и ли-
монная (9—13 мг/л) кислоты. Рост уровня молочной кислоты в крови, на-
пример, свидетельствует об усилении анаэробного гликолиза, что имеет
место в организме после интенсивной физической работы.
1.3.5. Белки плазмы крови
Белки плазмы крови (их около 200 видов) обеспечивают: 1) коллоидно-ос-
мотический и водный гомеостаз; 2) агрегатное состояние крови и ее реоло-
гические свойства (вязкость, свертываемость, суспензионные свойства);
3) кислотно-основный гомеостаз; 4) иммунный гомеостаз; 5) транспорт-
ную функцию крови; 6) питательную функцию крови как резерва амино-
кислот; 7) детоксикационную функцию.
Общее количество белков в плазме крови в норме составляет 57—81 г/л,
в том числе фибриногена — 2—4 г/л. С помощью электрофореза белки
плазмы разделяются на следующие фракции: преальбумин, альбумин, а(->
а2-, Р- и у-глобулины. Преальбумины выполняют в основном транспорт-
ную функцию, перенося гормон тироксин, ретинол (производное витами-
на А). В норме их количество составляет 0,18—0,37 г/л сыворотки. Недос-
таток белка или цинка в рационе человека нарушает синтез и уменьшает
концентрацию преальбумина в сыворотке крови.
Альбумины — это низкомолекулярные белки с молекулярной массой,
равной 69 000. Они, как и преальбумины, образуются в печени, поэтому
при ее заболевании количество этих белков в плазме крови уменьшается.
Примерно */3 общего количества альбуминов (200—300 г) в организме
взрослого человека находится в плазме крови (37—55 г/л сыворотки кро-
ви), а 2/з ~ вне сосудистого русла, во внеклеточной жидкости. Между плаз-
мой крови и внеклеточной жидкостью происходит непрерывный обмен
альбумина. Альбумин выполняет несколько функций в плазме крови и
тканях — участвует в формировании коллоидно-осмотического (онкотиче-
ского) давления (на его долю приходится 80 % величины этого показателя
в крови), что позволяет ему участвовать в поддержании транскапиллярного
обмена жидкости, тургора тканей и объема жидкости во внутрисосудистом
и внеклеточном пространствах. Альбумин легко соединяется с органиче-
скими (билирубин, свободные жирные кислоты) и неорганическими веще-
ствами (ионы Mg, Са и др.), стероидными гормонами (прогестерон и др.),
лекарственными средствами (антибиотики, сердечные гликозиды), достав-
ляя их с током крови в ткани, и, напротив, транспортирует многие про-
дукты метаболизма от тканей к районам их выведения из организма (на-
пример, билирубин, образовавшийся в ходе разрушения гемоглобина) — к
печени, почкам, легким, желудочно-кишечному тракту, способствуя деток-
сикационной функции организма. Альбумин является компонентом бу-
ферной системы плазмы крови, регулирующей кислотно-основное равно-
весие. Как легкоусвояемый белок альбумин легко расщепляется в тканях
до аминокислот, используемых клетками в качестве пластических и энер-
гетических ресурсов.
Глобулины имеют высокую молекулярную массу (105 000—900 000). Их
концентрация составляет около 30 % от общей массы белков в сыворотке
крови.
Отношение фракции альбумины/глобулины в сыворотке крови у взрос-
2 - 6095
34 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
лого человека колеблется от 1,3 до 2,3. На долю глобулинов приходится
15—18 % величины поддержания коллоидно-осмотического давления кро-
ви. В этой группе белков при электрофоретическом их разделении разли-
чают сц-глобулины (4 %), а2-глобулины (8,5 %), р-глобулины (12 %) и
у-глобулины (18 %).
а-Глобулины: гликопротеины, связанные с углеводами (2/3 всей глюкозы
плазмы циркулирует в составе гликопротеинов); они являются транспорт-
ными белками для гормонов, витаминов и микроэлементов; а-глобулины
осуществляют транспорт липидов, участвуя в образовании липопротеид-
ных комплексов, в составе которых переносятся триглицериды, фосфоли-
пиды, холестерин и сфингомиелины.
^-Глобулины — богатая липидами фракция белка, которая содержит 3/4
всех липидов плазмы крови, в том числе фосфолипиды, холестерин и
сфингомиелины.
Выполняя функцию белков-переносчиков различных соединений (на-
пример, а- и р-глобулины — так называемые транскобаламины — участву-
ют в переносе витамина В12 с кровью от кишечника к тканям; ^-глобу-
лин — трансферрин — съязыъаът: и переносит железо), а- и р- глобулины
участвуют в формировании «острофазных белков крови» (название связано
с выявлением этих белков в крови при остром начале заболевания; а,-
глобулины содержат aj-антитрипсин и агкислый гликопротеин; а2-глобу-
лины — содержат гаптоглобин, а2-макроглобулин, церулоплазмин, ргглобу-
лин — гемопексин). У взрослых людей физиологическая концентрация ост-
рофазных белков крови создает наряду с иммунной системой и грануло-
цитарно-моноцитарной линиями кроветворной ткани надежный барьер
против инфекций или поступления (образования) в организме токсичных
веществ (табл. 1.3). В последних случаях формирование острофазных бел-
ков в печени компенсаторно возрастает и их уровни в крови резко увели-
чиваются. Физиологические функции острофазных белков характеризуют-
ся следующими свойствами. С-реактивный белок связывает соматический
пневмококковый полисахарид, активирует комплемент по классическому
и альтернативному пути, активирует комплементзависимую цитотоксич-
ность, а также фагоцитарную активность нейтрофилов и макрофагов,
тормозит агрегацию тромбоцитов. Фибронектин обладает опсонизирую-
щей функцией, способствует фагоцитозу бактерий нейтрофилами и моно-
цитами/макрофагами. Амилоид А очищает плазму от токсичных субстан-
ций; И] антитрипсин и а2-макроглобулин — ингибиторы протеаз, регули-
рующие иммунологические и воспалительные реакции; а]-антитрипсин
тормозит протеолиз, особенно выражено угнетение активности трипсина,
хемотрипсина, плазмина, тромбина и протеаз, высвобождающихся при
разрушении лейкоцитов и клеток тканей. На долю сц-антитрипсина при-
ходится до 90 % всей антитрипсиновой активности сыворотки крови; аг
кислый гликопротеин модулирует функции Т- и В-лимфоцитов (синтез
антител, активность Т-киллеров, антителозависимую цитотоксичность).
Гаптоглобин связывает антигены макрофагов, участвует в угнетении ак-
тивности образующихся в избытке протеолитических ферментов. Церуло-
плазмин связывает и транспортирует ионы меди к тканям. На его долю
приходится 80 % антиокислительной активности крови, он угнетает пере-
кисное окисление липидов, предупреждая накопление их токсичных пе-
рекисей. Церулоплазмин ускоряет очищение и заживление ран, является
антиагрегантом. Таким образом, острофазные белки крови выполняют
важную функцию, связанную с повышением резистентности организма
человека к инфекции, токсичным соединениям, перекисному окислению
1. Жидкие среды организма • 35
Таблица 1.3. Физиологический уровень острофазных белков плазмы крови у лиц моло-
дого возраста (*, ** — по разным источникам)
Острофазные белки плазмы крови Содержание в плазме крови, г/л
арКислый гликопротеин 0,4-1,05* 0,6-1,42**
Гаптоглобин 0,4-2,9* 1,2-2,6**
Церулоплазмин 0,2-0,4* 0,2-0,6**
агАнтитрипсин 2,0-4,0* 1,4-4,0**
а2- Макроглобул ин 1,5-3,5
С-реактивный белок 0,47-1,34
липидов мембран клеток, усиливающемуся, например, при стрессе, что
является частью компенсационного интегрального ответа систем организ-
ма на действие чрезвычайных раздражителей, повышающего его выжи-
ваемость при болезнетворном воздействии среды. Снижение в крови од-
ного из острофазных белков — аj-антитрипсина, тормозящего протеолиз
при разрушении лейкоцитов и тканей, предрасполагает к бронхопульмо-
нальным заболеваниям (легочная эмфизема, хронический бронхит, брон-
хоэктатическая болезнь).
До 10 % глобулинов (значительная часть р-глобулинов) сыворотки кро-
ви поддерживают функцию системы комплемента — комплекс системы
белков крови, обеспечивающий защиту организма человека от инфекции,
инициирующий воспаление и участвующий в поражении многих бактерий
и вирусов. Система комплемента состоит из 11 в физиологических услови-
ях неактивных белков плазмы крови, обозначаемых Cl, С2 и т. д. Актива-
ция системы комплемента происходит при иммунологической реакции,
реже под влиянием полисахаридов и при обязательном участии ионов маг-
ния, ведет к лизису клетки.
К белковой защитной системе плазмы крови относят пропердиновую сис-
тему, состоящую из собственно пропердина (белок Р), фактора В (В-гли-
копротеид, богатый глицином) и профермента (протеаза D). Система акти-
вируется при обязательном участии ионов магния следующим образом:
вначале активируется пропердин (зимозаном дрожжей, эндотоксином бак-
терий и другими липополисахаридами, гормоном инсулином); пропердин
активирует фактор D, который в свою очередь активирует фактор В, а
фактор В — систему комплемента, разрушающую клетки.
Другие гуморальные факторы, защищающие плазму крови от инфек-
ции, — лейкины, плакины и р-лизины плазмы крови и тканевой жидкости.
Лейкины (выделяются лейкоцитами) и плакины (секретируются в плазму
крови тромбоцитами) оказывают отчетливое бактериологическое действие.
Например, нейтрофильные лейкоциты при дегрануляции выделяют в
кровь, интерстициальную жидкость противомикробные пептиды — дефенси-
ны и кателицидины, низкомолекулярные катионные пептиды, поражающие
грамположительные микроорганизмы, грибки, вирусы. Выраженным лити-
ческим эффектом на стафилококки и анаэробные микроорганизмы обла-
дают fy-лизины плазмы крови, секретируемые тромбоцитами. Ныне они
идентифицированы как тромбоцитарные антибактериальные катионные
белки. Ферментативную активность микроорганизмов и вирусов подавляют
2*
36 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Таблица 1.4. Иммуноглобулины, содержащиеся в сыворотке крови здорового человека
Свойства и функции иммуног- лобулинов IgGl IgG2 lgG3 IgG4 IgM IgAl lgA2 IgD IgE
Концентрация в сыворотке крови (мг/дл) 90 0 0 0,5 5 30 0,5 0,3 0,0005
Величина ежедневного синте- за (мг/кг массы тела в день) 3 3 3 3 3 4 4 0,4 0,002
Продолжительность полужиз- ни (сут) 1 0 7 1 0 6 6 3 2
Способность проникать в ор- ганизм плода через плаценту Способность связывать и ак- + + + + + -
тивировать тучные клетки и базофильные лейкоциты Способность связываться и - - - - - - +++
активировать клетки — моно- нуклеары + — + —
Молекулярная масса (кДа) 46 46 70 46 70 60 60 94 199
ингибиторы гиалуронидазы, фосфолипаз, коллагеназы, плазмина и интер-
ферон лейкоцитов.
Значительная часть белков плазмы (p-глобулины) представлена белками
свертывающей, антисвертывающей и фибринолитической систем крови,
являющихся составными частями системы гемостаза, т. е. системы, сохра-
няющей жидкое состояние крови, поддерживающей целостность стенки
сосудов и останавливающей кровопотерю образованием тромба в месте по-
вреждения сосуда (см. главу 7).
Главной функцией у-глобулинов является обеспечение гуморального
иммунитета. С помощью иммунологического метода у-глобулины разделя-
ют на 5 классов иммуноглобулинов (Ig): IgG, IgA, IgM, IgD, IgE, являю-
щихся антителами (т. e. специфическими веществами, образующимися
при поступлении в организм человека чужеродных веществ — бактерий,
вирусов, токсинов — и нейтрализующих их) (табл. 1.4).
Около 75 % всех иммуноглобулинов в крови представлены IgG. Их об-
щая концентрация в сыворотке крови взрослого человека составляет 6,32—
14,79 г/л. IgG образуют антитела против подавляющего числа возбудителей
инфекций и в свою очередь представлены еще четырьмя субклассами —
IgGl, IgG2, IgG3, IgG4 (см. табл. 1.4). Все субклассы IgG могут проникать
через плаценту к плоду из организма матери и обеспечивать новорожден-
ному иммунную защиту против инфекции.
Содержание IgA в сыворотке крови составляет 20 % от общей массы
иммуноглобулинов (0,89—3,14 г/л). IgA-антитела обильно представлены в
различных секреторных жидкостях организма — слюне, кишечном соке,
трахеобронхиальных и мочеполовых секретах, молоке — и обеспечивают
секреторный иммунитет.
IgM-антител содержится 0,59—2,91 г/л. Эти антитела могут активиро-
вать систему комплемента.
IgE-антитела участвуют в антипаразитарной защите организма, аллерги-
ческих реакциях. IgE прикрепляются к специфическим рецепторам на
мембране тучных клеток и базофильных лейкоцитов и, связываясь с анти-
генами (чужеродные вещества, на которые вырабатывались данные антите-
ла), вызывают высвобождение из этих клеток биологически активных ве-
ществ.
1. Жидкие среды организма • 37
IgD-антитела выполняют функцию мембранных рецепторов для антиге-
нов на поверхности В-лимфоцитов, участвуя таким образом в регуляции
иммунного ответа.
Поскольку продолжительность жизни иммуноглобулинов не велика
(особенно иммуноглобулинов A, D —см. табл. 1.4), то и в здоровом орга-
низме человека интенсивность их синтеза значительна. Например, имму-
нокомпетентными клетками кишечника за сутки синтезируется 3 г IgA.
В плазме крови имеются белковые молекулы, представляющие собой
«плавающие» рецепторы многих гемопоэтических гормонов и цитокинов
(эритропоэтина, тромбопоэтина, интерлейкинов, колониестимулирующих
факторов) — биологически активные вещества, секретируемые клетками
крови, фибробластами и др. и регулирующие различные функции организ-
ма (кроветворение, регенерацию тканей и др.). Клетки, продуцирующие
цитокиновые рецепторы, производят их в двух формах — в растворимой,
секретируемой в плазму крови, и в виде белковых компонентов, встраивае-
мых в мембрану клетки. Растворимые формы рецепторов действуют как
белки, соревнующиеся с рецепторами клеток за связывание лиганда, регу-
лируя таким образом действие цитокинов на клетки-мишени.
1.4. Факторы, обеспечивающие жидкое состояние крови
Жизнедеятельность клеток организма, транспорт питательных веществ,
продуктов обмена и газов кровью, протекающие в ней биохимические и
иммунные реакции могут осуществляться только при сохранении жидкого
состояния крови. Поэтому они нарушаются при агрегации (склеивании
друг с другом) клеток крови. Поддержание крови в жидком состоянии
обеспечивается системой гемостаза (сбалансированным соотношением
компонентов свертывающей, антисвертывающей и фибринолитической
систем крови — см. главу 7), электрохимическими свойствами клеток и
белков крови. Электрохимические заряды расположены на поверхности
или вокруг этих структур крови и сохраняют эритроциты, другие клетки
крови во взвешенном состоянии в кровеносном русле, обеспечивают ус-
тойчивость дисперсии белков в плазме крови. Поэтому поток крови в со-
судах представляет собой движение отрицательно заряженных частиц (кле-
ток крови, белков плазмы), что приводит к формированию электростати-
ческого распора, обеспечивающего расстояние между клетками и коллоид-
ными элементами крови, препятствующее их агрегации. Препятствуют аг-
регации клеток в потоке крови также силы поверхностного натяжения
мембран клеток, гидродинамические (механические) силы, перемещающие
клетки в направлении тока крови. Солидизации (соединению клеток крови
в агрегаты различной величины) способствуют снижение энергии движу-
щейся по сосудам крови, например в результате ухудшения работы сердца,
нарушение межмолекулярных отношений в мембранах эритроцитов, сни-
жение их деформабельности (способности клеток изменять и восстанавли-
вать форму), силы гравитации. При непродолжительном замедлении кро-
вотока агрегация эритроцитов обратима: при восстановлении кровотока
образовавшиеся агрегаты эритроцитов — «монетные столбики» — распада-
ются на функционально полноценные клетки.
Коллоидная стабильность плазмы крови обусловлена гидратацией белко-
вых молекул и наличием поверхностного электрокинетического «^-потен-
циала («^-потенциал — часть ср-потенциала, создаваемого двойным электри-
ческим слоем ионов на поверхности белковых молекул), т. е. потенциала
поверхности скольжения частицы в коллоидном растворе, который создает
38 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
одноименные заряды и силы электростатического отталкивания белковых
молекул и клеток крови. Это препятствует агрегации клеток и поддержива-
ет коллоидную устойчивость крови. Величина ^-потенциала выше у альбу-
минов, чем у других белков плазмы крови. Поэтому альбумины, которые ад-
сорбируются на поверхности эритроцитов, поддерживают клетки в плазме
крови во взвешенном состоянии (т. е. поддерживают суспензионные свойст-
ва крови). Крупнодисперсные белки (фибриноген, глобулины) снижают
суспензионные свойства крови. Эти свойства крови оценивают по скоро-
сти оседания эритроцитов (СОЭ) в ее неподвижном объеме, которая у
мужчин равна 4—10 мм/ч, у женщин — 5—12 мм/ч. У новорожденных ско-
рость оседания эритроцитов замедлена в сравнении с ее значениями у
взрослых людей: она равна 1 мм/ч. Объясняется это низким (по сравне-
нию со взрослыми) содержанием у-глобулинов в плазме крови новорож-
денных.
Скорость оседания эритроцитов возрастает при увеличении в крови
уровней фибриногена, а2,- 0- и у-глобулинов, получивших в силу этого
свойства название агломеринов. Агломерины уменьшают отрицательный
заряд эритроцитов (^-потенциал). Уменьшение последнего приводит к
формированию агломератов эритроцитов, которые оседают быстрее, чем
отдельные клетки крови. Альбумины плазмы крови, содержащийся в ней
лецитин замедляют СОЭ. Нарастание в плазме крови содержания холесте-
рола, снижение соотношения в ней фракции альбуминов и глобулинов
(белкового коэффициента альбумины/глобулины, в норме равного 1,3—
2,3) увеличивают СОЭ. На величину СОЭ влияет и масса формирующихся
клеточных агломератов. Она оказывается тем больше, чем больше площадь
поверхности эритроцита, поэтому макроциты (эритроциты с большим, чем
в норме, диаметром) увеличивают и СОЭ. Увеличение СОЭ имеет место
при гиперфибриногенемии, при воспалительных заболеваниях оно связано
с гиперглобулинемией.
Вязкость определяют как свойство жидкости оказывать сопротивление
при смещении ее слоев относительно друг друга.
Вязкость плазмы или цельной крови — это сопротивление течению
жидкости при взаимном перемещении ее слоев и частиц за счет внутрен-
него трения. Вязкость цельной крови создается взаимодействием воды
плазмы крови, макромолекулярных коллоидов (белков плазмы, липопро-
теидов и др.) и форменных элементов крови. Вязкость воды составляет
1,008 сантипуаз (сПз). Вязкость плазмы колеблется между 1,7 и 2,2 сПз,
цельной крови — от 4 до 5 сПз, т. е. вязкость плазмы в 1,8—2,5 раза, а
цельной крови — в 4—5 раз выше, чем воды. Повышению вязкости крови
способствуют: увеличение в ней содержания фибриногена, липопротеинов;
повышение концентрации углекислоты (поэтому вязкость артериальной
крови ниже, чем венозной); рост гематокрита (при повышении гематокри-
та до 60—70 вязкость крови возрастает в 10—20 раз по сравнению с вязко-
стью воды); снижение суспензионных свойств крови за счет агрегации
эритроцитов. При этом рост вязкости крови всегда усиливает агрегацию
эритроцитов. Величина, обратная вязкости, называется текучестью, поэто-
му с увеличением вязкости крови ее текучесть уменьшается.
Ток крови в сосудах с диаметром меньше 1,5 мм характеризуется мень-
шей вязкостью, чем в сосудах более крупного диаметра (эффект Фареуса—
Линдквиста). Этот эффект связан с перемещением эритроцитов к центру
движущейся струи крови («центровкой» эритроцитов) при прохождении ее
через сосуд с диаметром меньше 1,5 мм и с формированием пристеночных
слоев плазмы крови. Пристеночные же слои плазмы обладают низкой вяз-
1. Жидкие среды организма • 39
костью. Вязкость крови резко возрастает, как только снижается скорость
ее тока. Так, при снижении скорости тока крови в мелких сосудах ниже 1
мм/с, вязкость крови возрастает почти в 10 раз, что связано с увеличиваю-
щейся агрегацией эритроцитов и смещением их к стенке сосуда.
Удельный вес плазмы крови колеблется от 1024 до 1030. Удельный вес
эритроцитов составляет 1094—1107. Поэтому удельный вес цельной крови в
норме варьирует от 1052 до 1064, так как он зависит от количества эритро-
цитов, содержания в них гемоглобина и состава плазмы. Например, повы-
шение гематокрита увеличивает удельный вес крови.
1.5. Лимфа как внутренняя среда организма
Лимфа образуется из жидкости, оттекающей из интерстициального про-
странства в кровь по лимфатическим сосудам. В лимфатических сосудах
она проходит через лимфатические узлы, где ее состав меняется за счет
поступления в лимфу в основном лимфоцитов. Поэтому различают пери-
ферическую (до лимфатических узлов), промежуточную (прошедшую через
лимфатические узлы) и центральную лимфу грудного лимфатического про-
тока.
Основные функции лимфы: 1) поддержание постоянства состава и объе-
ма интерстициальной жидкости и микросреды клеток; 2) возврат белка из
интерстициальной жидкости в кровь; 3) участие в перераспределении жид-
кости в организме; 4) обеспечение гуморальной связи между тканями и ор-
ганами, лимфоидной системой и кровью; 5) всасывание и транспорт из же-
лудочно-кишечного тракта в кровь продуктов гидролиза пищи, особенно
липидов; 6) транспорт антигенов и антител, перенос из лимфоидных орга-
нов плазматических клеток, иммунных лимфоцитов и макрофагов в кровь.
Количество, состав и свойства лимфы. В теле человека в среднем цирку-
лирует 1,5—2,0 л лимфы. Лимфа состоит из лимфоплазмы и форменных эле-
ментов белой крови. В центральной лимфе отношение объема форменных
элементов к общему объему (лимфокрит) менее 1 %. Удельный вес лимфы
колеблется от 1010 до 1023; pH находится в диапазоне 8,4—9,2. Осмотиче-
ское и онкотическое давление и вязкость лимфы ниже благодаря меньше-
му, чем в плазме крови, содержанию белка и ионов Na+.
Химический состав периферической лимфы зависит от ее источников.
Лимфа, оттекающая от кишечника, содержит от 30 до 40 г/л белка, богата
жирами (до 40 г/л), оттекающая от печени — содержит больше белков (до
60 г/л) и углеводов (до 1,3 г/л).
Центральная лимфа имеет следующий белковый и ионный состав:
Белковые фракции
Альбумины (г/л)
Глобулины (г/л)
«[-Глобулины (%)
а2-Глобулины (%)
р-Глобулины (%)
у-Глобулины (%)
Фибриноген (г/л)
Протромбин (%)
Общий белок (г/л)
Содержание Ионы
15,0-40,0 Na+
10,0-16,1 К+
2,9-9,1 Са++
5,2-11,0 Mg+
6,7-17,7 10,0-23,8 1,5-4,6 30,0-78,7 25,0-56,1 сг
Концентрация (ммоль/л)
114,3-137,5
3,6-5,8
2,0-3,1
0,6-1,5
92,0-140,7
Альбуминоглобулиновый коэффициент лимфы приближается к 3, что
отражает значительно большую долю содержащихся в лимфе альбуминов,
чем глобулинов, по сравнению с плазмой крови — в последней альбуми-
ноглобулиновый коэффициент равен 1,3—2,3.
40 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Клетки лимфы представлены, прежде всего, лимфоцитами (от 1 до 22
на 109/л) и моноцитами. Гранулоцитов в лимфе мало, а эритроциты у
здорового человека в лимфе отсутствуют. Появление эритроцитов в лим-
фе — диагностический признак повышенной капиллярной проницае-
мости.
Соотношение отдельных видов лейкоцитов в лимфе получило название
лейкоцитарной формулы лимфы. Она выглядит следующим образом: лимфо-
цитов — 90 %, моноцитов — 5 %, сегментоядерных нейтрофилов — 1 %, эо-
зинофилов — 2 %, других клеток —2%. В лимфе могут формироваться
тромбы благодаря присутствию в ней тромбоцитов (5—35 • 109/л), фибри-
ногена и других белковых фракций. Время свертывания — 10—15 мин. Об-
разование фибриновых тромбов в лимфатических сосудах предупреждает
распространение по ним инфекции из очага воспаления.
1.6. Механизм образования лимфы
Большая часть жидкости, профильтровавшейся в артериальном конце ка-
пилляров в интерстициальное пространство, реабсорбируется в их веноз-
ном конце, перенося из интерстиция в кровь продукты конечного обмена,
ионы и др. Однако около */10 объема этой жидкости возвращается из ин-
терстициума в кровь через лимфатическую систему. Возможность возврата
интерстициальной жидкости по лимфатическим путям в кровь крайне
важна, так как содержащиеся в ней белки, в небольшом количестве посту-
пающие из артериальной части капилляров в межклеточное пространство,
не могут возвращаться в капилляр в его венозной части, но беспрепятст-
венно поступают в начальный отдел лимфатической системы — лимфати-
ческие капилляры. Лимфатические капилляры начинаются слепыми кон-
цами, «погруженными» в интерстиций (см. рис. 1.3). Выстилающие стенку
лимфатического капилляра эндотелиальные клетки прикреплены коллаге-
новыми волокнами, филаментом к окружающей капилляр соединительной
ткани, благодаря чему просвет лимфатического капилляра остается откры-
тым. «Конструкция» стенки лимфатического капилляра организована так,
что в местах соединения соседствующих эндотелиальных клеток край од-
ной клетки прикрывает свободно свисающий в просвет капилляра край
другой клетки, формируя маленький клапан. Под давлением интерстици-
альной жидкости клапан открывается внутрь капилляра и жидкость вместе
с содержащимися в ней молекулами белка, другими суспензированными в
ней частицами поступает в лимфатический капилляр. Размер пор, прикры-
ваемых краями таких клапанов, позволяет проходить через них крупным
молекулам белков плазмы, вышедшим из крови в интерстициальное про-
странство. Жидкость не может покидать капилляр через эту же щель между
эндотелиальными клетками, так как ее обратное движение закрывает
створку клапана. Лимфа из лимфатического капилляра оттекает в более
крупные лимфатические коллекторы, в которых также имеются клапаны,
располагающиеся в нескольких миллиметрах один от другого по току лим-
фы и препятствующие ее обратному движению.
На отток лимфы по лимфатическим капиллярам влияют: 1) величина
давления интерстициальной жидкости, 2) автоматические сокращения
гладких мышц стенок лимфатических коллекторов и больших лимфатиче-
ских сосудов, 3) ритмические сдавления лимфатических сосудов внешни-
ми тканями, 4) допускается также, что перекачиванию жидкости из лим-
фатических капилляров способствует сокращение их эндотелиальных кле-
ток, содержащих сократительный филамент актомиозин.
1. Жидкие среды организма • 41
На отток лимфы влияет величина давления интерстициальной жидко-
сти. Отток лимфы из капилляров, слабый при давлении интерстициальной
жидкости, меньшем атмосферного на 6 мм рт. ст. (в нормальных условиях
давление интерстициальной жидкости на 3 мм рт.ст. ниже атмосферного),
при нарастании интерстициального давления до 0 мм рт. ст. возрастает в
20 раз. Когда же давление интерстициальной жидкости становится на 1—2
мм рт. ст. выше атмосферного нарастание оттока лимфы из тканей прекра-
щается, поскольку возросшее давление интерстициальной жидкости сдав-
ливает лимфатические сосуды.
Поэтому факторы, увеличивающие давление интерстициальной жидко-
сти, такие как повышенное гидростатическое давление крови в капилляре,
уменьшенное онкотическое давление плазмы крови, повышенная концен-
трация белка в интерстициальной жидкости, увеличивая объем и давление
последней на лимфатические капилляры, одновременно усиливают отток
из них лимфы. Другие факторы, регулирующие отток лимфы, рассматри-
ваются в главе 9.
1.7. Трансцеллюлярные жидкости организма
Состав этих жидкостей имеет отличие от состава плазмы крови, что отра-
жает разные функциональные задачи, выполняемые этими жидкостями в
различных полостях организма, о чем можно судить на примере спинно-
мозговой и синовиальной жидкостей.
Спинномозговая жидкость (цереброспинальная жидкость, ликвор) обра-
зуется секрецией сосудистыми сплетениями головного мозга, мягкой моз-
говой оболочкой, глией, эпендимой желудочков мозга. За сутки ее форми-
руется до 600 мл. Однако количество спинномозговой жидкости у взросло-
го человека составляет 100—150 мл благодаря постоянному обратному вса-
сыванию жидкости арахноидальными грануляциями и оттоку ее по веноз-
ным синусам подпаутинного пространства.
Спинномозговая жидкость заполняет пространства между паутинной и
мягкой оболочками мозга, все желудочки мозга, четвертый из которых со-
единяется с центральным каналом спинного мозга. Спинномозговая жид-
кость бесцветна, прозрачна, на 90 % состоит из воды, плотность ее состав-
ляет 1,002-1,008; pH 7,35-7,8.
В составе ликвора представлены белки, аминокислоты, углеводы (глю-
коза, фруктоза), ионы натрия и хлора, гормоны, олигопептиды, ферменты,
СО2. Спинномозговая жидкость содержит белков в 200—400 раз меньше
(15—33 мг%), чем сыворотка крови (60—80 г%), благодаря низкой прони-
цаемости для белковых молекул гематоэнцефалического барьера. Увеличе-
ние содержания белка в спинномозговой жидкости указывает на органиче-
ское заболевание нервной системы (менингиты, компрессии спинного
мозга). Содержание глюкозы в ликворе несколько ниже, чем в крови, — 70
мг%. Ее увеличение в ликворе отмечено при диабете.
Спинномозговая жидкость защищает мозг от внешних механических
воздействий, играя роль своеобразной «подушки» (мозг оказывается как
бы взвешенным в ликворе, который играет по отношению к нему роль
гидравлического амортизатора). Ликвор поддерживает метаболизм нервной
ткани, питая ее глюкозой, аминокислотами, кислородом и устраняя мета-
болиты — СО2, избыток воды. В ликворе содержатся клетки крови, пред-
ставленные только лимфоцитами (не более 3 в 1 мкл), обеспечивающими
защиту ликвора и мозга от чужеродных веществ, микроорганизмов в слу-
чае их проникновения через гематоэнцефалический барьер. Увеличение
42 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
содержания лейкоцитов в ликворе (плеоцитоз) является признаком заболе-
вания центральной нервной системы (менингиты, полиомиелиты).
Синовиальная жидкость представляет собой водный раствор, заполняю-
щий полости суставов, синовиальные влагалища сухожилий, синовиальные
сумки и содержащий многие из входящих в состав плазмы крови веществ.
Но синовиальная жидкость, в отличие от плазмы крови, содержит и очень
важный для функции суставов биополимер — гликозаминогликан-гиалуро-
новую кислоту, входящий в состав белково-полисахаридного комплекса
суставной жидкости. Синовиальная жидкость вырабатывается синовиаль-
ными клетками — синовицитами. Ее количество в суставах (даже круп-
ных — коленном, тазобедренном) не превышает 4 мл. На 95 % эта жид-
кость состоит из воды, ее pH равен 7,7. Присутствие гиалуроновой кисло-
ты обеспечивает вязкоупругие свойства синовильной жидкости, формирует
защитный слой на поверхности хряща суставов, при движении предотвра-
щающий от повреждения их трущиеся поверхности. Уменьшение содержа-
ния гликозаминогликана в суставной жидкости, например при ревматиче-
ских заболеваниях суставов (ревматоидном артрите, деформирующем ос-
теоартрозе), ухудшает ее вязкоупругие и защищающие суставную поверх-
ность свойства, приводит к микротравматизации хрящей и их механиче-
скому разрушению. Синовиальная жидкость, благодаря содержащимся в
ней глюкозе, белкам, поддерживает трофику лишенных кровеносных сосу-
дов хрящей суставов. Содержащиеся в суставной жидкости небольшие ко-
личества лимфоцитов, моноцитов — макрофагов поддерживают ее гомео-
стаз. Количество лейкоцитов в синовиальной жидкости возрастает при за-
болевании суставов, в связи с чем имеет важное диагностическое значение.
1.8. Обмен жидкостей между водными секторами в организме
человека
Внеклеточные жидкости организма являются транспортной системой,
обеспечивающей постоянный обмен между различными водными сектора-
ми организма — из желудочно-кишечного тракта вода с растворенными в
ней веществами поступает в плазму крови, из нее — во внеклеточный и да-
лее — во внутриклеточный сектор. Из клеток вода с растворенными в ней
продуктами обмена, СО2 поступает во внеклеточный сектор, затем в кровь
и лимфу, которая доставляет ее к органам выделения — желудочно-кишеч-
ному тракту, почкам, легким, потовым железам кожи.
Исследования, проведенные с использованием «тяжелой» воды (D2O)
или воды, меченной по тритию, которые при введении в организм равно-
мерно распределяются во всех водных секторах организма, показали, что
интенсивность ее обмена может изменяться. Она увеличивается, например,
выше нормального уровня у человека, выполняющего физическую работу
на жаре: скорость обмена воды в организме, определенная у таких обсле-
дованных с помощью меченной по тритию воды, демонстрирует возраста-
ние скорости ее полного обмена с 27 сут, регистрируемой в норме, до 16
сут. В организме человека имеет место и непрерывное перемещение раз-
личных ионов с водой из плазмы крови в интерстициальную жидкость, из
нее — во внутриклеточную, а затем наоборот — из клеток в интерстициаль-
ную жидкость и далее — в плазму крови и лимфу, доставляющие их к орга-
нам выделения. Использование изотопов К, Mg, т. е. химических элемен-
тов, в наибольших концентрациях содержащихся во внутриклеточном сек-
торе (см. раздел 1.2.3 данной главы), обнаружило, что скорость их обмена
зависит от функционального состояния клеток. Например, в ответ на по-
1. Жидкие среды организма • 43
вышение возбудимости нервных клеток скорость обмена ионов К, Mg воз-
растает.
Обмен между внеклеточной и внутриклеточной жидкостями осуществ-
ляется через клеточные мембраны. В основе обмена воды и растворенных
в ней веществ через мембраны клеток лежат разные формы трансмембран-
ного транспорта — пассивного (простая диффузия, облегченная диффузия,
фильтрация) и активного.
Простая диффузия зависит от толщины мембраны клеток (их толщина
варьирует от 5 до 13,5 нм), размеров пор мембраны, разницы величины
электрического заряда между внутренней и наружной поверхностью мем-
браны, величины химического градиента для веществ по разные стороны
мембраны, величины гидрофобных участков мембраны (они обеспечивают
быстрое прохождение через мембрану более жирорастворимых веществ —
например, СО2 более жирорастворим, чем О2 и молекулы СО2 быстрее
проходят через мембраны клеток, чем О2). Величина пассивной диффузии
зависит от наличия положительных зарядов в гидрофильных порах мем-
бран клеток, которые облегчают прохождение через мембрану анионов и
ограничивают — катионов (например, через поры мембраны эритроцитов
легко проходят СГ, HCOj и много труднее — Na+). На величину пассивной
диффузии влияет гидростатическое давление жидкости на мембрану клет-
ки (например, в почках гидростатическое давление крови в капиллярах
клубочка на мембраны клеток, образующих стенку капсулы Боумона, яв-
ляется главной силой, обеспечивающей фильтрацию до 170 л воды в сутки
с растворенными в ней веществами из плазмы крови в просвет капсулы
Боумона).
Облегченная диффузия характеризуется большей, чем при простой диф-
фузии, скоростью прохождения молекул вещества через мембрану клетки с
помощью специальных белков — переносчиков. Например, молекулы глю-
козы в три раза быстрее проходят через мембрану эритроцита человека,
чем это следовало бы из механизма простой диффузии, связанной с разни-
цей концентрации глюкозы в плазме крови и эритроците. В мембране
эритроцита перенос глюкозы из плазмы крови внутрь этой клетки осуще-
ствляют около 800 тыс. участков, ответственных за ее перенос, и каждый
из них за секунду переносит 180 молекул глюкозы.
На направление движения воды между внеклеточным и внутриклеточ-
ным секторами влияет не общее осмотическое давление внеклеточной
жидкости, а ее эффективное осмотическое давление, которое создается в
ней концентрацией плохо проходящих через клеточную мембрану веществ.
Так, внеклеточная жидкость содержит большое количество Na+, поскольку
Na+ плохо проходят через клеточную мембрану в клетку из внеклеточной
жидкости. Кроме того, в нее благодаря непрерывной работе №+-К+-АТФ-
азе (т. е. с помощью активного транспорта) переносятся из клетки ионы
натрия.
Во внутриклеточной среде эффективное осмотическое давление созда-
ется внутриклеточными белками-протеинатами — анионами, которые, бла-
годаря эффекту Даннона (см. раздел 1.3.2), увеличивают концентрацию од-
новалентных катионов вокруг их молекул, создавая осмотическую силу для
движения воды в клетку. Увеличение во внеклеточной жидкости концен-
трации ионов натрия повышает ее эффективное осмотическое давление,
что приводит к выходу воды из клетки. Напротив, уменьшение эффектив-
ного осмотического давления внеклеточной жидкости, связанное с умень-
шением в ней концентрации Na+ вызывает перемещение воды в клетку.
Таким образом, Na+ является основным внеклеточным катионом, опре-
44 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
деляющим объем внеклеточной жидкости в организме человека. Поэтому
механизмы осморегуляции, контролирующие осмотическое давление вне-
клеточной жидкости, контролируют и объем внеклеточной жидкости в ор-
ганизме, поддерживая в ней концентрацию Na+ на постоянном уровне
(142 ммоль/л), позволяя отклоняться этой величине в норме в узких пре-
делах—от 138 до 146 ммоль/л. Так, увеличение концентрации Na+ выше
нормальной величины во внеклеточной жидкости, воспринимаемое осмо-
рецепторами, вызывает у человека чувство жажды, а почки концентрируют
мочу, уменьшая выведение воды из организма. Выпитая человеком вода
поступает во внеклеточное пространство и возвращает концентрацию Na+
к его нормальному содержанию во внеклеточной жидкости. Возбуждение
осморецепторов уменьшается, в результате чувство жажды у человека уст-
раняется, выведение воды с мочой становится нормальным.
ГЛАВА 2
Физиология возбудимых тканей
Действие внешней среды на биологические системы — организмы, органы,
ткани и клетки — всегда вызывает в них физико-химические изменения.
Способность живой системы реагировать на внешнее воздействие процес-
сом изменения физико-химических свойств является универсальным про-
явлением жизнедеятельности всех без исключения биологических систем и
называется раздражимостью. Некоторые живые ткани и клетки при дейст-
вии на них внешней среды отвечают проявлением определенной специфи-
ческой для данной живой системы функции, или возбуждением. Если жи-
вая система способна приходить в возбуждение при изменении состояния
внешней среды, она обладает возбудимостью, а ткани и клетки, имеющие
такое свойство, относятся к возбудимым.
Возбудимыми тканями организма человека являются нервная и мышеч-
ная, которые в ответ на воздействие внешней среды переходят из состоя-
ния покоя в состояние возбуждения (генерация нервного импульса, сокра-
щение и др.). Такой сложный физиологический процесс, как сокращение,
а равно вызывающий его нервный импульс, называются возбуждением.
Основной структурой возбудимых тканей (равно как секреторной или
эпителиальной) является клетка. Клеточные компоненты (цитоплазма, ор-
ганеллы) окружены единой клеточной мембраной, или плазмолеммой. Эта
мембрана представляет собой барьер между возбудимой клеткой и интер-
стициальной средой возбудимой ткани. Через мембрану возбудимой клет-
ки осуществляются все процессы, обеспечивающие ее жизнедеятельность,
и одновременно мембрана сохраняет состав внутриклеточной среды на от-
носительно постоянном уровне, несмотря на изменения в окружающей
интерстициальной среде. Клетки возбудимых тканей через свою мембрану
могут взаимодействовать с окружающей их средой и обе эти клеточные
функции — обмена и взаимодействия — обусловлены наличием белков в
пределах мембраны возбудимых клеток.
2.1. Строение и физиологические функции мембраны клеток
возбудимых тканей
Клетки возбудимых тканей отделены от интерстициальной среды клеточ-
ной мембраной. Мембрана нервной и мышечной клетки представляет со-
бой эластичную структуру, толщиной 6—8 нм, состоящую из липидов и
белков. Липиды мембраны возбудимых клеток представлены различными
фосфолипидами: фосфатидилхолин, фосфатидилэтаноламин, фосфатидил-
серин, сфингомиелин и фосфатидилинозит. Фосфолипиды образуют двой-
ной слой, который состоит из фосфолипидных молекул. Полярные голов-
ки этих молекул обращены наружу. К каждой полярной головке прикреп-
лены две углеводородные цепочки, которые погружены в толщу мембраны
(рис. 2.1). Обращенные наружу полярные головки фосфолипидных моле-
кул клеточных мембран придают им гидрофильные свойства, а удерживаю-
46 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Мембрана
Интегральные белки
6—8 нм
Ионный канал Ионный насос
Периферический белок
Углеводородные
связи
Внутренняя
поверхность
Наружная
поверхность
Полярные головки
фосфолипидных
молекул
Рис. 2.1. Схематическое изображение мембраны клетки возбудимых тканей.
Мембрана представляет собой двойной липидный слой, в который погружены интегральные
белки, функционирующие как ионные насосы и ионные каналы. Ионные насосы за счет
энергии АТФ перекачивают ионы К+, Na+, Са2+ против их концентрационного градиента. На-
личие градиентов концентрации указанных ионов на мембране возбудимых клеток является
основой возникновения на мембране клеток электрического потенциала и возбудимости нерв-
ной и мышечной ткани. Ионные каналы мембраны возбудимой клетки в зависимости от их
зактытого или открытого состояния обусловливают возникновение возбуждения в нервной и
мышечной ткани и его распространение по поверхности мембран клеток. Периферические
белки расположены, как правило, на внутренней поверхности мембраны и являются энзима-
ми, участвующими в регуляции метаболических процессов в возбудимых клетках.
щие их довольно жестко связи углеводородных цепей — гидрофобные. Бел-
ки мембран возбудимых клеток подразделяются на два типа: интегральные
и периферические. Молекулы интегральных белков погружены в толщу
мембраны на большую или меньшую глубину. Этот тип белков обеспечи-
вает взаимодействие (интеграцию) между клетками возбудимых тканей.
Интегральные белки выполняют функцию специальных ионных насосов в
мембране клеток. Например, с помощью ионных насосов различные ионы
(Na+, К+, Са2+ и др.) перемещаются через мембрану между интерстициаль-
ной и внутриклеточной средами, что лежит в основе формирования элек-
трических явлений в возбудимых клетках. Периферические белки располо-
жены, как правило, с внутренней стороны мембраны. Они являются ката-
лизаторами протекающих в клетке химических реакций, т. е. являются эн-
зимами. Периферические белки формируют цитоскелет клеток, который
придает мембране клеток прочность и одновременно гибкоэластические
свойства.
Мембрана клеток возбудимых тканей имеет функции избирательной
проницаемости для ионов и веществ, межклеточного взаимодействия, а
также экзо- и эндоцитоза, благодаря которым в нервной и мышечной тка-
ни осуществляются физиологические процессы возбуждения, проведения
возбуждения и сокращения.
2.1.1. Транспорт веществ через клеточную мембрану
Клеточная мембрана является разделительным барьером между цитоплаз-
мой и внеклеточной средой. При этом важнейшая функция подобного
барьера связана с транспортом воды, различных ионов (Na+, К+, СГ, Са2+
и др.) и питательных веществ (например, глюкозы) между этими средами с
целью обеспечения жизнедеятельности клеток.
2. Физиология возбудимых тканей • 47
2.1.1.1. Движение воды через мембрану клеток
Вода довольно свободно проникает через мембрану большинства клеток,
так как внутри клеточной мембраны имеются свободные пространства,
обусловленные временными дефектами или кинками в СН2 цепях фосфо-
липидов. Кинки и связанные с ними пространства свободно диффундируют
в пределах мембраны. Вода, заключенная в свободно диффундирующие
пространства, способна проникать с их помощью через биологические
мембраны. Благодаря кинкам и свободным пространствам внутри мембран
их «водная» проницаемость примерно в три раза выше, чем в мембранах,
организованных только из кристаллической липидной фазы. Однако дви-
жение воды через мембрану внутрь клетки и из клетки наружу регулирует-
ся осмотическим давлением растворов по обе ее стороны.
2.1.1.2. Осмос
Вода диффундирует через мембрану из области меньшей в область боль-
шей концентрации растворенного вещества. Этот процесс называется ос-
мосом. Наименьшее гидростатическое давление, которое необходимо при-
ложить к раствору для того, чтобы предотвратить перетекание раствори-
теля через полупроницаемую мембрану в раствор с большей концентра-
цией вещества называется осмотическим давлением. Концентрация осмо-
тически активных молекул в единице массы воды называется осмолярно-
стью или осмотическим коэффициентом. Осмотическое давление обуслов-
ливает поступление воды в клетку, формирует ее объем и упругость (тур-
гор) ткани.
Тоничность раствора представляет собой осмотическое давление, произ-
водимое раствором через проницаемую мембрану, отделяющую его от
плазмы крови. Терминами изо-, гипо- и гипертонический характеризуют
растворы, осмолярность которых соответственно равна, меньше или выше,
чем таковая в плазме крови.
2.1.1.3. Диффузия
Процесс свободного движения молекул вещества в пределах раствора на-
зывается диффузией. Большие молекулы движутся в растворе медленнее,
чем маленькие, а движение молекул вещества в воде происходит быстрее,
чем в среде с более высокой вязкостью. Молекулы биологически важных
веществ (медиаторы, гормоны), О2 и СО2 при температуре окружающей
среды 30 °C преодолевают расстояние 1 мкм в течение 1 мс, 100 мкм — в
течение 10 с, а расстояние 1 см — в течение 24 ч. В клетках возбудимых
тканей диффузия молекул в цитоплазме лимитирована их размером. В ин-
терстициальной среде расстояние диффузии молекул веществ ограничено
дистанцией между клетками либо между клетками и капиллярами. Так, в
среднем расстояние между капиллярами и клетками тканей у млекопитаю-
щих животных составляет 30 мкм. Поэтому небольшие по размерам моле-
кулы О2 и СО2 преодолевают это расстояние в течение двух-трех десятков
миллисекунд, что обеспечивает обмен газами между кровью и тканями ор-
ганизма.
Диффузия молекул вещества через проницаемую мембрану осуществля-
ется по концентрационному градиенту вещества и описывается первым за-
коном Фика. Подобная диффузия носит название простой и ее величина
тем больше, чем выше температура раствора. Первый закон диффузии Фи-
48 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
ка, который отражает скорость диффузии вещества через мембрану в зави-
симости от разницы концентраций вещества по обе ее стороны, выражает-
ся следующей формулой:
Скорость диффузии = —(С7~ Се),
х
где D — коэффициент диффузии, А — площадь мембраны (см2), х — тол-
щина мембраны (см), С — концентрация вещества по обе стороны мембра-
ны (i — внутри, е — снаружи). С помощью простой диффузии большинство
субстанций не проникают через мембрану клетки. Поэтому, наряду с про-
стой диффузией, имеет место движение веществ через мембрану с помо-
щью облегченной диффузии. Облегченная диффузия происходит при уча-
стии специфических белковых молекул-переносчиков. Скорость облегченной
диффузии лимитируется количеством транспортных белковых молекул и
их подвижностью в пределах мембраны. Этот вид мембранного транспорта
имеет более высокую скорость в сравнении с простой диффузией. Перенос
веществ через мембрану с помощью облегченной диффузии обусловлен
строением переносимых молекул, которые связываются с белком-перенос-
чиком. Ингибиторы способны подавлять этот вид транспорта. Для облег-
ченной диффузии характерна специфичность белков-переносчиков к типу
молекулы переносимого вещества и количеством транспортных белков в
мембране клетки. Например, перенос глюкозы или аминокислот осуществ-
ляется строго специфичными белками-переносчиками. Вещества, которые
транспортируются через мембрану одним и тем же белком-переносчиком,
конкурируют за переносчик {явление конкуренции). Если все белки-пере-
носчики заняты транспортируемыми молекулами или ионами, то увеличе-
ние градиента концентрации вещества по обе стороны мембраны не при-
ведет к росту диффузии — явление насыщения.
2.1.1.4. Первично-активный транспорт
Перенос ионов через клеточную мембрану может происходить против их
градиента концентрации за счет энергии макроэргических соединений
(АТФ). Транспорт ионов через мембрану возбудимых клеток против кон-
центрационного градиента, обусловленный функцией транспортных
АТФаз называется первично активным. Первично активный транспорт ха-
рактерен для переноса через мембрану возбудимых клеток ионов Na+, К+
или Са2+. Транспортные АТФазы — это интегральный тип белков клеточ-
ной мембраны, поэтому ионы могут активно транспортироваться, напри-
мер, из внеклеточной среды в цитоплазму (ионы К+) или наоборот (ионы
Na+).
В клетках возбудимых тканей первично активный транспорт осуществ-
ляется с помощью трех типов АТФаз: натрий-калиевой, кальциевой и про-
тонной помпы.
Транспорт ионов Na+ и К+ через мембрану. В мембранах всех клеток орга-
низма человека и животных локализована Иа+/К+-АТФаза, или натрий-ка-
лиевый насос (рис. 2.2). Функция этого насоса заключается в поддержании
градиента концентрации ионов Na+ и К+ между цитозолем клетки и вне-
клеточной средой. В возбудимых клетках создание и поддержание подоб-
ного градиента является основным условием возникновения потенциала
покоя на мембране клеток, а также последующей генерации и распростра-
нения потенциала действия по мембране нервного волокна и мышечной
клетки.
2. Физиология возбудимых тканей • 49
NaV^-АТФаза состоит из а-
и р-белковых субъединиц, а-
Субъединица содержит места свя-
зывания для АТФ, ионов Na+ и
К+. р-Субъединица обусловлива-
ет транспортные свойства
АТФазы и играет основную роль
во встраивании ее молекулы в
мембрану клетки. Схематически
цикл активности Na’/K1-АТФазы
представлен на рис. 2.3. Na+/
К+-АТФаза существует в двух
конформационных формах — Е, и
Е2. В конформации АТФ Е, участ-
ки насоса, связывающие катионы,
обращены в цитоплазму и способ-
ны связываться с ионами Na+. В
конформации АТФ Р-Е2 участки
связывания катионов обращены
во внеклеточную среду и преиму-
щественно связываются с ионами
калия. Места связывания локали-
зованы в специальном углублении
Рис. 2.2. Натрий-калиевый насос (Na+/
К+-АТФаза) в мембране возбудимой клет-
ки.
Натрий-калиевый насос за счет гидролиза од-
ной молекулы АТФ выводит 3 иона натрия из
клетки и аккумулирует 2 иона калия внутри
клетки, чем создается градиент концентрации
этих ионов по обе стороны возбудимой мем-
браны.
Рис. 2.3. Схематическое изображение цикла работы электрогенного натрий-калие-
вого насоса.
Конформация Е| насоса обращена в цитозоль и имеет на поверхности два отрицательно заря-
женных места, с которыми у внутриклеточных ионов Na+ и К+ имеется различное сродство.
Третье электрически нейтральное место связывания ионов натрия находится внутри молекулы
насоса. После связывания ионов с насосом они погружаются внутрь его молекулы. Фосфори-
лирование насоса с помощью АТФ вызывает «закупорку» ионов натрия (Na3E,-P). Переход к
конформации Е, открывает «карман» во внеклеточную среду. После связывания двух ионов
калия процесс повторяется в обратной последовательности, только ионы калия выводятся из
клетки.
50 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Внешняя сторона
АТФ ---► АДФ+Ф
АТФаза
Мембрана
саркоплазматического
ретикулума
+ + + 2 Са++
Внутренняя сторона
Рис. 2.4. Первично активный транс-
порт ионов кальция через мембрану.
При гидролизе одной молекулы АТФ каль-
циевый насосактивно переносит два иона
кальция через мембрану, например, сарко-
плазматического ретикулума против их кон-
центрационного градиента.
канала, поэтому во время транспор-
та ионов Na+ и К+ через мембрану
клетки они окружены молекулой
насоса и не могут взаимодейство-
вать с другими ионами. За один
цикл активности насоса три иона
натрия выводятся из клетки, а
внутрь транспортируются два иона
калия.
Транспорт ионов Са! через мем-
брану. В мембране саркоплазмати-
ческого ретикулума всех типов мы-
шечных клеток локализована Са2+-
АТФаза (рис. 2.4). Функция этого
насоса заключается в поддержании
низкой концентрации ионов каль-
ция в цитоплазме мышечных кле-
ток за счет депонирования ионов в
саркоплазматический ретикулум. Поддержание низкой концентрации ио-
нов Са2+ в саркоплазме является основным условием расслабления мы-
шечных клеток (менее 10“7 моль/л, против 10“3—10"2 моль/л в ретикулу-
ме). Са++-АТФ-аза функционирует следующим образом. В присутствии
АТФ после присоединения со стороны цитоплазмы иона кальция к каль-
ций-связывающему участку Са2+-АТФаза фермент изменяет свою конфор-
мацию, и область связывания ионов Са2+ оказывается внутри саркоплаз-
матического ретикулума. При этом сродство ионов к Са2+-АТФазе умень-
шается, и ионы Са2+ высвобождаются во внутриретикулярное простран-
ство. Под действием ионов Mg2+ саркоплазматического ретикулума фер-
мент Са2+-АТФаза дефосфорилируется и кальций-связывающий участок
вновь оказывается снаружи мембраны. В такой последовательности по-
вторяется цикл работы кальциевого насоса. В саркоплазме мышечных
клеток ионы кальция принимают участие в механизме мышечного сокра-
щения либо выполняют функцию вторичного посредника (описаны
ниже).
Транспорт протонов через мембрану. Протонная помпа (Н+-АТФаза)
транспортирует протоны через внутренние мембраны митохондрий. Про-
тонная помпа в митохондриях увеличивает градиент электрохимического
потенциала ионов водорода на мембране до порогового или критического
уровня, который необходим для синтеза АТФ. Протонная помпа увеличи-
вает электрическую и концентрационную составляющую электрохимиче-
ского потенциала на мембране митохондрий.
2.1.1.5. Вторично-активный транспорт
В возбудимых клетках имеет место активный транспорт веществ через
мембрану клеток против их концентрационного градиента, если эквива-
лентное количество энергии высвобождается в результате движения друго-
го вещества по его концентрационному градиенту. Этот вид трансмем-
бранного транспорта называется вторично активным. Вторично активный
транспорт называется также сопряженным транспортом, поскольку более
одного вещества транспортируется через мембрану клетки с помощью ин-
тегрального белка-переносчика. При этом виде транспорта молекулы ве-
ществ конкурируют между собой за белок-переносчик. Например, если ве-
2. Физиология возбудимых тканей • 51
щество А равномерно распределено по обе стороны мембраны, а вещество
Б создает более высокую концентрацию по одну стороны мембраны, то
конкуренция вещества Б за переносчик по сравнению с веществом А будет
тем больше, чем выше будет градиент концентрации вещества Б. При
этом, если белок-переносчик переносит две субстанции, то это называется
котранспортом. Если два вещества переносятся через мембрану одновре-
менно в одном направлении, то это называется симпортом. Если вещества
переносятся через мембрану одновременно, но в противоположных на-
правлениях, это называется антипортом. Как правило, натрий-калиевый
насос создает градиент концентрации ионов натрия и калия по обе сторо-
ны от мембраны клеток, который обеспечивает сопряженный или вторич-
но активный транспорт.
Этот вид активного транспорта широко представлен в физиологических
процессах различных органов. В скелетных мышцах, кардиомиоцитах и
гладкомышечных клетках с функцией №+/К+-АТФазы связан транспорт
ионов Са2+ через плазматическую мембрану при участии Na+/Ca2+-обменни-
ка. Этот белок транспортирует ионы Са2+ через мембрану клеток в обмен
на ионы натрия и за счет энергии №+/К+-АТФазы. Кроме того, в мембра-
нах клеток имеется белок, обменивающий внеклеточные ионы натрия на
внутриклеточные протоны, — Nа*/Н"-обменник, который поддерживает по-
стоянство pH внутриклеточной среды. Скорость обмена ионов при участии
Na+/Ca2+- и Ма+/Н+-обменников пропорциональна электрохимическому
градиенту ионов Na+ по обе стороны мембраны. В невозбудимых клетках,
например, слизистой оболочки тонкой кишки глюкоза транспортируется в
эпителиоциты лишь в том случае, если ионы Na+ одновременно с глюко-
зой связывается с белком-переносчиком.
2.1.1.6. Эндоцитоз и экзоцитоз
Молекулы некоторых веществ, например химических передатчиков возбу-
ждения в нервной системе (медиаторов) или гормонов, не способны про-
ходит через мембрану клеток. Поэтому их движение через клеточную мем-
брану осуществляется путем эндоцитоза и экзоцитоза. При эндоцитозе суб-
станции первоначально адсорбируются специальными рецепторами на
внешней стороне мембраны. С помощью мембранного белка клатрина,
расположенного на внутренней поверхности мембраны клетки, в мембране
формируется углубление, а затем образуется везикула и везикулярный ком-
плекс «рецептор—клатрин—белок» оказывается внутри клетки. Комплекс
утрачивает рецептор, белок клатрин вновь возвращается на внутреннюю
поверхность клеточной мембраны, а поступивший внутрь белок метаболи-
зируется в клетке.
Путем экзоцитоза из клетки выводятся крупные белковые молекулы,
которые инкапсулируются мембранами эндоплазматического ретикулума в
форме везикул. Подобные везикулы, а для многих клеток они представля-
ют собой секреторные гранулы, транспортируются вдоль внутриклеточной
микротрубулярной системы к мембране клеток, сливаются с клеточной
мембраной в строго определенных местах мембраны, содержащей специ-
альный белок синапсин. Процесс слияния секреторных гранул активирует-
ся ионами Са++. Мембрана секреторной везикулы соединяется с мембра-
ной клетки, и секретируемые субстанции оказываются во внеклеточной
среде.
52 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.1.1.7. Внутриклеточный транспорт молекул
Внутри клеток везикулы и
различные субстанции так-
же транспортируются с по-
мощью специальных бел-
ков. Например, в нервных
клетках имеет место аксон-
ный транспорт везикул как в
Аксон
Сома
нейрона
◄-------
Рис. 2.5. Внутриклеточное движение органелл с
помощью белков, связанных с микротрубочка-
ми: динеин перемещает органеллы нейронов в
направлении их сомы, а кинезин — в направле-
нии нервной терминали.
направлении нервных окон-
чаний (антероградно), так и
в направлении тела нейро-
нов (ретроградно). Внутри-
клеточные транспортные
процессы осуществляются
вдоль цитоплазматических
микротрубочек при участии
специфических белков кинезина и динеина (рис. 2.5). При этом кинезин
транспортирует везикулы по микротрубочкам аксона в направлении тер-
миналей нейронов, а динеин — в направлении их сомы.
2.2. Возбудимость как основное свойство нервной и мышечной
ткани
Возбудимые ткани организма человека в ответ на действие раздражителей
способны переходить из состояния покоя в состояние возбуждения. Так, жи-
вая нервная ткань при внешних воздействиях, достигающих определенного
уровня по силе раздражения, переходит в состояние возбуждения, генери-
руя нервный импульс. Возбудимая мышечная ткань под влиянием внешних
раздражителей или нервного импульса производит сокращение. Свойство
возбудимых тканей приходить в возбуждение при изменениях внешней или
внутренней среды организма называется возбудимостью. Возбудимые ткани
способны распространять возникшее в них возбуждение, например по по-
верхности мембран мышечных клеток или по нервным волокнам нейронов.
Это свойство возбудимых тканей называется проводимостью. Наконец, мы-
шечная ткань обладает проводимостью (как и нервная), а также сократимо-
стью, поскольку способна сокращаться в ответ на раздражение.
2.2.1. Понятие о раздражении и раздражителях
Процесс возбуждения нервной и мышечной ткани возникает при их раз-
дражении факторами, или раздражителями, внешней и внутренней среды
организма. Процесс воздействия раздражителя на возбудимые ткани назы-
вается раздражением. Раздражители различаются по их биологической зна-
чимости, а также по качественному и количественному признаку. По био-
логической значимости раздражители относят к адекватным и неадекват-
ным. Адекватным считается раздражитель, к восприятию которого данная
живая ткань приспособилась в процессе эволюции. Например, для органа
слуха адекватным раздражителем являются упругие механические колеба-
ния среды. К неадекватным раздражителям относят факторы внешней или
внутренней среды, которые в естественных условиях жизнедеятельности
организма не являются источником возбуждения живой ткани. Все типы
адекватных и неадекватных раздражителей по силе их действия на возбу-
2. Физиология возбудимых тканей • 53
димые ткани подразделяются на пороговые, подпороговые, максимальные,
субмаксимальные и супермаксимальные. При действии внешних раздра-
жителей на клетки возбудимых тканей они способны возбуждаться в том
случае, если сила раздражения достигает определенной, а именно порого-
вой, величины. Минимальная сила раздражителя, необходимая для воз-
никновения возбуждения нервной или мышечной ткани, называется поро-
гом возбуждения. Раздражители, сила которых ниже порога возбуждения,
являются подпороговыми. Если сила раздражения превосходит порог возбу-
ждения, то величина ответной реакции возбудимых тканей возрастает
вплоть до определенного для каждой возбудимой ткани предела. При этом
дальнейшее увеличение силы раздражителя не приводит к росту ответной
реакции ткани. На этом основании минимальная сила раздражителя, вы-
зывающего максимальный ответ возбудимой ткани, называется максималь-
ной силой раздражения. Раздражители, сила которых меньше или больше
максимальной, называются, соответственно, субмаксимальными и супер-
максимальными.
2.2.2. Зависимость возникновения возбуждения от длительности
и силы раздражения
Для возникновения возбуждения в нервной или мышечной ткани необхо-
димо, чтобы сила внешнего раздражения изменялась с достаточной скоро-
стью в единицу времени. Поэтому возникновение возбуждения обусловле-
но не действием абсолютной силы внешнего раздражителя, а скоростью ее
изменения от одной величины до другой. Это называется законом раздра-
жения Дюбуа-Реймона. Однако для возникновения возбуждения нервной и
мышечной ткани недостаточно только скорости изменения силы раздраже-
ния в единицу времени, нужно, чтобы время действия внешнего раздражи-
теля было не меньше определенной (пороговой) величины. Минимальное
время, которое необходимо для того, чтобы сила раздражителя вызвала
возбуждение в нервной или мышечной ткани, называется полезным време-
нем. Продолжительность полезного времени зависит от возбудимости жи-
вой ткани, которая обусловлена различными факторами. Например, к
этим факторам относятся тип мышц (гладкая или поперечно-полосатая),
интенсивность обмена веществ в нервной и мышечной ткани или вид жи-
вотного. Например, полезное время m. biceps человека составляет 1—2 мс,
а гл. gastrocnemius лягушки — около 3—7 мс. Более того, различные нервы
и иннервируемые ими мышцы, например мышцы-сгибатели или разгиба-
тели у одного животного или человека, имеют также разную возбудимость.
Поэтому факторы, которые обусловливают возбудимость живой ткани,
обусловливают величину полезного времени.
При действии внешнего раздражителя на возбудимую ткань она способ-
на возбуждаться в том случае, если сила раздражителя достигает опреде-
ленной, а именно — пороговой величины. Минимальная сила раздражите-
ля, необходимая для возникновения возбуждения нервной или мышечной
ткани, называется порогом возбуждения. Величина порога возбуждения яв-
ляется мерой возбудимости нервной и мышечной ткани. Когда возбуди-
мость нервной или мышечной ткани в результате каких-либо причин по-
нижается, то сила порогового раздражения возрастает, т. е. порог возбуж-
дения становится выше. Напротив, когда возбудимость нервной или мы-
шечной ткани в результате каких-либо причин повышается, то сила поро-
гового раздражения уменьшается и в этом случае, как говорят, порог воз-
буждения ткани становится ниже.
54 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Сила
раздражения
Рис. 2.6. Кривая зависимости силы раздражения возбудимой ткани от длительности
действия раздражителя (кривая сила—длительность).
Мембрана клеток возбудимых тканей является высокочувствительной к действию электриче-
ского тока. Для того, чтобы электрический ток вызвал возбуждение клетки, его сила должна
быть достаточной силы. Сила тока, которая вызывает возбуждение нервной и мышечной тка-
ни, называется реобазой. Время, необходимое для того, чтобы ток силой удвоенной реобазы,
вызвал возбуждение в ткани, называется хронаксией. Кривая сила—длительность отражает тот
факт, что для достижение порога возбуждения в возбудимых тканях необходима пороговая
скорость нарастания силы раздражения.
Хронаксия как мера возбудимости. Поскольку возникновение возбужде-
ния в нервной или мышечной ткани обусловлено силой и продолжитель-
ностью раздражения, то величина полезного времени не является точной
мерой функционального состояния возбудимых тканей. Это обусловлено
следующими обстоятельствами. Взаимодействие между силой и длительно-
стью стимула, вызывающего возбуждение нервной или мышечной ткани
характеризуется кривой «сила-длительность» (рис. 2.6). Для определения
величины полезного времени возбудимых тканей необходимо построение
подобной кривой, что требует многочисленных воздействий электриче-
ским током на возбудимые ткани и проведения многократных измерений
этого показателя. Однако при минимальной силе раздражения, способного
вызвать возбуждение в нервной или мышечной ткани, значение полезного
времени соответствует, как показано на рисунке, точке Р. Вправо от точки
Р кривая становится параллельной оси абсцисс, а следовательно, возбуж-
дение в нервной или мышечной ткани может возникать при любой дли-
тельности действующего раздражителя (закон раздражения) и становится
невозможным по полезному времени точно определить функциональное
состояние возбудимых тканей. Поэтому французским ученым Лапиком
экспериментально был выработан способ получения более точной характе-
ристики возбудимости нервной или мышечной ткани как функции време-
ни. Согласно этому предложению, минимальная сила электрического раз-
дражения (названа реобазой), которая способная вызвать возбуждение тка-
2. Физиология возбудимых тканей • 55
ни независимо от длительности действия электрического тока, должна
быть удвоена, чтобы установить полезное время. Минимальное время, в
течение которого ток силы, равной двум реобазам, должен действовать на
возбудимую ткань, чтобы вызвать минимальный эффект возбуждения, бы-
ло названо хронаксией. Хронаксия (обозначается буквой о) является мерой
возбудимости нервной или мышечной ткани и отражает скорость возник-
новения возбуждения в этих тканях. Значение хронаксии, как показано на
кривой рисунка, находится в области больших значений длительности раз-
дражения (восходящая часть кривой). Так, у мышц человека хронаксия со-
ставляет 0,1—0,7 мс. При этом хронаксия мышц-сгибателей (0,08—0,16 мс)
равна вдвое меньше, чем хронаксия мышц-рагибателей (0,16—0,32 мс).
Указанные величины продолжительности хронаксии мышц-сгибателей и
разгибателей у человека обусловлены возбудимостью нервных волокон,
иннервирующих эти мышцы. От возбудимости нервной или мышечной
ткани, как это будет изложено в разделах настоящей главы, зависит, на-
пример, скрытый период появления возбуждения в возбудимых тканях,
длительность и интенсивность возникающего в них возбуждения. Поэтому,
в наиболее общем виде, чем меньше хронаксия, тем сильнее в ткани про-
цесс возбуждения, тем меньше скрытый период возникновения возбужде-
ния. Определение хронаксии производится в клинической практике для
оценки функционального состояния мышц при их повреждении. При этом
чем больше хронаксия, тем хуже функциональное состояние поврежден-
ных мышц человека.
2.2.3. Возбудимость и возбуждение при действии постоянного
тока на нервную и мышечную ткань
Постоянный ток действует как раздражитель возбудимых тканей только
при замыкании и размыкании цепи электрического тока и в том месте, где
на ткани расположены катод и анод. Дюбуа-Реймоном в XIX в. было уста-
новлено, что в момент замыкания раздражение возбудимой ткани проис-
ходит на катоде, а в момент размыкания — на аноде. Эти явления называ-
ются полюсным действием постоянного тока на возбудимую ткань.
2.2.3.1. Физиологический электротон
Впервые Пфлюгер (1859) показал, что при раздражении нерва постоянным
током возбудимость на нерве или мышце при замыкании электрической це-
пи повышается в месте расположения катода и понижается в месте распо-
ложения анода. Эти изменения возбудимости на катоде называются катэ-
лектротоном, а на аноде — анэлектротоном. Наряду с изменением возбу-
димости в нервной и мышечной ткани под катодом повышается проведе-
ние возбуждения, а под анодом — понижается.
В момент размыкания электрической цепи постоянного тока возбуди-
мость и проводимость нервной и мышечной ткани меняются в обратном
направлении. Так, при размыкании цепи постоянного тока возбудимость
понижается под катодом, а на участке возбудимой ткани в области анода
она повышается. Соответственно проводимость понижается под катодом и
повышается под анодом.
При действии постоянного тока на нервную или мышечную ткань, на-
ряду с появлением кат- и анэлектротона, возбудимость ткани изменяется
за пределами участков электротонических изменений возбудимости. Так,
за пределами катэлектротона на участке нерва возбудимость ткани пони-
56 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
жается, а за пределами анэлектротона, напротив, повышается. Эти элек-
тротонические проявления были открыты Н. Е. Введенским и названы пе-
риэлектротоническими явлениями (периэлектротоном). Периэлектротониче-
ское изменение возбудимости всегда противоположно соответствующему
физиологическому электротону и выражено настолько, что может препят-
ствовать (катпериэлектротон) или способствовать (анпериэлектротон) про-
ведению возбуждения по нерву от места его возникновения к мышце.
При длительном действии постоянного тока подпороговой силы на
нерв в последнем развивается снижение возбудимости под катодом вплоть
до полного ее исчезновения. Это изменение возбудимости называется ка-
тодической депрессией.
2.2.3.2. Закон полярности раздражения нервной и мышечной ткани
Возбуждение от места его возникновения в нервном волокне способно
распространяться на мышцу, что вызывает ее сокращение. Особенностью
полюсного действия постоянного тока на возбудимую ткань является то,
что при одной и той же силе постоянного тока его замыкательное действие
на возбудимую ткань проявляется сильнее, чем размыкательное. Раздраже-
ние нерва замыканием или размыканием цепи постоянного тока вызывает
строго определенную реакцию иннервируемой мышцы в зависимости от
силы постоянного тока и расположения катода и анода на нерве относи-
тельно мышцы. В целом проявление действия постоянного тока на возбу-
димые ткани получало название закона полярности раздражения Пфлюгера
(табл. 2.1).
Очень слабый по силе (подпороговый) постоянный ток вызывает сокра-
щение мышцы только при замыкании, независимо от расположения като-
да и анода на нерве относительно мышцы. Постоянный ток пороговой
(средней) силы вызывает сокращение мышцы при замыкании и размыка-
нии также независимо от расположения катода и анода на нерве относи-
тельно мышцы. Эффекты подпорогового и порогового постоянного тока
на нерв и возникающее при этом сокращение мышцы обусловлены полюс-
ным действием постоянного тока. Проявления действия сверхпорогового
постоянного тока на нерв и возникающее при этом сокращение мышцы
обусловлены электротоническими изменениями возбудимости на катоде и
аноде при замыкании или размыкании цепи постоянного тока. Например,
если анод расположен на нерве между катодом и мышцей, то при замыка-
нии цепи постоянного тока мышца не ответит сокращением, поскольку
катодзамыкательное возбуждение нерва не способно распространиться че-
рез участок анэлектротонического понижения возбудимости. Напротив, в
этих условиях анодразмыкательное возбуждение нерва вызывает сокраще-
Таблица 2.1. Проявление закона полярности раздражения и физиологического электро-
тона
Сила постоянно- го тока Расположение электродов на нерве относительно мышцы
ближе к мышце катод ближе к мышце анод
замыкание размыкание замыкание размыкание
Подпороговая Сокращение Нет сокращения Сокращение Нет сокращения
Пороговая » Сокращение » Сокращение
Сверхпороговая » Нет сокращения Нет сокращения »
2. Физиология возбудимых тканей • 57
ние мышцы, возбуждение беспрепятственно распространяется от участка
нерва между анодом и мышцей. В этом случае участок катэлектротониче-
ского понижения возбудимости нерва не может препятствовать распро-
странению возбуждения по нерву.
В отношении мышечной ткани явления физиологического электротона
и закона сокращения Пфлюгера проявляются в описанной выше законо-
мерности.
2.2.3.3. Электродиагностический закон
При действии постоянного электрического тока на ткани человека ток
проходит не только по нерву, но и по окружающим нерв тканям. В этом
случае проявление закона полярности раздражения развивается по иному,
так называемому электродиагностическому, закону.
Суть этого закона заключается в возникновении возбуждения в нерве
под катодом и анодом в момент замыкания и размыкания в соответствии с
полюсным действием постоянного тока и явлением физиологического элек-
тротона. Однако при прохождении постоянного тока через нерв (физиче-
ский электротон) в нем возникает по обеим сторонам полюсов постоянного
тока поляризация осевого цилиндра нервного волокна (так называемый фи-
зиологический катод и анод) (рис. 2.7). Физиологические катод и анод при
пороговой величине поляризации нервных волокон также способны вызы-
вать в нерве возбуждение. Для электродиагностического закона характер-
ным является возникновение такой последовательности возбуждения в нер-
ве под катодом и анодом и появление сокращения в иннервируемой нервом
мышце: катодзамыкательное сокращение (действие катода) — анодзамыка-
тельное сокращение (действие физиологического катода) — анодразмыка-
тельное сокращение (действие анода) — катодразмыкательное сокращение
(действие физиологического анода). Возбуждение в нерве при действии фи-
зиологического катода и анода возникает при силе тока, как правило, боль-
шей, чем при действии на нерв постоянного тока под полюсами.
Указанные законы обосновали применение в медицине терапевтическо-
го эффекта анэлектротона для прерывания проведения по нерву импуль-
сов, в том числе болевых, при судорогах и невралгии у больных.
Рис. 2.7. Схематическое изображение распределения силовых линий между катодом
и анодом в области нерва, окруженного тканями.
Электродиагностический закон проявляется в том, что при замыкании цепи постоянного тока
сокращение мышцы возникает в области действия на нерв физического катода и физиологи-
ческого катода в области действия на нерв физического анода. При размыкании цепи посто-
янного тока сокращение мышцы возникает в области действия на нерв физического анода и
физиологического катода в области действия на нерв физического анода.
58 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.2.4. Понятие о функциональной подвижности возбудимых
тканей
Возбудимые ткани при их раздражении электрическим током отвечают оп-
ределенным числом возбуждений в единицу времени. Способность возбу-
димых тканей отвечать максимальным числом возбуждений в единицу вре-
мени на их электрическое раздражение называется функциональной по-
движностью, или лабильностью (Н. Е. Введенский). Лабильность возбуди-
мых тканей измеряется количеством волн возбуждения в единицу времени.
Лабильность и хронаксия возбудимой ткани взаимосвязаны между собой.
Эта взаимосвязь проявляется в том, что возбудимая ткань (нервная или
мышечная), имеющая высокую скорость возникновения возбуждения в
единицу времени, имеет и высокую скорость протекания процессов возбу-
ждения, и наоборот. Поэтому чем больше волн возбуждения может воспро-
извести нерв или мышца в единицу времени при их ритмическом раздра-
жении электрическим током, тем выше их лабильность. Эта взаимосвязь
лабильности и хронаксии возбудимой ткани позволяет в клинической
практике оценивать функциональную подвижность и контролировать, на-
пример, процесс восстановления лабильности поврежденных нервной и
мышечной тканей.
Лабильность нервного волокна специфическим образом снижается при
воздействии на нерв, например, анестетиков. Снижение лабильности нер-
ва начинается с уравнительной стадии, когда сильные и слабые по силе
раздражения нерва вызывают сходный по амплитуде сократительный ответ
мышцы. Дальнейшее снижение лабильности проявляется в парадоксальной
стадии ответа мышцы, когда слабое по силе раздражение нерва вызывает
большее по силе сокращение, и наоборот. Исчезновение лабильности нер-
ва свидетельствует об отсутствии в нем возбудимости и проводимости, что
характерно для тормозящей стадии, когда сокращение мышцы отсутствует
при любой силе раздражения нерва. Это явление было названо парабиозом
(Н. Е. Введенский). Оно объяснялось появлением сильного утратившего
способность распространяться возбуждения в участке нерва, находящемся
под действием длительного раздражения. С позиций современной физио-
логии утрата нервом возбудимости и проводимости при парабиозе обу-
словлена блокадой потенциалзависимых натриевых ионных каналов (см.
ниже).
2.3. Электрические явления в возбудимых клетках
Мембраны возбудимых клеток имеют ионные каналы, которые могут нахо-
диться в двух состояниях: закрытом и открытом. Когда ионный канал от-
крыт, то ионы свободно диффундируют через мембрану по градиенту их
концентрации. Проницаемость мембраны для иона, диффундирующего че-
рез определенный тип канала, взаимосвязана с величиной мембранного
потенциала. Процесс закрытия или открытия ионного канала обусловлен
потенциалзависимыми воротными эффектами, которые возникают в ион-
ном канале очень быстро и сохраняются кратковременно (1—2 мс). К та-
кому типу потенциалзависимых ионных каналов относятся калиевые ион-
ные каналы. Более медленные потенциалзависимые воротные эффекты
имеют место в ионных каналах, которые могут находиться в трех состоя-
ниях: открытом, инактивированном и закрытом. Механизм инактивирова-
ния ионного канала принципиально отличается от такового, который име-
ет место при открытии или закрытии (например, калиевого ионного кана-
2. Физиология возбудимых тканей • 59
ла), тем, что канал вначале открывается, а затем инактивируется, и остает-
ся в таком состоянии некоторое время. К такому типу потенциалзависи-
мых ионных каналов относятся натриевые ионные каналы мембраны воз-
будимых клеток, обусловливающие генерацию потенциала действия.
Потенциал действия нервной и мышечной клетки является основным
проявлением в них процесса возбуждения. В состоянии покоя потенциал-
зависимые ионные каналы мембраны клеток возбудимых тканей закрыты,
но сама мембрана находится в поляризованном состоянии, т. е. имеет лгел*-
бранный потенциал.
2.3.1. Мембранный потенциал покоя
Мембранный потенциал покоя, или потенциал покоя, является результатом
разности потенциалов между внутренней и наружной сторонами мембраны
клетки (Ет). В покое внутренняя сторона мембраны клетки заряжена от-
рицательно по отношению к наружной, т. е. клеточная мембрана поляризо-
вана. Поляризация мембраны клеток заключается в следующем (рис. 2.8).
Если мембрана клетки проницаема только для одного типа ионов, то в
случае, когда концентрация этого типа ионов по обе стороны мембраны
различна, они способны путем диффузии переходить через мембрану из
среды с большей концентрацией (С7) в среду с меньшей концентрацией
ионов (С2). В случае, если ионы заряжены положительно (см. рис. 2.8)
как, например, ионы калия, то на стороне С2 мембраны накапливаются
только положительно заряженные ионы. Положительный заряд ионов на
стороне С2 мембраны будет препятствовать дальнейшему проникновению
одноименных ионов, так как возникшее электрическое поле уравновеши-
вает движущую силу концентрационного градиента положительных ионов
на мембране. Электрические силы положительных ионов на стороне С2
мембраны будут притягивать отрицательные ионы со стороны С1 мембра-
ны, для которых она непроницаема. При достижении равновесия концен-
трационные потенциалы ионов по обе стороны мембраны будут равны по
величине, но противоположны по знаку их электрическому потенциалу. В
результате по обе стороны мембраны возникает разность потенциалов.
Возникновение на мембране разности потенциалов (Ет) описывается
уравнением Нернста, которое позволяет рассчитать равновесный электри-
ческий потенциал для любого иона:
Ет =
Zr с2
где С/ и С2 — концентрации ионов после достижения
равновесия по обе стороны мембраны, z — валент-
Рис. 2.8. Механизм возникновения потенциала Нернста.
Положительные ионы диффундируют из области высокой концен-
трации (С1) в область низкой концентрации (С2). В результате од-
на сторона мембраны заряжается положительно, что препятствует
дальнейшей диффузии положительных ионов на сторону С2. Меж-
ду сторонами С1 и С2 возникает электрическое поле, которое урав-
новешивает движущую силу концентрационного потенциала, и ус-
танавливается динамическое равновесие. Со стороны С1 на мем-
бране формируется отрицательныйй заряд, а со стороны С2 — по-
ложительный. В целом, на мембране возникает разность потен-
циалов.
Мембрана
60 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
ность ионов, F = 96 500 Кл/моль — постоянная Фарадея, R = 8,31 Дж/
мольбК — универсальная газовая постоянная, Т — абсолютная температура
системы, zF — заряд одного моля ионов, переносимых через мембрану.
Рассмотренный на рис. 2.8 пример свидетельствует о том, что потенци-
ал на мембране клетки обусловлен ионами в том случае, если она для них
проницаема. В отношении возбудимых клеток потенциал на их мембране
может быть обусловлен ионами Na+, К+ и СГ. В случае, если бы потенци-
ал на мембране определялся только ионами калия, то его величина, со-
гласно уравнению Нернста, должна равняться —90 мВ, принимая во вни-
мание значение температуры тела человека (57 °C), валентность иона (z —
— Г), постоянную Фарадея (F = 96 500 Кл/моль) и универсальную газовую
постоянную (R = 8,31Дж/мольКу.
Е, -И„ГЙ,
tCl
где К* и К+ви — наружная и внутренняя концентрация ионов калия вне и
внутри нейрона. В нейронах теплокровных животных отношение [Л„+] к
[А^] составляет порядка ’/28.
В случае, если бы мембрана клеток была пассивно проницаема только
для ионов Na+ или только для ионов С1", то на ней устанавливался бы по-
тенциал, равный +60 мВ и —80 мВ соответственно. При этом для хлора ве-
личина потенциала Нернста близка к реальной величине потенциала по-
коя нейрона, т. е. примерно —80 мВ. Однако в норме потенциал покоя на
мембране нейронов составляет порядка -60 мВ. Причина отклонения по-
тенциала на мембране нейрона от потенциала, рассчитанного по уравне-
нию Нернста для каждого из названных ионов, заключается в том, что че-
рез мембрану нейрона постоянно движутся встречные потоки ионов К+,
Na+ и СГ, каждый из которых имеет разный коэффициент проницаемости.
Потенциал покоя в возбудимых клетках. В возбудимых клетках между
цитозолем и внеклеточной средой существует концентрационный градиент
для ионов Na+ и К+, который поддерживается работой натрий-калиевого
насоса, функционирующего за счет гидролиза энергии АТФ. Активность
натрий-калиевого насоса создает разность потенциалов на мембране по-
рядка —4 мВ, поэтому его называют электрогенным насосом. Основной
функцией натрий-калиевого насоса является поддержание градиента кон-
центрации ионов Na+ и К+ на мембране нейрона. В результате активности
натрий-калиевого насоса концентрация ионов К+ с внутренней стороны
мембраны клеток в 20—50 раз выше, чем с внешней стороны мембраны, а
с внешней стороны мембраны в 8—10 раз выше концентрация ионов Na+.
Поэтому ионы калия стремятся выйти из клетки, а ионы натрия — проник-
нуть внутрь. В покое мембрана возбудимых клеток проницаема в разной
степени для нескольких ионов, т. е. ионов К+, Na+ и СГ. Это является
причиной отклонения потенциала на мембране от потенциала, рассчитан-
ного по формуле Нернста для каждого из этих ионов. В этом случае мем-
бранный потенциал может быть описан уравнением Гольдмана—Ходжки-
на—Катца:
Ет = ЯГ,„ Л1С] + PNa[Fa„+] + Л.,[СУ1
F Рк[£и] + Р^[Ка„/] + Рс/[ С1„]'
где Ет — мембранный потенциал; R — газовая постоянная; Т — абсолютная
температура; F— число Фарадея; Рк, PNn и Ра — константы проницаемости
2. Физиология возбудимых тканей • 61
мембраны для ионов К+, Na+ и СГ; [КГ], [K]J, [Na]], [Na].], [С1Н] и
fCla,J — концентрации ионов К+, Na+ и СГ снаружи (и) и внутри (вн)
клетки.
Согласно формуле Гольдмана—Ходжкина—Катца концентрационный
градиент каждого из ионов обусловливает величину мембранного потен-
циала. В покое величина мембранного потенциала пропорциональна про-
ницаемости мембраны для ионов К+, Na+ и СГ, которая составляет —
Рк : PNa : РС| = 1 : 0,04 : 0,45. Величина коэффициента проницаемости ио-
нов калия свидетельствует о том, что концентрационный градиент поло-
жительно заряженных ионов калия направлен из клетки во внеклеточную
среду. По своему характеру проницаемость ионов калия через мембрану
нейрона носит диффузионный характер (ток утечки ионов калия) и потен-
циал покоя определяется, в основном, этим ионом. Во внутриклеточной
среде возбудимых клеток содержатся различные анионы, которые не могут
проникать через мембрану (белки, аминокислоты, органические кислоты,
фосфаты, сульфаты). В результате избыток отрицательных ионов внутри
клетки создает отрицательный заряд на поверхности мембраны с ее внут-
ренней стороны. Выход ионов калия через мембрану из клетки создает из-
быток положительных ионов на внешней поверхности мембраны.
Концентрация ионов хлора по обе стороны мембраны возбудимых кле-
ток находится в равновесном состоянии. Поэтому ионы хлора могут дви-
гаться через мембрану в обоих направлениях с одинаковой скоростью. На
этом основании ионы хлора могут не входить в уравнение Гольдмана-
Ходжкина—Каца. Лишь при быстром нарушении равновесного состояния
ионы хлора могут оказывать влияние на величину потенциала на мембране
возбудимых клеток.
2.3.2. Потенциал действия возбудимых клеток
Потенциал действия возбудимых клеток представляет собой кратковремен-
ное фазное изменение знака потенциала на внутренней поверхности мем-
браны и проницаемости мембраны для ионов, вызванное внешним раздра-
жением мембраны возбудимых клеток. Потенциал действия состоит из фаз
деполяризации, реполяризации и следовых потенциалов.
Фаза деполяризации возникает в результате открытия в мембране потен-
циалзависимых натриевых ионных каналов (рис.2.9). При этом коэффици-
ент проницаемости мембраны для ионов Na+ возрастает в сотни раз, т. е. с
PNa+ = 0,04 до PNa+ = 20. Потенциал действия возникает по закону «все или
Рис. 2.9. Натриевый воротный
ионный канал. При деполяриза-
ции мембраны канал находится в
открытом или инактивированном
состояниях (закрыты h-ворота).
При поляризованной мембране,
т. е. когда внутренняя поверхность
мембраны имеет отрицательный за-
ряд относительно наружной, ион-
ный канал находится в закрытом
состоянии (m-ворота закрыты).
Д — деполяризация мембраны, П —
поляризация мембраны в состоянии
покоя.
62 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Рис. 2.10. Потенциал дейст-
вия.
Изменения мембранного потен-
циала (А), которые называются
потенциалом действия нервов и
мышц, являются результатом из-
менения ионной проницаемости
мембраны для Na’ и К' (Б). В за-
висимости от иона, участвующего
в генерации потенциала действия,
изменение ионной проницаемости
мембраны происходит в результате
открытия, закрытия или инакти-
вирования ионных каналов. Боль-
шинство потенциалзависимых на-
триевых ионных каналов открыва-
ется, когда деполяризация дости-
гает порогового уровня (Ek). По-
сле спонтанной инактивации этих
каналов открываются калиевые
ионные каналы, что вызывает ре-
поляризацию мембраны. В пери-
од, когда натриевые ионные кана-
лы находятся в инактивированном
состоянии, мембрана клетки явля-
ется рефрактерной. Натриевые
ионные каналы функционируют
примерно в 10 раз быстрее, чем
калиевые.
ничего». Согласно этому
закону величина потенциа-
ла действия остается посто-
янной, независимо от силы
вызвавшего его стимула.
Стимулом для генерации
потенциала действия явля-
ется быстрая кратковре-
менная деполяризация мембраны возбудимой клетки, которая называется
критическим уровнемдеполяризщии (£к). Величина потенциала на мембра-
не {Ет), необходимая для достижения критического уровня деполяриза-
ции, называется пороговым потенциалом {Ет—Ек), или порогом генерации
потенциала действия (рис. 2.10). Амплитуда критического уровня деполя-
ризациии мембраны нервного волокна и мышечной клетки в среднем
меньше величины потенциала покоя на 15—20 мВ. Пороговый сдвиг мем-
бранного потенциала возникает лишь в том случае, если натриевый ток
(4,), направленный внутрь клетки начинает превышать противоположно
направленный калиевый ток (ZJ. £сли уменьшение мембранного потен-
циала происходит медленно в течение нескольких миллисекунд, то инак-
тивационные ворота в натриевых ионных каналах инактивируются раньше,
чем произойдет открывание достаточного количества активационных во-
рот. В результате натриевый ток {1Па) будет недостаточным для открытия
потенциалзависимых натриевых каналов. Подобное явление называется
аккомодацией мембраны В этом случае не будет возникать потенциал дей-
ствия. Деполяризация мембраны начинается с активации инактивирован-
ных потенциалзависимых натриевых ионных каналов, и только после это-
го натриевые каналы открываются. Ионы ла * через открытые потенциал-
2. Физиология возбудимых тканей • 63
зависимые натриевые каналы устремляются по концентрационному гради-
енту внутрь клетки (натриевый ток /Л,а), заряжая внутреннюю поверхность
мембраны положительным зарядом. При этом отношение коэффициентов
проницаемостей мембраны для ионов калия и натрия составляет иную ве-
личину относительно состояния покоя: Рк : PNa = 1 : 20, а генерация по-
тенциала действия обусловлена высокой проницаемостью мембраны для
ионов Na+. Деполяризация мембраны клетки увеличивает число открытых
потенциалзависимых натриевых каналов, что существенно повышает на-
триевую проницаемость. В результате потенциал на мембране возбудимой
клетки за 1—2 мс стремится достичь величины натриевого равновесного
потенциала. При этом потенциал на внутренней поверхности мембраны
меняет знак и его величина достигает +40 мВ (явление реверсии заряда
мембраны).
Реполяризация. Во время реверсии заряда мембраны величина потенциа-
ла Нернста для ионов натрия приближается к их равновесному потенциа-
лу, что вызывает через 1—2 мс после начала деполяризации инактивацию
потенциалзависимых натриевых ионных каналов. При этом инактивацион-
ные ворота натриевых ионных каналов не открываются до тех пор, пока
мембранный потенциал покоя не восстанавливается до своего исходного
уровня. В результате прекращается входящий натриевый ток. Одновремен-
но под влиянием деполяризации мембраны открываются потенциалзави-
симые калиевые ионные каналы и увеличивается выходящий калиевый
ток, который стремится сместить мембранный потенциал в сторону калие-
вого равновесного потенциала и, таким образом, реполяризует мембрану
(рис. 2.11). Мембранный потенциал приближается к величине потенциала
покоя, но в мембране еще
остаются открытыми ка-
лиевые ионные каналы и
инактивироваными — на-
триевые ионные каналы.
Следовые потенциалы.
В возбудимых клетках фаза
реполяризации переходит в
следовые потенциалы, ко-
торые подразделяют на
следовую деполяризацию и
следовую гиперполяриза-
цию. Следовые потенциа-
лы обусловлены тем, что
натриевые и калиевые
ионные каналы не возвра-
тились к своему исходному
состоянию после быстрых
фаз деполяризации и репо-
ляризации. При этом чем
медленнее в фазу деполя-
ризации открываются по-
тенциалзависимые калие-
вые ионные каналы, тем
длительнее будет следовая
гиперполяризация и тем
медленнее в мембране
клетки будет восстанавли-
3 Na+ ik
Внешняя поверхность
мембраны клетки
Мембрана клетки
Внутренняя поверхность
мембраны клетки
Активный Диффузия
транспорт по градиенту
концентрации
Рис. 2.11. Генерация потенциала действия.
Ионы натрия по градиенту концентрации проникают
внутрь клетки (INo) и заряжают внутреннюю поверх-
ность мембраны положительным зарядом до величины
потенциала действия (ПД). После открывания натрие-
вых ионных каналов соотношение PNa : РК — 20 : 1.
Коэффициент проницаемости для ионов натрия меня-
ется с диффузионного на канальный. Открывание по-
тенциалзависимых калиевых ионных каналов (IK) вос-
станавливает отрицательный заряд на внутренней по-
верхности мембраны. Увеличение концентрации ионов
натрия внутри клетки активирует работу натрий-калие-
вого насоса.
64 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
ваться ее нормальная возбудимость. Величина потенциала покоя мембра-
ны нейронов составляет —60 мВ, а мембраны нервного волокна и скелет-
ной мышечной клетки —90мВ, что свидетельствует о более высокой возбу-
димости сомы нейронов, чем их аксонов и, тем более, скелетных мышеч-
ных волокон.
При этом амплитуда потенциала действия в нервных волокнах состав-
ляет 110 мВ, что на +20 мВ превышает величину потенциала покоя. В ске-
летных мышечных клетках во время фазы деполяризации величина потен-
циала на мембране достигает максимального значения +30—40 мВ. Поэто-
му амплитуда потенциала действия в клетках скелетных мышц составляет
120—130 мВ. Эти различия в амплитуде реверсии заряда мембраны нерв-
ных и мышечных волокон обусловливают большую скорость протекания
возбуждения в нервных волокнах, а следовательно, более высокую их ла-
бильность по сравнению с мышечными волокнами.
2.2.3. Рефрактерный период в возбудимых клетках
В фазу деполяризации потенциала действия потенциалзависимые натрие-
вые ионные каналы кратковременно открываются, но затем инактивиру-
ются h-ворота (см. рис. 2.9). В период инактивации натриевых ионных ка-
налов возбудимые клетки не способны реагировать повышением натрие-
вой проницаемости на повторный стимул. Поэтому во время фазы деполя-
ризации мембрана не может генерировать потенциал действия в ответ на
действие пороговых или сверхпороговых раздражителей. Это состояние на-
зывается абсолютной рефрактерностью, время которой составляет в нерв-
ных волокнах 0,5—1,0 мс, а в скелетных мышечных клетках —в среднем 2
мс. Период абсолютной рефрактерности заканчивается после того, как
снижается количество инактивированных натриевых каналов и постепенно
увеличивается количество натриевых каналов, находящихся в закрытом со-
стоянии. Происходят эти процессы во время фазы реполяризации, когда
уменьшению количества потенциалзависимых натриевых ионных каналов,
находящихся в состоянии инактивации, соответствует период относитель-
ной рефрактерности. Период относительной рефрактерности характеризу-
ется тем, что лишь некоторая часть потенциалзависимых натриевых ион-
ных каналов переходит в закрытое состояние, а в силу этого порог возбу-
димости мембраны клетки имеет более высокие значения, чем в исходном
состоянии. Поэтому возбудимые клетки в период относительной рефрак-
терности могут генерировать потенциалы действия, но при воздействии на
них раздражителей сверхпороговой силы. Однако из-за небольшого коли-
чества потенциалзависимых натриевых ионных каналов, находящихся в за-
крытом состоянии, амплитуда генерируемых при этом потенциалов дейст-
вия будет меньше, чем в условиях исходной возбудимости нервной или
мышечной клетки.
В клетках возбудимых тканей максимальное количество генерируемых
потенциалов действия в единицу времени обусловлено двумя факторами:
длительностью потенциала действия и длительностью периода абсолютной
рефрактерности после каждого импульса. На этом основании в физиоло-
гии формулируется современное понятие лабильности', чем меньше период
абсолютной рефрактерности при возбуждении возбудимой ткани, тем вы-
ше ее функциональная подвижность или лабильность, тем больше в ней
генерируется потенциалов действия в единицу времени.
При непрерывной стимуляции нерва электрическим током лабильность
нерва зависит от частоты и силы раздражения. В зависимости от частоты и
2. Физиология возбудимых тканей • 65
силы раздражения нерва сокращение иннервируемой им мышцы может
быть максимальной или минимальной амплитуды. Эти явления были на-
званы соответственно оптимумом и пессимумом (Н. Е. Введенский). Мак-
симальное (оптимально большое) сокращение мышцы возникает в том
случае, если каждый последующий электрический стимул действует на
нерв в периоде его состояния супернормальной возбудимости после пре-
дыдущего потенциала действия. Минимальное (или пессимальное) сокра-
щение мышцы возникает в том случае, если каждый последующий элек-
трический стимул действует на нерв, находящийся в периоде относитель-
ной рефрактерности после предыдущего потенциала действия. Поэтому
значения оптимальной частоты раздражения нерва всегда меньше, чем
значения пессимальной частоты раздражения.
2.3.4. Локальный ответ мембраны
возбудимых клеток
При действии электрического стимула подпороговой силы уменьшение
мембранного потенциала оказывается недостаточным для того, чтобы на-
триевый ток (JNa) стал больше, чем сумма калиевого выходящего тока или
тока утечки ионов калия. При этом мембранный потенциал уменьшается
на несколько милливольт, но в результате реполяризации восстанавливает-
ся до величины мембранного потенциала покоя. Подобные пассивные или
электротонические изменения потенциала на мембране, которые не дости-
гают критического уровня деполяризации под действием деполяризующего
электрического тока подпороговой величины, называются локальным отве-
том. Для локального ответа характерны следующие физиологические про-
явления на мембране нервной
клетки или нервного волокна: ам-
плитуда локального ответа не
имеет линейной зависимости от
силы подпорогового электриче-
ского раздражения; отсутствует
проявление закона «все или ниче-
го»; в период локального ответа
мембрана клетки или нервного
волокна не только сохраняет, но
и имеет повышенную (супернор-
мальную) возбудимость. Следова-
тельно, локальный ответ не имеет
четкого порога возникновения, а
в период его развития отсутствует
абсолютная рефрактерность нерв-
ного волокна. Во время локально-
го ответа мембрана нервной клет-
ки имеет высокую способность к
регенеративному увеличению
ионной натриевой проницаемо-
сти, поскольку амплитуда мем-
бранного потенциала снижается
электротонически. Локальные
электротонические изменения по-
тенциала на мембране нервного
волокна после прекращения дей-
Рис. 2.12. Влияние подпорогового элек-
трического раздражения возбудимой мем-
браны на степень ее деполяризации.
При раздражении мембраны возбудимых кле-
ток электрическим током подпороговой силы
происходит электротоническое уменьшение
потенциала на мембране и/или активация по-
тенциалзависимых натриевых каналов. Про-
цесс является обратимым, и мембранный по-
тенциал возвращается к исходному уровню, не
достигая критического уровня деполяризации
(Ek). В период локальных изменений мембран-
ного потенциала мембрана возбудимых клеток
имеет супернормальную возбудимость.
3 - 6095
66 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
ствия кратковременного импульса электрического тока подвергаются об-
ратному самостоятельному развитию. При действии на возбудимую мем-
брану серией подпороговых электрических стимулов происходит суммиро-
вание амплитуды локального ответа. Если амплитуда локального ответа
достигает порога возбудимости мембраны, то потенциалзависимые натрие-
вые ионные каналы открываются по закону «все или ничего» и происходит
генерация потенциала действия. На рис. 2.12 показано суммирование ам-
плитуды локального ответа по типу деполяризации при действии на мем-
брану нервного волокна серией подпороговых электрических стимулов.
Локальный ответ нервного волокна всегда сопровождается повышением
возбудимости мембраны клеток.
2.4. Проведение импульса по нервным волокнам
Потенциал действия или нервный импульс может возникать в любой точке
возбудимой мембраны нервного или мышечного волокна и способен рас-
пространяться вдоль ее поверхности. При этом роль потенциала действия
заключается в передаче информации по нервным волокнам от тела нейро-
на к нервному окончанию. Когда потенциалы действия достигают терми-
налей аксона, то информация передается на другие нейроны благодаря вы-
делению из нервных окончаний молекул медиаторов. В мышечных клетках
потенциалы действия распространяются по сарколемме и активируют ме-
ханизм сокращения мышц. Проведение нервного импульса от тела нейро-
на к окончанию аксона различается в немиелинизированных и миелини-
зированных нервных волокнах.
2.4.1. Демиелинизированные волокна
Проведение потенциала действия по немиелинированному нервному во-
локну происходит путем активации потенциалзависимых натриевых ион-
ных каналов участка мембраны волокна, прилегающего к тому месту, где
возник потенциал действия. При этом между возбужденным и невозбуж-
денным участками мембраны нервного волокна возникают локальные
электрические токи, которые вызывают деполяризацию мембраны невоз-
бужденного участка до критического уровня, и только после этого в мем-
------------►!«---------►-------------►
Рефрактерный Активный ' Нормальная
участок участок возбудимость
Рис. 2.13. Распространение потенциа-
ла действия по немиелинизированно-
му нервному волокну.
При деполяризации мембраны нервного во-
локна возникают продольные токи, которые
распространяются по аксоплазме и направ-
лены от активного участка генерации потен-
циала действия в обе стороны нервного во-
локна. Продольные токи (1а) вызывают от-
крытие натриевых потенциалзависимых
ионных каналов в участке мембраны, кото-
рый имеет нормальную возбудимость и вы-
зывают в этом месте генерацию потенциала
действия. Рефрактерный участок невозбу-
дим, поскольку натриевые ионные каналы
находятся в инактивированом состоянии. В
результате потенциал действия генерируется
на соседнем участке мембраны и движется
по нервному волокну в одном направлении.
2. Физиология возбудимых тканей • 67
бране нервного волокна мгновенно открываются потенциалзависимые на-
триевые каналы. Потенциал действия, таким образом, генерируется в по-
следующем участке нервного волокна (рис. 2.13).
2.4.2. Миелинизированные волокна
В миелинизированном нервном волокне потенциалы действия генериру-
ются только в области перехвата Ранвье, т. е. той части мембраны, которая
не покрыта шванновскими клетками. С одной стороны, это обусловлено
тем, что цитоплазма шванновской клетки содержит липид — сфингомиелин,
который уменьшает поток ионов через мембрану нервного волокна при-
мерно в 5000 раз и снижает ее емкость в 50 раз. С другой стороны, в об-
ласти перехвата Ранвье в мембране нервного волокна имеется наибольшее
число потенциалзависимых натриевых ионных каналов, а сама мембрана
обладает нормальным уровнем возбудимости. При нанесении точечного
раздражения на миелинизированное нервное волокно потенциал действия
генерируется в зоне перехвата Ранвье и возникают электрические токи, ко-
торые текут вдоль силовых линий от плюса к минусу потенциала на мем-
бране (рис. 2.14). Одновременно в аксоплазме нервного волокна возника-
ют продольные токи, которые направлены от места генерации потенциала
действия в обе стороны (1а). Продольные токи вызывают открывание по-
тенциалзависимых натриевых ионных каналов, а следовательно, движение
ионов Na+ через мембрану и генерацию потенциала действия в соседних
перехватах Ранвье, минуя часть нервного волокна, покрытого шванновски-
ми клетками. Поскольку потенциалы действия передаются (перепрыгива-
ют) от одного перехвата Ранвье к другому, то механизм проведения потен-
циалов действия в миелинизированных нервных волокнах получил назва-
' ' Миелин ч-------------►
Рефрактерный Активный
участок
Нормальная
возбудимость
Рис. 2.14. Распространение потенциала действия по миелиновому нервному во-
локну.
При деполяризации мембраны активного перехвата Ранвье продольные токи (1а) направлены
от активного участка в обе стороны волокна. Движение ионов, обусловленное продольными
токами, вызывает открытие потенциалзависмых натриевых ионных каналов мембраны пере-
хвата Ранвье, имеющей нормальную возбудимость. Аналогичный процесс не возникает на
мембране рефрактерного участка, натриевые каналы которого находятся в инактивированном
состоянии. В результате потенциал действия распространяется сальтаторно в одном направле-
нии. Стрелками обозначено направление токов из возбужденного перехвата вдоль нервного
волокна.
з-
68 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
ние сальтаторного. В миелинизированных нервных волокнах реполяриза-
ция мембраны после потенциала действия происходит с очень высокой
скоростью. В результате нервные волокна имеют высокую функциональ-
ную лабильность и способны проводить значительное число потенциалов
действия в единицу времени. Скорость распространения потенциалов дей-
ствия в зависимости от диаметра и типа миелинизированных нервных во-
локон чрезвычайно высока и варьирует от 6 до 120 м/с. В толстых миели-
низированных нервных волокнах скорость проведения возбуждения про-
порциональна диаметру волокна, а проводимость нервного волокна имеет
обратную зависимость от его диаметра.
2.4.3. Законы проведения возбуждения по нервному волокну
Проведение возбуждения по нервному волокну возможно при условии его
структурной целостности и физиологической непрерывности (закон физио-
логической непрерывности). Физиологическая непрерывность нерва может
быть нарушена, например, при сдавлении нерва без его структурного по-
вреждения, что препятствует проведению потенциалов действия. При про-
ведении возбуждения по нервному волокну потенциал действия не распро-
страняется с одного волокна на другое, например рядом расположенное
(закон изолированного проведения). Несмотря на то что нервные волокна
проводят потенциалы действия либо в направлении тела нейрона (центро-
стремительно), либо от тела нервной клетки к окончаниям аксона (цен-
тробежно), отдельное нервное волокно обладает двусторонней проводимо-
стью (закон двустороннего проведения). При искусственном электрическом
раздражении в любой точке по ходу нервного волокна может возникать
потенциал действия и распространяться как центростремительно, так и
центробежно.
2.5. Проведение возбуждения через синапс
Передача возбуждения с отростка одной нервной клетки на отросток или
тело другой нервной клетки возможна двумя способами; электрическим и
химическим. Электрический способ передачи возбуждения осуществляет-
ся с помощью щелевых контактов (нексусов) в том случае, если мембра-
ны клеток отделены щелью размером примерно 2 нм. Щелевые контакты
распространены, например, в миокарде и гладких мышцах пищеваритель-
ного тракта и представляют собой межклеточные тубулярные белковые
гидрофильные каналы, состоящие их шести молекул интегральных белков-
коннексонов. Передача возбуждения электрическим способом осуществля-
ется аналогично его проведению по нервным волокнам с помощью мест-
ных токов, возникающих между деполяризованным участком мембраны
нервного волокна и поляризованным участком мембраны нервной клетки.
Локальные токи деполяризуют мембрану нейрона до критического уров-
ня, после чего возникает спонтанный процесс регенеративной деполяри-
зации. Электрическая передача возбуждения осуществляется с высокой
скоростью, близкой к скорости проведения возбуждения по нервным во-
локнам.
Основным способом передачи возбуждения между нейронами, а также
между нервными и мышечными клетками является химический, который
реализуется с помощью синапсов. Синапс — это специализированное по
структуре и функции место контакта мембран между двумя возбудимыми
клетками, которое обеспечивает передачу импульсов от одной возбудимой
2. Физиология возбудимых тканей • 69
клетки к другой с помощью молекул химических веществ, или медиаторов.
Проведение возбуждения между нервом и мышцей осуществляется через
нервно-мышечный синапс, который представляет собой место контакта
окончания аксона нейрона с мембраной мышечного волокна. Функцией
такого соединения является передача сигналов возбуждения от «-мотоней-
ронов спинного мозга к скелетным мышечным волокнам, вызывающих со-
кращение мышцы.
2.5.1. Проведение возбуждения через нервно-мышечный синапс
Нервно-мышечный синапс образован мембраной терминали аксона, или
пресинаптического окончания мотонейрона спинного мозга, и мембраной
мышечного волокна, или постсинаптической мембраной, между которыми
имеется синаптическая щель (рис. 2.15). Постсинаптическая мембрана
нервно-мышечного синапса называется концевой пластинкой, поскольку
представляет собой специализированную часть мембраны мышечного во-
локна, имеющую складчатый вид, что существенно увеличивает площадь
мембраны, контактирующей с аксолеммой нервной терминали. Кроме то-
го, постсинаптическая мембрана содержит N-холинорецепторы и фермент
ацетилхолинестеразу, которая разрушает медиатор ацетилхолин. Исходя
из структурной организации нервно-мышечного синапса, передачу возбу-
ждения с нерва на мышцу можно разделить на два физиологических ме-
ханизма: пресинаптический и постсинаптический. Функция нервно-мы-
шечного синапса заключена в передаче очень небольших по амплитуде
потенциалов действия, достигающих двигательных нервных окончаний,
на мышечные волокна и инициации, таким образом, сокращения во всей
скелетной мышце.
Рис. 2.15. Схема нервно-мышечного синапса.
Функцией соединения является трансформация энергии нервного импульса в кванты ацетил-
холина, который выделяется из терминали в синаптическую щель с последующим превраще-
нием химического сигнала в электрический на мембране мышечного волокна. На складчатой
постсинаптической мембране находятся ионотропные холинорецепторы, которые, связываясь
с ацетилхолином, открывают натриевые ионные каналы постсинаптической мембраны. Депо-
ляризация постсинаптической мембраны электротонически открывает потенциалзависимые
натриевые ионные каналы сарколеммы и, таким образом, в ней генерируются распространяю-
щиеся потенциалы действия.
70 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.5.1.1. Пресинапти ческий механизм
Рис. 2.16. Высвобождение и рециркуляция
ацетилхолина в нервно-мышечном синапсе.
Пресинаптический механизм передачи возбуждения с нерва на мышцу
осуществляется в двигательных нервных окончаниях и его функция заклю-
чается в трансформации числа нервных импульсов в количество выделяю-
щегося из нервной терминали медиатора. Проведение возбуждения через
нервно-мышечный синапс связано с функцией активных зон пресинапти-
ческой мембраны, которые представляют собой комплекс рядами располо-
женных Са2+-каналов, синаптических везикул, специализированных бел-
ков экзоцитоза или выделения медиатора и элементов цитоскелета (акти-
новые филаменты и микротрубочки), объединенные вокруг так называе-
мых плотных тел синапса. Последние представляют собой участки утолще-
ния пресинаптической мембраны, в области которых происходит секреция
медиатора. В синаптическом окончании содержится примерно от 1 до 5
активных зон, которые имеют непосредственное отношение к выделению,
или экзоцитозу, медиатора в синаптическую щель.
В синаптических пузырьках
терминали аксона, иннерви-
рующего мышцу, содержится
медиатор ацетилхолин. Ацетил-
холин синтезируется непосред-
ственно в нервном окончании
из холина и ацетил-Ко-А при
участии фермента ацетил-
трансферазы. Наряду с медиа-
тором в везикулах содержатся
различные ионы, АТФ, фер-
менты и другие вещества, уча-
ствующие в выделении медиа-
тора в синаптическую щель. В
каждом синаптическом пузырь-
ке содержится примерно 10'4
молекул ацетилхолина. Количе-
ство ацетилхолина, высвобож-
даемое из одной везикулы,
принято называть квантом ме-
диатора. Без ресинтеза ацетил-
холина в двигательном нервном
окончании запаса его молекул
Ацетилхолин (АЦХ) синтезируется в нервных
окончаниях ацетилированием холина ферментом
ацетилхолинтрансферазой (АХТ) из ацетил-КоА
как источника ацетиловой группы и холина. Об-
разующийся ацетилхолин накапливается в вези-
кулах с помощью механизма активного транс-
порта с использованием АТФ и высвобождается
в синаптическую щель с помощью экзоцитоза во
время генерации потенциалов действия в нерв-
ном окончании. После связывания ацетилхолина
с холинорецептором концевой пластинки медиа-
тор подвергается гидролизу ферментом холин-
эстеразой до ацетата и холина до того, как медиа-
тор успеет связаться с другими холинорецептора-
ми. Примерно половина холина захватывается
нервным окончанием и используется в синтезе
ацетилхолина.
в синаптических пузырьках
достаточно для проведения
примерно 10 000 импульсов.
Везикулы пресинаптической
терминали находятся в актив-
ной и депонированной формах.
Активные везикулы непосред-
ственно участвуют в передаче
возбуждения через нервно-мы-
шечное соединение, т. е. в вы-
делении медиатора в синапти-
ческую щель. Молекулы медиа-
тора депонированной формы
везикул постоянно пополняют
2. Физиология возбудимых тканей • 71
пул активной формы ме-
диатора по мере их ис-
пользования в синаптиче-
ской передаче. Число ве-
зикул является постоян-
ной величиной пресинап-
тической терминали и
обусловливает функцио-
нальную лабильность си-
напса (рис. 2.16).
После того как нерв-
ный импульс достигает
пресинаптического окон-
чания в фазу деполяриза-
ции потенциала действия,
в мембране окончания от-
крываются потенциалза-
висимые кальциевые ион-
ные каналы, и ионы Са2+
пассивно по градиенту
концентрации поступают
внутрь нервного оконча-
ния из внеклеточной сре-
ды. Движение ионов Са2+
обусловлено тем, что в по-
Рис. 2.17. Схема формирования кальциевого мик-
родомена и секретосомы.
В фазу деполяризации потенциала действия в активных
зонах пресинаптической мембраны открываются потен-
циалзависимые кальциевые ионные каналы. Ионы каль-
ция по градиенту концентрации поступают внутрь тер-
минали и образуют облако ионов (кальциевый микродо-
мен). В зоне кальциевого микродомена происходит фор-
мирование секретосомы и связывание везикулы с актив-
ной зоной пресинаптической мембраны.
кое их концентрация в ци-
топлазме нервного окончания составляет порядка 10“7 М, а на внешней
поверхности его мембраны, благодаря активности Са2+-АТФазы, поддер-
живается более высокая концентрация ионов кальция, чем внутри. В мо-
мент открытия кальциевых ионных каналов в активных зонах пресинапти-
ческой мембраны формируются микродомены (рис. 2.17). Микродомен
представляет собой область высокой концентрации ионов Са2+ вблизи
устья кальциевого ионного канала. В момент появления микродомена бел-
ки мембраны везикулы способны связывать везикулярную мембрану с ак-
тивной зоной пресинаптического окончания. При этом образуется специ-
фический белковый комплекс, состоящий из везикулы и элементов актив-
ной зоны пресинаптической мембраны, которые связаны между собой
специализированными белками экзоцитоза. Этот комплекс называется сек-
ретосомой. Формирование секретосомы является основным процессом,
обеспечивающим экзоцитоз ацетилхолина в нервном окончании.
Белки экзоцитоза локализованы как на мембране везикул (синапсин,
синаптотагмин, синаптобревин), так и на пресинаптической мембране
(синтаксин, синапсоассоциированный белок). Белок синаптотагмин явля-
ется кальциевым рецептором. После связывания ионов Са2+ с синаптотаг-
мином и последующего взаимодействия синаптобревина с синаптосомас-
социированным белком и синтаксином пресинаптической мембраны про-
исходит связывание везикулы с активной зоной пресинаптической мем-
браны. Экзоцитоз медиатора обеспечивается за счет АТФазной активности
специфического цитозольного белка. При высвобождении ацетилхолина в
синаптическую щель мембрана везикулы полностью сливается с пресинап-
тической мембраной и медиатор выходит в синаптическую щель через так
называемую пору. Белки экзоцитоза в синапсах периферической нервной
системы являются мишенями для токсинов при их действии на организм
72 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
человека и животных. Например, ботулиновый, столбнячный и другие ток-
сины способны нарушать процессы формирования секретосомы, что обу-
словливает появление специфического симптомокомплекса при отрав-
лении.
2.5.1.2. Диффузия ацетилхолина через синаптическую щель
нервно-мышечного синапса
Синаптическая щель нервно-мышечного синапса заполнена аморфной се-
тью соединительной ткани, в которой находится фермент ацетилхолин-
эстераза. Молекулы этого фермента синтезируются в мышечной клетке и
фиксируются к ее сарколемме со стороны синаптической щели.
Ацетилхолин путем экзоцитоза поступает в синаптическую щель и за-
тем диффундирует к постсинаптической мембране в течение 1 мс. Во вре-
мя диффузии ацетилхолина через синаптическую щель примерно ’/3 от вы-
делившегося количества медиатора разрушается ферментом ацетилхолин-
эстеразой на ацетат и холин, а 2/3 молекул ацетилхолина достигает постси-
наптической мембраны и связывается с N-холинорецепторами. Ацетилхо-
лин после отделения от рецептора расщепляется ацетилхолинестеразой на
холин и ацетат. Синтез, хранение и высвобождение ацетилхолина в нерв-
но-мышечном соединении показаны на рис. 2.16. Активность ацетилхоли-
нэстеразы столь высока, что период полураспада молекул ацетилхолина не
превышает 1 мс. Инактивация ацетилхолина в синаптической щели ис-
ключает многократное связывание одних и тех же молекул медиатора с хо-
линорецепторами постсинаптической мембраны, что предотвращает воз-
можность непрерывного сокращения мышц или так называемой мышечной
тетании.
2.5.1.3. Постсинаптический механизм
Постсинаптический механизм передачи возбуждения через нервно-мышеч-
ный синапс представляет собой трансформацию количества выделившего-
ся ацетилхолина в число потенциалов действия, которые генерирует сарко-
лемма мышечного волокна. В постсинаптической мембране нервно-мы-
шечного соединения расположены ионотропные N-холинорецепторы (чув-
ствительны к никотину). Плотность этих рецепторов выше, чем в постси-
наптической мембране любого синапса нервной системы (1 • 104 на
1 мкм2). Молекулы ацетилхолина кратковременно (не более 2 мс) взаимо-
действуют с N-холинорецепторами, в результате в постсинаптической
мембране открываются натриевые и калиевые ионные каналы. Ионы Na+
по электрохимическому градиенту перемещаются из синаптической щели
внутрь мышечного волокна, а ионы К+ — в обратном направлении. Транс-
мембранный ток для этих ионов рассчитываются по закону Ома:
4 = g*.(Em ~ Ек); /Na = gNa(Em - £Na),
где 1к и INtt — трансмембранный ток ионов калия и натрия, gK и gNa — про-
ницаемость постсинаптической мембраны соответственно для ионов калия
и натрия, Ет — потенциал покоя постсинаптической мембраны, Ек и
ENa — равновесный калиевый и натриевый потенциалы.
Поскольку gNa = gK, а электродвижущая сила (Em — ENa) для натрия
больше (равновесный потенциал для натрия +60 мВ, а для калия — 90 мВ),
входящий ток ионов натрия преобладает над выходящим калиевым током,
в результате происходит деполяризация постсинаптической мембраны
2. Физиология возбудимых тканей • 73
нервно-мышечного синапса, которая называется потенциалом концевой
пластинки. Продолжительность потенциала очень мала — не более 1—2 мс,
а его амплитуда варьирует в зависимости от количества высвобождающего-
ся ацетилхолина. При этом между амплитудой потенциала концевой пла-
стинки и количеством высвобождающегося ацетилхолина имеется прямая
зависимость.
Потенциал концевой пластинки электротонически распространяется
вдоль мышечного волокна. При этом, если потенциал концевой пластинки
достигает по величине порога возбуждения мышечной мембраны (—15—
20 мВ), то он вызывает в ней генерацию распространяющихся потенциа-
лов действия. В результате возбуждение с нервного окончания передается
на мышечную клетку. Если пресинаптического окончания достигает один
потенциал действия, то в синаптическую щель нервно-мышечного синапса
высвобождается в среднем 100 квантов ацетилхолина. В этом случае возни-
кает потенциал концевой пластинки, величина которого достаточна для
того, чтобы вызвать генерацию потенциалов действия в мышечной мем-
бране, прилегающей к концевой пластинке. Даже при снижении амплиту-
ды потенциала концевой пластинки на 75 % он способен вызывать возбу-
ждение в мышечном волокне, что является фактором высокой надежности
процесса передачи возбуждения в нервно-мышечном синапсе. Надежность
функции нервно-мышечного синапса обусловлена также значительным ко-
личеством высвобождаемого из нервного окончания ацетилхолина и высо-
кой плотностью N-холинорецепторов на концевой пластинке.
Как только потенциалы действия, распространяясь по мембране мы-
шечного волокна, достигают t-системы, а ионы Са2+ высвобождаются из
саркоплазматического ретикулума, в мышце начинается сокращение.
2.5.1.4. Восстановительные процессы структуры мембраны и функции
нервно-мышечного синапса после передачи возбуждения
После завершения синаптической передачи в нервно-мышечном синапсе
ионы Са2+ с помощью Са2+-АТФ-азы первично активным транспортом вы-
водятся из цитоплазмы нервного окончания во внеклеточную среду. Про-
цесс восстановления исходного градиента концентрации ионов Са2+ между
аксоплазмой нервной терминали и внеклеточной средой осуществляется с
помощью вторично активного транспорта при участии белка Na+/Ca2+ об-
менника. Молекулярная структура пресинаптической мембраны восстанав-
ливается в результате эндоцитоза везикул, участвовавших в синаптической
передаче возбуждения. В нервном окончании образовавшиеся везикулы за-
полняются вновь синтезированными молекулами ацетилхолина. На пост-
синаптической мембране, благодаря работе Ыа+/К2+-АТФазы, восстанав-
ливаются ионные градиенты Na+ и К+, в результате возбудимость постси-
наптической мембраны возвращается к исходному уровню.
2.5.2. Проведение возбуждения через аксосоматический синапс
В нервной системе проведение возбуждения осуществляется между отрост-
ками нейронов и между отростками и телами нервных клеток. Так, прове-
дение возбуждения с аксона одного нейрона на аксон другого происходит
при участии аксо-аксональных синапсов. Передача импульсов от аксона
нейрона на на дендриты другой нервной клетки осуществляется аксоденд-
ритными синапсами, а от одних дендритов на другие — с помощью денд-
ро-дендритных синапсов. К наиболее часто встречающемуся в нервной
74 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Микротрубочки
Рис. 2.18. Схема строения и функции химического синапса.
Предшественник медиатора попадает в терминаль нейрона из крови или ликвора, под дейст-
вием специализированного фермента превращается в медиатор и активным транспортом нака-
пливается в синаптических везикулах. Медиатор после экзоцитоза или после выделения ко-
медиатора (КМ) взаимодействует с ионотропным (И) или метаботропным (М) рецептором
постсинаптической мембраны. В результате изменяется проницаемость этой мембраны для
ионов Na+, К+, Са2+, СГ и возникает локальная де- или гиперполяризация путем непосредст-
венного изменения ионной проницаемости, либо при участии вторичного посредника. Медиа-
тор разрушается ферментом в синаптической щели, захватывается активным транспортом в
пресинаптическое окончание либо в клетки нейроглии (пунктирные стрелки).
системе пути передачи импульсов между нейронами относится аксосома-
тический синапс, т. е. от аксона одного нейрона на тело другого. Количе-
ство синапсов на дендритах и теле одного нейрона достигает нескольких
тысяч. В качестве примера может быть рассмотрен аксосоматический си-
напс, т. е. синапс между окончаниями одной нервной клетки и телом дру-
гой (рис. 2.18).
Аксосоматический синапс образован пресинаптическим окончанием
аксона, синаптической щелью, за которой расположена постсинаптическая
мембрана. Передача импульсов в аксосоматических синапсах осуществля-
ется с помощью медиаторов, которые называются нейротрансмиттерами.
В химической передаче возбуждения между нейронами участвуют различ-
ные типы нейротрансмиттеров: глутамат, ацетилхолин, дофамин, норадре-
налин, адреналин, серотонин, гамма-аминомасляная кислота (ГАМК),
нейропептиды. При передаче возбуждения через аксосоматический синапс
в постсинаптическом нейроне возникает два типа ответных реакций: быст-
рые и медленные. Быстрые реакции в постсинаптических нейронах возни-
кают в случаях, если медиатор связывается с рецепторами ионных каналов
постсинаптической мембраны и, непосредственно вызывая их открытие, в
миллисекундный интервал передает возбуждение на постсинаптический
нейрон. При этом возникают быстрые реакции постсинаптической мем-
браны в виде увеличения ионной проницаемости и ее деполяризации. К
быстрым реакциям увеличения ионной проницаемости постсинаптической
2. Физиология возбудимых тканей • 75
мембраны аксосоматических синапсов относится открытие ионных кана-
лов постсинаптической мембраны после образования внутриклеточных
вторичных посредников (описаны ниже). При этом типе быстрой реакции
нейротрансмиттеры (адреналин, норадреналин, дофамин, гистамин, серо-
тонин) связываются на постсинаптической мембране с рецепторами систе-
мы G-белков. В результате активации системы G-белков в постсинаптиче-
ском нейроне образуются внутриклеточные вторичные посредники, кото-
рые через рецепторы ионных каналов открывают их и увеличивают таким
образом ионную проницаемость постсинаптической мембраны аксосома-
тических синапсов. При передаче возбуждения через аксосоматический
синапс возникают медленные или нейромодуляторные реакции постсинап-
тических нейронов. Они также обусловлены взаимодействием нейротранс-
миттеров с рецепторами системы G-белков постсинаптической мембраны,
но в этом случае при участии внутриклеточных вторичных посредников
возникают медленные, а именно — метаболические реакции постсинапти-
ческих нейронов. Данный способ передачи информации через синапсы
нервной системы присущ большой группе нейротрансмиттеров, которые
называются нейропептидами.
По аналогии с нервно-мышечным синапсом в аксосоматическом си-
напсе импульс предается с помощью пре- и постсинаптического меха-
низмов.
2.5.2.1. Функция пресинаптического окончания нейронов
В нервных терминалях локализовано два типа везикул — мелкие и круп-
ные. Мелкие везикулы терминалей нейронов центральной и перифериче-
ской нервной системы содержат разные группы медиаторов: амины (аце-
тилхолин, дофамин, норадреналин, серотонин и гистамин), аминокислоты
(глютамат, глицин, гамма-аминомасляная кислота) и пурины (АТФ, адено-
зин). Мелкие везикулы в пресинаптической терминали находятся в актив-
ной и депонированной формах. Активные везикулы непосредственно участ-
вуют в экзоцитозе медиатора, депонированная форма везикул пополняет
пул активной по мере ее использования в синаптической передаче
(рис. 2.19). Медиаторы накапливаются в везикулах с помощью механизма
активного транспорта. Крупные или пептидные везикулы нервных терми-
налей центральной и периферической нервной систем содержат нейроак-
тивные пептиды (субстанция Р, вазопрессин, окситоцин, кортиколиберин,
тиреолиберин, соматолиберин, гонадолиберин, энкефалины, ангиотензин-
2 и др.). В отличие от мелких везикул пептидные синтезируются в соме
нейронов, наполняются молекулами пептидов и транспортируются с по-
мощью аксотока в нервное окончание. Функции медиаторов мелких и
крупных везикул взаимосвязаны между собой в процессе синаптической
передачи: первые обеспечивают химическую передачу возбуждения через
синапс, а вторые — модулируют, т. е. усиливают или уменьшают этот про-
цесс. Генерация потенциала действия на мембране пресинаптической тер-
минали активирует кальцийзависимый процесс освобождения или экзоци-
тоза медиатора. Быструю, кратковременную (доли миллисекунды) и ло-
кальную передачу возбуждения между нейронами обеспечивает экзоцитоз
классических нейротрансмиттеров, содержащихся в мелких везикулах
нервной терминали. Экзоцитоз нейропептидов из нервных терминалей
может осуществляться в течение многих секунд и минут при активации
различных внутриклеточных энзимных систем. Благодаря этой особенно-
сти экзоцитоза нейропептидов, они могут модулировать экзоцитоз класси-
76 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Рис. 2.19. Везикулярный механизм эк-
зоцитоза медиатора из нервной терми-
нали.
Везикулы (1) формируются в теле нейрона,
транспортируются в нервное окончание, где
заполняются медиатором (2). При экзоци-
тозе, с помощью механизма полного слия-
ния и с последующим образованием покры-
тых везикул (3), запасы медиатора пополня-
ются через стадию эндосом. При неполном
слиянии и повторном заполнении медиато-
ром везикулы сразу могут участвовать в эк-
зопитозе (4). Крупные везикулы заполня-
ются пептидами в теле нейрона (5), их эк-
зопитоз происходит в других зонах нервно-
го окончания (6).
ческих нейротрансмиттеров в ней-
ронах на протяжении продолжи-
тельного интервала времени, уси-
ливая или замедляя передачу импульсов через аксосоматический синапс. В
аксосоматическом синапсе экзоцитоз классических медиаторов осуществ-
ляется в активных зонах пресинаптического окончания. Функциями ак-
тивных зон в нервных терминалях (как и в нервно-мышечном синапсе)
является мобилизация медиатора из депонированной формы в активную
при участии элементов цитоскелета. В пределах активной зоны пресинап-
тической мембраны происходит взаимодействие между везикулами и ме-
стом экзоцитоза медиаторов. При участии активных зон пресинаптической
мембраны экзоцитоз происходит только в момент повышения концентра-
ции ионов кальция в цитоплазме пресинаптической терминали. Наконец,
активные зоны нервной терминали участвуют в процессах восстановления
целостности синаптических пузырьков, заполнения их молекулами медиа-
торов, что обусловливает число везикул, находящихся в активной и депо-
нированной формах.
2.5.2.2. Пресинаптический механизм проведения возбуждения
Пресинаптический механизм проведения возбуждения осуществляет
трансформацию энергии нервных импульсов в кванты выделяющегося ме-
диатора. В фазу деполяризации потенциала действия в мембране нервной
терминали открываются потенциалзависимые кальциевые ионные каналы,
и ионы кальция по концентрационному градиенту поступают из внекле-
точной среды внутрь нервного окончания. Ионы Са2+ вблизи внутренней
поверхности пресинаптической мембраны образуют микродомен (см.
рис. 2.17) и связываются со специфическим везикулярным белком каль-
циевым сенсором — синаптотагмином. Последовательное взаимодействие
специфических белков экзоцитоза, локализованых на мембране везикул
(синапсин, синаптотагмин, синаптобревин) и на пресинаптической мем-
бране (синтаксин, синапсоассоциированный белок) связывает везикулу с
активной зоной пресинаптической мембраны, формируя секретосому. При
этом мембрана везикулы полностью сливается с пресинаптической мем-
браной, и медиатор освобождается в синаптическую щель через пору. В си-
напсах нервной системы экзоцитоз медиаторов может проходит через по-
стоянную или временную пору. В первом случае происходят полное слия-
2. Физиология возбудимых тканей • 77
ние мембран везикулы и пресинаптического окончания и высвобождение
всего количества молекул медиатора из везикул. Во втором случае не про-
исходит полного слияния мембран везикулы и пресинаптической термина-
ли. При образовании временной поры освобождается только часть медиа-
тора в синаптическую щель, а затем везикула отделяется от активной зоны
и возвращается в цитоплазму нервной терминали. Этот механизм позволя-
ет одной и той же везикуле без пополнения запаса медиатора неоднократ-
но участвовать в экзоцитозе.
После высвобождения молекулы нейротрансмиттера диффундируют от
мест экзоцитоза к постсинаптической мембране. Во время диффузии часть
медиатора может подвергаться ферментному разрушению в синаптической
щели или захватываться обратно внутрь нервного окончания. Постсинап-
тической мембраны могут достигать все или только часть молекул ней-
ротрансмиттеров, что обусловливает соответственно большее или меньшее
количество передаваемых через аксосоматический синапс импульсов.
2.5.2.3. Пресинаптическая регуляция экзоцитоза медиаторов
Пресинаптическая регуляция экзоцитоза медиаторов осуществляется с по-
мощью специфического типа ионного канала, который идентичен калие-
вым ионным каналам мембран возбудимых клеток, но открываются иона-
ми кальция. Такие ионные каналы пресинаптической мембраны называ-
ются кальцийзависимыми калиевыми каналами. Эти ионные каналы распо-
ложены в активных зонах пресинаптической мембраны нервной термина-
ли в непосредственной близости от кальциевых ионных каналов. Физиоло-
гические свойства подобного типа ионных каналов заключается в том, что
они быстро активируются при увеличении концентрации ионов кальция
на внутренней поверхности пресинаптической мембраны, т. е. в момент
формирования микродомена. При открытии кальцийзависимых калиевых
ионных каналов происходит выход из нервного окончания ионов калия по
концентрационному градиенту. Это локально реполяризует пресинаптиче-
скую мембрану нервной терминали, что ограничивает длительность фазы
деполяризации в тех участках пресинаптической мембраны, где происхо-
дит высвобождение медиатора. В результате уменьшения длительности фа-
зы деполяризации пресинаптической мембраны нервной терминали распо-
ложенные в ней кальциевые ионные каналы открываются на меньший от-
резок времени. Это ведет к уменьшению количества ионов кальция, посту-
пающих внутрь нервной терминали, а следовательно, уменьшаются коли-
чество высвобождаемого экзоцитозом медиатора и передача сигналов через
синапс.
Освобождение медиаторов из нервных терминалей изменяется (модули-
руется) субстанциями, содержащимися в интерстициальной жидкости, ок-
ружающей терминаль. Механизм регуляции экзоцитоза медиаторов из
нервных терминалей осуществляется через рецепторы, локализованные на
пресинаптической мембране. Эти рецепторы (N-холинергические, глюта-
матные, ГАМК-ергические и пуринергические Р2-рецепторы) ассоцииро-
ваны с ионными каналами и могут активироваться медиатором, который
выделяется из этого же нервного окончания, либо медиаторами, высвобо-
ждающимися из близлежащих нервных окончаний.
Вещества, которые оказывают модуляторный эффект на передачу им-
пульсов в синапсах нервной системы, в результате которой увеличивается
или уменьшается частота потенциалов действия постсинаптических нейро-
нах, называются нейромодуляторами.
78 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Экзоцитоз ацетилхолина из нервной терминали стимулируется связыва-
нием выделившегося в этом же синапсе ацетилхолина с N-холинорецепто-
рами пресинаптической мембраны аксосоматического синапса. Глютамат-
ные рецепторы пресинаптической мембраны аксосоматического синапса
(NMDA, агонист N-метил-О-аспартат; АМРА, агонист альфа-амино-3-
гидрокси-5-метил-4-изоксазолпропионовая кислота и каинатные — аго-
нист каиновая кислота) участвуют в разных синапсах в стимуляции или
торможении экзоцитоза других медиаторов. Например, NMDA-рецепторы
пресинаптической мембраны нервных терминалей, оканчивающихся на
нейронах коры головного мозга, стимулируют процесс экзоцитоза медиа-
тора ацетилхолина, а в нервных терминалях гиппокампа — медиатора но-
радреналина.
На пресинаптической мембране нервных терминалей нейронов цен-
тральной нервной системы распространен А, тип аденозиновых рецепто-
ров, которые связываются с нейромодулятором аденозином. Концентра-
ция аденозина во внеклеточной среде возрастает по мере активации пост-
синаптического нейрона. Функцией типа А] аденозиновых рецепторов
пресинаптической мембраны нервной терминали является уменьшение
экзоцитоза медиаторов (ацетилхолина, дофамина), что снижает актив-
ность постсинаптических нейронов, из которых в окружащую их внекле-
точную жидкость поступает аденозин. Тип А2А-аденозиновых рецепторов
локализован на пресинаптической мембране нейронов тех отделов мозга,
которые регулируют движения человека при участии медиатора дофамина
(хвостатое ядро и подушка таламуса). Активация А2а рецепторов преси-
наптической мембраны стимулирует высвобождение ацетилхолина из
нервных терминалей и вызывает усиление функции постсинаптического
нейрона.
2.5.2.4. Постсинаптический механизм проведения возбуждения
Постсинаптический механизм проведения возбуждения в аксосоматиче-
ском синапсе нервной системы осуществляет передачу нервного импульса
на постсинаптический нейрон (быстрые реакции). Передача нервного им-
пульса от одного нейрона к другому осуществляется в течение короткого
отрезка времени (мс). Этот процесс на этапе постсинаптического механиз-
ма передачи возбуждения осуществляется при участии ионотропных ре-
цепторов постсинаптической мембраны аксосоматического синапса. Ионо-
тропные рецепторы представляют собой часть молекулярной структуры хе-
мозависимых ионных каналов (натриевых, калиевых, кальциевых) постси-
наптической мембраны нейронов. Количество ионотропных рецепторов на
постсинаптической мембране нейронов является регулируемой величиной:
оно повышается в ответ на длительное воздействие на мембрану нейрона
малой концентрации молекул медиаторов (явление сенситизации). Напро-
тив, число рецепторов на постсинаптической мембране понижается в от-
вет на длительное воздействие на постсинаптическую мембрану медиато-
ров в высокой концентрации (явление десенситизации). Вещества, которые
связываются с рецепторами хемозависимых ионных каналов постсинапти-
ческой мембраны, называются лигандами. Лиганды подразделяются на аго-
нисты и антагонисты. Первые активируют, а вторые блокируют актив-
ность рецепторов хемозависимых ионных каналов постсинаптической
мембраны.
Результатом взаимодействия медиаторов с ионотропными рецепторами
постсинаптической мембраны является ее кратковременная деполяризация
2. Физиология возбудимых тканей • 79
или гиперполяризация постсинаптической мембраны. Деполяризация
постсинаптической мембраны в области одного синапса называется возбу-
ждающим постсинаптическим потенциалом (ВПСП). Химические синап-
сы, в которых при передаче возбуждения возникают ВПСП, называются
возбуждающими. По аналогии медиатор, вызывающий возникновение
ВПСП, называется возбуждающим медиатором. Возникновение ВПСП
обусловлено открытием преимущественно натриевых ионных каналов.
При ВПСП происходит деполяризация постсинаптической мембраны и
уменьшение порога ее возбудимости. Амплитуда ВПСП прямо зависит от
количества молекул медиатора взаимодействующих с рецепторами постси-
наптической мембраны, а также последовательностью выделения квантов
медиатора из пресинаптического окончания.
Результатом взаимодействия медиаторов с ионотропными рецепторами
хемозависимых ионных каналов постсинаптической мембраны может быть
ее кратковременная гиперполяризация. В результате понижается возбуди-
мость постсинаптического нейрона и в нем не происходит генерация по-
тенциала действия. Гиперполяризация постсинаптической мембраны в об-
ласти одного синапса называется тормозным постсинаптическим потен-
циалом (ТПСП), такие химические синапсы называются тормозными. По
аналогии медиаторы, вызывающие ТПСП, называются тормозными медиа-
торами. Взаимодействие тормозного медиатора с рецепторами постсинап-
тической мембраны открывает хлорные и калиевые ионные каналы, что
резко повышает ионную проводимость постсинаптической мембраны для
ионов хлора (/с/) или калия (7^). В соответствии с электрохимическими
градиентами ионов калия и хлора по обе стороны от постсинаптической
мембраны, ТПСП представляет собой сдвиг потенциала на постсинаптиче-
ской мембране в сторону увеличения его величины. В результате происхо-
дит гиперполяризация постсинаптической мембраны и увеличение порога ее
возбудимости.
При передаче возбуждения через аксосоматические синапсы на мембра-
не одного нейрона одновременно возникают ВПСП и ТПСП. При этом
происходит сложение (суммация) амплитуды ВПСП и ТПСП многих си-
напсов. Если в условиях суммации ВПСП и ТПСП суммарная амплитуда
ВПСП достигает критического уровня деполяризации, то в мембране ак-
сонного холмика нейрона электротонически открываются потенциалзави-
симые натриевые ионные каналы и генерируется потенциал действия. В
результате возбуждение передается от нервных терминалей пресинаптиче-
ского нейрона на постсинаптический нейрон.
При передаче возбуждения через аксосоматический синапс с пресинап-
тического нейрона на постсинаптический происходит синаптическая за-
держка скорости проведения электрического импульса. Синаптическая за-
держка в возбуждающих аксосоматических синапсах представляет собой
время от момента прихода потенциала действия к пресинаптическому
окончанию до начала деполяризации постсинаптической мембраны, т. е.
ВПСП. Время синаптической задержки в возбуждающих синапсах в сред-
нем составляет 0,5 мс, и оно прямо зависит от скорости процессов экзоци-
тоза медиатора из нервного окончания, диффузии медиатора через синап-
тическую щель к постсинаптической мембране, соединения медиатора с
рецепторами постсинаптической мембраны и последующего увеличения
ионной проницаемости постсинаптической мембраны. Время синаптиче-
ской задержки обусловливает функциональную подвижность или лабиль-
ность химического синапса. Лабильность химического синапса ниже, чем
нервного волокна, но выше, чем мышечного.
80 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.5.2.5. Функции метаботропных рецепторов постсинаптической мембраны
аксосоматического синапса
На постсинаптической мембране аксосоматических синапсов нервной сис-
темы локализованы хемозависимые метаботропные рецепторы. Функция
хемозависимых метаботропных рецепторов постсинаптической мембраны
состоит в регуляции метаболической активности постсинаптических ней-
ронов при передаче возбуждения через химический синапс (медленная ре-
акция). Эта функция метаботропных рецепторов постсинаптической мем-
браны обусловлена свойствами нескольких высокомолекулярных семейств
G-белков (Gs, Gi и Gq) либо низкомолекулярных семейств G-белков (ras,
rho, rab и др.), с которыми ассоциированы метаботропные рецепторы
постсинаптической мембраны (рис. 2.20).
Как показано на рис. 2.20, метаботропный рецептор постсинаптической
мембраны находится на стороне молекулы G-белка, обращенной в синап-
тическую щель. Соединение нейротрансмиттера с хемозависимым метабо-
тропным рецептором постсинаптической мембраны вызывает активацию
G-белков. При этом тример G-белков, состоящий из а-субъединицы, ас-
социированной с Ру-димером, диссоциирует на а-субъединицу и Ру-димер
с последующим образованием комплекса гуанинтрифосфат-Ga и Gpy-ди-
мер. Функция этих двух соединений состоит в активации внутриклеточных
энзимов постсинаптической мембраны, например фосфолипазы А2, фос-
фолипазы С, при участии которых в постсинаптических нейронах образу-
ются основные внутриклеточные регуляторы их метаболической активно-
сти — вторичные посредники.
Функция вторичных посредников в постсинаптическом нейроне. Основ-
ными вторичными посредниками в постсинаптических нейронах являют-
ся циклический аденозинмонофосфат, инозитол-3-фосфат/диацилглицерол и
ионы кальция. Циклический АМФ образуется в цитоплазме клеток при
участии активированной аденилатциклазы из АТФ. Посредник диффун-
дирует в пределах постсинаптической клетки, активирует протеинкиназы,
Рис. 2.20. Схема активации G-белка при взамодействии лиганда с метаботропным
рецептором постсинаптической мембраны.
Связывание лиганда с рецептором G-белка вызывает диссоциацию G-белка на гуанозинтри-
фосфат, связанный с a-субъединицей, и на ру-димер. a-Субъединица и py-димер могут взаи-
модействовать в клетке с эффекторными молекулами, такими как ионные каналы или связан-
ные с мембраной энзимы, и изменять их активность.
2. Физиология возбудимых тканей • 81
Мембрана клетки
Сигнальная
молекула Фосфолипид
ЭР
Рецептор,
связанный с
G-протеином
Активация
различных
протеинов
>ДАГ
'‘ИФЗ
Вторичные
посредники
Ga-белок
-> Са
Вторичный
ИФЗ-воротный посредник
кальциевый Реакции клетки
канал
Рис. 2.21. Схема образования вторичных посредников инозитол-3-фосфата/диа-
цилглицерола и ионов кальция при активации системы G-белка сигнальной моле-
кулой. Инозитол-З-фосфат/диацилглицерол, а также ионы кальция являются вто-
ричными посредниками сигнальных молекул в возбудимых клетках.
Фс — фосфолипаза С, ИФЗ — инозитол-3-фосфат, ЭР — эндоплазматический ретикулум, Ga-
белок — активированный G-белок, ДАГ — диацилглицерол.
вызывающие фосфорилирование внутриклеточных ферментов, которые
обусловливают метаболический эффект клеток. Системы внутриклеточ-
ных посредников на основе фосфоинозитида включают активацию энзи-
ма фосфолипазы С и образование двух метаболитов: инозитол-3-фосфата
и диацилглицерола (рис. 2.21). Диацилглицерол функционирует в преде-
лах клеточной мембраны и активирует киназы, открывающие ионные ка-
налы постсинаптической мембраны нейрона, а инозитол-3-фосфат взаи-
модействует с рецепторами кальциевых ионных каналов эндоплазматиче-
ского ретикулума и вызывает выход ионов Са2+ в цитоплазму клеток. Ио-
ны Ссг+ в качестве вторичного посредника активируют в нейронах мозга
процессы нейросекреции, а в возбудимых мышечных клетках — их сокра-
щение.
Примерно 60 % медикаментов, которые применяются в современной
медицине, оказывают свой терапевтический эффект на клетки, связываясь
с мембранными метаботропными рецепторами, ассоциированными с сис-
темой G-белков.
2.5.3. Проведение возбуждения в основных типах синапсов
центральной нервной системы
В нервных терминалях нейронов центральной и периферической нервной
системы в зависимости от типа нейротрансмиттера, с помощью которого
возбуждение проводится через синапс и/или регулируется метаболическая
активность постсинаптического нейрона, различают холинергический, до-
фаминергический, серотонинергический, гистаминергический, глутаматер-
гический, ГАМКергический и глицинергический синапсы центральной
нервной системы.
82 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.5.3.1. Холинергический синапс (рис. 2.22)
Ацетилхолин является нейротрансмиттером в нервной системе. Медиатор
синтезируется в нервных окончаниях из ацетил-коэнзима А при участии
фермента ацетилтрансферазы или холинацетилазы. Фермент синтезируется
в теле нейрона и транспортируется аксоплазматическим током в нервные
окончания. Холин накапливается в нервных окончаниях в результате его
захвата из интерстициальной жидкости. При этом количество холина, за-
хватываемого нервными окончаниями, обусловливает количество синтези-
руемого ацетилхолина в нервных окончаниях. Ацетилхолин накапливается
в везикулах нервных окончаний и высвобождается квантами в синаптиче-
скую щель во время генерации потенциалов действия в нервных оконча-
ниях. Физиологический эффект ацетилхолина на постсинаптический ней-
рон является результатом действия нейротрансмиттера на рецепторы пост-
синаптической мембраны. На постсинаптической мембране холинергиче-
ского синапса имеются N- и М-типы (подтипы М,—М4) рецепторов аце-
тилхолина. N-тип рецепторов чувствителен к никотину, а М-тип — к мус-
карину, что послужило основанием для обозначения буквами N и М этих
двух типов холинорецепторов постсинаптической мембраны.
Связывание ацетилхолина с ионоторопными N- холинорецепторами
открывает ионные каналы Са2+ и Na+, ионы поступают внутрь клетки,
вызывая быструю деполяризацию (ВПСП) постсинаптической мембраны.
Связывание ацетилхолина с ионотропаными М-холинорецепторами пост-
синаптической мембраны открывает кальциевые и калиевые ионные ка-
налы постсинаптической мембраны через систему вторичных посредни-
ков инозитол-3-фосфата и диацилглицерола (М,-тип рецепторов) и цик-
лического АМФ (М2-тип рецепторов) соответственно: ионы Са2+ поступа-
ют через кальциевые каналы постсинаптической мембраны холинергиче-
ского синапса внутрь нейрона, а ионы К+ выходят из него, вызывая ги-
Мускариновый
о
• Na
Na
Синаптическая
щель
/1) HI 11 111Ш1И 'll II» ' Постсинаптическая
I II II IllUHi НН» I 1 " •'-''-’•’ч-»'’
' мембрана
Никотиновый
постсинаптический
рецептор
Ацетилхолин
Ацетат
о
Холин
Мускариновый
постсинаптический
рецептор
Na
Na
+
Na
Na
Na
Рис. 2.22. Схематическое изображение холинергического синапса.
На постсинаптической мембране локализованы хемозависимые ионотропные и метаботроп-
ные рецепторы, при участии которых не только возбуждение проводится через синапс (ионо-
тропные рецепторы), но и может регулироваться метаболическая активность постсинаптиче-
ского нейрона (М-холинорецептор).
2. Физиология возбудимых тканей • 83
перполяризацию мембраны. Эффекты изменения ионной проницаемости
постсинаптической мембраны при участии ацетилхолина, М-холиноре-
цепторов и вторичных посредников во времени являются более длитель-
ными и отставлены, чем при активации ионотропных N-холинорецепто-
ров. Благодаря долговременному механизму действия ацетилхолина на
постсинаптические нейроны, этот нейротрансмиттер участвует в механиз-
мах памяти и обучения в нейронах гиппокампа и коры больших полуша-
рий мозга.
2.5.3.2. Адренергический синапс
Адренергический синапс осуществляет передачу возбуждения при участии
медиаторов адреналина или норадреналина как в центральной нервной
системе, так и в периферических тканях. В мозге норадренергические ней-
роны, в нервных терминалях которых высвобождается медиатор норадре-
налин, сконцентрированы, в основном, в голубом пятне моста. Ответвле-
ния аксонов этих нейронов встречаются во всех отделах ЦНС и норадре-
налин модулирует активность различных по функции нервных клеток. В
ЦНС и периферических тканях адренергические синапсы функционируют
при участии а (подтипы а,, а2) и р (подтипы (р1; р2 и р3) типов адренерги-
ческих рецепторов.
cq-Адренорецепторы ЦНС (гиппокамп, кора головного мозга) и пери-
ферической нервной системы осуществляют возбуждающую функцию,
увеличивая ионную проницаемость хемозависимых ионотропных кальцие-
вых каналов.
а2-Ддренорецепторы выполняют тормозную роль в ЦНС и перифериче-
ской нервной системе. В ЦНС а2-адренорецепторы расположены на пост-
синаптической мембране, представляющей собой мембрану нервной тер-
минали, из которых высвобождаются нейротрансмиттеры норадреналин и
серотонин. Торможение высвобождения в нервных терминалях норадрена-
лина и серотонина при участии а2-адренорецепторов осуществляется путем
модуляции ионной проницаемости кальциевых и калиевых каналов при
участии системы G-белков, аденилатциклазы/цАМФ. Прямое увеличение
проницаемости калиевых ионных каналов мембраны нервной терминали
гиперполяризует пресинаптическую мембрану и тормозит экзоцитоз ме-
диатора. Это, в свою очередь, уменьшает количество высвобождающегося
медиатора (норадреналина или серотонина) в адренергических или серото-
нинергических синапсах ЦНС, модулируя функцию постсинаптических
нейронов.
В центральной нервной системе представлены Р^адренорецепторы, ко-
торые идентифицированы на мембранах нейронов полосатых тел базаль-
ных ганглиев мозга. Взаимодействие норадреналина с Р^адренорецептора-
ми через систему вторичных посредников (система G-белков, аденилат-
циклаза/цАМФ) ведет к модуляции ионной проницаемости кальциевых и
калиевых ионных каналов постсинаптической мембраны. Активация сис-
темы G-белков непосредственно увеличивает проницаемость кальциевых
ионных каналов и повышает возбудимость и функцию нейронов полоса-
тых тел.
Наиболее важная функция норадреналина в ЦНС заключается в регуля-
ции пробуждения в цикле «сон—бодрствование» (совместно с серотони-
нергическими структурами ядер «шва» ствола мозга).
84 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.5.3.3. Дофаминергический синапс
Дофаминергический синапс нервной системы осуществляет передачу воз-
буждения от нервной терминали на постсинаптический нейрон с помощью
медиатора дофамина. Постсинаптическая мембрана синапсов, в которых
химическим передатчиком возбуждения является медиатор дофамин, со-
держит D,-D5 типы рецепторов, каждый из которых связан с системой
внутриклеточных посредников аденилатциклаза — цАМФ (рис. 2.23).
Взаимодействие дофамина с Dj рецепторами постсинаптической мем-
браны через стимулирующий Gs-белок активирует аденилатциклазу, кото-
рая приводит к повышению уровня цАМФ. Функция вторичного посред-
ника цАМФ заключается в открытии кальциевых ионных каналов постси-
наптической мембраны. Диффузия ионов кальция внутрь нейрона по их
концентрационному градиенту деполяризует постсинаптическую мембрану
и повышает возбудимость нейрона.
Взаимодействие дофамина с D2 рецепторами постсинаптической мем-
браны через тормозящий Gi-белок прямо или опосредованно путем умень-
шения внутриклеточного уровня цАМФ модулирует калиевые ионные ка-
налы и опосредованно — кальциевые ионные каналы постсинаптической
мембраны, вызывая ее гиперполяризацию и уменьшение возбудимости
нейрона. Например, при активации нейронов дофаминергической системы
мозга дофамин в нейронах передней доли гипофиза уменьшается содержа-
ние внутриклеточных вторичных посредников. Это вызывает уменьшение
внутриклеточной концентрации ионов кальция, что является основным
механизмом ограничения секреции из клеток передней доли гипофиза гор-
мона пролактина, а следовательно, лактации молока. Блокада D2 рецепто-
ров нейронов мозга специфическимии агонистами используется в медици-
проницаемости
проницаемости
кальциевых каналов
калиевых каналов
Пресинаптическая
мембрана
Синаптическая
щель
Постсинаптическая
мембрана
Рис. 2.23. Схематическое изображение дофаминергического синапса;
Дофамин через метаботропные рецепторы постсинаптической мембраны и систему G-протеи-
нов регулирует активность различных хемозависимых ионных каналов. Дофамин после взаи-
модействия с рецепторами постсинаптической мембраны транспортируется в пресинаптиче-
ско окончание. D,—D5 — различные типы дофаминовых рецепторов.
2. Физиология возбудимых тканей • 85
не при лечении шизофрении. Напротив, при нарушении функции дофами-
нергической системы мозга и ограничением синтеза в ее нейронах медиа-
тора дофамина, в связи с деградацией нейронов «черной субстанции»
среднего мозга, у человека возникают ограничение произвольных движе-
ний и дрожание конечностей в покое (синдром Паркинсона).
Рецепторы к дофамину располагаются на мембране пресинаптического
окончания, из которого выделяется этот нейротрансмиттер. После экзоци-
тоза дофамина в дофаминергическом синапсе его молекулы диффундируют
не только к постсинаптической мембране, но и действует на дофаминовые
рецепторы мембраны пресинаптического окончания. При участии вторич-
ного посредника цАМФ на пресинаптической мембране дофаминергическо-
го синапса открываются калиевые ионные каналы, что вызывает выход этих
ионов из цитоплазмы нервной терминали в интерстициальную среду. В ре-
зультате развивается гиперполяризация пресинаптической мембраны, что
тормозит генерацию потенциалов действия в нервном окончании и снижает
экзоцитоз дофамина из пресинаптической терминали, а следовательно,
уменьшает влияние медиатора на функцию постсинаптического нейрона.
2.5.3.4. Серотонинергический синапс
Серотонин является нейротрансмиттером как для центральной нервной
системы, так и для клеток периферических тканей. На постсинаптической
мембране серотонинергических синапсов нейронов мозга локализовано
семь типов рецепторов к серотонину — 5-НТ!_7. Основными являются ти-
пы 5-НТ]_4. Рецепторы на постсинаптической мембране к серотонину от-
носятся к ионотропному (5-НТ3) и метаботропному типам (остальные).
Время действия серотонина при передаче возбуждения через серотонинер-
гический синапс обусловлено обратным захватом медиатора в нервную
терминаль, где серотонин разрушается ферментом моноаминоксидазай.
Подтипы семейства рецепторов 5-HTj (5-HTIA, 5-НТ1В, 5-HT1D, 5-НТ1Е,
5-HT1F), ассоциированные с G-белками, широко представлены в нейронах
ЦНС (лимбическая система, лобная кора, базальные ганглии). При взаи-
модействии серотонина с 5-HTj рецепторами при участии аденилатцикла-
зы и вторичного посредника цАМФ повышается проводимость калиевых
ионных каналов постсинаптической мембраны и уменьшается ионная про-
ницаемость кальциевых каналов. В результате происходит гиперполяриза-
ция постсинаптической мембраны и понижение возбудимости нейронов и
торможение их функции.
Рецепторы 5-НТ2 представлены в периферических тканях и участвуют в
сокращении гладких мышц сосудов, кишечника, мочевыводящих путей,
агрегации тромбоцитов и увеличении проницаемости капилляров. Дейст-
вие серотонина на 5-НТ2 тип метаборецепторов постсинаптической мем-
браны обусловлено образованием через активацию системы G-белков
внутриклеточного вторичного посредника — инозитол-3-фосфата. Инози-
тол-3-фосфат вызывает открытие кальциевых ионных каналов саркоплаз-
матического ретикулума, что повышает внутриклеточную концентрацию
этих ионов. В результате ионы кальция вызывают мышечное сокращение.
5-НТ3 рецепторы имеются в ядрах продолговатого мозга (ядро одиноч-
ного пучка, ядра тройничного и блуждающего нервов), а также в нейронах
сенсорных и энтеральной нервных систем, на пре- и постганглионарных
нейронах вегетативной нервной системы. Действие серотонина на 5-НТ3
тип рецепторов избирательно повышает проницаемость катионных кана-
лов (Na+ и Са2+), вызывая деполяризацию постсинаптической мембраны
86 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
нейронов, повышение возбудимости нервных клеток и активацию их
функции. Это приводит к усилению сократительной функции миокарда,
расширению сосудов, стимуляции легочной и интестинальных функций,
возникновению болевых ощущений.
Рецепторы 5-НТ4 распространены в структурах мозга, в которых основ-
ным медиатором является дофамин (базальные ганглии, прилегающее ядро
лимбической системы). Рецепторы локализованы также на пресинаптиче-
ской мембране ГАМКергических интернейронов, в которых они облегчают
высвобождение основного тормозного медиатора центральной нервной
системы гамма-аминомасляной кислоты. Однако в нервных окончаниях
этот же тип рецепторов пресинаптической мембраны холинергических ин-
тернейронов тормозит высвобождение ацетилхолина.
В мозге серотонинергические нейроны концентрируются в участке
среднего мозга, который называется «шов». Аксоны серотонинергических
нейронов ядер «шва» имеют нервные терминали на нейронах переднего
мозга и мозжечка. Функцией этих связей является регуляция цикла «сон-
бодрствование».
2.5.3.5. Глутаматергический синапс
Глутамат является основным возбуждающим нейротрансмиттером в цен-
тральной нервной системе. После выделения из нервного окончания глу-
тамат действует на ионотропные и метаботропные рецепторы постсинап-
тической мембраны. Ионотропные рецепторы представлены двумя типами:
NMDA- и АМРА-рецепторами (рис. 2.24). Название рецепторов связано с
их способностью связываться с синтетическими аналогами глутамата —
Система
обратного /
транспорта
глутамата
1)№№!1№№1!№ВП(|
iaiattiaiaiiia Л
ннашшатвц
Метаботропный
пресинаптический
глутаматный рецептор
Са Na
Na
Na
NMDA
He-NMDA
MlBIKtlBIBWRnUiilil
йвйишшвшв1йнвжт
l№l»i№ltattSBt№t>№ail!f
Глутамат
fH'l ill
_Щ|1Ш VIHtdHIlilHIt) 1Щ
Mi'iiifiiitiiHiiiaiiiiiriii
baH^aiaiijau^ajMiaiiimi
Пресинаптическая
мембрана
Синаптическая
щель
Постсинаптическая
мембрана
Метаботропный + +
постсинаптический + Na Na ,
- |ча Na
глутаматный Ng+
рецептор
Ca
Ca
Са +
Na .
Са
Рис. 2.24. Схематическое изображение возбуждающего глутаматергического си-
напса.
На постсинаптической мембране локализованы хемозависимые NMDA и не-NMDA ионо-
тропные рецепторы, при участии которых возбуждение проводится через синапс. Метаболиче-
ская активность постсинаптического нейрона регулируется метаботропными рецепторами
постсинаптической мембраны. Медиатор после взаимодействия с рецептором постсинаптиче-
ской мембраны транспортируется в пресинаптическое окончание.
2. Физиология возбудимых тканей • 87
М-метил-О-аспартатом (NMDA) и альфа-амино-3-метил-изоксазол-про-
пионовой кислотой (АМРА). Оба типа рецепторов, как правило, располо-
жены на постсинаптической мембране одного нейрона и их основной
функцией является деполяризация постсинаптической мембраны. Глутамат
связывается с АМРА-рецепторами потсинаптической мембраны. Актива-
ция АМРА-рецепторов снимает блокирование ионами Mg2+ NMDA-рецеп-
торов, которые ассоциированы с кальциевым ионным каналом. Это при-
водит к открытию кальциевого ионного канала и поддерживает кальцие-
вый ток через постсинаптическую мембрану в течение нескольких милли-
секунд. Физиологически важной особенностью нейронов, имеющих
NMDA-рецепторы на мембране, является то, что глутаматные синапсы
способны поддерживать повышенную возбудимость нейронов {долговре-
менная потенциация) в течение длительного времени (дни, недели и меся-
цы), что является основой механизмов долговременной памяти.
2.5.3.6. ГАМКергический синапс
Гамма-аминомасляная кислота (ГАМК) является основным тормозным
медиатором в ЦНС. При действии ГАМК, на хлорные ионотропные рецеп-
торы (ГАМКа-тип) постсинаптической мембраны эти каналы открываются
и ионы хлора поступают внутрь нейрона, вызывая гиперполяризацию
постсинаптической мембраны и понижая, таким образом, возбудимость
нейрона. ГАМКб рецепторы постсинаптической мембраны выполняют
иную функцию. Комплекс ГАМКБ-рецептор функционирует через систему
G-белков, уменьшая концентрацию вторичного посредника цАМФ, что
приводит к снижению кальциевой ионной проводимости и увеличению
калиевой. В результате происходят гиперполяризация постсинаптической
мембраны, снижение возбудимости постсинаптического нейрона и тормо-
жение его функции.
В нейронах мозга ГАМКергические синапсы повсеместно взаимодейст-
вуют с нервными терминалями глутаматергических синапсов, выполняя
функцию торможения экзоцитоза глутамата из нервных окончаний. Это
взаимодействие прекращает возбуждающее действие глутамата на постси-
наптический нейрон, что обеспечивает точность исходящего количества
сигналов, которыми управляются другие нейроны ЦНС.
2.5.3.7. Глицинергический синапс
В мозге глицин является вторым после ГАМК тормозным медиатором.
Глициновые рецепторы локализованы на постсинаптической мембране
нейронов древних отделов мозга — ствол мозга, спинной мозг, сетчатка. В
этих участках ЦНС глицин и ГАМК выделяются из одних и тех же нерв-
ных терминалей. Глицин после экзоцитоза из нервной терминали взаимо-
действует (как и ГАМК) с хлорными ионотропными рецепторами постси-
наптической мембраны. В результате открытия хлорных ионных каналов
ионы хлора поступают внутрь постсинаптического нейрона, гиперполяри-
зуют постсинаптическую мембрану (возникает ТПСП) и, таким образом,
понижают возбудимость нейрона. Глицин понижает возбудимость постси-
наптического нейрона не только за счет генерации ТПСП, но и за счет
уменьшения сопротивления постсинаптической мембраны. Этот эффект
глицина в комплексе с возникновением ТПСП уменьшает амплитуду
ВПСП постсинаптического нейрона ниже порога возбуждения, что пре-
пятствует генерации в нейроне потенциалов действия.
88 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.6. Функции мышечной ткани
Мышцы в организме человека представлены двумя группами: поперечно-
полосатыми и гладкими. К поперечно-полосатому типу мышц относятся
скелетная и сердечная мышцы (сердце). Функцией скелетных поперечно-
полосатых мышц является движение скелета, а в целом — тела человека.
При этом скелетные поперечнополосатые мышцы называются произволь-
ными, поскольку человек по своей воле может управлять их действиями.
Сокращение сердечной мышцы не может возникать произвольно, т. е. по
воле человека, и в этом отношение сердце, как поперечнополосатая мыш-
ца, представляет исключение.
В гладких мышцах отсутствует поперечно-полосатая исчерченность. Эти
мышцы образуют основную массу стенок внутренних органов (пищевари-
тельный канал, кровеносные сосуды, внутренние полые органы, выводные
протоки желез внешней секреции и дыхательные пути). Гладкие мышцы
организма человека называются непроизвольными, поскольку человек по
своей воле не способен контролировать их сократительную способность и
функцию.
Скелетная, сердечная и гладкие мышцы имеют как структурные, так и
функциональные различия образующих их мышечных волокон и, в свою
очередь, подразделяются на следующие подтипы.
Скелетные мышцы подразделяются на белые и красные-, первые по функ-
ции являются быстро сокращающимися мышцами, а вторые — медленно
сокращающимися. Большинство мышц тела человека являются смешанны-
ми и их функция обусловлена быстрыми и медленными мышечными во-
локнами.
Гладкие мышцы подразделяются на два разных по строению и функции
типа: унитарные и мультиунитарные. Первые образованы большим числом
связанных между собой гладкомышечных клеток — так называемыми плот-
ными контактами , а вторые состоят из отдельных гладкомышечных воло-
кон. В унитарной гладкой мышце возбуждение отдельного гладкомышеч-
ного волокна способно распространяться на всю мышцу. Напротив, в
мультиунитарной мышце каждое гладкомышечное волокно имеет собст-
венную иннервацию, через которую управляется сократительная способ-
ность мышцы.
Сердечная мышца подразделяется на сократительные клетки, которые
образуют предсердия и желудочки (миокард), и клетки, которые не облада-
ют сократительной способностью, способны спонтанно генерировать элек-
трические импульсы и проводить их к сократительным клеткам сердца.
2.6.1. Скелетная мышца
Скелетная мышца представляет собой комплекс отдельных многоядерных
мышечных клеток или волокон, которые состоят из функциональных еди-
ниц — миофибрилл, осуществляющих механизм сокращения в результате
взаимодействия белковых филаментов миозина и актина. Основной харак-
теристикой скелетной мышечной ткани является ее способность под кон-
тролем со стороны нервной системы развивать силу сокращения и укороче-
ние. Степень механического напряжения и укорочения мышцы обусловле-
на активацией числа двигательных единиц, образующих целую мышцу.
Двигательной единицей скелетной мышцы называется количество двига-
тельных (моторных) мышечных волокон, которые иннервируются одним
двигательным нейроном спинного мозга.
2. Физиология возбудимых тканей • 89
Скелетное мышечное во-
локно представляет собой
вытянутую многоядерную
клетку, которая содержит
сократительные элементы,
или миофибриллы. В каж-
дом мышечном волокне со-
держится до 1000 миофиб-
рилл и более, толщиной 1 —
3 мкм. Отдельная миофиб-
рилла состоит, в свою оче-
редь, из множества толстых
и тонких миофиламентов —
нитей белка миозина и белка
актина соответственно.
Взаимодействие между тол-
стыми и тонкими миофила-
ментами является физиче-
ской основой мышечного
сокращения. Миофиламен-
ты окружены цитоплазмой,
называемой саркоплазмой.
Кроме того, в состав нитей
актина входят белки — тро-
понин и тропомиозин, кото-
рые необходимы для разви-
тия процессов сокращения
и расслабления мышцы. Ка-
ждая миофибрилла содер-
жит примерно 1500 миози-
новых и около 3000 актино-
вых миофиламентов.
По всей длине мышеч-
ной клетки миофиламенты
миофибриллы разделены на
структурно-функциональ-
ные единицы, которые на-
зываются саркомерами
(рис. 2.25). Саркомеры от-
делены друг от друга Z-мем-
Рис. 2.25. Схематическое изображение саркоме-
ра скелетной мышцы.
I. Саркомер является структурно-функциональной
единицей скелетного мышечного волокна, включает в
себя сократительные белки миозин и актин, располо-
женные между белковыми Z-мембранами. Саркомер
состоит из анизотропных (А) дисков, в пределах кото-
рых перекрываются между собой тонкие актиновые й
толстые миозиновые филаменты. Изотропные (И)
диски саркомера заключают в себе только филаменты
актина. В средней части анизотропного диска распо-
ложена полоса Н, в пределах которой находятся фила-
менты миозина. СПР — саркоплазматический ретику-
лум. Рр и ДР — рианодиновый и дигидропиридино-
вый рецепторы или кальциевые ионные каналы сар-
колеммы и СПР соответственно. II — пространствен-
ное гексамодальное расположение актиновых фила-
ментов в пределах изотропных дисков (а, г) и миози-
новых и актиновых филаментов в пределах анизотроп-
ного диска по обе стороны от полосы Н (б, в).
бранами, к которым прикреплены тонкие актиновые филаменты. Располо-
жены толстые и тонкие филаменты в саркомере гексамодально, т. е. вокруг
каждого толстого миозинового филамента расположено шесть тонких ак-
тивновых филаментов.
Нити миозина лежат в центре саркомера и выглядят в световом микро-
скопе как темная полоска, которая называется анизотропным А-диском.
По обе стороны от A-диска лежат тонкие нити актина, которые в световом
микроскопе образуют светлые изотропные диски. Нити актина, которые
образуют в саркомере два изотропных диска, тянутся до Z-мембран.
90 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.6.1.1. Функции миофиламентов
Сократительная способность скелетной мышцы обусловлена функцией
мышечных белков актина и миозина, взаимодействие которых между со-
бой является основой механизма сокращения скелетной мышцы.
Актиновые филаменты представляют собой двойные белковые спирали
F-актина, каждая из которых образована примерно 200 единицами (моно-
мерами) глобулярного G-актина. Каждый G-актин состоит из четырех суб-
доменов. Два больших субдомена (3-й и 4-й) актина выполняют функцию
связывания между собой белковой спирали F-актина, а два маленьких (1-й
и 2-й), локализованные на периферии глобулярного G-актина, являются
местами взаимодействия каждого мономера G-актина с толстым филамен-
Тропомиозин
Рис. 2.26. Схематическое изображение
пространственного расположения актина,
миозина, тропомиозина и тропонина.
Субъединица С тропонина является мио-
фибриллярной молекулой, при соедине-
нии с которой ионы кальция активируют
механизм актомиозинового взаимодей-
ствия.
А — пространственная организация сократи-
тельных белков в состоянии расслабления
мышцы. Б — организация сократительных бел-
ков после связывания двух ионов кальция с до-
меном NH2. После этого тормозная область и
домен СООН субъединицы I тропонина осво-
бождаются от соединения с актином. Это ведет
к взаимодействию головки поперечного мости-
ка миозина с актином. Благодаря конформаци-
онным изменениям толстого филамента разви-
вается усилие, и толстый филамент скользит к
центру саркомера.
том. Основную роль в связыва-
нии актина с миозином во время
сокращения мышцы выполняет
субдомен 1, содержащий NH2
СООН терминали актина. Этот
субдомен актина выполняет
функцию связывающего центра.
Поэтому каждый глобулярный
G-актин представляет собой ме-
сто связывания с миозином при
сокращении мышцы. В состоя-
нии покоя скелетной мышцы
центры связывания на каждом
глобулярном G-актине закрыты
белками тропонином и тропо-
миозином. Эти белки функцио-
нально связаны между собой и
образуют тропонин-тропомиози-
новый комплекс, который распо-
ложен вдоль всей двойной белко-
вой спирали F-актина (рис. 2.26).
Тропомиозин представляет со-
бой молекулу длиной 42 нм, об-
разованную двумя белковыми ?-
цепями по типу спирали. Тропо-
миозин связан электростатически
с актиновым филаментом. Одна-
ко элетростатическая связь между
актиновым филаментом и тропо-
миозином является плотной
только во время расслабления
скелетной мышцы. В присутст-
вии ионов Са2+ эта связь стано-
вится менее плотной, тропомио-
зин становится подвижным и
способен открывать места связы-
вания на актиновом филаменте
для поперечных мостиков миози-
на, что происходит во время со-
кращения.
В разных типах мышечной
2. Физиология возбудимых тканей • 91
ткани тропомиозин имеет различную подвижность относительно актина.
Например, в скелетной мышце тропомиозин более подвижен, чем в сердеч-
ной мышце, что обусловливает отличие в сократительных свойствах этих
двух типов поперечно-полосатых мышечных тканей. Подвижность тропо-
миозина относительно актинового филамента регулируется тропонином.
Тропонин состоит из трех белковых субъединиц: С, I и Т. Субъединица
С тропонина является своеобразным сенсором для ионов Са2+ в регуляции
сокращения как в скелетной, так и в сердечной мышцах. Тропонин С име-
ет две глобулярные области — NH2 и СООН терминали. В скелетном мы-
шечном волокне каждая из этих концевых областей тропонина С содержит
два возможных места связывания ионов Са2+. Функция субъединицы I тро-
понина заключается в связывании тропонина с актином. Субъединица Т
тропонина прочно удерживает вместе тропонин-тропомиозиновый комплекс
на нитях актина.
В присутствии ионов Са2+ связь тропонина Т и тропомиозина относи-
тельно тонкой нити актина становится подвижной. Это приводит к откры-
тию мест связывания на актиновом филаменте, с которым способны свя-
зываться поперечные мостики толстого филамента с началом процесса
мышечного сокращения.
Толстый филамент представляет собой биполярный полимер белка мио-
зина, который способен взаимодействовать с актином и развивать в ре-
зультате этого взаимодействия силу и укорочение мышцы.
Двигательный белок миозин состоит из двух тяжелых и четырех легких
полипептидных цепей. Тяжелые полипептидные цепи образуют спираль
миозина. Глобулярные концевые NH2 области называются головками, с по-
мощью которых происходит взаимодействие между центрами связывания
на актиновом филаменте с миозином во время сокращения мышцы. Одна
пара легких полипептидных цепей связана с каждой головкой. Основа тол-
стого филамента представлена миозиновым стержнем, который скручен из
множества белковых нитей. Образование миозинового стержня происходит
в период развития мышечной ткани, благодаря функции белка С саркоме-
ра, который является антителом для белка миозина.
В скелетных мышцах от миозинового стержня по его окружности отходят
с равным интервалом в 14,3 нм пары головок поперечных, а расстояние ме-
жду головками по одной линии толстого филамента составляет 43 нм. При
этом каждая пара головок на филаменте миозина ориентирована на 180° от-
носительно другой. Подобная «геометрия» расположения поперечных мос-
тиков на толстом филаменте обусловливает их взаимодействие с любой из
шести окружающих его тонких филаментов с интервалом в 14,3 нм.
Важным внутриклеточным элементом эластичности мышечных волокон
является высокоэластичный белок титин, который формирует своеобраз-
ную сеть вокруг нитей актина и миозина в пределах каждого саркомера и
придает ему механические свойства как в покое, так и при мышечном со-
кращении. Благодаря наличию белка титина в саркомере, в состоянии по-
коя сократительные элементы развивают напряжение при растяжении
мышцы.
2.6.1.2. Механизм сокращения скелетной мышцы
Во время укорочения мышцы уменьшается длина саркомеров. При этом
филаменты актина и миозина, входящие в состав саркомеров, не изменяют
своей длины. Уменьшение длины соркомеров происходит в результате
движения (скольжения) тонких филаментов с двух сторон саркомера к его
92 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Рис. 2.27. Миозиновые поперечные мос-
тики связываются с любым из шести ак-
тивновых филаментов, которые окружа-
ют миозиновый филамент. В результате
актомиозинового взаимодействия проис-
ходит «скольжение» активновых фила-
ментов относительно миозиновых. При
этом длина саркомера уменьшается за
счет сближения Z-мембран без измене-
ния длины сократительных белковых фи-
ламентов.
СПР — саркоплазматический ретикулум. И,
А — изотропный и анизотропный диски. Рр,
Др — дигидропиридиновый и рианодиновый
рецепторы или кальциевые ионные каналы.
центру относительно толстых мио-
зиновых филаментов (рис. 2.27).
Молекулы актина и миозина спо-
собны взаимодействовать между
собой как в условиях in vivo, так и
in vitro. В мышце эта связь миози-
новых головок с актиновым фила-
ментом является циклической и
представляет собой многочислен-
ные укорочения миозиновых по-
перечных мостиков, что поддер-
живает постоянство сокращения в
целой активной мышце.
Цикл поперечного мостика во
время сокращения мышцы — это
связывание, укорочение, высвобо-
ждение и вновь связывание попе-
речного мостика миозина с акти-
новым филаментом, что весьма
напоминает собой человека, кото-
рый на руках поднимается вверх
по канату. В саркомере каждый
миозиновый филамент находится
в окружении шести актиновых
филаментов. Миозиновые фила-
менты вращаются во время взаи-
модействия с окружающими их
шестью филаментами актина, а их
поперечные мостики связываются
лишь с тремя случайными тонки-
ми миофиламентами. Согласно
теории скольжения филаментов
мышечного сокращения попереч-
ные мостики миозина взаимодействуют с актином асинхронно, т. е. одни
находятся «в захвате», в то время как другие «изменяют свой захват». Мио-
зиновые головки, которые образуют поперечные мостики, расположены
только на концах миозиновых филаментов. Поэтому при максимальном
увеличении длины саркомеров, несмотря на раздражение мышцы, полно-
стью отсутствует сокращение мышцы.
Активность актомиозиновых мостиков, с помощью которых актиновые
филаменты прикрепляются к миозину, представляет собой процесс, в ходе
которого химическая энергия АТФ превращается в сокращение мышцы.
Каждый отдельный поперечный мостик имеет структурные особенности,
которые вовлечены в сокращение мышцы. Эти особенности включают
двойную головку миозина, которая содержит АТФазную активность попе-
речного мостика, и способность связываться плотно с актином, и в то же
время вращается наподобие шарнира относительно стержня поперечного
мостика. В свою очередь стержень поперечного мостика также наподобие
шарнира связан с миозиновым стержнем и обладает эластическими свой-
ствами, заключающими в себе энергию, необходимую при развитии на-
пряжения во время вращении головки поперечного мостика.
Поперечные мостики отходят от миозинового стержня таким образом,
что образуют спираль вдоль толстного филамента с интервалом 14,3 нм
2. Физиология возбудимых тканей • 93
Тяжелые
белковые
Шарнир спирали Актин
1 i Д—Головка поперечного
'' .мостика
| | х Шарнир •<-----------------
Рис. 2.28. Цикл «замыкание—сокращение—размыкание» поперечного мостика мио-
зина.
Стадия 1 — в саркоплазме низкая концентрация ионов кальция (менее 10"7 М), места связыва-
ния на филаменте актина блокированы тропонин-тропомиозиновым комплексом. Стадия 2 —
коцентрация ионов кальция в саркоплазме возрастает до порогового уровня, поперечный
мостик миозина связывается с актином. Связывание ускоряет гидролиз АТФ примерно в 200
раз с образованием АДФ и фосфата (Ф). Стадия 3 — энергия гидролиза АТФ вызывает движе-
ние головки поперечного мостика и одновременно растягивается эластичный компонент по-
перечного мостика. Стадия 4 — эластический компонент поперечного мостика укорачивается,
что вызывает перемещение актинового филамента — это является «шагом» укорочения сарко-
мера. Стадия 5 — с головкой поперечного мостика связывается новая молекула АТФ и попе-
речный мостик миозина высвобождается от связи с атином. При высокой концентрации ио-
нов кальция в саркомплазме цикл «замыкание—сокращение—размыкание» будет неоднократ-
но повторяться, а при удалении ионов кальция в саркоплазматический ретикулум скорость
гидролиза АТФ резко снижается и тормозится прочное актомиозиновое взаимодействие.
между мостиками. Этот интервал столь мал, что каждой мостик должен
многократно взаимодействовать с актином по мере продвижения вдоль ак-
тинового филамента, чтобы произошло существенное укорочение длины
всей мышцы.
В состоянии покоя поперечный мостик расположен параллельно мио-
зиновому стержню (рис.2.28), а его головка — перпедикулярно относитель-
но последнего, и не прикреплена к актиновому филаменту (стадия I). По-
сле того, как ионы Са2+ высвобождаются из саркоплазматического ретику-
лума (описано ниже) и активируют сокращение мышцы, активновый фи-
ламент может связываться с миозиновой головкой (стадия 2). После свя-
зывания головка наклоняется (стадия 3), используя энергию, заключенную
в миозин-АТФазном комплексе головки. Это является основной силой по-
перечного мостика, которая растягивает его эластический компонент для
того, чтобы произвести энергию движения вдоль активного филамента
(стадия 4). В результате головка мостика совершает движение на 40° к цен-
тру саркомера и перемещает при этом на 10 нм нить актина. В это время
от головки высвобождаются АДФ и монофосфат, что необходимо для за-
хвата головкой миозина новой молекулы АТФ. Только после этого головка
94 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
миозина отделяется от молекулы актина, необходимой для размыкания по-
перечного мостика с актином (положение 5).
Цикл будет продолжаться до тех пор, пока концентрация ионов Са++ в
межфибриллярном пространстве не снижается до подпороговой величины,
т. е. 10“* М, что приведет к расслаблению мышцы, или пока не будут ис-
черпаны запасы АТФ. В последнем случае в мышце возникает спазм или
мышечная контрактура (rigor), при которой в течение нескольких секунд
или минут отсутствует расслабление мышцы. При трупном окоченении (rig-
or mortis), возникающем после смерти человека, значительный процент
поперечных мостиков остается в связанном с актиновыми филаментами
положении, что придает мышцам необычно высокую плотность.
Таким образом, функция АТФ проявляется в двух точках процесса со-
кращения мышцы. Во-первых, АТФ является источником энергии для со-
кращения. Во-вторых, АТФ отсоединяет поперечной мостик для того, что-
бы повторялся цикл поперечных мостиков «замыкание—сокращение—раз-
мыкание».
2.6.1.3. Активация мышечного сокращения
Процесс, при котором потенциалы действия, генерируемые на сарколемме
в области нервно-мышечного синапса, активируют сокращение мышцы,
называется электромеханическим сопряжением (рис. 2.29). Центральным
процессом в электромеханическом сопряжении является взаимодействие
ионов Са2+ с тропонином, когда изменяется положение тропомиозина, за-
крывающего места связывания на активном филаменте для поперечных
мостиков миозина.
В состоянии покоя мышечного волокна Са2+-АТФаза саркоплазматиче-
ского ретикулума активно закачивает ионы Са2+ из саркоплазмы в его цис-
терны, где большинство ионов находится в обратимо связанном состоянии
с белком кальсеквестрином. В саркоплазме мышечного волокна концентра-
ция ионов Са2+ снижена до уровня порядка 0,1 мкМ/л или 10-7 М, при ко-
тором тропонин-тропомиозиновый комплекс закрывает места связывания
актина с головками поперечных мостиков миозина.
Тем не менее даже при столь низкой концентрации ионов Са2+ в сарко-
плазме отдельные места на концевых субдоменах актина могут осуществ-
лять слабое электростатическое взаимодействие с миозином. В этом случае
имеет место прямое связывание головки миозина с актином. Благодаря
этому в состоянии покоя мышечного волокна тонкие филаменты в сарко-
мере находится не в свободном, а в зафиксированном положении, что под-
держивает упорядоченное пространственное расположение тонких и тол-
стых миофиламентов между собой в саркомерах мышц.
Для того чтобы вызвать сокращение, потенциал действия мышцы, вы-
зваемый двигательным нервом, распространяется по сарколемме мышеч-
ной клетки и внутрь поперечных трубочек (t-система), открывая кальцие-
вые ионные каналы мембраны. В фазу деполяризации потенциала дейст-
вия сарколеммы в области t-системы открывается L-тип потенциалзависи-
мых кальциевых ионных каналов, которые, в свою очередь, вызывают от-
крытие кальциевых ионных каналов саркоплазматического ретикулума.
Это обусловлено тем, что L-тип потенциалзависимых кальциевых ионных
каналов сарколеммы в области t-системы, или дигидропиридиновый рецеп-
тор (агонист—дигидропиридин), прямо структурно связан с рианодиновым
рецептором (агонист — рианодин) или кальциевым ионным каналом сарко-
плазматического ретикулума. Функцией структурного взаимодействия этих
2. Физиология возбудимых тканей • 95
Т-трубочка мембраны мышечной клетки
Na+/K+- Na++/Ca++- Дигидропиридиновый
АТФаза обменник рецептор (Са++- канал)
Рис. 2.29. Схема механизма электромеханического сопряжения в скелетных мышеч-
ных клетках во время начала их сокращения.
А. Механизм представляет собой последовательную цепь деполяризации сарколеммы в облас-
ти Т-трубочки мышечного волокна, активации дигидропиридинового рецептора и сопряжен-
ного с ним рианодинового рецептора мембраны саркоплазматического ретикулума. Б. В ре-
зультате активации обоих ионных каналов концентрация ионов кальция в цитозоле повыша-
ется за счет поступления ионов кальция из внеклеточной среды, а также из саркоплазматиче-
ского ретикулума. В. Повышение концентрации ионов кальция в области сократительных бел-
ков и их связывание с субъединицей С тропонина вызывает мышечное сокращение. СПР —
саркоплазматический ретикулум. Рр — рианодиновый рецептор (высвобождающий ионы каль-
ция канал).
двух рецепторов является открытие кальциевых каналов саркоплазматиче-
ского ретикулума и выход ионов Са2+ в саркоплазму. Когда концентрация
ионов Са2+ в межфибриллярном пространстве, т. е. в саркоплазме между
нитями актина и миозина, увеличивается до 10 мкМ/л (10-5 М) и более,
ионы Са2+ начинают связываться с тропонином С. Активация тропонина С
при связывании с ионами Са2+ вызывает смещение тропонин-тропомиози-
нового комплекса примерно на 25° вокруг поверхности актинового фила-
мента и погружение молекул тропомиозина вглубь между белковыми цепя-
ми актина. В результате на актиновых филаментах открываются места свя-
зывания для головок поперечных мостиков миозина и начинается цикл их
сокращения. При этом начинается сокращение всей скелетной мышцы,
которое будет продолжаться до тех пор, пока концентрация ионов Са2+ в
саркоплазме не станет пороговой или сверхпороговой для поддержания со-
кратительного процесса.
96 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.6.1.4. Расслабление скелетной мышцы
Расслабление скелетной мышцы начинается только после того, как кон-
центрация ионов Са2+ в саркоплазме мышечных клеток снижается ниже
порогового уровня, т. е. ниже 10 мкМ/л, или 10-5 М. Расслабление мышцы
обусловливают три процесса: 1. Отсоединение (диссоциация) ионов Са2+
от субъединицы С тропонина. 2. Диффузия ионов Са2+ в сторону сарко-
плазматического ретикулума. 3. Активный транспорт ионов Са2+ внутрь
саркоплазматического ретикулума при участии Са2+-АТФазы.
В начальный момент расслабления длина мышцы остается постоянной.
Это происходит из-за того, что первоначально происходит расслабление
саркомеров, находящихся на концах мышц, в то время как центральные
саркомеры остаются укороченными. Это состояние характерно лишь для
начального момента мышечного расслабления, а именно фазы изометриче-
ского расслабления. Процесс расслабления во всех саркомерах усиливается
после того как сила сокращения мышцы уменьшается примерно на 20—
30 %. Это обусловлено тем, что во время расслабления мышцы уменьшает-
ся подвижность тропомиозина по поверхности белковых цепей актина
примерно с той же скоростью, что и сила сокращения. В результате чем
меньше сила сокращения мышцы, тем меньше открыто мест связывания
на актиновых филаментах для поперечных мостиков миозина
2.6.1.5. Типы мышечных сокращений
Скелетные мышцы развивают сокращение под влиянием нервных импуль-
сов, поступающих от двигательных нейронов, расположенных в передних
рогах спинного мозга. Мышечные волокна, иннервируемые одним мото-
нейроном спинного мозга, образуют двигательную единицу. В зависимости
от количества потенциалов действия, которые поступают к двигательным
единицам, в мышце могут возникать несколько типов сокращения: оди-
ночное мышечное сокращение, тетаническое, изометрическое, изотониче-
ское и смешанное. Сокращение мышцы в целом является результатом сум-
мации одиночных мышечных сокращений.
Одиночное мышечное сокращение. Сократительный ответ мышечного во-
локна или отдельной мышцы на одно раздражение называется одиночным
сокращением (рис. 2.30). Одиночное сокращение имеет латентный период,
фазу развития напряжения, или укорочения, и фазу удлинения, или расслаб-
ления. Длительность каждой фазы зависит от морфофункциональных
свойств мышечных волокон: у быстро сокращающихся волокон глазных
мышц фаза напряжения составляет 7—10 мс, а у волокон камбаловидной
мышцы — 50—100 мс. Фаза укорочения в одиночном мышечном сокраще-
нии всегда меньше по времени, чем фаза расслабления. Одиночное мышеч-
ное сокращение возникает в том случае, если интервал времени между по-
следовательными разрядами мотонейронов, вызывающими подобный тип
сокращения, равен или превышает время одиночного сокращения. Так, во-
локна глазодвигательных мышц имеют режим одиночного сокращения при
частоте импульсации мотонейрона менее 50 Гц, а волокна камбаловидной
мышцы человека — при частоте менее 10 Гц. В режиме одиночного сокра-
щения мышца способна работать длительное время без развития утомления.
Тетаническое мышечное сокращение. Тетаническим называется сокраще-
ние мышцы, при котором происходит суммация одиночных мышечных со-
кращений в результате ее раздражения частыми (тетаническими) стимула-
ми. Различают зубчатый и гладкий тетанусы. Зубчатый тетанус возникает
2. Физиология возбудимых тканей • 97
Зубчатый тетанус f
Гладкий тетанус I т т т т т
I I
Раздражение электрическим током
Рис. 2.30. Одиночное и суммированные типы мышечного сокращения.
Одиночное сокращение мышцы (А) подразделяется на латентный период, фазу укорочения и
расслабления. Суммированными типами мышечного сокращения (Б) являются зубчатый и
гладкий тетанусы, которые возникают в результате суперпозиции (наложения) одиночных мы-
шечных сокращений. Стрелками обозначены моменты нанесения электрического раздра-
жения.
в том случае, если мышца раздражается серией электрических стимулов с
частотой, при которой каждый из последующих импульсов приходится на
фазу расслабления. При тетаническом сокращении мышцы фаза напряже-
ния сменяется фазой неполного расслабления в течение всего периода раз-
дражения. Сила сокращения, развиваемая мышцей при зубчатом тетанусе,
больше, чем при одиночном мышечном сокращении. Гладкий тетанус воз-
никает в том случае, если нерв или мышцу раздражать серией электриче-
ских импульсов с частотой, при которой каждый последующий импульс
приходится на фазу напряжения. При гладком тетаническом сокращении
мышца не расслабляется и в ней поддерживается постоянное напряжение.
Это происходит в результате суперпозиции или наложения фазы укороче-
ния мышцы, возникающей в мышце после каждого последующего раздра-
жения. В результате при гладком тетанусе мышца развивает силу в 2—4 ра-
за больше, чем при одиночном мышечном сокращении.
В разных мышцах сокращение по типу гладкого тетануса возникает при
различной частоте раздражения. Гладкий тетанус быстрой глазодвигатель-
ной мышцы проявляется при частотах свыше 150—200 Гц, а у медленной
камбаловидной мышцы — при частоте около 30 Гц.
Изометрическое сокращение. Если мышца развивает напряжение в усло-
виях чрезмерной нагрузки, то ее длина не изменяется и такое сокращение
называется изометрическим. Согласно теории скольжения, при изометри-
ческом мышечном сокращении укорачивается длина саркомеров. Однако
общая длина мышцы не изменяется, поскольку на такую же величину уд-
линяется последовательно связанный с сократительными филаментами
эластический компонент мышцы.
Изотоническое сокращение. Если мышца до начала сокращения отягоще-
на грузом, а масса груза и развиваемое мышцей напряжение во время со-
кращения не изменяются, то такое сокращение называется изотоническим.
Во время перемещения тела в пространстве имеет место смешанный, или
4-6095
98 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
ауксотонический, или анизотонический тип сокращения, при котором мыш-
ца развивает напряжение и укорачивается. Изотонический и анизотониче-
ский типы сокращения лежат в основе динамической работы локомоторно-
го аппарата человека.
Если внешняя нагрузка на мышцу меньше, чем развиваемое во время
сокращения напряжение, то мышца укорачивается и совершается движе-
ние (концентрический тип сокращения). Если при чрезмерной внешней на-
грузке мышца, напрягаясь, все же растягивается (удлиняется), то такой тип
сокращения называется эксцентрическим.
2.6.1.6. Типы скелетных мышечных волокон
Сокращение в одних скелетных мышцах происходит быстро, и такие мыш-
цы быстро утомляются, а другие мышцы сокращаются медленно и так же
медленно утомляются. Эта физиологическая особенность сократительной
способности мышц зависит от двух типов мышечных волокон, образую-
щих мышцу: медленно и быстро сокращающихся (рис. 2.31).
Медленные неутомляемые мышечные волокна скелетных мышц, или тип I,
развивают небольшую силу при сокращении, поскольку в них мало мио-
фибрилл. Низкая скорость сокращения этого типа мышечных волокон
обусловлена невысокой активностью миозиновой АТФазы, медленным вы-
ходом ионов кальция из саркоплазматического ретикулума и, наконец, их
медленным связыванием с тропонином. Мышечные волокна медленных
двигательных единиц способны выполнять работу умеренной мощности
длительное время без утомления, если хорошо развита капиллярная сеть и
достаточно количество поступающего к ним О2. Например, мышцы туло-
вища, обеспечивающие поддержание положения тела в пространстве, со-
держат в основном медленные мышечные волокна. Поэтому данные мыш-
цы могут находиться в состоянии сокращения длительное время, поддер-
живая позу и положение тела человека в пространстве.
Быстрые мышечные волокна скелетных мышц (тип II) приспособлены
для выполнения мощной, но кратковременной работы, так как богаты
миофибриллами и имеют высокую скорость сокращения благодаря актив-
ности миозиновой АТФазы. Мышечные волокна этого типа мышц содер-
жат много гликолитических ферментов (быстро освобождается большое
количество энергии), в них мало митохондрий и миоглобина, они окруже-
ны небольшим количеством капилляров. Например, мышцы рук и ног
имеют в своем составе преимущественно быстрые мышечные волокна, что
обусловливает силу, быстроту и точность движения верхних и нижних ко-
нечностей человека.
У человека быстрые мышечные волокна подразделяют на тип ПА (легко
утомляемые) и ИХ (устойчивые к утомлению). Тип ИХ приспособлен для
выполнения мощной работы, обладает большой аэробной выносливостью
благодаря использованию энергии как аэробных, так и анаэробных про-
цессов.
Соотношение числа медленных и быстрых двигательных единиц в од-
ной мышце определено генетически. Например, у человека в четырехгла-
вой мышце бедра процент медленных волокон варьирует от 40 до 98. При-
чем если в мышце больше медленных волокон, то такая мышца приспо-
соблена к работе на выносливость. Если в составе мышцы выше процент
быстрых волокон, то такая мышца развивает большую скорость и силу со-
кращения. При нарастающей работе мышц активируются первоначально
медленные мышечные волокна, а затем быстрые. Это обусловлено тем, что
2. Физиология возбудимых тканей • 99
Время, мс
Эргограммы
__1___i__
100
НИИ
0 2 4 60 0 2 4 60 0 2 4 6 мин
Рис. 2.31. Двигательные единицы (ДЕ) скелетной мышцы и их типы.
1 — медленные, слабые, неутомляемые мышечные волокна. Низкий порог активации мото-
нейрона; 2 — промежуточный тип ДЕ; 3 — быстрые, сильные, быстроутомляемые мышечные
волокна. Высокий порог активации мотонейрона.
медленные неутомляемые мышечные волокна иннервированы мотонейро-
нами спинного мозга, которые имеют низкие пороги возбуждения и невы-
сокую, но постоянную на протяжении длительного времени частоту гене-
рации потенциалов действия (в среднем 6—10 Гц). Повышение силы со-
кращения медленных неутомляемых мышечных волокон лишь незначи-
тельно увеличивает частоту генерации потенциалов действия в мотонейро-
нах этого типа мышечных волокон. Кроме того, нервные терминали аксо-
нов этих мотонейронов оканчиваются на небольшой группе мышечных во-
локон. При силе мышечного сокращения более 20—25 % от максимальной
произвольной силы активируются быстрые мышечные волокна. Это обуслов-
лено тем, что иннервирующие их мотонейроны спинного мозга имеют вы-
сокий порог возбуждения, их частота генерации потенциалов действия на-
растает по мере увеличения силы сокращения мышечных волокон, но со-
храняется на постоянном уровне не продолжительное время.
Спортивная физическая нагрузка у человека не может изменить один
тип мышечных волокон на другой, а также число медленных и быстрых
мышечных волокон в мышце. В результате физических упражнений изме-
няются масса мышечной ткани, а также сила, выносливость и скоростные
качества мышц тела человека.
4*
100 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
2.6.1.7. Физиологические показатели сокращения скелетной мышцы
Основными физиологическими параметрами сокращения скелетных мышц
человека являются сила, скорость развития силы и укорочения мышцы,
мощность и работа
Скорость укорочения скелетной мышцы находится в обратной зависимо-
сти от величины нагрузки. Эта взаимосвязь описывается классическим
уравнением Хилла. Укорочение мышечных волокон обусловливает силу
мышцы во время сокращения. При этом чем больше укорочение мышцы,
тем меньшую она развивает силу. Уменьшение силы мышцы по мере
уменьшения ее длины во время сокращения связано с ростом концентра-
ции ионов Са2+ в саркоплазме. Чем ближе к максимальной становится ак-
тивация ионами Са2+ взаимодействия актина и миозина, тем меньшую си-
лу развивает мышца при сокращении. Основными причинами снижения
силы мышцы при ее укорочении является увеличение скорости отсоедине-
ния поперечных мостиков от актина. Этот фактор, в свою очередь, снижа-
ет сродство ионов Са2+ с местами связывания на тропонине С и высвобож-
дает ионы Са2+ из тропонина. В результате снижается активность тропо-
нин-тропомиозинового комплекса и тропомиозин блокирует места связы-
вания на активновых филаментах для поперечных мостиков миозина. В
результате снижается сила мышцы при сокращении. Скелетная мышца те-
ла человека способна поднять груз определенной величины. Максимальная
сила, которую мышца развивает при мышечном сокращении в случае, если
она не в состоянии поднять груз, называется абсолютной силой мышцы. Из-
меряется абсолютная сила мышцы величиной минимального груза, кото-
рый она не способна поднять.
Работа мышц равна произведению силы, развиваемой мышцей, на путь
перемещения груза, либо механическая работа мышц равна произведению
поднятого груза на величину укорочения мышцы. Между величиной на-
грузки мышцы и ее укорочением нет пропорциональной зависимости. При
постепенном увеличении нагрузки мышцы, которая при сокращении раз-
вивает максимальную силу, величина работы мышцы сначала растет, а за-
тем уменьшается. В основе изменения мышечной работы при увеличении
нагрузки лежат следующие физиологические процессы (рис. 2.32). При
максимальном исходном растяжении мышцы (максимальная длина сарко-
меров) в ней осутствует оптимальное число поперечных мостиков, которые
могут сокращаться (положение 1) и развивать мышечное напряжение. При
постепенном уменьшении исходного растяжения мышцы поперечные мос-
тики миозина во все большей степени перекрываются с тонкими филамен-
тами актина, что ведет к увеличению укорочения мышцы и росту работы
мышцы (положение 2). Оптимальная скорость укорочения скелетной
мышцы зависит от оптимальной исходной длины саркомеров перед началом
сокращения, которая в среднем составляет 1,7—2,7 мкм. Однако при ми-
нимальной исходной длине саркомеров, когда миозиновые филаменты за-
нимают всю их длину (положение 3), вновь отсутствует оптимальное число
поперечных мостиков, которые могут сокращаться, развивать мышечное
напряжение, а следовательно, совершать работу.
Величина мышечной работы зависит от количества мышечных волокон
в мышце. При этом чем больше поперечное сечение мышцы с параллель-
ным расположением мышечных волокон, тем большую она совершает ра-
боту. В мышцах человека не всегда мышечные волокна расположены по
оси мышцы. В мышцах перистого строения, что имеет место в большинст-
ве случаев расположения мышечных волокон относительно продольной
2. Физиология возбудимых тканей • 101
Рис. 2.32. Изменение изометрического тетанического напряжения скелетной мыш-
цы как функция от длины саркомера.
Максимальное напряжение развивается при длине мышцы L0 = 100 %. Затененная полоса по-
казывает предел изменения длины (от 70 до 130 %), который возможен в организме, если
мышцы связаны со скелетом.
оси мышцы, длина мышечных волокон нередко составляет лишь '/3— '/4
часть всей мышцы. В таких мышцах величина укорочения относительно
мала, но они развивают значительную абсолютную силу и выполняют
большую работу, чем мышцы с параллельным расположением мышечных
волокон. Это обусловлено тем, что в перистых мышцах величина работы
пропорциональна площади сечения, проходящего через все мышечные во-
локна, а не через анатомический поперечник мышцы в случае параллель-
ного расположения мышечных волокон.
Наконец, мощность мышечной работы как физиологический параметр
сокращения мышцы измеряется произведением величины силы мышцы на
скорость укорочения мышцы при сокращении.
2.6.2. Утомление скелетной мышцы
Утомлением называется временное снижение способности мышцы разви-
вать усилие в результате предшествующей физической активности. При
этом происходит снижение всех физиологических параметров сокращения:
скорости нарастания и снижения силы мышцы, скорости укорочения
мышцы, работы и мощности. Снижение мощности, наряду с другими па-
раметрами, является наиболее значимым для изменения количественных
параметров способности мышцы выполнять движение. Как показано на
рис. 2.33, во время сокращения скелетной мышцы максимальная изомет-
рическая сила непрерывно уменьшается (тонкая линия) от начала сокра-
щения, что отражает процесс утомления мышцы. Мощность мышцы (тол-
стая линия): после начала сокращения мышцы мощность первоначально
возрастает, а затем уменьшается, если мышца не способна поддерживать
заданную интенсивность работы.
102 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Сила
Рис. 2.33. Общие понятия утомления, истощения и прекращения возбуждения
мышцы при изометрическом и динамическом сокращениях.
Максимальную изометрическую силу мышца имеет при нулевой скорости укорочения. При
сокращении максимальная сила мышцы (тонкая линия) снижается в большей степени, чем
скорость ее максимального укорочения, а максимальная мощность мышцы (толстая линия)
снижается в большей степени, чем оба этих параметра. Во время периода сокращения макси-
мальная изометрическая сила постепенно уменьшается от начала сокращения и динамика
уменьшения силы отражает то, как мышца утомляется. Если мышца активируется, достигнув
точки, ниже которой она не способна больше развивать силу, то ее возбуждение прекраща-
ется.
Человек прекращает физическую работу в результате утомления, как
правило, еще до потери способности мышцы поддерживать сокращение.
Прекращение физической работы может происходить также в результате
дискомфортных или даже болевых ощущений, которые ассоциируются у
человека с утомлением. Эти ощущения возникают у высокотренированных
людей, когда утомление при чрезмерных нагрузках на мышцы является ре-
зультатом неспособности метаболических и сократительных процессов
поддерживать мышечное сокращение. У нетренированных людей при фи-
зической нагрузке развитие утомления не является результатом дефицита
метаболических и сократительных процессов.
По механизму возникновения утомления различают центральный и пе-
риферический процессы. Центральное утомление представляет собой про-
цессы, происходящие во время мышечной работы на различных уровнях
нервной регуляции движения, однако среди причин утомления оно состав-
ляет не более 10 %. При этом показана относительная неутомляемость
нервных волокон при проведении по ним потенциалов действия. Перифе-
рическое утомление обусловлено процессами, происходящими на уровне
нервно-мышечного синапса t-системы мышечных клеток, и ему принадле-
жит основная роль в снижении силы, скорости укорочения и расслабле-
ния, а также работу и мощности сокращающихся мышц.
Снижение возбудимости сарколеммы является основной причиной в
механизме периферического утомления. В области t-системы медленных и
быстрых скелетных мышечных волокон локализованы Са2+-АТФаза и Na+/
Са2+-обменник, а также Ма+/К+-АТФаза, энергия которой используется
для осуществления вторично активного транспорта ионов Са2+ через сар-
колемму. При мышечной работе в наибольшей степени снижается актив-
2. Физиология возбудимых тканей • 103
ность Na+/K+-Hacoca, что существенным образом уменьшает сократитель-
ные свойства мышечных волокон. Основными факторами, снижающими
активность Na+/K+-Hacoca, являются изменение концентрационного гра-
диента ионов Na+ и К+ по обе стороны мембраны мышечного волокна в
области t-системы сарколеммы в результате генерации потенциалов дейст-
вия на мембране. Так, во время мышечного сокращения увеличивается
внеклеточная и уменьшается внутриклеточная концентрация ионов К+.
Одновременно происходит снижение концентрации ионов Na+ на внеш-
ней поверхности мембраны мышечной клетки и повышение с внутренней
стороны. Это приводит к деполяризации сарколеммы до величины мем-
бранного потенциала, равной в среднем — 60 мВ. При величине мембран-
ного потенциала миоцитов порядка — 60 мВ происходит снижение силы
мышечного сокращения более чем на 20 %. Однако наиболее существен-
ное уменьшение силы мышечного сокращения происходит, когда мем-
бранный потенциал уменьшается до —60—55 мВ. При этой величине мем-
бранного потенциала мышечные волокна утрачивают возбудимость и в
мышечных клетках прекращается генерация потенциалов действия, а сле-
довательно, человек не способен выполнять физическую работу.
Во время физической работы у человека в плазме крови повышается
концентрация адреналина и норадреналина. Эти гормоны стимулируют ра-
боту Na+/K+-Hacoca, который эффективнее восстанавливает нормальную
величину градиентов ионов К+ и Na+ в области t-системы сарколеммы и,
таким образом, препятствует развитию утомления.
В развитии мышечного утомления у человека важное значение прида-
ется роли метаболизма как фактору, лимитирующему сокращение мышц
при физической работе из-за истощения энергетических субстратов и на-
копления продуктов метаболизма. Однако при развитии мышечного утом-
ления реально не обнаруживается недостатка АТФ. Так, в миоцитах даже
при максимальном произвольном мышечном сокращении запасы АТФ не
снижаются до нуля, как это имеет место у человека при мышечной кон-
трактуре или непосредственно после смерти человека при трупном око-
ченении.
2.7. Гладкая мышца
Функции гладкой мышцы связаны с деятельностью внутренних органов
(пищеварительный канал, кровеносные сосуды, внутренние полые органы,
выводные протоки желез внешней секреции и дыхательные пути), радуж-
ной оболочки глаза и кожи. В состав гладкомышечной клетки входят мио-
филаменты актин, миозин и тропомиозин (тропонин отсутствует), однако
нити актина и миозина более тонкие, чем в поперечно-полосатых мышеч-
ных клетках, они имеют несколько отличный химический состав от по-
следних, саркоплазматический ретикулум слабо развит, а t-система отсут-
ствует.
Эти факторы обусловливают низкую возбудимость гладкой мускулатуры
по сравнению с поперечно-полосатой, процесс возбуждения в гладкомы-
шечных клетках может продолжаться несколько секунд, они способны на-
ходиться в активном тоническом сокращении без видимого внешнего про-
цесса укорочения мышцы. Процесс сокращения гладкой мышцы (как и
поперечно-полосатой) зависит от диффузии ионов Са2+ через кальциевые
каналы мембраны гладкомышечных клеток из интерстициальной среды,
которая выполняет роль депо этих ионов. При этом кальциевые ионные
каналы локализованы только в местах погружений мембраны гладкомы-
104 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
шечных клеток, которые называются кавеолами. Эти структурные особен-
ности мембраны гладкомышечных клеток и интерстициальное депо ионов
Са2+ обусловливают относительно низкую скоростью развития сокращения
и расслабления.
Уникальные сократительные свойства гладкомышечных клеток обуслов-
лены структурами, которые называются плотными тельцами. Они образо-
ваны а-актином и представляют собой скопление молекул актина на кон-
цах тонких миофиламентов. Роль плотных телец идентична таковой Z-дис-
ка в саркомерах скелетного мышечного волокна, т. е. они удерживают ни-
ти актина в процессе их скольжения между толстыми филаментами и тем
самым обеспечивают укорочение мышечного волокна. Плотные тельца,
расположенные на мембране гладкомышечных клеток, присоединены к
микрофибриллам, которые выходят за пределы клеток, и соединяются с
коллагеновыми волокнами окружающей соединительной ткани. Таким об-
разом, отдельные гладкомышечные клетки объединяются в единую
мышцу.
Между актиновыми филаментами, связанными с их плотными тельца-
ми, разрозненно расположены толстые филаменты, которые в 2,5 раза тол-
ще, чем актиновые филаменты, но составляют примерно х/п часть от об-
щего числа нитей актина. Несмотря на то, что в гладкой мышце меньше
миозина, она способна развивать такое же напряжение, как поперечно-по-
лосатая идентичного поперечного сечения. Это обусловлено тем, что акти-
новые филаменты в гладкомышечных клетках длиннее, чем в клетках ске-
летной мышцы, и поэтому могут продвигаться на большую длину относи-
тельно миозиновых филаментов.
2.7.1. Типы гладких мышц
Организация гладкой мышцы в пределах органа существенным образом
предопределена выполняемой им функцией. На основании организации
гладкой мышцы в виде единичных глакдкомышечных клеток или пучков
из многочисленных гладкомышечных клеток, они классифицируются на
два типа: унитарные и мультиунитальные.
Унитарная гладкая мышца (рис. 2.34) называется висцеральной, так как
она окружает полые органы — желудок, кишечник, мочевой пузырь — и
некоторые сосуды. Гладкомышечные клетки унитарной мышцы взаимо-
действуют между собой с помощью различных молекулярных соединений
(нексусы, десмосомы, щелевые соединения, адгезия), однако большинст-
во клеток осуществляют межклеточное взаимодействие с помощью щеле-
вых соединений. Щелевые соединения выполняют две важные функции —
образуют единый гладкомышечный синтиций, в пределах которого по-
тенциалы действия через эти же контакты передаются от одной клетки к
другой, вызывая сокращение во всей гладкой мышце. Каждое щелевое
соединение состоит из двух белковых гексамеров, соединенных между
собой коннексонами. При этом один гексамер начинается в клетке и вы-
ходит в межклеточное пространство, где соединяется с гексамером дру-
гой клетки. Поэтому изменение потенциала на мембране одной гладко-
мышечной клетки посредством щелевого контакта электротонически вы-
зывает аналогичное изменение потенциала на мембране другой, что яв-
ляется основной распространения возбуждения в этом типе гладкой
мышцы.
В унитарной мышце только некоторые гладкомышечные клетки получа-
ют импульсы от нервного волокна и после возбуждения инициируют со-
2. Физиология возбудимых тканей • 105
Рис. 2.34. Схематическое изображение структурного взаимодействия гладкомышеч-
ных клеток унитарной мышцы.
Унитарная гладкая мышца представляет собой синцитий гладкомышечных клеток (А), объеди-
ненных между собой плотными контактами (Б). Функцией плотных контактов в унитарной
мышце является распространение медленных волн колебания мембранного потенциала и по-
тенциалов действия от одной гладкомышечной клетки к другой. Отдельная мышечная клетка,
иннервируемая вегетативным волокном в области варикозного расширения, является пейсме-
кером для рядом расположенных гладкомышечных клеток унитарной мышцы.
кращение в других гладкомышечных клетках. Такие клетки в унитарной
гладкой мышце называются пейсмекерами, или водителями ритма их со-
кращения.
Унитарные гладкие мышцы имеют два типа автоматической активно-
сти: ауторитмическую и тоническую. Ауторитмическую, или фазную, ак-
тивность имеют клетки пищеварительного тракта, а их ритмичность возбу-
ждения модулируется нервной активностью. Гладкие мышцы с ауторитми-
ческой активностью способны развивать быстрое, но проходящее (транзи-
торное) сокращение. Гладкие мышцы, имеющие тоническую активность,
способны поддерживать постоянный мышечный тонус на фоне их частич-
ного укорочения или расслабления. Тоническая активность унитарной
гладкой мышцы поддерживает мышечный тонус сфинктеров пищевари-
тельного тракта, которые находятся большую часть времени в состоянии
непрерывного сокращения и расслабляются лишь при эвакуации содержи-
мого из проксимального
отдела кишечника в дис-
тальный, например из же-
лудка в двенадцатиперст-
ную кишку.
Мультиунитарная глад-
кая мышца образована от-
дельными гладкомышеч-
ными клетками, каждая
из которых иннервирова-
на отдельным нервным
волокном вегетативной
нервной системы. Поэто-
му электрическая актив-
ность и сокращение муль-
тиунитарной мышцы воз-
никают под влиянием им-
пульсов, поступающих к
клетки
Рис. 2.35. Схематическое изображение гладкомы-
шечных клеток мультиунитарной мышцы.
Мультиунитарная мышца состоит из отдельных гладко-
мышечных клеток, каждая из которых иннервируется
нервным волокном. Медиаторы, которые выделяются из
варикозных расширений нервных волоки вызывают мед-
ленные волны деполяризации мембраны клеток, что
инициирует их сокрашение.
106 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
отдельным гладкомышечным клеткам (рис. 2.35). К такому типу гладких
мышц относятся цилиарная мышца глаза, мышца радужной оболочки,
гладкие мышцы сосудов.
2.7.2. Электрическая активность клеток гладкой мышцы
Мембранный потенциал в гладкомышечных клетках в условиях физиологи-
ческой нормы варьирует от —35 до -70 мВ. Подобное колебание величины
мембранного потенциала обусловлено физиологическими условиями тка-
ней, в которых производится измерение мембранного потенциала. Напри-
мер, мембранный потенциал продольных и циркулярных мышечных кле-
ток миометрия может существенно изменяться под действием гормонов во
время беременности и после родов. Кроме того, потенциал покоя различа-
ется на мембране гладкомышечных клеток, расположенных в разных слоях
гладких мышц одного и того же органа. Например, мембранный потенци-
ал в гладкомышечных клетках продольного слоя пищеварительного тракта,
как правило, на несколько милливольт меньше, чем циркулярного. Это
обусловливает более высокую возбудимость гладкомышечных клеток про-
дольного слоя и их сократительные свойства.
Гладкие мышцы сокращаются как при медленных изменениях величи-
ны мембранного потенциала, так и при генерации потенциала действия.
Амплитуда колебаний медленных волн мембранного потенциала составляет
—20—30 мВ. В том случае, если деполяризация мембраны в гладкомы-
шечной клетке достигает критического уровня деполяризации, в них ге-
нерируются единичные импульсы либо серии потенциалов действия, ко-
торые вызывают сокращение гладкой мышцы. В унитарных гладких
мышцах медленные волны колебания величины мембранного потенциала
обусловлены спонтанным изменением проницаемости мембраны ионов
Na+ и Са2+. Медленные волны мембранного потенциала сами по себе не
приводят к генерации потенциалов действия в этом типе гладких мышц.
Возбудимость гладких мышц к каждому типу раздражителей обусловлена
типом гладкой мышцы. Гладкомышечные клетки унитарной гладкой
мышцы генерируют потенциалы действия, которые распространяются по
всей мышце через плотные контакты (продольные гладкие мышечные
клетки пилорического отдела желудка, тощей кишки, пилорической об-
ласти уретры, миометрия, полой вены), либо потенциалы действия гене-
рируются в ответ на возбуждение периферических нервов (продольные
гладкомышечные клетки дна желудка, гладкие мышечные клетки сосудов,
а также трахеи). Поэтому унитарная гладкая мышца реагирует на раздра-
жение по закону «все или ничего», как поперечно-полосатая мышца. Се-
рия потенциалов действия, генерируемых в гладкомышечных клетках
унитарной мышцы, поддерживает в ней постоянное сокращение (рис.
2.36, А). Медленные изменения мембранного потенциала ведет к спон-
танным изменениям мембранной проницаемости ионов Na+ и Са2+, что
также вызывает генерацию потенциалов действия, инициирующих сокра-
щение мышцы (рис. 2.36, Б). В унитарной гладкой мышце медленные из-
менения мембранного потенциала сами по себе не вызывают сокраще-
ния. Такой тип активности занимает промежуточное положение между
медленными волнами мембранного потенциала и потенциалом действия,
имеющего плато (рис. 2.36, В). Это плато, которое продолжается около
одной секунды, ведет к тому, вызывает длительное сокращение, напри-
мер, в гладких мышцах матки.
Мембрана гладкомышечных клеток унитарных мышц способна деполя-
2. Физиология возбудимых тканей • 107
А
Мембранный потенциал (мВ)
0-
Напряжение
Мембранный потенциал (мВ)
Мембранный потенциал (мВ)
Мембранный потенциал (мВ)
В
Рис. 2.36. Мембранный потенциал и напряжение гладкой мышцы.
А — генерация потенциалов действия вызывает суммацию сокращения гладкой мышцы. Б —
медленные изменения мембранного потенциала гладкомышечной клетки приводят к генера-
ции потенциалов действия. В — плато потенциала действия способствует длительному сокра-
щению гладкомышечного волокна. Г — деполяризация мембраны клеток гладких мышц со-
провождается увеличением напряжения их волокон без генерации потенциалов действия.
ризоваться при ее растяжении. Возникающая при этом деполяризация мо-
жет комбинироваться с амплитудой медленных волн деполяризации, вызы-
вать генерацию серии потенциалов действия и, как следствие, мышечное
сокращение. Подобный тип активности характерен для пищеварительного
тракта при растяжении его стенок пищей.
В мультиунитарных гладких мышцах (образуют стенку многих крове-
носных сосудов), состоящей из отдельных гладкомышечных клеток,
электрическая активность не передается от одной клетки к другой. В
данном типе гладких мышц степень сокращения пропорциональна степе-
ни деполяризации мембраны гладкомышечной клетки даже в отсутствие
генерации потенциалов действия (рис. 2.36, Г). Отсутствие генерации
потенциалов действия в мультиунитарной мышце обусловлено тем, что
клетки имеют незначительный размер (20—200 мкм в длину и 5—10 мкм
в диаметре) и для генерации потенциалов необходимо вызвать деполяри-
зацию значительного количества клеток (50 и более), прежде чем в муль-
тиунитарной мышце возникнет распространяющийся потенциал дейст-
вия. Отсутствие потенциалов действия в мультиунитарной гладкой мыш-
це не исключает реакцию гладкомышечных клеток мультиунитарных
мышц на ацетилхолин или норадреналин. Эти нейротрансмиттеры при
связывании с рецепторами мембраны глакдомышечной клетки вызывают
ее локальную деполяризацию, которая распространяется электротониче-
ски на все волокно, вызывая его сокращение. Например, окончания нер-
108 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
вов, иннервирующих радужку, пиломоторные подкожные мышцы, секре-
тируют ацетилхолин или норадреналин, которые вызывают деполяриза-
цию мембраны гладкомышечных клеток и последующее их быстрое со-
кращение.
2.7.3. Нервно-мышечный синапс гладкой мышцы
Унитарные и мультиунитарные типы гладких мышц иннервируются тремя
категориями нервов: нервы вегетативной нервной системы (симатиче-
ские — артерии, и парасимпатические — цилиарная мышца и пищевари-
тельный тракт); нервы собственных сплетений в пределах гладкой мышцы
(преимущественно пищеварительный тракт); нервы афферентных чувстви-
тельных нейронов, которые рефлекторно активируют гладкие мышцы.
Значение иннервации гладкой мышцы иллюстрируется тем фактом, что
иннервация гладких мышц пищеварительного тракта осуществляется боль-
шим количеством нейронов, чем скелетной мускулатуры.
Нервные волокна распространяются по поверхности мышечного слоя
и образуют при этом не концевые пластинки (как в скелетной мышце), а
многочисленные расширения (варикозы) по ходу нервных волокон. В
среднем варикозы в нервных волокнах, иннервирующих, например, глад-
кими мышцами артерий и вен, имеют размеры порядка 2 мкм в длину и
1 мкм в диаметре и встречаются с интервалом 5—10 мкм. В местах вари-
козов нервное волокно не имеет слоя шванновских клеток, и химические
субстанции путем простой диффузии высвобождаются из варикозных рас-
ширений нервных волокон. В варикозах содержится два типа везикул,
наполненных медиаторами. Маленькие плотные везикулы, в которых со-
держится ацетилхолин, норадреналин и АТФ, и большие плотные везику-
лы, содержащие норадреналин, АТФ и нейропептиды (кальцитонин-ген-
связанный пептид, нейропептид Y и вазоактивный интестинальный пеп-
тид). Ацетилхолин и норадреналин являются медиаторами, высвобождаю-
щимися в гладкой мышце. Возбуждающий и тормозный эффект этих ме-
диаторов обусловлен типом адрено- и холинорецепторов на мембране
конкретной гладкой мышцы. Процесс высвобождения медиатора иденти-
чен таковому в нервно-мышечном синапсе, однако такой синапс в глад-
кой мышце нередко называется контактным соединением. Так, в муль-
тиунитарной гладкой мышце щель между мембраной гладкомышечной
клетки и нервом очень мал (порядка 20—30 мкм) и в эта мышца имеет
такой же латентный период реакции на действие медиатора, как в ске-
летной мышце. В случае, если расстояние для диффузии высвобождаемо-
го в варикозах медиатора является большим (диффузионный контакт), то
в ответ на возбуждение нерва сокращение гладкой мышцы начинается
через 0,2—30 с.
Рецепторы на мембране гладкой мышцы связываются с субстанциями,
которые циркулируют и крови или образуются локально в самих тканях.
В результате гладкая мышца способна реагировать открытием ионных ка-
налов в мембране гладкомышечных клеток, что вызывает эффекты сокра-
щения или расслабления гладких мышц, обусловливающие функции внут-
ренних органов: моторики и секреции в пищеварительном тракте, увеличе-
ние или уменьшение силы и частоты сокращения сердца, расслабление
или сокращение гладких мышц кровеносных сосудов.
2. Физиология возбудимых тканей • 109
2.7.4. Молекулярный механизм сокращения гладкой мышцы
Сокращение гладкой мышцы активируется так же, как и в скелетной
мышце, увеличением внутриклеточной концентрации ионов кальция. В
гладкомышечных клетках, в которых отсутствует саркоплазматический ре-
тикулум, ионы Са2+ диффундируют через кальциевые ионные каналы из
интерстициальной среды. Деполяризация мембраны гладкомышечных кле-
ток и/или генерация потенциалов действия открывает потенциалзависи-
мые натриевые и кальциевые ионные каналы, вызывая входящий внутрь
клетки натриевый и кальциевый ток. Большинство ионов Са2+ диффунди-
руют внутрь гладкомышечных клеток в результате открытия потенциалза-
висимых кальциевых ионных каналов саркоплазмы. Открытие каналов
происходит в течение 200—300 мс после деполяризации мембраны, поэто-
му в гладкой мышце длительный латентный период начала сокращения
после раздражения гладкой мышцы.
Некоторые гладкомышечные клетки содержат умеренно развитый сар-
коплазматический ретикулум, который соприкасается с углублениями, или
кавеолами, сарколеммы (рис. 2.37). В этом случае потенциалы действия
распространяются на мембрану кавеол, что вызывает выход ионов Са2+ из
цистерн саркоплазматического ретикулума и повышение концентрации
этих ионов в цитоплазме гладкомышечных клеток, что инициирует их со-
кращение. Высвобождение ионов Са2+ из саркоплазматического ретикулу-
ма гладкомышечных клеток может происходить в результате открытия
кальциевых ионных каналов при участии системы G-белков, фосфолипазы
С и вторичных посредников (инозитол-3-фосфата). В этом случае концен-
Рис. 2.37. Схематическое изображение кавеолы мембраны гладкомышечной клетки.
В гладкомышечных клетках, в которых саркоплазматический ретикулум слабо раз-
вит и представлен лишь небольшими трубочками, мембрана образует специфиче-
ские углубления, или кавеолы. Кавеолы наряду с небольшими размерами гладкомы-
шечных клеток уменьшают расстояние диффузии ионов кальция из внеклеточного
депо до сократительных филаментов, что связано с относительно медленной скоро-
стью процессов сокращения и расслабления гладкой мышцы.
В мембране кавеол имеются потенциалзависимые кальциевые ионные каналы (Пк), хемозави-
симые ионные каналы, через которые ионы кальция поступают внутрь гладкомышечных кле-
ток и вызывают их сокращение. Ионы кальция из саркоплазмы гладкомышечных клеток с по-
мощью вторично активного транспорта выводятся в интерстициальную среду, что сопровож-
дается расслабление гладкой мышцы. ИФЗ — инозитол-3-фосфат, Ац — ацетилхолин, М — хо-
линорецептор.
ПО • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
трация ионов Са2+ в саркоплазме увеличивается без увеличения проницае-
мости этих ионов через клеточную мембрану, что также сопровождается
сокращением гладкой мышцы.
В условиях покоя в цитоплазме гладкомышечных клеток концентрация
ионов кальция в среднем составляет незначительную величину — 100—
150 нМ/л. В покое концентрация ионов кальция в саркоплазме гладкомы-
шечных клеток зависит от ионного состава цитозольной и внеклеточной
сред, температуры тканей. Так, в цитозоле гладких мышц сосудов концен-
трация ионов кальция равна в среднем 79 нМ/л, а в сердечных — 155 нМ/л.
Напротив, минимальная концентрация ионов кальция в цитозоле гладких
мышечных клеток, при которой начинается мышечное сокращение, в
среднем составляет 200 нМ/л, а максимальное сокращение гладкие мыш-
цы развивают при концентрации в саркоплазме ионов Са2+ 0,8—1,0 мкМ/л.
В зависимости от указанных концентраций ионов кальция в цитозоле пря-
мо пропорционально изменяется сила сокращения гладкомышечных кле-
ток. При этом в гладких мышцах увеличение концентрации ионов Са2+ мо-
жет происходить под действием различных факторов (растяжение мембра-
ны, спонтанная электрическая активность, действие нейротрансмиттеров,
гормонов).
В гладкомышечных клетках отсутствует белок тропонин, с которым в
скелетных мышечных клетках связываются ионы Са2+, инициируя их со-
кращение. В гладких мышцах функцию, аналогичную тропонину, выпол-
няет белок кальмодулин. После повышения концентрации ионов Са2+ в
саркоплазме гладкомышечных клеток выше пороговой величины они свя-
зываются с белком кальмодулином, и комплекс «ионы Са2+—кальмодулин»
активирует фермент миозиновую киназу, который фосфорилирует одну из
легких цепей головок миозина. После этого головки поперечных мостиков
миозина приобретают способность циклически связываться с актином по
типу «замыкание—сокращение—размыкание» (как в скелетной мышце).
АТФ является источником энергии для актомиозинового взаимодействия в
гладкой мышце и необходима для отсоединения поперечных мостиков
миозина от актинового филамента. Это обусловливает повторение цикла
поперечных мостиков «замыкание—сокращение—размыкание».
Гладкая мышца в отличие от поперечно-полосатой имеет значительно
больший латентный период в связи с медленной диффузией ионов Са2+
через мембрану гладкомышечных клеток из интерстициальной среды. По-
этому после распространения потенциалов действия по сарколемме глад-
комышечных клеток они начинают сокращаться через 50—100 мс. Макси-
мум сокращения достигается примерно через 500 мс после его начала, а
затем в течение 1—2 с в гладких мышечных клетках медленно уменьшает-
ся сила сокращения, и они полностью расслабляются через 1—3 с. В раз-
ных группах гладких мышц период сокращения гладких мышц может
варьировать от 0,2 до 30 с, например в гладкой мышце матки. Медленное
начало и окончание мышечного сокращения в гладких мышцах обуслов-
лено низкой скоростью работы цикла «замыкание—размыкание» попереч-
ных мостиков и низкой скоростью процесса электромеханического сопря-
жения. Кроме того, цикл «замыкание—сокращение—размыкание» попе-
речных мостиков миозина в гладких мышечных клетках совершается с
частотой примерно в 10—300 раз меньшей, чем в скелетной мышце. На-
конец, поперечные мостики миозина могут находиться длительное время
в замкнутом состоянии с нитями актина, что обусловлено низкой АТФаз-
ной активностью головок поперечных мостиков миозина, медленным гид-
ролизом АТФ.
2. Физиология возбудимых тканей • 111
В гладкой мышце взаимодействие сократительных филаментов актина и
миозина имеет свою физиологическую особенность. Это проявляется в
том, что в гладкой мышце после развития максимального напряжения
происходит снижение степени активации сократительного аппарата, кото-
рое проявляется в замедлении процесса размыкания в циклической работе
поперечных мостиков миозина. Подобный феномен получил название ме-
ханизма «замка». Механизм «замка» дает возможность гладкой мышце не
снижая силу сокращения, осуществлять процесс сокращения при низких
энергетических затратах. Функциональное значение механизма «замка» за-
ключается в поддержании длительного, на протяжении многих часов, то-
нического сокращения гладкой мышцы, как при минимальных затратах
энергии, так и при незначительной активности нервов, иннервирующих
гладкомышечные клетки. Низкий расход АТФ в процессе сокращения
гладкомышечных клеток обусловлен тем, что миозиновых филаментов
примерно в 15 раз меньше, чем тонких актиновых нитей. Поэтому ско-
рость сокращения волокон гладких мышц и скорость расщепления в них
АТФ в 100—1000 раз меньше, чем в скелетных мышцах. Считается, что для
развития одного и того же по величине напряжения гладкой мышце требу-
ется от ’/io до ’/зоо части энергии скелетной мышцы. Это обусловлено низ-
кой скоростью работы поперечных мостиков в цикле «замыкание—размы-
кание», на один цикл которого независимо от его длительности расходует-
ся энергия одной молекулы АТФ. Этот механизм сокращения гладкой
мышцы чрезвычайно экономичен для энергетики организма, поскольку в
организме человека гладкие мышцу кишечника, мочевого пузыря и других
полых органов находятся в практически постоянном тоническом сокра-
щении.
2.7.5. Молекулярный механизм расслабления гладкой мышцы
Расслабление гладкомышечных клеток осуществляется при участии фер-
мента — миозиновой фосфатазы, от концентрации которой в саркоплазме
зависит скорость расслабления гладкой мышцы. Прекращение генерации
потенциалов действия на мембране гладкомышечной клетки вызывает
снижение концентрации ионов Са2+ в ее саркоплазме ниже порогового
уровня путем активного транспорта ионов кальция в саркоплазматический
ретикулум при участии Са2+-АТФазы и выведения ионов кальция из цито-
золя через сарколемму с помощью Са2+-АТФазы, а также путем облегчен-
ной диффузии при участии №+/Са2+-обменника. При низкой концентра-
ции ионов Са2+ в цитозоле фермент миозиновая фосфатаза дефосфорили-
рует легкую белковую цепь головки поперечного мостика миозина; в ре-
зультате прекращается цикл «замыкание—размыкание» и гладкая мышца
расслабляется до исходной длины мышечных волокон.
2.7.6. Физиологические параметры сокращения гладкой мышцы
По аналогии с поперечно-полосатой мышцей, основными физиологиче-
скими параметрами сокращения скелетных мышц человека являются сила,
скорость развития силы и укорочения мышцы, мощность и работа гладкой
мышцы.
При сокращении гладкая мышцы развивает максимальную силу, кото-
рая нередко больше (4—6 кг/см2), чем сила сокращения скелетной мышцы
(2—3 кг/см2). Гладкая мышца способна укорачиваться во время сокраще-
ния на 2/3 ее максимальной длины (скелетная мышца — не более ’/3—!/4
112 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
филаменты
Рис. 2.38. Особенности сократительных свойств гладкой мышцы. Гладкая мышца
может укорачиваться до меньших размеров относительно ее исходной длины, чем
скелетная мышца. Это обусловлено тем, что из-за разной длины актиновых фила-
ментов они могут продвигаться на большее расстояние относительно миозиновых
филаментов, чем в скелетной мышце. Благодаря этому свойству гладкая мышца
сфинктеров пищеварительного канала способна полностью «перекрывать» просвет
пищеварительной трубки.
А — исходная длина гладкомышечной клетки при расслаблении, Б — длина глакдомышечной
клетки существенно уменьшается при ее сокращении. СПР — саркоплазматический ретику-
лум.
своей длины). Эта особенность гладких мышечных клеток делает возмож-
ным гладкомышечным висцеральным органам изменять свой диаметр в
широком диапазоне. Объясняется это свойство тем, что в гладкомышной
клетке актиновые филаменты имеют разную длину. Поэтому во время со-
кращения актиновые и миозиновые филаменты имеют оптимальную длину
перекрывания, несмотря на изменение длины мышцы. Кроме того, широ-
кий диапазон изменения диаметра висцеральных органов обусловлен тем,
что актиновые филаменты в гладкомышечных клетках длиннее, чем в
клетках скелетной мышцы, и поэтому могут продвигаться на большую
длину относительно миозиновых филаментов (рис. 2.38). Гладкие мышцы
полых висцеральных органов способны восстанавливать в течение не-
скольких секунд или минут начальную силу своего сокращения после уко-
рочения или, наоборот, растяжения мышечных волокон. Это обусловлено
наличием феномена в полых висцеральных органах так называемой
стресс-релаксации. Суть феномена стресс-релаксации состоит в том, что
механизм «замка» первочально препятствует растяжению стенки полых
висцеральных органов. Затем в гладкомышечных волокнах совершается
новый цикл «размыкание—замыкание» поперечных мостиков миозина с
нитями актина. В результате длина гладкой мышцы может непрерывно из-
меняться, а напряжение стенки полых висцеральных органов остается по-
стоянным. Благодаря этому феномену внутри них давление сохраняется на
постоянном значении, несмотря на изменение их объема.
Например, увеличение объема жидкости в мочевом пузыре вызывает
2. Физиология возбудимых тканей • 113
растяжение его стенок и повышает внутрипузырное давление. В среднем
через 15—60 с путем стресс-релаксации происходит увеличение длины
гладкомышечных клеток и давление внутри мочевого пузыря восстанавли-
вается до начального уровня.
2.8. Функции мышечных клеток сердца
Клетки сердечной мышцы подразделяется на сократительные кардиомиоци-
ты, которые образуют предсердия и желудочки (миокард), и клетки синус-
ного узла и проводящей системы сердца, которые генерируют и проводят
электрические импульсы в сердце к кардиомиоцитам и не обладают сокра-
тительной способностью.
Рис. 2.39. Электромеханическое сопряжение в кардиомиоците. Плотность ионных
каналов в мембране t-трубочек кардиомиоцитов выше, чем в других участках сарко-
леммы.
Дигидропиридиновый рецептор сарколеммы и рианодиновый рецептор мембраны саркоплаз-
матического ретикулума представляют собой кальциевые ионные каналы и непосредственно
связаны между собой кальмодулином (СаМ). Деполяризация сарколеммы t-трубочек вызывает
последовательное открытие потенциалзависимых кальциевых ионных каналов сарколеммы и
при участии кальмодулина — саркоплазматического ретикулума. Открытие ионных каналов
ретикулума вызывает выход ионов Са2+ в саркоплазму, что является триггером механизма со-
кращения кардиомиоцитов.
114 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Сердечная мышца является поперечно-полосатой, однако ее сократи-
тельная способность, в отличие от скелетной мышцы, характеризуется
синхронностью сокращения и расслабления всего миокарда. Синхронность
сокращения и расслабления сердечных мышечных клеток обусловлена на-
личием специализированных плотных контактов между ними, или так на-
зываемых нексусов. Плотный контакт представляет собой участок мембра-
ны двух кардиомиоцитов, в пределах которого мембраны прилегающих
кардиомиоцитов сливаются между собой, эти плотные контакты является
местом низкого электрического сопротивления между клетками и пред-
ставляют, таким образом, путь электротонического распространения по-
тенциала действия от одного кардиомиоцита к другому. Эта структурная
особенность сердечной мышцы способствует быстрому распространению
электрической активности, т. е. потенциала действия в миокарде. Понятие
функциональный синцитий обычно относят к сердечной мышце, чтобы по-
казать, что сердце является единым в функциональном отношении орга-
ном в результате взаимодействия отдельных сердечных мышечных клеток.
Кардиомиоциты объединены в функциональный синцитий благодаря низ-
кому сопротивлению нексусов, с помощью которых осуществляются кон-
такт между миоцитами и передача потенциала действия от одного миоцита
к другому. В результате потенциал действия из центра синоатриального уз-
ла достигает всех сердечных мышечных клеток, вызывая сокращение пред-
сердий и желудочков
Кардиомиоциты имеют как сходные черты в своем строении со скелет-
ными мышечными клетками, так и отличия. Взаимное расположение фи-
ламентов актина и миозина в саркомерах придает поперечную исчерчен-
ность сердечным мышечным клеткам. Рядом расположенные миоциты от-
делены друг от друга на их концах вставочными дисками. Диски являются
местом проведения потенциала действия в сердечной мышце. Сердечная
мышечная клетка содержит одно ядро; t-система ее сарколеммы примерно
в два раза шире, чем в скелетном мышечном волокне, и t-трубочки лока-
лизованы ближе к Z-линии саркомеров, саркоплазматический ретикулум
контактирует с t-системой и с сарколеммой (рис. 2.39). Саркоплазматиче-
ский ретикулум представляет собой замкнутую систему трубочек, ветвя-
щихся в пределах кардиомиоцита и содержащих ионы кальция. Саркоплаз-
матический ретикулум кардиомиоцитов, как депо ионов Са2+, играет ос-
новную роль сопряжения процессов возбуждения и сокращения в сердеч-
ной мышце.
2.8.1. Электрическая активность клеток сердечной мышцы
2.8.1.1. Потенциал покоя
Потенциал покоя в разных типах сердечных мышечных клеток изменяется
в пределах от —60 до —90 мВ: он меньше в атипических клетках проводя-
щей возбуждение системы сердца и больше в сократительных сердечных
клетках предсердий и желудочков. Механизм происхождения мембранного
потенциала покоя в сердечных мышечных клетках является таким же, как
и для всех клеток организма, и зависит от высокой внутриклеточной и
низкой внеклеточной концентрации ионов калия, а также низкой прони-
цаемости саркоплазмы для ионов натрия. Мембранный потенциал сердеч-
ных мышечных клеток приближается к равновесному калиевому потенциа-
лу (Ej<). Однако потенциал покоя большинства кардиомиоцитов равен -80
мВ. Мышечные клетки синоатриального узла и клетки проводящей систе-
2. Физиология возбудимых тканей • 115
мы сердца имеют меньший, чем кардиомиоциты мембранный потенциал,
амплитуда которого находится в пределах -60—70 мВ. Поэтому клетки си-
ноатриального узла имеют более низкий порог возбудимости, чем кардио-
миоциты, что обусловлено так называемыми внутренними свойствами
мембраны синоатриальных клеток.
Электрогенный Na+/K+-Hacoc в кардиомиоцитах участвует в формиро-
вании 5—10 % нормальной величины потенциала покоя на мембране кле-
ток сердца, что составляет в среднем —10—11 мВ. Это обусловлено тем,
что плотность Ка+/К+-АТФазы в области t-трубочек мембраны кардиомио-
цитов выше, чем в других участках их сарколеммы. В отличие от скелет-
ных мышечных клеток, в кардиомиоцитах физиологическая роль Na+/K+-
насоса является ведущей в поддержании постоянной величины потенциала
покоя на мембранах клеток и, следовательно, физиологического уровня их
возбудимости. Данное обстоятельноство обусловлено тем, что инактивация
натриевых (а также кальциевых) ионных каналов и их способность участ-
вовать в генерации потенциала действия в сердечных мышечных клетках
зависит от величины максимального потенциала покоя сердечных клеток в
диастолу. В клетках сердца одной из важных функций Na+/K+-Hacoca яв-
ляется создание максимальной величины потенциала на мембране сердеч-
ных мышечных клеток в диастолу, что исключает колебания потенциала
покоя кардиомиоцитов и формирует у клеток постоянный исходный уро-
вень возбудимости.
2.8.1.2. Молекулярный механизм потенциала действия в типичных сердечных
мышечных клетках
Потенциал действия кардиомиоцитов имеет большую продолжительность:
до 200 мс в миоцитах предсердий, до 400 мс в клетках желудочков сердца.
Потенциал действия кардиомиоцитов состоит из фазы деполяризации, фа-
зы плато и фазы реполяризации (рис. 2.40). Изменение мембранного по-
тенциала сердечных мышечных клеток во время генерации в них потен-
циала действия обусловлено трансмембранными токами ионов Na+, Са2+
и К+.
Фаза деполяризации потенциала действия в кардиомиоците. Потенциал
действия на мембране кардиомиоцита возникает под влиянием возбужде-
ния атипичных мышечных клеток проводящей системы сердца. При этом
потенциал действия распространяется по проводящей системе сердца до
волокон Пуркинье. Между мембранами волокон Пуркинье и кардиомио-
цитами имеются плотные вставочные диски, или нексусы. Реверсия потен-
циала на мембране волокон Пуркинье электротонически при участии вста-
вочных дисков вызывает деполяризацию мембранного потенциала кардио-
миоцита до порога возбуждения (£к), который для кардиомиоцитов со-
ставляет —60 мВ. С этого момента по закону «все или ничего» в мембране
кардиомиоцитов открываются быстрые потенциалзависимые натриевые
ионные каналы. Ионы натрия через открытые каналы по концентрацион-
ному градиенту устремляются внутрь кардиомиоцита, сдвигая его мем-
бранный потенциал в сторону равновесного натриевого потенциала (Е^а
составляет +70 мВ) и происходит перезарядка мембраны сердечных сокра-
тительных клеток. Деполяризация сердечных мышечных клеток развивает-
ся чрезвычайно быстро, и на мембране кардиомиоцитов за 2 мс потенциал
достигает +30 мВ. В фазу деполяризации при открытых быстрых натрие-
вых ионных каналах мембраны кардиомиоциты утрачивают свою возбуди-
мость (абсолютная рефрактерность), и сократительные клетки миокарда не
116 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Мембранный
потенциал (мВ)
Время (мс)
Рис. 2.40. Потенциал действия кардиомиоцтита. В происхождении потенциала дей-
ствия кардиомиоцита участвуют натриевые, кальциевые и калиевые ионные токи.
В фазу деполяризации (1) открываются потенциалзависимые натриевые каналы и мембран-
ный потенциал резко уменьшается за счет входящего натриевого тока. Натриевые каналы че-
рез несколько миллисекунд закрываются и открываются потенциалзависимые калиевые ион-
ные каналы, что вызывает начало реполяризации мембраны кардиомиоцита (2). Поскольку
под влиянием деполяризации открываются потенциалзависимые кальциевые ионные каналы,
то входящий кальциевый ток в фазу реполяризации уравновешивает выходящий калиевый ток
и возникает фаза плато (3). После закрытия кальциевых ионных каналов выходящий калие-
вый ток реполяризует (4) мембранный потенциал до исходного уровня (Em).
способны реагировать даже на сверхпороговое раздражение. Деполяриза-
ция в сердечных мышечных клетках прекращается под влиянием двух про-
цессов: 1) быстрые натриевые каналы закрываются через несколько мил-
лисекунд после открытия; 2) открываются потенциалзависимые калиевые
ионные каналы. В результате на мембране кардиомиоцитов начинает до-
минировать калиевый ток (1к). Оба фактора — закрытие быстрых натрие-
вых ионных каналов и открытие калиевых каналов — вызывают уменьше-
ние положительного потенциала на мембране кардиомиоцитов, и начина-
ется фаза реполяризации мембранного потенциала. В кардиомиоцитах в
отличие от скелетных мышечных клеток фаза реполяризации имеет отли-
чительную особенность — она прерывается фазой плато потенциала дейст-
вия, длительность которой обусловливает общее время периода абсолют-
ной рефрактерности сердечной мышцы.
Фаза плато потенциала действия в кардиомиоцитах является уникаль-
ной особенностью потенциала действия кардиомиоцитов. Фаза плато по-
тенциала действия в кардиомиоцитах представляет собой задержку процес-
са реполяризации положительного потенциала, обусловленную появлени-
ем в этот момент генерации потенциала действия кардиомиоцита входяще-
го кальциевого тока. Основной причиной возникновения входящего каль-
циевого тока является деполяризация потенциала на мембране сердечных
сократительных клеток. Деполяризация потенциала на мембране кардио-
миоцитов открывает L-тип потенциалзависимых кальциевых ионных кана-
лов. В результате в мембране клеток увеличивается проницаемость для ио-
2. Физиология возбудимых тканей • 117
нов Са2+, которые по концентрационному градиенту входят внутрь кардио-
миоцитов. Кальциевые ионные каналы в мембране кардиомиоцитов оста-
ются открытыми в среднем 200 мс (варьирует от 150 до 400 мс) и, таким
образом, обусловливают длительность фазы плато. Мембранный потенци-
ал в фазу плато сохраняется на относительно постоянной величине, по-
скольку входящий кальциевый ток уравновешивается выходящим калие-
вым. Время фазы плато обусловливает продолжительность периода абсо-
лютной рефрактерности сердечной мышцы, которая у человека является
невозбудимой структурой на протяжении порядка 270 мс. Кардиомиоциты
имеют абсолютную рефрактерность на протяжении почти всей систолы
(0,3 с). Это создает идеальные физиологические условия для проявления
сократительных свойств всей сердечной мышцы, которая в этих условиях
сокращается по типу одиночного мышечного сокращения и осуществляет,
таким образом, выброс крови в аорту и легочные артерии из желудочков
сердца.
Фаза реполяризации потенциала действия в кардиомиоцитах. Фаза плато
оканчивается в результате закрытия кальциевых ионных каналов и увели-
чения калиевого тока. Поэтому выходящий калиевый ток (1К) снижает по-
тенциал на мембране кардиомиоцитов в течение порядка 50 мс до —90 мВ.
В фазу реполяризации в кардиомиоцитах уменьшается количество инакти-
вированных натриевых ионных каналов и они способны реагировать на
сверхпороговые раздражения. Это состояние сердечной мышцы называется
относительной рефрактерностью. Когда потенциал на мембране кардио-
миоцитов уменьшается до своей исходной величины, закрываются потен-
циалзависимые калиевые ионные каналы, а натриевые потенциалзависи-
мые ионные каналы переходят в закрытое состояние. С этого момента кар-
диомиоциты имеют исходный уровень возбудимости и способны реагиро-
вать на очередной импульс, исходящий от клеток проводящей системы
сердца.
2.8.1.3. Механизм возникновения пейсмекерной активности в клетках
синоатриального узла
Частота сокращения сердечных мышечных клеток предсердий и желудоч-
ков сердца обусловлена частотой генерации потенциалов действия клетка-
ми синусного узла. Синоатриальный узел расположен в задней стенке пра-
вого предсердия в месте его соединения с верхней полой веной и состоит
из небольших сердечных мышечных клеток, которые не способны сокра-
щаться.
Пейсмекерные потенциалы. Мембранный потенциал клеток синоатриаль-
ного узла равен примерно —60 мВ. В интервале времени между генерацией
потенциалов действия (в фазу диастолы сердца) мембранный потенциал
синоартриальных мышечных клеток постепенно уменьшается до критиче-
ского уровня деполяризации (порядка —40 мВ). Медленная деполяризация
мембранного потенциала синоатриальных мышечных клеток, поскольку
она совершается в фазу диастолы сердца, называется спонтанной диастоли-
ческой деполяризацией и заканчивается генерацией пейсмекерного потен-
циала при достижении критического уровня деполяризации этих клеток
(рис. 2.41).
Механизм возникновения пейсмекерного потенциала в синоатриальных
мышечных клетках связан с ролью натриевого, калиевого и кальциевого
ионных токов. Два ионных тока (/,Va+ и 1Са2+) направлены внутрь клеток си-
ноатриального узла и вызывают деполяризацию их мембраны. Наибольшее
118 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Рис. 2.41. Основные ионные токи калия, натрия и кальция, участвующие в генера-
ции пейсмекерного потенциала и потенциала действия клеток синоатриального
узла.
Внутренним свойством мембраны пейсмекерных клеток синоатриального узла является ее
спонтанная деполяризация, которая, достигая критического уровня (Ек), вызывает на ее мем-
бране генерацию потенциала действия. Стрелками обозначено направление входящих токов
(натрия, Кальция) и выходящего калиевого тока в период спонтанной деполяризации мембра-
ны и генерации потенциала действия клетки синоатриального узлаатри. Длина стрелок услов-
но отражает величину трансмембранных токов этих ионов. СДД — спонтанная диастолическая
деполяризация.
значение в появлении пейсмекерного потенциала имеет входящий натрие-
вый ток через специфические натриевые каналы, которые открываются ре-
поляризацией мембраны синоатриальных мышечных клеток. Финальная
фаза пейсмекерного потенциала, т. е. приближение его амплитуды к кри-
тическому уровню деполяризации мембраны в клетках синоатриального
узла, связана с открытием Т-типа потенциалзависимых кальциевых ион-
ных каналов и появлением входящего кальциевого тока. Входящие натрие-
вый и кальциевый токи (спонтанная деполяризация) противодействуют
выходящему калиевому току, который гиперполяризует мембрану синоа-
триальных мышечных клеток (см. рис. 2.41). В этих условиях под влияни-
ем токов ионов Na+ и Са2+ деполяризация мембраны синоатриальных мы-
шечных клеток достигает критического уровня деполяризации и в клетках
синоатриального узла генерируется очередной потенциал действия, кото-
рый распространяется по проводящей системе сердца и является раздра-
жителем для сократительных сердечных клеток сердца.
Частота генерации потенциалов действия в синоатриальных мышечных
клетках зависит от скорости деполяризации пейсмекерного потенциала. В
отсутствии влияний со стороны вегетативной нервной системы на клетки
2. Физиология возбудимых тканей • 119
синоатриального узла, частота генерации потенциалов действия составляет
в среднем 2 в секунду (120/мин).
Клетки, расположенные в центре синоатриального узла, имеют боль-
шую скорость развития пейсмекерного потенциала и высокую частоту ге-
нерации потенциалов действия. Клетки периферических областей узла
имеют более медленные изменения пейсмекерных потенциалов, но боль-
шее число миофиламентов. Таким образом, это обусловливает постепен-
ный переход в свойствах миоцитов от пейсмекерных клеток центральной
области синоатриального узла к свойствам, присущим сократительным
предсердным мышечным клеткам. Частота сокращения сердца в покое
(порядка 70 в минуту) обусловлена тормозным действием блуждающего
нерва на пейсмекерные клетки центральной области синоатриального узла.
2.8.2. Молекулярный механизм сокращения кардиомиоцитов
Механизм сокращения сердечных мышечных клеток в основном иденти-
чен таковому в скелетных мышечных клетках. Ионы Са2+ являются «клю-
чом» к сокращению миокарда, поскольку они выполняют связующую
функцию между деполяризацией мембраны кардиомиоцитов, вызванной
распространяющимся потенциалом действия по их поверхности, и укоро-
чением саркомеров в сократительных клетках сердца. Концентрация ионов
Са2+ в саркоплазме невозбужденного кардиомиоцита составляет 0,1 пМ/л
(менее 10-7 М/л). Во время деполяризации мембраны сарколеммы и t-сис-
темы кардиомиоцитов внутриклеточная концентрация ионов Са2+ достига-
ет 1—10 мкмМ/л (более 10-6 М/л) в результате появления входящего каль-
циевого тока.
Увеличение ионов кальция в саркоплазме кардиомиоцитов активирует
так называемую сократительную машину сердца. Взаимодействие ионов
Са2+ с тропонином, когда изменяется положение тропомиозина, закры-
вающего места связывания на актиновом филаменте для поперечных мос-
тиков миозина, является центральным процессом в электромеханическом
сопряжении, как в скелетной мышце, так и в сердечной. В кардиомиоцитах
ионы кальция связываются с субъединицей С тропонина тропомиозиново-
го комлекса. Связывание ионов Са2+ с тропонином С уменьшает плотность
электростатической связи тропонина Т и тропомиозина относительно тон-
ких филаментов и тропомиозиновый комплекс, смещаясь с перифериче-
ской части глобулярного актина в зону контакта двух белковых нитей спи-
рали актинового филамента, открывает на нем места связывания головок
поперечных мостиков миозина с актином. В отличие от скелетной мыш-
цы, в кардиомиоцитах концевая МН2-область тропонина С содержит толь-
ко одно место связывания ионов Са2+. Поэтому, даже при максимальной
концентрации ионов Са2+ в цитозоле кардиомиоцитов, не происходит пол-
ной активации ионами Са2+ тропонина С, что не приводит к открытию
всех мест связывания поперечных мостиков с актиновыми филаментами.
Это создает потенциальные условия в кардиомиоцитах для дополнитель-
ной активации сократительной способности сердечной мышцы уже непо-
средственно в процессе сокращения миокарда. Например, под влиянием
циклических процессов «замыкание—сокращение—размыкание» попереч-
ных мостиков миозина дополнительно увеличивается связывание ионов
Са2+ с тропонином С. Сила сокращения, генерируемая поперечными мос-
тиками миозина в кардиомиоцитах сердца, прямо зависит от внутрикле-
точной концентрации ионов Са2+ во время фазы плато потенциала дей-
ствия.
120 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
В электромеханическом сопряжении сердечных мышечных клеток уча-
ствуют два рецепторных белка: потенциалзависимый дигидропиридиновый
рецептор мембраны t-трубочек и рианодиновый рецептор мембраны сарко-
плазматического ретикулума. Оба белка являются медленными кальциевы-
ми ионными каналами. Деполяризация мембраны кардимиоцитов при рас-
пространении по их мембране потенциала действия открывают потенциал-
зависимые кальциевые ионные каналы мембраны t-трубочек. Ионы каль-
ция по концентрационному градиенту поступают внутрь кардиомиоцитов и
связываются с кальмодулином, расположенным на рианодиновом рецепторе
или кальциевом ионном канале саркоплазматического ретикулума. Белок
кальмодулин открывает кальциевые ионные каналы саркоплазматического
ретикулума, и концентрация ионов Са2+ в саркоплазме повышается до по-
роговой (примерно 1 мкМ/л). Кальмодулин осуществляет взаимодействие в
кардиомиоцитах между дигидропиридиновыми рецепторами мембраны t-
трубочек и рианодиновыми рецепторами мембраны саркоплазматического
ретикулума, за счет его активации внеклеточными ионами Са2+. Поэтому в
сократительных клетках сердца процесс элеткромеханического сопряжения
обусловлен ионами Са2+ внеклеточной среды. Электромеханическое сопря-
жение в кардиомиоцитах отсутствует при пониженной концентрации ионов
Са2+ в интерстициальной среде, несмотря на нормальное их содержание в
саркоплазматическом ретикулуме кардиомиоцитов.
Открытие кальциевых ионных каналов саркоплазматического ретикулу-
ма в кардиомиоцитах при участии белка кальмодулина вызывает выход
этих ионов в межфибриллярное пространство кардиомиоцитов.
Наибольшая активация сокращения кардиомиоцитов происходит при
концентрации ионов Са2+ в саркоплазме порядка 10 мкМ/л. В кардиомио-
цитах имеет место так называемое Са2+-индуцированное высвобождение ио-
нов Са2+, проявляющееся в том, что во время фазы плато потенциала дей-
ствия ионы Са2+ входят в саркоплазму кардиомиоцитов через медленный
кальциевый ионный канал дигидропиридинового рецептора. Далее ионы
Са2+ связываются с медленным рианодиновым рецептором мембраны сар-
коплазматического ретикулума. При этом открытие одного Са2+-канала яв-
ляется сигналом для открытия других кальциевых каналов эндоплазмати-
ческой сети. В результате из саркоплазматического ретикулума в цитоплаз-
му кардиомиоцитов поступает нарастающее количество ионов Са2+, что су-
щественно увеличивает силу сокращения сердечной мышцы.
2.8.3. Молекулярный механизм расслабления кардиомиоцитов
Их расслабление происходит во время фазы реполяризации потенциала
действия и вызывается снижением внутриклеточной концентрации ионов
Са2+. Процесс расслабления кардиомиоцитов обусловлен остановкой вхо-
дящего кальциевого тока и прекращением высвобождения ионов Са2+ из
саркоплазматического ретикулума. Основным механизмом расслабления
миокарда является первично активный транспорт ионов Са2+ из саркоплаз-
мы клеток в саркоплазматический ретикулум при участии Са2+-АТФазы
кардиомиоцитов. Ионы Са2+ выводятся из саркоплазмы кардиомиоцитов в
интерстициальную среду с помощью Ка+/Са2+-обменника. В результате их
концентрация в межфибриллярном пространстве становится ниже порога
активации тропонина С и тропонин-тропомиозиновый комплекс закрыва-
ет места связывания на актиновых филаментах для поперечных мостиков
миозина. Миокард расслабляется, что соответствует фазе диастолы сердеч-
ного цикла.
2. Физиология возбудимых тканей • 121
Ключевым регулятором расслабления миокарда является белок мембра-
ны саркоплазматического ретикулума фосфоламбан. Фосфорилирование
фосфоламбана при участии вторичного посредника цАМФ ускоряет гидро-
лиз АТФ, активирует кальциевый насос саркоплазматического ретикулума,
повышает сродство ионов Са2+ к Са2+-АТФазе, увеличивая скорость актив-
ного транспорта ионов Са2+ из саркоплазмы внутрь саркоплазматического
ретикулума. Благодаря функции фосфоламбана сродство ионов кальция и
кальциевого насоса в кардиомиоцитах идентично таковому в быстрых ске-
летных мышечных волокнах, в которых отсутствует этот белок. Фосфолам-
бан увеличивает кальциевую чувствительность Са2+-АТФазы саркоплазма-
тического ретикулума при взаимодействии адреналина или норадреналина
с 0,-адренорецепторами мембраны кардиомиоцитов. Положительный ино-
тропный эффект, возникающий при стимуляции этого типа рецепторов
адреналином или норадреналином, проявляется в активации как сокраще-
ния миокарда, так и расслабления. Сила сокращения сердечной мышцы
возрастает при увеличении скорости выхода ионов Са2+ из саркоплазмати-
ческого ретикулума в саркоплазму и связывания с тропонином С. Расслаб-
ление миокарда прямо зависит от скорости депонирования ионов Са2+ в
саркоплазматический ретикулум при участии фосфоламбана.
2.8.4. Медиаторный контроль сокращения кардиомиоцитов
В физиологических условиях механизмы сокращения сердечных мышеч-
ных клеток контролируются адрено- и холинорецепторами, расположен-
ными на мембране кардиомиоцитов.
Из трех типов щ-адренорецепторов (а1А, а1В и a1D) в сердце человека до-
минирует а1А тип, а также 0-адренорецепторы. Стимуляция (^-адреноре-
цепторов увеличивает сократимость кардиомиоцитов и в целом — силу
сердечных сокращений. Влияние стимуляции адренорецепторов на силу
сокращения сердечной мышцы называется инотропным эффектом (лат.
inos — сила). Молекулярный механизм инотропного эффекта норадренали-
на, вызванный стимуляцией cq-адренорецепторов, опосредован активацией
системы G-белков (семейство Gq/ц, обозначаемое как семейство Gh для
сердца человека) с последующим образованием в кардиомиоцитах инози-
тол-3-фосфата и диацилглицерола. Эти вторичные посредники через риа-
нодиновые рецепторы мембраны саркоплазматического ретикулума увели-
чивают активность кальциевых ионных каналов, что повышает концентра-
цию ионов кальция в саркоплазме кардиомиоцитов и усиливает силу их
сокращения. Кроме того, стимуляция а,-адренорецепторов увеличивает
кальциевый ток через сарколемму внутрь кардиомиоцитов из внеклеточ-
ной среды, а также чувствительность сократительных миофиламентов к
ионам Са2+. Функция а2-адренорецепторов в контроле физиологических
свойств кардиомиоцитов человека связана с пресинаптическим торможе-
нием выделения норадреналина из нервных окончаний, иннервирующих
сердце.
Из трех типов 0-адренорецепторов в сердце человека идентифицирова-
ны 0Г и 02-адренорецепторы. Молекулярный механизм стимуляции сокра-
щения миоцитов в сердце человека при участии 0Г и 02-адренорецепторов
осуществляется при участии семейства Gs-белка, который повышает внут-
риклеточное содержание аденилатциклазы и вторичного посредника
цАМФ. Это ведет к стимуляции фосфорилирования внутриклеточных бел-
ков при участии протеинканазы А, включая L-тип кальциевых ионных ка-
налов мембран t-системы. В результате открытия кальциевых ионных ка-
122 • БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
налов увеличивается содержание этих ионов в саркоплазме и, как следст-
вие, возрастает сократимость сердечной мышцы (положительный инотроп-
ный эффект). В различных тканях организма человека идентифицировано
пять генов, контролирующих синтез мускариновых М,—М5 рецепторов. При
этом в сердце человека, локализован М2 тип холинорецепторов. Взаимо-
действие ацетилхолина с М2 (а также М4) типом холинорецепторов через
семейство Gi/Go белков угнетает аденилатциклазу миоцитов и уменьшает
активность калиевых и кальциевых ионных каналов, активируя фосфоли-
пазу Д2, фосфолипазу D и протеинкиназы. В результате снижается ско-
рость развития пейсмекерного потенциала в клетках синоатриального узла
и уменьшается сократимость миокарда. Ацетилхолин через М2 холиноре-
цепторы уменьшает сократимость кардиомиоцитов, если исходный уро-
вень их сократимости был повышен под влиянием стимуляции р-адрено-
рецепторов.
II РЕГУЛИРУЮЩИЕ
И УПРАВЛЯЮЩИЕ
СИСТЕМЫ
124 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Организм человека представляет собой сложную саморегулирующуюся
иерархическую систему, которая обменивается с окружающей средой ве-
ществом, энергией и информацией. В ходе непрерывного взаимодействия
со средой организм проявляет себя как реактивная система, приспосабли-
вающая свои физиологические процессы к меняющимся условиям среды
для уравновешивания с ней (гомеостазис), и одновременно как активная
система, изменяющая среду в собственных целях и для удовлетворения
своих потребностей (поведение). Как в том, так и в другом случае необхо-
дима интеграция всех иерархических уровней организации живой системы:
молекулярного, субклеточного, клеточного, тканевого, органного. Согла-
сование биофизических, биохимических и физиологических процессов,
происходящих в тканях и органах, а также приспособление этих процессов
к изменяющимся условиям внешней среды осуществляют регулирующие и
управляющие системы организма: нервная и эндокринная.
Под регуляцией в физиологии понимается активное управление функ-
циями биологической системы (вплоть до организма в целом и его поведе-
ния) с целью поддержания оптимального уровня ее жизнедеятельности и
приспособления системы к меняющимся условиям внешней среды.
Изменение параметров функций при поддержании их в границах гомео-
стазиса происходит на каждом уровне организации или в любой иерархи-
ческой системе за счет саморегуляции, т. е. внутренних для системы меха-
низмов управления жизнедеятельностью. Так, например, уменьшение кро-
воснабжения ткани ведет к образованию в ней химических веществ, рас-
ширяющих артерии и восстанавливающих тем самым приток крови. Такие
механизмы саморегуляции получили название местных.
Для осуществления функций организма в целом необходимы взаимо-
связь и взаимозависимость функций составляющих его систем. Поэтому
наряду с внутренними механизмами саморегуляции систем в организме
должны существовать и внешние для каждой из них механизмы регуляции,
соподчиняющие и координирующие деятельность систем. Например, для
реализации функции перемещения тела человека в пространстве необходи-
мо изменение деятельности не только скелетных мышц, но и кровообра-
щения, дыхания, обмена веществ и т. п. Изменения параметров этих
функций реализуются под влиянием сформировавшейся в процессе эволю-
ции специализированной системы регуляции функций.
Однако и организму в целом присуща саморегуляция жизненных про-
цессов, позволяющая осуществлять приспособление к меняющимся усло-
виям существования. Организм является самоорганизующейся системой,
т. е. сам выбирает и поддерживает значения огромного числа параметров
функций, меняет их в зависимости от собственных потребностей, что по-
зволяет ему обеспечивать наиболее оптимальный характер функциониро-
вания. Так, например, при низких температурах внешней среды организм
снижает температуру поверхности тела (чтобы уменьшить теплоотдачу),
повышает скорость окислительных процессов во внутренних органах и
мышечную активность (чтобы увеличить теплообразование). Одновремен-
но реализуется и адаптивное поведение — человек утепляет жилище, меня-
ет одежду (для увеличения теплоизолирующих свойств), причем делает это
даже заранее, опережающе реагируя на изменения внешней среды.
Основой физиологической регуляции является передача и обработка ин-
формации. Под термином «информация» следует понимать все, что несет в
себе отражение фактов или событий, которые произошли, происходят или
могут произойти. Информация содержит количественные характеристики
определенных параметров внешней и внутренней среды организма, поэто-
2. Физиология возбудимых тканей • 125
му особую важность имеет ее объем. Примером способа количественного
выражения информации, принятого в информатике как науке и исполь-
зуемого в организме, является двоичная система. Единицей количества в
таком случае является бит, характеризующий информацию, получаемую
при выборе одного из двух одинаково вероятных состояний, например
да—нет, все—ничего, быть—не быть и т. п. Материальным носителем ин-
формации является сигнал, в форме которого и передается информация.
Это могут быть как физические, так и химические сигналы, например
электрические импульсы, форма молекулы, концентрация молекул и т. д.
В тех случаях, когда регуляторным сигналом являются электрические им-
пульсы (нервные потенциалы), регуляция функций осуществляется нерв-
ной системой. В случаях, когда регуляторный сигнал представлен химиче-
скими молекулами, находящимися в жидкостях внутренней среды, регуля-
цию называют гуморальной. Она осуществляется либо тканевыми метабо-
литами, либо специальной эндокринной системой.
Центральная нервная система обеспечивает согласованное взаимодейст-
вие органов и частей организма, участвующих в той или иной его реакции
на воздействие среды, и формирует поведение, направленное на удовле-
творение потребностей и преобразование среды. Центральная нервная сис-
тема постоянно получает информацию об изменениях среды и может не-
медленно запускать в ответ на них врожденный комплекс стандартных
приспособительных реакций — рефлексов, проявляющихся мышечными
сокращениями или секрецией желез.
Сокращениями скелетных мышц управляет соматическая нервная сис-
тема, посредством которой регулируются не только рефлекторные мышеч-
ные реакции, но и произвольные сокращения мышц, необходимые для ак-
тивного взаимодействия человека со средой. Вегетативная нервная система
иннервирует гладкие мышцы внутренних органов, сердце и железы внеш-
ней секреции; она осуществляет нервную регуляцию параметров внутрен-
ней среды организма. Регуляторная функция вегетативной нервной систе-
мы в отличие от соматической непосредственно не контролируется созна-
нием и осуществляется непроизвольно. Однако нервные центры головного
и спинного мозга координируют активность соматической и вегетативной
нервной системы и согласуют друг с другом их ответные реакции.
Эндокринные железы, или железы внутренней секреции, выделяют в
кровь гормоны — химические вещества, предназначенные для передачи
специфических управляющих сигналов другим клеткам. Специфичность
действия гормонов определяется рецепторами, расположенными на мем-
бранах клеток-мишеней и принимающих адресованные им сигналы, под
влиянием которых происходят изменения функциональной активности
управляемых клеток. Нервные клетки тоже используют химические веще-
ства для передачи сигналов, такие вещества называются нейромедиатора-
ми. Нейроны могут действовать только на такие клетки, которые образуют
с ними функциональные соединения — синапсы, а поэтому действие ме-
диаторов проявляется немедленно и только в синапсе. Гормоны поступают
в общий кровоток и поэтому способны влиять на большое количество кле-
ток, имеющих специфические рецепторы для связывания гормонов. Гор-
мональная регуляция осуществляется значительно медленнее, чем нервная,
однако гормональные влияния проявляются гораздо дольше, чем измене-
ния, происходящие под влиянием нейромедиаторов. Нервная и гормональ-
ная регуляция функций взаимно дополняют друг друга и осуществляются
согласованно.
Следует в то же время подчеркнуть, что как нервная, так и эндокринная
126 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
системы являются функциональными системами и находятся в организме
в ряду других систем (см. раздел III) с присущими им специфическими
функциями. И только чисто в дидактических целях — чтобы подчеркнуть
их особую значимость в общеинтегративных, регуляторных процессах ор-
ганизма — эти системы выделены в особый раздел управления и контроля
физиологических функций и процессов в организме.
ГЛАВА 3
Общие принципы и механизмы
регуляции физиологических функций
3.1. Общие принципы организации системы регуляции
Обработка информации осуществляется управляющей системой, или сис-
темой регуляции. Она состоит из отдельных структурных элементов, свя-
занных информационными каналами (рис. 3.1). Управляющее устройство
(например, центральная нервная система) получает информацию о дея-
тельности системы, обрабатывает ее в соответствии с имеющейся програм-
мой оптимизации функций (врожденной или приобретенной на основе
жизненного опыта) и посылает управляющие сигналы к исполнительным
органам и системам. Входные каналы связи (нервы, жидкости внутренней
среды) передают нервные импульсы или информационные молекулы ве-
ществ, сигнализирующие о состоянии внутренней среды и параметрах
функций, в управляющее устройство, а выходные каналы связи — от
управляющего устройства к исполнительным органам. Датчики (сенсорные
рецепторы) воспринимают информацию из внешней или внутренней сре-
ды на входе системы. Сигналы, идущие по выходным каналам связи из
управляющего устройства и предназначенные для изменения функции,
воспринимаются образованиями, располагающиеся на исполнительных
клетках (клеточные рецепторы). Часть управляющего устройства, служа-
щая для хранения информации, называется запоминающим устройством,
или аппаратом памяти. Характер переработки поступающих сигналов зави-
сит от той информации, которая записана в аппарате памяти управляюще-
го устройства системы регуляции.
3.1.1. Уровни организации системы регуляции
Система регуляции физиологических функций организма представляет со-
бой иерархическую структуру трех уровней.
Первый уровень системы регуляции состоит из относительно автоном-
ных местных систем, поддерживающих физиологические константы, зада-
ваемые собственными метаболическими потребностями. Так поддержива-
ются, в частности, осмотическое давление крови, вентиляционно-перфузи-
онные отношения в легких, тканевой кровоток. Например, при активации
деятельности скелетной мышцы возникает относительно недостаточное ее
кислородное обеспечение, в результате начинает накапливаться не успе-
вающая окисляться молочная кислота, способствующая расширению арте-
риол и увеличению кровоснабжения и кислородоснабжения работающей
мышцы. Для реализации механизмов этого уровня не обязательны сигналы
из управляющего устройства центральной нервной системы, они обеспечи-
ваются местными реакциями метаболической природы и носят поэтому
название «местная саморегуляция».
128 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 3.1. Схематическое изображение системы регуляции.
Жирным шрифтом и стрелками выделен выходной канал прямой регуляторной связи — канал управления.
Обычным шрифтом, рамками и сплошными стрелками обозначена регуляция по возмущению, включаемая в результате действия фактора среды
на сенсорные рецепторы. Обычным шрифтом, пунктирными стрелками и рамками выделен канал обратной связи, осуществляющий регуляцию по
отклонению при изменении параметров физиологической функции.
3. Общие принципы и механизмы регуляции физиологических функций • 129
Второй уровень системы регуляции осуществляет приспособительные
реакции в связи с изменениями внутренней среды. Этот уровень может
быть представлен низшими этажами центральной нервной системы (спин-
ной, продолговатый мозг) или отдельными эндокринными железами (око-
лощитовидные железы секретируют гормон паратирин при снижении
уровня кальция в крови, 0-клетки островков Лангерганса поджелудочной
железы секретируют гормон инсулин при повышении уровня сахара в кро-
ви). На этом уровне задается величина физиологических параметров, кото-
рые в дальнейшем могут поддерживаться системами первого уровня. Здесь
подбирается оптимальный режим работы физиологических систем для
адаптации организма к внешней среде. Например, выполнение физиче-
ской работы или даже подготовка к ней требует увеличенного снабжения
мышц кислородом, что обеспечивается усилением внешнего дыхания, по-
ступлением в кровь депонированных эритроцитов и повышением артери-
ального давления.
Третий уровень системы регуляции, представленный, как правило, об-
разованиями головного мозга (гипоталамус, лимбическая система, кора
больших полушарий), обеспечивает выработку критериев оценки состоя-
ния внутренней и внешней среды, настройку режимов работы первого и
второго уровней, гарантирующих в итоге изменение вегетативных фунций
и поведения организма с целью оптимизации его жизнедеятельности.
3.1.2. Типы и механизмы регуляции
На всех трех уровнях структурной организации системы регуляции воз-
можны 3 типа регуляции: 1) по возмущению, или адаптивная регуляция,
2) по отклонению, или гомеостатическая регуляция, 3) по упреждению или
опережающая регуляция. Регуляция по возмущению (саморегуляция по вхо-
ду) системы возможна для систем, имеющих связи с внешней для них сре-
дой (см. рис. 3.1). Этот тип регуляции включается в тех случаях, когда на
живую систему оказывает воздействие внешний для нее фактор, меняю-
щий условия ее существования. Например, регуляция дыхания обычно
обеспечивает оптимальную для метаболизма клеток взаимосвязь процессов
газообмена в легких, транспорта газов кровью и газообмена крови с клет-
ками в тканях. Физическая же нагрузка, не являющаяся частью структуры
приведенной системы (внешняя для нее), представляет собой возмущаю-
щее воздействие и, поскольку физическая нагрузка ставит новые условия в
виде повышенной потребности мышц в кислороде, реализуется регуляция
по возмущению, меняющая интенсивность составляющих дыхание процес-
сов. Или регуляция постоянства температуры тела подразумевает взаимо-
связь процессов образования тепла в организме и его отдачи в окружаю-
щую среду. Изменение температуры окружающей среды, например повы-
шение ее, является возмущающим раздражителем, воспринимается термо-
рецепторами кожи и слизистых оболочек. От рецепторов по входному ка-
налу связи (нервным проводникам) сигнал поступает в управляющее уст-
ройство (нервный центр), меняющее путем выходных сигналов процессы
образования и отдачи тепла так, чтобы поддерживать температуру тела на
постоянном уровне. Этот тип регуляции получил название адаптивной ре-
гуляции, так как она способствует быстрому приспособлению организма к
изменившимся условиям среды.
Регуляция по отклонению (саморегуляция по выходу системы) возникает
при отклонении параметров функции за пределы диапазона физиологиче-
ских колебаний (см. рис. 3.1). Регуляция обеспечивается сравнением
5-6095
130 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
имеющихся параметров реакции физиологических систем с требующимися
в конкретных условиях, определением степени рассогласования между ни-
ми и включением исполнительных устройств для устранения этого рассо-
гласования. Частным примером регуляции по отклонению является под-
держание физиологических констант внутренней среды. Стоит только от-
клониться от заданного уровня и повыситься в крови напряжению углеки-
слого газа из-за недостаточного его удаления через легкие или повышен-
ного образования в тканях, как начнут реализовываться регуляторные ме-
ханизмы, направленные на восстановление и поддержание гомеостазиса.
Этот тип регуляции поэтому называется гомеостатической регуляцией, ее
реакции направлены, например, на устранение описанного сдвига содер-
жания СО2: образование углекислоты и бикарбоната натрия, связывание
водородных ионов буферными системами, повышение выведения прото-
нов через почки, активация дыхания для выведения углекислого газа во
внешнюю среду.
Регуляция по отклонению требует наличия канала связи между выходом
системы регуляции и ее центральным аппаратом управления и даже между
выходом и входом системы регуляции. Этот канал связи получил название
механизма обратной связи (см. рис. 3.1). Именно обратная связь позволяет
регуляции по отклонению работать в двух режимах: компенсационном и
слежения. Компенсационный режим обеспечивает быструю корректировку
рассогласования реального и оптимального состояния физиологических
систем при внезапных влияниях среды, т. е. оптимизирует реакции орга-
низма. При режиме слежения регуляция осуществляется по заранее задан-
ным программам, а обратная связь контролирует соответствие параметров
деятельности физиологической системы заданной программе. Если возни-
кает отклонение — реализуется компенсационный режим.
По конечному эффекту регуляции механизм обратной связи может быть
положительным и отрицательным.
Механизм положительной обратной связи означает, что выходной сигнал
системы регуляции усиливает входной, активация какой-либо функции
вызывает усиление механизмов регуляции, еще больше ее активирующих.
Такая обратная связь усиливает процессы жизнедеятельности, например
прием пищи и поступление ее в желудок усиливают отделение желудочно-
го сока, необходимого для гидролиза веществ (рис. 3.2). Появляющиеся в
желудке и частично всасывающиеся в кровь продукты гидролиза в свою
очередь стимулируют сокоотделение, что ускоряет и усиливает дальнейшее
переваривание пищи. Однако положительная
обратная связь нередко приводит систему в
неустойчивое состояние, способствует фор-
мированию «порочных кругов», когда реакция
организма на вредное воздействие среды уси-
ливает следствия этого воздействия, что ле-
жит в основе многих патологических процес-
сов в организме.
Рис. 3.2. Пример механизма положительной обрат-
ной связи в регуляции секреции желудочного сока.
Прием пищи вызывает секрецию желудочного сока, рас-
щепление пищевого белка и всасывание продуктов его
гидролиза в кровь. Последние в свою очередь еще больше
стимулируют секрецию желудочного сока (положительная
обратная связь).
3. Общие принципы и механизмы регуляции физиологических функций • 131
Рис. 3.3. Пример механизма от-
рицательной обратной связи в
регуляции уровня глюкозы в
крови.
Прием сахара с пищей и последую-
щий его гидролиз до глюкозы ведут
к повышенному поступлению глю-
козы в кровь. Рост содержания глю-
козы в крови стимулирует секрецию
инсулина р-клетками островков
Лангерганса поджелудочной желе-
зы. Под влиянием инсулина растет
захват глюкозы из крови клетками
мышечной и жировой ткани, уро-
вень глюкозы в крови снижается,
что тормозит секрецию инсулина
(отрицательная обратная связь).
г — — ”1
I Подавление >
секреции
' инсулина ।
Механизм отрицательной обратной связи (рис. 3.3) означает, что выход-
ной сигнал уменьшает входной, активация какой-либо функции подавляет
механизмы регуляции, усиливающие эту функцию. Отрицательные обрат-
ные связи способствуют сохранению устойчивого, стационарного состоя-
ния системы. Благодаря им возникающее отклонение регулируемого пара-
метра уменьшается и система возвращается к первоначальному состоянию.
Например, под влиянием паратирина (гормона околощитовидных желез) в
крови возрастает содержание ионизированного кальция. Повышенный
уровень кальция тормозит секрецию паратирина, усиливает поступление в
кровь кальцитонина (гормона щитовидной железы), под влиянием которо-
го уровень кальция снижается и его содержание в крови нормализуется.
Отрицательные обратные связи способствуют сохранению стабильности
физиологических параметров внутренней среды при возмущающих воздей-
ствиях внешней среды, т. е. поддерживают гомеостазис. Они работают и в
обратном направлении, т. е. при уменьшении параметров включают систе-
мы регуляции, повышающие их и тем самым обеспечивающие восстанов-
ление гомеостазиса.
В условиях, когда воздействия среды относительно неизменны, возму-
щающие сигналы слабы или отсутствуют, основную роль играет система
гомеостатической регуляции, тогда как адаптационная регуляция мало ак-
тивна. Но стоит только в окружающей среде возникнуть изменению, т. е.
новому фактору, как сразу же активируется адаптационная регуляция и ос-
лабляется гомеостатическая.
Эффект обратной связи всегда запаздывает, так как она включает ком-
пенсационный режим уже после того как произошло рассогласование
имеющихся и должных параметров. Например, при низкой температуре
окружающей среды, включение механизмов поддержания температуры те-
ла после того, как снизилась температура крови, т. е. отклонилась гомео-
статическая константа, является слишком поздней регуляторной реакцией.
Если человек выходит раздетым на сильный мороз, включаются механиз-
мы поддержания постоянства температуры тела благодаря холодовому воз-
действию и адаптивной регуляции, но эффект низкой температуры может
привести к простуде. Необходимой в таких случаях является регуляция по
упреждению, в результате которой перед выходом на мороз человек надева-
5*
132 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
ет теплую одежду, а в его организме опережающе активируются механизмы
образования тепла. В центральном аппарате управления системы регуля-
ции заложен механизм контроля, позволяющий получать информацию не
об уже достигнутых параметрах деятельности, а осуществляющий сравне-
ние сигналов, посылаемых к исполнительным устройствам, с хранящими-
ся в памяти сигналами, требуемыми для заданной программы. Этот меха-
низм опережающей регуляции свойствен третьему уровню системы регуля-
ции.
3.1.3. Реактивность и эффект регуляции
Необходимым свойством живой системы, влияющим на эффективность
механизмов регуляции, является ее реактивность. Реактивность — это спо-
собность живой системы отвечать (реагировать) изменениями функции на
раздражители внешней или внутренней среды. Характер ответной реакции
любой живой системы определяется не только качественными и количест-
венными характеристиками раздражителя, но и реактивностью самой сис-
темы. Соответственно величина и характер эффектов регуляторных сигна-
лов (нервных импульсов, молекул химических веществ) зависят не только
от характеристик этих сигналов, но и от реактивности регулируемого объ-
екта, т. е. эффектора.
Одним из свойств реактивности является «правило исходного состоя-
ния», согласно которому величина и направленность эффекта регуляторно-
го сигнала зависят от особенностей метаболизма и функции, имевшихся в
регулируемой системе перед действием этого сигнала. Так, если функция
клетки, ткани, органа или физиологической системы находятся в активи-
рованном состоянии, то на стимулирующий регуляторный сигнал отмеча-
ется или слабый эффект, или отсутствие его, или даже противоположный
эффект, а на тормозящий регуляторный сигнал, напротив, возникает адек-
ватный или максимально возможный эффект. Если же в исходном состоя-
нии функция или метаболизм снижены, то стимулирующий регулятор вы-
зывает максимальный эффект, а действие подавляющего регулятора ослаб-
лено или даже приводит к стимуляции эффектора.
3.1.4. Механизмы регуляции жизнедеятельности
Различают нервные и гуморальные механизмы регуляции жизнедеятельности
организма. Первые используют для передачи и переработки информации
структуры нервной системы (нейроны, нервные волокна) и импульсы
электрических потенциалов, вторые — жидкости внутренней среды и моле-
кулы химических веществ.
Нервная регуляция осуществляет быструю и направленную передачу сиг-
налов, которые в виде нервных импульсов по нервным волокнам поступа-
ют к объекту регуляции. Быстрая передача сигналов (до 80—120 м/с) без
затухания и потери энергии обусловлена свойствами проводящих возбуж-
дение структур, преимущественно состоянием их мембран. Нервной регу-
ляцией обеспечены как соматические (деятельность скелетной мускулату-
ры), так и вегетативные (деятельность внутренних органов) функции.
Нервная регуляция функций включает в себя как произвольную, так и не-
произвольную, реализуемую в ответ на раздражение нервных сенсорных
рецепторов, т. е. рефлекторную.
Гуморальная регуляция — способ передачи информации к эффекторам
через жидкости внутренней среды организма с помощью молекул химиче-
3. Общие принципы и механизмы регуляции физиологических функций • 133
ских веществ, выделяемых клетками или специализированными для регу-
ляции эндокринными железами. Этот вид регуляции жизнедеятельности
осуществляется на двух уровнях: 1) обеспечивая относительно автономный
местный межклеточный обмен информацией об особенностях метаболизма
и функции клеток и тканей, 2) представляя системный эфферентный ка-
нал информационной связи, находящийся в большей или меньшей зависи-
мости от нервных процессов восприятия и обработки информации о со-
стоянии внешней и внутренней среды. Соответственно гуморальную регу-
ляцию подразделяют на местную клеточно-тканевую, и систему гормональ-
ной регуляции, обеспечивающую генерализованные эффекты в организме с
помощью гормонов. Местная гуморальная регуляция (тканевая саморегуля-
ция) практически не управляется нервной системой, тогда как система
гормональной регуляции составляет часть единой нейрогуморальной сис-
темы.
Деление механизмов регуляции жизнедеятельности организма на нерв-
ные и гуморальные условно и может использоваться только для дидактиче-
ских целей как способ изучения. В организме нервные и гуморальные ме-
ханизмы регуляции неразделимы и представляют собой единую нейрогу-
моральную систему регуляции. Так, во-первых, информация о состоянии
внешней и внутренней среды воспринимается почти всегда элементами
нервной системы (сенсорные рецепторы), обрабатывается в нервной систе-
ме, где может трансформироваться в сигналы исполнительных устройств
либо нервной, либо гуморальной природы. Следовательно, для второго и
третьего уровней системы регуляции физиологических функций управляю-
щим устройством является, как правило, нервная система. Во-вторых, сиг-
налы, поступающие по управляющим каналам нервной системы, переда-
ются в местах окончания нервных проводников в виде химических моле-
кул — нейромедиаторов, поступающих в микроокружение клеток, т. е. гу-
моральным путем, как было показано в главе 2. На мембранах одних и тех
же эффекторных клеток имеются рецепторы и к нейромедиаторам, и к
гормонам, и к тканевым факторам саморегуляции. Специализированные
для гормональной регуляции железы внутренней секреции управляются
нервной системой.
3.2. Рефлекторная регуляция функций организма
Рефлекс — стереотипная реакция организма в ответ на раздражитель, реа-
лизуемая с помощью нервной системы. Вызывающие рефлексы раздражи-
тели могут иметь как физическую (механические, электрические, темпера-
турные, звуковые, световые и т. п. раздражители), так и химическую при-
роду. Структурной основой рефлекса является рефлекторная дуга, пред-
ставляющая собой совокупность морфологически взаимосвязанных обра-
зований, обеспечивающих восприятие, передачу и переработку сигналов,
необходимых для реализации рефлекса.
Рефлекторная дуга по своему строению и назначению элементов пред-
ставляет собой описанный выше контур регуляции. Она включает следую-
щие звенья (рис. 3.4):
1) сенсорные рецепторы (датчики), воспринимающие стимулы внешней
или внутренней среды,
2) афферентные, или чувствительные, нервные проводники (каналы
сигналов входа),
3) нервные центры (аппарат управления), состоящие из афферентных,
промежуточных, или вставочных, и эфферентных нейронов, т. е. по-
134 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Двигательный
нейрон
Афферентный нейрон
спинального ганглия
Нервный центр
спинного мозга
Вставочный
нейрон
Эфферентный проводник
Эффектор (мышца)
Обратная связь
Рецептор
Афферентный проводник
Рис. 3.4. Схема рефлекторной дуги.
Раздражение рецептора ведет к поступлению нервного импульса по афферентному проводни-
ку к афференнтному нейрону спинального ганглия, а от него к вставочному и двигательному
нейронам спинного мозга (нервный центр рефлекса). От двигательного нейрона передних ро-
гов спинного мозга по эфферентному проводнику нервный импульс передается к эффекто-
ру — скелетной мышце. Сокращение последней вызывает раздражение расположенных в
мышце рецепторов и по афферентным проводникам информация о сокращении поступает об-
ратно в нервный центр (канал обратной связи)
думающих, обрабатывающих и выдающих информацию нервных кле-
ток,
4) эфферентные, или двигательные, нервные проводники (каналы вы-
хода),
5) эффекторы, или исполнительные органы (объекты управления).
Однако для оптимальности регуляции необходима информация о реак-
циях эффектора на управляющие сигналы, в связи с чем обязательным
звеном рефлекторного акта является канал обратной связи. Таким обра-
зом, структурную основу рефлекса лучше называть не рефлекторной дугой,
а рефлекторным кольцом.
Рецепторами называют специализированные образования, предназна-
ченные для восприятия клетками или нервной системой различных по
своей природе стимулов или раздражителей. Различают два типа рецепто-
ров — сенсорные, т. е. обеспечивающие восприятие нервной системой раз-
личных раздражителей внешней или внутренней среды, и клеточные хими-
ческие рецепторы — специальные структуры мембран, обеспечивающие
восприятие информации, переносимой молекулами химических веществ —
медиаторов, гормонов, антигенов и т. п.
3. Общие принципы и механизмы регуляции физиологических функций • 135
3.2.1. Сенсорные рецепторы
Сенсорные рецепторы в зависимости от их структурной организации и
функции могут быть первично или вторично чувствующими (рис. 3.5).
Первично чувствующие рецепторы — это нервные окончания отростков чув-
ствительных нейронов. Они имеются в коже и слизистых оболочках, ске-
летных мышцах, сухожилиях и надкостнице, а также барьерных структурах
внутренней среды — стенках кровеносных и лимфатических сосудов, ин-
терстициальном пространстве, в оболочках головного и спинного мозга,
ликворной системе. По характеру воспринимаемых раздражителей первич-
но чувствующие рецепторы делят на механорецепторы (восприятие растя-
жения или сдавления, линейного или радиального сдвига ткани), хеморе-
цепторы (восприятие химических раздражителей), терморецепторы (вос-
приятие температуры). В особую группу выделяют ноцицепторы, т. е. ре-
цепторы, воспринимающие боль, хотя их существование признается не
всеми.
Вторично чувствующие рецепторы — это специализированные на вос-
приятии определенных раздражителей рецепторные клетки, как правило
эпителиальной природы, входящие в состав органов чувств — зрения, слу-
ха, вкуса, равновесия. После восприятия раздражителя эти рецепторные
клетки передают информацию на окончания афферентных проводников
чувствительных нейронов. Таким образом, афферентные нейроны нервной
системы получают информацию о раздражителе, уже обработанную в ре-
цепторных клетках (что и определило название этих рецепторов).
Все виды рецепторов в зависимости от источника воспринимаемого
раздражения делят на экстероцепторы (воспринимающие стимулы из
внешней среды) и интероцепторы (предназначенные для раздражителей
внутренней среды). Среди интероцепторов выделяют проприоцепторы,
т. е. собственные рецепторы опорно-двигательного аппарата, ангиорецеп-
торы, расположенные в стенках сосудов, и тканевые рецепторы, локализо-
ванные в интерстициальном пространстве и клеточной микросреде. Осо-
бое место среди проприорецепто-
ров занимают мышечные верете-
на, представляющие собой образо-
вания, реагирующие на растяжение
мышцы и способные менять свою
чувствительность под влиянием
импульсов, приходящих к ним из
центральной нервной системы. Эти
рецепторы участвуют в регуляции
мышечного тонуса.
Общим функциональным свой-
ством всех видов сенсорных рецеп-
торов является способность преоб-
разовывать один вид энергии в
другой: механическую, тепловую,
света, звука и т. п. энергию раздра-
жителей в электрическую энергию
биопотенциалов, т. е. нервных им-
пульсов.
Поскольку рецепторы специа-
лизированы для восприятия опре-
деленного вида раздражителей, их
А. Тельце Пачини Б. Окончание Руффини
В. Тельце Мейснера Г. Диски Меркеля
Рис. 3.5. Виды рецепторов кожи.
А, Б, В — первично чувствующие рецепто-
ры — окончания чувствительных нервных
проводников; Г — вторично чувствующие ре-
цепторы — специализированные рецептор-
ные клетки.
136 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
чувствительность для таких раздражителей оказывается наибольшей. Вели-
чина минимальной силы раздражителя, способной вызвать возбуждение рецеп-
тора, носит название абсолютного порога раздражения. В связи с этим раз-
дражители, для которых рецептор обладает наивысшей чувствительностью,
т. е. минимальной величиной порога, получили название адекватных. В то
же время некоторые рецепторы могут реагировать и на несоответствующие
их специализации раздражители (например, рецепторы органа зрения на
механическое раздражение), порог для таких раздражителей, называемых
неадекватными, оказывается очень высоким и требуется значительная сила
раздражителя для возбуждения рецептора («искры из глаз» при ударе).
Если первично чувствующие рецепторы образованы разветвлениями от-
ростка одного сенсорного нейрона, они формируют рецептивное поле сен-
сорного нейрона. Обычно рецепторы образуют в тканях скопления различ-
ной плотности. В случаях, когда эти скопления дают начало определенно-
му рефлексу, их называют рецептивными полями рефлекса. Если в скопле-
нии представлены разные по характеру раздражителей рецепторы, дающие
начало разным рефлексам, их называют рефлексогенными зонами. Приме-
ром являются сосудистые рефлексогенные зоны (дуги аорты, каротидного
синуса), где расположены и механо- и хеморецепторы, раздражение кото-
рых вызывает различные рефлекторные реакции сердечно-сосудистой, ды-
хательной и других систем организма.
3.2.2. Афферентные и эфферентные нервные проводники
Основной функцией нервов является проведение сигналов к нервному
центру от рецепторов (афферентные проводники) или от нервного центра к
эффектору (эфферентные проводники). Собственно проводниками являются
нервные волокна, входящие в состав периферических нервов или белого
вещества головного и спинного мозга.
В зависимости от диаметра, наличия или отсутствия миелиновой обо-
лочки и скорости проведения нервных импульсов нервные волокна делят
на три класса: А (четырех типов), В и С (табл. 3.1). Миелинизированные
волокна более толстые, скорость проведения сигналов по ним существенно
больше. Так, волокна A-типов обеспечивают проведение сигналов при
Таблица 3.1. Характеристика нервных волокон периферических нервов
Тип воло- кон Диа- метр воло- кон (мкм) Ско- рость прове- дения (м/с) Назначение
А 13-22 70-120 Эфферентные волокна скелетных мышц, афферентные волок- на рецепторов — мышечных веретен
А 8-13 40-70 Афферентные волокна от рецепторов давления и прикоснове- ния
4-8 15-40 Эфферентные волокна рецепторов — мышечных веретен, часть афферентов от рецепторов давления и прикосновения
А 1-4 5-15 Афферентные волокна от кожных рецепторов температуры и боли, частично давления
В 1-3 3-14 Преганглионарные эфферентные волокна вегетативной нерв- ной системы
с 0,5-1.5 0,5-2 Постганглионарные эфферентные волокна вегетативной нерв- ной системы, афференты кожных рецепторов боли и тепла
3. Общие принципы и механизмы регуляции физиологических функций • 137
рефлекторной регуляции скелетных мышц. Немиелинизированные тонкие
волокна С-типа участвуют в проведении сигналов, регулирующих деятель-
ность внутренних органов, скорость проведения сигнала по ним — самая
низкая.
3.2.3. Возбуждение и торможение в рефлекторной дуге
При огромном количестве раздражителей, действующих одновременно на
многочисленные рецепторные образования организма, наличии множества
взаимосвязанных информационных каналов, в виде рефлекторных ответов
реализуются лишь некоторые из воздействий. Целесообразность такого
ограничения очевидна, поскольку в противном случае одновременно осу-
ществляемые противодействующие рефлексы сделали бы просто невоз-
можной не только регуляцию, но и саму жизнедеятельность. Поэтому, на-
ряду с процессом возбуждения, т. е. возникновения активной деятельности,
распространение которого лежит в основе всех рефлексов, существует вто-
рой процесс, ограничивающий и подавляющий возникновение и распро-
странение возбуждения в элементах нервной системы и тем самым не по-
зволяющий реализовываться рефлекторным актам. Этот второй основной
процесс в нервной системе называется торможение. Под торможением по-
нимают активный нервный процесс, возникающий при действии распро-
страняющихся нервных импульсов и проявляющийся в ослаблении или
подавлении возбуждения. Процесс торможения не способен распростра-
няться, он возникает и проявляется локально.
3.2.4. Механизмы связи между звеньями рефлекторной дуги
Передача возбуждения с отростка одной нервной клетки на отросток или
тело другой нервной клетки возможна двумя способами: электрическим
(электротоническим) и химическим. Электрический способ передачи воз-
буждения осуществляется благодаря тесным контактам передающей и
воспринимающей структур (щель между мембранами меньше 2 нм). Пере-
дача возбуждения в таком случае осуществляется аналогично его проведе-
нию по нервным волокнам с помощью местных токов. Электрическая пе-
редача возбуждения осуществляется с высокой скоростью, близкой скоро-
сти проведения возбуждения по нервным волокнам и, так же как нерв,
практически неутомляема.
Основным способом передачи информации между нервными клетками
является химический, реализуемый с помощью специальных образований,
получивших название синапсов. Синапсы — это специализированная фор-
ма контакта между отростками нейронов и любыми возбудимыми образо-
ваниями (нейронами, мышечными или секреторными клетками), обеспе-
чивающая передачу сигнала с помощью молекул химических веществ, на-
зываемых медиаторами. В нервной системе находятся центральные синап-
сы, которые образуются между отростками разных нейронов, а также меж-
ду отростками и телами клеток. Соответственно их называют аксо-аксо-
нальными, аксодендритными, аксосоматическими, дендросоматическими,
дендро-дендритными. Количество синапсов на нейроне очень большое и
достигает нескольких тысяч (рис. 3.6). В качестве примера может быть рас-
смотрен аксосоматический синапс (между аксоном одной нервной клетки
и телом другой), структура которого показана на рис. 3.7. Аксон, подходя к
телу другого нейрона, образует расширение, называемое пресинаптическим
окончанием, или терминалью. Мембрана такого окончания называется пре-
138 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 3.6. Множество синапсов на теле и
отростках нейрона.
1 — ядро, 2 — аксонный холмик, 3 — дендриты,
4 — миелиновая оболочка аксона, 5 — синап-
сы, 6 — тело нейрона.
синаптической. Под ней распола-
гается синаптическая щель, шири-
на которой составляет 20—40 нм.
За синаптической щелью лежит
мембрана тела нейрона, называе-
мая в области синапса постсинап-
тической.
Передача информации в си-
напсах осуществляется с помо-
щью молекул специальных хими-
ческих веществ — медиаторов,
т. е. посредников передачи, обра-
зуемых в терминали и выводимых
через пресинаптическую мембра-
ну в синаптическую щель. Медиа-
тор содержится в пресинаптиче-
ской терминали в пузырьках и его
выход происходит порциями
(квантами), равными содержимо-
му каждого пузырька.
Выделившиеся через преси-
наптическую мембрану кванты
медиатора диффундируют через
синаптическую щель к постси-
наптической мембране, где связываются со специальными химическими
клеточными рецепторами, специфическими для молекул медиатора. В за-
висимости от химической природы медиатора и особенностей клеточных
рецепторов на постсинаптическои
мембране возможны два эффекта
синаптической передачи сигнала:
возбуждение и торможение. Соот-
ветственно и синапсы делят на
два типа: возбуждающие и тор-
мозные. Передача информации
через синапсы осуществляется
значительно медленнее, чем по
нервам или через тесные контак-
ты, поскольку для процессов вы-
ведения медиатора, диффузии че-
рез синаптическую щель, связы-
вания с рецепторами постсинап-
тической мембраны требуется
больше времени. Прекращение
действия медиатора и соответст-
вующее завершение передачи
нервного импульса осуществляет-
ся за счет удаления медиатора из
синаптической щели. Это проис-
ходит в результате трех процес-
сов — обратного «захвата» медиа-
тора пресинаптическим оконча-
нием, разрушения медиатора спе-
циальными ферментами, находя-
Рис. 3.7. Структура аксосоматического
синапса.
1 — аксон, 2 — синаптическая пуговка, 3 —
пресинаптическая мембрана, 4 — постсинапти-
ческая мембрана, 5 — рецепторы постсинапти-
ческой мембраны, 6 — синаптические пузырь-
ки с медиатором, 7 — кванты медиатора в си-
наптической щели, 8 — митохондрии, СаСб —
кальцийсвязывающий белок.
3. Общие принципы и механизмы регуляции физиологических функций • 139
Рис. 3.8. Структура нервно-мышечного синапса.
1 — пресинаптическая концевая пластинка, 2 — синаптические пузырьки с медиатором, 3 —
митохондрии, 4 — синаптическая щель с квантами медиатора, 5 — складчатая постсинаптиче-
ская мембрана, 6 — рецепторы постсинаптической мембраны, 7 — саркоплазматический рети-
кулум, 8 — сократительный аппарат (миофибриллы) мышечной клетки.
щимися у рецепторов постсинаптической мембраны, и диффузии медиато-
ра из синапсов в микроокружение клетки.
Энергетическое и субстратно-ферментативное обеспечение синаптиче-
ской передачи сигналов осуществляется с помощью аксонного транспорта
из тела клетки в пресинаптическое окончание белков и ферментов, орга-
нелл, в частности митохондрий, низкомолекулярных веществ и уже синте-
зированных медиаторных веществ. Транспорт происходит с помощью ней-
рофибрилл или микротрубочек, тянущихся внутри аксона по всей его дли-
не. Аксонный транспорт требует обязательного участия ионов кальция, а
энергия для его реализации черпается из непрерывно ресинтезируемой в
аксоне АТФ. Помимо транспорта веществ по аксонам из тела нейрона к
синаптическому окончанию, существует и ретроградный аксонный транс-
порт, обеспечивающий поступление от пресинаптического окончания в те-
ло клетки веществ, регулирующих в ней синтез белка.
Периферические синапсы. Периферические синапсы расположены между
окончаниями эфферентного нерва и мембраной эффектора. Для соматиче-
ских рефлексов это синапсы между двигательным нервом и скелетной мыш-
цей, поэтому их еще называют нервно-мышечными синапсами (рис. 3.8).
140 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Благодаря форме синаптического образования они получили название кон-
цевых пластинок. Строение и основные свойства концевых пластинок в об-
щих чертах подобны центральным синапсам, однако постсинаптическая
мембрана у них имеет многочисленные складки, существенно увеличиваю-
щие поверхность взаимодействия медиатора с рецепторами.
3.2.5. Нервные центры и их свойства
Нервным центром называют функционально связанную совокупность ней-
ронов, расположенных в одной или нескольких структурах центральной
нервной системы и обеспечивающих регуляцию определенных функций
организма. В более узком понимании применительно к рассматриваемой
структуре рефлекторного акта нервный центр как аппарат управления
представляет собой функциональное объединение разных нейронов, обес-
печивающее реализацию конкретного рефлекса. Основные общие свойства
нервных центров определяются тремя главными факторами: 1) свойствами
нервных клеток, входящих в состав центра, 2) особенностями структурно-
функциональных связей нейронов, 3) свойствами синапсов между нейро-
нами центра.
Функциональные свойства нервных клеток зависят от их строения, ло-
кализации и связей, устанавливаемых отростками. С позиций функцио-
нального предназначения различают 3 типа нейронов: афферентные, или
чувствительные, эфферентные и интернейроны (промежуточные). Аффе-
рентные нейроны не входят в состав центра, по количеству отростков они
обычно униполярны, биполярны или псевдоуниполярны (т. е. имеют один
Т-образно делящийся отросток). Эти клетки получают сенсорную инфор-
мацию от рецепторов и осуществляют ее передачу нейронам центра. Эффе-
рентные нейроны обычно мультиполярны, т. е. имеют один аксон и не-
сколько дендритов, они осуществляют передачу информации из нервного
центра к эффекторам, например, мышцам. Наибольшую по количеству
группу нервных клеток в центральной нервной системе составляют интер-
нейроны, или ассоциативные нейроны, которые по своей форме обычно
мультиполярны, причем отростки клеток не покидают пределов централь-
ной нервной системы и их основной функцией является обеспечение взаи-
мосвязей между разными группами нейронов. Интернейроны могут быть
возбуждающими или тормозными, в зависимости от выделяемых на окон-
чаниях их отростков медиаторов. Тело и дендриты нервных клеток являют-
ся структурами, которыми осуществляется интеграция поступающих к
нейрону многочисленных сигналов.
Функция нервных клеток связана с особенностями их метаболизма, в
частности, с высоким уровнем энергетических и пластических процессов.
Наиболее существенным отличием нейронов от других видов клеток орга-
низма является интенсивный обмен нуклеиновых кислот и очень высокий
уровень транскрипции, т. е. синтеза молекул информационной РНК на
матрицах ДНК генома. Интенсивность транскрипции в нейронах превы-
шает более чем в 5 раз ее уровень в других клетках организма. Имеется
четкая взаимосвязь уровня синтеза белка в нейронах и характера их элек-
трической активности. Образуемые в нейронах специфические белки и
пептиды участвуют в регуляции эффективности синаптической передачи
сигналов и хранении информации, и, в конечном счете, реализации реф-
лекторных актов и особенностей поведения.
Схематизировано наиболее простой формой связи нейронов центра яв-
ляется нервная цепь (рис. 3.9, А), в которой возбуждение последовательно
3. Общие принципы и механизмы регуляции физиологических функций • 141
Рис. 3.9. Особенности распространения возбуждения в нейронных сетях.
А — нервная цепочка, Б — дивергенция возбуждения в нервной сети, В — конвергенция возбу-
ждения, Г — реверберация возбуждения в кольцевой цепи Лоренто де Но. Стрелки показыва-
ют направление распространения возбуждения.
передается от одного нейрона к другому. Нервная цепь может содержать
всего два нейрона с одним синапсом между ними, соответственно реф-
лекс, реализуемый через такой нервный центр, носит название моносинап-
тического. С увеличением числа нейронов и синапсов между ними время
рефлекторной реакции на раздражитель увеличивается, поскольку каждый
синапс имеет синаптическую задержку проведения. Такие рефлексы назы-
ваются полисинаптическими. Сигналы по нервным цепям распространяют-
ся в одну сторону — от афферентного входа к эфферентному выходу — бла-
годаря одностороннему проведению в синапсах.
Нейроны нервного центра за счет структурно-функциональных связей
(ветвления отростков и установления множества синапсов между разными
клетками) объединяются в нервные сети. Связи между нервными клетками
являются генетически детерминированными.
Различают 3 основных типа нервных сетей: иерархические, локальные и
дивергентные с одним входом. Иерархические сети обеспечивают посте-
пенное включение нейронных структур более высокого уровня. В тех слу-
чаях, когда афферентная информация поступает к увеличивающемуся чис-
лу нейронов, принято говорить о дивергенции возбуждения (рис. 3.9, Б).
Если, напротив, от многих возбужденных клеток информация (возбужде-
ние) сходится к меньшему числу нейронов, то такой принцип распростра-
нения сигналов называется конвергенцией (рис. 3.9, В). Так, например,
конвергенция в эффекторном пути двигательных спинальных рефлексов
ведет к тому, что малое число двигательных клеток (мотонейронов) спин-
ного мозга получает импульсы возбуждения от различных эфферентных
путей многих рефлекторных дуг. Изучая этот механизм на уровне спинно-
го мозга, Ч. Шеррингтон сформулировал принцип общего конечного пути,
согласно которому мотонейроны спинного мозга являются общим конеч-
ным путем многочисленных рефлексов (мотонейроны, управляющие сги-
бателями правой руки, участвуют в многочисленных двигательных рефлек-
торных реакциях — почесывании, жестикуляции при речи, переносе пищи
в рот и т. п.). Соответственно на уровне многочисленных синапсов кон-
вергентных путей возникает конкуренция за общий конечный путь. Одна-
ко конвергенция существует и для афферентной информации, когда на ог-
раниченном числе нейронов конвергируют сигналы, приходящие от раз-
ных сенсорных систем (слуха, обоняния, зрения и т. п.), что позволяет
формировать в нервных центрах полноценный образ воспринимаемого
объекта.
142 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Нервные сети обеспечивают реализацию принципа субординации, когда
деятельность ниже расположенных нейронных структур подчинена выше
расположенным. «Выше» и «ниже» здесь означает уровень расположения в
отделах мозга, например, нейроны спинного мозга подчинены влияниям
со стороны головного мозга.
Локальные сети нервных центров содержат нейроны с короткими аксо-
нами, осуществляющие взаимосвязи в пределах одного уровня. Они обес-
печивают задержку информации в пределах этого уровня, что играет роль в
механизмах памяти. Примером такой локальной сети являются кольцевые
нейронные цепочки Лоренто де Но, возбуждение в которых циркулирует
по замкнутому кругу (рис. 3.9, Г). Возврат возбуждения к «первому»нейро-
ну кольцевой цепи получил название реверберации возбуждения. Локаль-
ные сети обеспечивают надежность нервной регуляции за счет дублирова-
ния элементов, так как многие нейроны локальных сетей имеют одинако-
вые синаптические связи и функционируют попеременно, т. е. являются
взаимозаменяемыми.
Дивергентные сети с одним входом представляют собой нейронные ан-
самбли, в которых один нейрон образует выходные связи с большим коли-
чеством других клеток разных иерархических уровней и, главное, разных
нервных центров. Максимально выраженная дивергенция связей разных
нервных центров свидетельствует о том, что эти нервные сети не являются
специфическими для реализации определенных рефлексов, а обеспечивают
интеграцию разных рефлекторных актов и общее состояние активности
многочисленных нейронов разных отделов мозга.
Поскольку в нервных сетях включено огромное число интернейронов,
они могут, передавая нервные импульсы другим нейронам центра, реали-
зовывать как их возбуждение, так и торможение. Различают следующие
типы тормозных процессов в нейронных сетях:
• реципрокное торможение (рис. 3.10, В), т. е. процесс, основанный на
том, что сигналы по одним и тем же афферентным путям обеспечива-
ют возбуждение одной группы нейронов, а через вставочные тормоз-
ные клетки вызывают торможение другой группы нейронов. Этот тип
торможения называют иначе сопряженным. Проявляется реципрокное
торможение, например, на уровне мотонейронов спинного мозга, ин-
нервирующих мышцы-антагонисты (сгибатели-разгибатели) конечно-
стей. Передача возбуждения к мотонейронам мышц-сгибателей одно-
временно через вставочные тормозные нейроны тормозит мотонейро-
ны мышц-разгибателей, благодаря чему только и возможно осуществ-
ление сгибательного рефлекса;
• возвратное торможение (рис. 3.10, Г), т. е. процесс торможения эф-
ферентных нейронов сигналами, возвращающимися к ним по колла-
тералям аксонов через вставочные тормозные клетки. Возбуждение от
одних нейронов поступает через ответвления на аксонах к вставоч-
ным тормозным нейронам, а аксоны этих клеток в свою очередь об-
разуют тормозные синапсы на телах первых нейронов. Примером яв-
ляется торможение Реншоу в спинном мозге, где импульсы от двига-
тельных нейронов посылаются по аксонам не только к скелетной
мышце, но поступают через аксонные коллатерали к тормозным ин-
тернейронам Реншоу, а от них назад к двигательным нейронам, при-
водя к их торможению. Таким образом осуществляется короткая от-
рицательная обратная связь, не позволяющая возникнуть избыточно-
му возбуждению мотонейронов спинного мозга;
• латеральное торможение (рис. 3.10, Б), т. е. процесс торможения
3. Общие принципы и механизмы регуляции физиологических функций • 143
Рис. 3.10. Тормозные процессы в нейронных сетях центра.
А — окружающая тормозная зона, Б — латеральное торможение, В — реципрокное торможе-
ние в центрах спинного мозга, управляющих сокращениями мышц-сгибателей (С) и -разгиба-
телей (Р) нижних конечностей, Г — возвратное торможение (Реншоу) в спинном мозге. Знаки
(+) и (-) — соответственно возбужденное и заторможенное состояние нейронов, стрелки —
направление распространения возбуждения.
группы нейронов, расположенной рядом с группой возбужденных
клеток. Этот вид торможения распространен в сенсорных системах,
где ответвления аксонов возбужденных нейронов образуют синапсы
на вставочных тормозных нейронах, а аксоны последних оканчивают-
ся тормозными синапсами на соседних, рядом расположенных клет-
ках, получающих информацию от другого афферентного входа. Бла-
годаря явлению латерального торможения удается различать контраст
в освещенности объектов, звуковые тоны и другие различия воспри-
нимаемых раздражителей;
• тормозная зона, т. е. процесс торможения нескольких групп нейро-
нов, расположенных рядом с возбужденными клетками. Сигналы от
возбужденных нейронов по коллатералям аксонов поступают к вста-
вочным нейронам, а аксоны последних образуют тормозные синапсы
на группах клеток, получающих информацию по другим афферент-
ным входам (рис. 3.10, А). Если эта группа заторможенных клеток
располагается вокруг возбужденных нейронов, тормозную зону назы-
вают окружающей;
• нисходящее торможение, т. е. процесс торможения ниже расположен-
ных или эволюционно более древних структур нервной системы вы-
ше расположенными или эволюционно более новыми нейронными
структурами головного мозга. Реализация нисходящего торможения
способствует реализации принципа субординации нервных центров.
144 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
3.2.6. Взаимодействие различных рефлексов. Принципы
координации рефлекторной деятельности
Поскольку в организме одновременно или последовательно реализуется
обычно несколько рефлексов, простейшие связи между ними характеризу-
ются тремя вариантами взаимодействия. Во-первых, рефлексы могут вза-
имно содействовать друг другу, облегчая реализацию требуемого эффекта.
Такие рефлексы называют аллиированными, или союзными. Примерами ал-
лиированных рефлексов являются рефлексы мигания и слезоотделения,
слюноотделения и глотания. Во-вторых, рефлексы могут оказывать друг на
друга тормозящее влияние, и тогда их называют антагонистическими. На-
пример, рефлекс глотания тормозит рефлекс вдоха, рефлекс разгибания
тормозит рефлекс сгибания конечности. В-третьих, взаимодействие между
рефлексами может быть последовательным, когда один рефлекс, а вернее
его результат, вызывает возникновение другого рефлекса. Такие рефлексы
называют цепными, причем в цепи может быть взаимосвязано большое
число разных рефлексов. Простейшими цепными рефлексами являются,
например, шагательный рефлекс, когда сгибательный рефлекс (сгибание
ноги) становится причиной следующего за ним разгибательного, а он в
свою очередь вызывает сгибательный рефлекс и т. д.
При реализации сложных рефлекторных реакций, обеспечивающих ре-
гуляцию жизнедеятельности и приспособление организма к меняющимся
условиям среды, необходимым условием является координация рефлексов, в
основе которой лежит согласованное формирование и взаимодействие
процессов возбуждения и торможения в центральной нервной системе.
Основными принципами координации являются: принцип сопряженного
торможения (реципрокности), принцип доминанты, принцип общего ко-
нечного пути, принцип субординации нервных центров, принцип обрат-
ной афферентации.
• Принцип сопряженного торможения, или реципрокности, базируется на
описанном выше одноименном виде торможения и означает возник-
новение при возбуждении одного центра одновременного торможе-
ния другого центра для обеспечения цепных рефлексов и сложной
функции. Примером реципрокности может быть регуляция спинным
мозгом противоположных по функциональному назначению мышц
конечностей (рис. 3.11). Так, при возбуждении мотонейронов, иннер-
вирующих мышцы-сгибатели правой ноги, реципрокно тормозятся
мотонейроны мышц-разгибателей этой ноги и возбуждаются мото-
нейроны мышц-разгибателей левой ноги. Формирующийся цепной
характер рефлексов вслед за этим вызывает возбуждение мотонейро-
нов разгибателей правой ноги и реципрокно — торможение мотоней-
ронов сгибателей правой ноги и возбуждение мотонейронов сгибате-
лей левой ноги. Таким образом реципрокные взаимоотношения меж-
ду указанными рефлексами обеспечивают цепной шагательный реф-
лекс.
• Принцип доминанты основан на открытом А. А. Ухтомским свойстве
нервных центров. Доминантой называют общий принцип деятельно-
сти нервной системы, проявляющийся в виде господствующей в тече-
ние определенного времени системы рефлексов, реализуемых доми-
нирующими центрами, которые подчиняют себе или подавляют дея-
тельность других нервных центров и рефлексов. Нейроны доминини-
рующих центров становятся более возбудимыми и способны эффек-
тивнее осуществлять пространственную и временную суммацию
3. Общие принципы и механизмы регуляции физиологических функций • 145
Молекула гормона (первичный посредник)
J
Рецептор клеточной мембраны
I
Мембранный фермент
I
Вторичный посредник
Внутриклеточные эффекты
Рис. 3.11. Принцип реципрокности в рефлекторной регуляции мышц-антагонистов.
Возбуждение мотонейронов (1) мышцы-сгибателя тормозит через тормозные интернейроны
(2) мотонейроны мышцы-разгибателя (3). Знаки (+) и (—) — соответственно возбуждение и
торможение.
нервных импульсов. Синаптическое проведение к этим нейронам об-
легчено, и поэтому они могут возбуждаться и за счет «посторонних»
импульсов от не имеющих прямых связей с доминирующими центра-
ми информационных каналов. Вследствие суммации многочисленных
сигналов возбуждение нейронов, как и число возбужденных клеток в
доминирующем центре, нарастает и осуществляемые им рефлектор-
ные реакции легко реализуются. Преобладание рефлексов домини-
рующего центра над другими рефлекторными актами становится осо-
бенно выраженным, поскольку через систему вставочных нейронов
доминирующий центр сопряженно тормозит другие центры и теку-
щие рефлексы. Принцип доминанты позволяет концентрировать вни-
мание и строить поведение для достижения определенной намечен-
ной цели.
• Принцип общего конечного пути, описанный Ч. Шеррингтоном, рас-
сматривался при изложении процесса конвергенции возбуждения.
• Принцип субординации нервных центров (принцип подчинения) Прояв-
ляется в виде регулирующего влияния выше расположенных нервных
центров на ниже расположенные. Так, двигательные центры головно-
го мозга управляют спинальными мотонейронами. Примером такого
влияния является феномен центрального торможения спинальных
рефлексов, открытый И. М. Сеченовым и получивший название се-
ченовского торможения. В эксперименте И. М. Сеченова раздраже-
ние зрительных бугров лягушки с помощью кристаллика поваренной
соли (т. е. раздражение ретикулярной формации среднего мозга) при-
146 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
водило к торможению спинальных двигательных рефлексов, вызывае-
мых погружением лапки лягушки в слабый раствор кислоты. Следо-
вательно, торможение центров спинного мозга явилось следствием
возбуждения центров среднего мозга. Прекращение этого тормозного
контроля при перерыве цереброспинальных проводящих путей вызы-
вает резкое повышение возбудимости спинальных центров и гипер-
рефлексию.
• Принцип обратной афферентации заключается в рецепторном воспри-
ятии результатов рефлекторного акта и проведении информации об-
ратно в структуры нервного центра, где она обрабатывается и сравни-
вается с сохраняющимися параметрами возбуждения. Обратная аффе-
рентация реализуется в виде механизмов положительной или отрица-
тельной обратной связи. Таким образом, с помощью обратной аффе-
рентации нервные центры осуществляют непрерывный контроль эф-
фективности, целесообразности и оптимальности рефлекторной дея-
тельности.
3.2.7. Рефлекторная регуляция висцеральных функций
Висцеральными функциями называют физиологические процессы, осуще-
ствляемые внутренними органами, железами, сердцем, кровеносными и
лимфатическими сосудами, гладкой мускулатурой, клетками крови и на-
правленные на поддержание обмена веществ, роста, развития и размноже-
ния. Регуляция висцеральных функций осуществляется вегетативной нерв-
ной системой, реализующей эфферентные сигналы рефлексов, получив-
ших название вегетативных. Так, например, при растяжении желудка пи-
щей происходит раздражение механорецепторов, заложенных в его стенке
и являющихся окончаниями блуждающего нерва. По афферентным волок-
нам этого нерва импульсы от рецепторов поступают в нервный центр про-
долговатого мозга, а из него по эфферентным волокнам того же блуждаю-
щего Нерва посылаются к сердцу, вызывая замедление ритма его сокраще-
ний. Таким образом, структурной основой вегетативных рефлексов, как и
соматических, является рефлекторная дуга (или с обратной связью —
кольцо).
3.3. Произвольная (волевая) регуляция физиологических функций
В отличие от автоматической непроизвольной рефлекторной регуляции,
при которой регуляторный эффект является следствием действия раздра-
жителя, т. е. ответной реакцией организма на воздействия внешней или
внутренней среды, произвольная нервная регуляция осуществляет измене-
ние функции под влиянием волевой команды, т. е. нервных импульсов,
формирующихся в высших отделах головного мозга, в том числе в коре
больших полушарий. Так, например, помимо рефлекторной регуляции ды-
хания, обеспечивающей возникновение вдоха, его смену выдохом, измене-
ния частоты и глубины дыхания, человек может произвольно задерживать
дыхание, дышать чаще или реже. Наряду с автоматическим рефлекторным
разгибанием ноги в коленном суставе при ударе молоточком по сухожи-
лию коленной чашечки, человек может произвольно осуществлять то же
движение. Однако произвольная регуляция свойственна не всем физиоло-
гическим системам. Как правило, произвольная регуляция направлена на
осуществление поведения, т. е. психической деятельности, регулирующей
взаимоотношения организма и окружающей среды. В основе произволь-
3. Общие принципы и механизмы регуляции физиологических функций • 147
ной регуляции лежит потребность в изменении функции и образование
мотивации, т. е. побуждения к деятельности.
Многообразие потребностей организма определяет и множество моти-
ваций, среди которых выделяют 3 основные группы: биологические, соци-
альные и творческие. Биологические мотивации есть результат наличия по-
требностей в пищевых веществах, воде и солях, продолжении рода и т. п.
Значительная часть биологических мотиваций обусловлена необходимо-
стью поддержания гомеостазиса. Социальные мотивации — трудовые, по-
знавательные, эстетические, нравственные. Мотивации, возникающие в
результате ориентировочных потребностей, обеспечивают стремление к по-
знанию непонятных или новых явлений (познавательная мотивация), регу-
лирование своих действий в зависимости от эмоционального отношения к
ним других людей (мотивация эмоционального контакта), стремление со-
относить ценность своей личности с коллективными и общечеловеческими
ценностями (мотивация смысла жизни). Творческие мотивации, или иде-
альные потребности, возникают как в результате необходимости создания
эстетически или познавательно ценных новых вещей, так и как следствие
потребности в признании, успехе.
Благодаря мотивациям поведение направлено на достижение опреде-
ленной цели и может становиться приспособительным. Так, в условиях
низкой температуры окружающей среды наряду с рефлекторными автома-
тическими реакциями приспособления (ограничение теплоотдачи, усиле-
ние образования тепла в тканях) произвольно реализуется приспособи-
тельное поведение — человек утепляет жилище, надевает сохраняющую те-
пло одежду и т. п.
3.4. Гормональная регуляция функций организма
Генерализованные специализированные эффекты гуморальной регуляции
осуществляются с помощью особых химических регуляторов внутренней
среды — гормонов. Гормонами называют химические вещества, образую-
щиеся и выделяющиеся специализированными эндокринными клетками
(например, р-клетки островков Лангерганса секретируют гормон инсулин),
тканями и органами (щитовидной железой, надпочечниками и др.) во
внутреннюю среду для регуляции обмена веществ и физиологических
функций организма, гуморального обеспечения координации и интеграции
процессов жизнедеятельности. Гормоны отличают от других биологически
активных веществ, например метаболитов и медиаторов, по двум основ-
ным критериям: 1) гормоны образуются специализированными эндокрин-
ными клетками, 2) гормоны оказывают свое влияние через внутреннюю
среду на удаленные от секретирующей их ткани клетки-мишени, т. е. обла-
дают дистантным действием.
Гормоны являются чрезвычайно высокоактивными химическими соеди-
нениями, их концентрации в крови выражаются в нанограммах и даже в
пикограммах. Согласно расчетам, 1 г адреналина способен активировать
100 миллионов сердец лягушки, 1 г фолликулина может вызвать течку у 10
миллионов кастрированных мышей, 1 г инсулина — снизить уровень глю-
козы в крови у 125 тысяч кроликов. Гормоны оказывают выраженное по-
ложительное влияние на эмоциональную сферу, интеллектуальную и фи-
зическую активность, выносливость организма, половое поведение.
Эндокринные клетки, образующие гормоны, получили свое название
благодаря наличию у них специализированной функции внутренней секре-
ции (инкреции), т. е. активного выведения образовавшихся информацион-
148 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
ных молекул — гормонов — во внутреннюю среду. Эти специализирован-
ные клетки образуют эндокринную систему, т. е. функциональное объеди-
нение всех инкреторных клеток организма, осуществляющее гормональ-
ную регуляцию.
Гормональная регуляция, как и любая система регуляции, имеет аппарат
управления, каналы прямой и обратной передачи информации, сигналы,
которыми информация передается, исполнительные органы или объекты
управления. Эти элементы системы названы звеньями и составляют струк-
турно-функциональную организацию системы гормональной регуляции.
3.4.1. Общая характеристика звеньев гормональной системы
регуляции
Звенья управления. Деятельность эндокринных клеток управляется (регули-
руется) нервными центрами и гормонами других эндокринных образова-
ний, т. е. нейроэндокринным путем. Для некоторых эндокринных клеток
основным путем управления является местная саморегуляция за счет меха-
низма обратной связи (например, секреция сахаррегулирующих гормонов'
островками Лангерганса регулируется уровнем глюкозы в крови, секреция
кальцийрегулирующих гормонов — паратирина и кальцитонина — уровнем
кальция в крови), а звенья нейро гуморального управления обеспечивают
лишь усиление или ослабление эффекта местной саморегуляции.
Нервная система (рис. 3.12) осуществляет регуляцию деятельности эн-
докринных клеток двумя путями. Первый из них реализуется структурами
центральной нервной системы, непосредственно передающими нервные
импульсы к эндокринным структурам, синтезирующим и секретирующим
гормоны. Этот путь управления получил название нервного, или парагипо-
физарного, т. е. реализуемого мимо гипофиза. Так регулируется деятель-
ность практически всех эндокринных клеток. Второй путь управления эн-
докринными клетками нервная система реализует через гипофиз, обозна-
чаемый в этом случае как гуморальное звено управления; этот путь регуля-
ции получил название гипофизарного. Таким путем регулируется деятель-
Рис. 3.12. Основные пути регуляции эндокринной системы.
Пунктирные стрелки — обратные связи регуляции.
3. Общие принципы и механизмы регуляции физиологических функций • 149
ность тех желез, для которых в гипофизе секретируются специальные
тропные гормоны или тропины, например щитовидная железа или кора
надпочечников.
Центральной для регуляции эндокринных функций структурой нервной
системы является гипоталамус. Этот отдел осуществляет оба пути управле-
ния, т. е. и нервный, и гипофизарный. Управляющая функция гипотала-
муса связана с наличием здесь групп нейронов, обладающих способностью
синтезировать и секретировать специальные регуляторные пептиды — ней-
рогормоны. Таким образом, гипоталамус является одновременно и нерв-
ным, и эндокринным образованием, играя ключевую роль в интеграции
нервных и гуморальных механизмов регуляции, осуществляя нейрогумо-
ральное управление функциями. Свойство нейронов гипоталамуса синте-
зировать и секретировать регуляторные пептиды получило название нейро-
секреции. В принципе, этим свойством обладают все нервные клетки, по-
скольку нейроны транспортируют синтезированные в них белки, фермен-
ты и другие молекулы с помощью аксонального тока. У нейронов гипота-
ламуса эта способность приобрела специфические свойства регуляторов
эндокринной системы.
Нейросекрет, образующийся в теле гипоталамического нейрона, хра-
нится в нем в виде гранул и путем аксонального транспорта переносится в
структуры мозга, ликвор или гипофиз. Соответственно направлению
транспорта, в гипоталамусе человека выделены 3 нейроэндокринные сис-
темы.
1) Гипоталамо-экстрагипоталамная система представлена нейросекре-
торными клетками, аксоны которых уходят за переделы гипоталамуса в
другие структуры головного мозга — таламус, лимбику, продолговатый
мозг — и выделяют нейропептиды, выполняющие медиаторную и модуля-
торную роль: вазопрессин, эндогенные опиоиды, нейротензин, вещество
Р, соматостатин, киоторфин и др.
2) Гипоталамо-аденогипофизарная (переднегипофизарная) система обра-
зована пептид- и моноаминергическими нейросекреторными клетками
мелкоклеточных ядер заднего гипоталамуса. Аксоны этих клеток образуют
контакты в области срединного возвышения на капиллярах (аксовазальные
контакты) первичной сети портальной системы гипофиза, в которую и по-
ступают нейропептиды, стимулирующие (либерины) или подавляющие
{статины) синтез и секрецию тропных гормонов аденогипофиза. Таким
образом, связь гипоталамуса с аденогипофизом осуществляется нейроге-
мальным путем (через кровь).
3) Гипоталамо-нейрогипофизарная (заднегипофизарная) система пред-
ставлена нейросекреторными клетками крупноклеточных ядер переднего
гипоталамуса — супраоптического и паравентрикулярного. Аксоны этих
клеток опускаются в нейрогипофиз, по ним в заднюю долю гипофиза по-
ступают и депонируются нейропептиды вазопрессин и окситоцин, связан-
ные со специфическими белками нейрофизинами.
Гипоталамические нейропептиды в зависимости от места реализации
регуляторного эффекта делят на три группы: 1) висцерорецепторные ней-
рогормоны, обладающие преимущественным действием на висцеральные
органы (вазопрессин, окситоцин), 2) нейрорецепторные нейрогормоны
или нейромодуляторы и медиаторы, обладающие выраженными эффекта-
ми на функции нервной системы (эндорфины, энкефалины, нейротензин,
вазопрессин, ангиотензин и др.), 3) аденогипофизрецепторные нейрогор-
моны, регулирующие деятельность железистых клеток аденогипофиза
(кортиколиберин, соматостатин, тиреолиберин и др.).
150 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Звенья общего и гуморального управления имеют многочисленные об-
ратные связи, контролирующие процессы синтеза и секреции, уровень
гормонов в крови и реализацию их эффектов в органах и тканях.
Звено синтеза и секреции гормонов. Особенности синтеза гормонов в эн-
докринных клетках зависят от химической структуры гормонов. По хими-
ческой природе все гормоны подразделяют на три группы: 1) производные
аминокислот — тиреоидные гормоны, адреналин, гормоны эпифиза;
2) пептидные гормоны, простые (протеины) и сложные (гликопротеиды)
белки — гипоталамические нейропептиды, гормоны гипофиза, островково-
го аппарата поджелудочной железы, околощитовидных желез; 3) стероид-
ные гормоны, образующиеся из холестерина гормоны коры надпочечни-
ков, половых желез, гормон почечного происхождения кальцитриол.
Синтез гормонов эндокринными клетками происходит непрерывно, его
интенсивность зависит не только от регуляторных сигналов звена управле-
ния, но и от величины секреции. Известный в биохимии принцип тормо-
жения синтеза конечным продуктом обусловливает подавление образова-
ния гормонов при сниженном их удалении из клеток и, напротив, актива-
ция секреции повышает синтез гормонов. Таким образом, звенья синтеза и
секреции гормонов взаимосвязаны.
Звено депонирования связано с синтезом и секрецией гормонов, посколь-
ку гормоны обычно депонируются в тех же клетках тканей, где образуют-
ся. Депонирование гормонов эндокринной тканью может осуществляться в
специализированных гранулах (мозговое вещество надпочечников) или в
специализированных структурах железы (коллоид фолликулов щитовидной
железы). Гормоны депонируются в виде связанных форм с белками, мак-
роэргическими фосфатами, нуклеопротеидами или металлами. Однако не-
которые гормоны могут депонироваться и в несекреторных тканях, клетка-
ми которых они захватываются из крови. Так, например, могут депониро-
ваться катехоламины.
Звено транспорта представлено жидкостями внутренней среды, напри-
мер кровью, переносящими гормоны как в свободной, так и в связанной
формах. Связывание гормонов происходит как с мембранами клеток крови
(эритроциты, тромбоциты), так и с белками плазмы крови, при этом ак-
тивность таких связанных форм крайне низкая, поскольку они плохо про-
ходят через гистогематические барьеры и не могут взаимодействовать со
специфическими для них клеточными рецепторами. Свободные (т. е. не-
связанные) формы гормонов являются активными, поскольку проходят че-
рез барьеры, взаимодействуют с мембранными рецепторами и вызывают
физиологические эффекты. Вместе с тем физико-химическая связь гормо-
нов с клетками крови и белками плазмы является формой их депонирова-
ния во внутренней среде, поскольку удаление связанных гормонов во
внешнюю среду через органы выделения затруднено, а при необходимости
гормоны могут освобождаться из связанных форм, переходить в свободную
активную форму и вызывать регуляторные эффекты без дополнительной
активации их синтеза и секреции.
Транспорт гормонов кровью участвует в реализации механизма обрат-
ной связи со звеном управления, поскольку содержащиеся в крови гормо-
ны могут прямо воздействовать на гипоталамус или гипофиз.
Звено метаболизма гормонов требуется не только для разрушения обра-
зовавшихся гормонов, но и для уменьшения числа информационных моле-
кул и ослабления их регуляторного эффекта. Метаболические превраще-
ния гормонов приводят к образованию новых информационных молекул с
отличающимися от основного гормона свойствами, метаболическими и
3. Общие принципы и механизмы регуляции физиологических функций • 151
физиологическими эффектами. Метаболизм гормонов осуществляется под
влиянием ферментов в самих эндокринных тканях, печени, почках и в
тканях-эффекторах. Образование при метаболизме гормонов новых ин-
формационных молекул в тканях-эффекторах обеспечивает в них реализа-
цию и новых биохимических и физиологических эффектов. Так, напри-
мер, дейодирование тироксина в клетках-мишенях ведет к образованию
трийодтиронина, обладающего значительно более сильным физиологиче-
ским эффектом; метаболизм аланиновой боковой цепи тиреоидных гормо-
нов приводит к образованию тироуксусных кислот с более выраженным
эффектом на энергетический обмен клеток. В то же время снижение мета-
болической деградации молекул гормонов в печени влечет за собой избы-
точный эффект гормонов на ткани, несмотря на то что эндокринные клет-
ки не повышают секреции гормонов. Многие метаболиты гормонов выде-
ляются из организма с мочой и их концентрации в моче исследуются для
оценки функции железы.
Звено выделения гормонов представлено в организме почками, потовыми
железами, слюнными железами, желчью и пищеварительными соками.
Выведение информационных молекул гормонов и их метаболитов из крови
происходит в наибольшей степени через почки с мочой. Выведение гормо-
нов, наряду с депонированием и метаболическим разрушением, защищает
организм от избыточности гормональных эффектов.
В звене эффектора гормонов осуществляется реализация биохимических
и физиологических эффектов гормональной регуляции. Поступая через
жидкую внутреннюю среду к клеткам эффекторов, гормоны связываются
со специфическими для них клеточными рецепторами. В связи с этим
влияния гормонов не диффузные, предназначенные всем клеткам организ-
ма, а строго специфичные, адресованные конкретным клеткам, содержа-
щим рецепторы к определенному гормону. Соответственно, и чувствитель-
ность разных тканей к гормональному регулирующему воздействию неоди-
накова, так как определяется наличием и количеством специфических ре-
цепторов. Ткани, имеющие большое количество рецепторов с высоким
сродством к определенному гормону, называют тканями- или органами-
мишенями этого гормона. В зависимости от локализации в клетках рецеп-
торы делят на плазматические (расположенные на плазматической мем-
бране клеток), цитозольные (находящиеся в цитоплазме), и ядерные (лока-
лизованные в ядре).
3.4.2. Виды и пути действия гормонов
Различают пять видов действия гормонов на ткани-мишени: метаболиче-
ское, морфогенетическое, кинетическое, корригирующее и реактогенное.
• Метаболическое действие гормонов вызывает изменение обмена ве-
ществ в тканях. Оно происходит за счет трех основных гормональных
влияний. Во-первых, гормоны меняют проницаемость мембран клет-
ки и органоидов и транспорт через них субстратов, ферментов, ионов
и метаболитов. Во-вторых, гормоны меняют активность ферментов в
клетке. В-третьих, гормоны изменяют синтез ферментов, индуцируя
или подавляя их образование за счет влияния на генетический аппа-
рат ядра клетки, как прямо вмешиваясь в процессы синтеза нуклеи-
новых кислот и белка, так и опосредованно через энергетическое и
субстратно-ферментное обеспечение этих процессов. Сдвиги метабо-
лизма, вызываемые гормонами, лежат в основе изменения функций
клеток, ткани или органа.
152 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 3.13. Схема действия пептидных гормонов на клетки-эффекторы с участием
вторичных посредников.
Молекула гормона, называемая также первичным посредником регуляции функции) связыва-
ется со специфичным для нее рецептором клеточной мембраны. Образование гормон-рецеп-•
торного комплекса приводит к активации мембранного фермента (например, аденилатцикла-
зы), под влиянием которого в клетке происходит образование молекул вторичного посредника
(например, циклической АМФ). Вторичный посредник ведет к активации внутриклеточных
ферментов (протеинкиназ), способствующих фосфорилированию субстратов, что и обеспечи-
вает формирование внутриклеточных эффектов в виде изменения проницаемости клеточных
мембран, изменения метаболических процессов и активации генома. Следствием этих внутри-
клеточных эффектов вторичного посредника регуляторного сигнала является изменение
функции клетки.
• Морфогенетическое действие — это стимулирующее влияние гормо-
нов на процессы формообразования, дифференцировки и роста
структурных элементов. Осуществляются эти процессы за счет акти-
вации генетического аппарата клетки и обмена веществ. Примера-
ми может служить влияние соматотропина на рост тела и внутрен-
них органов, половых гормонов — на развитие вторичных половых
признаков.
• Кинетическое действие — способность гормонов запускать деятель-
ность эффектора, включать реализацию определенной функции. На-
пример, окситоцин вызывает сокращение мускулатуры беременной
матки; адреналин запускает распад гликогена в печени и выход глю-
козы в кровь, вазопрессин включает обратное всасывание воды в со-
бирательных трубочках нефрона, без него не происходящее.
• Корригирующее (исправляющее) действие — изменение деятельности
органов или процессов, которые осуществляются независимо от на-
личия гормона. Примером корригирующего действия гормонов явля-
ется влияние адреналина на частоту сердечных сокращений, актива-
3. Общие принципы и механизмы регуляции физиологических функций • 153
Молекула гормона
Рис. 3.14. Схема классического пути действия стероидных гормонов на клетки-эф-
фекторы.
Благодаря липофильности молекула гормона проникает через клеточную мембрану и в цито-
плазме взаимодействует с плазматическим рецептором. Гормон-рецепторный комплекс про-
никает в ядро клетки, где после связывания с ядерным рецептором гормон вызывает актива-
цию процессов транскрипции и трансляции, приводящих к стимуляции синтеза белков, в том
числе ферментов и, таким образом, изменяя метаболизм и функцию клетки.
ция окислительных процессов тироксином, уменьшение обратного
всасывания ионов калия в почках под влиянием альдостерона. Разно-
видностью корригирующего действия является нормализующий эф-
фект гормонов, когда их влияние направлено на восстановление из-
мененного или даже нарушенного процесса. Например, при исход-
ном превалировании анаболических процессов белкового обмена
глюкокортикоиды вызывают катаболический эффект, но если исход-
но преобладает распад белков, глюкокортикоиды стимулируют их
синтез.
• Реактогенное действие гормонов — способность гормона менять реак-
тивность ткани к действию того же гормона, других гормонов или
медиаторов нервных импульсов. Так, например, избыток кальцийре-
гулирующих гормонов снижает чувствительность дистальных отделов
нефрона к действию вазопрессина, тиреоидные гормоны усиливают
эффекты катехоламинов. Разновидностью реактогенного действия
гормонов является пермиссивное действие, означающее способность
одного гормона давать возможность реализоваться эффекту другого
гормона. Так, например, глюкокортикоиды обладают пермиссивным
действием по отношению к катехоламинам, т. е. для реализации эф-
фектов адреналина необходимо присутствие малых количеств корти-
154 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
зола, инсулин обладает пермиссивным действием для соматотропина
(гормона роста) и др. Особенностью гормональной регуляции являет-
ся то, что реактогенное действие гормоны могут реализовать и в тех
тканях и органах, где имеется лишь небольшое число рецепторов к
гормону.
Пути действия гормонов на эффекторы рассматриваются в виде двух
возможностей: 1) действия гормона (первичного посредника регуляции) с
поверхности клеточной мембраны после связывания со специфическим
рецептором, что ведет к активации мембранного фермента, образованию
химического вторичного посредника и запуска тем самым цепочки биохи-
мических превращений в мембране, цитоплазме и ядре клетки (преимуще-
ственно эффекты пептидных гормонов и катехоламинов) (рис. 3.13);
2) действия гормона, путем проникновения через мембрану благодаря рас-
творимости в липидах и связывания с рецептором цитоплазмы (рис. 3.14),
после чего гормон-рецепторный комплекс проникает в ядро и органоиды
клетки, где и реализует свой регуляторный эффект (преимущественно, сте-
роидные гормоны, гормоны щитовидной железы). Однако эти возможно-
сти не альтернативны, так как для многих гормонов существуют оба пути
действия на эффектор. В частности, для стероидных гормонов установле-
ны два пути действия на клетку: 1) классический, ядерный или геномный
(проникновение в ядро и активация процессов транскрипции, трансляции
и синтеза) и 2) альтернативный негеномный (связывание с мембранным
рецептором и активация вторичных посредников).
3.5. Местная гуморальная регуляция функций клеток
Физиологический контроль процессов пролиферации и дифференцировки
клеток, синтеза и продукции ими белков, гликопротеидов, липидов и дру-
гих веществ, жизнедеятельности клеток вплоть до их гибели осуществляет-
ся с помощью взаимодействия многочисленных внеклеточных регулятор-
ных молекул (называемых лигандами) с различными классами рецепторов
клеточных мембран.
Местная регуляция функций клеток обеспечивается тремя основными
способами, в основе которых лежит передача химических сигналов в пре-
делах одной ткани или органа с помощью: 1) креаторных связей клеток,
2) простейших метаболитов и 3) более сложных продуктов обмена — био-
логически активных веществ.
Понятие креаторные связи обозначает обмен между клетками макромо-
лекулами, несущими информацию, необходимую для направленного регу-
лирования внутриклеточного синтеза молекул белка и других процессов с
целью объединения клеток в ткань, обеспечения дифференцировки, роста
и развития и, в конечном счете функционирования отдельных клеток тка-
ни как единой многоклеточной системы. Межклеточное взаимодействие
через микрокружение клеток является одним из наиболее эволюционно
старых способов регуляции, возникших с появлением многоклеточных ор-
ганизмов.
Межклеточное взаимодействие обеспечивается, преимущественно, сле-
дующими факторами: 1) адгезивными молекулами клеточных мембран,
2) внеклеточным матриксом с содержащимися в нем макромолекулами,
3) растворимыми медиаторами межклеточного взаимодействия, 4) сверх-
слабым свечением (биохемилюминесценция) типа митогенетического из-
лучения в короткой ультрафиолетовой области, 5) онкогенами, 6) электри-
ческими потенциалами.
3. Общие принципы и механизмы регуляции физиологических функций • 155
Наиболее тесное взаимодействие между однотипными клетками осуще-
ствляется в области десмосом.
На поверхности клеток расположены многочисленные мембранные гли-
копротеины, обеспечивающие взаимодействие клеток. Ключевую роль во
взаимодействии клеток между собой и с экстрацеллюлярным матриксом
играют несколько классов мембранных рецепторных молекул: селектины,
иммуноглобулины, кадгерины, интегрины, адрессины. Описано 3 вида се-
лектинов'. L, Р и Е. Являясь рецепторами клеточной адгезии (прилипания),
они обеспечивают задержку лимфоцитов в лимфатических узлах (L-селек-
тин), адгезию тромбоцитов и лейкоцитов в места поврежденного эндоте-
лия и участки тромбообразования (Р-селектин) и адгезию активированных
клеток эндотелия к нейтрофилам (Е-селектин). Иммуноглобулиновые рецеп-
торы представлены в клеточных мембранах как минимум 11 видами моле-
кул, обеспечивающими адгезию лейкоцитов и эндотелия, рост и диффе-
ренцировку в эмбриогенезе эктодермы, особенно нервной ткани, в том
числе формирование синапсов, миелинизацию, морфогенез мезодермы и
ее производных, адгезию нейрон—нейрон и нейрон—глия, адгезию Т-лим-
фоцитов с клетками-мишенями и реакции иммунитета, адгезию клеток
мышц, красной пульпы селезенки, гемопоэтических клеток и т. д. Класс
кадгериновых рецепторов включает три вида (Е, Р и N). Эти молекулы обес-
печивают Са++-зависимый процесс межклеточной адгезии в процессах эм-
бриональной дифференцировки, морфогенезе, образовании клеточных
пластов в тканях мускулатуры, мозга, хрусталика, плаценты, эпителиаль-
ных и мезодермальных клетках развивающихся организмов. В зрелом воз-
» расте они представлены в эпителиальных тканях — базальном слое кожи,
эпителии урогенитального тракта, дыхательных путей, в лимфоидных орга-
нах и экзокринных железах.
Взаимодействие клеток с внеклеточным матриксом обеспечивается
мембранными рецепторами класса интегринов. К этому классу принадле-
I жат три группы поверхностных гликопротеинов, получивших название по
। особенностям с-субъединиц молекул (сг, с2- и с3-). Эти рецепторы обеспе-
I чивают реакции внутриклеточного цитоскелета на лиганды внеклеточного
матрикса. Они участвуют в агрегации тромбоцитов, иммунологических
процессах. Интегриновые рецепторы взаимодействуют с многочисленными
белками внеклеточного матрикса, обеспечивая реакции клеток в ответ на
измененное микроокружение. Вместе с тем интегрины могут существовать
в активной и неактивной формах в зависимости от внеклеточных сигна-
лов, а также менять, подобно другим мембранным рецепторам, сродство к
лигандам. Образование лиганд-интегринового комплекса ведет к передаче
информации внутрь клетки с изменением в ней не только метаболизма
(например, pH), но и процессов экспрессии генов.
К числу медиаторов межклеточного взаимодействия относят цитокины
(интерлейкины, опухольнекротизирующие факторы, хемокины, интерфе-
роны) и факторы роста, метаболиты арахидоновой кислоты (простагланди-
ны и лейкотриены), регуляторные метаболиты клеток (эндотелии, бради-
кинин, аденозин, оксид азота и др.), протеолитические ферменты и др.
Цитокины и факторы роста в мембраноассоциированной или диффузной
форме связываются либо с рецепторами на одноименных клетках-проду-
центах, вызывая эффект аутокринного регулирования, либо на соседних
клетках ткани (эффект паракринного регулирования), либо на клетках за
пределами мест синтеза (эффект «эндокринного» регулирования). Цитоки-
ны связываются и с внеклеточным матриксом, регулируя продукцию его
компонентов.
11
156 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
К числу медиаторов межклеточного взаимодействия относится и адено-
зин, выделяемый клетками, особенно в условиях функционального напря-
жения, и играющий роль местного регулятора их функций по принципу
обратной связи. Аденозиновая саморегуляция клеточных функций пред-
ставлена в тканях центральной нервной, сердечно-сосудистой, иммунной и
гемопоэтической систем, в желудочно-кишечном тракте и легких. Адено-
зин является регуляторным метаболитом, его высвобождение клетками за-
висит от реакций потребления и генерирования АТФ, т. е. от метаболиче-
ского статуса клеток.
К числу медиаторов аутокринного взаимодействия относят простаглан-
дины и лейкотриены, образующиеся практически во всех видах клеток,
выделяющиеся во внеклеточную микросреду и оказывающие регуляторное
влияние через мембранные рецепторы разных типов. Среди медиаторов та-
кого рода взаимодействия описаны также трансформирующий фактор рос-
та, тканевые ингибиторы металлопротеаз, инсулиноподобный фактор рос-
та и др.
В органах, имеющих клетки и ткани разных видов, существенную роль
в местной саморегуляции функций играют медиаторы гетерогенных клеток.
Так, например, в регуляции функционального состояния гладкомышечных
клеток кровеносных сосудов принимают участие гуморальные вещества,
выделяемые эндотелиальными клетками — эндотелиальные релаксирую-
щий и констрикторный факторы. Чем больше гетерогенность клеточных
популяций в органе, тем большее количество местных внутриорганных гу-
моральных факторов используется для регуляции функций. При этом
внутриорганные гуморальные регуляторы обеспечивают изменение не
только функции клеток, но и их метаболического обеспечения, меняя, на-
пример, условия кровоснабжения. Типичным примером наличия такого
рода многообразия гуморальных внутриорганных регуляторов является
почка, клеточными структурами которой образуются как гормональные
(ренин-ангиотензин, кальцитриол), так и местные метаболические (про-
стагландины, нейтральные липиды, кинины и т. п.) регуляторы. Мощное
гуморальное регуляторное взаимодействие существует между тканевыми
(в том числе эндотелиальными) клетками и клетками крови. Только в по-
чечном клубочке взаимодействие клеток крови, сосудистого эндотелия,
мезангиальных клеток и подоцитов обеспечивается следующим, далеко не
полным, перечнем гуморальных факторов: реактивно-оксилительные ве-
щества, интерлейкины, тромбоксан, простациклин, лейкотриены, эндоте-
лии, фактор активации тромбоцитов, тромбоцитарный фактор роста, мо-
ноцитарный гемоаттрактный белок, фактор роста фибробластов, эпидер-
мальный фактор роста, основной фактор роста фибробластов, фибронек-
тин, ламинин, тромбосподин, нидоген/энтактин, протеогликаны и т. п.
Простейшие метаболиты нередко являются регуляторами обменных
процессов и функции клеток органов по механизму обратной связи. На-
пример, образование избытка метаболитов с кислотными свойствами (мо-
лочной кислоты, пировиноградной кислоты) при интенсивной деятельно-
сти мышц, создающей относительный дефицит кислорода, активирует ме-
таболические пути их использования, ведет к расширению артериол и пре-
капилляров для увеличения притока крови и кислорода. В то же время в
связи с накоплением Н-ионов сократительная способность мышц ослабе-
вает. Регуляторные эффекты простейших метаболитов, как правило, не-
специфичны, поскольку для них не существует мембранных рецепторов.
Метаболиты более сложного химического строения носят название био-
логически активных веществ, или тканевых «гормонов». К их числу отно-
3. Общие принципы и механизмы регуляции физиологических функций • 157
сятся, например, биогенные амины (гистамин, серотонин), кинины и про-
стагландины. Эти вещества оказывают регулирующее влияние на функции
клеток и ткани в целом за счет изменения их биофизических свойств (про-
ницаемости мембран, величины мембранного заряда и т. п.), процессов
обеспечения функции — энергетического обмена, клеточной рецепции,
ферментативных реакций, образования вторичных посредников и сдвигов
кровоснабжения ткани. Биологически активные вещества меняют чувстви-
тельность клеток (количество и сродство клеточных химических рецепто-
ров) к нервным и гормональным влияниям, почему их называют также мо-
дуляторами регуляторных сигналов. Образуясь преимущественно в тканях,
большинство из биологически активных веществ могут проникать из кле-
точной микросреды в кровь, а некоторые (кинины) — даже образовываться
из предшественников в самой плазме крови. Таким образом, наряду с ос-
новным местным регуляторным действием эти вещества способны оказы-
вать и регионарное регуляторное влияние, и даже генерализованные эф-
фекты, подобно гормонам. Однако образование этих веществ, в отличие от
гормонов, осуществляется неспециализированными, т. е. неэндокринными
клетками.
3.6. Системный принцип организации механизмов регуляции
физиологических функций
Рассмотренные выше нервные и гуморальные принципы регуляции функ-
ционально и даже структурно (примером чему является гипоталамус) объе-
динены в единую систему нейрогуморальной регуляции. При этом началь-
ным звеном такого регуляторного механизма, как правило, является аффе-
рентный нервный сигнал на входе системы, а эффекторные каналы ин-
формационной связи являются либо нервными, либо гуморальными. Сле-
довательно, принципом реализации механизмов нейрогуморальной регуля-
ции является рефлекс, имеющий два типа путей передачи эфферентной
информации — нервный и гуморальный (рис. 3.15). Вместе с тем, каким
бы сложным ни был рефлекс, какие бы сложные информационные каналы
при этом ни использовались, он представляет собой только базисный, от-
носительно простой, уровень нейрогуморальной регуляции жизнедеятель-
ности. Даже в случаях более сложных рефлекторных механизмов, напри-
мер приобретенных условных рефлексов, регуляторные реакции охватыва-
ют лишь некоторые, причем весьма узкие стороны жизнедеятельности или
поведения. Однако, благодаря тому что организм является целостной
сложной системой, регуляция его жизнедеятельности носит системный ха-
рактер, т. е. не определяется простой арифметической суммой рефлектор-
ных регуляций составляющих его элементов, а проявляется перестройкой
всех взаимоотношений и взаимосвязей внутри системы для обеспечения
полноценного оптимального приспособления к меняющейся среде оби-
тания.
Сказанное отнюдь не отвергает большую биологическую значимость
рефлекторного принципа нейрогуморальной регуляции, напротив, рефлек-
торные реакции организма являются начальным звеном более сложного
целостного реагирования, конечной целью которого является приспособ-
ление живой системы к внешней среде, т. е. оптимизирование жизнедея-
тельности. Механизмы реализации системного принципа регуляции явля-
ются сложными и многообразными, особенно принимая во внимание ди-
намичность, т. е. постоянство изменчивости организации и функций жи-
' вой системы. В самом деле, ведь регуляция жизнедеятельности организма
158 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Афферентное звено Эфферентные звенья
Рис. 3.15. Схема рефлекторной нейрогуморальной регуляции.
Раздражение рецептора через афферентное звено (нервные проводники) вызывает активацию
нервного центра. Регуляторные влияния последнего передаются к эффектору по эфферентным
звеньям, которыми могут быть как нервные проводники (нервная регуляция), так и выделяе-
мые эндокринными железами гормоны (гуморальная регуляция). Происходящие изменения
функции эффектора (например, повышение тонуса мышцы) воспринимаются его рецептора-
ми (например, мышечными веретенами), от которых по механизму обратной связи через аф-
ферентное звено информация об эффекте поступает вновь в нервный центр.
в условиях, например, низкой температуры окружающей среды не может
быть сведена только к рефлекторным реакциям поддержания температуры
тела. Динамическая перестройка метаболизма и функций затрагивает все
без исключения органы и физиологические системы организма, меняется
и характер питания, и образ жизни, и поведение. Изменения затрагивают
не только биологическую, но и социальную сущность человека.
Системный характер регуляции физиологических функций проявляется
и во взаимосвязи между нейроэндокринными и иммунными процессами.
Так, на мембранах и в ядрах клеток, обеспечивающих реакции иммуните-
та, локализованы рецепторы ко многим гормонам (коры надпочечников,
щитовидной железы и др.). В результате взаимодействия с ними гормо-
нальный сигнал вовлекает в регуляторную реакцию иммунные клетки, вы-
деляющие во внутреннюю среду многочисленные гуморальные регулято-
ры — цитокины, мембранные рецепторы к которым существуют и у нерв-
ных, и у эндокринных клеток организма. Наличие в организме единой
нейрогуморально-иммунной системы регуляции функций способствует удале-
нию из организма генетически чужеродных агентов — антигенов.
Одним из установленных механизмов системной организации процес-
сов регуляции жизнедеятельности является общий адаптационный син-
дром, или стресс, представляющий собой совокупность неспецифических
и специфических реакций систем нейрогуморальной регуляции, метабо-
лизма и физиологических функций, приводящих к повышению устойчиво-
сти организма к действия неблагоприятных факторов среды. Основы уче-
ния о стрессе были заложены канадским физиологом и патологом Гансом
3. Общие принципы и механизмы регуляции физиологических функций • 159
Селье (Selye). Системный уровень нейрогуморальной регуляции жизнедея-
тельности проявляется при стрессе в виде увеличения энергообеспечения
клеток, что способствует повышению устойчивости организма в целом к
действию факторов окружающей среды, в том числе и вредных для орга-
низма.
Другой механизм системной организации регуляции функций был уста-
новлен отечественным физиологом П. К. Анохиным и сформулирован им
в виде учения о функциональной системе. По автору, классическое пред-
ставление о рефлексе всегда означает ответную реакцию организма на раз-
дражитель, т. е. действие. Например, рефлекс чиханья, сгибательный реф-
лекс, сосудодвигательный рефлекс и т. п. Но для организма важным явля-
ется не само действие, не процесс, а результат этого действия. Рефлекс чи-
ханья возникает как реакция приспособления не ради самого процесса, а
для удаления посторонних частиц, раздражающих слизистую оболочку но-
са. Удалены раздражающие частицы — результат достигнут, не удалены —
рефлекторное чиханье продолжается. Функциональной системой, по
П. К. Анохину, называют динамически складывающиеся единицы инте-
грации целостного организма, избирательно объединяющие специальные
центральные и периферические образования и направленные на достиже-
ние результатов приспособительной деятельности. Иначе говоря, функ-
циональная система есть временное объединение разных уровней органи-
зации (клеток, тканей, органов, физиологических систем) и механизмов их
регуляции для достижения конкретного, полезного для организма в целом,
результата. Системогенез — это процесс формирования функциональных
систем, обеспечивающих возможность приспособления организма к усло-
виям окружающей среды. Причиной, ведущей к образованию функцио-
нальной системы, или системообразующим фактором является потреб-
ность в конкретном результате приспособительной деятельности. При этом
параметры получаемого результата постоянно анализируются за счет об-
ратной афферентации.
ГЛАВА 4
Функции центральной нервной
системы
Нервная система регулирует совершаемые человеком движения, дея-
тельность его внутренних органов и обеспечивает осуществление психиче-
ских процессов. Она воспринимает изменения внешней среды и внутрен-
него состояния организма, организует ответные реакции на эти измене-
ния, координирует деятельность физиологических систем и органов и фор-
мирует целенаправленное поведение. Центральная нервная система обра-
зована спинным и головным мозгом, в периферическую нервную систему
включают периферические нервы и нервные сплетения, или ганглии.
В каждом отделе центральной нервной системы существуют два типа
клеток: нервные клетки, или нейроны, и вспомогательные клетки нейроглии,
образовавшиеся в процессе эмбриогенеза из общих для них стволовых кле-
ток. В головном мозге человека содержится приблизительно 10'1 различаю-
щихся по морфофункциональным признакам нейронов, клеток нейроглии
больше примерно в 10—50 раз и они занимают около половины объема
мозга. Нейроны вступают в контакт друг с другом в синапсах, численность
которых в мозгу человека равна 1014 (по некоторым данным — 1018). Спе-
цифическая функция нейронов заключается в их способности возбуждать-
ся, т. е. генерировать и проводить электрические сигналы. Таким способом
возбуждающие нейроны передают состояние возбуждения другим нейронам
или рабочим органам. Одновременно тормозные нейроны после своего воз-
буждения избирательно тормозят этот процесс и, тем самым, ограничива-
ют его распространение в пределах функционально необходимых контак-
тов между клетками.
Нервные клетки являются структурными и функциональными единица-
ми ЦНС, а их объединение в упорядоченные нервные сети осуществляют
постоянные синапсы между нейронами. Эффективность синаптических
переключений может изменяться, что лежит в основе пластичности нерв-
ной системы, т. е. способности перестраивать свои функциональные свой-
ства в зависимости от длительных изменений среды и характера деятельно-
сти. Модификация синаптических переключений происходит на протяже-
нии всей жизни, она позволяет приобретать индивидуальный поведенче-
ский опыт и приспосабливаться к изменяющимся условиям существо-
вания.
4.1. Основы функционирования нейронов и глии
4.1.1. Общая характеристика нейронов
Нервная ткань превосходит все другие ткани по количеству генов, вовле-
ченных в развитие клеток, и по количеству видов информационной РНК.
В мозге существует примерно 200 000 отличающихся друг от друга молекул
4. Функции центральной нервной системы • 161
Пресинап- Синапсы
возбуждающего Постсинаптические
нейрона нейроны
Рис. 4.1. Основные структуры типичного нейрона
Дендриты — ветвящиеся отростки нейрона, представляющие область формирования входных
сигналов. Аксон — отросток тела нейрона, служащий для проведения возбуждения от него,
имеет постоянный диаметр (большинство аксонов в ЦНС окружены миелиновой оболочкой)
и заканчивается пресинаптическими окончаниями. Аксонный холмик — конически расширенный
участок начала аксона, представляющий наиболее возбудимую часть плазматической мембра-
ны нейрона. Показаны окончания тормозного нейрона на теле клетки и возбуждающего ней-
рона на одном из дендритов.
иРНК, что в 10—20 раз больше, чем в печени или почках. Поэтому нерв-
ные клетки представлены многими популяциями нейронов, резко разли-
чающихся морфологическими признаками (строение тела, дендритов и ак-
сонов) и синтезируемыми в них нейромедиаторами. В то же время актив-
ность любого нейрона, независимо от выполняемой функции, проявляется
только в его возбуждении, т. е. генерации потенциалов действия, приводя-
щей к выделению в синапсе нейромедиатора.
Выполняемая нейроном функция всецело определяется специфично-
стью его синаптических соединений с другими нейронами. Некоторые по-
пуляции нейронов образуют компактные ядра, предназначенные для пере-
работки информации, которая передается из одного отдела мозга в другой.
Другие популяции нейронов представлены небольшими скоплениями кле-
ток, обладающих длинными разветвленными отростками, направленными
сразу к нескольким регионам мозга для регуляции их активности.
Четыре морфологически различаемые области нейрона (тело, дендриты,
аксон и пресинаптические окончания — рис. 4.1) отличаются друг от друга
и в функциональном отношении. Тело нейрона является центром происхо-
дящих в ней обменных процессов. В теле нейрона находится ядро, где
происходит переписывание генетической информации с ДНК на иРНК.
Дендриты представляют собой разветвленные отростки нервной клетки,
служащие для проведения нервных импульсов к телу нейрона. Степень
разветвленности и общее количество дендритов коррелируют с количест-
вом синапсов между нейроном и другими нервными клетками, способны-
ми повлиять на его активность и определяемыми как пресинаптические
нейроны. Так, например, на дендритах мотонейронов спинного мозга в
среднем существует около 8000 синапсов, а на дендритах клеток Пуркинье,
находящихся в коре мозжечка, — 150 000. Если одни пресинаптические
нейроны оказывают возбуждающее действие, а другие — тормозное, актив-
6 - 6095
162 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
кость постсинаптического нейрона будет определяться тем, какое из этих
влияний преобладает.
Нейроны имеют один аксон, в большинстве случаев самый длинный от-
росток клетки, предназначенный для проведения нервных импульсов от ее
тела. Обычно аксоны разделяются на несколько ветвей, или коллатералей,
позволяющих нейрону активировать или тормозить с помощью своего ме-
диатора сразу несколько клеток, с которыми он образует синапсы. В плаз-
матической мембране аксона содержится большое количество потенциал-
зависимых каналов для ионов натрия, необходимых для генерации и прове-
дения потенциалов действия. Наибольшая плотность таких каналов харак-
терна для аксонного холмика — суживающегося начального участка аксона,
отходящего от клеточного тела. Это наиболее возбудимый участок мембра-
ны центральных нейронов, где легче всего достигается критический уровень
деполяризации. Именно поэтому потенциалы действия обычно возникают в
аксонном холмике, представляющим собой наиболее чувствительную к
возбуждению область клетки.
Пресинаптические окончания аксонов свободны от миелинового покрытия
и предназначены для хранения и выделения медиаторов в химических си-
напсах. К выделению медиаторов приводит входящий ток ионов кальция
через быстро инактивирующиеся каналы, плотность которых в пресинап-
тических окончаниях высока. Аксоны возбуждающих нейронов в боль-
шинстве случаев образуют синапсы с дендритами и реже с телом постси-
наптических нейронов. Аксоны тормозных нейронов чаще всего образуют
синапсы на теле постсинаптического нейрона, их действие создает препят-
ствие проведению возбуждения от дендритов к аксонному холмику. Аксо-
аксоналъные синапсы, которые могут быть как возбуждающими, так и тор-
мозными, не влияют непосредственно на зону возникновения электриче-
ского сигнала. Их функция заключается в модуляции выходной активности
постсинаптического нейрона, т. е. в увеличении или уменьшении количе-
ства выделяемого в окончаниях его аксона медиатора.
4.1.2. Функциональная модель нейрона
Функция нейрона заключается в его способности возбуждаться и, действуя
через синапсы на другие клетки, генерировать возбуждение в них, что при-
водит к распространению этого процесса от одного нейрона к другому.
Возбуждение тормозного нейрона сопровождается подавлением процесса
возбуждения в клетках, на которые он действует. Процесс возбуждения
включает в себя возникновение нескольких типов электрических сигналов,
образование и распространение которых происходит по единым правилам
во всех нервных клетках: сенсорных, моторных, вставочных и нейросекре-
торных. Это позволяет рассмотреть закономерности возникновения и рас-
пространения электрических сигналов на основе единой функциональной
модели нейрона, пригодной для характеристики любого типа нервных
клеток.
Функциональная модель нейрона предусматривает выделение в нем че-
тырех функциональных областей, каждая из которых предназначена для
формирования одной из четырех разновидностей сигналов, характеризую-
щих процесс возбуждения: 1) входного (постсинаптический и рецепторный
потенциалы), 2) объединенного (потенциал действия), 3) проводящегося и
4) выходного (выделение медиатора) (рис. 4.2).
4. Функции центральной нервной системы • 163
4.1.2.1. Входные сигналы
Входные сигналы представляют собой изменения величины мембранного
потенциала в результате действия химических медиаторов на мембраны
постсинаптических нейронов (постсинаптические потенциалы), а в чувст-
вительных окончаниях сенсорных нейронов они возникают в ответ на дей-
ствие адекватных раздражителей (рецепторный потенциал). В возбуждаю-
щих синапсах ЦНС входной сигнал представляет собой деполяризацию
постсинаптической мембраны на 0,2—0,3 мВ, в тормозных синапсах —
столь же незначительную гиперполяризацию, соответственно этому различа-
ют возбуждающие постсинаптические потенциалы (ВПСП) и тормозные
постсинаптические потенциалы (ТПСП).
Постсинаптические потенциалы могут распространяться на очень малое
расстояние от места возникновения и в связи с этим определяются как ме-
стные, или локальные, потенциалы. Их амплитуда пропорциональна коли-
честву молекул нейромедиатора, связавшегося с постсинаптическими ре-
цепторами, такая зависимость является градуальной, а поэтому и входные
сигналы градуальны. Назначение возбуждающих входных сигналов состоит
в том, чтобы деполяризовать мембрану нейрона до критической величины,
достаточной для генерации потенциалов действия, а тормозные входные
сигналы предназначены для того, чтобы этому препятствовать.
Одиночные ВПСП электротонически, т. е. пассивно, распространяются
от места возникновения (дендриты или тело нейрона) к аксонному холми-
ку, служащему интегративной зоной или местом генерации потенциалов
действия. Изначально малая амплитуда одиночных ВПСП при пассивном
проведении становится еще меньше (рис. 4.3), поэтому для достижения
Функциональные
области:
Сенсорный
Типы нейронов
Объединенная Вид
модель сигнала
Рис. 4.2. Функциональная модель нейрона
Различные по выполняемой функции нейроны (сенсорные, моторные, интернейроны, нейро-
секретирующие клетки) имеют четыре функционально сопоставимые области: входную, инте-
гративную, проводящую и выходную. Эти области предназначены для возникновения или
проведения электрических сигналов, среди которых различают: входной (рецепторный или
постсииаптический потенциалы), интегративный (объединенный сигнал или потенциал дей-
ствия), проводящийся сигнал и выходной сигнал.
6’
164 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 4.3. Уменьшение амплитуды постсинаптического потенциала (ПСП) при рас-
пространении его по мембране клетки
А. В результате возбуждения окончаний пресинаптических нейронов и выделения из них ней-
ромедиатора на дендрите постсинаптического нейрона образуется возбуждающий постсинап-
тический потенциал (ВПСП). Амплитуда ВПСП уменьшается по мере его распространения от
дендрита к телу нейрона.
Б. Величина потенциала покоя постсинаптического нейрона составляет —65 мВ, порог возбу-
ждения (уровень критической деполяризации) на теле нейрона соответствует —35 мВ, а в области
аксонного холмика — около —50 мВ. Несмотря на уменьшение амплитуды распространяюще-
гося по мембране ВПСП, в области аксонного холмика деполяризующий сдвиг достигает кри-
тического значения.
критической деполяризации мембраны необходима пространственная или
последовательная (временная) суммация ВПСП.
Если постсинаптические потенциалы возникнут одновременно в разных
синапсах на дендритах и теле нейрона, то их общая сумма может оказаться
достаточной для деполяризации мембраны интегративной зоны до крити-
ческого уровня. Эта разновидность суммации называется пространствен-
ной, она возникает при одновременной возбуждающей активности группы
нейронов, аксоны которых конвергируют к одной общей постсинаптиче-
ской клетке (рис. 4.4). Средняя величина мембранного потенциала покоя у
4. Функции центральной нервной системы • 165
А. Пространственная суммация
ВПСП
Б. Последовательная суммация
Рис. 4.4. Суммация возбуждения в центральных синапсах
А. Пространственная суммация возникает при одновременном возбуждении нейронов, конвер-
гирующих к общей постсинаптической клетке.
Б. Последовательная суммация происходит вследствие ритмического возбуждения пресинапти-
ческого нейрона с достаточно высокой частотой генерации потенциалов действия (через каж-
дые 10 мс).
центральных нейронов составляет —65 мВ, а среднее значение критической
деполяризации —55 мВ, исходя из этого, для возбуждения постсинаптиче-
ского нейрона необходима сумма ВПСП, превышающая 10 мВ. Деполяри-
зующий сдвиг, превышающий это значение, приводит к возбуждению
постсинаптического нейрона.
Последовательная или временная суммация ВПСП происходит вследст-
вие продолжительного возбуждения пресинаптического нейрона, генери-
рующего потенциалы действия, которые следуют друг за другом через ко-
роткие промежутки времени. Если 50—100 нервных импульсов достигнут
пресинаптического окончания непосредственно друг за другом, то общего
количества выделившегося медиатора окажется достаточно для получения
амплитуды ВПСП, превышающей уровень критической деполяризации,
что вызовет возбуждение постсинаптического нейрона. В центральных си-
напсах ЦНС оба вида суммации, пространственная и последовательная,
обычно происходят одновременно, что способствует возбуждению постси-
наптических нейронов.
На дендритах, но чаще на теле клетки существуют тормозные синапсы,
расположенные между возбуждающими синапсами и интегративной зоной.
Их функция состоит в том, чтобы помешать деполяризующим ВПСП дос-
тичь интегративной зоны. В тормозных синапсах активируются хемозави-
симые каналы для ионов калия или хлора, что ведет к гиперполяризации
мембраны, представляющей тормозной постсинаптический потенциал. Ес-
ли на пути распространения деполяризующих ВПСП окажутся гиперполя-
ризованные действием тормозных медиаторов участки мембраны, то сум-
марная амплитуда ВПСП не сможет достичь значения критической депо-
ляризации, необходимой для возникновения потенциала действия
166 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
А. Постсинаптическое
1+2+3
ВПСП
Раздельное действие возбуждающих
и тормозного нейронов
Е кр.
Б. Пресинаптическое
Е кр.
ВПСП после действия
тормозного нейрона
Суммация ВПСП и ТПСП
Рис. 4.5. Торможение возбуждения в центральных синапсах
А. Постсинаптическое торможение обусловлено гиперполяризацией мембраны постсинапти-
ческой клетки, в результате которой возникающие в ее входной зоне ВПСП (возбуждающие
постсинаптические потенциалы) не могут достичь критического уровня деполяризации.
Б. Пресинаптическое торможение вызывается действием тормозного нейрона на аксон возбу-
ждающего пресинаптического нейрона. Мембрана аксона деполяризуется, а вследствие этого
уменьшается входящий ток ионов кальция в его окончание и выделение медиатора из этого
окончания.
1, 2, 3 — возбуждающие нейроны; Т — тормозной нейрон, Екр. — уровень критической депо-
ляризации.
(рис. 4.5). В этом случае способность нейрона к генерации потенциалов
действия в целом окажется заторможенной, несмотря на сохраняющуюся
активность возбуждающих пресинаптических клеток.
4.1.2.2. Объединенный сигнал — потенциал действия
Объединенным сигналом в нервной клетке является потенциал действия,
который возникает по принципу «все или ничего» при уменьшении мем-
бранного потенциала до порогового значения. Если же величина деполя-
ризующего сдвига мембранного потенциала продолжает увеличиваться над
критическим уровнем, то амплитуда потенциала действия остается неиз-
менной, составляя в среднем около 100 мВ. Объединенный сигнал возни-
кает в тех участках мембраны, где наиболее высока плотность потенциал-
зависимых каналов (аксонный холмик), и поэтому критический уровень
деполяризации здесь оказывается самым низким. Это интегративная, или
триггерная, зона, в которой суммируются все деполяризующие сдвиги
ВПСП. У нейронов с длинными дендритами на них имеются добавочные
триггерные зоны, наличие которых повышает эффективность возбуждаю-
щих синапсов, расположенных далеко от клеточного тела.
4. Функции центральной нервной системы • 167
Рис. 4.6. Изменение спайковой активности нейрона в результате его постсинапти-
ческого торможения
А. Исходная активность постсинаптического нейрона, регулярная генерация потенциалов дей-
ствия.
Б. Действие тормозного нейрона вызывает возникновение тормозных постсинаптических по-
тенциалов (ТПСП) и в это время генерация потенциалов действия прекращается.
Отдельные нейроны различаются характерной, специфической для них
частотой и ритмом генерации потенциалов действия. Эти особенности воз-
буждения определяются биофизическими свойствами нейронов: индивиду-
альным набором потенциалзависимых каналов и их распределением в раз-
ных областях клеточной мембраны. Характер спайковой активности ней-
рона в каждом конкретном случае зависит от соотношения деполяризую-
щих и гиперполяризующих входных сигналов (рис. 4.6) и биофизических
свойств самого нейрона. В результате изменений спайковой активности
нейрона изменяется и выходной сигнал, т. е. количество нейромедиатора,
выделяющегося в пресинаптическом окончании его аксона.
4.1.2.3. Проводящийся сигнал
Проведение возбуждения от интегративной зоны к окончаниям аксона
осуществляют проводящиеся сигналы. Это потенциалы действия, которые
последовательно возникают на мембране аксона в результате действия на
нее локальных электрических токов, образующихся между возбужденным
и невозбужденным участками. В отличие от дендритов, тела нейрона и ак-
сонного холмика, где представлено большое разнообразие ионселекгивных
каналов, в мембране аксона такие каналы однотипны во всех перехватах
Ранвье или на всем протяжении немиелинизированного аксона. Поэтому
при действии локального электрического тока на мембрану невозбужден-
ных участков в них открываются однотипные потенциалзависимые каналы
для ионов натрия, что вызывает стандартную саморегенерирующую депо-
ляризацию с возникновением типичного для данного нейрона потенциала
действия.
При таком способе проведения возбуждения амплитуда и длительность
потенциалов действия одинаковы на всем протяжении аксона вне зависи-
мости от его длины. Постоянство параметров потенциалов действия обес-
печивает передачу электрических сигналов другим нервным клеткам или
г
168 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
исполнительным органам без искажения передаваемой информации. Если
потенциалы действия возникают в триггерной зоне с определенной часто-
той, образуя «пачку», или серию, импульсов, то и к окончанию аксона они
распространяются без изменения рисунка спайковой активности, а это
значит, что цифровая информация о характере возбуждения нейрона при
проведении полностью сохраняется. По существу аксон является простым
и надежным проводником сигналов от своего начального сегмента до
окончания, а сами проводящиеся сигналы высоко стереотипны и принци-
пиально не различаются у разных нейронов. Только два признака проводя-
щихся сигналов определяют характер передаваемой информации: число
потенциалов действия и временные интервалы между ними.
4.1.2.4. Выходной сигнал
Выходным сигналом нейронов является выделение медиатора в окончани-
ях аксона, т. е. преобразование электрических процессов в другую форму
активности нервной клетки. Выходной сигнал градуален: его величина оп-
ределяется количеством выделяющегося медиатора, которое, в свою оче-
редь, зависит от общего числа и частоты потенциалов действия, пришед-
ших к окончанию аксона. Выходной сигнал адресуется одной или одно-
временно нескольким постсинаптическим клеткам, имеющим специфиче-
ские рецепторы для связывания медиатора.
Выделение медиатора происходит в результате повышения свободной
концентрации ионов кальция в окончании с 10-8—10-7 моль/л-1 до прибли-
зительно 10-5 моль/л-1. Даже при незначительной деполяризации мембра-
А. Пресинаптический Гиперполяризация Деполяризация
Время ►
Рис. 4.7. Изменения величины возбуждающего постсинаптического потенциала в
зависимости от значения мембранного потенциала пресинаптического аксона
А. Незначительная гиперполяризация пресинаптического окончания уменьшает входящий ток
ионов кальция. Соответственно этим изменениям уменьшается или увеличивается количество
нейромедиатора, выделяющегося из пресинаптического окончания.
Б. Амплитуда возбуждающего постсинаптического потенциала снижается при гиперполяриза-
ции пресинаптического окончания и возрастает при его деполяризации.
4. Функции центральной нервной системы • 169
пресинаптическом
окончании
Б. Пресинаптическое усиление
нейрон
нейрон
кальция в
пресинаптическом
окончании
Рис. 4.8. Пресинаптическое торможение и усиление
А. Пресинаптическое торможение в результате действия тормозного нейрона на окончание
пресинаптического нейрона в аксо-аксональном синапсе приводит к уменьшению входного
тока ионов кальция в пресинаптическое окончание. В результате уменьшается количество вы-
деляющегося из окончания медиатора и, соответственно, уменьшается амплитуда возбуждаю-
щего постсинаптического потенциала.
Б. Пресинаптическое усиление происходит под влиянием медиатора возбуждающего аксо-аксо-
нального синапса, что приводит к повышению входного тока кальция в пресинаптическое
окончание, увеличенному выделению из него нейромедиатора и увеличению амплитуды воз-
буждающего постсинаптического потенциала.
ны пресинаптического окончания в ней открываются потенциалзависимые
каналы для ионов кальция, и тогда его концентрация в окончании быстро
повышается, что вызывает выход медиаторов. И наоборот, даже небольшая
гиперполяризация пресинаптической мембраны приводит к уменьшению
тока ионов кальция в окончание аксона и снижает вероятность выхода ме-
диатора (рис. 4.7).
Величину выходных сигналов могут регулировать тормозные или возбу-
ждающие нейроны, действующие своими медиаторами на пресинаптиче-
ское окончание в аксо-аксональных синапсах (рис. 4.8). Медиаторы тор-
мозных нейронов гиперполяризуют мембрану пресинаптического оконча-
ния аксона, что уменьшает ток ионов кальция в это окончание, поэтому
выделение медиатора из него тоже уменьшается. Пресинаптическое тормо-
жение предназначено для временного уменьшения выходной активности
нейрона и не распространяется на интегративную зону, где происходит ге-
нерация потенциалов действия.
Если в образовании аксо-аксонального синапса участвует пресинапти-
ческий возбуждающий нейрон, то действие его медиатора приведет к депо-
170 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
ляризации мембраны аксона постсинаптического нейрона. Следствием
этого становится повышение тока ионов кальция в это окончание и, соот-
ветственно, увеличение количества выделяемого из него медиатора. В ре-
зультате управляемое окончание возбужденного нейрона станет эффектив-
нее действовать на постсинаптическую мишень. Этот эффект получил на-
звание пресинаптического усиления.
4.1.3. Функциональная характеристика нейроглии
Клетки нейроглии расположены между нервными клетками: они способст-
вуют объединению элементов нервной системы и, одновременно, изолиру-
ют друг от друга отдельные группы нейронов, а также большую часть их
аксонов.
4.1.3.1. Астроциты
Астроциты с помощью многочисленных ветвящихся отростков образуют
пространственную сеть, занимающую около половины межклеточного
пространства серого вещества. Отростки соседних астроцитов расширяют-
ся на концах и смыкаются друг с другом на поверхности кровеносных ка-
пилляров, изолируя снаружи более 90 % их поверхности. Это способствует
формированию гематоэнцефалического барьера, препятствующего проник-
новению из крови крупных молекул, большинства продуктов патологиче-
ских процессов и лекарств.
Остальные отростки астроцитов почти целиком обертывают нервные
клетки; вместе с синапсами, находящимися на теле и дендритах, они вы-
полняют функцию опоры и создают окружающую нейроны микросреду. Ас-
троциты поглощают избыток ионов калия, выходящих из нейронов при
возбуждении; тем самым они выполняют буферную функцию, обеспечи-
вающую постоянство ионного состава с наружной стороны нейронов. Ста-
бильная концентрация ионов калия на наружной мембране нейронов по-
зволяет сохранять возбудимость нейронов неизменной при разных услови-
ях их функционирования.
При поглощении калия астроциты деполяризуются и возникает ток этих
ионов через щелевые контакты между соседними астроцитами. Однако это
не приводит к возбуждению астроцитов, поскольку в их мембране для воз-
буждения недостаточно потенциалзависимых каналов. В состоянии покоя
мембрана астроцитов проницаема только для ионов калия, которые диф-
фундируют из клетки по градиенту концентрации, пока не установится
значение равновесного потенциала для калия (—75 мВ). Несмотря на то что
повышение концентрации ионов калия при поглощении их излишков ведет
к изменению некоторых свойств астроцитов, в переносе нервных импуль-
сов они не участвуют, эта функция возложена исключительно на нейроны.
Помимо опорной и буферной функции астроциты участвуют в регуля-
ции энергетического обмена нейронов, перекисного окисления липидов,
регуляции обмена нейромедиаторов; они секретируют нейротрофические
факторы, цитокины и иммуномодуляторы, а некоторые разновидности ас-
троцитов участвуют в фагоцитозе. Специфическую функцию выполняют
астроциты, сосредоточенные вокруг глутаматергических синапсов. Глута-
мат, самый распространенный возбуждающий нейромедиатор, в высокой
концентрации оказывается токсичным для многих нейронов, имеющих к
нему рецепторы. Повышенная концентрация глутамата стимулирует ток
кальция в клетки, а при высокой концентрации кальция повышается ак-
4. Функции центральной нервной системы • 171
тивность ферментов, способствующих усилению перекисного окисления
липидов (подобная ситуация возникает, например, вследствие кровоизлия-
ний и некоторых заболеваний мозга). Астроциты обладают высокой спо-
собностью к захвату молекул глутамата и содержат ключевой фермент (глу-
таминсинтетазу), с помощью которого захваченный глутамат преобразуется
в лишенный нейротоксической активности глутамин.
4.1.3.2. Олигодендроциты
Основной функцией олигодендроцитов является синтез миелина и образо-
вание миелиновых оболочек вокруг аксонов. Расположенные возле тел
нейронов и вдоль аксонов олигодендроциты своими отростками много-
кратно обертывают аксоны, обеспечивая им изолирующий миелиновый
футляр. Обычно отростки одного олигодендроцита участвуют в миелиниза-
ции нескольких соседних аксонов, однако они могут миелинизировать не
только аксоны, но некоторые нейроны, например клетки обонятельной лу-
ковицы, гранулярные клетки мозжечка. В периферической нервной систе-
ме миелинизацию аксонов осуществляют шванновские клетки, являющиеся
аналогами олигодендроцитов.
4.1.3.3. Эпендимная глия
Различают типичные эпендимные клетки, выстилающие поверхности желу-
дочков мозга и центральный канал спинного мозга, а также танициты,
расположенные преимущественно в области гипоталамуса и на дне IV же-
лудочка, в стенках водопровода и в спинном мозге. Эпендимные клетки
участвуют в трансцеллюлярном транспорте, с помощью своих ресничек
приводят в движение цереброспинальную жидкость и обладают секретор-
ной функцией. Танициты участвуют в транспорте веществ из спинномоз-
говой жидкости к аденогипофизу и в обратном направлении.
4.1.3.4. Микроглия
Клетки микроглии происходят от моноцитов, проникших из кровяного рус-
ла, они могут фагоцитировать значительные объемы разрушенных клеток
и нервных волокон, бактерии и вирусы. Это самые мелкие и самые немно-
гочисленные среди клеток глии (около 5 % глиальных клеток); диффузно
распределенные в сером и белом веществе мозга, они обладают подвижно-
стью и выраженной способностью захватывать и перерабатывать продукты
обмена. При повреждениях мозга наблюдается пролиферация микроглии, а
кроме того, их популяцию увеличивают вновь проникшие из крови моно-
циты. Клетки микроглии вырабатывают цитокины для активации астроци-
тов и Т-лимфоцитов.
4.2. Общие принципы функционального объединения нейронов
В соответствии с выполняемой деятельностью разные классы нейронов
объединены в функциональные системы', сенсорные, моторные или мотива-
ционные. Даже такая простая деятельность, как, например, перелистывание
страниц этого учебника, требует нескольких видов сенсорной информа-
ции: о расположении книги, о положении туловища, рук, пальцев, о кон-
такте кончиков пальцев с бумагой. Воспринимающие эту информацию
сенсорные системы перерабатывают ее и доставляют к ассоциативным об-
172 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Изменения внешней ___________ Поведение, направленное
или внутренней ------------ на уравновешивание
среды со средой
Рис. 4.9. Схема взаимодействия функциональных систем ЦНС.
Сенсорные системы осуществляют непрерывное слежение за состоянием внешней и внутрен-
ней среды и немедленно оповещают об изменениях параметров среды. Моторные системы
обеспечивают действия, направленные на уравновешивание с меняющимся состоянием среды.
Мотивационные системы получают сенсорную информацию, регистрируют прежний опыт
приспособления к условиям среды и побуждают к действиям, направленным на удовлетворе-
ние возникающих потребностей.
ластям коры, создающим замысел последующих действий. В соответствии
с этим замыслом моторные системы формируют программы движений.
Само решение о начале, как и об окончании, действий принимает мотива-
ционная система, влияющая на выходную моторную активность, а через
гипоталамус — и на состояние вегетативных функций.
Таким образом, сенсорные системы предоставляют информацию об из-
менениях внешней или внутренней среды, моторные системы обеспечива-
ют ответные действия, направленные на приспособление к изменившимся
условиям. Мотивационные системы активируются при возникновении ка-
ких-либо потребностей и побуждают к совершению действий, направлен-
ных на удовлетворение потребности. Функциональные системы постоянно
взаимодействуют друг с другом: сенсорные системы активируют мотиваци-
онные, а они побуждают моторные к созданию оптимальных для сложив-
шейся ситуации двигательных программ (рис. 4.9).
4.2.1. Общие принципы организации функциональных систем
мозга
В любой из функциональных систем соблюдаются единые принципы ее
организации.
4.2.1.1. Существование нескольких уровней переработки информации
Передача сигналов от одних нейронов к другим происходит в переключа-
тельных ядрах, находящихся в спинном и головном мозге. Каждое пере-
ключательное ядро представляет собой, во-первых, распределительную
4. Функции центральной нервной системы • 173
систему, образующую проекции на другие структуры мозга для передачи
им информации. Во-вторых, в любом переключательном ядре происходит
переработка информации, заключающаяся в изменениях параметров элек-
трических сигналов. Переключательные ядра образованы разными типами
нервных клеток, которые, в зависимости от выполняемой функции, под-
разделяются на два класса: проекционные и локальные интернейроны.
Проекционные нейроны имеют достаточно длинные аксоны, с помо-
щью которых переносят электрические сигналы от одного переключатель-
ного ядра к другому, передавая информацию на следующий уровень ее пе-
реработки. Короткие аксоны возбуждающих и тормозных локальных ин-
тернейронов не выходят за пределы переключательного ядра. Функция ло-
кальных интернейронов состоит в облегчении передачи одних сигналов и
подавлении других, что способствует передаче на следующий уровень наи-
более значимых сигналов и подавлению «информационного шума».
4.2.1.2. Топографическая упорядоченность проводящих путей
В переключательных ядрах и в проводящих путях сенсорных систем элек-
трические сигналы распространяются упорядоченно от каждого рецептив-
ного поля, например в сетчатке глаза или коже, к соответствующей именно
ему проекционной области коры. Благодаря этому проекционная кора пред-
ставляет собой своего рода нейронную карту, отражающую пространствен-
ные характеристики действующих стимулов. Нейроны моторной коры тоже
расположены топографически упорядоченно и могут влиять только на ие-
рархически подчиненные им нейроны головного и спинного мозга для пе-
редачи команд строго определенным мышцам.
4.2.1.3. Наличие параллельных проводящих путей
В каждой функциональной системе мозга существуют независимые парал-
лельные пути переработки специализированной информации. Например, в
зрительной сенсорной системе независимо и параллельно перерабатывает-
ся информация о форме, цвете и движении каждого объекта, оказавшегося
в зрительном поле. В двигательных системах существуют раздельные спе-
циализированные пути, по которым информация поступает от высших
двигательных центров к мотонейронам спинного мозга: одни пути служат
для контроля положения тела, рефлексов спинного мозга, другие — для
управления точными движениями кистей рук и пальцев.
4.2.2. Типы нейронных сетей
Специфические функциональные соединения нервных клеток друг с дру-
гом образуют нейронные сети, в зависимости от характера межнейронных
соединений различают три типа сетей: иерархические, локальные и дивер-
гентные.
Иерархический тип нейронной сети характерен для сенсорных и мотор-
ных систем, он включает несколько уровней переработки информации,
представленных переключательными ядрами. В сенсорных системах ин-
формация от первичных сенсорных нейронов передается вторичным сен-
сорным нейронам, от них — нейронам третьего порядка и далее до проек-
ционной зоны коры. В моторных системах существует нисходящая иерар-
хия: созданные в двигательных областях коры команды поступают к пере-
ключательным ядрам ствола, оттуда — в спинной мозг, а из него — к мыш-
174 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
цам. Передача сигналов в синапсах иерархических сетей происходит с по-
мощью возбуждающих нейромедиаторов. На каждом уровне иерархической
организации имеются локальные сети, образованные возбуждающими и
тормозными интернейронами. Функция нейронов локальной сети состоит
в сужении или расширении передаваемого потока информации и ее кон-
трастировании путем усиления информационно значимых сигналов и по-
давления шума.
Дивергентные сети образованы небольшими компактными группами
нейронов, тела которых находятся в нескольких ядрах ствола, а многочис-
ленные ветви их длинных аксонов распространяются диффузно ко многим
структурам мозга, включая кору. Области проекции таких нейронов пред-
назначены для выполнения разных функций, поэтому дивергентные сети
принято связывать с неспецифической деятельностью мозга. Наиболее
изученные дивергентные сети в головном мозге образованы норадренерги-
ческими, серотонинергическими, дофаминергическими и холинергически-
ми нейронами моста, продолговатого и среднего мозга. Действие медиато-
ров дивергентных систем модулирует передачу сигналов, т. е. облегчает
или затрудняет ее, что изменяет характер интегративной деятельности моз-
га (чередование сна и бодрствования, формирование внимания, возникно-
вение мотиваций и эмоций).
4.2.3. Нейрохимические классы нейронов
Разные популяции нейронов синтезируют различные нейромедиаторы:
синтез конкретного медиатора обусловлен имеющимся в клетке набором
ферментов, который, в свою очередь, определяется экспрессией тех или
иных генов. Многие нейроны используют комбинацию из одного низкомо-
лекулярного и одного или нескольких пептидных нейромедиаторов, способ-
ных усилить или ослабить действие низкомолекулярного медиатора. В пер-
вом случае они действуют на общую постсинаптическую мишень как си-
нергисты (например, ацетилхолин и вазоинтестинальный пептид), во вто-
ром как — антагонисты (глутамат и дайнорфин в гиппокампе, где глутамат
возбуждает, а опиоидный пептид дайнорфин уменьшает активность пост-
синаптической клетки).
Нейроны, синтезирующие определенный медиатор, образуют популя-
ции, распределение которых в разных регионах мозга строго упорядочено.
Эта упорядоченность позволяет выделить глутаматергическую, холинергиче-
скую, дофаминергическую, ГАМКергическую и т. п. системы нейронов.
4.2.3.1. Глутаматергическая система
Глутамат является медиатором большинства сенсорных нейронов, пира-
мидных клеток коры, многих нейронов гиппокампа, мозжечка, ствола и
спинного мозга. Это самый распространенный в головном мозге возбуж-
дающий медиатор, для которого существуют ионотропные и метаботропные
рецепторы. Ионотропные рецепторы подразделяют в зависимости от чувст-
вительности к NMDA (N-метил-D-аспартату), АМРА (аминометилпропио-
нату) и каиновой кислоте (рис. 4.10). Метаботропные глутаматные рецеп-
торы после присоединения к ним медиатора изменяют чувствительность к
глутамату у ионотропных рецепторов, расположенных на общей постси-
наптической мембране.
Самые быстрые возбуждающие процессы в глутаматергических синап-
сах происходят в результате активации АМРА-рецепторов. Они содержат
4. Функции центральной нервной системы • 175
бифосфат
Рис. 4.10. Схема постсинаптических рецепторов глутаматергической системы
А. Ионотропные АМРА-рецепторы: присоединение глутамата (Глу) приводит к открытию ка-
нала для ионов натрия; рецепторы чувствительны к бензодиазепинам (2,3-БДЗ).
Б. Ионотропные NMDA-рецепторы представляют собой белковую молекулу, содержащую ка-
нал для ионов кальция, который при значении мембранного потенциала покоя блокирован
ионами магния. Для открывания канала с последующим током через него ионов кальция не-
обходимо присоединение глутамата к рецептору и деполяризация мембраны, после которой
ионы магния освобождают канал. Рецепторы чувствительны к концентрации глицина (Гли) в
окружающей среде.
В. Ионотропные рецепторы для глутамата, одновременно чувствительные к каиновой кислоте
(КА); содержат канал для ионов натрия и калия.
Г. Метаботропные рецепторы для глутамата: после присоединения нейромедиатора происхо-
дит активация вторичных посредников. Рецепторы чувствительны к квискулату (КВИС).
катионные каналы для ионов калия и натрия и используются для получе-
ния сигналов от афферентных нейронов. Рецепторы NMDA-типа имеют
катионные каналы, пригодные для тока не только ионов калия и натрия,
но и кальция, однако ток ионов возможен только при условии предвари-
тельной деполяризации мембраны, поскольку при значении мембранного
потенциала покоя каналы блокированы ионами магния. При достаточно
длительной деполяризации постсинаптической мембраны, обусловленной
активацией нечувствительных к NMDA рецепторов, ионы магния уступа-
ют вход в канал ионам кальция, которые поступают в постсинаптический
нейрон.
При низкой активности пресинаптического нейрона и соответственно
малом выделении глутамата постсинаптический потенциал возникает пре-
имущественно за счет активации нечувствительных к NMDA ионотропных
рецепторов, например АМРА. Если же ритмическая активность пресинап-
тического нейрона будет продолжаться, то произойдет ток ионов кальция
через каналы NMDA рецепторов. Повышение концентрации ионов каль-
ция в постсинаптическом нейроне вызовет активацию нескольких систем
вторичных посредников: протеинкиназы С, кальций-кальмодулин-зависи-
мой протеинкиназы и тирозинкиназы. Активация вторичных посредников
приведет к длительному возбуждению постсинаптического нейрона, сохра-
няющемуся часами или сутками (долговременная потенциация).
Вторичные посредники обеспечивают открытое состояние каналов
NMDA рецепторов, и, кроме того, в ходе происходящих в постсинаптиче-
ском нейроне процессов в нем образуются и выделяются оксиды азота и
углерода. Эти низкомолекулярные вещества диффундируют к пресинапти-
176 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
вескому нейрону и вызывают в нем активацию посредников, способствую-
щих длительному выделению нейромедиатора. Длительная, а тем более по-
вторяющаяся долговременная потенциация в одних и тех же синапсах при-
водит к экспрессии генов и синтезу белков, необходимых для структурной
перестройки синапсов. Перестройка заключается в увеличении количества
дендритных шипиков постсинаптического нейрона и вступающих с ними в
контакт терминальных окончаний пресинаптического нейрона. Эти изме-
нения происходят в синапсах, которые активируются при часто повторяю-
щемся действии одних и тех же внешних раздражителей, а также при сов-
падении действующих стимулов друг с другом во времени. Возникшие в
синапсах преобразования являются материальной основой энграмм (сле-
дов) долговременной памяти.
4.2.3.2. Холинергическая система
Ацетилхолин в качестве нейромедиатора используется в ядрах черепно-моз-
говых нервов, мотонейронах спинного мозга и нейронах вегетативной
нервной системы. В головном мозге холинергические нейроны представлены
компактными популяциями в базальных ядрах, перегородке и ядрах диаго-
нального пучка Брока. Их аксоны образуют проекции к структурам перед-
него мозга, гиппокампу и коре, на которые ацетилхолин действует через
М-холинорецепторы. Ацетилхолин используется в качестве нейромодулято-
ра, повышающего эффективность переработки сенсорной информации и
ассоциативных процессов в коре. Большое число холинергических нейро-
нов имеется в полосатом теле, где они выполняют функцию локальных
интернейронов.
Помимо переднего мозга холинергические нейроны имеются в рост-
ральном отделе ствола: они связывают средний мозг с таламусом и корой
больших полушарий. Активация этих нейронов у спящих ведет к пробуж-
дению и одновременно обусловливает изменения ритма электроэнцефало-
граммы, который становится характерным для состояния бодрствования.
В коре преобладают холинорецепторы мускаринового типа, а в некоторых
подкорковых структурах (голубое пятно, супраоптическое ядро гипотала-
муса, отдельные ядра таламуса) существуют Н-холинорецепторы, с участием
которых осуществляются быстрые синаптические процессы. Таким обра-
зом, ацетилхолин в головном мозге может действовать через мускаринчув-
ствительные рецепторы в качестве нейромодулятора и выполняет функцию
классического нейромедиатора при действии на никотинчувствительные
рецепторы.
4.2.3.3. Системы нейронов, использующих биогенные амины
К биогенным аминам относят катехоламины {дофамин, норадреналин, адрена-
лин) и серотонин.
Синтезирующие дофамин нейроны находятся, в основном, в среднем и
промежуточном мозге, образуя нигростриатную, мезолимбическую (мезокор-
тикальную) и гипоталамо-гипофизарную дофаминэргические системы. Ниг-
ростриатная система представлена дофаминергическими нейронами ком-
пактной части черной субстанции среднего мозга, их аксоны достигают по-
лосатого тела; они необходимы для осуществления произвольных движений
человека. Мезолимбическую (мезокортикальную) систему представляют ло-
кализованные вне черной субстанции нейроны среднего мозга, аксоны ко-
торых оканчиваются в лимбических структурах мозга (миндалины, перего-
4. Функции центральной нервной системы • 177
Ионные каналы
Вторичные посредники
Рис. 4.11. Схема дофаминергического синапса.
На постсинаптической мембране находятся рецепторы Д, и Д5, связанные с G-белками, кото-
рые используются для активации или инактивации (+ или —) вторичных посредников.
На пресинаптической мембране расположены рецепторы Д2 и Д3, используемые в качестве ау-
торецепторов для регуляции количества медиатора, освобождающегося из пресинаптического
окончания.
родка, обонятельный бугорок), а также в лобных долях, поясных извилинах
и энторинальной области коры. Эта система участвует в формировании мо-
тиваций и эмоций, а ее чрезмерная активность рассматривается как один из
главных механизмов нарушения деятельности мозга при шизофрении. До-
фаминергические нейроны промежуточного мозга проецируют свои аксоны
на ядра гипоталамуса, влияя на синтез некоторых либеринов.
Метаботропные рецепторы для дофамина образуют два рецепторных се-
мейства: к первому относят рецепторы подтипов Д, и Д5, а ко второму —
Д2, Д3, и Д4 (рис. 4.11). Рецепторы Д, и Д5 связаны с Ов-белком, активи-
рующим цАМФ-зависимое фосфорилирование протеинкиназ А и С, они
сосредоточены преимущественно в полосатом теле, прилежащем ядре,
обонятельном бугорке, также в лобной и височной областях коры, гиппо-
кампе и некоторых ядрах таламуса. Рецепторы второго семейства связаны
с Gj-белком, их характерная особенность состоит в угнетении активности
аденилатциклазы вслед за присоединением медиатора. Высокая плотность
рецепторов Д2 и Д3 характерна для полосатого тела, черной субстанции,
лимбических структур, вентральной покрышки и гипофиза, где они ис-
пользуются в качестве ауторецепторов, т. е. участвуют в пресинаптической
регуляции высвобождения нейромедиатора. Действие многих психотроп-
ных препаратов (нейролептиков, психостимуляторов, антидепрессантов)
основано на модуляции дофаминергических синапсов.
Самое большое скопление норадренергических нейронов представлено
в голубых пятнах (locus coeruleus). Образующие многочисленные, широко
ветвящиеся коллатерали аксоны этих нейронов оканчиваются в спинном
мозге и стволе, мозжечке, таламусе, миндалинах, гиппокампе и образуют
диффузную дивергентную проекцию к коре больших полушарий. Норадре-
нергические нейроны голубого пятна совместно с дофаминергическими
178 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
нейронами среднего мозга своими аксонами образуют медиальный пучок
переднего мозга, с возбуждением которого связано возникновение эмоций.
Норадренергические нейроны голубого пятна активируются при стрессе,
причем их модулирующее действие направлено не на стимуляцию, а на
уменьшение активности иннервируемых ими областей мозга.
Серотонин (5-гидрокситриптамин) используется в качестве медиатора
нейронами ядер срединного шва продолговатого мозга, а также отдельны-
ми группами нейронов среднего мозга и гипоталамуса. Дивергентные про-
екции серотонинергических нейронов достигают гипоталамуса, таламуса,
гиппокампа, миндалин, полосатого тела и коры больших полушарий. Серо-
тонинергические нейроны регулируют восходящую активность ретикуляр-
ной формации, связанную с управлением цикла сон—бодрствование, их
влияние на переключательные ядра таламуса связано с распределением
внимания и фильтрацией сенсорных сигналов. Проекция серотонинергиче-
ских нейронов на структуры лимбической системы, полосатое тело и лоб-
ные области коры определяет эмоциональное состояние человека: сниженное
содержание серотонина ведет к развитию депрессии, страха и проявлению
немотивированной агрессивности, в особенности если сниженный уровень
серотонина сочетается с повышенной активностью катехоламинэргических
нейронов. При низкой активности серотонинергической системы увеличи-
вается риск возникновения алкоголизма. Коррекция угнетенного психиче-
ского состояния с помощью некоторых антидепрессантов основана на том,
что они повышают уровень серотонина в синаптической щели путем бло-
кады его обратного захвата пресинаптическими окончаниями.
Количество серотонинергических нейронов в мозге мало, в то же время
серотонинергическая система отличается разнообразием рецепторов, чув-
ствительных к серотонину: в настоящее время в ЦНС известно 16 типов
таких рецепторов, различающихся своим строением и свойствами. Серото-
нин препятствует проявлениям агрессивного поведения, оказывая тормоз-
но-модулирующее действие на рецепторы типа 1А и 1В нейронов лобной
коры, полосатого тела и лимбических структур. Нисходящая проекция се-
ротонинергических нейронов ядер шва на интернейроны задних рогов
спинного мозга служит для торможения передачи сигналов от болевых ре-
цепторов и является частью антиноцицептивной системы, понижающей по-
рог болевого восприятия.
Серотонинергические нейроны участвуют в регуляции вегетативных и
эндокринных процессов посредством модуляции активности соответствую-
щих центров гипоталамуса. Они регулируют нейронные переключения в
холинергических и норадренергических структурах ствола мозга, где дейст-
вие серотонина может быть опосредовано как метаботропными, так и ио-
нотропными рецепторами, управляющими постсинаптическими мембран-
ными каналами для ионов калия (постсинаптическое торможение). Дейст-
вие серотонина не обязательно является тормозным и при активации не-
которых разновидностей серотонинергических рецепторов происходит воз-
буждение постсинаптических структур: подобное происходит, например, в
гиппокампе и способствует регистрации новой информации и извлечению
следов памяти.
4.2.3.4. ГАМКергическая система
ГАМК (гамма-аминомасляная кислота) является важнейшим и самым рас-
пространенным тормозным медиатором нейронов коры, базальных гангли-
ев, мозжечка, гиппокампа, некоторых интернейронов спинного мозга. Су-
4. Функции центральной нервной системы • 179
Рис. 4.12. Схема ионотропного рецептора ГАМКа.
Рецептор образован пятью субъединицами, способными связывать ГАМК (все субъединицы),
а также бензодиазепины и барбитураты (альфа-субъединицы), которые усиливают связь ре-
цепторов с ГАМК. В результате связывания ГАМК с рецептором открывается ионный канал
для хлора, после вхождения ионов хлора в клетку возникает ее гиперполяризация.
шествует два типа ГАМКергических рецепторов: ионотропные ГАМКа (от-
крывают каналы для ионов хлора) и метаботропные ГАМКВ Метаботроп-
ные ГАМКВ рецепторы ассоциированы с G-белком, после присоединения
медиатора они активируют систему вторичных посредников, которые от-
крывают каналы для ионов калия. Эти рецепторы находятся на пресинап-
тических окончаниях неГАМКергических нейронов и служат для регуля-
ции количества выделяющихся из таких окончаний возбуждающих нейро-
медиаторов, например глутамата или катехоламинов.
У постсинаптических рецепторов ГАМК имеются отдельные центры
связывания (рис. 4.12) ряда фармакологических веществ (бензодиазепинов
и барбитуратов), являющихся агонистами ГАМК и усиливающих ее дейст-
вие. Такие вещества тормозят процесс возбуждения, сопровождающегося
реакцией страха и повышением тонуса мышц, они обладают снотворным и
успокаивающим действием. В окончаниях многих ГАМКергических ней-
ронов помимо основного медиатора происходит одновременное выделение
нейропептидов, обладающих нейромодулирующим действием. Некоторые
эндогенные белки и пептиды (кальретинин, сериновая протеаза-нексин),
модифицируя чувствительность тормозных ГАМК-рецепторов, способству-
ют улучшению процессов научения и памяти.
4.2.3.5. Пептидергические нейроны
Около 50 пептидов синтезируются в различных нервных клетках и затем
используются в качестве нейромедиаторов. Они образуются только в кле-
точном теле в ходе ферментативного преобразования крупных молекул
белков-предшественников в эндоплазматическом ретикулуме и аппарате
Голвджи, а затем в секреторных пузырьках доставляются в окончание ак-
сона быстрым аксонным транспортом. Разные пептиды служат в качестве
возбуждающих или тормозных медиаторов в соответствующих синапсах,
180 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
они также могут использоваться в качестве нейромодуляторов, усиливаю-
щих или ослабляющих действие низкомолекулярного медиатора. Некото-
рые нейроны способны одновременно синтезировать низкомолекулярный
медиатор и пептидный нейромодулятор для него. Нейропептиды могут
секретироваться в кровяное русло, и в таких случаях рассматриваются в
качестве нейрогормонов: классическим примером такой ситуации можно,
например, считать поступление в кровоток вазопрессина и окситоцина,
синтезируемых нейронами паравентрикулярных и супраоптических ядер
гипоталамуса.
По одинаковой последовательности аминокислот в достаточно длинных
участках молекул нейроактивных пептидов они разделены на несколько
семейств, среди которых наиболее изучены опиатные пептиды. Они состо-
ят из разного количества аминокислот, от 5 до 31, но обязательно включа-
ют участок с последовательностью аминокислот: Тир-Гли-Гли-Фен. К се-
мейству опиатных пептидов принадлежат лейцин-энкефалин, метионин-эн-
кефалин, альфа-эндорфин, гамма-эндорфин, бета-эндорфин, дайнорфин, аль-
фа-неоэндорфин. Указанные вещества взаимодействуют с опиатными ре-
цепторами, которые представлены не менее чем пятью разновидностями,
различающимися своими функциональными свойствами. Опиатные пепти-
ды модулируют переключения в синапсах, использующих в качестве ме-
диаторов дофамин, ГАМК, глутамат и ацетилхолин. Опиатные рецепторы
активируют систему вторичных посредников цАМФ-аденилатциклазы, эти
рецепторы кроме эндогенных пептидов могут связывать вводимые в орга-
низм для подавления боли производные морфина, а также налоксон — ис-
кусственно синтезированный антагонист морфина, применяемый для
борьбы с наркозависимостью.
Воздействие опиатных пептидов на нейронные переключения в гипота-
ламусе, гипофизе и лимбических структурах регулирует характер мотива-
ций и эмоций, пищевое и сексуальное поведение, ответные реакции орга-
низма на действие стрессоров. При повышенном образовании энкефалина
в полосатом теле и прилегающем ядре у человека возникает эйфория.
Опиаты участвуют в регуляции локомоторной активности и осуществлении
стереотипных поведенческих реакций. При участии опиоидных пептидов
изменяется порог болевой чувствительности, они существенно притупляют
ощущение боли, действуя на нейронные переключения в спинном мозге,
стволе, лимбических структурах, таламусе и соматосенсорной коре. Опио-
идные пептиды препятствуют выделению ГАМК из интернейронов гиппо-
кампа, что облегчает возникновение долговременной потенциации в этом
регионе и последующее образование следов долговременной памяти об ин-
дивидуальном поведенческом опыте.
Опиатные пептиды могут секретироваться в цереброспинальную жид-
кость, например, их концентрация в ликворе повышается при обезболива-
нии с помощью акупунктуры и при эффекте плацебо (положительный
клинический эффект от приема полученного под видом лекарства, но не
содержащего активного начала вещества). Образование бета-эндорфина
возрастает вследствие занятий марафонским бегом или аналогичными ви-
дами интенсивной физической деятельности, оно повышается во время бе-
ременности и достигает максимума за 3—4 нед перед родами, что необхо-
димо не только для уменьшения болевой чувствительности, но и для фор-
мирования так называемого материнского поведения.
Остальные семейства нейроактивных пептидов представляют тахикини-
ны (вещество Р, используемое в качестве медиатора при проведении боле-
вой чувствительности, а также бомбезин, физалемин, кассинин, упероле-
4. Функции центральной нервной системы • 181
ин, эледоизин, вещество К), пептиды нейрогипофиза (вазопрессин, оксито-
цин, нейрофизин), секретины (секретин, глюкагон, вазоактивный интести-
нальный пептид, соматолиберин), инсулины (инсулин, инсулиноподобные
ростковые факторы 1 и 2), семейство панкреатического полипептида (ней-
ропептид РР, нейропептид Y, пептид YY, участвующие в формировании
пищевого поведения, смене биологических ритмов и возникновении эмо-
ций), соматостатины (соматостатин, полипептид поджелудочной железы)
и гастрины (гастрин, холецистокинин).
4.3. Функции спинного мозга
Функции спинного мозга состоят в осуществлении двигательных и вегета-
тивных рефлексов, в проведении сенсорных сигналов от рецепторов кожи,
мышц и внутренних органов к головному мозгу, и в передаче информации
от головного мозга, служащей для координации деятельности мышц и
внутренних органов. Спинной мозг разделен на ряд участков — сегментов,
каждый сегмент связан только со своей областью иннервации (рис. 4.13).
В каждом сегменте спинного мозга центрально расположено серое ве-
щество, образованное нейронами, их дендритами и клетками нейроглии.
Две симметричные половины серого вещества соединены перешейком, в
центре которого находится спинномозговой канал. На периферии располо-
жено белое вещество, образованное преимущественно миелинизированны-
ми волокнами нисходящих и восходящих путей, обеспечивающих взаимо-
действие спинного и головного мозга. От каждого сегмента отходят два
дорсальных корешка, образованных центральными отростками первичных
афферентных нейронов спинальных ганглиев, и два вентральных корешка,
состоящих из волокон мотонейронов и преганглионарных вегетативных
нейронов. Таким образом, дорсальные или задние корешки состоят из чув-
ствительных афферентных волокон, а вентральные или передние — из эффе-
рентных', их функциональное взаимодействие обеспечивают интернейроны
серого вещества (рис. 4.14).
4.3.1. Функциональная организация спинного мозга
Компактные скопления нейронов серого вещества образуют ядра, как пра-
вило, вытянутые вдоль спинного мозга на протяжении нескольких сегмен-
тов и образующие столбы или колонны. Поэтому афферентные и эффе-
рентные волокна, связанные с одним ядром, входят и выходят в составе
нескольких корешков, принадлежащих соседним сегментам.
В верхушках дорсальных рогов сосредоточены клетки с короткими, густо
ветвящимися и переплетающимися отростками: они образуют студневидное
вещество, которое тянется непрерывно вдоль спинного мозга. Интерней-
роны студневидного вещества получают афференты первичных сенсорных
нейронов, проводящих сигналы от рецепторов кожи, а также от некоторых
висцерорецепторов и проприоцепторов. Переработку сенсорных сигналов
регулируют нисходящие к интернейронам из ствола волокна моноаминер-
гических нейронов голубого пятна, ядер шва и моста. Они образуют си-
напсы с интернейронами, а около 60 % окончаний норадренергических и
серотонинэргических нейронов имеют варикозные расширения, поэтому
выделяющийся из них медиатор распространяется диффузно на значитель-
ном протяжении и модулирует передаваемые от сенсорных нейронов сиг-
налы.
В основании заднего рога находятся переключательные релейные ней-
182 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Боковая проекция
спинного мозга:
С—шейный отдел
Т—грудной
L—поясничный
S—крестцовый
Рис. 4.13. Схема топографически организованной иннервации поверхности тела
(дерматомы).
На схеме показаны области иннервации кожи, соответствующие определенным сегментам
спинного мозга (С — шейные, Т — грудные, L — поясничные, S — крестцовые). Кожу лица
иннервирует тройничный нерв (на схеме не показан).
роны, на которых образуют синапсы аксоны сенсорных нейронов, пере-
дающих болевую и температурную чувствительность. Аксоны релейных
нейронов переходят на противоположную сторону, где образуют восходя-
щие спиноталамический и спиноретикулярный тракты, по которым аффе-
рентная информация поступает в головной мозг. Другая часть афферент-
ных окончаний сенсорных нейронов образует синапсы с интернейронами
студневидного вещества, причем во всех сенсорных волокнах в качестве
медиаторов используются глутамат и вещество Р. Интернейроны студне-
видного вещества способны тормозить передачу болевых импульсов с по-
мощью ГАМК и энкефалина, а их тормозная активность регулируется нис-
ходящими моноаминоергическими волокнами ствола.
Переработка сенсорной чувствительности разных модальностей проис-
ходит параллельно: в проведении тактильной, болевой и температурной
4. Функции центральной нервной системы • 183
Мотонейрон
Рис. 4.14. Поперечный разрез спинного мозга (схема).
Рефлекторную функцию спинного мозга обеспечивают сенсорные нейроны, интернейроны
серого вещества, образующие ядра спинного мозга, и мотонейроны. Стрелками показано на-
правление распространения возбуждения при осуществлении рефлекторной деятельности.
чувствительности участвуют нейроны студневидного вещества и собствен-
ного ядра заднего рога. Переработку висцеральной чувствительности осу-
ществляют нейроны промежуточной зоны, а для передачи проприоцептив-
ной чувствительности используется ядро Кларка, нейроны которого обра-
зуют спиномозжечковый путь, а также тонкое и клиновидное ядра, находя-
щиеся в продолговатом мозге.
В вентральных рогах спинного мозга расположены мотонейроны, тела
которых образуют моторные ядра, вытянутые вдоль спинного мозга в виде
колонн. Медиально сосредоточены мотонейроны, управляющие мышцами
туловища, необходимыми для сохранения равновесия и поддержания нуж-
ного положения тела. Активность этой группы мотонейронов определяет
нисходящий из головного мозга медиальный путь, предназначенный для
контроля за положением тела. В латеральной области передних рогов со-
средоточены мотонейроны, управляющие мышцами дистальных отделов
конечностей и контролирующие движения кистей и пальцев рук и ног. Их
активность регулируется посредством нисходящего из головного мозга ла-
терального пути. Топографическая упорядоченность распределения мото-
нейронов проявляется еще и в том, что мотонейроны, служащие для ин-
нервации мышц-разгибателей, расположены вентрально по отношению к
мотонейронам, контролирующим мышцы-сгибатели.
Все мотонейроны подразделяются на два типа: альфа-мотонейроны, ин-
нервирующие основную массу мышечных волокон, и гамма-мотонейроны,
служащие для иннервации особой разновидности мышечных волокон, на-
ходящихся внутри мышечных веретен и необходимых для изменения их
чувствительности к растяжению мышцы. У альфа-мотонейронов диаметр
тела достигает 80 мкм, от него отходят 10—15 первичных дендритов, кото-
рые благодаря многократному ветвлению заполняют окружающее мото-
нейрон пространство радиусом 2—3 мм. На всем этом протяжении мото-
нейрон формирует несколько тысяч синапсов с нервными окончаниями:
184 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Мышца
Клетка
Реншоу
Рис. 4.15. Схема возвратно-
го торможения в спинном
мозге.
Тормозной нейрон Реншоу воз-
буждается коллатералью мото-
нейрона и после этого умень-
шает его активность.
1) нейронов головного мозга, образующих
нисходящие двигательные пути; 2) интерней-
ронов спинного мозга, расположенных в про-
межуточном ядре между задним и передним
рогом; 3) первичных сенсорных нейронов, об-
разующих моносинаптические рефлекторные
дуги с мотонейронами. Таким образом, мото-
нейрон представляет собой общий конечный
путь, используемый как при осуществлении
элементарных рефлекторных реакций, вы-
званных раздражением различных рецепторов,
так и при выполнении произвольных движе-
ний, связанных с первоначальной активно-
стью моторных областей коры.
Активность мотонейронов регулируется
при участии возбуждающих и тормозных ин-
тернейронов спинного мозга, благодаря кото-
рым координируется работа мышц, участвую-
щих в совершении того или иного движения;
при этом одни мышцы должны сокращаться, а другие — расслабляться.
Особую популяцию тормозных нейронов представляют клетки Реншоу, ко-
торые активируются возвратными коллатералями аксонов мотонейронов,
после чего тормозят деятельность самих мотонейронов: такое торможение
называется возвратным (рис. 4.15). Возвратное торможение мотонейронов
посредством клеток Реншоу стабилизирует частоту потенциалов действия,
возникающих в мотонейронах. В норме она варьирует между 8 и 15 им-
пульсами в секунду, и лишь при самых сильных сокращениях мышц дос-
тигает 25 Гц.
Осуществление сильных или слабых сокращений одних и тех же мышц
происходит вследствие активации разных мотонейронов. Каждый альфа-
мотонейрон иннервирует в мышце определенное количество мышечных
волокон, образуя вместе с ними двигательную единицу. Двигательные еди-
ницы подразделяются, во-первых, на большие и малые в зависимости от
количества мышечных волокон, получающих иннервацию от одного мото-
нейрона. Во-вторых, они различаются в зависимости от типа мышечных
волокон как: 1) быстрые утомляемые; 2) медленные мало утомляемые;
3) быстрые мало утомляемые. Мотонейроны, иннервирующие мышечные
волокна первого типа, отличаются большим размером тела и высоким по-
рогом возбуждения. Мотонейроны для волокон второго типа, напротив,
являются мелкими и наиболее возбудимыми, а мотонейроны третьего типа
занимают промежуточное положение. При относительно слабом аффе-
рентном возбуждении активируются мотонейроны второго и третьего типа,
при этом сила мышечных сокращений оказывается достаточной, напри-
мер, для стояния или спокойной ходьбы. Сильные афферентные стимулы
возбуждают мотонейроны первого типа, при этом сила мышечных сокра-
щений возрастает настолько, что позволяет бежать или прыгать.
Группы дендритов соседних мотонейронов образуют между собой элек-
трические синапсы, что способствует синхронизации электрической актив-
ности этих клеток. Гамма-мотонейроны отличаются от альфа-мотонейро-
нов не только малой величиной, но и тем, что они иннервируют только
интрафузальные мышечные волокна, связанные с чувствительными к изме-
нениям длины мышцы проприоцепторами. Вместе с интрафузальными во-
локнами гамма-мотонейроны образуют так называемую фузимоторную сис-
4. Функции центральной нервной системы • 185
тему, тогда как альфа-мотонейроны вместе с экстрафузальными волокна-
ми составляют скелетомоторную систему, первая используется лишь для
регуляции мышечного тонуса, вторая — обеспечивает сократительную
функцию мышц.
На протяжении от верхнего шейного до II—III поясничного сегментов
располагается латеральное ядро промежуточной зоны, в котором содержат-
ся преганглионарные нейроны симпатического отдела вегетативной нерв-
ной системы, объединенные друг с другом внутриспинальными проводя-
щими путями. К этому ядру поступают сигналы от окончаний сенсорных
нейронов, несущих информацию, во-первых, от висцерорецепторов внут-
ренних органов (нейромедиаторами являются глутамат и вещество Р), во-
вторых, от интернейронов медиального ядра промежуточной зоны (медиа-
торы: холецистокинин, соматостатин и ВИП) и, в третьих, нисходящие во-
локна норадренергических нейронов голубого пятна и серотонинергиче-
ских нейронов ядер шва. Симпатические нейроны, контролирующие пото-
вые железы, сосуды кожи и скелетных мышц располагаются латерально по
отношению к нейронам, регулирующим деятельность внутренних органов.
Парасимпатические преганглионарные нейроны входят в состав медиально-
го ядра промежуточной зоны на уровне II поясничного — III крестцового
сегментов. Латерально расположены нейроны, иннервирующие мочевой
пузырь, медиально — иннервирующие толстый кишечник.
4.3.2. Рефлексы спинного мозга
Рефлексы спинного мозга осуществляются благодаря упорядоченному рас-
пространению возбуждения от рецепторов сенсорного нейрона до рабочего
органа по рефлекторной дуге. Представление о собственной рефлекторной
деятельности спинного мозга сложилось на основе изучения рефлекторных
реакций у животных, наблюдающихся после экспериментального отделе-
ния спинного мозга от головного хирургическим путем или фармакологи-
ческой блокадой проводящих путей между спинным и головным мозгом.
После разрыва спинного мозга возникает спинальный шок, проявляю-
щийся обратимым угнетением двигательных и вегетативных рефлексов.
Утративший связь с головным мозгом спинной мозг определяют как изоли-
рованный, а рефлексы, которые могут осуществляться с его помощью, на-
зывают спинномозговыми или спинальными. Изоляция спинного мозга или
его части может произойти в результате травматического разрыва у челове-
ка, и тогда сохраняющаяся рефлекторная деятельность сводится исключи-
тельно к спинальным рефлексам. Спинномозговые рефлексы являются
врожденными, их осуществление не требует осознания, но в естественных
условиях, т. е. при сохраненных связях спинного мозга с головным, спи-
нальные рефлексы включаются в более сложные программы поведения.
Нисходящие влияния головного мозга могут изменять или даже прекра-
щать те или иные виды рефлексов, дуга которых замыкается через спин-
ной мозг, например, болевой сгибательный рефлекс можно сознательно
подавить усилием воли.
4.3.2.1. Сухожильные рефлексы
Сухожильные рефлексы вызываются легким ударом по сухожилию и прояв-
ляются резким сокращением мышцы, прикрепленной к этому сухожилию.
Сенсорным стимулом для возникновения сухожильных рефлексов служит
растяжение мышцы в момент удара по ее сухожилию (рис. 4.16), при этом
186 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Растяжение
четырехглавой
мышцы бедра
III — IV сегменты поясничного
отдела спинного мозга
Афферентный
нейрон
Берцовые
кости
Рефлекторное сокращение
четырехгпавой мышцы бедра,
разгибание ноги в коленном суставе
Бедренная
кость
Рис. 4.16. Схема воспроизведения коленного рефлекса.
Удар молоточком по сухожилию четырехглавой мышцы бедра вызывает ее растяжение и ответ-
ное рефлекторное сокращение, приводящее к разгибанию ноги в коленном суставе. Моноси-
наптическая рефлекторная дуга замыкается на уровне III—IV сегментов поясничного отдела
спинного мозга.
Надколенник
Удар
молоточком
по сухо-
жилию
Эфферентный
нейрон
мышца не должна напрягаться, чтобы не препятствовать растяжению. Су-
хожильные рефлексы можно вызвать в разных мышцах, например трехгла-
вой мышце голени (ахиллов рефлекс), двуглавой и трехглавой мышцах
плеча, мышцах нижней челюсти и т. д. Воспроизведение сухожильных
рефлексов применяется в неврологической практике для диагностики
уровня поражения двигательных функций.
При растяжении мышцы в момент удара по ее сухожилию возбуждают-
ся проприоцепторы, находящиеся в небольших мышечных волокнах, окру-
женных капсулой из соединительной ткани. Такие капсулы называют мы-
шечными веретенами, а заключенные в них мышечные волокна — интрафу-
зальными (от лат. fusus — веретено). При увеличении длины мышцы интра-
фузальные волокна растягиваются, и тогда в проприоцепторах возникает
рецепторный потенциал, а вслед за ним и потенциал действия. Возникаю-
щие потенциалы действия распространяются вначале по периферической,
а затем по центральной ветви аксона первичного сенсорного нейрона, рас-
положенного в спинальном ганглии. В составе заднего корешка афферент-
ные глутаматергические волокна этого нейрона входят в спинной мозг и
оканчиваются непосредственно на мотонейронах, образуя моносинаптиче-
скую рефлекторную дугу. В дуге коленного рефлекса каждый мотонейрон
имеет от 2 до 6 синаптических контактов с единичным сенсорным нейро-
ном, а каждый сенсорный нейрон посредством многочисленных термина-
лей своего аксона вступает в контакт с 500—1000 гомонимных (т. е. относя-
щихся к той же мышце) мотонейронов, находящихся во II—IV пояснич-
ных сегментах. Возбуждение этих мотонейронов приводит к сокращению
четырехглавой мышцы.
4.3.2.2. Рефлекс растяжения мышцы
Сухожильные рефлексы представляют собой частный случай рефлексов
растяжения, являющихся физиологическим механизмом регуляции длины
мышц. При быстром растяжении мышцы возникает фазический рефлекс в
4. Функции центральной нервной системы • 187
Мышечные
Мышца
Интрафузальные
Сухожилие
Рецептор Гольджи
Расположение
проприоцепторов
Мышечное веретено
Рис. 4.17. Расположение проприоцепторов в скелетных мышцах.
В центральной части рисунка показана локализация интрафузальных рецепторов и сухожиль-
ных рецепторов Гольджи. Справа показано мышечное веретено с несколькими интрафузаль-
ными волокнами внутри него: эфферентный аксон принадлежит у-мотонейрону спинного
мозга; чувствительные окончания афферентного нейрона возбуждаются при растяжении цен-
тральной части интрафузальных волокон. Слева изображены рецепторы Гольджи, активация
которых происходит пропорционально росту напряжения мышцы и натяжению сухожильных
коллагеновых волокон, сдавливающих чувствительные окончания.
виде быстрого ответного сокращения мышцы, а при медленном растяже-
нии — тонический рефлекс, направленный на сохранение неизменной дли-
ны мышцы при постоянном растяжении. Тонические рефлексы необходи-
мы для поддержания мышечного тонуса, под которым понимают сопротив-
ление мышц растяжению какой-либо внешней силой. Мышечный тонус
сохраняет положение тела и обеспечивает сопротивление силе тяжести,
растягивающей мышцы-разгибатели. Изменения тонуса мышц позволяют
удерживать и перемещать груз, сохранять равновесие при отклонениях ту-
ловища вперед, назад или в сторону.
Служащие в качестве рецепторов растяжения интрафузальные мышеч-
ные волокна значительно тоньше и короче остальных, т. е. экстрафузаль-
ных, образующих основную массу мышцы и обеспечивающих ее сокраще-
ния (рис. 4.17). Мышечные веретена обоими концами прикреплены полос-
кой соединительной ткани к пучку экстрафузальных волокон, поэтому
растягиваются одновременно со всей мышцей. В средней части каждого
интрафузального волокна содержатся окончания первичного афферентно-
го нейрона, относящиеся к механорецепторам, которые реагируют на вы-
званную растяжением деформацию. Возбуждение сенсорных нейронов пе-
редается мотонейронам через моносинаптическую рефлекторную дугу, что
приводит к рефлекторному сокращению растянутой мышцы.
При растяжении мышцы внешней силой происходит одновременная ак-
тивация а-мотонейронов, иннервирующих основную массу мышечных во-
локон и у-мотонейронов, которые иннервируют только интрафузальные
188 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 4.18. Схема рефлекса растяжения.
А. Мышца-сгибатель растягивается внешней силой, что вызывает активацию рецепторов ин-
трафузальных волокон и возбуждение афферентного нейрона, действующего непосредственно
на мотонейроны сгибателей (синергистов) и уменьшающего активность мотонейрона мышцы-
разгибателя (антагониста) посредством тормозного интернейрона.
Б. Рефлекторный ответ на растяжение мышцы заключается в ее сокращении, приводящем к
сгибанию конечности в суставе.
волокна. Оба типа мотонейронов возбуждаются разными коллатералями
одного и того же сенсорного нейрона, поэтому сокращения обоих типов
мышечных волокон тоже происходит одновременно. Другие ветви аффе-
рентного аксона активируют тормозные интернейроны спинного мозга,
которые реципрокно подавляют активность мотонейронов для мышц-анта-
гонистов (рис. 4.18). Таким образом, в механизме рефлекса растяжения за-
программирована одновременная согласованная деятельность нескольких
мышц, необходимая для совершения или предотвращения движения в оп-
ределенном суставе.
Чувствительность интрафузальных волокон к растяжению изменяется
при повышении или понижении активности у-мотонейронов. Чем чаще
они возбуждаются, тем больше сократятся концевые участки интрафузаль-
ных волокон, в то время как их центральная часть, лишенная сократитель-
ных белков, будет вынуждена растягиваться. Небольшое ее растяжение су-
ществует в условиях физиологического покоя, что поддерживает возбужде-
ние рецепторов мышечных веретен и тоническое сокращение мышц. Из-
менения активности у-мотонейронов, приводящие к изменениям тонуса
4. Функции центральной нервной системы • 189
мышц, происходят под влиянием двигательных центров головного мозга. В
зависимости от конкретной функциональной задачи избирательно активи-
руются у-мотонейроны, иннервирующие разные группы мышц, например
сгибатели или разгибатели. Соответственно этому тонус одних групп
мышц будет повышаться, тогда как другие группы мышц останутся в
прежнем состоянии или даже расслабятся, если произойдет торможение
активности соответствующих у-мотонейронов.
4.3.2.3. Рефлекторная регуляция напряжения мышц
Сухожильные
рецепторы
Гольджи
Тормозной
интернейрон
Рост напряжения мышцы
без изменения ее длины
Уменьшение
возбуждения мотонейрона
Сокращения мышцы сопряжены с изменениями не только ее длины, но и
напряжения. Рефлекторная регуляция напряжения осуществляется с помо-
щью сухожильных рецепторов Гольджи — немиелинизированных оконча-
ний афферентного нейрона, которые расположены между коллагеновыми
пучками сухожильных волокон в области их прикрепления к мышечным
волокнам (см. рис. 4.17). Если интрафузальные волокна — датчики длины
лежат параллельно экстрафузальным волокнам, составляющим основу
мышцы, то сухожильные рецепторы соединены с ними последовательно.
При нарастании напряжения в мышце коллагеновые волокна сухожилий
натягиваются тем сильнее, чем большим становится напряжение. Пропор-
ционально ему деформируются сухожильные рецепторы и возрастает час-
тота потенциалов действия в афферентном нейроне. Окончания афферент-
ных нейронов образуют синапсы с тормозными интернейронами, дейст-
вующими на гомонимные (относящиеся к этой же мышце) мотонейроны
(рис. 4.19). Такой механизм регуляции по принципу отрицательной обрат-
ной связи предупреждает
неадекватное нарастание
мышечного напряжения.
Тормозные интернейро-
ны, контролирующие возбу-
ждение мотонейронов, а
значит, и силу мышечных
сокращений, получают аф-
ферентную информацию не
только от сухожильных ре-
цепторов Гольджи, но также
от суставных рецепторов,
высокочувствительных кож-
ных рецепторов и нисходя-
щих двигательных путей.
Конвергенция кожных аф-
ферентов на тормозных ин-
тернейронах требуется в тех
случаях, когда происходит
ощупывание хрупких или
мелких объектов и силу мы-
шечных сокращений нужно
согласовать с характером
тактильных ощущений.
Рис. 4.19. Схема рефлекторной регуляции на-
пряжения мышцы.
Нарастание напряжения в мышце при неизменной ее
длине приводит к раздражению сухожильных рецепто-
ров Гольджи и активации афферентного нейрона, ко-
торый уменьшает возбуждение мотонейрона с помо-
щью тормозного интернейрона.
190 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
4.3.2.4. Сгибательные и разгибательные рефлексы
Сгибательный рефлекс состоит в немедленном сгибании руки или ноги в
ответ на болевое раздражение кожи (укол, ожог). Он обеспечивает удале-
ние конечности от источника болевой стимуляции и по своему биологиче-
скому значению относится к категории защитных рефлексов. Сгибание ко-
нечности происходит одновременно в нескольких суставах, причем выра-
женность сгибательного рефлекса пропорциональна силе раздражителя.
Чем сильнее оказывается действующий стимул, тем больше мышц вовле-
кается в ответную реакцию.
Рефлекторные дуги всех сгибательных рефлексов являются полисинап-
тическишс. разветвленные окончания сенсорных нейронов проецируются
на возбуждающие и тормозные интернейроны, которые действуют на мо-
тонейроны для мышц сгибателей и разгибателей (рис. 4.20). Мотонейроны
мышц-сгибателей подвергаются действию возбуждающих интернейронов,
что приводит к сокращению этих мышц. Одновременно тормозные интер-
нейроны реципрокно угнетают активность мотонейронов, контролирую-
щих мышцы-разгибатели.
Рис. 4.20. Схема возникновения сгиба-
тельного рефлекса.
При действии болевого раздражителя, вы-
звавшего рефлекторное сгибание ноги, на
другой ноге повышается тонус мышц-разги-
бателей (+) и уменьшается тонус мышц-сги-
бателей (—), что позволяет опорной ноге
удерживать массу тела. Тормозные интерней-
роны на схеме темные, возбуждающие —
светлые.
Многие суставы способны про-
изводить движения в двух или даже
трех плоскостях, а для этого требу-
ются согласованные сокращения
одних групп мышц при одновре-
менном расслаблении других. Ме-
ханизм сгибательного рефлекса
включает взаимодействие возбуж-
дающих и тормозных интернейро-
нов спинного мозга с мотонейро-
нами мышц-синергистов и антаго-
нистов. Такое взаимодействие пре-
дусматривает решение трех задач:
1) непосредственный контроль ка-
ждой мышцы; 2) координация дея-
тельности мышц, управляющих од-
ним суставом; 3) координация дея-
тельности мышц, необходимых для
управления несколькими суста-
вами.
Рефлекторное сгибание конеч-
ности часто сопровождается одно-
временным разгибанием контрала-
теральной конечности, которое то-
же происходит рефлекторно {пере-
крестный разгибательный рефлекс).
Так, например, человек, наступив-
ший босой ногой на колючку, реф-
лекторно сгибает эту ногу, что
приводит к росту нагрузки на
опорную ногу. Немедленное реф-
лекторное повышение тонуса
мышц-разгибателей на опорной
ноге позволяет справиться с воз-
росшей нагрузкой на нее. Осуще-
4. Функции центральной нервной системы • 191
От нейронов моторной коры
(кортикоспинальный путь),
от двигательных ядер ствола
От сухожильных
рецепторов
Гольджи
От интрафузальных
рецепторов
мышцы-антагониста
От сенсорного нейрона,
рецепторы которого
находятся в интрафуза-
льных мышечных
волокнах
Клетка Реншоу
(возвратное
торможение)
К скелетной
мышце
Возбуждающие
нейроны
1 Тормозные
J нейроны
Рис. 4.21. Конвергенция возбуждающих и тормозных входов на мотонейроне.
Мотонейрон могут возбуждать сенсорные нейроны, реагирующие на растяжение мышцы, воз-
буждающие интернейроны и нисходящие моторные пути головного мозга. На него могут дей-
ствовать тормозные интернейроны, активированные рядом афферентных нейронов, а также
клетки Реншоу. Схема показывает лишь малую часть реальных влияний на мотонейрон.
ствление перекрестного разгибательного рефлекса обеспечивается контра-
латеральной проекцией аксонов, берущих начало от интернейронов, кото-
рые возбуждаются при поступлении в спинной мозг сигналов от болевых
рецепторов. В результате на противоположной стороне спинного мозга
возбуждаются мотонейроны мышц-разгибателей и тормозятся мотонейро-
ны сгибателей, т. е. наблюдается картина, прямо противоположная проис-
ходящему при сгибательном рефлексе.
Интернейроны, участвующие в образовании рефлекторных дуг сгиба-
тельных и разгибательных рефлексов, активируются окончаниями сенсор-
ных нейронов разных модальностей. К ним конвергируют афференты от
поверхности кожи, мышц, суставов, кроме того, они находятся под контро-
лем нисходящих двигательных путей головного мозга. В связи с конверген-
цией многих афферентов интернейроны спинного мозга, как и зависимые
от них мотонейроны, участвуют не только в замыкании сгибательных и раз-
гибательных рефлексов, но вовлекаются в осуществление двигательных
рефлексов ствола и выполнение произвольных движений (рис. 4.21).
192 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
4.3.2.5. Ритмические рефлексы
Ритмический чесательный рефлекс вызывается слабым раздражением кожи
и включает два компонента: 1) обнаружение раздраженного участка (пол-
зающие насекомые, зуд) и 2) ритмичные чесательные движения, заклю-
чающиеся в повторных сгибаниях-разгибаниях конечности, которая зани-
мает при этом определенное положение по отношению к кожной поверх-
ности за счет повышения тонуса приводящих мышц. Комплекс подобных
движений направлен на устранение нежелательных ощущений. Чесатель-
ный рефлекс сохраняется у собак с изолированным от головного спинным
мозгом, что свидетельствует о спинальном механизме этого рефлекса и его
относительной независимости от супраспинальных центров.
Правильные чередования активности сгибателей и разгибателей при
осуществлении ритмических рефлексов обеспечиваются с помощью интер-
нейронов, которые поочередно тормозят мотонейроны то сгибателей, то
разгибателей (рис. 4.22). Чередуемые движения регулирует двигательный
центр спинного мозга, составленный из двух «полуцентров»: сгибательного
и разгибательного, которые реципрокно тормозят друг друга. Продолжи-
тельность торможения в каждом выполняемом цикле зависит от свойств
самих тормозных синапсов, в которых происходит динамическое повыше-
ние порога возбуждения постсинаптических нейронов.
4.3.2.6. Участие спинного мозга в локомоции
Под локомоцией понимают совокупность координированных движений, с
помощью которых человек активно перемещается в пространстве. Его шаг
представляет собой цикл движений, в котором участвуют обе конечности,
и в каждой из них совершаются ритмические сокращения мышц-сгибате-
лей и разгибателей. Выполняемые каждой ногой попеременно движения
представляют собой чередование двух фаз: 1) мах, при котором нога сгиба-
ется и отрывается от пола; 2) упор, при котором нога касается пола и раз-
гибается. При этом движения ног синхронизированы так, что одна из них
находится в фазе маха, а другая — упора.
Мотонейрон
Интернейрон сгибателя
Интернейрон
Мотонейрон
разгибателя
Попеременная
активация мышц
Рис. 4.22. Схема осуществления ритмического рефлекса.
Активность мотонейронов, иннервирующих мышцы-антагонисты, проявляется попеременно
благодаря включению в рефлекторную дугу тормозных интернейронов, обеспечивающих ре-
ципрокное торможение.
4. Функции центральной нервной системы • 193
В управлении координированной деятельностью мышц при ходьбе и бе-
ге участвуют интернейроны и мотонейроны, находящихся в разных сег-
ментах спинного мозга. Согласованное чередование их циклической ак-
тивности обеспечивается топографически упорядоченными связями между
собой, сформировавшимися еще в процессе развития мозга и позволяю-
щими реализовать врожденную программу движений. Спинномозговые
центры локомоции контролируются двигательными ядрами ствола, мотор-
ной корой и мозжечком, для осуществления контроля используется сен-
сорная информация (проприоцептивная, вестибулярная, зрительная), пе-
реработка которой происходит в головном мозге. Активность спинальных
локомоторных центров у человека полностью подчинена супраспинальным
отделам и самостоятельно не проявляется.
4.3.2.7. Спинальные вегетативные рефлексы
В зависимости от локализации рецепторов и расположения рабочих орга-
нов различают висцеро-висцеральные, висцеродермальные и дермовисцераль-
ные рефлексы. Рефлекторные дуги всех этих рефлексов относятся к поли-
синаптическим, рефлекторные реакции происходят при участии гладких
мышц внутренних органов и кровеносных сосудов, сердечной мышцы и
желез внешней секреции, которые реагируют на раздражение интероре-
цепторов или рецепторов кожи.
Висцеро-висцеральные рефлексы вызываются раздражением рецепторов
внутренних органов и проявляются изменениями деятельности этих же
(например, кардио-кардиальные, кишечно-кишечные, эвакуаторные реф-
лексы) или других внутренних органов. Висцеродермальные рефлексы воз-
никают при раздражении рецепторов внутренних органов и проявляются
изменениями кожной чувствительности, потоотделения и электрического
сопротивления кожи. Область рефлекторного ответа определяется сегмен-
тарной организацией вегетативной иннервации: афферентные сигналы от
рецепторов внутренних органов поступают в определенные сегменты
спинного мозга, а кожные изменения обнаруживаются только в тех дерма-
томах, которые иннервируются из этих же сегментов. Изменения кожной
чувствительности или так называемая отраженная боль в ограниченных
участках кожи, получивших название зон Захарьина—Геда, указывают на
поражение определенного органа и имеют диагностическое значение.
Сегментарная организация рефлекторных дуг характерна и для дермо-
висцеральных рефлексов, которые вызываются раздражением рецепторов
кожи, после чего происходит рефлекторное изменение деятельности опре-
деленных внутренних органов и возникают реакции сосудов. На сегмен-
тарном принципе организации рефлекторных дуг дерматовисцеральных
рефлексов основано терапевтическое действие согревания, охлаждения или
иного раздражения кожи (точечный массаж, акупунктура, горчичники).
Нисходящий контроль рефлекторных дуг спинальных вегетативных
рефлексов осуществляют вегетативные центры ствола мозга и гипоталамус.
Окончания серотонинергических, норадренергических и пептидергических
нейронов головного мозга проецируются на преганглионарные нейроны и
интернейроны, участвующие в замыкании рефлекторных дуг вегетативных
рефлексов.
7-6095
I
194 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
4.3.3. Функциональная организация проводящих путей спинного
мозга
Проводящие пути спинного мозга образованы топографически упорядо-
ченными пучками длинных аксонов нейронов, входящих в состав задних,
передних и боковых канатиков белого вещества спинного мозга. В соот-
ветствии с источником и адресатом передаваемой информации они под-
разделяются на восходящие (чувствительные), нисходящие (двигательные) и
собственные, или проприоспинальные, проводящие пути. Собственные про-
водящие пути образованы аксонами интернейронов и служат для установ-
ления межсегментарных связей в пределах самого спинного мозга.
Восходящие пути задних столбов образованы длинными ветвями аксо-
нов чувствительных нейронов спинальных ганглиев, которые следуют к го-
ловному мозгу без перекреста и оканчиваются на нейронах ядер задних
столбов продолговатого мозга. Они передают сенсорные сигналы от мы-
шечных и сухожильных проприоцепторов и от тактильных рецепторов ко-
жи. Восходящие пути болевой и температурной чувствительности образо-
ваны аксонами нейронов задних рогов спинного мозга, которые возбужда-
ются соответствующими сенсорными нейронами. Аксоны клеток задних
рогов проходят через переднюю комиссуру на противоположную сторону и
направляются к головному мозгу в составе переднебокового канатика, в
котором различают спиноталамический и спиноретикулярный тракты. Спи-
ноталамический тракт служит для проведения сенсорных сигналов от ре-
цепторов в проекционную область коры, а спиноретикулярный тракт про-
водит нервные импульсы от рецепторов в ретикулярную формацию, кото-
рая неспецифически активирует обширные регионы коры. Дорсальный и
вентральный спиномозжечковые тракты, расположенные в боковых стол-
бах, передают информацию от механорецепторов кожи, мышц и суставов к
мозжечку: это канал обратной связи, необходимой мозжечку для коррек-
ции совершаемых движений.
Нисходящие пути спинного мозга, вестибулоспинальный, руброспиналь-
ный и ретикулоспинальный, предназначены для регуляции тонуса скелетных
мышц и координации рефлекторной деятельности спинного мозга. Они
образованы аксонами нейронов вестибулярного, красного ядер и ретику-
лярной формации. Берущий свое начало от нейронов покрышки среднего
мозга тектоспинальный путь служит для перераспределения тонуса скелет-
ных мышц при совершении ответных реакций на зрительные и слуховые
раздражители. Кортикоспинальный путь берет начало от нейронов мотор-
ных областей коры, он предназначен для регуляции произвольных дви-
жений.
4.4. Функции ствола мозга
К стволу мозга относят продолговатый мозг, мост и средний мозг, эти три
отдела мозга функционально связаны друг с другом. Ствол содержит ядра
II — XII черепных нервов (табл. 4.1), которые обеспечивают афферентную и
эфферентную иннервацию тканей головы и внутренних органов, в нем на-
ходятся нервные центры двигательных и вегетативных рефлексов. Нервные
центры ствола программируют циклическую деятельность (ходьба, жева-
ние, дыхание). Через ствол проходят сенсорные и двигательные пути,
обеспечивающие взаимодействие головного и спинного мозга, переключа-
тельные ядра ствола являются обязательным звеном для многих из них.
Ствол мозга содержит популяции нейронов, которые образуют восходящие
4. Функции центральной нервной системы • 195
и нисходящие проекции, оказывающие модулирующее влияние на осуще-
ствление рефлекторной деятельности, поведенческих реакций и психиче-
ских процессов.
4.4.1. Функциональная организация ствола мозга
Афферентные сигналы поступают к чувствительным ядрам ствола от сен-
сорных нейронов, тела которых расположены в чувствительных узлах че-
репно-мозговых нервов. Афферентные волокна образуют синапсы с интер-
нейронами чувствительных ядер, аксоны которых проецируются на двига-
тельные или вегетативные ядра ствола. Эти ядра образованы эфферентны-
ми нейронами, их аксоны иннервируют скелетные и гладкие мышцы голо-
вы, шеи и внутренних органов, экзокринные железы, которые являются
рабочими органами для стволовых рефлексов. В замыкании большинства
рефлекторных дуг принимает участие ретикулярная формация, занимающая
промежуточное положение между расположенными дорсально чувстви-
тельными ядрами и находящимися вентрально двигательными ядрами.
В стволе мозга расположены сегментарные нервные центры собствен-
ных рефлексов, которые возникают при активации чувствительных волокон
черепно-мозговых нервов и проявляются ответными сокращениями раз-
личных групп мышц головы или вегетативными реакциями (например,
слюноотделительный, зрачковый рефлексы). Наряду с ними в стволе име-
ются координационные центры системных рефлексов, возникающих в ре-
зультате возбуждения чувствительных волокон черепно-мозговых нервов,
затем их ядер, от которых оно распространяется по восходящим или нис-
ходящим путям, приводя к цепи моторных и вегетативных рефлексов ство-
ла. Системные рефлексы могут проявляться последовательными сокраще-
ниями мышц языка, глотки, гортани, секреторными реакциями, измене-
ниями частоты сокращений сердца, величины артериального давления,
частоты дыхания (например, глотание, кашель).
Комплексы ответных рефлекторных реакций на сенсорный стимул, ха-
рактеризующиеся запрограммированными (автоматическими) действиями,
определяются как центральные программы, например программа ходьбы,
жевания, дыхания. Под программой понимают заданную генетически по-
следовательность активации нервных центров, которая вызвана пусковым
влиянием какого-либо раздражителя. Запущенная центральная программа
реализуется автоматически, без дополнительного участия внешних стиму-
лов, однако порядок запрограммированных действий может изменяться в
зависимости от характера сенсорной информации (обратная связь).
4.4.1.1. Черепные нервы
В соответствии с основной выполняемой функцией выделяют группу вы-
соко специфических сенсорных черепно-мозговых нервов (И пара — зри-
тельный нерв, VIII пары — слуховой и вестибулярный нервы, I пара — обо-
нятельный нерв). Другую группу представляют смешанные черепно-мозго-
вые нервы, образованные соматическими и висцеральными афферентами
и эфферентами: V пара — тройничный нерв, VII пара — лицевой нерв, IX
пара — языкоглоточный нерв, X пара — блуждающий нерв. В третью груп-
пу включают черепно-мозговые нервы, иннервирующие мышцы головы, в
составе которых, помимо аксонов мотонейронов, содержится небольшое
количество афферентных волокон от проприоцепторов соответствующих
мышц.
Т
196 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
4.4.1.2. Функциональная специализация ядер ствола
В ядрах задних столбов продолговатого мозга происходит переключение
восходящих сенсорных путей тактильной и проприоцептивной чувстви-
тельности. Аксоны получивших афферентную информацию нейронов ядер
задних столбов переходят на противоположную сторону, образуя характер-
ные симметричные структуры продолговатого мозга, которые называются
лемнисками (от лат. lemnisci — ленты для почетных венков). От этого тер-
мина происходит название лемнисковой системы, предназначенной для пе-
редачи специфической информации соматосенсорной коре.
Восходящие пути температурной и болевой чувствительности относятся
к экстралемнисковой системе, они проходят через ствол без перекрещива-
ния к таламусу, отдавая возбуждающие коллатерали к нейронам ретику-
лярной формации ствола. Все виды сенсорной информации от специаль-
ных органов чувств, за исключением обоняния, проецируются на соответ-
ствующие ядра ствола, нейроны которых перерабатывают полученную ин-
формацию и передают ее следующему иерархическому уровню {проводни-
ковая функция), представленному ядрами таламуса. Помимо этого аффе-
рентные сигналы используются для осуществления собственных или сис-
темных рефлексов ствола {рефлекторная функция).
Моторные ядра черепных нервов участвуют в осуществлении рефлек-
торной деятельности ствола, но в то же время находятся под контролем
Таблица 4.1. Функциональная специализация черепных нервов
Па- ра Название Афферент- ный, эффе- рентный Специфическая функция
1 Обонятель- ный Афферентный Обоняние
II Зрительный Афферентный Зрение
III Глазодвига- Эфферентный Движения глаз, сужение зрачка, изменение формы
тельный Афферентный хрусталика Передача информации от глазодвигательных мышц
IV Блоковый Эфферентный Движения глаз
Афферентный Передача информации от глазодвигательных мышц
V Тройничный Эфферентный Жевание
Афферентный Чувствительность лица
VI Отводящий Эфферентный Движения глаз
Афферентный Передача информации от глазодвигательных мышц
VII Лицевой Эфферентный Движения мимических мышц лица, выделение
слюны и слез
Афферентный Вкусовая чувствительность передних 2/3 языка
VIII Слуховой Афферентный Слух
Вестибуляр- ный Афферентный Чувство равновесия
IX Языкогло- Афферентный Вкусовая чувствительность задней */3 языка, неба,
точный Эфферентный сведения о газовом составе артериальной крови Выделение слюны, глотание
X Блуждаю- Афферентный Передача сигналов от внутренних органов
щий Эфферентный Регуляция сокращений гладких мышц, сердца; сек- реция экзокринных желез
XI Добавочный Эфферентный Движения головы, шеи, плеч
Афферентный Передача информации от мышц шеи
XII Подъязыч- Эфферентный Движения языка
ный Афферентный Передача информации от мышц языка
4. Функции центральной нервной системы • 197
двигательных областей коры. Берущий начало в этих областях кортико-
бульбарный тракт регулирует выходную активность двигательных ядер че-
репно-мозговых нервов и участвует в управлении сокращениями лицевой
мускулатуры. Волокна кортикобульбарного тракта оканчиваются также на
нейронах координационных центров ствола: красного ядра, вестибулярных
ядер, ядер шва, голубого пятна. Эти ядра образуют нисходящие проекции к
интернейронам и мотонейронам спинного мозга, посредством которых
обеспечивают контроль рефлекторной деятельности спинного мозга и ре-
гулируют распределение тонуса скелетных мышц, необходимого для под-
держания позы и выполнения движений. Красное и вестибулярное ядра,
ядра шва и голубого пятна получают также коллатерали волокон корти-
коспинального тракта — прямого пути от двигательной коры к спинному
мозгу. Таким образом, моторные ядра ствола участвуют в его собственной
рефлекторной деятельности и одновременно являются иерархическим
уровнем моторной системы мозга: двигательная кора—ствол—спинной
мозг.
При осуществлении двигательной функции ЦНС ядра ствола взаимо-
действуют с мозжечком-, нейроны таких ядер активируются сенсорными
системами (вестибулярная, проприоцептивная, зрительная), возбуждающи-
мися в процессе движения, а затем передают информацию мозжечку, ко-
торый, в свою очередь, регулирует выходную активность координацион-
ных центров ствола, в первую очередь, вестибулярных ядер. У мозжечка
нет прямых связей с корой, и все его афферентные и эфферентные связи с
ней осуществляются посредством нескольких ядер ствола (ретикулярные
ядра продолговатого мозга, ядра нижней оливы и моста).
В регуляции жизненно важных вегетативных функций (кровообраще-
ние, дыхание, пищеварение) участвуют относительно небольшие, сложно
организованные группы нейронов ретикулярной формации, координирую-
щие активность симпатического и парасимпатического отделов вегетатив-
ной нервной системы. Большинство нейронов ретикулярной формации яв-
ляются полимодальными-. они получают разные виды сенсорной информа-
ции, а также проекции нейронов гипоталамуса и лимбической системы
мозга. Гипоталамус координирует деятельность вегетативных центров ство-
ла, а лимбическая система изменяет их активность при возникновении
эмоций.
Компактно сгруппированные в нескольких ядрах ствола моноаминерги-
ческие нейроны (ядра шва, голубого пятна, среднего мозга) образуют ди-
вергентные восходящие и нисходящие проекции, способные повлиять на про-
цесс передачи нервных импульсов в различных регионах мозга (таламус,
лимбические структуры, полосатое тело, кора). Нейромодулирующее дей-
ствие моноаминергических нейронов используется при регуляции цикли-
ческих процессов (чередование сна и бодрствования, распределение вни-
мания), возникновении мотиваций и эмоций.
4.4.2. Рефлекторная функция ствола мозга
Рефлексы ствола участвуют в осуществлении функций пищеварения (вы-
деление слюны, жевание, глотание), защиты от повреждающих воздейст-
вий (рвота, чиханье, кашель, слезоотделение, мигание) и приспособлении
к меняющимся условиям среды (ориентировочные, сторожевые рефлексы).
Слюноотделительный рефлекс возникает при раздражении вкусовых ре-
цепторов и механорецепторов полости рта, афферентная импульсация про-
водится по чувствительным волокнам лицевого и языкоглоточного, а так-
198 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
же тройничного и блуждающего нервов. Слюноотделительный центр про-
долговатого мозга представляет функциональное объединение нейронов,
расположенных в ретикулярной формации, которые активируются при
раздражении вкусовых и обонятельных рецепторов. Это координационный
центр, который связан не с одним, а с несколькими вегетативными ядра-
ми. В частности, рефлекторное выделение слюны происходит в результате
одновременной активации парасимпатических волокон лицевого и языко-
глоточного нервов, а также симпатических нейронов из II—IV грудных
сегментов спинного мозга. Стимуляция слюноотделения одним лишь запа-
хом, видом пищи или воспоминаниями о ней указывает на возможность
активации слюноотделительного центра нисходящими корковыми проек-
циями.
Рецептивные поля глотательного рефлекса находятся в корне языка, не-
бе, гортани и надгортаннике, где расположены механорецепторы тройнич-
ного и языкоглоточного нервов, а также верхнегортанной ветви блуждаю-
щего нерва. Они раздражаются пищевым комком, проглатываемой жидко-
стью или слюной, после чего афферентная импульсация поступает к ядрам
тройничного и солитарного трактов и достигает глотательного центра ре-
тикулярной формации, находящегося между ядром лицевого нерва и ниж-
ней оливой. Эфферентная импульсация, обеспечивающая координирован-
ные сокращения мышц рта, глотки и пищевода, поступает к ним по волок-
нам подъязычного и лицевого нервов, а также при посредстве мотонейро-
нов первых трех шейных сегментов спинного мозга. Глотательный центр
функционально связан с дыхательным центром, эта связь обеспечивает за-
держку дыхания при глотании и перекрытие надгортанником входа в дыха-
тельные пути во избежание поступления в них пищи.
Жевательные рефлексы (открывания и закрывания рта) осуществляются
в соответствии с центральной программой жевания. Она запускается стиму-
ляцией механорецепторов при изменениях давления на твердое небо, зубы,
десны. Афферентная информация поступает в продолговатый мозг по чув-
ствительным волокнам тройничного нерва, двигательное ядро которого
осуществляет эфферентную иннервацию жевательных мышц. Представле-
ние о жевании как о рефлексе описывает этот процесс в виде чередования
двух реципрокных фаз: открывания и закрывания рта. В современном
представлении центр жевания функционирует как центральный генератор
ритмических мышечных сокращений, который зависит от сенсорных сиг-
налов (поступление твердой пищи в ротовую полость) и от влияния цен-
тральных структур (жевание можно начать и прекратить произвольно).
Нисходящий контроль центра жевания осуществляют двигательные ядра
ствола, моторные области коры, мозжечок и базальные ганглии.
Рвотный рефлекс является результатом раздражения механо- и хеморе-
цепторов, расположенных в корне языка, глотке, слизистой оболочке же-
лудка, кишечника, брюшины, а также стимуляцией обонятельных, вкусо-
вых рецепторов (при резко неприятном запахе или вкусе пищи) и вестибу-
лярных рецепторов внутреннего уха (вследствие укачивания). Афферент-
ные импульсы поступают в ретикулярную формацию продолговатого мозга
по волокнам блуждающего, языкоглоточного и других чувствительных нер-
вов. Рвотный центр осуществляет сложную координацию соматических и
висцеральных двигательных реакций, приводящих к извержению содержи-
мого желудка, а иногда и кишечника, через рот. При расслаблении дна же-
лудка и сфинктера между желудком и пищеводом происходит непроиз-
вольное сокращение диафрагмы и брюшной стенки одновременно с сокра-
щением привратника, что приводит к выталкиванию содержимого желудка
4. Функции центральной нервной системы • 199
в пищевод. Повышение внутриплеврального давления способствует про-
движению содержимого желудка из пищевода в ротовую полость. Рефлек-
торный подъем мягкого неба предупреждает попадание рвотных масс в но-
совую часть глотки, а рефлекторное закрытие голосовой щели и прекраще-
ние дыхания предотвращают аспирацию содержимого желудка в легкие.
Эфферентные пути рвотного рефлекса представлены волокнами блуждаю-
щего и языкоглоточного нервов, а также мотонейронами, иннервирующи-
ми мышцы брюшной стенки и диафрагму. Рвотный центр находится под
контролем кортикальных центров мозга.
Чихательный рефлекс обеспечивает удаление инородных веществ, попав-
ших в полость носа, и возникает при механическом раздражении чувстви-
тельных окончаний тройничного нерва. Сначала происходит глубокий
вдох, а затем форсированный выдох при открытой голосовой щели. Под-
нимающийся к небу язык закрывает путь в ротовую полость, и поток вы-
ходящего воздуха идет через нос, освобождая его полость от инородных
частиц. Чихательный рефлекс представляет собой сложный моторный акт
с жестко запрограммированной последовательностью действий, которой
управляет центральный генератор, связанный с дыхательным центром.
Возбуждение чувствительных волокон передается нейронам дыхательного
центра и эфферентным волокнам языкоглоточного, блуждающего, подъ-
язычного нервов; в акте чиханья участвуют и мотонейроны дыхательных
мышц. Запущенный механизм чихательного рефлекса всегда приводит к
его завершению вне зависимости от поступления сенсорных сигналов.
Кашлевой рефлекс возникает при раздражении находящихся в носоглот-
ке, гортани и трахее чувствительных окончаний тройничного, языкогло-
точного, верхнего гортанного и блуждающего нервов. Механическими раз-
дражителями этих чувствительных окончаний являются вдыхаемые с воз-
духом твердые частицы или продукты воспалительных процессов, химиче-
скими раздражителями служат вдыхаемые газы. Вследствие раздражения
чувствительных окончаний происходит глубокий вдох, закрывается голосо-
вая щель, расслабляется диафрагма, а скелетные дыхательные мышцы со-
кращаются, что в итоге приводит к повышению внутригрудного давления,
которому противодействует закрытая голосовая щель. При открытии голо-
совой щели возникает быстрый выдох, способствующий удалению инород-
ных тел или воспалительной слизи. При кашлевом рефлексе происходит
согласование активности дыхательного центра с нейронами, управляющи-
ми деятельностью мышц гортани и скелетных мышц. Эфферентное звено
рефлекса включает возвратный гортанный нерв, регулирующий закрытие
голосовой щели, и спинномозговые нервы, контролирующие сокращения
грудных и брюшных мышц.
Слезный рефлекс обусловлен подсыханием роговицы или попаданием на
нее инородных тел, рефлекторный механизм защищает роговицу и конъ-
юнктиву от высыхания и от раздражения инородными телами, смываемы-
ми слезной жидкостью. Афферентная импульсация от роговицы осуществ-
ляется по волокнам слезной ветви тройничного нерва, в переключении
сигналов, следующих к слезным железам, участвуют чувствительные ядра
тройничного тракта и ретикулярная формация. Эфферентное звено пред-
ставляют парасимпатические нейроны, стимулирующие выделение слезной
жидкости. Преганглионарные парасимпатические нейроны расположены в
области моста, они могут возбуждаться и вызывать повышенное выделение
слезной жидкости при чиханье, кашле, рвоте, жевании, действии яркого
света, а также при переживании эмоций. Сопутствующее указанным реак-
циям слезотечение объясняется стимуляцией парасимпатических нейронов
200 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
из чувствительного ядра тройничного нерва, гипоталамуса и лимбической
системы. Активация симпатического отдела вегетативной нервной системы
приводит к уменьшению количества выделяемой слезной жидкости.
При раздражении роговицы помимо слезного возникает мигательный
рефлекс, связанный с раздражением чувствительных окончаний тройнич-
ного нерва. Рефлекторное опускание века происходит вследствие сокраще-
ния орбикулярной мышцы, получающей иннервацию от лицевого нерва.
При движении века инородное тело удаляется с роговицы и уносится слез-
ной жидкостью, а сама слезная жидкость равномерно распределяется по
роговице и конъюнктиве.
В верхнем отделе ствола, представленном структурами среднего мозга,
расположен центр зрачкового рефлекса, который служит для регуляции по-
тока света, поступающего в глаз: на ярком свету зрачок суживается, а в
темноте — расширяется. Афферентные импульсы от сетчатки глаза посту-
пают к верхним буграм четверохолмия, где находятся нейроны, осуществ-
ляющие замыкание рефлекторной дуги в зависимости от характера аффе-
рентной импульсации. При ярком освещении центр зрачкового рефлекса
активирует вегетативные ядра глазодвигательных нервов, которые иннер-
вируют мышцы, суживающие зрачок. В темноте характер афферентной
импульсации изменяется, в связи с этим рефлекторная дуга замыкается на
симпатические нейроны, находящиеся в нижнем шейном и верхнем груд-
ном сегментах спинного мозга. Происходит активация этих нейронов, что
ведет к сокращению расширяющих зрачок радиальных мышц.
Если верхние (передние) бугры четверохолмия получают афферентные
сигналы от сетчатки, то нижние (задние) — от слуховых рецепторов. При
действии новых зрительных или слуховых раздражителей здесь замыкаются
рефлекторные дуги ориентировочных рефлексов, проявляющихся поворотом
головы и глаз в сторону раздражителя и одновременным перераспределе-
нием тонуса скелетных мышц так, чтобы получилась поза, из которой лег-
че начать движение. Указанные реакции получили название сторожевых
или стартовых рефлексов. Физиологическая основа их возникновения опи-
сана в следующих разделах этой главы.
4.4.2.1. Статические и статокинетические рефлексы
Статические и статокинетические рефлексы обеспечивают степень тони-
ческого напряжения мышц, фиксирующих неподвижное положение суста-
вов, которое необходимо для поддержания позы и сохранения равновесия
и ориентации конечностей во время движения. Статические рефлексы
подразделяются на позные, или рефлексы положения, благодаря которым со-
храняется вертикальная поза, и установочные (выпрямления) рефлексы, про-
являющиеся при смене одной позы на другую, например при вставании из
положения сидя или лежа. Статокинетические рефлексы вызываются дей-
ствием на организм прямолинейного или углового ускорения.
Одним из источников афферентной импульсации, требующейся для
возникновения обеих разновидностей рефлексов, служат рецепторы вести-
булярного аппарата, реагирующие на изменения положения тела, наклоны
и повороты головы. Еще одну рефлексогенную зону образуют проприоцеп-
торы шейных мышц, возбуждающиеся в связи с наклонами головы. В соот-
ветствии с источником афферентной импульсации различают вестибуляр-
ные (или лабиринтные) и шейные тонические рефлексы. Определение реф-
лексов как тонических указывает на перераспределение тонуса мышц, тре-
буемого для сохранения равновесия и необходимой позы при смещении
4. Функции центральной нервной системы • 201
Шея
Лабиринт
голова вверх
нормальное
положение
головы
голова вниз
Отклонение
дорсально
Нормальное
отклонение
Отклонение
вентрально
А
В
Рис. 4.23. Схема лабиринтных и шейных рефлексов: распределение тонуса мышц
конечностей.
Тонус мышц, сгибающих или разгибающих конечности, изменяется рефлекторно в соответст-
вии с положением головы относительно горизонта (афферентные сигналы поступают от вес-
тибулорецепторов) и наклонами головы (афферентные сигналы поступают от проприоцепто-
ров мышц шеи). Окончательный вариант распределения тонуса мышц на конечностях зависит
от характера афферентной информации, поступившей из обоих источников.
центра тяжести тела. Для удержания равновесия необходимо повысить то-
нус мышц, противодействующих силе гравитации. К таким мышцам отно-
сятся разгибатели туловища и проксимальных отделов конечностей
(рис. 4.23).
Рефлекторные дуги вестибулярных и шейных рефлексов замыкаются в
соответствующих сенсорных ядрах продолговатого мозга, нейроны кото-
рых образуют проекции на стволовые центры нисходящих двигательных
путей, оканчивающихся в сером веществе спинного мозга. Двигательные
центры ствола представлены крупными нейронами красного ядра (его маг-
ноцеллюлярная часть), вестибулярными ядрами, медиальной частью ретику-
лярной формации и покрышкой среднего мозга. Нейроны этих ядер образуют
202 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
нисходящие проекции на интернейроны спинного мозга и гамма-мотоней-
роны, что позволяет им координировать активность мотонейронов, непо-
средственно управляющих мышечными сокращениями.
Позные рефлексы проявляются при подготовке к любому движению,
поскольку для его выполнения требуется определенная исходная поза: на-
пример, для того, чтобы встать из положения сидя, вначале необходимо
чуть наклонить вперед корпус и голову. Подъем с постели тоже начинается
с опережающего изменения положения головы, при котором возбуждаются
вестибулярные рецепторы, проприоцепторы шеи и происходит рефлектор-
ное перераспределение тонуса мышц туловища и конечностей, необходи-
мое для того, чтобы подняться.
Примером статокинетического рефлекса может служить сохранение
равновесия у стоящего в транспорте пассажира, когда это равновесие на-
рушается при резком начале движения или внезапной остановке. При сме-
щении центра тяжести рефлекторно повышается тонус разгибателей на той
стороне, в которую отклоняется тело, а выставленная рефлекторно в эту
сторону прямая нога помогает удержать равновесие. При вертикальных ус-
корениях имеют место лифтные рефлексы', в момент начала движения лиф-
та вверх у находящегося на лифтной площадке человека снижается тонус
разгибателей, и поэтому его ноги подгибаются, а при опускании площадки
тонус разгибателей увеличивается и ноги фиксируются в положении мак-
симального разгибания.
Во время ходьбы и бега возникает смещение центра тяжести тела впе-
ред. Если его уже нельзя вернуть в исходное положение, не оторвав конеч-
ность от опоры, то для сохранения равновесия приходится сделать шаг
вперед. В том случае, если поскользнувшийся человек начинает падать, на
стороне падения рефлекторно повышается тонус разгибателей. Этот эво-
люционно древний рефлекторный механизм приводит к повышению тону-
са разгибателей не только ноги, выставленной в сторону падения, но и ру-
ки, что нередко приводит к типичному перелому лучевой кости, когда при
падении на нее приходится вся сила удара. Спортсмены, чья деятельность
предполагает частые падения, учатся это делать безопасно, а овладение но-
вой техникой указывает на возможность перепрограммирования статоки-
нетического рефлекса, что связано с участием двигательных центров моз-
жечка и моторных областей коры.
В клинической практике статические и статокинетические рефлексы
называют постуральными и исследуют их электромиографически.
4.4.2.2. Нисходящие двигательные пути ствола мозга
Связь двигательных центров ствола с интернейронами и мотонейронами
спинного мозга обеспечивается двумя различающимися в функциональном
отношении нисходящими путями: медиальным и латеральным (рис. 4.24).
Медиальный путь образуют волокна вестибулоспинального, ретикулоспи-
нального и тектоспинального трактов, которые оканчиваются билатерально
на медиальных колоннах вентральных рогов спинного мозга, где находятся
интернейроны и мотонейроны, управляющие мышцами туловища и про-
ксимальных отделов конечностей. Основная функциональная задача этого
пути состоит в управлении деятельностью мышц-разгибателей, обеспечи-
вающих сохранение равновесия и вертикального положения тела. Еще од-
на его функция заключается в модулировании активности возбуждающих и
тормозных интернейронов спинного мозга, что проявляется торможением
или облегчением спинальных рефлексов. Специфической задачей текто-
4. Функции центральной нервной системы • 203
А. Латеральный
путь
Б. Медиальный
путь
Медиальный
путь
Вестибуло-
спинальный
путь
Рис. 4.24. Схема нисходящих двигательных путей ствола к спинному мозгу.
А. Латеральный путь начинается от крупноклеточных нейронов красного ядра и оканчивается
на противоположной стороне спинного мозга.
Б. Медиальный путь образован аксонами нейронов вестибулярного ядра, ретикулярной форма-
ции и покрышки среднего мозга; в спинном мозге представлен передним канатиком, оканчи-
вается билатерально.
спинального тракта является координация поворотов головы и следящих
движений глаз с изменениями позы.
Латеральный путь от ствола к спинному мозгу представлен руброспи-
нальным трактом, волокна которого переходят на противоположную сто-
рону и после прохождения ствола спускаются в спинной мозг, где образу-
ют синапсы с интернейронами и мотонейронами, расположенными лате-
рально и контролирующими деятельность дистальных мышц конечностей.
Эти мышцы предназначены для выполнения сложных движений, связан-
ных, например, с захватом предметов рукой и пальцами.
После перерезки руброспинального пути у экспериментальных живот-
ных возникает стойкое повышение тонуса разгибателей туловища и конеч-
ностей, что получило название децеребрационной ригидности. Она исчезает
после дополнительной перерезки вестибулоспинального и ретикулоспи-
нального путей. Сопоставление этих фактов дало повод к выводу о необхо-
димости красных ядер для противодействия стволовым антигравитацион-
ным центрам, т. е. вестибулярным ядрам и ретикулярной формации. Одна-
ко у человека после изолированного поражения красного ядра повышение
тонуса разгибателей выражено Незначительно, а иногда повышения их то-
204 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
нуса и вовсе не наблюдается. Только при массивных очагах повреждения в
области верхнего двухолмия, когда страдает и кортикоспинальный путь,
обнаруживается повышенный тонус разгибателей в нижних конечностях,
но одновременно с этим в верхних конечностях преобладает тонус сгибате-
лей. Возможно, это обстоятельство обусловлено изменением функциональ-
ного назначения верхних конечностей у человека по сравнению с четверо-
ногими.
4.4.2.3. Глазодвигательные центры ствола
Движения глазного яблока осуществляют четыре прямые мышцы (наруж-
ная, внутренняя, верхняя, нижняя) и две косые мышцы (верхняя и ниж-
няя). Они поворачивают глаз внутрь или наружу, вверх или вниз, чтобы
сфокусировать рассматриваемые объекты в центральной ямке сетчатки,
обеспечивающей максимальную остроту зрения; при этом движения глаз-
ных яблок происходят содружественно. Сокращения мышц определяются
влиянием глазодвигательных и блоковых нервов, ядра которых расположе-
ны в ростральной части среднего мозга, а также ядер отводящих нервов,
содержащихся в каудальной части моста. Различают очень быстрые движе-
ния глаз (саккады) и плавные следящие движения. Саккады, происходящие
около трех раз в секунду, обеспечивают перемещение изображения по ре-
цепторам сетчатки, которые не реагируют на стабильное изображение, а
отвечают на его движение. Плавные следящие движения глаз возникают в
связи с перемещением наблюдаемых объектов и при движении головы и
тела наблюдателя. Следящие движения позволяют удерживать изображе-
ние в пределах центральной ямки сетчатки.
Афферентная информация к мотонейронам глазодвигательных мышц
поступает от зрительных, вестибулярных, слуховых рецепторов, а также от
проприоцепторов мышц глаза. В зависимости от характера афферентной
информации дуги глазодвигательных рефлексов замыкаются на разных
уровнях ствола. Стволовые центры, осуществляющие замыкание рефлек-
торных дуг и обеспечивающие определенное положение взора, расположе-
ны в парамедианной ретикулярной формации и ретикулярной формации
среднего мозга. Уровень возбуждения глазодвигательных центров регули-
руют нейроны верхних бугров четверохолмия, вестибулярные ядра и моз-
жечок, кора больших полушарий.
4.5. Функции ретикулярной формации
Ретикулярная формация расположена во внутренней части ствола на про-
тяжении от нижних отделов промежуточного мозга до продолговатого моз-
га, где она проходит между двигательными и чувствительными ядрами.
Собственные ядра ретикулярной формации имеют низкую плотность ней-
ронов, относительно высокий процент клеток нейроглии и слабо выражен-
ные границы ядер, количество которых измеряется несколькими десятка-
ми. Большинство нейронов ретикулярной формации принимает коллатера-
ли от нескольких сенсорных путей (полисенсорная конвергенция) и обладает
тонической активностью покоя (5—10 импульсов в секунду), растущей в
связи с поступлением афферентных сигналов. Аксоны нейронов ретику-
лярной формации обычно делятся на две ветви, следующие в восходящем
и нисходящем направлениях. Восходящие влияния ретикулярной форма-
ции регулируют возбудимость нейронов коры в связи с циклом сон—бодр-
ствование, они необходимы для сохранения сознания и восприятия сенсор-
4. Функции центральной нервной системы • 205
ных сигналов. Нисходящие влияния осуществляются посредством ретику-
лоспинального тракта, входящего в состав медиального пути к спинному
мозгу и регулирующего его двигательную функцию. Координационные цен-
тры ретикулярной формации обеспечивают согласование афферентной и
эфферентной систем, участвующих в регуляции вегетативных функций
(кровообращение, дыхание, пищеварение).
4.5.1. Особенности нейронной организации ретикулярной
формации
Афферентные сенсорные входы представлены волокнами спиноретикулярно-
го тракта, образованного отростками нейронов задних рогов спинного
мозга, коллатералями от аксонов интернейронов чувствительных ядер
(тройничного, блуждающего, языкоглоточного, вестибулярного и слухово-
го нервов), а также небольшим количеством волокон от других сенсорных
проводящих путей, следующих через ствол. Остальные проекции на нейро-
ны ретикулярной формации представлены окончаниями нейронов ядер
мозжечка, гипоталамуса и коры, они регулируют активность нейронов ре-
тикулярной формации. Разные области и ядра ретикулярной формации об-
разуют синаптические контакты между собой, как, например, каудальная
область с ростральной, и могут реципрокно влиять друг на друга.
В дорсальном отделе продолговатого мозга находится ядро солитарного
или одиночного тракта, получающее сенсорную информацию от внутрен-
них органов. В составе волокон языкоглоточного и блуждающего нервов
сюда поступают сигналы от периферических баро- и хеморецепторов. От
нейронов ядра солитарного тракта отходят аксоны к парасимпатическим
нейронам черепно-мозговых нервов (дорсальные ядра блуждающих нервов
и обоюдные ядра) и к рострально-вентролатеральной области продолгова-
того мозга, регулирующей активность симпатических нейронов спинного
мозга.
Вегетативные центры ретикулярной формации продолговатого мозга
находятся под контролем гипоталамуса, который изменяет активность
симпатического и парасимпатического отделов соответственно возникаю-
щим задачам, например поддержанию температуры тела, осуществлению
оборонительного, пищевого или полового поведения. Гипоталамус обеспе-
чивает функциональное объединение вегетативных центров ствола и спин-
ного мозга в целостные системы, регулирующие функции кровообращения
и дыхания, температуру тела. Гипоталамус подвержен влиянию лимбиче-
ских структур мозга, обеспечивающих формирование эмоциональных ре-
акций, при возникновении которых происходит комплексное изменение
активности вегетативных центров, приспосабливающее внутренние про-
цессы организма к решению текущих поведенческих задач.
В ретикулярной формации моста и продолговатого мозга располагаются
структуры сосудодвигательного центра, состоящего из прессорного и депрес-
сорного отделов. Прессорный отдел находится в рострально-вентральной
области продолговатого мозга, где сосредоточены нейроны, имеющие ре-
шающее значение в повышении общей выходной активности симпатиче-
ского отдела вегетативной нервной системы. Депрессорный отдел располо-
жен в медиокаудальных отделах ствола, его активация сопровождается по-
нижением тонуса симпатических нервов и одновременным повышением
тонуса блуждающего нерва.
Поблизости от ядер одиночного тракта билатерально расположен дыха-
тельный центр, состоящий из дорсальной и вентральной групп нейронов.
206 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
В зависимости от выполняемой функции нейроны дыхательного центра
подразделяются на инспираторные, экспираторные, преинспираторные и
постинспираторные. Инспираторные нейроны генерируют ритм дыхания,
их нисходящие в спинной мозг аксоны возбуждают мотонейроны диафраг-
мы и наружных межреберных мышц, при сокращении которых происходит
вдох. Экспираторные нейроны регулируют активность мотонейронов внут-
ренних межреберных мышц, сокращения которых происходят при актив-
ном выдохе. Преинспираторные и постинспираторные нейроны регулиру-
ют активность инспираторных нейронов. Смена вдоха выдохом и наоборот
происходит при участии нейронов пневмотаксического центра, располо-
женного в передней части моста.
4.5.2. Нисходящие и восходящие влияния ретикулярной формации
Нисходящие влияния ретикулярной формации предназначены для регуля-
ции рефлекторной деятельности спинного мозга (см. раздел 4.2.2). Восхо-
дящие дивергентные проекции ретикулярной формации к другим отделам
головного мозга образованы ветвящимися аксонами моноаминергических
нейронов среднего мозга, голубого пятна и ядер шва, а также холинергиче-
ских нейронов. Метаботропное действие моноаминергических медиаторов
влияет на биохимические процессы нейронов-мишеней, это действие про-
является медленно по сравнению с распространением сигналов, передаю-
щихся через синапсы с помощью ионотропных рецепторов. Результатом
такого действия является модуляция функций многих нервных клеток,
участвующих в формировании мотиваций, эмоций и целенаправленного
поведения.
Норадренергические нейроны голубого пятна обладают фоновой актив-
ностью, изменяющейся в зависимости от сенсорной афферентации разных
модальностей, а также под влиянием других центров ретикулярной форма-
ции, гипоталамуса и мозжечка. Аксоны нейронов голубого пятна проеци-
руются почти во все области мозга и заканчиваются варикозными расши-
рениями, из которых выделяется норадреналин, диффундирует через меж-
клеточное пространство и через него действует на большие популяции
нейронов. Активность норадренергических нейронов необходима для под-
держания бодрствования, при физиологическом повышении этой активно-
сти формируются двигательные и вегетативные компоненты стресс-реак-
ции, чрезмерное повышение активности нейронов голубого пятна приво-
дит к развитию панического синдрома, а при сниженной активности воз-
никает депрессия.
Проекция нейронов черной субстанции среднего мозга к полосатому те-
лу (нигростриатный путь) необходима для регуляции его активности при
осуществлении произвольных движений. Дегенеративное поражение ней-
ронов черной субстанции приводит к развитию болезни Паркинсона, прояв-
ляющейся возникновением ригидности мышц и тремором. Еще одна про-
екция дофаминергических нейронов направлена к структурам лимбиче-
ской системы и фронтальной коре (мезолимбический и мезокортикальный
пути), участвующим в образовании мотиваций и эмоций. Влияние дофами-
на на эти структуры усиливается при удовлетворении биологических по-
требностей, а переживание в этот момент положительных эмоций побуж-
дает людей к повторению действий, приводящих к подобному результату.
Роль дофамина состоит в модуляции возбудимости нейронов-мишеней
лимбических структур и коры, чтобы их взаимодействие становилось опти-
мальным при создании программы поведения, а сами дофаминергические
4. Функции центральной нервной системы • 207
нейроны являются необходимым звеном образующихся поведенческих
программ. При избытке дофамина вследствие патологически повышенной
активности дофаминергических нейронов возникают грубые нарушения
поведения и мышления, проявляющиеся, например, при заболевании ши-
зофренией.
Нейроны ядер шва, синтезирующие серотонин, образуют проекции в
спинной мозг, кору мозжечка, ядра таламуса и кору больших полушарий,
что в совокупности создает восходящую и нисходящую системы, каждая из
которых принимает участие в регуляции циклических процессов сна и
бодрствования. Активация холинергических нейронов рострального отдела
среднего мозга и гигантоклеточной области покрышки моста, образующих
проекции к зрительному бугру и коре, необходима для поддержания созна-
ния, переработки сенсорной и ассоциативной информации в коре и обес-
печения познавательной интеллектуальной деятельности. Активность хо-
линергических, норадренергических и серотонинергических нейронов в
цикле сна и бодрствования периодически изменяется под влиянием супра-
хиазмальных и вентромедиальных ядер гипоталамуса, играющих роль основ-
ных водителей суточной ритмики физиологических процессов.
4.6. Функции мозжечка
Функция мозжечка состоит, в первую очередь, в формировании двигатель-
ных программ, необходимых для поддержания равновесия тела, регуляции
силы мышц и объема совершаемых с их участием движений. Он не является
составной частью иерархической сети двигательных систем, но имеет свя-
зи со всеми ее уровнями: моторной корой, двигательными центрами ство-
ла и спинным мозгом. Благодаря этим связям мозжечок может сравнивать
моторные программы коры с их исполнением, а затем корректировать дви-
жения, не соответствующие намеченной программе. Мозжечок способен
накапливать опыт двигательной активности, сохраняя его в долговремен-
ной моторной памяти.
4.6.1. Функциональная организация мозжечка
В мозжечке имеются три области, различающиеся спецификой своих со-
единений с головным и спинным мозгом и выполняемыми функциями:
1) вестибулоцеребеллум (червь с примыкающими к нему областями старой
коры); 2) спиноцеребеллум (примыкающие к червю медиальные области по-
лушарий мозжечка); 3) цереброцеребеллум, включающий в себя латеральные
области полушарий мозжечка (рис. 4.25). Вестибулоцеребеллум получает
афферентную информацию от вестибулярных ядер и к ним же посылает
эфферентные сигналы: его основная задача состоит в сохранении равнове-
сия при стоянии и ходьбе, а также в управлении движениями глаз. Спино-
церебеллум основную часть сенсорной информации получает от проводя-
щих афферентных путей спинного мозга — двух спиномозжечковых трак-
тов, несущих импульсы от проприоцепторов мышц и сухожилий. Эффе-
рентная информация от ядер этой части мозжечка поступает к двигатель-
ным центрам ствола, относящимся как к медиальному, так и к латерально-
му нисходящим путям. Спиноцеребеллум контролирует правильность на-
чинающихся движений ног и рук. Цереброцеребеллум получает при уча-
стии ядер моста информацию о планирующемся движении из сенсомотор-
ных областей коры и посылает эфферентные сигналы к первичной и вто-
ричной моторной коре, участвуя в планировании движений.
208 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Спиноцеребеллум
к медиальным
нисходящим
системам
к латеральным
нисходящим
системам
к моторной и
премоторной
коре
к вестибулярным
ядрам
От зри-
тельных и
слуховых
путей
От вес-
тибуляр-
ных
ядер
От спинного
мозга и ядра
тройничного
нерва
От коры
Управление
движениями
Планирование
движений
Сохранение
равновесия
и движения
глаз
Рис. 4.25. Схема афферентных и эфферентных связей трех функциональных облас-
тей мозжечка.
Вестибулоцеребеллум (функция сохранения равновесия и управления движениями глаз) полу-
чает зрительную и слуховую информацию, входные сигналы от вестибулярных ядер, ядер
тройничного нерва и спинного мозга. Основной эфферентный выход к вестибулярным ядрам
ствола.
Спиноцеребеллум (управление движениями конечностей) получает афферентную информацию
от проприоцепторов посредством спиномозжечковых трактов; эфферентный выход к первич-
ной и вторичной моторной коре.
Цереброцеребеллум (формирование двигательных программ) получает афферентную информа-
цию при посредничестве ядер моста от сенсомоторных областей коры; основной эфферент-
ный выход к первичной и вторичной моторной коре.
Таким образом, мозжечок постоянно получает информацию о плани-
рующихся движениях от коры, о положении головы и глаз, о тонусе
мышц, необходимом для совершения движения,— от двигательных цен-
тров ствола, а от спинного мозга к нему поступают сведения о характере
уже совершаемых движений. Располагая всей полнотой информации о
движении — от замысла до исполнения, мозжечок постоянно сравнивает
совпадение замысла с исполнением. При несоответствии хода движения
намеченному плану мозжечок немедленно исправляет возникающие ошиб-
ки. Он может вносить коррективы как в двигательную программу, благода-
ря своим связям с моторной корой, так и в исполнение движения, дейст-
вуя на двигательные центры ствола, на медиальные и латеральные нисхо-
дящие пути.
При выполнении запрограммированных произвольных движений вы-
ходная активность нейронов зубчатых ядер мозжечка регистрируется при-
близительно на 10 мс раньше, чем она обнаружится в моторной коре. Это
опережение имеет особенное значение при выполнении быстрых движе-
4. Функции центральной нервной системы • 209
ний, когда из-за дефицита времени кора не может исправить ошибку. В
таких случаях мозжечок влияет на другие двигательные центры, определяя
наиболее рациональную последовательность активации их нейронов, для
того чтобы необходимые движения были выполнены максимально точно, а
ошибочные, лишние движения исключались. Мозжечок не может иниции-
ровать движение, он лишь обеспечивает необходимый баланс действующих
мышц, причем его участие особенно необходимо в заключительной стадии
движений, когда замысел сопоставляется с результатом. При мозжечковом
поражении пациент, например, промахивается, когда его просят быстро
прикоснуться к кончику своего носа указательным пальцем.
4.6.2. Взаимодействие нейронов коры и ядер мозжечка
Афферентная информация, адресованная мозжечку, распределяется между
его ядрами и корой. Под влиянием афферентных сигналов нейроны ядер
мозжечка создают основную часть его выходной активности, и лишь самая
старая часть коры — клочок имеет непосредственную связь с вестибуляр-
ными ядрами. Функция коры мозжечка состоит в непрерывной модуляции
выходной активности ядер тормозными нейронами Пуркинье, использую-
щими в качестве медиатора ГАМК (рис. 4.26).
Во внутреннем слое трехслойной коры мозжечка расположены зерни-
стые клетки, являющиеся входными нейронами коры, а также тормозные
по отношению к ним клетки Гольджи. Афферентные моховидные волокна,
образованные нейронами не-
скольких ядер ствола и спинного
мозга, передают информацию
зернистым клеткам от проприо-
цепторов скелетных мышц, вести-
булярного аппарата и коры. Аксо-
ны зернистых клеток направлены
в наружный слой коры мозжечка,
где они Т-образно делятся на
длинные волокна, которые возбу-
ждают нейроны Пуркинье и одно-
временно тормозные интернейро-
ны, сдерживающие активность
клеток Пуркинье. Нигде в ЦНС
больше не встречается столь вы-
раженное, как в коре мозжечка,
преобладание торможения над
возбуждением, поэтому посту-
пающее в кору мозжечка возбуж-
дение прекращается уже через
100 мс: так быстро «стирается»
информация об осуществлении
актов движения — она не должна
мешать поступлению новых опе-
ративных данных.
Грушевидные нейроны Пурки-
нье формируют средний слой ко-
ры, они имеют аксоны, образую-
щие синапсы с нейронами ядер
мозжечка. Каждая клетка Пурки-
Рис. 4.26. Схема взаимодействия коры и
ядер мозжечка.
Афферентная информация, поступающая в
мозжечок, распределяется между его ядрами и
корой. Тормозные нейроны Пуркинье контро-
лируют эфферентные пути мозжечка, начи-
нающиеся от нейронов ядер, поэтому эффе-
рентная активность мозжечка зависит от возбу-
ждающих афферентных сигналов и тормозных
влияний клеток Пуркинье.
210 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
нье получает сигналы от приблизительно 200 000 параллельных волокон
зернистых клеток, а каждая зернистая клетка собирает афферентные входы
от десятка моховидных волокон. Еще один афферентный вход в мозжечок
представлен лазающими волокнами, которые образованы нейронами ниж-
него оливарного ядра продолговатого мозга и доставляют информацию от
коры и от спинного мозга. Лазающие волокна обертывают тела и дендриты
клеток Пуркинье, образуя с ними многочисленные синапсы, причем каж-
дое лазающее волокно контактирует с несколькими (от одного до десяти)
нейронами Пуркинье, но каждая клетка Пуркинье получает возбуждение
только от одного лазающего волокна. Синапс между окончанием лазающе-
го волокна и нейроном Пуркинье является одним из наиболее эффектив-
ных в ЦНС потому, что в ответ на одиночный потенциал действия, прове-
денный лазающим волокном, в нем возникает высокоамплитудный ВПСП.
Результатом этого становится высокочастотный залп потенциалов дейст-
вия клетки Пуркинье, направленный на нейроны ядер мозжечка и тормо-
зящий их активность. Этот тормозной выход из коры мозжечка является
конечным и единственным результатом ее деятельности.
Нейронная сеть коры мозжечка организована по принципу функцио-
нальных модулей, каждый из которых занимает сагиттальную зону шириной
1—2 мм. В каждый модуль входит лазающее волокно из определенной час-
ти оливы, а каждой клетке Пуркинье достается обособленная информация
от моховидных и лазающего волокон. Лазающие волокна модулируют ак-
тивность моховидных волокон, они могут усиливать их влияние, но могут
и ограничивать эффективность определенных входов. В функциональном
отношении это означает, что активность одного пути будет изменяться в
зависимости от активности другого пути. Соотношение активности отдель-
ных лазающих и моховидных волокон может меняться вследствие приоб-
ретаемого двигательного опыта — в результате подобной модуляции синап-
тических переключений повышается эффективность совершаемых дей-
ствий.
4.6.3. Эфферентные связи мозжечка с моторными структурами
мозга
Клетки ядер мозжечка представляют собой обычные переключательные
нейроны: в ответ на поступающую по коллатералям моховидных и лазаю-
щих волокон афферентную информацию они возбуждаются и отправляют
эфферентные сигналы двигательным ядрам ствола. Клетки Пуркинье огра-
ничивают их активность таким образом, что выполнение одних моторных
программ допускается, а других — отменяется в соответствии с характером
афферентной информации и организацией связей между корой мозжечка и
ядрами.
Эфферентное влияние зубчатых и промежуточных ядер мозжечка на
сенсомоторную кору больших полушарий осуществляется при участии мо-
торных вентролатеральных ядер таламуса, которые служат узловым пунк-
том нейронной сети, связывающей сенсомоторную кору не только с моз-
жечком, но и с базальными ганглиями. Аксоны таламических нейронов
способны изменять активность пирамидных клеток, образующих корти-
коспинальный путь, что позволяет мозжечку корректировать двигательные
программы, создаваемые в моторных областях коры.
Еще одним пунктом переключения моторных систем мозга являются
красные ядра, служащие источником образования руброспинальных путей.
Здесь оканчиваются аксоны возбуждающих нейронов промежуточных ядер
4. Функции центральной нервной системы • 211
мозжечка и моторной коры, оказывающей тормозное влияние. От ядер
шатра эфферентные волокна идут к вестибулярным ядрам и моторным яд-
рам ретикулярной формации ствола мозга, что позволяет мозжечку кон-
тролировать нисходящую активность вестибулоспинальных и ретикулоспи-
нальных путей. Действие мозжечка на стволовые двигательные центры по-
зволяет исправлять ошибки уже запущенной двигательной программы, ес-
ли они возникают в процессе ее осуществления.
4.7. Функции промежуточного мозга
К промежуточному мозгу относят две структуры: зрительные бугры (тала-
мус), расположенные с обеих сторон III желудочка мозга, и гипоталамус
(подбугорье), находящийся книзу от III желудочка. Функция таламуса со-
стоит в распределении потоков афферентной информации, следующей к
коре больших полушарий. Таламус участвует в восприятии сенсорных сиг-
налов и формировании целенаправленного поведения. Гипоталамус явля-
ется высшим центром регуляции вегетативных функций.
4.7.1. Функции таламуса
Таламус расположен между большими полушариями и мозговым стволом,
в нем происходят переключение и переработка афферентных сигналов, по-
ступающих к коре. В таламусе насчитывают до 120 ядер, среди которых
различают специфические, или релейные, ядра (подразделяются на сенсорные
и моторные), неспецифические ядра, а также ассоциативные ядра. Специфи-
ческие ядра таламуса содержат две основные группы клеток. Во-первых,
релейные нейроны с ветвящимися в пределах ядра дендритами и длинным
аксоном, образующим моносинаптическую проекцию на нейроны IV и III
слоев коры. Во-вторых, локальные возбуждающие и тормозные интернейро-
ны, аксоны которых не выходят за пределы ядра и участвуют в образова-
нии локальной сети. Ассоциативные ядра имеют такую же нейронную ор-
ганизацию, а в неспецифических ядрах основная масса нейронов похожа
на клетки ретикулярной формации ствола.
Основной характеристикой всех релейных ядер является специализация
афферентных входов от периферических рецепторов и чувствительных
ядер ствола и высокоточная передача нервных импульсов к проекционной
области коры с минимальным искажением входного сигнала. Так, напри-
мер, наружные коленчатые тела получают информацию от сетчатки и пе-
редают ее в первичную зрительную кору (затылочные доли). Медиальные ко-
ленчатые тела относятся к слуховой сенсорной системе и образуют проек-
ции на первичную слуховую кору (височные доли), а вентробазальные ядра,
получающие афферентную информацию от поверхности тела и от мышц,—
на соматосенсорную кору (постцентральные извилины).
При переключении сигналов в сенсорных ядрах сохраняется топографи-
ческая разграниченность проекций. Каждому рецептивному полю сетчатки
соответствуют определенные переключательные нейроны латерального ко-
ленчатого тела (ретинотопическая организация). Так, например, нейроны
вентральной части латерального коленчатого тела получают афферентную
информацию от наружных участков сетчатки, а дорсальные нейроны — от
внутренней области сетчатки. После этого таламические нейроны в том же
порядке передают информацию в проекционную зрительную кору, в кото-
рой пространственное распределение возбужденных нейронов повторяет
топографическую организацию сетчатки и действующих на нее зрительных
212 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
стимулов. Это соответствие создает в зрительной коре нейронную карту
зрительного поля.
Рецептивные поля разных участков тела соматотопически представлены
на нейронах вентробазального ядра. Каждый участок поверхности лица,
туловища, конечностей имеет сенсорный путь к определенным нейронам
этого ядра. По тому же принципу осуществляется проекция от разных уча-
стков кортиева органа внутреннего уха на определенные нейроны внутрен-
него коленчатого тела. Такая организация называется тонотопической, она
позволяет передавать независимо друг от друга сигналы от слуховых рецеп-
торов, различающихся по своей чувствительности к звукам разной высоты.
Передача информации о разных качествах одного стимула происходит
параллельно и осуществляется разными нейронами сенсорного ядра. Так,
например, магноцеллюлярный путь от больших рецептивных полей перифе-
рии сетчатки используется для передачи информации о положении объек-
тов в зрительном поле и об их движении, а парвоцеллюлярный путь от ма-
лых рецептивных полей центральной ямки сетчатки предназначен для пе-
редачи информации о форме и цвете объектов. В наружном коленчатом
теле переключение сигналов каждого параллельного пути происходит неза-
висимо. По такому же принципу, т. е. параллельно, с участием разных
нейронов вентробазального ядра передается соматосенсорная чувствитель-
ность от тактильных рецепторов кожи и проприоцепторов мышц, сухожи-
лий и суставов. Нейроны таламуса не объединяют эту информацию, пере-
правляя ее в разные регионы проекционной области, которой является со-
матосенсорная кора.
Моторные ядра таламуса, среди которых самым крупным является вен-
тролатеральное ядро, обеспечивают переключение афферентации от ба-
зальных ганглиев и мозжечка к коре, что необходимо для создания двига-
тельных программ. В оральном отделе вентролатерального ядра переклю-
чаются импульсы, следующие от внутренних областей бледного шара к
вторичной моторной коре, а в каудальном отделе проецируются афферен-
ты от ядер мозжечка и красного ядра, которые переключаются на релей-
ные нейроны, связанные с первичной моторной корой. В медиокаудальной
части вентролатерального ядра переключаются сигналы, следующие от вес-
тибулярных ядер и мышечных веретен во вторичную моторную кору, а в
ростральном отделе афференты от наружных областей бледного шара про-
ецируются на нейроны, имеющие выход в первичную и во вторичную мо-
торную кору.
Ассоциативные ядра отличаются от специфических релейных ядер тем,
что не относятся к какой-либо одной сенсорной системе, а большую часть
своих афферентов получают от других ядер таламуса, как релейных, так и
неспецифических. Функциональная связь проекционных и ассоциативных
ядер способствует интеграции всех этапов переработки сенсорной инфор-
мации, необходимой для комплексного восприятия сенсорных стимулов.
Интегративная функция ассоциативных ядер состоит в объединении дея-
тельности всех ядер таламуса с ассоциативными областями коры. Каждое
ассоциативное ядро имеет проекцию на определенную ассоциативную об-
ласть коры: медиодорсальное ядро образует проекцию на орбитофронталь-
ную кору, группа латеральных ядер — на теменную кору, а группа задних
ядер — на височную. Упорядоченное распределение информационных по-
токов отражает функциональную специализацию указанных областей
коры.
Характерным признаком ассоциативных ядер является полимодальная
(полисенсорная) конвергенция афферентных входов от двух и более сенсор-
4. Функции центральной нервной системы • 213
ных систем, а также от гипоталамуса и структур лимбической системы
мозга. Среди нейронов ассоциативных ядер имеются клетки, которые воз-
буждаются только в ответ на действие определенного сочетания сенсорных
стимулов. Полисенсорная конвергенция позволяет объединять в таламусе
разные виды сенсорной информации для комплексного восприятия окру-
жающего мира, а включение в этот процесс лимбических структур форми-
рует субъективное эмоциональное отношение к полученной информации.
Неспецифические ядра получают афферентные сигналы преимуществен-
но из двух источников: ретикулярной формации и спиноталамического
тракта. Помимо этого неспецифические ядра получают афферентные вхо-
ды от гипоталамуса, лимбических структур, базальных ганглиев, специфи-
ческих и ассоциативных ядер таламуса. В результате неспецифические яд-
ра оказываются морфологически и функционально связанными со многи-
ми системами, что определяет их активность, зависящую не столько от оп-
ределенной модальности, сколько от общей суммы афферентных сигналов.
При такой организации точность проекции неспецифических ядер на кору
не имеет значения, их проекции диффузно распространяются практически
на все области коры, а также на лимбическую систему и гипоталамус.
Влияние неспецифических ядер на кору состоит в модуляции активности
ее нейронов, т. е. прямо не возбуждает и не тормозит, но изменяет их воз-
будимость при получении специфической афферентации. Этот механизм
используется для регуляции функциональной активности мозга, которая
изменяется на протяжении суток.
Таламус, благодаря организации своих связей с другими регионами
мозга, вовлекается в осуществление различных функций: формирование
эмоций (совместно с лимбической системой), чувственное восприятие (со-
вместно с сенсорными областями коры), осуществление психической дея-
тельности (совместно с ассоциативными регионами коры).
4.7.2. Функции гипоталамуса
Гипоталамус является высшим центром регуляции вегетативных функций,
он координирует деятельность симпатического и парасимпатического от-
делов вегетативной нервной системы и согласует ее с двигательной актив-
ностью человека. Одновременно гипоталамус управляет секрецией гормо-
нов гипофиза, и тем самым контролирует эндокринную регуляцию внутрен-
них процессов. Ядра гипоталамуса являются центрами регуляции водно-
солевого баланса, температурного гомеостазиса, чувства голода и насыще-
ния, полового поведения, выполняют функцию координатора биологиче-
ских ритмов. Будучи важнейшей мотивационной структурой мозга, гипота-
ламус участвует в формировании биологических мотиваций, эмоций и орга-
низации целенаправленного поведения.
4.7.2.1. Роль гипоталамуса в регуляции вегетативных функций
Гипоталамус оперативно получает информацию от интерорецепторов внут-
ренних органов, сигналы от которых поступают через ствол мозга, кроме
того в гипоталамус поступает часть сенсорных сигналов от поверхности те-
ла и некоторых специализированных сенсорных систем (зрительная, обо-
нятельная, слуховая). Некоторые нейроны гипоталамуса имеют собствен-
ные осмо-, термо- и глюкорецепторы, которые воспринимают сдвиги ос-
мотического давления, температуры и уровня глюкозы в ликворе и крови.
В отличие от периферических рецепторов они называются центральными, а
214 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
их наличие именно в гипоталамусе связано со слабой выраженностью ге-
матоэнцефалического барьера.
Гипоталамус имеет двусторонние связи с лимбической системой мозга,
ретикулярной формацией и корой больших полушарий, что позволяет ему
координировать вегетативные функции с определенным поведением, на-
пример с переживанием эмоций. Гипоталамус образует проекции на веге-
тативные центры ствола и спинного мозга, что позволяет ему осуществлять
прямой контроль деятельности этих центров. Он регулирует ключевые ме-
ханизмы эндокринной регуляции вегетативных функций, осуществляя
управление секрецией гормонов гипофиза.
Необходимые для вегетативной регуляции переключения осуществляют
нейроны ядер гипоталамуса, в разных классификациях их насчитывают от
16 до 48. Искусственное раздражение гипоталамуса электрическим током
через вживленные электроды у экспериментальных животных сопровожда-
ется разными комбинациями ответных вегетативных и поведенческих ре-
акций в зависимости от того, какая область гипоталамуса подвергалась
раздражению. Так, например, при стимуляции задней области гипоталаму-
са происходит повышение кровяного давления, увеличение частоты сокра-
щений сердца, учащение и углубление дыхания, расширение зрачков. Од-
новременно с этим поднимается шерсть, выпускаются когти, изгибается
горбом спина и оскаливаются зубы. Вегетативные сдвиги свидетельствуют
об активации симпатического отдела, а поведение является аффективно-
оборонительным. Эта область гипоталамуса получила название эрготроп-
ной, т. е. способствующей мобилизации энергии.
Раздражение ростральных отделов гипоталамуса и преоптической облас-
ти вызывало у тех же животных пищевое поведение: они начинали погло-
щать пищу, даже если были досыта накормлены. У них увеличивалось вы-
деление слюны, усиливалась моторика желудка и кишечника, а частота
сердечных сокращений и дыхания уменьшалась, становился меньшим и
мышечный кровоток, что характерно для повышения парасимпатического
тонуса. Эта область гипоталамуса была названа трофотропной.
Таким образом, активация разных областей гипоталамуса запускает за-
программированные комплексы поведенческих и вегетативных реакций. За-
дача гипоталамуса заключается в том, чтобы оценить поступающую к нему
из разных источников информацию и на ее основе выбрать тот или иной
комплекс ответных реакций, объединяющий поведение с определенной
активностью обоих отделов вегетативной нервной системы. Следует обра-
тить внимание, что не только уже произошедшие отклонения параметров
гомеостазиса, но и любое потенциально угрожающее ему событие может
активировать деятельность гипоталамуса (опережающее реагирование).
Так, например, при внезапном возникновении опасной ситуации, угро-
жающей жизни, вегетативные сдвиги у человека (увеличение частоты со-
кращений сердца, повышение кровяного давления и т. п.) происходят бы-
стрее, чем он обратится в бегство, т. е. такие сдвиги уже учитывают харак-
тер последующей мышечной активности.
Непосредственный контроль тонуса вегетативных центров, а значит и
выходной активности вегетативной нервной системы, гипоталамус осуще-
ствляет с помощью эфферентных связей с тремя важнейшими областями
(рис. 4.27). Во-первых, он регулирует активность нейронов ядра солитар-
ного тракта, которое является главным адресатом сенсорной информации
от внутренних органов, участвует в контроле температуры, кровообраще-
ния, дыхания и взаимодействует с ядром блуждающего нерва и других па-
расимпатических нейронов. Во-вторых, гипоталамус определяет актив-
4. Функции центральной нервной системы • 215
Рис. 4.27. Схема гипоталамического контроля вегетативных функций.
1. Эфферентный путь от гипоталамуса к ядру солитарного тракта.
2. Эфферентный путь от гипоталамуса к ростральной вентральной области продолговатого
мозга.
3. Эфеерентный путь от гипоталамуса к вегетативным нейронам спинного мозга.
ность ростральной вентральной области продолговатого мозга, имеющей ре-
шающее значение в повышении общей выходной активности симпатиче-
ского отдела. Эта активность проявляется в повышении кровяного давле-
ния, увеличении частоты сокращений сердца, секреции потовых желез,
расширении зрачков и сокращении мышц, поднимающих волосы. В треть-
их, некоторые нейроны гипоталамуса образуют прямые проекции на веге-
тативные нейроны спинного мозга, что позволяет быстро изменять характер
их активности.
4.1.2.2. Роль гипоталамуса в регуляции эндокринных функций
Группы мелкоклеточных нейронов гипоталамуса способны действовать как
нейроэндокринные преобразователи, трансформирующие кратковременные
нервные импульсы в длительные гуморальные влияния. В области воронки
мозга секретируемые мелкоклеточными нейронами гипоталамуса пептиды
по системе портальных вен поступают в аденогипофиз (рис. 4.28) и регули-
руют образование его гормонов. Гипофизотропные нейроны гипоталамуса
получают информацию от разветвленной афферентной сети волокон, пере-
дающих сигналы от лимбической системы, ствола, зрительной, слуховой и
обонятельной сенсорных систем. В результате интеграции этой обширной
информации формируется выходной сигнал в виде необходимой порции
нейрогормонов.
Две разновидности крупноклеточных нейронов гипоталамуса, находя-
щиеся в супраоптическом и паравентрикулярном ядрах, синтезируют вазо-
прессин и окситоцин. Эти нейропептиды доставляются аксонным транспор-
том к окончаниям нейронов в задней доле гипофиза, где депонируются и
216 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
III
желудочек
Гипоталамус
Секретирующие
гормоны клетки
аденогипофиза
Верхняя
артерия
гипофиза
Портальные
вены
Капилляры
Поступление
нейросекретов
в капилляры
Нейро-
гипофиз
Нижняя
артерия
гипофиза
Адено-
1 гипофиз
Рис. 4.28. Схема гипоталамического контроля деятельности гипофиза.
1. Нейроны супраоптического и паравентрикулярного ядер гипоталамуса образуют вазопрес-
син и окситоцин, которые выделяются в задней доле гипофиза в кровь.
2 и 3. Нейроны, секретирующие либерины и статины в кровеносные капилляры; через пор-
тальные вены эти пептиды поступают к эндокринным клеткам аденогипофиза.
4 и 5. Нейроны, регулирующие деятельность нейросекреторных клеток гипоталамуса.
откуда выделяются в кровь, чтобы действовать в качестве гормонов. Круп-
ноклеточные нейроны гипоталамуса выполняют функцию нейроэндокрин-
ного преобразователя, в котором количество выделяемых нейрогормонов
увеличивается пропорционально электрической активности таких клеток.
4.8. Функции лимбической системы мозга
Лимбическая система участвует в формировании целенаправленного пове-
дения: она определяет возникновение мотиваций (побуждений к действию)
и эмоций, способствует научению и образованию следов долговременной
памяти. Структуры мозга, входящие в эту систему, объединены пучками
нервных волокон в кольцо, охватывающее верхнюю часть ствола
(рис. 4.29). Мамиллярные тела, миндалины, гиппокамп, свод и перегородка
соединены несколькими пучками аксонов так, что нервные импульсы рас-
пространяются от одной лимбической структуры к другой. Гипоталамус
соединен с гиппокампом и перегородкой волокнами свода, а терминальной
полоской (stria terminalis) и вентральным амигдалофугалъным путем — с
миндалиной. В лимбическую систему включают также обонятельный мозг,
гипоталамус и соседние области древней и старой коры мозга: прилегаю-
щие к гиппокампу грушевидные доли и поясные извилины, которые занимают
медиальную поверхность больших полушарий непосредственно над мозо-
листым телом и, как пояс, огибают таламус.
4. Функции центральной нервной системы • 217
Терминальная
полоска
Свод _Д_
Передняя
комиссура
Поясная
извилина
Обонятельная
луковица
Передние
ядра таламуса
От мамиллярного
тела к таламусу"”
Таламус
К медиальной части
переднего мозга
Продольный
— дорсальный
пучок
Мамиллярное
Гиппокамп тело
К гипоталамусу
Миндалина
Рис. 4.29. Схема взаимодействия отдельных структур лимбической системы.
Стрелки показывают преимущественное направление распространения возбуждения, возник-
шего в лимбической системе. Однако возбуждение может передаваться и в обратном направ-
лении, поскольку связи между лимбическими структурами являются бидирекциональными.
Входящие в лимбическую систему структуры имеют двусторонние связи
с вентральной покрышкой среднего мозга и благодаря этому способны из-
менять характер управления вегетативными и двигательными реакциями,
контролируемыми ретикулярной формацией, а ретикулярная формация, со
своей стороны, регулирует активность лимбической системы. С помощью
передних ядер таламуса лимбическая система имеет связь с лобной корой,
контролирующей активность лимбической системы. Гиппокамп и минда-
лины, расположенные в височных долях, непосредственно сообщаются с
височной корой, через которую получают переработанную в ней информа-
цию от зрительной, слуховой и соматосенсорной коры.
Ассоциативные области коры, функционально взаимодействующие с
лимбической системой, относят к лимбической коре и включают в нее ме-
диальные и вентральные поверхности лобных долей, часть медиальной по-
верхности затылочных долей, а также обращенную кпереди поверхность
височных долей. Лимбическая кора совместно с лимбической системой
организует мотивационные и эмоциональные аспекты поведения, участву-
ет в образовании долговременной памяти и формировании научения. Она
интегрирует информацию, полученную одновременно от лимбической
системы и регионов вторичной сенсорной коры, после чего оказывает
влияние на ассоциативные области коры, создающие план предстоящих
действий. Взаимодействие лимбических структур и коры является физио-
логической основой для формирования субъективных переживаний эмо-
ций, а связи между лимбической системой и стволом служат для возникно-
218 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
вения вегетативных и двигательных реакций, являющихся еще одним ком-
понентом эмоций.
Во время проведения стереотаксических операций некоторым категори-
ям больных была установлена зависимость между искусственной электри-
ческой стимуляцией различных лимбических структур и субъективным пе-
реживанием приятных ощущений (радость, комфорт, покой) или, напро-
тив, неприятных (тревога, беспокойство, уныние). К позитивным эмоцио-
генным зонам принадлежат латеральный и перивентрикулярный гипотала-
мус, перегородка, покрышка среднего мозга. Негативных эмоциогенных
зон в мозгу гораздо меньше, чем позитивных, а многие структуры (и в том
числе, по-видимому, гиппокамп) нейтральны — их стимуляция не сопро-
вождается приятным или неприятным ощущением. В мозгу человека боль-
ше областей, стимуляция которых вызывает приятные ощущения.
Естественное возбуждение эмоциогенных структур и соответствующее
ему возникновение эмоций происходит в результате модуляции их дея-
тельности дофаминергическими нейронами вентральной покрышки сред-
него мозга. Волокна дофаминергических нейронов идут в составе медиаль-
ного пучка переднего мозга и образуют диффузные проекции на перегород-
ку, обонятельные луковицы, миндалины, прилегающее ядро переднего
мозга и фронтальную лимбическую кору. Субъективные ощущения, пере-
живаемые человеком как эмоции, фиксируются в памяти, и поэтому он
стремится к повторению своих действий, если они вели к переживанию
положительных эмоций, и избегает совершать действия, связанные с воз-
никновением отрицательных эмоций.
4.8.1. Функции миндалин
Миндалевидный комплекс ядер находится в глубине передней части ви-
сочной доли, его базолатералъные ядра получают афферентные проекции
из двух источников: 1) от сенсорных ядер таламуса и 2) от височной и ор-
битофронтальной коры (рис. 4.30). Эмоциогенные стимулы, действующие
на организм, активирует миндалины с помощью короткого прямого пути
передачи сигналов от сенсорных ядер таламуса, лишь после этого к ним
поступает информация о том же раздражителе, предварительно перерабо-
танная в коре. Миндалины сопоставляют сенсорную информацию, посту-
пившую от таламуса, с информацией, пришедшей из коры, чтобы выде-
лить сигналы опасности. Сличение информации происходит в базолате-
ральном ядре миндалины, а затем в ее центральном ядре формируется эф-
ферентный ответ. Нейроны центрального ядра образуют прямые и непря-
мые проекции на другие лимбические структуры, гипоталамус, ствол, по-
ясную извилину, орбитофронтальную кору и ассоциативные области
коры.
В угрожающей ситуации нейроны базолатеральных ядер миндалин не-
медленно активируются, что происходит, например, при вызывающем
боль действии электрического тока. Если пришедшая чуть позже информа-
ция из коры подтверждает наличие опасности, нейроны центрального ядра
генерируют эфферентный ответ миндалин. Миндалины сохраняют следы
долговременной памяти об эмоциональном значении стимула, и поэтому
после их удаления или искусственной блокады синапсов исчезают не толь-
ко врожденные реакции на угрожающие стимулы, но и приобретенная ра-
нее способность оценивать ситуацию как угрожающую по косвенным при-
знакам.
После двустороннего удаления части височных долей, включая минда-
4. Функции центральной нервной системы • 219
Лемнисковый Экстра-
путь слуховой лемнисковый
системы путь
Рис. 4.30. Схема вовлечения миндалин мозга в ответную реакцию на угрожающие
слуховые раздражители.
Миндалины получают прежде всего афферентную информацию от сенсорных ядер таламуса, а
затем к ним поступают сигналы о том же раздражителе, но после переработки информации о
нем в коре. В базолатеральном ядре происходит сличение информации, поступившей из обоих
источников, на этой основе нейроны центрального ядра создают выходные сигналы.
лины и гиппокамп, у обезьян возникает комплекс поведенческих рас-
стройств, определяемый как синдром Клювера—Бьюси и характеризующий-
ся прожорливостью, или гиперфагией, гиперсексуальностью и исчезнове-
нием чувства страха. Все проявления этого синдрома обусловлены наруше-
нием способности оценивать значение информации, поступающей из окру-
жающей среды, и связывать ее со своим собственным состоянием. После
двустороннего удаления только миндалин нарушается социальное поведе-
ние обезьян, что проявляется замкнутостью, покорностью, постоянным
стремлением к уединению.
Миндалины участвуют и в возникновении положительных эмоций, на-
пример удовольствия, связанного с определенной пищей (ее видом, вкусом
и запахом), они определяют предпочтение той или иной жидкости для пи-
тья, определенного места обитания, выбор полового партнера. Некоторые
нейроны миндалин чувствительны к действию половых гормонов, и, бла-
годаря своим связям с обонятельными луковицами, способны изменять
активность под влиянием феромонов — веществ, выделяемых особями про-
тивоположного пола во внешнюю среду. При возникновении конкурирую-
220 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Анатомическая структура
Эффект, обусловленный
раздражением миндалин
Внешние проявления
Безусловный
угрожающий
раздражитель
Парабрахиальмор ядро
Миндалина
Каудальная часть ретику-
Ядро тройничного нерва
Покрышка,
Голубое пятно
Условный
угрожающий
раздражитель
Цегиральное серое вещество
вокруг водопровода
Дорсальное ядро
блуждающего нерва
Двойное ядро
Усиленное дыхание
Активация дофаминергических.
J* норадренергических и
/ холинергических структур
Повышение тонуса мышц
Паравентрикулярное ядро
гипоталамуса
Выделение кортикотропина
(АКТГ)
Парасимпатическая активация
Брадикардия, мочеиспускание,
дефекация, язвы желудка
Активное бодрствование,
десинхронизация ритма ЭЭГ
.? Оцепенение, мышечная дрожь
Выделение кортикостероидов
(стресс)
Рис. 4.31. Схема эфферентных путей от миндалин к гипоталамусу и стволу, при ак-
тивации которых наблюдаются внешние проявления страха.
При активации миндалин возникает комплекс поведенческих реакций, возникновение каждой
реакции обусловлено проведением электрических сигналов от миндалин к нескольким струк-
турам мозга, каждая из которых определяет разные проявления поведения.
щих мотиваций миндалины обеспечивают выбор доминирующей потребно-
сти, определяя ее на основе зафиксированной предшествующим опытом
эмоциональной оценки событий. В результате такой оценки миндалины
могут переключать цели поведения с второстепенных на наиболее значи-
мую. После удаления миндалин такое переключение становится почти не-
возможным.
Выходная активность миндалин адресована лимбической коре, а также
гипоталамусу и стволу мозга. Активация эфферентного пути от миндалин
к орбитофронтальной коре и поясной извилине приводит к осознанию пе-
реживаемых эмоций. Одновременная передача информации от миндалин к
латеральному гипоталамусу и стволу мозга активирует центры регуляции
вегетативных и эндокринных реакций, сопровождающих, например, воз-
никновение страха: увеличение частоты сокращений сердца, учащение ды-
хания и т. д. (рис. 4.31). Эти периферические реакции способствуют моби-
лизации энергии для реакций борьбы или бегства.
4.8.2. Функции гиппокампа
Гиппокамп расположен в медиальной части височной доли, афферентную
информацию он получает от близлежащей энторинальной области коры, в
которой происходит переключение сигналов, приходящих из нескольких
источников. Это, во-первых, сенсорные и ассоциативные области коры, в
которых перерабатывается информация обо всех действующих сенсорных
стимулах. Во-вторых, это мотивационные структуры мозга, прежде всего
гипоталамус, перегородка и миндалины. В третьих, это фронтальная ассо-
циативная кора, определяющая стратегию поведения. В четвертых, ретику-
лярная формация, регулирующая активность корковых нейронов и связан-
4. Функции центральной нервной системы • 221
ная с гиппокампом волокнами свода. Функция гиппокампа состоит в регу-
ляции распределения всех поступающих информационных потоков, класси-
фикации таких потоков и создании условий для образования энграмм (сле-
дов) памяти.
Двустороннее удаление гиппокампа у человека приводит к антероград-
ной амнезии — утрате способности образовывать новую долговременную
память при сохранении памяти о событиях, предшествовавших операции.
Двустороннее удаление гиппокампа у экспериментальных животных не по-
зволяет им вырабатывать новые поведенческие навыки. После кратковре-
менной стимуляции входящих в гиппокамп волокон обонятельного тракта
его пирамидные нейроны начинают разряжаться необычно долго, на про-
тяжении нескольких суток и даже недель, что было названо долговременной
потенциацией. Она возникает в глутаматергических синапсах при участии
NMDA-рецепторов, связанных с мембранными каналами для ионов каль-
ция. Ток этих ионов в постсинаптический нейрон вызывает активацию
вторичных посредников и последующее изменение метаболической актив-
ности не только постсинаптического, но и пресинаптического нейрона.
При повторяющейся активации одних и тех же афферентных входов, кон-
вергирующих к гиппокампу, в таких синапсах происходит структурная пе-
рестройка, которая повышает эффективность синаптической передачи и
является основой долговременной памяти.
В то же время гиппокамп не является единственным регионом мозга,
где происходит долговременная потенциация, как не является и единст-
венным местом хранения следов о предшествующем индивидуальном опы-
те. Память о прежних событиях сохраняется и после удаления гиппокампа,
а некоторые разновидности долговременной памяти могут образоваться и
без него.
4.9. Функции базальных ганглиев (стриопаллидарная система)
К базальным ганглиям относят хвостатое ядро и скорлупу, вместе образую-
щие полосатое тело (corpus striatum, а также neostriatum), бледный шар (glo-
bus pallidus, pallidum) и субталамическое ядро (рис. 4.32). С базальными
ганглиями функционально связана черная субстанция среднего мозга, со-
стоящая из ретикулярной части, сходной по происхождению с внутренним
сегментом бледного шара, и компактной части, образующей дофаминерги-
ческую проекцию на полосатое тело. Наиболее изученная функция базаль-
ных ганглиев заключается в их участии в планировании и инициации про-
извольных движений. Наряду с этим базальные ганглии участвуют в позна-
вательной деятельности мозга, а также в формировании эмоций.
4.9.1. Взаимодействие базальных ганглиев с другими
структурами мозга
Афферентная информация поступает к базальным ганглиям из нескольких
регионов коры', префронтальной ассоциативной коры, первичной и вто-
ричной моторной коры, соматосенсорной коры, теменных и височных об-
ластей ассоциативной коры. Все афферентные волокна оканчиваются на
нейронах полосатого тела, которые объединяются в функциональные моду-
ли, однотипные по способу переработки информации. Соматотопически
организованные волокна нейронов моторных и сенсорных регионов коры
направляются к соответствующим им модулям, в результате пучки волокон
распределяются в виде отдельных полосок, из-за которых полосатое тело
Ill • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Первичная моторная кора
Соматосенсорная кора
Заднетеменная кора
Добавочный
моторный ареал
Премоторная
кора
Г лутамат(+)
Таламус
офамин
Полосатое
тело
Бледный шар
внутр, наруж.
К стволу и
спинному
мозгу
нальный
путь
Субталамическое
ядро
АХ+
компактная ретикулярная
Черная субстанция
Кортикоспи-
*АМК(-
Рис. 4.32. Схема циркуляции возбуждения между системой базальных ганглиев и
корой больших полушарий.
Базальные ганглии включают хвостатое ядро и скорлупу, вместе образующие полосатое тело,
бледный шар и субталамическое ядро; с базальными ганглиями взаимодействует черная суб-
станция среднего мозга. Базальные ганглии взаимодействуют с корой при планировании и
инициации произвольных движений, при этом образуются две цепи нейронных переключе-
ний: 1) кора—полосатое тело—бледный шар—таламус—кора; 2) кора—полосатое тело—блед-
ный шар—таламус—полосатое тело.
получило свое название. Окончания волокон кортикальных нейронов об-
разуют с нейронами полосатого тела возбуждающие глутаматергические
синапсы.
В базальных ганглиях входящие сигналы распределяется независимо
друг от друга по параллельным путям переработки информации. Эти пути
образуют несколько самостоятельных функциональных кругов, включаю-
щих в себя помимо базальных ганглиев разные регионы коры. В зависимо-
сти от того, какая область коры включена в функциональный круг, опре-
деляется и специфическая роль каждой структуры базальных ганглиев.
Скорлупа участвует преимущественно в координации движений тела, одни
регионы хвостатого ядра необходимы для регуляции движений глаз, дру-
гие — для формирования побуждений к конкретным действиям, выбора
стратегии поведения или осуществления когнитивных (познавательных)
4. Функции центральной нервной системы • 223
функций. Вентральные отделы полосатого тела участвуют в формировании
эмоций.
Скелетомоторный круг нейронных переключений используется при
планировании и осуществлении контроля произвольной двигательной ак-
тивности человека. Вначале активируются премоторная и моторная лоб-
ные области коры, которые возбуждают топографически соответствующие
им модули полосатого тела. Выходная активность полосатого тела распре-
деляется между внутренней частью бледного шара и ретикулярной частью
черной субстанции среднего мозга, для которых характерна спонтанная
фоновая активность. Под влиянием преобладающих в полосатом теле эф-
ферентных тормозных ГАМКергических нейронов эта активность умень-
шается. Значительно меньшая часть выходных нейронов полосатого тела
использует в качестве медиатора вещество Р, оказывающее возбуждающее
действие. Одновременно с этим многие нейроны полосатого тела синтези-
руют энкефалин, действующий в качестве нейромодулятора, и суммарный
итог действия полосатого тела на бледный шар и черную субстанцию ока-
зывается тормозно-модулирующим.
Находящаяся под контролем полосатого тела выходная активность ней-
ронов бледного шара и черной субстанции адресуется клеткам моторных
вентролатеральных ядер таламуса. Эти нейроны образуют проекцию на
премоторные области коры, замыкая большой круг нейронных переключе-
ний: премоторные и моторные области коры—полосатое тело—бледный
шар или ретикулярная часть черной субстанции—таламус—премоторная
кора. Нейроны моторных ядер таламуса кроме премоторной коры образу-
ют проекцию на клетки полосатого тела: так замыкается малый круг пере-
ключений между полосатым телом — бледным шаром или черной субстан-
цией — таламусом — и вновь полосатым телом. Субталамическое ядро по-
лучает топографически организованные сигналы сигналы от премоторной
и моторной коры, в соответствии с которыми оно регулирует выходную
активность бледного шара.
Распространение возбуждения по этим кругам нейронных переключе-
ний происходит до того момента, пока базальные ганглии не выберут из
предложенного корой ряда моторных программ наиболее приемлемую для
конкретной ситуации, а затем инициируют ее реализацию. Конечным ре-
зультатом такого взаимодействия нейронов является создание в моторной
коре двигательных программ, предназначенных для исполнения двигатель-
ными центрами ствола и нейронами спинного мозга.
Окуломоторный круг, образующийся при участии базальных ганглиев,
используется для регуляции произвольных движений глаз, связанных с при-
влечением внимания к одному из объектов, находящихся в зрительном по-
ле. Этот круг объединяет афферентную информацию от заднетеменной об-
ласти коры (поле 7) и небольшого по площади фронтального глазного по-
ля (поле 8 по Бродману). В поле 7 коры создается нейронная карта окру-
жающего зрительного пространства, а поле 8 необходимо для регуляции
движений глаз, фиксирующих определенную часть этого пространства.
Входные сигналы от обоих полей поступают к соответствующим им моду-
лям хвостатого ядра, которое в зависимости от характера сигналов опреде-
ляет активность нейронов бледного шара и ретикулярной части черной
субстанции. Выходная активность последних адресована тем нейронам та-
ламуса, которые связаны с фронтальным глазным полем и влияют на ак-
тивность его нейронов.
Активность одиночных нейронов базальных ганглиев повышается еще
до начала произвольных движений контралатеральных конечностей, что
224 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
указывает на их участие в планировании и формировании начальной стадии
произвольных движений. Базальные ганглии не имеют непосредственного
выхода к моторным ядрам ствола и спинного мозга и поэтому не влияют
на исполнение уже начавшихся движений, а их электрическая активность
не коррелирует с активностью и силой сокращения отдельных мышц. На-
личие в составе нейронных кругов переключения возбуждающих и тормоз-
ных нейронов позволяет сбалансировать выходную активность двигатель-
ных систем так, чтобы обеспечить необходимую плавность движений. Ба-
зальные ганглии облегчают одни и угнетают другие движения, что способ-
ствует чередованию тонических и фазных компонентов движения, т. е. сна-
чала формированию позы, требующейся для совершения конкретного дви-
жения, а затем и самого движения.
Одним из источников афферентной информации, доставляемой к ба-
зальным ганглиям, являются нейроны лобных ассоциативных областей
коры: префронтальной и орбитофронтальной, причастных к программиро-
ванию поведения и эмоционально окрашенному прогнозу возможных ре-
зультатов предстоящих действий. Их глутаматергические аксоны оканчи-
ваются на нейронах полосатого тела, далее формируется большой круг
нейронных переключений, в образовании которого участвуют бледный
шар и ретикулярная часть черной субстанции, а затем ассоциативные ядра
таламуса, замыкающие круг в ассоциативной коре. С этим кругом ней-
ронных переключений связаны формирование мотиваций поведения, про-
гнозирование его результатов, познавательная деятельность и возникнове-
ние эмоций.
4.9.2. Модуляции нейронных переключений в базальных ганглиях
Помимо кортикостриатной проекции полосатое тело получает афферент-
ную информацию от нескольких ядер таламуса, черной субстанции сред-
него мозга, миндалин, ядер шва продолговатого мозга и нейронов голубого
пятна. В результате в каждом модуле полосатого тела создается сложная
сеть переключений из возбуждающих глутаматергичеких окончаний корко-
вых нейронов, модулирующих дофаминергических окончаний нейронов
компактной части черной субстанции, серотонинергических окончаний
нейронов ядер шва и норадренергических окончаний нейронов голубого
пятна. В переработке информации участвуют также локальные интерней-
роны полосатого тела, синтезирующие ацетилхолин и несколько пептид-
ных нейротрансмиттеров.
Дофаминергические нейроны компактной дорсальной части ретикуляр-
ной формации образуют диффузную нигростриатную проекцию на нейроны
полосатого тела. Эта проекция топографически не организована и предна-
значена для модуляции кортикостриатных глутаматергических переключе-
ний. При дегенерации дофаминергических нейронов черной субстанции
нарушается необходимый баланс активности нейронов полосатого тела,
что приводит к характерным двигательным нарушениям.
Дисфункция, вызванная потерей дофаминергических нейронов, прояв-
ляется в виде болезни Паркинсона и характеризуется мышечной ригидностью
(значительным повышением тонуса мышц), резким ограничением сопутст-
вующих движений, или акинезией, а также появлением крупноразмашисто-
го тремора — дрожания конечностей во время покоя. Указанные наруше-
ния двигательной функции возникают в связи с уменьшением влияния до-
фаминергических нейронов и изменением баланса активности в пользу ос-
тальных нейронов полосатого тела. Если же этот баланс нарушается вслед-
4. Функции центральной нервной системы • 225
ствие поражения холинергических или ГАМКергических нейронов полоса-
того тела, то возникает другая разновидность двигательных расстройств,
проявляющихся появлением непроизвольных и нежелательных для челове-
ка насильственных движений — гиперкинезов.
Модулирующее влияние на нейронные переключения в базальных ганг-
лиях оказывают не только дофамин, но и другие биогенные амины (серо-
тонин, норадреналин), а также многочисленные пептидные нейротранс-
миттеры. Столь сложный контроль деятельности базальных ганглиев отра-
жает важность выполняемых ими функций.
4.10. Функции коры больших полушарий
Складчатая наружная поверхность полушарий достигает 1700—2500 см2,
толщина серого вещества варьирует между 1,5 и 5 мм, а находящиеся в
нем нейроны образуют шесть слоев, различающихся между собой по со-
ставу клеток. В зависимости от выполняемых функций области коры под-
разделяются на сенсорные, моторные и ассоциативные (рис. 4.33). К сен-
сорным областям относят: соматосенсорную кору, занимающую задние
центральные извилины, зрительную кору, находящуюся в затылочных до-
лях, и слуховую кору, занимающую часть височных долей. Моторная кора
находится в передних центральных извилинах и в примыкающих к ним
спереди регионах лобных долей. Ассоциативная кора занимает всю осталь-
Первичная
Рис. 4.33. Схема функциональных областей коры.
На рисунке обозначены примерные границы функциональных областей коры на боковой по-
верхности левого полушария.
8-6095
226 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
ную поверхность мозга и подразделяется на префронтальную кору лобных
долей, теменно-височно-затылочную (парието-темпорально-окципиталь-
ную) и лимбическую, к которой относят внутренние и нижние поверхно-
сти лобных долей, внутренние поверхности затылочных долей и нижние
отделы височных долей.
Различные области коры взаимодействуют друг с другом посредством
внутрикортикальных ассоциативных связей или с помощью подкорковых
структур (таламус, базальные ганглии). Регионы коры, находящиеся в раз-
ных полушариях, связаны друг с другом волокнами мозолистого тела, что
позволяет передавать информацию из одного полушария в другое.
4.10.1. Функциональное распределение нейронов в коре
Дендриты пирамидных нейронов коры образуют синапсы с очень большим
количеством афферентов, а их аксоны направляются в белое вещество и
своими разветвленными коллатералями образуют 3/4 всех синапсов корти-
кальных нейронов. Большинство пирамидных нейронов используют в ка-
честве медиаторов возбуждающие аминокислоты (глутамат или аспартат),
но одновременно могут секретировать несколько комедиаторов (нейропеп-
тиды, АТФ), модулирующих синаптический перенос сигналов. Непирамид-
ные нейроны (корзинчатые и звездчатые клетки) в большинстве случаев
синтезируют ГАМК в разных комбинациях с нейропептидами и обладают
тормозным действием, однако часть непирамидных клеток использует воз-
буждающие медиаторы. В отличие от пирамидных нейронов все виды не-
пирамидных клеток образуют не выходящие за пределы коры аксоны и иг-
рают роль внутрикортикальных интернейронов.
Нейроны расположены в коре упорядоченными слоями, специфика ко-
торых отражает их роль в переработке информации. В наружном молеку-
лярном слое проходят соединительные нервные волокна, осуществляющие
внутрикорковые связи соседних нейронов. В наружном зернистом II слое
и в наружном пирамидном III слое содержатся мелкие и средней величи-
ны пирамидные нейроны, аксоны которых служат для связи региона с
другими областями коры. Во внутреннем зернистом IV слое имеются не-
большие звездчатые клетки, получающие афферентные волокна таламиче-
ских нейронов и выполняющие функцию входных нейронов. Наряду с
ними в этом слое содержится много пирамидных нейронов, использую-
щихся для передачи эфферентных сигналов. Во внутреннем пирамидном
V слое сосредоточены крупные пирамидные нейроны (клетки Беца), аксо-
ны которых служат для передачи информации к базальным ганглиям,
стволу и спинному мозгу (кортикоспинальный тракт). Во внутреннем по-
лиморфном VI слое, граничащим с белым веществом, встречаются разной
формы нейроны, а в подслое Via обнаружен самый большой процент пи-
рамидных нейронов (97 %), которые осуществляют передачу информации
к таламусу.
4.10.2. Модульная организация коры
Каждый релейный нейрон таламуса возбуждается в ответ на раздражение
собственного рецептивного поля, после чего активирует звездчатые клетки
IV слоя коры в кортикальной колонке, получающей проекцию от этого та-
ламического нейрона. Каждая кортикальная колонка представляет ориен-
тированный перпендикулярно поверхности цилиндр диаметром 300—500
мкм. Нейроны одной колонки получают сходные афферентные сигналы и
4. Функции центральной нервной системы • 227
образуют синапсы преимущественно друг с другом. В колонках проекци-
онной коры имеются две разновидности пирамидных нейронов, которые
получили название простых и сложных.
Простые нейроны расположены поблизости от входного IV слоя и вы-
полняют функцию детекторов элементарных признаков раздражителя. На-
пример, простые нейроны зрительной коры сильнее всего реагируют на
линейный зрительный стимул в виде полосы или грани между светлой и
темной плоскостью, т. е. реагируют на контраст. Друг от друга они отлича-
ются по чувствительности к разным углам наклона линейного стимула. Ак-
соны нескольких однотипных простых нейронов конвергируют к сложным,
или комплексным, нейронам, поэтому комплексные нейроны реагируют на
стимул позже простых клеток. Рецептивное поле сложных нейронов состо-
ит из небольших рецептивных полей простых нейронов, а эффективными
раздражителями для комплексных нейронов становится уже не линейный
раздражитель определенной ориентации, а характер и направление его
движения через рецептивное поле. Так осуществляется переход от детек-
ции простых сигналов к абстрагированию, обобщению. В этом процессе
участвуют интернейроны локальной сети кортикальной колонки, а итогом
преобразования становится уровень активности выходных нейронов, аксоны
которых покидают колонку, чтобы обеспечить ее взаимодействие с други-
ми колонками.
Аналогичная нейронная организация имеет место в слуховой и мотор-
ной коре, что послужило поводом для гипотезы о модульном строении как
общем принципе организации нейронных ансамблей в коре. Подобный
нейронный ансамбль представляет собой функциональную единицу коры,
для которой используют термины «кортикальная колонка» или «микромо-
дуль». С помощью горизонтальных связей они взаимодействуют с соседни-
ми колонками (микромодулями), образуя вместе с ними гиперколонку, или
модуль. Если микромодули предназначены для распознания элементарных
признаков раздражителя или образа, то модуль служит для комплексного
объединения элементарных признаков.
Разные модули могут быть связаны друг с другом ассоциативными во-
локнами, образованными преимущественно пирамидными нейронами II—
III слоев. Если модули находятся на большом расстоянии друг от друга, та-
кие волокна проходят в составе белого вещества. Модули, расположенные
в разных полушариях, могут передавать информацию посредством волокон
мозолистого тела. Волокна пирамидных нейронов обычно группируются, и
аксоны направляются к клеткам-мишеням в виде пучка волокон. В моду-
лях ассоциативной коры отсутствуют клетки, реагирующие на элементар-
ные раздражители, но имеются нейроны, разряжающиеся только в ответ
на определенный сложный комплекс стимулов. Например, в нижней ви-
сочной доле существуют нейроны, которые активируется только при зри-
тельном восприятии человеческого лица.
4.10.3. Электрическая активность коры
Нейроны коры обладают спонтанной ритмической фоновой активностью,
характер которой изменяется в ответ на стимуляцию периферических ре-
цепторов (вызванные потенциалы). Обе эти разновидности электрической
активности можно зарегистрировать с помощью электродов, помещенных
на поверхности коры (электрокортикограмма) или на коже головы (элек-
троэнцефалограмма, ЭЭГ). ЭЭГ отражает суммарную активность большого
количества одновременно возбуждающихся и тормозимых нейронов, нахо-
8'
228 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Бета-ритм
Альфа-ритм
50 мкВ
/Зрльта-ритм
Рис. 4.34. Основные ритмы электроэнцефалограммы.
Бета-ритм — электрические колебания малой амплитуды с частотой 13—26 Гц, наиболее выра-
женные в лобных областях и характерные для активного бодрствования. Альфа-ритм — харак-
теризуется близкими к синусоидальным волнами с амплитудой 50—100 мкВ и частотой 8—
12 Гц, наблюдается при пассивном бодрствовании и монотонной деятельности. Тета-ритм от-
личается волнами, имеющими амплитуду до 100 мкВ и более с частотой 4—7 Гц, возникает
при переходе ко сну, а также при некоторых формах измененного сознания. Дельта-ритм
представлен волнами с большой амплитудой и малой частотой, 0,3—0,5 Гц, он характерен для
глубокого сна.
дящихся в поверхностных и глубоких слоях коры (рис. 4.34). Регистрируе-
мые разности потенциалов примерно в сто раз слабее, чем в ЭКГ, а час-
тотные характеристики сигналов легли в основу классификации ритмов
ЭЭГ, где различают альфа-волны, следующие с частотой 8—12 Гц, бета-
волны (13—26 Гц), тета-волны (4—7 Гц) и дельта-волны (0,3—0,5 Гц). Из-
менения ритмов ЭЭГ связаны с активностью таламуса и восходящих про-
екций некоторых ядер ствола. После перерезки ствола мозга между верх-
ними и нижними буграми четверохолмия у кошки на электроэнцефало-
грамме постоянно регистрируются низкочастотные волны, обычно харак-
терные для картины сна. Замедленная фоновая активность ЭЭГ имеет ме-
сто у человека во время комы, характеризующейся утратой сознания
вследствие поражения верхних отделов моста или диффузных поражений
коры. Активация холинергических нейронов гигантоклеточной области по-
крышки моста и ростральных отделов ретикулярной формации среднего
мозга приводит к десинхронизации ЭЭГ, т. е. переходу от низкочастотных
волн к бета-ритму. Блокада альфа-ритма происходит в связи с привлече-
нием внимания к новому раздражителю, а смена альфа-волн ЭЭГ в такой
ситуации на бета-ритм обусловлена восходящим активирующим влиянием
норадренергических нейронов голубого пятна.
4. Функции центральной нервной системы • 229
4.10.4. Функции сенсорных областей коры
Сенсорные области коры являются пунктом назначения передающейся в
сенсорных системах информации. Все сенсорные системы организованы
по единому иерархическому принципу. В их рецепторах энергия внешних
стимулов преобразуется в электрохимическую энергию нервных импуль-
сов, которые от чувствительных нейронов передаются нейронам второго
порядка, находящимся в ЦНС. После преобразования сигналов на низшем
переключательном уровне с помощью релейных нейронов информация со-
общается следующему уровню переключения и переработки, пока не посту-
пит в первичную проекционную кору. Почти все проводящие пути вклю-
чают проекционные ядра таламуса, распределяющего информационные
потоки к разным проекционным областям коры (табл. 4.2).
Топографическая упорядоченность нейронных переключений обеспечи-
вает пространственное представительство или проекцию рецептивных по-
лей в коре в виде нейронной карты. Например, каждый квадратный милли-
метр поверхности кожи любого из пальцев имеет собственное представи-
тельство в строго определенной области постцентральной извилины про-
тивоположного полушария. Принцип строго упорядоченной топографиче-
ской организации в соматосенсорной системе называется соматотопией, в
зрительной системе — ретинотопией, в слуховой системе — тонотопией.
Сенсорные системы создают ощущения и восприятие окружающего мира,
контролируют правильность совершаемых человеком движений, использу-
ются в качестве обратной связи для регуляции физиологических процессов
и поддерживают необходимую для бодрствования активность мозга.
В формировании восприятия участвуют первичная и вторичная сенсор-
ная кора, а также ассоциативные области. Первичная сенсорная кора полу-
чает афферентную информацию преимущественно от мономодальных ней-
ронов специфических ядер таламуса. Проекционный нейрон таламуса, по-
лучивший информацию от своего рецептивного поля, возбуждает входные
звездчатые нейроны преимущественно одной кортикальной колонки: этот
Таблица 4.2. Уровни обработки информации в сенсорных системах
Модаль- ность Локализация рецеп- торов Первое пере- ключение Повторные переключения Проекционные области коры
Осяза- ние Кожа Продолгова- тый мозг Таламус Постцентраль- ная извилина
Зрение Палочки и кол- бочки сетчатки Сетчатка гла- за Таламус, верхнее четве- рохолмие Затылочные до- ли, поле 17
Слух Волосковые клет- ки улитки Улитка, спи- ральный ганглий Мост, оливы, нижнее четверохолмие, таламус Поперечная ви- сочная извили- на, поле 41
Равнове- Волосковые клет- Вестибуляр- Глазодвигательные ядра, Постцентраль-
сие ки вестибулярно- го аппарата ные ядра ствол, спинной мозг, мозжечок, таламус ная извилина
Вкус Вкусовые почки языка Продолгова- тый мозг Таламус Постцентраль- ная извилина
Обоня- ние Биполярные клет- ки носовой пазухи Обонятель- ная луковица Пириформная кора Лимбическая система
Боль Ноцицепторы ко- жи Задние рога спинного мозга Таламус, ретикулярная формация Постцентраль- ная извилина, передняя пояс- ная извилина
230 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
принцип соединений обеспечивает топографическое соответствие рецеп-
тивных полей определенным кортикальным колонкам. Большинство ней-
ронов первичных областей коры имеют очень небольшую спонтанную ак-
тивность и возбуждаются сильнее всего в ответ на действие элементарного
и специфического именно для них сенсорного стимула.
Вторичная сенсорная кора отличается от первичной тем, что большую
часть афферентной информации получает не напрямую из таламуса, а из
первичной сенсорной коры, а также из других вторичных областей этой же
сенсорной системы. Благодаря конвергенции входных сигналов из разных
источников здесь происходит поэтапная интеграция информации, посту-
пившей по параллельным путям ее переработки, каждый из которых акти-
вируется каким-то одним, элементарным признаком комплексного раздра-
жителя. Во вторичной коре нейроны сильнее всего реагируют на опреде-
ленный комплекс признаков, а не на один из них, что присуще нейронам
первичной коры.
4.10.4.1. Функция соматосенсорной коры
Центральные отростки первичных сенсорных нейронов, передающих сиг-
налы от тактильных рецепторов кожи и от проприоцепторов мышц, сухо-
жилий и суставов, входят в спинной мозг через задние корешки и образу-
ют коллатерали, которые в составе задних столбов спинного мозга дости-
гают продолговатого мозга. Поступающие сигналы принимают нейроны
ядер задних столбов, аксоны которых немедленно переходят на противопо-
ложную сторону продолговатого мозга в составе медиальной петли или ме-
диального лемниска (благодаря этому термину весь путь получил название
лемнискового) и направляются к
Рис. 4.35. Пространственное представи-
тельство тела в соматосенсорной коре.
Соматотопическая проекция тела в постцен-
тральной извилине отражает плотность чувст-
вительной иннервации периферии: чем выше
она на периферии, тем больше представитель-
ство этой области в соматосенсорной коре.
заднему вентролатеральному ядру
таламуса. К этому же ядру таламу-
са поступает информация от кожи
противоположной стороны лица,
переданная по ветвям тройнично-
го нерва. Нейроны вентролате-
ральных ядер таламуса образуют
проекцию на заднюю централь-
ную извилину, представляющую
собой проекционную соматосен-
сорную область S,. Часть инфор-
мации поступает также во вторич-
ную соматосенсорную область Slb
которая расположена на верхней
стенке боковой борозды, разде-
ляющей теменную и височную
доли, и имеет билатеральное
представительство поверхности
тела. Она участвует в координа-
ции одновременных движений
обеих рук (ног).
Карта соматосенсорного пред-
ставительства в коре — сенсорный
гомункулус, т. е. человечек (рис.
4.35), не соответствует пропорци-
ям человеческого тела, она отра-
4. Функции центральной нервной системы • 231
Поперечный разрез постцентральной извилины
Рис. 4.36. Регионы соматосенсорной коры в области представительства руки.
Формирование ощущений, а затем и восприятия происходит в несколько этапов. Четыре ре-
гиона постцентральной извилины (За, 36, 1 и 2) получают афферентную информацию от раз-
ных видов рецепторов, расположенных в одной области тела: За — от интрафузальных рецеп-
торов, 36 — от поверхностных рецепторов кожи, 1 — от быстро адаптирующихся рецепторов,
2 — от рецепторов, реагирующих на сильное давление. В полях За и 36 осуществляется первый
элементарный этап переработки информации, в полях 1 и 2 — следующий этап с участием
комплексных нейронов, реагирующих на несколько элементарных признаков. В полях 5 и 7
заднетеменной области осуществляется дальнейшая переработка информации, в результате
которой возникает целостное восприятие действующего раздражителя.
жает относительную плотность чувствительной иннервации противополож-
ной половины тела. Соматосенсорная кора разделяется на четыре области
(рис. 4.36), занимающие поля За, 36, 1 и 2. В поле За поступает информа-
ция от рецепторов мышц и суставов, в поле 36 — от поверхностных рецеп-
торов кожи: эта информация содержит самые элементарные характеристи-
ки стимула. В поле 1 происходит дальнейшая переработка информации,
поступившей от рецепторов кожи, а в поле 2 она комбинируется с той, ко-
торая содержит сведения о мышцах и суставах. Таким образом, если в по-
лях За и 36 формируются элементарные представления о стимуле, то в по-
лях 1 и 2 — комплексные.
Большую часть входов во вторичную сенсорную кору образуют клетки
232 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
областей S, от обоих полушарий, а поэтому в областях Sn представлены обе
половины тела. Выходы из первичной сенсорной коры, а также из вторич-
ной — Sn, направлены к прилежащим регионам теменной коры — это ассо-
циативные области, которые интегрируют сенсорную информацию. Кроме
того, от поля 2 есть выход к первичной моторной коре, он имеет большое
значение для осуществления точных движений.
Представительство различных частей тела в соматосенсорной коре ос-
новано на генетически предопределенном характере нейронных переклю-
чений на разных иерархических уровнях. Однако при изменении характера
поступающей информации с периферии представительство разных частей
тела в коре может измениться у взрослых животных и у людей. Замена
коркового представительства руки после ее ампутации была обнаружена у
людей, продолжавших ощущать отсутствующую конечность и даже чувст-
вовать в ней боль: такие боли называют фантомными. В одних случаях ме-
сто руки на пространственной карте соматосенсорной коры заняли отдель-
ные участки лица. Вследствие этого прикосновение к таким участкам вос-
принималось, как прикосновение к отсутствующим пальцам. В других слу-
чаях представительство кисти руки занимали другие смежные области:
предплечье, плечо, верхняя часть туловища. Подобные перестройки сома-
тотопического представительства определяются как перекартирование.
Карту представительства руки в коре у людей можно с точностью до
1 мм исследовать с помощью магнитоэнцефалограммы. У людей этим мето-
дом обнаруживаются четкие границы полей всех пяти пальцев. При врож-
денном сращении пальцев (такой дефект обозначается термином синдак-
тилия) их отдельное представительство в коре не обнаруживается. Но
спустя несколько недель после хирургического разделения пальцев начи-
нают выявляться промежутки между отдельными представительствами
пальцев в коре.
4.10.4.2. Функция зрительной коры
Информация от сетчатки глаз передается по волокнам зрительных нервов
и поступает к нейронам латеральных коленчатых тел, относящихся к тала-
мусу. Релейные нейроны латеральных коленчатых тел образуют ретиното-
пическую проекцию на кортикальные колонки 17-го поля (в литературе
оно чаще обозначается как V[). Это поле занимает около четверти поверх-
ности затылочной области преимущественно на внутренней стороне каж-
дого полушария. 17-е поле является первичной зрительной корой, в которой
каждый участок соответствует определенной точке сетчатки, причем об-
ласть центральной ямки представлена с наибольшим разрешением и зани-
мает около половины всей поверхности первичной зрительной коры.
В зрительной коре существует около 20 популяций простых нейронов,
каждая из которых возбуждается линейным стимулом, отличающимся по
своему наклону от остальных минимум на 10°. Комплексные нейроны от-
личаются от простых большей величиной рецептивного поля и обычно
возбуждаются в момент движения стимула в строго определенном направ-
лении, тогда как линейная ориентация имеет для них меньшее значение,
чем для простых нейронов. Многие комплексные нейроны бинокулярны,
т. е. способны реагировать на стимуляцию обоих глаз, в то же время им
присуща глазодоминантностъ, т. е. большая реакция на стимуляцию одного
из глаз.
Простые и комплексные нейроны, сходные по своей чувствительности
к стимулу определенной линейной ориентации, образуют вертикальную
4. Функции центральной нервной системы • 233
Рис. 4.37. Схема формирования кортикальных колонок первичной зрительной
коры.
Ориентационные колонки имеют вид узких полосок шириной 25—30 мкм, в каждой колонке
содержатся нейроны с одинаковой ориентацией рецептивных полей.
А. Группа ориентационных колонок, различающихся наибольшей чувствительностью к опре-
деленной ориентации линейных стимулов и поступлению информации от определенного
глаза.
Б. Каждая группа ориентационных колонок содержит регулярно повторяющиеся скопления
клеток в виде капель (blobs), которые отсутствуют в IV слое коры. Эти клетки осуществляют
переработку информации о цвете наблюдаемых объектов.
ориентационную колонку. Соседние колонки различаются систематическим
изменением оси линейной ориентации стимула, которая от одной колонки
к другой смещается приблизительно на 10° (рис. 4.37). На площади при-
мерно в 1 мм2 размещается комплект ориентационных колонок, рассчи-
танный на регистрацию наклона линейных стимулов в пределах 360°. По-
мимо этого в комплект входят две глазодоминантные колонки, одна из кото-
рых получает информацию только от левого, а другая — только от правого
глаза. Еще одним обязательным компонентом такого набора колонок яв-
ляется несколько скоплений клеток, предназначенных для восприятия
цвета и расположенных в виде вставок или капель {blobs) между ориентаци-
онными колонками. Полный комплект, включающий ориентационные,
глазодоминантные колонки и воспринимающие цвет вставки-blobs, полу-
чил название гиперколонки, которая является элементарным модулем пере-
работки зрительной информации.
Вторичная зрительная кора примыкает к первичной зрительной коре и
занимает 18-е и 19-е поля затылочных долей. В них, однако, процесс пере-
работки зрительной информации не завершается, и у человека в нем уча-
ствует свыше 30 регионов, связанных между собой приблизительно 300 со-
единительными путями, образующими сложную сеть нейронных переклю-
234 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
чений. Эти регионы различаются
между собой по выполняемой
функции, например, регион Vv,
или МТ (медиотемпоральный)
осуществляет переработку инфор-
мации о движении объекта в зри-
тельном поле, а регион Viv, рас-
положенный на стыке теменной и
височной областей, обеспечивает
восприятие формы и цвета объ-
екта.
Существуют два пути, начи-
нающихся в первичной зритель-
ной коре и предназначенных для
раздельной переработки зритель-
ной информации: вентральный и
дорсальный (рис. 4.38). Вентраль-
ный путь проходит к нижней ви-
сочной извилине, где обнаружены
нейроны, имеющие очень боль-
шие рецептивные поля. В этой
области отсутствует ретинотопи-
ческая организация, в ней проис-
ходит опознание зрительных сти-
мулов, устанавливается их фор-
ма, величина и цвет. Здесь име-
ются лицеспецифические нейроны, избирательно реагирующие на появле-
ние в зрительном поле лица человека, причем одни нейроны активируют-
ся, если лицо повернуто в профиль, а другие реагируют на поворот в фас.
При поражении этой области может возникнуть прозопагнозия, при кото-
рой человек перестает узнавать знакомые ему лица. В средней, а также в
верхней височной извилинах находятся нейроны, необходимые для вос-
приятия движущихся объектов и для фиксации внимания на неподвижных
объектах.
Дорсальный путь из первичной зрительной коры ведет к заднетеменным
областям. Его функциональное значение заключается в определении вза-
имного пространственного расположения всех одновременно действующих
зрительных стимулов. Повреждение этой области коры приводит к дефекту
пространственных ощущений и нарушению зрительно-моторной интегра-
ции: человек видит предмет и может правильно описать его форму и цвет,
но при попытке взять этот предмет рукой промахивается. Для удобства за-
поминания вентральный путь принято ассоциировать с вопросом, «что»
представляет собой объект, а дорсальный путь — с вопросом, «где» этот
объект находится.
Дорсальный
путь ("где")
Первичная
зрительная
кора
Вентральный
путь ("что")
Рис. 4.38. Схема двух основных направле-
ний переработки зрительной информации
за пределами первичной сенсорной об-
ласти.
Вентральный путь из зрительной коры прохо-
дит к нижней и средней извилинам височной
коры, объединяя структуры, обеспечивающие
восприятие формы и цвета наблюдаемых зри-
тельных объектов.
Дорсальный путь ведет из зрительной коры к
структурам, функция которых состоит в фор-
мировании пространственного представления
о наблюдаемом объекте.
4.10.4.3. Функция слуховой коры
Первичная слуховая кора получает сенсорную информацию от релейных
нейронов таламуса, она расположена в глубине латеральной борозды (поле
41), отделяющей височную долю от лобной и передних отделов теменной
доли. Вторичная слуховая кора получает большую часть афферентных сиг-
налов от первичной коры и располагается вокруг нее. К первичной слухо-
вой коре поступает тонотопически организованная информация от слухо-
4. Функции центральной нервной системы • 235
вых рецепторов, предварительно переработанная в переключательных яд-
рах слухового тракта. В передних областях слуховой коры находятся ко-
лонки, предназначенные для получения и переработки высокочастотных
акустических сигналов, позади них — колонки, получающие информацию
о низкочастотных звуковых раздражителях. Параллельно с обработкой ин-
формации о высоте тона происходит оценка интенсивности звука и вре-
менных интервалов между отдельными звуками. Для некоторых нейронов
слуховой коры наиболее сильными стимулами являются звуки определен-
ной длительности, для других — повторяющиеся звуки, для третьих — шу-
мы, имеющие широкий диапазон частот.
Нейроны такого рода относятся к категории простых, наряду с ними су-
ществуют комплексные нейроны, реагирующие на определенные частот-
ные или амплитудные модуляции звуков. Наличие простых и комплексных
нейронов свидетельствует о соблюдении общего принципа переработки
информации в сенсорной коре, когда этап регистрации элементарных при-
знаков раздражителя и определения различных составляющих звукового
сигнала предшествует следующему этапу, на котором возникает комплекс-
ный образ.
4.10.5. Функции ассоциативных областей коры
Ассоциативная кора организована подобно сенсорным областям: ее нейро-
ны объединены в вертикальные колонки. Она интегрирует активность сен-
сорных областей коры и осуществляет их функциональное объединение с
моторной корой при выполнении целенаправленных и осознаваемых дей-
ствий человека. Три ассоциативные области коры участвуют в реализации
разных когнитивных (познавательных) функций.
4.10.5.1. Функции теменно-височно-затылочной коры
Здесь происходит интеграция сенсорных функций, прежде всего сомато-
сенсорной, зрительной и слуховой, эта ассоциативная область связана с
когнитивными процессами — мышлением и речью, хотя и то, и другое тре-
бует совместной деятельности многих регионов коры, а не одних лишь ас-
социативных полей.
Задняя часть коры теменных долей (поля 5 и 7) получает информацию от
соматосенсорной, зрительной (дорсальный путь) и слуховой коры
(рис. 4.39). Объединение информации дает пространственное ощущение ок-
ружающего мира, позволяющее ориентироваться в нем и соотносить его с
собственным телом, а также с отдельными частями тела, при этом функ-
циональное значение левой и правой заднетеменных областей не одинако-
во. Если повреждение левого полушария у большинства людей приводит к
нарушениям речи и письма, то при правостороннем повреждении наруше-
ний речи, как правило, нет, но почти утрачивается сенсорная связь с ле-
вой половиной тела, хотя там и сохраняется чувствительность. Такие боль-
ные обычно игнорируют свою левую половину, например, когда одеваются
или когда умываются, иногда они не в состоянии воспринимать левую ру-
ку или ногу как собственную. Причина игнорирования левой половины тела
у людей с повреждением правой заднетеменной области заключается в ут-
рате сознательного контроля и характерном изменении памяти, когда
больной пренебрегает не только реальными предметами слева, но и воспо-
минаниями об этих предметах.
Распределение внимания у здоровых людей связано с активацией задне-
236 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Вторичная
моторная -<
кора
Добавочный
моторный
ареал
Область, ------
контролирующая
следящие
движения
глаза
Первичная
моторная
кора
Соматосенсорная
кора
Заднетеменная
ассоциативная
кора
Первичная
слуховая
кора
Первичная
зрительная
кора
Премоторная
кора
Дорсолатеральная
префронтальная
ассоциативная
кора
Рис. 4.39. Последовательность переработки информации в коре при формировании
комплексного восприятия окружающего мира и определении стратегии поведения.
Ассоциативная кора теменной, затылочной и височной областей интегрирует полученные из
сенсорных зон сигналы, используя их для осуществления когнитивной деятельности. Пре-
фронтальная кора получает информацию от теменно-височно-затылочной ассоциативной об-
ласти и использует ее для создания поведенческих программ.
теменной, а также фронтальной коры. Однако каждая из этих областей ре-
шает разные задачи. Теменная область активируется, когда происходит
только переключение внимания с одних сенсорных сигналов на другие. В от-
личие от этого, лобная кора становится активной только в том случае, ес-
ли переключение внимания сопровождается связанным с ним движением.
Правое полушарие позволяет фиксировать внимание на объектах, нахо-
дящихся как в левом, так и в правом поле зрения, тогда как левое полуша-
рие способно это делать только в правом поле зрения. Таким образом, при
удержании внимания на объектах, расположенных в правом поле зрения,
активны оба полушария, а при переносе его на объекты, представленные в
левое поле зрения, сознательный контроль осуществляет одно лишь правое
полушарие. В правой заднетеменной области имеются два отдельных ре-
гиона для пространственного восприятия, а в левой — только один.
Височные доли участвуют в восприятии зрительных (в нижних извили-
нах) и слуховых (в верхних извилинах) стимулов: они сопоставляют новую
информацию, полученную от первичных сенсорных областей, со старой,
поступавшей прежде и зафиксированной в долговременной памяти. По-
вреждения нижних областей височных долей затрудняют процесс научения
при решении задач, связанных со зрительным восприятием, а вследствие
повреждения верхних областей височных долей нарушается запоминание
информации, предоставляемой слуховой сенсорной системой. Переработка
информации в височной коре связана с использованием памяти о про-
шлом опыте. После двустороннего удаления некоторых областей височной
коры и гиппокампа (с целью устранения эпилепсии) у пациентов утрачи-
вается способность к образованию долговременной памяти (за исключени-
4. Функции центральной нервной системы • 237
ем неосознаваемой моторной памяти, образование которой не требует уча-
стия височных долей). Функция левой и правой височной областей не со-
всем одинаковы, что свидетельствует о функциональной асимметрии боль-
ших полушарий мозга. При повреждении только левой височной области
больные хуже, чем до операции, могли вспомнить предъявленный им спи-
сок существительных, а при повреждении правой височной области вер-
бальная память почти не изменялась, но хуже запоминались образцы ри-
сунков, геометрических фигур, человеческих лиц.
4.10.5.2. Функции префронтальной ассоциативной коры
Префронтальная ассоциативная кора занимает дорсолатеральную поверх-
ность лобных долей, ее основная функция состоит в создании плана произ-
вольных действий. Большую часть информации префронтальная область
получает от заднетеменной ассоциативной коры, с которой она связана
многочисленными внутрикортикальными и субкортикальными связями.
Благодаря этим связям префронтальная кора получает полную пространст-
венную карту находящихся в поле зрения предметов. Сведения о внешнем
пространстве здесь объединяются с информацией о положении тела и от-
дельных его частей, причем префронтальная кора включает все эти данные
в кратковременную рабочую память. На этой основе создается план пред-
стоящих действий, т. е. из множества возможных вариантов деятельности
выбираются необходимые и в наиболее рациональной последовательности.
Прежде всего, программируется положение глаз, направляемых на нужный
объект, Предусматривается координация действий обеих рук и т. д. Боль-
шая часть выходящих из префронтальной коры сигналов поступает в пре-
моторную кору, создающую конкретные программы моторных действий.
Префронтальная область коры отличается изобилием заканчивающихся
в ней дофаминергических окончаний. Дофамин здесь выполняет роль моду-
лятора, необходимого для сохранения кратковременной рабочей памяти.
После локальной инъекции в префронтальную область обезьяны вещества,
избирательно нарушающего дофаминергическую передачу, ухудшается вы-
бор правильных действий, которые она должна совершать, чтобы добрать-
ся до корма. С нарушениями в дофаминэргической системе связывают
развитие шизофрении', у большинства шизофреников величина лобных до-
лей меньше, чем у здоровых людей.
4.10.5.3. Функции лимбической коры
Лимбическая кора связана с лимбической системой мозга и занимает меди-
альные и вентральные поверхности лобных долей, часть медиальной по-
верхности затылочных долей, поясные извилины, находящиеся в глубине
межполушарной щели, а также передние полюса височных долей. Лимби-
ческая кора получает информацию от вторичных сенсорных областей и иг-
рает важную роль в формировании мотиваций, эмоций и образовании дол-
говременной памяти, она влияет на создание плана будущих действий. Ор-
битофронтальная кора имеет прямые связи с миндалинами, от эффектив-
ности использования этой связи в значительной мере зависят эмоциональ-
ные аспекты поведения. Взаимодействие орбитофронтальной и дорсолате-
ральной областей лобных долей необходимо при планировании предстоя-
щих действий, которое определяется полученными сведениями об окру-
жающем мире, с одной стороны, и субъективным отношением к нему — с
другой. При поражении лобных долей часто нарушается формирование
238 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
мотиваций и затрудняется прогнозирование результатов действий, такие
больные отличаются вспыльчивостью, грубы в общении с другими людьми
и одновременно легкомысленны, не способны критически оценивать свое
состояние.
4.10.6. Функции моторных областей коры
Моторные области коры занимают часть корковой поверхности лобных
долей и подразделяются на первичную и вторичную моторную кору. Первич-
ная моторная кора (прецентральные извилины мозга или поле 4) получает
афферентную информацию преимущественно от соматосенсорной коры и
непосредственно управляет двигательными центрами ствола и спинного
мозга, контролирующими сокращения скелетных мышц. Вторичная мотор-
ная кора (поле 6) получает большую часть афферентной информации от
ассоциативной коры и на ее основе создает программы предстоящих дви-
жений, которые отправляет для исполнения первичной моторной коре. В
функциональной иерархии моторных систем вторичная моторная кора гла-
венствует над первичной.
4.10.6.1. Функция первичной моторной коры
Соматотопическое распределение ее нейронов соответствует регулируемым
мышцам лица, тела и конечностей пропорционально объему движений,
совершаемых с их помощью (рис. 4.40). Пропорции двигательного гомунку-
луса, искаженные, по сравнению с реальным человеком, отражают не объ-
Рис. 4.40. Схема соматотопической орга-
низации первичной моторной коры.
Первичная моторная кора занимает область
передних центральных извилин и организована
соматотопически. Непропорциональное пред-
ставительство отдельных частей тела отражает
степень сложности движений, осуществляемых
под контролем соответствующей соматотопи-
ческой области.
ем регулируемых мышц, а объем
и степень сложности движений,
которые они выполняют, поэтому
кисти рук, пальцы, лицо (мими-
ческие мышцы) и язык имеют
большее представительство, чем,
например, мышцы спины. Отно-
сящиеся к каждому участку тела
кортикальные колонки содержат
популяции нейронов, активация
которых вызывает изменение по-
ложения управляемого сустава в
одном определенном направлении
в результате согласованных сокра-
щений мышц-синергистов при
одновременном расслаблении ан-
тагонистов. Для изменения поло-
жения этого сустава в противопо-
ложном направлении необходима
активность других колонок.
Каждая соматотопическая об-
ласть моторной коры имеет об-
ратную связь с управляемой ча-
стью тела. Связь обеспечивается
поступлением информации от
проприоцепторов управляемых
мышц и от тактильных рецепто-
ров кожи в соматосенсорную ко-
4. Функции центральной нервной системы • 239
ру, а из нее — в моторную. Например, в область представительства рук
сенсорная информация поступает от рецепторов кожи, что позволяет тон-
ко регулировать движения пальцев при ощупывании предметов руками
или пальцами. С другой стороны, сами движения пальцев или рук по по-
верхности предмета активируют находящиеся в них проприоцепторы, а по-
ступающая от этих рецепторов информация помогает установить форму
предмета на ощупь. Такая деятельность рук называется стереогностиче-
ской.
4.10.6.2. Функция вторичной моторной коры
Вторичная моторная кора создает план предстоящих движений, исполне-
ние которого осуществляет первичная моторная кора. При электрической
стимуляции первичной моторной коры возникают сравнительно простые
сокращения мышц, а при стимуляции вторичной коры — целые комплек-
сы движений, но чтобы их вызвать, необходима более сильная и продол-
жительная электрическая стимуляция, чем та, которая способна активиро-
вать первичную моторную кору. При повреждениях вторичной моторной
коры нарушается выполнение комплексов автоматизированных, заученных
движений.
Вторичная моторная кора состоит из двух соматотопически организо-
ванных областей: премоторной коры и добавочного моторного ареала, распо-
ложенного на верхней стороне лобных долей. Каждая из этих областей по-
лучает афферентную информацию от префронтальной ассоциативной ко-
ры, а выходную активность адресует первичной моторной коре и мотор-
ным ядрам ствола. Некоторые аксоны ее нейронов входят в состав волокон
кортикоспинального пути, спускающегося в спинной мозг и осуществляю-
щего управление мотонейронами мышц туловища и конечностей.
Кроме того, премоторная область получает от заднетеменных областей
переработанную сенсорную информацию о комплексе зрительных ощуще-
ний. Эфферентный выход из премоторной области к стволу осуществляет-
ся с участием медиального пути, в основном ретикулоспинального тракта.
С помощью медиального пути премоторная область коры управляет мыш-
цами туловища и проксимальных отделов конечностей, которые сокраща-
ются в начальной фазе выпрямления тела или движения руки к намечен-
ной цели.
Добавочный моторный ареал принимает непосредственное участие в
переработке соматосенсорной информации, в соответствии с которой про-
граммирует последовательность движений, которые выполняются билате-
рально. При экспериментальном повреждении этой области коры у обезьян
отдельные движения могут выполняться правильно, но координировать их
в общем потоке действий животные уже не могут, особенно в тех случаях,
когда необходимы согласованные движения обеих рук.
4.11. Регуляция движений
Каждое из движений, выполняемых человеком, содержит два регулируе-
мых компонента: тонический, который обеспечивает необходимую для на-
чала движения позу и на время фиксирует в стабильном положении суста-
вы, и фазный, определяющий направление и скорость движения. В зависи-
мости от степени сложности выполняемых движений и участия сознания в
их регуляции они подразделяются на рефлекторные, ритмические и произ-
вольные.
240 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рефлекторные движения представляют собой наиболее простые мотор-
ные действия, которые выполняются быстро, стереотипно и не нуждаются
в сознательном контроле. Примером могут быть защитные сгибательные
рефлексы, осуществляемые при участии спинного мозга в ответ на болевое
раздражение кожи. Статические и статокинетические рефлексы ствола осу-
ществляются тоже стереотипно в соответствии с полученной сенсорной
информацией от вестибулярных, проприоцептивных, зрительных и слухо-
вых рецепторов.
Двигательные структуры спинного мозга и ствола участвуют в формиро-
вании ритмических движений, таких как ходьба, бег, жевание. Эти движе-
ния относительно стереотипны и могут повторяться почти автоматически,
без осознаваемого контроля. Осознание и произвольный контроль необхо-
димы лишь в самом начале, т. е. при запуске ритмических движений, и в
конце — при остановке движений.
Произвольные движения — это сложные комбинированные действия, не-
обходимые, например, для набора текста на клавиатуре компьютера или
игры на фортепьяно. Их отличительными признаками являются направле-
ние движений к определенной, заранее намеченной цели и совершенство-
вание координации в связи с приобретаемым опытом. Спинной мозг и
ствол необходимы при всякой моторной деятельности, при совершении
произвольных движений они участвуют в качестве исполнителей двига-
тельных команд моторных областей коры.
4.11.1. Иерархическая организация моторных систем
Любое движение человека возникает при сокращении одних мышц (синер-
гистов) и одновременном расслаблении других мышц (антагонистов), что
зависит только от активности управляющих ими мотонейронов. Порядок
возбуждения мотонейронов определяют действующие на них возбуждаю-
щие и тормозные интернейроны. Функциональное объединение мотоней-
ронов с соседними интернейронами образует низшую моторную систему
или локальный моторный аппарат, предназначенный для управления опре-
деленной частью тела. В любой низшей моторной системе у взрослого че-
ловека запрограммированы все варианты последовательности возбуждения
мотонейронов, представляющие программы возможных движений управ-
ляемой части тела.
Возбуждение мотонейронов в ходе стереотипной рефлекторной реакции
происходит в результате действия на них возбужденных афферентных ней-
ронов и интернейронов, объединенных синапсами в общую рефлекторную
ДУГУ- Двигательные центры головного мозга могут усилить или затормозить
осуществление рефлекторного ответа, и тогда выполняемое движение бу-
дет исполнено с большей или меньшей амплитудой либо совсем не воз-
никнет (рис. 4.41). Выбор конкретной двигательной программы зависит от
наиболее значимой сенсорной информации и чаще всего состоит в пред-
почтении наиболее эффективных действий.
Командные двигательные центры расположены в стволе мозга и мотор-
ных областях коры, их нисходящие пути могут оканчиваться непосредст-
венно на мотонейронах спинного мозга, но чаще регулируют их актив-
ность опосредованно, с помощью соседних интернейронов. Существует
несколько параллельных нисходящих путей, которые участвуют в решении
разных функциональных задач. Так, например, намеренное движение руки
к находящемуся на уровне головы предмету может напоминать по своей
траектории нечаянный взмах при попытке сохранить равновесие. При
4. Функции центральной нервной системы • 241
Рис. 4.41. Иерархическая организация двигательных структур мозга.
Иерархия моторных центров отражает эволюционное развитие двигательной системы (спинной
мозг—ствол—моторная кора), способствующее решению все более сложных задач и одновре-
менной передачи управления от низших отделов к высшим. Иерархические уровни соединяют-
ся несколькими параллельными путями, каждый уровень получает необходимую сенсорную
информацию. ВМК — вторичная моторная кора (премоторная и дополнительная области).
внешнем сходстве этих движений, осуществляющие их механизмы оказы-
ваются разными, так как в них участвуют разные двигательные центры и
нисходящие пути.
Все двигательные центры, на каком бы уровне иерархии они ни находи-
лись, организованы соматотопически. Они специализируются на управле-
нии только определенными группами мышц, от которых постоянно полу-
чают сенсорную информацию. Непрерывное поступление сенсорной ин-
формации на всех уровнях организации моторных систем своевременно
обеспечивает каждую двигательную структуру оперативной обратной свя-
зью, т. е. сведениями о том, как выполняется то или иное движение, дос-
тигается или нет намеченная цель: в соответствии с этими данными вы-
полняемые движения постоянно корректируются.
242 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Иерархия двигательных центров состоит в том, что высшие из них могут
отменить команды низших или поручить им выполнение собственной ко-
манды. Так, например, стволовые двигательные центры способны подчи-
нять себе активность низших моторных систем спинного мозга, но в то же
время сами подчиняются моторным областям коры. Вторичная и первич-
ная моторная кора создают конкретную программу движений, т. е. выбира-
ют из множества существующих вариантов наиболее подходящие сочета-
ния. Ассоциативная кора готовит общий план действий. Многочисленные
обратные связи между разными иерархическими уровнями моторной сис-
темы предоставляют им возможность взаимодействовать и в рамках отно-
шений субординации, и в качестве партнеров, согласованно решающих об-
щие задачи.
Важную роль как в формировании движений, так и в контроле их ис-
полнения играют мозжечок и базальные ганглии. Они не имеют прямого
выхода к мотонейронам и поэтому их нельзя отнести к какому-то опреде-
ленному иерархическому уровню. Мозжечок и базальные ганглии взаимо-
действуют с несколькими уровнями организации моторной системы и ко-
ординируют их активность.
4.11.2. Нисходящие пути моторной коры
Моторные области коры определяют наиболее сложную моторную дея-
тельность и регулируют ее посредством двух нисходящих путей: кортико-
спинального и кортикобулъбарного, объединяемых под названием пирамид-
ный тракт. Кортикоспинальный путь, необходимый для управления мыш-
цами туловища и конечностей, заканчивается либо прямо на мотонейро-
нах, либо на интернейронах спинного мозга, а часть аксонов этого пути
образует синапсы с нейронами красных ядер среднего мозга (рис. 4.42).
Кортикобульбарный путь осуществляет контроль двигательных ядер череп-
но-мозговых нервов, управляющих мышцами лица и движениями глаз.
Помимо этого он проецируется на сеть двигательных ядер ствола, связан-
ных со спинным мозгом.
Прямой кортикоспинальный тракт является самым большим нисходя-
щим моторным путем, он образован приблизительно одним миллионом
аксонов, больше половины которых принадлежат нейронам V слоя первич-
ной моторной коры (поле 4). Приблизительно */3 волокон кортикоспиналь-
ного пути происходит от нейронов премоторной коры (поле 6), а остальные
начинаются в соматосенсорной коре (поля 3,2, 1) и осуществляют перенос
сенсорной информации.
Кортикоспинальный путь проходит через внутреннюю капсулу мозга, а
в продолговатом мозге образует характерные утолщения на вентральной
поверхности — они называются пирамидами (от этого термина произошло
название: пирамидный тракт). На границе между продолговатым и спин-
ным мозгом большая часть волокон, приблизительно 3/4, переходит на
противоположную сторону. Вместе с ними на противоположную сторону
переходят волокна руброспинального пути, в результате этого объединения
образуется дорсолатеральный нисходящий путь. Он заканчивается на интер-
нейронах и мотонейронах спинного мозга, контролирующих дистальные
мышцы конечностей на противоположной, по отношению к моторной ко-
ре, стороне тела.
Не перешедшая на другую сторону часть волокон кортикоспинального
пути совместно с медиальными путями двигательных центров ствола обра-
зует вентромедиальный путь, оканчивающийся синапсами с медиальным
4. Функции центральной нервной системы • 243
А. Латеральный кортико-
спинальный путь
Премоторная
кора
Первичная
моторная
кора
Сомато-
сенсорная
кора
Перекрест в
пирамидах
продолговатого
мозга
Рубро-
спинальный
путь
Латеральный
кортикоспи-
нальный
путь
,2,1
Красное
ядро
Вентромедиальный
кортикоспинальный
путь
Б. Вентромедиальный
кортикоспинальный путь
Медиальный
путь
ствола
Рис. 4.42. Схема нисходящих двигательных путей коры к спинному мозгу.
А. Латеральный кортикоспинальный путь предназначен для управления дистальными группами
мышц конечностей, он начинается от моторных полей 4 и 6, сенсорных полей 3, 2, 1 и окан-
чивается в медиальной части заднего рога спинного мозга на противоположной стороне. Пе-
реход волокон этого пути на противоположную сторону происходит на уровне пирамид про-
долговатого мозга; ниже они идут вместе с волокнами руброспинального пути.
Б. Вентромедиальный кортикоспинальный путь предназначен для управления мышцами тулови-
ща и проксимальными мышцами конечностей, он образован аксонами пирамидных нейронов
моторной и премоторной коры, которые оканчиваются в спинном мозгу билатерально. Пере-
креста волокон не происходит, в спинном мозге они проходят параллельно медиальным путям
двигательных центров ствола.
столбом нейронов спинного мозга, которые контролируют мышцы тулови-
ща и проксимальных отделов конечностей, участвующие в формировании
позы и приспособлении ее к совершаемым движениям. Такое управление
осуществляется билатерально, т. е. относится к обеим половинам спинного
мозга и, соответственно, к обеим половинам тела.
244 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
4.11.3. Контроль выполняемых движений
Контроль совершаемых движений осуществляют одновременно спинной
мозг, ствол и моторная кора. Каждая из этих структур получает свою пор-
цию сенсорных сигналов {обратная связь) и может непосредственно влиять
на локальные моторные аппараты (прямая связь), каждая из них способна
осуществлять независимый контроль, однако высшие центры могут огра-
ничивать выбор программ низшими центрами или предлагать им свои ко-
манды. Сенсорная информация о положении тела и отдельных его частей
в пространстве используется для определения силы сокращения мышц, ре-
гуляции точности и скорости движений. Контролирующая роль двигатель-
ных центров состоит в том, чтобы обеспечить: 1) точность и своевремен-
ность доставки команд одновременно к нескольким (многим) группам
мышц; 2) адекватное совершаемым движениям положение тела; 3) дина-
мическую оценку механических свойств мышц, костей и суставов.
4.12. Межполушарная функциональная асимметрия
Каждое из больших полушарий мозга получает большую часть информа-
ции от тактильных рецепторов и проприоцепторов противоположной по-
ловины тела и одновременно контролирует двигательные функции проти-
воположной половины тела. Полушария соединены несколькими пучками
нервных волокон, самый крупный из них — мозолистое тело, функция ко-
торого состоит в обмене информации между полушариями. Сравнение
сенсорных и двигательных функций обоих полушарий свидетельствует, на
первый взгляд, об их симметрии. Однако речевая деятельность у большин-
ства людей определяется левым полушарием, асимметричной является и
способность к пространственному восприятию, в большей степени завися-
щая от работы правого полушария. Асимметрично представительство ряда
других функций, что выражается, например, в функциональном превос-
ходстве одной руки, ноги, глаза, уха.
4.12.1. Функциональные возможности изолированных полушарий
Полушария изолируются друг от друга после комиссуротомии — хирургиче-
ского пересечения мозолистого тела (к такому вмешательству прибегали,
чтобы ограничить распространение возбуждения у больных эпилепсией
при локализации патологического очага в одном из полушарий). Мозг че-
ловека с искусственно разделенными полушариями в литературе определя-
ют как расщепленный мозг. Функциональные возможности каждого из раз-
деленных полушарий были определены с помощью тахистоскопии — мето-
да раздельного введения зрительной информации в каждое из полушарий
(рис. 4.43). Люди с сохраненным мозолистым телом при подобных иссле-
дованиях способны называть предметы и читать демонстрируемые им сло-
ва независимо от того, в каком поле зрения они появляются, поскольку их
полушария через мозолистое тело обмениваются информацией.
При демонстрации изображения предмета или обозначающего его слова
в правом поле зрения испытуемый с расщепленным мозгом воспринимает
информацию только левым полушарием. Он правильно называет предмет,
а слово — читает: изолированное левое полушарие справляется с постав-
ленными задачами точно так же, как и целый мозг. Когда же изображение
появляется в левом зрительном поле, испытуемый может левой рукой вы-
брать на ощупь среди нескольких предметов продемонстрированный ему,
4. Функции центральной нервной системы • 245
Рис. 4.43. Схема тахистоскопического исследования людей, перенесших комиссуро-
томию.
Слева: пациентка перед экраном тахистоскопа, в левом и правом полях которого появляются
изображения предметов или слова; за экраном находятся предметы, опознать которые можно
только на ощупь.
Справа показана схема переработки зрительной информации: если пациентка смотрит на точ-
ку в центре экрана тахистоскопа, то информация из правого зрительного поля поступает в ле-
вое полушарие мозга, а из левого зрительного поля — в правое.
но не способен его назвать. Правое полушарие может воспринимать толь-
ко относительно простые слова. Если, например, испытуемому показывали
для обработки в правом полушарии слово D-O-G (собака) или A-P-P-L-E
(яблоко), то он правильно выбирал из предъявленных предметов игрушеч-
ную собачку или яблоко, но назвать слово не мог. Правое полушарие спо-
собно перерабатывать простую входящую лингвистическую информацию,
но не может обеспечить языковый выход.
В других видах деятельности, например в формировании пространст-
венного ощущения, правое полушарие превосходит левое, и его нельзя
считать лишь копией левого минус речевая деятельность.
4.12.2. Выявление функций неразделенных полушарий
После кратковременной наркотизации только одного из полушарий (путем
введения амитала натрия в левую или правую сонную артерию) можно оп-
ределить, в каком из них расположены центры речи. Приблизительно у
96 % правшей речевые центры находятся в левом полушарии и только у
4 % — в правом, однако и у большинства левшей, приблизительно у 70 %,
речевые центры также находятся в левом полушарии и лишь у 15 % лев-
шей — в правом, а еще у 15 % — в обоих полушариях. Если в левое и пра-
246 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
вое ухо через стереофонические наушники поступает различающаяся рече-
вая информация (произносимые диктором слова или числа), то правши в
большинстве случаев лучше воспринимают информацию, поступившую в
правое ухо. Этот феномен получил название «эффекта правого уха», кото-
рый принято объяснять более эффективным проведением информации в
левое «речевое» полушарие от контрлатерального уха.
На функциональную специализацию регионов мозга указывает уровень
их функциональной активности во время речевой деятельности. Так, на-
пример, при чтении вслух функционально активны речевые центры доми-
нантного левого полушария, что удалось обнаружить с помощью позитрон-
но-эмиссионной томографии (ПЭТ) и ядерно-магнитного резонанса (ЯМР).
При проведении ПЭТ в сонную артерию вводят меченную короткоживу-
щим изотопом фтора 2-дезоксиглюкозу, которая включается в обменные
процессы наиболее активных нейронов. Ее присутствие в функционально
активных участках мозга можно зарегистрировать по радиомаркеру с помо-
щью компьютерной томографии. Другой вариант ПЭТ основан на том, что
чем активнее функционируют нейроны, тем большим становится кровоток
в этом регионе, а для оценки кровотока вводят помеченную короткоживу-
щим изотопом (кислород-15) воду. Поскольку применяемые при ПЭТ изо-
топы существуют очень недолго, их надо получать непосредственно перед
проведением исследования с помощью циклотрона, что делает всю проце-
дуру ПЭТ очень дорогой. В отличие от ПЭТ исследование с помощью
ЯМР не требует предварительной инъекции пациенту. Оно регистрирует
электромагнитное излучение, возникающее в связи с влиянием внешнего
магнитного поля на вращающиеся ядра атомов, например ядра атома водо-
рода в составе воды. Электромагнитное излучение можно регистрировать в
любой плоскости и на этой основе получать представление о пространст-
венной характеристике слоя толщиной 5—10 мм. При повышенном по-
треблении нейронами кислорода картина ЯМР характерным способом из-
меняется, что и представляет основу для применения этого метода с целью
определения функциональной активности отдельных регионов мозга.
4.12.3. Функциональная специализация полушарий мозга
Левое полушарие доминирует в формальных лингвистических операциях,
обеспечивает фонетическое представление и синтаксический анализ, одна-
ко правое полушарие может узнавать звучащие слова и ассоциативное зна-
чение отдельных слов, а также определяет эмоциональную окраску речи
(табл. 4.3). Правое полушарие значительно лучше левого обеспечивает
пространственное ориентирование человека и пространственное располо-
жение зрительных сигналов, ориентацию линий и их кривизну, много-
угольники неправильной формы. Однако с задачей обнаружения мини-
мальных различий в таких фигурах лучше справляется левое полушарие.
Встретив хорошо знакомого человека, мы узнаем его мгновенно благодаря
правому полушарию, воспринимающему целостный образ («гештальт»).
Левое полушарие в таких случаях последовательно оценивает все характер-
ные признаки: волосы, лоб, глаза, нос, уши, подбородок, губы (вербальное
кодирование).
Два полушария могут функционировать независимо и поочередно до-
минировать в зависимости от характера решаемых задач. Например, в вос-
приятии или узнавании знакомых мелодий доминирует правое полушарие,
воспринимающее мелодию как целостный образ, с его гармонией и тем-
бром. Но при сравнении двух мелодий, отличающихся всего одной нотой,
4. Функции центральной нервной системы • 247
Таблица 4.3. Латерализация функций у правшей
Доминирование левого полушария Доминирование правого полушария
Восприятие устной речи и ритма му- зыки Понимание значения букв и слов, чтение и письмо Арифметический счет Вербальное мышление Аналитическое и последовательное восприятие Контроль сложных движений Конструирование временных отноше- ний Восприятие интонации речи, гармонии и тем- бра музыки Восприятие сложных геометрических фигур, уз- навание лиц Пространственная ориентация Невербальное мышление Целостное и одновременное восприятие («геш- тальт»-образ) Передвижение в пространстве Конструирование пространственных отношений
компетентнее оказывается левое полушарие; оно же оценивает ритмиче-
ский рисунок мелодии, причем эта функция хорошо коррелирует с рече-
вой деятельностью.
Левое полушарие упорядочивает и программирует события во времени,
тогда как правое представляет и конструирует пространственные отноше-
ния. При этом не имеет значения, являются ли сами стимулы акустико-
временными или же зрительно-пространственными. Левое полушарие дей-
ствует логически и аналитически, т. е. подобно компьютеру: оно последо-
вательно выделяет детали и снабжает их словесными определениями. Пра-
вое полушарие функционирует синтетически: определяет общую конфигу-
рацию стимула и перерабатывает информацию как единое целое. Накоп-
лен большой фактический материал, свидетельствующий о доминировании
правого полушария в эмоциональной сфере. Пациенты с расщепленным
мозгом при демонстрации им в левом зрительном поле картин эмоцио-
нального содержания реагировали на них так же, как и здоровые люди, что
демонстрирует возможность невербального возникновения эмоций. Выра-
жение левой половины чужого лица (информация обрабатывается в пра-
вом полушарии) оказывается более важным для здоровых людей при оцен-
ке эмоционального значения этого выражения. Эмоциональная окраска
голоса, плач или смех, эмоциональная окраска музыкальных фрагментов
воспринимаются лучше, если звуковые сигналы попадают в левое ухо: при
этом перекрещивающиеся и проводящие сигналы в правое полушарие
нервные волокна играют более важную роль, чем неперекрещивающиеся.
Большая способность правого полушария к объединению сложных сигна-
лов в целостный образ играет решающую роль в эмоциональных пережи-
ваниях, в то время как преимущество левого полушария при анализе вре-
менных отношений и отдельных деталей дает ему возможность контроли-
ровать и ослаблять эмоциональные реакции. Таким образом, эмоциональ-
ное преимущество правого полушария и когнитивное — левого уравнове-
шивают и дополняют друг друга.
ГЛАВА 5
Вегетативная нервная система*
5.1. Строение вегетативной нервной системы
Вегетативная нервная система [синонимы: автономная, висцеральная нерв-
ная система] представляет собой комплекс центральных и периферических
нервных структур, участвующих в регуляции деятельности внутренних ор-
ганов, кровеносных и лимфатических сосудов, гладкой и отчасти попереч-
но-полосатой мускулатуры.
В стволе головного мозга и спинном мозге расположены четыре центра
вегетативной нервной системы:
1) мезэнцефалический, из которого вегетативные нервные волокна вы-
ходят в составе глазодвигательного нерва;
2) бульбарный, из которого нервные волокна выходят в составе лицево-
го, языкоглоточного и блуждающего черепных нервов;
3) тораколюмбальный — ядра боковых рогов CV111—Lln сегментов;
4) сакральный — ядра боковых рогов SH—S1V сегментов.
Из двух последних центров волокна выходят в составе передних кореш-
ков спинномозговых нервов.
Эффекторные импульсы, исходящие из стволовых и спинномозговых
центров, достигают внутренних органов и сосудов по двухнейронному пу-
ти. Первые нейроны располагаются в головном или спинном мозге, их от-
ростки направляются к периферическим узлам вегетативной нервной сис-
темы (ганглиям) и оканчиваются на телах вторых нейронов, отростки кото-
рых разветвляются в иннервируемых органах (рис. 5.1). Отростки первых
нейронов называются преганглионарными, а отростки вторых — постганг-
лионарными волокнами.
По морфологическим и функциональным свойствам в вегетативной
нервной системе различают симпатический и парасимпатический отделы.
Тораколюмбальные спинномозговые центры, выходящие от них преганг-
лионарные волокна, соответствующие им ганглии и постганглионарные
волокна составляют симпатический отдел вегетативной нервной системы.
По выходе из позвоночного канала в составе передних корешков преганг-
лионарные волокна вступают в виде белых соединительных ветвей (гг.
communicantes albi), которые расположены двумя цепочками, лежащими по
обе стороны позвоночного столба, и образуют симпатические стволы
(trunci sympathici).
Симпатические ганглии содержат нейроны, отростки которых (пост-
ганглионарные волокна) направляются непосредственно к внутренним орга-
нам и кровеносным сосудам либо входят в состав спинномозговых нервов в
виде серых соединительных ветвей (rr. communicantes grisei), вместе с ними
достигая кровеносных сосудов, гладких мышц и желез тела и конечностей.
Центры, расположенные в стволе головного мозга и крестцовых сегмен-
*Глава написана доцентом Л. В. Надежкиным.
5. Вегетативная нервная система • 249
Б
Рис. 5.1. Схема строения вегетативной нервной системы.
Центры вегетативной нервной системы в стволе головного мозга и спинном мозге: М — ме-
зэнцефалический (преганглионарные нервные волокна выходят в составе глазодвигательного
нерва); Б — бульбарный (преганглионарные нервные волокна выходят в составе лицевого,
языкоглоточного и блуждающего нервов); Т — торако-люмбальный — ядра боковых рогов
CVIII-LHI сегментов спинного мозга; С — сакральный — ядра боковых рогов SII-SIV сегмен-
тов спинного мозга.
Из Т и С центров преганглионарные волокна выходят в составе передних корешков спинно-
мозговых нервов.
max спинного мозга, выходящие из них преганглионарные волокна, а также
ганглии с постганглионарными волокнами составляют парасимпатический
отдел вегетативной нервной системы. Парасимпатические ганглии распо-
лагаются вблизи или в стенке иннервируемых органов, поэтому постганг-
лионарные парасимпатические волокна короткие.
Ганглии и нервные волокна в стенках полых органов (в составе подсе-
розных, межмышечных, подслизистых сплетений) составляют энтеральную
часть вегетативной нервной системы.
5.2. Функции вегетативной нервной системы
Функция вегетативной нервной системы — регуляция физиологических
процессов, обеспечивающих поддержание гомеостазиса в организме. Для
обеспечения этой основной функции вегетативной нервной системой осу-
ществляется:
1. Управление деятельностью висцеральных органов:
а) пусковая функция — активация специфической для органа функции
(например, сокращения гладкой мышцы, секреции железы);
б) корригирующая функция — усиление (или ослабление) деятельности
органа в соответствии с меняющимися потребностями организма.
2. Влияние на процессы обмена веществ в органах (трофическая функ-
ция), особенно во время приспособления их к изменяющимся усло-
виям деятельности:
250 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
а) усиление функциональных ответов на раздражители за счет ис-
пользования энергетических ресурсов (эрготропная функция) ',
б) коррекция и поддержание процессов восстановления и сохранения
резерва энергии для дальнейшей деятельности органов (трофо-
тропная функция).
3. Участие в регуляции деятельности всех органов путем изменения кро-
воснабжения.
При возбуждении симпатических эфферентов усиливаются сокращения
сердца и увеличивается их ритм, возрастает скорость проведения возбуж-
дения по мышце сердца, возникает констрикция кровеносных сосудов, по-
вышающая артериальное давление, расширяются бронхи и зрачки, увели-
чивается секреция мозгового слоя надпочечников, снижается тонус глад-
ких мышц желудочно-кишечного тракта, повышаются скорость свертыва-
ния крови, активность ферментов, увеличивается в крови содержание глю-
козы и свободных жирных кислот.
Активность парасимпатических нервов уменьшает силу и частоту сокра-
щений сердца и за счет этого понижает артериальное давление, увеличива-
ет секрецию инсулина, снижает концентрацию глюкозы в крови, приводит
к усилению моторной и секреторной функций желудочно-кишечного трак-
та (табл. 5.1).
Вегетативная нервная система способна изменять функциональную реак-
тивность или адаптационные свойства тканей иннервируемых органов пу-
тем изменения биохимических характеристик их клеток. Например, ске-
летная мышца, доведенная до утомления длительным возбуждением ее мо-
торного нерва, под влиянием дополнительного возбуждения симпатиче-
ских волокон начинает вновь отвечать интенсивными сокращениями (фе-
номен Орбели—Гинецинского). Возбуждение симпатических волокон не вы-
зывает сокращение скелетной мышцы, но при утомлении мышцы повыша-
ет ее возбудимость и ускоряет протекающие в ней биохимические процес-
сы, оказывая адаптационно-трофическое влияние.
Адаптационно-трофические влияния вегетативной нервной системы су-
ществуют помимо ее воздействия на тонус и проницаемость стенок сосу-
дов. За счет такого влияния симпатической и парасимпатической иннерва-
ции создаются условия для улучшения функциональных свойств не только
нервно-мышечных приборов, но и рецепторов, а также всей центральной
нервной системы.
Специальных нервов, оказывающих только адаптационно-трофические
влияния на иннервируемые ткани, не найдено. Эти влияния осуществля-
ются не импульсами передачи возбуждения в синапсах, а гуморальными
факторами, например действием не разрушившегося в синапсе медиатора
или продуктов его обмена. В механизме адаптационно-трофического влия-
ния на иннервируемые ткани существенная роль принадлежит непрерыв-
ному проксимально-дистальному медленному (со скоростью около 1 мм в
сутки) току аксоплазмы по нервному волокну, благодаря которому в аксо-
нальные терминали доставляются нуклеотиды и продукты метаболизма
нуклеиновых кислот, которые синтезированы в ядре нервной клетки. При
диффузии из аксональных терминалей эти биологически активные вещест-
ва достигают клеток эффекторных органов и оказывают на них адаптаци-
онно-трофическое воздействие.
5. Вегетативная нервная система • 251
5.3. Функции периферических отделов вегетативной нервной
системы
5.3.1. Симпатический и парасимпатический отделы
Практически ко всем органам подходят постганглионарные симпатиче-
ские и преганглионарные парасимпатические волокна, при этом влияния
отделов вегетативной нервной системы имеют свои особенности: парасим-
патический отдел осуществляет в основном мощные локальные воздейст-
вия на иннервируемые органы, а симпатический вовлекает в реакцию сра-
зу несколько органов и систем.
В вегетативном ганглии клеток нейронов в несколько раз больше, чем
подходящих к нему преганглионарных волокон. Разветвляясь внутри ганг-
лия, каждое из этих волокон образует синапсы на телах до 30 ганглионар-
ных нейронов, а на каждом из них оканчиваются синапсами несколько
преганглионарных волокон. В связи с этим возбуждение преганглионарно-
го волокна в ганглии способно оказывать влияние на большое число пост-
ганглионарных нейронов, и, следовательно, на еще большее число эффек-
торных клеток иннервируемого органа. Поэтому возбуждения, поступаю-
щие, например, из спинного мозга по симпатическим волокнам, при пере-
даче в ганглиях получают значительное пространственное распространение.
Передача возбуждения в синапсах вегетативной нервной системы осу-
ществляется исключительно с помощью биологически активных ве-
ществ — медиаторов. В передаче возбуждения с преганглионарных на пост-
ганглионарные нейроны в ганглиях симпатического и парасимпатического
отделов медиатором служит ацетилхолин, который связывается с И-(нико-
тиночувствительными) холинорецепторами.
Медиатором передачи возбуждения с постганглионарных парасимпати-
ческих нейронов на иннервируемую ткань является ацетилхолин, а с пост-
ганглионарных симпатических — норадреналин. Принято называть нервные
клетки и волокна вегетативной нервной системы по выделяемому их тер-
миналями медиатору. Поэтому все парасимпатические и преганглионар-
ные симпатические нейроны, с медиатором ацетилхолином называют хо-
линергическими, а симпатические постганглионарные нейроны, у которых
основной медиатор норадреналин, — адренергическими.
Не каждая клетка внутренних органов непосредственно контактирует с
волокнами вегетативной иннервации. В скелетных мышцах, например, не
все гладкомышечные клетки кровеносных сосудов имеют прямой контакт
с волокнами симпатических нервов. Однако симпатический нерв вызывает
констрикторную реакцию сосудов, поскольку выделяющиеся при его воз-
буждении медиаторы, кроме влияния на иннервируемые клетки, широко
диффундируют и воздействуют также и на соседние группы гладкомышеч-
ных клеток, вызывая их сокращение.
Передача возбуждения с постганглионарных нейронов на эффекторы
происходит в контактах, которые отличаются от «классических» синапсов
в соматической нервной системе. Так, симпатические постганглионарные
нейроны образуют синапсы на эффекторных клетках не только пресинап-
тическими окончаниями, но и имеют особые синаптические контакты в
варикозах (расширениях) периферических участков симпатических воло-
кон в области иннервируемых тканей. Количество варикозов в этих участ-
ках достигает 15—30 на 100 мкм длины пресинаптической терминали и на-
растает по мере уменьшения ее диаметра. В варикозах имеются скопления
везикул, которые содержат медиатор норадреналин.
252 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
В синаптических контактах варикоз щель очень широкая. Из-за относи-
тельной «открытости» этих контактов осуществляется обмен медиатором
терминали с окружающей тканевой жидкостью. Медиатор, выделившийся
в одной варикозе, путем диффузии достигает нескольких эффекторных
клеток, а медиатор разных варикоз может воздействовать на одну и ту же
клетку. Кроме того, в вегетативных синапсах выражен механизм так назы-
ваемого обратного захвата медиатора (re-uptake) Благодаря ему, напри-
мер, симпатические терминали поглощают катехоламины из экстрацеллю-
лярного пространства и крови, восстанавливая запасы медиатора за счет
циркулирующих в крови катехоламинов, выделенных мозговым слоем над-
почечников
Передача возбуждения симпатических нервов на эффекторы осуществ-
ляется, как правило, катехоламинами: адреналином и в большей степени
норадреналином. Тела нервных клеток и нетерминальная часть их аксонов
содержат от 10 до 100 мкг норадреналина на 1 г ткани, а пресинаптические
терминали — до 10 000 мкг на 1 г. Катехоламины синтезируются в теле
нервной клетки, переходят в состав цитоплазматических гранул и в этой
форме медленно транспортируются по аксону в направлении концевых
разветвлений.
В пресинаптических терминалях катехоламины депонированы в везику-
лах или синаптических пузырьках. В терминалях норадреналин существует
в двух пулах (депо): малом, лабильном, легко высвобождающем медиатор
под действием нервного импульса, и большом, стабильном, прочно свя-
занном с белком. В цитоплазме аксона катехоламины быстро разрушаются
ферментом моноаминоксидазой, а длительно сохраняться они могут толь-
ко внутри гранул. Для механизма переноса катехоламинов из цитоплазмы
в гранулы необходимы АТФ и ионы Mg2+.
В ответ на нервный импульс катехоламины лабильного пула освобожда-
ются из гранул и выходят в синаптическую щель, где часть медиатора раз-
рушается ферментом катехол-метил-трансферазой, другая — реабсорбиру-
ется аксонной терминалью, а третья
вступает в контакт с белками ре-
цепторами постсинаптической
мембраны, участвуя в передаче
нервного импульса эффектору
(рис. 5.2).
На постсинаптических мембра-
нах норадреналин связывается с
Рис. 5.2. Симпатический синапс и
его регуляция.
1 — варикоза симпатической терминали,
2 — синаптическая везикула, 3 — синапти-
ческая щель, 4 — постсинаптическая мем-
брана, 5 — рядом расположенный холинер-
гический синапс. АХ — ацетилхолин, НА —
норадреналин, МАО — моноаминоксидаза,
КОМТ — катехол-О-метилтрансфераза (раз-
рушающие норадреналин ферменты), вто-
ричные посредники а-рецепторов: ФЛ-ИФ3
(фосфолипаза-С—инозитол-3-фосфат); вто-
ричные посредники p-адренорецепторов:
АЦ-цАМФ (аденилатциклаза—цикличе-
ский аденозинмонофосфат), а, а2, р — ад-
ренорецепторы, (+) — стимуляция, (—) —
подавление освобождения медиатора.
5. Вегетативная нервная система • 253
альфа- и /3-адренорецепторами, с образованием медиатор-рецепторных
комплексов.
а-Рецепторный комплекс активирует метаболизм мембраны клеток, что
приводит к появлению внутриклеточных вторичных посредников медиа-
торного эффекта, которыми являются инозитол-3-фосфат и ионизирован-
ный кальций. Под их воздействием повышается проницаемость для ионов
постсинаптической мембраны и происходит ее локальная деполяриза-
ция — возбуждающий постсинаптический потенциал (ВПСП).
о^-Адренорецепторы — находятся в гладких мышцах органов. Их возбуж-
дение приводит к сужению сосудов, расслаблению кишечника, расшире-
нию зрачка и сокращению миометрия.
а2-Адренорецепторы расположены на пресинаптической мембране вари-
коз, их возбуждение приводит к уменьшению дальнейшего выделения ме-
диатора.
Стимуляция p-адренорецепторов активирует другую систему вторичных
посредников. Действие катехоламинов на p-адренорецепторы увеличивает
активность фермента аденилатциклазы с образованием из АТФ цикличе-
ского 3',5'-аденозинмонофосфата (цАМФ). Последний активирует проте-
инкиназу, что в свою очередь приводит к росту концентрации фосфопро-
теидов в мембране с последующим увеличением ее проницаемости.
/3,-Адренорецепторы находятся в сердечной мышце (их возбуждение уве-
личивает силу и частоту сердечных сокращений) и в жировой ткани (уве-
личение липолиза жира и калоригенеза).
/32-Адренорецепторы имеются в гладких мышцах артериальных сосудов,
особенно скелетных мышц, коронарных артерий, бронхов, матки, мочево-
го пузыря. Их стимуляция вызывает тормозной эффект в виде расслабле-
ния гладких мышц.
В вегетативных синапсах выделившиеся в ответ на нервные импульсы
медиаторы активируют рецепторы не только постсинаптической, но и пре-
синаптической мембраны, оказывая влияние на дальнейшее высвобожде-
ние медиаторов из нервных окончаний. При действии норадреналина на
а-адренорецепторы выделение медиатора снижается, а на р-адренорецеп-
торы — возрастает.
Поэтому при сильном возбуждении постганглионарных нейронов зна-
чительное повышение концентрации норадреналина в синаптической ще-
ли приводит к торможению выделения медиатора вследствие активации
пресинаптических а-адренорецепторов {отрицательная обратная связь).
Напротив, при низкой концентрации норадреналина (в условиях слабого
возбуждения нейронов) выброс этого медиатора увеличивается в результате
возбуждения p-адренорецепторов {положительная обратная связь).
В парасимпатических постганглионарных синапсах ацетилхолин содер-
жится в аксоплазме и синаптических пузырьках пресинаптических оконча-
ний в трех основных пулах или фондах. Это: 1) стабильный пул медиатора,
прочно связанный с белком и не готовый к освобождению; 2) мобилизиро-
ванный пул, менее прочно связанный с белком и пригодный к освобожде-
нию; 3) готовый к освобождению спонтанно или активно выделяемый пул. В
пресинаптическом окончании постоянно происходит перемещение пулов и
пополнение активного пула путем продвижения синаптических пузырьков
по направлению к пресинаптической мембране, так как медиатор активно-
го пула содержится в тех пузырьках, которые непосредственно прилежат к
этой мембране.
В отсутствие импульсного возбуждения из пресинаптической терминали
спонтанно освобождаются единичные кванты медиатора. Но с приходом
254 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 5.3. Парасимпатический синапс и
его регуляция.
1 — пресинаптическое окончание, 2 — си-
наптическая везикула, 3 — синаптическая
щель, 4 — постсинаптическая мембрана эф-
фекторной клетки, 5 — рядом расположен-
ный адренергический синапс. НА — норад-
реналин, АХ — ацетилхолин, М — мускари-
новый холинорецептор, Н — никотиновый
холинорецептор, ХЭ — холинэстераза, вто-
ричные посредники: ГЦ-цГМФ (гуанилат-
циклаза — циклический гуанозинмонофос-
фат), (+) — стимуляция, (—) — подавление
освобождения медиатора.
импульсов, деполяризующих пре-
синаптическую мембрану, спон-
танное фоновое выделение еди-
ничных квантов сменяется актив-
ным выбросом групп квантов.
Процесс освобождения медиатора является кальцийзависимым. Вызванная
импульсом деполяризация пресинаптической мембраны открывает в ней
каналы, по которым ионы кальция входят внутрь окончания, обеспечивая
выделение медиатора в синаптическую щель.
Высвобождение ацетилхолина в синаптическую щель зависит от:
1) связывания ацетилхолина с холинорецепторами пресинаптической
мембраны, относящимися к М- (мускариночувтвителыюму) типу,
что оказывает тормозящее влияние на дальнейший выход медиато-
ра — отрицательная обратная связь (рис. 5.3);
2) взаимодействия ацетилхолина с Н-холинорецептором пресинаптиче-
ской мембраны, что усиливает освобождение ацетилхолина — поло-
жительная обратная связь',
3) поступления в синаптическую щель парасимпатического синапса
норадреналина из рядом расположенного симпатического синапса,
что тормозит освобождение ацетилхолина;
4) выделения в синаптическую щель под влиянием ацетилхолина из
постсинаптической клетки большего числа молекул АТФ, которые
связываются с пуринергическими рецепторами пресинаптической
мембраны и подавляют освобождение медиатора — ретро-ингибиро-
вание.
Выделившийся в синаптическую щель ацетилхолин удаляется из нее не-
сколькими путями. Во-первых, часть медиатора связывается с холиноре-
цепторами пре- и постсинаптической мембраны; во-вторых, он разрушает-
ся ацетилхолинэстеразой с образованием холина и уксусной кислоты, ко-
торые подвергаются обратному захвату пресинаптической мембраной и
вновь используются для синтеза ацетилхолина; в-третьих, медиатор путем
диффузии выносится в межклеточное пространство и кровь, причем этот
процесс происходит после связывания медиатора с рецептором. Послед-
ним путем инактивируется почти половина выделившегося ацетилхолина.
На постсинаптической мембране ацетилхолин связывается с М-холино-
рецепторами нескольких типов, что и определяет различия в характере ре-
акций органов на этот медиатор. По чувствительности к различным фар-
макологическим препаратам выделяют М^-М^-холинорецепторы, которые
локализуются:
5. Вегетативная нервная система • 255
М^холинорецепторы — в вегетативных ганглиях и ЦНС;
М2-холинорецепторы — в сердце, в гладких мышцах желудочно-кишеч-
ного тракта;
М3- холинорецепторы — в гладких мышцах, в большинстве экзокринных
желез.
М4-холинорецепторы изучены мало.
На постсинаптической мембране ацетилхолин образует с М-холиноре-
цепторами медиатор-рецепторный комплекс, который активирует натрие-
вые каналы и возбуждает гладкомышечные и секреторные клетки желудоч-
но-кишечного тракта, клетки гладких мышц бронхов, мочевого пузыря и
мочеточника.
Возбуждение М-холинорецепторов вызывает: сужение бронхов, увеличе-
ние слюноотделения и слезоотделения, увеличение моторики и секреции
соков в желудочно-кишечном тракте, а также сужение зрачка. Этому спо-
собствует активация вторичных посредников — инозитол-3-фосфата и ио-
низированного кальция.
В клетках проводящей системы сердца и гладких мышц сосудов поло-
вых органов ацетилхолин активирует калиевые каналы и выходящий ток
калия, что приводит к гиперполяризации постсинаптических мембран. В
результате этого происходят торможение ритма сердца, снижение проводи-
мости и возбудимости в миокарде, расширение артерий половых органов.
Одновременно в клетках активируется система вторичных посредников —
цГМФ. М-холинорецепторы блокирует атропин, который тормозит пара-
симпатическую стимуляцию сокращения гладких мышц и парасимпатиче-
ское торможение деятельности сердца.
Как правило, передача возбуждения с симпатических нервов на эффек-
торные органы осуществляется медиаторами катехоламинового ряда. Од-
нако имеются и симпатические холинергические волокна, например, ин-
нервирующие сосуды скелетных мышц, потовые железы. Освобождающий-
ся в синаптических структурах этих волокон ацетилхолин расслабляет
гладкие мышцы сосудов, действуя через так называемый эндотелиальный
релаксирующий фактор (NO), а также вызывает секрецию потовых желез.
К медиаторам в вегетативной нервной системе относят также серото-
нин. При этом различают три типа его рецепторов: D, М, Т.
D-серотонинергические рецепторы локализуются в гладких мышцах.
Действуя на эти рецепторы, серотонин вызывает сокращение гладких
мышц.
М-серотонинергические рецепторы расположены в основном в вегета-
тивных ганглиях. Влияя на эти рецепторы, серотонин облегчает синапти-
ческую передачу в ганглии за счет повышения возбудимости Н-холинерги-
ческих структур.
Т-серотонинергические рецепторы обнаружены в сердечной и легочной
рефлексогенных зонах.
Эффективность синаптической передачи нервных импульсов на эффек-
тор зависит от количества активных рецепторов на постсинаптической
мембране. Число мембранных рецепторов возрастает при интенсивной ра-
боте синапса. Перерезка вегетативного нерва в эксперименте на животных
(прекращение выделения медиатора) увеличивает число мембранных ре-
цепторов, что повышает чувствительность денервированных структур к ме-
диатору (явление сенситизации).
В органах, имеющих двойную вегетативную иннервацию (табл. 5.1), эф-
фекты от изолированного раздражения парасимпатических или симпатиче-
ских нервов могут быть противоположными. Так, под влиянием парасим-
256 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
патических нервов замедляется ритм и уменьшается сила сокращений к
сердца, тогда как симпатические нервы учащают ритм сердца и увеличива- f
ют силу его сокращений. Вместе с тем взаимосвязь влияний симпатиче-
ской и парасимпатической иннервации в процессе регуляции функций 8
имеет сопряженный характер благодаря близкому расположению симпати-
ческих и парасимпатических синапсов. Во-первых, медиатор одной систе-
мы подавляет через рецепторы пресинаптической мембраны выделение
медиатора другой системы. Во-вторых, в постсинаптических структурах
взаимодействие медиаторов осуществляется за счет вторичных посредни-
ков цАМФ, цГМФ и кальция. При этом, например, тормозной эффект
возбуждения парасимпатической иннервации в миокарде тем сильнее, чем
выше уровень симпатической активности (так называемый акцентирован-
ный антагонизм).
Противоположный характер влияний симпатического и парасимпатиче-
ского отделов нервной системы проявляется еще и в том, что один из них j
может иннервировать железу внутренней секреции, вызывающую измене-
ния состояния организма в одном направлении, а второй иннервирует дру-
гую железу, которая изменяет состояние организма в противоположном I
направлении. Так, симпатические нервы стимулируют мозговой слой над-
почечника и увеличивают секрецию адреналина, что приводит к увеличе-
нию сахара в крови — гипергликемии, а парасимпатические блуждающие
нервы, иннервируя островки поджелудочной железы, увеличивают продук-
цию инсулина, который вызывает снижение содержание сахара в крови —
гипогликемию.
Однако взаимодействие симпатической и парасимпатической систем
может осуществляться и по пути синергизма. Последний проявляется во
влиянии этих отделов вегетативной нервной системы на трофику тканей и '
особенно при адаптивных реакциях, когда симпатическая нервная система
обеспечивает быструю «аварийную» мобилизацию энергетических ресурсов
и активирует функциональные ответы на раздражители, а парасимпатиче- (
ская — поддерживает гомеостазис, обеспечивая резервы для аварийной ре- t
гуляции, т. е. симпатические влияния обеспечивают эрготропную регуля-
цию приспособления, а парасимпатические — трофотропную.
5.3.2. Энтеральная нервная система
Органы, обладающие собственным миогенным моторным ритмом (сердце,
желудок и кишечник, желчный пузырь и желчевыводящие протоки, матка,
мочевой пузырь и мочеточники), продолжают осуществлять свои мотор-
ные функции и после перерезки подходящих к ним симпатических или па-
расимпатических нервных волокон и даже после извлечения этих органов
из организма. Относительную функциональную автономию этих органов
обеспечивает энтеральная часть вегетативной нервной системы.
Энтеральная нервная система содержит вегетативные ганглии, располо-
женные в стенках внутренних органов (интрамурально). Под ее контролем
находятся гладкие мышцы, всасывающий и секретирующий эпителий, ло-
кальный кровоток, местные эндокринные элементы, иммунные структуры.
Ганглии энтеральной нервной системы по своей структурной организа-
ции сходны с ЦНС. В этих ганглиях имеется большинство медиаторов
ЦНС: ацетилхолин, норадреналин, дофамин, гистамин, ГАМК, субстанция
Р, простагландины, гастроинтестинальные гормоны, пуриновые нуклео-
тиды.
Ганглии энтеральной нервной системы содержат весь набор структур,
5. Вегетативная нервная система • 257
которые характеризуют интегративную функцию нервной системы: сен-
сорные элементы, промежуточные нейроны, моторные нейроны, а также
собственные нейрогенные водители ритма.
Нейроны ганглиев имеют синаптические контакты с волокнами симпа-
тического и парасимпатического отделов нервной системы. Действуя на
структуры энтеральной системы, эти волокна модулируют активность же-
лудочно-кишечного тракта. Тормозное влияние волокон симпатических
нервов на моторные функции кишечника, в частности, осуществляется че-
рез аксосоматические и аксо-аксональные синапсы, образованные этими
волокнами внутри интрамуральных ганглиев на клетках и терминалях
постганглионарных парасимпатических нейронов.
Сенсорные элементы ганглия энтеральной нервной системы дают ин-
формацию о состоянии стенки органа или химическом составе содержимо-
го его полости. Получая эту сенсорную информацию, связующие внутри-
ганглионарные интегративные цепи нейронов перерабатывают ее и гене-
рируют возбуждающий (или тормозный) поток импульсов к эффекторным
нейронам и далее к гладкой мускулатуре, железам, сосудам стенки органа.
Ганглии энтеральной нервной системы выполняют функции низших цен-
тров интеграции висцеральных функций. Поэтому внутренние органы с
разрушенными (или выключенными с помощью ганглиоблокаторов) ганг-
лиями утрачивают способность к координации ритмической моторной
функции.
5.4. Рефлексы вегетативной нервной системы
Вегетативные рефлексы, возникающие при возбуждении рецепторов внут-
ренних органов, подразделяются на сегментарные, реализуемые спинным
мозгом и стволовыми структурами, и надсегментарные, которые обеспечи-
ваются высшими центрами вегетативной регуляции.
Передачу импульсов от интерорецепторов в головной мозг осуществля-
ют аксоны вставочных нейронов спинного мозга, на которые передается
возбуждение и от висцеральных (спинальных и интрамуральных) сигналы
и от соматических (кожных, мышечных) афферентов.
Совмещение афферентной импульсации, поступающей от рецепторов
внутренних органов и кожи, является основой для появления так называе-
мых отраженных болей. При некоторых процессах во внутренних органах
болевые ощущения как бы проецируются в участки наружных покровов.
Вместе с тем возбуждение от рецепторов внутренних органов в любом сег-
менте спинного мозга передается не только на вегетативные, но и на сома-
тические эфферентные нейроны. Поэтому раздражение внутренних орга-
нов может вызвать возникновение двигательных реакций (напряжение
скелетных мышц, вынужденные позы).
Поскольку рефлекторные процессы в вегетативной и соматической
нервной системе тесно связаны, в организме существуют сопряженные
рефлексы, когда рецептор и эффектор расположены в разных органах и
даже разных системах.
Такие рефлексы разделяют на висцеро-висцеральные, висцеросоматиче-
ские, соматовисцеральные, висцеросенсорные.
Висцеро-висцеральный рефлекс — раздражение рецепторов в одном внут-
реннем органе приводит к изменению деятельности другого внутреннего
органа. Так, механическое раздражение брыжейки вызывает уменьшение
частоты сердечных сокращений (рефлекс Гольца); раздражение рецепторов
аортальной или каротидной зон влечет за собой изменение глубины и час-
9-6095
258 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
тоты дыхания, уровня артериального давления и частоты сокращений
сердца; надавливание на глазные яблоки вызывает снижение ритма сердца
(рефлекс Данини—Ашнера).
Висцеросоматический рефлекс — возбуждение рецепторов некоторых
внутренних органов способно изменить тонус скелетной мускулатуры. Раз-
дражение, например, рецепторов желудочно-кишечного тракта (в частно-
сти — брюшины) вызывает сокращение мышц передней брюшной стенки.
Соматовисцеральный рефлекс — раздражение экстерорецепторов приво-
дит к изменению деятельности внутренних органов. Так, действие ряда
раздражителей (тепло, холод, некоторые физиотерапевтические процеду-
ры) на рецепторы кожи вызывает сосудистые реакции в органах грудной
или брюшной полости.
Висцеросенсорный рефлекс — раздражение рецепторов внутренних орга-
нов приводит к изменениям чувствительности кожи. Зона повышенного
восприятия в этом случае ограничивается участком кожи, иннервируемым
сегментом, к которому поступают импульсы от раздражаемого внутреннего
органа. Эти зоны называют зонами Захарьина—Геда. При недостаточности
коронарного кровотока (приступ стенокардии) характерна иррадиация бо-
лей в левое плечо, левую руку, левую половину шеи и головы, нижнюю че-
люсть, межлопаточное пространство, а иногда в верхнюю часть живота.
Иррадиация болей в этом случае обусловлена повышенной чувствительно-
стью кожи к болевым ощущениям в зонах, которые иннервируются VII
шейным и I—V грудными сегментами спинного мозга. Раздражение желче-
выводящих путей вызывает повышенную чувствительность (гиперестезию)
кожи в области проекции желчного пузыря, в эпигастральной области, в
плечевой зоне, в точке лопаточного угла, в паравертебральных точках
справа от VIII до XI грудного позвонка, в точке диафрагмального нерва —
болезненность при надавливании между передними ножками правой гру-
дино-ключично-сосцевидной мышцы (положительный френикус-симптом,
симптом Мюсси—Георгиевского).
5.5. Высшие центры вегетативной регуляции
Группа центров головного мозга, расположенных в продолговатом мозге и
подбугорной области, осуществляет регуляцию гомеостазиса в условиях,
которые характеризуются некоторым постоянством, например в покое. В
стволе мозга располагаются сердечно-сосудистый и дыхательный центры,
центры глотания, слюноотделения, чиханья, рвоты.
Ретикулярная формация, способная осуществлять как активирующие,
так и тормозящие влияния на различные отделы центральной нервной
системы, регулирует уровень активности также и нервных центров вегета-
тивных функций организма. Специализированным образованием ретику-
лярной формации является бульбарный сосудодвигательный центр, кото-
рый поддерживает тонус и интегрирует деятельность сосудодвигательных
центров спинного мозга.
Главным центром поддержания постоянства внутренней среды организ-
ма, является гипоталамус, в котором расположены центры регуляции тем-
пературы тела и водного баланса, а также пищевого и полового поведения.
Более 40 пар ядер гипоталамуса через симпатический и парасимпатиче-
ский отделы вегетативной нервной системы обеспечивают регуляцию
большинства висцеральных функций. Возбуждение заднегипоталамических
ядер повышает системное артериальное давление, учащает ритм сердца,
увеличивает содержание сахара в крови. При возбуждении переднегипота-
5. Вегетативная нервная система • 259
ламических ядер происходит снижение артериального давления, замедле-
ние ритма сердца, а также усиление перистальтики кишечника. Ядра сред-
ней части гипоталамуса {латеральные и вентромедиальные) регулируют ап-
петит и пищевое поведение. Раздражение ядер заднего и среднего гипота-
ламуса способно вызывать агрессивное поведение и эмоции удовольствия.
Благодаря нервным и гуморальным связям с передней и задней долями
гипофиза, гипоталамус также участвует в регуляции эндокринных функций
и тем самым влияет практически на все виды обмена веществ.
Регулируя функции симпатической и парасимпатической нервной сис-
темы и эндокринных желез, гипоталамус обеспечивает вегетативный ком-
понент всех реакций организма. В свою очередь функции гипоталамуса
контролируются высшими отделами центральной нервной системы — под-
корковыми ядрами и корой больших полушарий, с которыми гипоталамус
связан как прямыми нервными путями, так и через посредство ретикуляр-
ной формации мозгового ствола.
На функции органов, иннервируемых вегетативной системой, влияет
также мозжечок. Последствия его удаления не ограничиваются только рас-
стройствами движений скелетных мышц, но охватывают и ряд вегетатив-
ных функций. У животных с удаленным мозжечком обнаружены различ-
ные нарушения функций внутренних органов: угнетение моторной функ-
ции и секреции желез желудка и кишечника, изменения возбудимости со-
судодвигательного центра. После удаления мозжечка все основные вегета-
тивные функции сохраняются, но протекают менее совершенно (напри-
мер, механизмы терморегуляции, предохраняющие организм от перегрева-
ния, включаются позже, чем у нормальных животных; уровень глюкозы в
крови оперированного животного в меньшей степени снижается после
введения инсулина).
В регуляции деятельности внутренних органов существенна роль струк-
тур переднего мозга, входящих в состав так называемой лимбической сис-
темы, или «висцерального мозга»', гиппокамп, поясная извилина, миндалевид-
ные ядра. Лимбическая система во многом определяет поддержание гомео-
стазиса в организме, оказывая регулирующее влияние на функции орга-
нов, иннервируемых вегетативной нервной системой. Она также участвует
в организации мотиваций, эмоций и поведенческих реакций с выраженны-
ми вегетативными компонентами.
Об участии коры больших полушарий головного мозга в регуляции вис-
церальных функций свидетельствует выработка условных рефлексов с ре-
цепторов внутренних органов при изменении их деятельности. В образова-
нии таких условных рефлексов важная роль принадлежит передним корко-
вым отделам, в первую очередь области сигмовидной и коронарной изви-
лин. После удаления у животных в эксперименте этих участков коры воз-
никают нарушения условных рефлексов, выработанных с рецепторов орга-
нов сердечно-сосудистой системы, желудочно-кишечного тракта, почек, а
также других висцеральных систем.
9*
ГЛАВА 6
Эндокринная нервная система —
регулятор функций и процессов
в организме
Эндокринная система представляет собой функциональное объедине-
ние специализированных для внутренней секреции клеток, тканей и орга-
нов. Основной их функцией является синтез и секреция во внутреннюю
среду организма (инкреция) молекул гормонов. Таким образом, эндокрин-
ная система осуществляет гормональную регуляцию процессов жизнедея-
тельности. Эндокринной функцией обладают: 1) органы или железы внут-
ренней секреции, 2) эндокринная ткань в органе, функция которого не
сводится лишь к внутренней секреции, 3) клетки, обладающие наряду с
эндокринной и неэндокринными функциями. Эти три вида структур и об-
разуемые ими гормоны приведены в табл. 6.1.
6.1. Химическая природа и общие механизмы действия гормонов
Гормоны подразделяют по химической природе на три группы: 1) произ-
водные аминокислот — тиреоидные гормоны, адреналин, гормоны эпифи-
за; 2) пептидные гормоны, простые (протеины) и сложные (гликопротеи-
ды) белки — гипоталамические нейропептиды, гормоны гипофиза, остров-
кового аппарата поджелудочной железы, околощитовидных желез; 3) сте-
роидные гормоны, образующиеся из холестерина гормоны коры надпочеч-
ников, половых желез, гормон почечного происхождения кальцитриол.
6.1.1. Механизмы действия пептидных, белковых гормонов
и катехоламинов
Молекулу гормона обычно называют первичным посредником регулятор-
ного эффекта, или лигандом. Молекулы большинства гормонов связывают-
ся со специфическими для них рецепторами плазматических мембран кле-
ток мишеней, образуя лиганд-рецепторный комплекс. Для пептидных, белко-
вых гормонов и катехоламинов его образование является основным началь-
ным звеном механизма действия и приводит к активации мембранных
ферментов и образованию различных вторичных посредников гормонально-
го регуляторного эффекта, реализующих свое действие в цитоплазме, орга-
ноидах и ядре клетки. Среди ферментов, активируемых лиганд-рецептор-
ным комплексом, описаны: аденилатциклаза, гуанилатциклаза, фосфолипа-
зы С, D и А2, тирозинкиназы, фосфаттирозинфосфатазы, фосфоинозитид-3-
ОН-киназа, серинтреонин-киназа, синтаза NO и др. Вторичными посредни-
ками, образующимися под влиянием этих мембранных ферментов, явля-
ются: 1) циклический аденозинмонофосфат (цАМФ); 2) циклический гуано-
зинмонофосфат (цГМФ); 3) инозитол-3-фосфат (ИФЗ); 4) диацилглицерол',
6. Эндокринная нервная система — регулятор функций и процессов в организме • 261
Таблица 6.1. Органы, ткани и клетки с эндокринной функцией
№ Органы Ткань, клетки Гормоны
1. ГИПОФИЗ а) аденогипо- 1. Эндокринные желе Кортикотрофы зы Кортикотропин
физ б) нейроги- Гонадотрофы Тиреотрофы Соматотрофы Лактотрофы Питуициты Меланотропин Фоллитропин Лютропин Тиреотропин Соматотропин Пролактин Вазопрессин
2. пофиз Надпочечники а) корковое Клубочковая зона Окситоцин Эндорфины Минералокортикоиды
вещество Пучковая зона Глюкокортикоиды
б) мозговое Сетчатая зона Хромаффинные клетки Половые стероиды Адреналин (Норадреналин)
3. вещество Щитовидная же- Фолликулярные тиреоциты Адреномедудлин Трийодтиронин
4. леза Околощитовид- К-клетки Главные клетки Тетрайодтиронин Кальцитонин Паратирин
ные железы К-клетки Кальцитонин
5. Эпифиз Пинеоциты Мелатонин
6. Поджелудочная 2. Органы с эндокринной Островки Лангерганса тканью
железа альфа-клетки Глюкагон
7. Половые железы а) семенники бета-клетки дельта-клетки Клетки Лейдига Инсулин Соматостатин Тестостерон
б)яичники Клетки Сертолли Клетки гранулезы Эстеро гены Ингибин Эстрадиол
8. Желудочно-ки- Желтое тело 3. Органы с инкреторной функ Эндокринные и энтерохром- Эстрон Прогестерон Прогестерон цией клеток Регуляторные пептиды
9. шечный тракт Плацента аффинные клетки желудка и тонкого кишечника Синцитиотрофобласт Хорионический гонадотропин
10. Тимус Цитотрофобласт Тимоциты Пролактин Эстриол Прогестерон Тимозин, Тимопоэтин, Тимулин
11. Почка ЮГА Ренин
12. Сердце Перитубуляерные клетки Канальцы Миоциты предсердий Эритропоэтин Кальцитриол Атриопептид
13. Кровеносные Эндотелиоциты Соматостатин Ангиотензин-П Эндотел ины
сосуды NO Гиперполяризуюгций фактор Простагландины Регуляторы адгезии
262 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
5) олиго (А) (2,5-олигоизоаденилат)', 6) Са2+ (ионизированный кальций);
7) фосфатидная кислота; 8) циклическая аденозиндифосфатрибоза; 9) NO
(оксид азота). Многие гормоны, образуя лиганд-рецепторные комплексы,
вызывают активацию одновременно нескольких мембранных ферментов и,
соответственно, вторичных посредников.
Значительная часть гормонов и биологически активных веществ взаи-
модействуют с семейством рецепторов, связанных с G-белками плазмати-
ческой мембраны (андреналин, норадреналин, аденозин, ангиотензин, эн-
дотелии и др.).
6.1.1.1. Основные системы вторичных посредников
1. Система аденилатциклаза—цАМФ (рис. 6.1). Мембранный фермент
аденилатциклаза может находиться в двух формах — активированной и не-
активированной. Активация аденилатциклазы происходит под влиянием
гормон-рецепторного комплекса, образование которого приводит к связы-
ванию гуанилового нуклеотида (ГТФ) с особым регуляторным стимули-
рующим белком (GS-белок), после чего GS-белок вызывает присоединение
Mg к аденилатциклазе и ее активацию. Так действуют активирующие аде-
нилатциклазу гормоны — глюкагон, тиротропин, паратирин, вазопрессин
(через V-2-рецепторы), гонадотропин и др. Ряд гормонов, напротив, по-
давляет аденилатциклазу — соматостатин, ангиотензин-II и др. Гормон-ре-
цепторные комплексы этих гормонов взаимодействуют в мембране клетки
с другим регуляторным ингибирующим белком (GI-белок), который вызы-
вает гидролиз гуанозинтрифосфата (ГТФ) до гуанозиндифосфата (ГДФ) и,
соответственно, подавление активности аденилатциклазы. Адреналин через
0-адренорецепторы активирует аденилатциклазу, а через а,-адренорецепто-
ры ее подавляет, что во многом и определяет различия эффектов стимуля-
ции разных типов рецепторов.
Под влиянием аденилатциклазы из АТФ синтезируется цАМФ, вызы-
вающий активацию двух типов протеинкиназ в цитоплазме клетки, веду-
щих к фосфорилированию многочисленных внутриклеточных белков. Это
повышает или снижает проницаемость мембран, активность и количество
ферментов, т. е. вызывает типичные для гормона метаболические и, соот-
ветственно, функциональные сдвиги жизнедеятельности клетки.
В табл. 6.2 приведены основные эффекты активации цАМФ-зависимых
протеинкиназ.
Кроме активации протеинкиназ внутриклеточные эффекты цАМФ реа-
лизуются также через другие механизмы: систему кальций—кальмодулин,
трансметилазную систему, аденозин-5-монофосфат — аденозин, тирозин-
киназы. О роли системы кальций—кальмодулин сказано ниже.
Таблица 6.2. Некоторые эффекты фосфорилирования белков клетки цАМФ-зависимой
протеинкиназой
Виды белков Эффекты фосфорилирования
Компоненты мембран Ферменты, лимитирующие скорость метаболического процесса Белки рибосом Ядерные белки Белки микротрубочек Изменения проницаемости Активация или подавление Активирование или подавление трансляции Активация или подавление транскрипции Секреторный, двигательный эффекты или изме- нение конфигурации клетки
6. Эндокринная нервная система — регулятор функций и процессов в организме • 263
Рис. 6.1. Опосредование гормонального сигнала системой аденилатциклаза—
цАМФ.
Образование гормон-рецепторного комплекса приводит к связыванию гуанилового нуклеоти-
да (ГТФ) с регуляторным стимулирующим белком (GS-белок), последний вызывает присоеди-
нение Mg к аденилатциклазе и ее активацию (слева вверху). Так действуют активирующие аде-
нилатциклазу гормоны — глюкагон, тиротропин, паратирин и др. Гормон-рецепторные ком-
плексы соматостатина, ангиотензина-11 и других подавляющих аденилатциклазу гормонов
взаимодействуют в мембране клетки с другим регуляторным ингибирующим белком (GI-бе-
лок), который вызывает гидролиз ГТФ до ГДФ и, соответственно, подавление активности аде-
нилатциклазы (вверху справа). Под влиянием аденилатциклазы из АТФ синтезируется цАМФ,
вызывающий активацию протеинкиназ в цитоплазме клетки и фосфорилирование многочис-
ленных внутриклеточных белков. Это меняет проницаемость мембран, активность и количест-
во ферментов, т. е. вызывает типичные для гормона метаболические и, соответственно, функ-
циональные сдвиги. Внутриклеточные эффекты цАМФ реализуются также через другие меха-
низмы: систему кальций-кальмодулин, трансметилазную систему, аденозин-5-монофосфат —
аденозин, тирозинкиназы.
264 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Трансметилазная система обеспечивает метилирование ДНК, всех типов
РНК, белков хроматина и мембран, ряда гормонов на уровне тканей, фос-
фолипидов мембран. Это способствует реализации многих гормональных
влияний на процессы пролиферации, дифференцировки, состояние про-
ницаемости мембран и свойства их ионных каналов и, что важно подчерк-
нуть особо, влияет на доступность мембранных рецепторных белков моле-
кулам гормонов.
Прекращение гормонального эффекта, реализуемого через систему аде-
нилатциклаза—цАМФ, осуществляется с помощью специального фермента
фосфодиэстеразы цАМФ, вызывающей гидролиз этого вторичного посред-
ника с образованием аденозин-5-монофосфата. Однако этот продукт гид-
ролиза превращается в клетке в аденозин, также обладающий эффектами
вторичного посредника, так как подавляет в клетке процессы метилиро-
вания.
2. Система гуанилатциклаза-цГМФ. Активация мембранной гуанилат-
циклазы происходит не под непосредственным влиянием гормон-рецеп-
торного комплекса, а опосредованно через ионизированный кальций и ок-
сидантные системы мембран. Определяющая эффекты ацетилхолина сти-
муляция активности гуанилатциклазы также осуществляется опосредован-
но через Са2+. Через активацию гуанилатциклазы реализует эффект и на-
трийуретический гормон предсердий — атриопептид. Путем активации пе-
рекисного окисления стимулирует гуанилатциклазу гормон эндотелия со-
судистой стенки оксид азота — расслабляющий эндотелиальный фактор.
Под влиянием гуанилатциклазы из ГТФ синтезируется цГМФ, активирую-
щий цГМФ-зависимые протеинкиназы, которые уменьшают скорость фос-
форилирования легких цепей миозина в гладких мышцах стенок сосудов,
приводя к их расслаблению. В большинстве тканей биохимические и фи-
зиологические эффекты цАМФ и цГМФ противоположны. Примерами
могут служить стимуляция сокращений сердца под влиянием цАМФ и тор-
можение их цГМФ, стимуляция сокращения гладких мышц кишечника
цГМФ и подавление цАМФ. цГМФ обеспечивает гиперполяризацию ре-
цепторов сетчатки глаза под влиянием фотонов света. Ферментативный
гидролиз цГМФ, а следовательно, и прекращение гормонального эффекта,
осуществляется с помощью специфической фосфодиэстеразы.
3. Система фосфолипаза С — инозитол-3-фосфат (рис. 6.2). Гормонре-
цепторный комплекс с участием регуляторного G-белка ведет к активации
мембранного фермента фосфолипазы С, вызывающей гидролиз фосфоли-
пидов мембраны с образованием двух вторичных посредников: инозитол-
3-фосфата и диацилглицерола. Инозитол-З-фосфат вызывает выход Са2+ из
внутриклеточных депо, в основном из эндоплазматического ретикулума,
ионизированный кальций связывается со специализированным белком
кальмодулином, что обеспечивает активацию протеинкиназ и фосфорили-
рование внутриклеточных структурных белков и ферментов. В свою оче-
редь диацилглицерол способствует резкому повышению сродства протеин-
киназы С к ионизированному кальцию, последний без участия кальмоду-
лина ее активирует, что также завершается процессами фосфорилирования
белков. Диацилглицерол одновременно реализует и другой путь опосредо-
вания гормонального эффекта за счет активирования фосфолипазы А-2.
Под влиянием последней из мембранных фосфолипидов образуется арахи-
доновая кислота, являющаяся источником мощных по метаболическим и
физиологическим эффектам веществ — простагландинов и лейкотриенов. В
разных клетках организма превалирует один или другой путь образования
вторичных посредников, что в конечном счете и определяет физиологиче-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 265
Рис. 6.2. Опосредование гормонального сигнала системой фосфолипаза С—инози-
тол-3-фосфат.
Образование гормон-рецепторного комплекса при участии регуляторного G-белка активирует
мембранную фосфолипазу С, вызывающую гидролиз фосфолипидов мембраны с образовани-
ем двух вторичных посредников: инозитол-3-фосфата и диацилглицерола. Инозитол-З-фосфат
ведет к выходу Са2+ из внутриклеточных депо. Связывание ионизированного кальция со спе-
циализированным белком кальмодулином активирует протеинкиназы и вызывает фосфорили-
рование внутриклеточных структурных белков и ферментов. Диацилглицерол повышает срод-
ство протеинкиназы С к Са2+, способствуя ее активации, что также завершается процессами
фосфорилирования белков. Диацилглицерол одновременно реализует другой путь опосредова-
ния гормонального эффекта, активируя фосфолипазу А-2 и образование простаноидов.
ский эффект гормона. Через рассмотренную систему вторичных посредни-
ков реализуются эффекты адреналина (при связи с альфа-адренорецепто-
ром), вазопрессина (при связи с V-1-рецептором), ангиотензина-II, сома-
тостатина, окситоцина.
4. Система кальций—кальмодулин. Ионизированный кальций поступает
в клетку после образования гормон-рецепторного комплекса либо из вне-
клеточной среды за счет активирования медленных кальциевых каналов
мембраны (как это происходит, например, в миокарде), либо из внутри-
клеточных депо под влиянием инозитол-3-фосфата. В цитоплазме немы-
шечных клеток кальций связывается со специальным белком-кальмодули-
ном, а в мышечных клетках роль кальмодулина выполняет тропонин С.
Связанный с кальцием кальмодулин изменяет свою пространственную ор-
266 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
ганизацию и активирует многочисленные протеинкиназы, обеспечиваю-
щие фосфорилирование, а следовательно изменение структуры и свойств
белков. Кроме того комплекс кальций—кальмодулин активирует фосфо-
диэстеразу цАМФ, что подавляет эффект циклического соединения как
вторичного посредника. Вызываемое гормональным стимулом кратковре-
менное увеличение в клетке кальция и его связывание с кальмодулином
является пусковым стимулом для многочисленных физиологических про-
цессов — сокращения мышц, секреции гормонов и выделения медиаторов,
синтеза ДНК, изменения подвижности клеток, транспорта веществ через
мембраны, изменения активности ферментов.
6.1.1.2. Взаимосвязи вторичных посредников
В клетках организма присутствуют или могут образовываться одновремен-
но несколько вторичных посредников. В связи с этим между вторичными
посредниками устанавливаются различные взаимоотношения: 1) равно-
значное участие, когда разные посредники необходимы для полноценного
гормонального эффекта; 2) один из посредников является основным, а
другой лишь способствует реализации эффектов первого; 3) посредники
действуют последовательно (например, инозитол-3-фосфат обеспечивает
освобождение кальция, диацилглицерол облегчает взаимодействие кальция
с протеинкиназой С); 4) посредники дублируют друг друга для обеспече-
ния избыточности с целью надежности регуляции; 5) посредники являются
антагонистами, т. е. один из них включает реакцию, а другой — тормозит
(например, в гладких мышцах сосудов инозитол-3-фосфат и кальций реа-
лизуют их сокращение, а цАМФ — расслабление).
6.1.2. Механизм действия стероидных гормонов
Стероидные гормоны (рис. 6.3) обладают двумя путями действия на клет-
ки: 1) классическим геномным или медленным и 2) быстрым негеномным.
6.1.2.1. Геномный механизм действия
Геномный механизм действия на клетки-мишени начинается трансмем-
бранным переносом молекул стероидных гормонов в клетку (благодаря их
растворимости в липидном бислое клеточной мембраны), с последующим
связыванием гормона с цитоплазменным белком-рецептором. Эта связь с
рецепторным белком необходима для поступления стероидного гормона в
ядро, где происходит его взаимодействие с ядерным рецептором. После-
дующее взаимодействие комплекса гормон—ядерный рецептор с хромати-
новым акцептором, специфическим кислым белком и ДНК влечет за со-
бой: активацию транскрипции специфических мРНК, синтез транспорт-
ных и рибосомных РНК, процессинг первичных РНК-транскриптов и
транспорт мРНК в цитоплазму, трансляцию мРНК при достаточном уров-
не транспортных РНК с синтезом белков и ферментов в рибосомах. Все
эти явления требуют длительного (часы, сутки) присутствия гормон-рецеп-
торного комплекса в ядре.
6.1.2.2. Негеномный механизм действия
Эффекты стероидных гормонов проявляются не только спустя несколько
часов, что требуется для ядерного влияния, часть из них проявляется очень
6. Эндокринная нервная система — регулятор функций и процессов в организме • 267
быстро, в течение нескольких минут. Это такие эффекты, как повышение
проницаемости мембран, усиление транспорта глюкозы и аминокислот,
освобождение лизосомальных ферментов, сдвиги энергетики митохондрий.
К числу быстрых негеномных эффектов стероидных гормонов относятся,
например, увеличение в течение 5 мин после введения человеку альдосте-
рона общего периферического сосудистого сопротивления и артериального
давления, изменение транспорта натрия через мембрану эритроцитов (во-
обще лишенных ядра) под влиянием альдостерона в опытах in vitro, быст-
рый вход Са2+ в клетки эндометрия под влиянием эстрогенов и др. Меха-
низм негеномного действия стероидных гормонов заключается в связыва-
нии на плазматической мембране клетки со специфическими рецепторами
и активации каскадных реакций систем вторичных посредников, например
фосфолипазы С, инозитол-3-фосфата, ионизированного Са2+, протеинки-
назы С. Под влиянием стероидных гормонов в клетке может увеличиваться
содержание цАМФ и цГМФ. Негеномный эффект стероидных гормонов
Рис. 6.3. Схема путей действия стероидных гормонов.
1 — классический геномный путь действия (гормон проникает через клеточную мембрану и
цитоплазму в ядро, где после взаимодействия с ядерным рецептором воздействует на гены-ми-
шени, активируя их). 2а и 26 — негеномные пути действия через мембранные рецепторы: 2а —
пути, связанные с мембранным ферментом и образованием вторичного посредника, ведущего
к активации протеинкиназ. Последние через фосфорилирование в ядре белка-коактиватора
(БКА) активируют гены-мишени; 26 — пути, связанные с ионными каналами клеточной мем-
браны, в результате чего гормон-рецепторный комплекс активирует ионные каналы, меняя
возбудимость клетки. 3 — альтернативный негеномный путь действия (молекула гормона, про-
никая через мембрану в цитоплазму, взаимодействует с цитозольным рецептором, что приво-
дит к активации цитозольных киназ.
268 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
может быть реализован и после их связывания с цитоплазматическими ре-
цепторами. Часть негеномных эффектов стероидных гормонов осуществ-
ляется благодаря их взаимодействию с рецепторами, связанными с ворот-
ным механизмом ионных каналов мембран нервных клеток, являясь тем
самым модуляторами, например, глицин-, серотонин- или гамма-аминобу-
тиратергических нейронов. Наконец, растворяясь в липидном бислое мем-
браны, стероидные гормоны могут менять физические свойства мембраны,
такие как ее текучесть или проницаемость для гидрофильных молекул, что
также является негеномным эффектом.
Таким образом, механизмы действия гормонов разной химической
структуры имеют не только различия, но и общие черты. Как и стероиды,
пептидные гормоны обладают способностью избирательно влиять на
транскрипцию генов в ядре клетки. Этот эффект пептидных гормонов мо-
жет быть реализован не только с поверхности клетки при образовании вто-
ричных посредников, но и путем поступления пептидных гормонов внутрь
клетки за счет интернализации гормон-рецепторного комплекса.
6.1.3. Саморегуляция чувствительности эффектора
к гормональному сигналу
Изменение характера и интенсивности регуляторных сигналов, поступаю-
щих к клетке, ведет к адаптивной перестройке ее рецепторных структур за
счет специальных механизмов саморегуляции, изменяющих чувствитель-
ность клетки к этому сигналу. Так, при избыточности внеклеточного гумо-
рального сигнала формируется десенситизация соответствующего рецепто-
ра, т. е. уменьшение его чувствительности к гуморальному регулятору из-за
ослабления сродства рецептора к молекуле лиганда или числа рецепторных
молекул. Механизм десенситизации рецепторов, реализующих регулятор-
ный эффект через системы вторичных посредников, представлен на
рис. 6.4. При наличии во внеклеточной среде избыточного гуморального
сигнала и/или увеличении времени связывания лиганда с рецептором ак-
тивируемая лиганд-рецепторным взаимодействием протеинкиназа плазма-
тической мембраны фосфорилирует рецептор, в результате его аффинность
к лиганду уменьшается в 2—5 раз, что предотвращает чрезмерность регуля-
торного эффекта. Если этого снижения чувствительности рецептора недос-
таточно, рецептор фосфорилируется другой протеинкиназой, активируе-
мой соответствующим вторичным посредником. Следствием такого фос-
форилирования является нарушение сопряжения рецепторов с G-белками
и нарушение эффектов G-белков (активирующих или ингибирующих) на
системы вторичных посредников и ионные каналы. Наконец, в случаях
недостаточной эффективности двух указанных механизмов десенситизации
гормон-рецепторные комплексы интернализуются внутрь клетки с образо-
ванием рецепторосомы (вакуоль с рецептором). В последующем, если уро-
вень гуморального регулятора во внеклеточной микросреде снизится в те-
чение 2—3 ч, интернализированный рецептор может вновь встроиться в
мембрану, а если повышенная концентрация регулятора сохраняется — ре-
цепторосомы сливаются с лизосомами и рецепторы разрушаются. Десенси-
тизация рецепторов может быть гомологической, проявляющейся в сниже-
нии чувствительности только к тому гуморальному регулятору, избыток
которого ее вызвал, и гетерологической, заключающейся в снижении чув-
ствительности и к другим гуморальным факторам регуляции, но также реа-
лизующим эффект через связанные с G-белком рецепторы и те же вторич-
ные посредники.
6. Эндокринная нервная система — регулятор функций и процессов в организме • 269
Рис. 6.4. Схема трех путей десенситизации клетки к избыточному внешнему регуля-
торному сигналу.
1. Снижение сродства рецептора к лиганду из-за фосфорилирования рецептора мембранной
протеинкиназой. 2. Нарушение связи молекулы рецептора с G-белком при фосфорилирова-
нии рецептора цитоплазматическими протеинкиназами. 3. Интернализация лиганд-рецептор-
ного комплекса и разрушение рецептора ферментами лизосом. Стрелками со знаком (—) обо-
значено подавление процесса взаимодействия.
Чувствительность клеток к внешним регуляторным сигналам зависит
также и от числа так называемых свободных, или резервных, рецепторов,
т. е. доступных для лиганда. Увеличивая синтез этих мембранных структур
или их «всплывание» на поверхность мембраны, клетка способна повы-
шать чувствительность к регуляторному сигналу (сенситизация}, например
в случаях недостаточного количества молекул гуморального регулятора во
внеклеточной микросреде. Повышение чувствительности к регуляторному
сигналу является также следствием возросшего сопряжения рецепторов с
G-белком и системой вторичных посредников.
Клетки организма, как правило, используют не один, а несколько путей
реализации одного гуморального регуляторного сигнала, в связи с чем для
каждого лиганда (например, молекул одного гормона) на мембранах суще-
ствуют несколько типов рецепторов. Так, для ацетилхолина, гистамина,
катехоламинов, опиоидных пептидов, серотонина, вазопрессина иденти-
270 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
фицировано два (вазопрессин), три (опиоиды), четыре (мускариновые для
ацетилхолина) и более (катехоламины, серотонин) мембранных рецепто-
ров. Соответственно, меняя за счет саморегуляции чувствительность ре-
цепторов одного типа, клетка изменяет характер функционирования и
способна компенсировать неадекватные сдвиги в регуляторных влияниях.
Одним из регуляторных эффектов, реализуемых внутри клетки с помо-
щью систем вторичных посредников мембранного лиганд-рецепторного
взаимодействия, является изменение функционального состояния генома
клетки и биосинтеза белка. Но и ядро клетки способно менять состояние
плазматической мембраны, ее чувствительность к сдвигам внеклеточной
среды, в том числе и изменению в ней количества и характера регулятор-
ных сигналов. В клетках под контролем генома происходит синтез специ-
альных белков — инверторов, оказывающих различное влияние на функ-
циональное состояние мембраны в зависимости от адаптивно-компенса-
торных потребностей клетки — изменение активности рецепторов, состоя-
ния ионных каналов и насосов. Эти белки могут вести к гиперполяриза-
ции клеточной мембраны, ослаблению функциональной активности клет-
ки, с одновременным повышением синтеза белка и процессов регенерации
внутриклеточных структур.
6.2. Регуляторные функции гормонов гипофиза
Гипофиз представляет собой эндокринный орган, объединяющий три желе-
зы, соответстветствующие его отделам, или долям. Передняя доля гипофи-
за получила название аденогипофиза. По морфологическим критериям это
железа эпителиального происхождения, содержащая несколько типов эн-
докринных клеток, составляет 3/4 общей массы гипофиза. Задняя доля ги-
пофиза, или нейрогипофиз, образуется в эмбриогенезе как выпячивание
вентрального гипоталамуса и имеет общее с ним нейроэктодермальное
происхождение. В нейрогипофизе локализованы веретенообразные клет-
ки — питуициты и аксоны гипоталамических нейронов. Третья, или про-
межуточная, доля гипофиза, как и передняя, — эпителиального происхож-
дения, у человека практически отсутствует, но отчетливо выражена у мно-
гих животных. У человека ее функцию выполняет небольшая группа кле-
ток передней части задней доли, эмбриологически и функционально свя-
занных с аденогипофизом.
Кровоснабжение аденогипофиза и нейрогипофиза. Передняя доля гипофи-
за прямого артериального снабжения не получает, а кровь в нее поступает
из срединного возвышения через воротную систему гипофиза. Эта особен-
ность кровоснабжения играет основную роль в регуляции функций перед-
ней доли, поскольку аксоны нейросекреторных клеток гипоталамуса в об-
ласти срединного возвышения образуют аксовазальные контакты и нейро-
секрет с регуляторными пептидами через воротные сосуды поступает в
аденогипофиз. Задняя доля гипофиза получает артериальную кровь из
нижней гипофизарной артерии. Наиболее интенсивный кровоток имеет
место в аденогипофизе, при этом его уровень (0,8 мл/г/мин) выше, чем в
большинстве других тканей организма.
Венозный отток из гипофиза осуществляется в пещеристый венозный
синус твердой мозговой оболочки (меньшая часть). Большая часть крови
оттекает ретроградно в срединное возвышение, что, благодаря транспорту
кровью гипофизарных гормонов, играет определяющую роль в реализации
механизмов обратной связи между гипофизом и гипоталамусом.
6. Эндокринная нервная система — регулятор функций и процессов в организме • 271
6.2.1. Гормоны аденогипофиза и их эффекты в организме
Структура передней доли гипофиза представлена 8 типами клеток, из ко-
торых секреторная функция присуща хромафильным клеткам. Выделяют
5 типов хромафильных клеток: 1) ацидофильные красные клетки с мелки-
ми гранулами или соматотрофы — вырабатывают соматотропин (СТГ, гор-
мон роста); 2) ацидофильные желтые клетки с крупными гранулами или
лактотрофы — вырабатывают пролактин-, 3) базофильные тиреотрофы —
вырабатывают тиреотропин (тиреотропный гормон — ТТГ); 4) базофиль-
ные гонадотрофы — вырабатывают гонадотропины-, фоллитропин (фоллику-
лостимулирующий гормон — ФСГ) и лютропин (лютеинизирующий гор-
мон — ЛГ); 5) базофильные кортикотрофы — вырабатывают кортикотро-
пин (адренокортикотропный гормон — АКТГ). Кроме того, так же как и в
клетках промежуточной доли, в базофильных кортикотрофах образуются
бета-эндорфин и меланотропин, поскольку все эти вещества происходят из
единой общей молекулы предшественника липотропинов.
Кортико-
либерин
Тирео-
либерин
Г онадо- Пролактостатин?
либерин (дофамин)
।
I
♦
Кортикотропин
Тиреотропин
Половые железы
Пролактин
Кора надпо-
чечников
Глюкокортикоиды
Половые гормоны
Щитовидная
железа
♦ ▼
Ткани органов-мишеней
Андрогены
Эстрогены
Прогестерон
И | J
Рост тела Ткани органов-мишеней Молочные
железы
+
Рис. 6.5. Основные гормоны аденогипофиза (кортикотропин, тиреотропин, сомато-
тропин, гонадотропины, пролактин), регуляция их секреции гипоталамическими
нейропептидами и основные эффекты.
Сплошные стрелки — стимуляция, пунктирные стрелки — подавление секреции.
272 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Таким образом в аденогипофизе синтезируются и секретируются пять
гормонов: кортикотропин, гонадотропины (фоллитропин и лютропин), ти-
реотропин, пролактин и соматотропин. Первые три из них обеспечивают
гипофизарную регуляцию периферических эндокринных желез (коры над-
почечников, половых желез и щитовидной железы), т. е. участвуют в реа-
лизации гипофизарного пути управления. Для двух других гормонов (сома-
тотропина и пролактина) гипофиз является периферической эндокринной
железой, поскольку эти гормоны сами действуют на ткани-мишени
(рис. 6.5). Регуляция секреции аденогипофизарных гормонов осуществля-
ется с помощью гипоталамических нейро пептидов (либеринов и статинов),
приносимых кровью воротной системы гипофиза.
6.2.1.1. Регуляция секреции и физиологические эффекты кортикотропина
Кортикотропин является продуктом расщепления крупного (239 аминокис-
лот) гликопротеина проопиомеланокортина, образующегося базофильны-
ми кортикотрофами. Этот белок делится на две части, одна из которых
при расщеплении служит источником кортикотропина и меланотропина, а
вторая, называемая липотропином,— расщепляясь, дает, кроме меланотро-
пина, пептид эндорфин, обладающий морфиноподобным действием и иг-
рающий важнейшую роль в антиноцицептивной (антиболевой) системе
мозга и в модуляции секреции гормонов аденогипофиза.
Секреция кортикотропина происходит пульсирующими вспышками с
четкой суточной ритмичностью. Наивысшая концентрация гормона в кро-
ви отмечается в утренние часы (6—8 часов), а наиболее низкая — с 18 до 2
часов ночи. Механизм регуляции секреции представлен прямыми и обрат-
ными связями. Прямые связи реализуются кортиколиберином гипоталамуса
и вазопрессином. Усиливают эффекты кортиколиберина адреналин и ан-
гиотензин-П, ацетилхолин, норадреналин, адреналин, серотонин, холеци-
стокинин, бомбезин, атриопептид, а ослабляют — опиоидные пептиды,
гамма-аминомасляная кислота. Глюкокортикоиды, циркулирующие в кро-
ви (кортизол), в гипоталамусе тормозят секрецию кортиколиберина, а в
гипофизе — секрецию кортикотропина (обратная связь). Обратная связь
имеет отрицательную направленность и замыкается как на уровне гипота-
ламуса (подавление секреции кортиколиберина), так и гипофиза (тормо-
жение секреции кортикотропина). Продукция кортикотропина резко воз-
растает при действии на организм стрессорных раздражителей, например
холода, боли, физической нагрузки, повышения температуры тела, эмо-
ций, а также под влиянием гипогликемии (снижение сахара в крови), при
этом суточная ритмика секреции изчезает.
Физиологические эффекты кортикотропина принято делить на надпо-
чечниковые и вненадпочечниковые. Надпочечниковое действие гормона яв-
ляется основным и заключается в стимуляции (через системы аденилат-
циклаза—цАМФ и Са2+ с последующей активацией протеинкиназы А) кле-
ток пучковой зоны коры надпочечников, секретирующей глюкокортикои-
ды (кортизол и кортикостерон). Значительно меньший эффект кортико-
тропин оказывает на продукцию минералокортикоидов и половых стерои-
дов клетками клубочковой и пучковой зон коры надпочечников. Под
влиянием кортикотропина не только усиливается стероидогенез (синтез
гормонов) за счет повышения образования и активности ферментов, но и
синтез белков ткани за счет избирательной активации транскрипции ге-
нов, что при избытке гормона вызывает гипертрофию и гиперплазию тка-
ни коры надпочечников. При этом происходит и перестройка структуры
6. Эндокринная нервная система — регулятор функций и процессов в организме • 273
Рис. 6.6. Гипоталамо-аденогипофизарно-надпочечниковая ось регуляции.
Сплошные стрелки — стимуляция, пунктирные стрелки — отрицательная обратная связь. Ос-
новным стимулятором секреции кортикотропина аденогипофизом является гипоталамический
нейропептид — кортиколиберин, слабый стимулирующий эффект на секрецию кортикотропи-
на оказывает вазопрессин. Кортикотропин стимулирует секрецию глюкортикоидов (кортизо-
ла) корой надпочечников. Содержание в крови кортизола по механизму отрицательной обрат-
ной связи подавляет секрецию кортикотропина аденогипофизом и кортиколиберина гипота-
ламусом.
коры, с преимущественным увеличением массы клеток пучковой зоны, в
ткани уменьшается содержание холестерина и аскорбиновой кислоты,
опосредующей синтез гормонов. Таким образом, кортикотропин является
основным звеном гипоталамо-аденогипофизарно-надпочечниковой регуля-
торной оси (рис. 6.6).
Вненадпочечниковое действие кортикотропина заключается в прямых эф-
фектах гормона: 1) липолитическом действии на жировую ткань, 2) повы-
шении секреции инсулина и соматотропина, 3) гипогликемии из-за стиму-
ляции секреции инсулина, 4) увеличении захвата аминокислот и глюкозы
мышечной тканью, 5) повышенном отложении меланина с гиперпигмента-
цией из-за родства молекулы гормона с меланотропином.
Избыток секреции кортикотропина сопровождается развитием гипер-
кортицизма с преимущественным увеличением секреции надпочечниками
кортизола и носит название болезни Иценко—Кушинга. Дефицит корти-
котропина ведет к недостаточности синтеза и секреции в кровь глюкокор-
тикоидов с выраженными метаболическими сдвигами и снижением устой-
чивости организма к стрессорным влияниям среды.
6.2.1.2. Регуляция секреции и физиологические эффекты гонадотропинов
Секреция гонадотропинов из специфических гранул аденогипофизарных
клеток имеет четко выраженную суточную и месячную цикличность как у
мужчин, так и, особенно, у женщин. Молекулы гонадотропинов секрети-
руются с прикрепленными на конце углеводных цепей гликопротеида сиа-
ловыми кислотами, что защищает их от разрушения в печени. Фоллитро-
пин и лютропин образуются и секретируются одними и теми же клетками,
и активация их секреции обеспечивается единым гонадолиберином гипота-
ламуса. Эффект последнего на секрецию фоллитропина и лютропина зави-
274 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
сит от циклических изменений содержания в крови половых гормонов —
эстрогенов, прогестерона и тестостерона (отрицательная обратная связь).
Секреция гонадолиберина стимулируется катехоламинами, а подавляется
эндогенными опиоидными пептидами, гастрином, холецистокинином и
соматостатином. Главный тормозной эффект на продукцию фоллитропина
оказывает по механизму обратной связи гормон семенников — ингибин.
Тормозят секрецию гонадотропинов гормон аденогипофиза пролактин и
мелатонин эпифиза; секрецию лютропина угнетают и глюкокортикоиды.
Основное действие на половые железы гонадотропины реализуют через
систему аденилатциклаза—цАМФ, причем они влияют не только на обра-
зование и секрецию половых гормонов, но и на функции яичников и се-
менников. Фоллитропин связывается с рецепторами клеток примордиаль-
ного фолликула в яичниках и клеток Сертоли в семенниках. Следствием
является рост фолликулов яичника и пролиферация клеток гранулезы у
женщин, рост яичек, пролиферация клеток Сертоли и сперматогенез у
мужчин. В продукции половых гормонов фоллитропин оказывает вспомо-
гательный эффект, готовя секреторные структуры к действию лютропина и
стимулируя ферменты биосинтеза половых стероидов. Лютропин вызывает
овуляцию и рост желтого тела в яичниках, стимулирует клетки Лейдига в
семенниках. Он является ключевым гормоном стимуляции образования и
секреции половых гормонов: эстрогенов и прогестерона в яичниках, анд-
рогенов в семенниках. Для оптимального развития гонад и секреции поло-
вых гормонов необходимо синергичное действие фоллитропина и лютро-
пина, поэтому их часто объединяют единым названием гонадотропины.
Таким образом, гонадотропины гипофиза являются основным звеном ги-
поталамо-аденогипофизарно-гонадной регуляторной оси.
6.2.1.3. Регуляция секреции и физиологические эффекты тиреотропина
Тиреотропин — гликопротеидный гормон аденогипофиза — секретируется
непрерывно, с ритмичными колебаниями в течение суток, при этом мак-
симум содержания в крови приходится на вечерние часы. Секреция тирео-
тропина стимулируется тиреолиберином гипоталамуса, а подавляется сома-
тостатином. По механизму отрицательной обратной связи повышение со-
держания в крови гормонов щитовидной железы (трийодтиронина и тет-
райодтиронина), секрецию которых тиреотропин усиливает, ведет к подав-
лению секреции тиреотропина. Замыкание обратной связи возможно и на
уровне гипоталамуса в виде подавления продукции тиреолиберина. Таким
образом, тиреотропин является ведущим звеном гипоталамо-аденогипофи-
зарно-тиреоидной регуляторной оси. Тормозят секрецию тиреотропина до-
фамин, глюкокортикоиды и соматотропин, а стимулируют — вазопрессин
и антагонисты дофаминовых рецепторов. Тиреотропин секретируется в по-
вышенных количествах при действии на организм низкой температуры,
другие же воздействия — травма, боль, наркоз — секрецию гормона подав-
ляют.
Тиреотропин связывается со специфическим рецептором фолликуляр-
ных клеток щитовидной железы и вызывает метаболические реакции с по-
мощью четырех вторичных посредников: цАМФ, инозитол-3-фосфата,
диацилглицерола и комплекса Са2+—кальмодулин. Под влиянием тирео-
тропина в клетках фолликулов щитовидной железы меняются все виды об-
мена веществ, ускоряется захват иода и осуществляется синтез тиреоглобу-
лина и тиреоидных гормонов. Тиреотропин увеличивает секрецию гормо-
нов щитовидной железы активацией гидролиза тиреоглобулина. Благодаря
6. Эндокринная нервная система — регулятор функций и процессов в организме • 275
увеличению синтеза пурина и пиримидина, РНК, белка и фосфолипидов
тиреотропин вызывает увеличение массы щитовидной железы.
Внетиреоидное действие тиреотропина проявляется повышением обра-
зования гликозаминогликанов в коже и подкожной клетчатке, что ведет к
так называемому слизистому отеку, или микседеме. Это происходит, на-
пример, при дефиците йода и недостаточной продукции гормонов щито-
видной железы, когда по механизму обратной связи возрастает секреция
тиреотропина.
Избыточная секреция тиреотропина приводит к развитию зоба, гипер-
функции щитовидной железы с эффектами избытка тиреоидных гормонов
(тиреотоксикоз), пучеглазию (экзофтальм), что в совокупности называют
«базедова болезнь».
6.2.1.4. Регуляция секреции и физиологические эффекты соматотропина
Соматотропин секретируется аденогипофизарными клетками непрерывно
и «вспышками» через 20—30 мин и 3—5 ч с отчетливой суточной ритми-
кой — повышение секреции соматотропина происходит во время глубокого
сна, на ранних его стадиях (народная мудрость гласит: «человек растет, ко-
гда спит»). Секреция соматотропина стимулируется гипоталамическим
нейропептидом соматолиберином, в механизме действия которого основ-
ным вторичным посредником является Са2+. Подавляется секреция сома-
тотропина гипоталамическим соматостатином, приводящим к снижению
концентрации ионов кальция в соматотрофах аденогипофиза. Увеличива-
ется секреция гормона после мышечных нагрузок, под влиянием травм,
инфекций, голодания. Стимулируют продукцию соматотропина вазопрес-
син, глюкагон, эстрогены (увеличивая число рецепторов соматолиберина
на соматотрофах), дофамин, норадреналин, эндорфин и серотонин, при-
чем последний обеспечивает рост секреции гормона в начале глубокого
сна, а также изменения обмена веществ. Так, гипогликемия активирует
секрецию соматолиберина и соматотропина, а гипергликемия — тормозит;
избыток аминокислот и снижение свободных жирных кислот в крови ак-
, тивируют их секрецию. Эти влияния реализуются через специальные ре-
цепторные нейроны гипоталамуса, воспринимающие сдвиги биохимиче-
ского состава крови и участвующие в регуляции обмена веществ. Угнетает
синтез и секрецию гормона повышение в крови содержания прогестерона
I и свободных жирных кислот. Механизм отрицательной обратной связи в
саморегуляции уровня гормона в крови реализуется стимуляцией сомато-
тропином нейросекреции соматостатина.
Механизм действия соматотропина на клетки-эффекторы заключается в
его связывании с двумя молекулами мембранных рецепторов, активации
1 тирозинкиназы — протеинкиназы С, фосфорилировании и активации ци-
топлазматических белков. Гормон-рецепторный комплекс активирует и
мембранную фосфолипазу С, что ведет к образованию диацилглицерола,
мобилизации внутриклеточного кальция и активации протеинкиназы С.
Следствием является фосфорилирование и активация цитоплазматических
белков, стимулирование транскрипции генов и синтез новых белков. Спе-
цифические рецепторы к гормону имеются на клетках жировой, мышеч-
ной, хрящевой и лимфоидной ткани, клетках печени, поджелудочной же-
лезы, кишечника, половых органов, мозга, легких, сердца и почек.
Соматотропин участвует в регуляции роста и развития организма. По-
вышая синтез хрящевой ткани в эпифизарных отделах костей, гормон в
детском возрасте стимулирует рост тела в длину, а активирование пери-
276 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
остального роста увеличивает толщину и ширину костей. Возрастание мас-
сы тканевых структур происходит в мышечной и соединительной ткани,
растет масса и внутренних органов. Основные эффекты соматотропина
связаны с его влияниями на обмен веществ, приводящими к: 1) усилению
липолиза и уменьшению массы жировой ткани; 2) повышению усвоения
аминокислот и синтеза белков, в результате чего масса тела возрастает за
счет нежировой ткани; 3) увеличению глюконеогенеза и повышению уров-
ня сахара в крови. Вместе с тем большинство ростовых эффектов гормона
опосредуется специальными гуморальными факторами (гормонами) пече-
ни, почек и костной ткани, получившими название соматомедины
(рис. 6.7). Поскольку эффекты соматомединов на обмен веществ во мно-
гом сходны с эффектами инсулина, а их структура имеет сходство с моле-
кулой проинсулина, их еще называют инсулиноподобные факторы роста
(ИФР). Химическая структура и основные эффекты установлены для двух
факторов (ИФР-1 и ИФР-2). ИФР-1 обладает большим, чем ИФР-2, влия-
нием на рост, а также является основным фактором, реализующим отри-
цательную обратную связь в виде угнетения секреции соматолиберина и
соматотропина, увеличения продукции соматостатина. Действие на хряще-
вую ткань инсулиноподобных факторов проявляется в виде стимуляции
включения сульфата в синтезируемые протеогликаны, стимуляции включе-
ния тимидина в образуемую ДНК, активации синтеза РНК и белка. Эти
эффекты выражены у ИФР-1 и ИФР-2 в 100 раз больше, чем у инсулина, а
влияние на обмен глюкозы у них в 50 раз слабее, чем у инсулина. В то же
время дифференцировка прехондроцитов, повышение транспорта амино-
кислот через их клеточную мембрану обеспечиваются не соматомединами,
а самим соматотропином. Несмотря на то что соматомедины называют ин-
сулиноподобными факторами роста, рецепторы клеточной мембраны для
них отличаются от рецепторов инсулина. Рецепторы инсулиноподобных
факторов находятся не только в хрящевой ткани, но и в мышечной и со-
единительной тканях, где эти регуляторы также стимулируют митогенез и
синтез белка.
При длительной и чрезмерной секреции соматотропина хотя и сохраня-
ется действие соматомединов на хрящевую ткань, но в целом эффекты со-
матотропина приобретают четкие контринсулярные черты. Они проявля-
ются в изменениях углеводного и жирового обмена в тканях. Так, сомато-
тропин вызывает гипергликемию из-за распада гликогена в печени и мыш-
цах и угнетения утилизации глюкозы в тканях, благодаря повышению сек-
реции глюкагона островками Лангерганса поджелудочной железы. Сомато-
тропин увеличивает и секрецию инсулина островками Лангерганса, как за
счет прямого стимулирующего действия, так и благодаря гипергликемии.
Но в то же время соматотропин активирует инсулиназу печени — фермент,
разрушающий инсулин, и вызывает инсулинорезистентность тканей. По-
добное сочетание стимуляции секреции инсулина с его разрушением и по-
давлением эффекта в тканях может вести к сахарному диабету, который по
происхождению называют гипофизарным. Как антагонист инсулина сома-
тотропин проявляет свои эффекты и в метаболизме липидов. Гормон ока-
зывает пермиссивное (облегчающее) действие по отношению к влияниям
катехоламинов и глюкокортикоидов, следствием чего являются стимуля-
ция липолиза жировой ткани, повышение уровня свободных жирных ки-
слот в крови, избыточное образование кетоновых тел в печени (кетоген-
ный эффект) и даже жировая инфильтрация печени. Инсулинорезистент-
ность тканей может быть связана и с этими сдвигами жирового обмена.
Избыточная секреция соматотропина и, соответственно, возросший под
6. Эндокринная нервная система — регулятор функций и процессов в организме • 277
Соматотропин
Соматотропин
Рецептор соматотропина
Соматомедины
(ИПФ-1 и ИПФ-2)
Усиление транспорта
аминокислот и ускорение
синтеза белка
Тирозинкиназа
Фосфолипаза С
Диацилглицерол
Протеинкиназа С
Фосфорилирование
МАПК
Ядро
Метаболические
эффекты
Фосфорилирование
белков
ранскрипция
генов
Синтез новых белков
Рис. 6.7. Механизм действия со-
матотропина. Секретируемый аде-
ногипофизом соматотропин ока-
зывает эффект на рост, стимули-
руя транспорт в клетку аминокис-
лот и синтез белка. Эффект сома-
тотропина опосредуется сомато-
мединами или инсулиноподобны-
ми факторами роста (ИПФ-1, в
большей степени, и ИПФ-2), об-
разуемыми в печени.
В нижнем квадрате показан клеточ-
ный механизм действия гормона. Свя-
зывание соматотропина с одной из
двух молекул специфического мем-
бранного рецептора вызывает актива-
цию ассоциированной с ней тирозин-
киназы, что ведет к фосфорилирова-
нию и активации митогенактивируе-
мой протеинкиназы (МАПК), вызы-
вающей транскрипцию генов и синтез
новых белков. Вторая молекула рецептора после связывания с соматотропином через G-белок
активирует мембранную фосфолипазу С, ускорение метаболизма фосфоинозитидов ведет к
образованию диацилглицерола, мобилизации внутриклеточного кальция, активации протеин-
киназы С. Последняя вызывает фосфорилирование белков, активацию ферментов и метаболи-
ческие эффекты, в том числе активирует транспорт аминокислот в клетку. Протеинкиназа С
также вызывает фосфорилирование и активацию МАПК и других индукторов транскрипции
генов, способствуя синтезу белка.
его влиянием уровень ИФР-1, возникающие в раннем детстве, ведут к раз-
витию гигантизма с пропорциональным развитием конечностей и тулови-
ща. В юношеском и зрелом возрастах избыток гормона усиливает рост
эпифизарных участков костей скелета, зон с незавершенным окостенени-
ем, что получило название акромегалия. Растут кисти и стопы, нос, подбо-
родок и т. д. Увеличиваются в размерах и внутренние органы, что называ-
ют спланхомегалия. Появляются утолщение кожи, повышенная потливость,
ущемление нервов, резистентность к инсулину. При врожденном дефиците
соматотропина, особенно при нечувствительности тканей к нему (при
278 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
этом в организме имеет место низкий уровень ИФР-1 при высоком уровне
соматотропина), формируется карликовость, называемая «гипофизарный
нанизм». После выхода в 1726 г. романа Дж. Свифта «Путешествия Гулли-
вера» таких карликов стали называть лилипутами. Приобретенный дефи-
цит гормона в зрелом возрасте выраженного морфогенетического эффекта
не вызывает.
6.2.1.5. Регуляция секреции и физиологические эффекты пролактина
Синтез и секреция аденогипофизом пролактина в основном регулируется
гипоталамическим ингибитором дофамином (пролактостатин), а также сти-
муляторами пролактолиберином и тиреолиберином (рис. 6.8). Образование
дофамина происходит в тубероинфундибулярных нейронах дугообразного
и перивентрикулярного ядер гипоталамуса. На лактотрофах аденогипофиза
выявлено 2 типа дофаминовых рецепторов: D, и D2 Первые стимулируют
аденилатциклазу, а вторые, напротив, ингибируют ее активность. Пролак -
толиберин образуется в нейронах переднего гипоталамуса и срединного та-
ламуса. Пролактолибериновой активностью обладают также окситоцин,
серотонин и вазоинтестинальный пептид. Секреция пролактина зависит и
от уровня в крови эстрогенов, глюкокортикоидов и тиреоидных гормонов,
изменяющих число рецепторов пролактолиберина и тиреолиберина на
Тиреолиберин
(пролактолиберин)
олочная
железа Гипофиз —
Г ипоталамус
Инстинкт материнства ' +
Иммунорегуляция
Рост молочных желез,
лактация
Пролактин
Дофамин
Беременность
Синтез прогестерона
+ "^Послеродовое ожирение
Эритропоэз
Водно-солевой обмен
Рис. 6.8. Регуляция секреции и основные эффекты пролактина. Раздражение рецеп-
торов матки при беременности и сосков молочных желез при кормлении (обозначе-
ны пунтиром) вызывает рефлекторное повышение секреции пролактина. Гипотала-
мическая регуляция секреции пролактина осуществляется стимуляцией тиреолибе-
рином (пролактолиберином), что обозначено знаком «+», и подавлением секреции
дофамином (обозначено знаком «-»). Внизу показаны основные эффекты пролак-
тина в организме.
6. Эндокринная нервная система — регулятор функций и процессов в организме • 279
лактотрофах. Повышается секреция пролактина при беременности, корм-
лении грудью, во время сна, физической нагрузки, стресса.
Одним из органов-мишеней пролактина является молочная железа, где
гормон стимулирует развитие секреторной ткани, рост желез и лактацию,
оказывая свой эффект после связывания со специфическим рецептором и
образования вторичного посредника цАМФ. В молочных железах пролак-
тин влияет на процессы образования молока, а не на его выделение. При
этом гормон стимулирует синтез белка — лактальбумина, а также жиров и
углеводов молока. В регуляции роста и развития молочных желез синерги-
стами пролактина являются эстрогены, но при начавшейся лактации эст-
рогены выступают в роли антагонистов пролактина. Эффект пролактина
на лактацию во многом опосредуется образованием в печени лактогенного
фактора.
Пролактин способствует поддержанию секреторной активности желтого
тела в яичниках и образованию прогестерона. Он является одним из регу-
ляторов водно-солевого обмена организма, уменьшая экскрецию воды и
электролитов, повышает в крови содержание альдостерона и вазопрессина,
стимулирует рост внутренних органов, эритропоэз, способствует появле-
нию инстинкта материнства (см. рис. 6.8). Помимо усиления синтеза бел-
ка, пролактин увеличивает образование жира из углеводов, способствуя
послеродовому ожирению.
6.2.2. Гормоны нейрогипофиза и их эффекты в организме
В нейрогипофизе не образуются, а лишь накапливаются и секретируются в
кровь нейрогормоны супраоптического и паравентрикулярного ядер гипо-
таламуса — вазопрессин и окситоцин. Оба гормона находятся в гранулах в
связанном состоянии со специальными белками — нейрофизинами. В про-
цессе секреции содержимое гранул путем экзоцитоза поступает в кровь.
6.2.2.1. Регуляция секреции и физиологические эффекты вазопрессина
Секреция вазопрессина зависит от его синтеза в гипоталамических нейро-
нах и регулируется тремя типами стимулов: 1) сдвигами осмотического
давления и содержания натрия в крови, воспринимаемыми интероцепто-
рами сосудов и сердца (осмо-, натрио-, волюмо- и механорецепторы), а
также непосредственно гипоталамическими нейронами (центральные ос-
морецепторы); 2) активацией гипоталамических ядер при эмоциональном
и болевом стрессе, физической нагрузке, 3) гормонами плаценты и ангио-
тензином-П, как содержащимся в крови, так и образуемым в мозге.
В крови вазопрессин не связывается белками плазмы, но ассоциирован
с тромбоцитами, выполняющими по отношению к гормону транспортную
функцию.
Эффекты вазопрессина реализуются за счет связывания пептида в тка-
нях-мишенях с двумя типами мембранных рецепторов — V! и V2.
Стимуляция Vj-рецепторов, локализованных на мембране эндотелиаль-
ных и гладкомышечных клеток стенки кровеносных сосудов, через вторич-
ные посредники инозитол-3-фосфат и кальций-кальмодулин вызывает су-
жение сосудов, что соответствует названию «вазопрессин». Это влияние в
физиологических условиях выражено слабо из-за низких концентраций
гормона в крови, но играет существенную роль в изменениях кровообра-
щения при стрессе, шоке, артериальной гипертензии. Через У^рецепторы
вазопрессин повышает чувствительность механорецепторов в каротидных
280 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 6.9. Механизм действия вазопрессина на клетку эпителия собирательной тру-
бочки нефрона.
Связывание вазопрессина с У2-рецептором на базолатеральной мембране через мембранный
G-белок активирует аденилатциклазу (АЦ), что ведет к образованию вторичного посредни-
ка — цАМФ. Последний через активацию протеинкиназы А вызывает фосфорилирование мо-
лекул аквапоринов в агрефорах, их взаимодействие с белками микротубул и путем экзоцитоза
встраивание аквапоринов в апикальную мембрану. Протеинкиназа А путем активации генома
повышает синтез агрефор с аквапоринами. Аквапорины из апикальной мембраны путем эндо-
цитоза интернализуются и повергаются рециркуляции или деградации, особенно в
отсутствие вазопрессина, а также экскретируются с мочой.
синусах к изменениям артериального давления и этим способствует баро-
рефлекторной регуляции артериального давления.
Стимуляция У2-рецепторов базолатеральной мембраны клеток дисталь-
ных отделов почечных канальцев через вторичный посредник цАМФ вы-
зывает повышение проницаемости стенки канальцев для воды, ее реаб-
сорбцию и концентрирование мочи, что соответствует второму названию
вазопрессина — «антидиуретический гормон». Вазопрессин является един-
ственным гормоном, способным стимулировать канальцевую реабсорбцию
воды без задержки натрия. Эффект вазопрессина на транспорт воды связан
с особыми транспортными белками «аквапоринами». Только аквапорины
2-го типа являются вазопрессинозависимыми. При наличии в крови гор-
мона вазопрессина (рис. 6.9), он связывается на базолатеральной мембране
клеток эпителия почечного канальца с У2-рецептором, следствием чего яв-
ляется активация аденилатциклазы, образование цАМФ, активация проте-
инкиназы А. Последняя вызывает фосфорилирование молекул аквапори-
на-2 в цитоплазматических пузырьках (агрефорах), их транспорт с помо-
щью микротубулярных белков динеина, динактина и миозина-1 к апикаль-
ной мембране, где специальные рецепторные молекулы (синтаксин-4, ре-
цепторы-мишени пузырьков и др.) обеспечивают встраивание молекул ак-
вапорина-2 в мембрану и формирование водных каналов. Протеинкиназа
А является также регулятором синтеза белка аквапорина 2 в ядре клеток
эпителия. Поступающая в клетки через водные каналы молекул аквапори-
на-2 вода по микротубулярной системе клеток перемещается к базолате-
ральной мембране, где постоянно встроены вазопресин-независимые бел-
ки аквапорины 3-го и 4-го типа. Через них вода выходит в интерстициаль-
ную жидкость по осмотическому градиенту (рис. 6.10). В отсутствие вазо-
прессина молекулы аквапорина-2 подвергаются эндоцитозу (интернализа-
ция) в цитоплазму, где вновь способны к рециркуляции, т. е. новому цик-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 281
Вазопрессин
Рис. 6.10. Схема механизма действия вазопрессина на транспорт воды через стенку
собирательной трубочки нефрона.
Взаимодействуя с Vj-рецептором, вазопрессин обеспечивает как трансцеллюлярный транспорт
воды (из внутриканальцевой жидкости через водные каналы аквапоринов-2 апикальной мем-
браны, микротубулярную систему клеток и через аквапорины-3 и -4 базолатеральной мемеб-
раны в интерстициальное перетубулярное пространство), так и парацеллюлярный транспорт
воды (через межклеточные плотные соединения)
лу активации, транспорта и встраивания в мембрану, или подвергаются
разрушению. Вазопрессин стимулирует всасывание воды и в железах
внешней секреции, в желчном пузыре.
Нейропептид вазопрессин поступает по аксонам экстрагипоталамиче-
ской системы в другие отделы мозга (лимбика, средний мозг) и участвует в
формировании жажды и питьевого поведения, механизмах терморегуля-
ции, в нейрохимических механизмах памяти, формировании биологиче-
ских ритмов и эмоционального поведения.
Вазопрессин стимулирует секрецию кортикотропина в аденогипофизе,
подавляет выделение лютропина при стрессе. Метаболические эффекты
вазопрессина заключаются в стимуляции гликогенолиза в печени, стиму-
ляции секреции инсулина, повышении синтеза в печени антигемофиличе-
ского глобулина А, продукции фактора Виллебрандта.
Недостаток вазопрессина проявляется резко повышенным выделением
мочи низкого удельного веса, что называют «несахарным диабетом», а из-
быток гормона ведет к задержке воды в организме.
6.2.2.2. Регуляция секреции и физиологические эффекты окситоцина
Синтез окситоцина в гипоталамических нейронах и его секреция нейроги-
пофизом в кровь стимулируется рефлекторным путем при раздражении ре-
цепторов растяжения матки и механорецепторов сосков молочных желез.
282 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Усиливают секрецию гормона эстрогены. Болевой стресс резко повышает
секрецию окситоцина, а этиловый спирт ее угнетает. Возрастает секреция
окситоцина и при повышении осмотического давления внеклеточной сре-
ды. Несмотря на то что мембранный рецептор окситоцина в клетках-ми-
шенях относится к той же группе, что и У2-рецептор вазопрессина, связы-
вание гормона со специфическим рецептором вызывает снижение в клет-
ках уровня цАМФ. Образование окситоцин-рецепторного комплекса по-
вышает в клетках содержание Са2+ (вторичный посредник) и активирует
ионные каналы мембраны, приводя к ее деполяризации.
Основные эффекты окситоцина состоят в стимуляции сокращения мат-
ки при родах (чему способствуют высокие концентрации эстрогенов в кро-
ви), сокращении гладких мышц протоков молочных желез, что вызывает
выделение молока, а также в регуляции водно-солевого обмена и питьево-
го поведения. Окситоцин является одним из дополнительных факторов ре-
гуляции секреции гормонов аденогипофиза, наряду с либеринами. В
структурах мозга окситоцин может выступать в роли медиатора или моду-
лятора синаптических процессов, участвовать в механизмах памяти, стиму-
лируя процессы забывания. Окситоцин активирует клеточный иммунитет,
оказывает инсулиноподобное действие на жировую ткань. Повышенные
количества окситоцина в крови могут вызывать снижение артериального
давления.
6.2.3. Гормоны промежуточной доли
Меланотропин у взрослого человека, в отличие от животных с обильным
волосяным покровом, практически не синтезируется. Функции этого гор-
мона, заключающиеся в синтезе меланина, его дисперсии в отростках ме-
ланоцитов кожи, увеличении свободного пигмента в эпидермисе и, в ко-
нечном счете, повышении пигментации кожи и волос, выполняют у чело-
века кортикотропин и липотропин. Эти гормоны, как и меланотропин, об-
разуются из единого предшественника, в связи с чем при избыточной
секреции кортикотропина усиливается пигментация кожи. Этому способ-
ствуют и близкие гормону пептиды плаценты. Меланотропин тем не ме-
нее играет роль как мозговой пептид в нейрохимических процессах па-
мяти.
6.2.4. Эндогенные опиаты
Гипоталамо-гипофизарная система является основным местом синтеза и
секреции эндогенных опиатов — полипептидов эндогенного происхожде-
ния, обладающих морфиноподобным аналгезирующим действием. Выделя-
ют четыре семейства эндогенных опиатов: эндорфины (a-, {3- и у-), динорфи-
ны А и В, неоэндорфины а и /3, метионин- и лейцин-энкефалины. В аденоги-
пофизе образуются эндорфины, динорфины и неоэндорфины, тогда как
энкефалины синтезируются нейронными структурами разных отделов моз-
га, прежде всего таламуса и гипоталамуса, и клетками слизистой оболочки
кишечника. Опиатные пептиды в центральной нервной системе выполня-
ют роль медиаторов и модуляторов синаптической передачи, обеспечивают
функцию противоболевой системы мозга. На мембранах клеток выявлены
специфические опиатные рецепторы трех основных типов: ц, 5 и к. Мор-
фин и бета-эндорфин взаимодействуют преимущественно с ц-рецептора-
ми, энкефалины — с 5-рецепторами, а динорфины — с к-рецепторами, од-
нако существует и перекрестное взаимодействие. Помимо подавления бо-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 283
левых ощущений эндогенные опиаты участвуют в регуляции адаптивного
поведения и реакций организма на стресс, модулируют секрецию гормонов
адено- и нейрогипофиза (соматотропина, пролактина, гонадотропинов, ва-
зопрессина). Опиоидные пептиды регулируют процесс всасывания в желу-
дочно-кишечном тракте, угнетают секрецию панкреатического сока и со-
ляной кислоты в желудке. Опиоидные рецепторы выявлены на окончаниях
аксонов в мозговом слое надпочечников и симпатических ганглиях, где
опиоидные пептиды выполняют роль модуляторов синаптической пере-
дачи.
6.3. Регуляторные функции гормонов надпочечников
Надпочечники являются парной железой внутренней секреции, морфоло-
гически и функционально состоящей из двух разных по эмбриональному
происхождению тканей — коркового и мозгового вещества. Кора надпочеч-
ников — производное мезодермы, синтезирует три вида гормонов стероид-
ной природы, или кортикостероидов. Мозговое вещество называют также
хромаффинной тканью, которая имеет общее с нервной системой происхо-
ждение и развивается из эмбриональных симпатических клеток. Кроме
надпочечника хромаффинная ткань, образующая катехоламины, имеется в
составе симпатического пограничного ствола, брюшном и солнечном
сплетении.
Кровоснабжение надпочечника осуществляется тремя группами артерий,
. отходящих от диафрагмальной и почечной артерий и аорты, а также не-
* скольких добавочных артерий. Ветви этих сосудов образуют раздельные
корковые и мозговые артерии, дающие обильные капиллярные сети. Ве-
нозный отток крови осуществляется по многочисленным венам в систему
нижней полой вены и бассейн воротной вены печени. Как в корковом, так
< и в мозговом веществе имеется большое количество нервных волокон.
Особое значение имеют ветви чревного нерва, несущие пре- и постганг-
лионарные волокна к хромаффинным клеткам мозгового вещества и обес-
печивающие поступление к ним регуляторных стимулов.
6.3.1. Гормоны коры надпочечников и их эффекты в организме
Кора надпочечников, занимающая по объему 80 % всей железы, состоит
из трех клеточных зон: наружной клубочковой зоны, образующей минера-
локортикоиды, средней пучковой зоны, образующей глюкокортикоиды, и
внутренней сетчатой зоны, в небольшом количестве продуцирующей поло-
вые стероиды. Все кортикостероиды образуются из холестерина крови и
синтезируемого в самих корковых клетках. При синтезе кортистероидов
1 образуется порядка 50 различных соединений, однако секретируются в
• кровь в физиологических условиях лишь 7—9 из них.
! 6.3.1.1. Регуляция секреции и физиологические эффекты
минералокорт икоидов
У человека единственным минералокортикоидом, поступающим в кровь,
! является альдостерон. Регуляция синтеза и секреции альдостерона осуще-
। ствляется преимущественно ангиотензином-П, что дало основание считать
альдостерон частью ренин-ангиотензин-альдостероновой системы или регу-
ляторной оси (рис. 6.11), обеспечивающей регуляцию водно-солевого об-
мена и гемодинамики. Регуляции секреции альдостерона может осуществ-
284 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 6.11. Ренин-ангиотензин-альдостероновая система.
Секреция юкстагломерулярными клетками почек в кровь фермента ренина вызывает отщепле-
ние пептида ангиотензина-1 от белка плазмы крови ангиотензиногена, образуемого в печени.
В сосудистом русле почек, печени, легких, мозга ангиотензин-1 подвергается воздействию
превращающего фермента, вызывающего образование из ангиотензина-1 ангиотензина-2. Ан-
гиотензин-2 стимулирует секрецию альдостерона клубочковой зоной коры надпочечников.
Пунктирной стрелкой обозначена отрицательная обратная связь — подавление секреции рени-
на ангиотензином-2.
ляться и под влиянием собственной адренокортикальной ренин-ангиотен-
зиновой системы, что объясняет частое несоответствие уровней активно-
сти ренина в плазме крови и секреции альдостерона. Поскольку альдосте-
рон регулирует содержание в крови ионов Na+ и К+, обратная связь в регу-
ляции его секреции реализуется прямым влиянием ионов К+ на клубочко-
вую зону коры надпочечников. В ренин-ангиотензин-альдостероновой
системе обратные связи включаются при сдвигах содержания Na+ в моче
дистальных канальцев, объема и давления крови.
Механизм действия альдостерона, как и всех стероидных гормонов,
состоит в прямом влиянии на генетический аппарат ядра клеток со сти-
муляцией синтеза соответствующих РНК, активации синтеза транспорти-
рующих катионы белков и ферментов, а также повышении проницаемо-
сти мембран для аминокислот. Негеномные эффекты гормона реализуют-
ся через системы вторичных посредников. Механизм действия альдосте-
рона на клетки почечных канальцев представлен на рис. 6.12. Стимуля-
ция всасывания натрия под влиянием альдостерона происходит не только
в нефроне, но и в желудочно-кишечном тракте, протоках желез внешней
секреции, желчном пузыре. Негеномные эффекты альдостерона обуслов-
лены стимуляцией мембранного антипорта Na+/H+ в клетках разных ти-
пов (гладкие мышцы матки, эпителий дистальных канальцев почек, глад-
кие мышцы артерий и артериол, клетки крипт кишечника). Эти эффекты
обусловлены образованием вторичного посредника диацилглицерола и
активацией протеинкиназы С. Повышение уровня внутриклеточного
кальция в эндотелиальных и гладкомышечных клетках сосудов под влия-
нием альдостерона обусловлено активацией вторичного посредника ИФЗ.
Альдостерон вызывает в клетках и двукратное повышение уровня цАМФ,
модулируя геномные эффекты стероидных гормонов. Быстрые негеном-
ные эффекты альдостерона проявляются и со стороны сердечно-сосуди-
стой системы в виде: повышения сосудистого сопротивления и артери-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 285
Рис. 6.12. Геномный и внегеномный механизмы действия альдостерона на клетку
почечного канальца.
Геномный механизм: проникновение молекулы гормона через мембрану внутрь клетки, свя-
зывание с цитоплазматическим рецептором, транспорт в ядро, связывание с ядерным рецеп-
тором, активация синтеза белков (Na-транспортирующего белка-переносчика) и №+-К+-анти-
порта через люминальную мембрану. Внегеномный механизм: связывание молекулы гормона
с мембранным рецептором, образование вторичных посредников (ИФЗ), фосфорилирование и
активация №+-протонного антипорта через люминальную мембрану.
ального давления при снижении сердечного выброса, противодействия
повышению в гладких мышцах сосудов уровня цАМФ и увеличения чув-
ствительности к прессорным эффектам катехоламинов и ангиотензина-П,
что дало основание считать альдостерон циркуляторным гормоном
стресса.
Альдостерон поддерживает оптимальный водно-солевой обмен между
внешней и внутренней средой организма. Одним из главных органов-ми-
шеней гормона являются почки, где альдостерон вызывает усиленную ре-
абсорбцию натрия в дистальных канальцах с его задержкой в организме и
повышение экскреции калия с мочой. Под влиянием альдостерона проис-
ходит задержка в организме хлоридов и воды, усиленное выведение Н-ио-
нов и аммония, увеличивается объем циркулирующей крови, формируется
сдвиг кислотно-основного состояния в сторону алкалоза. Действуя на
клетки сосудов и тканей, гормон способствует транспорту натрия и воды
во внутриклеточное пространство.
Минералокортикоиды являются жизненно важными гормонами, гибель
организма после удаления надпочечников можно предотвратить, вводя
гормоны извне. Минералокортикоиды усиливают воспаление и реакции
иммунной системы. Избыточная их продукция ведет к задержке в организ-
ме натрия и воды, отекам и повышению артериального давления, потере
калия и водородных ионов, вследствие чего возникают нарушения возбу-
286 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
димости нервной системы и миокарда. Недостаток альдостерона у челове-
ка сопровождается уменьшением объема крови, гиперкалиемией, гипотен-
зией, угнетением возбудимости нервной системы.
6.3.1.2. Регуляция секреции и физиологические эффекты глюкокортикоидов
Клетки пучковой зоны секретируют в кровь у здорового человека два ос-
новных глюкокортикоида: кортизол и кортикостерон, причем кортизола
примерно в 10 раз больше. Секреция глюкокортикоидов регулируется кор-
тикотропином аденогипофиза. Избыток кортизола в крови по механизму
обратной связи угнетает секрецию кортиколиберина в гипоталамусе и кор-
тикотропина в гипофизе. Секреция глюкокортикоидов происходит непре-
рывно с отчетливой суточной ритмикой, повторяющей ритмику секреции
кортикотропина: максимальные уровни гормона в крови у человека отме-
чаются в утренние часы, а минимальные — вечером и ночью (рис. 6.13).
Поступающие в кровь гормоны транспортируются к тканям в свободной и
связанной (до 95 %) с а2-глобулином плазмы (транскортин) формах. Ме-
ханизм действия глюкокортикоидов на клетки-мишени представлен на
рис. 6.14.
Благодаря растворимости в липидах кортизол проникает через мембра-
ну клетки-мишени и взаимодействует с цитоплазматическим рецептором,
образуя лиганд-рецепторный комплекс, что обеспечивает транспорт моле-
кулы гормона в ядро, где кортизол связывается с ядерным рецептором, ак-
тивируя синтез новых белков и ферментов, тем самым обеспечивая мета-
болические эффекты. Молекула кортизола может образовывать лиганд-ре-
цепторный комплекс и с мембранными рецепторами. Хотя роль этого про-
цесса в реализации эффектов гормона еще изучается, тем не менее извест-
ны быстрые негеномные эффекты гормона на возбудимость нервных кле-
ток, связанные с изменением ионного трансмембранного транспорта, обу-
словливающие изменение поведения.
Среди синтезированных под влиянием кортизола белков в клетке-ми-
шени важнейшую роль для реализации опосредованных эффектов гормона
играет семейство липокортинов. Последние, выходя из клетки, связывают-
ся со специфическими липокортиновыми рецепторами клеточной мембра-
ны (аутокринный путь влияния), что вызывает подавление активности
фосфолипазы-А. Липокортины способны ингибировать фосфолипазу-А и
непосредственно, следствием чего является подавление синтеза в клетках
простагландинов и лейкотриенов, ослабление их метаболических и регуля-
торных эффектов. Уменьшение проницаемости клеточных мембран и про-
тивовоспалительный эффект кортизола обусловлены ингибированием син-
теза лейкотриенов.
Глюкокортикоиды прямо или опосредованно регулируют практически
все виды обмена веществ и физиологические функции. Метаболические
эффекты глюкокортикоидов проявляются, прежде всего, со стороны угле-
водного, белкового и жирового обменов. Обобщенно эти сдвиги можно
свести к распаду белков и липидов в тканях, после чего метаболиты посту-
пают в печень, где из них синтезируется глюкоза, использующаяся как ис-
точник энергии. Эффекты на углеводный обмен в целом противоположны
инсулину, поэтому глюкокортикоиды называют контринсулярными гормо-
нами. Гипергликемия под влиянием гормонов возникает за счет усиленно-
го образования глюкозы в печени из аминокислот — глюконеогенеза и по-
давления утилизации ее тканями. Гипергликемия является причиной акти-
вации секреции инсулина. Чувствительность тканей к инсулину глюкокор-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 287
Рис. 6.13. Суточный ритм секреции кортикотропина и кортизола. Максимум кон-
центрации гормонов в крови приходится на утренние часы, при этом прирост уров-
ня кортикотропина опережает рост содержания кортизола.
тикоиды снижают, а контр инсулярные метаболические эффекты могут
вести к развитию стероидного сахарного диабета. На белковый обмен гор-
моны оказывают катаболический и антианаболический эффекты, приводя
к отрицательному азотистому балансу. Распад белка происходит в мышеч-
ной, соединительной и костной тканях, снижается уровень альбумина в
крови, уменьшается проницаемость клеточных мембран для аминокислот.
Однако синтез некоторых белков в печени, например а2-глобулинов, глю-
кокортикоиды повышают. Со стороны жирового обмена имеют место липо-
литический эффект в тканях, гиперлипидемия и гиперхолестеринемия, ак-
тивация кетогенеза в печени, угнетение липогенеза в печени, стимуляция
липогенеза и перераспределения жира в жировой ткани центральной оси
туловища и лица, стимуляция аппетита и потребления жира
Влияние глюкокортикоидов на реактивность тканей проявляется не
только в виде подавления чувствительности к инсулину, но и в повыше-
нии чувствительности адренорецепторов к катехоламинам. Глюкокорти-
коиды вызывают снижение в крови количества лимфоцитов, эозинофилов
288 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 6.14. Схема механизма действия кортизола на клетку-мишень.
Проникая внутрь клетки через мембрану, молекула гормона последовательно взаимодействет с
цитозольным, а затем с ядерным рецептором. Следствие геномного влияния — активация син-
теза новых белков, в том числе являющихся внутриклеточными ферментами, что вызывает из-
менения обмена веществ. К числу синтезируемых под влиянием кортизола белков относятся
липокортины. Последние либо выводятся из клетки и взаимодействуют со специфическими
для них мембранными рецепторами, либо действуют внутриклеточно. Основной эффект липо-
кортинов — ингибирование мембранного фермента фосфолипазы-А и образования из арахи-
доновой кислоты простагландинов и лейкотриенов.
и базофилов, повышение чувствительности сенсорных структур и возбуди-
мости нервной системы, обеспечение оптимальной симпатической регуля-
ции сердечно-сосудистой системы. Почечные эффекты глюкокортикоидов
состоят в стимуляции диуреза путем снижения реабсорбции воды и повы-
шении клубочковой фильтрации; подобно минералокортикоидам, они мо-
гут вызывать задержку натрия при потере калия. Глюкокортикоиды повы-
шают синтез ангиотензиногена в печени и тем самым способствуют боль-
шему образованию в крови ангиотензина-П и секреции альдостерона, уве-
личивают синтез катехоламинов в мозговом веществе надпочечников. Гор-
моны повышают устойчивость организма к действию чрезмерных раздра-
жителей, подавляют сосудистую проницаемость и воспаление (поэтому их
называют адаптивными и противовоспалительными), из-за катаболизма
белка в лимфоидной ткани и угнетения иммунных реакций они оказывают
антиаллергические эффекты. Физиологическое влияние кортизола на им-
мунокомпетентные клетки носит защитный, регуляторный характер
(рис. 6.15).
Избыток гормонов в крови вызывает активацию желудочной секреции,
выделения НС1, уменьшение числа мукоцитов и продукции слизи, что
способствует возникновению язвы желудка — ульцерогенный эффект.
6. Эндокринная нервная система — регулятор функций и процессов в организме • 289
Кортиколиберин
Вазопрессин
Кортикотропин
Кора надпочечников
Интерлейкины (ИЛ-1)
<>
Кортизол
Кортикотропин
Макрофаг
Иммунологический стимул
ФИО и другие токсичные соединения
Рис. 6.15. Физиологическое значение влияния кортизола на иммунокомпетентные
клетки (на примере макрофага).
Кортизол тормозит выделение макрофагом фактора некроза опухолей (ФИО) и других токсич-
ных для клеток организма макрофагальных цитокинов. Под влиянием иммунологического
стимула макрофаг выделяет кортикотропин, что усиливает продукцию кортизола надпочечни-
ками и ослабляет выделение макрофагом цитотоксические соединений. Кроме того выделяе-
мые макрофагом интерлейкины, в частности интерлейкин-1, стимулируют гипоталамо-адено-
гипофизарно-надпочечниковую ось регуляции, что также повышает продукцию кортизола,
снижающего цитотоксические эффекты макрофага.
Повышенная секреция глюкокортикоидов, не связанная с описанной
выше усиленной секрецией кортикотропина, вызывает развитие синдрома
Иценко—Кушинга. Его основные проявления близки болезни Иценко-Ку-
шинга, однако, благодаря механизму обратной связи, избыток глюкокор-
тикоидов угнетает секрецию кортикотропина и усиленную пигментацию.
Среди проявлений повышенной секреции гормонов: антиростовые эффек-
ты (подавление действия кальцитриола и митоза фибробластов, деградация
коллагена вплоть до остеопороза; подавление заживления ран из-за угнете-
ния пролиферации фибробластов и редукции соединительной ткани; по-
давление секреции соматотропина и синтеза ИФР-1; мышечная атрофия и
слабость), склонность к сахарному диабету, гипертензия, нарушения поло-
вых функций, лимфопения, пептические язвы желудка, изменения психи-
ки, предрасположенность к инфекциям. Дефицит глюкокортикоидов вы-
зывает гипогликемию, снижение чувствительности сердечно-сосудистой
системы к симпатической регуляции, замедление сердечного ритма, гипо-
тензию, нейтропению, эозинофилию и лимфоцитоз, снижение сопротив-
ляемости организма инфекциям.
10-6095
290 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
6.3.1.3. Регуляция секреции и физиологические эффекты половых стероидов
коры надпочечников
Клетками сетчатой зоны у человека секретируются в кровь мужские по-
ловые гормоны или андрогены', андростендион, дегидроэпиандростерон и
существенно меньше — 11-бета-гидроксиандростендион. Наиболее высо-
кий уровень этих гормонов отмечается в 6 часов утра, а наиболее низ-
кий — в 19 часов. Секреция андрогенов регулируется кортикотропином
гипофиза.
Андрогены надпочечника стимулируют окостенение эпифизарных хря-
щей, повышают синтез белка (анаболический эффект) в коже, мышечной
и костной ткани, а также формируют половое поведение у женщин. Гор-
моны являются предшественниками основного андрогена семенников —
тестостерона и могут превращаться в него при метаболизме в тканях-ми-
шенях. Андрогены надпочечников способствуют развитию оволосения по
мужскому типу, а их избыток у женщин — к вирилизации, т. е. появлению
мужских черт. Ранний избыток андрогенов ведет к преждевременному
проявлению вторичных половых признаков у мальчиков и вирилизации
женских половых органов. Нарушения секреции андрогенов коры надпо-
чечников получили название адреногенитальных синдромов.
6.3.2. Гормоны мозгового вещества надпочечников и их эффекты
в организме
Мозговое вещество надпочечников содержит хромаффинные клетки, на-
званные так из-за избирательной окраски хромом. По происхождению и
функции они являются постганглионарными нейронами симпатической
нервной системы, однако, в отличие от типичных нейронов, клетки надпо-
чечников: 1) синтезируют больше адреналина, а не норадреналина (отноше-
ние у человека между ними 6:1); 2) накапливая секрет в гранулах, после
поступления нервного стимула они немедленно выбрасывают гормоны в
кровь. Регуляция секреции гормонов мозгового вещества надпочечников
осуществляется благодаря наличию гипоталамо-симпатоадреналовой оси,
при этом симпатические нервы стимулируют хромаффинные клетки через
холинорецепторы, выделяя медиатор ацетилхолин.
Хромаффинные клетки являются частью общей системы нейроэндо-
кринных клеток организма, или APUD-системы (Amine and amine Precur-
sors Uptake and Decarboxylation), т. e. системы поглощения и декарбокси-
лирования аминов и их предшественников. К этой системе относятся ней-
росекреторные клетки гипоталамуса, клетки желудочно-кишечного тракта
(энтериноциты), продуцирующие кишечные гормоны, клетки островков
Лангерганса поджелудочной железы и К-клетки щитовидной железы.
Гормоны мозгового вещества — катехоламины — образуются из амино-
кислоты тирозина поэтапно: тирозин—ДОФА—дофамин-норадреналин—
адреналин. Хотя надпочечник и секретирует значительно больше адрена-
лина, тем не менее в состоянии покоя в крови содержится в четыре раза
больше норадреналина, так как он поступает в кровь и из симпатических
окончаний. Секреция катехоламинов в кровь хромаффинными клетками
осуществляется с обязательным участием Са2+, кальмодулина и особого
белка синексина, обеспечивающего агрегацию отдельных гранул и их связь
с фосфолипидами мембраны клетки.
Катехоламины называют гормонами срочного приспособления к дейст-
вию сверхпороговых раздражителей среды.
6. Эндокринная нервная система — регулятор функций и процессов в организме • 291
Физиологические эффекты катехоламинов обусловлены различиями в
адренорецепторах (альфа и бета) клеточных мембран, при этом адреналин
обладает большим сродством к бета-адренорецепторам, а норадреналин —
к альфа. Чувствительность адренорецепторов к адреналину увеличивают
гормоны щитовидной железы и глюкокортикоиды. Основные функцио-
нальные эффекты адреналина проявляются в виде: 1) учащения и усиле-
ния сердечных сокращений, 2) сужения сосудов кожи и органов брюш-
ной полости, 3) повышения теплообразования в тканях, 4) ослабления
сокращений желудка и кишечника, 5) расслабления бронхиальной муску-
латуры, 6) стимуляции секреции ренина почкой, 7) уменьшения образо-
вания мочи, 8) повышения возбудимости нервной системы, скорости
рефлекторных процессов и эффективности приспособительных реакций.
Адреналин вызывает мощные метаболические эффекты в виде усиленно-
го расщепления гликогена в печени и мышцах из-за активации фосфо-
рилазы, а также подавление синтеза гликогена, угнетение потребления
глюкозы тканями, что в целом ведет к гипергликемии. Адреналин вызы-
вает активацию распада жира, мобилизацию в кровь жирных кислот и их
окисление. Все эти эффекты противоположны действию инсулина, по-
этому адреналин называют контринсулярным гормоном. Адреналин усили-
вает окислительные процессы в тканях и повышает потребление ими ки-
слорода.
Таким образом, как кортикостероиды, так и катехоламины обеспечива-
ют активацию приспособительных защитных реакций организма и их
энергоснабжение, повышая устойчивость организма к неблагоприятным
влияниям среды.
В мозговом веществе надпочечников, кроме катехоламинов, образуется
и пептидный гормон адреномедуллин. Кроме мозгового вещества надпочеч-
ников и плазмы крови он выявлен в тканях легких, почек и сердца, а так-
же эндотелиальных клетках сосудов. Этот пептид состоит у человека из 52
аминокислот. Основное действие гормона заключается в мощном сосудо-
расширяющем эффекте, в связи с чем его называют гипотензивным пепти-
дом. Второй физиологический эффект гормона заключается в подавлении
продукции альдостерона клетками клубочковой зоны коры надпочечников.
При этом пептид подавляет не только базальный, фоновый уровень обра-
зования гормона, но и его секрецию, стимулированную высоким уровнем
калия в плазме крови или действием ангиотензина-II.
6.4. Регуляторные функции гормонов щитовидной железы
Щитовидная железа — орган эпителиального происхождения, который за-
кладывается в эмбриогенезе вначале как типичная экзокринная железа, и
лишь в процессе дальнейшего эмбрионального развития она становится
эндокринной. Кровоснабжение щитовидной железы очень интенсивное,
осуществляется через 2 пары артерий, берущих начало от выходящих из
аорты крупных артериальных стволов. Капилляры окружают фолликулы,
тесно прилегают к эпителиальным клеткам. Иннервация железы осуществ-
ляется ветвями симпатических шейных узлов и блуждающего нерва. Ин-
тенсивный лимфоотток, наряду с венозным оттоком, обеспечивает транс-
порт тиреоидных гормонов в общую циркуляцию. Эндокринные функции
присущи А-клеткам или тироцитам, образующим фолликулы и способ-
ным захватывать йод и синтезировать йодсодержащие тиреоидные гормо-
ны, и парафолликулярным К-клеткам, образующим кальций-регулирую-
щий гормон кальцитонин.
10*
292 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
6.4.1. Регуляция секреции и физиологические эффекты
йодсодержащих тиреоидных гормонов
Тироциты образуют фолликулы, заполненные коллоидной массой тирео-
глобулина. Базальная мембрана тироцитов тесно прилежит к кровеносным
капиллярам, и из крови эти клетки получают не только необходимые для
энергетики и синтеза белка субстраты, но и активно захватывают соедине-
ния йода — йодиды. В тироцитах происходит синтез тиреоглобулина, окис-
ление иодидов до образования атомарного йода. Тиреоглобулин содержит
на поверхности молекулы значительное количество остатков аминокисло-
ты тирозина (тиронины), которые и подвергаются йодированию. Через
апикальную мембрану тироцита тиреоглобулин выделяется в просвет фол-
ликула. При секреции гормонов в кровь ворсинки апикальной мембраны
окружают и поглощают путем эндоцитоза капельки коллоида, которые в
цитоплазме подвергаются гидролизу лизосомальными ферментами, и два
продукта гидролиза — трийодтиронин (Т3) и тетрайодтиронин {тироксин,
Т4) секретируются через базальную мембрану в кровь и лимфу. Все опи-
санные процессы регулируются тиреотропином аденогипофиза. Наличие
столь многочисленных процессов, регулируемых одним тиреотропином,
обеспечивается за счет включения многих внутриклеточных вторичных по-
средников. Существует и прямая нервная регуляция щитовидной железы
вегетативными нервами, хотя для активации секреции гормонов она играет
меньшую роль, чем эффекты тиреотропина. Механизм отрицательной об-
ратной связи в регуляции функции щитовидной железы реализуется уров-
нем тиреоидных гормонов в крови, что подавляет секрецию тиреолиберина
гипоталамусом и тиреотропина гипофизом. Интенсивность секреции ти-
реоидных гормонов влияет на объем их синтеза в железе (механизм мест-
ной положительная обратная связь).
Транспорт Т3 и Т4 в крови осуществляется с помощью специальных бел-
ков, однако, в такой связанной с белком форме гормоны не способны
проникать в клетки-эффекторы. Значительная часть тироксина депониру-
ется и транспортируется эритроцитами. Дестабилизация их мембран, на-
пример под воздействием ультрафиолетового облучения, ведет к выходу
тироксина в плазму крови. При взаимодействии гормона с рецептором на
поверхности клеточной мембраны происходит диссоциация гормон-белко-
вого комплекса, после чего гормон проникает внутрь клетки. Внутрикле-
точными мишенями тиреоидных гормонов являются ядро и органоиды
(митохондрии). Механизм действия тиреоидных гормонов представлен на
рис. 6.16. Т3 в несколько раз активнее Т4, и в тканях происходит превраще-
ние Т4 в Т3 В связи с этим основная часть эффектов тиреоидных гормонов
обеспечивается Т3.
Основными метаболическими эффектами тиреоидных гормонов являют-
ся: 1) усиление поглощения кислорода клетками и митохондриями с акти-
вацией окислительных процессов и увеличением основного обмена,
2) стимуляция синтеза белка за счет повышения проницаемости мембран
клетки для аминокислот и активации генетического аппарата клетки,
3) липолитический эффект и окисление жирных кислот с уменьшением их
уровня в крови, 4) активация синтеза холестерина в печени и его экскре-
ции с желчью, 5) гипергликемия за счет активации распада гликогена в пе-
чени и повышения всасывания глюкозы в кишечнике, 6) повышение по-
требления и окисления глюкозы клетками, 7) активация инсулиназы пече-
ни и ускорение инактивации инсулина, 8) стимуляция секреции инсулина
за счет гипергликемии. Таким образом, избыточные количества тиреоид-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 293
Трийодтиронин (Т3) Эффекты на мембранный Т4
Рис. 6.16. Геномный и внегеномный механизмы действия тиреоидных гормонов на
клетку.
Эффекты гормонов реализуются как после проникновения гормонов внутрь клетки (влияние
на транскрипцию в ядре и синтез белка, влияние на окислительно-восстановительные реак-
ции и освобождение энергии в митохондриях), так и после связывания гормона с мембран-
ным рецептором (образование вторичных посредников, увеличение транспорта в клетку суб-
стратов, в частности аминокислот, необходимых для синтеза белка).
ных гормонов, стимулируя секрецию инсулина и одновременно вызывая
контринсулярные эффекты, могут также способствовать развитию сахарно-
го диабета.
Основные физиологические эффекты тиреоидных гормонов, обусловлен-
ные перечисленными выше сдвигами обмена веществ, проявляются в сле-
дующем: 1) обеспечении нормальных процессов роста, развития и диффе-
ренцировки тканей и органов, особенно центральной нервной системы, а
также процессов физиологической регенерации тканей, 2) активации сим-
патических эффектов (тахикардия, потливость, сужение сосудов и т. п.),
как за счет повышения чувствительности адренорецепторов, так и в ре-
зультате подавления ферментов (моноаминоксидаза), разрушающих норад-
реналин, 3) повышении энергообразования в митохондриях и сократимо-
сти миокарда, 4) повышении теплообразования и температуры тела, 5) по-
вышении возбудимости центральной нервной системы и активации психи-
ческих процессов, 6) предотвращении стрессорных повреждений миокарда
и язвообразования в желудке, 7) увеличении почечного кровотока, клубоч-
ковой фильтрации и диуреза при угнетении канальцевой реабсорбции в
почках, 8) поддержании репродуктивной функции.
Избыточная продукция тиреоидных гормонов носит название гиперти-
реоза. При этом проявляются характерные метаболические (повышение ос-
новного обмена, гипергликемия, гипертермия, похудание) и функциональ-
ные признаки повышенного симпатического тонуса (тахикардия, потли-
вость, повышение артериального давления и др.). Врожденная недостаточ-
ность тиреоидных гормонов, из-за наследственных дефектов или дефицита
294 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 6.17. Баланс йода в организме.
В сутки поступает в организм с пищей и водой 500 мкг йода. Всасываясь в кровь, йодиды дос-
тавляются к щитовидной железе, где депонирован основной тиреоидный пул йода. Его расхо-
дование при секреции тиреоидных гормонов восполняется из резервного пула крови. Основ-
ное количество йода выделяется через почки с мочой (485 мкг), часть теряется с калом (15
мкг), следовательно, экскреция йода равна его поступлению в организм, что и составляет
внешний баланс.
йода в организме матери, нарушает рост, развитие и дифференцировку
скелета, тканей и органов, особенно центральной нервной системы, что
ведет к умственной отсталости («кретинизм»). Приобретенная недостаточ-
ность щитовидной железы (дефицит йода в воде и пище, нарушение про-
дукции гипофизом тиреотропина, повреждения ткани щитовидной желе-
зы — механические или химическими веществами) проявляется в замедле-
нии окислительных процессов и снижении основного обмена, гипоглике-
мии, падении возбудимости нервной системы и психической деятельности,
снижении температуры тела, накоплении гликозаминогликанов и воды в
подкожно-жировой клетчатке и коже (гипотиреоз, микседема или слизи-
стый отек).
Одной из наиболее частых причин врожденной или приобретенной ти-
реоидной недостаточности является дефицит йода в пище и воде или нару-
шения его захвата из крови и утилизации железой. Основные показатели
обмена йода приведены на рис. 6.17. При снижении количества йодида в
крови щитовидная железа быстро компенсирует этот дефицит, усиливая
захват йода. Запасы тиреоидных гормонов в щитовидной железе значи-
тельно превышают суточную потребность в них организма, поэтому крат-
ковременные и нерезкие уменьшения потребления йода не вызывают на-
рушений функции железы. Если дефицит йода более выражен и длителен,
синтез и секреция тиреоидных гормонов начинает отставать от потребно-
стей организма, активируется секреция тиреолиберина гипоталамуса и ти-
реотропина аденогипофиза, развивается гипертрофия и гиперплазия желе-
зы, получившая название зоба. В таких случаях нередко увеличенная масса
железы более эффективно поглощает и утилизирует йодид, что компенси-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 295
рует йодную недостаточность и поддерживает нормальную секрецию ти-
реоидных гормонов. Недостаточность этой компенсаторной реакции ведет
к тому, что несмотря на развитие зоба железа не в состоянии синтезиро-
вать и секретировать адекватные потребностям количества тиреоидных
гормонов и возникает тиреоидная недостаточность. Особенно часто начало
ее развития совпадает с повышением потребности организма в тиреоидных
гормонах, например при охлаждении.
6.4.2. Регуляция секреции и физиологические эффекты
кальцитонина
Кальцитонин является пептидным гормоном парафолликулярных К-кле-
ток щитовидной железы, но образуется также в тимусе и в легких. В орга-
низме существует ряд близких по химической структуре гормонов, поэтому
они получили собирательное название гормонов семейства кальцитонина.
К ним относятся также катакальцин и мозговой пептид, родственный гену
кальцитонина, причем последний, наряду с близкими кальцитонину эф-
фектами, рассматривается в роли возможного медиатора сосудистых нерв-
ных регуляторных влияний, так как показано его выделение на окончани-
ях сосудодвигательных нервов. Обнаружен гормон и в спинном мозге, где
также, вероятно, играет роль медиатора или модулятора синаптической пе-
редачи.
Кальцитонин относится к кальцийрегулирующим гормонам, и регуля-
ция его секреции осуществляется уровнем ионизированного кальция плаз-
мы крови по механизму обратной связи. Стимуляция секреции кальцито-
нина происходит при значительном повышении кальция в крови, тогда
как обычные физиологические колебания концентрации кальция мало
сказываются на секреции кальцитонина. Мощным регулирующим секре-
цию кальцитонина эффектом обладают нейропептиды и пептидные гормо-
ны желудочно-кишечного тракта, особенно гастрин. Повышение секреции
Рис. 6.18. Основные эффекты кальцитонина на органы-мишени.
Гипокальциемический гормон кальцитонин усиливает выведение почками из крови (угнетая
реабсорбцию) кальция и фосфата, одновременно с увеличением реабсорбции магния повыша-
ет поступление кальция и фосфата в кости, увеличивая их минерализацию. Возможен слабый
эффект кальцитонина, угнетающий всасывание кальция.
296 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
кальцитонина после перорального приема кальция обусловлено выделени-
ем в кровь гормона желудка — гастрина.
Кальцитонин оказывает эффекты в результате взаимодействия с рецеп-
торами органов-мишеней (почка, желудочно-кишечный тракт, костная
ткань) через вторичные посредники цАМФ и цГМФ. Гормон снижает уро-
вень кальция в крови {гипокальциемический гормон) за счет облегчения ми-
нерализации и подавления резорбции костной ткани, а также путем сни-
жения реабсорбции кальция в почках (рис. 6.18). Кальцитонин вызывает
фосфатурию в результате подавления реабсорбции фосфата в почечных ка-
нальцах. Гормон оказывает слабое диуретическое и натрийуретическое
действие, он способен тормозить секрецию гастрина в желудке и снижать
кислотность желудочного сока (эффект механизма отрицательной обрат-
ной связи).
6.5. Регуляторные функции гормона околощитовидных желез
Околощитовидные железы (у человека в среднем 4 железы) эпителиального
происхождения, кровоснабжаются из щитовидных артерий и, так же как
щитовидная железа, иннервируются симпатическими и парасимпатически-
ми волокнами. Гормон околощитовидных желез — паратирин — является
кальцийрегулирующим гормоном, повышающим уровень кальция в плазме
крови, в связи с чем его называют гиперкальциемическим гормоном. Регу-
ляция секреции паратирина происходит по механизму отрицательной об-
ратной связи уровнем ионизированного кальция плазмы крови. Низкая
концентрация кальция стимулирует секрецию паратирина при одновре-
менном повышении уровня цАМФ в клетках. Стимулируют продукцию па-
ратирина и симпатические влияния на клетки околощитовидных желез че-
рез бета-адренорецепторы, также приводящие к возрастанию в клетках же-
лезы содержания цАМФ. Подавляют секрецию паратирина высокий уро-
вень кальция в крови и почечный гормон кальцитриол.
Основные эффекты паратирина проявляются со стороны органов-ми-
шеней гормона — костной ткани, почек и желудочно-кишечного тракта
(рис. 6.19). Реализация действия паратирина осуществляется через цАМФ,
и повышение уровня этого вторичного посредника в моче является важ-
ным диагностическим критерием избыточной секреции паратирина. Эф-
фект гормона на костную ткань обусловлен стимуляцией активности и
увеличением количества остеокластов, резорбирующих кость. Под влияни-
ем паратирина в костной ткани накапливаются лимонная и молочная ки-
слоты, вызывающие местный ацидоз. Кислая реакция среды в костной
ткани тормозит активность щелочной фосфатазы — фермента, необходи-
мого для образования основного минерального вещества кости — фосфор-
нокислого кальция. Избыток лимонной и молочной кислот ведет к образо-
ванию растворимых в воде солей кальция — цитрата и лактата, вымыванию
их в кровь, что приводит к деминерализации кости. Избыток цитрата вы-
водится с мочой, что является важным диагностическим признаком повы-
шенного уровня секреции паратирина. В почках гормон снижает реабсорб-
цию кальция в проксимальных канальцах, но резко усиливает ее в дис-
тальных канальцах, что предотвращает потери кальция с мочой и способ-
ствует гиперкальциемии. Реабсорбция фосфата в почках под влиянием па-
ратирина угнетается, это приводит к фосфатурии и снижению содержания
фосфата в крови — гипофосфатемии. Почечные эффекты паратирина про-
являются также в диуретическом и натрийуретическом действии, угнете-
нии канальцевой реабсорбции воды, снижении эффективности действия
6. Эндокринная нервная система — регулятор функций и процессов в организме • 297
Рис. 6.19. Регуляция секреции и основные эффекты паратирина.
Снижение уровня кальция в крови является стимулом для околощитовидных желез (ОЩЖ),
секретирующих паратирин. Основные эффекты паратирина проявляются со стороны органов-
мишеней: почек (подавление реабсорбции кальция и его выведение с мочой, повышение реаб-
сорбции фосфата), кишечника (стимуляция всасывания кальция, в основном через стимуля-
цию синтеза в почках кальцитриола) и костной ткани (вымывание кальция в кровь). Эти эф-
фекты ведут к повышению в крови уровня кальция (паратирин является гиперкальциемиче-
ским гормоном).
на канальцы вазопрессина. В кишечнике паратирин прямо, но главным
образом опосредованно через кальцитриол, стимулирует всасывание каль-
ция, что также способствует гиперкальциемии.
Паратирин повышает поступление кальция во внутриклеточную среду
и транспорт иона из цитозоля во внутриклеточные депо, увеличивает уда-
ление свободного кальция из клеток. Благодаря этому изменяется возбу-
димость и реактивность клеток к нейрогенным и гуморальным регулятор-
ным стимулам. Паратирин вызывает повышение образования в почках
кальци-триола, стимулирует секрецию соляной кислоты и пепсина в же-
лудке.
Повышенная секреция паратирина при гиперплазии или аденоме око-
лощитовидных желез сопровождается деминерализацией скелета с дефор-
мацией длинных трубчатых костей, образованием почечных камней, мы-
шечной слабостью, депрессией, нарушениями памяти и концентрации
внимания.
6.6. Регуляторные функции гормонов эпифиза
Эпифиз представляет собой структуру эпиталамуса межуточного мозга и
расположен по срединной плоскости глубоко между полушариями. Будучи
железой нейроглиального происхождения, эпифиз связан с многослойным
участком эпендимы задней стенки III желудочка, называемым субкомиссу-
ральным органом. Кровоснабжение эпифиза осуществляется из ветвей пе-
редней, средней и задней мозговых артерий. Основными секреторными
клетками эпифиза являются пинеалоциты. Ими образуется и секретирует-
298 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Рис. 6.20. Схема механизма регуляции секреции мелатонина эпифизом и основные
эффекты гормона.
Свет, воспринимаемый глазом, тормозит секрецию мелатонина, а в темноте нервные импуль-
сы через ретикулогипоталамический тракт, гипоталамус, верхний шейный симпатический
ганглий приводят к освобождению на симпатических терминалях в эпифизе медиатора норад-
реналина, стимулирующего секрецию гормона эпифизом.
ся в кровь и цереброспинальную жидкость гормон мелатонин (название
получил благодаря способности менять окраску кожи и чешуи у земновод-
ных и рыб, у человека на пигментацию не влияет). Мелатонин является
производным аминокислоты триптофана, он обеспечивает регуляцию био-
ритмов эндокринных функций и метаболизма для приспособления орга-
низма к разным условиям освещенности. Эпифиз имеет нервные связи со
структурами лимбики, однако основная регуляторная информация посту-
пает в эпифиз из верхнего шейного узла пограничного ствола по симпати-
ческим волокнам, которые формируют шишковидный нерв.
Синтез и секреция мелатонина зависят от освещенности — избыток све-
та тормозит его образование. Путь регуляции секреции (рис. 6.20) начина-
ется от сетчатки глаза ретиогипоталамическим трактом, из межуточного
мозга по преганглионарным волокнам информация поступает в верхний
шейный симпатический ганглий, затем отростки постганглионарных кле-
ток возвращаются в мозг и доходят до эпифиза. Снижение освещенности
повышает выделение на окончаниях симпатического шишковидного нерва
6. Эндокринная нервная система — регулятор функций и процессов в организме • 299
норадреналина и, соответственно, синтез и секрецию мелатонина. У чело-
века на ночные часы приходится 70 % суточной продукции гормона.
Основной физиологический эффект мелатонина заключается в тормо-
жении секреции гонадотропинов как на уровне аденогипофиза, так и опо-
средованно через угнетение нейросекреции либеринов гипоталамуса. Кро-
ме того, снижается, но в меньшей степени, секреция и других гормонов
аденогипофиза — кортикотропина, тиреотропина, соматотропина. Секре-
ция мелатонина подчинена суточному ритму (циркадианный ритм), опре-
деляющему ритмичность гонадотропных эффектов и половой функции, в
том числе продолжительность менструального цикла у женщин. Деятель-
ность эпифиза называют «биологическими часами» организма, так как же-
леза обеспечивает процессы адаптации организма к смене часовых поясов.
Введение мелатонина человеку вызывает легкую эйфорию и сон. В экспе-
риментальных условиях экстракты эпифиза оказывают инсулиноподобный
(гипогликемический) и паратиреоподобный (гиперкальциемический) эф-
фекты, что, по-видимому, связано не только с мелатонином, но и с други-
ми биологически активными веществами эпифиза — серотонином, адре-
ногломерулотропином, гиперкалиемическим фактором и др. Проявляется
и диуретическое влияние экстрактов эпифиза, что позволяет считать его
ответственным за ритмическую регуляцию водно-солевого обмена. Пока-
зана способность экстрактов эпифиза тормозить апоптоз (естественную
гибель клеток) и старение, выявлен противоопухолевый эффект эпифизар-
ных гормонов.
6.7. Регуляторные функции гормонов эндокринных тканей
в органах, обладающих неэндокринными функциями
6.7.1. Регуляторные функции гормонов поджелудочной железы
Эндокринную функцию в поджелудочной железе выполняют скопления
клеток эпителиального происхождения, получившие название островков
Лангерганса и составляющие всего 1 —2 % массы поджелудочной железы —
экзокринного органа, образующего панкреатический пищеварительный
сок. Количество островков в железе взрослого человека очень велико и со-
ставляет от 200 тысяч до полутора миллионов. В островках различают не-
сколько типов клеток, продуцирующих гормоны: альфа-клетки образуют
глюкагон, бета-клетки — инсулин, дельта-клетки — соматостатин, джи-
клетки — гастрин и РР- или F-клетки — панкреатический полипептид. По-
мимо инсулина в бета-клетках синтезируется гормон амилин, обладающий
противоположными инсулину эффектами. Кровоснабжение островков бо-
лее интенсивно, чем основной паренхимы железы. Иннервация осуществ-
ляется постганлионарными симпатическими и парасимпатическими нерва-
ми, причем среди клеток островков расположены нервные клетки, обра-
зующие нейроинсулярные комплексы.
Инсулин синтезируется в эндоплазматическом ретикулуме бета-клеток
вначале в виде пре-проинсулина, затем от него отщепляется 23-аминокис-
лотная цепь и остающаяся молекула носит название проинсулина. В ком-
плексе Гольджи проинсулин упаковывается в гранулы, в них осуществля-
ется расщепление проинсулина на инсулин и соединительный пептид (С-
пептид). В гранулах инсулин депонируется в виде полимера и частично в
комплексе с цинком. Количество депонированного в гранулах инсулина
почти в 10 раз превышает суточную потребность в гормоне. Секреция ин-
сулина происходит путем экзоцитоза гранул, при этом в кровь поступает
300 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Ацетилхолин
Са2+, аминокислоты, желудочно-
Рис. 6.21. Функциональная организация островков Лангерганса как «мини-органа».
Сплошные стрелки — стимуляция, пунктирные — подавление гормональных секретов. Веду-
щий регулятор — глюкоза — при участии кальция стимулирует секрецию инсулина р-клетками
и, напротив, тормозит секрецию глюкагона а-клетками. Всасывающиеся в желудке и кишеч-
нике аминокислоты являются стимуляторами функции всех клеточных элементов «мини-орга-
на». Ведущий «внутриорганный» ингибитор секреции инсулина и глюкагона — соматостатин,
активация его секреции происходит под влиянием всасывающихся в кишечнике аминокислот
и гастроинтестинальных гормонов при участии ионов Са2+. Глюкагон является стимулятором
секреции как соматостатина, так и инсулина.
эквимолярное количество инсулина и С-пептида. Определение содержания
последнего в крови является важным диагностическим тестом оценки сек-
реторной способности р-клеток.
Секреция инсулина является кальцийзависимым процессом. Под влия-
нием стимула — повышенного уровня глюкозы в крови — мембрана бета-
клеток деполяризуется, ионы кальция входят в клетки, что запускает про-
цесс сокращения внутриклеточной микротубулярной системы и перемеще-
ние гранул к плазматической мембране с последующим их экзоцитозом.
Секреторная функция разных клеток островков взаимосвязана, зависит
от эффектов образуемых ими гормонов, в связи с чем островки рассматри-
ваются как своеобразный «мини-орган» (рис. 6.21). Выделяют два вида
секреции инсулина: базальную и стимулированную. Базальная секреция
инсулина осуществляется постоянно, даже при голодании и уровне глюко-
зы крови ниже 4 ммоль/л.
Стимулированная секреция инсулина представляет собой ответ бета-
клеток островков на повышенный уровень D-глюкозы в притекающей к
бета-клеткам крови. Под влиянием глюкозы активируется энергетический
рецептор бета-клеток, что увеличивает транспорт в клетку ионов кальция,
активирует аденилатциклазу и пул (фонд) цАМФ. Через эти посредники
глюкоза стимулирует выброс инсулина в кровь из специфических секре-
торных гранул. Усиливает ответ бета-клеток на действие глюкозы гормон
двенадцатиперстной кишки — желудочный ингибиторный пептид (ЖИП).
В регуляции секреции инсулина определенную роль играет и вегетативная
нервная система. Блуждающий нерв и ацетилхолин стимулируют секрецию
инсулина, а симпатические нервы и норадреналин через альфа-адреноре-
цепторы подавляют секрецию инсулина и стимулируют выброс глюкагона.
Специфическим ингибитором продукции инсулина является гормон дель-
та-клеток островков — соматостатин. Этот гормон образуется и в кишеч-
6. Эндокринная нервная система — регулятор функций л процессов в организме • 301
нике, где тормозит всасывание глюкозы и тем а мым уменьшает ответную
реакцию бета-клеток на глюкозный стимул. Образование в поджелудочной
железе и кишечнике пептидов, аналогичных мосговым, например сомато-
статина, подтверждает существование в организк е единой APUD-системы.
Секреция глюкагона стимулируется снижением уровня глюкозы в крови,
гормонами желудочно-кишечного тракта (ЖИП гастрин, секретин, холе-
цистокинин-панкреозимин) и при уменьшении в крови ионов Са2+. По-
давляют секрецию глюкагона инсулин, соматостатин, глюкоза крови и
Са2+. В эндокринных клетках кишечника образуется глюкагоноподобный
пептид-1, стимулирующий всасывание глюкозы и секрецию инсулина по-
сле приема пищи. Клетки желудочно-кишечного тракта, продуцирующие
гормоны, являются своеобразными «приборами раннего оповещения» кле-
ток панкреатических островков о поступлении пищевых веществ в орга-
низм, требующих для утилизации и рас пределе шя участия панкреатиче-
ских гормонов. Эта функциональная взаимосвязь нашла отражение в тер-
мине «гастро-энтеро-панкреатическая система».
6.7.1.1. Физиологические эффекты инсулина
Действие инсулина на клетки-мишени начинается после его связывания со
специфическими димерными мембранными рецепторами (рис. 6.22), при
этом внутриклеточный домен рецептора обладае г тирозинкиназной актив-
ностью. Инсулин-рецепторный комплекс не только передает сигнал внутрь
клетки, но и частично путем эндоцитоза поступает внутрь клетки к лизо-
сомам. Под влиянием лизосомальной протеазы инсулин отщепляется от
рецептора, при этом последний либо разрушается, либо возвращается к
мембране и вновь встраивается в нее. Многократное перемещение рецеп-
тора от мембраны к лизосомам и обратно к мембране носит название ре-
циклизация рецептора. Процесс рециклизации вакен для регуляции колич-
ства инсулиновых рецепторов, в частности обес течения обратной зависи-
мости между концентрацией инсулина и количеством мембранных рецеп-
торов к нему.
Образование инсулин-рецепторного комплекса активирует тирозинки-
назу, запускающую процессы фосфорилирования внутриклеточных бел-
ков. Происходящее при этом аутофосфорилирование рецептора ведет к
усилению первичного сигнала. Инсулин-рецепторный комплекс вызывает
активирование фосфолипазы С, образование вторичных посредников
инозитолтрифосфата и диацилглицерола, активацию протеинкиназы С,
ингибирование цАМФ. Участие нескольких систем вторичных посредни-
ков объясняет многообразие и различия эффс ктов инсулина в разных
тканях.
Инсулин оказывает влияние на все виды обмена веществ, способствует
анаболическим процессам, увеличивая синтез пикогена, жиров и белков,
тормозя эффекты многочисленных контринсулярных гормонов (глюкаго-
на, катехоламинов, глюкокортикоидов и соматотзопина). Все эффекты ин-
сулина по скорости их реализации подразделяют на 4 группы: очень быст-
рые (через несколько секунд) — гиперполяризация мембран клеток (за ис-
ключением гепатоцитов), повышение проницаемости для глюкозы, актива-
ция Na-K-АТФазы, входа К+ и откачивания Na , подавление Са-насоса и
задержка Са2+; быстрые эффекты (в течение нес кольких минут) — актива-
ция и торможение различных ферментов, подав; яющих катаболизм и уси-
ливающих анаболические процессы; медленные процессы (в течение не-
скольких часов) — повышенное поглощение ами гокислот, изменение син-
302 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Внеклеточный домен реце птора
Трансмембранный домек-----
—рецептора
Внутриклеточный домен рецептора
Молекула инсулина
Активация тирозинкиназы
Молекулы
ГЛЮТ-4
Фосфорилирование
Аутофосфорилирование
♦
Активация внутри! неточных белков, биологические эффекты
Синтез липидов Синтез гликогена Рост и экспрессия Синтез белков Транспорт
генов глюкозы
Рис. 6.22. Схема механизма х ействия инсулина на клетку-мишень.
Взаимодействие инсулина со сп< цифическим рецептором вызывает активацию тирозинкина-
зы, связанной с внутриклеточны и доменом рецептора. Следствием является фосфорилирова-
ние внутриклеточных белков, в г ом числе и аутофосфорилирование внутриклеточного домена
рецептора инсулина, усиливают :е тирозинкиназную активность. Фосфорилирование внутри-
клеточных белков ведет к актива) (ии ферментов, увеличению синтеза новых белков, в том чис-
ле и переносчиков ГЛЮТ-4, обеспечивающих транспорт в клетку молекул глюкозы.
теза РНК и белков-ферме зтов; очень медленные эффекты (от часов до су-
ток) — активация митогенсза и размножения клеток.
Важнейшим эффектом инсулина в организме является увеличение в
20—50 раз транспорта глю <озы через мембраны мышечных и жировых кле-
ток путем облегченной диффузии по градиенту концентрации с помощью
чувствительных к гормон) мембранных белковых переносчиков, называе-
мых ГЛЮТ. В мембранах разных видов клеток выявлены 6 типов ГЛЮТ
(рис. 6.23), но только один из них — ГЛЮТ-4 — является инсулинозависи-
мым и находится в мембранах клеток скелетных мышц, миокарда, жиро-
вой ткани.
Инсулин влияет на угле водный обмен, что проявляется:
1) активацией утилизации глюкозы клетками,
2) усилением процессов фосфорилирования;
3) подавлением распаде и стимуляцией синтеза гликогена;
4) угнетением глюконеогенеза;
5) активацией процессов гликолиза;
6) гипогликемией.
Действие инсулина на белковый обмен состоит в: 1) повышении прони-
цаемости мембран для ам знокислот; 2) усилении синтеза иРНК; 3) акти-
вации в печени синтеза av инокислот; 4) повышении синтеза и подавлении
распада белка.
Рис. 6.23. Схема транспорта глюкозы через клеточные мембраны. Переносчики имеют общее название ГЛЮТ-1, 2, 3, 4, 5, 6.
Только ГЛЮТ-4 является инсулинозависимым.
6. Эндокринная нервная система — регулятор функци 1 и процессов в организме • 303
304 • РЕГУЛИРУЮЩИЕ И УП ’АВЛЯЮЩИЕ СИСТЕМЫ
Основные эффекты инс/лина на липидный обмен:
• стимуляция синтеза свободных жирных кислот из глюкозы;
• стимуляция синтеза липопротеиновой липазы в клетках эндотелия
сосудов и благодаря этому активация гидролиза связанных с липо-
протеинами крови триглицеридов и поступления жирных кислот в
клетки жировой ткаш ;
• стимуляция синтеза триглицеридов;
• подавление распада жара;
• активация окисления кетоновых тел в печени.
Благодаря влиянию на тдеточную мембрану инсулин поддерживает вы-
сокую внутриклеточную концентрацию ионов калия, что необходимо для
обеспечения нормальной в эзбудимости клеток.
Широкий спектр метаболических эффектов инсулина в организме сви-
детельствует о том, что гормон необходим для осуществления функциони-
рования всех тканей, оргаь ов и физиологических систем, реализации эмо-
циональных и поведенческих актов, поддержания гомеостазиса, осуществ-
ления механизмов приспособления и защиты организма от неблагоприят-
ных факторов среды.
Недостаток инсулина (о гносительный дефицит по сравнению с уровнем
контринсулярных гормонов, прежде всего глюкагона) приводит к сахарно-
му диабету. Избыток инсу; ина в крови, например при передозировке, вы-
зывает гипогликемию с эезкими нарушениями функций центральной
нервной системы, использующей глюкозу как основной источник энергии
независимо от инсулина.
6.7.1.2. Физиологические э(2 >фекты глюкагона
Глюкагон является мощный контринсулярным гормоном и его эффекты
реализуются в тканях череэ систему вторичного посредника аденилатцик-
лаза—цАМФ. В отличие о' инсулина, глюкагон повышает уровень сахара
крови, в связи с чем его называют гипергликемическим гормоном. Основ-
ные эффекты глюкагона проявляются в следующих сдвигах метаболизма в
организме:
• активация гликогенолиза в печени и мышцах;
• активация глюконеоп неза;
• активация липолиза и подавление синтеза жира в адипоцитах;
• повышение синтеза кетоновых тел в печени и угнетение их окисле-
ния;
• стимуляция катаболи: ма белков в клетках тканей, прежде всего пече-
ни, и увеличение син еза в ней мочевины.
Образующиеся в острсвках Лангерганса гастрин и панкреатический
полипептид основную рол > играют в регуляции процессов пищеварения,
их эффекты и физиологг ческая роль рассмотрены в соответствующей
главе.
6.7.2. Регуляторные ф ункции гормонов половых желез
Половые железы (семенники и яичники), наряду с функцией гаметообра-
зования, содержат клетки, синтезирующие и секретирующие половые гор-
моны. При этом эндокринная функция присуща и специализированным
для внутренней секреции клеткам (клетки Лейдига семенников, клетки
желтого тела яичников), и клеткам, участвующим в процессах гаметогенеза
(клетки Сертоли семенниюв, клетки гранулезы яичников). Как семенни-
6. Эндокринная нервная система — регулятор функций i процессов в организме • 305
ки, так и яичники синтезируют и мужские горк оны (андрогены), и жен-
ские половые гормоны (эстрогены), являющиеся стероидами — производ-
ными холестерина.
6.7.2.1. Гормоны семенников и их эффекты в организме
Основной структурой семенника, где происходгт образование и созрева-
ние гамет — сперматозоидов, — являются извт ые семенные канальцы.
Базальная мембрана изнутри покрыта отростчатыми клетками Сертоли и
располагающимися между ними клетками сперматогенного эпителия,
внутренний слой которых составляют сперматогонии. Клетки Сертоли,
наряду с обеспечением процесса созревания сперматид, поглощения ос-
татков их цитоплазмы при превращении сперматиды в сперматозоид, об-
ладают секреторной и инкреторной функцией. Их секреторная функция
заключается в выделении в просвет канальца жидкости, в которой плава-
ют сперматозоиды. Инкреторная функция сводт тся к двум процессам: 1)
образованию и секреции с жидкостью в просвет канальца гормона ингиби-
на — основного тормозящего продукцию фоллит ропина механизма обрат-
ной связи с гипофизом и 2) образованию и секэеции в периканальцевую
лимфу эстрогенов. Кровеносные капилляры не проникают в просвет ка-
нальцев, а ветвятся между их петлями. Рядом с кровеносными капилляра-
ми расположены скопления клеток мезенхимнсго происхождения, назы-
ваемых клетками Лейдига. Эти клетки отделены от семенных канальцев
лимфатическими пространствами. Клетки Лейдига являются основными
продуцентами мужских половых гормонов, главным образом тестостеро-
на. Регуляция продукции гормонов семенниками осуществляется преиму-
щественно лютропином аденогипофиза, специфи тески регулирующим сек-
реторную активность клеток Лейдига и продукц по тестостерона, и отчас-
ти фоллитропином, меняющим активность клетж Сертоли и продукцию
ими эстрогенов и ингибина. Функциональная активность клеток Сертоли
и Лейдига регулируется влияниями через лимф} секретируемых ими гор-
монов.
Основные метаболические и функциональные эффекты тестостерона:
• обеспечение процессов половой дифференцировки в эмбриогенезе;
• развитие первичных и вторичных половых признаков;
• формирование структур центральной нервной системы, обеспечиваю-
щих половое поведение и функции;
• генерализованное анаболическое действие, обеспечивающее рост ске-
лета, мускулатуры, распределение подкожш го жира;
• регуляция сперматогенеза;
• задержка в организме азота, калия, фосфате, кальция;
• активация синтеза РНК;
• стимуляция эритропоэза.
6.7.2.2. Гормоны яичников и их эффекты в организме
Гормонопродуцирующие клетки гранулезы фолликулов являются по проис-
хождению и функциям аналогом клеток Сертоли семенников, но их функ-
ция регулируется не только гипофизарным фоллитропином, но в большей
мере лютропином. Основным гормоном гранулемы является эстрадиол, об-
разуемый из предшественника тестостерона. В меньшем количестве грану-
леза образует эстрон, из которого в печени и пл; центе образуется эстриол.
Клетки гранулезы образуют в малых количества? и прогестерон, необходи-
306 ‘ РЕГУЛИРУЮЩИЕ И УГ РАВНЯЮЩИЕ СИСТЕМЫ
мый для овуляции, но главным источником прогестерона служат клетки
желтого тела, регулируем ле гипофизарным лютропином.
Секреторная активность этих эндокринных клеток характеризуется вы-
раженной цикличностью, связанной с женским половым циклом. Послед-
ний обеспечивает интеграцию во времени различных процессов, необходи-
мых для осуществления репродуктивной функции — периодическую подго-
товку эндометрия к имплантации оплодотворенной яйцеклетки, созрева-
ние яйцеклетки и овуляцию, изменение вторичных половых признаков.
Эстрогены необходимы для процессов половой дифференцировки в эм-
бриогенезе, полового созревания и развития женских половых признаков,
установления женского полового цикла, роста мышцы и железистого эпи-
телия матки, развития мог очных желез. В итоге эстрогены регулируют по-
ловое поведение, овогенез, процессы оплодотворения и имплантации яй-
цеклетки, развитие и дифференцировку плода, нормальный родовой акт.
Благодаря геномному механизму действия эстрогены подавляют резорб-
цию кости, оказывают общее анаболическое действие, хотя и более слабое,
чем андрогены. Негеномгый механизм действия эстрогенов ведет к за-
держке в организме азота воды и солей. Одним из важных негеномных
эффектов эстрогенов является активация под их влиянием NO-синтетазы
и образование оксида азе та в коронарных сосудах, что профилактирует
развитие у женщин ишемической болезни сердца. Эстрогены модулируют
секрецию инсулина и внутриклеточный гомеостазис кальция. Распростра-
ненность рецепторов эстрогенов на мембранах разных типов клеток орга-
низма (костные, мышечные, секреторные, нервные, соединительной ткани
и крови) и, соответственна, множественность эффектов гормонов в орга-
низме, объясняют причин г многочисленных функциональных изменений,
происходящих в женском организме при климаксе, когда секреция эстроге-
нов резко снижается. Ввехение в организм женщины в этот период экзо-
генных веществ с эстрогеноподобным действием, в том числе и раститель-
ного происхождения, позголяет существенно уменьшить выраженность и
характер функциональных проявлений климактерического периода.
Прогестерон является п рмоном сохранения беременности (гестагеном),
так как ослабляет готовность мускулатуры матки к сокращению. Малые
концентрации гормона необходимы и для овуляции. Большие количества
прогестерона, образующиеся желтым телом, подавляют секрецию гипофи-
зарных гонадотропинов. Прогестерон обладает выраженным антиальдосте-
роновым эффектом, поэтому стимулирует натриурез.
6.8. Регуляторные функции гормонов клеток, сочетающих
выработку гормонов и неэндокринные функции
6.8.1. Регуляторные функции гормонов плаценты
Плацента тесно анатомически и функционально связана с организмами
матери и плода, поэтому принято говорить о комплексе «мать—плацента-
плод», или «фетоплацента эном комплексе». Синтез в плаценте эстриола
происходит не только из эстрадиола матери, но и из дегидроэпиандросте-
рона, образуемого надпочечниками плода. Поэтому по экскреции с мочой
матери эстриола можно судить о жизнеспособности плода. В плаценте об-
разуется прогестерон, действующий преимущественно на мускулатуру мат-
ки. С плацентарным прогестероном, например, связан временной интер-
вал между рождениями пледов при двойне.
Основная часть гормон эв плаценты у человека по своим свойствам и
6. Эндокринная нервная система — регулятор функц) 'й и процессов в организме • 307
даже строению напоминает гипофизарные го тадотропин и пролактин. В
наибольших количествах при беременности плацентой продуцируется хо-
рионический гонадотропин, оказывающий регул 1торные эффекты не только
на процессы дифференцировки и развития п. ода, но и на метаболизм в
организме матери. Гормон обеспечивает в орг шизме матери задержку со-
лей и воды, необходимых для растущего плоде , стимулирует секрецию ва-
зопрессина, активирует механизмы иммунитет; у матери.
6.8.2. Регуляторные функции гормонов п имуса
Тимус (вилочковая железа) является центральным органом иммунитета,
обеспечивающим продукцию специфических Т-лимфоцитов. Тимоциты
секретируют в кровь гормональные факторь, оказывающие не только
влияние на дифференцировку Т-клеток, но и вызывающие ряд общих ре-
гуляторных эффектов в организме. Основные эффекты гормонов тимуса
(тимозина, тимопоэтина, тимулина) описаны < главе 8 «Иммунная систе-
ма». Гормоны тимуса влияют на процессы синтеза клеточных рецепторов к
медиаторам и гормонам, стимулируют разруше лие ацетилхолина в нервно-
мышечных синапсах, регулируют состояние угг еводного и белкового обме-
на, а также обмена кальция, функции щитовидной и половых желез, моду-
лируют эффекты глюкокортикоидов, тироксин i (антагонизм) и соматотро-
пина (синергизм). В целом вилочковая железе рассматривается как орган
интеграции иммунной и эндокринной систем с рганизма.
6.8.3. Регуляторные функции гормонов п )чек
В почках отсутствует специализированная эндокринная ткань, однако ряд
клеток обладает способностью к синтезу и сек эеции многих биологически
активных веществ, обладающих всеми свойствами классических гормонов.
Установленными гормонами почек являются: 1) кальцитриол — третий
кальцийрегулирующий гормон, 2) ренин — на1 альное звено ренин-ангио-
тензин-альдостероновой системы, 3) эритропоэтин.
6.8.3.1. Синтез, секреция и физиологические эффекты кальцитриола
Кальцитриол является активным метаболитом витамина D3 и в отличие от
двух других кальцийрегулирующих гормонов - паратирина и кальцитони-
на—имеет стероидную природу. Синтез каль щтриола происходит в три
этапа (рис. 6.24). Первый этап протекает в кож?, где под влиянием ультра-
фиолетовых лучей из провитамина образуется штамин D3 или холекалъци-
ферол. Второй — связан с печенью, куда холека льциферол транспортирует-
ся кровью и где в эндоплазматическом ретикул 'ме гепатоцитов происходит
его гидроксилирование по 25-му атому углерод i с образованием 25(OH)D3.
Этот метаболит поступает в кровь и циркулирует в связи с альфа-глобулй-
ном. Его физиологические концентрации не влияют на обмен кальция.
Третий этап осуществляется в почках, где в мттохондриях клеток прокси-
мальных канальцев происходит второе гидроксилирование и образуются
два соединения: 1,25-(OH)2-D и 24,25-(OH)2-D. Первое — является наибо-
лее активной формой витамина D3, обладает мощным регуляторным влия-
нием на обмен кальция в организме и называется кальцитриолом. Образо-
вание в почках этого гормона регулируется паратирином, который стиму-
лирует гидроксилирование по первому атому углерода. Таким же эффек-
том обладает и гипокальциемия. При избытке кальция в крови гидрокси-
308 • РЕГУЛИРУЮЩИЕ И УПР/ ВЛЯЮЩИЕ СИСТЕМЫ
Инсоляция
Рис. 6.24. Схема образования кальцитриола или активной формы витамина D3.
Под воздействием ультрафиолетовых лучей в коже из холестерина образуется витамин D3; по-
ступая с кровью в печень, он подвс м ается первому гидроксилированию по 25-му атому угле-
рода, затем из печени с кровью noi адает в почки, где подвергается второму гидроксилирова-
нию по 1-му атому углерода, что и i едет к образованию дважды гидроксилированного витами-
на или кальцитриола.
Рис. 6.25. Основные эффекты f альцитриола.
Под влиянием паратирина почка се <ротирует кальцитриол, основные эффекты которого (жир-
ные стрелки) заключаются в стиму пяции всасывания в кишечнике в кровь ионов кальция и
фосфата и усилении их захвата кост ной тканью. Эффекты кальцитриола на почку (стимуляция
реабсорбции кальция и фосфора) в сражены слабее.
6. Эндокринная нервная система — регулятор функций и процессов в организме • 309
Рис. 6.26. Эффекты трех кальцийрегулирующих горм >нов на органы-мишени.
В почке паратирин и кальцитриол активируют реабсорбции кальция, а кальцитонин ее угне-
тает. И паратирин, и кальцитонин подавляют реабсорбцию < юсфата. В кишечнике кальцитри-
ол и паратирин активируют всасывание кальция и фосфата Кальцитонин и кальцитриол спо-
собствуют отложению кальция в костях, а паратирин акти шрует резорбцию кости и выход
кальция в кровь.
лирование происходит по 24-му атому углерода и синтезируется второе со-
единение — 24,25-(OH)2-D, которое обладает си )собностью угнетать секре-
цию паратирина по принципу обратной связи. Инактивация кальцитриола
происходит в печени.
Основной эффект кальцитриола (рис. 6.25) заключается в активации
всасывания кальция в кишечнике. Гормон стал улирует все три этапа вса-
сывания: захват ворсинчатой поверхностью клетки, внутриклеточный
транспорт, выброс кальция через базолатеральн 'Ю мембрану во внеклеточ-
ную среду. Действие кальцитриола на эпителиальные клетки кишечника
состоит в индуцировании синтеза энтероцитаии специальных кальций-
связывающих и транспортирующих белков — к зльбайндинов. Кальцитриол
повышает в кишечнике и всасывание фосфато г Почечные эффекты гор-
мона заключаются в стимуляции реабсорбции < эосфата и кальция каналь-
цевым эпителием. Эффекты кальцитриола на костную ткань связаны с
прямой стимуляцией остеобластов и обеспечением костной ткани усилен-
но всасывающимся в кишечнике кальцием, что активирует рост и минера-
лизацию кости. Эффекты кальцитриола, как и зсех стероидных гормонов,
делятся на геномные и негеномные. Геномные эффекты обусловливают
синтез кальбайндинов, активирование остеоблш тов и синтеза костной тка-
ни. Увеличение транспорта кальция внутрь клегок, например в скелетных
и сердечной мышцах, в остеобластах, энтерош тах, гепатоцитах и клетках
310 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
околощитовидных желез, происходит быстро и обусловлено негеномным
действием гормона. Под влиянием гормонрецепторного мембранного ком-
плекса в клетках происх< щит образование вторичного посредника диацил-
глицерола и активация протеинкиназы С. Кальцитриол меняет в клетке и
уровень цАМФ и цГМФ, что ведет к модификации геномного эффекта.
Наличие специфических рецепторов к гормону у многих клеток тканей (в
молочной железе, эндокринных железах, нервной системе), способность
кальцитриола активиров; ть транспорт кальция в большинстве из них сви-
детельствуют о широком спектре эффектов этого гормона.
Участие трех кальцийрегулирующих гормонов в гомеостазисе кальция и
фосфора показано на рис. 6.26.
Недостаточность кальцитриола проявляется в виде рахита, т. е. наруше-
ния созревания и кальцификации хрящей и кости у детей, либо остеомаля-
ции, т. е. падения минер ишзации костей после завершения роста скелета.
При этом сдвиги уровня кальция в крови и клетках обусловливают угнете-
ние нейромышечной возбудимости и мышечную слабость.
6.8.3.2. Образование ренина и основные функции
ренин-ангиотензи н-алъдостероновой системы
Ренин образуется в виде г роренина и секретируется в юкстагломерулярном
аппарате (ЮГА) (от латинских слов juxta — около, glomerulus — клубочек)
почек миоэпителиоидны ии клетками приносящей артериолы клубочка,
получившими название юкстагломерулярных (ЮГК). Структура ЮГА при-
ведена на рис. 6.27. В ЮГА кроме ЮГК также входит прилегающая к при-
носящим артериолам час ь дистального канальца нефрона, многослойный
эпителий которого образ} ет здесь плотное пятно — macula densa. Секреция
ренина в ЮГК регулируется четырьмя основными влияниями. Во-первых,
величиной давления крови в приносящей артериоле, т. е. степенью ее рас-
тяжения. Снижение растяжения активирует, а увеличение — подавляет
секрецию ренина. Во-втоэых, регуляция секреции ренина зависит от кон-
центрации натрия в моче дистального канальца, которая воспринимается
macula densa — своеобразным Na-рецептором. Чем больше натрия оказы-
вается в моче дистального канальца, тем выше уровень секреции ренина.
В-третьих, секреция ренина регулируется симпатическими нервами, ветви
которых заканчиваются на ЮГК, медиатор норадреналин через бета-адре-
норецепторы стимулирует секрецию ренина. В-четвертых, регуляция сек-
реции ренина осуществляется по механизму отрицательной обратной свя-
зи, включаемой уровнем I крови других компонентов системы — ангиотен-
зина и альдостерона, а т юсе их эффектами — содержанием в крови на-
трия, калия, артериальным давлением, концентрацией простагландинов в
почке, образующихся под влиянием ангиотензина.
Кроме почек образование ренина происходит в эндотелии кровеносных
сосудов многих тканей, м чокарде, головном мозге, слюнных железах, клу-
бочковой зоне коры надпочечников.
Секретированный в кр >вь ренин вызывает расщепление альфа-глобули-
на плазмы крови — ангиотензиногена, образующегося в печени. При этом в
крови образуется (рис. 6.'.’.8) малоактивный декапептид ангиотензин-1, ко-
торый в сосудах почек, легких и других тканей подвергается действию пре-
вращающего фермента (карбоксикатепсин, кининаза-2), отщепляющего от
ангиотензина-1 две аминокислоты. Образующийся октапептид ангиотен-
зин-11 обладает большим числом различных физиологических эффектов, в
том числе стимуляцией К1убочковой зоны коры надпочечников, секрети-
6. Эндокринная нервная система — регулятор функщ й и процессов в организме • 311
Юкстагломерулярные
клетки
Приносящая артериола
Капилляры
Дистальный,
каналец
Мезангиальные
клетки
Рис. 6.27. Схема юкстагломерулярного аппарата по1 ек, включающего юкстагломе-
рулярные клетки стенки приносящей артериолы, г летки плотного пятна (macula
densa) стенки дистального канальца и мезангиальные клетки. Основное место выра-
ботки ренина — юкстагломерулярные клетки принос пцей артериолы клубочка.
Симпатическая активация
АД —* ЮГК -*—Macula —NaCl
—-i— densa
Ренин
Ангиотензиноген 1 2 3 4 5 6 7 8 ' 10 U11 12 13 14
Asp-Arg-Val-Tyr-lle-His-Pro-Phe-Hs-Leu-Leu-Val-Tyr-Ser--R
Ангиотензин-l 1 23 4567 8 9 10
Asp-Arg-Val-Tyr-lle-His-Pro-Phe -Н'3-Leu
r I— Дипептидкарбоксилаза (ПФ)
Ангиотензин-ll 1 23 4567 8
Asp-Arg-Val-Tyr-lle-His-Pro-Phe
Рис. 6.28. Активация секреции ренина и образование в крови ангиотензина-II.
Показаны три вида стимулов для секреции ренина юкст; гломерулярными клетками почек:
снижение АД в приносящей артериоле клубочка, повыи ение симпатической активности,
влияния macula densa, вызванные сдвигами уровня натрия. 1од влиянием фермента ренина от
молекулы белка ангиотензиногена отщепляется декапеш ид —ангиотензин-I. Этот пептид
подвергается воздействию превращающегося фермента (1Ф) дипептидкарбоксилазы клеток
эндотелия сосудов легких, почек и др., отщепляющей две ;г инокислоты. Образующийся окта-
пептид является ангиотензином-П.
312 • РЕГУЛИРУЮЩИЕ И УПР ХВЛЯЮЩИЕ СИСТЕМЫ
рующей альдостерон, что и гало основание называть эту систему ренин-ан-
гиотензин-альдостероновой. Хнгиотензин-П, кроме стимуляции продукции
альдостерона, обладает след тощими эффектами:
• вызывает сужение арте риальных сосудов,
• активирует симпатиче< кую нервную систему как на уровне центров,
так и способствуя сип гезу и освобождению норадреналина в синап-
сах,
• повышает сократимост ь миокарда,
• увеличивает реабсорбт ню натрия и ослабляет клубочковую фильтра-
цию в почках,
• способствует формиро !анию чувства жажды и питьевого поведения.
Таким образом, ренин-а тгиотензин-альдостероновая система участвует
в регуляции системного и почечного кровообращения, объема циркули-
рующей крови, водно-солев эго обмена и поведения.
6.8.4. Регуляторные эффекты гормонов сердца
Миоцитами предсердий (преимущественно правого) образуется пептидный
гормон с установленной химической структурой, получивший название
предсердный натрийуретический гормон, или атриопептид. Гормон накап-
ливается в специфических i ранулах саркоплазмы миоцитов и секретирует-
ся в кровь под влиянием ряда регуляторных стимулов: растяжения пред-
сердий объемом крови, урозня натрия в крови, эффектов блуждающего и
симпатических нервов, соде ржания в крови вазопрессина.
Физиологические эффе) гы атриопептида многообразны, так как во
многих органах и тканях обнаружены специфические для него мембран-
ные рецепторы (рис. 6.29). Сосудистые эффекты гормона состоят в рас-
слаблении гладких мышц с >судов и вазодилатации (через цАМФ), сниже-
нии артериального давления. Кроме того, гормон повышает проницае-
Рис. 6.29. Основные эффекты атриопептида — вазодилатирующего и натрийурети-
ческого гормона. Атриопептид является полным антагонистом ренин-ангиотензин-
альдостероновой системы, так как подавляет и образование каждого элемента сис-
темы, и вызываемые ими эфф( кты.
6. Эндокринная нервная система — регулятор функций и процессов в организме • 313
мость гистогематических барьеров и увеличивает транспорт воды из крови
в тканевую жидкость.
Почечные эффекты атриопептида включают: 1) повышение экскреции
натрия (до 90 раз) и хлора (до 50 раз) в связи с подавлением их реабсорб-
ции в канальцах. Гормон — в 1000 раз более эффективный натрийуретик,
чем фуросемид; 2) выраженное диуретическое действие за счет увеличения
клубочковой фильтрации и подавления реабсорбции воды; 3) подавление
секреции ренина, ингибирование эффектов ангиотензина-П и альдостеро-
на, т. е. гормон является полным антагонистом РААС.
Атриопептид, кроме того, расслабляет гладкую мускулатуру кишечника,
уменьшает величину внутриглазного давления, объема и давления ликвора
в желудочках мозга.
Близкий по натрийуретическому эффекту гормон выявляется и в ткани
головного мозга.
Помимо атриопептида в предсердиях образуются атриопептины, повы-
шающие артериальное давление и обладающие антидиуретическим эффек-
том. Атриопептины участвуют в регуляции питьевого поведения, солевого
аппетита и жажды. В малых количествах в сердце образуются соматоста-
тин, ангиотензин-II и релаксин, обладающие хронотропным влиянием на
миокард.
6.8.5. Регуляторная функция гормонов сосудистого эндотелия
Клетки сосудистого эндотелия синтезируют и выделяют через апикальную
и базальную мембраны три группы гормонов: сосудосуживающие (эндоте-
лины, тромбоксаны), сосудорасширяющие (оксид азота, гиперполяризую-
щий фактор, простагландины) и факторы адгезии и агрегации клеточных
элементов.
Эндотелины (ЭТ) являются крупными полипептидами (21 аминокисло-
та), образуются путем частичного гидролиза молекулы предшественника
или «большого эндотелина», состоящей из 38 аминокислот, под влиянием
связанного с мембраной клетки и находящегося в цитоплазматических ве-
зикулах эндотелинпревращающего фермента. Этот фермент локализован в
эндотелии сосудов легких, сердца, почек, плаценты, поджелудолчной же-
лезы, надпочечников, головного мозга и даже в сосудистых гладких мыш-
цах. Наличие двух форм эндотелинпревращающего фермента (мембрано-
связанной и внутриклеточной) определяет и два места образования эндоте-
линов — в цитоплазме и на поверхности клеточной мембраны. Эндотелин-
превращающий фермент ведет не только к образованию молекул эндоте-
лина, но способен вызывать гидролиз и инактивацию на поверхности эн-
дотелиоцитов ряда регуляторных пептидов и гормонов (инсулина, бради-
кинина, нейротензина и др.). Активация эндотелинпревращающего фер-
мента происходит под влиянием цитокинов.
Эффекты эндотелинов обусловлены их взаимодействием со специфиче-
скими мембранными рецепторами двух типов — ЭТ-А и ЭТ-В. Следствием
специфического связывания эндотелина с рецептором является активация
систем вторичных посредников (фосфолипаза С, ИФЗ, диацилглицерол,
цГМФ и цАМФ, фосфолипазы D и А2). Различают прямые и опосредован-
ные сосудистые эффекты эндотелинов. Прямые эффекты заключаются в
действии на гладкие мышцы сосудов. Связывание эндотелина с рецептора-
ми гладких мышц сосудов вызывает их сокращение и вазоконстрикцию
(через образование ИФЗ и повышение внутриклеточного Са2+), а также
стимуляцию митогенеза и пролиферации клеток (Через активацию тиро-
314 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
зинкиназы и фосфорилирование тирозина). Взаимодействие эндотелинов с
рецепторами клеток эндотелия вызывает реализацию опосредованных эф-
фектов, в виде высвобождения из эндотелия вазоактивных факторов, при-
водящих к расширению сосудов (NO, гиперполяризующий фактор, про-
стагландины). Кроме сосудистых эффектов эндотелины изменяют секре-
цию гормонов гипофиза и надпочечников, стимулируют секрецию атрио-
пептида миокардом, угнетают эффекты вазопрессина в почках, способст-
вуя диурезу и натрийурезу, увеличивают реакции сердца на симпатические
стимулы.
Среди сосудорасширяющих гормонов эндотелия основное место по вы-
раженности и распространенности эффекта занимает оксид азота (NO),
постоянно образующийся из L-аргинина под влиянием фермента NO-син-
тетазы. Одним из стимулов, активирующих фермент и образование оксида
азота, является механическое растяжение стенки сосудов. Активация фер-
мента и синтез оксида азота происходят при действии на мембранные ре-
цепторы эндотелиоцитов ацетилхолина, адреномедуллина, гистамина, бра-
дикинина, АТФ, а также в результате повышения в клетке эндотелия кон-
центрации ионизированного Са2+. NO-синтетаза помимо образования ок-
сида азота стимулирует синтез некоторых цитокинов: интерлейкина — 1р,
альфа-интерферона, тогда как другие цитокины: ИЛ-4, ИЛ-8, ИЛ-10, на-
против, подавляют активацию фермента.
Вазодилатирующий эффект NO опосредуется активацией образования в
гладкомышечных клетках цГМФ. Оксид азота также подавляет вазоконст-
рикторное действие ангиотензина-П. Синтезированный клетками эндоте-
лия оксид азота выделяется не только через базальную мембрану в сторону
гладкомышечных клеток кровеносных сосудов, но и через апикальную
мембрану, где тормозит адгезию тромбоцитов и лейкоцитов крови к эндо-
телиальной выстилке стенки сосуда. Антиагрегантное действие оксида азо-
та отчасти опосредовано простациклином, образующимся в эндотелии.
В нервной системе NO является модулятором синаптической передачи,
так как выявлено его поступление в синаптическую щель и показано инги-
бирующее влияние на выделение медиаторных аминокислот.
Эндотелиальный гиперполяризующий фактор также вызывает дилатацию
артериальных и венозных сосудов, но отличается от NO механизмом дей-
ствия — способностью активировать ионные каналы (К+,СГ) и снижать
возбудимость эндотелиальных клеток.
К числу факторов регуляции адгезии и агрегации клеток относятся мно-
гочисленные интегрины и селектины, образуемые клетками эндотелия.
6.8.6. Регуляторная функция гормонов желудочно-кишечного
тракта
Клетки эпителия желудка и двенадцатиперстной кишки обладают способ-
ностью секретировать большое количество пептидных соединений, многие
из которых выявляются также в головном мозге. Это дало основание рас-
сматривать продуцирующие пептиды клетки в качестве единой APUD-сис-
темы организма. Основной функцией гастроинтестинальных гормонов яв-
ляется регуляция процессов пищеварения, а мозговых пептидов — пищево-
го поведения. В связи с этим гастроинтестинальные гормоны, их природа
и регуляторные эффекты представлены в главе 15 «Функции пищевари-
тельной системы».
6. Эндокринная нервная система — регулятор функций и процессов в организме • 315
6.9. Роль эндокринной системы в неспецифических
приспособительных реакциях
6.9.1. Гормональное обеспечение общего адаптационного
синдрома, или стресса
Примером неспецифического участия эндокринной системы в приспосо-
бительных реакциях организма являются изменения ее деятельности при
стрессе. Состояние стресса возникает как следствие действия на организм
любых сильных, в том числе экстремальных и повреждающих, раздражите-
лей. При действии на организм экстремальных факторов неспецифические
реакции стресса прежде всего направлены на стимуляцию энергетического
обеспечения приспособительных процессов. Ведущую роль в этих неспе-
цифических реакциях играют катехоламины и глюкокортикоиды, в значи-
тельных количествах мобилизуемые в кровь (рис. 6.30). Активируя катабо-
лические процессы, эти гормоны ведут к гипергликемии — одной из на-
чальных реакций субстратного энергообеспечения. Как следствие гипер-
гликемии на некоторое время повышается в крови уровень инсулина. Ме-
таболические перестройки при таком гипергормональном профиле связа-
ны, прежде всего, с активацией в печени фосфорилазы и гликогенолизом,
а поступающая в кровь глюкоза под влиянием инсулина интенсивно ути-
лизируется тканями, прежде всего скелетными мышцами, что увеличивает
их работоспособность и по-
вышает теплообразование в
организме. Жиромобили-
зующий эффект глюкокор-
тикоидов и катехоламинов
способствует повышению в
крови второго важнейшего
энергетического субстрата —
свободных жирных кислот.
Однако подобный «форси-
рованный» режим функцио-
Рис. 6.30. Схема участия эндо-
кринной системы в компенса-
торных реакциях при стрессе.
Чрезмерные, часто повреждающие
воздействия среды, раздражая ре-
цепторы, вызывают мощный поток
афферентных импульсов в цен-
тральную нервную систему, что ве-
дет к активации гипоталамических
центров. Быстрым следствием этих
процессов является симпатическая
активация и поступление в кровь из
надпочечников катехол-аминов, что
вызывает срочные адаптивные реак-
ции. Одновременно возрастает ней-
росекреция кортиколиберина, обу-
словливающая повышение активно-
сти гипоталамо-аденогипофизарно-
надпочечниковой оси регуляции,
способствующей реализации ком-
пенсаторных реакций за счет акти-
вации их энергообеспечения.
316 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
нирования эндокринной системы из-за ограниченности функциональных
резервов не может длиться долго, вскоре содержание инсулина в крови
уменьшается, что носит название «функциональный транзиторный диа-
бет». Это необходимое условие для усиления жиромобилизующего эффекта
глюкокортикоидов и активации глюконеогенеза. Важнейшим источником
глюкозы как энергетического материала в этот период становится глюко-
неогенез, но на образование глюкозы расходуется дефицитный пластиче-
ский материал — аминокислоты. Необходимым условием длительно повы-
шенного энергоснабжения является переключение энергетического обмена
с углеводного типа на липидный, тем более что углеводные резервы в виде
гликогена достаточно быстро оказываются практически исчерпанными.
Постепенно снижается продукция глюкокортикоидов, устанавливается но-
вое гормональное соотношение: нерезко повышенный уровень глюкокор-
тикоидов при более значительном снижении уровня инсулина. Этот новый
уровень функционирования эндокринной системы способствует восста-
новлению равновесия между катаболическими и анаболическими процес-
сами, расходы белка на энергетические нужды снижаются. Жиромобили-
зующий эффект гормональной перестройки и образование транспортной
формы эндогенного жира — липопротеинов очень низкой плотности —
приводят к тому, что растет использование клетками липидов как источ-
ников энергии. Жирные кислоты интенсивно окисляются в скелетных
мышцах, миокарде и печени. Образующиеся при этом кетоновые тела уси-
ленно окисляются в мышечной ткани, почках, а также сердце и мозге.
Особенно резко ограничивается потребление углеводов мышечной и жиро-
вой тканью, что экономит глюкозу для углеводзависимых тканей — голов-
ного мозга, кроветворной ткани и эритроцитов, в некоторой степени —
миокарда. Таким образом, происходящие при стрессе гормональные и ме-
таболические перестройки обеспечивают длительное неспецифическое по-
вышение энергообеспечения приспособительных процессов.
Стресс как неспецифическая реакция приспособления и компенсации
нарушенных функций может при резкой выраженности сопровождаться и
активацией гипоталамо-гипофизарно-тиреоидной системы, приводя к рос-
ту в крови уровня тиреоидных гормонов. Физиологический стресс, напри-
мер эмоциональное напряжение, напротив, характеризуется снижением
гормональной активности щитовидной железы. Гиперфункция щитовид-
ной железы в ответ на повреждение тканей является неспецифической ре-
акцией компенсации, поскольку тиреоидные гормоны, благодаря стимуля-
ции синтеза белков, способствуют клеточным регенеративным процессам
и репарации поврежденных структур. Активация гипоталамо-гипофизарно-
тиреоидной системы может носить и черты специфической компенсации,
например в условиях холодового стресса, как компенсация термического
угнетения метаболизма. Стресс при экстремальных состояниях сопровож-
дается активацией и других гормональных систем — гипоталамо-гипофи-
зарно-андрогенной и гипоталамо-нейрогипофизарной. Повышение актив-
ности системы гонадолиберины—гонадотропины—андрогены (одним из
проявлений является повышение либидо) и избыточно секретируемые при
этом андрогены за счет анаболического эффекта способствуют репаратив-
ным процессам.
При хирургическом и эмоциональном стрессе повышается секреция ва-
зопрессина. Компенсаторное значение активации секреции вазопрессина
заключается в облегчении консолидации процессов памяти, формирования
аналгезии, потенцировании эффектов кортиколиберина на секрецию кор-
тикотропина, восстановлении нарушенной при кровопотере гемодинами-
6. Эндокринная нервная система — регулятор функций и процессов в организме • 317
ки. Избыточная секреция вазопрессина при травмах является примером
опережающей перестройки эндокринных механизмов компенсации, реали-
зующей избыточный гормональный сигнал для предотвращения возмож-
ности потери организмом воды при кровотечении. Реакция гипоталамо-
нейрогипофизарной системы может быть и специфической компенсацией
возникших в организме нарушений водно-солевого и осмотического го-
меостазиса (осмотический стресс).
6.9.2. Гормональная регуляция местных компенсаторных реакций
Происходящие при стрессе гормональные перестройки, помимо активации
энергетического обеспечения тканей и, соответственно, функциональной
активации органов, способствуют местным тканевым компенсаторным ре-
акциям — регенерации и репарации, равно как и развертыванию воспали-
тельной реакции. Изменения кровообращения в участках повреждения,
мобилизация в поврежденные ткани пластического материала, регуляция
процессов клеточного деления и пролиферации, утилизации материала по-
врежденных клеток — далеко не полный перечень компенсаторных про-
цессов, обеспечиваемых неспецифическими эффектами гормонов. Даже
используемые как липидный источник энергии липопротеиды низкой и
очень низкой плотности, проникая в клетки с помощью специального ре-
цепторного аппарата, подвергаются в них лизосомному расщеплению с об-
разованием аминокислот, холестерина, фосфолипидов и др., идущих на
пластические цели и способствующих регенерации. Важнейшую роль в ор-
ганизации тканевых компенсаторных реакций (регенерации, воспаления)
играют местные «тканевые гормоны», образование которых во многом за-
висит от эндокринных влияний на клетки. Среди таких «тканевых гормо-
нов» важнейшую роль играют продукты перекисного окисления арахидо-
новой кислоты, биогенные амины и кинины.
Образование арахидоновой кислоты происходит из мембранных фосфо-
липидов при активации фосфолипазы А2, чему способствуют многие пеп-
тидные гормоны через вторичный посредник диацилглицерол, а также
нейромедиаторы, химические и физические повреждающие факторы. Од-
ним из наиболее мощных ингибиторов фосфолипазы является кортизол.
Из арахидоновой кислоты под влиянием специфических ферментов проис-
ходит образование трех основных групп соединений: лейкотриенов (при
действии липоксигеназы), эпоксидов (при действии эпоксигеназы) и про-
стагландинов (под влиянием циклооксигеназы). В дальнейшем, в зависи-
мости от наличия в тканях определенного типа ферментов, происходит об-
разование простагландинов (PGE, PGF, тромбоксана, простациклина и др.)
и лейкотриенов (LTB, LTC, LTD, LTE). В механизме действия простаглан-
динов на клетки важное место занимает изменение активности аденилат-
циклазы и уровня цАМФ. Простагландины участвуют в регуляции многих
физиологических процессов — тонуса артериальных сосудов, сокращения
гладких мышц матки, овуляции, агрегации тромбоцитов и свертывания
крови, моторики кишечника и секреции соляной кислоты в желудке, тер-
морегуляции, проницаемости сосудов, модуляции функций лейкоцитов,
тонуса гладких мышц бронхов и др. Простагландины способны индуциро-
вать и усиливать воспалительную реакцию тканей. Благодаря реализации
эффектов через цАМФ и Са2+, простагландины способны менять секрецию
гормонов, а в органах-мишенях модулировать их эффекты. Так, увеличи-
вая уровень цАМФ после связи с собственным мембранным рецептором,
простагландины имитируют и модулируют эффекты тиротропина, корти-
318 • РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
котропина, лютропина, паратирина, а снижая уровень цАМФ в других тка-
нях — ослабляют гормональные эффекты, например липолитическое дей-
ствие гормонов на жировую ткань, антидиуретическое действие вазопрес-
сина в почечных канальцах. Очевидно, что эти эффекты простагландинов
могут обусловливать как приспособительные реакции к действию неблаго-
приятных факторов среды, так и компенсаторные неспецифические реак-
ции в условиях патологии. Приток в мышцы жирных кислот повышает
синтез простагландинов, что благодаря активации аденилатциклазы повы-
шает содержание цАМФ в клетках. Результатом является стимуляция
РНК-полимеразной активности и повышение синтеза РНК в ядрах мы-
шечных клеток, а соответственно и синтеза белков, что обеспечивает фи-
зиологическую регенерацию сократительного аппарата клеток и их гипер-
трофию. Простагландины ускоряют включение аминокислот в белки, ос-
нований в РНК и ДНК, активируют синтез коллагена и стимулируют за-
живление ран.
В клетках, например, эндотелия артериол при оксидативном превраще-
нии арахидоновой кислоты с участием цитохрома Р-450 и эпоксигеназы
происходит образование эпоксида, получившего название эндотелиального
гиперполяризующего фактора, способствующего активации калиевых и
хлорных каналов гладкомышечных клеток и падению их возбудимости.
Нередко повреждение тканей (прежде всего иммунной природы) сопро-
вождается образованием лейкотриенов. При этом лейкотриены не только
способствуют развертыванию патологического процесса, но и ведут к реа-
лизации клеточных, тканевых и внутриорганных компенсаторных реакций.
Так, например, снижение коронарного кровотока под влиянием лейко-
триенов сочетается с отрицательным инотропным эффектом, причем
уменьшение сократимости миокарда выражено сильнее, чем ограничение
коронарного кровотока, что не позволяет развиться кислородному голода-
нию миокарда (кислородэкономящая реакция). Лейкотриены являются
мощными стимуляторами лейкоцитов, усиливающими их хемотаксис, ат-
тракцию, дегрануляцию и высвобождение лизосомальных ферментов, т. е.
способствуют реализации защитной функции лейкоцитов в очаге воспале-
ния. Выделяемые клетками и образуемые в очагах повреждения биогенные
амины (гистамин, серотонин) и кинины ведут к сосудистым, гемореологи-
ческим и метаболическим эффектам, играющим неспецифическую роль в
компенсации структурных и функциональных нарушений.
Ill ФУНКЦИИ СИСТЕМ
ЖИЗНЕОБЕСПЕЧЕНИЯ
ОРГАНИЗМА
320 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
В основе всех проявлений жизнедеятельности организма лежат процес-
сы обмена веществ и энергии в клетках органов и тканей. Интенсивность
этих процессов поддерживается на определенном уровне, необходимом для
осуществления многообразных функций организма в различных условиях
его существования. Для поддержания жизнедеятельности клеток на нор-
мальном уровне организм обеспечивает относительное постоянство (гомео-
стазис) внутренней среды (ее состав, осмотическое давление, кислотно-ос-
новное равновесие, температуру).
Способность поддерживать постоянство количественного и качествен-
ного состава внутренней среды организма является необходимым условием
его нормальной жизнедеятельности. Гомеостазис организма нарушается
под действием факторов внешней и внутренней среды и быстро восстанав-
ливается благодаря функционированию и надежному регулированию фи-
зиологических систем. Под физиологической системой понимают генети-
чески детерминированную совокупность клеток, тканей и органов, кото-
рые выполняют общие функции в организме (например, системы кровооб-
ращения, дыхания, пищеварения, выделения и т. д.). В результате физио-
логические системы позволяют клеткам организма существовать и выпол-
нять свои функции в относительно постоянных (стабильных) условиях.
При изменениях параметров внешней среды, например при перепадах тем-
пературы, влажности, атмосферного давления, освещения, при недостаточ-
ном поступлении питательных веществ и т. д., поддержание гомеостазиса
осуществляется за счет включения механизмов регуляции соответствующих
физиологических систем.
Обеспечение метаболических потребностей клеток осуществляется бла-
годаря функции кровообращения. Движущаяся по сосудам кровь доставляет
тканям поступившие из желудочно-кишечного тракта органические и не-
органические вещества, а из легких — кислород, без которого не могут про-
ходить окислительные процессы в клетках. Одновременно кровью к орга-
нам выделения переносятся продукты обмена, подлежащие удалению из
внутренней среды организма. С кровью к тканям доставляются также гу-
моральные регуляторы функции клеток — гормоны, олигопептиды, медиа-
торы, биологически активные промежуточные продукты обмена.
Для обеспечения защиты внутренней среды организма от проникнове-
ния чужеродных веществ и болезнетворных агентов (микробов и их токси-
нов) система крови находится в тесном функциональном взаимодействии с
иммунной системой, продуцирующей клеточные и гуморальные факторы
иммунитета.
Указанные физиологические процессы должны протекать при постоян-
стве температуры организма, постоянстве pH крови и отделов желудочно-
кишечного тракта, так как их отклонение вызовет нарушение гомеостазиса
и, следовательно, необходимость корригирующего управления со стороны
одной или нескольких физиологических систем.
Огромную роль в обеспечении жизнедеятельности организма играют
сенсорные системы, поскольку зрение, слух, все виды чувствительности
организма, обоняние, вкус являются непременными контролирующими
деятельность организма или способствующими его адаптации к факторам
окружающей среды системами.
Что касается репродуктивной функции человека, то она определяет и
продолжение рода, и получение удовольствия от физического общения с
партнером.
Взаимодействие компонентов сложных процессов жизнеобеспечения
физиологических систем крови, дыхания, пищеварения, выделения, вод-
• 321
но-солевого обмена, кислотно-основного равновесия, теплопродукции и
теплоотдачи, иммунной, сенсорной систем, в конечном счете обеспечивает
метаболические потребности организма.
Из дидактических соображений функции каждой анатомо-физиологи-
ческой системы и механизма ее регуляции рассматриваются отдельно. За-
кономерности интеграции деятельности различных систем на организмен-
ном уровне изложены в IV разделе учебника.
ГЛАВА 7
Функции клеток крови. Гемостаз.
Регуляция кроветворения. Основы
трансфузиологии
Цельная кровь состоит из жидкой части (плазмы) (см. главу 1) и форменных
элементов, к которым относят эритроциты, лейкоциты и кровяные пла-
стинки — тромбоциты.
Функции крови: 1) транспортная — перенос газов (О2 и СОД, пластиче-
ских (аминокислот, нуклеозидов, витаминов, минеральных веществ), энер-
гетических (глюкоза, жиры) ресурсов к тканям, а конечных продуктов об-
мена — к органам выделения (желудочно-кишечный тракт, легкие, почки,
потовые железы, кожа); 2) гомеостатическая — поддержание температуры
тела, кислотно-основного состояния организма, водно-солевого обмена,
тканевого гомеостаза и регенерации тканей; 3) защитная — обеспечение
иммунных реакций, кровяного и тканевого барьеров против инфекции;
4) регуляторная — гуморальной и гормональной регуляции функций раз-
личных систем и тканей; 5) секреторная — образование клетками крови
биологически активных веществ.
7.1. Функции эритроцитов
7.1.1. Функции и свойства эритроцитов
Эритроциты переносят О2 содержащимся в них гемоглобином от легких к
тканям и СО2 от тканей к альвеолам легких. Функции эритроцитов обу-
словлены высоким содержанием гемоглобина (95 % массы эритроцита),
деформируемостью цитоскелета, благодаря чему эритроциты легко прони-
кают через капилляры с диаметром меньше 3 мкм, хотя имеют диаметр от
7 до 8 мкм. Глюкоза является основным источником энергии в эритроци-
те. Восстановление формы деформированного в капилляре эритроцита, ак-
тивный мембранный транспорт катионов через мембрану эритроцита, син-
тез глютатиона обеспечиваются за счет энергии анаэробного гликолиза в
цикле Эмбдена—Мейергофа. В ходе метаболизма глюкозы, протекающего
в эритроците по побочному пути гликолиза, контролируемого ферментом
дифосфоглицератмутазой, в эритроците образуется 2,3-дифосфоглицерат
(2,3-ДФГ). Основное значение 2,3-ДФГ заключается в уменьшении срод-
ства гемоглобина к кислороду.
В цикле Эмбдена—Мейергофа расходуется 90 % потребляемой эритро-
цитами глюкозы. Торможение гликолиза, возникающее, например, при
старении эритроцита и уменьшающее в эритроците концентрацию АТФ,
приводит к накоплению в ней ионов натрия и воды, ионов кальция, по-
вреждению мембраны, что понижает механическую и осмотическую устой-
чивость эритроцита, и стареющий эритроцит разрушается. Энергия глюко-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 323
зы в эритроците используется
также в реакциях восстановления,
защищающих компоненты эрит-
роцита от окислительной денату-
рации, которая нарушает их
функцию. Благодаря реакциям
восстановления атомы железа ге-
моглобина поддерживаются в вос-
становленной, т. е. двухвалент-
ной форме, что препятствует пре-
вращению гемоглобина в метге-
моглобин, в котором железо
окислено до трехвалентного,
вследствие чего метгемоглобин
неспособен к транспорту кисло-
рода. Восстановление окисленно-
го железа метгемоглобина до
двухвалентного обеспечивается
ферментом — метгемоглобинре-
дуктазой. В восстановленном со-
стоянии поддерживаются и серу-
содержащие группы, входящие в
мембрану эритроцита, гемогло-
бин, ферменты, что сохраняет
функциональные свойства этих
структур.
Эритроциты имеют дисковид-
ную, двояковогнутую форму, их
поверхность — около 145 мкм2, а
объем достигает 85—90 мкм3. Та-
кое соотношение площади к объ-
ему способствует деформабильно-
сти (под последней понимают
способность эритроцитов к обра-
тимым изменениям размеров и
формы) эритроцитов при их про-
хождении через капилляры. Фор-
ма и деформабильность эритро-
цитов поддерживаются липидами
мембран — фосфолипидами (гли-
церофосфолипидами, сфинголи-
пидами, фосфотидилэтанолами-
ном, фосфатидилсирином и др.), гликолипидами и холестерином, а также
белками их цитоскелета. В состав цитоскелета мембраны эритроцита вхо-
дят белки — спектрин (основной белок цитоскелета), анкирин, актин, бел-
ки полосы 4.1, 4.2, 4.9, тропомиозин, тропомодулин, аддуцин. Основой
мембраны эритроцита является липидный бислой, пронизанный инте-
гральными белками цитоскелета — гликопротеинами и белком полосы 3.
Последние связаны с частью белковой сети цитоскелета — комплексом
Рис. 7.1. Схема модели изменений цито-
скелета мембраны эритроцита во время
его обратимой деформации.
Обратимая деформация эритроцита изменяет
лишь пространственную конфигурацию (сте-
реометрию) эритроцита, следующую за изме-
нением пространственного расположения мо-
лекул цитоскелета. При этих изменениях фор-
мы эритроцита площадь поверхности эритро-
цита остается неизменной.
а — положение молекул цитоскелета мембраны
эритроцита при отсутствии его деформации.
Молекулы спектрина находятся в свернутом
состоянии;
б — деформация эритроцита, вызванная изме-
нением положения молекул его цитоскелета.
Молекулы спектрина раскручены и растянуты,
но площадь мембраны эритроцита остается
прежней (пояснения в тексте).
Обозначения:
~ Молекулы спектрина, обозначенные
двойными извитыми лентами.
0 — Спектрин-спектриновые взаимодействия
+ — Участки взаимодействия белка полосы
4.1, актина и спектрина.
\\\ — Внутренняя поверхность мембраны
эритроцита.
спектрин—актин—белок полосы 4.1, локализованным на цитоплазматиче-
ской поверхности липидного бислоя мембраны эритроцита (рис. 7.1).
Взаимодействие белкового цитоскелета с липидным бислоем мембраны
обеспечивает стабильность структуры эритроцита, поведение эритроцита
и*
324 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
как упругого твердого тела при его деформации. Нековалентные межмоле-
кулярные взаимодействия белков цитоскелета легко обеспечивают измене-
ние размеров и формы эритроцитов (их деформацию) при прохождении
этих клеток через микроциркуляторное русло, при выходе ретикулоцитов
из костного мозга в кровь — благодаря изменению расположения молекул
спектрина на внутренней поверхности липидного бислоя. Генетические
аномалии белков цитоскелета у человека сопровождаются появлением де-
фектов мембраны эритроцитов. В результате последние приобретают изме-
ненную форму (так называемые сфероциты, элиптоциты и др.) и имеют
повышенную склонность к гемолизу. Увеличение соотношения холесте-
рин—фосфолипиды в мембране увеличивает ее вязкость, уменьшает теку-
честь и эластичность мембраны эритроцита. В результате снижается де-
формируемость эритроцита. Усиление окисления ненасыщенных жирных
кислот фосфолипидов мембраны перекисью водорода или супероксидны-
ми радикалами вызывает гемолиз эритроцитов (разрушение эритроцитов с
выходом гемоглобина в окружающую среду), повреждение молекулы гемо-
глобина эритроцита. Постоянно образующийся в эритроците глютатион, а
также антиоксиданты (а-токоферол), ферменты — глутатионредуктаза, су-
перо ксиддисмутаза и др. защищают компоненты эритроцита от этого по-
вреждения.
До 52 % массы мембраны эритроцитов составляют белки гликопротеи-
ны, которые с олигосахаридами образуют антигены групп крови. Глико-
протеины мембраны содержат сиаловую кислоту, которая придает отрица-
тельный заряд эритроцитам, отталкивающий их друг от друга.
Энзимы мембраны — №+/К+-зависимая АТФаза обеспечивает актив-
ный транспорт Na+ из эритроцита и К+ в его цитоплазму. Са2+-зависимая
АТФаза выводит Са2+ из эритроцита. Фермент эритроцита карбоангидраза
катализирует реакцию: Са2+ Н2О <-> Н2СО3 <-> Н+ + НСО7, поэтому эритро-
цит транспортирует часть углекислого газа от тканей к легким в виде би-
карбоната, до 30 % СО2 переносится гемоглобином эритроцитов в форме
карбаминового соединения с радикалом NH2 глобина.
7.1.2. Гемоглобин
Гемоглобин — это гемопротеин, с молекулярной массой около 60 тыс., ок-
рашивающий эритроцит в красный цвет после связывания молекулы О2 с
ионом железа (Fe++). У мужчин в 1 л крови содержится 157 (140—175) г ге-
моглобина, у женщин— 138 (123—153) г. Молекула гемоглобина состоит
из четырех субъединиц гема, связанных с белковой частью молекулы —
глобином, сформированной из полипептидных цепей. Синтез гема проте-
кает в митохондриях эритробластов. Синтез цепей глобина осуществляется
на полирибосомах и контролируется генами 11-й и 16-й хромосом. Схема
синтеза гемоглобина у человека представлена на рис. 7.2. Гемоглобин, со-
держащий две а- и две p-цепи, называется A-тип (от adult — взрослый). 1 г
гемоглобина A-типа связывает 1,34 мл О2. В первые три месяца жизни
плода человека в крови содержатся эмбриональные гемоглобины типа
Gower I (4 эпсилон цепи) и Gower II (2а и 28 цепи). Затем формируется
гемоглобин F (от faetus — плод). Его глобин представлен двумя цепями а и
двумя р. Гемоглобин F обладает на 20—30 % большим сродством к О2, чем
гемоглобин А, что способствует лучшему снабжению плода кислородом.
При рождении ребенка до 50—80 % гемоглобина у него представлены ге-
моглобином F и 15—40 % — типом А, а к 3 годам уровень гемоглобина F
снижается до 2 %.
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 325
' Глицин + янтарная кислота - коэнзим А
-*— АЛК-синтетаза
У
а-Амино-р-кето-адипиновая кислота
s
о.
Ct
X
о
X
о
8-Аминолевулиновая кислота
- *— АЛК-дегидрогеназа
У
Порфобилиноген
- *— ПБГ-дезаминаза
У
Уропорфириноген III
- *— Декарбоксилаза
У
Копропорфириноген III
<— КПГ-оксидаза
У
Протопорфириноген IX
- *— Оксидаза
У
Протопорфирин IX
к- Гемсинтетаза
иРНК <- Хромосома 16
(ген а-цепи)
иРНК <- Хромосома 11
(ген р, у, 8-цепей)
Полирибосомы
Полирибосомы
Fe+ -> ГЕМ
2 а-цепи
2 р-цепи
Глобин (2 а-цепи и 2 Р-цепи) -*
t
Гемоглобин А
Рис. 7.2. Схема синтеза гемоглобина у человека в эритроидной клетке.
Указаны ферменты, контролирующие этапы синтеза молекулы гема в митохондриях, а также
хромосомы, ответственные за синтез цепей глобина А (р-цепи) и гемоглобинов плода (у- и 5-
цепи). Пояснения в тексте.
Соединение гемоглобина с молекулой О2 называется оксигемоглобином.
Сродство гемоглобина к кислороду и диссоциация оксигемоглобина (отсо-
единения молекул кислорода от оксигемоглобина) зависят от напряжения
кислорода (РО2), углекислого газа (РСО2) в крови, pH крови, ее температу-
ры и концентрации 2,3-ДФГ в эритроцитах. Так, сродство повышают уве-
личение РО2 или снижение РСО2 в крови, нарушение образования 2,3-
ДФГ в эритроцитах. Напротив, повышение концентрации 2,3-ДФГ, сни-
жение РО2 крови, сдвиг pH в кислую сторону, повышение РСО2 и темпе-
ратуры крови — уменьшают сродство гемоглобина к кислороду, тем самым
облегчая ее отдачу тканям. 2,3-ДФГ связывается с p-цепями гемоглобина,
облегчая отсоединение О2 от молекулы гемоглобина. Увеличение концен-
трации 2,3-ДФГ наблюдается у людей, тренированных к длительной физи-
ческой работе, адаптированных к длительному пребыванию в горах. Окси-
гемоглобин, отдавший кислород, называется восстановленным, или дезок-
сигемоглобином. В состоянии физиологического покоя у человека гемогло-
бин в артериальной крови на 97 % насыщен кислородом, в венозной — на
70 %. Чем выраженней потребление кислорода тканями, тем ниже насы-
326 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
щение венозной крови кислородом. Например, при интенсивной физиче-
ской работе потребление кислорода мышечной тканью увеличивается в не-
сколько десятков раз и насыщение кислородом оттекающей от мышц ве-
нозной крови снижается до 15 %. Содержание гемоглобина в отдельном
эритроците составляет 27,5—33,2 пикограмма. Снижение этой величины
свидетельствует о гипохромном (т. е. пониженном), увеличение — о гипер-
хромном (т. е. повышенном) содержании гемоглобина в эритроцитах. Этот
показатель имеет диагностическое значение. Например, гиперхромия эрит-
роцитов характерна для В,2-дефицитной анемии, гипохромия — для желе-
зодефицитной анемии.
7.1.3. Старение и разрушение эритроцитов в организме
Эритроциты у человека функционируют в крови максимум 120 дней, в
среднем 60—90 дней. Старение эритроцитов связано с уменьшением обра-
зования в эритроците количества АТФ в ходе метаболизма глюкозы в этой
клетке крови. Уменьшенное образование АТФ, ее дефицит нарушает в
эритроците процессы, обеспечиваемые ее энергией, — восстановление
формы эритроцитов, транспорт катионов через его мембрану и защиту
компонентов эритроцитов от окисления, их мембрана теряет сиаловые ки-
слоты. Старение эритроцитов вызывает изменения мембраны эритроцитов:
из дискоцитов они превращаются в эхиноциты, т. е. эритроциты, на по-
верхности мембраны которых образуются многочисленные выступы, вы-
росты (рис. 7.3).
Причиной формирования эхиноцитов помимо уменьшения воспроиз-
водства молекул АТФ в эритроците при старении клетки является усилен-
ное образование лизолецитина в плазме крови, повышенное содержание в
ней жирных кислот. Под влиянием перечисленных факторов изменяется
соотношение поверхности внешнего и внутреннего слоев мембраны эрит-
роцита за счет увеличения поверхности внешнего слоя, что и приводит к
появлению выростов на мембране.
По степени выраженности изменений мембраны и формы эритроцитов
различают эхиноциты I, II, III классов и сфероэхиноциты I и II классов.
При старении эритроцит последовательно проходит этапы превращения в
эхиноцит III класса, теряет способность изменять и восстанавливать дис-
ковидную форму, превращается в сфероэхиноцит и разрушается. Устране-
ние дефицита глюкозы в эритроците легко возвращает эхиноциты I—II
классов к форме дискоцита. Эхиноциты начинают появляться, например,
в консервированной крови, сохраняемой в течение нескольких недель при
4 °C, или в течение 24 ч, но при температуре 37 °C. Это связано с уменьше-
нием образования АТФ внутри клетки, с появлением в плазме крови лизо-
лецитина, образующегося под влиянием лецитин-холестерол-ацетилтранс-
ферразы, ускоряющих старение клетки. Отмывание эхиноцитов в свежей
плазме от содержащегося в ней лизолецитина или активация в них глико-
лиза, восстанавливающей уровень АТФ в клетке, уже через несколько ми-
нут возвращает им форму дискоцитов.
Стареющие эритроциты становятся менее эластичными, вследствие чего
разрушаются внутри сосудов (внутрисосудистый гемолиз) или же становят-
ся добычей захватывающих и разрушающих их макрофагов в селезенке,
купферовских клетках печени и в костном мозге (внесосудистый или внут-
риклеточный гемолиз). Внутриклеточным гемолизом в сутки разрушается
80—90 % старых эритроцитов, содержащих 6—7 г гемоглобина, из которых
освобождается в макрофагах до 30 мг железа. После отщепления от гемо-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 327
Рис. 7.3. Схема формирования эхиноцитов и стоматоцитов из дискоцита (нормоци-
та) при разных значениях pH, создаваемых in vitro.
I — сферостоматоцит, II — стоматоцит, III — дискоцит, IV — эхиноцит, V — сфероэхиноцит.
глобина гем превращается в желчный пигмент билирубин, который посту-
пает с желчью в кишечник и под влиянием микрофлоры кишечника по-
следовательно превращается в уробилиноген, а затем в стеркобилиноген.
Оба соединения выводятся из организма с калом и мочой, под влиянием
света и воздуха превращаясь в стеркобилин и уробилин. При метаболизме
1 г гемоглобина образуется 33 мг билирубина.
Внутрисосудистым гемолизом разрушается 10—20 % эритроцитов. При
этом гемоглобин поступает в плазму, образует с плазменным гликопротеи-
ном гаптоглобином комплекс гемоглобин—гаптоглобин. В течение 10 мин
50 % комплекса поглощается из плазмы паренхиматозными клетками пе-
чени, что предупреждает поступление свободного гемоглобина в почки и
тромбирование им их нефронов. У здорового человека в плазме содержит-
ся около 1 г/л плазмы гаптоглобина, что оставляет несвязанным с ним в
плазме крови не более 3—10 мг гемоглобина. Молекулы гема, высвобож-
дающиеся из связи с глобином при внутрисосудистом гемолизе, связыва-
ются белком плазмы — гемопексином, транспортируются им в печень и
также поглощаются паренхиматозными клетками печени, где подвергаются
ферментному разрушению до билирубина.
7.1.4. Роль ионов железа в эритропоэзе
В организме содержится 4—5 г железа в виде резервного ('/<) и функциональ-
но-активного железа (3/4). 62—70 % железа находится в гемоглобине эрит-
роцитов, 5—10 % — в миоглобине, остальное — в тканях, где оно участвует
во многих метаболических процессах: в составе металсодержащих энзи-
мов—цитохромов обеспечивают митохондриальный транспорт электронов,
синтез ДНК и деление клеток, метаболизм гормонов мозгового вещества
надпочечников, детоксикационные механизмы, снижающие активность
токсичных веществ под влиянием цитохрома Р450, в состав которого входит
железо; поддерживают активность глицерол-3-фосфатдегидрогеназы в по-
перечно-полосатых мышцах, что сохраняет физическую работоспособность
328 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
человека. Например, у людей, страдающих дефицитом железа, физическая
работоспособность резко снижается, но восстанавливается после приема
препаратов железа.
Резервное железо постоянно переходит в функциональное и обратно.
Так, за счет повторного использования костный мозг получает ежесуточно
20—25 мг железа. Ионы железа (Fe++) ежедневно всасываются в кишечни-
ке из пищи. Fe2+ в кишечном содержимом вначале связывается с белком-
рецептором на поверхности эпителия слизистой оболочки кишки (р3-ин-
тегрином), что необходимо для перехода его через мембрану клетки слизи-
стой в цитозоль. Далее, образовав комплекс с цитозольным транспортным
белком- мобилферрином, Fe2+ пересекает клетку эпителия слизистой ки-
шечника оставаясь в этом комплексе вплоть до поступления Fe2+ в капил-
лярную сеть кишечника. С выходом в кровоток двухвалентное железо
окисляется до трехвалентного белком плазмы крови церрулоплазмином и
присоединяется к трансферрину, гликопротеину плазмы крови. Трансфер-
рином Fe3+ доставляется к тканям и используется в митохондриях эритроб-
ластов для синтеза гема, депонируется в макрофагах в виде резерва. Плаз-
ма содержит от 1,8 до 2,6 мг/л трансферрина, 1 мг которого связывает
1,25 мкг Fe. В общем объеме плазмы содержится около 3 мг Fe2+. В норме
лишь 73 трансферрина плазмы насыщена железом. Количество железа, ко-
торое может быть связано трансферрином, называется общей железосвязы-
вающей способностью крови и в норме составляет 250—400 мкг % (45—
72 мкмоль/л). Концентрация железа в сыворотке крови у взрослых людей
обоего пола колеблется от 50 до 160 мкг % (9—29 мкмоль/л). С мочой вы-
водится за сутки 60—100 мкг железа.
Комплекс трансферрин—железо фиксируется на рецепторах мембран
эритробластов. Количество этих рецепторов уменьшается за счет торможе-
ния их синтеза в ходе созревания эритроидных клеток и исчезает после со-
зревания ретикулоцитов, поэтому в зрелые эритроциты железо не включа-
ется. Освобождение железа из комплекса трансферрин — железо обеспечи-
вается энергией молекул АТФ, образуемых в эритробластах. Молекула
трансферрина, отдавшая железо, смещается с мембранного участка моле-
кулами трансферрина, связанными с железом, поскольку их сродство к ре-
цепторам более сильное. Железо, поступившее в эритробласт, использует-
ся в митохондриях для синтеза гема и депонируется в эритробласте в виде
резерва. В макрофагах печени и костного мозга резервное железо депони-
руется в молекуле ферритина. Внутри лизосом молекулы ферритина обра-
зуют большие аморфные нерастворимые агрегаты — гемосидерин. Таким
образом, ферритин и гемосидерин — это формы резервного железа в клет-
ках. Из клеточного резерва железо освобождается в двухвалентном состоя-
нии (благодаря энзиму ксантиноксидазе, аскорбиновой кислоте и др.), за-
тем церулоплазмин окисляет Fe2+ до трехвалентного состояния, Fe3+ соеди-
няется с трансферрином и транспортируется с плазмой крови к эритробла-
стам.
Абсорбции железа эпителиальными клетками желудочно-кишечного
тракта способствуют сниженное насыщение трансферрина железом и по-
вышенная эритропоэтическая активность крови. Абсорбция снижается при
увеличении концентрации железа в клетках слизистой оболочки кишечни-
ка. В кишечнике более эффективна абсорбция Fe2+, чем Fe3+. Поэтому ас-
корбиновая кислота, фруктоза, аминокислоты (цистеин, метионин) под-
держивают двухвалентную форму железа и ускоряют его абсорбцию. В ки-
шечнике лучше абсорбируется биодоступное железо, входящее в состав ге-
ма (мясные продукты, кровяная колбаса), чем железо из пищи раститель-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 329
ного происхождения. Абсорбция Fe2+ в желудочно-кишечном тракте зави-
сит от возраста человека, функционального сотояния его организма. Она
наиболее высокая у детей первых месяцев жизни и достигает 57 %, у 7—
10-летних — 7,75—17,75 %, у взрослых мужчин и женщин — 1,1—11,2 % и
у беременных женщин — 20 % от поступившего в желудочно-кишечный
тракт железа. Отсюда ежедневная потребность в железе, абсорбируемом из
пищи в организме человека, составляет: у детей от 0 до 4 мес жизни — 0,5
мг/день, от 1 года до 12 лет — 1,0 мг/день, женщины детородного перио-
да — 2,8 мг/день (во время менструации потери железа составляют от 5 до
45 мг), молодые мужчины — 1 мг/день, старики — 0,9 мг/день, беременные
женщины — 3—3,5 мг/день. В организм последних за время беременности
должно поступать до 1 г железа (500 мг железа используется для синтеза
дополнительного количества гемоглобина в организме беременной, 300 мг
Fe2+ необходимо формирующемуся плоду и 200 мг Fe2+ компенсируют ес-
тественные потери металла организмом беременной женщины). Дефицит
железа в организме человека приводит к развитию гипохромной анемии.
7.1.5. Эритропоэз
Эритропоэз — процесс образования эритроцитов в костном мозге. Первой
клеткой эритроидного ряда, образующейся из колониеобразующей клетки
эритроцитарной (КОК-Э) — клетки-предшественницы эритроидного ряда,
является проэритробласт, из которого в ходе 4—5 последующих удвоений и
созревания образуется 16—32 зрелых эритроцита. Схема процесса: 1 про-
эритробласт (удвоение) два базофильных эритробласта I порядка 4
базофильных эритробласта II порядка 8 полихроматофильных эритро-
бластов I порядка 16 полихроматофильных эритробластов II поряд-
ка -> 32 полихроматофильных нормобласта 32 оксифильных нормобла-
Рис. 7.4. Эритробластический островок костного мозга человека.
1 — эритробласт, 2 — цитоплазма макрофага.
330 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ста -> денуклеация нормобластов -> 32 ретикулоцита -> 32 эритроцита.
Эритропоэз до формирования ретикулоцита занимает 5 дней.
Эритропоэз у человека и животных (от проэритробласта до ретикулоци-
та) протекает в эритробластических островках костного мозга, которых в
норме содержится до 137 на 1 мг ткани костного мозга (рис. 7.4). Макро-
фаги эритроцитарных островков играют основную роль в физиологии
эритроидных клеток, влияя на их пролиферацию и созревание. Макрофаги
фагоцитируют вытолкнутые из нормобластов ядра, обеспечивают эритро-
бласты ферритином и пластическими веществами, секретируют эритропо-
этин и гликозаминогликаны, последние повышают концентрацию ростко-
вых факторов в островках. Эти благоприятные условия для развития эрит-
робластов макрофаги создают благодаря наличию рецепторов к эритроид-
ным клеткам-предшественницам.
Из костного мозга ретикулоциты выходят в кровь и в течение суток со-
зревают в эритроциты. По количеству ретикулоцитов в крови судят об
эритроцитарной продукции костного мозга и интенсивности эритропоэза.
У человека их количество составляет 5—10%о. За сутки в 1 мкл крови по-
ступает 60—80 тыс. эритроцитов. В 1 мкл крови у мужчин содержится 5,21
(4,52—5,9) млн, а у женщин — 4,6 (4,1—5,1) млн эритроцитов. Уменьшение
количества эритроцитов в единице объема крови называется анемией, уве-
личение — эритроцитозом. Последний может носить физиологический,
приспособленный для организма человека характер (например, при подъе-
ме человека в горы, на высоту более 3000 м над уровнем моря).
7.1.6. Регуляция эритропоэза
Эритропоэтин, секретируемый тубулярными и перитубулярными клетками
почек, является гуморальным регулятором эритропоэза. Небольшое коли-
чество гормона (10—15 %) вырабатывается в макрофагах костного мозга,
купферовских клетках и гепатоцитах. В почках синтез и секреция эритро-
поэтина определяются уровнем обеспечения кислородом их тканей.
К уровню кислорода в почечной ткани чувствителен гемсодержащий бе-
лок- цитохром b , входящий составной частью в НАДФ-зависимую окси-
дазу пери- и тубулярных клеток. При нормальном уровне рО2 в ткани поч-
ки радикалы кислорода, продукцируемые оксидазой, прежде всего пере-
кись водорода, препятствуют формированию в почечной ткани «индуци-
руемого гипоксией фактора-1 (ИГФ-1)», стимулирующего транскрипцию
эритропоэтиновой иРНК и синтез эритропоэтина. При снижении кисло-
родного обеспечения ткани почек (рО2 до 20—40 мм рт. ст.) продукция ок-
сидазой перекиси водорода уменьшается. Нарастает активация ИГФ-1 в
цитозоле и его перемещение в ядро клетки, где ИГФ-1 специфически свя-
зывается с ДНК, вызывая экспрессию гена эритропоэтина. Гипоксия по-
чечных структур активирует также ферменты (фосфолипаза А2), ответст-
венные за синтез простагландинов (Е! и Е2), которые через систему «аде-
нилатциклаза—цАМФ» также усиливают синтез эритропоэтина в пери- и
тубулярных клетках почек. Адреналин и норадреналин через [^-адреноре-
цепторы синтезирующих эритропоэтин клеток почек и систему вторичных
посредников в них — цАМФ и цГМФ — вызывают усиление синтеза и сек-
реции эритропоэтина в кровь. При гипоксии почечной ткани количество
эритропоэтина возрастает в 1000 раз и более при норме 0,01—0,08 ME (ме-
ждународных единиц)/мл плазмы. При гематокрите, равном 40—45, коли-
чество эритропоэтина составляет 5—80 мМЕ/мл, а при гематокрите, рав-
ном 10—20 (его уменьшение может быть вызвано острой кровопотерей, ге-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 331
молизом эритроцитов),— 1—8 МЕ/мл плазмы. Эритропоэтин тормозит
апоптоз, регулирует пролиферацию и дифференциацию КОК-Э, про- и
эритробластов, ускоряет синтез гемоглобина в эритроидных клетках и ре-
тикулоцитах, «запускает» в чувствительных к нему клетках синтез эритро-
поэтиновой иРНК и энзимов, участвующих в формировании гема и глоби-
на, цитоскелета эритроцитов, увеличивает кровоток в эритропоэтической
ткани костного мозга и выход в кровь ретикулоцитов. Наконец, катехола-
мины через [3-адренорецепторы КОК-Э также усиливают пролиферацию
эритроидных клеток-предшественниц. Продукция эритропоэтина почками
угнетается при повышенном образовании в организме человека опухоль-
некротизирующего фактора а, интерлейкинов-1а и 13, интерферонов а, [3
и у, что имеет место, например, у больных с хроническими паразитарной и
бактериальной инфекциями, при ревматоидном артрите, так как перечис-
ленные вещества тормозят синтез эритропоэтина в клетках почек. В ре-
зультате у больных развивается анемия вследствие сниженной продукции
почками эритропоэтина.
Половые гормоны и эритропоэз. Андрогены, а точнее продукты их 5-0-ре-
дукгазного превращения — 5-[3-Н-метаболиты, увеличивают чувствитель-
ность клеток-предшественниц эритроидного ряда к эритропоэтину, что де-
лает эритропоэз более интенсивным. Эстрогены обладают противополож-
ным действием на эритропоэз, снижая его интенсивность. Поэтому после
полового созревания у мужчин устанавливаются более высокие значения
эритроцитов и гемоглобина, чем у женщин.
7.2. Лейкоциты
Лейкоциты формируют мощный кровяной и тканевый барьеры против
микробной, вирусной и паразитарной (гельминтной) инфекции, поддер-
живают тканевый гомеостазис и регенерацию тканей. У взрослого человека
в крови содержится 4—10-109/ л лейкоцитов. Увеличение их количества
называется лейкоцитозом, уменьшение — лейкопенией. Лейкоциты, имею-
щие в цитоплазме зернистость, называются гранулоцитами, а не содержа-
щие зернистости — агранулоцитами. Процентное отношение в крови лей-
коцитов, происшедших из разных линий кроветворения, называется лей-
коцитарной формулой (табл. 7.1).
7.2.1. Функции нейтрофильных гранулоцитов
Функцией зрелых нейтрофильных лейкоцитов является уничтожение про-
никших в организм инфекционных агентов. Осуществляют ее они, тесно
взаимодействуя с макрофагами, Т- и В-лимфоцитами. На важность функ-
Таблица 7.1. Лейкоцитарная формула*
Общее чис- ло лейко- цитов в 1 л крови Показа- тели Гранулоциты Агранулоциты
базофилы эозино- филы нейтрофильные лимфоциты МОНО- ЦИТЫ
палочко- ядерные сегменто- ядерные
4-10-109 в 1 мкл (%) 1-75 0,25-0,75 100-250 1-4 180-400 1-5 3065-5600 55-68 1200-2800 25-39 200-700 1-9
•Количество и процентное содержание лейкоцитов в 1 мкл крови.
332 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ционального вклада нейтрофилов в защиту организма от инфекции указы-
вает, например, тяжесть течения инфекционных заболеваний у больных,
страдающих сниженной продукцией или качественными нарушениями
этих клеток. Нейтрофилы фагоцитируют поврежденные клетки собствен-
ного организма, секретируют бактерицидные вещества и способствуют ре-
генерации тканей, удаляя из них поврежденные клетки, а также секрети-
руя стимулирующие регенерацию тканей вещества (нейтрофилокины).
Зрелый нейтрофил содержит сегментированное на 2—5 долей ядро, уплот-
ненный хроматин, а в цитоплазме — многочисленные мелкие секреторные
гранулы трех типов. Часть гранул, дающих положительную окраску на
фермент миелопероксидазу, представлена лизосомами, которые содержат
лизоцим (повреждает стенку бактерий), низкомолекулярные катионные
белки — дефенсины и кателицидины (нарушают дыхание и рост микроорга-
низмов и грибков; дефенсины повышают проницаемость сосудов микро-
циркуляторного русла), протеазы и кислые гидролазы — позволяют ней-
трофилам легко переваривать фагоцитированные объекты. Гранулы 2-го
типа содержат лактоферрин, обладающий бактериостатическим действием,
транскобаламины I и III — переносчики витамина В12 в крови и лизоцим.
Вещества гранул 3-го типа содержат кислые гликозаминогликаны, участ-
вующие в процессах размножения, роста и регенерации тканей, благодаря
их способности концентрировать факторы роста в тканях костного мозга.
Гранулы 2-го и 3-го типов, содержащиеся в цитоплазме нейтрофилов, по-
стоянно секретируют биологически активные вещества даже вне фагоцито-
за. К ним относятся опухольнекротизирующий фактор а, интерлейкин-1,
интерлейкин-6, интерлейкин-11. Вместе с такими же цитокинами, секре-
тируемыми макрофагами в острой фазе заболевания, вызванного инфекци-
ей, травмой (например, ожоговой), они стимулируют синтез и секрецию в
кровь из печени так называемых белков острой фазы (с-реактивный белок,
сц-антитрипсин, сц-кислый гликопротеин, церулоплазмин), повышающих
устойчивость организма к инфекции, токсичным соединениям, образуе-
мым микроорганизмами (см. главу 1).
Нейтрофилы осуществляют свои функции благодаря способности быст-
ро мигрировать и накапливаться в инфицированном или поврежденном
участках организма, фагоцитировать, т. е. захватывать и разрушать в фаго-
цитарных вакуолях внутри клетки поглощенные бактерии и поврежденные
клетки. Их способность к миграции связана с хорошо развитым аппаратом
движения, выбор направления их движения к воспаленным или инфици-
рованным тканям обусловлен появлением в этих тканях вазоактивных и
хемотаксических факторов. Вазоактивные факторы (дефенсины, гистамин,
секретируемый под влиянием дефенсинов тучными клетками тканей) по-
вышают проницаемость капилляров, что способствует миграции нейтро-
филов в ткань. Хемотаксические факторы взаимодействуют с рецепторами
на поверхности гранулоцитов, образуя лиганд-рецепторный комплекс, оп-
ределяющий движение нейтрофилов к воспаленному участку. Мощным хе-
мотаксическим эффектом обладают N-формилпепетиды бактериального
происхождения и лейкотриены, производные метаболизма арахидоновой
кислоты в мембране клеток. Лейкотриены секретируются активированны-
ми Т-лимфоцитами и макрофагами после воздействия на них бактериаль-
ных веществ. Помимо лейкотриенов эти клетки секретируют другие хемо-
аттрактанты — эндотоксины. Важными хемотакическими факторами явля-
ются продукты активации комплемента — фрагменты его молекул С3в и
С5а. Некоторые из этих факторов, особенно С3в, функционируют как опсо-
нины, т. е. вещества, облегчающие фагоцитоз бактерий (от греческого ор-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 333
выход в кровь
КОК-Г
J б
миелобласты
промиелоциты
КСФг' миелоциты
I
метамиелоциты
палочкоядерные
сегментоядерные
Количество делений — 4
Продолжительность развития клеток — 5—7 дней
Деление клеток отсутствует
Продолжительность развития клеток — 5—7 дней
Продолжительность жизни клеток в крови — 1—16
дней
Рис. 7.5. Развитие нейтрофильных гранулоцитов.
КОК-Г — колониеобразующая единица гранулоцитарная; КСФг — колониестимулирующий
фактор гранулоцитарный. Выделены отделы способных к делению (КОК-Г-миелоциты) и не-
размножающихся клеток (метамиелоциты—сегментоядерные) нейтрофильной линии лейкоци-
тов. КСФг активирует деление и усиливает функциональную активность нейтрофилов.
sonein — делать съедобным). Недостаточность хемотаксической активности
нейтрофилов препятствует накоплению этих лейкоцитов в очаге воспале-
ния, что способствует распространению инфекции в организме. К таким
последствиям приводит, например, нарушение хемотаксиса нейтрофилов у
больных при ожоговой болезни, диабете, при низком содержании белка в
рационе человека.
Бактерицидный эффект нейтрофилов. Через 30—60 с после контакта
мембраны нейтрофила с бактерией в нем возникает «вспышка метаболиз-
ма» с образованием перекиси водорода и суперокисных ионов, которые
поражают поглощенные нейтрофилом бактерии, окисляя галогены (СГ, Г)
мембраны бактерий. Одновременно нейтрофилы секретируют лизоцим,
лактоферрин, катионные белки, кислые и нейтральные гидролазы, пора-
жающие фагоцитированные бактерии.
Нейтрофильный гранулопоэз в костном мозге представлен одновремен-
но пролиферирующими и созревающими клетками — от миелобластов до
миелоцитов включительно, и только созревающими (недедящимися) клет-
ками — от метамиелоцитов до сегментоядерных нейтрофилов (рис. 7.5).
Зрелые сегментоядерные нейтрофилы поступают из костного мозга в кровь
и составляют в ней до 50—70 % всех лейкоцитов.
В небольшом количестве (1—5 %) в кровь поступают и палочкоядерные
нейтрофилы. Их увеличение в крови — важный признак нарастания интен-
сивности нейтрофильного гранулопоэза, а также остроты воспалительного
процесса. Под гранулоцитозом понимают увеличение количества нейтро-
фильных лейкоцитов в крови выше 10* 109/л, под нейтропенией — сниже-
ние их числа ниже 1,5 • 109/л крови.
В костном мозге в 20—25 раз больше зрелых сегментоядерных нейтро-
филов, чем в крови. Это костномозговой резерв нейтрофилов. У взрослого
334 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
человека он составляет 7,7 ± 1,2 • 109/кг массы тела. Гранулоцитарный ко-
лониестимулирующий фактор (КСФ-Г), бактериальный эндотоксин,
N-формилпептиды бактериального происхождения, лейкотриены, катехол-
амины вызывают выброс из костного мозга резервных гранулоцитов в
кровь. В крови часть гранулоцитов циркулирует, а часть — оседает у сосу-
дистой стенки малых вен и капилляров, образуя пристеночный нециркули-
рующий резерв. Выброс адреналина и КСФ-Г в кровь возвращает присте-
ночные гранулоциты в циркулирующую кровь. За адгезию нейтрофилов и
моноцитов к поверхности эндотелиальных клеток ответственны поверхно-
стные рецепторы этих лейкоцитов — интегрины СД18, СДПа, СДПв,
СДПс, а также синтезируемые в эндотелиоцитах селектины СД62 (селек-
тин Р) и СД62Е (селектин Е). Последние представляют собой молекулы
адгезии на поверхности эндотелиальных клеток, с которыми связываются
интегрины нейтрофилов. При нарушении функций этих рецепторов ней-
трофилы оказываются неспособными прикрепляться к эндотелию капил-
ляра, а затем мигрировать из сосуда в инфицированную микроорганизма-
ми ткань. Указанный дефект нейтрофилов способствует развитию тяжелых
гнойных инфекций, что наблюдается, например, при врожденном заболе-
вании — «синдроме недостаточности лейкоцитарной адгезии», связанном
со слабым представительством (или отсутствием) интегрина СД18 на ней-
трофилах. Пристеночный резерв нейтрофилов равен 0,17 ± 0,08* 109 ней-
трофилов/кг массы тела, циркулирующий — 0,22 ± 0,05 • 109 кг/массы.
Средний полупериод жизни циркулирующих гранулоцитов составляет 6—
8 ч, а полный — не более 30 ч.
7.2.2. Функции базофильных гранулоцитов
Функции базофильных гранулоцитов крови и тканей: поддержание крово-
тока в мелких сосудах; трофика тканей и рост новых капилляров; обеспе-
чение миграции других лейкоцитов в ткани; защита кишечника, кожи и
слизистых оболочек при инфицировании гельминтами и клещами; участие
в формировании аллергических реакций. Базофильные гранулоциты спо-
собны к фагоцитозу, миграции из кровеносного русла в ткани и передви-
жению в них.
Цитоплазма зрелых базофилов содержит гранулы неравных размеров, ок-
рашивающиеся в филолетово-розовые тона при окраске по Романовскому-
Гимзе. Базофилы могут синтезировать и накапливать в гранулах биологиче-
ски активные вещества, очищая от них ткани, а затем и секретировать их.
Постоянно присутствуют в клетках: а) кислые гликозаминогликаны — хон-
дроитинсульфат, дерматансульфат, гепарансульфат и гепарин — основной
антикогуляционный фактор; б) гистамин — антагонист гепарина, активатор
внутрисосудистого тромбообразования и стимулятор фагоцитоза, оказывает
про воспалительное действие на ткань. В каждом базофиле содержится: 1—
2 пг гистамина; «фактор, активирующий тромбоциты» (вещество, вызываю-
щее агрегацию тромбоцитов и освобождение их содержимого); «эозино-
фильный хемотаксический фактор анафилаксии» и лейкотриен С4, способ-
ствующие выходу эозинофилов из сосудов в места скопления базофилов.
При повышении чувствительности организма к аллергенам в базофилах об-
разуется «медленно реагирующая субстанция анафилаксии», лейкотриен С4,
вызывающие спазм гладкой мускулатуры мелких бронхов, что наблюдается,
например, при бронхиальной астме. Базофилы способны мигрировать из
крови в ткани, активировать микроциркуляцию инфицированного района
ткани, выход других лейкоцитов через стенку капилляров в зону накопле-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 335
ния паразитарного антигена. Так, в местах локализации гельминтов (в ки-
шечнике) или клещей (в коже) наблюдается инфильтрация тканей базофи-
лами, способствующими уничтожению и изгнанию этих паразитов.
Основные хемотаксические факторы движения базофилов — лимфоки-
ны, секретируемые лимфоцитами в присутствии аллергена, а также кал-
ликреин, фактор комплемента С567. Базофилы и тучные клетки, окружаю-
щие мелкие сосуды печени и легких, интенсивно секретируют гепарин,
препятствующий образованию тромбов, что поддерживает нормальный
кровоток в сосудах этих органов. Активаторами выброса веществ из гранул
базофилов являются иммуноглобулин Е и аллергены — вещества антиген-
ной природы.
Базофильные гранулоциты образуются из КОК базофильной, а тучные
клетки из КОК тучноклеточной. Продукция базофилов активируется ин-
терлейкинами -3 и -4, стимулирующими пролиферацию и дифференциа-
цию КОК базофильной. В лейкоцитарной формуле доля базофилов состав-
ляет 0,25—0,75 %, или около 0,04 • Юул крови.
7.2.3. Функции эозинофильных лейкоцитов
Это защита организма от паразитарной инфекции (шистосомы, трихинел-
лы, гельминты, аскариды и др.); инактивация биологически активных со-
единений, образующихся при аллергических реакциях; препятствуют дли-
тельному действию биологически активных веществ, секретируемых туч-
ными клетками и базофилами; обладают фагоцитарной и бактериоцидной
активностью. Зрелый эозинофил имеет 2- или 3-дольчатое ядро и два типа
гранул в цитоплазме. Большие гранулы содержат специфический основной
белок, токсичный для паразитов; вещества, нейтрализующие гепарин, ме-
диаторы воспаления и секретируют ферменты — p-глюкуронидазу, рибо-
нуклеазу, фосфолипазу Д. Так, фосфолипаза Д инактивирует «фактор, ак-
тивирующий тромбоциты», синтезируемый базофилами, что предупрежда-
ет агрегацию тромбоцитов. Маленькие гранулы содержат кислую фосфата-
зу и арилсульфатазу В, нейтрализующую «медленно реагирующую анафи-
лактическую субстанцию».
Хемотаксический фактор «эозинофильный хемотаксический фактор
анафилаксии» кислый пептид (МВ 500), секретируемый тучными клетками
и базофилами, обусловливает выход эозинофилов в места скопления туч-
ных клеток и базофилов. Хемотаксис эозинофилов усиливают фрагменты
молекул комплемента СЗа, С5а и С567, гистамин и активированный пара-
зитарным антигеном секрет лимфоцитов. Хемотаксис позволяет эозинофи-
лам накапливаться в инфицированном участке ткани и участвовать в анти-
паразитарной защите организма. Например, эозинофилы повреждают
шистосомулу (молодая форма шистосомы), секретируя в нее токсичные в
микроконцентрациях белки своих гранул: спецефический основной белок,
эозинофильный катионный белок и эозинофильный нейротоксин, мигри-
руют в ее интерстициальную ткань и вызывают гибель паразита. Однако
при аллергических заболеваниях, например при бронхиальной астме, эти
токсичные белки эозинофилов накапливаются в тканях, участвующих в ал-
лергических реакциях (перибронхиальная ткань легких при бронхиальной
астме) поражают бронхиальные эпителиальные клетки, вызывая их слущи-
вание, что делает более тяжелым течение заболевания.
В костном мозге эозинофилы образуются из КОК эозинофильной и
представлены пулом пролиферирующих и созревающих клеток — от эози-
нофильного миелобласта до миелоцита, а также пулом созревающих кле-
336 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ток, начиная от метамиелоцита. Продолжительность развития первого со-
ставляет 5,5 дня, второго — 2,5 дня. Стимулируют эозинофилопоэз в кост-
ном мозге ИЛ-5, а также ИЛ-3 и КСФ-ГМ.
В крови человека содержится 1—4 % эозинофилов или 0,15—0,25 • 109/л
крови. Увеличение их количества (эозинофилия) свидетельствует о возмож-
ной паразитарной инфекции или аллергическом заболевании. Эозинофилы
накапливаются в тканях, которые контактируют с внешней средой (легкие,
желудочно-кишечный тракт, кожа, урогенитальный тракт), формируя тка-
невый барьер против паразитарной инфекции. В этих тканях их количест-
во в 100—300 раз превышает содержание в крови. По параметрам плотно-
сти и функциональной активности эозинофилы в крови и тканях подраз-
деляются на три последовательно формирующиеся субпопуляции: неакти-
вированные эозинофилы нормальной плотности; активированные, нор-
мальной плотности эозинофилы; активированные, низкой плотности эози-
нофилы, что является следствием фенотипических изменений зрелых эо-
зинофилов, возникающих под влиянием цитокинов (ИЛ-3, ИЛ-5, КСФ-
ГМ). Наиболее активно секретирует токсичные белки субпопуляция акти-
вированных эозинофилов низкой плотности. Именно накопление в тканях
этой субпопуляции эозинофилов делает успешной защиту организма при
паразитарной инфекции, а при аллергических заболеваниях обусловливает
повреждение тканей легких, кожи, слизистых оболочек.
7.2.4. Функции моноцитов-макрофагов
Функции моноцитов-макрофагов (система фагоцитирующих мононуклеа-
ров): фагоцитарная защита организма против микробной инфекции; ток-
сический эффект метаболитов макрофагов на паразитов в организме чело-
века; участие в иммунном ответе организма и воспалении; регенерация
тканей и противоопухолевая защита; регуляция гемопоэза; фагоцитоз ста-
рых и поврежденных клеток крови, регуляция продукции острофазных
белков печенью.
В тканях моноциты дифференцируются в тканевые макрофаги. Моно-
циты-макрофаги имеют аэробный гликолиз, обеспечивающий энергией их
фагоцитарную активность. Макрофаги функционируют также в анаэроб-
ных условиях (например, в полости абсцесса), используя для генерации
энергии гликолитический путь. Макрофаги распознают микроорганизмы,
поврежденные клетки, медиаторы, гормоны, лимфокины с помощью ре-
цепторов своей плазменной мембраны.
Цитотоксическое действие на опухолевые клетки, токсоплазмы (кокци-
дии, паразитические простейшие), лейшмании и возбудителей малярии
макрофаги оказывают супероксидом, перекисью водорода, гидроксильным
радикалом и др.
Макрофаги человека секретируют в кровь и тканевую жидкость более
100 биологически активных веществ с молекулярной массой от 32 (анион
супероксида) до 440 000 (фибронектин). Функция этих веществ: стимуля-
ция пролиферации остеобластов и лимфоцитов, продукции фибробластами
КСФ-ГМ (интерлейкин-1); регуляция гемопоэза и механизмов воспаления
(КСФ-ГМ, КСФ-Г, эритропоэтин, простагландины, лейкотриены В, U, С,
D, Е, тромбоксан); цитотоксический и цитостатический эффекты на опу-
холевые клетки (опухольнекротизирующий фактор — ОНФ). Наконец, ин-
терлейкин-1 и ОНФ повышают температуру тела через терморегуляторные
центры гипоталамуса.
В костном мозге взрослого человека число моноцитов составляет около
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 337
7,3 • 109 клеток. В крови человека циркулирует 1,7 • 109 моноцитов, которые
распределяются на циркулирующий (25 %) и краевой пулы (75 %). Из кро-
ви в ткани выходит 1,6 • 107 клеток в час. Средний полупериод пребывания
моноцитов в крови колеблется от 36 до 104 ч. Продолжительность жизни
моноцитов-макрофагов в тканях человека составляет не менее 3 нед. Под
моноцитозом понимают увеличение абсолютного их количества свыше 800
клеток/мкл.
7.2.5. Регуляция грануло- и моноцитопоэза
Гранулоцитопоэз стимулируют гранулоцитарные колониестимулирующие
факторы (КСФ-Г, КСФ-ГМ) макрофагов и Т-лимфоцитов, а угнетают —
секретируемые нейтрофилами кейлоны (тканевоспецифические ингибито-
ры синтеза ДНК в клетках предшественницах) и лактоферрин — ингиби-
тор, угнетающий секрецию КСФг, а также простагландины серии Е, обра-
зуемые моноцитами и макрофагами.
Моноцитопоэз стимулируют моноцитарный колониестимулирующий фак-
тор (КСФ-М) и катехоламины через а-адренорецепторы KOK-ГМ. Моно-
цитопоэз тормозят простагландины Е, а- и 0-интерфероны. Продукцию
КСФ-М ослабляет также лактоферрин нейтрофилов. Выходу моноцитов из
костного мозга в кровь препятствуют большие дозы гидрокортизона.
7.3. Функции тромбоцитов
Кровяные пластинки препятствуют внезапной потере крови, мгновенно
закупоривая место повреждения кровеносных сосудов вначале временной,
а затем постоянной тромбоцитной пробкой. Установлена также антипара-
зитарная активность тромбоцитов. При инфецировании организма челове-
ка шистосомами, трематодами (долгоживущие паразиты, вызывающие по-
ражение внутренних органов человека — легких, печени и др.) тромбоциты
оказывают на них цитотоксический эффект и поражают их.
7.3.1. Структура и функции тромбоцитов
Тромбоциты имеют дисковидную форму, диаметр от 2 до 5 мкм, объем 5—
10 мкм3 (рис. 7.6). В тромбоците выделяют несколько зон: перифериче-
скую, золя-геля, внутриклеточ-
ных органелл. На наружной по-
верхности периферической зо-
ны располагается покров тол-
щиной до 50 нм, содержащий
Рис. 7.6. Ультраструктурная органи-
зация тромбоцита. Сечение тромбо-
цита, параллельное горизонтальной
плоскости.
ЕС — периферическая зона тромбоцита,
СМ — трехслойная мембрана, SMF —
субмембранный филамент, МТ — мик-
ротрубочки, Gly — гликоген. Зона орга-
нелл: М — митохондрии, G — гранулы,
DB — плотные гранулы, DTS — система
плотных трубочек, CS — система откры-
тых канальцев.
338 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
плазматические факторы свертывания крови, энзимы, рецепторы, необхо-
димые для активации тромбоцитов, их адгезии (приклеивания к суб-эндо-
телию) и агрегации (приклеивания друг к другу).
Периферическая зона. Мембрана тромбоцитов содержит «мембранный
фосфолипидный фактор 3» — «фосфолипидную матрицу», формирующую
активные коагуляционные комплексы с плазменными факторами сверты-
вания крови. Мембрана богата арахидоновой кислотой, из которой фер-
мент мембраны — фосфолипаза А2 образует свободную арахидоновую ки-
слоту для синтеза простагландинов. Метаболиты простагландинов форми-
руют короткоживущий агент — тромбоксан А2, вызывающий мощную агре-
гацию тромбоцитов. Фосфолипаза А2 активируется при контакте с адгезив-
ным белком субэндотелия — коллагеном, обнажающемся при повреждении
эндотелия, и с фактором Виллебранда, адгезирующемуся к субэндотелию
после повреждении эндотелия сосудов.
В липидный бислой мембраны тромбоцита «встроены» гликопротеины I
(субъединицы la, lb, Ic), II (субъединицы Па, ПЬ), III (субъединицы Ша,
Illb), TV, V, VI, которые обусловливают адгезивные и агрегационные
функции тромбоцитов.
Функции гликопротеинов. 1а — рецептор адгезии (прилипание, приклеи-
вание) тромбоцитов к коллагену субэндотелия. Комплекс «1Ь — фактор
свертывания крови IX» обеспечивает рецепцию фактора Виллебранда, что
необходимо для адгезии пластинок на поврежденный субэндотелий. 1с —
взаимодействуя с адгезивным белком субэндотелия фибронектином, обес-
печивает адгезию пластинки к субэндотелию. Па и ПЬ — необходимы для
всех видов агрегации тромбоцитов. Ша с гликопротеином ПЬ образуют
Са2+-зависимый комплекс, связывающий на тромбоцитах фибриноген, что
обеспечивает дальнейшую агрегацию тромбоцитов и ретракцию (сокраще-
ние) сгустка. V — гидролизуется тромбином, поддерживает агрегацию
тромбоцитов. Недостаток в мембране тромбоцитов различных субъединиц
гликопротеинов I—V вызывает повышенную кровоточивость.
Зона золя-геля гиалоплазмы прилегает к нижнему краю периферической
зоны тромбоцита и, в свою очередь, отделяет зону внутриклеточных орга-
нелл. В указанной зоне вдоль края клетки располагается сократительный
аппарат тромбоцита — краевое кольцо микротрубочек, контактирующее с
микрофиламентом. При стимуляции тромбоцита кольцо микротрубочек,
сокращаясь, смещает гранулы к центру тромбоцита («централизация гра-
нул»), сжимает их, вызывая секрецию содержимого гранул через систему
открытых канальцев в плазму крови. Сокращение кольца микротрубочек
позволяет тромбоциту образовывать псевдоподии (выросты цитоплазмы),
что увеличивает его способность к агрегации.
Зона плотных и а-гранул I и II типа. Плотные гранулы содержат АДФ,
АТФ, кальций, серотонин, норадреналин и адреналин. Кальций участвует
в регуляции адгезии, сокращения, секреции тромбоцитов, активации его
фосфолипаз и, следовательно, продукции в мембране тромбоцитов про-
стагландинов, необходимых для образования тромбоксана А2. При адгезии
тромбоцитов к поврежденной стенке сосуда плотные гранулы начинают
секретировать в больших количествах АДФ, которая усиливает приклеива-
ние циркулирующих тромбоцитов к адгезированным, т. е. агрегацию тром-
боцитов, поддерживая тем самым и рост тромбоцитарного агрегата. Реак-
ция «освобождения гранул» усиливает секрецию серотонина, вызывающего
вазоконстрикцию в районе поврежденного участка ткани, что уменьшает
кровотечение из нарушенных травмой сосудов.
а-Гранулы I типа содержат и секретируют антигепариновый фактор
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 339
тромбоцитов 4, тромбоцитарный ростовой фактор, тромбоспондин (глико-
протеин G). АДФ, тромбин, адреналин вызывают секрецию антигепарино-
вого фактора тромбоцитов 4, что усиливает агрегацию тромбоцитов. Тром-
боспондин, взаимодействуя с фибриногеном, образует на поверхности ак-
тивированных тромбоцитов комплекс, необходимый для формирования
тромбоцитарных агрегатов. Тромбоцитарный ростовой фактор — полипеп-
тид, стимулирующий рост гладких мышц сосудов и фибробластов, восста-
новление сосудистой стенки и соединительной ткани. Благодаря его свой-
ствам тромбоциты поддерживают целостность сосудистой стенки. У боль-
ных тромбоцитопенией имеется сниженная устойчивость стенки капилля-
ра, поэтому петехии (точечные кровоизлияния в коже) появляются вслед
за легкими травмами или изменениями давления крови. Петехии вызыва-
ются слущиванием эндотелия капилляров. В нормальных условиях возни-
кающий дефект в стенке капилляров устраняется тромбоцитами, секрети-
рующими тромбоцитарный ростовой фактор.
а-Гранулы II типа содержат лизосомальные энзимы (кислые гидрола-
зы). После адгезии или агрегации большая часть гранул в тромбоците ис-
чезает. Данный феномен получил название «реакции освобождения гра-
нул». Он имеет место после активации тромбоцитов тромбоксаном А2,
АДФ, адреналином, тромбином, протеолитическими энзимами, бактери-
альными эндотоксинами, коллагеном.
7.3.2. Тромбоцитопоэз и его регуляция
Тромбоцитопоэз (образование тромбоцитов в организме) протекает в кост-
ном мозге и включает следующие этапы: колониеобразующая клетка мега-
кариоцитарная (КОК-мег) -> промегакариобласт -> мегакариобласт ->
промегакариоцит -> зрелый мегакариоцит -> тромбоцитогенный мегака-
риоцит -> тромбоциты (рис. 7.7). Истинные митозы, т. е. деление клеток,
присущи только КОК-мег. Для промегакариобластов и мегакариобластов
характерен эндомитоз, т. е. удвоение ДНК в клетке без ее деления. После
остановки эндомитоза, в основном после 8-, 16-, 32-, 64-кратного удвое-
ния ДНК, мегакариобласт начинает дифференциацию до тромбоцитарного
мегакариоцита, образующего тромбоциты
Митоз и дифференциацию КОК-мег активирует гемопоэтический цито-
кин — тромбопоэтин (тромбоцитопоэтин) при взаимодействии с интерлей-
кином-3. Этот гуморальный фактор стимулирует также эндомитоз мегака-
риоцитов, он необходим для нормального созревания цитоплазмы мегака-
риоцита и формирования в ней тромбоцитов. Стимулируют образование
тромбоцитопоэтина уменьшение мегакариоцитов и их предшественников в
костном мозге, а также тромбоцитопения, вызванная усиленным исполь-
зованием тромбоцитов при формировании тромба (воспаление, необрати-
мая агрегация тромбоцитов). Активированные тромбоциты и селезенка вы-
деляют в кровь гуморальный ингибитор пролиферации КОК-мег, а также
немитотической стадии развития мегакариоцитов (эндомитоза) и созрева-
ния цитоплазмы мегакариоцитов. Это гликопротеин, массой 12—17 кДа.
В костном мозге тромбоцитарные мегакариоциты локализованы на по-
верхности синусного эндотелия. Часть их цитоплазматических отростков
на 1—2 мкм проникает в просвет синусоида костного мозга через эндоте-
лий и фиксирует мегакариоцит на эндотелии, выполняя функцию «якоря».
Вторая часть отростков, представленных цитоплазматическими лентами до
120 мкм в длину и числом 6—8, проникает через эндотелий в просвет си-
нусоида костного мозга. Эти отростки называются протромбоцитами. В
340 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
СКК—► КОКгммэ—► КОКмег —►КОКмег-1 —►КОКмег-2
(митозы)
I
0,07 0,48 0,74
III
Промегакариобласт: 2N
I
Мегакариобласты: 4N—► 8N —*-16N—*-32N—*-64N—*-128N
Illi
(эндомитоз)
Illi
Промегакариоциты
I
Зрелые мегакариоциты
I
Тромбоцитогенный мегакариоцит
I
Протромбоциты —► Тромбоциты
Рис. 7.7. Схема дифференцировки клеток мегакариоцитарной серии.
СКК — стволовая кроветворная клетка, КОКгммэ — КОКмегакариоцитарная (КОКмег-1 ме-
нее дифференцированная, а КОКмег-2 — более дифференцированная клетка).
Цифрами 0.07; 0.48; 0.74 обозначена вероятность вовлечения клетки-предшественницы в мега-
кариоцитарную дифференциацию, т. е. по мере снижения пролиферативного потенциала
КОКмег возрастает вероятность ее дифференциации в мегакариобласт.
просвете синусоида цитоплазма протромбоцита после локальных сокраще-
ний разрывается и он образует до 1000 тромбоцитов, поступающих с кро-
вью из просвета синусоидов в русло циркулирующей крови. Однако сами
протромбоциты или их фрагменты, содержащие до 100 тромбоцитов, также
могут выходить из синусоида костного мозга в кровь. Они достигают мик-
роциркуляторного русла легких, где из них освобождаются тромбоциты.
Поэтому количество тромбоцитов оказывается более высоким в легочных
венах, чем в легочной артерии. Количество тромбоцитов, образовавшихся
в легких, может достигать 7—17 % от массы тромбоцитов в крови.
Костный мозг человека содержит около 15 • 106 мегакариоцитов на 1 кг
массы тела. Дневная продукция тромбоцитов у человека 66 000 ± 14 600 в
1 мкл крови. В среднем мегакариоцит высвобождает до 3000 тромбоцитов.
Количество тромбоцитов в крови взрослого человека достигает 150—
375 • 109/л; у детей — 150—250 • 109/л. Общая популяция тромбоцитов пред-
ставлена циркулирующими в крови (70 %) и находящимися в селезенке
(30 %). Накопление тромбоцитов в селезенке возникает из-за их медлен-
ного движения через извилистые селезеночные корды, занимающему до 8
мин. Сокращение селезенки (например, вызванное адреналином) освобож-
дает депонированные тромбоциты в общий кровоток. Удаление селезенки
у человека, устраняющее депонирование тромбоцитов, повышает число
тромбоцитов в крови у спленэктомированных индивидуумов. Напротив,
увеличение селезенки (спленомегалия) вызывает рост селезеночного депо
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 341
тромбоцитов у больных и, как следствие этого, тяжелую тромбоцитопе-
нию. В результате у таких больных имеет место повышенная кровоточи-
вость. Тромбоцитопенией называют содержание тромбоцитов в крови
взрослого ниже 150 • 109/л. Продолжительность жизни тромбоцитов 6,9—
9,9 дня. Стареющие клетки разрушаются макрофагами в костном мозге и,
в меньшей степени, в селезенке и печени.
7.4. Механизмы свертывания крови (гемостаза)
7.4.1. Тромбоцитарный гемостаз
Тромбоцитарная реакция, т. е. реакция тромбоцитов на нарушение целост-
ности сосудистой стенки, формируется параллельно реакции самих сосу-
дов на повреждение — их сокращение в месте повреждения, вызывающее
шунтирование крови выше поврежденного участка сосуда. Данная сосуди-
сто-тромбоцитарная реакция на повреждение сосудистой стенки первой
останавливает кровотечение из микрососуда и поэтому называется сосуди-
сто-тромбоцитарным, или первичным, гемостазом. Формирование и закре-
пление тромбов с помощью плазменных факторов свертывания называют
вторичным коагуляционным гемостазом. Первая волна агрегации тромбоци-
тов начинается с приклеивания тромбоцитов рецепторами гликопротеинов
I и II к фактору Виллебранда, фибронектину и коллагену субэндотелия по-
врежденных тканей. Вторая волна агрегации тромбоцитов вызвана высво-
бождением АДФ из плотных гранул тромбоцитов, образованием тромбок-
сана А2 в их мембране, взаимодействием мембранных гликопротеинов ПЬ
и Ша с фибриногеном, а V — с тромбином. Но в основном вторую волну
агрегации тромбоцитов обеспечивают тромбоксан А2 и тромбин. Мощным
стимулом для образования тромбоксана А2 в мембране тромбоцита явля-
ются коллаген и фактор Виллебранда, активирующий фосфолипазу А2.
Сильный агрегационный эффект оказывает тромбин, который образуется
на мембране тромбоцитов при участии фактора V и гликопротеина V. Ад-
гезированные тромбоциты являются основой для агрегации, т. е. приклеи-
вания к ним тромбоцитов, приносимых с кровью к участку поврежденного
сосуда. В ходе агрегации тромбоцитарный тромб уплотняется и сокращает-
ся. Его формирование усиливается с образованием фибрина (фибринового
тромба) вследствие активации плазменной системы свертывания крови
(рис. 7.8).
7.4.2. Система свертывания крови
Останавливает окончательно кровотечение из поврежденных сосудов обра-
зование фибриновых тромбов, закрывающих их просвет. В плазме крови
содержатся факторы свертывания в виде неактивных форм ферментов,
обозначаемых римскими цифрами: I, II, VIII, IX, X, XI, XII, XIII
(табл. 7.2). Повреждение тканей, эндотелия сосуда или клеток крови вызы-
вает каскадную реакцию активации этих ферментов, которая приводит к
образованию фибриновых нитей, формирующих сеть тромба (рис. 7.9).
Начало каскадной реакции связано с контактом неактивных форм факто-
ров свертывания с поврежденными тканями, окружающими сосуды,
(внешний путь активации свертывания крови), а также при контакте крови
с поврежденными тканями сосудистой стенки или с поврежденными сами-
ми клетками крови (внутренний путь активации свертывания крови).
Внешний путь. Мембраны поврежденных клеток тканей выделяют в
342 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 7.8. Схема формирования коагуляционного гемостаза.
1 — активация тромбином ф.У; 2 — активация тромбином ф.УШ, высвобождаемого из связи с
ф.Вилленбранта; 3 — активация тромбином ф.Х1. Контакт крови с поверхностью субэндоте-
лия активирует «внутренний» путь свертывания крови; контакт крови с поврежденными клет-
ками ткани активирует «внешний» путь активации свертывания крови.
плазму крови тканевый фактор — трансмембранный белок. Тканевой фак-
тор с активированным им фактором свертывания крови VII активируют
фактор X. Фактор Ха (а-активированный) в присутствии ионов кальция
немедленно соединяется с тканевыми фосфолипидами и фактором V. Об-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 343
Таблица 7.2. Факторы свертывания крови
Фактор Период полу- жизни Молекуляр- ная масса (Да) Нормальная кон- центрация в плазме (цг/мл)
I — фибриноген 3,7 дня 340.000 1500-4000
II — протромбин 2,8 дня 72.500 150
III — тромбопластин — — 0
IV — ионы кальция — — 0,9—1,2 ммоль/л
V — проакцелерин 15-24 ч 330.000 10
VII — проконвертин 1,2-6 48.000 менее 1
VIIIc — антигемофильный фактор А 5-12 ч 1.000.000 менее 5
VIII ФВ — фактор Виллебранда 24-40 ч
IX — антигемофильный фактор В 20-24 ч 57.000 5
X — фактор Стюарта—Прауэра 32-48 ч 59.000 8
XI — антигемофильный фактор В 40-48 ч 160.000 5
XII — фактор Хагемана 48-52 76.000 35
XIII — фибрин-стабилизирующий фактор 5—12 дней 320.000 20
Плазминоген 2,2 дня 90.000 150
Прекалликреин — 85.000 30
Высокомолекулярный кининоген — 150.000 80
разевавшийся комплекс через несколько секунд после его формирования
превращает часть протромбина в тромбин. Тромбин начинает действовать
как протеолитический фермент на фибриноген, а также активировать фак-
тор V, тем самым дополнительно ускоряя превращение протромбина в
тромбин.
Внутренний путь. Разрушение тромбоцитов и эритроцитов или контакт
этих клеток с субэндотелием поврежденного сосуда активирует фактор XII.
Фактор ХПа (а-активированный), взаимодействуя с высокомолекулярным
кининогеном плазмы, превращает фактор XI в фактор Х1а. Реакция уско-
ряется прекалликреином плазмы. Х1а активирует фактор IX (тромбопла-
стин плазмы), образующийся фактор 1Ха формирует комплекс: «фактор
1Ха + фактор VIII (антигемофильный фактор) + тромбоцитарный фосфо-
липид (тромбоцитарный фактор 3) + ионы кальция». Этот комплекс акти-
вирует фактор X. Факторы Ха и Va, взаимодействуя с фосфолипидной по-
верхностью клетки в присутствии Са2+, образуют новый комплекс, назы-
ваемый кровяной протромбиназой, превращающей протромбин в тромбин.
Особую роль в повышении активности протромбиназы играет тромбоци-
тарный фактор 3. Его отсутствие снижает в 1000 раз активность протром-
биназы!
В крови содержатся неферментные белки — акселераторы или факторы
V и VII, которые при взаимодействии с фосфолипидными поверхностями
тромбоцитов и участками мембран других поврежденных клеток ускоряют
во много тысяч раз реакцию свертывания крови. Дефицит тромбоцитарно-
го фактора III приводит к геморрагическому диатезу, дефицит фактора IX
вызывает гемофилию В, а дефицит фактора VIII — гемофилию А.
Фактор VIII циркулирует в крови, будучи связанным со своим белком-
носителем — фактором Виллебранда. Последний стабилизирует молекулу
фактора VIII, увеличивает ее период жизни внутри сосуда и способствует
переносу фактора VIII к месту повреждения сосуда. Активированный фак-
тор VIII может проявить свое действие, лишь отсоединившись от белка-
носителя. Эта «операция» с комплексом «фактор VIII — фактор Вилле-
бранда» происходит под воздействием тромбина, следовые количества ко-
344 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Повреждение сосуда
Рис. 7.9. Последовательность формирования и закрепления тромба с помощью
плазменных факторов свертывания при «внутреннем» пути активации свертывания
крови. Время указывает на продолжительность процесса после контакта крови с
субэндотелием.
торого постоянно образуются в крови как следствие реакции на разруше-
ние стареющих клеток крови.
Внешний путь активации свертывания крови занимает около 15 с, а
внутренний — 2—10 мин. Оба они завершаются превращением протромби-
на в тромбин. Протромбин синтезируется в печени, для его образования,
как и для образования факторов IX, X, VII и II, необходим витамин К, по-
ступающий в организм с пищей, депонирующийся в печени и активирую-
щий в ней синтез вышеуказанных факторов свертывания крови. Поэтому
поражение печени или возникающий в организме дефицит витамина К со-
провождаются кровоточивостью. Количество тромбина, образующегося
при активации свертывания крови, прямо пропорционально количеству
активирующих его комплексов.
Тромбин — активный протеолитический фермент, отщепляет от молеку-
лы фибриногена 4 пептида мономера. Каждый из мономеров имеет 4 сво-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 345
бодные связи. Соединяясь ими друг с другом: конец к концу, бок к боку, —
мономеры в течение нескольких секунд формируют сеть волокон фибрина.
Под влиянием фибринстабилизирующего фактора (фактор XIII) в фибрине
образуются дополнительные дисульфидные связи, и сеть фибриновых во-
локон становится прочной. В этой сети задерживаются тромбоциты, лейко-
циты, эритроциты и белки плазмы, формируя фибриновый тромб.
После образования сгустка через 30—60 мин начинается его сокраще-
ние, или ретракция. Ретракция происходит за счет сокращения нитей ак-
тина и миозина тромбоцитов, а также нитей фибрина под влиянием тром-
бина и ионов Са2+. В результате ретракции фибриновый сгусток сжимается
в плотную массу, тромб уплотняется и становится непроницаемым как для
клеток, так и для плазмы крови. Продолжению свертывания крови в кро-
веносном русле после формирования фибринового тромба препятствует
противосвертывающая система крови.
7.4.3. Противосвертывающие механизмы крови
Физиологические антикоагулянты поддерживают кровь в жидком состоя-
нии и ограничивают процесс тромбообразования. К ним относятся анти-
тромбин III, гепарин, протеины С и S, а-2-макроглобулин, нити фибрина.
Антитромбин III является а2-глобулином и создает 75 % всей антикоагу-
лянтной активности плазмы. Он является основным плазменным кофакто-
ром гепарина, ингибирует активность тромбина, факторов Ха, IXa, VII,
ХПа. Его концентрация в плазме достигает 240 мкг/мл.
Гепарин — сульфатированный полисахарид — трансформирует антитром-
бин III в антикоагулянт немедленного действия, в 1000 раз усиливая его
эффекты.
Протеины С и S— синтезируются в печени. Их синтез активирует вита-
мин К. Протеин С высвобождает активатор плазминогена из стенки сосу-
да, инактивирует активированные факторы VIII и V. Протеин S снижает
способность тромбина активировать факторы VIII и V. Нити фибрина об-
ладают антитромбинным действием, так как адсорбируют до 80—85 %
тромбина крови. В результате тромбин концентрируется в формирующем-
ся сгустке и не распространяется по току крови (рис. 7.10).
Регуляция агрегации тромбоцитов сосудистой стенкой. Адгезии тромбо-
цитов к неповрежденной сосудистой стенке препятствуют: эндотелиальные
клетки; гепариноподобные соединения, секретируемые тучными клетками
соединительной ткани; синтезируемые эндотелиальными и гладкомышеч-
ными клетками сосудов (рис. 7.11) — простациклин 12, оксид азота (NO),
тромбомодулин, тканевый активатор плазминогена и эктоэнзимы
(АДФаза), ингибитор тканевого фактора (ингибитор внешнего пути свер-
тывания крови).
Простациклин 12 — мощный ингибитор агрегации тромбоцитов, образу-
ется в венозных и артериальных эндотелиальных клетках из арахидоновой
кислоты. Между антиагрегационной способностью простациклина и про-
агрегационной субстанцией — тромбоксаном А2 тромбоцитов в нормаль-
ных условиях имеет место динамическое равновесие, регулирующее агрега-
цию тромбоцитов. При преобладании эффекта простациклина над тром-
боксаном А2 агрегации томоцитов не происходит. Напротив, сниженная
или утраченная продукция простациклина участком эндотелия может быть
одной из причин агрегации кровяных пластинок к стенке сосуда и форми-
рования тромба. Синтез простациклинов в эндотелии усиливается при
стрессе под влиянием тромбина.
346 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Повреждение сосудистой стенки
Адгезия Агрегация тромбоцитов
тканевого обратимая
. V .... Выброс в кровь
Активация XII —►тромбопластина
фактора (Хагемана) (фактор III)
f , , • Антитромбин III +
Xlla + XI + ТФЗ * Гепарин
+ УП + Са
необратимая
Протромбин (II)
III
IXa + VIII + ТФЗ + Са
Ха + Va + ТФЗ + Са
Тромбин
Армирование тромба
Фибринстабилизирующиий
фактор XIII
Фибрин ◄— Растворимые комплексы ◄—Фибриноген
фибрин мономера
V
Консолидация тромба
Продукты деградации
фибрина
Плазминоген
Плазмин
Тканевый активизатор Активация фактора XII
плазминогена
Белок С |
Выброс
Тканевый активатор
Повреждение сосудистой стенки
плазминогена
Рис. 7.10. Схема действия компонентов противосвертывающей и фибринолитиче-
ской системы крови на разных этапах образования фибринового сгустка.
Пояснения в тексте. ТФЗ-тромбоцитарный фактор 3.
Тромбомодулин — рецептор тромбина на эндотелии сосудов — взаимо-
действует с тромбином и активирует белок С, обладающий способностью
высвобождать тканевый активатор плазминогена из стенки сосуда. Дефи-
цит белка С повышает свертываемость крови.
NO образуется в эндотелиальных клетках и угнетает адгезию и рекрути-
рование тромбоцитов. Его эффект усиливается взаимодействием с проста-
циклином. Атеросклеротические повреждения сосуда, гиперхолестерине-
мия понижают способность эндотелия к продукции оксида азота, повышая
риск формирования тромбов.
Тканевый активатор плазминогена — это белок, воспроизводимый и по-
стоянно секретируемый эндотелием сосудов. Обеспечивает прямую ло-
кальную тромболитическую активность в отношении образовавшегося
Л
Антикоагулянты
Простагландины I
расслабляющий
фактор (NO)
—► экто-АДФаза
Тромбоциты •*
Прокоагулянты
Фактор Вилленбранда
л Эндотелиальные
Г Тромбомодулин
Антитромбин
Гепариноподобные
соединения
[—►Па, Ха, Xia, IXa
тканевого
фактора
Т каневый активатор
плазминогена------
Формирование
фибринового
сгустка
Тромбин
Простациклин
Белок S
Белок С-------
IXa <- IX
Протромбин
Фибринолиз
Тканевый
фактор
Villa
Ингибитор активатора
ллазминогена-1
V
Рис. 7.11. Роль эндотелия кровеносного сосуда в свертывании крови.
V
Под надписью «Антикоагулянты» указаны факторы эндотелия, обладающие антикоагулянтным действием, благодаря ингибиции агрегации тром-
боцитов, формирования фибринового сгустка и активации фибринолиза.
Под названием «Прокоагулянты» указаны факторы эндотелия, участвующие в образовании тромбоцитарного тромба, фибринового сгустка и по-
давляющие фибринолиз (-<—| |— показаны ингибирующие действия факторов).
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 347
348 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
тромба. В крови поддерживается постоянный уровень этого фактора, что
обеспечивает системную тромболитическую активность крови.
Эктоэнзимы — это образуемые эндотелием АДФаза, АТФаза и адено-
зинконвертирующий фермент. Эндотелиальная АДФаза быстро расщепля-
ет проагрегант — АДФ, секретируемый активированными тромбоцитами.
Ютетки эндотелия сосудов синтезируют и протромботические факторы:
тканевый фактор, ингибиторы активатора плазминогена, фактор Вилле-
бранда.
Тканевый фактор — это сложный белок мембраны клеток массой
46 кДа. Часть его молекулы при повреждении клетки плотно связывается с
фактором коагуляции Vila, поддерживая его функцию ускорителя во
внешнем пути свертывания крови.
Ингибитор активатора плазминогена-1 — это белок массой 52 кДа, со-
держащейся в циркулирующей крови. Тесно связываясь с активатором
плазминогена, он инактивирует его, участвуя таким образом в регуляции
фибринолиза в организме.
Фактор Виллебранда — это многомерная молекула массой 1—20 млн Да,
синтезируется эндотелием и хранится в эндотелиальных секреторных гра-
нулах. Высвобождаясь из них, выполняет функцию адгезивной молекулы
для тромбоцитов, поддерживает их агрегацию. Увеличенное высвобожде-
ние фактора Виллебранда из эндотелия индуцируется тромбином.
Свертывание крови в сосуде предупреждает и гладкая поверхность эн-
дотелия, препятствующая включению внутреннего пути формирования ак-
тивной протромбиназы. Мономолекулярный слой белка, адсорбированный
на поверхности эндотелия, отталкивает факторы свертывания и тромбоци-
ты, также предупреждает свертывание крови.
Антикоагулянты применяются в клинической практике. Например, для
понижения повышенной свертываемости крови у больных с ишемической
болезнью сердца, для поддержания крови в жидком состоянии при ис-
пользовании аппарата искуственного кровообращения, вызывающих трав-
матизацию клеток крови, в результате чего активируется внутренний путь
свертывания крови.
7.4.4. Фибринолиз
Это процесс разрушения (лизиса) сгустка крови и восстановление просвета
сосудов, закупоренных тромбами. Лизис сгустка крови осуществляет сис-
тема ферментов, компонентами которой являются плазминоген, плазмин,
активатры плазминогена и их ингибиторы.
В плазме крови содержится неактивный белок плазминоген. Плазмино-
ген переходит в активный плазмин под влиянием: а) белкового тканевого
активатора плазминогена, высвобождающегося из эндотелиальных клеток
на участке формирующегося кровяного сгустка, особенно активно он об-
разуется эндотелием капилляров почек, печени, легких; б) активированно-
го фактора свертывания крови ХПа в комплексе с калликреином, высоко-
молекулярным кининогеном; в) лизосомальных ферментов поврежденной
ткани; г) урокиназы почек, которая обеспечивает 15 % общей фибриноли-
тической активности крови, и д) стрептокиназы бактерий. Стрептокиназа
бактерий в инфицированных тканях растворяет плазменный сгусток в
лимфе и тканевой жидкости, что способствует распространению ин-
фекции.
Активный плазмин расщепляет нити фибрина. Тормозят агрегацию
тромбоцитов и формирование волокон фибрина продукты фибринолиза.
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 349
Лизис кровяных сгустков продолжается в течение нескольких дней. Вы-
брос тканевых активаторов фибринолиза происходит в случае выполнения
физических нагрузок, появления в крови адреналина и норадреналина.
Плазмин инактивируют антиплазмины: а2-антиплазмин (а2-глобулин),
нейтрализующий 2/з плазмина, и а2-макроглобулин. Возможность регуля-
ции активности компонентов фибринолитической системы позволяет пре-
дупреждать нежелательные следствия фибринолиза. Например, исходно
высокая локальная активность тканевых активаторов плазминогена в лег-
ких и печени, не в полной мере регулируемая антиплазминами, может со-
провождаться отрывом тромбов от стенки поврежденного сосуда, вызы-
вающим опасные для жизни человека осложнения — массивные кровоте-
чения, тромбоэмболию сосудов, т. е. закупорку сосудов кусочками тромба.
7.5. Общие закономерности кроветворения
7.5.1. Кроветворные клетки-предшественницы
Ежечасно в крови человека разрушаются, заканчивая свой жизненный
цикл, и вновь образуются 20 млрд, тромбоцитов, 10 млрд, эритроцитов и 5
млрд, лейкоцитов. В результате количество этих клеток остается в крови на
постоянном уровне. Примерно каждые два года в организме человека про-
изводится масса клеток крови, равная массе его тела. Пролиферативный
потенциал кроветворной ткани заключен в стволовых кроветворных клет-
ках (СКК)-предшественницах. Согласно одной точке зрения, эти клетки
способны к самообновлению, т. е. производству дочерних СКК, на протя-
жении всей жизни человека. Другая точка зрения допускает, что человек
рождается с готовым «запасом» СКК, которые до дифференциации в опре-
деленные клетки-предшественницы костного мозга находятся в состоянии
фазы клеточного покоя — Go. Выход СКК из состояния покоя Go сопрово-
ждается производством дочерних СКК и их дифференциацией в направле-
нии: 1) клетки-предшественницы всех линий миелопоэза — гранулопоэза,
моноцитопоэза, мегакариоцитопоэза и эритропоэза; 2) клетки-предшест-
венницы Т-лимфоцитов; 3) клетки-предшественницы В-лимфоцитов. Все
клетки-предшественницы получили название колониеобразующцх клеток
(КОК). В костном мозге имеются: 1) гранулоцитарно-моноцитарно-мега-
кариоцитарно-эритроцитарная колониеобразующая единица (КОК-
ГММЭ), формирующая гранулоциты, макрофаги, мегакариоциты и эрит-
роциты; 2) пре-КОК-Т, представленная Т-клеточными субпопуляциями; 3)
пре-КОК-В, которая состоит из В-лимфоцитов. КОК-ГММЭ дифферен-
цируется в би- и унипотентные КОК, которые также классифицируют по
произведенному ими потомству при культивировании in vitro. Нейтро-
фильные гранулоциты и макрофаги образуются из общей КОК, которая
называется КОК гранулицитарно-моноцитарная (KOK-ГМ), а из КОК-
ЭМег образуются колонии из эритроидных клеток и мегакариоцитов. Би-
потентные клетки дифференцируются в направлении каких-либо двух ли-
ний гемопоэза. Формирующиеся из них унипотентные КОК образуют
клетки только одной линии гемопоэза. В связи с этим возникли их назва-
ния — КОК-эритроцитарные, эозинофильные, базофильные, тучно-кле-
точные, нейтрофильные, мегакариоцитарные.
Клетки-предшественницы несут на мембране различные антигенные
структуры, по которым эти клетки можно дифференцировать и с помощью
моноклональных антител. Так, определяемая на основании данного подхо-
да, самая ранняя СКК человека (стадия развития СКК, предшествующая
КСФтучн.кл.
«
u
КОКэр. -<--Эритроциты
I Кейлоны,
-= ингибитор :---1
эритропоэза
'кОКмег.
Рис. 7.12. Регуляция формирования кроветворных клеток.
СКК — стволовая кроветворная клетка; КОК-лимф. — колониеобра-
зующая клетка лимфоцитарная; пре-КОК-Т — колониеобразующая
клетка- предшественница Т-клеток;
пре-КОК-В — колониеобразующая клетка-предшественница В-клеток;
КОК-ГММЭ — колониеобразующая клетка гранулоцитарно-мегака-
риоцитарно-моноцитарно-эритроцитарная;
КОКнейтр.эоз. — колониеоброзующая клетка нейтрофильно-эозино-
фильная;
КОК-ГМ — колониеобразующая клетка гранулоцитарно-моноцитар-
ная;
КОКэр.эоз. — колониеобразующая клетка эритроцитарно-эозино-
фильная;
КОКгр.эр. — колониеобразующая клетка гранулоцитарно-эритроци-
тарная;
КОКэр.мег. — колониеобразующая клетка эритроцитарно-мегакарио-
цитарная;
КОКбаз. — колониеобразующая клетка базофильная;
КОКтучн.кл. — колониеобразующая клетка тучноклеточная;
КОК-мег. — колониеобразующая клетка мегакариоцитарная;
КОК-нейтр. — колониеобразующая клетка нейтрофильная;
КОК-эоз. — колониеобразующая клетка эозинофильная;
КОК-мон. — колониеобразующая клетка моноцитарная;
КОК-эр. — колониеоброазующая клетка эритроцитарная;
ФБ — фибробласт;
ФСК — фактор стволовой клетки; его связанная и растворимая фрак-
ции синтезируются фибробластом;
C-kit — рецептор ФСК на мембране стволовой кроветворной клетки,
контролируемый C-kit геном (взаимодействие СКК со связаной фрак-
цией ФСК делает ее чувствительной к действию растворимой фракции
ФСК, ИЛ-1 и ИЛ-6, вызывающих ее дифференциацию в КОК-
ГММЭ);
ЭЦМ — экстрацеллюлярный матрикс;
ИЛ-1 — интерлейкин-1;
ИЛ-3 — интерлейкин-3;
ИЛ-4 — интерлейкин-4;
ИЛ-5 — интерлейкин-5;
ИЛ-6 — интерлейкин-6;
ИЛ-7 — интерлейкин-7;
ИЛ-11 — интерлейкин-11;
КСФ-ГМ — колониестимулирующий фактор гранулоцитарно-моноци-
тарный;
КСФ-Г — колониестимулирующий фактор гранулоцитарный;
КСФ-мон — колониестимулирующий фактор моноцитарный;
ТРФ-р — трансформирующий рост фактор- (секрет а-гранул тромбо-
цитов).
— регуляция на основе отрицательной обратной связи (направле-
ние угнетающего эффекта);
— ► — направление дифференциации клеток;
— ► — регулирующее влияние.
352 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
КОК-ГММЭ) несет дифференцировочный маркер CD 133 и такие клетки
обозначаются как CD 133+ (положительные); из CD 133+ образуются
клетки, предшествующие КОК-ГММЭ, несущие маркер CD 34, и обозна-
чаемые как CD 34+. У человека CD 133+ СКК обнаруживают наибольшую
пролиферативную активность из всех распознаваемых с помощью моно-
клональных антител СКК — их пролиферативная активность в несколько
раз выше, чем у CD 34+ клеток. КОКгммэ способна совершать до 21 ми-
тоза. Бипотентные КОК также обладают значительной способностью к
размножению. Например, КОК-ЭМег, будучи стимулированной ростковы-
ми факторами, способна совершать до 13 митозов и образовывать затем
сотни КОК-Э, из которых формируются тысячи эритроидных клеток. Ко-
личество КОК-ГММЭ составляет 2—5 • 105 содержащих ядро клеток кост-
ного мозга, количество КОК-ЭМег достигает 40—120-105 клеток в кост-
ном мозге, а содержание КОК-Э — 200—600 • 105 КОКЭ способна к 2—4
делениям. Из каждой КОК-Э образуется до 50 эритроцитов. KOK-ГМ спо-
собна совершать 5—6 делений, образующиеся из нее КОК-Г и КОК-М —
еще 5—6 делений каждая. Это позволяет одной KOK-ГМ формировать ты-
сячи зрелых клеток-потомков — гранулоцитов и моноцитов (рис. 7.12).
Дифференциация клеток-предшественниц КОК-ГММЭ до унипотент-
ных КОК сопровождается формированием рецепторов к гемопоэтическим
цитокинам (гормонам) — интерлейкинам (ИЛ), колониестимулирующим
факторам (КСФ), эритропоэтину, тромбоцитопоэтину, к нейромедиато-
рам, катехол-аминам, тиреотропному гормону, производным тестостерона,
поэтому перечисленные гормоны регулируют пролиферацию и дифферен-
циацию клеток крови.
7.5.2. Регуляция пролиферации и дифференциации КОК
Рост различных КОК в культурах удается получить, добавляя стимулирую-
щие его факторы. Отсюда факторы, стимулирующие образование грануло-
цитарных колоний, получили название «колониестимулирующие факторы
гранулоцитарные» — КСФ-Г, макрофагальные — КСФ-М, гранулоцитар-
но-макрофагальные — КСФ-ГМ, эозинофильные — КСФэоз (ИЛ-5), мега-
кариоцитарные — КСФмег (тромбопоэтин), стимулирующие развитие ко-
лоний из КОК-ГММЭ — мульти-КСФ или интерлейкин-3 (ИЛ-З).
ИЛ-3 и КСФ-ГМ стимулируют пролиферацию и дифференциацию не
только КОК-ГММЭ, но и бипотентных КОК—KOK-ГМ, КОК-нейтро-
фильно-эозинофильных и др. В связи с этим ИЛ-3 и КСФ-ГМ рассматри-
ваются как неспецифические факторы, поддерживающие самообновление
и пролиферацию КОК-ГММЭ и бипотентных КОК. Все КСФ гликопро-
теины относятся к полипептидным гормонам, которые регулируют Гемо-
поэз. Источниками ИЛ-3 и КСФэоз (ИЛ-5) являются Т-лимфоциты и
спленоциты, КСФ-ГМ — Т-лимфоциты, моноциты, эндотелиальные клет-
ки и фибробласты, КСФ-Г, КСФ-М-моноциты и фибробласты, а эритро-
поэтина — перитубулярные и тубулярные клетки почек, купферовские
клетки печени. Эти вещества получили общее название «гемопоэтические
цитокины».
Гены, контролирующие синтез ИЛ-3, КСФ-Г и КСФ-М у человека, на-
ходятся на дистальной части длинного плеча 5-й хромосомы, КСФ-Г — ло-
кализованы у человека на 17-й хромосоме, эритропоэтины — на 7-й хро-
мосоме.
Тяжелые нарушения в системе крови могут возникать при повреждении
участков генома, ответственных за синтез молекулярных регуляторов гемо-
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 353
поэза, за синтез их рецепторов на КОК. Так, например, причиной тяжелой
наследственной гранулоцитопении (нейтропении детей), приводящей их к
ранней гибели из-за легкой поражаемости микробными и вирусными ин-
фекциями, является нарушенное воспроизводство КСФ-Г в организме
больных. Гранулоцитопоэз и уровень нейтрофилов в крови быстро восста-
навливается при введении КСФ-Г этим больным. Точечная мутация над-
мембранного района рецепторов эритропоэтина вызывает их резкую акти-
визацию даже при малых количествах эритропоэтина в крови, что приво-
дит к усиленной пролиферации эритроидной ткани и развитию эритроци-
тоза. Продукцию КСФ усиливают различные стимулы: гипоксия — эритро-
поэтина; тромбоцитопения — тромбопоэтина; микробная инфекция —
КСФ-ГМ, КСФ-Г, КСФ-М; гельминтная инфекция — КСФэоз (ИЛ-5).
Каждый из перечисленных стимулов одновременно вызывает продукцию
неспецифических ростковых факторов ИЛ-3 и КСФ-ГМ. При этом ИЛ-3 и
КСФ-ГМ стимулируют как пролиферацию КОК-ГММЭ и КОК-бипотент-
ных, так и формирование на их мембране рецепторов к КСФ, действие ко-
торых направлено на унипотентные КОК. Поэтому высокие концентрации
КСФ-Г, КСФ-М и т. д. стимулируют пролиферацию и дальнейшую диф-
ференциацию унипотентных клеток-предшественниц гранулоцитарного,
моноцитарного и других рядов.
На интенсивность продукции КСФ оказывает регулирующее влияние
вегетативная нервная система. Так, введение животным а- и [3-адренобло-
каторов значительно уменьшает уровень КСФ в крови. Стимуляторами ге-
мопоэза являются Т-лимфоциты (Т-хелперы). Действие на организм возбу-
ждающих гемопоэз факторов (кровопотеря, высотная гипоксия и др.) вы-
зывает миграцию лимфоцитов в костный мозг и активацию ими КОК.
КСФ регулируют функции не только КОК, но и зрелых клеток крови. Так,
КСФ-ГМ усиливает фагоцитарную активность, метаболизм, миграцию в
ткани зрелых нейтрофилов и моноцитов-макрофагов. КСФ-Г стимулирует
бактерицидную, фагоцитарную и цитотоксическую активность этих кле-
ток. ИЛ-3 также усиливает цитотоксическую активность макрофагов, уве-
личивает эозинофильную фагоцитарную активность, а КСФ-М — бактери-
цидную и тумороцидную (разрушающую опухолевые клетки) функции мо-
ноцитов и макрофагов. Ответ клеточных линий кроветворной ткани на
стимул внешней среды реализуется через рецепцию не только гемопоэти-
ческих цитокинов, но и медиаторов нервных окончаний, находящихся в
костномозговой ткани. Так, в паренхиме костного мозга представлены
симпатические и пуринергические нервные окончания. Первые из них
секретируют адреналин, норадреналин, дофамин, вторые — нейрокини-
ны — субстанцию Р и др. Рецепторы этих соединений представлены на
КОК-ГММЭ, KOK-ГМ, КОК-Г, КОК-ЭРМЕГ, КОК-Э. Нейрокинины,
связываясь с рецепторами на КОК, могут как усиливать, так и тормозить
эффект гемопоэтических цитокинов на кроветворные клетки-предшест-
венницы.
Например, связывание субстанции Р с рецепторами клеток-предшест-
венниц активирует взаимодействие различных гемопоэтических цитокинов
(ИЛ-3, ИЛ-6, КСФ-ГМ, ФСК и др.) с рецепторами этих цитокинов на
указанных клетках-мишенях, вызывая сигнал трансдукции, направленный
к геному клетки с последующей активацией пролиферации и дифферен-
циации клеток. Напротив, нейрокинин А, секретируемый клетками стро-
мы, связываясь с другими рецепторами этих же кроветворных клеток-
предшественниц, взаимодействует с трансформирующим ростовым факто-
ром р, снижая пролиферативный эффект кроветворных клеток. Это свиде-
12-6095
354 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
тельствует, что регуляция гемопоэза осуществляется в интересах целого
организма и приспособительные реакции кроветворной ткани обеспечива-
ются не только цитокинами, но и передачей к ней нервными волокнами
регулирующих сигналов.
В регуляцию гемопоэза «включен» механизм ингибирования гумораль-
ной обратной связью клеток-предшественниц продуктами, образуемыми
клетками разных линий на последних этапах их созревания. Кислый изо-
ферритин, продуцируемый зрелыми нейтрофилами, тормозит их новооб-
разование из КОК-Г; образуемый а-гранулами тромбоцитов «трансформи-
рующий рост фактор-p» угнетает развитие КОКмег; простагландины се-
рии Е, а- и p-интерфероны, продуцируемые макрофагами, тормозят про-
лиферацию КОКмон; вещества ядер, вытолкнутых нормобластами, кейло-
ны, образуемые зрелыми эритроцитами, тормозят дифференциацию КОК-
Э. К цитокинам, тормозящим гемопоэз, относят опухольнекротизирую-
щий фактор (ОНФ). Его рецепторы представлены на мембране различных
КОК. Активация рецепторов ОНФ, в частности fas-рецептора опухольнек-
ротизирующего фактора, на КОК вызывает выраженное угнетение гемо-
поэза и «запускает» апоптоз-запрограммированную смерть клетки-пред-
шественницы. Fas-опосредованный апоптоз стволовых кроветворных кле-
ток лежит в основе ряда наследуемых форм недостаточности костного
мозга.
7.5.3. Роль стромы гемопоэтических органов в регуляции
кроветворения
Строма костного мозга [макрофаги, соединительнотканная оболочка, вы-
стилающая костномозговую полость (эндост), костномозговые синусоиды,
жировые клетки, соединительная ткань и нервные окончания] создает «ге-
мопоэтическое индуцирующее микроокружение», необходимое для пролифе-
рации, дифференциации и фиксации СКК в костном мозге, размножения
и созревания миелоидных клеток. Оно включает ростковые факторы, а
также экстрацеллюлярный матрикс, содержащий фибронектин, ламинин,
коллаген и гликозаминогликаны.
Делящиеся КОК взаимодействуют преимущественно с эндостом, яв-
ляющимся источником ростовых факторов. Более зрелые клетки распола-
гаются ближе к костномозговым синусоидам, что облегчает выход созре-
вающих клеток крови через стенку синусоида в кровь.
СКК, КОК-ГММЭ и малодифференцированые бипотенциальные клет-
ки постоянно выходят из костного мозга в кровь и транспортируются в
другие участки кроветворной ткани. Это обеспечивает постоянный обмен
клетками между анатомически разделенными участками кроветворной тка-
ни. Однако пролиферировать и дифференцироваться в клетки крови кро-
ветворные клетки-предшественницы способны лишь в ткани костного
мозга, и необходимые для этого условия создает ГИМ. Так, адгезивный
гликопротеин — фибронектин выполняет «якорную», фиксирующую функ-
цию в отношении КОК-ГММЭ и эритроидных КОК и одновременно уве-
личивает их пролиферацию. Особые же микроворсинки ретикулярных и
эндотелиальных клеток стромы фиксируют СКК и КОК-ГМ.
Строма костного мозга продуцирует КСФгм, КСФг, КСФмон, ИЛ-6, а
также особый фактор — модулятор, «фактор стволовой клетки», который
не вызывает пролиферации КОК, но резко усиливает пролиферативный
эффект КСФ на эти клетки. При дефекте его воспроизводства стромой
возникает нарушение кроветворения.
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 355
Нейтральные и кислые гликозаминогликаны увеличивают концентра-
цию цитокинов в непосредственной близости от клеток-мишеней, а также
проницаемость мембран гемопоэтических клеток для кальция. Цитокины
активируют в КОК формирование гемопоэтических вторичных мессендже-
ров, получивших название «сигнальные трансдукторы и активаторы транс-
крипции» (STAT), «ядерные факторы» (NF-E1, NF-E2, NF-kb/REL). Вто-
ричные мессенджеры обеспечивают быстрое распространение сигнала от
возбужденного цитокином рецептора гемопоэтической клетки к ее гено-
му, активируют транскрипцию его участков, ответственных за специализа-
цию клетки-предшественницы, т. е. дифференциацию ее в направлении
определенной клеточной линии — эритроидной, моноцитарной, нейтро-
фильной и т. д. Нарушение формирования вторичных мессенджеров в
КОК, не позволяющее цитокину реализовать регулирующий эффект на
кроветворную клетку-предшественницу, приводит к нарушениям крове-
творения. Так, нарушение, вызванное мутацией в участке генома, ответст-
венного за синтез одного из транскрипционных факторов — NF-E1 обры-
вает развитие и влечет гибель эритроидных клеток на стадии проэритроб-
ластов.
7.5.4. Регуляция выхода форменных элементов крови из костного
мозга в кровеносное русло
Сосудистая сеть костного мозга начинается от артерии, проникающей в
кроветворную ткань через костный канал. Ответвления артерии формиру-
ют элементарную морфофункциональную единицу костного мозга — «сину-
соидальное дерево». Стенка синусоидов состоит из эндотелиальных клеток и
лежащих на них со стороны гемопоэтической ткани широких мононукле-
арных адвентициальных клеток. Гемопоэтическая ткань распространяется
между синусоидами. Созревающие эритроидные и гранулоцитарные клет-
ки, мегакариоциты и макрофаги прилегают к наружной поверхности сосу-
дистых синусов.
Зрелые клетки из костного мозга попадают в кровь через отверстия ме-
жду эндотелиальными клетками костно-мозговых синусоидов. Диаметр
этих отверстий — 2,3 мкм, т. е. много меньше проходящих через них в
кровь клеток. Поэтому мигрирующие клетки должны обладать хорошей
деформируемостью, чтобы преодолеть барьер и выйти в кровь. Плохо де-
формирующиеся ригидные ядра эритробластов, миелоцитов, промиелоци-
тов не позволяют им мигрировать через отверстия в синусоиде. Напротив,
хорошо деформирующиеся ядра зрелых гранулоцитов, моноцитов и лим-
фоцитов дают им возможность пересекать эндотелий.
Важным условием миграции клеток в кровь является способность рети-
кулоцитов и клеток белой крови к движению. Они пересекают узкие от-
верстия синусоидов, выпуская в них сначала цитоплазматические отрост-
ки-псевдоподии. В костномозговых синусоидах края эндотелиальных
клеток тесно прилегают друг к другу и не позволяют клеткам проходить
через отверстия между ними. Однако миграция клеток через них облегча-
ется ритмичными расслаблениями синусоидов, которые уменьшают пере-
крытие этими клетками пор. Поверхность эндотелиальных клеток частич-
но покрыта адвентициальными клетками, которые легко скользят по ней
благодаря сократительному аппарату, имеющемуся у адвентициальных кле-
ток. Сокращаясь, адвентициальные клетки уменьшают покрываемую ими
поверхность костномозговых синусоидов и увеличивают возможность ми-
грации клеток через стенку синусоидов в кровоток.
12*
356 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Выход клеток в просвет синусоида возрастает под влиянием молеку-
лярных регуляторов гемопоэза. Эритропоэтин стимулирует быстрый вы-
ход ретикулоцитов, КСФ и бактериальный эндотоксин — нейтрофилов.
Эритропоэтин ускоряет «сборку» цитоскелета ретикулоцитов, находящих-
ся в костном мозге, увеличивает их деформабельность, что облегчает их
выход через отверстия стенки синусоидов в кровь. Регуляторы облегчают
формирование отверстий в эндотелии, уменьшая внешнюю поверхность
эндотелия костномозгового синуса, покрытого адвентициальными клетка-
ми. Например, инъекция животному эритропоэтина резко уменьшает его
адвентициальный покров и ретикулоциты легко пересекают стенку сину-
соида.
В норме небольшая часть клеток не достигает стадии созревания, поги-
бает в костном мозге и подвергается фагоцитозу макрофагами, распола-
гающимися на наружной поверхности костномозговых синусоидов. При-
менительно к эритроидному ряду это явление называется неэффективным
эритропоэзом, применительно к гранулоцитарному — неэффективным гра-
нулопоэзом. Неэффективный гемопоэз охватывает от 2 до 10 % эритробла-
стов и от 10 до 15 % костномозговых гранулоцитов. Их мембраны теряют
сиаловые кислоты, в результате уменьшается отрицательный заряд мембра-
ны и макрофаги легко фагоцитируют эти клетки. Неполноценные клетки в
кровоток не поступают.
7.5.5. Особенности метаболизма кроветворной ткани
Масса костного мозга у взрослого человека составляет 4,6 % от массы тела,
или 3,4 кг, в том числе масса красного костного мозга — 1,7 кг. Общее ко-
личество ядросодержащих клеток достигает в среднем 8,1 • 109 на кг от мас-
сы тела, а занимаемый ими объем колеблется от 1320 до 4192 мл.
Клетки костного мозга представляют одну из наиболее пролиферирую-
щих тканей организма. Для осуществления митоза клеток используется
энергия макроэргических соединений, образующихся в процессе окисли-
тельного фосфорилирования. Энергообмен в ткани костного мозга усили-
вается при повышении пролиферативной активности. Например, усиление
эритропоэза после кровопотери активирует в костном мозге процессы
аэробного окисления, увеличивает скорость потребления кислорода клет-
ками, интенсивность дыхания и окислительного фосфорилирования в их
митохондриях. Одновременно раскрываются нефункционирующие сину-
соиды и увеличивается кровоток в костном мозге, что укорачивает путь
диффузии кислорода от отдельного сосуда к клетке. При активации гемо-
поэза в костном мозге возрастает интенсивность синтеза ядерных и мито-
хондриальных ДНК, РНК и белков, увеличивается количество и площадь
поверхности митохондрий, растет общее число рибосом и потребление
свободных аминокислот. Например, усиление регенерации эритроцитов
увеличивает потребление в ткани костного мозга лизина, серосодержащих,
ароматических аминокислот, глутаминовой кислоты. При их дефиците ре-
генерация красной крови замедляется. Усиление пролиферации гемопо-
этических клеток требует повышения проницаемости их мембран, что
обеспечивается нарастанием интенсивности перекисного окисления липи-
дов. Активации гранулопоэза и, особенно, эритропоэза предшествует ре-
зорбция жировой ткани костного мозга. Высвобождающиеся из нее гемо-
поэтические цитокины- КСФгм, КСФг активируют гемопоэз, а полинена-
сыщенные жирные кислоты используются в ходе гемопоэза для формиро-
вания клеток крови.
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 357
7.6. Роль витаминов и микроэлементов в кроветворении
Витамин В12 и фолиевая кислота (витамин В9) необходимы для синтеза
нуклеопротеинов в разных тканях организма, созревания и деления ядер
эритроидных клеток в кроветворной ткани. При дефиците витаминов В12 и
В9 в наиболее интенсивно делящейся ткани организма — эритроидной —
раньше, чем в других, возникают нарушения, вызывающие анемию. При
дефиците витамина В12 в костном мозге появляются большие ядросодержа-
щие эритроидные клетки — мегалобласты, которые образуют с замедлен-
ной скоростью большие эритроциты — мегалоциты с резко укороченным
периодом жизни. Замедление поступления эритроцитов в кровь и быстрое
их разрушение ведет к анемии. Дефицит витамина В12 возникает в орга-
низме при утрате париетальными клетками желудка способности продуци-
ровать «внутренний фактор» — гликопротеин (молекулярная масса 60 000).
Фактор связывает витамин В12, поступающий с пищей, и предохраняет его
от расщепления пищеварительными ферментами. Эти нарушения возника-
ют при атрофии слизистой оболочки желудка, эпителия двенадцатиперст-
ной кишки, часто наблюдающейся, например, у стариков. И хотя запаса
витамина В12 в печени достаточно взрослому человеку на 1—5 лет, посте-
пенное его истощение приводит к заболеванию. Суточная потребность в
витамине В12— 5 мкг, содержание в плазме крови — 150—450 мкг/л. В ки-
шечнике комплекс гликопротеин—витамин В12 фиксируется специальны-
ми рецепторами слизистой оболочки тонкого кишечника, далее витамин
поступает в интестинальные клетки, затем в кровь и переносится с помо-
щью особых молекул — транскобаламинов (I, II, III типов). Транскобала-
мины I и III типа продуцируются лейкоцитами, II — макрофагами. Поэто-
му при выраженном лейкоцитозе отмечается гипервитаминоз В12. Витамин
В12 содержится в больших количествах в печени, почках, куриных яйцах.
Фолиевая кислота (витамин В9) поддерживает синтез ДНК в клетках ко-
стного мозга, благодаря обеспечению этого процесса одним из нуклеоти-
дов — диокситимидилатом, образующимся в результате митилирования ди-
оксиуридиловой кислоты в присутствии тетрагидрофолата (одной из реду-
цированных форм фолиевой кислоты). При дефиците витамина В9 в пище
у человека уже через 1—6 мес нарушается синтез ДНК и деление эритро-
идных клеток, ускоряется разрушение эритроцитов, что ведет к анемии.
Суточная потребность организма человека в фолиевой кислоте 500—700
мкг. Ее резерв в организме равен 5—10 мг, '/3 которого находится в пече-
ни. Фолиевой кислотой богаты овощи (шпинат), дрожжи, молоко.
1,2,5- дигидроксивитамин D3 и ретиноловая кислота (производное вита-
мина А) участвуют в организме человека в дифференциации кроветворных
клеток до их зрелых форм. Витаминами А и D богаты печень трески, тун-
ца, сельдь, коровье молоко и сливочное масло.
Витамин В6 (пиридоксин) является кофактором (т. е. дополнительным
фактором активности) фермента — синтетазы 8-аминолевулиновой кисло-
ты (АЛК-синтетазы), участвующей в образовании гема в эритроидных
клетках в ткани костного мозга (см. рис. 7.2). Дефицит витамина В6 в орга-
низме человека нарушает синтез гемоглобина и вызывает анемию. Витами-
ном В6 богаты зерна злаков, капуста, картофель, молоко.
Витамин С поддерживает основные этапы эритропоэза, способствуя ме-
таболизму фолиевой кислоты в эритробластах. Он участвует в метаболизме
железа, повышая как его абсорбцию в желудочно-кишечном тракте, так и
мобилизацию депонированного в клетках железа.
Витамин Е (а-токоферол) и витамин РР защищают фосфатидилэтанол-
358 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
амин мембраны эритроцитов от усиливающего их гемолиз перекисного
окисления.
Защиту гемоглобина и мембраны эритроцитов от окисления осуществ-
ляет и витамин РР, являющийся одной из составляющих пиридиннуклео-
тидов НАД и НАДФ.
Дефицит витамина В2, участвующего в окислительно-восстановитель-
ных реакциях, вызывает у человека анемию вследствие замедленного обра-
зования эритроцитов в костном мозге.
Микроэлементы: медь способствует всасыванию ионов железа в кишеч-
нике и мобилизации его резерва из печени и макрофагов; никель и кобальт
способствуют утилизации железа, поддерживая синтез гемоглобина в эрит-
роидных клетках и гемсодержащих молекул в тканях. Кобальт входит в со-
став витамина В12. При дефиците кобальта в организме человека возникает
мегалобластическая анемия. Никель индуцирует синтез фосфатидилсери-
на, стабилизирующего мембрану эритроцитов. При дефиците никеля уси-
ливается выведение ионов железа из организме человека. Селен, тесно воз-
действуя с витамином Е, защищает мембрану эритроцита от повреждения
свободными радикалами. Почти 75 % всего цинка в организме человека
находится в эритроцитах в составе фермента карбоангидразы, цинк входит
в состав транскрипционных факторов, регулирующих активность генома
гемопоэтических клеток.
7.7. Основы трансфузиологии
Трансфузиология (от лат. transfusio — переливание) — отрасль медицинской
науки, изучающая способы и средства управления функциями организма
путем воздействия на него переливания цельной крови, ее компонентов
(эритроцитов, лейкоцитов, тромбоцитов, стволовых кроветворных клеток,
плазмы крови) и кровезаменителей.
Переливание крови или ее компонентов от человека человеку основы-
вается на знании антигенных (иммунных) свойств клеток и белков крови.
7.7.1. Группы крови
Эритроцитарные антигены. Часть из более чем 300 антигенов мембраны
эритроцитов человека объединена в 23 генетически контролируемые систе-
мы групп крови (ABO, Rh-Hr, Дафи, М, N, S, Леви, Диего). Система анти-
генов эритроцитов АВО содержит в сыворотке крови естественные анти-А
и анти-В антитела. Генетический локус, контролирующий образование ан-
тигенов этой системы, расположен в длинном плече 9-й хромосомы и
представлен генами Н, А, В и 0. Гены А, В, Н контролируют синтез фер-
ментов, которые формируют особые моносахариды или антигены мембра-
ны эритроцита — А, В и Н. Образование антигенов начинается с гена Н,
который через контролируемый им энзим гликолизилтранферазу формиру-
ет из особого вещества-предшественника — церамидпентасахарида — анти-
ген Н эритроцитов. Далее гены А и В через активность контролируемых
ими энзимов формируют из Н-антигена, являющегося для них исходным
материалом, антигены А или В. Ген «0» не контролирует трансферазу, и Н-
антиген остается неизменным, формируя группу крови 0(1). Таким обра-
зом, на мембране эритроцитов человека присутствуют антигены А, В и Н.
У 20 % людей антиген А имеет антигенные отличия (А! и А2). Антитела
против антигенов А, А„ А2 и В начинают формироваться после рождения
человека иммунной системой в ответ на стимуляцию ее антигенами пищи
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 359
Таблица 7.3. Группы крови системы АВО
Ген Антигены на мем- бране эритроцита Антитела (в плазме крови) Группа крови сис- темы АВО
Н(О) Н Анти-А+ Анти-А2+ Анти-В 1(0)
А1 А, Анти-В П(А])
А2 а2+н Анти-А] у 1 % обследуемых + Анти-В П(А2)
В В Анти-А+ Анти-А, Ш(В)
А„ В А,+В Отсутствуют IV(A„ В)
а2, в а2+в Анти-А] у 25 % обследуемых IV(A2,B)
Таблица 7.4. Определение группы крови системы АВО
Группа крови ис- следуемых эритро- цитов Антитела (антисыворотки), добавляемые к исследуемым эритроцитам
анти-А анти-В
0(1) — —
А (II) + —
В (III) — +
А, В (IV) + +
«—» — агглютинация отсутствует, «+» — агглютинация эритроцитов.
и бактерий, поступающих, например, в организм с вдыхаемым воздухом.
Максимум продукции анти-А и анти-В антител приходится на 8—10-лет-
ний возраст. При этом в плазме крови накапливается анти-А больше, чем
анти-В. Антитела анти-А и анти-В называются изоантителами, или агглю-
тининами, а соответствующие антигены мембраны — агглютиногенами. Ха-
рактеристика групп крови системы АВО представлена в табл. 7.3.
Естественные анти-А- и анти-В-антитела принадлежат к иммуноглобу-
линам класса М. Выработанные в процессе иммунизации А или В антиге-
ном анти-А- и анти-В-антитела являются иммунными и относятся к имму-
ноглобулинам класса G. Иммуноглобулины склеивают эритроциты (явле-
ние агглютинации) и вызывают их гемолиз. При несовместимости группы
крови донора (т. е. человека, у которого берут кровь для переливания) и ре-
ципиента (т. е. человека, которому переливают кровь) переливание крови
вызывает гемоконфликт, связанный с агглютинацией и гемолизом эритро-
цитов, заканчивающийся гибелью реципиента. Для исключения гемокон-
фликтов человеку переливают лишь одногруппную кровь. Для определения
группы крови по системе АВО смешивают антитела анти- А и анти- В с ис-
следуемыми эритроцитами и по наличию или отсутствию агглютинации
эритроцитов определяют группу крови (табл. 7.4).
Rh-Hr антигены эритроцитов. Rh-антигены представлены на мембране
эритроцитов тремя связанными участками: антигенами С (rh') или с (hr"),
Е (rh") или е (hr") и D (Rho) или d. Из этих антигенов сильным является
D, он способен иммунизировать человека, у которого антиген D отсутству-
ет. Люди, имеющие D-антиген, называются «резус-положительными» (Rh+),
среди европейцев их 85 %, а не имеющие его — «резус-отрицательными»
(Rh-) (15 %). У некоторых народов, например эвенков, отмечается 100 %
Rh+ принадлежность.
Резус-положительная кровь донора образует иммунные антитела (анти-
D) у резус-отрицательного реципиента. Повторное переливание резус-по-
ложительной крови может вызвать гемоконфликт. Подобная же ситуация
360 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
возникает у резус-отрицательной женщины, беременной резус-положи-
тельным плодом. Во время родов (или аборта) эритроциты плода поступа-
ют в кровь матери и иммунизируют ее организм (вырабатывают анти-D-
антитела). При последующих беременностях резус-положительным плодом
анти-D-антитела проникают через плацентарный барьер, повреждают тка-
ни и эритроциты плода, вызывая выкидыш, а при рождении ребенка — ре-
зусную болезнь, одним из проявлений которой является гемолитическая
анемия. Для профилактики иммунизации резус-отрицательной женщины
D-антигёнами плода во время родов или абортов ей вводят концентриро-
ванные анти-Э-антитела. Они агглютинируют резус-положительные эрит-
роциты плода, поступающие в ее организм, и имммунизации не наступает.
Слабые резусные антигены С и Е при их значительном поступлении в ор-
ганизм резус-положительного человека могут вызвать антигенные реакции.
Чаще всего антитела к антигенам эритроцитов системы резус являются им-
муноглобулинами класса G. Для выявления этих антител используют анти-
глобулиновую сыворотку крови, содержащую анти-^С-антитела.
Антигенные свойства лейкоцитов и тромбоцитов. В настоящее время
вместе с эритроцитарными антигенами открыто более 500 антигенов кле-
ток крови и белков плазмы, которые создают свыше 40 различных анти-
генных систем крови, сочетания же различных антигенных систем образу-
ют в человеческой популяции множество антигенных комбинаций. Поэто-
му, несмотря на подбор донора и реципиента по системам АВО эритроци-
тов, всегда имеется несовместимость в других антигенных структурах их
крови, приводящая к иммунизации организма реципиента.
На мембранах лейкоцитов помимо антигенов системы ABO, MN, Левис
содержатся антигены гистосовместимости HLA (от Human Leucocyte Anti-
gens), представленные более чем 150 антигенами, а также антигены не-
скольких других генетических систем — NA, NB, NC, ND, NE и др. Анти-
гены гистосовместимости HLA представлены и на поверхности тромбоци-
тов. Несовместимость по антигенам HLA комплекса у донора и реципиен-
та при переливании цельной крови и ее компонентов (лейкоцитарной или
тромбоцитарной массы) приводит к иммунизации реципиента. Минималь-
ное количество лейкоцитов, вызывающее иммунизацию реципиента, со-
ставляет 1 • 10б. Образовавшиеся в организме реципиента антитела против
антигенов системы HLA или антигенов лейкоцитов NA-NE при повторном
переливании крови реципиенту вызывают различные осложнения (лихо-
радку, возникающую в результате освобождения пирогенных веществ из
поврежденных антителами лейкоцитов; антитела вызывают разрушение до-
норских лейкоцитов и тромбоцитов и др.). Такие реакции могут развивать-
ся у реципиентов после повторного переливания цельной крови, эритро-
цитарной или тромбоцитарной массы, плазмы крови, так как лейкоцитар-
ные антигены находятся в растворимой форме в плазме крови, присутству-
ют на поверхности других клеток крови. Из сказанного следует, что транс-
фузия цельной крови от донора реципиенту, даже имеющим одну и ту же
группу крови по системе АВО и Rh-Hr, тем не менее, может привести к ос-
ложнениям, связанным с поступлением в организм реципиента лейкоци-
тов и тромбоцитов, имеющих разные с лейкоцитами и тромбоцитами ре-
ципиента антигенные характеристики, что вызывает иммунизацию и по-
слетрансфузионную реакцию у реципиента. Поэтому цельную кровь для
переливания используют в клинике крайне редко, лишь по экстренным
жизненным показаниям, например с целью полного восполнения крово-
потери у детей, весьма чувствительных к гипоксии, вызванной острой кро-
вопотерей. Кроме того, переливание цельной свежезаготовленной крови
7. Функции клеток крови. Гемостаз. Регуляция кроветворения. Трансфузиология • 361
несет опасность передачи инфекции реципиенту, если донор находился в
латентном (скрытом), не поддающемся диагностике периоде заболевания
гепатитом В, С, D, СПИДом, сифилисом и малярией. Поэтому больным с
целью восстановления газотранспортной функции крови переливают эрит-
роцитарную массу, отмытую от плазмы крови, обедненную или лишенную
лейкоцитов и тромбоцитов. Так, 5-кратное отмывание эритроцитов изото-
ническим раствором NaCl удаляет из эритроцитарной массы до 80 % лей-
коцитов и почти все тромбоциты и эффективно предупреждает иммуниза-
цию организма реципиента.
7.7.2. Влияние переливаемой крови и ее компонентов на организм
человека
Перелитая пациенту кровь или ее компоненты позволяют восстановить
объем потерянной крови, коллоидно-осмотическое давление крови, газо-
транспортную, кроветворную, защитную (иммунную), питательную, регу-
ляторную, гемостатическую функции крови, стимулируют гомеостатиче-
ские механизмы, поддерживающие объем жидкостей различных секторов
организма.
После кровопотери взрослый человек может выжить без переливания
крови или кровезаменителей, если потеря циркулировавших в сосудистом
русле эритроцитов не превысит 2/3, а плазма крови — */3 от их исходного
количества. Большая потеря объема циркулирующих эритроцитов и объе-
ма циркулирующей плазмы без возмещения их потери приводит организм
человека к гибели. Для получения лечебного эффекта дефицит потерянных
с кровопотерей эритроцитов восполняют на 80 %, а плазмы —на 150—
170 %, т. е. объем плазмы крови, переливаемой больному, должен быть
больше утраченного с кровопотерей. Введение избыточного объема плазмы
улучшает реалогические характеристики циркулирующей крови, способст-
вует вымыванию эритроцитов из депо в циркуляцию. Например, с целью
восстановления газотранспортной функции крови больным, перенесшим
кровопотерю или страдающим анемией с уменьшением гемоглобина ниже
80 г/л крови, переливают одногруппную (по системе AB0) эритроцитарную
массу, полученную от одного донора, и изотонический раствор NaCl, уст-
раняя дефицит эритроцитов в организме больного на 80 %, а плазмы кро-
ви — на 130—150 % от нормального уровня.
Переливание одногруппной крови (по системе AB0) от разных доноров
одному реципиенту может вызвать сильное иммунологическое воздействие
на организм реципиента и синдром «гомологичной крови», связанный с
появлением микроагрегатов клеток крови (до 150—200 мкм), вызывающих
закупорку мелких легочных и мозговых сосудов реципиента, других, опас-
ных для жизни осложнений.
С целью восстановления тромбоцитарного звена гемостаза (например,
при тромбоцитопении или функциональной недостаточности тромбоци-
тов) переливают тромбоцитарный концентрат. Так, при снижении тромбо-
цитов у человека ниже 20 • 109/л крови могут возникать самопроизвольные,
внезапные кровотечения в желудочно-кишечном тракте, внутричерепные,
что требует переливания таким больным тромбоцитарного концентрата.
Лечебная доза тромбоцитарного концентрата, обеспечивающая прекраще-
ние кровоточивости, содержит не менее 2,8—3,0 • 10” тромбоцитов.
Трансфузия концентратов лейкоцитов производится с целью повыше-
ния иммунных и защитных сил организма больных (например, с гнойно-
деструктивными заболеваниями легких, при стафилококковом сепсисе,
362 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
гнойном менингите и др.). Концентрат лейкоцитов (состоящий из грану-
лоцитов и лимфоцитов) донора должен быть совместим с лейкоцитами ре-
ципиента по системам АВО, Rh-Hr и лейкоцитарным антигенам. Лечебная
доза концентрата, переливаемая больному, содержит от 10 • 109 до 40 • 109
лейкоцитов. При использовании концентрата лейкоцитов или тромбоци-
тов должно строго выполняться правило «один донор — один реципиент».
Переливание плазмы крови производится с целью поддержания плазмен-
но-коагуляционного гомеостаза (например, для остановки кровотечения
больному с гемофилией, страдающему дефицитом VIII фактора); с целью
создания пассивного гуморального иммунитета (например, при инфекции
организма синегнойной палочкой или стафилококками больным перелива-
ют антисинегнойную плазму донора, содержащую антитела против синег-
нойной палочки, антистафилококковую — с антителами против стафило-
кокков, повышая устойчивость организма больных против этих инфек-
ций); с целью заместительной терапии — например, при ожоговой болез-
ни, когда имеет место выраженная потеря плазмы, связанная с выходом
плазмы крови из сосудов, вызывающая сгущение крови и ухудшение кро-
воснабжения тканей. Так, при ожоговом поражении до 10—15 % площади
тела плазмопотеря достигает 6—8 л в сутки и ее замещение производят
плазмой крови и кровезаменителями. Средняя доза вводимой плазмы —
500 мл.
Переливаемая плазма должна быть одной группы с кровью реципиента
по системе АВО. В чрезвычайных ситуациях, когда отсутствует одногруп-
пная плазма, переливают плазму донора группы А(П) или В(Ш) реципиен-
ту с 0 (I) группой крови и плазму АВ (IV) группы реципиентам любой
группы крови.
При дефиците донорской крови и ее компонентов для лечения острой
кровопотери, травматического шока, плазмопотери (при ожоговой травме)
используют кровезаменители, которые восстанавливают объем циркули-
рующей крови, нарушенную гемодинамику (повышают до нормального
значения уровень артериального давления, нормализуют микроциркуля-
цию), коллоидно-осмотическое давление крови.
В качестве кровезаменителей, восстанавливающих нарушенное крово-
потерей кровообращение, используют, например, декстран — водораство-
римый высокомолекулярный (мол. масса 80 000) полимер глюкозы — и
созданные на его основе полиглюкин, полиглюсоль, реополиглюкин. Кро-
везаменители, благодаря молекулярной массе, препятствующей выходу мо-
лекул через стенку сосудов в интерстиций, поддерживают высокое колло-
идно-осмотическое давление плазмы крови, вследствие чего способствуют
движению тканевой жидкости в кровоток, поддерживают нормальный объ-
ем циркулирующей крови.
ГЛАВА 8
Иммунная система *
Во внутренней среде организма присутствуют клетки и молекулы, кото-
рые обеспечивают защитную функцию (иммунитет). Часть из них относят-
ся к факторам врожденного иммунитета, т. е. присутствуют в организме
еще до встречи с каким-либо болезнетворным микроорганизмом или чу-
жеродной молекулой. К таким факторам защиты относятся фагоцитирую-
щие клетки крови и тканей, а также большие гранулярные лимфоциты, по-
лучившие название «натуральные (естественные) киллеры». Врожденными
факторами защиты организма являются также молекулы, продуцируемые и
секретируемые вышеназванными клетками, а также лимфоцитами, стро-
мальными и другими клетками. К таким молекулам относятся: белки сис-
темы комплемента, медиаторы межклеточного взаимодействия — цитокины
и др. Белки системы комплемента продуцируются постоянно и циркулиру-
ют в крови, большинство цитокинов начинают продуцироваться и секре-
тироваться клетками только в ответ на сигнал активации (например, на
контакт с микроорганизмом).
Более эффективным способом защиты внутренней среды организма от
проникающих в нее чужеродных агентов (антигенов) является специфиче-
ский иммунный ответ, в результате которого организм приобретает допол-
нительные защитные механизмы: активированные клетки и продуцируе-
мые ими молекулы. Защитное действие этих механизмов строго избира-
тельно (специфично) в отношении того конкретного антигена (например,
патогенного микроорганизма), контакт с которым вызвал иммунный ответ.
Специфический иммунный ответ является функцией клеток и органов им-
мунной системы.
Рис. 8.1. Рециркуляция лимфоцитов.
Из центральных органов иммунной системы (костного мозга и тимуса) созревающие лимфо-
циты поступают в кровь и заселяют периферические органы и ткани иммунной системы, от-
куда снова поступают в лимфоток, в кровоток, продолжая рециркуляцию.
’Глава написана членом-корреспондентом РАМН И. С. Фрейдлин.
364 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Главными клетками иммунной системы являются Т- и В-лимфоциты,
которые рециркулируют в системе кровотока и лимфотока, постоянно пе-
ремещаясь из одних органов иммунной системы в другие, обладают спо-
собностью выходить в ткани для выполнения защитных функций
(рис. 8.1).
В защитных реакциях специфического иммунитета кроме Т- и В-лим-
фоцитов участвуют фагоцитирующие клетки (гранулоциты, моноциты,
макрофаги), «естественные киллеры, тучные клетки, эндотелиальные и
эпителиальные клетки, которые играют роль вспомогательных, взаимодей-
ствуя с Т- и В-лимфоцитами.
8.1. Происхождение и функции клеток иммунной системы
Лимфоциты, как и другие клетки крови, ведут свое происхождение из еди-
ной стволовой гемопоэтической клетки. Лимфопоэз имеет черты сходства с
общим гемопоэзом: зависит от контактов клеток-предшественниц со стро-
мальными клетками и от влияния цитокинов. Вместе с тем лимфопоэзу
присущи некоторые особенности: возникновение в процессе пролифера-
ции клеток-предшественниц широкого разнообразия Т- и В-лимфоцитов с
разными антигенраспознающими рецепторами; гибель путем апоптоза зна-
чительной части пролиферирующих клеток-предшественниц; избиратель-
ное накопление в результате пролиферации клеток — потомков одной
клетки, первоначально активированной при контакте с соответствующим
антигеном, «клона» клеток, все клетки которого несут одинаковые по спе-
цифичности антигенраспознающие рецепторы.
Лимфоциты — это единственные клетки организма, способные специ-
фически распознавать собственные и чужеродные антигены и отвечать ак-
тивацией на контакт с конкретным антигеном. При весьма сходной мор-
фологии малые лимфоциты делятся на две популяции, имеющие различ-
ные функции и продуцирующие разные белки (табл. 8.1).
Одна из популяций получила название В-лимфоциты, от названия орга-
на «бурса Фабрициуса», где было впервые обнаружено созревание этих
клеток у птиц. У человека В-лимфоциты созревают в костном мозге. В-
лимфоциты распознают антигены специфическими рецепторами иммуно-
глобулиновой природы, которые по мере созревания В-лимфоцитов появля-
ются на их мембранах. Взаимодействие антигена с такими рецепторами яв-
ляется сигналом активации В-лимфоцитов и их дифференцировки в плаз-
Таблица 8.1. Классификация лимфоцитов человека
Свойства клеток В-лимфоциты Т-лимфоциты NK-клетки
Органы, где происхо- дит развитие клеток Морфологические особенности Антигенраспознаю- щий рецептор Содержание в крови (% от общего количе- ства лимфоцитов) Функции Костный мозг Малые лимфоциты Иммуноглобулин 8-20 % Предшественники антителопродуцен- тов (плазмоцитов) Костный мозг, ти- мус Малые лимфоциты Т-клеточный рецеп- тор — димер 65-80 % Предшественники эффекторных и ре- гуляторных клеток Костный мозг, се- лезенка Большие грануляр- ные лимфоциты Отсутствует 5-20 % Естественные (на- туральные) киллеры
8. Иммунная система • 365
матические клетки, продуцирующие и секретирующие специфические для
данного антигена антитела — иммуноглобулины.
Другая популяция получила название Т-лимфоциты в связи с диффе-
ренцировкой их предшественников в тимусе. Эта популяция включает раз-
ные субпопуляции, среди которых наиболее отчетливо различаются Т-хел-
перы (CD4+) и цитотоксические Т-лимфоциты (CD8+). В отличие от В-лим-
фоцитов, способных распознавать и связывать белковые, полисахаридные
и липопротеидные растворимые антигены, Т-лимфоциты могут распозна-
вать только короткие пептидные фрагменты белковых антигенов на по-
верхности вспомогательных антигенпредставляющих клеток (дендритных,
макрофагов или В-лимфоцитов) в комплексах с собственными молекулами
главного комплекса гистосовместимости. Связывание антигена с антиген-
распознающим рецептором на клеточной мембране индуцирует активацию
Т-лимфоцита, которая проявляется продукцией и секрецией цитокинов,
усиливающих процессы пролиферации и дифференцировки самих Т-лим-
фоцитов, В-лимфоцитов и макрофагов.
Т-лимфоциты выполняют важнейшую функцию специфического распо-
знавания и связывания антигена, что ведет к их активации, пролиферации
и дифференцировке разных субпопуляций Т-лимфоцитов, участвующих
далее во всех формах иммунного ответа.
Главной функцией В-лимфоцитов также является специфическое рас-
познавание антигена, которое ведет к их активации, пролиферации и диф-
ференцировке в плазматические клетки — продуценты специфических ан-
тител — иммуноглобулинов, т. е. к гуморальному иммунному ответу. Чаще
всего В-лимфоциты для развития гуморального иммунного ответа нужда-
ются в помощи Т-лимфоцитов в виде продукции активирующих цито-
кинов.
Имеется еще один тип лимфоцитов — большие гранулярные лимфоци-
ты, отличающиеся от меньших по величине Т- и В-лимфоцитов не только
особенностями структуры, но и отсутствием антигенраспознающего рецеп-
тора. Эти клетки получили название «естественные киллеры»: они способ-
ны убивать инфицированные разными вирусами клетки-мишени или опу-
холевые клетки (см. табл. 8.1).
Все процессы транспортировки лимфоцитов из первичных во вторич-
ные органы иммунной системы, из вторичных органов иммунной системы
в ткани зависят от взаимодействия особого типа цитокинов — хемокинов с
соответствующими рецепторами на поверхности клеток. Кроме хемокинов
миграция лимфоцитов зависит от присутствия на их поверхности адгезион-
ных молекул, которым соответствуют молекулы-лиганды на поверхности
эндотелиальных клеток сосудов, на поверхности макрофагов и дендритных
клеток, на элементах внеклеточного матрикса.
8.1.1. Т-лимфоциты
8.1.1.1. Характеристики Т-лимфоцитов
Решающее событие в развитии Т-лимфоцитов — формирование антиген-
распознающего Т-клеточного рецептора — происходит только в тимусе.
Для обеспечения возможности распознавания любого антигена нужны
миллионы различных по специфичности антигенраспознающих рецепто-
ров. Формирование огромного разнообразия антигенраспознающих рецеп-
торов возможно благодаря перестройке генов в процессе пролиферации и
дифференцировки клеток-предшественниц.
366 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 8.2. Типы молекул на поверхности Т-лимфоцитов
Молекулы Функции
Антигенраспознающий рецеп- тор: Т-клеточный рецептор Распознавание и связывание комплекса: антигенный пептид + собственная молекула главного комплекса гистосовместимости
Корецепторы: CD4, CD8 Участвуют в связывании молекулы главного комплек- са гистосовместимости
Адгезионные молекулы Адгезия лимфоцитов к эндотелиальным клеткам, к антигенпредставляющим клеткам, к элементам вне- клеточного матрикса
Костимулирующие молекулы Участвуют в активации Т-лимфоцитов после взаимо- действия с антигеном
Рецепторы иммуноглобулинов Рецепторы цитокинов Связывают иммунные комплексы Связывают цитокины
По мере созревания Т-лимфоцитов на их поверхности появляются ан-
тигенраспознающие рецепторы и другие молекулы, опосредующие их
взаимодействие с антигенпредставляющими клетками. Так, в распознава-
нии собственных молекул главного комплекса гистосовместимости (см.
раздел 8.3.2) наряду с Т-клеточным рецептором участвуют молекулы CD4
или CD8. Межклеточные контакты обеспечиваются наборами поверхност-
ных адгезионных молекул, каждой из которых соответствует молекула —
лиганд на поверхности другой клетки. Как правило, взаимодействие Т-
лимфоцита с антигенпредставляющей клеткой не ограничивается распо-
знаванием антигенного комплекса Т-клеточным рецептором, а сопровож-
дается связыванием других попарно комплементарных поверхностных
«костимулирующих» молекул.
Сочетание поверхностных молекул лимфоцитов, которые принято обо-
значать порядковыми номерами «кластеров дифференцировки» (clusters of
differentiation — CD), обозначается как «поверхностный фенотип клетки», а
отдельные поверхностные молекулы называют «маркерами», так как они
служат метками конкретных субпопуляций и стадий дифференцировки Т-
лимфоцитов. Так, например, на поздних этапах дифференцировки одни Т-
лимфоциты утрачивают молекулу CD8 и сохраняют только CD4, а другие
утрачивают CD4, а сохраняют CD8. Поэтому среди зрелых Т-лимфоцитов
различают CD4+ (Т-хелперы) и CD8+ (цитотоксические Т-лимфоциты).
Среди циркулирующих в крови Т-лимфоцитов клеток с маркером CD4
примерно в два раза больше, чем клеток с маркером CD8.
Зрелые Т-лимфоциты несут на поверхности рецепторы для разных ци-
токинов и рецепторы для иммуноглобулинов (табл. 8.2).
При распознавании Т-клеточным рецептором антигена Т-лимфоциты
получают сигналы активации, пролиферации и дифференцировки в на-
правлении клеток-эффекторов, т. е. клеток, способных непосредственно
участвовать в защитных или повреждающих эффектах. Для этого на их по-
верхности резко возрастает количество адгезионных и костимулирующих
молекул, а также рецепторов для цитокинов. Активированные Т-лимфоци-
ты начинают продуцировать и секретировать цитокины, активирующие
макрофаги, другие Т-лимфоциты и В-лимфоциты.
После завершения инфекции, сопряженной с усиленной продукцией,
дифференцировкой и активацией Т-эффекторов соответствующего клона,
в течение нескольких дней 90 % эффекторных клеток погибают, поскольку
не получают дополнительных сигналов активации. В организме остаются
8. Иммунная система • 367
долгоживущие клетки памяти, несущие соответствующие по специфично-
сти рецепторы и способные ответить пролиферацией и активацией на по-
вторную встречу с тем же антигеном.
8.1.1.2. Субпопуляции Т-лимфоцитов
Основная функция CD4+ Т-лимфоцитов — продуцировать цитокины (см.
раздел 8.4.2), активирующие функции других клеток, в связи с этим дан-
ные клетки называют помощниками (helper) и обозначают соответственно
Th. Основной функцией СГ)8+Т-лимфоцитов является цитотоксичность,
т. е. способность убивать клетки-мишени, инфицированные соответствую-
щим по специфичности вирусом, поэтому их называют цитотоксическими
Т-лимфоцитами и обозначают CTL (табл. 8.3).
Таблица 8.3. Характеристики трех типов эффекторных Т-лимфоцитов
Типы клеток CD8+ Т-клетки CD4+ Т-клетки CD4+ Т-клетки
Обозначение Функция Основные продуци- руемые молекулы CTL Цитотоксическое дейст- вие на клетки-мишени Перфорины, грензимы ТЫ Активация макро- фагов ИФНу, ГМ-КСФ, ТНФа, CD40 L ТЬ2 Активация В-лимфоцитов ИЛ-4, ИЛ-5, ИЛ-6, CD40 L
С04+Т-лимфоциты после распознавания антигена начинают секретиро-
вать интерлейкины (ИЛ-2, ИЛ-4), гамма-интерферон и дифференцируются
в двух направлениях. Одни — Th 1 — приобретают способность активиро-
вать макрофаги и участвовать в клеточном иммунном ответе. Другое на-
правление дифференцировки ведет к формированию Th2, способных акти-
Рис. 8.2. Цитокиновый баланс, от которого зависит преобладание клеточного или
гуморального иммунного ответа. Превалирование Th2 приводит к усилению гумо-
рального ответа и подавлению ТЫ. Превалирование ТЫ приводит к усилению кле-
точного иммунного ответа и подавлению Th2. Эти альтернативные эффекты опо-
средованы разными группами цитокинов.
368 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
вировать В-лимфоциты к продукции соответствующих по специфичности
антител, т. е. участвуют в гуморальном иммунном ответе. Каждая из двух
субпопуляций Th продуцирует аутокринно и паракринно действующие ци-
токины, способные помешать дифференцировке и активации альтернатив-
ной субпопуляции (рис. 8.2).
8.1.1.3. Функции Т-лимфоцитов
При развитии специфического иммунного ответа Т-лимфоциты в лимфа-
тических узлах, селезенке и мукозноассоцииросванных лимфоидных тка-
нях выполняют секреторные и эффекторные функции.
Активированные Т-лимфоциты продуцируют и секретируют молекулы
цитокинов. Цитокины связываются со специфическими рецепторами на
поверхности клеток-мишеней. Соединение цитокина с его рецептором по-
рождает сигнал активации, который передается соответствующими факто-
рами трансдукции к ядру клетки-мишени, где начинают функционировать
определенные гены, контролирующие функции клеток. Таким образом, Т-
лимфоциты своими продуктами цитокинами контролируют процессы про-
лиферации, дифференцировки и функциональную активность макрофагов,
дендритных клеток, других Т-лимфоцитов, В-лимфоцитов.
Некоторые цитокины непосредственно участвуют в защите организма:
• интерфероны обладают противовирусным действием,
• туморнекротизирующий фактор (ТНФ) оказывает цитотоксическое
действие на некоторые опухолевые клетки.
Активированные CD8+ Т-лимфоциты выполняют эффекторную функ-
цию цитотоксических Т-лимфоцитов (CTL): распознают своими рецепто-
рами и убивают клетки-мишени, несущие на своей поверхности соответст-
вующий по специфичности антигенный пептид. При непосредственном
контакте CTL с клеткой-мишенью содержимое гранул CTL (цитотоксины:
перфорины и грензимы) проникают в клетку-мишень и вызывают ее ги-
бель.
8.1.2. В-лимфоциты
8.1.2.1. Характеристики В-лимфоцитов
Предшественники В-лимфоцитов из эмбриональной печени человека на
8—9-й неделе развития плода перемещаются в костный мозг, где они со-
зревают под влиянием интерлейкина 7, который секретируют стромальные
клетки.
В ранний период созревания в костном мозге формируется и фиксиру-
ется на поверхностной мембране В-лимфоцита иммуноглобулиновый анти-
генраспознающий рецептор. Незрелый В-лимфоцит уже несет мембранные В-
клеточные рецепторы (IgM) определенной специфичности, т. е. способные
распознать и связать конкретный антиген. При встрече незрелых В-лим-
фоцитов с аутоантигенами связывание последних с мембранными IgM ве-
дет к остановке дифференцировки этих В-лимфоцитов, с последующей их
гибелью путем апоптоза. На этой стадии погибают до 90 % незрелых В-
клеток. Часть незрелых В-клеток после контакта с растворимым аутоанти-
геном не погибают, а переходят в состояние анергии по отношению к дан-
ному антигену. Этим объясняется врожденное состояние иммунологической
толерантности (неспособности ответить) на собственные аутоантигены.
Выжившие в этом процессе отбора В-клетки по мере созревания поки-
8. Иммунная система • 369
дают костный мозг, переходя в кровяное русло через венозные синусы, и
поступают в периферические органы иммунной системы, где превращают-
ся в зрелые В-лимфоциты. В кровяном русле В-лимфоциты включаются в
рециркуляцию и заселяют В-зоны периферических органов и тканей им-
мунной системы.
Около 2 • 107 В-лимфоцитов покидают костный мозг ежедневно, что со-
ставляет 5—10 % общей популяции В-клеток периферической крови. Это-
го достаточно, чтобы полностью обновить популяцию периферических
В-лимфоцитов за 4—5 дней. Среди сохранившихся и вступивших в рецир-
куляцию относительно долгоживущих В-лимфоцитов (с полупериодом
жизни 1—2 мес) значительную часть составляют клетки памяти, способ-
ные быстро ответить пролиферацией, активацией, продукцией и секрецией
специфических антител (иммуноглобулинов) на повторную встречу с тем
же антигеном.
8.1.2.2. Функции В-лимфоцитов
В-лимфоциты с помощью своих иммуноглобулиновых рецепторов могут
распознавать не только пептиды, но и более крупные молекулы белков,
нуклеиновых кислот, углеводов, липидов, полисахаридные и липопротеид-
ные растворимые антигены. Зрелые В-лимфоциты способны самостоятель-
но распознавать растворимые антигены, в отличие от Т-лимфоцитов они
не нуждаются в презентации чужеродных антигенов. Более того, В-лимфо-
циты сами могут выполнять функцию антигенпредставляющих клеток при
взаимодействии с Th2. Для этого В-лимфоциты имеют на своей поверхно-
сти адгезионные и костимулирующие молекулы, рецепторы для иммуно-
глобулинов, компонентов системы комплемента и для цитокинов
(табл. 8.4).
Распознавание антигена В-клеточным рецептором служит одним из
сигналов активации В-лимфоцитов, которая проявляется их пролифераци-
ей и дифференцировкой в продуцирующие иммуноглобулины плазматиче-
ские клетки. Дополнительные сигналы активации В-клетки получают при
взаимодействии с Th2, которые продуцируют и секретируют соответствую-
щие цитокины: интерлейкины 4, 5, 6, 10.
Синтез и секреция антител (иммуноглобулинов) В-лимфоцитами явля-
ется завершающей стадией специфического гуморального иммунного отве-
та на конкретный антиген. В динамике иммунного ответа происходит пере-
ключение синтеза антител, который начинается с IgM, на другие изотипы:
Таблица 8.4. Типы молекул на поверхности В-лимфоцитов
Молекулы Функции
Антигенраспознающий рецептор им- муноглобулиновой природы Адгезионные молекулы Костимулирующие молекулы Рецепторы иммуноглобулинов Рецепторы компонентов комплемента Рецепторы цитокинов Молекулы главного комплекса гисто- совместимости Распознавание и связывание антигена Адгезия лимфоцитов к эндотелиальным клет- кам, к элементам внеклеточного матрикса Участвуют в активации В-лимфоцитов после взаимодействия с антигеном Связывают иммунные комплексы Участвуют в связывании иммунных комплексов Связывают цитокины Участвуют в презентации антигенов
370 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
IgG, IgA или IgE. Переключение индуцируется разными цитокинами и
взаимодействием костимулирующих молекул (CD40—CD40L), присутст-
вующих на поверхности В-лимфоцитов и Th2.
8.1.3. Антигенпредставляющие клетки
Способностью представлять (презентировать) антигенные пептиды Т-лим-
фоцитам обладают антигенпредставляющие клетки: дендритные клетки,
макрофаги и В-лимфоциты. Презентации антигенных пептидов предшест-
вуют стадии: 1) захвата поступившего в организм антигена, 2) его перера-
ботки (дезинтеграции), 3) формирования комплексов накопившихся анти-
генных пептидов с собственными молекулами главного комплекса гисто-
совместимости, постоянно синтезирующиеся в этих клетках (см. раздел
8.3.2), 4) транспортировки образовавшихся комплексов на мембрану анти-
генпрезентирующей клетки, 5) доставки во вторичные лимфоидные орга-
ны, где и происходит встреча с Т-лимфоцитами и распознавание образо-
вавшегося комплекса Т-клеточным рецептором.
Дендритные клетки, как и макрофаги и лимфоциты, имеют гемопоэти-
ческое происхождение. Дендритные клетки локализованы в эпителии ки-
шечника, урогенитального тракта, воздухоносных путей, легких, в эпидер-
мисе кожи (клетки Лангерганса) и интерстициальных пространствах.
8.2. Структура и функции органов иммунной системы
К органам иммунной системы относятся:
— центральные (первичные): костный мозг и тимус,
— периферические (вторичные): селезенка, лимфатические узлы, ассо-
циированная со слизистыми оболочками (мукозно-ассоциированная)
лимфоидная ткань.
В центральных органах иммунной системы постоянно идут процессы
пролиферации клеток-предшественниц Т- и В-лимфоцитов, их созревания
(дифференцировки), их отбора (селекции), сопровождающиеся их частич-
ной гибелью или транспортировкой созревающих клеток через кровь в пе-
риферические органы.
Периферические органы иммунной системы являются местом встречи Т-
и В-лимфоцитов с поступающими туда антигенами, местом распознавания
антигенов и развития последовательных стадий специфического иммунного
ответа на данный антиген. Распознавание антигена лимфоцитом служит сиг-
налом его усиленной пролиферации, ускоренной дифференцировки и акти-
вации. В-лимфоциты после активации в периферических органах иммунной
системы дифференцируются в плазматические клетки, продуцирующие и
секретирующие антитела — иммуноглобулины. Активированные Т- и В-
лимфоциты в периферических органах иммунной системы продуцируют и
секретируют межклеточные медиаторы — цитокины, влияющие на иммун-
ный ответ. Там же, в периферических органах иммунной системы, накап-
ливаются и сохраняются долгоживущие Т- и В-лимфоциты, ответственные
за поддержание «иммунологической памяти» о встрече с данным антигеном.
8.2.1. Костный мозг
Продолжая функцию эмбриональной печени, костный мозг является ме-
стом гемопоэза (см. главу 7), в том числе лимфопоэза (рис. 8.3). Единая
гемопоэтическая стволовая клетка может дифференцироваться в сторону
8. Иммунная система • 371
ГМ-КСФ, ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-11
О
Кроветворная
стволовая клетка
ИЛ-1, ИЛ-6
Миеломоно-
цитарная
---------. клетка-пред-
ГМ-КСФ шественница
Г-КСФ, ’
ИЛ-1
Миелобласт
ИЛ-3
Дендритная
клетка
ИЛ-1, ИЛ-2,
ИЛ-4, ИЛ-6,
ИЛ-7
ИЛ-3, ИЛ-9,
ГМ-КСФ,
М-КСФ,
ТНФ-а
ГМ-КСФ,
ИЛ-4,
ТНФ-а
Естественный
киллер (ЕК)
Лимфоцитарная
клетка-
предшественница
ИЛ-1, ИЛ-2,
ИЛ-4, ИЛ-5,
ИЛ-6, ИЛ-7
Тимоцит Пре-В-лимфоцит
Монобласт
Т-лимфоцит
В-лимфоцит
Макрофаг
Т-хелпер
CTL
Плазматическая клетка
Рис. 8.3. Миеломоноцитопоэз и лимфопоэз в общей системе гемопоэза.
Единая стволовая гемопоэтическая клетка является общей клеткой-предшественницей всех
ростков кроветворения. Миеломоноцитарные клетки-предшественницы под контролем ука-
занных групп цитокинов созревают и дифференцируются в гранулоциты и моноциты/макро-
фаги. Клетки-предшественницы лимфоцитов под влиянием указанных групп цитокинов со-
зревают и дифференцируются в Т- и В-лимфоциты, или в естественные киллеры. Миелоид-
ные и лимфоидные клетки-предшественницы могут дифференцироваться в дендритные клет-
ки в присутствии указанных цитокинов.
I
372 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
общей клетки-предшественницы лимфоцитов. Эта клетка дает начало
клеткам-предшественницам В-лимфоцитов, Т-лимфоцитов и естественных
киллеров. Дальнейшая дифференцировка лимфоцитов зависит от влияния
колониестимулирующих факторов, продуцируемых стромальными клетка-
ми костного мозга. Созревающие активированные лимфоциты начинают
продуцировать цитокины, аутокринно влияющие на их пролиферацию и
дифференцировку. Так, интерлейкин-1 и интерлейкин-6 служат синергиста-
ми колониестимулирующих факторов в стимуляции пролиферации клеток-
предшественниц; интерлейкин-2 является ростовым фактором Т-лимфоци-
тов; интерлейкины-4, -6, -7 способствуют выживанию, пролиферации и
дифференцировке ранних предшественниц лимфоцитов; туморнекротизи-
рующий фактор (ТНФ), гамма-интерферон, трансформирующий ростовой
фактор-бета (ТРФ-бета), напротив, ингибируют процессы пролиферации
и дифференцировки клеток-предшественниц.
Созревшие в костном мозге клетки-предшественницы Т-лимфоцитов
через кровоток попадают в тимус для дальнейшей дифференцировки. В от-
личие от Т-лимфоцитов, местом дальнейшего созревания В-лимфоцитов у
млекопитающих является костный мозг. Один из продуктов стромальных
клеток костного мозга — интерлейкин- 7 преимущественно стимулирует
процесс созревания В-лимфоцитов из костномозговых предшественников.
Созревшие В-лимфоциты выходят в кровь, заселяют лимфоидные органы,
участвуют в рециркуляции, проявляя способность к распознаванию кон-
кретного антигена. Распознавание антигена служит одним из сигналов ак-
тивации В-лимфоцитов и их дальнейшей дифференцировки в плазматиче-
ские клетки, продуцирующие и секретирующие антитела (иммуноглобули-
ны). Иммуноглобулины продуцируются в костном мозге и в перифериче-
ских органах иммунной системы, откуда поступают в кровоток.
Таким образом, костный мозг в качестве одного из центральных орга-
нов иммунной системы выполняет следующие функции: является местом
начальной дифференцировки и пролиферации ранних клеток-предшест-
венниц лимфоцитов; является местом дальнейшей дифференцировки В-
лимфоцитов вплоть до их выхода в кровоток и заселения периферических
органов иммунной системы; является местом продукции и секреции коло-
ниестимулирующих факторов и цитокинов, влияющих на процессы проли-
ферации, дифференцировки и транспортировки Т- и В-лимфоцитов; явля-
ется одним из мест продукции и секреции антител (иммуноглоблинов).
8.2.2. Тимус (вилочковая железа)
Тимус в качестве одного из центральных органов иммунной системы явля-
ется местом созревания Т-лимфоцитов из клеток-предшественниц и фор-
мирования огромного разнообразия зрелых Т-лимфоциов, способных рас-
познать своими рецепторами любой антиген.
Лимфоциты, находящиеся в тимусе, называют тимоцитами. В тимусе
идут параллельно несколько процессов:
• пролиферация Т-лимфоцитов,
• их созревание (дифференцировка),
• отбор пригодных для данного организма клеток, которому сопутству-
ет гибель значительной части непригодных клеток.
Ранние клетки-предшественницы Т-лимфоцитов из костного мозга по-
ступают в корковый слой тимуса. Постепенно они мигрируют из корково-
го слоя тимуса в мозговой, контактируя с тимическими эпителиальными
клетками, макрофагами и дендритными клетками. На тимоциты влияют
8. Иммунная система • 373
Позитивная селекция
Негативная селекция
Рис. 8.4. Этапы отбора тимоцитов.
А. Тимоцит, способный распознать своим рецептором (ТКР) собственные антигены главного
комплекса гистосовместимости (1) на поверхности тимических эпителиальных клеток (ТЭК),
вступает в дальнейшую дифференцировку.
Б. Тимоцит, не способный распознать своим ТКР собственные антигены главного комплекса
гистосовместимости (1) на поверхности ТЭК, получает сигнал апоптоза.
В. Тимоцит, способный распознать своим ТКР пептидный фрагмент аутоантигена (2) на по-
верхности антигенпредставляющей клетки в комплексе с собственными молекулами главного
комплекса гистосовместимости (1), получает сигнал апоптоза.
продуцируемые тимическими эпителиальными клетками гормоны: тиму-
лин, al- и р4-тимозины, тимопоэтин, контролирующие их дифференци-
ровку. В процессе продвижения в мозговой слой тимоциты созревают
(дифференцируются), поэтапно приобретая характерные для зрелых Т-
лимфоцитов поверхностные рецепторы и антигены.
От стромальных клеток тимуса и продуцируемых ими молекул каждый
тимоцит получает сигналы: усиления пролиферации, изменений поверхно-
стного фенотипа (сочетания молекул, присутствующих на поверхности
клеток), перестройки («реаранжировки») генов, ответственных за широкое
разнообразие специфических антигенраспознающих рецепторов.
Основная функция зрелых Т-лимфоцитов — это распознавание чуже-
родных антигенных пептидов в комплексе с собственными молекулами
главного комплекса гистосовместимости на поверхности антигенпредстав-
ляющих клеток. Для выполнения этой функции Т-лимфоциты располага-
ют специфическими антигенраспознающими рецепторами.
Отбор тимоцитов идет в два этапа (рис. 8.4). После того как на тимоци-
те формируется Т-клеточный рецептор определенной специфичности, в
следующие этапы развития вступают только те тимоциты, которые прояви-
ли способность распознавать собственные молекулы главного комплекса
гистосовместимости, присутствующие на эпителиальных клетках коры ти-
муса данного организма. В отличие от зрелого Т-лимфоцита, который при
встрече с антигенным пептидом, специфичным для его Т-клеточного ре-
цептора, получает сигнал активации, незрелые тимоциты в тимусе при
распознавании специфичных для их Т-клеточных рецепторов аутоантиген-
374 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ных пептидов получают сигнал генетически программированной гибели —
апоптоза. Таким образом, идет «выбраковка» аутореактивных Т-лимфоци-
тов, 95—98 % тимоцитов ежедневно погибают в тимусе этим путем. Только
2—5 % тимоцитов (1 • 10б) в виде зрелых Т-лимфоцитов ежедневно покида-
ют тимус, поступая в кровь и расселяясь в лимфоидных органах.
В результате в кровоток и в лимфоидные органы поступают только те
Т-лимфоциты, которые несут рецепторы, распознающие чужеродные пеп-
тиды в сочетании с собственными молекулами главного комплекса гисто-
совместимости.
Тимус представляет собой единственный орган иммунной системы,
подвергающийся быстрой возрастной инволюции, которая у людей начи-
нается после первого года жизни. Человек ежегодно теряет до 3 % истинно
тимической ткани, которая постепенно замещается жировой и соедини-
тельной тканью. Соответственно снижается и продукция Т-лимфоцитов.
Высокая продукция Т-лимфоцитов сохраняется до двух лет, а затем быстро
снижается. Однако количество Т-лимфоцитов в организме сохраняется на
достигнутом уровне благодаря тому, что значительную часть популяции Т-
лимфоцитов составляют долгоживущие клетки, которые не нуждаются в
постоянном обновлении. Более того, зрелые Т-лимфоциты, распознавшие
«свой» антиген в периферических органах иммунной системы, получают
сигналы активации и избирательной пролиферации, за счет чего их чис-
ленность возрастает. Перенесенная взрослым человеком тимэктомия не
приводит к катастрофическому снижению количества циркулирующих Т-
лимфоцитов.
В качестве одного из центральных органов иммунной системы тимус
выполняет следующие функции: а) контролирует пролиферацию, диффе-
ренцировку, отбор и окончательное созревание Т-лимфоцитов, б) проду-
цирует тимические гормоны, влияющие на функции Т-лимфоцитов.
8.2.3. Селезенка
Селезенка выполняет функции фильтра крови, удаляющего из крови попа-
дающие туда чужеродные частицы и молекулы, а также состарившиеся
эритроциты.
В селезенке имеются Т- и В-зоны, заселенные преимущественно Т- или
В-лимфоцитами. В качестве одного из периферических органов иммунной
Рис. 8.5. Периферические органы иммунной системы, обеспечивающие специфиче-
ский иммунный ответ на антигены, поступающие в организм разными путями.
На антиген, попадающий в кровяное русло (1), специфический иммунный ответ развивается в
селезенке. На антиген, проникающий через кожу (2), отвечают преимущественно лимфоциты
в регионарном лимфатическом узле. Местом ответа на антигены, проникающие через слизи-
стые оболочки (3), является мукозно-ассоциированная лимфоидная ткань.
8. Иммунная система • 375
системы селезенка является местом: 1) созревания естественных киллеров,
2) распознавания антигена, 3) антигензависимой пролиферации и диффе-
ренцировки Т- и В-лимфоцитов, 4) активации Т- и В-лимфоцитов, 5) про-
дукции цитокинов, 6) продукции и секреции специфических антител —
иммуноглобулинов.
Селезенка является местом специфического иммунного ответа на анти-
гены, циркулирующие в крови, а в лимфатических узлах разыгрываются
процессы специфического иммунного ответе на антигены, попадающие
через лимфу (рис. 8.5).
8.2.4. Лимфатические узлы
Лимфатические узлы функционируют в качестве своеобразных фильтров
лимфы, задерживая микроорганизмы, попавшие в лимфу. В качестве пери-
ферических органов иммунной системы лимфатические узлы являются ме-
стом: 1) распознавания антигена, 2) антигензависимой пролиферации и
дифференцировки Т- и В-лимфоцитов, 3) активации Т- и В-лимфоцитов,
4) продукции цитокинов, 5) продукции и секреции специфических анти-
тел — иммуноглобулинов.
Один лимфатический узел имеет массу около 1 г. Каждый час из лим-
фоузла выходит в лимфу количество лимфоцитов, эквивалентное его утро-
енной массе. Большая часть (90 %) клеток в этой эфферентной лимфе
представляют собой лимфоциты, покинувшие кровяное русло на террито-
рии этого лимфатического узла. Среди клеток лимфатического узла около
10 % составляют макрофаги и около 1 % — дендритные клетки.
В кортикальном (наружном) слое лимфатического узла преобладают
скопления лимфоцитов. В мозговом (внутреннем) слое лимфатического уз-
ла лимфоциты содержатся в сочетании с макрофагами (около 10 %) и ден-
дритными клетками (около 1 %), которые выполняют функцию представ-
ления антигенов Т-лимфоцитам и продуцируют цитокины. В пределах
лимфатического узла происходит свободная циркуляции лимфоцитов меж-
ду лимфой, кровью и лимфоидной тканью.
При развитии специфического иммунного ответа в мозговом слое лим-
фатических узлов накапливаются образовавшиеся из В-лимфоцитов плаз-
матические клетки, продуцирующие и секретирующие соответствующие
антигену по специфичности антитела — иммуноглобулины. Строго опреде-
ленное взаимное расположение Т- и В-лимфоцитов, макрофагов, дендрит-
ных клеток в лимфатических узлах обеспечивает последовательное разви-
тие стадий иммунного ответа: представления антигена, его распознавания
Т-клеточными рецепторами, активации процессов пролиферации и диф-
ференцировки (созревания) лимфоцитов. В результате в лимфатических
узлах формируются зрелые Т-лимфоциты, которые принимают участие в
защитных реакциях. Там же формируются клоны долгоживущих Т- и В-
лимфоцитов, обеспечивающих поддержание иммунологической памяти о
встрече с конкретным антигеном.
8.2.5. Ассоциированная со слизистыми оболочками лимфоидная
ткань (мукозно-ассоциированная лимфоидная ткань)
Непосредственно под мукозным эпителием слизистых оболочек в тесной
связи с эпителиальными клетками располагаются лимфоциты пейеровых
бляшек кишечника, лимфоидных фолликулов аппендикса, миндалин глот-
ки, лимфоидных фолликулов подслизистого слоя верхних дыхательных пу-
376 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
тей и бронхов, мочеполового тракта. Все эти лимфоидные скопления по-
лучили собирательное название — мукозно-ассоциированная лимфоидная
ткань.
В лимфоидных тканях, ассоциированных со слизистыми оболочками,
проходят все стадии специфического иммунного ответа в тех случаях, ко-
гда антиген проникает в организм через слизистые оболочки. В мукозно-
ассоциированных лимфоидных тканях активированные В-лимфоциты
дифференцируются в плазматические клетки, продуцирующие специфиче-
ские антитела, относящиеся к классу иммуноглобулинов A (IgA). Имму-
ноглобулины А, пройдя через эпителиальные клетки, где они присоединя-
ют секреторный компонент, выходят на поверхность слизистых оболочек в
форме секреторного иммуноглобулина A (SIgA), который обеспечивает мест-
ную антибактериальную и антивирусную защиту.
В качестве одного из периферических органов иммунной системы му-
козно-ассоциированная лимфоидная ткань является местом: 1) распозна-
вания антигена, 2) антигензависимой пролиферации и дифференцировки
Т- и В-лимфоцитов, 3) активации Т- и В-лимфоцитов, 4) продукции цито-
кинов, 5) продукции и секреции специфических антител — иммуноглобу-
линов.
8.3. Стадии и формы иммунного ответа
8.3.1. Ранний защитный воспалительный ответ
Ранний защитный воспалительный ответ призван препятствовать внедре-
нию и распространению возбудителя, по возможности быстро удалять его
из организма. Ранний защитный воспалительный ответ разыгрывается в
течение первых 4 сут после внедрения возбудителя. Ранний защитный от-
вет против внедрившихся в организм патогенных микроорганизмов обес-
печивается факторами врожденного иммунитета, к которым относятся фа-
гоцитирующие клетки крови и тканей, естественные киллеры, циркули-
рующие в крови белковые молекулы, обладающие защитными свойствами
(компоненты системы комплемента и др.), а также межклеточные медиа-
торы — цитокины. Ранний воспалительный ответ стимулирует последую-
щий специфический иммунный ответ, влияет на его форму, способствуя
развитию наиболее эффективного против конкретного микроорганизма
специфического иммунного ответа.
Ранний воспалительный ответ начинается с привлечения лейкоцитов из
кровяного русла в очаг инфекции с последующей их активацией для удале-
ния возбудителя (рис. 8.6). Мобилизация лейкоцитов в очаг инфекции сти-
мулируется провоспалителъными цитокинами, а опосредуется адгезионными
молекулами на поверхности лейкоцитов и эндотелиальных клеток, а также
молекулами хемокинов и их рецепторами. Провоспалительные цитокины
(туморнекротизирующий фактор—ТНФ, интерлейкин-1 — ИЛ-1) продуци-
руются и секретируются макрофагами при фагоцитозе возбудителя. Под
действием этих цитокинов на поверхности эндотелиальных клеток, появ-
ляются адгезионные молекулы вначале для нейтрофилов, а затем для мо-
ноцитов и лимфоцитов, ответственные за прилипание лейкоцитов к эндо-
телию сосудов. Кроме того, активированные цитокинами эндотелиальные
клетки продуцируют хемокины, достигающие высокой концентрации на
уровне очага инфекции (рис. 8.7). Этим обеспечивается прочная адгезия
лейкоцитов к эндотелиальным клеткам с последующей их трансэндотели-
8. Иммунная система • 377
Рис. 8.6. Этапы взаимодействия лейкоцитов крови с эндотелиальными клетками
при воспалении: адгезия и трансэндотелиальная миграция обеспечивают выход лей-
коцитов из кровяного русла и движение к очагу инфекции.
Рис. 8.7. Взаимное влияние макрофагов и эндотелиальных клеток в процессах вос-
паления и иммунного ответа. Продукты активированных макрофагов — провоспа-
лительные цитокины — активируют эндотелиальные клетки, индуцируя появление
на поверхности эндотелиальных клеток адгезионных молекул для лейкоцитов и
продукцию эндотелиальными клетками хемокинов, усиливающих процессы мигра-
ции лейкоцитов.
альной миграцией и выходом в ткани в направлении очага инфекции (см.
рис. 8.6).
Таким образом, ранний воспалительный ответ проявляется инфильтра-
цией очага инфекции фагоцитирующими клетками, где эти клетки получа-
ют дополнительные сигналы активации от микробных продуктов и компо-
нентов (липополисахарид клеточной стенки бактерий), от компонентов ак-
тивированной системы комплемента и от провоспалительных цитокинов, в
том числе, от гамма-интерферона, продуцируемого и секретируемого акти-
вированными естественными киллерами. Активированные фагоцитирую-
щие клетки приобретают повышенную способность убивать захваченных
микробов, что способствует удалению возбудителя. Активированные под
378 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
влиянием провоспалительных цитокинов эндотелиальные клетки продуци-
руют молекулы вазодилататоров, под действием которых усиливается мест-
ный кровоток, повышается проницаемость сосудов для макромолекул
фибриногена, который, покидая сосуды, превращается в фибрин, что спо-
собствует ограничению очага инфекции. В случае попадания в организм
небольшого количества низковирулентных возбудителей ранний воспали-
тельный ответ подавляет очаг инфекции.
Удаление из кровяного русла попавших в кровь единичных бактериаль-
ных клеток является функцией системы комплемента. Большая часть ком-
понентов комплемента синтезируются гепатоцитами и мононуклеарными
фагоцитами. Компоненты комплемента (Cl, С2, СЗ, С4, С5, Сб, С7, С8,
С9, факторы В и D) содержатся в крови в неактивной форме. При попада-
нии в кровяное русло бактерий на их поверхности каскад ферментативных
реакций ведет к последовательной активации компонентов системы ком-
племента («альтернативный путь активации») с формированием мембран-
атакующего комплекса (С5—С9), вызывающего лизис бактерий. В процес-
се активации системы комплемента накапливаются фрагменты, которые
опосредуют разные биологические эффекты: привлечение лейкоцитов в
очаг инфекции или воспаления (хемотаксис) — фрагмент С5а, усиление
фагоцитоза (опсонизацию) — СЗЬ, индукцию синтеза и секреции медиато-
ров воспаления — СЗа, С5а. (При развитии специфического иммунного от-
вета и появлении в кровяном русле комплексов антиген—антитело другой
каскад ферментативных реакций ведет к активации системы комплемен-
та — «классический путь активации», см. раздел 8.3.5.)
8.3.2. Представление и распознавание антигена
В случае неэффективности раннего защитного ответа и факторов врожден-
ного иммунитета в организме накапливаются микроорганизмы, содержа-
щие чужеродные антигены. Контакт этих антигенов с клетками иммунной
системы приводит к развитию специфического иммунного ответа, который
начинается с этапа представления и распознавания антигена.
Растворимые антигены в нативной форме связываются антителами и
антигенраспознающими иммуноглобулиновыми рецепторами В-клеток.
Т-лимфоциты своими антигенраспознающими Т-клеточными рецепто-
рами связывают короткие пептидные фрагменты белковых антигенов в
комплексах с собственными молекулами главного комплекса гистосовме-
стимости I или II классов на мембранах антигенпредставляющих клеток.
Молекулы главного комплекса гистосовместимости на поверхности
лейкоцитов человека, получили название «человеческие лейкоцитарные
антигены» (Human Leukocyte Antigens — HLA). Молекулы главного ком-
плекса гистосовместимости I класса присутствуют на поверхности всех ти-
пов клеток, кроме эритроцитов и клеток трофобласта. Молекулы главного
комплекса гистосовместимости II класса находятся на поверхности анти-
генпредставляющих клеток: дендритных клеток, макрофагов, В-лимфо-
цитов.
Антигенпредставляющие клетки играют важную роль на начальном
(пусковом) этапе специфического иммунного ответа: 1) макрофаги, как
правило, представляют антигены бактериального происхождения — про-
дукты захвата и внутриклеточной переработки ими бактерий, 2) В-лимфо-
циты представляют микробные антигены, антигены токсинов, связанные
их поверхностными иммуноглобулиновыми рецепторами, 3) наиболее уни-
версальными антигенпредставляющими клетками являются дендритные
8. Иммунная система • 379
Рис. 8.8. Взаимодействие Т-хелперов (Thl) с антигенпредставляющими дендритны-
ми клетками (ДК) опосредовано цитокинами (ИЛ-12, гамма-интерферон) и кости-
мулирующими молекулами (CD40, CD40L).
В центре «иммунологического синапса» находятся: Т-клеточный антиген распознающий ре-
цептор (1) и распознаваемый им комплекс антигенного пептида с молекулой главного ком-
плекса гистосовместимости (2).
клетки, которые необходимы для запуска первичного иммунного ответа,
представляют многие, в том числе опухолевые, антигены.
После захвата чужеродного белка — антигена этими клетками и его раз-
ложения до коротких пептидов внутри клеток происходит комплексирова-
ние отдельных молекул главного комплекса гистосовместимости с кон-
кретными пептидами. Такие комплексы транспортируются на поверхность
антигенпредставляющих клеток (рис. 8.8).
При встрече антигенпредставляющей клетки с Т-лимфоцитом, несущим
на своей поверхности подходящий по структуре антигенраспознающий ре-
цептор, замыкается лиганд-рецепторная связь, т. е. происходит распозна-
вание антигена.
CD4+ Т-лимфоциты способны распознавать антигенные пептиды в ком-
плексах с собственными молекулами главного комплекса гистосовмести-
мости II класса, a CD8+ Т-лимфоциты — в комплексах с собственными мо-
лекулами главного комплекса гистосовместимости I класса. Связывание
антигена Т-клеточным рецептором служит сигналом усиления пролифера-
ции данного Т-лимфоцита с последующей ускоренной дифференцировкой
и активацией его клеток-потомков. В результате формируется клон Т-лим-
фоцитов, способных при встрече с тем же антигеном быстро активировать-
ся для участия в удалении патогенного агента.
8.3.3. Активация Т- и В-лимфоцитов в иммунном ответе
Передача сигнала активации от антигенраспознающего рецептора в ядро
клетки приводит к активации конкретных генов, ответственных за митоти-
ческую активность клеток, контролирующих продукцию клеткой биологи-
чески активных молекул, участвующих в антимикробной защите (антитела,
цитокины).
В-лимфоцит получает одновременно два сигнала активации: от анти-
генраспознающего рецептора при его соединении с антигеном и от связы-
вания его поверхностных костимулирующих молекул с соответствующими
лигандами на Т-лимфоцитах. После этого В-лимфоцит пролиферирует и
потомки его превращаются в зрелые антителопродуцирующие плазматиче-
ские клетки.
380 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 8.9. Основные формы специфического иммунного ответа.
В зависимости от направления дифференцировки ThO в сторону Thl или Th2 развивается, со-
ответственно, клеточный или гуморальный иммунный ответ с активацией либо макрофагов,
либо В-лимфоцитов под влиянием указанных цитокинов. Как Т-, так и В-лимфоциты могут
дать начало долгоживущим клеткам, обеспечивающим поддержание «иммунологической па-
мяти».
Т-лимфоцит в ответ на контакт с антигеном начинает пролиферировать,
потомки его приобретают способность продуцировать определенные цито-
кины или превращаются в зрелые цитотоксические клетки. В зависимости
от того, какие дополнительные сигналы активации (цитокины, костимули-
рующие молекулы) получает Т-лимфоцит в момент контакта с антигеном,
его потомки дифференцируются в двух разных направлениях, превращаясь
в Т-хелперы, продуцирующие гамма-интерферон (Thl), или в Т-хелперы,
продуцирующие интерлейкины-4, -5, -6, -10, -13 (Th2). Количественное
преобладание Thl над Th2 является условием развития клеточного (кле-
точно-опосредованного) иммунного ответа. В случае преобладания Th2
развивается гуморальный иммунный ответ, проявляющийся продукцией
специфических антител (рис. 8.9).
8.3.4. Клеточный иммунный ответ
Клеточный иммунный ответ направлен против внутриклеточно паразити-
рующих микроорганизмов, основная защитная роль в нем принадлежит
активированным макрофагам и цитотоксическим лимфоцитам (CD8+
CTL).
Макрофаги, инфицированные микроорганизмами, получают от Thl в
качестве сигналов активации цитокины: гамма-интерферон и туморнекро-
тизирующий фактор (ТНФ), которые действуют через свои рецепторы, вы-
зывая усиленную продукцию макрофагами супероксидных и нитроксид-
ных радикалов, убивающих внутриклеточные паразиты.
8. Иммунная система • 381
Цитотоксические CD8+ CTL способны убивать зараженные вирусами
клетки при непосредственном контакте с ними. В месте контакта из CTL в
мембрану клетки-мишени проникают порообразующие белки — перфори-
ны, формирующие в мембране микроканалы, через которые в клетку-ми-
шень проникают ферменты — фрагментины, вызывающие разрушение яд-
ра клетки и ее гибель.
8.3.5. Гуморальный иммунный ответ
Гуморальный иммунный ответ защищает преимущественно против внекле-
точно паразитирующих микроорганизмов, которые доступны действию
специфических антител. Продуцентами антител (иммуноглобулинов) явля-
ются потомки активированных В-лимфоцитов — плазматические клетки.
В-лимфоцит получает сигнал активации при соединении его антиген-
распознающего рецептора с антигеном. Для активной пролиферации и
дифференцировки в плазматические клетки потомки В-лимфоцита получа-
ют дополнительные сигналы активации — продуцируемые Th2 цитокины:
интерлейкины-4, -5, -6, -10, -13. Те же цитокины усиливают продукцию и
секрецию специфических для данного антигена антител зрелыми плазма-
тическими клетками.
Специфические антитела — иммуноглобулины против конкретных ан-
тигенов бактерий (стафилококки, стрептококки, возбудители дифтерии,
кишечных инфекций, клостридии и др.), связываясь с бактериальными
токсинами, вызывают их нейтрализацию, т. е. утрату токсического дейст-
вия на организм. Сами бактерии, связавшиеся со специфическими антите-
лами, быстрее и легче захватываются и убиваются фагоцитирующими
клетками или лизируются активированной системой комплемента.
Иммуноглобулины делятся на пять классов (изотипов): IgG, IgM, IgA,
IgE, IgD. В нормальной сыворотке крови 80 % всех иммуноглобулинов со-
ставляют IgG, на долю IgM приходится 6 %, на долю IgA — 13 %, а на до-
лю IgE и IgD — сотые или тысячные доли процента. Главными защитными
иммуноглобулинами являются IgG. У иммуноглобулинов разных изотипов
различаются защитные свойства (табл. 8.5).
Иммуноглобулин G (IgG) — мономер, доминирующий среди других изо-
типов иммуноглобулинов у взрослых в кровяном русле, легко диффунди-
рующий из крови в ткани, единственный из иммуноглобулинов способен
преодолевать плацентарный барьер и обеспечивать гуморальный иммуни-
тет новорожденных первых месяцев жизни. Дополнительные порции мате-
ринского, содержащегося в молозиве IgG, поступают в кровоток новорож-
денного через слизистую оболочку кишечника. IgG составляют основную
массу антител при иммунном ответе на повторную встречу с тем же анти-
Таблица 8.5. Защитные функции иммуноглобулинов (антител) разных изотипов
Изотипы (классы) иммуноглобулинов Локализация Защитные функции
Иммуноглобулин G Иммуноглобулин М Иммуноглобулин А (секреторный) Кровяное русло и ткани Только кровяное русло Секреты на слизистых оболочках, грудное мо- локо Нейтрализация токсинов и вирусов, активация системы комплемента, уси- ление фагоцитоза Активация системы комплемента Препятствие адгезии бактерий и виру- сов на слизистых оболочках, их связы- вание
382 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
геном. IgG обеспечивает наиболее эффективную антибактериальную и ан-
титоксическую защиту организма, действуя и в крови, и в тканях.
Иммуноглобулин М (IgM) — пентамер, состоящий из пяти четырехцепо-
чечных структур, (называют еще макроглобулином из-за высокой молеку-
лярной массы). IgM синтезируется раньше других классов в онтогенезе,
может продуцироваться в организме плода в ответ на внутриутробную ин-
фекцию. К этому классу иммуноглобулинов относятся так называемые
нормальные антитела — изогемагглютинины групп крови: анти-А, анти-В.
У взрослых иммуноглобулины этого изотипа первыми появляются на са-
мых ранних стадиях гуморального иммунного ответа на инфекцию. Из-за
больших размеров молекулы IgM обычно не покидают кровяного русла, но
играют ведущую роль в антибактериальной защите кровяного русла от по-
падающих туда бактерий.
Иммуноглобулин A (IgA) циркулирует в сыворотке крови в виде моно-
меров или димеров. Димер IgA может связываться с полиглобулиновым
рецептором на базолатеральной поверхности эпителиальных клеток и в
комплексе с этим рецептором проникать в эпителиальные клетки. Внутри
эпителиальной клетки такой комплекс подвергается протеолизу и через
апикальную поверхность эпителиальной клетки секретируется образовав-
шийся комплекс димера IgA с фрагментом полиглобулинового рецептора,
который получил название «секреторный компонент». Основными места-
ми синтеза и секреции IgA являются: эпителий желудочно-кишечного
тракта, респираторного тракта, лактирующие молочные железы, слюнные
и слезные железы. В организм ребенка IgA попадают с молоком матери
при грудном вскармливании. В составе секретов оболочек слизистых сек-
реторный IgA — SIgA обеспечивает местный иммунитет слизистых оболо-
чек. Защитное действие SIgA от проникновения через слизистые оболочки
бактерий и вирусов зависит от его способности связывать микроорганизмы
и препятствовать их адгезии или адсорбции на рецепторах слизистой обо-
лочки.
В сыворотке крови здоровых людей иммуноглобулин Е (IgE), как прави-
ло, не содержится.
Защитное действие специфических антител реализуется с помощью не-
скольких механизмов: 1) усиление фагоцитоза бактерий, 2) нейтрализация
бактериальных экзотоксинов и вирусов; 3) активация системы комплемен-
та с последующим бактериолитическим действием ее мембранатакующего
комплекса, 4) препятствие колонизации слизистых оболочек патогенными
бактериями и адсорбции вирусов.
В результате гуморального иммунного ответа на бактериальную инфек-
цию в сыворотке крови накапливаются специфические антитела классов
IgG и IgM. При взаимодействии этих антител с антигенами на поверхно-
сти бактерий создаются условия активации системы комплемента по клас-
сическому пути, результатом которого становится лизис бактерий (бакте-
риолиз). Классический путь активации системы комплемента начинается
со стадии соединения Clq с определенным участком молекулы иммуно-
глобулина, который становится доступным только после взаимодействия
иммуноглобулина — антитела со своим антигеном. Clq при этом активиру-
ется, приобретая активность сериновой протеиназы (эстеразы), которая за-
пускает каскадный процесс расщепления и присоединения последующих
фракций: С4, С2, СЗ. После активации СЗ запускается дальнейший каскад
формирования мембранатакующего комплекса (С5—С9), что ведет к лизи-
су бактерий. Некоторые защитные функции активированных компонентов
системы комплемента были описаны выше (см. раздел-8.3.1).
8. Иммунная система • 383
8.3.6. Иммунологическая память как форма специфического
иммунного ответа
После первой встречи организма с чужеродным антигеном в лимфоидных
органах сохраняются долгоживущие потомки Т- и В-лимфоцитов, проли-
ферировавших в ответ на сигнал активации, полученный от антигенрас-
познающих рецепторов. На мембране этих клеток-потомков сохраняются
специфические для данного антигена рецепторы, способные связаться с
ним при его повторном попадании в организм.
Способность иммунной системы организма ускоренно и усиленно отве-
чать активацией на повторную встречу с тем же антигеном характеризуется
как «иммунологическая память».
Особенностями долгоживущих В-клеток памяти являются: 1) способ-
ность быстро отвечать пролиферацией и дифференцировкой в плазматиче-
ские клетки на повторную встречу с антигеном, 2) способность быстро пе-
реключаться с синтеза IgM на синтез IgG и IgA, 3) способность быстро
продуцировать и секретировать большое количество специфических анти-
тел с выраженными защитными свойствами.
Некоторые из ответивших пролиферацией на контакт с антигеном Т-лим-
фоцитов не претерпевают дальнейшей дифференцировки, а после несколь-
ких делений превращаются в клетки, способные сохраняться в органах им-
мунной системы более 20 лет и поддерживать состояние иммунологической
памяти в отношении конкретного антигена. Особенностями Т-клеток памя-
ти являются: 1) повышенная активность связывания антигена антигенрас-
познающими рецепторами, 2) повышенное количество рецепторов интер-
лейкина-2, 3) готовность быстро ответить на повторный контакт с антигеном
активацией, пролиферацией и дифференцировкой в клетки-эффекторы.
8.3.7. Иммунологическая толерантность
Иммунологической толерантностью называют отсутствие специфического
иммунного ответа на собственные антигены организма (аутоантигены).
В период внутриутробного развития фрагменты аутоантигенов могут за-
носиться в тимус с током крови. В тимусе происходит встреча функцио-
нально незрелых тимоцитов, уже имеющих антигенраспознающие рецеп-
торы, с антигенпредставляющими клетками, несущими на поверхности ау-
тологичные пептиды. Для незрелого тимоцита связывание его антигенрас-
познающего рецептора с аутологичным пептидом служит сигналом апо-
птоза (гибели) или превращения в «анергичную» клетку, не способную в
дальнейшем активироваться при контакте с данным антигеном. Приобре-
тенная во внутриутробный период развития организма иммунологическая
толерантность сохраняется на протяжении всей жизни.
8.4. Механизмы, контролирующие иммунную систему
Количественные характеристики популяций и субпопуляций клеток им-
мунной системы и их функциональная активность находятся под гормо-
нальным и цитокиновым контролем.
8.4.1. Гормональный контроль
Влияние нейроэндокринной системы на иммунную систему может быть
прямым или опосредованным. Прямой контроль осуществляется путем свя-
384 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
зывания нейромедиаторов или гормонов с соответствующими рецепторами
на клетках иммунной системы. Контроль может быть опосредован влияни-
ем нейромедиаторов и гормонов на стромальные клетки органов иммун-
ной системы. Клетки иммунной системы имеют рецепторы, позволяющие
им получать сигналы от многих гормонов и нейромедиаторов: кортикосте- ;
роидов, инсулина, гормона роста (соматотропин), эстрадиола, тестостеро-
на, p-адренергических агентов, ацетилхолина, эндорфинов и энкефалинов.
Секреция глюкокортикоидов в ответ на стресс, который нередко сопут-
ствует иммунному ответу, играет роль регуляции по механизму обратной свя-
зи. Провоспалительные цитокины ИЛ-1, ИЛ-6, ТНФ-альфа, секреция ко-
торых сопутствует клеточному иммунному ответу, стимулируют продукцию
глюкокортикоидов через гипоталамус-гипофизарно-надпочечниковую сис-
тему. Это ведет к подавлению активности клеточного иммунного ответа.
Глюкокортикоиды, являющиеся физиологическими регуляторами им-
мунного ответа, влияют на разные функции лимфоцитов и антигенпрезен-
тирующих клеток: на рециркуляцию лимфоцитов, на адгезию лейкоцитов
к эндотелиальным клеткам, на продукцию цитокинов и количество цито-
киновых рецепторов на клетках.
Глюкокортикоиды действуют на клетки, формируя комплексы с соот-
ветствующими внутриклеточными рецепторами, которые препятствуют
транскрипции отдельных генов, в том числе генов, ответственных за про-
дукцию цитокинов. В концентрациях, превосходящих физиологический
уровень, глюкокортикоиды блокируют транскрипцию многих цитокинов:
туморнекротизирующего фактора, гамма-интерферона, интерлейкинов-1,
-2, -5, -6, -12. В то же время глюкокортикоиды индуцируют продукцию Т-
лимфоцитами ингибирующих цитокинов: трансформирующего ростового
фактора-бета и интерлейкина-10. С нарушениями продукции и рецепции
цитокинов связаны противовоспалительные и иммуномодулирующие эф- Р
фекты глюкокортикоидов.
Глюкокортикоиды способны влиять на характер иммунного ответа: уси-
ливать синтез цитокинов, характерных для Th2, потенцировать эффекты
интерлейкина-4. Разная чувствительность субпопуляций Thl и Th2 к глю-
кокортикоидам связана с различиями количества гормональных рецепто-
ров у этих клеток. Под влиянием ИЛ-4 в сочетании с ИЛ-2 у лимфоцитов
повышается количество рецепторов для глюкокортикоидов, а под влияни-
ем гамма-интерферона количество рецепторов для глюкокортикоидов сни-
жается. Так, дифференцировка лимфоцитов в отсутствие ИЛ-4, в присут-
ствии высокой концентрации гамма-интерферона приводит к созреванию
Thl, резистентных к глюкокортикоидам.
У отдельных индивидуумов генетически детерминирован высокий уро-
вень стресс-индуцированной продукции глюкокортикостероидов. Повы-
шенная чувствительность этих индивидуумов к внутриклеточно паразити-
рующим патогенным агентам объясняется низким уровнем клеточного им-
мунного ответа, подавленного в присутствии повышенного уровня глюко-
кортикостероидов .
Стимулирующим действием эстрогенов на иммунную систему объясня-
ют более высокую устойчивость женщин к инфекциям и более высокую
частоту аутоиммунных заболеваний среди женщин по сравнению с мужчи-
нами. С возрастным снижением продукции андрогенного гормона дегид-
роэпиандростерона, сопряженным с повышением продукции цитокина
ИЛ-6, связывают состояние возрастного иммунодефицита у людей старше
60 лет.
Тироксин стимулирует процессы пролиферации и дифференцировки
8. Иммунная система • 385
Таблица 8.6. Влияние гормонов и нейромедиаторов на иммунную
систему
Угнетение Стимуляция
Кортизол Кортикотропин Адреналин Андрогены Гестагены Медиаторы симпатического отдела нервной системы Эстрогены Соматотропин Тироксин Инсулин
иммунокомпетентных клеток. Соматотропин оказывает прямое митоген-
ное действие на Т-лимфоциты.
Эффекты многих гормонов опосредованы их влиянием на секрецию
гормонов тимуса: глюкокортикоиды, андрогены и эстрогены подавляют ее,
а пролактин, СТГ и прогестерон — стимулируют (табл. 8.6).
Лимфоциты имеют 0-адренергические рецепторы и получают сигналы
от соответствующих медиаторов: норадреналина и нейроксина, которые
угнетают пролиферацию лимфоцитов, стимулируя их дифференцировку.
Ацетилхолин и холинергические стимулы, напротив, усиливают пролифе-
рацию лимфоцитов. Холинергические влияния в тимусе способствуют на-
коплению и эмиграции тимоцитов. Таким образом, проявляется альтерна-
тивный характер адренергических и холинергических эффектов нервной
системы в отношении иммунной системы.
Свидетельствами нейроэндокринной регуляции иммунной системы мо-
гут служить: зависимость иммунологических функций от циркадных рит-
мов, иммуномодулирующие эффекты гипноза, возможность условно-реф-
лекторной стимуляции или угнетения иммунного ответа.
8.4.3. Цитокиновый контроль
Цитокинами называют белковые и полипептидные молекулы, которые
продуцируются клетками иммунной системы (лимфоцитами, мононукле-
арными фагоцитами, дендритными клетками), а также стромальными
клетками. Цитокины действуют на клетки, соединяясь со своими поверх-
ностными рецепторами. Связывание цитокина с рецептором служит ис-
точником сигнала активации генов, который передается в ядро клетки.
Чувствительность клеток-мишеней к действию цитокинов изменяется в за-
висимости от количества цитокиновых рецепторов на их поверхности.
Время синтеза цитокина, как правило, краткосрочно: лимитирующим фак-
тором служит нестабильность молекул мРНК. Отдельные цитокины (на-
пример, ростовые факторы) продуцируются спонтанно. Синтез цитокинов
индуцируется: чаще всего, микробными компонентами и продуктами (на-
пример, бактериальным эндотоксином). Кроме того, один цитокин может
служить индуктором синтеза других цитокинов (например, интерлейкин 1
индуцирует продукцию интерлейкинов-6, -8, -12), чем обеспечивается кас-
кадный характер цитокинового контроля. Для биологических эффектов
цитокинов характерно то, что один и тот же цитокин проявляет разную
биологическую активность, в то же время одну и ту же функцию могут вы-
полнять разные цитокины. Этим обеспечивается запас прочности системы
цитокинового контроля. При совместном влиянии на клетки цитокины
могут выступать в качестве синергистов или антагонистов.
13-6095
386 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Большинство цитокинов оказывают наиболее выраженное влияние в
ближайшем микроокружении клеток-продуцентов. Местное действие ци-
токинов может быть как аутокринное, так и паракринное, в зависимости от
того, связывается ли цитокин с рецептором на самой клетке-продуценте,
или — на соседней клетке. Системное (эндокринное) влияние цитокинов
возможно только в случае интенсивного накопления их в циркулирующей
крови (например, при эндотоксическом шоке). Обычно в крови присутст-
вуют следовые количества цитокинов, недостаточные для проявления сис-
темных эффектов.
В костном мозге, в тимусе и лимфоидных органах постоянно происхо-
дит спонтанный синтез цитокинов, выполняющих функции ростовых фак-
торов, которые связываются с рецепторами на клетках-предшественницах,
стимулируя процессы их пролиферации и дифференцировки.
В очаге инфекции бактериальные компоненты и продукты индуцируют
синтез макрофагами так называемых провоспалителъных цитокинов (интер-
лейкины-1, -6, -8, туморнекротизирующий фактор), которые связываются
с рецепторами на других макрофагах, лимфоцитах и эндотелиальных клет-
ках. Их основные функции: усилить приток лейкоцитов из кровяного рус-
ла в очаг инфекции, активировать синтез фагоцитирующими клетками
бактерицидных молекул — супероксидных и нитроксидных радикалов.
В динамике процесса воспаления те же макрофаги начинают продуци-
ровать противовоспалительные цитокины—интерлейкины-10, -13, транс-
формирующий ростовой фактор, которые связываются со своими рецепто-
рами на клетках, посылая к ядру сигналы ингибиции функций фагоцити-
рующих клеток, в том числе продукции провоспалительных цитокинов.
Цитокины, контролирующие силу и форму специфического иммунного
ответа, продуцируются Thl (гамма-интерферон и туморнекротизирующий
фактор бета) или Th2 (интерлейкины-4, -5, -6, -10, -13). Первая группа
цитокинов обеспечивает перевес клеточного иммунного ответа над гумо-
ральным, а вторая обеспечивает перевес гуморального иммунного ответа
над клеточным. Например, продукт Th2 интерлейкин-4 ингибирует боль-
шинство функций макрофагов, активированных гамма-интерфероном. Ин-
терлейкин-10 ингибирует представление антигена, продукцию провоспали-
тельных цитокинов, является синергистом ИЛ-4. Продукт Thl гамма-ин-
терферон ингибирует функции В-лимфоцитов, участвующих в гумораль-
ном ответе. Таким образом, под влиянием отдельных цитокинов и их соче-
таний может изменяться характер специфического иммунного ответа. Это
значит, что от баланса цитокинов зависит эффективность противоинфек-
ционной защиты, поскольку от внутриклеточно паразитирующих микроор-
ганизмов эффективна клеточная защита, а против внеклеточно паразити-
рующих микроорганизмов эффективнее специфический гуморальный от-
вет. Цитокины выполняют функции межклеточных медиаторов, обеспечи-
вающих передачу сигналов активации или ингибиции от одних клеток к
другим.
ГЛАВА 9
Функции систем кровообращения
и лимфообращения
9.1. Система кровообращения
Основное назначение сердечно-сосудистой системы — обеспечение крово-
обращения, т. е. постоянной циркуляции крови из сердца в сосуды и из
них вновь к сердцу. Движущей силой кровотока является энергия, задавае-
мая сердцем потоку крови в сосудах, и градиент давления — разница давле-
ний между последовательными отделами сосудистого русла: кровь течет от
области высокого давления к области низкого давления. Поэтому из аорты
(где среднее давление составляет 100 мм рт. ст.) кровь поступает через сис-
тему магистральных артерий (80 мм рт. ст.) и артериол (40—60 мм рт. ст.) в
капилляры (15—25 мм рт. ст.), откуда продолжает движение в венулы (12—
15 мм рт. ст.), венозные коллекторы — более крупные вены (3—5 мм рт.
ст.) и полые вены (1—3 мм рт. ст.).
Центральное венозное давление — давление в устье полых вен — состав-
ляет около 0 мм рт. ст. В легочной артерии (в которой течет венозная
кровь) кровяное давление равно 18—25 мм рт. ст., в легочной вене — 3—
4 мм рт. ст. и в левом предсердии — 2—3 мм рт. ст.
Благодаря постоянному движению крови в сосудах выполняются основ-
ные функции системы кровообращения: 1) транспорт веществ, необходи-
мых для обеспечения специфической деятельности клеток организма;
2) доставка к клеткам организма химических веществ, регулирующих их
обмен; 3) отвод от клеток продуктов метаболизма; 4) гуморальная, т. е.
осуществляемая через жидкость, связь органов и тканей между собой;
5) доставка тканям средств защиты; 6) удаление вредных веществ из орга-
низма; 7) обмен тепла в организме. Следовательно, система кровообраще-
ния выполняет одновременно две задачи: обеспечивает циркуляцию крови
в системе и нутритивную (питательную) функцию клеток всех органов и
тканей. При этом к тканям доставляются не только питательные вещества,
но также кислород, физиологически активные вещества, в том числе гор-
моны, вода, соли, а из тканей выводятся углекислота и другие продукты
обмена веществ.
Кровоток в организме теплокровных животных осуществляется по двум
кругам, соединенным между собой через сердце. Малый (или легочный)
круг кровообращения осуществляет прямой контакт с внешней средой, а
большой — обеспечивает контакт с органами и тканями.
9.1.1. Функциональные классификации системы кровообращения
Распространено и обосновано деление сердечно-сосудистой системы по
уровню кровяного давления: область высокого и область низкого давления.
К области высокого давления относят левый желудочек сердца, артерии
13'
388 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
крупного, среднего и малого калибра, артериолы; к области низкого давле-
ния — остальные отделы системы (от капилляров до левого предсердия).
В функциональной классификации шведского физиолога Б. Фолкова
предусмотрено деление системы кровообращения на «последовательно со-
единенные звенья».
1. Сердце — насос, ритмически выбрасывающий кровь в сосуды.
2. Упруго-растяжимые сосуды, которые превращают периодичный вы-
брос крови из сердца в равномерный кровоток (аорта с ее отделами,
легочная артерия).
3. Резистивные сосуды (сосуды сопротивления) — прекапиллярный (в ос-
новном артериолы) и посткапиллярный отделы (венулы), которые
вместе создают общее сопротивление кровотоку в сосудах органов.
4. Прекапиллярные сфинктеры — специализированный отдел мельчайших
артериальных сосудов, сокращение гладкомышечных клеток этих
сфинктеров может приводить к перекрытию просвета мелких сосудов.
Эти сосуды регулируют объем кровотока в капиллярном русле.
5. Обменные сосуды, или истинные капилляры, где кровь контактирует с
тканью благодаря огромным поверхностям капиллярного ложа. Здесь
реализуется основная функция сердечно-сосудистой системы — об-
мен между кровью и тканями.
6. Шунтирующие сосуды (артериовенозные анастомозы), наличие кото-
рых доказано не для всех тканей.
7. Емкостные сосуды, в которых изменения просвета, даже столь неболь-
шие, что не оказывают существенного влияния на общее сопротивле-
ние, вызывают выраженные изменения распределения крови и вели-
чины притока ее к сердцу (венозный отдел системы).
Однако разделение на «резистивные» и «емкостные» сосуды весьма ус-
ловно, поскольку сопротивлением обладают как артериальные, так и ве-
нозные сосуды, хотя в количественном плане эта функция различна для
указанных отделов. С другой стороны, емкостью обладают как венозные
сосуды, так и артериальные. Весьма расплывчатым является и понятие
«емкостные сосуды», поскольку одни авторы относят к ним все венозное
ложе, другие — только венулы и мелкие вены. Неудачно выделены в клас-
сификации и «прекапиллярные» сфинктеры, поскольку в венозном русле
также существуют сосуды с расположением гладкомышечных волокон типа
сфинктеров или запирательных образований.
Функциональное назначение различных отделов сердечно-сосудистой
системы отражает следующая классификация (Б. И. Ткаченко):
1. Генератор давления и расхода крови — сердце, подающее кровь в аорту
и легочную артерию во время систолы.
2. Сосуды высокого давления — аорта и крупные артериальные сосуды, в
которых поддерживается характерный для индивидуума уровень кро-
вяного давления.
3. Сосуды — стабилизаторы давления — мелкие артерии и артериолы, ко-
торые путем сопротивления кровотоку и во взаимоотношении с сер-
дечным выбросом поддерживают оптимальный для системы уровень
артериального давления.
4. Распределители капиллярного кровотока — терминальные сосуды, глад-
комышечные образования которых при сокращении прекращают
кровоток в капилляре или возобновляют его (при расслаблении),
обеспечивая необходимое в данной ситуации число функционирую-
щих и нефункционирующих капилляров.
5. Обменные сосуды — капилляры и частично посткапиллярные участки
9. Функции систем кровообращения и лимфообращения • 389
венул, функция которых состоит в обеспечении обмена между кро-
вью и тканями.
6. Аккумулирующие сосуды — венулы и мелкие вены, активные или пас-
сивные изменения просвета которых ведут к накоплению крови (с
возможностью ее последующего использования) или к экстренному
выбросу ее в циркуляцию. Функция этих сосудов в основном емкост-
ная, но они обладают и резистивной функцией, хотя и намного мень-
шей, чем стабилизаторы давления.
7. Сосуды возврата крови — крупные венозные коллекторы и полые ве-
ны, через которые обеспечивается подача крови к сердцу.
8. Шунтирующие сосуды — различного типа анастомозы, соединяющие
между собой артериолы и венулы и обеспечивающие ненутритивный
кровоток.
9. Резорбтивные сосуды — лимфатический отдел системы кровообраще-
ния, в котором главная функция лимфатических капилляров состоит в
резорбции из тканей белков и жидкости, а лимфатических сосудов —
в транспортировке резорбированного материала обратно в кровь.
9.1.2. Общая характеристика движения крови по сосудам
Отличительной особенностью характеристики сердечно-сосудистой систе-
мы на современном этапе является требование выражать все составляющие
ее параметры количественно. Геометрические (табл. 9.1) и гидродинамиче-
ские (табл. 9.2) характеристики системы кровообращения свидетельствуют
о том, что аорта представляет собой трубку диаметром 1,6—3,2 см с пло-
щадью поперечного сечения 2,0—3,5 см2, постепенно разветвляющуюся на
109 капилляров, площадь поперечного сечения каждого из которых равна
5 • 10~7 см2.
Радиус усредненного капилляра может составлять 3 мкм, длина — около
750 мкм (хотя диапазон реальных значений довольно велик). Площадь по-
верхности стенки каждого усредненного капилляра равна 15 000 мкм2, а
площадь поперечного сечения — 30 мкм2. Поскольку доказано, что обмен
происходит и в посткапиллярных венулах, можно допускать, что общая об-
менная поверхность мельчайшего сосуда большого круга составляет
25 000 мкм2. Общее число функционирующих капилляров у человека мас-
сой 70 кг должно быть порядка 40 000 млн., тогда общая обменная пло-
щадь поверхности капилляров должна составлять около 1000 м2.
В сосудах различают скорость кровотока объемную и линейную.
Объемная скорость кровотока — количество крови, протекающее через
поперечное сечение сосуда в единицу времени. Объемная скорость крово-
Таблица 9.1. Геометрические характеристики сосудистого русла большого круга крово-
обращения
Сосуд Диаметр, см Общее число в организме Длина, см
Аорта 1,6-3,2 1 80
Большие артерии 0,6-0,1 103 20-40
Малые артерии, артериолы 0,1-0,02 108 0,2-5
Капилляры 0,0005-0,001 109 0,1
Венулы, малые вены 0,02-0,2 109 0,2-1,0
Большие вены 0,5-1,0 103 10-30
Полые вены 2,0 2 50
390 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 9.2. Гидродинамические характеристики сосудистого русла большого круга
кровообращения
Сосуд Давление, мм рт. ст. Объем, см3 Скорость крово- тока, см • с-1 Сопротивление, дин • с • см-5
Аорта 100-120 30 50 64
Магистральные артерии 100-120 60 13 3,9 • 103
Ветвящиеся артерии 80-90 50 8 1,6- 10s
Терминальные артерии 80-90 25 6 1,2- 10s
Артериолы 40-60 25 0,3 2 1010
Капилляры 15-25 60 0,07 3,9-10"
Венулы 12-18 НО 0,07 4 • 109
Терминальные вены 10-12 130 1,3 3,2- 103
Ветвящиеся вены 5-8 270 1,5 0,5 • 104
Венозные коллекторы 3-5 220 3,6 250
Полые вены 1-3 100 33 26
тока через сосуд прямо пропорциональна давлению крови в нем и обратно
пропорциональна сопротивлению току крови в этом сосуде.
Линейная скорость кровотока отражает скорость продвижения частиц
крови вдоль сосуда и равна объемной скорости, деленной на площадь се-
чения кровеносного сосуда. Линейная скорость различна для частиц кро-
ви, продвигающихся в центре потока и у сосудистой стенки. В центре со-
суда линейная скорость максимальна, а около стенки сосуда она мини-
мальна в связи с тем, что здесь особенно велико трение частиц крови о
стенку.
Под сердечным выбросом понимают количество крови, выбрасываемой
сердцем в сосуды в единицу времени.
Исходя из величины сердечного выброса в покое и средней скорости
кровотока в капилляре (см. табл. 9.2) подсчитано, что площадь поперечно-
го сечения капиллярного ложа должна в 700 раз превышать площадь попе-
речного сечения аорты. В покое функционирует только 25—35 % капилля-
ров и общая площадь их обменной поверхности составляет 250—350 м2.
Давление и скорость кровотока в системе кровообращения уменьшают-
ся от аорты до венул (см. табл. 9.2), а кровеносные сосуды становятся все
более мелкими и многочисленными. В капиллярах скорость кровотока за-
медляется наиболее выражение, что благоприятствует отдаче кровью ве-
ществ тканям. Для венозного отдела характерны низкий уровень давления
и более медленная по сравнению с артериальным руслом скорость крово-
тока.
Сопоставление величин давления, кровотока и сопротивления сосудов в
различных отделах сосудистого русла (табл. 9.2) свидетельствует о том, что
внутрисосудистое давление от аорты до полых вен резко снижается, а объ-
ем крови в венозном русле, наоборот, возрастает. Следовательно, артери-
альное русло характеризуется высоким давлением и сравнительно неболь-
шим объемом крови, а венозное — большим объемом крови и низким дав-
лением.
Считается, что в венозном русле содержится 75—80 % крови, а в арте-
риальном — 15—17 % и в капиллярах — около 5 % (в диапазоне 3—10 %).
Исходя из этого в функциональной схеме сердечно-сосудистой системы
(рис. 9.1) выделены 3 области: высокого давления, транскапиллярного об-
мена и большого объема.
При функциональном единстве, согласованности и взаимообусловлен-
9. Функции систем кровообращения и лимфообращения • 391
ОБЛАСТИ
Большого объема Транскапиллярного Высокогодавления
(венозная) обмена (капилляры) (артериальная)
70—80 % 5—10 % 15—20 %
ОБЪЕМ СОДЕРЖАЩЕЙСЯ КРОВИ
Рис. 9.1. Сердечно-сосудистая
система (функциональная схе-
ма).
Цифры в скобках — величина кро-
вотока в покое (в % к минутному
объему), цифры внизу рисунка —
содержание крови (в % к общему
объему).
Артериальная часть сердечно-сосу-
дистой системы (светлая часть схе-
мы) содержит всего 15—20 % обще-
го объема крови и характеризуется
высоким (относительно остальных
отделов системы) давлением. В
центре схемы находится область
транскапиллярного обмена, т. е.
капиллярных (обменных) сосудов,
для обеспечения оптимальной
функции которых служит, в основ-
ном, сердечно-сосудистая система.
При этом в виде точек обозначено
большое число капилляров в орга-
низме и огромная площадь их воз-
можной поверхности во время
функционирования органа или тка-
ни, хотя цифры внизу указывают
на сравнительно небольшой объем
содержащейся в них крови в усло-
виях покоя. Наибольшее количест-
во крови содержится в области
большого объема, которая обозна-
чена штриховкой. Эта область со-
держит в 3—4 раза больше крови,
чем область высокого давления, в
связи с чем и площадь, обозначен-
ная на схеме штриховкой, больше
площади светлой части схемы.
ности подразделов сердеч-
но-сосудистой системы и
характеризующих их пара-
метров в ней условно выде-
ляют три уровня:
а) системная гемодинамика — обеспечивающая процессы циркуляции
крови (кругооборота) в системе;
б) органное кровообращение — кровоснабжение органов и тканей в зави-
симости от их функциональной потребности;
в) микрогемодинамика (микроциркуляция) — обеспечение транскапил-
лярного обмена, т. е. нутритивной (питательной) функции сосудов.
9.1.3. Системная гемодинамика
Основными параметрами, характеризующими системную гемодинамику,
являются: системное артериальное давление, общее периферическое со-
противление сосудов, сердечный выброс, работа сердца, венозный возврат
крови к сердцу, центральное венозное давление, объем циркулирующей
крови к сердцу.
392 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
9.1.3.1. Системное артериальное давление
Внутрисосудистое давление крови является одним из основных парамет-
ров, по которому судят о функционировании сердечно-сосудистой систе-
мы. Артериальное давление есть интегральная величина, составляющими и
определяющими которой являются объемная скорость кровотока (Q) и со-
противление (R) сосудов. Поэтому системное артериальное давление
(САД) является результирующей величиной сердечного выброса (СВ) и об-
щего периферического сопротивления сосудов (ОПСС):
САД = СВ • ОПСС.
Давление в крупных ветвях аорты (собственно артериальное) определя-
ется как:
АД = Q • R.
Применительно к артериальному давлению различают систолическое,
диастолическое, пульсовое и среднее давления. Систолическое — возникает
в артериях в период систолы левого желудочка сердца, диастолическое — в
период его диастолы, разница между величиной систолического и диасто-
лического давлений характеризует пульсовое давление (рис. 9.2). Выделяют
также среднее давление, которое представляет собой среднюю (не арифме-
тическую) между систолическим и диастолическим давлениями величину,
которая была бы способна при отсутствии пульсовых колебаний давления
крови дать такой же гемодинамический эффект, какой имеет место при ес-
тественном, колеблющемся движении крови. Среднее давление выражает
энергию непрерывного движения крови. Поскольку продолжительность
диастолического давления больше, чем систолического, то среднее давле-
ние ближе к величине диастолического давления и вычисляется как сумма
диастолического давления плюс '/3 пульсового.
Величина внутрисосудистого давления при прочих равных условиях оп-
Рис. 9.2. Систолическое (3), диастолическое (1), среднее (2) и пульсовое (1—3) дав-
ление в сосудах.
9. Функции систем кровообращения и лимфообращения • 393
ределяется расстоянием места его измерения от сердца. Различают поэтому
аортальное давление, артериальное давление, артериолярное, капиллярное, ве-
нозное (в мелких и крупных венах) и центральное венозное (в устье полых
вен) давление.
В биологических и медицинских исследованиях артериальное давление
выражают в миллиметрах ртутного столба (мм рт. ст.), а венозного — в
миллиметрах водного столба (мм водн. ст.).
У человека в покое наиболее усредненным из всех средних величин
считается систолическое давление 120—125 мм рт. ст., диастолическое 70—
75 мм рт. ст. Эти величины зависят от пола, возраста, конституции челове-
ка, условий его работы, географического пояса проживания и т. д.
Уровень АД не позволяет, однако, судить о степени кровоснабжения
органов и тканей или величине объемной скорости кровотока в сосудах.
Выраженные перераспределительные сдвиги в системе кровообращения
могут происходить при неизменном уровне АД, поскольку изменения
ОПСС могут компенсироваться противоположными сдвигами СВ, а суже-
ние сосудов в одних регионах — сопровождаться их расширением в других.
Одним из важнейших факторов, определяющих интенсивность кровоснаб-
жения тканей, является величина просвета сосудов, определяющая их со-
противление кровотоку.
9.1.3.2. Общее периферическое сопротивление сосудов
Под этим термином понимают общее сопротивление всей сосудистой сис-
темы выбрасываемому сердцем потоку крови. Это соотношение описыва-
ется уравнением:
ОПСС =
Как следует из этого уравнения, для расчета ОПСС необходимо опреде-
лить величину системного артериального давления и сердечного выброса.
Прямых бескровных методов измерения общего периферического со-
противления не разработано, и его величина определяется из уравнения
Пуазейля для гидродинамики:
R = 8-Щ,
пг
где R — гидравлическое сопротивление, / — длина сосуда, v — вязкость
крови, г — радиус сосудов.
Поскольку при исследовании сосудистой системы животного или чело-
века радиус сосудов, их длина и вязкость крови остаются обычно неизвест-
ными, Франк, используя формальную аналогию между гидравлической и
электрической цепями, привел уравнение Пуазейля к следующему виду:
R = P,~QP2 • 1332,
где Р1—Р2 — разность давлений в начале и в конце участка сосудистой сис-
темы, Q — величина кровотока через этот участок, 1332 — коэффициент
перевода единиц сопротивления в систему CGS.
Уравнение Франка широко используется на практике для определения
сопротивления сосудов, хотя оно не всегда отражает истинные физиологи-
ческие взаимоотношения между объемным кровотоком, АД и сопротивле-
394 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
|— 20 мм рт. ст.
Аортальное давление
Перфузионное давление в плечеголовной артерии
Перфузионное давление в грудной аорте
Отметка стимуляции
Рис. 9.3. Более выраженная величина повышения сопротивления сосудов бассейна
грудной аорты по сравнению с его изменениями в бассейне плечеголовной артерии
при прессорном рефлексе.
нием сосудов кровотоку у теплокровных. Эти три параметра системы дей-
ствительно связаны приведенным соотношением, но у разных объектов, в
разных гемодинамических ситуациях и в разное время их изменения могут
быть в разной мере взаимозависимыми. Так, в конкретных случаях уро-
вень САД может определяться преимущественно величиной ОПСС или в
основном СВ.
В обычных физиологических условиях ОПСС составляет от 1200 до
1700 дин • с см , при гипертонической болезни эта величина может воз-
растать в два раза против нормы и быть равной 2200—3000 дин • с • см-5.
Величина ОПСС состоит из сумм (не арифметических) сопротивлений
регионарных сосудистых отделов. При этом в зависимости от большей или
меньшей выраженности изменений регионарного сопротивления сосудов в
них соответственно будет поступать меньший или больший объем крови,
выбрасываемый сердцем. На рис. 9.3 показан пример более выраженной
степени повышения сопротивления сосудов бассейна нисходящей грудной
аорты по сравнению с его изменениями в плечеголовной артерии. Поэтому
прирост кровотока в плечеголовной артерии будет больше, чем в грудной
аорте. На этом механизме базируется эффект «централизации» кровообра-
щения у теплокровных, обеспечивающий в тяжелых или угрожающих орга-
низму условиях (шок, кровопотеря и др.) перераспределение крови, преж-
де всего, к головному мозгу и миокарду.
9.1.3.3. Сердечный выброс
В клинической литературе чаще используют понятие «минутный объем
кровообращения» (МОК).
Минутный объем кровообращения характеризует общее количество крови,
перекачиваемое правым и левым отделом сердца в течение одной минуты
в сердечно-сосудистой системе. Размерность минутного объема кровообра-
9. Функции систем кровообращения и лимфообращения • 395
щения — л/мин или мл/мин. Чтобы нивелировать влияние индивидуаль-
ных антропометрических различий на величину МОК, его выражают в ви-
де сердечного индекса. Сердечный индекс — это величина минутного объема
кровообращения, деленная на площадь поверхности тела в м. Размерность
сердечного индекса — л/(мин • м2).
В системе транспорта кислорода аппарат кровообращения является ли-
митирующим звеном, поэтому соотношение максимальной величины
МОК, проявляющейся при максимально напряженной мышечной работе,
с его значением в условиях основного обмена дает представление о функ-
циональном резерве сердечно-сосудистой системы. Это же соотношение
отражает и функциональный резерв сердца в его гемодинамической функ-
ции. Гемодинамический функциональный резерв сердца у здоровых людей
составляет 300—400 %. Это означает, что МОК покоя может быть увеличен
в 3—4 раза. У физически тренированных лиц функциональный резерв вы-
ше — он достигает 500—700 %.
Для условий физического покоя и горизонтального положения тела ис-
пытуемого нормальные величины МОК соответствуют диапазону 4—6 л/
мин (чаще приводятся величины 5—5,5 л/мин). Средние величины сердеч-
ного индекса колеблются от 2 до 4 л/(мин • м2) — чаще приводятся величи-
ны порядка 3—3,5 л/(мин • м2).
Поскольку объем крови у человека составляет только 5—6 л, полный
кругооборот всего объема крови происходит примерно за 1 мин. В период
тяжелой работы МОК у здорового человека может увеличиваться до 25—
30 л/мин, а у спортсменов — до 30—40 л/мин.
Факторами, определяющими величину МОК, являются систолический
объем крови, частота сердечных сокращений и венозный возврат крови к
сердцу.
Систолический объем крови. Объем крови, нагнетаемый каждым желу-
дочком в магистральный сосуд (аорту или легочную артерию) при одном
сокращении сердца, обозначают как систолический, или ударный, объем
крови.
В покое объем крови, выбрасываемый из желудочка, составляет в норме
от трети до половины общего количества крови, содержащейся в этой ка-
мере сердца к концу диастолы. Оставшийся в сердце после систолы ре-
зервный объем крови является своеобразным депо, обеспечивающим уве-
личение сердечного выброса при ситуациях, в которых требуется быстрая
интенсификация гемодинамики (например, при физической нагрузке,
эмоциональном стрессе и др.).
Величина систолического (ударного) объема крови во многом пред-
определена конечным диастолическим объемом желудочков. В условиях
покоя диастолическая емкость желудочков сердца подразделяется на три
фракции: ударного объема, базального резервного объема и остаточного
объема. Все эти три фракции суммарно составляют конечно-диастоличе-
ский объем крови, содержащийся в желудочках (рис. 9.4).
После выброса в аорту систолического объема крови оставшейся в же-
лудочке объем крови — это конечно-систолический объем. Он подразделя-
ется на базальный резервный объем и остаточный объем. Базальный ре-
зервный объем — это количество крови, которое может быть дополнитель-
но выброшено из желудочка при увеличении силы сокращений миокарда
(например, при физической нагрузке организма). Остаточный объем — это
то количество крови, которое не может быть вытолкнуто из желудочка да-
же при самом мощном сердечном сокращении (см. рис. 9.4).
Величина резервного объема крови является одной из главных детерми-
396 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
«симальная диастолическая емкость желудочка Максимальный ударный объем Дополнительный резервный объем
Ударный объем
Базальный резервный объем
си Остаточный объем
Рис. 9.4. Фракции диастолической емкости левого желудочка.
нант функционального резерва сердца по его специфической функции —
перемещению крови в системе. При увеличении резервного объема, соот-
ветственно, увеличивается максимальный систолический объем, который
может быть выброшен из сердца в условиях его интенсивной деятельности.
Регуляторные влияния на сердце реализуются в изменении систоличе-
ского объема путем воздействия на сократительную силу миокарда. При
уменьшении мощности сердечного сокращения систолический объем сни-
жается.
У человека при горизонтальном положении тела в условиях покоя сис-
толический объем составляет от 60 до 90 мл (табл. 9.3).
Таблица 9.3. Некоторые параметры системной гемодинамики и насосной функции
сердца у человека (в условиях основного обмена)
Сердечный выброс, л/мин Частота сердечных сокращений, мин-1 Конечно-диастолический объем, мл Конечно-систолический объем, мл Ударный объем сердца, мл 4,0-6,0 60-80 65-130 40-60 60-90
Давление, мм рт. ст. Аорта Легочный ствол
систолическое 110-120 25-30
диастолическое 60-75 6-12
среднее 70-80 9-17
Общее периферическое сопротивление сосу- Большой круг Малый круг
дов, дин • с • см-5 1200-1700 150-350
Давление, мм рт. ст. Левое предсердие Правое предсердие
конечно-систолическое 10-12 3-7
конечно-диастолическое 0-6 0-2
Давление, мм рт. ст. Левый желудочек Правый желудочек
конечно-систолическое 110-120 25-30
конечно-диастолическое 9-12 6-10
Максимальная скорость прироста давления (dP/dt,nax), мм рт. ст./с 2000-2500 350-400
Работа за систолу, Дж 0,9-1,0 0,2
9. Функции систем кровообращения и лимфообращения • 397
9.1.3.4. Частота сердечных сокращений (пульс)
Частота сердечных сокращений (пульс) в покое составляет от 60 до 80 уда-
ров в минуту. Регуляторные влияния, вызывающие изменения частоты
сердечных сокращений, называются хронотропными, а изменения силы
сокращений сердца — инотропными.
Повышение частоты сердечных сокращений является важным адапта-
ционным механизмом увеличения МОК, осуществляющим быстрое при-
способление его величины к требованиям организма. При некоторых экс-
тремальных воздействиях на организм сердечный ритм может повышаться
в 3—3,5 раза по отношению к исходному.
Важным показателем системной гемодинамики и сердечной деятельно-
сти является работа сердца за систолу.
9.1.3.5. Работа сердца
Миокард желудочков в период изометрического сокращения сообщает на-
ходящейся в них крови потенциальную энергию, которая в период изгна-
ния крови переходит в кинетическую энергию движущейся крови. Боль-
шая часть последней переходит в сосудистой системе в потенциальную
энергию растянутых эластических стенок аорты и легочной артерии, и
лишь небольшое количество ее (2—5 %) сохраняется в виде кинетической
энергии движущейся крови.
Поскольку вся полезная работа сердца переходит в энергию крови, пол-
ную работу сердца за систолу можно рассчитать как сумму потенциальной
и кинетической энергии крови. Потенциальная часть работы сердца вы-
числяется как произведение ударного объема сердца на среднее артериаль-
ное давление. Кинетическая часть работы сердца рассчитывается как поло-
вина произведения массы выбрасываемой из сердца крови за систолу на
квадрат средней скорости изгнания крови из сердца. В покое кинетическая
часть работы сердца составляет лишь 2—5 % от полной работы сердца, по-
этому ее обычно не учитывают при расчетах. Однако при физических на-
грузках кинетическая часть работы сердца у человека может составлять до
30 % от полной работы сердца за систолу. Рассчитанная таким образом ра-
бота характеризует деятельность левой половины сердца. Работа правых
отделов сердца составляет около 25 % от этой величины.
9.1.3.6. Сократимость
Сократимость, т. е. способность сокращаться, характерная для всех разно-
видностей мышечной ткани, реализуется в миокарде благодаря трем спе-
цифическим свойствам сердечной мышцы: автоматизм — способность кле-
ток водителей ритма генерировать импульсы без каких-либо внешних воз-
действий; проводимость — способность элементов проводящей системы к
элекгротонической передаче возбуждения; возбудимость — способность
кардиомиоцитов возбуждаться в естественных условиях под влиянием им-
пульсов, передаваемых по волокнам Пуркинье (рис. 9.5). Важной особен-
ностью возбудимости сердечной мышцы является длительный рефрактор-
ный период (полное исчезновение или резкое снижение возбудимости
кардиомиоцитов после их предыдущего сокращения), гарантирующий рит-
мический характер последующего сокращения.
398 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
9.1.3.6.1. Автоматизм и проводимость миокарда
В области правого предсердия, а также на границе предсердий и желудоч-
ков располагаются участки, ответственные за возбуждение сердечной
мышцы. Автоматизм сердца имеет миогенную природу и обусловлен спон-
танной активностью части клеток его атипической ткани.
Указанные клетки образуют скопления в определенных участках мио-
карда. Наиболее важным в функциональном отношении из них является
синусный, или синоатриальный, узел, расположенный между местом впаде-
ния верхней полой вены и ушком правого предсердия. В нижней части
межпредсердной перегородки, непосредственно над местом прикрепления
септальной створки трехстворчатого клапана, располагается атриовентри-
кулярный узел. От него отходит пучок атипических мышечных волойон, ко-
торый пронизывает фиброзную перегородку между предсердиями и пере-
ходит в узкий длинный мышечный тяж, заключенный в межжелудочковую
перегородку. Он называется атриовентрикулярным пучком, или пучком Ги-
са. Пучок Гиса разветвляется, образуя две ножки, от которых приблизи-
тельно на уровне середины перегородки отходят волокна Пуркинье, также
образованные атипической тканью и формирующие субэндокардиальную
сеть в стенках обоих желудочков (см. рис. 9.5).
Функция проводимости в сердце имеет электротоническую природу.
Она обеспечивается низким электрическим сопротивлением щелевидных
контактов (нексусов) между элементами атипического и рабочего миокар-
да, а также в области вставочных пластинок, разделяющих кардиомиоци-
ты. В результате сверхпороговое раздражение любого участка вызывает ге-
нерализованное возбуждение всего миокарда. Это позволяет считать ткань
сердечной мышцы, морфологически разделенную на отдельные клетки,
функциональным синцитием.
Возбуждение миокарда зарождается в синоатриальном узле, который
называют водителем ритма, или пейсмекером первого порядка, и далее
Пучок Гиса
Правая ножка
Левая ножка
Передняя ветвь
Задняя ветвь
Волокна Пуркинье
Синоатриальный узел
Атриовентрикулярный узел
Правое
предсердие
Рис. 9.5. Проводящая система сердца.
9. Функции систем кровообращения и лимфообращения • 399
распространяется на мускулатуру предсердий с последующим возбуждени-
ем атриовентрикулярного узла, который является водителем ритма второго
порядка. Скорость распространения возбуждения в предсердиях составляет
1 м/с. При переходе возбуждения на атриовентрикулярный узел имеет ме-
сто так называемая атриовентрикулярная задержка, составляющая 0,04—
0,06 с. Механизм атриовентрикулярной задержки состоит в том, что про-
водящие ткани синоатриального и атриовентрикулярного узлов контакти-
руют не непосредственно, а через волокна рабочего миокарда, для которых
характерна более низкая скорость проведения возбуждения. Последнее
распространяется далее по ножкам пучка Гиса и волокнам Пуркинье, пе-
редаваясь на мускулатуру желудочков, которую оно охватывает со скоро-
стью 0,75—4,0 м/с. В силу особенностей расположения волокон Пуркинье
возбуждение сосочковых мышц происходит несколько раньше, чем оно
охватывает стенки желудочков. Благодаря этому нити, удерживающие
трехстворчатый и митральный клапаны, оказываются натянутыми раньше,
чем на них начинает действовать сила сокращения желудочков. По той же
причине наружная часть стенки желудочков у верхушки сердца возбужда-
ется несколько раньше участков стенки, прилежащих к ее основанию. Та-
ким образом, волна возбуждения последовательно охватывает различные
отделы сердца в направлении от правого предсердия к верхушке. Однако
указанные сдвиги во времени крайне невелики и обычно принимается, что
весь миокард желудочков охватывается возбуждением одновременно.
9.1.3.6.2. Мембранная природа автоматии сердца
Возбудимость клеток проводящей системы и рабочего миокарда имеет ту
же биоэлектрическую природу, что и в поперечно-полосатых мышцах. На-
личие заряда на мембране здесь также обеспечивается разностью концен-
траций ионов калия и натрия возле ее внешней и внутренней поверхности
и избирательной проницаемостью мембраны для этих ионов. В покое мем-
брана кардиомиоцитов проницаема для ионов калия и почти непроницае-
ма для ионов натрия. В результате диффузии ионы калия выходят из клет-
ки и создают положительный заряд на ее поверхности. Внутренняя сторо-
на мембраны становится электроотрицательной по отношению к на-
ружной.
В клетках атипического миокарда, обладающих автоматией, мембран-
ный потенциал способен спонтанно уменьшаться до критического уровня,
что приводит к генерации потенциала действия. В норме ритм сердечных
сокращений задается всего несколькими наиболее возбудимыми клетками
синоатриального узла, которые называются истинными водителями ритма,
или пейсмекерными клетками. В этих клетках во время диастолы мембран-
ный потенциал, достигнув максимального значения, соответствующего ве-
личине потенциала покоя (60—70 мВ), начинает постепенно снижаться.
Этот процесс называют медленной спонтанной диастолической деполяризаци-
ей. Она продолжается до того момента, когда мембранный потенциал дос-
тигает критического уровня (40—50 мВ), после чего возникает потенциал
действия.
Для потенциала действия пейсмекерных клеток синоатриального узла
характерны малая крутизна подъема, отсутствие фазы ранней быстрой ре-
поляризации, а также слабая выраженность «овершута» и фазы «плато».
Медленная реполяризация плавно сменяется быстрой. Во время этой фазы
мембранный потенциал достигает максимальной величины, после чего
вновь возникает фаза медленной спонтанной деполяризации (рис. 9.6).
400 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 9.6. Развитие потенциала действия истинного водителя ритма автоматии
сердца.
Во время диастолы спонтанная деполяризация уменьшает мембранный потенциал (Етах) до
критического уровня (Екр) и вызывает потенциал действия.
Рис. 9.7. Развитие потенциала действия истинного и латентного водителей ритма
автоматии сердца.
Скорость медленной диастолической деполяризации истинного водителя ритма больше, чем у
латентного.
Частота возбуждения пейсмекерных клеток у человека составляет в по-
кое 70—80 в минуту при амплитуде потенциала действия 70—80 мВ. Во
всех остальных клетках проводящей системы потенциал действия в норме
возникает под влиянием возбуждения, приходящего из синоатриального
узла. Такие клетки называют латентными водителями ритма. Потенциал
действия в них возникает раньше, чем их собственная медленная спонтан-
9. Функции систем кровообращения и лимфообращения • 401
ная диастолическая деполяризация достигает критического уровня. Ла-
тентные водители ритма принимают на себя ведущую функцию только при
условии разобщения с синоатриальным узлом. Частота спонтанной депо-
ляризации таких клеток у человека составляет 30—40 в минуту (рис. 9.7),
Спонтанная медленная диастолическая деполяризация обусловлена со-
вокупностью ионных процессов, связанных с функциями плазматических
мембран. Среди них ведущую роль играют медленное уменьшение калие-
вой и повышение натриевой и кальциевой проводимости мембраны во
время диастолы, параллельно чему происходит падение активности элек-
трогенного натриевого насоса. К началу диастолы проницаемость мембра-
ны для калия на короткое время повышается, и мембранный потенциал
покоя приближается к равновесному калиевому потенциалу, достигая мак-
симального диастолического значения. Затем проницаемость мембраны
для калия уменьшается, что и приводит к медленному снижению мембран-
ного потенциала до критического уровня. Одновременное увеличение про-
ницаемости мембраны для натрия и кальция приводит к поступлению этих
ионов в клетку, что также способствует возникновению потенциала дейст-
вия. Снижение активности электрогенного насоса дополнительно умень-
шает выход натрия из клетки и, тем самым, облегчает деполяризацию мем-
браны и возникновение возбуждения.
9.1.3.6.3. Возбудимость сердечной мышцы
Клетки миокарда обладают возбудимостью, но им не присуща автоматия.
В период диастолы мембранный потенциал покоя этих клеток стабилен, и
его величина выше (80—90 мВ), чем в клетках водителей ритма. Потенциал
действия в этих клетках возникает под влиянием возбуждения клеток во-
дителей ритма, которое достигает кардиомиоцитов, вызывая деполяриза-
цию их мембран.
Потенциал действия клеток рабочего миокарда состоит из фазы быст-
рой деполяризации, начальной быстрой реполяризации, переходящей в
фазу медленной реполяризации (фаза плато), и фазы быстрой конечной
Рис. 9.8. Потенциал действия клетки рабочего миокарда.
Быстрое развитие деполяризации и продолжительная реполяризация. Замедленная реполяри-
зация (плато) переходит в быструю реполяризацию.
402 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
.... Возбудимость---------Исходный уровень
Рис. 9.9. Сопоставление потенциала действия и сокращения миокарда с фазами из-
менения возбудимости.
1 — фаза деполяризации; 2 — фаза начальной быстрой реполяризации; 3 — фаза медленной ре-
поляризации (фаза плато); 4 — фаза конечной быстрой реполяризации; 5 — фаза абсолютной
рефрактерности; 6 — фаза относительной рефрактерности; 7 — фаза супернормальной возбу-
димости.
Рефрактерность миокарда практически совпадает не только с возбуждением, но и с периодом
сокращения.
реполяризации (рис. 9.8). Фаза быстрой деполяризации создается резким
повышением проницаемости мембраны для ионов натрия, что приводит к
возникновению быстрого входящего натриевого тока. Последний, однако,
при достижении мембранного потенциала 30—40 мВ инактивируется и в
последующем, вплоть до инверсии потенциала (около +30 мВ) и в фазу
«плато», ведущее значение имеют кальциевые ионные токи. Деполяриза-
9. Функции систем кровообращения и лимфообращения • 403
ция мембраны вызывает активацию кальциевых каналов, в результате чего
возникает дополнительный деполяризирующий входящий кальциевый ток.
Конечная реполяризация в клетках миокарда обусловлена постепенным
уменьшением проницаемости мембраны для кальция и повышением про-
ницаемости для калия. В результате входящий ток кальция уменьшается, а
выходящий ток калия возрастает, что обеспечивает быстрое восстановле-
ние мембранного потенциала покоя. Длительность потенциала действия
кардиомиоцитов составляет 300—400 мс, что соответствует длительности
сокращения миокарда (рис. 9.9).
9.1.3.6.4. Сопряжение возбуждения и сокращения
миокарда
Инициатором сокращения миокарда, как и в скелетной мышце, является
потенциал действия, распространяющийся вдоль поверхностной мембраны
кардиомиоцита. Поверхностная мембрана волокон миокарда образует впя-
чивания, так называемые поперечные трубочки (Т-система), к которым
примыкают продольные трубочки (цистерны) саркоплазматического рети-
кулума, являющиеся внутриклеточным резервуаром кальция (рис. 9.10).
Саркоплазматический ретикулум в миокарде выражен в меньшей степени,
чем в скелетной мышце. Нередко к поперечной Т-трубочке примыкают не
две продольные трубочки, а одна (система диад, а не триад, как в скелет-
ной мышце). Считается, что потенциал действия распространяется с по-
верхностной мембраны кардиомиоцита вдоль Т-трубочки в глубь волокна
и вызывает деполяризацию цистерны саркоплазматического ретикулума,
что приводит к освобождению из цистерны ионов кальция.
Следующим этапом электромеханического сопряжения является пере-
мещение ионов кальция к сократительным протофибриллам. Сократитель-
ная система сердца представлена сократительными белками — актином и
миозином, и модуляторными белками — тропомиозином и тропонином.
Молекулы миозина формируют толстые нити саркомера, молекулы акти-
на—тонкие нити. В состоянии диастолы тонкие актиновые нити входят
своими концами в промежутки между толстыми и более короткими миози-
новыми нитями. На толстых нитях миозина располагаются поперечные
мостики, содержащие АТФ, а на нитях актина — модуляторные белки —
тропомиозин и тропонин. Эти белки образуют единый комплекс, блоки-
рующий активные центры актина, предназначенные для связывания мио-
зина и стимуляции его АТФазной активности. Сокращение волокон мио-
карда начинается со связывания тропонином вышедшего из саркоплазма-
тического ретикулюма в межфибриллярное пространство кальция. Связы-
вание кальция вызывает изменения конформации тропонин-тропомиози-
нового комплекса. В результате этого открываются активные центры и
происходит взаимодействие актиновых и миозиновых нитей. При этом
стимулируется АТФазная активность миозиновых мостиков, происходит
распад АТФ и выделяющаяся энергия используется на скольжение нитей
друг относительно друга, приводящее к сокращению миофибрилл. В отсут-
ствие ионов кальция тропонин препятствует образованию актомиозиново-
го комплекса и усилению АТФазной активности миозина. Морфологиче-
ские и функциональные особенности миокарда свидетельствуют о тесной
связи между внутриклеточным депо кальция и внутриклеточной средой.
Так как запасы кальция во внутриклеточных депо невелики, большое зна-
чение имеет вход кальция в клетку во время генерации потенциала дейст-
вия (см. рис. 9.10). Потенциал действия и сокращение миокарда совпадают
404 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Триггерный эффект
Рис. 9.10. Схема соотношений между возбуждением, током Са2+ и активацией со-
кратительного аппарата. Начало сокращения связано с выходом Са2+ из продольных
трубочек при деполяризации мембраны. Са2+, входящий через мембраны кардио-
миоцита в фазу плато потенциала действия, пополняет запасы Са2+ в продольных
трубочках.
по времени. Поступление кальция из наружной среды в клетку создает ус-
ловия для регуляции силы сокращения миокарда. Большая часть входяще-
го в клетку кальция, очевидно, пополняет его запасы в цистернах сарко-
плазматического ретикулума, обеспечивая последующие сокращения.
Удаление кальция из клеточного пространства приводит к разобщению
процессов возбуждения и сокращения миокарда. Потенциалы действия
при этом регистрируются почти в неизменном виде, но сокращения мио-
карда не происходит. Вещества, блокирующие вход кальция во время гене-
рации потенциала действия, вызывают аналогичный эффект. Вещества, уг-
нетающие кальциевый ток, уменьшают длительность фазы плато и потен-
циала действия и понижают способность миокарда к сокращению. При
повышении содержания кальция в межклеточной среде и при введении ве-
ществ, усиливающих вход этого иона в клетку, сила сердечных сокраще-
ний увеличивается. Таким образом, потенциал действия играет роль пус-
кового механизма, вызывая освобождение кальция из цистерн саркоплаз-
матического ретикулума, регулирует сократимость миокарда, а также по-
полняет запасы кальция во внутриклеточных депо.
9. Функции систем кровообращения и лимфообращения • 405
9.1.3.6.5. Сердечный цикл и его фазовая структура
Работа сердца представляет собой непрерывное чередование периодов со-
кращения (систола) и расслабления (диастола). Сменяющие друг друга
систола и диастола составляют сердечный цикл. Поскольку в покое часто-
та сокращений сердца составляет 60—80 циклов в минуту, то каждый из
них продолжается около 0,8 с. При этом 0,1 с занимает систола предсер-
дий, 0,3 с — систола желудочков, а остальное время — общая диастола
сердца.
К началу систолы миокард расслаблен, а сердечные камеры заполнены
кровью, поступающей из вен. Атриовентрикулярные клапаны в это время
раскрыты и давление в предсердиях и желудочках практически одинаково.
Генерация возбуждения в синоатриальном узле приводит к систоле пред-
сердий, во время которой за счет разности давлений конечно-диастоличе-
ский объем желудочков возрастает приблизительно на 15 %. С окончанием
систолы предсердий давление в них понижается.
Поскольку клапаны между магистральными венами и предсердиями от-
сутствуют, во время систолы предсердий происходит сокращение кольце-
вой мускулатуры, окружающей устья полых и легочных вен, что препятст-
вует оттоку крови из предсердий обратно в вены. В то же время систола
предсердий сопровождается некоторым повышением давления в полых ве-
нах. Большое значение имеет обеспечение турбулентного характера потока
крови, поступающего из предсердий в желудочки, что способствует захло-
пыванию атриовентрикулярных клапанов. Максимальное и среднее давле-
ние в левом предсердии во время систолы составляют соответственно 8—
15 и 5—7 мм рт. ст., в правом предсердии — 3—8 и 2—4 мм рт. ст.
(рис. 9.11).
С переходом возбуждения на атриовентрикулярный узел и проводящую
систему желудочков начинается систола последних. Ее начальный этап
(период напряжения) продолжается 0,08 с и состоит из двух фаз. Фаза
асинхронного сокращения (0,05 с) представляет собой процесс распростране-
ния возбуждения и сокращения по миокарду. Давление в желудочках при
этом практически не меняется. В процессе начинающегося синхронного
сокращения миокарда желудочков, когда давление в них возрастает до ве-
личины, достаточной для закрытия атриовентрикулярных клапанов, но не-
достаточной для открытия полулунных, наступает фаза изоволюмического,
или изометрического, сокращения.
Дальнейшее повышение давления приводит к раскрытию полулунных
клапанов и началу периода изгнания крови из сердца, общая длительность
которого составляет 0,25 с. Этот период состоит из фазы быстрого изгнания
(0,13 с), во время которой давление в желудочках продолжает расти и дос-
тигает максимальных значений, и фазы медленного изгнания (0,13 с), во
время которой давление в желудочках начинает снижаться, а после окон-
чания сокращения оно резко падает. В магистральных артериях давление
снижается значительно медленнее, что обеспечивает захлопывание полу-
лунных клапанов и предотвращает обратный ток крови. Промежуток вре-
мени от начала расслабления желудочков до закрытия полулунных клапа-
нов называется протодиастолическим периодом.
После окончания систолы желудочков возникает диастолический пери-
од желудочков сердца (диастола), длящаяся 0,47 с. Он включает в себя
следующие периоды и фазы (при частоте сердечных сокращений 75 в ми-
нуту):
Период расслабления (0,12 с), состоящий из:
406 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Систола
Давление в аорте
Давление влевом
предсердии
Давление влевом
желудочке
Объем левого
желудочка
Рис. 9.11. Изменения объема левого желудочка и колебания давления в левом пред-
сердии, левом желудочке и аорте в течение сердечного цикла.
I — начало систолы предсердий; II — начало систолы желудочков; III — момент раскрытия по-
лулунных клапанов; IV — конец систолы желудочков и момент закрытия полулунных клапа-
нов; V — раскрытие атриовентрикулярных клапанов.
Опускание линии, показывающей объем желудочков, соответствует динамике их опорож-
нения.
— протодиастолического интервала — 0,04 с (время от начала расслабле-
ния миокарда желудочков до закрытия полулунных клапанов);
— фазы изометрического (изоволюмического) расслабления — 0,08 с
(время от закрытия полулунных клапанов до открытия атриовентри-
кулярных).
Период наполнения (0,35 с) состоящий из:
— фазы быстрого наполнения — 0,08 с (с момента открытия атриовен-
трикулярных клапанов);
— фазы медленного наполнения — 0,18 с;
— фазы наполнения желудочков, обусловленной систолой предсер-
дий — 0,09 с.
К концу систолы желудочков и началу диастолы (с момента закрытия
полулунных клапанов) в желудочках содержится остаточный, или резерв-
ный, объем крови (конечно-систолический объем). В это же время начи-
нается резкое падение давления в желудочках (фаза изоволюмического,
или изометрического, расслабления). Способность миокарда быстро рас-
слабляться является важнейшим условием для наполнения сердца кровью.
Когда давление в желудочках (начальное диастолическое) становится
9. Функции систем кровообращения и лимфообращения • 407
меньше давления в предсердиях, открываются атриовентрикулярные кла-
паны и начинается фаза быстрого наполнения, во время которой кровь с
ускорением поступает из предсердий в желудочки. Во время этой фазы в
желудочки поступает до 85 % их диастолического объема. По мере запол-
нения желудочков скорость их наполнения кровью снижается (фаза мед-
ленного наполнения). В конце диастолы желудочков начинается систола
предсердий, в результате чего в желудочки поступает еще 15 % их диасто-
лического объема. Таким образом, в конце диастолы в желудочках создает-
ся конечно-диастолический объем, которому соответствует определенный
уровень конечно-диастолического давления в желудочках. Конечно-диа-
столический объем и конечно-диастолическое давление составляет так на-
зываемую преднагрузку сердца, которая является определяющим условием
для растяжения волокон миокарда, т. е. реализации закона Франка—Стар-
линга (см. «Миогенные механизмы регуляции сердца» в разделе 9.1.3.6.7).
Частота генерации возбуждения клетками проводящей системы и соот-
ветственно сокращений миокарда определяется длительностью рефрактер-
ной фазы, возникающей после каждой систолы. Как и в других возбудимых
тканях, в миокарде рефрактерность обусловлена инактивацией натриевых
ионных каналов, возникающей в результате деполяризации (см. рис. 9.9).
Для восстановления входящего натриевого тока необходим уровень репо-
ляризации около 40 мВ. До этого момента имеет место период абсолютной
рефрактерности, который продолжается около 0,27 с. Далее следует период
относительной рефрактерности (см. рис. 9.9), в течение которого возбуди-
мость клетки постепенно восстанавливается, но остается еще сниженной
(длительность 0,03 с). В этот период сердечная мышца может ответить до-
полнительным сокращением, если стимулировать ее очень сильным раз-
дражителем. За периодом относительной рефрактерности следует короткий
период супернормальной возбудимости (см. рис. 9.9). В этот период возбуди-
мость миокарда высока и можно получить дополнительный ответ в виде
сокращения мышцы, нанося на нее подпороговый раздражитель.
Длительный рефрактерный период имеет для сердца важное биологиче-
ское значение, так как он предохраняет миокард от быстрого или повтор-
ного возбуждения и сокращения. Этим исключается возможность тетани-
ческого сокращения миокарда и нарушения нагнетательной функции
сердца.
К тетаническому сокращению и утомлению в физиологическом пони-
мании этого термина миокард не способен. При раздражении сердечная
ткань ведет себя как функциональный синцитий, и сила каждого сокраще-
ния определяется по закону «все или ничего», согласно которому при возбу-
ждении, превышающем пороговую величину, сокращающиеся волокна
миокарда развивают максимальную силу, не зависящую от величины над-
порогового раздражителя.
Преждевременное сокращение всего сердца или его частей в результате
дополнительного возбуждения миокарда вызывает экстрасистолу. По мес-
ту возникновения дополнительного возбуждения различают синусовые,
предсердные, атриовентрикулярные и желудочковые экстрасистолы.
9.1.3.6.6. Механические, электрические и физические
проявления деятельности сердца
Запись сокращений сердца, выполненная каким-либо инструментальным
способом, называется кардиограммой.
При сокращении сердце изменяет свое положение в грудной клетке.
408 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Оно несколько поворачивается вокруг своей оси слева направо, плотнее
прижимаясь изнутри к грудной стенке. Регистрация сердечного толчка оп-
ределяет механокардиограмму (апекс-кардиограмму), которая находит весь-
ма ограниченное использование на практике.
Более широко в клинике и в научных исследованиях используются раз-
личные модификации электрокардиографии. Последняя представляет собой
метод исследования сердца, основанный на регистрации и анализе элек-
трических потенциалов, возникающих при деятельности сердца.
Электрокардиограмма. Метод электрокардиографии основан на том,
что в процессе распространения возбуждения по миокарду поверхность не-
возбужденных (поляризованных) кардиомиоцитов несет положительный
заряд, а возбужденных (деполяризованных) — отрицательный. При этом
возникает электрическое поле, которое можно зарегистрировать с поверх-
ности тела. Поскольку между различными тканями тела создается в этом
случае разность потенциалов, изменяющаяся в соответствии с колебания-
ми величины и направления электрического поля сердца, регистрируемые
изменения разности потенциала во времени и составляют суть метода
электрокардиографии. Кривая изменений этой разницы потенциалов, оп-
ределяемая с помощью высокочувствительного вольтметра, называется
электрокардиограммой (ЭКГ), а соответствующий прибор для записи этой
кривой — электрокардиографом. Важно подчеркнуть, что ЭКГ отражает
возбуждение сердца, но не его сокращение.
Для регистрации ЭКГ используют различные схемы наложения элек-
тродов — отведения ЭКГ. К обязательно регистрируемым в клинике отно-
сятся следующие 12 отведений: 3 стандартных (двухполюсные от конечно-
стей), 3 усиленных (однополюсные от конечностей), 6 грудных (однопо-
люсные от грудной клетки).
При использовании двухполюсных (биполярных) отведений электроды
регистрируют разность потенциалов между двумя точками тела, потенциал
каждой из которых меняется в течение сердечного цикла. Электроды по
этой схеме накладываются на обе руки и левую ногу, образуя три так на-
зываемых стандартных отведения, обозначаемых римскими цифрами I, II,
III (рис. 9.12).
I отведение: правая рука (—) — левая рука (+);
II отведение: правая рука (—) — левая нога (+);
III отведение: левая рука (—) — левая нога (+).
Правую руку всегда соединяют с отрицательным, а левую ногу — с поло-
жительным полюсом прибора. Левую руку в I стандартном отведении соеди-
няют с положительным полюсом, а в III стандартном — с отрицательным.
При регистрации ЭКГ в однополюсных (униполярных) отведениях один
из электродов — активный — накладывают на участок тела с меняющимся
электрическим потенциалом и подключают к положительному полюсу из-
мерительного прибора. Потенциал второго электрода, называемого индиф-
ферентным, остается практически постоянным и условно принимается за
нулевой. Этот электрод подключают к отрицательному полюсу измеритель-
ного прибора.
На теле человека трудно найти участок с постоянным электрическим
потенциалом, поэтому для получения индифферентного электрода исполь-
зуют искусственные приемы. Один из них состоит в том, что соединяются
вместе провода от трех электродов, наложенных на обе руки и левую ногу.
Полученный таким способом условный электрод называют объединенным,
а производимые с его помощью однополюсные отведения обозначают ла-
тинской буквой V (от англ. Voltage). Этот электрод применяют для регист-
9. Функции систем кровообращения и лимфообращения • 409
I отведение
Рис. 9.12. Двухполюсные (стандартные) отведения электрокардиограммы.
Концы стрелок соответствуют конечностям, соединяемым с кардиографом в I (вверху), II (по-
средине) и III (внизу) отведениях. Справа —левые конечности, слева — правые. В правой час-
ти — схематическое изображение электрокардиограммы в каждом из этих отведений.
рации однополюсных грудных отведений (V,— V6).
Другой способ получения индифферентного электрода используется
при регистрации однополюсных отведений от конечностей. В этом случае
его получают, соединяя электроды только от двух конечностей — тех, на
которых не находится активный электрод, и присоединяют к отрицатель-
ному полюсу прибора. Амплитуда ЭКГ при этом способе в 1,5 раза боль-
ше, чем в предыдущем случае. Поэтому эти однополюсные отведения от
конечностей получили название «усиленных» и обозначаются символами
aVR, aVL, aVF (от англ, augmented — усиленный, right — правый, left — ле-
вый, foot — нога).
При графической записи электрокардиограммы в любом отведении в
каждом цикле отмечается совокупность характерных зубцов, которые при-
нято обозначать буквами Р, Q, R, S и Т (см. рис. 9.12). Считается, что зу-
бец Р отражает процессы деполяризации в области предсердия, интервал
P—Q характеризует процесс распространения возбуждения в предсердиях и
атриовентрикулярном узле, комплекс зубцов QRS — процессы деполяриза-
ции в желудочках, а сегмент 5— Ти зубец Т— процессы реполяризации в
желудочках. Таким образом, комплекс зубцов QRST характеризует распро-
странение электрических процессов в миокарде или электрическую систо-
лу. Важное диагностическое значение имеют временные и амплитудные
характеристики составляющих электрокардиограммы. Во втором стандарт-
ном отведении в норме амплитуда зубца R составляет 0,8—1,2 мВ, а ам-
плитуда Q не должна превышать '/4 этой величины. Длительность интерва-
ла P—Q в норме составляет 0,12—0,20 с, комплекса QRS— не более 0,08 с,
а сегмент S—T — 0,36—0,44 с.
410 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 9.13. Нормальная фонокардиограмма.
Римские цифры — тоны сердца; ЭКГ — синхронно регистрируемая электрокардиограмма.
Тоны сердца. Каждый сердечный цикл сопровождается несколькими
раздельными звуками, которые называются тонами сердца. Первый тон
низкий, он возникает одновременно с началом систолы желудочков и обу-
словлен вибрацией створок атриовентрикулярных клапанов, включая их
сухожильные струны, сокращением мускулатуры желудочков и механиче-
скими колебаниями начальных отделов аорты и легочной артерии. Первый
тон называют систолическим, его общая продолжительность составляет
приблизительно 0,12 с, что соответствует фазе напряжения и началу пе-
риода изгнания крови.
Второй тон высокий и продолжается около 0,08 с, его возникновение
связано с захлопыванием полулунных клапанов и происходящей при этом
вибрацией их стенок. Этот тон называют диастолическим. Интенсивность
первого тона зависит от крутизны нарастания давления в желудочках во
время систолы, а второго — от давления в аорте и легочной артерии.
Графическая запись тонов сердца называется фонокардиограммой. Фоно-
кардиография позволяет выявить третий и четвертый тоны сердца: менее
интенсивные, чем первый и второй, и поэтому неслышные при обычной
аускультации. Третий тон отражает вибрацию стенок желудочков вследст-
вие быстрого поступления крови в начале фазы наполнения. Четвертый
тон возникает во время систолы предсердий и продолжается до начала их
расслабления (рис. 9.13).
Сфигмо- и флебография. Сократительный процесс, протекающий в ходе
сердечного цикла, находит отражение в ритмических колебаниях стенок
9. Функции систем кровообращения и лимфообращения • 411
Рис. 9.14. Графическая запись пульсо-
вого колебания давления крови в арте-
рии.
А — анакрота; К — катакрота; ДП — дикро-
тический подъем.
крупных артерий и вен. Методика
графической регистрации артериаль-
ного пульса называется сфигмогра-
фией (от греч. sphygmos — пульс), а
венозного пульса — флебографией
(от греч. phleps — вена). Централь-
ный артериальный пульс чаще реги-
стрируют над сонными артериями
(каротидная сфигмография), а цен-
тральный венозный пульс — над
яремными венами (югулярная фле-
бография).
Сфигмограмма сонной артерии
(рис. 9.14) характеризуется высоко-
амплитудной главной волной, крутой
восходящий участок которой назы-
вается анакротой (А). Этот участок
отражает ускоренное поступление
крови в артерии из левого желудочка
в начале фазы быстрого изгнания,
что приводит к увеличению давления
в артериях и их растяжению. Верши-
на этой волны, отражающая примерное равенство между притоком крови в
магистральной артерии и ее оттоком в периферические сосуды, переходит
в нисходящее колено — катакроту (К). Последняя соответствует по време-
ни фазе медленного изгнания, когда отток крови из растянутых эластиче-
ских артерий начинает преобладать над притоком. В период катакроты
проявляется остроконечный, направленный вниз зубец, который называет-
ся инцизурой (вырезкой) и соответствует окончанию систолы левого желу-
дочка, когда давление в нем становится ниже, чем в аорте. Самая низкая
точка инцизуры соответствует полному закрытию аортального клапана.
Диастолическая часть сфигмограммы начинается дикротической волной
(дикротическим подъемом — ДП), которая возникает в результате отраже-
ния гидравлической волны от замкнутых кармашков аортального клапана.
Последующий плавный спуск кривой соответствует равномерному оттоку
крови из центральных сосудов в
дистальные во время диастолы.
Косвенная характеристика со-
кратительных процессов, проис-
ходящих в правых отделах сердца,
может быть получена при помощи
югулярной флебографии, отра-
жающей динамику оттока крови
из полых вен в правое предсер-
дие. Каждый сердечный цикл на
югулярной флебограмме (рис.
9.15) представлен тремя положи-
тельными (а, с, v) и двумя отри-
цательными (х, у) волнами.
Систола правого предсердия
вызывает замедление оттока кро-
ви из вен, что приводит к возник-
новению положительной волны
Рис. 9.15. Графическая запись венозного
пульса (флебограмма).
Объяснение в тексте.
412 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
флебограммы, обозначаемой символом «а» (от лат. atrium). Следующая по-
ложительная волна возникает в начале систолы правого желудочка при за-
крытии трехстворчатого клапана и обозначается символом «с» (от лат. саг-
otis), так как ее появление совпадает с началом подъема каротидной сфиг-
мограммы. Далее следует отрицательная волна «х», которая отражает уско-
ренный отток крови из центральных вен в расслабляющееся предсердие.
Самая глубокая точка этой волны совпадает по времени с закрытием полу-
лунных клапанов.
Наполнение правого предсердия при закрытом трехстворчатом клапане
приводит к повышению в нем давления и затруднению оттока крови из
вен, что отражается на флебограмме положительной волной «V» (от лат.
ventriculus). Последующее быстрое поступление крови из правого предсер-
дия в желудочек в период диастолы сердца проявляется в виде отрицатель-
ной волны флебограммы, которая называется волной диастолического
коллапса и обозначается символом «у».
9.1.3.6.7. Общие принципы регуляции сердечного
выброса
От величины сердечного выброса зависят два условия выполнения адек-
ватной текущим задачам нутритивной функции системы кровообращения:
обеспечение оптимального количества циркулирующей крови и поддержа-
ние (совместно с сосудами) определенного уровня среднего артериального
давления (70—90 мм рт. ст.), необходимого для удержания физиологиче-
ских констант в капиллярах (25—30 мм рт. ст.). При этом обязательным
условием нормальной работы сердца является равенство притока крови по
венам и ее выброса в артерии. Решение этой задачи обеспечивается, в ос-
новном, механизмами, обусловленными свойствами самой сердечной
мышцы. Проявление этих механизмов называют миогенной ауторегуляци-
ей насосной функции сердца. Существуют два способа ее реализации: ге-
терометрическая — осуществляется в ответ на изменения исходной длины
волокон миокарда, гомеометрическая — происходит при их сокращениях в
изометрическом режиме.
Миогенные механизмы регуляции деятельности сердца. Изучение зави-
симости силы сокращений сердца от растяжения его камер показало, что
сила каждого сердечного сокращения зависит от величины венозного при-
тока и определяется конечной диастолической длиной волокон миокарда.
Эта зависимость получила название гетерометрическая регуляция сердца и
известна как закон Франка—Старлинга: «Сила сокращения желудочков
сердца, измеренная любым способом, является функцией длины мышеч-
ных волокон перед сокращением», т. е. чем больше наполнение камер
сердца кровью, тем больше сердечный выброс (рис. 9.16). Установлена
ультраструктурная основа этого закона, заключающаяся в том, что количе-
ство актомиозиновых мостиков является максимальным при растяжении
каждого саркомера до 2,2 мкм.
Увеличение силы сокращения при растяжении волокон миокарда не со-
провождается увеличением длительности сокращения, поэтому указанный
эффект одновременно означает увеличение скорости нарастания давления
в камерах сердца во время систолы.
Инотропные влияния на сердце, обусловленные эффектом Франка-
Старлинга, играют ведущую роль в увеличении сердечной деятельности
при усиленной мышечной работе, когда сокращающиеся скелетные мыш-
цы вызывают периодическое сжатие вен конечностей, что приводит к уве-
9. Функции систем кровообращения и лимфообращения • 413
Центральное венозное давление
Рис. 9.16. Увеличение силы сокращений сердца (амплитуда колебаний верхней кри-
вой) при его растяжении. Опыт на сердечно-легочном препарате.
личению венозного притока за счет мобилизации резерва депонированной
в них крови. Отрицательные инотропные влияния по указанному механиз-
му играют существенную роль в изменениях кровообращения при переходе
в вертикальное положение (ортостатическая проба). Эти механизмы имеют
большое значение для согласования изменений сердечного выброса и при-
тока крови по венам малого круга, что предотвращает опасность развития
отека легких.
Термином «гомеометрическая регуляция» обозначают миогенные механиз-
мы, для реализации которых не имеет значения степень конечно-диастоли-
ческого растяжения волокон миокарда. Среди них наиболее важным явля-
ется зависимость силы сокращения сердца от давления в аорте (эффект
414 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Анрепа) и хроно-инотропная зависимость. Этот эффект состоит в том, что
при увеличении давления «на выходе» из сердца сила и скорость сердеч-
ных сокращений возрастают, что позволяет сердцу преодолевать возросшее
сопротивление в аорте и поддерживать оптимальным сердечный выброс.
Иннервация сердца. Сердце — обильно иннервированный орган. Среди
чувствительных образований сердца основное значение имеют две популя-
ции механорецепторов, сосредоточенных, главным образом, в предсердиях
и левом желудочке: А-рецепторы реагируют на изменение напряжения
сердечной стенки, а В-рецепторы возбуждаются при ее пассивном растя-
жении. Афферентные волокна, связанные с этими рецепторами, идут в со-
ставе блуждающих нервов. Свободные чувствительные нервные оконча-
ния, расположенные непосредственно под эндокардом, представляют со-
бой терминали афферентных волокон, проходящих в составе симпатиче-
ских нервов. Эфферентная иннервация сердца осуществляется при участии
обоих отделов вегетативной нервной системы. Тела симпатических пре-
ганглионарных нейронов, участвующих в иннервации сердца, располага-
ются в сером веществе боковых рогов трех верхних грудных сегментов
спинного мозга. Преганглионарные волокна направляются к нейронам
верхнего грудного (звездчатого) симпатического ганглия. Постганглионар-
ные волокна этих нейронов вместе с парасимпатическими волокнами блу-
ждающего нерва образуют верхний, средний и нижний сердечные нервы,
Симпатические волокна пронизывают весь орган и иннервируют не только
миокард, но и элементы проводящей системы.
Тела парасимпатических преганглионарных нейронов, участвующих в
иннервации сердца, располагаются в продолговатом мозге. Их аксоны идут
в составе блуждающих нервов. После вхождения блуждающего нерва в
грудную полость от него отходят веточки, которые включаются в состав
сердечных нервов.
Отростки блуждающего нерва, проходящие в составе сердечных нервов,
представляют собой парасимпатические преганглионарные волокна. С них
возбуждение передается на интрамуральные нейроны и далее — преимуще-
ственно на элементы проводящей системы. Влияния, опосредованные пра-
вым блуждающим нервом, адресованы, в основном, клеткам синоатриаль-
ного, а левым — клеткам атриовентрикулярного узла. Прямого влияния на
желудочки сердца блуждающие нервы не оказывают.
Иннервируя ткань водителей ритма, вегетативные нервы способны ме-
нять их возбудимость, тем самым вызывая изменения частоты генерации
потенциалов действия и сокращений сердца (хронотропный эффект). Нерв-
ные влияния изменяют скорость электротонической передачи возбуждения
и, следовательно, длительности фаз сердечного цикла. Такие эффекты на-
зывают дромотропными.
Поскольку действие медиаторов вегетативной нервной системы заклю-
чается в изменении уровня циклических нуклеотидов и энергетического
обмена, вегетативные нервы в целом способны влиять и на силу сердечных
сокращений (инотропный эффект). В лабораторных условиях получен эф-
фект изменения величины порога возбуждения кардиомиоцитов под дей-
ствием нейромедиаторов, его обозначают как батмотропный.
Перечисленные пути воздействия нервной системы на сократительную
активность миокарда и насосную функцию сердца представляют собой хо-
тя и исключительно важные, но вторичные по отношению к миогенным
механизмам модулирующие влияния.
9. Функции систем кровообращения и лимфообращения • 415
9.1.3.6.8. Нейрогенная регуляция деятельности сердца
Влияние на сердце блуждающих нервов. Результатом стимуляции этих нер-
вов является отрицательный хронотропный эффект сердца (рис. 9.17), на
фоне которого проявляются также отрицательные и дромотропный ино-
тропный эффекты. Существуют постоянные тонические влияния на сердце
со стороны бульбарных ядер блуждающего нерва: при его двусторонней
перерезке частота сердцебиений возрастает 1,5—2,5 раза. При длительном
сильном раздражении влияние блуждающих нервов на сердце постепенно
ослабевает или прекращается, что получило название «эффекта ускольза-
ния» сердца из-под влияния блуждающего нерва.
Различные отделы сердца по-разному реагируют на возбуждение пара-
симпатических нервов. Так, холинергические влияния на предсердия вы-
зывают значительное угнетение автоматии клеток синусного узла и спон-
танно возбудимой ткани предсердий. Сократимость рабочего миокарда
предсердий в ответ на стимуляцию блуждающего нерва снижается. Реф-
рактерный период предсердий при этом также уменьшается в результате
значительного укорочения длительности потенциала действия предсердных
кардиомиоцитов. С другой стороны, рефрактерность кардиомиоцитов же-
лудочков под влиянием блуждающего нерва, напротив, значительно воз-
растает, а отрицательный парасимпатический инотропный эффект на же-
лудочки выражен в меньшей степени, чем на предсердия.
Электрическая стимуляция блуждающего нерва вызывает урежение или
прекращение сердечной деятельности вследствие торможения автоматиче-
ской функции водителей ритма синоатриального узла. Выраженность этого
Рис. 9.17. Электрическое раздражение эфферентных нервов сердца.
Вверху — уменьшение частоты сокращений при раздражении блуждающего нерва; внизу —
увеличение частоты и силы сокращений при раздражении симпатического нерва.
Стрелками отмечены начало и конец раздражения.
416 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
эффекта зависит от силы и частоты раздражения блуждающего нерва. По
мере увеличения силы раздражения отмечается переход от небольшого за-
медления синусного ритма до полной остановки сердца.
Отрицательный хронотропный эффект раздражения блуждающего нерва
связан с угнетением (замедлением) генерации импульсов в водителе ритма
сердца синусного узла. Поскольку при раздражении блуждающего нерва в
его окончаниях выделяется медиатор — ацетилхолин, при его взаимодейст-
вии с мускариночувствительными рецепторами сердца повышается прони-
цаемость поверхностной мембраны клеток водителей ритма для ионов ка-
лия. Вследствие этого возникает гиперполяризация мембраны, которая за-
медляет (подавляет) развитие медленной спонтанной диастолической де-
поляризации, и поэтому мембранный потенциал позже достигает критиче-
ского уровня. Это приводит к урежению ритма сокращений сердца.
При сильных раздражениях блуждающего нерва диастолическая деполя-
ризация подавляется, возникают гиперполяризация водителя ритма и пол-
ная остановка сердца. Развитие гиперполяризации в клетках водителей
ритма снижает их возбудимость, затрудняет возникновение очередного ав-
томатического потенциала действия и тем самым приводит к замедлению
или даже остановке сердца. Стимуляция блуждающего нерва, усиливая вы-
ход калия из клетки, увеличивает мембранный потенциал, ускоряет про-
цесс реполяризации и при достаточной силе раздражающего тока укорачи-
вает длительность потенциала действия клеток водителя ритма.
При вагусных воздействиях имеет место уменьшение амплитуды и дли-
тельности потенциала действия кардиомиоцитов предсердия. Отрицатель-
ный инотропный эффект связан с тем, что уменьшенный по амплитуде и
укороченный потенциал действия не способен возбудить достаточное ко-
личество кардиомиоцитов. Кроме того, вызванное ацетилхолином повы-
шение калиевой проводимости противодействует потенциалзависимому
входящему току кальция и проникновению его ионов внутрь кардиомио-
цита. Холинергический медиатор ацетилхолин может также угнетать АТФ-
азную активность миозина и, таким образом, уменьшать величину сокра-
тимости кардиомиоцитов. Возбуждение блуждающего нерва приводит к
повышению порога раздражения предсердий, подавлению автоматии и за-
медлению проводимости атриовентрикулярного узла. Указанное замедле-
ние проводимости при холинергических влияниях может вызвать частич-
ную или полную атриовентрикулярную блокаду.
Влияние симпатических нервов на сердце проявлется в виде положитель-
ного хронотропного и положительного инотропного эффекта. Сведения о
наличии тонических влияний симпатической нервной системы на миокард
основываются в основном на хронотропных эффектах.
Электрическая стимуляция волокон, отходящих от звездчатого ганглия,
вызывает увеличение ритма сердца и силы сокращений миокарда (см. рис.
9.17). Под влиянием возбуждения симпатических нервов скорость медлен-
ной диастолической деполяризации повышается, снижается критический
уровень деполяризации клеток водителей ритма синоатриального узла,
уменьшается величина мембранного потенциала покоя. Подобные измене-
ния увеличивают скорость возникновения потенциала действия в клетках
водителей ритма сердца, повышают его возбудимость и проводимость. Эти
изменения электрической активности связаны с тем, что выделяющийся
из окончаний симпатических волокон медиатор норадреналин взаимодей-
ствует с р1 -адренорецепторами поверхностной мембраны клеток, что при-
водит к повышению проницаемости мембран для ионов натрия и кальция,
а также уменьшению проницаемости для ионов калия.
9. Функции систем кровообращения и лимфообращения • 417
Ускорение медленной спонтанной диастолической деполяризации кле-
ток водителей ритма, увеличение скорости проведения в предсердиях, ат-
риовентрикулярном узле и желудочках приводит к улучшению синхронно-
сти возбуждения и сокращения мышечных волокон и к увеличению силы
сокращения миокарда желудочков. Положительный инотропный эффект
связан также с повышением проницаемости мембраны для ионов кальция.
При увеличении входящего тока кальция возрастает степень электромеха-
нического сопряжения, в результате чего увеличивается сократимость мио-
карда.
Менее изучено участие в регуляции сердечной деятельности интракар-
диальных ганглиозных нервных элементов. Известно, что они обеспечива-
ют передачу возбуждения с волокон блуждающего нерва на клетки синоат-
риального и атриовентрикулярного узлов, выполняя функцию парасимпа-
тических ганглиев. Описаны инотропные, хронотропные и дромотропные
эффекты, полученные при стимуляции этих образований в условиях экс-
перимента на изолированном сердце. Значение этих эффектов в естествен-
ных условиях остается неясным.
9.1.3.6.9. Механизмы адренергической
и холинергической регуляции деятельности
сердца
Симпатические и парасимпатические нервы оказывают влияние на сердце
посредством высвобождения нейромедиаторов, которые взаимодействуют с
рецепторами, расположенными на наружной мембране миокардиальных
клеток. К основным медиаторам, высвобождающимся из симпатических и
парасимпатических терминалей, относят соответственно норадреналин и
ацетилхолин. Норадреналин взаимодействует с адренорецепторами мио-
карда, а ацетилхолин — с холинорецепторами.
Адренергические механизмы. В сердце человека содержатся преимущест-
ренцо [31- и в меньшей степени [32-адренорецепторы, а также al- и а2-ад-
ренорецепторы. Поскольку [3-адренорецепторы расположены на поверхно-
сти миокардиальных клеток, это делает их легкодоступными как для но-
радреналина, высвобождающегося из симпатических нервных окончаний,
так и для циркулирующего в крови адреналина.
При повышении активности симпатической нервной системы или кон-
центрации катехоламинов в крови количество [3-адренорецепторов на по-
верхности миокардиальной клетки уменьшается, тогда как в случае сниже-
ния симпатической активности их количество возрастает. Йодсодержащие
гормоны щитовидной железы, особенно тироксин, а также гормон коры
надпочечников кортизол увеличивают количество p-адренорецепторов в
миокарде, что приводит к усилению эффектов катехоламинов на сердце.
Возбуждение p-адренорецепторов сердца приводит к росту частоты со-
кращений сердца, сократимости миокарда, значительному увеличению по-
требления им кислорода и уменьшению длительности диастолы.
Физиологические механизмы, связанные с активацией а,-адренорецеп-
торов миокарда, известны в меньшей степени. а2-Адренорецепторы распо-
лагаются на пресинаптических мембранах симпатических и парасимпати-
ческих нервных окончаний, их возбуждение тормозит выделение норадре-
налина и ацетилхолина из нервных терминалей.
Холинергические механизмы. На наружной мембране кардиомиоцитов
представлены, в основном, мускаринчувствительные (М-) холинорецепто-
ры. Доказано наличие в миокарде и никотинчувствительных (N-) холино-
14 -6095
418 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
рецепторов, однако их значение в парасимпатических влияниях на сердце
менее ясно. Плотность мускариновых рецепторов в миокарде зависит от
концентрации мускариновых агонистов в тканевой жидкости.
Возбуждение мускариновых рецепторов тормозит активность пейсме-
керных клеток синусного узла и в то же время увеличивает возбудимость
предсердных кардиомиоцитов. Эти два процесса могут привести к возник-
новению предсердных экстрасистол в случае повышения тонуса блуждаю-
щего нерва, например ночью во время сна. Таким образом, возбуждение
М-холинорецепторов вызывает снижение частоты и силы сокращений
предсердий, но повышает их возбудимость.
Ацетилхолин угнетает проводимость в атриовентрикулярном узле. Это
связано с тем, что под влиянием ацетилхолина возникает гиперполяриза-
ция клеток атриовентрикулярного узла вследствие усиления выходящего
калиевого тока.
Таким образом, возбуждение мускариновых холинорецепторов оказыва-
ет противоположное, по сравнению с активацией [3-адренорецепторов,
действие на сердце. При этом снижается частота сердечных сокращений,
угнетается проводимость и сократимость миокарда, а также потребление
миокардом кислорода. Возбудимость предсердий в ответ на применение
ацетилхолина возрастает, тогда как возбудимость желудочков, напротив,
уменьшается.
Рефлекторные влияния на сердце. Выделены три категории кардиальных
рефлексов: собственные, вызываемые раздражением рецепторов сер-
дечно-сосудистой системы; сопряженные, обусловленные активностью
любых других рефлексогенных зон; неспецифические, которые вос-
производятся в ответ на неспецифические влияния (в условиях физиологи-
ческого эксперимента, а также в патологии).
Наибольшее физиологическое значение имеют собственные рефлексы
сердечно-сосудистой системы, которые возникают чаще всего при раздра-
жении барорецепторов магистральных артерий в результате изменения
системного давления. Так, при повышении давления в аорте и каротидном
синусе происходит рефлекторное урежение частоты сердцебиения.
Особую группу собственных кардиальных рефлексов представляют те
из них, которые возникают в ответ на раздражение артериальных хемо-
рецепторов изменением напряжения кислорода в крови. В условиях ги-
поксемии развивается рефлекторная тахикардия, а при дыхании чистым
кислородом — брадикардия. Эти реакции отличаются исключительно вы-
сокой чувствительностью: у человека увеличение частоты сердцебиений
наблюдается уже при снижении напряжения кислорода всего на 3 %, ко-
гда никаких признаков гипоксии в организме обнаружить еще невоз-
можно.
Собственные рефлексы сердца проявляются и в ответ на механическое
раздражение сердечных камер, в стенках которых находится большое ко-
личество барорецепторов. К их числу относят рефлекс Бейнбриджа, прояв-
ляющийся в виде тахикардии в ответ на быстрое внутривенное введение
определенного объема крови. Считается, что эта реакция сердца является
рефлекторным ответом на раздражение барорецепторов полых вен и пред-
сердия, поскольку она устраняется при денервации сердца. Отрицательные
хронотропные и инотропные реакции сердца рефлекторной природы воз-
никают в ответ на раздражение механорецепторов как правых, так и левых
отделов сердца. Значение интракардиальных рефлексов состоит в том, что
увеличение исходной длины волокон миокарда приводит к усилению со-
кращений не только растягиваемого отдела сердца (в соответствии с зако-
9. Функции систем кровообращения и лимфообращения • 419
Рис. 9.18. Эфферентная иннервация сердца.
Гф — гипофиз; Гт — гипоталамус; Пм — продолговатый мозг; Цсд — бульбарный центр сер-
дечно-сосудистой системы; К — кора больших полушарий; Гл — симпатические ганглии;
См — спинной мозг; Th — грудные сегменты.
ном Франка—Старлинга), но и к усилению сокращений других отделов
сердца, не подвергающихся растяжению.
Рефлексы с сердца изменяют функцию других висцеральных систем. К
их числу относят, например, кардиоренальный рефлекс Генри—Гауэра, ко-
торый представляет собой увеличение диуреза в ответ на растяжение стен-
ки левого предсердия.
Собственные кардиальные рефлексы составляют основу нейрогенной
регуляции деятельности сердца, хотя реализация его насосной функции
возможна без участия нервной системы.
Сопряженные кардиальные рефлексы представляют собой эффекты раз-
дражения рефлексогенных зон, не принимающих прямого участия в регу-
ляции кровообращения. К числу таких рефлексов относят рефлекс Гольца,
который проявляется в форме брадикардии (до полной остановки сердца) в
ответ на раздражение механорецепторов брюшины или органов брюшной
полости. Возможность проявления такой реакции учитывается при прове-
дении оперативных вмешательств на брюшной полости, при нокауте у
боксеров и т. д. При раздражении некоторых экстерорецепторов (резкое
охлаждение кожи области живота) может иметь место рефлекторная оста-
14-
420 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
новка сердца. Именно такую природу имеют несчастные случаи при ныря-
нии в холодную воду. Сопряженным соматовисцеральным кардиальным
рефлексом является рефлекс Данини—Ашнера, который проявляется в ви-
де брадикардии при надавливании на глазные яблоки. Таким образом, со-
пряженные рефлексы сердца, не являясь составной частью общей схемы
нейрогенной регуляции, могут оказывать влияние на его деятельность.
Замыкание большинства кардиорефлекторных дуг происходит на уровне
продолговатого мозга, где находятся: 1) ядро солитарного тракта, к которо-
му подходят афферентные пути рефлексогенных зон сердечно-сосудистой
системы; 2) ядра блуждающего нерва и 3) вставочные нейроны бульбарно-
го кардиоваскулярного центра. В то же время реализация рефлекторных
влияний на сердце в естественных условиях всегда происходит при участии
вышележащих отделов центральной нервной системы (рис. 9.18). Сущест-
вуют различные по знаку инотропные и хронотропные влияния на сердце
со стороны мезэнцефальных адренергических ядер (голубое пятно, черная
субстанция), гипоталамуса (паравентрикулярное и супраоптические ядра,
мамиллярные тела) и лимбической системы. Имеют место и кортикальные
влияния на сердечную деятельность, среди которых особое значение при-
дают условным рефлексам — таким, например, как положительный хроно-
тропный эффект при предстартовом состоянии. Достоверных данных о
возможности произвольного управления человеком сердечной деятельно-
стью не получено.
9.1.3.6.10. Гуморальные влияния на сердце
Прямое и опосредованное действие на сердце оказывают практически все
биологически активные вещества, содержащиеся в плазме крови. В то же
время круг фармакологических агентов, осуществляющих гуморальную ре-
гуляцию сердца, в подлинном смысле этого слова, достаточно узок. Таки-
ми веществами являются катехоламины, выделяемые мозговым веществом
надпочечников — адреналин, норадреналин и дофамин. Действие этих гор-
монов опосредуется p-адренорецепторами кардиомиоцитов, что и опреде-
ляет конечный результат их влияния на миокард. Он аналогичен симпати-
ческой стимуляции и заключается в активации фермента аденилатциклазы
и усилении синтеза циклического АМФ (3,5-циклического аденозинмоно-
фосфата), с последующей активацией фосфорилазы и повышением уровня
энергетического обмена. Такое действие на пейсмекерную ткань вызывает
положительный хронотропный, а на клетки рабочего миокарда — положи-
тельный инотропный эффекты. Усиливающим инотропный эффект дейст-
вием катехоламинов является повышение проницаемости мембран кардио-
миоцитов к ионам кальция.
Действие других гормонов (глюкагон, йодсодержащие гормоны щито-
видной железы) на миокард неспецифическое и реализуется опосредован-
но, например через влияние на активность симпатоадреналовой системы.
Положительное инотропное действие на сердце оказывают также гормоны
коры надпочечников (кортикостероиды), вазопрессин и ангиотензин. Вме-
сте с тем прямые положительные инотропные эффекты последних на серд-
це скрываются за их непрямыми эффектами, обусловленными повышени-
ем давления в аорте и увеличением объема циркулирующей крови.
В регуляции деятельности сердца принимают участие и местные гумо-
ральные факторы, образующиеся в миокарде. К таким веществам относят-
ся аденозин, гистамин и простагландины.
Аденозин, взаимодействуя с аденозиновыми рецепторами кардиомио-
9. Функции систем кровообращения и лимфообращения • 421
цитов, уменьшает пейсмекерную активность клеток синоатриального узла
и снижает скорость проведения возбуждения в атриовентрикулярном узле
и в проводящей системе сердца. Это свойство аденозина используется в
клинической кардиологии — для лечения пароксизмальных желудочковых
тахикардий применяются соли аденозинтрифосфорной кислоты.
В миокарде человека имеются два типа гистаминовых рецепторов: Ht и
Н2. Активация этих рецепторов вызывает повышение сократимости мио-
карда. При возбуждении Н|-рецепторов продолжительность потенциала
действия кардиомиоцитов желудочков увеличивается, тогда как при возбу-
ждении Н2-рецепторов она, напротив, уменьшается. Однако в случае мас-
сивного выброса в кровь гистамина (например, при анафилактическом
шоке) происходит резкое снижение ОПСС, падение АД и, следовательно,
постнагрузки сердца (давление в аорте, против которого происходит изгна-
ние крови желудочками в систолу). В результате этих гемодинамических
сдвигов сократимость миокарда снижается.
В миокарде синтезируются простагландины, которые могут уменьшать
симпатические влияния на сердце и коронарные артерии. Синтез проста-
гландинов увеличивается при ишемии миокарда и имеет в этом случае за-
щитный характер.
Сердце проявляет чувствительность и к ионному составу протекающей
крови. Катионы кальция повышают возбудимость клеток миокарда как за
счет участия в сопряжении возбуждения и сокращения, так и за счет акти-
вации фосфорилазы. Повышение концентрации ионов калия по отноше-
нию к норме, составляющей 4 ммоль/л, приводит к снижению величины
потенциала покоя и увеличению проницаемости мембран для этих ионов.
Возбудимость миокарда и скорость проведения возбуждения при этом воз-
растают. Обратные явления, часто сопровождающиеся нарушениями рит-
ма, имеют место при недостатке в крови калия, в частности в результате
применения некоторых диуретических препаратов. Такие соотношения ха-
рактерны для сравнительно небольших изменений концентрации катионов
калия, при ее увеличении более чем в два раза возбудимость и проводи-
мость миокарда резко снижаются. На этом эффекте основано действие
кардиоплегических растворов, которые используются в кардиохирургии
для временной остановки сердца. Угнетение сердечной деятельности на-
блюдается и при повышении кислотности внеклеточной среды.
Гормональная функция сердца. Вокруг миофибрилл в клетках миокарда
предсердий обнаружены гранулы, подобные тем, которые имеются в щито-
видной железе или аденогипофизе. В этих гранулах образуется группа гор-
монов, которые высвобождаются при растяжении предсердий, стойком по-
вышении давления в аорте, нагрузке организма натрием, повышении ак-
тивности блуждающих нервов. Отмечены следующие эффекты предсерд-
ных гормонов: а) снижение ОПСС, МОК и АД, б) увеличение гематокри-
та, в) увеличение клубочковой фильтрации и диуреза, г) угнетение секре-
ции ренина, альдостерона, кортизола и вазопрессина, д) снижение кон-
центрации в крови адреналина, е) уменьшение освобождения норадрена-
лина при возбуждении симпатических нервов. Подробнее см. главу 6.
9.1.3.7. Венозный возврат крови к сердцу
Этим термином обозначают объем венозной крови, протекающей по верх-
ней и нижней (у животных, соответственно, по передней и задней) полым
венам.
Количество крови, протекающей за единицу времени через артерии и
422 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 9.4. Комплекс факторов, участвующих в формировании величины венозного
возврата
вены, в устойчивом режиме функционирования системы кровообращения
остается постоянным, поэтому в норме величина венозного возврата равна
величине минутного объема крови, т. е. 4—6 л/мин у человека. Однако
вследствие перераспределения массы крови от одной области к другой это
равенство может временно нарушаться при переходных процессах в систе-
ме кровообращения, вызываемых различными воздействиями на организм
как в норме (например, при мышечных нагрузках или перемене положе-
ния тела), так и при развитии патологии сердечно-сосудистой системы
(например, недостаточности правых отделов сердца).
Величина общего или суммарного венозного возврата между полыми
венами как у животных, так и у человека состоит примерно из ’/3 объемно-
го кровотока по верхней (или передней) полой вене и 2/3 — по нижней
(или задней) полой вене. Величина кровотока у людей в верхней полой ве-
не составляет примерно 42 %, а в нижней полой вене — 58 % общей вели-
чины венозного возврата.
Факторы, участвующие в формировании величины венозного возврата,
условно делят на две группы (табл. 9.4) в соответствии с направлением
действия сил, способствующих продвижению крови по сосудам большого
круга кровообращения.
Первую группу представляет сила «vis a tergo» (т. е. действующая сзади),
сообщаемая крови сердцем; она продвигает кровь по артериальным сосу-
дам и участвует в обеспечении ее возврата к сердцу. Если в артериальном
русле эта сила соответствует давлению 100 мм рт. ст., то в начале венул об-
щее количество энергии, которой обладает кровь, прошедшая через капил-
лярное русло, составляет около 13 % от ее начальной энергии. Именно по-
следняя величина энергии и образует «vis a tergo» и расходуется на приток
венозной крови к сердцу. К силе, действующей «vis a tergo», относят также
9. Функции систем кровообращения и лимфообращения • 423
ряд других факторов, способствующих продвижению крови к сердцу: со-
кращения скелетной мускулатуры (так называемый мышечный насос),
способствующие «выжиманию» крови из вен; функционирование веноз-
ных клапанов (препятствующих обратному току крови); влияние уровня
гидростатического давления в системе кровообращения (особенно в верти-
кальном положении тела).
Ко второй группе факторов, участвующих в венозном возврате, относят
силы, действующие на кровоток в полых венах «vis a fronte» (т. е. спереди)
и включающие, прежде всего, присасывающую функцию грудной клетки и
сердца. Присасывающая функция грудной клетки обеспечивает поступле-
ние крови из периферических вен в грудные вследствие существования от-
рицательного давления в плевральной полости: во время вдоха отрицатель-
ное давление в последней еще более снижается, что приводит к ускорению
кровотока в нижней полой вене, а во время выдоха давление, напротив,
относительно исходного несколько возрастает и кровоток в этой вене за-
медляется. Для присасывающей функции правых отделов сердца характер-
но то, что силы, способствующие поступлению в него крови, развиваются
не только во время диастолы сердца (вследствие понижения давления в
правом предсердии), но также и во время систолы (в результате смещения
атриовентрикулярного кольца увеличивается объем предсердия и быстрое
падение в нем давления способствует наполнению сердца кровью из полых
вен). Однако не все исследователи разделяют мнение о важной роли при-
сасывающей функции грудной клетки и правой половины сердца в форми-
ровании величины венозного возврата.
Наряду с этим важное значение имеют взаимоотношения встречных по-
токов по полым венам, которые при переходных процессах в системе мо-
гут изменяться неоднократно, а также констрикторные реакции венозных
сосудов, проявляющиеся при действии на систему кровообращения нейро-
генных или гуморальных стимулов; изменения транскапиллярного обмена
жидкости, обеспечивающие ее переход из интерстиция в кровоток вен.
Повышение артериального давления сопровождается возрастанием ве-
личины венозного возврата, что проявляется при прессорных рефлексах
(синокаротидном — вызываемом снижением давления в каротидных сину-
сах, при стимуляции афферентных волокон соматических нервов), увели-
чении объема циркулирующей крови, внутривенном введении вазоактив-
ных веществ (адреналин, норадреналин, простагландин Р2, ангиотензин
II). Гормон задней доли гипофиза вазопрессин вызывает на фоне повыше-
ния артериального давления уменьшение венозного возврата.
Депрессорные системные изменения могут сопровождаться как умень-
шением венозного возврата, так и возрастанием его величины. Совпадение
направленности системной реакции с изменениями венозного возврата
имеет место при депрессорном синокаротидном рефлексе (повышении дав-
ления в каротидных синусах), в ответ на ишемию миокарда, при уменьше-
нии объема циркулирующей крови. Системная депрессорная реакция может
сопровождаться и возрастанием притока крови к сердцу по полым венам,
как это наблюдается, например, при гипоксии (дыхание газовой смесью с
пониженным до 6—10 % содержанием в ней О2), гиперкапнии (6 % СО2),
введении в сосудистое русло ацетилхолина, стимулятора р-адренорецепто-
ров изопротеренола, местного гормона брадикинина, проста-гландина Е,.
Степень увеличения суммарного венозного возврата крови к сердцу при
применении различных препаратов (или нервных влияний на систему) оп-
ределяется не только величиной, но и направленностью изменений крово-
тока в каждой из полых вен. Кровоток по передней полой вене у животных
424 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
100 п
Рис. 9.19. Разнонаправленные изменения венозного возврата по передней и задней
полым венам при прессорном рефлексе.
Сверху вниз: системное артериальное давление (мм рт. ст.), кровоток по передней полой вене,
кровоток по задней полой вене, отметка времени (10 с), отметка раздражения. Исходная вели-
чина кровотока по передней полой вене — 52 мл/мин, по задней полой вене — 92,7 мл/мин.
в ответ на введение вазоактивных веществ (любой направленности дейст-
вия) или при нейрогенных стимулах всегда увеличивается. Различная на-
правленность изменений кровотока отмечена только в задней полой вене
(рис. 9.19). Эта разнонаправленность изменений кровотока в полых венах
и является фактором, обусловливающим относительно небольшое увеличе-
ние общего венозного возврата сравнительно с его изменениями при одно-
направленных сдвигах кровотоков в полых венах.
Механизм разнонаправленных сдвигов кровотока в полых венах состоит
в том, что в результате преобладающего влияния вазоконстрикторных пре-
паратов на артериолы имеет место большая степень увеличения сопротив-
ления сосудов бассейна брюшной аорты по сравнению с изменениями со-
противления сосудов бассейна плечеголовной артерии. Это приводит к пе-
рераспределению сердечного выброса между указанными сосудистыми
руслами (увеличение доли сердечного выброса в направлении сосудов бас-
сейна плечеголовной артерии и уменьшение — в направлении бассейна
брюшной аорты) и вызывает соответствующие разнонаправленные измене-
ния кровотока в полых венах.
Помимо вариабельности кровотока в задней полой вене, зависящей от
гемодинамических факторов, на его величину оказывают существенное
влияние другие системы организма (дыхательная, мышечная, нервная).
Так, перевод животного на искусственное дыхание почти в 2 раза умень-
шает кровоток по задней полой вене, а наркоз и открытая грудная клетка
еще в большей степени снижают его величину.
Спланхническое сосудистое русло в результате изменений находящегося в
нем объема крови вносит наибольший вклад (по сравнению с другими ре-
гионами системы кровообращения) в величину венозного возврата. Изме-
нения емкости отдельных сосудистых регионов спланхнического русла не-
одинаковы, и их вклад в обеспечение венозного возврата различен. Напри-
мер, при прессорном синокаротидном рефлексе имеет место уменьшение
объема селезенки на 2,5 мл/кг массы тела, объема печени — на 1,1 мл/кг, а
кишечника — лишь на 0,2 мл/кг (в целом спланхнический объем уменьша-
ется на 3,8 мл/кг). Во время умеренной геморрагии (9 мл/кг) выброс крови
из селезенки составляет 3,2 мл/кг (35 %), из печени — 1,3 мл/кг (14 %) и
9. Функции систем кровообращения и лимфообращения • 425
из кишечника — 0,6 мл/кг (7 %), что в сумме составляет 56 % величины
изменений общего объема крови в организме.
Указанные изменения емкостной функции сосудов органов и тканей
организма участвуют в величине венозного возврата крови к сердцу за счет
сдвигов кровотока в нижней полой вене и, тем самым, в преднагрузке
сердца (т. е. конечно-диастолическое давление), и в результате оказывают
влияние на формирование величины сердечного выброса и уровня систем-
ного артериального давления.
9.1.3.8. Центральное венозное давление
Под центральным венозным давлением обычно понимают величину давле-
ния в правом предсердии, которая в норме близка к 0 мм рт. ст. Однако в
действительности центральному венозному давлению соответствует вели-
чина давления в устье полых вен. Между давлением в правом предсердии и
в устье полых вен далеко не всегда существует соответствие, особенно в фа-
зу переходных процессов в системе кровообращения, когда давление в пра-
вом предсердии нормализуется быстрее, чем центральное венозное давле-
ние. Уровень центрального венозного давления (ЦВД) оказывает сущест-
венное влияние на величину венозного возврата крови к сердцу. При пони-
жении давления в правом предсердии от 0 до 4 мм рт. ст. приток венозной
крови возрастает на 20—30 %, но когда давление в нем становится ниже —
4 мм рт. ст., дальнейшее снижение давления не вызывает уже увеличения
притока венозной крови, что обусловлено спадением вен, впадающих в
грудную клетку, вызванным резким падением давления крови в этих венах.
Если снижение ЦВД увеличивает приток венозной крови к сердцу по по-
лым венам, то его повышение на 1 мм рт. ст. снижает венозный возврат
примерно на 14 %. Исходя из сказанного, повышение давления в правом
предсердии до 7 мм рт. ст. должно снизить приток венозной крови к сердцу
до нуля, что привело бы к катастрофическим нарушениям гемодинамики.
Однако в исследованиях, в которых сердечно-сосудистые рефлексы
функционировали, а давление в правом предсердии повышалось медленно,
приток венозной крови к сердцу продолжался и при повышении давления
в правом предсердии до 12—14 мм рт. ст. (рис. 9.20). Снижение притока
Рис. 9.20. Венозный возврат крови к сердцу при медленном подъеме давления в
правом предсердии (когда успевают развиться компенсаторные механизмы).
426 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
крови к сердцу в этих условиях приводит к проявлению в системе компен-
саторных рефлекторных реакций, возникающих при раздражении бароре-
цепторов артериального русла, а также возбуждению сосудодвигательных
центров в условиях развивающейся ишемии центральной нервной систе-
мы. Это вызывает увеличение потока импульсов, генерируемых в симпати-
ческих сосудосуживающих центрах и поступающих к гладким мышцам со-
судов, что определяет повышение тонуса, уменьшение емкости перифери-
ческого сосудистого русла и, следовательно, увеличение количества крови,
подаваемой к сердцу, несмотря на рост ЦВД до уровня, когда теоретиче-
ски венозный возврат должен быть близким к 0.
Определены минимальные и максимальные пределы изменений ЦВД,
ограничивающие область устойчивой работы сердца. Минимально допусти-
мое среднее давление в правом предсердии составляет 5—10, а максималь-
ное — 100—120 мм водн. ст., при выходе за эти пределы ЦВД зависимость
энергии сокращения сердца от величины притока крови не наблюдается
из-за необратимого ухудшения функционального состояния миокарда.
Средняя величина ЦВД у здоровых людей составляет в состоянии мы-
шечного покоя около 40 мм водн. ст. и в течение дня меняется, нарастая
днем и особенно к вечеру на 10—30 мм водн. ст., что связано с ходьбой и
мышечными движениями. Увеличение внутриплеврального давления, со-
провождаемое сокращением мышц брюшной полости (кашель, натужива-
ние), приводит к кратковременному резкому возрастанию ЦВД, а задержка
дыхания на вдохе — к его временному падению.
При вдохе ЦВД уменьшается за счет падения плеврального давления,
что вызывает дополнительное растяжение правого предсердия и более пол-
ное заполнение его кровью. При этом возрастает скорость венозного кро-
вотока и увеличивается градиент давления в венах, что приводит к допол-
нительному падению ЦВД. Так как давление в венах, лежащих вблизи
грудной полости (например, в яремных венах), в момент вдоха является
отрицательным, их ранение опасно для жизни, поскольку при вдохе в этом
случае возможно проникновение в вены воздуха, пузырьки которого, раз-
носясь с кровью, могут закупорить кровеносное русло (развитие воздуш-
ной эмболии).
При выдохе ЦВД растет, а венозный возврат крови к сердцу уменьшает-
ся. Это является результатом повышения плеврального давления, увеличи-
вающего венозное сопротивление вследствие спадения грудных вен и сдав-
ливающего правое предсердие, что затрудняет его кровенаполнение.
Для увеличения производительности сердца в клинике часто использу-
ют искусственное повышение венозного возврата за счет увеличения объе-
ма циркулирующей крови, которое достигается путем внутривенных вли-
ваний кровезаменителей. Однако вызываемое этим повышение давления в
правом предсердии эффективно только в пределах соответствующих вели-
чин средних давлений, приведенных выше. Чрезмерное повышение веноз-
ного притока таким способом и, следовательно, ЦВД не только не способ-
ствует улучшению деятельности сердца, но может принести и вред, созда-
вая перегрузки в системе и приводя, в конечном итоге, к чрезмерному рас-
ширению правой половины сердца.
9.1.3.9. Объем циркулирующей крови
Объем крови у мужчины массой 70 кг составляет примерно 5,5 л (75—
80 мл/кг), у взрослой женщины он несколько меньше (около 70 мл/кг).
Объем крови колеблется в зависимости от пола, возраста, телосложения,
9. Функции систем кровообращения и лимфообращения • 427
условий жизни, степени физического развития и тренированности орга-
низма и составляет от 50 до 80 мл на 1 кг массы тела. У здорового челове-
ка, находящегося в лежачем положении 1—2 нед, объем крови может сни-
зиться на 9—15 % от исходного.
Из 5,5 л крови у взрослого мужчины 55—60 %, т. е. 3,0—3,5 л, прихо-
дится на долю плазмы, остальное количество — на долю эритроцитов.
В течение суток по сосудам циркулирует около 8000—9000 л крови. Из
этого количества приблизительно 20 л выходит в течение суток из капил-
ляров в ткань в результате фильтрации и возвращается вновь (путем аб-
сорбции) через капилляры (16—18 л) и с лимфой (2—4 л). Объем жидкой
части крови, т. е. плазмы (3—3,5 л), существенно меньше, чем объем жид-
кости во внесосудистом интерстициальном пространстве (9—12 л) и во
внутриклеточном пространстве тела (27—30 л); с жидкостью этих «про-
странств» плазма находится в динамическом осмотическом равновесии.
Общий объем циркулирующей крови (ОЦК) состоит из активно циркули-
рующей крови по сосудам и той ее части, которая не участвует в данный
момент в кровообращении, т. е. депонированной (в селезенке, печени,
почке, легких и др.). Последняя может быстро включаться в циркуляцию
при соответствующих гемодинамических ситуациях. Количество депони-
руемой крови более чем в два раза превышает объем циркулирующей. Де-
понированная кровь не находится в состоянии полного застоя, некоторая
ее часть все время включается в циркуляцию, а соответствующая часть бы-
стро движущейся крови переходит в состояние депонирования.
Уменьшение или увеличение объема циркулирующей крови на 5—10 % у
человека с нормальным объемом крови компенсируется изменением емко-
сти венозного русла и не вызывает сдвигов ЦВД. Более значительное уве-
личение ОЦК обычно сопряжено с увеличением венозного возврата и при
сохранении эффективной сократимости сердца приводит к увеличению
сердечного выброса.
Важнейшими факторами, от которых зависит объем крови, являются:
1) регуляция объема жидкости между плазмой и интерстициальным про-
странством, 2) регуляция обмена жидкости между плазмой и внешней сре-
дой (осуществляется, главным образом, почками), 3) регуляция объема
эритроцитной массы. Нервная регуляция этих трех механизмов осуществ-
ляется с помощью предсердных рецепторов типа А, реагирующих на изме-
нение давления и, следовательно, являющихся барорецепторами, и типа
В — реагирующих на растяжение предсердий и весьма чувствительных к
изменению объема в них крови.
Существенное влияние на объем крови оказывает инфузия различных
растворов. Вливание в вену изотонического раствора хлорида натрия не
повышает длительно объем плазмы на фоне нормального объема крови,
так как образующийся в организме избыток жидкости быстро выводится
путем усиления диуреза. При дегидратации и дефиците солей в организме
указанный раствор, введенный в кровь в адекватных количествах, быстро
восстанавливает нарушенное равновесие. Внутривенное введение раство-
ров высокомолекулярных декстранов на длительный период (до 12—24 ч)
повышает объем циркулирующей крови.
9.1.3.10. Соотношение основных параметров системной гемодинамики
Параметры системной гемодинамики — системное артериальное давле-
ние, периферическое сопротивление сосудов, сердечный выброс, работа
сердца, венозный возврат, центральное венозное давление, объем цирку-
428 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 9.5. Первичные и компенсаторные изменения в системе кровообращения чело-
века при переходе из горизонтального положения в вертикальное
Первичные изменения Компенсаторные изменения
Дилатация сосудистого русла нижней по- ловины тела как результат повышения внутрисосудистого давления. Уменьшение венозного притока к право- му предсердию. Уменьшение сердечного выброса Снижение общего периферического со- противления. Снижение систолического, диастоличе- ского, пульсового и среднего артериаль- ного давления. Снижение мозгового кровотока Рефлекторная веноконстрикция, ведущая к уменьшению емкости вен и увеличению ве- нозного притока к сердцу. Рефлекторное увеличение частоты сердеч- ных сокращений, ведущее к увеличению сердечного выброса. Повышение тканевого давления в нижних конечностях и насосного действия мышц ног, рефлекторная гипервентиляция и уве- личение напряжения брюшных мышц: уве- личение венозного притока к сердцу. Снижение сопротивления мозговых сосудов. Увеличение секреции норадреналина, альдо- стерона, антидиуретического гормона, вызы- вающего как увеличение сосудистого сопро- тивления, так и гиперволемию.
лирующей крови — находятся в сложных тонко регулируемых взаимоотно-
шениях, что позволяет системе обеспечивать выполнение своих функций.
Так, снижение давления в синокаротидной зоне вызывает повышение
системного артериального давления, учащение сердечного ритма, увеличе-
ние общего периферического сопротивления сосудов, работы сердца и ве-
нозного возврата крови к сердцу. Минутный и систолический объем кро-
ви могут меняться при этом неоднозначно. Повышение давления в сино-
каротидной зоне вызывает снижение системного артериального давления,
замедление частоты сердечных сокращений, снижение общего сосудистого
сопротивления и венозного возврата, уменьшение работы сердца. Измене-
ния сердечного выброса при этом выражены, но неоднозначны по на-
правленности.
Переход из горизонтального положения человека в вертикальное сопро-
вождается последовательным развитием характерных изменений систем-
ной гемодинамики. Эти сдвиги включают в себя как первичные, так и вто-
ричные компенсаторные изменения в системе кровообращения, которые
схематически представлены в табл. 9.5.
Важным является поддержание постоянного соотношения между объе-
мом крови, содержащейся в большом круге кровообращения, и объемом
крови, находящейся в органах грудной клетки (легкие, полости сердца). В
сосудах легких содержится до 15 %, а в полостях сердца (в фазе диасто-
лы)—до 10 % всей массы крови; исходя из сказанного, центральный
(внутригрудной) объем крови может составлять до 25 % общего количества
крови в организме.
Растяжимость сосудов малого круга, в особенности легочных вен, по-
зволяет аккумулировать в этой области значительный объем крови при
увеличении венозного возврата к правой половине сердца. Аккумуляция
крови в малом круге имеет место у людей во время перехода тела из верти-
кального положения в горизонтальное, при этом в сосуды грудной полости
из нижних конечностей может перемещаться до 600 мл крови, из которых
примерно половина скапливается в легких. Напротив, при переходе тела в
9. Функции систем кровообращения и лимфообращения • 429
вертикальное положение этот объем крови переходит в сосуды нижних ко-
нечностей.
Резерв крови в легких используется, когда необходима срочная мобили-
зация дополнительного количества крови для поддержания должной вели-
чины сердечного выброса. Это особенно важно в начале интенсивной мы-
шечной работы, когда, несмотря на включение мышечного насоса, веноз-
ный возврат к сердцу еще не достигает уровня, обеспечивающего сердеч-
ный выброс в соответствии с кислородным запросом организма.
Одним из источников, обеспечивающих резерв сердечного выброса, яв-
ляется также остаточный объем крови в полости желудочков. В горизон-
тальном положении человека остаточный объем левого желудочка состав-
ляет в среднем 100 мл, а в вертикальном — 45 мл. Близкие к этим величины
характерны и для правого желудочка. Увеличение ударного объема, наблю-
даемое при мышечной работе или действии катехоламинов, не сопровож-
дающееся увеличением размеров сердца, происходит за счет мобилизации,
главным образом, части остаточного объема крови в полости желудочков.
Таким образом, наряду с изменениями венозного возврата к сердцу, к
числу факторов, определяющих динамику сердечного выброса, относятся:
объем крови в легочном резервуаре, реактивность сосудов легких и оста-
точный объем крови в желудочках сердца.
Совместное проявление гетеро- и гомеометрического типов регуляции
сердечного выброса выражается в такой последовательности: а) увеличение
венозного возврата к сердцу, обусловленное констрикцией артериальных и
особенно венозных сосудов в системе циркуляции, ведет к увеличению
сердечного выброса; б) последнее, наряду с ростом общего перифериче-
ского сопротивления сосудов, повышает системное АД; в) это соответст-
венно ведет к увеличению давления в аорте и, следовательно, кровотока в
коронарных сосудах; г) гомеометрическая регуляция сердца, основанная на
последнем механизме, обеспечивает преодоление сердечным выбросом
возросшего сопротивления в аорте и поддержание сердечного выброса на
повышенном уровне; д) увеличение сократительной функции сердца вызы-
вает рефлекторное снижение периферического сопротивления сосудов (од-
новременно с проявлением рефлекторных влияний на периферические со-
суды с барорецепторов синокаротидных зон), что способствует уменьше-
нию работы сердца, затрачиваемой на обеспечение необходимого кровото-
ка и давления в капиллярах.
Увеличение количества циркулирующей крови в организме изменяет
минутный объем крови, главным образом, вследствие повышения степени
наполнения кровью сосудистой системы. Это вызывает усиление притока
крови к сердцу, увеличение его кровенаполнения, повышение центрально-
го венозного давления и, следовательно, интенсивности работы сердца.
Изменение количества крови в организме влияет на величину минутного
объема крови также путем изменения сопротивления притока венозной
крови к сердцу, которое находится в обратно пропорциональной зависи-
мости от объема крови, притекающей к сердцу. Между объемом циркули-
рующей крови и величиной среднего системного давления существует пря-
мая пропорциональная зависимость. Однако повышение последнего, воз-
никающее при остром увеличении объема крови, продолжается около
1 мин, после чего оно начинает снижаться и устанавливается на уровне,
лишь немного превосходящем норму. Если объем циркулирующей крови
уменьшается, величина среднего давления временно падает и возникаю-
щий эффект в сердечно-сосудистой системе прямо противоположен повы-
шению среднего давления при увеличении объема крови.
430 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Возвращение величины среднего давления к исходному уровню являет-
ся результатом включения компенсаторных механизмов. Известны три из
них, которые выравнивают сдвиги, возникающие при изменении объема
циркулирующей крови в сердечно-сосудистой системе: 1) рефлекторные
компенсаторные механизмы; 2) непосредственные реакции сосудистой
стенки; 3) нормализация объема крови в системе.
Рефлекторные механизмы связаны с изменением уровня системного ар-
териального давления, обусловленным влиянием с барорецепторов сосуди-
стых рефлексогенных зон. Однако удельный вес этих механизмов сравни-
тельно небольшой. В то же время при сильном кровотечении возникают
другие очень мощные нервные влияния, которые могут привести к ком-
пенсаторным сдвигам этих реакций в результате ишемии центральной
нервной системы. Так, снижение системного артериального давления ни-
же 55 мм рт. ст. вызывает изменения гемодинамики, которые в 6 раз пре-
вышают сдвиги, возникающие при максимальной стимуляции симпатиче-
ской нервной системы через сосудистые рефлексогенные зоны. Таким об-
разом, нервные влияния, возникающие при ишемии центральной нервной
системы, могут играть чрезвычайно важную роль в качестве «последней
линии обороны», предотвращающей резкое снижение минутного объема
крови в терминальных состояниях организма после массивной кровопоте-
ри и значительного падения артериального давления.
Компенсаторные реакции самой сосудистой системы возникают вслед-
ствие ее способности растягиваться при повышении давления крови и спа-
даться, когда давление крови снижается. В наибольшей мере этот эффект
присущ венозным сосудам. Считается, что указанный механизм является
более действенным, нежели нервный, особенно при сравнительно неболь-
ших изменениях давления крови. Главное различие этих механизмов со-
стоит в том, что рефлекторные компенсаторные реакции включаются в
действие через 4—5 с и достигают максимума через 30—40 с, в то время
как расслабление самой сосудистой стенки, возникающее в ответ на усиле-
ние ее напряжения, лишь начинается в этот период, достигая максимума
через минуты или десятки минут.
Нормализация объема крови в системе в случае ее изменений достига-
ется следующим образом. После переливания больших объемов крови дав-
ление во всех сегментах сердечно-сосудистой системы, включая капилля-
ры, повышается, что приводит к фильтрации жидкости через стенки ка-
пилляров в интерстициальные пространства и через капилляры клубочков
почек в мочу. При этом величины системного давления, периферического
сопротивления и минутного объема крови возвращаются к исходным зна-
чениям.
В случае кровопотери возникают противоположные сдвиги. При этом
большое количество белка из межклеточной жидкости поступает через
лимфатическую систему в сосудистое русло, повышая уровень белков
плазмы крови. Кроме того, значительно возрастает количество белков, об-
разующихся в печени, что также приводит к восстановлению уровня бел-
ков плазмы крови. Одновременно нормализуется объем плазмы, компен-
сирующий сдвиги, возникающие вследствие кровопотери. Восстановление
объема крови до нормы является медленным процессом, но тем не менее
через 24—48 ч у человека объем крови становится нормальным, в результа-
те нормализуется и гемодинамика. Долгосрочная нормализация объема
циркулирующей крови осуществляется гормонами эндокринной системы
(ренин-альдостероновая, АДГ).
Следует особо подчеркнуть, что ряд параметров системной гемодинами-
9. Функции систем кровообращения и лимфообращения • 431
Время, с
О 50 100 150 200 250 300
Время, с
Время, с
Рис. 9.21. Соотношение параметров системной гемодинамики при внутривенном
введении норадреналина (10 мкг/кг).
АД — артериальное давление; ВВ — суммарный венозный возврат; ОПС — общее перифериче-
ское сопротивление; ПГА — кровоток по плечеголовной артерии; ППВ — кровоток по перед-
ней полой вене; ЦВД — центральное венозное давление; СВ — сердечный выброс; УО — удар-
ный объем сердца; НГА — кровоток по грудной аорте; ЗПВ — кровоток по задней полой вене.
ки или их взаимоотношений у человека в настоящее время практически
невозможно исследовать, особенно в динамике развития реакций в сердеч-
но-сосудистой системе. Это связано с тем, что человек не может быть объ-
ектом экспериментирования, а число датчиков для регистрации величин
указанных параметров, даже в условиях торакальной хирургии, явно не-
достаточно для выяснения этих вопросов, тем более в условиях нормаль-
432 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ного функционирования системы. Поэтому изучение сдвигов всего ком-
плекса параметров системной гемодинамики и механизмов их регуляции
возможно в настоящее время только у животных.
В результате сложнейших технических подходов, использования специ-
альных датчиков, применения физических, математических и кибернети-
ческих приемов сегодня можно представить изменения параметров систем-
ной гемодинамики количественно, в динамике развития процесса у одного
и того же животного (рис. 9.21). Как следует из рисунка, однократное
внутривенное введение норадреналина вызывает значительное повышение
артериального давления, не соответствующее ему по длительности — крат-
ковременное повышение общего периферического сопротивления сосудов
и соответствующее ему увеличение центрального венозного давления. Сер-
дечный выброс и ударный объем сердца при этом в момент повышения
периферического сопротивления снижаются, а затем — резко возрастают,
соответствуя во второй фазе сдвигам артериального давления. Кровоток в
плечеголовной и грудной аорте изменяется соответственно сердечному вы-
бросу, хотя в последней эти сдвиги более выражены (очевидно, в связи с
высоким исходным кровотоком). Венозный возврат крови к сердцу, есте-
ственно, соответствует по фазам сердечному выбросу, однако в передней
полой вене он увеличивается, а в задней — вначале снижается, потом не-
сколько возрастает. Вот эти сложные, взаимоподкрепленные сдвиги пара-
метров системной гемодинамики и обусловливают повышение ее инте-
грального показателя — артериального давления.
Изучение соотношения венозного возврата и сердечного выброса при
применении прессорных вазоактивных веществ (адреналин, норадреналин,
ангиотензин) показало, что при качественно единообразном изменении
венозного возврата, который в этих случаях, как правило, увеличивается,
характер сдвигов сердечного выброса варьирует: он может как увеличи-
ваться, так и уменьшаться. При этом имеют место два основных варианта
различий между этими параметрами: дефицит величины сердечного вы-
броса по сравнению с величиной притока крови к сердцу по полым венам
и избыток сердечного выброса над величиной венозного возврата. На ос-
новании данных о депонирующей функции сосудов малого круга кровооб-
ращения можно полагать, что первый вариант различий между сердечным
выбросом и венозным возвратом обусловлен депонированием крови в ма-
лом круге, а второй — дополнительным выбросом крови из малого в боль-
шой круг кровообращения.
9.1.4. Общие закономерности органного кровообращения
9.1.4.1. Функционирование органных сосудов
Под тонусом органа, ткани, сосуда или клетки понимают состояние дли-
тельно поддерживаемого возбуждения, выражающегося специфической
для этого образования деятельностью, без развития утомления.
Сосудистый тонус создается в своей основе периферическими механиз-
мами, а нервные импульсы корригируют его, обеспечивая перераспределе-
ние крови между различными сосудистыми областями.
Регионарное кровообращение — термин, принятый для характеристики
движения крови в органах и системе органов, относящихся к области тела
(региону). На уровне органа или региона могут быть определены такие па-
раметры, как величина и скорость кровотока; давление крови в артерии,
капилляре, венуле; сопротивление кровотоку в различных отделах орган-
9. Функции систем кровообращения и лимфообращения • 433
ного сосудистого русла; объем крови в органе. Именно эти параметры, ха-
рактеризующие движение крови по сосудам органа, и подразумеваются,
когда используют термин «органное кровообращение».
Согласно формуле Пуазейля скорость кровотока в сосудах определяется
(помимо нервных и гуморальных влияний) соотношением пяти местных
факторов: упомянутого в начале главы градиента давления (который зави-
сит от артериального давления и венозного давления) и рассмотренного
выше сопротивления сосудов (которое зависит от радиуса сосуда, длины
сосуда, вязкости крови).
Повышение давления в артерии органа ведет к увеличению
градиента давления и, следовательно, к увеличению кровотока в нем. Сни-
жение артериального давления вызывает замедление кровотока. Повыше-
ние давления в вене влечет за собой уменьшение градиента давле-
ния, в результате чего кровоток уменьшается. При снижении венозного
давления градиент давления увеличится, что будет способствовать увеличе-
нию кровотока.
Изменения радиуса сосудов могут происходить активно и пассив-
но. Изменения радиуса сосуда, которые возникают не в результате измене-
ний сократительной активности их гладких мышц, являются пассивными.
Последние могут быть следствием как интраваскулярных, так и экстравас-
кулярных факторов.
Интраваскулярным фактором, вызывающим в организме пассивные из-
менения просвета сосуда, является внутрисосудистое давление. Повыше-
ние артериального давления вызывает пассивное расширение просвета со-
судов, которое может даже нивелировать активную констрикторную реак-
цию артериол в случае их малой выраженности. Аналогичные пассивные
реакции могут возникать в венах при изменении венозного давления.
Экстраваскулярные факторы, способные вызвать пассивные изменения
просвета сосудов, присущи не всем сосудистым областям и зависят от
специфической функции органа. Так, сосуды сердца могут пассивно изме-
нять свой просвет в результате: а) изменений частоты сердечных сокраще-
ний, б) степени напряжения сердечной мышцы при ее сокращениях, в)
изменений внутрижелудочкового давления. Бронхомоторные реакции
влияют на просвет легочных сосудов, а двигательная или тоническая ак-
тивность отделов желудочно-кишечного тракта или скелетной мускулату-
ры изменит просвет сосудов этих областей. Следовательно, степень сжа-
тия сосудов внесосудистыми элементами может определить величину их
просвета.
Активными реакциями сосудов обозначаются те из них, которые возни-
кают в результате сокращения гладкой мускулатуры стенки сосуда. Многие
стимулы вызывают активные изменения просвета сосудов. К их числу от-
носятся, прежде всего, физические, нервные и химические влияния.
Одним из физических факторов является внутрисосудистое давление, из-
менения которого сказываются на степени напряжения (сокращения)
гладкой мускулатуры сосудов. Так, повышение внутрисосудистого давле-
ния влечет за собой увеличение сокращения гладких мышц сосудов, и на-
оборот, его снижение вызывает уменьшение напряжения сосудистых
мышц (эффект Остроумова-Бейлисса). Этот механизм обеспечивает ауто-
регуляцию кровотока в сосудах.
Под ауторегуляцией, кровотока понимают тенденцию к сохранению его
величины в органных сосудах. Это, конечно, не значит, что при резких из-
менениях артериального давления (от 70 до 200 мм рт. ст.) органный кро-
воток сохраняется постоянным. Речь идет о том, что указанные сдвиги ар-
434 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 9.6. Регионарные особенности ауторегуляции кровотока и постокклюзионной
(реактивной) гиперемии
Регион Ауторегуляция кровотока при изменениях артери- ального давления Реактивная гиперемия
порого- вая дли- тель- ность окклюзии максимальная крат- ность увеличения кровотока основной фактор раз- вития
Мозг Хорошо выражена, Дс « 80-160 3-5 с 1,5-2 Механизм реагиро- вания на растяже- ние
Мио- кард Хорошо выражена, дс « 75-140 2-20 с 2-3 Аденозин, ионы ка- лия и др.
Скелет- ные мышцы Выражена при высоком исходном тонусе сосу- дов, Дс ~ 50—100 1-2 с 1,5-4 Механизм реагиро- вания на растяже- ние, метаболиче- ские факторы, не- достаток О2
Кишеч- ник По общему кровотоку не столь четко выражена. В слизистой оболочке выражена полнее, Дс = 40-125 30-120 с 1,5-2 Метаболиты. Мест- ные гормоны
Печень Не обнаружена. Не изу- чена Слабо выражена. Гиперемия, вторая фаза реакции на окклюзию артерии. Простагландины
Кожа ? 0,5- 6 мин 1,5-4 Метаболиты
Примечание. Д. — диапазон величин артериального давления (мм рт.ст.), в котором
стабилизируется кровоток.
териального давления вызывают меньшие изменения кровотока, чем они
могли бы быть в пассивно-эластической трубке.
Ауторегуляция кровотока высокоэффективна в сосудах почек и мозга
(изменения давления в этих сосудах почти не вызывают сдвигов кровото-
ка), несколько меньше — в сосудах кишечника, умеренно эффективна — в
миокарде, относительно неэффективна — в сосудах скелетных мышц и
весьма слабо эффективна — в легких (табл. 9.6).
Несколько теорий объясняют механизм ауторегуляции кровотока:
а) миогенная, признающая за основу передачу возбуждения по гладкомы-
шечным клеткам; б) нейрогенная, предполагающая взаимодействие между
гладкомышечными клетками и рецепторами в сосудистой стенке, чувстви-
тельными к изменению внутрисосудистого давления; в) теория тканевого
давления, основывающаяся на данных о сдвигах капиллярной фильтрации
жидкости при изменении давления в сосуде; г) обменная теория, предпола-
гающая зависимость степени сокращения гладких мышц сосудов от обмен-
ных процессов (сосудоактивных веществ, выделяющихся в кровоток в про-
цессе метаболизма).
Физическим фактором, также способным изменять просвет сосудов, яв-
ляется температура. На повышение температуры крови сосуды внутрен-
них органов отвечают расширением, но на повышение температуры окру-
жающей среды — сужением, сосуды же кожи при этом расширяются.
Длина сосуда в большинстве регионов относительно постоянна, од-
9. Функции систем кровообращения и лимфообращения • 435
нако в органах, выполняющих периодическую или ритмичную деятель-
ность (легкие, сердце, желудочно-кишечный тракт), длина сосуда может
играть роль в изменениях сопротивления сосудов и кровотока в них. Так,
например, увеличение объема легких (на вдохе) вызывает повышение со-
противления легочных сосудов как в результате их сужения, так и удлине-
ния. В этом случае изменения длины сосуда участвуют в дыхательных ва-
риациях легочного кровотока.
Вязкость крови также влияет на кровоток в сосудах. При высоком
показателе гематокрита сопротивление кровотоку может быть значи-
тельным.
Сосуды, лишенные нервных и гуморальных влияний, как оказалось, со-
храняют (хотя и в меньшей мере) способность оказывать сопротивление
кровотоку. Денервация сосудов скелетных мышц, например, увеличивает
кровоток в них примерно в два раза, но последующее введение ацетилхо-
лина в кровоток этой сосудистой области может вызвать дальнейшее деся-
тикратное увеличение в ней кровотока, свидетельствующее о сохраняю-
щейся в этом случае способности сосудов к вазодилатации. Для обозначе-
ния этой особенности денервированных сосудов оказывать сопротивление
кровотоку введено понятие «базальный тонус сосудов».
Базальный тонус сосудов определяется структурными и миогенными
факторами. Структурная часть его создается жесткой сосудистой «сумкой»,
образованной коллагеновыми волокнами, которая определяет сопротивле-
ние сосудов, если активность их гладких мышц полностью исключена.
Миогенная часть базального тонуса обеспечивается напряжением гладких
мышц сосудов в ответ на растягивающее усилие артериального давления.
Следовательно, изменения сопротивления сосудов под влиянием нерв-
ных или гуморальных факторов наслаиваются на базальный тонус, кото-
рый для определенной сосудистой области более или менее постоянен. Ес-
ли нервные и гуморальные влияния отсутствуют, а нейрогенный компо-
нент сопротивления сосудов равен нулю, сопротивление их кровотоку оп-
ределяется базальным тонусом.
Поскольку одной из биофизических особенностей сосудов является их
способность к растяжению, то при активной констрикторной реакции со-
судов изменения их просвета находятся в зависимости от двух противопо-
ложно направленных влияний: сокращающихся гладких мышц сосудов,
которые уменьшают их просвет, и повышенного давления в сосудах, кото-
рое их растягивает. Растяжимость сосудов различных органов значительно
отличается. При повышении артериального давления только на 10 мм рт.
ст. (со ПО до 120 мм рт. ст.) кровоток в сосудах кишечника увеличивается
на 5 мл/мин, а в сосудах миокарда в 8 раз больше — на 40 мл/мин.
На величине реакций сосудов могут сказываться и различия их исход-
ного просвета. При этом имеет значение отношение толщины стенки сосу-
да к его просвету. Чем выше указанное отношение (стенка/просвет), т. е.
чем больше масса стенки находится внутри «линии силы» укорочения
гладких мышц, тем более выражено сужение просвета сосудов. В этом слу-
чае при одной и той же величине сокращения гладких мышц в артериаль-
ных и венозных сосудах уменьшение просвета всегда будет более выражено
в артериальных сосудах, так как структурные «возможности» уменьшения
просвета в большей степени присущи сосудам с высоким отношением
стенка/просвет. На этой основе строится одна из теорий развития гиперто-
нической болезни у человека.
Изменения трансмурального давления (разность внутри- и внесосудисто-
го давлений) влияют на просвет кровеносных сосудов и, следовательно, на
436 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
их сопротивление кровотоку и содержание в них крови, что особенно ска-
зывается в венозном отделе, где растяжимость сосудов велика и значитель-
ные изменения объема содержащейся в них крови могут иметь место при
небольших сдвигах давления. Поэтому изменения просвета венозных сосу-
дов будут -вызывать соответствующие изменения трансмурального давле-
ния, что может привести к пассивно-эластической отдаче кро-
ви из этой области.
Следовательно, выброс крови из вен, возникающий при усилении им-
пульсации в вазомоторных нервах, может быть обусловлен как активным
сокращением гладкомышечных клеток венозных сосудов, так и их пассив-
но-эластической отдачей. Относительная величина пассивного выброса
крови в этой ситуации будет зависеть от исходного давления в венах. Если
исходное давление в них низкое, дальнейшее его уменьшение может вы-
звать спадение вен, ведущее к весьма выраженному пассивному выбросу
крови. Нейрогенная констрикция вен в этой ситуации не вызовет сколько-
нибудь значительного выброса из них крови и в результате может быть
сделано ошибочное заключение, что нервная регуляция этого отдела не-
значительна. Напротив, если исходное трансмуральное давление в венах
высокое, то уменьшение этого давления не повлечет за собой спадение вен
и пассивно-эластическая их отдача будет минимальной. В этом случае ак-
тивная констрикция вен вызовет значительно больший выброс крови и по-
кажет истинное значение нейрогенной регуляции венозных сосудов.
Пассивный компонент мобилизации крови из вен при низком давлении
в них очень выражен, но становится весьма малым при давлении 5—10 мм
рт. ст. В этом случае вены имеют циркулярную форму и выброс крови из
них при нейрогенных влияниях обусловлен активными реакциями указан-
ных сосудов. Однако при подъеме венозного давления выше 20 мм рт. ст.
величина активного выброса крови вновь уменьшается, что является след-
ствием «перенапряжения» гладкомышечных элементов венозных стенок.
Во время спокойного стояния человека объем вен, расположенных ни-
же уровня сердца, увеличивается примерно на 500 мл и даже больше, если
расширены кожные вены. Именно это может быть причиной головокруже-
ния или даже обморока при продолжительном стоянии, особенно в тех
случаях, когда при высокой температуре окружающей среды имеет место
расширение сосудов кожи. Недостаточность венозного возврата при этом
обусловлена не тем, что «кровь должна подниматься вверх», а повышен-
ным трансмуральным давлением и обусловленным этим растяжением вен,
а также застоем в них крови. Гидростатическое давление в венах тыльной
поверхности стопы в этом случае может достигать 80—100 мм рт. ст. Одна-
ко уже первый шаг создает наружное давление скелетных мышц на вены,
и кровь устремляется к сердцу, так как клапаны вен препятствуют обрат-
ному току крови. Это приводит к опорожнению вен в скелетных мышцах
конечностей и снижению в них венозного давления, которое возвращается
к первоначальному уровню со скоростью, зависящей от кровотока в этой
конечности. Растяжение вен органов брюшной полости в положении стоя
сводится к минимуму в результате того, что при переходе в вертикальное
положение давление внутри брюшной полости повышается.
К числу основных феноменов, присущих органному кровообращению,
помимо ауторегуляции кровотока, зависимости реакции сосудов от их ис-
ходного тонуса, от силы раздражителя, относятся функциональная (рабо-
чая) гиперемия, а также реактивная (постокклюзионная) гиперемия. Эти
феномены свойственны регионарному кровообращению во всех областях.
Рабочая (или функциональная) гиперемия — увеличение органного кро-
9. Функции систем кровообращения и лимфообращения • 437
вотока, сопровождающее усиление функциональной активности органа.
Так, в сокращающейся скелетной мышце увеличиваются кровоток и кро-
венаполнение, саливация также сопровождается резким увеличением кро-
вотока по расширенным сосудам слюнной железы. Гиперемия возникает в
поджелудочной железе в момент пищеварения, а также в кишечной стенке
в период усиления моторики и секреции. Увеличение сократительной ак-
тивности миокарда ведет к росту коронарного кровотока, активация зон
головного мозга сопровождается усилением их кровоснабжения, усиленное
кровоснабжение ткани почки регистрируется при увеличении натрийуреза.
Реактивная (или постокклюзионная) гиперемия — увеличение кровотока
в сосудах органа после временного прекращения кровотока. Она проявля-
ется в скелетных мышцах и в конечности человека и животных, хорошо
выражена в почке и в головном мозге, имеет место в коже и кишечнике.
Установлена связь изменений кровотока в органе с химическим соста-
вом среды, окружающей внутриорганные сосуды. Выражением этой связи
являются местные вазодилататорные реакции в ответ на искусственное
введение в сосуды продуктов тканевого обмена (СО2, лактат) и веществ,
изменения концентрации которых в межклеточной среде сопутствуют
сдвигам функции клеток (ионы, аденозин и др.). Отмечена органная спе-
цифичность этих реакций: особая активность СО2, ионов К в церебраль-
ных сосудах, аденозина — в коронарных.
Ауторегуляторная реакция на понижение давления, в принципе, соот-
ветствует «реактивной» гиперемии, вызванной временной окклюзией арте-
рии. В соответствии с этим данные табл. 9.6 свидетельствуют, что наиболее
кратковременные пороговые окклюзии артерий регистрируются в тех же
самых регионах, где эффективна ауторегуляция.
Функциональная гиперемия органов является веским доказательством ос-
новного постулата физиологии кровообращения, согласно которому регу-
ляция кровообращения необходима для осуществления нутритивной функ-
ции движения крови по сосудам. Данные табл. 9.7 свидетельствуют о том,
что усиление деятельности практически каждого органа сопровождается
увеличением кровотока по его сосудам.
В большей части сосудистых регионов (миокард, скелетные мышцы,
кишечник, пищеварительные железы) функциональная гиперемия выявля-
ется как существенное увеличение общего кровотока (максимально до 4—
10-кратного) при усилении функции органа. К этой группе относится и
мозг, хотя общее увеличение его кровоснабжения при усилении активно-
сти «всего мозга» не установлено, но локальный кровоток в зонах повы-
шенной нейрональной активности существенно возрастает. Функциональ-
ная гиперемия не обнаружена в печени — главном химическом реакторе
тела. Возможно, это связано с тем, что печень не бывает в функциональ-
ном «покое», либо с тем, что она и без того обильно снабжается кровью из
печеночной артерии и воротной вены. В другом химически активном «ор-
гане» — жировой ткани — функциональная гиперемия выражена. Функ-
циональная гиперемия проявляется также и в мозговом веществе почки,
работающей «безостановочно», где кровоснабжение коррелирует со скоро-
стью реабсорбции натрия, хотя диапазон изменений кровотока невелик.
Применительно к коже понятие «функциональная гиперемия» не исполь-
зуется, хотя обусловленные ею изменения кровоснабжения происходят
здесь постоянно. Основная функция теплообмена организма со средой
обеспечивается кровоснабжением кожи, но и другие (не только нагрева-
ние) виды стимуляции кожи (ультрафиолетовое облучение, механические
воздействия) обязательно сопровождаются гиперемией.
438 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 9.7. Регионарные особенности функциональной гиперемии
Орган Показатель усиления функциональной ак- тивности Изменение кровотока Основной фактдр (факторы) ме- ханизма
Мозг Локальная нейрон- ная активность моз- говых зон. Общая Локальное увеличе- ние на 20—60 %. В коре увеличение в Начальный «быстрый» фактор (нервный или химический: ка- лий, аденозин и др.). После-
активность коры 1,5—2 раза дующий «медленный» фактор (РСО22, pH и др.)
Миокард Увеличение часто- ты и силы сокраще- ний сердца Увеличение до 6-кратного Аденозин, гиперосмия, ионы калия и другие гистомеханиче- ские влияния
Скелет- Сокращения мы- Увеличение до Ионы калия, водорода. Гисто-
ные мыш- цы шечных волокон 10-кратного в двух режимах механические влияния
Кишеч- Усиление секре- Увеличение до 2— РО2, метаболиты, интестиналь-
ник ции, моторики И всасывания 4-кратного ные гормоны, серотонин, мест- ный рефлекс
Поджелу- дочная железа Усиление экзосек- реции Увеличение Метаболиты, интестинальные гормоны, кинины
Слюнные Усиление слюноот- Увеличение до Влияние импульсации парасим-
железы деления 5-кратного патических волокон, кинины, гистомеханические влияния
Печень Усиление обмен- ных реакций Локальное увеличе- ние (?) Мало исследовано
Почка (мозговое вещество) Увеличение реаб- сорбции натрия Увеличение до 2-кратного Брадикинин,гиперосмия
Костный мозг Стимуляция эри- тропоэза Увеличение Аденозин
Кость Ритмическая дефор- мация кости Увеличение до 2-кратного Механические влияния
Жир Нейрогенное усиле- ние липолиза через циклический АМФ Увеличение Аденозин, адренергические влияния
Кожа Повышение темпе- ратуры, УФ-облуче- ние, механическая стимуляция Изменение до 5-кратного Уменьшение констрикторной импульсации, метаболиты, ак- тивные вещества из дегранули- рованных тучных клеток, ос- лабление чувствительности к симпатической импульсации
Табл. 9.7 показывает также, что все известные механизмы регуляции ре-
гионарного кровотока (нервные, гуморальные, местные) могут быть прича-
стны к механизмам функциональных гиперемий, причем в разной комби-
нации для различных органов.
9.1.4.2. Нервные и гуморальные влияния на органные сосуды
Нейрогенное сужение сосудов осуществляется путем возбуждения адренер-
гических волокон, которые действуют на гладкие мышцы сосудов путем
высвобождения в области нервных окончаний медиатора адреналина. Тор-
можение импульсов в симпатических нервных волокнах влияет на гладкие
мышцы сосудов путем снижения их тонуса.
Парасимпатические вазодилататорные волокна холинергической приро-
9. Функции систем кровообращения и лимфообращения • 439
ды доказаны для группы волокон сакрального отдела, идущих в составе п.
pelvicus. В блуждающих нервах отсутствуют сосудорасширяющие волокна
для органов брюшной полости.
В скелетных мышцах доказано наличие симпатических вазодилататор-
ных нервных волокон, которые являются холинергическими. Внутрицен-
тральный путь этих волокон начинается в моторной зоне коры мозга. Тот
। факт, что эти волокна могут возбуждаться при стимуляции двигательной
области коры мозга, позволяет предположить, что они вовлекаются в сис-
темную реакцию, способствующую увеличению кровотока в скелетных
мышцах в начале их работы. Гипоталамическое представительство этой
системы волокон указывает на их участие в эмоциональных реакциях орга-
' низма.
У теплокровных отсутствует «дилататорный» центр с особой системой
«дилататорных» волокон. Вазомоторные сдвиги бульбоспинального уровня
осуществляются исключительно путем изменения числа возбужденных
констрикторных волокон и частоты их разрядов, т. е. сосудодвигательные
эффекты возникают только путем возбуждения или торможения констрик-
торных волокон симпатических нервов.
Адренергические волокна при электрической стимуляции могут переда-
вать импульсацию с частотой 80—100 в 1 с. Однако в физиологическом по-
кое частота импульсов в них составляет 1—3 в 1 с и может увеличиваться
при прессорном рефлексе только до 12—15 имп/с. Из сказанного ясно, что
практически весь диапазон величин сосудистых реакций, которые можно
I получить при электрической стимуляции нервов, соответствует увеличе-
нию частоты импульсов всего лишь на1—12в1си что вегетативная нерв-
ная система в норме функционирует при частоте разрядов значительно
меньшей 10 имп/с.
Электрическая стимуляция соответствующих симпатических волокон
। приводит к достаточно сильному повышению сопротивления сосудов ске-
летных мышц, кишечника, селезенки, кожи, печени, почки, жира; эффект
выражен слабее в сосудах мозга, сердца. В сердце и почке этой вазоконст-
, рикции противостоят местные вазодилататорные влияния, опосредованные
активацией функций основных или специальных клеток ткани, одновре-
менно запускаемые нейрогенным адренергическим механизмом. В резуль-
тате такой суперпозиции двух механизмов выявление адренергической
, нейрогенной вазоконстрикции в сердце и почке составляет более слож-
1 ную, чем для других органов, задачу. Общая закономерность все же состо-
ит в том, что во всех органах стимуляция симпатических волокон вызывает
активацию гладких мышц сосудов, иногда маскируемую одновременными
или вторичными тормозными эффектами.
При рефлекторном возбуждении симпатических нервных волокон, как
правило, имеет место повышение сопротивления сосудов всех изученных
областей (рис. 9.22). При торможении симпатической нервной системы
(рефлексы с полостей сердца, депрессорный синокаротидный рефлекс) на-
блюдается обратный эффект. Различия между рефлекторными вазомотор-
ными реакциями органов в основном количественные, качественные — об-
наруживаются значительно реже. Одновременная параллельная регистра-
ция сопротивления в различных сосудистых областях свидетельствует о ка-
I чественно однозначном характере активных реакций сосудов при нервных
влияниях.
Учитывая небольшую величину рефлекторных констрикторных реакций
сосудов сердца и мозга, можно полагать, что в естественных условиях кро-
воснабжения этих органов симпатические вазоконстрикторные влияния на
440 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 9.22. Величины изменений сопротивления сосудов (активные реакции) в раз-
личных областях системы кровообращения при прессорном рефлексе у кошки.
По оси ординат — изменения сопротивления (в процентах к исходному); по оси абсцисс: 1 —
коронарные сосуды; 2 —мозговые; 3 — печеночные; 4—таза и задних конечностей; 5 — зад-
ней конечности; 6 — обеих задних конечностей; 7 — мышц таза; 8 — почки; 9 — толстой киш-
ки; 10 — селезенки; 11 — передней конечности; 12 — желудка; 13 — подвздошной кишки; 14 —
печени.
них нивелируются метаболическими и общими гемодинамическими фак-
торами, в результате чего конечным эффектом может быть расширение со-
судов сердца и мозга. Этот суммарный дилататорный эффект обусловлен
сложным комплексом влияний на указанные сосуды, а не только нейро-
генных. Кроме того, эти отделы сосудистой системы обеспечивают обмен
веществ в жизненно важных органах, поэтому слабость вазоконстриктор-
ных рефлексов в этих органах обычно интерпретируют тем, что выражен-
ные симпатические констрикторные влияния на сосуды мозга и сердца
биологически нецелесообразно, так как это значительно уменьшало бы их
кровоснабжение.
Сосуды легких, обеспечивающих кислородом органы и ткани и выведе-
ние из них углекислоты, т. е. выполняющих функцию, жизненная важ-
ность которой бесспорна, на том же основании не должны подвергаться
выраженным констрикторным влияниям симпатической нервной системы,
поскольку это вело бы к нарушению соответствия их основному физиоло-
гическому значению. Такое суждение относится и к сосудам печени и по-
чек, функционирование которых определяет жизнеобеспечения организма
менее «экстренно», но не менее ответственно.
В то же время при вазомоторных рефлексах сужение сосудов скелетных
мышц и органов брюшной полости значительно больше, чем рефлектор-
ные реакции сосудов сердца, мозга и легких (см. рис. 9.22). Аналогичная
величина вазоконстрикторных реакций в скелетных мышцах больше, чем в
чревной области, а увеличение сопротивления сосудов задних конечностей
больше, чем сосудов передних конечностей.
Причинами неодинаковой выраженности нейрогенных реакций отдель-
ных сосудистых зон могут быть: различная степень симпатической иннер-
вированности; количество, распределение в тканях и сосудах и чувстви-
9. Функции систем кровообращения и лимфообращения • 441
тельность а- и 0-адренорецепторов; местные факторы (особенно метаболи-
ты); биофизические особенности сосудов; неодинаковая интенсивность
импульсов к различным сосудистым областям.
Применение катехоламинов показывает, что во всех органах активация
а-адренорецепторов сопровождается констрикцией артерий и вен. Актива-
ция p-адренорецепторов (обычно связь их с симпатическими волокнами
существенно менее тесная, чем у а-адренорецепторов) приводит к вазоди-
латации; для кровеносных сосудов некоторых органов р-адренорецепция
не обнаружена. Следовательно, в качественном отношении регионарные
адренергические изменения сопротивления кровеносных сосудов первично
однотипны.
Большое количество химических веществ вызывает активные измене-
ния просвета сосудов. Концентрация этих веществ определяет выражен-
ность вазомоторных реакций. Небольшое увеличение концентрации ионов
калия в крови вызывает дилатацию сосудов, а при более высоком уровне —
они суживаются, ионы кальция вызывают артериальную констрикцию,
ионы натрия и магния — являются дилататорами, равно как и ионы ртути
и кадмия. Ацетаты и цитраты также являются активными вазодилататора-
ми, значительно меньшим эффектом обладают хлориды, бифосфаты, суль-
фаты, лактаты, нитраты, бикарбонаты. Ионы соляной, азотной и других
кислот вызывают обычно расширение сосудов. Прямое действие адренали-
на и норадреналина на сосуды вызывает преимущественно их констрик-
цию, а гистамина, ацетилхолина, АДФ и АТФ — дилатацию. Ангиотензин
и вазопрессин — сильные местные констрикторы сосудов. Влияние же се-
ротонина на сосуды зависит от их исходного тонуса: если последний вы-
сок — серотонин расширяет сосуды и, наоборот, при низком тонусе — дей-
ствует сосудосуживающе. Кислород может быть высокоактивным в органах
с интенсивным обменом веществ (мозг, сердце) и значительно меньшее
действие оказывать на другие сосудистые области (например, конечности).
То же относится и к углекислоте. Снижение концентрации кислорода в
крови и соответственно увеличение углекислоты ведет к расширению со-
судов.
Простагландины (ПГ) относятся к физиологически активным вещест-
вам, связанным происхождением с арахидоновой кислотой, и эту группу
веществ называют эйкозаноидами. ПГ выделяются в процессе тканевого
обмена и отличаются от остальных эйкозаноидов тем, что способны вли-
ять на тонус сосудов. При этом важно отметить, что вазоактивны такие
же их концентрации, которые образуются во внеклеточной среде, окру-
жающей кровеносные сосуды в условиях их нормального функционирова-
ния. Дистанционность действия на сосуды веществ рассматриваемой груп-
пы невелика. ПГ типа Е способны в очень малых концентрациях вызы-
вать снижение сопротивления кровотоку сосудов практически всех орга-
нов. Содержание ПГ в венозной крови интенсивно работающих мышц
повышается, и это дает основание предполагать участие данного фактора
в развитии рабочей гиперемии. Доказана роль ПГ в регуляции выхода
трансмиттера (норадреналин) из нервных окончаний. Так, ПГ группы Е
тормозят как количество освобождающего норадреналина, так и действие
его на рецепторы, а ПГ группы F производят противоположный эффект.
Следовательно, ПГ принимают участие и в нервном контроле тонуса со-
судов.
Кинины (действующее начало калликреин-кининовой системы) подоб-
но ПГ относят к биологически активным веществам местного действия.
Активным компонентом этой системы является брадикинин. При введе-
442 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
нии в кровоток брадикинин расширяет сосуды большинства органов. Он
вызывает увеличение кровотока в скелетных мышцах в 15 раз более силь-
ное, чем ацетилхолин в такой же концентрации.
9.1.4.3. Роль эндотелия сосудов в регуляции их просвета
Эндотелий сосудов обладает способностью синтезировать и выделять фак-
торы, вызывающие расслабление или сокращение гладких мышц сосудов в
ответ на разного рода стимулы. Общая масса эндотелиоцитов, монослойно
выстилающих кровеносные сосуды изнутри (интима), у человека прибли-
жается к 500 г. Общая масса, высокая секреторная способность эндотели-
альных клеток позволяют рассматривать эту «ткань» как своеобразный эн-
докринный орган (железу). Распределенный по сосудистой системе эндо-
телий, очевидно, предназначен для вынесения своей функции непосредст-
венно к гладкомышечным образованиям сосудов. Период полужизни вы-
деляемого эндотелиоцитами инкрета очень мал — 6—25 с (вследствие бы-
строго перехода его в нитраты и нитриты), но он способен сокращать и
расслаблять гладкие мышцы сосудов, не оказывая влияния на эффектор-
ные образования других органов (кишечник, бронхи, матка).
Выделяемые эндотелием сосудов расслабляющие факторы (ЭРФ) — не-
стабильные соединения, одним из которых является оксид азота (NO). В
эндотелиальных клетках сосудов NO образуется из а-аргинина при уча-
стии фермента — синтетазы окиси азота.
NO рассматривается как некоторый общий путь передачи сигнала от
эндотелия к гладким мышцам сосудов. Выделение из эндотелия NO инги-
бируется гемоглобином и потенцируется ферментом — дисмутазой.
Участие эндотелия в регуляции тонуса сосудов общепризнанно. Для
всех магистральных артерий показана чувствительность эндотелиоцитов к
скорости кровотока, выражающаяся в выделении ими расслабляющего
гладкие мышцы сосудов фактора, приводящего к увеличению просвета
этих артерий. Таким образом, артерии непрерывно регулируют свой про-
свет соответственно скорости течения по ним крови, что обеспечивает ста-
билизацию давления в артериях в физиологическом диапазоне изменений
величин кровотока. Этот феномен имеет большое значение в условиях раз-
вития рабочей гиперемии органов и тканей, когда происходит значитель-
ное увеличение кровотока, а также при повышении вязкости крови, вызы-
вающей рост сопротивления кровотоку в сосудистой сети. Повреждение
механочувствительности сосудистых эндотелиоцитов может быть одним из
этиологических (патогенетических) факторов развития облитерирующего
эндоартериита и гипертонической болезни.
9.1.5. Особенности кровоснабжения органов и тканей
9.1.5.1. Головной мозг
Интенсивность кровотока в сосудах мозга высока и составляет в состоя-
нии покоя человека 55—60 мл/100 г/мин, т. е. около 15 % сердечного вы-
броса. При относительно небольшой массе (2 % от массы тела) мозг по-
требляет до 20 % всего кислорода и 17 % глюкозы, которые поступают в
организм человека. Интенсивность потребления кислорода мозгом состав-
ляет в среднем 3—4 мл/100 г/мин. В отличие от других органов мозг прак-
тически не располагает запасами кислорода.
При относительном постоянстве общего мозгового кровотока локаль-
9. Функции систем кровообращения и лимфообращения • 443
ный кровоток в различных отделах мозга не постоянен и зависит от интен-
сивности их функционирования. Так, при напряженной умственной рабо-
те локальный кровоток в коре головного мозга человека может возрастать
в 2—3 раза по сравнению с состоянием покоя.
В условиях герметичности и жесткости черепа общее сопротивление со-
судистой системы головного мозга мало зависит от изменений давления в
его артериях. Так, при повышении АД происходит расширение мозговых
артерий, что ведет к повышению давления ликвора, сжатию вен мозга и
оттоку ликвора в спинальную полость. При этом сопротивление артерий
падает, а вен — возрастает, так что общее сопротивление сосудистой систе-
мы мозга в целом практически не меняется.
Миогенная регуляция мозгового кровотока осуществляется за счет
реакции гладких мышц артериальных сосудов мозга на изменение давле-
ния в них. Повышение АД приводит к возрастанию тонуса миоцитов и су-
жению артерий, снижение АД — к снижению тонуса и расширению арте-
рий. Миогенная регуляция мозгового кровообращения считается централь-
ным звеном системы ауторегуляции кровообращения в мозге.
Гуморальная регуляция. Мощным регулятором мозгового крово-
тока является уровень напряжения углекислого газа в артериальной крови
и связанный с этим уровень pH спинномозговой жидкости. На каждый
миллиметр изменения напряжения СО2 величина мозгового кровотока из-
меняется примерно на 6 %. Возрастание напряжения СО2 в крови (гипер-
капния) сопровождается расширением мозговых сосудов, а гипокапния —
их сужением, столь значительным, что достигается порог кислородной не-
достаточности мозга (одышка, судороги, потеря сознания). Увеличение
мозгового кровотока при гиперкапнии обеспечивает быстрое «вымывание»
углекислоты и возвращение уровня напряжения СО2 и концентрации во-
дородных ионов к исходной величине.
Метаболическая реакция клеток мозга играет существенную роль при
локальных перераспределениях крови между областями мозга, имеющими
разный уровень функциональной активности в каждой конкретной ситуа-
ции. Локальное повышение функциональной активности нервных клеток
приводит к росту в межклеточной среде концентрации аденозина и ионов
калия, что ведет к местному расширению сосудов и усилению в них крово-
тока.
Поскольку уровень концентрации ионов калия в межклеточной среде
головного мозга может меняться в течение долей секунды от момента уси-
ления функциональной активности нейронов, описанный механизм счита-
ется быстрым контуром регуляции. Более медленный контур регуляции
мозгового кровотока связан с повышением напряжения СО2 в мозговом
веществе вследствие активного потребления кислорода работающими
клетками. Это вызывает локальное снижение pH в межклеточной среде и
приводит к расширению сосудов активно функционирующей области
мозга.
Биологически активные вещества и гормоны могут оказывать как пря-
мое, так и опосредованное влияние на сосуды мозга. К внутрисосудистым
вазоконстрикторам относятся: вазопрессин, ангиотензин, простагландины
группы F, катехоламины. Сосудорасширяющий эффект оказывают: аце-
тилхолин, гистамин (средние и крупные артерии), брадикинин (мелкие ар-
терии).
Нейрогенная регуляция сосудов головного мозга менее эффек-
тивна, чем метаболическая. Основной зоной приложения нейрогенных
влияний являются мелкие артериальные мозговые сосуды диаметром до
444 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
25—30 мкм. Доказано существование адренергических, холинергических,
серотонинергических и пептидергических нервных волокон, обеспечиваю-
щих регуляцию тонуса мозговых сосудов, Нервные влияния на стенку со-
судов головного мозга опосредуются через а- и 0-адренорецепторы (норад-
реналин), М-холинорецепторы (ацетилхолин, вазоинтестинальный пеп-
тид), D-рецепторы (серотонин).
Основной источник нервных влияний на сосуды мозга — постганглио-
нарные симпатические волокна, начинающиеся в верхних шейных гангли-
ях. Существование парасимпатических влияний на мозговые сосуды не до-
казано.
Нейрогенные влияния на кровоснабжение мозга во многом зависят от
выраженности ауторегуляции, исходного тонуса сосудов, напряжения СО2,
О2, состава и концентрации ионов, присутствия биологически активных
веществ в спинномозговой жидкости и тканях мозга. Именно поэтому ко-
нечный эффект нейрогенных влияний на мозговой кровоток не одно-
значен.
9.1.5.2. Миокард
В состоянии покоя у человека через коронарные сосуды протекает 200—
250 мл крови в минуту (60 мл/100 г/мин), это составляет 4—5 % минутного
объема крови. При интенсивной мышечной работе коронарный кровоток
может возрастать до 400 мл/мин на 100 г ткани.
В период систолы желудочков, расположенные в них сосуды частично
пережимаются, кровоток к миокарду резко ослабевает (до 15 %). Во время
диастолы напряжение в стенке миокарда падает и кровоток увеличивается
до 85 %.
Несмотря на резкое снижение кровотока в миокарде во время систолы,
коронарное кровообращение полностью удовлетворяет высокие метаболи-
ческие потребности миокарда. Это достигается большой объемной скоро-
стью кровотока, растяжимостью сосудов сердца, фазными колебаниями
кровотока в коронарных венах (во время систолы ускоряется отток крови
из коронарного синуса, в диастолу он понижается), наличием густой ка-
пиллярной сети (сердечная мышца богато снабжена капиллярами — на ка-
ждое мышечное волокно приходится капилляр; малое диффузное расстоя-
ние от капилляра до кардиомиоцита облегчает доставку кислорода и пита-
тельных веществ к кардиомиоцитам), высокой экстракцией кислорода
миокардом [в состоянии покоя миоглобин сердечной мышцы извлекает из
артериальной крови 60—75 % кислорода (в мозге — 25—30 %)]. Миокард
потребляет в покое кислорода до 10 мл/100 г/мин. При интенсивной мы-
шечной работе экстракция кислорода в миокарде возрастает примерно в
6 раз.
Миогенная регуляция. Для коронарных сосудов характерен высо-
кий тонус, величина которого находится в обратно пропорциональной за-
висимости от интенсивности обменных процессов миокарда.
Гуморальная регуляция. Наиболее мощным регулятором коронар-
ного кровотока является напряжение кислорода в крови: расширение ко-
ронарных сосудов проявляется при снижении содержания кислорода в
крови на 5 %. Взаимосвязь между потреблением кислорода и уровнем кро-
вотока осуществляется через метаболические механизмы. «Аденозиновая
теория» метаболической регуляции коронарного кровообращения является
общепризнанной. Аденозин блокирует захват Са2+ гладкими мышцами.
При физической нагрузке имеет место увеличение концентрации ионов
9. Функции систем кровообращения и лимфообращения • 445
калия, водородных ионов, молочной кислоты, повышение концентрации
двуокиси углерода, появление в межклеточной жидкости вазоактивных ве-
ществ (гистамин, кинины, простагландины группы Е, пептиды) что ведет к
расширению сосудов и увеличению кровотока в миокарде. Ацетилхолин
через М-холинорецепторы расширяет коронарные артерии. Адреналин и
норадреналин через а-адренорецепторы вызывают сужение, через [3-адре-
норецепторы — расширение коронарных артерий и вен. Ангиотензин и ва-
зопрессин только в больших дозах приводят к сужению коронарных арте-
рий. Тироксин, кортизол и другие гормоны усиливают кровоснабжение
миокарда через метаболические посредники.
Нервная регуляция. Нейрогенный тонус сосудов сердца невелик
(около 20 %). Симпатические нервы содержат сосудосуживающие волокна,
тормозные процессы в них вызывают коронародилатацию. Нервные влия-
ния находятся в конкурентных отношениях с метаболическими, которые
представляют более мощный контур регуляции.
9.1.5.3. Легкие
Легочная артерия и ее ветви, имеющие диаметр более 1 мм, относятся к
артериям эластичного типа, они демпфируют (смягчают) пульсовые толчки
крови, выбрасываемой в момент систолы правого желудочка. Артериолы в
легких тесно связаны с окружающей альвеолярной паренхимой, это опре-
деляет непосредственную зависимость уровня кровоснабжения легких от
режима вентиляции.
В отличие от большого круга кровообращения, капилляры которого
имеют диаметр около 7—8 мкм, в легких имеются два типа капилляров —
широкие (20—40 мкм) и узкие (6—12 мкм). Общая площадь капиллярного
русла легких у человека составляет 35—40 м2. Стенка капилляров легких и
стенка альвеол представляют в совокупности функциональное целое, обо-
значаемое как альвеоло-капиллярная мембрана.
Если функциональное значение сосудов малого круга кровообращения
заключается, главным образом, в поддержании адекватного легочного га-
зообмена, то бронхиальные сосуды обеспечивают питание тканей самих
легких. Венозная бронхиальная сеть дренирует кровь как в систему боль-
шого круга кровообращения (верхняя непарная вена, правое предсердие),
так и малого — в легочные вены и левое предсердие. Только 30 % крови,
поступающей в бронхиальные артерии по системе большого круга крово-
обращения, достигает правого желудочка, основная же часть кровотока на-
правляется через капиллярные и венозные анастомозы в легочные вены.
Указанная особенность бронхиального кровотока формирует так называе-
мый физиологический дефицит напряжения кислорода в артериальной
крови большого круга. Примесь бронхиальной венозной крови к артериа-
лизированной крови легочных вен понижает на 6—10 мм рт.ст. напряже-
ние кислорода по сравнению с его напряжением в крови легочных капил-
ляров, что практически не сказывается на кислородном режиме в процессе
обычной жизнедеятельности организма.
Основным условием, определяющим степень оксигенации крови в лег-
ких, являются величины легочной вентиляции и кровотока, а также сте-
пень их соответствия друг другу. Минутный объем кровообращения через
легкие соответствует МОК в большом круге и составляет в условиях покоя
5—6 л/мин. Сопротивление сосудистого русла малого круга при этом при-
близительно в 8—10 раз меньше, чем в системе большого круга кровообра-
щения. Легочные сосуды характеризуются высокой растяжимостью, по-
446 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
скольку их сосудистая стенка значительно тоньше, чем у соответствующих
по калибру сосудов скелетной мускулатуры и спланхнической области. Это
определяет роль легочных сосудов как депо крови.
Важной особенностью кровоснабжения легких является то, что сосуды
малого круга кровообращения — это система низкого давления. Среднее
давление в легочной артерии у человека составляет 15—25 мм рт. ст., а дав-
ление в легочных венах — 6—8 мм рт. ст. Таким образом, градиент давле-
ния, определяющий движение крови по сосудам малого круга, составляет
9—15 мм рт. ст., что значительно меньше градиента давления в большом
круге кровообращения. Отсюда понятен физиологический смысл высокой
растяжимости легочных сосудов: значительное увеличение кровотока в
системе малого круга (например, при физической нагрузке) не будет со-
провождаться повышением давления крови в силу указанных свойств сосу-
дов легких.
Другим следствием низкого градиента давления в малом круге является
неравномерность кровоснабжения легких от их верхушки к основанию.
В вертикальном положении тела кровоснабжение верхних долей несколько
меньше, чем нижних.
Нервная регуляция. Легочные сосуды имеют двойную иннервацию:
вагусную (афферентную) и симпатическую (эфферентную). Основным ис-
точником афферентной иннервации легочных сосудов являются блуждаю-
щие нервы (волокна, идущие от чувствительных клеток узлового ганглия).
Главными источниками эфферентной иннервации являются шейные и
верхние грудные симпатические узлы.
Влияние нервной системы на легочные сосуды, в отличие от сосудов
большого круга кровообращения, выражено намного меньше.
Крупные легочные сосуды (особенно легочная артерия и область ее би-
фуркации) являются рефлексогенной зоной, обеспечивающей рефлектор-
ные реакции сосудов малого круга. Так, повышение давления в легочных
сосудах приводит к рефлекторному падению системного артериального
давления, замедлению ритма сердечных сокращений, увеличению кровена-
полнения селезенки и вазодилатации в скелетных мышцах. Расширение
периферических сосудов уменьшает приток крови в малый круг кровооб-
ращения и, тем самым, «разгружает» легочные капилляры и предохраняет
их от отека. Описанный комплекс рефлекторных реакций с барорецепто-
ров малого круга получил в литературе обозначение как рефлекс Швигка—
Парина.
Рецепторный аппарат сосудов в малом круге представлен преимущест-
венно p-адренорецепторами (хотя плотность их распределения значитель-
но меньше, чем сосудов большого круга), Д-серотониновыми, Нггистами-
новыми рецепторами и в меньшей степени М-холинорецепторами.
Гуморальная регуляция. В реализации гуморального контроля ле-
гочного кровообращения катехоламины и ацетилхолин играют значитель-
но меньшую роль, чем в большом круге кровообращения. Повышение
концентрации ацетилхолина в крови сопровождается умеренной дилатаци-
ей легочных сосудов. Гуморальная регуляция легочного кровотока опреде-
ляется серотонином, гистамином, ангиотензином-П, простагландином F.
9.1.5.4. Желудочно-кишечный тракт (ЖКТ)
В состоянии покоя на ЖКТ приходится до 20 % сердечного выброса. Кро-
воток в различных отделах ЖКТ неодинаков: в желудке человека он дости-
гает 40 мл/100 г/мин, в тонкой кишке — 35 мл/100 г/мин, в толстой —
9. Функции систем кровообращения и лимфообращения • 447
20 мл/100 г/мин. При максимальной
дилатации сосудов кишечника кро-
воток в нем увеличивается в 8—
10 раз, причем более 90 % дополни-
тельной крови поступает в слизи-
сто-подслизистую сеть.
В первые минуты после еды про-
исходит увеличение кровотока в со-
судах ЖКТ (отчасти за счет вы-
прямления спиралевидных арте-
рий), причем лишь в отделах, функ-
циональная активность которых
усилена. Эта фаза реакции сосуди-
стой системы ЖКТ на прием пищи
продолжается 5—30 мин. Через
30 мин после приема пищи крово-
ток в брыжеечной артерии значи-
тельно возрастает (на 30—130 % по
сравнению с состоянием «натощак»)
и сохраняется на таком уровне в те-
80
Рис. 9.23. Различная величина измене-
ний сопротивления в сосудах селезен-
ки и тонкой кишки при прессорном
синокаротидном рефлексе.
Сверху вниз: системное артериальное давле-
ние, перфузионное давление в сосудах селе-
зенки, тощей кишки, отметка раздражения,
отметка времени (5 с). Шкалы — в мм рт. ст.
чение 3—7 ч. Степень увеличения кровотока после приема пищи опреде-
ляется ее химическим составом и исходным функциональным состоянием
органов ЖКТ.
Нервная регуляция сосудов ЖКТ осуществляется симпатическими вазо-
констрикторными волокнами. Возбуждение этих волокон вызывает суже-
ние артериальных (рис. 9.23) и венозных сосудов, а также прекапиллярных
сфинктеров, что приводит к увеличению регионарного сопротивления и
уменьшению кровотока в органе. При снижении частоты импульсов в сим-
патических волокнах возникает обратный эффект — расширение сосудов
ЖКТ.
В артериальных сосудах кишечника широко представлены а- и р-адре-
норецепторы. В венозных сосудах [3-адренорецепторов значительно мень-
ше. В сосудистом русле ЖКТ при выделении катехоламинов р-адреноре-
цепторы ограничивают констрикторный эффект, возникающий при возбу-
ждении а-адренорецепторов. Это служит механизмом обеспечения необхо-
димого кровоснабжения кишечника при активации симпатоадреналовой
системы. Холинергические сосудорасширяющие волокна в кишечнике и в
желудке не обнаружены. При длительном возбуждении симпатических ва-
зоконстрикторных нервов в кишечнике происходит «ауторегуляторное ус-
кользание» из-под влияния сосудосуживающих волокон нервной системы.
Значение его в поддержании постоянного кровотока и защите тканей ЖКТ
(например, от ишемии при стрессорных воздействиях) велико.
Гуморальная регуляция. Объектом гуморальных влияний агентов
в сосудах ЖКТ являются, главным образом, артериолы и прекапиллярные
сфинктеры. Велика роль в регуляции кровотока в ЖКТ таких метаболитов,
как СО2, Н+ и др. Эти вещества, как и недостаток кислорода, оказывают
вазодилататорное действие. Метаболиты снижают тонус гладких мышц ар-
териол и тем самым понижают сопротивление в системе брыжеечной арте-
рии, увеличивая кровоток в сосудах ЖКТ. В ЖКТ существует ряд специ-
фических механизмов вазодилатации, не связанных с изменением содер-
жания в тканях истинных метаболитов. Так, функциональная гиперемия
ЖКТ после приема пищи обусловлена действием на сосуды ряда гастроин-
тестинальных гормонов, например гастрина и гистамина. Противополож-
448 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ное действие на них оказывают вазопрессин, гастрон, простагландин Е.
Большое значение отводится веществам, высвобождающимся в активно
работающем органе. В тонкой кишке расширение артериальных сосудов и
прекапиллярных сфинктеров вызывают секретин и холицистокинин.
Миогенная регуляция. Сосуды ЖКТ отличаются высокой способ-
ностью к ауторегуляции кровотока. В тонкой кишке проявляется вено-ар-
териолярный рефлекс, который состоит в том, что при локальном увеличе-
нии венозного давления в этом участке повышается тонус прекапиллярных
сфинктеров, что уменьшает местный капиллярный кровоток и создает воз-
можности для «разгрузки» вен и снижения давления на них.
9.1.5.5. Главные пищеварительные железы
Слюнные железы при активации вырабатывают за 2 мин количество
слюны, равное по массе самим железам. Возбуждение парасимпатических
нервов вызывает гиперемию слюнных желез. При этом артериолы и прека-
пиллярные сфинктеры расслабляются, а проницаемость капилляров воз-
растает, что создает благоприятные условия для транспорта веществ в же-
лезистые клетки. При активной работе желез имеющиеся в них артерио-
венозные шунты не функционируют и вся кровь проходит через капилля-
ры. В создании функциональной гиперемии участвуют: 1) вазодилататор-
ные волокна, которые способствуют выработке сосудорасширяющих кини-
нов, 2) метаболиты, 3) гистомеханический фактор — снижение трансму-
рального давления на стенку сосуда вследствие фильтрации части слюны в
интерстиций. Симпатические влияния на сосуды слюнных желез вызыва-
ют их сужение. Вазоконстрикторный эффект осуществляется через /7-адре-
норецепторы.
Поджелудочная железа. Кровоснабжение эндокринных и экзо-
кринных тканей железы осуществляется через одни и те же сосуды. Капил-
ляры снабжают кровью а-, а затем 0-клетки островков Лангерганса и далее
эндокринные ткани. Железа иннервирована симпатическими и парасимпа-
тическими нервами, сосуды железы — только симпатическими. Возбужде-
ние симпатических нервов приводит к сужению сосудов и торможению
секреции. Увеличение кровотока в железе вызывают вещества, расслаб-
ляющие гладкие мышцы сосудов (АТФ, АДФ, брандикинин, холицистоки-
нин и др.).
Функциональная гиперемия железы, возникающая при приеме пищи,
связана с активацией центров вагуса, выделением соляной кислоты в же-
лудке и эвакуацией пищи в двенадцатиперстную кишку. Высвобождаю-
щиеся в дуоденум секретин и холицистокинин стимулируют выделение
местных метаболитов, увеличивающих панкреатический кровоток. Допус-
кают возможность выделения под влиянием парасимпатических нервов
кининов, которые способствуют увеличению кровотока и проницаемости
капилляров в поджелудочной железе.
9.1.5.6. Печень
В печени кровь протекает по печеночной артерии (25—30 %) и воротной
вене (70—75 %). По прохождении капиллярной сети кровь дренируется в
систему печеночных вен, которые впадают в нижнюю полую вену. Важной
особенностью сосудистого русла печени является наличие большого коли-
чества анастомозов между сосудами систем воротной вены, печеночной
артерии и печеночных вен.
9. Функции систем кровообращения и лимфообращения • 449
Давление в печеночной артерии соответствует давлению в других маги-
стральных сосудах — 100—120 мм рт. ст. В воротной вене оно в 10 раз
меньше и составляет около 10 мм рт. ст., в синусоидах — 3—5 мм рт. ст., в
печеночных венах — 2—3 мм рт. ст. Такая небольшая разница между пор-
тальным давлением и давлением в печеночных венах оказывается доста-
точной для портального кровотока вследствие низкого сопротивления пор-
тальных сосудов.
Величина кровотока через печень человека составляет около 100 мл/
100 г/мин, т. е. 20—30 % от величины сердечного выброса. На долю пор-
тального кровотока приходится 70—80 % этого объема, а на долю кровото-
ка в печеночной артерии 20—30 %. При максимальной вазодилатации кро-
воток в печени может возрастать до 5000 мл/мин.
Печень является одним из органов, выполняющих функцию депо крови
в организме (в норме в печени содержится свыше 500 мл крови). За счет
этого может поддерживаться определенный объем циркулирующей крови
(например, при кровопотере) и обеспечиваться необходимая каждой кон-
кретной гемодинамической ситуации величина венозного возврата крови к
сердцу.
Миогенная регуляция обеспечивает высокую степень ауторегуля-
ции кровотока в печени. Даже небольшое увеличение объемной скорости
портального кровотока приводит к сокращению гладких мышц воротной
вены, что ведет к уменьшению ее диаметра, а также включает миогенную
артериальную констрикцию в печеночной артерии. Оба этих механизма
направлены на обеспечение постоянства кровотока и давления в сину-
соидах.
Гуморальная регуляция. Адреналин вызывает сужение воротной
вены, активируя расположенные в ней а-адренорецепторы. Действие адре-
налина на артерии печени сводится, преимущественно, к вазодилатации
вследствие стимуляции преобладающих в печеночной артерии 0-адреноре-
цепторов. Норадреналин при действии как на артериальную, так и на ве-
нозную систему печени приводит к сужению сосудов и повышению сосу-
дистого сопротивления в обоих руслах, что ведет к уменьшению кровотока
в печени. Ангиотензин суживает как портальные, так и артериальные сосу-
ды печени, значительно уменьшая при этом кровоток в них. Ацетилхолин
расширяет артериальные сосуды, увеличивая приток артериальной крови к
печени, но сокращает печеночные венулы, ограничивая отток венозной
крови из органа, что приводит к увеличению портального давления и уве-
личению объема крови в печени.
Метаболиты и тканевые гормоны (двуокись углерода, аденозин, гиста-
мин, брадикинин, простагландины) вызывают сужение портальных венул,
уменьшая портальный кровоток, но расширяют печеночные артериолы,
усиливая приток артериальной крови к печени (артериализация печеноч-
ного кровотока). Другие гормоны (глюкортикостероиды, инсулин, глюка-
гон, тироксин) вызывают увеличение кровотока через печень вследствие
усиления метаболических процессов в печеночных клетках.
Нервная регуляция выражена сравнительно слабо. Вегетативные
нервы печени идут от левого блуждающего нерва (парасимпатические) и от
чревного сплетения (симпатические). Электрическая стимуляция блуждаю-
щего нерва существенно не влияет ни на скорость печеночного кровотока,
ни на его распределение в органе. Стимуляция печеночных симпатических
нервов повышает сосудистое сопротивление в печеночной артерии и в во-
ротной вене.
15-6095
450 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
9.1.5.7. Кожа
Кожа в большей степени, чем другие органы, подвержена прямому дейст-
вию высоких и низких температур, ультрафиолетовых лучей, механических
факторов и т. д. Кровоток по ее сосудам значительно превышает собствен-
ные нутритивные потребности. Это объясняется тем, что выполнение важ-
нейшей функции кожи человека — участие в терморегуляции — определя-
ется не активностью метаболических процессов в ней, а теплопереносящей
функцией кровотока.
В покое при нейтральной температуре внешней среды кожа получает от
5 до 10 % сердечного выброса. Суммарный кожный кровоток взрослого че-
ловека при этом составляет 200—500 мл/мин. В различных частях поверх-
ности тела кожный кровоток значительно отличается. Например, в коже
спины он составляет 9,5 мл/100 г/мин, на передней поверхности тела
15,5 мл/100 г/мин. Наиболее интенсивный кровоток отмечается в коже
пальцев рук и ног, где находится большое количество артериовенозных
анастомозов.
Диапазон возможного возрастания кровотока в коже велик: отношение
объемной скорости кровотока в покое к максимальной его величине со-
ставляет 1:8. Максимальной величины кожный кровоток у человека дости-
гает при тепловом стрессе. В условиях высокой внешней температуры он
может возрастать с 200—500 мл/мин до 2,5—3 л/мин, а при продолжитель-
ном нагревании организма человека (температура кожи 42 °C) увеличивает-
ся до 8 л/мин, составляя 50—70 % сердечного выброса.
Нервная регуляция кровоснабжения кожи обеспечивается широко
представленной иннервацией ее сосудов (особенно артериовенозных ана-
стомозов) симпатическими адренергическими сосудосуживающими волок-
нами. Повышение их активности обусловливает сужение кожных сосудов,
а торможение — приводит к вазодилатации.
Главным фактором в регуляции кожного кровотока является температу-
ра тела, снижение которой приводит к рефлекторному сужению как арте-
риальных, так и венозных сосудов кожи, что способствует перемещению
крови в глубокие вены и сохранению тепла. При общем охлаждении сни-
жается кровоток как через артериовенозные анастомозы, так и через ка-
пилляры кожи. Эта реакция опосредована через гипоталамус, а эффектор-
ными ее путями являются адренергические нервные волокна. При общем
воздействии на организм высоких температур происходит увеличение кож-
ного кровотока за счет, главным образом, раскрытия артериовенозных
анастомозов, кровоток через которые увеличивается в 3—3,5 раза. Раскры-
тие анастомозов является следствием угнетения сосудосуживающей им-
пульсации к кожным сосудам по симпатическим адренергическим волок-
нам, обусловливающим стимуляцию p-адренорецепторов. Медиаторами
активной кожной вазодилатации являются гистамин и допамин.
Гуморальная регуляция. В коже имеется большое количество туч-
ных клеток — источника вазоактивных веществ. Дегрануляция тучных кле-
ток и выделение вазоактивных веществ (гистамина, серотонина и др.) про-
исходит при непосредственном воздействии на кожу ультрафиолетового
облучения, механических и других факторов. В сосудах кожи имеются Н,-
и Н2-гистаминовые рецепторы, опосредующие вазодилататор ное действие
эндогенного и экзогенного гистамина. Расширение сосудов кожи вызывает
субстанция Р, оказывая при этом как прямое влияние на гладкую мышцу
сосудов, так и опосредованное — через гистамин, выделяющийся из туч-
ных клеток. В коже происходит биосинтез простагландинов. Внутрикож-
9. Функции систем кровообращения и лимфообращения • 451
ное введение простагландинов Е2 и Н2 вызывает расширение кожных сосу-
дов, а простагландина F2a — сужение их.
Температура самой крови является фактором, играющим важную роль в
локально действующих механизмах контроля сосудистых функций в коже.
При локальном нагревании кожи имеет место увеличение капиллярного
кровотока без существенных изменений кровотока через артериовенозные
анастомозы. В механизме вазодилатации при локальном нагревании кожи
большую роль играет освобождение вазоактивных веществ (АТФ, субстан-
ция Р, гистамин) и накопление метаболитов. Однако большее значение в
развитии гиперемии в этом случае имеет прямое действие тепла на гладко-
мышечные элементы кожных сосудов. При повышении температуры крови
снижается миогенный тонус и уменьшаются реакции гладких мышц сосу-
дов кожи на симпатическую импульсацию и вазоконстрикторные вещест-
ва, в частности на норадреналин. Снижение адренореактивности гладких
мышц кожных сосудов под влиянием гипертермии связано с уменьшением
чувствительности их а-адренорецепторов.
При локальном действии на кожу низких температур имеют место вазо-
констрикции и снижение кожного кровотока, что обусловлено как повы-
шением сосудистого тонуса, так и увеличением вязкости крови.
9.1.5.8. Почка
Почка является одним из наиболее снабжаемых кровью органов — 400 мл/
100 г/мин, что составляет 20—25 % сердечного выброса. Удельное крово-
снабжение коркового вещества значительно превышает кровоснабжение
мозгового вещества почки. У человека через корковое вещество почки
протекает 80—90 % общего почечного кровотока. Медуллярный кровоток
мал только в сравнении с корковым, однако если сравнивать его с другими
тканями, то он, например, в 15 раз выше, чем в покоящейся скелетной
мышце.
Гидростатическое давление крови в капиллярах клубочков значительно
выше, чем в соматических капиллярах и составляет 50—70 мм рт. ст. Это
обусловлено близким расположением почек к аорте и различием диамет-
ров афферентных и эфферентных сосудов корковых нефронов.
Метаболизм в почках протекает более интенсивно, чем в других орга-
нах, включая печень, головной мозг и миокард. Интенсивность его опре-
деляется величиной кровоснабжения почек. Эта особенность характерна
именно для почек, поскольку в других органах (мозг, сердце, скелетные
мышцы) наоборот — интенсивность метаболизма определяет величину
кровотока.
Гуморальная регуляция. Ангиотензин II (ATII) является мощным
констриктором для сосудов почек, он влияет на почечный кровоток также
опосредованно, стимулируя выброс медиатора из симпатических нервных
окончаний. АТП стимулирует также выработку альдостерона и антидиуре-
тического гормона, которые, в свою очередь, усиливают констрикторный
эффект в сосудах почек.
Простагландины оказывают на сосуды почки выраженное дилататорное
действие. В состоянии покоя простагландины практически не участвуют в
регуляции почечного кровотока, однако их активность резко возрастает
при любых вазоконстрикторных эффектах, что и обусловливает ауторегу-
ляцию почечного кровотока.
Кинины являются местным гуморальным фактором регуляции корково-
го кровотока в почках. Действуя на кининовые рецепторы в сосудах, они
15*
452 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
вызывают вазодилатацию, увеличивая почечный кровоток и активируя на-
трийурез.
Катехоламины через а-адренорецепторы сосудов почек вызывают их
констрикцию, преимущественно в корковом слое. Допамин в малых дозах,
воздействуя на допаминовые рецепторы, вызывает в почках вазодилата-
цию. При больших концентрациях допамин, влияя на /?-адренорецепторы,
вызывает вазоконстрикцию, наиболее выраженную в корковом слое почки.
Вазопрессин, наряду со специфическим влиянием на канальцы почек,
вызывает констрикцию артериол, усиливает действие катехоламинов, пере-
распределяет кровоток в почке, повышая корковый и снижая мозговой
кровоток. Вазопрессин подавляет секрецию ренина и стимулирует синтез
простагландинов. Ацетилхолин, воздействуя на гладкие мышцы артериол и
повышая активность внутрипочечных холинергических нервов, увеличива-
ет почечный кровоток. Секретин также вызывает увеличение общего по-
чечного кровотока. Накопление продуктов метаболизма (СО2), состояние
гипоксии ведет к снижению кортикального кровотока без изменения ме-
дуллярного кровотока. Аденозин вызывает вазоконстрикцию, уменьшая
общий почечный кровоток, при этом происходит перераспределение кро-
вотока в пользу мозгового вещества почки.
Нервная регуляция. По сравнению с гуморальными факторами
нервная регуляция почечного кровотока выражена меньше. Постганглио-
нарные симпатические нервные волокна локализованы в перивазальной
ткани основной, междолевых, междольковых артерий и достигают арте-
риол коркового слоя, реализуя констрикторные эффекты через а-адрено-
рецепторы. Сосуды почки, особенно мозгового слоя, иннервируются сим-
патическими холинергическими нервными волокнами, которые оказывают
вазодилататорный эффект.
Миогенная регуляция обеспечивает в основном ауторегуляцию
кровоснабжения почек. Миогенными реакциями прегломерулярных сосу-
дов обусловливается относительно постоянный уровень коркового крово-
тока в диапазоне колебаний системного артериального давления крови в
пределах от 70 до 180 мм рт. ст.
9.1.5.9. Скелетные мышцы
Большая масса скелетных мышц (около 40 % массы тела) обусловливает
необходимость обеспечения значительного кровотока в мышцах при их ра-
боте. В покое интенсивность кровотока в скелетных мышцах колеблется от
2 до 5 мл/100 г/мин, что составляет 15—20 % величины сердечного выбро-
са. Функциональный резерв для увеличения кровотока в мышцах при фи-
зической работе обеспечивается высоким исходным базальным тонусом
сосудов скелетных мышц. Кровоток в мышцах при их интенсивной работе
может возрасти более чем в 30 раз, достигая величины 100—120 мл/100 г/
мин (80—90 % сердечного выброса). Потребление кислорода мышцами
при этом увеличивается с 0,3 до 6 мл/100 г/мин. Возросшая интенсивность
метаболических процессов обеспечивается значительным увеличением
числа функционирующих капилляров. В покое открыто 20—30 % имею-
щихся в мышце капилляров. При работе скелетных мышц число функцио-
нирующих в них капилляров увеличивается в 2—3 раза.
Миогенная регуляция. Высокий исходный тонус сосудов скелет-
ных мышцах обусловлен, главным образом, миогенной активностью сосу-
дистой стенки и в меньшей степени — влиянием симпатических вазокон-
стрикторов (15—20 % тонуса в покое нейрогенного происхождения).
9. Функции систем кровообращения и лимфообращения • 453
Нервная регуляция сосудов скелетных мышц осуществляется через
симпатические адренергические вазоконстрикторы. В артериях скелетных
мышц имеются а- и p-адренорецепторы, в венах — только a-адренорецеп-
торы. Активизация a-адренорецепторов приводит к сокращению миоцитов
и сужению сосудов, активизация p-адренорецепторов — к расслаблению
миоцитов и расширению сосудов. Сосуды скелетных мышц иннервируют-
ся также симпатическими холинергическими нервными волокнами, возбу-
ждение которых оказывает дилататорный эффект. В покое сосуды скелет-
ных мышц находятся под тоническим влиянием симпатических вазоконст-
рикторов.
Гуморальная регуляция. Наиболее мощным фактором гумораль-
ной регуляции тонуса сосудов являются метаболиты, накапливающиеся в
работающей мышце. В межклеточной жидкости и в оттекающей от мышцы
венозной крови при этом резко падает содержание кислорода, растет кон-
центрация угольной и молочной кислот, аденозина. Среди факторов, обес-
печивающих снижение тонуса сосудов в мышце при ее работе, ведущими
являются быстрое повышение внеклеточной концентрации ионов калия,
гиперосмолярность, а также снижение pH тканевой жидкости.
Серотонин, брадикинин, гистамин оказывают сосудорасширяющее дей-
ствие в скелетных мышцах. Адреналин при взаимодействии с а-адреноре-
цепторами вызывает констрикцию, с p-адренорецепторами — дилатацию
мышечных сосудов, норадреналин обладает сосудосуживающим действием
через a-адренорецепторы. Ацетилхолин и АТФ приводят к выраженной
дилатации сосудов скелетных мышц.
9.1.5.10. Сопряженные функции сосудов
Органным сосудам присущи три сопряженные функции: резистивная, ем-
костная и обменная. Резистивная функция присуща сосудам всех органов
и тканей и определяет величину сопротивления кровотоку. В естественных
условиях циркуляции изменения резистивной функции гладких мышц со-
судов в регионах определяет степень перераспределения кровотока между
ними.
В артериальном отделе сосудистого русла, играющем наиболее сущест-
венную роль в резистивной функции сосудов, содержится только 15—18 %
циркулирующей крови. В то же время в области большого объема (или, по
другой классификации, — низкого давления) находится 70—75 % крови.
В связи с этим для понимания механизмов и закономерностей органного
кровообращения немаловажное значение имеет емкостная функция со-
судов.
Главной отличительной особенностью венозных сосудов по сравнению
с артериальными является то, что они имеют по крайней мере две четко
выраженные функции: емкостную и резистивную, тогда как артериальным
сосудам присуща лишь резистивная функция, а емкостная практически
сведена к минимуму. Емкостная функция венозных сосудов направлена на
формирование венозного возврата крови к сердцу и объема циркулирую-
щей крови.
9.1.6. Микроциркуляция (микрогемодинамика)
К микроциркуляторному руслу относят сосуды: распределители капилляр-
ного кровотока (терминальные артериолы, метартериолы, артериовенуляр-
ные анастомозы, прекапиллярные сфинктеры) и обменные сосуды (капил-
454 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Эндотелиальные клетки
Капиллярная мембрана
Рис. 9.24. Проницаемость капилляров для жидкости, белков и клеток крови.
ляры и посткапиллярные венулы). В месте отхождения капилляров от ме-
тартериол имеются одиночные гладкомышечные клетки, получившие
функциональное название «прекапиллярные сфинктеры».
Стенки капилляров гладкомышечных элементов не содержат. В капил-
лярах наиболее благоприятные условия для обмена между кровью и ткане-
вой жидкостью: высокая проницаемость стенки капилляров для воды и
растворенных в плазме веществ; большая обменная поверхность капилля-
ров; гидростатическое давление, способствующее фильтрации на артери-
альном и реабсорбции на венозном концах капилляра; медленная линей-
ная скорость кровотока, обеспечивающая достаточный контакт крови с об-
менной поверхностью капилляров.
Стенки капилляров образованы расположенными в один слой эндотели-
альными клетками (рис. 9.24). В зависимости от ультраструктуры стенок
выделяют три типа капилляров: соматический, висцеральный, синусоидный.
Стенка капилляров соматического типа образована сплошным слоем эндо-
телиальных клеток, в мембране которых имеется большое количество
мельчайших пор диаметром 4—5 нм. Этот тип капилляров характерен для
кожи, скелетных и гладких мышц, миокарда, легких. Стенка капилляров
соматического типа легко пропускает воду, растворенные в ней кристал-
лоиды, но мало проницаема для белка. Клетки капилляров висцерального
типа имеют в мембране эндотелия «окошки» — фенестры, которые пред-
ставляют собой пронизывающие цитоплазму эндотелиальных клеток от-
верстия, диаметром 40—60 нм, затянутые тончайшей мембраной. Такой
тип капилляров представлен в почках, кишечнике, эндокринных железах,
т. е. органах, в которых всасывается большое количество воды с раство-
ренными в ней веществами. Капилляры синусоидного типа имеют преры-
вистую стенку с большими просветами. Диаметр капилляров — синусоид —
колеблется от 1 до 4 мкм. Такой тип капилляров обеспечивает высокую
9. Функции систем кровообращения и лимфообращения • 455
проницаемость не только для жидкости, но и для белка и клеток крови.
Он имеется в селезенке, печени, костном мозге.
Состояние капиллярного русла характеризуется отношением числа
функционирующих капилляров к нефункционирующим. При увеличении
числа функционирующих капилляров возрастает величина их обменной
поверхности, снижается диффузионное расстояние между капиллярами и
клетками и улучшается кровоснабжение ткани. Несмотря на небольшую
толщину стенки капилляров (0,7—1,5 мкм), растяжимость их мала. Это
обусловлено механическими свойствами окружающей капилляр соедини-
тельной ткани органа.
Гидростатическое давление на артериальном конце «усредненного» ка-
пилляра равно примерно 30 мм рт. ст., на венозном — 10—15 мм рт. ст.
Этот показатель варьирует в различных органах и тканях и зависит от со-
отношения пре- и посткапиллярного сопротивления, которое и определяет
его величину. Так, в капиллярах почек он может достигать 70 мм рт. ст., а
в легких — только 6—8 мм рт. ст.
Транскапиллярный обмен веществ обеспечивается путем диффузии,
фильтрации-абсорбции и микропиноцитоза. Скорость диффузии высока:
60 л/мин. Легко осуществляется диффузия жирорастворимых веществ
(СО2, О2), водорастворимые вещества попадают в интерстиций через поры,
крупные вещества — путем пиноцитоза.
Второй механизм, обеспечивающий обмен жидкости и растворенных в
ней веществ между плазмой и межклеточной жидкостью,— фильтрация-аб-
сорбция. Давление крови на артериальном конце капилляра способствует
переходу воды из плазмы в тканевую жидкость. Белки плазмы, создавая
онкотическое давление, равное примерно 25 мм рт. ст., задерживают выход
воды. Гидростатическое давление тканевой жидкости около 3 мм рт. ст.,
онкотическое — 4 мм рт. ст. На артериальном конце капилляра обеспечи-
вается фильтрация, на венозном — абсорбция. Между объемом жидкости,
фильтрующейся на артериальном конце капилляра и абсорбирующейся в
венозном конце, существует динамическое равновесие.
Линейная скорость кровотока в сосудах микроциркуляторного русла ма-
ла—от 0,1 до 0,5 мм/с. Низкая скорость кровотока обеспечивает относи-
тельно длительный контакт крови с обменной поверхностью капилляров и
создает оптимальные условия для обменных процессов.
Отсутствие мышечных клеток в стенке капилляров указывает на невоз-
можность активного сокращения капилляров. Пассивное сужение и рас-
ширение капилляров, величина кровотока и количество функционирую-
щих капилляров зависят от тонуса гладкомышечных структур терминаль-
ных артериол, метартериол и прекапиллярных сфинктеров.
Процессы транскапиллярного обмена жидкости в соответствии с урав-
нением Старлинга (рис. 9.25) определяется силами, действующими в об-
ласти капилляров: капиллярным гидростатическим давлением (Рс) и гид-
ростатическим давлением интерстициальной жидкости (Pi), разность кото-
рых (Pc — Pi) способствует фильтрации, т. е. переходу жидкости из внутри-
сосудистого пространства в интерстициальное; коллоидно-осмотическим
давлением крови (Пс) и интерстициальной жидкости (П1), разность кото-
рых (Пс — ni) способствует абсорбции, т. е. движению жидкости из тканей
во внутрисосудистое пространство, ст — осмотический коэффициент отра-
жения капиллярной мембраны, который характеризует реальную прони-
цаемость мембраны не только для воды, но и для растворенных в ней ве-
ществ, а также белков. Если фильтрация и абсорбция сбалансированы, то
наступает «старлинговое равновесие».
456 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
AV/At = ± CFG {(Рс - Р) - ст (тгс - л,)}
Рис. 9.25. Транскапиллярный обмен
жидкости (по Старлингу).
ИП — интерстициальное пространство.
Остальные обозначения — в тексте.
Своеобразие строения терми-
нального сосудистого русла различ-
ных органов и тканей отражает и
зависит от их функциональных осо-
бенностей, прежде всего от уровня
обмена кислорода, интенсивности
процессов метаболизма. Так, в раз-
личных тканях и органах капилля-
ры образуют сеть определенной
плотности в зависимости от их ме-
таболической активности. На осно-
вании этих данных введено понятие
«критическая толщина тканевого
слоя» — наибольшая толщина тка-
ни между двумя капиллярами, ко-
торая обеспечивает оптимальный
транспорт кислорода и эвакуацию
продуктов метаболизма. Чем интен-
сивнее обменные процессы в орга-
не, тем меньше критическая тол-
щина ткани, т. е. между этими по-
казателями существует обратно
пропорциональная зависимость. В
большинстве паренхиматозных ор-
ганов величина этого показателя
составляет всего 10—30 мкм, а в ор-
ганах с замедленными процессами обмена она возрастает до 1000 мкм.
Для оценки функциональной активности шунтирующих сосудов
(артериовенозных анастомозов) используют возможность перехода частиц,
превышающих по размерам диаметр капилляров, из артериального отдела
сосудистого русла в венозный.
Рассчитано, что кровоток через анастомозы во много раз превышает кро-
воток по капиллярам. Так, через анастомоз диаметром 40 мкм может про-
брасываться в 250 раз больше крови, чем через капилляр такой же длины,
но диаметром 10 мкм. Диаметр артериовенозных анастомозов в разных ор-
ганах колеблется в широких пределах (например, в сердце — 70—170 мкм, в
почках — 30—440 мкм, в печени — 100—370 мкм, в тонком кишечнике —
20—180 мкм, в легких — 28—500 мкм, в скелетных мышцах — 20—40 мкм).
9.1.7. Центральная регуляция кровообращения
Нервная регуляция сердечно-сосудистой системы является мощным и бы-
стрым путем изменения кровообращения. Нервные влияния на гладкомы-
шечные клетки сосудов являются высшими по сравнению с влиянием на
сосуды гуморальных веществ, содержащихся в крови, поскольку первые
обеспечивают не только быструю, но и, если необходимо, локализованную
регуляцию.
Нормальное функционирование системы кровообращения в условиях
постоянно меняющихся внешних воздействий осуществляется только при
участии механизмов регуляции (управления, контроля). Непременным ус-
ловием рассматриваемого вопроса является цель регуляции (управления).
В физиологии принято употреблять выражение не «цель», а «физиологиче-
ское значение (роль)» регуляторного механизма.
9. Функции систем кровообращения и лимфообращения • 457
Две части системы управления — управляющее устройство и объект
управления — образуют вместе контур управления. В кровообращении в ка-
честве объекта управления можно выделить капиллярное русло и течение
крови по нему, обеспечивающие питание тканей, тогда управляющими
устройствами являются отделы сердечно-сосудистой системы и свойства
крови, обеспечивающие необходимый кровоток в капиллярном русле в хо-
де быстротекущих событий. Однако большинство авторов в качестве объ-
екта управления в системе кровообращения рассматривают системное ар-
териальное давление. Поддержание артериального давления на постоян-
ном уровне является залогом обеспечения нутритивной функции систе-
мой. Другие авторы объектом управления в системе считают сердечный
выброс. В любом из этих случаев на первый план выступает рефлекторная
регуляция кровообращения.
9.1.7. /. Рефлекторная регуляция кровообращения
Роль каротидных синусов в регуляции кровообращения была доказана тем,
что при раздражении ветви языкоглоточного нерва — нерв каротидного си-
нуса, имело место рефлекторное замедление частоты сердечных сокраще-
ний и независимое от него снижение артериального давления. В дальней-
шем была открыта барорецепторная область не только в каротидном сину-
се, но и в дуге аорты, чувствительные волокна от которой проходят в со-
ставе аортального нерва.
При двустороннем повышении давления в изолированных каротидных
синусах на 15 мм рт. ст. происходят снижение системного артериального
давления и замедление частоты сокращений сердца. Снижение давления в
обоих каротидных синусах до 30 мм рт. ст. (пережатие сонных артерий) со-
провождается значительным повышением АД и ростом частоты сокраще-
ний сердца. Степень вовлечения артериальных сосудов различных регио-
нарных областей в рефлекторную барорецепторную реакцию различна.
Наиболее мощные вазомоторные эффекты отмечены в скелетных мышцах,
менее выражены реакции сосудов брыжейки, кожи и почек. Изменения
сердечного выброса при синокаротидных барорефлексах невелики.
Указанные рефлексогенные зоны чувствительны и к изменениям на-
пряжения в крови О2, СО2 и pH. Стимулятором хеморецепторов этих об-
ластей являются также изменения температуры крови, кровопотеря, а так-
же некоторые вещества (цианиды, никотин, лобелии, серотонин и др.).
При раздражении каротидных хеморецепторов повышается сопротивле-
ние сосудов, замедляется частота сердечных сокращений и снижается на-
сосная функция сердца, повышается артериальное давление, кровоток в
коронарных сосудах увеличивается. Наименьшее сужение сосудов при
этом рефлексе отмечено в сосудах почек и мозга, наибольшее — в сосудах
бассейна подвздошной артерии и конечностей.
В ответ на раздражение аортальных хеморецепторов увеличивается час-
тота сердечных сокращений, повышается АД, суживаются сосуды, причем
более выражено в брыжеечной и почечной областях, менее — в бедренной
артерии.
9.1.7.2. Спинальный уровень регуляции
Эффекторными клетками, осуществляющими регуляцию сердца и сосудов
на уровне спинного мозга, являются симпатические преганглионарные
нейроны. Они обладают спонтанной фоновой импульсной активностью,
458 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
частота которой коррелирует с изменениями частоты сокращений сердца и
пульсовыми колебаниями артериального давления. Спинальные нейроны
представляют собой важное промежуточное звено по пути нисходящих
нейрогенных влияний, адресованных эффекторам в сердечно-сосудистой
системе. Ядра боковых рогов спинного мозга могут обеспечивать преобра-
зование нервных импульсов по дивергентному и конвергентному типу.
Перерезка в эксперименте на животных спинного мозга на уровне шей-
ных и верхних грудных сегментов приводит к значительному снижению
(до 60 мм рт. ст.) артериального давления. Однако если перерезка выпол-
нена в щадящих условиях препаровки (например, при помощи ультразву-
кового ножа), уже через несколько часов давление восстанавливается
практически до нормальных величин. В этих же условиях восстанавлива-
ются и сосудодвигательные рефлексы. Эти факты свидетельствуют о том,
что спинной мозг при определенных условиях может являться уровнем за-
мыкания рефлексов сердечно-сосудистой системы. В то же время в естест-
венных условиях рефлексы сердечно-сосудистой системы осуществляются
при обязательном участии бульбарных и гипоталамических образований
головного мозга.
9.1.7.3. Бульбарный уровень регуляции
Современные представления о бульбарных механизмах регуляции кровооб-
ращения основаны на исследованиях, в которых показано влияние как пе-
ререзок на разных уровнях, так и стимуляции различных бульбарных
структур на артериальное давление и частоту сердечных сокращений, а
также на импульсации в симпатических и парасимпатических нервах. Раз-
дражение продолговатого мозга и варолиева моста оказывает либо прес-
сорные, либо депрессорные изменения АД в зависимости от локализации
раздражающих электродов: прессорные зоны располагаются, как правило,
ростральнее и латеральнее депрессорных. При перерезке на уровне ядер
лицевого нерва отделяется расположенная ростральнее часть прессорной
области, тогда как депрессорная остается интактной. В этом случае снижа-
ется артериальное давление и уменьшается импульсация в эфферентных
вегетативных нервах. Перерезка продолговатого мозга на уровне задвижки
ведет к еще большему снижению давления и полному прекращению им-
пульсации по вегетативным нервам. Бульбарная медиальная депрессорная
область оказывает угнетающее тоническое влияние на спинальные пре-
ганглионарные нейроны, способные к спонтанной импульсивной актив-
ности.
Латеральные участки бульбарной ретикулярной формации содержат
скопления спонтанно активных нейронов, которые через нисходящие пути
спинного мозга оказывают стимулирующее влияние на спинальные пре-
ганглионарные симпатические нейроны. Эти нейроны образуют латераль-
ную «прессорную» область продолговатого мозга. Функции этих нейронов
осуществляются в реципрокном взаимодействии с нейронами медиальной
«депрессорной» зоны.
Указанный медуллярный комплекс, реализующий влияния на сердечно-
сосудистую систему через симпатические нервы, называют бульбарным со-
судодвигательным центром. Функционально он включает в себя и структу-
ры дорсального ядра блуждающего нерва, обеспечивающие тонические
кардиоингибирующие влияния. Поэтому говорят о едином бульбарном
центре сердечно-сосудистой системы, получающем информацию от раз-
личных рецепторных зон и обеспечивающем поддержание циркуляторного
9. Функции систем кровообращения и лимфообращения • 459
гомеостазиса. Это достигается благодаря реципрокному взаимодействию
симпатических и парасимпатических рефлекторных влияний на сердце, а
также дозировкой тонических констрикторных влияний симпатических
нервов на сосуды.
В вентролатеральных отделах продолговатого мозга сосредоточены об-
разования, соответствующие по своим характеристикам тем представлени-
ям, которые вкладывают в понятие «вазомоторный центр». Здесь сконцен-
трированы нервные элементы, играющие ключевую роль в тонической и
рефлекторной регуляции кровообращения. В вентральных отделах продол-
говатого мозга расположены нейроны, изменение тонической активности
которых ведет к активации симпатических преганглионарных нейронов.
Структуры этих отделов мозга контролируют выброс вазопрессина клетка-
ми супраоптического и паравентрикулярного ядер гипоталамуса.
Доказаны проекции нейронов каудальной части вентральных отделов
продолговатого мозга к клеткам его ростральной части, что свидетельству-
ет о возможности тонического угнетения активности этих клеток. Функ-
ционально значимы связи структур вентральных отделов продолговатого
мозга с ядром солитарного тракта, которое играет ключевую роль в обра-
ботке афферентации от хемо- и барорецепторов сосудов.
К вентральным отделам продолговатого мозга информация поступает
как от его структур — ядер солитарного тракта, area postrema, вестибуляр-
ных ядер, ядер шва, обоюдного ядра, так и от клеток спинного мозга и
супрабульбарных структур центральной нервной системы. Наиболее значи-
мыми в функциональном плане для структур ростральной части вентраль-
ных отделов являются их связи с симпатическими преганглионарными
нейронами спинного мозга, а для структур каудальных вентролатеральных
отделов — восходящие проекции к нейросекреторным клеткам гипотала-
муса.
Структуры ростральной части вентральных отделов продолговатого моз-
га играют ключевую роль в тоническом контроле уровня артериального
давления. Стимуляция нейронов рострального вентролатерального ядра
вызывает подъем АД, тахикардию, торможение барорецепторных рефлек-
сов, высвобождение адреналина из надпочечников и аргинин-вазопресси-
на из нейросекреторных клеток гипоталамуса. Возбуждение нейронов кау-
дальной части вентральных отделов ведет к снижению системного АД.
Структуры вентральных отделов участвуют в контроле сердечной деятель-
ности и через обоюдное ядро блуждающего нерва.
Нейроны ростральной части вентральных отделов продолговатого мозга
являются звеном дуг собственных и сопряженных сосудистых рефлексов, а
нейроны каудальной части — модулируют реализацию на сосуды указан-
ных рефлексов.
Сердечно-сосудистый центр продолговатого мозга оказывает тониче-
ское возбуждающее влияние на преганглионарные симпатические нейро-
ны тораколюмбального отдела спинного мозга. Однако его влияние явля-
ется не единственными связями, конвергирующими на спинальных веге-
тативных нейронах. В медуллярной области происходит переключение пу-
тей нисходящих возбуждающих и тормозных кардиоваскулярных влияний,
исходящих из вышележащих отделов центральной нервной системы. Соб-
ственная тоническая активность спинальных нейронов в норме зависит от
нисходящих бульбарных и супрабульбарных влияний, эти нисходящие
влияния полностью доминируют над их спонтанной активностью. Не-
смотря на то что сегментарные спинальные вазомоторные рефлексы (на-
пример, побледнение кожи над областью внутрибрюшинной воспалитель-
460 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ной реакции) могут иметь место, замыкание собственных сердечно-сосу-
дистых рефлексов происходит не ниже, чем на бульбарном уровне. Одна-
ко и бульбарный уровень регуляции в естественных условиях модулирует-
ся сложной совокупностью нисходящих к нему влияний, среди которых
ведущую роль играют нейрогуморальные влияния гипоталамического про-
исхождения.
9.1.7.4. Гипоталамические влияния
Традиционно гипоталамус делят на передний — депрессорный (парасимпа-
тический, трофотропный) и задний — прессорный (симпатический, эрго-
тропный), однако это деление весьма условно. Выраженные прессорные
реакции могут быть вызваны и с передних отделов гипоталамуса (супраоп-
тическое ядро). Депрессорные или прессорные эффекты можно получить в
зависимости от частоты подаваемых на структуру импульсов (депрессор-
ные — при низких частотах — 20—40 имп/с, прессорные — 60—100 имп/с),
функционального состояния центров (вид, глубина наркоза) и от исходно-
го тонуса сосудов.
Опыты на животных с перерезками, коагуляцией и раздражением гипо-
таламических структур и нисходящих путей показали, что имеются два ос-
новных вазоконстрикторных пути: от переднего гипоталамуса (от супраоп-
тического ядра по срединному пучку переднего мозга и по периферии вен-
тромедиального ядра) и от заднего гипоталамуса (от периферии III желу-
дочка над маммилярными телами и дорсолатеральнее субталомической об-
ласти: неопределенная зона и поля Фореля 1 —2 через перивентрикулярные
безмякотные волокна). Каудальнее оба пути проходят в центральном сером
веществе сильвиева водопровода, в покрышке мозга и в продольном дор-
зальном пучке спускаются к дну IV желудочка. Здесь образуются синапсы
с клетками ядра блуждающего нерва и вазомоторного центра. На протяже-
нии этих путей отходят многочисленные коллатерали в ретикулярную фор-
мацию ствола мозга. Таким образом, первое симпатическое переключение
имеет место в ретикулярной формации продолговатого мозга, второе — на
вазомоторных нейронах спинного мозга (общий конечный путь сосудодви-
гательных реакций).
Наиболее универсальным механизмом реакции сердечно-сосудистой
системы на возбуждение гипоталамуса является активация симпатической
системы. Типичным результатом возбуждения гипоталамических структур
в этом случае является альфа-адренергическая констрикция перифериче-
ских сосудов в сочетании с положительными хронотропными и инотроп-
ными влияниями на сердце.
Симпатические эффекторы, обусловленные гипоталамическими влия-
ниями, вызывают и дилатацию периферических сосудов. Это достигается
симпатической холинергической или p-адренергической дилатацией, а
также результатом центрального нисходящего гипоталамического торможе-
ния бульбарных сердечно-сосудистых центров.
Гипоталамические влияния на кровообращение реализуются и гумо-
ральным путем. Возбуждение гипоталамуса сопровождается выделением
вазопрессина в гипофизе и адреналина в надпочечниках с их последую-
щим действием на сердце и сосуды, а также выделением ренина почками в
результате прямых нейрогенных влияний на их юкстагломерулярный аппа-
рат. Конечным результатом этого процесса является повышение артери-
ального давления за счет усиленного образования и нарастания в крови
ангиотензина II.
9. Функции систем кровообращения и лимфообращения • 461
Гипоталамической отдел регуляции кровообращения оказывает диффе-
ренцированное влияние на сердце и некоторые сосудистые бассейны. Так,
раздражение гипоталамических центров «защитных» реакций организма
вызывает увеличение сердечного выброса, повышение артериального дав-
ления, сужение сосудов внутренних органов, но при этом имеет место рас-
ширение артериальных сосудов скелетных мышц. Стимуляция пищевых
центров в латеральном гипоталамусе приводит, наоборот, к расширению
сосудов желудочно-кишечного тракта и их сужению в скелетных мышцах.
Гипоталамические нейроны получают сигналы практически от всех
экстеро- и интероцепторов организма, включая артериальные барорецеп-
торы, импульсы от которых поступают в передний гипоталамус. Кроме то-
го, гипоталамические нейроны получают информацию об изменениях
внутренней среды организма (температуры, кислотности, осмолярности
и др.). Следовательно, существует еще одна рефлекторная функция гипо-
таламуса, физиологический смысл которой заключается в обработке и ин-
теграции этой информации, и включения в общую реакцию сердечно-со-
судистой системы, направленную на поддержание гомеостазиса. Эта
функция гипоталамуса включает в себя и контроль деятельности нижеле-
жащих бульбарных и спинальных структур, ответственных за регуляцию
кровообращения.
Важная роль гипоталамуса заключается также в координации кровооб-
ращения с другими висцеральными и соматическими функциями. Так,
эмоциональным состояниям человека, связанным с активацией гипотала-
мических структур, соответствуют и определенные изменения гемодинами-
ки. Поэтому считается, что гипоталамус осуществляет координацию сома-
томоторных и вегетативных проявлений эмоционального поведения.
9.1.7.5. Участие лимбических структур
Лимбическая система, или «висцеральный мозг», представляет собой ком-
плекс структур на медиальной поверхности полушарий мозга. При элек-
трическом раздражении передних отделов системы имеет место умеренная
гипотензия и брадикардия. Стимуляция переднего отдела лимбической ко-
ры вызывает депрессорную реакцию с верхних участков, а с базальных
(нижних) участков провоцируются прессорные ответы.
Выраженная гипотензия и брадикардия зарегистрированы при раздра-
жении прозрачной перегородки, функционально относящейся к лимбиче-
ской системе. Эта реакция аналогична эффектам раздражения сердечно-
тормозного центра гипоталамуса и является одним из элементов централь-
ного комплекса экстренного снижения сосудистого тонуса и частоты со-
кращений сердца.
Для стимуляции гиппокампа характерны «парасимпатические ответы»
(снимаемые атропином гипотензия и брадикардия). Функционально к ро-
стральной части гиппокампа примыкает миндалевидно-ядерный (амигда-
лярный) комплекс, имеющий тесные связи с заднелатеральным гипотала-
мусом. Амигдалярные влияния модулируют чрезмерные гипоталамические
реакции. Для раздражения самого миндалевидного комплекса характерны
дифференцированные влияния на сердце и тонус сосудов: брадикардия
предшествует позже развивающейся гипертензии, т. е. изменения ритма
сердца не являются вторичными, обусловленными барорецептивными
рефлексами. Через амигдалу и передние отделы гипоталамуса реализуются
рефлекторные изменения кровообращения при раздражении афферентных
волокон блуждающего нерва.
462 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
9.1.7.6. Кортикальные влияния
Изучению роли коры больших полушарий в регуляции кровообращения
уделялось пристальное внимание, но результаты этих исследований до на-
стоящего времени весьма противоречивы. В экспериментах на животных
удаление обоих полушарий не вызывает каких-либо существенных измене-
ний кровообращения. Электрическая стимуляция участков лобной и те-
менной областей коры ведет к изменениям артериального давления в ре-
зультате сужения или расширения сосудов, причем, в зависимости от пара-
метров раздражения стимуляция одних и тех же точек, может вызвать как
вазоконстрикцию, так и вазодилатацию.
Влияние коры больших полушарий на сердце связано с высшей нерв-
ной деятельностью человека — реализацией эмоциональных, поведенче-
ских реакций и условных рефлексов. Последние могут образовываться на
основе сопряженных кардиальных рефлексов. Изменения кровообраще-
ния, обусловленные собственными рефлексами сердечно-сосудистой сис-
темы, воспроизвести условнорефлекторным путем не удалось. Условно-
рефлекторные реакции сердца являются составной частью сложных пове-
денческих рефлексов. У человека эти реакции могут быть вызваны и сло-
весными раздражителями (речевыми сигналами). Однако произвольно из-
менить параметры сердечной деятельности возможно только косвенно че-
рез заведомо управляемые системы: дыхание или скелетную мускулатуру, а
также в некоторых случаях — изменяя эмоциональное состояние орга-
низма.
Большее значение имеют кортикальные механизмы в реализации изме-
нений кровообращения при стрессорных и невротических состояниях че-
ловека.
9.1.7.7. Общая схема центральной регуляции
На рис. 9.26 представлена общая схема организации центральных звеньев
нервной регуляции кровообращения. Сердечно-сосудистый центр продол-
говатого мозга состоит из двух зон (А и Б) одна из которых определяет су-
жение сосудов (А'), ускорение и усиление сердцебиения (АП), тогда как
другая — расширение сосудов (Б') и замедление частоты сокращений сердца
(БП). Тонически активные нейроны зоны А непосредственно или через
промежуточные нейроны возбуждают преганглионарные и ганглионарные
симпатические нейроны, иннервирующие сердце и сосуды, и в то же время
тормозят нейроны зоны Б. Последние активируются сигналами барорецеп-
торов сердечно-сосудистой системы. Через короткие внутрицентральные
пути нейроны зоны Б тормозят нейроны зоны А, а посредством нисходящих
тормозных путей — и преганглионарные симпатические нейроны. Кроме
того, через моторное ядро блуждающих нервов нейроны зоны Б влияют на
ганглионарные парасимпатические нейроны сердца. В результате обе зоны
(А и Б), получая основную сигнализацию от рецепторов сердечно-сосуди-
стой системы, являются основой поддержания кровообращения, сочетанно
подвергая сердце тоническому контролю тормозных вагусных и возбуждаю-
щих симпатических волокон, тогда как постоянный контроль сосудов обес-
печивается только симпатическими вазоконстрикторными волокнами.
Афферентная нервная система является источником сигналов собствен-
ных и сопряженных рефлексов. Вход первых ограничен продолговатым
мозгом, причем барорецепторы сердечно-сосудистой системы связаны с
зоной Б, а хеморецепторы — с зоной А.
9. Функции систем кровообращения и лимфообращения • 463
Кора
центральных звеньев
Рис. 9.26. Схема организации
сосудистой системы (объяснение в тексте).
нервной регуляции сердечно-
Поскольку сердечно-сосудистый центр продолговатого мозга регулирует
основные параметры системной гемодинамики, даже будучи отключенным
от связей с вышележащими нервными структурами, он функционирует как
истинный «центр», обладающий значительной независимостью. Следова-
тельно, супрабульбарные структуры признаются несущественными ни для
прямой тонической регуляции кровообращения, ни для осуществления
сердечно-сосудистых рефлексов. В то же время при общих реакциях орга-
низма (поведенческих, эмоциональных, стрессорных и др.) кора мозга, ги-
поталамус и другие супрабульбарные структуры могут вмешиваться в дея-
тельность сердечно-сосудистой системы, изменяя путем нисходящих (мо-
дулирующих) влияний возбудимость бульбарного центра.
Вазомоторные центры спинного мозга самостоятельного значения в ре-
гуляции кровообращения не имеют. Действующими элементами спиналь-
ного уровня являются лишь симпатические преганглионарные нейроны,
которые входят в состав эфферентного звена бульбарных рефлекторных
дуг.
9.2. Лимфообращение
Основной функцией лимфатической системы является резорбция из ин-
терстиция белков и других веществ, вышедших в него из кровеносного
русла и неспособных вновь вернуться в кровоток через кровеносные ка-
пилляры, с последующей транспортировкой этих веществ и жидкости по
лимфатическим сосудам в венозную систему.
464 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Поскольку в указанной системе транспорт жидкости и включенных в
нее веществ осуществляется по пути: кровеносное русло—интерстиций—
лимфатические сосуды (лимфообразование и лимфоток)—кровеносное
русло, это является основанием для использования термина «лимфообра-
щение».
Физиологическая характеристика интерстиция и лимфообразования
представлена в главе 2, поэтому здесь рассматриваются только вопросы
лимфотока.
У взрослого человека за сутки из кровеносного русла в интерстиций вы-
ходит около 20 л жидкости, из которой 1,5—2,5 л в виде лимфы возвраща-
ется в кровеносную систему по лимфатическим сосудам. Вместе с жидко-
стью из крови в интерстиций за сутки мигрирует 50—100 % всех циркули-
рующих в плазме белков. Значительная часть их должна возвратиться в
кровеносное русло, что и достигается благодаря функционированию лим-
фатической системы, регулирующей экстраваскулярное обращение плаз-
менных белков. Увеличение объема интерстициальной жидкости ведет к
повышению резорбирующей функции лимфатической системы. При этом
деятельность ее направлена на нормализацию объема интерстициальной
жидкости как путем ускорения вывода избытка ее из тканей в кровь, так и
частичного депонирования в лимфатической системе.
Находящаяся в интерстиции жидкость движется по направлению к
лимфатическим капиллярам по так называемым соединительнотканевым
пре- или паралимфатическим транспортным путям. Примером такого пу-
ти могут служить пространства Диссе в печени, с одной стороны ограни-
ченные кровеносными синусоидами, а с другой — «балочками» из клеток
печени. Лимфатические капилляры окружают «кольцом» фрагменты сети
кровеносных капилляров и топографически связаны с посткапиллярными
венулами. Это создает предпосылки для возникновения градиентов дав-
ления на границе раздела интерстициальная жидкость — терминальная
лимфа, определяющих в интерстициальном пространстве направление
перемещения жидкости и белка и поступление их в корни лимфатиче-
ской системы. Последние сообщаются с интерстициальным пространст-
вом через отверстия в эндотелиальной выстилке лимфатических капил-
ляров.
Основными путями попадания крупно- и жидкодисперсных частиц в
просвет лимфатических капилляров являются: а) места соединения эндоте-
лиальных клеток, б) пиноцитозные пузырьки, в) цитоплазма эндотелиаль-
ной клетки. Когда гидростатическое давление в тканях становится выше,
чем в лимфатическом капилляре, проникающая в него жидкость растяги-
вает межэндотелиальные соединения. При этом эндотелиальная клетка, не
связанная с фиксирующими волокнами и контактирующая в обычных ус-
ловиях с другой клеткой, свободно отгибается в просвет сосуда, открывая
для крупных молекул доступ в лимфатический капилляр. Этому содейству-
ет приводящее к накоплению интерстициальной жидкости повышение ос-
мотического давления в интерстиции, при котором набухание тканей вы-
зывает натяжение фиксирующих волокон и расширение лимфатических
капилляров.
Макромолекулы и частицы диаметром 3—50 мкм проникают в просвет
лимфатических капилляров через эндотелиальные клетки с помощью пи-
ноцитозных пузырьков или везикул. Этим путем в лимфатические капил-
ляры поступают белки, хиломикроны и ионы.
9. Функции систем кровообращения и лимфообращения • 465
9.2.1. Лимфатические сосуды
Лимфатическая система состоит из мелких внутриорганных лимфатиче-
ских сосудов, отводящих лимфу из лимфатических капилляров; внеорган-
ных лимфатических сосудов, отводящих лимфу из органов в лимфатиче-
ские узлы; внеорганных лимфатических сосудов, отводящих лимфу из
лимфатических узлов; крупных лимфатических сосудов — стволов, прото-
ков, отводящих лимфу в венозную систему.
Форма лимфатических сосудов преимущественно цилиндрическая. Она
отличается от вида артерий и вен чередованием многочисленных, сменяю-
щих друг друга расширений и сужений, придающих им сходство с четка-
ми, что объясняется наличием в этих сосудах многочисленных клапанов.
Клапаны лимфатических сосудов препятствуют обратному току лимфы.
Благодаря им при сокращении стенки лимфатического сосуда лимфа течет
только в центрипетальном направлении. Клапаны и стенка лимфатическо-
го сосуда в структурном и функциональном отношении составляют единое
целое. Клапаны имеют створки — парные, расположенные друг против
друга полулунные складки интимы. Створки имеют два края, один из них
прикреплен к стенке сосуда в месте его сужения, другой — свободно свиса-
ет в просвет сосуда.
Часть лимфатического сосуда между двумя клапанами называется лим-
фангион, или клапанный сегмент. В лимфангионе различают мышцесодер-
жащую часть, или мышечную манжетку, и область прикрепления клапана,
в которой мускулатура развита слабо или отсутствует.
Поскольку средняя и адвентициальная оболочки не имеют нервных
окончаний, активность лимфангионов считается миогенной. Лимфатиче-
ским сосудам свойственны: фазные ритмические сокращения, медленные
волны, тонус. Фазное ритмическое сокращение представляет собой быстрое
сужение отдельного участка сосуда, сменяемое быстрым расслаблением.
Эта активность может быть спонтанной или вызванной (растяжением, по-
вышением температуры, гуморальными воздействиями). Фазные ритмиче-
ские сокращения следуют с частотой 10—20 в 1 мин.
Медленные волны представляют собой колебания просвета сосуда неоди-
наковой продолжительности и амплитуды. Продолжительность медленной
волны может составлять от 2 до 5 мин. Эти волны лимфатических сосудов
непостоянны, появляются спонтанно или в ответ на действие вазоактив-
ных веществ.
Тонус лимфатических сосудов является отражением активности их тони-
ческих клеток, модулируемой местными, гуморальными или нервными
факторами. В естественных условиях тонус гладких мышц лимфатических
сосудов обусловливает определенную жесткость их стенок, препятствуя пе-
рерастяжению последних, создает исходный фон для фазных сокращений,
поддерживает внутрисосудистое давление, необходимое для реализации
фазной активности. Изменения тонуса лимфатических сосудов лежат в ос-
нове регуляции объема лимфатической системы.
9.2.2. Лимфатические узлы
На объем и состав лимфы влияет наличие в системе лимфатических узлов.
У человека их примерно 460. Функции лимфатических узлов — гемопоэти-
ческая, иммунопоэтическая, защитно-фильтрационная, обменная, резерву-
арная, пропульсивная. Лимфатические узлы играют роль не только меха-
нического, но и биологического фильтра, задерживающего поступление в
466 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
кровь инородных частиц, бактерий, клеток злокачественных опухолей,
токсинов, чужеродных белков. К узлу обычно подходят 2—4 приносящих
сосуда, а выходят 1—2 выносящих, поэтому количество послеузловой лим-
фы в среднем в 3 раза меньше, чем доузловой. Сопротивление узлов току
перфузата в несколько раз больше, чем в лимфатических сосудах; возмож-
но, поэтому входящие в узел лимфатические сосуды обладают более мощ-
ным слоем гладкомышечных клеток, чем выходящие. Роль узла заключает-
ся не только в создании сопротивления току лимфы и замедления ее дви-
жения, но и в депонировании лимфы, перераспределении жидкости между
кровью и лимфой. При возникновении венозного застоя лимфатические
узлы увеличиваются на 40—50 %.
Лимфатические узлы содержат гладкомышечные элементы и могут по-
этому сокращаться при нейрогуморальных или местных влияниях: они об-
ладают сходной с лимфатическими сосудами сократительной активностью.
Лимфатические узлы, кровоснабжение которых очень обильно, содер-
жат фагоцитарные клетки, разрушающие чужеродные вещества, которые
поступают в узлы с лимфой. Они также вырабатывают лимфоциты и плаз-
матические клетки и синтезируют антитела. Лимфатические узлы играют
роль мощных защитных барьеров против бактерий, попадающих в орга-
низм.
9.2.3. Лимфоток
Содержимое двух больших терминальных каналов — правого и левого
грудных протоков — поступает, соответственно, в правую и левую подклю-
чичные вены у их соединения с яремными венами. Скорость тока лимфы
варьирует в широких пределах в различных магистральных и органных
лимфатических сосудах. Так, лимфоток в грудном протоке собаки состав-
ляет 0,03—0,4 мл/мин, в кишечнике — 0,06—0,2 мл/мин, в печени — 0,01 —
0,2 мл/мин, в миокарде — 0,01—0,6 мл/мин, в почке и конечности — 0,01—
0,7 мл/мин. У человека лимфоток в грудном протоке колеблется от 0,4 до
1,3 мл/кг/мин.
Скорость движения лимфы по сосудам зависит от ряда факторов: вне-
сосудистых — сокращений скелетных мышц, перистальтики кишечника,
дыхательных экскурсий грудной клетки, пульсаций рядом лежащих арте-
рий; внутрисосудистых — сил vis a tergo, лимфообразования, сократитель-
ной активности стенок лимфатических сосудов.
Уменьшение онкотического давления белков плазмы или повышение
венозного (и, соответственно, капиллярного) давления ведет к увеличению
объема оттекающей лимфы. Любые водные нагрузки, вызывающие увели-
чение объема циркулирующей крови, приводят к увеличению лимфотока.
Повышение венозного давления в портальной системе на 1—2 см водн. ст.
увеличивает лимфоток из печени в 10—12 раз.
9.2.4. Нервные и гуморальные влияния
Существование лимфатических сосудов только у позвоночных является
свидетельством наибольшего совершенства их интерстиция, а сам аппарат
лимфатических сосудов — наисовершеннейшей системой интерстиция, ко-
торая не обязательна для примитивной жизни тканей (она отсутствует у
беспозвоночных).
Нервные волокна достигают лимфатических сосудов по ходу vasa vaso-
rum; степень развития гладких мышц лимфатических сосудов коррелирует
9. Функции систем кровообращения и лимфообращения • 467
с их иннервацией. Там, где мускулатура стенки лимфатических сосудов
достигает большей степени развития, она содержит мышечные и адвенти-
циальные вегетативные нервные сплетения.
Лимфатические сосуды снабжены адренергическими и холинергически-
ми нервными волокнами. Эти волокна из периадвентициальных сплетений
артерий распространяются на сопровождающие их вены и близлежащие
лимфатические сосуды, образуя из них сплетения. Интенсивность иннер-
вации лимфатических сосудов в 2—2,5 раза слабее, чем артерий. Адренер-
гические и холинергические волокна концентрируются в местах перехода
лимфатических сосудов малого диаметра в более крупные, а также в местах
расположения клапанов.
Возбуждение симпатических нервов вызывает сокращение лимфатиче-
ских сосудов, возбуждение парасимпатических — как сокращение, так и
расслабление (зависит от исходного тонуса и ритмической активности со-
суда). Грудной проток, брыжеечные лимфатические сосуды имеют двой-
ную иннервацию — симпатическую и парасимпатическую (волокна блуж-
дающего нерва), в то время как крупные лимфатические сосуды конечно-
стей иннервируются симпатическим отделом нервной системы. В магист-
ральных и периферических лимфатических сосудах учащение ритма фаз-
ных сокращений достигается активацией а-адренорецепторов. Торможе-
ние ритма спонтанных сокращений лимфатических сосудов осуществляет-
ся двойным тормозным механизмом: посредством выделения АТФ и путем
активации p-адренорецепторов.
Адреналин вызывает усиление тока лимфы и повышение давления в
грудном протоке, увеличивает частоту и амплитуду спонтанных сокраще-
ний лимфатических сосудов брыжейки. Внутривенное введение гистамина
увеличивает ток лимфы в грудном протоке и кишечном стволе, не только
влияя на моторику сосудов, но и повышая проницаемость лимфатических
капилляров. Малые концентрации гистамина стимулируют спонтанный
ритм и повышают тонус лимфатических сосудов, высокие концентрации —
тормозят фазную сократительную активность и увеличивают тоническое
сокращение. Аналогично действует на лимфатические сосуды гепарин.
АТФ тормозит ритмические сокращения грудного протока и брыжеечных
лимфатических сосудов. Серотонин вызывает сокращение просвета груд-
ного протока и брыжеечных сосудов, величина их сокращения зависит от
дозы серотонина. Спонтанные ритмические сокращения лимфатических
сосудов прекращаются в бескальциевой среде или при блокаде кальциевых
каналов. В отличие от фазных, тонические сокращения этих сосудов в ана-
логичных условиях практически не уменьшаются. В низких концентрациях
ионы калия вызывают увеличение частоты фазных сокращений лимфати-
ческих сосудов, в более высоких — увеличение тонических сокращений.
Гиперкальциевый раствор увеличивает амплитуду спонтанных сокраще-
ний, гипокальциевый — подавляет амплитуду и ритм фазных сокращений.
Уменьшение в среде ионов натрия вызывает увеличение частоты сокраще-
ний и снижение амплитуды спонтанных фазных сокращений лимфатиче-
ских сосудов. Наркоз подавляет их ритмическую активность.
Лимфу в экспериментальных исследованиях на животных или в клини-
ческой лимфологии у человека забирают не из капилляров, что технически
очень сложно или невозможно, а из крупных доузловых или послеузловых
сосудов грудного протока, через который проходит лимфа, дренируемая от
3/4 тела. Лимфа из лимфатических сосудов существенно отличается от ка-
пиллярной в силу проницаемости стенки лимфатических сосудов для низ-
комолекулярных веществ и воды. Проницаемость лимфатических сосудов
468 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
уменьшается от периферии к центру, что связано с наличием у них базаль-
ной мембраны и гладкой мускулатуры, увеличением количества эластиче-
ских и коллагеновых волокон, уплотнением межэндотелиальных щелей.
Итак, основной движущей силой лимфотока является собственная со-
кратительная активность лимфатических сосудов и узлов. При этом сред-
нее внутрилимфатическое давление колеблется в небольших пределах и
обусловлено фазами сокращений лимфатических ососудов по типу пери-
стальтической волны. Нейрогуморальные влияния, физические и фармако-
логические воздействия влияют на сократительную активность лимфатиче-
ских сосудов и узлов, на их емкость и пропускную способность, а также на
обменные процессы в лимфатических узлах.
ГЛАВА 10
Функции дыхательной системы
Дыхание в организме человека и животных представляет собой процесс ис-
пользования кислорода клетками тканей в биологическом окислении с об-
разованием энергии и конечного продукта дыхания — углекислого газа.
Дыхательная система человека обеспечивает газообмен между атмо-
сферным воздухом и легкими, в результате которого кислород из легких
поступает в кровь и переносится кровью к тканям организма, а углекис-
лый газ транспортируется от тканей в противоположном направлении.
В состоянии покоя тканями организма взрослого человека потребляется
примерно 0,3 л кислорода в 1 мин и в них образуется несколько меньшее
количество углекислого газа. Отношение количества образующегося в его
тканях СО2 к потребляемому организмом количеству О2 называется дыха-
тельным коэффициентом, величина которого в обычных условиях равна
0,9. Поддержание нормального уровня газового гомеостазиса О2 и СО2 ор-
ганизма в соответствии со скоростью тканевого метаболизма (дыхания) яв-
ляется основной функцией дыхательной системы организма человека. Эта
система состоит из единого комплекса костной, хрящевой, соединитель-
ной и мышечной тканей грудной клетки, дыхательных путей (воздухонос-
ный отдел легких), обеспечивающих движение воздуха между внешней
средой и воздушным пространством альвеол, а также легочной ткани (рес-
пираторный отдел легких), которая обладает высокой эластичностью и
растяжимостью. В состав дыхательной системы входит собственный нерв-
ный аппарат, управляющий дыхательными мышцами грудной клетки, чув-
ствительные и двигательные волокна нейронов вегетативной нервной сис-
темы, имеющие терминали в тканях органов дыхания.
Местом газообмена между организмом человека и внешней средой яв-
ляются альвеолы легких, общая площадь которых достигает в среднем
100 м2. Альвеолы (порядка 3 • 108) находятся на конце мелких дыхательных
путей легких, имеют диаметр примерно 0,3 мм и плотно контактируют с
легочными капиллярами. Циркуляция крови между клетками тканей орга-
низма человека, потребляющих О2 и продуцирующих СО2, и легкими, где
эти газы обмениваются с атмосферным воздухом, осуществляется систе-
мой кровообращения.
В организме человека дыхательная система выполняет дыхательную и не-
дыхательную функции. Дыхательная функция системы поддерживает газо-
вый гомеостазис внутренней среды организма в соответствии с уровнем ме-
таболизма его тканей. С вдыхаемым воздухом в легкие попадают микрочас-
тицы пыли, которые задерживаются слизистой оболочкой дыхательных пу-
тей и затем удаляются из легких с помощью защитных рефлексов (кашель,
чиханье) и механизмов мукоцилиарного очищения (защитная функция).
Недыхательные функции системы обусловлены такими процессами, как
синтез (сурфактанта, гепарина, лейкотриенов, простагландинов), актива-
ция (ангиотензина II) и инактивация (серотонина, простагландинов, нор-
адреналина) биологически активных веществ, при участии альвеолоцитов,
470 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
тучных клеток и эндотелия капилляров легких (метаболическая функция).
Эпителий слизистой оболочки дыхательных путей содержит иммунокомпе-
тентные клетки (Т- и В-лимфоциты, макрофаги) и тучные клетки (синтез
гистамина), обеспечивающие защитную функцию организма. Через легкие
из организма выводятся с выдыхаемым воздухом пары воды и молекулы
летучих веществ (выделительная функция), а также незначительная часть
тепла из организма (терморегулирующая функция). Дыхательные мышцы
грудной клетки участвуют в поддержании положения тела в пространстве
(позно-тоническая функция). Наконец, нервный аппарат дыхательной сис-
темы, мышцы голосовой щели и верхних дыхательных путей, а также
мышцы грудной клетки участвуют в речевой деятельности человека (функ-
ция речеобразования).
Основная дыхательная функция системы дыхания реализуется в процес-
сах внешнего дыхания, которые представляют собой обмен газов (О2, СО2
и N2) между альвеолами и внешней средой, диффузию газов (О2 и СО2) ме-
жду альвеолами легких и кровью (газообмен). Наряду с внешним дыхани-
ем в организме осуществляется транспорт дыхательных газов кровью, а
также газообмен О2 и СО2 между кровью и тканями, который называется
нередко внутренним (тканевым) дыханием.
10.1. Внешнее дыхание
Газообмен между атмосферным воздухом и альвеолярным пространством
легких происходит в результате циклических изменений объема легких в
течение фаз дыхательного цикла. В фазу вдоха объем легких увеличивает-
ся, воздух из внешней среды поступает в дыхательные пути и затем дости-
гает альвеол. Напротив, в фазу выдоха происходит уменьшение объема лег-
ких и воздух из альвеол через дыхательные пути выходит во внешнюю сре-
ду. Увеличение и уменьшение объема легких обусловлены биомеханиче-
скими процессами изменения объема грудной полости при вдохе и выдохе.
10.1.1. Биомеханика дыхания
10.1.1.1. Биомеханика вдоха
Увеличение объема грудной полости при вдохе происходит в результате со-
кращения инспираторных мышц: диафрагмы и наружных межреберных.
Основной дыхательной мышцей является диафрагма, которая находится в
нижней трети грудной полости и разделяет
грудную и брюшную полости. При сокраще-
нии диафрагмальной мышцы диафрагма
движется вниз и смещает органы брюшной
полости вниз и кпереди, увеличивая объем
грудной полости преимущественно по вер-
тикали (рис. 10.1).
Рис. 10.1. Влияние сокращения диафрагмальной
мышцы на объем грудной полости. Сокращение
диафрагмальной мышцы при вдохе (пунктирная
линия) вызывает опускание диафрагмы вниз,
смещение органов брюшной полости вниз и впе-
ред. В результате увеличивается объем грудной
полости.
10. Функции дыхательной системы • 471
Увеличению объема грудной
полости при вдохе способствует
сокращение наружных межребер-
ных мышц, которые поднимают
грудную клетку вверх, увеличи-
вая объем грудной полости. Этот
эффект сокращения наружных
межреберных мышц обусловлен
особенностями прикрепления
мышечных волокон к ребрам —
волокна идут сверху вниз и сзади
кпереди (рис. 10.2). При подоб-
ном направлении мышечных во-
локон наружных межреберных
мышц их сокращение поворачи-
вает каждое ребро вокруг оси,
проходящей через точки сочлене-
ния головки ребра с телом и по-
перечным отростком позвонка. В
результате этого движения каждая
нижележащая реберная дуга под-
мышцы оси вращения
а реберной дуги б
Рис. 10.2. Направление волокон наруж-
ных межреберных мышц и увеличение
объема грудной полости при вдохе.
а — сокращение наружных межреберных мышц
при вдохе поднимает нижнее ребро больше,
чем опускает вниз верхнее. В результате ребер-
ные дуги поднимаются вверх и увеличивают (б)
объем грудной полости в сагиттальной и фрон-
тальной плоскости.
нимается вверх больше, чем опус-
кается вышерасположенная. Одновременное движение вверх всех ребер-
ных дуг приводит к тому, что грудина поднимается вверх и кпереди, а объ-
ем грудной клетки увеличивается в сагиттальной и фронтальной плоско-
стях. Сокращение наружных межреберных мышц не только увеличивает
объем грудной полости, но и препятствует опусканию грудной клетки
вниз. Например, у детей, имеющих неразвитые межреберные мышцы,
грудная клетка уменьшается в размере во время сокращения диафрагмы
(парадоксальное движение).
При глубоком дыхании в биомеханизме вдоха, как правило, участвует
вспомогательная дыхательная мускулатура — грудино-ключично-сосцевидные
и передние лестничные мышцы, и их сокращение дополнительно увеличи-
вает объем грудной клетки. В частности, лестничные мышцы поднимают
верхние два ребра, а грудино-ключично-сосцевидные — поднимают груди-
ну. Вдох является активным процессом и требует расхода энергии при со-
кращении инспираторных мышц, которая затрачивается на преодоление
эластического сопротивления относительно ригидных тканей грудной
клетки, эластического сопротивления легко растяжимой легочной ткани,
аэродинамического сопротивления дыхательных путей потоку воздуха, а
также на повышение внутриабдоминального давления и возникающего
при этом смещения органов брюшной полости книзу.
10.1.1.2. Биомеханизм выдоха
Выдох в покое у человека осуществляется пассивно под действием эласти-
ческой тяги легких, которая возвращает объем легких к исходной величине.
Тем не менее при глубоком дыхании, а также при кашле и чиханье, выдох
может быть активным, и уменьшение объема грудной полости происходит
за счет сокращения внутренних межреберных мышц и мышц живота. Мы-
шечные волокна внутренних межреберных мышц идут относительно точек
их прикрепления к ребрам снизу вверх и сзади кпереди. При их сокраще-
нии ребра поворачиваются вокруг оси, проходящей через точки их сочлене-
472 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ния с позвонком, и каждая вышерасположенная реберная дуга опускается
вниз больше, чем нижерасположенная поднимается вверх. В результате все
реберные дуги вместе с грудиной опускаются вниз, уменьшая объем груд-
ной полости в сагиттальной и фронтальной плоскостях. При глубоком ды-
хании человека сокращение мышц живота в фазу выдоха увеличивает дав-
ление в брюшной полости, что способствует смещению купола диафрагмы
вверх и уменьшает объем грудной полости в вертикальном направлении.
10.1.2. Изменение объема легких во время вдоха и выдоха
10.1.2.1. Функция внутриплеврального давления
Сокращение дыхательных мышц грудной клетки и диафрагмы при вдохе
вызывает увеличение объема легких, а при их расслаблении во время вы-
доха легкие спадаются до исходного объема. Объем легких как при вдохе,
так и при выдохе изменяется пассивно, поскольку благодаря своей высо-
кой эластичности и растяжимости легкие следуют за изменениями объема
грудной полости, вызванными сокращением дыхательных мышц. Это по-
ложение иллюстрирует следующая модель пассивного увеличения объема
легких (рис. 10.3). В этой модели легкие могут быть рассмотрены в качест-
ве эластичного баллона, помещенного внутрь емкости, выполненной из
ригидных стенок и гибкой диафрагмы. Пространство между эластичным
баллоном и стенками емкости является герметичным. Эта модель позволя-
ет изменять давление внутри емкости при движении вниз гибкой диафраг-
мы. При увеличении объема емкости, вызванном движением вниз гибкой
диафрагмы, давление внутри емкости, т. е. вне баллона, становится ниже
атмосферного в соответствии с законом идеального газа. Баллон раздувает-
ся, поскольку давление внутри него
давление в емкости вокруг баллона.
В приложении к легким человека,
грудной полости, их поверхность и
Рис. 10.3. Схема модели, демонстри-
рующей пассивное раздувание легких
при опускании диафрагмы. При опус-
кании вниз диафрагмы давление возду-
ха внутри емкости становится ниже ат-
мосферного давления, что вызывает
раздувание эластичного баллона.
Р — атмосферное давление.
(атмосферное) становится выше, чем
которые полностью заполняют объем
внутренняя поверхность грудной по-
лости покрыты плевральной мем-
браной. Плевральная мембрана по-
верхности легких (висцеральная
плевра) физически не соприкасает-
ся с плевральной мембраной, по-
крывающей грудную стенку (парие-
тальная плевра), так как между эти-
ми мембранами имеется плевральное
пространство (синоним — внутри-
плевральное пространство), запол-
ненное тонким слоем жидкости —
плевральной жидкости. Эта жид-
кость увлажняет поверхность долей
легких и способствует их скольже-
нию относительно друг друга во
время раздувания легких, а также
облегчает трение между париеталь-
ным и висцеральным листками
плевры. Жидкость несжимаема и ее
объем не увеличивается при умень-
шении давления в плевральной по-
лости. Поэтому высокоэластичные
10. Функции дыхательной системы • 473
Рис. 10.4. Давление в альвеолах и внутриплевральное давление в фазу вдоха и выдо-
ха дыхательного цикла.
В отсутствии потока воздуха в дыхательных путях давление в них равно атмосферному (А), а
эластическая тяга легких создает в альвеолах давление Е. В этих условиях величина внутри-
плеврального давления равна разнице А — Е. При вдохе сокращение диафрагмы увеличивает
величину отрицательного давления в плевральной полости до —10 см водн. ст., которое спо-
собствует преодолению сопротивления потоку воздуха в дыхательных путях, и воздух движется
из внешней среды в альвеолы. Величина внутриплеврального давления обусловлена разницей
между давлениями А — R — Е. При выдохе диафрагма расслабляется и внутриплевральное дав-
ление становится менее отрицательным относительно атмосферного давления (—5 см
водн. ст.). Альвеолы вследствие своей эластичности уменьшают свой диаметр, в них повыша-
ется давление Е. Градиент давлений между альвеолами и внешней средой сопособствует выве-
дению воздуха из альвеол по дыхательным путям во внешнюю среду. Величина внутриплев-
рального давленния обусловлена суммой A+R за вычетом давления внутри альвеол, т. е.
А + R — Е. А — атмосферное давление, Е —давление в альвеолах, возникающее вследствие
эластической тяги легких, R — давление, обеспечивающее преодоление сопротивления потоку
воздуха в дыхательных путях, Р — внутриплевральное давление.
легкие в точности повторяют изменение объема грудной полости во время
вдоха. Бронхи, кровеносные сосуды, нервы и лимфатические сосуды фор-
мируют корень легкого, с помощью которого легкие фиксированы в облас-
ти средостения. Механические свойства этих тканей обусловливают основ-
ную степень усилия, которое должны развивать дыхательные мышцы при
сокращении, чтобы вызывать увеличение объема легких. В обычных усло-
виях эластическая тяга легких создает незначительную величину отрица-
тельного давления в тонком слое жидкости внутриплеврального простран-
ства относительно атмосферного давления. Отрицательное внутриплев-
ральное давление варьирует в соответствии с фазами дыхательного цикла
от —5 (выдох) до —10 см водн. ст. (вдох) ниже атмосферного давления
(рис. 10.4). Отрицательное внутриплевральное давление способно вызвать
уменьшение (коллапс) объема грудной полости, которому ткани грудной
клетки противодействуют своей чрезвычайно ригидной структурой. Диа-
фрагма по сравнению с грудной клеткой, является более эластичной, и ее
купол поднимается вверх под влиянием градиента давления, существующе-
го между плевральной и брюшной полостями.
В состоянии, когда легкие не расширяются и не спадаются (пауза соот-
ветственно после вдоха или выдоха), в дыхательных путях отсутствует по-
ток воздуха и давление в альвеолах равно атмосферному. В этом случае
градиент между атмосферным и внутриплевральным давлением будет точ-
но уравновешивать давление, развиваемое эластической тягой легких (см.
рис. 10.4). В этих условиях величина внутриплеврального давления равна
474 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
разности между давлением в дыхательных путях и давлением, развиваемым
эластической тягой легких. Поэтому чем больше растянуты легкие, тем
сильнее будет эластическая тяга легких и более отрицательным относи-
тельно атмосферного является величина внутриплеврального давления.
Так происходит во время вдоха, когда диафрагма опускается вниз и эла-
стическая тяга легких противодействует раздуванию легких, а величина
внутриплеврального давления становится более отрицательной. При вдохе
это отрицательное давление способствует продвижению воздуха по дыха-
тельным путям в сторону альвеол, преодолевая сопротивление дыхатель-
ных путей. В результате воздух поступает из внешней среды в альвеолы.
При выдохе диафрагма расслабляется и величина внутриплеврального
давления становится менее отрицательной. В этих условиях альвеолы в
связи с высокой эластичностью их стенок начинают уменьшаться в разме-
ре и выталкивают воздух из легких через дыхательные пути. Сопротивле-
ние дыхательных путей потоку воздуха поддерживает положительное дав-
ление в альвеолах и препятствует их быстрому спадению. Таким образом, в
спокойном состоянии при выдохе поток воздуха в дыхательных путях обу-
словлен только эластической тягой легких.
Пневмоторакс. Если воздух входит во внутриплевральное пространство,
например через раневое отверстие, в легких возникает коллапс, грудная
клетка незначительно увеличивается в объеме, а диафрагма опускается
вниз, как только внутриплевральное давление становится равным атмо-
сферному давлению. Это состояние называется пневмотораксом, при кото-
ром легкие утрачивают способность следовать за изменением объема груд-
ной полости во время дыхательных движений. Более того, во время вдоха
воздух через раневое отверстие входит в грудную полость и выходит во
время выдоха без изменения объема легких во время дыхательных движе-
ний, что делает невозможным газообмен между внешней средой и орга-
низмом.
10.1.2.2. Легочные объемы воздуха в течение фаз дыхательного цикла
Процесс внешнего дыхания обусловлен изменением объема воздуха в лег-
ких в течение фаз вдоха и выдоха дыхательного цикла. При спокойном ды-
хании соотношение длительности вдоха к выдоху в дыхательном цикле
равняется в среднем 1:1,3. Внешнее дыхание человека характеризуется час-
тотой и глубиной дыхательных движений. Частота дыхания человека из-
меряется количеством дыхательных циклов в течение 1 мин и ее величина
в покое у взрослого человека варьирует от 12 до 20 в 1 мин. Этот показа-
тель внешнего дыхания возрастает при физической работе, повышении
температуры окружающей среды, а также изменяется с возрастом. Напри-
мер, у новорожденных частота дыхания равна 60—70 в 1 мин, а у людей в
возрасте 25—30 лет — в среднем 16 в 1 мин. Глубина дыхания определяется
по объему вдыхаемого и выдыхаемого воздуха в течение одного дыхатель-
ного цикла. Произведение частоты дыхательных движений на их глубину
характеризует основную величину внешнего дыхания — вентиляцию лег-
ких. Количественной мерой вентиляции легких является минутный объем
дыхания — это объем воздуха, который человек вдыхает и выдыхает за 1
мин. Величина минутного объема дыхания человека в покое варьирует в
пределах 6—8 л. При физической работе у человека минутный объем дыха-
ния может возрастать в 7—10 раз.
Легочные объемы воздуха. В физиологии дыхания принята единая но-
менклатура легочных объемов у человека, которые заполняют легкие при
10. Функции дыхательной системы • 475
Сопрограмма
Глубокий
вдох
Резервный
объем вдоха
Спокойное
дыхание
Дыхательный
объем
Глубокий
выдох
_±_
Резервный
объем
выдоха
И
ФОЕ
о
5
ш
Остаточный объем
Рис. 10.5. Объемы и емкости воздуха в лег-
ких человека и кривая (спирограмма) изме-
нения объема воздуха в легких при спокой-
ном дыхании, глубоком вдохе и выдохе.
ФОЕ — функциональная остаточная емкость.
спокойном и глубоком дыхании
в фазу вдоха и выдоха дыха-
тельного цикла (рис. 10.5). Ле-
гочный объем, который вдыха-
ется или выдыхается челове-
ком при спокойном дыхании,
называется дыхательным объе-
мом. Его величина при спокой-
ном дыхании составляет в сред-
нем 500 мл. Максимальное ко-
личество воздуха, которое мо-
жет вдохнуть человек сверх ды-
хательного объема, называется
резервным объемом вдоха (в
среднем 3000 мл). Максималь-
ное количество воздуха, кото-
рое может выдохнуть человек
после спокойного выдоха, на-
зывается резервным объемом вы-
доха (в среднем 1100 мл). Нако-
нец, количество воздуха, кото-
рое остается в легких после
максимального выдоха, называ-
ется остаточным объемом, его
величина равна примерно
1200 мл.
Сумма величин двух легочных объемов и более называется легочной ем-
костью. Объем воздуха в легких человека характеризуется инспираторной
емкостью легких, жизненной емкостью легких и функциональной остаточ-
ной емкостью легких. Инспираторная емкость легких (3500 мл) представля-
ет собой сумму дыхательного объема и резервного объема вдоха. Жизненная
емкость легких (4600 мл) включает в себя дыхательный объем и резервные
объемы вдоха и выдоха. Функциональная остаточная емкость легких
(1600 мл) представляет собой сумму резервного объема выдоха и остаточ-
ного объема легких. Сумма жизненной емкости легких и остаточного объе-
ма называется общей емкостью легких, величина которой у человека в сред-
нем равна 5700 мл.
При вдохе легкие человека за счет сокращения диафрагмы и наружных
межреберных мышц начинают увеличивать свой объем с уровня функцио-
нальной остаточной емкости, и его величина при спокойном дыхании со-
ставляет дыхательный объем, а при глубоком дыхании — достигает различ-
ных величин резервного объема вдоха. При выдохе объем легких вновь
возвращается к исходному уровню функциональной остаточной емкости
пассивно, за счет эластической тяги легких. Если в объем выдыхаемого
воздуха начинает входит воздух функциональной остаточной емкости, что
имеет место при глубоком дыхании, а также при кашле или чиханье, то
выдох осуществляться за счет сокращения мышц брюшной стенки. В этом
случае величина внутриплеврального давления, как правило, становится
выше атмосферного давления, что обусловливает наибольшую скорость
потока воздуха в дыхательных путях.
т
476 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
10.1.3. Факторы, влияющие на легочный объем в фазу вдоха
При вдохе увеличению объема грудной полости препятствуют эластическая
тяга легких, движение ригидной грудной клетки, органы брюшной полости
и, наконец, сопротивление дыхательных путей движению воздуха в на-
правлении альвеол. Первый фактор, а именно эластическая тяга легких, в
наибольшей степени препятствует увеличению объема легких во время ин-
спирации.
10.1.3.1. Растяжимость легочной ткани
В нормальных физиологических условиях глубина вдоха может быть огра-
ничена только физическими свойствами легочной ткани и грудной клетки.
Сопротивление раздуванию легких, которое возникает при поступлении в
них воздуха, обусловлено растяжимостью их соединительной ткани и со-
противлением дыхательных путей потоку воздуха. Мерой эластических
свойств легочной ткани является растяжимость легких, которая характери-
зует степень увеличения объема легких в зависимости от степени уменьше-
ния внутриплеврального давления:
С = AV/ДР,
где С — растяжимость (англ. — compliance), AV — изменение легочного объ-
ема (мл), а ДР — изменение внутриплеврального давления (см водн. ст.).
Растяжимость характеризует количественно степень изменения объема лег-
ких у человека в зависимости от степени изменения при вдохе внутриплев-
рального давления. Грудная клетка также обладает эластическими свойства-
ми, поэтому растяжимость тканей легких и тканей грудной клетки обуслов-
ливает эластические свойства всего аппарата внешнего дыхания человека.
На рис. 10.6 показано изменение легочного объема, которое возникает
при изменениях внутриплеврального давления. Восходящая и нисходящая
линии характеризуют раздувание и спадение легких соответственно. Фраг-
мент рис. 10.6 слева демонстрирует, каким образом могут быть измерены
значения легочного объема и внутриплеврального давления, вынесенные
на график. Объем легких не уменьшается до нуля, когда величина внутри-
плеврального давления становится равной нулю. Раздувание легких с уров-
ня их минимального объема требует усилия для того, чтобы раскрыть
спавшиеся стенки альвеол в силу значительного поверхностного натяже-
ния жидкости, покрывающей как их поверхность, так и поверхность дыха-
тельных путей. Поэтому кривые, полученные при раздувании и спадении
легких, не совпадают друг с другом, и их нелинейное изменение называет-
ся гистерезисом.
Легкие при спокойном дыхании никогда не спадаются полностью, по-
этому кривая спадения описывает изменения легочного объема при изме-
нении величины внутриплеврального давления в диапазоне от —2 см. водн.
ст. до —10 см водн. ст. В норме легкие человека имеют высокую растяжи-
мость (200 мл/см водн. ст.). Эластичность легочной ткани обусловлена
свойствами соединительных волокон легочной ткани. С возрастом эти во-
локна, как правило, снижают тонус, что сопровождается увеличением рас-
тяжимости и уменьшением эластической тяги легких. При повреждении
легочной ткани или при избыточном развитии в ней соединительной тка-
ни (фиброз) легкие становятся плохо растяжимыми, величина их растяжи-
мости снижается, что затрудняет выполнение вдоха и требует значительно
большего усилия дыхательных мышц, чем в норме.
10. Функции дыхательной системы • 477
клетка
Внутриплевральное
давление (см водн. ст.)
Рис. 10.6. Кривая комплайенса легких человека.
Кривые справа показывают изменение дыхательного объема легких и общей емкости легких,
возникающие при изменениях внутриплеврального давления без учета влияния тканей груд-
ной клетки. Легкие полностью не спадаются, если внутриплевральное давление становится
равным нулю (точка 1). Кривые комплайенса совпадают в точке 2 при большом объеме в лег-
ких, когда легочная ткань достигает предела эластического растяжения. Вд — внутриплевраль-
ное давление. Слева — схема регистрации изменений внутриплеврального давления и дыха-
тельного объема легких.
Легочная растяжимость обусловлена не только эластичностью легочной
ткани, но и поверхностным натяжением слоя жидкости, покрывающей
альвеолы. По сравнению с эластической тягой легких, влияние на величи-
ну легочной растяжимости во время дыхания фактора поверхностного на-
тяжения слоя жидкости, покрывающей альвеолы, имеет более сложную
природу.
10.1.3.2. Поверхностное натяжение слоя жидкости в альвеолах
Тонкий слой жидкости покрывает поверхность альвеол легких. Переходная
граница между воздушной средой и жидкостью имеет поверхностное натя-
жение, которое формируется межмолекулярными силами и которое будет
уменьшать площадь покрываемой молекулами поверхности. Однако мил-
лионы альвеол легких, покрытых мономолекулярным слоем жидкости, не
спадаются, поскольку эта жидкость содержит субстанции, которые в целом
называются сурфактантом (поверхностно активный агент). Поверхностно
активные агенты обладают свойством снижать поверхностное натяжение
слоя жидкости в альвеолах легких на границе фаз воздух—жидкость, благо-
даря которому легкие становятся легко растяжимыми.
Альвеолярный эпителий состоит из плотно контактирующих между со-
бой альвеолоцитов (пневмоцитов) I и II типа и покрыт мономолекулярным
слоем сурфактанта, состоящего из фосфолипидов, белков и полисахари-
дов (глицерофосфолипиды 80 %, глицерол 10 %, белки 10 %). Синтез сур-
фактанта осуществляется альвеолоцитами 11 типа из компонентов плазмы
крови. Основным компонентом сурфактанта является дипальмитоилфос-
фатидилхолин (более 50 % фосфолипидов сурфактанта), который адсорби-
руется на границе фаз жидкость—воздух с помощью белков сурфактанта
SP-B и SP-C. Эти белки и глицерофосфолипиды уменьшают поверхност-
ное натяжение слоя жидкости в миллионах альвеол и обеспечивают легоч-
478 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 10.7. Приложение закона Лапласа к изменению
поверхностного натяжения слоя жидкости, покрываю-
щего поверхность альвеол.
Изменение радиуса альвеол изменяет в прямой зависимости
величину поверхностного натяжения в альвеолах (Т). Давле-
ние (Р) внутри альвеол также варьирует при изменении их ра-
диуса: уменьшается при вдохе и увеличивается при выдохе.
ной ткани свойство высокой растяжимости. По-
верхностное натяжение слоя жидкости, покры-
вающей альвеолы, изменяется в прямой зависи-
мости от их радиуса (рис. 10.7). В легких сурфактант изменяет степень по-
верхностного натяжения поверхностного слоя жидкости в альвеолах при
изменении их площади. Это обусловлено тем, что во время дыхательных
движений количество сурфактанта в альвеолах остается постоянным. По-
этому при растяжении альвеол во время вдоха слой сурфактанта становит-
ся тоньше, что вызывает снижение его действия на поверхностное натяже-
ние в альвеолах. При уменьшении объема альвеол во время выдоха моле-
кулы сурфактанта начинают более плотно прилегать друг к другу и, увели-
чивая поверхностное давление, снижают поверхностное натяжение на гра-
нице фаз воздух—жидкость. Это препятствует спадению (коллапсу) альвеол
во время экспирации, независимо от ее глубины. Сурфактант легких влия-
ет на поверхностное натяжение слоя жидкости в альвеолах в зависимости
не только от ее площади, но и от направления, в котором происходит из-
менение площади поверхностного слоя жидкости в альвеолах. Этот эффект
сурфактанта называется гистерезисом (рис. 10.8). Физиологический смысл
эффекта заключается в
следующем. При вдохе по
мере увеличения объема
легких под влиянием сур-
фактанта увеличивается
натяжение поверхностно-
го слоя жидкости в альве-
олах, что препятствует
растяжению легочной
ткани и ограничивает глу-
бину инспирации. Напро-
тив, при выдохе поверх-
ностное натяжение жид-
кости в альвеолах под
влиянием сурфактанта
Внутриплевральное давление (см водн. ст.)
Рис. 10.8. Эффект поверхностного натяжения слоя
жидкости на изменение объема легких в зависимо-
сти от внутриплеврального давления при раздува-
нии легких солевым раствором и воздухом. Когда
объем легких увеличивается за счет их наполнения
солевым раствором, то в них отсутствуют поверх-
ностное натяжение и феномен гистерезиса. Отно-
сительно интактных легких — площадь петли гис-
терезиса свидетельствует об увеличении поверхно-
уменыпается, но не исче-
зает полностью. Поэтому
даже при самом глубо-
ком выдохе в легких не
происходит спадения,
т. е. коллапса альвеол.
В составе сурфактанта
имеются белки типа SP-A
и SP-D, благодаря кото-
рым сурфактант участву-
стного натяжения слоя жидкости в альвеолах при
вдохе и снижении этой величины при выдохе.
ют в местных иммунных
реакциях, опосредуя фа-
10. Функции дыхательной системы • 479
гоцитоз, поскольку на мембранах альвеолоцитов II типа и макрофагов
имеются рецепторы SP-A. Бактериостатическая активность сурфактанта
проявляется в том, что это вещество опсонизирует бактерии, которые за-
тем легче фагоцитируются альвеолярными макрофагами. Кроме того, сур-
фактант активирует макрофаги и влияет на скорость их миграции в альве-
олы из межальвеолярных перегородок. Сурфактант выполняет защитную
роль в легких, предотвращая непосредственный контакт альвеолярного
эпителия с частицами пыли, агентами инфекционного начала, которые
достигают альвеол с вдыхаемым воздухом. Сурфактант способен обволаки-
вать инородные частицы, которые затем транспортируются из респиратор-
ной зоны легкого в крупные дыхательные пути и удаляются из них со сли-
зью. Наконец, сурфактант снижает поверхностное натяжение в альвеолах
до близких к нулевым величинам и тем самым создает возможность рас-
правления легких при первом вдохе новорожденного.
10.1.3.3. Сопротивление дыхательных путей
Растяжимость легких количественно характеризует растяжимость легочной
ткани в любой момент изменения их объема в течение фазы вдоха и выдо-
ха. Поэтому растяжимость представляет собой статическую характеристику
эластических свойств легочной ткани. Однако во время дыхания возникает
сопротивление движению аппарата внешнего дыхания, обусловливающее
его динамические характеристики, среди которых наибольшее значение
имеет сопротивление потоку воздуха при его движении через дыхательные
пути легких. На движение воздуха из внешней среды через дыхательные
пути к альвеолам и в обратном направлении оказывает влияние градиент
давления: при этом воздух движется из области высокого давления в об-
ласть низкого давления. При вдохе давление воздуха в альвеолярном про-
странстве меньше, чем атмосферное, а при выдохе — наоборот. Сопротив-
ление дыхательных путей потоку воздуха зависит от градиента давления
между полостью рта и альвеолярным пространством. Поток воздуха через
дыхательные пути может быть ламинарным, турбулентным и переходным
между этими типами. Воздух движется в дыхательных путях, в основном,
ламинарным потоком, скорость которого выше в центре этих трубок и
меньше вблизи их стенок. При ламинарном потоке воздуха его скорость
линейно зависит от градиента давления вдоль дыхательных путей. В местах
деления дыхательных путей (бифуркации) ламинарный поток воздуха пе-
реходит в турбулентный. При возникновении турбулентного потока в ды-
хательных путях возникает дыхательный шум, который может выслуши-
ваться в легких с помощью стетоскопа. Сопротивление ламинарному пото-
ку газа в трубе обусловлено ее диаметром. Поэтому, согласно закону Пуа-
зейля величина сопротивления дыхательных путей потоку воздуха пропор-
циональна их диаметру, возведенному в четвертую степень. Поскольку со-
противление дыхательных путей находится в обратной зависимости от их
диаметра в четвертой степени, то этот показатель самым существенным
образом зависит от изменений диаметра воздухоносных путей, вызванных,
например, выделением в них слизи из слизистой оболочки или сужением
просвета бронхов. Общий диаметр сечения дыхательных путей возрастает в
направлении от трахеи к периферии легкого и становится максимально
большим в терминальных дыхательных путях, что вызывает резкое сниже-
ние сопротивления потоку воздуха и его скорости в этих отделах легких.
Так, линейная скорость потока вдыхаемого воздуха в трахее и главных
бронхах равна примерно 100 см/с. На границе воздухопроводящей и пере-
480 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ходной зон дыхательных путей линейная скорость воздушного потока со-
ставляет около 1 см/с, в дыхательных бронхах она снижается до 0,2 см/с, а
в альвеолярных ходах и мешочках — до 0,02 см/с. Столь низкая скорость
воздушного потока в альвеолярных ходах и мешочках обусловливает в них
незначительное сопротивление движущемуся воздуху и не сопровождается ।
значимыми затратами энергии мышечного сокращения. Напротив, наи-
большее сопротивление дыхательных путей потоку воздуха возникает на
уровне сегментарных бронхов в связи с наличием в их слизистой оболочке
секреторного эпителия и хорошо развитого гладкомышечного слоя, т. е.
факторов, которые в наибольшей степени влияют как на диаметр воздухо-
носных путей, так и на сопротивление в них потоку воздуха. В преодоле-
нии этого сопротивления заключается одна из функций дыхательных
мышц.
10.1.3.4. Зависимость «поток—объем» в легких
В легких большинство дыхательных путей представляют собой эластичные
трубки, за исключением трахеи и бронхов, стенки которых «укреплены»
хрящевой тканью. Бронхиолы имеют высокоэластичные стенки, и диаметр
их просвета может изменяться пассивно во время дыхательных движений.
В обычных физиологических условиях при вдохе (как спокойном, так и
глубоком) растяжение легочной ткани вызывает растяжение стенки мелких
дыхательных путей. Согласно закону Пуазейля, незначительное увеличение
радиуса дыхательных путей резко снижает в них сопротивление потоку
воздуха. Поэтому при вдохе сопротивление дыхательных путей потоку воз-
духа не оказывает существенного влияния на силу сокращения дыхатель-
ных мышц. Напротив, при выдохе, особенно при глубоком и усиленном
(форсированном) выдохе, диаметр мелких дыхательных путей уменьшает- (
ся, что вызывает значительное увеличение сопротивления потоку воздуха в
них. Влияние объема легких при выдохе на поток воздуха в дыхательных »
путях количественно характеризуется зависимостью «поток—объем». В ?
клинической физиологии дыхания оценка этой зависимости является ос-
новным критерием типа и степени нарушения функции дыхательных
путей. ‘
Зависимость «поток—объем» следующим образом характеризует влия-
ние большого объема воздуха в легких на экспираторный поток воздуха в
дыхательных путях (рис. 10.9). В момент, предшествующий началу выдоха,
после глубокой инспирации в дыхательных путях отсутствует поток возду-
ха, а внутриплевральное давление равно —10 см водн. ст. С началом фор-
сированной экспирации внутриплевральное давление возрастает примерно
до +30 см водн. ст. относительно атмосферного давления, вызывая умень-
шение радиуса как альвеол, так и мелких дыхательных путей. В этих усло-
виях давление газов внутри альвеол становится выше, чем в плевральной
полости, благодаря действию на стенки альвеол эластической тяги легких.
В результате поток воздуха выходит из альвеолярного пространства по ды-
хательным путям во внешнюю среду по градиенту давления, который по-
степенно уменьшается в дыхательных путях по мере приближения к тра-
хее. Спадению эластичных стенок бронхиол препятствует градиент давле-
ния воздуха между дыхательными путями и внутриплевральным давлени-
ем. Однако в некоторой точке дыхательных путей (как правило, в области
бронхиол) этот градиент давления становится равным нулю (эквипотенци-
альная точка давления) и стенки дыхательных путей могут частично или
полностью спадаться. В этих условиях продвижение воздуха по дыхатель-
10. Функции дыхательной системы • 481
О-----Полость рта
РА = +1
Р = 0
Плевральная
полость
Р = -5
О-----Полость рта
Трахея
ЭТД
Диафрагма
Г рудная
клетка
Альвеолярное
пространство
РА = +36
Р = +34
Р = +6
Р = +30
Р = +30
Рис. 10.9. Давление в дыхательных путях при выдохе.
Вертикальными стрелками показаны величины давления, возникающие в дыхательных путях
под влиянием комплайенса легких и грудной клетки. Горизонтальными стрелками в области
дыхательных путей показано, что давление, оказываемое на стенки дыхательных путей, может
увеличивать их просвет при спокойном выдохе (а) либо уменьшать их диаметр при глубоком
выдохе (б) в том участке общей площади поперечного сечения мелких дыхательных путей, где
сравниваются величины внутриплеврального и альвол и давления в дыхательных путях (экви-
потенциальная точка — ЭПТ). Р — давление (см водн. ст.), РА — давление в альвеолах.
ным путям может обеспечиваться только за счет увеличения силы сокра-
щения (работы) внутренних межреберных мышц и мышц живота.
Снижение эластической тяги легких, например при эмфиземе легких,
вызывает смещение ближе к альвеолярному пространству эквипотенциаль-
ной точки давления в дыхательных путях при выдохе, и, таким образом,
блокируется выход воздуха непосредственно из альвеол. Дыхательные шу-
мы, которые возникают в легких у больных, обусловлены прохождением
воздуха через спавшиеся мелкие дыхательные пути. Увеличение экспира-
торного усилия у таких пациентов повышает риск спадения мелких дыха-
тельных путей и еще больше затрудняет выдох. При бронхиальной астме у
пациентов дыхательные пути уменьшают свой просвет в результате сокра-
щения гладких мышц стенки бронхиол. В этом случае увеличение сопро-
тивления потоку воздуха в мелких дыхательных путях вызывает рост гради-
ента давления вдоль дыхательных путей при вдохе и смещает эквипотенци-
альную точку ближе к альвеолярному пространству, вызывая коллапс ды-
хательных путей при выдохе. Усиление сокращения экспираторных мышц
в фазу выдоха еще больше затрудняет выдох у пациентов вследствие
уменьшения просвета мелких дыхательных путей.
10.1.4. Работа дыхательных мышц в течение дыхательного цикла
Сокращение дыхательных мышц создает градиент давления по ходу дыха-
тельных путей. При этом преодолевается эластическое сопротивление лег-
ких и грудной клетки, а также сопротивление дыхательных путей потоку
воздуха. Наряду с этим последние два показателя позволяют измерять ра-
боту дыхательных мышц во время дыхательного цикла. Если принять, что
величина работы (W) представляет собой произведение силы (F) на путь
16-6095
482 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Внутриплевральное давление (см водн. ст.)
Рис. 10.10. Работа дыхательных мышц при
спокойном дыхании. Изменения дыхательно-
го объема (вертикальная ось) при вдохе и вы-
дохе сопровождаются изменениями внутри-
плеврального давления. При одновременной
регистрации этих величин во время дыхатель-
ного цикла общая площадь петель дыхатель-
ный объем — внутриплевральное давление от-
ражает количественно работу дыхательных
мышц. Работа дыхательных мышц при вдохе
больше, поскольку она затрачивается на пре-
одоление эластического сопротивления лег-
ких. При выдохе работа дыхания минималь-
ная, поскольку совершается за счет энергии
эластической тяги легких, т. е. пассивно.
Стрелками показаны изменения внутриплев-
рального давления в течение фаз дыхательно-
го цикла. Чем больше площадь петли, тем
больше работа дыхательных мышц.
(х), то получим: W = F х х. В
дыхательной системе, в кото-
рой измеряемыми величина-
ми являются дыхательный
объем и внутриплевральное
давление, сила сокращения
дыхательных мышц приравни-
вается к развиваемому ими
давлению (Р), которое они
оказывают на площадь (А).
Поэтому, подставляя выраже-
ние F - Р А ъ формулу ра-
боты дыхательных мышц в те-
чение дыхательного цикла, по-
лучим: W = Р х А хх По-
скольку величина А, умножен-
ная на путь (х), в дыхательной
системе представляет собой
аналог дыхательного объема
(V), то общая формула работы
дыхательных мышц имеет вид:
W = Р х V.
Работа дыхательных мышц
при спокойном дыхании. При
спокойном дыхании объем
вдоха достигает максимум 1 л,
а инспираторные мышцы со-
вершают минимальную рабо-
ту (рис. 10.10). Сокращение
инспираторных мышц обеспе-
чивает вдох, а выдох осущест-
вляется пассивно за счет эла-
стической тяги легких. В этих
условиях сопротивление дыха-
тельных путей при вдохе и вы-
дохе не оказывает лимитирую-
щего влияния на процесс
внешнего дыхания.
По мере увеличения глубины дыхания дыхательный объем формируется за
счет объема функциональной остаточной емкости и резервного объема
вдоха, а работа дыхания совершается против существенного нарастания
поверхностного натяжения жидкости на поверхности альвеол. Поэтому
чем глубже инспирация, тем большую работу совершают инспираторные
мышцы. Во время выдоха, когда глубина дыхательных движений осуществ-
ляется в пределах объема жизненной емкости легких, объем легких возвра-
щается пассивно к уровню функциональной остаточной емкости за счет
эластической тяги легких, а в пределе функциональной остаточной емко-
сти выдох происходит активно в результате сокращения мышц живота, ко-
торые при этом совершают работу.
Работа дыхательных мышц при глубоком дыхании. При глубоком дыха-
нии на силу сокращения дыхательных мышц начинает оказывать влияние
изменение диаметра дыхательных путей. Глубокий вдох вызывает расшире-
ние дыхательных путей и снижение сопротивления в них потоку вдыхае-
10. Функции дыхательной системы • 483
мого воздуха, поэтому работа инспираторных мышц обусловлена только
величинами комплайенса легких и тканей грудной клетки. При глубоком
выдохе, при котором в выдыхаемом воздухе оказывается объем воздуха
функциональной остаточной емкости, возникает сдавление мелких дыха-
тельных путей высоким градиентом давления между дыхательными путями
и внутриплевральным давлением. Существенное увеличение потока газов
через дыхательные пути приводит к росту их сопротивления потоку возду-
ха, которое становится основным фактором, обусловливающим величину
работы дыхания. Однако при глубоком дыхании механизмы регуляции
диаметра дыхательных путей при участии вегетативной нервной системы
способны минимизировать величину работы, которые выполняют дыха-
тельные мышцы. Так, при глубоком дыхании за счет регулирующих влия-
ний вегетативной нервной системы на гладкие мышцы дыхательных путей
увеличивается их диаметр. В результате на сокращение дыхательных мышц
затрачивается минимальное количество энергии. Например, при астме ды-
хание у пациентов становится медленным и глубоким, что снижает затраты
энергии на преодоление сопротивления дыхательных путей потоку воздуха
и уменьшает работу дыхательных мышц.
10.2. Вентиляция и перфузия кровью легких
10.2.1. Вентиляция легких
Вентиляцией легких обозначают процесс обмена воздуха между легкими и
атмосферой. Количественным показателем вентиляции легких служит ми-
нутный объем дыхания, определяемый как количество воздуха, которое про-
ходит (или вентилируется) через легкие в 1 мин. В покое у человека ми-
нутный объем дыхания составляет 6—8 л/мин. Только часть воздуха, кото-
рым вентилируются легкие, достигает альвеолярного пространства и непо-
средственно участвует в газообмене с кровью. Эта часть вентиляции легких
называется альвеолярной вентиляцией. В покое альвеолярная вентиляция
равна в среднем 3,5—4,5 л/мин. Основная функция альвеолярной вентиля-
ции заключается в поддержании необходимой для газообмена концентра-
ции О2 и СО2 в воздухе альвеол.
Легкие состоят из воздухопроводящей (дыхательные пути) и респиратор-
ной зон (альвеолы). Дыхательные пути, начиная от трахеи и до альвеол, де-
лятся по типу дихотомии и образуют 23 генерации элементов дыхательного
тракта (рис. 10.11). В воздухопроводящей или кондуктивной зонах легких
(16 генераций) отсутствует газообмен между воздухом и кровью, поскольку
в этих отделах дыхательные пути не имеют достаточной для этого процесса
сосудистой сети, а стенки дыхательных путей, из-за их значительной тол-
щины, препятствуют обмену газов через них. Этот отдел воздухоносных
путей называется анатомическим мертвым пространством, объем которого
составляет в среднем 175 мл. На рис. 10.12 показано, каким образом воз-
дух, заполняющий анатомическое мертвое пространство в конце выдоха,
смешивается с «полезным», т. е. атмосферным воздухом и вновь поступает
в альвеолярное пространство легких.
Дыхательные бронхиолы 17—19-й генераций относят к переходной
(транзиторной) зоне, в которой начинается газообмен в малочисленных
альвеолах (2 % от общего числа альвеол). Альвеолярные ходы и альвеоляр-
ные мешочки, непосредственно переходящие в альвеолы, образуют альвео-
лярное пространство, в области которого происходит в легких газообмен
О2 и СО2 с кровью. Однако у здоровых людей и, особенно, у пациентов с
Рис. 10.11. Схема дыхательных путей
легких человека. Дыхательные пути
от уровня трахеи (1-я генерация) до
долевых бронхов (2—4-я генерации
деления) поддерживают свой просвет
благодаря хрящевым кольцам в их
стенке. Дыхательные пути от сегмен-
тарных бронхов (5—11-я генерации)
до терминальных бронхиол (12— 16-я
генерации) стабилизируют свой про-
свет с помощью тонуса гладких
мышц их стенок. 1—16-я генерации
дыхательных путей образуют возду-
хопроводящую зону легких, в кото-
рой не происходит газообмена. Рес-
пираторная зона легких имеет длину
порядка 5 мм и включает первичные
дольки или ацинусы: дыхательные
бронхиолы (17—19-я генерации) и
альвеолярные протоки (20—22-я ге-
нерации). Альвеолярные мешочки
состоят из многочисленных альвеол
(23-я генерация), альвеолярная мем-
брана которых является идеальным
местом для диффузии О2 и СО2.
Рис. 10.12. Эффект воздуха мертвого (вредного) пространства на вдыхаемый воздух
в легкие. В конце выдоха анатомическое мертвое пространство заполняется выды-
хаемым воздухом, в котором пониженное количество кислорода и высокое про-
центное содержание углекислого газа. При вдохе «вредный» воздух анатомического
мертвого пространства смешивается с «полезным» атмосферным воздухом. Эта га-
зовая смесь, в которой меньше, чем в атмосферном воздухе, кислорода и больше уг-
лекислого газа, поступает в респираторную зону легких. Поэтому газообмен в лег-
ких происходит между кровью и альвеолярным пространством, заполненным не ат-
мосферным воздухом, а смесью «полезного» и «вредного» воздуха.
10. Функции дыхательной системы • 485
заболеваниями легких часть альвеолярного пространства может вентилиро-
ваться, но при этом не участвовать в газообмене, поскольку эти отделы
легких не перфузируются кровью. Сумму объемов таких областей легких и
анатомического мертвого пространства обозначают как физиологическое
мертвое пространство. Увеличение физиологического мертвого простран-
ства в легких приводит к недостаточному снабжению тканей организма
кислородом и к увеличению содержания в крови углекислого газа, что на-
рушает в ней газовый гомеостазис.
10.2.2. Перфузия легких кровью
Эффективность выполнения основной функции дыхательной системы за-
висит от соответствия перфузии (кровотока) в регионах легких с их вен-
тиляцией. Так, хороший кровоток будет недостаточным для газообмена в
регионах легких, если они слабо вентилируются воздухом с низким со-
держанием кислорода, при этом незначительный объем вентиляции лег-
ких не позволит удалить из крови углекислый газ. Соответственно, при
слабом кровотоке в регионах легких возрастает объем функционального
мертвого пространства, и перфузия легких кровью будет недостаточной
для транспорта в ней нормального количества газов. Наконец, перфузия
кровью регионов легких с недостаточной вентиляцией этих же отделов
называется шунтированием, и это состояние является неадекватным для
нормального газообмена. При шунтировании венозная кровь в легких не
обогащается кислородом, что снижает его содержание в крови организ-
ма. В нормальных физиологических условиях фактор гравитации оказы-
вает наиболее выраженный эффект на вентиляцию и перфузию регионов
легких кровью.
10.2.3. Эффект гравитации на вентиляцию и перфузию легких
кровью
Легкие окружены плевральным пространством, отрицательное давление в
котором изменяется от —5 до —10 см водн. ст. в различные фазы дыхатель-
ного цикла. Этот фактор взаимодействует с эффектом гравитации на жид-
кие среды, прежде всего кровь, содержащуюся в артериальных и венозных
сосудах тканей легких. В результате под действием силы тяжести на ткань
легких величина внутриплеврального давления на уровне основания лег-
ких у человека в положении стоя менее отрицательная относительно атмо-
сферного, чем в области верхушек легкого. Поэтому альвеолы верхушек
легких имеют большие размеры, а стенка их растянута и более напряжена,
чем у альвеол нижних участков легких. Альвеолы на уровне основания лег-
ких растянуты в незначительной степени и имеют значительно больше по-
тенциальные возможности для растягивания и вентиляции, чем в области
верхушек. Поэтому растянутые альвеолы верхушки легких вентилируются
меньше, чем альвеолы основания (рис. 10.13). Эти различия в вентиляции
отделов легких приводят к тому, что вдыхаемый воздух неравномерно рас-
пределяется в отделах альвеолярного пространства. Особенности распреде-
ления воздуха, вдыхаемого в легкие, дополняется разницей в величине
кровотока на уровне верхушек и основания легких. Относительно положе-
ния тела в пространстве кровоток в верхних и нижних отделах легкого раз-
личается под влиянием фактора гравитации.
У человека в вертикальном положении тела величина легочного крово-
тока на единицу объема ткани легкого линейно убывает в направлении
486 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 10.13. Влияние внутриплеврального давления и фактора гравитации на разме-
ры альвеол верхних и нижних отделов легких. Между верхушками и основанием
легких имеется градиент внутриплеврального давления, возникающий под влияни-
ем гравитации на массу жидких сред и ткань легких.
В результате размеры альвеол в верхушках легких больше, чем в основании (А). Альвеолы в
нижних отделах легких имеют большие потенциальные возможности для увеличения в них
вентиляции легких при вдохе, чем альвеолы в области верхних отделов легких (Б).
снизу вверх, и меньше всего снабжаются кровью верхушки легких. Соот-
ветственно в положении тела человека на спине кровоток в нижних (дор-
сальных) отделах легких становится выше, чем в верхних (вентральных).
Это обусловлено тем, что артериальная кровь, поступающая в легкие из
правого желудочка, проходит по сосудам легких из областей низкого внут-
риплеврального давления в области тонкостенных капилляров, которые
окружены альвеолами, содержащими воздух под давлением, близким к ат-
мосферному. Поэтому в зависимости от соотношения давления в альвео-
лах (РА), мелких артериях (Ра) и мелких легочных венах (Pv) легкие разде-
лены на функциональные зоны Веста (рис. 10.14).
В верхушках легких (зона 1) могут возникнуть области с давлением в ле-
гочных капиллярах (особенно в фазу диастолы) ниже альвеолярного
(Ра > Ра > Pv)- Капилляры в таких зонах могут спадаться, и кровоток че-
рез них становится невозможным. Такие участки легких вентилируются,
но не участвуют в газообмене и формируют альвеолярное мертвое про-
странство.
В средних отделах легких (зона 2) под действием гравитации давление в
альвеолах, как правило, превышает венозное (Ра > РА > Pv)- Поэтому вели-
чину кровотока в зоне 2 по Весту определяет разность между артериаль-
ным и альвеолярным давлениями. В зоне 2 практически не возникает аль-
веолярное мертвое пространство.
В нижних отделах легких (зона 3) давление в легочных венах выше аль-
веолярного (Ра > Pv > РА) и величина кровотока, как и в обычных сосудах,
определяется разницей между артериальным и венозным давлениями.
Величина зон Веста динамично изменяется в зависимости от положе-
ния тела в пространстве или глубины дыхания. При выдохе на уровне
функциональной остаточной емкости примерно 2/3 объема легких может
занимать зона 2. После глубокой экспирации (на уровне остаточного объе-
ма) большая часть легких по соотношению перфузии кровью и вентиляции
соответствует зоне 3 Веста. Относительная однонаправленность изменения
градиента внутриплеврального давления и влияния гравитации на крово-
ток в легких от верхних отделов легких к нижним теме не менее не сопря-
жены в каждом отдельном регионе легких.
10. Функции дыхательной системы • 487
10.2.3. Коэффициент вентиляционно-перфузионных отношений
в легких
Коэффициент представляет собой отношение величины вентиляции лег-
ких (V) к величине их перфузии кровью (Q). При адекватности вентиляци-
онно-перфузионных отношений (V/Q) величина коэффициента близка к 1.
В обычных физиологических условиях в разных регионах легких, как пра-
вило, коэффициент варьирует: 1<V/Q >1. Региональные отличия значения
этого коэффициента обусловлены действием гравитации либо в случае по-
явления в каком-либо регионе легких феномена шунтирования
(рис. 10.15). Однако в целом 97—98 % от общего количества О2 и СО2, уча-
ствующих в газообмене в легких, обменивается между альвеолярным про-
странством и кровью легочных капилляров в условиях полного соответст-
вия величины вентиляции и перфузии легких кровью (V/Q =1).
10.3. Газообмен в легких
Обмен газов между кровью и воздухом относится к основной функции
легких. Воздух, поступающий в легкие при вдохе, нагревается и насыщает-
ся водяными парами при движении в дыхательных путях и достигает аль-
веолярного пространства, имея температуру 37 °C. Парциальное давление
Рис. 10.14. Модель, связывающая неравномерность распределения легочного кро-
вотока при вертикальном расположении тела человека с величиной давления, дей-
ствующего на капилляры.
В зоне 1 (верхушки легких) альвеолярное давление (РА) превышает давление в артериолах (Ра)
и кровоток ограничен. В средней зоне легких (зона 2), где Ра > РА, кровоток больше, чем в зо-
не 1. В основаниях легких (зона 3) кровоток усилен и определяется разностью давления в ар-
териолах (Ра) и венулах (Pv). В центре схемы легкого — легочные капилляры; вертикальные
трубочки по сторонам легкого — манометры.
488 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Кровоток------► Q = 1 Кровоток------► Q = 1
Смешанная венозная Кровь,насыщенная
кровь
кислородом
! V = 1
Г Альвеолярное ;
I пространство ;
... О" 0 = 0
Отсутствует кровоток
Рис. 10.15. Соотношение вентиляции и перфузии кровью -легких.
При прекращении вентиляции в каком-либо регионе легких увеличивается их функциональ-
ное мертвое пространство (а). При этом венозная кровь перфузирует этот отдел легких и, не
обогащаясь кислородом, поступает в большой круг кровообращения. Нормальное вентиляци-
онно-перфузионное отношение формируется, когда вентиляция регионов легких соответству-
ет величине их перфузии кровью (б). При отсутствии кровотока в каком-либо регионе легких
(в) вентиляция также не обеспечивает нормальное вентиляционно-перфузионное отношение.
V — вентиляция легких, Q — кровоток в легких.
водяных паров в альвеолярном воздухе при этой температуре составляет
47 мм рт. ст. Поэтому согласно закону парциальных давлений Дальтона
вдыхаемый воздух находится в разведенном водяными парами состоянии и
парциальное давление кислорода в нем меньше, чем в атмосферном воз-
духе.
Обмен кислорода и углекислого газа в легких происходит в результате
разницы парциального давления этих газов в воздухе альвеолярного про-
странства и их напряжения в крови легочных капилляров. Процесс движе-
ния газа из области высокой концентрации в область с низкой его концен-
трацией обусловлен диффузией. Кровь легочных капилляров отделена от
воздуха, заполняющего альвеолы, альвеолярной мембраной, через которую
газообмен происходит путем пассивной диффузии. Процесс перехода газов
между альвеолярным пространством и кровью легких объясняется диффу-
зионной теорией.
10.3.1. Состав альвеолярного воздуха
Газовый состав альвеолярного воздуха обусловлен альвеолярной вентиля-
цией и скоростью диффузии О2 и СО2 через альвеолярную мембрану.
В обычных условиях у человека количество О2, поступающего в единицу
времени в альвеолы из атмосферного воздуха, равно количеству О2, диф-
фундирующего из альвеол в кровь легочных капилляров. Равным обра-
зом количество СО2, поступающего в альвеолы из венозной крови, равно
количеству СО2, которое выводится из альвеол в атмосферу. Поэтому в
норме парциальное давление О2 и СО2 в альвеолярном воздухе остается
практически постоянным, что поддерживает процесс газообмена между
альвеолярным воздухом и кровью капилляров легких. Газовый состав аль-
веолярного воздуха отличается от атмосферного воздуха тем, что в нем
10. Функции дыхательной системы • 489
Таблица 10.1. Парциальное давление газов в воздушной среде легких
Газы Атмосферный воздух, мм рт. ст. (%) Альвеолярный воздух, мм рт. ст. (%) Выдыхаемый воздух, мм рт. ст. (%)
n2 597,0 (78,62 %) 573,0 (75 %) 566,0 (74 %)
О2 159,0 (20,84 %) 100,0 (13,5 %) 120,0 (16 %)
со2 0,3 (0,04 %) 40,0 (5,5 %) 27,0 (4 %)
Н2О 3,7 (0,5 %) 47,0 (6 %) 47,0 (6 %)
Итого... 760,0 (100,0 %) 760,0 (100,0 %) 760,0 (100,0 %)
меньше процентное содержание кислорода и выше процент углекислого
газа. Состав альвеолярного воздуха отличается от выдыхаемого воздуха
большим содержанием углекислого газа и меньшим содержанием кисло-
рода (табл. 10.1).
10.3.2. Напряжение газов в крови капилляров легких
Диффузия газов через альвеолярную мембрану происходит между альвео-
лярным воздухом и венозной, а также артериальной кровью легочных ка-
пилляров. В табл. 10.2 приведены стандартные величины напряжения ды-
хательных газов в артериальной и венозной крови легочных капилляров.
Градиенты парциального давления кислорода и углекислого газа обу-
словливают процесс пассивной диффузии через альвеолярную мембрану
кислорода из альвеол в венозную кровь (градиент 60 мм рт. ст.), а углеки-
слого газа — из венозной крови в альвеолы (градиент 6 мм рт. ст.). Парци-
альное давление азота по обе стороны альвеолярной мембраны остается
постоянным, поскольку этот газ не потребляется и не продуцируется тка-
нями организма. При этом сумма парциального давления всех газов, рас-
творенных в тканях организма, меньше, чем величина атмосферного дав-
ления, благодаря чему газы в тканях не находятся в газообразной форме.
Если величина атмосферного давления будет меньше, чем парциальное
давление газов в тканях и в крови, то газы начинают выделяться из крови
в виде пузырьков, вызывая тяжелые нарушения в кровоснабжении тканей
организма (кессонная болезнь).
10.3.3. Скорость диффузии О2 и СО2 в легких
Скорость диффузии (M/t) кислорода и углекислого газа через альвеоляр-
ную мембрану количественно характеризуется законом диффузии Фика.
Согласно этому закону газообмен (M/t) в легких прямо пропорционален
градиенту (ДР) концентрации О2 и СО2 по обе стороны от альвеолярной
мембраны, площади ее поверхности (S), коэффициентам (к) растворимо-
Таб л и ца 10.2. Напряжение дыхательных газов в артериальной и венозной крови легоч-
ных капилляров
Дыхательные газы Артериальная кровь (мм рт. ст.) Венозная кровь (мм рт. ст.)
Кислород 100 40
Углекислый газ 40 46
Азот 573 573
Вода 47 47
490 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Альвеолярное Альвеолярная Просвет
пространство
Базальная мембрана
АльвеолоцитI порядка
Слой сурфактанта
О2 = 100 мм рт. ст.
СО2 = 40 мм рт. ст.
капилляра
Мембрана эритроцита
Эндотелиоцит
О2 = 40 мм рт. ст.
СО2 = 46 мм рт. ст.
Рис. 10.16. Диффузия газов через альвеолярную мембрану. Диффузия газов в легких
осуществляется по градиентам концентрации О2 и СО2 между альвеолярным про-
странством и кровью капилляров легких, которые разделены альвеолярной мембра-
ной. При этом диффузия тем эффективнее, чем тоньше альвеолярная мембрана и
области контакта альвеолоцитов и эндотелиоцитов. Поэтому альвеолярная мембра-
на образована уплощенными частями альвеолоцитов I порядка (0,2 мкм) и эндоте-
лиоцитов капилляров легких (0, 2 мкм), между которыми находится тонкая общая
базальная мембрана (0,1 мкм) этих клеток. В состав мембраны входит также моно-
молекулярный слой сурфактант а. Мембрана эритроцитов является препятствием
для диффузии газов в легких.
сти О2 и СО2 в биологических средах альвеолярной мембраны и обратно
пропорционален толщине альвеолярной мембраны (L), а также молекуляр-
ной массе газов (М). Формула этой зависимости имеет следующий вид:
М = др S • к
t L ’ JM '
Структура легких образует максимальное по величине поле для диффу-
зии газов через альвеолярную стенку, которая имеет минимальную толщи-
ну (рис. 10.16). Так, количество альвеол в одном легком человека прибли-
зительно равно 300 млн. Суммарная площадь альвеолярной мембраны, че-
рез которую происходит обмен газов между альвеолярным воздухом и ве-
нозной кровью, имеет огромные размеры (порядка 100 м2), а толщина аль-
веолярной мембраны составляет лишь — 0,3—2,0 мкм.
В обычных условиях диффузия газов через альвеолярную мембрану
происходит в течение очень короткого отрезка времени (не более 3/4 с),
пока кровь проходит через капилляры легких. Даже при физической рабо-
те, когда эритроциты проходят капилляры легкого в среднем за ’/4 с, ука-
занные выше структурные особенности альвеолярной мембраны создают
оптимальные условия для формирования равновесия парциальных давле-
ний О2 и СО2 между альвеолярным воздухом и кровью капилляров легких
(рис. 10.17). В уравнении Фика константы диффузии (к) пропорциональны
растворимости газа в альвеолярной мембране. Углекислый газ имеет при-
мерно в 20 раз большую растворимость в альвеолярной мембране, чем ки-
слород. Поэтому, несмотря на существенное различие в градиентах парци-
альных давлений О2 и СО2 по обе стороны от альвеолярной мембраны,
10. Функции дыхательной системы • 491
Альвеолярный газ
(РО2 = 106 мм рт. ст.)
Альвеолярный газ
(РСО2 = 106 мм рт. ст.)
РаО2
97 мм рт. ст.
Рис. 10.17. Градиенты парциального давления дыхательных газов в смешанной ве-
нозной крови легочной артерии, альвеолярном воздухе и артериальной крови. Рав-
новесие парциальных давлений углекислого газа и кислорода между альвеолярным
воздухом и кровью легочных капилляров достигается в течение короткого времени
(‘/4—3/4 с) движения плазмы крови и эритроцитов в капиллярах легких.
РаСО2
39 мм рт. ст.
диффузия этих газов совершается за очень короткий отрезок времени дви-
жения эритроцитов крови через легочные капилляры.
Газообмен через альвеолярную мембрану количественно оценивается
диффузионной способностью легких, которая измеряется количеством газа
(мл), проходящего через эту мембрану за 1 мин при разнице давления газа
по обе стороны мембраны в 1 мм рт. ст.
Наибольшее сопротивление диффузии О2 в легких создают альвеоляр-
ная мембрана и мембрана эритроцитов, в меньшей степени — плазма кро-
ви в капиллярах. У взрослого человека в покое диффузионная способность
легких О2 равна 20—25 мл • мин-1 • мм рт. ст.-1. СО2, как полярная молекула
(О=С=О), диффундирует через указанные мембраны чрезвычайно быстро,
благодаря высокой растворимости этого газа в альвеолярной мембране.
Диффузионная способность легких СО2 равна 400—450 мл • мин-’ • мм
рт. ст.-1.
10.4. Транспорт газов кровью
Кровообращение выполняет одну из важнейших функций переноса кисло-
рода от легких к тканям, а углекислого газа — от тканей к легким. Потреб-
ление кислорода клетками тканей может изменяться в значительных пре-
делах, например при переходе от состояния покоя к физической нагрузке
и наоборот. В связи с этим кровь должна обладать большими резервами,
необходимыми для увеличения ее способности переносить кислород от
легких к тканям, а углекислый газ в обратном направлении.
492 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
10.4.1. Транспорт кислорода
При 37 °C растворимость О2 в жидкости составляет 0,225 мл • л-1 • кПа~'
(0,03 мл/л/мм рт. ст.). В условиях нормального парциального давления ки-
слорода в альвеолярном воздухе, т. е. 13,3 кПа или 100 мм рт.ст., 1 л плаз-
мы крови может переносить только 3 мл О2, что недостаточно для жизне-
деятельности организма в целом. В покое в организме человека за минуту
потребляется примерно 250 мл кислорода. Чтобы тканям получить такое
количество кислорода в физически растворенном состоянии, сердце долж-
но перекачивать за минуту огромное количество крови. В эволюции живых
существ проблема транспорта кислорода была более эффективно решена
за счет обратимой химической реакции с гемоглобином эритроцитов. Ки-
слород переносится кровью от легких к тканям организма молекулами ге-
моглобина, которые содержатся в эритроцитах. Гемоглобин способен за-
хватывать кислород из альвеолярного воздуха (соединение называется ок-
сигемоглобином) и освобождать необходимое количество кислорода в тка-
нях. Особенностью химической реакции кислорода с гемоглобином явля-
ется то, что количество связанного кислорода ограничено количеством мо-
лекул гемоглобина в эритроцитах крови. Молекула гемоглобина имеет
4 места связывания с кислородом, которые взаимодействуют таким обра-
зом, что зависимость между парциальным давлением кислорода и количе-
ством переносимого кислорода с кровью имеет S-образную форму, кото-
рая носит название кривой насыщения или диссоциации оксигемоглобина
(рис. 10.18). При парциальном давлении кислорода 10 мм рт. ст. насыще-
ние гемоглобина кислородом составляет примерно 10 %, а при РО2 30 мм
рт. ст. — 50—60 %. При дальнейшем увеличении парциального давления
кислорода от 40 мм рт. ст. до 60 мм рт. ст. происходит уменьшение крутиз-
ны кривой диссоциации оксигемоглобина и процент его насыщения ки-
Рис. 10.18. Кривая диссоциации оксигемоглобина. Пределы колебания кривой при
РСО2 = 40 мм рт. ст. (артериальная кровь) и РСО2 = 46 мм рт. ст. (венозная кровь)
показывают изменение сродства гемоглобина к кислороду (эффект Ходена).
10. Функции дыхательной системы • 493
слородом возрастает в диапазоне от 70—75 до 90 % соответственно. Затем
кривая диссоциации оксигемоглобина начинает занимать практически го-
ризонтальное положение, поскольку увеличение парциального давления
кислорода с 60 до 80 мм рт. ст. вызывает прирост насыщения гемоглобина
кислородом на 6 %. В диапазоне от 80 до 100 мм рт. ст. процент образова-
ния оксигемоглобина составляет порядка 2. В результате кривая диссоциа-
ции оксигемоглобина переходит в горизонтальную линию и процент насы-
щения гемоглобина кислородом достигает предела, т. е. 100. Насыщение
гемоглобина кислородом под влиянием РО2 характеризует своеобразный
молекулярный «аппетит» этого соединения к кислороду.
Значительная крутизна кривой насыщения гемоглобина кислородом в
диапазоне парциального давления от 20 до 40 мм рт. ст. способствует тому,
что в ткани организма значительное количество кислорода может диффун-
дировать из крови в условиях градиента его парциального давления между
кровью и клетками тканей (не менее 20 мм рт. ст.). Незначительный про-
цент насыщения гемоглобина кислородом в диапазоне его парциального
давления от 80 до 100 мм рт. ст. способствует тому, что человек без риска
снижения насыщения артериальной крови кислородом может перемещать-
ся в диапазоне высот над уровнем моря до 2000 м.
Общие запасы кислорода в организме обусловлены его количеством,
находящимся в связанном состоянии с ионами Fe2+ в составе органиче-
ских молекул гемоглобина эритроцитов и миоглобина мышечных клеток.
Один грамм гемоглобина связывает 1,34 мл О2. Поэтому в норме при
концентрации гемоглобина 150 г/л каждые 100 мл крови могут переносить
20,0 мл О2.
Количество О2, которое может связаться с гемоглобином эритроцитов
крови при насыщении 100 % его количества, называется кислородной емко-
стью гемоглобина. Другим показателем дыхательной функции крови явля-
ется содержание О2 в крови (кислородная емкость крови), которое отража-
ет его истинное количество, как связанного с гемоглобином, так и физиче-
ски растворенного в плазме. Поскольку в норме артериальная кровь насы-
щена кислородом на 97 %, то в 100 мл артериальной крови содержится
примерно 19,4 мл О2.
10.4.1.1. Изменение сродства гемоглобина к кислороду
Молекула гемоглобина может находиться в двух формах — напряженной и
расслабленной. Расслабленная форма гемоглобина имеет свойство насыщать-
ся кислородом в 70 раз быстрее, чем напряженная. Изменение фракций
напряженной и расслабленной формы в общем количестве гемоглобина в
крови обусловливает S-образный вид кривой диссоциации оксигемоглоби-
на, а следовательно, так называемое сродство гемоглобина к кислороду.
Если вероятность перехода от напряженной формы гемоглобина к расслаб-
ленной больше, то возрастает сродство гемоглобина к кислороду, и наобо-
рот. Вероятность образования указанных фракций гемоглобина изменяется
в большую или меньшую сторону под влиянием нескольких факторов. Ос-
новной фактор — это связывание кислорода с геминовой группой молеку-
лы гемоглобина. При этом чем больше геминовых групп гемоглобина свя-
зывают кислород в эритроцитах, тем более легким становится переход мо-
лекулы гемоглобина к расслабленной форме и тем выше их сродство к ки-
слороду. Поэтому при низком РО2, что имеет место в метаболически ак-
тивных тканях, сродство гемоглобина к кислороду ниже, а при высоком
РО2 — выше. Как только гемоглобин захватывает кислород, повышается
494 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
его сродство к кислороду и молекула гемоглобина становится насыщенной
при связывании с четырьмя молекулами кислорода.
Когда эритроциты, содержащие гемоглобин, достигают тканей, то ки-
слород из эритроцитов диффундирует в клетки. В мышцах он поступает в
своеобразного депо кислорода — в молекулы миоглобина, из которого ки-
слород используется в биологическом окислении мышц.
Диффузия кислорода из гемоглобина эритроцитов в ткани обусловлена
низким РО2 в тканях — 35 мм рт. ст. Внутри клеток тканей напряжение ки-
слорода, необходимое для поддержания нормального метаболизма, состав-
ляет еще меньшую величину — не более 1 кПа. Поэтому кислород путем
диффузии из капилляров достигает метаболически активных клеток. Неко-
торые ткани приспособлены к низкому содержанию РО2 в капиллярах кро-
ви, что компенсируется высокой плотностью капилляров на единицу объе-
ма тканей. Например, в скелетной и сердечной мышцах РО2 в капиллярах
может снизиться чрезвычайно быстро во время сокращения. В мышечных
клетках содержится белок миоглобин, который имеет более высокое срод-
ство к кислороду, чем гемоглобин. Миоглобин интенсивно насыщается
кислородом и способствует его диффузии из крови в скелетную и сердеч-
ную мышцы, где он обусловливает процессы биологического окисления.
Эти ткани способны экстрагировать до 70 % кислорода из крови, проходя-
щей через них, что обусловлено снижением сродства гемоглобина к кисло-
роду под влиянием температуры тканей и pH.
Эффект pH и температуры на сродство гемоглобина к кислороду. Моле-
кулы гемоглобина способны реагировать с ионами водорода, в результате
этой реакции происходит снижение сродства гемоглобина к кислороду.
При насыщении гемоглобина менее 100 % низкое pH понижает связыва-
ние кислорода с гемоглобином — кривая диссоциации оксигемоглобина
смещается вправо по оси х. Это изменение свойства гемоглобина под
влиянием ионов водорода называется эффектом Бора. Метаболически ак-
тивные ткани продуцируют кислоты, такую как молочная, и СО2. Если pH
плазмы крови снижается от 7,4 в норме до 7,2, что имеет место при сокра-
щении мыщц, то концентрация кислорода в ней будет возрастать вследст-
вие эффекта Бора. Например, при постоянном pH 7,4 кровь отдавала бы
порядка 45 % кислорода, т. е. насыщение гемоглобина кислородом снижа-
лось до 55 %. Однако когда pH снижается до 7,2, кривая диссоциации сме-
щается по оси х вправо. В результате насыщение гемоглобина кислородом
падает до 40 %, т. е. кровь может отдавать в тканях до 60 % кислорода, что
на */3 больше, чем при постоянном pH.
Метаболически активные ткани повышают продукцию тепла. Повыше-
ние температуры тканей при физической работе изменяет соотношение
фракций гемоглобина в эритроцитах и вызывает смещение кривой диссо-
циации оксигемоглобина вправо вдоль оси х. В результате большее коли-
чество кислорода будет освобождаться из гемоглобина эритроцитов и по-
ступать в ткани.
Эффект 2,3-дифосфоглицерата (2,3-ДФГ) на сродство гемоглобина к ки-
слороду. При некоторых физиологических состояниях, например при пони-
жении РО2 в крови ниже нормы (гипоксия) в результате пребывания чело-
века на большой высоте над уровнем моря, снабжение тканей кислородом
становится недостаточным. При гипоксии может понижаться сродство ге-
моглобина к кислороду вследствие увеличения содержания в эритроцитах
2,3-ДФГ. В отличие от эффекта Бора, уменьшение сродства гемоглобина к
кислороду под влиянием 2,3-ДФГ не является обратимым в капиллярах
легких. Однако при движении крови через капилляры легких эффект 2,3-
10. Функции дыхательной системы • 495
ДФГ на снижение образования оксигемоглобина в эритроцитах (плоская
часть кривой диссоциации оксигемоглобина) выражен в меньшей степени,
чем отдача кислорода под влиянием 2,3-ДФГ в тканях (наклонная часть
кривой), что обусловливает нормальное кислородное снабжение тканей.
10.4.2. Транспорт углекислого газа
СО2 является продуктом метаболизма клеток тканей и поэтому переносит-
ся кровью от тканей к легким. Углекислый газ выполняет жизненно важ-
ную роль в поддержании во внутренних средах организма уровня pH меха-
низмами кислотно-основного равновесия. Поэтому транспорт углекислого
газа кровью тесно взаимосвязан с этими механизмами (см. глава 15).
В плазме крови небольшое количество СО2 находится в растворенном
состоянии; при РСО2= 40 мм рт. ст. переносится 2,5 мл/100 мл крови СО2,
или 5 %. Количество растворенного в плазме углекислого газа в линейной
зависимости возрастает от уровня РСО2.
В плазме крови СО2 реагирует с водой с образованием Н+ и НСО^. Уве-
личение напряжения углекислого газа в плазме крови вызывает уменьше-
ние величины ее pH. Напряжение СО2 в плазме крови может быть измене-
но функцией внешнего дыхания, а количество ионов водорода или pH —
буферными системами крови и HCOJ, например путем их выведения через
почки с мочой. Величина pH плазмы крови зависит от соотношения кон-
центрации растворенного в ней СО2 и ионов бикарбоната. В виде бикарбо-
ната плазмой крови, т. е. в химически связанном состоянии, переносится
основное количество углекислого газа — порядка 45 мл/100 мл крови, или
до 90 %. Эритроцитами в виде карбаминового соединения с белками гемо-
глобина транспортируется примерно 2,5 мл/100 мл крови СО2, или 5 %.
Транспорт углекислого газа кровью от тканей к легким в указанных формах
не связан с явлением насыщения, как при транспорте кислорода, т. е. чем
больше образуется углекислого газа, тем большее его количество транспор-
тируется от тканей к легким. Однако между парциальным давлением СО2 в
крови и количеством переносимого кровью углекислого газа имеется кри-
волинейная зависимость: кривая диссоциации углекислого газа.
10.4.2.1. Роль эритроцитов в транспорте СО2
В крови капилляров тканей организма напряжение СО2 составляет 5,3 кПа
(40 мм рт. ст.), а в самих тканях — 8,0—10,7 кПа (60—80 мм рт. ст.). В ре-
зультате СО2 диффундирует из тканей в плазму крови, а из нее — в эритро-
циты по градиенту парциального давления СО2. В эритроцитах СО2 обра-
зует с водой угольную кислоту, которая диссоциирует на Н+ и НСО^.
(СО2 + Н2О = Н2СО3 = Н+ + НСОр. Эта реакция протекает быстро, по-
скольку СО2 + Н2О = Н2СО3 катализируется ферментом карбоангидразой
мембраны эритроцитов, которая содержится в них в высокой концентра-
ции (рис. 10.19). Эта реакция протекает по закону действия масс и в норме
выражается в логарифмической форме, известной как уравнение Гендерсо-
на—Гассельбаха (см. главу 15).
В эритроцитах диссоциация СО2 продолжается постоянно по мере обра-
зования продуктов этой реакции, поскольку молекулы гемоглобина дейст-
вуют как буферное соединение, связывая положительно заряженные ионы
водорода. В эритроцитах по мере освобождения кислорода из гемоглобина
его молекулы будут связываться с ионами водорода (СО2 + Н2О = Н2СО3 =
= Н+ + НСО7), образуя соединение (НЬ-Н+). В целом это называется эф-
496 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
а
Эритроцит Плазма Клетки
Рис. 10.19. Роль эритроцитов в транспорте углекислого газа от тканей к легким.
А. Процессы газообмена СО2 между клетками тканей и плазмой крови. СО2 поступает путем
диффузии в плазму крови из тканей и дифференцирует внутрь эритроцитов. В эритроцитах
СО2 превращается при участии фермента карбоангидразы мембраны эритроцитов в НСО, и в
виде этого иона вновь активно транспортируется в плазму крови, в которой в составе бикар-
бонатов (NaHCO3) переносится к капиллярам легких. Водородные ионы, образующиеся при
реакции дегидратации Н2СО3, захватываются молекулами гемоглобина и также транспортиру-
ются кровью от тканей к легким. При транспорте из эритроцитов НСО3-ионное постоянство
внутри эритроцитов поддерживается хлорным сдвигом. Кроме того, СО2 в эритроцитах непо-
средственно соединяется с Nh2 группами белков гемоглобина.
Б. Процессы газообмена СО2 между плазмой крови и альвеолами легких. В капиллярах легких
в эритроцитах под влиянием эффекта Бора происходит диссоциация карбаминовых соедине-
ний и СО2 диффундирует через мембрану эритроцитов и альвеолярную мембрану в просвет
альвеол легких.
фектом Холдена, который приводит к сдвигу кривой диссоциации оксиге-
моглобина вправо по оси х, что снижает сродство гемоглобина к кислоро-
ду и способствует более интенсивному освобождению его из эритроцитов в
ткани. При этом в составе соединения НЬ-Н+ транспортируется примерно
200 мл СО2 в одном литре крови от тканей к легким.
Диссоциация СО2 в эритроцитах может быть лимитирована только бу-
ферной емкостью молекул гемоглобина. Образующиеся внутри эритроци-
тов в результате диссоциации СО2 ионы НСО^ с помощью специального
белка-переносчика мембраны эритроцитов выводятся из эритроцитов в
плазму, а на их место из плазмы крови закачиваются ионы СГ (феномен
«хлорного» сдвига) (рис. 10.19). Основная роль реакции СО2 внутри эрит-
роцитов заключается в обмене ионами СГ и HCOJ между плазмой и внут-
ренней средой эритроцитов. В результате этого обмена продукты диссо-
циации углекислого газа Н+ и НСО3 будут транспортироваться внутри
эритроцитов в виде соединения (НЬ-Н+), а плазмой крови — в виде бикар-
бонатов.
Эритроциты участвуют в транспорте углекислого газа от тканей к лег-
ким, поскольку СО2 образует прямую комбинацию с — 1ЧН2-группами бел-
ковых субъединиц гемоглобина: СО2 + НЬ -> НЬСО2 или карбаминовое со-
единение. Транспорт кровью СО2 в виде карбаминового соединения и ио-
нов водорода гемоглобином зависит от свойств молекул последнего; обе
реакции обусловлены величиной парциального давления кислорода в плаз-
ме крови на основе эффекта Холдена.
В количественном отношении транспорт СО2 в растворенной форме и в
форме карбаминового соединения является незначительным, по сравне-
10. Функции дыхательной системы • 497
нию с его переносом СО2 кровью в виде бикарбонатов. Однако при газо-
обмене СО2 в легких между кровью и альвеолярным воздухом эти две фор-
мы приобретают основное значение.
Когда венозная кровь возвращается от тканей к легким, СО2 диффунди-
рует из крови в альвеолы и РСО2 в крови снижается с 46 мм рт. ст. (веноз-
ная кровь) до 40 мм рт.ст. (артериальная кровь). При этом в величине об-
щего количества СО2 (6 мл/100 мл крови), диффундирующего из крови в
альвеолы, доля растворенной формы СО2 и карбаминовых соединений ста-
новится более значительной относительно бикарбонатной. Так, доля рас-
творенной формы составляет 0,6 мл/100 мл крови, или 10 %, карбамино-
вых соединений — 1,8 мл/100 мл крови, или 30 %, а бикарбонатов —
3,6 мл/100 мл крови, или 60 %.
В эритроцитах капилляров легких по мере насыщения молекул гемогло-
бина кислородом начинают освобождаться ионы водорода, диссоциировать
карбаминовые соединения и НСО/ вновь превращается в СО2 (Н+ + НСО/ =
= Н2СО3 = СО2 +Н2О), который путем диффузии выводится через легкие
по градиенту его парциальных давлений между венозной кровью и альвео-
лярным пространством. Таким образом, гемоглобин эритроцитов играет
основную роль в транспорте кислорода от легких к тканям, и углекислого
газа в обратном направлении, поскольку способен связываться с О2 и Н+.
В состоянии покоя через легкие из организма человека за минуту удаляет-
ся примерно 300 мл СО2: 6 мл/100 мл крови х 5000 мл/мин минутного
объема кровообращения.
10.5. Регуляция дыхания
Дыхательная система должна обеспечивать полное соответствие между ко-
личеством кислорода, поступающего в кровь через легкие, и скоростью его
потребления в тканях, а так же соответствие между продукцией в тканях
СО2 и количеством удаляемого из организма через легкие СО2. Эта функ-
ция дыхательной системы достигается за счет приспособления (регуляции)
объема легочной вентиляции к уровню метаболизма в организме. Приспо-
собление легочной вентиляции к процессам метаболически обусловленно-
го газообмена в организме человека осуществляется сложной иерархиче-
| н + ро2 |
Периферические
хеморецепторы
Центральные
хеморецепторы
Механорецепторы
легких
Рецепторы Дыхательные
мышц fr. ^ыш1|Ы-
> Дыхательный ____^Гх
центр
Дыхательные
мотонейроны
спинного мозга
'Мышцы “
верхних
дыхательных
.путей_______
4___Вентиляция
легких
Гладкие
мышцы
дыхательных
путей
Рис. 10.20. Регуляция вентиляции легких осуществляется сложной иерархической
системой, включающей механизмы обратных связей при участии хеморецепторных
(центральные и периферические) и механорецепторных (механорецепторы легких и
дыхательных мышц) рефлексов. Местом генерации дыхательного ритма является
дыхательный центр продолговатого мозга.
тг
498 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ской системой регуляции дыхания, состоящей из центральных и перифе-
рических хеморецепторных рефлексов (общий контроль), а также механо-
рецепторных рефлексов (локальный контроль) (рис. 10.20). В этой системе
дыхательный центр генерирует дыхательный ритм, интегрирует афферент-
ные сигналы, посылает импульсы к дыхательным мышцам грудной клетки
и к гладким мышцам дыхательных путей, обеспечивая приспособление ле-
гочной вентиляции к условиям жизнедеятельности организма.
10.5.1. Дыхательный центр
Величина легочной вентиляции обусловлена частотой и глубиной дыха-
тельных движений (дыхательного ритма), происхождение которых связано
с функцией дыхательного центра центральной нервной системой. Под ды-
хательным центром понимают ограниченный участок ЦНС, где происхо-
дит формирование дыхательного импульса, вызывающего координирован-
ную деятельность дыхательных мышц, обеспечивающих для организма не-
обходимую величину газообмена в легких. В центральной нервной системе
местом генерации дыхательного ритма, вызывающего ритмические сокра-
щения дыхательных мышц при вдохе и выдохе, является продолговатый
мозг, в котором расположен дыхательный центр. Дыхательный центр со-
стоит из нервных клеток (дыхательных нейронов), для которых характерна
периодическая электрическая активность в одну из фаз дыхания. Нейроны
дыхательного центра локализованы двусторонне в продолговатом мозге в
виде двух вытянутых столбов вблизи obex — точки, где центральный канал
спинного мозга впадает в четвертый желудочек. Эти два образования дыха-
тельных нейронов в соответствии с их положением относительно дорсаль-
ной и вентральной поверхности продолговатого мозга обозначают как дор-
сальная и вентральная дыхательные группы (рис. 10.21).
Дорсальная дыхательная группа нейронов образует вентролатеральную
часть ядра одиночного тракта. Дыхательные нейроны вентральной дыха-
тельной группы расположены в области n. ambiguus каудальнее уровня obex,
n. retroambigualis непосредственно ростральнее obex и представлены ком-
плексом Бетзингера, который находится непосредственно вблизи n. retrofa-
cialis вентролатеральных отделов продолговатого мозга. В состав дыхатель-
ного центра входят нейроны двигательных ядер черепно-мозговых нервов
(обоюдное ядро, ядро подъязычного нерва), которые иннервируют мышцы
гортани и глотки.
Основным критерием классификации нейронов дыхательного центра
является фаза дыхательного цикла, в которую они активны, т. е. инспира-
ция или экспирация. По этому критерию дыхательные нейроны подразде-
ляют на инспираторные и экспираторные. Дорсальная дыхательная группа
состоит полностью из инспираторных нейронов. Вентральная дыхательная
группа образована инспираторными и экспираторными нейронами, а ком-
плекс Бетзингера образуют только экспираторные нейроны. По паттерну
электрической активности нейронов в пределах фаз дыхательного цикла
инспираторные и экспираторные нейроны подразделяют на нейроны с на-
растающим, постоянным или декрементным типом активности
(рис. 10.22).
По проекции аксонов дыхательные нейроны разделяют на премоторные
или бульбоспинальные нейроны и проприобульбарные. Аксоны премотор-
ных дыхательных нейронов переходят на противоположную сторону про-
долговатого мозга, а затем направляются к мотонейронам спинного мозга.
Функция инспираторных премоторных дыхательных нейронов заключается
10. Функции дыхательной системы • 499
IV желудочек
Рис. 10.21. Схема положения нейронов дорсальной (ДРГ) и вентральной дыхатель-
ных групп (ВДГ) дыхательного центра продолговатого мозга.
Слева — проекция отделов дыхательного центра на дорсальную поверхность продолговатого
мозга. Справа — расположение отделов дыхательного центра на поперечном плане продолго-
ватого мозга. Диафрагмальные мотонейроны и мотонейроны межреберных мышц являются
«общим конечным путем» моторных команд дыхательного центра, обусловливающих сокра-
щение дыхательных мышц и вентиляцию легких. NA — n. ambiguus.
1
2
3
4
5
6
iwrnjjL—_
L_J_UWt
Вдох
Выдох
у Потенциал
; действия
Разряд
потенциалов
действия
Сопрограмма
Рис. 10.22. Образцы электрической активности основных типов нейронов дыха-
тельного центра. Наиболее общая классификация типов дыхательных нейронов ос-
нована на связи их электрической активности с фазами вдоха и выдоха дыхательно-
го цикла и паттерном разряда потенциалов действия.
По этому критерию классифицируют: полные инспираторные с нарастающей активностью (1),
экспираторные нейроны с нарастающей активностью (2), ранние инспираторные нейроны с
декрементной активностью (3), поздние инспираторные нейроны с нарастающей активностью
(4), экспираторные или постинспираторные нейроны с декрементной активностью (5), пол-
ные инспираторные нейроны с постоянной активностью (6).
500 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
в управлении электрической активностью инспираторных мотонейронов
диафрагмы и наружных межреберных мышц во время их сокращения при
вдохе. В обычных условиях экспирация осуществляется пассивно, поэтому
функция экспираторных премоторных дыхательных нейронов реализуется
только при увеличении глубины дыхательных движений. Премоторные
нейроны комплекса Бетзингера выполняют уникальную функцию — они
тормозят все типы инспираторных нейронов дыхательного центра и диа-
фрагмальные мотонейроны. Поэтому их аксоны распределяются билате-
рально, т. е. направляются к соответствующим нейронам, расположенным
как ипсилатерально, так и контралатерально.
Аксоны проприобульбарных дыхательных нейронов (ранние инспира-
торные, постинспираторные, поздние инспираторные, экспираторные ней-
роны комплекса Бетзингера) оканчиваются на мембране нейронов самого
дыхательного центра, расположенных в вентральной дыхательной группе.
Функция большинства проприробульбарных нейронов заключается в гене-
рации дыхательного ритма.
10.5.1.1. Происхождение дыхательного ритма
У человека дыхательные движения впервые начинаются у плода (в начале
III триместра беременности). Эти дыхательные движения носят периоди-
ческий и нерегулярный характер. Современные исследования свидетельст-
вуют о том, что дыхательные движения плода инициируются проприобуль-
барными инспираторными пейсмекерными клетками вентральной дыха-
тельной группы, расположенными вблизи нейронов комплекса Бетзингера
(пребетзингерова область). Эти нейроны обладают двумя уникальными
свойствами. Мембрана нейронов способна к спонтанной деполяризации и,
когда их мембранный потенциал достигает порога возбуждения, нейроны
генерируют короткий залп потенциалов действия. Вторым свойством мем-
браны пейсмекерных нейронов дыхательного центра является механизм
аккомодации, т. е. самоограничения электрической активности. Этот меха-
низм представляет собой гиперполяризацию мембранного потенциала ней-
рона под влиянием нарастающего входящего тока ионов хлора и формиро-
вание декрементного паттерна электрической активности. В результате в
течение короткого отрезка времени нейроны способны самостоятельно
прекращать свою электрическую активность. Разряды электрических им-
пульсов по аксонам дыхательных нейронов распространяется к мотонейро-
нам спинного мозга и вызывают через них сокращение диафрагмы и на-
ружных межреберных мышц у плода. Внутриутробные ритмические сокра-
щения инспираторных мышц у плода способствуют развитию у него аппа-
рата внешнего дыхания к моменту рождения. Первый вдох новорожденно-
го и дыхание в первые сутки после рождения по-прежнему обусловлены
спонтанной пейсмекерной активностью проприобульбарных нейронов
вентральной дыхательной группы дыхательного центра.
У взрослых млекопитающих животных происхождение дыхательного
ритма объясняют двумя теориями, которые основаны на пейсмекерной ги-
потезе и гипотезе нейронной сети. Согласно первой, дыхательный ритм
возникает в пейсмекерных проприобульбарных дыхательных нейронах
пребетзингеровой области. Затем залпы нервных импульсов от пейсмекер-
ных нейронов передаются другим типам нейронов дыхательного центра,
при участии которых возникает соответствующий ритм дыхательных дви-
жений.
Происхождение дыхательного ритма при участии нейронной сети дыха-
10. Функции дыхательной системы • 501
тельного центра объясняют функцией синаптических связей между различ-
ными типами дыхательных нейронов. Основным проявлением синаптиче-
ских связей является взаимное торможение между группами нейронов
противоположных фаз дыхательного цикла. Вторым свойством ритмгене-
рирующей нейронной сети является наличие в ней хотя бы одного типа
дыхательных нейронов, возбудимая мембрана которых обладает механиз-
мом аккомодации, т. е. самоограничения электрической активности. В ды-
хательном центре взрослых животных имеется два типа дыхательных ней-
ронов, обладающих аккомодативным механизмом: ранние инспираторные
и постсинспираторные (рис. 10.22). Оба типа нейронов взаимно тормозят
друг друга и все другие типы нейронов дыхательного центра в период, ко-
гда активны либо ранние инспираторные нейроны, либо постинспиратор-
ные нейроны. Ключевая роль в дыхательном ритмогенезе ранних инспира-
торных нейронов с декрементным типом активности заключается в ини-
циации дыхательного ритма, а именно вдоха. С момента начала разряда
этого типа нейронов, благодаря механизму самоограничения активности
ранних инспираторных нейронов, прекращается их тормозное действие на
другие типы инспираторных нейронов дыхательного центра. В результате
происходит активация инспираторных нейронов (полных и поздних) и
возникает фаза вдоха. Как только вдох выключается (функция центров
моста и ядер блуждающего нерва), активируются постинспираторные ней-
роны с декрементным типом активности и начинается фаза выдоха. Благо-
даря механизму самоограничения активности постинспираторные нейроны
постепенно прекращают тормозное действие на экспираторные нейроны с
нарастающей активностью и на ранние инспираторные нейроны. С мо-
мента активации последних прекращается фаза выдоха и начинается оче-
редная фаза вдоха.
10.5.2. Влияние нервных центров виролиеви мости
ни дыхительный ритм
В верхних отделах моста расположены две области — n. parabrachealis medi-
alis и n. Kolliker—Fuse, которые влияют на дыхательный центр продолгова-
того мозга и образуют пневмотаксический центр. Нейроны этого центра ре-
ципрокно связаны с инспираторными нейронами дорсальной дыхательной
группы. Функцией пневмотаксического центра моста является уменьшение
периода активности инспираторных нейронов дыхательного центра путем
выключения фазы вдоха и более раннее появление в дыхательном цикле
фазы выдоха. В результате пневмотаксический центр вызывает в дыхатель-
ном центре генерацию большего числа небольших по длительности инспи-
раций, т. е. увеличивает частоту дыхания.
На уровне нижней трети моста имеется область, определяемая как ап-
нейстический центр. В обычных условиях активность этого центра затор-
можена со стороны пневмотаксического центра. Название центра обуслов-
лено тем, что его отделение от пневмотаксического центра и/или от тор-
мозных афферентных влияний блуждающего нерва (описано ниже), вызы-
вает остановку дыхания на вдохе (апнейзис). Эта область оказывает возбу-
ждающее влияние на нейроны дорсальной дыхательной группы дыхатель-
ного центра и ее роль заключается в увеличении времени фазы выдоха, а,
следовательно, глубины дыхательных движений.
502 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
10.5.3. Функция спинальных дыхательных мотонейронов
При спокойном дыхании в течение фазы вдоха электрические импульсы
инспираторных бульбоспинальных нейронов дыхательного центра переда-
ются к мотонейронам сегментов спинного мозга: Сш—Cv и Тг—Тхп. Аксо-
ны мотонейронов СП1—Cv образуют диафрагмальные нервы, а Т(—Тхп —
межреберные нервы, которые иннервируют соответственно диафрагму и
наружные межреберные мышцы. Активация электрической активности
этих мотонейронов под влиянием нисходящих команд дыхательного цен-
тра вызывает сокращение диафрагмы и наружных межреберных мышц, что
определяет уровень вентиляции легких. При глубоком дыхании фаза выдо-
ха также становится активной, и нисходящие команды дыхательного цен-
тра начинают включать импульсы к мотонейронам мышц живота и внут-
ренних межреберных мышц (сегменты Т—L). Активация электрической
активности мотонейронов, которые иннервируют экспираторные мышцы,
обусловливает глубину выдоха.
10.5.4. Рефлекторная регуляция дыхания
10.5.4.1. Хеморецепторный контроль дыхания
Хеморецепторный контроль дыхания осуществляется при участии цен-
тральных и периферических хеморецепторов. Центральные (медуллярные)
хеморецепторы расположены непосредственно в в ростральных отделах
вентральной дыхательной группы, в структурах голубого пятна (locus
coeruleus), в ретикулярных ядрах шва ствола мозга и реагируют на водо-
родные ионы в окружающей их межклеточной жидкости мозга (рис.
10.23). Центральные хеморецепторы представляют собой нейроны, кото-
рые в определенной степени являются рецепторами углекислого газа, по-
скольку величина pH обусловлена парциальным давлением СО2, согласно
уравнению Гендерсона—Гасельбаха, а также тем, что концентрация ионов
Артериальное РСО2 (мм рт. ст.)
Рис. 10.23. Зависимость вентиляции
легких от степени стимуляции цен-
тральных хеморецепторов измене-
ниями [Н+]/РСО2 в артериальной
крови. Увеличение парциального
давления СО2 в артериальной крови
выше порога (РСО2 = 40 мм рт. ст.)
линейно увеличивает объем вентиля-
ции легких.
водорода в межклеточной жидкости
мозга зависит от парциального давле-
ния углекислого газа в артериальной
крови. Увеличение вентиляции легких
при стимуляции центральных хеморе-
цепторов ионами водорода называется
центральным хеморефлексом, который
оказывает выраженное влияние на
дыхание. Так, в ответ на уменьшение
pH внеклеточной жидкости мозга в
области локализации рецепторов на
0,01 легочная вентиляция возрастает в
среднем на 4,0 л/мин. Однако цен-
тральные хеморецепторы медленно
реагируют на изменения СО2 в арте-
риальной крови, что обусловлено их
локализацией в ткани мозга. У чело-
века центральные хеморецепторы сти-
мулируют линейное увеличение вен-
тиляции легких при увеличении СО2 в
артериальной крови выше порог, рав-
ного 40 мм рт. ст.
10. Функции дыхательной системы • 503
Периферические (артериальные) хе-
морецепторы расположены в каротид-
ных тельцах в области бифуркации
общих сонных артерий и в аорталь-
ных тельцах в области дуги аорты.
Периферические хеморецепторы реа-
гируют как на изменение концентра-
ции водородных ионов, так и парци-
ального давления кислорода в артери-
альной крови. Рецепторы чувстви-
тельны к анаэробным метаболитам,
которые образуются в ткани каротид-
ных телец при недостатке кислорода.
Недостаток кислорода в тканях каро-
тидных телец может возникнуть, на-
пример, при гиповентиляции, веду-
щей к гипоксии, а также при гипо-
тензии, вызывающей снижение кро-
вотока в сосудах каротидных телец.
При гипоксии (низкое парциальное
давление кислорода) периферические
хеморецепторы активируются под
влиянием увеличения концентрации в
артериальной крови, прежде всего,
ионов водорода и РСО2. Действие на
периферические хеморецепторы этих
раздражителей усиливается по мере
снижения в крови РО2 (мультиплика-
тивное взаимодействие). Гипоксия
Артериальное РСО2 (мм рт. ст.)
Рис. 10.24. Зависимость вентиляции
легких от степени стимуляции пери-
ферических хеморецепторов гипок-
сическим стимулом.
При стимуляции периферических хеморе-
цепторов гипоксией имеет место мульти-
пликативное взаимодействие парциально-
го давления СО2 в артериальной крови и
гипоксии, в результате которого происхо-
дит максимальное увеличение вентиляции
легких. Напротив, при высоком Парци-
альном давлении кислорода в артериаль-
ной крови периферические хеморецепто-
ры слабо реагируют на увеличение РСО2.
Если в артериальной крови парциальное
давление СО2 становится ниже порога
(40 мм рт.ст.), то периферические хеморе-
цепторы также слабо реагируют на гипок-
сию.
увеличивает чувствительность пери-
ферических хеморецепторов к [Н+] и СО2. Это состояние называется ас-
фиксией и возникает при прекращении вентиляции легких. Поэтому пери-
ферические хеморецепторы называются часто рецепторами асфиксии. Им-
пульсы от периферических хеморецепторов по волокнам синокаротидного
нерва (нерв Геринга — часть языкоглоточного нерва) и аортальной ветви
блуждающего нерва достигают чувствительных нейронов ядра одиночного
тракта продолговатого мозга, а затем переключаются на нейроны дыха-
тельного центра. Возбуждение последнего вызывают прирост вентиляции
легких. Вентиляция легких увеличивается линейно в соответствии с вели-
чиной [Н+] и РСО2 выше порога (40 мм рт. ст.) в артериальной крови, про-
текающей через каротидные и аортальные тельца (рис. 10.24). Наклон кри-
вой на рисунке, который отражает чувствительность периферических хе-
морецепторов к [Н+] и РСО2, варьирует в зависимости от степени ги-
поксии.
10.5.4.2. Механорецепторный контроль дыхания
Механорецепторный контроль дыхания осуществляется рефлексами, кото-
рые возникают при раздражении механорецепторов дыхательных путей
легких. В тканях дыхательных путей расположено два основных типа меха-
норецепторов, импульсы от которых поступают к нейронам дыхательного
центра: быстро адаптирующиеся, или ирритантные, рецепторы и рецепто-
ры растяжения.
504 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Быстроадаптирующиеся рецепторы расположены в эпителии и субэпите-
лиальном слое, начиная от верхних дыхательных путей вплоть до альвеол.
Название рецепторов свидетельствует о том, что они активируются при
раздражении не продолжительно и быстро снижают свою активность при
сохранении действия стимула. Поэтому быстроадаптирующиеся рецепторы
реагируют на изменение силы раздражения. Эти рецепторы инициируют
такие сложные рефлексы, как нюхательный или кашлевой. Они возбужда-
ются при попадании на слизистую оболочку трахеи и бронхов механиче-
ских или химических раздражителей (пыль, слизь, табачный дым, пары ед-
ких веществ — аммиак, эфир). В зависимости от местоположения ирри-
тантных рецепторов в дыхательных путях возникают специфические реф-
лекторные реакции дыхания. Раздражение рецепторов слизистой оболочки
носовой полости при участии тройничного нерва вызывает рефлекс чиха-
нья; рецепторов эпифарингеальной области — через волокна языкоглоточ-
ного нерва — нюхательный или аспирационный рефлекс; рецепторов сли-
зистой оболочки гортани и трахеи — через волокна блуждающего нерва —
рефлекс чиханья; рецепторов слизистой оболочки от уровня трахеи и до
бронхиол — при участии блуждающих нервов — парадоксальный рефлекс
Геда (при раздувании легких) и рефлекс выдоха и, наконец, рецепторов
стенки альвеол в месте их контакта со стенкой легочных капилляров — че-
рез волокна блуждающего нерва — вызывает рефлекторную реакцию в виде
частого и поверхностного дыхания.
Медленно адаптирующиеся рецепторы растяжения легких локализованы в
гладких мышцах главных дыхательных путей бронхиального дерева (брон-
хи и трахея) и раздражаются в результате увеличения объема легких (разду-
вание). Рецепторы связаны с нейронами дорсальной дыхательной группы
дыхательного центра миелинизированными афферентными волокнами
блуждающего нерва. Стимуляция этих рецепторов вызывает рефлекс Ге-
ринга—Брейера, который у животных проявляется в том, что раздувание
легких вызывает рефлекторное переключение фазы вдоха на фазу выдоха.
У человека в состоянии бодрствования этот рефлекторный эффект возни-
кает при величине дыхательного объема, которая превышает примерно в
три раза его нормальную величину при спокойном дыхании. Во время сна
рефлекторное выключение вдоха с помощью рефлекса Геринга—Брейера
обусловливает смену фаз дыхательного цикла.
Легочные J-рецепторы локализованы в пределах стенок альвеол в месте
их контакта с капиллярами и способны реагировать на стимулы как со
стороны легких, так и со стороны легочного кровообращения. Рецепторы
связаны с дыхательным центром немиелинизированными афферентными
С-волокнами. Эти рецепторы повышают свою активность при увеличении
в плазме крови концентрации ионов водорода, при сдавливании легочной
ткани или легочном отеке. Наибольшую активность легочные J-рецепторы
имеют во время физической активности большой мощности и при подъе-
ме на большую высоту над уровнем моря. В этих случаях одним из прояв-
лений изменения давления в сосудах малого круга кровообращения может
быть отек легких разной выраженности. Возникающее при этом раздраже-
ние J-рецепторов вызывает частое, поверхностное дыхание, рефлекторную
бронхоконстрикцию и одышку.
Проприорецепторы. Дыхательный центр непрерывно получает афферент-
ные входы от прориорецепторов мышц (мышечные веретена и сухожильные
рецепторы Гольджи) по восходящим спинальным трактам. Эти афферент-
ные входы являются как неспецифическими (рецепторы расположены в
мышцах и суставах конечностей), так и специфическими (рецепторы рас-
10. Функции дыхательной системы • 505
положены в дыхательных мышцах). Импульсация от проприорецепторов
распространяется преимущественно к спинальным центрам дыхательных
мышц, а также к центрам головного мозга, контролирующим тонус скелет-
ной мускулатуры. Активация проприорецепторов в момент начала физиче-
ской нагрузки является основной причиной увеличения активности дыха-
тельного центра и повышения вентиляции легких. Проприорецепторы
межреберных мышц и диафрагмы рефлекторно регулируют ритмическую
активность дыхательного центра продолговатого мозга в зависимости от
положения грудной клетки в различные фазы дыхательного цикла, а на
сегментарном уровне — тонус и силу сокращения дыхательных мышц.
10.6. Дыхание при физической нагрузке
При физической нагрузке регуляция дыхания не обусловлена исключи-
тельно ролью хеморецепторов, поскольку парциальное напряжение О2 в
альвеолах повышено относительно нормы в связи с увеличенной вентиля-
цией, или гиперпноэ, а прирост СО2 недостаточен для хеморецепторной
стимуляции внешнего дыхания. При физической нагрузке в мышцах воз-
растает продукция молочной кислоты, которая стимулирует перифериче-
ские хеморецепторы дыхания, но основное значение в увеличении венти-
ляции имеют супрабульбарные входы, которые изменяют активность дыха-
тельного центра продолговатого мозга.
Увеличение дыхания при физической нагрузке проявляется в виде трех
фаз:
1) первая фаза гиперпноэ возникает в первые 20 с под влиянием нисхо-
дящих двигательных команд от нейронов моторной коры и входов от
проприорецепторов сокращающихся мышц;
2) вторая фаза характеризуется медленным (экспотенциальным) прирос-
том вентиляции в результате активации под влиянием нисходящих
центральных команд центров варолиева моста, регулирующих дыха-
ние (например, пневмотаксического);
3) третья фаза проявляется относительно постоянным уровнем актива-
ции механизмов регуляции легочной вентиляции, которые включают
процессы температурного и хеморецепторного контроля внутренней
среды организма при физической нагрузке.
Начальная стадия увеличения вентиляции при физической нагрузке
обусловлена так называемыми нейрогенными стимулами. Механизм нейро-
генной стимуляции вентиляции легких обусловлен афферентными импуль-
сами от рецепторов работающих мышц, центральных двигательных команд
моторной коры и двигательных центров ствола мозга. Двигательные ко-
манды моторной коры обусловливают специфику вентиляции легких при
разных видах физической активности. Дыхание человека в определенной
степени может находиться под произвольным (корковым) контролем; че-
ловек может прекратить дыхательные движения или, наоборот, увеличить
вентиляцию легких (гипервентиляция). Кора больших полушарий головно-
го мозга регулирует паттерн дыхательных движений при речи, пении, заня-
тиях физическими упражнениями, например при плавании, при игре на
духовых музыкальных инструментах. Нисходящие двигательные команды
от соответствующих областей коры поступают к нейронам дыхательного
центра, а по пирамидным трактам в составе боковых столбов — непосред-
ственно к дыхательным мотонейронам сегментов спинного мозга. Непро-
извольная стимуляция вентиляции легких при физической нагрузке проис-
ходит под влиянием двигательных центров ствола мозга, нисходящие влия-
506 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Мощность физической нагрузки
ния которых в составе бульбоспи-
нальных трактов адресованы двига-
тельным нейронам дыхательных
мышц в соответствующие сегменты
спинного мозга, с помощью которых
непосредственно осуществляются ре-
гуляция тонуса и сокращение скелет-
ных мышц организма человека. На-
ряду с корковыми влияниями на
прирост вентиляции во время физи-
ческой нагрузки важную роль выпол-
няет таламический «генератор двига-
тельного паттерна», с помощью кото-
рого частота и амплитуда дыхатель-
ных движений могут быть включены
в определенный ритм движения, на-
пример, при плавании или беге.
Физическая нагрузка низкой и сред-
ней интенсивности. При физической
нагрузке низкой и средней интенсив-
ности, когда потребление О2 (VO2)
составляет менее 55 % от максималь-
ной скорости этого процесса в орга-
низме, VO2 и вентиляция легких
взаимосвязаны между собой линей-
но (рис. 10.25). Для обеспечения по-
требления 1 л О2 в организме при
физической нагрузке необходимо
увеличение вентиляции легких (VE)
гвается дыхательным эквивалентом и
Рис. 10.25. Зависимость вентиляциии
легких относительно аэробной и ана-
эробной мощности физической на-
грузки.
При аэробной физической нагрузке венти-
ляция легких линейно связана с мощно-
стью физической нагрузки. В этих услови-
ях активация дыхательного центра и при-
рост вентиляции легких обусловлены ней-
рогенными стимулами. При анаэробной
физической нагрузке прирост вентиляции
легких становится не линейным относи-
тельно метаболизма в мышцах. В этих ус-
ловиях активация дыхательного центра и
прирост вентиляции легких обусловлены
стимуляцией центральных и перифериче-
ских хеморецепторов.
на 20—25 л. Это соотношение н;
оно равно 25 : 1 при физической нагрузке низкой и средней интенсивно-
сти. В этих условиях у человека из крови в ткани экстрагируется 20—25 %
О2, поскольку 25 л вдыхаемого воздуха содержит 5 л кислорода. Для физи-
чески тренированных людей максимальная вентиляция легких должна со-
ставить порядка 120 л/мин, чтобы обеспечить необходимое потребление О2
в организме, равное порядка 5 л/мин.
Физическая нагрузка высокой интенсивности. Начиная с так называемо-
го анаэробного порога, происходит увеличение дыхательного эквивалента
(35—40:1). Интенсивность физической нагрузки, при которой достигается
это изменение, представляет собой так называемый дыхательный порог.
Дыхательный порог проявляется значительным ростом вентиляции легких,
что обусловлено увеличением содержания СО2 в плазме крови в результате
его высвобождения из бикарбонатного буфера ионами водорода, что соот-
ветствует так называемому лактатному порогу. Лактатный порог представ-
ляет собой количество потребляемого О2, при котором возрастает концен-
трация молочной кислоты в плазме крови (более чем на 1 мМ выше уров-
ня покоя), а продукция молочной кислоты в мышцах начинает превышать
буферную емкость крови. С этого момента, который соответствует дыха-
тельной компенсаторной точке, у человека начинается второй прирост ле-
гочной вентиляции.
При анаэробной физической нагрузке увеличение вентиляции легких
вызывают не только центральные двигательные команды и афферентные
обратные связи от работающих мышц, но и стимуляция периферических
10. Функции дыхательной системы • 507
хеморецепторов снижением pH артериальной крови {молочный ацидоз),
увеличение температуры тела (стимулирует дыхательный центр через центр
терморегуляции гипоталамуса) и, наконец, увеличение в плазме крови со-
держания катехоламинов.
Энергетическая стоимость дыхания. В покое дыхательные мышцы чело-
века потребляют примерно 4 % от общего количества О2, потребляемого
тканями организма. При физической нагрузке VO2 дыхательных мышц
возрастает до 8—11 % и более. В этих условиях дыхательные мышцы со-
вершают работу, равную 50 % от их максимальной способности развивать
силу сокращения. Физическая тренировка вызывает приспособление вен-
тиляции; при этом максимальная нагрузка линейно увеличивает вентиля-
цию легких в зависимости от роста VO2 у человека. В результате физиче-
ской тренированности человека у него снижается относительная энергети-
ческая стоимость дыхания за счет увеличения лактатного порога, причем
чем выше физическая тренированность человека, тем меньше в работаю-
щих мышцах продуцируется ионов Н+, которые инициируют высвобожде-
ние СО2 из бикарбонатного буфера. Соответственно у тренированных лю-
дей меньше выражена хеморецепторная стимуляция дыхания при физиче-
ской нагрузке.
После прекращения физической нагрузки в первые 20—30 с резко
уменьшается вентиляция легких, что обусловлено прекращением двига-
тельных команд со стороны коры головного мозга и двигательных центров
ствола мозга. Быстрое начальное уменьшение вентиляции легких перехо-
дит в стадию медленного восстановления исходных показателей частоты,
глубины и объема легочной вентиляции. В эту стадию в мышцах снижают-
ся потребление кислорода и продукция углекислого газа. Благодаря венти-
ляции легких происходит нормализация содержания газов в артериальной
крови до физиологических величин состояния покоя.
10.7. Дыхание человека при измененном барометрическом
давлении воздуха
10.7.1. Дыхание человека при пониженном давлении воздуха
Снижение парциального давления кислорода в окружающем воздухе, что в
естественных условиях имеет место при восхождении человека на высокие
горы, вызывает недостаток О2 в крови, что называется артериальной гипок-
семией. Так, на высоте 3000 м над уровнем моря барометрическое давление
воздуха уменьшается на '/3, а на высоте 8500 м — на 2/3. При подъеме чело-
века на высоту до 3—3,5 км над уровнем моря артериальная кровь насы-
щена кислородом в пределах нормы — на 90—95 % (табл. 10.3).
Поскольку насыщение артериальной крови кислородом при подъеме на
высоту до 3000 м над уровнем моря составляет не менее 90 %, то в этих ус-
ловиях незначительное снижение напряжения О2 в артериальной крови че-
ловека происходит за счет уменьшения содержания в крови физически
растворенного кислорода. Это, тем не менее, обусловливает появление
слабо выраженной гипоксемии и сопровождается незначительным увели-
чением вентиляции легких. Восхождение человека на высокую гору всегда
сопряжено с усиленной мышечной деятельностью, повышением темпера-
туры тела, увеличением в плазме крови содержания катехоламинов. Эти
факторы оказывают комплексное стимулирующее влияние на дыхание че-
ловека при восхождении на горную высоту. В результате слабо выраженная
артериальная гипоксемия при участии периферических хеморецепторов
508 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 10.3. Парциальное давление кислорода во вдыхаемом и альвеолярном воздухе
и процент насыщения артериальной крови кислородом на различной высоте над уровнем
моря
Высота над уровнем моря (м) Барометриче- ское давление (мм рт. ст.) Парциальное давление О2 (мм рт. ст) Насыщение артериальной крови О2 (%)
вдыхаемый воздух альвеолярный воздух
0 760 159 105 95
1000 680 140 90 94
2000 600 125 70 92
3000 530 110 62 90
4000 460 98 50 85
5000 405 85 45 75
6000 355 74 40 70
7000 310 65 35 60
8000 270 56 30 50
9000 230 48 <25 >40
увеличивает степень гиперпноэ у человека, обусловленное работой мышц.
Повышенное содержание катехоламинов в плазме крови повышает чувст-
вительность периферических хеморецепторов к гипоксемии и, усиливая
активность периферических хеморецепторов, ведет к росту параметров
внешнего дыхания. Наконец, повышение температуры тела человека при
восхождении на высокие горы в результате мышечной деятельности также
повышает чувствительность периферических хеморецепторов к гипоксе-
мии. Повышение температуры тела при физической работе может стиму-
лировать дыхание через усиление скорости метаболизма в организме, через
периферические хеморецепторы и нейроны дыхательного центра. При
этом периферические хеморецепторы являются основными источниками
стимуляции вентиляции легких у человека при гипоксии. Поэтому при
восхождении человека на высокую гору до высоты 3—3,5 км над уровнем
моря усиление вентиляции обусловлено активацией механизмов гумораль-
ной и нервной регуляции дыхания в пределах физиологической нормы.
Начиная с высоты 3,5 км над уровнем моря в артериальной крови чело-
века уменьшение напряжения О2 в плазме крови происходит как за счет
падения фракции физически растворенного О2, так и за счет понижения
насыщения гемоглобина кислородом. При снижении насыщения артери-
альной крови кислородом до 80—85 % у человека появляются признаки
недостаточности дыхания, которые проявляются головокружением, увели-
чением частоты сердцебиений, слабостью, вплоть до потери способности
мышечной деятельности, или обморочным состоянием. Явления дыхатель-
ной недостаточности, как правило, возникают внезапно и исчезают при
отдыхе, но могут вновь появиться при возобновлении восхождения на вы-
сокую гору в результате мышечной деятельности. Указанный комплекс
симптомов получил название горной болезни. При выраженной гипоксии
(4000 м и выше над уровнем моря), когда напряжение кислорода в артери-
альной крови достигает 50 мм рт. ст. и менее, возникает метаболический
ацидоз ткани мозга в результате активации анаэробного гликолиза. Как
следствие, уменьшается pH внеклеточной жидкости мозга. Повышение
концентрации водородных ионов во внеклеточной жидкости мозга стиму-
лирует центральные хеморецепторы, что обусловливает первоначальное су-
щественное увеличение вентиляции легких. Однако выраженная гипоксия
оказывает угнетающий эффект на вентиляцию легких двумя механизмами.
10. Функции дыхательной системы • 509
Во-первых, гипоксия может прямо угнетать нейронную активность дыха-
тельного центра и таким образом усугублять степень дыхательной недоста-
точности у человека, восходящего на высокую гору. Во-вторых, гипоксия
вызывает усиление кровотока в сосудах мозга под влиянием низкого на-
пряжения О2 в крови. Этот механизм способствует повышению кислород-
ного снабжения ткани мозга человека в условиях выраженной гипоксемии.
Однако усиление мозгового кровотока повышает скорость газообмена СО2
между межклеточной жидкостью мозга и кровью и вызывает снижение на-
пряжения СО2 в ткани мозга, а следовательно, уменьшение стимулирую-
щего влияния центральных хеморецепторов на дыхательный центр и сни-
жение вентиляции легких. Этот механизм вызывает нарастание степени
артериальной гипоксемии, и в условиях пониженного парциального давле-
ния О2 в атмосферном воздухе у человека увеличивается дыхательная не-
достаточность. При высокой степени развития горной болезни у человека
ее проявления становятся опасными для жизни. Проявление крайней фи-
зической слабости и угнетение функциональной активности ЦНС могут
быть настолько велики, что пострадавший без посторонней помощи не в
состоянии принять меры защиты против грозящей ему смертельной опас-
ности. Аналогичные явления дыхательной недостаточности могут возник-
нуть у людей при воздушных полетах на большой высоте, если происходит
разгерметизация летательного аппарата. При понижении барометрического
давления до величин, вызывающих снижение насыщения артериальной
крови кислородом до 50—45 %, у человека под влиянием гипоксии насту-
пает смерть.
10.7.2. Дыхание человека при повышенном давлении воздуха
Дыхание человека при повышенном давлении воздуха имеет место на зна-
чительной глубине под водой при работе водолазов или при кессонных ра-
ботах. Поскольку давление одной атмосферы соответствует давлению стол-
ба воды высотой 10 м, то в соответствии с глубиной погружения человека
под воду в скафандре водолаза или в кессоне поддерживается давление
воздуха по этому расчету. Человек, находясь в атмосфере повышенного
давления воздуха, не испытывает каких-либо дыхательных расстройств.
При повышенном давлении атмосферного воздуха человек может дышать в
том случае, если в его дыхательные пути поступает воздух под таким же
давлением. При этом растворимость газов в жидкости прямо пропорцио-
нальна его парциальному давлению. Поэтому при дыхании воздухом на
уровне моря в 1 мл крови содержится 0,011 мл физически растворенного
азота. При давлении воздуха, которым дышит человек, например, 5 атмо-
сфер, в 1 мл крови будет содержаться в 5 раз больше физически растворен-
ного азота. При переходе человека к дыханию при более низком давлении
воздуха (подъем кессона на поверхность или всплытие водолаза) кровь и
ткани тела могут удержать только 0,011 мл М2/мл крови. Остальное коли-
чество азота переходит из раствора в газообразное состояние. Переход че-
ловека из зоны повышенного давления вдыхаемого воздуха к более низко-
му его давлению должен происходить достаточно медленно, чтобы освобо-
ждающийся азот успел выделиться через легкие. Если азот, переходя в га-
зообразное состояние, не успевает полностью выделиться через легкие, что
имеет место при быстром подъеме кессона или нарушении режима всплы-
тия водолаза, пузырьки азота в крови могут закупорить мелкие сосуды тка-
ней организма. Это состояние называется газовая эмболия. В зависимости
от локализации газовой эмболии (сосуды кожи, мышц, центральной нерв-
510 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ной системы, сердца и др.) у человека возникают различные расстройства
(боли в суставах и мышцах, потеря сознания), которые в целом называют-
ся «кессонной болезнью». Развитие кессонной болезни предотвращается
определенной скоростью декомпрессии, т. е. скоростью перехода человека
от дыхания при повышенном давлении воздуха к дыханию воздухом при
нормальном атмосферном давлении на уровне моря. Например, переход
человека от дыхания при одной добавленной атмосфере к дыханию атмо-
сферным воздухом на уровне моря должен происходить 5 мин, от двух до-
бавленных атмосфер — 30 мин, а от четырех — 60 мин. В случае возникно-
вения кессонной болезни пострадавшего немедленно помещают в барока-
меру, в которой быстро повышают давление воздуха, которое обеспечивает
растворение мелких пузырьков азота в тканях организма. Это приводит к
исчезновению проявлений у человека кессонной болезни. Последующая
декомпрессия давления воздуха в барокамере производится по специаль-
ным нормам времени под наблюдением медицинского персонала за по-
страдавшим человеком.
ГЛАВА 11
Функции пищеварительной системы
Пищеварительная система включает в себя пищеварительный (желудочно-
кишечный) тракт, имеющий трубчатое строение (пищевод, желудок, тон-
кая и толстая кишка), слюнные железы, печень и поджелудочную железу.
Благодаря нервным и гуморальным механизмам регуляции функций орга-
ны пищеварения объединены в единую систему.
Для поддержания обмена веществ и энергии и осуществления жизне-
деятельности организма необходимо поступление из внешней среды орга-
нических и неорганических веществ. Содержащиеся в пище белки, жиры,
углеводы и другие сложные органические вещества не могут быть усвоены
организмом человека и животных без предварительной физико-химиче-
ской обработки в желудочно-кишечном тракте, в результате которой про-
исходит деполимеризация молекул питательных веществ. Образующиеся
под влиянием гидролитических ферментов пищеварительных соков олиго-
меры и мономеры, лишенные видовой специфичности, поступают в кровь,
лимфу и тканевую жидкость и включаются в метаболизм клеток. Комплекс
процессов механической, физико-химической и химической обработки пищи, а
также всасывание в пищеварительном тракте конечных продуктов гидролиза
и называется пищеварением. Пищеварение осуществляется благодаря реа-
лизации функций пищеварительной системы (моторной, секреторной и
всасывательной).
11.1. Состояние голода и насыщения
Пищеварительная система является частью более сложной системы —
функциональной системы питания, обеспечивающей сложное пищедобыва-
тельное поведение и поддерживающей относительно постоянный уровень
содержания питательных веществ в организме.
В процессе метаболизма клеток происходит постоянное потребление ими
питательных веществ. Снижение концентрации питательных веществ в кро-
ви приводит к возникновению у животных и человека неприятного чувства
голода, которое является субъективным выражением потребности организма
в пище. Физиологической основой для чувства голода является возбуждение
центра голода, локализованного в латеральных ядрах гипоталамуса. Чувство
голода является побудительной причиной (мотивацией) целенаправленной
пищедобывательной деятельности (поиска и приема пищи). При электро-
стимуляции через вживленные электроды (в опытах на животных) латераль-
ных ядер гипоталамуса возникает чрезмерная потребность в пище (гиперфа-
гия), а при их разрушении — отказ от приема пищи (афагия). Мощным сти-
мулятором центра голода является кровь с пониженным содержанием глю-
козы, аминокислот, жирных кислот и глицеридов, продуктов метаболизма
цикла Кребса. Она возбуждает ядра латерального гипоталамуса через хемо-
рецепторы сосудов и рецепторы самого гипоталамуса, избирательно чувст-
вительные к недостатку в крови определенных питательных веществ.
512 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
После приема пищи у животного и человека возникает субъективно
приятное чувство насыщения, сменяющее чувство голода и прекращающее
потребление пищи. Чувство насыщения является следствием возбуждения
центра насыщения, расположенного в вентромедиальных ядрах гипоталаму-
са. Между центрами голода и насыщения имеются реципрокные отноше-
ния (возбуждение одного сопровождается торможением другого).
Чувство насыщения, прекращающее прием пищи, имеет нейрогенную
природу и обусловлено поступлением афферентных импульсов от раздра-
жаемых пищей рецепторов проксимальных отделов пищеварительного
тракта (слизистых оболочек и мускулатуры полости рта, пищевода и же-
лудка). Эта афферентация возбуждает центр насыщения и тормозит центр
голода. Она является причиной сенсорного насыщения (первичного) и пред-
шествует увеличению питательных веществ в крови.
Через 1,5—2 ч после приема пищи исходный уровень содержания пита-
тельных веществ в крови восстанавливается за счет их поступления из ор-
ганов, в которых они находятся в депонированном состоянии.
Так, превращение гликогена печени в глюкозу приводит к возрастанию
ее концентрации в крови. Это обусловливает гуморальное возбуждение
центра насыщения, что является причиной метаболического (обменного,
вторичного) насыщения.
Поддержание уровня концентрации питательных веществ в крови осу-
ществляется за счет поступления из желудочно-кишечного тракта продук-
тов гидролиза пищевых веществ.
11.2. Общая характеристика функций пищеварительной системы
и механизмов ее регуляции
11.2.1. Секреторная функция
Секреторная функция — деятельность пищеварительных желез, вырабаты-
вающих секрет (пищеварительный сок), с помощью ферментов которого в
желудочно-кишечном тракте осуществляется физико-химическое преобра-
зование принятой пищи.
Секреция — процесс образования из веществ, поступивших из крови в
секреторные клетки (гландулоциты), секрета определенного функциональ-
ного назначения и выделения его из железистых клеток в протоки пищева-
рительных желез.
Секреторный цикл железистой клетки состоит из трех последователь-
ных и взаимосвязанных этапов — поглощения веществ из крови, синтеза
из них секреторного продукта и секретовыделения. Клетки пищеваритель-
ных желез по характеру продуцируемого секрета подразделяются на бе-
лок-, мукоид- и минералсекретирующие. Пищеварительные железы отли-
чаются обильной васкуляризацией. Из крови, протекающей по сосудам
железы, секреторные клетки поглощают воду, неорганические и органиче-
ские низкомолекулярные вещества (аминокислоты, моносахариды, жир-
ные кислоты). Этот процесс осуществляется за счет активности ионных
каналов, базальных мембран эндотелиоцитов капилляров, мембран самих
секреторных клеток. Из поглощенных веществ на рибосомах гранулярного
эндоплазматического ретикулума синтезируется первичный секреторный
продукт, который подвергается дальнейшим биохимическим превращени-
ям в аппарате Гольджи и накапливается в конденсирующих вакуолях глан-
дулоцитов. Вакуоли превращаются в гранулы зимогена (профермента), по-
крытые липопротеиновой оболочкой, с помощью которой окончательный
11. Функции пищеварительной системы • 513
секреторный продукт транспортируется через мембрану гландулоцита в
протоки железы.
Гранулы зимогена выводятся из секреторной клетки по механизму экзо-
цитоза: после перемещения гранулы к апикальной части гландулоцита
происходит слияние двух мембран (гранулы и клетки), и через образовав-
шиеся отверстия содержимое гранул поступает в ходы и протоки железы.
По характеру выделения секрета этот тип клеток относят к мерокри-
новым.
Для голокриновых клеток (клеток поверхностного эпителия желудка) ха-
рактерно превращение всей массы клетки в секрет в результате ее фермен-
тативной деструкции. Апокриновые клетки выделяют секрет с апикальной
(верхушечной) частью своей цитоплазмы (клетки протоков слюнных желез
человека в период эмбриогенеза).
Секреты пищеварительных желез состоят из воды, неорганических и
органических веществ. Наибольшее значение для химической трансформа-
ции пищевых веществ имеют ферменты (вещества белковой природы), яв-
ляющиеся катализаторами биохимических реакций. Они относятся к груп-
пе гидролаз, способных присоединять к перевариваемому субстрату Н+ и
ОН”, превращая высокомолекулярные вещества в низкомолекулярные. В
зависимости от способности расщеплять определенные вещества фермен-
ты подразделяются на 3 группы: глюколитические (гидролизующие углево-
ды до ди- и моносахаридов), протеолитические (гидролизующие белки до
пептидов, пептонов и аминокислот) и липолитические (гидролизующие жи-
ры до глицерина и жирных кислот). Гидролитическая активность фермен-
тов возрастает в известных пределах при повышении температуры перева-
риваемого субстрата и наличия в ней активаторов, их активность снижает-
ся под влиянием ингибиторов. Максимальная гидролитическая активность
ферментов слюны, желудочного и кишечного соков обнаруживается при
разном оптимуме pH среды.
Принято различать несколько типов и подтипов пищеварения.
Гидролиз пищевых веществ за счет ферментов, вырабатываемых пище-
варительными железами самого организма, характерен для собственного
типа пищеварения. В результате осуществления собственного типа пище-
варения образуется основное количество олигомеров, поступающих в
кровь и лимфу. Расщепление компонентов пищи ферментами, синтезируе-
мыми микроорганизмами, которые обитают в пищеварительном тракте,
называют симбионтным типом пищеварения (поскольку он является след-
ствием симбиоза организмов хозяина и микробов). Так переваривается
клетчатка в толстом кишечнике человека.
Гидролиз пищевых веществ ферментами, поступающими в пищевари-
тельный тракт вместе с пищей, относят к аутолитическому типу пищеваре-
ния, так как происходит самопереваривание. Аутолитическое пищеварение
играет важную роль у новорожденного, потому что компоненты грудного
молока перевариваются ферментами, входящими в его состав.
В зависимости от локализации процесса гидролиза пищевых веществ
различают два типа пищеварения — внутриклеточное и внеклеточное.
Внутриклеточное пищеварение — расщепление мельчайших частичек
пищевых веществ, поступивших в энтероцит путем эндоцитоза, за счет
клеточных ферментов. Этот тип пищеварения играет важную роль в ки-
шечном пищеварении в раннем постнатальном периоде развития. По мере
формирования функций пищеварительного тракта у ребенка значение
внутриклеточного пищеварения уменьшается. Внеклеточное пищеварение
А. М. Уголев предложил делить на 2 подтипа — дистанционное и присте-
17-6095
514 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
В Г
Рис. 11.1. Схема внеклеточного, внутриклеточного и мембранного гидролиза пище-
вых веществ.
1 — внеклеточная среда; 2 — перевариваемый субстрат и продукты его гидролиза; 3 — фермен-
ты; 4 — внутриклеточная среда; 5 — мембрана энтероцита; 6 — ядро; 7 — внутриклеточная пи-
щеварительная вакуоль, 8 — мезосома.
А — внеклеточное (дистантное) пищеварение. Полимеры и олигомеры пищевых веществ под
влиянием ферментов пищеварительных соков в полости кишки гидролизуются до мономеров,
которые через мембрану энтероцита транспортируются в его цитоплазму. Б — внутриклеточ-
ное цитоплазматическое пищеварение. Олигомеры пищевых веществ проникают через мем-
брану энтероцита в его цитоплазму и под влиянием ферментов, находящихся в цитоплазме,
превращаются в мономеры. В — внутриклеточное вакуольное (внеплазматическое) пищеваре-
ние, связанное с эндоцитозом. В мембране энтероцита образуется выпячивание, которое за-
полняется перевариваемым субстратом и превращается в вакуоль. Вакуоль соединяется с мезо-
сомой, заполненной ферментами, которые расщепляют субстрат до конечных продуктов гид-
ролиза, поступающих через мембрану вакуоли в цитоплазму энтероцита. Г — мембранное пи-
щеварение. Адсорбированные на внешней поверхности мембраны энтероцита ферменты рас-
щепляют олигомеры пищевых веществ до мономеров, которые затем поступают в цитоплазму
клетки.
ночное. Дистанционное (полостное) пищеварение осуществляется в полос-
тях пищеварительного тракта, удаленных от мест выработки ферментов.
В процессе полостного пищеварения деполимеризация молекул пищевых
веществ совершается в основном до олигомеров. Пристеночное пищеваре-
ние (контактное, мембранное) совершается в тонком кишечнике — в при-
стеночном слое слизи, на поверхности ворсинок и микроворсинок, в гли-
кокаликсе (мукополисахаридных нитях, связанных с мембраной микровор-
синок). В слизи и гликокаликсе содержится много адсорбированных фер-
ментов пищеварительных соков, выделенных в полость кишки и располо-
женных на огромной площади соприкосновения с перевариваемым суб-
стратом. Поэтому в процессе пристеночного пищеварения значительно
увеличивается скорость гидролиза пищевых веществ, что приводит к воз-
растанию объема всасывания продуктов гидролиза.
Схема внеклеточного и мембранного пищеварения представлена на
рис. 11.1. Из этой схемы следует, что при внеклеточном пищеварении (А)
ферменты расщепляют субстрат в полости пищеварительного тракта до ко-
нечных продуктов гидролиза, которые затем проникают в цитоплазму эн-
тероцита.
В процессе внутриклеточного цитоплазматического пищеварения (Б)
крупные осколки молекул пищевых веществ проникают через мембрану
энтероцита в его цитоплазму и расщепляются ее ферментами до мономе-
ров. При внутриклеточном вакуольном пищеварении (В) мельчайшие час-
11. Функции пищеварительной системы • 515
Рис. 11.2. Схема деполимеризации
частичек пищевых веществ на по-
верхности микроворсинок энтероци-
та, апикальном и латеральном глико-
каликсе.
1—3 — перевариваемые субстраты в по-
дсети кишки; 4 — апикальный гликока-
ликс; 5 — латеральный гликокаликс; 6 —
мембрана микроворсинки; 7 — микровор-
синки энтероцита.
Крупные осколки пищевых веществ в по-
лости кишки под влиянием ферментов
пищеварительных соков расщепляются до
олигометров (1—3). Их гидролиз на по-
верхности микроворсинок и нитей глико-
каликса (4—6) завершается образованием
монометров, молекулы которых проника-
ют в микроворсинку, а затем — в цито-
плазму энтероцита (8).
тички субстрата захватываются из по-
лости кишки мембраной энтероцита,
образующей выпячивание, которое
превращается в вакуоль. Вакуоль, со-
держащая субстрат, объединяется с
вакуолью (мезосомой), наполненной
ферментом, который и осуществляет
гидролиз субстрата. Мембранное пи-
щеварение (Г) характеризуется тем,
что интенсивный процесс гидролиза
крупных осколков молекулы пита-
тельных веществ до мономеров про-
исходит на поверхности энтероцита за
счет ферментов, адсорбированных на
его мембране.
На рис. 11.2 схематически показа-
но, что на поверхности микроворси-
нок энтероцитов и нитей апикального
и латерального гликокаликса проис-
ходит гидролиз частичек пищевых ве-
ществ за счет адсорбированных фер-
ментов. Поскольку суммарная пло-
щадь, на которой происходит гидро-
лиз пищевых веществ, очень боль-
шая, то это обусловливает высокую
эффективность мембранного пищева-
рения
Заключительные стадии гидролиза
пищевых веществ осуществляют фер-
менты, синтезируемые энтероцитами
и встроенные в структуры их мем-
бран. Образующиеся на поверхности
мембраны энтероцита мономеры вса-
сываются за счет активности ее ион-
ных каналов.
Таблица 11.1. Гормоны желудочно-кишечного тракта, место их образования и вызы-
ваемые ими эффекты
Название гормона Место выработ- ки гормона Типы эндо- крин- ных клеток Эффект действия гормонов
Соматоста- тин Вазоактив- ный инте- стинальный Желудок, про- ксимальный от- дел тонкой кишки, подже- лудочная железа Во всех отделах желудочно-ки- шечного тракта D- клетки Dr клетки Тормозит выделение инсулина и глюкагона, большинства известных желудочно-кишеч- ных гормонов (секретина, ГИПа, мотилина, гастрина); тормозит активность париеталь- ных клеток желудка и ацинарных клеток поджелудочной железы Тормозит действие холецистокинина, секре- цию соляной кислоты и пепсина желудком, стимулированную гистамином, расслабляет
’17*
516 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Продолжение
Название гормона Место выработ- ки гормона Типы эндо- крин- ных клеток Эффект действия гормонов
(ВИП) пеп- тид Панкреати- Поджелудочная D2- гладкие мышцы кровеносных сосудов, желч- ного пузыря Антагонист ХЦК-ПЗ, усиливает пролифера-
ческий по- железа клетки цию слизистой оболочки тонкой кишки,
липептид (ПП) Гастрин Антральная G- поджелудочной железы и печени; участвует в регуляции обмена углеводов и липидов Стимулирует секрецию и выделение пепсина
Гастрон часть желудка, поджелудочная железа, прокси- мальный отдел тонкой кишки Антральный от- клетки G- желудочными железами, возбуждает мотори- ку расслабленного желудка и двенадцатипер- стной кишки, а также желчного пузыря Снижает объем желудочной секреции и вы-
Бульбогас- дел желудка Антральный от- клетки G- ход кислоты в желудочном соке Тормозит секрецию и моторику желудка
трон Дуокринин дел желудка Антральный от- клетки G- Стимулирует выделение секрета бруннеро-
Бомбезин дел желудка Желудок и про- клетки Р-клет- вых желез двенадцатиперстной кишки Стимулирует высвобождение гастрина, уси-
(гастринвы- ксимальный от- ки ливает сокращение желчного пузыря и выде-
свобождаю- щий пептид) Секретин дел тонкой кишки Тонкий кишеч- S-клет- ление ферментов поджелудочной железой, усиливает выделение энтероглюкагона Стимулирует секрецию бикарбонатов и воды
Холецисто- НИК Тонкий кишеч- КИ 1-клет- поджелудочной железой, печенью, железами Бруннера, пепсина; тормозит секрецию в желудке Возбуждает выход ферментов и в слабой сте-
кинин-пан- ник ки пени стимулирует выход бикарбонатов под-
креозимин (ХЦК-ПЗ) Энтероглю- Тонкий кишеч- ЕС,- желудочной железой, тормозит секрецию со- ляной кислоты в желудке, усиливает сокра- щение желчного пузыря и желчевыделение, усиливает моторику тонкой кишки Тормозит секреторную активность желудка,
кагон ник клетки снижает в желудочном соке содержание К+ и
Мот илин Проксималь- ес2- повышает содержание Са2+, тормозит мото- рику желудка и тонкой кишки Возбуждает секрецию пепсина желудком и
Гастроинги- ный отдел тон- кой кишки Тонкий кишеч- клетки К-клет- секрецию поджелудочной железы, ускоряет эвакуацию содержимого желудка Тормозит выделение соляной кислоты и
бирующий ник ки пепсина, высвобождение гастрина, моторику
пептид (ГИП) Нейротен- Дистальный от- N- желудка, возбуждает секрецию толстой кишки Тормозит секрецию соляной кислоты желе-
ЗИН дел тонкой клетки зами желудка, усиливает высвобождение
Энкефали- кишки Проксималь- L-клет- глюкагона Тормозит секрецию ферментов поджелудоч-
ны (эндор- ный отдел тон- КИ ной железой, усиливает высвобождение гаст-
финн) Субстанция кой кишки и поджелудочная железа Тонкая кишка ЕС,- рина, возбуждает моторику желудка Усиливает моторику кишечника, слюноотде-
Р клетки ление, тормозит высвобождение инсулина
11. Функции пищеварительной системы • 517
Продолжение
Название гормона Место выработ- ки гормона Типы эндо- крин- ных клеток Эффект действия гормонов
Вилликинин Двенадцатипер- стная кишка ЕС,- клетки Стимулирует ритмические сокращения вор- синок тонкой кишки
Энтерогас- Двенадцатипер- ЕС,- Тормозит секреторную активность и мотори-
трон стная кишка клетки ку желудка
Серотонин Желудочно-ки- шечный тракт ЕС,, ес2- клетки Тормозит выделение соляной кислоты в же- лудке, стимулирует выделение пепсина, ак- тивирует секрецию поджелудочной железы, желчевыделение, кишечную секрецию
Гистамин Желудочно-ки- шечный тракт ес2- клетки Стимулирует выделение секрета желудка и поджелудочной железы, расширяет крове- носные капилляры, оказывает активирующее влияние на моторику желудка и кишечника
Инсулин Поджелудочная железа Бета- клетки Стимулирует транспорт веществ через кле- точные мембраны, способствует утилизации глюкозы и образованию гликогена, тормозит липолиз, активирует липогенез, повышает интенсивность синтеза белка
Глюкагон Поджелудочная железа Альфа- клетки Мобилизует углеводы, тормозит секрецию желудка и поджелудочной железы, тормозит моторику желудка и кишечника
Регуляция секреторной деятельности пищеварительных желез осуществ-
ляется за счет нервных и гуморальных механизмов. Основными стимули-
рующими секрецию нервными волокнами являются парасимпатические.
Они представляют собой аксоны постганглионарных нейронов. Симпати-
ческие нервные волокна тормозят стимулированную секрецию пищевари-
тельных желез и оказывают на железы трофические влияния, усиливая
синтез компонентов секрета.
Стимуляторами, ингибиторами и модуляторами секреции пищевари-
тельных желез являются гастроинтестинальные регуляторные пептиды
(табл. 11.1).
11.2.2. Моторная функция
Процесс пищеварения во всех отделах пищеварительного тракта осуществ-
ляется при участии двигательной активности его мускулатуры. Сокраще-
ния мышц обеспечивают: прием и измельчение пищи в процессе жевания
в ротовой полости, глотание и продвижение порции пищи по пищеводу,
накопление ее в желудке и эвакуацию его содержимого в кишечник, со-
кращение и расслабление желчного пузыря, перемешивание и продвиже-
ние кишечного содержимого, движение ворсинок, переход химуса из тон-
кой кишки в толстую, его перемещение по толстой кишке, сокращение и
расслабление сфинктеров, перистальтику выводных протоков пищевари-
тельных желез и выведение экскрементов.
Гладкая мускулатура пищеварительного тракта состоит из гладкомы-
шечных клеток (миоцитов). Они собраны в пучки и соединены друг с дру-
гом нексусами. Пучок получает нервные терминали, артериолу и выполня-
ет роль функциональной единицы гладкой мышцы. Миоциты обладают
518 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
способностью к спонтанному ритмическому возбуждению за счет периоди-
ческой деполяризации их мембраны. Это возбуждение распространяется
благодаря нексусам от клетки к клетке (как по синцитию). Пучки миоци-
тов образуют гладкомышечные слои пищеварительной трубки — циркуляр-
ный (внутренний), продольный (наружный) и подслизистый (косой).
Растяжение мышц содержимым желудочно-кишечного тракта является
для них адекватным раздражителем, вызывающим деполяризацию мембран
их клеток и сокращение мышечных волокон. Частота и сила сокращений
миоцитов изменяются в широком диапазоне под влиянием нервных им-
пульсов эфферентных терминалей вегетативных нервных волокон, гормо-
нов и гастроинтестинальных регуляторных пептидов. Комплексная нерв-
но-гуморальная регуляция миоцитов обеспечивает соответствие уровня ак-
тивности мускулатуры объему и составу содержимого желудка и кишеч-
ника.
Характер сократительной деятельности мускулатуры пищеварительного
тракта зависит от активности водителей ритма, расположенных в желудке
и кишечнике (см. раздел 11.5.2). Они представляют собой гладкомышеч-
ные клетки, более чувствительные к биологически активным веществам и
имеющие более обильную иннервацию, чем другие пучки миоцитов.
На протяжении пищеварительного тракта у человека имеется около 35
сфинктеров. Они состоят из мышечных пучков, расположенных циркуляр-
но (в основном), спирально и продольно. Сокращение циркулярных пуч-
ков приводит к смыканию сфинктера, а сокращение спиральных и про-
дольных пучков увеличивает его просвет, что способствует переходу содер-
жимого пищеварительного тракта в нижележащий отдел. Сфинктеры обес-
печивают движение содержимого пищеварительной трубки в каудальном
направлении и временное разобщение функционально различных частей
пищеварительного тракта. Основные из них — кардиальный (на входе в
желудок), пилорический (на выходе из желудка), в основании баугиниевой
заслонки (на входе в слепую кишку), внутренний и наружный анальный
(на выходе из прямой кишки).
Координация сократительной деятельности различных участков муску-
латуры пищеварительной трубки осуществляется за счет регулирующих
влияний периферической и центральной нервной системы. Парасимпати-
ческие нервные волокна в основном усиливают моторику желудка и ки-
шечника. Однако в составе блуждающих нервов имеются и волокна, тор-
мозящие моторику миоцитов. Симпатические нервы оказывают преимуще-
ственно тормозные влияния на мышечные пучки.
Миоциты обладают способностью генерировать медленные ритмиче-
ские колебания мембранного потенциала. В фазе деполяризации мембра-
ны происходит накопление ионов Са2+ в клетке, что активирует кальций-
зависимые калиевые каналы. Это вызывает выход К+ из клетки и приводит
к деполяризации мембраны. Если деполяризация достигает критического
уровня, то на гребне медленной волны возникают потенциалы действия,
которые вызывают открытие быстрых потенциалозависимых кальциевых
каналов, что приводит к сокращению миоцита (рис. 11.3). На этом рисун-
ке приведена синхронная запись медленных и быстрых колебаний мем-
бранного потенциала миоцита тонкой кишки при внутриклеточной и вне-
клеточной регистрации. С появлением серии быстрых потенциалов возни-
кает сокращение мышечного волокна.
Медленные волны деполяризации возникают в миоцитах наружной час-
ти циркулярного слоя и распространяются как на круговой, так и на про-
дольный мышечные слои (по мышечным мостикам). Это определяет по-
11. Функции пищеварительной системы • 519
следовательность сокращения мы-
шечных пучков, расположенных в
разных мышечных слоях. Мем-
брана миоцитов в области распо-
ложения водителей ритма облада-
ет способностью генерировать
медленные волны с более высо-
кой частотой, чем в соседних уча-
стках. Это способствует «навязы-
ванию» частоты медленных волн
клеток водителя ритма миоцитам
соседних участков.
Продольный мышечный слой
имеет выраженную холинергиче-
скую иннервацию, а циркулярный
слой испытывает тонические вли-
яния интрамуральных тормозных
нейронов через их многочислен-
ные пептидергические окончания.
Нейромедиатор окончаний пара-
симпатических нервных волокон
(ацетилхолин), действуя на М-хо-
линорецепторы, вызывает повы-
шение проницаемости клеточной
мембраны к ионам Са2+, Na+ и К+.
тенциалов действия, длительность
кой мышцы, что лежит в основе
Рис. 11.3. Связь между медленными и
быстрыми электрическими волнами мио-
цита и сокращением мышечного волокна
тонкой кишки.
а — внутриклеточная регистрация медленных и
быстрых потенциалов миоцита (потенциалы
действия накладываются на плато волны депо-
ляризации); б — внеклеточная регистрация по-
тенциалов миоцита (серия спайковых потен-
циалов совпадает с медленной волной, регист-
рируемой при внутриклеточном отведении);
в — локальное сокращение гладкого мышечно-
го волокна.
При этом увеличивается количество по-
и амплитуда фазного сокращения глад-
усиления моторики желудка и кишеч-
ника.
При сильном возбуждении парасимпатических постганглионарных ней-
ронов повышается концентрация ацетилхолина во внеклеточной среде.
Это приводит к непрерывной генерации миоцитами потенциалов дейст-
вия, которые сопровождаются слиянием фазных сократительных эффектов
в слитное сокращение.
Катехоламины влияют на миоциты через адренорецепторы двух типов
(альфа и бета), которые подразделяются на четыре подтипа (at и а2, р] и
р2). Воздействие норадреналина на а- и р-адренорецепторы окончаний хо-
линергических нейронов миэнтерального и подслизистого сплетений угне-
тает выход ацетилхолина из холинергических окончаний, что ослабляет па-
расимпатические влияния на миоциты и способствует торможению глад-
кой мускулатуры.
При электрическом раздражении симпатических нервных волокон име-
ет место торможение сократительной активности кишечника и усиление
тонуса циркулярных пучков сфинктеров.
11.2.3. Функция всасывания
Всасывание — это совокупность физиологических и физико-химических
процессов транспорта питательных веществ, минеральных соединений и
витаминов из полости пищеварительного тракта во внутреннюю среду ор-
ганизма (кровь, лимфу, тканевую жидкость). Всасывание веществ осущест-
вляется на всем протяжении пищеварительного тракта. Но интенсивность
этого процесса в разных ее отделах не одинакова. В ротовой полости вса-
сывание компонентов пищи осуществляется в ничтожно малых объемах.
520 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Гликокаликс
Филаменты
актина
Клеточная
мембрана
Филаменты
миозина
Практическое значение имеет
всасывание лишь некоторых ле-
карственных веществ (например,
нитроглицерина, валидола). В же-
лудке всасывается небольшое ко-
личество воды, минеральных со-
лей, аминокислот, глюкозы. В
значительном количестве из же-
лудка всасывается алкоголь. Ос-
новным местом всасывания пита-
тельных веществ, минеральных
солей и воды является слизистая
оболочка тонкого кишечника. В
толстом кишечнике всасываются
вода, некоторые минеральные со-
ли и продукты микробного гидро-
лиза компонентов пищи. Слизи-
стая оболочка тонкого кишечника
представляет собой специализи-
рованный орган всасывания. За
счет складок, ворсинок и микро-
Рис. 11.4. Роль сократительных белков
щеточной каймы энтероцита.
Филаменты актина, расположенные вдоль
микроворсинки, у ее основания приобретают
направление расположения филаментов мио-
зина. Взаимодействие актина и миозина вызы-
вает движение микроворсинки, что способст-
вует транспорту в цитоплазму энтероцита про-
никших через мембрану микроворсинки моно-
меров пищевых веществ.
ворсинок ее всасывательная по-
верхность возрастает в 300—
500 раз (в сравнении с ее площа-
дью без учета перечисленных ана-
томо-гистологических образова-
ний) и составляет у человека око-
ло 200 м2. На 1 мм2 слизистой
оболочки приходится от 30 до 40
ворсинок. На апикальной мем-
бране энтероцита, обращенной в
полость кишки, обнаружено от
1700 до 4000 микроворсинок. У взрослого человека имеется около 1010 эн-
тероцитов. Следовательно, на 1 мм2 слизистой оболочки кишки приходит-
ся 50—100 млн. микроворсинок. Высокая интенсивность всасывания из
тонкой кишки тесно сопряжена с высокой эффективностью гидролиза пи-
щевых веществ, обусловленной механизмом мембранного пищеварения и
пространственной близостью встроенных в мембрану энтероцита молекул
ферментов и транспортных систем продуктов гидролиза.
Процессу всасывания способствует взаимодействие филаментов белка
актина микроворсинки с филаментами белка миозина щеточной каймы
энтероцита (рис. 11.4).
В процессе гидролиза высокомолекулярных веществ и последующего
всасывания продуктов гидролиза принимает участие гликокаликс на по-
верхности мембраны микроворсинки. Гликокаликс состоит из мукополи-
сахаридных нитей, образующих слой толщиной около 0,1 мкм. Нити свя-
заны друг с другом кальциевыми мостиками и образуют сеть, которая вы-
полняет роль молекулярного сита, препятствующего проникновению к
мембране микроворсинки высокомолекулярных веществ. Гликокаликс
удерживает на поверхности кишечного эпителия слой слизи и образует
единый комплекс, который адсорбирует из содержимого кишки гидроли-
тические ферменты, продолжающие полостной гидролиз на поверхности
энтероцита. На мембране микроворсинки процесс деполимеризации моле-
]]. Функции пищеварительной системы • 521
кул пищевых веществ завершается. Образовавшиеся мономеры через мем-
брану микроворсинки поступают в энтероцит.
В транспорте питательных веществ в энтероцит важную роль играют
микроциркуляторная система ворсинок и их сократительная деятельность.
Сеть капилляров располагается непосредственно под базальной мембраной
энтероцитов. Это способствует транспорту веществ через мембрану энте-
роцита в кровь. Эндотелий капилляров имеет большое количество фенестр
значительного размера (45—67 нм), через которые из межклеточных про-
странств в кровь проникают крупные молекулы и надмолекулярные струк-
туры. При сокращении мускулатуры ворсинки из нее выжимается лимфа в
более крупные лимфатические сосуды, а во время ее расслабления создает-
ся присасывающий эффект, так как возврату лимфы препятствуют клапа-
ны лимфатических сосудов. Снижение давления в лимфатическом сосуде
ворсинки способствует транспорту веществ из энтероцитов и межклеточ-
ных пространств между ними.
Всасывание макромолекул. Крупные молекулы и их агрегаты всасывают-
ся в кишечнике по механизму трансцитоза. В энтероцит они поступают
путем эндоцитоза. В везикуле, образовавшейся из участка мембраны клет-
ки, вещество транспортируется через цитоплазму энтероцита и выделяется
из него в межклеточное пространство путем экзоцитоза. При этом мембра-
на везикулы (вакуоли), содержащей макромолекулы транспортируемых ве-
ществ, «встраивается» в мембрану энтероцита. Сокращения ворсинок спо-
собствуют эндоцитозу. Так в кишечнике транспортируются иммуноглобу-
лины, витамины, ферменты, а у новорожденных — белки грудного молока.
Всасывание микромолекул энтероцитами осуществляется по механизму
пассивного транспорта (диффузии и осмоса, облегченной диффузии и
фильтрации), а также активного транспорта.
Движение молекул через полупроницаемые биологические мембраны
энтероцитов в процессе диффузии и осмоса происходит вследствие концен-
трационных градиентов веществ в цитоплазме клетки и внеклеточной среде.
Для облегченной диффузии необходимо наличие мембранных переносчиков.
Процесс фильтрации растворенных в воде веществ осуществляется в си-
лу разности давления жидкости над мембраной, выполняющей роль
фильтра, и под ней. О существовании механизма фильтрации при всасыва-
нии веществ в кишечнике свидетельствует увеличение скорости всасыва-
ния изотонического раствора натрия хлорида при повышении внутрики-
шечного давления в процессе осуществления моторики кишки.
Двигательная активность кишечника способствует всасыванию продук-
тов гидролиза пищевых веществ еще и потому, что она обеспечивает пере-
мешивание пристеночного слоя содержимого кишечника.
Процесс всасывания регулируется с помощью нервных и гуморальных
механизмов. При механическом раздражении ворсинок тонкой кишки и
под влиянием продуктов гидролиза пищевых веществ (пептидов, амино-
кислот, глюкозы) они резко усиливают и учащают свои сокращения. Этот
эффект сохраняется и после перерезки парасимпатических и симпатиче-
ских нервов кишечника в опытах на Животных, что говорит о его реализа-
ции через энтеральную нервную систему.
Редкая и слабая сократительная активность ворсинок кишки голодного
животного значительно оживляется при переливании ему крови сытого
животного. Это свидетельствует о наличии гуморальных регуляторов вса-
сывания. Установлено, что мощным гуморальным стимулятором активно-
сти ворсинок является гормон вилликин, выделяемый эндокринными
клетками слизистой оболочки двенадцатиперстной кишки.
522 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Вода
Алкоголь
Кальций
Магний
Железо
Водорастворимые
витамины
П ищеварительн ый
тракт
Жирорастворимые
витамины
елудок
Желчный проток
Белки и аминокислоты
одвздошная
Жиры, жирные кислоты,
моноглицериды
Проток
поджелудочной
железы
Моносахариды,
глюкоза
венадцати-
перстная
кишка
Рис. 11.5. Топография всасывания различных веществ в разных последовательных
отделах пищеварительного тракта.
Стрелки указывают, что перечисленные вещества поступают из полости пищеварительного
тракта во внутреннюю среду организма.
Обобщенная схема, отражающая топографию всасывания различных ве-
ществ, представлена на рис. 11.5.
Из данного рисунка следует, что зоны всасывания различных веществ
частично или полностью перекрываются. В желудке всасываются в основ-
ном вода, соединения меди, алкоголь. Продукты гидролиза белков, жиров
11. Функции пищеварительной системы • 523
и углеводов всасываются в тех же отделах пищеварительного тракта, где
осуществляется их переваривание, т. е. в двенадцатиперстной и тощей
кишке. Жирорастворимые и водорастворимые витамины (кроме витамина
В12) всасываются в тощей кишке. В подвздошной кишке осуществляется
всасывание солей желчных кислот и витамина В)2. Двухзарядные катионы
(цинка, кальция, магния и железа) всасываются в основном в двенадцати-
перстной кишке, а вода, хлориды, основания, жирные кислоты и газы — в
толстой кишке.
11.2.4. Общая характеристика механизмов регуляции функций
пищеварительной системы
Вне периода пищеварения железы и гладкие мышцы желудочно-кишечно-
го тракта находятся у человека в состоянии относительного покоя, кото-
рый на короткие промежутки времени прерывается периодической («го-
лодной») активностью (см. раздел 11.3). Прием пищи вызывает рефлектор-
ное усиление секреции слюнных, желудочных и поджелудочных желез, вы-
деление желчи из общего желчного протока (что обусловлено поступлени-
ем нервных импульсов из парасимпатических центров регуляции), кратко-
временное расслабление мускулатуры желудка (пищевая релаксация) и ос-
лабление моторики проксимального отдела тонкой кишки (что является
следствием возбуждения симпатических нервных волокон).
Содержимое желудка и кишечника поддерживают вызванную рефлек-
торным путем секторную и двигательную активность вплоть до заверше-
ния пищеварения в кишечнике и всасывания продуктов гидролиза пи-
щевых веществ. Этот эффект является следствием влияния на рецепто-
ры и эндокринные элементы слизистой оболочки желудка и кишечника
объема химуса, его консистенции, осмотического давления, pH, темпе-
ратуры, продуктов гидролиза пищевых веществ и экстрактивных ве-
ществ.
Нервная регуляция секреции пищеварительных соков и моторики же-
лудка и кишечника осуществляется с помощью центральных, перифериче-
ских и местных рефлексов. Примером центрального рефлекса является мо-
торный пищеводно-кишечный эффект, реализуемый через ядро блуждаю-
щего нерва продолговатого мозга (рис. 11.6),
примером периферического — антрофундальная
тормозная реакция, рефлекторная дуга которой
замыкается в симпатическом ганглии солнечно-
го сплетения (рис. 11.7), примером местного —
изменения моторики желудка через нейроны
миэнтерального сплетения (рис. 11.8).
Начальные отделы пищеварительного тракта
(слюнные железы, мышцы, осуществляющие
жевание и глотание, пищевода, желудка и
Рис. 11.6. Рефлекторная дуга центрального пи-
щеводно-кишечного моторного рефлекса.
1 — механорецепторы пищевода; 2 — афферентный нейрон
ганглия; 3 — чувствительное ядро продолговатого мозга; 4 —
ядро блуждающего нерва (тело преганглионарного нейрона);
5 — преганглионарное парасимпатическое волокно; 6 — ганг-
лионарный парасимпатический нейрон; 7 — гладкомышеч-
ные клетки тонкой кишки.
524 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 11.7. Рефлекторная дуга перифери-
ческого антрофундального тормозного
рефлекса.
1 — механорецепторы антрального отдела же-
лудка; 2 — сенсорный нейрон (клетка Догеля
II типа); 3 — симпатический ганглий солнеч-
ного сплетения; 4 — ганглионарный симпати-
ческий нейрон; 5 — гладкомышечные клетки
фундального отдела желудка.
сфинктера Одди, гландулоциты же-
лудка и поджелудочной железы) в
наибольшей степени подвержены
влияниям УНС. Ее роль в регуля-
ции тонкого и толстого кишечника
снижается, но значение интраор-
ганной (энтеральной) нервной сис-
темы возрастает. Окончания аксо-
нов ее нейронов выделяют различ-
ные медиаторы. Возбуждающие
влияния на миоциты и гландуло-
циты оказывают холинергические
нейроны, а тормозные — оконча-
ния аксонов постганглионарных
симпатических нейронов. Тормоз-
ной эффект осуществляется также
за счет угнетения терминалей хо-
линергических волокон, располо-
женными на них окончаниями ак-
сонов симпатических нейронов.
Торможение миоцитов и гландуло-
цитов может быть достигнуто за
счет влияния тормозных медиаторов пептидергических нейронов — вазоак-
тивного интестинального пептида (ВИП) и АТФ.
Центральные, периферические и местные рефлексы осуществляются в
тесном взаимодействии с гуморальным механизмом регуляции миоцитов,
гландулоцитов и нервных клеток.
В слизистой оболочке желудочно-кишечного тракта и в поджелудочной
железе имеются эндокринные клетки, которые вырабатывают гастроинте-
стинальные гормоны (регуляторные пептиды, энтерины). Эти гормоны че-
рез кровоток и местно (паракринно, диффундируя через межклеточную
жидкость) оказывают влияние на миоциты, гландулоциты, интрамураль-
ные нейроны и эндокринные клетки. Их выработка запускается рефлек-
торно (через блуждающий нерв) во время приема пищи и длительное вре-
Рис. 11.8. Рефлекторная дуга местного моторного рефлекса желудка.
1 — механорецепторы; 2 —продольный мышечный слой; 3, 4 — нейроны миэнтерального
сплетения (3 — сенсорный, 4 — эфферентный холинергический); 5 — циркулярный мышеч-
ный слой.
11. Функции пищеварительной системы • 525
мя поддерживается за счет раздражающего влияния продуктов гидролиза
пищевых веществ и экстрактивных веществ.
Место выработки основных гастроинтестинальных гормонов, вызывае-
мые ими эффекты и продуцирующие их клетки представлены в табл. 11.1.
В настоящее время обнаружено около 30 регуляторных пептидов. Как сле-
дует из представленной таблицы, они оказывают стимулирующее, тормоз-
ное и модулирующее влияние на секрецию пищеварительных соков, мото-
рику гладкой мускулатуры желудочно-кишечного тракта, всасывание, вы-
деление энтеринов эндокринными элементами слизистой оболочки желуд-
ка, кишечника и поджелудочной железы.
Выделение гастроинтестинальных гормонов имеет каскадный характер.
Например, под влиянием гастрина обкладочные клетки желез желудка уве-
личивают выработку соляной кислоты, которая в слизистой оболочке тон-
кой кишки стимулирует выделение S- и J-клетками секретина и холеци-
стокинна — панкреозимина. Секретин усиливает секрецию воды и бикар-
бонатов поджелудочной железой и печенью, а холецистокинин — панкрео-
зимин — возбуждает выделение ферментов поджелудочной железой и тор-
мозит секрецию соляной кислоты обкладочными клетками, усиливает мо-
торику тонкой кишки и желчного пузыря.
Регуляторные пептиды, поступая в кровоток, быстро разрушаются в пе-
чени и почках и тем самым создают условия для осуществления эффектов
других гастроинтестинальных гормонов.
Выработка некоторых энтеринов носит циклический характер и может
осуществляться и при отсутствии пищевого раздражителя. Например, мо-
талин, вырабатываемый ЕС2-клетками проксимального отдела тонкой
кишки, вызывает сокращения мышц желудка и кишечника, совпадающие
с периодами «голодной» активности пищеварительного тракта (см. раздел
11.3).
Обобщенная схема механизмов регуляции функций пищеварительного
тракта представлена на рис. 11.9. Эта схема демонстрирует взаимодействие
нервных и гуморальных механизмов регуляции миоцитов, гландулоцитов и
эндокринных клеток пищеварительного тракта. В ответ на внешние раз-
дражители, адресованные экстерорецепторам, структуры пищеварительно-
го тракта через вегетативные нервные волокна получают регулирующие
влияния, причем по парасимпатическим волокнам — возбуждающие, а по
симпатическим — тормозные. При раздражении интерорецепторов слизи-
стой оболочки желудочно-кишечного тракта компонентами химуса (про-
дуктами гидролиза пищи, экстрактивными веществами, крупными фраг-
ментами пищи) рефлекторно изменяется активность мышечных, секретор-
ных и эндокринных элементов. Рефлекс с интерорецепторов удлиняет во
времени секреторную и моторную реакцию, вызванную раздражением экс-
терорецепторов. Эндокринное звено этого рефлекса подкрепляется отве-
том эндокриноцитов на непосредственные химические раздражители, диф-
фундирующие в слизистую оболочку пищеварительного тракта из химуса,
заполняющего его полость. Гастроинтестинальные гормоны через крово-
ток и интерстициальные пространства достигают регулируемых секретор-
ных, мышечных и эндокринных элементов.
Нервные импульсы от рецепторов желудочно-кишечного тракта по аф-
ферентным путям достигают пищевого центра, доставляя информацию о
состоянии процесса пищеварения. Она дополняется гуморальными влия-
ниями, обусловленными поступлением в кровь продуктов гидролиза пище-
вых веществ и гастроинтестинальных гормонов. Информация, поступаю-
щая в пищевой центр по каналам нервной и гуморальной обратной связи,
526 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Экстерорецепторы
Внешние стимулирующие
Секреты Стимулирующие и тормозные
Полость желудочно-кишечного тракта влияния из полости желудочно-
кишечного тракта
Рис. 11.9. Схема механизмов регуляции функций желудочно-кишечного тракта.
ПЖ — пищеварительные железы, ГМ — гладкие мышцы, ЭК — эндокринные клетки, Р — ре-
цепторы, АН — афферентные нейроны, ЭН — эфферентные нейроны, С и П — аксоны симпа-
тических и парасимпатических преганглионарных нейронов, ПК — паракринные влияния,
ЦНС — центральная нервная система
вызывает его ответную реакцию, направленную на оптимизацию процесса
пищеварения и пищевого поведения животного и человека (см. раздел
11.1).
11.3. Периодическая деятельность пищеварительной системы
Мембрана гладких мышечных волокон пищеварительного тракта обладает
способностью генерировать медленные электрические волны деполяриза-
ции (от 3 до 12 в минуту). Во время деполяризации повышается проницае-
мость мембраны к ионам натрия. Если снижение мембранного потенциала
11. Функции пищеварительной системы • 527
достигает критического уровня, то повышается проницаемость к ионам
кальция, и на высоте медленной волны возникает серия потенциалов дей-
ствия, которые вызывают сокращение миоцитов, лежащей в основе пери-
стальтической волны. Этот ритм электрической активности называется ба-
зальным.
Ритмические колебания мембранного потенциала миоцитов желудка со-
вершаются с частотой около 3 циклов в минуту. Они обусловлены часто-
той электрической активности водителя ритма, расположенного в области
малой кривизны рядом с кардиальным сфинктером желудка. В двенадца-
типерстной кишке частота ритма колеблется от 10 до 12 в минуту и зави-
сит от работы водителя ритма, расположенного в области впадения общего
желчного протока в кишку. Частота медленных волн в подвздошной киш-
ке уменьшается до 8—6 в минуту, а в толстой кишке — до 6— 1.
На фоне базального ритма (медленных колебаний мембранного потен-
циала миоцитов) у человека вне периода пищеварения (в состоянии отно-
сительного покоя) через каждые 45—90 мин возникает так называемая го-
лодная периодическая деятельность пищеварительного тракта, которая про-
должается 20—50 мин. Она проявляется вначале в нерегулярных сокраще-
ниях гладкой мускулатуры, переходящих в регулярные. Голодная моторика
с желудка и двенадцатиперстной кишки перемещается до конечного отдела
подвздошной кишки. После завершения сокращений подвздошной кишки
начинается новый цикл голодной моторики в желудке и двенадцатиперст-
ной кишке (мигрирующий моторный комплекс).
В настоящее время установлено, что вне периода пищеварения (в со-
стоянии физиологического голода) концентрация глюкозы в крови снижа-
ется, что возбуждает хеморецепторы кровеносных сосудов и желудочно-
кишечного тракта, а также глюкорецепторы гипоталамуса. Это вызывает
возбуждение гипоталамического пищевого центра, что инициирует актив-
ность подкорковых и корковых структур, являющихся субстратом пищево-
го поведения, а также ядер блуждающих нервов, стимулирующих пищева-
рительный тракт.
Поступление эфферентных нервных импульсов по парасимпатическим
нервам к миоцитам пищеварительного тракта и вызывают его периодиче-
скую голодную перистальтику.
Центральный механизм возникновения голодной моторики дополняет-
ся периферическим, лежащим в основе осуществления деятельности энте-
рильной нервной системы. Об этом говорит тот факт, что после перерезки
вегетативных нервов желудка и кишечника периодическая моторика пище-
варительного тракта не прекращается.
Вероятно, ферменты пищеварительных соков поджелудочной железы и
кишечных желез, поступающие в кишечник, готовят его к осуществлению
полостного и пристеночного пищеварения.
Высказывается мнение, что во время периодической перистальтики пи-
щеварительного тракта из него удаляются остатки химуса и экскретируе-
мые вещества, а также предотвращается проникновение в тонкий кишеч-
ник микроорганизмов из толстой кишки.
В желудочно-кишечный тракт вне периода пищеварения в составе сек-
ретов и слущивающихся эпителиоцитов поступает значительное количест-
во органических веществ, которые подвергаются гидролизу выделяющими-
ся во время периодической деятельности ферментами. Продукты гидроли-
за усваиваются в кишечнике и тем самым обеспечивают эндогенное пита-
ние организма в условиях физиологического голода (при отсутствии пище-
вых веществ в пищеварительном тракте).
528 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
11.4. Пищеварение в ротовой полости и функция глотания
11.4.1. Ротовая полость
Ротовая полость является начальным отделом пищеварительного тракта,
где осуществляется анализ вкусовых свойств веществ и разделение их на
пищевые и отвергаемые, защита пищеварительного тракта от попадания
некачественных пищевых веществ и экзогенной микрофлоры, измельче-
ние, смачивание слюной пищи, начальный гидролиз углеводов и формиро-
вание пищевого комка. Кроме того происходит раздражение механо-, хе-
мо-, терморецепторов, вызывающее рефлекторное возбуждение деятельно-
сти слюнных желез, желез желудка, поджелудочной железы, печени, желез
двенадцатиперстной кишки.
Защита организма от патогенной микрофлоры, попадающей в ротовую
полость, осуществляется благодаря наличию в слюне бактерицидного ве-
щества лизоцима (муромидазы), антивирусному действию нуклеазы слю-
ны, способности иммуноглобулина А слюны связывать экзотоксины, а
также в результате фагоцитоза лейкоцитов, содержащихся в слюне.
Пища находится в ротовой полости 16—18 с, и за это время слюна сма-
чивает сухие вещества пищи, растворяет растворимые и обволакивает твер-
дые ее частички, нейтрализует раздражающие жидкости или уменьшает их
концентрацию, облегчает удаление несъедобных (отвергаемых) веществ,
смывая их со слизистой оболочки ротовой полости.
11.4.2. Слюноотделение
У человека имеется три пары больших слюнных желез {околоушные, подъя-
зычные, подчелюстные) и большое количество мелких желез, локализован-
ных в слизистой оболочке рта. Слюнные железы состоят из слизистых и
серозных клеток. Первые выделяют мукоидный секрет густой консистен-
ции, вторые — жидкий, серозный или белковый. Околоушные слюнные
железы содержат только серозные клетки. Такие же клетки находятся и на
боковых поверхностях языка. Подчелюстные и подъязычные содержат как
серозные, так и слизистые клетки. Подобные железы расположены и в
слизистой оболочке губ, щек, на кончике языка. Подъязычные и мелкие
железы слизистой оболочки выделяют секрет постоянно, а околоушные и
подчелюстные — при их стимуляции.
Ежедневно у человека продуцируется от 0,5 до 2,0 л слюны. Ее pH ко-
леблется от 5,25 до 8,0, а скорость секреции слюны у человека при «спо-
койном» состоянии слюнных желез составляет 0,24 мл/мин. Однако ско-
рость секреции может колебаться даже в состоянии покоя от 0,01 до
18,0 мл/мин, что обусловлено раздражением рецепторов слизистой обо-
лочки ротовой полости и возбуждением слюноотделительного центра под
влиянием условных раздражителей. Слюноотделение при жевании пищи
возрастает до 200 мл/мин.
Количество и состав секрета слюнных желез меняется в зависимости от
характера раздражителя. Слюна человека представляет собой вязкую, опа-
лесцирующую, слегка мутную (благодаря присутствию клеточных элемен-
тов) жидкость с удельным весом 1,001—1,017 и вязкостью 1,10—1,33.
Секрет смешанных всех слюнных желез человека содержит 99,4—99,5 %
воды и 0,5—0,6 % плотного остатка, который состоит из неорганических и
органических веществ (табл. 11.2). Неорганические компоненты в слюне
представлены ионами калия, натрия, кальция, магния, железа, меди, хло-
11. Функции пищеварительной системы • 529
Таблица 11.2. Состав смешанного секрета всех слюнных желез человека
Вещество Содержание, г/л Вещество Содержание, ммоль/л
Вода 994 Соли натрия 6-23
Белки 1,4-6,4 Соли калия 14-41
Муцин 0,9-6,0 Соли кальция 1,2-2,7
Холестерин 0,02-0,50 Соли магния 0,1-0,5
Глюкоза 0,1-0,3 Хлориды 5-31
Аммоний 0,01-0,12 Гидрокарбонаты 2-13
Мочевая кислота 0,005-0,030 Мочевина 140-750
ра, фтора, йода, роданистых соединений, фосфата, сульфата, бикарбоната
и составляют примерно ’/3 часть плотного остатка, а 2/3 приходится на ор-
ганические вещества. Минеральные вещества слюны поддерживают опти-
мальные условия среды, в которой осуществляется гидролиз пищевых ве-
ществ ферментами слюны (осмотическое давление, близкое к нормально-
му, необходимый уровень pH). Значительная часть минеральных компо-
нентов слюны всасывается в кровь слизистой оболочки желудка и кишеч-
ника (см. раздел 11.2.3). Это говорит об участии слюнных желез в поддер-
жании постоянства внутренней среды организма.
Органические вещества плотного остатка — это белки (альбумины, гло-
булины, свободные аминокислоты), азотсодержащие соединения небелко-
вой природы (мочевина, аммиак, креатин), лизоцим и ферменты (альфа-
амилаза и мальтаза). Альфа-амилаза является гидролитическим ферментом
и расщепляет 1,4-глюкозидные связи в молекулах крахмала и гликогена с
образованием декстринов, а затем мальтозы и сахарозы. Мальтаза (глюко-
зидаза) расщепляет мальтозу и сахарозу до моносахаридов. Вязкость
и ослизняющие свойства слюны обусловлены наличием в ней мукополиса-
харидов (муцина). Слизь слюны склеивает частички пищи в пищевой ко-
мок; обволакивает слизистую оболочку ротовой полости и пищевода, она
защищает ее от микротравм и проникновения патогенных микробов. Дру-
гие органические компоненты слюны, например холестерин, мочевая ки-
слота, мочевина, являются экскретами, подлежащими удалению из орга-
низма.
Слюна образуется как в ацинусах, так и в протоках слюнных желез.
В цитоплазме железистых клеток содержатся секреторные гранулы, распо-
лагающиеся преимущественно в околоядерной и апикальной частях кле-
ток, вблизи аппарата Гольджи. В ходе секреции размер, количество и рас-
положение гранул изменяются. По мере созревания секреторных гранул
они смещаются от аппарата Гольджи к вершине клетки. В гранулах осуще-
ствляется синтез органических веществ, которые двигаются с водой через
клетку по эндоплазматической сети. В ходе секреции слюны количество
коллоидного материала, находящегося в виде секреторных гранул, посте-
пенно уменьшается по мере его расходования и возобновляется в период
покоя в процессе его синтеза.
В ацинусах слюнных желез осуществляется первый этап образования
слюны. В первичном секрете содержится альфа-амилаза и муцин, которые
синтезируются гландулоцитами. Содержание ионов в первичном секрете
незначительно отличается от их концентрации во внеклеточных жидко-
стях, что говорит о переходе этих компонентов секрета из плазмы крови.
В слюнных протоках состав слюны существенно изменяется по сравнению
с первичным секретом: ионы натрия активно реабсорбируются, а ионы ка-
530 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
лия активно секретируются, но с меньшей скоростью, чем всасываются
ионы натрия. В результате концентрация натрия в слюне снижается, тогда
как концентрация ионов калия возрастает. Существенное преобладание
реабсорбции ионов натрия над секрецией ионов калия увеличивает элек-
тронегативность мембран клеток слюнных протоков (до 70 мВ), что вызы-
вает пассивную реабсорбцию ионов хлора. Одновременно усиливается сек-
реция ионов бикарбоната эпителием протоков, что обеспечивает ощелачи-
вание слюны.
Отделение слюны является сложным рефлекторным актом, вызванным
раздражением рецепторов ротовой полости пищей или другими вещества-
ми (безусловно-рефлекторными раздражителями), а также раздражением
зрительных и обонятельных рецепторов внешним видом и запахом пищи,
видом обстановки, в которой происходит прием пищи (условно-рефлектор-
ными раздражителями).
Возбуждение, возникающее при раздражении механо-, хемо- и терморе-
цепторов ротовой полости, достигает центра слюноотделения в продолго-
ватом мозге по афферентным волокнам V, VII, IX, X пар черепно-мозго-
вых нервов. Эфферентные влияния к слюнным железам поступают по па-
расимпатическим и симпатическим нервным волокнам. Преганлионарные
парасимпатические волокна к подъязычным и подчелюстным слюнным
железам находятся в составе барабанной струны (ветвь VII пары) к подъя-
зычному и подчелюстному ганглиям, расположенным в теле соответствую-
щих желез, а постганглионарные — от указанных ганглиев к секреторным
клеткам и сосудам желез. К околоушным железам преганглионарные пара-
симпатические волокна идут от нижнего слюноотделительного ядра про-
долговатого мозга в составе IX пары черепно-мозговых нервов к ушному
узлу, от которого постганглионарные волокна направляются к секретор-
ным клеткам и сосудам.
Прегангионарные симпатические волокна, иннервирующие слюнные
железы, являются аксонами нейронов боковых рогов II—VI грудных сег-
ментов спинного мозга и заканчиваются в верхнем шейном ганглии. От-
сюда постганглионарные симпатические волокна направляются к слюн-
ным железам. Электрическое раздражение парасимпатических нервов в
экспериментах на животных вызывает обильную секрецию жидкой слюны,
содержащей небольшие количества органических веществ, а в гландулоци-
тах уменьшается количество секреторных гранул. При стимуляции симпа-
тических нервов из слюнных протоков выделяется небольшое количество
густой и вязкой слюны с высоким содержанием ферментов и муцина, а в
гландулоцитах увеличивается количество секреторных гранул. В связи с
этим парасимпатические нервы называют секреторными, а симпатиче-
ские — трофическими. При раздражении рецепторов слизистой оболочки
рта пищевыми веществами у животных и человека парасимпатические
влияния на слюнные железы преобладают над симпатическими, что вызы-
вает обильную секрецию жидкой слюны.
Во время приема пищи раздражаются тактильные, температурные и
вкусовые рецепторы слизистой оболочки полости рта. Афферентные им-
пульсы от них по чувствительным волокнам тройничного, языкоглоточно-
го, лицевого и блуждающего нервов достигают слюноотделительного цен-
тра продолговатого мозга. При возбуждении верхнего слюноотделительно-
го ядра центра эфферентные импульсы по преганглионарным парасимпа-
тическим волокнам барабанной струны достигают подъязычного и нижне-
челюстного ганглиев, где они переключаются на постганглионарные, пара-
симпатические волокна подъязычного нерва, иннервирующего подъязыч-
11. Функции пищеварительной системы • 531
ные и подчелюстные слюнные железы. При возбуждении нижнего слюно-
отделительного ядра центра эфферентные импульсы по преганглионарным
парасимпатическим волокнам языкоглоточного нерва достигают ушного
ганглия, где они переключаются на постганглионарные волокна ушнови-
сочного нерва, иннервирующего околоушные слюнные железы.
Под влиянием эфферентных импульсов в окончаниях парасимпатиче-
ских постганглионарных волокон выделяется ацетилхолин, возбуждающий
гландулоциты и расширяющий кровеносные сосуды железы. Поэтому сек-
реторный эффект сопровождается повышением уровня кровоснабжения
железы.
Поступающие в продолговатый мозг афферентные импульсы возбужда-
ют также чувствительные нейроны одиночного пучка, по аксонам которых
сенсорные импульсы достигают ядер таламуса, где они переключаются на
таламокортикальное пути и достигают коркового представительства вкусо-
вой сенсорной системы (в области роландовой борозды). В коре больших
полушарий сенсорная информация переключается на эфферентные корко-
вые нейроны, аксоны которых передают эфферентные импульсы на пара-
симпатические и симпатические ядра гипоталамуса. Нисходящие влияния
от парасимпатических ядер активируют бульбарный слюноотделительный
центр, а от симпатических ядер — активируют преганглионарные симпати-
ческие нейроны II—V грудных сегментов спинного мозга, где возбуждение
переключается на постганглионарные волокна, в окончаниях которых вы-
деляется норадреналин.
Центры слюноотделения рефлекторно могут не только возбуждаться, но
и тормозиться. Например, при болевом раздражении, во время отрицатель-
ных эмоций и при умственном напряжении слюноотделение резко ослаб-
ляется или прекращается («пересыхает во рту»).
Фармакологические вещества, относящиеся к группе холиномиметиков
(например, пилокарпин, прозерин), усиливают слюноотделение, так как
способствуют выделению ацетилхолина, а холинолитики — блокируют (на-
пример, атропин). Обильное отделение слюны наблюдается при асфиксии
вследствие раздражения слюноотделительного центра угольной кислотой.
11.4.3. Жевание
Жевание — физиологический акт, заключающийся в измельчении с помо-
щью зубов пищевых веществ и формировании пищевого комка. Жевание
обеспечивает качество механической обработки пищи и определяет время
ее пребывания в полости рта, оказывает рефлекторное возбуждающее
влияние на секреторную и моторную деятельность желудка и кишечника.
В жевании участвуют верхняя и нижняя челюсти, жевательная и мимиче-
ская мускулатура лица, язык, мягкое небо. Механическая обработка пищи
между верхними и нижними рядами зубов осуществляется благодаря пере-
мещению нижней челюсти относительно верхней. У взрослого человека в
ряду справа и слева имеются зубы разного функционального назначения —
два резца и один клык (откусывающие пищу), два малых и три больших
коренных, которые раздавливают и растирают пищу, — всего 32 зуба. Про-
цесс жевания имеет четыре фазы — введения пищи в рот, ориентировоч-
ную, основную и формирования пищевого комка (рис. 11.10).
Регуляция жевания осуществляется рефлекторно. Возбуждение от рецеп-
торов слизистой оболочки рта (механо-, хемо- и терморецепторов) переда-
ется по афферентным волокнам II, III ветви тройничного, языкоглоточно-
го, верхнего гортанного нерва и барабанной струны в центр жевания, ко-
532 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
1 1 11 111 1 zi 6 IV 1 Л V 1
। । . _а _ Я. Qi _______- I-
Рис. 11.10. Кимограмма жевательного периода.
I — покой жевательной мускулатуры; II — фаза введения пищи в рот; III — ориентировочная
фаза; IV — основная фаза; V — фаза формирования пищевого комка; а—б — опускание ниж-
ней челюсти, б—в — подъем нижней челюсти; о—О| — момент размалывания пищи. Под кимо-
граммой — отметка времени — 1 с.
торый находится в продолговатом мозге. Возбуждение от центра к жева-
тельным мышцам передается по эфферентным волокнам тройничного, ли-
цевого и подъязычного нервов. Возбуждение от чувствительных ядер ство-
ла мозга по афферентному пути через специфические ядра таламуса пере-
ключается на корковый отдел вкусовой сенсорной системы, где осуществ-
ляется анализ и синтез информации, поступающей от рецепторов слизи-
стой оболочки ротовой полости.
На уровне коры больших полушарий происходит переключение сен-
сорных импульсов на эфферентные нейроны, которые по нисходящим пу-
тям посылают регулирующие влияния к центру жевания продолговатого
мозга.
11.4.4. Глотание
Глотание — рефлекторный акт, при помощи которого пища переводится из
ротовой полости в желудок. Акт глотания состоит из трех фаз: ротовой
(произвольной), глоточной (непроизвольной, быстрой) и пищеводной (не-
произвольной, медленной).
Пищевой комок (объемом 5—15 см3) скоординированными движениями
мускулатуры щек и языка продвигается к его корню (за передние дужки
глоточного кольца). Так завершается первая фаза глотания и начинается
вторая. С этого момента акт глотания становится непроизвольным. Раз-
дражение пищевым комком рецепторов слизистой оболочки мягкого не-
ба и глотки передается по языкоглоточным нервам к центру глотания в
продолговатом мозге. Эфферентные импульсы от него идут к мышцам
полости рта, глотки, гортани и пищевода по волокнам подъязычных,
тройничных, языкоглоточных и блуждающих нервов. Этот центр обеспе-
чивает координированные сокращения мышц языка и мускулатуры, при-
поднимающей мягкое небо. Благодаря этому вход в полость носа со сто-
роны глотки закрывается мягким небом, и язык перемещает пищевой
комок в глотку. Одновременно происходит сокращение мышц, подни-
мающих нижнюю челюсть. Это приводит к смыканию зубов и прекраще-
нию жевания, а сокращение челюстно-подъязычной мышцы — к подня-
тию гортани. В результате осуществляется закрытие входа в гортань над-
гортанником. Этим предотвращается попадание пищи в дыхательные пу-
ти. В это же время открывается верхний пищеводный сфинктер, образо-
ванный волокнами циркулярного направления в верхней половине шей-
ной части пищевода, и пищевой комок поступает в пищевод. Так начи-
нается третья фаза. Верхний пищеводный сфинктер сокращается после
перехода пищевого комка в пищевод, предотвращая пищеводно-глоточ-
ный рефлюкс (т. е. обратное поступление пищи в глотку). Затем осуще-
11. Функции пищеварительной системы • 533
ствляется прохождение пищи по пищеводу и переход ее в желудок. Пи-
щевод является мощной рефлексогенной зоной. Рецепторный аппарат
представлен здесь в основном механорецепторами. Вследствие раздраже-
ния последних пищевым комком происходит рефлекторное сокращение
мускулатуры пищевода. При этом последовательно сокращаются кольце-
вые мышцы (с одновременным расслаблением нижележащих). Волны пе-
ристальтических сокращений распространяются в сторону желудка, пере-
двигая пищевой комок. Скорость их распространения 2—5 см/с. Сокра-
щение мускулатуры пищевода связано с поступлением из продолговатого
мозга эфферентной импульсации по волокнам возвратного и блуждаю-
щего нервов.
Движение пищи по пищеводу обусловлено рядом факторов: во-первых,
перепадом давления между полостью глотки и началом пищевода — от
45 мм рт. ст. в полости глотки (в начале глотания) до 30 мм рт. ст. (в пи-
щеводе); во-вторых, наличием перистальтических сокращений мышц пи-
щевода, в-третьих, тонусом мускулатуры пищевода, который в торакаль-
ном отделе почти в три раза ниже, чем в шейном, и, в-четвертых, силой
тяжести пищевого комка. Скорость прохождения пищи по пищеводу зави-
сит от консистенции пищи: плотная проходит за 3—9 с, жидкая —за 1 —
2 с.
Центр глотания через ретикулярную формацию связан с другими цен-
трами продолговатого и спинного мозга. Его возбуждение в момент глота-
ния вызывает торможение деятельности дыхательного центра и снижение
тонуса блуждающего нерва. Последнее вызывает задержку дыхания и уча-
щение сердечных сокращений. Задержка дыхания предотвращает попада-
ние пищи в дыхательные пути.
При отсутствии глотательных сокращений вход из пищевода в желу-
док закрыт, так как мышцы кардиального отдела желудка находятся в
состоянии тонического сокращения. Когда перистальтическая волна и
комок пищи достигают конечной части пищевода, тонус мышц карди-
альной части желудка рефлекторно снижается, и комок пищи поступает
в желудок. При наполнении желудка пищей тонус мышц кардиального
отдела желудка повышается и препятствует обратному поступлению же-
лудочного содержимого из желудка в пищевод (желудочно-пищеводный
рефлюкс).
11.5. Пищеварение в желудке
В желудке пища, смешанная со слюной и слизью, задерживается от 3 до 10
ч для ее механической и химической обработки. Желудок осуществляет
следующие функции: 1) депонирование пищи-, 2) секрецию желудочного сока,
обеспечивающего химическую обработку пищи; 3) перемешивание пищи с
пищеварительными соками; 4) ее эвакуацию — передвижение порциями в
двенадцатиперстную кишку; 5) всасывание в кровь небольшого количества
веществ, поступивших с пищей; 6) выделение (экскрецию) вместе с желу-
дочным соком в полость желудка метаболитов (мочевины, мочевой кисло-
ты, креатина, креатинина), веществ, поступивших в организм извне (солей
тяжелых металлов, йода, фармакологических препаратов); 7) образование
активных веществ (инкрецию), принимающих участие в регуляции деятель-
ности желудочных и других пищеварительных желез (гастрина, гистамина,
соматостатина, мотилина и др.); 8) бактерицидное и бактериостатическое
действие желудочного сока; 9) удаление недоброкачественной пищи, преду-
преждающее ее попадание в кишечник.
534 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
11.5.1. Секреторная функция желудка
Секреторная функция желудка осуществляется желудочными железами,
продуцирующими желудочный сок. Они состоят из трех видов клеток:
главных, принимающих участие в выработке ферментов; обкладочных, (па-
риетальных), участвующих в выработке хлористоводородной (соляной) ки-
слоты, и добавочных, выделяющих мукоидный секрет (слизь). В его состав
входит также внутренний фактор кастла (гастромукопротен), участвующий
в регуляции кроветворения. Натощак слизь выделяется также цилиндриче-
ским эпителием, которым покрыта слизистая оболочка желудка. Железы
кардиального отдела желудка секретируют в основном слизь. В железах
пилорического отдела отсутствуют обкладочные клетки. Поэтому в секрете
желез этого отдела отсутствует соляная кислота и его pH равен 7,8—8,4.
Основную роль в желудочном пищеварении играют железы фундального
отдела, включающего три секректорные зоны: дна, малой кривизны и тела
желудка (рис. 11.11). Эти железы имеют все три типа клеток и выделяют
основное количество желудочного сока.
Состав желудочного сока. В состоянии покоя (натощак) из желудка че-
ловека можно извлечь около 50 мл желудочного содержимого нейтральной
или слабокислой реакции (pH 6,0). Это смесь слюны и желудочного сока.
Общее количество желудочного сока, отделяющегося у человека при
обычном пищевом режиме, составляет 2,0—2,5 л в сутки. Это бесцветная,
Рис. 11.11. Образование соляной кислоты желудочного сока. Пояснения в тексте.
Символ ® означает активность ферментных транспортных систем мембраны кислотопроду-
цирующих клеток. Стрелками показано направление движения ионов и воды.
11. Функции пищеварительной системы • 535
прозрачная, слегка опалесцирующая жидкость с удельным весом 1,002—
1,007. В соке могут быть хлопья слизи. Желудочный сок имеет кислую ре-
акцию (pH 0,8—1,5) вследствие высокого содержания в нем хлористоводо-
родной (соляной) кислоты (0,3—0,5 %). Содержание воды в соке — 99,0—
99,5 %, а плотных веществ — 1,0—0,5 %. Плотный остаток представлен ор-
ганическими и неорганическими веществами: хлоридами (5—6 г/л), суль-
фатами (10 мг/л), фосфатами (10—60 мг/л), гидрокарбонатами (0—1,2 г/л)
натрия, калия, кальция и магния, аммиалом (20—80 мг/л). Значительная
часть минеральных веществ всасывается в желудке и кишечнике в кровь и
участвует в поддержании постоянства внутренней среды. Основной неорга-
нический компонент желудочного сока — соляная кислота (см. ниже). Ор-
ганическая часть плотного остатка состоит из ферментов и мукоидов (см.
ниже). В небольшом количестве находятся в остатке азотсодержащие ве-
щества небелковой природы (мочевина, мочевая кислота, молочная кисло-
та и др.), подлежащие удалению из организма.
Механизм секреции соляной кислоты. Хлористоводородная кислота вы-
рабатывается париетальными (обкладочными) клетками желез желудка.
Эти клетки характеризуются богатством митохондрий, расположенных
вдоль внутриклеточных канальцев. Площадь мембраны канальцев и апи-
кальной поверхности клеток во время стимуляции на высоте секреции рез-
ко возрастает за счет встроенных в мембрану тубовезикул (трубочек-пу-
зырьков), что сопровождается значительным увеличением клеточных ка-
нальцев, проникающих вплоть до базальной мембраны. Это значительно
увеличивает возможности синтеза гландулоцитом соляной кислоты. Вдоль
канальцев располагается множество митохондрий, площадь внутренней
мембраны которых возрастает в процессе биосинтеза НС1. Соответственно
увеличивается площадь контакта канальцев и апикальной мембраны клет-
ки. Таким образом, увеличение секреторной активности париетальных
клеток обусловлено увеличением площади секреторной мембраны.
Секреция НС1 является ярко выраженным цАМФ-зависимым процес-
сом, активация которого протекает на фоне усиления гликогенолитиче-
ской и гликолитической активности, что сопровождается продукцией пи-
рувата. Окислительное декарбоксилирование пирувата до ацетил-КоА. СО2
осуществляется пируватдегидрогеназным комплексом и сопровождается
накоплением в цитоплазме НАД • Н2. Последний используется для генери-
рования Н+ в процессе секреции НС1. Расщепление триглицеридов в сли-
зистой оболочке желудка под влиянием триглицеридлипазы и последую-
щая утилизация жирных кислот создает в 3—4 раза больший приток вос-
становительных эквивалентов в митохондриальную цепь переноса электро-
нов. Как аэробный гликолиз, так и окисление жирных кислот запускаются
посредством цАМФ-зависимого фосфорилирования соответствующих фер-
ментов, обеспечивающих генерирование ацетил-КоА в цикле Кребса и
восстановительных эквивалентов для электронпереносящей цепи митохон-
дрий. Са2+ является необходимым элементом секреторной системы НС1.
Процесс цАМФ-зависимого фосфорилирования обеспечивает актива-
цию желудочной карбоангидразы, которая является регулятором кислотно-
основного равновесия в кислотопродуцирующих клетках. Работа этих кле-
ток сопровождается длительной и массовой потерей ионов Н+, что приво-
дит к накоплению в клетке ОН-, способных оказать повреждающее дейст-
вие на клеточные структуры. Нейтрализация гидроксильных ионов и явля-
ется главной функцией карбоангидразы. Образующиеся бикарбонатные
ионы посредством электронейтрального механизма выводятся в кровь, а
ионы СГ входят в клетку.
536 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Кислотопродуцирующие клетки на наружных мембранах имеют две
мембранные ферментные системы, участвующие в механизмах продукции
Н+ и секреции НС1. Ими являются Ка+-К+-АТФаза и Н+-К+-АТФаза.
№+-К+-АТФаза, расположенная в базолатеральных мембранах клеток, пе-
реносит К+ из крови в обмен на Na+, а Н+-К+-АТФаза, локализованная в
секреторной мембране, транспортирует калий из первичного секрета в об-
мен на выводимые в желудочный сок ионы Н+. Процесс образования со-
ляной кислоты кислотопродуцирующими клетками схематически пред-
ставлен на рис. 11.11.
В период секреции митохондрии всей массой охватывают в виде муфты
секреторные канальцы, и их мембраны сливаются, образуя митохондри-
ально-секреторный комплекс, где ионы Н+ непосредственно акцептируют-
ся Н+-К+-АТФазой секреторной мембраны и транспортируются из клетки.
Таким образом, кислотообразующая функция обкладочных клеток осу-
ществляется благодаря процессу фосфорилирования — дефосфорилирова-
ния, наличию митохондриальной окислительной цепи, транспортирующей
ионы Н+ из матриксного пространства, а также активности Н+-К+-АТФазы
секреторной мембраны, перекачивающей протоны из клетки за счет энер-
гии АТФ.
Вода поступает в канальцы клетки путем осмоса. Конечный секрет, по-
ступающий в канальцы, содержит НС1 в концентрации 155 ммоль/л, хло-
ристый калий в концентрации 15 ммоль/л и очень малое количество хло-
ристого натрия.
Роль соляной кислоты в пищеварении. В полости желудка хлористоводо-
родная кислота: 1) стимулирует секреторную активность желез желудка;
2) способствует превращению пепсиногена в пепсин путем отщепления
ингибирующего белкового комплекса; 3) создает оптимальную кислот-
ность для действия протеолитических ферментов желудочного сока; 4) вы-
зывает денатурацию и набухание белков (что способствует их расщепле-
нию ферментами); 5) обеспечивает антибактериальный эффект секрета;
6) участвует в осуществлении механизма перехода пищи из желудка в две-
надцатиперстную кишку, раздражая хеморецепторы ее слизистой оболоч-
ки; 7) участвует в регуляции секреции желудочных и поджелудочных же-
лез, стимулируя образование гастроинтестинальных гормонов (гастрина,
секретина); 8) возбуждает секрецию фермента энтерокиназы энтероцитами
слизистой оболочки двенадцатиперстной кишки; 9) участвует в створажи-
вании молока; 10) стимулирует моторную активность желудка.
Ферменты желудочного сока и их роль в пищеварении. В полости желуд-
ка под влиянием протеолитических ферментов осуществляется начальный
гидролиз белков до альбумоз и пептонов. Протеолитические ферменты же-
лудочного сока обладают активностью в широком диапазоне колебаний pH
с оптимумом действия при pH 1,5—2,0 и 3,2—4,0. Это обеспечивает гидро-
лиз белков в условиях значительных колебаний концентрации соляной ки-
слоты в желудочном соке, в слоях пищи, прилежащих к слизистой оболоч-
ке желудка, и в глубине содержимого желудка.
В желудочном соке представлены семь видов пепсиногенов, объединен-
ных общим названием пепсины. Образование пепсинов осуществляется из
неактивных предшественников — пепсиногенов, находящихся в клетках же-
лудочных желез в виде гранул зимогена. В просвете желудка пепсиноген
активируется НС1 путем отщепления от него ингибирующего белкового
комплекса. В дальнейшем в ходе секреции желудочного сока активация
пепсиногена осуществляется аутокаталитически под действием уже образо-
вавшегося пепсина.
11. Функции пищеварительной системы • 537
При оптимальной величине pH среды пепсин осуществляет гидролиз
белков, разрывая в белковой молекуле пептидные связи, образованные
группами фениламина, тирозина, триптофана и других аминокислот. В ре-
зультате этого белковая молекула распадается на пептоны и пептиды. Пеп-
син обеспечивает гидролиз основных белковых веществ, особенно коллаге-
на — основного компонента волокон соединительной ткани.
К основным пепсинам желудочного сока относятся следующие.
Пепсин А — группа ферментов, гидролизирующих белки при оптимуме
pH 1,5—2,0. Часть пепсиногена (около 1 %) переходит в кровеносное рус-
ло, откуда вследствие небольшого размера молекулы фермента проходит
через клубочковый фильтр в почках и выделяется с мочой {уропепсиноген).
Определение содержания уропепсина в моче используется в лабораторной
практике для характеристики протеолитической активности желудочного
сока.
Гастриксин (пепсин С), гидролизирующий белки при оптимуме pH 3,2—
3,5. Пепсин В (парапепсин) расщепляет желатину и белки соединительной
ткани. При pH 5,6 и выше протеолитическое действие фермента ослабля-
ется.
Реннин (пепсин Д, химозин) расщепляет казеин молока в присутствии ио-
нов Са2+.
Желудочный сок содержит ряд непротеолитических ферментов. Среди
них — желудочная липаза, расщепляющая жиры, которые находятся в пище
в эмульгированном состоянии (жиры молока), на глицерин и жирные ки-
слоты при pH 5,9—7,9. У грудных детей желудочная липаза расщепляет до
59 % жира молока. В желудочном соке взрослых людей липазы мало. По-
этому основное количество жиров переваривается в тонком кишечнике.
Клетками поверхностного эпителия слизистой оболочки желудка выра-
батывается лизоцим (муромидаза). Лизоцим обусловливает бактерицидные
свойства желудочного сока.
Уреаза расщепляет мочевину в желудке при pH 8,0. Освобождающийся
при этом аммиак нейтрализует соляную кислоту и предотвращает избыточ-
ную кислотность химуса, поступающего из желудка в двенадцатиперстную
кишку.
Желудочная слизь и ее значение. Органическим компонентом желудоч-
ного сока является слизь.
Нерастворимая слизь (муцин) является продуктом секреторной актив-
ности добавочных клеток и клеток поверхностного эпителия. Муцин выде-
ляется через апикальную мембрану мукоцита, образует слой слизи толщи-
ной 0,5—1,5 мм, он обволакивает слизистую оболочку желудка и препятст-
вует повреждающему воздействию соляной кислоты и пепсинов на клетки
слизистой оболочки и раздражающих веществ, поступивших с пищей. Эти-
ми же клетками одновременно с муцином продуцируется и бикарбонат.
Образующийся при взаимодействии муцина и бикарбоната мукозо-бикар-
бонатный барьер предохраняет слизистую оболочку от аутолиза под воз-
действием соляной кислоты и пепсинов.
Слой слизи является преградой для обратной диффизии ионов водорода
из полости желудка; он нейтрализует соляную кислоту благодаря буфер-
ным свойствам из-за наличия гидрокарбонатов, а также адсорбирует фер-
менты. Под влиянием длительного воздействия желчных кислот (при за-
брасывании их из двенадцатиперстной кишки), салицилатов, масляной и
пропионовой кислот, алкоголя происходит нарушение слизистого барьера.
Это приводит к обратной диффизии ионов водорода из полости желудка,
контакту слизистой оболочки с пепсинами и ее повреждению в результате
538 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
аутолиза. Так формируются пептические язвы желудка. Возникновению
язвенного процесса способствуют продукты жизнедеятельности микроор-
ганизма Helicobacter pylori, которые усиливают секрецию соляной ки-
слоты.
11.5.2. Регуляция секреции желудочного сока
Железы желудка в состоянии относительного покоя (в условиях отсутствия
процесса пищеварения) выделяют небольшое количество сока нейтральной
или слабощелочной реакции (фоновая секреция). Под влиянием пищевого
раздражения во время приема пищи железы желудка секретируют значи-
тельный объем желудочного сока, богатого протеолитическими фермента-
ми. Эта реакция желез является рефлекторным ответом на раздражение
пищей рецепторов слизистой оболочки ротовой полости, глотки и желудка
(безусловный рефлекс) и воздействие комплекса раздражителей, влияющих
на другие рецепторы, предшествующих и сопутствующих приему пищи
(условный рефлекс).
Афферентные импульсы с раздражаемых пищей рецепторов (тактиль-
ных, температурных и вкусовых) по афферентным волокнам V, VII, IX и X
черепно-мозговых нервов передаются в бульбарный, таламический, гипо-
таламический и корковый отделы пищевого центра. Нисходящие влияния
коркового представительства пищевого центра активируют парасимпатиче-
ские и симпатические ядра гипоталамуса. При возбуждении парасимпати-
ческих ядер их эфферентные влияния активируют клетки бульбарного от-
дела пищевого центра, импульсы от которых по преганглионарным волок-
нам блуждающего нерва передаются на постганглионарные нейроны ин-
трамуральных ганглиев желудка (клетки Догеля I типа). Выделяемый в
окончаниях их аксонов медиатор ацетилхолин через М-холинорецепторы
мембраны гландулоцитов стимулирует секреторную деятельность главных,
обкладочных и мукоидных клеток желез желудка. Поэтому после перерез-
ки блуждающих нервов у собак не выделяется желудочный сок при раздра-
жении пищей рецепторов полости рта, на ее вид и запах.
Ацетилхолин активирует также эндокринные У-клетки слизистой обо-
лочки антрального отдела желудка, вырабатывающие гастрин, который че-
рез кровоток достигает обкладочных клеток желез и через специальные ре-
цепторы их мембраны вызывает выработку соляной кислоты.
Ацетилхолин и гастрин в слизистой оболочке фундального отдела же-
лудка стимулируют активность эндокринных клеток, выделяющих гиста-
мин, который, диффундируя по межклеточным пространствам, через Н2-
рецепторы мембраны обкладочных клеток стимулирует выработку большо-
го количества кислого желудочного сока, бедного ферментами и мукоида-
ми. После перерезки блуждающих нервов (в экспериментах на животных)
сокогонный эффект ацетилхолина, гастрина и гистамина резко ослабля-
ется.
Поступление пищи в желудок во время еды усиливает безусловно-реф-
лекторное отделение желудочного сока, что обусловлено раздражением ме-
ханорецепторов при растяжении его стенок (особенно антрального отдела)
и хеморецепторов пептидами и экстрактивными веществами пищи. При
этом рефлекс осуществляется с участием бульбарного отдела пищевого
центра, дополняется местной реакцией, реализуемой через холинергиче-
ские нейроны интраорганной нервной системы.
Увеличение концентрации соляной кислоты в желудочном соке приво-
дит к угнетению выработки гастрина У-клетками и последующим умень-
11. Функции пищеварительной системы • 539
шением кислотности сока. Этот механизм саморегуляции предотвращает
избыточное выделение соляной кислоты. Подавляют секрецию соляной
кислоты соматостатин, ВИП и серотонин (см. табл. 11.1).
При возбуждении симпатических ядер гипоталамуса его нисходящие
влияния стимулируют преганглионарные симпатические нейроны боковых
рогов торакальных сегментов (IV—X) спинного мозга. Их эфферентные
влияния стимулируют постганглионарные нейроны симпатических гангли-
ев, аксоны которых в составе чревных нервов достигают желез желудка.
Выделяющийся в их окончаниях медиатор норадреналин тормозит секре-
цию желудочного сока, но приводит к накоплению секреторного материа-
ла в гландулоцитах, содержащих пепсиногены и мукоиды.
Поступление в двенадцатиперстную кишку не полностью переваренно-
го химуса из желудка вызывает дополнительное возбуждающее и тормоз-
ное влияние на железы желудка. Гастрин, вырабатываемый У-клетками
слизистой оболочки кишки, бомбезин (гастрин-рилизинг-гормон), выде-
ляемый Р-клетками, продукты гидролиза белков гуморально стимулируют
выработку соляной кислоты.
Выделяемые эндокринными клетками двенадцатиперстной кишки холе-
цистокинин, гастроингибирующий пептид, нейротензин и другие пептиды
(см. табл. 11.1) под влиянием продуктов гидролиза жиров в двенадцатипер-
стной кишке оказывают тормозной эффект на выработку соляной кислоты
железами желудка.
11.5.2.1. Фазы желудочной секреции
В естественных условиях приема пищи сокоотделение в желудке начинает-
ся под влиянием условных раздражителей — вида и запаха пищи, привыч-
ной обстановки. При попадании пищи в ротовую полость в процессе же-
вания и глотания раздражаются рецепторы слизистой оболочки рта, глотки
и пищевода, что вызывает безусловно-рефлекторную секрецию желудочно-
го сока. Этот комплекс условных и безусловных секреторных рефлексов
составляет сложнорефлекторную первую фазу желудочного сокоотделения,
которая продолжается около двух часов. Выделяющийся при этом сок от-
личается высоким содержанием ферментов, что способствует возникнове-
нию второй фазы сокоотделения — нейрогуморальной. Она обусловлена
раздражением рецепторов желудка и кишечника компонентами химуса,
влиянием на железы желудка гастроинтестинальных гормонов (см. табл.
11.1) и продуктов гидролиза пищевых веществ, поступающих в кровоток.
Поэтому секреция желудочного сока (в опытах на собаках) продолжается
6—10 ч.
Нейрогуморалъная фаза секреции желудочного сока «наслаивается» на
сложнорефлекторную и состоит из желудочной и кишечной фаз. Желудоч-
ная фаза сокоотделения обусловлена влиянием содержимого желудка на
рецепторы его слизистой оболочки и эндокринные клетки (см. табл. 11.1).
Так, механическое раздражение пищей приводит к выделению гастрина.
Его выделение усиливается под влиянием продуктов гидролиза белков и
экстрактивных веществ.
Кишечная фаза секреции желудочного сока обусловлена раздражением
рецепторов слизистой оболочки двенадцатиперстной кишки кислым желу-
дочным содержимым, поступающим из желудка, всасыванием в кровь про-
дуктов гидролиза белков и гастроинтестинальных гормонов, выделяемых
эндокринными элементами слизистой оболочки кишечника (см. табл.
11.1). При осуществлении кишечной фазы возникают и тормозные влия-
540 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Часы
12345678123456789 10 123456
~1 I I I I 1 I I I I I I I I ) I I I I I I I Г"
| 14 -
2 12 -
Мясо Хлеб Молоко
Рис. 11.12. Динамика желудочной секреции на различные пищевые вещества (в
опытах на собаках).
Желудочный сок собирался через фистульную трубку, вставленную в маленький желудочек,
выкроенный из желудка по методике И. П. Павлова (с сохраненной иннервацией).
На белковую пищу (мясо) выделяется наибольшее количество желудочного сока. На хлеб по-
сле всплеска секреции в сложнорефлекторную фазу сокоотделение продолжается длительное
время (около 10 ч) на низких цифрах. После кормления животного молоком секреция сока
медленно достигает максимума (к концу 3-го часа).
ния на железы желудка, вызванные продуктами гидролиза жиров, крахмала
и регуляторными пептидами.
Желудочная секреция при переваривании различных пищевых веществ.
Адаптация секреторного аппарата желудка к характеру пищи обусловлена
ее качеством, количеством, режимом питания. Классическим примером
приспособительных реакций желудочных желез является изученная
И. П. Павловым реакция пищеварительных желез в ответ на прием пищи,
содержащей преимущественно углеводы (хлеб), белки (мясо), жиры (мо-
локо).
Наиболее эффективным возбудителем секреции является белковая пи-
ща (рис. 11.12). Белки и продукты их переваривания обладают выражен-
ным сокогонным действием. После приема мяса развивается довольно
энергичная секреция желудочного сока с максимумом на 2-м часу. Она
продолжается около 7 ч. Длительная мясная диета (на протяжении 30—40
сут) приводит к усилению желудочной секреции на все пищевые раздра-
жители, повышению кислотности и переваривающей силы желудочного
сока. Это говорит о том, что под влиянием сильных возбудителей секре-
ции происходит устойчивая перестройка в деятельности желез желудка и
механизмов их регуляции.
Углеводная пища (хлеб) — более слабый возбудитель секреции желудоч-
ного сока. Хлеб беден химическими возбудителями секреции, поэтому по-
сле его приема развивается ответная секреторная реакция с максимумом
на 1-м часе (рефлекторное отделение сока), а затем резко уменьшается и
11. Функции пищеварительной системы • 541
на невысоком уровне удерживается продолжительное время (из-за слабой
мобилизации гуморальных механизмов возбуждения гландулоцитов). При
длительном питании человека углеводами кислотность и переваривающая
сила сока снижаются, что является следствием приспособления желез же-
лудка к пище, содержащей небольшое количество белка, продукты непол-
ного гидролиза которого стимулируют выработку соляной кислоты и пеп-
синогена.
Жиры молока вызывают желудочную секрецию в две стадии: тормозную
и возбуждающую. Этим объясняется тот факт, что после приема жирной
пищи максимум секреции сока наблюдается только к концу 3-го часа.
Первые порции молока, попав из желудка в двенадцатиперстную кишку,
оказывают тормозное влияние на секрецию желудочного сока. В результа-
те длительного питания жирной пищей происходит усиление желудочной
секреции на все пищевые раздражители за счет второй половины секре-
торного периода. Переваривающая сила сока при использовании в пище
жиров ниже по сравнению с соком, выделяющимся на мясо, но выше, чем
при питании углеводной пищей.
Количество отделяющегося желудочного сока, его кислотность и проте-
олитическая активность зависят также от количества и консистенции при-
нятой пищи. По мере увеличения объема пищи секреция желудочного
сока.
11.5.3. Сократительная деятельность мускулатуры желудка
Во время приема пищи происходит расслабление мускулатуры фундально-
го отдела желудка, что является рефлекторной реакцией в ответ на раздра-
жение рецепторов слизистой оболочки полости рта, глотки и пищевода.
Это расслабление получило название рецептивной релаксации, которая спо-
собствует заполнению пищей фундального отдела. Через 5—30 мин после
завершения приема пищи происходит усиление моторики желудка. Внача-
ле автоматическое возбуждение миоцитов возникает в водителе ритма, ко-
торый располагается в области кардии, откуда волна перистальтического
сокращения распространяется в дистальном направлении — на тело желуд-
ка, его антральную часть и пилорический сфинктер. За 1 мин возникает
около трех волн. Скорость распространения перистальтической волны по
мере ее приближения к пилорическому отделу желудка возрастает с 1 до
3—4 см/с. Волна перистальтики представляет собой сокращение участков
(полос) циркулярного слоя мышц по всей окружности желудка. После со-
кращения участка циркулярных мышц происходит их расслабление, а вол-
на сокращения переходит на другой участок.
Перистальтические волны смещают в дистальном направлении слои хи-
муса, прилежащие к слизистой оболочке желудка, в наибольшей степени
обработанные желудочным соком. На фоне перистальтических волн воз-
никают сильные и продолжительные сокращения (тонические волны), ко-
торые смещают химус из фундального отдела в пилорический. По мере на-
копления содержимого желудка в пилорическом отделе возникают силь-
ные систолические сокращения его мускулатуры. Они создают значительное
возрастание давления химуса в желудке, что при открытом пилорическом
сфинктере вызывает эвакуацию порции желудочного содержимого в двена-
дцатиперстную кишку.
Если с помощью введенного в желудок человека баллона регистриро-
вать колебания давления желудочного содержимого, то можно выявлять
три типа волн сокращения (рис. 11.13): фазные волны с амплитудой до
542 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 11.13. Волны сокращений мускулатуры желудка здорового человека.
I — перистальтические сокращения первого типа, II — волны второго типа, III — волны
третьего типа: на фоне длительного сокращения (в течение 5 мин), повышающего исходное
давление содержимого желудка до 10 мм рт. ст., возникают волны II типа с большей ампли-
тудой.
5 мм рт. ст., продолжительностью около 20 с, частотой 3 в 1 мин (I тип),
фазные волны той же продолжительности и частоты, но большей амплиту-
ды (10—50 мм рт. ст. — II тип). На фоне волн I и II типов происходит воз-
растание исходного тонуса мускулатуры, что повышает давление на 5—
10 мм рт. ст. в течение 5—6 мин (III тип).
11.5.3.1. Регуляция сократительной деятельности желудка
Регулирующие влияния на мускулатуру желудка передаются из вегетатив-
ных центров по блуждающему и чревному нервам. Возбуждение холинер-
гических волокон блуждающего нерва за счет выделения в его окончаниях
ацетилхолина усиливает моторику желудка, что выражается в увеличении
силы и частоты перистальтических волн, а также скорости их распростра-
нения. Если через парасимпатические волокна блуждающего нерва возбу-
ждаются тормозные нейроны интрамуральных ганглиев, то наблюдается
эффект релаксации мышц желудка и расслабления пилорического сфинк-
тера. Это является следствием выделения в окончаниях аксонов этих ней-
ронов тормозных медиаторов (ВИП и АТФ).
При возбуждении симпатических (адренергических) волокон чревного
нерва наступает торможение мускулатуры желудка (уменьшение силы и
частоты сокращений, уменьшение скорости их распространения), но акти-
вация а- и p-адренорецепторов постсинаптических мембран миоцитов пи-
лорического сфинктера вызывает повышение его тонуса.
Координация парасимпатических и симпатических влияний осуществ-
ляется благодаря взаимодействию надсегментарных вегетативных центров
гипоталамуса и корковых представительств пищевого центра. При элекро-
стимуляции ядер передних и средних отделов гипоталамуса через вживлен-
ное электроды (в опытах на животных), как правило, возникает эффект
стимуляции моторики желудка, а при раздражении задних — торможение
его двигательной активности.
И. Функции пищеварительной системы • 543
11.5.3.2. Эвакуация содержимого желудка в двенадцатиперстную кишку
Желудочное содержимое переходит в двенадцатиперстную кишку порция-
ми. Этот процесс осуществляется благодаря согласованной деятельности
мускулатуры антрального отдела желудка, пилорического сфинктера и две-
надцатиперстной кишки. Скорость эвакуации содержимого желудка зави-
сит от ряда условий.
1. Чем больше давление в желудке превышает давление в двенадцати-
перстной кишке и чем ниже тонус мышечных волокон пилорическо-
го сфинктера, тем больше скорость эвакуации. Градиент давлений
достигает 20—30 см водн. ст.
2. Жидкая часть желудочного содержимого начинает поступать в кишку
вскоре после приема пищи. Скорость перехода кашицеобразного хи-
муса из желудка постепенно замедляется. Твердые компоненты пищи
задерживаются в желудке до их измельчения до частичек размером в
2—3 мм. Если человеком была принята средняя по объему порция
смешанной пищи, то она задерживается в желудке на 3,5—4,5 ч.
3. С возрастанием объема принятой пищи продолжительность эвакуа-
ции возрастает. Так, удвоение объема твердой углеводной пищи удли-
няет процесс эвакуации на 17 %, а белково-жировой — на 43 %.
4. При прочих равных условиях быстрее всего покидает желудок угле-
водная пища, дольше задерживается в желудке белковая пища и еще
дольше — жирная.
5. Высокое осмотическое давление желудочного содержимого замедляет
процесс его эвакуации.
Важным регулятором сократительной активности пилорического
сфинктера является pH эвакуируемых порций желудочного содержимого.
Раздражение кислым химусом хеморецепторов слизистой оболочки пило-
рического отдела рефлекторно способствует открытию сфинктера и пере-
ходу порции химуса в двенадцатиперстную кишку. Раздражение соляной
кислотой хеморецепторов кишки вызывает рефлекторное торможение мус-
кулатуры желудка (энтерогастральный рефлекс) и сокращение пилориче-
ского сфинктера. Такую же направленность имеет рефлекс с механорецеп-
торов двенадцатиперстной кишки. По мере ощелачивания поступившей в
кишку порции желудочного содержимого панкреатическим и кишечным
соками, а также желчью, описанные рефлекторные реакции с хеморецеп-
торов слизистой оболочки кишки прекращаются и создаются условия для
эвакуации из пилорического отдела желудка другой порции химуса.
Рефлекторная дуга этих рефлексов замыкается как в центрах мозга, так
и на периферии (в экстраорганных и интраорганных ганглиях).
Рефлекторная регуляция эвакуаторной функции подкрепляется гумо-
ральными факторами. Так, выделяющиеся эндокринными клетками киш-
ки под влиянием соляной кислоты секретин и холецистокинин тормозят
моторику желудка и эвакуацию. Но в тоже время они ускоряют ощелачи-
вание химуса в двенадцатиперстной кишке, стимулируя выработку пан-
креатического сока. Другие гастроинтестинальные гормоны (энкефалины
и ГИП) тормозят моторику желудка и эвакуаторную функцию.
11.6. Пищеварение в двенадцатиперстной кишке
В двенадцатиперстной кишке продолжается процесс гидролиза пищевых
веществ, начатый в желудке. Но его объем значительно возрастает, так как
в полость кишки выделяются пищеварительные соки поджелудочной желе-
544 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
зы и кишечных желез, содержащие ферменты для гидролиза белков, жиров
и углеводов. Эти ферменты наиболее активны в щелочной среде, которая
создается пищеварительными соками поджелудочной железы, бруннеро-
вых и либеркюнновых желез, а также желчью.
11.6.1. Пищеварительные функции поджелудочной железы
За сутки поджелудочная железа человека выделяет от 1,5 до 2,5 л сока, ко-
торый вырабатывается ацинарными, центроацинарными и эпителиальны-
ми клетками протоков железы. В состоянии относительного покоя (нато-
щак) железа выделяет небольшое количество сока, а при поступлении же-
лудочного содержимого в двенадцатиперстную кишку скорость сокоотде-
ления возрастает до 4,7 мл/мин.
11.6.1.1. Состав и свойства панкреатического сока
Панкреатический сок имеет высокую концентрацию бикарбонатов, кото-
рые обусловливают его щелочную реакцию. Его pH колеблется от 7,5 до
8,8. В соке содержатся хлориды натрия, калия и кальция, сульфаты и фос-
фаты. Вода и электролиты выделяются в основном центроацинарными и
эпителиальными клетками выводах протоков. В состав сока входит и
слизь, которая вырабатывается бокаловидными клетками главного протока
поджелудочной железы.
Панкреатический сок богат ферментами, осуществляющими гидролиз
белков, жиров и углеводов. Они вырабатываются ацинарными панкреаци-
тами.
Протеолитические ферменты (трипсин, химотрипсин, эластаза, карбок-
сипептидазы А и В) выделяются панкреацитами в неактивном состоянии,
что предотвращает самопереваривание клеток. Трипсиноген превращается
в трипсин в полости двенадцатиперстной кишки под влиянием фермента
энтерокиназы, который вырабатывается слизистой оболочкой кишки. Вы-
деление энторокинизы обусловлено влиянием желчных кислот. С появле-
нием трипсина наступает аутокаталитический процесс активации всех про-
теолитических ферментов, выделяющихся в зимогенной форме.
Трипсин, химотрипсин и эластаза расщепляют внутренние пептидные
связи белковой молекулы и высокомолекулярных полипептидов. Процесс
гидролиза завершается образованием низкомолекулярных пептидов и ами-
нокислот. Образовавшиеся пептиды подвергаются заключительному гидро-
лизу карбоксипептидазами А и В, которые расщепляют С-концевые связи
молекул белков и пептидов с образованием аминокислот.
Содержащаяся в панкреатическом соке а-амилаза расщепляет крахмал
на декстрины, мальтозу и мальтотриозу. Ионы кальция, входящие в состав
а-амилазы, обеспечивают устойчивость фермента при изменении pH сре-
ды и ее температуры, а также препятствуют его гидролизу под влиянием
протеолитических ферментов.
Панкреатическая липаза секретируется в активной форме. Но ее актив-
ность значительно возрастает под влиянием колипазы после ее активации в
двенадцатиперстной кишке трипсином. Колипаза образует комплекс с
панкреатической липазой. В образовании этого комплекса участвуют соли
жирных кислот. Липаза гидролизует жир на моноглицериды и жирные ки-
слоты. Эффективность гидролиза жира резко возрастает после его эмуль-
гирования желчными кислотами и их солями.
Под влиянием холестеразы холестериды расщепляются до холестерина и
11. Функции пищеварительной системы • 545
жирных кислот. Фосфолипиды подвергаются гидролизу с помощью пан-
креатической фосфолипазы А2, которая активируется трипсином. Конечны-
ми продуктами гидролиза являются жирная кислота и изолецетин. Рибо-
нуклеазы и дезоксирибонуклеазы панкреатического сока расщепляют РНК и
ДНК пищевых веществ до нуклеотидов.
11.6.1.2. Нервная и гуморальная регуляция секреторной функции
поджелудочной железы
Слабое сокоотделение натощак резко усиливается во время (через 2—
3 мин) и после приема пищи. В привычных условиях приема пищи пан-
креатическое сокоотделение начинается уже на вид, запах пищи и другие
раздражители, сопутствующие приему пищи, что свидетельствует об услов-
но-рефлекторном механизме секреции. При раздражении пищевыми веще-
ствами рецепторов слизистой оболочки ротовой полости, глотки и пище-
вода возникает безусловно-рефлекторное отделение панкреатического со-
ка. Нервные импульсы от раздражаемых рецепторов по афферентным пу-
тям достигают бульбарного центра панкреатической секреции, где они пе-
реключаются на преганглионарные нейроны ядер блуждающего нерва, по
эфферентным волокнам которого достигают постганглионарных нейронов.
Их аксоны образуют синаптические окончания на базальных мембранах
панкреацитов. Выделяющийся при возбуждении этих окончаний ацетилхо-
лин активирует М-холинорецепторы постсинаптической мембраны. При
этом освобождаются вторичные посредники (Са++ и ГЦ-цГМФ), которые
и вызывают секреторную деятельность панкреоцитов и эпителиальных
клеток протоков.
При раздражении чревных нервов, осуществляющих симпатическую
иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет
активации p-адренорецепторов медиатором норадреналином). Но тормо-
жение секреции сопровождается накоплением секреторного материала в
панкреацитах.
Поступление пищи в желудок во время приема пищи вызывает раздра-
жение хемо- и механорецепторов желудка, что приводит к продолжению
безусловно-рефлекторного сокоотделения поджелудочной железой, вы-
званного стимуляцией рецепторов ротовой полости.
Поступление в двенадцатиперстную кишку желудочного содержимого
вызывает наряду с дуоденопанкреатическим секреторным рефлексом (с ре-
цепторов слизистой оболочки) выделение из эндокринных клеток гастро-
интестинальных гормонов, стимулирующих или тормозящих сокоотделе-
ние поджелудочной железой. Роль основных гуморальных регуляторов вы-
полняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками
слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усили-
вают влияние друг друга на панкреоциты, особенно на фоне выделения
ацетилхолина в синаптических окончаниях холинергических нервных во-
локон, иннервирующих железу. Важным гуморальным возбудителем пан-
креатической секреции является гастрин, выделяемый У-клетками слизи-
стой оболочки антрального отдела желудка. Возбуждающее влияние этих
гормонов подкрепляется бомбезином, серотонином и инсулином.
Тормозят выделение сока соматостатин, глюкагон, энкефалины, вещест-
во Р, ГИП, ПП (см. табл. 11.1), а также кальцитонн и АКТГ. Но их роль в
комплексной гуморальной регуляции поджелудочной железы изучена не-
достаточно.
Секреция сока поджелудочной железой осуществляется в три фазы.
18-6095
546 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Первая называется сложнорефлекторной. Она обусловлена комплексом ус-
ловных и безусловных раздражителей, предшествующих и сопутствующих
приему пищи. В первую фазу выделяется около 10—15 % от общего объема
сока за три фазы, а ферментов — около 25 %. Во вторую желудочную фазу
вырабатывается около 10 % сока от общего его объема (с высоким содер-
жанием ферментов). В третью кишечную фазу выделяется основное количе-
ство сока (около 75 % от общего объема), но менее богатого ферментами.
Его состав отличается большим содержанием бикарбонатов, необходимых
для ощелачивания содержимого двенадцатиперстной кишки.
11.6.2. Пищеварительные функции печени
В пищеварении печень принимает участие благодаря образованию желчи,
которая из общего протока через сфинктер Одди поступает в полость две-
надцатиперстной кишки. Желчь — результат секреторной деятельности ге-
патоцитов и эпителиальных клеток желчных протоков. Секреторные клет-
ки печени исходный материал для образования желчи получают из крови,
протекающей по капиллярам дольки печени и под базальной мембраной
эпителиальных клеток протоков.
Кровь поступает в печень по воротной вене от пищеварительного трак-
та и печеночной артерии (около 1,2 л за 1 мин).
11.6.2.1. Механизм образования желчи
Желчные кислоты синтезируются гепатоцитами из холестерина. Кроме то-
го, из синусоидного кровеносного капилляра гепатоциты активно извлека-
ют желчные кислоты, которые с помощью переносчиков выделяются в
желчные капилляры. При этом за желчными кислотами по осмотическому
градиенту следует вода.
Через мембрану гепатоцита осуществляется активный транспорт Na+ в
желчные капилляры; за Na+ по электрическому градиенту перемещаются
анионы СГ и HCOJ. Возникающий осмотический градиент привлекает в
желчный капилляр воду. Механизм транспорта других катионов и анио-
нов аналогичен описанному. Гепатоциты выделяют также в желчные ка-
пилляры фосфолипиды, холестерин и билирубин. Билирубин образуется
в крови из гемоглобина гемолизированных эритроцитов и в комплексе с
альбумином плазмы поступает в гепатоцит, где образуется прямой били-
рубин (после его соединения с глюкуроновой кислотой). Прямой били-
рубин через мембрану гепатоцита транспортируется в желчные капил-
ляры.
В желчных протоках продолжается процесс формирования желчи за
счет деятельности эпителиальных клеток, которые осуществляют реабсорб-
цию электролитов из содержимого протоков, выводят HCOj и воду.
11.6.2.2. Состав и свойства желчи
За сутки у человека образуется 0,6—1,5 л желчи. Печеночная желчь, запол-
няющая желчные протоки, поступая в желчный пузырь, изменяется по
своему составу.
Эпителиальные клетки слизистой оболочки желчного пузыря осуществ-
ляют активную реабилитацию Na+ из его содержимого, что является при-
чиной реабсорбции анионов СГ, HCOj и воды. Это приводит к сгущению
пузырной желчи и уменьшению ее pH (с 7,3—8,0 до 6,5).
11. Функции пищеварительной системы • 547
Печеночная и пузырная желчь, поступив в двенадцатиперстную кишку,
принимает участие в пищеварении. Это выражается в следующем.
Снижая кислотность поступившего в кишку желудочного содержимого,
желчь прекращает действие пепсинов и создает среду для проявления ак-
тивности ферментов поджелудочного сока.
За счет солей желчных кислот происходит эмульгирование жиров, круп-
ные капли которых распадаются на мелкие капельки, резко увеличиваю-
щие площадь соприкосновения с липазой панкреатического сока и эффек-
тивность гидролиза жиров. Около 7—20 % желчных кислот выводится из
организма с калом, большая часть всасывается в подвздошной кишке в
кровь воротной вены, откуда гепатоциты повторно извлекают желчные ки-
слоты.
Желчные кислоты способствуют всасыванию жирных кислот и жирора-
створимых витаминов (D, Е, К).
Желчь является возбудителем моторики кишечника и кишечных ворси-
нок, стимулирует пролиферацию энтероцитов, угнетает развитие кишеч-
ной микрофлоры и предотвращает гнилостные процессы в толстом кишеч-
нике.
11.6.2.3. Регуляция желчеобразования и желчевыведения
Желчеобразование в печени происходит непрерывно. Под влиянием услов-
ных и безусловных раздражителей во время приема пищи образование
желчи усиливается. Продолжительность латентного периода секретного
рефлекса варьирует от 3 до 12 мин. После приема пищи темпы желчеобра-
зования возрастают и достигают максимума при переваривании углеводис-
той пищи через 2—3 ч, белковой — через 3 ч, жирной — через 5—7 ч. Это
зависит от продолжительности пребывания пищевых веществ в желудке,
кислотности порций желудочного содержимого, поступающего в двенадца-
типерстную кишку, образования эндокринными клетками слизистой обо-
лочки желудка и кишки гастроинтестинальных гормонов (гастрина, секре-
тина, холецистокинина-панкреозимина, глюкагона), которые стимулируют
желчеобразование.
Рефлекторные стимулирующие влияния на желчеобразование реализу-
ются через блуждающий нерв в ответ на раздражение рецепторов всех от-
делов пищеварительного тракта, а тормозные — через симпатические
нервы.
При отсутствии процесса пищеварения (натощак) желчь поступает в
желчный пузырь, потому что сфинктеры Мирицци (препятствующий по-
ступлению желчи из общего желчного протока в проток желчного пузыря)
и Люткинса (в шейке желчного пузыря) находятся в расслабленном со-
стоянии, а сфинктер Одди — в сокращенном. Емкость желчного пузыря у
взрослого человека равна 50—60 мл, но за счет сгущения желчи резервиру-
ется ее объем, выделенный печенью за 12—14 ч.
Начинается желчевыведение в ответ на комплекс условных и безуслов-
ных раздражителей, связанных с приемом пищи. Реализуется этот рефлекс
через эфферентные волокна блуждающего нерва, возбуждение которых
стимулирует моторику желчного пузыря и общего желчного протока, но
расслабляет сфинктер Одди. Продолжается желчевыведение от 3 до 6 ч.
Оно обусловлено не только раздражением рецепторов желудка и двенадца-
типерстной кишки химусом, но и влиянием на мускулатуру желчевыводя-
щего аппарата гастроинтестинальных гормонов (гастрина, секретина, холе-
цистокинина-панкреозимина, бомбезина), вырабатываемых эндокринны-
18*
548 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ми клетками желудка и двенадцатиперстной кишки под влиянием химуса.
Наибольший сокогонный эффект наблюдается после приема молока, яич-
ных желтков, жиров и мяса.
Раздражение симпатических нервов вызывает расслабление мышц
желчного пузыря, общего желчного протока и сокращение сфинктера Од-
ди, что приводит к уменьшению и прекращению выделения желчи в две-
надцатиперстную кишку. Торможение желчевыведения наблюдается также
под влиянием ВИП, глюкагона и кальцитонина.
11.6.3. Непищеварительные функции печени
Деятельность печени в большой степени влияет на состояние обмена ве-
ществ организма. В печени синтезируется основное количество белков
плазмы крови — альбумины, глобулины, факторы свертывания крови (про-
тромбин, фибриноген и др.). В гепатоцитах синтезируется и депонируется
гликоген, из которого образуется глюкоза при возрастании энергетических
затрат организма (например, во время мышечной деятельности) для под-
держания константного уровня ее концентрации в крови. Это происходит
под влиянием нервных импульсов по симпатическим нервам, адреналина
и глюкагона. Гепатоциты также расщепляют жиры до жирных кислот. Пе-
чень оказывает косвенное влияние на обмен веществ, осуществляя дезак-
тивацию гормонов. Белково-пептидные гормоны дезактивируются протеи-
назами, стероидные — гидроксилазами, катехоламины — моноаминокси-
дазой.
Печень защищает организм от ядовитых продуктов пищеварения, по-
ступающих из кишечника (индола, скатола, фенола) и чужеродных ве-
ществ. Обезвреживание токсичных веществ гепатоциты осуществляют за
счет их окисления, восстановления, соединения с глюкуроновой и серной
кислотами, глицином и глутамином, что приводит к образованию неядо-
витых продуктов.
При дезаминировании промежуточных продуктов белкового обмена в
гепатоцитах образуется токсичный аммиак, который инактивируется за
счет синтеза из него мочевины.
11.7. Пищеварение в тонком кишечнике
В тонкой кишке осуществляются завершающие стадии гидролиза пище-
вых веществ, начатого с помощью ферментов слюны, желудочного и под-
желудочного соков. В кишечном соке имеется полный набор ферментов,
расщепляющий поли- и олигомеры пищевых веществ до мономеров, ко-
торые по мере своего образования всасываются слизистой оболочкой тон-
кой кишки в кровь и лимфу (см. раздел 11.7.3). Начинается гидролиз пи-
щевых веществ в тонкой кишке в процессе полостного пищеварения, а за-
вершается при осуществлении пристеночного (мембранного пищева-
рения).
11.7.1. Секреторная функция тонкой кишки
Кишечный сок является продуктом секреторной активности всей слизи-
стой оболочки тонкой кишки. В проксимальном отделе двенадцатиперст-
ной кишки сок вырабатывается бруннеровыми железами. Секрет этих желез
содержит значительное количество слизи, которая защищает слизистую
оболочку двенадцатиперстной кишки от механического и химического раз-
11. Функции пищеварительной системы • 549
дражения. Основная часть кишечного сока выделяется либеркюновыми же-
лезами. Они заложены в криптах у основания ворсинок. В образовании со-
ка принимают участие все энтероциты. Они обладают высокой пролифера-
тивной активностью, в результате чего происходит их перемещение от ос-
нования ворсинки до ее вершины, где они подвергаются слущиванию (де-
сквамации) и становятся компонентом кишечного сока. В полости кишки
спущенные эпителиальные клетки распадаются, освобождая содержащиеся
в них ферменты.
Мембраны апикальной поверхности эпителиоцитов покрыты большим
числом микроворсинок, связанных с гликокаликсом. Они создают боль-
шую площадь, на которой фиксированы молекулы ферментов. Эти фер-
менты транспортируются из эпителиоцита и адсорбируются из содержимо-
го тонкой кишки. Они обусловливают мембранное пищеварение. Слизи-
стая оболочка кишки покрыта слоем слизи. Она вырабатывается бокало-
видными клетками. Слизь не только защищает слизистую оболочку от хи-
мического и механического повреждения, но и является субстратом, на ко-
тором абсорбируются ферменты.
Слизистая оболочка тонкой кишки взрослого человека выделяет за су-
тки около 2,5 л сока. Он состоит из жидкой и плотной частей. Жидкая
часть содержит небольшое количество ферментов, катионы (Na+, К+, Са2+
и др.), анионы (в основном НСО^, СГ), белки, аминокислоты, мочевину,
молочную кислоту и др. Ее pH 7,2—7,5. В плотной части кишечного сока
содержится основное количество ферментов, слущенные эпителиоциты и
их фрагменты, лейкоциты и слизь.
В кишечном соке и в энтероцитах имеется около 20 ферментов, осуще-
ствляющих заключительный гидролиз пищевых веществ. Ферменты, выра-
батываемые эпителиальными клетками, вначале проявляют свою актив-
ность в процессе мембранного пищеварения (на поверхности микроворси-
нок и нитей гликокаликса, а после слущивания энтероцитов принимают
участие в полостном пищеварении. К этим ферментам относятся лейцин-
аминопептидиза, нуклеаза, нуклеотидаза, липаза, фосфолипаза, щелочная
фосфатаза, холинэстераза.
В кишечном соке содержится энтерокиназа, которая выходит из десква-
мированных эпителиальных клеток и активирует трипсиноген поджелудоч-
ного сока. Лейцинаминопептидаза расщепляет пептиды до аминокислот.
Фосфатазы осуществляют гидролиз моноэфиров ортофосфорной кислоты.
Нуклеаза деполимеризует нуклеиновые кислоты, а нуклеотидиза дефосфо-
рилизует мононуклеотиды.
Ферменты, связанные с мембраной энтероцитов, имеют высокую ак-
тивность. Олигосахаридазы и дисахаридазы завершают гидролиз углеводов
превращая их в моносахариды. Аминопептидазы и дипептидазы осуществ-
ляют расщепление пептидов до аминокислот. Моноглицеридлипазы завер-
шают гидролиз жиров.
Таким образом, в процессе полостного пищеварения в тонкой кишке об-
разуются олигомеры молекул пищевых веществ. Из полости кишки олиго-
меры разной степени сложности проникают через слой слизи на гликока-
ликс и подвергаются дополнительному гидролизу (за счет ферментов, ад-
сорбированных в слизи и на глико кал иксе). Заключительный гидролиз
алигомеров происходит на мембране микроворсинок апикальной части эн-
тероцитов (мембранное пищеварение). Мембранное пищеварение характери-
зуется высокой эффективностью, так как происходит на очень большой
площади.
550 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
11.7.1.1 . Регуляция секреторной функции тонкой кишки
Основную роль в регуляции сокоотделения слизистой оболочки кишки иг-
рают местные рефлексы, осуществляемые энтеральной нервной системой.
Механическое раздражение усиливает секрецию жидкой части кишечного
сока и не изменяет содержания в нем ферментов. Стимуляция хеморецеп-
торов продуктами переваривания белков и жиров вызывает секрецию сока,
богатого ферментами. При раздражении блуждающего нерва увеличивается
содержание ферментов в кишечном соке. Такой же эффект оказывают аце-
тилхолин и холиномиметики. Раздражение чревного нерва угнетает отделе-
ние сока.
Во время приема пищи наблюдается рефлекторное усиление секреции
бруннеровых желез двенадцатиперстной кишки и рефлекторное торможе-
ние остального железистого аппарата, что предотвращает избыточную вы-
работку сока и его ферментов (сверх того количества секрета, которое обу-
словлено местным раздражением химуса рецепторов кишки).
Гастроинтестинальные гормоны, вырабатываемые эндокринными эле-
ментами слизистой оболочки кишки под влиянием химуса (дуокринин,
энтерокринин, ГИП, ВИП и мотилин), а также гормоны коры надпочеч-
ников (кортизон, дезоксикортикостерон) стимулируют кишечную секре-
цию, подкрепляя функцию местных рефлекторных механизмов, и только
один гормон — соматостатин — оказывает тормозное влияние на кишечное
сокоотделение.
11.7.2. Двигательная функция тонкой кишки
Благодаря сократительной деятельности гладкой мускулатуры тонкой киш-
ки ее содержимое перемешивается с пищеварительными соками и переме-
щается в дистальном направлении. При этом пристеночные слои химуса
смещаются раньше остального содержимого, которое приходит в сопри-
косновение со слизистой оболочкой. Моторика тонкой кишки обеспечива-
ет высокую эффективность гидролиза пищевых веществ и способствует
всасыванию их мономеров.
Миоциты кишки обладают способностью к автоматическому возбужде-
нию, что является причиной сокращения мышечных пучков. Их деятель-
ность приобретает упорядоченный характер благодаря энтеральной нерв-
ной системе, которая обеспечивает несколько видов скоординированных
сокращений (ритмическую сегментацию, маятникообразные, перистальти-
ческие и тонические сокращения).
Ритмическая сегментация представляет собой сокращение пучков цир-
кулярных мышц, которые вызывают в разных участках кишки глубокие
перетяжки шириной 1—2 см.
Содержимое кишки из зоны перетяжки перемещается в соседние участки
(шириной 15—20 см), мускулатура которых находится в расслабленном со-
стоянии. Через несколько минут сокращенные участки расслабляются, а пе-
ретяжки возникают в ранее расслабленных участках. Чередование сокращен-
ных и расслабленных участков осуществляется 8—10 раз в 1 мин. Ритмиче-
ская сегментация обеспечивает перемешивание химуса в полости кишки.
Маятникообразные сокращения — это ритмические сокращения в основ-
ном продольных пучков мышечных волокон участка кишки, который, уко-
рачиваясь, приводит к перемещению химуса «вперед—назад», что способ-
ствует его перемешиванию. Маятникообразные сокращения в одном и том
же участке кишки чередуются с ритмической сегментацией.
11. Функции пищеварительной системы • 551
Перистальтические сокращения подобны волне, распространяющейся в
каудальном направлении и продвигающей химус вдоль по кишке. Эта
волна возникает выше порции химуса в результате сокращения циркуляр-
ных мышц и сокращения продольных мышц ниже образовавшегося пере-
хвата.
Сужение участка кишки, вызванное сокращением циркулярных мышц,
и расширение рядом расположенного участка, обусловленное сокращени-
ем продольных мышц, создает градиент давления, который и вызывает пе-
ремещение химуса в каудальном направлении. Скорость распространения
перистальтической волны в тонкой кишке 1—2 см/с. Такие волны могут
возникать в любых частях тонкой кишки. Но чаще они начинаются в две-
надцатиперстной кишке. При этом их возникновение совпадает с началом
эвакуации желудочного содержимого в кишку. В процессе кишечного пи-
щеварения одновременно осуществляется несколько волн перистальтики,
что обеспечивает равномерное перемещение химуса всей тонкой кишки в
каудальном направлении. Слабые перистальтические волны перемещают
только пристеночные слои химуса.
В конце пищеварительного периода возникают сильные перистальтиче-
ские сокращения, которые распространяются вдоль всей тонкой кишки.
Они освобождают ее от химуса, который через илиоцекальный сфинктер
переходит в слепую кишку. Скорость распространения таких волн возрас-
тает в каудальном направлении и достигает в подвздошной кишке 7—
21 см/с. Такая перистальтика называется пропульсивной.
Тонические сокращения возникают на фоне исходного (базального) тону-
са гладких мыщц тонкой кишки, могут распространяться с малой скоро-
стью и вызывать повышение тонуса кишки, суживая ее на большом протя-
жении. Тонические сокращения мускулатуры кишки приводят в соответст-
вие ее емкость с объемом содержимого и создают давление химуса около
8—9 см водн. ст. Другие виды моторики кишки совершаются на фоне то-
нических сокращений. Тонические сокращения характерны также для
гладкомышечных волокон сфинктеров.
Функционально значимыми являются также сокращения и расслабле-
ния ворсинок тонкой кишки, которые осуществляются в течение всего
процесса пищеварения. Благодаря ритмичным сокращениям ворсинок они
постоянно контактируют с новыми порциями химуса, что способствует
всасыванию продуктов гидролиза пищевых веществ и оттоку лимфы по
лимфатическим сосудам.
11.7.2.1. Регуляция моторики тонкой кишки
Моторная активность тонкой кишки зависит от механических свойств и
химического состава ее содержимого. Если пища содержит грубые частицы
(изделия из муки грубого помола, отруби, овощи, фрукты с высоким со-
держанием клетчатки) или жиры, то двигательная активность кишки воз-
растает. На моторику кишечника оказывают также стимулирующее влия-
ние слабые растворы кислот, щелочи и соли.
Регуляция моторики тонкой кишки обеспечивается миогенными меха-
низмами, нервными интрамуральными влияниями на миоциты, централь-
ной нервной системой, гормонами энтерального и неэнтерального проис-
хождения.
В основе миогенного механизма регуляции моторики кишки лежит спо-
собность миоцитов реагировать сокращением на растяжение. Так, при со-
кращении продольных мышечных пучков происходит растяжение цирку-
552 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
лярных мышечных волокон, которые реагируют на механическое воздейст-
вие сокращением. Возбуждение с одного миоцита на другой легко переда-
ется с помощью нексусов. Нексусы — участки тесного контакта мембран
рядом находящихся миоцитов, имеющие низкое электрическое сопротив-
ление. Благодаря этому механизму распространения возбуждения на часто-
ту генерации «спонтанных» медленных волн и потенциалов действия мио-
цитов оказывает влияние электрическая активность мышечных волокон
водителей ритма.
Энтеральный механизм нервной регуляции моторики кишки реализует-
ся за счет взаимодействия нейронов интрамуральных ганглиев (сенсорных,
промежуточных и эфферентных), являющихся структурной основой мест-
ных рефлексов. Так, при локальном раздражении механо- и хеморецепто-
ров слизистой оболочки кишки возникает сокращение циркулярных мышц
выше места стимуляции и их расслабление ниже места раздражения. Это
способствует перемещению химуса в каудальном направлении. При этом
возбуждение от раздраженных рецепторов передается к сенсорным нейро-
нам подслизистого сплетения, а по их аксонам — к промежуточным нейро-
нам межмышечного сплетения. Интернейроны передают возбуждение к
исполнительным (эфферентным) нервным клеткам. Проксимальнее от
места раздражения возбуждаются холинергические нейроны, вызывающее
сокращение циркулярных мышц, а дистальнее от места стимуляции приво-
дятся в действие пептидергические тормозные нервные клетки, которые
тормозят циркулярные мышечные пучки. В окончаниях аксонов этих ней-
ронов выделяются тормозные медиаторы (ВИП и АТФ), которые вызыва-
ют гиперполяризацию мембраны миоцитов.
Моторика тонкой кишки регулируется также центральной нервной сис-
темой через парасимпатические и симпатические нервы. Возбуждение хо-
линергических волокон блуждающего нерва вызывает, как правило, усиле-
ние сократительной активности мускулатуры кишки, а возбуждение адре-
нергических волокон чревных нервов — торможение. Электростимуляция
передних и средних ядер гипоталамуса, сигмовидной извилины коры боль-
ших полушарий усиливает моторику кишки, а раздражение задних ядер ги-
поталамуса и орбитальной извилины коры — тормозит. Электростимуля-
ция переднего отдела поясной извилины и миндалевидного комплекса в
зависимости от функционального состояния тонкой кишки либо активи-
рует, либо тормозит ее моторику. Центры регуляции сократительной ак-
тивности тонкой кишки располагаются также в продолговатом и спинном
мозге.
В естественных условиях функционирования пищеварительной системы
центральные структуры регуляции включаются в ответ на раздражение раз-
личных рефлексогенных зон, что и определяет характер вызываемых реф-
лексов. Различают две группы рефлексов — моторные и тормозные.
При осуществлении пищеводно-кишечного моторного рефлекса раздраже-
ние механорецепторов пищевода во время глотания вызывает повышение
тонуса и силы волн перистальтики тонкой кишки. Центр этого рефлекса
расположен в продолговатом мозге. Свои влияния на кишку он реализует
через блуждающий нерв. Гастродуоденальный, гастроеюнальный и гастро-
илеоцекальный моторные рефлексы возникают в ответ на раздражение меха-
норецепторов наполненного пищей желудка. Эти рефлексы имеют две
компоненты: одна из них реализуется через энтеральную нервную систему,
другая — через центр блуждающего нерва. Раздражение механо- и хеморе-
цепторов слизистой оболочки тонкой кишки порцией химуса вызывает ки-
шечно-кишечный моторный рефлекс, который выражается в усилении
11. Функции пищеварительной системы • 553
двигательной активности участников кишки, расположенных каудальнее
места раздражения. Кишечно-кишечный моторный рефлекс тоже имеет
две компоненты — местную и центральную, осуществляемую через ядро
блуждающего нерва. Группа моторных рефлексов обеспечивает ускорение
темпов пищеварения в тонкой кишке.
Тормозные рефлексы выражаются в угнетении моторики тонкой кишки
и других отделов желудочно-кишечного тракта. Так, при сильном раздра-
жении механорецепторов кишечника возникает ослабление моторики же-
лудочно-кишечного тракта за счет возбуждения спинальных центров чрев-
ного нерва {кишечно-кишечный тормозной рефлекс). В физиологических ус-
ловиях он предотвращает поступление в нижележащие отделы кишечника
недостаточно переработанных порций химуса.
Аналогичный характер имеет прямокишечно-кишечный тормозный
рефлекс, который выражается в угнетении моторики тонкой и толстой
кишки в ответ на раздражение механорецепторов прямой кишки при ее
переполнении каловыми массами. Реализуется этот рефлекс через спи-
нальный центр чревного нерва. Значение описанного рефлекса очевидно:
он уменьшает поступление химуса в конечный отдел толстой кишки до ее
освобождения от каловых масс.
Гуморальная регуляция моторики тонкой кишки осуществляется за счет
комплекса гормонов энтерального и неэнтерального происхождения. Так,
гастрин, мотилин, гистамин, ХЦК, серотонин, вещество Р (см. табл. 11.1),
вазопрессин, брадикинин и окситоцин усиливают электрическую и двига-
тельную активность миоцитов, а секретин, ВИП и ГИП — тормозят.
Гастроинтестинальные гормоны вырабатываются эндокринными эле-
ментами двенадцатиперстной и тонкой кишки под влиянием рефлекторно-
го и химического раздражения химуса и действуют не только на мембран-
ные рецепторы миоцитов, но и на нейроны энтеральной нервной системы.
11.7.3. Функция всасывания тонкой кишки
Общая характеристика процессов всасывания в пищеварительном тракте
изложена в разделе 11.2.3.
Тонкая кишка является основным отделом пищеварительного тракта,
где осуществляется всасывание продуктов гидролиза пищевых веществ, ви-
таминов, минеральных веществ и воды. Высокая скорость всасывания и
большой объем транспорта веществ через слизистую оболочку кишки объ-
ясняются большой площадью ее соприкосновения с химусом за счет нали-
чия макро- и микроворсинок и их сократительной активности, густой сети
капилляров, расположенных под базальной мембраной энтероцитов и
имеющих большое количество широких пор (фенестров), через которые
могут проникать крупные молекулы.
Через поры клеточных мембран энтероцитов слизистой оболочки две-
надцатиперстной и тощей кишки вода легко проникает из химуса в кровь и
из крови в химус, так как ширина этих пор равна 0,8 нм, что значительно
превышает ширину пор в других отделах кишечника. Поэтому содержимое
кишки изотонично плазме крови. По этой же причине в верхних отделах
тонкого кишечника всасывается основное количество воды. При этом вода
следует за осмотически активными молекулами и ионами. К их числу от-
носятся ионы минеральных солей, молекул моносахаридов, аминокислот и
олигопептидов.
С наибольшей скоростью всасываются ионы Na+ (около 500 м/моль за
сутки). Существует два пути транспорта ионов Na+ — через мембрану энте-
554 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Панкреатические
протеазы
Рис. 11.14. Схема переваривания и всасывания белков.
Дипептидазы и аминопептидазы мембраны микроворсинок энтероцита расщепляют олигопеп-
тиды до аминокислот и мелких осколков белковой молекулы, которые транспортируются в
цитоплазму клетки, где цитоплазматические пептидазы завершают процесс гидролиза. Амино-
кислоты через базальную мембрану энтероцита поступают в межклеточное пространство, а за-
тем — в кровь.
роцитов и по межклеточным каналам. В цитоплазму энтероцитов они по-
ступают в соответствии с электрохимическим градиентом. А из энтероцита
в интерстиций и кровь Na+ транспортируется с помощью Na+/K+-Hacoca,
локализованного в базолатеральной части мембраны энтероцитов. Помимо
Na+ по межклеточным каналам по механизму диффузии всасываются ионы
11. Функции пищеварительной системы • 555
Капля жира
Просвет кишечника
Мицелла
Микроворсинка-
Гладкий —
эндоплаз-
матический
ретикулум
Аппарат____
Гольджи >
Межклеточное
пространство
ш
11
Панкреатическая
липаза
Триглицерид
Диглицерид
Моноглицерид
Жирная кислота
Соли желчных кислот
Глицерин
Хиломикрон
Центральный
лимфатический
сосуд
Рис. 11.15. Схема переноса продуктов гидролиза жиров из просвета кишки в цито-
плазму энтероцита и в межклеточное пространство.
Из продуктов гидролиза жиров (моноглицеридов, жирных кислот и глицерина) в гладком эн-
доплазматическом ретикулуме ресинтезируются триглицериды, а в гранулярном эндоплазма-
тическом ретикулуме и аппарате Гольджи формируются хиломикроны. Хиломикроны через
латеральные участки мембраны энтероцита поступают в межклеточное пространство, а за-
тем — в лимфатический сосуд.
К+ и СГ. Высокая скорость всасывания СГ обусловлена тем, что они сле-
дуют за ионами Na+.
Транспорт HCOj сопряжен с транспортом Na+. В процессе его всасыва-
ния в обмен на Na+ энтероцит секретирует в полость кишки Н+, который,
взаимодействуя с HCOj, образует Н2СО3. Н2СО3 под влиянием фермента
556 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
карбоангидразы превращается в молекулу воды и СО2. Двуокись углерода
всасывается в кровь и удаляется из организма с выдыхаемым воздухом.
Всасывание ионов Са2+ осуществляет специальная транспортная система,
которая включает Са2+-связываюгций белок щеточной каймы энтероцита и
кальциевый насос базолатеральной части мембраны. Этим и объясняется от-
носительно высокая скорость всасывания Са2+ (в сравнении с другими двух-
валентными ионами). При значительной концентрации Са2+ в химусе объем
его всасывания возрастает за счет механизма диффузии. Всасывание Са2+
усиливается под влиянием паратгормона, витамина D и желчных кислот.
Всасывание Fe2+ осуществляется с участием переносчика. В энтероците
Fe2+ вступает в соединение с апоферритином, образуя ферритин. В составе
ферритина железо и используется в организме. Всасывание ионов Zn2+ и
М^+ происходит по законам диффузии.
При высокой концентрации моносахаридов (глюкозы, фруктозы, галак-
тозы, пентозы) в химусе, заполняющем тонкую кишку, они всасываются
по механизму простой и облеченной диффузии. Механизм всасывания глюкозы
и галактозы является активным натрий-зависимым. Поэтому при отсутст-
вии Na+ скорость всасывания этих моносахаридов замедляется в 100 раз.
Продукты гидролиза белков (аминокислоты и трипептиды) всасываются
в кровь в основном в верхнем отделе тонкого кишечника — двенадцати-
перстной и тощей кишке (около 80—90 %). Главный механизм всасывания
аминокислот — активный натрийзависимый транспорт. Меньшая часть ами-
нокислот всасывается по механизму диффузии. Процессы гидролиза и вса-
сывания продуктов расщепления белковой молекулы тесно связаны (рис.
11.14). Небольшое количество белка всасывается без расщепления до мо-
номеров — путем пиноцитоза (см. раздел 11.2.3). Так из полости кишки по-
ступают в организм иммуноглобулины, ферменты, а у новорожденного —
белки, содержащиеся в грудном молоке.
Процесс всасывания продуктов гидролиза жиров (моноглицериды, гли-
церин и жирные кислоты) осуществляется в основном в двенадцатиперст-
ной и тощей кишке и отличается существенными особенностями.
Моноглицериды, глицерин и жирные кислоты взаимодействуют с фос-
фолипидами, холестерином и солями желчных кислот, образуя мицеллы.
На поверхности микроворсинок энтероцита липидные компоненты мицел-
лы легко растворяются в мембране и проникают в его цитоплазму, а соли
желчных кислот остаются в полости кишки. В гладком эндоплазматиче-
ском ретикулуме энтероцита происходит ресинтез триглицеридов, из кото-
рых в гранулярном эндоплазматическом ретикулуме и аппарате Гольджи с
участием фосфолипидов, холестерина и гликопротеинов образуются мель-
чайшие капельки жира (хиломикроны), диаметр которых равен 60—75 нм.
Хиломикроны скапливаются в секреторных везикулах. Их мембрана
«встраивается» в латеральную мембрану энтероцита, и через образовавшее-
ся отверстие хиломикроны поступают межклеточные пространства, а затем
в лимфатический сосуд (рис. 11.15).
11.8. Пищеварение в толстом кишечнике
В толстом кишечнике завершается гидролиз остатков пищевых веществ за
счет ферментов, поступающих с химусом из подвздошной кишки, и фер-
ментов, вырабатываемых слизистой оболочкой толстой кишки, и энзимов,
выделяемых микрофлорой, населяющей толстую кишку; осуществляется
всасывание воды, продуктов гидролиза пищевых веществ и микробного раз-
ложения остатков пищи; происходит также формирование каловых масс.
И. Функции пищеварительной системы • 557
11.8.1. Перемещение химуса из тощей кишки в слепую
Перемещение химуса из тощей кишки в слепую осуществляется благодаря
ритмической деятельности илеоцекального сфинктера, который представ-
ляет собой утолщение циркулярных пучков мышечных волокон в конеч-
ном отделе подвздошной кишки. Воронкообразная складка слизистой обо-
лочки кишки в области сфинктера выполняет роль клапана, пропускающе-
го химус только в дистальном направлении.
При отсутствии пищеварительной деятельности желудочно-кишечного
тракта сфинктер находится в состоянии тонического напряжения и пере-
крывает выход из тощей кишки. Через 1—4 мин после приема пищи
сфинктер начинает ритмически сокращаться и расслабляться. Его расслаб-
ление связано с перистальтикой подвздошной кишки, осуществляемой по
механизму местного рефлекса. Когда перистальтическая волна достигает
конечного участка подвздошной кишки, давление химуса в нем возрастает.
При этом сфинктер расслабляется, и химус переходит в слепую кишку до
тех пор, пока не освободится конечный отдел подвздошной кишки. За
1 мин в слепую кишку переходит около 15 мл химуса.
Установлено, что илеоцекальный сфинктер работает синхронно с пило-
рическим сфинктером за счет осуществления бисфинктерного рефлекса. Это
обеспечивает согласование уровней наполнения химусом начальных и ко-
нечных отделов тонкого кишечника.
11.8.2. Сокоотделение в толстом кишечнике
Сокоотделение в толстом кишечнике является в основном реакцией в от-
вет на местное механическое раздражение слизистой оболочки химусом.
Сок толстой кишки состоит из плотной и жидкой компонент. Плотная
компонента включает в себя слизистые комочки, состоящие из спущенных
эпителиоцитов, лимфоидных клеток и слизи. Жидкая компонента имеет
pH 8,5—9,0. Ферменты сока содержатся в основном в спущенных эпите-
лиоцитах, при распаде которых их ферменты (пептидазы, амилаза, липаза,
нуклеаза, катепсины, щелочная фосфатаза) поступают в жидкую компо-
ненту. Содержание ферментов в соке толстой кишки и их активность зна-
чительно ниже, чем в соке тонкого кишечника. Но имеющихся ферментов
достаточно для завершения гидролиза в проксимальных отделах толстой
кишки остатков непереваренных пищевых веществ.
Регуляция сокоотделения слизистой оболочки толстого кишечника осу-
ществляется в основном за счет энтеральных местных нервных механиз-
мов.
11.8.3. Двигательная активность толстого кишечника
Двигательная активность толстого кишечника имеет особенности, которые
обеспечивают накопление химуса, его сгущение за счет всасывания воды,
формирование каловых масс и их удаление из организма во время дефе-
кации.
О временных характеристиках процесса передвижения содержимого по
отделам жулудочно-кишечного тракта судят по перемещению рентгено-
контрастного вещества (например, сернокислого бария). После приема
оно начинает поступать в слепую кишку через 3—3,5 ч. В течение 24 ч
происходит заполнение толстой кишки, которая освобождается от контра-
стной массы через 48—72 ч.
558 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Начальным отделам толстой кишки свойственны очень медленные ма-
лые маятникообразные сокращения. С их помощью осуществляется переме-
шивание химуса, что ускоряет всасывание воды. В поперечной ободочной
и сигмовидной кишке наблюдаются большие маятникообразные сокращения,
вызванные возбуждением большого количества продольных и циркуляр-
ных мышечных пучков. Медленное перемещение содержимого толстой
кишки в дистальном направлении осуществляется благодаря редким пери-
стальтическим волнам. Задержке химуса в толстой кишке способствуют ан-
типеристальтические сокращения, которые перемещают содержимое в рет-
роградном направлении и тем самым способствуют всасыванию воды. Сгу-
щенный обезвоженный химус накапливается в дистальном отделе толстой
кишки. Этот участок кишки отделяется от вышележащего, заполненного
жидким химусом, перетяжкой, вызванной сокращением циркулярных мы-
шечных волокон, что является выражением сегментации.
При заполнении поперечной ободочной кишки сгущенным плотным
содержимым усиливается раздражение механорецепторов ее слизистой
оболочки на значительной площади, что способствует возникновению
мощных рефлекторных пропульсивных сокращений, перемещающих боль-
шой объем содержимого в сигмовидную и прямую кишку. Поэтому подоб-
ного рода сокращения называются масс-сокращениями. Прием пищи ус-
коряет возникновение пропульсивных сокращений за счет осуществления
желудочно-ободочного рефлекса.
Перечисленные фазные сокращения толстой кишки осуществляются на
фоне тонических сокращений, которые в норме продолжаются от 15 с от
5 мин.
В основе моторики толстой кишки, как и тонкой, лежит способность
мембраны гладкомышечных элементов к спонтанной деполяризации. Ха-
рактер же сокращений и их координация зависят от влияний эфферентных
нейронов интраорганной нервной системы и вегетативного отдела ЦНС.
Местные рефлексы при раздражении механорецепторов толстой кишки
усиливают ее моторику; раздражение же механорецепторов только прямой
кишки приводит к торможению двигательной активности толстой и тон-
кой кишки, предотвращая ее переполнение каловыми массами. Торможе-
ние миогенной активности миоцитов толстой кишки через интраорганную
нервную систему реализуется с помощью тормозных медиаторов (ВИП и
АТФ).
Стимуляция парасимпатических нервов (блуждающего и тазового) уве-
личивает частоту и амплитуду сокращений мышц толстой кишки, а раздра-
жение симпатических нервных волокон, выходящих из верхнего и нижнего
брыжеечного сплетений, оказывает противоположный эффект.
Рефлекторные влияния, реализуемые через экстраорганные нервы, сум-
мируются с гуморальными. Так, гастрин, ХЦК и кортизон стимулируют
моторику толстой кишки, а адреналин, серотонин, секретин и глюкагон —
тормозят.
11.8.4. Роль микрофлоры толстой кишки в процессе пищеварения
и формировании иммунологической реактивности
организма
Толстая кишка является местом обитания большого числа микроорганиз-
мов. Они формируют эндоэкологический микробный биоценоз (сообщест-
во). Микрофлора толстого кишечника состоит из трех групп микроорга-
низмов: главной (бифидобактерии и бактероиды — почти 90 % от всех
11. Функции пищеварительной системы • 559
Рис. 11.16. Функции нормальной микрофлоры кишечника.
микробов), сопутствующей (лактобактерии, эшерехии, энтерококки —
около 10 %) и остаточной (цитробактер, энтеробактер, протеи, дрожжи,
клостридии, стафилококки и др. — около 1 %). В толстой кишке находится
максимальное количество микроорганизмов (по сравнению с другими от-
делами пищеварительного тракта). На 1 г кала приходится 1010—1013 мик-
роорганизмов.
Нормальная микрофлора здорового человека участвует в формировании
иммунологической реактивности организма человека, предотвращает раз-
витие в кишечнике патогенных микробов, синтезирует витамины (фолие-
вую кислоту, цианокобаламин, филлохиноны) и физиологически активные
амины, осуществляет гидролиз токсичных продуктов метаболизма белков,
жиров и углеводов, предотвращая эндотоксинемию (рис. 11.16).
В процессе жизнедеятельности микроорганизмов, относящихся к нор-
мальной микрофлоре, образуются органические кислоты, которые снижа-
ют pH среды и тем самым препятствуют размножению патогенных, гнило-
стных и газообразующих микроорганизмов.
Бифидобактерии, лактобактерии, эубактерии, пропионбактерии и бак-
тероиды усиливают гидролиз белков, сбраживают углеводы, омыляют жи-
ры, растворяют клетчатку и стимулируют перистальтику кишечника. Би-
фидо- и эубактерии, а также эшерихии за счет своих ферментных систем
участвуют в синтезе и всасывании витаминов, а также незаменимых ами-
нокислот. Бактериальные модулины бифидо- и лактобактерий стимулиру-
ют лимфоидный аппарат кишечника, увеличивают синтез иммуноглобули-
нов, интерферона и цитокинов, подавляя развитие патогенных микробов.
Кроме того, модулины усиливают активность лизоцима. Анаэробные бак-
терии вырабатывают биологически активные вещества (бета-аланин,
5-аминовалериановую и гамма-аминомасляную кислоты), медиаторы, ока-
зывающие влияние на функции пищеварительной и сердечно-сосудистой
систем, а также на органы кроветворения.
На состав микробного сообщества толстого кишечника оказывают
влияние многие эндогенные и экзогенные факторы. Так, растительная пи-
ща приводит к увеличению энтерококков и эубактерий, животные белки и
жиры способствуют размножению клостридий и бактероидов, но снижают
количество бифидобактерий и энтерококков, молочная пища приводит к
увеличению числа бифидобактерий. Естественным регулятором микрофло-
ры кишечника являются антимикробные вещества, продуцируемые слизи-
стой оболочкой кишечника и содержащиеся в пищеварительных секретах
560 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
(лизоцим, лактоферрин, дефенины, секреторный иммуноглобулин А).
Нормальная перистальтика кишечника, продвигающая химус в дистальном
направлении, оказывает большое влияние на уровень заселенности микро-
бами каждого отдела кишечного тракта, препятствуя их распространению в
проксимальном направлении. Поэтому нарушения двигательной активно-
сти кишечника способствуют возникновению дисбактериоза (изменению
количественных соотношений и состава микрофлоры).
11.8.5. Акт дефекации
Опорожнение нижних отделов толстой кишки от экскрементов осуществ-
ляется с помощью акта дефекации. Позыв к дефекации вызывает раздра-
жение рецепторов прямой кишки при ее заполнении каловыми массами и
повышение давления в ней до 40—50 мм водн. ст. Возникает дефекация
благодаря моторной деятельности прямой кишки и ее двух сфинктеров —
внутреннего гладкомышечного и наружного, образованного поперечно-по-
лосатой мышцей. Как внутренний, так и наружный сфинктеры вне акта
дефекации находятся в состоянии тонического сокращения, что препятст-
вует выпадению каловых масс. Регуляция непроизвольного процесса опо-
рожнения прямой кишки осуществляется интрамуральной нервной систе-
мой, парасимпатическими и соматическими нервными центрами крестцо-
вых сегментов спинного мозга, образующими центр дефекации (S,—S,v).
Афферентные импульсы от механорецепторов слизистой оболочки пря-
мой кишки по срамным и тазовым нервам поступают в спинальный центр,
откуда по эфферентным парасимпатическим волокнам этих же нервов пе-
редаются к гладкой мускулатуре прямой кишки и ее внутреннего сфинкте-
ра. Тонус наружного анального сфинктера в начале осуществления реф-
лекса дефекации повышается, а при достижении надпороговой силы раз-
дражения механорецепторов прямой кишки тормозится, что сопровожда-
ется дефекацией.
Произвольный акт дефекации осуществляется при участии центров
продолговатого мозга, гипоталамуса и коры больших полушарий головного
мозга. Произвольное управление актом дефекации формируется в течение
первого года жизни. Центр в продолговатом мозге, принимающий участие
в регуляции этого акта, находится вблизи дыхательного и рвотного. Близо-
стью центров объясняется усиление дыхания и торможение рвотного реф-
лекса при растяжении анальных сфинктеров и непроизвольная дефекация
при остановке дыхания.
Естественный акт дефекации является отчасти волевым, отчасти непро-
извольным. При значительном раздражении прямой кишки происходит ее
сокращение и расслабление внутреннего анального сфинктера. Произволь-
ная часть акта дефекации включает расслабление наружного сфинктера,
сокращение мышц диафрагмы и брюшных мышц. Все это ведет к умень-
шению объема брюшной полости и повышению внутрибрюшного давле-
ния (до 220 см водн. ст.). Рефлекс дефекации полностью исчезает после
разрушения крестцовых сегментов — спинного мозга, где расположен
центр дефекации. При разрушении спинного мозга выше этих сегментов
спинальные рефлексы дефекации сохраняются, однако произвольный ком-
понент рефлекса дефекации не осуществляется из-за отсутствия нисходя-
щих влияний центров головного мозга.
Дефекация оказывает рефлекторные влияния на различные органы и
системы. Так, во время дефекации систолическое артериальное давление
повышается примерно на 60 мм рт. ст., а диастолическое — на 20 мм рт.
11. Функции пищеварительной системы • 561
ст., пульс учащается на 20 ударов в минуту. Это обусловлено сокращением
большой мышечной массы и задержкой дыхания при натуживании. В нор-
ме у 70 % здоровых людей акт дефекации совершается 1 раз в сутки, у
30 % — 2 раза и чаще.
11.8.6. Иммунная система пищеварительного тракта
Помимо неспецифических факторов противодействия болезнетворным ан-
тигентам содержимого желудочно-кишечного тракта организм располагает
и специфической системой защиты. Она представляет собой совокупность
структур и элементов лимфоидной ткани, расположенных на протяжении
всего пищеварительного тракта и являющихся частью иммунной системы
организма. Функции иммунной защиты выполняют: лимфоидные фолли-
кулы, которые образуют скопления (пейеровы бляшки тонкой кишки,
миндалины глоточного кольца), Т-лимфоидные клетки и плазматические
элементы слизистой оболочки, а также малые лимфоидные клетки. Имму-
нокомпетентные клетки пищеварительного тракта осуществляют распозна-
ние антигенов химуса, обеспечивают местный иммунный ответ и форми-
руют иммунную память (чтобы облегчить реакцию на антиген при повтор-
ном контакте с ним).
Миндалины выделяют на слизистую оболочку глотки иммуноглобули-
ны, лизоцим, интерферон, простагландины, лимфоциты и макрофаги.
Плазматические клетки, находящиеся в строме ворсинок, продуцируют
иммуноглобулины всех классов (I, М, A, D, Е). Особую роль играет имму-
ноглобулин А, который не разрушается протеолитическими ферментами
пищеварительных соков. Выделившись на поверхность эпителиальных
клеток слизистой оболочки, он соединяется с антигенами химуса и микро-
бов и приобретает способность отталкиваться от эпителиоцитов. Вместе со
слизью этот комплекс включается в состав химуса, подвергается разруше-
нию протеолитическими ферментами. Степень выраженности и эффектив-
ность местного иммунного ответа зависят от состояния общей иммунной,
нервной и эндокринной систем.
Иммунная система пищеварительного тракта выполняет роль энтероге-
матического барьера, предотвращая поступление в кровь патогенных мик-
робов, их токсинов и ядовитых веществ пищи.
При ослаблении механизмов неспецифической и специфической, мест-
ной и общей защиты организма от болезнетворного влияния антигенов и
ядовитых компонентов химуса экзогенного и эндогенного происхождения
возникают патологические нарушения функций пищеварительной сис-
темы.
11.8.7. Тошнота и рвота
Тошнота — неприятное ощущение, формирующееся при значительном по-
вышении возбудимости нейронов ретикулярной формации области рвот-
ного центра. Это ощущение вызвано раздражением рецепторов желудочно-
кишечного тракта токсинами или метаболитами внутренней среды, а также
возбуждением высших отделов центральной нервной системы (коркового
отдела зрительной и обонятельной сенсорных систем) на фоне повышен-
ной возбудимости рвотного центра. Тошнота предшествует рвоте и сопро-
вождается учащением дыхания, тахикардией, саливацией, появлением мел-
ких глотательных движений, направленных на освобождение полости рта
от избыточного количества слюны, усилением потоотделения и сужением
562 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
кожных сосудов. Возникновение этого комплекса реакций объясняется ир-
радиацией возбуждения из рвотного центра на центры других рефлексов.
Рвота — защитная сложнокоординированная рефлекторная реакция
гладкой мускулатуры желудочно-кишечного тракта и скелетных мышц,
осуществляющих выбрасывание содержимого желудка и тонкой кишки на-
ружу через рот. Эта реакция возникает при возбуждении нейронов рвотно-
го центра ретикулярной формации продолговатого мозга в ответ на раздра-
жение рецепторов желудочно-кишечного тракта токсичными метаболита-
ми внутренней среды или вредными веществами, введенными извне, раз-
дражением рецепторов вестибулярного аппарата. Рвота может быть обу-
словлена обонятельными, зрительными, вкусовыми раздражениями, вызы-
вающими чувство отвращения, а также раздражением нейронов рвотного
центра при повышении внутричерепного давления. Импульсы от рецепто-
ров желудочно-кишечного тракта поступают в рвотный центр по сенсор-
ным волокнам блуждающего и языкоглоточного нервов. Эфферентные
влияния по центробежным нервным волокнам в составе блуждающего и
чревного нервов передаются к кишечнику, желудку, пищеводу, а также по
моторным нервам, иннервирующим мышцы брюшной стенки, диафрагмы
и наружных межреберных мышц.
Рвота сопровождается урежением и углублением дыхания, поднятием
подъязычной кости и гортани, что способствует открытию верхнего пище-
водного сфинктера, закрытию глотки, поднятию мягкого неба с закрытием
хоан. Вслед за этим начинается сильное сокращение диафрагмы при одно-
временном сокращении всех мышц брюшной стенки. При этом нижний
пищеводный сфинктер расслабляется, позволяя содержимому желудка
пройти через пищевод наружу.
Акту рвоты предшествует возникновение антиперистальтики тонкой
кишки, забрасывающей ее содержимое в желудок.
ГЛАВА 12
Обмен веществ и энергии. Питание
Обмен веществ и энергии лежит в основе всех проявлений жизнедеятельно-
сти и представляет собой совокупность процессов превращения веществ и
энергии в живом организме и обмен веществами и энергией между орга-
низмом и окружающей средой.
Для поддержания жизнедеятельности в процессе обмена веществ и
энергии обеспечиваются пластические и энергетические потребности орга-
низма. Пластические потребности удовлетворяются за счет веществ, ис-
пользуемых для построения биологических структур, а энергетические —
путем преобразования химической энергии поступающих в организм пита-
тельных веществ в энергию макроэргических (АТФ и другие молекулы) и
восстановленных (НАДФ • Н — никотин-амид-адениндинуклеотидфосфат)
соединений. Их энергия используется организмом для синтеза белков,
нуклеиновых кислот, липидов, а также компонентов клеточных мембран и
органелл клетки, для выполнения деятельности клеток, связанной с ис-
пользованием химической, электрической и механической энергии.
Обмен веществ и энергии (метаболизм) в организме человека — сово-
купность взаимосвязанных, но разнонаправленных процессов: анаболизма
(ассимиляции) и катаболизма (диссимиляции).
Анаболизм — это совокупность процессов биосинтеза органических ве-
ществ, компонентов клетки и других структур органов и тканей. Анабо-
лизм обеспечивает рост, развитие, обновление биологических структур, а
также непрерывный ресинтез макроэргических соединений и их накоп-
ление.
Катаболизм — это совокупность процессов расщепления сложных моле-
кул, компонентов клеток, органов и тканей до простых веществ (с исполь-
зованием части из них в качестве предшественников биосинтеза) и до ко-
нечных продуктов метаболизма (с образованием макроэргических и вос-
становленных соединений).
Взаимосвязь процессов катаболизма и анаболизма основывается на
единстве биохимических превращений, обеспечивающих энергией все про-
цессы жизнедеятельности и постоянное обновление тканей организма. Со-
пряжение анаболических и катаболических процессов в организме могут
осуществлять различные вещества, но главную роль в этом сопряжении иг-
рают АТФ, НАДФ • Н. В отличие от других посредников метаболических
превращений АТФ циклически рефосфорилируется, а НАДФ • Н — восста-
навливается, что обеспечивает непрерывность процессов катаболизма и
анаболизма.
Обеспечение энергией процессов жизнедеятельности осуществляется за
счет анаэробного (бескислородного) и аэробного (с использованием кислорода)
катаболизма поступающих в организм с пищей белков, жиров и углеводов.
В ходе анаэробного расщепления глюкозы (гликолиза) или ее резервного
субстрата гликогена (гликогенолиза) превращение 1 моля глюкозы в 2 мо-
ля лактата приводит к образованию 2 молей АТФ. Лактат — промежуточ-
564 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ный продукт обмена. В химических связях его молекулы аккумулировано
значительное количество энергии. Энергии, образующейся в ходе анаэроб-
ного обмена, недостаточно для осуществления процессов жизнедеятельно-
сти животных организмов. За счет анаэробного гликолиза могут удовлетво-
ряться лишь относительно кратковременные энергетические потребности
клетки.
В организме животных и человека в процессе аэробного обмена органи-
ческие вещества, в том числе продукты анаэробного обмена, окисляются
до конечных продуктов — СО2 и Н2О. Общее количество молекул АТФ, об-
разующихся при окислении 1 моля глюкозы до СО2 и Н2О, составляет 25,5
моля. При окислении молекулы жиров образуется большее количество мо-
лей АТФ, чем при окислении молекулы углеводов. Так, при окислении 1
моля пальмитиновой кислоты образуется 91,8 моля АТФ. Количество мо-
лей АТФ, образующихся при полном окислении аминокислот и углеводов,
примерно одинаково. АТФ играет в организме роль внутренней «энергети-
ческой валюты» и аккумулятора химической энергии клеток.
Основным источником энергии восстановления для реакции биосинте-
за жирных кислот, холестерина, аминокислот, стероидных гормонов, пред-
шественников синтеза нуклеотидов и нуклеиновых кислот является
НАДФ • Н. Образование этого вещества осуществляется в цитоплазме клет-
ки в процессе фосфоглюконатного пути катаболизма глюкозы. При таком
расщеплении из 1 моля глюкозы образуется 12 молей НАДФ • Н.
Процессы анаболизма и катаболизма находятся в организме в состоя-
нии динамического равновесия или временного превалирования одного из
них. Преобладание анаболических процессов над катаболическими приво-
дит к росту, накоплению массы тканей, а катаболических — к частичному
разрушению тканевых структур, выделению энергии. Состояние равновес-
ного или неравновесного соотношения анаболизма и катаболизма зависит
от возраста. В детском возрасте преобладают процессы анаболизма, а в
старческом — катаболизма. У взрослых людей эти процессы находятся в
равновесии. Их соотношение зависит также от состояния здоровья, выпол-
няемой человеком физической или психоэмоциональной деятельности.
12.1. Роль белков, жиров, углеводов, минеральных веществ
и витаминов в метаболизме
Потребность организма в пластических веществах может быть удовле-
творена тем минимальным уровнем их поступления с пищей, который
уравновешивает потери структурных белков, липидов и углеводов. Эти по-
требности индивидуальны и зависят от таких факторов, как возраст чело-
века, состояние здоровья, интенсивность и вид труда.
Человек получает в составе пищевых продуктов заключенные в них пла-
стические вещества, минеральные вещества и витамины. Суточная потреб-
ность человека в белках, жирах и углеводах представлена в разделе 12.5.
12,1.1. Белки и их роль в организме
Белки в организме находятся в состоянии непрерывного обмена и обнов-
ления. У здорового взрослого человека количество распавшегося за сутки
белка равно количеству вновь синтезированного. Животные существа мо-
гут усваивать азот только в составе аминокислот, поступающих в организм
с белками пищи. Десять аминокислот из 20 (валин, лейцин, изолейцин,
лизин, метионин, триптофан, треонин, фенилаланин, аргинин и гистидин)
12. Обмен веществ и энергии. Питание • 565
в случае их недостаточного поступления с пищей не могут быть синтезиро-
ваны в организме. Эти аминокислоты называют незаменимыми. Другие де-
сять аминокислот (заменимые) не менее важны для жизнедеятельности,
чем незаменимые, но в случае недостаточного поступления с пищей заме-
нимых аминокислот они могут синтезироваться в организме. Важным фак-
тором обмена белков организма является повторное использование (реути-
лизация) аминокислот, образовавшихся при распаде одних белковых моле-
кул, для синтеза других.
Скорость распада и обновления белков организма различна. Полупери-
од распада гормонов пептидной природы составляет минуты или часы,
белков плазмы крови и печени —около 10 сут, белков мышц — около 180
сут. В среднем все белки организма человека обновляются за 80 сут. О
суммарном количестве белка, подвергшегося распаду за сутки, судят по
количеству азота, выводимого из организма человека. В белке содержится
около 16 % азота (т. е. в 100 г белка — 16 г азота). Таким образом, выделе-
ние организмом 1 г азота соответствует распаду 6,25 г белка. За сутки из
организма взрослого человека выделяется около 3,7 г азота. Из этих дан-
ных следует, что масса белка, подвергшегося за сутки полному разруше-
нию, составляет 3,7 х 6,25 = 23 г, или 0,028—0,075 г азота на 1 кг массы
тела в сутки {коэффициент изнашивания по Рубнеру).
Если количество азота, поступающего в организм с пищей, равно коли-
честву азота, выводимого из организма, принято считать, что организм на-
ходится в состоянии азотистого равновесия. В случаях, когда в организм
поступает азота больше, чем его выделяется, говорят о положительном азо-
тистом балансе (задержке, ретенции азота). Такие состояния бывают у че-
ловека при увеличении массы мышечной ткани, в период роста организма,
беременности, выздоровления после тяжелого истощающего заболевания.
Состояние, при котором количество выводимого из организма азота
превышает его поступление в организм, называют отрицательным азоти-
стым балансом. Оно имеет место при питании неполноценными белками,
когда в организм не поступают какие-либо из незаменимых аминокислот,
при белковом голодании или при полном голодании.
Белки, использующиеся в организме в первую очередь в качестве пла-
стических веществ, в процессе их разрушения освобождают энергию для
синтеза в клетках АТФ и образования тепла.
Всемирной организацией здравоохранения рекомендуется потребление
белка не менее 0,75 г/кг в сутки, или для взрослого здорового человека
массой 70 кг не менее 52,5 г легкоусвояемого полноценного белка.
12.1.2. Липиды и их роль в организме
Липиды организма человека — это, главным образом, нейтральные слож-
ные эфиры глицерина и высших жирных кислот — триглицериды, фосфо-
липиды и стерины. Высшие жирные кислоты, входящие в состав сложных
липидных молекул в виде углеводородных радикалов, бывают насыщенны-
ми и ненасыщенными, содержащими одну и более двойных связей. Липи-
ды играют в организме энергетическую и пластическую роль. По сравне-
нию с молекулами углеводов и белков молекула липидов является более
энергоемкой. Поэтому при окислении липидов в организме образуется
больше молекул АТФ и тепла. За счет окисления жиров обеспечивается
около 50 % потребности в энергии взрослого организма. Запасы нейтраль-
ных жиров-триглицеридов в жировых депо человека в среднем составляют
10—20 % массы его тела. Из них около половины локализуется в подкож-
566 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ной жировой клетчатке. Кроме того, значительные запасы нейтрального
жира откладываются в большом сальнике, околопочечной клетчатке, в об-
ласти гениталий и между мышцами. Жиры, откладываясь в жировых депо,
служат долгосрочным резервом питания организма.
Жиры являются источником образования эндогенной воды. При окис-
лении 100 г нейтрального жира в организме образуется около 107 г воды.
Если в удовлетворении энергетических потребностей организма основную
роль играют нейтральные молекулы жира (триглицериды), то пластическая
функция липидов в организме осуществляется, главным образом, за счет
фосфолипидов, холестерина, жирных кислот. Эти липидные молекулы яв-
ляются структурными компонентами клеточных мембран (липопротеинов)
и предшественниками синтеза стероидных гормонов, желчных кислот и
простагландинов.
12.1.2.1. Клеточные липиды
В состав клеточных липидов входят фосфолипиды и холестерин, являющие-
ся необходимыми структурными компонентами поверхностной и внутри-
клеточных мембран. Триглицериды откладываются в клетках в виде жиро-
вых капель, формируя жировые депо. Последние являются не инертной
массой, а активной динамической тканью, в которой запасенные жиры
подвергаются постоянному расщеплению и ресинтезу. При действии на
организм холода, в состоянии голода, при физической или психоэмоцио-
нальной нагрузке происходит интенсивное расщепление (липолиз) запа-
сенных триглицеридов. Образующиеся при этом неэстерифицированные
жирные кислоты используются в организме как энергодающие или как
пластические вещества, необходимые для синтеза сложных липидных мо-
лекул. В условиях покоя после приема пищи происходят ресинтез и отло-
жение нейтральных липидов в подкожной жировой клетчатке, брюшной
полости, мышцах.
12.1.2.2. Бурый жир
Такой оттенок жировой ткани придают окончания симпатических нервных
волокон, а также многочисленные митохондрии, содержащиеся в клетках
этой ткани. Поэтому бурый жир легко мобилизуется для обеспечения
энергетических потребностей организма. Он располагается в межлопаточ-
ной области, вдоль крупных сосудов грудной и брюшной полостей, в заты-
лочной области шеи. Масса бурой жировой ткани достигает у взрослого
0,1 % массы тела. В митохондриях жировых клеток имеется полипептид
молекулярной массой 32 000, способный разобщать идущие здесь процес-
сы окисления и образования АТФ. Результатом такого разобщения являет-
ся образование в бурой жировой ткани в ходе метаболизма жира значи-
тельно большего количества тепла, чем в белой жировой ткани. Бурая жи-
ровая ткань участвует в теплопродукции.
12.1.2.3. Липиды плазмы крови
Липидные молекулы, обладая гидрофобными свойствами, не могут раство-
ряться в водной среде или образовывать молекулярную взвесь. Попадая в
значительных количествах в кровь при переломе крупных трубчатых кос-
тей или при переливании неустойчивых жировых эмульсий, они формиру-
ют липидные капельки и могут вызвать закупорку сосудов — жировую эм-
12. Обмен веществ и энергии. Питание • 567
болию. В физиологических условиях в эпителиальных клетках тонкого ки-
шечника и клетках печени из белковых молекул (апопротеинов), фосфоли-
пидов, холестерина, эфиров холестерина и триглицеридов формируются
транспортные частицы, называемые липопротеинами. По показателям мо-
лекулярной массы, размеров и плотности липопротеины крови подразде-
ляют на классы: хиломикроны, липопротеины очень низкой плотности,
липопротеины низкой плотности и липопротеины высокой плотности.
Липидные молекулы, поступившие из кишечника в эпителиоциты, упа-
ковываются в транспортные частицы экзогенных липидов, главным обра-
зом в хиломикроны. Хиломикроны через лимфатические сосуды и протоки
поступают в кровоток. Под действием липопротеидлипазы эндотелия ка-
пилляров главный компонент хиломикронов — нейтральные триглицери-
ды — расщепляются до глицерина и свободных жирных кислот. Часть жир-
ных кислот может связываться с альбумином, а глицерин и свободные
жирные кислоты поступают в жировые клетки и реэстерифицируются в
триглицериды. Остатки хиломикронов крови захватываются гепатоцитами
по механизму экзоцитоза и разрушаются в лизосомах.
В печени формируются липопротеины для транспорта синтезированных
в ней липидных молекул. Это липопротеины очень низкой и липопротеины
низкой плотности, которые транспортируются из печени к другим тканям.
Как простые, так и сложные липидные молекулы могут синтезироваться
в организме. Исключением являются полиненасыщенные линолевая, ли-
ноленовая и арахидоновая жирные кислоты, синтез которых в организме
не осуществляется, и они должны поступать с пищей. Эти кислоты, полу-
чившие название незаменимых, входят в состав молекул фосфолипидов. Из
арахидоновой кислоты, отщепляемой от молекулы мембранных фосфоли-
пидов, образуются простагландины, простациклины, тромбоксаны, лей-
котриены. Отсутствие в пищевых продуктах или недостаточное поступле-
ние в организм незаменимых жирных кислот приводит к задержке роста,
нарушению функции почек, заболеваниям кожи, бесплодию.
12.1.3. Углеводы и их роль в организме
Организм человека получает углеводы, главным образом, в виде раститель-
ного полисахарида крахмала и в небольшом количестве в виде животного
полисахарида гликогена. В желудочно-кишечном тракте осуществляется их
расщепление до уровня моносахаридов (глюкозы, фруктозы, лактозы, га-
лактозы). Моносахариды, основным из которых является глюкоза, всасыва-
ются в кровь и через воротную вену поступают в печеночные клетки. Здесь
фруктоза и галактоза превращается в глюкозу. Внутриклеточная концен-
трация глюкозы в гепатоцитах близка к ее концентрации в крови. При из-
быточном поступлении в печень глюкозы она фосфорилируется и превра-
щается в резервную форму ее хранения — гликоген. Количество гликогена
может составлять у взрослого человека 150—200 г. В случае ограничения
потребления пищи или по мере снижения концентрации глюкозы в крови
происходит расщепление гликогена и поступление глюкозы в кровь. Кон-
центрация глюкозы в крови через 3—4 ч после приема пищи поддержива-
ется на уровне 0,8—1,0 г/л.
Такой же процесс перехода глюкозы в кровь осуществляется, когда ее
концентрация в клетке повышается за счет глюконеогенеза — синтеза глю-
козы из лактата или аминокислот. В течение первых 12 ч и более после
приема пищи поддержание концентрации глюкозы в крови и обеспечение
потребности организма в углеводах реализуются за счет распада гликогена
568 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
в печени. Вслед за истощением запасов гликогена усиливается синтез фер-
ментов, обеспечивающих глюконеогенез.
Глюкоза, являющаяся источником энергии для процессов жизнедея-
тельности и, в частности, главным источником энергии для клеток мозга,
выполняет в организме и пластические функции. Так, в ходе ее окисления
образуются промежуточные продукты — пентозы, которые являются со-
ставной частью молекулы нуклеотидов и нуклеиновых кислот. Глюкоза не-
обходима для синтеза некоторых аминокислот, синтеза и окисления липи-
дов, полисахаридов.
Следовательно, обмен углеводов является необъемным компонентом
единого метаболизма.
12.1.4. Минеральные вещества и их роль в организме
Минеральные вещества растворены в жидкостях, составляющих внутрен-
нюю среду организма, а также в цитоплазме клеток. Образуемый ими по-
лиэлектролит создает необходимые условия для осуществления многочис-
ленных химических реакций. Минеральные вещества являются кофактора-
ми ферментативных реакций, создают необходимый уровень осмотическо-
го давления, обеспечивают кислотно-основное равновесие, участвуют в
процессах свертывания крови, создают мембранный потенциал и потенци-
ал действия возбудимых клеток.
Данные о физиологической роли, суточной потребности и пищевых ис-
точниках минеральных веществ приведены в табл. 12.1.
Таблица 12.1. Физиологическая роль, суточная потребность организма и источник по-
ступления минеральных веществ
Вещества Физиологическая роль, суточная потребность Источник
Натрий Содержится в больших количествах во внекле- точной жидкости и плазме крови. Играет важ- нейшую роль в процессах возбуждения, поддер- жании осмотического давления, распределении и выведении воды из организма; участвует в формировании бикарбонатной буферной систе- мы. Суточная потребность — 2—3 г, а в виде NaCl — 5 г Поваренная соль, в соста- ве растительной и живот- ной пищи, в жидкостях, потребляемых при питье
Кальций Один из наиболее важных минеральных элемен- тов организма. Выполняет функцию структурно- го компонента в тканях зубов и костей. В этих тканях содержится около 99 % от общего коли- чества Са2+ в организме. Необходим для осуще- ствления процессов свертывания крови, возбуж- дения клеток, синаптической передачи, сокра- щения мыщц, вторичный посредник в регуля- ции внутриклеточного метаболизма и др. Суточ- ная потребность — 0,8 г Молоко и молочные про- дукты, овощи, зеленые листья
Калий Содержится преимущественно внутри клеток, а также в жидкостях внутренней среды. Играет важную роль в процессах реполяризации после возбуждения в нервных волокнах, сокращении мышц, в том числе миокарда. Суточная потреб- ность — 2—3 г Потребность при нор- мальном питании удовле- творяется за счет пищево- го калия. Наиболее бога- ты мясо, овощи, орехи сухофрукты
Хлор Содержится как во внеклеточной, так и во внут- риклеточной жидкости. Играет роль в процессах Поваренная соль, в соста- ве растительной и живот-
12. Обмен веществ и энергии. Питание • 569
Продолжение
Вещества Физиологическая роль, суточная потребность Источник
возбуждения и торможения, в синаптической передаче, образовании соляной кислоты желу- дочного сока. Суточная потребность — 3—5 г ной пищи; в жидкостях, потребляемых при питье
Фосфор Около 80 % в виде минеральных веществ содер- жится в костях и зубах. В составе .фосфолипидов входит в структуру клеточных мембран, липо- протеидов. В составе АТФ и ее производных иг- рает большую роль в метаболизме, осуществле- нии важнейших физиологических процессов. Суточная потребность около 0,7—0,8 г Пищевые продукты, в особенности молоко, мя- со, яйца, рыба, орехи, злаки
Железо Около 65 % содержится в гемоглобине крови, находится в скелетных мышцах, печени, селе- зенке, костном мозге, в составе ферментов. Ос- новная функция — связывание кислорода. Су- точная потребность — 10—15 мг Пищевые продукты, в особенности мясо, пе- чень, свежая рыба, яйца, сухофрукты, орехи
Йод Важнейший компонент гормонов и предшест- венников гормонов щитовидной железы. Суточ- ная потребность —0,15—0,3 мг Йодированная поварен- ная соль, морепродукты, рыбий жир, овощи, выра- щенные на обогащенных йодом почвах
Медь Содержится в печени, селезенке. Играет роль в процессах всасывания железа, образовании ге- моглобина, пигментации. Суточная потреб- ность — 2—5 мг Пищевые продукты, в особенности яйца, пе- чень, почки, рыба, шпи- нат, сухие овощи, вино- град
Фтор Содержится в зубных тканях и необходим для сохранения их целостности. Суточная потреб- ность — 1 мг. При пятикратной передозировке токсичен Пищевые продукты, фто- рированная NaCl, фтори- рованные зубные пасты и растворы
Магний Содержится в костной ткани, необходим для ее образования, а также нормального осуществле- ния функции мышечной и нервной тканей. Не- обходим для синтеза многих коферментов. Су- точная потребность — 250—350 мг Мясо, молоко, целые зерна
Сера Входит в состав аминокислот, белков (инсулин) и витаминов (В,, Н), суточная потребность предположительно равна 1 г Пищевые продукты, в особенности мясо, пе- чень, рыба, яйца Пищевые продукты: кра- бы, мясо, бобы, яичный желток
Цинк Важный компонент ряда ферментов. Необходим для нормального роста. Суточная потребность — 10—15 мг
Кобальт Входит в состав витамина В12 и необходим для нормального осуществления эритропоэза. Су- точная потребность точно не известна, предпо- ложительно 100—200 мкг Печень
Йод, железо, медь, марганец, цинк, фтор, хром, кобальт относятся к
группе микроэлементов; они содержатся в пище и воде в чрезвычайно ма-
лых количествах, но необходимы для осуществления метаболических про-
цессов, так как входят в состав молекулы ферментов, гормонов и витами-
нов. Сведения, приведенные в табл. 12.1, необходимы для анализа пище-
вого рациона человека с целью научно обоснованной его коррекции.
570 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
12.1.5. Вода и ее роль в организме — см. раздел 14.3. Водно-солевой
обмен
12.1.6. Витамины и их роль в организме
Витамины — группы разнородных по химической природе веществ, не син-
тезируемых или синтезируемых в недостаточных количествах в организме,
но необходимых для нормального осуществления обмена веществ, роста,
развития организма и поддержания здоровья. Эти вещества не являются
непосредственными источниками энергии и не выполняют пластических
функций. Они являются составными компонентами ферментных систем и
играют роль катализаторов в обменных процессах. Сведения об источни-
ках витаминов, их суточной потребности для взрослого человека и значе-
нии в осуществлении физиологических функций приведены в табл. 12.2.
Таблица 12.2. Физиологическая роль, потребность организма и источник поступления
витаминов
Вита- мин Суточная потреб- ность взросло- го чело- века Основные ис- точники Физиологическая роль Признаки недостаточности
А* (ре- А,- Животные жи- Необходим для синтеза Нарушаются функции
тинол) 0,9 мг, бета-ка- ротин — 1,8 мг ры, мясо, ры- ба, яйца, мо- локо зрительного пигмента родопсина; оказывает влияние на процессы роста, размножения, пролиферации и оро- говения эпителия сумеречного зрения, рос- та, развития и размноже- ния. Развивается сухость поверхности конъюнкти- вы и роговицы, изъязвле- ние роговицы
D (каль- цифе- рол) 2,5 мкг Печень и мя- со млекопи- тающих, пе- чень рыб, яйца Необходим для всасы- вания из кишечника ионов кальция и для обмена в организме кальция и фосфора Недостаточное поступле- ние в детском возрасте приводит к развитию ра- хита, что проявляется на- рушением окостенения и роста костей, их декаль- цификацией и размягче- нием
РР** (нико- тино- вая ки- слота) 150 мг Мясо, печень, почки, рыба, дрожжи Участвует в процессах клеточного дыхания (переносе водорода и электронов); регуляции секреторной и мотор- ной функции желудоч- но-кишечного тракта Воспаление кожи (пелла- гра), расстройства желу- дочно-кишечного тракта (понос)
К (фил- лохино- ны) До 1 мг Зеленые ли- стья овощей, печень Участвует в синтезе факторов свертывания крови, протромбина и др. Замедленное свертыва- ние крови, спонтанные кровотечения
Е (то- коферо- лы) 10—12 мг Растительные масла, зеле- ные листья овощей, яйца Антиоксидант (инги- битор окисления) Четко определенных симптомов недостаточ- ности у человека не опи- сано
С (ас- 50- Свежие фрук- Участвует в гидрокси- Развивается цинга, про-
корби- новая 100 мг ты и растения (особенно ши- лировании, образова- нии коллагена, вклю- явлением которой явля- ются кровоточивость де-
12. Обмен веществ и энергии. Питание • 571
Продолжение
Вита- мин Суточная потреб- ность взросло- го чело- века Основные ис- точники Физиологическая роль Признаки недостаточности
кисло- та) повник, чер- ная смороди- на, цитрусо- вые) чении железа в ферри- тин. Повышает устой- чивость организма к инфекциям сен, мелкие кровоизлия- ния в коже, поражение стенок кровеносных со- судов
В, (тиа- 1,4- Целые зерна, Участвует в энергети- Развивается заболевание
мин) 2,4 мг бобы, печень, почки, отру- би, дрожжи ческом обмене (про- цессах декарбоксили- рования), является ко- ферментом пируват - карбоксилазы бери-бери, сопровож- дающееся полиневри- том, нарушением сердеч- ной деятельности и функций желудочно-ки- шечного тракта
В2 (рибо- флавин) 2—3 мг Зерновые, бо- бы, печень, молоко, дрож- жи, яйца Входит в состав фла- виновых ферментов. Осуществляет перенос водорода и электронов Поражение глаз (свето- боязнь), поражение сли- зистой оболочки полости рта и языка
В3 (пан- тотено- вая ки- слота) 10 мг Зерновые, бо- бы, карто- фель, печень, яйца, рыба Перенос ацетильной группы (КоА) при син- тезе жирных кислот, стероидов и других со- единений Общая слабость, голово- кружение, нейромотор- ные нарушения, воспале- ния кожи, поражения слизистых оболочек
В6 (пи- ридок- син) 1,5—3 мг Зерно, бобы, мясо, печень, дрожжи, ры- ба. Синтезиру- ется микро- флорой ки- шечника Кофермент трансами- назы, декарбоксилазы, дегидратазы, десульфо- гидразы Повышенная раздражи- тельность, судороги, ги- похромная анемия. Игра- ет важную роль в обмене аминокислот, белков и жиров, а также в процес- сах кроветворения
В12 (ци- аноко- бала- мин) 2 мкг Печень, синте- зируется мик- роорганизма- ми кишечника Компонент ферментов метаболизма нуклеино- вых кислот и метили- рования. Необходим для гемопоэза Злокачественная анемия
Фолие- вая ки- слота 400 мг Зеленые ли- стья, овощи, мясо, молоко, дрожжи. Син- тезируется микроорганиз- мами кишеч- ника Необходима для синте- за пуринов и метиони- на и метаболизма од- ноуглеродных фраг- ментов молекул. Сти- мулирует процесс кро- ветворения Анемия
150- Молоко, яич- Кофермент дезаминаз, карбоксилаз, трансфе- раз, осуществляет пе- ренос СО2 Дерматит (воспаление
(био- тин) 200 мкг ный желток, печень, синте- зируется мик- роорганизма- ми кишечника кожи) с гиперфункцией сальных желез
‘Проявления передозировок: головные боли, эйфория, анемия, изменения со стороны ко-
жи, слизистых оболочек, костной ткани.
“Проявление передозировки: нарушение функций ЦНС и почек; вымывание Са2+ из кос-
тей и повышение его уровня в крови.
“‘Гиповитаминоз может развиваться при потреблении больших количеств сырого яичного
белка, связывающего биотин.
572 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Основными источниками водорастворимых витаминов (группы В, вита-
мин С) являются, как правило, пищевые продукты растительного происхо-
ждения и в меньшей мере животного происхождения. Эти витамины легко
всасываются из желудочно-кишечного тракта в кровь и лимфу.
Основными источниками жирорастворимых витаминов (витамины A, D,
Е, К) являются продукты животного происхождения. Для удовлетворения
потребностей организма в витаминах имеет значение не только достаточ-
ное содержание в пищевом рационе богатых витаминами продуктов расти-
тельного и животного происхождения, но и нормальное осуществление
процессов пищеварения и всасывания веществ в желудочно-кишечном
тракте. Так, при нарушениях пищеварения в тонком кишечнике, связан-
ных с недостаточным поступлением в двенадцатиперстную кишку желчи
или панкреатической липазы, может наблюдаться недостаточное всасыва-
ние из желудочно-кишечного тракта витаминов при их нормальном содер-
жании в пище.
Дополнительным источником витаминов К, В6, и В12 является микро-
флора толстой кишки. Микроорганизмы синтезируют эти витамины (наря-
ду с другими веществами), которые частично усваиваются организмом.
Длительное голодание, питание пищевыми продуктами, не содержащи-
ми или содержащими малое количество витаминов, употребление в пищу
продуктов после их длительного хранения или неправильной переработки,
нарушение пищеварительных функций могут приводить к недостаточному
поступлению витаминов в организм (гиповитаминозу). Гиповитаминоз или
полное прекращение поступления витамина в организм {авитаминоз) при-
водят как к неспецифическим изменениям функций (снижению умствен-
ной и физической работоспособности), так и к специфическим нарушени-
ям в организме, характерным для гипо- и авитаминоза (см. табл. 12.2). Из-
быточный прием витаминов может приводить к гипервитаминозу. При по-
ступлении водорастворимых витаминов в дозах, превышающих суточную
потребность, эти вещества могут быстро выводиться из организма с мочой.
При этом обычно признаков гипервитаминоза не отмечается. Однако, на-
пример, потребление больших количеств витамина В6 может сопровож-
даться нарушением функции периферических нервов. Изменения в орга-
низме, возникающие при гипервитаминозах A, D, РР, приведены в
табл. 12.2.
12.2. Роль обмена веществ в обеспечении энергетических
потребностей организма
Количество энергии, поступающей в организм с пищей, должно обеспечи-
вать подержание равновесного энергетического баланса на фоне неизмен-
ной массы тела, физической активности и соответствующих скоростях
роста и обновления структур организма. Организм человека получает энер-
гию в виде потенциальной химической энергии питательных веществ. Эта
энергия аккумулирована в химических связях молекул жиров, белков и уг-
леводов, которые в процессе катаболизма преобразуются в конечные про-
дукты обмена с более низким содержанием энергии. Высвобождающаяся в
процессе биологического окисления энергия используется, прежде всего,
для синтеза АТФ, которая как универсальный источник энергии необходи-
ма в организме для осуществления механической работы, химического
синтеза и обновления биологических структур, транспорта веществ, осмо-
тической и электрической работы. Схема процессов превращения энергии
в клетке представлена на рис. 12.1.
12. Обмен веществ и энергии. Питание • 573
Количество синтезированных молей АТФ на моль окисленного субстра-
та зависит от его вида (белка, жира, углевода) и от величины коэффициен-
та фосфорилирования. Этот коэффициент, обозначаемый как P/О, равен
количеству синтезированных молекул АТФ в расчете на один атом кисло-
рода, потребленный при окислении восстановленных органических соеди-
нений в процессе дыхания. При переносе каждой пары электронов по ды-
хательной цепи от НАД • Н до О2 величина Р/О = 2. Для субстратов, окис-
ляемых НАД • Н2-зависимыми ферментами, Р/О = 1,3. Эти соотношения
P/О отражают энергетические затраты клетки на синтез АТФ в митохонд-
риях и транспорт макроэрга против химического градиента из митохонд-
рий к местам потребления.
Таким образом, одна часть аккумулированной в химических связях мо-
лекул жиров, белков и углеводов энергии в процессе биологического окис-
ления используется для синтеза АТФ, другая часть этой энергии превраща-
ется в теплоту. Эта теплота, выделяющаяся сразу же в процессе биологиче-
ского окисления питательных веществ, получила название первичной. Ка-
кая часть энергии будет использована на синтез АТФ и будет вновь акку-
мулирована в ее химических макроэргических связях, зависит от величины
Р/О и эффективности сопряжения в митохондриях процессов дыхания и
фосфорилирования. Разобщение дыхания и фосфорилирования под дейст-
вием гормонов щитовидной железы, ненасыщенных жирных кислот, липо-
протеидов низкой плотности, динитрофенола ведет к уменьшению коэф-
фициента Р/О, превращению в первичную теплоту большей, чем в услови-
ях нормального сопряжения дыхания и фосфорилирования, части энергии
химических связей окисляемого вещества. При этом снижается коэффици-
ент полезного действия синтеза АТФ, количество синтезированных моле-
кул АТФ уменьшается.
При полном окислении 1 г смеси углеводов пищи выделяется 4 ккал те-
СО2
Рис. 12.1. Обмен энергии в клетке.
В процессе биологического окисления аминокислот, моносахаридов и жирных кислот освобо-
ждающаяся химическая энергия используется для синтеза макроэргического соединения
(АТФ). При расщеплении АТФ его энергия реализуется для осуществления всех видов работы
клетки (химической, электрической, осмотической и механической)
574 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
пла. В процессе окисления в организме 1 г углеводов синтезируется
0,13 моля АТФ. Если считать, что энергия пирофосфатной связи в АТФ
равна 7 ккал/моль, то при окислении 1 г углеводов лишь 0,91 (0,13 х 7)
ккал энергии будет запасено в организме в синтезированной АТФ. Осталь-
ные 3,09 ккал будут рассеяны в виде тепла (первичная теплота). Отсюда
можно рассчитать коэффициент полезного действия синтеза АТФ и акку-
мулирования в ней энергии химических связей глюкозы:
к.п.д. = (0,91 : 4,0) х Ю0 = 22,7 %.
Из приведенного расчета видно, что только 22,7 % энергии химических
связей глюкозы в процессе ее биологического окисления используется на
синтез АТФ и вновь запасается в виде химической макроэргической связи,
77,3 % энергии химических связей глюкозы превращается в первичную те-
плоту и рассеивается в тканях.
Аккумулированная в АТФ энергия в последующем используется для
осуществления в организме химических, транспортных, электрических
процессов, производства механической работы и в конечном итоге тоже
превращается в теплоту, получившую название вторичной.
В названиях первичная и вторичная теплота отражено представление о
двухступенчатости полного превращения всей энергии химических связей
питательных веществ в тепло (первая ступень — образование первичной
теплоты в процессе биологического окисления, вторая ступень — образова-
ние вторичной теплоты в процессе затраты энергии макроэргов на произ-
водство различных видов работы). Таким образом, если измерить все коли-
чество тепла, образовавшегося в организме за час или сутки, то это тепло
станет мерой суммарной энергии химических связей питательных веществ,
подвергшихся за время измерения биологическому окислению. По количе-
ству образовавшегося в организме тепла можно судить о величине энерге-
тических затрат, произведенных на осуществление процессов жизнедея-
тельности.
Основным источником энергии для осуществления в организме процес-
сов жизнедеятельности является биологическое окисление питательных ве-
ществ. На это окисление расходуется кислород. Следовательно, измерив
количество потребленного организмом кислорода за минуту, час, сутки,
можно судить о величине энергозатрат организма за время измерения.
Между количеством потребленного за единицу времени организмом ки-
слорода и количеством образовавшегося в нем за это же время тепла суще-
ствует связь, выражающаяся через калорический эквивалент кислорода
(КЭО2). Под КЭО2 понимают количество тепла, образующегося в организ-
ме при потреблении им 1 л кислорода.
12.2.1. Способы оценки энергетических затрат организма
Прямая калориметрия основана на измерении количества тепла, непосред-
ственно рассеянного организмом в теплоизолированной камере. При пря-
мой калориметрии достигается высокая точность оценки энергозатрат ор-
ганизма, однако из-за громоздкости и сложности способ используется
только для специальных целей.
Непрямая калориметрия основана на измерении количества потреблен-
ного организмом кислорода и последующем расчете энергозатрат с ис-
пользованием данных о величинах дыхательного коэффициента (ДК) и
КЭО2. Под дыхательным коэффициентом понимают отношение объема
выделенного углекислого газа к объему поглощенного кислорода.
12. Обмен веществ и энергии. Питание • 575
Таблица 12.3. Значение ДК и КЭ при окислении в организме различных питательных
веществ
Вещества Энергетическая ценность (ккал/г) О2 (л/г) СО2 (л/г) ДК КЭО2 (ккал/л)
Углеводы 4,0 0,81 0,81 1,0 5,05
Белки 4,0 0,94 0,75 0,80 4,46
Жиры 9,0 1,96 1,39 0,70 4,69
Сущность непрямой калориметрии видна на примере окисления глюко-
зы. Этот процесс описывается следующими превращениями:
С6Н12О6 + 6О2 — 6СО2 + 6Н2О + 675 ккал.
При окислении 1 г глюкозы количество выделяющейся энергии состав-
ляет 675:180 (масса 1 моля глюкозы) = 3,75 ккал. На окисление 1 моля
глюкозы затрачивается 6 молей О2 или 134,4 (6 х 22,4) л. Калорический
эквивалент 1 л О2, использованного на окисление глюкозы, равен 675 ккал :
: 134,4 л = 5,02 ккал/л. Так как смесь углеводов пищевых продуктов имеет
более высокую, чем чистая глюкоза, энергетическую ценность, то при
окислении их в организме КЭО2 = 5,05 ккал/л. Из приведенного уравне-
ния реакции окисления глюкозы видно, что объем выделенного в процессе
окисления углекислого газа равен объему затраченного кислорода. Следо-
вательно, при окислении глюкозы:
ДК= 6СО2 : 6О2 = 1.
В случае окисления жиров, в которых на 1 атом углерода приходится
меньше атомов кислорода, чем в углеводах и белках, величина ДК имеет
значение 0,7. При окислении белковой, а также смешанной пищи величи-
на ДК принимает промежуточное значение между 1,0 и 0,7. Поскольку в
организме все питательные вещества одновременно подвергаются окисле-
нию, то, определив величину ДК, можно условно судить о преимуществен-
ном окислении в организме того или иного вида питательных веществ. Так
как для каждого питательного вещества характерна своя энергетическая
ценность, то по величине дыхательного коэффициента можно рассчитать
значение калорического эквивалента кислорода. Величины этих значений
представлены в табл. 12.3.
В условиях интенсивной физической нагрузки и при состояниях, когда
в организме имеет место накопление углекислоты, ДК может принимать
значение > 1,0. В этих случаях по нему нельзя судить о природе окисляе-
мых веществ.
12.3. Обмен веществ и энергии при различных уровнях
функциональной активности организма
12.3.1. Основной обмен
Под основным обменом понимают минимальный уровень энергозатрат, не-
обходимых для поддержания жизнедеятельности организма в условиях от-
носительно полного физического, эмоционального и психического покоя.
В состоянии относительного покоя энергия затрачивается на осуществле-
ние функций нервной системы, постоянно идущий синтез веществ, работу
ионных насосов, поддержание температуры тела, деятельность дыхатель-
ной мускулатуры, гладких мышц, работу сердца и почек.
576 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 12.4. Уравнения для расчета величины основного обмена.
Пол Возраст, лет Уравнения для расчета основного обмена (ккал/сутки)
М 10-18 16,6 мт* + 77Р + 572
Ж 7,4 мт + 482Р + 217
м 18-30 15,4 мт - 27Р + 717
Ж 13,3 мт + 334Р + 35
м 30-60 11,3 мт + 16Р + 901
Ж 8,7 мт - 25Р + 865
м 60 8,8 мт + 1128Р- 1071
Ж 9,2 мт + 637Р- 302
*Мт — масса тела (кг), Р — рост (м).
Энергозатраты организма возрастают при физической и умственной ра-
боте, психоэмоциональном напряжении, после приема пищи, при пониже-
нии температуры среды. Для того, чтобы исключить влияние перечислен-
ных факторов на величину энергозатрат, определение основного обмена
проводят в стандартных строго контролируемых условиях: утром, в поло-
жении лежа, при максимальном расслаблении мышц, в состоянии бодрст-
вования, в условиях температурного комфорта (около 22 °C), натощак (че-
рез 12—14 ч после приема пищи). Полученные в таких условиях величины
основного обмена характеризуют исходный «базальный» уровень энергоза-
трат организма.
Для взрослого человека среднее значение величины основного обмена
равно 1 ккал/кг/ч (4,19 кДж). Следовательно, для взрослого мужчины мас-
сой 70 кг величина энергозатрат составляет около 1700 ккал/сут (7117
кДж), для женщин — около 1500 ккал/сут. Интенсивность основного обме-
на тесно связана с размерами поверхности тела, что обусловлено прямой
зависимостью величины отдачи тепла от площади поверхности тела. У теп-
локровных организмов, имеющих разные размеры тела, с 1 м2 поверхности
тела в окружающую среду рассеивается одинаковое количество тепла. На
этом основании сформулирован закон поверхности тела, согласно которо-
му энергетические затраты теплокровного организма пропорциональны ве-
личине поверхности тела.
Величины основного обмена определяют методами прямой или непря-
мой калориметрии. Его должные величины можно рассчитать по уравне-
ниям с учетом пола, возраста, роста и массы тела (табл. 12.4).
Нормальные величины основного обмена у взрослого человека можно
рассчитать также по формуле Дрейера:
Н = W/K • А,
где W — масса тела (г), А —возраст, К —константа (0,1015 для мужчин и
0,1129 — для женщин).
Величина основного обмена зависит от соотношения в организме про-
цессов анаболизма и катаболизма. Преобладание в детском возрасте про-
цессов анаболической направленности в обмене веществ над процессами
катаболизма обусловливает более высокие значения величин основного
обмена у детей (1,8 и 1,3 ккал/кг/ч у детей 7 и 12 лет соответственно) по
сравнению со взрослыми людьми (1 ккал/кг/ч), у которых уравновешены в
состоянии здоровья процессы анаболизма и катаболизма.
Так как показатели теплопродукции, артериального давления и пульса
12. Обмен веществ и энергии. Питание • 577
взаимосвязаны, то можно рассчитать величину основного обмена и его от-
клонения от нормы по следующей формуле:
ПО = 0,75 • (ЧСС + ПД • 0,74) - 72,
где ПО — процент отклонения от нормы, ЧСС — частота сердечных сокра-
щений, ПД — пульсовое давление.
Для каждой возрастной группы людей установлены и приняты в каче-
стве стандартов величины основного обмена. Это дает возможность при
необходимости измерить его величину у человека и сравнить полученные
у него показатели с нормативными. Отклонение величины основного об-
мена от стандартной не более чем на ±10 % считается в пределах нормы.
Более значительные отклонения основного обмена могут служить диаг-
ностическими признаками таких состояний организма, как нарушение
функции щитовидной железы; выздоровление после тяжелых и длитель-
ных заболеваний, сопровождающееся активацией метаболических про-
цессов; интоксикация и шок, сопровождающиеся угнетением метабо-
лизма.
Интенсивность основного обмена в различных органах и тканях неоди-
накова. По мере уменьшения энерготрат в покое их можно расположить в
таком порядке: внутренние органы—мышцы—жировая ткань.
12.3.2. Энергетические затраты организма в условиях физической
нагрузки
Интенсивность обменных процессов в организме значительно возрастает в
условиях физической нагрузки. Объективным критерием для оценки энерго-
затрат, связанных с двигательной активностью разных профессиональных
групп, является коэффициент физической активности. Он представляет со-
бой отношение общих энергозатрат к величине основного обмена. Прямая
зависимость величины энергозатрат от тяжести нагрузки позволяет ис-
Таблица 12.5. Энергетические затраты организма при различной интенсивности физи-
ческой работы
Группа Вид деятельности Пол Уровень энергетиче- ских затрат (ккал/сутки) Коэффи- циент фи- зической активности
I В условиях определения основного обмена м 1700
ж 1500
Выполнение работы, не требующей физиче- и 2300 1,4
ских усилий (врачи-терапевты, педагоги, дис- петчеры, секретари и др.) ж 2000
11 Физическая нагрузка: легкой тяжести (работ- м 2800 1,6
ники сферы обслуживания, конвейерных про- изводств, агрономы, медсестры) ж 2500
III умеренно тяжелая (продавцы продовольствен- м 3300 1,9
ных магазинов, станочники, слесари-наладчи- ки, врачи-хирурги, водители транспорта) ж 3000
V тяжелая (строительные и сельскохозяйствен- м 3800 2,2
ные рабочие, механизаторы, работники неф- тяной и газовой промышленности) ж 3700
V очень тяжелая (шахтеры, сталевары, камен- щики, грузчики) м 4800 2,5
19-6095
578 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
пользовать уровень энергозатрат в качестве одного из показателей интен-
сивности выполняемой работы (табл. 12.5).
Разница между величинами энергозатрат организма на выполнение раз-
личных видов работ и энергозатрат на основной обмен составляет так на-
зываемую рабочую прибавку (к минимальному уровню энергозатрат). Пре-
дельно допустимая по тяжести работа, выполняемая на протяжении ряда
лет, не должна превышать по энергозатратам уровень основного обмена
для данного индивидуума более чем в 3 раза.
Умственный труд не требует столь значительных энергозатрат, как фи-
зический. Энергозатраты организма возрастают при умственной работе в
среднем лишь на 2—3 %. Умственный труд, сопровождающийся легкой
мышечной деятельностью, психоэмоциональным напряжением, приводит
к повышению энергозатрат уже на 11—19 % и более.
Специфически-динамическое действие пищи — это усиление интенсивно-
сти обмена веществ под влиянием приема пищи и увеличение энергетиче-
ских затрат организма относительно уровней обмена и энергозатрат, имев-
ших место до приема пищи. Специфически-динамическое действие пищи
обусловлено затратами энергии на переваривание пищи, всасывание в
кровь и лимфу питательных веществ из желудочно-кишечного тракта, ре-
синтез белковых, сложных липидных и других молекул; влиянием на мета-
болизм биологически активных веществ, поступающих в организм в соста-
ве пищи (в особенности белковой) и образующихся в нем в процессе пи-
щеварения.
Увеличение энергозатрат организма выше уровня, имевшего место до
приема пищи, проявляется примерно через час после приема пищи, дости-
гает максимума через три часа, что обусловлено развитием к этому време-
ни высокой интенсивности процессов пищеварения, всасывания и ресин-
теза поступающих в организм веществ. Специфически-динамическое дей-
ствие пищи может продолжаться 12—18 ч. Оно наиболее выражено при
приеме белковой пищи, повышающей интенсивность обмена веществ до
30 %, и менее значительно при приеме смешанной пищи, повышающей
интенсивность обмена на 6—15 %.
Уровень общих энергозатрат, как и основного обмена, зависит от воз-
раста: суточный расход энергии возрастает у детей с 800 ккал (6 мес—
1 год) до 2850 ккал (11—14 лет). Резкий прирост энергозатрат имеет место
у подростков-юношей 14—17 лет (3150 ккал). После 40 лет энергозатраты
снижаются и к 80 годам составляют около 2000—2200 ккал/сут.
В повседневной жизни уровень энергозатрат у взрослого человека зави-
сит не только от особенностей выполняемой работы, но и от общего уров-
ня двигательной активности, характера отдыха и социальных условий
жизни.
12.4. Регуляция обмена веществ и энергии
Регуляция обмена организма с окружающей средой теплом обсуждается в
главе 13. Вопросы регуляции водно-солевого обмена описаны в главе 14.
Поэтому в данной главе представлены общие вопросы нейрогумораль-
ной регуляции обмена веществ и энергии в организме и, главным образом,
регуляция метаболизма. Конечной целью регуляции обмена веществ и
энергии является обеспечение потребностей организма, его органов, тка-
ней и отдельных клеток в энергии и в разнообразных веществах в соответ-
ствии с уровнем функциональной активности. В целостном организме по-
стоянно существует необходимость согласования общих метаболических
12. Обмен веществ и энергии. Питание • 579
потребностей с потребностями клетки органа, ткани. Такое согласование
достигается посредством распределения между органами и тканями ве-
ществ, поступающих из окружающей среды и синтезированных внутри ор-
ганизма. *
Обмен веществ, протекающий внутри организма, не связан непосредст-
венно с окружающей средой. Питательные вещества, прежде чем они смо-
гут вступить в обменные процессы, должны быть получены из пищи в же-
лудочно-кишечном тракте в молекулярной форме. Кислород, необходимый
для биологического окисления, должен быть получен из воздуха в легких,
доставлен в кровь, связан с гемоглобином и перенесен кровью к тканям.
Скелетные мышцы, являясь в организме одним из мощных потребителей
энергии, также обслуживают обмен веществ и энергии, обеспечивая поиск,
прием и обработку пищи. Непосредственное отношение к обмену веществ
и энергии имеет выделительная система. Таким образом, регуляция обмена
веществ и энергии является мультипараметрической, включающей в себя
регулирующие системы множества функций организма (например, дыха-
ния, кровообращения, выделения, теплообмена и др.).
Роль центра в регуляции обмена веществ и энергии играют ядра гипо-
таламуса. Они имеют непосредственное отношение к генерации чувства
голода и насыщения, теплообмену, осморегуляции. В гипоталамусе имеют-
ся полисенсорные нейроны, реагирующие на изменения концентрации
глюкозы, водородных ионов, температуры тела, осмотического давления,
т. е. важнейших гомеостатических констант внутренней среды организма.
В ядрах гипоталамуса осуществляется анализ состояния внутренней среды
и формируются управляющие сигналы, которые посредством эфферентных
систем приспосабливают ход метаболизма к потребностям организма.
В качестве звеньев эфферентной системы регуляции обмена используют-
ся симпатический и парасимпатический отделы вегетативной нервной сис-
темы. Выделяющиеся их нервными окончаниями медиаторы оказывают
прямое или опосредованное вторичными посредниками влияние на функ-
цию и метаболизм тканей. Под управляющим влиянием гипоталамуса на-
ходится и используется в качестве эфферентной системы регуляции обме-
на веществ и энергии — эндокринная система. Гормоны гипоталамуса, ги-
пофиза и других эндокринных желез оказывают прямое влияние на рост,
размножение, дифференцировку, развитие и другие функции клеток. Гор-
моны принимают участие в поддержании в крови необходимого уровня та-
ких веществ, как глюкоза, свободные жирные кислоты, минеральные ве-
щества.
Химическая энергия питательных веществ используется для ресинтеза
АТФ, выполнения всех видов работы (см. рис. 12.1) и процессы, проте-
кающие внутри клетки. Поэтому важнейшим эффектором, через который
оказывается регулирующее воздействие на обмен веществ и энергии, явля-
ются клетки органов и тканей. Регуляция обмена веществ заключается в
воздействии на скорость биохимических реакций, протекающих в клетках.
Наиболее частыми эффектами регуляторных воздействий на клетку яв-
ляются изменения каталитической активности ферментов и их концентра-
ции, сродства фермента и субстрата, свойств микросреды, в которой функ-
ционируют ферменты. Регуляция активности ферментов может осуществ-
ляться различными способами. «Тонкая настройка» каталитической актив-
ности ферментов достигается посредством влияния веществ — модулято-
ров, которыми нередко являются сами метаболиты.
Метаболизм клетки в целом невозможен без интеграции многих биохи-
мических превращений. Эта интеграция обеспечивается, главным образом,
19*
580 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
с помощью аденилатов, участвующих в регуляции любых метаболических
превращений клетки.
Интеграция обмена белков, жиров и углеводов клетки осуществляется
посредством общих для них источников энергии. При биосинтезе любых
простых и сложных органических соединений, макромолекул и надмоле-
кулярных структур в качестве общих источников энергии используется
АТФ, которая поставляет энергию для процессов фосфорилирования, или
НАД • Н, НАДФ • Н, поставляющих энергию для восстановления окислен-
ных соединений других веществ. За общий энергетический запас клетки,
полученный в ходе катаболизма, конкурируют все анаболические процес-
сы, протекающие с затратой энергии. Так, например, при осуществлении
печенью синтеза глюкозы из лактата и аминокислот (глюконеогенез) она не
может одновременно синтезировать жиры и белки. Глюконеогенез сопро-
вождается расщеплением в печени белков и жиров и окислением обра-
зующихся при этом жирных кислот, что ведет к освобождению энергии,
необходимой для синтеза АТФ и НАД - Н, необходимых для глюконеоге-
неза.
Еще одним проявлением интеграции метаболических превращений бел-
ков, жиров и углеводов в клетке является существование общих предшест-
венников и общих промежуточных продуктов обмена веществ. Общим про-
межуточным продуктом обмена является ацетил-КоА. Важнейшими конеч-
ными путями превращений веществ в клетке являются цикл лимонной ки-
слоты и реакции дыхательной цепи, протекающие в митохондриях. Цикл
лимонной кислоты — главный источник СО2 для последующих реакций
глюконеогенеза, синтеза жирных кислот и мочевины.
Одним из механизмов согласования общих метаболических потребно-
стей организма с потребностями клетки являются нервные и гормональные
влияния на ключевые ферменты. Характерными особенностями этих фер-
ментов являются: положение в начале того метаболического пути, к кото-
рому принадлежит фермент; приближенность расположения или ассоции-
рованность со своим субстратом; реагирование не только на действие
внутриклеточных регуляторов метаболизма, но и на внеклеточные нервные
и гормональные воздействия.
Примерами ключевых ферментов являются гликогенфосфорилаза, фос-
фофруктокиназа, липаза. Их роль в процессах регуляции метаболизма вид-
на, в частности, при подготовке организма к «борьбе или бегству». При
повышении в этих условиях в крови уровня адреналина до 1СГ9 М он свя-
зывается с адренорецепторами плазматической мембраны, активирует аде-
нилатциклазу, которая катализирует превращение АТФ в циклический
АМФ. Последний активирует гликогенфосфорилазу, многократно усили-
вающую расщепление гликогена в печени.
Процесс гликогенолиза в мышцах может одновременно активироваться
нервной системой и катехоламинами. Этот эффект достигается с участием
ионов Са2+, который связывается с кальмодулином, являющимся субъеди-
ницей фосфорилазы. Она при этом активируется и приводит к мобилиза-
ции гликогена. Нервный механизм мобилизации гликогена осуществляется
через меньшее число промежуточных этапов, чем гормональный. Этим
достигается его быстродействие.
Удовлетворение энергетических потребностей организма посредством
ускорения внутриклеточных процессов расщепления триглицеридов в жи-
ровой клетчатке достигается активацией гормончувствительной липазы.
Повышение активности этого фермента (адреналином, норадреналином,
глюкагоном) приводит к мобилизации свободных жирных кислот, являю-
12. Обмен веществ и энергии. Питание • 581
щихся основным энергетическим субстратом окисления в мышцах при вы-
полнении ими интенсивной и длительной работы.
Переход органов и тканей с одного уровня функциональной активности
на другой всегда сопровождается соответствующими изменениями их тро-
фики (питания). Например, при рефлекторном сокращении скелетных
мышц нервная система осуществляет не только пусковое действие, но и
трофическое влияние путем усиления в них местного кровотока и интен-
сивности обмена веществ. Увеличение силы сокращений миокарда под
влиянием симпатической нервной системы обеспечивается одновремен-
ным усилением коронарного кровотока и метаболизма в мышце сердца.
О влиянии нервной системы на трофику скелетных мышц свидетельствует
тот факт, что денервация мышцы приводит к постепенной атрофии мы-
шечных волокон. Важнейшее значение в осуществлении трофической
функции нервной системы играет ее симпатический отдел. Через симпато-
адреналовую систему достигается не только активация обмена веществ и
энергии в клетке.
Норадреналин и адреналин, выброс которых в кровоток возрастает при
возбуждении симпатической нервной системы, вызывают увеличение глу-
бины дыхания, расширяют мускулатуру бронхов, что способствует достав-
ке кислорода в кровь. Адреналин, оказывая положительное инотропное и
хронотропное действие на сердце, увеличивает минутный объем крови, по-
вышает систолическое артериальное давление. В результате активации ды-
хания и кровообращения возрастает доставка кислорода к тканям.
Одним из интегральных показателей внутренней среды, отражающим
обмен в организме углеводов, белков и жиров, является концентрация в
крови глюкозы. Она является не только источником энергии для синтеза
жиров и белков, но и субстратом для их синтеза. В печени происходит но-
вообразование углеводов из жирных кислот и аминокислот.
Нормальное функционирование клеток нервной системы, поперечно-
полосатых и гладких мышц, для которых глюкоза является важнейшим
энергосубстратом, возможно при условии, что приток к ним глюкозы обес-
печит их энергетические потребности. Это достигается при содержании в
литре крови у человека в среднем 1 г (0,8—1,2 г) глюкозы (рис. 12.2). Из
схемы на этом рисунке следует, что при нормальном уровне содержания
глюкозы в крови происходит образование гликогена в печени и мышцах,
синтез жиров, ее потребление клетками мозга, мышцами и другими тканя-
ми. В условиях гипергликемии избыточное количество глюкозы удаляется
из крови через почки, увеличивается синтез гликогена. При гипогликемии
усиливается гликогенолиз под влиянием адреналина и глюкагона.
Сдвиги в концентрации глюкозы в крови от «заданного» (константного)
значения воспринимаются глюкорецепторами гипоталамуса, который реа-
лизует свои регулирующие влияния на клетки через симпатический и па-
расимпатические отделы вегетативной нервной системы. Эти влияния обу-
словливают срочное повышение или снижение выработки инсулина, глю-
кагона и адреналина эндокринным аппаратом поджелудочной железы и
надпочечников. Более медленный эффект гипоталамических влияний осу-
ществляется через гормоны гипофиза. Для поддержания константного
уровня концентрации глюкозы существует и более короткая петля обрат-
ной связи — влияние глюкозы, циркулирующей в крови, непосредственно
на бета-клетки островков Лангерганса поджелудочной железы, вырабаты-
вающих гормон инсулин.
При снижении содержания глюкозы в литре крови до уровня менее
0,5 г, вызванном голоданием, передозировкой инсулина, имеет место не-
582 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
центральная нервная система
Рис. 12.2. Схема регуляции концентрации глюкозы в крови.
Положительная направленность физиологических эффектов в ответ на: Ц — гипергликемию;
Д — гипогликемию; отрицательная направленность физиологических эффектов в ответ на:
а — гипергликемию; Q — гипогликемию
12. Обмен веществ и энергии. Питание • 583
достаточность снабжения энергией клеток мозга. Нарушение их функций
проявляется учащением сердцебиения, слабостью и тремором мышц, голо-
вокружением, усилением потоотделения, ощущением голода. При даль-
нейшем снижении концентрации глюкозы в крови указанное состояние,
именуемое гипогликемией, может перейти в гипогликемическую кому, харак-
теризующуюся угнетением функций мозга вплоть до потери сознания.
Введение в кровь глюкозы, прием сахарозы, инъекция глюкагона преду-
преждают или ослабляют эти проявления гипогликемии. Кратковременное
повышение уровня глюкозы в крови (гипергликемия) не представляет угро-
зы для здоровья человека.
В крови организма человека обычно содержится около 5 г глюкозы.
При среднесуточном потреблении с пищей взрослым человеком, занимаю-
щимся физическим трудом, 430 г углеводов в условиях относительного по-
коя, тканями ежеминутно потребляется около 0,3 г глюкозы. При этом за-
пасов глюкозы в циркулирующей крови достаточно для питания тканей на
3—5 мин и без ее восполнения неминуема гипогликемия. Потребление глю-
козы возрастает при физической и психоэмоциональной нагрузках. Так
как периодический (несколько раз в день) прием углеводов с пищей не
обеспечивает постоянного и равномерного притока глюкозы из кишечника
в кровь, в организме существуют механизмы, восполняющие убыль глюко-
зы из крови в количествах, эквивалентных ее потреблению тканями. При
достаточном уровне концентрации глюкозы в крови она частично превра-
щается в запасаемую форму — гликоген. При уровне более 1,8 г в литре
крови происходит выведение ее из организма с мочой.
Избыток глюкозы, поступившей из кишечника в кровь воротной вены,
поглощается гепатоцитами. При повышении в них концентрации глюкозы
активируются ферменты углеводного обмена печени, превращающие глю-
козу в гликоген. В ответ на повышение уровня сахара в крови, протекаю-
щей через поджелудочную железу, возрастает секреторная активность бета-
клеток островков Лангерганса. В кровь выделяется большее количество
инсулина — единственного гормона, обладающего резким понижающим
концентрацию сахара в крови действием. Под влиянием инсулина повы-
шается проницаемость для глюкозы плазматических мембран клеток мы-
шечной и жировой тканей. Инсулин активирует в печени и мышцах про-
цессы превращения глюкозы в гликоген, улучшает ее поглощение и усвое-
ние скелетными, гладкими и сердечной мышцами. Под влиянием инсули-
на в клетках жировой ткани из глюкозы синтезируются жиры. Одновре-
менно выделяющийся в больших количествах инсулин тормозит распад
гликогена печени и глюконеогенез.
Содержание глюкозы в крови оценивается глюкорецепторами переднего
гипоталамуса, а также его полисенсорными нейронами. В ответ на повы-
шение уровня глюкозы в крови выше «заданного значения» (> 1,2 г/л) воз-
растает активность нейронов гипоталамуса, которые посредством влияния
парасимпатической нервной системы на поджелудочную железу усиливают
секрецию инсулина.
При понижении уровня глюкозы в крови уменьшается ее поглощение
гепатоцитами. В поджелудочной железе снижается секреторная активность
бета-клеток, уменьшается секреция инсулина. Тормозятся процессы пре-
вращения глюкозы в гликоген в печени и мышцах, уменьшается поглоще-
ние и усвоение глюкозы скелетными и гладкими мышцами, жировыми
клетками. При участии этих механизмов замедляется или предотвращается
дальнейшее понижение уровня глюкозы в крови, которое могло бы при-
вести к развитию гипогликемии.
584 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
При уменьшении концентрации глюкозы в крови имеет место повыше-
ние тонуса симпатической нервной системы. Под ее влиянием усиливается
секреция в мозговом веществе надпочечников адреналина и норадренали-
на. Адреналин, стимулируя распад гликогена в печени и мышцах, вызыва-
ет повышение концентрации сахара в крови. Норадреналин обладает сла-
бовыраженной способностью повышать уровень глюкозы в крови.
Под влиянием симпатической нервной системы стимулируется выра-
ботка альфа-клетками поджелудочной железы глюкагона, который активи-
рует распад гликогена печени, стимулирует глюконеогенез и приводит к
повышению уровня глюкозы в крови.
Понижение в крови концентрации глюкозы, являющейся для организма
одним из наиболее важных энергетических субстратов, вызывает развитие
стресса. В ответ на снижение уровня сахара крови глюкорецепторные ней-
роны гипоталамуса через рилизинг-гормоны стимулируют секрецию гипо-
физом в кровь гормона роста и адренокортикотропного гормона.
Под влиянием гормона роста уменьшается проницаемость клеточных
мембран для глюкозы, усиливается глюконеогенез, активируется секреция
глюкагона, в результате чего уровень сахара в крови увеличивается.
Секретируемые под действием адренокортикотропного гормона в коре
надпочечников глюкокортикоиды активируют ферменты глюконеогенеза и
этим способствуют увеличению содержания сахара в крови.
Регуляция обмена веществ и энергии в организме находится под кон-
тролем нервной системы и ее высших отделов. Об этом свидетельствуют
факты условно-рефлекторного изменения интенсивности метаболизма у
спортсменов в предстартовом состоянии, у рабочих перед началом выпол-
нения тяжелой физической работы, у водолазов перед их погружением в
воду. В этих случаях увеличивается скорость потребления организмом ки-
слорода, возрастает минутный объем дыхания, минутный объем кровотока,
усиливается энергообмен.
Развивающееся при снижении в крови содержания глюкозы, свободных
жирных кислот, аминокислот чувство голода обусловливает поведенческую
реакцию, направленную на поиск и прием пищи и восполнение в организ-
ме питательных веществ.
12.5. Питание
Питание — поступление в организм и усвоение им веществ, необходимых
для роста, жизнедеятельности и воспроизводства. От качества и режима
питания зависят его здоровье, работоспособность и продолжительность
жизни.
Недостаточность или избыточность питания приводят к нарушениям
обмена веществ.
Удовлетворение пластических и энергетических потребностей организ-
ма служит критерием для формирования норм питания. В свою очередь,
нормы питания, определяющие величины потребления пищевых веществ,
основываются на данных научных исследований обмена жиров, белков, уг-
леводов, воды, минеральных веществ, витаминов у различных групп насе-
ления.
При определении физиологических норм питания с учетом удовлетво-
рения потребностей организма в пластических веществах исходят из того,
что большинство из них может синтезироваться в организме; другие веще-
ства (незаменимые аминокислоты, незаменимые жирные кислоты, все ми-
неральные вещества и микроэлементы, витамины) в организме человека не
12. Обмен веществ и энергии. Питание • 585
синтезируются и должны поступать с пищей. Так, источником аминокис-
лот являются белки пищи, резервом белка или аминокислот организм не
располагает. Это обусловливает необходимость поступления в организм
белка из расчета 0,75—1 г на 1 кг массы тела взрослого человека в сутки.
При этом 55—60 % суточной потребности белка должно обеспечиваться
белками животного происхождения (молоко, молочные продукты, яйца,
мясо, рыба).
Такие необходимые организму вещества, как витамин К и витамины
группы В, организм получает не только с пищей, но и в составе продуктов
жизнедеятельности микрофлоры кишечника (см. раздел 11.8.4).
Соотношение в пищевом рационе белков, жиров и углеводов должно
быть 1:1,2:4,6. В состав пищевого рациона должны входить продукты жи-
вотного и растительного происхождения (например, жиров растительного
происхождения должно быть не менее 30 % от общего количества жиров).
Необходимо включение в пищевые рационы свежих натуральных продук-
тов питания, являющихся источниками витаминов, ненасыщенных жир-
ных кислот и минеральных веществ.
При небольших отклонениях в течении короткого времени от рекомен-
дуемых соотношений количества жиров и углеводов (при условии поступ-
ления в организм белков из расчета 0,75 г/кг/сут) нарушений метаболизма
у человека не происходит. Жиры и углеводы могут заменять друг друга как
энергетические субстраты в соответствии с правилом изодинамии. При
энергетической ценности 1 г жиров, равной 9,0 ккал, и 1 г углеводов — 4,0
ккал, грамм жиров заменяет при окислении в организме 2,25 г углеводов.
Однако прием жиров в количестве, превышающем потребность организма,
ведет к ожирению и риску сердечно-сосудистых заболеваний. Поступление
жиров в организм в количествах ниже его потребности ограничивает вса-
сывание жирорастворимых витаминов и может быть причиной развития
авитаминозов. Особенно неблагоприятным для пластических процессов
является недостаточное поступление в организм незаменимых жирных ки-
слот (линолевой, арахидоновой) (см. раздел 12.1.2).
Движущей силой обмена веществ в организме и выполнения любых ви-
дов работы является энергия, освобождаемая при осуществлении катаболи-
ческих процессов. Ее источником служит энергия химических связей пита-
тельных веществ, поступающих с пищей. Поэтому при определении физио-
логических норм питания необходимо соблюдать соответствие энергетиче-
ской ценности (калорийности) пищевого рациона энергозатратам конкрет-
ного организма. Они складываются из затрат энергии основного обмена,
энергозатрат, связанных со специфически-динамическим действием пищи
и особенностями трудовой деятельности. Взрослое трудоспособное населе-
ние в возрасте 18—60 лет может быть отнесено к 5 группам, дифференци-
рованным в зависимости от величин энергозатрат. Для этих групп рассчи-
таны средние величины энергозатрат и потребления питательных веществ.
Рекомендуемые нормы питания для этих групп приведены в табл. 12.6.
Как следует из данных этой таблицы, у взрослых мужчин и женщин су-
точная потребность в энергии возрастает в зависимости от коэффициента
физической активности. Так, при коэффициенте 1,9 (III группа) в возрасте
18—29 лет требуется в сутки 3300 ккал мужчинам и 2600 ккал женщинам, а
при коэффициенте 2,2 (IV группа) — соответственно 3850 и 3050 ккал, что
обеспечивается за счет увеличения в пищевом рационе белков, жиров и уг-
леводов. С возрастом у той же группы населения (например, III) потреб-
ность в энергии снижается (у мужчин до 2950 ккал, у женщин —до
2500 ккал).
586 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Табл ица 12.6. Нормы суточной потребности в пищевых веществах для взрослого чело-
века
Группа Коэффициент физической активности Воз- раст, годы Энер- ГИЯ, ккал Белки,г Жи- ры, г Угле- воды, г
общее количество в т.ч. животные
Мужчины
I 1,4 18-29 2450 72 40 81 358
30-39 2300 68 37 77 335
40-59 2100 65 36 70 303
II 1,6 18-29 2800 80 44 93 411
30-39 2650 77 42 88 387
40-59 2500 72 40 83 366
III 1,9 18-29 3300 94 52 110 484
30-39 3150 89 49 105 462
40-59 2950 84 46 98 432
IV 2,2 18-29 3850 108 59 128 565
30-39 3600 102 56 120 528
40-59 3400 96 53 113 499
V 2,5 18-29 4200 117 64 154 586
30-39 3950 111 61 144 550
40-59 3750 104 57 137 524
Женщины
I 1,4 18-29 2000 61 34 67 289
30-39 1900 59 33 63 274
40-59 1800 58 32 60 257
11 1,6 18-29 2200 66 36 73 318
30-39 2150 65 36 72 311
40-59 2100 63 35 70 305
III 1,9 18-29 2600 76 42 87 378
30-39 2500 74 41 85 372
40-59 2500 72 40 83 366
IV 2,2 18-29 3050 87 48 102 462
30-39 2950 84 46 98 432
40-59 2850 417
Примечание. В I группу входят работники преимущественно умственного труда; II груп-
па — работники легкого физического труда; III группа — работники среднего по тяжести фи-
зического труда; IV группа — работники тяжелого физического труда; V группа — работники
особо тяжелого физического труда.
Хотя подразделение трудоспособного населения на группы, основанное
на особенностях трудовой деятельности, носит во многом условный харак-
тер, выделяют группы лиц, занятых преимущественно умственным или
физическим трудом. У людей преимущественно умственного труда в про-
цессе этой деятельности развиваются свойственный данному индивидууму
уровень психоэмоционального напряжения, гипокинезия, может увеличи-
ваться масса тела. Эти состояния являются факторами риска развития
многих заболеваний. Для предупреждения подобных осложнений лица, за-
нятые преимущественно умственным трудом, должны выполнять разум-
ный объем физической нагрузки и в случае увеличения массы тела умерен-
но ограничить питание. Ограничение питания должно идти лишь по пока-
зателю его энергетической ценности (преимущественно за счет уменьше-
ния приема углеводов) и не в ущерб его пластической ценности. Умерен-
ное ограничение питания следует сочетать с введением в пищевой рацион
широкого ассортимента продуктов питания растительного происхождения.
12. Обмен веществ и энергии. Питание • 587
Жиры являются не только энергетическими и пластическими веществами,
но и поставщиками таких необходимых организму компонентов, как поли-
ненасыщенные жирные кислоты, фосфолипиды, токоферолы, витамины А
и D и др. Для поддержания высокого уровня умственной работоспособно-
сти с пищевым рационом в организм должны поступать в соответствии с
суточной потребностью минеральные вещества, витамины, микроэле-
менты.
При выполнении преимущественно физического труда в пищевом рацио-
не соотношение белков, жиров и углеводов должно составлять примерно
1:1,3:5,1. Пищевой рацион должен содержать разнообразные калорийные
продукты питания, удельный вес животного белка должен составлять в нем
55 % от суточной нормы белка, а жиры растительного происхождения —
30 % от суточной нормы жиров. Чем тяжелее и продолжительнее труд, тем
более витаминизированными должны быть пищевые продукты.
Для восстановления здоровья после заболеваний, профилактики забо-
леваний, сохранения высокой работоспособности разработаны особые ре-
жимы и рационы лечебно-профилактического питания. Они при необходи-
мости рекомендуются работникам как физического, так и умственного
труда.
Уровни энергетических затрат и потребностей организма в пластиче-
ских веществах зависят не только от интенсивности труда, но и от множе-
ства других факторов, в частности от возраста, массы тела, физической ак-
тивности, функционального состояния организма. Для беременных и кормя-
щих женщин содержание белка в пищевом рационе должно быть увеличено
до 2 г/кг в сутки. Увеличение белка необходимо для обеспечения роста
тканей развивающегося организма, а у кормящих женщин — для образова-
ния молока. Количество белка в рационе детского питания должно состав-
лять 1,2—1,5 г/кг в сутки. Большее количество белка необходимо вводить в
рацион питания лицам тяжелого физического труда, у которых потери бел-
ка больше, чем у лиц, выполняющих более легкую физическую нагрузку.
Для скорейшего выздоровления, восстановления массы тканей организма
после тяжелых истощающих заболеваний, перенесенных операций, обшир-
ных ожогов также требуется пищевой рацион с более высоким, чем для
здорового, содержанием белка (1,5—2,0 г/кг в сутки). Общая калорийность
пищевого рациона в расчете на 1 кг массы тела выше у растущего детского
организма в сравнении со взрослым и ниже у стариков (см. табл. 12.7).
Табл ица 12.7. Рекомендуемые средние величины потребления пищевых веществ в зави-
симости от возраста
Возраст Энергия, ккал Белки, г/сут Жиры, г/сут Углеводы, г/сут
всего в т.ч. жи- вотные всего в т.ч. расти- тельные
1—3 года 1540 53 37 53 5 212
4—6 лет 1970 68 44 68 10 272
7—10 лет 2300 79 47 79 16 315
11—13 лет 2700 (2450)' 93 (85) 56 (51) 93 (85) 19 (17) 370 (340)
14—17 лет 2900 (2600) 100 (90) 60 (54) 100 (90) 20 (18) 400 (360)
60—74 года 2300 (2100) 69 (63) 38 (35) 77 (70) 26 (23) 333 (305)
75 лет и старше 2000 (1900) 60 (57) 33 (31) 67 (63) 23 (21) 290 (275)
*В скобках приведены величины, рекомендуемые для женщин.
588 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
При ограниченном поступлении в организм человека питательных ве-
ществ имеет место повышенная утомляемость, снижаются как физическая,
так и умственная работоспособность, замедляется рост и развитие детей,
уменьшается масса тела, могут появляться отеки (при белковой недоста-
точности), снижается устойчивость организма к инфекционным заболева-
ниям. Переедание приводит к возникновению нарушений функций желу-
дочно-кишечного тракта, сонливости, ожирению, снижению физической
активности и трудоспособности. Увеличение массы тела и ожирение явля-
ются факторами риска сердечно-сосудистых заболеваний, сахарного диабе-
та и приводит к уменьшению продолжительности жизни.
К важнейшим физиологическим принципам, которые необходимо со-
блюдать при составлении пищевых рационов, относится режим питания,
т. е. приспособление характера питания, частоты и периодичности приема
пищи к суточным ритмам труда и отдыха, к физиологическим закономер-
ностям деятельности желудочно-кишечного тракта. Принято считать, что
наиболее рациональным является четырехразовый прием пищи в одни и те
же часы суток. Интервал между приемами пищи должен составлять 4—5 ч.
Этим достигается более равномерная функциональная нагрузка на пище-
варительный аппарат, ЧТО способствует созданию оптимальных условий
для полной обработки пищи. Рекомендуется вечерний прием легкоусвояе-
мой пищи не позднее, чем за 3 ч до отхода ко сну.
Общую калорийность суточного пищевого рациона целесообразно рас-
пределять следующим образом: на завтрак —25 %, второй завтрак — 15 %,
обед —35 %, ужин — 25 %. В случае невозможности осуществления четы-
рехразового питания оно может быть трехразовым (30 % калорий суточно-
го пищевого рациона на завтрак, 45 % — на обед, 25 % — на ужин).
Опасность для здоровья человека могут представлять вещества, которые
содержатся в пищевых продуктах, выращенных или переработанных без
соблюдения санитарно-гигиенических требований к сельскохозяйствен-
ным или промышленным технологиям. Это пестициды, нитраты, радио-
нуклиды, лекарственные средства, металлы, пищевые добавки, консерван-
ты. При попадании в организм они могут оказывать на ткани токсическое
воздействие (металлы, радионуклиды), вызывать аллергические реакции
(пищевые добавки, консерванты, лекарственные вещества). Пестициды
могут накапливаться в жировой ткани и, медленно выводясь из организма,
оказывать длительное токсическое влияние.
Условием для эффективного всасывания и усвоения питательных ве-
ществ из желудочно-кишечного тракта является переваривание пищевых
веществ до мономеров при полостном и пристеночном пищеварении.
Часть веществ пищи не подвергается в желудочно-кишечном тракте гидро-
лизу (например, растительный полисахарид целлюлоза) или расщепляется
не полностью. Степень переваривания пищевых веществ зависит от их
предварительной обработки, в процессе приготовления пищи или механи-
ческой обработки при жевании. Таким образом, пищевые продукты не
полностью усваиваются организмом, и при питании смешанной пищей
животного и растительного происхождения ее усвояемость по калориче-
ской ценности составляет около 90—95 %.
Неусваиваемыми веществами пищи являются ее грубоволокнистые ком-
поненты — пищевые волокна (целлюлоза, гемицеллюлоза, пектин и лег-
нин). Хотя эти вещества снижают калорическую ценность пищевого ра-
циона, они стимулируют перистальтику кишечника, ускоряют продвиже-
ние в желудочно-кишечном тракте пищевых масс, способствуют формиро-
ванию оптимальной для выведения из организма консистенции каловых
12. Обмен веществ и энергии. Питание • 589
масс, способствуют выведению из организма избытка пищевого холесте-
рина.
Потребность конкретного человека в различных компонентах пищи, ко-
личестве и соотношениях питательных веществ не только индивидуальны,
но и зависят от возраста, выполняемой физической или умственной на-
грузки, состояния покоя или психоэмоционального напряжения. Поэтому
определение норм и характера питания, хотя и должно учитывать общие
физиологические требования и рекомендации, может быть лишь строго
индивидуализированным.
Суточная потребность организма в белках, жирах и углеводах приведена
в табл. 12.6 и 12.7, а его потребность в минеральных веществах, микроэле-
ментах и витаминах, а также источники их поступления указаны в
табл. 12.1 и 12.2.
12.5.1. Рациональное питание как фактор сохранения и укрепления
здоровья
Рациональным является питание, которое основано на достижениях науч-
ных данных (физиологии, биохимии и гигиены) относительно качества и
количества принимаемой пищи, возможности усвоения ее организмом и
режима ее приема.
От характера питания (биологической полноценности пищевых продук-
тов, количества принимаемой пищи и режима ее потребления) в большой
степени зависит состояние здоровья человека и продолжительность его
жизни. Рациональное питание — мощный фактор укрепления здоровья,
неправильное питание — способ его разрушения.
При формировании здорового образа жизни необходимо учитывать че-
тыре принципа рационального питания: 1) калорийность суточного рацио-
на питания должна соответствовать энергетическим затратам человека;
2) химический состав пищи должен удовлетворять потребности организма
в белках, жирах, углеводах, минеральных веществах, витаминах, биологи-
чески активных веществах и «балластных» компонентах пищи; 3) разнооб-
разие пищевых продуктов должно обеспечивать поступление в организм
всех ингредиентов, которые не синтезируются в организме; 4) прием пищи
в течение суток должен подчиняться оптимальному режиму (как по энер-
гоценности, так и по объему).
Нарушение этих принципов имеет отрицательные последствия для здо-
ровья. В практике последних лет нередко приходится иметь дело с послед-
ствием избыточного питания — ожирением, являющимся фактором риска
возникновения различных заболеваний. Увеличение массы тела по отно-
шению к должной ее величине на 6—14 % считается показателем избыточ-
ного питания. Для стандартизованной оценки массы тела используют ин-
декс массы тела (ИМТ), рассчитываемый по формуле m/рост2 (т —масса
в килограммах, рост — в метрах). При ИМТ более 25 массу тела расцени-
вают как повышенную, требующую коррекции. При наличии избыточной
массы необходимо сокращение общей калорийности пищи за счет жиров и
углеводов (прежде всего — моносахаридов), а также увеличение двигатель-
ной активности.
Правильный режим приема пищи соответствует фазам суточных био-
ритмов и легко «вписывается» в распорядок дня. Более того — этот фактор
в значительной мере определяет режим жизни: человек в определенное
время прерывает работу, чтобы пообедать, ужин ассоциируется со време-
нем отдыха, вечерний стакан молока — с предстоящим сном и т. д. Вот по-
590 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
чему большой вред здоровью может наносить привычка есть урывками, во
время работы и вообще по принципу «когда придется», вредны и так назы-
ваемые перекусы. При этом нарушается не только «схема» упорядоченно-
сти нервно-психической деятельности, но и нормальное функционирова-
ние механизмов регуляции пищеварительной системы (что является фак-
тором риска заболеваний органов желудочно-кишечного тракта).
Пищевой рацион должен обеспечивать здоровое, сбалансированное пи-
тание взрослого человека. Сбалансированным является питание, которое
включает все необходимые компоненты пищи в оптимальных количествах
и соотношениях. Для обеспечения такого питания необходимо учитывать
следующие требования к составлению рациона питания.
Рацион питания должен включать незаменимые компоненты пищи. По-
линенасыщенных жирных кислот в суточном рационе должно быть около
3—5 г; богатым источником их содержания являются такие продукты, как
подсолнечное и кукурузное масла (53—55 %).
Помимо углеводов, являющихся источником энергии, в пищевом ра-
ционе должны содержаться сложные полисахариды — пищевые волокна;
они обладают способностью связывать воду и набухать, стимулировать пе-
ристальтику кишечника и ускорять транзит кишечного содержимого, ад-
сорбировать токсичные вещества и выводить их из организма, связывать
желчные кислоты и стерины, снижать содержание холестерина и нормали-
зовывать микрофлору кишечника. Пищевые волокна содержатся в ржаных
и пшеничных отрубях, овощах и фруктах. Они не являются балластом, а
активно участвуют в процессах пищеварения. В сутки организм взрослого
человека нуждается в 30 г пищевых волокон.
Сбалансированное питание в ряде случаев становится мерой профилак-
тики серьезных заболеваний. Диетические мероприятия, направленные на
снижение уровня холестерина в крови, следующие: уменьшение в рационе
питания количества насыщенных жиров и холестерина, являющихся ос-
новными факторами формирования гиперхолестеринемии, для чего часть
насыщенных жиров, содержащихся в продуктах животного происхожде-
ния, заменяют ненасыщенными, содержащимися в растительных маслах;
увеличение в рационе сложных углеводов, содержащихся в овощах и фрук-
тах; снижение общей калорийности рациона, избыточной массы тела;
уменьшение количества холестерина в пище до уровня ниже 300 мг в сутки
(для взрослого человека).
Необходимо обратить внимание на бесполезность, а нередко и вред-
ность различных модных «строгих» диет. Эти диеты обычно бывают несба-
лансированными, кроме того, их очень трудно выдержать длительное вре-
мя, что в конечном счете сводит на нет огромные нервно-психические
усилия, затраченные на привыкание к такой диете. Выбирая диету, не сле-
дует забывать, что прием пищи должен приносить удовольствие. Из обыч-
ных, традиционно используемых продуктов всегда можно составить такую
диету, которая будет низкокалорийной, содержащей умеренное количество
насыщенных жиров и холестерина. ,
Учитывая традиционные привычки питания, комитет экспертов ВОЗ
выдвинул рекомендации по диете. Согласно им, в диету следует включать
продукты растительного происхождения (бобовые, зерновые злаки, овощи,
фрукты) — в них содержатся белки, мало жира, холестерина, калорий и
много минеральных солей, витаминов, клетчатки; рыбу, птицу и тощие
сорта мяса, которые лучше употреблять небольшими порциями, — эти
продукты содержат полноценные белки и минеральные соли при низком
содержании насыщенных жиров, холестерина, калорий.
12. Обмен веществ и энергии. Питание • 591
При приготовлении пищи и приправ ВОЗ рекомендует меньше исполь-
зовать животные жиры, отдавая предпочтение растительным. Необходимо
ограничивать в рационе: жирные сорта мяса, в которых много насыщен-
ных жиров, холестерина и калорий; жирные молочные продукты (цельное
молоко, сметана, сыры); яйца, если они не являются основным источни-
ком белков (желтки яиц содержат много холестерина); кондитерские изде-
лия (они содержат много насыщенных жиров); алкогольные напитки, так
как они высококалорийны и содержат мало питательных веществ (при
«сгорании» в организме 1 г 96 % алкоголя выделяется 6,93 ккал).
Эти рекомендации касаются взрослых людей. Потребности в питатель-
ных веществах изменяются в период роста детей и подростков, во время
беременности, кормления, что необходимо учитывать при составлении
диеты.
При ожирении для ограничения энергетической ценности пищи и соз-
дания отрицательного энергетического баланса можно использовать дли-
тельную ограничительную диету, разгрузочные дни и сочетание этих двух
методов. Тактика диетотерапии зависит от возраста индивида. Диетотера-
пию у старых и пожилых людей следует применять с известной долей ос-
торожности, а темпы снижения массы тела должны быть не столь высоки-
ми, как у молодых.
Существует распространенное мнение, что абсолютное воздержание от
еды освобождает организм от вредных веществ, кишечник очищается от
остатков непереваренной пищи и слизи. В процессе голодания — доказы-
вают приверженцы этого метода — организм «сжигает» для процессов жиз-
недеятельности менее ценные белки, освобождаясь прежде всего от избы-
точных запасов жира и «шлаков». Однако необходимо учитывать, что с пе-
реходом организма на так называемое эндогенное питание в качестве ис-
точника энергии используются собственные «материалы», в то время как
организм нуждается в поступлении определенного минимума белка (50 г/
сут). При полном голодании организм вынужден кроме жира расходовать
структурные белки клеток и тканей, в первую очередь — мышц. Помимо
этого, при полном голодании неизбежна витаминная недостаточность. На-
конец, голодание является серьезным стрессом, приводящим к усугубле-
нию и без того нарушенного обмена веществ у лиц с избыточной массой
тела; многие люди, похудев в результате голодания, затем быстро восста-
навливают прежнюю массу тела.
ГЛАВА 13
Температура тела и ее регуляция
Основу жизнедеятельности организма составляет обмен веществ и энер-
гии, сопровождающийся выработкой тепла при биологическом окислении
молекул белков, жиров и углеводов. Существует непосредственная прямая
взаимосвязь между интенсивностью обмена веществ и количеством обра-
зующегося тепла в организме: увеличение скорости обменных процессов
вызывает рост теплообразования, а повышение температуры тела ускоряет
биологическое окисление в организме. Поскольку прирост температуры
тела сопровождается усилением отдачи организмом тепла в окружающую
среду, это препятствует лавинообразному росту скорости обменных про-
цессов и величины температуры.
Температура тела млекопитающих и человека поддерживается на отно-
сительно постоянном уровне независимо от колебаний температуры окру-
жающей среды. Такое постоянство температуры тела носит название изо-
термин. Изотермия свойственна только гомойотермным, или теплокровным,
животным, постоянство температуры внутренней среды которых является
необходимым условием для функционирования ферментов, влияющих на
скорость течения метаболических процессов. Температура тела гомойо-
термных организмов обусловливается интенсивностью теплообразования
за счет протекающих внутри них обменных процессов, поэтому они явля-
ются также эндотермными.
Изотермия отсутствует у пойкилотермных, или холоднокровных, живот-
ных, температура тела которых переменна и мало отличается от температу-
ры окружающей среды (низшие позвоночные и беспозвоночные). Для них
характерен более низкий по сравнению с теплокровными организмами
уровень энергетического обмена. Температура окружающей среды опреде-
ляет интенсивность энергетических превращений и уровень активности
холоднокровных животных, поэтому их называют эктотермными. В усло-
виях пониженной температуры среды их обитания пойкилотермные жи-
вотные впадают в состояние анабиоза, характеризующееся низкой активно-
стью ферментов и минимальной интенсивностью обменных процессов.
Тем не менее такие организмы обладают способностью противодейство-
вать сдвигам температуры тела при колебаниях температуры окружающей
среды за счет принятия определенной позы, выбора (если есть возмож-
ность) более холодной или более теплой среды, целенаправленного избега-
ния воздействия более высоких или низких температур.
Кроме гомойотермных и пойкилотермных в природе существует проме-
жуточная группа — гетеротермные организмы. При благоприятных услови-
ях существования они обладают способностью к изотермии, а при внезап-
ном понижении температуры внешней среды, недостатке пищи и воды эти
организмы теряют такую способность и становятся холоднокровными. К
этой группе относятся как млекопитающие (грызуны, летучая мышь), так
и некоторые виды птиц (колибри).
13. Температура тела и ее регуляция • 593
13.1. Нормальная температура тела
Способность теплокровных животных и человека поддерживать температу-
ру тела на относительно постоянном уровне в изменяющихся условиях
внешней и внутренней среды обеспечивается за счет непрерывной деятель-
ности физиологической системы терморегуляции. Эта система включает в
себя: 1) температурные рецепторы, реагирующие на изменение температу-
ры внешней и внутренней среды; 2) центр терморегуляции, расположен-
ный в гипоталамусе; 3) эффекторное (исполнительное) звено терморегуля-
ции. Основная функция системы терморегуляции — поддерживать опти-
мальную для метаболизма организма, или нормальную, температуру тела.
Полезным для организма приспособительным результатом работы этой
системы является определенная величина температуры крови, обеспечи-
вающая нормальное течение обменных процессов в организме, с одной
стороны, и определяющаяся интенсивностью этих процессов — с другой.
Обладая высокой теплоемкостью, кровь переносит тепло от тканей с высо-
ким уровнем теплообразования к тканям с более низким уровнем и, таким
образом, содействует выравниванию уровня температуры в различных час-
тях тела.
Температура глубоких тканей тела за счет теплопереноса кровью рас-
пределена более равномерно и составляет около 36,7—37,0 °C. Ее суточные
колебания в условиях относительного покоя организма находятся в преде-
лах 1 °C, поэтому говорят о гомойотермном «ядре» тела человека. В это по-
нятие включают ткани человеческого тела, расположенные на глубине 1 см
от поверхности кожи и глубже. В тканях печени, мозга, почек температура
несколько выше благодаря их метаболической активности, чем в тканях
других внутренних органов. Температура
поверхности тела и дистальных отделов
конечностей ниже, чем глубоких тканей и
проксимальных отделов конечностей. Она
зависит от интенсивности кровотока в со-
судах органов и тканей и от охлаждающе-
го или согревающего действия температу-
ры внешней среды, поэтому говорят о
пойкилотермной «оболочке» тела человека.
Относительное постоянство температуры
сохраняется в большей массе глубоких
тканей человека, если организм находится
в среде с температурой 25—26 °C. Это зна-
чение температуры для легко одетого че-
ловека называют термонейтральной зоной,
или температурой комфорта, потому что
в таком температурном диапазоне темпе-
ратура организма поддерживается посто-
янной без дополнительного участия тер-
морегуляторных механизмов. При охлаж-
дающем действии внешней среды темпе-
ратура глубоких тканей уменьшается, а
при согревании организма — возрастает
(рис. 13.1).
Температура тела человека изменяется
в течение суток (рис. 13.2), что является
проявлением суточных (циркадианных)
Рис. 13.1. Температура в различ-
ных областях тела в условиях хо-
лода (А) и тепла (Б).
Колебания температуры тела, вы-
званные изменениями внешней тем-
пературы, больше выражены вблизи
поверхности тела и в области дис-
тальных отделов конечностей (в
«оболочке» тела). В условиях холод-
ной внешней среды граница гомойо-
термного «ядра» температурой 37 °C
отодвигается в глубь тела.
594 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
В полости рта
Активные дети
Эмоции, работа
Норма
Раннее утро
40
39
38
37
36
°C
— Норма
— Раннее утро
Тяжелые физические
нагрузки
Активные дети
— Эмоции, работа
В прямой кишке
Рис. 13.2. Температура тела человека, измеренная в полости рта и прямой кишке в
различных физиологических состояниях.
В течение дня температура тела человека изменяется, при этом она минимальна во время сна
в предутренние часы (4—6 часов утра), так как в это время отмечается самый низкий уровень
обменных процессов (а значит и теплопродукции) в организме. Под влиянием физических и
психоэмоциональных нагрузок обмен веществ в организме интенсифицируется, увеличивается
теплопродукция, что приводит к увеличению температуры тела человека. Рост температуры
при этом находится в прямой зависимости от интенсивности нагрузок.
ритмов (см. главу 23). Суточные колебания температуры тела происходят
под влиянием эндогенных ритмов («биологических часов»), которые син-
хронизированы с внешними сигналами, например с вращением Земли.
Кроме того, температура тела человека зависит от его физиологического
состояния (сон или бодрствование, покой или физические и психо-эмо-
циональные нагрузки и т. д.). Максимального значения температура тела
человека достигает в 18—20 часов и снижается до своего минимума в пред-
утренние часы, к 4—6 часам утра. Амплитуда этих суточных колебаний не
превышает 1 °C.
Среднее значение температуры «ядра» тела отражает температура крови
в полостях сердца, аорте и других крупных сосудах. Однако измерение
температуры в этих частях тела человека практически невозможно, поэто-
му для клинических целей в качестве показателя температуры глубоких
тканей тела используют такие относительно доступные для ее измерения
значения, как ректальная температура, подъязычная и подмышечная тем-
пература, температура в наружном слуховом проходе у барабанной пере-
понки. Очевидно, что подобные измерения в каждом из перечисленных
участков тела имеют свои особенности и ограничения, а полученные вели-
чины температур лишь в большей или меньшей степени отражают темпе-
ратуру глубоких тканей (см. рис. 13.2).
13.2. Теплопродукция и теплоотдача
У человека и теплокровных животных температура «ядра» тела поддержи-
вается на относительно постоянном уровне. Это достигается с помощью
процессов эндогенной терморегуляции, результатом которой является устой-
чивое равновесие между количеством продуцируемого в организме в еди-
ницу времени тепла (теплопродукцией) и количеством тепла, рассеиваемого
организмом за то же время в окружающую среду (теплоотдачей). Другим
важнейшим процессом в регуляции температуры тела является поведение-
13. Температура тела и ее регуляция • 595
ская терморегуляция, обусловленная изменением характера поведения че-
ловека и животных для поддержания температурного баланса (например,
ношение человеком теплой одежды в холодное время года или сворачива-
ние животного в «клубок»),
13.2.1. Теплопродукция
Суммарная теплопродукция (теплообразование) в организме состоит из
первичной и вторичной теплоты. Первичная теплота выделяется в ходе по-
стоянно протекающих во всех органах и тканях реакций обмена веществ.
Вторичная теплота образуется при расходовании энергии макроэргиче-
ских соединений на выполнение человеком определенной мышечной ра-
боты. Уровень теплообразования в организме зависит от величины основ-
ного обмена, «специфически динамического действия» принимаемой пи-
щи, мышечной активности и интенсивности метаболизма в тканях.
Метаболические процессы осуществляются с неодинаковой интенсив-
ностью в различных органах и тканях, поэтому вклад в общую теплопро-
дукцию организма отдельных органов и тканей неравнозначен. Наиболь-
шее количество тепла образуется в скелетных мышцах при их тоническом
напряжении или сокращении. Образование тепла, наблюдающееся в мыш-
цах при этих условиях, получило название сократительного термогенеза
(сократительной теплопродукции), который является наиболее значимым
механизмом теплообразования у взрослого человека.
У новорожденных, а также у мелких млекопитающих животных имеется
Мембрана
клетки
Цитозоль
Митохондрия
Рис. 13.3. Схематичное изображение стимулирующего влияния норадреналина (НА)
на окислительное фосфорилирование жирных кислот в митохондриях бурой жиро-
вой ткани в механизме недрожательного термогенеза.
Норадреналин, высвобождаемый симпатическими нервными окончаниями в условиях холода,
воздействует на p-адренорецепторы (Р) на мембране клеток бурой жировой ткани и через сис-
тему вторичного посредника — циклический аденозинмонофосфат (цАМФ) стимулирует обра-
зование свободных жирных кислот из триацилглицеролов, которые окисляются в митохондри-
ях. ГЛ — гормончувствительная липаза, НП — несвязанный протеин, Н+ — протоны водорода,
АТФ — аденозинтрифосфат.
596 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
механизм ускоренного теплообразования за счет возрастания метаболиче-
ской активности в других тканях и, прежде всего, в буром жире. Бурую ок-
раску этой ткани придает большое количество окончаний симпатических
нейронов, содержащих медиатор норадреналин. В условиях холодового
воздействия на организм под влиянием выделяющегося из симпатических
нервных окончаний норадреналина происходит интенсивное окисление
жирных кислот. Бурый жир характеризуется избытком митохондрий, кото-
рые окружают мелкие капельки жира в цитоплазме. Окисление жирных
кислот в митохондриях бурой жировой ткани осуществляется без значимо-
го синтеза макроэргов и, таким образом, с максимально возможным обра-
зованием теплоты. Этот механизм получил название несократителъного
термогенеза (несократительной теплопродукции) (рис. 13.3). Посредством
механизмов несократительного термогенеза уровень теплопродукции у че-
ловека может быть увеличен примерно в три раза по сравнению с уровнем
основного обмена.
13.2.2. Теплоотдача
Существуют следующие пути отдачи тепла организмом в окружающую сре-
ду: излучение, теплопроведение, конвекция и испарение.
Излучение — это способ отдачи тепла в окружающую среду поверхно-
стью тела человека в виде электромагнитных волн инфракрасного диапазо-
на (а = 5—20 мкм). Количество тепла, рассеиваемого организмом в окру-
жающую среду излучением, пропорционально площади поверхности излу-
чения и разности средних значений температур кожи и окружающей сре-
ды. Площадь поверхности излучения — это суммарная площадь поверхно-
сти тех частей тела, которые соприкасаются с воздухом. При температуре
окружающей среды 20 °C и относительной влажности воздуха 40—60 % ор-
ганизм взрослого человека рассеивает путем излучения около 40—50 %
всего отдаваемого тепла. Теплоотдача путем излучения возрастает при по-
нижении температуры окружающей среды и уменьшается при ее повыше-
нии. В условиях постоянной температуры окружающей среды излучение с
поверхности тела возрастает при повышении температуры кожи и умень-
шается при ее понижении. Если средние температуры поверхности кожи и
окружающей среды выравниваются (разность температур становится рав-
ной нулю), отдача тепла излучением становится невозможной. Снизить те-
плоотдачу организма излучением можно за счет уменьшения площади по-
верхности излучения («сворачивания тела в клубок»). Если температура
окружающей среды превышает среднюю температуру кожи, тело человека,
поглощая инфракрасные лучи, излучаемые окружающими предметами, со-
гревается.
Теплопроведение — способ отдачи тепла, имеющий место при контакте,
соприкосновении тела человека с другими физическими телами. Количест-
во тепла, отдаваемого организмом в окружающую среду этим способом,
пропорционально разнице средних температур контактирующих тел, пло-
щади контактирующих поверхностей, времени теплового контакта и теп-
лопроводности контактирующего тела. Сухой воздух, жировая ткань харак-
теризуются низкой теплопроводностью и являются теплоизоляторами. Ис-
пользование одежды из тканей, содержащих большое число маленьких не-
подвижных «пузырьков» воздуха между волокнами (например, шерстяные
ткани), дает возможность организму человека уменьшить рассеяние тепла
путем теплопроводности. Влажный, насыщенный водяными парами воз-
дух, вода характеризуются высокой теплопроводностью. Поэтому пребыва-
13. Температура тела и ее регуляция • 597
Рис. 13.4. Виды теплоотдачи. Пути отдачи тепла организмом во внешнюю среду
можно условно подразделить на «влажную» теплоотдачу, связанную с испарением
пота и влаги с кожи и слизистых оболочек, и на «сухую» теплоотдачу, которая не
связана с потерей жидкости.
ние человека в среде с высокой влажностью при низкой температуре со-
провождается усилением теплопотерь организма. Влажная одежда также
теряет свои теплоизолирующие свойства.
Конвекция — способ теплоотдачи организма, осуществляемый путем пе-
реноса тепла движущимися частицами воздуха (воды). Для рассеяния тепла
конвекцией требуется обтекание поверхности тела потоком воздуха с более
низкой температурой, чем температура кожи. При этом контактирующий с
кожей слой воздуха нагревается, снижает свою плотность, поднимается и
замещается более холодным и более плотным воздухом. В условиях, когда
температура воздуха равна 20 °C, а относительная влажность — 40—60 %,
тело взрослого человека рассеивает в окружающую среду путем теплопро-
ведения и конвекции около 25—30 % тепла (базисная конвекция). При
увеличении скорости движения воздушных потоков (ветер, вентиляция)
значительно возрастает и интенсивность теплоотдачи (форсированная кон-
векция).
Отдача тепла организмом путем теплопроведения, конвекции и излуче-
ния, называемых вместе «сухой» теплоотдачей, становится неэффективной
при выравнивании средних температур поверхности тела и окружающей
среды.
Теплоотдача путем испарения — это способ рассеяния организмом тепла
в окружающую среду за счет его затраты на испарение пота или влаги с
поверхности кожи и влаги со слизистых оболочек дыхательных путей
(«влажная» теплоотдача). У человека постоянно осуществляется выделе-
ние пота потовыми железами кожи («ощутимая», или железистая, потеря
воды), увлажняются слизистые оболочки дыхательных путей («неощутимая»
потеря воды) (рис. 13.4). При этом «ощутимая» потеря воды организмом
оказывает более существенное влияние на общее количество отдаваемого
путем испарения тепла, чем «неощутимая».
При температуре внешней среды около 20 °C испарение влаги составля-
ет около 36 г/ч. Поскольку на испарение 1 г воды у человека затрачивается
0,58 ккал тепловой энергии, нетрудно подсчитать, что путем испарения ор-
ганизм взрослого человека отдает в этих условиях в окружающую среду
около 20 % всего рассеиваемого тепла. Повышение внешней температуры,
выполнение физической работы, длительное пребывание в теплоизоли-
598 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
рующей одежде усиливают потоотделение и оно может возрасти до 500—
2000 г/ч. Если внешняя температура превышает среднее значение темпера-
туры кожи, то организм не может отдавать во внешнюю среду тепло излу-
чением, конвекцией и теплопроведением. Организм в этих условиях начи-
нает поглощать тепло извне, и единственным способом рассеяния тепла
становится усиление испарения влаги с поверхности тела. Такое испарение
возможно до тех пор, пока влажность воздуха окружающей среды остается
меньше 100 %. При интенсивном потоотделении, высокой влажности и
малой скорости движения воздуха, когда капли пота, не успевая испарить-
ся, сливаются и стекают с поверхности тела, теплоотдача путем испарения
становится менее эффективной.
13.2.3. Поведенческая терморегуляция
Наряду с эндогенными процессами для поддержания нормальной темпера-
туры тела важнейшим механизмом является изменение характера поведе-
ния, или поведенческая терморегуляция. Для холоднокровных животных
этот механизм является определяющим. Поддерживающими постоянную
температуру факторами являются изменение позы, поиск укрытия, по воз-
можности выбор более теплой или холодной среды и т. п. Человек для
поддержания оптимальной температуры тела нередко прибегает к усилен-
ным мышечным движениям, особенно для согревания на холоде. При
ходьбе теплопродукция увеличивается в 2 раза, а при беге или интенсив-
ной работе — в 4—5 раз. Повышение температуры тела при этом даже на
несколько десятых градуса способствует ускорению окислительных про-
цессов, в частности — окислению продуктов белкового катаболизма. Кроме
того, для человека не менее важными факторами поддержания оптималь-
ной температуры тела является ношение одежды, соответствующей темпе-
ратуре окружающей среды, и оборудование жилища (утепление жилища
зимой и использование кондиционеров в жаркое время года).
13.3. Регуляция температуры тела
13.3.1. Восприятие организмом температурных воздействий
(терморецепция)
Изменение температуры внутренней среды («ядра») и поверхностных отде-
лов («оболочки») тела человека воспринимается организмом с помощью
терморецепторов. Температурная рецепция осуществляется окончаниями
тонких чувствительных нервных волокон типа С и А (8), которые пред-
ставлены в коже, слизистых оболочках, мышцах, сосудах, во внутренних
органах (периферические терморецепторы). Холодо- и теплочувствительные
нейроны располагаются в медиальной преоптической области переднего
гипоталамуса (центральные терморецепторы).
Восприятие температурных раздражений из внешней среды и формиро-
вание температурных ощущений у человека осуществляется с помощью
терморецепторов кожи и слизистых оболочек, среди которых имеются хо-
лодовые рецепторы (повышают частоту передачи нервных импульсов по
афферентным нервным волокнам к терморегуляторному центру при их ох-
лаждении и снижают эту частоту при их нагревании) и тепловые рецепто-
ры (реагируют на изменение температуры тела противоположным обра-
зом). В коже и на слизистых оболочках человека больше холодовых рецеп-
торов (около 250 000), чем тепловых (около 30 000). Кроме того, холодовые
13. Температура тела и ее регуляция • 599
рецепторы кожи расположены более поверхностно, на глубине 0,17 мм, а
тепловые — более глубоко, на глубине 0,3 мм. Эта особенность расположе-
ния терморецепторов обусловливает более раннее восприятие организмом
человека холода, чем тепла. Другая особенность терморецепторов — их не-
равномерное распределение в коже по площади, что определяет различный
уровень чувствительности к холоду и теплу разных участков тела. Наиболь-
шей чувствительностью обладает кожа лица, наименьшей — кожа нижних
конечностей.
Афферентный поток нервных импульсов от периферических терморе-
цепторов поступает через задние корешки спинного мозга к вставочным
нейронам задних рогов. Затем по спиноталамическому тракту этот поток
импульсов достигает передних ядер таламуса и далее проводится в сомато-
сенсорную кору больших полушарий головного мозга. Поступление нерв-
ных импульсов от периферических терморецепторов в соматосенсорную
кору обеспечивает возникновение и топическую локализацию субъектив-
ных температурных ощущений, таких как «тепло», «холодно», «прохладно»,
«жарко», «температурный комфорт» или «дискомфорт». На их основе фор-
мируются поведенческие терморегуляторные реакции. Значительная часть
афферентных импульсов от периферических рецепторов кожи и внутрен-
них органов поступает из спинного мозга по волокнам спиноталамическо-
го тракта к нейронам гипоталамического центра терморегуляции.
13.3.2. Центральное звено системы терморегуляции
Регуляция теплообмена, а следовательно, и температуры тела человека осу-
ществляется центром терморегуляции, который расположен в медиальной
преоптической области переднего отдела гипоталамуса и в заднем отделе
гипоталамуса. Разрушение этих отделов гипоталамуса или нарушение их
нервных связей посредством перерезки на уровне среднего мозга в экспе-
риментах на животных приводит к нарушению контроля за температурой
Термочувствительные нейроны
преоптической области гипоталамуса
Нейроны переднего Эффекторные нейроны
гипоталамуса
заднего гипоталамуса
Рис. 13.5. Схема взаимодействия различных типов нейронов терморегуляторного
центра гипоталамуса между собой и с кожными терморецепторами.
Стимуляция тепловых рецепторов кожи (Рт) и гипоталамуса активирует процессы теплоотдачи
в организме человека, а холодовых рецепторов (Рх) кожи и гипоталамуса — теплопродукции.
Ин — интернейроны гипоталамуса.
600 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 13.6. Схема механизмов регуляции теплообмена в организме человека.
Поддержание относительного постоянства температуры тела достигается с помощью баланса
между количеством продуцируемого в единицу времени тепла в организме человека и количе-
ством тепла, которое организм отдает за то же время в окружающую среду. Тепловой баланс
регулируется нейрогуморальными механизмами, которые активируются в результате измене-
ния импульсной активности эффекторных нейронов терморегуляторного центра гипоталаму-
са. В гипоталамический терморегуляторный центр поступает афферентная информация об из-
менениях внешней температуры от периферических терморецепторов и об изменения темпе-
ратуры «ядра» — от центральных терморецепторов (пояснения в тексте).
тела у гомойотермных организмов. Кроме того, местное нагревание перед-
ней гипоталамической области вызывает усиление потоотделения и учаще-
ние дыхания у экспериментальных животных, охлаждение — возникнове-
ние дрожи и «свертывание в клубок». Регистрация активности отдельных
нейронов гипоталамуса с помощью микроэлектродов показала ее измене-
ние как в ответ на локальные колебания температуры в самом гипоталаму-
се, так и при воздействии раздражителей на терморецепторы кожи, внут-
ренних органов и сосудов. Вышеперечисленные факты доказывают, что
центр терморегуляции расположен в гипоталамусе.
В терморегуляторном центре гипоталамуса обнаружены различные по
функциям группы нервных клеток: 1) термочувствительные нейроны пре-
оптической области; 2) клетки, «задающие» уровень поддерживаемой в ор-
ганизме температуры тела («установочная точка» терморегуляции) в перед-
нем гипоталамусе; 3) вставочные нейроны (интернейроны) гипоталамуса;
13. Температура тела и ее регуляция • 601
4) эффекторные нейроны, управляющие процессами теплопродукции и те-
плоотдачи, в заднем гипоталамусе (рис. 13.5).
Термочувствительные нервные клетки преоптической области гипотала-
муса непосредственно «измеряют» температуру артериальной крови, проте-
кающей через мозг, и обладают высокой чувствительностью к температур-
ным изменениям (способны различать разницу температуры крови в
0,011 °C). Отношение холодо- и теплочувствительных нейронов в гипотала-
мусе составляет 1:6, поэтому центральные терморецепторы преимущест-
венно активируются при повышении температуры «ядра» тела человека.
На основе анализа и интеграции информации о значении температуры
крови и периферических тканей, в преоптической области гипоталамуса
непрерывно определяется среднее (интегральное) значение температуры те-
ла. Эти данные передаются через вставочные нейроны в группу нейронов
переднего отдела гипоталамуса, задающих в организме определенный уро-
вень температуры тела — «установочную точку» терморегуляции. На основе
анализа и сравнений значений средней температуры тела и заданной вели-
чины температуры, подлежащей регулированию, механизмы «установоч-
ной точки» через эффекторные нейроны заднего гипоталамуса воздейству-
ют на процессы теплоотдачи или теплопродукции, чтобы привести в соот-
ветствие фактическую и заданную температуру. Таким образом, за счет
функции центра терморегуляции устанавливается равновесие между тепло-
продукцией и теплоотдачей, позволяющее поддерживать температуру тела
в оптимальных для жизнедеятельности организма пределах (рис. 13.6).
В механизме формирования «установочной точки» имеет значение уро-
вень спонтанной активности вставочных нейронов гипоталамуса. Напри-
мер, если уровень спонтанной активности интернейрона является высо-
ким, то для усиления термогенеза требуется более высокая активность
кожных холодовых рецепторов, а значение пороговой температуры для ре-
гулируемой теплопродукции является более низким. И наоборот, если
вставочный нейрон проявляет низкую спонтанную активность, то даже не-
значительная афферентация от кожных холодовых рецепторов может ока-
заться достаточной для запуска дополнительного теплообразования в орга-
низме. Уровень спонтанной активности вставочных нейронов зависит от
соотношения концентрации ионов натрия и кальция в гипоталамусе и не-
которых других нетемпературных факторов.
13.3.3. Эффекторное (исполнительное) звено системы
терморегуляции
Система терморегуляции не имеет собственных специфических эффектор-
ных органов, она использует эффекторные пути других физиологических
систем (сердечно-сосудистой, дыхательной, скелетной мускулатуры, выде-
лительной и др.). Эти эффекторные механизмы усиливают либо ослабляют
процессы теплопродукции и теплоотдачи в организме в зависимости от
температурных условий окружающей среды.
В термонейтральных условиях внешней среды баланс теплопродукции и
теплоотдачи в организме человека для поддержания оптимальной температу-
ры тела достигается преимущественно за счет изменения просвета сосудов
поверхности тела под влиянием симпатического отдела вегетативной нервной
системы. Увеличение симпатического тонуса вызывает сужение кровеносных
сосудов, а его снижение — расширение сосудов. Это приводит соответствен-
но к уменьшению или увеличению переноса тепла кровью от «ядра» тела к
«оболочке» и его рассеивания во внешнюю среду физическими способами.
602 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
В условиях высокой внешней температуры для поддержания оптимально-
го температурного баланса в организме человека включения механизма со-
судодвигательных реакций может быть недостаточно. Если уровень сред-
ней интегральной температуры тела, несмотря на расширение поверхност-
ных сосудов, превышает величину установочной температуры, происходит
резкое усиление потоотделения. Эта реакция также контролируется симпа-
тической нервной системой через выделение из окончаний нервных воло-
кон ацетилхолина. Испарение влаги с поверхности тела и поведенческие
реакции (например, обмахивание веером, включение вентилятора или кон-
диционера) приобретают в усилении теплоотдачи ведущее значение.
В условиях низкой внешней температуры основную роль для поддержа-
ния оптимальной температуры тела играет активизация процессов тепло-
продукции, особенно когда, несмотря на сужение поверхностных сосудов
и минимальное потоотделение, уровень средней интегральной температу-
ры тела человека становится ниже, чем величина «установочной точки».
Уровень теплопродукции в организме контролируется нейронами заднего
отдела гипоталамуса и осуществляется посредством соматических и симпа-
тических нервных волокон, а также при участии ряда гормонов и биологи-
чески активных веществ. Так, при увеличении притока афферентных нерв-
ных импульсов от холодовых рецепторов кожи в гипоталамус первоначаль-
но усиливается сократительный термогенез. Для этого от нейронов дорсо-
медиальной области гипоталамуса через ядра двигательной системы сред-
него и продолговатого мозга поток эфферентных нервных импульсов по-
ступает к мотонейронам спинного мозга. Последние осуществляют ритми-
ческую посылку эффекторных нервных импульсов к скелетным мышцам
шеи, туловища, проксимальных отделов конечностей. Первоначально это
проявляется в увеличении амплитуды и частоты электромиографической
активности этих мышц, росте их тонического напряжения, однако види-
мых сокращений данные мышцы при этом не совершает. При этом в тер-
морегуляционный тонус последовательно вовлекаются мышцы подбород-
ка, шеи, верхнего плечевого пояса, туловища, сгибатели конечностей. По-
вышенный тонус мышц придает телу характерную позу «сворачивание в
клубок», которая уменьшает площадь поверхности тела, контактирующей с
внешней средой, и снижает интенсивность теплоотдачи.
При продолжающемся охлаждении организма, когда начинается сниже-
ние его внутренней температуры (температуры «ядра»), повышение тонуса
скелетных мышц переходит в качественно новое состояние — возникают
непроизвольные периодические сокращения скелетной мускулатуры, полу-
чившие название холодовой дрожи. В этом случае совершается сравнитель-
но небольшая механическая работа скелетных мышц, и почти вся их мета-
болическая энергия освобождается в виде тепла. Скорость метаболизма и
теплообразования в мышцах при холодовой дрожи может возрастать в
5 раз по сравнению с метаболизмом и теплообразованием в них в условиях
относительного покоя. В условиях холода благодаря активизации симпати-
ческой нервной системы через ее медиатор норадреналин стимулируется
липолиз в жировой ткани. В кровоток выделяются и в последующем окис-
ляются с образованием большого количества тепла свободные жирные ки-
слоты. Под влиянием норадреналина и адреналина происходит быстрое,
но непродолжительное повышение теплопродукции в организме человека.
Более продолжительное усиление обменных процессов достигается под
влиянием гормонов щитовидной железы — тироксина и трийодтиронина
(см. главу 6).
Если, несмотря на активацию обмена веществ, величина теплопродук-
13. Температура тела и ее регуляция • 603
ции организма становится меньше величины теплоотдачи, возникает по-
нижение температурь! тела, получившее название гипотермии. Противопо-
ложное состояние организма, сопровождающееся повышением температу-
ры тела,— гипертермия, имеет место в том случае, когда интенсивность те-
плопродукции превышает способность организма отдавать тепло в окру-
жающую среду посредством имеющихся у него способов теплоотдачи.
13.4. Гипертермия и гипотермия
Состояние организма человека, при котором температура его тела подни-
мается выше 37 °C, принято называть гипертермией. Она легко развивается
в условиях действия на организм внешней температуры, превышающей
37 °C при 100 % влажности воздуха, когда испарение пота или влаги с по-
верхности тела становится невозможным. В случае продолжительной ги-
пертермии может развиваться «тепловой удар». Это состояние организма
характеризуется покраснением кожи в результате расширения перифериче-
ских сосудов, отсутствием потоотделения, признаками нарушения функ-
ций центральной нервной системы (нарушение ориентации, бред, судоро-
ги). В более легких случаях гипертермия может проявиться в виде теплово-
го обморока, когда в результате расширения периферических кожных сосу-
дов происходит резкое падение артериального давления.
В процессе эволюции теплокровных организмов у них выработана осо-
бая ответная реакция на попадание во внутреннюю среду чужеродных па-
тогенных микроорганизмов — лихорадка — состояние организма, при кото-
ром центр терморегуляции стимулирует повышение температуры тела. Это
достигается перестройкой механизма «установочной точки» на более высо-
кую, чем в норме, заданную температуру регуляции. Переход «установоч-
ной точки» на более высокий уровень происходит в результате действия на
соответствующую группу нейронов преоптической области гипоталамуса
эндогенных пирогенов — веществ, вызывающих подъем температуры тела.
Эндогенными пирогенами являются пептиды: интерлейкин-1 в формах а и
р, фактор некроза опухолей, интерлейкин-6, а-интерферон и другие. На-
личие в организме ряда дублирующих друг друга эндопирогенных факто-
ров свидетельствует о том, что вызываемая ими лихорадка играет для орга-
низма важную защитную роль.
Состояние организма человека, при котором температура его тела ста-
новится ниже 35 °C, называют гипотермией. Наиболее быстро гипотермия
возникает при погружении организма в холодную воду. При этом происхо-
дит возбуждение симпатического отдела вегетативной нервной системы,
рефлекторно ограничивается теплоотдача и усиливается теплопродукция в
первую очередь за счет мышечной дрожи. Если человек находится в воде в
неподвижном состоянии, то через некоторое время температура тела начи-
нает падать, снижается интенсивность обмена веществ, замедляется дыха-
ние, урежаются сердечные сокращения, снижается сердечный выброс, по-
нижается артериальное давление (при температуре тела 24—25 °C оно мо-
жет составлять 15—20 % от исходного). Если же человек плавает, то в ре-
зультате работы скелетных мышц включается механизм сократительного
термогенеза, который компенсирует усиленную теплоотдачу в холодной
воде. В хирургической практике используется искусственная гипотермия с
охлаждением тела до 24—28 °C при операциях на сердце и ЦНС. Искусст-
венная гипотермия значительно снижает интенсивность обмена веществ в
ЦНС и, как следствие, потребность ее в кислороде. Больные легче перено-
сят временное выключение сердечной деятельности и остановку дыхания.
604 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Как при гипотермии, так и при гипертермии в организме человека име-
ет место нарушение основного условия поддержания постоянства темпера-
туры его тела — баланса теплопродукции и теплоотдачи. Изменение темпе-
ратуры тела при этих состояниях осуществляется вопреки «усилиям» цен-
тра терморегуляции и других механизмов системы терморегуляции удер-
жать нормальную температуру тела.
13.5. Взаимодействие системы терморегуляции с другими
физиологическими системами организма
Система терморегуляции использует для осуществления своих функций
эффекторные компоненты других физиологических систем. Это обстоя-
тельство обусловливает необходимость постоянного взаимодействия, со-
пряжения или конкуренции механизмов, регулирующих теплообмен и дру-
гие гомеостатические функции. Такое сопряжение регуляции теплообмена
и других гомеостатических функций имеет место, прежде всего, в гипота-
ламусе, где термочувствительные нейроны преоптической области гипота-
ламуса являются одновременно чувствительными к изменению осмотиче-
ского давления, артериального давления крови, концентрации ионов Н+,
Na+, Са2+, СО2, глюкозы. Эти нейроны изменяют свою биоэлектрическую
активность при сдвигах температуры тела, под действием эндопирогенов,
половых гормонов, некоторых нейромедиаторов. Таким образом, центр
терморегуляции в гипоталамусе постоянно взаимодействует с другими рас-
положенными здесь же центрами регуляции гомеостаза.
Используемые организмом в системе регуляции теплообмена нейроме-
диаторы, гуморальные вещества также одновременно принимают участие в
регуляции других функций и показателей гомеостаза. Их примерами могут
быть катехоламины, которые параллельно выполняют функции медиато-
ров в центральной и симпатической нервных системах, функции сосудоак-
тивных веществ, активаторов обменных процессов.
13.5.1. Сердечно-сосудистая система и терморегуляция
В качестве эффекторов в реакциях теплообмена используются сосуды по-
верхности тела, посредством которых регулируется кровоток в коже, ее
температура и интенсивность теплоотдачи. В термонейтральных условиях,
при действии на организм умеренно низких температур или неглубокой
гипотермии изменение кровотока в поверхностных тканях не оказывает
существенного влияния на деятельность сердца и системную гемодинами-
ку. При действии же на организм высоких температур, гипертермии, лихо-
радке резкое расширение сосудов поверхности тела, влияние высокой тем-
пературы на центральные механизмы регуляции кровообращения могут
привести к падению давления крови, развитию коллаптоидного состояния.
Использование при гипертермии многочисленных поверхностных сосудов,
как общих эффекторов сердечно-сосудистой и терморегулирующей систем,
соподчинено более важной в этот момент времени гомеостатической по-
требности организма — поддержанию системного кровотока.
13.5.2. Водно-солевой баланс и терморегуляция
Когда температура поверхности тела достигает величин, равных температу-
ре окружающей среды, ведущее значение в Механизмах теплоотдачи при-
обретает уже не повышение кровотока в поверхностных тканях, а потоот-
13. Температура тела и ее регуляция • 605
деление и испарение пота и влаги с поверхности тела. Более существенную
роль начинают играть учащение дыхания и испарение влаги с поверхности
дыхательных путей. Включение в реакции теплообмена потоотделения яв-
ляется примером использования общих эффекторов для системы терморе-
гуляции и систем регуляции водно-солевого баланса и осмотического дав-
ления.
Если при подъеме температуры тела в силу потери жидкости за счет
усиленного потоотделения уменьшается объем циркулирующей крови и
повышается ее осмотическое давление, организм стремится сохранить
водный гомеостаз, даже если это идет в ущерб терморегуляторным реак-
циям. С развитием гипогидратации и повышением осмотического давле-
ния в организме отдача тепла за счет потоотделения уменьшается и тем-
пература тела устанавливается на еще более высоком уровне. Развивается
чувство жажды, уменьшается диурез. В конкуренции за общие эффектор-
ные механизмы начинают преобладать системы осмо- и волюморегуля-
ции, как более древние и в экстремальных условиях более важные для со-
хранения гомеостаза. Сопряжение осмо- и терморегуляции достигается в
нервных центрах медиальной преоптической области гипоталамуса, где те-
пло- и холодочувствительные нейроны наделены одновременно высокой
осмочувствительностью. Подтверждением сопряженного протекания в ор-
ганизме процессов термо- и осморегуляции являются изменения водного
обмена противоположной направленности — при охлаждении организма.
При действии на организм низкой температуры имеют место уменьшение
потребления воды, усиление диуреза и повышение осмолярности плазмы
крови.
Если дегидратация при действии на организм высокой внешней темпе-
ратуры приводит к торможению терморегуляторных реакций, то при дей-
ствии на него низкой температуры дегидратация ведет к торможению теп-
лочувствительных нейронов гипоталамуса и в результате — к снижению те-
плоотдачи.
13.5.3. Дыхание и терморегуляция
При действии на организм высокой внешней температуры активация по-
тоотделения и дыхания ведет к усиленному выделению из организма СО2,
некоторых минеральных ионов. Как при гипер-, так и при гипотермии
могут наблюдаться сдвиги кислотно-основного состояния. За счет полип-
ноэ и интенсификации потоотделения развивается дыхательный алкалоз,
сопровождающиейся увеличением pH и снижением рСО2 в крови. При
нарастании гипертермии в связи с ухудшением доставки к тканям кисло-
рода в них развивается метаболический ацидоз. Смена щелочной реак-
ции крови на кислую при выраженной гипертермии вновь начинает иг-
рать положительную регуляторную роль, как для усиления теплоотдачи,
так и для предупреждения дальнейшего закисления крови и улучшения
оксигенации тканей. Это достигается стимуляцией дыхательного центра
посредством избытка Н2+, увеличения минутного объема дыхания, усиле-
ния испарения влаги с поверхности дыхательных путей, что ведет к сни-
жению рСО2 и увеличению рО2. Обратные взаимоотношения между про-
цессами регуляции теплообмена и дыхания прослеживаются при гипотер-
мии. Развивающаяся при этом гиповентиляция является общим эффек-
торным механизмом, обеспечивающим снижение теплопотерь, поддержа-
ние на более низком уровне pH крови соответственно сниженной темпе-
ратуре тела.
ГЛАВА 14
Выделение. Функции почек.
Водно-солевой обмен
Выделение — часть обмена веществ, осуществляемая путем выведения из
внутренней среды организма во внешнюю среду конечных и промежуточ-
ных продуктов метаболизма, чужеродных и излишних веществ для обеспе-
чения оптимального состава внутренней среды и нормальной жизнедея-
тельности организма. Процессы выделения являются неотъемлемым при-
знаком жизни, поэтому их нарушение неизбежно приводит к нарушениям
гомеостазиса, обмена веществ и функций организма, вплоть до его гибели.
Выделение неразрывно связано с обменом воды, поскольку основная часть
предназначенных для выведения из организма веществ выделяется в рас-
творенном виде. Основным органом выделения являются почки, образую-
щие и выделяющие мочу и вместе с ней вещества, подлежащие удалению
из организма. Почки являются также основным органом обеспечения вод-
но-солевого обмена, поэтому в этой главе и рассматриваются функции по-
чек, выделение и водно-солевой обмен.
14.1. Органы и процессы выделения
Функция выделения веществ из внутренней среды организма осуществля-
ется почками, желудочно-кишечным трактом, легкими, кожей и слизисты-
ми оболочками, слюнными железами. Реализуемые ими процессы выделе-
ния находятся в координированной взаимосвязи и поэтому функциональ-
но эти органы объединяют понятием выделительная система организма
(рис. 14.1). Между органами выделения существуют функциональные и ре-
гуляторные взаимосвязи, в результате чего сдвиг функционального состоя-
ния одного из органов выделения меняет активность другого в пределах
единой выделительной системы. Так, например, при избыточном выведе-
нии жидкости через кожу путем потоотделения при высокой температу-
ре — снижается объем мочеобразования, при уменьшении экскреции азо-
тистых соединений с мочой — увеличивается их выведение через желудоч-
но-кишечный тракт, легкие и кожу.
14.1.1. Выделительная функция кожи
Выделительная функция кожи, непосредственно контактирующей с внеш-
ней средой большой площадью поверхности, преимущественно обеспечи-
вается деятельностью потовых и, в меньшей степени, сальных желез.
В среднем у человека за сутки выделяется от 300 до 1000 мл пота, что за-
висит от температуры окружающей среды и интенсивности энергетическо-
го метаболизма. Составы пота и плазмы крови отличаются, поскольку пот
является не простым фильтратом плазмы, а секретом потовых желез. С по-
том из организма выводится в покое до ‘/3 общего количества экскретируе-
14. Выделение. Функции почек. Водно-солевой обмен • 607
Внешняя среда
Моча
Рис. 14.1. Выделительная система организма.
Органы выделения (кожа, легкие, слюнные железы, печень и желудочно-кишечный тракт,
почки) выводят из внутренней среды организма во внешнюю среду метаболиты и ксенобиоти-
ки в составе секретов и экскретов.
мой воды, 5—7 % всей мочевины, мочевая кислота, креатин, хлориды, на-
трий, калий, кальций, органические вещества, липиды, микроэлементы.
Через кожу может выделяться даже больше кальция, чем выводится с мо-
чой. При недостаточности функции почек или печени возрастает выделе-
ние через кожу веществ, обычно экскретируемых с мочой, — мочевины,
ацетона, желчных пигментов и др. С потом выделяются пепсиноген, ами-
лаза и щелочная фосфатаза, отражая тем самым функциональное состоя-
ние органов пищеварения.
Регуляция потоотделения осуществляется симпатическими холинерги-
ческими влияниями, а также гормонами — вазопрессином, альдостероном,
гормонами щитовидной железы и половыми стероидами.
Секрет сальных желез на 2/3 состоит из воды, а */3 составляют неомыляе-
мые соединения — холестерин, сквален (алифатический углеводород), ана-
608 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
логи казеина, продукты обмена половых гормонов, кортикостероидов, ви-
таминов и ферментов. В выделительной системе сальные железы не имеют
большой значимости, так как за сутки выделяется всего лишь около 20 г
секрета. Регуляция сальных желез осуществляется в основном половыми и
надпочечниковыми стероидами.
14.1.2. Выделительная функция печени и пищеварительного
тракта
Выделительная функция печени реализуется за счет образования и секре-
ции в ней желчи. За сутки печень секретирует от 500 до 2000 мл желчи, но
большая часть ее объема затем реабсорбируется в желчном пузыре и ки-
шечнике. С желчью из организма экскретируются конечные продукты об-
мена гемоглобина и других порфиринов в виде желчных пигментов, конеч-
ные продукты обмена холестерина — в виде желчных кислот. Несмотря на
всасывание в кишечнике и обратный транспорт в печень с кровью ворот-
ной вены, часть этих веществ покидает организм с фекальными массами.
В составе желчи из организма выделяются тироксин, мочевина, кальций и
фосфор, а также вещества, поступающие в организм: лекарственные пре-
параты, ядохимикаты и др. В желчном пузыре происходит обратное всасы-
вание в кровь воды и растворенных в ней веществ, прежде всего электро-
литов. Этот процесс приводит к концентрированию желчи и регулируется
гормоном вазопрессином, повышающим проницаемость стенки желчного
пузыря.
Выделительная функция желудка обеспечивает выведение в составе же-
лудочного сока продуктов метаболизма (мочевины, мочевой кислоты), ле-
карственных и ядовитых веществ (ртуть, йод, салицилаты, хинин).
Выделительная функция кишечника состоит, во-первых, в выделении
продуктов распада пищевых веществ, не подвергшихся всасыванию в
кровь и представляющих излишние или вредные для организма соедине-
ния. Во-вторых,, кишечник экскретирует вещества, поступившие в его про-
свет с пищеварительными соками (желудочным, поджелудочным) и жел-
чью. При этом многие из них в кишечнике подвергаются метаболизму и с
калом выделяются не сами вещества, а их метаболиты, например метабо-
литы билирубина желчи. В-третьих, стенка кишечника способна экскрети-
ровать из крови ряд веществ, среди которых особое значение имеет экс-
креция плазменных белков. При чрезмерности этого процесса возникает
избыточная потеря организмом белка, ведущая к патологии. Из крови ки-
шечный эпителий экскретирует соли тяжелых металлов, магний, почти по-
ловину всего выделяемого организмом кальция. Вместе с экскрементами
кишечником выделяется и некоторое количество воды (в среднем около
100 мл/сут).
14.1.3. Выделительная функция легких и верхних дыхательных
путей
Процессы газообмена, происходящие в легких, обеспечивают удаление из
внутренней среды организма летучих метаболитов и экзогенных веществ —
углекислого газа, аммиака, ацетона, этанола, метилмеркаптана и др. Кро-
ме того, за счет мерцательного эпителия удаляются продукты обмена ве-
ществ самой легочной ткани и эпителия воздухоносных путей, например
продукты деградации сурфактанта. Легкие выделяют в просвет дыхатель-
ных путей небольшие количества белка, в том числе гамма-глобулинов,
14. Выделение. Функции почек. Водно-солевой обмен • 609
обладающих сродством к легочной ткани, а также входящих в состав сек-
рета желез бронхиального дерева. Через слизистую оболочку дыхательных
путей испаряется значительное количество воды (от 400 мл в покое до 1 л
при усиленном дыхании), а при повышении проницаемости аэрогематиче-
ского барьера из крови могут в избытке выделяться пурины, аденозин- и
гуанозинмонофосфаты. Гиперсекреция желез слизистой оболочки верхних
дыхательных путей имеет место при нарушениях выделительной функции
почек, в этом случае через слизистую оболочку выделяется много мочеви-
ны, которая, разлагаясь, образует аммиак, определяющий соответствую-
щий запах изо рта.
14.2. Функции почек
Функции почек многообразны, при этом часть из них связана с процес-
сами выделения, в которых почки играют ведущую роль, другая же часть
подразумевает невыделительные функции почек. Почки участвуют в регу-
ляции:
1) водного баланса организма и, соответственно, объемов вне- и внут-
риклеточных водных пространств, поскольку меняют количество вы-
водимой с мочой воды;
2) ионного баланса и состава жидкостей внутренней среды путем изби-
рательного изменения экскреции ионов с мочой;
3) постоянства осмотического давления жидкостей внутренней среды за
счет изменения количества выводимых осмотически активных ве-
ществ (солей, мочевины, глюкозы и др.);
4) кислотно-основного баланса, путем изменения экскреции водород-
ных ионов, нелетучих кислот и оснований;
5) метаболизма белков, липидов, углеводов, нуклеиновых кислот и дру-
гих органических соединений, во-первых, за счет изменений экскре-
ции продуктов метаболизма и избытка соединений, поступивших с
пищей или образовавшихся в организме, во-вторых, благодаря собст-
венной метаболической функции (синтез аммиака и мочевины, но-
вообразование глюкозы, гидролиз белков и липидов, синтез фермен-
тов, простаноидов и т. п.);
6) циркуляторного гомеостазиса, путем регуляции обмена электроли-
тов, объема циркулирующей крови, внутренней секреции гормонов,
регулирующих функции сердечно-сосудистой системы, — ренина,
кальцитриола и др., а также экскреции катехоламинов и других гор-
мональных регуляторов системы кровообращения;
7) эритропоэза, за счет внутренней секреции эритропоэтина — гумо-
рального регулятора эритрона;
8) гемостаза, путем образования гуморальных регуляторов свертывания
крови и фибринолиза (урокиназы, тромбопластина, тромбоксана и
простациклина) и участвуя в обмене физиологических антикоагулян-
тов (гепарина).
Экскретируя из внутренней среды чужеродные и вредные вещества,
почки выполняют защитную функцию. Таким образом, выделяют следую-
щие функции почек: экскреторную, гомеостатическую, метаболическую,
инкреторную и защитную. Основной функцией почек, обеспечивающей ве-
дущую роль в выделительной системе организма, является образование и
выделение мочи.
20-6095
610 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
14.2.1. Механизмы мочеобразования
Моча образуется в почках из плазмы крови, причем почка относится к
наиболее интенсивно кровоснабжаемым органам — ежеминутно через поч-
ку проходит ’/4 всего объема крови, выбрасываемой сердцем, при этом
объем кровотока в коре почки, где происходит фильтрация плазмы крови
и образование первичной мочи, составляет свыше 90 % общего почечного
кровотока. Основной структурно-функциональной единицей почки, обес-
печивающей образование мочи, является нефрон. В почке человека нахо-
дится около 1,2 млн. нефронов. Однако не все нефроны функционируют в
почке одновременно, существует определенная периодичность активности
отдельных нефронов, когда часть из них функционирует, а другие нет. Эта
периодичность обеспечивает надежность деятельности почки за счет функ-
ционального дублирования. В связи с этим важным показателем функцио-
нальной активности почки является масса действующих нефронов в кон-
кретный момент времени.
Нефрон состоит из нескольких последовательно соединенных отделов
(рис. 14.2), располагающихся в корковом и мозговом веществе почки.
Сосудистый клубочек, или мальпигиево тельце, является структурой, где
происходит процесс ультрафильтрации плазмы крови через фильтрацион-
ный барьер и образование первичной мочи (рис. 14.3). Он расположен в
корковом веществе, имеет около 50 капиллярных петель, связанных друг с
другом и подвешенных как на брыжейке с помощью мезангия, состоящего
из волокнистых структур и мезангиальных клеток. Снаружи клубочки по-
крыты двухслойной капсулой Боумена—Шумлянского. Висцеральный лис-
ток этой капсулы покрывает капилляры клубочка и состоит из эпителиаль-
ных отростчатых клеток — подоцитов. Отростки подоцитов (большие и ма-
лые), называемые педикулами, покрывают всю поверхность капилляров,
тесно переплетаясь друг с другом и оставляя межпедикулярные простран-
ства не более 30 нм. Пространства заполнены фибриллярными структура-
ми, образующими щелевую диафрагму, формирующую решетку или сито с
диаметром пор около 10 нм. Наружный или париетальный листок капсулы
состоит из базальной мембраны, покрытой кубическими эпителиальными
клетками, переходящими в эпителий канальцев. Между двумя листками
капсулы, расположенными наподобие чаши, имеется щель или полость
капсулы, в которую происходит ультрафильтрация плазмы крови. Полость
капсулы переходит в просвет главного или проксимального отдела ка-
нальцев.
Стенка всех канальцев нефрона, где по мере продвижения мочи проис-
ходит обратное всасывание в кровь воды и различных веществ (реабсорб-
ция), а также секреция веществ из крови в мочу, состоит из эпителиаль-
ных клеток, расположенных на базальной мембране (рис. 14.4). По строе-
нию и функции у этих клеток выделяют апикальную, или люминальную,
мембрану, обращенную в просвет канальца, и базолатеральную мембрану.
Канальцевый аппарат нефрона подразделяют на несколько отделов.
Главный, или проксимальный, отдел канальцев, начинающийся от полос-
ти капсулы извитой частью, которая затем переходит в прямую часть ка-
нальца. Клетки проксимального отдела на апикальной мембране имеют
щеточную каемку из микроворсин, покрытых гликокаликсом. Прокси-
мальный отдел расположен в корковом веществе, где переходит в петлю
Генле.
Тонкий нисходящий отдел петли Генле, который покрыт плоскими клет-
ками с щелевидными пространствами в цитоплазме шириной до 7 нм,
14. Выделение. Функции почек. Водно-солевой обмен • 611
Рис. 14.2. Типы и структура нефронов.
1 — клубочек интракортикального нефрона; 2 — клубочек юкстамедуллярного нефрона; 3 —
петля Генле интракортикального нефрона; 4 — петля Генле юкстамедуллярного нефрона; 5 —
проксимальные извитые канальцы; 6 —дистальные извитые канальцы; 7 — собирательные
трубочки; 8 — капиллярная сеть интракортикального нефрона; 9 — прямые капиллярные сосу-
ды юкстамедуллярного нефрона; 10 — артерии и артериолы; 11 — венулы и вены.
Интракортикальные нефроны имеют короткие петли Генле, выносящая артериола клубочка
образует густую капиллярную сеть вокруг канальцев. Юкстамедуллярные нефроны имеют
длинные петли Генле, спускающиеся вглубь мозгового вещества к почечному сосочку и обра-
зующие канальцевую противоточную систему почки, а выносящие артериолы клубочка фор-
мируют в мозговом веществе почки прямые нисходящие и восходящие капиллярные сосуды,
образующие сосудистую противоточную систему.
спускающийся в мозговое вещество почки, где поворачивает на 180° и пе-
реходит в тонкую восходящую часть, соединяющуюся с дистальным отде-
лом канальцев.
Дистальный отдел канальцев, состоящий из толстой восходящей части
петли Генле или прямого отдела и извитой части. Восходящая часть по-
крыта клетками, напоминающими клетки проксимального отдела, но ли-
шенными щеточной каемки. Восходящая часть дистального канальца
вновь входит в кору почки, подходит к клубочку и обязательно соприкаса-
ется с его полюсом между приносящей и выносящей артериолами. Здесь
эпителий канальца становится цилиндрическим, ядра клеток — гипер-
хромными, этот участок выглядит темным, плотным, что и дало ему назва-
ние macula densa —• плотное пятно. Поскольку непрерывная базальная
мембрана здесь отсутствует и клетки эпителия канальца имеют тесный
20-
612 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
— Отрицательно заряженные молекулы
эндотелия
Рис. 14.3. Схема строения клубочка.
А — схематическое изображение клубочка в целом, Б — фрагмент трехслойного фильтрацион-
ного барьера, В — увеличенный участок фильтрационного барьера. Отчетливо выявляются три
слоя барьера: эндотелий капилляра клубочка, базальная мембрана и клетки висцерального ли-
стка капсулы Боумена—Шумлянского (подоциты). Фильтрация воды с растворенными в ней
веществами происходит из плазмы крови капилляра клубочка через фенестры эндотелия, по-
ры базальной мембраны и щелевые диафрагмы между ножками подоцитов. Все эти структуры
фильтрационного барьера имеют отрицательный заряд.
14. Выделение. Функции почек. Водно-солевой обмен • 613
Рис. 14.4. Схема строения почечного канальца.
Из просвета канальца через люминальную мембрану клеток эпителия канальца происходит
процесс реабсорбции воды и веществ, содержащихся в первичной моче. Поступившие в клет-
ки вещества далее транспортируются через базолатеральные мембраны. Выведение через базо-
латеральные мембраны натрия в интерстициальную жидкость способствует повышению ее ос-
мотического давления и обеспечивает осмотический транспорт воды. Из интерстициальной
жидкости вода и вещества всасываются в кровь перитубулярных капилляров. Клетки эпителия
канальцев обладают также способностью захватывать вещества из кровеносных перитубуляр-
ных капилляров и затем секретировать их в просвет канальца через люминальные мембраны.
контакт с гранулированными миоэпителиоидными клетками артериолы
клубочка (юкстагломерулярными клетками), плотное пятно является
структурой юкстагломерулярного аппарата почки. Дистальные извитые ка-
нальцы через короткий связующий отдел впадают в коре почек в следую-
щий отдел нефрона — собирательные трубки.
Собирательные трубки спускаются из коры почек вглубь мозгового ве-
щества, где их эпителий из кубического становится цилиндрическим. Тем-
ные цилиндрические эпителиальные клетки дистальных отделов собира-
тельных трубок богаты карбоангидразой и обеспечивают секрецию ионов
водорода. В глубине мозгового вещества в области вершин пирамид соби-
рательные трубки сливаются в выводные протоки, открывающиеся в по-
лость лоханки.
По особенностям локализации клубочков в коре почек, строения ка-
нальцев и особенностям кровоснабжения различают три типа нефронов:
суперфициалъные, интракортикальные и юкстамедуллярные (см. рис. 14.2),
Суперфициальные нефроны имеют поверхностно расположенные в ко-
ре клубочки, наиболее короткую петлю Генле, их 20—30 %. Интракорти-
кальные нефроны, клубочки которых расположены в средней части коры
почки, наиболее многочисленны (60—70 %) и выполняют основную роль в
процессах ультрафильтрации мочи. Диаметр их приносящей артериолы
614 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
больше, чем у выносящей, ветви последней дают густую сеть капилляров в
корковом и мозговом веществе. Юкстамедуллярных нефронов значительно
меньше (10—15 %), клубочки их расположены у границы коркового и моз-
гового вещества почки, выносящие артериолы шире приносящих, петли
Генле самые длинные и спускаются почти до вершины сосочка пирамид.
Выносящие артериолы образуют прямые капиллярные нисходящие и вос-
ходящие сосуды, идущие в глубину мозгового вещества параллельно пет-
лям Генле. Юкстамедуллярные нефроны играют ведущую роль в процессах
концентрирования и разведения мочи.
Механизм мочеобразования складывается из трех основных процессов:
• клубочковой ультрафильтрации из плазмы крови воды и низкомоле-
кулярных компонентов с образованием первичной мочи;
• канальцевой реабсорбции (обратного всасывания в кровь) воды и не-
обходимых для организма веществ из первичной мочи;
• канальцевой секреции ионов, органических веществ эндогенной и
экзогенной природы.
14.2.1.1. Клубочковая ультрафильтрация и ее регуляция
Процесс клубочковой ультрафильтрации (далее просто фильтрация) осу-
ществляется под влиянием физико-химических и биологических факторов
через структуры гломерулярного фильтра, находящегося на пути выхода
жидкости из просвета капилляров клубочка в полость капсулы Боумена—
Шумлянского. Гломерулярный фильтр состоит из 3 слоев: эндотелия капил-
ляров, базальной мембраны и эпителия висцерального листка капсулы или
подоцитов (см. рис. 14.3). Эндотелий капилляров пронизан отверстиями
диаметром до 100 нм. На поверхности эндотелия находится особая выстил-
ка отрицательно заряженными молекулами гликопротеинов, мешающая
доступу форменных элементов и крупных молекул, в том числе и белков, к
лежащей под эндотелием базальной мембране. Базальная мембрана являет-
ся основной частью фильтра, препятствующей проникновению из плазмы
крови крупномолекулярных соединений (белков). При этом не только раз-
мер пор мембраны (около 2,9 нм), но и их отрицательный заряд противо-
действуют прохождению молекул с отрицательным зарядом, например аль-
буминов. Базальная мембрана довольно быстро «изнашивается» за счет не-
прерывного процесса фильтрации, и ее элементы постоянно восстанавли-
ваются с помощью мезангиальных клеток, при этом в течение года проис-
ходит полная замена ее основного вещества. Третий слой фильтра образо-
ван отростками подоцитов, между которыми остаются щелевые диафрагмы
с диаметром пор около 10 нм, поры покрыты гликокаликсом, оставляю-
щим отверстия радиусом около 3 нм. Эта часть фильтра также несет отри-
цательный заряд.
Поскольку подоциты содержат внутри отростков — педикул актомиози-
новые миофибриллы, они могут сокращаться и расслабляться, действуя
как микронасосы, откачивающие фильтрат в полость капсулы. Эта актив-
ность подоцитов составляет один из биологических факторов обеспечения
процесса фильтрации, к числу которых относится также сокращение и
расслабление мезангиальных клеток, изменяющих тем самым площадь по-
верхности клубочкового фильтра.
Физико-химические факторы обеспечения фильтрации представлены от-
рицательным зарядом структур фильтра и фильтрационным давлением, яв-
ляющимся основной причиной фильтрационного процесса.
Фильтрационное давление — это сила, обеспечивающая движение жидко-
14. Выделение. Функции почек. Водно-солевой обмен • 615
Приносящая Капилляры Выносящая
артериола клубочка артериола
Норма
Сужение
выносящей
артериолы
| Рг
Сужение
приносящей
артериолы
Рис. 14.5. Зависимость величины гидростатического давления в капиллярах клубоч-
ка (Рг) от соотношения просветов приносящей и выносящей артериол.
При сужении выносящей артериолы гидростатическое давление растет и скорость клубочко-
вой фильтрации (СКФ) повышается, а при сужении приносящей артериолы гидростатическое
давление и СКФ падают.
сти с растворенными в ней веществами из плазмы крови капилляров клу-
бочка в просвет капсулы. Эта сила создается гидростатическим давлением
крови в капилляре клубочка. Препятствующими фильтрации силами явля-
ются онкотическое давление белков плазмы крови (так как белки почти не
проходят через фильтр) и давление жидкости (первичной мочи) в полости
капсулы клубочка. Таким образом, фильтрационное давление (ФД) пред-
ставляет собой разность между гидростатическим давлением крови в ка-
пиллярах (Рг) и суммой онкотического давления плазмы крови (Ро) и дав-
ления первичной мочи (Рм) в капсуле: ФД = Рг — (Ро + Рм). По ходу ка-
пилляров клубочка от приносящего к выносящему отделу гидростатиче-
ское давление снижается за счет сосудистого сопротивления, а онкотиче-
ское давление плазмы, благодаря потере фильтрующейся воды и сгуще-
нию, возрастает. Гидростатическое давление крови в приносящей части
капилляров клубочка высокое, примерно 50—60 мм рт. ст., т. е. выше, чем
в капиллярах других тканей. Это связано, во-первых, с тем, что капилляры
клубочка находятся близко к аорте (короткие почечные и внутрипочечные
артерии), и, во-вторых,—диаметр приносящих артериол клубочка больше,
чем у выносящих. Гидростатическое давление в выносящей части капилля-
ров ниже на 2—5 мм рт. ст. Гидростатическое давление увеличивается или
снижается при изменении соотношения диаметров приносящей и вынося-
щей артериол, что является ведущим механизмом регуляции процесса
фильтрации (рис. 14.5). Онкотическое давление белков плазмы крови в
приносящей части капилляров клубочка около 25 мм рт. ст., а в вынося-
щей части капилляров, благодаря фильтрации из плазмы воды, оно возрас-
тает до 35—40 мм рт.ст. Давление первичной мочи в капсуле Боумена—
Шумлянского примерно равно 15—20 мм рт. ст. Таким образом, ФД в
1
616 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
приносящей части капилляров клубочка составляет в среднем: 60 — (25 +
15) = 20 мм рт. ст. В выносящей части капилляров фильтрации практиче-
ски не происходит, так как ФД равно: 58 — (40 + 15) = 3 мм рт. ст.
Основной количественной характеристикой процесса фильтрации явля-
ется скорость клубочковой фильтрации (СКФ). СКФ —это объем ультра-
фильтрата или первичной мочи, образующийся в почках за единицу време-
ни. Эта величина зависит от нескольких факторов: 1) от объема крови,
точнее плазмы, проходящей через корковое вещество почек в единицу вре-
мени, т. е. почечного плазмотока, составляющего в среднем у здорового
человека массой 70 кг около 600 мл/мин; 2) фильтрационного давления,
обеспечивающего сам процесс фильтрации; 3) фильтрационной поверхно-
сти, которая равна примерно 2—3 % от общей поверхности капилляров
клубочка (1,6 м2) и может изменяться при сокращении подоцитов и мезан-
гиальных клеток; 4) массы действующих нефронов, т. е. числа клубочков,
осуществляющих процесс фильтрации в определенный промежуток вре-
мени.
В физиологических условиях СКФ поддерживается на довольно посто-
янном уровне (несмотря на изменения системного артериального давле-
ния) за счет механизмов ауторегуляции. К их числу относятся: 1) миоген-
ная ауторегуляция тонуса приносящих артериол по принципу феномена
Остроумова—Бейлиса; 2) канальцево-клубочковая обратная связь, приво-
дящая к изменению соотношения тонуса приносящих и выносящих арте-
риол клубочка. Вовлечение в регуляцию механизма обратной связи обу-
словлено изменением доставки с фильтратом в область плотного пятна
(macula densa) ионов натрия и хлора, что ведет к изменению продукции в
ЮГА гуморальных регуляторов: аденозина (суживающий афферентные ар-
териолы фактор), NO (дилатирующий артериолы фактор), ренина и ангио-
тензина-П, кининов и простагландинов (рис. 14.6); 3) изменения числа
функционирующих нефронов. Первые два механизма поддерживают по-
стоянство кровотока в клубочках и фильтрационное давление, гумораль-
ные регуляторы могут менять площадь фильтрационной поверхности и
функции подоцитов, третий механизм определяет конечный эффект ауто-
регуляции СКФ в органе, что в итоге обеспечивает постоянство объема об-
разуемой первичной мочи.
СКФ определяют в результате сопоставления концентрации определен-
ного вещества в плазме крови и моче. При этом используемое вещество
должно выделяться вместе с водой только путем фильтрации и не всасы-
ваться в нефроне обратно в кровь. Таким условиям больше всего соответ-
ствует полисахарид фруктозы инулин. Исходя из концентрации инулина в
плазме [Пин], и, определив его концентрацию в определенном объеме (V)
конечной мочи [Мин], рассчитывают, какой объем первичной мочи соот-
ветствует найденной концентрации инулина. Насколько выросла концен-
трация инулина в конечной моче по сравнению с его концентрацией в
плазме, во столько раз больше объем профильтровавшейся плазмы (т. е.
первичной мочи) объема конечной мочи. Этот показатель получил назва-
ние «клиренса» инулина или коэффициента очищения и рассчитывается
по формуле:
Син = Мин X Умечи = скф (мл/мин)
Пин
показывающей, в каком объеме плазмы крови в единицу времени выдели-
лось в мочу найденное количество инулина или какой объем плазмы «очи-
стился» от инулина. По мере прохождения мочи по канальцам вода всасы-
14. Выделение. Функции почек. Водно-солевой обмен • 617
Увеличение гидростатического
давления в капиллярах — -
У
Увеличение сокрости
клубочковой фильтрации
Т
Увеличение объема
первичной мочи и скорости
ее движения по канальцам _ —
Клубочек Macula densa
Снижение сокрости
клубочковой фильтрации
Сужение приносящей
артериолы
Снижение гидростатического
давления в капиллярах
'Увеличение синтеза и
секреции в кровь аденозина
У
Увеличение загрузки канальцев
натрием и хлоридом________
У
Увеличение реабсорбции
натрия и хлорида клетками
macula densa
Рис. 14.6. Канальцево-клубочковая обратная связь как механизм ауторегуляции
клубочковой фильтрации.
Сплошные стрелки указывают последовательность реакций, штриховые стрелки — участки
нефрона, где происходит соответствующий процесс. В качестве примера приведен случай пер-
вичного повышения гидростатического давления в капиллярах клубочка при расширении при-
носящей артериолы. В результате повышенного гидростатического давления растут скорость
клубочковой фильтрации, объем образующейся первичной мочи и скорость ее продвижения
по канальцам. При повышенной скорости тока мочи в проксимальных канальцах не успевает
реабсорбироваться должная часть профильтровавшихся натрия и хлорида, соответственно,
увеличивается загрузка дистальных отделов канальцев натрием и хлоридом и их реабсорбция
клетками macula densa. Последние увеличивают синтез и секрецию в кровь аденозина, побуж-
дают юкстагломерулярные клетки к активации синтеза и секреции ренина. Поступающий в
кровь аденозин, равно как и образующийся в ней под влиянием ренина ангиотензин, вызыва-
ют сужение приносящей артериолы клубочка, снижение гидростатического давления и восста-
новление скорости клубочковой фильтрации.
вается обратно в кровь и концентрация инулина растет, что и находят в
конечной моче.
Поскольку инулин в организме отсутствует, для определения СКФ его
необходимо капельно вводить в кровоток, создавая постоянную концен-
трацию. Это затрудняет исследование, поэтому в клинике обычно исполь-
зуют эндогенное вещество креатинин, концентрация которого в крови до-
вольно стабильна.
618 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 14.1. Основные гуморальные влияния на процессы мочеобразовання
Процесс Увеличивают Уменьшают
Клубочковая Простагландины — PGE2 и простациклин Норадреналин и адреналин
фильтрация Атриопептид Оксид азота Прогестерон Глюкокортикоиды Окситоцин Глюкагон Т-3 и Т-4 Паратирин Хорионический гонадотропин Допамин Брадикинин Ангиотензин-П Аденозин Вазопрессин Тромбоксан А2 Лейкотриены Эндотелии
Канальцевая Вазопрессин Простагландины
реабсорбция Пролактин Атриопептид
воды Ангиотензин-П Инсулин Эстрогены Хорионический гонадотропин Кинины Паратирин Кальцитриол Т-3 и Т-4 Эпифизарный экстракт
Сравнивая клиренс инулина с клиренсом других веществ, определяют
процессы, участвующие в выделении этих веществ с мочой. Если клиренс
определенного вещества равен клиренсу инулина, значит вещество выде-
ляется почками только путем фильтрации в клубочках. Если клиренс ве-
щества больше клиренса инулина, следовательно, вещество выделяется не
только за счет фильтрации, но и секрецией эпителием канальцев. Если
клиренс вещества меньше, чем у инулина,— вещество после фильтрации
реабсорбируется в канальцах.
В норме СКФ составляет у мужчин около 125 мл/мин, а у женщин —
ПО мл/мин. В сутки образуется около 180 л первичной мочи, а за 25 мин
фильтруется примерно 3 л плазмы крови, т. е. весь циркулирующий ее
объем. За сутки этот объем плазмы крови фильтруется, т. е. очищается,
примерно 60 раз. Так как объем конечной мочи составляет около 1,5 л в
сутки, очевидно, что из объема первичной мочи за это время всасывается в
канальцах обратно в кровь примерно 178,5 л жидкости.
Поскольку первичная моча (клубочковый ультрафильтрат) образуется
из плазмы крови, по своему составу она близка плазме, почти полностью
лишенной белков. Так, в ультрафильтрате такое же, как в плазме крови,
количество аминокислот, глюкозы, мочевины, креатинина, свободных ио-
нов и низкомолекулярных комплексов. В связи с тем что белки-анионы не
проникают через клубочковый фильтр, для сохранения мембранного рав-
новесия Доннана (равенства произведений концентрации противоположно
заряженных ионов электролитов, находящихся по обе стороны мембраны)
в первичной моче оказывается на 5 % больше концентрация анионов хло-
ра и бикарбоната и пропорционально меньше концентрация катионов на-
трия и калия, меньше концентрация одновалентных катионов (натрия и
калия). В первичную мочу проходит небольшое количество наиболее мел-
ких молекул белка — менее 3 % гемоглобина и 0,01 % альбуминов.
Регуляция СКФ осуществляется за счет нервных и гуморальных меха-
низмов. Независимо от природы, регулирующие факторы влияют на СКФ
за счет изменения: 1) тонуса артериол клубочков и, соответственно, объем-
14. Выделение. Функции почек. Водно-солевой обмен • 619
ного кровотока (плазмотока) через них и величины фильтрационного дав-
ления; 2) тонуса мезангиальных клеток и фильтрационной поверхности;
3) активности подоцитов и их «отсасывающей» функции. Нервные влия-
ния реализуются вазомоторными ветвями почечных нервов, преимущест-
венно симпатической природы, обеспечивающими изменение соотноше-
ния тонуса приносящих и выносящих артериол клубочков. Кроме того,
симпатические влияния на юкстагломерулярные клетки через бета-адрено-
рецепторы стимулируют секрецию ренина и тем самым реализуют ангио-
тензинный механизм регуляции фильтрации (спазм выносящих и/или
приносящих артериол). Гуморальные факторы (табл. 14.1) могут как увели-
чивать, так и уменьшать клубочковую фильтрацию через три описанных
выше механизма, причем эффекты вазопрессина реализуются через Vrpe-
цепторы. Важнейшую роль в обеспечении постоянства СКФ играет мест-
ная ауторегуляция коркового кровотока в почке.
14.2.1.2. Канальцевая реабсорбция и ее регуляция
При сравнении состава и количества первичной и конечной мочи выяв-
ляется, что в канальцах нефрона происходит процесс обратного всасыва-
ния воды и веществ, профильтровавшихся в клубочках, что необходимо
для поддержания их внешнего баланса. Этот процесс называется каналь-
цевой реабсорбцией и в зависимости от отдела канальцев, где он происхо-
дит, различают реабсорбцию проксимальную и дистальную. В процессе ре-
абсорбции вода и вещества из просвета канальцев через люминальную
мембрану поступают в цитоплазму клеток эпителия, затем через базолате-
ральную мембрану выносятся из клеток эпителия в интерстициальное
пространство, после чего поступают в перитубулярные (околоканальце-
вые) капилляры. Такой путь реабсорбции носит название трансцеллюляр-
ного, в его основе лежат общие механизмы транспорта веществ через
плазматические мембраны. Кроме того, возможен путь реабсорбции че-
рез плотные соединения между клетками эпителия посредством простой
диффузии или переносом вещества вместе с растворителем, что носит на-
звание парацеллюлярного пути реабсорбции. Реабсорбция представляет со-
бой транспорт веществ из мочи в лимфу и кровь, и в зависимости от ме-
ханизма выделяют пассивный, первично и вторично активный транспорт
(см. главу 2).
Проксимальная реабсорбция обеспечивает полное всасывание ряда ве-
ществ первичной мочи — глюкозы, белка, аминокислот и витаминов.
В проксимальном отделе канальцев всасывается 2/3 профильтровавшихся
воды и ионов натрия (рис. 14.7), большие количества ионов калия, двухва-
лентных катионов, анионов хлора, бикарбоната, фосфата, а также мочевая
кислота и мочевина. К концу проксимального отдела в его просвете оста-
ется только 73 объема ультрафильтрата, и, хотя его состав из-за неодина-
ковой реабсорбции разных компонентов уже существенно отличается от
плазмы крови, осмотическое давление первичной мочи остается таким же,
как в плазме.
Эпителий проксимального канальца хорошо проницаем для воды, бла-
годаря наличию в апикальной мембране водных каналов, образованных
особыми белковыми молекулами аквапоринами. В структурах нефрона опи-
сано 6 типов аквапоринов, первый из них (AQP-1) имеется в мембранах
клеток проксимальных канальцев (рис. 14.8). Всасывание воды происходит
пассивно путем простой диффузии по осмотическому градиенту и прямо
зависит от реабсорбции ионов натрия хлорида, других осмотически актив-
620 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
С КФ = 180 л/день
[Na] плазмы = 140 ммоль/л
Фильтрационная загрузка Na+ = 25 200 ммоль/день
16 800 ммоль/день
67 % фильтр, загрузки
6300 ммоль/день
25 % ф. з.
1200 ммоль/день
5 % ф. з.
750 ммоль/день
3 % ф. з. ~*~
V мочи = 1500 мл/день
[Na] мочи = 100 ммоль/л
Поступление Na+ в организм = 155 ммоль/день
Экскреция Na = 150 ммоль/день
0,6 % ф. з.
Рис. 14.7. Почечные процессы, обеспечивающие поддержание баланса натрия.
Прямые стрелки и цифры — реабсорбция и экскреция натрия в разных отделах нефрона,
СКФ — скорость клубочковой фильтрации. При поступлени в сутки 155 ммоль натрия в орга-
низм, фильтруется в мочу за сутки 25 200 ммоль, из которых реабсорбируется 25 050 ммоль, в
Том числе 2/3 (67 %) в проксимальном отделе канальцев, а ‘/3 (33 %) — в дистальном отделе.
В результате за сутки с мочой выделяется 150 ммоль натрия. Оставшиеся 5 ммоль от поступив-
шего количества натрия экскретируются из организма с калом и потом.
ных веществ. Благодаря этому содержимое проксимального отдела остает-
ся изоосмотичным плазме крови.
Реабсорбция ионов натрия в проксимальном отделе осуществляется не-
сколькими механизмами активного и пассивного транспорта (рис. 14.9).
Во-первых, реабсорбция натрия осуществляется первично активным транс-
портом. Ионы натрия входит в клетки эпителия через апикальную мембра-
ну пассивно через натриевые каналы по концентрационному градиенту,
его выведение через базолатеральные мембраны эпителиальных клеток
происходит активно с помощью натрий-калиевых насосов, использующих
энергию АТФ. Именно деятельность этих насосов обеспечивает градиент
концентрации ионов натрия между внутриканальцевой и внутриклеточной
средами. Во-вторых, на апикальной мембране имеется электронейтраль-
ный переносчик, обеспечивающий активный обмен Na+ и Н+, при этом
ион натрия поступает в клетку в обмен на удаляемый из клетки Н-ион. Та-
кой механизм транспорта носит название антипорта. Этот переносчик
обеспечивает и всасывание бикарбонатного аниона. Профильтровавшийся
бикарбонатный анион вместе с Н-ионом образуют угольную кислоту:
HCOj + Н+ = Н2СО3. Располагающаяся на щеточной каемке эпителия ка-
нальца карбоангидраза катализирует разложение в канальцевой жидкости
угольной кислоты: Н2СО3 <-> Н2О + СО2, после чего СО2 диффундирует в
клетку по градиенту концентрации. В клетке под влинием цитоплазменной
14. Выделение. Функции почек. Водно-солевой обмен • 621
Дистальный извитой каналец
Проксимальный
каналец
Нисходящая
тонкая часть
петли Генле
Клубочек
AQP1
Вода
Вода
Вода
Собирательная трубка
AQP1
AQP1
Восходящая тонкая
часть петли Генле
Вода
AQP2
AQP4—► Вода
Вазопрессин
AQP3—► Вода
Вода
AQP2
Вазопрессин
Собирательная
трубка
AQP3 _
AQP4 Вода
Кора
Внешняя
медулла
со
ф
о.
£
X
со
5
s
(X
Конечная моча
Рис. 14.8. Локализация в нефроне и участие в транспорте воды через эпителий по-
чечных канальцев аквапоринов (водных каналов) разных типов (AQP 1—4).
AQP1 — водные каналы, постоянно локализованные в проксимальных извитых канальцах и
нисходящем отделе петли Генле, обеспечивают трансэпителиальный транспорт воды из про-
света канальцев, т. е. проксимальную реабсорбцию воды.
AQP2 — вазопрессинозависимый тип аквапоринов. Эти водные каналы встраиваются в люми-
нальную мембрану главных клеток собирательных трубочек только при наличии вазопрессина,
обеспечивая зависимую от вазопрессина дистальную реабсорбцию воды и концентрирование
мочи.
AQP3 и AQP4— водные каналы, постоянно локализованные в базолатеральных мембранах
главных клеток эпителия собирательных трубочек, обеспечивают транспорт воды из эпители-
альных клеток в интерстиций мозгового вещества.
карбоангидразы протекает обратная реакция: СО2 + Н2О <-> Н2СО3, уголь-
ная кислота диссоциирует: Н2СО3 <-> Н+ + HCOj. Бикарбонатный анион
(HCOJ) пассивно переносится в перитубулярную жидкость по электрохи-
мическому градиенту, создаваемому активным переносом натрия через ба-
золатеральную мембрану, а Н-ион через апикальную мембрану с помощью
антипорта Na+-H+ выводится в просвет канальца. Таким образом, сопрово-
ждающим всасывающийся ион натрия в начальных отделах проксимально-
го канальца анионом является бикарбонат. Анионы хлора всасываются в
начальных отделах плохо из-за низкой проницаемости стенки. Объем мочи
в канальце уменьшается из-за пассивной реабсорбции воды, и концентра-
ция хлоридов в его содержимом растет. В конечных участках проксималь-
ных канальцев межклеточные контакты уже проницаемы для хлоридов
(концентрация которых повысилась) и они пассивно по градиенту концен-
трации всасываются из мочи путем парацеллюлярной диффузии, создавая
электрохимический градиент для натрия. В-третьих, ион натрия реабсор-
622 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Просвет канальца
Аминокислоты
Na
Na
Глюкоза
Na*, Cl-
Люминальная
мембрана
Na
Na
Н
Na
НРО;2
Интерстиций
Аминокислоты,
глюкоза
НРО;2
Базолатеральная
мембрана
Рис. 14.9. Основные системы транспорта натрия в проксимальном канальце неф-
рона.
Жирной стрелкой выделено поступление натрия в клетку через натриевый канал по концен-
трационному градиенту (пассивный транпорт путем простой диффузии). На люминальной
мембране черными кружками обозначены переносчики, осуществляющие вторично активный
котранспорт путем облегченной диффузии (Na* и глюкоза, Na+ и аминокислоты), либо пас-
сивный котранспорт (Na+ и фосфат), либо антипорт (Na+ и Н+). На базолатеральной мембране
расположены насосы, обеспечивающие активный транспорт натрия из клетки (Na+-K+-Hacoc).
Черный квадрат — основное вещество плотного межклеточного соединения, при деполимери-
зации которого возможен пассивный межклеточный транспорт Na.
бируется пассивно, по электрохимическому градиенту, вслед за анионом
хлора. Такой пассивный транспорт одного иона (натрия) вместе с пассив-
ным транспортом другого (хлорида) носит название копгранспорта. В-чет-
вертых, на апикальной мембране расположены переносчики-котранспор-
теры натрия и органических веществ (глюкозы, аминокислот), натрия и
фосфата или сульфата.
Проксимальная реабсорбция глюкозы и аминокислот осуществляется с
помощью специальных переносчиков щеточной каемки апикальной мем-
браны эпителиальных клеток. Эти переносчики транспортируют глюкозу
или аминокислоту, только если одновременно связывают и переносят на-
трий. Пассивное перемещение натрия по градиенту концентрации внутрь
клеток ведет к транспорту через мембрану и переносчика с глюкозой или
аминокислотой. Для реализации этого процесса необходима низкая кон-
центрация натрия в эпителиальной клетке, создающая градиент концен-
14. Выделение. Функции почек. Водно-солевой обмен • 623
трации между внешней и внутриклеточной средой, что обеспечивается
энергозависимой работой натрий-калиевого насоса базальной мембраны.
Поскольку перенос глюкозы или аминокислоты связан с натрием, а его
транспорт определяется активным удалением натрия из клетки, такой вид
транспорта называют вторично активным, или симпортом, т. е. совместным
пассивным транспортом одного вещества (глюкоза) из-за активного транс-
порта другого (натрия) с помощью одного переносчика.
Ввиду того что для реабсорбции глюкозы необходимо связывание каж-
дой ее молекулы с молекулой переносчика, при избытке глюкозы в пер-
вичной моче может произойти полная загрузка всех молекул переносчиков
и глюкоза уже не сможет всасываться в кровь. Эта ситуация характеризует-
ся понятием «максимальный канальцевый транспорт вещества» (Тм глюко-
зы), которое отражает максимальную загрузку канальцевых переносчиков
при определенной концентрации вещества в первичной моче и, соответст-
венно, в крови. Эта величина составляет от 303 мг/мин у женщин до 375
мг/мин у мужчин. Величине максимального канальцевого транспорта со-
ответствует понятие «почечный порог выведения».
Почечным порогом выведения называют ту концентрацию вещества в кро-
ви и, соответственно, в первичной моче, при которой оно уже не может
быть полностью реабсорбировано в канальцах и появляется в конечной
моче. Такие вещества, для которых может быть найден порог выведения,
т. е. реабсорбирующиеся при низких концентрациях в крови полностью, а
при повышенных концентрациях — не полностью, носят название порого-
вых. Примером является глюкоза, которая полностью всасывается из пер-
вичной мочи при концентрациях в плазме крови ниже 10 ммоль/л, но по-
является в конечной моче, т. е. полностью не реабсорбируется, при содер-
жании ее в плазме крови выше 10 ммоль/л. Следовательно, для глюкозы
порог выведения составляет 10 ммоль/л.
Вещества, которые вообще не реабсорбируются в канальцах (инулин,
маннитол) или мало реабсорбируются и выделяются пропорционально на-
коплению в крови (мочевина, сульфаты и др.), называются непороговыми,
так как для них порога выведения не существует.
Переносчики апикальной мембраны для симпорта натрия и аминокис-
лот лишь относительно специфичны, каждый из них способен транспор-
тировать несколько видов аминокислот. Так, для глутамата и аспартата су-
ществует один вид переносчика, для аргинина, лизина — другой.
Реабсорбция пептидов и белков осуществляется практически полностью в
проксимальных канальцах. Количество профильтровавшегося белка отно-
сительно невелико и составляет около 1,8 г в сутки. Некоторую его часть
составляют альбумины, но фильтрационный барьер клубочков проходят и
полипептиды меньшего размера, и в первичную мочу фильтруются, напри-
мер, соматотропин, а также лизоцим и др. В конечную мочу поступает не
более 0,15 г белка в сутки (рис. 14.10). Молекулы альбуминов после связы-
вания с рецепторами на люминальной мембране клеток эпителия каналь-
цев подвергаются эндоцитозу, внутриклеточные пузырьки поглощенного
белка сливаются с лизосомами и белковые молекулы гидролизуются пеп-
тидазами (аналог внутриклеточного пищеварения). Продукты гидролиза, в
основном аминокислоты, выводятся в интерстициальную жидкость и по-
ступают в перитубулярные капилляры. Пептиды, особенно с короткой це-
пью, подвергаются гидролизу ферментами щеточной каемки (аналог мем-
бранного пищеварения), образующиеся аминокислоты реабсорбируются из
просвета канальцев.
Дистальная реабсорбция ионов и воды по объему значительно меньше
624 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Альбумины, лизоцим,
Основные белки мочи уромукоид и др.
Экскреция белка (г/сут)
< 0,15
Рис. 14.10. Фильтрация белка и его реабсорбция в проксимальном отделе каналь-
цев.
Гломерулярный фильтр пропускает в первичную мочу лишь мелкие молекулы белков и пепти-
дов. В проксимальных канальцах эти молекулы поглощаются клетками эпителия и гидролизу-
ются. Аминокислоты и мелкие пептиды реабсорбируются далее в кровь. С конечной мочой из
организма выделяется не более 0,15г белка в сутки.
проксимальной. Однако, существенно меняясь под влиянием регулирую-
щих ее, преимущественно гормональных, воздействий, она определяет со-
став конечной мочи и способность почки выделять либо концентрирован-
ную, либо разведенную мочу (в зависимости от водного баланса организ-
ма). В дистальном отделе нефрона происходит активная реабсорбция ио-
нов натрия. Хотя здесь всасывается всего 10 % от профильтровавшегося
количества катиона, этот процесс обеспечивает выраженное уменьшение
его концентрации в моче и, напротив, повышение концентрации в интер-
стициальной жидкости, что создает значительный градиент осмотического
давления между мочой и интерстицием, необходимый для всасывания из
14. Выделение. Функции почек. Водно-солевой обмен • 625
мочи воды. Анион хлора всасывается в толстой восходящей части петли
Генле благодаря наличию в апикальной мембране клеток эпителия пере-
носчика-котранспортера ионов хлора, натрия и калия, а в дистальных из-
витых канальцах и собирательных трубочках анион хлора реабсорбируется
пассивно по электрохимическому градиенту вслед за активным транспор-
том натрия. Способность эпителия дистальных канальцев секретировать в
мочу Н-ионы связана с реабсорбцией ионов натрия, этот вид транспорта в
виде обмена натрия на протон получил название «антипорт». Активно вса-
сывается в дистальном отделе канальцев ионы калия, кальция и фосфатов.
Стенка дистального извитого канальца из-за отсутствия аквапоринов имеет
низкую проницаемость для воды и, несмотря на реабсорбцию здесь ионов
натрия и хлорида, вода почти не всасывается и в собирательные трубки
поступает гипотоническая моча.
Способность почки образовывать концентрированную или разведенную
мочу обеспечивается деятельностью противоточно-множительной канальце-
вой системы почки, которая представлена параллельно расположенными
коленами петли Генле и собирательными трубочками (рис. 14.11). Моча
двигается в этих канальцах в противоположных направлениях (почему сис-
тема и названа противоточной), а процессы транспорта веществ в одном
колене системы усиливаются («умножаются») за счет деятельности другого
колена. Определяющую роль в работе противоточного механизма играет
восходящее колено петли Генле. Его эпителий активно реабсорбирует в
окружающее интерстициальное пространство ионы натрия, поддерживая
гиперосмолярность интерстициальной жидкости. Транспорт натрия сопря-
жен с транспортом калия и хлора (Na, К, 2С1-котранспорт) и антипортом с
ионами водорода, осуществляется посредством соответствующих перенос-
чиков. Проницаемость для воды здесь очень низкая из-за отсутствия аква-
поринов. В результате интерстициальная жидкость становится гиперосмо-
тичной по отношению к содержимому нисходящего колена петли и по на-
правлению к вершине петли осмотическое давление в окружающей ткани
растет. Клетки эпителия нисходящего колена, содержащие в апикальной
мембране аквапорины 1-го типа (см. рис. 14.8), проницаемы для воды, ко-
торая пассивно уходит из просвета в гиперосмотический интерстиций. Та-
ким образом, в нисходящем колене моча из-за всасывания воды становит-
ся все более и более гиперосмотичной, т. е. устанавливается осмотическое
равновесие с интерстициальной жидкостью. В восходящем колене из-за
всасывания натрия моча становится все менее осмотичной и в корковый
отдел дистального канальца восходит уже гипотоничная моча. Однако ее
количество из-за всасывания воды и солей в петле Генле существенно
уменьшилось.
Собирательная трубочка, в которую затем поступает моча, тоже образу-
ет с восходящим коленом петли Генле противоточную систему. Если в со-
бирательных трубочках вода всасывается мало, а натрий, хлориды, мочеви-
на продолжают реабсорбироваться, то нефрон выделяет большие количест-
ва гипотоничной слабо концентрированной мочи. Этот процесс носит на-
звание разведения мочи. Он реализуется при необходимости выделения
избытка воды из внутренней среды.
Стенка собирательной трубочки становится проницаемой для воды
только в присутствии вазопрессина. Этот эффект вазопрессина обусловлен
тем, что гормон вазопрессин обеспечивает встраивание в апикальную мем-
брану эпителия собирательных клеток молекул аквапорина 2-го типа (см.
рис. 14.8), образующих водные каналы, а в отсутствии вазопрессина аква-
порины-2 путем эндоцитоза поглощаются внутрь клеток, что носит назва-
626 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 14.11. Противоточно-множительная канальцевая система мозгового вещества
почки.
Цифрами обозначены величины осмотического давления интерстициальной жидкости и мочи.
В собирательных трубочках цифрами в скобках обозначено осмотическое давление мочи в от-
сутствие вазопрессина (разведение мочи), а цифрами без скобок — осмотическое давление мо-
чи в условиях действия вазопрессина (концентрирование мочи).
Первичная моча, образовавшаяся путем фильтрации плазмы крови в клубочке, поступает в
проксимальный извитой каналец с осмотическим давлением, изотоничным плазме крови.
Здесь реабсорбируются вода и соли и осмотическое давление остается без изменений. В нис-
ходящем колене петли Генле вода реабсорбируется в гиперосмотичный интерстиций и по мере
продвижения к вершине моча становится все более гипертоничной. Восходящее колено петли
Генле реабсорбирует в интерстиций ионы натрия, тем самым создавая в нем высокое осмоти-
ческое давление, а в моче, теряющей натрий, осмотическое давление снижается. Гипотонич-
ная моча поступает в извитые дистальные отделы и собирательные трубочки, где реабсорбиру-
ются натрий и, в присутствии вазопрессина, вода. Если действует вазопрессин и реасорбиру-
ется вода, осмотическое давление мочи растет, она концентрируется и ее выделяется мало. Ес-
ли вазопрессина нет, вода не всасывается, а за счет реабсорбции натрия осмотическое давле-
ние становится все ниже и ниже, происходит разведение мочи, ее выделяется много с низким
удельным весом.
ние интернализации водного канала. На базолатеральных мембранах кле-
ток собирательных трубочек постоянно встроены независимые от вазо-
прессина аквапорины 3-го и 4-го типов, свободно пропускающие воду в
интерстициальное пространство. По мере продвижения мочи по собира-
тельным трубочкам вглубь мозгового вещества вода пассивно уходит в ги-
перосмотичный интерстиций и моча становится все более концентриро-
ванной.
Под влиянием вазопрессина реализуется еще один важный для концен-
трирования мочи механизм: активация переносчика, усиливающая выход
мочевины из собирательных трубочек, расположенных в глубине мозгового
вещества, в окружающий интерстиций. Всасывание воды в верхних отде-
14. Выделение. Функции почек. Водно-солевой обмен 627
Рис. 14.12. Противоточная со-
судистая система мозгового ве-
щества почки.
Цифрами обозначены величины
осмотического давления плазмы
крови (в просвете капилляров) и
интерстициальной жидкости. Пас-
сивная диффузия воды (сплошные
стрелки) происходит по градиенту
осмотического давления, в резуль-
тате кровь в нисходящем колене
капилляра теряет воду, натрий и
мочевина пассивно по градиенту
концентрации диффундируют из
интерстиция в кровь и она стано-
вится более высоко осмотичной, а
в восходящем колене капилляра из
крови в интерстиций происходит
диффузия натрия и мочевины, а из
интерстиция в кровь — диффузия
воды. В результате к концу восхо-
дящего колена капилляра осмоти-
ческое давление крови восстанав-
ливается. Таким образом сосуди-
стая противоточная система сохра-
няет высокие концентрации на-
трия и мочевины в интерстиции и
его высокое осмотическое дав-
ление.
—► Пассивная диффузия воды
лах собирательных трубо- — Пассивная диффузия натрия и мочевины
чек ведет к нарастанию
концентрации мочевины в
моче, а в самых нижних их отделах, расположенных в глубине мозгового
вещества, вазопрессин повышает проницаемость для мочевины и она пас-
сивно диффундирует в интерстиций, резко повышая его осмотическое дав-
ление. Таким образом, интерстиций мозгового вещества становится наибо-
лее высоко осмотичным в области вершины почечных пирамид, где и про-
исходит увеличение всасывания воды из просвета канальцев в интерсти-
ций и концентрирование мочи.
Мочевина интерстициальной жидкости по концентрационному гради-
енту диффундирует в просвет тонкой восходящей части петли Генле и
вновь поступает с током мочи в дистальные канальцы и собирательные
трубочки. Так осуществляется кругооборот мочевины в канальцах, сохра-
няющий высокий уровень ее концентрации в мозговом веществе. Описан-
ные процессы транспорта мочевины осуществляются, в основном, в юк-
стамедуллярных нефронах, имеющих наиболее длинные петли Генле, спус-
кающиеся глубоко внутрь мозгового вещества почки.
В мозговом веществе почки имеется и другая — сосудистая противоточ-
ная система, образованная кровеносными капиллярами. Кровеносная сеть
юкстамедуллярных нефронов образует длинные параллельные прямые
нисходящие и восходящие капиллярные сосуды (рис. 14.12), спускающие-
ся вглубь мозгового вещества. Двигающаяся по нисходящему прямому ка-
пилляру кровь постепенно отдает воду в окружающее интерстициальное
пространство в силу нарастающего осмотического давления в ткани и, на-
против, обогащается натрием и мочевиной, сгущается и замедляет свое
движение. В восходящем капиллярном отделе сосуда по мере движения
628 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
крови в ткани с постепенно снижающимся осмотическим давлением про-
исходят обратные процессы — натрий и мочевина по концентрационному
градиенту диффундируют обратно в ткань, а вода всасывается в кровь (см.
рис. 14.12). Таким образом, сосудистая противоточная система не создает,
а лишь способствует поддержанию высокого осмотического давления в
глубоких слоях ткани мозгового вещества почек, обеспечивая удаление во-
ды и удержание натрия и мочевины в интерстиции.
Деятельность описанных противоточных систем находится в зависимо-
сти от скорости движения находящихся в них жидкостей (мочи или кро-
ви). Чем быстрее будет двигаться моча по трубкам противоточной системы
канальцев, тем меньшие количества натрия, мочевины и воды успеют ре-
абсорбироваться в интерстиций и большие количества менее концентриро-
ванной мочи будут выделяться почкой. Чем выше будет скорость кровото-
ка по прямым капиллярным сосудам мозгового вещества почки, тем боль-
ше натрия и мочевины унесет кровь из почечного интерстиция, так как
они не успеют диффундировать из крови обратно в ткань. Этот эффект на-
зывают «вымыванием» осмотически активных веществ из интерстиция, в
результате его осмолярность падает, концентрирование мочи уменьшается
и почкой выделяется больше мочи низкого удельного веса (разведение мо-
чи). Чем медленнее происходит движение мочи или крови в мозговом ве-
ществе почек, тем больше осмотически активных веществ накапливается в
интерстиции и выше способность почки концентрировать мочу.
Регуляция канальцевой реабсорбции осуществляется как нервным, так и, в
большей мере, гуморальным путем.
Нервные влияния реализуются преимущественно симпатическими про-
водниками и медиаторами через бета-адренорецепторы мембран клеток
проксимальных и дистальных канальцев. Симпатические эффекты прояв-
ляются в виде активации процессов реабсорбции глюкозы, ионов натрия,
воды и анионов фосфатов и осуществляются через систему вторичных по-
средников (аденилатциклаза — цАМФ). Нервная регуляция кровообраще-
ния в мозговом веществе почки увеличивает или уменьшает эффектив-
ность сосудистой противоточной системы и концентрирование мочи. Со-
судистые эффекты нервной регуляции также опосредуются через внутри-
почечные системы гуморальных регуляторов — ренин-ангиотензиновую,
кининовую, простагландины и др.
Основным фактором регуляции реабсорбции воды в дистальных отделах
нефрона является гормон вазопрессин, называвшийся ранее антидиурети-
ческим гормоном. Этот гормон образуется в супраоптическом и паравен-
трикулярных ядрах гипоталамуса, по аксонам нейронов транспортируется
в нейрогипофиз, откуда и поступает в кровь. Влияние вазопрессина на
проницаемость эпителия канальцев обусловлено наличием рецепторов к
гормону, относящихся к У2-типу, на поверхности базолатеральной мембра-
ны клеток эпителия. Образование гормон-рецепторного комплекса влечет
за собой через посредство GS-белка и гуанилового нуклеотида активацию
аденилатциклазы и образование цАМФ, активацию синтеза и встраивания
аквапоринов 2-го типа («водных каналов») в апикальную мембрану клеток
эпителия собирательных трубочек. Перестройка ультраструктур мембраны
и цитоплазмы клетки ведет к образованию внутриклеточных специализи-
рованных структур, переносящих большие потоки воды по осмотическому
градиенту от апикальной к базолатеральной мембране, не позволяя транс-
портируемой воде смешиваться с цитоплазмой и препятствуя набуханию
клетки. Такой трансцеллюлярный транспорт воды через клетки эпителия
реализуется вазопрессином в собирательных трубочках. Кроме того, в дис-
14. Выделение. Функции почек. Водно-солевой обмен • 629
тальных канальцах вазопрессин обусловливает активацию и выход из кле-
ток гиалуронидаз, вызывающих расщепление гликозаминогликанов основ-
ного межклеточного вещества, тем самым способствуя межклеточному
пассивному транспорту воды по осмотическому градиенту.
Канальцевая реабсорбция воды регулируется и другими гормонами
(табл. 14.1). По механизму действия все гормоны, регулирующие реабсорб-
цию воды, делятся на шесть групп:
• повышающие проницаемость мембран дистальных отделов нефрона
для воды (вазопрессин, пролактин, хорионический гонадотропин);
• меняющие чувствительность клеточных рецепторов к вазопрессину
(паратирин, кальцитонин, кальцитриол, простагландины, альдосте-
рон);
• меняющие осмотический градиент интерстиция мозгового слоя почки
и, соответственно, пассивный осмотический транспорт воды (парати-
рин, кальцитриол, тиреоидные гормоны, инсулин, вазопрессин);
• меняющие активный транспорт натрия и хлорида, а за счет этого и
пассивный транспорт воды (альдостерон, вазопрессин, атриопептид,
прогестерон, глюкагон, кальцитонин, простагландины);
• повышающие осмотическое давление канальцевой мочи за счет нере-
абсорбированных осмотически активных веществ, например глюкозы
(контринсулярные гормоны);
• меняющие кровоток по прямым сосудам мозгового вещества и, тем
самым, накопление или «вымывание» осмотически активных веществ
из интерстиция (ангиотензин-П, кинины, простагландины, парати-
рин, вазопрессин, атриопептид).
Канальцевая реабсорбция электролитов, так же как и воды, регулирует-
ся преимущественно гормональными, а не нервными влияниями.
Реабсорбция ионов натрия в проксимальных канальцах активируется
альдостероном и угнетается паратирином, в толстой части восходящего ко-
лена петли Генле реабсорбция натрия активируется вазопрессином, глюка-
гоном, кальцитонином, а угнетается — простагландинами Е. В дистальном
отделе канальцев главными регуляторами транспорта натрия являются аль-
достерон (активация), простагландины и атриопептид (угнетение). Основ-
ной активатор реабсорбции натрия — альдостерон — обеспечивает образо-
вание и активацию всех структур, необходимых для транспорта натрия: ком-
понентов натрий-калиевого насоса базолатеральной мембраны и ферментов
его энергетического обеспечения, структур натриевых каналов апикальной
мембраны и переносчиков иона. Наиболее отчетливо выражена активация
реабсорбции ионов натрия под влиянием альдостерона в корковом отделе
собирательных трубочек. Действие альдостерона имеет место не только в
почках, но также и в желудочно-кишечном тракте, железах внешней секре-
ции — везде этот гормон способствует всасыванию натрия в кровь. В почках
альдостерон стимулирует также секрецию ионов калия в мочу.
Регуляция канальцевого транспорта ионов кальция, фосфата и частично
магния обеспечивается, в основном, кальций-регулирующими гормонами.
Влияния паратирина отличаются в разных отделах канальцевого аппарата
почки. В проксимальных канальцах (прямой отдел) всасывание кальция
происходит параллельно с транспортом натрия и воды. Угнетение реаб-
сорбции ионов натрия в этом отделе под влиянием паратирина сопровож-
дается параллельным снижением реабсорбции ионов кальция. За предела-
ми проксимального канальца паратирин избирательно усиливает реабсорб-
цию кальция, особенно в дистальном извитом канальце и корковой части
собирательных трубочек. Реабсорбция ионов кальция активируется также
630 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
калыштриолом, а подавляется кальцитонином. Всасывание анионов фос-
фата в канальцах почки угнетается и паратирином (проксимальная реаб-
сорбция), и кальцитонином (дистальная реабсорбция), а усиливается каль-
цитриолом и соматотропином. Паратирин активирует реабсорбцию ионов
магния в корковой части восходящего колена петли Генле и тормозит про-
ксимальную реабсорбцию аниона бикарбоната.
14.2.1.3 . Канальцевая секреция и ее регуляция
Канальцевой секрецией называют активный транспорт в мочу веществ, со-
держащихся в крови или образуемых в самих клетках канальцевого эпите-
лия, например аммиака. Секреция осуществляется, как правило, против
концентрационного или электрохимического градиента с затратами энер-
гии. Путем канальцевой секреции из крови выделяются как ионы К+, Н+,
органические кислоты и основания эндогенного происхождения, так и по-
ступившие в организм чужеродные вещества, в том числе органического
происхождения. Для ряда чужеродных организму веществ органической
природы (антибиотиков, красителей и рентгеноконтрастных препаратов)
скорость и интенсивность выделения из крови путем канальцевой секре-
ции значительно превышает их выведение путем клубочковой фильтрации.
Таким образом канальцевая секреция является одним из механизмов обес-
печения гомеостазиса в организме.
Способностью к секреции обладают клетки эпителия проксимального и
дистальных отделов канальцев. При этом клетки проксимальных каналь-
цев секретируют органические соединения с помощью специальных пере-
носчиков: один из которых обеспечивает секрецию органических кислот
(парааминогиппуровой кислоты, диодраста, фенолрота, пенициллина
и др.), а другой — секрецию органических оснований (гуанидина, пипери-
дина, тиамина, холина, серотонина, хинина, морфина и т. п.). Молекулы
переносчиков, расположенные в базолатеральной мембране эпителиальных
клеток канальцев, обеспечивают поступление органических веществ из
околоканальцевой жидкости и крови в цитоплазму клеток. Перемещаясь
внутри клеток к апикальной мембране, секретируемые вещества затем про-
ходят через нее в просвет канальца с помощью облегченной диффузии. В
базальной мембране клеток имеются переносчики — котранспортеры, ис-
пользующие энергию движения натрия по градиенту концентрации.
Секреция водородных ионов происходит в проксимальных канальцах в
большей мере, чем в дистальных. Однако дистальная секреция водородных
ионов играет основную роль в регуляции кислотно-основного состояния
внутренней среды.
Секреция ионов калия происходит в дистальных канальцах и собира-
тельных трубочках. Поскольку почти весь профильтровавшийся калий ре-
абсорбируется в канальцах до дистальной извитой их части, то выделяе-
мый с мочой калий является результатом его секреции в дистальных изви-
тых канальцах и собирательных трубочках. Регуляция выделения ионов ка-
лия осуществляется альдостероном, усиливающим секрецию К+ и подав-
ляющим его реабсорбцию. Секреция аммиака, образующегося в самих
клетках эпителия, происходит в проксимальном и в дистальном отделах.
Регуляция канальцевой секреции осуществляется с помощью гормонов
и симпатической нервной системы. Эффекты нервной регуляции реализу-
ются за счет изменений кровотока в постгломерулярных капиллярах поч-
ки, т. е. транспорта веществ кровью к секретирующим клеткам, и влияния
на энергетический обмен в клетках эпителия канальцев. Гормонами, уси-
14. Выделение. Функции почек. Водно-солевой обмен • 631
ливающими проксимальную канальцевую секрецию органических веществ
за счет метаболических эффектов, являются соматотропин аденогипофиза,
йодсодержащие гормоны щитовидной железы и андрогены.
Процесс секреции некоторых веществ в проксимальных канальцах идет
настолько интенсивно, что за одно прохождение крови через корковое ве-
щество почек из нее удаляются полностью путем секреции такие вещества,
как, например, парааминогиппуровая кислота или рентгеноконтрастные
препараты. Следовательно, определяя клиренс этих веществ, можно рас-
считать объем плазмы крови, проходящей в единицу времени через кору
почек, или величину эффективного (т. е. участвующего в мочеобразовании)
почечного плазмотока.
14.2.1.4 . Состав и свойства конечной мочи
В сутки у человека образуется и выделяется от 0,7 до 2 л мочи. Эта величина
носит название суточного диуреза и зависит от количества выпитой жидко-
сти, так как здоровым человеком выделяется 65—80 % ее объема с мочой.
Основное количество мочи образуется днем, тогда как ночью оно составля-
ет не более половины дневного объема. Удельный вес мочи колеблется в
широком диапазоне — от 1005 до 1025, обратно пропорционально объему
принятой жидкости и образовавшейся мочи. Реакция суточной мочи обыч-
но слегка кислая, однако pH колеблется в зависимости от характера пита-
ния. При растительной пище моча приобретает щелочную реакцию, а при
белковой — становится более кислой. Моча обычно прозрачна, но имеет не-
большой осадок, получаемый при центрифугировании и состоящий из ма-
лого количества эритроцитов, лейкоцитов и эпителиальных клеток. В осад-
ке мочи, собранной за 12 ночных часов, содержится от 0 до 400 000 эритро-
цитов, от 300 000 до 1,8 миллионов лейкоцитов. Здесь также могут присут-
ствовать кристаллы мочевой кислоты, уратов и оксалата кальция (в кислой
моче) или кристаллы мочекислого аммония, фосфорнокислого и углекисло-
го кальция (в щелочной моче). Белок и глюкоза в конечной моче практиче-
ски отсутствуют, содержание аминокислот не превышает 0,5 г за сутки. По-
скольку в канальцах нефрона происходит обратное всасывание основной
части профильтровавшейся воды, солей и других веществ, то выделяется их
с мочой от 45 % (мочевина) до 0,04 % (бикарбонат) от профильтровавшего-
ся количества. Однако за счет всасывания воды и процессов концентриро-
вания мочи, а также секреции в канальцах, содержание в конечной моче ря-
да веществ превышает их концентрацию в плазме крови: мочевины в 67 раз,
калия в 7, сульфатов в 90, фосфатов в 16 раз, тем самым обеспечивается эф-
фективность их экскреции из организма. В небольших количествах в мочу
поступают производные продуктов гниения белков в кишечнике — индола,
скатола, фенола. В моче содержится широкий спектр органических кислот,
небольшие концентрации витаминов (кроме жирорастворимых), биогенные
амины и их метаболиты, стероидные гормоны и их метаболиты, ферменты
и пигменты, определяющие цвет мочи. С мочой в разных концентрациях,
зависящих от ее количества, выделяются практически все неорганические
катионы и анионы, в том числе и широкий спектр микроэлементов.
14.2.1.5 . Механизмы выведения мочи и мочеиспускания
Образовавшаяся в структурах нефрона моча поступает в почечные лохан-
ки. По мере их заполнения и растяжения достигается порог раздражения
механорецепторов, приводящий к рефлекторному сокращению мускулату-
632 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ры лоханки и раскрытию мочеточника. За счет перистальтических сокра-
щений их гладкой мускулатуры моча поступает в мочевой пузырь. Гладкие
мышцы лоханки и мочеточников обладают значительной степенью автома-
тии, в связи с чем их перистальтика вызывается растяжением объемом по-
ступающей мочи.
Заполняющая мочевой пузырь моча по мере накопления начинает рас-
тягивать его стенки, но при этом напряжение стенок пузыря не повышает-
ся до определенной величины растяжения, обычно соответствующей объе-
му мочи в пузыре около 400 мл. Появление напряжения стенки мочевого
пузыря вызывает позывы к мочеиспусканию, так как раздражение механо-
рецепторов ведет к поступлению афферентной информации в крестцовые
отделы спинного мозга и формированию сложного рефлекторного акта.
В этом акте участвуют не только спинальные, но и расположенные в го-
ловном мозге центральные структуры, позволяющие осуществлять произ-
вольную задержку мочеиспускания или его начало, а также обеспечиваю-
щие сенсорно-эмоциональную реакцию. Акт мочеиспускания реализуется
благодаря тому, что эфферентные имупльсы из спинального центра по па-
расимпатическим нервным волокнам достигают мочевого пузыря и моче-
испускательного канала, одновременно обеспечивая сокращение гладкой
мышцы стенки мочевого пузыря и расслабление двух сфинктеров — шейки
мочевого пузыря и мочеиспускательного канала.
14.2.2 . Экскреторная функция почек
Экскреторная функция почек состоит в выделении из внутренней среды ор-
ганизма с помощью процессов мочеобразования конечных и промежуточ-
ных продуктов обмена (метаболитов), экзогенных веществ, а также избыт-
ка воды и физиологически ценных минеральных и органических соедине-
ний. Особое значение для жизнедеятельности организма имеет при этом
выделение продуктов азотистого метаболизма (мочевина, мочевая кислота,
креатинин и др.), Н+-ионов, индолов, фенолов, гуанидинов, аминов и аце-
тоновых тел. Их экскреция осуществляется преимущественно почками, а
накопление этих веществ в крови при нарушении экскреторной функции
почек ведет к развитию токсического состояния, называемого уремией.
14.2.3 . Метаболическая функция почек
Состоит в обеспечении гомеостазиса обменных процессов в организме,
поддержании во внутренней среде определенного уровня и состава компо-
нентов метаболизма. При этом участие почки в процессах обмена веществ
в организме обеспечивается не только экскрецией субстратов и метаболи-
тов, но и протекающими в ней биохимическими процессами. Почка мета-
болизирует фильтрующиеся с мочой пептиды малой молекулярной массы
и денатурированные белки, возвращая в кровь аминокислоты и поддержи-
вая в крови уровень этих пептидов, в том числе и гормонов. Ткань коры
почки обладает способностью к новообразованию глюкозы — глюконеоге-
незу, причем в расчете на единицу массы органа эта способность у почки
выше, чем у печени. При длительном голодании примерно половина по-
ступающей в кровь глюкозы образуется почками. Почка является основ-
ным органом окислительного катаболизма инозитола, здесь синтезируются
важный компонент клеточных мембран фосфатидилинозитол, глюкуроно-
вая кислота, триацилглицерины и фосфолипиды, поступающие в крово-
ток, а также простагландины и кинины.
14. Выделение. Функции почек. Водно-солевой обмен • 633
14.2.4 . Роль почек в регуляции артериального давления
Почки участвуют в регуляции артериального давления благодаря несколь-
ким механизмам.
1. В почках образуется ренин, являющийся частью ренин-ангиотензин-
альдостероновой системы (РААС), которая обеспечивает регуляцию тонуса
кровеносных сосудов, поддержание баланса натрия в организме и объема
циркулирующей крови, активацию адренергических механизмов регуляции
насосной функции сердца и сосудистого тонуса. Уменьшение уровня дав-
ления крови в приносящей артериоле клубочка, повышение симпатическо-
го тонуса и концентрации натрия в моче дистального канальца активирует
секрецию ренина, что с помощью ангиотензина-П и альдостерона способ-
ствует нормализации сниженной величины артериального давления. Не-
адекватно избыточная секреция ренина и активация РААС может быть
причиной повышенного артериального давления.
2. В почке образуются вещества депрессорного действия, т. е. снижаю-
щие тонус сосудов и артериальное давление. Их образование получило на-
звание «антигипертензивной» функции почек, поскольку ее нарушение
может приводить к артериальной гипертензии.
Антигипертензивные гуморальные факторы почек представлены не-
сколькими соединениями, образующимися в основном интерстициальны-
ми клетками мозгового вещества: 1) простагландинами — ПГА, ПГЕ, ПГф,
ПГ^ 2) алкиловыми эфирами фосфатидилхолина (активирующий тромбо-
циты фактор); 3) нейтральным липидом мозгового вещества; а также обра-
зующимися в корковом веществе кининами. Активация синтеза почечных
простагландинов происходит при артериальной гипертензии, после ише-
мизации почки, под влиянием норадреналина, вазопрессина, ангиотензи-
на-П, почечных кининов и стимуляции почечных симпатических нервов.
Противогипертензивное действие простагландинов заключается как в вазо-
дилатирующем эффекте, так и в стимуляции почечной экскреции ионов
натрия и воды. Большая часть поступающих в кровь простагландинов раз-
рушается в легких, поэтому в артериальные сосуды не попадает, за исклю-
чением ПП2, почти не деградирущего в малом круге и обладающего мощ-
ным системным вазодилатирующим эффектом. Важное значение для про-
тивогипертензивного действия имеет торможение синаптической передачи
в адренергических синапсах под влиянием ПГЕ. Наличием обратной связи
между простагландинами и ренин-ангиотензин-альдостероновой системой
обусловлен стимулирующий эффект простагландинов на синтез ренина в
юкстагломерулярном аппарате почек.
Алкиловые эфиры фосфатидилхолина и нейтральный липид мозгового
вещества почек в отличие от большинства простагландинов оказывают, в
основном, системное вазодилатирующеее действие и являются поэтому ос-
новными гемодинамическими антигипертензивными факторами почек.
Кининовая система почек проявляет антигипертензивное действие как за
счет системного вазодилатирующего эффекта, так и, в большей мере, бла-
годаря внутрипочечной вазодилатации, повышению почечного кровотока,
диуретическому и натрийуретическому эффектам.
3. Почка экскретирует воду и электролиты, а их содержание в крови,
вне- и внутриклеточной среде является важным для поддержания уровня
артериального давления. Задержка ионов натрия и воды во внутренней
среде обеспечивает прирост объема циркулирующей крови. Однако боль-
шую роль играет содержание ионов натрия, калия и кальция во вне- и
внутриклеточной среде, поскольку оно определяет сократимость миокарда
634 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
и сосудистый тонус, а также реактивность сердца и сосудов к регулятор-
ным нейрогуморальным влияниям.
4. Почки участвуют в регуляции артериального давления с помощью
феномена «давление—натрийурез—диурез», суть которого заключается в
том, что повышение артериального давления вызывает увеличение экскре-
ции с мочой ионов натрия, возрастание объема диуреза и, в итоге, восста-
новление уровня артериального давления. Четыре основных механизма ле-
жат в его основе. Во-первых, повышенное артериальное давление тормозит
секрецию ренина, образование в почечной циркуляции ангиотензина-П, и
в результате ослабляется стимуляция ангиотензином реабсорбции ионов
натрия. Во-вторых, повышенное артериальное давление увеличивает обра-
зование в почках натрийуретических веществ: окиси азота и простагланди-
нов Е2, продукции последнего способствует и снижение уровня ангиотен-
зина-II. В-третьих, повышенное артериальное давление способствует росту
гидростатического давления в почечном интерстиции, что уменьшает реаб-
сорбцию воды и натрия. В-четвертых, повышение артериального давления
ускоряет кровоток по прямым сосудам мозгового вещества почки. Это
приводит к вымыванию осмотического градиента натрия и мочевины, что
снижает реабсорбцию воды, а следовательно, ослабляет концентрацион-
ную способность почки. Таким образом происходит увеличение натрий-
уреза и диуреза, следствием которых является уменьшение объема цирку-
лирующей крови, снижение прессорной активности сосудов, и в результа-
те нормализуется артериальное давление.
14.3. Водно-солевой обмен
Водно-солевым обменом называют совокупность процессов поступления во-
ды и электролитов в организм, распределения их во внутренней среде и
выделения из организма. Жизнедеятельность биологических систем, яв-
ляющихся с позиций термодинамики открытыми, требует равновесия или
баланса, общего поступления веществ и энергии (как экзо-, так и эндоген-
ных) с общим их выведением из системы. Наличие у большинства живых
систем основного циркадианного (суточного) ритма обусловливает баланс
основных веществ и энергии во временном промежутке, равном 24 ч. От-
сутствие в системе равенства суточного поступления и выведения веществ
позволяет говорить о нарушении баланса или дисбалансе — основном при-
знаке нарушения гомеостазиса. При этом обычно выделяют два полярных
состояния — положительный баланс (когда поступление вещества вместе с
его эндогенным образованием превышает выведение из системы) и отри-
цательный баланс (выведение превышает сумму поступления и образова-
ния).
14.3.1. Внешний водный баланс организма
Баланс между поступлением веществ из внешней среды и их выделением
из внутренней среды носит название «внешнего баланса». У здорового чело-
века поддерживается равенство объемов выделяющейся из организма и по-
ступившей в него за сутки воды, что называют внешним водным балансом
организма. Средние показатели внешнего водного баланса здорового чело-
века в состоянии покоя показаны в табл. 14.2. Принято рассматривать так-
же и внешний баланс электролитов — натрия, калия, кальция и других ве-
ществ (табл. 14.3).
Вода является важнейшим неорганическим компонентом организма,
14. Выделение. Функции почек. Водно-солевой обмен • 635
обеспечивающим связь внешней и внутренней среды, транспорт веществ
между клетками и органами. Являясь растворителем органических и неор-
ганических веществ, вода представляет собой основную среду развертыва-
ния метаболических процессов. Она входит в состав различных систем ор-
ганических веществ. Каждый грамм гликогена, например, содержит 1,5 мл
воды, каждый грамм белка — 3 мл воды. При ее участии формируются кле-
точные мембраны, транспортные частицы крови, макромолекулярные и
надмолекулярные образования.
В процессе обмена веществ и окислении водорода, отделенного от суб-
страта, образуется эндогенная «вода окисления», причем ее количество за-
висит от вида распадающихся субстратов и уровня обмена веществ. Так, в
покое при окислении 100 г жира образуется более 100 мл воды, 100 г бел-
ка — около 40 мл воды, 100 г углеводов — 55 мл воды. Повышение катабо-
лизма и энергетического обмена ведет к увеличению объема образуемой
эндогенной воды.
Однако эндогенной воды у человека недостаточно для обеспечения вод-
ной среды метаболических процессов, особенно выведения в растворен-
ном виде продуктов метаболизма. В частности, повышение потребления
белков и, соответственно, конечное превращение их в мочевину, удаляе-
мую из организма с мочой, ведет к абсолютной необходимости возраста-
ния потерь воды в почках, что требует повышенного ее поступления в ор-
ганизм. При питании преимущественно углеводной, жировой пищей и не-
большом поступлении в организм NaCl потребность организма в поступле-
нии воды меньше, так как и эндогенной воды образуется больше, и для
экскреции веществ с мочой воды требуется меньше. У здорового взрослого
человека суточная потребность в воде колеблется от 1 до 3 л.
Общее количество воды в организме составляет у человека от 44 до
70 % массы тела, или примерно 38—40 л. Содержание ее в разных тканях
варьирует от 10 % в жировой ткани до 83—87 % в почках и крови.
Таблица 14.2. Средние величины параметров внешнего водного баланса организма че-
ловека
Потребление и образование воды Количест- во, мл/сут Выделение воды Количест- во, мл/сут
Питье и жидкая пища 1200 С мочой 1500
С твердой пищей 1000 С потом 500
Эндогенная вода «окисления» 300 С выдыхаемым воздухом 400
С калом 100
Поступление 2500 Выделение 2500
Таблица 14.3. Среднесуточный баланс обмена некоторых веществ у человека
Вещества Поступление Выделение
пища метаболизм моча фекалии пот и воздух
Натрий (ммоль) 155 150 2,5 2,5
Калий (ммоль) 75 70 5,0 —
Хлориды (ммоль) 155 150 2,5 2,5
Азот (г) 10 9 1
Кислоты (мэкв)
нелетучие — 50 50 —
летучие — 14 000 — — 14 000
636 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
14.3.2. Внутренний водный баланс организма
Вода организма образует два водных пространства', внутриклеточное (2/3 об-
щей воды) и внеклеточное ('/3 общей воды). Небольшое количество воды
входит в жидкости полостей тела: брюшной, плевральной и т. д., а также в
цереброспинальную, внутриглазную, внутрисуставную жидкости. Внекле-
точное водное пространство включает два сектора (рис. 14.13): 1) внутри-
сосудистый водный сектор, т. е. плазму крови, объем которой составляет
около 8 % общей воды организма, и 2) интерстициальный водный сектор,
содержащий */4 всей воды организма (15 % массы тела) и являющийся
наиболее подвижным, меняющим объем при избытке или недостатке воды
в теле. Вся вода организма обновляется примерно через месяц, а внекле-
точное водное пространство — за неделю.
Таким образом, в организме человека вода распределена неодинаково в
разных водных средах, тканях, клетках и внеклеточном пространстве. Кон-
центрации ионов различаются в водных пространствах и секторах, причем
перемещения через биологические мембраны клеток одних ионов вызыва-
ют движение, часто в противоположном направлении, других (Na+ и К+; К+
и Н+; и др.). Соответственно, соотношение содержания воды и веществ в
разных средах внутренней среды получило название «внутреннего баланса».
Система регуляции водного баланса обеспечивает два основных гомео-
статических процесса: во-первых, поддержание постоянства общего объема
жидкости в организме и, во-вторых, оптимальное распределение воды ме-
жду водными пространствами и секторами организма. К числу факторов
поддержания водного гомеостазиса относятся осмотическое и онкотиче-
ское давление жидкостей водных пространств, гидростатическое и гидро-
динамическое давление крови, проницаемость гистогематических барьеров
и других мембран, активный транспорт электролитов и неэлектролитов,
нейроэндокринные механизмы регуляции деятельности почек и других ор-
ганов выделения, а также питьевое поведение и жажда.
14.3.3. Электролитный, или солевой, баланс организма
Водный баланс организма тесно связан с обменом электролитов. Суммар-
ная концентрация минеральных
— Внутриклеточная жидкость
и других ионов создает величину осмоти-
ческого давления водных пространств
организма. Концентрация отдельных
минеральных ионов в жидкостях внут-
ренней среды определяет функциональ-
ное состояние возбудимых и невозбу-
димых тканей, а также состояние про-
ницаемости биологических мембран,—
поэтому принято говорить о водно-элек-
тролитном (или солевом) обмене. По-
скольку синтез минеральных ионов в
Рис. 14.13. Распределение воды по секторам
водных пространств в % от общего содержа-
ния воды в организме. Наибольший объем
воды приходится на внутриклеточное про-
странство, а наименьший — на плазму крови
или внутрисосудистый сектор внеклеточно-
го водного пространства.
14. Выделение. Функции почек. Водно-солевой обмен • 637
организме не осуществляется, они должны поступать в организм с пищей
и питьем. Для поддержания электролитного баланса и, соответственно,
жизнедеятельности организм в сутки должен получать примерно 130
ммоль натрия и хлора, 75 ммоль калия, 26 ммоль фосфора, 20 ммоль каль-
ция и других элементов.
Основным катионом внеклеточного водного пространства является на-
трий, а анионом — хлор. Во внутриклеточном пространстве основной ка-
тион — калий, а анионами являются фосфат и белки.
Для обеспечения физиологических процессов важна не столько общая
концентрация каждого электролита в водных пространствах, сколько их
активность или эффективная концентрация свободных ионов, поскольку
часть ионов находится в связанном состоянии (Са2+ и Mg2+ с протеинами,
Na+ в ячейках клеточных органелл и и т. п.). Роль электролитов в жизне-
деятельности организма многообразна и неоднозначна.
Натрий поддерживает осмотическое давления внеклеточной жидкости,
причем его дефицит не может быть восполнен другими катионами. Изме-
нение уровня натрия в жидкостях организма неизбежно влечет за собой
сдвиг осмотического давления и в результате — объема жидкостей. Умень-
шение концентрации натрия во внеклеточной жидкости способствует пе-
ремещению воды в клетки, а увеличение содержания натрия — способству-
ет выходу воды из клеток. Количество натрия в клеточной микросреде оп-
ределяет величину мембранного потенциала и, соответственно,— возбуди-
мость клеток.
Основное количество калия (98 %) находится внутри клеток в виде
непрочных соединений с белками, углеводами и фосфором. Часть калия
содержится в клетках в ионизированном виде и обеспечивает их мем-
бранный потенциал. Во внеклеточной среде небольшое количество калия
находится преимущественно в ионизированном виде. Обычно выход ка-
лия из клеток зависит от увеличения их биологической активности, рас-
пада белка и гликогена, недостатка кислорода. Концентрация калия уве-
личивается при ацидозе и снижается при алкалозе. Уровень калия в
клетках и внеклеточной среде играет важнейшую роль в деятельности
сердечно-сосудистой, мышечной и нервной систем, в секреторной и мо-
торной функциях пищеварительного тракта, экскреторной функции
почек.
Кальций участвует в физиологических процессах только в ионизирован-
ном виде. Этот катион необходим для обеспечения возбудимости нервно-
мышечной системы, проницаемости мембран, свертывания крови. Иони-
зация кальция в крови зависит от ее pH. При ацидозе содержание ионизи-
рованного кальция повышается, а при алкалозе — падает. Алкалоз и сни-
жение уровня кальция во внеклеточной среде ведут к резкому снижению
порога возбуждения, повышению нейромышечной возбудимости и тетани-
ческим судорогам. Влияет на уровень свободного ионизированного каль-
ция и концентрация белков в плазме крови. Содержание кальция в крови
поддерживается в норме в диапазоне 2,3—2,6 ммоль/л. Внутриклеточный
ионизированный кальций является важнейшим вторичным посредником
нервно-гуморальных регуляторных влияний на клетки, обеспечивает про-
цессы освобождения медиаторов в синапсах и секрецию гормонов, энерге-
тику клетки. Основное депо кальция — костная ткань, в которой содер-
жится 90 % катиона в связанном виде.
Магний, как и калий, является основным внутриклеточным катионом,
его концентрация в клетках значительно выше, чем во внеклеточной среде.
Половина всего количества магния находится в костях, 49 % в клетках
638 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
мягких тканей и лишь 1 % во внеклеточном водном пространстве. Уровень
магния в крови составляет 0,7—1,0 ммоль/л, при этом более 60 % катиона
находится в ионизированном виде. Магний находится в составе более 300
разных ферментных комплексов, обеспечивая их активность. Он способст-
вует синтезу белков, необходим для поддержания состояния клеточных
мембран. Увеличивая потенциал покоя и порог возбуждения клеток, кати-
он уменьшает возбудимость нервно-мышечной системы, сократительную
способность миокарда и гладких мышц сосудов, оказывает депрессивное
действие на психические функции.
Главным анионом внеклеточной жидкости является хлор. Его концен-
трация в плазме крови в норме колеблется от 90 до 105 ммоль/л. Поступая
в клетку через хлорные каналы, анион участвует в формировании потен-
циала покоя возбудимых клеток, а способствуя гиперполяризации
мембран — определяет формирование процессов синаптического и преси-
наптического торможения. Избыток хлора ведет к ацидозу. Анион необхо-
дим для образования соляной кислоты в желудке.
Фосфаты являются основными внутриклеточными анионами, концен-
трация их в клетках выше, чем во внеклеточной среде, в 40 раз. Содержа-
ние неорганического фосфата в крови составляет 0,94—1,44 ммоль/л, но
50 % неорганического фосфата находится в костях, где он вместе с кальци-
ем образуют основное минеральное вещество костной ткани. Фосфаты —
необходимый компонент клеточных мембран, играют ключевую роль в ме-
таболических процессах, входя в состав многих коферментов, нуклеиновых
кислот и фосфопротеинов, вторичных посредников и макроэргических со-
единений.
Сульфаты в большем количестве содержатся во внутриклеточном про-
странстве, входят в состав многих биологически активных веществ. В плаз-
ме крови неорганических сульфатов содержится 0,3—1,5 ммоль/л. Они
участвуют в обезвреживании токсичных соединений в печени.
Для гомеостаза электролитов необходимо взаимодействие нескольких
процессов: поступление в организм, перераспределение и депонирование в
клетках и их микроокружении, выделение из организма. Поступление их в
организм зависит от состава и свойств пищевых продуктов и воды, особен-
ностей их всасывания в желудочно-кишечном тракте и состояния энте-
рального барьера. Однако, несмотря на широкие колебания количества и
состава пищевых веществ и воды, внешний водно-солевой баланс в здоро-
вом организме неуклонно поддерживается за счет изменений экскреции с
помощью органов выделения. Основную роль в этом гомеостатическом ре-
гулировании выполняют почки.
14.3.4. Общие принципы регуляции водно-солевого обмени
Регуляция водно-солевого обмена включает афферентное, центральное и
эфферентное звенья. Афферентное звено представлено массой рецептор-
ных аппаратов сосудистого русла, тканей и органов, воспринимающих
сдвиги осмотического давления, объема жидкостей и их ионного состава.
В результате в нервных центрах промежуточного мозга (гипоталамус, эпи-
таламус), лимбической системы и коры больших полушарий создается ин-
тегрированная картина состояния водно-солевого баланса в организме.
Следствием центрального анализа является изменение питьевого и пище-
вого поведения, перестройка работы желудочно-кишечного тракта и систе-
мы выделения (прежде всего функции почек), реализуемая через эффе-
рентные звенья регуляции. Последние представлены вегетативной нервной
14. Выделение. Функции почек. Водно-солевой обмен • 639
системой и, в большей мере, гормонами нейрогипофиза (вазопрессин, ок-
ситоцин), миокарда (атриопептид), коры надпочечников (альдостерон),
других эндокринных желез.
Важнейшим элементом регуляции водно-солевого обмена является осу-
ществляемое осморегулирующими системами организма поддержание ос-
мотического гомеостазиса. Его основными показателями являются осмоти-
ческие давления крови, интерстициальной жидкости, внутрцклеточной
среды. Осмотическое давление любой жидкости внутренней среды, являю-
щейся сложным раствором, равно сумме парциальных осмотических дав-
лений содержащихся в ней ингредиентов. Осмотическое давление, созда-
ваемое каждым из ингредиентов, зависит от его концентрации. Осмотиче-
ское давление принято выражать в миллиосмолях. Для электролитов вели-
чина парциального осмотического давления определяется согласно их ва-
лентности: одновалентные ионы создают число миллиосмолей, равное их
концентрации в миллимолях, а для двухвалентных ионов количество мил-
лиосмолей составляет половину их концентрации в миллимолях. Осмоти-
ческое давление плазмы крови у здорового человека относится к числу же-
стких гомеостатических констант и находится в пределах 275—305 моем/
кг, при этом электролиты создают осмотическое давление порядка 270—
300 мосм/кг, а остальная незначительная часть общего осмотического дав-
ления приходится на неэлектролиты — сахар, мочевину, липиды и т. д. В
условиях патологии осмотическое давление плазмы крови может выходить
за пределы нормального диапазона колебаний. Границы осмотического
давления, совместимого с жизнью человека, находятся в пределах 203—
375 мосм/кг.
14.4. Интегративные механизмы регуляции водно-солевого обмена
и гомеостатическая функция почек
Водно-солевой гомеостазис является одной из важнейших констант внут-
ренней среды организма. От состояния водно-солевого обмена зависят
фундаментальные свойства клеток и тканей (энергетический и пластиче-
ский метаболизм, возбудимость, проводимость и сократимость, секретор-
ная способность и т. п.) и, соответственно, функции практически всех фи-
зиологических систем. Основные характеристики водно-солевого обме-
на — количество поступающих в организм жидкости и солей, объемы вне-
клеточных и внутриклеточных водных пространств, содержание в них от-
дельных электролитов и осмотическое давление, наконец, выделение воды
и солей почками — имеют взаимосвязанные комплексные механизмы регу-
ляции. Эти механизмы легко проследить на примерах изменения приема
жидкости или ее потерь, сдвигов баланса ионов натрия и калия, измене-
ния объема крови.
Нарушения баланса веществ могут проявляться в виде положительного
или отрицательного дисбаланса. Положительный водный дисбаланс полу-
чил название гипергидратации, а отрицательный — дегидратации. Измене-
ния водного баланса могут сопровождаться или не сопровождаться изме-
нениями электролитного баланса и, соответственно, осмотического давле-
ния. В зависимости от осмолярности виды водного дисбаланса подразделя-
ют на изоосмотические, гиперосмотические и гипоосмотические. Эти по-
нятия относят к внеклеточной среде организма, но благодаря механизмам
саморегуляции поддерживается осмотическое равновесие между вне- и
внутриклеточной средой, поэтому изменения осмолярности внеклеточной
среды неизбежно сопровождаются аналогичными сдвигами осмотического
640 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
давления и внутри клеток. Водный дисбаланс может быть и следствием из-
менения коллоидно-осмотического (онкотического) давления плазмы
крови.
14.4.1. Гомеостатические механизмы при гиперосмотической
дегидратации
Такой вид водного дисбаланса возникает при дефиците поступления воды
в организм, а также в случаях, когда потери воды превышают потери солей
или при потерях свободной от электролитов воды. Примером этого типа
дисбаланса является пребывание человека в пустыне, когда недостаток во-
ды для питья сочетается с повышенными перспирационными (через кожу
и дыхательные пути) потерями воды. Недостаточное поступление воды в
организм может быть следствием отсутствия чувства жажды при патологии
диэнцефальной области головного мозга, нарушений режима кормления и
ухода за детьми грудного возраста, больными при потере сознания, нару-
шений глотания.
Ограничение приема воды (рис. 14.14) приводит к дегидратации внутрен-
ней среды организма, при этом имеет место повышение осмотического
давления плазмы крови и внеклеточной жидкости, что дало основание
именовать этот вид дегидратации гиперосмотической, или истинной. Повы-
шение осмотического давления плазмы крови вызывает перераспределение
жидкости и электролитов между кровью, внеклеточной жидкостью и лим-
фой, а также между внеклеточной и внутриклеточной средой, направлен-
ное на нормализацию сдвига осмотического давления. Начальные меха-
низмы регуляции гомеостазиса при этом типе дегидратации заключаются в
буферировании белками и эритроцитами катионов, образовании с ними
сложных химических соединений, с измененными осмотическими свойст-
вами и гидрофильностью, диффузии осмотически активных веществ из
крови в тканевую жидкость и лимфу, осмотическом токе воды в противо-
положном направлении, усилении всасывания воды в кишечнике. При не-
больших физиологических уровнях дегидратации этих механизмов может
быть достаточно для обеспечения водно-солевого гомеостазиса.
Повышенное осмотическое давление жидкостей внутренней среды раз-
дражает осморецепторы, приводя к формированию осморегулирующих
рефлексов и жажды.
Осморегулирующие рефлексы начинаются от периферических и централь-
ных осморецепторов (рис. 14.15). Периферические осморецепторы распо-
ложены в интерстициальном пространстве тканей и кровеносных сосудах
печени (система воротной вены), сердца, пищеварительного тракта, почек,
селезенки, а также в рефлексогенной зоне каротидного синуса. Поскольку
осмотическое давление связано преимущественно с ионами натрия, часть
осморецепторов являются специализированными натриорецепторами.
Наиболее мощными рецептивными полями для натрия являются интер-
стициальные пространства печени и предсердий. Имеются также тканевые
рецепторы, воспринимающие концентрацию ионов калия, кальция и маг-
ния. Центральные осморецепторы — клетки супраоптического ядра гипота-
ламуса, непосредственно воспринимающие сдвиги осмотического давле-
ния притекающей в межуточный мозг крови и концентрацию в ней натрия
(натриорецепторы).
Афферентная информация от тканевых осмо- и натриорецепторов по-
ступает по волокнам блуждающего нерва и задних корешков спинного
мозга в центральную нервную систему, где направляется в центр осморегу-
14. Выделение. Функции почек. Водно-солевой обмен • 641
Рис. 14.14. Схема компенсаторных реакций при ограничении приема или потерях
воды, т. е. при гиперосмотической дегидратации. Основные реакции направлены на
восполнение объемов воды (жажда, повышение реабсорбции воды в почках) и вы-
ведение избытка натрия с мочой (угнетение реабсорбции катиона в почках).
ляции, локализованный в ядрах переднего гипоталамуса — супраоптиче-
ском и паравентрикулярном. Эфферентное звено осморегулирующих и на-
трийрегулирующих рефлексов включает как вегетативные нервы, так и
гормональные сигналы. Осмо- и натрийрегулирующие рефлексы увеличи-
вают нейросекрецию и содержание в крови вазопрессина. Избыток ионов
натрия в крови стимулирует секрецию натрийуретического атриопептида.
Вазопрессин повышает в почках проницаемость дистальных канальцев и
собирательных трубочек для воды, и она пассивно реабсорбируется по ос-
мотическому градиенту, т. е. реализуется механизм концентрирования мо-
чи. Вазопрессин повышает всасывание воды и в других органах — кишеч-
нике, желчном пузыре, слюнных и пищеварительных железах. Натрийуре-
тический атриопептид, как и, в меньшей степени, вазопрессин, ведут к по-
вышенному выведению натрия из крови, что устраняет увеличение осмо-
тического давления.
21 -6095
642 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
I
Вазопрессин
Активация реабсорбции воды, концентрирование мочи, снижение осмолярности крови
Рис. 14.15. Осморегулирующий рефлекс.
Повышение осмотического давления крови при ограниченном поступлении воды в организм
или ее чрезмерных потерях вызывает раздражение сосудистых и центральных (гипоталамиче-
ских) осморецепторов, следствием чего является увеличение секреции в кровь вазопрессина,
активация обратного всасывания воды в канальцах почек и ее поступление в кровь, что ведет
к нормализации осмотического давления крови.
Продукция вазопрессина возрастает и при значительном снижении объ-
ема циркулирующей крови и артериального давления. Это происходит бла-
годаря механорецептивному рефлексу, берущему начало от рефлексоген-
ных зон сосудов и сердца.
Важнейшей приспособительной реакцией организма при дегидратации
организма является жажда. Жажда — это субъективное ощущение челове-
ка, возникающее при абсолютном или относительном (к содержанию на-
трия) дефиците воды в организме и приводящее к поведенческой реакции
по приему воды. Таким образом, жажда является одной из основных био-
логических мотиваций (т. е. побуждений к деятельности), обеспечивающих
поддержание жизнедеятельности человека. Формирование чувства жажды
связано с возбуждением питьевого центра, локализованного в структурах
гипоталамуса и лимбики (рис. 14.16). Возбуждение этого центра обуслов-
лено несколькими причинами. Во-цервых, при повышении осмотического
давления крови и внеклеточной жидкости, уровня натрия в них происхо-
дит раздражение центральных и периферических осморецепторов и реф-
лекторное возбуждение центра. Во-вторых, возбуждение центра и возник-
новение жажды имеет место и при уменьшении объема циркулирующей
крови без сдвига осмотического давления. В этом случае ее формирование
обусловлено двумя механизмами: 1) волюморефлексом от волюморецепто-
ров правого предсердия и полых вен, возникающим в ответ на уменьшение
растяжения этих областей низкого давления сниженным объемом крови;
14. Выделение. Функции почек. Водно-солевой обмен • 643
Рис. 14.16. Механизм развития жажды.
Стимуляция гипоталамического питьевого центра и возникновение чувства жажды происходит
под влиянием повышенного осмотического давления крови, образования в крови и ткани моз-
га ангиотензина-П, снижении объема крови и артериального давления.
2) уменьшенный объем крови ведет к активации юкстагломерулярного ап-
парата почек и секреции ренина, что увеличивает в крови уровень ангио-
тензина-П. Ангиотензин-П — один из мощных дипсогенных (от греческого
dipsa —жажда) факторов, стимулирующий структуры субфорникального
органа (СФО) межуточного мозга, который является частью питьевого
центра. В-третьих, возбуждение питьевого центра и жажду вызывают ней-
ропептиды, образуемые в самой мозговой ткани. Так, повышение концен-
трации натрия в крови при дегидратации влечет образование в мозговой
ткани натрийуретического пептида, подобного атриопептиду, также отно-
сящегося к дипсогенным факторам. Дипсогенными являются и образуе-
мые в мозговой ткани ангиотензин-П, окситоцин и вазопрессин.
Питьевой центр может возбуждаться и после еды. Механизм жажды в
этих случаях обусловлен гормональными эффектами (рис. 14.17).
Выраженное и продолжительное ограничение приема воды — водное го-
лодание — ведет к выходу жидкости из клеток во внеклеточную среду и
формированию состояния дегидратации клеток, что вызывает тяжелые
расстройства, особенно со стороны центральной нервной системы, клетки
которой чувствительны к сдвигам водно-солевого гомеостазиса. Как пра-
вило, вместе с водой клетки теряют и К+, что вызывает сдвиги мембранно-
го потенциала и еще более усугубляет расстройства деятельности нервной
системы и других возбудимых тканей.
Избыточные потери воды (при гипервентиляции легких, обильном по-
тоотделении при тяжелой физической работе и высокой температуре сре-
ды) также ведут к гипертонической дегидратации, поскольку при этом по-
тери воды намного превышают потерю электролитов (главным образом,
натрия). Формирующиеся при этом интегративные механизмы регуляции
водно-солевого гомеостазиса аналогичны описанным выше при дефиците
приема жидкости.
2Г
644 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 14.17. Механизм жажды после еды.
При поступлении пищи в кишечник двенадцатиперстная кишка и поджелудочная железа сек-
ретируют во внутреннюю среду гормон кальцитонин. Бета-клетки поджелудочной железы,
особенно после сладкой пищи, секретируют гормон амилин. Под влиянием этих гормонов ак-
тивируются расположенные в межуточном мозге вокруг III мозгового желудочка структуры,
получившие название циркумвентрикулярного органа, входящего в питьевой центр.
Осмотическое давление плазмы крови (мосм/кг) в норме:
275----280-----285-----288-----290-----295-----300-----305
203*
Порог роста
секреции
вазопрессина
Водный
баланс
Порог
жажды
Нестерпимая
жажда
Границы выживания
► 375
Рис. 14.18. Пороги стимуляции секреции вазопрессина и жажды как факторы регу-
ляции осмотического гомеостазиса.
При повышении осмотического давления плазмы крови первой регуляторной приспособи-
тельной реакцией является увеличение секреции вазопрессина, а жажда появляется при более
выраженном повышении осмолярности.
Таким образом, основными физиологическими механизмами, препятст-
вующими повышению осмотического давления жидкостей внутренней сре-
д^>1, являются повышение секреции вазопрессина и жажда (рис. 14.18).
При этом порог реакции увеличения секреции вазопрессина расположен
на уровне более низких значений осмотического давления, чем порог жаж-
ды; следовательно, в процесс нормализации повышенного осмотического
давления вначале вовлекается прирост секреции вазопрессина, а затем уже
жажда.
14. Выделение. Функции почек. Водно-солевой обмен • 645
14.4.2. Гомеостатические механизмы при изоосмотической
дегидратации
При рвоте и поносах у человека происходит потеря жидкости; изотонич-
ной плазме крови, т. е. теряется не только вода, но и содержащиеся в ней
электролиты, преимущественно ионы натрия и хлорида. Такая дегидрата-
ция носит название изоосмотической и для нее характерен истинный де-
фицит натрия в организме. Изотоническая дегидратация сопровождается
уменьшением объема циркулирующей плазмы и крови, что определяет
формирование в организме приспособительных реакций: повышается ней-
росекреция вазопрессина за счет рефлекса с волюморецепторов предсер-
дий и полых вен (что активирует реабсорбцию воды в почках), усиливается
секреция ренина и повышается в крови содержание ангиотензина и альдо-
стерона (что поддерживает сосудистый тонус и вызывает активацию реаб-
сорбции натрия в дистальных канальцах почек). Поскольку при изоосмо-
тической дегидратации осмотическое давление внеклеточной и внутрикле-
точной жидкости не меняется, жажда может быть мало выраженной.
При этом типе дегидратации происходит потеря и Na+, и воды, а белки
крови и эритроциты сохраняются, поэтому в крови отмечается увеличение
концентрации белка и гематокрита. При выраженной степени дегидрата-
ции и недостаточности компенсаторных механизмов из-за начального
уменьшения объема плазмы крови быстро наступают циркуляторные нару-
шения как системной гемодинамики, так и микроциркуляции.
14.4.3. Гомеостатические механизмы при гипоосмотической
дегидратации
При этом виде дисбаланса в организме возникает истинный дефицит Na+,
как правило, связанный с избыточной потерей хлорида натрия при чрез-
мерном потоотделении, осмотическом диурезе. Классическим клиниче-
ским примером гипоосмотической дегидратации является потеря натрия
организмом при надпочечниковой недостаточности (дефицит альдосте-
рона).
Поскольку NaCl содержится, в основном, во внеклеточном пространст-
ве, именно здесь и снижается прежде всего осмотическое давление и вода
начинает перемещаться во внутриклеточное пространство с относительно
более высоким осмотическим давлением. Следствием компенсаторного пе-
рераспределения воды является одновременное уменьшение объема вне-
клеточного протсранства (дегидратация) и увеличение объема внутрикле-
точного пространства (гипергидратация), а осмолярность в обоих про-
странствах оказывается сниженной.
Компенсаторные механизмы при гипоосмотической дегидратации не
всегда эффективны, так как из-за клеточной гипергидратации и уменьше-
ния осмолярности плазмы крови чувство жажды отсутствует. Если причи-
ной этого вида дегидратации является не патология надпочечников, а из-
быточное потоотделение или осмотический диурез, ведущей компенсатор-
ной реакцией является повышение секреции альдостерона, вызванное как
снижением Na+ в плазме крови, так и активацией ренин-ангиотензин-аль-
достероновой системы при уменьшении артериального давления.
646 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
14.4.4. Гомеостатические механизмы при гипоосмотической
гипергидратации
Гипоосмотическая гипердегидратация возникает при значительном посту-
плении воды в кровь в результате ее всасывания в желудочно-кишечном
тракте или введении в сосудистое русло бессолевых растворов (например,
5 % раствора глюкозы). Возникает этот тип дисбаланса и при почечной не-
достаточности, и при циррозе печени. При умеренных физиологических
объемах выпиваемой воды (рис. 14.19), приспособительные реакции начи-
наются еще до всасывания воды из желудочно-кишечного тракта в кровь
системной циркуляции, так как снижение осмотического давления слюны
и химуса воспринимаются рецепторами ротовой полости и пищеваритель-
ного тракта, а позднее всасывающаяся в кровь воротной вены вода умень-
шает концентрацию ионов натрия и осмотическое давление крови, посту-
пающей в печень, где этот сдвиг воспринимается соответствующими осмо-
и натриорецепторами. Возникающее при их раздражении рефлекторное (в
том числе и условно-рефлекторное) подавление нейросекреции вазопрес-
сина и нарастание мочеобразования происходят вскоре после приема жид-
Рис. 14.19. Схема компенсаторных реакций, возникающих при гипоосмотической
гипергидратации.
Как при физиологических (прием воды), так и при искусственных условиях (введение избытка
жидкости) в основе компенсации гидратации лежит подавление образования и секреции в
кровь вазопрессина, вследствие чего возрастает выведение избытка воды через почки.
14. Выделение. Функции почек. Водно-солевой обмен • 647
кости. Таким образом, вовлечение систем интегративного контроля водно-
солевого баланса начинается еще до сдвигов уровня натрия и осмотиче-
ского давления в крови системной циркуляции, поэтому в обычных усло-
виях рефлекторная стимуляция почечной экскреции воды и натрия огра-
ничивает увеличение объема крови из-за всасывания выпитой воды.
Чрезмерный прием воды или введение гипотонических растворов в сосу-
дистую систему (см. рис. 14.19) вызывают выраженную гипоосмотическую
гипергидратацию, ведут к уменьшению осмотического давления и концен-
трации ионов натрия в плазме. Следствием этого является подавление
нейросекреции вазопрессина, стенки дистальных канальцев и собиратель-
ных трубочек в почках остаются непроницаемыми для воды. Поскольку
реабсорбция ионов натрия сохраняется, почки выделяют большие количе-
ства гипотоничной мочи и избыток воды удаляется из крови. В отсутствие
вазопрессина мочевина не реабсорбируется в собирательных трубочках,
что снижает осмолярность интерстиция мозгового вещества почек и тем
самым также ослабляет реабсорбцию воды. Если описанные механизмы
регуляции не обеспечивают выведение объемов воды, адекватных ее посту-
пившему количеству, устанавливается положительный водный баланс, во-
да задерживается в организме. Задержка воды в крови (гидремия) снижает
ее осмотическое давление и вода выходит в ткани — формируются отеки.
14.4.5. Гомеостатические механизмы при изоосмотической
гипер гидратации
Это состояние является наиболее частым проявлением у человека положи-
тельного водного баланса с одновременной задержкой NaCl (недостаточ-
ность кровообращения, токсикозы беременности, болезнь Кушинга). При
этом, поскольку ионы Na+ и СГ являются почти исключительно внекле-
точными, имеет место увеличение лишь объема изотонической жидкости
во внеклеточной среде и перемещений воды между вне- и внутриклеточ-
ным пространствами не отмечается. Существенно увеличивается объем ин-
терстициального сектора, где депонируется изотоническая жидкость и
формируются отеки, увеличивается масса тела. Внутриклеточное простран-
ство остается без изменений. Общее количество воды и натрия в организ-
ме возрастает, но концентрация ионов Na+ остается в пределах нормы. Не-
смотря на гипергидратацию возникает потребность в осмотически свобод-
ной воде и нередко появляется жажда. Избыток изотонической жидкости в
системе гемоциркуляции может вести к острой сердечно-сосудистой не-
достаточности, острой почечной недостаточности при наличии патологии
почек. Наиболее простым примером этого типа гипергидратации является
внутривенное вливание больших количеств изотонического раствора NaCl.
Следствием гипергидратации внутрисосудистого сектора является умень-
шение гематокрита и концентрации плазменных белков. Избыток изоос-
мотической жидкости выводится почками путем увеличения диуреза. Не-
соответствие скорости мочеобразования степени гипергидратации ведет к
отекам. В условиях патологии компенсация гипергидратации этого типа
отличается малой эффективностью.
14.4.6. Гомеостатические механизмы при гиперосмотической
гипергидратации
Этот вид водного дисбаланса возникает при первичном альдостеронизме
(синдром Кона), остром гломерулонефрите, введении в организм больших
648 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
количеств гипертонических растворов хлорида натрия, гидрокарбоната в
послеоперационный период, особенно при почечной недостаточности.
Объем и осмолярность внеклеточного пространства увеличиваются из-за
прироста концентрации Na+ и С1+, вода перемещается из внутриклеточно-
го пространства во внеклеточное до равенства в них осмотического давле-
ния. Вследствие этого объем внутриклеточного пространства уменьшается,
а осмолярность становится повышенной в обоих пространствах. Если не
происходит потерь белка и эритроцитов из плазмы крови, как это бывает
при гломерулонефрите, в крови повышаются гематокрит и концентрация
белка. Компенсация нарушений водно-солевого гомеостазиса при этом ти-
пе дисбаланса осуществляется путем почечного выведения избытка жидко-
сти и натрия и требует значительного времени. Без устранения причины,
вызвавшей гиперосмотическую гипергидратацию, компенсация нарушен-
ного гомеостазиса в большинстве случаев не эффективна.
14.4.7. Нарушения баланса электролитов
Нарушения баланса электролитов в организме являются, как правило,
следствием водного дисбаланса. Это прежде всего относится к дисбалансу
ионов натрия, поскольку гиперосмотическая дегидратация и гипоосмоти-
ческая гипергидратация, являясь следствием водного дисбаланса, неизбеж-
но сопровождаются гипер- или гипонатриемией. Гипер- или дегидратация
внутриклеточного пространства ведут, соответственно, к уменьшению или
повышению содержания внутриклеточных ионов, прежде всего, калия.
Как правило, сдвиги содержания электролитов в плазме крови являются
следствием трех основных механизмов: 1) изменения всасывания в желу-
дочно-кишечном тракте, 2) перераспределения между вне- и внутрикле-
точной средами, в том числе и депонирования, 3) изменения экскреции из
организма, прежде всего через почки. Электролитный гомеостазис необхо-
дим для обеспечения фундаментальных свойств клеток (метаболизм, реге-
нерация, возбудимость, специфические функции), а также их регуляции и
саморегуляции, поэтому его нарушения ведут к тяжелейшим расстрой-
ствам жизнедеятельности.
ГЛАВА 15
Кислотно-основное состояние
Кислотно-основное состояние (кислотно-щелочное равновесие; кислотно-
щелочной баланс; кислотно-основное равновесие) — одно из важнейших
гомеостатических свойств внутренней среды организма, характеризующее-
ся относительным постоянством соотношения водородных и гидроксиль-
ных ионов и определяющее оптимальный характер обменных процессов и
физиологических функций.
Основой внутренней среды является вода, молекулы которой при дис-
социации образуют Н+ и ОН" ионы. Соотношение их концентрации опре-
деляет актуальную реакцию жидкостей, прежде всего крови.
Актуальной реакцией называют существующую в организме в конкрет-
ных условиях кислотность или щелочность внутренней среды. От актуаль-
ной реакции среды зависят условия функционирования большинства бел-
ков, так как электронейтральность белковой молекулы определяется ее
изоэлектрической точкой, что напрямую связано с кислотностью среды.
Актуальная реакция среды определяет активность ферментов, для боль-
шинства из которых свойственны свои оптимальные значения реакции
среды. Кислотно-основное состояние внутренней среды определяет в тка-
нях направление и интенсивность процессов окисления и восстановления,
расщепления и синтеза белков, углеводов и жиров, нуклеиновых кислот,
активность витаминов и микроэлементов. Изменения актуальной реакции
среды помимо метаболизма влияют и на функции клеток, органов и сис-
тем (сердца и сосудов, нервной системы и мышц, легких и почек), опреде-
ляют биофизические свойства клеток и молекул, например проницаемость
мембран и возбудимость, степень дисперсности коллоидов и пр.
В свою очередь, кислотно-основное состояние является интегральным
параметром внутренней среды организма и зависит от состояния клеточ-
ного метаболизма, газотранспортной функции крови, процессов питания и
внешнего дыхания, водно-солевого обмена.
Внутриклеточная среда для большинства клеток нейтральна, так как это
обеспечивает оптимальную возможность образования субстрат-фермент-
ных комплексов. Внеклеточная жидкость и кровь имеют слабую щелочную
реакцию, что способствует нейтрализации и удалению из клеток кислых
метаболитов и Н-ионов.
15.1. Кислоты и основания внутренней среды
Кислотами называют вещества, способные при диссоциации в растворах
выделять ион водорода (донаторы протона), а основаниями или щелоча-
ми — вещества, способные связывать ион водорода (акцепторы протона).
Вода в слабой степени диссоциирована на Н+ и ОН- ионы. При ней-
тральной реакции воды в ней имеется одинаковое количество Н+ и ОН-
ионов, при 22 °C составляющее 10-7 каждого. Произведение Н+ и ОН- ио-
нов в водном растворе независимо от того, нейтральный, кислый он или
650 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 15.1. Соотношение величины pH и концентрации водородных ионов ([Н*])
pH [Н+] нмоль/л
7,7 20
7,4 (Плазма) 40
7,3 (Цереброспинальная жидкость) 50
7,1 (Внутриклеточная жидкость) 80
7,00 100
6,9 126
Таблица 15.2. Актуальная реакция жидкостей организма
Наименование жидкостей PH
Артериальная кровь 7,36-7,42
Венозная кровь 7,26-7,36
Цереброспинальная жидкость 7,24-7,5
Межклеточная жидкость тканей 7,26-7,38
Тканевая жидкость мышц 6,7-6,9
Поджелудочный сок 7,8-8,4
Желчь печеночная 7,3-8,0
Желчь пузырная 6,0-7,0
Слюна 5,8-7,8
Желудочный сок 1,4-1,8
Сок тонкой кишки 7,5-8,6
Сок толстой кишки 8,0-9,0
щелочной, всегда одинаково и равно: 10-7 • 10-7 - 10-14. Эта величина на-
зывается константой диссоциации воды. Очевидно, что если концентрация
водородных ионов — [Н+] — будет расти, то число ОН- ионов — [ОН-] —
будет уменьшаться.
Поскольку актуальная реакция водного раствора зависит от соотноше-
ния [Н+] и [ОН-], оценивать ее можно по одной из них, т. е., как это при-
нято, по [Н+]. Таким образом, раствор с нейтральной реакцией имеет
[Н+] = 10 , с кислой реакцией — [Н+] >10-7, а со щелочной реакцией —
[Н+] <10-7. Для оценки [Н+] используется отрицательный десятичный лога-
рифм концентрации, который обозначается pH и называется водородным
показателем. Таким образом, pH = —log[H+]. Следовательно, для нейтраль-
ного раствора pH = 7,0; для кислотного — <7, а для щелочного — >7. Со-
отношения величины pH и [Н+] приведены в табл. 15.1.
В норме величина pH в различных жидких средах организма неодина-
кова, но ее колебания невелики, pH внутриклеточной, тканевой жидкости
и крови относится к числу жестких гомеостатических констант. В табл.
15.2 приведены значения pH различных сред организма.
Кислоты образуются из принятой пищи и в результате межуточного об-
мена веществ. Процессы окисления в клетках обусловливают появление
органических и минеральных кислот. Так, катаболизм серосодержащих
аминокислот вызывает образование серной кислоты; фосфопротеины и
нуклеопротеины ведут к образованию фосфорной и мочевой кислоты; ли-
пиды формируют свободные жирные кислоты; углеводы при окислении и
гликолизе поставляют в кровь пировиноградную и молочную кислоту и
т. п. Углекислый газ, образуемый как конечный продукт процессов окис-
ления, также является кислотой, поскольку вместе с другим конечным
15. Кислотно-основное состояние • 651
продуктом окисления — водой — дает реакцию: СО2 + Н2О -> Н2СО3 ->
Н+ + НС Оу
Углекислота является слабой кислотой, но служит в свою очередь ис-
точником углекислого газа, удаляемого через легкие, поэтому она еще на-
зывается летучей кислотой. Другие органические и неорганические кисло-
ты называют нелетучими. Летучей кислоты образуется в организме намного
больше (до 20 000 ммоль СО2), чем нелетучих кислот (около 100 ммоль).
Кислоты, как промежуточный этап метаболизма, являются биохимиче-
ской необходимостью, характеризующей процессы жизнедеятельности
клетки. При этом интенсивность образования кислот при метаболизме яв-
ляется одним из основных факторов, обеспечивающих поступление прото-
нов во внутреннюю среду. Протоны, поступающие с пищей, играют в pH
внутренней среды существенно меньшую роль. Содержание свободных во-
дородных ионов в жидкостях зависит от интенсивности их связывания ос-
нованиями и выделения во внешнюю среду.
Основания поступают в организм преимущественно в растительной пи-
ще, богатой щелочными и щелочноземельными солями. Образуются щело-
чи и клетками организма, например внешнесекреторными клетками под-
желудочной железы, секрет которых богат ОН-.
Интенсивность образования кислот прямо связана с окислительно-вос-
становительными реакциями клеток, обеспечением их кислородом. При
ограничении обеспечения клеток кислородом или наличии кислородного
долга из-за чрезмерной потребности в нем усиливаются или активизиру-
ются процессы анаэробного гликолиза. Появляющиеся в клетках в избыт-
ке пировиноградная и молочная кислоты переходят в тканевую жидкость и
кровь. Так происходит, например, при интенсивной мышечной работе за
счет отставания возможности кислородного обеспечения от потребности в
нем работающей мышцы, в результате для целей энергообеспечения ско-
рость гликолиза повышается в сотни раз. Образующаяся при этом в избыт-
ке молочная кислота является причиной более низкого pH в тканевой
жидкости скелетных мышц, чем в других тканях.
Однако, несмотря на поступление в кровь пировиноградной и молоч-
ной кислот, так же как и других нелетучих и летучих кислот, pH крови
практически не меняется. Это свидетельствует о наличии в организме
мощных гомеостатических систем, поддерживающих константу pH. К их
числу относят физико-химические (буферные системы внутренней среды,
тканевые обменные процессы) и физиологические гомеостатические систе-
мы (легкие, почки, желудочно-кишечный тракт, костная ткань).
15.2. Физико-химические гомеостатические механизмы
К физико-химическим механизмам кислотно-основного гомеостазиса отно-
сятся буферные системы внутренней среды организма и тканевые гомео-
статические обменные процессы.
15.2.1. Буферные системы внутренней среды организма
Основными буферными системами внутриклеточной, межклеточной жидко-
сти и крови являются бикарбонатная, фосфатная и белковая буферная систе-
ма, причем из последней для крови особо выделяют гемоглобиноный буфер.
Основное значение для поддержания pH межклеточной жидкости и
плазмы крови имеет бикарбонатная буферная система. Угольная кислота в
плазме и межклеточной жидкости присутствует в четырех формах: физиче-
652 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ски растворенного углекислого газа (СОД, угольной кислоты (Н2СО3),
аниона карбоната (СО^) и аниона бикарбоната (гидрокарбонатный анион)
(HCOj). В условиях физиологического диапазона pH в плазме крови боль-
ше всего содержание бикарбоната, примерно в 20 раз меньше содержание
растворенного углекислого газа и угольной кислоты, а ион карбоната
практически отсутствует. Бикарбонат представлен в виде натриевых и ка-
лиевых солей. Константа диссоциации (К) есть отношение:
КН2СО) = а для бикарбоната — KNaHCO} =
[712'^L/V 6?/7 U Czj J
Анион НСО3 является общим как для кислоты, так и для соли, а соль
диссоциирует сильнее, поэтому данный анион, образуясь из бикарбоната,
будет подавлять диссоциацию угольной кислоты, т. е. практически все ко-
личество аниона HCOJ в бикарбонатном буфере происходит из NaHCO3
Следовательно:
„„ [/Гпясо;] го+1 К[Н2со-л
КН2СО} = I -а — ц, а отсюда: [FT] = -L ;-„3J-.
[//2ьО3] [J\aHCO3]
(формула Гендерсона, где К —константа диссоциации угольной кислоты).
В связи с использованием отрицательного логарифма концентрации фор-
мула, названная уравнением Гендерсона—Гассельбаха, для бикарбонатного
буфера приняла выражение:
^PK=-lSK.
При физиологических значениях pH отношение концентрации углеки-
слоты к бикарбонату составляет 1:20 (рис. 15.1).
В условиях взаимодействия бикарбонатного буфера с кислотами проис-
ходит их нейтрализация с образованием слабой угольной кислоты. Угле-
кислый газ, появляющийся при ее разложении, удаляется через легкие. Из-
быток оснований, взаимодействуя с бикарбонатным буфером, связывается
с угольной кислотой и приводит в конечном счете к образованию бикарбо-
ната, излишки которого удаляются, в свою очередь, из крови через почки.
Другой буферной системой плазмы крови, а в большей мере внутрикле-
точной среды, является фосфатная буферная система. Она образована од-
но- и двузамещенными слоями фосфорной кислоты, где однозамещенные
соли являются слабыми кислотами, а двузамещенные — имеют щелочные
свойства. Уравнение для фосфатного буфера следующее:
/ [NaH2POA
Двухосновной фосфатной соли содержится в плазме в 4 раза больше,
чем кислой одноосновной. Общим анионом в этой системе является
НРО4. Ее буферная емкость меньше, чем бикарбонатной, так как и фосфа-
тов в крови содержится меньше, чем бикарбонатов. Принцип действия
фосфатного буфера аналогичен бикарбонатному, хотя роль его в плазме
крови мала и, в основном, сводится к поддержанию концентрации бикар-
боната при реакции буфера с избытком угольной кислоты. В то же время в
клетках значение фосфатного буфера велико.
Третьей буферной системой клеток, плазмы крови и межклеточной жид-
кости является белковая. Белки выполняют буферную роль из-за их амфо-
15. Кислотно-основное состояние • 653
Рис. 15.1. Кислотно-основное равновесие. Стрелка весов указывает на значения pH
в норме и при смещениях в сторону ацидоза или алкалоза. На чашах весов помеще-
ны Н2СО3 (дыхательная компонента) и NaCO3 (недыхательная компонента), что от-
ражает соотношение между ними в норме 1:20.
терности, а характер их диссоциации зависит от природы белка и актуаль-
ной реакции внутренней среды. При этом глобулины обладают более выра-
женной кислой диссоциацией, т. е. отщепляют больше протонов, чем гид-
роксильных ионов, и соответственно играют большую роль в нейтрализа-
ции щелочей. Белки, содержащие много диаминокислот, диссоциируют
больше как щелочи и поэтому в большей мере нейтрализуют кислоты. Бу-
ферная емкость белков плазмы крови сравнительно с бикарбонатной систе-
мой невелика, однако в клетках тканей ее роль может быть очень высокой.
Наибольшую буферную емкость крови обеспечивает гемоглобиновая бу-
ферная система. Содержащаяся в гемоглобине человека аминокислота гис-
тидин (до 8,1 %) имеет в своей структуре как кислые (СООН), так и ос-
новные (NH2) группы. Константа диссоциации у гемоглобина ниже, чем
pH крови, поэтому гемоглобин диссоциирует как кислота. Оксигемогло-
бин является более сильной кислотой, чем редуцированный гемоглобин.
При диссоциации оксигемоглобина в капиллярах тканей с отдачей кисло-
рода появляется большее количество щелочно-реагирующих солей гемо-
глобина, способных связывать Н-ионы, поступающие из кислот тканевой
жидкости, например угольной кислоты. Оксигемоглобин обычно представ-
ляет собой калиевую соль. При взаимодействии кислот с калиевой солью
оксигемоглобина образуется соответствующая калиевая соль кислоты и
свободный гемоглобин со свойствами очень слабой кислоты. Гемоглобин в
тканевых капиллярах связывает углекислый газ за счет аминогрупп, обра-
зуя карбогемоглобин:
Hb- NH2+CO2 Hb- NHCOOH.
654 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 15.3. Относительная эффективность буферных систем крови
Плазма крови % Эритроциты %
Бикарбонатная 35 Гемоглобиновый 35
Белковая 7 Бикарбонатный 18
Фосфатная 1 Фосфатный 4
Общая эффективность буферных Общая эффективность буферных
систем 43 систем 57
Для кислотно-основного гомеостазиса важное значение имеет также об-
мен анионов СГ и НСО7 между плазмой и эритроцитами. Если в плазме
крови увеличивается концентрация углекислоты, то образующийся при
диссоциации NaCl анион С1 входит в эритроциты, где образует КС1, а ион
Na+, для которого мембрана эритроцита непроницаема, соединяется с из-
бытком НСОу, образуя бикарбонат натрия, восполняя его убыль в бикар-
бонатом буфере. При снижении концентрации углекислоты в бикарбо-
натом буфере происходит обратный процесс — анионы СГ выходят из
эритроцитов и соединяются с избытком Na+, освободившимся из бикарбо-
ната, этим, следовательно, предотвращается ощелачивание плазмы.
Буферные системы плазмы крови и эритроцитов имеют разную относи-
тельную эффективность. Так, эффективность буферных систем эритроци-
тов выше (за счет гемоглобинового буфера), чем плазмы крови (табл. 15.3).
Концентрация Н-ионов уменьшается в направлении: клетка — межкле-
точная среда — кровь. Это свидетельствует о том, что наибольшую буфер-
ную емкость имеет кровь, а наименьшую — внутриклеточная среда. Обра-
зуемые в клетках при метаболизме кислоты поступают в межклеточную
жидкость тем легче, чем больше их образуется в клетках, так как избыток
Н-ионов повышает проницаемость клеточной мембраны. В буферных
свойствах межклеточной среды играет роль соединительная ткань, особен-
но коллагеновые волокна, известные как «ацидофильные». На минимальное
накопление кислот они реагируют набуханием, быстро поглощая кислоту и
освобождая от Н-ионов межклеточную жидкость. Эта способность колла-
гена объясняется свойством абсорбции.
15.2.2. Тканевые гомеостатические обменные процессы
Кислотно-основное состояние поддерживается в пределах физиологиче-
ских значений pH и путем метаболических превращений в тканях. Это
достигается за счет совокупности биохимических и физико-химических
процессов, которые обеспечивают: 1) потерю кислотных и щелочных
свойств продуктов обмена веществ, 2) их связывание в средах, препятст-
вующих диссоциации, 3) образование новых, более легко нейтрализуемых
и выводимых из организма соединений.
Так, например, органические кислоты могут соединяться с продуктами
белкового обмена (бензойная кислота с глицином) и тем самым терять
кислые свойства. Избыток молочной кислоты ресинтезируется в гликоген,
кетоновых тел — в высшие жирные кислоты и жиры. Неорганические ки-
слоты нейтрализуются солями калия, натрия, освобождающимися при дез-
аминировании аминокислот аммиаком, образующим аммонийные соли.
После внутривенного введения животным кислоты 43 % ее количества
нейтрализуется бикарбонатом плазмы крови, 36 % нейтрализуются за счет
клеточного натрия, а 15 % — за счет выходящего из клеток калия. Основа-
15. Кислотно-основное состояние • 655
ния нейтрализуются преимущественно молочной кислотой, образуемой из
гликогена при ощелачивании микроокружения клеток.
В поддержании внутриклеточного pH существенную играет роль обмен
производных имидазола и его изомера пиразола. Особенности пятичленно-
го кольца этих соединений определяют их амфотерные свойства, т. е. спо-
собность быть одновременно и донатором, и акцептором протонов. Ими-
задол способен очень быстро образовывать соли с сильными кислотами и
щелочными металлами. Наиболее распространенным соединением имида-
зола является а-аминокислота гистидин, участвующая в кислотном и ос-
новном катализе.
Сильные кислоты и щелочи могут растворяться в липидах, обладающих
низкой диэлектрической константой, что препятствует их диссоциации.
Наконец, органические кислоты могут подвергаться окислению с образо-
ванием летучей слабой угольной кислоты.
15.3. Физиологические гомеостатические механизмы
Эти механизмы, включающие функции легких, почек, желудочно-кишеч-
ного тракта, печени и костной ткани, обеспечивают нейтрализацию, свя-
зывание и выведение из организма Н+ и ОН- ионов в составе различных
соединений, в зависимости от особенностей метаболизма и состояния бу-
ферных систем внутренней среды.
15.3.1. Легкие и кислотно-основное состояние
Процессы газообмена между кровью и окружающим воздухом, происходя-
щие в легких, обеспечивают регуляцию кислотно-основного состояния, по-
скольку внешнее дыхание — это основной путь удаления из крови летучей
угольной кислоты. При физиологических условиях в покое в организме
вырабатывается 10 ммоль/мин угольной кислоты, а выделяется через лег-
кие 13 000—15 000 ммолей СО2 в сутки. Благодаря бикарбонатному буферу
образующиеся при метаболизме кислоты не вызывают закисления крови, а
приводят к увеличению содержания СО2 в крови с последующим удалени-
ем через легкие.
Удаленный из плазмы углекислый газ немедленно восполняется за счет
двух механизмов, происходящих в крови легочных капилляров. Во-первых,
образование оксигемоглобина ведет к распаду карбаминовой связи гемо-
глобина и освобождению СО2 из эритроцитов; во-вторых, под влиянием
карбангидразы, фермента, катализирующего реакцию: Н2СО3 Н2О +
СО2
Количество выводимого из организма углекислого газа зависит от глу-
бины и частоты дыхания, т. е. от альвеолярной вентиляции. Величина же
альвеолярной вентиляции регулируется дыхательным центром, деятель-
ность которого зависит или даже определяется парциальным напряжением
СО2 в крови и pH внеклеточной жидкости мозга. Таким образом, усиление
дыхания и альвеолярной вентиляции происходит при увеличении напря-
жения в крови СО2 и уменьшении pH. Нарастающее при этом удаление уг-
лекислоты через легкие приводит ее содержание в крови в соответствие с
концентрацией бикарбоната. Следовательно, легкие выступают в роли фи-
зиологической гомеостатической системы, не только выводящей избыток
летучей кислоты, но и восстанавливающей емкость буферных систем, пре-
жде всего бикарбонатной и гемоглобиновой.
При быстром поступлении во внутреннюю среду оснований вентиляция
656 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
легких уменьшается и количество выделяемого СО2 существенно снижает-
ся. Концентрация углекислоты возрастает и pH поддерживается на исход-
ном уровне.
15.3.2. Почки и кислотно-основное состояние
Почки осуществляют выведение из внутренней среды организма нелетучих
органических и неорганических кислот. Избыток кислот при этом может
выводиться как в свободном состоянии, так и в виде нейтральных солей.
Сильные неорганические кислоты выделяются из внутренней среды только
через почки и только в связанной форме.
В физиологических условиях почки выделяют кислую мочу, pH которой
колеблется от 5 до 7. Однако, в зависимости от кислотно-основного со-
стояния внутренней среды, моча может быть более кислой (pH 4,0) или
даже щелочной (pH 8,0).
Участие почек в регуляции кислотно-основного состояния внутренней
среды обеспечивается совокупностью взаимосвязанных процессов, проис-
ходящих в структурах нефрона. К их числу относятся: 1) секреция водо-
родных ионов в мочу клетками канальцевого эпителия, 2) реабсорбция
(обратное всасывание) профильтровавшихся в мочу оснований, прежде
всего бикарбоната, пополняющих резерв в крови, 3) образование в клетках
эпителия канальцев ионов бикарбоната, больше или меньше всасываемого
в кровь, 4) образование и диффузия в мочу аммиака, создающего катион
аммония, присоединяя Н-ион, 5) фильтрация в первичную мочу из плаз-
мы крови соединений, обладающих кислыми или щелочными свойствами,
6) обмен ионов СГ, Na+, К+, фосфатов, сульфатов и двухвалентных катио-
нов (Са2+, Mg2+).
Важнейшую роль в способности почек к выделению кислой мочи игра-
ет секреция водородных ионов клетками эпителия проксимальных и дисталь-
ных канальцев (рис. 15.2). Эта способность обусловлена наличием в клет-
ках эпителия фермента карбоангидразы, обеспечивающего образование из
СО2 и Н2О угольной кислоты, с последующей диссоциацией:
Н2О + СО2 -> Н2СО3 Н+ + нсо;.
Ион водорода активно секретируется с затратой энергии через апикаль-
ную мембрану клетки эпителия в просвет канальца, а из канальцевой мочи
в клетку диффундирует ион натрия, обеспечивая равновесие электриче-
ских зарядов. Из клетки, уже через базальную мембрану, Na+ активно с
помощью Na-K-насоса удаляется в интерстиций и кровь, туда же пассивно
по градиенту концентрации поступает HCOJ образуя бикарбонат натрия.
Следовательно, секреция Н-ионов обеспечивает обратное всасывание би-
карбоната и натрия, причем основная часть секретируемых ионов водорода
тратится именно на обеспечение реабсорбции бикарбоната. Так, секрети-
руемый Н-ион в просвете проксимального канальца взаимодействует с
профильтровавшимся в мочу из плазмы ионом бикарбоната (в сутки около
5000 ммоль), образуя угольную кислоту. Под влиянием фермента карбоан-
гидразы, встроенного в мембрану щеточной каемки клеток эпителия ка-
нальца, угольная кислота разлагается на СО2 и воду:
карбоангидраза
Н+ + НСО7 -> Н2СО3---------------> СО2 + Н2О.
Молекулы углекислого газа легко диффундируют в клетки, где реак-
ция катализируется в обратном направлении. Таким образом, на суточ-
15. Кислотно-основное состояние • 657
ПРОСВЕТ
КАНАЛЬЦА
КЛЕТКА ПЕРИКАНАЛЬЦЕВАЯ
ЭПИТЕЛИЯ ЖИДКОСТЬ
Рис. 15.2. Основные механизмы участия клеток почечных канальцев в поддержании
кислотно-основного состояния внутренней среды организма.
Кружки обозначают мембранные транспортные системы.
Первый фрагмент (сверху) — секреция Н+-ионов в мочу клетками эпителия проксимальных
канальцев обеспечивает реабсорбцию профильтровавшегося в первичную мочу натрия и
транспорт во внутреннюю среду бикарбоната натрия, что способствует сохранению нормаль-
ного значения величины актуального бикарбоната крови. Второй (средний) фрагмент — секре-
ция водородных ионов в мочу ведет к реабсорбции натрия из диссоциирующих солей неорга-
нических кислот и появлению титруемых кислот мочи. Третий фрагмент — аммониогенез как
путь выведения избытка Н+-ионов. Секретируемые эпителием в просвет канальца аммиак и
Н-ионы взаимодействуют между собой и с ионом хлора, образуя экскретируемый хлорид ам-
мония.
ную реабсорбцию до 5000 ммоль бикарбоната расходуется эквивалентное
количество секретированных ионов водорода. Выделяется же с мочой
только 60 ммоль/сут ионов водорода, преимущественно в связанном
виде.
Наибольшая часть секретированных канальцами Н+ (65—75 %) участву-
ют в процессе аммониогенеза, обеспечивающего выведение из плазмы кро-
ви анионов сильных неорганических и органических кислот в виде аммо-
нийных солей. Клетки проксимальных и дистальных почечных канальцев,
благодаря процессам дезаминирования и дезамидирования аминокислот,
образуют аммиак. Основным источником является глутамин, который де-
заминируется под влиянием фермента глутаминазы, 40 % аммиака образу-
ется из аминокислот аланина и глицина. Аммиак, благодаря высокой рас-
творимости в липидах, легко диффундирует в просвет канальца, где при-
соединяет секретированные Н+ и образует ион аммония; NH3 + Н+ -> NH^.
Ионы аммония вытесняют натрий из соединений с анионами сильных ки-
слот, натрий реабсорбируется, а анионы кислот в виде аммонийных солей
выделяются с мочой.
Содержащиеся в крови кислые и щелочные компоненты буферных сис-
тем, например одно- и двухосновный фосфаты, бикарбонат, а также сла-
бые органические кислоты — молочная, лимонная, р-оксимаслянная и др.,
так же как и анионы сильных неорганических кислот (СГ, SO^), фильтру-
ются из плазмы крови в первичную мочу. Часть из них реабсорбируется в
658 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
проксимальных канальцах, особенно существенно бикарбонат (до 80—
90 %), мочевая кислота (более 90 %). Но наряду с обратным всасыванием
многие из указанных веществ, особенно органические кислоты и основа-
ния (холин), активно секретируются клетками канальцевого эпителия. Та-
ким образом, уровень этих веществ в моче определяется соотношением
трех основных процессов: клубочковой фильтрации, канальцевых реаб-
сорбции и секреции.
Секретированные в мочу водородные ионы участвуют в образовании
титруемых кислот мочи. Основную роль при этом играют фосфаты. По-
скольку в плазме крови в 4 раза больше двухосновных фосфатов, поэтому
и в первичную мочу их фильтруется соответственно больше. Под воздейст-
вием секретированного эпителием канальца Н-иона происходит перевод
двухосновного фосфата в одноосновный, путем ионного обмена протона
на натрий: Na2HPO4 + Н+ -> NaH2PO4 + Na+
Освобождаемый ион Na+ реабсорбируется в канальцах в кровь. Таким
образом, этот процесс способствует сохранению во внутренней среде на-
трия и удалению из нее избытка Н+-ионов.
Многие органические кислоты содержатся в плазме крови в виде солей
и, фильтруясь в клубочках, поступают в первичную мочу. Под воздействи-
ем секретированных канальцами Н-ионов в моче происходит образование
свободных слабых, т. е. плохо диссоциирующих, кислот, также составляю-
щих титруемую кислотность мочи. Титруемой кислотностью мочи называ-
ют сумму Н-ионов слабых кислот и солей, определяемую путем титрова-
ния мочи щелочью до величины pH, равной pH крови. Эта величина пока-
зывает, насколько больше водородных ионов содержится в моче по срав-
нению с плазмой крови, т. е. насколько эффективно почки поддерживают
постоянство pH.
В клетках эпителия дистальных отделов нефрона кроме описанной вы-
ше секреции водородных ионов происходит секреция ионов К+, при этом
ионы Н+ и К+ конкурируют в обмене на Na+. В связи с этим выведение ка-
лия и протона может изменяться противоположно друг другу. Так, при из-
бытке водородных ионов секреция калия снижается, а при недостатке —
возрастает. Напротив, избыток К+ снижает секрецию водородных ионов, а
недостаток К+ Повышает ее. Недостаток калия стимулирует и синтез ам-
миака в эпителиальных клетках канальцев для выведения избытка Н-ио-
нов в виде аммонийных солей. Однако секреция протонов и калия связана
с транспортом натрия и меняется пропорционально сдвигам реабсорбции
натрия. При снижении дистальной реабсорбции натрия соответственно
снижается секреция Н+ и К+ ионов, напротив, усиление всасывания на-
трия в дистальных канальцах приводит к повышению секреции калия и
водородных ионов.
При поступлении в кровь избытка кислот или оснований компенсация
сдвига кислотно-основного состояния осуществляется прежде всего буфер-
ными системами крови, дыхательная компенсация формируется через 16—
18 ч, а почечная компенсация развивается еще медленнее — спустя 2—
3 сут. При этом почечные механизмы обладают значительной инерционно-
стью и даже после удаления из организма избытка кислот или оснований
возвращение к исходному кислотно-основному состоянию продолжается
также около 2—3 сут.
15. Кислотно-основное состояние • 659
15.3.3. Желудочно-кишечный тракт, печень, костная ткань
и кислотно-основное состояние
Характер питания человека и особенности пищеварения в значительной ме-
ре сказываются на кислотно-основном состоянии его внутренней среды,
В пищевых продуктах растительного происхождения содержится большое
количество солей органических кислот (щавелевой, яблочной, лимонной,
янтарной и др.), после окисления которых и выведения кислотных валент-
ностей с углекислотой через легкие остаются и накапливаются связанные с
катионами щелочные валентности. Поэтому растительная пища способст-
вует ощелачиванию внутренней среды, соответственно, основания в боль-
шем количестве выводятся с мочой, что увеличивает pH мочи и она может
приобретать щелочную реакцию. Белковая пища животного происхожде-
ния способствует закислению внутренней среды, так как в составе живот-
ных белков содержатся сера и фосфор, что приводит к образованию силь-
ных неорганических кислот. Повышенное выведение их с мочой приводит
к снижению pH мочи и она приобретает резко кислую реакцию. Всасыва-
ние в желудочно-кишечном тракте кислот или щелочей, содержащихся в
различных напитках, в том числе алкогольных и минеральных водах, также
приводит к изменению кислотно-основного состояния внутренней среды.
Сравнительно небольшую роль в регуляции кислотно-основного со-
стояния играют пищеварительные функции желудка и кишечника. Парие-
тальные клетки слизистой оболочки желудка секретируют соляную кисло-
ту. Водородные ионы образуются в клетках под влиянием карбоангидразы,
анион хлора поступает в клетки из хлористого натрия крови путем обмена
на бикарбонат. Однако при этом ощелачивания крови не происходит, так
как СГ желудочного сока в кишечнике всасывается обратно в кровь. Зна-
чительное количество бикарбонатов поступает в кишечник в составе сока
поджелудочной железы и кишечного сока. При этом ионы Н+ всасываются
в кровь. Обратное всасывание претерпевают и бикарбонатные ионы, по-
этому сдвигов кислотно-основного состояния крови не происходит. При
кислой диете увеличивается выделение двухвалентных катионов Са2+ и
Mg2+ а при щелочной — и одновалентных катионов. Хотя значение желу-
дочно-кишечного тракта в физиологической регуляции кислотно-основно-
го состояния невелико, тем не менее при изменениях, а тем более наруше-
ниях функций желудка и кишечника неизбежно происходят сдвиги и ки-
слотно-основного состояния. Стойкое и резкое повышение кислотности
желудочного сока может вести к увеличение щелочного резерва крови из-
за потерь Н-ионов и избыточного всасывания бикарбоната. Частая рвота,
особенно при непроходимости кишечника, вызывая потерю Н+-ионов и
хлоридов, вызывает гипохлоремию и защелачивание внутренней среды,
что требует обязательной врачебной коррекции.
В печени происходит утилизация молочной кислоты с образованием
гликогена, здесь разрушаются кетоновые тела (ацетоуксусная и бета-окси-
масляная кислоты), происходят метаболические превращения органиче-
ских кислот и оснований. Образуя желчь с изменяемой степенью щелоч-
ной реакции, печень способствует удалению из внутренней среды разных
количеств оснований.
В нейтрализации кислот внутренней среды принимает участие костная
ткань, поскольку в ней связано огромное количество катионов и фосфата.
При длительном накоплении в крови кислот и снижении pH внеклеточной
жидкости происходит остеолиз и вымывание из скелета фосфата кальция.
В результате реакции с угольной кислотой образуются бикарбонатный и
660 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
однозамещенный фосфатный анионы, нейтрализующие избыток Н-ионов.
Благодаря огромным запасам в скелете фосфата кальция этот механизм
обладает высокой эффективностью, выход из скелета 1 моль фосфата каль-
ция нейтрализует и способствует в дальнейшем выведению из организма
через почки 4 эквивалентов кислоты. Однако потеря костной тканью фос-
фата кальция вызывает деминерализацию скелета и потерю его прочности.
В костной ткани находится до 45 % всего натрия, содержащегося в орга-
низме, однако выход натрия из костной ткани возможен только при ре-
зорбции кости, например, в условиях кислой реакции среды. В таких слу-
чаях натрий костей, поступая во внеклеточную среду, способствует нейтра-
лизации кислотного сдвига.
15.4. Основные физиологические показатели кислотно-основного
состояния
Кислотно-основное состояние внутренней среды характеризуют следую-
щие показатели: актуальный pH; парциальное напряжение углекислоты;
актуальный бикарбонат крови; стандартный бикарбонат крови; буферные
основания крови; избыток или дефицит буферных оснований крови.
Актуальный pH — фактическая величина отрицательного логарифма
концентрации водородных ионов плазмы артериальной крови, измеренная
при 38 °C. Физиологические пределы колебаний от 7,35 до 7,45. Значение
pH крови соответствует pH внеклеточной жидкости.
Парциальное напряжение углекислоты (рСО^ — равно парциальному дав-
лению над кровью углекислого газа, находящегося в равновесии с раство-
ренным СО2 в артериальной крови при 38 °C. В физиологических условиях
рСО2 в покое составляет в среднем 40 мм рт. ст., колеблясь от 35 до 45 мм
рт. ст. Крайние отклонения при патологии составляют диапазон 10—130
мм рт. ст. При произвольной задержке дыхания напряжение углекислоты
может достигать 90 мм, а при произвольной гипервентиляции — снижаться
до 20 мм рт. ст.
Актуальный бикарбонат крови — истинная концентрация бикарбонатно-
го аниона (НСО^) при фактическом состоянии плазмы артериальной кро-
ви. Этот показатель называют также «щелочной резерв» крови. В физиоло-
гических условиях равен 22—25 ммоль/л.
Стандартный бикарбонат крови — содержание бикарбонатного аниона в
плазме крови при стандартных условиях, т. е. полном насыщении гемогло-
бина кислородом, уравновешивании с газовой средой, имеющей рСО2 40
мм рт. ст., и 38 °C. Этот показатель важен для клинической диагностики,
так как отражает только метаболические, не связанные с дыханием, сдвиги
кислотно-основного состояния крови. У здоровых людей стандартный би-
карбонат не отличается от актуального.
Буферные основания (БО) крови — это общая сумма концентрации всех
анионов цельной крови, обладающих буферными свойствами при условии
полного насыщения крови кислоррдом, при температуре крови 38 °C и
рСО2 40 мм рт. ст. В физиологических условиях величина БО составляет в
среднем 41,7 + 0,043 НЬ ммоль/л, где НЬ — концентрация гемоглобина в г/
л. Пределы колебаний показателя в норме от 46 до 52 ммоль/л.
Дефицит или избыток буферных оснований (ДИБО) — является наиболее
важным для диагностики метаболическим параметром кислотно-основного
состояния крови. Этот показатель характеризует разницу между фактиче-
ской величиной буферных оснований крови, найденных у исследуемого
человека, и значением БО, определенным в стандартных условиях. Устано-
15. Кислотно-основное состояние • 661
вить значение этого параметра можно путем титрования крови. Ввиду тех-
нической сложности и трудоемкости такого титрования, на практике этот
параметр определяют по специальным номограммам. Если число буфер-
ных оснований в исследуемой крови выше, чем стандартный показатель
БО, параметр ДИБО обозначается со знаком плюс, а если ниже — со зна-
ком минус. Физиологический диапазон колебания ДИБО в артериальной
крови составляет от —2 до +2. В условиях патологии пределы сдвига ДИБО
значительно шире: от —30 при резких кислотных сдвигах до +30 при сдви-
гах в щелочную сторону.
15.5. Основные изменения кислотно-основного состояния
и их компенсация
Кислотно-основное состояние внутренней среды характеризуют по содер-
жанию в ней количества водородных ионов, зависящего от их поступления
и выделения, т. е. баланса. В связи с этим выделяют две формы сдвига ки-
слотно-основного состояния: ацидоз и алкалоз. Ацидоз — это сдвиг кислот-
но-основного состояния в связи с положительным балансом водородных
ионов, т. е. при накоплении Н-ионов в крови. Алкалоз — это сдвиг кислот-
но-основного состояния в связи с отрицательным балансом водородных
ионов, т. е. при уменьшении Н-ионов в крови.
Концентрация водородных ионов в крови определяется, во-первых,
[Н2СО3] или рСО2, выводимого в большей или меньшей степени через лег-
кие, — дыхательная, или респираторная, компонента. Во-вторых, содержа-
ние Н+ определяется концентрацией бикарбоната (HCOj), не выделяемого
через легкие,— недыхательная, или нереспираторная, компонента.
Таким образом, как ацидозы, так и алкалозы можно представить в двух
видах: 1) дыхательные или респираторные и 2) недыхательные или нерес-
пираторные (см. рис. 15.1). При дыхательных сдвигах кислотно-основного
состояния изменяется концентрация углекислоты (повышается при ацидо-
зе и снижается при алкалозе), а при недыхательных — бикарбоната, т. е.
основания (снижается при ацидозе, повышается при алкалозе). Однако
нарушения баланса водородных ионов не обязательно приводят к сдвигу
уровня свободных Н+, т. е. pH, поскольку буферные системы и физиоло-
гические гомеостатические системы компенсируют изменения баланса во-
дородных ионов. Компенсацией называют процесс выравнивания наруше-
ния путем изменения в той системе, которая не была нарушена. Таким
образом, дыхательные нарушения компенсируются сдвигами в недыха-
тельной компоненте и, наоборот, сдвиги уровня бикарбоната компенсиру-
ются изменениями дыхательной компоненты, т. е. выведения углеки-
слоты.
В зависимости от степени эффективности компенсации возможны три
варианта сдвигов кислотно-основного состояния: некомпенсированные
(сдвиг рН<7,29 или >7,56), частично компенсированные (рН<7,35 или
>7,45, отмечается избыток иди дефицит буферных оснований), компенси-
рованные (сдвиг pH отсутствует /7,35—7,45/, благодаря полной компенса-
ции изменившегося баланса водородных ионов) (см. рис. 15.1).
Определение характера сдвига кислотно-щелочного состояния в случаях
компенсированных ацидоза или алкалоза требует специального расширен-
ного исследования. Выявление частично компенсированных и некомпен-
сированных сдвигов может быть осуществлено путем сопоставления трех
параметров: pH, дыхательной и недыхательной компонент баланса, как это
показано в табл. 15.4.
662 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Таблица 15.4. Сдвиги основных параметров кислотно-основного состояния при ацидо-
зах и алкалозах
Вид нарушения кислотно-основного состояния Параметры
PH ДИБО pCO
Ацидоз недыхательный Частично компенсированный г 4'4*
Некомпенсированный и u =
Ацидоз дыхательный Частично компенсированный I t tt
Некомпенсированный и = tt
Алкалоз недыхательный Частично компенсированный t tt t
Некомпенсированный tt tt =
Алкалоз дыхательный Частично компенсированный t г 44
Некомпенсированный tt = г;
У здоровых людей дыхательный ацидоз может возникать при длитель-
ном пребывании в среде с повышенным содержанием углекислого газа,
например в замкнутых помещениях малого объема, шахтах, подводных
лодках. Недыхательный ацидоз у здоровых людей бывает при длительном
употреблении кислой пищи, углеводном голодании, усиленной мышечной
работе, особенно у нетренированных людей.
Дыхательный алкалоз формируется у здоровых людей при нахождении в
условиях сниженного атмосферного давления и, соответственно, парци-
ального давления СО2, например при пребывании высоко в горах, полетах
в негерметизированных летательных аппаратах, что создает условия для
избыточных потерь СО2 кровью. Гипервентиляция легких также способст-
вует потере двуокиси углерода и дыхательному алкалозу.
Недыхательный алкалоз развивается у здоровых людей при длительном
приеме щелочной пищи или минеральной воды типа боржоми.
15.5.1. Функциональное значение ацидозов и алкалозов
• Физиологические свойства ферментов, транспорт и связывание бел-
ков, обусловленные диссоциацией кислых или основных аминокис-
лот (особенно гистидина), зависят от pH.
• Ферменты гликолиза, особенно фосфофрукто киназа, зависят от pH.
Внутриклеточный алкалоз стимулирует, а ацидоз подавляет гликолиз
и образование молочной кислоты. Алкалоз подавляет глюконеогенез
и повышает содержание субстратов цикла Кребса, например цитрата.
Ацидоз стимулирует деградацию глюкозы через пентозо-фосфатный
цикл.
• Для синтеза ДНК и пролиферации клеток pH не должен быть кис-
лым. Легкий алкалоз всегда имеет место при клеточной пролифера-
ции. Множество регуляторов роста активируют Na/H-обменник мем-
браны и приводят к защелачиванию внутриклеточной среды. Щелоч-
ной pH характерен для опухолевых клеток.
• К+-каналы большинства клеток активируются алкалозом и подавля-
ются ацидозом, что способствует изменению потенциала покоя и воз-
будимости клеток.
• Ащидоз ведет к уменьшению входа Са2+ через Са-каналы, приводя,
например, к снижению сократимости миокарда.
• В легочных артериолах Н+-ионы вызывают вазоконстрикцию, а в со-
судах большого круга они способствуют вазодилатации, что в услови-
15. Кислотно-основное состояние • 663
ях кислородного голодания может вести к снижению артериального
давления. Алкалоз способствует вазоконстрикции, особенно мозговых
сосудов.
• Ацидоз снижает, а алкалоз повышает проводимость в плотных меж-
клеточных контактах, в частности в миокарде, следствием чего могут
быть аритмии сердца.
• Ацидоз снижает, а алкалоз повышает сродство гемоглобина к кисло-
роду (эффект Бора).
• Алкалоз стимулирует диссоциацию плазменных белков, которые за-
тем связывают больше Са2+ и снижают его активность в плазме. На-
против, ацидоз снижает кальций-связывающую активность белков
плазмы. С другой стороны, бикарбонат соединяется с Са2+ и умень-
шает его активность.
Все случаи возникновения сдвигов кислотно-основного состояния у
здоровых людей обычно полностью компенсированы. В условиях патоло-
гии ацидозы и алкалозы у человека бывают значительно чаще и, соответст-
венно, чаще являются частично компенсированными или даже некомпен-
сированными, что требует искусственной коррекции.
15.5.2. Дыхательный ацидоз
Причиной развития дыхательного ацидоза у человека является гиперкап-
ния, т. е. повышение содержания и напряжения СО2 в крови (выше 45 мм
рт. ст.) в результате того, что скорость образования СО2 в тканях превыша-
ет скорость его выделения легкими. Это одно из наиболее тяжелых нару-
шений кислотно-основного состояния. В зависимости от скорости возник-
новения и тяжести, выделяют острый и хронический дыхательный ацидоз.
Острое повышение напряжения СО2 в плазме крови вызывает рост со-
держания Н+-ионов до почти двукратного их увеличения, а также повыше-
ние концентрации НСО3 за счет реакции, катализируемой карбоангидра-
зой:
СО2 +Н2О -> Н2СО3 -> Н+ + НСО3
Острая гиперкапния вызывает повышение концентрации Н+-ионов в
соотношении: на каждый мм рт. ст. прироста РСО2 увеличение [Н+] на
0,75 нмоль/л.
Компенсаторные реакции организма, возникающие немедленно вслед
за ростом концентрации Н+-ионов, состоят в связывании избытка водо-
родных ионов протеинами, гемоглобином и фосфатной буферной систе-
мой. Спустя несколько часов включаются почечные компенсаторные про-
цессы — в канальцах почек активируется обмен Н+-ионов на Na+, повы-
шается реабсорбция HCOJ, избыток Н+-ионов выводится с солями аммо-
ния, прежде всего в виде NH4Cl. Избыточное выведение хлорида аммония
с мочой влечет за собой усиление потерь СГ и истощение его запасов в
организме, что замедляет процессы почечной компенсации ацидоза и тре-
бует экзогенного восполнения хлоридного аниона. Содержание НСО3 в
жидкостях внутренней среды в результате активации реабсорбции в поч-
ках повышается, актуальный бикарбонат крови становится выше стан-
дартного бикарбоната при неизмененном количестве буферных оснований
крови и, если причина гиперкапнии устранена (например, при искусст-
венной вентиляции легких), на смену дыхательному ацидозу в результате
роста избытка буферных оснований может формироваться недыхательный
«постгиперкапнический» алкалоз. Избыток Н+-ионов во внеклеточной
664 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
среде приводит к поступлению в клетки не только Н+-ионов, но и ионов
Na+, что вызывает выход из них ионов К+ и развитие гиперкалиемии. По-
вышается и концентрация фосфатов в плазме. В то же время при компен-
сации ацидоза накопление в плазме НСО2 снижает содержание калия в
плазме крови, а «постгиперкапнический» алкалоз может вести к гипока-
лиемии.
15.5.3. Недыхательный ацидоз
Ацидоз возникает при превышении образования в организме над возмож-
ностями выведения из него нелетучих органических и неорганических ки-
слот, при повышенной потере организмом оснований и относительном из-
бытке кислот, а также при поступлении кислот в организм из внешней
среды. Соответственно этому, выделяют четыре типа недыхательного аци-
доза: 1) метаболический — возникающий при повышенном образовании
нелетучих кислот в случаях шока, гипоксии, лихорадки, гипертиреоза, не-
достаточности кровообращения, диабета, голодания и усиленной мышеч-
ной работы; 2) выделительный — формирующийся в результате сниженного
выделения нелетучих кислот почками (уремия, дистальный канальцевый
ацидоз, пиелонефрит) или повышенных потерь HCOJ с содержимым ки-
шечника (кишечные и желчные свищи, диарея); 3) экзогенный — прояв-
ляющийся при введении в организм с лечебной целью нелетучих кислот
(прием салицилатов, хлорида аммония, парентеральное вливание 0,1 моль/
л раствора хлористоводородной кислоты); 4) дилюционный — причиной ко-
торого является разведение внеклеточной жидкости, ведущее к уменьше-
нию концентрации НСО3 (при быстрых и чрезмерных внутривенных вли-
ваниях изотонического раствора хлорида натрия, в первой фазе действия
осмотических диуретиков типа маннита).
Общие компенсаторные реакции организма при недыхательных типах
ацидоза состоят в буферировании кровью избытка Н+-ионов, прежде всего
бикарбонатной буферной системой, и выведении избытка освобождающе-
гося из нее СО2 через легкие. При ацидозе метаболического типа важней-
шим компенсаторным механизмом является выделение почками избытка
Н+. К числу компенсаторных реакций при недыхательном ацидозе (глав-
ным образом, метаболического типа) также относятся: 1) активация сим-
патоадреналовой системы, повышающая производительность сердца и вы-
зывающая вазодилатацию за счет стимуляции в условиях ацидоза преиму-
щественно бета-адренорецепторов сосудов; усиливающийся при этом тка-
невый кровоток повышает доставку кислорода и уменьшает тканевый аци-
догенез; 2) смещение кривой диссоциации оксигемоглобина вправо, что
облегчает отдачу кислорода тканям; 3) возбуждение дыхательного центра,
приводящее к гипервентиляции, облегчающей выведение СО2.
Наиболее распространенным метаболическим ацидозом у человека яв-
ляется лактат-ацидоз. При физиологических условиях обмена углеводов
во всех клетках и тканях организма образуется молочная кислота, посту-
пающая во внеклеточную жидкость. Извлекается при этом лактат из крови
в значительных количествах преимущественно печенью и корковым веще-
ством почек, где основная его часть превращается в глюкозу, а остаток ме-
таболизирует до СО2 и воды. При гипоксии в тканях образуется больше
лактата, чем может превратиться в пируват, и отношение лактат/пируват, в
норме составляющее 10:1, возрастает, т. е. развивается избыток лактата в
крови. К появлению избытка лактата и лактат-ацидозу ведет и чрезмерная
активация симпатоадреналовой системы при стрессе, поскольку адреналин
15. Кислотно-основное состояние • 665
снижает поступление пирувата в цикл трикарбоновых кислот и, следова-
тельно, повышает образование лактата. Кроме того, возникающая при
симпатической стимуляции констрикция сосудов печени снижает утилиза-
цию ею избытка лактата. Основными поставщиками лактата в плазму кро-
ви являются эритроциты (до 25 % всего лактата) и активно работающие
скелетные мышцы.
Таким образом, накопление лактата в крови является следствием изме-
нения как его продукции, так и утилизации. Нормальное содержание лак-
тата в плазме крови составляет около 1 ммоль/л, но лишь возрастание
концентрации молочной кислоты свыше 4 ммоль/л является патологиче-
ским и ведет к лактат-ацидозу.
Одним из компенсаторных процессов, реализуемых при этом на уровне
образования лактата, является метаболическая ауторегуляция по механиз-
му обратной связи: ацидоз подавляет активность ключевого фермента гли-
колиза в эритроцитах фосфофруктокиназы, в результате скорость гликоли-
за падает и образование молочной кислоты снижается.
15.5.4. Дыхательный алкалоз
Дыхательный алкалоз возникает как следствие гипервентиляции, в резуль-
тате которой выведение СО2 из крови превышает его образование в орга-
низме. В крови при этом снижается РСО2, т. е. возникает гипокапния и
уменьшается концентрация водородных ионов.
Остро возникающий дыхательный алкалоз, как правило, в начальный
период декомпенсирован, и pH может повышаться до критических преде-
лов (7,6—7,8).
Компенсаторные реакции организма при дыхательном алкалозе начина-
ются прежде всего в системе крови с буферирования сдвига pH, что осу-
ществляется пополнением недостатка Н+ путем диссоциации протеинов и
фосфатов, при этом содержание не гидрокарбонатных анионов возрастает.
В эритроцитах при алкалозе увеличивается активность фосфофруктокина-
зы, и образуются повышенные количества молочной кислоты, восполняю-
щей низкую концентрацию Н+-ионов.
Почечная компенсация дыхательного алкалоза формируется медленно,
в течение 2—4 сут. Она проявляется торможением в дистальных отделах
нефрона обмена протонов на Na+, подавлением реабсорбции НСО3 и сек-
реции Н+. При этом повышается экскреция с мочой НСО7, в крови умень-
шается избыток буферных оснований.
Сниженные концентрации СО2 в плазме крови ведут к сужению внут-
риорганных сосудов, гипоксии и активации гликолиза в клетках, что
приводит к увеличению продукции ими молочной кислоты и компенса-
ции алкалоза. Ослабление тонуса сосудодвигательного центра при недос-
татке возбуждающих его Н+ и, как следствие, уменьшение симпатических
эффектов на сосуды способствует артериальной гипотензии, снижению
кровотока в тканях и активации образования в них молочной кислоты, а
снижение почечного и печеночного кровотока ослабляет ее утилизацию
из крови, способствуя тем самым восстановлению [Н+]. Кислородному
голоданию тканей и, соответственно, росту образования в них лактата
способствует и сдвиг при алкалозе кривой диссоциации оксигемоглобина
влево, уменьшающий отдачу кислорода. В то же время в эритроцитах
растет содержание 2,3-дифосфоглицерата, что ограничивает величину
сдвига кривой влево и не позволяет резко снизиться диссоциации оксиге-
моглобина.
666 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Следствием алкалоза является изменение внутреннего баланса электро-
литов: перемещение К+ из плазмы крови в клетки и гипокалиемия; выход
Na+ из клеток в плазму крови и увеличение его экскреции с мочой; умень-
шается в плазме крови и содержание фосфата.
15.5.5. Недыхательный алкалоз
Различают три основных типа недыхательного алкалоза: 1) выделительный
(при избыточных потерях Н+-ионов в сочетании с потерями СГ или К+);
2) экзогенный или нагрузочный (при поступлении в организм НСОу или его
предшественников); 3) контракционный (при уменьшении объема внекле-
точной жидкости без соответствующего снижения концентрации НСО^).
Выделительный алкалоз может быть следствием: потери ионов Н+ и Cl-
при частой рвоте; выведения Н+-ионов с мочой при подавлении реабсорб-
ции ионов СГ в почках при назначении диуретиков-хлоруретиков; избыт-
ка глюкокортикоидов; потери Н+-ионов в комбинации с ионами К+ при
альдостеронизме; повышении секреции Н+-ионов канальцами почек; поте-
ри Н+-ионов с калом.
Недыхательный, особенно выделительный, алкалоз сопровождается, как
правило, гипокалиемией, причиной которой является перемещение калия
из плазмы крови в клетки, а также общий дефицит калия в организме.
Наиболее выражен дефицит калия при алкалозе, связанном с гиперкорти-
цизмом. Обмен калия и Н+-ионов имеет положительную обратную связь:
алкалоз усиливает гипокалиемию, но гипокалиемия усиливает недыхатель-
ный алкалоз. Гипокалиемический алкалоз нередко бывает хлоррезистив-
ным, т. е. введение экзогенного СГ не устраняет алкалоз. Дефицит калия
вызывает активацию канальцевой реабсорбции НСО3 в почках, гипокалие-
мия стимулирует продукцию NH3 в почках и снижение реабсорбции СГ,
вследствие чего при искусственной компенсации алкалоза введением в ор-
ганизм кислых валентностей необходимым условием является восстановле-
ние содержания калия.
Недыхательный алкалоз редко бывает компенсированным, чаще он суб-
компенсирован, при этом pH > 7,45. Компенсаторные реакции состоят в
буферировании слабыми кислотами негидрокарбонатных буферных сис-
тем. Как и при дыхательном алкалозе, в этом случае увеличивается образо-
вание молочной кислоты, прежде всего в эритроцитах, благодаря актива-
ции гликолиза.
Компенсация недыхательного алкалоза со стороны легких заключается
в уменьшении вентиляции, что связано со снижением возбудимости дыха-
тельного центра в условиях уменьшения в плазме крови величины [Н+] и
[СО2]. Следствием гиповентиляции является повышение РСО2 в плазме
крови. Почечная компенсация недыхательного алкалоза осуществляется за
счет усиленного выведения НСО7 и падения экскреции титруемых кислот.
Контракционный алкалоз усиливается за счет изменений функции по-
чек, так как уменьшение при нем объема внеклеточной жидкости ведет к
снижению почечной экскреции НСО3 и увеличивает выраженность алка-
лоза. Таким образом, искусственная компенсация контракционного алка-
лоза достигается уже при восполнении дефицита внеклеточного объема
вводимой жидкостью, что не только снижает концентрацию HCOj в кро-
ви, но и усиливает экскрецию бикарбоната почками, способствуя тем са-
мым устранению алкалоза. Однако для более эффективного устранения
контракционного алкалоза необходимо искусственно восполнять имею-
щийся в организме дефицит С Г.
15. Кислотно-основное состояние • 667
15.5.6. Общие закономерности компенсации нарушений кислотно-
основного состояния
Как правило, компенсация изменений кислотно-основного состояния в
организме человека является вторичным физиологическим процессом в
ответ на первичное нарушение одной из систем поддержания pH. При
этом отклонение pH восстанавливается благодаря компенсаторным реак-
циям ненарушенных систем регуляции pH.
Основные компенсаторные реакции у человека:
• реакция буферных систем внеклеточной жидкости и перемещение
Н+-ионов через клеточные мембраны в направлении, нормализую-
щем сдвиги pH крови;
• дыхательная компенсация, вызванная изменениями pH крови и влия-
нием Н+-ионов на дыхательный центр, что в результате увеличения
или уменьшения альвеолярной вентиляции меняет РСО2 в альвеоляр-
ном воздухе и артериальной крови и таким образом противодействует
сдвигам pH. При этом повышение РаСО2, является потенциальной
причиной роста [Н+], так как реакция с участием карбангидразы:
СО2 + Н2О -> Н2СО3 -> Н+ + НСО; ведет к росту [Н+]. Эта дыхатель-
ная компенсация первичного накопления либо дефицита некарбоно-
вых кислот или оснований, как правило, не восстанавливает pH кро-
ви до нормы;
• компенсация костной тканью: бикарбонатный ион при ацидозе осво-
бождается из кости для нейтрализации некарбоновых кислот крови, а
Н+ связывается костной тканью;
• почечная компенсация путем увеличения экскреции некарбоновых ки-
слот или оснований при дыхательных ацидозах или алкалозах, веду-
щих к изменениям РСО2 в плазме крови, уменьшения или увеличе-
ния выведения Н+ и НСОу из организма.
ГЛАВА 16
Репродуктивная функция человека
Отличительной особенностью репродуктивной функции человека от дру-
гих физиологических функций организма является то, что ее нормальное
функционирование проводит к слиянию половых клеток мужского и жен-
ского организмов в процессе половой репродукции. Ооциты и спермато-
зоиды называются женскими и мужскими половыми клетками, или гаме-
тами. Мужские и женские гаметы в зрелой форме содержат гаплоидное
число хромосом, т. е. половину нормального числа. Гаплоидное число хро-
мосом в гаметах формируется в процессе сперматогенеза и оогенеза
(рис. 16.1). В мужском организме мейотическое деление сперматогенных
клеток происходит постоянно на протяжении всей жизни после начала пе-
риода полового созревания (пубертатный период). Напротив, в ооците гап-
лоидное число хромосом образуется непосредственно перед овуляцией яй-
цеклетки из фолликула. В результате способности ооцита и сперматозоида
Разрыв фолликула
Сперматиды
(гаплоиды)
Второе
мейотическое
деление
2^ "I Вторичные
сперматоциты
J (гаплоиды)
Первое
мейотическое
-1 деление
(II)
+Х+Х
Второе мейотическое
деление во время
фертилизации
(I)
Вторичный
ооцит (гаплоид)
Первое мейотическое
деление во время
овуляции
Первичный
ооцит (диплоид)
Зигота
44+ХХ
ли+ХУ
Оогенез
Митоз
Сперматозоид (гаплоид]
22 +Х или +Y
44
44
Первичные сперматоциты
(диплоиды)
Сперматогенез
Рис. 16.1. Схема сперматогенеза и оогенеза.
Первичные сперматоциты образуются путем митоза из сперматогоний и содержат диплоидное
число хромосом. В результате первого мейотического деления образуются сперматоциты, со-
держащие гаплоидное число двухроматидных хромосом (22 единичные соматические хромосо-
мы и одна половая — X или Y). После второго мейотического деления образуются спермати-
ды, содержащие гаплоидный набор монохроматидных хромосом. Оогенез начинается со ста-
дии первичного ооцита, содержащего диплоидное число хромосом. Первичный ооцит путем
первого мейотического деления превращается во вторичный ооцит, содержащий гаплоидное
число двухроматидных хромосом (22 единичные соматические хросомы и одна половая Х-хро-
мосома). В процессе оплодотворения ооцита сперматозоидом образуется зигота, содержащая
22 пары соматических хромосом и одну пару половых (XX или XY). Таким образом, в зиготе
восстанавливается диплоидный набор хромосом.
16. Репродуктивная функция человека • 669
соединяться друг с другом во время оплодотворения в женском половом
тракте происходит образование зиготы. Этот процесс называется фертили-
зацией. В зиготе содержится диплоидное число хромосом, как в любой со-
матической клетке организма человека и животных. Две хромосомы из ди-
плоидного числа в зиготе, а именно половые X- и Y-хромосомы, обуслов-
ливают мужской или женский пол будущей особи в новом поколении.
Женская половая клетка содержит только Х-хромосомы, а мужская — X- и
Y-хромосомы. Хромосомы заключаю? в себе гены, которые передают гене-
тические особенности одного поколения другому.
Способность человека как вида передавать одну половину генетической
информации будущего поколения от отца к матери обеспечивается физио-
логическими особенностями репродуктивной функции мужского организ-
ма. Репродуктивная функция женского организма обеспечивает процесс
фертилизации, внутриутробное развитие плода, рождение ребенка и его
вскармливание грудным молоком.
16.1. Половая дифференциация человека
При фертилизации, в зиготе, находящейся в репродуктивном тракте жен-
ского организма, на основе генетической программы первоначально фор-
мируется хромосомный, или генетический, пол человека. Затем в ходе раз-
вития эмбриона формируется гонадный пол, когда недифференцирован-
ные гонады развиваются либо в семенники, либо в яичники. В зависимо-
сти от гонадного пола у плода происходит дифференциация мужских или
женских внутренних и наружных гениталий (формируется фенотипиче-
ский пол). В целом указанные превращения называется половой дифферен-
циацией человека (рис. 16.2).
Стадия индифферентных гонад
Семенники
Хромосомный пол
Гонадный пол
XY
Яичники
Рис. 16.2. Половая дифференциация человека.
Эмбрион имеет мужской (XY) и женский (XX) хромосомный пол. Хромосомный пол сменяет-
ся развитием индифферентных гонад в яичники или семенники. В результате формируется го-
надный пол. Семенники секретируют тестостерон, который обусловливает развитие мужского
фенотипического пола, а отсутствие секреции тестостерона обусловливает развитие женского
фенотипического пола.
670 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
16.1.1. Генетический пол
Половые хромосомы (X и Y) являются единственными из 23 хромосом, ко-
торые определяют пол организма. Половые клетки, продуцируемые жен-
ским организмом, содержат Х-хромосому, тогда как сперматозоиды — X-
или Y-хромосому. Остальные 22 хромосомы предопределяют характеристи-
ки организма, не связанные с полом, и поэтому называются аутосомами.
Во время фертилизации образуется зигота, содержащая 23 пары хромо-
сом. В зиготе, из которой будет развиваться плод женского пола, содер-
жится две Х-хромосомы, т. е. по одной из каждой женской и мужской га-
меты. В зиготе, из которой будет развиваться плод мужского пола, содер-
жится X- и Y-хромосомы. Развитие гонад и гениталий в мужском организ-
ме обусловлено Y-хромосомой. При отсутствии в генах эмбриона Y-хромо-
сомы у эмбриона развиваются яичники. Y-хромосома ответственна за на-
чальный половой диморфизм, т. е. две различные половые формы, мужская
и женская, одного и того же индивидуума.
16.1.2. Гонадный пол
Примерно до 6—7-й недели внутриутробной жизни эмбриона его гонады
развиваются идентично как у мужчин, так и у женщин. Это так называе-
мая индифферентная стадия развития эмбриона, когда половая система
обоих полов развивается по одной генетической программе. В период фор-
мирования гонадного пола у плода развиваются мужской (вольфов) и жен-
ский (мюллеров> протоки. Первоначально развитие этих протоков начина-
ется унипотенциально, т. е. независимо от будущего пола, и только один
из них может развиваться в половой тракт, связанный с конкретным гене-
тическим полом плода. В частности, вольфов проток превращается в
структуры мужского полового тракта, а мюллеров — женского (табл. 16.1).
Закладка индифферентных гонад происходит у эмбриона в виде гонад-
ных валиков. В эпителии гонадных валиков обособляются крупные пер-
вичные половые клетки, которые в дальнейшем дают начало развитию по-
ловых клеток. Первичные половые клетки дифференцируются на 5-й день
после фертилизации. По прошествии 3 нед они определяются среди клеток
внезародышевой энтодермы в стенке желточного мешка (часть первичной
кишки, вынесенная за пределы зародыша). От поверхности внутрь поло-
вых валиков прорастают тяжи, состоящие из первичных половых клеток
фолликулярного эпителия, который выполняет для первичных половых
клеток трофическую роль. В течение 3 нед жизни эмбриона в половых ва-
ликах эти клеточные элементы развивается в ткань, которая будет основой
первичных гонад. Первичные половые клетки из вольфова тела мигрируют
в эту ткань, в которой из них будут образовываться половые клетки или
гаметы. В индифферентный период развития первичных гонад в их стро-
мальной ткани содержится два типа клеток. Один тип клеток на гонадной
Таблица 16.1. Особенности формирования гонадного пола у плода
Генетический статус пола Развиваю- щиеся гона- ды Источники развития гонад Наружные половые ор- ганы
женский проток мужской проток
XY хромосомы (мужской) Семенники Регрессия Развитие Мужские
XX хромосомы (женский) Яичники Развитие Регрессия Женские
16. Репродуктивная функция человека • 671
стадии дифференциации пола развивается в гранулезные клетки яичников
или в клетки Сертоли семенных трубочек в яичках. Второй тип клеток на
стадии гонад дифференцируются в клетки прозрачной оболочки (theca pel-
lucida) в яичниках или в клетки Лейдига в семенниках.
У эмбриона мужского пола на 6—7-й неделе эмбриональной жизни по-
сле миграции первичных половых клеток в первичные гонады в присутст-
вии Y-хромосомы, содержащей SRY-ген, происходит дифференциация
клеток Сертоли. В процессе дифференциации клетки Сертоли располага-
ются вокруг первичных половых клеток, в результате начинается развитие
трубочек семенников в первичных гонадах. Дифференциация мезенхи-
мальных (стромальных) клеток гонад в интерстициальные клетки Лейдига,
которые в последующем будут секретировать мужской половой гормон
тестостерон, начинается с 8—9-й недели и заканчивается на 10-й неделе
развития плода.
У женского эмбриона дифференцирование первичных гонад в яичники
начинается с 9-й недели, когда активируются Х-хромосомы. Их активация
приводит к пролиферации первичных половых клеток, деление которых
осуществляется путем митоза. К моменту рождения плода женского пола в
яичниках содержатся первичные ооциты, которые образуются в период
развития плода из первичных половых клеток путем мейотического деле-
ния. Однако уже в периоде внутриутробного развития мейоз первичных
половых клеток блокируется (стадия поздней профазы) после образования
первичного ооцита. В первичном ооците мейоз на стадии поздней профа-
зы вновь возобновится спустя много лет в одну из будущих овуляций, под
влиянием гонадотропинов гипофиза и гормонов половых желез, в периоде
пубертатного развития женского организма.
16.1.3. Фенотипический пол
Развитие внутренних мужских половых органов у плода происходит под
влиянием тестостерона. Отсутствие его секреции в фетальных клетках Лей-
дига приведет к образованию у плода женских половых путей. Секреция
тестостерона клетками Лейдига у плода мужского пола начинается при-
мерно на 9-й неделе развития, под влиянием хорионического гонадотро-
пина плаценты, секреция которого стимулируется гормоном роста. Под
влиянием тестостерона вольфовы протоки трансформируется в своем раз-
витии в придатки яичек, семявыносящие протки и семенные пузырьки.
Кроме того, на формирование фенотипического пола оказывает влияние
мюллеров ингибирующий фактор (синоним — анти-мюллеров гормон),
секреция которого начинается в яичках примерно с 7-й недели развития
эмбриона и который вызывает регрессию мюллеровых протоков у плода
мужского пола. Механизм регрессии мюллеровых протоков связан с тем,
что мюллеров ингибирующий фактор связывается с рецепторами мембра-
ны мезенхимальных клеток, окружающих эти протоки, а те в свою очередь
играют роль посредников в этом процессе.
У плода, развивающегося по женскому типу, гранулезные клетки и
клетки прозрачной оболочки не секретируют мюллеров тормозящий гор-
мон и тестостерон. При отсутствии аинти-мюллерова гормона мюллеровы
протоки превращается во внутренние женские репродуктивные органу
(маточные трубы, матка, верхняя часть влагалища), и одновременно про-
исходит регрессия вольфовых протоков в связи с отсутствием секреции
тестостерона у плода.
Развитие наружных половых органов. В отличие от внутренних органов
672 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
мужского и женского репродуктивного тракта, первичные наружные гени-
талии у плода могут развиваться либо по мужскому, либо по женскому ти-
пу. У плода, развивающегося по женскому типу, в отсутствии секреции
тестостерона одновременно с уретральными складками и генитальными
выпуклостями развиваются малые и большие половые губы, клитор. У
плода, развивающегося по мужскому типу, клетки Лейдига секретируют
тестостерон, который с этого периода способен превращаться в более ак-
тивный метаболит — дигидротестостерон. Этот гормон регулирует нор-
мальное развитие мужских половых органов, их слияние с уретральными
складками, а также с генитальными выпуклостями. В результате образуется
пенис. Кроме того, слияние генитальных выпуклостей по средней линии
приводит к образованию мошонки, в которую позднее у плода опускаются
яички, а генитальные бугорки развиваются в головку пениса. Клетки Лей-
дига секретируют тестостерон под влиянием лютропина аденогипофиза, а
регуляция секреции клеток Лейдига осуществляется при участии гипотала-
муса. Нейроны гипоталамуса примерно на 4-й неделе развития эмбриона
начинают секретировать гонадолиберин, а на 10-й неделе — это стимули-
рует в гипофизе секрецию лютропина. Другие фенотипические признаки
женского или мужского пола у индивидуума развиваются в пубертатном
периоде. В периоде детства концентрация гонадотропина, фоллитропина и
лютропина сохраняется на низком уровне, а также не функционирует ме-
ханизм отрицательной обратной связи их взаимной регуляции из-за низ-
кой чувствительности нейронов гипоталамуса к циркулирующим в крови
половым стероидам.
16.2. Репродуктивная функция мужского организма
В пубертатный период в мужском организме происходит развитие и ста-
новление репродуктивной функции, характерной для организма взрослого
мужчины. При этом во время пубертатного периода в коре надпочечников
начинается секреция адреналовых мужских половых гормонов — андроге-
нов. В среднем в И-летнем возрасте у мальчиков в семенниках начинается
увеличение количества клеток Лейдига и благодаря их функции уровень
секреции тестостерона постепенно достигает плато взрослого мужчины
(порядка 6 нг/мл плазмы крови). В течение пубертатного периода происхо-
дит рост и развитие тела, изменение в регуляции многих функций орга-
низма, в том числе ЦНС и гипоталамуса. Секреция гонадолиберина в ги-
поталамусе и гонадотропинов (фоллитропина и лютропина) в гипофизе
постепенно достигает уровня взрослого мужчины и сохраняется на относи-
тельно постоянном уровне в среднем до 50 лет, обусловливая характер
синтеза и секреции тестестерона в мужских половых железах (рис. 16.3).
16.2.1. Функции семенников
Основными функциями мужских гонад являются сперматогенез и секре-
ция мужских половых гормонов. Активность мужской репродуктивной
функции является непрерывной и не имеет циклических проявлений, как
в женском организме. Семенники развиваются в брюшной полости плода,
но примерно за 2—3 мес до рождения под влиянием нарастающей концен-
трации тестостерона в плазме крови семенники опускаются в мошонку. В
мошонке температура составляет примерно 34 °C и ниже, что является аде-
кватной температурной средой для нормального течения сперматогенеза.
Оптимальная для сперматогенеза температура в яичках регулируется в про-
16. Репродуктивная функция человека • 673
Содержание
Месяцы Годы
Рис. 16.3. Изменение концентрации тестостерона в плазме крови мужского орга-
низма в различные периоды жизни.
На протяжении жизни мужского организма секреторная функция клеток Лейдига существен-
но изменяется. Высокая концентрация тестостерона отмечается в поздний период внутриут-
робного развития (4 нг/мл), когда он необходим для регуляции интенсивного развития наруж-
ных половых органов. Низкий уровень тестостерона в плазме крови поддерживается до пред-
пубертатного периода, поскольку отсутствуют клетки Лейдига. Следующий подъем секреции
тестостерона отмечается в 11 лет и удерживается в течение пубертатного периода. По оконча-
нии пубертатного периода секреция тестостерона достигает уровня взрослого мужчины и со-
храняется вплоть до 50 лет жизни.
цессе противоточного температурного обмена между притекающей артери-
альной кровью и оттекающей от тканей яичек венозной кровью, а также
при сокращении или расслаблении гладкой мышцы, поднимающей яичко,
что изменяет положение последнего относительно области центральной
температуры тела.
Сперматогенез осуществляется в семенных канальцах, стенки которых
выстланы сперматогенным эпителием, содержащим сперматозоиды и
предшествующие им клетки — сперматогонии, сперматоциты I и II поряд-
ка, а также сперматиды, которые расположены между поддерживающими
клетками Сертоли (рис. 16.4). Плотные контакты между клетками Сертоли
создают надежный барьер с кровью (гематотестикулярный), который пре-
пятствует развитию аутоиммунных реакций организма на антигены, распо-
ложенные на мембране развивающихся половых клеток, а также действию
на сперматогенный эпителий токсичных веществ, попадающих в организм
из внешней среды. Клетки Сертоли выполняют трофическую функцию в
отношении развивающегося сперматозоида, фагоцитируют половые клет-
ки, утратившие способность к делению, а также остатки цитоплазмы деля-
щихся половых клеток. Клетки Сертоли синтезируют белок, способный
связывать тестостерон. Это поддерживает высокую концентрацию в спер-
матогенном эпителии гормона, необходимого для стимуляции сперматоге-
неза.
Клетки Лейдига синтезируют мужские половые гормоны андрогены и
находятся в соединительной ткани между семенными канальцами. Высо-
кая активность этих клеток и соответственно высокая секреция тестосте-
рона имеет место в поздний период развития плода, что необходимо для
22 - 6095
674 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Ядро клетки
Сертоли
Базальная
мембрана ►
Клетка Сертоли
Митохондрия
Просвет
семявыносящей
трубочки
Сперматозоиды
Сперматида
Сперматоцит II
Клетка Сертоли
Сперматоцит I
Плотный контакт
Сперматогония
Рис. 16.4. Расположение сперматогенных клеток в эпителии семявыносящей тру-
бочки.
Семявыносящая трубочка окружена базальной мембраной, на которой находятся клетки Сер-
толи и сперматогенные клетки (сперматогонии, сперматоциты I и II порядка, сперматиды).
Между клетками Сертоли расположены сперматогенные клетки, находящиеся на различной
стадии деления (митоз — сперматогонии и мейоз — сперматоциты I и II порядка) и морфоло-
гической дифференциации (сперматиды). Плотные контакты между клетками Сертоли пред-
ставляют эффективный барьер между кровью и внутренней средой семявыносящей трубочки,
препятствуя попаданию вырабатываемых половых клеток в кровь и развитию иммунной реак-
ции мужского организма на эти клетки.
развития у него наружных половых органов. После рождения, к концу
первого года жизни происходит интенсивная регрессия количества клеток
Лейдига и снижение секреции в гонадах ребенка тестостерона в связи со
снижением секреции лютропина в гипофизе. В результате до начала пу-
бертатного периода в организме ребенка мужского пола не развиваются
первичные и вторичные половые признаки, которые характерны для нор-
мального взрослого организма мужчины. Начало пубертатного периода в
развитии мужского организма связано с началом секреции гонадолиберина
в гипоталамусе и лютропина в аденогипофизе. Под влиянием этих гормо-
нов, клетки Лейдига дифференцируются во взрослый тип и в них начина-
ется секреция тестостерона и дигидротестостерона, обусловливающих по-
ловое развитие мужского организма (табл. 16.2).
16.2.2. Сперматогенез
В пубертатный период развития мужского организма активируется продук-
ция сперматозоидов, которая сохраняется на протяжении репродуктивного
периода жизни мужчины вплоть до глубокой старости. При этом ежеднев-
но продуцируется 200—400 млн. сперматозоидов. Сперматогенез, или об-
разование зрелых сперматозоидов, происходит в семенных канальцах на
протяжении стадий размножения, развития и созревания. Стадия размно-
жения начинается с митотического деления сперматогоний (см. рис. 16.1).
Несколько митотических делений сменяются двумя мейозами, во время
16. Репродуктивная функция человека • 675
Таблица 16.2. Роль основных андрогенов в мужской репродуктивной функции
Гормон Период развития мужского организма
внутриутробный пубертатный взрослый мужчина
Тесто- Стимуляция диф- Стимуляция пубертатного Сперматогенез
стерон ференциации: роста организма Секреция гонадотропинов
Дигид- ротесто- стерон семявынося- щие протоки семенные пу- зырьки придатки яи- чек Рост пениса, семенных тру- бочек Сперматогенез Секреция гонадотропинов Регуляция по механизму об- ратной связи: гипоталамус- гипофиз—половые железы Формирование вторичных половых признаков: разви- тие голосового аппарата, мышц, распределение жира Рост волос по мужскому ти- пу Развитие сальных желез, простаты Половые стимулы Половое поведение Регуляция по механизму об- ратной связи: гипоталамус- гипофиз—половые железы Ткани пениса, семенных пу- зырьков Вторичные половые призна- ки: голос, мышцы, распре- деление жира Количество эритроцитов к крови и содержание в плаз- ме крови липидов Сперматогенез Секреция гонадотропинов Половые стимулы Половое поведение Наружные половые органы Семенные пузырьки Регуляция по механизму об- ратной связи: гипоталамус- гипофиз—половые железы
которых диплоидное число хромосом (первичные сперматоциты) уменьша-
ется наполовину, т. е. становится гаплоидным (сперматоциты I и II поряд-
ка). В стадии развития сперматоциты остаются связанными между собой
цитоплазматическими мостиками, которые исчезают только на финальной
стадии сперматогенеза. Одна первичная сперматогония образует в резуль-
тате деления 16 сперматид. В стадии созревания (спермиогенез) одна спер-
матида продуцирует один сперматозоид. Зрелый сперматозоид обладает
способностью транспортировать хромосомы к ооциту, после того как он
окажется в женском репродуктивном тракте. Период превращения сперма-
тогоний в зрелые сперматозоиды продолжается 70—80 сут.
Зрелый сперматозоид состоит из следующих структурных компонентов
(рис.16.5). Головка сперматозоида (2—5 мкм в диаметре) содержит ядро,
две центриоли и акросому. Ядро занимает большую часть головки сперма-
тозоида и содержит гаплоидный набор хромосом. Акросома головки содер-
жит ионы Са2+ и энзимы, необходимые для проникновения сперматозоида
в ооцит. Жгутик (хвост) сперматозоида имеет длину порядка 100 мкм и со-
держит комплекс микротрубочек, необходимых для движения спермато-
зоида. Митохондрии продуцируют энергию для сократительных белков,
осуществляющих волнообразные движения жгутика сперматозоида
16.2.3. Гормональная регуляция сперматогенеза
Сперматогенез стимулирует лютропин аденогипофиза. Под влиянием лю-
тропина интерстициальные клетки Лейдига секретируют мужской половой
гормон тестостерон. Гормон диффундирует через базальную мембрану в
22'
676 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рис. 16.5. Схематическое изображение
строения зрелого сперматозоида.
Головка сперматозоида содержит ядро с гаплоид-
ным набором хромосом и имеет акросому. Акро-
сома в процессе акросомальной реакции выделяет
энзимы, с помощью которых сперматозоид про-
никает через прозрачную оболочку внутрь ооцита
во время его оплодотворения. Средний отдел
сперматозоида содержит митохондрии, которые
производят энергию для двигательных филамент
жгутика. Зрелый сперматозоид способен достав-
лять в женском репродуктивном тракте хромосо-
мы к ооциту благодаря двигательной активности
жгутика.
семенные канальцы, связывает-
ся со специфическим белком,
который секретируется клетка-
ми Сертоли, и, таким образом,
накапливается в сперматоген-
ном эпителии, стимулируя
сперматогенез. Концентрация
тестостерона во внутренней
среде семенных канальцев в 100
раз выше, чем в плазме крови.
Тестостерон, образующийся в
клетках Лейдига, поступает в
клетки Сертоли и в них превра-
щается в более активный гор-
мон — дигидротестостерон, ко-
торый транспортируется в жид-
кость канальцев, и в присутст-
вии которого происходит деле-
ние сперматогоний.
Тестостерон стимулирует
формирование метаботропных
рецепторов фоллитропина в
клетках Сертоли. Фоллитропин
через эти рецепторы активирует
клетки Сертоли, благодаря тро-
фической и барьерной функции
которых осуществляются про-
цессы митотических и мейоти-
ческих делений, а также спер-
миогенез мужских половых
клеток. Гормон роста аденоги-
пофиза через метаботропные
рецепторы мембраны клеток
Сертоли стимулирует их мета-
болическую активность, а в от-
ношении сперматогоний — активирует начало их митотического деления.
Под влиянием фоллитропина в клетках Сертоли тестостерон ароматизиру-
ется в эстрадиол-17р, который поддерживает нормальное течение спермио-
генеза мужских половых клеток. Фоллитропин и тестостерон являются си-
нергистами стимуляции в клетках Лейдига синтеза белка, который транс-
портирует эстрогены и тестостерон от клеток Лейдига в жидкость, запол-
няющую семенные канатики, в которых эти гормоны активируют сперма-
тонгенез. На рис. 16.6 представлена общая схема стероидогенеза, происхо-
дящего в клетках Лейдига и Сертоли.
Лютропин контролирует метаболическую активность клеток Лейдига,
связываясь, с мембранными рецепторами системы G-белков, которые ак-
тивируют энзимы аденилатциклазу и фосфолипазу С. Вторичным посред-
ником действия лютропина на клетки Лейдига является цАМФ. При этом
в клетках Лейдига в первые минуты действия гормона возникает быстрый
стероидогенный эффект. Эффект проявляется в резком увеличении содер-
жания цАМФ и росте продукции тестостерона за счет активации переме-
щения холестерина из цитозоля клеток на внутреннюю мембрану митохон-
дрий. Лютропин оказывает метаботропный {трофический) долговременный
16. Репродуктивная функция человека • 677
Жидкость просвета канальцев
Рис. 16.6. Схема стероидогенеза в клетках мужских половых желез.
Биосинтез основного мужского полового гормона тестостерона происходит в клетках Лейдига.
Импульсная секреция лютропина (Лт) в аденогипофизе стимулирует в клетках Лейдига секре-
цию тестостерона, концентрация которого в плазме крови также имеет незначительные коле-
бания в течение суток. Лютропин стимулирует секрецию клеток Лейдига путем связывания с
метаботропными рецепторами на их мембране (РЛт). Превращение тестостерона в эстрадиол -
17b и дигидротестостерон происходит в клетках Сертоли. Гормон эстрон синтезируется в клет-
ках Сертоли из андростендиона, образующегося в клетках Лейдига. Тестостерон и другие муж-
ские половые гормоны транспортируются в кровь и жидкость семенных канальцев, оказывая
физиологические эффекты на ткани мужского организма.
эффект на структуру и функцию клетки Лейдига. Тестостерон, который
синтезируется в клетках Лейдига, под влиянием лютропина диффундирует
через базальную мембрану в семенные канатики, где его нормальная кон-
центрация необходима для процесса сперматогенеза. Гормон аденогипо-
физа пролактин и фактор роста ингибин, секретируемые в клетках Сертоли
в качестве модуляторов, повышают стимулирующее действие лютропина
на функцию клеток Лейдига.
В клетках Сертоли секретируются так называемые факторы роста инги-
бин и активин, которые тормозят по механизму отрицательной обратной
связи секрецию гонадотропинов в аденогипофизе, а следовательно, спер-
матогенез.
Функция эстрогенов. Концентрация эстрогенов (эстрон и эстрадиол- 17р)
в плазме крови мужского организма поддерживается за счет их поступле-
ния в кровь из клеток Сертоли (ароматизируются из андрогенов), клеток
пучковой и сетчатой зон коры надпочечников.
Эстрогены по механизму отрицательной обратной связи тормозят син-
тез тестостерона в клетках Лейдига, в результате уменьшается активность
всех стадий сперматогенеза, а также количество рецепторов фоллитропина
на мембране клеток Лейдига и синтез в них адроген-транспортного белка.
678 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Кроме того, эстрогены через мембранные и ядерные эстрогенные рецепто-
ры p-типа оказывают влияние на зрелые клетки Лейдига семенников
взрослых мужчин, а также на сперматогонии, сперматоциты и развиваю-
щиеся сперматиды. Однако эстрогены не способны оказывать свое физио-
логическое действие на зрелые сперматозоиды, поскольку в их ядре нет эс-
трогеновых рецепторов. С p-типом эстрогенных рецепторов на мембране
клеток Лейдига способны связываться такие внеклеточные гуморальные
субстанции, как пептидные факторы роста и цитокины, и, таким образом,
стимулировать секреторную функцию этих клеток. Роль эстрогенов заклю-
чается в модуляции образования рецепторов к аквапорину-I на мембране
клеток проксимальных отделов протоков придатков яичек. Под влиянием
аквапорина-1 увеличивается реабсорбция воды из жидкости семенных ка-
нальцев и, таким образом, регулируется нормальное осмотическое давле-
ние в канальцевой среде, окружающей зрелые сперматозоиды.
16.2.4. Мужской половой акт
Половое влечение является биологической мотивацией, формирующейся
на основе наследственно обусловленных механизмов гормональной регуля-
ции полового поведения и научения в процессе индивидуального психо-
физиологического развития человека. Наследственно обусловленные меха-
низмы полового влечения связаны с функцией структур гипоталамуса как
первичного центра половой мотивации и лимбической системы. Синхро-
низация во времени активности ядер лимбической системы (комплекс
ядер: миндалина—гипоталамус—мезенцефальные структуры—миндалина)
формирует функциональное состояние, характерное для сексуального по-
ведения. Ассоциативные области коры головного мозга и, прежде всего,
префронтальный неокортекс обеспечивают психофизиологическую регуля-
цию полового поведения (выбор объекта полового влечения, морально-
этический опыт общения половых партнеров). В мужском организме фи-
зиологические механизмы регуляции репродуктивной функции поддержи-
вают исходный уровень возбудимости физиологических систем или так на-
зываемое состояние предварительной нейрогуморальной готовности, кото-
рое обусловливает половое поведение.
16.2.4.1. Стадии мужского полового акта
Мужской половой акт подразделяется на стадии возбуждения, оргазма и
разрешения. Психические стимулы существенно увеличивают способность
субъекта выполнять половой акт. Например, эротические переживания
или даже сновидения сексуального характера могут сопровождаться семя-
извержением.
Основным эффектом стадии возбуждения мужчины является эрекция.
Эрекция — это появление механической твердости пениса в результате уве-
личения его кровенаполнения, обеспечивающая реализацию мужского по-
лового акта. Степень эрекции прямо зависит от психических и физических
сексуальных стимулов, действующих на механизмы ее регуляции.
Основным физиологическим механизмом, вызывающим у мужчины
эрекцию пениса, является расслабление гладких мышц артерий пещеристых
тел и губчатого тела уретры с усилением их кровенаполнения. Регуляция
эрекции осуществляется центральными и периферическими нервными ме-
ханизмами.
Центральная нервная регуляция эрекции. Центры, регулирующие возник-
16. Репродуктивная функция человека • 679
новение эрекции, находятся в паравентрикулярном ядре и в преоптиче-
ской области гипоталамуса. Глутаматергические нейроны медиальной пре-
оптической области гипоталамуса, имеющие на своей мембране NMDA ре-
цепторы, при участии возбуждающего медиатора глутамата инициируют
эрекцию пениса через парасимпатические центры эрекции II—IV крестцо-
вых сегментов спинного мозга. Функция NMDA рецепторов на мембране
нейронов гипоталамических центров эрекции усиливается (модулируется)
оксидом азота. Оксид азота увеличивает активность нейронов медиальной
преоптической области и паравентирикулярного ядра гипоталамуса, имею-
щих на своей мембране D2 тип дофаминовых рецепторов. Терминали ней-
ронов дофаминергической системы мозга, проецирующиеся к гипоталамиче-
ским центрам эрекции, выделяют нейротрансмиттер дофамин и активиру-
ют указанные гипоталамические центры. Дофаминергические нейроны ги-
поталамуса иннервируют нейроны пояснично-крестцовых сегментов и че-
рез эти нейроны активируют вегетативные и соматические рефлексы сек-
суального поведения, в частности, усиливают эрекцию.
Торможение эрекции происходит при участии нейронов паравентрику-
лярного ядра гипоталамуса. На мембране нейронов паравентрикулярного
ядра гипоталамуса локализованы рецепторы, агонистом которых является
ГАМК. Активация ГАМКа типа мембранных рецепторов нейронов гипота-
ламического центра эрекции снижает их активность. Нейроны паравентри-
кулярного ядра через нисходящие проекции аксонов и при участии
ГАМКВ типа рецепторов мембраны нейронов пояснично-крестцовых сег-
ментов спинного мозга тормозят эрекцию.
Центральная нервная регуляция эрекции осуществляется медиаторными
системами мозга, которые оказывают нисходящее активирующее влияние
на спинальные центры эрекции — парасимпатические нейроны крестцо-
вых сегментов спинного мозга. Так, серотонинергическая система мозга,
включающая нейроны вентральной ретикулярной формации продолговато-
го мозга, проецирует аксоны своих нейронов к нейронам поясничных и
крестцовых сегментов спинного мозга, которые иннервируют органы ма-
лого таза. Серотонин выделяется из окончаний серотонинергических ней-
ронов и связывается с рецепторами типа 5-НТ2С, расположенных на мем-
бранах нейронов спинальных центров эрекции. Снижение активности се-
ротонинергических нейронов ядер ретикулярной формации продолговато-
го мозга уменьшает экзоцитоз серотонина в терминалях нейронов, в ре-
зультате увеличивается активность парасимпатических нейронов пояснич-
ных и крестцовых сегментов спинного мозга, что усиливает эрекцию поло-
вого члена. Норадренергические нейроны области голубого пятна ствола моз-
га проецируют свои терминали к нейронам крестцовых сегментов спинно-
го мозга, которые непосредственно регулируют кровенаполнение сосудов
пениса и вызывают его эрекцию.
Периферическая регуляция эрекции осуществляется через парасимпатиче-
ские центры II—IV крестцовых сегментов спинного мозга. Парасимпатиче-
ские волокна богато иннервируют гладкие мышцы тканей пениса. При
эрекции под влиянием гипоталамических центров ее регуляции и указан-
ных выше медиаторных систем мозга возрастает активность парасимпати-
ческих центров эрекции спинного мозга. Холинергическая иннервация
оказывает непрямой расслабляющий эффект на гладкие мышцы каверноз-
ных тел и пещеристых тел уретры, увеличивая их кровенаполнение. Аце-
тилхолин выделяется из нервных окончаний и связывается с М2-холиноре-
цепторами пресинаптических окончаний адренергических нервов, которые
иннервируют гладкие мышцы указанных структур пениса. Активация М2-
680 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Гуанозинтрифосфат
Циклический
гуанозинмонофосфат
цГМФ-зависимые
протеинкиназы
Фосфодиэстеразы
Регнуляция активности
ионных каналов
холинорецепторов тормозит
высвобождение адреналина
из окончаний адренергиче-
ских нервов.
Функцией адренергиче-
ской иннервации тканей пе-
ниса является уменьшение
его кровенаполнения при
отсутствии половой мотива-
ции у мужчины. Высвобож-
дение норадреналина стиму-
лирует преимущественно а,-
адренорецепторы мембраны
гладкомышечных клеток
стенки спиральных сосудов
пениса, вызывая их сокра-
щение. В кавернозных телах
адреналин вызывает сокра-
щение трабекулярных глад-
комышечных волокон. В ре-
зультате активации адренер-
гических нервных волокон
Рис. 16.7. Молекулярный механизм действия
NO на клетки гладких мышц сосудов каверноз-
ных тел пениса.
Растворимая гуанилатциклаза, представленная в свя-
занной (с мембраной) и растворимой изоформах, со-
держится практически во всех клетках. В гладких
мышцах кавернозных тел растворимая гуанилатцик-
лаза является основным рецептором для оксида азота.
Активность фермента повышается примерно в 400
раз после присоединения оксида азота. Энзим ката-
лизирует превращение гуанозинтрифосфата в цикли-
ческий гуанозинмонофосфат. Циклический гуано-
зинмонофосфат в эукариотических клетках путем из-
менения активности ионных каналов, фосфодиэсте-
разы и протеинканазы вызывает расслабление глад-
ких мышц сосудов кавернозных тел. В результате
происходит их кровенаполнение и возникает эрекция
пениса.
повышается тонус гладких
мышц тканей пениса и по-
нижается его кровенаполне-
ние.
Торможение высвобожде-
ния адреналина из оконча-
ний адренергических воло-
кон под влиянием ацетилхо-
лина вызывает расслабление
трабекулярных гладкомы-
шечных волокон, увеличение
кровенаполнения каверноз-
ных тел и эрекцию полового
члена.
Ацетилхолин при высво-
бождении из терминалей хо-
линергических волокон уси-
ливает эрекцию полового
члена за счет активации М3-холинорецепторов мембраны эндотелия сосу-
дов пениса и образования в них оксида азота. Основная роль оксида азота
в регуляции эрекции заключается в расслаблении гладких мышц каверноз-
ных тел и сосудов пениса. Механизм стимуляции эрекции пениса при уча-
стии оксида азота заключается в следующем (рис. 16.7). Фермент гуани-
латциклаза гладких мышечных клеток кавернозных тел является основным
рецептором оксида азота, поскольку активность фермента после связыва-
ния с оксидом азота возрастает примерно в 400 раз. Энзим катализирует
превращение гуанозинтрифосфата в циклический гуанозин монофосфат,
который вызывает расслабление гладкомышечных клеток кавернозных тел,
открывает в их мембране кальций-зависимые калиевые ионные каналы.
Выход ионов калия из гладкомышечных клеток гиперполяризует их мем-
бранный потенциал, что вызывает закрытие L-типа потенциалзависимых
16. Репродуктивная функция человека • 681
кальциевых ионных каналов. В результате прекращается диффузия ионов
Са2+ из внеклеточной среды в саркоплазму гладкомышечных клеток и про-
исходит их расслабление, вызывающее кровенаполнение кавернозных тел
пениса. АТФ в качестве активатора открытия калиевых ионных каналов
мембраны гладкомышечных клеток кавернозных тел также регулирует то-
нус их гладких мышц, увеличивая или уменьшая эрекцию пениса.
Во время полового акта у мужчин происходит раздражение механоре-
цепторов головки полового члена. Парасимпатические импульсы от нейро-
нов сакральных сегментов спинного мозга, наряду с эрекцией, вызывают
секрецию слизи в уретральных и бульбоуретральных железах. Эта слизь по-
ступает по мочеиспускательному каналу пениса во влагалище и способст-
вует увлажнению (любрикации) коитуса.
Импульсы от механорецепторов головки полового члена по афферент-
ным волокнам достигают крестцовых сегментов спинного мозга и усилива-
ют возбуждение парасимпатических нейронов. При пороговом уровне аф-
ферентных импульсов от механорецепторов головки полового члена реф-
лекторно возбуждаются сегментарные центры спинного мозга (Ц-Ьц), вы-
зывающие выброс спермы из придатков яичек, или эякуляцию.
16.2.4.2. Регуляция эякуляции
Эякуляция (семяизвержение) представляют собой заключительную
часть мужского полового акта. Эякуляции предшествует процесс эмиссии.
Он начинается в результате сокращения гладких мышц семявыносящих
протоков. Возбуждение нейронов сегментарных центров эякуляции акти-
вирует преганглионарные симпатические волокна нейронов поясничных
сегментов, которые прерываются в нижнем брыжеечном узле и подчрев-
ном сплетении. Постганглионарные симпатические волокна оканчиваются
на гладких мышцах семенных пузырьков и семявыносящих протоков. Из
терминалей симпатических волокон высвобождается медиатор норадрена-
лин, который активирует а-адренорецепторы мембраны гладкомышечных
волокон и при участии вторичных посредников открывает кальциевые
ионные каналы, что повышает внутриклеточный уровень ионов Са2+ и вы-
зывая сокращение гладких мышц семявыносящих протоков придатков.
Сокращение этих мышц выбрасывает сперму из протоков во внутреннюю,
или предстательную, часть уретры. Сокращение гладких мышц этих прото-
ков усиливается окситоцином, концентрация которого во время эякуляции
в плазме крови у мужчин повышается примерно в пять раз. Окситоцин
действует на гладкие мышцы семявыносящих протоков через окситоцино-
вые рецепторы на мембране гладкомышечных клеток. Внутренняя часть
уретры является рефлексогенной зоной. Раздражение этой зоны спермой
возбуждает парасимпатические нейроны сегментов Sn—S1V, иннервирую-
щие гладкие мышцы предстательной железы. В результате секрет предста-
тельной железы начинает также поступать в уретру и смешиваться со спер-
мой, образуя там семенную жидкость. Процесс до момента образования се-
менной жидкости является эмиссией. Раздражение спермой рефлексоген-
ной зоны предстательной части уретры рефлекторно через крестцовые сег-
менты спинного мозга вызывает сокращения поперечно-полосатой муску-
латуры луковично-губчатых и седалищно-пещерестых мышц, а также
мышц тазового дна. Ритмическое сокращение этих мышц сдавливает про-
ксимальные отделы кавернозных тел и губчатое тело мочеиспускательного
канала, способствуя выбросу семенной жидкости из уретры во влагалище
женщины. Этот процесс называется эякуляцией.
682 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
16.2.4.3. Оргазм
Оргазм — эмоционально-психическое ощущение полового наслаждения у
мужчин, которое появляется во время семяизвержения. Интенсивность ор-
газма у мужчин напрямую коррелирует с концентрацией окситоцина в
плазме крови. Окситоцин сенсетизирует нейроны крестцового отдела
спинного мозга и головного мозга, участвующих в формировании сексу-
ального чувства у мужчин. Стадия оргазма мужского полового акта сменя-
ется стадией разрешения, во время которой в течение 1—2 мин прекраща-
ется эрекция, состоянием рефрактерности в центральной и перифериче-
ской регуляции эрекции. Основным проявлением стадии разрешения яв-
ляется снижение кровенаполнения кавернозных тел пениса, уменьшение
его объема и механической твердости, а также снижение у мужчины сексу-
ального чувства (период рефрактерности).
16.3. Репродуктивная функция женского организма
Женская репродуктивная функция осуществляется в виде двух больших
стадий подготовки организма женщины к оплодотворению яйцеклетки и
гестации. Последняя традиционно обозначается как беременность.
Подготовка женского организма к оплодотворению яйцеклетки осуще-
ствляется в среднем в течение 28 дней, но может варьировать в пределах
21—33 дня. В этот период
Динамика
секреции
гонадотропинов
Доминирует
секреция
фоллитропина
|6 Дес
|g мсь| |
Доминирует
секреция
лютропина
Доминирует
секреция
фоллитропина
46 и более"!
Старость
16-45
1-11 ,
I _ J Pei
1— ( Детство Пубертат
_ I Грудной возраст
Плод
12-1^ | Старосп
4_Репродуктивный
период
Годы
жизни
Рис. 16.8. Динамика секреции концентрации
фоллитропина и лютропина в плазме крови в
различные периоды жизни женского организма.
От момента начала секреции гонадотропинов у плода
и до окончания пубертатного периода секреция фол-
литропина и лютропина в женском организме имеет
сходную динамику с таковой в мужском организме.
В репродуктивный период жизни женщины секреция
гонадотропных гормонов имеет циклический характер
в соответствии с овариальным и менструальным цик-
лами.
женская репродуктивная
функция обеспечивает со-
зревание яйцеклетки в яич-
никах. Созревание яйце-
клетки происходит в тече-
ние овариального цикла.
Подготовка женского орга-
низма к гестации характери-
зуется циклическими фи-
зиологическими процесса-
ми, происходящими в ре-
продуктивном тракте жен-
ского организма, прежде
всего в эндометрии матки, и
называются маточным, или
менструальным, циклом.
Принципиальной особенно-
стью функциональной ак-
тивности женской репро-
дуктивной функции являет-
ся ежемесячная циклич-
ность секреции гонадотро-
пинов (рис. 16.8), которая
обусловливает нормальное
течение овариального и
менструального циклов и их
длительность.
16. Репродуктивная функция человека • 683
16.3.1. Овариальный цикл и оогенез
Овариальный цикл состоит из трех физиологических фаз: фолликулярной,
овуляторной и лютеальной, или фазы желтого тела. Фолликулярная фаза
овариального цикла начинается у женщины с момента начала менструаль-
ного кровотечения. Эта фаза варьирует по времени от 9 до 23 дней, но яв-
ляется относительно постоянной у каждой женщины. Овуляторная фаза
продолжается примерно 1—3 дня й заканчивается овуляцией. Заключи-
тельная фаза овариального цикла, лютеальная, в течение которой сохраня-
ется гормональная активность желтого тела, длится примерно 14 дней.
16.3.1.1. Фолликулярная фаза
В течение каждого овариального цикла начинается медленное развитие
фолликулов, количество которых нарастает в течение двух последующих
циклов. В этот период примерно 20 фолликулов достигают размера 2—
4 мм и в последующем цикле, благодаря появлению на мембране фоллику-
лярных клеток рецепторов к фоллитропину, они начинают развиваться под
влиянием этого гормона. Примерно за одну неделю в течение фолликуляр-
Чрогесгероь
Капилллр
Эстрон—> Эстрадиол-17
---► Холестерин
Прегненолон
-----Прогестерон
Клетка
theca interna
Андростедин —> Тестостерон
Ацетат |
Эстрон
РФт
Гранулезная
клетка
Эстрадиол-17
Андростендион “►Тестостерон
Рис. 16.9. Схема стероидогенеза в клетках женских половых желез.
Эффективность развития фолликулов в яичниках зависит от регулирующего влияния на них
гонадотропинов аденогипофиза. Рецепторы, на мембране клеток внутренней оболочки — к
лютропину (РЛт) и на мембране гранулезных клеток — к фоллитропину (РФт), появляются в
преантральной и ранней антральной стадиях развития фолликула. Связывание гормонов с ре-
цепторами к лютропину и фоллитропину стимулирует в клетках синтез стероидных гормонов.
Лютропин в текальных клетках стимулирует превращение ацетата и холестерина в андрогены
(70 % от циркулирующих в плазме крови). Под влиянием лютропина текальные клетки синте-
зируют небольшое количество эстрогенов. Эстрогены и андрогены в небольшом количестве
диффундируют в гранулезные клетки и под влиянием фоллитропина эти гормоны ароматизи-
руются в эстрогены. Основными эстрогенами являются эстрадиол-17(3 и эстрон.
684 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
МЕ/л
100—1
Овуляция
80-
60-
40-
0
Менструация Фоллитропин
Дни
Рис. 16.10. Динамика содержания лютропина и фоллитропина в плазме крови в те-
чение менструального цикла.
Успешное развитие овуляции во время короткой овуляторной фазы зависит от резкого и бы-
стро проходящего пика секреции лютропина и фоллитропина. Пик секреции гонадотропинов
нарастает в течение примерно 15 ч, достигает относительно постоянного уровня (следующие
15 ч), а затем снижается до исходного уровня в течение последующих 20 ч.
ной фазы один из фолликулов, на мембране которого выше, чем у других
фолликулов, плотность рецепторов к фоллитропину, достигает размеров в
среднем 11 мм и становится доминантным (вторичный фолликул). Это
обусловлено тем, что в нем синтезируется больше, чем в других фоллику-
лах, эстрадиола-17р. При высокой плотности рецепторов к фоллитропину
на мембране доминантного фолликула, в нем сохраняется способность
синтезировать эстрадиол-17р во время циклического снижения секреции
фоллитропина в аденогипофизе женщины. В этих условиях другие фолли-
кулы, имеющие низкую плотность рецепторов к фоллитропину на своей
мембране, синтезируют незначительное количество эстрадиола-17р и под-
вергаются атрезии. Функция фоллитропина в развитии доминантного фол-
ликула заключается в следующем. Этот гормон связывается с рецепторами
мембраны гранулезных клеток и стимулирует в них синтез ароматазы, пре-
вращающей тестостерон в эстрадиол^17р. Тестостерон синтезируется во
внутренних клетках наружной оболочки фолликула (theca) и диффундиру-
ет в гранулезные клетки, где гормон превращается в эстрогены (рис. 16.9).
Количество андрогенов фолликулярного происхождения у женщины со-
ставляет примерно 70 % от их общей концентрации в плазме крови. Гра-
нулезные клетки содержат эстрогеновые рецепторы, с которыми связыва-
ется образующийся в них эстрадиол и стимулирует пролиферацию этих
клеток, увеличивая размер фолликула. Одновременно эстрадиол-17(3 через
р-тип эстрогенных рецепторов активирует в гранулезных клетках абразова-
16. Репродуктивная функция человека • 685
ние новых рецепторов к фоллитропину. Поэтому чем больше образуется
гранулезных клеток в фолликуле, тем больше андрогенов ароматизируется
в эстрогены, которые стимулируют продукцию гранулезными клетками
еще большего количества эстрадиола-17₽ (механизм положительной обрат-
ной связи). Нарастающий синтез эстрогенов в гранулезных клетках приво-
дит к повышению концентрации женских половых гормонов в плазме кро-
ви. Эстрогены вместе с фоллитропином стимулируют образование на мем-
бране гранулезных клеток рецепторов к лютропину, который является ос-
новным регулятором следующей стадии развития антрального фолликула
(третичный фолликул). Лютропин связывается с рецепторами на мембране
гранулезных клеток и клеток внутреннего слоя наружной мембраны (theca)
и стимулирует накопление в них липидов, желтого пигмента, а также пре-
овуляторное увеличение образования прогестерона, который инициирует
овуляцию. В конце фолликулярной фазы под влиянием нарастающей сек-
реции эстрогенов в фолликуле и увеличения в плазме крови концентрации
яичникового цитокина ингибина, тормозится секреция фоллитропина в
аденогипофизе по механизму отрицательной обратной связи. При этом
высокий уровень эстрогенов в плазме крови вызывает торможение секре-
ции гонадолиберина в гипоталамусе и фоллитропина в гипофизе. Напро-
тив, эти же гормоны (эстрогены и ингибин) по механизму положительной
обратной связи стимулируют резкое повышение концентрации лютропина
в плазме крови за 24—36 ч до начала овуляции (рис. 16.10). Нормальная
овуляция во время короткой овуляторной фазы зависит от выброса, в виде
быстро проходящего пика концентрации в плазме крови гонадотропинов,
прежде всего лютропина.
16.3.1.2. Овуляторная фаза
Овуляция происходит примерно через 15 ч после достижения концентра-
цией лютропина в плазме крови максимальных значений (пик концентра-
ции). Лютропин связывается с рецепторами мембраны фолликулярных
клеток и активирует завершение первого мейотического деления первич-
ного ооцита (см. рис. 16.1). В результате образуется вторичный ооцит,
имеющий гаплоидное число хромосом, которые во время оплодотворения
яйцеклетки сливаются с гаплоидным числом хромосом сперматозоида.
Кроме того, лютропин, связываясь с рецепторами системы G-белков
фолликулярных клеток, увеличивает в них содержание вторичного посред-
ника цАМФ. Под влиянием цАМФ в фолликулярных клетках непосредст-
венно перед овуляцией прекращается фолликулогенез, связанный с обра-
зованием в гранулезных клетках стероидогенных энзимов, рецепторов
фоллитропина, p-эстрогеновых рецепторов и циклина D2, являющегося
регулятором митоза в фолликулярных клетках.
Под влиянием высокой концентрации лютропина в плазме крови воз-
растает кровоток в капиллярах ткани фолликула за счет роста продукции
NO-синтазы и образования вазодилататора оксида азота в гладких мышцах
сосудов, питающих фолликул. Это увеличивает размер фолликула, стиму-
лируя накопление фолликулярной жидкости, которая своим гидростатиче-
ским давлением истончает слой гранулезных клеток и клеток наружной
оболочки (theca) настолько, что лишь эпителиальный слой, покрывающий
поверхность яичника, отделяет стенку фолликула от перитонеального про-
странства. В предовуляторную фазу лютропин стимулирует синтез рецеп-
торов прогестерона, пептида, активирующего синтез аденилатциклазы в
гипофизе, регуляторного фактора роста, которые вызывают овуляцию.
686 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Преовуляторное увеличение концентрации прогестерона в плазме крови
женщины активирует синтез в фолликуле протеолитических энзимов и
простагландинов (Е и F). Под действием простагландинов, а также тром-
боксанов и лейкотриенов происходит истончение, растяжение и разруше-
ние стенки фолликула. В результате зрелая яйцеклетка выходит через стен-
ку лопнувшего фолликула в жидкость перитонеального простраства. Дви-
гательная активность цилий ампулярного расширения фаллопиевой трубы
направляет движение ооцита в просвет фаллопиевой трубы.
16.3.1.3. Лютеальная фаза
После овуляции на месте лопнувшего фолликула образуется желтое тело,
функцией которого является секреция эстрадиола-17р, эстрона и прогесте-
рона, регулирующих подготовку эндометрия матки к имплантации яйце-
клетки. Если не происходит оплодотворения яйцеклетки, то желтое тело
функционирует в лютеальную фазу овариального цикла (меструальное
желтое тело). При оплодотворении яйцеклетки желтое тело функциониру-
ет первую половину беременности (желтое тело беременности), а затем его
функция прекращается. Преовуляторное резкое повышение концентрации
лютропина в плазме крови женщины приводит к лютеинизации гранулез-
ных клеток и клеток theca и изменяет в них стероидогенез таким образом,
что после овуляции в плазме крови из женских половых гормонов домини-
рует концентрация прогестерона. Связывание лютропина с рецепторами
системы G-белков на мембране лютеальных клеток увеличивает содержа-
ние в них вторичных посредников, которые стимулируют транспорт холе-
стерина из плазмы крови в клетки желтого тела, где он используется для
синтеза прогестерона. Концентрация этого гормона в плазме крови жен-
щины в лютеальную фазу зависит от массы стероидогенной ткани в жел-
том теле, интенсивности кровотока и способности стероидогенной ткани
синтезировать прогестерон под влиянием лютропина. Стероидогенез в
клетках желтого тела постепенно увеличивается и достигает пика пример-
но на 8—9-й день после овуляции. В этот период концентрация прогесте-
рона в плазме крови женщины в 5 раз выше, чем в другие фазы овариаль-
ного цикла. Повышение в плазме крови эстрогенов и прогестерона в люте-
альную фазу по механизму отрицательной обратной связи тормозит секре-
ции гонадотропинов в аденогипофизе, увеличивая с 1 до 2—3 ч интервал
времени между периодами импульсного выброса этих гормонов кровь. В
результате не начинается развитие очередного фолликула в яичниках. Сек-
реция прогестерона в лютеальную фазу контролирует подготовку (секре-
торная фаза маточного цикла) эндометрия матки к имплантации яйцеклет-
ки. В эту фазу происходит функциональное созревание клеток эндометрия,
который полностью подготавливается к имплантации яйцеклетки. Если
яйцеклетка не оплодотворяется, то желтое тело сохраняется около 14 дней
и затем подвергается лютеолизу.
16.3.1.4. Лютеолиз желтого тела
Лизис, или структурное разрушение, желтого тела называется лютеолизом.
В процессе лютеолиза утрачивается способность клеток желтого тела син-
тезировать и секретировать прогестерон. Физиологическое значение пре-
кращения эндокринной функции желтого тела заключается в завершении
овариального цикл, а также менструального цикла, поскольку не произош-
ло оплодотворения яйцеклетки.
16. Репродуктивная функция человека • 687
Лютеолиз регулируется простагландином ПГР2а, который образуется в
клетках основной оболочки (theca) желтого тела. Простагландин ПГР2а
связывается со специфическими метаботропными рецепторами, располо-
женными на мембране гранулезных стероидогенных клеток желтого тела.
Взаимодействие простагландина ПГР^ с рецептором мембраны стероидо-
генных клеток через систему G-белков и вторичные посредники инозитол-
3-фосфат и диацилглицерол активирует фосфолипазу С, которая вызывает
в позднюю фазу развития желтого тела гидролиз мембранных фосфолипи-
дов с высвобождением арахидоновой кислоты. Арахидоновая кислота, в
свою очередь, является источником для синтеза простагландина ПГР^, что
приводит к дополнительному росту концентрации простагландина и акти-
вации лютеолиза. Последний усиливается под влиянием эндотелина-1,
секретируемого эндотелиальными клетками кровеносных сосудов желтого
тела. Эндотелии-1 вызывает спазм сосудов желтого тела, прекращение в
лютеальной ткани кровотока и, как следствие, разрушение стероидогенных
клеток из-за недостатка в них кислорода и питательных веществ.
Удаление разрушенных клеток желтого тела осуществляется клетками
крови (лейкоцитами, эозинофилами и макрофагами). Т-лимфоциты в жел-
том теле секретируют у-интерферон, с помощью которого иммунокомпе-
тентные клетки распознают поврежденные клетки желтого тела и удаляют
их из ткани яичника. Макрофаги фагоцитируют дегенеративные лютеаль-
ные клетки, тормозят с помощью цитокинов секрецию прогестерона в
клетках желтого тела и активируют секрецию основного активатора лютео-
лиза простагландина ПГР2а. Макрофаги продуцируют a-фактор некроза
опухоли, который тормозит секрецию прогестерона клетками желтого тела.
Продукцию прогестерона ПГР2а активирует интерлейкин-I, который вы-
свобождается макрофагами, фибробластами и эндотелиальными клетками
капилляров желтого тела, что также усиливает лютеолиз. При участии им-
мунных клеток крови и цитокинов во время лютеолиза регулируется син-
тез простагландина ПГР2а в клетках желтого тела, в них тормозится сте-
роидогенез и активируется фагоцитоз клеток в период его структурной ин-
волюции.
16.3.2. Менструальный цикл (маточный цикл)
Подготовка женского организма к гестации характеризуется циклическими
изменениями эндометрия матки, которые состоят из трех последователь-
ных фаз: менструальной, пролиферативной и секреторной — и называются
маточным, или менструальным, циклом.
16.3.2.1. Менструальная фаза
Менструальная фаза при длительности маточного цикла 28 дней продол-
жается в среднем 5 дней. Эта фаза представляет собой кровотечение из по-
лости матки, которое возникает в конце овариального цикла, если не про-
исходит фертилизации и имплантации яйцеклетки. Менструация — это
процесс отторжения слоя эндометрия. Пролиферативная и секреторная
фазы менструального цикла включают процессы восстановления эндомет-
рия для возможной имплантации яйцеклетки в течение следующего овари-
ального цикла.
688 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
16.3.2.2. Пролиферативная фаза
Пролиферативная фаза варьирует по продолжительности от 7 до 11 дней.
Эта фаза совпадает с фолликулярной и овуляторной фазами яичникового
цикла, в течение которого возрастает уровень эстрогенов, в основном эст-
радиола- 17Р, в плазме крови. Основная функция эстрогенов в пролифера-
тивную фазу менструального цикла — стимуляция клеточной пролифера-
ции тканей органов репродуктивной системы с восстановлением функцио-
нального слоя эндометрия и развитием эпителиальной выстилки слизи-
стой оболочки матки. В эту фазу под влиянием эстрогенов происходит
утолщение эндометрия матки, увеличение в размере его желез, секрети-
рующих слизь, растет длина спиральных артерий. Эстрогены вызывают
пролиферацию эпителия влагалища, усиливают секрецию слизи в шейке
матки. Секреция становится обильной, в ее составе увеличивается количе-
ство воды, что облегчает движение в ней сперматозоидов. Стимуляция
пролиферативных процессов в эндометрии связана с увеличением количе-
ства рецепторов прогестерона на мембране клеток эндометрия, что усили-
вает в нем пролиферативные процессы под влиянием этого гормона. На-
конец, увеличение концентрации эстрогенов в плазме крови стимулирует
сокращение гладких мышц и микроворсин фаллопиевых труб, что способ-
ствует продвижению сперматозоидов в направлении ампулярного отдела
фаллопиевых труб, где должно произойти оплодотворение яйцеклетки.
16.3.2.3. Секреторная фаза
Секреторная фаза менструального цикла начинается непосредственно после
овуляции и продолжается до начала менструации (в среднем 12—16 дней).
Основная роль секреторной фазы состоит в подготовке слизистой оболочки
матки к имплантации яйцеклетки. В эту фазу активно функционирует жел-
тое тело, стероидогенные клетки которого секретируют этсрадиол-17р, эс-
трон и прогестерон. Основным условием нормального течения секреторной
фазы является высокая концентрация прогестерона в плазме крови. Прогес-
терон совместно с эстрогенами действует на клетки желез слизистой обо-
лочки матки, вызывая в них секрецию. В секрете желез повышается уровень
гликогена, гликопротеинов, гликолипидов, которые необходимы для под-
держания метаболизма оплодотворенной яйцеклетки и способствуют ее
внедрению в слизистую оболочку стенки матки. Если не происходит опло-
дотворения яйцеклетки и маточный цикл не завершается беременностью,
менструальное желтое тело подвергается лютеолизу. Это вызывает сниже-
ние секреции клетками желтого тела эстрогенов и прогестерона и, как след-
ствие, уменьшение концентрации в плазме крови женщины содержания
этих гормонов. Понижение концентрации эстрогенов и прогестерона в
плазме крови вызывает в слизистой оболочке функционального слоя мио-
метрия повышение тонуса гладкомышечных клеток спиральных артерий, их
скручивание и уменьшение просвета. В результате ишемии эндометрия воз-
никает его некроз с последующим менструальным кровотечением.
16.3.3. Женский половой акт
Женский половой акт, как и мужской, состоит из стадии возбуждения,
эрекции и разрешения. По аналогии с мужчиной, у женщины способность
к осуществлению полового акта зависит от психической сексуальной сти-
муляции и от стимуляции эрогенных зон.
16. Репродуктивная функция человека • 689
Половое возбуждение у женщины возникает при эротических пережи-
ваниях, вызванных ментальными процессами или просматриванием эроти-
ческих картин или фильмов. Степень сексуального возбуждения варьирует
на протяжении овариального цикла; половое влечение выше накануне ову-
ляции, когда в крови увеличивается концентрация эстрогенов. Половое
возбуждение у женщины тесно связано с рецептивной направленностью
женской сексуальности, зависящей, от ситуации общения и эротической
стимуляции со стороны мужчины (словесные, а также тактильные ласки,
особенно эрогенных зон). Стимуляция эрогенных зон у женщин, связан-
ная с раздражением рецепторов наружных половых органов (половых губ,
преддверия влагалища и, особенно, клитора), в том числе во время поло-
вого акта, активирует многочисленные механорецепторы, сигналы от кото-
рых по срамным нервам поступают в парасимпатические центры крестцо-
вых сегментов спинного мозга. Возбуждение парасимпатических эффе-
рентных сосудорасширяющих нервов, а также симпатических вазоактив-
ных волокон, идущих от нейронов грудных и поясничных сегментов к со-
судам наружных и внутренних половых органов женщины, вызывает их
кровенаполнение.
Основными проявлениями стадии возбуждения являются эрекция кли-
тора и секреция слизи. Строение клитора идентично строению ткани муж-
ского полового члена. Поэтому активация иннервирующих клитор параси-
мпатических волокон вызывает расслабление гладких мышц артерий при
участии оксида азота, расширение просвета артерий и увеличение кровена-
полнения ткани клитора. Импульсы по парасимпатическим нервам дости-
гают также бартолиниевых желез, локализованных в глубине малых поло-
вых губ, и вызывает секрецию слизи. Слизь, секретируемая бартолиниевы-
ми железами и слизистой оболочкой влагалища, а также слизь, выделяю-
щаяся из желез мужской уретры во влагалище во время полового акта,
способствует увлажнению влагалища и формированию оптимального сек-
суального чувства во время движений пениса во влагалище. Во время по-
лового акта усиление кровенаполнения органов малого таза по мере нарас-
тания нервного возбуждения изменяет размеры и положение матки, кото-
рая отклоняется к задней стенке влагалища, образуя в области шейки мат-
ки так называемую семяпринимающую полость. Одновременно в нижней
части влагалища в результате застоя крови может формироваться оргасти-
ческая манжетка.
Оргазм. У женщин ощущение оргазма формируется в результате рефлек-
торных сокращений внутренних (например, ритмические сокращения ор-
гастической манжетки влагалища под влиянием симпатических рефлектор-
ных механизмов) либо наружных половых органов (клитор). При этом ин-
тенсивность мышечных сокращений во время оргазма прямо взаимосвяза-
на с увеличением концентрации в плазме крови окситоцина. Секреция ок-
ситоцина из клеток нейрогипофиза во время оргазма регулируется минда-
левидным комплексом и гипоталамусом. Ритмические сокращения мышц
промежности у женщины во время оргазма регулируются также рефлек-
торно центрами спинного мозга крестцовых сегментов. У мужчин нейроны
этих же крестцовых сегментов рефлекторно регулируют эякуляцию. Во
время оргазма у женщины рефлекторные сокращения гладких мышц внут-
ренних половых органов, а также расслабление шейки матки способствуют
продвижению спермы через матку и фаллопиевы трубы.
Интенсивное сексуальное чувство, которое возникает у женщины во
время оргазма, сопряжено с активацией двигательных систем головного
мозга, что сопровождается повышением тонуса скелетных мышц. Во время
690 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
оргазма увеличиваются частота дыхания, системное артериальное давление
и частота сердцебиений. Стадия оргазма у женщины, так же как и у муж-
чин, сменяется стадией разрешения, во время которой нормализуются па-
раметры систем кровообращения и дыхания. Однако у женщин отсутствует
период рефрактерности после оргазма.
16.4. Оплодотворение (фертилизация)
Вероятность фертилизации зависит от времени полового сношения отно-
сительно момента овуляции. Это обусловлено тем, что после эякуляции
сперма сохраняет жизнеспособность 3—5 дней, а ооцит — порядка 24 ча-
сов. Оптимальным местом фертилизации являются фаллопиевы трубы, по-
скольку это увеличивает возможность имплантации зиготы в наиболее бла-
гоприятное для ее развития место матки — на задней стенке в верхней ее
трети. Сперматозоиды, попавшие во влагалище женщины во время поло-
вого акта, примерно через 30 мин после эякуляции достигают устьев фал-
лопиевых труб, а еще через 15 мин —их ампулярной части, где обычно
происходит оплодотворение яйцеклетки.
В репродуктивном тракте мужчины сперматозоиды имеют низкую опло-
дотворяющую способность. Нормальная оплодотворяющая способность
сперматозоидов формируется после эякуляции в женском репродуктивном
тракте. Формирование нормальной оплодотворяющей способности спер-
матозоидов, или капацитация, происходит в результате образования спер-
мы, т. е. смешивания сперматозоидов с семенной жидкостью во влагали-
ще, а также во время продвижения сперматозоидов через слизь шейки
матки. Продвижению сперматозоидов в просвете фаллопиевых труб спо-
собствуют собственная двигательная активность сперматозоидов и направ-
ленные в сторону каудального отдела труб волны сокращения гладких
мышц стенки яйцевода. Двигательная активность сперматозиодов, находя-
щихся в каудальной части фаллопиевых труб, возрастает через несколько
минут после овуляции. Это свидетельствует о том, что ооцит или фоллику-
лярные клетки высвобождают сигнальные факторы, которые активируют
подвижность сперматозоидов (хемотаксис) и направляют их в зону оплодо-
творения. Только небольшая часть сперматозоидов человека (2—12 %) об-
ладает хемотаксисом, т. е. реагирует на хемостимуляцию фолликулярными
факторами. Поэтому в оплодотворении ооцита избирательно участвуют
только капацитированные сперматозоиды. Следующим физиологическим
процессом, предшествующим фертилизации, является акросомальная ре-
акция сперматозодида.
Физиологическая роль акросомальной реакции заключается в слиянии
акросомальной мембраны сперматозоида с наружной мембраной его го-
ловки и формировании в обеих мембранах пор, через которые способны
выходить из сперматозоида акросомальные энзимы. Эти энзимы способны
разрушать контакты между гранулезными клетками холмика (cumulus
oophorus), внутри которого находится^ ооцит, формируя путь проникнове-
ния к нему сперматозоида через толстую прозрачную оболочку (zona pellu-
cida). Основным физиологическим активатором акросомальной реакции
сперматозоида является zona pellucida, гликопротеины которой связывают-
ся с олигосахаридами полипептидов мембраны сперматозоида. Физиологи-
ческим эффектом связывания рецепторов zona pellucida с мембраной го-
ловки сперматозоида является увеличение концентрации ионов Са2+ спер-
матозоида. Молекулярный механизм этого процесса связан с открытием Т-
типа потенциалзависимых кальциевых ионных каналов в мембране голов-
16. Репродуктивная функция человека • 691
Рис. 16.11. Акросомальная реакция сперматозоида.
Zona pellucida ооцита связывается с рецепторами мембраны головки сперматозоида (напри-
мер, спермадгезином — СА). Активированные рецепторы мембраны сперматозоида при уча-
стии Gi-белка стимулируют мембранный белок, транспортирующий Н+-ионы из цитоплазмы
сперматозоидов, что увеличивает в ней pH (1). Активация рецепторов мембраны сперматозои-
да стимулирует образование вторичных посредников инозитол-3-фосфата (ИФЗ), а также цик-
лического аденозин-монофосфата. ИФЗ открывает кальциевые ионные каналы акросомы и
мембраны сперматозоида. В результате в цитоплазме сперматозоида повышается концентра-
ция ионов Са2+. Вторичные посредники стимулируют образование энзимов, вызывающих раз-
рушение мембраны акросомы и головки сперматозоида. Акросомальная реакция модулируется
прогестероном и у-аминомасляной кислотой. При этом прогестерон является основным регу-
лятором проницаемости кальциевых ионных каналов мембраны головки сперматозоида. От-
крывание калиевых потенциалзависимых ионных каналов (2) гиперполяризует мембрану го-
ловки сперматозоида и инактивирует потенциалзависимые кальциевые ионные каналы. АР —
акросома, ФИ2 — фосфатидилинозитол, ДАГ — диацилглицерол.
ки сперматозоида в результате ее деполяризации (непрямой эффект связы-
вания рецепторов zona pellucida с рецепторами мембраны сперматозоида,
действие прогестерона). Прогестерон в наибольшей степени влияет на уве-
личение концентрации ионов Са2+ внутри сперматозоида человека, так как
активирует натриевые ионные каналы и, таким образом, вызывает деполя-
ризацию мембраны сперматозоидов. Прогестерон увеличивает взаимодей-
ствие у-аминомасляной кислоты с ГАМКА-рецепторами мембраны сперма-
тозоидов, что увеличивает выход ионов хлора из цитоплазмы и также уси-
ливает деполяризацию их мембраны.
Через открытые каналы ионы Са2+ диффундируют внутрь сперматозоди-
да по концентрационному градиенту из внеклеточной среды. Увеличение
концентрации ионов Са2+ внутри сперматозоида происходит за счет высво-
бождения ионов из депо акросомы при участии системы семейства Gi-бел-
ков, вторичного посредника инозитол-3-фосфата, цАМФ и при условии
высокого внутриклеточного pH (рис. 16.11). Инозитол-З-фосфат и цАМФ
изменяют активность соответственно протеинкиназы С и протеинкиназы
А, что приводит к фосфорилированию различных протеинов (протеолити-
ческий фермент акрозин, нейраминидаза и гиалуронидаза), которые вызы-
вают образование пор в мембране сперматозоида, выходят во внеклеточ-
ную среду и растворяют толстую оболочку ооцита. Проникновение голов-
ки сперматозоида через отверстие в zona pellucida вызывает диффузию ио-
692 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
нов Са2+ из головки сперматозоида во внеклеточную среду, окружающую
ооцит. В присутствии ионов Са2+ из ооцита высвобождаются субстанции,
которые изменяют свойства поверхностных гликопротеинов zona pellucida,
что препятствует проникновению через нее других сперматозоидов. Изме-
нение свойств поверхностных гликопротеинов zona pellucida называется
зональной реакцией и свидетельствует о том, что ооцит оплодотворен одним
сперматозоидом. В момент зональной реакции из ооцита высвобождается
проядро ооцита (второе полярное тельце) (см. рис. 16.1) с гаплоидным
числом хромосом и происходит его слияние с проядром сперматозоида,
также содержащего гаплоидное число хромосом. В результате образуется
зигота с диплоидным числом хромосом.
16.5. Имплантация оплодотворенной яйцеклетки
После начального периода митотической сегментации зиготы, благодаря
слабым движениям ресничек реснитчатых клеток слизистой оболочки ма-
точных труб в направлении матки, оплодотворенная яйцеклетка транспор-
тируется к матке, и оказывается в ней через 5 дней после овуляции (18—
19-й день менструального цикла). К этому времени эндометрий матки под
влиянием эстрогенов и прогестерона полностью сформирован для имплан-
тации оплодотворенной яйцеклетки.
Оптимальные условия для имплантации яйцеклетки имеют место в се-
редине лютеальной фазы овариального цикла, когда под влиянием прогес-
терона желтого тела полностью сформировался функциональный слой эн-
дометрия. Процесс имплантации происходит после контакта бластоцисты
со стенкой матки, как правило, в ее верхней задней области. В течение од-
ного часа после начала имплантации в матке происходят изменения в
стромальной ткани матки, называемые децидуализацией, течение которой
зависит от эстрадиола-17р и прогестерона.
Децидуализация характеризуются ростом проницаемости стенки сосу-
дов функционального слоя эндометрия, в результате которого возникает
местный тканевой отек, сопровождающийся изменением состава межкле-
точного матрикса, структуры стромальных клеток и увеличением прони-
цаемости капилляров. В этих условиях трофобластные клетки бластоцисты
полностью погружаются в глубину слизистой оболочки эндометрия. Этот
процесс называется интерстициальной имплантацией. Имплантация яйце-
клетки в эндометрий матки регулируется хорионическим гонадотропином,
который продуцируется в трофобластных клетках бластоцисты после нача-
ла его внедрения в слизистую оболочку матки. С другой стороны, имплан-
тация зародыша вызывает усиление афферентных импульсов от рецепто-
ров слизистой оболочки матки в гипоталамус. Постоянный поток этих аф-
ферентных импульсов активирует секрецию либерина в гипоталамусе и
приводит к непрерывной секреции в кровь лютропина аденогипофиза. Не-
прерывный характер секреции лютропина тормозит следующий менстру-
альный цикл у беременной женщины;
16.6. Беременность
По мере того как эмбрион внедряется в слизистую оболочку матки (в сред-
нем 48 ч), материнские сосуды слизистой оболочки повреждаются, а вновь
развивающиеся фетальные кровеносные сосуды оказываются погруженны-
ми непосредственно в материнскую кровь. В результате образуется гемохо-
риальная плацента, которая разделяет, по крайней мере, одним слоем эн-
16. Репродуктивная функция человека • 693
дотелиальных клеток капилляров на протяжении всего периода беременно-
сти кровь плода и матери. Это позволяет поддерживать между организмом
матери и плода газообмен и обмен веществ.
16.6.1. Функции плаценты
Плацента является местом диффузии и активного транспорта метаболитов,
дыхательных газов и электролитов между матерью и плодом, за исключе-
нием клеток и больших молекул белков. Проникновение веществ из крови
матери в кровь плода и в обратном направлении регулируется функцией
плацентарного барьера, образованного эпителием ворсинок и эндотелием
погруженных в них капилляров. Функциями плаценты является дыхатель-
ная (обмен О2 из крови матери в кровь плода и СО2 в обратном направле-
нии), питательная (расщепление белков, углеводов и жиров энзимами
плаценты и синтез питательных веществ, поступающих в кровь плода),
экскреторная (выведение продуктов метаболизма из крови плода в кровь
матери), детоксикационная (например, лекарственных препаратов, прини-
маемых во время беременности женщиной) и эндокринная. Плацента вы-
полняет во время беременности роль эндокринного органа, синтезирую-
щего стероидные, пептидные и белковые гормоны. Функциями гормонов
плаценты являются сохранение беременности, регуляция обмена веществ в
организме матери и плода, структурная и физиологическая подготовка мо-
лочных желез беременной женщины к лактации. В отношении продукции
гормонов плод и плацента функционируют совместно и представляют со-
бой фетоплацентраную единицу (рис. 16.12).
Диффузия кислорода через плацентарную мембрану является жизненно
необходимым процессом для развития плода. В среднем напряжение О2 в
синусах плаценты составляет 50 мм рт. ст., в крови плода — примерно
30 мм рт. ст. При столь низком градиенте О2 эффективность кислород-
транспортной функции поддерживается за счет фетального гемоглобина,
его высокой концентрации в крови плода (на 50 % выше, чем в крови ма-
тери), а также эффекта Бора. Физиологическая роль последнего состоит в
том, что повышенная концентрация углекислого газа в крови матери по-
нижает сродство кислорода с гемоглобином и облегчает отдачу О2 из эрит-
роцитов при газообмене в плаценте и способствует кислородному снабже-
нию организма плода при низком его содержании в крови матери. Угле-
кислый газ непрерывно образуется в тканях плода и выводится из его ор-
ганизма через плаценту в кровь матери. В крови плацентарных синусов со-
держание углекислого газа не превышает 40 мм рт. ст. Напряжение СО2 в
крови плода на 2—3 мм рт. ст. выше, чем в крови матери. Это достаточно
низкий градиент концентрации СО2, но он не препятствует эффективному
выведению из организма плода углекислого газа, поскольку проницаемость
углекислого газа через гемохориальную плаценту в 20 раз выше, чем у ки-
слорода. В крови небеременной женщины напряжение углекислого газа
выше (46 мм рт. ст.), чем в крови беременной. Понижение содержания уг-
лекислого газа в крови беременной женщины вызвано стимуляцией дыха-
тельного центра половыми стероидами; эстрогеном и прогестероном. В ре-
зультате в крови плода также поддерживается пониженное содержание
СО2 — не выше 40 мм рт. ст.
Основным источником углеводов, используемых плодом, является глю-
коза. Перенос через плацентарную мембрану глюкозы осуществляется пу-
тем облегченной диффузии. В мембране трофобластных клеток имеется
белок — переносчик глюкозы, однако в крови плода ее содержание на
694 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Дегидроэпиандростерон
Кровообра- Холестерол Прогестерон
щениев .
организме
мвтери ______ ________
Эстрогены Эстрадиол Эстриол (основной
. а гормон беременности)
Плацента Холестерол—v/►Прогестерон^егидро"-^?стР°геиы—► Эстрадиол
„ _____ эпиандро-Х f 16-гидс
1 /—► Эстриол
16-гидрокси-дегидро-
стерон Андростендион эпиандростерон сульфат
Прегненолон
Плод
Прегненолон
> Дегидро-------► Дегидро------► 16-гидрокси-
эпиандростерон эпиандростерон дегидроэпиандростерон
сульфат (синтезируется в организме
плода)
Рис. 16.12. Схема продукции стероидных гормонов в плаценте и в организме плода.
В начале П триместра берменности, когда желтое тело прекращает свою функцию, основным
источником эстрогенов и прогестерона становится плацента. Оптимальный синтез эстрогена
осуществляется в условиях нормальной функции надпочечников плода. Дигидроэпиандросте-
рон, который синтезируется в надпочечниках матери и плода, транспортируется с кровью в
плаценту, где он ароматизируется в эстрогены. Примерно 50 % дигидроэпиандростерона син-
тезируется в надпочечниках плода. Основным эстрогеном, продуцируемым плацентой, являет-
ся эстриол; в меньшем количестве продуцируются эстриол-170 и эстрон-170. Напротив, во
время менструального цикла основным эстрогеном является эстадиол-17р. Синтез в плаценте
эстриола происходит за счет 16а-гидрокси-дегидроэпиандростерона, образующегося в орга-
низме плода. Напротив, эстрадиол и эстрон синтезируются за счет дигидроэпиандростерона
как организма матери, так и плода. В конце беременности концентрация эстриола в плазме
крови равна 400 нМ/л, а эстрадиола — 50 нМ/л.
20—30 % ниже, чем в крови матери. Аминокислоты транспортируются че-
рез плаценту механизмом активного транспорта, и их концентрация в
крови плода выше, чем в крови матери. Жирные кислоты с короткой це-
пью и глицерол диффундируют через плацентарный барьер свободно по
концентрационному градиенту. В плаценте или в организме плода они яв-
ляются источником синтеза более сложных липидов. Жирорастворимые
витамины свободно диффундируют через плаценту, а водорастворимые —
с помощью механизма активного транспорта. Белки, исключая материн-
ские белковые и пептидные гормоны, не проникают из крови матери в
кровь плода через плаценту. Исключение составляет иммуноглобулин IgG,
который транспортируется через плацентарный барьер в кровь плода с по-
мощью специфического белка переносчика. Большинство минеральных
веществ и электролитов свободно диффундирует через плацентарный
барьер. Однако развивающийся плод испытывает повышенные потребно-
сти в минеральных веществах. Поэтому в его крови поддерживается поло-
жительный минеральный баланс (например, ионов Са2+, Fe2+, Na+, К+) за
счет работы специфических мембранных ионных насосов клеток плацен-
тарного барьера.
Выведение продуктов метаболизма* образующихся в организме плода,
осуществляется путем диффузии: прежде всего, это касается мочевины,
мочевой кислоты и креатинина. Уровень мочевины в крови плода ниже,
чем в крови матери, так как это вещество диффундирует через плацентар-
ную мембрану достаточно легко. Напротив, креатинин диффундирует ме-
нее легко, но его диффузия происходит достаточно эффективно из-за вы-
сокого градиента концентрации между кровью плода и матери.
16. Репродуктивная функция человека • 695
16.6.2. Плацентарные гормоны
Клетки трофобласта во время его имплантации в функциональный слой
эндометрия дифференцируются в цитотрофобласт и синцитиотрофобласт
Оба типа клеток секретируют белковые и пептидные гормоны, а син-
цитиотрофобластные — стероидные. Функция гормонов плаценты — это
регуляция нормального течения беременности и развития плода. В плацен-
те синтезируются хорионический гонадотропин, хорионический сомато-
маммотропин, плацентарный лактогенный гормон, а также стероиды — эс-
трогены и прогестерон
Человеческий хорионический гонадотропин. Секреция этого гормона на-
чинается на ранних этапах беременности — примерно с 9-го дня гестации.
В течение I триместра беременности концентрация гормона в плазме кро-
ви беременной женщины интенсивно нарастает до максимума на 9—12-й
неделях (рис. 16.13). Появление гормона в моче женщины является тестом
на наличие беременности. Функция хорионического гонадотропина ис-
ключительно важна для продолжения беременности. В течение овариаль-
ного цикла, если оплодотворения не происходит, прогестерон желтого тела
и эстрогены тормозят секрецию лютропина в аденогипофизе, поддерживая
сохранение желтого тела. В период снижения концентрации прогестерона
и эстрогенов в плазме крови беременной женщины происходит меструа-
ция. Во время беременности стабильный уровень концентрации эстроге-
нов и прогестерона в плазме крови поддерживает нормальное функцио-
нальное состояние эндометрия. Это обеспечивается регулирующим эффек-
том хорионического гонадотропина, который во время беременности дей-
ствует как плацентарный гонадотропин. С начала II триместра беременно-
сти плацента способна самостоятельно продуцировать достаточное количе-
ство эстрогенов и прогестерона, при участии которых эндометрий поддер-
живается в нормальном функциональном состоянии, независимо от секре-
ции гормонов в желтом теле. С этого периода концентрация хориониче-
Рис. 16.13. Динамика содержания в плазме крови уровня человеческого хориониче-
ского гонадотропина, прогестерона и эстрогенов в различные сроки беременности.
Во время беременности постоянно возрастает уровень эстрогена в плазме крови. В I триместре
его основным источником являются клетки желтого тела. В этот же период беременности уве-
личивается концентрация прогестерона в плазме крови, который продуцируется в плаценте. С
началом II триместра, когда желтое тело прекращает функционировать, основным источни-
ком эстрогенов и прогестерона является плацента.
696 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ского гонадотропина в плазме крови беременной женщины постепенно
снижается, а желтое тело может функционировать без регулирующего
влияния хорионического гонадотропина. В этот период развития плода
физиологическая роль гормона заключается в стимуляции секреции клет-
ками Лейдига фетального тестостерона, а клетками надпочечников — де-
гидроэпиандростерона. Эти гормоны необходимы для нормального разви-
тия наружных половых органов у плода, т. е. становлению фенотипическо-
го пола.
Эстрогены. На протяжении беременности концентрация эстрогенов в
плазме крови беременной женщины постоянно увеличивается (см. рис.
16.13). Основным эстрогеном, синтезируемым в плаценте, является эстри-
ол, в меньшем количестве синтезируются эстрадиол-17р и эстрон-17р. На-
пример, в поздний период беременности концентрация эстриола в плазме
крови женщины составляет 400 нМ/л, а эстрадиола — порядка 50 нМ/л.
Плацентарный синтез эстриола обеспечивается 16а-гидрокси-дегидроэпи-
андростероном, который поступает только от плода, в то время как источ-
ником эстрадиола-17р и эстрона-17р является дегидроэпиандростерон ма-
тери и плода. Поэтому нормальная концентрация эстрогенов в моче бере-
менной женщины отражает нормальное функционирование фетоплацен-
тарной единицы. Если снижается функционирование организма плода, то
существенно понижается экскреция с мочой эстриола в общем количестве
выводимых из организма беременной женщины эстрогенов.
Прогестерон. В течение беременности лютропин аденогипофиза стиму-
лирует рост и гормональную активность материнского желтого тела. Жел-
тое тело секретирует прогестерон, который является наиболее важным гор-
моном в отношении сохранения жизнеспособности плода во время его
развития в полости матки. Примерно с 12-й недели беременности в плазме
крови беременной женщины сохраняется постоянный повышенный уро-
вень прогестерона, который достигает концентрации порядка 500 нМоль/л
во II и III триместрах беременности (см. рис. 16.13). Гормон существенно
снижает сократительные свойства миометрия во время беременности, по-
скольку тормозит функцию всей системы окситоциновых рецепторов в
матке.
К концу беременности, которая продолжается в среднем 9 мес, в крови
беременной женщины уровень прогестерона снижается. Происходит «сме-
щение» содержания в плазме крови половых гормонов в сторону эстроге-
нов, которые резко увеличивают чувствительность миометрия к окситоци-
ну. Под действием окситоцина повышается сократительная активность
мускулатуры матки. К началу родов возрастает концентрация в амниотиче-
ской жидкости некоторых простагландинов (F2, Е, и Е2), которые также
стимулируют родовую деятельность. Эффекты окситоцина и простагланди-
нов на миометрий реализуется через цАМФ, при этом происходит деполя-
ризация мембранного потенциала клеток за счет повышения проницаемо-
сти мембран миоцитов для ионов натрия. Повышается также проницае-
мость мембран миоцитов для ионов кайьция, мобилизация которых из вне-
клеточной среды необходима для начала активной родовой деятельности.
Человеческий хорионический соматомаммотропин. Нормальная концен-
трация этого гормона в плазме крови беременной женщины является ос-
новным индикатором нормального развития плода. Имеется прямая зави-
симость нарастания концентрации гормона в плазме крови беременной
женщины по мере увеличения массы плаценты, однако этот феномен не
получил объяснения. По структуре и функции гормон идентичен пролак-
тину аденогипофиза и поэтому имеет другое название — человеческий пла-
16. Репродуктивная функция человека • 697
центарный лактоген. Действие гормона на ткани идентично гормону роста,
поскольку они имеют одинаковую молекулярную структуру. Гормон сти-
мулирует захват протеинов клетками тканей и усиливает рост тела плода.
Этот гормон обладает также липолитическим действием, что позволяет ор-
ганизму беременной женщины использовать запасы жира в ее организме
как альтернативный источник энергии при дефиците белков и углеводов.
Хорионический соматомаммотропин оказывает антиинсулиновый эффект,
что сохраняет уровень глюкозы в крови на необходимом для жизнедея-
тельности организма плода и матери уровне. На ранних этапах беременно-
сти недостаточная секреция гормона в плаценте может стать причиной
спонтанного аборта, а на поздних — служит показателем недостаточной
секреторной функции плаценты.
Пролактин. В период беременности секреция пролактина в гипофизе
женщины прогрессивно увеличивается, превышая в 10 раз его содержание
в крови небеременной женщины. Функция гормона проявляется в совме-
стном действии эстрогена и прогестерона на развитие секреторных отделов
молочных желез у беременной женщины. Во время всего периода беремен-
ности высокая концентрация в плазме крови беременной женщины эстро-
генов и прогестерона тормозит в ее молочных железах продукцию молока
под влиянием пролактина, хотя лактогенная функция молочных желез
формируется на четвертом месяце гестации. Пролактин резко усиливает
продукцию молока лишь после снижения в плазме крови эстрогенов и
прогестерона, что имеет место непосредственно перед родами. Высокий же
уровень пролактина в плазме крови у кормящей женщины тормозит в ран-
ний период после родов начало овариального цикла, что обусловливает
аменорею послеродового периода.
Релаксин в I триместре беременности секретируется клетками желтого
тела, а в поздние сроки — децидуальными клетками эндометрия. Функция
этого гормона в наибольшей степени проявляется в подготовке родовых
путей к родам: расслабление тазовых связок и, особенно, шейки матки. В
течение беременности гормон снижает возбудимость клеток миометрия и
их сократительную способность.
16.7. Роды и лактация
16.7.1. Роды
Физиологический процесс, возникающий в конце беременности, когда
плод через родовые пути женщины изгоняется из матки, называется рода-
ми. Два процесса обеспечивают изгнание плода из матки: координирован-
ные сокращения миометрия и раскрытие шейки матки. Биомеханика ро-
дов состоит из фазы раскрытия шейки матки, изгнания плода и послеродово-
го периода. Ритмические сокращения миометрия, или схватки, свидетель-
ствуют о начале родовой деятельности. Во время схваток волна мышечного
сокращения распространяется со дна на тело и далее в сторону шейки мат-
ки. Это укорачивает и расширяет шейку матки, а также повышает давле-
ние в плодном пузыре. Шейка матки постепенно расширяется и формиру-
ется единый родовой канал, через который плод изгоняется из матки. Обо-
лочка плодного пузыря разрывается под влиянием внутриматочного давле-
ния. Изгнанию плода способствуют потуги — ритмические схватки, скоор-
динированные с сокращением мышц брюшного пресса. После рождения
ребенка начинается послеродовый период, в течение которого под влиянием
сокращений матки отслаивается плацента.
698 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
На протяжении беременности гладкие мышечные клетки миометрия
повышают чувствительность к стимулирующему действию эстрогенов: уве-
личиваются размер и масса гладких мышечных клеток матки, передача
возбуждения между клетками осуществляется через щелевые контакты,
или нексусы. Во время беременности физиологическое действие прогесте-
рона на мембрану гладкомышечных клеток миометрия проявляется в ги-
перполяризации мембранного потенциала. Гиперполяризация блокирует
потенциалзависимые кальциевые ионные каналы в мембране гладкомы-
шечных клеток и, таким образом, исключается возможность их сокраще-
ния во время беременности.
Гладкие мышцы матки во время родов являются главной тканью-мише-
нью для действия гормона нейрогипофиза — окситоцина. Окситоцин син-
тезируется во время беременности в тканях матки — амнионе, хорионе и
децидуальной оболочке. Образующийся в клетках этих тканей окситоцин
достигает паракринно с амниотической жидкостью миометрия и способен
стимулировать его сокращение.
Окситоцин на фоне его нормального содержания в плазме крови бере-
менной женщины вызывает сокращение миометрия во время родов в ре-
зультате резкого увеличения концентрации окситоциновых рецепторов в
хориодецидуальной ткани, вызывающего повышение чувствительности
миометрия к действию окситоцина. Физиологический механизм повыше-
ния чувствительности миометрия к окситоцину заключается в следующем.
Во время III триместра беременности концентрация эстрогенов и прогес-
терона в плазме крови беременной женщины достигает плато. Однако в
соотношении эстрогены/прогестерон последний доминирует и, таким об-
разом, тормозит сокращение матки. Синтез окситоцина в децидуальной
оболочке эндометрия стимулирует высвобождение простагландина ПГР2а,
который вызывает лютеолиз желтого тела. Прекращение функции желтого
тела приводит к снижению концентрации прогестерона в плазме крови бе-
ременной женщины к концу III триместра беременности. Примерно за
24 ч до начала родов в плазме крови беременной женщины в соотношении
концентрации эстрогены/прогестерон начинают доминировать эстрогены.
Понижение концентрации прогестерона в плазме крови восстанавливает
возбудимость гладкомышечных клеток матки. Эстрогены активируют про-
дукцию в миометрии простагландинов, увеличение концентрации которых
стимулирует образование в гладкомышечных клетках матки рецепторов ок-
ситоцина. В результате в течение нескольких часов, предшествующих на-
чалу родов, плотность окситоциновых рецепторов на мембране гладкомы-
шечных клеток матки увеличивается примерно в 200 раз, возрастает коли-
чество нексусов между гладкими мышечными клетками матки. Эти про-
цессы являются основой механизма повышения чувствительности миомет-
рия к окситоцину. Пик плотности окситоциновых рецепторов на мембране
гладкомышечных клеток матки приходится на ранний период родовой дея-
тельности, поэтому с началом родовой деятельности окситоцин начинает
стимулировать сокращение матки при такой его концентрации в плазме
крови, которой недостаточно для активации ее сокращений вне периода
беременности. Во время родовой деятельности потенциалы действия в
клетках миометрия распространяются по гладкомышечному синцитию, что
является источником координированных сокращений отделов матки во
время родов.
После родов концентрация окситоциновых рецепторов в миометрии
интенсивно снижается, только за первые 24 ч концентрация мРНК, кон-
тролирующей синтез окситоциновых рецепторов в гладкомышечных клет-
16. Репродуктивная функция человека • 699
ках матки, снижается примерно в семь раз. Это обусловлено тем, что от-
торжение плаценты во время родов вызывает резкое снижение концентра-
ции эстриола в плазме крови женщины, в результате снижаются продук-
ция простагландинов и уровень мРНК в клетках миометрия, контролирую-
щей образование окситциновых рецепторов. Снижение количества оксито-
циновых рецепторов в тканях матки в послеродовый период исключает ак-
тивацию ее сокращений во время лактации, когда рефлекторно возрастает
уровень окситоцина в плазме крови/
16.7.2. Лактация
Во время беременности молочные железы развиваются, что позволяет ма-
тери после родов адекватно обеспечивать питанием новорожденного. Раз-
витие молочных желез во время беременности регулируется гормонами
пролактином, эстрогенами и прогестероном, под действием которых сти-
мулируется развитие протоков, долей и альвеол молочных желез, а следо-
вательно, их подготовка к периоду вскармливания ребенка после родов ма-
теринским молоком. Физиологические эффекты пролактина на молочные
железы включают их рост и развитие (маммогенез), синтез молока (лакто-
генез) и поддержание секреции молока (галактопоэз). Пролактин влияет
на морфогенез молочных желез двумя путями: до первой беременности
гормон контролирует ветвление протоков, во время беременности — непо-
средственно контролирует развитие долек и альвеол. Лактогенез и галакто-
генез контролируются пролактином, который секретируется лактотрофами
аденогипофиза. Однако его физиологические действие на молочные желе-
зы осуществляется только в комплексе с другими гормонами и факторами
роста. Например, развитие молочных желез до первой беременности про-
исходит под влиянием на молочные железы пролактина, эстрогенов, про-
гестерона и глюкокортикоидов. В ранние сроки беременности гипертро-
фия структурных элементов протоков, долей и альвеол в молочных железах
происходит под влиянием эстрадиола и прогестерона. Пролактин и инсу-
лин стимулируют рост тканей. В ткани молочных желез во время беремен-
ности синтезируются специфические факторы роста, в частности эпидер-
мальный фактор роста, трасформирующийся в a-фактор роста. Молочные
железы способны продуцировать молоко с четвертого месяца беременно-
сти, поскольку к этому времени полностью завершается развитие протоков
и долей и происходит дифференциация альвеол. Последние выстланы ку-
бическим секреторным эпителием, способным к синтезу и секреции моло-
ка при участии миоэпителиальных клеток, которые окружают снаружи
стенку альвеол и выводные протоки секреторных отделов молочной желе-
зы. Основная функция пролактина в процессе лактогенеза — стимуляция
транспорта из плазмы крови в цитоплазму секреторных клеток альвеол мо-
лочной железы аминокислот, синтез секреторными клетками белков моло-
ка (казеин, а-лактальбумина), транспорт глюкозы внутрь альвеолярных
клеток и синтез молочного сахара лактозы. В период беременности про-
лактин не стимулирует секрецию молока в молочных железах, поскольку
эстрогены и, особенно, прогестерон тормозят действие пролактина на сек-
реторные клетки молочных желез (рис. 16.14). После родов в плазме крови
матери происходит снижение концентрации прогестерона, и пролактин
начинает стимулировать секрецию молока в альвеолярных клетках. Во вре-
мя лактации в секреторных отделах секретируются вода, электролиты, жи-
ры, а-лактоальбумин, казеин, сывороточный альбумин, лактоферрин, ли-
зоцим, лактоза, антитела IgA.
700 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Оплодотворение Роды Прекращение
вскармливания
молоком
Лактация
Базальный уровень
эстрадиола до начала
менструальных циклов
Рис. 16.14. Секреция материнских гормонов, вызывающая лактацию.
До рождения плода эстрогены и прогестерон тормозят секрецию молока в молочных железах,
несмотря на высокую концентрацию этого гормона в плазме крови беременной женщины.
После родов уровень пролактина в плазме крови снижается, но остается более высоким, чем у
небеременной женщины. Этот уровень концентрации пролактина в плазме крови поддержива-
ется только в течение 3—4 нед, если не начинается грудное вскармливание ребенка, а затем
снижается. После рождения ребенка нормальная секреция пролактина в гипофизе матери
поддерживается за счет нейроэндокринного рефлекса, возникающего у кормящей женщины
во время кормления ребенка грудью.
*
Во время кормления ребенка грудью рефлекторно возникает сокраще-
ние миоэпитеальных клеток альвеол молочных желез у кормящей женщи-
ны, и молоко выдавливается из них в протоки молочных желез.
Нейроэндокринный рефлекс выделения молока. Рефлекторное выделение
молока у матери из секреторных отделов молочных желез во время корм-
ления ребенка грудью называется нейроэндокринным рефлексом
(рис. 16.15). Рефлекс возникает, когда ребенок начинает сосать сосок. При
этом происходит стимуляция тактильных рецепторов кожи сосковой об-
ласти. Нервные импульсы от рецепторов поступают в спинной мозг, а за-
тем к нейронам супраоптического и паравентрикулярного ядер гипотала-
муса, которые секретируют окситоцин. Эти нейроны гипоталамуса генери-
руют электрическую активность в среднем каждые 5— 15 мин в виде корот-
кого, длительностью 3—4 с, высокочастотного разряда потенциалов дейст-
16. Репродуктивная функция человека • 701
Миоэпителиальные
клетки
клетки
Рецепторы кожц
сосков
Молочные железы
Супраоптическое и
паравентрикулярнце
ядра гипоталамуса
Сегменты
спинного мозга
Хиазма ▼
Рис. 16.15. Регуляция секреции окситоцина в нейрогипофизе и пролактина в адено-
гипофизе кормящей женщины во время кормления ребенка грудью (нейроэндо-
кринный рефлекс). Раздражение рецепторов кожи сосков во время кормления вы-
зывает активацию нейронов супраоптического и паравентрикулярного ядер гипота-
ламуса, повышает концентрацию окситоцина и пролактина в плазме крови.
Аденогипофиз '
Медиальная пучок
переднеголюзга
вия. Во время каждого залпа активности нейросекреторных клеток гипота-
ламуса потенциалы действия достигают терминалей нейронов, локализо-
ванных в нейрогипофизе, и в плазму крови кормящей ребенка женщины
выделяется окситоцин, который с кровью транспортируется к молочным
железам. В мембранах миоэпителиальных клеток молочных желез локали-
зованы рецепторы к окситоцину. Этот гормон при взаимодействии с ре-
цепторами мембраны миоэпителиальных клеток секреторных отделов мо-
лочных желез вызывает внутри клеток образование вторичного посредника
инозитол-3-фосфата, который открывает кальциевые ионные каналы сар-
коплазматического ретикулума этих клеток. Высвобождение из депо ионов
Са2+ вызывает сокращение гладкомышечных клеток и продвижение молока
из альвеолярных клеток в протоки и синусы молочных желез. При кормле-
нии ребенка грудью из синусов молочной железы молоко поступает в ро-
товую полость ребенка. У кормящей женщины, в среднем через 30—60 с
после начала кормления ребенка грудью, молоко начинает активно выде-
ляться в протоки молочной железы.
Для нормальной секреции молока в молочных железах кормящей жен-
щине необходимо придерживаться постоянного режима кормления ребен-
ка, что сохраняет секрецию пролактина и препятствует снижению лакта-
ции. Это поддерживает в грудном молоке содержание иммуноглобулинов,
особенно IgA, неспецифических антимикробных факторов (лизоцим, лак-
тоферрин), которые обеспечивают неспецифические защитные реакции в
организме новорожденного в период формирования у него механизмов
клеточного иммунитета.
ГЛАВА 17
Сенсорные системы
Сенсорные системы человека являются частью его нервной системы, спо-
собной воспринимать внешнюю для мозга информацию, передавать ее в
мозг и анализировать. Получение информации от окружающей среды и
собственного тела является обязательным и необходимым условием суще-
ствования человека. Термин «сенсорные (лат. sensus — чувство) системы»
сменил название «органы чувств», сохранившееся только для обозначения
анатомически обособленных периферических отделов некоторых сенсор-
ных систем (как, например, глаз или ухо). В отечественной литературе в
качестве синонима сенсорной системы применяется предложенное
И. П. Павловым понятие «анализатор», указывающее на функцию сенсор-
ной системы.
Все сенсорные системы состоят из периферических рецепторов, проводя-
щих путей и переключательных ядер, первичных проекционных областей коры
и вторичной сенсорной коры. Сенсорные системы организованы иерархиче-
ски, т. е. включают несколько уровней последовательной переработки ин-
формации. Низший уровень такой переработки обеспечивают первичные
сенсорные нейроны, которые расположены в специализированных органах
чувств или в чувствительных ганглиях и предназначены для проведения
возбуждения от периферических рецепторов в центральную нервную сис-
тему. Периферические рецепторы — это чувствительные высокоспециали-
зированные образования, способные воспринять, трансформировать и пе-
редать энергию внешнего стимула первичным сенсорным нейронам.
Центральные отростки первичных сенсорных нейронов оканчиваются в
головном или спинном мозге на нейронах второго порядка, тела которых
расположены в переключательном ядре. В нем имеются не только возбуж-
дающие, но и тормозные нейроны, участвующие в переработке передавае-
мой информации. Представляя более высокий иерархический уровень,
нейроны переключательного ядра могут регулировать передачу информа-
ции путем усиления одних и торможения или подавления других сигналов.
Аксоны нейронов второго порядка образуют проводящие пути к следую-
щему переключательному ядру, общее число которых обусловлено специ-
фическими особенностями разных сенсорных систем. Окончательная пе-
реработка информации о действующем стимуле происходит в сенсорных
областях коры.
Сенсорные системы человека обеспечивают: 1) формирование ощущений
и восприятие действующих стимулов; 2) контроль произвольных движений;
3) контроль деятельности внутренних органов; 4) необходимый для бодрст-
вования человека уровень активности мозга. Ощущение представляет со-
бой субъективную чувственную реакцию на действующий сенсорный сти-
мул (например, ощущение света, тепла или холода, прикосновения и т. п.).
Однородные сенсорные стимулы активируют одну из сенсорных систем и
вызывают субъективно одинаковые ощущения, совокупность которых обо-
значается термином модальность. Самостоятельными модальностями явля-
17. Сенсорные системы • 703
ются осязание, зрение, слух, обоняние, вкус, чувство холода или тепла, бо-
ли, вибрации, ощущение положения конечностей и мышечной нагрузки.
Внутри модальностей могут существовать разные качества, или субмодаль-
ности', например, во вкусовой модальности различают сладкий, соленый,
кислый и горький вкус. На основе совокупности ощущений формируется
чувственное восприятие, т. е. осмысление ощущений и готовность их опи-
сать. Восприятие не является простым отражением действующего стимула,
оно зависит от распределения внимания в момент его действия, памяти о
прошлом сенсорном опыте и субъективного отношения к происходящему,
выражающегося в эмоциональных переживаниях.
Сенсорное восприятие включает следующие этапы: 1) действие раздра-
жителя на периферические рецепторы; 2) преобразование энергии стимула
в электрические сигналы — потенциалы действия, возникающие в первич-
ном сенсорном нейроне; 3) последующую переработку передаваемых сиг-
налов на всех иерархических уровнях сенсорной системы; 4) возникнове-
ние субъективной реакции на раздражитель, представляющей собой вос-
приятие или внутреннее представительство действующего стимула в виде
образов или словесных символов. Указанная последовательность соблюда-
ется во всех сенсорных системах, отражая иерархический принцип их ор-
ганизации.
17.1. Общая физиология сенсорных систем
Человек воспринимает не все изменения окружающей среды, он не спосо-
бен, например, ощущать действие ультразвука, рентгеновских лучей или
радиоволн. Диапазон сенсорного восприятия человека ограничен имеющи-
мися у него сенсорными системами, каждая из которых перерабатывает
информацию о стимулах определенной физической природы. Сенсорная
специфичность каждой системы определяется, прежде всего, свойствами
входящих в нее рецепторов.
17.1.1. Классификации рецепторов
Рецепторы различают по специфической чувствительности к разным сти-
мулам, по строению и местоположению. Специфическая чувствительность
рецепторов к раздражителям различной природы (механическим, химиче-
ским, температурным и т. д.) обусловлена разными механизмами управле-
ния ионными каналами плазматических мембран, состояние которых опре-
деляет возникновение рецепторного потенциала и переход от физиологиче-
ского покоя к возбуждению. Стимулы, к которым рецепторы наиболее чув-
ствительны, называются адекватными (лат. adaequatus — приравненный).
Механорецепторы возбуждаются сильнее всего вследствие деформации
их клеточной мебраны при давлении или растяжении, к ним относятся
тактильные рецепторы кожи, проприоцепторы мышц и сухожилий, слухо-
вые и вестибулярные рецепторы во внутреннем ухе, барорецепторы и во-
люморецепторы, находящиеся во внутренних органах и кровеносных сосу-
дах. Хеморецепторы возбуждаются вследствие присоединения к ним опре-
деленных химических молекул, они представлены обонятельными и вкусо-
выми рецепторами, а также хемочувствительными рецепторами внутрен-
них органов и кровеносных сосудов. Для расположенных в сетчатке глаза
фоторецепторов адекватным раздражителем являются поглощенные ими
кванты света, для терморецепторов (холодовых и тепловых) — изменения
температуры.
704 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Большинство рецепторов возбуждаются в ответ на действие стимулов
только одной физической природы и поэтому относятся к мономодалъным.
Их можно возбудить и некоторыми неадекватными раздражителями, на-
пример фоторецепторы — сильным давлением на глазное яблоко, а вкусо-
вые рецепторы — прикосновением языка к контактам гальванической ба-
тареи, но получить качественно различаемые ощущения в таких случаях
невозможно. Наряду с мономодальными существуют полимодалъные рецеп-
торы, адекватными стимулами которых могут служить раздражители раз-
ной природы. К такому типу рецепторов принадлежат некоторые болевые
рецепторы, или ноцицепторы (лат. nocens — вредный), которые можно воз-
будить механическими, термическими и химическими стимулами. Поли-
модальность имеется у терморецепторов, реагирующих на повышение кон-
центрации калия во внеклеточном пространстве так же, как на повышение
температуры.
В зависимости от строения рецепторов их подразделяют на первичные,
или первичночувствующие, которые являются специализированными окон-
чаниями чувствительного нейрона, и вторичные, или вторичночувствую-
щие, представляющие собой клетки эпителиального происхождения, спо-
собные к образованию рецепторного потенциала в ответ на действие аде-
кватного стимула. Первичночувствующие рецепторы могут сами генери-
ровать потенциалы действия в ответ на раздражение адекватным стиму-
лом, если величина их рецепторного потенциала достигнет пороговой ве-
личины. К ним относятся обонятельные рецепторы, большинство меха-
норецепторов кожи, терморецепторы, болевые рецепторы или ноцицепто-
ры, проприоцепторы и большинство интерорецепторов внутренних ор-
ганов.
Вторичночувствующие рецепторы отвечают на действие раздражителя
лишь возникновением рецепторного потенциала, от величины которого
зависит количество выделяемого этими клетками медиатора. С его помо-
щью вторичные рецепторы действуют на нервные окончания чувствитель-
ных нейронов, генерирующих потенциалы действия в зависимости от ко-
личества медиатора, выделившегося из вторичночувствующих рецепторов.
Вторичные рецепторы представлены вкусовыми, слуховыми и вестибуляр-
ными рецепторами, а также хемочувствительными клетками синокаротид-
ного клубочка. Фоторецепторы сетчатки, имеющие общее происхождение
с нервными клетками, чаще относят к первичным рецепторам, но отсутст-
вие у них способности генерировать потенциалы действия указывает на их
сходство с вторичными рецепторами.
В зависимости от источника адекватных стимулов рецепторы подразде-
ляют на наружные и внутренние, или экстерорецепторы и интерорецепто-
ръг, первые стимулируются при действии раздражителей внешней среды
(электромагнитные и звуковые волны, давление, действие пахучих моле-
кул), а вторые — внутренней (к этому типу рецепторов относят не только
висцерорецепторы внутренних органов, но также проприоцепторы и вес-
тибулярные рецепторы). В зависимости от того, действует стимул на рас-
стоянии или непосредственно на рецепторы, их подразделяют еще на дис-
тантные и контактные.
17.1.2. Преобразование энергии раздражителя в рецепторах
В результате действия адекватного раздражителя у большинства рецепто-
ров увеличивается проницаемость клеточной мембраны для катионов, что
приводит к ее деполяризации. Исключением из общего правила являются
17. Сенсорные системы • 705
фоторецепторы, где после поглощения энергии квантов света в связи осо-
бенностями управления ионными каналами (см. раздел 17.3.2.2) происхо-
дит гиперполяризация мембраны. Изменение величины мембранного по-
тенциала рецепторов в ответ на действие стимула представляет собой ре-
цепторный потенциал — входной сигнал первичных сенсорных нейронов.
Если величина рецепторного потенциала достигнет критического уровня
деполяризации или превысит его, генерируются потенциалы действия, с
помощью которых сенсорные нейройы передают в центральную нервную
систему информацию о действующих стимулах.
Генерация потенциалов действия происходит в ближайшем к рецепто-
рам перехвате Ранвье миелинизированных волокон или ближайшей к ре-
цепторам части мембраны безмиелинового волокна. Минимальная сила
адекватного стимула, достаточная для генерации потенциалов действия в
первичном сенсорном нейроне, определяется как его абсолютный порог.
Минимальный прирост силы стимула, сопровождающийся значимым из-
менением реакции сенсорного нейрона, представляет собой дифференци-
альный порог его чувствительности.
Информация о силе действующего на рецепторы стимула кодируется
двумя способами: частотой потенциалов действия, возникающих в сенсор-
ном нейроне (частотное кодирование), и числом сенсорных нейронов, воз-
будившихся в ответ на действие стимула. При увеличении силы действую-
щего на рецепторы раздражителя повышается амплитуда рецепторного по-
тенциала, что, как правило, сопровождается увеличением частоты потен-
циалов действия в сенсорном нейроне первого порядка. Чем шире имею-
щийся частотный диапазон потенциалов действия у сенсорных нейронов,
тем большее число промежуточных значений силы раздражителя способна
различать сенсорная система. Первичные сенсорные нейроны одинаковой
модальности различаются порогом возбуждения, поэтому при действии
слабых стимулов возбуждаются только наиболее чувствительные нейроны,
но с увеличением силы раздражителя на него реагируют и менее чувстви-
тельные нейроны, имеющие более высокий порог раздражения. Чем боль-
ше первичных сенсорных нейронов возбудится одновременно, тем сильнее
будет их совместное действие на общий нейрон второго порядка, что в
итоге отразится на субъективной оценке интенсивности действующего раз-
дражителя.
Длительность ощущения зависит от реального времени между началом и
прекращением воздействия на рецепторы, а также от их способности
уменьшать или даже прекращать генерацию нервных импульсов при про-
должительном действии адекватного стимула. При длительном действии
стимула порог чувствительности рецепторов к нему может повышаться,
что определяется как адаптация рецепторов. Механизмы адаптации не
одинаковы в рецепторах разных модальностей, среди них различают быст-
ро адаптирующиеся (например, тактильные рецепторы кожи) и медленно
адаптирующиеся рецепторы (например, проприоцепторы мышц и сухожи-
лий). Быстро адаптирующиеся рецепторы сильнее возбуждаются в ответ на
быстрое нарастание интенсивности стимула (фазический ответ), а их быст-
рая адаптация способствует освобождению восприятия от биологически
незначительной информации (например, контакт между кожей и одеж-
дой). Возбуждение медленно адаптирующихся рецепторов мало зависит от
скорости изменения стимула и сохраняется при его длительном действии
(тонический ответ), поэтому, например, медленная адаптация проприо-
цепторов позволяет человеку получать нужную ему для сохранения позы
информацию в течение всего необходимого времени.
23-6095
706 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Существуют сенсорные нейроны, генерирующие потенциалы действия
спонтанно, т. е. при отсутствии раздражения (например, сенсорные нейро-
ны вестибулярной системы), такая активность называется фоновой. Частота
нервных импульсов в этих нейронах может увеличиваться или уменьшать-
ся в зависимости от интенсивности действующего на вторичные рецепто-
ры стимула, кроме того, она может определяться направлением, в котором
отклоняются чувствительные волоски механорецепторов. Например, от-
клонение волосков вторичных механорецепторов в одну сторону сопрово-
ждается повышением фоновой активности сенсорного нейрона, которому
они принадлежат, а в противоположную сторону — понижением его фоно-
вой активности. Указанный способ рецепции позволяет получать инфор-
мацию и об интенсивности стимула, и о направлении, в котором он дейст-
вует.
17.1.3. Рецептивные поля
Рецептивным полем называется область, занимаемая совокупностью всех
рецепторов, стимуляция которых приводит к возбуждению сенсорного
нейрона (рис. 17.1). Максимальная величина рецептивного поля первич-
ного сенсорного нейрона определяется пространством, которое занимают
все ветви его периферического отростка, а число рецепторов, имеющихся в
этом пространстве, указывает на плотность иннервации. Высокая плот-
ность иннервации сочетается, как правило, с малыми размерами рецептив-
ных полей и, соответственно, высоким пространственным разрешением,
позволяющим различать стимулы, действующие на соседние рецептивные
поля. Малые рецептивные поля типичны, например, для центральной ям-
Рецептивные поля
ного ядра)
Рис. 17.1. Рецептивные поля первичных сенсорных нейронов и сенсорных нейро-
нов второго порядка.
А. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствитель-
ных окончаний. Рецептивное поле переключательного нейрона образуется из суммы рецеп-
тивных полей конвергирующих к нему первичных сенсорных нейронов.
Б. Раздражение центральной или периферической области рецептивного поля сенсорного
нейрона второго и следующих порядков сопровождается противоположным эффектом. Как
видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного
нейрона, а раздражение периферической области — торможение с помощью интернейронов
переключательного ядра (латеральное торможение). В результате контраста, создаваемого ме-
жду центром и периферией рецептивного поля, выделяется информация для передачи на сле-
дующий иерархический уровень.
17. Сенсорные системы • 707
ки сетчатки и для пальцев рук, где плотность рецепторов значительно вы-
ше, чем на периферии сетчатки или в коже спины, для которых характер-
ны большая величина рецептивных полей и меньшее пространственное
разрешение. Рецептивные поля соседних сенсорных нейронов могут час-
тично перекрывать друг друга, поэтому информация о действующих на них
стимулах передается не по одному, а по нескольким параллельным аксо-
нам, что повышает надежность ее передачи.
Величина рецептивных полей сенсорных нейронов второго и следую-
щих порядков больше, чем у первичных сенсорных нейронов, поскольку
центральные нейроны получают информацию от нескольких конвергирую-
щих к ним нейронов предшествующего уровня. От центра рецептивного
поля информация передается непосредственно к сенсорным нейронам сле-
дующего порядка, а от периферии — к тормозным интернейронам пере-
ключательного ядра, поэтому центр и периферия рецептивного поля явля-
ются реципрокными по отношению друг к другу. В результате сигналы от
центра рецептивного поля беспрепятственно достигают следующего иерар-
хического уровня сенсорной системы, тогда как сигналы, поступающие от
периферии рецептивного поля, тормозятся (в другом варианте организа-
ции рецептивного поля легче пропускаются сигналы от периферии, а не от
центра). Такая функциональная организация рецептивных полей обеспе-
чивает выделение наиболее значимых сигналов, легко различаемых на кон-
трастном с ними фоне.
17.1.4. Переработка информации в переключательных ядрах
и проводящих путях сенсорной системы
Сенсорные нейроны низшего иерархического уровня способны передавать
электрические сигналы одновременно нескольким нейронам следующего
уровня благодаря дивергенции к ним своих аксонов. Это повышает надеж-
ность передачи информации от одного иерархического уровня к другому и
позволяет сохранять чувствительность сенсорной системы при утрате от-
дельных нейронов. Возбуждение нейронов более высокого иерархического
уровня определяется не только действием на них нейронов предыдущего
уровня, но и механизмом латерального торможения, существующего в пе-
реключательных ядрах (рис. 17.2).
Латеральное торможение происходит вследствие активации тормозных
интернейронов переключательного ядра коллатералями возбуждающих
нейронов. Чем сильнее возбужден релейный сенсорный нейрон, тем боль-
ше он активирует тормозные нейроны, которые подавляют активность со-
седних релейных нейронов. Релейные нейроны, передающие сигналы от
центра рецептивного поля, сильнее других повышают активность тормоз-
ных нейронов, а их влияние на соседние релейные нейроны делает еще
большим контраст между возбужденными и тормозимыми нейронами, вы-
деляя линию передачи избранных сигналов.
Высшие уровни сенсорной системы регулируют переработку информа-
ции в низших по отношению к ним переключательных ядрах посредством
нисходящего торможения или усиления передаваемых сигналов. Нисходящее
торможение (рис.17.3) происходит вследствие активации тормозных ней-
ронов переключательного ядра, деятельность которых определяют нисхо-
дящие пути, начинающиеся на более высоком иерархическом уровне сен-
сорной системы. В результате нисходящего торможения повышается порог
афферентной синаптической передачи в переключательном ядре низшего
уровня. Нисходящее торможение представляет собой регуляцию сенсорно-
23*
708 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Действие раздражителя
Рецепторы первичных
сенсорных нейронов
Первые переклю-
чательные нейроны
Рис. 17.2. Схема латерального торможения в сенсорных системах.
Тормозные интернейроны (их тела выделены черным цветом) возбуждаются возвратными
коллатералями переключательных сенсорных нейронов: чем сильнее возбужден переключа-
тельный нейрон, тем большее торможение соседних переключательных нейронов он вызовет.
Степень возбуждения переключательных нейронов определяется интенсивностью раздраже-
ния центра или периферии его рецептивного поля. В результате возбуждение будет переда-
ваться от одной части рецептивного поля на фоне торможения или прерывания сигналов от
другой его части, что создает эффект контрастирования, способствующий различению дейст-
вующих стимулов.
го восприятия по механизму отрицательной обратной связи. Тормозные
интернейроны переключательных ядер уменьшают частоту передаваемых
на следующий уровень сигналов, в связи с чем сенсорное ощущение ос-
лабляется.
Нисходящее усиление происходит с помощью возбуждающих интерней-
ронов переключательного ядра, которые активируются аксонами нейронов
более высокого иерархического уровня. Вследствие этого в управляемом
по механизму положительной обратной связи переключательном ядре пони-
жается порог синаптической передачи, что делает возможным прохожде-
ние относительно слабых сигналов на следующий иерархический уровень.
Одновременное использование механизмов отрицательной и положитель-
ной обратной связи позволяет освобождаться от избыточной информации,
подавлять «шум», т. е. нейронную активность, не содержащую физиологи-
чески значимых сообщений, и одновременно выделять и усиливать сигна-
лы, на которых будет сосредоточено внимание.
Большинство раздражителей, воспринимаемых сенсорными системами,
оказывает на них комплексное действие, поскольку одновременно возбуж-
дает разные типы рецепторов. Например, прикасающийся к ограниченно-
му участку кожи предмет может быть гладким или шершавым, теплым или
холодным, сухим или влажным, а его давление на кожу может усиливаться
или ослабевать, он может передвигаться в том или ином направлении.
17. Сенсорные системы • 709
Таламус
Латеральное
торможение
Тормозные
интернейроны
Проекционный
нейрон
Переклю-
чательное
ядро
сенсорной
системы
Афферентные
сигналы
К другим
областям ЦНС
Рис. 17.3. Схема нисходящего торможения в сенсорных системах.
Передачу афферентных сигналов от одного иерархического уровня сенсорной системы к дру-
гому в переключательных ядрах контролируют высшие иерархические уровни. На схеме пока-
зан принцип нисходящего торможения коры, которое осуществляется с помощью тормозных
интернейронов (выделены черным цветом) переключательного ядра. Возбуждение тормозных
интернейронов под влиянием коры препятствует передаче афферентных сигналов в переклю-
чательном ядре.
Зрительные ощущения представляют собой сочетание определенной фор-
мы воспринимаемого предмета, его движения и комбинации различных
цветов, заполняющих ту или иную деталь формы. Сенсорные системы ор-
ганизованы так, что информация о каждом компоненте комплексного раз-
дражителя передается одновременно по нескольким параллельным путям,
каждый из которых относится к определенной субмодальности, как, на-
пример, форма, движение или цвет наблюдаемого зрительного объекта.
В каждом из таких путей независимо друг от друга перерабатываются сиг-
налы, несущие информацию о разных качествах подействовавшего на ре-
цепторы комплексного раздражителя. При этом разные каналы, используе-
мые для передачи сенсорной информации, не дублируют друг друга, в чем
проявляется принцип многоканальности, присущий всем сенсорным систе-
мам. Высокая упорядоченность нейронных переключений на каждом ие-
рархическом уровне обеспечивает поступление информации от каждого
рецептивного поля к соответствующим именно ему кортикальным колон-
кам проекционной коры. Необходимое для целостного восприятия раздра-
жителя объединение информации, относящейся к разным субмодально-
стям, происходит в сенсорных регионах коры.
710 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
17.1.5. Субъективное сенсорное восприятие
Каждая сенсорная система воспринимает действие адекватного стимула в
ограниченном диапазоне значений его силы. Наименьший по интенсивно-
сти стимул, способный вызвать ощущение, называется абсолютным поро-
гом ощущения. Его величина устанавливается опытным путем, она не оди-
накова у разных людей и может изменяться у одного и того же человека в
зависимости от функционального состояния, различаясь, например, при
оптимальной работоспособности и утомлении. Величина абсолютного по-
рога ощущения выше абсолютного порога первичных сенсорных нейро-
нов, так как возбуждение отдельных чувствительных нейронов или их не-
большой группы не обязательно приводит к возбуждению высших уровней
сенсорной системы вследствие тормозных процессов на предыдущих ие-
рархических уровнях. Поэтому не каждый стимул, возбуждающий рецепто-
ры первичного сенсорного нейрона, субъективно ощущается и осознается.
Величина, на которую один стимул надпорогового диапазона должен
отличаться от другого, чтобы их разницу можно было субъективно разли-
чить, получила название дифференциального порога или порога различения.
Ощущаемый прирост интенсивности раздражения (дифференциальный по-
рог) должен превышать ранее действовавший стимул на определенную и
постоянную величину. Например, человек легко различает по весу гирьки
весом 14 и 15 граммов (разница в 1 грамм), но гирьки весом 29 и 30 грам-
мов кажутся ему одинаковыми, и различия между ними выявляются, если
разница окажется не меньше 2 граммов. Если же гирька составит 60 грам-
мов, то субъективное ощущение большего груза возможно только тогда,
когда его прирост будет не меньше 4 граммов. Таким образом, ощущаемый
прирост силы раздражителя возможен при ее увеличении на постоянную
величину, что сформулировано в законе Вебера как 6S/S = к, где S — раз-
дражение, 6S — его ощущаемый прирост, к — постоянная величина.
Закон Вебера справедлив для разных модальностей, но лишь в границах
раздражения средней силы, а при слабых и сильных стимулах способность
к различению силы стимулов у человека снижается. Предложена также
ступенчатая шкала интенсивности ощущений, в которой за ноль принима-
ется величина абсолютного порога, а зависимость между интенсивностью
стимула и ощущением выражается как: Е = k х log(S/S0), где Е — интен-
сивность ощущения, к — постоянная величина, S — интенсивность стиму-
ла, So — абсолютный порог. Эта зависимость, получившая название закона
Вебера—Фехнера, показывает, что линейное увеличение интенсивности
ощущения отражает логарифмический рост интенсивности стимула. Закон
Вебера—Фехнера характеризует субъективную способность человека разли-
чать действующие стимулы (шкала различения), но не оценивает саму ин-
тенсивность ощущений, поскольку этот закон основан на допущении ра-
венства любых ее приростов, как к слабым, так и к сильным стимулам.
Субъективная оценка интенсивности раздражителя (качественное раз-
личие) возрастает при увеличении разницы между пороговой и действую-
щей силой стимула (количественное различие). Однако зависимость между
силой стимула и ощущением не одинакова при разной интенсивности сти-
мулов, а потому имеет не линейный, а степенной характер. Для оценки
интенсивности ощущений всего диапазона используется шкала Стивенса,
устанавливающая зависимость ощущений от силы стимула в виде степен-
ной функции: Е = k(S — S0)n, где Е — интенсивность ощущения, S — дей-
ствующая сила стимула, So — абсолютный порог, к — константа шкалы,
п — показатель степени, который зависит от сенсорной модальности (на-
17. Сенсорные системы • 711
пример, для восприятия светового ощущения он составляет 0,33, громко-
сти звука — 0,6, а для проприоцептивных ощущений — 1,7).
Пространственные характеристики действующих стимулов, необходи-
мые для их различения, зависят от специфических особенностей каждой
сенсорной системы и величины рецептивных полей. Прикосновение к ко-
же дистальной фаланги пальца руки двух ножек циркуля с расстоянием
между ними 2 мм ощущается раздельно, но чтобы ощутить раздельное
прикосновение к коже спины, ножки циркуля необходимо раздвинуть до
60 мм. Пространственное восприятие этих тактильных стимулов зависит от
размеров соответствующих рецептивных полей: раздельное ощущение воз-
можно только при условии раздражения каждой ножкой циркуля незави-
симого рецептивного поля. Лишь тогда информация о каждом стимуле бу-
дет перерабатываться раздельно на каждом уровне организации сенсорной
системы, включая проекционную область коры. Аналогичная ситуация
имеет место при восприятии двух точек зрительного поля: они не сливают-
ся в одну, если отражаемые ими световые лучи попадут на разные рецеп-
тивные поля сетчатки. Имеет значение и степень контраста между дейст-
вующим стимулом и его фоном: хорошо контрастируемые объекты (напри-
мер, черное на белом) различаются легче, чем мало контрастируемые (чер-
ное на сером).
Временная характеристика восприятия действующих стимулов у челове-
ка имеет абсолютный порог различения коротких временных отрезков, ко-
торый соответствует примерно ’/18 секунды. Например, 18 зрительных изо-
бражений, предъявленных в течение 1 секунды, сливаются в непрерывное
движение, 18 прикосновений к коже за 1 секунду воспринимаются как од-
но, а 18 звуковых колебаний в секунду воспринимаются как один очень
низкий звук. Разрешающая способность сенсорных систем для восприятия
действующих через малые промежутки времени стимулов ограничена реф-
рактерным периодом, во время которого система не способна реагировать
на предъявленный стимул.
17.2. Соматовисцералыгая сенсорная система
Соматовисцеральная система перерабатывает сенсорную информацию не-
скольких модальностей, получая ее от кожи, мышц, суставов, внутренних
органов и кровеносных сосудов. Она содержит разные типы рецепторов:
механорецепторы, терморецепторы, хеморецепторы, осморецепторы, но-
цицепторы. Поступающая информация передается по афферентным во-
локнам в составе чувствительных и смешанных нервов, перерабатывается
раздельными проводящими путями и переключательными ядрами, а затем
доставляется в соматосенсорную кору, занимающую постцентральные из-
вилины мозга и организованную топически.
17.2.1. Тактильная чувствительность
Часть соматовисцеральной системы, обеспечивающая чувство осязания,
включает несколько разновидностей механорецепторов кожи, представлен-
ных свободными нервными окончаниями либо инкапсулированными, т. е. за-
ключенными в капсулу из соединительной ткани или видоизмененных
клеток эпидермиса (рис. 17.4), Свободные нервные окончания иннервиру-
ют волосяные фолликулы пушковых волос, покрывающих большую часть
тела человека, а также грубых волос, растущих на голове, в подмышечных
впадинах, на лобке, а у мужчин еще и на лице. Свободные нервные окон-
712 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Свободные
Тельце Диски нервные Тельце
Руффини Меркеля окончания Пачини
Волосяной
фолликул
Рис. 17.4. Схема распределения механорецепторов
в коже человека.
Свободные нервные окончания имеют высокий порог
раздражения и слабо реагируют на изменение интенсив-
ности стимула. Быстро адаптирующиеся рецепторы
(тельца Пачини, рецепторы волосяных фолликулов) слу-
жат датчиками скорости действующих стимулов, а мед-
ленно адаптирующиеся рецепторы (диски Меркеля,
тельца Руффини) являются датчиками интенсивности
действующего раздражителя. Наличие нескольких раз-
новидностей рецепторов позволяет передавать аффе-
рентные сигналы о разных свойствах одного и того же
раздражителя.
чания волосяных фолли-
кулов являются механоре-
цепторами и возбуждают-
ся при смещении волос
или их подергивании.
Другая разновидность сво-
бодных нервных оконча-
ний имеется в эпидермисе
и в сосочковом слое дер-
мы, большинство из них
являются ноцицепторами
или терморецепторами, но
некоторые принадлежат к
механорецепторам, кото-
рые специфически реаги-
руют на слабое околопо-
роговое раздражение.
Предполагается, что при
раздражении этой разно-
видности рецепторов воз-
никают ощущения щекот-
ки и зуда.
Среди инкапсулирован-
ных окончаний различа-
ют тельца Пачини, Мейс-
снера, Руффини, диски
Меркеля, тактильные тель-
ца Пинкуса—Игго, колбы
Краузе. В зависимости от
строения и формы капсу-
лы нервные окончания
подвержены наиболее
сильному воздействию ли-
бо в результате давления
действующим перпендику-
лярно раздражителем, ли-
бо вследствие бокового
смещения капсулы, кото-
рая играет роль механиче-
ского преобразователя
энергии внешних стимулов. Большинство инкапсулированных рецепторов
содержится в лишенной волос коже пальцев рук и ног, ладоней и подошв,
лица, губ, языка, сосков и половых органов, где они распределены с раз-
личной плотностью и на разной глубине. Тельца Пачини имеются также в
сухожилиях, связках и брыжейке.
Механорецепторы кожи различаются по скорости адаптации к дейст-
вующему раздражителю. Быстроадаптирующиеся (фазные) рецепторы воз-
буждаются только в момент смещения кожи и волос и служат датчиками
скорости воздействия стимула. Это свойство присуще тельцам Мейснера,
рецепторам волосяных фолликулов и особенно тельцам Пачини, способ-
ным реагировать на изменения скорости продолжающего свое действие
стимула. Медленно адаптирующиеся (тонические) рецепторы не прекраща-
ют генерировать потенциалы действия при продолжительном действии
17. Сенсорные системы • 713
раздражителя, если он оказывает давление
на кожу: такие рецепторы служат датчиками
интенсивности действующего стимула
(тельца Руффини, диски Меркеля).
Площадь рецептивных полей сенсорных
нейронов, иннервирующих тельца Мейсне-
ра и диски Меркеля, составляет в среднем
около 12 мм2, а у нейронов с окончаниями
в виде телец Пачини и Руффини она на по-
рядок больше. Рецептивные поля различаю-
щихся своими рецепторами сенсорных ней-
ронов перекрываются, поэтому при дейст-
вии на кожу комплекса стимулов одновре-
менно возбуждаются разные виды рецепто-
ров, что позволяет ощущать все динамиче-
ские и статические свойства такого ком-
плекса. Обработка и анализ информации
Таблица 17.1. Пространствен-
ный двухточечный порог в раз-
ных участках тела
Поверхность тела Порог (мм)
Кончик языка 1
Кончик пальца 2
Кайма губ 4,5
Кончик носа 6,8
Щека п,з
Скула 15,8
Тыл кисти 31,6
Надколенник 36,1
Предплечье, голень 40,6
Область грудины 45,1
Плечо 60
Затылок, спина 67,7
сигналов от различных рецепторов проис-
ходит на высших уровнях сенсорной системы, формирующих комплексное
восприятие действующих на поверхность тела стимулов. Плотность меха-
норецепторов в разных участках кожи не одинакова, чем определяются
разные показатели пространственного дифференциального порога, т. е. наи-
меньшего расстояния между двумя точками, раздражение каждой из кото-
рых ощущается раздельно (табл. 17.1). Приведенные в таблице данные не
следует считать эталоном, поскольку дифференциальная чувствительность
различается у разных людей.
Инкапсулированные рецепторы иннервируются миелинизированными
волокнами первичных сенсорных нейронов, которые проводят нервные
импульсы в ЦНС со скоростью около 30—70 м/с. Немиелинизированные
волокна передают потенциалы действия от свободных нервных окончаний
со значительно меньшей скоростью — около 1 м/с, поэтому ощущение
действующего на них стимула возникает относительно позже. Централь-
ные отростки первичных сенсорных нейронов входят в спинной мозг в со-
ставе задних корешков и разделяются в задних рогах спинного мозга на
коллатерали. Восходящие коллатерали достигают переключательных ядер
заднего столба продолговатого мозга, откуда специфическая информация
передается на противоположную сторону мозга по лемнисковому пути, по-
ступает к проекционным ядрам таламуса, а затем в соматосенсорную кору,
с участием которой формируется чувство осязания.
17.2.2. Проприоцептивная чувствительность
Проприоцептивные ощущения дают человеку возможность воспринимать из-
менения положения отдельных частей тела в покое и во время совершае-
мых движений. Информация, поступающая от проприоцепторов, позволя-
ет ему постоянно контролировать позу и точность произвольных движе-
ний, дозировать силу мышечных сокращений при противодействии внеш-
нему сопротивлению, например при подъеме или перемещении груза.
Проприоцепторы представлены интрафузалъными волокнами мышц (датчи-
ки длины мышцы), сухожильными рецепторами Гольджи (датчики напря-
жения мышцы) и механорецепторами суставных капсул, отвечающими из-
менениями фоновой активности на сгибание или разгибание в суставе и
морфологически сходными с тельцами Руффини и рецепторами Гольджи.
714 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Афферентная информация от проприоцепторов в составе задних ко-
решков поступает в спинной мозг и без переключения в нем передается по
восходящим путям к ядрам задних столбов продолговатого мозга. Располо-
женные там сенсорные нейроны второго порядка передают информацию
по лемнисковому пути на противоположную сторону мозга к проекцион-
ным ядрам таламуса. Нейроны переключательных ядер таламуса образуют
соматотопическую проекцию на соматосенсорную кору, с помощью кото-
рой формируются проприоцептивные ощущения. Сенсорные сигналы от
проприоцепторов мышц лица и головы следуют в ЦНС по чувствительным
волокнам черепных нервов, а в соматосенсорную кору информация посту-
пает тем же способом, что и сигналы от мускулатуры тела, т. е. по лемни-
сковому пути.
Проприоцептивное восприятие положения тела и движений происходит
в результате объединения в соматосенсорной коре информации от всех
разновидностей проприоцепторов. Порог воспринимаемых изменений уг-
лового положения проксимальных суставов ниже, чем дистальных: 0,2—
0,4’ в плечевом суставе и около Г в суставах пальца руки. Порог возраста-
ет при увеличении угловой скорости движений, в связи с чем уменьшается
точность выполняемых движений. Способность человека дифференциро-
вать силу произвольных сокращений мышц подчиняется закону Вебера—
Фехнера, эта способность уменьшается при самых слабых и очень сильных
мышечных усилиях, необходимых для выполнения работы.
17.2.3. Температурная чувствительность
Ощущения тепла или холода, вызванные прикосновением к телу человека
нагретых или холодных предметов, теплого или охлажденного воздуха, воз-
никают вследствие раздражения одной из двух разновидностей температур-
ных рецепторов, представленных в коже свободными нервными оконча-
ниями. Среди них различают тепловые и холодовые рецепторы, которым
соответствуют две субмодальности температурного восприятия внешних
воздействий. Температурные рецепторы распределены в коже неравномер-
но и образуют специфические тепловые и холодовые точки площадью око-
ло 1 мм2 и менее, с наибольшей плотностью на лице и ладонях, причем
холодовых точек примерно в десять раз больше, чем тепловых.
Импульсная активность терморецепторов возрастает в зависимости от
направления и скорости изменений температуры поверхности кожи, со-
ставляющей в условиях температурного комфорта около 33 °C. При ее сни-
жении растет активность холодовых рецепторов с максимумом в диапазоне
от 26 до 17 °C, что субъективно расценивается как действие на кожу холо-
да. При повышении температуры кожи от 34 до 45 °C увеличивается им-
пульсная активность тепловых рецепторов, что субъективно ощущается
как возрастающее по интенсивности действие на кожу тепла. При умень-
шении температуры кожи ниже 17 °C и увеличении выше 45 °C к темпера-
турным ощущениям прибавляются болевые.
Тоническая активность температурных рецепторов при неизменной тем-
пературе сохраняется в течение некоторого времени, а затем постепенно
уменьшается. В таком случае, как и при медленных изменениях действую-
щей на рецепторы температуры, происходит их адаптация, в связи с кото-
рой постепенно ослабевают ощущения тепла или холода. Но в ответ на
резкие ступенчатые изменения действующей температуры (температурный
контраст) возникает залповая фазическая активность терморецепторов, ко-
торая отражает произошедшие изменения температуры. Наличие адапта-
17. Сенсорные системы • 715
ции терморецепторов наряду с их способностью реагировать залповой ак-
тивностью на контрастные изменения действующей температуры проявля-
ются в уменьшении порога тепловой чувствительности при пониженной
температуре кожи и в уменьшении порога холодовой чувствительности при
повышении ее температуры.
Кожные терморецепторы позволяют человеку распознавать нагретые
или холодные предметы и окружающий воздух, прикасающиеся к коже.
Однако субъективное восприятие, определяемое как «мне холодно» или
«мне жарко», возникает в результате суммарной оценки информации, по-
ступившей не только от кожных рецепторов, но также и от терморецепто-
ров спинного мозга, внутренних органов и центральных рецепторов перед-
него гипоталамуса, где расположен центр терморегуляции. Нервные им-
пульсы от кожных терморецепторов поступают в спинной мозг, в задних
рогах которого расположены сенсорные нейроны второго порядка. Их ак-
соны переходят на противоположную сторону спинного мозга, входя в со-
став спиноретикулярного и спиноталамического путей.
Спиноталамический путь ведет к задним вентролатеральным ядрам та-
ламуса, на нейронах которых конвергируют проводящие пути температур-
ной и тактильной чувствительности. Благодаря такой конвергенции у че-
ловека проявляется способность устанавливать, на какую часть поверхно-
сти тела подействовал температурный стимул. Спиноретикулярный тракт
служит для проведения возбуждения от терморецепторов к ретикулярной
формации и принадлежит к неспецифической части сенсорной системы.
Ретикулярная формация вместе с неспецифическими ядрами таламуса ак-
тивирует одновременно многие регионы коры, определяя общую актив-
ность мозга и уровень бодрствования. Поэтому повышение притока аффе-
рентной информации к ней от терморецепторов, например во время прие-
ма холодного душа, оказывает на человека тонизирующее влияние.
17.2.4. Болевая чувствительность
Боль определяется как неприятное сенсорное и эмоциональное пережива-
ние, связанное с истинным или потенциальным повреждением ткани или
описываемое в терминах такого повреждения. В отличие от других сенсор-
ных модальностей боль всегда субъективно неприятна и служит не столько
источником информации об окружающем мире, сколько сигналом повре-
ждения или болезни. Болевая чувствительность побуждает к прекращению
контактов с повреждающими факторами среды.
Болевые рецепторы или ноцицепторы представляют собой свободные
нервные окончания, расположенные в коже, слизистых оболочках, мыш-
цах, суставах, надкостнице и во внутренних органах. Чувствительные
окончания принадлежат либо безмякотным, либо тонким миелинизиро-
ванным волокнам, что определяет скорость проведения сигналов в ЦНС и
дает повод к различению ранней боли, короткой и острой, возникающей
при проведении импульсов с большей скоростью по миелиновым волок-
нам, а также поздней, тупой и длительной боли, в случае проведения сигна-
лов по безмякотным волокнам. Ноцицепторы относятся к полимодальным
рецепторам, поскольку могут активироваться стимулами разной природы:
механическими (удар, порез, укол, щипок), термическими (действие горя-
чих или холодных предметов), химическими (изменение концентрации ио-
нов водорода, действие гистамина, брадикинина и ряда других биологиче-
ски активных веществ). Порог чувствительности ноцицепторов высок, по-
этому лишь достаточно сильные стимулы вызывают возбуждение первич-
716 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ных сенсорных нейронов: например, порог болевой чувствительности для
механических стимулов примерно в тысячу раз превышает порог тактиль-
ной чувствительности.
Центральные отростки первичных сенсорных нейронов входят в спин-
ной мозг в составе задних корешков и образуют синапсы с нейронами вто-
рого порядка, расположенными в задних рогах спинного мозга. Аксоны
нейронов второго порядка переходят на противоположную сторону спин-
ного мозга, где образуют спиноталамический и спиноретикулярный тракты.
Спиноталамический тракт оканчивается на нейронах нижнего заднелате-
рального ядра таламуса, где происходит конвергенция проводящих путей
болевой и тактильной чувствительности. Нейроны таламуса образуют про-
екцию на соматосенсорную кору: этот путь обеспечивает осознаваемое
восприятие боли, позволяет определять интенсивность стимула и его лока-
лизацию.
Волокна спиноретикулярного тракта оканчиваются на нейронах ретику-
лярной формации, взаимодействующих с медиальными ядрами таламуса.
При болевом раздражении нейроны медиальных ядер таламуса оказывают
модулирующее влияние на обширные регионы коры и структуры лимбиче-
ской системы, что приводит к повышению поведенческой активности че-
ловека и сопровождается эмоциональными и вегетативными реакциями.
Если спиноталамический путь служит для определения сенсорных качеств
боли, то спиноретикулярный путь предназначен играть роль сигнала об-
щей тревоги, оказывать на человека общее возбуждающее действие.
Субъективную оценку боли определяет соотношение нейронной активно-
сти обоих путей и зависимая от нее активация антиноцицептивных нисхо-
дящих путей, способных изменить характер проведения сигналов от ноци-
цепторов. В сенсорную систему болевой чувствительности встроен эндо-
генный механизм ее уменьшения путем регуляции порога синаптических
переключений в задних рогах спинного мозга («ворота боли»). На передачу
возбуждения в этих синапсах влияют нисходящие волокна нейронов серо-
го вещества вокруг водопровода, голубого пятна и некоторых ядер средин-
ного шва. Медиаторы этих нейронов (энкефалин, серотонин, норадрена-
лин) тормозят активность нейронов второго порядка в задних рогах спин-
ного мозга, чем уменьшают проведение афферентных сигналов от ноци-
цепторов.
Аналгезирующим (обезболивающим) действием обладают опиатные пеп-
тиды (дайнорфин, эндорфины), синтезируемые нейронами гипоталамуса,
которые имеют длинные отростки, проникающие в другие отделы голов-
ного мозга. Опиатные пептиды присоединяются к специфическим рецеп-
торам нейронов лимбической системы и медиальной области таламуса, их
образование увеличивается при некоторых эмоциональных состояниях,
стрессе, продолжительных физических нагрузках, у беременных женщин
незадолго перед родами, а также в результате психотерапевтического воз-
действия или акупунктуры. В результате повышенного образования опиат-
ных пептидов активируются антиноцицептивные механизмы и повышается
порог болевой чувствительности. Баланс между ощущением боли и ее
субъективной оценкой устанавливается с помощью лобных областей мозга,
участвующих в процессе восприятия болевых стимулов. При поражении
лобных долей (например, в результате травмы или опухоли) порог болевой
чувствительности не изменяется и поэтому сенсорный компонент болевого
восприятия сохраняется неизменным, однако субъективная эмоциональная
оценка боли становится иной: она начинает восприниматься лишь как
сенсорное ощущение, а не как страдание.
17. Сенсорные системы • 717
17.2.5. Висцеральная чувствительность
Большая часть афферентной информации от висцерорецепторов (visceralis —
внутренностный) не осознается и служит для рефлекторной регуляции ве-
гетативных процессов в организме человека. Висцерорецепторы располо-
жены во внутренних органах и тканях организма и представляют собой как
свободные, так и инкапсулированные нервные окончания. В зависимости
от чувствительности к адекватным стимулам они подразделяются на меха-
норецепторы, хеморецепторы, терморецепторы, осморецепторы и ноци-
цепторы. Информация от висцерорецепторов поступает в ЦНС двумя пу-
тями: от сенсорных нейронов чувствительных ганглиев и по чувствитель-
ным афферентным волокнам, имеющимся в составе эфферентных симпа-
тических и парасимпатических нервов.
Висцеральные механорецепторы раздражаются вследствие изменения
давления в полых органах и кровеносных сосудах. Осознаваемые ощуще-
ния, вызванные раздражением такого типа рецепторов, чаще всего обу-
словлены изменениями в пищеварительном тракте, поскольку из всех
внутренних органов только он имеет общее эмбриологическое происхож-
дение с поверхностью тела. Растяжение стенок желудка пищей вызывает
ощущение сытости, растяжение стенок кишечника газами — ощущение
дискомфорта и тяжести в области живота, а растяжение прямой кишки ка-
ловыми массами осознается как позыв к дефекации. Подобно этому им-
пульсация от механорецепторов, расположенных в стенке мочевого пузы-
ря, ощущается субъективно как позыв к его опорожнению.
Висцеральные хеморецепторы обладают специфической чувствительно-
стью к действию определенных химических молекул; например, хеморецеп-
торы, расположенные в дуге аорты и синокаротидной области, раздражают-
ся изменениями концентрации углекислого газа, кислорода и водородных
ионов. Изменения активности таких рецепторов ощущаются не прямо, а
опосредованно, проявляясь, например, головокружением или удушьем. По-
вышение осмотического давления вызывает раздражение периферических
осморецепторов, расположенных в бассейне воротной вены, что, наряду с
информацией от центральных осморецепторов промежуточного мозга, спо-
собствует появлению жажды. Афферентная информация от висцеральных
терморецепторов служит для возникновения ощущений жара или озноба.
Боль, возникающая при раздражении ноцицепторов внутренних орга-
нов, называется висцеральной, в отличие от соматической боли, вызван-
ной раздражением болевых рецепторов в коже, мышцах, суставах и костях.
Острую висцеральную боль может вызвать сильное растяжение полых ор-
ганов вследствие закупорки выходных протоков желчного пузыря, моче-
точников, в результате непроходимости кишечника, а также спастических
сокращений гладких мышц. Плотность ноцицепторов особенно высока в
брюшине, при воспалении которой или при ее сильном механическом раз-
дражении возникает острая и интенсивная боль. При воспалительных про-
цессах во внутренних органах выделяются биологически активные вещест-
ва, которые служат химическими раздражителями ноцицепторов и причи-
ной болевых ощущений.
17.3. Зрительная сенсорная система
Зрительное восприятие оставляет в памяти человека наибольшую часть его
чувственных впечатлений об окружающем мире. Оно происходит в резуль-
тате поглощения фоторецепторами сетчатки отраженной от окружающих
718 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
предметов энергии световых лучей или электромагнитных волн в диапазо-
не от 400 до 700 нм. Энергия поглощенных квантов света (адекватный раз-
дражитель) преобразуется сетчаткой в нервные импульсы, поступающие по
зрительным нервам к латеральным коленчатым телам, а от них — в проек-
ционную зрительную кору. В дальнейшей переработке зрительной инфор-
мации у человека участвуют свыше тридцати отделов мозга, представляю-
щих вторичные сенсорные и ассоциативные области коры.
17.3.1. Проецирование световых лучей на сетчатку глаза
Прежде чем попасть на сетчатку, световые лучи последовательно проходят
через роговицу, жидкость передней камеры глаза, хрусталик и стекловид-
ное тело, вместе образующие оптическую систему глаза (рис. 17.5). На ка-
ждом из этапов этого пути свет преломляется и в результате на сетчатке
возникает уменьшенное и перевернутое изображение наблюдаемого пред-
мета, этот процесс называется рефракцией. Преломляющая сила оптиче-
ской системы глаза составляет около 58,6 диоптрий при рассматривании
удаленных предметов и возрастает до приблизительно 70,5 диоптрий при
фокусировании на сетчатку световых лучей, отраженных от близко распо-
ложенных предметов (1 диоптрия соответствует преломляющей силе линзы
с фокусным расстоянием 1 м).
17.3.1.1. Аккомодация
Чтобы фокусировать на сетчатке световые лучи, отраженные от близко
расположенных предметов, оптическая система глаза должна преломлять
их тем сильней, чем ближе расположен наблюдаемый объект. Механизм, с
помощью которого глаз настраивается на рассмотрение удаленных или
Рис. 17.5. Оптическая система глаза и
проекция световых лучей на сетчатку.
Световые лучи, отраженные от рассматриваемой части наблюдаемого объекта (точка фикса-
ции), преломляются оптическими средами глаза (роговица, передняя камера, хрусталик, стек-
ловидное тело) и фокусируются в центральной ямке сетчатки. Проекция световых лучей на
поверхность центральной ямки обеспечивает максимальную остроту зрения благодаря малым
размерам рецептивных полей и отсутствию ганглиозных и биполярных клеток на пути прохо-
ждения световых лучей к фоторецепторам.
17. Сенсорные системы • 719
близких предметов и в обоих случаях фокусирует их изображение на сет-
чатку, называется аккомодацией. Аккомодацию обеспечивают изменения
кривизны хрусталика, зависимой от степени натяжения его тонкой и про-
зрачной капсулы, которая переходит по краям в циннову связку, прикреп-
ляющуюся к ресничному телу. Гладкие мышцы ресничного тела, управляе-
мые парасимпатическими нейронами, регулируют натяжение цинновой
связки: при полном расслаблении мышц связка натягивает капсулу хруста-
лика, заставляя его принимать максимально уплощенную форму, необхо-
димую для рассмотрения далеких предметов. При сокращении ресничных
мышц натяжение цинновой связки уменьшается, хрусталик в силу своей
эластичности принимает более выпуклую форму и поэтому сильнее пре-
ломляет световые лучи, что, например, происходит во время чтения этого
текста.
При максимальном сокращении ресничных мышц кривизна хрусталика
становится максимальной, что позволяет фокусировать на сетчатке свето-
вые лучи, отраженные от ближайшей точки ясного видения. У детей эта точ-
ка расположена примерно в семи сантиметрах от глаза, но с возрастом
эластичность хрусталика уменьшается, что ограничивает возможность уве-
личения его кривизны и соответственно преломляющей силы. В связи с
возрастным уменьшением диапазона аккомодации (интервала изменения
преломляющей силы хрусталика) ближайшая точка ясного видения посте-
пенно удаляется: в 20 лет она располагается на расстоянии около 10 см от
глаза, в 30 лет — 14 см, в 40 лет — 22 см, в 50 лет — 40 см и т. д. Возрастное
уменьшение аккомодационной способности {пресбиопия, или возрастная
дальнозоркость) принуждает человека использовать при чтении очки с
двояковыпуклыми линзами.
17.3.1.2. Аномалии рефракции
У большинства людей глаз имеет шарообразную форму, максимально при-
способленную для того, чтобы световые лучи фокусировались на сетчатке
при любой преломляющей силе хрусталика, изменяющейся в процессе ак-
комодации. При нормальной рефракции на сетчатке одинаково четко про-
ецируются изображения далеких и близких предметов, а проявление такой
универсальной рефракционной способности определяют термином эмме-
тропия. Аномалии рефракции возникают при изменениях формы глаза,
проявляющихся в удлинении его оптической оси {близорукость, или мио-
пия) или в ее укорочении {дальнозоркость, или гиперметропия).
При удлиненной форме глаза, характерной для миопии, обычная пре-
ломляющая способность его оптической системы оказывается избыточной,
в связи с чем изображение фокусируется перед сетчаткой и субъективно
воспринимается нечетким. Дефект зрительного восприятия ощущается
только при рассматривании удаленных предметов, тогда как световые лу-
чи, отраженные от близко находящихся предметов, фокусируются точно на
сетчатке благодаря ограничению механизма аккомодации. Например, при
чтении близоруким людям нужна меньшая, чем людям с нормальной реф-
ракцией, преломляющая сила хрусталика, поэтому нагрузка на ресничные
мышцы у них снижается. Но для ясного видения далеких предметов близо-
рукому человеку приходится носить очки с двояковогнутыми рассеиваю-
щими линзами, уменьшающими преломление световых лучей. При харак-
терной для гиперметропии укороченной оптической оси глаза световые лу-
чи фокусируются за сетчаткой, что принуждает ресничные мышцы к со-
кращению даже при рассмотрении далеких предметов. Для компенсации
720 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
этой аномалии рефракции необходимы очки с двояковыпуклыми линзами,
преломляющими световые лучи так, чтобы они фокусировались на сет-
чатке.
Наибольшей преломляющей способностью в оптической системе глаза
обладает роговица (около 48,8 диоптрий), которая не участвует в аккомо-
дации. Поверхность роговицы не идеально сферична почти у всех людей,
ее кривизна по вертикали выражена больше, чем по горизонтали. При зна-
чительном отклонении формы роговицы от идеальной пропорции возни-
кает аномалия рефракции, которая определяется как астигматизм. В ре-
зультате такой аномалии световые лучи, отраженные от наблюдаемых объ-
ектов, преломляются не одинаково, поэтому часть лучей фокусируется на
сетчатке, а остальные — за ее пределами, что ведет к искажению зритель-
ного восприятия. Для исправления этого дефекта применяются индивиду-
ально подобранные линзы очков с различной кривизной по вертикальному
и горизонтальному меридианам.
17.3.1.3. Регуляция интенсивности светового потока
Приспособление к изменяющимся условиям освещения происходит с по-
мощью зрачкового рефлекса. Ширина зрачка уменьшается при ярком свете
благодаря сокращению кольцевых мышц, управляемых парасимпатически-
ми волокнами глазодвигательного нерва, а при слабом освещении зрачок
расширяется с помощью радиальных мышц, получающих симпатическую
иннервацию. Уменьшая просвет зрачка, глаз защищается от избытка света,
а увеличивая ширину зрачка, он повышает чувствительность зрительной
системы к воспринимаемым стимулам. Сужение зрачков повышает глуби-
ну резкости, что позволяет лучше видеть удаленные предметы. При расши-
рении зрачков глубина резкости снижается, а вместе с ней снижается ост-
рота зрения, которая характеризуется максимальной способностью глаза
различать две соседние точки зрительного пространства как отдельные.
В норме глаз различает две точки, видимые под углом в одну минуту при
достаточно ярком освещении.
17.3.1.4. Проекция зрительного поля на сетчатку
Благодаря механизму аккомодации глаз представляет собой самофокуси-
рующуюся оптическую систему, позволяющую получать на сетчатке четкое
изображение приближенных и удаленных предметов. Кроме того, глаз по-
движен, что позволяет ему следить за перемещающимися объектами. Дви-
жения глаза обеспечиваются сокращениями глазодвигательных мышц (двух
косых и четырех прямых), которые позволяют повернуть глаз внутрь или
наружу, вверх или вниз. Оба глаза поворачиваются бинокулярно, т. е. содру-
жественно, и при рассмотрении близких предметов глазные оси сходятся
(конвергенция глаз), а при рассмотрении далеких предметов — расходятся
(дивергенция). Благодаря бинокулярному зрению каждая точка зрительного
поля проецируется на соответствующие друг другу участки сетчатки обоих
глаз. Изображение одного и того же предмета на сетчатках левого и право-
го глаза немного различается, поскольку глаза занимают разную позицию
по отношению к зрительному полю. Разница изображений наблюдаемого
предмета на сетчатке левого и правого глаза обозначается термином попе-
речная диспарация, она способствует последующему преобразованию двух-
мерного изображения на сетчатке в трехмерное изображение, создаваемое
в процессе переработки зрительной информации в коре.
17. Сенсорные системы • 721
Рис. 17.6. Ретинотопическая проекция сетчатки в первичную зрительную кору.
Пространственное соотношение между объектами зрительного поля соблюдается в его проек-
ции на соответствующие друг другу (корреспондирующие) области сетчатки каждого глаза.
Афферентные сигналы от носовых половин сетчатки благодаря частичному перекресту воло-
кон зрительных нервов поступают в противоположное полушарие. Афферентные сигналы от
височных половин сетчаток попадают с помощью неперекрещивающихся волокон зрительных
нервов в ипсилатеральное полушарие. В связи с этим в каждое полушарие поступает зритель-
ная информация о противоположной ему половине зрительного поля и с сохранением его
пространственных координат (принцип ретинотопии). При направленном на определенные
объекты внимании световые лучи, отраженные от них, всегда фокусируются в центральной
ямке. Поэтому центральная ямка и прилегающие к ней области сетчатки имеют наибольшую
площадь представительства в первичной зрительной коре.
Зрительное поле представляет собой область, видимую обоими глаза-
ми в тот момент, когда они и голова неподвижны. При таком условии
левая половина зрительного поля проецируется на носовую половину
сетчатки левого глаза и височную половину сетчатки правого глаза. Со-
ответственно, правая половина зрительного поля проецируется на носо-
вую половину сетчатки правого глаза и височную половину сетчатки ле-
вого глаза. Поскольку оптическая система глаза перевертывает изображе-
ние, верхняя половина зрительного поля проецируется на нижнюю поло-
вину сетчатки, а нижняя половина зрительного поля — на верхнюю по-
ловину сетчатки. При дальнейшей переработке информации в наружном
коленчатом теле и первичной зрительной коре сохраняются все про-
странственные координаты зрительного поля: каждая область сетчатки
связана с соответствующим представительством в коре, что называется
ретинотопией (рис. 17.6).
722 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
17.3.1.5. Движения глаз
При наблюдении за движущимися в зрительном поле объектами, а также
при движении человека относительно окружающего мира происходят сле-
дящие движения глаз, благодаря которым изображение в одной и той же
области сетчатки сохраняется неизменным. При зрительном восприятии
неподвижных объектов, имеющих многочисленные детали формы, а также
во время чтения происходят быстрые движения глаз, предназначенные для
фиксации наиболее информативных деталей объекта. Например, при вос-
приятии человеческого лица фиксируются его контуры, но с наибольшим
вниманием рассматриваются глаза и рот, изображения которых глаз про-
ецирует не вообще на сетчатку, а на ту ее область, которая обладает макси-
мальной разрешающей способностью. Это центральная ямка, или fovea,
представляющая собой небольшое углубление диаметром около 3 мм в
центре сетчатки.
При рассматривании любых объектов глаза ежесекундно совершают
около трех очень быстрых непроизвольных и субъективно не ощущаемых
движений, которые называются саккадами. Благодаря таким движениям
изображение на сетчатке регулярно смещается, вызывая раздражение раз-
ных фоторецепторов. Необходимость саккад объясняется свойством зри-
тельной системы сильнее реагировать на изменяющееся раздражение (по-
явление или исчезновение стимула), тогда как на постоянную стимуляцию
она отвечает слабо. При проецировании на сетчатку постоянного изобра-
жения с помощью миниатюрного проектора, прикрепленного к контакт-
ной линзе, это изображение перестает восприниматься спустя несколько
секунд, и вместо него в зрительном ощущении появляется сплошное серое
поле. По истечении нескольких секунд восприятие проецируемого на сет-
чатку изображения восстанавливается, но затем снова исчезает и т. д.
17.3.2. Преобразование энергии света в сетчатке
Задняя стенка глаза выстлана эпителием, содержащим черный пигмент
меланин, который препятствует отражению света и поглощает его излиш-
ки. К пигментному эпителию примыкает слой фоторецепторов, а перед
ним расположены слои горизонтальных, биполярных, амакриновых и
ганглиозных клеток, чьи аксоны образуют зрительный нерв. В связи с ин-
версией сетчатки решаются две задачи: 1) обеспечение максимальной про-
зрачности клеток, расположенных на пути прохождения света к фоторе-
цепторам, 2) выделение в слое фоторецепторов промежутка для прохожде-
ния волокон зрительного нерва.
Решение первой задачи заключается в анатомическом устройстве цен-
тральной ямки, где биполярные и ганглиозные клетки сетчатки распола-
гаются вокруг нее, а их относительно прозрачные безмиелиновые волок-
на не мешают прохождению света к фоторецепторам. В других областях
сетчатки световые лучи отчасти рассеиваются при прохождении сквозь
тела клеток, расположенных перед фоторецепторами, но в центральной
ямке кванты света без помех попадают на фоторецепторы, представлен-
ные здесь преимущественно колбочками. При направленном на опреде-
ленный объект зрительного поля внимании его изображение всегда фо-
кусируется в центральной ямке для обеспечения максимальной остроты
зрения.
Сосок зрительного нерва образован аксонами ганглиозных клеток, от-
сутствие в этой области фоторецепторов делает ее не чувствительной к
17. Сенсорные системы • 723
свету, в связи с чем она получила название слепого пятна. На соски право-
го и левого глаза благодаря диспарации попадают неодинаковые части
проецируемого изображения, поэтому в условиях естественного зрительно-
го восприятия пробел изображения на сетчатке не осознается. Кроме того,
быстрые движения глаз позволяют получать нужную информацию о любой
области рассматриваемого изображения, и отсутствие сигналов от слепого
пятна замещается информацией, поступившей от ближайших к нему фото-
рецепторов.
17.3.2.1. Скотопическая и фотопическая системы сетчатки
Фоторецепторный слой сетчатки человека образован примерно 130 мил-
лионами клеток, из которых около семи миллионов являются колбочками,
основная масса которых сосредоточена в области центральной ямки, а все
остальные фоторецепторы представлены палочками. У обеих разновидно-
стей фоторецепторов существуют три функциональные области: 1) наруж-
ный, или внешний, сегмент, ориентированный в направлении эпителиаль-
ного пигментного слоя и содержащий зрительный пигмент', 2) внутренний
сегмент, в котором расположено клеточное ядро и происходят биохимиче-
ские процессы, связанные с жизнедеятельностью клетки; 3) синаптические
окончания, предназначенные для передачи информации от фоторецепто-
ров к биполярным клеткам с помощью медиатора глутамата.
Зрительный пигмент палочек родопсин состоит из двух компонентов: это
молекула ретиналя, образующаяся из витамина А и способная поглощать
свет, а также крупная белковая молекула опсина, не поглощающая свет.
Молекула опсина представляет собой извитую цепь из 348 аминокислот,
которая семь раз проходит через мембрану зрительного диска, образован-
ного из клеточной мембраны фоторецептора. В наружном сегменте фото-
рецептора имеется большое количество таких дисков, расположенных по-
добно стопке поставленных друг на друга монет. Ретиналь существует в
темноте как 11-цис-ретиналь, такая форма изомера идеально соответствует
упорядоченному расположению аминокислот в опсине. Энергия погло-
щенных фотонов превращает ретиналь в 11-транс-изомер, что приводит к
конформационным изменениям молекулы опсина и превращению родоп-
сина в нестабильный метародопсин, который сразу же распадается на рети-
наль и опсин. Таким образом, действие света уменьшает концентрацию
родопсина в фоторецепторе, что приводит к изменениям активности вто-
ричных посредников и величины мембранного потенциала фоторецептора.
В темноте происходит ферментативный ресинтез расщепленного родопси-
на, для которого используется витамин А, поступающий в организм чело-
века с пищей.
Родопсин наиболее чувствителен к электромагнитным волнам длиной
около 500 нм, но хорошо поглощает и другие волны в диапазоне от 400 до
600 нм. Способность родопсина поглощать волны почти всего светового
диапазона позволяет палочкам обеспечить только ахроматическое, т. е.
черно-белое, зрение и лишает их возможности различать цвет. Высокое со-
держание зрительного пигмента в палочках и его способность суммировать
фотоны, поглощенные в течение около 100 мс, делает палочки наиболее
чувствительными фоторецепторами сетчатки. При слабом сумеречном ос-
вещении зрение обеспечивают только палочки, способные возбуждаться
вследствие поглощения всего лишь одного фотона. Палочки образуют ско-
топическую систему, или систему ночного зрения.
Зрительные пигменты колбочек подобны родопсину палочек и состоят
724 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
С Р 3 к
Рис. 17.7. Спектры поглощения четырех разновидностей фоторецепторов.
Зрительный пигмент палочек родопсин (Р) имеет максимум поглощения световых волн дли-
ною 496 нм, но способен также к поглощению коротких и длинных волн светового диапазона.
Зрительный пигмент колбочек, чувствительных к синему цвету (С), имеет максимум поглоще-
ния 419 нм и не поглощает длинные волны оптического диапазона. Пигмент колбочек, чувст-
вительных к зеленому цвету (3), имеет максимум поглощения при 531 нм, а пигмент чувстви-
тельных к красному цвету колбочек (К) максимально поглощает волны длиной 596 нм.
из светопоглощающей молекулы ретиналя и опсина, который отличается
составом аминокислот от белковой части родопсина. Кроме того, колбоч-
ки содержат меньшее, чем палочки, количество зрительного пигмента, и
для их возбуждения требуется энергия нескольких сотен фотонов. Поэто-
му колбочки активируются лишь при дневном или достаточно ярком ис-
кусственном освещении, они образуют фотопическую систему, или систе-
му дневного зрения.
В сетчатке человека существуют три типа колбочек, различающихся ме-
жду собой по составу аминокислот в опсине зрительного пигмента. Разли-
чия в белковой части молекулы определяют особенности взаимодействия
каждой из трех форм опсина с ретиналем и специфическую чувствитель-
ность к световым волнам разной длины (рис. 17.7). Колбочки одного из
трех типов максимально поглощают короткие световые волны с длиной
419 нм, что необходимо для восприятия синего цвета. Другой тип зритель-
ного пигмента наиболее чувствителен к волнам средней длины и имеет
максимум поглощения при 531 нм, он служит для восприятия зеленого
цвета. Третий тип зрительного пигмента максимально поглощает длинные
волны с максимумом при 559 нм, что позволяет воспринимать красный
цвет. Наличие трех типов колбочек обеспечивает человеку восприятие всей
цветовой палитры, в которой существует свыше семи миллионов цветовых
градаций, тогда как скотопическая система палочек позволяет различать
лишь около пятисот черно-белых градаций.
17. Сенсорные системы • 725
17.3.2.2. Рецепторный потенциал палочек и колбочек
Специфической особенностью фоторецепторов является темновой ток ка-
тионов через открытые мембранные каналы внешних сегментов (рис. 17.8).
Эти каналы открываются при высокой концентрации циклического гуано-
зинмонофосфата, который является вторичным посредником рецепторного
белка (зрительного пигмента). Темновой ток катионов деполяризует мем-
брану фоторецептора до приблизительно —40 мВ, что приводит к выделе-
нию медиатора в его синаптическом окончании. Активированные поглоще-
нием света молекулы зрительного пигмента стимулируют активность фос-
фодиэстеразы — фермента, расщепляющего цГМФ, поэтому при действии
света на фоторецепторы в них уменьшается концентрация цГМФ. В резуль-
тате управляемые этим посредником катионные каналы закрываются, и ток
катионов в клетку прекращается. Вследствие непрерывного выхода ионов
калия из клеток, мембрана фото-
рецепторов гиперполяризуется
приблизительно до —70 мВ, эта ги-
перполяризация мембраны являет-
ся рецепторным потенциалом. При
возникновении рецепторного по-
тенциала прекращается выделение
глутамата в синаптических оконча-
ниях фоторецептора.
Фоторецепторы образуют си-
напсы с биполярными клетками
двух типов, различающихся по
способу управления хемозависи-
мыми натриевыми каналами в си-
напсах. Действие глутамата приво-
дит к открытию каналов для ионов
натрия и деполяризации мембра-
ны одних биполярных клеток и к
закрытию натриевых каналов и ги-
перполяризации биполярных кле-
ток другого типа. Наличие двух
типов биполярных клеток необхо-
димо для формирования антаго-
низма между центром и перифери-
ей рецептивных полей ганглиоз-
ных клеток.
ЦГМФ-управляемые
Высокая
Низкая
концентрация
цГМФ, каналы
открыты
концентрация
цГМФ, каналы
закрыты
17.3.2.3. Адаптация
фоторецепторов
к изменениям
освещенности
Временное ослепление при быст-
ром переходе от темноты к яркому
освещению исчезает спустя не-
сколько секунд благодаря процес-
су световой адаптации. Одним из
механизмов световой адаптации
является рефлекторное сужение
Рис. 17.8. Ток ионов через мембрану фо-
торецептора в темноте и на свету.
Ионы натрия поступают в фоторецепторы че-
рез катионные каналы внешнего сегмента, ко-
торые открываются при высокой концентра-
ции цГМФ в темноте (темновой ток). Поступ-
ление в клетку ионов натрия вызывает депо-
ляризацию мембраны фоторецептора до —40
мВ. На свету концентрация цГМФ уменьша-
ется, что приводит к закрытию катионных ка-
налов. Непрекращающийся выход ионов ка-
лия ведет к гиперполяризации мембраны, ко-
торая представляет собой рецепторный потен-
циал.
726 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
зрачков, другой зависит от концентрации ионов кальция в колбочках. При
поглощении света в мембранах фоторецепторов закрываются катионные
каналы, что прекращает вхождение ионов натрия и кальция и уменьшает
их внутриклеточную концентрацию. Высокая концентрация ионов кальция
в темноте подавляет активность гуанилатциклазы — фермента, определяю-
щего образование цГМФ из гуанозинтрифосфата. Вследствие снижения
концентрации кальция, обусловленного поглощением света, активность
гуанилатциклазы повышается, что ведет к дополнительному синтезу
цГМФ. Повышение концентрации этого вещества приводит к открытию
катионных каналов, восстановлению тока катионов в клетку и, соответст-
венно, способности колбочек отвечать на световые раздражители как
обычно. Низкая Концентрация ионов кальция способствует десенситизации
колбочек, т. е. уменьшению их чувствительности к свету. Десенситизация
обусловлена изменением свойств фосфодиэстеразы и белков катионных
каналов, становящихся менее чувствительными к концентрации цГМФ.
Способность различать окружающие предметы исчезает на некоторое
время при быстром переходе от яркого света к темноте. Она постепенно
восстанавливается в ходе темновой адаптации, обусловленной расширени-
ем зрачков и переключением зрительного восприятия с фотопической сис-
темы на скотопическую. Темновую адаптацию палочек определяют мед-
ленные изменения функциональной активности белков, приводящие к по-
вышению их чувствительности. В механизме темновой адаптации участву-
ют и горизонтальные клетки, способствующие увеличению центральной
части рецептивных полей в условиях низкой освещенности.
17.3.3. Рецептивные поля клеток сетчатки
Выходные сигналы, передаваемые в ЦНС от сетчатки, возникают только в
ганглиозных клетках, импульсная активность которых зависит от возбужде-
ния фоторецепторов, а затем биполярных клеток, входящих в округлое ре-
цептивное поле ганглиозной клетки. Размер рецептивных полей и количе-
ство фоторецепторов, относящихся к одному рецептивному полю, варьи-
руют от минимального в области центральной ямки до наибольшего на пе-
риферии сетчатки. Малые рецептивные поля служат для различения мел-
ких деталей наблюдаемых объектов в тех случаях, когда соседние детали
воспринимаются под углом в несколько угловых минут. Большие рецеп-
тивные поля вмещают изображение целого объекта, воспринимаемого под
углом в несколько угловых градусов (Г соответствует рецептивному полю
на поверхности сетчатки с диаметром около 0,25 мм).
Существуют два пути для передачи сигналов от фоторецепторов к ганг-
лиозной клетке: прямой и непрямой. Прямой путь начинается от фоторе-
цепторов, расположенных в центре рецептивного поля и образующих си-
напс с биполярной клеткой, которая через другой синапс действует на ганг-
лиозную клетку. Непрямой путь берет начало от фоторецепторов перифе-
рии рецептивного поля, которая с центром состоит в реципрокных отно-
шениях, обусловленных тормозным действием горизонтальных и амакрино-
вых клеток (латеральное торможение).
17.3.3.1. Рецептивные поля с on-центрами и off-центрами
В сетчатке человека имеются два типа ганглиозных клеток, отличающихся
реакцией на точечные световые стимулы, воздействовавшие на центр или
периферию их рецептивного поля (рис. 17.9). Примерно половина гангли-
/ 7. Сенсорные системы • 727
Биполярные Фоторецепторы
К зрительному клетки (палочки)
нерву
Рис. 17.9. Рецептивные поля ганглиозных клеток сетчатки с on- и off-центрами.
А. Рецептивное поле ганглиозной клетки образуют все фоторецепторы и биполярные клетки,
имеющие с нею синаптические контакты. Ганглиозные клетки постоянно генерируют потен-
циалы действия, частота возникновения которых зависит от активности фоторецепторов и би-
полярных клеток, входящих в ее рецептивное поле.
Б. Ганглиозная клетка on-типа увеличивает частоту электрических разрядов в ответ на свето-
вое раздражение центра рецептивного поля и снижает свою электрическую активность при
действии светового раздражителя на периферию рецептивного поля.
В. Ганглиозная клетка off-типа тормозится при действии света на центр ее рецептивного поля
и увеличивает частоту нервных импульсов в ответ на раздражение периферии рецептивного
поля.
озных клеток возбуждается действием света на центр рецептивного поля и
тормозится при действии светового стимула на периферию рецептивного
поля. Такие клетки принято называть оп-нейронами. Другая половина ганг-
лиозных клеток возбуждается действием светового раздражителя на пери-
ферию рецептивного поля и тормозится в ответ на световую стимуляцию
центра рецептивного поля — они получили название off-нейронов.
Рецептивные поля ганглиозных клеток обоих типов в сетчатке пред-
ставлены поровну, чередуясь друг с другом. Оба типа клеток очень слабо
отвечают на равномерную диффузную засветку всего рецептивного поля, а
наиболее сильным раздражителем для них является световой контраст,
т. е. различная интенсивность засветки центра и периферии. Именно кон-
трастирование деталей изображения дает необходимую информацию для
зрительного восприятия в целом, тогда как абсолютная интенсивность от-
раженного от наблюдаемого объекта света не столь важна. Восприятие гра-
ней, т. е. восприятие контраста между соседними поверхностями с разной
освещенностью, является наиболее информативным признаком изображе-
ния, определяющим протяженность и позиции разных объектов.
728 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
17.3.3.2. Рецептивные поля цветового восприятия
Восприятие цвета основано на существовании шести первичных цветов, об-
разующих три антагонистичные, или цветооппонентных, пары\ красный —
зеленый, синий — желтый, белый — черный. Ганглиозные клетки, пере-
дающие в центральную нервную систему информацию о цвете, различают-
ся организацией своих рецептивных полей, состоящих из комбинаций трех
существующих типов колбочек. Каждая колбочка предназначена для по-
глощения электромагнитных волн определенной длины, однако сами они
не кодируют информацию о длине волны и способны реагировать на
очень яркий белый свет. И только наличие в рецептивном поле ганглиоз-
ной клетки антагонистичных фоторецепторов создает нейронный канал
для передачи информации об определенном цвете. При наличии только
одного типа колбочек (монохромазия) человек не способен различить ни
один цвет и воспринимает окружающий мир в черно-белой градации, как
при скотопическом зрении. При наличии только двух типов колбочек (ди-
хромазия) цветовое восприятие ограничено, и лишь существование трех ти-
пов колбочек (трихромазия) обеспечивает полноту цветового восприятия.
Возникновение монохромазии и дихромазии у человека обусловлено гене-
тическими дефектами Х-хромосомы.
Концентрические широкополосные ганглиозные клетки имеют округлые
рецептивные поля on- или off-типа, которые образованы колбочками, но
предназначены для фотопического черно-белого зрения. Белый свет, попа-
дающий в центр или на периферию такого рецептивного поля, возбуждает
или тормозит активность соответствующей ганглиозной клетки, которая в
итоге передает информацию об освещенности. Концентрические широко-
полосные клетки суммируют сигналы от колбочек, поглощающих красный
и зеленый цвет и расположенных в центре и на периферии рецептивного
поля. Поступление сигналов от колбочек обоих типов происходит незави-
симо друг от друга, а потому не создает цветового антагонизма и не позво-
ляет широкополосным клеткам дифференцировать цвет (рис. 17.10).
Наиболее сильным раздражителем для концентрических противоцветных
ганглиозных клеток сетчатки является действие антагонистических цветов
на центр и периферию рецептивного поля. Одну разновидность противо-
цветных ганглиозных клеток возбуждает действие красного цвета на центр
ее рецептивного поля, в котором сосредоточены колбочки, чувствительные
к красной части спектра, и зеленого цвета — на периферию, где имеются
колбочки, чувствительные к нему. У другой разновидности концентриче-
ских противоцветных клеток в центре рецептивного поля расположены
колбочки, чувствительные к зеленой части спектра, а на периферии — к
красной. Эти две разновидности концентрических противоцветных клеток
различаются ответными реакциями на действие красного или зеленого цве-
та на центр или периферию рецептивного поля подобно тому, как on- и off-
нейроны различаются в зависимости от воздействия света на центр или пе-
риферию рецептивного поля. Каждая из двух разновидностей противоцвет-
ных клеток представляет собой нейронный канал, передающий информа-
цию о действии красного или зеленого цвета, причем передача информа-
ции тормозится действием антагонистического или оппонентного цвета.
Оппонентные отношения при восприятии синего и желтого цветов
обеспечиваются в результате объединения в рецептивном поле колбочек,
поглощающих короткие волны (синий цвет) с комбинацией из колбочек,
реагирующих на зеленый и красный цвет, что при смешении дает воспри-
ятие желтого цвета. Синий и желтый цвет оппонентны по отношению друг
17. Сенсорные системы • 729
Концентрические
широкополосные
клетки (ахроматические)
Концентрические
простые
противоцветные
клетки
Концентрические
простые
противоцветные
клетки
Простые
противоцветные
клетки
Рис. 17.10. Рецептивные поля ганглиозных клеток, образованные колбочками трех
типов.
А. Концентрические широкополосные клетки имеют в своем рецептивном поле колбочки для
восприятия красного (К) и зеленого (3) цветов. Колбочки обоих типов расположены в рецеп-
тивном поле так, что их входные сигналы суммируются независимо друг от друга, поэтому
широкополосные ганглиозные клетки не передают информацию о цвете, а реагируют лишь на
световой контраст между центром и периферией рецептивного поля как on- или off-нейроны.
Б. Простые противоцветные клетки двух разновидностей, отвечающие на стимуляцию зеле-
ным цветом центра или периферии рецептивного поля (красный цвет действует как антаго-
нист зеленому).
В. Простые противоцветные клетки двух разновидностей, различающиеся по характеру ответа
на действие красного цвета в центре или на периферии рецептивного поля (зеленый цвет дей-
ствует как антагонист красному).
Г. Колбочки, поглощающие короткие волны (синий цвет), находятся в антагонистических от-
ношениях со входящими в общее рецептивное поле колбочками, поглощающими средние и
длинные волны светового диапазона (зеленый и красный цвет).
к другу, и сочетание в рецептивном поле колбочек, поглощающих эти цве-
та, позволяет противоцветной ганглиозной клетке передавать информацию
о действии одного из них. Каким именно окажется этот нейронный канал,
т. е. передающим информацию о синем или желтом цвете, определяет рас-
положение колбочек внутри рецептивного поля концентрической противо-
цветной клетки. В зависимости от этого нейронный канал возбуждается
синим или желтым цветом и тормозится оппопентным цветом.
17.3.3.3. М- и P-типы ганглиозных клеток сетчатки
Зрительное восприятие происходит в результате согласования друг с дру-
гом различных сведений о наблюдаемых объектах. Но на низших иерархи-
ческих уровнях зрительной системы, начиная с сетчатки глаза, осуществ-
ляется независимая переработка информации о форме и глубине объекта,
о его цвете и его движении. Паралелльная переработка информации об
этих качествах зрительных объектов обеспечивается специализацией ганг-
лиозных клеток сетчатки, которые подразделяются на магноцеллюлярные
(М-клетки) и парвоцеллюлярные (Р-клетки).
730 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
В большом рецептивном поле относительно крупных М-клеток, состоя-
щем преимущественно из палочек, может проецироваться цельное изобра-
жение крупных объектов: М-клетки регистрируют грубые признаки таких
объектов и их движение в зрительном поле, отвечая на раздражение всего
рецептивного поля непродолжительной импульсной активностью. Клетки
P-типа имеют малые рецептивные поля, состоящие преимущественно из
колбочек и предназначенные для восприятия мелких деталей формы объ-
екта или для восприятия цвета. Среди ганглиозных клеток каждого типа
имеются как оп-нейроны, так и off-нейроны, дающие наиболее сильный
ответ на раздражение центра или периферии рецептивного поля. Сущест-
вование М- и P-типов ганглиозных клеток позволяет разделить информа-
цию о разных качествах наблюдаемого объекта, которая перерабатывается
независимо в параллельных путях зрительной системы: о тонких деталях
объекта и о его цвете (пути начинаются от соответствующих рецептивных
полей клеток P-типа) и о движении объектов в зрительном поле (путь от
клеток М-типа).
17.3.4. Проводящие пути и переключительные центры зрительной
системы
Аксоны ганглиозных клеток образуют зрительный нерв, состоящий прибли-
зительно из одного миллиона волокон. Войдя через зрительные отверстия
в череп, правый и левый зрительные нервы сходятся друг с другом и час-
тично перекрещиваются: волокна от носовых половин сетчаток переходят
на противоположную сторону, волокна от височных половин сетчаток сле-
дуют ипсилатерально. В результате частичного перекреста образуются пра-
вый и левый зрительные тракты, передающие информацию от противопо-
ложной половины зрительного поля.
Окончания аксонов ганглиозных клеток на нейронах претектальной об-
ласти среднего мозга служат для осуществления зрачкового рефлекса. Аксо-
ны, следующие к верхним буграм четверохолмия, переносят информацию,
необходимую для осуществления глазодвигательных рефлексов и ориентиро-
вочных реакций. Проводящий путь от сетчатки к гипоталамусу предназна-
чен для синхронизации эндогенного циркадианного 24-часового ритма с ес-
тественным чередованием светлого и темного времени суток. Собственно
сенсорная функция, т. е. восприятие зрительной информации, обеспечивает-
ся при участии латерального коленчатого тела, принимающего информа-
цию от сетчатки и передающего ее в зрительную кору.
17.3.4.1. Функциональная организация латерального коленчатого тела
Аксоны ганглиозных клеток образуют топографически организованные со-
единения с нейронами латерального коленчатого тела, которые представ-
лены шестью слоями клеток. Два первых слоя, расположенные вентрально,
состоят из магноцеллюлярных клеток, имеющих синапсы с М-клетками
сетчатки, причем первый слой получает сигналы от носовой половины
сетчатки контралатерального глаза, а второй — от височной половины ип-
силатерального глаза. Остальные четыре слоя клеток, расположенные дор-
сальнее, получают сигналы от Р-клеток сетчатки: четвертый и шестой — от
носовой половины сетчатки контралатерального, а третий и пятый — от
височной половины сетчатки ипсилатерального глаза. В результате такой
организации афферентных входов в каждом латеральном коленчатом теле,
т. е. левом и правом, формируются шесть расположенных точно одна над
17. Сенсорные системы • 731
другой нейронных карт противоположной половины зрительного поля. Ней-
ронные карты организованы ретинотопически, в каждой из них около 25 %
клеток получают информацию от фоторецепторов центральной ямки.
Рецептивные поля нейронов латерального коленчатого тела имеют
округлую форму с центрами on- или off-типа и антагонистичной по отно-
шению к центру периферией. К каждому нейрону конвергирует небольшое
количество аксонов ганглиозных клеток, и потому характер передающейся
зрительной коре информации здесь почти не изменяется. Сигналы от пар-
воцеллюлярных и магноцеллюлярных клеток сетчатки перерабатываются
независимо друг от друга и передаются в зрительную кору параллельными
путями. Нейроны латерального коленчатого тела получают от сетчатки не
более 20 % афферентных входов, а остальные афференты образованы в ос-
новном нейронами ретикулярной формации и коры. Эти входы в латераль-
ное коленчатое тело регулируют передачу сигналов от сетчатки к коре.
17.3.5. Переработка зрительной сенсорной информации в коре
Проекционная зрительная кора расположена в затылочных долях каждого
полушария (поле 17 или VI) и получает организованные ретинотопически
афферентные входы от соответствующего, т. е. левого или правого, лате-
рального коленчатого тела. Каждый участок проекционной коры соответ-
ствует определенным точкам сетчатки, на которые проецируются изобра-
жения предметов, расположенных в противоположной полушарию полови-
не зрительного поля. Проекция центральной ямки и ближайших к ней
участков сетчатки занимает около половины первичной зрительной коры.
Информация о форме наблюдаемых объектов, об их цвете и их движении
поступает по параллельным путям в разные слои первичной зрительной ко-
ры. Объединение информации обо всех качествах наблюдаемых объектов
происходит в процессе ее переработки во вторичной зрительной и ассоциа-
тивной коре.
Аксоны нескольких нейронов латерального коленчатого тела передают
информацию входным звездчатым клеткам коры, отростки которых кон-
вергируют к одному из простых пирамидных нейронов первичной зритель-
ной коры. Поэтому рецептивное поле последнего имеет форму вытянутого
в длину прямоугольника, составленного из нескольких округлых рецептив-
ных полей нейронов предыдущего уровня (рис. 17.11). Центральная часть
такого рецептивного поля представляет собой on- либо off-зону, антагони-
стичную по отношению к периферии. Наиболее эффективным раздражи-
телем для простых нейронов является действующая вдоль их рецептивного
поля световая грань (светлая полоса на темном фоне, темная линия на
светлом фоне, контраст между темной и светлой плоскостью). В первич-
ной зрительной коре имеется около 20 популяций простых нейронов, раз-
личающихся чувствительностью к определенному углу наклона световой
грани.
Наряду с простыми нейронами в первичной зрительной коре имеются
комплексные нейроны, получающие афферентную информацию от несколь-
ких простых нейронов и от входных клеток коры. Рецептивные поля ком-
плексных нейронов больше, чем у простых нейронов, а линейная ориента-
ция стимулов уже не имеет решающего значения для их возбуждения: са-
мым значимым для них раздражителем служит движение грани или свето-
вого контура через рецептивное поле. Более половины комплексных ней-
ронов глазодоминантны, т. е. сильнее реагируют на раздражение одного
глаза, каждый из которых поставляет чуть отличающуюся информацию в
732 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Рецептивные поля нейронов
латерального коленчатого тела
IV слоя первичной
зрительной коры
Рецептивные поля простых
нейронов зрительной коры
Звездчатые клетки
Простой нейрон
зрительной коры
Рис. 17.11. Рецептивные поля нейронов разных иерархических уровней зрительной
системы.
А. Нейроны латерального коленчатого тела имеют округлые рецептивные поля с on- или off-
центрами, подобные рецептивным полям ганглиозных клеток. Конвергенция нейронов лате-
рального коленчатого тела к простым нейронам проекционной зрительной коры (опосредо-
ванная звездчатыми клетками) формирует их рецептивные поля прямоугольной формы и мак-
симальной чувствительности к определенному углу наклона линейного стимула.
Б. Конвергенция нейронов зрительной коры к комплексным нейронам создает рецептивное
поле последних, в котором угол наклона линейных стимулов не имеет значения.
связи с поперечной диспарацией (см. раздел 17.3.1.4). Глазодоминантные
нейроны обеспечивают бинокулярное зрение, необходимое для определе-
ния пространственной глубины наблюдаемых объектов.
Ни простые, ни комплексные нейроны не реагируют на монотонные
участки внутри объекта или фона — такие участки не дают информации, и
восприятие внутренних участков зрительного объекта полностью опреде-
ляется их границами.
Анализ информации о цвете в первичной зрительной коре осуществля-
ют сосредоточенные там в виде так называемых капель (blobs) двойные про-
тивоцветные клетки, получающие афферентную информацию от простых
противоцветных клеток сетчатки. Двойные противоцветные клетки возбу-
ждаются (или тормозятся) при попадании одного цвета в центр рецептив-
ного поля, а антагонистичного цвета — на периферию. Первичная зритель-
ная кора не может обеспечить цветовое восприятие, которое осуществляет-
ся на следующей ступени преобразования поступившей информации в ре-
гионах V2 и V4 (последний расположен на стыке теменной и височной об-
ластей).
17. Сенсорные системы • 733
17.3.5.1. Зрительное восприятие
Целостное восприятие окружающего мира происходит в результате объеди-
нения всей информации, поступившей по параллельным путям зрительной
системы. Магноцеллюлярный путь обеспечивает регистрацию движения
объектов, восприятие пространственной глубины и определение простран-
ственных отношений между объектами. В объединении этой информации
помимо первичной зрительной коры участвуют регионы V2 (поле 18), V3
(поле 19), V5 или МТ (медиотемпоральный), MST (область коры, проходя-
щая вдоль верхней височной борозды) и заднетеменной коры (поля 5 и 7).
Парвоцеллюлярный путь, берущий начало от on- и off-ганглиозных клеток
сетчатки и служащий для анализа информации о контурах объекта и ли-
нейной ориентации его деталей, после переработки в регионе VI продол-
жается в направлении нижних височных долей коры, при участии которых
происходит восприятие формы наблюдаемых объектов. Параллельный пар-
воцеллюлярный путь начинается от концентрических противоцветных ганг-
лиозных клеток сетчатки и предназначается для формирования зрительно-
го представления о цвете объекта. Необходимая информация последова-
тельно перерабатывается в регионах VI, V2 и V4, где происходит объедине-
ние зрительного восприятия формы и цвета. В регионе V4 расположены
нейроны, воспринимающие определенный цвет, а не длину волн, соответ-
ствующих тому или иному цвету. С активностью этих нейронов связан фе-
номен цветового постоянства, т. е. способность воспринимать цвет одних
и тех же объектов неизменным, несмотря на естественные изменения ос-
вещенности на протяжении светового дня.
Восприятие образуется из элементарных зрительных ощущений подобно
тому, как музыкальная мелодия складывается из отдельных звуков опреде-
ленной высоты и узнается независимо от того, в какой тональности она
сыграна. Процесс зрительного восприятия состоит из нескольких этапов.
Первый из них осуществляется очень быстро и состоит в определении важ-
нейших контуров объектов, их границ и текстуры поверхности, когда вни-
мание еще не фиксировано на каких-либо деталях. После этого происходит
фокусирование внимания на деталях, которые, в зависимости от их важно-
сти для целостного восприятия, могут быть просмотрены несколько раз, то-
гда как другие, менее информативные остаются незамеченными. Например,
при рассматривании человеческого лица внимание смотрящего помимо об-
щего контура более всего сосредоточено на таких деталях, как глаза и губы,
тогда как щеки обычно привлекают внимание в меньшей мере. Зрительное
восприятие не сводится к элементарному отражению зрительного поля, оно
складывается и в результате конструктивной и высокопроизводительной ак-
тивности всех участвующих в этом процессе регионов мозга: благодаря это-
му человек может с первого взгляда узнавать лица, вещи или пейзажи.
17.4. Слуховая сенсорная система
Функция слуховой системы состоит в формировании слуховых ощущений
человека в ответ на действие звуковых волн, представляющих собой распро-
страняющиеся колебания молекул воздуха (упругой среды). Перифериче-
ская часть слуховой системы включает наружное, среднее и внутреннее ухо,
в котором расположены слуховые рецепторы. Ее центральную часть образу-
ют проводящие пути, переключательные ядра и слуховая кора, расположенная
в обоих полушариях в глубине латеральной борозды, отделяющей височную
долю от лобной и передних отделов теменной доли (поля 41 и 42).
734 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
17.4.1. Психофизические характеристики звуковых сигналов
Звуковые волны представляют собой передаваемые от источника звука ме-
ханические смещения молекул воздуха (или иной упругой среды). Ско-
рость распространения звуковых волн в воздухе около 343 м/с при 20 °C (в
воде и металлах она значительно выше). Правильно чередующиеся участки
сжатия и разрежения молекул упругой среды можно представить в виде си-
нусоид, которые различаются частотой и амплитудой. При суперпозиции
звуковых волн с различными частотами и амплитудами они наслаиваются
друг на друга, образуя комплексные волны. Физическим понятиям амплиту-
ды, частоты и комплексности соответствуют ощущения громкости, высоты
и тембра звука (рис. 17.12). Звук, образованный синусоидальными колеба-
ниями только одной частоты, вызывает ощущение определенной его
высоты и определяется как тон. Комплексные тоны состоят из основного
тона (самая низкая частота колебаний) и определяющих тембр обертонов,
или гармоник, представляющих более высокие частоты, кратные основной.
В повседневной жизни тоны всегда бывают комплексными, т. е. состав-
ленными из нескольких синусоид. Индивидуальное сочетание комплекс-
ных волн определяет характерный тембр человеческого голоса или музы-
кального инструмента. Слуховая система человека способна различать вы-
соту звука лишь у периодических звуковых сигналов, тогда как звуковые
раздражители, состоящие из беспорядочного сочетания частотных и ам-
плитудных компонентов, воспринимаются как шум.
Физический
параметр
Амплитуда
Ощущение
Звуковые волны
Громкость
Громко
Тихо
Частота
Высота
Низко
Высоко
Чистый ; Комплексный
тон тон
Рис. 17.12. Соотношение физических параметров звуковых раздражителей с субъек-
тивными ощущениями человека.
Уменьшение амплитуды звуковых волн субъективно воспринимается как уменьшение громко-
сти звука. Увеличение частоты звуковых волн субъективно оценивается как повышение высо-
ты звука. Комплексные звуковые волны, образованные суперпозицией нескольких волн, фор-
мируют субъективное восприятие тембра звука.
17. Сенсорные системы • 735
17.4.1.1. Диапазон частотного восприятия
Дети воспринимают звуковые волны в диапазоне от 16 до 20 000 Гц, но
приблизительно с 15—20 лет диапазон частотного восприятия начинает су-
живаться в связи с утратой чувствительности слуховой системы к самым
высоким звукам. В норме независимо от возраста человек легче всего вос-
принимает звуковые волны в диапазоне от 100 до 2000 Гц, что имеет для
него особенное значение, поскольку человеческая речь и звучание музы-
кальных инструментов обеспечиваются передачей звуковых волн именно в
этом диапазоне (табл. 17.2).
Чувствительность слуховой системы к минимальному изменению высо-
ты звука определяется как разностный порог частоты. В оптимальном для
восприятия частотном диапазоне, приближающемся к 1000 Гц, порог раз-
личения частот составляет около 3 Гц. Это значит, что изменение частоты
звуковых волн на 3 Гц в большую или меньшую сторону человек замечает
как повышение или понижение звука.
Таблица 17.2. Частотная и волновая характеристики некоторых источников звука
Источники звука Частота (Гц) Длина волны (м)
Нижняя нота рояля 27,5 12,4
Низкий певческий бас 80 2,7
Нота «ля» для настройки 440 78 • 1(Г2
Высокий тенор 500 69 • Ю’2
Альт 650 53 • 10’2
Верхнее «до» 1048 33 • 10’2
Колоратурное сопрано 1350 25 • 10’2
Самая высокая нота рояля 4186 8,2 • 10’2
17.4.1.2. Громкость звука
Амплитуда звуковых волн определяет величину звукового давления, под ко-
торым понимают силу сжатия, действующую на перпендикулярно располо-
женную к ней площадь. Акустическим эталоном, близким к абсолютному
порогу слухового восприятия, принято считать 2 • 10-5 Н/м2, а сравнитель-
Таблица 17.3. Интенсивность некоторых источников звука
Интенсив- ность (Вт/м2) Уровень интен- сивности (дБ) Качественная субъ- ективная оценка Источник звука
ю-“ 10 Едва слышно Спокойное дыхание
Ю-ю 20 Очень тихо Шелест листвы
кг’ 30 Тихо Перелистывание книги
ю-8 40 Умеренно Тихая контора
1(Г7 50 Умеренно Домашняя обстановка
10~6 60 Умеренно Обычный разговор
10"! 70 Умеренно Лектор
10"4 80 Шумно Уличный транспорт
Ю’3 90 Очень шумно Близко идущий поезд
КГ2 100 Очень шумно Пожарная сирена
кг1 110 Невыносимо Взлет авиалайнера
10 120 Болевое ощущение Разрыв артиллерийского снаряда
736 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
ной единицей измерения громкости, выражаемой в логарифмической шка-
ле, служит децибел (дБ). Громкость измеряется в децибелах как 201g(Px/Po),
где Рх — действующее звуковое давление, а Ро — эталонное давление. Также
в децибелах принято измерять интенсивность различных источников звука,
понимая под интенсивностью звука мощность или плотность звуковых волн
в единицу времени. Принимая за эталонную интенсивность 10“12 Вт/м2 (10),
количество децибел для измеряемой интенсивности (1Х) определяют по фор-
муле lOlgCVIJ. Интенсивность звука пропорциональна квадрату звукового
давления, поэтому 101g(Ix/Io) = 201g(Px/Po). Сравнительная характеристика
интенсивности некоторых источников звука представлена в табл. 17.3.
Субъективно воспринимаемая громкость звучания зависит не только от
уровня звукового давления, но и от частоты звукового стимула. Чувстви-
тельность слуховой системы максимальна для раздражителей с частотами
от 500 до 4000 Гц, при других частотах она снижается.
17.4.2. Периферическая часть слуховой системы
В наружном, среднем и внутреннем ухе происходят необходимые для слу-
хового восприятия подготовительные процессы, смысл которых состоит в
оптимизации параметров передаваемых звуковых колебаний при одновре-
менном сохранении характера сигналов. Во внутреннем ухе происходит
преобразование энергии звуковых волн в рецепторные потенциалы волос-
ковых клеток.
17.4.2.1. Функция наружного уха
Движения ушных раковин млекопитающих в направлении источника звука
помогают обнаружить его пространственное расположение, эта функция
определяется как ототопика. Большинство людей не могут изменять поло-
жение ушных раковин, а сохранность такой способности у человека квали-
фицируется как атавизм. Функцию ототопики у человека выполняют рель-
еф ушных раковин и их расположение на противоположных сторонах го-
ловы, позволяющее различать поступление звука спереди или сзади. На-
ружный слуховой проход ведет к барабанной перепонке, представляющей во-
гнутую в полость среднего уха перегородку, которая приводится в колеба-
ния распространяющимися звуковыми волнами. Ориентация коллагеновых
волокон барабанной перепонки позволяет ей колебаться с частотой дейст-
вующих звуковых волн относительно оси, расположенной вблизи ее верх-
него края.
17.4.2.2. Функция среднего уха
Воздушная полость среднего уха соединяется евстахиевой трубой с носо-
глоткой, что позволяет выравнивать давление в среднем ухе по атмосфер-
ному давлению (соприкасающиеся стенки евстахиевой трубы раскрывают-
ся при глотательных движениях). В полости среднего уха имеются три под-
вижно сочлененные слуховые косточки (молоточек, наковальня и стремеч-
ко), служащие для передачи колебаний от барабанной перепонки к оваль-
ному окну, которое ведет в вестибулярную часть внутреннего уха. Рукоятка
молоточка прикреплена к барабанной перепонке, а основание стремечка
закрывает овальное окно, подвижную связь между ними обеспечивает на-
ковальня (рис. 17.13).
Колебания барабанной перепонки сообщаются молоточку, рукоятка ко-
17. Сенсорные системы • 737
Рис. 17.13. Среднее и внутреннее ухо.
А. Строение среднего и внутреннего уха: колебания барабанной перепонки сообщаются слухо-
вым косточкам, которые передают их во внутреннее ухо через овальное окно.
Б. Улитка показана в развернутом виде: колебания перилимфы вестибулярной лестницы сооб-
щаются через геликотрему перилимфе барабанной лестницы, заставляя колебаться основную
мембрану.
В. Поперечный разрез кортиева органа: 1) вестибулярная лестница; 2) барабанная лестница;
3) средняя лестница (перепончатый канал улитки); 4) вестибулярная мембрана; 5) основная
мембрана; 6) покровная пластинка; 7) волосковые клетки; 8) первичные сенсорные нейроны.
торого в полтора раза длиннее отростка наковальни; благодаря этому соз-
дается рычаг, повышающий силу колебаний стремечка. Увеличение силы
колебаний необходимо для их передачи из воздушной среды среднего уха в
заполненную жидкостью полость внутреннего уха. Решению этой задачи
способствует и большая площадь барабанной перепонки по сравнению с
площадью овального окна, соотносящихся между собой как 20:1.
При высоких значениях звукового давления амплитуда колебаний слу-
ховых косточек уменьшается вследствие рефлекторного сокращения двух
мышц, прикрепленных к рукоятке молоточка и стремечку. При сокраще-
нии одной из них (ш. tensor tympani) увеличивается натяжение барабанной
перепонки, что ведет к уменьшению амплитуды ее колебаний, а сокраще-
ние другой мышцы (ш. stapedius) ограничивает колебания стремечка. Эти
мышцы участвуют в приспособлении слуховой системы к звукам высокой
интенсивности и начинают сокращаться примерно через 10 мс после нача-
ла действия звука, превышающего 40 дБ.
24 - 6095
738 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
17.4.2.3. Внутреннее ухо
В пирамиде височной кости имеется сложной формы полость (костный ла-
биринт), составными частями которой являются преддверие, улитка и полу-
кружные каналы (см. рис. 17.13). Внутри костного лабиринта как в футляре
размещен перепончатый лабиринт, по форме соответствующий костному.
Полость трехслойного (эпителий, базальная мембрана, соединительная
ткань) перепончатого лабиринта заполнена жидкостью (эндолимфа), а сам
он взвешен в наружной жидкости (перилимфа), отличающейся по своему
составу от внутренней.
Слуховой частью лабиринта является улитка, которая представляет со-
бой спираль из двух с половиной завитков, закрученных вокруг полого ко-
стного веретена. Внутри костного веретена расположены тела чувствитель-
ных нейронов, образующие спиральный ганглий. Периферические отростки
нейронов спирального ганглия образуют синапсы со вторичными рецепто-
рами улитки (волосковыми клетками), а центральные — составляют слухо-
вой нерв. Перепончатый лабиринт улитки имеет трехгранную форму, его
дно образовано основной, или базилярной, мембраной, крыша — вестибуляр-
ной, или рейснеровой, мембраной. Заполненная перилимфой полость под ба-
зилярной мембраной называется барабанной лестницей, а полость над вес-
тибулярной мембраной — вестибулярной лестницей', обе полости сообщают-
ся у вершины улитки посредством тонкого прохода — геликотремы. Собст-
венная полость перепончатого лабиринта заполнена эндолимфой и назы-
вается средней лестницей.
На базилярной мембране расположен кортиев орган, состоящий из не-
скольких рядов волосковых рецепторных клеток, поддерживаемых опор-
ным эпителием. Около 3500 волосковых клеток образуют внутренний ряд
(внутренние волосковые клетки), а приблизительно 12—20 тысяч наружных
волосковых клеток образуют три, а в области верхушки улитки пять про-
дольных рядов. На обращенной внутрь средней лестницы поверхности во-
лосковых клеток имеются покрытые плазматической мембраной чувстви-
тельные волоски — стереоцилии. Волоски соединены с цитоскелетом, их
механическая деформация ведет к открытию катионных каналов мембраны
и возникновению рецепторного потенциала волосковых клеток. Над кор-
тиевым органом имеется желеобразная покровная, или текториальная, пла-
стинка, образованная гликопротеином и коллагеновыми волокнами и при-
крепленная к внутренней стенке лабиринта. Верхушки стереоцилий на-
ружных волосковых клеток погружены в вещество покровной пластинки.
17.4.2.4. Функция внутреннего уха
Вызванные звуковыми волнами колебания барабанной перепонки и слухо-
вых косточек сообщаются через овальное окно перилимфе вестибулярной
лестницы и распространяются через геликотрему на барабанную лестницу,
которую отделяет от полости среднего уха круглое окно, закрытое тонкой
и упругой мембраной, повторяющей колебания перилимфы. Колебания
стремечка вызывают распространение следующих одна за другой бегущих
волн, которые перемещаются по основной мембране от основания улитки
к геликотреме.
Звуковая волна распространяется по вестибулярной лестнице к гелико-
треме практически мгновенно (примерно за 2 • 10-5 секунды). Вызванное
этой волной гидростатическое давление смещает весь улитковый ход в на-
правлении барабанной лестницы, одновременно с этим покровная пла-
17. Сенсорные системы • 739
стинка сдвигается относительно поверхности кортиева органа. Ось враще-
ния покровной пластинки расположена выше оси вращения основной
мембраны, и поэтому в области амплитудного максимума бегущей волны
возникает срезающее усилие. В результате покровная пластинка деформи-
рует пучки стереоцилий волосковых клеток, что приводит к их возбужде-
нию, передающемуся окончаниям первичных сенсорных нейронов.
Помимо воздушной передачи колебаний с участием барабанной пере-
понки движение жидкости внутреннего уха можно вызвать посредством
костного проведения звуковых волн. Так, например, звуковые колебания
камертона можно слышать и при закрытом слуховом проходе, если поста-
вить его ножку на сосцевидный отросток височной кости или на темя. Ис-
пользование костных телефонов, установленных за ушными раковинами,
тоже основано на костном проведении звука.
17.4.2.5. Биоэлектрические процессы в кортиевом органе
Различия ионного состава эндолимфы, содержащей около 155 ммоль/л ио-
нов калия, т. е. близкой к внутриклеточной, и перилимфы с высокой кон-
центрацией ионов натрия (около 140 ммоль/л) создают между ними раз-
ность потенциалов около +80 мВ, которая называется эндокохлеарным по-
тенциалом. Мембранный потенциал покоя волосковых клеток составляет
около —80 мВ, поэтому между эндолимфой и внутриклеточной средой ре-
цепторных клеток существует разность потенциалов около 160 мВ. Столь
значительная разность потенциалов способствует возбуждению волосковых
клеток даже при действии слабых звуковых сигналов, вызывающих незна-
чительные колебания основной мембраны. При деформации стереоцилий
волосковых клеток в них возникает рецепторный потенциал, что приводит
к выделению медиатора глутамата, действующего на окончания слуховых
нервов и тем самым возбуждающего их.
17.4.2.6. Частотное кодирование
Амплитуда и скорость распространяющихся по основной мембране бегу-
щих волн зависят от механических свойств этой мембраны. Вблизи оваль-
ного окна жесткость основной мембраны приблизительно в 100 раз выше,
чем в завитках улитки, поэтому возле овального окна волны распространя-
ются с максимальной скоростью и с минимальной амплитудой. Амплитуда
бегущих волн возрастает в менее натянутых участках основной мембраны,
но затем гасится сопротивлением жидкости неподалеку от геликотремы.
Между точкой появления и точкой затухания бегущей волны существует
амплитудный максимум, место возникновения которого зависит от частоты
звуковых колебаний. Это место расположено тем ближе к овальному окну,
чем большей окажется частота звуковых колебаний, и наоборот: чем ниже
частота колебаний, тем дальше от овального окна возникнет амплитудный
максимум.
Звуковые волны разной частоты вызывают наиболее сильные колебания
основной мембраны в разных ее участках, каждый из которых соответству-
ет определенной частоте слухового диапазона. Ближе к овальному окну
располагаются участки мембраны, настроенные на восприятие высоких
звуков, область вершины улитки предназначена для возбуждения низкими
частотами. Такая структурно-функциональная организация рецепторной
поверхности основной мембраны определяется как тонотопическая. Во-
лосковые клетки сильнее всего возбуждаются в области амплитудного мак-
24*
740 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
симума, при действии комплексных звуковых волн одновременно активи-
руются рецепторы нескольких участков основной мембраны. Каждый из
них обладает специфической чувствительностью к определенной частоте
звуковых колебаний, на которую он «настроен», и не возбуждается при от-
сутствии такой частоты в комплексной звуковой волне. Рецепторные по-
тенциалы волосковых клеток отражают не только значение амплитуды, но
также крутизну нарастания волн и интервалы между ними, что предостав-
ляет дополнительные характеристики действующего звукового сигнала.
17.4.2.7. Кодирование сенсорной информации в окончаниях слухового нерва
Информацию о характере колебаний основной мембраны проводят при-
близительно 30—40 тысяч волокон слухового нерва, среди которых 90 %
несут сигналы от внутренних волосковых клеток. Разрушение этих клеток
у экспериментальных животных повышает порог слуховой чувствительно-
сти на 80—100 дБ, что фактически означает глухоту, тогда как разрушение
всех наружных волосковых клеток увеличивает порог только на 40—45 дБ.
Каждый нейрон спирального ганглия контактирует только с одной внут-
ренней волосковой клеткой, которая, в свою очередь, имеет синапсы при-
близительно с 10—20 сенсорными нейронами, образующими нейронный ка-
нал для передачи информации о собственной характеристической частоте
звука. Такие каналы не активируются другими частотами, поэтому диффе-
ренцировка звуковых раздражителей по частоте осуществляется уже на
уровне преобразования механических процессов в электрическую актив-
ность.
В восприятии каждой характеристической частоты звукового стимула
участвуют приблизительно 150 наружных волосковых клеток, передающих
возбуждение 10—15 сенсорным нейронам. Наружные волосковые клетки
являются не только рецепторами, но также эффекторами, получающими в
составе слухового нерва эфферентные волокна от нейронов верхнеоливарно-
го ядра. Эфферентная активация наружных волосковых клеток происходит
в ответ на действие звукового стимула и состоит в сокращениях актиновых
филаментов этих клеток, происходящих с частотой звукового сигнала. По-
вторяя частоту колебаний слабых звуковых сигналов, наружные волоско-
вые клетки их усиливают и одновременно создают звуковое излучение
кортиева органа (эндокохлеарная эмиссия), которое можно зарегистрировать
с помощью микрофона.
Волокна слухового нерва обладают фоновой активностью в пределах
50—100 импульсов в 1 с, которая увеличивается при повышении уровня
звука, тем самым кодируя информацию об интенсивности звукового дав-
ления. Кроме этого, кодирование информации об уровне звука происходит
за счет возбуждения дополнительного числа сенсорных нейронов, имею-
щих относительно высокий порог чувствительности и находившихся ранее
в состоянии фоновой активности. При действии звуковых раздражителей с
частотой до 5000 Гц импульсная активность нейронов отражает повыше-
ние звукового давления в каждой фазе повторяющихся циклов звуковых
колебаний. Максимальная активность возникает во время фазы подъема
звуковой волны, а затем она уменьшается, чем передается информация о
временной структуре сигналов. При продолжительной слуховой стимуляции
частота нервных импульсов постепенно уменьшается вследствие развития
адаптации к действию звуковых стимулов.
17. Сенсорные системы • 741
17.4.3. Проводящие пути и переключательные ядра слуховой
системы
Центральные отростки первичных слуховых нейронов спирального ганглия
образуют две ветви: к вентральному и дорсальному улитковым (кохлеар-
ным) ядрам продолговатого мозга (рис. 17.14). В каждом ядре существует
тонотопически организованное представительство кортиева органа. Нейро-
ны дорсального улиткового ядра образуют проекцию на верхнее ядро оливы
противоположной стороны, а нейроны вентрального ядра — на ипсилате-
ральные и контралатеральные верхние ядра олив. Перекрест нервных во-
локон обеспечивает передачу акустической информации на противополож-
ную каждому уху сторону мозга.
В ядрах верхних олив большинство нейронов возбуждается бинаурально,
т. е. в ответ на сигналы, поступающие как от ипсилатерального, так и от
контралатерального уха. Однако часть имеющихся в этих ядрах нейронов
реагирует на сигналы, поступившие только от одного уха, и тормозится
сигналами от противоположного уха. Существование таких нейронов обес-
печивает сравнительный
анализ звуковых сигна-
лов, возникающих с ле-
вой или правой от чело-
века стороны, что необ-
ходимо для его простран-
ственной ориентации.
Некоторые нейроны ядер
верхней оливы макси-
Рис. 17.14. Упрощенная схе-
ма проводящих путей и пе-
реключательных ядер слухо-
вой сенсорной системы.
Сенсорные нейроны второго
порядка расположены в улитко-
вых (кохлеарных) ядрах, в каж-
дом ядре имеется собственное
представительство кортиева ор-
гана. Нейроны улитковых ядер
образуют ипсилатеральные и
контралатеральные проекции
на ядра верхней оливы, здесь
происходит бинауральный ана-
лиз звуковых сигналов и сохра-
няется тонотопия. В нижних
буграх четверохолмия существу-
ют нейронные переключения,
необходимые для ориентиро-
вочных реакций на звуковые
сигналы. В медиальном колен-
чатом теле фоновая активность
нейронов изменяется в соответ-
ствии с афферентными сигна-
лами и происходит анализ их
временных характеристик. Слу-
ховая кора расположена в полях
41 и 42, кортикальные колонки
в ней упорядочены тонотопиче-
ски.
742 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
мально активны при расхождении времени поступления сигналов от правого
и левого уха, другие нейроны наиболее сильно реагируют на различную ин-
тенсивность сигналов.
Такие нейроны образуют проекцию на верхние бугры четверохолмия, ку-
да одновременно поступает зрительная информация, что позволяет нейро-
нам четверохолмия создавать трехмерную карту слухового пространства и
определять пространственное положение источника звука. Благодаря бинау-
ральному слуху сенсорная система человека определяет источники звука,
находящиеся в стороне от средней линии, поскольку звуковые волны
раньше действуют на ближнее к этому источнику ухо. Слуховая система
реагирует на 1 дБ различий звукового давления, действующего на правое и
левое ухо, и фиксирует временное запаздывание действия звука на отда-
ленное от него ухо всего в 3 • 10-5 с.
Большая часть аксонов верхних оливарных ядер проходит в составе ла-
терального лемнискового пути к нижним буграм четверохолмия, нейроны
которого участвуют в формировании рефлекторной дуги ориентировочного
рефлекса, а также осуществляют анализ поступающей информации. Он
обеспечивается наличием в нижних буграх четверохолмия нейронов, кото-
рые возбуждаются только частотными модуляциями тона, т. е. повышени-
ем или понижением высоты действующего звукового сигнала. Другие ней-
роны этих переключательных ядер активируются только изменениями ам-
плитуды звуковой волны, т. е. увеличением или уменьшением громкости
звукового сигнала. Большинство переключательных нейронов нижнего
двухолмия образуют проекцию на медиальное коленчатое тело, представ-
ляющее следующий иерархический уровень слуховой системы.
Медиальное коленчатое тело, относящееся к таламусу, является послед-
ним переключательным ядром слуховой системы на пути к коре. Его ней-
роны расположены тонотопически и образуют проекцию в слуховую кору.
Некоторые нейроны медиального коленчатого тела активируются в ответ
на возникновение либо на окончание сигнала, другие реагируют только на
частотные или амплитудные его модуляции. Во внутреннем коленчатом те-
ле имеются нейроны, способные постепенно увеличивать активность при
неоднократном повторении одного и того же сигнала. Большинство нейро-
нов медиального коленчатого тела обладают спонтанной фоновой активно-
стью, проявляющейся чередованием серий потенциалов действия и пауз
между ними.
17.4.4. Переработка сенсорной информации в слуховой коре
В первичной слуховой коре (поле 41) кортикальные колонки расположены
тонотопически для раздельной переработки информации о звуках раз-
личной частоты слухового диапазона. Они также содержат нейроны, кото-
рые избирательно реагируют на звуки различной продолжительности, на
повторяющиеся звуки, на шумы с широким частотным диапазоном и т. п.
В слуховой коре происходит объединение информации о высоте тона и его
интенсивности, о временных интервалах между отдельными звуками.
Вслед за этапом регистрации и объединения элементарных признаков
звукового раздражителя, который осуществляют простые нейроны, в пере-
работку информации включаются комплексные нейроны, избирательно реа-
гирующие только на узкий диапазон частотных или амплитудных модуля-
ций звука. Подобная специализация нейронов позволяет слуховой системе
создавать целостные слуховые образы, с характерными только для них со-
четаниями элементарных компонентов слухового раздражителя. Такие со-
17. Сенсорные системы • 743
четания могут быть зафиксированы энграммами памяти, что в дальнейшем
позволяет сравнивать новые акустические стимулы с прежними. Некото-
рые комплексные нейроны слуховой коры возбуждаются сильнее всего в
ответ на действие звуков человеческой речи.
17.5. Вестибулярная сенсорная система
Функция вестибулярной сенсорной системы состоит в обеспечении мозга
информацией о положении головы в пространстве, о действии гравитации
и сил, вызывающих линейные или угловые ускорения. Эта функция необ-
ходима для поддержания равновесия, т. е. устойчивого положения тела в
пространстве, и для пространственной ориентации человека. Вестибуляр-
ная система включает в себя периферический отдел, состоящий из распо-
ложенного во внутреннем ухе вестибулярного аппарата, проводящие пути,
переключательные центры, представленные вестибулярными ядрами про-
долговатого мозга и таламусом, и проекционную область коры в постцен-
тральной извилине. Адекватными раздражителями вестибулярной системы
являются гравитация и силы, сообщающие телу линейное или угловое ус-
корение. Специфическая особенность вестибулярной системы состоит в
том, что значительная часть перерабатываемой в ней сенсорной информа-
ции используется для автоматической регуляции функций, осуществляемой
без сознательного контроля. Вестибулярная система взаимодействует на
нескольких уровнях своей иерархической организации со зрительной и со-
матосенсорной системами; три эти системы дополняют друг друга в пре-
доставлении человеку информации, необходимой для его пространствен-
ной ориентации.
17.5.1. Вестибулярный аппарат
Внутри костных лабиринтов, расположенных в пирамидах височных кос-
тей, имеются прикрепленные к ним соединительнотканными тяжами пере-
пончатые лабиринты, заполненные вязкой жидкостью эндолимфы
(рис. 17.15). Лабиринт образован двумя отолитовыми органами и тремя по-
лукружными каналами, расположенными в трех плоскостях: горизонтальный
канал, передний вертикальный канал — во фронтальной плоскости и задний
вертикальный канал — в сагиттальной плоскости. Все три канала соедине-
ны в полости преддверия, от латинского определения которого (vestibulum)
происходит само название вестибулярного аппарата. В месте соединения с
преддверием каналы расширены в виде ампул, в которых содержится ре-
цепторный эпителий, выступающий внутрь в форме гребня или кристы.
Каждая криста покрыта купулой, представляющей собой аморфное желеоб-
разное вещество, скрепленное множеством фибриллярных волокон. При
угловых ускорениях, когда в силу инерции происходит сдвиг эндолимфы,
купула тоже смещается, что приводит к деформации погруженных в нее
вторичных рецепторных клеток с последующим возникновением в них ре-
цепторного потенциала.
В полости преддверия имеются два расширения: мешочек (sacculus) и
маточка (utriculus), сообщающиеся между собой с помощью У-образного
протока и представляющие собой отолитовые органы. Рецепторный эпите-
лий маточки и мешочка расположен на небольших возвышениях — маку-
лах, покрытых отолитовой мембраной, которая имеет слоистое строение и
содержит множество мелких, но тяжелых кристаллов карбоната кальция
{отолиты или отокинии). Макула маточки расположена в горизонтальной
744 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Б
ИмпульсацИя в афферентных Окончаниях
вестибулярного нерва I
11 11 11 । iiiHiHiuiuiiiiiii I—1.......। 1
В покое ; Возбуждение ; Торможение
Рис. 17.15. Вестибулярная сенсорная система.
А. Схема перепончатого лабиринта, рецепторные области выделены черным цветом: 1) гори-
зонтальный канал; 2) задний вертикальный канал; 3) передний вертикальный канал; 4) маточ-
ка; 5) мешочек; 6) канал улитки; 7) эндолимфатический мешочек.
Б. Волосковые рецепторные клетки вестибулярного аппарата: показана зависимость между на-
правлением смещения волосков и активностью клеток.
плоскости (при вертикальном положении головы), а макула мешочка ори-
ентирована вертикально. При действии силы тяжести или линейного уско-
рения отолитовые мембраны сдвигаются относительно макул, а вследствие
этого раздражаются имеющиеся в них вторичные рецепторные клетки.
Таким образом, устройство вестибулярного аппарата обеспечивает воз-
буждение вторичных рецепторных клеток благодаря действию силы тяже-
сти и прямолинейного ускорения (макулы мешочка и маточки) и вследст-
вие угловых ускорений (купулы полукружных каналов).
17.5.1.1. Свойства рецепторных клеток вестибулярного аппарата
Рецепторы макул и купул представлены волосковыми клетками, являющи-
мися вторичными механорецепторами и образующими синапсы с перифе-
рическими окончаниями нейронов вестибулярного ганглия (первичные сен-
сорные нейроны). Каждый рецептор имеет пучок из 40—80 волосков —
стереоцилий, достигающих в длину 50 мкм, а также расположенный экс-
17. Сенсорные системы • 745
центрично по отношению к стереоцилиям один длинный волосок — кино-
цилию. Если пучок стереоцилий наклоняется под влиянием механического
стимула в сторону киноцилии, рецептор деполяризуется, а при отклонении
стереоцилий от киноцилии происходит гиперполяризация мембраны ре-
цептора. Вследствие этого при сгибании пучка стереоцилий в одном на-
правлении волосковая клетка возбуждается, а при сгибании этого же пучка
в противоположном направлении — тормозится, т. е. у каждой волосковой
клетки выявляются два функциональных полюса. Направление функцио-
нальной поляризации изменяется от одной клетки к другой, а рецептор-
ный эпителий в целом содержит полный комплект клеток для регистрации
стимулов, действующих в любом возможном направлении.
Нейроны вестибулярного ганглия, образующие синапсы на основании
рецепторов, обладают спонтанной фоновой активностью, характер которой
меняется под влиянием медиаторов волосковых клеток, которыми предпо-
ложительно могут быть глутамат и/или ГАМК. Рецептивные поля нейро-
нов вестибулярного ганглия включают в среднем три волосковых клетки
ампул полукружных каналов либо 4—6 рецепторов макул маточки или ме-
шочка.
17.5.1.2. Адекватные раздражители рецепторов отолитовых органов
Рецепторы маточки и мешочка служат датчиками гравитации и линейных
ускорений. При вертикальном положении головы человека макула маточки
расположена в горизонтальной плоскости, при наклоне головы покрываю-
щая макулу отолитовая мембрана смещается, подчиняясь силе тяжести.
Смещение отолитовой мембраны сгибает стереоцилии рецепторных кле-
ток, отвечающих на деформацию образованием рецепторного потенциала.
В зависимости от направления и степени наклона головы сильнее дру-
гих возбуждаются такие рецепторы, функциональная поляризация которых
соответствует именно этому направлению, прочие рецепторы возбуждают-
ся слабее или же тормозятся. Изменившееся соотношение возбужденных и
заторможенных рецепторных клеток вызывает в зависимых от них нейро-
нах вестибулярного ганглия адекватное изменение фоновой активности.
Нейроны вестибулярного ганглия передают полученную от рецепторных
клеток информацию в центральную нервную систему. Указанные процес-
сы возникают не только при наклоне головы, но и при любом отклонении
положения всего тела от вертикальной оси, например при спортивной или
профессиональной деятельности, случайном падении, использовании ат-
тракционов (качели, американские горки).
Макула мешочка при вертикальном положении тела и головы располо-
жена в вертикальной плоскости, и ее отолитовая мембрана сдвигается при
действии линейных ускорений, вызывая раздражение рецепторов. В зави-
симости от направления, в котором происходит линейное ускорение, воз-
буждаются наиболее чувствительные именно к нему рецепторы. Наличие
нескольких популяций рецепторов, различающихся своей функциональной
поляризацией, позволяет им в целом передавать сенсорным нейронам ин-
формацию о линейных перемещениях в любом направлении. Чувствитель-
ность этих рецепторов позволяет человеку ощутить прямолинейное уско-
рение, превысившее 2 см/с2 и наклон головы в сторону всего на Г. Наряду
с этим рецепторный аппарат мешочка высокочувствителен к действию
вибрации.
746 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
17.5.1.3. Адекватные раздражители рецепторов полукружных каналов
Угловые ускорения возникают при вращении тела вокруг одной из трех
пространственных осей, расположенных перпендикулярно друг другу, они
возникают также при поворотах головы и ее наклонах. При вращении во-
круг вертикальной оси кресла с сидящим человеком у него раздражаются
рецепторы горизонтального канала. Раздражение возникает в самом начале
вращения, когда инертная эндолимфа остается неподвижной, что создает
усилие, смещающее купулу в противоположную вращению сторону. При
остановке вращения движение эндолимфы по инерции продолжается, что
вызывает смещение купулы в сторону уже прекратившегося движения.
В результате смещения купулы стереоцилии горизонтального канала сги-
баются сначала в одном направлении, что сопровождается деполяризацией
волосковых клеток, а затем — в противоположном направлении, что вызы-
вает гиперполяризацию рецепторов. Соответственно этому волосковая
клетка увеличивает или уменьшает выделение медиатора, действующего на
окончание нейрона вестибулярного ганглия, что повышает или понижает
его фоновую активность.
При вращении вокруг осей, перпендикулярных фронтальной или сагит-
тальной плоскостям, аналогичные вышеописанным изменения активности
рецепторов происходят в переднем или заднем вертикальных полукружных
каналах. Вращение вокруг какой-либо диагональной оси вызывает движе-
ние эндолимфы в двух каналах одновременно, соответственно реагируют
рецепторы, расположенные в купулах обоих каналов. Наличие трех полу-
кружных каналов обеспечивает человеку восприятие вращения и поворо-
тов головы в любой плоскости трехмерного пространства. Сенсорная чув-
ствительность вестибулярной системы позволяет ощущать угловые ускоре-
ния, достигшие значения 2°/с2.
17.5.2. Центральная часть вестибулярной системы
Центральные аксоны первичных сенсорных нейронов вестибулярного
ганглия оканчиваются на нейронах вестибулярных ядер: верхнего (ядро
Бехтерева), нижнего (ядро Роллера), латерального (ядро Дейтерса) и меди-
ального (ядро Швальбе). Эти ядра представляют собой единый функцио-
нальный комплекс, в котором объединяется афферентная информация от
вестибулярных ганглиев и от проприоцепторов; эта афферентация опреде-
ляет характер активности нейронов вестибулярных ядер. Вестибулярные
ядра являются подкорковыми центрами познотонических и статокинетиче-
ских рефлексов (регуляция равновесия), глазодвигательных рефлексов
(зрительное наблюдение окружающего мира) и опосредованных через ги-
поталамус вестибуловисцеральных реакций, с механизмом которых связа-
ны проявления кинетозов — морской болезни, проявляющейся головокру-
жением и тошнотой.
Осознаваемое восприятие изменений положения головы происходит в
результате последовательной переработки информации сначала в вестибу-
лярных ядрах, затем в задних вентральных ядрах таламуса, образующих
проекцию к постцентральным извилинам. Дополнительная информация
поступает в проекционную кору непрямым путем: от вестибулярных ядер в
мозжечок, а из него к вентролатеральным ядрам таламуса и проекционной
коре. Первичная проекционная область вестибулярной чувствительности
размещена в задней центральной извилине преимущественно той стороны
тела, на которой расположен вестибулярный аппарат. Еще одна проекция,
17. Сенсорные системы • 747
отличающаяся двусторонним представительством вестибулярной чувстви-
тельности, имеется во вторичной моторной коре. Осознание пространст-
венного расположения и схемы тела происходит с участием заднетеменных
регионов коры (поля 5 и 7), где осуществляется интеграция вестибуляр-
ной, зрительной и соматосенсорной чувствительности человека.
17.6. Вкусовая сенсорная система
Функция вкусовой сенсорной системы состоит в оценке качества пищи
или отвергаемых веществ. Адекватным раздражителем вкусовой системы
служат молекулы органических или неорганических веществ, которые по-
ступают в полость рта при приеме пищи и присоединяются к хеморецеп-
торным клеткам языка и ротовой полости. Эти клетки являются вторичны-
ми рецепторами и обладают высокой химической избирательностью, а
присоединение химических веществ вызывает в них образование рецептор-
ного потенциала, что сопровождается выделением медиатора, действующе-
го на чувствительные окончания первичных сенсорных нейронов. В пере-
даче сигналов от рецепторов в цен-
тральную нервную систему участву-
ют афферентные волокна IX, VII и
X черепных нервов (рис. 17.16).
Центральный отдел вкусовой сен-
сорной системы образуют ядра оди-
ночных пучков продолговатого мозга
и вентральные ядра таламуса, обра-
зующие проекцию в первичную про-
екционную кору. Проекционная
вкусовая кора соответствует области
представительства языка в постцен-
тральных извилинах.
17.6.1. Вкусовая рецепция
Выпячивания слизистой оболочки
языка (сосочки) содержат у человека
около 2000 вкусовых почек, каждая из
которых образована 30—60 клетка-
ми, расположенными в почке напо-
добие долек апельсина. Рецепторные
клетки составляют около 5—7 %
всех клеток вкусовой почки, они от-
Рис. 17.16. Рецепторы и афферентные
пути вкусовой системы.
Рецепторные клетки вкусовой почки имеют
ворсинки, выступающие во вкусовую пору.
Они реагируют на присоединение химических
веществ изменением количества медиатора,
действующего на афферентные волокна ней-
ронов первого порядка. Афферентные сигна-
лы поступают в ЦНС ипсилатерально по во-
локнам лицевого, языкоглоточного и блуж-
дающего нервов к нейронам второго порядка,
расположенным в ядре одиночного пучка.
Блуждающий
нерв
К таламусу
Полость рта
Язык
Лицевой нерв
Языко-
глоточный
нерв
Ядро одиночного
тракта
Вкусовая
почка
Эпителий языка
Вкусовая
пора
Афферентные з
волокна ----=
Рецепторные клетки
748 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
личаются от остальных клеток (опорных, базальных) наличием микроворси-
нок, выступающих во вкусовую пору — отверстие на вершине вкусовой поч-
ки. Продолжительность жизни рецепторной клетки составляет 10—12
дней, разрушенные клетки заменяются новыми, образующимися из деля-
щихся базальных клеток и устанавливающими точно такие же, как у своей
предшественницы, синаптические контакты с 1—6 отростками первичного
сенсорного нейрона, поэтому специфичность вкусовой чувствительности
рецепторов после регенерации не изменяется.
17.6.1.1. Рецепторные потенциалы вкусовых клеток
Мембрана микроворсинок вкусовых клеток содержит специфические уча-
стки (рецепторы), предназначенные для связывания растворенных в жид-
кой среде полости рта химических молекул. Существует четыре разновид-
ности вкусовых ощущений, или четыре вкусовые модальности: сладкое, ки-
слое, соленое и горькое. Строгой зависимости между химической природой
вещества и вкусовым ощущением нет: например, сладким вкусом облада-
ют не только сахара, но и некоторые неорганические соединения (соли
свинца, бериллия), а самым сладким веществом является не усваиваемый
организмом сахарин. Большинство вкусовых клеток полимодальны, т. е.
могут реагировать на стимулы всех четырех вкусовых модальностей.
Присоединение к специфическим рецепторам молекул, обладающих
сладким вкусом, активирует систему вторичных посредников аденилатцик-
лазы — циклического аденозинмонофосфата, которые закрывают мембран-
ные каналы ионов калия, и поэтому мембрана рецепторной клетки депо-
ляризуется. Вещества, обладающие горьким вкусом, активируют одну из
двух систем вторичных посредников: 1) фосфолипазу С — инозитол-3-фос-
фат, что приводит к выходу из внутриклеточного депо ионов кальция с по-
следующим выделением медиатора из рецепторной клетки; 2) специфиче-
ский G-белок гастдуцин, регулирующий внутриклеточную концентрацию
цАМФ, которая управляет катионными каналами мембраны и этим опре-
деляет возникновение рецепторного потенциала. Действие на рецепторы
молекул, имеющих соленый вкус, сопровождается открытием управляемых
натриевых каналов и деполяризацией вкусовой клетки. Вещества, обла-
дающие кислым вкусом, закрывают мембранные каналы для ионов калия,
что ведет к деполяризации рецепторной клетки.
Величина рецепторного потенциала зависит от вкусового качества и кон-
центрации химического вещества, действующего на клетку. Возникновение
рецепторного потенциала приводит к выделению вкусовой клеткой медиа-
тора, действующего через синапс на афферентное волокно первичного сен-
сорного нейрона, в котором через 40—50 мс от начала действия стимула по-
вышается частота потенциалов действия. Возникшие в афферентных волок-
нах нервные импульсы проводятся к ядрам одиночных пучков продолгова-
того мозга. При повышении концентрации действующего вещества общее
количество реагирующих чувствительных волокон возрастает за счет вовле-
чения высокопороговых афферентов в передачу информации от рецепторов.
17.6.1.2. Вкусовая чувствительность
Пороги вкусовой чувствительности выявляются поочередным нанесением
на поверхность языка растворов веществ, обладающих разными вкусовыми
качествами (табл. 17.4). Абсолютным порогом чувствительности считают
появление определенного вкусового ощущения, отличающегося от вкуса
17. Сенсорные системы • 749
Таблица 17.4. Абсолютные пороги восприятия веществ с характерным вкусом
Вкусовое качество Вещества Порог восприятия (ммоль/л)
Горькое Сульфат хинина 0,000008
Кислое Лимонная кислота 0,0023
Сладкое Глюкоза 0,08
Сахарин 0,000023
Соленое Хлорид натрия 0,01
дистиллированной воды. Вкус одного и того же вещества может восприни-
маться по-разному в зависимости от его концентрации в растворе; напри-
мер, при малой концентрации хлорида натрия он ощущается сладким, а
при большей концентрации — соленым. Максимальная способность разли-
чать концентрацию растворов одного и того же вещества и, соответствен-
но, самый низкий дифференциальный порог вкусовой чувствительности ха-
рактерны для среднего диапазона концентраций, а при высоких концен-
трациях вещества дифференциальный порог повышается.
Абсолютные пороги вкусовой чувствительности индивидуально разли-
чаются, но у подавляющего большинства людей самым низким оказывает-
ся порог определения веществ с горьким вкусом. Эта особенность воспри-
ятия возникла в эволюции, она способствует отказу от употребления в пи-
щу веществ горького вкуса, к которым принадлежат алкалоиды многих
ядовитых растений. Вкусовые пороги различаются у одного и того же че-
ловека в зависимости от его потребности в тех или иных веществах, они
повышаются вследствие длительного употребления веществ с характерным
вкусом (например, сладостей или соленостей) или курения, потребления
алкоголя, обжигающих напитков. Разные области языка различаются вку-
совой чувствительностью к различным веществам, что обусловлено осо-
бенностями распределения вкусовых рецепторов. Кончик языка более дру-
гих областей чувствителен к сладкому, боковые стороны языка — к кисло-
му и соленому, а корень языка — к горькому. Вкусовые ощущения в боль-
шинстве случаев мультимодальны и основаны не только на избирательной
химической чувствительности вкусовых рецепторных клеток, но и на раз-
дражении пищей терморецепторов и механорецепторов ротовой полости, а
также действии летучих компонентов пищи на обонятельные рецепторы.
17.6.2. Центральный отдел вкусовой системы
Афферентную иннервацию вкусовых почек передних 2/3 языка осуществля-
ют чувствительные волокна лицевых нервов (барабанная струна), вкусовые
рецепторы задней трети языка и твердого неба образуют синапсы с чувст-
вительными волокнами языкоглоточных нервов, а передачу сигналов от не-
большого количества вкусовых рецепторов в глотке и надгортаннике осу-
ществляют чувствительные волокна блуждающих нервов. Волокна барабан-
ной струны полимодальны и реагируют повышением частоты нервных им-
пульсов на действие веществ с любым вкусовым качеством. Однако если
одни волокна максимально возбуждаются при действии веществ со слад-
ким вкусом, то наибольшая активность других волокон возникает в ответ
на действие веществ соленого вкуса, либо кислого или горького вкуса. Если
определить активность одиночного волокна к веществам каждой из четырех
вкусовых субмодальностей, то можно получить «вкусовой профиль» такого
волокна, который будет отличаться от вкусовых профилей других волокон.
750 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Сенсорные нейроны второго порядка содержатся в составе ядра одиноч-
ного пучка, вытянутого вдоль продолговатого мозга. На этих нейронах
оканчиваются чувствительные волокна лицевого, языкоглоточного и блуж-
дающего нервов, а их аксоны переходят на противоположную сторону в
составе медиального лемниска и оканчиваются на нейронах вентрального
ядра таламуса. Нейроны таламуса образуют проекцию на латеральную
часть постцентральной извилины — корковую проекционную область вку-
совой чувствительности.
Электрическая активность нейронов ядра одиночного пучка повышает-
ся в ответ на действие вкусовых раздражителей разных модальностей, а
также вследствие тактильного и температурного раздражения языка и ро-
товой полости. Эти нейроны так же, как и нейроны первого порядка,
мультимодальны, однако каждый такой нейрон отличается своей индиви-
дуальной активностью. Различия состоят в том, что максимальная или ми-
нимальная активность нейрона проявляется в ответ на действие веществ,
обладающих разным вкусом, что формирует индивидуальный профиль вку-
совой активности данного нейрона. Указанные индивидуальные различия
сохраняются у нейронов вентрального ядра таламуса, и лишь в коре суще-
ствуют клетки, реагирующие только на действие вещества с определенным
вкусовым качеством, т. е. обладающие специфической вкусовой чувстви-
тельностью. Наряду с ними в проекционной коре имеются нейроны, реа-
гирующие не только на действие вкусовых раздражителей, но и на так-
тильное и температурное раздражение языка, что способствует целостному
восприятию всех качеств пищи.
17.6.3. Вкусовое восприятие
Вкусовые ощущения отчетливо проявляются уже у новорожденных детей,
а грудной ребенок 4—5 мес хорошо различает вкусовые изменения пищи.
Вкусовое восприятие человека формируется в результате соединения муль-
тимодальных сенсорных ощущений и механизмов памяти, хранящей инди-
видуальный опыт и связанной с усвоенными традициями питания. Изме-
нения вкусового восприятия часто связаны с нарушениями обоняния, а не
собственно вкусовой чувствительности. Ротовая полость сообщается с но-
совой, и пахучие вещества пищи достигают обонятельных рецепторов, по-
этому представление о вкусе пищи почти всегда основано не только на
вкусовом, но и на обонятельном восприятии. Гастрономические предпоч-
тения людей связаны с комплексным действием тех или иных блюд, вклю-
чающем не только вкусовые и обонятельные, но также тактильные темпе-
ратурные и зрительные ощущения.
17.7. Обонятельная сенсорная система
Обонятельные ощущения возникают в результате действия пахучих химиче-
ских веществ, попадающих из внешней среды в полость носа вместе с воз-
духом во время вдоха или из ротовой полости во время еды. Пахучие ве-
щества действуют на хеморецепторные клетки обонятельного нейроэпите-
лия (первичные рецепторы) носовой полости. В дальнейшей переработке
сенсорной информации участвуют обонятельные луковицы, лимбические
структуры мозга и кора больших полушарий. В отличие от большинства
млекопитающих, относящихся к макросматикам с высоко развитым обо-
нянием, человек принадлежит к микросматикам, для которых роль обоня-
ния в организации поведения значительно меньше.
17. Сенсорные системы • 751
17.7.1. Классификация запахов
Все пахучие вещества должны быть летучими, чтобы поступать в носовую
полость с воздухом, и растворимыми, чтобы проникать к рецепторным
клеткам через слой обонятельной слизи, покрывающей эпителий носовых
раковин. Таким требованиям удовлетворяет огромное количество веществ,
а человек способен различать тысячи всевозможных запахов, при этом от-
сутствует строгое соответствие между структурой химической молекулы и
ее запахом. Большинство имеющихся теорий запахов основано на произ-
вольном выделении нескольких запахов в качестве основных (по аналогии с
четырьмя вкусовыми модальностями) и объяснении всех остальных запа-
хов их различными комбинациями. И только стереохимическая теория за-
пахов основана на выявлении соответствия между геометрическим сходст-
вом молекул пахучих веществ и присущим им запахом.
Построение трехмерных моделей пахучих молекул на основе их предва-
рительного изучения с помощью дифракции рентгеновских лучей и ин-
фракрасной стереоскопии показало, что не только природные, но и искус-
ственно синтезированные молекулы обладают запахом, соответствующим
определенной форме и отличным от запаха, присущего другой форме мо-
лекул. В связи с этим проверяется гипотеза существования семи разновид-
ностей хеморецепторов, способных присоединять вещества, которые сте-
реохимически им соответствуют. Среди нескольких сотен эксперименталь-
но исследованных пахучих молекул удалось выявить семь классов, в кото-
рых расположились вещества со сходной стереохимической конфигураци-
ей молекул и сходным запахом (табл. 17.5). Семь указанных запахов назва-
ны первичными, а все остальные запахи объясняются различными сочета-
ниями первичных запахов.
17.7.2. Периферический отдел обонятельной системы
Обонятельный эпителий человека расположен в верхних и отчасти в сред-
них раковинах носовой полости, он состоит из биполярных хеморецепторных
клеток, а также опорных и базальных клеток. Биполярные клетки являются
первичночувствующими рецепторами (рис. 17.17), их количество у челове-
ка приближается к 10 миллионам (у макросматиков оно на порядок выше).
Биполярные сенсорные нейроны существуют около 60 сут и после их есте-
ственной гибели замещаются новыми рецепторами, образующимися из ба-
зальных клеток. Регенерировавшие сенсорные нейроны восстанавливают
прежние синаптические контакты с центральными отделами обонятельной
системы, а опорные клетки фагоцитируют разрушенные рецепторы.
Дендриты биполярных клеток имеют 10—20 ресничек, выступающих из
Таблица 17.5. Классификация первичных запахов (по Эймуру)
Первичный запах Известные вещества с таким запахом (количество) Примеры веществ с первичным запахом
Камфарный 106 Камфара, эвкалипт
Едкий 95 Уксус, муравьиная кислота
Эфирный 53 Эфир, груши
Цветочный 71 Розы
Мятный 77 Мята, ментол
Мускусный 69 Железы ондатры, кабарги
Гнилостный 49 Тухлые яйца
752 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Медиальное
Рис. 17.17. Обонятельная сенсорная система.
Пахучие молекулы присоединяются к ресничкам рецепторных клеток обонятельного эпителия
(первичные сенсорные нейроны). Сенсорные нейроны второго порядка расположены в обоня-
тельных луковицах, образующих диффузные проекции к нескольким структурам лимбической
системы. Передача информации от лимбической системы к орбитофронтальной коре (ОФК)
осуществляется с помощью медиального дорсального ядра таламуса.
эпителия в слой обонятельной слизи, они увеличивают рецепторную по-
верхность плазматической мембраны и содержат специфические хеморе-
цептивные белки, а также G-белки, активирующие вторичные посредники.
Пахучие вещества вначале абсорбируются на обонятельной слизи, выде-
ляемой боуменовыми железами, и доставляются к рецепторам неспецифи-
ческими белками-переносчиками. Нарушения транспорта пахучих веществ
к рецепторам возникают вследствие набухания слизистой оболочки и из-
менения секреции обонятельной слизи при воспалительных или аллерги-
ческих поражениях носовой полости.
17.7.2.1. Механизм возбуждения обонятельных клеток
Присоединение пахучих молекул к хеморецептивным белкам запускает
каскад биохимических реакций, приводящих к возникновению потенциа-
лов действия рецепторных клеток. После присоединения пахучих молекул
к рецепторам происходит активация G-белков, а затем аденилатциклазы,
что ведет к повышению внутриклеточной концентрации циклического аде-
нозинмонофосфата, который активирует протеинкиназу А. Ее фосфорили-
рующее действие на белки мембранных каналов для катионов приводит к
открытию каналов, току катионов в клетку и деполяризации мембраны,
являющейся рецепторным потенциалом первичного сенсорного нейрона.
При деполяризации мембраны до критического уровня в сенсорном ней-
роне возникают потенциалы действия, служащие для передачи информа-
ции на следующий иерархический уровень обонятельной системы.
17. Сенсорные системы • 753
Еще одна разновидность G-белка в мембране хеморецепторных клеток
связана с другим вторичным посредником — фосфолипазой С. При ее ак-
тивации образуется инозитол-3-фосфат, способствующий выходу из внут-
риклеточного депо ионов кальция, и диацилглицерат, активирующий про-
теинкиназу С. Протеинкиназа С фосфорилирует белки катионных кана-
лов, определяя тем самым деполяризацию мембраны, а высокая концен-
трация ионов кальция повышает выделение химического медиатора сен-
сорным нейроном первого порядка.
Все первичные обонятельные нейроны имеют широкий спектр чувстви-
тельности и реагируют на многие пахучие вещества, в то же время разные
рецепторные клетки отличаются индивидуальной чувствительностью к раз-
личным веществам (обонятельный профиль сенсорного нейрона). Каждый
запах возбуждает одновременно большое количество рецепторов, индиви-
дуальные особенности которых в совокупности создают неповторимую
комбинацию, характерную именно для этого запаха. На этом основана
способность человека к различению сложных запахов, принадлежащих, на-
пример, двум разновидностям духов.
17.7.3. Центральный отдел обонятельной системы
Безмякотные аксоны биполярных рецепторных клеток объединяются в
15—20 пучков, каждый из которых содержит несколько сотен или тысяч
волокон. Такие пучки входят в полость черепа через отверстия решетчатой
кости и объединяются в обонятельные нервы (I пара черепных нервов).
Окончания первичных сенсорных нейронов образуют синапсы с дендрита-
ми митральных клеток обонятельных луковиц. Обонятельные луковицы
расположены в передней черепной ямке над решетчатой костью, их на-
ружный слой образован сплетениями волокон обонятельных нервов. Чуть
глубже, в клубочковом слое, информация передается митральным клеткам,
причем каждая митральная клетка (сенсорный нейрон второго порядка)
получает в одном клубочке сигналы приблизительно тысячи аксонов ре-
цепторных клеток. Митральным клеткам свойственна ритмическая актив-
ность, изменения которой обусловлены вдыханием пахучих веществ.
Локальные интернейроны обонятельных луковиц (перигломерулярные и
зернистые клетки) осуществляют латеральное торможение, способствую-
щее контрастированию передаваемых сигналов. На локальных интерней-
ронах оканчиваются эфферентные волокна противоположной обонятель-
ной луковицы, лимбических структур и ретикулярной формации ствола:
эфферентное влияние позволяет контролировать характер выходных сиг-
налов обонятельной луковицы. Аксоны митральных клеток вместе с при-
легающими к ним аксонами пучковых клеток, которые получают аффе-
рентную информацию от нескольких клубочков одновременно, образуют
латеральный обонятельный тракт к центрам высшего порядка.
Латеральный обонятельный тракт разделяется на несколько частей,
оканчивающихся в лимбических структурах переднего мозга: переднем
обонятельном ядре, перегородке, пириформной и парагиппокампальной
извилинах. Нейроны этих структур возбуждаются при поступлении аффе-
рентной информации от обонятельных рецепторов и передают ее гиппо-
кампу, миндалинам, орбитофронтальной коре и ретикулярной формации
среднего мозга. Большинство областей проекции обонятельного тракта не
принимают непосредственного участия в восприятии запахов, их физиоло-
гическая роль состоит в образовании ассоциативных связей обонятельной
системы с другими сенсорными системами при формировании пищевого,
754 • ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
полового и оборонительного поведения. Активация лимбической системы
создает эмоциональный компонент обонятельного восприятия, который оп-
ределяет субъективное отношение к запахам.
17.7.4. Физиологическая роль обоняния у человека
Абсолютные пороги обонятельной чувствительности у человека определяют-
ся с помощью ольфактометров, позволяющих втягивать носом воздух из
емкостей с дозированной концентрацией пахучего вещества. Дифференци-
альные пороги, характеризующие способность оценивать изменения интен-
сивности запаха, у человека довольно высоки: концентрацию вдыхаемых
веществ необходимо повысить на 30 %, а для некоторых веществ на 60 %,
чтобы почувствовать усиление запаха. При продолжительном действии па-
хучих молекул на обонятельные рецепторы происходит их адаптация, в ре-
зультате которой человек перестает воспринимать запах не только дейст-
вующего вещества, но и всех других веществ со сходным запахом (гетеро-
генная адаптация). Нарушения обоняния обусловлены чаще всего затруд-
ненным доступом пахучих веществ к обонятельному эпителию, а так же
поражениями самого эпителия или проводящих путей. Утрата обонятель-
ной чувствительности определяется термином аносмия.
17.7.4.1. Физиологические реакции на запахи
Обонятельные ощущения могут сопровождаться возникновением положи-
тельных или отрицательных эмоций, возникающих в результате активации
лимбической системы. На этом свойстве запахов основано применение
благовоний, ароматических эссенций, нюхательных солей, духов, а также
различных поглотителей неприятных запахов. Ароматерапия учитывает
свойства отдельных запахов (например, лимона, жасмина, лаванды, розма-
рина и т. п.) и вырабатывает рекомендации их использования для повыше-
ния чувствительности сенсорных систем и работоспособности. Неприят-
ные запахи (например, сероводорода, сернистого аммония, продуктов
гниения) действуют неблагоприятно на ряд физиологических функций:
они могут вызывать тошноту, раздражительность, изменения артериально-
го давления и ритма сокращений сердца.
17.7.4.2. Способность к восприятию феромонов у человека
Феромонами называют химические вещества, которые вырабатываются
' специальными железами, выделяются во внешнюю среду и влияют на пове-
дение других представителей вида. Дистантная хеморецепция играет суще-
ственную роль в формировании полового и агрессивного поведения многих
млекопитающих, но, как считали долгое время, не у человека. В конце XX
века было установлено, что вомероназальный орган, или сошник, описанный
ранее только у новорожденных, сохраняется и во взрослом состоянии чело-
века. Он расположен у основания носовой перегородки и содержит рецеп-
торные клетки с ворсинками, способными присоединять половые феромо-
ны, которые выделяются в окружающую среду. Центральные отростки ре-
цепторных клеток сошника образуют вомероназальный нерв, переносящий
сигналы к дополнительным обонятельным луковицам. Присутствие феро-
монов во вдыхаемом воздухе большинством людей не осознается, сущест-
вуют большие различия индивидуальной чувствительности к ним, но мно-
гие люди способны определить пол другого человека по его запаху.
IV ИНТЕГРАТИВНЫЕ
ФУНКЦИИ ОРГАНИЗМА
756 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Организм человека является открытой термодинамической системой,
которая постоянно обменивается с окружающей средой веществом и энер-
гией в соответствии с характером выполняемой работы и изменениями па-
раметров среды. Центральная нервная система отвечает на изменения
внешней и внутренней среды координированными врожденными приспо-
собительными реакциями, в осуществлении которых участвуют все физио-
логические системы организма. Одновременно с этим физические и соци-
альные факторы окружающего мира определяют поведение, направляя его
на поиск контактов с одними внешними объектами и избегание контактов
с другими. По ходу такой деятельности происходит интеграция физиологи-
ческих процессов, сопряженная с их адаптацией — приспособлением к из-
менениям среды.
Поведение человека складывается из последовательности действий, на-
правляемых на удовлетворение возникающих у него потребностей в пище,
воде для питья, температурном комфорте, безопасности продолжения ро-
да, социального общения, интеллектуальной деятельности. Направленное
на удовлетворение потребности поведение определяется как мотивация, а
субъективная оценка вероятности удовлетворения потребностей порождает
чувственные переживания, или эмоции.
Определение объектов внешнего мира осуществляется с помощью сен-
сорных систем организма, трансформирующих энергию внешних стимулов
в нервные импульсы. Вследствие активации нейрофизиологических про-
цессов происходит восприятие объектов и явлений внешнего мира, заклю-
чающееся в создании внутреннего образа реальных событий. Необходи-
мым условием для восприятия является наличие сознания, которым харак-
теризуется активное состояние человека во время бодрствования. Сознание
отключается во время сна, образующим вместе с бодрствованием повто-
ряющийся суточный цикл.
Познавательная деятельность человека направлена на установление свя-
зей между воспринимаемыми объектами и явлениями. Процесс познания
сопровождается приобретением индивидуального поведенческого опыта, в
ходе которого активируются физиологические механизмы памяти, заклю-
чающиеся в регистрации приобретенного опыта, его хранении и воспроиз-
ведении. На основе памяти возникают новые формы поведения, которые
представляют как относительно простой результат ассоциативного науче-
ния (условные рефлексы), так и более сложные его формы, связанные с
биографическими сведениями и познанием окружающего мира. На ис-
пользовании следов памяти основан и процесс мышления, в ходе которого
создаются символически обобщенные представления объектов и явлений
внешнего мира и формируется стратегия поведения. Результат мышления
может быть передан с помощью речи, являющейся средством общения ме-
жду людьми и вовлекающей их в социальную среду. Специфической осо-
бенностью существования человека является его трудовая деятельность,
при осуществлении которой интегрируются механизмы гомеостатического
регулирования и поведения.
ГЛАВА 18
Высшая нервная деятельность
(по И. П. Павлову)
Академик Иван Петрович Павлов (1849—1936) в 1904 г. был удостоен. Но-
белевской премии по медицине за фундаментальные исследования по фи-
зиологии пищеварения. Примерно в то же время он начал изучать в опы-
тах на собаках условные рефлексы, или индивидуально приобретаемые ре-
акции организма на различные внешние раздражения. Это научное на-
правление явилось результатом его наблюдения за слюноотделительным
рефлексом у собак, который нередко возникал у животных на предшест-
вующие еде раздражители. Научные исследования в этой области физиоло-
гии сделали И. П. Павлова основоположником научного направления, ко-
торое было названо им высшей нервной деятельностью. Высшая нервная
деятельность — это приспособление организма к меняющимся условиям
внешней среды на основе безусловных (врожденных) и условных, или при-
обретенных в течение жизни, рефлексов. Высшую нервную деятельность
И. П. Павлов объяснял с позиций созданной им условно-рефлекторной
теории. Согласно классической рефлекторной теории нервная деятель-
ность животных и человека проявляется в виде врожденных рефлексов,
т. е. безусловных. Однако И. П. Павлов считал, что полноценная жизнь
организма требует формирования у него приспособительных реакций к
внешней среде. Согласно условно-рефлекторной теории формирование
приспособительных поведенческих реакций организма состоит не только в
том, что на него действует бесчисленное количество сигнальных раздражи-
телей внешней среды, но и в том, что при определенных условиях эти раз-
дражители, будучи индифферентными, меняют свое физиологическое зна-
чение и становятся условными. Поэтому высшая нервная деятельность
представляет собой взаимодействие врожденных и индивидуально приоб-
ретенных форм реагирования на внешние воздействия с целью приспособ-
ления организма к окружающей среде.
В основе высшей нервной деятельности лежит научение. Научением на-
зывается относительно постоянное изменение поведения, которое является
результатом индивидуального опыта. Научение имеет место только в том
случае, если оно приводит к изменению поведения, а результаты поведе-
ния могут быть как положительными, так и отрицательными. Наиболее
простыми формами научения являются привыкание и сенситизация. При-
выканием, или габитуацией, называется снижение ответной реакции нерв-
ной системы при повторном действии одного и того же раздражителя. Фе-
номен угасания впервые был открыт И. П. Павловым, и его физиологиче-
ский смысл будет описан ниже. Сенситизация проявляется как состояние
повышенной реактивности организма на действие стимула, возникающее
при повторном его предъявлении. Более сложной формой научения явля-
ется ассоциативное, которое представлено классическими условными реф-
лексами по И. П. Павлову и инструментальными; или оперантными, ус-
758 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ловными рефлексами (о них речь пойдет ниже). С помощью классических
условных рефлексах животное научается соотносить, или ассоциировать,
действие на него двух стимулов внешней среды.
18.1. Классические условные рефлексы
Условным рефлексом называется рефлекс, вырабатываемый в организме на
основе временной нервной связи в отделах центральной нервной системы.
Классический условный рефлекс представляет собой научение животного
ассоциировать (связывать) стимул с подкреплением. При изучении услов-
но-рефлекторной деятельности у животных в лаборатории И. П. Павлова
«сосредоточились»* на пищевом, в частности секреторном, и оборонитель-
ном рефлексах. Секреторный рефлекс оказался более удобным в исследо-
ваниях, учитывая технические возможности физиологической лаборатории
того времени, поскольку имелась возможность определить его интенсив-
ность по количеству капель слюны у собаки. Для этого животных подвер-
гали предварительной операции — устье протока околоушной слюнной же-
лезы, вместе с кусочком слизистой оболочки рта, выводили через разрез в
стенке рта и подшивали к коже щеки. В результате можно было опреде-
лять количество вытекающей слюны, например через устье специальной
воронки, прикрепленной к коже в месте выхода протока слюнной железы
(рис. 18.1). Если такому животному дается пища, то уже через 1—2 с у него
происходит выделение слюны. Пища является безусловным стимулом, а вы-
зываемая ею слюноотделительная реакция называется безусловно-рефлек-
торным ответом, или безусловным рефлексом.
Безусловный рефлекс является закономерной реакцией организма на
внешнее раздражение, которая осуществляется при помощи определенного
отдела центральной нервной системы. В экспериментах И. П. Павлова со-
бака до того, как получала пищу, всегда слышала звук (условный стимул).
В результате, после нескольких сочетанных действий условного и безус-
ловного стимулов, у собаки выделение слюны начиналось только на
предъявление одного условного стимула (рис. 18.2), т. е. образовался ус-
ловный рефлекс. В отличие от безусловных рефлексов, т. е. врожденных,
условные рефлексы указанного типа образуются в процессе индивидуаль-
ной жизни животных.
Образование классического условного рефлекса происходит при сочета-
нии двух стимулов — условного и безусловного, из которых безусловный
вызывает безусловно-рефлекторный ответ. Стандартной процедурой соче-
танного действия условного и безусловного стимулов при выработке ус-
ловного рефлекса является так называемая задержка начала действия ус-
ловного стимула относительно начала действия безусловного. Сочетанное
действие условного (например, звук) и безусловного (например, пища)
стимулов формирует условный слюноотделительный рефлекс:
УС + БС => УС -> Условнр-рефлекторный ответ.
При неоднократном сочетанном предъявлении животному двух стиму-
лов «звук—пища» в ответ на действие только условного стимула, т. е. зву-
ка, появляется безусловно-рефлекторная реакция слюноотделения. Пища
при ассоциативном научении выполняет роль подкрепления. В результате у
подопытного животного выделяется слюна, когда оно только слышит зво-
*В кавычках курсивом приведены слова и фразы из книги И. П. Павлова «Двадцати-
летний опыт исследования высшей нервной деятельности».
нок. Выделение слюны в данном
случае является условно-рефлек-
торным ответом. Этот «удиви-
тельный результат», по мнению
И. П. Павлова, был связан с
формированием в ЦНС связи ме-
жду центрами, воспринимающи-
ми действие на животное двух
стимулов, которую ученый назвал
временной.
В наиболее общем виде счита-
ется, что временная связь может
формироваться в ЦНС следую-
щим образом. Звук, как услов-
ный стимул, вызывает так назы-
ваемую репрезентацию безуслов-
ного стимула в соответствующем
мозговом центре, т. е. на одну и
ту же структуру мозга конверги-
руют импульсы, вызванные дей-
ствием условного и безусловного
стимулов. При этом в нервном
Рис. 18.1. Схематическое изображение по-
следствия операции выведения протока
околоушной слюнной железы у собаки на
кожу щеки. Подсчет количества капель
слюны позволяет контролировать проце-
дуру выработки классического условного
пищевого рефлекса.
Рис. 18.2. Схематическое изображение экспериментальной установки для выработ-
ки классических условных рефлексов по И. П. Павлову. Раздражение электриче-
ским током участков кожи, рецепторы которых посылают импульсы в сенсомотор-
ную кору, позволило выявить распространение процессов возбуждения и торможе-
ния в коре головного мозга животного.
1 — воронка для сбора и отведения слюны; 2 — система для измерения количества выделяю-
щейся слюны. На коже собаки закреплены раздражающие электроды: со знаком плюс — поло-
жительный условный стимул, знак минус — отрицательный (тормозный) условный стимул.
760 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
центре на действие условного стимула возникает такое же возбуждение,
как и на действие безусловного стимула, и это приводит к появлению ус-
ловно-рефлекторной реакции слюноотделения. В этом случае животное
«знает», что звонок предполагает получение пищи. Поэтому модель клас-
сического условного рефлекса — сочетанное действие двух раздражителей,
всегда приводит к научению путем образования временнбй связи. С помо-
щью условных рефлексов животные различных видов научаются прогнози-
ровать по стимулу либо опасность для организма, либо получение пищи,
либо иные события, обусловливающие жизнедеятельность организма.
18.1.1. Условия, влияющие на ассоциативное научение
Для безусловных рефлексов характерен строго определенный латентный пе-
риод ответной реакции, независимо от внешних условий, при которых дей-
ствуют на организм животного раздражители. Напротив, в условных реф-
лексах как латентный период ответной реакции, так и особенности ее про-
явления (например, количество капель слюны у собаки в опытах И. П. Пав-
лова) полностью зависят от «большого числа условий». Важнейшим
параметром при научении по типу классических условных рефлексов явля-
ется интервал времени между началом действия условного и безусловного
стимулов. Временная связь между двумя стимулами легко возникает в том
случае, если условный стимул предъявляется непосредственно (минимум за
100 мс) перед началом действия безусловного. Однако временная связь об-
разуется медленно, если условный и безусловный стимулы предъявляются
одновременно и, тем более, если предъявление безусловного стимула начи-
нается через минуту после условного. Функцией ассоциативного научения
в этом случае является предвосхищение будущих событий, поэтому, как
указывал И. П. Павлов, условный раздражитель должен несколько предше-
ствовать действию безусловного. Например, свет (условный раздражитель)
включается до начала подачи пищи (безусловный раздражитель). В резуль-
тате условный раздражитель становится заменителем безусловного и спосо-
бен вызывать не свойственную ему биологическую реакцию в организме.
Другим параметром выработки условного рефлекса является сила безус-
ловного стимула, или подкрепление. Временная связь формируется быстрее,
если сила подкрепления больше, и наоборот. Данная особенность в форми-
ровании временнбй связи позволяет животному предвосхищать более зна-
чимые события среди многих других. Причем важным является не физиче-
ская сила стимула, а его биологическая значимость. Например, условный
слюноотделительный рефлекс легче образуется у голодного животного.
Третьим параметром, оказывающим влияние на ассоциативное науче-
ние, является сила условного стимула. Эта зависимость проявляется в том,
что при предъявлении сильного условного стимула формируется более
прочная временная связь. Среди всего многообразия раздражителей внеш-
ней среды разные виды животных в эволюции приспособились к воспри-
ятию определенных по специфичности стимулов. Следовательно, в ассо-
циативном научении имеет значение предрасположенность животных од-
ного вида реагировать на те или иные стимулы внешней среды. Поэтому у
разных видов животных не все раздражители внешней среды могут участ-
вовать в научении в качестве условных стимулов. Кроме этого, с фактором
силы условного стимула связана его новизна для животного. Если на орга-
низм животного впервые действует какой-либо условный стимул, то его
организм реагирует на новый раздражитель сильнее (явление сенситиза-
ция), чем ранее встречавшийся.
18. Высшая нервная деятельность (по И. П. Павлову) • 761
18.1.2. Рефлекторная дуга классического условного рефлекса
И. П. Павлов полагал, что рефлекторная дуга условного рефлекса состоит
из афферентного, центрального и эфферентного звеньев, т. е. имеет все
составляющие классической рефлекторной дуги. И. П. Павлов считал, что
образование временной связи полностью происходит в коре, а именно ме-
жду корковым полем восприятия условного раздражителя и корковым
представительством безусловного рефлекса (рис. 18.3). Доказывалось это
тем, что после удаления у животного коры головного мозга безусловные
рефлексы (слюноотделительные, двигательные и др.) становились более
грубыми и инертными, менее точными и совершенными, хуже приспособ-
ленными к качеству, силе и продолжительности раздражителей. В своих
трудах И. П. Павлов писал, что в основе физиологического механизма об-
разования временной связи лежит суммационный рефлекс, явление прото-
рения пути между центрами, воспринимающими действие условного и без-
условного раздражителей или облегчения проведения возбуждения между
нейронами, образующими временную связь, а также доминанта. Эти пред-
положения И. П. Павлова отчасти остаются гипотетическими.
Классические условные рефлексы является филогенетически наиболее
ранним образцом простого ассоциативного научения, который представля-
ет собой пример недекларативной (имплицитной) памяти (см. глава 16).
Имплицитная память формируется в пределах различных по иерархической
сложности подкорковых нейронных сетей мозга. В современной нейробио-
логии показано, что в двух наиболее широко изучаемых формах павловско-
го ассоциативного научения, к которым относится условный рефлекс стра-
ха и защитный мигательный условный рефлекс, временная связь образуется
соответственно в миндалевидном комплексе и в структурах мозжечка.
И. П. Павлов считал, что так называемый суммационный рефлекс (сум-
мационная реакция) является простейшей формой индивидуально-приспо-
собительной деятельности организма, в основе которой лежит феномен
повышения возбуждения в нейронах коры головного мозга при неодно-
кратном совпадении во времени условного и безусловного раздражения.
Современные исследова-
ния свидетельствуют о
том, что механизм дли-
тельной циркуляции воз-
буждения в нейронных
сетях мозга (явление дол-
говременной потенциа-
ции) при участии глута-
матергических рецепторов
нейронов миндалевидно-
го комплекса и гиппокам-
па является основным в
формировании времен-
ной связи классического
условного рефлекса стра-
ха. Многократные сочета-
ния «условный—безуслов-
ный стимулы» повышают
эффективность синапти-
ческой передачи между
нейронами за счет увели-
Рис. 18.3. Схематическое изображение представле-
ний И. П. Павлова о рефлекторной дуге классиче-
ского условного слюноотделительного рефлекса.
Ур — корковый центр, воспринимающий действие услов-
ного раздражителя. Бр — центр коры, воспринимающий
действие безусловного раздражителя. 1 — подкорковый
центр ориентировочного рефлекса; 2 — центр пищевого
рефлекса.
762 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
чения пресинаптического высвобождения медиаторов. Повышение эффек-
тивности синаптической передачи при выработке условного рефлекса ле-
жит в основе феномена проторения или облегчения проведения возбужде-
ния, о котором говорил И. П. Павлов.
18.1.3. Стадии формирования условного рефлекса
Выработка классического условного рефлекса происходит в условиях акти-
вации различных центров подкорковых образований, как показано в по-
следующих исследованиях, и состоит из стадий генерализации и специали-
зации. В стадии генерализации все близкие по физическим параметрам ус-
ловные раздражители вызывают одинаковую условную реакцию.
И. П. Павлов полагал, что так называемая генерализация стимула является
следствием распространения возбуждения по коре больших полушарий го-
ловного мозга. Однако современные исследования свидетельствуют о том,
что условный раздражитель заключает в себе несколько параметров. На-
пример, звук имеет определенную частоту, интенсивность и длительность.
Каждый из этих параметров может быть условным раздражителем. Поэто-
му многие компоненты условного раздражителя, на который был вырабо-
тан условный рефлекс, способны вызывать условно-рефлекторный ответ.
Например, реакция привыкания к определенному стимулу может сме-
ниться генерализацией при появлении нового раздражителя.
После выработки стойкого положительного условного рефлекса услов-
но-рефлекторный ответ становится дифференцированным, т. е. возникает
только на строго определенный параметр условного раздражителя. В ре-
зультате близкие к условному стимулу по физическим параметрам раздра-
жители, действующие в пределах одной модальности, становятся неэффек-
тивными и не вызывают условного рефлекса. Это происходит на стадии
специализации условного рефлекса, в основе которой лежит явление диф-
ференцировки, которое будет описано далее.
18.1.4. Условные рефлексы высшего порядка
Классический условный рефлекс, выработанный на основе сочетанного
действия условного и безусловного стимулов, И. П. Павлов назвал услов-
ным рефлексом первого порядка. На основе условного рефлекса первого
порядка может образовываться условный рефлекс второго порядка. Проце-
дура выработки условного рефлекса второго порядка состоит в следующем.
Вначале вырабатывается классический условный рефлекс первого порядка,
как это описано в разделе 18.1 настоящей главы. Затем новый условный
стимул (например, свет) сочетается с условным стимулом рефлекса перво-
го порядка (звонок). На этой стадии звонок выполняет функцию подкреп-
ления. В результате неоднократного сочетанного действия «свет—звонок»,
свет, как новый условный стимул, начинает вызывать условно-рефлектор-
ную реакцию (условный рефлекс второго порядка). Во время выработки
условного рефлекса второго порядка временная связь образуется между
новым условным стимулом и реакцией, которую вызывает условный сти-
мул рефлекса первого порядка.
По аналогии со сказанным, условный рефлекс, выработанный на базе
условного рефлекса второго порядка, называется условным рефлексом
третьего порядка. Условные рефлексы этого типа И. П. Павлов назвал ус-
ловными рефлексами высшего порядка. В лаборатории И. П. Павлова было
установлено, что условные рефлексы высшего порядка не могут иметь
18. Высшая нервная деятельность (по И. П. Павлову) • 763
множественный порядок. Так, например, на основе условного пищевого
рефлекса можно выработать условный рефлекс только второго прядка. На
базе условных, оборонительных рефлексов можно выработать условный реф-
лекс максимум третьего порядка.
18.1.5. Виды классических условных рефлексов
Разнообразие условий внешней среды, к которым происходит приспособ-
ление организмов, предполагает формирование множества соответствую-
щих условных рефлексов. Тем не менее все многообразие условных реф-
лексов можно классифицировать на несколько групп.
В зависимости от вида сенсорного (безусловного) раздражения различа-
ют: экстероцептивные, интероцептивные и проприоцептивные условные
рефлексы. По эффекторному признаку выделяют вегетативные и сомато-
двигательные условные рефлексы. По соотношению во времени действия
условного и безусловного стимулов различают совпадающие и следовые
условные рефлексы (рис. 18.4). Совпадающие условные рефлексы форми-
руются при совпадении во времени действия условного и безусловного
стимулов. Следовые условные рефлексы формируются в ситуации, когда
условный и безусловный стимулы следуют во времени один за другим с
некоторым интервалом. В свою очередь совпадающие условные рефлексы
по отставленности начала действия условного стимула и подкрепления от-
носят к строго совпадающим (задержка порядка 500 мс), отставленным
(5—30 с) и запаздывающим условным рефлексам (30 с—3 мин). Условные
рефлексы могут вырабатываться на время. Такие условные рефлексы обра-
зуются при повторении безусловного стимула через строго определенные
интервалы времени. При этом только по истечении интервала времени
проявляется условно-рефлекторная реакция.
18.2. Торможение условных
рефлексов
Деятельность коры больших полу-
шарий обусловлена не только про-
цессами возбуждения, но и тормо- а
жения. В лаборатории И. П. Павло-
ва было установлено два типа тор-
можения условных рефлексов:
1) внешнее, врожденное или безус-
ловное; 2) внутреннее, приобретен-
ное индивидуально или условное.
Рис. 18.4. Схематическое изображение Б
последовательности предъявления сти-
мулов при выработке различных видов
классического условного рефлекса по
И. П. Павлову.
УС — условный стимул, БС — безусловный
стимул. Стрелками обозначена длительность
предъявления условного и безусловного сти-
мулов. А — совпадающий условный рефлекс, в
Б и В —следовые условные рефлексы. УС—
БС — интервал времени между началом дей-
ствия условного и безусловного стимулов.
Время УС—БС •
УС
БС
БС
УС
УС
Время
УС- БС
БС
764 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
18.2.1. Внешнее торможение
Внешним безусловным торможением называется снижение или исчезнове-
ние условно-рефлекторной реакции при действии на организм сильного
или даже чрезвычайно сильного безусловного раздражения. Внешнее тор-
можение возникает без предшествующей выработки. Снижение или даже
прекращение активности нервного центра условного рефлекса происходит
под влиянием другого нервного центра, воспринимающего действие силь-
ного раздражителя. В естественном поведении животных имеют место час-
тые проявления внешнего торможения, возникающее в ответ на действие
сильного внешнего раздражения. Однако при действии на организм жи-
вотного сильных или даже чрезвычайно сильных раздражителей у них мо-
жет проявиться либо агрессивная реакция, либо пассивно-оборонительная.
Только во втором случае речь идет о внешнем торможении. Внешнее тор-
можение условных рефлексов носит, как правило, временный характер. Ес-
ли животное встречается с действием сильного внешнего раздражителя по-
вторно, которое не сопровождается «-для животного никаким дальнейшим
существенным результатом», то они становятся индифферентными для не-
го, а торможение условного рефлекса прекращается.
По мнению И. П. Павлова, любой новый раздражитель вызывает у жи-
вотного исследовательский рефлекс, и животное ориентирует на источник
раздражения свои органы чувств (зрение, слух). В результате появления
ориентировочного рефлекса тормозится условный рефлекс. Безусловный
ориентировочный рефлекс, который И. П. Павлов обозначал термином
рефлекс «что такое?», проявляется в повороте головы, глаз и ушных рако-
вин животного в сторону источника зрительного или звукового раздра-
жения.
18.2.2. Внутреннее торможение
Внутренним или условным торможением называется изменение проявле-
ния условно-рефлекторной реакции вплоть до ее исчезновения при изме-
нении условий, вызывавших конкретный вид ассоциативного научения
(условного рефлекса). Внутреннее торможение возникает в том случае, ес-
ли предъявление условного раздражителя не подкрепляется безусловным
рефлексом. Основными видами внутреннего торможения, открытыми и
исследованными в лаборатории И. П. Павлова, являются: угасательное, ус-
ловный тормоз, запаздывающее и дифференцировочное.
18.2.2.1. Угасательное торможение
Угасанием называется стирание или подавление приобретенной временнбй
связи между условным стимулом и условно-рефлекторной реакцией при
его повторном предъявлении без сочетания с безусловным подкреплением.
Способность мозга утрачивать научение и временную связь лежит в ос-
нове адаптации организма к меняющимся условиям внешней среды. Уга-
сание прежде выработанной временнбй связи происходит в результате по-
вторяющегося предъявления условного стимула без подкреплений. На-
чальной поведенческой реакцией животного на отмену подкрепления яв-
ляется активация ориентировочной реакции, которая постепенно снижает
условно-рефлекторную реакцию на не подкрепляемый условный стимул.
По мере развития угасательного торможения при действии на животное
условного стимула без подкрепления постепенно увеличивается латентный
18. Высшая нервная деятельность (по И. П. Павлову) • 765
период условного рефлекса и уменьшается условно-рефлекторная реакция
вплоть до ее полного исчезновения. Скорость развития угасательного тор-
можения возрастает при однообразии и постоянстве действия условного
раздражителя, а также при возникновении внешнего торможения. Напро-
тив, угасательное торможение слюноотделительного условного рефлекса
при натуральных условных рефлексах происходит медленно, особенно ес-
ли пища находится вблизи животного. Развитие процесса угасательного
торможения в ЦНС зависит от особенностей типологических особенно-
стей центральной нервной системы животных — «у одних животных услов-
ные рефлексы угасают быстро, у других очень медленно». Таким образом, ес-
ли действие условного стимула связывается с отсутствием подкрепления,
то условный рефлекс тормозится и животное «научается» тому, что за ус-
ловным стимулом не последует его подкрепление.
У животных может иметь место феномен спонтанного восстановления
условного рефлекса после его угасания. И. П. Павлов называл это явление
растормаживанием условного рефлекса, которое свидетельствует о том, что
в основе ассоциативного научения лежат механизмы памяти. Растормажи-
вание проявляется в том случае, когда животное вновь начинает научаться
прежде выработанной временной связи. В этом случае восстановление ус-
ловного рефлекса происходит очень быстро, в течение буквально несколь-
ких сочетаний условного и безусловного стимулов. Показателем глубины
угасания условного рефлекса является период времени, в течение которого
может произойти полное восстановление прежнего эффекта условного
рефлекса.
Сходной с угасанием формой научения является привыкание к неодно-
кратному действию условного стимула. Например, у животных довольно
быстро исчезает ориентировочная реакция на несъедобную пищу. Птицы
довольно скоро привыкают к звуковым средствам их отпугивания в аэро-
портах и т. п.
18.2.2.2. Запаздывающее торможение
Запаздывающим торможением условного рефлекса называется задержка
проявления условно-рефлекторной реакции на положительный условный
стимул в результате увеличения интервала времени между началом предъ-
явления условного стимула и безусловного подкрепления. Запаздывающее
торможение при наличных условных рефлексах проявляется в том, что за-
держка условно-рефлекторной реакции у животного может формироваться
по мере увеличения интервала времени между началом действия условного
раздражители и началом сочетанного безусловного подкрепления. При
следовых условных рефлексах запаздывающее торможение формируется по
мере увеличения интервала времени между окончанием действия условно-
го раздражители и началом сочетанного безусловного подкрепления (см.
рис. 18.4, Б, В). У собак запаздывающее торможение в указанных типах
условных рефлексов возникает в интервале времени задержки не более
3 мин.
Минимальный интервал времени задержки между началом предъявле-
ния условного и безусловного стимулов при наличных и следовых услов-
ных рефлексов составляет порядка 100 мс. Если после выработки запазды-
вающего или следового условного рефлексов постепенно увеличивать ин-
тервал времени между началом действия условного и безусловного стиму-
лов (на 5 с каждый последующий день), то, например, условно-рефлектор-
ная реакция организма начинает запаздывать, т. е. «отодвигаться от нача-
766 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ла условного раздражителя». Физиологические значение запаздывающего
торможения условных рефлексов заключается в приспособлении условно-
рефлекторных реакций организма животных к меняющимся условиям со-
четанного действия условных и безусловных раздражителей внешней
среды.
18.2.2.3. Дифференцировочное торможение
Дифференцировочным торможением называется исчезновение условно-
рефлекторной реакции на физические параметры положительного услов-
ного стимула, которые предъявляются животному без сочетанного дейст-
вия с безусловным подкреплением.
При выработке нового условного рефлекса условный раздражитель,
действующий на организм животного, «носит обобщенный характер». На-
пример, если звук с частотой 1000 Гц в процессе выработки условного
рефлекса становится положительным условным раздражителем, то и дру-
гие частоты, т. е. близкие по значению к 1000 Гц, также первоначально
оказывают условное действие. Однако это действие будет тем меньше, чем
больше звук отличается по частоте в ту или иную сторону от условного
раздражителя. «Специализация условного раздражителя», или его дифферен-
цирование, происходит в условиях многократного сочетания или повторе-
ния подкрепляемого условного раздражителя с близким к нему по физиче-
ским свойствам условным неподкрепляемым раздражителем. В результате,
например, частота звука 1000 Гц сохраняет свое положительное сигнальное
значение, а звуки, отличающиеся по частоте в ту или иную сторону от ус-
ловного раздражителя, без подкрепления утрачивают свое первоначальное
сигнальное значение для животного. Таким образом, при помощи диффе-
ренцировочного торможения условных рефлексов в ЦНС осуществляется
различение биологической значимости различных параметров одного и то-
го же внешнего раздражителя.
18.2.2.4. Условное торможение
Условным торможением называется исчезновение условно-рефлекторной
реакции на положительный условный раздражитель, если он предъявляет-
ся животному в комплексе с новым условным стимулом без сочетанного
их действия с безусловным подкреплением.
После выработки условного рефлекса индифферентный для животного
раздражитель становится условным по отношению к выработанной услов-
но-рефлекторной реакции. Новый условный стимул может предъявляться
животному в комплексе с положительным условным стимулом, но без со-
четанного их действия с безусловным подкреплением. В этом случае но-
вый условный раздражитель приобретает тормозные свойства, а его соче-
танное предъявление животному с положительным условным стимулом не
вызовет условно-рефлекторного ответа. Этот феномен И. П. Павлов на-
звал условным тормозом, значение которого заключается в том, что в ассо-
циативном научении животного один и тот же раздражитель может быть
как условным, так и безусловным. Это способствует более адекватному
приспособлению высшей нервной деятельности животного к условиям
внешней среды.
18. Высшая нервная деятельность (по И. П. Павлову) • 767
18.3. Оперантный условный рефлекс
В то время как И. П. Павловым
было начато исследование класси-
ческих условных рефлексов, в зару-
бежных лабораториях параллельно
изучались так называемые инстру-
ментальные условные рефлексы,
или оперантные рефлексы (рис.
18.5). Оперантные условные реф-
лексы представляют собой науче-
ние методом «проб и ошибок». На-
пример, голодное животное поме-
щается в специальный ящик, выход
из которого оно совершенно слу-
чайно, тем не менее, может обнару-
жить. Как правило, животное со-
вершает множество движений по
всему внутреннему пространству
Рис. 18.5. Скиннеровский ящик для
изучения оперантных условных рефлек-
сов. Случайное нажатие на ключ мага-
зина с пищей позволяет голубю полу-
чить подкрепление за выполненное
действие. Подкрепление является сти-
мулом для следующего «произвольного»
действия.
ящика, пытаясь выбраться из него,
прежде чем случайно не нажимает
на запирающий дверь механизм.
При следующих пробах с тем же
ящиком животное концентрирует
свое поведение все ближе к запи-
рающему механизму, пока не нау-
чается вести себя весьма целена-
правленно, выбираясь из ящика с
минимальными затратами усилий.
Главное отличие между оперантными и классическими павловскими ус-
ловными рефлексами заключается в активном, т. е. «произвольном» ответе
животного. Поведение животных на основе оперантных условных рефлек-
сов может быть усилено через положительное и отрицательное подкрепле-
ние, а также наказанием и угасанием (табл. 18.1).
На основе инструментальных условных рефлексов могут формироваться
новые двигательные реакции. Знания об оперантном научении имеют
большое значение в различных аспектах деятельности человека, в частно-
сти, для содержания и воспитания домашних животных и приручения ди-
Таблица 18.1. Особенности оператного поведения
Характер подкрепления Последствие Результат Пример
Положительное Положительное нака- зание Отрицательное Отсутствие Реакция приводит к же- лаемым последствиям Ответ приводит к избега- нию последствия Ответ блокируется или предупреждается нежела- тельное последствие Ответ блокируется или предупреждаются жела- тельные последствия Усиливается в поведении Уменьшает- ся в поведе- нии Увеличивает- ся в поведе- нии Уменьшает- ся в поведе- нии Собака сидит, полу- чает еду Собаку наказывают, она убегает Собака демонстриру- ет подчинение и не получает наказание Если собака ошиб- лась, то не получает награду
768 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ких животных. Особенности оперантного научения проявляются в разной
степени у человека. Например, различные группы учащихся реагируют по-
разному на тип поощрения, т. е. обратной связи на свое поведение. В ча-
стности, информирование детей средних классов об успехе их ответа на
уроке повышает успеваемость, но оказывается малоэффективным для дру-
гих групп детей.
18.4. Аналитико-синтетическая деятельность коры больших
полушарий
Способность к анализу поступающей в мозг информации о явлениях или
событиях окружающей среды является важным свойством различных уров-
ней центральной нервной системы и, особенно, коры головного мозга.
При этом огромное количество информации в процессе ее обработки в
мозге разделяется на составляющие ее части. Процесс разложения целого
на составляющие его части по нескольким составляющим называется ана-
лизом.
Вместе с тем для высшей нервной деятельности значимыми являются
не отдельные и конкретные раздражители, а их комплексы или комбина-
ция. Поэтому в центральной нервной системе происходят процессы синте-
за, т. е. объединения отдельных частей некогда целого процесса по сущест-
венным признакам в единое целое. Поэтому способность центральной
нервной системы и, прежде всего, коры головного мозга объединять сиг-
нальное значение различных раздражителей в единое целое и на этой ос-
нове формировать приспособительные реакции называется синтезом. Про-
цессы анализа и синтеза в центральной нервной системе осуществляются
взаимосвязанно, и на этом основании И. П. Павлов эту деятельность мозга
назвал аналитико-синтетической.
Анализ в нервной системе начинается на этапе возбуждения рецепто-
ров, с помощью которых осуществляется восприятие различных стимулов
окружающей среды. В зависимости от способности рецепторов реагировать
на простые и сложные стимулы их роль в аналитико-синтетической дея-
тельности мозга относится к категории элементарного анализа и синтеза.
Более сложные процессы анализа и синтеза осуществляются на различных
уровнях (ядрах) сенсорных систем или анализаторов по И. П. Павлову. На
каждом уровне переключения при передаче сигналов от рецепторов до ко-
ры головного мозга происходит обработка передаваемой информации.
Высший уровень анализа и синтеза осуществляется в коре головного моз-
га, структурная организация которой приспособлена к анализу простых и
сложных комплексов раздражений и формированию на этой основе вре-
менных связей.
18.5. Динамический стереотип
Временная связь условного рефлекса является результатом синтетической
деятельности центральной нервной системы. Эта деятельность проявляется
в формировании динамического стереотипа. Динамический стереотип —
это зафиксированная в долговременной памяти мозга система индивиду-
ально выработанных условных рефлексов при определенной последова-
тельности воздействия условных раздражителей внешней среды на орга-
низм.
В лаборатории И. П. Павлова было установлено, что стереотипное по
последовательности и временнбму интервалу предъявление разных услов-
18. Высшая нервная деятельность (по И. П. Павлову) • 769
ных раздражителей фиксируется в деятельности коры в виде системы ус-
ловно-рефлекторных реакций или «динамического стереотипа». После
формирования динамического стереотипа, предъявление животному толь-
ко одного из условных стимулов в той же стереотипной последовательно-
сти вызывает условно-рефлекторные реакции, свойственные предъявле-
нию других раздражителей.
В мозговой деятельности животных и человека формируется система за-
фиксированных условных рефлексов под влиянием внешних и внутренних
стереотипно повторяющихся воздействий на организм различных последо-
вательностей условных стимулов. Многократные повторения определенной
системы действующих на центральную нервную систему условных стиму-
лов будет фиксироваться в долговременной памяти мозга в виде системы
условных рефлексов, которые затем воспроизводятся в строгой последова-
тельности. Таким образом, внешний стереотип предъявления раздражите-
лей обусловливает стереотипный комплекс приобретенных на них услов-
но-рефлекторных реакций организма.
Формирование динамического стереотипа происходит в течение опреде-
ленного интервала времени, но при сформированном динамическом сте-
реотипе центральная нервная система у животных и человека работает
экономно на фоне автоматически повторяющихся цепей условных рефлек-
сов. Например, образование динамического стереотипа лежит в основе вы-
работки профессионального навыка. Динамический стереотип формирует-
ся в течение определенного периода жизни и не может мгновенно изме-
ниться в случае изменения условий внешней среды.
18.6. Фазовые явления в коре больших полушарий головного мозга
И. П. Павлов считал, что высшая нервная деятельность обусловлена дина-
микой внутренних процессов возбуждения и торможения в популяциях
корковых нейронов, которая возникает в результате воздействия на орга-
низм стимулов внешней и внутренней среды. Процессы возбуждения и
торможения могут возникать в отделах коры, которые не связаны непо-
средственно с анализом и синтезом условного или безусловного раздраже-
ния, т. е. они способны иррадиировать из участков коры, где они возника-
ют, на другие нейронные популяции коры. Например, на иррадиации воз-
буждения в коре головного мозга основана выработка временнбй связи и
стадия генерализации в формировании условного рефлекса. По мнению
И. П. Павлова, «деятельность больших полушарий управляется законами:
законом иррадиирования и концентрирования процессов возбуждения и
торможения, и законом их взаимной индукции».
Явление иррадиации процесса торможения в коре головного мозга было
обнаружено в лаборатории И. П. Павлова при выработке дифференциро-
вочного торможения условного слюноотделительного рефлекса на так на-
зываемое электрокожное раздражение. Условное электрическое раздраже-
ние на участки кожи наносили через близко расположенные между собой
электроды (см. рис. 18.2). В соответствии с соматотопической организаци-
ей сенсомоторной коры, сигналы от рецепторов ограниченного участка ко-
жи поступают в рядом расположенные центры сенсомоторной коры жи-
вотного. Предъявление животному условного электрического раздражения
через один электрод без сочетания с безусловным подкреплением вызыва-
ет торможение в соответствующем центре коры. Последующее электриче-
ское раздражение через рядом расположенный на коже электрод в сочета-
нии с безусловным пищевым подкреплением сопровождалось незначитель-
25-6095
770 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ной по сравнению с фоном условной слюноотделительной реакцией, кото-
рая до применения «тормозного» электрокожного раздражения имела по-
стоянную величину слюноотделения.
Процесс концентрации торможения, как было установлено И. П. Пав-
ловым, взаимосвязан с иррадиацией торможения на другие отделы коры
головного мозга. Иррадиация нервных процессов обусловлена их силой.
В лаборатории И. П. Павлова было показано, что иррадиация процессов
возбуждения и торможения в пределах ограниченных зон коры больших
полушарий происходит, как правило, при их слабой выраженности. На-
против, при достаточной силе как процесса возбуждения, так и торможе-
ния, доминирует явление их концентрации в центрах коры, воспринимаю-
щих действие положительного и отрицательного условного раздражений.
При чрезмерной силе процессов возбуждения и торможения доминируют
явления их иррадиации с распространением на обширные отделы коры го-
ловного мозга. Следовательно, нервным процессам возбуждения и тормо-
жения в коре головного мозга характерна определенная динамика, которая
обусловлена многими факторами.
Взаимная индукция нервных процессов возбуждения и торможения. Дина-
мика процессов возбуждения и торможения в коре головного мозга, про-
являющаяся в возникновении возбуждения вслед за торможением и наобо-
рот или в их усилении в одном и том же участке коры головного мозга,
была названа И. П. Павловым индукцией. Взаимная индукция процессов
возбуждения и торможения в коре головного мозга связана со сменой од-
ного процесса другим в ограниченном участке коры головного мозга. Вза-
имная индукция процессов обусловлена выраженностью (силой) возбужде-
ния и торможения, развитием их во времени, с большей или меньшей сте-
пенью их распространения в пределах нервных центров коры, восприни-
мающих действие условных и безусловных стимулов, наконец, с типологи-
ческими особенностями центральной нервной системы (описаны ниже).
По И. П. Павлову, взаимная индукция процессов возбуждения и торможе-
ния бывает двух видов: последовательная, или временная, и одновремен-
ная, или пространственная, индукция. Последовательная индукция характе-
ризуется сменой возбуждения торможением или наоборот либо их усиле-
нием в пределах одного нервного центра коры головного мозга. Простран-
ственная индукция нервных процессов проявляется в том, что в участках
коры головного мозга, расположенных рядом с корковыми зонами возбуж-
дения или торможения, развивается противоположный нервный процесс.
С учетом влияния одного нервного процесса на другой явление индукции
классифицируют по знаку. Резкое усиление условного рефлекса при воз-
действии на центр коры головного мозга положительным условным стиму-
лом, находившийся под влиянием процесса торможения (например, диф-
ференцировки), называется положительной индукцией. Напротив, появле-
ние и усиление процесса торможения в нейронах коры головного мозга,
окружающих корковый процесс возбуждения, называется отрицательной
индукцией.
Положительная индукция в участке коры головного мозга, как правило,
имеет место после дифференцировочного торможения условного рефлекса.
Примером проявления отрицательной индукции является внешнее тормо-
жение условных рефлексов. Отрицательная индукция ограничивает рас-
пространение (иррадиацию) возбуждения по коре больших полушарий го-
ловного мозга. В лаборатории И. П. Павлова было установлено, что ин-
дукция нервных процессов, по аналогии с иррадиацией, взаимосвязана с
силой возбуждения и торможения в участках коры головного мозга. При
18. Высшая нервная деятельность (по И. П. Павлову) • 771
слабых и чрезмерно сильных процессах возбуждения и торможения явле-
ние индукции в коре головного мозга не проявляется. При нормальной ак-
тивации коры головного мозга, а следовательно, достаточной выраженно-
сти процессов возбуждения и торможения явление индукции не только
возникает, но и может возрастать в своем проявлении. На этом основании
И. П. Павлов пришел к выводу о наличии в коре головного мозга тесного
взаимодействия иррадиации и взаимной индукции нервных процессов воз-
буждения и торможения. Экспериментальные модели изучения условно-
рефлекторной деятельности животных позволили проследить определен-
ную динамику подобного взаимодействия. В наиболее общем виде это
проявлялось в том, что в коре головного мозга первоначально формирует-
ся зона либо возбуждения, либо торможения в зависимости от сигнальных
особенностей раздражителя, и начинает развиваться положительная или
отрицательная индукция. Дальнейшим этапом в динамике возбуждения и
торможения является иррадиация первично вызванного нервного процесса
в другие отделы коры головного мозга с последующим обратным его дви-
жением в начальный пункт. При этом в нейронах коры, в которых исчеза-
ют процессы возбуждения или торможения, индуцированные их же ирра-
диацией, также могут происходить явления взаимной индукции.
И. П. Павлов считал, что «закон иррадиации и концентрации и закон индук-
ции тесно связаны друг с другом, взаимно ограничивая, уравновешивая, и укре-
пляя друг друга и, таким образом обусловливая точное соотношение деятель-
ности организма к условиям внешней среды».
18.7. Типология высшей нервной деятельности
И. П. Павлов, исследуя высшую нервную деятельность животных, устано-
вил, что «нервные задачи», которым подвергали животных, сопровождались
проявлением различий в их высшей нервной деятельности. Эти различия у
одних животных в деятельности коры больших полушарий были выражены
больше, у других меньше, а у некоторых — вообще не наблюдали их воз-
никновения. Особенности индивидуального проявления у животных выс-
шей нервной деятельности И. П. Павлов связывал с типами их нервной
системы. Так по описанию И. П. Павлова, один тип животных — это «в
высшей степени суетливые», «быстро реагирующие на малейшие звуки», «на-
вязчивые в общении с людьми». Такие животные энергичны при быстрой
смене раздражений, но при их однообразии животные становятся бездея-
тельными и вялыми. Другой тип собак — медленно осваивающиеся с об-
становкой опытов, малоподвижные, но как только обстановка опытов ста-
новится однообразной, у них легко вырабатываются условные рефлексы,
особенно тормозные.
Индивидуальные особенности высшей нервной деятельности животных
проявляются в различной скорости образования и закрепления условных
рефлексов, в различной скорости выработки внутреннего торможения. Ти-
пологические особенности высшей нервной деятельности проявляются во
взаимодействии нервных процессов возбуждения и торможения в коре го-
ловного мозга. В основе классификации типов высшей нервной деятельно-
сти по И. П. Павлову лежат врожденные индивидуальные различия основ-
ных свойств нервных процессов возбуждения и торможения. Такими ос-
новными свойствами нервных процессов возбуждения и торможения в ко-
ре головного мозга являются их сила и слабость. Сила нервных процес-
сов—это свойство нервной системы отвечать на сильные раздражители
сильной реакцией, а также способность длительное время поддерживать
25*
Ill • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
процесс возбуждения. По И. П. Павлову, в зависимости от врожденных
особенностей нейронов коры головного мозга нервные процессы могут
быть сильными или слабыми; в зависимости от соотношения силы возбуди-
тельного и тормозного процессов они могут быть уравновешенными или не-
уравновешенными. Наконец, у нервных процессов есть скорость их возник-
новения и прекращения, легкость перехода от одного процесса к другому.
По данному критерию И. П. Павлов разделял нервные процессы на по-
движные и инертные.
И. П. Павлов на основании исследований из достаточно большого числа
сочетания свойств процессов возбуждения и торможения выделил только
четыре типа высшей нервной деятельности у животных: 1) сильный, неурав-
новешенный, с резким преобладанием процесса возбуждения над торможе-
нием, «безудержный» тип; 2) сильный, уравновешенный, подвижный (ла-
бильный) тип; 3) сильный, уравновешенный (инертный) тип; 4) слабый тип.
И. П. Павлов проводил параллель между четырьмя типами высшей нервной
деятельности и их проявлением в поведении, т. е. темпераментом по Гип-
пократу. При этом холерический, сангвинический, меланхолический и
флегматический темпераменты обусловлены наличием у животного соот-
ветствующего (порядок указан выше) типа высшей нервной деятельности.
Особенности высшей нервной деятельности человека. У человека, по срав-
нению с животными, уровень развития аналитико-синтетических процес-
сов в коре головного мозга обусловил специфику его высшей нервной дея-
тельности. По словам И. П. Павлова, эволюционной «прибавкой» к меха-
низмам корковой деятельности у человека стало слово, что «составило вто-
рую сигнальную систему действительности». В целом же особенностью выс-
шей нервной деятельности человека по И. П. Павлову является наличие
двух сигнальных систем. Первая сигнальная система является системой
мозгового обеспечения восприятия непосредственных раздражителей
внешней среды. Вторую сигнальную систему, которая является специфиче-
ски человеческой, составляет слово. На физиологическом уровне вторую
сигнальную систему можно определить как систему мозгового обеспечения
вербальной (словесной) деятельности. Вторая сигнальная система является
основой абстрактного мышления, благодаря которому человек имеет воз-
можность структурировать среду без непосредственного контакта с ней пу-
тем операций с мысленными образами и символами объектов. В результате
у человека аналитико-синтетическая деятельность коры больших полуша-
рий осуществляется как путем непосредственных ощущений и представле-
ний, так и путем оперирования словом. Слово, «будучи сигналом первых
сигналов», создает неограниченные возможности для отвлечения процессов
анализа и синтеза от непосредственного восприятия внешнего мира.
И. П. Павлов предполагал, что по соотношению первой и второй сиг-
нальных систем у людей можно выделить художественный, мыслительный и
промежуточный типы людей. У первых особенно ярко выражена деятель-
ность первой сигнальной системы. Для мыслительного типа заметно пре-
обладает деятельность второй сигнальной системы и, наконец, средний
тип, у которого деятельность первой и второй сигнальных систем проявля-
ются примерно в равной степени. Однако эта точка зрения И. П. Павлова
не получила в последующем научного подтверждения. И. П. Павлов вы-
сказывал также мнение, что типологические особенности высшей нервной
деятельности человека, обусловленные соотношением деятельности двух
сигнальных систем, формируются в процессе индивидуальной жизни под
влиянием многих факторов внешней среды, в том числе воспитания и об-
разования.
ГЛАВА 19
Мотивации и эмоции
19.1. Мотивации
Слово мотивация происходит от латинского глагола movere, что значит
двигаться, а смысл самого понятия мотивация литературным языком мож-
но выразить словами «нечто, что движет другим». Мотивации обеспечива-
ют поведение человека при удовлетворении биологических потребностей.
К биологическим потребностям человека относятся, в основном, потреб-
ность в пище, воде, сне, сексе и т. д. Биологической потребностью называ-
ется изменение гомеостатических параметров внутренней среды организ-
ма — снижение концентрации глюкозы в плазме крови, повышение в ней
осмотического давления, увеличение содержания в плазме крови половых
гормонов либо норадреналина, серотонина или ацетилхолина. В условиях
голода (снижение в плазме крови питательных веществ) или жажды в ЦНС
возникает возбуждение (стимул, драйв — от английского слова управление),
которое инициирует, направляет, интенсифицирует и обусловливает по-
стоянство целенаправленного поведения человека на удовлетворение био-
логических потребностей и которое называется мотивацией. Целенаправ-
ленное поведение человека в процессе удовлетворения биологической по-
требности также представляет собой мотивацию.
19.1.1. Понятие о первичных и вторичных мотивациях
Мотивации, которые возникают в организме человека на основе таких фи-
зиологических состояний, как изменение гомеостатических параметров
внутренней среды, называются первичными. Наиболее важными видами
первичных мотиваций, обусловливающих целенаправленное удовлетворе-
ние биологических потребностей в течение суток, являются пищевая и
цикл сон—бодрствование. Первичные мотивации обеспечивают выжива-
ние индивидуума, например в результате восполнения в организме пита-
тельных веществ.
Мотивации, которые могут возникать только в процессе научения чело-
века, через его индивидуальный жизненный опыт, называются вторичны-
ми. Вторичные мотивации (синоним — мотив) обусловлены познаватель-
ными и социальными потребностями человека. Примерами вторичных мо-
тиваций человека являются мотивация достижения цели и изменения по-
ведения. Так, мотивация достижения цели является «повседневной».
В процессе удовлетворения этой мотивации человек стремится к совер-
шенствованию профессиональной компетенции, либо к овладению навы-
ками новой профессии, либо к получению положительной оценки за ре-
зультаты своей профессиональной деятельности со стороны окружающих.
Мотивация изменения поведения может быть направлена на получение мате-
риальной выгоды либо на расширение количества знаний и профессио-
нальных навыков (так называемый потенциал мастерства).
774 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Вторичные мотивации человека представлены также внутренней и внеш-
ней мотивациями. Вторичная внутренняя мотивация возникает у человека
из потребности сделать что-либо для себя. Внутренняя мотивация у чело-
века связана с высокими успехами в учебе, психологической совместимо-
стью с одноклассниками, сверстниками и студентами. У взрослых внутрен-
няя мотивация является источником активной, продуктивной работы, иг-
ры, творчества. Вторичная внешняя мотивация человека направлена на по-
лучение вознаграждения либо обусловлена стремлением избежать нака-
зания.
19.1.2. Понятие о мотивациях влечения и избегания
На основе аффективной (чувственной) природы сенсорных ощущений
(приятные или неприятные), которые возникают у человека при первич-
ных и вторичных мотивациях, они могут быть разделены на две категории:
влечения и избегания. Мотивации влечения обусловливают поведение чело-
века, направленное на получение пищи, взаимодействие с половым партне-
ром и достижение успехов в работе (достижение победы), и вызывающих у
человека чувство удовольствия. Мотивации влечения обусловлены получе-
нием вознаграждения (положительные подкрепления, положительные эмо-
ции) и их проявление связывают с функцией систем удовольствия мозга
человека. Мотивации избегания обусловливают поведение человека, при
котором он стремится избежать неприятного состояния, как правило, это
чувство боли или страха (отрицательное подкрепление, наказание, отрица-
тельные эмоции).
19.1.3. Пищевая мотивация человека
Питание является обязательным условием выживания индивидуума. Под-
держание высокой скорости метаболических процессов в организме воз-
можно при условии постоянного восполнения энергетических запасов.
Чтобы обеспечить адекватное питание, в мозге человека имеются нейрон-
ные сети, которые регулируют содержание питательных веществ (жиров,
углеводов и белков) в плазме крови и в так называемых депо организма.
Пищевая мотивация проявляется под влиянием гомеостатических стиму-
лов, которые возникают у человека при дискомфорте, ассоциирующемся с
чувством голода. Наряду с гомеостатическими стимулами пищевая мотива-
ция у человека обусловлена подкрепляющей ролью пищи (вознаграждени-
ем), поскольку питание всегда сопряжено с возникновением у человека
чувства удовольствия.
19.1.3.1. Гомеостатические механизмы регуляции пищевой мотивации
у человека
Основным фактором формирования пищевой мотивации у человека явля-
ется химический состав плазмы крови: концентрация глюкозы, жирных
кислот и аминокислот. Уровень глюкозы в плазме крови контролируется
глюкорецепторами нейронов латеральных отделов гипоталамуса (глюкоста-
тическая теория регуляции голода). Согласно этой теории, ощущение го-
лода у человека возникает при понижении концентрации глюкозы в плаз-
ме крови, и наоборот. При увеличении содержания глюкозы в плазме кро-
ви увеличивается активность глюкорецепторных нейронов вентромедиаль-
ного гипоталамуса и одновременно снижается чувствительных к глюкозе
19. Мотивации и эмоции • 775
нейронов так называемого центра голода латерального гипоталамуса. При
потреблении пищи увеличивается высвобождение инсулина в плазму крови
из р-клеток островков Лангерганса поджелудочной железы. Инсулин, сти-
мулируя транспорт глюкозы из плазмы крови через мембраны клеток тка-
ней, вызывает уменьшение концентрации глюкозы в плазме крови и уве-
личивает проявление у человека чувства голода. Уменьшение концентра-
ции аминокислот и липидов в плазме крови увеличивает электрическую
активность нейронов латерального' гипоталамуса (аминостатическая или
липостатическая теории регуляции пищевой мотивации), что обусловлива-
ет активацию пищевого поведения.
Эти теории были основаны на представлении о мозговых механизмах
регуляции питания при участии двух противоположных по функции групп
клеток — латерального гипоталамуса, регулирующего питание, и вентроме-
диального ядра, тормозящего латеральный гипоталамус в случае пищевого
насыщения. Однако гомеостатические механизмы регуляции пищевой мо-
тивации в связи с открытием роли гормонов лептина, грелина и ряда пеп-
тидов (меланокортины, нейропептид Y, орексины и др.) в регуляции ини-
циации и прекращения пищевого поведения включают, наряду с гумораль-
ными, гормональные и центральные гипоталамические механизмы пище-
вой мотивации у человека.
Роль лептина в регуляции пищевой мотивации. Гормон лептин синтезиру-
ется жировыми клетками в организме человека и поступает в плазму кро-
ви. Его концентрация в плазме крови зависит от массы жировой ткани у
человека. Циркулирующий в крови лептин связывается с типом 1 цитоки-
нового рецептора (синоним — лептиновые рецепторы) на мембране нейро-
нов вентрального отдела гипоталамуса (дуговое, вентромедиальное, дорсо-
медиальное и вентральное премаммилярное ядра). Нейроны медиальной
части дугового ядра гипоталамуса, которые способны связываться с лепти-
ном, содержат нейропептид Y, про-опиомеланокортин и его производные,
включая а-меланоцитостимулирующий гормон. При этом нейроны, синте-
зирующие нейропептид Y, образуют гипоталамическую систему потребле-
ния пищи, а нейроны, содержащие про-опиомеланокортин/производные
кокаина и амфетамина, — систему пищевого насыщения. При активации
системы потребления пищи заторможена активность системы пищевого
насыщения и наоборот.
Физиологический механизм действия лептина заключается в торможе-
нии высвобождения нейропептида Y в нейронах паравентрикулярных ядер
гипоталамуса. В результате уменьшения секреции нейропептида Y в ней-
ронах системы потребления пищи, активируется нейроны системы насы-
щения, синтезирующие про-опиомеланокортини, что вызывает у человека
снижение ощущения голода.
Считается, что гипоталамус регулирует постоянство массы тела челове-
ка, или так называемую установочную точку массы тела, которая обуслов-
лена размерами и количеством жировых клеток в организме человека, и,
следовательно, содержанием гормона лептина в плазме крови. При этом
чем больше количество жировых клеток в организме, тем выше установоч-
ная точка массы тела человека и наоборот. Установочная точка массы тела
обусловлена генетическими факторами, поскольку каждый человек имеет
индивидуальную скорость метаболизма, при которой пища превращается в
энергию и используется в тканях организма. Например, люди, имеющие
высокую скорость метаболизма, чаще потребляют пищу без существенного
изменения массы тела. Напротив, люди, имеющие низкую скорость мета-
болизма, потребляют пищу относительно редко, но имеют избыточный вес.
776 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Роль грелина в регуляции пищевой мотивации. Гормон грелин синтезирует-
ся в клетках слизистой оболочки желудка и его концентрация в плазме
крови человека становится максимальной при пищевой мотивации непо-
средственно перед приемом пищи. Увеличение его содержания в плазме
крови и одновременное снижение в ней уровня лептина являются стиму-
лами инициации у человека чувства голода. Мишенью для грелина явля-
ются грелиновые рецепторы на мембране нейронов вентромедиального и
дугового ядер гипоталамуса, которые синтезируют нейропептид Y. Увели-
чение концентрации грелина в плазме крови вызывает рост синтеза нейро-
пептида Y в нейронах гипоталамуса, что активирует систему регуляции по-
требления пищи у человека. Блокада рецепторов этих нейронов гипотала-
муса антагонистами нейропептида Y непосредственно препятствует дейст-
вию на них грелина и тормозит пищевую мотивацию. У пациентов, поте-
рявших примерно 17 % массы тела в результате интенсивной диеты, имеет
место чрезвычайно высокая концентрация грелина в плазме крови непо-
средственно перед началом еды. У людей с ожирением и чрезмерным ап-
петитом в плазме крови более высокое содержание грелина, чем у субъек-
тов с ожирением и обычным уровнем питания.
Регуляция пищевой мотивации при участии факторов химического со-
става плазмы крови (глюкоза, аминокислоты, жирные кислоты и гормоны)
обеспечивает поддержание постоянства запасов питательных веществ в ор-
ганизме человека между приемами пиши.
Наряду с этим гастроинтестинальные стимулы выполняют еще одну
роль, — они регулируют оптимальное количество пищи, которое может
принять человек за один прием (завтрак, обед или ужин).
Нейрогормональная регуляция пищевого насыщения. Структуры мозга, ре-
гулирующие пищевую мотивацию человека, получают сенсорные сигналы
от рецепторов желудочно-кишечного тракта через дорсальный комплекс
блуждающего нерва продолговатого мозга. Импульсы, которые передаются
в гипоталамус от механорецепторов афферентных волокон блуждающего
нерва, сигнализируют о степени растяжения стенки желудка пищей, а так-
же об уровне глюкозы и липидов в печени. Эта информация поступает
первоначально в медиальные и дорсомедиальные ядра одиночного тракта
продолговатого мозга, а далее, через нейроны парабрахиальных ядер мос-
та, — к паравентрикулярным, дорсомедиальным и дуговым ядрам гипота-
ламуса, а также к латеральному гипоталамусу, обусловливая момент пре-
кращения приема пищи.
Сенсорные сигналы, поступающие от нейронов ядер одиночного тракта
продолговатого мозга через парабрахиальные ядра моста к висцеральным
сенсорным ядрам таламуса, а далее к нейронам висцеральных сенсорных
отделов коры головного мозга, обусловливают формирование у человека
сознательного контроля степени наполнения желудка пищей.
Наконец, сенсорные сигналы, поступающие от нейронов ядер одиноч-
ного тракта продолговатого мозга через парабрахиальные ядра моста к
центральному ядру миндалины, обусловливают возникновение чувства
удовольствия при восприятии вкуса пищи.
Роль гастроинтестинальных гормонов в регуляции насыщения. Прекраще-
ние питания регулируется гормонами гастроинтестинального тракта при
участии нейронов area postrema. Area postrema, или так называемый циркум-
вентрикулярный орган, расположен над ядрами одиночного тракта продол-
говатого мозга. Нейроны area postrema не имеют гематоэнцефалического
барьера, поэтому они непосредственно реагируют на циркулирующие в
крови гастроинтестинальные гормоны холецистокинин и глюкагонсвязан-
19. Мотивации и эмоции • 777
ный пептид-1. Повышение содержания этих гормонов в плазме крови про-
исходит при приеме пищи и поступлении химуса в двенадцатиперстную
кишку, что вызывает их высвобождение из клеток ее слизистой оболочки.
Гормоны активируют нейроны area postrema, от которых сигналы передают-
ся к ядрам одиночного тракта, а затем через нейроны парабрахиальных
ядер моста в гипоталамус, что вызывает прекращение приема пищи.
Эндокринные клетки тонкой кишки человека секретируют в плазму
крови пептид YY. Этот гормон тормозит нейроны дугового ядра гипотала-
муса, секретирующие нейропептид Y. Торможение секреции в нейронах
гипоталамуса нейропептида Y активирует нейроны, синтезирующие про-
опиомеланокортин. В результате у человека наступает насыщение и он
прекращает принимать пищу.
Роль механорецепторов стенки пищеварительного канала в регуляции на-
сыщения. При растяжении пищей стенок желудка и двенадцатиперстной
кишки происходит раздражение чувствительных окончаний блуждающего
нерва. Импульсы достигают нейронов ядер латерального гипоталамуса и,
вызывая торможение их активности, приводят к уменьшению степени пи-
щевой мотивации.
19.1.3.2. Роль структур продолговатого мозга в регуляции пищевой мотивации
Стволовые центры непосредственно регулируют процесс приема пищи че-
ловеком за один присест. При этом осуществляются секреция слюны, вку-
совая апробация пищи, а также двигательные акты жевания и глотания.
Сигналы от вкусовых рецепторов слизистой оболочки языка и глоточ-
ной области поступают в ядра одиночного тракта продолговатого мозга: от
рецепторов передней 2/3 языка по чувствительным волокнам лицевого нер-
ва', вкусовые ощущения, возникающие при раздражении рецепторов со-
сочков корня языка и других задних областей слизистой оболочки рта, —
по волокнам языкоглоточного нерва', от рецепторов слизистой оболочки ос-
нования языка и глоточной области — по волокнам блуждающего нерва. От
нейронов ядра одиночного пучка волокна направляются к медиальному
ядру таламуса, а затем к структурам сильвиевой борозды: постцентральная
извилина теменной коры и островковой коры. Корковые проекции вкусо-
вой рецепции обусловливают так называемое вкусовое предпочтение, ко-
торое заключается в том, что человек отдает предпочтение некоторым ви-
дам пищи среди многих. Во вкусовом предпочтении основную роль вы-
полняют структуры мезолимбической системы мозга, при участии которой у
человека возникают приятные или неприятные ощущения после приема
того или иного типа пищевых веществ. В частности, если человек испыты-
вает неприятные (например, болезненные) ощущения после приема опре-
деленного типа продукта, то у него возникает так называемое отрицатель-
ное пищевое предпочтение, или вкусовая неприязнь.
Нейроны ядра одиночного тракта синаптически связаны с верхними и
нижними слюноотделительными ядрами ствола мозга, которые регулируют
слюноотделение околоушной, подъязычной и подчелюстной слюнных же-
лез. Одной из функций слюны является растворение молекул пищевых ве-
ществ, что способствует их доставке к рецепторам вкуса в слизистой обо-
лочке языка. Нейроны ядер одиночного тракта рефлекторно возбуждают
двигательные центры этого же отдела мозга, которые регулируют процессы
жевания и глотания (двигательные ядра V, VII, X, XI, XII черепных нервов
и CI—СШ). Активация стволовых нейронных механизмов регуляции жева-
ния, слюноотделения, глотания и рецепции вкуса вызывает торможение
778 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
нейронов гипоталамической системы потребления пиши, что также обу-
словливает момент прекращения приема пищи. Однако этот процесс осу-
ществляется медленно (порядка 20—40 мин), и тормозной эффект выра-
жен в меньшей степени, чем при действии гастроинтестинальных стимулов
на вентромедиальный отдел гипоталамуса.
19.1.3.3. Роль латерального гипоталамуса в возникновении пищевой
мотивации
Регуляция пищевой мотивации у человека осуществляется нейронными се-
тями латерального, вентромедиального и паравентрикулярных ядер гипо-
таламуса. Двустороннее повреждение ядер латерального гипоталамуса вы-
зывает отказ от приема пищи у человека, электрическая стимуляция, на-
против,— приводит к чрезмерному потреблению пищи.
Роль ядер латерального гипоталамуса в мотивации голода обусловлена
началом потребления пищи в результате активации нейронов этого отдела
мозга. Считают, что эта роль латерального гипоталамуса связана с нейро-
нами, содержащими пептиды — меланинконцентрирующий гормон и орекси-
ны (синоним — гипокретины). Эти нейроны инициируют начало приема
пищи, активируя двигательные системы ствола мозга, регулирующие жева-
ние, облизывание и глотание при участии нейронов черепных нервов:
тройничного, лицевого и подъязычного ядер.
Нейроны латерального гипоталамуса иннервируют симпатические и па-
расимпатические преганглионарные ядра продолговатого и спинного моз-
га, которые регулируют процессы слюноотделения, пищеводной и желу-
дочной моторики, секреции соляной кислоты желудка и регуляцию экзо-
кринной (панкреатический сок) и эндокринной (инсулин и глюкагон) сек-
реции поджелудочной железы. Нейроны латерального гипоталамуса, сек-
ретирующие меланин-концентрирующий гормон и орексины, через проек-
ции к моноаминергическим нейронам голубого пятна, серотонинергиче-
ским нейронам дорсального и срединного ядер шва, гистаминергическим
нейронам бугорково-маммилярной области, активирует восходящую рети-
кулярную систему мозга. В результате при пищевой мотивации усиливают-
ся возбуждение организма и его двигательная активность.
Наконец, нейроны латерального гипоталамуса, содержащие меланин-
концентрирующий гормон, проецируют терминали своих аксонов к нейро-
нам прилегающего ядра (и. accumbens). Увеличение активности нейронов
прилегающего ядра под влиянием нейронов латерального гипоталамуса
обусловливает инициацию и усиление у человека чувства удовольствия
при еде.
При участии нейротрансмиттеров паравентрикулярные ядра гипотала-
муса регулируют так называемое вкусовое предпочтение человека по отно-
шению к определенному типу питательных веществ. Например, ней-
ротрансмиттеры нейропептид Y, ГАМК и норадреналин увеличивают вку-
совое предпочтение пищи, насыщенной углеводами, а серотонин, напро-
тив, уменьшает.
19.1.3.4. Роль меланокортиновой системы гипоталамуса в прекращении
пищевой мотивации
Электрическая стимуляция вентромедиальных ядер гипоталамуса вызывает
чувство насыщения и отказ от приема пищи, разрушение, напротив, при-
водит к непрерывному и чрезмерному потреблению пищи даже в том слу-
19. Мотивации и эмоции • 779
чае, если масса животного в несколько раз превышает норму. Роль нейро-
нов ядер вентромедиального гипоталамуса заключается в инициации мо-
мента прекращения потребления пищи. Например, при опухолях вентро-
медиального отдела гипоталамуса резко возрастает потребление пищи у че-
ловека. Электрическое раздражение вентромедиального гипоталамуса ведет
либо к прерыванию, либо к полному прекращению потребления пищи.
Прекращение приема пищи осуществляется так называемой меланокорти-
новой системой дугового ядра гипоталамуса, нейроны которого синтезиру-
ют про-опиомеланокортин и производные кокаина и амфетамина. Так, у
человека взаимодействие а-меланоцитостимулирующего гормона с рецепто-
рами меланокортина на мембране нейронов этого ядра вызывает анорексию
(отказ от пищи) и резкое снижение массы тела в результате прекращения
питания. Рецепторы меланокортина имеются также на мембране нейронов
паравентрикулярного и латерального гипоталамуса. Молекулярный меха-
низм действия а-меланоцитостимулирующего гормона на нейроны паравен-
трикулярных ядер осуществляется через ГАМК-ергические интернейроны.
Нейроны меланокортиновой системы дугового ядра гипоталамуса имеют
прямые синаптические связи с преганглионарными симпатическими ней-
ронами в спинном мозге. Изменение активности этого пути под влиянием
лептина или а-меланоцитостимулирующего гормона при взаимодействии
этих гормонов с лептиновыми и меланокортиновыми рецепторами нейро-
нов меланокортиновой системы может непосредственно регулировать (уве-
личивать и уменьшать соответственно) артериальное давление, расход
энергии и секрецию инсулина при пищевой мотивации или при ее тормо-
жении.
19.1.3.5. Роль лимбической системы в регуляции пищевой мотивации
у человека
Если бы пищевая мотивация регулировалась только гомеостатическими
механизмами, то большинство людей имели бы идеальную массу тела. Од-
нако еда, особенно вкусная, является для человека своеобразным возна-
граждением, как деньги за выполненную работу. Эта особенность пищи
человека при пищевой мотивации модулируется такими качествами пищи,
как ее вкус и запах. Различение вкусовых качеств пищи на молекулярном
уровне осуществляется вкусовыми рецепторами языка, большинство из ко-
торых способны воспринимать более одного первичного вкуса, а многие —
три или все четыре одновременно. Импульсы от вкусовых рецепторов по-
ступают к нейронам ядра одиночного тракта, затем к парабрахиальным яд-
рам моста (второй синаптический уровень) и через релейные ядра таламуса
в латеральные области лобной коры, к центральному ядру миндалины и к
латеральному гипоталамусу. Подкрепляющая роль пищи в пищевом пове-
дении человека обусловлена нейронами подкорковых областей мозга,
имеющих на мембране. Так, нейроны прилегающего ядра, содержащие на
своей мембране опиоидные рецепторы и дофаминергические входы к этим
нейронам, непосредственно образуют систему пищевого вознаграждения
пищевой мотивации человека. Активация опиоидных и глютаматергиче-
ских рецепторов нейронов прилегающего ядра и дофаминергических вхо-
дов к его нейронам увеличивают потребление вкусной пищи: сладкой, со-
леной или жирной, а также жидкости. Нейроны прилегающего ядра лим-
бической системы взаимосвязаны синаптически с нейронами латеральных,
вентромедиальных и паравентрикулярных ядер гипоталамуса и через них
регулируют выраженность пищевой мотивации у человека в отношении
780 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
специфического типа пищи. Считается, что активность этой нейронной
сети мозга человека обусловливает аппетит. Чувство аппетита ориентирует
человека на определенный состав пищи, которую он употребляет при еде.
Регуляция аппетита происходит при участии обонятельной сенсорной сис-
темы, к которой относится значительная часть миндалевидного комплекса.
У человека двустороннее повреждение миндалины приводит к утрате вы-
бора типа и качества пищи (потеря аппетита).
Прилегающие ядра при участии ГАМК-ергических нейронов этого об-
разования реципрокно взаимодействуют с нейронами латерального гипота-
ламуса. Роль прилегающего ядра в инициации пищевого поведения заклю-
чается в растормаживании нейронов латерального гипоталамуса. Актив-
ность дофаминергических входов к нейронам прилегающего ядра обуслов-
ливает регуляцию таких процессов, как внимание и/или ожидание получе-
ния пищи при пищевой мотивации.
19.1.4. Половая мотивация человека
Половая мотивация человека имеет многофакторную природу. Эта мотива-
ция не связана с изменением гомеостатических параметров внутренней
среды организма человека, а является в значительной степени результатом
научения человека, его индивидуального жизненного опыта. Основными
факторами, которые инициируют возникновение у человека половой мо-
тивации, являются генетически обусловленный половой диморфизм (от гре-
ческого dimorphos — имеющий две формы) центральной нервной системы,
приобретенный в процессе индивидуальной жизни сексуальный опыт об-
щения с половым партнером и общая культура человека. Половая мотива-
ция не относится к первичным мотивациям, поскольку она не является
необходимой для выживания организма индивидуума. У человека половая
мотивация не обусловлена физиологическим состоянием внутренней сре-
ды организма человека (уровень половых гормонов в плазме крови) и не
имеет прямой зависимости с депривацией (длительное отсутствие полового
партнера). В отличие от животных, у человека нормальное содержание по-
ловых гормонов в плазме крови лишь усиливает степень проявления поло-
вой мотивации, но не инициирует ее возникновение, а длительное отсутст-
вие полового партнера не заключает в себе угрозу жизни субъекта, как это
имеет место при пищевой депривации.
19.1.4.1. Генетические, социальные и психологические факторы возникновения
половой мотивации у человека
У человека половая мотивация возникает под влиянием полового партне-
ра, зрительных стимулов (картина или фильм эротического содержания),
запахов, исходящих от тела человека, или изделий парфюмерии, звуков
(например, ассоциирующихся с половым поведением) илй'“других стиму-
лов (например, сексуальные фантазии), которые способны по типу ассо-
циативного научения вызвать половое возбуждение у людей. Сексуальные
фантазии (например, воображение интимного общения с половым партне-
ром и др.) вызывают у людей обоего пола возникновение полового возбуж-
дения. Так, фактором половой мотивации могут быть мысли о сексуальной
привлекательности потенциального или реального полового партнера, что
характерно как для мужчин, так и для женщин. Половая самостимуляция
{мастурбация) относится к одному из факторов нередко практикуемой по-
ловой активности у людей.
19. Мотивации и эмоции • 781
Половая ориентация человека. К фактору, оказывающему влияние на
возникновение половой мотивации у человека, относится половая (сексу-
альная) ориентация. Половая ориентация человека обусловлена генетиче-
скими, гормональными и социальными факторами. Основную роль в воз-
никновении половой ориентации у человека играет генетический фактор.
В частности, сексуальная ориентация человека взаимосвязана с половым
диморфизмом преоптического поля переднего гипоталамуса. У человека это
поле состоит из четырех групп нейронов и называется интертициальными
ядрами переднего гипоталамуса (intestinial nuclei of the anterior hypothalamus —
INAH). Половой диморфизм связан с различием в нейронной организации
одной из ядерных групп NAH-3. Считается, что гетеросексуальная половая
ориентация мужчин обусловлена большим, примерно наполовину, количе-
ством нейронов TNAH-3, чем это имеет место у гетеросексуальных жен-
щин и гомосексуальных мужчин. Однако генетическая предопределенность
половой ориентации у мужчин и женщин относится к категории факторов,
предрасполагающих к гетеросексуальности, бисексуальности или гомосек-
суальности.
Половая ориентация человека проявляется в его желании вступить в
сексуальные взаимоотношения с половым партнером и отражает направ-
ленность индивидуального полового влечения в отношении конкретного
полового партнера. Большинство индивидуумов имеют гетеросексуальную
ориентацию, которая проявляется в половом влечении к человеку проти-
воположного пола. Гомосексуальная ориентация человека выражается во
влечении индивидуума к партнеру того же пола, а бисексуальная — к парт-
нерам обоего пола.
Гетеросексуальность. Примерно половина женщин до вступления в брак
в период с 15 до 19 лет имеет гетеросексуальный опыт. Такой же опыт
имеет половина мужчин в возрасте до 18 лет, т. е. до вступления в брак, а
в возрасте 20 лет — гетеросексуальный опыт имеют порядка 88 % мужчин.
В период супружеской жизни возникновение половой мотивации у людей
имеет различную частоту. Так, 43 % супружеских пар занимаются сексом
несколько раз в месяц, 36 % — 2—3 раза в неделю. По мере увеличения
стажа супружеской жизни половая активность гетеросексуальных партне-
ров постепенно снижается. Частота возникновения половой мотивации и
занятие сексом женатых мужчин и замужних женщин обусловлено также
внебрачными половыми связями. Тем не менее примерно 85 % замужних
женщин и 75 % женатых мужчин сохраняют верность своим супругам, и
возникновение у них половой мотивации обусловлено одним и тем же по-
ловым партнером.
Получение удовольствия. Удовольствие относится к важному фактору,
инициирующему половую мотивацию у людей. Например, дети в возрасте
от 2 до 5 лет спонтанно дотрагиваются до своих гениталий, хотя в этом
возрасте у детей отсутствует какое-либо половое поведение. Тактильные
раздражения наружных половых органов у человека (при мастурбации или
половым партнером) являются для него источником приятных ощущений.
Самостимуляция органов чувств в области гениталий не связана с репро-
дуктивной функцией человека, а направлена на получение только чувства
удовольствия. Так, взрослые женщины довольно часто достигают состоя-
ния оргазма не в результате движений пениса во влагалище, а за счет сти-
муляции чувствительных окончаний тканей клитора, имеющей место при
половом акте.
Познавательный аспект. Мужчины и женщины выполняют различные
гендерные (половые) роли и поэтому по-разному объясняют одни и те же
782 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
факторы, инициирующие у них половую мотивацию. В частности, в по-
ловой мотивации Левушек доминирует романтический интерес, в то вре-
мя как у юношей половая мотивация обусловлена преимущественно воз-
можностью осуществления секса. Например, просмотр эротического
фильма, предназначенного для мужчин, вызывает у мужчин и женщин
примерно идентичное физиологическое возбуждение. Однако для мужчин
этот просмотр вызывает чувство полового возбуждения и положительные
эмоции, а у женщин, напротив, чувство отвращения и отсутствие полово-
го возбуждения. Если же эротический фильм предназначен для женщин,
то женщинам свойственны те же физиологические реакции, что при про-
смотре «мужской эротики», но они к тому же испытывают чувство поло-
вого возбуждения, интерес к фильму и положительные эмоции. Поэтому
возникновение у человека половой мотивации обусловлено тем, как муж-
чина и женщина понимают эротический «смысл» воспринимаемых раз-
дражителей.
Влечение к другим людям — это еще один элемент половой мотивации,
обусловленный биологической природой человека и его воспитанием. Че-
ловек от рождения испытывает влечение к окружающим. Способность
взрослого человека выбрать среди окружающих его людей полового парт-
нера обусловлена его общей культурой и научением.
Научение половому поведению оказывает существенное влияние на поло-
вую мотивацию человека, поскольку человек запоминает и повторяет то,
что ему нравится и доставляет удовольствие. Для большинства людей по-
ловая мотивация ассоциируется с вознаграждением (чувством удовольст-
вия). С другой стороны, человек стремиться избегать сексуального поведе-
ния, которое приносит ему отрицательные результаты. Поэтому ассоциа-
тивное научение напрямую влияет на половую мотивацию человека, вызы-
вая усиление или уменьшение ее проявлений. Так, горящая свеча может
стать причиной возникновения половой мотивации у человека, если этому
предшествовала сексуальная встреча, например в виде приятного романти-
ческого ужина при свечах. У человека чрезвычайно выражено влияние
научения на половую мотивацию. Научение относится к ключевому компо-
ненту, предопределяющему половое влечение, познавательный интерес к
половому поведению, половую ориентацию и, наконец, уровень половой
мотивации человека.
Общая культура человека предопределяет присущее мужчине и женщине
поведение в обществе, а также, какое поведение подразумевает проявление
полового возбуждения. Культура человека приобретается в процессе науче-
ния, поскольку без соответствующего научения индивидуум не сможет ос-
воить нормы культурного поведения мужчины и женщины. Например, по-
ловое воспитание школьников и социальное равноправие мужчин и жен-
щин в обществе обусловливают примерно идентичное понимание предста-
вителями обоего пола внешних проявлений половой мотивации людей.
Однако нормальное половое поведение человека, воспитанного на одной
культуре, может не оказывать возбуждающего влияния на человека другой
культуры. Что касается полового поведения, то отношение человека к его
проявлениям со стороны полового партнера обусловлено культурным
уровнем людей.
Роль феромонов в половой мотивации у человека. Феромонами (запахи)
называются химические стимулы, которые обусловливают физиологиче-
ские и поведенческие реакции человека (и, в большей степени, животных).
Роль феромонов заключается в повышении степени проявления половой
мотивации человека.
19. Мотивации и эмоции • 783
У человека феромоны продуцируются апокриновыми железами кожи,
их источником также являются микрофлора кожи в отдельных регионах
тела: подмышечные впадины, ротовая полость, ступни ног и гениталии.
Апокриновые железы кожи расположены преимущественно в области ге-
ниталий, вокруг пупка, на грудной клетке, молочных железах, особенно
вокруг сосков. Различные типы желез расположены также в коже головы
вокруг волос. К мужским феромонам, содержащимся в секретах апокрино-
вых желез, относятся андростенон й андростенол. У человека имеет место
так называемый обонятельный половой диморфизм, который проявляется в
том, что женщины воспринимают андростенон при более низком пороге
концентрации в окружающем воздухе, чем мужчины. Однако восприятие
этого феромона женщинами уменьшает для них сексуальную привлека-
тельность мужчины. Напротив, восприятие андростенола женщинами вы-
зывает у них положительные эмоции, в результате чего увеличивается сек-
суальная привлекательность мужчины. Восприятие феромонов обонятель-
ной системой увеличивает выброс гонадолиберина из нейронов гипотала-
муса, что в свою очередь, повышая импульсную секрецию лютропина/
фоллитропина или тестостерона, может усиливать инициацию полового
возбуждения человека.
На пике овуляции у женщин возникает максимум остроты и чувстви-
тельности восприятия мужских феромонов. Это обусловлено тем, что обо-
нятельная система женщин в этот период под влиянием повышенной кон-
центрации в плазме крови эстрогенов и лютропина становится наиболее
чувствительной к феромонам. У женщин феромоны содержатся в ваги-
нальном секрете. Однако запах этого секрета вызывает как у мужчин, так и
у женщин, как правило, неприятные субъективные ощущения и может не-
гативно влиять на половую мотивацию. В целом феромоны обусловливают
индивидуальный запах тела человека, который позитивно либо негативно
может влиять на половое поведение людей.
Зрительные стимулы, возникающие при зрительном восприятии полово-
го партнера, рассматривании эротической картины или просмотре фильма
аналогичного содержания, вызывают наиболее сильные проявления поло-
вой мотивации у человека, особенно у мужчин. У мужчин более выражен-
ная активация структур миндалевидного комплекса, как ключевой структу-
ры системы удовольствия мозга человека, возникает при рассматривании
красивого женского лица, чем при рассматривании изображения мужского
лица. Мужчины чаще, чем женщины, просматривают порнографические
материалы. Просмотр фильмов с сексуальной тематикой активирует у
мужчин области ассоциативной коры головного мозга, которые выполня-
ют основную функцию в возникновении половой мотивации у человека:
нижняя височная область ассоциативной зрительной коры, правая орби-
тофронтальная кора, левая передняя поясная кора.
У женщин половая мотивация формируется преимущественно под
влиянием психологических (психогенных) стимулов: романтические слова,
картины и темы из фильмов или историй, следовательно, при участии зри-
тельных и слуховых ассоциативных областей коры головного мозга.
Основную триггерную роль в увеличении степени полового возбужде-
ния и инициации полового поведения человека (особенно у женщин) иг-
рает стимуляция рецепторов кожи, прежде всего так называемых эроген-
ных зон (рецептивные зоны тканей гениталий) как у мужчин, так и у жен-
щин при непосредственном общении с половым партнером.
784 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
19.1.4.2. Роль половых гормонов в модуляции половой мотивации человека
Количество женских и мужских половых гормонов в плазме крови увели-
чивает или уменьшает у человека степень половой мотивации, формирую-
щейся под влиянием познавательных и социальных факторов (см. ранее).
У женщин половое влечение увеличивается в предменструальный период,
что обусловлено увеличением концентрации эстрогенов и прогестерона в
плазме крови. Изменения в степени субъективного полового возбуждения
полностью отсутствует у женщин, принимающих оральные контрацепти-
вы, поскольку в этих условиях в их плазме крови поддерживается относи-
тельно постоянный уровень женских половых гормонов.
У мужчин степень проявления половой мотивации в большей степени,
чем у женщин, зависит от содержания мужских половых гормонов в плаз-
ме крови. Основную роль в усилении половой мотивации у мужчин вы-
полняет мужской половой гормон тестостерон. Физиологический эффект
тестостерона на половую мотивацию у мужчин зависит от пороговой кон-
центрации гормона в плазме крови, выше которой он не вызывает допол-
нительное увеличение степени половой мотивации. Механизм усиления
половой мотивации у мужчин под влиянием тестостерона в меньшей сте-
пени обусловлен стимулирующим действием гормона на нейроны структур
гипоталамуса, а в большей — стимулирующим эффектом гормона на
нейроны центра эрекции спинного мозга в сегментах SH—S[V (рис. 19.1).
Стероидные гормоны повышают активность нейронов мозга двумя путями:
1. Гормоны увеличивают возбудимость мембраны нейронов и ее чувстви-
тельность к нейротрансмиттерам, а также стимулируют высвобождение
нейротрансмиттеров. Это обусловлено тем, что стероидные гормоны, пря-
мо связываясь с мембранными рецепторами, увеличивают активность ион-
ных каналов и рецепторов к нейротрансмиттерам (например, тормозных
Непрямое действие Прямое действие
Рис. 19.1. Пути увеличения возбудимости и метаболической активности нейронов
гипоталамуса и сакральных сегментов спинного мозга стероидными гормонами.
Стероидные гормоны изменяют синтез медиаторов и их высвобождение из пресинаптических
окончаний, а также могут связываться с рецепторами медиаторов на постсинаптической мем-
бране (прямое действие). В результате прямого действия стероидных гормонов на нейроны
мозга изменяется их возбудимость и функциональная активность, например в гипоталамусе и
сегментах спинного мозга (S,,—S,v), что обусловливает формирование физиологического уров-
ня половой мотивации. Непрямое действие стероидных гормонов на нейроны мозга человека
осуществляется через цитоплазматические или ядерные рецепторы. При этом функции нейро-
нов мозга регулируются стероидными гормонами в течение длительного времени (минуты или
часы). У женщин варьирование концентрации половых гормонов в плазме крови способно из-
менять степень половой мотивации на протяжении менструального цикла, а у мужчин эффект
проявляется в постоянстве половой мотивации, если концентрация тестостерона в плазме
крови достигает порового уровня.
19. Мотивации и эмоции • 785
ГАМКа рецепторов). 2. Стероидные гормоны, проникая внутрь нейронов и
связываясь со специфическими типами стероидных рецепторов цитоплаз-
мы и ядра, могут усиливать или тормозить транскрипцию специфических
генов в ядре нейронов, обусловливая их метаболическую реакцию на про-
тяжении многих минут и часов. В результате кастрации у мужчин и вы-
званного этим снижения содержания тестостерона ниже порогового уров-
ня примерно наполовину уменьшаются длина дендритов и размер поля
нейронов в спинальном центре эрекции, что приводит к нарушению ин-
нервации гладких мышц артерий кавернозных тел пениса и прекращению
эрекции. Снижение концентрации тестостерона в плазме крови у мужчин
в результате кастрации вызывает периодическое уменьшение и даже исчез-
новение половой мотивации, однако сексуальный интерес к женскому по-
лу у таких мужчин сохраняется постоянно.
Усиление половой мотивации у человека под влиянием тестостерона не
связано с полом. Так, у женщин после хирургического удаления яичников
уменьшается степень половой мотивации. Однако введение тестостерона
таким женщинам увеличивает степень полового возбуждения, инициирует
сексуальные фантазии, т. е. усиливает половую мотивацию.
19.1.4.3. Стадии полового возбуждения у человека при половой мотивации
Проявления полового возбуждения у мужчин и женщин в основном иден-
тичны и зависят от приемов половой стимуляции при мастурбации, взаим-
ных ласках или половом акте. Сексуальный ответ мужского и женского ор-
ганизмов при половой мотивации состоит из стадий возбуждения, плато,
оргазма и разрешения.
Стадия возбуждения в мужском и женском организме характеризуется
быстрым увеличением полового возбуждения, при котором возрастают
частота сердцебиений, кровяное давление и параметры дыхания. У мужчин
стадия полового возбуждения инициируется в основном под влиянием
сексуальных зрительных стимулов, которые активируют структуры зри-
тельной ассоциативной зоны нижней височной коры. Мотивационные и
эмоциональные феномены при половом возбуждении (равно как и при
других мотивациях) при зрительном восприятии сексуальных раздражите-
лей активируют правую глазолобную кору. Аффективные (чувственные)
реакции организма на сексуальные стимулы обусловлены активностью ле-
вой передней поясной коры, которая активирует проявление эндокринных
и вегетативных реакций организма при половом возбуждении. Зрительное
и соматосенсорное восприятие проявлений полового возбуждения у муж-
чин (частое сердцебиение, эрекция пениса и др.) обусловлены активацией
нейронов островковой коры правого полушария. У мужчин в стадию воз-
буждения активируются парасимпатические нейроны сегментов Sn—S)v,
сигналы от которых достигают нервных терминалей волокон и при уча-
стии оксида азота вызывают расслабление гладких мышц артерий кавер-
нозных тел и губчатого вещества уретры, обусловливая эрекцию пениса.
Парасимпатические импульсы стимулируют секрецию уретральными и
бульбоуретральными железами слизи, которая способствует движению пе-
ниса во влагалище.
У женщин тактильное раздражение рецепторов тканей клитора и акти-
вация соматосенсорной системы вызывает более быстрое нарастание сте-
пени полового возбуждения, чем тактильное раздражение слизистой обо-
лочки влагалища или рецепторов других областей тела. Половые реакции в
организме женщин в стадию возбуждения проявляются в некотором увели-
786 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
чении молочных желез, набухании сосков и половых губ, увеличении раз-
меров клитора, увлажнении (любрикация) слизистой оболочки влагалища.
Любрикация влагалища обусловливается усилением кровотока в его тканях
и увеличением секреции слизи батролиниевыми железами за счет расслаб-
ления гладких мышц артерий вазоактивным интенстинальным пептидом.
Этот гормон вызывает расслабление гладких мышц артерий кавернозных
тел клитора и его эрекцию.
Стадия плато. При продолжающейся непрерывной половой стимуля-
ции (мастурбация, половой акт) у мужчин и женщин быстрое увеличение
степени полового возбуждения сменяется более медленным ростом часто-
ты сердцебиений, кровяного давления и параметров дыхания (стадия пла-
то). У женщин в фазу плато головка клитора выступает над окружающими
тканями, что усиливает тактильное раздражение его рецепторов мастурба-
цией или движениями пениса. Стенка влагалища из-за нарастающего кро-
венаполнения становится более плотной.
Прекращение половой стимуляции в стадии возбуждения и плато
уменьшает степень полового возбуждения как у женщин, так и у мужчин.
Стадия оргазма. У мужчин и женщин стадия оргазма длится несколько
секунд, во время нее половое возбуждение достигает максимума и затем
резко прекращается. Основной физиологической реакцией у мужчин в фа-
зу оргазма является эякуляция 3—4 мл спермы, содержащей 300—500 млн.
сперматозоидов, из которых только несколько тысяч достигнут фаллопие-
вых труб и лишь один сперматозоид будет участвовать в оплодотворении
яйцеклетки. В женском организме стадия оргазма, которая проявляется в
виде ритмических сокращений гладких мышц влагалища, характеризуется
значительной вариабельностью по интенсивности и длительности. Так, не-
которые женщины способны на протяжении относительно длительного
времени продолжающегося полового акта неоднократно достигать стадии
оргазма. Напротив, мужчины способны достигать следующей стадии оргаз-
ма лишь по прошествии некоторого времени после предыдущего.
При половом возбуждении, особенно в стадию оргазма, у людей в плаз-
ме крови в 3—4 раза повышается концентрация окситоцина, которая со-
храняется и после оргазма. У мужчин гормон стимулирует процесс семяиз-
вержения, а у женщин — сокращение гладких мышц фаллопиевых труб,
что способствует продвижению спермы в направлении яичников. Эффект
окситоцина на половое возбуждение человека зависит от его количества в
плазме крови. При невысокой концентрации окситоцина в плазме крови
гормон увеличивает половое возбуждение, а при высокой концентрации,
напротив, угнетает его. Механизм действия гормона связан с активацией
окситоциновых рецепторов на мембране нейронов ядер вентромедиально-
го гипоталамуса.
Оргазм является кульминацией полового возбуждения человека. В этот
момент человек испытывает ощущение полового наслаждения, которое
обусловлено активацией нейронных кругов систем удовольствия мозга
(описано ниже). Наиболее интенсивное сексуальное чувство возникает во
время семяизвержения (эякуляции) у мужчин и сокращения гладких мышц
матки у женщин. Одновременно у человека увеличивается напряжение
скелетных мышц и возникает их спазм, увеличиваются частота сердцебие-
ний, артериальное давление, уровень в плазме крови норадреналина (все
физиологические реакции обусловлены увеличением активности симпати-
ческой нервной системы). Во время оргазма в плазме крови повышается
содержание пролактина, концентрация которого сохраняется на повышен-
ном уровне в течение 30—60 мин, а также лютропина и тестостерона. Счи-
19. Мотивации и элюции • 787
тают, что механизм стимуляции секреции гормонов, в частности пролакти-
на, обусловлен стимуляцией рецепторов тканей половых органов, импуль-
сы от которых направляются в преоптическое поле гипоталамуса. Увеличе-
ние содержания стероидных гормонов в плазме крови повышает возбуди-
мость мембраны нейронов гипоталамуса и спинальных центров эрекции.
Например, у мужчин в результате повышения возбудимости нейронов ме-
диального преоптического поля гипоталамуса уменьшается латентный пе-
риод времени наступления эякуляцйи у мужчин во время полового акта.
Стадия разрешения. После оргазма у человека снижается половое возбу-
ждение, при этом нормализуются частота сердцебиений и параметры дыха-
ния. У мужчин прекращается эрекция пениса, и в течение некоторого вре-
мени не возникает повторное половое возбуждение на половую стимуля-
цию (рефрактерный период). У женщин в стадию разрешения также норма-
лизуются параметры частоты сердцебиения, дыхания, но отсутствует реф-
рактерный период инициации повторного полового возбуждения.
19.1.4.4. Нервная регуляция половой мотивации у человека
Проявление половой мотивации человека регулируется тремя нейронными
сетями ЦНС, которые интегрируют гормональную, сенсорную и познава-
тельную информацию.
1. Роль спинного мозга и ствола мозга в проявлении полового возбуждения
у человека. Активация парасимпатических и симпатических структур спин-
ного мозга при половом возбуждении регулирует у мужчин вегетативные
половые реакции во время стадий возбуждения и оргазма (эрекция пени-
са, выброс жидкости из бульбоуретральных желез, эякуляция), а у жен-
щин — набухание сосков, эрекция клитора, усиление кровенаполнения
тканей полового тракта и любрикация влагалища при участии бартоли-
ниевых желез.
Активность спинальных нейронных сетей при половом возбуждении ре-
гулируется нейронами центрального серого вещества вокруг сильвиева во-
допровода, которые являются основной непроизвольной двигательной сис-
темой ствола мозга. Нейронами центрального серого вещества вокруг
сильвиева водопровода активируют нейроны позадилицевых ядер продол-
говатого мозга, а далее симпатические преганглионарные нейроны боко-
вых рогов спинного мозга, обусловливая сердечно-сосудистые реакции при
половом возбуждении (увеличение артериального давления и частоты
сердцебиений). Центральное вещество через нейроны позадиобоюдного
ядра продолговатого мозга регулирует активность мотонейронов, иннерви-
рующих мышцы гортани, глотки, мягкого неба и экспираторные мышцы,
обусловливая регуляцию артикуляции при половом возбуждении. Нейроны
центрального серого вещества вокруг сильвиева водопровода через вен-
тральные каудальные отделы моста (норадренергические ядра моста) повы-
шают в целом активность симпатической нервной системы при половом
возбуждении. При участии дофаминергической системы мозга (медиальная
покрышка ствола мозга) и центрального серого вещества при половом воз-
буждении у человека снижается возбудимость нейронов дорсальных рогов
спинного мозга, при участии которых в ЦНС передаются болевые сиг-
налы.
Наконец, нейроны центрального серого вещества вокруг сильвиева во-
допровода среднего мозга проецируют свои аксоны к нейронам ядер рети-
кулярной формации ствола мозга, которые, в свою очередь, синаптически
активируют мотонейроны спинного мозга через ретикулоспинальный
788 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
тракт, обусловливая проявление локомоторных реакций при половом воз-
буждении.
2. Роль миндалины в проявлении полового возбуждения человека. При поло-
вом возбуждении (мастурбация, взаимные ласки, слуховые и обонятельные
стимулы, половой акт) нейроны базолатеральной области миндалины по-
лучают сенсорные сигналы всех модальностей (зрительной, соматосенсор-
ной, слуховой, обонятельной), что непосредственно обусловливает степень
полового возбуждения человека. Пресинаптические окончания на нейро-
нах центрального ядра миндалины являются тормозными и высвобождают
ГАМК. Эфферентным выходом миндалины является ее центральное ядро,
нейроны которого проецируют свои аксоны к нейронам центрального се-
рого вещества вокруг сильвиева водопровода.
Миндалина в регуляции половой мотивации человека обеспечивает
инициацию и степень проявления полового возбуждения через структуры
гипоталамуса и ствола мозга. Медиальное преоптическое — переднее гипо-
таламическое поле регулирует латентный период времени проявления эя-
куляции у мужчин во время полового акта. Так, билатеральное разрушение
этого отдела гипоталамуса у животных увеличивает латентный период эя-
куляции, а электрическая стимуляция, напротив, уменьшает количество
движений (интромиссий) пениса во влагалище, в результате которых воз-
никает семяизвержение.
Нейроны вентромедиального гипоталамического ядра имеют на мем-
бране высокую концентрацию стероидных рецепторов. Это обусловливает
степень полового возбуждения индивидуумов под влиянием повышенного
содержания стероидных гормонов в плазме крови при половом поведении.
3. Роль лимбической системы мозга в регуляции половой мотивации. Выс-
ший уровень регуляции половой мотивации, как и других форм мотиваци-
онного поведения, осуществляется при участии системы удовольствия моз-
га: мезолимбическая система мозга, или лимбическая кора (описана выше).
При этом мотивационные эффекты эмоционально значимых стимулов
осуществляются при участии вентральной части полосатых тел, а именно
нейронных кругов прилегающего ядра (n. accumbens). Структуры приле-
гающего ядра при любом виде мотивации, в том числе и половой, осуще-
ствляют взаимодействие между лимбической и двигательной системами
мозга, что обеспечивает проявление сексуально ориентированного поведе-
ния, ассоциирующегося с чувством удовольствия. Ключевой структурой
прилегающего ядра, обусловливающей появление у человека чувства удо-
вольствия при половом возбуждении (стимуляция рецепторов эрогенных
зон, стадия оргазма полового возбуждения), являются нейроны ракушки,
имеющие на своей мембране опиоидные рецепторы. Опиоид-зависимая
активация нейронов ракушки прилегающего ядра при половом возбужде-
нии происходит под влиянием импульсов, поступающих из отделов корко-
во-стриато-паллидо-таламо-корковой нейронной «петли», гиппокампа и
глазолобной коры. Проекции дофаминергической системы мозга к нейро-
нам ракушки прилегающего ядра играете основную роль в усилении внима-
ния к сексуальной стимуляции различной модальности.
Передняя поясная кора, как часть лимбической системы мозга, располо-
жена на медиальной поверхности лобных долей (префронтальная кора).
Структуры передней поясной коры интегрирует информацию от различ-
ных входов — мотивации, познавательные процессы и эмоции. В комплек-
се с другими отделами мозга осуществляет направленность произвольного
внимания человека на сексуальные стимулы различной модальности (зри-
тельные, слуховые, тактильные, обонятельные), что обусловливает ини-
19. Мотивации и эмоции • 789
циацию и увеличение степени полового возбуждения человека. У человека
передняя поясная кора всегда активируется при эмоциях (см. ниже), кото-
рые увеличивают степень полового возбуждения человека.
19.2. Эмоции
Слово эмоция происходит от латинского глагола movere, означающего дви-
гаться. Это означает, что люди всегда реагируют на происходящие вокруг
них события, например, усилением сердцебиений, возбуждением организ-
ма, прекращением деятельности (упадок сил), потоотделением, например
на коже ладоней, напряжением или расслаблением скелетных мышц; по-
краснением или моментальной бледностью лица, нахмуренными бровями
или улыбкой. Наряду с этим подавляющее большинство людей испытыва-
ли чувство счастья, например после успешной сдачи экзамена, либо стра-
дания — при потере любимого человека, или гнева — при несправедливо-
сти. Люди испытывают удовольствие от общения с друзьями, занятия
спортом, в том числе его экстремальными видами, либо испытывают сму-
щение при поломке взятой у кого-то вещи. Независимо от причин всех
этих чувств, субъективное переживание смысла событий относятся к кате-
гории эмоций.
Чувства, которые посредством физиологических и/или познавательных
процессов изменяют целенаправленное поведение людей, называются эмо-
циями. Эмоции проявляются у человека на самых ранних этапах его жиз-
ни, а именно до развития речи. Так, плачущий ребенок быстро обращает
на себя внимание взрослых людей, и они оказывают ему необходимую по-
мощь и заботу. Жизнерадостный и улыбающийся ребенок вызывает у
взрослых людей чувство заботы и любви. Субъективные проявления эмо-
ций в интонации голоса человека, выражениях его лица, жестах или позе
изменяет поведение других людей. Так, крик человека при страхе может
вызвать панику у окружающих его людей, а улыбка, напротив, успокоить
испуганных людей. Эти реакции человека обозначают общим понятием
эмоции.
19.2.1. Виды эмоций
Французский ученый, философ-дуалист Р. Декарт (XVII век) описал у че-
ловека шесть базовых, или фундаментальных, эмоций: восхищение, любовь,
ненависть, страсть, радость и страдание. Все эти эмоции Р. Декарт считал
положительными, включив в перечень любовь. В XX веке классифициро-
валось семь базовых эмоций: страх, гнев, счастье, ярость, страдание, от-
вращение, удивление. В психиатрии и социальной психологии распростра-
нена классификация социальных эмоций, включающая эмоции вины, гор-
дости, смущения, стыда, материнской любви, сексуальной любви, увлечен-
ности, восхищения и ревности.
С учетом реакций мимической мускулатуры лица человека или мимиче-
ских выражений лица, возникающих при эмоциях, их перечень был рас-
ширен до десяти: интерес, наслаждение, удивление, страдание, гнев, отвра-
щение, стыд, страх, презрение и вина. Поскольку каждая из перечисленных
эмоций характеризуется однотипным выражением лица у людей, эмоции
были названы базовыми, или врожденными, эмоциями (рис. 19.2). Лицо
человека уникально тем, что из 44 мышц этой области тела — двигательная
функция 40 мышц обусловливает выражение лица при базовых эмоциях, и
только 4 — участвуют в жевательной функции.
790 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Рис. 19.2. Примеры базовых эмоций человека.
Реакции мимической мускулатуры лица при эмоциях у всех людей проявляется сходными вы-
ражениями лица при одной и той же эмоции: нейтральное выражение лица (1), эмоция сча-
стья (2), испуг (3), отвращение (4), гнев (5), страх (6), печаль (7).
Эмоции характеризуются интенсивностью, направленностью, качеством
и осознанием. Под интенсивностью эмоций подразумевается, прежде все-
го, их выраженность или сила; направленность эмоций связана с их зна-
ком, т. е. положительная или отрицательная эмоция. К отрицательным
эмоциям относят гнев, страх, страдание и отвращение. Отрицательные
эмоции могут оказывать негативное влияние на отношения между людьми,
препятствовать принятию правильного решения при целенаправленном
поведении. Например, эмоция страха, вызываемая одними и теми же фак-
торами внешней среды, может стать причиной так называемой фобии (си-
ноним — боязнь) у человека. Качество эмоций обусловлено степенью ин-
теллектуального развития человека, т. е. его умственными способностями,
а осознанность эмоций подразумевает, осознает или нет субъект наличие у
него самого или у другого человека эмоций. Эти проявления эмоций раз-
личаются по степени, с которой они воздействуют на индивидуума.
19.2.2. Роль эмоций в поведении человека
В наиболее общей форме роль эмоций характеризуется следующим обра-
зом. Эмоции активируют функции вегетативной и эндокринной систем и
этим способствуют реализации целенаправленного поведения человека
(активационная роль). Отрицательные эмоции, которые возникают при не-
приятных болевых ощущениях, приводят к тому, что человек стремится их
предотвратить или избежать возникновения (регулирующая роль). Подкреп-
ляющая роль эмоций в процессе накопления человеком индивидуального
жизненного опыта усиливает (положительные эмоции) либо снижает (от-
19. Мотивации и эмоции • 791
рицательные эмоции) социальную активность человека. Эмоции человека
проявляются в жестах, мимике лица либо интонации слов и, таким обра-
зом, через эти внешние проявления могут изменить поведение окружаю-
щих его людей (коммуникативная роль). Социальная роль эмоций регулирует
социальные взаимодействия между людьми, способствуя формированию
таковых с одними людьми и игнорирование взаимодействия с другими.
Например, положительные эмоции влияют на привязанность родителей к
своим детям и наоборот, и поэтому они являются важным механизмом
развития внутрисемейных отношений. Познавательная роль эмоций заклю-
чается в том, что эмоции повышают эффективность памяти человека. При
этом мозг человека запоминает не только поведенческие действия, но и
сопутствующие им эмоции.
19.2.3. Нейрофизиологические механизмы проявления эмоций
Проявление эмоций обусловлено активностью, прежде всего, лимбической
системы мозга.
19.2.3.1. Гипоталамус как центр регуляции вегетативных и эндокринных
реакций организма при эмоциях
Гипоталамическая регуляция вегетативных и эндокринных проявлений
при эмоциях осуществляется двумя областями: перифорникальной обла-
стью и медиальной частью латерального гипоталамуса, которая нередко на-
зывается гипоталамическим полем, регулирующим эмоциональные реак-
ции; паравентрикулярными ядрами гипоталамуса, которые состоят из
взаимодействующих между собой популяции нейронов, и также регулиру-
ют вегетативные и эндокринные реакции организма человека при эмоциях
При эмоциях интегративная функция латерального гипоталамуса в регу-
ляции вегетативных и эндокринных реакций организма обусловлена тем,
что гипоталамус состоит из специфических нейронных сетей, каждая из
которых обусловливает возникновение одного из конкретных видов целе-
направленного мотивационного поведения — пищевого, питьевого или ре-
продуктивного. Функциональные состояния голода, жажды и половой ак-
тивности возникают и усиливаются под влиянием пептидных гормонов,
которые выделяются из окончаний нервных волокон нейронов гипотала-
муса. Гипоталамические нейроны передают сигналы к вентральным кау-
дальным отделам моста и медиальной покрышке ствола мозга, которые
обусловливают общий уровень активации симпатических преганглионар-
ных нейронов боковых рогов спинного мозга. В результате активации сим-
патической нервной системы под влиянием гипоталамуса постганглионар-
ные симпатические нервные волокна высвобождают биогенные амины —
норадреналин и адреналин. Эти гормоны оказывает диффузное и длитель-
ное активирующее влияние на функцию различных систем организма, вы-
зывая увеличение частоты сердцебиений, рост артериального давления,
увеличение частоты и глубины дыхательных движений, расширение зрач-
ков, увеличение уровня сахара в крови, снижение произвольного контроля
мочеиспускания и дефекации, блокаду слюноотделения и выброс гормонов
в кровь при эмоциях.
Паравентрикулярные ядра гипоталамуса при эмоциях регулируют секре-
цию гормонов гипофиза, щитовидной железы, паращитовидных желез, ко-
ры надпочечников и клеток островков Лангерганса поджелудочной желе-
зы. Так, при отрицательных эмоциях у человека при участии гипоталамо-
792 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
гипофизарно-надпочечниковой оси активируется секреция кортизола, обу-
словленная избыточной секрецией кортикотропина или кортиколиберина.
Эти эндокринные реакции возникают в результате понижения секреции
тиреолиберина и тиреотропина, а также снижения чувствительности аг-ад-
ренорецепторов нейронов гипоталамуса к гормону роста, что характерно
для состояния депрессии (эмоциональной подавленности) у человека.
19.2.3.2. Роль миндалины в осуществлении базовых эмоций
Нейроны центрального ядра миндалины непосредственно активируют ядра
латерального гипоталамуса. В результате в организме человека возникают
как вегетативные (симпатические), так и эндокринные реакции при эмо-
циях.
Миндалина занимает довольно ограниченный участок мозга в височных
долях переднего мозга. Однако при ее повреждении у человека ухудшается
восприятие эмоций, снижается запоминание эмоций, возникает скудность
эмоциональных выражений лица, снижается память на эмоциональные со-
бытия.
Миндалина является входом в сложную нейронную сеть, состоящую из
ядер различных отделов головного мозга, которые обусловливают возник-
новение эмоций. Миндалина получает поток афферентных нервных им-
пульсов от каждой из основных сенсорных систем ЦНС и от ассоциатив-
ных областей коры головного мозга (таламо-кортико-миндалевидные пу-
ти). Сенсорные сигналы миндалина получает непосредственно от ядер та-
ламуса (таламо-миндалевидные пути). Поэтому нейроны миндалины чело-
века активируются на различные стимулы: при вознаграждении и наказа-
нии в ходе целенаправленного поведения человека, при восприятии им со-
циально значимых стимулов, при прослушивании приятной музыки, рас-
сматривании картины, при слуховом восприятии смеха или плача.
Таламо-миндалевидные пути ответственны за формирование эмоций
через короткий промежуток времени после действия на организм раздра-
жителей внешней среды, таких как громкий звук, свет. Таламо-кортико-
миндалевидные пути, которые являются полисинаптическими и мультимо-
дальными, обусловливают медленное развитие эмоций. Сложный поток
информации, который поступает в миндалину по этим путям, играет важ-
ную роль в понимании человеком эмоций, возникающих под влиянием
различных стимулов внешней среды. Поэтому сложные раздражители, воз-
никающие, например, при рассматривании картины, вызывают эмоции
под влиянием информации, поступающей в миндалину по таламо-корти-
ко-миндалевидному пути, а простые индифферентные стимулы вызывают
эмоции путем прямой активации миндалины через ядра таламуса.
Миндалина состоит из нескольких субъядерных популяций нейронов,
которые сгруппированы в цитоархитектонические и функциональные еди-
ницы. Две такие единицы миндалины участвуют в контроле эмоций и
представлены базолатеральной областью миндалины, включающей лате-
ральное, базальное и добавочное ядро, а также центральным ядром.
Большинство сенсорных входов от ядер таламуса конвергирует на ней-
ронах латерального ядра миндалины, а от высших отделов головного моз-
га — на нейронах базального ядра (рис. 19.3). Эти два ядра представляют
собой входные ворота для сенсорной и когнитивной (познавательной) ин-
формации, под воздействием которой возникают эмоции. При этом базо-
латеральная область миндалины имеет обширные реципрокные проекции
с лобной корой, мультимодальными отделами коры головного мозга. Ней-
19. Мотивации и эмоции • 793
Сенсорные
входы
Гипоталамус,
средний мозг,
мост,
продолговатый
мозг
Гипоталамус
Первичная
зрительная
кора
Первичная
слуховая
кора <
Сомато-
сенсорная
кора
Ассоциативная кора
(височные доли) -
Ядра вкусовой и
висцеральной
чувствительности
продолговатого
мозга
ентральное
► %
Гиппокамп
Вентральный
отдел
стриарных тел
Обонятельный
мозг
едиально
ядро
обавочное
базальное
ядро
Базо-
латеральное
ядро
> Серое вещество вокруг
сильвиеаа водопровода
Дорсомедиальное
ядро таламуса
Рис. 19.3. Афферентные входы и выходы, а также связи между отделами минда-
лины.
Базовые эмоции могут формироваться при участии нескольких нейронных сетей миндалины.
Это достигается за счет последовательной и параллельной организации нейронных сетей и
аналогичной (последовательной и параллельной) обработки сенсорной информации (слева от
рисунка). Эфферентным выходом миндалины является центральное ядро, при участии кото-
рого обеспечивается взаимодействие нейронных сетей миндалины с различными структурами
мозга (справа от рисунка), которые непосредственно или опосредованно регулируют вегета-
тивные и эндокринные реакции при эмоциях.
роны латеральных и базальных ядер миндалины посылают сигналы, преж-
де всего, к нейронам центрального ядра. Центральное ядро миндалины
имеет стриарное строение, благодаря которому от строго определенных
популяций нейронов начинается большинство эфферентных проекций
миндалины. Эти проекции активируют структуры головного мозга, обу-
словливающие формирование в организме вегетативных и двигательных
реакций при эмоциях.
В нейронных сетях миндалины на основе оценки поступающей сенсор-
ной информации возникают специфическая базовая эмоция и характерные
для нее вегетативные и двигательные реакции. Так, в случае «опасности»
для жизнедеятельности организма воздействий внешней среды возникает
эмоция страха — это либо затаивание и мышечная дрожь, либо бегство от
опасности для жизни человека.
От центрального ядра миндалины начинаются проекции нейронов, ко-
торые достигают ядер ствола головного мозга, в частности нейронов цен-
трального серого вещества вокруг сильвиева водопровода (рис. 19.4). Цен-
тральное серое вещество относится к основной моторной системе мозга,
которая контролирует непроизвольные двигательные реакции при эмоци-
ях, например реакции крика, смеха и гнева, формируя так называемую
эмоциональную двигательную систему.
Нейроны центрального серого вещества контролируют двигательные ре-
акции при агрессивном (эмоция гнева) и защитном поведении (эмоция
Серое вещество вокруг сильвиева водопровода
Вентральная часть каудального отдела моста и
медиальная покрышка продолговатого мозга
N. subretrofacialis
N. retroambiguus
Симпатические преганглионарные
нейроны боковых рогов спинного
мозга
Сенсорные нейроны
дорсальных рогов
спинного мозга
Общий уровень симпатической
активности
Ноцицептивный
контроль
Мотонейроны и
премоторные
интернейроны
вентральных рогов и
промежуточной зоны
сегментов спинного
мозга
Симпатические
преганглионарные
нейроны боковых
рогов спинного мозга
Сердечно-сосудистые
эффекты
Локомоция
Мотонейроны мышц
гортани, глотки, мягкого
неба, экспираторных
мышц
Речь
Парасимпатические Сегменты спинного мозга Грудные сегменты Т,—Т„
преганглионарные Мотонейроны VII и VII
нейроны сакральных пластины по Рекседу
сегментов спинного мозга |
Защитная поза
Мочеиспускание
Рис. 19.4. Роль нейронов центрального серого вещества вокруг сильвиева водопровода в регуляции вегетативных и моторных ре-
акций при эмоциях.
Нейроны центрального серого вещества получают информацию от ядер гипоталамуса, ядра дна терминальных полосок и миндалины, обусловли-
вая регуляцию базовых реакций, таких как агрессивное, защитное или репродуктивное поведение. Действиям сопутствуют эмоции, которые состо-
ят из специфических соматических и вегетативных реакций. Так, при агрессивном поведении (эмоция ярости) активируется речевая функция (че-
рез n. retroambiguus), увеличивается артериальное давление (n. subretrofacialis), понижается порог болевой чувствительности (вентральная покрыш-
ка моста и продолговатого мозга), расширяются зрачки (преганглионарные нейроны Т,—Тп).
Преганглионарные
симпатические нейроны
Расширен^ зрачков
794 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
19. Мотивации и эмоции • 795
страха), которые чрезвычайно важны для сохранения жизни индивидуума.
Каждая из этих реакций состоит из комплекса специфической двигатель-
ной активности и регулируется определенной нейронной сетью централь-
ного серого вещества.
Активация при эмоциях нейронов вентролатеральной части централь-
ного серого вещества вокруг сильвиева водопровода вызывает снижение
двигательной активности (пассивность или неподвижность), гипотензию,
брадикардию и опиоид-зависимую аналгезию. Активация системы обезбо-
ливания мозга сигналами, поступающими в центральное серое вещество,
тормозит болевые импульсы, входящие в ЦНС через периферические нер-
вы, и снижает степень воздействия боли на человека, а также реакции ор-
ганизма на болевые воздействия.
При защитной реакции, такой как бегство (эмоция страха), активиру-
ются нейроны латерального отдела серого вещества вокруг сильвиева водо-
провода. При этом происходит повышение артериального давления, воз-
никает тахикардия, способствующие усилению кровообращения скелетных
мышц при бегстве.
Нейроны центрального серого вещества вокруг сильвиева водопровода
регулируют двигательную активность через интернейронные группы кле-
ток (n. reteofaciaiis, n. retroambiguus). Так, нейроны задней части обоюдных
ядер продолговатого мозга, иннервируют мышцы верхних дыхательных пу-
тей и гортани и, таким образом, влияют на артикуляцию человека при
эмоциях. Под влиянием нейронов центрального серого вещества вокруг
сильвиева водопровода активируется рефлекс мочеиспускания, например
при эмоции страха.
Эфферентный выход центрального ядра миндалины проецируется к та-
ким структурам ствола мозга, как пневмотаксический центр моста (увели-
чение частоты дыхания), нейроны голубого пятна (увеличивают насторо-
женность), дофаминергическое поле вентральной покрышки мозга (пове-
дение, обусловленное мотивациями), дорсальное двигательное ядро блуж-
дающего нерва (изменение парасимпатической активности), ядра ретику-
лярной формации ствола мозга (общая активация мозга), а также нейроны
тройничного нерва (мимическое выражение лица при эмоциях).
Роль миндалины в формировании базовых эмоций осуществляется при
участии последовательного и параллельного механизмов взаимодействия ее
структур с афферентными входами и эфферентными выходами. Последо-
вательный механизм осуществляется путем взаимодействия сенсорных вхо-
дов первоначально с нейронами базолатерального ядра миндалины. Затем
сигналы от нейронов базолатерального ядра миндалины поступают к ней-
ронам центрального ядра, а далее — к структурам-мишеням ствола мозга.
Параллельные механизмы функционирования миндалины при возникно-
вении базовых эмоций осуществляются нейронными сетями базолатераль-
ного и центрального ядер миндалины. Нейронные сети базолатерального и
центрального ядра миндалины участвует в формировании эмоций при по-
мощи раздельных афферентных и эфферентных путей. При этом базолате-
ральное ядро миндалины получает сенсорные входы непосредственно из
таламуса и коры головного мозга. Далее проекции нейронов направляются
к структурам вентрального отдела стриарных тел базальных ганглиев (регу-
ляция эмоционального выражения лица), к префронтальной коре (перед-
няя поясная извилина, латеральная и медиальная орбитальная кора, лим-
бическая кора), а также к зрительной затылочной и височной коре, обу-
словливая эмоции при познавательной деятельности человека. Централь-
ное ядро получает прямые сенсорные входы из таламуса и коры головного
796 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
мозга и независимо от функции базолатерального ядра миндадины увели-
чивает через ядра латерального гипоталамуса активность симпатической
нервной системы, структур промежуточного мозга и ствола мозга, обуслов-
ливая вегетативные и соматические проявление базовых эмоций.
В целом, каждая фундаментальная эмоция осуществляются в соответст-
вии с жесткой программой конкретной нейронной сети миндалины, реа-
лизация которой обусловливает эмоциональное выражение лица человека.
19.2.3.3. Регуляция положительных эмоций у человека
Под положительной эмоцией понимают осознаваемое человеком чувство
удовольствия. Выражение лица детей при восприятии сладкого вкуса явля-
ется типичным примером положительной чувственной реакции на прият-
ное сенсорное раздражение: расслабление мышц середины лица, а в ряде
случаев —появление улыбки. Хотя у человека на приятные для него раз-
дражения (вкусовые, обонятельные, тактильные и др.) активируются об-
ласти глазолобной коры, инициация положительных эмоций обусловлена
ролью подкорковых мозговых структур.
Впервые «центры удовольствия мозга» были открыты полвека назад в
опытах на животных с самораздражением электрическим током через
вживленные электроды в латеральный гипоталамус и в область перегород-
ки переднего мозга. Места самораздражения включали также вентральную
покрышку среднего мозга, вентральный отдел бледного шара, вентральный
отдел таламуса, прилегающее ядро (n. accumbens) и предлобную кору. По-
скольку в отношении животных невозможно говорить о чувственной при-
роде их поведения, то электрическая самостимуляция «центров удовольст-
вия» или «нейронных кругов удовольствия» рассматривается как получение
вознаграждения (синоним — подкрепление по И. П. Павлову) подопыт-
ным животным при выполнении самораздражения. Лабораторные живот-
ные могли нажимать на рычаг, соединенный с электродами, вживленными
в соответствующие центры удовольствия, с чрезвычайно высокой частотой
(боле 6000 раз в час). В результате экспериментов с самораздражением ос-
новные (первичные) центры удовольствия у животных были локализованы в
латеральном гипоталамусе, а вторичные — в перегородке, миндалине, тала-
мусе, базальных ганглиях и базальной покрышке промежуточного мозга.
У человека «нейронные круги удовольствия», при участии которых ини-
циируются положительные эмоции, включают ракушку прилегающего ядра
(n. accumbens), латеральное поле перегородки, каудальное вентромедиаль-
ное и базальное ядра Мейнерта лимбической системы, вентральный отдел
бледного шара и парабрахиальные ядра ствола мозга. Каудальная область
ракушки прилегающего ядра (n. accumbens) содержит специфические
опиоидные нейронные круги, активация которых морфином лежит в осно-
ве усиления чувства удовольствия, например при еде. В частности, нейро-
ны ракушки представляют собой единственную область прилегающего яд-
ра, которая активируется при положительных чувственных (аффективных)
реакциях человека на приятные вкусовые раздражители. Поэтому нейроны
ракушки прилегающего ядра представляют собой морфологическую основу
возникновения у человека чувства удовольствия на потенциально прият-
ные раздражения — вкусовые, тактильные или сексуальные.
Нейроны ракушки прилегающего ядра синаптически связаны с меди-
альной частью вентрального бледного шара, а затем сигналы направляются
к срединному дорсальному ядру таламуса, а далее непосредственно в об-
ласти глазолобной коры коры. Активация этой нейронной сети обусловли-
19. Мотивации и эмоции • 797
Рис. 19.5. Нейронные круги положительных эмоций.
Нейроны ракушки прилегающего ядра имеют на мембране опиоидные рецепторы, активация
которых вызывает у человека положительные эмоции (чувство удовольствия). Прилегающее
ядро является частью мезолимбической системы мозга (парабрахиальные ядра моста, бледный
шар базальных ганглиев, таламус, лобная часть передней поясной извилины, и глазолобная
кора), нейронные сети которой обусловливают возникновение положительных эмоций и чув-
ства удовольствия при восприятии вкусной пищи, приятных запахов, родительской заботе о
детях, спортивной игре или занятии сексом.
вает появление у человека приятных аффективных или чувственных ре-
акций.
В целом эти синаптические связи представляют собой один потенци-
альный путь формирования у человека положительных эмоций, вызванно-
го опиоид-зависимой активацией ракушки прилегающего ядра (рис. 19.5).
Отделы глазолобной коры у человека, расположенные в области пре-
фронтальной коры непосредственно над глазными яблоками, выполняют
ключевую функцию в восприятии приятных сенсорных раздражений. Ней-
роны глазолобной коры активируется на приятные вкусовые, обонятель-
ные, тактильные раздражения, прослушивание приятной музыки и даже
при выигрыше денег.
Приятные раздражения могут вызвать положительные эмоции у челове-
ка при участии нейронных кругов ракушки прилегающего ядра, минуя
корковый уровень. Эту роль выполняют прямые восходящие чувствитель-
ные пути к нейронам ракушки прилегающего ядра от нейронов вкусовых
ядер продолговатого мозга — ядер одиночного тракта. Активация нейронов
ядер одиночного тракта происходит при стимуляции рецепторов вкуса на
слизистой оболочке языка.
Нейроны ракушки прилегающих ядер имеют прямые синаптические
связи с восходящей мезолимбической дофаминергической системой мозга.
Электрическое раздражение структур дофаминергической системы мозга
вызывает у человека физиологические эффекты с чувством выраженного
удовольствия, например необычно сильный оргазм.
Нейроны ракушки прилегающего ядра образуют нейронный круг сис-
тем удовольствия мозга с парабрахиальными ядрами верхнего отдела мос-
та. Нейроны парабрахиальных ядер моста образуют бензодиазепин/ГАМК-
ергический круг нижнего отдела ствола мозга, активация которого усили-
вает проявление мотиваций влечения при воздействии на организм челове-
798 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ка приятных сенсорных раздражений. Функция парабрахиальных ядер
моста обусловлена двумя восходящими путями: один достигает первичной
вкусовой коры головного мозга через переключательные ядра таламуса, а
другой — прямо идет к нейронам в прилегающие ядра. Бензодиазепино-
вый/ГАМК-ергический нейронный круг парабрахиальных ядер в составе
обширных нейронных кругов системы удовольствия мозга, обусловливает
возникновение приятного вкусового сенсорного ощущения у человека (по-
ложительной эмоции) при участии нижнего отдела ствола мозга.
В целом, системы удовольствия мозга человека функционируют на ос-
нове ключевой роли прилегающего ядра и представляют собой параллель-
ные нейронные пути, которые входят в состав корковых и подкорковых
нейронных кругов. Считают, что в мозге человека функционирует не-
сколько нейронных кругов положительных эмоций, активирующихся при
пищевой и половой мотивациях, при родительской заботе о детях, видео-
играх, приеме наркотиков и др.
19.2.3.4. Регуляция отрицательных эмоций у человека
У здоровых субъектов нейроны миндалины в наибольшей степени активи-
руются при восприятии эмоционального выражения лица при страхе и
гневе, чем радости. Наряду с активацией миндалины при эмоции страха
активируются подкорковые области мозга (таламическая и бугорковая), в
меньшей степени корковые поля. Повреждение миндалины у человека
снижает восприятие эмоционального выражения лица, особенно выраже-
ния эмоции страха, ухудшается также распознавание эмоциональной инто-
нации голоса. Повреждение миндалины обусловливает уменьшение им-
плицитной эмоциональной памяти (см. ниже)
У человека эмоцию страха вызывают стимулы различной модальности.
Основной структурой-мишенью для афферентных входов, вызывающих от-
рицательные эмоции у человека, является латеральная область миндалины
(см. рис. 19.3). Нейроны этой области активируются слуховыми, ноцицеп-
тивными сигналами, нейронами таламических полей, получающих аффе-
рентные импульсы от спиноталамического тракта. Структуры латеральной
области миндалины активируются соматосенсорными и ноцицептивными
стимулами, поступающими от сенсомоторных областей коры. Ноцицеп-
тивные стимулы, поступающие от парабрахиальных ядер моста, непосред-
ственно активируют нейроны центрального ядра миндалины. Аналогичные
стимулы, поступающие от задних отделов таламуса, вызывают активацию
дополнительной базальной области миндалины.
Основным эфферентным выходом миндалины являются нейроны цен-
трального ядра, которые интегрируют сигналы от нейронов афферентных
отделов миндалины. Активация нейронных сетей центрального ядра вызы-
вает характерные реакции в организме человека при эмоции страха. Так,
через двигательные ядра ствола мозга инициируется эмоциональное выра-
жение страха на лице человека. Посредством структур латерального гипо-
таламуса регулируются реакции со стороны системного артериального дав-
ления в организме. Нейроны центрального ядра миндалины, активируя
нейроны центрального серого вещества вокруг сильвиева водопровода, вы-
зывают в организме мышечную дрожь, при возбуждении ядра дна и терми-
нальных полосок происходит высвобождение стрессовых гормонов оси ги-
пофиз—кора надпочечников.
Наиболее выраженные отрицательные эмоции у человека возникают
при восприятии боли. Возникновение неприятных ощущений у человека
19. Мотивации и эмоции • 799
при ноцицептивных раздражениях обусловлено ролью в этом процессе
нейронов лобного отдела передней поясной извилины, расположенной на ме-
диальной поверхности префронтальной коры. Активация этого участка
мозга при болевой стимуляции посредством структур миндалины, цен-
трального серого вещества вокруг сильвиева водопровода, прилегающего
ядра, гипоталамуса инициирует у человека отрицательные эмоции (страх,
страдание), мотивации избегания и соответствующие им проявления ак-
тивности вегетативной и эндокринной систем (описаны выше).
ГЛАВА 20
Физиологические основы
познавательной деятельности
человека
Познавательной деятельностью человека называется получение и сохране-
ние знаний. Эта деятельность осуществляется на основе базовых познава-
тельных процессов, к которым относят внимание, восприятие, память,
научение и мышление. Основным результатом познавательной деятельно-
сти человека является знание. В наиболее общей форме знание определяет-
ся как объекты, концепции и взаимосвязи, которые находятся в поле инте-
реса (деятельности) человека. Знание человека имеет многочисленные
проявления: умение, отношение к чему-либо или к кому-либо, информа-
ция, словарный запас, лексические правила, практические навыки и др.
Физиологической основой познавательной деятельности человека явля-
ются функции ассоциативных областей коры головного мозга. Эти области
коры занимают большую ее площадь и имеют сложную синаптическую
многоуровневую иерархию ядер (рис. 20.1). Иерархия ядер ассоциативных
отделов коры включает в себя первичные сенсорные проекционные поля,
вторичные унимодальные (имеют отношение к определенному виду чувст-
вительности), гетеромодальные поля коры. В гетеромодальных полях много-
численные ядра, входящие в их состав, функционируют на основе разных
видов информации. Уникальной функцией этих полей является связыва-
ние многочисленных унимодальных и трансмодальных полей в распреде-
ленные и интегрированные мультимодальные репрезентации, на основе ко-
Рис. 20.1. Схема расположения первичных проекционных и ассоциативных полей
коры головного мозга человека (А) и шимпанзе (Б). У человека по сравнению с
высшими приматами преимущественное развитие получили ассоциативные области
коры, при участии которых осуществляются базовые познавательные процессы вос-
приятия, научения и мышления.
Первичные проекционные области коры: 1 — двигательная, 2 —слуховая, 3 — обонятельная,
4 — зрительная, 5 — соматосенсорная, 6 — ассоциативная.
20. Физиологические основы познавательной деятельности человека ' 801
торого в мозге происходит превращение ощущения в познавательный про-
цесс. Под репрезентацией (синоним — представление) понимают актива-
цию нейронной сети ядер как унимодальных, так и гетеромодальных полей
под влиянием поступающих в них сигналов. В познавательной деятельно-
сти человека функцией унимодальных ассоциативных полей коры голов-
ного мозга является восприятие базовых особенностей ощущений цвета,
движения, формы и пространственной локализации стимулов внешней
среды. Более сложное восприятие таких сенсорных признаков, как объек-
ты, лица, словесные формы, пространственные локализации стимулов и
последовательности звуков, осуществляется на более высоких уровнях си-
наптического переключения (4-й и 5-й уровни) унимодальных полей. Выс-
шие синаптические уровни обработки сенсорной информации при позна-
вательной деятельности происходят в гетеромодальных ассоциативных по-
лях коры головного мозга, а также в паралимбической и лимбической ко-
ре. Эти поля коры головного мозга называются также трансмодальными.
Трансмодальные поля коры головного мозга (средние отделы височной ко-
ры, зона Вернике, гиппокампально-энториальный комплекс, задняя те-
менная кора) выполняют в познавательной деятельности функцию транс-
формации: восприятия — в распознавание, словесных форм — в мысли,
сцен и событий — в индивидуальный жизненный опыт человека.
Префронтальная ассоциативная кора в познавательной деятельности вы-
полняет высшие функции мозга: вероятностное прогнозирование и програм-
мирование познавательной деятельности, анализ поступающей в этот отдел
коры больших полушарий информации с учетом выполняемых результатов
деятельности. Функции префронтальной коры в познавательной деятель-
ности человека осуществляются на основе эксплицитной рабочей памяти.
Механизмы этого вида памяти, для которой необходима точная и ясно вы-
раженная или четко наблюдаемая человеком «информация», позволяют
структурам префронтальной коры неоднократно обрабатывать в течение
короткого времени одну и ту же осознаваемую человеком информацию и
на этой основе включать ее элементы в содержание знания.
Познавательная деятельность человека обусловлена его сознанием. Соз-
нание человека представляет собой психический процесс. Тем не менее
сознание человека коррелирует с проявлениями активности (электриче-
ской или метаболической) в различных расположенных на расстоянии друг
от друга ассоциативных полях коры головного мозга при базовых познава-
тельных процессах внимания, восприятия, памяти и мышления.
20.1. Внимание
Сенсорные системы мозга человека одновременно воспринимают большое
число влияний внешней среды. Одни из них имеют отношение к поведе-
нию человека в текущий момент времени, а другие — не влияют на тако-
вое. Мозг человека не может воспринимать одновременно все многообра-
зие воздействий внешней среды на организм. Поэтому в ЦНС происходят
процессы, которые вычленяют лишь некоторые из стимулов внешней сре-
ды и «фильтруют» их от многих других. Это связано с тем, что организм
человека способен эффективно функционировать во внешней среде, если
происходит отбор или селекция тех стимулов, которые необходимы для
осуществления его приспособления. Процессы отбора информации в мозге
человека обозначаются понятием «внимание». Избирательная или селек-
тивная реакция организма человека на определенный вид сенсорной ин-
формации внешней среды называется вниманием. Внимание человека обу-
26 - 6095
802 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
словлено функциями слуховой, соматовисцеральной, обонятельной, вкусо-
вой и вестибулярной сенсорных систем. Внимание человека ответственно
за выбор во внешней среде объектов и их локализации, о которых затем
необходимо получить более подробную информацию и, одновременно, ис-
ключить из восприятия объекты другой локализации. Внимание человека
одновременно не способно ориентироваться на несколько стимулов внеш-
ней среды или несколько мыслительных процессов.
20.1.1. Формы внимания
Избирательный отбор одного стимула из множества раздражений внешней
среды происходит у человека на основе непроизвольного (пассивного, или
рефлекторного) и активного, или произвольного, внимания.
Непроизвольное внимание представляет собой автоматическую, рефлек-
торную реакцию организма человека (как правило, поворот глаз и/или го-
ловы в направлении источника раздражения) в ответ на действие стимулов
внешней среды. При этом стимулы внешней среды, которые способны вы-
звать внимание человека, должны быть очень сильными либо значитель-
ными, либо резкими и неожиданными, и иметь конкретную природу —
световые, звуковые, обонятельные или ноцицептивные (например, внеш-
няя или внутренняя боль). Непроизвольное внимание возникает неосоз-
нанно и не требует умственных усилий со стороны человека для направле-
ния взгляда или поворота головы в сторону источника раздражения. Как
правило, непроизвольное внимание обусловлено такими качествами сти-
мулов, как интенсивность или неожиданность их возникновения для чело-
века, или их новизна.
Произвольное внимание у человека представляет осознаваемое человеком
ориентирование органов чувств, как правило, органа зрения, на воспри-
ятие конкретного объекта или явления внешней среды. Основным видом
произвольного внимания является зрительное произвольное внимание, ко-
торое подразделяется на пространственное и объект-ориентированное.
Пространственное зрительное внимание представляет собой процесс изме-
нения зрительного внимания с одного объекта на другой. Направленность
зрительного внимания на конкретный объект называется объект-ориенти-
рованным зрительным произвольным вниманием (синоним — фокусное
внимание).
Произвольное внимание возникает в том случае, если человек осознает
действие (поворот глаз и/или головы), направленное на рассматривание,
выслушивание или осязание какого-то объекта или явления внешней сре-
Таблица 20.1. Две формы зрительного внимания человека
Свойства внимания Формы внимания
непроизволное произвольное
Пространственная спе- цифичность Детальная специфич- ность Продолжительность Произвольный контроль Зависимость от задания Через поле зрения Действует в любое время и во всех пространственных направлениях Временное Отсутствует Не зависит Пространственно обу- словленное (фокусное) Может избирать специ- фичный атрибут Устойчивое (с произ- вольным усилием) Обязателен Зависит
20. Физиологические основы познавательной деятельности человека • 803
ды. Произвольное внимание у человека обусловлено предшествующим
научением (ассоциативное научение, инструкция, задание, внутреннее же-
лание). Произвольное внимание человек может направлять на мыслитель-
ные процессы, обусловливающие, например, принятие решения и после-
дующие за этим действия. По сравнению с произвольным вниманием, не-
произвольное внимание возникает независимо от инструкций или внут-
реннего намерения человека (табл. 20.1).
20.1.2. Нейрофизиологические механизмы внимания
Физиологическая основа внимания состоит в том, что оно обусловлено
увеличением и синхронизацией нейронной активности в тех отделах ЦНС,
которые воспринимают сенсорные стимулы, и увеличением активности
структур теменной и лобной коры мозга человека.
Наиболее полное проявление избирательности, или селективности,
внимания связано с функцией зрения у человека. Зрительное пространство
содержит множество стимулов, специфическая локализация которых явля-
ется избирательной для процесса зрительного внимания. Емкость зритель-
ного внимания ограничена. Своеобразной ценой избирательности внима-
ния является «слепота» к зрительным стимулам, на которые человек не об-
ращает внимания. При этом зрительное внимание способно выбирать наи-
более интересный объект в поле зрения или изменять «фокус» внимания с
одного объекта на новый. Наиболее важной функцией непроизвольного и
произвольного зрительного внимания является быстрое направление
взгляда человека на интересующий объект в окружающем зрительном про-
странстве. Физиологические механизмы непроизвольного и произвольного
внимания человека тесно взаимосвязаны между собой. У человека нейрон-
ные структуры внимания представлены отделами верхних бугров четверо-
холмия, подушки таламуса, зрительной коры, лобного глазного поля, до-
полнительного глазного поля лобной коры, задней теменной коры, ниж-
ней височной и префронтальной коры. Все отделы зрительных областей
коры головного мозга играют важную роль в зрительном внимании. Одна-
ко наибольший дефицит зрительного внимания у человека возникает при
повреждениях теменных долей коры. Это проявляется в игнорировании
контралатерального зрительного пространства, которое возникает чаще
при повреждении правой теменной области коры головного мозга и ре-
же — левой. Поэтому правая теменная область коры головного мозга вы-
полняет доминирующую функцию во внимании человека. Физиологиче-
ские механизмы внимания обусловлены функцией следующих подкорко-
вых и корковых центров внимания.
20.1.2.1. Функции среднего мозга и моста в контроле внимания
Функция состоит в контроле движения глаз и/или головы человека в на-
правлении источника раздражения. Этот механизм лежит в основе обеих
форм внимания человека, т. е. непроизвольного (ориентировочный реф-
лекс по И. П. Павлову) и произвольного.
Ориентированное движение глаз при внимании осуществляется синер-
гичными сокращениями трех пар глазодвигательных мышц. Нейроны, ко-
торые иннервируют мышечные волокна этих мышц, образуют III (глазо-
двигательный), IV (блоковой) и VI (отводящий) черепные нервы (рис.
20.2).
В стволе мозга, на уровне моста, расположена нейронная сеть ретику-
26‘
804 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Таламус
Интерстициальное
ядро Кахала
Ростральное интерстициальное
ядро продольного пучка
Верхние бугры четверохолмия
Ретикулярная формация
среднего мозга
Мозжечок
Мост
Ядро подъязычного нерва
Ретикулярная формация
продолговатого мозга
Парамедианная ретикулярная
формация моста
Рис. 20.2. Нейронный контроль саккадических движений глаз на уровне структур
среднего мозга.
Направление движения глаз (саккад) управляется мотонейронами двух центров взгляда рети-
кулярной формации. В парамедианной ретикулярной формации моста расположен центр го-
ризонтального направления движения вгляда, а в ростральном интерстициальном ядре ретику-
лярной формации находится центр вертикального направления движения взгляда. HI, IV,
VI — ядра глазодвигательного, блокового и отводящего нервов, которые иннервируют мышцы
глазных яблок человека. Стрелками показано направление регулирующих влияний структур
верхних бугров четверохолмия.
лярной формации, функция которой состоит в регуляции многих аспектов
внимания: генерация быстрых (саккадических) движений глаз (парамеди-
анная ретикулярная формация); участие в произвольной регуляции на-
правления взгляда человека (ретикулярные нейроны вблизи ядра отводя-
щего нерва); генерация горизонтального направления движения взгляда
(нейроны ретикулярной формации моста и прилегающие к ядру подъязыч-
ного нерва); содружественное горизонтальное направление движения
взгляда обоих глаз. Последний комплекс ядер в соответствии с его функ-
цией получил название центр горизонтального направления движения взгля-
да человека. Аналогичная нейронная сеть, или центр вертикального на-
правления движения взгляда, расположен в области интерстициального ядра
Кахала.
Центры горизонтального и вертикального направления движения взгля-
да человека регулируют поворот глазных яблок через мотонейроны ядер
III, IV и VI черепных нервов и иннервируемые ими мышцы.
Функция верхних бугров четверохолмия. Верхние бугры четверохолмия
представляют собой область ствола мозга, которая образует основный вы-
ход к ретикулярным нейронным сетям моста и среднего мозга, при уча-
стии которых возникает движение или остановка взгляда человека при
внимании.
Зрительные сигналы от сетчатки гдаз поступают к нейронам верхних
слоев верхних бугров четверохолмия. Аксоны этих нейронов направляются
к нейронам глубоких слоев верхних бугров четверохолмия, а также к тала-
мусу и стволовым ядрам. Функция этих «нисходящих» связей заключается
в том, что нейроны глубоких слоев верхних бугров четверохолмия стано-
вятся высокоактивными перед началом движения глаз и головы в направ-
лении источника раздражения во внешней среде. Высокочастотная актив-
ность нейронов глубоких слоев верхних бугров четверохолмия резко акти-
вирует глазодвигательный механизм человека через нейроны ретикулярных
20. Физиологические основы познавательной деятельности человека • 805
ядер ствола мозга при возникновении реакции внимания (поворот глаз и/
или головы человека). Поэтому глаза человека способны в течение не-
скольких десятков миллисекунд ориентироваться на объект после раздра-
жения рецепторов сетчатки.
Функция нейронов глубоких слоев верхних бугров четверохолмия явля-
ется мультимодальной при внимании, поскольку эти отделы получают
сигналы от зрительной коры, от слуховых, соматосенсорных релейных
ядер и других ядер ствола мозга. Глубокие слои нейронов верхних бугров
четверохолмия имеют строгую топографическую организацию в виде ней-
ронных карт. Это проявляется в том, что при возбуждении нейронов оп-
ределенной локализации участков глубоких слоев верхних бугров четверо-
холмия возникают строго координированные движения глаз и поворот го-
ловы.
20.1.2.2. Функции корковых центров внимания
Простое произвольное изменение внимания с одной точки зрительного
пространства на другую с помощью движений глаз активирует у человека
большую нейронную сеть коры головного мозга, которая включает ком-
плекс зрительная кора — средняя теменная область, лобные глазные поля,
дополнительные глазные поля, часть передней поясной извилины, заднюю
затылочную кору (рис. 20.3). Функции этой обширной нейронной сети
при внимании заключаются в контроле движения глаз и головы человека
через верхние бугры четверохолмия. Структуры верхних бугров четверохол-
мия и корковых центров внимания функционируют параллельно. В обыч-
ных физиологических условиях возникновение активности в нейронах
верхних бугров четверохолмия находится под контролем лобных зритель-
ных полей, задней теменной коры и нейронов ретикулярной части черной
субстанции ствола мозга.
Функции комплекса структур стриарной зрительной коры и средней те-
менной области (зона МТ) в механизмах внимания состоят в проведении
зрительной информации о направлении и скорости движения стимулов в
зрительном поле к структурам задней теменной коры головного мозга. Из
структур экстрастриарной коры информация передается в лобную кору, а
именно в лобные и дополнительные глазные поля, которые выполняют ос-
новную функцию в планировании и выполнении ориентированных движе-
ний глаз. Структуры первичной зрительной коры выполняют основную
функцию в объект-ориентированном внимании человека и в выборе или
селекции объекта глазодвигательной системой мозга человека с того мо-
мента, как только цель оказалась в сенсорном поле. В ответ на зрительный
стимул стриарная затылочная кора человека активируется довольно быст-
ро, примерно через 40 мс.
Роль теменной коры человека в механизмах внимания. При внимании те-
менные отделы мозга человека, а именно структуры внутритеменной бо-
розды и нижней теменной доли, повышают уровень метаболической ак-
тивности. Эта область коры головного мозга является центром, который
вовлекается в поддержание внимания специфической пространственной
локализации или, как правило, в генерацию сигналов в глазодвигательной
системе мозга при изменении внимания с одной локализации на другую.
Эту функцию отделы задней теменной коры выполняют через структуры
верхних бугров четверохолмия.
Нейроны задней теменной коры реагируют строго топографически на
зрительные стимулы и участвуют в произвольном изменении пространст-
806 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Произвольное зрительное внимание
(произвольное движение глаз)
Непроизвольное зрительное
внимание
Пространственное зрительное
внимание
Двигательные
программы
ЗТК
кора
Мозжечок
Зрительная
информация
Генератор
саккад
. Верхние бугры
четверохолмия
Интеграция
пространственной
зрительной
информации
Ретикулярная
формация
Структуры
ствола
мозга
Рис. 20.3. Функции высших центров внимания, расположенных в коре головного
мозга человека.
Центры внимания коры головного мозга человека включают зрительную кору (ЗК), которая
направляет зрительную информацию в заднюю теменную кору (ЗТК). Этот корковый центр
внимания регулирует механизм непроизвольного зрительного внимания, осуществляет инте-
грацию пространственной зрительной информации. Задняя теменная кора (преимущественно
правая) направляет сигналы в лобное глазное поле (ЛГП) и дополнительное глазное поле
(ДТП). ЛГП функционирует как центр произвольного зрительного внимания человека и вы-
полняет двигательные программы при участии ДГП. Префронтальная кора включена в произ-
вольную регуляцию внимания путем коррекции функции центров лобного и дополнительного
зрительного полей. Человек при участии структур префронтальной коры способен тормозить
непроизвольное внимание. Верхние бугры четверохолмия являются основным подкорковым
центром регуляции непроизвольного (рефлекторного) и произвольного внимания при участии
различных сенсорных систем и под влиянием двигательных команд центров внимания коры
головного мозга.
венного внимания. Это проявляется прежде всего в том, что при изменении
направления взгляда на новое положение рецептивные поля нейронов зад-
ней теменной коры также перемещаются на новое положение. Более того,
рецептивные поля задней теменной коры способны занимать новое поло-
жение до начала движения глаз и изменения направления взгляда на но-
вый объект, а следовательно, нейроны этого коркового центра внимания
получают сигналы из других отделов коры относительно предполагаемого
движения глаз в новом направлении. Поэтому активность нейронов задней
теменной коры как основного высшего центра внимания всегда нарастает
при ориентировании взгляда на объекте, если человек ожидает появления
объекта в зрительном поле или думает о появлении в нем такого объекта.
После начала действия зрительного стимула зрительная информация дос-
тигает теменных отделов коры примерно через 60 мс и передается к этим
структурам от стриарной коры. При этом активация структур стриарной и
20. Физиологические основы познавательной деятельности человека • 807
Рис. 20.4. Функция ретикулярной части
черной субстанции и базальных ганглиев в
контроле внимания.
Ретикулярная часть черной субстанции (ЧС)
ствола мозга получает вход от хвостатого ядра
(ХЯ) базальных ганглиев и функционирует как
«ворота» при произвольном контроле движения
глаз. При отсутствии зрительных стимулов в по-
ле зрения ЧС тонически тормозит структуры
верхних бугров четверохолмия, что препятствует
возникновению нежелаемых движений глаз и
изменению направления взгляда. Непосредст-
венно перед началом произвольного изменения
взгляда в черную субстанцию из базальных ганг-
лиев поступают импульсы, которые снижают
торможение верхних бугров четверохолмия. Ак-
тивация базальных ганглиев происходит под
влиянием двигательных команд из центров вни-
мания коры головного мозга. В результате ста-
новится возможным произвольное изменение
направления взгляда. Знак плюс обозначают ак-
тивацию центров регуляции направления взгля-
да со стороны верхних бугров четверохолмия и
торможение, а знак минус — торможение верх-
них бугров четверохолмия со стороны ЧС.
Центры горизонтального и вертикального
направления движения взгляда
экстрастриарной коры под влиянием зрительного стимула не осознается
человеком.
Функция лобных глазных полей заключается в планировании движения
глаз при изменении направления взгляда. Лобные глазные поля располо-
жены в прецентральной борозде лобных долей мозга человека. Реакции
нейронов этой области коры головного мозга организованы топографиче-
ски: нейроны активируются только в том случае, если объект находится в
определенной части зрительного поля. Функция лобных глазных полей
осуществляется при помощи проекций аксонов нейронов этого отдела ко-
ры к структурам верхних бугров четверохолмия и стволовым премоторным
(ретикулярным) нейронам. Лобные зрительные поля являются корковым
премоторным центром в регуляции внимания и обеспечивают выбор целей
для движений глаз и изменения направления взгляда среди многих зри-
тельных целей в окружающем человека пространстве. Функцией этой об-
ласти коры головного мозга является торможение непроизвольных движе-
ний глаз, а также генерация произвольных, не связанных со зрительными
стимулами движений глаз (например, при мышлении).
Функцией дополнительных глазных полей коры является выполнение дви-
жения глаз в направлении объекта внимания. Дополнительные глазные по-
ля расположены медиальнее лобных глазных полей, вдоль межполушарной
борозды на дорсомедиальной поверхности лобных долей. Нейроны этого
коркового центра внимания обладают пространственной чувствительно-
стью и регулируют движение глаз таким образом, чтобы объект или его ка-
кая-то часть находились в центре поля зрения. Благодаря функции струк-
тур дополнительного глазного поля у человека осуществляется объект-ори-
ентированное внимание.
Функция базальных ганглиев. В механизме произвольного внимания
функция базальных ганглиев состоит в активации структур ретикулярной
части черной субстанции среднего мозга (рис. 20.4). Нейроны ретикуляр-
808 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ной части черной субстанции функционируют подобно «воротам» относи-
тельно функции структур верхних бугров четверохолмия при внимании.
Так, при отсутствии в окружающем пространстве вызывающих внимание
зрительных стимулов нейроны черной субстанции тормозят нейроны верх-
них бугров четверохолмия и препятствуют, таким образом, случайному из-
менению направления взгляда. Под влиянием произвольных двигательных
команд из лобной коры, поступающих к нейронам черной субстанции че-
рез структуры хвостатого ядра базальных ганглиев, тоническое торможение
нейронов верхних бугров четверохолмия снижается, и они приобретают
способность регулировать произвольные движения глаз в направлении
объекта в поле зрения.
Дорсолатеральные области префронтальной коры обеспечивают произволь-
ный контроль различных форм поведения, в том числе ориентированные
движения глаз и головы на интересующий человека объект в окружающем
пространстве. При участии префронтальной коры осуществляется произ-
вольная задержка внимания, в результате которой человек осознанно не
производит объект-ориентированное движение глаз и головы в направле-
нии источника сильного сенсорного раздражения.
20.1.3. Внимание при различных модальностях
Внимание у человека проявляется в отношении стимулов разной модаль-
ности, т. е. зрительных, слуховых, соматосенсорных, обонятельных и вес-
тибулярных. Слуховые центры ствола мозга взаимодействуют с системой
регуляции зрительного внимания на уровне структур верхних бугров четве-
рохолмия (ориентировочный рефлекс), а на уровне коры — с теменными
областями коры, преимущественно со структурами нижней теменной доли
в правом полушарии мозга. Активация структур этой области коры голов-
ного мозга происходит при бинауральном и дихотическом звуковом воз-
действиях. Соматосенсорная система на подкорковом уровне (через верх-
ние бугры четверохолмия) и на уровне коры при участии структур сомато-
сенсорной коры также взаимодействуют с системой регуляции внимания
через отделы задней теменной коры.
Наибольшее внимание вызывает стимул одной модальности (зритель-
ной, слуховой или соматосенсорной), т. е. внимание эффективнее осуще-
ствляется при участии конкретной сенсорной системы. Однако, несмотря
на наличие внимания к различным внешним стимулам, в коре головного
мозга человека имеется одна единая надмодальная (или мультимодальная)
система регуляции внимания. Основным мультимодальным центром вни-
мания является задняя теменная кора головного мозга. Функция задней те-
менной коры при внимании проявляется в том, что пространственное слу-
ховое или пространственное зрительное внимание у человека всегда на-
правлены на один и тот же объект в окружающем пространстве. При про-
извольном изменении пространственного слухового внимания в направле-
нии наилучшего восприятия звукового стимула в эту же область направля-
ется пространственное зрительное внимание, и наоборот. При участии зад-
ней теменной коры каждая сенсорная система вызывает эквивалентное из-
менение пространственного внимания у человека, что способствует увели-
чению процессов локализации внимания на определенной цели в условиях
активации различных сенсорных систем мозга.
20. Физиологические основы познавательной деятельности человека • 809
20.2. Восприятие
Восприятие — это процесс трансформирования физического раздражения
в физиологическую информацию. Процессы восприятия сенсорной ин-
формации различной модальности осуществляется в первичных и вторич-
ных проекционных областях коры головного мозга. Наиболее изученными
видами восприятия у человека являются зрительное и слуховое, в меньшей
степени — соматосенсорное. Механизмы восприятия запаха и вкуса изло-
жены в главе 17 «Сенсорные системы».
20.2.1. Зрительное восприятие
Человек распознает новый объект, появившийся в поле зрения, в течение
долей секунды. При этом зрительное восприятие включает оценку контра-
ста, лица, объектов, зрительного пространства, а также единого изображе-
ния, воспринимаемого обоими глазами (бинокулярное зрение).
Структуры ЦНС, которые участвуют в «зрительном поведении» челове-
ка, широко представлены в коре головного мозга, однако не все из них во-
влечены в процесс восприятия. Например, двустороннее повреждение ви-
сочных долей коры вызывает выраженные нарушения в зрительном распо-
знавании и памяти, но при этом остается неизмененным зрительное вос-
приятие. Напротив, повреждение у человека затылочных долей вызывает
слепоту. Небольшой латентный период и длительность реакций нейронов
первичной зрительной коры (порядка 40 мс) на зрительные стимулы сви-
детельствуют об основной функции этого отдела зрительной коры в вос-
приятии зрительной информации.
20.2.1.1. Функции стриарной коры в зрительном восприятии
Первичная зрительная кора (стриарная кора, поле VI) является первым
корковым уровнем, на котором конвергируют сигналы от сетчатой оболоч-
ки обоих глаз человека, и последним полем в зрительной системе, нейроны
которого реагируют непосредственно на зрительные стимулы (рис. 20.5).
Первичная зрительная кора представляет собой единственную структуру
коры головного мозга человека, которая получает сигналы непосредственно
от нейронов латеральных коленчатых
ядер таламуса и создает точную ретино-
топическую карту зрительных полей, ко-
торые представляют собой нервный суб-
страт зрительного восприятия. Комплек-
сы нейронов стриарной коры формируют
разнонаправленные мозаичные столбы,
слои и модули, при участии которых про-
исходит восприятие цвета, стереопсиса
Рис. 20.5. Схема передачи импульсов от сет-
чатки к нейронам зрительной коры (поле VI).
Пути от латеральных коленчатых тел (Л КТ) в пра-
вую и левую области зрительной коры состоят из
относительно равных проекций от сетчатки обоих
глаз. Нейроны первичной зрительной коры активи-
руются на зрительные стимулы и обеспечивают
функцию зрительного восприятия.
810 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
(глубины), ориентационных стимулов, контуров объектов, движения, фор-
мируется глазодоминантность. Нейроны стриарной коры имеют высокую
чувствительность к световому контрасту, а их реакции прямо взаимосвяза-
ны с порогами светового контраста (описаны в главе 17 «Сенсорные систе-
мы»). Восприятие освещенности предмета также является функцией стри-
арной коры. Структуры полей V1/V2 являются высшими зрительными цен-
трами восприятия, в которых начинается восприятие бинокулярного зре-
ния, хотя в этих центрах зрительного восприятия расположено меньше
нейронов с подобными свойства, чем в последующих зрительных полях.
Восприятие стереоопсиса происходит в результате взаимодействия на ней-
ронах стриарной коры монокулярной зрительной информации, возникаю-
щей в результате перекрытия одного объекта другим в поле зрения.
К своеобразной форме зрительного восприятия относится воображение
человека в отсутствие какой-либо световой стимуляции сетчатки глаз. При
этом у человека активируются ретинотопические поля зрительной коры, а
не ее ассоциативные поля.
20.2.1.2. Зрительное восприятие при участии отделов экстрастриарной коры
Нейроны зрительного поля VI моносинаптически связаны с нейронами
других ретинотопических зрительных областей коры — V2, V3/VP, V3A,
V4v и МТ фис. 20.6).
При произвольном или непроизвольном внимании активируются выс-
шие зрительные корковые поля, преимущественно поля V2 и V4.
Лобная кора
Двигательная кора
Соматосенсорная кора
Теменная кора
Рука)
’Лицо
Восприятие движения
Восприятие скорости
движения
. /V7
ТА/ЗА
jbV3
[l;X-V2
Височная кора
Боль
V8 V4v
Восприятие цвета
1 \ Восприятие лица
Запах Слуховая кора
Первичная
зрительная
кора (поле V1)
Рис. 20.6. Схематическое изображение локализации функций познавательной дея-
тельности человека в коре головного мозга: восприятие (по данным позитронно-
эмиссионной томографии и функционального магнитного резонансного исследова-
ния).
На рисунке отмечены участки коры головного мозга человека, в которых возрастает локально
мозговой кровоток при выполнении человеком заданий на восприятие. VI, V2, V3, V3A, V/P,
V4v — классические ретинотопические поля зрительной коры человека, функцией которых яв-
ляется восприятие относительно простых зрительных стимулов, поля V7 и V8 — имеют слабо
выраженную ретинотопическую организацию. За пределами этих полей расположены области
зрительной коры, не имеющие ретинотопической организации и поэтому выполняющие «гло-
бальные» функции восприятия движения, пространства, цвета и формы объектов внешней
среды. МТ — средняя височная область.
20. Физиологические основы познавательной деятельности человека • 811
После восприятия зрительного раздражения структурами поля VI, далее,
в каждом последующем ретинотопическом поле, происходит копирование
'Д карты поля VI. Поэтому при повреждении полей V2, V3/VP и V4v у па-
циентов возникает выпадение восприятия */4 части зрительного поля (квад-
ранопсия), а поля VI и V3A —половины зрительного поля (гемианопсия).
Структуры полей V2, V3/VP, V3A, V4v и МТ зрительной коры образуют вто-
рой синаптической уровень и относятся к восходящим ассоциативным зри-
тельным полям, функцией которых является распознавание относительно
простых параметров зрительных сигналов. Кпереди от промежуточных по-
лей зрительной коры (поля V7 и V8), которые имеют слабо выраженную ре-
тинотопическую карту, расположены структуры зрительной коры, которые
не имеют ретинотопической организации и поэтому выполняют «глобаль-
ные» функции зрительного восприятия движения, цвета, лиц, участков зри-
тельного поля и кинетику движения граничащих между собой элементов.
В процессе зрительного восприятия у человека функции этих отделов
зрительной коры количественно разделены на теменные и височные ин-
формационные процессы («потоки»). Структуры средней теменной облас-
ти (МТ), верхней теменной области, полей V3A и V7 формируют у челове-
ка теменной (дорсальный) путь или поток, который обеспечивает воспри-
ятие движения и пространства.
Вентральный (височный) «поток» информации (структуры полей V8,
лицевого поля веретенообразной извилины) выполняют функцию воспри-
ятия более сложно организованных особенностей зрительной информа-
ции, таких как цвет и форма объектов.
20.2.1.3. Особенности зрительного восприятия индивидуальных лиц и объектов
Зрительное поле, специализирующееся на восприятии лица, расположено
в пределах средней части веретенообразной извилины (поля 37 и 20 по Брод-
ману). Нейроны веретенообразной извилины активируются в большей сте-
пени при рассматривании анатомически нормальных и вертикально распо-
ложенных лиц, чем перевернутых или хаотично расположенных в поле
зрения объектов. В то же время степень активации «лицевого» поля коры
не различается при рассматривании знакомых и незнакомых лиц. Нейро-
ны «лицевого» поля распознают лица на категориальном или генетическом
уровне, т. е. до стадии индивидуального распознавания лица.
Непосредственно латеральнее веретенообразной извилины расположено
поле, нейроны которого специализируются на распознавании словесных
форм и слов, связанных в последовательный ряд при их написании. У че-
ловека письменный язык является поздним филогенетическим приобрете-
нием, и в онтогенезе он развивается относительно медленно. Поэтому ор-
ганизация «словесного» поля коры не запрограммирована генетически или
эпигенетически. Четвертый синаптический уровень зрительного воспри-
ятия содержит нейроны, которые избирательно активируются при рассмат-
ривании зданий, предметов домашнего быта (стул, стол).
При восприятии цвета активируются нейроны задней части продольной
и веретеноообразной извилины. Односторонее разрушение этого участка
мозга ведет к контралатеральной потере восприятия цвета (полушарная ах-
роматопсия), без нарушения восприятия остроты зрения, движения или
идентификации объекта в поле зрения. Дорсальная теменно-затылочная
область (место соединения полей 19 и 37 по Бродману) участвует в воспри-
ятии зрительной информации движения, которая позволяет идентифици-
ровать целенаправленность перемещения объекта или человека.
812 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Нейронные популяции зрительных ассоциативных полей обеспечивают
восприятие лиц и объектов путем группового кодирования: один нейрон
способен реагировать на различные лица и в то же время одно лицо акти-
вирует несколько нейронов. Нейроны «лицевого» поля нижней височной
коры избирательно реагируют на категорийно-специфические особенно-
сти, такие как расстояние между глазами на лице, прическа, выражение
глаз, направление взгляда. Нейроны, реагирующие на идентичные особен-
ности объекта, образуют вертикальные колонки диаметром примерно 400
мкм. Несколько рядом расположенных колонок, реагирующих сходным
образом на воспринимаемую зрительную информацию, взаимосвязаны ме-
жду собой и образуют модули размером несколько миллиметров. Группы
подобных взаимосвязанных, но распределенных ансамблей настроены на
восприятие строго определенных особенностей, которые определяют класс
объекта. Восприятие и основанное на этой информации зрительное распо-
знавание может быть градуальным и основанным на частичной информа-
ции, так как одна и та же информация может проходит через многие ассо-
циативные нейронные сети.
20.2.2. Слуховое восприятие
Слуховое восприятие у человека представляет собой процесс восприятия
звука как смыслового феномена. Благодаря процессу слухового восприятия
человек может идентифицировать, объяснить и понять смысл звуков. Зву-
ковое восприятие осуществляется структурами слуховой коры (см.
рис. 20.6). Проекции слуховой сенсорной системы в кору головного мозга
имеют тонотопическую организацию. Первичная слуховая кора верхнего
отдела височной части коры состоит из трех тонотопических карт: Al, R и
СМ. Нейроны полей А1 и R получают большее количество афферентных
волокон от слуховых релейных ядер таламуса, вентральных медиальных
коленчатых тел и реагируют непосредственно на тональные стимулы.
Нейроны поля А1 настроены на восприятие чистых тонов и силы звука.
Средняя и передняя части верхней височной извилины в большей степени
воспринимают специфические фонетические параметры разговорного язы-
ка. Нейроны верхней височной извилины настроены на восприятие сег-
ментов и последовательности фонем, так же как и на их когерентность при
восприятии многослоговых и сложных слов. Эти отделы мозга распознают
речь на пресемантическом уровне. Средняя височная извилина у человека
(поле 21 по Бродману) содержит нисходящие слуховые ассоциативные по-
ля. Примерно половина нейронов этих полей обладает высоко избиратель-
ными реакциями на звуковые стимулы в основном в форме подавления на
понимаемую речь. По аналогии со зрительным восприятием, структуры
восходящих слуховых полей воспринимают элементарные особенности
звука — частоту и силу, а нисходящих слуховых полей — кодируют более
сложную информацию, связанную с идентификацией слов, локализацией
источника звука и характеристикой индивидуального голоса человека.
20.2.3. Соматосенсорное восприятие
Соматосенсорное восприятие обеспечивает восприятие у человека тактиль-
ных, проприоцептивных и висцеральных ощущений. Предметы в значи-
тельной степени отличаются друг от друга по форме и тактильным свойст-
вам. При восприятии с помощью тактильных рецепторов формы и поверх-
ностных свойств предметов возникает соответствующая активация полей
20. Физиологические основы познавательной деятельности человека • 813
Рис. 20.7. Активация участков первичной со-
матосенсорной коры при восприятии так-
тильных раздражителей.
На схеме показаны поля 1 и 36 (моторные поля 6 и
4 по Бродману), структуры которых воспринимают
все виды тактильных стимулов, и структуры поля 2
(моторное поле 44 по Бродману), которые воспри-
нимают прикосновение к участку кожной поверх-
ности. Структуры передней части внутритеменной
извилины и надкраевой извилины активируются
при тактильном восприятии формы предметов.
сенсомотороной коры головного мозга
человека (рис. 20.7). Форма предметов
воспринимается как при активном, так
и при пассивном исследовании предметов при участии структур передней
части внутритеменной и надкраевой извилин. Все виды тактильных раз-
дражений воспринимаются структурами поля 36 и поля 1. Нейроны сома-
тосенсорной коры, локализованные в пределах поля 2, воспринимают пре-
имущественно структуру поверхности исследуемых человеком предметов,
другие — скорость движения предмета по поверхности кожи пальцев руки.
Более тонкое отличие формы и поверхностной структуры предметов вос-
принимается структурами вторичной сенсомоторной коры и распознается
при участии нейронных сетей премоторных полей коры головного мозга и
префронтальной коры.
20.3. Сознание
Сознание — специфически человеческая форма идеального отражения и ду-
ховного освоения действительности, она включает в себя и осознание че-
ловеком своей психической деятельности.
Слово «сознание» имеет латинский корень conscio, образованный при-
ставкой сит, обозначающей «с», и глагола scio, обозначающиего «знать». В
этом оригинальном смысле слова быть сознающим — это значит знать о
чем-то или о ком-то, либо о самом себе.
В нейрологии сознание ассоциируют с состоянием бодрствования, спо-
собностью воспринимать, взаимодействовать и общаться с окружающей
средой и с другими людьми. По объективным критериям состояние созна-
ния человека определяют как суженное, уменьшенное или восстанавли-
вающееся; сознание может быть потерянным, подавленным и вновь вос-
становленным. В наиболее общей форме быть в сознании — это значит
быть бодрым, активным, бдительным и настороженным.
При нормальном состоянии бодрствования сознание — это, прежде все-
го, проявление поведения человека. Сознание является содержанием жиз-
ненного опыта в каждый момент времени. В этом смысле сознание обра-
щено внутрь человека и касается такой качественной стороны, как субъек-
тивные размеры жизненного опыта. Наряду с этим сознание является все-
гда персональным и требует от субъекта определенной точки зрения. Со-
держание сознания является стабильным на протяжении коротких перио-
дов жизни, но, как правило, варьирует в течение длительных интервалов.
Сознание является центральной для человека величиной: продолжение че-
ловеческой жизни, когда его сознание утрачивается необратимо, в клини-
ческой практике рассматривается как напрасное усилие.
814 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Осознание человеком своей психической деятельности относится к ка-
тегории самосознания. Самосознание проявляется в склонности к замеша-
тельству, осведомленности об осведомленности и знании о самом себе.
20.3.1. Нейрофизиологические корреляты сознания
Любым произвольным (субъективным, осознанным) действиям человека
соответствует определенный паттерн нейронной активности в различных
областях мозга. Сознательный жизненный опыт является результатом
взаимодействия значительного количества распределенных областей мозга,
которые вовлечены в процессы внимания, восприятия, памяти и целена-
правленного действия. Поэтому под нервными коррелятами сознания пони-
мают совпадение паттернов активности (электрической или метаболиче-
ской) между разделенными и расположенными на расстоянии друг от дру-
га полями коры головного мозга при различных состояниях сознания. На-
пример, сознание тесно коррелирует с состоянием сна и бодрствования,
зрительным осознанием (синоним осведомленность) человека, а также вни-
манием, восприятием, памятью и мыслительными функциями мозга
(рис. 20.8).
20.3.1.1. Электрическая активность мозга человека
Электрическая активность мозга может быть физиологической основой
осознаваемого жизненного опыта, но может и не иметь отношения к тако-
вому. По данным современной нейрофизиологии мозга, поведенческие и
электрофизиологические проявления в деятельности мозга человека в цик-
ле сон—бодрствование коррелируют с тремя главными состояниями созна-
ния: бодрствование, быстрое движение глаз и медленные волны сна (см.
главу 21 «Сон и бодрствование»). Состояние сознания во время бодрство-
вания и быстрых движений глаз характеризуется десинхронизацией ЭЭГ
мозга человека, а в стадии медленных волн сна — электрической синхро-
низацией мозга. Однако в у-диапазоне ЭЭГ (35—45 Гц) у человека синхро-
низация электрической активности мозга имеет место как при бодрствова-
нии, так и при быстрых движениях глаз. Электрофизиологическими корре-
лятами сознания при этих состояниях являются сенсорные вызванные по-
тенциалы в сенсомоторных областях коры и огромное разнообразие «ког-
нитивных» потенциалов (потенциалы готовности), возникающих в различ-
ных участках двигательных полей коры головного мозга примерно за 1 с до
начала выполнения движения. Поэтому все осознаваемые человеком по-
знавательные процессы имеют определенные электрические корреляты.
20.3.1.2. Активация мозга человека как нейрофизиологическая основа
проявлений состояний сознания
Представление о «восходящей активирующей системе» базируется на веду-
щей роли ретикулярных структур ствола мозга в регуляции состояний соз-
нания. В регуляции состояний бодрствования, а следовательно, проявле-
ний сознания у человека участвуют: холинергические ядра верхнего отдела
ствола мозга и базального отдела переднего мозга; норадренергические яд-
ра и особенно голубое пятно (locus coeruleus); гистаминергические проек-
ции от заднего гипоталамуса; дофаминергические и серотонинергические
пути, начинающиеся в стволе мозга.
В состоянии бодрствования таламокортикальные нейроны тонически
Рис. 20.8. «Нервные корреляты сознания». Совпадение паттернов активности (электрической или метаболической) между распре-
деленными полями коры головного мозга коррелирует с состоянием сознания человека и обусловливает индивидуальную форму
отражения действительности (сознание) и способность осознавать психическую деятельность (самосознание).
20. Физиологические основы познавательной деятельности человека • 815
816 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
деполяризованы под влиянием холинергических, норадренергических и
гистаминергических входов от структур ствола мозга и гипоталамуса, бло-
кирующих калиевую гиперполяризацию мембраны нейронов. Это обуслов-
ливает спайковый тип активности таламокортикальных нейронов, что
обеспечивает беспрепятственную передачу сигналов в таламус и далее в
кору головного мозга. Снижение числа деполяризующих входов к нейро-
нам таламуса вызывает в них гиперполяризацию и инициирует в них зал-
повый тип активности, что имеет место при медленных волнах сна. Залпо-
вые разряды таламических нейронов на фоне их гиперполяризации обу-
словлены низкопороговой кальциевой мембранной проницаемостью, но в
целом этот тип активности таламических нейронов ограничивает поступ-
ление сигналов к таламусу.
Одновременное растормаживание ретикулярных ядер таламуса в начале
сна как следствие снижения торможения холинергических входов от
структур ствола мозга, приводит к синхронизированному ГАМК-ергиче-
скому торможению таламокртикальных нейронов, которое ведет к увели-
чению «веретен» ЭЭГ во вторую стадию сна. Дальнейшая гиперполяриза-
ция мембраны таламокортикальных нейронов по мере увеличения глубины
сна обусловливает их участие в медленных волнах сна на ЭЭГ. В возник-
новении этого состояния участвуют как индивидуальные, так и сетевые
свойства таламокортикальных, кортикоталамических нейронов и нейронов
ядер ретикулярной формации. Различия между спайковыми и залповыми
реакциями таламокортикальных нейронов совпадают с поведенческими
различиями человека, т. е. способностью реагировать при состоянии бодр-
ствования и отсутствием таковой во время сна.
Во время бодрствования изменение уровня активации мозга также кор-
релирует с активностью структур покрышки ствола мозга и таламуса, кото-
рые регулируют проявления сознания. Так, при переходе от состояния по-
коя к выполнению зрительных и соматосенсорных тестов происходит ак-
тивация ядер покрышки среднего мозга и интраламинарных ядер таламуса.
Снижение степени активации структур среднего мозга и таламуса ассоции-
руется со снижением способности человека выполнять длительное время
распознавание звуковых тестов.
Электрическая активность клеток холинергических и норадренергиче-
ских ядер ствола мозга полностью зависит от функционального состояния
мозга и коррелирует с состоянием бодрствования. При этом чем длитель-
нее период бодрствования, тем сильнее выражено у человека желание
спать. Важнейшую роль в качестве «гипнотоксина» может играть увеличе-
ние внеклеточной концентрации аденозина во время бодрствования. Это
приводит к торможению активных холинергических ядер верхнего отдела
ствола мозга и структур основания переднего мозга, что обусловливает
снижение степени проявления сознания у человека (например, возникает
дремота).
Уровень физиологической активности нейротрансмиттерных систем
влияет на познавательную деятельность человека, находящегося в состоя-
нии сознания. Так, повреждение норадренергической системы снижает
уровень внимания, повреждение холинергической системы снижает точ-
ность действий человека, нарушение дофаминергической системы увели-
чивает длительность латентного периода реакций и длительность самих ре-
акций, нарушение серотонинергической системы ведет к импульсивным
реакциям человека. Потеря сознания у человека под влиянием некоторых
анестетиков обусловлена избирательным подавлением функций структур
таламуса, которые связаны с механизмами обезболивания и сна.
20. Физиологические основы познавательной деятельности человека • 817
20.3.1.3. Осознание зрительного восприятия (зрительная осведомленность)
Процессы, лежащие в основе содержания элементарного сознания, тесно
связаны, прежде всего, с функцией зрения человека. При этом у человека
между уровнем мозговой активности и зрительным жизненным опытом
имеется высокая степень корреляции. Выше были изложены представле-
ния о функции ретинотопической карты зрительной коры в восприятии
простых зрительных стимулов.
Стриарная кора окружена многочисленными анатомо-функциональны-
ми зрительными полями, к основным из которых относятся затылочно-ви-
сочные и затылочно-теменные. «Потоки» зрительной информации (вен-
тральный и дорсальный соответственно) через эти поля обеспечивают вос-
приятие человеком формы, цвета, глубины и движения. При этом актив-
ность зрительных вентральных путей («вентральный поток») является обя-
зательным условием осознания «зрительного жизненного опыта». Поле в
области червеобразной извилины (вентральный поток) избирательно реа-
гирует на различные выражения человеческого лица, а структуры парагип-
покампального поля воспринимают локализацию объектов в окружающем
зрительном пространстве.
20.3.1.4. Внимание и сознание
Внимание имеет общее значение для человека. Так, педагог обращается к
своим студентам с фразой «будьте внимательными», когда он просит их
сконцентрироваться, чтобы выслушать его учебное задание. Эта общая
форма внимания зависит от состояния бодрствования и бдительности че-
ловека, когда зрение и слух «настраиваются» на лучшее восприятие, а ум-
ственная деятельность ориентируется на выполнение задания. При выпол-
нении какого-либо задания на внимание абстрактное знание начинает
управлять вниманием человека. Это называется произвольно контролируе-
мым вниманием. При этой форме внимания оно направляется в опреде-
ленную точку пространства и поэтому обозначается как фокусное внимание.
Фокусное внимание является часовым на воротах осознанного воспри-
ятия. Фокусное внимание, ориентированное на характерные особенности
объекта, всегда является активным, т. е. произвольным. Наиболее важным
нервным коррелятом произвольного внимания является изменение ней-
ронной активности в специфических областях коры головного мозга при
движении объекта к фокусу зрительного внимания и от него. Этот процесс
называется нейронной экспрессией внимания и в определенной степени отра-
жает нейронное «сопровождение» сознания. Активность веретенообразной
(восприятие лица) и гиппокампальной (восприятие объектов) областей ко-
ры головного мозга человека с высокой достоверностью изменяется при
переключении внимания с лиц на виды при их одновременной демонстра-
ции испытуемым. Феномен нейронной экспрессии внимания выражен в наи-
большей степени, если человек осведомлен о содержании воспринимаемого
зрительного стимула.
20.4. Память и научение
Памятью называется способность мозга человека приобретать, сохранять в
мозге полученную в процессе индивидуальной жизни информацию, а так-
же неоднократно использовать эту информации) в дальнейшем поведении.
Научением называется относительно устойчивое изменение в поведении
818 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
или знании человека, которое возникает в результате жизненного опыта
или тренировки. В результате научения у человека возникает новая форма
поведения, которая позволяет ему полнее приспосабливаться к условиям
окружающей среды. Научение основано на механизмах памяти мозга чело-
века, поэтому их развитие в процессе индивидуальной жизни человека не-
разрывно связаны между собой.
Мозг должен «уметь учиться», добавлять к врожденным механизмам по-
ведения индивидуальный опыт. Это «умение» основано на пластичности
ЦНС, т. е. на способности к перестройке функциональных свойств и свя-
зей нейронных сетей мозга человека под влиянием внешних воздействий.
Такая перестройка должна обеспечивать адаптивное изменение поведения
благодаря сохранению памяти о приобретаемом опыте. Для образования
энграмм (следов) памяти необходимо изменять эффективность синаптиче-
ских переключений между определенными классами нейронов.
У человека врожденное и приобретенное поведение основано на реали-
зации имеющихся в распоряжении его клеток генетических программ. Раз-
ница состоит в том, что программы врожденного поведения формируются
еще до рождения человека и обеспечивают стабильный минимум поведен-
ческих реакций, а программы приобретаемых форм поведения мозг реали-
зует в соответствии с индивидуальным опытом, приобретаемым в процессе
жизни, т. е. в условиях непрерывного научения.
20.4.1. Формы памяти и научения
Внимание человека привлекает любой новый раздражитель. Непроизволь-
ный механизм внимания обусловлен врожденным ориентировочным реф-
лексом (реакция «что такое?», как определял ее И. П. Павлов). Но если та-
кой раздражитель не представляет опасности и не связан с какой-либо
биологической потребностью, то он постепенно утрачивает новизну и ори-
ентировочная реакция исчезает, что получило название привыкания, или
габитуации. Примером привыкания может служить отсутствие реакции на
обычный городской шум за окнами квартиры или учебной аудитории.
Противоположную привыканию форму научения представляет собой сен-
ситизация, проявляющаяся в усилении обычных ответов на прежде ней-
тральные раздражители. Так, например, пациент, испытавший боль на
приеме у зубного врача, повышенно реагирует даже на манипуляции, не
вызывающие боли. Привыкание и сенситизация являются простейшими
формами научения, существующими благодаря специальным механизмам
памяти.
Привыкание, сенситизация и условные рефлексы образуются на основе
имплицитной (от англ, implicit — неявный, скрытый) памяти, не требую-
щей обязательного участия сознания в процессе научения. Благодаря та-
кой памяти у человека образуется большинство двигательных навыков, на-
пример умение ездить на велосипеде или танцевать, при ее участии все де-
ти овладевают основами родного языка. Другая форма памяти — экспли-
цитная (от англ, explicit — явный, высказанный до конца) — относится к
познанию окружающего мира, других людей; она образуется при активном
участии сознания и подразделяется на эпизодическую и семантическую.
Эпизодическая память содержит биографические сведения, включающие
данные о конкретных людях, времени и окружающем пространстве или
местности, а семантическая включает в себя общие знания об устройстве
мира и общества. Имплицитную память иначе называют процедурной, а
эксплицитную — декларативной. Например, приобретенное умение ездить
20. Физиологические основы познавательной деятельности человека • 819
Рис. 20.9. Основные формы памяти. Наиболее простыми формами памяти являются
габитуация (привыкание) и сенситизация, на основе которых у человека развивают-
ся различные виды, например, двигательной памяти (развитие речи или спортив-
ный навык). Познание окружающего мира осуществляется на основе эксплицитной
памяти. Формирование памяти — это последовательные во времени процессы сме-
ны кратковременной памяти на долговременную.
Рис. 20.10. Схематическое изображение последовательности процессов образования
памяти при ее восприятии, хранении, а также извлечении из долговременной па-
мяти.
на велосипеде базируется на процедурной, или имплицитной, памяти, а
все остальные знания о велосипеде, равно как и очередное его изобрете-
ние, требуют привлечения декларативной, или эксплицитной, памяти. Раз-
личные формы памяти можно представить в виде следующей схемы
(рис. 20.9).
Образование памяти у человека при восприятии сенсорной информа-
ции представляет собой ступенчатый процесс преобразования одного фи-
зиологического процесса в другой: активация сенсорных систем при по-
ступлении информации в мозг, реверберация возбуждения в нейронных
кругах мозга (кратковременная память), консолидация следов памяти и об-
разование энграмм (долговременная память), хранение энграмм памяти в
структурах мозга и процесс извлечения энграмм памяти при научении
(рис. 20.10). Новая информация в течение небольшого отрезка времени
(около 1 мин) сохраняется в кратковременной памяти, емкость которой не-
значительна: в среднем 7 ± 2 элементов информации, т. е. семь слов, букв,
цифр или символов (но не более двенадцати). Приблизительно 20—30 се-
кунд мозг обрабатывает эту информацию, решая, насколько она важна и
820 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
стоит ли сохранять ее в дальнейшем. Повторное поступление той же ин-
формации повышает вероятность ее преобразования в долговременную па-
мять, емкость которой практически не ограничена, а длительность хране-
ния варьирует от нескольких минут до десятилетий.
20.4.2. Нейронные механизмы имплицитной памяти
Поступающее в центральную нервную систему возбуждение способно не-
которое время циркулировать в замкнутых цепях нейронов. При этом по-
ступившая в мозг информация может циркулировать в структурах мозга и
после того, как прекратилось действие раздражения на органы чувств.
Процесс циркуляции возбуждения в замкнутых нейронных сетях мозга че-
ловека называется реверберацией (рис. 20.11). При реверберации потенциа-
лы действия возвращаются к одним и тем же нейронам, сохраняя их воз-
бужденное состояние в течение некоторого времени, этим обеспечивается
кратковременная память. В возбужденных нейронах стимулируется фосфо-
рилирование ферментов, связанных с окислением субстратов, что способ-
ствует кратковременному сохранению активности таких клеток. Если дей-
ствие раздражителя на органы чувств повторяется, то одни и те же синап-
сы мозга, куда поступает это возбуждение, используются многократно, и в
результате эффективность их деятельности постепенно повышается. Повы-
шение эффективности синап-
тической передачи лежит в ос-
нове механизма долговремен-
ной памяти.
20.4.2.1. Габитуация
и сенситизация
При габитуации и сенситиза-
ции проявляются две ступени
образования памяти: кратко-
временная и долговременная.
Однократное действие серии
слабых стимулов приводит к
непродолжительной габитуа-
ции, а однократно применен-
ный сильный раздражитель
(однократное действие серото-
нина) вызывает кратковремен-
ную сенситизацию, отмечае-
мую на протяжении несколь-
ких минут. Четыре-пять таких
тренировок, следующих друг за
другом, приводят уже к долго-
временной сенситизации, со-
храняющейся в течение суток,
а при дальнейшем повторении
тренировок сенситизация со-
храняется на протяжении 3 нед
и больше.
При повторяющихся трени-
ровках (повторяющемся дейст-
Замкнутые нейронные сети
вие, 20.11. Схема реверберации возбуждения
в замкнутых нейронных сетях как основы
кратковременной памяти.
Нервные импульсы от органов чувств, поступая в
замкнутые нейронные сети отделов мозга, способ-
ны циркулировать в них определенное время. По-
вторяющиеся раздражения органов чувств поддер-
живают циркуляцию (реверберацию) возбуждения
в этих нейронных сетях мозга. Время ревербера-
ции возбуждения соответствует длительности
кратковременной памяти. Результатом длительной
реверберации возбуждения может быть повыше-
ние эффективности синаптической передачи за
счет облегчения экзоцитоза медиаторов и увеличе-
ния количества рецепторов на постсинаптических
мембранах нейронов (компоненты механизма дол-
говременной памяти). Стрелками на схеме обозна-
чено направление циркуляции возбуждения в
замкнутых нейронных сетях мозга.
20. Физиологические основы познавательной деятельности человека • 821
Рис. 20.12. Изменения в синапсах при длительной габитуации и сенситизации.
Количество терминалей пресинаптического аксона уменьшается при продолжительной габи-
туации и возрастает при длительной сенситизации. В первом случае эффективность работы
синапса уменьшается, во втором — увеличивается. Происходящие изменения структуры си-
напса являются основой долговременной памяти.
вии серотонина) в клеточное ядро сенсорного нейрона постепенно переме-
щается некоторое количество цАМФ-зависимой протеинкиназы, что вы-
зывает фосфорилирование регуляторных белков, управляющих процессом
транскрипции — считывания информации с молекулы ДНК. В результате
активируются гены, руководящие синтезом белков, необходимых для до-
полнительной активации компонентов все той же цАМФ-зависимой про-
теинкиназы, и происходит образование новых синапсов на шипиках денд-
ритов нейронов. Последнее увеличивает число синаптических связей ней-
рона с другими нервными клетками, участвующими в научении. Этот про-
цесс является основой постепенного формирования долговременной памя-
ти в мозге. Сенсорный нейрон может увеличивать количество своих преси-
наптических окончаний, одновременно имеет место увеличение числа ак-
тивных зон, из которых непосредственно выделяется медиатор. При этом
образуются дополнительные шипики на дендритах мотонейрона, которые
вступают в синаптические контакты с дополнительными пресинаптически-
ми окончаниями аксона сенсорного нейрона, и таким образом облегчается
проведение возбуждения по нейронной сети, участвующей в научении
(рис. 20.12). При длительной габитуации имеют место противоположные
изменения: количество синаптических связей между сенсорным и мотор-
ным нейронами уменьшается приблизительно на '/3 с одновременным
уменьшением числа активных зон в синапсах. Это является структурным
компонентом снижения физиологической активности нейронной сети и
уменьшения проявления ее реакции на действие внешнего раздражения,
что не приводит к запоминанию информации.
Привыкание и сенситизация являются неассоциативными формами
научения. Изучение механизмов привыкания и сенситизации позволило
822 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
показать, что уменьшение или увеличение тока кальция в пресинаптиче-
ском окончании может быть одним из механизмов кратковременной памя-
ти. Образование же долговременной памяти оказалось связанным с ис-
пользованием вторичных посредников, экспрессией генов и возникнове-
нием дополнительных синапсов. Аналогичные механизмы в принципе мо-
гут быть использованы почти во всех синапсах мозга, поэтому нет основа-
ний считать, что эти механизмы специфичны лишь для каких-то особен-
ных «нейронов памяти».
20.4.2.2. Ассоциативное научение (условные рефлексы)
Гипотеза, объясняющая нейронные основы ассоциативного научения, со-
стоит в том, что между разными видами специфических нейронов в цен-
тральной нервной системе могут существовать малоэффективные и даже
нефункционирующие синапсы. Так, например, естественная реакция на
звуковой раздражитель не связана со слюноотделением, поскольку возбуж-
дение клеток слуховой коры не передается к нейронам, регулирующим пи-
щеварительную деятельность, из-за низкой эффективности синаптической
передачи между этими нервными центрами. При частом сочетании звуко-
вого сигнала с вкусовыми ощущениями малоэффективные синапсы начи-
нают функционировать, благодаря чему один лишь звуковой сигнал вызы-
вает слюноотделение. Изменения в синапсах могут быть следствием био-
химической модификации или увеличения контактирующих участков меж-
ду пре- и постсинаптическими нейронами (рис. 20.13).
Классический условный рефлекс, представляющий собой более слож-
ную, чем габитуация или сенситизация, форму научения, отличается необ-
ходимостью запомнить значение совпадения во времени двух стимулов:
слабого, обычно не вызывающего никакого ответа вследствие стойкого
привыкания к нему, с сильным, всегда вызывающим закономерную реф-
лекторную реакцию. Формирующаяся при образовании условного рефлек-
са память оказывается более стойкой, чем при габитуации или сенсити-
зации.
Генетические исследования подтверждают участие связанных с цАМФ
вторичных посредников в процессах научения. Установлено также, что из-
бирательная блокада одного из регуляторных белков, участвующих в экс-
прессии гена для цАМФ, приводит к утрате способности образовывать
долговременную память, но не затрагивает кратковременную. Однако было
бы ошибкой считать, что в основе процессов Научения и памяти лежат
только связанные с цАМФ обменные процессы. При других формах науче-
Рис. 20.13. Модификация синаптического
переключения в малоэффективном синап-
се (модель Хебба).
1. Нейрон А всегда возбуждает нейрон X.
2. Нейрон Б не способен возбудить нейрон X.
3. Нейроны А и Б в течение некоторого времени
действуют совместно, нейрон X возбуждается.
4. После этого эффективность синаптической
передачи между нейронами Б и X повышается и
нейрон Б приобретает способность возбуждать
нейрон X. Ек — критический уровень деполяри-
зации мембраны нейрона X.
МП — мембранный потенциал.
20. Физиологические основы познавательной деятельности человека • 823
ния могут использоваться иные системы вторичных посредников, а опи-
санные выше процессы простых форм научения могут комбинироваться с
другими молекулярными процессами, приводя в итоге к образованию ком-
плексных форм памяти. Имплицитная и эксплицитная формы памяти у
человека нередко оказываются связанными друг с другом.
20.4.3. Механизм образования эксплицитной памяти
В головном мозге человека отсутствует единое место хранения памяти.
Эксплицитная память формируется в тех регионах мозга, которые необхо-
димы для ее приобретения и участвуют в ее образовании. Такими структу-
рами памяти могут быть гиппокамп, обонятельный мозг, миндалины, ба-
зальный передний мозг и, конечно, кора больших полушарий. Из проек-
ционных областей коры больших полушарий в сохранении памяти участ-
вуют вторичные проекционные области и ассоциативная кора, осуществ-
ляющие распознавание наиболее сложной информации и мыслительные
функции человека. При этом вторичные зрительные области хранят зри-
тельную память, слуховая кора — слуховую, сенсомоторные области, наря-
ду с мозжечком и базальными ядрами, используются для фиксации следов
памяти о выработанных двигательных навыках. Префронтальная кора иг-
рает важную роль в хронологической организации событий памяти.
Структурным коррелятом долговременной памяти является энграмма
памяти. Поводом к возникновению процессов, приводящих к образова-
нию энграмм памяти, могут быть новые, неожиданные воздействия на ор-
ганизм человека стимулов внешней среды или, напротив, исчезновение
привычных либо ожидаемых событий. Место мозга, в котором происходит
образование энграммы памяти, зависит от модальности сенсорных воздей-
ствий и их сложности, что требует участия в запоминании разных вторич-
ных сенсорных и ассоциативных областей коры. Место образования новой
энграммы памяти обусловлено участками мозга, где хранятся образован-
ные ранее энграммы. Новая энграмма памяти образуется при участии
структур, выполняющих функцию внимания в текущий момент времени.
Наконец, энграмма памяти образуется при участии лимбических структур
мозга, от которых зависит эмоциональный фон, ускоряющий или замед-
ляющий процесс научения.
Однако знание места хранения долговременной памяти в мозге не по-
зволяет объяснить механизм воспоминания или извлечения следов памяти.
Эту ситуацию можно сравнить с памятью, хранимой на видеокассете: ее
можно извлечь лишь с помощью видеомагнитофона и телевизора. Поэтому
энграмма памяти, хранящаяся в определенных структурах мозга, может
быть извлечена при участии структур других регионов мозга, непосредст-
венно не участвующих в хранении информации. У человека извлечение
энграммы эксплицитной памяти (семантической или биографической)
связано с процессами осознания информации и речевой деятельностью.
Нейронные механизмы поддержания необходимого уровня бодрствования
и речевой деятельности у человека взаимодействуют с теми регионами
мозга, где зафиксированы следы памяти.
Показано, что удаление гиппокампа приводит к нарушению образова-
ния эксплицитной долговременной памяти, но не влияет на формирование
имплицитной долговременной памяти, что указывает на разные места об-
разования этих форм памяти. В пользу этого свидетельствуют и наблюде-
ния, демонстрирующие избирательное нарушение только имплицитной
памяти (например, при поражениях мозжечка).
824 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
В то же время отмечено, что в условиях указанного отсутствия структур
гиппокампа выработанные прежде навыки не исчезают, однако приобрете-
ние новых становится практически невозможным. Эти результаты свиде-
тельствует о том, что гиппокамп не является местом хранения долговре-
менной памяти, но он необходим для преобразования кратковременной
памяти в долговременную.
Память связана не с синтезом каких-либо уникальных белков, специ-
фичных для каждого ее вида. Она зависит не столько от конкретных био-
химических процессов, сколько от того, в каких нейронах и синапсах про-
исходят эти процессы, где находятся такие нейроны и синапсы и как они
связаны с другими клетками. Если указанные синапсы не активируются на
протяжении длительного времени, то происходит естественное разрушение
белковых молекул, синтезированных для образования энграмм памяти, и
таком случае происходит забывание.
20.5. Речь
Речь представляет собой исторически сложившуюся форму общения лю-
дей, опосредованную языком. Язык является системой звуковых, словар-
ных и грамматических средств для общения, мышления, передачи инфор-
мации от человека человеку, от одного поколения — другому. В повседнев-
ном обиходе эти два термина понимаются как способность говорить. Этой
уникальной способностью человек отличается от всех живых существ. Речь
в узком смысле слова понимают как использование в общении между
людьми голосовых, слуховых, зрительных и мануальных навыков, к кото-
рым относят способность произносить слова, воспроизводить письменные
знаки, улавливать речь на слух и распознавать зрительно печатный или ру-
кописный текст.
Различают речь экспрессивную (моторную), предназначенную для комму-
никации, или социального общения людей, и внутреннюю речь, «немую»,
используемую для решения мыслительных задач и внутреннего програм-
мирования. Внутренняя речь характеризуется своей фрагментарностью и
краткостью по сравнению с экспрессивной внешней речью. Перевод чело-
веком внутренней речи во внешнюю происходит путем синтаксического
переструктурирования, т. е. перевода в синтаксически расчлененную, раз-
вернутую и понятную другим людям форму. Особо выделяют письменную
речь, отличающуюся, во-первых, использованием письменной графики, а
во-вторых, специфическими для нее синтаксическими конструкциями и
функциональными стилями. Речь неразрывно связана с мышлением, памя-
тью, сознанием, эмоциями, вместе с которыми она включается в целостный
поведенческий акт речевой деятельности.
20.5.1. Свойства языка
Среди всех возможных средств коммуникации у людей различают вербаль-
ные, т. е. словесные, невербальные (геометрические фигуры, математиче-
ские знаки, рисунки, фотографии), а также паралингвистические (жесты,
мимика, музыкальные мелодии). Представляя собой типично человече-
скую форму общения, язык отличается от прочих форм коммуникации
своими свойствами. Во-первых, языку присуща креативность (созидатель-
ность). Уже на стадии овладения родным языком человек не заучивает
наизусть все услышанное, но через неосознаваемые (имплицитные) про-
цессы переходит к общим правилам построения осмысленных выражений.
20. Физиологические основы познавательной деятельности человека • 825
Словарный запас одного человека содержит большее или меньшее, но все-
таки ограниченное количество слов, из которых он может построить неог-
раниченное количество предложений, в том числе и такие, которые рань-
ше никогда не слышал и не говорил. Во-вторых, язык отличается своей
уникальной формой-, из сравнительно небольшого числа используемых в
нем звуков создается бесконечное множество комбинаций. Звуки, или фо-
немы, являются самыми малыми звуковыми единицами, изменение кото-
рых может привести к изменению значения слова, например: дом — том,
волос — голос и т. п. В-третьих, для языка человека характерна определен-
ная структура, которая регламентируется грамматикой, включающей пра-
вила комбинации фонем в словах (морфология) и правила комбинации
слов в предложениях и оборотах речи (синтаксис). В-четвертых, в каждом
языке существует возможность абстрагирования, т. е. передачи информа-
ции, не связанной непосредственно с конкретными событиями. Такая воз-
можность отсутствует у простых символических систем коммуникации: на-
пример, с помощью жестов можно передавать только специфическую ин-
формацию, связанную с текущими обстоятельствами. Но в то же время
речь посредством жестов, мимики, интонации и положения тела может
приобретать нужную или желаемую эмоциональную выразительность. В-пя-
тых, являясь основным средством социального общения, язык наряду с
коммуникацией способствует образованию индивидуального эмоциональ-
ного опыта людей. Он служит способом выражения сознания и самосознания
человека и его индивидуальности. Язык непосредственно связан с мышле-
нием человека.
20.5.2. Речевой аппарат
В создании звуков человеческой речи участвуют мышцы гортани, языка,
губ, мягкого неба, глотки и дыхательная мускулатура, которые иннервиру-
ются подъязычным, блуждающим, лицевым, диафрагмальным нервами, а
также мотонейронами спинного мозга, регулирующими сокращения дыха-
тельных мышц. Звуки возникают во время выдоха, когда поток воздуха
проходит через гортань, а ее голосовые связки вибрируют, изменяя тем са-
мым величину подсвязочного давления. Характер дыхания должен приспо-
сабливаться к речи, что проявляется в регуляции скорости и продолжи-
тельности выдоха и, соответственно, регуляции глубины вдоха, чтобы на-
бранного в легкие воздуха было достаточно для произнесения фразы опре-
деленной длины. Произвольное управление дыханием особенно наглядно
проявляется при пении, когда быстрый вдох сменяется продолжительным
контролируемым выдохом, соответствующим длине музыкальной фразы.
Строение лицевого черепа и челюстей человека обеспечивает такое по-
ложение языка, при котором его задняя часть оттеснена в глотку, а гор-
тань — вниз. Только такая позиция позволяет языку изменять форму рас-
положенной над гортанью полости и тем самым модулировать звуки, воз-
никающие в гортани. Во время прохождения воздуха через голосовую щель
происходят правильные вибрации голосовых связок, благодаря которым
возникают гармонические колебания воздушного потока, соответствующие
частоте вибрации. В зависимости от частоты колебаний воздуха возникают
звуки определенной высоты. Частоту вибрации, а следовательно, и высоту
звука, можно произвольно изменять посредством сокращений мышц гор-
тани, регулирующих натяжение голосовых связок. Рождающиеся при этом
звуки соответствуют частотам с правильным периодом, которые можно
разложить на ряд чистых тонов, а сами звуки можно пропеть; в составе
826 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
языка они называются гласными. Воспроизведение таких звуков гортанью
обозначается термином фонация.
Произведенные гортанью тоны преобразуются в звуки речи располо-
женными выше полостями носа и рта, выполняющими функцию резонато-
ров. Этот процесс управляется сокращениями мышц языка, губ, мягкого
неба и глотки и определяется как артикуляция. Согласные звуки возника-
ют в основном с помощью артикуляции; если гортань не участвует в их
произнесении, то получаются глухие согласные, например [т], а при уча-
стии гортани — звонкие согласные [д]. В отличие от гласных согласные
звуки невозможно пропеть.
Самая низкая частота колебаний воздушного столба в гортани характе-
ризуется наибольшей амплитудой, воспринимается на слух как самая
громкая и называется основной. Одновременно с этой частотой голосовые
связки, а затем и расположенный над ними воздушный столб, колеблются
с большими частотами, кратными частоте основного тона. Они обусловле-
ны колебаниями независимых сегментов воздушного столба, что образует
обертоны. Сочетание основного тона с обертонами создает индивидуально
узнаваемый тембр голоса. Полости носа и рта в речевой функции выпол-
няют роль резонаторов и фильтров, поскольку в большой мере поглощают
некоторые частоты, что определяется размером и формой этих полостей в
каждый конкретный момент. В результате при произнесении гласных со-
храняется лишь часть акустической энергии гортани, соответствующая не-
поглощенным частотам, которые называются формантами. Формантные
частоты различаются у одной и той же гласной в зависимости от того, с
какой согласной она образовала слог. Распознание речи на слух основано
на различении формант, и, следовательно, наименьшими смысловыми
единицами речи являются слоги.
20.5.3. Речевые структуры мозга
Речевая деятельность присуща только человеку, что почти исключает экс-
периментальное изучение ее физиологических основ на животных. Поэто-
му в основу представлений о нейрофизиологических механизмах речи лег-
ли клинические наблюдения: это весьма распространенный путь к понима-
нию функции, который состоит в анализе причин ее нарушения. Значи-
тельно позже к опыту клинических наблюдений добавились методы пози-
тронно-эмиссионной томографии (ПЭТ) и ядерно-магнитного резонанса
(ЯМР), позволяющие регистрировать динамику активности различных ре-
гионов мозга в процессе речевой деятельности.
20.5.3.1. Нарушения речи при очаговых повреждениях мозга
Утрата экспрессивной речи и/или способности понимать устную речь
вследствие приобретенных поражений мозга обозначается как афазия, для
менее выраженных речевых расстройств используется термин дисфазия. В
подавляющем большинстве случаев поражения мозга, вызывающие афа-
зию у правшей, имели место в левом полушарии мозга.
При поражении левой нижней лобной извилины (зона Брока, соответст-
вующая полям 44—45 Бродмана) развивается моторная афазия, проявляю-
щаяся нарушением разговорной речи, а нередко и письма. Выраженность
нарушений речи пропорциональна размерам очага поражения, но в любом
случае имеют место неудовлетворительные артикуляция и мелодичность ре-
чи, которая становится крайне немногословной (телеграфной).
20. Физиологические основы познавательной деятельности человека • 827
Центральная борозда
Рис. 20.14. Речевые структуры мозга человека в левом полушарии.
При участии зоны Вернике (поле 22) происходит программирование смысла речи человека.
Импульсы из зоны Вернике по дугоообразному пучку передаются в зону Брока (поле 45).
Функцией зоны Брока является создание двигательной программы речи. Эта программа вы-
полняется через первичную двигательную зону (поле 3). Цифры на рисунке соответствуют ну-
мерации полей по классификации Бродмана.
Сенсорная афазия отмечается у людей при поражении зоны Вернике, за-
нимающей заднюю часть левой верхней височной извилины (поле 22 по
Бродману). При сенсорной афазии у человека нарушается понимание обра-
щенной к нему речи. При попытке говорить больной правильно произно-
сит звуки, но допускает ошибки в понимании и использовании слов, мо-
жет произносить грамматически правильные, но бессмысленные фразы.
Отсюда был сделан вывод о том, что для восприятия и программирования
речи используются разные регионы левого полушария мозга.
Из этого представления возникла первая модель формирования речи, ко-
торая различала сенсорную деятельность, связанную с пониманием смысла
воспринимаемых слов и обеспечиваемую задней третью верхней височной
извилины (зона Вернике), а также моторную деятельность, формирующую
программы движений для мышц артикуляции и находящуюся в зоне Бро-
ка. Две эти анатомические области связаны между собой дугообразным пуч-
ком нервных волокон, что обеспечивает их взаимодействие (рис. 20.14).
20.5.3.2. Модель речевой деятельности Вернике—Гешвинда
На протяжении первой половины XX столетия доминировало представле-
ние об эквипотенциальности, т. е. функциональной равноценности раз-
личных областей коры, поэтому считалось, что речь не имеет анатомиче-
ских коррелят. Однако в дальнейшем были показаны различия в деятель-
ности речевых центров левого полушария при восприятии устной речи и
чтении написанного текста (рис. 20.15). Звуки голоса воспринимаются
слуховыми рецепторами внутреннего уха человека, где происходит транс-
формация энергии звуковой волны в потенциалы действия слуховых нер-
вов. По слуховым нервам они поступают через медиальное коленчатое те-
ло таламуса в первичную слуховую кору (поле 41), затем во вторичную
слуховую кору (поле 42), а оттуда в зону Вернике (поле 22), обеспечиваю-
828 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Рис. 20.15. Модель речевой деятельности Вернике—Гешвинда.
Устная речь формируется при поступлении информации в слуховую кору и при декодирова-
нии ее в зоне Вернике, где происходит программирование ответной речи. Из зоны Вернике
информация передается по дугообразному пучку в зону Брока, в которой готовится программа
артикуляции, поступающая в первичную моторную кору. Моторная кора осуществляет регуля-
цию деятельности артикуляционных мышц. При письменной речи информация поступает
вначале в зрительную кору, а затем через угловую извилину передается также в зону Вернике,
далее в зону Брока, затем в двигательную кору, регулирующую артикуляцию.
щую понимание значения услышанных слов и конструирование ответных.
Из зоны Вернике информация передается по дугообразному пучку в зону
Брока, где локализована память об артикуляции, на основе которой созда-
ется программа собственной речи. Электрическая активность нейронов зо-
ны Брока передается в первичную двигательную область коры, которая ре-
гулирует в соответствии в поступающей в нее программой деятельность
мышц артикуляции. Речь человека, которая возникает при этом, содержит
смысловые составляющие, которые являются источником информации для
другого человека.
При чтении текста соответствующие сигналы направляются от сетчатки
глаз по зрительным нервам к наружному коленчатому телу таламуса, отту-
да — в первичную, а затем во вторичную зрительную кору. Далее перерабо-
танная информация попадает в угловую извилину теменной доли (поле
39), где перерабатывается зрительная, слуховая и тактильная афферентная
информация, а оттуда — в зону Вернике. Если текст следует прочесть
вслух, то для этого требуется отправить надлежащую информацию по дуго-
образному пучку в зону Брока, а затем активировать первичную моторную
кору.
20.5.3.3. Современная модель речевой деятельности человека
Наблюдения клиницистов показывают, что как при моторной, так и при
сенсорной афазии очаги поражения мозга обычно захватывают довольно
большую территорию, а не ограничиваются лишь зоной Брока или Верни-
ке. В то же время встречающиеся иногда в клинике ограниченные повреж-
дения этих областей, как правило, не приводят к типичным афазиям Бро-
ка или Вернике.
20. Физиологические основы познавательной деятельности человека • 829
Применение специально разработанных когнитивных тестов в комплек-
се с ПЭТ и ЯМР привело к появлению новой модели речевой деятельно-
сти. Согласно этой модели информация, воспринимаемая на слух и при
чтении, перерабатывается в мозге человека и достигает зоны Вернике не
по общим, а по раздельным (параллельным) нейронным сетям. Поэтому
зрительные и слуховые репрезентации слов в структурах мозга нередко
происходят при участии полей коры головного мозга специфической мо-
дальности, откуда информация непосредственно поступает в зону Брока.
Наконец, слуховая информация может перерабатываться при участии не-
скольких параллельных нейронных сетей мозга. Например, восприятие
обычных слов, имеющих значение для понимания содержания речи, и вос-
приятие бессмысленных в контексте воспринимаемой речи слов происхо-
дят посредством переработки информации разными и независимыми путя-
ми, образующими отдельные входы в зону Брока. Это свидетельствует о
том, что информация о звуке как о средстве языка и информация о смы-
словом значении этого звука в воспринимаемой человеком речи перераба-
тывается разными нейронными сетями мозга (рис. 20.16).
20.5.3.4. Литерализация речи
Речевая деятельность, как правило, обеспечивается только одним, домини-
рующим относительно этой функции полушарием, обычно левым. У чело-
века левое полушарие в среднем на 1 см3 больше, и следовательно в левом
полушарии содержится больше нейронов. Анатомические различия между
полушариями мозга у человека подтверждаются при сравнении величины
объема зоны Вернике, которая слева больше примерно у 70 % взрослых
людей, а также у плодов, что указывает на врожденное, генетически запро-
граммированное происхождение асимметрии мозга. Указанная асимметрия
выявляется и при сопоставлении борозд и извилин лобных долей: в левом
полушарии обычно имеется четкое разделение всех трех лобных борозд,
каждая из которых хорошо выражена и ветвиста, тогда как в правом полу-
шарии чаще отмечается атипичное расположение борозд и извилин, неред-
ко прерывающихся и не имеющих общего ствола. Объем зоны Брока в ле-
вом полушарии значительно больше, чем в правом, а количество пирамид-
ных нейронов в этой зоне слева больше, чем справа, причем различия от-
мечены не только у взрослых людей, но и у новорожденных.
Функциональное доминирование левого полушария окончательно уста-
навливается лишь после изучения родного языка. Маленькие дети, пере-
несшие тяжелые повреждения речевых областей левого полушария, в по-
следующем способны овладеть языком благодаря вовлечению в эту функ-
цию правого полушария. Но взрослые люди после необратимых обширных
повреждений речевых областей левого полушария уже не могут компенси-
ровать утрату речи с помощью правого полушария. Из этого следует, что
анатомическая асимметрия является только необходимой основой, но не
гарантией языковой специализации полушарий, которая происходит на
ранних стадиях развития, когда специфическая функциональная актив-
ность способствует развитию и совершенствованию нейронных переклю-
чений между речевыми структурами левого полушария мозга.
Однако речи человека присущи не только когнитивные, но и аффектив-
ные компоненты: интонация, регулируемая высота голоса, ритм речи.
Считается, что они контролируются зеркально организованными в правом
полушарии структурами мозга. При повреждении правого полушария
больные говорят одинаково ровно, независимо от того, рассказывают они
830 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Речь Письмо
Рис. 20.16. Современная модель нейронных переключений, необходимых для рече-
вой деятельности.
Восприятие устной и письменной речи у человека осуществляется при участии слуховой и
зрительной коры (первый уровень переработки поступающей в мозг речевой информации). На
втором уровне в структурах зоны Вернике и вторичной зрительной коре происходят процессы
фонологического и зрительного кодирования информации соответственно. Второй уровень
переработки речевой информации представлен гетеромодальными структурами, которые уча-
ствуют в кодировании как устной, так и письменной информации (3). Речевая (устная или
письменная) информация из зоны Вернике и ассоциативной зрительной коры поступает не-
посредственно в премоторное поле Брока и одновременно в лобную кору, откуда также осу-
ществляется контроль формирования программ артикуляции. Результатом последовательной и
параллельной переработки речевой информации является содержание речи и письма.
о радостном событии или печальном; они часто не способны понимать
эмоциональную окраску обращенной к ним речи.
Глухие от рождения дети не могут услышать чужую речь, они усваивают
язык с помощью той или иной системы жестикуляции, в которых тоже су-
ществуют грамматические правила. Овладевающие языком жестов дети
проходят точно такие же этапы развития, что и дети, которые осваивают
обычный разговорный язык. Для овладения языком жестов используются
те же речевые структуры мозга, что и для овладения речью: при левосто-
роннем поражении речевых структур мозга глухие перестают понимать
20. Физиологические основы познавательной деятельности человека • 831
язык жестов, тогда как при правостороннем поражении эта способность
сохраняется.
Хотя существование речевых структур мозга определено генетически,
это отнюдь не гарантирует последующее овладение языком, поскольку его
познание возможно только в условиях социального межличностного обще-
ния ребенка со взрослыми людьми. Предложена также гипотеза о сущест-
вовании критического периода развития, после завершения которого овла-
дение языком происходит трудно. Доказательства в пользу этой гипотезы
основаны на результатах наблюдения за людьми, изучающими второй
язык. При изучении второго языка критический возраст длится от третьего
года жизни до пубертатного периода. После вхождения в пубертатный воз-
раст возможность выучить второй язык стремительно уменьшается, и под-
ростки усваивают его не быстрее и не лучше, чем взрослые люди 30—40
лет, а потому и не овладевают естественным произношением. По другим
данным, второй язык становится родным лишь в том случае, если дети на-
чали овладевать им до семилетнего возраста, а затем результаты ухудшают-
ся по мере прибавления возраста первого знакомства с новым языком.
Аналогичная зависимость имеет место и у глухих детей: чем позже глухой
ребенок начинает осваивать язык жестов, тем больше дефектов примене-
ния этого языка обнаруживается у него в последующей практике. Считает-
ся, что окончание критического возраста совпадает с завершением созре-
вания нейронных соединений речевых структур мозга.
20.6. Мышление
Мышление — психический процесс познавательной деятельности, позво-
ляющий человеку структурировать внешнюю среду без непосредственного
контакта с ней путем операций с мысленными образами и символами объ-
ектов.
Фундаментальным абстрактным символом, с помощью которого чело-
век может передавать информацию об окружающем мире или о себе друго-
му человеку, является слово. С помощью слова у человека осуществляются
словесно-логические мыслительные процессы, результатом которых явля-
ются различные виды смысловых понятий, категорий или гипотез. Наряду
со словесно-логическим человек обладает наглядно-образным мышлением.
Мысли человека, которые являются результатом обоих видов мышления,
характеризуются рядом особенностей. Прежде всего, мысли человека (как
правило, существенные) обладают относительной стабильностью, несмот-
ря на изменение сознания. Так, после бессознательного состояния, вы-
званного действием на мозг человека наркоза, мысли стабилизируются мо-
ментально и самопроизвольно. Человек способен запоминать мысли. При
этом он запоминает стабильные или существенные проявления какого-ли-
бо калейдоскопа событий. В результате человек запоминает наиболее су-
щественные мысли, а не промежуточные результаты мыслительного про-
цесса. Это можно подтвердить тем, что человек обычно не запоминает ле-
тящую птицу, а только сидящую. Мысли человека характеризуются
мультимодальностью. Обычно в мышлении человека используются различ-
ные модальности, т. е. мысли человека могут включать словесные, зри-
тельные или иные компоненты (зрительное воображение или внутреннюю
речь). В мозге человека мысли и обусловливающие их мыслительные про-
цессы возникают последовательно в разное время, а не одновременно не-
сколько мыслей в одно и то же время. Наконец, мысли человека имеют ог-
раниченную емкость. Под этим понимается, что только какая-то и, как пра-
832 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
вило, незначительная часть от общего количества информации, которой
обладает мозг, может быть представлена в мысли человека. Мыслительная
деятельность человека обусловлена функциями ассоциативных областей
коры головного мозга.
20.6.1. Нейрофизиологические основы мыслительной деятельности
человека
Мышление осуществляется преимущественно в ассоциативных отделах ко-
ры больших полушарий, которые являются местом интеграции информации,
поступающей из первичных и вторичных проекционных зон различных
сенсорных систем (рис. 20.17). В ассоциативных областях коры больших
полушарий текущая информация объединяется с таковой, содержащейся в
долговременной памяти (при участии височной коры и гиппокампа). Ин-
теграция всех сенсорных сигналов со следами долговременной памяти дает
возможность человеку оценивать, например, положение тела или головы в
пространстве. Ассоциативные отделы лобной коры больших полушарий
имеют особое значение в интерпретации явлений и событий. Благодаря
двусторонним связям лобной коры и лимбической системы в систему
оценки ситуации включаются эмоции. Лобная кора ответственна также за
выбор целей и прогнозирование событий. Наконец, использование язы-
ка — основа многих мыслительных процессов человека — возможно только
при совместной работе лобных и височных долей (зоны Брока и зоны Вер-
нике). Сохранение и манипулирование информацией является основным
моментом при процессах мышления человека. Эта функция мозга человека
осуществляется на основе эксплицитной рабочей памяти при участии ассо-
циативной префронтальной коры. Указанный отдел коры больших полу-
Предчувствие
Математические
аппроксимации
Первичная
зрительная
кора (поле V1)
Нога
Лобная кора
Аналитические и
образные рассуждения
Пространственная
рабочая память
Математические
аппроксимации
Предчувствие
боли
Объектная
рабочая
память
Точный Запах /
математический /
счет Приятные
прикосновения
Двигательная кора
Соматосенсорная кора
Аналитические рассуждения
Теменная кора
Понимание разговорной речи
Смысловое различение "зрительных" слов
Предчувствие боли
Разговорная речь
Височная кора
Рис. 20.17. Схема локализации функций познавательной деятельности человека в
коре головного мозга: мышление (по данным позитронно-эмиссионной томогра-
фии и функционального магнитно-резонансного исследования). На рисунке отме-
чены участки коры головного мозга человека, в которых возрастает локально мозго-
вой кровоток при выполнении человеком познавательных заданий.
20. Физиологические основы познавательной деятельности человека • 833
шарий мозга человека осуществляет умственные процессы контроля вни-
мания, выбора между альтернативными реакциями, а также репрезентацию
и сохранение инструкций мыслительного задания. Мышление как процесс
распознавания информации осуществляется при участии ассоциативных ви-
сочных и лобных отделов коры больших полушарий. Мышление как поиск
осуществляется при участии задних ассоциативных (теменно-затылочных)
отделов коры мозга. Соответствие решения выработанному критерию
(стратегия) реализуется при участии ассоциативных лобных, височных и
лимбических отделов коры головного мозга.
20.6.1.1. Нейрофизиологические основы абстрактного мышления (рассуждение
человека)
Рассуждение человека (синоним — абстрактное мышление) — это способ-
ность делать некоторые выводы (мысли), что связано с высшими уровнями
человеческого мышления, предполагающего достижение знания и его ис-
пользование в повседневном опыте.
Существуют индуктивная и дедуктивная формы абстрактного мышле-
ния. При дедуктивной форме рассуждения человека, когда происходит вы-
деление части из целого, активируются структуры левого полушария: в ча-
стности, средняя и нижняя лобные извилины (поля 45 и 47 по Бродману)
и область левой верхней затылочной извилины (поле 19 по Бродману).
При индуктивной форме абстрактного мышления, когда часть события рас-
сматривается в его движении, мыслительный процесс связан с активацией
структур левой медиальной лобной извилины, левой поясной извилины и
левой верхней лобной извилины (поля 8, 9, 24, 32 по Бродману). Кроме
того, индуктивное мышление сопровождается активацией структур левой
верхней затылочной извилины и небольшой области латеральной нижней
височной извилины.
В целом, рассуждение человека обусловлено функцией левого полуша-
рия мозга. Области дорсолатеральных лобных извилин имеют отношение к
рабочей памяти человека, а медиальная префронтальная кора — к функци-
ям произвольного внимания.
Функции полушарий мозга человека различаются при других видах
мышления. В частности, распознавание координатной информации, для
которого необходима специфическая информация о расстоянии и локали-
зации, осуществляется структурами правого полушария. Напротив, катего-
риальное распознавание, связанное с восприятием пространства и выража-
емое с помощью речи, осуществляется левым полушарием мозга.
Распознавание пространственного положения объектов является функ-
цией структур задней теменной коры при участии дорсальных зрительных
путей.
20.6.1.2. Нейрофизиологические основы умственных арифметических операций
У человека умственные арифметические операции обусловлены функцией
структур теменной коры. Левая и правая внутритеменные области активи-
руются в случае, если человек производит счет. Степень активации этих
отделов коры головного мозга пропорциональна сложности арифметиче-
ского задания (размер чисел, число операций в единицу времени). В осно-
ве физиологического механизма активации (например, теменной коры)
могут лежать процессы, способствующие выполнению математического за-
дания, такие как манипуляции с клавиатурой и пространственная зритель-
27 - 6095
834 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ная рабочая память. В области внутритеменной извилины у человека нахо-
дятся нейроны, которые воспринимают числа. Так, в результате пораже-
ния в области внутритеменной извилины возникает нарушение счета.
20.6.1.3. Нейрофизиологические основы мышления при чтении
Чтение является важнейшим культурным изобретением человека. При чте-
нии, даже отдельных слов, у человека активируются структуры областей
мозга, которые участвуют в речевой функции. В левой затылочно-темен-
ной борозде на латеральном краю червеобразной извилины расположено
поле зрительной формы слова, которое имеет высокий уровень активности
при написании человеком слов. Структуры расположены несколько лате-
ральнее структур веретенообразной извилины, которые активируются при
восприятии выражения лица человека, и ниже ретинотопических карт зри-
тельной коры и поля, участвующего в восприятии преимущественно объ-
ектов внешней среды. Поле зрительной формы слова является ближайшей
областью к стриарной зрительной коре, которая не имеет ретинотопиче-
ской организации. Функцией структур этого поля является извлечение аб-
страктной, инвариантной репрезентации последовательности (цепочки)
букв. Структуры левого поля зрительной формы слова включены в «вен-
тральный поток» зрительной информации и активируются у человека на
зрительно воспринимаемые как левым, так и правым глазом слова, а не
слова при речи. У человека в процессе развития мышления всегда взаимо-
связана активация структур поля зрительной формы слова с навыками чте-
ния. При нарушении чтения (дислексия) эта область мозга не реагирует при
попытке распознать буквы и слова.
20.6.2. Функции левого и правого полушарий мозга человека
при мышлении
Мышление человека осуществляется в условиях билатеральной (полушар-
ной) организации мозга. При этом языковые и речевые процессы у человека
очень редко представлены в обоих полушариях. Лексическое же знание
(словарный запас человека) широко представлено в коре головного мозга.
При участии структур левого полушария осуществляются все аспекты про-
дукции и понимания языка. Поэтому общий словарь человека «располо-
жен» в левом полушарии. Правое полушарие осуществляет только лексиче-
ские процессы с весьма ограниченной грамматикой.
Синтаксические мыслительные операции, связанные с пониманием глаго-
лов, множественными числами, притяжательными местоимениями, явля-
ются функцией правого полушария. Грамматические рассуждения челове-
ка также обусловлены функцией правого полушария мозга. Несмотря на
то, что при участии только правого полушария люди не могут использо-
вать синтаксис для понимания смысловых высказываний, они, тем не ме-
нее, в состоянии оценить, какое произношение является грамматическим,
а какое не является таковым. Таким образом, как в отношении словаря,
так и грамматики мыслительной способностью правого полушария являет-
ся пассивное распознавание звуковых слов без его продуктивного приме-
нения. Последнее проявляется в том, что люди не могут с помощью право-
го полушария организовать звуки в слова.
Мозг человека имеет неисчислимую емкость. Однако его способность к
интерпретации генерируемых человеком объяснений и гипотез является
функцией только левого полушария головного мозга. Этот мыслительный
20. Физиологические основы познавательной деятельности человека • 835
процесс осуществляется благодаря тому, что в левом полушарии сохраня-
ется информация, понимаемая человеком в целом. Осуществление подоб-
ного процесса в левом полушарии мозга основано на аккуратности распо-
знавания воспринимаемой информации, как словесной, так и зрительной,
при участии правого полушария. Правое полушарие обеспечивает точней-
шую запись событий, которые используются для построения выводов от-
носительно представленного в него материала. Оба полушария участвуют в
проблемном решении умственных задач, но разными путями: правое полу-
шарие — на основе простой информации, а левое — на основе образования
вырабатываемых гипотез. Различие участия двух полушарий мозга в мыш-
лении позволяет человеку видеть мир адаптивно во всем его многообразии.
ГЛАВА 21
Сон и бодрствование
Сон — периодически возникающее функциональное состояние организма
человека, которое проявляется отключением сознания, отсутствием реак-
ций на внешние раздражители, физической пассивностью и относительной
неподвижностью спящего. Важнейшими признаками бодрствования явля-
ются сознание, мышление и двигательная активность. В течение каждых
суток сон и бодрствование поочередно сменяют друг друга, образуя гене-
тически детерминированный суточный цикл сна—бодрствования.
Регулярное периодическое повторение во времени характера и интен-
сивности жизненных процессов, отдельных состояний или событий обра-
зует биологические ритмы, присущие всем живым организмам, включая че-
ловека. Связанный с естественным чередованием дня и ночи 24-часовой
ритм называется циркадным или циркадианным (от лат. circa — около,
dies —день). Циркадная ритмика характерна для большинства физиологи-
ческих функций. Это суточные колебания температуры тела, от минимума
около 6 часов утра до максимума около 6 часов вечера, ритмические изме-
нения интенсивности обменных процессов, ритмическая секреция гормо-
нов и т. д. В суточном цикле сна и бодрствования циркадная ритмика про-
является особенно заметно. Цикл сна—бодрствования синхронизирован с
другими биологическими ритмами, однако он не является причиной их
возникновения.
21.1. Физиологическое значение сна
Несмотря на более чем вековую историю научных исследований физиоло-
гических механизмов и роли сна, его суть остается непонятой. Определить,
в чем заключается функция сна, не менее сложно, чем ответить на вопрос:
в чем состоит функция бодрствования? Восстановительная теория сна объ-
ясняет его физиологическое значение необходимостью восстановления па-
раметров гомеостазиса, измененных в связи с активной деятельностью при
бодрствовании. Согласно циркадианной теории сна он появился в процессе
эволюции в качестве механизма гомеостатической регуляции, предупреж-
дающего появление физиологического дефицита функций. Основу цирка-
дианной теории составляет представление о существовании нервных цен-
тров, выполняющих функцию так называемых биологических часов, кото-
рые ритмически изменяют характер поведенческих реакций и мозговой
деятельности человека на протяжении суток.
21.1.1. Восстановительная теория сна
Восстановительная теория возникла из наблюдений за последствиями дли-
тельного лишения сна (депривации), исчезающими после его предоставле-
ния. В результате депривации сна снижается работоспособность, ухудшается
настроение, повышаются пороги чувствительности к сенсорным стимулам,
21. Сон и бодрствование • 837
а предоставление сна нормализует все указанные функции. Во время сна
происходит снижение мышечного тонуса, интенсивности обменных процес-
сов и температуры тела, уменьшение частоты сокращений сердца, частоты
и глубины дыхания, что способствует уменьшению потребления энергии,
необходимой для этих процессов. Одновременно с этим во время сна уве-
личивается секреция гормона роста, активируются анаболические процессы
и происходит репаративное восстановление белковых молекул в клетках, что
соответствует представлению о восстановительном значении сна.
И. П. Павлов рассматривал сон как распространяющийся в коре боль-
ших полушарий процесс охранительного торможения, цель которого состо-
ит в предупреждении или уменьшении утомления клеток коры, по мнению
автора, неизбежно развивающегося при их деятельности. Эта концепция
является одним из вариантов восстановительной теории сна, акцентирую-
щим внимание на необходимости восстановительных процессов, прежде
всего, в нервных клетках и, в первую очередь, в нейронах коры больших
полушарий. И. П. Павлов полагал, что торможение и сон присущи каждой
нервной клетке («мы видим сон каждой клетки») и специфических цен-
тров сна не существует. Он объяснял механизм сна особой разновидно-
стью торможения, которое начинается с отдельных, наиболее утомленных
клеток, после чего активно иррадиирует и распространяется на большую
часть полушарий мозга.
Однако при регистрации электрической активности нейронов установ-
лено, что большинство из них во время сна не менее активны, чем во вре-
мя бодрствования, и сон нельзя считать результатом только торможения
клеток коры и, соответственно, состоянием нейронного покоя. Электриче-
ская активность нейронов на протяжении сна неоднократно и ритмически
изменяется, поэтому сон не является однородным состоянием, каким его
традиционно представляла восстановительная теория. Ритмические изме-
нения электрической активности мозга во время ночного сна позволяют
выделить различающиеся между собой фазы медленноволнового и быстро-
волнового сна, которые вместе образуют один цикл сна. Цикл продолжается
около 90 мин и затем повторяется 4—6 раз на протяжении ночного сна.
Правильный эндогенный ритм с такой длительностью периода называется
улътрадианным (внутрисуточным).
Восстановительная теория сна не подтверждается при сопоставлении его
продолжительности у разных видов млекопитающих с их физической ак-
тивностью и скоростью обменных процессов. Если необходимость сна дей-
ствительно зависела бы только от накопления продуктов обмена, то у более
активных во время бодрствования млекопитающих продолжительность сна
должна быть большей. Однако животные с малой интенсивностью обмен-
ных процессов и подвижностью (например, ленивцы) спят значительно
дольше, чем гораздо более активные хищники или приматы, и дольше, чем
грызуны, с их высоким уровнем обменных процессов. С позиций восстано-
вительной теории трудно объяснить, почему кошки спят до 14 час в сутки,
а лошади довольствуются всего лишь двумя. Отмечено также, что для дея-
тельных и энергичных людей зачастую характерна меньшая продолжитель-
ность сна по сравнению с теми, кто ведет малоподвижный образ жизни.
21.1.2. Циркадианная теория сна
Эта теория рассматривает цикл сна и бодрствования как результат управ-
ления циркадного ритма с помощью эндогенного механизма, который не
зависит от внешних обстоятельств и определяется как внутренние биологи-
838 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ческие часы. Эндогенный ритм жизненных процессов синхронизирован с
естественным чередованием дня и ночи, и после перемещении человека в
другой часовой пояс он приспосабливается к местному времени. В услови-
ях неизменной освещенности сохраняется продолжительность обычного
24-часового цикла, который может лишь немного удлиняться или укорачи-
ваться. Сохранение правильного характера циркадных ритмов отмечено в
ходе наблюдений за людьми, пребывающими в условиях полярного дня
или ночи. Сходные результаты получены при исследовании добровольцев,
находящихся длительное время в темной пещере или ее искусственном
аналоге без возможности узнавать время суток.
Большинство имеющихся данных указывает на то, что главным коорди-
натором биоритмологических процессов является гипоталамус. Его роль в
происхождении сна стали исследовать после Первой мировой войны, ко-
гда выяснилось что распространившийся в Европе вирусный летаргиче-
ский энцефалит, сопровождающийся непомерной сонливостью, обуслов-
лен повреждением нейронов промежуточного мозга. Позже было обнару-
жено, что водителем циркадных ритмов, включая цикл сна—бодрствова-
ния, являются супрахиазмальные ядра (СХЯ) гипоталамуса, расположенные
над зрительным перекрестом. У приматов, в частности у человека, сущест-
вуют два первичных синхронизатора биологических ритмов. Один из них
представляют СХЯ, инициирующие возникновение медленноволнового
сна, регулирующие интенсивность секреции гормона роста и скорость вы-
ведения кальция из организма. Другой синхронизатор биологических рит-
мов находится в одной из областей вентромедиального ядра (ВМЯ) гипота-
ламуса и служит в качестве регулятора быстроволнового сна, интенсивно-
сти секреции кортикостероидов, температуры тела и выведения калия из
организма. Активность обоих компонентов биологических часов согласует-
ся между собой в естественном цикле сна—бодрствования.
Циркадианную и восстановительную теорию в настоящее время приня-
то рассматривать не как альтернативные, а как взаимодополняющие. В со-
ответствии с таким подходом сон следует считать, с одной стороны, след-
ствием длительного предшествующего бодрствования, а с другой — прояв-
лением активности эндогенного генератора циркадного ритма, синхрони-
зированного с чередованием дня и ночи.
21.2. Периодичность физиологических процессов во время сна
Об изменениях глубины сна проще всего судить по тому, как легко разбу-
дить спящего человека. Иногда для этого достаточно слабых раздражите-
лей, например негромкого голоса или легкого прикосновения, но по мере
углубления сна порог чувствительности спящего повышается, и чтобы раз-
будить его, требуются более сильные раздражители. На протяжении ночно-
го сна порог пробуждения у спящего человека изменяется неоднократно.
Изменение порогов чувствительности к внешним воздействиям дает повод
характеризовать сон как поверхностный или глубокий. Объективный метод
наблюдения за спящими людьми на протяжении всего ночного сна от за-
сыпания до пробуждения предоставляет большую информацию об особен-
ностях нормального сна и его нарушениях, он позволяет выявить отдель-
ные стадии сна, отражающие ультрадианную ритмику.
Любой отдельно рассмотренный признак сна не является достаточным
для его характеристики, поскольку может встречаться и при других состоя-
ниях человека. Поэтому для изучения сна используется полисомнография,
т. е. одновременная длительная регистрация нескольких физиологических
21. Сон и бодрствование • 839
функций у спящего человека. Полисомнографическое исследование вклю-
чает регистрацию электрической активности мозга (электроэнцефалогра-
фия, ЭЭГ), электрической активности скелетных мышц (электромиогра-
фия, ЭМГ), движений глаз (электроокулография, ЭОГ) с одновременным
измерением частоты и ритма сокращений сердца, артериального давления,
частоты дыхания и газового состава выдыхаемого воздуха, а также насыще-
ния крови кислородом. Датчики для слежения за указанными процессами
фиксируются на теле человека перед отходом ко сну, проявления которого
исследуют в специально оборудованных центрах изучения сна. Ритмичное
чередование физиологических проявлений сна является его главной харак-
теристикой, позволяющей отличить нормальный сон от других состояний,
имитирующих отдельные его стадии.
21.2.1. Стадии сна
Наиболее распространенная классификации отдельных стадий сна (по Де-
менту и Клейтману) различает их по изменениям частоты и глубины волн
ЭЭГ (рис. 21.1). Во время активного бодрствования на электроэнцефало-
грамме преобладает так
называемый p-ритм, ко-
торый характеризуется
низкой амплитудой коле-
баний регистрируемых
Активное бодрствование, 0-ритм
Расслабленное бодрствование
потенциалов при сравни-
тельно высокой частоте
электрических разря-
дов—от 13 до 26 Гц и
более. При расслаблен-
ном, или пассивном,
бодрствовании, когда че-
ловек лежит с закрытыми
глазами, p-ритм чередует-
ся с a-ритмом, отличаю-
щимся большей амплиту-
дой и меньшей частотой
а-волны
1-я стадия сна
Рис. 21.1. Изменения элек-
троэнцефалограммы во вре-
мя ночного сна.
Активное бодрствование харак-
теризуется низкоамплитудным
высокочастотным 0-ритмом
(13—26 Гц), на фоне которого
периодически возникают а-вол-
ны (8—12 Гц), особенно харак-
терные для расслабленного
бодрствования. При переходе ко
сну происходит синхронизация
волн ЭЭГ и повышение их ам-
плитуды. Характерным проявле-
нием глубокого сна является по-
явление высокоамплитудных 8-
волн, которые в 4-й стадии сна
составляют не менее половины
всех волн ЭЭГ.
4 -я стадия сна, преобладание 8-волн
840 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Таблица 21.1. Характеристики отдельных стадий сна
Ста- дии сна Особенности ЭЭГ Мышечная актив- ность Вегетативные функции Когнитивная активность Порог пробуж- дения
1-я Низкоампли- тудные, высоко- частотные вол- ны, отдельные а- и 8-волны Эпизодические движения, тонус мышц начинает уменьшаться Замедление работы сердца, дыхания, сни- жение АД и температуры Мимолетные мысли и обра- зы, переход «точки без воз- врата» — засы- пание Низкий
2-я Появление К- комплексов и сонных веретен Продолжается сни- жение тонуса мышц Углубление происходя- щих измене- ний Характерных примет нет Чуть вы- ше, чем в 1-й ста- дии
3-я 8-Волны состав- ляют не менее 20 % ЭЭГ Тонус мышц низ- кий, случайные движения Примерно то же, что и во 2-й стадии Разбуженные бывают дезори- ентированы Еще выше
4-я Минимум 50 % всех волн ЭЭГ представляют 8-волны Низкий тонус мышц, поза эм- бриона Минимизация функций, ста- бильная кар- тина Абсолютная дезориентация при пробужде- нии Высокий
БДГ Десинхрониза- ция ритма ЭЭГ Гиперполяризация мотонейронов, максимальное рас- слабление мышц, раскинутые конеч- ности, БДГ Повышение ЧСС, АД, час- тоты дыхания, эрекция поло- вого члена или клитора Примерно 80 % разбуженных рассказывают о ярких сюжет- ных сновиде- ниях Высокий
(8—12 Гц). Во время медленноволнового сна частота волн ЭЭГ становится
еще меньше, а их амплитуда увеличивается, что объясняется ритмичным
одновременным и согласованным возбуждением большого количества
нервных клеток, т. е. синхронизацией их электрических разрядов. Синхро-
низация обусловлена ритмичной восходящей активностью стволовых нерв-
ных центров сна, когда же такая активность прекращается, возбуждение
нейронов коры начинает происходить несогласованно. В это время на ЭЭГ
возникают, как и во время бодрствования, высокочастотные и низкоам-
плитудные волны, что обозначают термином десинхронизация.
После засыпания ритм электроэнцефалограммы последовательно изме-
няется, что позволило выделить четыре сменяющие друг друга стадии мед-
ленноволнового сна, для которого характерно нарастание синхронной дея-
тельности нейронов подкорковых центров и коры. Первая стадия сна ха-
рактеризуется низкоамплитудной и относительно быстрой электрической
активностью, перемежающейся короткими группами a-волн и 0-волн (4—
7 Гц), а также медленными движениями глаз, выявляемыми с помощью
ЭОГ. Во второй стадии сна амплитуда волн ЭЭГ увеличивается, а их час-
тота уменьшается, одновременно с этим периодически возникают харак-
терные именно для этой стадии сна К-комплексы и сонные веретена. К-
комплекс представляет собой острую высокоамплитудную волну (до 300
мкВ), а сонные веретена — серию низко- и среднеамплитудных колебаний
с частотой 12—14 Гц.
Для третьей стадии сна типичны медленные высокоамплитудные фо-
новые колебания в 0-ритме (4—7 Гц) и периодически появляющиеся на
этом фоне глубокие 5-волны. В этой стадии сна высокоамплитудные 5-
волны составляют от 20 до 50 % всех регистрируемых проявлений ЭЭГ. В
21. Сон и бодрствование • 841
четвертой стадии преобладают высокоамплитудные (75 мкВ и больше) и
медленные (0,5—2 Гц) 5-волны, составляющие более 50 % всех регистри-
руемых колебаний ЭЭГ. Во время первой и второй стадий сна спящего
легко разбудить, что послужило поводом называть такой сон легким, или
поверхностным. Разбудить спящего человека во время третьей стадии зна-
чительно труднее, но самым глубоким сон оказывается во время четвертой
стадии, когда на ЭЭГ преобладают 5-волны, такой сон часто называют
дельта-сном.
Медленноволновой сон внезапно сменяется быстроволновым сном,
проявляющимся десинхронизацией волн ЭЭГ, амплитуда которых резко
уменьшается, а частота увеличивается до 4—8 Гц. Наряду с этим возника-
ют быстрые периодические движения закрытых глаз, что послужило пово-
дом назвать эту фазу сном с быстрыми движениями глаз (БДГ-сон', англ.
REM — rapid eye movements). Еще одним характерным признаком БДГ-сна
является уменьшение тонуса скелетных мышц до минимального и исчез-
новение сухожильных рефлексов, что обусловлено гиперполяризацией мо-
тонейронов спинного мозга. Расслабленное состояние скелетных мышц
одновременно с высокой активностью нейронов головного мозга побудило
назвать быстроволновой сон парадоксальным в отличие от ортодоксального
медленноволнового сна. Типичные для сменяющих друг друга стадий сна
изменения функций представлены в табл. 21.1.
21.2.2. Структура сна
После засыпания происходит последовательная смена 1, 2, 3 и 4-1 стадий
медленноволнового сна, после чего спящий возвращается в 3-ю, потом во
2-ю и 1-ю стадию, которую сменяет фаза БДГ-сна, наступающая прибли-
зительно через 90—100 мин от момента засыпания. Все эти события со-
ставляют один цикл сна, такие циклы повторяются на протяжении ночного
сна 4—6 раз (рис. 21.2). Суммарная продолжительность 1-й стадии у взрос-
Рис. 21.2. Циклическая смена стадий сна на протяжении ночи.
Первый цикл длится около 100 мин, в течение которых спящий человек последовательно пе-
реходит из 1-й стадии медленноволнового сна во 2, 3 и 4-ю стадии, а затем в 3-ю и 2-ю, после
чего фазу медленноволнового сна сменяет фаза парадоксального сна, завершающего этот
цикл. В последующих циклах уменьшается продолжительность глубокого сна, не возникает
4-я, а затем и 3-я стадии медленноволнового сна, но удлиняются периоды парадоксального
сна с характерной для него утратой мышечного тонуса. В утренние часы возникают эпизоди-
ческие пробуждения.
842 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Таблица 21.2. Возрастные особенности сна
Возраст Длительность сна БДГ-сон (%)
Новорожденные 16-18 ч 50
1 год 13-14 ч 35
3 года Около 10 ч 28-30
После 20 лет 7-8 ч 24
70 лет Меньше 6 ч Около 20
лых людей составляет примерно 5—10 % ночного сна, продолжительность
2-й стадии —около 50 %, 3-й —примерно 15 %, 4-й —около 5 %, БДГ-
сна —20—25 %. Продолжительность эпизодов БДГ-сна увеличивается от
одного цикла к другому и становится наибольшей перед пробуждением. В
то же время 4-я стадия в каждом новом цикле становится все короче или
вообще не достигается. Чем ближе к концу ночного сна, тем чаще проис-
ходят переходы к 1-й стадии, а также эпизодические пробуждения.
Средняя продолжительность ночного сна и его структура зависят от
возраста (табл. 21.2). Начиная приблизительно с пятидесяти лет, люди спят
меньше, чем в молодом возрасте, причем сокращение сна происходит пре-
имущественно за счет 3-й и 4-й стадий медленноволнового сна, а также
БДГ-сна.
21.2.3. Фаза медленноволнового сна
При засыпании и последующем переходе от поверхностного к глубокому
сну уменьшается тонус большинства скелетных мышц, на электромиограм-
ме регистрируется постепенное понижение амплитуды и частоты потен-
циалов действия скелетных мышц. Тонус мышц, закрывающих веки, на-
против, повышается при переходе ко сну и остается высоким на всем его
протяжении. Совершаемые в это время случайные движения обратно про-
порциональны глубине сна, в среднем за ночь человек 8—10 раз меняет
позу, которая во время глубокого сна похожа на эмбриональную.
Реципрокные отношения между симпатическим и парасимпатическим
отделами вегетативной нервной системы во время сна сглажены, а внутри
каждого отдела выявляется функциональная дифференциация, что не по-
зволяет четко классифицировать происходящие во время сна изменения
как следствие только ваготонии или симпатикотонии. Динамика показате-
лей вегетативных функций во время медленноволнового сна указывает на
повышение активности парасимпатических центров, и в первую очередь
блуждающего нерва, без отчетливого снижения симпатической активности.
По мере перехода от поверхностного сна к глубокому сужаются зрачки,
уменьшается частота сокращений сердца и вариативность сердечного рит-
ма, снижается артериальное давление. Самое значительное снижение кро-
вяного давления отмечено в первых циклах ночного сна, для которых ха-
рактерна наибольшая продолжительность стадий дельта-сна. Одновремен-
но с этим уменьшаются объем циркулирующей крови, мозговой кровоток
и потребление глюкозы клетками мозга, понижаются интенсивность об-
менных процессов и выделение катехоламинов мозговым веществом над-
почечников. Средняя частота дыхания в стадиях медленного сна постепен-
но снижается и начинает возрастать только в ранние утренние часы в со-
ответствии с фазой циркадного ритма. Температура мозга в 3-й и 4-й ста-
диях медленноволнового сна снижается одновременно с уменьшением по-
21. Сон и бодрствование • 843
требления мозгом кислорода. В то же время сохраняется гомойотермная
регуляция температуры тела, и ее снижение во время медленноволнового
сна сопровождается уменьшением кожного кровотока и снижением темпе-
ратуры кожи.
Моторика желудка уменьшается по мере углубления медленноволнового
сна, но в ранние утренние часы она снова возрастает, что совпадает с уве-
личением продолжительности эпизодов БДГ-сна в это время. Секреция
слюнных желез и выделение соляной кислоты в желудке испытывают наи-
большее торможение в фазе парадоксального сна, а во время медленновол-
нового сна секреторная активность хотя и понижена по сравнению с бодр-
ствованием, но в меньшей степени, чем во время БДГ-сна. Во время мед-
ленноволнового сна увеличиваются образование и секреция гормона роста,
способствующего анаболическим и репаративным процессам в клетках,
восстановлению их работоспособности.
Совокупность физиологических изменений, происходящих во время
медленноволнового сна одновременно с уменьшением кортикальной ак-
тивности, побуждает многих исследователей рассматривать эту фазу сна
как состояние, необходимое для восстановления гомеостазиса мозговой
ткани и оптимизации управления внутренними органами.
21.2.4. Фаза парадоксального сна
Для парадоксального сна характерны отсутствие движений и резко сни-
женная чувствительность к внешним воздействиям на фоне высокой ак-
тивности большинства клеток мозга. В связи с атонией мышц, обусловлен-
ной гиперполяризацией мотонейронов, поза спящего человека характери-
зуется распростертыми, «раскинутыми» конечностями. Рефлекторные ре-
акции резко ослаблены, сухожильные рефлексы не воспроизводятся. По-
роги всех видов чувствительности, как и порог пробуждения, во время
БДГ-сна выше, чем в остальных стадиях сна. Одновременно с этим проис-
ходит десинхронизация электроэнцефалограммы, которая становится поч-
ти такой же, как и во время бодрствования. Повторяющиеся эпизоды бы-
стрых движений глаз включают от 5 до 50 движений и в целом занимают
около 10 % всей фазы парадоксального сна.
Большинство разбуженных в этой фазе сна, около 80 %, сообщают об
увиденных сновидениях, причем эмоционально насыщенным сновидениям
часто соответствуют наиболее интенсивные быстрые движения глаз. Одна-
ко у некоторых людей сновидения могут возникнуть в любой стадии сна, а
быстрые движения глаз не обязательно сопутствуют сновидениям. Актива-
ция нейронов коры во время парадоксального сна сочетается с блокадой
афферентных сигналов к ним от органов чувств и эфферентных сигналов
от них к мышцам, что происходит на фоне облегченной передачи аффе-
рентной информации от внутренних органов. Считается, что в это время
интенсивно перерабатывается полученная в предшествующем бодрствова-
нии информация, создаются энграммы памяти и формируются будущие
поведенческие программы (информационная теория сна).
Динамика вегетативных показателей во время парадоксального сна по
контрасту с картиной медленноволнового сна оценивается как «вегета-
тивная буря». Такое определение отражает крайне переменчивый и про-
тиворечивый характер происходящих изменений. Так, например, в начале
фазы БДГ-сна возникает тахикардия, обусловленная еще ббльшим, чем в
фазе медленного сна, снижением активности блуждающего нерва и отсут-
ствующая при его блокаде. Затем она сменяется брадикардией, которую
844 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
некоторые исследователи связывают с еще большим усилением парасим-
патической активности и одновременным понижением симпатического
тонуса. По другим данным, в фазе парадоксального сна происходят пе-
риодические повышения симпатической активности с одновременным
увеличением содержания адреналина в периферической крови. Повыше-
ние частоты сокращений сердца часто коррелирует с приятными сновиде-
ниями, тогда как эмоционально напряженные сновидения сопровождают-
ся высокой вариабельностью сердечного ритма. Изменения артериального
давления в фазе парадоксального сна характеризуются выраженными ин-
дивидуальными особенностями с тенденцией к незначительному повы-
шению.
Мозговой кровоток в фазе БДГ-сна увеличивается на 3—12 % по срав-
нению с активным бодрствованием, наибольшее увеличение кровотока за-
регистрировано в прецентральных регионах коры, ромбовидном мозге и
гипоталамусе: считается, что повышение кровотока определяют местные
механизмы его регуляции. Одновременно с этим мозг увеличивает потреб-
ление кислорода, а его температура повышается на несколько десятых гра-
дуса. В это же время прекращается гомойотермная регуляция, о чем свиде-
тельствует исчезновение ответных реакций на перепады внешней темпера-
туры (прекращаются потение, мышечная дрожь и обусловленные наруж-
ной температурой изменения кровотока через кожу).
В фазе парадоксального сна возрастает средняя частота дыхания, кото-
рое становится нерегулярным как по частоте, так и по глубине. При воз-
никновении быстрых движений глаз возможно появление периодического
дыхания и апноэ, причем подобные изменения дыхания совпадают с наи-
большими вариациями сердечного ритма, артериального давления и со-
провождаются нестабильностью показателей газового состава крови. Ряд
исследований указывают на связь между эмоциональным характером сно-
видений и возникновением нерегулярного дыхания.
Секреция пищеварительных желез (саливация, секреция соляной ки-
слоты в желудке) в фазе БДГ-сна угнетена, а моторные проявления в пи-
щеварительном тракте усиливаются, но вне связи с быстрыми движения-
ми глаз. У мужчин в этой стадии сна происходит эрекция полового чле-
на, у женщин — клитора, что не связано непосредственно с половой ак-
тивностью или сексуальным содержанием сновидений. Эрекция во время
парадоксального сна регулярно возникает у детей и подростков еще до
появления полового влечения. Она обусловлена активацией парасимпати-
ческих структур крестцового отдела спинного мозга, возбуждение кото-
рых определяется нисходящим влиянием лимбической системы и гипота-
ламуса.
Характерные изменения вегетативных функций во время парадоксаль-
ного сна не связаны с изменениями окружающей среды, ограничиваются
во времени именно этой фазой сна, а следовательно, определяются внут-
ренними факторами и отражают направленность происходящих процессов
на осуществление до сих пор не понятых функций сна. Остается дискусси-
онным и вопрос о том, что объединяет регулирующие механизмы столь
разных процессов, приводящих к угнетению моторной деятельности и сен-
сорного восприятия одновременно с высокой активностью коры и разно-
направленными изменениями вегетативных функций. Предполагается, что
все происходящее необходимо для деятельности мозга — самого активного
функционально органа в фазе БДГ-сна. В развитие информационной тео-
рии высказываются предположения о том, что во время быстрого сна мозг
настраивается на восприятие и переработку информации, поступающей от
21. Сон и бодрствование • 845
внутренних органов, производит реорганизацию ранее воспринятой ин-
формации, согласует и оптимизирует деятельность всех структур мозга,
участвующих в этих процессах.
21.3. Нейрофизиологические механизмы сна
Переход от бодрствования ко сну определяется, с одной стороны, сменой
фаз эндогенного циркадного ритма, с другой — возникновением потребно-
сти в отдыхе, возрастающей пропорционально продолжительности бодрст-
вования. В связи с этим механизмы перехода ко сну включают активные
внутренние процессы в центральной нервной системе и ее пассивные изме-
нения, обусловленные интенсивной предыдущей деятельностью. В реали-
зации этих механизмов участвуют несколько структур ствола мозга, регу-
лирующих взаимодействие таламуса и коры в цикле сна—бодрствования и
способных ритмически изменять свою активность, которая управляется
гипоталамусом и одновременно зависит от нисходящего влияния коры и
лимбической системы.
21.3.1. Участие стволовых центров в регуляции цикла
сна—бодрствования
Нервные центры ствола, регулирующие цикл сна—бодрствования, по-раз-
ному проявляют свою активность в обеих фазах этого цикла, что, однако,
не позволяет их классифицировать в качестве специфических центров
только сна или только бодрствования. Скорее это разные звенья единой
системы, образованной несколькими ядрами ретикулярной формации,
нейроны которых образуют восходящие и нисходящие дивергентные про-
екции к различным структурам мозга, посредством которых ритмически
изменяют их активность во время сна и бодрствования.
В ростральном отделе ретикулярной формации расположены небольшие
группы нейронов, постоянно активирующие кору во время бодрствования
и получившие название восходящей активирующей ретикулярной системы
(БАРС). К ней принадлежат холинергические нейроны покрышки среднего
мозга и гигантоклеточной области моста, а также норадренергические ней-
роны, самое большое скопление которых в области моста известно как го-
лубое пятно (рис. 21.3). В результате активирующего действия ВАРС на
неспецифические ядра таламуса и кору во время бодрствования происхо-
дит деполяризация большинства кортикальных нейронов приблизительно
на 10—15 мВ от исходного значения мембранного потенциала покоя (65—
70 мВ). Такая деполяризация получила название тонической, она снижает
порог возбуждения кортикальных нейронов и тем самым облегчает их от-
ветную реакцию на поступающую афферентную информацию. Спонтанная
активность ВАРС на протяжении бодрствования не одинакова, она повы-
шается, например, при действии новых сенсорных стимулов, вызывающих
ориентировочные реакции, или при переживании эмоций ярости и гнева,
когда особенно возрастает активность норадренэргических нейронов.
В каудальной области ретикулярной формации расположены серотони-
нергические нейроны, скопления которых образуют ядра шва. Проекции
этих нейронов распространяются на обширные области мозга: кору, где
имеется около 500 000 серотонинергических синапсов, таламус, лимбиче-
скую систему, гипоталамус, мост и продолговатый мозг. Серотонинергиче-
ские нейроны тормозят активность ВАРС, изменяют активность вегетатив-
ных центров ствола и характер нейронных переключений в моторных и
846 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Базальный
передний
мозг
Холинергические
нейроны РФ
среднего мозга и
покрышки моста
Норадренергические
нейроны голубого
пятна
Серотонин-
ергические
нейроны
ядер шва
Рис. 21.3. Нейронные популяции, участвующие в регуляции цикла сна—бодрство-
вания.
Нейронные популяции, указанные на схеме, обладают спонтанной активностью, характер ко-
торой закономерно изменяется на протяжении ночного сна и во время перехода к бодрствова-
нию. Активность указанных нейронов во время сна изменяется под влиянием супрахиазмаль-
ных ядер гипоталамуса (СХЯ).
сенсорных ядрах ствола. Повышение их фоновой активности приводит к
гиперполяризации нейронов рострального отдела ретикулярной формации,
в результате снижается активирующее влияние ВАРС на кору больших по-
лушарий и происходит переход от бодрствования к медленноволновому
сну. Одновременное действие серотонинергических нейронов на вегета-
тивные центры и переключательные ядра ствола вызывает характерные для
медленноволнового сна изменения вегетативных процессов, сенсорной
чувствительности и моторной деятельности.
В результате торможения ВАРС изменяется характер возбуждения зави-
симых от нее неспецифических ядер таламуса, нейроны которых гиперпо-
ляризуются и в них возникают длительные тормозные постсинаптические
потенциалы. Они прерываются пачками или сериями нейронных разрядов,
при этом большинство нейронов постепенно синхронизирует ритм своей
активности, а затем навязывает его коре, нейроны которой тоже переходят
от характерного для бодрствования состояния тонической деполяризации
к состоянию гиперполяризации. Синхронизированная активность талами-
ческих и корковых нейронов коррелирует с постепенным торможением
психической активности.
Переход из фазы медленноволнового сна в фазу парадоксального сна
инициируют вентромедиальные ядра гипоталамуса, которые тормозят ак-
тивность серотонинергических нейронов и тем самым прекращают их тор-
мозное действие на ростральную область ретикулярной формации. Вслед-
ствие этого возобновляется активность холинергических нейронов гиган-
токлеточной области покрышки моста, вызывающих тоническую деполя-
ризацию нейронов таламуса и коры. В результате в таламусе и коре исчеза-
ет характерная для фазы медленноволнового сна синхронизированная
электрическая активность нейронов, которые начинают разряжаться неза-
висимо друг от друга, что проявляется десинхронизацией электроэнцефа-
лограммы.
Десинхронизация в фазе парадоксального сна и при бодрствовании от-
21. Сон и бодрствование • 847
личаются мало, но в остальных проявлениях мозговой деятельности между
этими состояниями существуют большие различия. Во время парадоксаль-
ного сна резко заторможена активность всех моноаминергических нейро-
нов ствола, т. е. серотонинергических, норадренергических и дофаминер-
гических, высокоактивных при бодрствовании. Считается, что активность
холинергических и моноаминергических нейронов регулируется во время
парадоксального сна реципрокно, и чем активнее холинергические нейро-
ны, тем сильнее тормозятся моноаМинергические. Из всех восходящих ди-
вергентных систем ствола, модулирующих во время бодрствования харак-
тер нейронных переключений в других регионах мозга, включая кору, во
время парадоксального сна активны только холинергические нейроны.
Выключение моноаминергической активности сказывается на переклю-
чательной способности сенсорных и моторных ядер, на активности вегета-
тивных центров ствола, лимбических структур и коры, поскольку диффуз-
ные моноаминергические проекции при бодрствовании модулируют ней-
ронные переключения в этих регионах. Вследствие тормозного нисходяще-
го влияния орального ядра ретикулярной формации моста возникает мы-
шечная атония, характерная для парадоксального сна, во время которого
высокоактивные клетки коры лишаются как афферентной информации от
органов чувств, так и возможности передать информацию мышцам. Сно-
видения, возникающие в этой фазе сна, отражают противоречивую ситуа-
цию, когда высокая активность коры сочетается с ограниченными возмож-
ностями обмена информацией с периферией.
21.3.2. Регуляция циркадного ритма
Эндогенный суточный цикл сна—бодрствования синхронизирован с есте-
ственным чередованием светлого и темного времени суток. Афферентная
информация о характере освещения поступает по волокнам ретиногипота-
ламического тракта от сетчатки к супрахиазмальным ядрам гипоталамуса
(СХЯ), расположенным по обеим сторонам III желудочка мозга непосред-
ственно над зрительным перекрестом. Активность нейронов СХЯ ритмич-
но изменяется с периодичностью 24 ч, такой ритм определен генетически
и сохраняется в условиях не меняющегося освещения. Функция супрахиаз-
мальных ядер состоит в согласовании эволюционно древних ритмов актив-
ности—покоя и реализуется при эфферентном действии СХЯ на гипотала-
мические и стволовые центры, регулирующие специфические физиологи-
ческие процессы, включая сон и бодрствование, терморегуляцию, секре-
цию гормонов, экскрецию метаболитов и т. п. При этом бодрствование
должно соответствовать периодам активности, а сон — покоя. Афферент-
ная информация от сетчатки служит средством согласования этой врож-
денной ритмики с реальным временем суток (рис. 21.4).
Нейроны вентромедиальной области СХЯ являются их входной зоной и
кроме афферентных сигналов от сетчатки получают информацию из ствола
мозга от ядер шва, а также из других ядер гипоталамуса (функция обрат-
ной связи). Нейроны дорсомедиальной области СХЯ образуют ипсилате-
ральные эфферентные связи с другими ядрами гипоталамуса, в особенно-
сти с вентромедиальным ядром, они имеют также эфферентный выход к
нервным центрам ствола, лимбической системе и полосатому телу. По-
средством своих эфферентных путей СХЯ синхронизируют суточные рит-
мы активности физиологических процессов.
В управлении циркадным ритмом вместе с супрахиазмальными ядрами
участвует эпифиз, в котором в светлое время суток повышается образова-
848 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Аргинин-вазотоцин? Мелатонин?
Рис. 21.4. Схема организации циркадной ритмики.
Супрахиазмальные ядра гипоталамуса (СХЯ) состоят из комплекса нейронных осцилляторов,
связанных иерархически и параллельно. Они получают информацию о характере освещенно-
сти непосредственно от сетчатки по волокнам ретиногипоталамического тракта (РГТ) и после
переработки такой информации в латеральном коленчатом теле (ЛКТ). Эта информация необ-
ходима для согласования эндогенного ритма активности СХЯ с периодической сменой дня и
ночи. Активность СХЯ адресована другим ядрам гипоталамуса, стволу и эпифизу, которые
корректируют активность СХЯ по механизму отрицательных обратных связей (пунктирные
линии).
ние серотонина, а ночью из него синтезируется гормон мелатонин. Синтез
мелатонина происходит благодаря повышению в ночные часы активности
ключевого фермента N-ацетилтрансферазы. Ритмику этого процесса регу-
лируют СХЯ, но не прямо, а опосредованно, с помощью нескольких ней-
ронных переключений, активирующих симпатические нейроны верхнего
шейного ганглия, которые непосредственно иннервируют эпифиз.
Нейроны СХЯ имеют рецепторы для присоединения мелатонина, а его
содержание в супрахиазмальных ядрах растет в ночные часы. Поэтому ме-
латонин рассматривается рядом исследователей как ингибитор активности
СХЯ в ночное время, осуществляющий обратную связь между двумя
звеньями циркадной системы: эпифизом и СХЯ. В соответствии с другой
точкой зрения у человека мелатонин играет роль локального гормона эпи-
физа и влияет на синтез пептидов, которые обладают функциональной ак-
тивностью и способны проникать в кровь и ликвор. Одним из таких пеп-
тидов является аргинин-вазотоцин, попадающий с током цереброспиналь-
ной жидкости в супрахиазмальные ядра для модуляции активности их ней-
ронов.
21.3.3. Участие коры и лимбической системы в регуляции цикла
сна—бодрствования
Переход от бодрствования ко сну у человека определяет не только эндо-
генный ритм, синхронизированный с реальным временем суток, но и ха-
рактер поведенческой активности, связанной с переработкой сенсорной
информации, особенностями трудовой или иной деятельности, пережива-
нием эмоций и т. п. Не связанный с естественным чередованием дня и но-
чи переход ко сну, как и ситуативная бессонница, могут произойти в ре-
зультате нисходящего влияния коры или действия лимбической системы
21. Сон и бодрствование • 849
на гипоталамус и стволовые нервные центры, от активности которых не-
посредственно зависит поддержание бодрствования или переход ко сну.
В состоянии бодрствования восходящая активность рострального отдела
ретикулярной формации обеспечивает тоническую деполяризацию корко-
вых нейронов. В таком состоянии фронтальные отделы коры тормозят ак-
тивность подкорковых центров, индуцирующих переход к медленноволно-
вому сну. Предполагается, что нисходящая активность фронтальной коры
распространяется при бодрствовании на лимбические структуры (гиппо-
камп, миндалины, перегородку), а они подавляют активность гипоталами-
ческих центров сна. По-видимому, нельзя исключить и нисходящее тормо-
жение каудального отдела ретикулярной формации, где расположены ядра
шва, индуцирующие медленноволновой сон.
Спящего человека можно разбудить, обратившись к нему по имени, он
просыпается от плача своего ребенка, что в таких случаях свидетельствует
о первоначальной активации коры, без участия которой невозможно рас-
познать эти слабые и сложные стимулы. Механизм пробуждения включает
торможение центров сна с последующей активацией восходящей ретику-
лярной формации. Напротив, бессонница, возникающая иногда из-за мыс-
лей, которые не удается «отключить», вызвана торможением подкорковых
центров сна активно функционирующей корой.
Активность коры при бодрствовании поддерживается тонической сти-
муляцией со стороны ретикулярной формации и зависит от характера по-
ступающей сенсорной информации: возрастает при действии разнообраз-
ных сенсорных стимулов и уменьшается при ограничении сенсорного при-
тока или при действии однообразных, монотонно повторяющихся раздра-
жителей. При интенсивной сенсорной стимуляции торможение подкорко-
вых центров сна со стороны коры будет проявляться сильнее, а при огра-
ниченном или монотонном притоке сенсорной информации торможение
уменьшается, что ведет к повышению активности центров сна. Возникно-
вение сонливости при низком уровне сенсорной стимуляции (например,
при монотонной работе) обусловлено снижением активности коры и по-
следующим «растормаживанием» подкорковых центров, индуцирующих
медленноволновой сон.
Лимбическая система определяет субъективное отношение человека к
действующим стимулам и возникновение эмоций, связанное с его пове-
денческой активностью. Сенсорная информация и результаты деятельно-
сти, вызывающие переживание эмоций, способствуют сохранению или да-
же возрастанию восходящей активности ретикулярной формации и, тем
самым, поддержанию бодрствования. Отсутствие новизны в сенсорной ин-
формации, монотонная деятельность, скука приводят к снижению восхо-
дящей активности ретикулярной формации и, соответственно, снижению
тормозящего влияния фронтальной коры на подкорковые центры сна.
21.3.4. Гуморальные индукторы и регуляторы сна
Известен ряд гуморальных веществ, индуцирующих сон, и существует ги-
потеза, допускающая наличие не только гипногенных, но и препятствую-
щих возникновению сна гуморальных факторов. Такие вещества были из-
влечены из жидких сред организма (ликвор, кровь) экспериментальных
животных и человека в условиях длительного лишения сна или, напротив,
активного бодрствования. Большинство веществ, причастных к регуляции
цикла сна—бодрствования, относится к пептидам.
Путем фракционирования ликвора, полученного от длительно лишае-
850 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
мых сна животных, выделен состоящий из пяти аминокислот пептид, ко-
торый обладает снотворным эффектом. Он получил название фактора S
(от англ, sleep — сон), его удалось выделить и у человека. При введении
экспериментальным животным он вызывает медленноволновой сон с по-
явлением характерных изменений на ЭЭГ. Обнаружены и другие пептиды
эндогенного происхождения, обладающие способностью индуцировать
медленноволновой сон. К ним относятся: вызывающая сон субстанция с
молекулярной массой 70—200 (SPS), дипептид мураминовой кислоты, нона-
пептид, индуцирующий дельта-сон (DSIP). Эти пептиды способны преодо-
левать гематоэнцефалический барьер и влиять на метаболизм нейромедиа-
торов, однако после внутривенного введения они быстро разрушаются под
воздействием ферментов. При введении в желудочки мозга DSIP повышает
в головном мозге концентрацию серотонина и ГАМК, а их накопление в
стадии медленноволнового сна способствует уменьшению обменных про-
цессов и потребления кислорода мозгом.
Наряду с пептидами — индукторами медленноволнового сна, описан
выделенный из ликвора в период парадоксального сна неидентифициро-
ванный эндогенный пептид, вызывающий БДГ-сон при его введении экс-
периментальным животным. К этому пептиду получены антитела, введе-
ние которых угнетает парадоксальный сон. У человека сразу после оконча-
ния фазы БДГ-сна в спинномозговой жидкости в несколько раз повышает-
ся уровень аргинина-вазотоцина (АВТ). Его введение даже в очень малом
количестве (около 600 молекул) вызывает сон у кошек, снотворное дейст-
вие проявляется и при введении его людям, у которых при этом возникает
фаза БДГ-сна. Содержание АВТ у пожилых людей с нарушениями сна
уменьшено, высказано предположение о регулирующей роли этого пепти-
да в организации сна у человека. Имеются сведения о том, что возникно-
вению БДГ-сна способствует также вазоинтестинальный пептид (ВИП).
К пептидам, участвующим в регуляции сна, относят также субстанцию
Р (нормализует сон при стрессе), бета-эндорфин, ангиотензин-П, есть дан-
ные об участии в регуляции сна соматостатина и тиреотропина. Участие
нейропептидов в регуляции цикла сна—бодрствования возможно как в ка-
честве комедиаторов нейронных переключений, так и в качестве нейрогор-
монов. Мягкое модулирующее действие пептидов состоит в медленных из-
менениях нейронной активности мозговых структур, взаимодействующих
во время сна.
Наряду с нейропептидами снотворное действие на человека может ока-
зывать простагландин Д2, синтез которого стимулируется при воспалении
паутинной оболочки мозга, сопровождающемся повышенной сонливостью.
Мурамилпептиды бактериального происхождения даже в очень малой кон-
центрации увеличивают продолжительность медленноволнового сна и уг-
нетают фазу БДГ-сна; одновременно с этим они вызывают повышение
температуры тела у человека. Эти факты дают возможность приблизиться к
пониманию механизма повышенной продолжительности сна, наблюдаю-
щейся у человека при воспалительных и инфекционных заболеваниях.
21.4. Сновидения и физиологическая роль БДГ-сна
Большинство разбуженных во время парадоксального сна людей сообщают
о сюжетно связанных сновидениях, причем это относится и к людям, ут-
верждавшим, что они никогда не видят сны. Скорее всего, они просто за-
бывают о сновидениях, если не просыпаются сразу после фазы БДГ-сна.
Установлено, что люди не менее часто видят сновидения и во время мед-
21. Сон и бодрствование • 851
ленноволнового сна: разбуженные во 2-й фазе сна говорят об этом в 60 %
случаев, реже вспоминают сны в случае пробуждения в 3-й или 4-й стади-
ях. Но обычно в таких случаях разбуженные рассказывают лишь о несвяз-
ных, обрывочных образах, тогда как после БДГ-сна вспоминаются связные
эпизоды, кажущиеся реальными.
Зигмунд Фрейд создал теорию толкования сновидений, которые он объ-
яснял прорывом из подсознания вытесненных туда запретных по соображе-
ниям морали сексуальных и агрессивных желаний. В настоящее время
сновидения принято объяснять теорией активационного синтеза, основан-
ной на том, что во время фазы БДГ-сна одновременно активируются мно-
гие нейронные переключения в центрах ствола мозга, сигналы от которых
поступают в кору, а она осуществляет синтез этих нередко противоречивых
сигналов, пытаясь объединить их и придать им смысл. При этом кора ак-
тивно извлекает из памяти поступившую прежде информацию, которой не
приходится, как при бодрствовании, конкурировать с новой информацией,
предоставляемой сенсорными системами. Поэтому интерпретация инфор-
мации происходит иначе, чем при сохраненном сознании, она отражает
реальные потребности и конфликты, но не учитывает реальные временные
и пространственные обстоятельства.
На фоне ограниченного поступления информации о внешних раздра-
жителях освобождается доступ к коре афферентной информации от внут-
ренних органов. Это обстоятельство имеет особое значение при развитии
патологических процессов, при которых кора начинает получать необыч-
ную информацию, становящуюся причиной столь же необычных сновиде-
ний. Активация сенсорных областей во время БДГ-сна приводит к зри-
тельным, слуховым, тактильным компонентам сна, а повышение активно-
сти моторных структур ствола сопровождается представлениями о якобы
выполняемых движениях, хотя реальные движения не происходят в связи с
торможением активности мотонейронов ретикулярной формацией. Таким
образом, сновидения представляют собой не случайный, а биологически
значимый и полезный процесс, хотя содержательная сторона сновидений в
большинстве случаев не поддается достоверному истолкованию.
У детей продолжительность БДГ-сна больше, чем у взрослых: считается,
что в этой стадии сна совершенствуется эффективность переключений ме-
жду определенными нейронами, представляющая основной механизм об-
разования долговременной памяти. Возможно, именно поэтому в периоды
решения наиболее сложных жизненных задач или творческих проблем
продолжительность БДГ-сна увеличивается и у взрослых, причем в боль-
шинстве сновидений появляются эпизоды из предыдущей жизни и про-
фессиональной деятельности.
Отдельную разновидность сновидений представляют люцидные сны (от
лат. lucidus — ясный, очевидный), когда спящий понимает, что он спит, но
видит сон и может управлять событиями сна. Содержание таких снов при-
ближается к реальной действительности, а электрическая активность ней-
ронов коры практически не отличается от таковой в состоянии бодрство-
вания.
21.5. Продолжительность сна и последствия его лишения
Индивидуальная продолжительность сна у здоровых взрослых людей варь-
ирует в диапазоне от 4 до 10 ч, в связи с чем различают мало и много спя-
щих, тогда как средняя продолжительность ночного сна составляет 7,5 ч.
Около трети своей жизни человек проводит во сне. Существуют две проти-
852 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
воположные точки зрения по поводу необходимой длительности сна у че-
ловека. Сторонники первой из них демонстрируют примеры пагубных по-
следствий недосыпания, которые могут отражаться на функциональном со-
стоянии человека, вызывать снижение уровня внимания и сосредоточенно-
сти при выполнении работы, что, в свою очередь, может приводить к ава-
риям и катастрофам по вине недосыпающих людей, а также к психиче-
ским нарушениям у них. Альтернативный взгляд на этот вопрос основан
на представлении о том, что многие люди спят гораздо дольше, чем это им
необходимо, и в буквальном смысле слова просыпают свою жизнь, а един-
ственным следствием недосыпания является лишь повышенная сонли-
вость.
Во время длительного вынужденного бодрствования у человека регист-
рируют регулярные и очень короткие, продолжительностью в 1—2 с, пе-
риоды так называемого микросна, когда «слипаются» веки и понижается
реакция на внешние стимулы. Вследствие длительного лишения сна воз-
никают нарастающее утомление, ослабление внимания, раздражительность
и уменьшение поведенческой активности, поскольку потребность во сне
начинает доминировать по отношению ко всем остальным биологическим
потребностям. После длительного бодрствования изменяется структура
следующего за ним сна: вскоре после засыпания возникает глубокий мед-
ленноволновой сон и на протяжении всей ночи преобладают 3-я и 4-я ста-
дии сна. Восстановление обычной структуры сна происходит лишь на сле-
дующую ночь, что послужило поводом к представлению о первоочередной
потребности в медленноволновом сне.
В экспериментах с избирательным лишением БДГ-сна испытуемых бу-
дят, как только они перейдут в эту фазу, а затем предоставляют им воз-
можность заснуть вновь, в результате БДГ-сон у них появляется чаще, чем
обычно. Длительное лишение БДГ-сна у человека приводит к возникнове-
нию раздражительности, появлению зрительных или слуховых галлюцина-
ций и навязчивых идей. Компенсаторное увеличение продолжительности
только этой фазы в ответ на избирательное лишение БДГ-сна указывает на
самостоятельный механизм его управления, различающийся с механизмом
управления медленноволновым сном.
Эксперименты, в которых добровольцы уменьшали продолжительность
ежесуточного сна на 1,5—2 ч в течение нескольких недель не выявили у
них каких-либо выраженных нарушений психики и здоровья, за исключе-
нием сниженной способности длительного сосредоточения внимания.
Изучение возможности человека перейти от 24-часового ритма к 18-, 21-,
28- или 48-часовому ритму проводилось в пещерах или в условиях поляр-
ного дня, чтобы исключить естественное чередование дня и ночи. Оказа-
лось, что организм человека трудно приспосабливается к искусственным
ритмам, поскольку собственный эндогенный ритм весьма консервативен.
Обстоятельства, нарушающие этот естественный ритм, приводят к так
называемому десинхронозу, проявляющемуся в снижении умственной и фи-
зической работоспособности, настроения, а порой и невротическими рас-
стройствами. Причинами десинхроноза в повседневной жизни чаще всего
бывают дальние трансмеридиональные перелеты со сменой часовых поя-
сов, а также нерегулярное чередование дневных и ночных рабочих смен.
До 30 % взрослых людей имеют некоторые проблемы в связи со сном,
причем от 8 до 15 % жалуются на плохой или недостаточный сон, а среди
пожилых людей этот процент значительно выше; от 9 до 11 % взрослых
людей регулярно принимают снотворные средства. Следует, однако,
учесть, что жалобы многих людей на плохой сон, его недостаточность или
21. Сон и бодрствование • 853
многочисленные пробуждения часто не подтверждаются объективными ис-
следованиями. Так, например, многие люди, жалующиеся на бессонницу,
фактически спят около 6 ч, но считают, что должны спать 8 ч. Поэтому
они проводят в постели больше времени, чем им необходимо для сна, и,
соответственно, испытывают определенные трудности при засыпании. По-
степенно эта проблема усугубляется из-за боязни снова не заснуть быстро,
и чем больше человек думает об этом, тем труднее ему перейти ко сну. В
таких случаях порой помогают очень простые рецепты: соблюдать посто-
янный режим сна и ложиться спать лишь тогда, когда возникает усталость.
К основным проявлениям нарушений сна относятся бессонница, или
инсомния, и патологическая сонливость, или гиперсомния. Бессонница у
здоровых людей нередко появляется как результат ситуативной реакции
эмоциональной природы в ответ на различные житейские коллизии. Дру-
гие формы бессонницы связаны с нарушением функции дыхания во время
сна или возникновением ночных гиперкинезов — насильственных и болез-
ненных мышечных сокращений, появляющихся чаще всего в нижних ко-
нечностях.
Гиперсомнии проявляются повышенной сонливостью в дневное время.
При одной из форм гиперсомнии — нарколепсии — бодрствование внезапно
прерывается на 10—15 мин периодом БДГ-сна с характерной для него ут-
ратой мышечного тонуса, причем произойти это может в самый неподхо-
дящий момент: во время разговора, еды, вождения автомобиля и даже по-
лового акта. «Сонные атаки» могут повторяться через несколько часов, то-
гда как общая продолжительность ночного сна увеличена незначительно,
хотя он и нарушается частым пробуждением. Еще одна форма гиперсом-
нии — катаплексия, при которой кратковременные эпизоды потери мы-
шечного тонуса не сопровождаются утратой сознания. Эти состояния час-
то связаны с эмоциональными реакциями, например смехом, страхом или
злостью, когда переживающий эмоции больной теряет мышечный тонус, а
поэтому падает и затем лежит 1—2 мин.
21.6. Бодрствование и сознание
Пробуждение, или переход от состояния сна к бодрствованию, обусловле-
но активирующим влиянием на кору больших полушарий со стороны диф-
фузной системы нейронов верхних отделов мозгового ствола и зрительных
бугров, и, в первую очередь, нейронов, расположенных между ростральны-
ми отделами моста и каудальными отделами промежуточного мозга. Боль-
шинство этих нейронов являются холинергическими, и ацетилхолин явля-
ется, по-видимому, важнейшим нейромедиатором, необходимым для про-
буждения от сна. Наряду с ним пробуждению' способствует и активация
норадренэргических нейронов голубых пятен.
Активность ретикулярной формации периодически изменяется на про-
тяжении суток, подчиняясь, во-первых, генетически предопределенному
суточному ритму эндогенных осцилляторов, синхронно с которым проис-
ходят изменения концентрации гуморальных регуляторов активности (се-
ротонин, катехоламины, нейропептиды) в крови или ликворе. Во-вторых,
высокой активности ретикулярной формации во время бодрствования спо-
собствует поступление новой сенсорной информации. Если же ее приток
уменьшается (темнота, тишина, отсутствие новых тактильных, температур-
ных и других раздражителей), то и восходящая активность ретикулярной
системы снижается, что приводит к развитию сонливости. В третьих, ак-
тивность ретикулярной формации зависит от характера выполняемой дея-
854 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
тельности и субъективного интереса к ней, что, в свою очередь, определяет
эмоциональный подъем или, напротив, состояние скуки: первое способст-
вует активному бодрствованию, второе — развитию сонливости. Подобные
перепады восходящей активности ретикулярной формации определяются
ее двусторонними связями с корой больших полушарий и лимбической
системой.
Состояние бодрствования у человека характеризуется наличием у него
сознания. Как бодрствование, так и сознание не связаны с активностью ка-
кой-либо одной специфической области мозга, они являются продуктом
взаимодействия обоих его полушарий и ствола мозга и определяются сум-
марным количеством активных нейронов и эффективностью нейронных
переключений между ними. Утрата сознания и невосприимчивость к
внешним раздражителям может быть вызвана как метаболическими нару-
шениями мозга, так и поражением верхних отделов ствола: такое состоя-
ние обозначается термином кома. Сознание называют спутанным, если че-
ловек оказывается неспособным четко и последовательно мыслить — в та-
ких случаях могут появиться искаженные зрительные, слуховые или осяза-
тельные ощущения, а также галлюцинации.
21.7. Различные уровни бодрствования
Сознание человека после пробуждения и его сознание во время активной
деятельности различаются быстротой реагирования, способностью сосре-
доточить внимание на тех или иных объектах, мобилизовать ресурсы памя-
ти. Повышенное эмоциональное возбуждение, как и повышенная сонли-
вость, негативно сказываются на характере поведенческих реакций. В свя-
зи с этим выделяют несколько уровней бодрствования и сознания, кото-
рые отражают различную степень активности мозга (табл. 21.3).
При низкой активности мозга (кома, наркоз) сознание отсутствует, но и
при чрезмерно высокой активности (эпилептический приступ) оно тоже
исчезает. Таким образом, необходимым условием сохранения сознания яв-
ляется оптимальный, а не самый высокий уровень активности мозга
(рис. 21.5). Для активного бодрствования характерна способность концен-
трировать внимание, т. е. сосредоточить познавательную деятельность на
каком-либо одном объекте, наиболее значимом для человека в данный мо-
мент времени, но одновременно с этим обычно ухудшается восприятие
Таблица 21.3. Характеристика различных уровней бодрствования
Физиологи- ческое со- стояние Уровень бодрствования и внимание Поведенческие реакции Особенности ЭЭГ
Сильные эмоции, воз- буждение Активное бодрствова- ние Сонливость Засыпание Суженное сознание, рас- щепленное внимание, растерянность Концентрация внима- ния, предугадывание ре- зультата действий Снижение внимания, от- влеченные мысли, неяс- ные мечты Сильно сниженный уро- вень сознания, повышен- ный порог раздражения Дезорганизация, дефи- цит контроля, реакции страна или паники Четкая организация, оптимальная работо- способность Нарушенная коорди- нация, низкая работо- способность Реакция только на сильные или ожидае- мые стимулы Выраженная десин- хронизация ритма Избирательная десин- хронизация, наличие групп а-волн Варьирует; преоблада- ет а-ритм а- и 0-волны, эпизо- дически сонные вере- тена
21. Сон и бодрствование • 855
Рис. 21.5. Зависимость эффективности поведения от активации мозга.
Целенаправленная деятельность наиболее успешна при оптимальной активности мозга, соот-
ветствующей уровню активного бодрствования. При низких уровнях активации готовность к
действиям снижается и развивается сонливость, при усилении активации растет количество
совершаемых ошибок и может произойти дезорганизация поведения.
других объектов. Находясь на шумной вечеринке, человек способен фоку-
сировать внимание на словах своего собеседника, в то время как все ос-
тальные голоса сливаются в единый фон и не запоминаются. Избиратель-
ность внимания, направляемого на отдельные объекты, выделяемые из об-
щего фона, сопряжена с ограниченным объемом оперативной памяти, не
способной вместить всю входящую сенсорную информацию.
Появление нового раздражителя привлекает внимание человека по ме-
ханизму ориентировочного рефлекса (поворот головы, глаз в сторону но-
вого объекта), а последующее восприятие такого раздражителя вызывает
изменение ритма электроэнцефалограммы в специфической сенсорной об-
ласти коры (зрительной, слуховой, соматосенсорной), где характерный для
пассивного бодрствования ос-ритм сменяется p-ритмом — такая десинхро-
низация получила название блокады a-ритма. Избирательное внимание че-
ловека, направленное на конкретный объект, проявляется активацией не
только первичных, но также вторичных сенсорных и ассоциативных облас-
тей коры, что увеличивает ресурсы для анализа наблюдаемого объекта.
ГЛАВА 22
Физиологические основы труда
Физиология труда представляет собой раздел физиологии человека, изу-
чающий функции скелетных мышц и физиологических систем, обеспечи-
вающих их работу, а также механизмы их регуляции при трудовой деятель-
ности с целью обоснования рациональной организации труда человека и
поддержания высокой работоспособности организма.
В наиболее общем виде труд человека подразделяется на физический и
умственный в зависимости от степени вовлеченности в трудовой процесс
соответственно скелетных мышц и нервно-психических функций. Оба ви-
да труда характеризуются величиной трудовой нагрузки, отражающей уро-
вень физиологического напряжения работающего человека. Характер тру-
довой нагрузки определяется элементами трудового процесса (мышечная
работа, рабочая поза, нагрузка на зрительную систему, психоэмоциональ-
ное напряжение, монотонность рабочих действий, гипокинезия), которые
оказывают влияние на функции физиологических систем человека. Вели-
чина трудовой нагрузки зависит от характера трудового процесса, от режи-
ма труда и отдыха, от факторов производственной среды и т. д.
Организм человека адаптируется к физическим и нервно-психическим
нагрузкам в зависимости от двух основных факторов. Во-первых, от рабо-
тоспособности человека — это способность выполнять работу максимально
возможное время без снижения ее эффективности. Работоспособность за-
висит от степени тренированности человека, его склонности к данному
виду деятельности, состояния его здоровья, мотивации, состояния окру-
жающей среды. Во-вторых, адаптация к нагрузкам зависит от коэффициен-
та полезного действия, который определяется как отношение величины за-
трат энергии на внешнюю (физическую) работу к величине общих энерго-
затрат организма человека. Чем ниже коэффициент полезного действия,
тем меньше эффективность работы и тем больше напряжение организма.
Физическая работа может быть динамической и статической. Динамиче-
ская работа выполняется в том случае, когда в физическом смысле преодо-
левается сопротивление на определенном расстоянии (например, при
подъеме по лестнице или в гору, при беге или езде на велосипеде) и работа
может быть выражена в физических единицах. При положительной дина-
мической работе скелетные мышцы укорачиваются и действуют как «дви-
гатель», а при отрицательной динамической работе напряжение скелетной
мышцы играет роль «тормоза» и уступает весу груза (например, при спуске
с горы). Статическая работа производится при изометрическом мышеч-
ном сокращении (без изменения длины мышцы), при этом не преодолева-
ется никакое расстояние и в физическом смысле это не работа. Однако ор-
ганизм реагирует на нее изменениями физиологических функций, соответ-
ствующими нагрузке. Проделанная статическая работа измеряется произ-
ведением силы сокращения скелетной мышцы и его продолжительности.
Умственная работа включает мыслительный и эмоциональный компонен-
ты. Мыслительный компонент преобладает при работе, требующей прояв-
22. Физиологические основы труда • 857
ления интеллектуальных способностей человека. Примерами служат реше-
ние математических задач, обдумывание конкретных ситуаций и концен-
трация внимания, обработка сигналов при работе за компьютером или при
вождении автомобиля. Умственная работа с преобладанием эмоционально-
го компонента связана с реакциями вегетативной нервной системы и нахо-
дит выражение в вегетативных компонентах эмоций человека (например,
ощущение радости, гнева, печали и т. д.).
Наиболее часто в повседневной жизни человек выполняет комбиниро-
ванную работу, когда сочетаются элементы физического и умственного
труда {сенсомоторная деятельность). Для этого вида деятельности характер-
на не тяжелая мышечная работа, а определенный навык и тренировка, как,
например, при выполнении хирургических операций, сборке машин на
конвейере и т. д.
22.1. Образование энергии в скелетных мышцах при физической
работе
В мышечных клетках человека, как и в любых других видах клеток, проис-
ходит энергетический обмен, при котором богатые энергией питательные
вещества усваиваются и химически преобразуются, а конечные продукты
обмена с более низким содержанием энергии выделяются из клетки. Вы-
свобождающаяся при этом энергия в мышцах обеспечивает их сокраще-
ние. Основным источником энергии для мышечного сокращения при фи-
зической работе является гидролиз АТФ. Макроэргические фосфатные
связи молекулы АТФ нестойкие и концевые фосфатные группы легко от-
щепляются, при этом освобождается энергия (7—10 ккал/моль АТФ). При-
соединение молекул АТФ к головкам миозиновых мостиков и последую-
щий гидролиз АТФ являются ключевым моментом в процессе преобразо-
вания химической энергии фосфатных связей в механическую энергию
мышечного сокращения. В нормальных физиологических условиях мы-
шечная активность как низкой, так и высокой мощности не сопровождает-
ся истощением запасов энергии макроэргических соединений. Это связано
с процессами ресинтеза АТФ. Различают аэробный и анаэробный пути ре-
синтеза АТФ.
22.1.1. Анаэробный путь ресинтеза АТФ
При кратковременной физической работе большой мощности ресинтез
АТФ происходит анаэробным путем, в котором выделяют три процесса:
фосфогенный, миозинкиназный и гликолиз.
Фосфогенный анаэробный процесс (креатинфосфокиназная реакция).
В мышцах наряду с АТФ содержится другое макроэргическое фосфорное
соединение — креатинфосфат, являющееся вторым после АТФ по значи-
мости макроэргическим фосфатным соединением. Креатинфосфат адсор-
бируется на сократительных белках миофибрилл и на мембране саркоплаз-
матического ретикулума. С тонким филаментом — актином связан фер-
мент креатинфосфокиназа, который катализирует реакцию перефосфори-
лирования между креатинфосфатом и аденозидифосфатом (АДФ), обра-
зующимся при гидролизе АТФ.
Креатинфосфат + АДФ------------------> АТФ + Креатин
Креатинфосфокиназная реакция первой включается в процесс ресинте-
за АТФ в момент начала мышечной работы и протекает с максимальной
858 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
скоростью до тех пор, пока запасы креатинфосфата в мышцах не будут ис-
черпаны примерно на 30 % (в течение 5—6 с). Затем скорость креатинфос-
фокиназной реакции начинает уменьшаться и в процесс ресинтеза АТФ
вступают гликолиз и аэробный механизм.
Фосфогенный аэробный процесс ресинтеза АТФ играет решающую
роль в энергетическом обеспечении кратковременной физической работы
максимальной мощности (например, подъем и перенос тяжелых грузов), в
процессе быстрого перехода от покоя к работе, во время финишного уско-
рения у спортсменов.
Анаэробный гликолиз. Как только в процессе мышечной работы креатин-
фосфокиназная реакция перестает обеспечивать необходимую скорость ре-
синтеза АТФ и в саркоплазме мышечных волокон увеличивается концен-
трация молекул АДФ, происходит активация ресинтеза АТФ за счет ана-
эробного гликолиза. Источником энергии для ресинтеза АТФ являются за-
пасы гликогена в миоцитах и глюкоза плазмы крови. Повышение концен-
трации АДФ в саркоплазме, а также увеличение содержания ионов Са++
активируют ферменты фосфорилазу и гексокиназу. Молекулы этих фер-
ментов локализованы на мембране саркоплазматического ретикулума, а
также на других внутриклеточных компонентах.
В процессе гликолиза образуется до 2,5 кДж/кг/мин. Гликолиз как ис-
точник энергии мышечного сокращения эффективно обеспечивает сокра-
тительный процесс с 20-30-й секунды после начала сокращения и спосо-
бен обеспечивать мышечную активность высокой мощности до 2—2,5 мин.
Скорость гликолиза угнетается под влиянием повышения концентрации
молочной кислоты в миоцитах и уменьшения pH в саркоплазме. Накопле-
ние молочной кислоты при анаэробной работе находится в прямой зависи-
мости от мощности и общей продолжительности физической работы.
Увеличение количества молочной кислоты в саркоплазматическом про-
странстве мышц вызывает рост осмотического давления в саркоплазме,
при этом вода из межклеточной среды поступает внутрь мышечных воло-
кон и вызывает их набухание. Значительные изменения осмотического
давления в мышцах могут быть причиной болевых ощущений.
Миокиназная реакция. Физическая активность высокой мощности, а
также развитие утомления (фаза истощения) приводят к повышению в
саркоплазме концентрации молекул АДФ, которые являются источником
ресинтеза АТФ:
Аденилатциклаза
АДФ + АДФ--------------------> АТФ + АМФ.
Миокиназная реакция, как и креатинфосфокиназная, легко обратима и
используется в организме для уменьшения резких перепадов в скорости
образования и использования АТФ. В случае появления в клетке избытка
АТФ он быстро устраняется с помощью миокиназной реакции.
22.1.2. Аэробный гликолиз
Примерно 90 % запасов АТФ в миоцитах образуется в процессе аэробного
ресинтеза, т. е. при участии кислорода. Ферментные системы аэробного
обмена расположены в основном в митохондриях клеток. К числу суб-
стратных циклов окисления относятся: гликолитическое расщепление уг-
леводов, заканчивающееся образованием пировиноградной кислоты, окис-
лительное декарбоксилирование пировиноградной кислоты, цикл превра-
щений трикарбоновых кислот, окислительное дезаминирование аминокис-
22. Физиологические основы труда • 859
лот, [3-окисление жирных кислот. Суть химических превращений в суб-
стратных циклах заключается в постепенном преобразовании исходного
субстрата в форму, доступную действию специфических дегидрогеназ, с
последующим высвобождением энергии в ходе окислительно-восстанови-
тельных реакций, где участвуют дыхательные ферменты. Энергия окисле-
ния, выделяющаяся в реакциях дегидрогенирования, сохранятся в соеди-
нениях водорода с коферментами НАД или ФАД. Для ресинтеза АТФ она
используется при переносе водорода от коферментов НАД и ФАД на ки-
слород по системе дыхательных ферментов, которая расположена на внут-
ренней мембране митохондрий.
В дыхательной цепи имеется три пункта сопряжения, где за счет энер-
гии, высвобождающейся при переносе электронов, может синтезироваться
АТФ. Первый пункт сопряжения находится на участке переноса водорода
от НАД к ФАД. Количества свободной энергии, высвобождаемой в этой
реакции, достаточно для синтеза 1 моля АТФ. Второй пункт сопряжения
локализуется на участке переноса электронов от кофермента Q через цито-
хром к цитохрому с. Третья молекула АТФ синтезируется на завершающем
этапе дыхательной цепи в цитохромоксидазной реакции, где происходит
перенос электронов с системы цитохромов на кислород. В этом пункте со-
пряжения энергии освобождается больше, чем при любой другой реакции
дыхательной цепи. Этой энергии вполне могло бы хватить для образования
нескольких молей АТФ, но, тем не менее, в этом пункте сопряжения, как
и в первых двух, синтезируется только 1 моль АТФ. Излишек энергии, вы-
деляемой в цитохромоксидазной реакции, служит главной движущей си-
лой для всего процесса переноса электронов по дыхательной цепи. Кисло-
род, акцептирующий электроны, которые поставляет цитохромоксидазная
реакция, служит резервуаром, обеспечивающим постоянный отток элек-
тронов из дыхательной цепи и поддерживающим ее компоненты в окис-
ленном состоянии, благодаря чему они могут принимать водород от раз-
личных субстратов окисления.
Наряду с обычным путем окисления субстратов на внутренней мембра-
не мышечной клетки существует также путь окисления, локализованный
на внешней мембране, в котором принимают участие цитохром с, система
ФП5 — цитохром Ь5 и цитохромоксидаза. Активация этого пути приводит
к быстрому окислению внемитохондриального НАД-Н, но он не связан с
синтезом АТФ и ведет к рассеиванию энергии в виде тепла. Этот путь ис-
пользуется в качестве буферной системы, поддерживающей необходимую
концентрацию окисленной формы НАД в саркоплазме и устраняющей из-
быток молочной кислоты, образующейся при гликолизе.
Общая схема, иллюстрирующая взаимосвязь аэробных и анаэробных
превращений в энергетическом обмене скелетной мышцы, представлена
на рис. 22.1.
Общий выход энергии при аэробном процессе более чем в 10 раз пре-
вышает изменение свободной энергии при гликолитическом распаде угле-
водов в анаэробных условиях. В качестве субстратов аэробных превраще-
ний в работающих мышцах могут быть использованы не только внутримы-
шечные запасы гликогена, но и внемышечные резервы углеводов (напри-
мер, гликоген печени), жиров, а в отдельных случаях и белков. Поэтому
суммарная емкость аэробного процесса очень велика и трудно поддается
точной оценке. В отличие от гликолиза, метаболическая емкость которого
в значительной степени зависит от изменений внутренней среды вследст-
вие накопления в организме избытка молочной кислоты, конечные про-
дукты аэробных превращений — СО2 и Н2О — не вызывают каких-либо
860 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Рис. 22.1. Общая схема образования энергии в мышечных клетках.
Источниками образования АТФ в мышечных клетках являются окислительное фосфорилиро-
вание, а также фосфогенная система «гликоген—молочная кислота». Аэробное дыхание явля-
ется наиболее эффективным, так как из каждой молекулы глюкозы образуется 38 молекул
АТФ, а при анаэробном дыхании — только две.
значительных изменений внутренней среды и легко удаляются из орга-
низма.
Образование 1 моля АТФ в процессе окислительного фосфорилирова-
ния эквивалентно потреблению 3,45 л О2. Столько же кислорода в покое
потребляется в течение 10—15 мин, а при напряженной мышечной дея-
тельности (например, во время бега на марафонскую дистанцию) — за
1 мин. Однако в самих работающих мышцах запасы кислорода крайне не-
велики. Небольшое его количество находится в растворенном состоянии
во внутриклеточной саркоплазме и в связанном состоянии с миоглобином
мышц. Основное же количество кислорода, потребляемого в мышцах для
ресинтеза АТФ, доставляется в ткани через систему легочного дыхания и
кровообращения.
Для бесперебойной работы дыхательной цепи и механизма окислитель-
22. Физиологические основы труда • 861
ного фосфорилирования напряжение О2 в клетках должно поддерживаться
на уровне не ниже 5—10 мм рт. ст. Чтобы обеспечить его, напряжение О2
снаружи (в мышечных капиллярах) должно быть примерно 15—20 мм рт.
ст., поскольку кислород поступает в клетки путем диффузии.
Поддержание критического напряжения О2 на наружной клеточной
мембране независимо от изменений скорости расхода кислорода в тканях
осуществляет сложная система регуляции, в которую наряду с внутрикле-
точными механизмами метаболического контроля входят также нервная и
гормональная регуляция внешнего дыхания, центрального и перифериче-
ского кровообращения.
Максимальная мощность аэробного процесса в равной мере зависит как
от скорости утилизации О2 в клетках (а она, в свою очередь, от общего
числа митохондрий в клетке, количества и активности ферментов аэробно-
го окисления), так и от скорости поставки О2 в ткани.
22.1.3. «Кислородный каскад» и эффективность транспорта
кислорода к работающим мышцам
Скорость доставки кислорода к тканям является одним из важнейших
факторов, определяющих возможность обеспечения энергией работаю-
щих мышц. В митохондриях скелетных мышц, где образуется до 90 %
всей необходимой энергии, скорость ресинтеза АТФ находится в зависи-
мости от достигнутой концентрации или напряжения кислорода в клетке.
Общая скорость ресинтеза АТФ, обеспечиваемая различными источника-
ми энергии, является функцией напряжения кислорода в клетке. При
низком уровне метаболических превращений в клетке, как это имеет ме-
сто в покоящейся мышце, изменения в скорости доставки О2 в ткани не
оказывают влияния на скорость ресинтеза АТФ (зона насыщения). Одна-
ко при снижении рО2 в клетке ниже некоторого критического уровня под-
держание скорости ресинтеза АТФ на необходимом уровне возможно
только за счет адаптивных сдвигов клеточного метаболизма, связанных с
природой используемых субстратов, размерами их внутриклеточных запа-
сов, с состоянием фосфорилирования и величиной внутриклеточного pH,
что с неизбежностью потребует увеличения скорости доставки О2 в ткани.
Максимальная скорость потребления О2 митохондриями скелетных мышц
может быть поддержана только до определенного критического значения
рО2 в клетке, которое составляет от 0,5 до 3,5 мм рт. ст. Если уровень ме-
таболической активности при мышечной работе превысит значение мак-
симально возможного усиления аэробного ресинтеза АТФ, то возрастаю-
щая энергетическая потребность в этих условиях может быть компенсиро-
вана за счет анаэробного ресинтеза АТФ. Однако этот диапазон анаэроб-
ной метаболической компенсации весьма узок, и выше значения макси-
мального увеличения скорости ресинтеза АТФ, обеспечиваемого сочетан-
ной метаболической активностью аэробного и анаэробных процессов в
работающей мышце, дальнейшее исполнение ее сократительной функции
становится невозможным. Диапазоны метаболической активности, в пре-
делах которых текущая поставка О2 недостаточна для поддержания необ-
ходимого уровня ресинтеза АТФ, обычно обозначаются как гипоксиче-
ские состояния (т. е. состояния кислородной недостаточности) разной тя-
жести.
Для того чтобы поддерживать напряжение О2 в митохондриях на уров-
не, превышающем критическое значение, при котором еще сохраняются
условия для адаптивной регуляции клеточного обмена, напряжение О2 на
862 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Рис. 22.2. «Кислородный каскад» орга-
низма.
В обычных условиях процессы биологическо-
го окисления в клетках тканей на уровне ми-
тохондрий обеспечиваются при сохранении
постоянства парциального давления кислоро-
да на этапах внешнего дыхания (альвеоляр-
ный воздух), транспорта газов кровью (арте-
риальная и смешанная венозная кровь), а так-
же в тканях — на мембранах клеток и внутри
митохондрий.
наружной клеточной мембране
должно составлять не менее 15—20
мм рт. ст. Для поддержания этого
критического напряжения О2, при
котором возможно нормальное
функционирование мышц, напряже-
ние О2 в артериолах, доставляющих
кровь непосредственно к работающим мышцам, должно составлять при-
близительно 40 мм рт. ст., а в магистральных артериях —до 80—90 мм рт.
ст. В легочных альвеолах, где осуществляется газообмен между кровью и
атмосферным воздухом, напряжение О2 составляет около 100 мм рт. ст., во
вдыхаемом воздухе оно равняется 150 мм рт. ст.
На рис. 22.2 вышеуказанные значения критических напряжений О2 на
различных этапах транспорта кислорода в организме обозначены в виде
нисходящих ступеней с постепенно понижающимися уровнями рО2 и ско-
ростью доставки О2 к тканям. Этот «кислородный каскад» обеспечивает по-
стоянную «накачку» О2 в работающие ткани в точном соответствии с их
метаболической потребностью. Сохранение критических напряжений ки-
слорода на каждом из обозначенных на схеме этапов транспорта кислорода
к тканям обеспечивается совокупной деятельностью всех вегетативных
систем организма.
2
2
I
О.
О
о
§
§
ф
о
150—1
104 -
96 -
39-
20-
Вдыхаемый воздух
Альвеолярный воздух
Артериальная кровь
Смешанная венозная кровь
Мембрана клеток
Мембрана митохондрий
о
§
л
5 -
0
22.1.4. Потребление кислорода, кислородный дефицит,
кислородный долг и кислородный запрос при мышечной
работе
При переходе от состояния покоя к интенсивной мышечной деятельности
потребность организма в кислороде возрастает во много раз, однако сразу
она не может быть удовлетворена. Нужно время, чтобы усилилась деятель-
ность систем дыхания и кровообращения и чтобы кровь, обогащенная ки-
слородом, могла дойти до работающих мышц. По мере усиления активно-
сти системы вегетативного обеспечения мышечной деятельности постепен-
но увеличивается потребление кислорода работающими мышцами. При
умеренной работе, если ЧСС не превышает 150 уд/мин, скорость потреб-
ления О2 возрастает до тех пор, пока не наступит устойчивое состояние
метаболических процессов, при котором О2-потребление достигает посто-
янного уровня и в каждый данный момент времени точно соответствует
потребности организма в нем. Такое устойчивое состояние называется ис-
тинным. Уровень потребления О2 в устойчивом состоянии зависит от
мощности выполняемой работы (рис. 22.3).
При более интенсивной работе (ЧСС 150—180 уд/мин) устойчивое со-
стояние не устанавливается и потребление О2 может возрастать до конца
22. Физиологические основы труда • 863
Рис. 22.3. Потребление кислорода в организме человека при легкой и тяжелой фи-
зической работе. Потребление кислорода резко возрастает в тканях организма с мо-
мента начала физической работы.
Однако для приспособления кровотока к возросшим метаболическим потребностям тканей
требуется некоторое время, что приводит к формированию кислородного дефицита (Кд). При
легкой работе величина кислородного дефицита остается постоянной после достижения ус-
тойчивого состояния. При тяжелой физической работе величина кислородного дефицита воз-
растает до момента ее окончания. Поэтому по окончании как легкой, так и тяжелой физиче-
ской работы скорость потребления кислорода остается некоторое время выше уровня покоя,
что обеспечивает ликвидацию («выплату») кислородного долга. По окончании тяжелой физи-
ческой работы происходит ликвидация не только «алактатного», но и «лактатного» кислород-
ного долга. Лр — легкая и Тр — тяжелая физическая работа.
работы или до достижения максимально возможного уровня. В последнем
случае может наблюдаться «ложное устойчивое состояние», когда потреб-
ление О2 некоторое время (6—10 мин) поддерживается на постоянном
максимальном уровне не потому, что потребность организма в кислороде
полностью удовлетворяется, а потому, что исчерпаны возможности сердеч-
но-сосудистой системы по доставке его к тканям. Именно эти возможно-
сти в большинстве случаев ограничивают потребление О2 мышечными
клетками, хотя иногда лимитирующим фактором становится окислитель-
ная способность самих работающих мышечных клеток, в частности актив-
ность дыхательных ферментов в митохондриях.
Максимальный уровень потребления О2 не может поддерживаться дол-
го. Во время длительной работы он снижается из-за утомления.
Количество кислорода, которое необходимо организму, чтобы полно-
стью удовлетворить энергетические потребности за счет аэробных процес-
сов, называется кислородным запросом работы. При интенсивной работе ре-
альное потребление кислорода — кислородный приход — составляет только
часть кислородного запроса. Разность между кислородным запросом рабо-
ты и реально потребляемым кислородом составляет кислородный дефицит
организма (см. рис. 22.3). В условиях кислородного дефицита активируют-
ся анаэробные реакции ресинтеза АТФ, что приводит к накоплению в ор-
ганизме недоокисленных продуктов анаэробного распада. Во время рабо-
ты, при которой возможно установление устойчивого состояния, часть
анаэробных метаболитов может ликвидироваться по ходу работы за счет
усиления аэробных реакций, другая их часть устраняется после работы.
Если устойчивого состояния не наступает или при ложном устойчивом со-
864 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
стоянии количество недоокисленных продуктов все время увеличивается
по ходу работы, они устраняются в восстановительном периоде.
Для этих процессов требуется дополнительное количество кислорода,
поэтому некоторое время после окончания работы потребление его про-
должает оставаться повышенным по сравнению с уровнем покоя. Этот
восстановительный излишек кислородного потребления получил название
кислородного долга. Кислородный долг всегда больше кислородного дефи-
цита, и чем больше интенсивность и продолжительность работы, тем зна-
чительнее это различие.
В период восстановления после мышечной работы, когда в организме
имеется достаточное количество субстратов биологического окисления и
доставка кислорода к митохондриям клеток не ограничена, уровень по-
требления кислорода зависит от количества свободной АТФ, осуществляю-
щей дыхательный контроль в митохондриях. Субстратами окислительных
энергетических превращений являются накопившиеся во время работы
молочная кислота, янтарная кислота, альфа-глицерофосфат, а на поздних
стадиях восстановления — и жирные кислоты.
Некоторая часть избыточно потребленного в период отдыха кислорода
идет на восстановление его запасов в миоглобиновом и гемоглобиновом
депо. При утомительной работе эффективность использования кислорода
снижается вследствие частичного разобщения процессов окисления и об-
разования АТФ. Это разобщение может сохраняться и после работы, что
обусловливает увеличение потребления кислорода по сравнению с уровнем
покоя.
При работе различного характера разные факторы становятся ведущими
в образовании О2-долга. Например, при выполнении кратковременной ра-
боты решающую роль в образовании О2-долга играет ресинтез КрФ и
АТФ. С увеличением длительности нагрузки возрастает значение окисле-
ния молочной кислоты, восстановления pH, ионного равновесия и других
процессов.
После работы, в которой возможно установление устойчивого состоя-
ния, снижение потребления О2 происходит быстро. О2-долг наполовину
оплачивается за 27—30 с, а полностью — за 3—5 мин. При более интенсив-
ной работе в кривой снижения потребления О2 выявляются две фазы — на-
чальный быстрый спад и более медленный затяжной процесс возвращения
к уровню покоя. Быстрый компонент О2-долга (так называемый алактат-
ный) позволяет приблизительно оценить вклад креатинфосфатного меха-
низма в энергетическое обеспечение работы. Медленный компонент ки-
слородного долга (лактатный) при сравнительно кратковременной работе
может служить отражением развития анаэробного гликолитического про-
цесса. При длительной работе в его образовании значительна доля и дру-
гих процессов, оценить которую точно очень трудно. Медленный компо-
нент кислородного долга уменьшается наполовину за 15—20 мин, а ликви-
дируется полностью за 1,5—2 ч (см. рис. 22.3).
22.2. Физиологические основы тренировки двигательного навыка
22.2.1. Развитие силовых качеств мышц
Тренировка двигательного навыка увеличивает произвольную силу мышцы
или группы мышц. Силу мышечного сокращения можно контролировать
по массе груза, который поднимает человек. У нетренированного человека
произвольная активация мышцы может быть недостаточной для того, что-
22. Физиологические основы труда • 865
бы развить максимальную силу сокращения. Развитие силы мышц при
тренировке является результатом так называемой периферической и цен-
тральной адаптации мышцы к физической нагрузке, или «нервным трени-
рующим эффектом». Начальный этап развития мышечной силы включает
формирование правильной последовательности активации мышц. Под
этим подразумевается точность выполнения специфического произвольно-
го задания, например выполнение движения рукой, когда сустав должен
быть согнут под определенным углом и движение выполняется с опреде-
ленной скоростью.
Поперечное сечение мышцы. При физической тренировке рост макси-
мальной произвольной силы мышцы прямо зависит от увеличения площа-
ди ее поперечного сечения. Однако анатомическое поперечное сечение
мышцы, представляющее собой проходящую через середину мышцы пер-
пендикулярно ее волокнам плоскость, при физической тренировке не уве-
личивается в той же степени, как максимальная произвольная сила мыш-
цы. Способность мышцы развивать силу зависит от ее физиологического
поперечного сечения и напряжения. Так, физиологическое поперечное се-
чение (плоскость, пересекающая максимальное количество волокон мыш-
цы) в мышцах ноги человека в 2—8 раз больше их анатомического попе-
речного сечения. На зависимость между силой мышцы и ее физиологиче-
ским поперечным сечением влияют такие факторы, как специфическое
напряжение, изменения типа мышечных волокон, архитектуры мышечных
волокон и сухожилий и т. д.
Максимальная сила мышцы. При физической тренировке максимальная
произвольная сила мышечного сокращения возрастает примерно в три
раза больше, чем максимальная тетаническая сила сокращения мышцы,
которую можно вызвать искусственным электрическим раздражением дви-
гательного нерва у нетренированного человека. Максимальная произволь-
ная сила двустороннего сокращения мышц-синергистов меньше, чем сила,
развиваемая при одностороннем максимальном произвольном сокращении
одной группы мышц. При этом в норме отношение билатеральной макси-
мальной силы к суммарной силе, развиваемой, например, мышцами пра-
вой и левой руки, составляет порядка 90 %. Физическая тренировка сни-
жает билатеральный дефицит в показателях максимальной произвольной
силы.
22.2.2. Физиологические механизмы формирования трудовых
навыков
В процессе трудовой деятельности человека формируются различные дви-
гательные умения и навыки, составляющие основу его поведения.
Двигательные умения — способность на двигательном уровне справлять-
ся с новыми задачами поведения. Двигательные навыки — это освоенные и
упроченные действия, которые могут осуществляться человеком без уча-
стия сознания (автоматически) и обеспечивают оптимальное решение дви-
гательной задачи.
Любые навыки: бытовые, профессиональные, спортивные — не являют-
ся врожденными движениями. Они приобретаются в ходе индивидуального
развития человека. Комплекс нейронов, обеспечивающих формирование
двигательных навыков у человека, располагается в различных отделах
нервной системы, становясь доминантой, т. е. господствующим очагом
возбуждения в центральной нервной системе. Он подавляет деятельность
других нервных центров и, соответственно, «лишних» скелетных мышц.
28 - 6095
866 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
В результате движения выполняются все более экономно, при включении
лишь самых необходимых мышечных групп, и лишь в те моменты, кото-
рые нужны для его осуществления. Происходит экономизация энерготрат.
Порядок возбуждения в доминирующих нервных центрах закрепляется
в виде системы условных и безусловных рефлексов и сопровождающих их
вегетативных реакций, образуя двигательный динамический стереотип. Ка-
ждый предшествующий двигательный акт в этой системе запускает сле-
дующий, облегчает выполнение целостного комплекса двигательных актов
и освобождает сознание человека от контроля за каждым его элементом.
Процесс обучения двигательному навыку у человека начинается с побу-
ждения к действию, которое задается подкорковыми и корковыми мотива-
ционными зонами. У человека это, главным образом, стремление к удовле-
творению определенной социальной потребности (желание заниматься
данным видом труда, преуспеть в нем и пр.).
На первом этапе формирования двигательного навыка возникает замы-
сел действия, осуществляемый ассоциативными зонами коры больших по-
лушарий (переднелобными и нижнетеменными). Вначале это лишь общее
представление о двигательной задаче, которое возникает либо при показе
движения другим лицом, либо после словесной инструкции, самоинструк-
ции, речевого описания. В сознании человека создается определенный эта-
лон требуемого действия. Формирование такой наглядно-образной модели
складывается из образа ситуации в целом (задаваемые пространственные и
временные характеристики двигательной задачи) и образа тех мышечных
действий, которые необходимы для достижения цели. Имея представление
о требуемой модели движения, человек может осуществить ее разными
мышечными группами. Так, например, подпись человека имеет характер-
ные черты, независимо от мышечных групп, выполняющих ее (пальцы,
кисть, предплечье).
На втором этапе обучения двигательному навыку начинается непосред-
ственное выполнение разучиваемого упражнения. При этом отмечаются
три стадии формирования двигательного навыка: 1) стадия генерализации
(иррадиации возбуждения); 2) стадия концентрации; 3) стадия стабилиза-
ции и автоматизации.
На первой стадии созданная модель становится основной для перевода
внешнего образа во внутренние процессы формирования программы соб-
ственных действий. На ранних этапах онтогенеза, когда речевая регуляция
движений (внешней речью постороннего лица или внутренней собствен-
ной речью) еще не развита, особое значение имеют процессы подражания,
общие у человека и животных. Наблюдая за действиями другого лица и
имея некоторый опыт управления своими мышцами, ребенок превращает
свои наблюдения в программы собственных движений.
Некоторые особенности программирования движений у человека отра-
жаются в межцентральных взаимосвязях электрической активности мозга.
Можно видеть, например, что при наблюдении за выполнением бега по-
сторонним лицом в коре больших полушарий у наблюдающего человека
появляются потенциалы в темпе этого бега (своеобразная модель наблю-
даемого движения). При представлении и при мысленном выполнении
движений пространственные взаимосвязи мозговой активности начинают
отличаться от состояния покоя и приближаться к таковым при реальной
работе, появляется небольшая биоэлектрическая активность в нужных для
выполнения реального движения мышцах. Этим объясняется тренирующее
воздействие на формирование двигательного навыка идеомоторного (мыс-
ленного) выполнения упражнений. В создании двигательных программ
22. Физиологические основы труда • 867
принимают участие многие нейроны коры, мозжечка, таламуса, подкорко-
вых ядер и ствола мозга. Обширное вовлечение множества мозговых эле-
ментов необходимо для поиска наиболее нужных из них. Этот процесс
обеспечивается широкой иррадиацией возбуждения по различным зонам
мозга и сопровождается обобщенным характером (генерализацией) пери-
ферических реакций со стороны скелетных мышц. Стадия генерализации
характеризуется напряжением большого числа активированных скелетных
мышц, их продолжительным сокращением, одновременным вовлечением в
движение мышц-антагонистов. Все это нарушает координацию движений,
делает их закрепощенными, приводит к значительным энерготратам и, со-
ответственно, излишне выраженным вегетативным реакциям. Например,
подобные реакции можно наблюдать у человека, который впервые сел за
руль автомобиля.
На второй стадии формирования двигательного навыка происходит
концентрация возбуждения в необходимых для его осуществления корко-
вых зонах. В других зонах коры активность подавляется одним из видов
условного внутреннего торможения — дифференцировочным торможени-
ем. При формировании сложных двигательных программ концентрация
возбуждения происходит в сложной системе различных корковых зон, за-
интересованных в управлении движениями. Между ними устанавливается
высокий уровень пространственной синхронизации электрической актив-
ности (синхронность и синфазность колебаний потенциалов), который от-
ражает их функциональные взаимосвязи. На этой стадии двигательный на-
вык уже сформирован. Включаются лишь необходимые мышечные группы
и только в нужные моменты движения. В результате рабочие энерготраты
снижаются. Однако сформированный навык еще очень непрочен и нару-
шается при любых новых раздражениях (вождение автомобиля в условиях
интенсивного городского движения сразу после автодрома, появление вне-
запной опасности на дороге и т. д.).
На третьей стадии в результате многократного повторения навыка в
разнообразных условиях появляются стабильность и надежность двига-
тельного навыка, снижается сознательный контроль над его элементами,
т. е. возникает автоматизация навыка. Внешние раздражения на этой ста-
дии лишь подкрепляют рабочую доминанту данного двигательного навыка,
не разрушая ее. Большая часть посторонних афферентных потоков не про-
пускается в спинной и головной мозг: специальные команды из вышеле-
жащих центров вызывают пресинаптическое торможение импульсов от пе-
риферических рецепторов, препятствуя их доступу в спинной мозг и вы-
шележащие центры. Этим обеспечивается защита сформированных двига-
тельных программ от случайных влияний и повышается надежность сфор-
мированных навыков.
Процесс автоматизации не означает выключения коркового контроля за
выполнением движения. Однако в этой системе центров снижается уча-
стие лобных ассоциативных отделов коры, что, по-видимому, и отражает
снижение его осознаваемости.
Особое значение в отработке моторных программ имеют обратные свя-
зи между вовлеченными в исполнение двигательных навыков скелетными
мышцами и управляющими ими нервными центрами. Информация, по-
ступающая в нервные центры при движении, служит для сравнения полу-
ченного результата с имеющимся в этих центрах эталоном. При их несов-
падении в мозговых аппаратах сравнения (лобных долях, подкорковом
хвостатом ядре) возникают импульсы рассогласования и в программу вно-
сятся поправки — сенсорные коррекции. При кратковременных движениях
28‘
868 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
рабочие фазы настолько малы (составляют сотые и тысячные доли секун-
ды), что сенсорные коррекции по ходу движения вносить невозможно.
В этих случаях вся программа действия должна быть готова до начала дви-
гательного акта, а поправки могут вноситься при его повторениях.
В системе обратных связей различают «внутренний контур» регуляции
движений, передающий информацию в нервные центры от двигательного
аппарата и внутренних органов (в первую очередь — от рецепторов мышц,
сухожилий и суставных сумок), и «внешний контур», несущий сигналы от
экстерорецепторов (главным образом, зрительных и слуховых). При пер-
вых попытках выполнения движений, благодаря множественному и неоп-
ределенному характеру мышечной афферентации, основную роль в систе-
ме обратных связей играют сигналы «внешнего контура» — зрительный и
слуховой контроль. Поэтому на начальных этапах освоения двигательных
навыков для облегчения процесса обучения так важно использовать зри-
тельные ориентиры и звуковые сигналы. По мере освоения навыка «внут-
ренний контур» регуляции движений приобретает все большее значение,
обеспечивая автоматизацию навыка, а значимость «внешнего контура»
снижается.
Большую роль в процессе моторного научения играет речевая регуляция
движений (словесные указания педагога, внутренняя речь обучаемого).
В высших отделах мозга (кора больших полушарий и подкорковые образо-
вания) человека обнаружены специальные «командные» нейроны, которые
реагируют на словесные приказы и запускают нужные действия.
22.2.3. Работоспособность
Работоспособность — это свойство человека на протяжении длительного
времени и с определенной эффективностью выполнять максимальное ко-
личество физической или умственной работы. На работоспособность чело-
века влияют внешние (характер труда, условия окружающей среды, режи-
мы труда, отдыха и т. д.) и внутренние факторы (мотивация, степень со-
вершенства трудовых навыков, функциональные резервы человека).
Работоспособность фазно изменяется на протяжении рабочей смены.
Так, фаза врабатывания характеризуется повышением активности ЦНС и
уровня обменных процессов в организме человека, усилением деятельно-
сти сердечно-сосудистой и дыхательной систем. Фаза врабатывания короче
при физическом труде, чем при умственном.
Фаза относительно устойчивой работоспособности характеризуется опти-
мальным уровнем функционирования систем организма и максимальной
эффективностью труда. Время устойчивой работоспособности тем меньше,
чем больше физическая тяжесть и нервная напряженность труда.
Фаза снижения работоспособности связана с развитием утомления.
В конце рабочего дня возникает фаза вторичного повышения работоспособ-
ности, которая обусловлена условно-рефлекторными механизмами и свя-
зана с предстоящим концом работы и отдыхом.
22.3. Функции физиологических систем организма человека
при физической работе
Физическое напряжение вызывает изменения функций практически всех
физиологических систем организма человека, особенно сердечно-сосуди-
стой и дыхательной. Перестройка функций физиологических систем дает
возможность организму человека адаптироваться к физическим нагруз-
22. Физиологические основы труда • 869
кам и успешно справляться с выполнением трудовой задачи. Выражен-
ность адаптивных сдвигов в деятельности физиологических систем слу-
жит непосредственно мерой реакции организма на физическую нагрузку
и лежит в основе классификации физического труда по степени его тя-
жести.
22.3.1. Кровообращение
При физической работе активируется деятельность сердечно-сосудистой
системы и, таким образом, более полно удовлетворяется увеличенная по-
требность работающих мышц в кислороде, а образующееся тепло с током
крови отводится от работающей мышцы в те участки организма, где про-
исходит его отдача. Через 3—6 мин после начала легкой работы возникает
стационарное (устойчивое) повышение частоты сердечных сокращений,
которое обусловлено иррадиацией возбуждения из моторной зоны коры на
сердечно-сосудистый центр продолговатого мозга и поступлением активи-
рующих импульсов к этому центру от хеморецепторов работающих мышц.
Активация мышечного аппарата усиливает кровоснабжение в работающих
мышцах, которое достигает максимума уже через 60—90 с после начала ра-
боты. При легкой работе формируется соответствие между кровотоком и
метаболическими потребностями мышцы. По ходу легкой динамической
работы начинает доминировать аэробный путь ресинтеза АТФ с использо-
ванием в качестве энергетических субстратов глюкозы, жирных кислот и
глицерина. При тяжелой динамической работе частота сердечных сокра-
щений увеличивается до максимума по мере развития утомления. Крово-
ток в работающих мышцах возрастает в 20—40 раз. Однако доставка к
мышцам О2 отстает от потребностей мышечного метаболизма, и часть
энергии образуется за счет анаэробных процессов.
Интенсивность трудовой деятельности можно классифицировать по по-
казателю ответных изменений частоты сердечных сокращений. Различают
легкую работу (без утомления) с устойчивой величиной частоты сердечных
сокращений и тяжелую с развитием утомления на фоне постоянного роста
частоты сердечных сокращений.
При динамической работе минутный объем кровообращения и систоли-
ческое артериальное давление (диастолическое давление меняется мало)
возрастают пропорционально выполняемой работе. Увеличение систоличе-
ского давления (до 225 мм рт. ст. и выше) происходит в результате сокра-
щения времени сердечного выброса. Диастолическое давление отражает
баланс между возросшим сердечным выбросом и пониженным перифери-
ческим сосудистым сопротивлением. При статической работе происходит
значительное повышение и систолического, и диастолического давления,
что способствует лучшему прохождению крови через сжатые сокращаю-
щейся мышцей сосуды и имеет большое значение в увеличении крово-
снабжения работающих мышц. Изменения артериального давления при
физической работе могут быть вызваны тремя основными причинами, обу-
словленными возрастанием активности симпатоадреналовой системы. Во-
первых, благодаря повышению частоты сердечных сокращений и систоли-
ческого объема происходит увеличение минутного объема кровообраще-
ния. Во-вторых, сужение сосудов в неработающих мышцах увеличивает со-
противление кровотоку. В-третьих, увеличение объема циркулирующей
крови за счет сужения венозных сосудов способствует повышению артери-
ального давления. Таким образом, в регуляции системного кровообраще-
ния при физической работе ведущую роль играет повышение активности
870 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
центров симпатической нервной системы в результате воздействия им-
пульсов, поступающих из моторной зоны коры и от рецепторов работаю-
щих мышц.
22.3.2. Кровь
Во время легкой физической работы в артериальной крови происходят не-
значительные изменения парциального давления кислорода и углекислого
газа. Тяжелая работа вызывает отчетливое снижение РСО2 без заметных из-
менений РО2, что отражает нарастание вентиляции из-за ацидоза (который
возникает в результате накопления молочной кислоты в крови и мышцах).
Насыщение кислородом венозной крови уменьшается при нарастании фи-
зической нагрузки, при этом артериовенозная разница по кислороду увели-
чивается в результате улучшения извлечения кислорода из крови работаю-
щей мышцей и ограничения кровотока в неработающих отделах.
При физической работе уменьшается объем циркулирующей плазмы
вследствие ее усиленной фильтрации через стенку капилляров и происхо-
дит выброс эритроцитов из кроветворных органов в кровь (миогенный
эритроцитоз). В результате увеличивается показатель гематокрита. Количе-
ство лейкоцитов в крови также зависит от мощности выполняемой челове-
ком работы и может достигать 15 000—30 000 клеток в 1 мкл (миогенный
лейкоцитоз). Это увеличение обусловлено преимущественно нейтрофиль-
ными гранулоцитами, причем происходит изменение соотношения клеток
различных типов. Число тромбоцитов также увеличивается пропорцио-
нально интенсивности работы (миогенный тромбоцитоз), повышается кон-
центрация тромбопластина и фибриногена в плазме крови. В результате
время свертывания крови и время кровотечения сокращаются, что пред-
ставляет собой защитную реакцию организма.
22.3.3. Дыхание
При тяжелой динамической работе вентиляция легких прямо зависит от
уровня метаболической активности организма. Это связано с накоплением
в крови молочной кислоты (метаболический недыхательный ацидоз кро-
ви), оказывающей рефлекторное стимулирующее воздействие на дыхатель-
ный центр и систему дыхания.
Кислотно-основное состояние изменяется только при тяжелой работе,
при которой развивается метаболический недыхательный ацидоз. Выра-
женность ацидоза пропорциональна анаэробной мощности и, следователь-
но, скорости образования молочной кислоты.
Потребление О2 организмом человека возрастает как линейная функция
от мощности работы, зависит от тренированности человека и коэффици-
ента полезного действия данного вида работы. Если при легкой работе по-
требление О2 соответствует метаболическим потребностям для аэробных
процессов ресинтеза АТФ, то при тяжелой мышечной работе потребление
О2 возрастает до максимально возможного для человека уровня и после
этого больше не увеличивается. Поэтому тяжелая работа не может продол-
жаться длительное время.
22.3.4. Эндокринная система
Центральная моторная команда, которая распространяется по пирамидно-
му тракту от моторной зоны коры больших полушарий к мотонейронам
22. Физиологические основы труда • 871
спинного мозга, передается также на эндокринные железы. Нервная им-
пульсация, сопровождающая центральную моторную команду, достигает
промежуточного мозга (гипоталамуса). В ответ усиливается активность
центральных нервных структур, управляющих симпатоадреналовой систе-
мой, а также нейросекреторными клетками гипоталамуса. Последние реа-
гируют секрецией нейрогормонов (либеринов), поступающих через специ-
альную систему кровеносных сосудов в гипофиз и стимулирующих про-
дукцию тропных гормонов, в первую очередь кортикотропина. Общим ре-
зультатом этих регуляторных изменений является то, что уже через 10 с от
начала мышечной работы в крови увеличивается концентрация норадрена-
лина и адреналина, а чуть позже — и кортикотропина. В течение минуты
могут повышаться уровни кортизола и эндорфинов в крови. В дальнейшем
концентрации этих, а также других гормонов устанавливаются на необхо-
димом уровне под влиянием обратных нервных связей, берущих начало от
проприорецепторов и направляемых к центральным нервным структурам.
Важное значение имеют смещение pH, ионные сдвиги, а также накопле-
ние различных метаболитов. Под влиянием этих воздействий, а также под
прямым воздействием метаболических изменений, отражающихся на ак-
тивности эндокринных желез, постепенно нарастают уровни соматотропи-
на, альдостерона и вазопрессина в крови. Более медленно активируется и
щитовидная железа.
В отличие от других гормонов уровень инсулина в крови снижается.
Это изменение наступает через определенный период: снижение уровня
инсулина наблюдается лишь через 10—20 мин работы. В ряде случаев не-
большой латентный период выявляется также до нарастания уровня сома-
тотропина. Продолжительный латентный период (иногда до 60 мин) пред-
шествует увеличению концентрации глюкагона и кальцитонина.
Активность желез может по-разному изменяться во время мышечной
работы. Большие физические нагрузки, как правило, угнетают продукцию
эстрогенов. При хорошей тренированности угнетение мало выражено или
совсем отсутствует. У мужчин при кратковременной физической работе
уровень тестостерона в крови увеличивается, а при длительной работе, на-
оборот, снижается. Силовые упражнения приводят к повышенной продук-
ции андрогенов.
Окончательное формирование гормональных реакций во время мышеч-
ной работы зависит от ряда модулирующих факторов. Важна обеспечен-
ность тканей организма углеводами. При ее снижении особенно увеличи-
вается секреция адреналина, глюкагона, соматотропина и кортизона, а
уровень инсулина в крови значительно понижается. Диета, богатая углево-
дами, или прием глюкозы во время физической нагрузки вызывают повы-
шение уровня инсулина в крови вместо его снижения, что сопровождается
угнетением усиленного выброса адреналина, глюкагона и соматотропина.
Гормональные изменения модулируются также температурой среды, со-
держанием кислорода во вдыхаемом воздухе, суточным и сезонным ритма-
ми и особо значительно — эмоциональным напряжением. При воздейст-
вии гипоксии или эмоционального напряжения подпороговые интенсив-
ности работы могут вызвать гормональные изменения. В состоянии утом-
ления активность гормональных механизмов, ответственных за мобилиза-
цию энергетических и пластических ресурсов организма, падает. В частно-
сти, это выражается в уменьшении уровня адреналина, норадреналина и
глюкокортикоидов в крови. Эти изменения не связаны с истощением кле-
ток, продуцирующих гормоны, а представляют собой регуляторно обуслов-
ленную защитную реакцию, направленную на предотвращение чрезмерных
872 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
расходов ресурсов организма. При очень длительной мышечной работе эта
защитная реакция может заменяться новым усилением адренокортикаль-
ной активности.
22.4. Физиологические функции при умственном труде
При умственном труде изменения происходят прежде всего в функциональ-
ном состоянии ЦНС, что связано с возрастанием нервного напряжения (ак-
тивируются обменные процессы в нервной ткани). Нервное напряжение
при умственном труде могут вызвать следующие факторы: 1) активация
механизмов внимания, памяти и мышления; 2) восприятие и переработка
большого объема разнообразной информации; 3) лимит времени для пере-
работки информации и принятия решения; 4) выполнение работ в ночное
время; 5) возникновение нештатной и аварийной ситуации, дефицит вре-
мени, отпущенного на ее устранение; 6) степень ответственности за при-
нимаемые решения.
Нервное напряжение сопровождается активацией как специфических,
так и неспецифических образований мозга. При этом значительная роль в
осуществлении психических функций принадлежит лобным долям мозга.
В лобных долях объединяется обширная информация об эмоциональном
состоянии человека, поступающая из внешнего мира, от внутренней сре-
ды организма, а также нижележащих структур мозга и центров старой
коры.
Кровоснабжение мозга. Активация различных структур головного мозга
при умственной работе повышает уровень обменных процессов и кровото-
ка в этих структурах. Однако общее кровоснабжение головного мозга и по-
требление энергии нервной тканью мало меняется при различных видах
умственной деятельности, так как увеличение поступления О2 и энергети-
ческих веществ усиленно работающими зонами мозга обеспечивается за
счет внутреннего перераспределения поступающего в мозг потока крови.
Так, во время умеренной мышечной работы не меняется общее кровоснаб-
жение мозга, но кровоток в моторной зоне возрастает на 50 % по отноше-
нию к уровню покоя. При работе, требующей напряжения зрительного
анализатора, усиливается кровоснабжение зрительной области коры.
Эндокринная система. При умственной работе, связанной с нервно-эмо-
циональным напряжением, повышается активность симпатоадреналовой и
гипоталамо-гипофизарно-адреналовой систем, что приводит к активации
минерального, углеводного и жирового обменов в организме человека, по-
вышению возбудимости ЦНС, усилению деятельности сердечно-сосуди-
стой и дыхательной систем.
В механизме возникновения нервно-психического напряжения прини-
мают участие следующие медиаторные системы: 1) катехоламины — адре-
налин, норадреналин, дофамин; 2) биогенные амины — серотонин; 3) аце-
тилхолин (продуцируется в ЦНС, синапсах парасимпатической нервной
системы); 4) аминокислоты — ГАМК, глицин, глутамат, пролин и т. д.;
5) гипоталамические нейропептиды (рилизинг-гормоны); 6) гипофизарные
пептиды — АКТГ, пролактин и т. д.; 7) медиаторные олигопептиды — суб-
станция Р, эндорфин, брадикинин и др. Серотонин, эндогенные олиго-
пептиды и ГАМК относятся к стресс-лимитирующей системе, обеспечи-
вающей устойчивость к эмоциональному стрессу.
Нервно-психическое напряжение при умственной деятельности увели-
чивает частоту сердечных сокращений, артериальное давление, частоту ды-
хания. В условиях психоэмоциональной нагрузки у человека нередко воз-
22. Физиологические основы труда • 873
никают трудноподавляемые двигательные акты (жестикуляция, хождение и
т. д.). При произвольном подавлении этих реакций увеличивается тониче-
ское напряжение скелетной мускулатуры.
22.5. Работа в условиях зрительного напряжения
Под контролем зрения совершается до 80—90 % трудовых операций. Зри-
тельное напряжение является важным элементом трудового процесса, оп-
ределяющим физиологическую нагрузку на организм работающих при раз-
личных видах труда. В ряде случаев зрительное напряжение (труд операто-
ров ЭВМ) приводит к развитию зрительного утомления, при котором на-
блюдается изменение рефракции и аккомодационной способности глаз,
уменьшение контрастной чувствительности, снижение устойчивости хро-
матического и ахроматического видения, уменьшение границ поля зрения,
снижение остроты зрения, скорости восприятия и переработки информа-
ции. Чрезмерная интенсивность зрительного напряжения может привести
к развитию перенапряжения зрительной функции и патологии зрительного
аппарата. Перенапряжение понижает способность глаз фокусировать и
фиксировать объект наблюдения, уменьшает объем аккомодации и ухуд-
шает бинокулярное зрение. Субъективно человек ощущает затуманенность
объекта наблюдения, размывание его очертаний.
Для предотвращения развития зрительного утомления и перенапряже-
ния следует соблюдать следующие рекомендации: 1) не допускать резких
изменений уровня освещенности рабочего места, воздействий на зрение
сильных световых и цветовых контрастов, источников яркого света;
2) подбор оптимального светового и цветового режимов работы мониторов
ЭВМ, оптимальной яркости и контрастности изображения, возможность
индивидуального выбора для оператора высоты экрана, расстояния от эк-
рана до глаз, высоты пульта с клавиатурой и т. д.; 3) соблюдение правиль-
ного режима труда и отдыха, использование сеансов релаксации и выпол-
нение специальных упражнений для глаз; 4) проведение профессионально-
го отбора при приеме на работу, требующую зрительного напряжения.
22.6. Утомление при работе
Под влиянием продолжительной или интенсивной работы у человека вре-
менно возникает особое функциональное состояние, приводящее к сниже-
нию его работоспособности и называемое утомлением. Во время работы
утомление проявляется в уменьшении силы сокращения скелетных мышц,
в возрастании энергозатрат при выполнении одной и той же внешней ра-
боты, в снижении скорости переработки информации, ухудшении памяти
и мыслительных функций, в затруднении процессов сосредоточения и пе-
реключения внимания с одного вида деятельности на другой. Утомление
может быть мышечным (физическим) и центральным (нервно-психическим).
Тяжелая физическая работа приводит в первую очередь к мышечному
утомлению, а интенсивная умственная деятельность вызывает утомление
центрального происхождения. При различных видах комбинированного
труда обе формы утомления сочетаются и их нельзя отделить друг от друга.
Субъективно утомление проявляется в виде ощущения усталости, обу-
словленного потребностью во сне и вызывающего желание прекратить ра-
боту.
874 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
22.6.1. Утомление человека при физической работе
В процессе выполнения физической работы у человека происходит посте-
пенное снижение максимальной произвольной силы мышц и максималь-
ной произвольной скорости укорочения мышц или их мощности. Макси-
мальная сила, которую способны развивать мышцы человека при их про-
извольной активации, называется максимальной произвольной силой
мышц. Под произвольной активацией работающих мышц понимают двига-
тельные команды от нейронов моторной коры человека, которые поступа-
ют к мотонейронам спинного мозга, а от последних — к иннервируемым
скелетным мышцам. При физической работе начало уменьшения показа-
телей максимальной произвольной силы мышц и их мощности обусловле-
но развитием утомления. Начиная с ранних физиологических исследова-
ний, было установлено, что у человека снижение максимальных произ-
вольных изометрических сокращений мышц при физической работе, т. е.
развитие утомления не сопровождается уменьшением возбудимости мышц
(рис. 22.4). Поэтому снижение у человека способности произвольной ак-
тивации сокращений мышц во время выполнения физической работы на-
зывается центральным утомлением. Наряду с центральным утомлением при
физической работе у человека причиной снижения способности поддержи-
вать начальную максимальную силу сокращения мышц может быть пери-
ферическое утомление, вызванное изменениями физиологических свойств
самих работающих мышц (см. главу 2).
Физиологический механизм центрального утомления при физической
работе человека обусловлен процессами, возникающими на уровне спи-
нерва
Рис. 22.4. Примеры утомления мышцы-сгибателя указательного пальца при макси-
мальных изотонических сокращениях.
Электрическое раздражение нерва, иннервирующего эту мышцу тотчас после ее утомления,
вновь вызывает сокращения, что свидетельствует о центральной природе явления утомления.
В период мышечных сокращенй, вызванных электрическим раздражением двигательного нер-
ва, двигательные центры головного мозга находятся в состоянии покоя (отдыха). В результате
кратковременного отдыха двигательных центров головного мозга возможны произвольные со-
кращения мышцы-сгибателя указательного пальца тотчас после прекращения электрического
раздражения двигательного нерва этой мышцы.
22. Физиологические основы труда • 875
нальных механизмов регуляции мышечных сокращений. Спинальные ме-
ханизмы центрального утомления различаются при статической и динами-
ческой физической работе.
22.6.1.1. Утомление человека при статической физической работе
Спинальные механизмы «центрального» утомления человека при статиче-
ской физической работе обусловлены снижением функции мотонейронов
спинного мозга. Снижение функции мотонейронов спинного мозга при
физической работе происходит в результате уменьшения возбудимости а-
и у-мотонейронов передних рогов сегментов спинного мозга. Этот фено-
мен называется адаптацией. При развитии адаптации мотонейроны посте-
пенно уменьшают частоту генерации потенциалов действия в процессе вы-
полнения физической работы, что приводит к снижению физиологических
показателей работоспособности мышц во время статической физической
работы. Различают раннюю и позднюю адаптацию мотонейронов. Ранняя
адаптация мотонейронов возникает в первые секунды после начала мышеч-
ных сокращений, а поздняя — в процессе выполнения физической работы
по мере развития утомления.
Физиологический механизм адаптации при утомлении человека имеет
место в мотонейронах, иннервирующих мышцы как верхних, так и нижних
конечностей. Именно снижение частоты разрядов спинальных мотонейро-
нов является основной причиной уменьшения силы и мощности мышеч-
ного сокращения. Одновременно при развитии утомления увеличивается
период времени мышечного расслабления. Адаптация мотонейронов мо-
жет привести к полному прекращению активности отдельных двигатель-
ных единиц. Это характерно, прежде всего, для низкопороговых мотоней-
ронов спинного мозга и механизм развивается при участии тормозного ме-
диатора гамма-аминомасляной кислоты. При этом частота генерации им-
пульсов в мотонейронах снижается тем быстрее, чем выше скорость мы-
шечного сокращения, т. е. прямо зависит от мощности выполняемой
мышцами работы.
При развитии физического утомления у человека кроме снижения элек-
трической активности мотонейронов спинного мозга, у нейронов начинает
варьировать частота генерации импульсов. Эти изменения электрической
активности мотонейронов спинного мозга, как правило, проявляется с мо-
мента снижения показателей произвольной силы мышечного сокращения.
В основе физиологического механизма феномена адаптации мотонейро-
нов при утомлении у человека лежат такие процессы, как возвратное тор-
можение при участии клеток Реншоу, рефлекторное уменьшение возбуди-
мости мотонейронов под влиянием импульсов от рецепторов сокращаю-
щихся мышц, пресинаптическое торможение и, наконец, уменьшение воз-
будимости мотонейронов под влиянием нейромодуляторов.
Клетки Реншоу вызывают классическое торможение мотонейронов
спинного мозга, которые иннервируют мышцы-антагонисты с одной сто-
роны туловища или мышцы-синергисты обеих конечностей, и, таким об-
разом, усиливают развитие феномена адаптации мотонейронов.
Тормозные интернейроны сегментов спинного мозга, которые получают
нисходящие эфферентные сигналы от двигательных центров ствола мозга
и афферентные волокна от проприорецепторов скелетных мышц, также
уменьшают возбудимость мотонейронов спинного мозга.
В процессе мышечных сокращений к тормозным интернейронам спин-
ного мозга поступает нарастающий поток импульсов от проприорецепто-
876 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Возбуждающий
интернейрон
Тормозной
интернейрон
П реси наптическое
торможение
Рефлекторные
входы афферентных
волокон I—IV групп
Группы афферентных волокон
II III—IV
la 1Ь
а-Мотонейрон
у-Мотонейрон
Экстрафузальные
мышечные волокна
Нисходящие
кортикоспинальные,
проприоспинальные
и другие двигательные
команды
+— Клетка Реншоу
Интрафузальные
мышечные волокна
Рис. 22.5. Афферентные входы к а- и у-мотонейронам мышц антагонистов. Аффе-
рентные входы от проприорецепторов мышц и нисходящие супраспинальные влия-
ния оказывают активирующее, а также пре- и постсинаптическое торможение при
участии клеток Реншоу на а- и у-мотонейроны спинного мозга. Тормозные влияния
на а- и у-мотонейроны спинного мозга являются основным механизмом адаптации
и снижения электрической активности этих нейронов. В результате происходит
уменьшение физиологических показателей работающих мышц (сила и мощность) и
развитие утомления.
Темные кружки — тормозные входы, светлые — возбуждающие.
ров. В результате как клетки Реншоу, так и тормозные интернейроны сег-
ментов спинного мозга постепенно повышают свою электрическую актив-
ность и при участии тормозного медиатора гамма-аминомасляной кислоты
гиперполяризуют мембрану а- и у-мотонейронов, снижая их возбудимость.
В результате у мотонейронов снижается частота генерации потенциалов
действия мотонейронов, что обусловливает уменьшение силы и мощности
сокращений мыщц.
Во время мышечного сокращения растяжение сухожильных органов
Гольджи, расположенных на границе перехода соединительнотканных во-
локон в мышечные, вызывает генерацию потенциалов действия в чувстви-
тельных нервах. Степень возбуждения сухожильных рецепторов Гольджи
прямо зависит от силы как изометрических, так и изотонических сокраще-
ний скелетных мышц. Нервные импульсы от сухожильных органов Голь-
джи поступают в спинной мозг по 1b нервным волокнам и конвергируют
на те же мотонейроны сегментов спинного мозга, что 1а тип афферентных
волокон, через которые передаются к мотонейронам импульсы от мышеч-
ных веретен. Классическое действие 1b афферентных волокон на мотоней-
роны спинного мозга заключается в торможении мышц одной стороны ту-
ловища и одновременно в обширном торможении мыщц-синергистов. При
этом сухожильные органы Гольджи в процессе развития утомления у чело-
века оказывают влияние на снижение частоты разрядов как а-, так и у-мо-
тонейронов при участии механизма их пресинаптического торможения.
Свободные нервные окончания, расположенные в скелетных мышцах,
22. Физиологические основы труда • 877
возбуждаются при их растяжении, укорочении, ишемии и гипоксемии мы-
шечной ткани, что имеет место при физической работе. Нервные импуль-
сы по афферентным немиелинизированным волокнам III и IV групп пере-
даются в спинной мозг. Афферентные волокна III группы обильно иннер-
вируют также сухожилия мышц. Механизм взаимодействия афферентных
волокон III и IV групп, и 1b типа афферентных волокон на мотонейроны
спинного мозга показан на рис. 22.5. Центральный механизм развития
утомления при участии афферентных волокон la, 1b и мышечных веретен
(группа II) осуществляется по типу пресинаптического торможения элек-
трической активности мотонейронов спинного мозга.
Супраспинальные механизмы утомления человека при статической фи-
зической работе, в частности роль нейронов двигательной коры больших
полушарий головного мозга, не является первичной причиной развития
«центрального» утомления. Только при высокой активности нисходящих
двигательных команд имеет место феномен адаптации спинальных мото-
нейронов как причина утомления человека
22.6.1.2. Утомление человека при динамической мышечной работе
Динамическая мышечная работа сопровождается активацией сердечно-со-
судистой, дыхательной, эндокринной систем, двигательных центров голов-
ного и спинного мозга, а также функции многих нейротрансмиттерных
систем ЦНС. Физиологические проявления центрального действия этих
систем при динамической физической работе человека могут быть компо-
нентами супраспинальных механизмов «центрального» утомления. При
физической динамической работе происходит активация функции серото-
нинергической и норадренергической систем мозга, которые могут изме-
нять уровень активации мозга, степень мотивации выполнения физиче-
ской работы, уровень внимания и устойчивость к дискомфорту и действию
стрессовых факторов. Физическая работа как стрессовый фактор активиру-
ет иммунную систему, включая продукцию таких факторов, как кортизол и
цитокины. Роль этих процессов в качестве «пусковых» факторов супраспи-
нальных механизмов «центрального» утомления человека требует уточне-
ния. При этом наиболее распространенная теория супраспинальных меха-
низмов «центрального утомления» объясняет его природу нарушением ме-
таболизма аминокислот в нейронах серотонинергической системы мозга.
Так, нарушение метаболизма аминокислот в нейронах этой медиаторной
системы мозга является основной причиной изменений в цикле сна-
бодрствования, гипоталамо-гипофизарной оси регуляции функций орга-
низма, а также регуляции двигательной функции. При физической работе
возрастает проникновение через гематоэнцефалический барьер в мозг ами-
нокислоты триптофана. Этому способствует активация использования в
работающих мышцах аминокислот и, как следствие, увеличение концен-
трации в плазме крови жирных кислот. В результате в плазме крови воз-
растает отношение между содержанием свободного триптофана и амино-
кислот с короткой цепью. Триптофан в нейронах ЦНС метаболизируется в
серотонин, что повышает его концентрацию в структурах базальных ганг-
лиев, гиппокампа и ствола мозга. Так, агонисты серотонина снижают ус-
тойчивость организма к утомлению, а антагонисты, напротив, задержива-
ют развитие этого процесса. У человека блокада обратного захвата серото-
нина, выделяющегося из нервных терминалей, что исключает его повтор-
ное использование в синаптической передаче, увеличивает время физиче-
ской работоспособности человека (например, при езде на велосипеде или
878 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
во время бега). Аналогичный эффект на сохранение физической работо-
спосбности оказывает уменьшение соотношения свободного триптофана к
аминокислотам в плазме крови путем специальной диеты.
Напротив, активация дофаминергической системы мозга при физиче-
ской динамической работе тормозит синтез в нейронах мозга серотонина и
снижает скорость развития «центрального утомления» при участии супра-
спинальных механизмов. Однако при длительной динамической физиче-
ской работе происходит снижение уровня двигательной активности чело-
века за счет периферического утомления. Это ведет к снижению активно-
сти дофаминергической системы мозга и, как следствие, увеличивается ак-
тивность серотонинергической системы мозга. Серотонин, уменьшая ак-
тивность супраспинальных двигательных центров головного мозга, стиму-
лирует развитие «центрального утомления».
ГЛАВА 23
Адаптация человека к условиям
внешней среды
23.1. Общие принципы и механизмы адаптации
23.1.1. Адаптация
Адаптация — процесс приспособления организма к меняющимся условиям
среды, который создает возможность приспособления человека к новым
для него природным, производственным или социальным условиям. Адап-
тация человека к неадекватным условиям среды обеспечивает ему работо-
способность, воспроизведение здорового потомства, нормальный темп ста-
рения организма, максимальную продолжительность жизни, характерную
для популяции, к которой относится индивидуум. Процесс адаптации
обеспечивают компенсаторные механизмы — адаптивные реакции, направ-
ленные на устранение или ослабление функциональных сдвигов в организ-
ме, вызванных неадекватными факторами среды. Это динамические, быст-
ро возникающие физиологические средства аварийного обеспечения орга-
низма. Они мобилизуются, как только организм попадает в неадекватные
условия, и постепенно затухают по мере развития адаптационного процесса.
Например, под воздействием холода усиливаются процессы производства
и сохранения тепловой энергии, повышается обмен веществ, в результате
рефлекторного сужения периферических сосудов (особенно кожи) умень-
шается теплоотдача. Компенсаторные механизмы служат составной частью
резервных сил организма. Обладая высокой эффективностью, они могут
поддерживать относительно стабильный гомеостазис достаточно долго для
развития устойчивых форм адаптационного процесса, приспособления
строения и функций организма, его органов и систем к изменению окру-
жающей среды. Все живые системы обладают способностью к приспособ-
лению, т. е. обладают адаптивностью, или приспособляемостью. Мерой
соответствия адаптации организма к измененным условиям среды является
оценка степени приспособления к ним его жизнедеятельности. Например,
оценка степени приспособления человека к жизни в измененной среде
обитания положительна, если для него характерна высокая продолжитель-
ность жизни, рождение здорового потомства, а в плане социальной оценки
достигнутой адаптации организмом человека к новой среде обитания —
высокая производительность труда. Адаптацию организма человека к но-
вым климатическим условиям и биологическому окружению обозначают
понятием акклиматизация
Процесс, обратный адаптации, который возникает при воздействии на
организм человека факторов среды, количественно превышающих возмож-
ности адаптирующейся системы, и вызывает нарушение адаптивных реак-
ций организма, называется дизадаптацией. Дизадаптация приводит к дис-
функции, т. е. нарушению функций системы, органа или тканей организма,
880 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Время,ч
Рис. 23.1. Динамика температуры ядра
тела и метаболической продукции тепла
у австралийских аборигенов и европей-
цев в ночные часы в пустыне.
------австралийские аборигены;-----ев-
ропейцы.
Метаболическая продукция тепла в ваттах х
м2 поверхности тела.
характеризующемуся неадекватно-
стью ответа данных структур орга-
низма на действие раздражителей.
Организм способен возвращать
вовлеченные в процесс адаптации
системы в исходное положение по-
сле прекращения действия фактора
среды, повлекшего адаптацию к не-
му организма, т. е. возможна деа-
даптация организма.
Организм всегда сохраняет след,
память об адаптации к неадекват-
ным условиям среды (так называе-
мая вегетативная память), что дела-
ет более быстрым процесс повтор-
ной адаптации (реадаптации) к
этим условиям среды. Например,
такая память о сформировавшейся
адаптации к жаркому, влажному
климату в организме жителя сред-
них широт, вернувшегося из тропи-
ков в привычную среду обитания
умеренного климата, сохраняется
от нескольких недель до несколь-
ких месяцев, облегчая адаптацию
человека при возвращении в тро-
пики. В основе «вегетативной па-
мяти» лежат возникшие в ходе
адаптации организма к новым ус-
ловиям среды изменения в нейро-
нах гипоталамических ядер голов-
ного мозга, характеризуемые увели-
чением количества РНК и белков в
их цитоплазме.
Адаптация организма к воздействию факторов внешней среды может
идти по пассивному пути, по типу толерантности (выносливость, терпели-
вость), который характеризуется снижением, экономизацией энергетиче-
ских трат организма, связанных с поддержанием гомеостазиса. Другой, ак-
тивный путь — связан с возрастанием энергетических затрат организма на
поддержание гомеостазиса, на компенсацию изменений во внутренней
среде организма, вызванных воздействием факторов внешней среды. На-
пример, на снижение температуры внешней среды в ночное время суток
организм австралийского аборигена отвечает снижением метаболизма и
температуры ядра тела (пассивный путь адаптации), а организм европей-
ца — усилением метаболизма и теплопродукции (активный путь адапта-
ции) (рис. 23.1).
Организм человека обладает устойчивостью, или сопротивляемостью, к
воздействию раздражителей большой силы (стрессоров) — механическая
травма, боль, инфекция, физическая работа большого объема и др., а так-
же их разновидности — экстремальных факторов, т. е. угрожающих жизне-
деятельности организма факторов окружающей среды (холод, жара, изме-
нения атмосферного давления). Это свойство организмов называют рези-
стентностью. Различают специфическую (устойчивость по отношению к
23. Адаптация человека к условиям внешней среды • 881
определенному фактору) и неспецифическую (по отношению к различным
факторам) резистентность. Резистентность организма обеспечивается его
адаптивными реакциями на раздражитель. Адаптационные реакции орга-
низма также являются неспецифическими, если физиологические системы
организма (нервная, эндокринная, кровообращения и др.) сходно реагиру-
ют при действии на организм человека различных по качеству и силе раз-
дражителей. Так, при действии на организм сильных, чрезвычайных раз-
дражителей в ЦНС развивается резкое возбуждение, сменяющееся запре-
дельным торможением — крайней мерой защиты, проявляющейся в сниже-
нии возбудимости и чувствительности структур ЦНС к сильному раздра-
жителю, так как адекватный ответ на этот раздражитель мог бы привести
организм к гибели.
23.1.2. Неспецифические адаптивные реакции организма
При адаптации организма человека к действию на него различной силы
раздражителей внешней среды различают по характеру проявлений и меха-
низмам развития несколько неспецифических адаптивных реакций организ-
ма, «организуемых» нейрогормональными системами: симпатоадреналовую
реакцию, стресс-реакцию, реакцию тренировки и реакцию активации. Все
они повышают неспецифическую резистентность организма к нарушаю-
щим его жизнедеятельность факторам внешней среды.
На действие любых раздражителей большой силы, или стрессоров, орга-
низм отвечает развитием неспецифических адаптивных реакций — симпа-
тоадреналовой и стресс-реакции. Под термином «стресс» (напряжение) по-
нимают неспецифические психофизиологические проявления адаптацион-
ной активности организма и его систем при действии на него стрессоров.
При воздействии стрессора на организм происходит быстрая активация
симпатоадреналовой системы («запускающая» симпатоадреналовую реак-
цию) и системы гипофиз—кора надпочечников («запускающая» стресс-ре-
акцию), благодаря одновременному возбуждению нервных центров голов-
ного мозга, регулирующих функцию этих систем.
Центр, активация которого возбуждает симпатоадреналовую систему,
расположен в продолговатом мозге — в голубом пятне (locus coeruleus) и
является самым крупным скоплением норадренергических нейронов в го-
ловном мозге человека (см. главу 5).
Центр, активирующий систему гипофиз—кора надпочечников, локали-
зован в медиодорсальной области паравентрикулярных ядер гипотоламуса
и представлен самым крупным в головном мозге человека скоплением
нейронов, секретирующих кортикотропин-рилизинг-гормон. При дейст-
вии на организм стрессора нейроны паравентрикулярных ядер гипотоламу-
са секретируют кортикотропин-рилизинг-гормон, активирующий систему
гипофиз—кора надпочечников, а активация симпатоадреналовой системы
осуществляется сигнализацией с терминалей секретирующих кортикотро-
пин-рилизинг-гормон нейронов паравентрикулярных ядер гипотоламуса
на катехоламинергические нейроны голубого пятна. Активирующий эф-
фект на катехоламинергические нейроны голубого пятна оказывает и не-
посредственно кортикотропин-рилизинг-гормон.
Существует несколько механизмов, активирующих нейроны медиодор-
сальной области паравентрикулярных ядер гипотоламуса и секрецию ими
кортикотропин-рилизинг-гормона. Так, стрессоры (психоэмоциональное
напряжение, боль, травма, кровопотеря и др.) вызывают сдвиги величин
показателей внутренней среды организма. Сигнализация об этих наруше-
882 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ниях гомеостазиса, поступающая от центральных и периферических рецеп-
торов, контролирующих показатели внутренней среды организма к гипота-
ламусу, к коре головного мозга, ретикулярной формации, лимбической
системе (гиппокамп, миндалевидный комплекс), стимулирует образование
в тканях головного мозга нейропептида Y (соединение, относимое к классу
тахикининов). Нейропептид Y активирует нейроны медиодорсальной об-
ласти паравентрикулярных ядер гипотоламуса и секрецию ими кортико-
тропин-рилизинг-гормона. При стрессе же, вызванном инфекцией, макро-
фаги, фагоцитируя проникшие в организм микроорганизмы, взаимодейст-
вуя с их липополисахаридным комплексом резко усиливают секрецию ин-
терлейкина-1 в тканевую жидкость и кровь (см. главу 7). Интерлейкин-1
проникает в ткань головного мозга человека из плазмы крови через гема-
тоэнцефалический барьер благодаря специфическим механизмам транс-
порта для данного цитокина. Интерлейкин-1 стимулирует экспрессию
и-РНК кортикотропин-рилизинг-гормона, его синтез и секрецию в нейро-
нах медиодорсальной области паравентрикулярных ядер гипотоламуса. В
свою очередь кортикотропин-рилизинг-гормон стимулирует секрецию
АКТГ в гипофизе, который усиливает образование и секрецию в кровь
глюкокортикоидных гормонов корой надпочечников. Рассмотрим эффек-
ты симпатоадреналовой и стресс-реакции, повышающие устойчивость ор-
ганизма человека к действию на него стрессоров.
23.1.3. Симпатоадреналовая реакция
Симпатоадреналовая реакция развивается в результате рефлекторного
возбуждения расположенных в области голубого пятна норадренергиче-
ских нейронов, активирующих симпатический отдел вегетативной нерв-
ной системы, при действии стрессора на организм. Активация симпати-
ческого отдела вегетативной нервной системы стимулирует секрецию в
кровь катехоламинов — норадреналина и адреналина — мозговым вещест-
вом надпочечников. Под влиянием адреналина повышается обеспечение
энергетическими ресурсами и кислородом работающих мышц, сердца,
ЦНС: адреналин стимулирует образование глюкозы из гликогена, свобод-
ных жирных кислот из триглицеридов жировой ткани; активирует функ-
цию систем транспорта кислорода: кровообращения — увеличивая число
и силу сердечных сокращений, сердечный выброс и кровяное давление;
дыхания — увеличивая минутный объем дыхания, уменьшая газовое со-
противление вентиляции легких (за счет расслабления гладких мышц
бронхиол и увеличения их просвета); эритрона — увеличивая кислород-
ную емкость крови за счет выброса в нее депонированных эритроцитов.
Под влиянием адреналина повышается свертываемость крови, что преду-
преждает значительную потерю крови при повреждении кровеносных со-
судов. Одновременно адреналин стимулирует деятельность ЦНС, облегча-
ет восприятие сенсорной информации индивидуумом (расширяются зрач-
ки, увеличивая поля зрения), что способствует развитию адаптивной по-
веденческой реакции, направленной на избегание или борьбу с угрожаю-
щей организму опасностью. Активация функций перечисленных выше
систем и метаболизма под влиянием адреналина осуществляется быст-
ро — уже в первые минуты действия стрессора на организм, т. е. гормон
способствует быстрой мобилизации приспособительных реакций организ-
ма, направленных на восстановление нарушенного действием стрессора
гомеостазиса.
23. Адаптация человека к условиям внешней среды • 883
23.1.4. Стресс-реакция
Ее развитие связано с усилением секреции нейронами медиодорсальной
области паравентрикулярных ядер гипотоламуса кортикотропин рили-
зинг-гормона, резко усиливающего секрецию АКТЕ передней долей гипо-
физа и активации под влиянием АКТЕ секреции кортикостероидов (кор-
тизол, кортикостерон, альдестерон) надпочечниками в кровь. Эти гормо-
ны усиливают эффекты катехоламинов, направленные на энергообеспече-
ние тканей: активируют глюконеогенез, стимулируют разрушение части
лимфоидной ткани, в результате которого высвобождаются аминокислоты,
используемые в глюконеогенезе; глюкокортикоиды препятствуют сниже-
нию проницаемости кровеносных сосудов, возникающему под влиянием
биологически активных веществ (лейкотоксин, эксудин и др.), образую-
щихся в поврежденных тканях; подавляют воспаление; понижают прони-
цаемость гематоэнцефалического барьера, тем самым препятствуя поступ-
лению в ЦНС токсичных веществ, эндогенной и экзогенной природы
(наркотиков и др.); усиливают миграцию Т-хелперов в костный мозг, ак-
тивирующих гранулоцитопоэз, что увеличивает число нейтрофильных лей-
коцитов в крови и повышает резистентность организма к инфекции, по-
вреждению тканей. Одновременно АКТЕ и кортизон угнетают выработку
антител, что предупреждает аутосенсибилизацию организма (т. е. иммуни-
зацию организма антигенами собственных поврежденных тканей, в ре-
зультате чего антитела начинают повреждать здоровые ткани собственного
организма).
При стрессе могут возникать не только приспособительные, адаптивные
изменения в организме человека, но и выраженные расстройства его жиз-
недеятельности, характеризующиеся нарушением функций различных сис-
тем организма, с переходом даже в болезнь. Например, в современном об-
ществе одной из распространенных форм стресса стал эмоциональный
стресс, под которым понимают психоэмоциональное напряжение челове-
ка, развивающееся на стрессор — конфликтную ситуацию. Физиологиче-
ский эмоциональный стресс — это нормальная реакция человека и живот-
ных на конфликтную ситуацию. Механизмы данной реакции развились и
генетически закрепились в ходе эволюции, помогая выживанию организ-
ма, делая его устойчивым к психоэмоциональному напряжению в кон-
фликтных ситуациях. Однако эмоциональное перенапряжение, длительно
и часто повторяющийся эмоциональный стресс могут нарушать вырабо-
танные в эволюции адаптивные механизмы и приводить к развитию пси-
хосоматических заболеваний — психозов, неврозов; сердечно-сосуди-
стых — аритмий, инфаркта миокарда, гипертонической болезни; для тяже-
лого стресса характерны эрозивные поражения слизистой оболочки желуд-
ка, вызывающие формирование кровоточащих язв. Повышенная секреция
кортикотропин-рилизнг-гормона, при стрессе тормозит секрецию половых
гормонов: под его влиянием снижается секреция люлиберина фолликулин-
стимулирующего и лютеинизирующего гормонов. Поэтому длительный
стресс, снижая уровень продукции половых гормонов, может вызвать у
мужчин нарушение сперматогенеза и, как следствие, бесплодие. Поэтому
различают положительные, физиологические (эустресс — от греч. «эу» —
хороший) и отрицательные, патологические (дисстресс — от лат. «дис» —
плохой) формы стресса. Переходу эустресса в дисстресс препятствуют ан-
тистрессовые механизмы, «нейтрализующие, смягчающие» отрицательные
проявления стресс-реакции в тканях и системах организма.
К антистрессовым механизмам относят секрецию эпифизом гормона
884 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
мелатонина; синтезирующиеся в эмоциогенных зонах гипоталамуса эндо-
генные олигопептиды, субстанцию Р, опиатные гормоны (р-эндорфин
и др.), пептид, вызывающий 5-сон; ГАМК-ергическую систему; антиокси-
дантные механизмы (они защищают организм от длительной активации
перекисного окисления липидов); оксид азота, активирующий в тканях
синтез протекторных белков (так называемых белков теплового шока, защи-
щающих органы от стрессорных повреждений); систему простагландинов
(тормозят взаимодействие катехоламинов с адренорецепторами). Перечис-
ленные антистрессовые механизмы «включаются» в организме человека
одновременно с началом развития стресс-реакции. Так, молекулы нейро-
пептида Y, усиленно образующиеся в ткани головного мозга под влиянием
стрессора, одновременно активируют как секрецию кортикотропин-рили-
зинг-гормона нейронами медиодорсальной области паравентрикулярных
ядер гипотоламуса, так и секрецию мелатонина эпифизом. Мелатонин, в
свою очередь, угнетает выработку кортикотропин-рилизинг-гормона в ги-
поталамусе, АК.ТГ — в гипофизе и стероидных гормонов — в коре надпо-
чечников, снижая интенсивность стресс-реакции; повышает продукцию
субстанции Р, также угнетающую секрецию кортикотропин-рилизинг-гор-
мона в гипотоламусе; активирует ГАМК-ергическую систему и выработку
эндогенных опиоидных гормонов в тканях головного мозга. Образующий-
ся из ГАМК тормозной метаболит — гаммаоксимаслянная кислота
(ГОМК) — понижает возбудимость нервных центров, ответственных за
развитие стресс-реакции. Опиоидные гормоны оказывают аналгезирую-
щий эффект.
Если стрессор действует на организм длительно, если сила раздражите-
ля велика и к его действию полная адаптация организма не происходит
или же активность антистрессовых механизмов оказывается недостаточной
для устранения отрицательных следствий стресса, то эустресс переходит в
дисстресс. Дисстресс резко повышает риск развития заболевания гиперто-
нической болезнью, ишемической болезнью сердца, психоневрологических
заболеваний, иммунодефицитного состояния организма, в свою очередь
увеличивающего риск развития злокачественных опухолей у человека.
Чрезмерная выраженность стресс-реакции, т. е. дисстресс, вызванный
микробной инфекцией, вызывает мощную секрецию нейтрофильными
лейкоцитами, макрофагами и моноцитами в кровь и тканевую жидкость
супероксидных радикалов. Последние резко усиливают перекисное окис-
ление липидов мембран Т- и В-лимфоцитов, влекущие снижение активно-
сти их мембранных ферментов (Na+, К+-АТФазы, Са++-АТФазы, аденилат-
циклазы), повышение проницаемости клеточных и субклеточных мембран,
снижение чувствительности лимфоцитов к гормональной и нервной регу-
ляции. В результате функция данных иммунокомпетентных клеток нару-
шается, как следствие — угнетается иммунный ответ организма на инфек-
цию, у больных имеет место развитие «суперинфекции», делающий крайне
тяжелым течение инфекционного заболевания. При физиологическом же
стрессе, вызванном умеренной микробной инфекцией, также имеющее ме-
сто перекисное окисление липидов мембран иммунокомпетентных клеток
поддерживает обновление мембран, клеточное деление и синтез биоактив-
ных веществ в лимфоцитах, активирует эти клетки, что способствует раз-
витию адекватной иммунной реакции организма человека на инфекцию и
последняя уничтожается антителами и клетками иммунной системы (см.
главу 8).
Совокупность защитных реакций организма, возникающих в организме
при состоянии стресса, получила название общего адаптационного синдро-
23. Адаптация человека к условиям внешней среды • 885
Рис. 23.2. Изменение резистентности организма в разные стадии общего адаптив-
ного синдрома.
N — границы нормального уровня резистентности организма; I — стадия тревоги; II — стадия
резистентности; III — стадия истощения; а —фаза шока; б — фаза противотока (пояснение в
тексте).
ма, который характеризуется последовательно развивающимися тремя ста-
диями (рис. 23.2).
Первая стадия (реакция тревоги) может развиваться до 24—48 ч после
действия стрессора на организм. В ее развитии выделяют две фазы — шока
и противотока. Фаза шока характеризуется нарушением функций ряда
систем организма (нервной системы, кровообращения, дыхания, водно-со-
левого обмена, теплорегуляции, метаболизма и др.), вызывающих наруше-
ние гомеостазиса. Поэтому фаза шока характеризуется гипотонией (пони-
жением кровяного давления), гипотермией (понижением температуры «яд-
ра» тела), депрессией центральной нервной системы, повышением прони-
цаемости клеточных мембран, явлениями катаболизма, вызывающими ис-
худание, отрицательный азотистый баланс, гипогликемию, лейкопению,
лимфо- и эозинопению, уменьшение массы вилочковой железы и лимфа-
тических узлов, кровоизлияние в слизистую оболочку тонкого кишечника
и желудка, появление язв в слизистой оболочке этих органов. В результате
резистентность организма к стрессору снижается, что может вызвать нару-
шение его жизнедеятельности. Поэтому организм не может длительное
время находиться в фазе шока и погибает в первые часы или дни действия
стрессора на организм, если не развивается фаза противошока, характери-
зующаяся мобилизацией приспособительных ответов физиологических
систем организма в ходе симпатоадреналовой реакции и активации систе-
мы гипофиз—кора надпочечников. Фаза противошока характеризуется по-
вышенным содержанием адреналина, АКТГ и кортизола в крови, увеличе-
нием под влиянием АКТГ массы коры надпочечников, усилением в них
(особенно в пучковой зоне надпочечников) митотической и пролифера-
тивной активности. В результате действия гормонов надпочечников фаза
противошока характеризуется сдвигами, идущими в направлении, обрат-
ном фазе шока: повышается кровяное давление, повышается температура
тела, понижается проницаемость клеточных мембран, развивается гиперг-
ликемия и т. д., которые ведут к восстановлению нарушенного гомеостази-
са и обусловливают переход фазы противошока во вторую стадию — ста-
дию резистентности, или устойчивости, приспособления, повышения ус-
тойчивости организма к действующему на него стрессору. Стадия рези-
стентности характеризуется преобладанием анаболических процессов (т. е.
886 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
процессов биосинтеза) в различных тканях организма, что способствует
восстановлению поврежденных под влиянием стрессора тканей. В стадии
резистентности для организма характерно повышение устойчивости не
только к стрессору, вызвавшему развитие общего адаптационного синдро-
ма, но и к другим сильным раздражителям, т. е. имеют место так называе-
мая перекрестная резистентность. Стадия резистентности приводит к под-
держанию нормального существования организма в новых для него усло-
виях.
Если же стрессор продолжает действовать на организм, а его антистрес-
сорные механизмы оказываются недостаточно эффективными, то может
развиться третья стадия — стадия истощения, которая может привести к
заболеваниям организма, к раннему его постарению и даже гибели.
23.1.5. Реакция тренировки и реакция активации
При действии на организм слабых, пороговых раздражителей (системати-
ческие занятия физической культурой, утренняя физическая зарядка, вод-
ные процедуры) организм человека отвечает формированием адаптивной
реакции, получившей название реакции тренировки, в ходе которой в ЦНС
развивается возбуждение, быстро сменяющееся охранительным торможе-
нием, что обеспечивает снижение ее возбудимости, реактивности по отно-
шению к слабому раздражителю; повышается активность защитных систем
организма (иммунной системы, неспецифических факторов резистентно-
сти); в результате повышается устойчивость организма человека к простуд-
ным заболеваниям. При реакции тренировки умеренно активируется сек-
реторная функция передней доли гипофиза и контролируемых ее гормона-
ми периферических эндокринных желез (половых, щитовидной железы),
что повышает половую функцию, усиливает обмен веществ в организме
человека. Реакция тренировки повышает противоопухолевую резистент-
ность организма. Например, при развитии у экспериментальных животных
реакции тренировки у них быстро рассасываются перививаемые опухоли —
саркома и рак молочной железы.
При действии на организм раздражителей средней силы происходит
развитие «реакции активации» защитных систем организма, которая, одна-
ко, не носит характера их патологической гиперфункции. Реакция актива-
ции возникает, например, под влиянием «адаптогенов» — в результате
приема человеком настойки женьшеня, элеутерококка колючего, золотого
корня, эфедрина и др. Ее главной чертой является резкая активация тими-
ко-лимфатической системы (в противоположность инволюции этой систе-
мы, возникающей в стадию тревоги, вызванной действием стрессора),
следствием которой является гиперплазия лимфоидной ткани, активация
клеточного и гуморального иммунитета (см. главу 8), системы фагоцити-
рующих мононуклеаров (см. главу 7), что повышает устойчивость к инфек-
ции и препятствует развитию опухолей в организме человека. Реакция ак-
тивации повышает детоксикационную функцию организма, что увеличива-
ет его устойчивость к токсичным веществам (рис. 23.3).
23.1.6. Срочная и долговременная адаптация
При кратковременном воздействии экстремальных факторов на организм
они компенсируются резервными возможностями человека (например, в
ответ на жару возрастает секреция пота, усиливающая испарительный тер-
молиз). При длительном воздействии экстремальных факторов на организм
23. Адаптация человека к условиям внешней среды • 887
Рис. 23.3. Изменения резистентности организма при развитии реакции тренировки
и реакции активации (пояснение в тексте).
человека функциональные перестройки определяются совокупным вклю-
чением процессов восстановления нарушенного гомеостазиса, их силой и
продолжительностью. Например, при длительном действии холода на орга-
низм человека повышается продукция катехоламинов и тироксина, усили-
вающих жировой обмен, в результате резко активируется несократитель-
ный термогенез, температура ядра тела повышается.
Адаптационные реакции организма осуществляются в виде срочной и
долговременной адаптации.
Срочная адаптация возникает в организме человека как реакция физио-
логических систем на действие раздражителя большой силы. Она развива-
ется на основе ранее сформировавшихся приспособительных механизмов
физиологических систем организма (кровообращения, дыхания, крови
и др.) к среднему по интенсивности действия на организм средовому фак-
тору, например постоянно выполняемой человеком умеренной по интен-
сивности физической работы. Под действием раздражителя большой силы
(например, работы большой интенсивности и объема) органы и системы,
не адаптированные к такой физической нагрузке, функционируют на пре-
деле их физиологических возможностей, почти при полной мобилизации
функциональных резервов. Однако срочная адаптация не обеспечивает ор-
ганизму человека оптимальный адаптационный эффект, так как при ней
не успевают развиться приспособительные изменения в клетках тканей ор-
ганов (сердце, скелетные мышцы), обеспечивающих организму выполне-
ние работы большого объема. Так, например, при беге на длительную дис-
танцию нетренированного человека у него могут быть зарегистрированы
близкие к максимуму показатели работы физиологических систем — кро-
вообращения и дыхания. Однако при отсутствии адаптации к данной фи-
зической нагрузке физическая работоспособность нетренированного чело-
века лимитируется недостаточно быстрым ходом окислительного фосфо-
рилирования в митохондриях скелетных мышц, не обеспечивающим обра-
888 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
зования количества молекул АТФ, необходимого для выполнения скелет-
ными мышцами работы, связанной с длительным бегом.
Долговременная адаптация организма человека к действию раздражителя
большой силы развивается постепенно, в результате длительного, постоян-
ного или многократно повторяющегося действия на организм факторов
среды большой интенсивности. В ходе ее развития активируются морфо-
генетические, биосинтетические процессы в тканях органов, наиболее ин-
тенсивно функционирующих при адаптации организма к действию опреде-
ленного сильного раздражителя. Например, интенсивные физические тре-
нировки вызывают рабочую гипертрофию сердца, в его ткани увеличивает-
ся масса миоглобина, количество митохондрий, усиливающих процессы
окисления и образования энергии («сердце атлета»), увеличивается кисло-
родная емкость крови (за счет формирования большей массы новых эрит-
роцитов и гемоглобина костным мозгом). Увеличение рабочей мощности
систем транспорта кислорода облегчает организму выполнение работы
большого объема на основе возросших функциональных возможностей его
систем. По существу долговременная адаптация к факторам среды боль-
шой интенсивности развивается на основе многократной реализации срочной
адаптации к этим факторам. В результате организм приобретает новое ка-
чество — из неадаптированного превращается в адаптированный, что дела-
ет его способным, например, к выполнению физической работы в объеме
и продолжительности, невозможной для осуществления не тренированным
организмом человека.
23.1.7. Норма адаптивной реакции и дизадаптации
Норма адаптивной реакции — это пределы изменения ответов систем орга-
низма человека, возникающих под влиянием действующих на организм
факторов внешней среды, при которых не нарушаются структурно-функ-
циональные связи систем организма и их взаимодействие со средой. Пре-
вышение нормы адаптации организма вызывают явления дизадаптации,
которая может вызывать патологические изменения в организме. Напри-
мер, у спортсменов, адаптированных к большим физическим нагрузкам
(марафонцы), нередко могут регистрироваться признаки очагового кар-
диосклероза (т. е. развития участков соединительной ткани в миокарде),
которые могут вызвать нарушение работы сердца у спортсмена во время
бега.
23.1.8. Генотипическая и фенотипическая адаптация.
Покрывающие адаптации
Под генотипической адаптацией имеется в виду совокупность врожденных
морфофизиологических особенностей организма и адаптивного поведения
особи, направленных на поддержание гомеостаза и позволяющих организ-
му существовать в экстремальных условиях среды. Фенотипическая адап-
тация предусматривает формирование приобретенных адаптивных реак-
ций под влиянием среды (к ним относится, например, адаптация организ-
ма нетренированного человека к большим физическим нагрузкам).
Генотипическую и фенотипическую адаптацию подразделяют на эволю-
ционную (филогенетическую) и онтогенетическую, или индивидуальную. Эво-
люционная адаптация человеческой популяции прослеживается при наблю-
дении генотипических и фенотипических различий у рас и народов. На-
пример, различия между расами касаются пигментации кожи. Приспособ-
23. Адаптация человека к условиям внешней среды • 889
лению древних людей к условиям севера, где мало ультрафиолетовых лу-
чей, способствовала светлая кожа, которая лучше их пропускает.
Онтогенетическая адаптация, протекающая с момента рождения орга-
низма и до конца жизни, позволяет организму индивидуально приспосаб-
ливаться к меняющимся факторам среды. Например, у жителя средних
широт основной обмен повышается с понижением температуры воздуха
зимой, и снижается при повышении температуры внешней среды летом.
В жизни человека часто имеют место случаи так называемых покрываю-
щих форм адаптации, когда организм, адаптированный к одному экстре-
мальному фактору, подвергается воздействию другого экстремального фак-
тора, к которому человек не был адаптирован. В этом случае уровень пред-
шествующей адаптации организма к первому из экстремальных факторов
существенно влияет на устойчивость организма к воздействию второго из
них. Например, у людей, адаптированных к физической работе большого
объема, повышена устойчивость к высотной гипоксии, благодаря увеличе-
нию рабочей мощности систем транспорта кислорода, развившихся в ходе
адаптации организма к физическим нагрузкам.
23.1.9. Обратимость процессов адаптации
Прекращение действия экстремальных факторов вызывает в организме че-
ловека развитие деадаптации, т. е. процесс исчезновения развившихся в
тканях различных систем организма адаптивных морфологических и био-
синтетических изменений, самой адаптации с возвратом функций к услов-
ной норме. Например, при спуске альпинистов с гор на равнину возросшая
у них под влиянием высотной гипоксии кислородная емкость крови (за
счет повышенной продукции эритроцитов и массы гемоглобина, стимули-
рованной интенсивным образованием эритропоэтина почками под влияни-
ем гипоксии) возвращается к величинам, регистрируемым у них до подъема
в горы (следствие снижения продукции эритропоэтина до нормальных ве-
личин почками благодаря устранению действия гипоксии на организм).
23.2. Адаптация человека к климатическим факторам
При адаптации к определенным климатическим факторам организм чело-
века в естественных условиях обитания постоянно подвержен влиянию
сложного комплекса факторов. В результате в организме человека при адап-
тации к естественным биоклиматическим факторам (пребывание в горах,
высоких широтах, например Заполярье, в жарком климате) формируются
особенности функционирования различных физиологических систем, на-
правленных на поддержание гомеостазиса. Рассмотрим особенности адап-
тации организма человека к климатическим факторам на примере его
адаптации к жаркому климату.
23.2.1. Биоклиматические факторы жаркого климата
Почти половина населения Земли часть года находится в жарких естест-
венных условиях. Жаркий и влажный пояс, простирающийся до 10° южной
и северной широт, — это равномерно влажная среда с максимальными
температурами, колеблющимися в течение всего года от +24 до +28 °C.
Ночью относительная влажность в этих районах приближается к 100 %.
Большие пустыни находятся в основном между 10 и 30° северной и южной
широт. Лето здесь сухое, исключительно жаркое, температура достигает
890 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
+57—+58 °C, а зимой снижается до +10—+15 °C, солнечная радиация со-
ставляет 180—220 ккал/с м2/год. В пустынях субтропических поясов летом
температура воздуха достигает +22—+35 °C, зимой — от +5 до -5 °C, сол-
нечная радиация составляет 150—180 ккал/см2/год.
В пустынях сочетается действие на организм высокой температуры,
ультрафиолетового и теплового излучения, крайняя сухость воздуха, к ко-
торым добавляется ветер с пылью. Этот комплекс факторов оказывает тя-
желое, нередко губительное действие на неакклиматизированных людей.
Поступление тепла в организм человека в пустыне под воздействием горя-
чего воздуха и излучения тепла нагретыми камнями, песком и др. может
превысить теплообразование в организме в 3,5 раза. В дневные же часы
при дополнительном воздействии солнечных лучей поступление тепла в
тело человека в пустыне может в 10 раз превысить его образование в орга-
низме человека, находящегося в состоянии физического покоя. Поэтому
народы, населяющие зоны пустынь, используют одежду из плотной ткани,
предохраняющую тело от радиационного и конвенкционного нагревания.
При этом термическая нагрузка для одетого человека в пустыне на солнце
в 2,4 раза больше, чем во влажных тропиках
Несмотря на жесткие климатические условия тропиков, у их жителей
выработались адаптивные реакции, позволившие им успешно приспосо-
биться к жизни в этой среде. Однако следствие воздействий тропической
среды обитания на жителей не сводится лишь к приобретению их организ-
мом повышенной устойчивости к перегреванию. Для влажных тропиков,
где проходило формирование вида Ното, характерно мощное воздействие
на организм человека различных антигенов (широкое распространение
гельминтных инвазий, протозойных и вирусных инфекций, постоянные
укусы кровососущих насекомых), распространенность алиментарного бел-
кового дефицита и дефицита железа в пище, особенность геохимической
среды тропиков (низкое содержание солей и ряда микроэлементов в грун-
товых водах). Сочетание этих воздействий тропической среды на живущие
здесь популяции за многие тысячелетия сформировали и закрепили гене-
тические механизмы, определившие устойчивость организма к гипертер-
мии, тропической биологической агрессии, особенностям пищевого ра-
циона. В свою очередь, они повлекли значительные функциональные из-
менения систем терморегуляции, кровообращения, крови, обмена веществ,
иммунитета и их регуляции.
В умеренной зоне, между 30 и 50° широты, температура летом также ино-
гда повышается до больших величин, например на Южном Урале, на юге
Сибири —до 40 °C. Тепловую нагрузку на организм в умеренной зоне мо-
жет создавать и климат специальных помещений — микроклимат. Значи-
тельным тепловым нагрузкам подвергаются рабочие в металлургической
промышленности, на стекольных заводах, где высоки уровень радиации и
температура воздуха. Например, летом на металлургических заводах Юж-
ного Урала температура воздуха на рабочем месте горнового (доменный
цех) достигает 38 °C при относительной влажности воздуха от 53 до 91 %, а
излучение расплавленного чугуна достигает 2—3 кал/см2/мин. В мартенов-
ских цехах температура на рабочих местах колеблется от 22 до 43 °C, но
может достигать 53°C, при наличии инфракрасного излучения до Il-
ls кал/см2/мин. Для ткацких и бумажных фабрик, прачечных, глубоких
шахт характерно сочетание тепла и высокой влажности, двигатели многих
средств передвижения также создают повышенную тепловую нагрузку.
Имеются выраженные различия реакции организма на жару у адаптиро-
ванного и неадаптированного к ней жителя средних широт.
23. Адаптация человека к условиям внешней среды • 891
23.2.2. Адаптивные реакции организма человека к жаркой среде
обитания
У находящегося в покое человека высокая тепловая нагрузка за 30 мин вы-
зывает подъем температуры кожи до 40 °C, а ректальной температуры — до
39 °C. Под действием атмосферного тепла раздражаются температурные ре-
цепторы кожи, которые вместе с повышающейся температурой циркули-
рующей крови активируют центральные термочувствительные структуры (в
гипоталамусе). Активация последних вызывает изменение посылки симпа-
тической адренергической импульсации к сосудистым регионам оболочки
тела и внутренних органов. При этом симпатическая адренергическая ак-
тивность снижается в вазоконстрикторных волокнах кожи, что вызывает
раскрытие артериовенозных анастомозов, снижение тонуса вен, а также
подавляет нейрогенный тонус артериальных сосудов кожи (т. е. развивает-
ся пассивная вазодилатация). При дальнейшем повышении температуры
тела включаются механизмы активной нейрогенной дилатации сосудов ко-
жи, обусловленные активацией специализированных вазодилататорных
или судомоторных нервов. Активная дилатация сосудов на 90 % увеличи-
вает кровоток во время тепловой стресса и обеспечивает более эффектив-
ную теплоотдачу в коже, чем пассивная вазодилатация. В результате смены
пассивной вазодилатации на активную возросший кровоток в артериаль-
ных и венозных сосудах кожи, слизистых оболочках верхних дыхательных
путей увеличивает теплоотдачу во внешнюю среду путем теплопроведения,
излучения и снабжения потовых желез кровью. Кожный кровоток при
этом увеличивается до 8 л/мин по сравнению с 0,5 л/мин при термоней-
тральном диапазоне (рис. 23.4). Вазодилатация в пальцах и кистях рук на-
чинается уже при температуре воздуха, равной +22 °C, в стопе — при
+30 °C. При этом кровоток в верхних конечностях повышается в большей
мере, чем в нижних. Так, в сауне у человека кровоток через кисть может
возрастать в 30 раз, а через дистальные фаланги пальцев рук — в 70—
100 раз по сравнению с термонейтральным диапазоном среды.
При тепловом стрессе увеличивается посылка симпатической адренер-
гической активности от термочувствительных структур гипоталамуса к
сердцу и сосудам внутренних органов, оказывая положительный хроно- и
инотропный эффект, опосредуемый через Р2_алренорецепторы на сердеч-
ную мышцу, и вызывая сокращение а-адренореактивных артериальных и
венозных сосудов чревной области (кишечника, желудка, печени и др.), а
также артериальных сосудов почек. Усиление резистивной функции сосу-
дов в этих регионах при гипертермии снижает в них кровоток и компенси-
рует сдвиг последнего в места теплоотдачи.
Рост кровотока в коже обеспечивается, в основном, увеличением сер-
дечного выброса и, в меньшей степени, его перераспределением между
«ядром» и «оболочкой» тела. Так, при тепловой нагрузке на организм че-
ловека, находящегося в состоянии физического покоя, снижение кровото-
ка в сосудах чревной области, почках и скелетных мышцах «освобождает»
до 1 л крови в 1 мин, остальная же часть увеличивающегося кожного кро-
вотока — до 6—7 л/мин обеспечивается приростом сердечного выброса.
Его увеличение у людей при тепловом воздействии идет параллельно по-
вышению температуры тела и подчиняется требованиям терморегуляции и
интенсивности реакций, контролирующих транспорт тепла. В основе роста
величины сердечного выброса лежит, главным образом, увеличение числа
сердечных сокращений, являющееся следствием положительных хроно- и
инотропных эффектов симпатических нервов на сердце, аккумуляции в
892 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Рис. 23.4. Распределение минутного объема крови в тканях.
I — в условиях покоя в термонейтральном диапазоне, II — в условиях покоя, но в жаркой
среде, III — в жаркой среде, во время физической работы; кровоток: 1 — спланхнический, 2 —
почечный, 3 — церебральный, 4 — коронарный, 5 — в скелетных мышцах, 6 — кожный, 7 — в
других тканях.
миокарде адреналина во время гипертермии. У человека, находящегося в
покое, в условиях высокой внешней температуры прирост числа сердечных
сокращений может достигать 50—80 %. Солнечная радиация, повышающая
температуру кожи, резко усиливает влияние высокой температуры воздуха
на прирост числа сердечных сокращений. Увеличение числа сердечных со-
кращений в механизме повышения сердечного выброса является ведущим,
так как ударный объем сердца вначале не меняется или возрастает лишь в
первые минуты действия тепла. При дальнейшем перегревании человека
ударный объем сердца уменьшается, что связано со снижением сократимо-
сти миокарда под влиянием прямого действия повышенной температуры
крови на сердечную мышцу. Вызвано это тем, что увеличивающаяся в на-
чале перегревания интенсификация аэробных процессов (окислительного
фосфорилирования и гликолиза) в миокарде в последующем отстает от на-
растающей потребности сердечной мышцы в кислороде и в ней возникают
явления гипоксии. Снижение ударного объема связано не только с метабо-
лическими нарушениями в миокарде, но и с уменьшением венозного воз-
врата крови к сердцу, что снижает величину давления наполнения сердца.
Высокая температура воздуха, действующая на неадаптированного к жа-
ре человека, вызывает фазные изменения артериального давления. В пер-
23. Адаптация человека к условиям внешней среды • 893
вую из них, уже при небольшом увеличении температуры тела, снижаются
систолическое и диастолическое давление. Основной причиной снижения
диастолического давления является уменьшение общего периферического
сопротивления сосудов. Снижение последнего начинается при температуре
воздуха, равной +28 °C. Наиболее значимо общее периферическое сопро-
тивление сосудов снижается при повышении температуры тела до +38 °C,
составляя 40—55 % от исходной величины. Дальнейшее повышение темпе-
ратуры тела (вторая фаза), напротив, может характеризоваться повышени-
ем общего периферического сопротивления сосудов и диастолического
давления при выраженном снижении систолического давления. Повыше-
ние общего периферического сопротивления сосудов и диастолического
давления является следствием компенсаторных сосудистых реакций, на-
правленных на поддержание уровня системного артериального давления и
необходимого возврата крови к сердцу. Снижение систолического давле-
ния является следствием понижения ударного объема сердца.
В условиях высокой внешней температуры наиболее ранней реакцией,
вызывающей перераспределение регионарных объемов крови, является
расширение кожных вен, влекущее функциональное депонирование значи-
тельного количества крови, которое практически выключается из общей
циркуляции. Это связано с тем, что в венозных сплетениях кожи кровь
движется в 5—20 раз медленнее, чем в магистральных венах. Увеличение
объема венозного русла кожи компенсируется снижением у людей цен-
трального и легочного объемов крови (на 11—22 %). Однако в результате
депонирования крови в кожных венах снижается эффективный объем цир-
кулирующей крови — возникает относительная гиповолемия. Ее компенса-
ция достигается за счет перемещения части внеклеточной жидкости в со-
судистое русло. Так, уже в первые 30 мин теплового воздействия на орга-
низм человека у него увеличивается объем плазмы крови. Вода поступает в
кровь из межклеточного пространства внутренних органов — печени, лег-
ких и др. Повышенный объем плазмы при гипертермии может сохраняться
долго, если человек не ограничивает потребление воды. Важную роль в
этом процессе играет увеличение коллоидно-осмотического давления плаз-
мы крови, задерживающего воду в сосудистом русле, а также уменьшение
выведения воды с мочой.
Симпатическое возбуждение, возникающее при действии высокой тем-
пературы на организм, вызывает сужение почечных сосудов и, как следст-
вие, уменьшение почечного кровотока и скорости клубочковой фильтра-
ции. В результате выделительная функция почек снижается. Так, диурез
уменьшается в жару до 300—600 мл в сутки (при норме около 1,5 л). Одна-
ко на снижение выделительной функции почек влияет и повышенная сек-
реция гормонов — вазопрессина и альдостерона. Увеличение секреции
первого связано: 1) с рефлекторным ответом нейрогипофиза на снижение
центрального венозного давления; 2) с раздражением осморецепторов, вы-
званным увеличением осмотической концентрации крови и потерями воды
с потом. Возросшая секреция вазопрессина усиливает реабсорбцию воды в
почках, что повышает объем циркулирующей крови. Симпатическое возбу-
ждение при гипертермии снижает кровоток в почках, активирует ренин-
ангиотензин-альдостероновую систему. Так, у человека 20 мин пребыва-
ния в сауне увеличивает в плазме крови содержание ренина на 95 %, ан-
гиотензина — на 195 % и альдостерона — в 5 раз. Активация ренин-ангио-
тензин-альдостероновой системы, в свою очередь, усиливает почечную и
чревную вазоконстрикцию, дополняя эффект симпатической активации,
адресованной этим сосудистым зонам. Секрецию альдостерона стимулиру-
894 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ет и уменьшение хлористого натрия в крови, вызванное потерями послед-
него с потом.
Увеличение кровотока в коже обеспечивает потовые железы кровью в
количестве, необходимом для повышенного образования ими секрета. У не
адаптированного к жаре человека, находящегося в состоянии физического
покоя, воздействие внешнего тепла, повышая температуру «ядра» тела до
38 °C, повышает секрецию пота до 10—12 мл/мин (при комнатной темпе-
ратуре она составляет 600 мл/сут). У адаптированного к жаре человека,
также находящегося в состоянии физического покоя, потоотделение дос-
тигает 20 мл/мин. Это связано с более быстрым усилением кровотока в ко-
же и с повысившейся способностью к потоотделению потовых желез у
адаптированных к жаре людей. Неакклиматизированные к жаре люди в
первые дни действия тепла на организм теряют 15—30 г солей в день с по-
том и мочой, но после 4—6 нед их акклиматизации потери солей уменьша-
ются до 3—5 г в день. Лучшее сохранение хлористого натрия в организме
связано с повышенной секрецией альдостерона.
При высокой температуре окружающей среды дыхание учащается, ле-
гочная вентиляция увеличивается, в основном за счет глубины дыхания.
Значительная гипервентиляция легких, способствуя вымыванию угольной
кислоты из крови, понижает в ней напряжение СО2, что, в свою очередь,
смещает pH крови в щелочную сторону. Поэтому гипокапния и дыхатель-
ный алкалоз являются постоянными признаками перегревания организма
человека. Способность крови связывать СО2 снижается и под влиянием
роста ее температуры.
Уменьшение кровоснабжения желудочно-кишечного тракта, вызванное
перераспределением крови к кожным сосудам при повышении температу-
ры тела, ведет к снижению секреторной, всасывательной и моторной
функций органов пищеварения, понижению аппетита. Так, при повыше-
нии температуры тела уменьшается секреция слюны, в ней повышается со-
держание плотного остатка, что связано с потерей воды организмом при
испарительном термолизисе. При потере воды, равной 8 % от массы тела,
слюноотделение прекращается полностью. Гипертермия снижает объем
секреции желудочного сока и содержание в нем соляной кислоты и пепси-
на. Это, в свою очередь, уменьшает переваривающую силу желудочно-ки-
шечного сока. Повышение температуры тела у человека до 37,5 °C угнетает
моторику желудка. В результате замедляется эвакуация из него пищи. Под
влиянием высокой внешней температуры на организм у неадаптированных
людей уменьшается также объем секретируемого панкреатического сока, в
нем снижается содержание протеолитических ферментов. Эвакуация жел-
чи из желчного пузыря уменьшается вследствие снижения тонуса его сте-
нок. Это приводит к застою желчи. По-видимому, вследствие указанных
сдвигов у людей при тепловых воздействиях ухудшается аппетит, исчезает
желание принимать пищу животного происхождения. Существующий у
жителей жарких стран обычай выпивать перед едой некоторое количество
жидкости (чая и др.), сопутствующий приему пищи отдых, уменьшение те-
пловой нагрузки, по-видимому, не только уменьшают дегидратацию орга-
низма, но и способствуют перемещению части крови к желудочно-кишеч-
ному тракту.
Тепловой стресс, активируя симпатоадреналовую и гипофизарно-адре-
нокортикальную системы, вызывает усиление секреции катехоламинов
мозговым веществом надпочечников и глюкокортикоидов их корой. Под
влиянием этих гормонов в печени усиливается образование глюкозы в ходе
гликолиза, стимулируется глюконеогенез.
23. Адаптация человека к условиям внешней среды • 895
У акклиматизированных к жаре людей реакция нейроэндокринной систе-
мы на тепловые воздействия снижается. В результате уменьшается и кон-
центрация в плазме катехоламинов, глюкокортикоидов, тироксина. Поэто-
му у адаптированных к жаре людей замедляется глюконеогенез, снижается
скорость гликолиза, восстанавливаются запасы гликогена в печени и ске-
летных мышцах. В целом перегревание организма приводит к понижению
энергетического обмена.
Снижение кровотока во внутренних органах в условиях нарастающей
гипертермии приводит к уменьшению отношения доставки кислорода к
его потреблению в тонком кишечнике, печени, почках, скелетных мыш-
цах. В результате, в этих органах возникает тканевая гипоксия, что активи-
рует образование энергии в ходе анаэробного гликолиза и приводит к уве-
личению концентрации недоокисленных продуктов обмена (молочной ки-
слоты и др.) в этих тканях и крови.
Система крови отвечает на перегревание организма значительными из-
менениями. Тучные клетки и базофильные лейкоциты реагируют на повы-
шение температуры крови быстрой дегрануляцией, секретируя в кровь ге-
парин, гистамин и другие биологически активные вещества. При повыше-
нии температуры «ядра» тела уровень гистамина в крови увеличивается в
несколько раз. У адаптированных к жаре людей активируются и механиз-
мы, препятствующие росту гистамина в крови, нарастает содержание гис-
топексина и активность гистаминазы, понижающих концентрацию в ней
гистамина. Это снижает проницаемость кровеносных сосудов, повышае-
мую гистамином, снижает его вазодилататорный эффект на артериолы
брюшной полости и других регионов.
Под действием тепла в клетках тканей внутренних органов, скелетных
мышцах повышается проницаемость мембран лизосом, что вызывает вы-
ход лизосомальных протеиназ в кровь. Однако с адаптацией организма к
жаре в плазме крови повышается уровень а]-антитрипсина — мощного ин-
гибитора протеолитических ферментов, что предохраняет белки и клеточ-
ные элементы крови от избыточного протеолиза.
У людей — выходцев из умеренной зоны Земли, но живущих в жарких
влажных тропиках, — гематологические сдвиги (лейкопения, относитель-
ная и абсолютная нейтропения, лимфоцитоз) в целом сходны с измене-
ниями крови у аборигенов тропиков. У европейцев, адаптированных к тро-
пикам, и у рабочих, многие годы подвергавшихся тепловым воздействиям,
возникают изменения в эритроне: нарастает гемоглобинизация и гидрата-
ция отдельного эритроцита, увеличивается содержание гемоглобина в кро-
ви, эритроцитарный уровень удерживается на значениях, приближающих-
ся к нижней границе европейской нормы. Уровень эритропоэтина в крови
снижается, и это объясняет, почему у адаптированных к жаркому климату
людей происходит замедление регенерации красной крови, например по-
сле кровопотери.
23.2.3. Особенности адаптации человека к работе в жаркой среде
Потребление кислорода и температура «ядра» тела закономерно нарастают
во время работы возрастающей мощности. Так, работа мощностью до 50 %
от величины максимального потребления кислорода (МПК), производи-
мая в комфортной температурной среде, увеличивает температуру ядра до
37,4 °C, а работа с 75 % потреблением кислорода от величины МПК повы-
шает ее до 38,5 °C. Температура кожи при этом остается относительно не-
изменной, что обеспечивает эффективную отдачу тепла. Однако при нали-
896 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
чии внешней тепловой нагрузки тепло, образующееся в работающих мыш-
цах, может еще более увеличивать температуру тела, если нарушится эф-
фективность охлаждающего испарительного механизма и обеспечивающих
его сосудистых компенсаторных реакций. Работа человека на жаре требует
от системы кровообращения выполнения двух задач: 1) обеспечить мышцы
кислородом, чтобы поддержать в них возросший расход энергии; 2) обес-
печить перенос кровью метаболического тепла от глубоких тканей к обо-
лочке тела. При этом кровоток в мышцах повышается до 13 л/мин, кож-
ный — до 2,2 л/мин, а сердечный выброс — до 18 л/мин (см. рис. 23.3), что
обеспечивается за счет уменьшения кровотока в висцеральных органах.
Например, в желудочно-кишечном тракте и почках кровоток снижается в
этом случае соответственно до 300 и 450 мл/мин. Вазоконстрикция в вис-
церальных органах повышает общее периферическое сопротивление сосу-
дов, что поддерживает на оптимальном уровне артериальное давление кро-
ви во время работы на жаре, обеспечивает достаточную перфузию сосудов
головного мозга, препятствует развитию церебральной ишемии и артери-
альной гипотензии. Коронарный кровоток при работе на жаре может воз-
растать в четыре раза по сравнению с покоем.
Физическая нагрузка, даже хорошо переносимая в жару, выполняется с
нарастанием доли анаэробного метаболизма во внутренних органах, что
приводит к быстрому уменьшению запасов гликогена в этих тканях, нарас-
танию в них молочной кислоты. Этому способствует сниженная экстрак-
ция кислорода из крови в мышцах, так как, несмотря на возрастающий в
них кровоток, большое количество крови из мышц шунтируется, не отдав
кислород, к сосудам оболочки тела для рассеивания тепла. Росту уровня
молочной кислоты в крови способствует и сниженное поступление крови в
печень из-за значительного уменьшения печеночного кровотока. Накапли-
вающиеся в крови кислые метаболиты истощают буферные основания,
создавая условия для развития метаболического ацидоза.
Потери воды при работе в жару, например при очень жаркой погоде в
пустыне, могут достигать 8—12 л/сут с потом, при этом интенсивное пото-
отделение в течение нескольких часов может привести к «усталости» пото-
вых желез, вызванной истощением их секреторной функции. Это резко на-
рушает способность регулировать температуру «ядра» тела. Потери жидко-
сти при потоотделении не столь эффективно охлаждают кожу во время ра-
боты в жару при высокой влажности, когда испарительное охлаждение ко-
жи снижено высоким давлением водяных паров в окружающем воздухе.
Поэтому интенсивное потоотделение в этих условиях хотя и приводит к
большим потерям жидкости, но сопровождается минимальным испарени-
ем и охлаждением поверхности тела, повышая риск перегревания орга-
низма.
Значительная потеря жидкости с потом уменьшает объем плазмы и
циркулирующей крови, что снижает величины конечного диастолического
объема желудочков и систолического объема сердца, уменьшает кровоток в
коже и, следовательно, потоотделение. Минутный объем не уменьшается
вследствие компенсаторно нарастающего числа сердечных сокращений.
В результате снижения эффективности механизмов, обеспечивающих вы-
ведение тепла из организма, увеличивается температура «ядра» тела. Сле-
довательно, дегидратация организма уменьшает возможность систем кро-
вообращения и терморегуляции компенсировать тепловой стресс при фи-
зической нагрузке. Поэтому поддержание на оптимальном уровне объемов
плазмы и внеклеточной жидкости у человека в условиях работы на жаре
сохраняет функции кровообращения и продукцию пота. А это, в свою оче-
23. Адаптация человека к условиям внешней среды • 897
редь, достигается сбалансированием потери и восполнения воды. У чело-
века, получающего достаточное количество жидкости, работоспособность в
жару сохраняется на более высоком уровне, чем у работающего в состоя-
нии дегидратации. Употребление дополнительно 400—600 мл воды за 10—
20 мин до мышечной работы на жаре, задерживает развитие дегидратации,
увеличивает потоотделение во время работы и препятствует значительному
повышению температуры «ядра» трла.
Для рабочего, выполняющего работу средней тяжести на открытом воз-
духе в условиях Средней Азии, «норма» выпиваемой воды за смену состав-
ляет 4—4,5 л. Большинство людей устойчивы к потерям 3—4 % воды (от
массы тела). При потере же ее 5—8 % возникает чувство усталости в поло-
жении «сидя», сонливость, апатия. Потеря свыше 10 % воды вызывает у
человека физические и психические нарушения. При тяжелой дегидрата-
ции необходимы срочные врачебные меры: вводят внутривенно 5 % рас-
твор глюкозы, изотонический раствор хлорида натрия (до 1 л/ч в начале
лечения), но не более 4 л в сутки.
Потери пота при работе на жаре могут истощать солевой запас организ-
ма на 13—15 г/сут (2,3—3,4 г/л пота), у неакклиматизированных людей по-
тери солей могут быть больше. Это значительно превышает их величину в
ежедневном рационе, поэтому необходима добавка в пищу соли (*/3 чай-
ной ложки поваренной соли на литр воды). Потери калия, кальция и маг-
ния при выделении пота невелики и могут быть восполнены богатой ими
диетой. Например, стакан томатного сока возмещает почти весь калий,
кальций и магний, содержащиеся в 2—3 л пота. При значительных потерях
натрия внутривенно вводят изотонический раствор хлорида натрия. При
интенсивных потерях жидкости, связанных с жарой, возникает гиповита-
миноз водорастворимых витаминов С и В,.
Повышение устойчивости организма к действию тепла называется теп-
ловой акклиматизацией. Обычно акклиматизация организма заканчивается
к концу второй недели теплового воздействия. Для этого требуется от 2 до
4 ч ежедневных тепловых экспозиций. Однако первые тепловые воздейст-
вия должны быть тренирующими, продолжительностью около 15—20 мин.
Их продолжительность и интенсивность должны возрастать постепенно.
Акклиматизация достигается легче, если человек во время работы на жаре
имеет возможность полностью покрывать потребность в воде. Адаптиро-
ванные к жаре люди легче переносят ее воздействие, сохраняют большую
работоспособность, другие виды социальной активности.
23.2.4. Предупреждение тепловых поражений организма
Наиболее эффективный путь профилактики поражений организма пере-
греванием — это оценка окружающей среды с позиций опасности ее тепло-
вого влияния на организм. Она может быть получена, например, при рас-
чете величины индекса теплового стресса (ИТС). Этот показатель учитыва-
ет температуру окружающей среды, относительную влажность и уровень
радиации тепла:
ИТС (в °C) = 0,1 • ТСТ+0,7 • ТВТ+0,2 • ТШТ,
где ТСТ — температура «сухого» термометра, т. е. температура воздуха, ре-
гистрируемая обычным ртутным термометром; ТВТ — температура «влаж-
ного» термометра, т. е. температура воздуха, замеренная термометром, ре-
зервуар с ртутью которого покрыт влажной марлей (уровень относитель-
ной влажности влияет на его испарительное охлаждение); ТШТ — темпе-
29 - 6095
898 • ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
ратура шарового термометра, резервуар с ртутью которого заключен в по-
лый металлический шар с черной матовой поверхностью (адсорбирует из-
лучаемое тепло из окружающей среды).
На основании величины расчета индекса теплового стресса разработаны
рекомендации для степени физической активности на открытом воздухе.
Так, величина этого индекса, равная 26,5—28,8 °C, должна насторожить
неакклиматизированных людей в отношении перегревания. При значениях
индекса теплового стресса 29,5—30,5 °C адаптированным к жаре лицам
следует избегать большой физической активности на солнце. При индексе
теплового стресса 31,2 °C необходимо резко ограничить физические на-
грузки. Признаком границы предельно допустимой тепловой нагрузки на
организм в условиях физического покоя является увеличение частоты сер-
дечных сокращений до 140 уд/мин, повышение ректальной температуры
до 38,4—38,6°C. При физической работе на жаре эти показатели составля-
ют соответственно 170—180 уд/мин и 38,5—38,8 °C.
При длительном действии тепла может возникать нарушение терморегу-
ляции, проявляющееся в различных формах тепловых поражений организ-
ма: тепловые судороги, тепловое истощение организма, тепловой обморок
и тепловой удар.
Тепловые судороги — внезапно наступающее непроизвольное сокращение
мышц (обычно икроножных) во время или после интенсивной физической
работы на жаре. Они связаны с несбалансированностью электролитов в
организме, как результат потери солей (хлорида натрия) с потом. Если эти
потери не восстановлены, то возникает мышечная боль с судорогами. При
тепловых судорогах температура тела может не повышаться. Для их профи-
лактики достаточно употребления обильного количества воды с увеличе-
нием ежедневных приемов соли за несколько дней, предшествующих пе-
риоду теплового стресса, и во время работы в условиях длительного пере-
гревания.
Тепловое истощение обычно характеризуется общей слабостью, быстрым
пульсом, низким артериальным давлением в вертикальном положении те-?
ла, головокружением, головной болью. Тепловое истощение организма
развивается у неакклиматизированных людей в первые дни летней жары
или во время работы в жаркий день при недостаточно компенсируемых
потерях воды и солей с потом. Тепловое истощение вызвано неэффектив-
ной регуляцией кровообращения, сочетающейся с уменьшением внекле-
точной жидкости вследствие чрезмерного потоотделения. Кровь при этом
скапливается в дилатированных периферических сосудах, что резко умень-
шает венозный возврат, необходимый для поддержания производительной
работы сердца. Температура тела обычно не повышается до опасных пре-
делов. Человек с этими симптомами должен переместиться в прохладную
или холодную среду; явления дегидратации устраняются потреблением
жидкости.
Тепловой обморок (или тепловой коллапс) — внезапная кратковременная
потеря сознания, сопровождающаяся резкой бледностью, ослаблением ды-
хания и кровообращения. Обморок часто возникает после перехода тела в
вертикальное положение, влекущего ортостатическую гипотензию. Орто-
статическая гипотензия связана с генерализованной вазодилатацией пери-
ферических сосудов, особенно скелетных мышц и кожи нижних конечно-
стей. Скапливающаяся в них кровь резко уменьшает венозный возврат к
сердцу, в результате снижается сердечный выброс и уменьшается крово-
снабжение головного мозга, возникает его ишемия и кратковременная по-
теря сознания. Потере сознания обычно предшествует головокружение.
23. Адаптация человека к условиям внешней среды • 899
Тепловой обморок может наблюдаться у человека во время активной мы-
шечной работы в очень жаркой среде, когда температура «ядра» тела дос-
тигает 39 °C. Помощь при обмороке: нужно уложить пострадавшего в тень,
в хорошо проветриваемом месте, приведя его в сознание, дать ему про-
хладное питье в умеренном количестве. Обморок, как правило, наблюдает-
ся у людей, не акклиматизированных к жаре.
Тепловой удар — болезненное состояние, вызванное общим перегревани-
ем организма вследствие действия на него тепловых факторов внешней
среды. Тепловой удар возникает вследствие недостаточности деятельности
гипоталамических центров терморегуляции, неадекватности их регулирую-
щих влияний нарастающей тепловой нагрузке на организм. Это нарушение
терморегуляции связано, прежде всего, с недостаточностью механизмов
образования пота и выведения с ним тепла из организма. Вместе с тем
усиленное потоотделение при перегревании тела вызывает значительную
потерю воды и солей, что, в свою очередь, ведет к уменьшению объема
циркулирующей плазмы крови, увеличению ее вязкости. В результате на-
рушается кровообращение, возникает кислородное голодание тканей, за-
трудняется перенос тепла от «ядра» к «оболочке» тела. Высокая температу-
ра «ядра» на фоне гипоксии тканей приводит к разрушению клеток, дена-
турацию белков, вызывая необратимые изменения в тканях.
В зависимости от тяжести проявлений перегревания организма человека
различают легкую, среднюю и тяжелую формы теплового удара. При лег-
кой форме теплового удара пострадавший жалуется на головную боль, тош-
ноту, зрачки расширены, дыхание и пульс учащены, кожа влажная, темпе-
ратура тела повышена (до 38 °C, иногда и более). Если пострадавшего пе-
ренести в тень, все симптомы теплового удара проходят.
При средней тяжести теплового удара у пострадавшего усиливаются
признаки нервных и ментальных нарушений: головная боль становится
очень сильной, она сопровождается рвотой, нарушается координация дви-
жений, возникают кратковременные обмороки. Кожа горячая, влажная,
потоотделение усилено. Температура тела повышается до 39—40 °C.
Тяжелая форма теплового удара развивается, если температура тела по-
вышается до 41—42 °C. У пострадавшего возникает психомоторное возбуж-
дение, нарушение сознания (бред, галлюцинации), он впадает в кому.
Пульс нитевидный, 120—140 уд/мин, дыхание поверхностное, частое. Кожа
становится сухой и горячей. Без оказания немедленной медицинской по-
мощи пострадавший при средней и тяжелой форме теплового удара поги-
бает. Помощь: пострадавшего необходимо поместить в тень, желательно в
открытое для ветра место, раздеть до пояса, на голову и шею наложить
смоченную холодной водой ткань или пузыри с холодной водой. Если по-
зволяют условия, тело обкладывают пакетами со льдом или погружают в
ванну с холодной водой до понижения ректальной температуры ниже
39 °C.
29*
Предметный указатель
Авитаминоз 572
Агглютинация 359
Агранулоциты 331
Адаптация 875, 880
— генотипическая 888
— долговременная 888
— онтогенетическая 889
— световая 725
— срочная 887
— эволюционная 888
Адгезия 155
Аденилаты 580
Аденогипофиз 270
Аденозин 75
Аденозинмонофосфат цик-
лический 80
Адреномедуллин 291
Адренорецепторы а, р 83,
253
Аквапорины 280, 619, 625
Акинезия 224
Акклиматизация 879
— тепловая 897
Аккомодация 62, 719
Акромегалия 277
Акт дефекации 560
— полевой женский 688
мужской, стадии 678
Активатор плазминогена
тканевый 346
Активность пейсмекерная
клеток гладкомышеч-
ных 117
------синоатриального
узла 117
Алкалоз 661
— дыхательный 665
— значение функциональ-
ное 662
— недыхательный 666
Альбумины 33
Альвеолы 469
Альдостерон 283
Амилин 299
Амилоид А 34
Аминокислоты 75
Аминокислоты незамени-
мые 565, 567
Амины 75
Амины биогенные 176
Аммониогенез 657
Анаболиз 563
Ангиотензин I, II 75, 310
Андрогены 290, 684
Анемия 330
Анпериэлектротон 56
Антагонизм акцентирован-
ный 256
Антиген(ы) лейкоцитов
360
— распознавания 378
— тромбоцитов 360
— эритроцитов 358
Анти порт 51, 620
Антитромбин III 345
Анэлектротон 55
Апнейзис 501
Аппарат вестибулярный
743
— памяти 127
— речевой 825
Ароматерапия 754
Астроциты 170
Атриопептид 312
Аутоантигены 383
Афагия 511
Афашия Брока 830
— Вернике 830
— моторная 826
— сенсорная 827
Ахромотопсия полушар-
ная 811
Ацетилхолин 75, 82
Ацетилхолин, высвобожде-
ние 254
Ацетилхолинэстераза 72
Ацидоз 661
— дыхательный 663
— значение функциональ-
ное 662
— недыхательный 664
Баланс азотистый 565
— организма водный
внешний 634
------внутренний 636
---электролитный 636
Барьер(ы) гематоэнцефа-
лический 170
— гистогематические 22
---изолирующие 22
---неизолирующие 22
---частично изолирую-
щие 22
Белки 564
Белки-коннексоны 68
Белки острофазные крови
34
— кальциевый сенсор 76
— С-реактивный 34
Бикарбонат крови актуаль-
ный 669
----- стандартный 660
Биомеханика вдоха 470
— выдоха 471
Бледный шар 221
Бодрствование 853
— уровни 854
Болезнь горная 508
— кессоная 510
Боли фантомные 232
Боль 715
Вазопрессин 75, 149, 452
Варикозы 251
Везикулы 75
Вентиляция легких 483
---альвеолярная 483
Веретена мышечные 186
Вестибулоцеребеллум 207
Вещества(о) биологически
активные 156
— минеральные 568, 569
— студневидное 181
Вирилизация 290
Висцерфецепторы 717
Витамины 570, 585
— источники 572
— роль в кроветворении
357
Влечение 782
Внимание 801
— зрительное 803
— механизмы нейрофи-
зиологические 803
— пространственное 806
— фокусное 817
— формы 802
— центры корковые 805
— центр мультимодаль-
ный 808
— экспрессия нейронная
817
Вода 21
Водитель ритма 398
---латентный 400
Возбудимость 45
Возбуждение подпорого-
вое 53
Предметный указатель • 901
— порог 53
— проведение аксосомати-
ческое 73
— постсинаптическое 72
---пресинаптическое 76
— сигнал выходной 168
---проводящийся 167
— сила раздражения мак-
симальная 53
Волны звуковые 734
Волокна афферентные 181
— интрафузальные 186
— коллагеновые 25
— миелинизированные 67
— мышечные интрафу-
зальные 187
---скелетные 98
---экстрафузальные 187
— немиелинизированные
66
— нервные нервов пери-
ферических 136
— эфферентные 181
Восприятие 809
— вкусовое 750
— зрительное 717, 809
---функции стриарной
коры 809
— слуховое 812
— соматосенсорное 812
— цветовое
Выделение 606
Габитуация 818, 820
Газ углекислый 655
Газообмен в легких 487
Галактогенез 699
Гаметы 668
Ганглии базальные 221
Гастрин(ы) 181, 304
Гематокрит 28
Гемианопсия 811
Гемоглобин 492
— емкость кислородная
493
— сродство к О2 494
— форма расслабления 493
- А324
- а, р 324
- F324
— Gower I, II 324
Гемодинамика системная
391
Гемолиз 31
— внесосудистый 326
— внутриклеточный 326
— внутрисосудистый 326
Гемосидерин 328
Гемостаз вторичный 341
— первичный 341
Гепарин 345
Гепатит С хронический
377
Гетеросексуальность 781
Гипервитаминоз 572
Гипергидратация гиперос-
мотическая 647
— гипоосмртическая 646
— изоосмотическая 647
Гиперемия ауторегулятор-
ная 437
— рабочая 436
— реактивная 437
— функциональная 437
Гиперкапния 663
Гиперпноэ 505
Гиперполяризация 79
Гипертермия 603
Гипертиреоз 293
Гиперфагия 511
Гиповитаминоз 572
Гипогликемия 583
Гипоксемия артериальная
507
Гипоталамус 149
— область трофотропная
214
----эрготропная 214
— регуляция мотиваций
213
----поведения 213
----пищевого 214
----эмоций 213
— ядра вентромедиальные
207
----хиазмальные 207
Гипотермия 603
Гипофиз 270
Гипофосфатемия 296
Гиппокамп 83
— функции 221
Гистамин 75
Гистерезис 476, 478
Гликоген 567, 583
Гликолиз анаэробный 858
— аэробный 858
Гликопротеины, функции
338
Глицин 75, 87
Глия эпендимная 171
Глобулины 33
— функции 36
Глотание, фазы 532
Глубина дыхания 474
Глутамат 86, 170
Глюкагон 299, 304
Глюкоза 567, 581
Глюкокортикоиды, секре-
ция 286
— эффекты 286
Глюконеогенез 580
ГЛ ЮТ 302
Голодание водное 643
Голубое пятно 206
Гольдмана—Ходжкина-
Катца уравнение 60
Гомеостазис 21
— водно-солевой 639
— поддержание 249
Гомункулус сенсорный 230
Гонадолиберин 75
Гонадотропин(ы) 273
— хорионический 307
--- человеческий 695
Гормон(ы) 147
— аденогипофиза 271
— белковые 260
Гормоны гастроинтести-
нальные 524, 550, 553
— гипокальциемический
296
— гипофиза 270
— действие кинетическое
152
---корригирующее 152
---метаболическое 151
---морфогенетическое
152
---пермиссивное 153
---реактогенное 153
— доли промежуточной
282
— железы околощитовид-
ной 131
---щитовидной 131
— желудочно-кишечного
тракта 314, 515, 517
— избыток в крови 288
— коры надпочечников
283
— мозгового вещества над-
почечников 290
— надпочечников 283
— нейрогипофиза 279
— пептидные 260
— плацентарные 695
— половые 331
— почек 307
— предсердный натрийу-
ретический 312
— семенников 305
— сердца 312
— стероидные 260, 266
---механизм действия
геномный 266
---------негеномный 266
— тимуса 307
— тиреоидные йодсодер-
жащие 292
---эффекты метаболиче-
ские 292
------физиологические
293
— тканевые 156
902 • Предметный указатель
— эндотелия сосудистого
313
— яичников 305
Градиент концентрацион-
ный для Na+ и К+ 60
Гранулопоэз неэффектив-
ный 356
Гранулоцитоз 333
Гранулоциты 331
— базофильные, функции
334
— нейтрофильные 331
Громкость звука 735, 736
Давление артериальное
диастолическое 392
---пульсовое 392
---систолическое 392
---среднее 392
— венозное 433
---центральное 425
— внутриплевральное 472
— внутрисосудистое 433
— звуковое 735
— осмотическое 47
---эффективное 43
— парциальное газов 489,
492
— трансмуральное 435
— фильтрационное 30, 614
Движение(я) контроль,
связь обратная 244
------прямая 244
— произвольные 239
— рефлекторные 239
— ритмические 239
Деадаптация 880, 889
Дегидратация гиперосмо-
тическая 640
— гипоосмотическая 645
— изотоническая 645
Дезоксигемоглобин 325
Действие пищи специфи-
чески-динамическое 578
Деполяризация, критиче-
ский уровень 62
— спонтанная диастоличе-
ская 117
Депрессия католическая 56
Депривация 780, 836
Десенситизация 268
Дефенсины 35, 332
Дефицит организма кисло-
родный 863
Деятельность нервная выс-
шая 757
------типология 771
------человека, особен-
ности 772
— сенсомоторная 857
— человека познаватель-
ная 756, 800
----трудовая 756
Диацилглицерол 81
Дивергенция возбуждения
141
Дизадаптация 879, 888
Диморфизм половой 670,
780
Динеин 52
Диски Меркеля 712
Диспарация 720
Дифференциация половая
669
Диффузия пассивная 488
— явление конкуренции 48
----насыщения 48
Долг кислородный 864
Дофамин 75
Дуга рефлекторная 133
----возбуждение 137
----моносинаптическая
186
----полисинаптические
190
----связь между звенья-
ми 137
----торможение 137
Дыхание 469, 603
— внешнее 470
— при физической нагруз-
ке интенсивности высо-
кой 506
-----------низкой и
средней 506
--------фазы 505
— регуляция 497
Дюбуа—Реймона закон
раздражения 53
Единица фетоплацентар-
ная 693
Емкость легких жизнен-
ная 475
----инспираторная 475
----общая 475
----остаточная респира-
торная 475
— легочная 475
Жажда 642
Жевание 531
Железа(ы) бруннеровы 548
— вилочковая см. Тимус
— желудочные 534
— молочная, влияние про-
лактина 279
— околощитовидные 296
— сальные 607
— слюнные 528
---состав смешанного
секрета 529
— щитовидная 291
Железо, потребность еже-
дневная 329
— резервное 327
— функционально-актив-
ное 327
Желудок, деятельность
мускулатуры сократи-
тельная 541
---------регуляция 542
— функции 533
— эвакуация содержимого
543
Желчь, механизм образо-
вания 546
— регуляция образования
и выделения 547
— состав и своства 546
Жидкость внеклеточная,
величины средние 20
---отклонения нормаль-
ные 20
— внутриклеточная, пока-
затели физико-химиче-
ские 25
— интерстициальная 25
— обмен в организме 42
Жидкость семенная 681
— синовиальная 42
— спинномозговая 41
— тканевая см. Жидкость
интерстициальная
Жизнедеятельность, меха-
низмы регуляции 132
Жир бурый 566
Зависимость градуальная
163
— «поток—объем» в лег-
ких 480
Задержка синаптическая
79
Закон Вебера 710
— Вебера—Фехнера 710
— двустороннего проведе-
ния 68
— диффузии Фика 489
Закон Дюбуа—Реймона 53
Закон изолированного
проведения 68
— поверхности тела 576
Закон полярности раздра-
жения Пфлюгера 56
Закон физиологической
непрерывности 68
Закон Фика первый 47
Закон Франка—Старлинга
419
Предметный указатель • 903
Закон электродиагностиче-
ский 57
Знание 800
Зоб 294
Зона(ы) Брока 828
— Вернике 827
— Веста 486
— золя-геля гиалоплазмы
338
— легких 483
— плотных а-гранул 338
— рефлексогенные 136
Излучение 596
Изотермия 592
Иммуноглобулин А 382
- Е382
- G 381
- М 382
Ингибин 677, 685
Индукция нервных про-
цессов 770
Инозитол-З-фосфат 80
Инкряция 261
Инсулин 181, 299
— эффекты физиологиче-
ские 301
Интегрины 155
Интернейроны 140
— локальные 173
— проекционные 173
Интерорецепторы 704
Инсулин 616
Испарение 597
Истощение тепловое 898
Кавеолы 109
Кальбайндины 309
Кальсеквестрин 94
Кальмодулин 110, 120
Кальцитриол 297, 307
Канал(ы) калиевые каль-
цийзависимые 77
---спинномозговой 181
Капацитация 690
Капилляры, типы 454
Кардиомиоциты 113
— высвобождение 120
— потенциал действия,
фаза плато 115
--------реполяризации
117
— расслабление, меха-
низм молекулярный 120
— сокращение, контроль
медиаторный 121
---механизм молекуляр-
ный 119
«Каскад кислородный» ор-
ганизма 861
Катаболизм 563
Катакальцин 295
Кателицидины 35, 332
Катехоламины 290, 452
Катпериэлектротон 56
Квадранопсия 811
Квант медиатора 70
Кинезин 52
Кинетоды 746
Кинины 157, 451
Кинки 47
Кислота(ы) 650
— арахидоновая 317
— гамма-аминомасляная
75, 87
— ретиноловая 357
— соляная, механизм сек-
реции 535
---в пищеварении 536
— фолиевая 357
Кислотность мочи титруе-
мая 658
Кишечник толстый, актив-
ность двигательная 557
---перемещение химуса
557
---микрофлоры в пище-
варении 558
---сокоотделение 557
Кишка тонкая, регуляция
моторики 551
---------гуморальная 553
---функция всасывания
553
------двигательная 550
Классификация запахов
751
Клетки апокриновые 513
— вкусовые 748
— волосковые 736
— ганглиозные 726, 728
---сетчатки, типы 729
— голокриновые 513
— колониеобразующие 349
— Лангерганса 370
— Лейдига 304, 673, 676
— мерокриновые 513
— микросреда 27
— пейсмекерные 399
Клетки-предшественницы
349
— противоцветные двой-
ные 732
— Реншоу 184, 875
— Сертоли 304, 673
— функции, взаимодейст-
вие межклеточное 154
---регуляция местная
154
---связи креаторные 154
— шванновские 171
— эпендимные типичные
171
Клетки-эффекторы 366
— юкстагломерулярные
310
Кожа, кровоток 450
Колбы Краузе 712
Колориметрия непрямая
574
— прямая 574
Кома гипогликемическая
583
Комплекс тропонин-тро-
помиозиновый 91
Конвекция 596
Конвергенция 141
— полисенсорная 204
Коннексоны 104
Константа(ы) внутренней
среды 19
— диссоциации воды 650
Контрактура мышечная 94
---запасы АТФ 103
Контроль дыхания механо-
рецепторный 503
---хеморецепторный 502
Кора, активность электри-
ческая 227
— ассоциативная пре-
фронтальная 237
— зрительная вторичная
233
---первичная 233
---проекционная 731
— лимбическая 237
— моторная вторичная 239
---первичная 238
---путь вентральный 242
------ кортикобульбар-
ный 242
------кортикоспиналь-
ный 242
------нисходящий дор-
солатеральный 242
— области сенсорные 229
— слуховая вторичная 234
---первичная 234
— соматосенсорная, функ-
ция 230
Корти кол ибер ин 75, 272
Кортикостероиды 283
Кортикотропин 272
Котранспорт 51, 622
Коэффициент вентиляци-
он но- гравитацион ных
отношений в легких 487
— дыхательный 574
— изнашивания по Рубне-
ру 565
— осмотический 47
— физической активности
577
904 • Предметный указатель
— фосфорилирования 573
Креативность языка 824
Кретинизм 294
Кровезаменители 362
Кровообращение 491
Кровообращение, круг
большой 387
---малый 387
— объем минутный 394
---резервный 395
--- систолический 395
---ударный 395
— при работе 869
— регионарное 432
Кровоток, ауторегуляция
433
Кровь, вязкость 38, 435
— группы 359
— давление гидростатиче-
ское 30
---фильтрационное 30
— изменения при работе
870
— иммуноглобулины 36
— переливание 361
— резус-отрицательная 359
резус-положительная 359
— система свертывания
341
------путь внешний 341
---------внутренний 343
— свойства суспензион-
ные 38
— способность общая же-
лезосвязывающая 328
— текучесть 38
— факторы 343
— функция гомеостатиче-
ская 322
---регуляторная 322
---секреторная 322
---транспортная 322
— циркулирующая, объем
28
------гиперволемия 28
------нормоволемия 28
Ксенотрансплантаты I, 340
Культура человека общая
782
Лабильность 58,64
---стадия парадоксаль-
ная 58
---тормозящая 58
---уравнительная 58
Лактат-ацидоз 664
Лактация 699
Лактоген плацентарный
человеческий 696
Лактогенез 699
Лактоферрин 332
Легкие, кровоток 445
Лейкопения 331
Лейкоцитоз 331
Лейкотриены 318, 332
— миогенный 870
Лейкоциты 331
— эозинофильные, функ-
ции 335
Либерины 149
Лиганд 260
Лимфа, белки 39
— образование 40
— периферическая 39
— центральная 39
— формула, лейкоцитар-
ная 40
Лимфангион 465
Лимфообращение 463
Лимфопоэз 364
Лимфоток 466
Лимфоциты В 368
---синтез антител 369
---функции 369
-Т365
---цитотоксические 365
---функции 368
Липиды 565
— клеточные 566
— плазмы крови 566
Липокортины 286
Липопротеины 567
— плотности высокой 32
---низкой 32
---очень низкой 32
Лихорадка 603
Локализация речи 829
Локомоция 192
Лютеолиз 686
Лютропин 672, 675, 685
Маммогенез 699
Мастурбация 780
Медиатор(ы) 137
— возбуждающий 79
— клеток гетерогенных 156
— тормозные 79
Мелатонин 298
Меланотропин 282
Мембрана постсинаптиче-
ская 138
— пресинаптическая 138
Менструация, 687
Метаболизм 563
Метаболиты 156
Механизмы компенсатор-
ные 879
— мочеобразования 610,
614
Механорецепторы 703
Механорецепторы кожи
711
---плотность 713
Микроглия 171
Микродинамика см. Мик-
роциркуляция
Микродомен 71
Микросон 852
Микроциркуляция 453
Миндалины, выбор конку-
рирующих мотиваций
---положительных эмо-
ций 219
— функции 218
---сигналы опасности
218
— ядра базолатеральные
218
— ядро центральное 218
Минералокортикоиды 283
Миокард, кровоток 444
— проводимость 398
— сокращение 397, 403
Миофибриллы 88
Миофиламенты 89
Миоциты пищеваритель-
ного тракта 518
Модель речевой деятель-
ности человека Верни-
ке—Гешвинда 827
--------современная 828
Модуляторы 579
Мозг головной, кора,
функция 83
— — кровоток 442
— костный 370
---островки эритробла-
стические 330
---строма 354
— мост 194
— продолговатый, ядро со-
литарного тракта 205
— промежуточный, гипо-
таламус 211
---таламус 211
— расщепленный 244
— система лимбическая
216
— спинной, мотонейроны
альфа 183
------гамма 183
--- нейроны зоны про-
межуточной 183
---путь(и) вестибулос-
пинальный 194
------кортикоспиналь-
ный 194
------латеральный 183
------медиальный 183
—-----проводящие восхо-
дящие 194
--------нисходящие 194
--------собственные 194
Предметный указатель • 905
------рекутилоспиналь-
ный 194
------руброспинальный
194
------тектоспинальный
194
---рога дорсальные 181
--- ствол 194
---тракт спиноретику-
лярный 194
------спиноталамиче-
ский 194
---участие в локомоции
192
---функции 181
---ядра(о) 181
------медиальное 185
------моторные 183
— средний 194
---ростральный 207
---покрышка 201
— ствол, путь латераль-
ный 202
------медиальный 202
---центры глазодвига-
тельные 204
---ядра вегетативные
195
------двигательные 195
------ — чувствительные
195
— структуры лимбические
176
Мозжечок 259
— функции 207
---объем движений 207
---равновесие тела 207
---сила мышц 207
Молекулы, транспорт
внутриклеточный 52
Моноциты-макрофаги 336
Мотивациия(и) 147, 206,
773
— биологические 147
— влечения 774
— вторичные 773
— избегания 774
— ориентировочных по-
требностей 147
— первичные 773
— пищевая человека 774
— познавательные 147
— половая человека 780
------нервная регуля-
ция 787
— смысла жизни 147
— творческие 147
Мотонейроны 876
— геномные 186
— дыхательные спиналь-
ные 502
Моча первичная 614
Мускулатура дыхательная
471
Мышление 756, 824
— абстрактное 833
----формы 833
— — функции полушарий
мозга 834
— основы нейрофизиоло-
гические 832
------при чтении 834
Мышцы(а) гладкие 88, 103
----активность тониче-
ская 105
------электрическая 106
----мультиунитарная 105
— — расслабление, меха-
низм молекулярный 111
----синапс нервно-мы-
шечный 108
----сокращение, меха-
низм молекулярный 109
------параметры физио-
логические 111
----унитарная 104
— сердечная 88
----активность электри-
ческая 114
----функции 113
— скелетные 88, 452
----белые 88
----двигательная едини-
ца 88
----длина исходная 101
----красные 88
----неутомляемость от-
носительная 102
----расслабление 96
----регуляция сосудов гу-
моральная 453
---------миогенная 452
----нервная 453
----сила абсолютная 100
----сокращение 91
------изотоническое 97
------концентрическая
98
----укорочение, ско-
рость 100
----утомление 101
------периферическое
102
------центральное 102
— сокращение непроиз-
вольные 88
----произвольные 88
Навыки двигательные 865
----этапы формирования
866
Нагрузка физическая 577
Надпочечник, кровоснаб-
жение 283
Напряжение углекислоты
парциальное 660
Насос натрий-калиевый 48
— электрогенный 60
Научение 757, 817
— ассоциативное 760, 822
— половому поведению
782
Нейрогипофиз 270
Нейроглия 160
Нейрогормоны 149
Нейромодуляторы 77
Нейрон(ы) 160
— ассоциативные 140
— афферентные 140
— возбуждающие 160
— глазодоминантные 232
— группы дыхательные 498
— классы нейрохимиче-
ские 174
— коры мозжечка 209
---коры моторной 173
— окончание пресинапти-
ческое, функция 75
— парасимпатические пре-
ганглионарные 185
— пептидергические 179
— полимодальные 197
— простые 731, 742
— Пуркинье 209
— система глутаматергиче-
ская 174
---ГАМКергическая 178
---мотивационные 171
---моторные 171
---сенсорные 171
---холинергическая 176
— стриарной коры 810
— тормозные 160
— холинергические 176
---мозга головного 176
— черной субстанции 206
------путь мезокорти-
кальный 206
--------мезолимбиче-
ский 206
--------нигростриат-
ный 206
— эфферентные 140
Нейропептиды 75
— гипоталамические 149
Нейросекреция 149
Нейротрансмиттеры 74
Нейтрофилы, эффект бак-
терицидный 333
Нексус 68, 114
Неприязнь вкусовая 777
Нервы обонятельные 753
— черепные 195
Нефрон 610
906 • Предметный указатель
— типы 613
Норадреналин 75, 83
---эффект инотропный
121
Норма адативной реакции
888
Ноцицепторы 704
Обмен веществ транска-
пиллярный 455
--- и энергии 563, 592
— водно-солевой 634
Обмен водный 21
— жидкости транскапил-
лярный 455
— основной 575
---формула расчетная
547
— энергии в клетке 573
Обморок тепловой 603, 898
Обоняние 750
— роль физиологическая у
человека 754
Объем(ы) вдоха резерв-
ный 475
— воздуха легочные 474
— выдоха резервный 475
— грудной полости 470
— дыхания минутный 483
— дыхательный 475
---объем циркулирую-
щей крови 426
Окисление питательных
веществ биологическое
574
Оксигемоглобин 325
Оксид азота 314
Окситоцин 75, 180, 281
Олигодендроциты 171
Оогенез 668, 683
Опиаты эндогенные 282
---динорфины А, В 282
---лейцин-энкефалин
282
---метионин-энкефа-
лин 282
---неоэндорфины а, р
282
---эндорфины 282
Оплодотворение 690
Оргазм 682, 689, 786
Орган(ы) кортиев 738
---процессы биоэлек-
трические в нем 739
— половые наружные 671
Организм, среда внутрен-
няя 19
Осведомленность зритель-
ная 817
Осмолярность 47
Основные крови буфер-
ное 660
Основания 650
Островки Лангерганса 299
Ответ иммунный 376
---активация Т- и В-
лимфоцитов 379
---гуморальный 381
---клеточный 380
Ототопика 736
Ощущения проприоцеп-
тивные 713
Память 817, 824
— имплицитная 818
---механизмы нейрон-
ные 820
— кратковременная 819,
820
— формы 818
— эксплицитная 818
---механизм образова-
ния 823
— энграмм 818, 823
Парабиоз 58
Паратирин 131, 296
Пейсмекеры 105
Пепсины 536, 537
Пептид(ы) желудочный
ингибиторный 300
— мозговой 295
— нейроактивные 75
— нейрогипофиза 181
— опиатные 180, 716
Перехват Ранвье 67
Периэлекгротон 56
Перфузия легких кровью
485
Петля Генле 625, 627
Печень, кровоток 448
Пиноцитоз 556
Питание 584, 659
— рациональное 589
Пищеварение 511, 659
— в желудке 533
---тонком кишечнике
548
— типы 513, 514
Плазма крови 28, 322
---белки 33
---давление онктиче-
ское 29
------осмотические 29
---«плавающие» рецеп-
торы 37
---состав электролит-
ный 28, 32
---стабильность колло-
идная 37
---удельный вес 39
---функция защитная 28
------ транспортная 28
Плазмолемма 45
Плазмоток почечный эф-
фективный 631
Плацента гемохориальная
692
— функции 693
Пневмоторакс 474
Пол генетический 670
— гонадный 670
— фенотипический 671
Поле(я) зрительное 721
— рецептивные 706
Полипептид панкреатиче-
ский 304
Половое возбуждение, ста-
дии 785
Полосатое тело 221
Полость ротовая 528
Полушария 225
— большие 225
---кора ассоциативная
225
------зрительная 225
------моторная 225
------сенсорная 225
------соматосенсорная
225
— левое 246
— мозолистое тело 244
— правое 246
— речевая деятельность
244
Порог возбуждения 53
— выведения почечный 62
— генерации потенциала
действия 62
— дыхательный 506
— лактатный 507
— ощущения абсолютный
710
Потенциал(ы) вызванные
227
— действия 59, 705
---зона интегративная
163
------триггерная 166
---кардиомиоцита 114
— концевой пластинки 73
---мембранный 107
— Нернста 59
— пейсмекерные 117
— покоя возбудимых кле-
ток 60
---мембранный 164
---постсинаптические
возбуждающие 79, 163
------локальные 163
------тормозные 163
— пороговый 62
— рецепторный 163, 705
— следовые 63
Предметный указатель • 907
Потенциация долговре-
менная 175
Почка 451
— регуляция кровотока гу-
моральная 451
------миогенная 452
------нервная 452
Пресбиопия 719
Пресинаптическое тормо-
жение 169
— усиление 170
Привыкание 757
Принцип обратной аффе-
рентации 146
— общего конечного пути
Шеррингтана 141
— реципрокности 144
— субординации нервных
центров 145
— торможения сопряжен-
ного 144
Проводники нервные аф-
ферентные 136
----эфферентные 136
Прогестерон 305, 696
Прозопагнозия 234
Пролактин 278, 677, 697,
699
Проприорецепторы 504
— шейных мышц 200
Простагландин(ы) 157, 317
- Е2 451
- F2a 451, 687
- Н2 451
Простациклин 12 345
Пространство внеклеточ-
ное, жидкости 19
— мертвое анатомическое
483
----физиологическое 485
— плевральное 472
Протеины С и S 345
Протромбиназа кровяная
343
Процессы бодрствования
207
Процессы познавательные
базовые 800
— сна 207
------Пуазейля формула
433
Пурины 75
Путь(и) лемнисковый 230,
713
— проводящие параллель-
ные 173
— спинномозжечковый
183
Пучок Гиса 398
Пятно голубое 177
Работа дыхательных мышц
481
— запрос кислородный 863
— комбинированная 857
— мышечная, мощность
101
— умственная 856
---изменение физиоло-
гических функций при
ней 872
— физическая 856
---формирование гор-
мональных реакций 871
Работоспособность 868
— человека 856
Раздражитель адекватный
52, 136
— неадекватный 52, 136
Раствор, тоничность 47
Растяжимость легочной
ткани 476
Рацион питания 590
Рвота 562
Реабсорбция аминокислот
и глюкозы 622
— ионов натрия 620, 624,
629
— канальцевая 619
---воды 629
---регуляция 628
— пептидов и белков 623
— проксимальная 619
Реактивность 132
Реакция(и) жидкостей ак-
туальная 650
— организма адаптивные
неспецифические 881
------симпатоадренало-
вая 882
Реверберация возбужде-
ния 142
Регенерация 317
Регулирование аутокрин-
ное 155
— паракринное 155
— эндокринное 155
Регуляция адаптивная 129
— гомеостатическая 130
— гуморальная 132
---местная клеточно-
тканевая 133
Регуляция кровообраще-
ния, влияния гипотала-
мические 460
------кортикальные 462
---рефлекторная 457
---центральная 456
------схема общая 462
---уровень спинальный
457
------бульбарный 458
— нейрогуморальная реф-
лекторная 158
— нервная 132
— обмена водно-солевого
578, 605, 638
---организма с окру-
жающей средой 578
— по возмущению 129
---отклонению 129
---упреждению 131
Режим питания 588
Резистентность осмотиче-
ская максимальная 31
---минимальная 31
— организма 880
Рекомендации ВОЗ по
диете 590
Релаксин 697
Ренин 310
Реншоу: торможение 142
Реобаза 54
Репарация 317
Реполяризация 63
Репрезентации слов 829
Ретинопия 721
Рефлекс(ы) аллиирован-
ные 144
— антагонистические 144
— безусловный 758
— Бейнбриджа 418
— вегетативные 257
---надсегментарные 257
---сегментарные 257
— вестибулярные 200
— висцеро-висцеральный
193, 257
— висцероДермальный 193
— висцеросенсорный 258
— висцеросоматический
258
— выпрямления 200
— Геда парадоксальный
504
— Геда—Брейера 504
— Генри—Гауэра 419
— Геринга—Брейера 504
— глотательный 198
— Гольца 419
— Данини—Ашнера 420
— дермовисцеральные 193
— жевательные 198
— зрачковый 200, 720, 730
— кардиальные 418
— кашлевой 199
— коленный 186
— координация 144
— лабиринтные 200
— лифтные 202
— мигательный 200
— моносинаптический 141
— ориентировочные 200
— осморегулирующие 640
908 • Предметный указатель
— позные 200
— полисинаптические 141
— положения 200
— постуральные 202
— поля рецептивные 136
— разгибательный 190
— растяжения мышцы 186
— рвотный 198
— ритмические 192
— сгибательный 190
— слезный 199
— слюноотделительный
197
— соматовисцеральный
258
— спинальные вегетатив-
ные 193
— спинномозговые 185
— стартовые 200
— статические 200
— статокинетические 200
— суммационный 761
— сухожильные 185
— тонический 187
— условный 756, 758
--высшего порядка 762
--классический 762,
822
------виды 767
-- оперантный 767
--стадии формирова-
ния 762
--торможение 763
------внешнее 764
------дифференцировоч-
ное 766
------запаздывающее 765
— — — угасательное 764
------условное 766
— установочные 200
— физический 186
— цепные 144
— чихательный 199
— шейные тонические 200
— энтерогастральный 543
Рефрактерность абсолют-
ная 64
— относительная 64
Рефракция 718
— аномалии 719
Рецепторы 702
— адаптация 705
— аппарата вестибулярно-
го 200
— болевые 715
— возбуждение, порог раз-
дражения абсолютный
136
— глютаматные 78
— Гольджи 189
— Гольджи сухожильные
504
— дигидропиридиновый 94
— иммуноглобулиновые
155
— ионотропные 78, 86
— кадгериновые 155
— классификация 703
— клеточные 127
--химические 134
— метаботропные 80, 177
— мускариновые М,—М5
122
— отолитовых органов 745
— полилюдальные 715
— растяжения легких бы-
строадаптирующиеся
504
-------медленно адапти-
рующиеся 504
— рианодиновый 94
— сенсорные 133
— — интероцепторы 135
— серотонинергические D,
М, Т 255
— слуховые 733
— сухожильные Гольджи
713
— температурные 714
--активность тониче-
ская 714
--экстероцептеры 135
Рецепторы сенсорные
---чувствующие вторич-
но 135
— первично 135
------- механорецепто-
ры 135
------- ноцицепторы
135
-------термоцепторы
135
- V1; V2 279
J-рецепторы легочные 504
Рецепция вкусовая 747
Ригидность децеребраци-
онная 203
— мышечная 224
Ритм дыхательный 500
Роды 697
Русло микроциркулятор-
ное 453
— сосудистое спланхниче-
ское 424
Саккады 204
Саркомер 89Секретосома
71, 76
Секреция канальцевая 630
Селезенка, функции 374
Селектины 155
Семенники 672
Сенситизация 78, 757, 812,
820
Сердечный выброс 412
Сердце, возврат венозной
крови 421
— иннервация 414
— работа, цикл 405
---экстрасистолы 407
— сокращение, эффектт
батмотропный 414
------дромотропный 414
------инотропный 414
------ хронотропный 414
— функция гормональная
421
— Серотонин 75, 85, 178
Сети нейронные дивер-
гентные 174
--- иерархические 173
---локальные 173
---торможение возврат-
ное 142
------латеральное 142
------нисходящее 143
------реципрокное 142
------тормозная зона 143
— нервные дивергентные
141
--- иерархические 141
---локальные 141
---принципы суборди-
нации 142
Сила мышц максимальная
865
Симпорт 51
Симптом Мюсси—Георги-
евского 258
Синапс(ы) 68, 137
— адренергический 83
— аксо-аксональные 137,
162
— аксодендритные 137
— аксосоматические 137
---реакции быстрые 74
------медленные 75
— ГАМКергический 81, 87
— глицинергический 81,
87
— глутаматергический 86
— дендро-дендритные 137
— дендросоматические 137
— дофаминергический 84
— нервно-мышечный 69,
139
---механизм постсинап-
тический 72
------ пресинаптический
70
— периферические 139
— серотонинергический 85
— тормозные 79
Предметный указатель • 909
----каналы хемозависи-
мые 165
— химический 74
— холинергический 82
Синапсин 51
Синдактилия 232
— адаптационный общий,
стадии 885
— Иценко—Кушинга 289
— Клювера—Бьюси 219
Синусоидальное дерево
355
Синцитий функциональ-
ный 114
Система(ы) аденилатцик-
лаза —цАМФ 262, 330
— антиноцицептивная 178
— буферная белковая 652
----бикарбонатная
----гемоглобиновая 653
----фосфатная 652
— гипоталамо-аденогипо-
физарная 149
— гипоталамо-нейрогипо-
физарная 149
— гипоталамо-экстрагипо-
таламная 149
— глаза оптическая 718
— гомеостатические 651
— гормональная, звено
выделения гормонов 151
------ депонирования 150
------метаболизма 150
------синтеза и секре-
ции 150
------транспорта 150
------эффектора 151
----регуляция, путь гипо-
физарный 148
--------парагипофизар-
ный 148
— гуанилатциклаза —
цГМФ 264
— дофаминергичская ме-
золплебическая 176
----нигростриатная 176
— дыхательная 469
----функции 470
— зрения дневного 724
----ночного 723
— иммунная 363
----контроль гормональ-
ный 383
------цитокиновый 385
----органы 370
— — пищеварительного
тракта 561
— кальций—кальмодулин
265
— мозга дофаминергиче-
ская 679
----серотонинергическая
679
— моторные мышцы-анта-
гонисты 240
----мышцы-синергисты
240
— нейрогуморально-им-
мунная 158
— нервная вегетативная
248
------функция корриги-
рующая 249
---------пусковая 249
---------трофическая 249
---------трофотропная
250
---------эрготропная 250
------центр бульбарный
248
--------мезэнцефаличе-
ский 248
---------сакральный 248
---------тораколюмбаль-
ный 248
----энтеральная 256
------ганглии 256
— организма выделитель-
ная 606
— питания функциональ-
ная 511
— пищеварительная, дея-
тельность периодиче-
ская 526
----механизмы регуля-
ции функций 523, 525
----строение 511
----функция всасывания
519
---------макро- и мик-
ромолекул 521
------моторная 517
------секреторная 512
— пищевого насыщения
775
— потребления пищи 775
— пропердиновая 35
— почки канальцевая про-
тивоточно- множител ь-
ная 625
— противоточная сосуди-
стая 627
Система регуляции, меха-
низмы 129
----обмена эфферентная
579
----типы 129
Система регуляции, уров-
ни 129
— ренин-ангиотензин-аль-
достероновая 283, 310
— сенсорная 173, 702, 756
----вестибулярная 743
----вкусовая 747
— — зрительная 717, 733
---- обонятельная 750
------отдел перифериче-
ский 751
---------центральный
753
----переработка инфор-
мации 707
----пути проводящие 173
----слуховая 733
------диапазон частот-
ного восприятия 735
----физиология 703
— сердечно-сосудистая 387
----отделы, классифика-
ция 388
— соматовисцеральная 711
— стриопаллидарная 221
— фосфолипаза С — ино-
зитол-3-фосфат 264
— фузимоторная 185
— функциональная 159
— экстралемнисковая
— эндокринная 148
Системогенез 159
Скорость клубочковой
фильтрации 616
------регуляция 618
— кровотока линейная 455
Слизь желудочная 537
Слюноотделение 528
Сновидения 850
Сознание 813, 824, 854
— корреляты нейрофизио-
логические 814
Сок желудочный, регуля-
ция секреции 538
---- состав 534
----фазы секреции 539
-кишечный 548
---- состав 549
— панкреатический, регу-
ляция секреции 545
----состав и свойства 544
— толстой кишки, состав
557
Сокращение мышечное,
активация 94
----одиночное 96
----сопряжение электро-
механическое 94
----тетаническое 96
----типы 96
Соматолиберин 75, 275
Соматомаммотропин хо-
рионический человече-
ский 696
Соматомедины 276
Соматостатины 181, 274
Соматотропин 275
Сон 836
910 • Предметный указатель
— быстроволновый 837,
841
— индукторы гумораль-
ные 849
— медленноволновый 837,
842, 846
---стадии 840
— механизмы нейрофи-
зиологические 845
— парадоксальный 843,
846
— последствия его лише-
ния 852
— продолжительность 851
— регуляторы 850
— регуляция циркадного
ритма 847
— стадии 839
— структура 841
— теория восстановитель-
ная 836
---информационная 843
---циркадианная 837
Сопротивление дыхатель-
ных путей 479
Сопряжение электромеха-
ническое, мышца глад-
кая 119
------сердечная 120
------ скелетная 119
Состояние кислотно-ос-
новное 650
---изменение при рабо-
те 870
---и желудочно-кишеч-
ный тракт 659
------ легкие 655
------печень 659
------почки 656
------компенсация нару-
шений 667
Сосуды лимфатические
465
---проницаемость 467
— сопротивление общее
периферическое 393
— шунтирующие 456
Сперматогенез 668, 672,
675
Сперматозоиды 673, 690
Спиноцеребеллум 207
Спланхомегалия 277
Способность легких диф-
фузионная 491
Среда организма внутрен-
няя, вода 21
Средства коммуникации у
людей 824
Статины 149
Ствол мозга 194
Стероиды половые 283
Стоимость дыхания энер-
гетическая 507
Стресс-реакция 883
Стресс-релаксация 112
---механизм «замка» 112
— тепловой 891
— участие эндокринной
системы 315
Субстанция Р 75
Судороги тепловые 898
Сурфактант 477, 478
Сфингомиелин 67
Таламус, интернейроны
возбуждающие локаль-
ные 211
---тормозные 211
— кора моторная вторич-
ная 212
------первичная 212
— нейроны релейные 211
— путь передачи магно-
целлюлярный 212
------парвоцеллюляр-
ный 212
— ядро(а) ассоциативные
211
---вентролатеральное
212
---моторные 212
---неспецифические
211, 213
------специфические 211
Танициты 171
Тахикинины 180
Тахистоскопия 244
Тело желтое 306, 683, 686
— полосатое 224
Тельца Мейсснера 712
— Пачини 712
— Руффини 712
— тактильные Пинкуса—
Игго 712
Тембр голоса 826
Температура комфорта
593— тела 592
---колебания суточные
594
---нормальная 593
Теория диффузионная 488
Теплоотдача 594
— пути 596
Теплопроведение 596
Теплопродукция 594, 595
Теплота вторичная 574
— первичная 573
Терминаль 137
Термогенез несократитель-
ный 596
— сократительный 595
Терморегуляция 605
— поведенческая 595, 598
— эндогенная 594
Терморецепторы 598, 703
Тестостерон 675
Тетанус гладкий 96
— зубчатый 96
Тетрайодтиронин 292
Тимоциты 373
Тимус 372
Тиреолиберин 75, 274
Тиреотропин 274, 292
Тиронины 292
Тироциты 292
Титин 91
Ткань(и) возбудимые 45
свойства 52
— лимфоидная мукозно-
ассоциированная 376
Ткани-мишени 151
— мышечная 88
— раздражимость 45
Ток постоянный, дейст-
вие полюсное 55
Тонотопия 212
Точка массы тела устано-
вочная 775
Тошнота 561
Тракт(ы) вестибулоспи-
нальный 202
— желудочно-кишечный,
кровоток 446
— зрительные 730
— ретикулоспинальный
202
— руброспинальный 203
— спиноретикулярный 182
— спиноталамический 182
— тектоспинальный 202
Транскобаламины 34
Транскортин 286
Транспорт веществ через
мембрану 46
--------движение воды
47
--------диффузия 47
--------осмос 47
— вторично-активный 50
— кровью кислорода 492
----углекислого газа 495
— парацеллюлярный 22
— первично-активный 48
— трансцеллюлярный 22
----активный 23
----пассивный 22
Трансферрин 34
Трансфузиология 358
Трансцитоз 521
Трийодтиронин 292
Тромбин 344
Тромбомодулин 346
Тромбопоэтин 339
Предметный указатель • 911
Тромбоцитоз миогенный
870
Тромбоцитопоэз 339
Тромбоциты, функции 337
Тропомиозин 89
Тропонин 90
Углеводы 567
Удар тепловой, формы 899
Узел синоатриальный,
клетки, активность
пейсмекерная 117
Узлы лимфатические 375,
465
Ультрафильтрация клубоч-
ковая 614
Умения двигательные 865
Уравнение Гендерсона—
Гассельбаха 652
— Нернста 59
— Хилла 100
Утомление зрительное 873
— центральное 874
— человека при работе
мышечной динамиче-
ской 877
---------физической ста-
тической 875
Фактор(ы) Виллебранда
348
— колониестимулирую-
щий гранулоцитарный
337, 352
--моноцитарный 337
— опухольнекротизирую-
щий 336, 354
— роста инсулиноподоб-
ные 276
— тканевый 348
— туморнекротизирую-
щий 380
Феномен Орбели—Гине-
цинского 250
— Остроумова—Бейлиса
616
— эффекта правого уха 246
Ферменты желудочного
сока 536
— пищеварительных же-
лез 513
— протеолитические 544
Феромоны 219, 754, 783
— роль в половой мотива-
ции у человека 782
Фертилизация 669
Фибринолиз 348
Фибронектин 34
Физиология труда 856
Филамент(ы) актиновые 90
— толстый 91
Фонокардиограмма 410
Формация ретикулярная
201
----входы афферентные
сенсорные 205
----стресс-реакция 206
----центр(ы) вегетатив-
ные 205
------дыхательный 205
------пневмотаксиче-
ский 206
Фосфатаза миозиновая 111
Фосфатурия 296
Фосфоламбан 121
Фосфолипаза Д 335
Фосфолипиды 45
Франка уравнение 393
Френикус-симптом 258
Функции(я) висцераль-
ные, регуляция рефлек-
торная 146
— выделительная верхних
дыхательных путей 609
----желудка 608
----кишечника 608
----легких 608
---- печени 608
— двигательная 205
— печени непищевари-
тельные 548
----пищеварительные 546
— поджелудочной железы
544
— почек 609
----метаболическая 632
----экскреторная 632
— репродуктивная жен-
ская 682
— сосудов емкостная 453
— уха внутреннего 738
----наружного 736
----среднего 736
— физиологические, регу-
ляция произвольная 146
Хеморефлекс централь-
ный 502
Хеморецепторы перифери-
ческие (артериальные)
503, 508
— центральные (медулляр-
ные) 502
Хемотаксис 690
Хиломикроны 567
Холекальциферол 307
Холин 82
Холинорецепторы М 254
Хромосомы 668
Хронаксия 54
Ценность энергетическая
585
Центр(ы) апнейстический
501
— голода 511
— дефекации 560
— дыхательный 498
— двигательные 201
---командные 240
— жевания 531
— насыщения 512
— нервные 140
---свойства 140
— пневмотаксический 501
— речи 245
— слюноотделения 530,
531
— терморегуляции 599
Цепочки нейронные Ло-
ренто де Но 142
Цереброцеребеллум 207
Цикл менструальный 687
---фаза менструальная
688
------пролиферативная
688
------ секреторная 688
— овариальный 682, 683
---фаза лютеальная 686
------овуляторная 685
------фолликулярная 683
Цикл сон—бодрствование
204
Частота дыхания 474
Часы биологические 836,
838
Черная субстанция 206,
221
Число Авогадро 29
Чувствительность вкусо-
вая 748
Шкала Стивенса 710
Шок спинальный 185
Шунтирование 485
Эквивалент кислорода ка-
лорический 574
Экзоцитоз 51, 77, 521
— медиаторы, регуляция
пресинаптическая 77
Экстерорецепторы 704
Эктоэнзимы 348
Электрокардиограмма 408
Электротон физиологиче-
ский 55
— Эмбдена—Мейергофа
цикл 322
912 • Предметный указатель
Эмболия газовая 509
— жировая 567
Эмбрион 692
Эмоции 206, 789, 824
— механизмы проявления
791
— отрицательные, регуля-
ция 798
— положительные, регуля-
ция 796
— роль в поведении чело-
века 790
Энграммы долговремен-
ной памяти 176
Эндотелины 313
Эндоцитоз 51, 521
Энкефалины 75
Эозинофилия 336
Эпифиз 297
Эпоксид 318
Эритропоэз 329, 456
— неэффективный 356
Эритропоэтин 330
Эритроцитоз 330
— миогенный 870
Эритроциты 494
— роль в транспорте СО2
495
— свойства 322
— форма 323
Эстрадиол 305
Эстриол 305
Эстрогены 677, 686, 688,
696
Эстрон 305
Эффект Анрепа 413
— Бора 494, 693
— Остроумова—Бейлисса
438
— равновесия Даннона 30
— ускользания сердца 415
— Фареуса—Линдквиста 38
— Холдена 496
Эхиноциты 326
Эякуляция 681
Ядра(о) вестибулярные 201
— красное 201
— мозжечка 210
---зубчатые 210
---промежуточные 210
— переключательные 172
— субталамическое 321
— хвостатое 221
— шатра 211
— шва 207
Язык жестов 830
— свойства 825
Оглавление
К читателю (предисловие к 1-му изданию)..................................... 5
Предисловие ко 2-му изданию................................................. 8
Список сокращений............................................................ 10
Введение. Физиология как предмет и характеризующие его понятия............. 12
I БАЗИСНЫЕ ОСНОВЫ ФИЗИОЛОГИИ ЧЕЛОВЕКА
Глава 1 Жидкие среды организма............................................. 19
1.1. Внутренняя среда организма.................................... 19
1.2. Биологические свойства жидкостей, составляющих внутреннюю сре-
ду организма....................................................... 21
1.2.1. Вода как составная часть жидкостей организма .......... 21
1.2.2. Гистогематические барьеры ............................. 22
1.2.3. Внутриклеточная жидкость............................... 24
1.2.4. Интерстициальная, или тканевая, жидкость............... 25
1.3. Плазма крови как внутренняя среда организма................... 28
1.3.1. Электролитный состав плазмы крови...................... 28
1.3.2. Осмотическое и онкотическое давление плазмы крови .... 29
1.3.3. Обмен воды между плазмой крови и интерстициальной жидко-
стью ........................................................ 30
1.3.4. Продукты белкового обмена, углеводы и липиды плазмы
крови........................................................ 32
1.3.5. Белки плазмы крови.................................... 33
1.4. Факторы, обеспечивающие жидкое состояние крови................ 37
1.5. Лимфа как внутренняя среда организма.......................... 39
1.6. Механизм образования лимфы ................................... 40
1.7. Трансцеллюлярные жидкости организма........................... 41
1.8. Обмен жидкостей между водными секторами в организме человека . 42
Глава 2 Физиология возбудимых тканей..................................... 45
2.1. Строение и физиологические функции мембраны клеток возбудимых
тканей............................................................. 45
2.1.1. Транспорт веществ через клеточную мембрану............ 46
2.1.1.1. Движение воды через мембрану клеток........... 47
2.1.1.2. Осмос ........................................ 47
2.1.1.3. Диффузия...................................... 47
2.1.1.4. Первично-активный транспорт................... 48
2.1.1.5. Вторично-активный транспорт .................. 50
2.1.1.6. Эндоцитоз и экзоцитоз......................... 51
2.1.1.7. Внутриклеточный транспорт молекул............. 52
2.2. Возбудимость как основное свойство нервной и мышечной ткани . . 52
2.2.1. Понятие о раздражении и раздражителях................. 52
2.2.2. Зависимость возникновения возбуждения от длительности и
силы раздражения............................................. 53
2.2.3. Возбудимость и возбуждение при действии постоянного тока
на нервную и мышечную ткань.................................. 55
2.2.З.1. Физиологический электротон.................... 55
2.2.3.2. Закон полярности раздражения нервной и мышечной ткани . 56
2.2.3.3. Электродиагностический закон.................. 57
2.2.4. Понятие о функциональной подвижности возбудимых тканей 58
2.3. Электрические явления в возбудимых клетках.................... 58
2.3.1. Мембранный потенциал покоя ............................ 59
2.3.2. Потенциал действия возбудимых клеток................... 61
2.2.3. Рефрактерный период в возбудимых клетках............... 64
2.3.4. Локальный ответ мембраны возбудимых клеток............. 65
914 • Оглавление
2.4. Проведение импульса по нервным волокнам........................ 66
2.4.1. Немиелинизированные волокна............................. 66
2.4.2. Миелинизированные волокна............................... 67
2.4.3. Законы проведения возбуждения по нервному волокну .... 68
2.5. Проведение возбуждения через синапс............................. 68
2.5.1. Проведение возбуждения через нервно-мышечный синапс . . 69
2.5.1.1. Пресинаптический механизм....................... 70
2.5.1.2. Диффузия ацетилхолина через синаптическую щель нерв-
но-мышечного синапса..................................... 72
2.5.1.3. Постсииаптический механизм...................... 72
2.5.1.4. Восстановительные процессы структуры мембраны и функции
нервно-мышечного синапса после передачи возбуждения . . 73
2.5.2. Проведение возбуждения через аксосоматический синапс ... 73
2.5.2.1. Функция пресинаптического окончания нейронов.... 75
2.5.2.2. Пресинаптический механизм проведения возбуждения .... 76
2.5.2.3. Пресинаптическая регуляция экзоцитоза медиаторов .... 77
2.5.2.4. Постсинаптический механизм проведения возбуждения ... 78
2.5.2.5. Функции метаботропных рецепторов постсинаптической мем-
браны аксосоматического синапса.......................... 80
2.5.3. Проведение возбуждения в основных типах синапсов цен-
тральной нервной системы....................................... 81
2.5.3.1. Холинергический синапс (рис. 2.22).............. 82
2.5.3.2. Адренергический синапс.......................... 83
2.5.3.3. Дофаминергический синапс........................ 84
2.5.3.4. Серотонинергический синапс...................... 85
2.5.3.5. Глутаматергический синапс....................... 86
2.5.3.6. ГАМКергический синапс........................... 87
2.5.3.7. Глицинергический синапс......................... 87
2.6. Функции мышечной ткани.......................................... 88
2.6.1. Скелетная мышца......................................... 88
2.6.1.1. Функции миофиламентов........................... 90
2.6.1.2. Механизм сокращения скелетной мышцы............. 91
2.6.1.3. Активация мышечного сокращения.................. 94
2.6.1.4. Расслабление скелетной мышцы.................... 96
2.6.1.5. Типы мышечных сокращений........................ 96
2.6.1.6. Типы скелетных мышечных волокон ................ 98
2.6.1.7. Физиологические показатели сокращения скелетной мышцы 100
2.6.2. Утомление скелетной мышцы...............................101
2.7. Гладкая мышца...................................................103
2.7.1. Типы гладких мышц.......................................104
2.7.2. Электрическая активность клеток гладкой мышцы...........106
2.7.3. Нервно-мышечный синапс гладкой мышцы....................108
2.7.4. Молекулярный механизм сокращения гладкой мышцы .... 109
2.7.5. Молекулярный механизм расслабления гладкой мышцы ... 111
2.7.6. Физиологические параметры сокращения гладкой мышцы . . 111
2.8. Функции мышечных клеток сердца...................................ИЗ
2.8.1. Электрическая активность клеток сердечной мышцы.........114
2.8.1.1. Потенциал покоя.................................114
2.8.1.2. Молекулярный механизм потенциала действия в типичных
сердечных мышечных клетках...............................115
2.8.1.3. Механизм возникновения пейсмекерной активности в клетках
синоатриального узла.....................................117
2.8.2. Молекулярный механизм сокращения кардиомиоцитов .... 119
2.8.3. Молекулярный механизм расслабления кардиомиоцитов ... 120
2.8.4. Медиаторный контроль сокращения кардиомиоцитов..........121
II РЕГУЛИРУЮЩИЕ И УПРАВЛЯЮЩИЕ СИСТЕМЫ
Глава 3 Общие принципы и механизмы регуляции физиологических функций .... 127
3.1. Общие принципы организации системы регуляции...........127
3.1.1. Уровни организации системы регуляции............127
3.1.2. Типы и механизмы регуляции......................129
3.1.3. Реактивность и эффект регуляции.................132
3.1.4. Механизмы регуляции жизнедеятельности...........132
Оглавление • 915
3.2. Рефлекторная регуляция функций организма........................133
3.2.1. Сенсорные рецепторы .....................................135
3.2.2. Афферентные и эфферентные нервные проводники............136
3.2.3. Возбуждение и торможение в рефлекторной дуге ............137
3.2.4. Механизмы связи между звеньями рефлекторной дуги .... 137
3.2.5. Нервные центры и их свойства.............................140
3.2.6. Взаимодействие различных рефлексов. Принципы координа-
ции рефлекторной деятельности...................................144
3.2.7. Рефлекторная регуляция висцеральных функций .............146
3.3. Произвольная (волевая) регуляция физиологических функций .... 146
3.4. Гормональная регуляция функций организма........................147
3.4.1. Общая характеристика звеньев гормональной системы регуля-
ции ............................................................148
3.4.2. Виды и пути действия гормонов............................151
3.5. Местная гуморальная регуляция функций клеток....................154
3.6. Системный принцип организации механизмов регуляции физиологи-
ческих функций........................................................157
Глава 4 Функции центральной нервной системы...................................160
4.1. Основы функционирования нейронов и глии.......................160
4.1.1. Общая характеристика нейронов ...........................160
4.1.2. Функциональная модель нейрона............................162
4.1.2.1. Входные сигналы.................-................163
4.1.2.2. Объединенный сигнал — потенциал действия.........166
4.1.2.3. Проводящийся сигнал..............................167
4.1.2.4. Выходной сигнал..................................168
4.1.3. Функциональная характеристика нейроглии..................170
4.1.3.1. Астроциты........................................170
4.1.3.2. Олигодендроциты..................................171
4.1.3.3. Эпендимная глия..................................171
4.1.3.4. Микроглия........................................171
4.2. Общие принципы функционального объединения нейронов............171
4.2.1. Общие принципы организации функциональных систем мозга 172
4.2.1.1. Существование нескольких уровней переработки информации 172
4.2.1.2. Топографическая упорядоченность проводящих путей .... 173
4.2.1.3. Наличие параллельных проводящих путей............173
4.2.2. Типы нейронных сетей.....................................173
4.2.3. Нейрохимические классы нейронов..........................174
4.2.3.1. Глутаматергическая система.......................174
4.2.3.2. Холинергическая система..........................176
4.2.3.3. Системы нейронов, использующих биогенные амины .... 176
4.2.3.4. ГАМКергическая система...........................178
4.2.3.5. Пептидергические нейроны.........................179
4.3. Функции спинного мозга .........................................181
4.3.1. Функциональная организация спинного мозга................181
4.3.2. Рефлексы спинного мозга..................................185
4.3.2.1. Сухожильные рефлексы.............................185
4.3.2.2. Рефлекс растяжения мышцы ........................186
4.3.2.3. Рефлекторная регуляция напряжения мышц...........189
4.3.2.4. Сгибательные и разгибательные рефлексы...........190
4.3.2.5. Ритмические рефлексы.............................192
4.3.2.6. Участие спинного мозга в локомоции...............192
4.3.2.7. Спинальные вегетативные рефлексы.................193
4.3.3. Функциональная организация проводящих путей спинного
мозга............................................................194
4.4. Функции ствола мозга............................................194
4.4.1. Функциональная организация ствола мозга..................195
4.4.1.1. Черепные нервы ..................................195
4.4.1.2. Функциональная специализация ядер ствола.........196
4.4.2. Рефлекторная функция ствола мозга........................197
4.4.2.1. Статические и статокинетические рефлексы.........200
4.4.2.2. Нисходящие двигательные пути ствола мозга........202
4.4.2.3. Глазодвигательные центры ствола..................204
4.5. Функции ретикулярной формации...................................204
4.5.1. Особенности нейронной организации ретикулярной формации 205
916 • Оглавление
4.5.2. Нисходящие и восходящие влияния ретикулярной формации 206
4.6. Функции мозжечка...............................................207
4.6.1. Функциональная организация мозжечка......................207
4.6.2. Взаимодействие нейронов коры и ядер мозжечка.............209
4.6.3. Эфферентные связи мозжечка с моторными структурами мозга 210
4.7. Функции промежуточного мозга ..................................211
4.7.1. Функции таламуса ........................................211
4.7.2. Функции гипоталамуса.....................................213
4.7.2.1. Роль гипоталамуса в регуляции вегетативных функций . ... 213
4.7.2.2. Роль гипоталамуса в регуляции эндокринных функций ... 215
4.8. Функции лимбической системы мозга..............................216
4.8.1. Функции миндалин.........................................218
4.8.2. Функции гиппокампа.......................................220
4.9. Функции базальных ганглиев (стриопаллидарная система)..........221
4.9.1. Взаимодействие базальных ганглиев с другими структурами
мозга...........................................................221
4.9.2. Модуляции нейронных переключений в базальных ганглиях . 224
4.10. Функции коры больших полушарий ...............................225
4.10.1. Функциональное распределение нейронов в коре............226
4.10.2. Модульная организация коры..............................226
4.10.3. Электрическая активность коры...........................227
4.10.4. Функции сенсорных областей коры.........................229
4.10.4.1. Функция соматосенсорной коры ..................230
4.10.4.2. Функция зрительной коры .......................232
4.10.4.3. Функция слуховой коры..........................234
4.10.5. Функции ассоциативных областей коры ....................235
4.10.5.1. Функции теменно-височно-затылочной коры....235
4.10.5.2. Функции префронтальной ассоциативной коры......237
4.10.5.3. Функции лимбической коры ......................237
4.10.6. Функции моторных областей коры..........................238
4.10.6.1. Функция первичной моторной коры................238
4.10.6.2. Функция вторичной моторной коры................239
4.11. Регуляция движений............................................239
4.11.1. Иерархическая организация моторных систем........240
4.11.2. Нисходящие пути моторной коры...........................242
4.11.3. Контроль выполняемых движений...........................244
4.12. Межполушарная функциональная асимметрия.......................244
4.12.1. Функциональные возможности изолированных полушарий . . 244
4.12.2. Выявление функций неразделенных полушарий...............245
4.12.3. Функциональная специализация полушарий мозга ...........246
Глава 5 Вегетативная нервная система.........................................248
5.1. Строение вегетативной нервной системы..........................248
5.2. Функции вегетативной нервной системы...........................249
5.3. Функции периферических отделов вегетативной нервной системы . . 251
5.3.1. Симпатический и парасимпатический отделы.................251
5.3.2. Энтеральная нервная система..............................256
5.4. Рефлексы вегетативной нервной системы .........................257
5.5. Высшие центры вегетативной регуляции...........................258
Глава 6 Эндокринная нервная система — регулятор функций и процессов в организме 260
6.1. Химическая природа и общие механизмы действия гормонов .... 260
6.1.1. Механизмы действия пептидных, белковых гормонов и катехо-
ламинов .......................................................260
6.1.1.1.Основные системы вторичных посредников..................262
6.1.1.2. Взаимосвязи вторичных посредников...............266
6.1.2. Механизм действия стероидных гормонов....................266
6.1.2.1. Геномный механизм действия......................266
6.1.2.2. Негеномный механизм действия....................266
6.1.3. Саморегуляция чувствительности эффектора к гормональному
сигналу..................................................268
6.2. Регуляторные функции гормонов гипофиза.........................270
6.2.1. Гормоны аденогипофиза и их эффекты в организме...........271
Оглавление • 917
6.2.1.1. Регуляция секреции и физиологические эффекты кортикотро-
пина ....................................................272
6.2.1.2. Регуляция секреции и физиологические эффекты гонадотро-
пинов ...................................................273
6.2.1.3. Регуляция секреции и физиологические эффекты тиреотропи-
на ......................................................274
6.2.1.4. Регуляция секреции и физиологические эффекты соматотро-
пина ....................................................275
6.2.1.5. Регуляция секреции и физиологические эффекты пролактина 278
6.2.2. Гормоны нейрогипофиза и их эффекты в организме..........279
6.2.2.1. Регуляция секреции и физиологические эффекты вазопресси-
на ......................................................279
6.2.2.2. Регуляция секреции и физиологические эффекты окситоцина 281
6.2.3. Гормоны промежуточной доли..............................282
6.2.4. Эндогенные опиаты.......................................282
6.3. Регуляторные функции гормонов надпочечников...................283
6.3.1. Гормоны коры надпочечников и их эффекты в организме . . 283
6.3.1.1. Регуляция секреции и физиологические эффекты минерало-
кортикоидов .............................................283
6.3.1.2. Регуляция секреции и физиологические эффекты глюкокорти-
коидов ..................................................286
6.3.1.3. Регуляция секреции и физиологические эффекты половых сте-
роидов коры надпочечников................................290
6.3.2. Гормоны мозгового вещества надпочечников и их эффекты в
организме.....................................................290
6.4. Регуляторные функции гормонов щитовидной железы...............291
6.4.1. Регуляция секреции и физиологические эффекты йодсодержа-
щих тиреоидных гормонов...............................292
6.4.2. Регуляция секреции и физиологические эффекты кальцитони-
на ...........................................................295
6.5. Регуляторные функции гормона околощитовидных желез ..........296
6.6. Регуляторные функции гормонов эпифиза........................297
6.7. Регуляторные функции гормонов эндокринных тканей в органах, об-
ладающих неэндокринными функциями...................................299
6.7.1. Регуляторные функции гормонов поджелудочной железы . . . 299
6.7.1.1. Физиологические эффекты инсулина...............301
6.7.1.2. Физиологические эффекты глюкагона..............304
6.7.2. Регуляторные функции гормонов половых желез.............304
6.7.2.1. Гормоны семенников и их эффекты в организме ...305
6.7.2.2. Гормоны яичников и их эффекты в организме......305
6.8. Регуляторные функции гормонов клеток, сочетающих выработку гор-
монов и неэндокринные функции ......................................306
6.8.1. Регуляторные функции гормонов плаценты .................306
6.8.2. Регуляторные функции гормонов тимуса....................307
6.8.3. Регуляторные функции гормонов почек.....................307
6.8.3.1. Синтез, секреция и физиологические эффекты кальцитриола 307
6.8.3.2. Образование ренина и основные функции ренин-ангиотен-
зин-альдостероновой системы..............................310
6.8.4. Регуляторные эффекты гормонов сердца....................312
6.8.5. Регуляторная функция гормонов сосудистого эндотелия ... 313
6.8.6. Регуляторная функция гормонов желудочно-кишечного тракта
314
6.9. Роль эндокринной системы в неспецифических приспособительных
реакциях............................................................315
6.9.1. Гормональное обеспечение общего адаптационного синдрома,
или стресса...................................................315
6.9.2. Гормональная регуляция местных компенсаторных реакций . 317
III ФУНКЦИИ СИСТЕМ ЖИЗНЕОБЕСПЕЧЕНИЯ ОРГАНИЗМА
Глава 7 Функции клеток крови. Гемостаз. Регуляция кроветворения. Основы транс-
фузиологии .............................................................322
7.1. Функции эритроцитов...........................................322
7.1.1. Функции и свойства эритроцитов..........................322
918 • Оглавление
7.1.2. Гемоглобин...........................................324
7.1.3. Старение и разрушение эритроцитов в организме........326
7.1.4. Роль ионов железа в эритропоэзе .....................327
7.1.5. Эритропоэз...........................................329
7.1.6. Регуляция эритропоэза................................330
7.2. Лейкоциты ..................................................331
7.2.1. Функции нейтрофильных гранулоцитов...................331
7.2.2. Функции базофильных гранулоцитов.....................334
7.2.3. Функции эозинофильных лейкоцитов.....................335
7.2.4. Функции моноцитов-макрофагов.........................336
7.2.5. Регуляция грануло- и моноцитопоэза...................337
7.3. Функции тромбоцитов.........................................337
7.3.1. Структура и функции тромбоцитов......................337
7.3.2. ТроМбоцитопоэз и его регуляция ......................339
7.4. Механизмы свертывания крови (гемостаза) ....................341
7.4.1. Тромбоцитарный гемостаз..............................341
7.4.2. Система свертывания крови............................341
7.4.3. Противосвертывающие механизмы крови..................345
7.4.4. Фибринолиз...........................................348
7.5. Общие закономерности кроветворения..........................349
7.5.1. Кроветворные клетки-предшественницы..................349
7.5.2. Регуляция пролиферации и дифференциации КОК..........352
7.5.3. Роль стромы гемопоэтических органов в регуляции кроветво-
рения ......................................................354
7.5.4. Регуляция выхода форменных элементов крови из костного
мозга в кровеносное русло.............................355
7.5.5. Особенности метаболизма кроветворной ткани ..........356
7.6. Роль витаминов и микроэлементов в кроветворении ............357
7.7. Основы трансфузиологии .....................................358
7.7.1. Группы крови.........................................358
7.7.2. Влияние переливаемой крови и ее компонентов на организм
человека..............................................361
Глава 8 Иммунная система ................................................363
8.1. Происхождение и функции клеток иммунной системы.............364
8.1.1. Т-лимфоциты..........................................365
8.1.1.1. Характеристики Т-лимфоцитов..................365
8.1.1.2. Субпопуляции Т-лимфоцитов....................367
8.1.1.3. Функции Т-лимфоцитов ........................368
8.1.2. В-лимфоциты..........................................368
8.1.2.1. Характеристики В-лимфоцитов..................368
8.1.2.2. Функции В-лимфоцитов ........................369
8.1.3. Антигенпредставляющие клетки.........................370
8.2. Структура и функции органов иммунной системы................370
8.2.1. Костный мозг.........................................370
8.2.2. Тимус (вилочковая железа)............................372
8.2.3. Селезенка............................................374
8.2.4. Лимфатические узлы...................................375
8.2.5. Ассоциированная со слизистыми оболочками лимфоидная
ткань (мукозно-ассоциированная лимфоидная ткань)............375
8.3. Стадии и формы иммунного ответа.............................376
8.3.1. Ранний защитный воспалительный ответ.................376
8.3.2. Представление и распознавание антигена...............378
8.3.3. Активация Т- и В-лимфоцитов в иммунном ответе ..... 379
8.3.4. Клеточный иммунный ответ.............................380
8.3.5. Гуморальный иммунный ответ...........................381
8.3.6. Иммунологическая память как форма специфического иммун-
ного ответа ................................................383
8.3.7. Иммунологическая толерантность.......................383
8.4. Механизмы, контролирующие иммунную систему..................383
8.4.1. Гормональный контроль................................383
8.4.3. Цитокиновый контроль.................................385
Оглавление • 919
Глава 9 Функции систем кровообращения и лимфообращения............................387
9.1. Система кровообращения..............................................387
9.1.1. Функциональные классификации системы кровообращения . 387
9.1.2. Общая характеристика движения крови по сосудам...............389
9.1.3. Системная гемодинамика ......................................391
9.1.3.1. Системное артериальное давление.....................392
9.1.3.2. Общее периферическое сопротивление сосудов..........393
9.1.3.3. Сердечный выброс....................................394
9.1.3.4. Частота сердечных сокращений (пульс)................397
9.1.3.5. Работа сердца.......................................397
9.1.3.6. Сократимость........................................397
9.1.3.6.1. Автоматизм и проводимость миокарда........398
9.1.3.6.2. Мембранная природа автоматии сердца.......399
9.1.3.6.3. Возбудимость сердечной мышцы .............401
9.1.3.6.4. Сопряжение возбуждения и сокращения миокарда . 403
9.1.3.6.5. Сердечный цикл и его фазовая структура....405
9.1.3.6.6. Механические, электрические и физические прояв-
ления деятельности сердца ............................407
9.1.3.6.7. Общие принципы регуляции сердечного выброса . 412
9.1.3.6.8. Нейрогенная регуляция деятельности сердца . . . . 415
9.1.3.6.9. Механизмы адренергической и холинергической ре-
гуляции деятельности сердца...........................417
9.1.3.6.10. Гуморальные влияния на сердце............420
9.1.3.7. Венозный возврат крови к сердцу ...........................421
9.1.3.8. Центральное венозное давление.......................425
9.1.3.9. Объем циркулирующей крови...........................426
9.1.3.10. Соотношение основных параметров системной гемодинамики 427
9.1.4. Общие закономерности органного кровообращения................432
9.1.4.1. Функционирование органных сосудов...................432
9.1.4.2. Нервные и гуморальные влияния на органные сосуды .... 438
9.1.4.3. Роль эндотелия сосудов в регуляции их просвета......442
9.1.5. Особенности кровоснабжения органов и тканей..................442
9.1.5.1. Головной мозг.......................................442
9.1.5.2. Миокард.............................................444
9.1.5.3. Легкие..............................................445
9.1.5.4. Желудочно-кишечный тракт (ЖКТ)......................446
9.1.5.5. Главные пищеварительные железы .....................448
9.1.5.6. Печень..............................................448
9.1.5.7. Кожа................................................450
9.1.5.8. Почка...............................................451
9.1.5.9. Скелетные мышцы.....................................452
9.1.5.10. Сопряженные функции сосудов........................453
9.1.6. Микроциркуляция (микрогемодинамика)..........................453
9.1.7. Центральная регуляция кровообращения.........................456
9.1.7.1. Рефлекторная регуляция кровообращения...............457
9.1.7.2. Спинальный уровень регуляции........................457
9.1.7.3. Бульбарный уровень регуляции........................458
9.1.7.4. Гипоталамические влияния............................460
9.1.7.5. Участие лимбических структур .......................461
9.1.7.6. Кортикальные влияния................................462
9.1.7.7. Общая схема центральной регуляции...................462
9.2. Лимфообращение.....................................................463
9.2.1. Лимфатические сосуды........................................465
9.2.2. Лимфатические узлы...........................................465
9.2.3. Лимфоток.....................................................466
9.2.4. Нервные и гуморальные влияния................................466
Глава 10 Функции дыхательной системы..............................................469
10.1. Внешнее дыхание....................................................470
10.1.1. Биомеханика дыхания ........................................470
10.1.1.1. Биомеханика вдоха..................................470
10.1.1.2. Биомеханизм выдоха.................................471
10.1.2. Изменение объема легких во время вдоха и выдоха.............472
10.1.2.1. Функция внутриплеврального давления................472
10.1.2.2. Легочные объемы воздуха в течение фаз дыхательного цикла . 474
10.1.3. Факторы, влияющие на легочный объем в фазу вдоха............476
10.1.3.1. Растяжимость легочной ткани........................476
920 • Оглавление
10.1.3.2. Поверхностное натяжение слоя жидкости в альвеолах .... 477
10.1.3.3. Сопротивление дыхательных путей ...............479
10.1.3.4. Зависимость «поток—объем» в легких.............480
10.1.4. Работа дыхательных мышц в течение дыхательного цикла ... 481
10.2. Вентиляция и перфузия кровью легких..........................483
10.2.1. Вентиляция легких.....................................483
10.2.2. Перфузия легких кровью................................485
10.2.3. Эффект гравитации на вентиляцию и перфузию легких кровью 485
10.2.3. Коэффициент вентиляционно-перфузионных отношений в
легких.........................................................487
10.3. Газообмен в легких...........................................487
10.3.1. Состав альвеолярного воздуха..........................488
10.3.2. Напряжение газов в крови капилляров легких............489
10.3.3. Скорость диффузии 02 и СО2 в легких...................489
10.4. Транспорт газов кровью.......................................491
10.4.1. Транспорт кислорода...................................492
10.4.1.1. Изменение сродства гемоглобина к кислороду.....493
10.4.2. Транспорт углекислого газа ...........................495
10.4.2.1. Роль эритроцитов в транспорте СО2..............495
10.5. Регуляция дыхания............................................497
10.5.1. Дыхательный центр.....................................498
10.5.1.1. Происхождение дыхательного ритма...............500
10.5.2. Влияние нервных центров варолиева моста на дыхательный
ритм...........................................................501
10.5.3. Функция спинальных дыхательных мотонейронов...........502
10.5.4. Рефлекторная регуляция дыхания........................502
10.5.4.1. Хеморецепторный контроль дыхания...............502
10.5.4.2. Механорецепторный контроль дыхания.............503
10.6. Дыхание при физической нагрузке..............................505
10.7. Дыхание человека при измененном барометрическом давлении возду-
ха .................................................................507
10.7.1. Дыхание человека при пониженном давлении воздуха.....507
10.7.2. Дыхание человека при повышенном давлении воздуха .... 509
Глава 11 Функции пищеварительной системы...................................511
11.1. Состояние голода и насыщения.................................511
11.2. Общая характеристика функций пищеварительной системы и меха-
низмов ее регуляции...............................................512
11.2.1. Секреторная функция ..................................512
11.2.2. Моторная функция......................................517
11.2.3. Функция всасывания....................................519
11.2.4. Общая характеристика механизмов регуляции функций пище-
варительной системы...........................................523
11.3. Периодическая деятельность пищеварительной системы...........526
11.4. Пищеварение в ротовой полости и функция глотания.............528
11.4.1. Ротовая полость.......................................528
11.4.2. Слюноотделение .......................................528
11.4.3. Жевание...............................................531
11.4.4. Глотание..............................................532
11.5. Пищеварение в желудке........................................533
11.5.1. Секреторная функция желудка...........................534
11.5.2. Регуляция секреции желудочного сока...................538
11.5.2.1. Фазы желудочной секреции.......................539
11.5.3. Сократительная деятельность мускулатуры желудка.541
11.5.3.1. Регуляция сократительной деятельности желудка .542
11.5.3.2. Эвакуация содержимого желудка в двенадцатиперстную кишку 543
11.6. Пищеварение в двенадцатиперстной кишке.......................543
11.6.1. Пищеварительные функции поджелудочной железы..........544
11.6.1.1. Состав и свойства панкреатического сока........544
11.6.1.2. Нервная и гуморальная регуляция секреторной функции под-
желудочной железы......................................545
11.6.2. Пищеварительные функции печени........................546
11.6.2.1. Механизм образования желчи.....................546
Оглавление • 921
11.6.2.2. Состав и свойства желчи .......................546
11.6.2.3. Регуляция желчеобразования и желчевыведения....547
11.6.3. Непищеварительные функции печени.......................548
11.7. Пищеварение в тонком кишечнике................................548
11.7.1. Секреторная функция тонкой кишки.......................548
11.7.1.1. Регуляция секреторной функции тонкой кишки.....550
11.7.2. Двигательная функция тонкой кишки......................550
11.7.2.1. Регуляция моторики тонкой кишки................551
11.7.3. Функция всасывания тонкой кишки .......................553
11.8. Пищеварение в толстом кишечнике...............................556
11.8.1. Перемещение химуса из тощей кишки в слепую.............557
11.8.2. Сокоотделение в толстом кишечнике......................557
11.8.3. Двигательная активность толстого кишечника.............557
11.8.4. Роль микрофлоры толстой кишки в процессе пищеварения и
формировании иммунологической реактивности организма . . 558
11.8.5. Акт дефекации .........................................560
11.8.6. Иммунная система пищеварительного тракта...............561
11.8.7. Тошнота и рвота .......................................561
Глава 12 Обмен веществ и энергии. Питание....................................563
12.1. Роль белков, жиров, углеводов, минеральных веществ и витаминов в
метаболизме..........................................................564
12.1.1. Белки и их роль в организме............................564
12.1.2. Липиды и их роль в организме...........................565
12.1.2.1. Клеточные липиды...............................566
12.1.2.2. Бурый жир......................................566
12.1.2.3. Липиды плазмы крови ...........................566
12.1.3. Углеводы и их роль в организме.........................567
12.1.4. Минеральные вещества и их роль в организме.............568
12.1.5. Вода и ее роль в организме — см. раздел 14.3. Водно-солевой
обмен ..........................................................570
12.1.6. Витамины и их роль в организме.........................570
12.2. Роль обмена веществ в обеспечении энергетических потребностей ор-
ганизма .............................................................572
12.2.1. Способы оценки энергетических затрат организма.........574
12.3. Обмен веществ и энергии при различных уровнях функциональной
активности организма...............................................575
12.3.1. Основной обмен ........................................575
12.3.2. Энергетические затраты организма в условиях физической на-
грузки .........................................................577
12.4. Регуляция обмена веществ и энергии............................578
12.5. Питание.......................................................584
12.5.1. Рациональное питание как фактор сохранения и укрепления
здоровья........................................................589
Глава 13 Температура тела и ее регуляция.....................................592
13.1. Нормальная температура тела...................................593
13.2. Теплопродукция и теплоотдача..................................594
13.2.1. Теплопродукция.........................................595
13.2.2. Теплоотдача............................................596
13.2.3. Поведенческая терморегуляция...........................598
13.3. Регуляция температуры тела....................................598
13.3.1. Восприятие организмом температурных воздействий (терморе-
цепция) ........................................................598
13.3.2. Центральное звено системы терморегуляции...............599
13.3.3. Эффекторное (исполнительное) звено системы терморегуля-
ции ............................................................601
13.4. Гипертермия и гипотермия .....................................603
13.5. Взаимодействие системы терморегуляции с другими физиологически-
ми системами организма...............................................604
13.5.1. Сердечно-сосудистая система и терморегуляция...........604
13.5.2. Водно-солевой баланс и терморегуляция..................604
13.5.3. Дыхание и терморегуляция...............................605
922 • Оглавление
Глава 14 Выделение. Функции почек. Водно-солевой обмен.....................606
14.1. Органы и процессы выделения .................................606
14.1.1. Выделительная функция кожи............................606
14.1.2. Выделительная функция печени и пищеварительного тракта . 608
14.1.3. Выделительная функция легких и верхних дыхательных путей 608
14.2. Функции почек................................................609
14.2.1. Механизмы мочеобразования ............................610
14.2.1.1. Клубочковая ультрафильтрация и ее регуляция ..614
14.2.1.2. Канальцевая реабсорбция и ее регуляция........619
14.2.1.3. Канальцевая секреция и ее регуляция...........630
14.2.1.4. Состав и свойства конечной мочи...............631
14.2.1.5. Механизмы выведения мочи и мочеиспускания.....631
14.2.2. Экскреторная функция почек............................632
14.2.3. Метаболическая функция почек..........................632
14.2.4. Роль почек в регуляции артериального давления.........633
14.3. Водно-солевой обмен .........................................634
14.3.1. Внешний водный баланс организма.......................634
14.3.2. Внутренний водный баланс организма....................636
14.3.3. Электролитный, или солевой, баланс организма..........636
14.3.4. Общие принципы регуляции водно-солевого обмена........638
14.4. Интегративные механизмы регуляции водно-солевого обмена и го-
меостатическая функция почек......................................639
14.4.1. Гомеостатические механизмы при гиперосмотической дегид-
ратации ................................................640
14.4.2. Гомеостатические механизмы при изоосмотической дегидрата-
ции ..........................................................645
14.4.3. Гомеостатические механизмы при гипоосмотической дегидра-
тации ........................................................645
14.4.4. Гомеостатические механизмы при гипоосмотической гипер-
гидратации ...................................................646
14.4.5. Гомеостатические механизмы при изоосмотической гипергид-
ратации ......................................................647
14.4.6. Гомеостатические механизмы при гиперосмотической гипер-
гидратации ...................................................647
14.4.7. Нарушения баланса электролитов........................648
Глава 15 Кислотно-основное состояние.......................................649
15.1. Кислоты и основания внутренней среды.........................649
15.2. Физико-химические гомеостатические механизмы.................651
15.2.1. Буферные системы внутренней среды организма...........651
15.2.2. Тканевые гомеостатические обменные процессы...........654
15.3. Физиологические гомеостатические механизмы...................655
15.3.1. Легкие и кислотно-основное состояние..................655
15.3.2. Почки и кислотно-основное состояние...................656
15.3.3. Желудочно-кишечный тракт, печень, костная ткань и кислот-
но-основное состояние.........................................659
15.4. Основные физиологические показатели кислотно-основного состоя-
ния ..............................................................660
15.5. Основные изменения кислотно-основного состояния и их компенса-
ция ..............................................................661
15.5.1. Функциональное значение ацидозов и алкалозов .........662
15.5.2. Дыхательный ацидоз....................................663
15.5.3. Недыхательный ацидоз..................................664
15.5.4. Дыхательный алкалоз...................................665
15.5.5. Недыхательный алкалоз ................................666
15.5.6. Общие закономерности компенсации нарушений кислотно-
основного состояния...........................................667
Глава 16 Репродуктивная функция человека...................................668
16.1. Половая дифференциация человека..............................669
16.1.1. Генетический пол......................................670
16.1.2. Гонадный пол..........................................670
16.1.3. Фенотипический пол....................................671
Оглавление • 923
16.2. Репродуктивная функция мужского организма.......................672
16.2.1. Функции семенников.......................................672
16.2.2. Сперматогенез............................................674
16.2.3. Гормональная регуляция сперматогенеза....................675
16.2.4. Мужской половой акт .....................................678
16.2.4.1. Стадии мужского полового акта....................678
16.2.4.2. Регуляция эякуляции..............................681
16.2.4.3. Оргазм...........................................682
16.3. Репродуктивная функция женского организма.......................682
16.3.1. Овариальный цикл и оогенез ..............................683
16.3.1.1. Фолликулярная фаза...............................683
16.3.1.2. Овуляторная фаза.................................685
16.3.1.3. Лютеальная фаза..................................686
16.3.1.4. Лютеолиз желтого тела............................686
16.3.2. Менструальный цикл (маточный цикл).......................687
16.3.2.1. Менструальная фаза...............................687
16.3.2.2. Пролиферативная фаза.............................688
16.3.2.3. Секреторная фаза.................................688
16.3.3. Женский половой акт......................................688
16.4. Оплодотворение (фертилизация) ..................................690
16.5. Имплантация оплодотворенной яйцеклетки..........................692
16.6. Беременность....................................................692
16.6.1. Функции плаценты.........................................693
16.6.2. Плацентарные гормоны ....................................695
16.7. Роды и лактация.................................................697
16.7.1. Роды.....................................................697
16.7.2. Лактация.................................................699
Глава 17 Сенсорные системы.....................................................702
17.1. Общая физиология сенсорных систем ..............................703
17.1.1. Классификации рецепторов.................................703
17.1.2. Преобразование энергии раздражителя в рецепторах ........704
17.1.3. Рецептивные поля ........................................706
17.1.4. Переработка информации в переключательных ядрах и прово-
дящих путях сенсорной системы....................................707
17.1.5. Субъективное сенсорное восприятие........................710
17.2. Соматовисцеральная сенсорная система............................711
17.2.1. Тактильная чувствительность .............................711
17.2.2. Проприоцептивная чувствительность........................713
17.2.3. Температурная чувствительность ..........................714
17.2.4. Болевая чувствительность ................................715
17.2.5. Висцеральная чувствительность............................717
17.3. Зрительная сенсорная система....................................717
17.3.1. Проецирование световых лучей на сетчатку глаза...........718
17.3.1.1. Аккомодация......................................718
17.3.1.2. Аномалии рефракции...............................719
17.3.1.3. Регуляция интенсивности светового потока.........720
17.3.1.4. Проекция зрительного поля на сетчатку............720
17.3.1.5. Движения глаз....................................722
17.3.2. Преобразование энергии света в сетчатке..................722
17.3.2.1. Скотопическая и фотопическая системы сетчатки....723
17.3.2.2. Рецепторный потенциал палочек и колбочек ........725
17.3.2.3. Адаптация фоторецепторов к изменениям освещенности . . . 725
17.3.3. Рецептивные поля клеток сетчатки.........................726
17.3.3.1. Рецептивные поля с on-центрами и off-центрами ...726
17.3.3.2. Рецептивные поля цветового восприятия............728
17.3.3.3. М- и P-типы ганглиозных клеток сетчатки .........729
17.3.4. Проводящие пути и переключательные центры зрительной
системы..........................................................730
17.3.4.1. Функциональная организация латерального коленчатого тела 730
17.3.5. Переработка зрительной сенсорной информации в коре ... 731
17.3.5.1. Зрительное восприятие............................733
17.4. Слуховая сенсорная система......................................733
17.4.1. Психофизические характеристики звуковых сигналов.........734
924 • Оглавление
17.4.1.1. Диапазон частотного восприятия.................735
17.4.1.2. Громкость звука................................735
17.4.2. Периферическая часть слуховой системы..................736
17.4.2.1. Функция наружного уха..........................736
17.4.2.2. Функция среднего уха...........................736
17.4.2.3. Внутреннее ухо.................................738
17.4.2.4. Функция внутреннего уха........................738
17.4.2.5. Биоэлектрические процессы в кортиевом органе..739
17.4.2.6. Частотное кодирование..........................739
17.4.2.7. Кодирование сенсорной информации в окончаниях слухового
нерва....................................................740
17.4.3. Проводящие пути и переключательные ядра слуховой системы 741
17.4.4. Переработка сенсорной информации в слуховой коре.......742
17.5. Вестибулярная сенсорная система...............................743
17.5.1. Вестибулярный аппарат .................................743
17.5.1.1. Свойства рецепторных клеток вестибулярного аппарата . . . 744
17.5.1.2. Адекватные раздражители рецепторов отолитовых органов . . 745
17.5.1.3. Адекватные раздражители рецепторов полукружных каналов . 746
17.5.2. Центральная часть вестибулярной системы................746
17.6. Вкусовая сенсорная система....................................747
17.6.1. Вкусовая рецепция......................................747
17.6.1.1. Рецепторные потенциалы вкусовых клеток ........748
17.6.1.2. Вкусовая чувствительность .....................748
17.6.2. Центральный отдел вкусовой системы ....................749
17.6.3. Вкусовое восприятие....................................750
17.7. Обонятельная сенсорная система................................750
17.7.1. Классификация запахов .................................751
17.7.2. Периферический отдел обонятельной системы..............751
17.7.2.1. Механизм возбуждения обонятельных клеток.......752
17.7.3. Центральный отдел обонятельной системы.................753
17.7.4. Физиологическая роль обоняния у человека...............754
17.7.4.1. Физиологические реакции на запахи .............754
17.7.4.2. Способность к восприятию феромонов у человека..754
IV ИНТЕГРАТИВНЫЕ ФУНКЦИИ ОРГАНИЗМА
Глава 18 Высшая нервная деятельность (по И. П. Павлову)......................757
18.1. Классические условные рефлексы................................758
18.1.1. Условия, влияющие на ассоциативное научение............760
18.1.2. Рефлекторная дуга классического условного рефлекса.....761
18.1.3. Стадии формирования условного рефлекса.................762
18.1.4. Условные рефлексы высшего порядка......................762
18.1.5. Виды классических условных рефлексов...................763
18.2. Торможение условных рефлексов.................................763
18.2.1. Внешнее торможение.....................................764
18.2.2. Внутреннее торможение..................................764
18.2.2.1. Угасательное торможение........................764
18.2.2.2. Запаздывающее торможение.......................765
18.2.2.3. Дифференцировочное торможение .......766
18.2.2.4. Условное торможение............................766
18.3. Оперантный условный рефлекс...................................767
18.4. Аналитико-синтетическая деятельность коры больших полушарий . . 768
18.5. Динамический стереотип .......................................768
18.6. Фазовые явления в коре больших полушарий головного мозга .... 769
18.7. Типология высшей нервной деятельности.........................771
Глава 19 Мотивации и эмоции..................................................773
19.1. Мотивации.....................................................773
19.1.1. Понятие о первичных и вторичных мотивациях.............773
19.1.2. Понятие о мотивациях влечения и избегания .............774
19.1.3. Пищевая мотивация человека.............................774
19.1.3.1. Гомеостатические механизмы регуляции пищевой мотивации у
человека................................................774
19.1.3.2. Роль структур продолговатого мозга в регуляции пищевой мо-
тивации ................................................777
Оглавление • 925
19.1.3.3. Роль латерального гипоталамуса в возникновении пищевой
мотивации ..................................................778
19.1.3.4. Роль меланокортииовой системы гипоталамуса в прекращении
пищевой мотивации..........................................778
19.1.3.5. Роль лимбической системы в регуляции пищевой мотивации у
человека...................................................779
19.1.4. Половая мотивация человека ...............................780
19.1.4.1. Генетические, социальные и психологические факторы воз-
никновения половой мотивации у человека.....................780
19.1.4.2. Роль половых гормонов в модуляции половой мотивации че-
ловека .....................................................784
19.1.4.3. Стадии полового возбуждения у человека при половой мотива-
ции ........................................................785
19.1.4.4. Нервная регуляция половой мотивации у человека...787
19.2. Эмоции...........................................................789
19.2.1. Виды эмоций...............................................789
19.2.2. Роль эмоций в поведении человека..........................790
19.2.3. Нейрофизиологические механизмы проявления эмоций .... 791
19.2.3.1. Гипоталамус как центр регуляции вегетативных и эндокрин-
ных реакций организма при эмоциях..........................791
19.2.3.2. Роль миндалины в осуществлении базовых эмоций ...792
19.2.3.3. Регуляция положительных эмоций у человека........796
19.2.3.4. Регуляция отрицательных эмоций у человека........798
Глава 20 Физиологические основы познавательной деятельности человека............800
20.1. Внимание.........................................................801
20.1.1. Формы внимания............................................802
20.1.2. Нейрофизиологические механизмы внимания...................803
20.1.2.1. Функции среднего мозга и моста в контроле внимания . . . 803
20.1.2.2. Функции корковых центров внимания................805
20.1.3. Внимание при различных модальностях ......................808
20.2. Восприятие.......................................................809
20.2.1. Зрительное восприятие.....................................809
20.2.1.1. Функции стриарной коры в зрительном восприятии...809
20.2.1.2. Зрительное восприятие при участии отделов экстрастриарной
коры.......................................................810
20.2.1.3. Особенности зрительного восприятия индивидуальных лиц и
объектов...................................................811
20.2.2. Слуховое восприятие.......................................812
20.2.3. Соматосенсорное восприятие................................812
20.3. Сознание.........................................................813
20.3.1. Нейрофизиологические корреляты сознания...................814
20.3.1.1. Электрическая активность мозга человека..........814
20.3.1.2. Активация мозга человека как нейрофизиологическая основа
проявлений состояний сознания..............................814
20.3.1.3. Осознание зрительного восприятия (зрительная осведомлен-
ность) .....................................................817
20.3.1.4. Внимание и сознание..............................817
20.4. Память и научение................................................817
20.4.1. Формы памяти и научения...................................818
20.4.2. Нейронные механизмы имплицитной памяти ...................820
20.4.2.1. Габитуация и сенситизация........................820
20.4.2.2. Ассоциативное научение (условные рефлексы).......822
20.4.3. Механизм образования эксплицитной памяти...............823
20.5. Речь.............................................................824
20.5.1. Свойства языка............................................824
20.5.2. Речевой аппарат...........................................825
20.5.3. Речевые структуры мозга...................................826
20.5.3.1. Нарушения речи при очаговых повреждениях мозга...826
20.5.3.2. Модель речевой деятельности Вернике—Гешвинда.....827
20.5.3.3. Современная модель речевой деятельности человека.828
20.5.3.4. Латерализация речи...............................829
20.6. Мышление ........................................................831
20.6.1. Нейрофизиологические основы мыслительной деятельности
человека..........................................................832
926 • Оглавление
20.6.1.1. Нейрофизиологические основы абстрактного мышления (рас-
суждение человека)......................................833
20.6.1.2. Нейрофизиологические основы умственных арифметических
операций................................................833
20.6.1.3. Нейрофизиологические основы мышления при чтении . . . 834
20.6.2. Функции левого и правого полушарий мозга человека при
мышлении.....................................................834
Глава 21 Сон и бодрствование...............................................836
21.1. Физиологическое значение сна.................................836
21.1.1. Восстановительная теория сна..........................836
21.1.2. Циркадианная теория сна...............................837
21.2. Периодичность физиологических процессов во время сна.........838
21.2.1. Стадии сна ...........................................839
21.2.2. Структура сна.........................................841
21.2.3. Фаза медленноволнового сна ...........................842
21.2.4. Фаза парадоксального сна..............................843
21.3. Нейрофизиологические механизмы сна...........................845
21.3.1. Участие стволовых центров в регуляции цикла сна—бодрство-
вания .......................................................845
21.3.2. Регуляция циркадного ритма............................847
21.3.3. Участие коры и лимбической системы в регуляции цикла сна-
бодрствования ...............................................848
21.3.4. Гуморальные индукторы и регуляторы сна................849
21.4. Сновидения и физиологическая роль БДГ-сна....................850
21.5. Продолжительность сна и последствия его лишения .............851
21.6. Бодрствование и сознание.....................................853
21.7. Различные уровни бодрствования...............................854
Глава 22 Физиологические основы труда......................................856
22.1. Образование энергии в скелетных мышцах при физической работе . 857
22.1.1. Анаэробный путь ресинтеза АТФ.........................857
22.1.2. Аэробный гликолиз.....................................858
22.1.3. «Кислородный каскад» и эффективность транспорта кислоро-
да к работающим мышцам.......................................861
22.1.4. Потребление кислорода, кислородный дефицит, кислородный
долг и кислородный запрос при мышечной работе................862
22.2. Физиологические основы тренировки двигательного навыка.......864
22.2.1. Развитие силовых качеств мышц ........................864
22.2.2. Физиологические механизмы формирования трудовых навы-
ков ..........................................................865
22.2.3. Работоспособность.....................................868
22.3. Функции физиологических систем организма человека при физиче-
ской работе........................................................868
22.3.1. Кровообращение........................................869
22.3.2. Кровь ................................................870
22.3.3. Дыхание...............................................870
22.3.4. Эндокринная система...................................870
22.4. Физиологические функции при умственном труде..............872
22.5. Работа в условиях зрительного напряжения .................873
22.6. Утомление при работе.........................................873
22.6.1. Утомление человека при физической работе..............874
22.6.1.1. Утомление человека при статической физической работе . . 875
22.6.1.2. Утомление человека при динамической мышечной работе . . 877
Глава 23 Адаптация человека к условиям внешней среды.......................879
23.1. Общие принципы и механизмы адаптации.........................879
23.1.1. Адаптация.............................................879
23.1.2. Неспецифические адаптивные реакции организма..........881
23.1.3. Симпатоадреналовая реакция............................882
23.1.4. Стресс-реакция........................................883
23.1.5. Реакция тренировки и реакция активации................886
23.1.6. Срочная и долговременная адаптация....................886
23.1.7. Норма адаптивной реакции и дизадаптации...............888
Оглавление • 927
23.1.8. Генотипическая и фенотипическая адаптация. Покрывающие
адаптации..................................................888
23.1.9. Обратимость процессов адаптации ...................889
23.2. Адаптация человека к климатическим факторам...............889
23.2.1. Биоклиматические факторы жаркого климата...........889
23.2.2. Адаптивные реакции организма человека к жаркой среде оби-
тания .....................................................891
23.2.3. Особенности адаптации человека к работе в жаркой среде . . 895
23.2.4. Предупреждение тепловых поражений организма........897
Предметный указатель ...................................................900